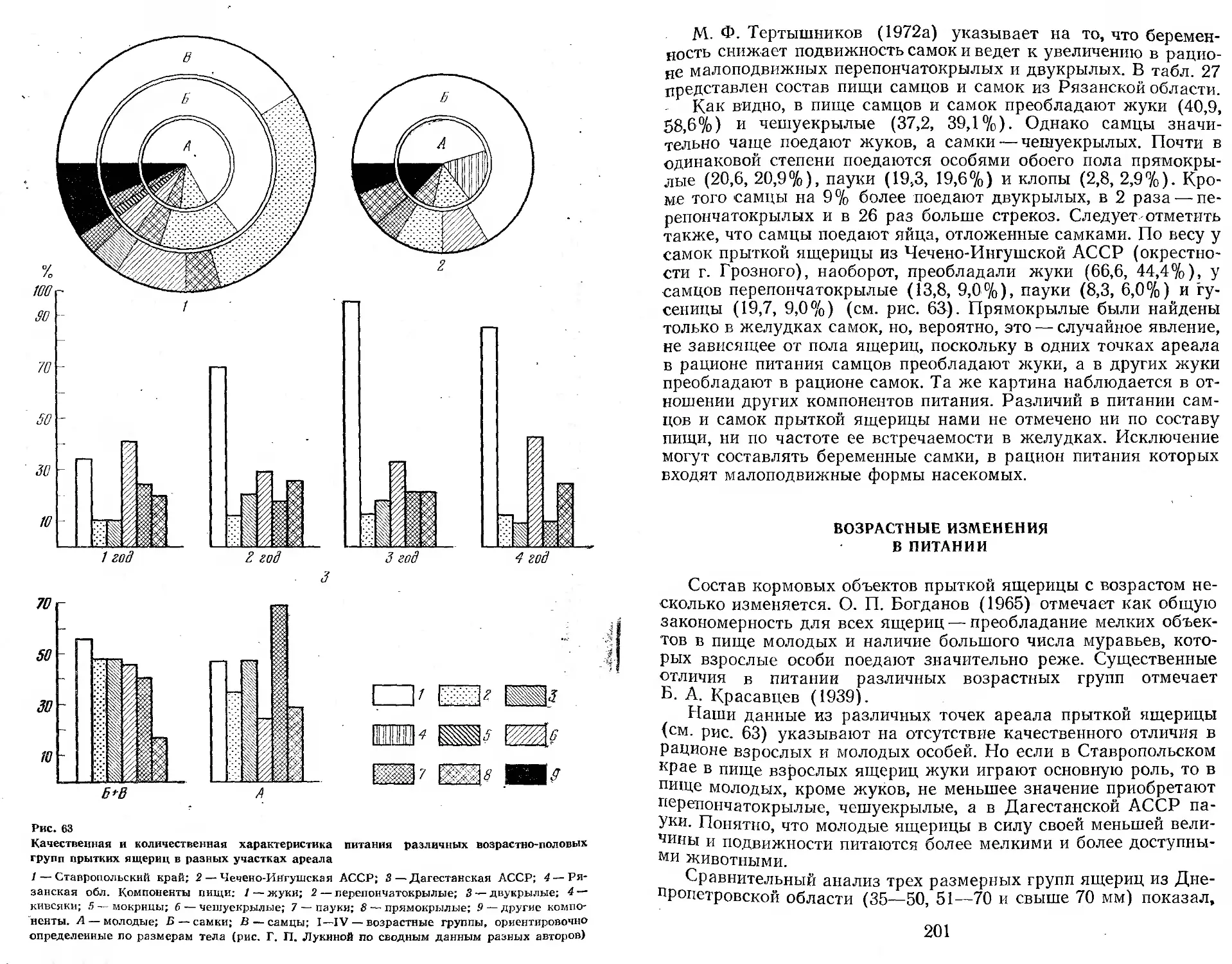
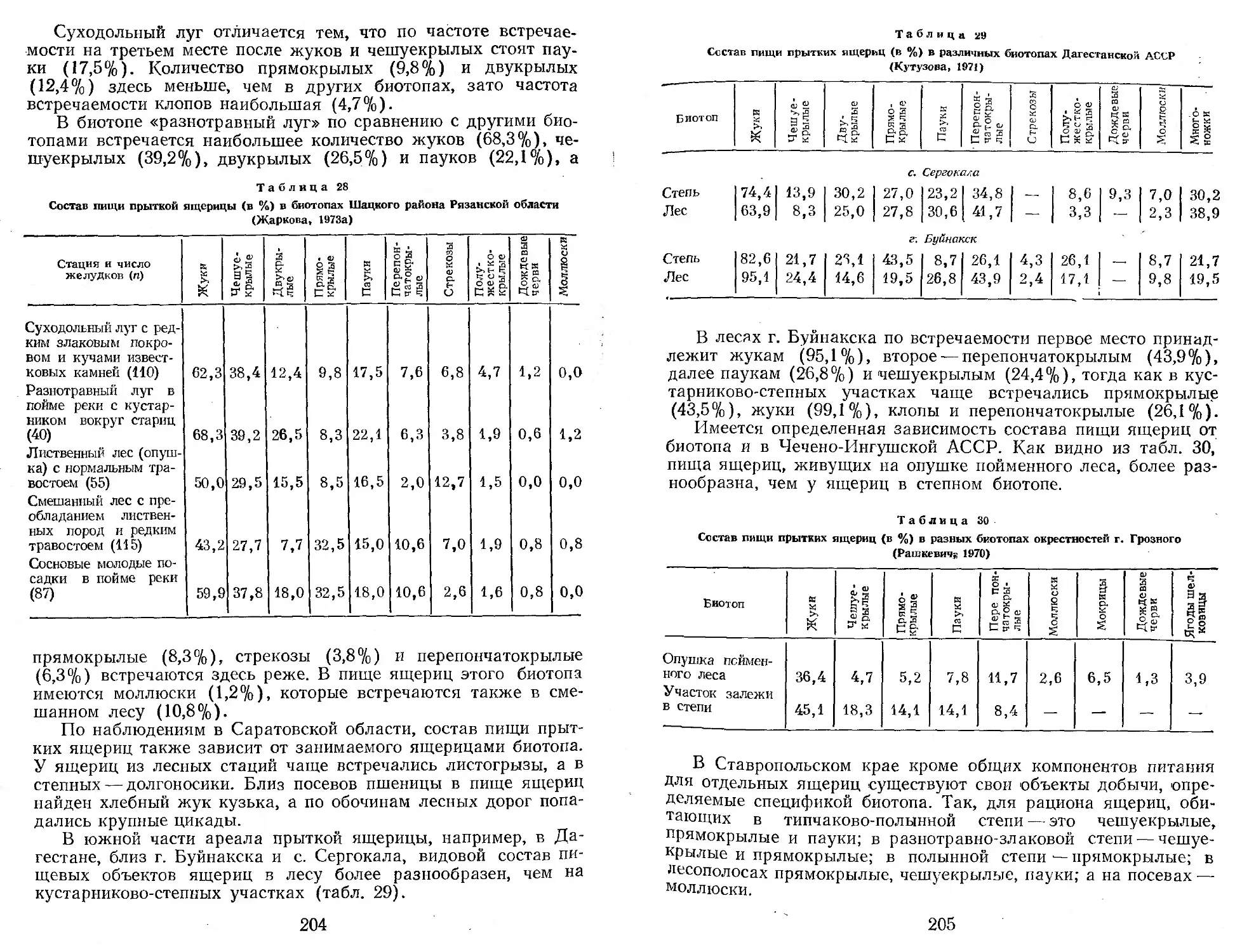
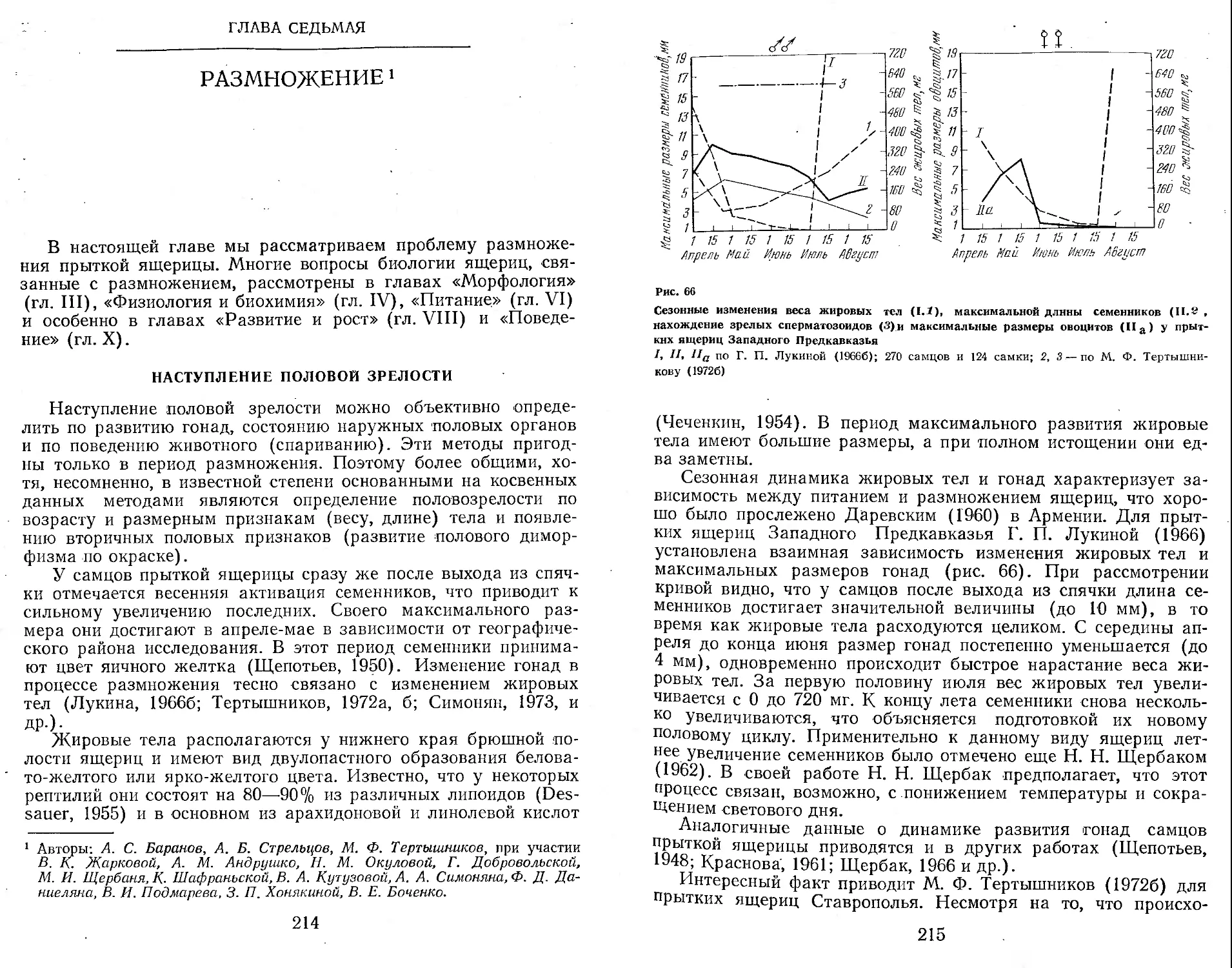
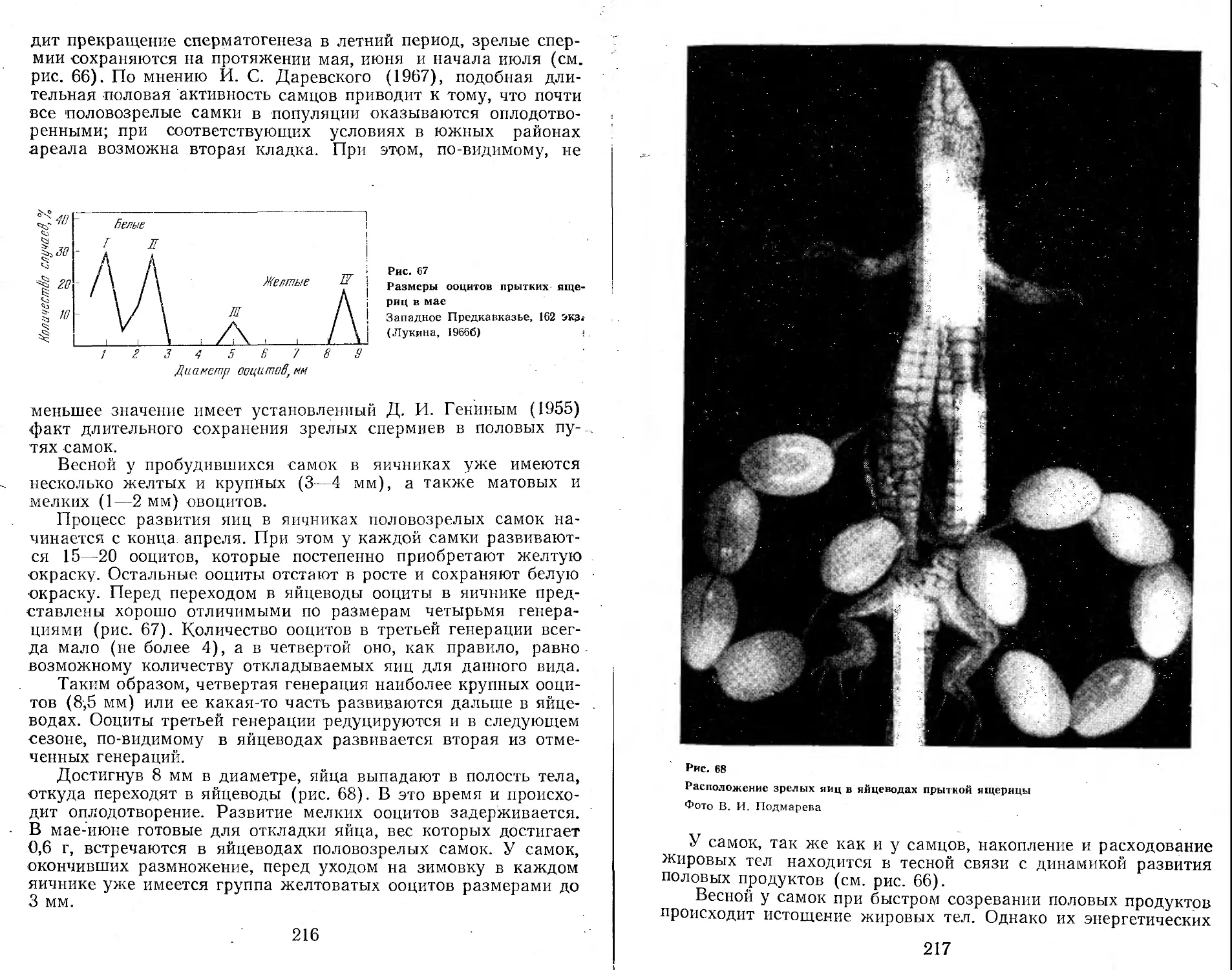
/






























































































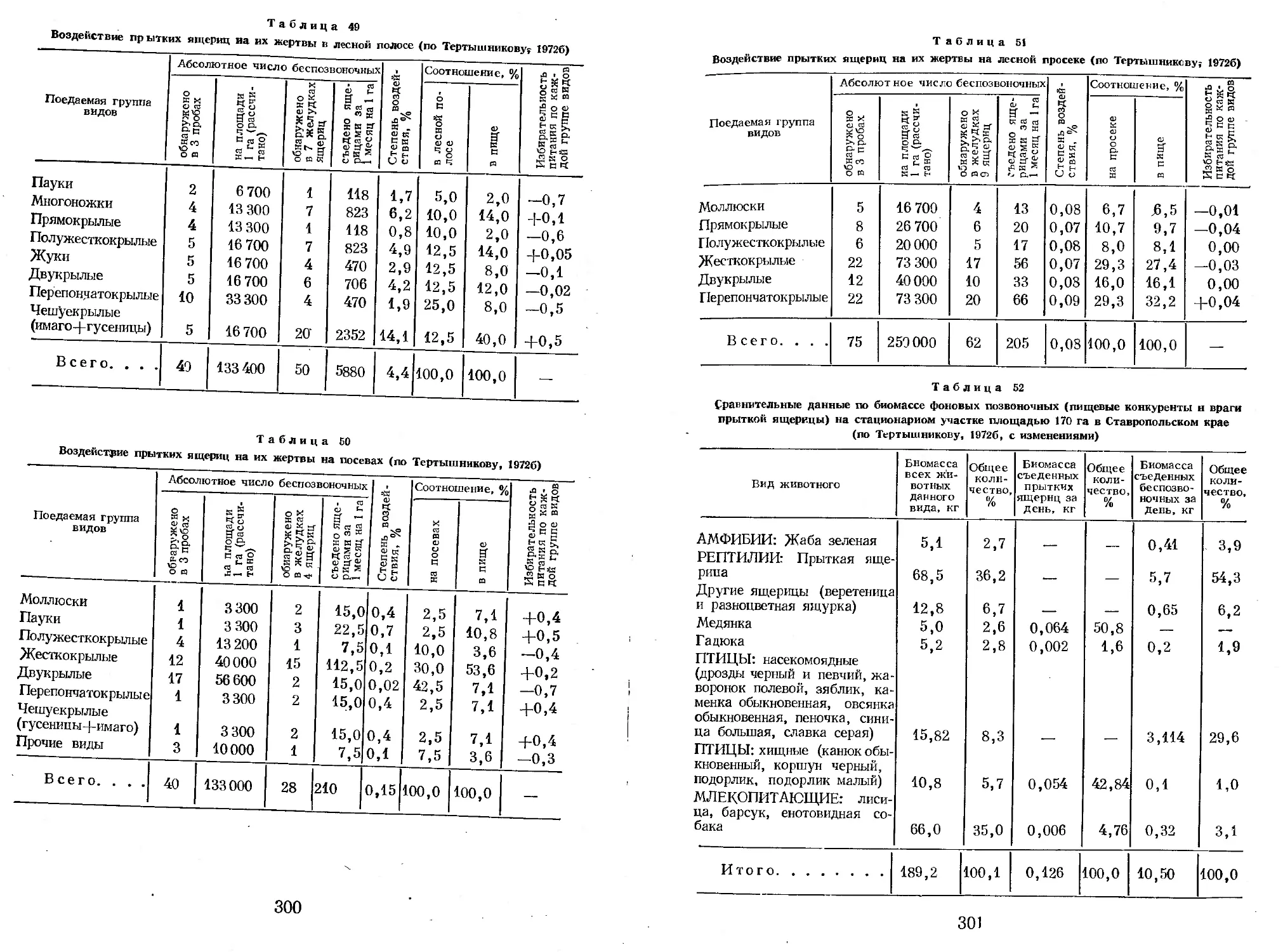


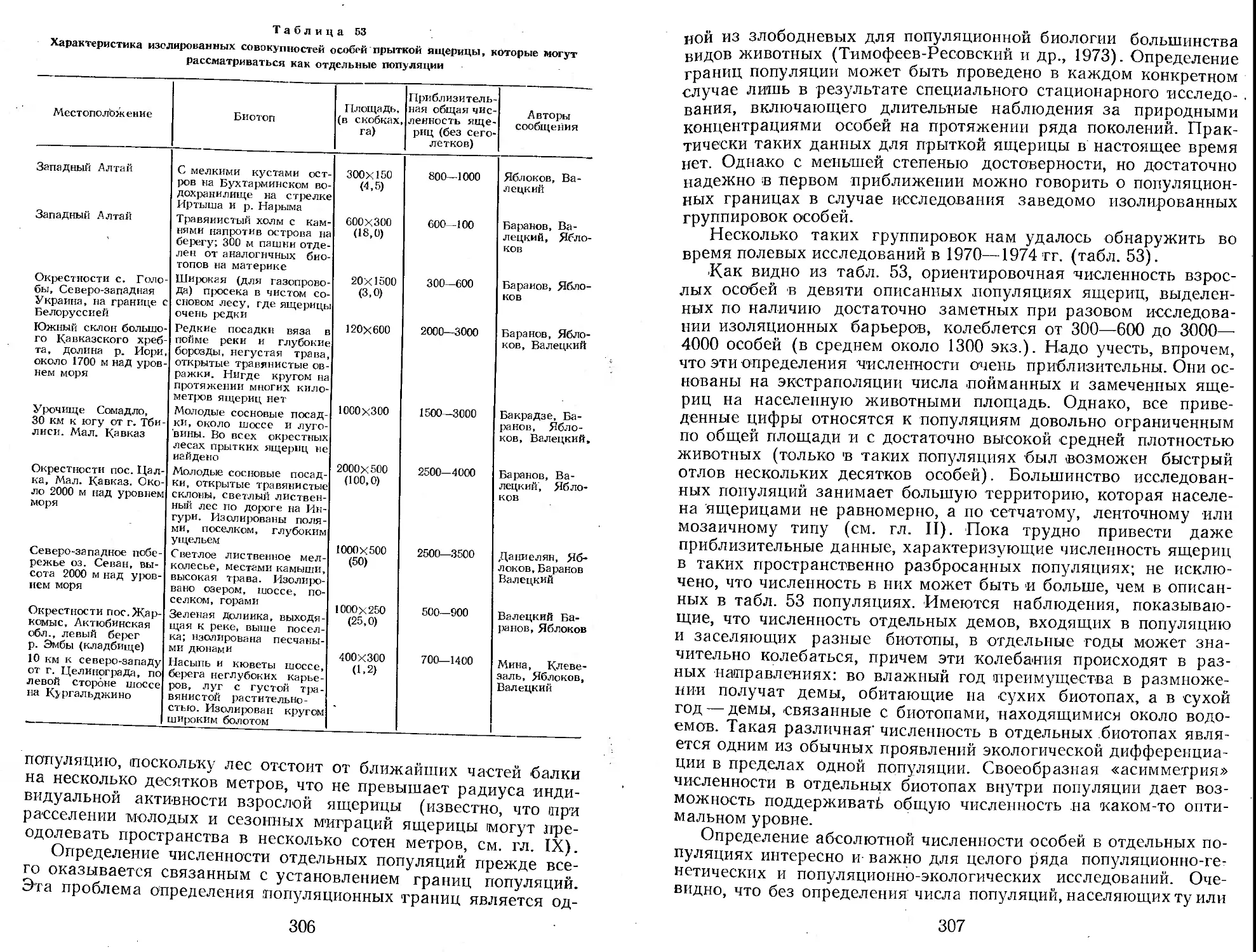


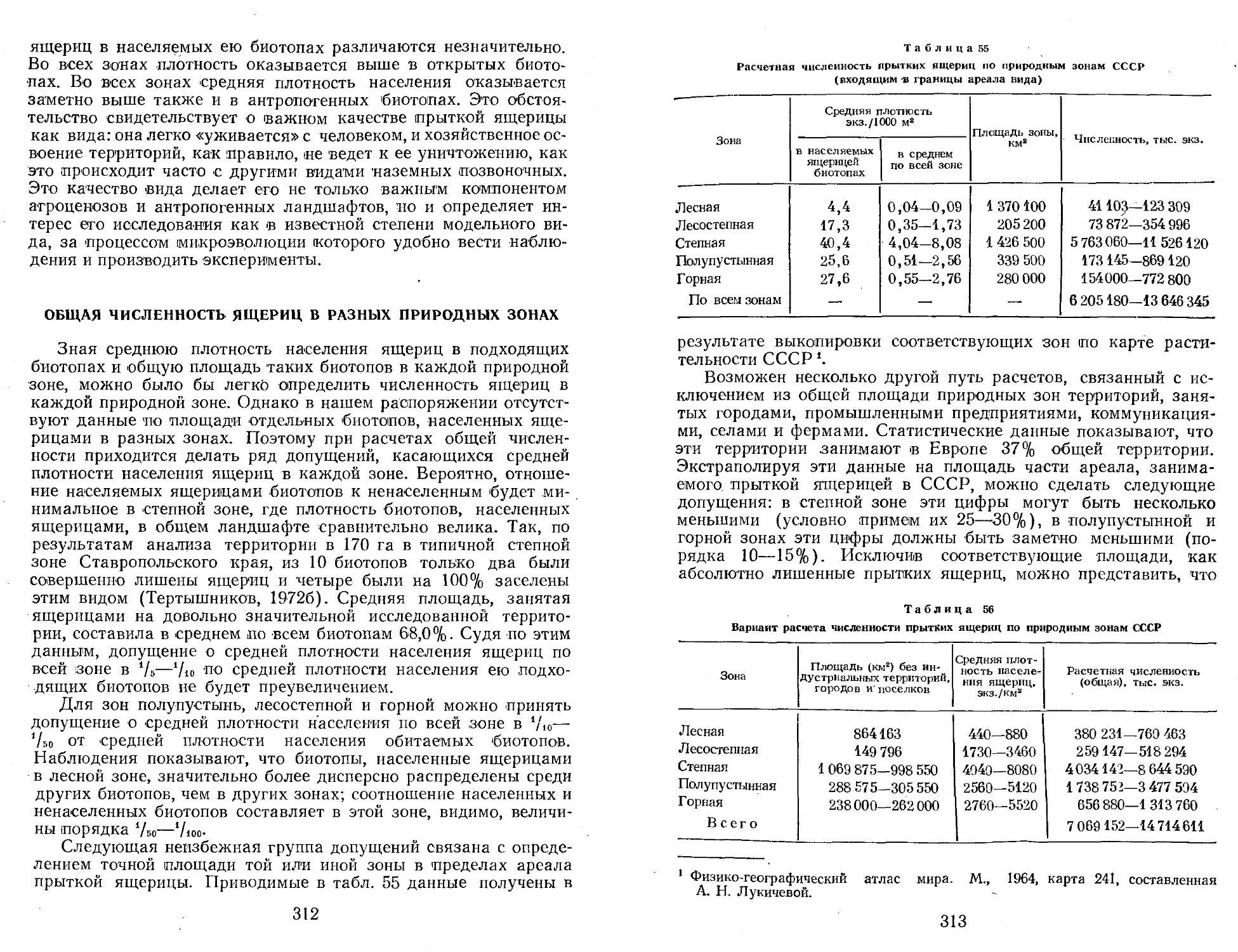

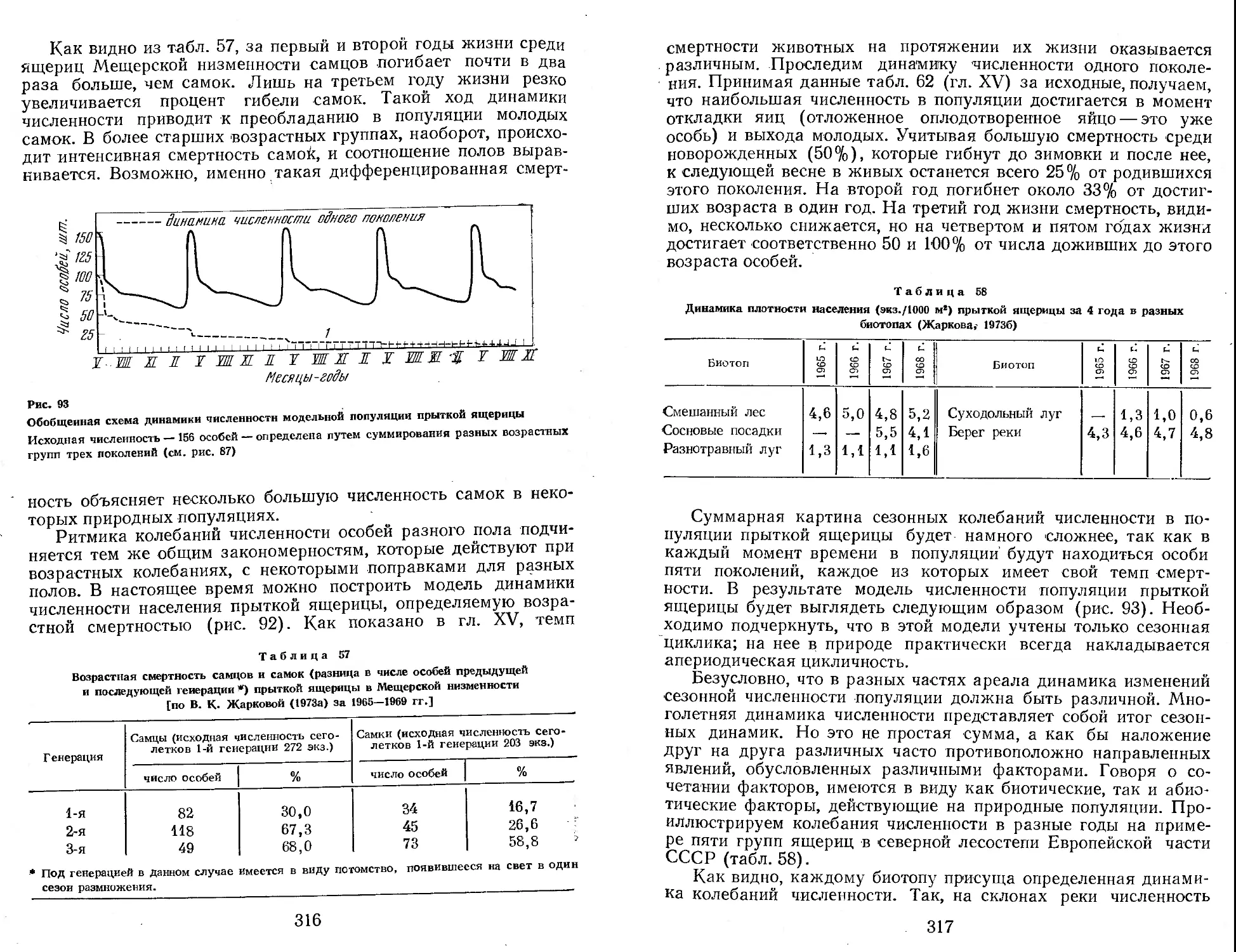



















Author: Яблоков А.В.
Tags: reptilia рептилии пресмыкающиеся герпетология биология зоология
Year: 1976




Text
АКАДЕМИЯ НАУК СССР
ИНСТИТУТ БИОЛОГИИ РАЗВИТИЯ ИМ. Н. К. КОЛЬЦОВА
ИНСТИТУТ ЭВОЛЮЦИОННОЙ МОРФОЛОГИИ И ЭКОЛОГИИ ЖИВОТНЫХ им. А. Н. СЕВЕРЦОВА
ПРЫТКАЯ ЯЩЕРИЦА
Монографическое описание вида
ИЗДАТЕЛЬСТВ О «Н А У К А» М О С К В А
1976
УДК 598-1
ПРЫТКАЯ ЯЩЕРИЦА. Монографическое описание вида. М., «Наука», 1976 г. 376 с.
Коллективная - монография посвящена описанию различных сторон биологии самого распространенного вида рептилий фауны СССР—прыткой ящерицы: особенностям распространения, систематике, морфологии, особенностям образа жизни, месту вида в биогеоценозах и цепях питания, численности, практическому значению. Затрагиваются многие вопросы, связанные с исследованием рептилий и позвоночных животных.
Рассчитана на герпетологов, биогеоценологов, систематиков, зоогеографов.
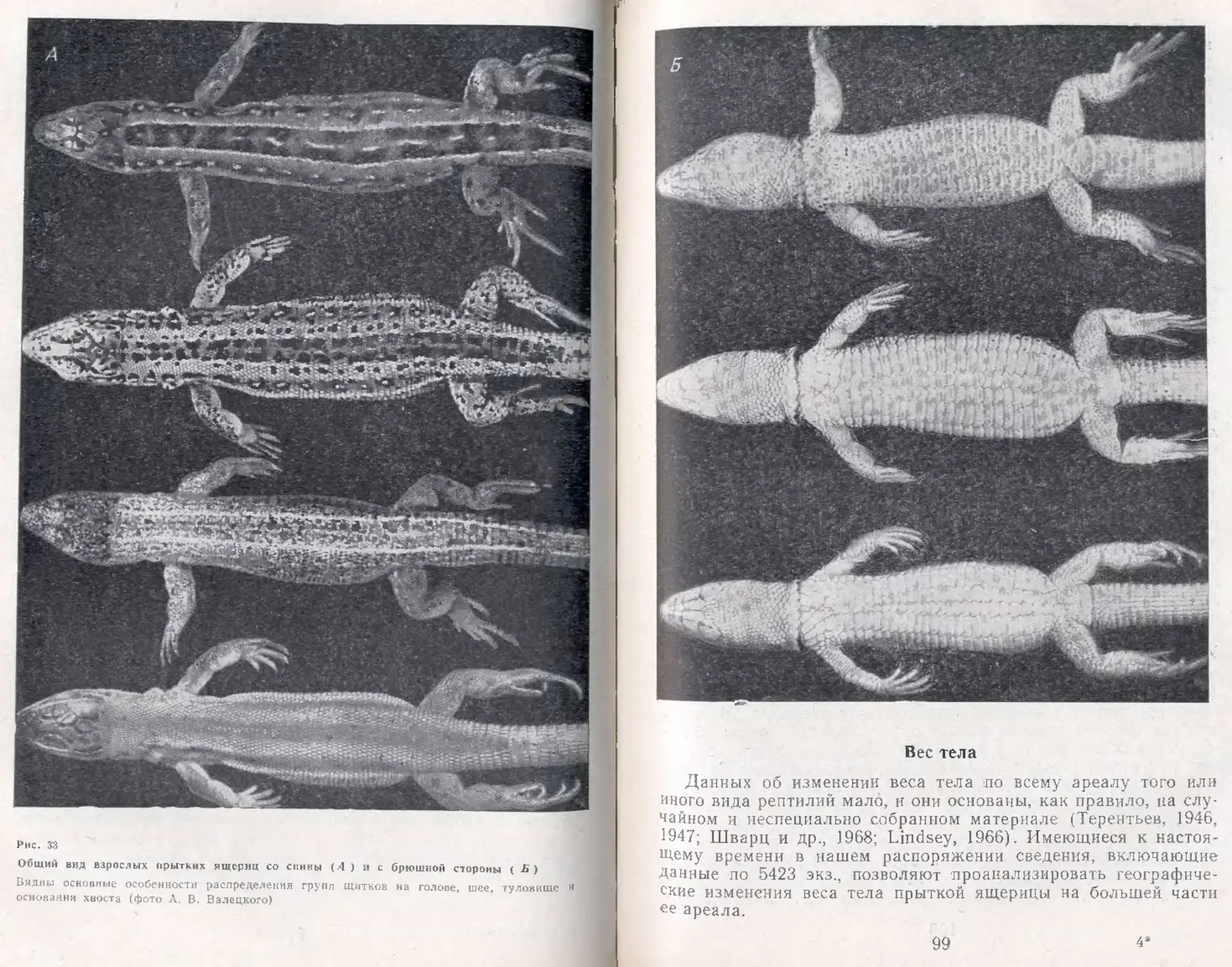
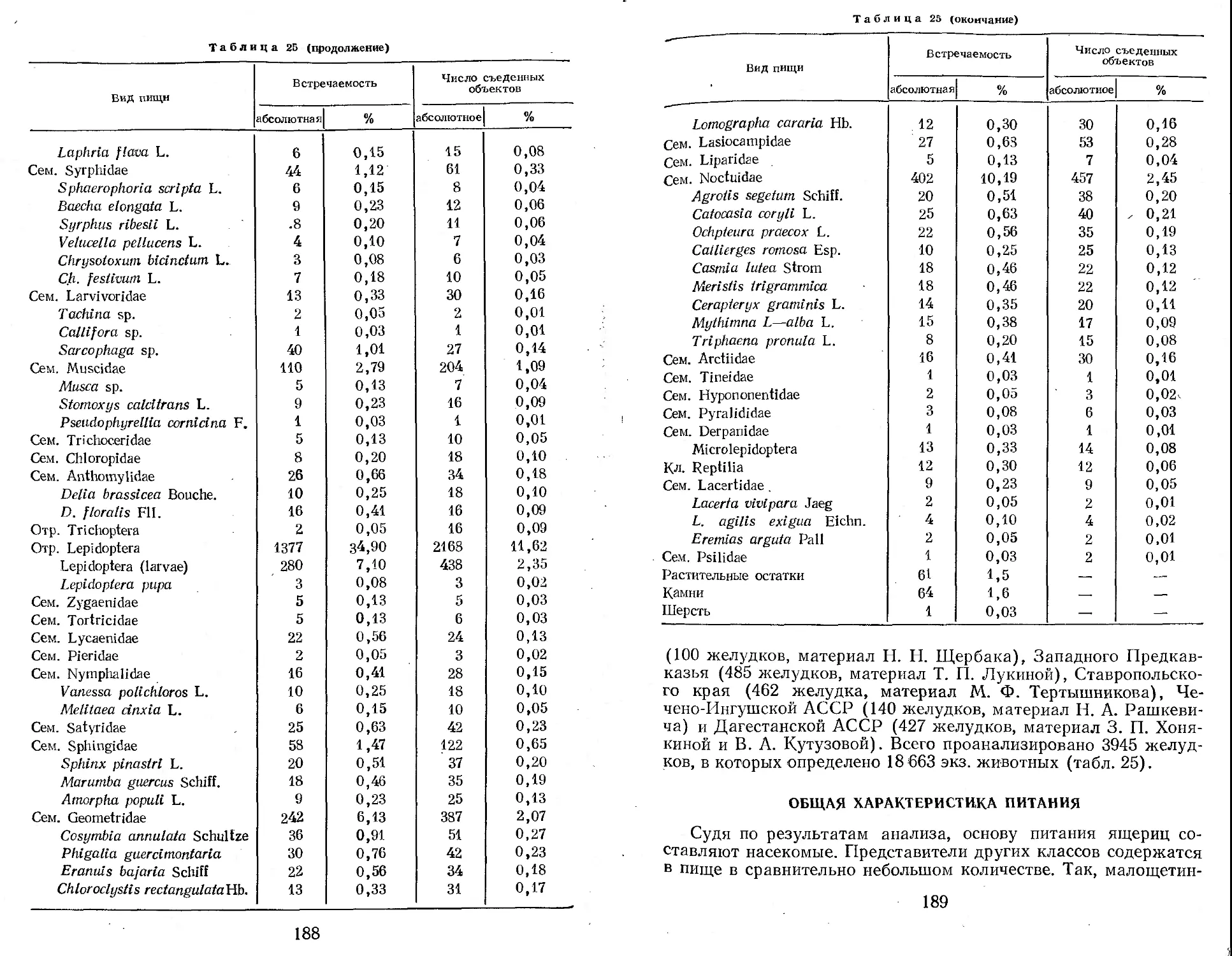
Табл. 63, илл. 97, список лит. — 647 назв.
Ответственный редактор доктор биологических наук А. В. ЯБЛОКОВ
р21'0<?8—g65^1Fi-7fij кн_ 2 055(.02)-76
© Издательство «Наука», 1976 г.
ПРЕДИСЛОВИЕ
Изучение животного мира любого крупного района нашей планеты проходит обычно несколько этапов. На первых этапах исследования уточняется состав фауны того или иного региона, выясняются общие особенности образа жизни каждого из входящих в данную фауну видов животных, их взаимоотношения в цепях питания с - другими видами, определяются возможности хозяйственного использования видов. Этот обычно весьма длительный этап зоологических исследований завершается созданием региональных многотомных фаунистических сводок типа «Животный мир Южной Америки», «Животный мир Средней Азии», «Фауна Индии» и т. п.
Параллельно проводящиеся фаунистические и систематические исследования отдельных крупных групп животных, связанных с углубленным исследованием какой-либо группы, приводит к созданию промежуточных по своему характеру сводок типа «Птицы СССР», «Млекопитающие Северной Америки», «Звери СССР и -прилежащих стран» и т. д.
Оценивая степень зоологической изученности нашей планеты, можно сказать, что сейчас по основным зоогеографическим районам такие фаунистические и обзорно-систематические сводки созданы или будут созданы в ближайшие годы. Одним из следующих шагов в углубленном изучении животного мира является теперь создание монографических сводок по отдельным видам. Такие сводки прежде всего, естественно, -могут быть созданы для тех видов, изучение которых’ привлекает особое внимание (например, видам, важным с хозяйственной точки зрения, видам, редким и исчезающим, и т. п.)_
Создание видовых монографий, несомненно, соответствует требованиям, которые современный этап освоения биосферы ставит перед зоологией. Объединение всех данных по какому-либо конкретному виду — особенностей распределения вида на всем пространстве его ареала и внутривидовой систематики, морфологии, экологии, поведения, выяснению места вида в биогеоцено- -зах и его положения в цепях питания и потоках энергии в разных участках ареала,— все это требует специального, по существу, монографического рассмотрения каждого вида.
3
Для такого монографического рассмотрения вида имеются и другие предпосылки. Вид представляет собой генетическое единство, систему связанных между собой популяций, обменивающихся время от времени -генетическим материалом посредством скрещивания особей из разных популяций. Такой поток генетической информации от популяции к популяции внутри вида поддерживает морфо-физиологическое единство вида как главнейшего этапа эволюционного процесса. Вид представляет собой генетически «закрытую» систему, обладающую собственной эволюционной судьбой. Человечество во все больших масштабах сейчас берет под сознательный (или чаще бессознательный) контроль развитие все более крупных участков биосферы, и при этом возникает реальная угроза исчезновения или необратимого изменения тех или иных видов. При таком давлении человека любой вид в живой -природе выступает как интегрированная система популяций, как единое целое, принципиально отличающееся от любой фаунистической общности (например, биоценоза), состоящей из многих популяций разных видов. Понимание такого особенного генетико-эволюционного положения вида, как единицы организации живой материи в биосфере Земли, также ведет к необходимости все более тщательного и глубокого специального исследования именно видового уровня организации.
Однако в наше время требуется не простое исследование вида в одной или немногих точках его ареала. Каждая из популяций, составляющих вид, занимает определенное место в биосфере, входит в разные биогеоценозы и экосистемы, отличается от других популяций внутри вида множеством специфических признаков и свойств. Поэтому для глубокого познания какого-либо вида необходимо исследование его на всем пространстве ареала или хотя бы (что практически более осуществимо) во многих точках ареала.
Монографическое описание отдельных видов важно и еще с одной точки зрения. При таком описании особенно четко заметны недостаточно исследованные проблемы, ранее не привлекшие достаточного внимания исследователей. Поэтому любая видовая монография должна являться, с одной стороны, итогом исследования данного вида, сводкой знаний об этом виде, а с другой стороны — программой дальнейших исследований.
Наконец, -неверно было бы считать, что каждая видовая монография относится исключительно к данному виду. При описании любых особенностей биологии данного вида, как правило, проводятся сравнительные данные по систематически близким видам; кроме того, многие проблемы оказываются общими для исследования многих видов данной группы животных. Поэтому, например, настоящая монография по прыткой ящерице, без сомнения, окажется чрезвычайно полезной для всех герпетологов, а также для специалистов, интересующихся общими проблемами зоологии позвоночных и биогеоцеиологии.
4
Отдельные видовые монографии появились уже много лет назад. Прекрасный пример этому — монография Т. Г. Гексли «Рак, Введение в изучение зоологии», в предисловии к которой автор, один из самых авторитетных зоологов того времени, писал: «...я старался показать, как внимательное изучение самого обыкновенного и самого незначительного животного доводит нас шаг за шагом от обыденного знания до широчайших обобщений и до труднейших проблем зоологии и'биологических наук вообще». В классической мировой литературе известны также монографии об одном из видов древоточцев, черепахах, майском жуке. В отечественной зоологической литературе также есть несколько примеров создания видовых монографий. Среди них книги А. В. Федюшина «Речной бобр, его история, жизнь и опыты по размножению», «Белуха. Опыт монографического исследования вида» С, Е. Клейненберга и др., А. А. Берзина «Кашалот», Н. Г. Коломиец и др. «Рыжий сосновый пилильщик».
Начиная с 1970 г. в вашей стране развернута программа исследований видов в пределах ареала, которая координировалась в рамках Советского национального комитета по Международ-
5
пой биологической программе, а затем — в рамках программы «Человек и биосфера».
Проведенное в 1974 г. в Ленинграде I Всесоюзное совещание по изучению вида в пределах ареала также единодушно высказалось в пользу своевременности и целесообразности создания серии видовых монографий. Все это позволяет надеяться, что в ближайшие годы в отечественной зоологической литературе появятся новые видовые монографий, касающиеся самых разных видов животных. Появление такой серии работ послужило бы хорошим примером для исследователей в других странах и привело бы к дальнейшему существенному прогрессу в зоологических исследованиях.
Настоящая работа была задумана в 1967—1968 гг. в процессе выбора объектов для всестороннего микроэволюпионного исследования. Прыткая ящерица (Lacerta ag'dis L.) — один из самых распространенных и многочисленных видов позвоночных фауны Евразии — представляет собой удобную модель для изучения общих особенностей процесса эволюции внутри вида (значения изоляции, естественного отбора и других факторов). Однако прежде чем перейти к такому эволюционному исследованию необходимым оказалось собрать достаточно полные данные по морфологии, распределению, популяционной структуре и другим биологическим параметрам вида в природе, проверить ряд вопросов, связанных с наследованием тех или иных признаков в эксперименте и т. д.
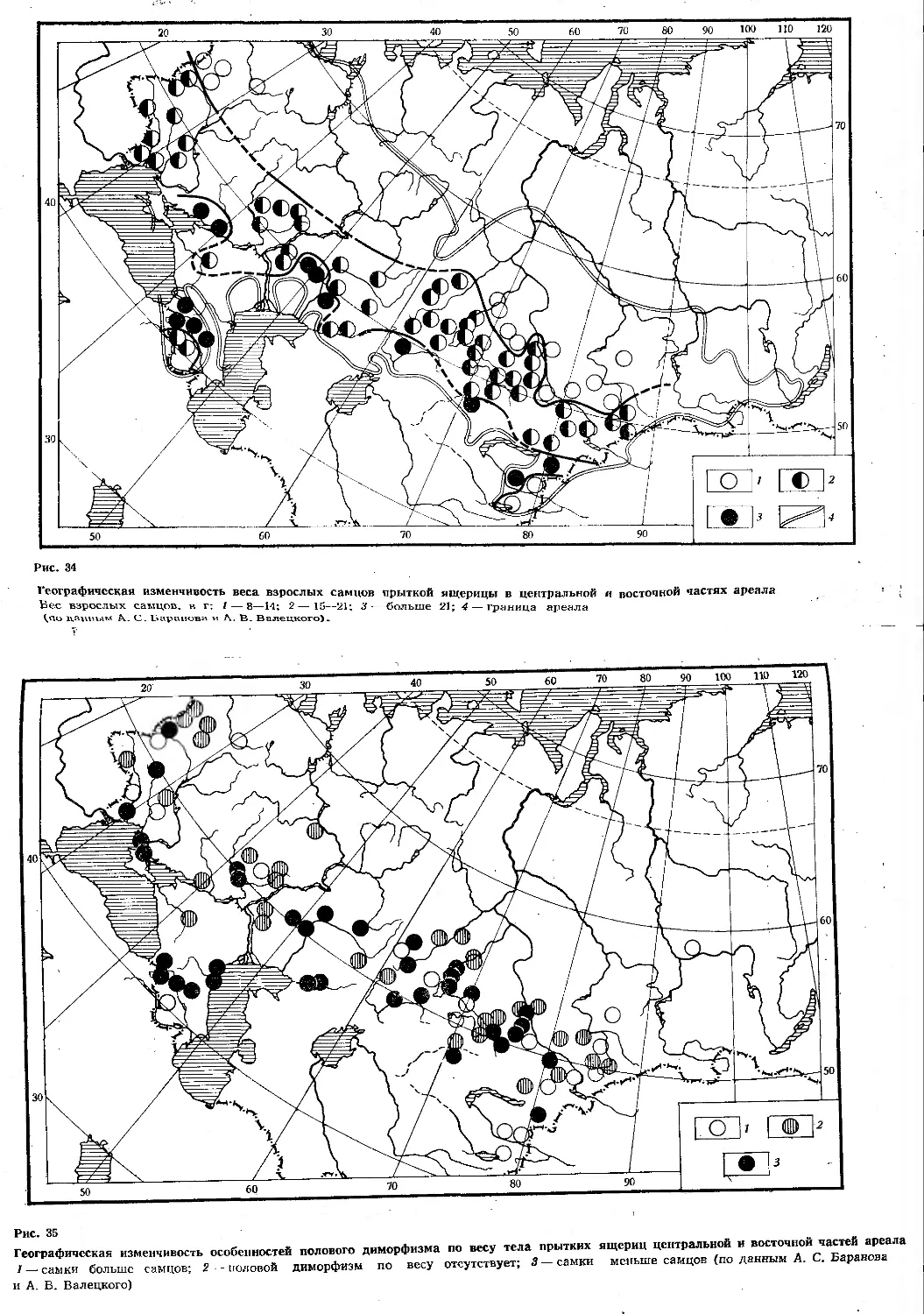
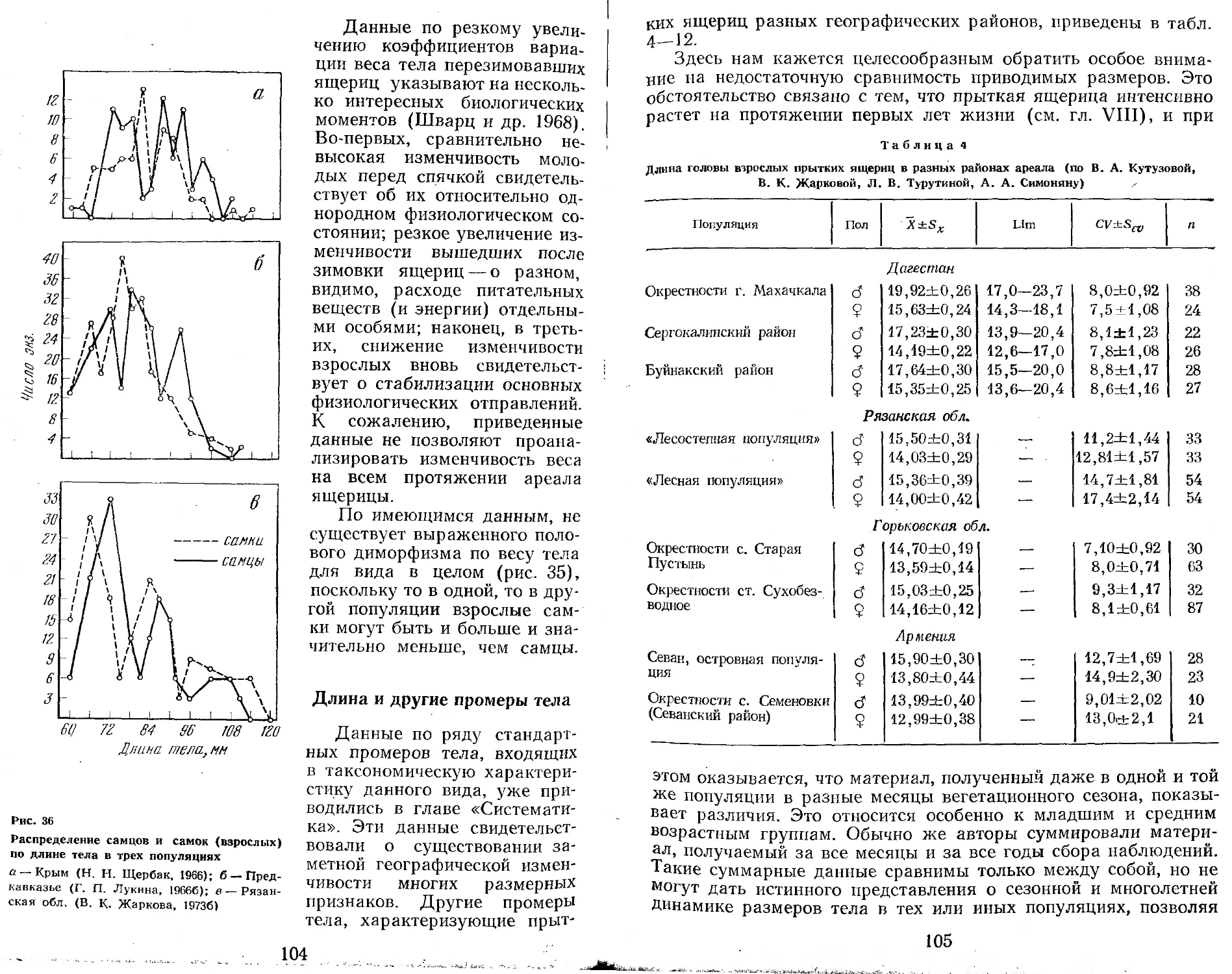
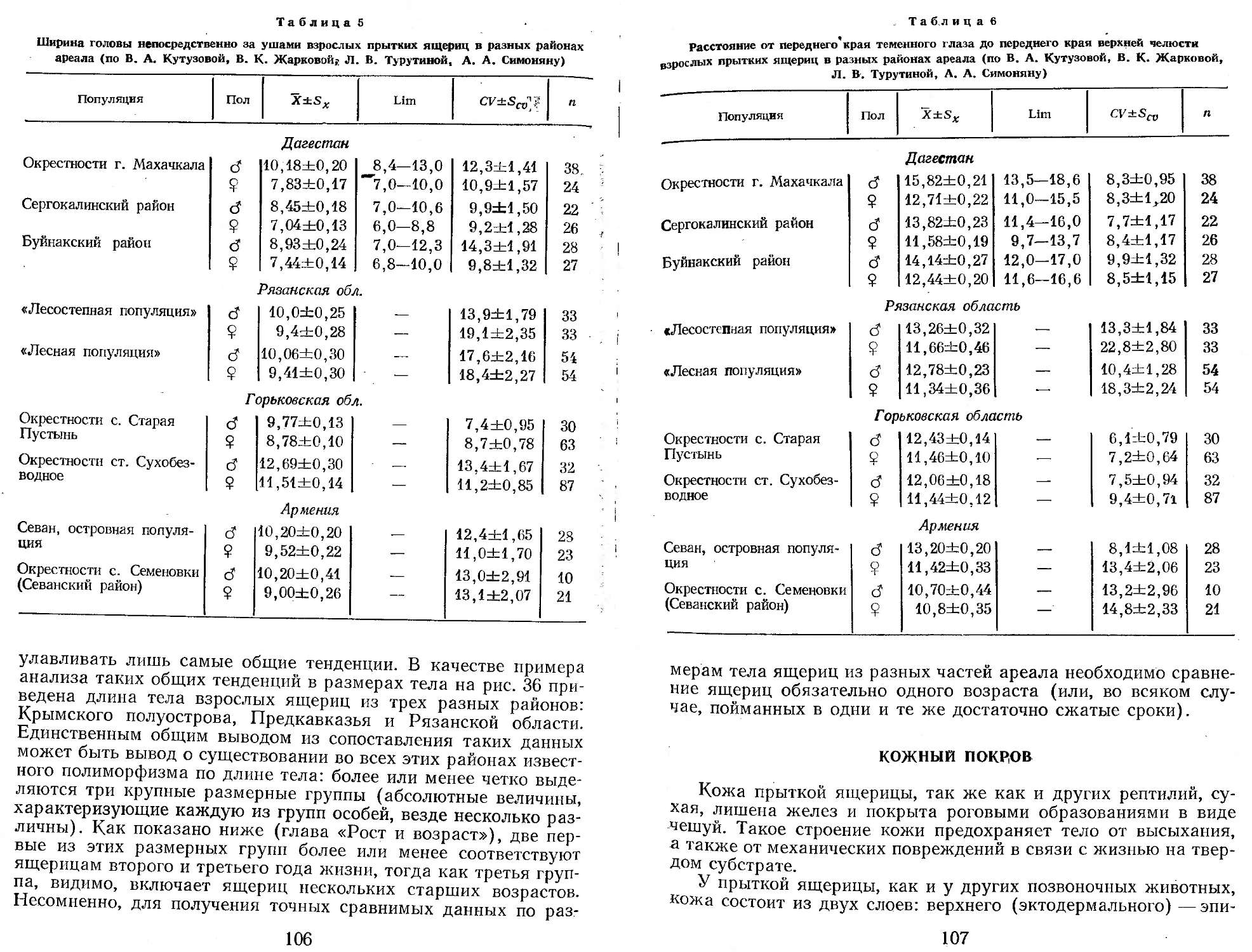
Первоначально казалось, что прыткая ящерица, изученная к настоящему времени едва ли не лучше любого другого вида рептилий нашей фауны (судя по числу выполненных исследований по биологии вида1), не требует дальнейшего общего исследования. Однако вскоре выяснилось, что большинство работ по биологии прыткой ящерицы относится к Европейской части СССР и Закавказью, тогда как огромные пространства ареала от Урала до Байкала остаются весьма слабо изученными. Недостаточно изученными оказались также и некоторые фундаментальные вопросы биологии, такие, например, как особенности роста, численность и биомасса в отдельных зонах и т. д.
Все это привело нас к необходимости организации специальных экспедиций в основные районы обитания этого вида. Первая из них была организована в 1969 г., и затем на протяжении следующих трех лет экспедиционными маршрутами была покрыта
1 Только специальному исследованию прыткой ящерицы были посвящены диссертации Н. В. Щепотьева (1948), М. Ф. Тертышникова (1972), В. К. Марковой (1973а); в целом ряде диссертационных работ прыткой ящерице уделялось большое внимание, как например, в работах 3. П. Хонякиной (1964), Э. А. Шебзуховой (1969), В. Ф. Орловой (1975) и др. Общее число публикаций, посвященных исключительно прыткой ящерице, составляет около 200< (см. список литературы).
6
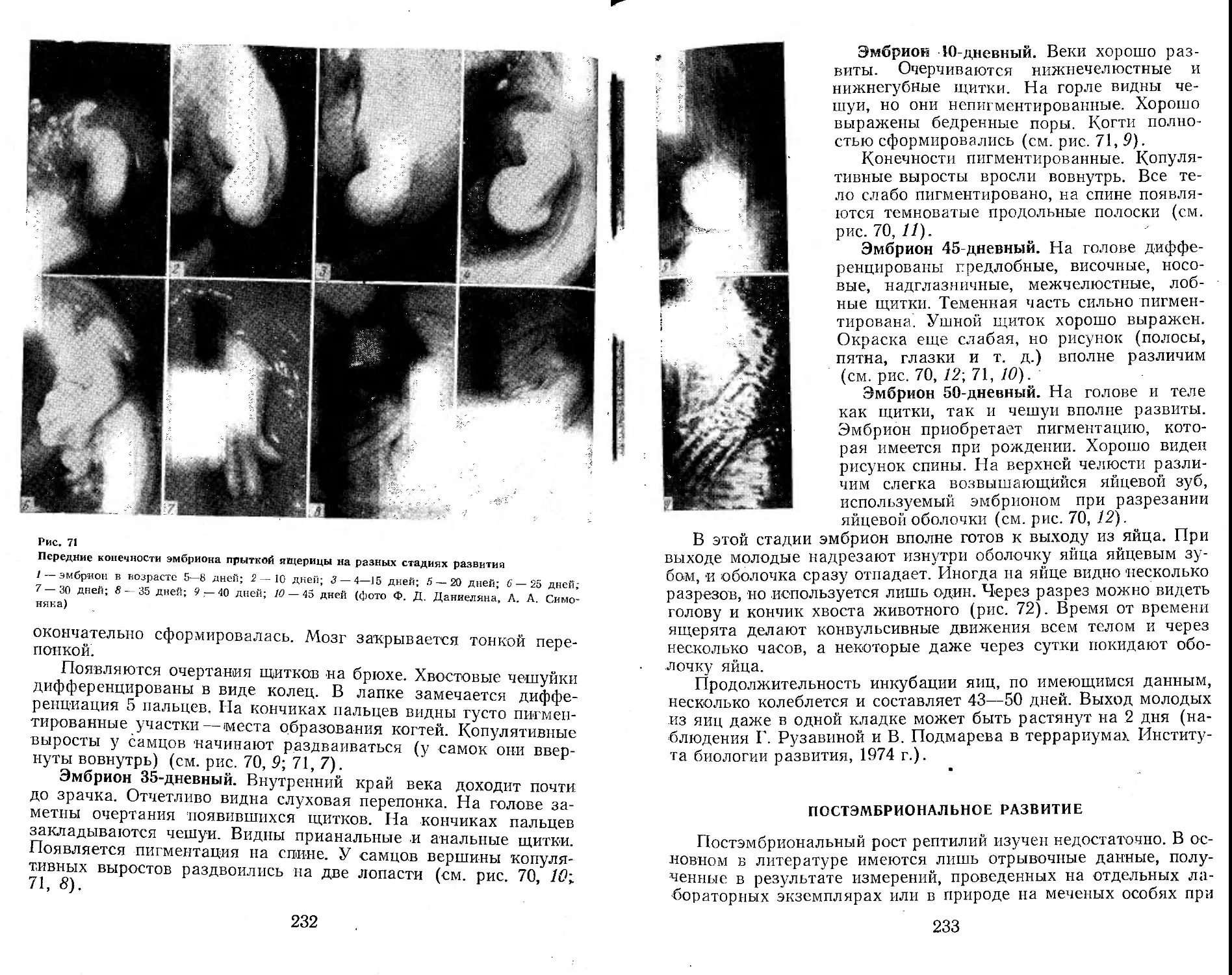
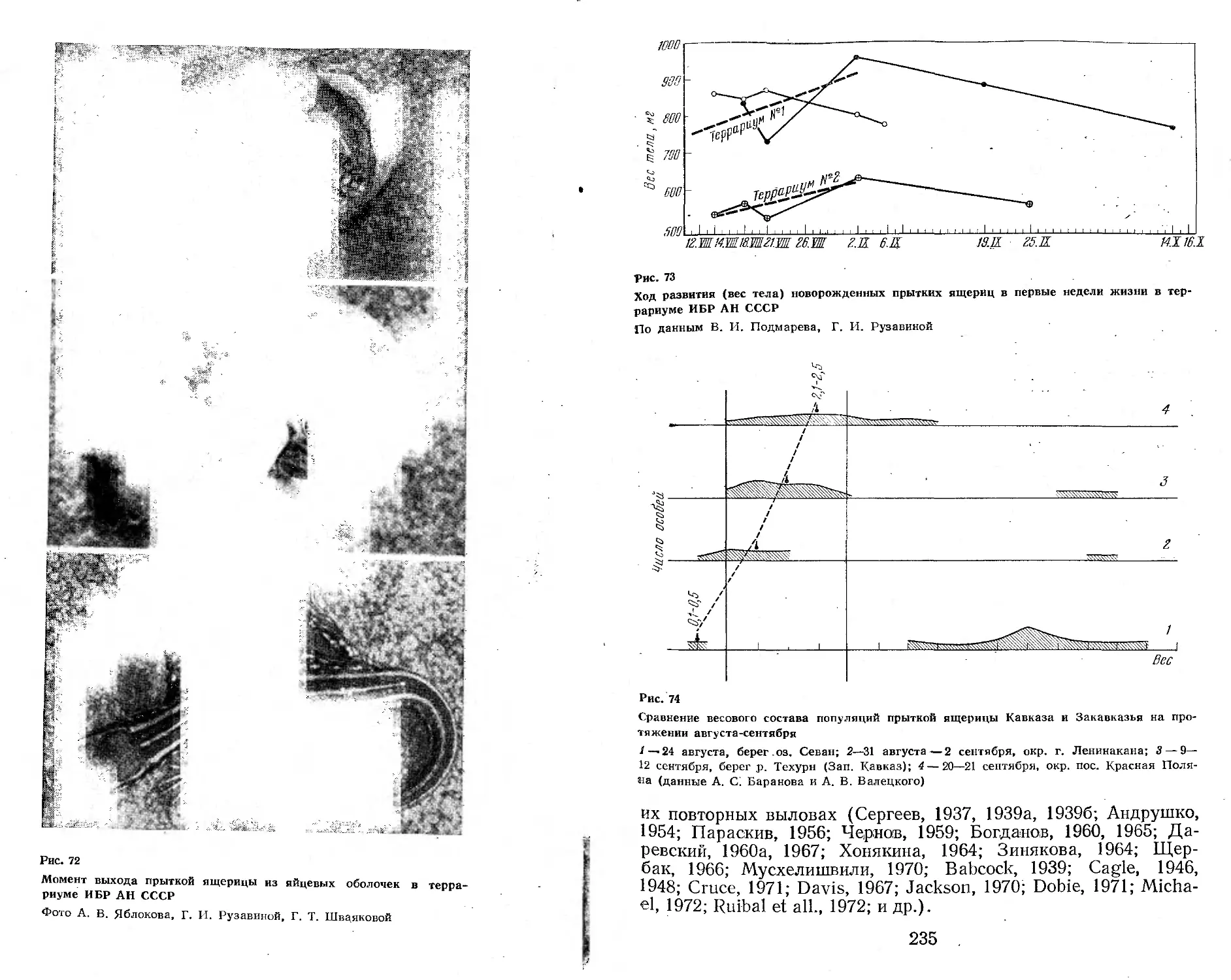
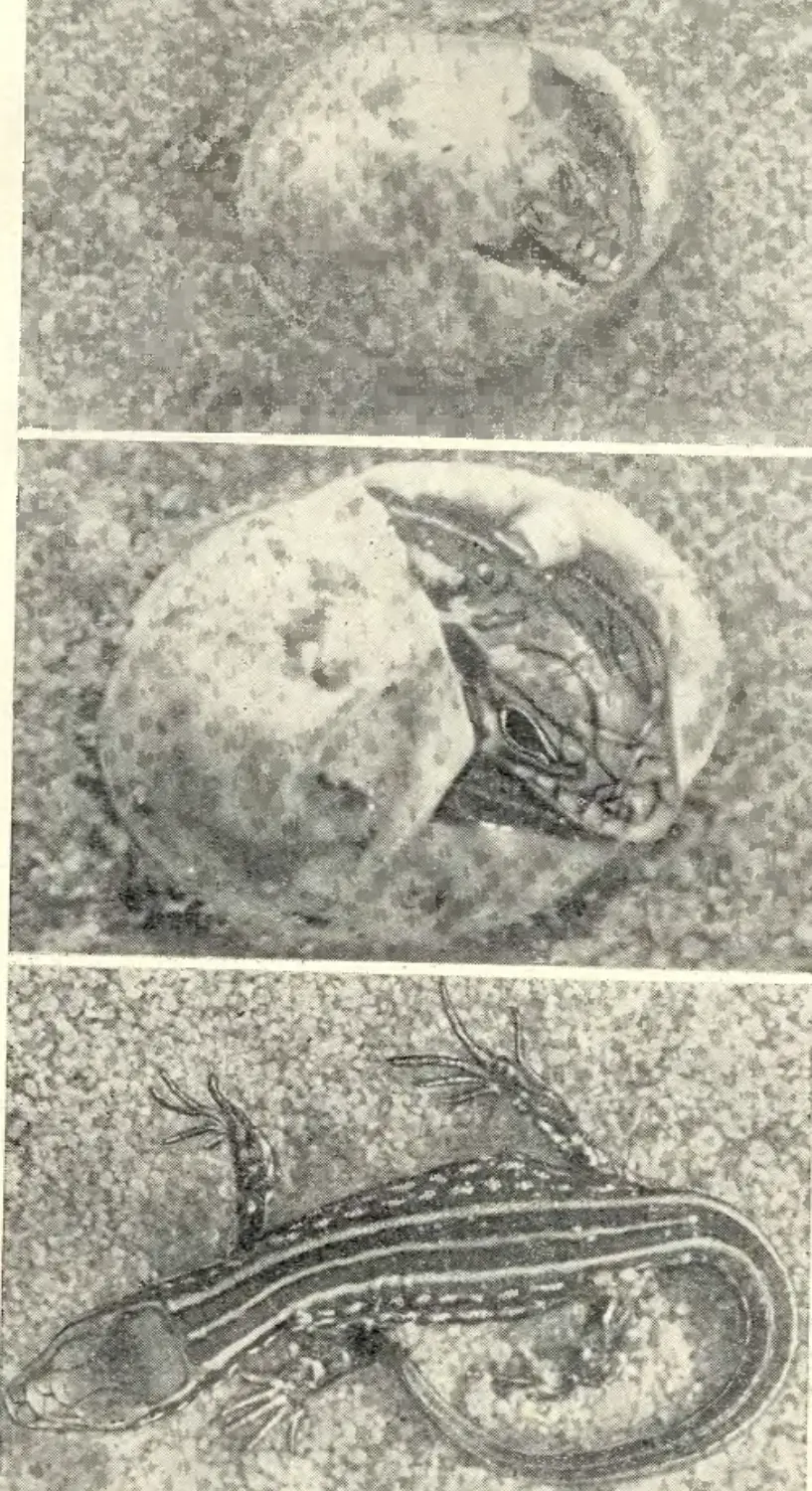
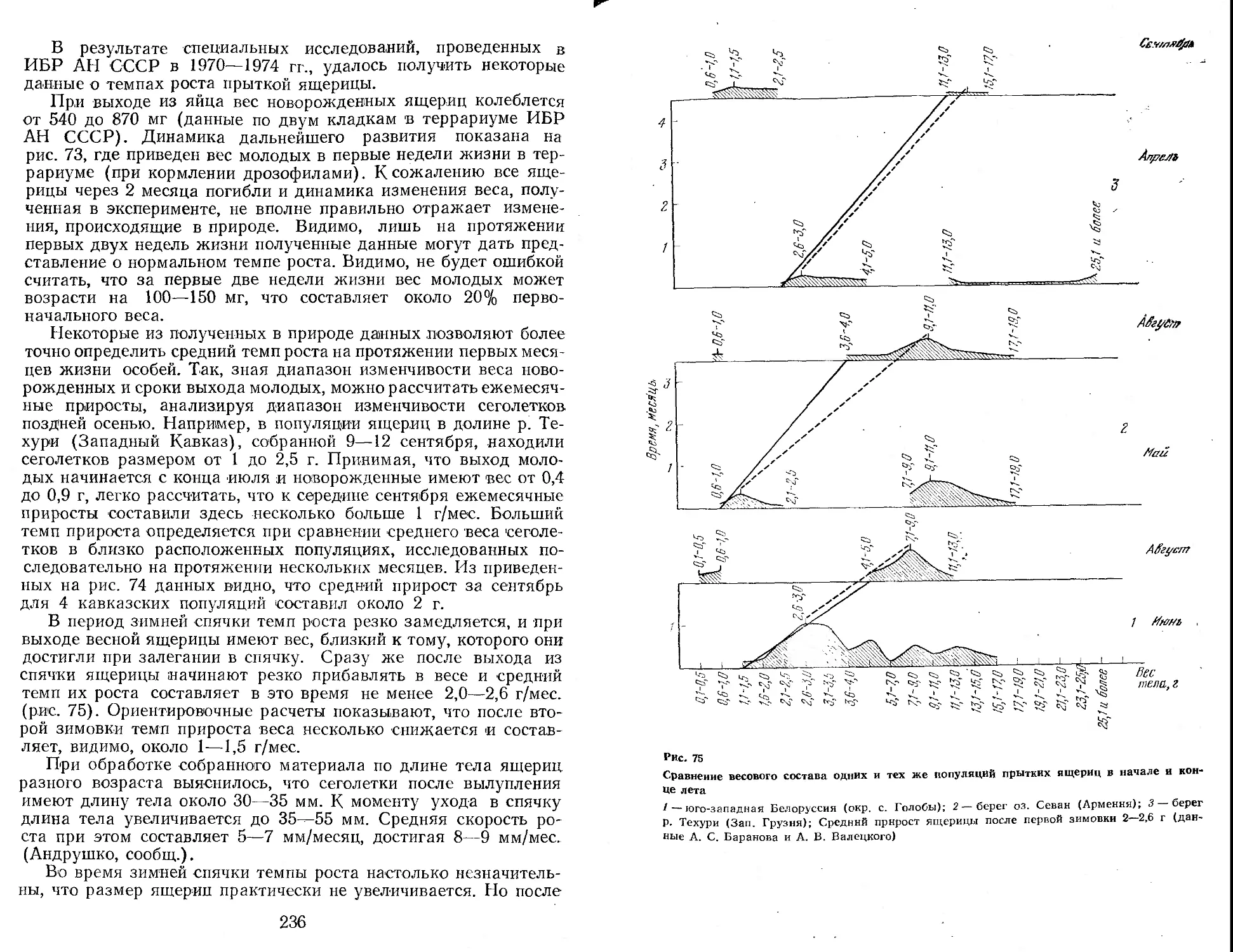
территория юга, запада и востока Европейской части страны, Закавказье, Кавказ, Северный, Южный, Центральный и Восточный Казахстан, Прииссыккулье, Западное Прибайкалье, некоторые районы Западной Сибири и Алтая (рис. 1). Непременными участниками этих экспедиций были А. С. Баранов, А. В. Валец-кий, А. В. Яблоков, В. И. Борисов. К ним присоединялись и другие сотрудники лаборатории постнатального онтогенеза Института биологии развития АН СССР (Г. А. Клевезаль, Э. М. Смп-рииа, М. С. Владимирская), а также сотрудники других учреждений (М. А. Бакрадзе, М. В. Мина, Ф. Д. Даниелян, Л. М. Смеловский, А. В. Шилейко и др.). Во время работы экспедиций был получен большой коллекционный материал (около 7000 экз. ящериц разного пола и возраста более чем из 100 популяций вида), проведено множество наблюдений за ящерицами в природных условиях. Одновременно в террариумах лаборатории шла отработка методики круглогодичного содержания и разведения ящериц (в этой работе приняли деятельное участие , А. Б. Стрельцов, Ф. Д. Даниелян, А. С. Баранов, В. И. Подмарев, Г. И. Рузавина).
Материал для настоящей монографии был собран также благодаря активной помощи ряда исследователей, объединенных в рамках программы «Изучение вида в пределах ареала», развернутой Советским национальным комитетом по МБП и продолжающейся сейчас в рамках программы «Человек и биосфера». Участие в этой программе разных исследователей выражалось по-разному. Одни, как например Н. В. Щепотьев, представили свои ценнейшие неопубликованные материалы по биологии прыткой ящерицы, собранные на протяжении целого ряда прошлых лет, другие помогали в сборе нового материала в доступных им районах ареала, третьи не только собирали, но и обрабатывали этот материал по заранее согласованным планам. При этом в некоторых высших учебных заведениях страны (например, в Горьковском, Дагестанском, Ереванском университетах, Даугавпил-ском и других пединститутах) были выполнены курсовые и дипломные работы, содержащие оригинальный материал, широко использованный в настоящей сводке (Л. В. Турутина, В. А. Кутузова, А. А. Симонян, С. А. Шарыгин, Т. М. Зорина, В. Д. Бахарев, А. С. Розанов и др.).
Живо откликнулись на нашу просьбу помочь в исследовании прыткой ящерицы зарубежные исследователи: В. А. Беш-ков (Зоологический институт НРБ, София), С. Ванци (Ясский университет, СРР), Г. Петерс (Зоологический музей, ГДР, Берлин), И. Фун (Зоологический институт АН СРР, Бухарест), Г. Добровольска и К- Шафраньска (Зоологический институт Варшавского университета). В ряде разделов нами использованы материалы, любезно предоставленные в наше распоряжение музеями Вены, Берна, Лондона, Стокгольма, Боппа и других европейских городов.
7
Более подробно вклад каждого исследователя, принявшего непосредственное участие в работе над монографией, отмечен перед началом отдельных глав и в оглавлении. Здесь же приведен общий список лиц (с указанием учреждений), принявших активное участие в работе по прыткой ящерице:
А. М. Андрушко — Ленинградский университет.
Ю. П. Антипчук — Мелитопольский пединститут.
М. А. Бакрадзе — Тбилисский университет.
А. С. Баранов — Институт биологии развития АН СССР, Москва.
В. А. Бахарев — Даугавпилский пединститут.
В. И. Борисов — Институт биологии развития АН СССР, Москва.
А. В. Валецкий — Институт биологии развития АН СССР, Москва.
В. Л. Булахов — Днепропетровский университет.
В. Е. Боченко — Криворожский пединститут.
В. И. Ведмедеря — Харьковский университет.
В. И. Гаранин — Биологический институт АН СССР, Казань.
В. Ф. Гинтовт — Гродненский пединститут.
И. С. Даревский — Зоологический институт АН СССР, Ленинград.
Ф. Д. Даниелян — Ереванский университет.
А. М. Дорофеев — Витебский пединститут.
В. К. Жаркова — Рязанский пединститут.
Е. Н. Инщикова — Уманский пединститут.
Н. Ф. Константинова — Днепропетровский университет.
Г. П. Лукина — Ростовский университет.
Н. М. Окулова — Институт полиомиелита АМН СССР, Москва.
В. И. Подмарёв — Институт биологии развития АН СССР, Москва.
В. Ф. Орлова — Московский университет.
Н. А. Рашкевпч — Чечено-Ингушский университет, Грозный.
А. А. Симонян — Институт зоологии АН АрмССР, Ереван.
Л. М. Смеловский — Волгоградский пединститут.
Э. М. Смирина — Институт биологии развития АН СССР, Москва.
А. Б. Стрельцов — Калужский областной краеведческий музей.
М. Ф. Тертышников — Ставропольский пединститут.
Л. В. Турутина — Горьковский университет.
3. П. Хонякина — Дагестанский университет, Махачкала.
В. П. Шарпило — Институт зоологии АН УССР, Киев.
С. П. Чащин — Пермский университет.
В. К. Щепотьев — Саратов ВНИИ «Микроб».
Н. Н. Щербак — Институт зоологии АН УССР, Киев.
М. И. Щербань — Ужгородский университет.
А. В. Яблоков — Институт биологии развития АН СССР, Москва.
В заключение от имени авторского коллектива мне приятно выразить благодарность за постоянную поддержку и помощь в исследовании прыткой ящерицы акад. Б. Л. Астаурову и члену-корр. АН СССР Т. М. Турпаеву, председателю Межсекционной рабочей группы Советского национального комитета по МБП Р. С. Вольскису, руководителю биологической части Советской национальной программы «Человек и биосфера» акад. В. Е. Соколову, а также Т. Н. Щербиновской, сотрудникам ИБР АН СССР Г. И. Рузавиной, Г. М. Шваяковой, Е. Г. Якутовской, Л. И. Суховской, М. С. Владимирской и всем другим лицам, активно помогавшим авторам на разных этапах проведения настоящей работы.
А. В. Яблоков-
ГЛАВА ПЕРВАЯ
АРЕАЛ1
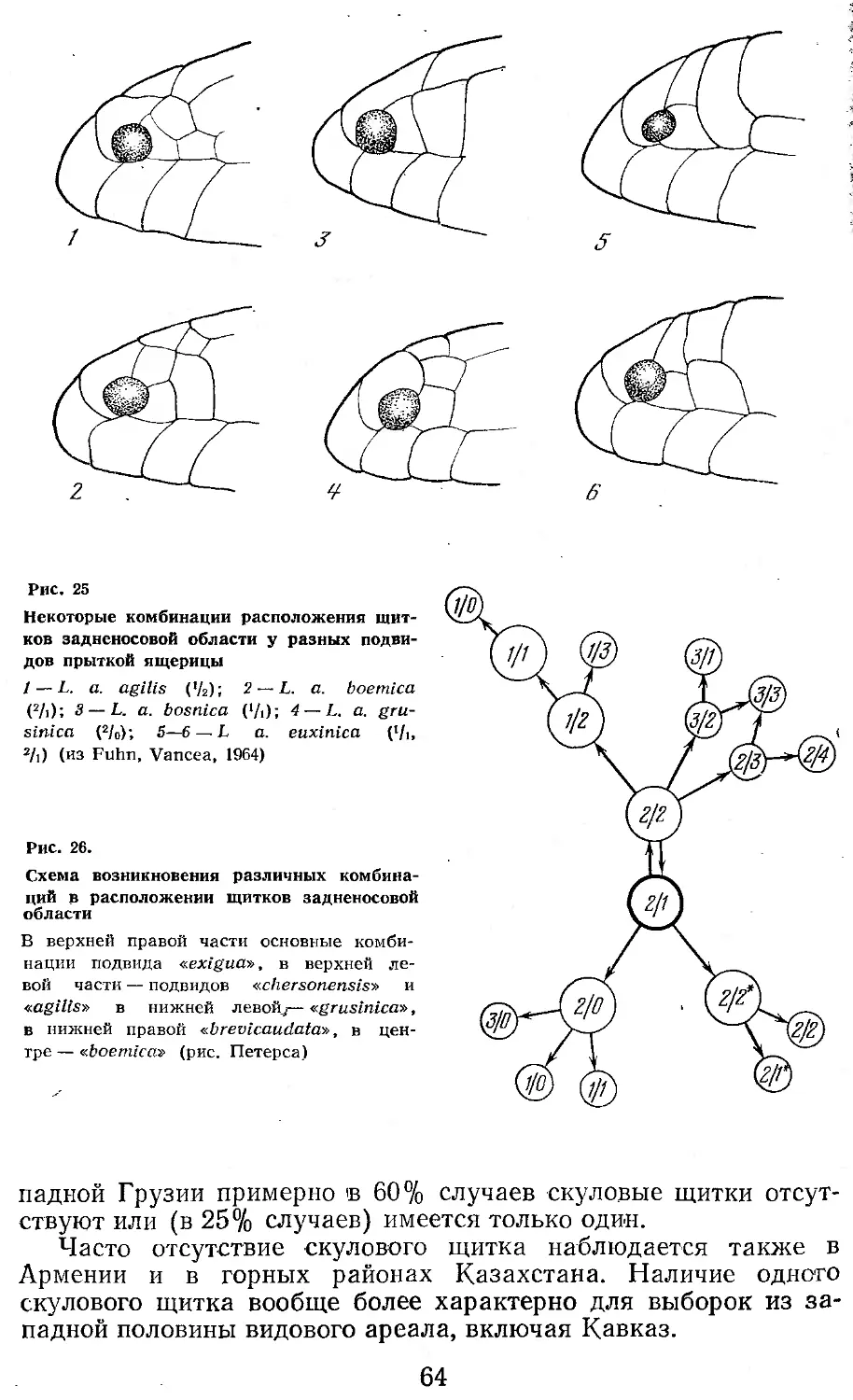
Прыткая ящерица — вид, широко распространенный, эври-топный, с большим диапазоном изменчивости. Свидетельством последнего является более чем два десятка синонимов, под которыми описывалось данное пресмыкающееся (Mertens, Wermuth, 1960). Этим объясняется то обстоятельство, что спустя почти полтора века после описания этого обычного в Европе вида отсутствовал достаточно достоверно составленный ареал. Одно из наиболее ранних и близких к действительности описаний ареала L. agilis принадлежит Буланже (Boulenger, 1887). Выяснение и уточнение границ ареала было закончено в начале текущего века и обобщено в серии работ А. М. Никольского (1902, 1905, 1907, 1915). В последующие годы происходило уточнение распространения прыткой ящерицы, которое позволило составить графическое изображение ее ареала как на территории СССР (Терентьев, Чернов, 1949; Банников, Даревский, Рустамов, 1971), так и всей Евразии (Fuhn, Vancea, 1961; Щербак, 1966; и др.). Однако все указанные карты ареалов или описания содержат много ошибок и очень приблизительны.
В последние годы в различных музеях накопилось много коллекционных материалов по прыткой ящерице, появились региональные сводки с указанием мест находок данного вида (Параскив, 1956; Яковлева, 1964; Fuhn, Vancea, 1961; и др.), для уточнения распространения L. agilis проводились специальные исследования сотрудниками Института биологии развития. Все эти материалы позволили предпринять попытку картирования всех ныне известных мест находок прыткой ящерицы и на этой базе, при помощи метода кадастров, уточнить границы и «кружево» ареала, а также рассмотреть факторы, ограничивающие распространение данного вида.
Выполнение настоящей работы показало различную степень изученности отдельных районов ареала и возможность новых
1 Авторы: И. И. Щербак, И. Г. Остатке, при участии И. С. Даревского, А. С. Баранова, А. М. Андрушко, М. А. Бакрадзе, В. И. Ведмедеря, В. И. Гаранина, В Г. Ищенко, Г. П. Лукиной, И. М. Окуловой, Н. А. Вашкевича, А1. Ф. Тертышникова, Л. Я. Топорковой, 3. П. Хонякиной, Ю. Г. Швецова, -М. И. Щербаня.
9
интересных находок (например, сообщение Г. А. Воронова, В. В. Демидова (1973) отодвинуло границу ареала на 400 км к северо-востоку). Часть известных мест находок, к сожалению, вследствие недостаточно точной этикетировки, переименования населенных пунктов или отсутствия их на доступных нам картах не была обозначена или обозначена весьма приблизительно.
При выполнении данной работы были обработаны фондовые коллекции музеев ЗИН АН СССР, ИЗАНУ, МГУ, ИБР АН СССР. Кроме того, для выяснения коллекционных материалов по L. agilis и уточнения деталей ее распространения нами были разосланы запросы во многие зарубежные музеи. На большинство из них получены ответы, содержащие много ценной информации. Ответы получены из следующих музеев и учреждений: Венский естественноисторический музей (I. Eiselt), Австрия; Уэльский национальный музей (Cardiff, I. A. Bateman); Шотландский королевский музей (Edinburgh, A. S. Clarke); Манчестерский музей (Е. L. Seyd); Норвичский замок-музей (Т. G. Goldsmith); Оксфордский универсальный музей (Т. S. Кешр); Шеффильдский городской музей (Т. Н. Riley); Хантингдонская опытная станция (Huntingdon Mouks Wood Experimentai Station Aiberts Ripton, F. H. Perring), (Англия); Бельгийский королевский институт естественных наук (Brussels, G. F. Witte); Венгерский естественноисторический музей (Budapest, I. Szabo); Веспремский музей (Veszprem, Bakonyi Museum, Bankovics Attila); Сегедский музей им. Мора Ференца (Szeged, Mora Ferenc Museum, M. Marian) (Венгрия); Государственный музей естествознания в Герлитце (W. Dunger); Государственный зоологический музей и исследовательский центр в Дрездене (F. I. Obst); Музей естествознания при Берлинском университете им. Александра Гумбольдта (G. Peters) (ГДР); Зоологический исследовательский институт и музей Александра Кёнига в Бонне (W. Bohme); Музей Ремера-Пелицеюса в Гильдесгайме (Н. Kayser); Государственный естественноисторический музей в Брауншвейге (V. Frisch); Зоологическое отделение Гессенского земельного музея в Дармштадте (Н. Feus-tel); Земельный музей естествознания в Карлсруэ (R. Angst); Государственный музей естествознания в Штутгарте (Н. Wermuth); Естественнонаучный музей в Висбадене (R. Mentzel); Дорнбирский музей (Vorarlberger Naturschau, М. Krieg (ФРГ); Тегеранский университет (М. Chojai) (Иран); Зоологический музей Амстердамского университета (D. Hillenius) и Государственный музей естественной истории в Лейдене (М. S. Hoogmo-ed) (Нидерланды); Ставангерский музей (Н. Holgersen); Музей королевского общества наук и литературы в Тронхейме (D. Dolmen) (Норвегия); Зоологический музей Хельсинского университета (G. Bergman); Музей в Куопио (Eino Salainen) (Финляндия); Лионский музей естественной истории (L. David); Музей естественной истории Этьена (G. de-la-Comole) (Франция);
10
Братиславский зоопарк (I. Lac); Восточно-Чешский музей в Пардубице (I. Sklenar); Музей Восточно-Словацкого края в Кошицах (О. Mosansky); Пражский природоведческий музей (Liri Cihar); Музей Орлицких гор в Рихнове над Кнежной (Zbynek Rocek) (Чехословакия); Музей естественной истории в Ля-Шоде-Фон (R. Forissier); Музей естественной истории в Женеве (Volker Mahnert); из Лампенберга (Urs Regenass); Ольтенский естественноисторический музей (Н. Furer) (Швейцария); Гетеборгский естественноисторический музей (S. Mat-hiasson); Хельсингборгский музей естественной истории (Н. Merker), Музей Мальмо (Н. Hallander); Зоологический институт Упсальского университета (A. Holm) (Швеция); -Словенский природоведческий музей в Люблянах (A. Polenes) (Югославия).
Всем перечисленным выше лицам мы выражаем глубокую благодарность.
Места добычи животных группировались по административному признаку. В каждой административной группе нумерация начинается с первого номера. Это упрощает работу при поступлении новых данных о находках на этой территории.
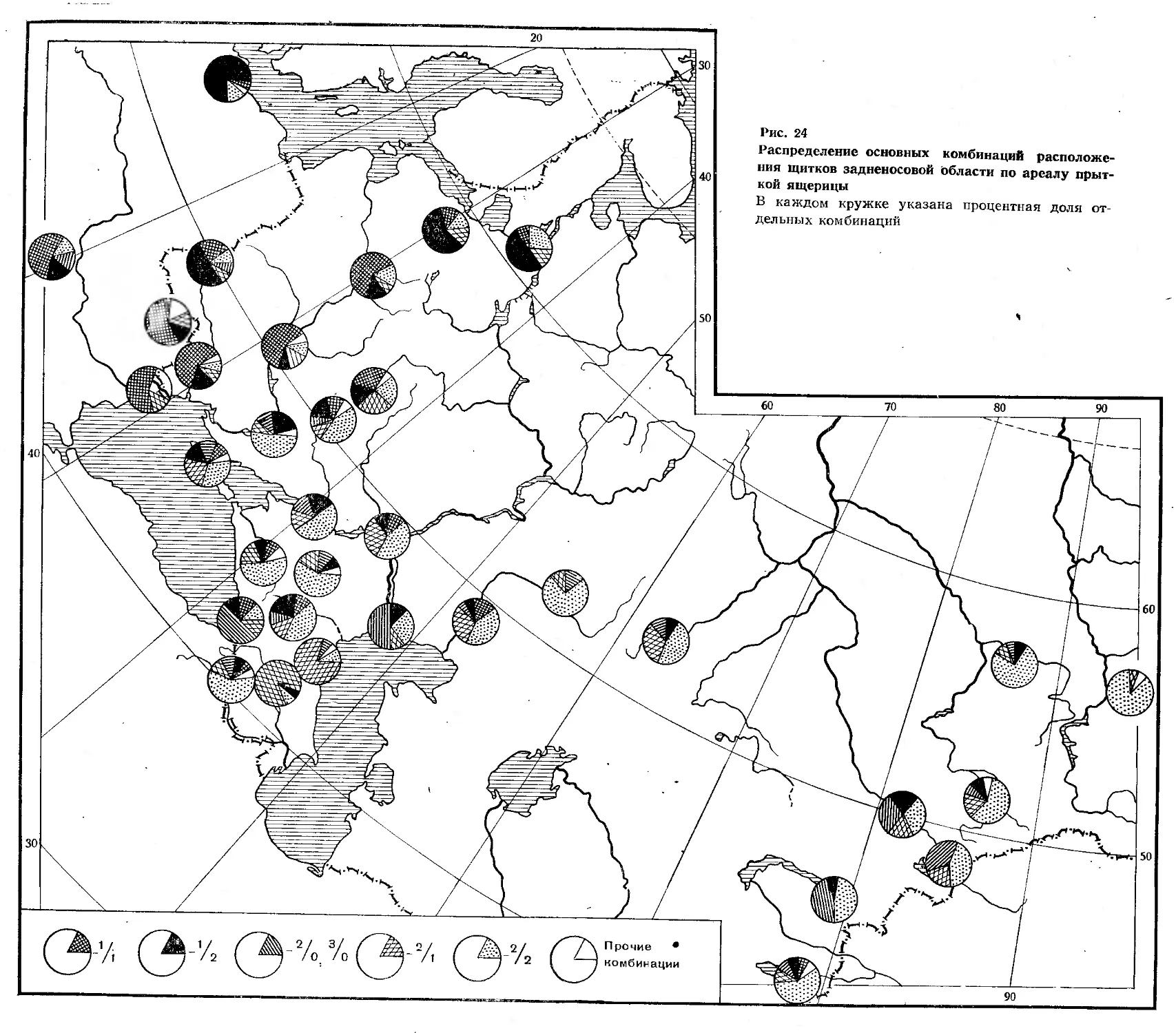
В настоящей работе весь ареал прыткой ящерицы разбит на 14 следующих районов:
I — Литовская, Латвийская, Эстонская, Белорусская ССР, Калининградская, Ленинградская, Псковская, Новгородская области.
II — Украинская, Молдавская ССР.
III—Грузинская, Армянская, Азербайджанская ССР, Автономные республики Северного Кавказа, Краснодарский, Ставропольский края, Северо-Восточная Турция.
IV—Вологодская, Московская, Калининская, Смоленская, Брянская, Калужская, Орловская, Тульская, Ивановская, Липецкая, Курская, Тамбовская, Ярославская, Костромская, Рязанская, Горьковская, Белгородская области.
V— Калмыцкая АССР, Ростовская, Астраханская, Волгоградская, Воронежская, Саратовская, Ульяновская, Куйбышевская, Пензенская области, Мордовская и Чувашская АССР.
VI— Татарская, Удмуртская и Марийская АССР, Кировская, Пермская, Свердловская области.
VII— Челябинская, Курганская, Оренбургская области, Башкирская АССР.
VIII — часть Казахской ССР восточнее 64° в. д.
IX— Тюменская, Томская, Омская, Новосибирская, Кемеровская, Иркутская области, Тувинская АССР, Алтайский и Красноярский края.
£Ср— часть Казахской ССР западнее 64° в. д., Киргизская
XI— Дания, Швеция.
XII— Великобритания, Бельгия, Люксембург, Нидерланды, Франция.
11
20 30
30
Рис. 2.
Места находок прытких ящериц в Литовской, Латвийской, Эстонской, Белорусской ССР, Калининградской, Ленинградской, Псковской, Новгородской областях
XIII— ФРГ, ГДР, Польша, Чехословакия, Австрия, Швейцария.
XIV— Венгрия, Румыния, Болгария, Югославия.
Если на одно и то же место находки встречаются ссылки у разных авторов и есть коллекционные экземпляры животных, то предпочтение отдавалось музейным материалам или автору последней региональной сводки или ревизии.
МЕСТА НАХОДОК ПРЫТКОЙ ЯЩЕРИЦЫ
Литовская, Латвийская, Эстонская, Белорусская ССР, Ленинградская, Псковская, Новгородская
и Калининградская обл.
(рис. 2)
1. БССР, окр. г. Минска; 2. Ленинградская обл., берег р. Свирь; 3. БССР, окр. г. Гродно; 4. БССР, окр. г. Бобруйска; 5. Ленинградская обл., Лужский р-н; 6. БССР, Беловежская Пуща; 7. Ленинградская обл., с. Толмачево; 9. БССР, окр. г. Полоцка; 10. Ленинградская обл., окр. г. Луги; 11. БССР, окр. с. Дмит-ровичи; 12. Могилевская обл., с. Гиженка; 13. Литовская ССР, окр. г. Друскининкай, (ЗИН); 14. БССР, Гомельская обл., окр.
12
г. Мозырь; 15. Окр. г. Калининграда; 16. БССР, окр. г. Луги; 17. Псковская обл., г. Гдов (МГУ); 19. БССР, Брестская обл., шоссе Кобрин — Малорита; 20. Гродненская обл., шоссе Сло-ним — Волковыск; 21. Гродненская обл., пос. Зельва; 22. БССР, г. Гродно; 23. БССР, окр. г. Барановичи; 24. БССР, шоссе Брест — Минск; 25. Витебская обл., окр. с. Заольша; 26. Новгородская обл., Крестецкий р-н, д. Болотница; 27. Карельская АССР, окр. г. Питкяранта; 28. Латвийская ССР, окр. г. Даугавпилс; 29. Латвийская ССР, окр. г. Илга (ИБР); 30. Карельская АССР, окр. г. Олонец (Ивантер); 32. Псковская обл., Плюсский р-н, окр. с. Окрино (ЗИН); 34. Окр. г. Петрозаводска (?), (Никольский, 1915); 35. БССР, Витебская обл., окр. г. Яновичи (ЗИН); 8, 18, 31, 33 — по письменному (карта) сообщению И. С. Даревского, а также Г. Петерса (1959).
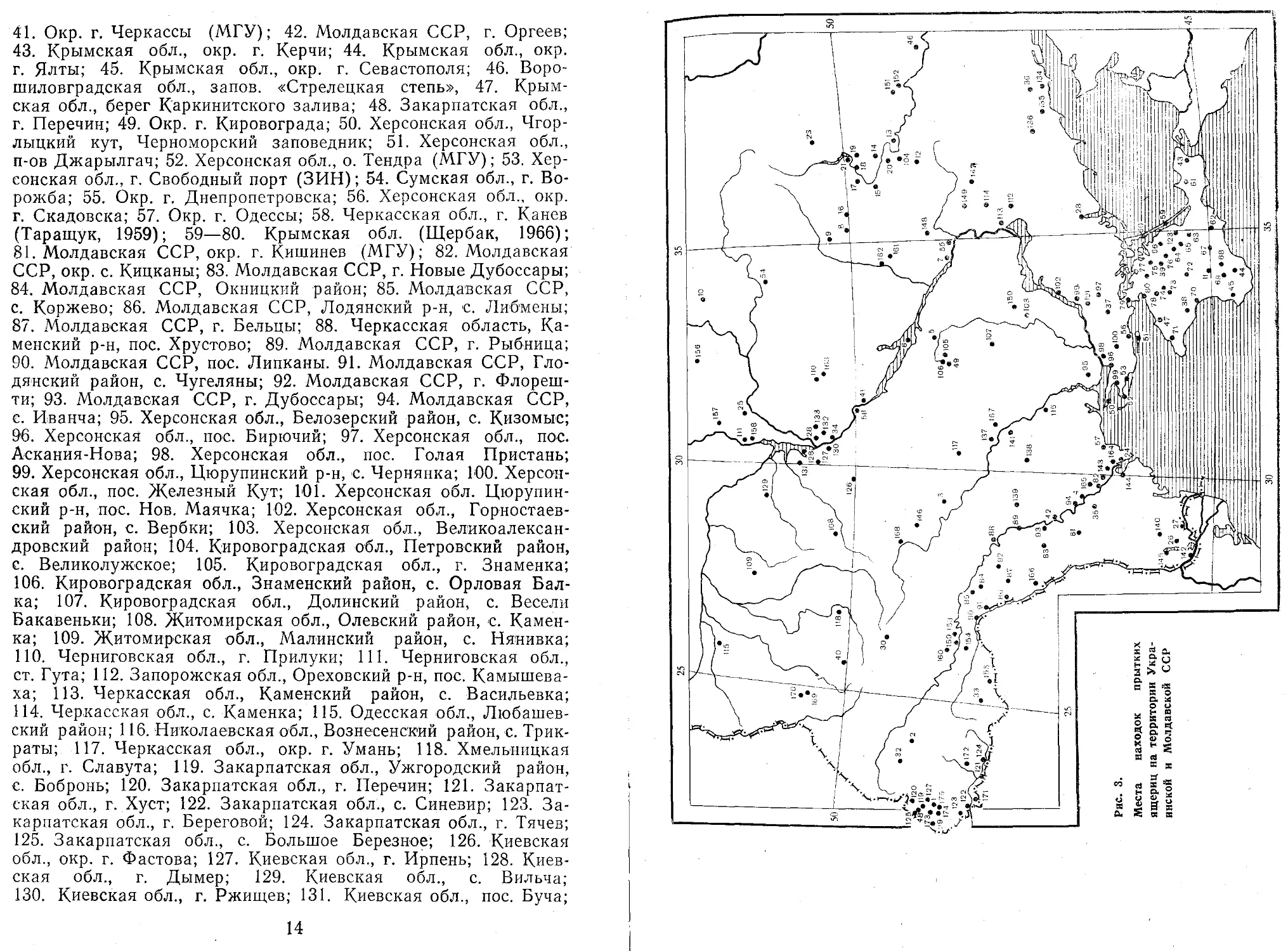
Украинская и Молдавская ССР (рис. 3)
1. Закарпатская обл., с. Воловец (В. И. Ведмедеря);
2. Львовская обл., Стрыйский р-н, с. Тунля; 3. Винницкая обл., г. Гайсин, с. Губник; 4. Молдавская ССР, окр. г. Тирасполя; 5. Кировоградская обл., окр. г. Александрии; 6. Полтавская обл., остров в Кременчугском водохранилище; 7. Днепропетровская обл., окр. г. Днепродзержинска; 8. Харьковская обл., Крас-нокутский район, с. Чернищина; 9. Сумская обл., Ахтырский район; 10. Сумская обл., Середино-Будский район; 11. Крымская обл., окр. г. Симферополя; 12. Харьковская обл., Близ-нюковский район; 13. Харьковская обл., Изюм; 14. Харьковская обл., ст. Сезонная; 15. Харьковская обл., Змиёвский район; 16. Харьковская обл., Богодуховский район, с. Шаровка; 17. Окр. Харькова; 18. Харьковская обл., Чугуевский район, пос. Эскар; 19. Харьковская обл., с. Покровка; 20. Харьковская обл., Балаклейский район; 21. Харьковская обл., Чугуевский район, с. Б. Бабка; (ХГУ); 22. Запорожская обл., Мелитопольский район, берег р. Молочная (ИЗАНУ); 23. Харьковская обл., Волчанский район, с. Землянки (ХГУ); 24. Одесская обл., окр. г. Белгород-Днестровского; 25. Окр. Чернигова; 26. Одесская обл., окр. г. Измаила; 27. Одесская обл., с. Вилково; 28. Окр. Киева, Быковня; 29. Закарпатская обл., окр. г. Ужгорода; 30. Хмельницкая обл., с. Волочиск; 31. Черниговская обл., с. Козелец; 32. Львовская обл., с. Трускавец; 33. Черновицкая обл., Вижницкий р-н; 34. Киевская обл., Триполье; 35. Молдавская ССР, окр. г. Бендеры (ЗИН); 36. Донецкая обл., Ждановский район, берег реки Грузской Еланчик (ИЗАНУ); 37. Херсонская обл., с. Колончак; 38. Крымская обл., окр. г. Евпатории; 39. Крымская обл., окр. г. Джанкоя (ЗИН); 40. Тернопольская обл.., г. Кременец (сообщ. И. В. Марисовой);
13
41. Окр. г. Черкассы (МГУ); 42. Молдавская ССР, г. Оргеев; 43. Крымская обл., окр. г. Керчи; 44. Крымская обл., окр. г. Ялты; 45. Крымская обл., окр. г. Севастополя; 46. Воро-шиловградская обл., запов. «Стрелецкая степь», 47. Крымская обл., берег К'аркинитского залива; 48. Закарпатская обл., г. Перечни; 49. Окр. г. Кировограда; 50. Херсонская обл., Чгор-лыцкий кут, Черноморский заповедник; 51. Херсонская обл., п-ов Джарылгач; 52. Херсонская обл., о. Тендра (МГУ); 53. Херсонская обл., г. Свободный порт (ЗИН); 54. Сумская обл., г. Ворожба; 55. Окр. г. Днепропетровска; 56. Херсонская обл., окр. г. Скадовска; 57. Окр. г. Одессы; 58. Черкасская обл., г. Канев (Таращук, 1959); 59—80. Крымская обл. (Щербак, 1966); 81. Молдавская ССР, окр. г. Кишинев (МГУ); 82. Молдавская ССР, окр. с. Кицканы; 83. Молдавская ССР, г. Новые Дубоссары; 84. Молдавская ССР, Окницкий район; 85. Молдавская ССР, с. Коржево; 86. Молдавская ССР, Лодянокий р-н, с. Либмены; 87. Молдавская ССР, г. Бельцы; 88. Черкасская область, Каменский р-н, пос. Хрустово; 89. Молдавская ССР, г. Рыбница; 90. Молдавская ССР, пос. Липканы. 91. Молдавская ССР, Гло-дянский район, с. Чугеляны; 92. Молдавская ССР, г. Флореш-ти; 93. Молдавская ССР, г. Дубоссары; 94. Молдавская ССР, с. Иванча; 95. Херсонская обл., Белозерский район, с. Кизомыс; 96. Херсонская обл., пос. Бирючий; 97. Херсонская обл., пос. Аскания-Нова; 98. Херсонская обл., пос. Голая Пристань; 99. Херсонская обл., Цюрупинский р-н, с. Чернянка; 100. Херсонская обл., пос. Железный Кут; 101. Херсонская обл. Цюрупинский р-н, пос. Нов. Маячка; 102. Херсонская обл., Горностаев-ский район, с. Вербки; 103. Херсонская обл., Великоалександровский район; 104. Кировоградская обл., Петровский район, с. Великолужское; 105. Кировоградская обл., г. Знаменка; 106. Кировоградская обл., Знаменский район, с. Орловая Балка; 107. Кировоградская обл., Долинский район, с. Весели Бакавеньки; 108. Житомирская обл., Олевский район, с. Каменка; 109. Житомирская обл., Малинский район, с. Нянивка; ПО. Черниговская обл., г. Прилуки; 111. Черниговская обл., ст. Гута; 112. Запорожская обл., Ореховский р-н, пос. Камышева-ха; 113. Черкасская обл., Каменский район, с. Васильевка; 114. Черкасская обл., с. Каменка; 115. Одесская обл., Любашев-ский район; 116. Николаевская обл., Вознесенский район, с. Трик-раты; 117. Черкасская обл., окр. г. Умань; 118. Хмельницкая обл., г. Славута; 119. Закарпатская обл., Ужгородский район, с. Бобронь; 120. Закарпатская обл., г. Перечни; 121. Закарпатская обл., г. Хуст; 122. Закарпатская обл., с. Синевир; 123. Закарпатская обл., г. Береговой; 124. Закарпатская обл., г. Тячев; 125. Закарпатская обл., с. Большое Березное; 126. Киевская обл., окр. г. Фастова; 127. Киевская обл., г. Ирпень; 128. Киевская обл., г. Дымер; 129. Киевская обл., с. Вильча; 130. Киевская обл., г. Ржищев; 131. Киевская обл., пос. Буча;
14
t
132. Киевская обл., г. Борисполь; 133. Киевская обл., г. Бровары; 134. Донецкая обл., Новоазовский район, з-х «Хомутовская степь»; 135. Донецкая обл., окр. г. Жданова; 136. Запорожская обл., Куйбышевский район, пос. Розовка; 137. Николаевская обл., окр. г. Первомайска; 138. Одесская обл., г. Троицк; 139. Одесская обл., г. Котовск; 140. Одесская обл., Измаильский район, пос. Суворове; 141. Одесская обл., с. Покровское; 142. Одесская обл., г. Рени; 143. Одесская обл., с. Беляевка; 144. Одесская обл., с. Кринички; 145. Одесская обл., с. Стар'оказачье; 146. Винницкая обл., Немировский район, с. Алексеевка; 147. Днепропетровская обл., с. Ва-сильковка; 148. Днепропетровская обл., окр. г. Новомосковска; 149. Днепропетровская обл., окр. г. Синельникова; 150. Днепропетровская обл., Апостоловский район, с. Шоколо-во; 151. Ворошиловградская обл., с Кременное; 152. Ворошилов-градская обл., с. Кузьмино; 153. Черновицкая обл., г. Хотин; 154. Черновицкая обл., с. Книжковцы; 155. Черновицкая обл., Сторожинецкий район; 156. Черниговская обл., Семеновский район, с. Барановка; 157. Черниговская обл., Репнинский район, с. Вирь; 158. Черниговская обл., с. Любеча; 159. Тернопольская обл., Залещицкий район; 160. Тернопольская обл., с. Бабинцы; 161. Полтавская обл., с. Жуковки; 164. Окр. г. Полтавы; 163. Полтавская обл., Пирятинскпй район; 164. Одесская обл., Беляевский район, с. Троицкое; 165. Молдавская ССР, Тираспольский район, с. Ближний хутор (ИЗАНУ); 166. Молдавская ССР, Унгенский район, г. Корнешты; 167. Николаевская обл., Первомайский район, с. Мигия; 168. Винницкая обл., берег р. Деснянки; 169. Волынская обл., шоссе Радехов—-Луцк; 170. Волынская обл., Ковельский район, с. Голобы (ИБР); 171. Закарпатская обл., Виноградовский район, с. Черный поток; 172. Закарпатская обл., Виноградовский район, с. Вербовец; 173. Закарпатская обл., Великоберезнянский р-н, с. Кострпно; 174. Закарпатская обл., Виноградовский район, с. Черна; 175. Закарпатская обл., Воловецкий район (М. И. Щербань).
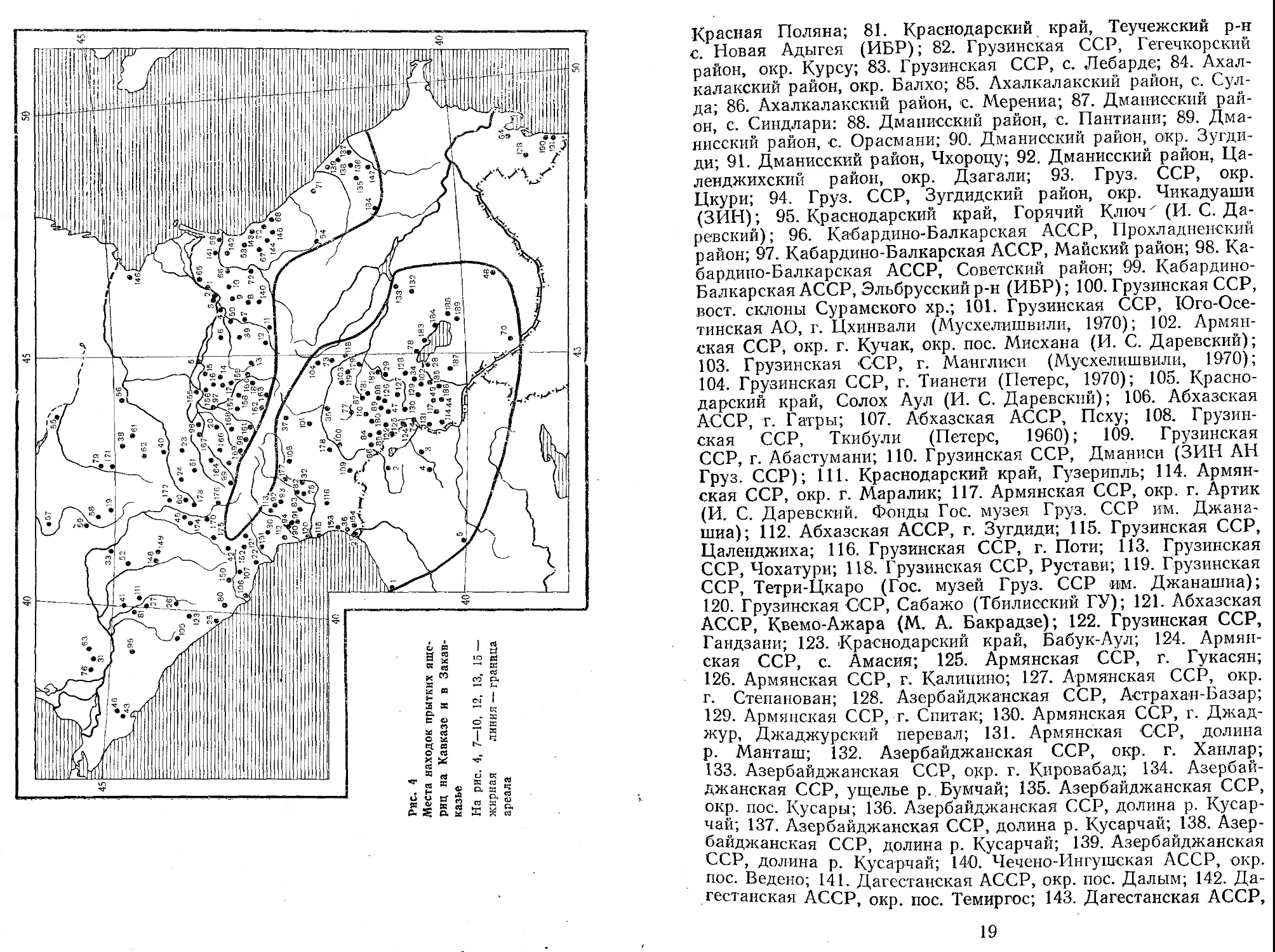
Северный Кавказ, Закавказье, Северо-Восточная Турция (рис. 4)
1—4. Чечено-Ингушская АССР, район ст. Старогладковской, Шелковской, Новощедринской; 5—6. Чечено-Ингушская АССР, Кеньюрт — Толстой Юрт; 7—8. Чечено-Ингушская АССР, Урус — Мартсы — Шам; 9—10. Чечено-Ингушская АССР, район Гудермес; И—13. Чечено-Ингушская АССР, Балсу — Мурже-ни —Сурхахи; 14—15. Чечено-Ингушская АССР, Серноводск — Гарагорски; 16—17. Чечено-Ингушская АССР, Карабулик— Пседах (Н. А. Рашкевич); 18. Северо-Осетинская АССР, окр.
16
г. Орджоникидзе; 19. Ставропольский край, окр. г. Ставрополя; 20. Кабардино-Балкарская АССР, окр. г. Нальчика; 21. Аджарская АССР, окр. г. Батуми; 22. Абхазская АССР, окр. г. Сухуми; 23. Ставропольский край, окр. г. Кисловодска; 24. Ставропольский край, окр. г. Ессентуки; 25. Краснодарский край, окр. г. Адлера; 26. Краснодарский край, Красная Поляна; 27. Кавказский заповедник (ЗИН); 28. Армянская ССР, п-ов Севан (И. С. Даревский); 29. Армянская ССР, Семеновка; 30. Абхазская АССР, окр. г. Ткварчели; 31. Окр. г. Краснодара; 32. Грузинская ССР, окр. г. Кутаиси; 33. Краснодарский край, окр. Армавира; 34. Армянская ССР, окр. Кировакана; 35. Грузинская ССР, окр. г. Гори; 36. Аджарская АССР, с. Кобулети (ЗИН); 37. Грузинская ССР, Юго-Осетинская АО, пос. Джава (И. С. Даревский); 38. Ставропольский край, ст. Александровская; 39. Чечено-Ингушская АССР, берег р. Асса (ЗИН); 40. Ставропольский край, окр. г. Пятигорска; 41. Краснодарский край, окр. г. Майкопа; 42. Ставропольский край, с. Те-бердинет; 43. Краснодарский край, окр. г. Новороссийска; 44. Армянская ССР, окр. Степанаван; 45. Ставропольский край, окр. Усть-Джегутинской; 46. Краснодарский край, Ахтырский перевал; 47. Армянская ССР, Гукасянский район, берег о. Арпалнч; 48. Армянская ССР, берег р. Акера, окр. г. Апаран; 49. Армянская ССР, подножье горы Арагац (МГУ); 50. Чечено-Ингушская АССР, окр. г. Грозного (Хозацкий, 1948, 1950); 51. Ставропольский край, окр. г. Минеральные Воды (ХГУ, по сообщ. В. И. Ведмедеря); 52. Краснодарский край, г. Лабинск (Никольский, 1913); 53. Дагестанская АССР, окр. г. Махачкалы (В. И. Ведмедеря). 54. Дагестанская АССР, Кумух (по сообщ. Г. П. Лукиной); 55. Ставропольский край, с. Дивное; 56. Ставропольский край, т. Прикумск; 57. Ставропольский край, с. Красногвардейское; 58. Ставропольский край, Изобильный; 59. Ставропольский край, Новоалександровская; 60. Ставропольский край, Невинномыск; 61. Ставропольский край, с. Благодарное; 62. Ставропольский край, Александровское (М. Ф. Тертышников); 63. Краснодарский край, станица Дин-ская (Г. П. Лукин); 64. Азербайджанская ССР, о-в Сара (А. М. Алекперов); 65. Дагестанская АССР, Кизляр; 66. Дагестанская АССР, т. Хасавюрт; 67. Дагестанская АССР, г. Буй-накс; 68. Дагестанская АССР, Урари, 69. Дагестанская АССР, устье р. Сулак (3. П. Хонякина); 70. Нахичеванская АССР, оз. Эдильаг (И. С. Даревский); Низовья р. Самур (3. И. Хонякина); 72. Дагестанская АССР, окр. г. Ботлих; 73. Грузинская ССР, окр. г. Тбилиси; 74. Армянская ССР, окр. г. Ленинакан; 75. Грузинская ССР, окр. г. Чегечкори (дорога на Ле-барде) (ИБР); 76. Краснодарский край, ст. Елизаветинская (И. С. Даревский); 77. Грузинская ССР, г. Цалка; 78. Армянская ССР, Севанский район, с. Еленовка; 79. Шоссе Ставцополь — Элиста; 80. Краснодарский край, шоссе Адлер —
Й7) Т г' A J
Красная Поляна; 81. Краснодарский край, Теучежский р-н с. Новая Адыгея (ИБР); 82. Грузинская ССР, Гегечкорский район, окр. Курсу; 83. Грузинская ССР, с. Лебарде; 84. Ахал-калакский район, окр. Балхо; 85. Ахалкалакский район, с. Сул-да; 86. Ахалкалакский район, с. Мерениа; 87. Дманисский район, с. Синдлари: 88. Дманисский район, с. Пантиани; 89. Дманисский район, с. Орасмани; 90. Дманисский район, окр. Зугдиди; 91. Дманисский район, Чхороцу; 92. Дманисский район, Ца-ленджихский район, окр. Дзагали; 93. Груз. ССР, окр. Цкури; 94. Груз. ССР, Зугдидский район, окр. Чикадуаши (ЗИН); 95. Краснодарский край, Горячий Ключ'ДИ. С. Даревский); 96. Кабардино-Балкарская АССР, Прохладненский район; 97. Кабардино-Балкарская АССР, Майский район; 98. Кабардино-Балкарская АССР, Советский район; 99. Кабардино-Балкарская АССР, Эльбрусский р-н (ИБР); 100. Грузинская ССР, вост, склоны Сурамского хр.; 101. Грузинская ССР, Юго-Осетинская АО, г. Цхинвали (Мусхелишвили, 1970); 102. Армянская ССР, окр. г. Кучак, окр. пос. Мисхана (И. С. Даревский); 103. Грузинская ССР, г. Манглиси (Мусхелишвили, 1970); 104. Грузинская ССР, г. Тианети (Петерс, 1970); 105. Краснодарский край, Солох Аул (И. С. Даревский); 106. Абхазская АССР, г. Гагры; 107. Абхазская АССР, Псху; 108. Грузинская ССР, Ткибули (Петерс, 1960); 109. Грузинская
ССР, г. Абастумани; ПО. Грузинская ССР, Дманиси (ЗИН АН Груз. ССР); 111. Краснодарский край, Гузерипль; 114. Армянская ССР, окр. г. Маралик; 117. Армянская ССР, окр. г. Артик (И. С. Даревский. Фонды Гос. музея Груз. ССР им. Джана-шиа); 112. Абхазская АССР, г. Зугдиди; 115. Грузинская ССР, Цаленджиха; 116. Грузинская ССР, г. Поти; 113. Грузинская ССР, Чохатури; 118. Грузинская ССР, Рустави; 119. Грузинская ССР, Тетри-Цкаро (Гос. музей Груз. ССР им. Джанашиа); 120. Грузинская ССР, Сабажо (Тбилисский ГУ); 121. Абхазская АССР, Квемо-Ажара (М. А. Бакрадзе); 122. Грузинская ССР, Гандзани; 123. Краснодарский край, Бабук-Аул; 124. Армянская ССР, с. Амасия; 125. Армянская ССР, г. Гукасян; 126. Армянская ССР, г. Калинино; 127. Армянская ССР, окр. г. Степанован; 128. Азербайджанская ССР, Астрахан-Базар; 129. Армянская ССР, г. Спитак; 130. Армянская ССР, г. Джад-жур, Джаджурский перевал; 131. Армянская ССР, долина р. Манташ; 132. Азербайджанская ССР, окр. г. Ханлар; 133. Азербайджанская ССР, окр. г. Кировабад; 134. Азербайджанская ССР, ущелье р. Бумчай; 135. Азербайджанская ССР, окр. пос. Кусары; 136. Азербайджанская ССР, долина р. Кусар-чай; 137. Азербайджанская ССР, долина р. Кусарчай; 138. Азербайджанская ССР, долина р. Кусарчай; 139. Азербайджанская ССР, долина р. Кусарчай; 140. Чечено-Ингушская АССР, окр. пос. Ведено; 141. Дагестанская АССР, окр. пос. Далым; 142. Дагестанская АССР, окр. пос. Темиргос; 143. Дагестанская АССР,
19
окр. пос. Манас-Аул; 144. Дагестанская АССР, окр. пос. Кути-ша; 145. Дагестанская АССР, окр. пос. Тинит; 146. Калмыцкая АССР, низовья р. Кумы; 147. Азербайджанская ССР, окр. г. Шемаха; 148. Краснодарский крап, Псебай; 149. Краснодарский край, долина р. Б. Лаба; 150. Абхазская АССР, перевал Анчха; 151. Абхазская АССР, ущелье р. Амткел; 152. Абхазская АССР, ущелье р. Лата; 153. Аджарская АССР, Чаква; 154. Аджарская АССР, Кеда; 155. Северо-Осетинская АССР, Моздок; 156. Северо-Осетинская АССР, Чиколя, долина р. Урух; 158. Северо-Осетинская АССР, Ардон; 159. Северо-Осетинская АССР, Оль-гинское; 160. Северо-Осетинская АССР, Тарское; 161. Северо-Осетинская АССР, Ахсарисар; 162. Северо-Осетинская АССР, долина р. Фиагдон; 163. Северо-Осетинская АССР, Балта, ущелье' р. Терек; 164. Кабардино-Балкарская АССР, верх, р. Малки; 165. Кабардино-Балкарская АССР, Терскол, ущелье Баксана; 166. Кабардино-Балкарская АССР, Тырны—Ауз, ущелье Баксана; 167. Кабардино-Балкарская АССР, Баксан; 168. Кабардино-Балкарская АССР, г. Ндрткала (Докшуки-ко); 169. Кабардино-Балкарская АССР, ущелье р. Черек; 170. Ставропольский край, окр. Зеленчукскип; 171. Ставропольский край, долина р. Калаус; 172. Ставропольский край, окр. г. Черкесск; 173. Ставропольский край, окр. г. Учкекен; 174. Ставропольский край, ст. Кардаманская; 175. Ставропольский край, г. Карачаевск; 176. Ставропольский край, Учкелен, верховья р. Кубань; 177. Грузинская ССР, окр. г. Риони; 178. Грузинская ССР, Сурамский перевал; 179. Грузинская ССР, Коджори; 180. Грузинская ССР, окр. пос. Богдановка; 181. Грузинская ССР, окр. пос. Ахалкалаки; 182. Армянская ССР, склоны г. Лальвар, окр. г. Калинино; 183. Армянская । АССР, окр. пос. Шорджа; 184. Армянская АССР, берег оз. Севан; 185. Армянская ССР, окр. г. Раздан; 186. Армянская ССР, окр. г. Фантан; 187. Армянская СССР, верховья р. Ведичай; 188. Армянская СССР, окр. пос. Басартечар; 189. Армянская ' ССР, окр. пос. Загалу; 190. Азербайджанская ССР, окр. Ленкорани (?); 191. Азербайджанская ССР, Шах-Агач (?) (И. С. Даревский) .
Северо-Восточная Турция: 1. Трабзон; 2. Ардаган; 3. Сары-камыш; 4. Карс; 5. Эрзерум (И. С. Даревский).
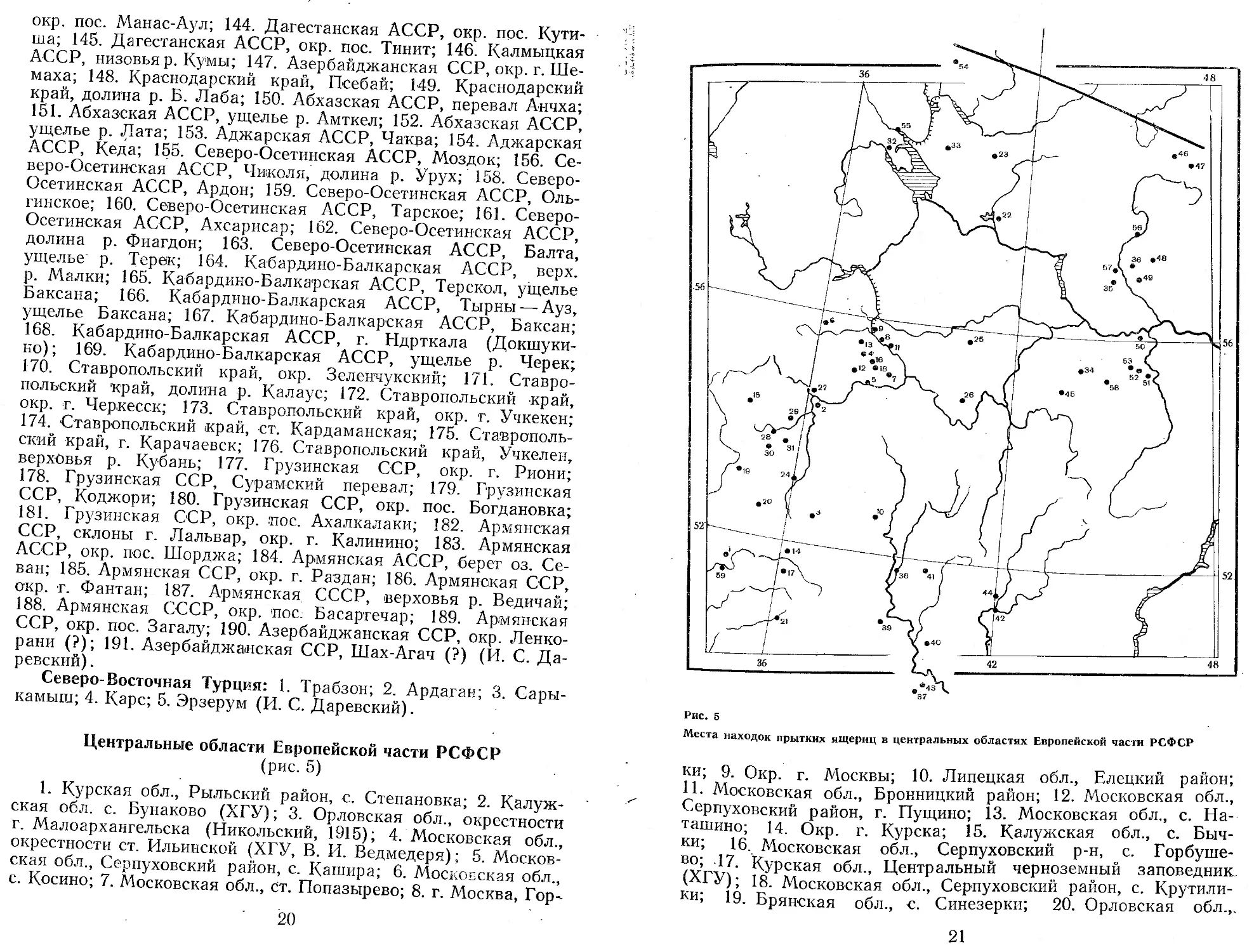
Центральные области Европейской части РСФСР (рис. 5)
1. Курская обл., Рыльский район, с. Степановка; 2. Калужская обл. с. Бунаково (ХГУ); 3. Орловская обл., окрестности । г. Малоархангельска (Никольский, 1915); 4. Московская обл., окрестности ст. Ильинской (ХГУ, В. И. Ведмедеря); 5. Московская обл., Серпуховский район, с. Кашира; 6. Московская обл., с. Косино; 7. Московская обл., Ст. Попазырево; 8. г. Москва, Гор-
20
Рис. 5
Места находок прытких ящериц в центральных областях Европейской части РСФСР
ки; 9. Окр. г. Москвы; 10. Липецкая обл., Елецкий район; 11. Московская обл., Бронницкий район; 12. Московская обл., Серпуховский район, г. Пущино; 13. Московская обл., с. Наташино; 14. Окр. г. Курска; 15. Калужская обл., с. Бычки; 16. Московская обл., Серпуховский р-н, с. Горбуше-во; 17. Курская обл., Центральный черноземный заповедник (ХГУ); 18. Московская обл., Серпуховский район, с. Крутили-ки; 19. Брянская обл., с. Синезерки; 20. Орловская обл.,.
21
•с. Дмитровичи; 21. Белгородская обл., с. Борисовка; 22. Окрестности г. Костромы; 23. Вологодская обл., с. Липин Бор (ЗИН); 24. Окрестности г. Орла (Никольский, 1915); 25. Владимирская обл. (Жаркова, 1973); 26. Рязанская обл., Окский заповедник (Стрельцов, Воронин, 1973); 27. Окрестности г. Калуги; 28. Калужская обл., Думиничский район; 29. Калужская обл., Козельский район; 30. Калужская обл., Хвастовичский район; 31. Калужская обл., Ульяновский район; 32. Вологодская обл., дер. Борок; 33. Вологодская обл., Вожегодский р-н, с. Бекетовское; 34. Горьковская обл., Арзамаский р-н, с. Старая пустынь; 35. Горьковская обл., с. Сухобездное; 36. Горьковская обл., с. Ше-маниха (ИБР); 37. Воронежская обл., Таловский р-н; 38. Окр. г. Воронежа; 39. Воронежская обл., Острогожский р-н; 40. Воронежская обл., Воронцовский колхоз (ЗИН); 41. Воронежская обл., с. Перелешино (МГУ); 42. Воронежская обл., Борисоглебский р-н; 43. Воронежская обл., г. Верхний Мамон; 44. Воронежская обл., Борисоглебский р-н (ИБР); 45. Горьковская обл., Ардатовский р-н; 46. Костромская обл., Шарьинский р-н; 47. Костромская обл., с. Нонадырево; 48. Горьковская обл., с. Воскресенское; 49. Горьковская обл., Семеновский р-н; 50. Горьковская обл., Воротынский р-н, пос. Васильсурск; 51. Горьковская обл., Пильненский р-н; 52. Горьковская обл., Сергачский р-н; 53. Горьковская обл., Чкаловский р-н, пос. Каунки (В. И. Гаранин) ; 54. Вологодская обл., с. Огибалово; 55. Вологодская обл..
I с. Суда; 56. Горьковская обл., с. Урень; 57. Горьковская обл. с. Красн. Баки; 58. Горьковская обл., окр. г. Лукоянов (ИБР), 59. Курская обл., заповедник «Лес на Ворскле» (А. М. Андрушко) .
Нижнее и Среднее Поволжье и Ростовская обл.
(рис. 6)
1. Окр. г. Астрахани; 2. Саратовская обл., с. Балашове (ЗИН); 3. Ростовская обл., пос. Зерновой (И. С. Даревский); 4. Окр. г. Новочеркасска; 5. Жигулевский заповедник; 6. Окр. г. Ростова-на-Дону; 7. Пензенская обл., заповедник «Сосновый -бор»; 8. Ростовская обл., с. Романовское (ЗИН); 9. Волгоградская обл., Михайловский р-н, лесхоз Глинище, правый и левый берег р. Медведицы (Марков, Косарева, Кубанец, 1969); 10. Ростовская обл., Сальский р-н, с. Федосеевка; И. Астраханский заповедник; 12. Ростовская обл., Сальский р-н, г. Атаманск; 13. Окр. г. Таганрога; 1^. Мордовская АССР, Мордовский запо-। веднпк; 15. Пензенская обл., Городищенский р-н (МГУ); 16. Ростовская обл., с. Журавлевка (И. С. Даревский); 17. Окр. г. Ульяновска (МГУ); 18. Волгоградская обл., бер. Волго-Донского канала (Марков, Косарева, Кубанцев, 1969); 19. Куйбышевская обл., окр. г. Сызрань; 20. Пензенская обл., Кузнецкий р-н; 21. Кал-1 мыцкая АССР, с. Смушковое (ЗИН); 22. Астраханская обл.,
22
с. Черный Яр (В. И. Ведмедеря); 23. Ростовская обл., с. Богородское; 24. Ростовская обл., с. Красное поле (Г. П. Лукина); 25. Ростовская обл., с. Аксае (ЗИН); 26. Волгоградская обл., Палласовскнй р-н; 27. Волгоградская обл., Палласовский р-н, с. Тараново; 28. Саратовская обл., г. Петровский; 29. Пензенская обл., с. Владыкино; 30. Саратовская обл., с. Котоврас (ИБР); 31. Камыш-Самарские озера (Чернов, 1954); 32. Куйбышевская обл., Сызранский р-н, берег р. Уса; 33. Куйбышевская обл., г. Тольяти; 3^. Мордовская АССР, окр. г. Саранск; 35. Мордовская АССР, Ардатовский р-н; 36. Куйбышевская обл., Динель-Черкасский р-н; 37. Куйбышевская обл., окр. с. Сергеевен; 38. Окр. г. Куйбышева; 39. Ульяновская обл., Чердаклпн-ский р-н; 40. Ульяновская обл., Сенгилейский р-н; 41. Ульяновская обл., Теренгульский р-н; 42. Ульяновская обл., Карсунский р-н; 43. Ульяновская обл., Ново-Спасский р-н; 44. Ульяновская обл., окр. г. Мелекесс; 45. Пензенская обл., Никольский р-н; 46. Пензенская обл., Кадодинское лесничество; 47. Саратовская обл., Хвалынский р-н; 48. Саратовская обл., Пугачевский р-н; 49. Саратовская обл., с. Воскресенское (В. И. Гаранин); 50. Саратовская обл., г. Красноармейск (б. Серента) (Никольский, 1915); 51. Окр. г. Саратова (Н. Н. Щербак).
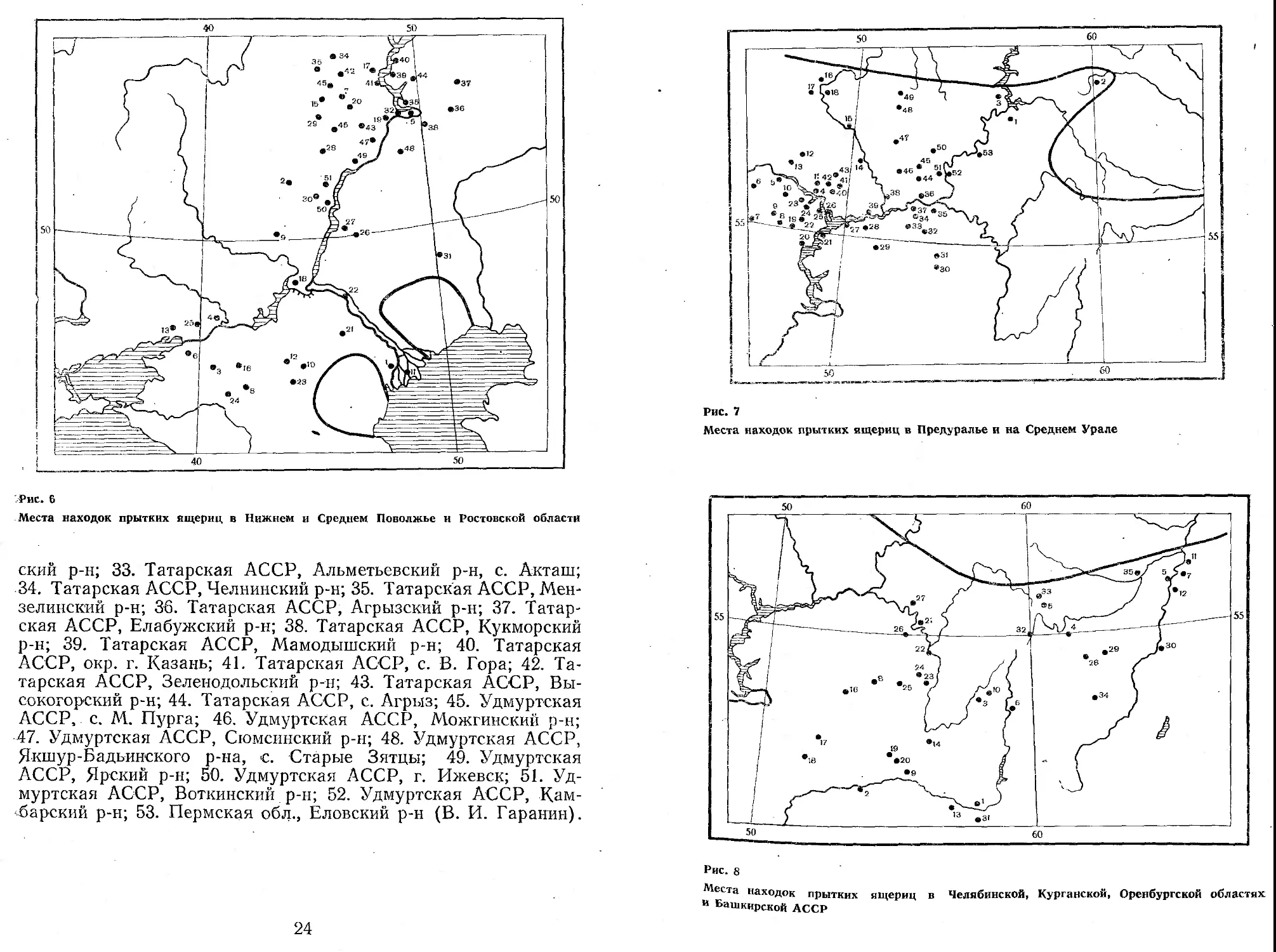
Предуралье и Средний Урал (рис. 7)
1. Окр. г. Пермь, южнее 50° с. ш. (МГУ, Болотников, Хазиева, Литвинов, Чолцин, 1973); 2. Свердловская обл., окр. г. Н. Тагил (Никольский, 1945); 3. Пермская обл., Краснокамский р-н (В. И. Гаранин); 4. Татарская АССР, Зеленодольский р-н (Гаранин, 1971); 5. Чувашская АССР, с. Большой Сундырь;. 6. Чувашская АССР, с. Ядрин; 7. Чувашская АССР, Алатырский р-н; 8. Чувашская АССР, Шимерлинский р-н; 9. Чувашская АССР,; Комсомольский р-н; 10. Чувашская АССР, Урмарский р-н; 11. Марийская АССР, Волжский р-н; 12. Марийская АССР, Йошкар-Олинский р-н; 13. Марийская АССР, Медведевский р-н; 14. Кировская обл., Малмыжекский р-н; 15. Кировская обл., с. Медвежий Бор; 16. Кировская обл., окр. г. Халтурина; 17. Кировская обл., Котельничский р-н; 18. Кировская обл., Оричев-ский р-н; 19. Татарская АССР, Гивинский р-н; 20. Татарская АССР, Тетюшский р-н; 21. Татарская АССР, Куйбышевский р-н;. 22. Татарская АССР, Аплетовский р-н; 23. Татарская АССР, Зеленодольский р-н; 24. Татарская АССР, окр. г. Верхний Ус-л°н; 25. Татарская АССР, Камско-Устьинскпй р-н; 26. Татарская АССР, Лайшевский р-н; 27. Татарская АССР, Алексеевский р-Н; 28. Татарская АССР, Чистопольский р-н, с. Кутлушки-Дсг^' барская АССР, Черепшанский р-н; 30. Татарская АССР, Бавлинский р-н, с. Урдалы-Тай; 31. Татарская АССР, Азнакаевский р-н, с. Четыр-Тан; 32. Татарская АССР, Муслюмов-
23
Рис. 6
Места находок прытких ящериц в Нижнем и Среднем Поволжье и Ростовской области
ский р-н; 33. Татарская АССР, Альметьевский р-н, с. Акташ; 34. Татарская АССР, Челнинский р-н; 35. Татарская АССР, Мен-зелинский р-н; 36. Татарская АССР, Агрызский р-н; 37. Татарская АССР, Елабужский р-н; 38. Татарская АССР, Кукморский р-н; 39. Татарская АССР, Мамодышский р-н; 40. Татарская АССР, окр. г. Казань; 41. Татарская АССР, с. В. Гора; 42. Татарская АССР, Зеленодольский р-н; 43. Татарская АССР, Высокогорский р-н; 44. Татарская АССР, с. Агрыз; 45. Удмуртская АССР, с. М. Пурга; 46. Удмуртская АССР, Можгинский р-н; 47. Удмуртская АССР, Сюмспнский р-н; 48. Удмуртская АССР, Якшур-Бадьинского р-на, с. Старые Зятцы; 49. Удмуртская АССР, Ярский р-н; 50. Удмуртская АССР, г. Ижевск; 51. Удмуртская АССР, Воткинский р-н; 52. Удмуртская АССР, Кам-барский р-н; 53. Пермская обл., Еловский р-н (В. И. Гаранин).
24
Рис. 7
Места находок прытких ящериц в Предуралье и на Среднем Урале
Рис. 8
Места находок прытких ящериц в Челябинской, Курганской, Оренбургской областях и Башкирской АССР
риц в Средней и Центральной Азии и в Центральном и Восточном Казахстане (восточнее 64° в. д.)
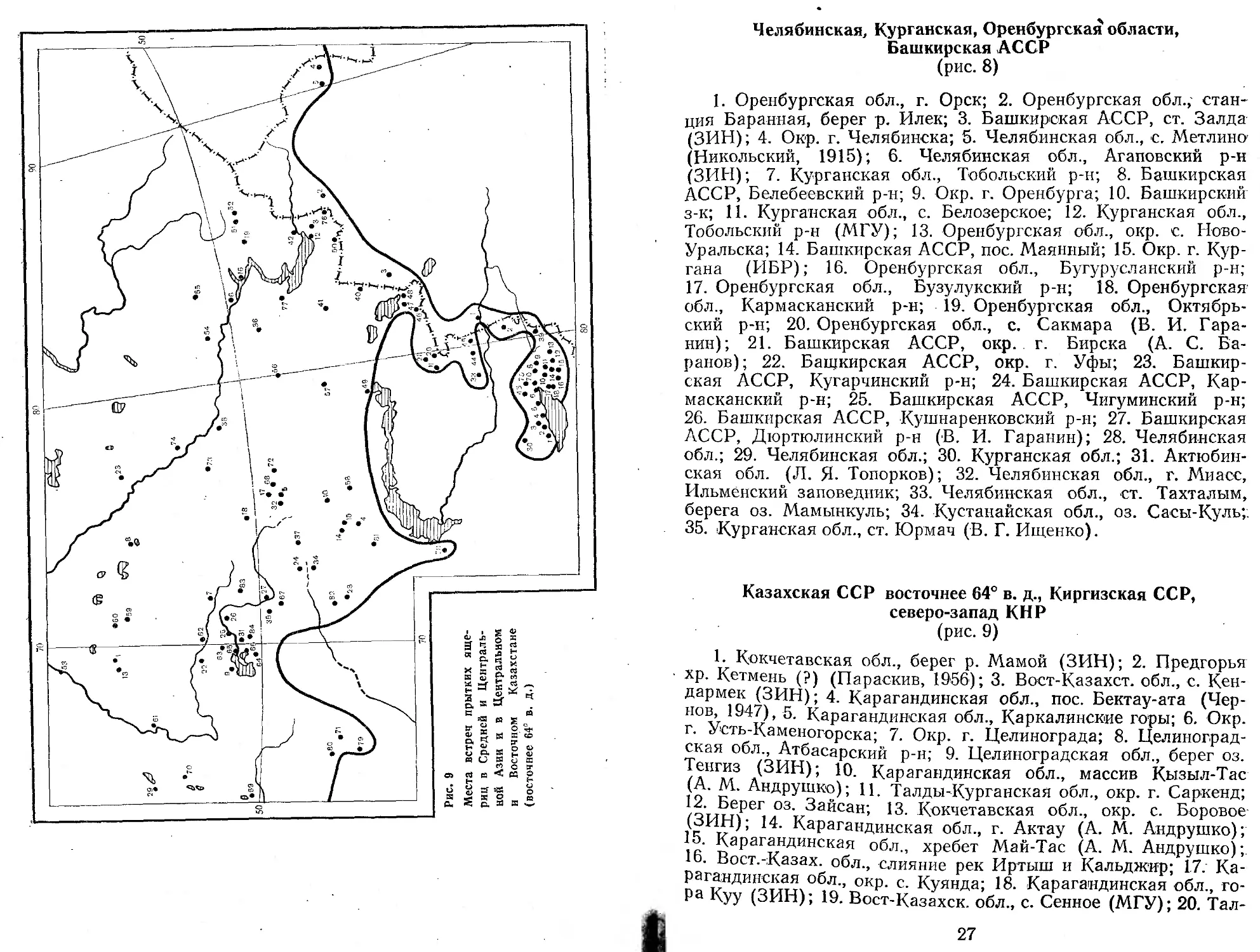
Челябинская, Курганская, Оренбургская области, Башкирская АССР (рис. 8)
1. Оренбургская обл., г. Орск; 2. Оренбургская обл., станция Баранная, берег р. Илек; 3. Башкирская АССР, ст. Залда (ЗИН); 4. Окр. г. Челябинска; 5. Челябинская обл., с. Метлино (Никольский, 1915); 6. Челябинская обл., Агаповский р-н (ЗИН); 7. Курганская обл., Тобольский р-н; 8. Башкирская АССР, Белебеевский р-н; 9. Окр. г. Оренбурга; 10. Башкирский з-к; 11. Курганская обл., с. Белозерское; 12. Курганская обл., Тобольский р-н (МГУ); 13. Оренбургская обл., окр. с. Ново-Уральска; 14. Башкирская АССР, пос. Маянный; 15. Окр. г. Кургана (ИБР); 16. Оренбургская обл., Бугурусланский р-н; 17. Оренбургская обл., Бузулукский р-н; 18. Оренбургская обл., Кармасканский р-н; 19. Оренбургская обл., Октябрьский р-н; 20. Оренбургская обл., с. Сакмара (В. И. Гаранин); 21. Башкирская АССР, окр. г. Бирска (А. С. Баранов); 22. Башкирская АССР, окр. г. Уфы; 23. Башкирская АССР, Кугарчинский р-н; 24. Башкирская АССР, Кармасканский р-н; 25. Башкирская АССР, Чигуминский р-н; 26. Башкирская АССР, Кушнаренковский р-н; 27. Башкирская АССР, Дюртюлинский р-н (В. И. Гаранин); 28. Челябинская обл.; 29. Челябинская обл.; 30. Курганская обл.; 31. Актюбинская обл. (Л. Я. Топорков); 32. Челябинская обл., г. Миасс, Ильменский заповедник; 33. Челябинская обл., ст. Тахталым, берега оз. Мамынкуль; 34. Кустанайская обл., оз. Сасы-Куль;. 35. Курганская обл., ст. Юрмач (В. Г. Ищенко).
Казахская ССР восточнее 64° в. д., Киргизская ССР, северо-запад КНР (рис. 9)
1. Кокчетавская обл., берег р. Мамой (ЗИН); 2. Предгорья хр. Кетмень (?) (Параскив, 1956); 3. Вост-Казахст. обл., с. Кен-дармек (ЗИН); 4. Карагандинская обл., пос. Бектау-ата (Чернов, 1947), 5. Карагандинская обл., Каркалинские горы; 6. Окр. г. Усть-Каменогорска; 7. Окр. г. Целинограда; 8. Целиноградская обл., Атбасарский р-н; 9. Целиноградская обл., берег оз.
/ЗИН); 10. Карагандинская обл., массив Кызыл-Тас 19 К Андрушко); И. Талды-Курганская обл., окр. г. Саркенд; !'чыг.п)еГ °3', Ззйсан; 13. Кокчетавская обл., окр. с. Боровое iKTz ’ КаРаганДинская обл., г. Актау (А. М. Андрушко); 1R Карагандинская обл., хребет Май-Тас (А. М. Андрушко); о. Вост.-Казах. обл., слияние рек Иртыш и Кальджир; 1.7. Карагандинская обл., окр. с. Куянда; 18. Карагандинская обл., гора Куу (ЗИН); 19. Вост-Казахск. обл., с. Сенное (МГУ); 20. Тал-
27
ды-Курганская обл., отроги Джунгарского Алатау (ЗИН); 21. Талды-Курганская обл., окр. г. Текелн (ИЗАНУ); 22. Целиноград, обл., окр. с. Майдан; 23. Павлодарская обл., Кулундин-ская степь (МГУ); 24. Карагандинская обл., окр. с. Жарык (А. С. Баранов); 25. Целиноградская обл., берег р. Ишим; 26. Целиноградская обл., с. Сабунды; 27. Окр. с. Казгородок, берег р. Нуры (ЗИН); 28. Карагандинская обл., окр. пос. Атасу (А. С. Баранов); 29. Кустанайская обл., с. Семиозерное; 30. Алма-Атинская обл., берег р. Алма-Атинка (ЗИН); 31. Целиноградская обл., берег оз. Кургальджин (МГУ); 32. Карагандинская обл., шоссе Караганда-Каркаралинск, не доезжая с. Петровка (А. С. Баранов); 33. Талды-Курганская обл., отроги хр. Алтынэмель; 34. Карагандинская обл., окр. пос. Успенский; 35. Карагандинская обл., берег р. Кулантпес (ЗИН); 36. Семипалатинская обл., ст. Георгиевка; 37. Карагандинская обл., окр. г. Узынбулак (МГУ); 38. Павлодарская обл., берег р. Ши-дерты (А. С. Баранов); 39. Алма-Атинская обл., с. Нарынкол; 40. Семипалатинская обл., окр. ст. Бахгы; 41. Семипалатинская обл., окр. дер. Базарка; 42. Вост.-Казах. обл., долина р. Черный Иртыш близ оз. Зайсан (Кащенко, 1909); 43. Алма-Атинская обл., верховье р. Чилик; 44. Талды-Курганская обл., отроги Джунгарского Алатау; 45. Талды-Курганская обл., отроги Джунгарского Алатау; 46. Талды-Курганская обл., оз. Баскан; 47. Талды-Курганская обл., берег оз. Алаколь; 48. Талды-Курганская обл., берег оз. Алаколь; 49. Северо-восточный берег оз. Балхаш; 50. Семипалатинская обл., отроги хребта Тарбагатай; 51. Вост.-Казах. обл., отроги Алтая; 52. Вост.-Казах. обл., отроги Алтая; 53. Северо-Казах. обл., окр. г. Петропавловска; 54. Вост.-Казах. обл.; 55. Вост.-Казах. обл.; 56. Семипалатинская обл., 57. Семипалатинская обл., хребет Чингеу-Тау; 58. Карагандинская обл., 59. Кокчетавская обл.; 60. Кокчетавская обл.; 61. Кокчетавская обл., окр. с. Рузаевка; 62—67. Пункты по карте К. П. Параскива (1956) точно не установлены; 68. Карагандинская обл., берег р. Тундык, шоссе на Егиндыбу-лак (А. С. Баранов); 69. Кустанайская обл., оз. Сарыкопа (Ди-несман, 1953); 70. Кустанайская обл., Наурзумскпй заповедник; 71. Карагандинская обл., р. Бала-Джезде (Чельцов-Бебутов, 1953); 72. Карагандинская обл., шоссе Егиндыбулак-Киикашкоп; 73. Павлодарская обл., 39 км по шоссе Баян — Аул—Павлодар; 74. Павлодарская обл., окр. г. Ермак (А. С. Баранов); 75. Тюп-ский район, хр. Кунгей-Алатау, Курменту; 76. Семипалатинская обл., хр. Саур; 77. Вост.-Казах. обл., хр. Калбинский Алтай; 78. Восточная Бетпак-Дала 45° 20' с. ш.; 79. Карагандинская обл., окр. г. Байканур; 80. Карагандинская обл., окр. г. Бай-канур; 81. Карагандинская обл., окр. ст. Мопнты; 82. Карагандинская обл., близ г. Актау; 83. Целинный край; 84. Целинный край (Параскив, 1956).
Кроме того, не установлены местонахождения следующих
28
пунктов по К- П. Параскиву (1956): верх р. Джиланчик, гора Булат-Тау и лесной массив Кара-Агач.
Киргизская ССР: 1—17 по И. Д. Яковлевой (1964), в том числе 4 окр. с. Григорьевка; 7-—перевал Курменты; 14—окр. г. Пржевальска; 17—окр. с. Покровка; 18—окр. с. Безбешик.
КНР: 1. Окр. г. Кульджа (ИЗАНУ); 2. К западу от озера Улюнгур; 3. Хр. Барлык; 4. Хр. Монгольский Алтай, берег р. Чин-гиль (Кащенко, 1909*), 5. Верховья р. Урунгу (Никольский, 1915).
Тюменская, Томская, Омская, Новосибирская, Кемеровская, Иркутская обл.,
Красноярский край, Алтайский край, Тувинская АССР (рис. 10)
1. Красноярский край, Сев.-Енис. р-н, с. Брянка (И. С. Даревский); 3. Окр. г. Иркутска (МГУ); 4. Иркутская обл. (Ю. Г. Швецов); 5. Алтайский край, с. Кашино; 6. Красноярский край, окр. г. Минусинска; 7. Алтайский край, окр. г. Павловска; 8. Алтайский край, окр. г. Горноалтайска; 9. Алтайский край, Троицкий р-н, с. Бедово (МГУ); 10. Алтайский край, окр. г. Барнаула; 11. Тувинская АССР, с. Донгул; 12. Окр. Омска; 13. Окр. г. Змеиногорска; 14. Алтайский край, Чуйская степь; 15. Окр. г. Томска; 16. Алтайский край, с. Ниж. Уймон; 17. Красноярский край, берег р. Мана; 18. Алтайский край, дер. Балэ, берег Телецкого озера; 19. Алтайский край, Змеино-горский р-н, с. Сростинское; 20. Алтайский край, с. Чемал; 21. Алтайский край, окр. г. Чуйска (ЗИН); 22. Иркутская обл., берег р. Иркут (Ю. Г. Швецов, И. С. Даревский); 23. Новосибирская обл., с. Ельцовка; 24. Красноярский край, ст. Иланская; 25. Алтайский край, берег Телецкого озера (ЗИН); 26. Иркутская обл., Жигаловский р-н, с. Якимовка (Воронов Г. А.); 27. Красноярский край, окр. Мотыгпно; 28. 50 км севернее г. Иркутска; 29. Иркутская обл., ст. Зима; 30. Иркутская обл., •с. Япды (Демидов, 1973); 31. Красноярский край, окр. г. Канска (Никольский, 1915); 32. Алтайский край, с. Целинное (ИБР); 33. Тувинская АССР, Каа-Хемский р-н, с. Ильинка (ИЗАНУ); 34. .Алтайский край, окр. г. Алейска (ИБР); 36. Алтайский край, хр. Айгулак (Кащенко, 1909); 37. Устье р. Иртыш (ЗИН); 38. Окр. г. Тюмени (Никольский, 1915).
Указывает прыткую ящерицу в Западной Монголии, однако в настоящее время эти пункты поимки животных находятся на территории КНР. Местонахождение 2 пунктов (Каирты, Курту), указанных Н. Ф. Кащенко, не удалось установить.
29
ноярском крае
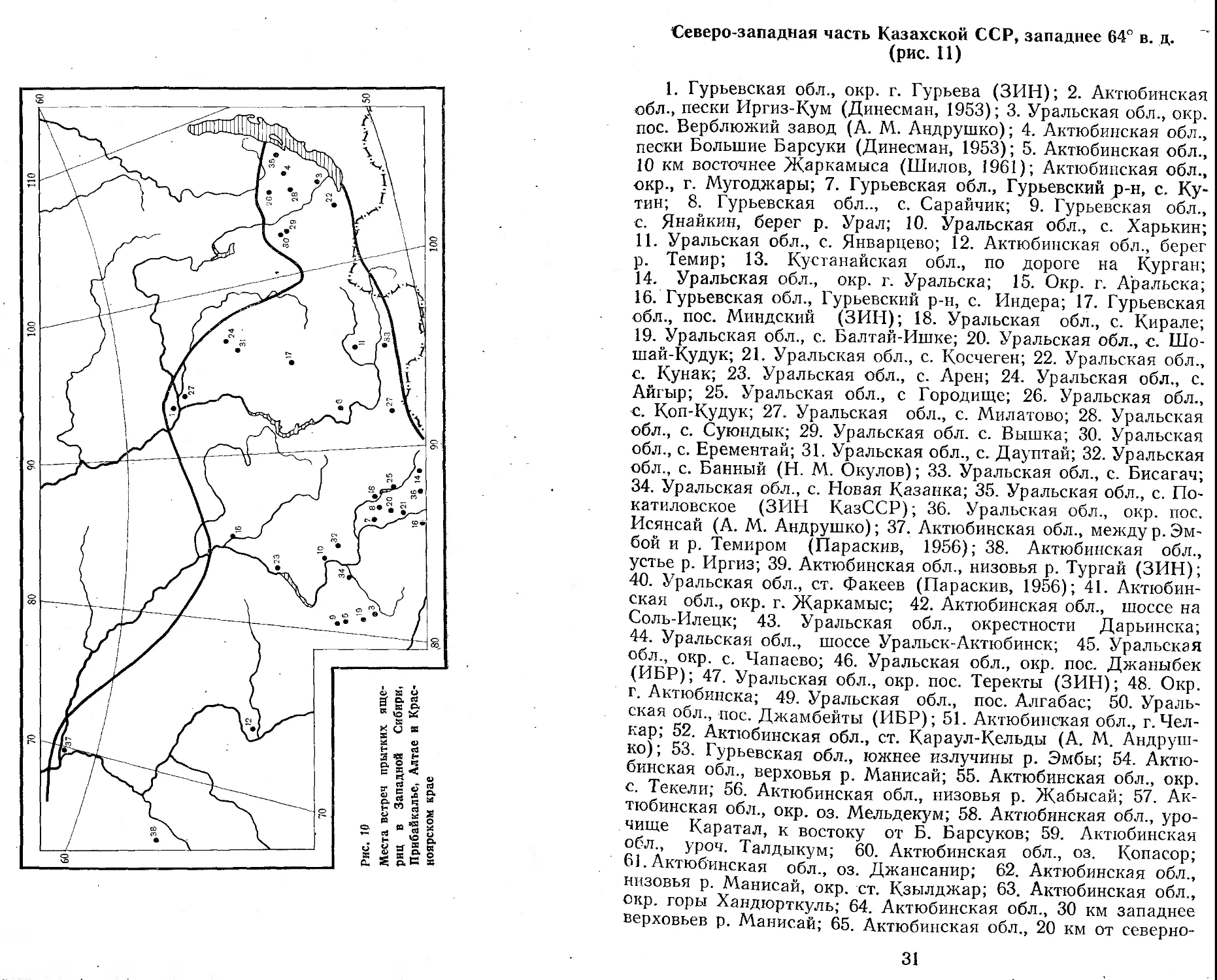
Северо-западная часть Казахской ССР, западнее 64° в. д. (рис. II)
1. Гурьевская обл., окр. г. Гурьева (ЗИН); 2. Актюбинская обл., пески Иргиз-Кум (Динесман, 1953); 3. Уральская обл., окр. пос. Верблюжий завод (А. М. Андрушко); 4. Актюбинская обл., пески Большие Барсуки (Динесман, 1953); 5. Актюбинская обл., 10 км восточнее Жаркамыса (Шилов, 1961); Актюбинская обл., окр., г. Мугоджары; 7. Гурьевская обл., Гурьевский j>-h, с. Кутин; 8. Гурьевская обл.., с. Сарайчик; 9. Гурьевская обл., с. Янайкин, берег р. Урал; 10. Уральская обл., с. Харькин; 11. Уральская обл., с. Январцево; 12. Актюбинская обл., берег р. Темир; 13. Кустанайская обл., по дороге на Курган; 14. Уральская обл., окр. г. Уральска; 15. Окр. г. Аральска; 16. Гурьевская обл., Гурьевский р-н, с. Индера; 17. Гурьевская обл., пос. Миндский (ЗИН); 18. Уральская обл., с. Кирале; 19. Уральская обл., с. Балтай-Ишке; 20. Уральская обл., с. Шо-шай-Кудук; 21. Уральская обл., с. Косчеген; 22. Уральская обл., с. Кунак; 23. Уральская обл., с. Арен; 24. Уральская обл., с. Айгыр; 25. Уральская обл., с Городище; 26. Уральская обл., с. Коп-Кудук; 27. Уральская обл., с. Милатово; 28. Уральская обл., с. Суюпдык; 29. Уральская обл. с. Вышка; 30. Уральская обл., с. Ерементай; 31. Уральская обл., с. Дауптай; 32. Уральская обл., с. Банный (Н. М. Окулов); 33. Уральская обл., с. Бисагач; 34. Уральская обл., с. Новая Казанка; 35. Уральская обл., с. По-катиловское (ЗИН КазССР); 36. Уральская обл., окр. пос. Исянсай (А. М. Андрушко); 37. Актюбинская обл., между р. Эм-бой и р. Темиром (Параскив, 1956); 38. Актюбинская обл., устье р. Иргиз; 39. Актюбинская обл., низовья р. Тургай (ЗИН); 40. Уральская обл., ст. Факеев (Параскив, 1956); 41. Актюбинская обл., окр. г. Жаркамыс; 42. Актюбинская обл., шоссе на Соль-Илецк; 43. Уральская обл., окрестности Дарышска; 44. Уральская обл., шоссе Уральск-Актюбинск; 45. Уральская обл., окр. с. Чапаеве; 46. Уральская обл., окр. пос. Джапыбек (ИБР); 47. Уральская обл., окр. пос. Теректы (ЗИН); 48. Окр. г. Актюбинска; 49. Уральская обл., пос. Алгабас; 50. Уральская обл., пос. Джамбейты (ИБР); 51. Актюбинская обл., г. Чел-кар; 52. Актюбинская обл., ст. Караул-Кельды (А. М. Андрушко); 53. 1урьевская обл., южнее излучины р. Эмбы; 54. Актюбинская обл., верховья р. Манисай; 55. Актюбинская обл., окр. с. Текели; 56. Актюбинская обл., низовья р. Жабысай; 57. Актюбинская обл., окр. оз. Мельдекум; 58. Актюбинская обл., урочище Каратал, к востоку от Б. Барсуков; 59. Актюбинская обл., уроч. Талдыкум; 60. Актюбинская обл., оз. Копасор; 6].Актюбинская обл., оз. Джансанир; 62. Актюбинская обл., низовья р. Манисай, окр. ст. Кзылджар; 63. Актюбинская обл., окр. горы Хандюрткуль; 64. Актюбинская обл., 30 км западнее верховьев р. Манисай; 65. Актюбинская обл., 20 км от северно-
31
Рис. 11
Места встреч прыткой ящерицы в северо-западном Казахстане (западнее 64° в. д.)
го чинка Устюрта, ур. Донгузтау; 66—67. Актюбинская обл., долина р. Чакан; 68. Актюбинская обл., горы Чушкакуль, южнее Карабутака; 69. Актюбинская обл., горы Чукуль, севернее Карабутака; 70. Актюбинская обл., верхнее течение р. Мани-сай, с. Шайтас, с. Байсуат; 71. Актюбинская обл., ст. Тобускен, 50 км юго-западнее Жаркамыса; 72. Актюбинская обл., окр. Жар-камыса, пойма ср. теч. Эмбы (Шилов, 1961); 73. Уральская обл., окр. пос. Донецкого; 74. Уральская обл., окр. пос. Владимировка (А. М. Андрушко); 75. Уральская обл., берег р. Улькаян (Ди-несман, 1953).
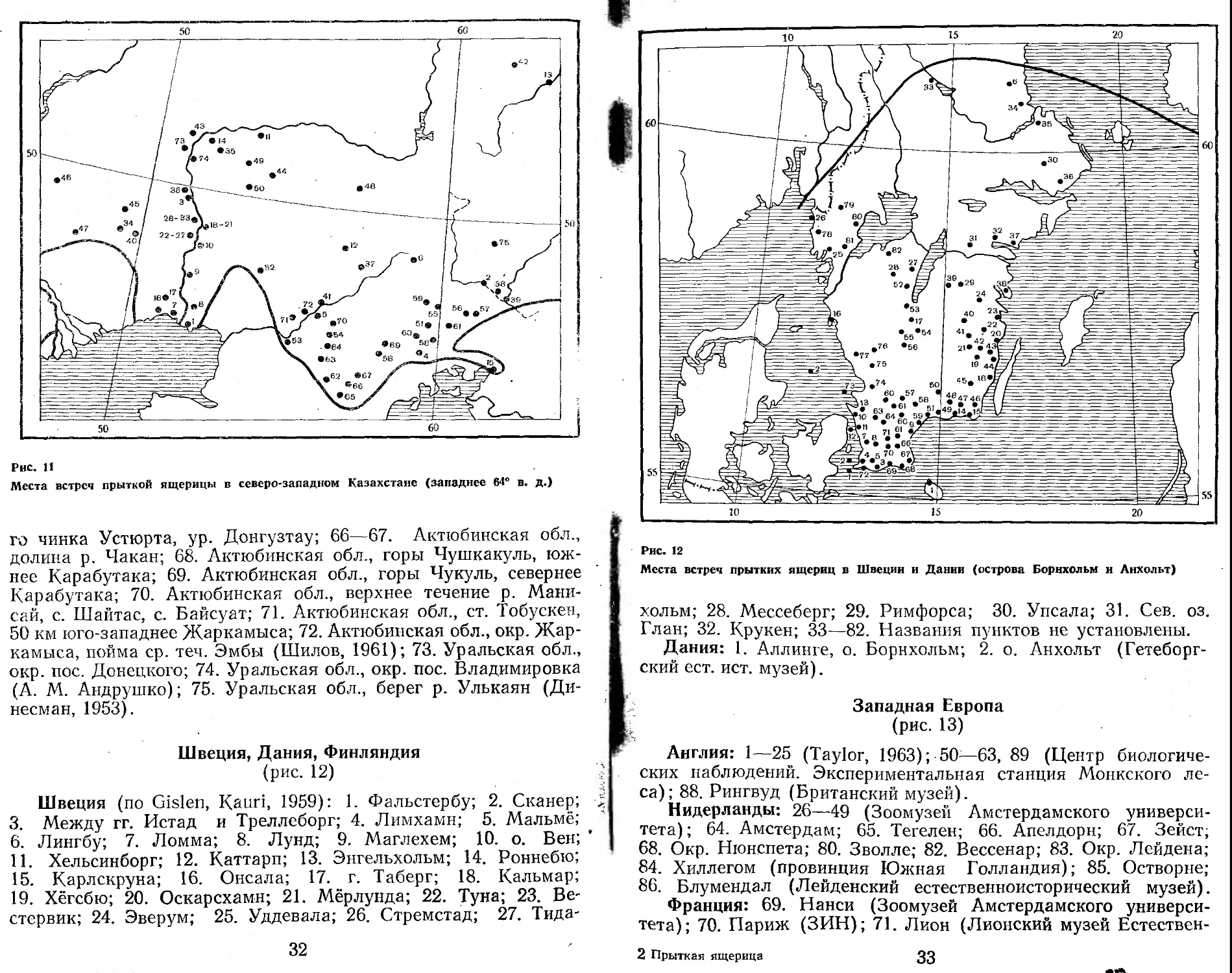
Швеция, Дания, Финляндия (рис. 12)
Швеция (по Gislen, Kauri, 1959): 1. Фальстербу; 2. Сканер; > 3. Между гг. Истад и Треллеборг; 4. Лимхамн; 5. Мальмё;
6. Лингбу; 7. Ломма; 8. Лунд; 9. Маглехем; 10. о. Вен; * 11. Хельсинборг; 12. Каттарп; 13. Энгельхольм; 14. Роннебю;
15. Карлскруна; 16. Онсала; 17. г. Таберг; 18. Кальмар;
19. Хёгсбю; 20. Оскарсхамн; 21. Мёрлунда; 22. Туна; 23. Ве-стервик; 24. Эверум; 25. Уддевала; 26. Стремстад; 27. Тида-
32
Рис. 12
Места встреч прытких ящериц в Швеции и Дании (острова Борнхольм и Лнхольт)
хольм; 28. Мессеберг; 29. Римфорса; 30. Упсала; 31. Сев. оз.
Глан; 32. Крукен; 33—82. Названия пунктов не установлены.
Дания: 1. Аллинге, о. Борнхольм; 2. о. Анхольт (Гетеборгский ест. ист. музей).
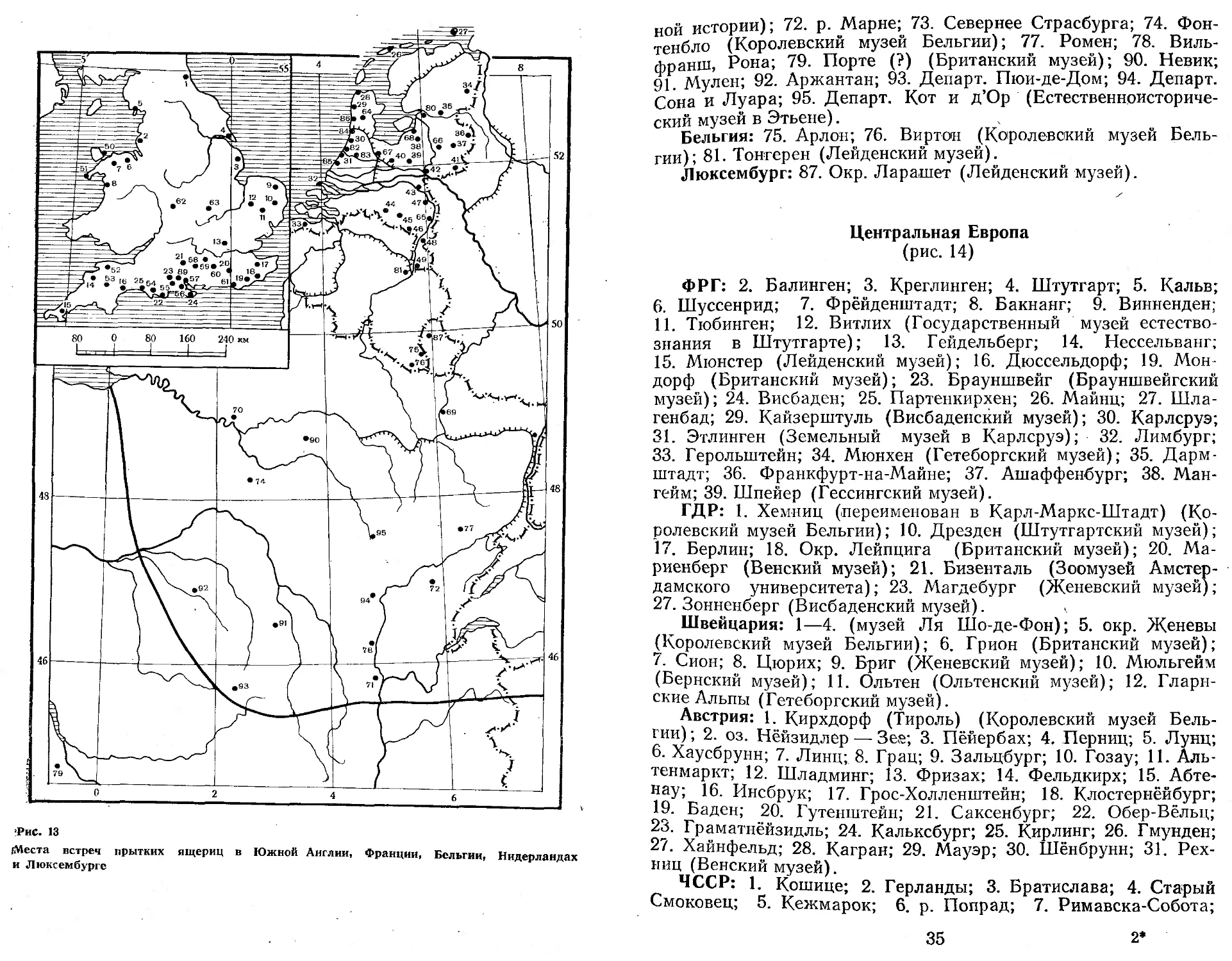
Западная Европа (рис. 13)
Англия: 1—25 (Taylor, 1963); 50-—63, 89 (Центр биологических наблюдений. Экспериментальная станция Монкского леса); 88. Рингвуд (Британский музей).
Нидерланды: 26—49 (Зоомузей Амстердамского университета) ; 64. Амстердам; 65. Тегелен; 66. Апелдорн; 67. Зейст, 68. Окр. Нюнспета; 80. Зволле; 82. Вессенар; 83. Окр. Лейдена; 84. Хиллегом (провинция Южная Голландия); 85. Остворне; 86. Блумендал (Лейденский естественноисторический музей).
Франция: 69. Нанси (Зоомузей Амстердамского университета); 70. Париж (ЗИН); 71. Лион (Лионский музей Естествен-
2 Прыткая ящерица 33
Рис. 13
/Места встреч прытких ящериц в Южной Англии, Франции, Бельгии, Нидерландах и Люксембурге
ной истории); 72. р. Марне; 73. Севернее Страсбурга; 74. Фонтенбло (Королевский музей Бельгии); 77. Ромен; 78. Виль-франш, Рона; 79. Порте (?) (Британский музей); 90. Невик; 91. Мулен; 92. Аржантан; 93. Департ. Пюи-де-Дом; 94. Департ. Сона и Луара; 95. Департ. Кот и д’Ор (Естественноисторический музей в Этьене).
Бельгия: 75. Арлон; 76. Виртои (Королевский музей Бельгии); 81. Тонгерен (Лейденский музей).
Люксембург: 87. Окр. Ларашет (Лейденский музей).
Центральная Европа (рис. 14)
ФРГ: 2. Балинген; 3. Креглинген; 4. Штутгарт; 5. Кальв; 6. Шуссенрид; 7. Фрёйденштадт; 8. Бакнанг; 9. Винненден; 11. Тюбинген; 12. Витлих (Государственный музей естествознания в Штутгарте); 13. Гейдельберг; 14. Нессельванг; 15. Мюнстер (Лейденский музей); 16. Дюссельдорф; 19. Мон-дорф (Британский музей); 23. Брауншвейг (Брауншвейгский музей); 24. Висбаден; 25. Партенкирхен; 26. Майнц; 27. Шла-генбад; 29. Кайзерштуль (Висбаденский музей); 30. Карлсруэ; 31. Этлинген (Земельный музей в Карлсруэ); 32. Лимбург; 33. Герольштейн; 34. Мюнхен (Гетеборгский музей); 35. Дармштадт; 36. Франкфурт-на-Майне; 37. Ашаффенбург; 38. Мангейм; 39. Шпейер (Гессингский музей).
ГДР: 1. Хемниц (переименован в Карл-Маркс-Штадт) (Королевский музей Бельгии); 10. Дрезден (Штутгартский музей); 17. Берлин; 18. Окр. Лейпцига (Британский музей); 20. Мариенберг (Венский музей); 21. Визенталь (Зоомузей Амстердамского университета); 23. Магдебург (Женевский музей); 27. Зонненберг (Висбаденский музей).
Швейцария: 1—4. (музей Ля Шо-де-Фон); 5. окр. Женевы (Королевский музей Бельгии); 6. Грион (Британский музей); 7. Сион; 8. Цюрих; 9. Бриг (Женевский музей); 10. Мюльгейм (Бернский музей); И. Ольтен (Ольтенский музей); 12. Гларн-ские Альпы (Гетеборгский музей).
Австрия: 1. Кирхдорф (Тироль) (Королевский музей Бельгии) ; 2. оз. Нёйзидлер — Зее; 3. Пёйербах; 4. Перниц; 5. Лунц; 6. Хаусбрунн; 7. Линц; 8. Грац; 9. Зальцбург; 10. Гозау; 11. Аль-тенмаркт; 12. Шладминг; 13. Фризах; 14. Фельдкирх; 15. Абте-нау; 16. Инсбрук; 17. Грос-Холленштейн; 18. Клостернёйбург; 19. Баден; 20. Гутенштейн; 21. Саксенбург; 22. Обер-Вёльц; 23- Граматнёйзидль; 24. Кальксбург; 25. Кирлинг; 26. Гмунден; 27. Хайнфельд; 28. Кагран; 29. Мауэр; 30. Шёнбрунн; 31. Рех-ниц (Венский музей).
ЧССР: 1. Кошице; 2. Герланды; 3. Братислава; 4. Старый Смоковец; 5. Кежмарок; 6. р. Попрад; 7. Римавска-Собота;
35
2*
Рис. 14 Места встреч прытких ящ< риц в Центральной Европе
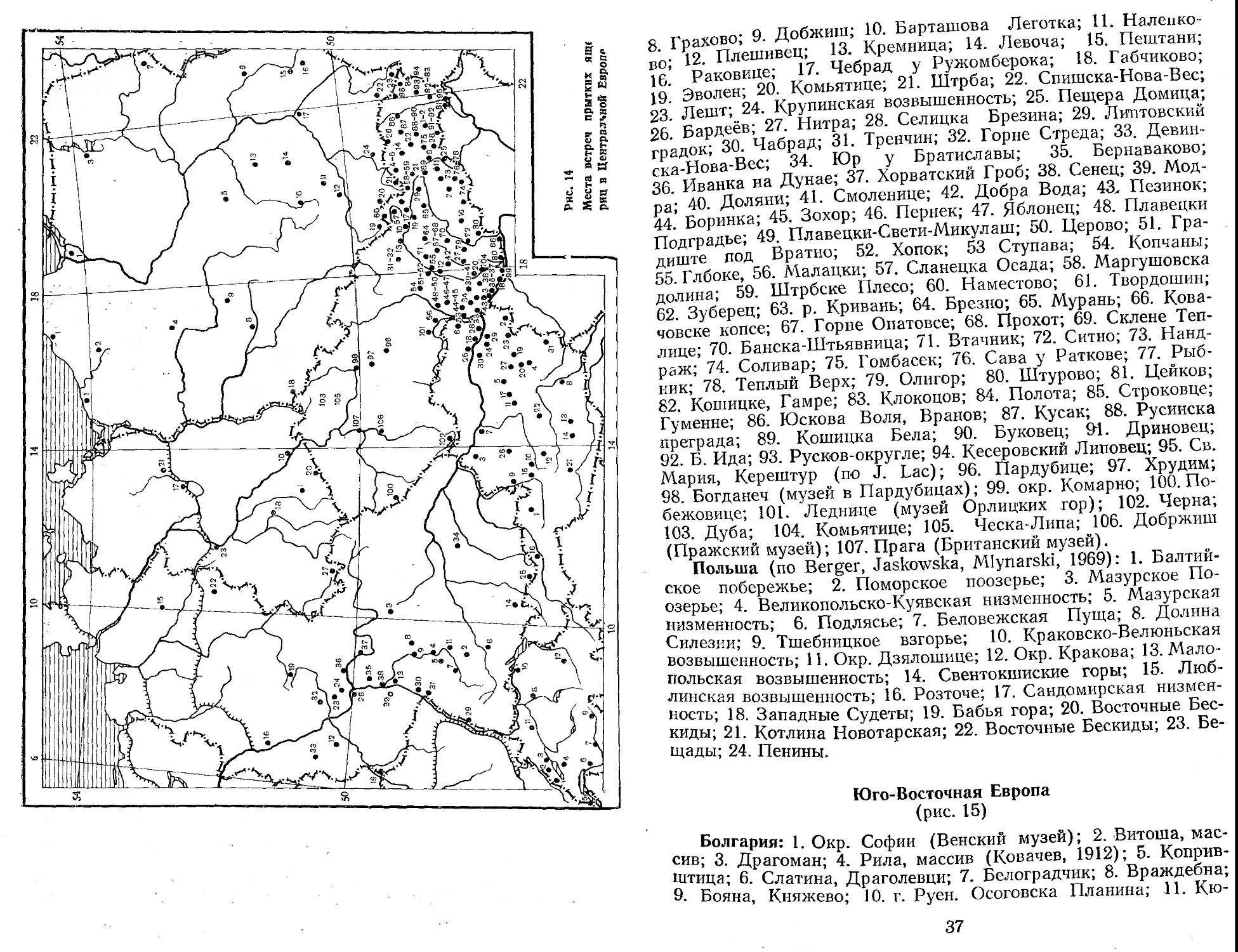
я Гоахово; 9. Добжиш; 10. Барташова Леготка; 11. Налеико-Ь’ • 12 Плешивей; 13. Кремница; 14. Левоча; 15. Пештани; ?б’ Раковице; 17. Чебрад у Ружомберока; 18. Габчиково; 19* Эволен; 20. Комьятице; 21. Штрба; 22. Спишска-Нова-Вес; 23 Лешт; 24. Крупинская возвышенность; 25. Пещера Домица; 26 Бардеёв; 27. Нитра; 28. Селицка Брезина; 29. Литовский грядок; 30. Чабрад; 31. Тренчин; 32. Горне Стреда; 33. Девин-ска-Нова-Вес; 34. Юр у Братиславы; 35. Бернаваково; 36 Иванка на Дунае; 37. Хорватский Гроб; 38. Сенец; 39. Мод-ра; 40. Доляни; 41. Смоленице; 42. Добра Вода; 43. Пезинок; 44.' Боринка; 45. Зохор; 46. Пернек; 47. Яблонец; 48. Плавецки Подградье; 49. Плавецки-Свети-Микулаш; 50. Церово; 51. Гра-диште под Вратно; 52. Хопок; 53 Ступава; 54. Копчаны; 55. Глбоке, 56. Малацки; 57. Сланецка Осада; 58. Маргушовска долина; 59. Штрбске Плесо; 60. Наместово; 61. Твордошин; 62. Зуберец; 63. р. Кривань; 64. Брезно; 65. Мурань; 66. Кова-човске копсе; 67. Горне Опатовсе; 68. Прохот; 69. Склене Теплице; 70. Банска-Штьявница; 71. Втачник; 72. Ситно; 73. Нанд-раж; 74. Соливар; 75. Гомбасек; 76. Сава у Раткове; 77. Рыбник; 78. Теплый Верх; 79. Олигор; 80. Штурово; 81. Цейков; 82. Кошицке, Гамре; 83. Клокоцов; 84. Полота; 85. Строковце; Гуменне; 86. Юскова Воля, Вранов; 87. Кусак; 88. Русинска преграда; 89. Кошицка Бела; 90. Буковец; 91. Дриновец; 92. Б. Ида; 93. Русков-округле; 94. Кесеровский Липовец; 95. Св. Мария, Керештур (по J. Lac); 96. Пардубице; 97. Хрудим; 98. Богданеч (музей в Пардубицах); 99. окр. Комарно; 100. По-бежовице; 101. Леднице (музей Орлицких гор); 102. Черна; 103. Дуба; 104. Комьятице; 105. Ческа-Липа; 106. Добржиш (Пражский музей); 107. Прага (Британский музей).
Польша (по Berger, Jaskowska, Mlynarski, 1969): 1. Балтийское побережье; 2. Поморское поозерье; 3. Мазурское Поозерье; 4. Великопольско-Куявская низменность; 5. Мазурская низменность; 6. Подлясье; 7. Беловежская Пуща; 8. Долина Силезии; 9. Тшебницкое взгорье; 10. Краковско-Велюньская возвышенность; 11. Окр. Дзялошице; 12. Окр. Кракова; 13. Малопольская возвышенность; 14. Свентокшиские горы; 15. Люблинская возвышенность; 16. Розточе; 17. Сандомирская низменность; 18. Западные Судеты; 19. Бабья гора; 20. Восточные Бес-киды; 21. Котлина Новотарская; 22. Восточные Бескиды; 23. Бе-щады; 24. Пенины.
Юго-Восточная Европа (рис. 15)
Болгария: 1. Окр. Софии (Венский музей); 2. Витоша, массив; 3. Драгоман; 4. Рила, массив (Ковалев, 1912); 5. Коприв-штица; 6. Слатина, Драголевци; 7. Белоградчик; 8. Враждебна; У- Бояна, Княжево; 10. г. Руен. Осоговска Планина; 11. Кю-
37
тендил; 12. Разлог; 13. Куклен; 14. Костенец; 15. Батак; *16 Р Ядепица’ с- Голямо-Белово; 17. Позарджик; 18. Чепеларе; 19 Бальчик; 20. Малко-Тырново; 21. Странджа (Буреш, Цонков, 1933)
Югославия: 1—40 (Буреш, Цонков, 1933); 41. Прозор; 42 Госпич; 43. Калиновик; 44. Мостар; 45. Огулин; 46. Травник; 47 Баня-Лука; 48. Вуковар; 49. Карловац; 50. Бойишта (Венский музей); 51. 80 км. сев. Белграда (Лейденский музей); 52. 44 км сев.-зап. Белграда (Зоомузей Амстердамского университета) .
Венгрия: 1. окр. Будапешта; 2. Бекешчаба; 3. Виллань; 4. Калочка (Венский музей); 5. Хевиз (Лейденский музей); 6. Зирц; 7. Баконьбель; 8—16 (пункты не названы) (Баконь-ский музей в Зирце).
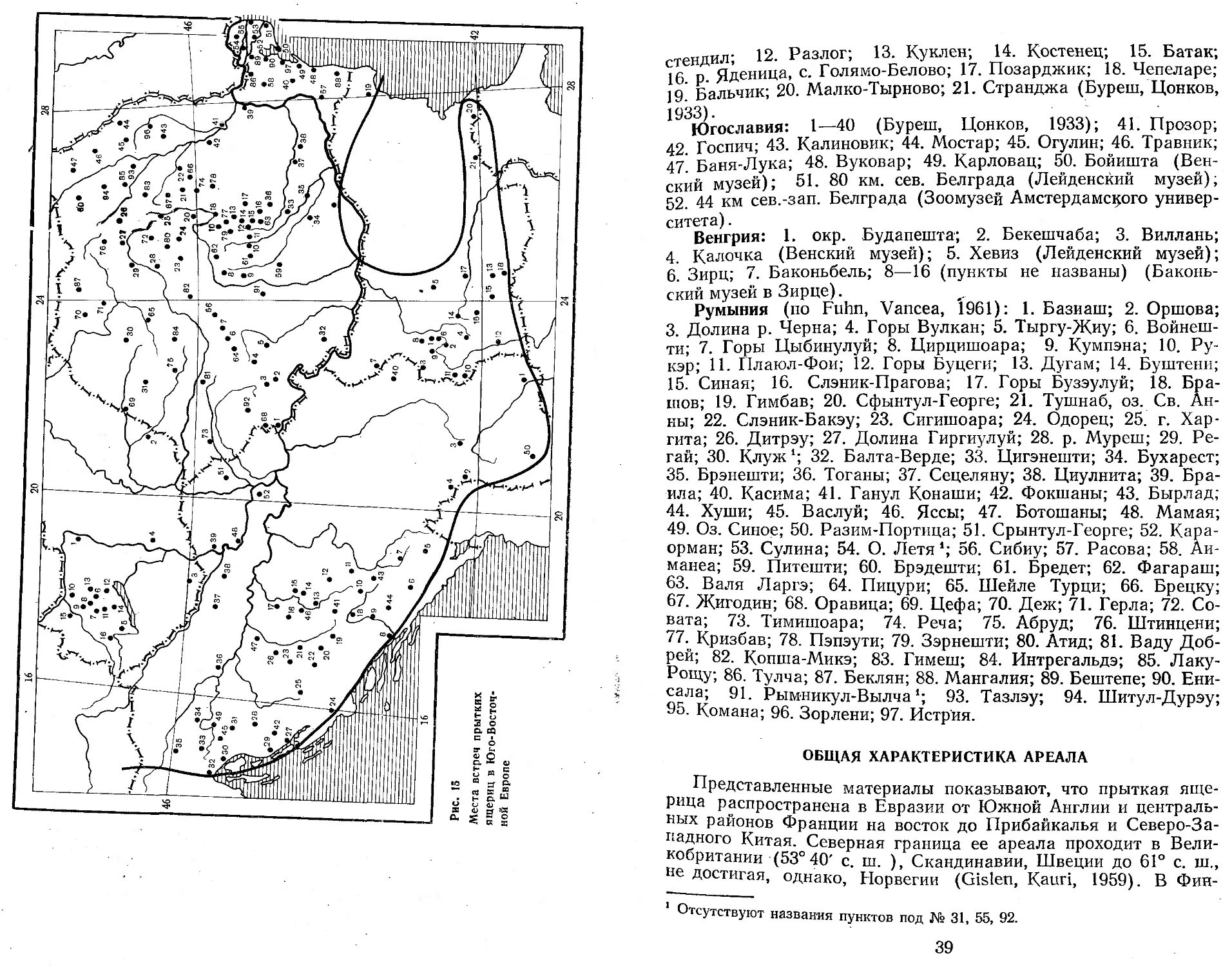
Румыния (по Fuhn, Vancea, 1961): 1. Базиаш; 2. Оршова; 3. Долина р. Черна; 4. Горы Вулкан; 5. Тыргу-Жиу; 6. Войнеш-ти; 7. Горы Цыбинулуй; 8. Цирцишоара; 9. Кумпэна; 10. Ру-кэр; И. Плаюл-Фои; 12. Горы Буцеги; 13. Дугам; 14. Буштени; 15. Синая; 16. Слэник-Прагова; 17. Горы Бузэулуй; 18. Бра-шов; 19. Гимбав; 20. Сфынтул-Георге; 21. Тушнаб, оз. Св. Анны; 22. Слэник-Бакэу; 23. Сигишоара; 24. Одорец; 25. г. Хар-гита; 26. Дитрэу; 27. Долина Гиргиулуй; 28. р. Муреш; 29. Ре-гай; 30. Клуж1; 32. Балта-Верде; 33. Цигэнешти; 34. Бухарест; 35. Брэнешти; 36. Тоганы; 37. Сецеляну; 38. Циулнита; 39. Бра-ила; 40. Касима; 41. Ганул Конаши; 42. Фокшаны; 43. Бырлад; 44. Хуши; 45. Васлуй; 46. Яссы; 47. Ботошаны; 48. Мамая; 49. Оз. Синое; 50. Разим-Портпца; 51. Срынтул-Георге; 52. Кара-орман; 53. Сулина; 54. О. Летя *; 56. Сибиу; 57. Расова; 58. Аи-манеа; 59. Питешти; 60. Брэдешти; 61. Бредет; 62. Фагараш; 63. Валя Ларгэ; 64. Пицури; 65. Шейле Турци; 66. Брецку; 67. Жигодин; 68. Оравица; 69. Цефа; 70. Деж; 71. Герла; 72. Со-вата; 73. Тимишоара; 74. Реча; 75. Абруд; 76. Штинцени; 77. Кризбав; 78. Пэпэути; 79. Зэрнешти; 80. Атид; 81. Ваду Добрей; 82. Копша-Микэ; 83. Гимеш; 84. Интрегальдэ; 85. Лаку-Рощу; 86. Тулча; 87. Беклян; 88. Мангалия; 89. Бештепе; 90. Ени-сала; 91. Рымникул-Вылча *; 93. Тазлэу; 94. Шитул-Дурэу; 95. Комана; 96. Зорлени; 97. Истрия.
ОБЩАЯ ХАРАКТЕРИСТИКА АРЕАЛА
Представленные материалы показывают, что прыткая ящерица распространена в Евразии от Южной Англии и центральных районов Франции на восток до Прибайкалья и Северо-Западного Китая. Северная граница ее ареала проходит в Великобритании (53° 40' с. ш. ), Скандинавии, Швеции до 61° с. ш., е достигая, однако, Норвегии (Gislen, Kauri, 1959). В Фин-
1 Отсутствуют названия пунктов под № 31, 55, 92.
39
ляндии L. agilis отсутствует. He встречена она и на западном берегу Ладожского озера. На восточном берегу упомянутого озера отлавливалась южнее Петрозаводска. Затем граница ареала спускается на юг, проходя через Вологодскую и Кировскую области РСФСР, пересекает Урал по северу Свердловской области, в Западно-Сибирской низменности поднимается до 60° с. ш., постепенно двигаясь на юг и в Прибайкалье достигает 54°30' с. ш. (Гагина, Скалой, 1965). Южная граница ареала в направлении на запад проходит по Тувинской АССР, севернее хребта Танну-Ола, Северо-Западному Китаю (Кульд-жа), откуда заходит узким клином в Иссыккульскую котловину (с востока, приблизительно, до середины озера) и далее через отроги Джунгарского Алатау обходит оз. Балхаш с востока. Данные о находках L. agilis в предгорьях Туркестанского и Зеравшанского хребтов в Восточном Узбекистане (Даль, 1936; Дубинин, 1954) ошибочны (сообщение О. П. Богданова). В Казахстане граница ареала прыткой ящерицы проходит от северного берега оз. Балхаш до устья р. Эмбы. Находки в окрестностях г. Аральска требуют подтверждения. В. С. Лобачев и др. (1973) приводят этот вид в долине р. Тургай как очень редкий, что связано, по-видимому, с близостью южной границы ареала. На Северном Кавказе, по данным И. С. Даревского, прыткая ящерица встречается почти повсеместно (исключая Восточное Предкавказье), кроме сплошных горных лесов и высоко* горья. Здесь она широко распространена на предгорных равнинах в пределах Краснодарского и Ставропольского краев, Кабардино-Балкарии, Северной Осетии и отчасти Чечено-Ингушетии и Дагестана. Южнее в предгорьях и на северных склонах Большого Кавказского хребта ее ареал разбивается на ряд языков, приуроченных к долинам и ущельям системы Кубани, Куры, Терека, Сулака, Самура и более мелких рек, стекающих в Каспий. Так в ущелье Кубани она поднимается до' высоты 2200 м (Учкулан), ущелье Зеленчука до 1800 м, ущелье Баксан 2200 м (Терскол), ущелье Фиагдон до 2000 м и т. д. (Стрельников, 1944, и др-).
Сходный характер имеет распространение этого вида и на Черноморском побережье Кавказа, где прыткая ящерица далеко проникает в горы по ущельям таких впадающих в море рек, как Шахе (окр. Псху), Кодори (окр. Лата), Риони (окр. Кутаиси и Тквибули) и отчасти Чороха и других рек. Далее к востоку на южных склонах Большого хребта она спорадически распространена в долинах верхнего течения некоторых левобережных притоков Куры — Б. Лиахви (окр. Джава). Пори (окр. Тианети) и Пирсагата (окр. Шемахи).
На большей части территории Закавказья прыткая ящерица является горным видом, приуроченным к горно-степной и горно-лесной зонам, и почти полностью отсутствует в Кура-Ара-ксинской низменности. Исключением является лишь небольшая,
40
видимо, остаточная популяция на юге Ыахичеваньской АССР /око оз. Эдильага).
1 Интересно отметить, что в целом закавказская популяция того вида в настоящее время полностью изолирована от основной части его ареала на Северном и Западном Кавказе, что находит свое отражение и в систематической принадлежности закавказских особей. Большое сомнение вызывают сообщения о находках прыткой ящерицы в Талыше. В этой местности на поисутствие L. agilis указывают Менетрие (Menetries, 1832), Го-генакер (Hohenacker, 1837), О. А. Гримм (1876). А. М. Никольский (1915) приводит в списке коллекций Зоологического музея Академии наук 2 экземпляра, добытые под Ленкоранью. Р. Д. Джафаров (1949) указывает этот вид для с. Гемушам. К этому следует добавить, что проводившие в Талыше сборы рептилий Н. И. Соболевский (19296), а позже И. С. Даревский, Н. Н. Щербак и В. И. Ведмедеря не находили здесь прыткую ящерицу. Вот что пишет по этому поводу В. М. Ведмедеря: «Довольно странным является то, что Менетрие (Menetries, 1832), на которого ссылается большинство последующих авторов, указывая Lacerta agilis (между прочим в его списке после этого названия стоит L. muralis?), не приводит в своих списках полосатую или, по тем временам, зеленую ящерицу, очень широко распространенную в Талыше. Еще более непонятна ссылка И. И. Соболевского (19296) на указания О. А. Гримма (1876), якобы добывавшего ее на острове Сара. О. А. Гримм приводит в своей работе Lacerta stirpium (без автора), тем более, что в его коллекции пресмыкающихся, хранившейся в зоологическом кабинете Петербургского университета, с о-ва Сара имелась лишь L. viridis strigata Eichn. (С. Царевский, 1914). Натуралисты, собравшие наиболее полные коллекции герпетофауны Та-лыша, такие, как Г. Ледер (Boettger, 1886), Э. М. Кириченко (1910), А. Н. Казнаков, А. Б. Шелковников. Р. Г. Шмидт (Никольский, 1913, 1915), Н. И. Соболевский (19296) не добыли ни одного экземпляра этого вида. К. А. Сатунин (1912, 1913), детально исследовавший фауну юго-восточной части Закавказья, не приводит для этих районов прыткой ящерицы. С. Н. Чернявская (1965) тоже не включает это животное в список герпетофауны Кызыл-Агачского заповедника, хотя О. А. Гримм (1876) якобы находил его именно на о-ве Сара. Очевидно, указания на нахождение прыткой ящерицы в Талыше основаны на неправильном определении, тем более, что пластические признаки L. agilis и L. strigata Eich., за исключением длины хвоста (L. strigata более длиннохвостая) во многом совпадают. Возможно, что за прыткую ящерицу были приняты полосатые с регенерировавшим (более коротким) хвостом.
Сообщение П. В. Терентьева и С. А. Чернова (1949) о том, что южная граница прыткой ящерицы проходит по северо-западному Ирану нашими материалами не подтверждается.
41
Распространение в Северо-Восточной Турции не выяснено. Немногие известные местонахождения (окрестности Карса, Эрзурума, Сарыкамыша и Ардагана) позволяют предполагать, что прыткая ящерица достаточно широко распространена по всему Армянскому нагорью. На Черноморском побережье она известна до ближайших окрестностей Трабзона. Заселяя северное побережье Черного моря, прыткая ящерица проникает в Крым, но не переходит его главную гряду гор (встречается на Яйле, но отсутствует на южном берегу). В Румынии прыткая ящерица распространена повсеместно, на юг — до Дуная, проникает на северо-востоке в Болгарию (Бешков, сообщ.). В Болгарию она заходит и на западе из Югославии, где встречается спорадично в Динарских горах. Здесь ее южная граница идет в Черногорию и Македонию, оставляя в стороне Адриатическое побережье. Далее следует через Австрию и Северную Швейцарию. Здесь прыткая ящерица практически не проникает через Альпы.
По общепринятому в настоящее время мнению, этот вид обитает в северо-восточных и центральных районах Франции, однако недостаток материала не позволяет нам более точно очертить здесь его границу, и она нуждается в уточнении.
Таким образом, на основании наших данных, можно заключить, что граница ареала прыткой ящерицы удовлетворительно выяснена на северо-западе (Англия, Скандинавия, Европейская часть СССР). Возможны ее существенные изменения в районах, расположенных восточнее Урала и на крайнем юго-востоке—в пределах КНР. Следует считать ошибочными сообщения о находках этого вида в Монголии, Кашгарии и Узбекистане, Талы-ше и южном берегу Крыма. Нуждается в уточнении граница ареала на территории Югославии и Франции. Судя по количеству данных, ареал L. agilis изучен неравномерно. Много сведений о находках этого вида собрано на Украине, в Молдавии, Кавказе, очень мало — в Прибалтике, на севере Центральной части РСФСР и Сибири.
ФАКТОРЫ, ОПРЕДЕЛЯЮЩИЕ РАСПРОСТРАНЕНИЕ ПРЫТКОЙ ЯЩЕРИЦЫ
Наиболее важными факторами, воздействующими на распространение и жизнь пресмыкающихся, являются: 1) абиотические (климат, температура, влажность, почва, рельеф, солнечная радиация и др.); 2) биотические (растительность, биоцено-логические взаимоотношения), 3) исторические и 4) антропиче-ские.
Климат
Ареал прыткой ящерицы находится в умеренном и частично субтропическом поясах [климатологическое районирование по Б. П. Алисову и др. (1954) и Б. П. Алисову (1956)].
42
Он занимает несколько климатических областей умеренного П°ЯАтлантико-континентальная лесная (и лесостепная) область.
Климат слагается под влиянием циклонических и антицикло-нических вхождений атлантического воздуха и последующей трансформации его в воздух континентальный. Зимой преобладают юго-западные ветры, летом — западные и северо-западные Температура января на западе Западной Европы -—5°, на востоке—10°, Европейской части СССР — на севере —20°, на юге —15°. Температура июля на севере 16, на западе и юге 20°. Годовое количество осадков на западе до 1000, на востоке 450 мм. Снежный покров на западе до 20—30, на востоке 40—во см. Осадки и испарение уравновешивают друг друга.
Континентальная лесная западносибирская область.
Климат слагается под действием арктического фронта. Зимой преобладают юго-западные ветры, летом — северные.
Температура января на юге —20, на севере —30°. Температура июля на севере — 21,5°, на юге — 20°; годовое количество осадков 400 мм. Высота снежного покрова на севере 80, на юге 50 см.
Континентальная лесная восточносибирская область.
Климат слагается под действием радиационных факторов, влияние которых усиливается благодаря слабым ветрам и пересеченному характеру рельефа. Температура января в Прибайкалье —20, температура июля 18°. Годовое количество осадков 200—400 мм.
Континентально-европейская и атлантико-континентальная степные области.
Климат формируется под влиянием радиационных факторов, характеризуется большей или меньшей засушливостью. Ветры несут континентальный воздух, снежный покров мал. Радиационный баланс превышает затрату тепла на испарение выпадающих осадков. Температура июля на западе 20°, января 0 —5°, годовое количество осадков 500—1000 мм.
Континентальная степная западносибирская область.
Климат слагается под влиянием радиационных факторов и характеризуется возрастающей засушливостью к югу. Занимает степи Казахстана и Западной Сибири. Южная граница проходит по северной границе зоны пустыни. Температура января — —20; температура июля 20—-25°; годовое количество осадков 200—300 мм. Снежный покров мал. Радиационный баланс превышает затраты тепла на испарение выпадающих осадков.
Горная область Алтая и Саян.
Климат слагается под действием циркулярных процессов Западной Сибири и влиянием высотной поясности. Температура января до —30°, июля 14—20°. Снежный покров распространяется неравномерно. Радиационный баланс, снижается. Годовое количество осадков 700 мм.
43
Континентальная пустынная область Средней Азии (частично).
Климат слагается под действием радиационных факторов. Зимой преобладают северо-восточные, летом—северо-западные ветры. Температура января на юге 0, на севере —15°; температура июля 25—30°; годовое количество осадков 200—300 мм. Относительная влажность летом 30%.
Горная область Северного Кавказа.
Климат слагается под действием циркулярных процессов и влиянием высотной поясности. Температура января —1,3—4,8°, июля 21,8—24,6, годовое количество осадков около 500 мм, снежный покров мал и неустойчив (в среднем около 2 месяцев). Радиационный баланс снижается.
Горная область Тянь-Шаня.
Климат слагается под действием циркулярных процессов и влиянием высотной поясности. Занимает горную часть Казахстана и Северной Киргизии. Температура января —5—16, июля — около 20°. Годовое количество осадков 100—500 мм.
Ареал L. agilis также находится в следующих климатических областях субтропического пояса.
Западная Закавказская область.
Климат слагается под действием циклонической деятельности малоазиатского фронта, непосредственной близости Черного моря и закрытости положения, в силу которого зимой затрудняется проникновение холодных масс с севера, летом выход влажного воздуха. В горах накладывается влияние высотной поясности. Годовое количество осадков 200'0 мм. Температура января 5°, июля 23°.
Горная область Закавказского нагорья.
Климат слагается под действием циркулярных процессов, влиянием высотной поясности и форм рельефа. Годовое количество осадков около 400 мм, температура января —2 —10°, июля 20—25°.
Суммируя полученные данные, можно отметить, что ареал прыткой ящерицы расположен в климатических поясах, характеризующихся температурой января 0 — —30°, также годовым количеством осадков 100—2000 мм, что свидетельствует о ее значительной эвритопности (учитывая, конечно, и микроклиматические условия). Кроме того, следует заметить, что южная граница распространения этого вида в Казахстане почти полностью совпадает с южной границей континентальной степной западносибирской климатической области. Находки прыткой ящерицы из мест, расположенных южнее этой границы (северный берег оз. Балхаш, долины рек Урал, Эмбы, Тургай и др.), объясняются специфическими условиями микроклимата отдельных интрозональных ландшафтов (например, ' долины рек, горных хребтов).
44
Температура воздуха и солнечная радиация
Во многих местах, особенно в Западной Европе, современная граница ареала прыткой ящерицы устанавливается с трудом, так как здесь этот вид в течение многих веков подвергался активному воздействию антропического фактора. Северная и северо-восточная граница в пределах Сибири, как указывалось, ёще недостаточно изучена. Таким образом, все это затрудняет решение вопроса о влиянии тех или иных факторов среды на ареал данного вида и современные выводы в дальнейшем в какой-то степени могут быть уточнены или даже оспариваться. Различная степень влияния одних и тех же факторов в разных частях ареала вида зависит от усиливающего или ослабляющего действия других факторов, а также определяется тем обстоятельством, что отдельные популяции могут различаться по экологическим требованиям.
Сравнение границы ареала прыткой ящерицы с различными изотермами дает следующие результаты.
1. Западная и юго-западная границы видового ареала в значительной степени совпадают с январской изотермой +4° (рис. 16), и ареал не выходит за линию средней длительности снегового покрова в течение 10 дней в году.
2. Зависимость границы ареала от температурных условий в Европейской части СССР не выявлена.
3. Границу распространения прыткой ящерицы с 50° в. д. и далее на восток можно сопоставить с изотермой июля 18° С, а в Сибири, начиная с 75° в. д., северная граница определяется январской изотермой —20°, а южная — изотермой января —14°.
4. В Казахстане распространение L. agilis ограничивается изолинией числа дней в году со средней суточной температурой воздуха выше 20°, равной 95 дням, и изолинией числа дней в году со среднесуточной температурой воздуха ниже —10°, равной 60 дням. Кроме того, южная граница распространения прыткой ящерицы совпадает с изолинией даты перехода среднесуточной температуры воздуха через 5° весной (11 апреля) и изолинией даты перехода среднесуточной температуры воздуха через 5° осенью (21 октября). В Западной Сибири северная граница ареала также совпадает с изолинией числа дней в году со среднесуточной температурой воздуха выше 20°, равной 20 дням, с изолинией даты перехода среднесуточной температуры воздуха через 5° весной (1 мая) и с изолинией даты перехода среднесуточной температурой воздуха через 5° осенью (1 октября). Число дней в году со среднесуточной температурой воздуха выше 20° и даты перехода среднесуточной температуры через 5° весной и осенью связано с минимальным количеством теплых дней, по-видимому, необходимых для процесса воспроизводства вида. Когда дата перехода среднесуточной
45
температуры через 5° весной соответствует 1 мая и дата перехода средней суточной температуры через 5° осенью происходит не позднее 1 октября, это условие соблюдается.
5. В Европейской части СССР распространение прыткой ящерицы на севере ограничивается изолинией суммарной солнечной радиации в июле, равной 14 ккал/см2, хотя в Западной и Восточной Сибири этот вид может и переходить в данную линию. В Киргизии, Казахстане и Закавказье распространение ограничивается изолинией суммарной солнечной радиации в июле 17 ккал/см2 и изотермой июля, равной 24°.
6. С минимальным количеством теплых дней, необходимых для процесса воспроизводства вида, связано, вероятно, и совпадение северной границы ареала с некоторыми фенологическими данными, в частности, изолинией срока зацветания черемухи 31 мая, которая является наиболее эффективным комплексным показателем весеннего развития природы (начало апогея весны), позволяющим сопоставить природные условия различных местностей (Атлас СССР, 1962).
Осадки
Среднегодовое количество осадков на большей части ареала прыткой ящерицы, по-видимому, не оказывает существенного влияния на ее распространение. Этот вид в Западной Европе обитает на территории со среднегодовыми атмосферными осадками от 400 до 2000 мм, в Азиатской части ареала — со среднегодовыми осадками от 200 до 700 мм. Некоторая зависимость границы ареала L. agilis от осадков наблюдается на юге. Так, по наблюдениям в Крыму (Щербак, 1966), прыткая । ящерица не встречается в местах с относительной влажностью ниже 55%. Здесь она не проникает в зону Южного берега Крыма, который характеризуется годовыми осадками от 400 до 600 мм, но обитает на Яйле при количестве осадков, годовая норма которых превышает 100 мм. В Казахстане южная граница ареала близка к изогиете 150 мм/год. Суммируя сказанное выше, следует отметить, что западные популяции прыткой ящерицы обитают в более влажных районах, чем восточные.
Почвы
Северная граница распространения прыткой ящерицы в Европейской части ареала проходит по зоне подзолистых почв. В ряде мест она совпадает с северной границей дерново-подзолистых почв. Южнее этот вид встречается на дерново-карбо-натных почвах, различных черноземных, бурых лесных, серых лесных, на осушенных участках подзолисто-болотных, пойменных, темно-каштановых, но избегает темно-каштановые солон-
47
цеватые почвы [в Крыму встречается в зоне солонцеватых, но только в лесополосах (Щербак, 1966)].
На Кавказе этот вид встречается на красноземах, желтоземах, пойменных почвах, горных черноземах, горных луговостепных, горных лесных дерново-карбонатных, но избегает сероземы, солончаки, серо-коричневые. На Северном Кавказе встречается на желтоземах, горно-луговых черноземах, горных черноземах, горных, лесных, дерново-карбонатных, темно-каштановых и даже на южных солонцеватых черноземах.
В Западной Сибири прыткая ящерица встречается на горных черноземах, горных лесных (Урал), подзолисто-дерновых, лесных серых, подзолисто-болотных, пойменных выщелоченных черноземах, черноземах, горно-лесных, горно-луговых, горно-таежно-подзолпстых, лугово-черноземных, а в Прибайкалье, Туве и Западной Сибири — на серых лесных, горно-таежноподзолистых, пойменных, черных лесных.
В Западном Казахстане прыткая ящерица обычна на пойменных, серо-бурых пустынных почвах, в Центральном Казахстане— на черноземах, южных черноземах, светло-каштановых, горных черноземах, в Восточном Казахстане — на лугово-черноземных, горно-луговых, горных черноземных, пойменных, в Киргизии —на горно-луговых, горных черноземах, сероземах, лугово-сероземах, а в Узбекистане — на горно-лугово-степных почвах.
Этот общий обзор почв, на которых обитает прыткая ящерица в разных районах ареала, позволяет сделать следующие выводы.
1. L. agilis повсеместно избегает солодовых, солонцеватых и солончаковых почв.
2. На крайнем северо-западе ареала, в Великобритании, прыткая ящерица обитает исключительно на береговых дюнах и песчаных почвах (Corbett, 1969), о чем свидетельствует также ее английское название (sand lizard). В умеренной зоне Европы этот вид не избегает песчаных почв с травянистой растительностью. Например, даже в степных районах Крыма прыткую ящерицу можно встретить у кромки приморских пляжей. В Восточном Предкавказье этот вид, однако, уже избегает песчаные участки с проэкционным покрытием травяной растительности менее 50% (Тертышников, 19726) и совсем не встречается в песках Калмыкии (наши наблюдения), междуречья Волги и Урала (сообщ. Н. М. Окуловой) и далее в Азиатской части ареала. Здесь граница ареала L. agilis жестко лимитируется песками зоны пустынь и полупустынь; ящерица встречается, однако, в глинистых полупустынях. Таким образом, в направлении с востока на запад у прытких ящериц заметно выражена смена местообитаний и возрастает степень предпочтения песчапых почв.
48
Рельеф
Граница ареала прыткой ящерицы наиболее зависима от „ельефа. На юге это объясняется скорее только лишь большим количеством горных систем Евразии. Северная граница ареала проходит по предгорьям Скандинавских гор (по высотам 200—300 м), а также обходит высокогорье Урала. На востоке, в Сибири, этот вид проникает через Саяны в Туву до Байкала и обитает здесь в межгорных понижениях и долинах. Более выражена зависимость ареала прыткой ящерицы от рельефа, как указывалось, на юге. Так, юго-западйая граница на территории Франции не переходит Центральный Французский массив: ящерица здесь избегает высоты 1000 м, затем обходит горы Юра, но полностью отсутствует в Центральных Альпах. Таким образом, распространение этого вида на юге Европы четко ограничено высокогорьем Альп, которые не позволяют L. agilis проникнуть на Ломбардскую низменность и Аппенинский полуостров. Восточнее Альп ящерицы обитают на Чешско-Моравских высотах, Западных и Восточных Карпатах, на высотах, не превышающих, как правило, 850—1000 м. Несколько иная картина наблюдается на Балканах. Прыткая ящерица здесь более или менее обычна севернее рек Савы и Дуная. В Динарских горах встречается локально и только в высокогорье, на высоте от 1000 до 2000 м, а на Адриатическом побережье полностью отсутствует (сообщение д-ра Антона Поленца из Югославии). То же наблюдается и в Западных Родопах в Болгарии, где ящерицы данного вида обитают в высокогорье и совсем не встречаются в низменных районах долины Марицы (Beskov, Вегой, 1964). Нечто подобное наблюдается и в Крыму, где L. agilis обычна на яйлах (до высоты 1400 м) и отсутствует на Южном берегу. На Северном Кавказе она изредка по долинам рек проникает до высоты 2200 м (Учку-лан, Баксан), как правило, не поднимаясь в Западном Предкавказье выше 600—1000 м, а в Восточном выше 200—400 м. В высокогорье Главного Кавказского хребта и Кура-Араксин-ской низменности прыткая ящерица отсутствует. Закавказская популяция, как указывалось, полностью изолирована от основной части видового ареала на Северном и Западном Кавказе и также обитает в условиях высокогорья Малого Кавказа на высоте 1500—2000 м. В Центральном Казахстане южная граница видового ареала не выходит в районы, расположенные ниже 400—600 м, в Восточном Казахстане и Киргизии прыткие ящерицы встречаются на высоте 1500—2500 м.
Суммируя приведенные выше сведения, можно заключить, что: 1) восточные и южные популяции прыткой ящерицы лучше приспособлены к высокогорным условиям, чем западные (первые могут обитать до высоты 2000—2500 м, вторые избегают высоты 1000 м), 2) зависимость распределения прыткой ящерицы от рельефа более выражена на юге ареала.
49
Растительность, ландшафтная структура
В Великобритании прыткие ящерицы обитают на береговых дюнах и сухих вересковых пустошах (Corbett, 1969). Граница ареала здесь и на территории Франции не зависит от зоны растительности и проходит через зону дубовых лесов при-атлантической Европы и по дубово-буковым лесам (Добрынин, 1948). Северная граница ареала L. agilis совпадает с северной границей широколиственно-елового леса только на юге Скандинавии. Южная граница вида в Западной Европе ограничена в Альпах зоной еловых горных лесов, на Балканском полуострове основное место обитания прыткой ящерицы — горные луга. Горные степи и луга являются также ее обычными биотопами в Крыму, на Кавказе и в Восточном Казахстане. В Карпатах L. agilis встречается на равнинных и предгорных биотопах и практически не проникает в зону хвойного леса у субальпийских лугов. В Европейской части СССР прыткая ящерица предпочитает лесостепные биотопы. Однако северная граница ее распространения здесь и в Западной Сибири примерно до 90° в. д. в значительной степени совпадает с северной границей зоны южнотаежных темнохвойных лесов (Атлас СССР, 1962). Участки остепненных лугов являются характерными местообитаниями этого вида и в Восточной Сибири. В Тувинской котловине добывали L. agilis в горной лесостепи. Южная граница распространения прыткой ящерицы в Казахстане почти полностью совпадает с южной границей зоны полупустынь (злаково-полынных степей). В Семиречье и Киргизии L. agilis распространена в зоне горных и предгорных степей, населяет высокотравные, злаково-разнотравные луга, придерживается долин рек и ручьев, не избегая склонов гор и плато, обычна в ар-чевниках, поднимаясь до альпийских лугов. В междуречье Волги и Урала, по сведениям Н. М.. Окуловой, в глинистой полупустыне данный вид придерживается понижений, занятых злаково-полынными или эфемерово-злаковыми ассоциациями. По данным Н. А. Рашкевича на Северном Кавказе в Чечено-Ингушетии, граница ареала L. agilis проходит по зоне лесостепи, а в Кабардино-Балкарии (Шебзухова, 1969) она обитает в остепненных участках пояса буковых и буково-грабовых лесов.
Приведенные данные позволяют сделать следующие выводы.
1. На большей части ареала прыткой ящерицы предпочитае-' мыми биотопами являются участки лесостепи.
2. Характер местообитания данного вида в разных частях ареала свидетельствует о том, что в направлении с севера и запада на юг и восток происходит смена его стаций: в первом случае L. agilis обитает на сухих солнечных местах (дюны, вересковые пустоши, поляны сосновых лесов и др.), во втором — на сырых прохладных местах (поймы рек, понижения, горные степи и луга).
50
3. Зависимость границы ареала L. agilis от характера растительности наиболее хорошо выражена на юге Скандинавии, в Европейской части СССР и Западной Сибири, а также в Казахстане.
АНТРОПОГЕННЫЙ ФАКТОР И ДИНАМИКА АРЕАЛА
В Великобритании прыткие ящерицы вымирают вследствие разрушения их естественных биотопов в результате хозяйственной деятельности человека (Corbett, 1969, рис. 17). Имеют место также удачные случаи интродукции этого вида за пределы естественного ареала на о-в Колл (50° 40' с. ш.), в Камберленде и Западном Сусексе (F. Н. Perring, сообщ.). В Западной и Центральной Европе вследствие распашки целинных земель, уничтожения лесов, осушения болот и создания обширных водохранилищ в значительной степени изменена картина естественного распространения вида, а в некоторых случаях и границы его ареала. В степных и лесостепных районах прыткие ящерицы оказались вытесненными в лесополосы, кюветы дорог и кое-где сохранились в нераспахиваемых поймах рек и оврагов. В то же время искусственные лесополосы в степных районах могут способствовать проникновению ящериц в те районы, в которых они отсутствовали. Так, в Крыму, данный вид попал на Керченский полуостров и в район Евпатории сравнительно недавно, после строительства железной дороги и создания лесополосы.
В лесных районах ареала наблюдается другая картина. Те
перь даже полоса сплошных лесов под влиянием антропогенного фактора превращается в лесостепь (примерно до 60° с. ш., т. е. северной границы видового ареала). Это наглядно показано на современных картах лесов (Добрынин, 1948; Атлас СССР, 1962). Данное обстоятельство способствует расселению вида. По сообщению М. И. Щербаня, L. agilis в Карпаты про-
никает через горные перевалы вдоль дорог, не встречаясь на этой же высоте в лесах. Как свидетельствуют северные пункты находок прыткой ящерицы в Сибири (карта IX), они расположены чаще в поймах рек по вырубкам (Воронов, Демидов, 1973; Гагина, Скалой, 1965), т. е. в местах, где тайга исчезает в первую очередь в связи с удобством лесосплава. Влиянию хозяйственной деятельности человека наименее подвержен участок южной границы ареала в пределах Казахстана, хотя и там этот вид кое-где исчезает в результате перевыпаса степей домашними животными и вызванной этим эрозии, а в других местах расселяется вдоль оросительных каналов и лесополос. Все сказанное выше позволяет сделать следующие выводы.
1. Антропогенный фактор имеет ведущее значение в изменении «кружева ареала». Особенно большое влияние он ока-
зывает на численность прытких ящериц в западных и южных районах ареала.
51
Рис. 17
Особенности распространения прыткой ящерицы в Англии
/ — места обитания, известные до I960 г.; II— места обитания, обнаруженные в 1960— 1969 гг.; III — места обитания, обнаруженные в 1970—1973 гг.- (треугольником обозначены пункты интродукции по данным, представленным для настоящей монографии)
2. Изменение лесных ландшафтов под влиянием хозяйственной деятельности человека на севере и востоке видового ареала может способствовать продвижению L. agilis на север.
Таким образом, антропогенный фактор в настоящее время оказывает наибольшее влияние на северную границу ареала прыткой ящерицы.
* * &
Обзор эколого-географических факторов, определяющих ареал Lacerta agilis, подтвердил мнение о высокой эвритопно-сти вида в целом и экологической разнородности отдельных популяций. Зависимость ареала от температурных условий более четко выражена в восточной части ареала. Западные популяции прыткой ящерицы обитают в более влажных условиях, чем восточные. L. agilis избегает солодовых, солонцеватых и солончаковых почв; на северо-западе ею предпочитаются пески, на юго-востоке она их избегает. Восточные и южные популяции лучше приспособлены к высокогорным условиям. Большая часть ареала прыткой ящерицы расположена в первичной или вторичной лесостепи. Изменение лесных ландшафтов под влиянием хозяйственной деятельности человека может способствовать продвижению Lacerta agilis на север.
I
ГЛАВА ВТОРАЯ
СИСТЕМАТИКА
И ВНУТРИВИДОВАЯ СТРУКТУРА1
ОБЪЕМ И АНАЛИЗ МАТЕРИАЛА
Как было показано выше, прыткая ящерица имеет очень широкий видовой ареал, охватывающий различные природные зоны. Данное обстоятельство в значительной мере определяет высокую степень географической изменчивости рассматриваемого вида, распадающегося на ряд в разной степени дифференцированных подвидовых форм. В этом отношении прыткая ящерица представляет собой классический пример политипического вида, у которого при наличии непрерывного ареала подвиды переходят друг в друга постепенно, и в длинной цепи замещающих популяций каждая из них представляет собой промежуточную стадию между двумя соседними, соединяя таким образом крайние.
К настоящему времени в руках специалистов, и прежде всего в СССР, сосредоточены достаточно обширные выборки из многочисленных популяций, собранные практически на всем протяжении видового ареала прыткой ящерицы. К сожалению, только меньшая часть из этих сборов обработана биометрически и может быть на должном уровне использована для таксономических исследований. Тем не менее критический разбор имеющейся литературы, наряду с изучением дополнительных коллекционных материалов, дают нам необходимую основу для выяснения закономерностей географической изменчивости Lacerta agilis и построения объективной внутривидовой системы этого вида.
Ниже приводится перечень использованных с этой целью выборок из популяций, охватывающих в общей сложности 3658 биометрически обработанных особей. В скобках указано число исследованных экземпляров (и).
Южная Англия (15); Франция (11); ГДР и ФРГ (72); Польша (14); Болгария (15); Румыния: Валахия (50), Молдова (78), Добруджа (43); Советский Союз: Могилев (14); Ле-нинградская обл.: Свирь (23), Луга (46); Львов (14), Киев (49); 1 Авторы: И. С. Даревский, И. Н. Щербак, Г. Петерс, при участии А. С. Баранова,^ В. К. Булахова, Н. Ф. Константиновой, В. К. Жарковой, Л. В. Ту-
Окуловой, Г. П. Лукиной, С. Ванци, В. А. Кутузовой,
53
Рио. 18
Средняя длина тела (Л1±2т) в различных популяциях прыткой ящерицы (л см. в перечне изученного материала)
Молдавия (70); Рязанская обл.: лес (54), степь (33); Калужская обл. (70); Горьковская обл.: Пустынь (93), Шеманиха (116); Белгородская обл.: «Лес на Ворскле» (352); Курская обл. (69); Днепропетровская обл.: Хощеватье (28); юго-восточные районы УССР (240); Крым (253); Западное Предкавказье (617); Ростов (29); Волгоград 21); Астрахань (16); Красно-
54
I —Англия;
2 — Франция;
3 -ГДР и ФРГ;
4__Швейцария;
5 — Австрия;
6 — Польша;
7 —- Венгрия;
g___Румыния (Валахия);
ga — Румыния (Молдова);
8б — Румыния (Добруджа);
у — Могилев;
д? — Львов;
11 — Луга;
12 — Свирь;
13 — Молдавия;
14 — Киев;
15 — Белгородская обл.;
16 — Днепропетровская обл.;
17 — Крым;
18 — Ростов;
19 — Волгоград;
20 — Астрахань;
21 — Гурьев;
22 — Ставрополь;
23 — Верхняя Кубань;
24 — Нижняя Кубань;
25 — Курск;
26 — Нальчик;
27 — Дагестан;
28 —• Пятигорск;
29 — Западное Предкавказье;
30 — Западная Грузия;
31 —Центральная Грузия (Нори);
32 — Восточная Грузия;
33 — Северная Армения;
33 а — оз. Севан;
336 — Спитак;
ЗЗв — Кировакан;
34 — Западный Казахстан;
35 — Оренбург;
36 — Акмолинск;
37 — Калужская обл.;
38 —Рязанская обл.;
32 — Горьковская обл.;
40 ~ Тургай;
47 — Курган;
^3 Горный Алтай;
43 — Томск;
44 — Иланский;
45 —оз. Зайсан;
45с — Семиречье;
456 Тихий Иртыш;
45 — 03. Иссык-Куль;
47 Иркутск (по номерам, отсутствующим на рисунке, нет данных)
дарский край: Новая Адыгея (96); Ставрополь (43); Пятигорск, Кисловодск (56); Нальчик (76); Орджоникидзе (24); Дагестан: Махачкала (61), Буйнакск (55), Сергокала (48); Западная Грузия (43); Восточная Грузия: долина р. Иори (59); Армения (254); Казахстан: долина р. Урал (48), Тургай (13), Курган (24); Джунгарский Алатау (12), оз. Зайсан-Семиречье (32); Горный Алтай (16); Томск (29), Красноярский край: Иланский (15); Киргизия: оз. Иссык-Куль (46); Северо-Восточная Турция (2).
При таксономических исследованиях подрода собственно Lacerta, к которому относится и L. agilis, специалисты оперируют обычно с традиционным набором следующих основных признаков:
Длина туловища, мм (Longitudo corporis) L.
Длина хвоста, мм (Longitudo cauda-lis) L. cd.
Число задненосовых щитков (Nasa-lia) Na
Число скуловых щитков (Loreale) Lor. Число ресничных зернышек (Grann-lae) Gran.
Число горловых чешуи (Squamae gu-lares) G.
Число чешуи вокруг середины туловища, не считая брюшных (Sguamac dorsalis), Sg.
Число поперечных рядов брюшных щитков (Ventralia), Ventr.
Число бедренных пор (Pori fcmorales) Р. f.
Соотношение ширины и длины аналь-Д./с
ного щитка — •
Характер расположения преанальных щитков (Praenalia) Рг. ап.
Специфика рисунка и прижизненной окраски тела.
55
Характеристика наиболее важных из перечисленных признаков рассматривается ниже.
АБСОЛЮТНЫЕ И ОТНОСИТЕЛЬНЫЕ РАЗМЕРЫ ТЕЛА
Как видно из рис. 18, максимальные размеры тела полово-зрелых особей наблюдаются в южных—-оптимальных районах ареала прыткой ящерицы: в Крыму, на Северном Кавказе, в некоторых районах Закавказья и на юге Казахстана. По направлению к востоку (Алтай, Иркутск, п. Иланский и оз. Иссык-Куль), на север (Курск, Могилев, Рязанская, Калужская и Ленинградская обл.) и на запад [СССР (Молдавия), Румыния, Центральная Европа, Англия] средняя длина туловища постепенно убывает. Наблюдающиеся исключения (относительно крупные ящерицы в Восточной Франции и в дельте Дуная и мелкие — в Абхазии), возможно, объясняются климатическими особенностями этих частей ареала. Как правило, самки прытких ящериц крупнее самцов, причем обнаруживается тен-, денция к нарастанию этих различий от Кавказа к востоку и
56
за'паду. Отметим, что изменения веса половозрелых особей прыткой ящерицы обнаруживают сходную тенденцию, т. е. нарастание по мере продвижения от северной границы ареала к южной (Баранов, Валецкий, 1975). Об изменчивости абсолютной длины хвоста (L. cd.) можно судить по величине индекса Т cd
-—— (рис. 19). Картина географической изменчивости этого признака подтверждает зоогеографическое правило Аллена для теплокровных животных, т. е. коррелирует с направлением отмеченной выше изменчивости длины тела. Самые длиннохвостые особи, как и обладающие максимальными размерами туловища, обитают на юге. Наблюдающиеся исключения (уменьшение данного индекса в Западной и Северной Европе, Сибири и горных районах Армении) можно объяснить тем, что рост хвоста находится в прямой зависимости от длительности зимней спячки, которая наиболее продолжительна на севере ареала и в горах.
ЧИСЛО ЧЕШУИ ВОКРУГ СЕРЕДИНЫ ТУЛОВИЩА (Sq.)
Географическая изменчивость данного признака показана на рис. 20.
Можно видеть, что абсолютный максимум числа туловищных чешуй обнаруживается на Кавказском перешейке, и в частности у особей из Западной Грузии. Четкое снижение этого показателя прослеживается в направлении на запад и более умеренное — на восток. При сравнении самцов и самок раздельно на достаточно большом материале обнаруживается, что число чешуи у первых достоверно выше, чем у вторых, несмотря на то, что самки обычно толще самцов. Для вида в целом характерно в среднем 38—49 туловищных чешуй при крайних вариантах этого признака 33 и 54 чешуи.
ЧИСЛО БЕДРЕННЫХ ПОР (Р. f.)
Г. Ф. Сухов (1928), изучив характер изменчивости данного признака у прыткой ящерицы, пришел к заключению, что количество бедренных пор достигает максимума в Крыму и радиально снижается по направлению к северу, западу и востоку. В соответствии с современными данными, полученными при обработке более обширного материала, максимум числа бедренных пор наблюдается на Кавказе, а минимум в Венгрии и на Трансильванском плато. Резкое снижение числа бедренных пор прослеживается от Кавказа «а запад и более умеренное — на восток (рис. 21).
57
Рис. 20
Среднее число чешуй вокруг середины тела в различных популяциях прыткой ящерицы (для самцов и самок вместе)
А их распределение по ареалу; Б — значение ЛГ±2т в изученных выборках. Условные обозначения те же, что и на рис. 18 ►
ЧИСЛО ПОПЕРЕЧНЫХ РЯДОВ БРЮШНЫХ ЩИТКОВ (Ventr.)
Картина изменчивости данного признака обнаруживает широкую область его «депрессии» севернее Черного моря (Румыния и СССР: Молдавия, Киев, Львов, Могилев), простирающуюся на юг до Крыма, Кубани и Северного Кавказа. Севернее, западнее и восточнее этих районов число брюшных щитков постепенно увеличивается. Интересно отметить, что если максимальное значение ряда других меристических признаков находится в центрах их радиальной изменчивости, то в случае с брюшными щитками картина обратная: по направлению от центра число их увеличивается. Как и у некоторых других видов рода Lacerta, число рядов брюшных щитков у самок достоверно превышает таковое у самцов. Как видно из рис. 22, у обоих полов максимум числового значения данного признака наблюдается в западной и восточной окраинах ареала.
58
Число чешуи
СООТНОШЕНИЕ ШИРИНЫ И ДЛИНЫ АНАЛЬНОГО ЩИТКА
L- ta \
Ч J
В данном случае рассматривается так называемый «анальный» индекс, т. е. отношение ширины анального щитка к его длине. В своем численном значении он отражает форму анального щитка: большие величины свидетельствуют о широком вытянутом в поперечном направлении щитке, тогда как малые (ниже 2,00) указывают на его округлую или треугольную форму.
У самцов анальный щиток всегда шире, чем у самок, что определяется общим утолщением основания хвоста, связанным с залеганием в нем парных копулятивных органов. Изучение клинальной изменчивости рассматриваемого индекса показывает, что максимальных значений он достигает на Главном Кавказском хребте, в Армении, в южнорусских, северокавказ-
59
Рис. 21
Среднее число бедренных пор по ареалу прыткой ящерицы (Д ) и значение М+2т (Б в изученных выборках для самцов и самок вместе
Условные обозначения мест сбора те же, что и на рис. 18 ►
ских, поволжских и казахстанских степях, т. е. в районах, где обитают максимально крупные особи. Однако корреляция этого признака с величиной тела все же не устанавливается. Абсолютный максимум по «анальному» индексу показывают среди самцов армянские, а среди самок — иланские ящерицы, хотя и те и другие не относятся к наиболее крупным. У особей из сибирских популяций (Томск, Алтай и др.) данный индекс выше, чем у западноевропейских, хотя по длине туловища они почти не разнятся между собой. Отметим также, что центр радиации признака находится в Кавказско-Причерноморском районе.
РАСПОЛОЖЕНИЕ ПРЕАНАЛЬНЫХ ЩИТКОВ
(Рг. ап.)
В качестве одного из диагностических признаков при изучении внутривидовой изменчивости вида L. agilis часто используется расположение преанальных щитков, образующих один или два полукруга впереди анального. Рассмотрение этого признака свидетельствует, однако, об отсутствии ' кажущейся альтернативной изменчивости, поскольку передний полукруг
60
может быть в той или иной степени не замкнутым (рис. 23). Один полукруг имеется, как правило, лишь у ящериц из самых западных популяций, причем процент особей с двумя полукругами быстро нарастает при продвижении на восток. Так, в Молдавии, на Украине,-в Белоруссии и Ленинградской области около 50% ящериц обладают двумя полукругами преанальных щитков, а восточнее Крыма, Днепропетровска и Ростова такие особи составляют уже 100% популяции. Не обнаруживается и четкой закономерности при рассмотрении такого признака, как величина центральных преанальных щитков, которые бывают увеличены по сравнению с соседними. Выяснилось, что более или менее равные по величине преанальные щитки наблюдаются в популяциях из Центральной Европы и СССР (Ленинградской области). Однако большинство популяций Кавказского перешейка обладают почти 100%-ным увеличением
преанальных щитков, причем этот признак обнаруживает картину радиальной изменчивости по направлению к западу, северу и востоку.
ЧИСЛО РЕСНИЧНЫХ ЗЕРНЫШЕК (Gran.)
Определенный процент особей с так называемыми «зернышками»— мелкими округлыми чешуйками между верхнересничными и надглазничными щитками, встречается почти на всем протяжении ареала прыткой ящерицы. Максимальное значение этого признака как по числу самих зернышек, так и проценту особей, у которых они имеются, наблюдается в некоторых кавказских популяциях, в частности на северо-востоке Кавказского перешейка (96,0+2,7%).
Особи с «зернышками» довольно обычны в Крыму, на Украине, во Франции, в Центральной Европе и в восточной половине ареала.
61
Рис. 22
Среднее число поперечных рядов брюшных щитков (Af±2m) в изученных выборках
А — самцы; Б — самки. Условные обозначения мест сбора те же, что и на рис. 18
ЧИСЛО И РАСПОЛОЖЕНИЕ ЗАДНЕНОСОВЫХ ЩИТКОВ (Na.)
Географическая изменчивость характера щиткования задненосовой области (числа и расположения задненосовых и скуловых щитков) отличается у прыткой ящерицы значительным разнообразием. Число и расположение этих щитков удобнее всего представить -в виде дроби, где в числителе указывается число задненосовых, а в знаменателе — число скуловых щитков.
62
Рис. 23
Расположение анального и преанальных щитков у некоторых подвидовых форм прыткой ящерицы
Сверху — L. a. exigua, снизу L. a. agilis (рис. Г. Петерса)
Из-за часто наблюдающейся асимметрии каждую сторону головы приходится учитывать отдельно. Скуловые и носовые щитки связаны общностью происхождения и нередко бывает трудно решить, какая из формул более справедлива, например ’/г или 2А. Формально задненосовым щитком всегда считается тот, который хотя бы в одном месте касается ноздри. В пределах ареала прыткой ящерицы в целом наиболее часто встречаются следующие пять комбинаций: Vi, V2, 7о, 7. и2/2 (рис. 24).
Как видно на рис. 25, каждая из указанных комбинаций имеет свои районы преобладания, по мере удаления от которых количество особей с данной комбинацией (в процентном отношении) постепенно снижается, уступая место особям с комбинациями, наблюдающимися по соседству. Перечень таких районов преобладания приводится ниже.
71 — Болгария, восточная Югославия, Румыния, СССР: Молдавия, Киев, Могилев.
*/2 — Центральная (Перлеберг) и, вероятно, вся Западная Европа.
7о — Западная Грузия, Астрахань, Восточный Казахстан.
71 — Нальчик, Орджоникидзе, Кубань, Крым, Волгоград, Западный Казахстан.
2/2 — Северный Кавказ, Оренбург, Томск, Иланский, Семиречье, оз. Иссык-Куль.
Комбинации 71, 7г и 71 обнаруживают радиальную изменчивость от своих центров преобладания. Остальные комбинации зачастую преобладают на значительных участках ареала и могут характеризоваться несколькими районами максимума. Местами примешиваются и другие комбинации (7о, 7з, 7з, 2Л, 7s, 7в, 7о, 71, 7г, 74, 7e, Vi, 7г, 7г), например в окрестностях Киева, Курска, в Воронежской и Калужской областях, в котловине оз. Иссык-Куль, Армении, однако доля их нигде не превышает 5—8%. По ареалу в целом вырисовывается следующая картина: наличие одного задненосового щитка характерно для западных популяций прыткой ящерицы (2 задненосовых встречается у них редко), а наличие двух носовых более характерно для восточных, хотя число особей с двумя задненосовыми и здесь местами достигает 30%. Интересно, что у особей из За-
63
Рис. 25
Некоторые комбинации расположения щитков задненосовой области у разных подвидов прыткой ящерицы
1 — L. a. agilis (’/2); 2 — L. a. boemica (2/j); 5 — L. a. bosnica (*/i); 4 — L. a. gru-sinica (2/o); 5—6— L a. euxinica (l/i. 2/i) (из Fuhn, Vancea, 1964)
Рис. 26.
Схема возникновения различных комбинаций в расположении щитков задненосовой области
В верхней правой части основные комбинации подвида «exigua», в верхней левой части — подвидов «chersonensis» и «agilis» в нижней левой^—«grusinica», в нижней правой «brevicaudata», в центре — «.boemica» (рис. Петерса)
ладной Грузии примерно б 60% случаев скуловые щитки отсутствуют или (в 25% случаев) имеется только один.
Часто отсутствие скулового щитка наблюдается также в Армении и в горных районах Казахстана. Наличие одного скулового щитка вообще более характерно для выборок из западной половины видового ареала, включая Кавказ.
64
20
Рис. 24
Распределение основных комбинаций расположения щитков задненосовой области по ареалу прыткой ящерицы
В каждом кружке указана процентная доля отдельных комбинаций
90
Описанная выше специфика изменчивости характера щит-кования задненосовой области, по-видимому, объясняется тем, что одни и те же комбинации могут образовываться из различных исходных соотношений. Это поясняет схема, предложенная Г. Петерсом (рис. 26). В качестве исходной здесь принимается соотношение 2/i прежде всего потому, что подобная комбинация характерна для всех остальных видов ящериц подрода собственно Lacerta. Кроме того, соотношение 2/, особенно часто-встречается на Кавказском перешейке, где, как мы видели выше, находятся центры радиальной изменчивости ряда других признаков чешуйчатого покрова. Наконец, указанная комбинация в более или менее значительном числе случаев наблюдается по всему ареалу прыткой ящерицы, возможно, за исключением лишь Центральной и Западной Европы.
ОСОБЕННОСТИ ОКРАСКИ И РИСУНКА
Специфика окраски и рисунка тела имеет важное значение” при изучении географической и внутривидовой изменчивости-вида Lacerta agilis. Обычно различают два основных типа спинного рисунка, характерные для западной и восточной групп популяций. Первый из них характеризуется проходящей вдоль середины спины относительно узкой, слагающейся из темных пятен, вертебральной полосой, ограниченной с боков светлыми теменными линиями, разбивающимися иногда на отдельные черточки (рис. 27, а). Продольный ряд таких черточек, образованных светлой затылочной линией, бывает заметен и посередине вертебральной полосы. Для рисунка второго типа характерно наличие широкой спинной полосы окаймленной светлыми верхневисочными и разделенной по хребту светлой же затылочной линией (рис. 27, б). Таким образом затылочная полоса фактически слагается здесь из сдвоенного ряда темных, неправильной формы пятен. Первый из описанных рисунков мы будем называть в дальнейшем типом «agilis», а второй — типом «exigua». В различных районах ареала встречаются также в той или иной мере многочисленные особи с промежуточным типом спинной полосы, что, как мы увидим ниже, в значительной мере обусловлено явлением вторичной интерградации.
Отметим, что характеризующийся тремя светлыми продольными линиями рисунок типа «exigua» как бы повторяет в несколько измененном виде продольную полосатость, свойственную другим крупным представителям рода Lacerta: L. trilinea-ta и L. strigata.
Помимо описанных выше, различают случаи, когда спинные полосы не выражены вовсе (aberr. erythronotus Fith). (рис. 27,6), или рисунок на одноцветно окрашенном теле отсутствует полностью (aberr. immaculata Dtirigen). Примесь таких аберраций,
3 Прыткая ящерица
65
Рис. 27
Рисунок верхней стороны тела
а — L. a. chersonensis; б — то же abernot erythrorus;
местами весьма значительная, наблюдается, как правило, лишь в западной части ареала.
Говоря о различиях типов «agilis» и «exigua», следует учитывать также, что рисунок тела прытких ящериц, как это было впервые отмечено Г. Петерсом, связан с количеством и формой спинных чешуй. Если узкая спинная полоса первого типа за-1 нимает 8—14 продольных рядов резко килеватых чешуй, то широкая сдвоенная полоса второго типа распространяется на 13—17 рядов, причем чешуи здесь более широкие и сравнительно слабо килеваты. У всех спинных полос обоего типа килева-гость туловищных чешуй резко уменьшена.
66
I
в — L. a. exigua (по Сухову, 1948)
Наряду с рисунком большое значение имеет также общая прижизненная. окраска верхней и нижней сторон тела раздельно у обеих полов. Специальное изучение этого вопроса (Баранов, 1970) показало, что зеленые самки наряду с коричневыми и бурыми особенно часто встречаются в некоторых популяциях Румынии, СССР — Молдавии, Правобережной Украины, Междуречья Волги и Дона, Предкавказья, Западного и Малого Кавказа и изредка также и в Волжско-Уральском междуречье и по средней Волге. Совсем нет зеленых самок в Закарпатье, Западной Украине, в районе Тирасполя и Одессы, на южных, склонах Главного Кавказского хребта и в Северо-Западном Казахстане. В окраске нижней стороны тела самок, исключая большинство популяций Молдавии и Украины, преобладают зелено-голубые тона, а восточнее Волги коричнево-желтые тона почти исчезают. В Закарпатье, северо-западной Украине, Белоруссии, среднем Приуралье окраска верхней стороны тела самцов только коричневая, в популяциях же Румынии, Молдавии, южной Украины, ниж
него Дона, Южного и Среднего Урала, а также всего Кавказа, как правило, зеленая. Доля самцов с коричнево-желтыми тонами в окраске брюха заметно выше в междуречьях Волги и Дона, в северо-западной Украине, Белоруссии и в некоторых популяциях Кавказа. В Закарпатье, Молдавии, южной Украине и некоторых районах Кавказа, а также восточнее Волги эти тона н окраске брюха самцов не участвуют вовсе. Не исключено, что наличие зеленых тонов в окраске верхней стороны тела обоих полов связано с повышенным режимом влажности местообитаний, т. е. мы встречаемся здесь с проявлением известного зоогеографического правила Глогера.
67
3*
ВНУТРИВИДОВАЯ СТРУКТУРА ВИДА LACERTA AGILIS
Анализируя изложенные выше данные об изменчивости размеров тела, признаков чешуйчатого покрова и особенно прижизненной окраски и рисунка прыткой ящерицы, можно видеть, что рассматриваемый вид в целом распадается на две достаточно хорошо дифференцированные группы географических форм: западную и восточную. Именно объективное существование названных группировок привело в свое время Г. Ф. Сухова (1948) к ошибочному выводу о наличии двух самостоятельных видов Lacerta agilis L. и Lacerta exigua Eichw. Ниже мы постараемся показать, что существование достаточно хорошо различающихся восточной и западной групп популяций прыткой ящерицы связано с имевшим место вторичным разрывом некогда единого видового ареала исходного типа Lacerta agilis. Переходя к рассмотрению внутривидовой структуры вида, важно подчеркнуть, что все признаваемые в настоящее время подвидовые формы выделяются на основании прежде всего определенных комплексов окраски и чешуйчатого покрова, имеющих диагностическое значение и указывающих на их видовую самостоятельность. В своей совокупности подвидовые признаки, дополняя друг друга, позволяют точно диагностировать почти каждый .подвид так, что достаточно бывает просмотреть серию из нескольких особей или даже единичные экземпляры, чтобы определить их подвидовую принадлежность.
В настоящее время можно считать доказанным существование у прыткой ящерицы 9 следующих подвидовых форм, относящихся к восточной и западной географическим группам.
Восточная группа
1. Прыткая ящерица грузинская L. agilis grusinica Peters, 1960.
2. Прыткая ящерица дагестанская L. a. boemica Suchow, 1929.
3. Прыткая ящерица короткохвостая L. a. brevicaudata Peters, 1958.
4. Прыткая ящерица йорская L. a. iorensis Peters et Musk-helischwili, 1968.
5. Прыткая ящерица восточная L. a. exigua (Eichwald, 1831).
Западная группа
; 6. Прыткая ящерица западная L. a. agilis Linnaeus, 1758.
7. Прыткая ящерица южная L. a. chersonensis Andrzejowski, 1832.
8. Прыткая ящерица добруджинская L. a. euxinica Fuhn et Vancea, 1964.
68
9. Прыткая ящерица боснийская L. a. bosnica Schreiber, 1912.
Характеристика вида в целом и описание каждой из подвидовых форм приводятся ниже.
Прыткая ящерица — Lacerta agilis Linnaeus, 1758
Terra typica (restricta) — Южная Швеция.
1758 — Lacerta agilis Linnaeus, part., Syst. Nat. Ed. 10. 1:203.
1768 — Seps argus Laurenti, Synops. Rept.: 61 (Вена).
1768 — Seps caerulescens Laurenti, Synops. Aept.: 62 (Вена).
1768 — Seps ruber Laurenti, Synops. Rept.: 62 (Вена).
1789 — Lacerta punctata Hablizl, Phys. Beschr. Taur. Statth.: 342 (Крым). 1789 — Lacerta pardus Razumowsky, Hist. nat. Jorat, 1:107 (Швейцария). 1798 — Seps stellatus Schrank, Fauna boica, Г : 286 (Бавария).
1802 — Lacerta stripium Daudin, Hist. nat. Rept., 3:155 (Париж).
1802 — Lacerta laurentii Daudin, Hist. nat. Rept., 3 : 227.
1802 — Lacerta arenicola Daudin, Hist. nat. Rept., 3 :230.
1804 — Lacerta anguiformes Sheppard, Transact. Linn. Soc., London. 7:51 (Англия),
1814 — Lacerta europaea Pallas, part. Zoogr. rosso — asiat., 3:29 (Европа). 1828 — Lacerta sepium Bory (ex errore) Res. Erpetol. Hist. nat. Rept.,: 105. 1831—Lacerta exigua Eichwald, Zool. spec. Ross. Polon., 3: 188 (Урал).
1831 — Lacerta muralis Eichwald (part.), Zool. spec. 3 : 189.
1832 — Lacerta chersonensis Andrzejowski, Nouv. Mem. Soc. Natural. Moscou, (2)2:327 (окр. Херсона).
1834 — Lacerta sylvicola Eversmann, Nouv. Mem. Soc. Natural. Moscou (2)3: 344 (между Симбирском и Сызранью).
1837 — Lacerta viridis Rathke, Mem. Ac. Petersb., Ill : 299.
1841 —Lacerta viridis var. colchica Eichwald, Nouv. Mem. Soc. Natural. Moscou, 7 : 83 (Черноморское побережье Грузии).
1851 —Lacerta sericea Glflckselig, Lotos, Prag, 1 : 113 (Прага).
1856 — Lacerta dilepis Lichtenstein, Nomenci. Rept. Amph. Mus. zool. berolin : 14 (киргизские степи).
1868 — Lacerta kochi Gistel, Die Lurche Europas (in: Blicke in das Leben der Natur) : 146 (Италия).
1868 — Lacerta italica Gistel, то же. 147.
1874 — Lacerta doniensis Bedriaga, Entst. Fabr. Eidechs,: 14 (Дон у Воронежа и Волга у Самары).
1886 — Lacerta paradoxa Bedriaga, Abh. senckenberg. naturf. Ges., Frankfurt a/M, 14,3:170 (Сухуми, окр. Киева, Крым, Константинополь (?)). 1929 — Lacerta boemica Suchow, 36, праць зоомузею, УАН, Труды ф!з-мат. в!д., Ки1в, 13 : 116 (г. Орджоникидзе).
Описание1. L.min-max 30,0—114,0 L. cd.mln_max 36,0—190,5. Самые крупные особи встречаются в Крыму и на Кавказе. Размеры туловища и хвоста убывают в северном, восточном и западном направлениях.
L cd
—nun-max 1,24—2,03. Для дифференциации прытких ящериц от всех других видов зеленых ящериц -весьма надежным признаком является относительная коротконогость первых. Так, У самцов прытких ящериц длина задних ног составляет 32—52%,
1 Более детальные сведения приведены в описаниях подвидов.
69
а у самок 34—47% длины туловища. Самые длинноногие особи встречены в Грузии.
Sq.mm-max 33—54, самое высокое число у кавказских особей (49,19±0,45 у особей из Сухуми), уменьшается у западных и восточных популяций. В среднем число чешуй вокруг середины туловища более высокое у самцов.
Ventr.min-max 23—34. Самые низкие показатели этого признака в области Причерноморья — от Молдавии до Нальчика. Севернее, западнее и восточнее этого района число брюшных щитков возрастает. У самок количество брюшных щитков несколько выше, чем у самцов.
Р. f-min-max 9—18. У самцов бедренных пор относительно (на 0,2—0,3) больше. Максимум этого признака на Кавказе, минимум у венгерско-трансильванской популяции.
G.mln_max 14—25; зависимость от пола не выражена.
Количество зернышек между верхнересничными и надглазничными щитками обычно незначительно, у многих восточных популяций особи с зернышками отсутствуют. Максимально этот признак выражен у L. a. boemica (у 96,0±2,7%). Задненосовых щитков 1—3, у. западных популяций чаще 1, восточных —2. Скуловых 0—6, у некоторых западных популяций 1, чаще 2. Анальный щиток у самцов относительно более широкий, чем у самок. Максимальных значений отношение его ширины к длине достигает там, где обитают более крупные особи, т. е. у восточных популяций. Абсолютный максимум этого индекса (2,7) найден у самцов севанской популяции. Данный индекс у сибирских популяций выше, чем у западноевропейских (при отсутствии различий в размерах туловища). Анальный щиток спереди окружен одним (чаще на западе ареала) или двумя (у восточных популяций) полукругами более мелких преанальных щитков. Пара центральных преанальных щитков увеличена у 100% особей с Кавказа и постепенно количество особей с этим признаком уменьшается, причем совсем отсутствуют увеличенные преанальные у особей западноевропейских популяций.
Окраска и рисунок. У особей западных популяций преобладают бурые тона (у самцов в брачный период зеленеют бока). У ящериц с/юга и востока ареала самцы (а иногда и самки) зеленые. Количество зеленых особей возрастает обычно у более старых возрастных категорий. Горло и брюхо варьирует от белого до зеленого цвета (особенно у особей из Причерноморья и некоторых других восточных популяций). Для подавляющего большинства прытких ящериц (особенно молодых и самок) характерно наличие темной продольной полосы посередине спины, более узкой у западных популяций- широкой — на востоке ареала. Эта темная спинная полоса, как правило, ограничена по сторонам светлыми теменными линиями, а у восточных ящериц к ним прибавляется еще одна светлая линия — хребтовая, которая делит широкую спинную полосу; в этом случае на спине
70
ящерицы четко выделяются три светлые линии. Первый вариант рисунка (с двумя линиями), как уже говорилось выше, относят к типу «agilis», второй (с тремя линиями) —к типу «exigua». Учитывая, что между отдельными формами прыткой ящерицы существуют широкие зоны интерградации, становится понятным то большое разнообразие рисунка, исходными для которого являются указанные выше типы.
На темной спинной полосе могут быть темно-бурые или черные пятна (от точек до крупных пятен, закрывающих почти сплошь спинную полосу). На боках туловища хорошо выделяются у большинства особей 2—3 ряда светлых «глазков», отороченных темным. Элементы светлых и темных полос продолжаются и на хвосте. Светлые глазки и темные точки покрывают сверху также и конечности.
Прыткая ящерица грузинская —
Lacerta agilis grasinicaPeters, 1960
Terra typica: Сухуми, Абхазия.
1841 — Lacerta viridis var. colchica Eichwald, (Part) Nouv. Mem. Soc. Natural. Moscou, 7—83 (Абхазия).
1886 — Lacerta paradoxa Bedriaga. Abh. Senck. Ges., XIV, 170; 114 (Сухуми, Трабзон).
1948 — Lacerta exigua paradoxa, Сухов, Труды Зоол. ин-та АН СССР, VII. 1960 — Lacerta agilis grusinica Peters, Zool. Anzeiger, Bd. 86: 179.
Описание1. L.mln_mai ad 57—105; L. & & (n=28) 57—92 (75,8+0,25), L. 9? (n=21) 59—105 (76,0+0,24); L. cd. ml„_max. ad 140—170; Л? (n=6) 1,68—1,86 (1,79+0,03); 99
(n=7) 1,51—1,71 (1,6+0,03); Sq.m)D_max ad. 44—54, cM (n=23) 44—54 (49,44 + 0,54); 99 (n=22) 44—54 (48,86+0,43); Ven-tr. mlo_max 24—30, (n=23) 24—28 (25,96+0,27); 99 (n=22)
27—30 (28,09+0,16); P. f. ml„-max 12—18, d'd (n=46) 12—18 (14,89+0,14), 99 (n=43) 12—17 (15,07+0,18); G. m)n_max 15—23.
Между верхнересничными и надглазничными щитами у 7% особей (ц=43) расположено 1—2 зернышка. Наиболее частое расположение щитков задненосовой области 2/0—39,5%, 1h— 16,3%, */2—12,8%, 7г —7%, 7о — 2,3%. Примерно у 60% особей скуловой щиток отсутствует. Анальный индекс (п=17) 1,7—2,29 (1,98 + 0,04) у самцов и 1,37—2,12 (1,79 ±0,05) при п~ 15, у самок.
Окраска и рисунок. Молодые сверху с тремя светлыми продольными линиями, ограничивающими темную сдвоенную спинную полосу типа «.exigua». По мере роста животного спинные полосы светлеют и постепенно исчезают, так и не разделившись
1 По Петерсу (Peters, 1960, 1962) с добавлениями.
71
на характерные или у других форм прытких ящериц темные, неправильной формы пятна. Взрослые самцы и самки в подавляющем большинстве случаев однотонно зеленые (реже оливково-зеленые или коричневатые), с мелкими коричневатыми пятнышками и крапинами на боках. Спинные полосы у них полностью отсутствуют или же очертания их вдоль хребта едва угадываются. Светлые глазки по бокам тела, если они имеются, выражены лишь в шейной области. Общий тип окраски в целом очень напоминает описанную выше аберрацию erythrono-tus.
Распространение. Встречается в Западной Грузии и на юго-западе Краснодарского края, где наиболее северные местонахождения известны из района Сочи. Вне СССР населяет районы Северо-Восточной Турции по крайней мере до Трапзона на юго-востоке. На юго-западе Краснодарского края граничит с подвидом L. a. exigua, где между ними имеется, по-видимому, зона интерградации (рис. 28).
{-rtt^rsonensis
* Прыткая ящерица дагестанская —
Lacerta agilis boemicaSuchow, 1929
Terra typica: Орджоникидзе, Северная Осетия.
1929 — Lacerta boemica, Сухов, Труды физ.-мат. вшд. Всеукр. Акад, наук XIII: 116.
1936 — Lacerta agilis boemia, Терентьев, Чернов, Кратк. опред. земн. и пресн. СССР, Л.: 34.
'Описание. L.mtn-max ad 84—107; L dd (n=156) 84—107 (87,6± ±1,29); L. 99 (n= 129) 60—97(81,21+1,19); L.cdmln_max 106—170; L.cd. dd (n=152) 106—170,6 (141,6+4,87), L.cd. $ 9 (n=126) L.cd.
86—156 (126,7+3,33); -y— dd (n=82) 1,58—2,05 (1,74+0,03) 99 (n=67) 1,39—1,70 (l,67±0,02); Sq.mln_max 38—49, dd-j-9 9 (Нальчик — Орджоникидзе, n=90) 39—49 (46,l±0,2); (Дагестан), dd (n=137) 38—49 (45,2+0,29), 99 (n=109) 38—48 (45,01+0,35); Ventr.mln_max 28—35 (Нальчик — Орджоникидзе), dd (n=65) 27—33 (26,9+0,2), 99 (n=67) 28—35 (28,7+0,1); (Дагестан); dd (n=132) 28—35 (30,5+0,15), 99 (n=109), 29—35 (32,0±0,2); P. 13—19 (Нальчик — Орджони-
кидзе), dd 13—18 (16,2+0,1), 99 13—18 (15,7+0,1); (Дагестан) dd 13—18 15,41+0,15), 99 13—19 (15,26+0,12). G-min-max 18—25 (20,69 ± 0,20).
Между верхнересничными и надглазничными щитками У 95% особей +=272) имеется 1—12 зернышек, образующих прерванный, реже сплошной ряд. Наиболее частое (87%) расположение щитков задненосовой области 2/i- В незначительной примеси встречаются также комбинации ®Л, 3/а и исключительно редко 2/0.
73
Анальный щиток, как правило, окружен двумя рядами пре-анальных (3—10 во внутреннем и 9—16 во внешнем). Центральные щитки внутреннего ряда увеличены у 55% особей (п = 212). Нередко (8—10%) впереди анального лежит один сильно расширенный преанальный. Индекс анального щитка 2,0+0,04 у самцов (« = 57) и 1,70+0,06 у самок («=46).
Окраска и рисунок. Верхняя сторона тела самцов зеленая, салатовая или оливково-бурая. Самки коричневато-бурые или коричневые, реже зеленые. Сдвоенная спинная затылочная линия «exigua» обычно хорошо выражена. Реже светлая затылочная линия простирается лишь до середины или задней трети спины, или же совсем отсутствует. В последнем случае, однако, слагающие ее пятна остаются разделенными на два параллельных ряда. На боках тела два или три продольных ряда светлых глазков, причем нередки случаи когда глазки среднего или нижнего ряда сливаются в пунктирную или даже сплошную полосу. У части особей рисунки отсутствуют вовсе (aberr. im-maculata) и тело одноцветно-зеленое или коричневато-бурое. У самцов в этом случае верхняя поверхность головы покрыта многочисленными червеобразными пятнышками и крапинами. У половозрелых самцов, а иногда и у старых самок в период размножения горло, бока и низ головы, включительно до груди, синие или фиолетово-синие. Брюхо желтоватое, зеленоватое или синевато-лиловое, часто с голубыми, синими или фиолетовыми пятнами на брюшных щитках, сливающимися иногда у самцов в яркие продольные полосы. Голубые, синие и фиолетовые тона особенно ярко выражены в период размножения, а также накануне ухода на зимовку осенью.
Распространение. Восточное Предкавказье от Дагестана на востоке через Чечено-Ингушетию и Северную Осетию, до центральной Кабардино-Балкарии на западе (см. рис. 28). В западной части ареала имеется зона интерградации с L. a. exigua.
Прыткая ящерица короткохвостая — Lacerta agilis brevicaudata Peters, 1958
Terra typica: Степанаван (Армения).
1878 — Lacerta agilis var. orientalis Кесслер (part.), Труды СПб., об-ва Еств. 8,2 : 151 (берег оз. Севан, Армения).
1939 — Lacerta agilis exigua, Чернов, Зоол. сборн. Арм. фил. АН СССР, 1 : 103. 1958 — Lacerta agilis brevicaudata Peters, Zool. Ib. Syst 86: 128 (Степанаван).
Описание. L.mm-max ad 76—96; L. dd («=110) 76—93 (81,2+ ±1,19); L.99 («=151) 82—96 (84,0+1,23); L. cd.mIn_mox 68—92; T cd
—j—- c? d («=96) 1,48—1,54 (1,52+0,01), 99 («=112) 1,32—1,45 (1,34+0,01); Sq.mln_max d*d+9 9 («=254) 40—54 (45,7+0,1), Ventr.mln_max25—32,dd («=110) 25—30 (27,2+0,1); 99 («=151) 25—32 (28,1+0,4); P.f.mln_maJ dd+99 11—19, dd («=110)
74
12-19 (14,5+0,1), 99 (n=147) 11-18 (14,2+0,1); G.ml„ (n=98) 15—23 (18,6 + 0,21).
Между верхнересничными и надглазничными щитками у 5% (ц=230) имеется от 1 до 3 зернышек. Наиболее частое (65%) расположение щитков задненосовой области 2/2. Реже наблюдается 7, (12%), 7о и 7о (до 10%), */2 и другие комбинации (5%). В целом для подвида очень характерно своеобразное расположение верхнего скулового щитка, выступающего на поверхность цилеуса и контактирующего с заднескуловым, предлобным, ло-боносовым и носовыми щитками. Подобная комбинация наблюдается в 78,2+1,8% случаев (рис. 29). Анальный щиток окружен двумя рядами преанальных, причем средние анальные внутреннего ряда обычно заметно увеличены. Индекс анального щитка 2,36+0,04 у самцов («=73) и 1,77+0,02 («=109) у самок. Таким образом, самцы по анальному индексу показывают максимум для вида.
Окраска и рисунок. Молодые сверху с тремя светлыми продольными ювенильными полосками. По мере роста животного они постепенно становятся менее четкими, а ограниченное ими вдоль хребта темное пространство разбивается на два продольных ряда крупных темно-бурых или черных пятен, образующих сдвоенную спинную полосу типа «exigua». Ряды темных пятен, особенно у самок, появляются и на боках тела, где они местами окантованы белым (рнс. 30). Общая окраска тела у самцов зеленая и салатовая, у самок коричневая, коричневато-бурая или реже, как и у самцов, зеленая. В последнем случае спинные полосы и боковые пятна полностью или частично исчезают (aberr. erythronotus). Встречаются и одноцветно-зеленые, без всякого рисунка самцы (aberr. immaculata). Нижняя сторона тела зеленоватая или белая, обычно с многочисленными темными пятнышками.
Распространение. Армянское нагорье в пределах Северной Армении и Южной Грузии, южные склоны центральной части Большого Кавказского хребта в Южной Осетии, а вне СССР — Северо-Восточная Турция. В Закавказье западнее Сурамского хребта не известна (см. рис. 28).
Прыткая ящерица морская — Lacerta agilis iorensis Peters et Muskhelischwili, 1968
Terra typica: долина p. Пори близ г. Тианети (Грузия). 1968 — Lacerta agilis iorensis Peters et Muskhelischwili, Zool. Ib. Sust. 95:214.
Описание1. L. min_max ad 74 — 90,5; L dtf (n=37) 74 — 90,5 (85+0,08); L. 99 («=22) 74—89 (81+0,10); L. cd.mln_max, 85—126; («=4) 1,42.-1,43; 99 («=2) 1,28—1,29;
1 По Петерсу и Мусхелишвили (Peters, Muskhelischwili, 1968).
75
Рис. 29
Различные комбинации расположения щитков задненосовой области у L. a. brericaudata а-—д) по сравнению с типичной комбинацией у L. a. exigua (е) (no Peters, 1958)
Sq.mm-max- d'd'4-99 (n=59) 38—46 (41,5+0,24); Ventr.raIn_mal 25—32, dd (n=37) 25—30 (27,6+0,16); 99 (n=22) 25—32 (29,0=0,33); P.f.mln-max cfcr+9 9 13-18, dd(n=37) 13—18 (16,0+0,13); 9 9 (n=22) 13—18 (15,4+0,18); G.mln_max
(n = 22) 15—24.
Между верхнересничными и надглазничными щитками у 73% особей (п=59) имеется одно (24% случаев), два (22%), три (17%) или совсем редко четыре зернышка. Наиболее частое расположение щитков задненосовой области — 2Л (наблюдается у 91,5% особей); остальные комбинации (4/2, 2/г, ‘А) встречаются в единичных случаях. Анальный окружен спереди двумя рядами преанальных щитков; средние щитки внутреннего ряда заметно расширены. Индекс анального щитка v самцов (п=31), 1,42—2,48 (1,94-0,04), у самок (п= 11) (1,51+0,07).
Окраска и рисунок. Сдвоенная спинная полоса типа «exigua» у самцов и самок в той или иной степени всегда выражена; слагающие ее пятна сильно варьируют по величине, иногда очень крупные в светлой окантовке. Светлые продольные линии в средней части и по краям спинной полосы имеются или отсутствуют. На боках ряды различных по величине темных пятен. Общая окраска верхней стороны туловища светло-зеленая у самцов и серовато-коричневая у самок. Брюхо зеленоватое у самцов и беловатое у самок. Зеленые самки по всей видимости отсутствуют.
76
Распространение. Известен только из типовой территории в долине верхнего течения р. Иори (близ г. Тианети) в северной Грузии (см. рис. 28).
Прыткая ящерица восточная — Lacerta agilis exigua Eichwald, 1831
Terra typica: Уральские горы.
1785 — Lacerta viridis Таблиц, Физ. опис. Таврич. обл. по ее местоположению и по всем трем царствам природы. СПб. (Крым).
1785 — Lacerta punctata Таблиц, там же (Крым).
1831 —Lacerta exigua Eichwald. Zool. spec. Ross. Polon., 3: 188. (Урал). 1831—Lacerta europaea Pallas, Zoograph. Rosso-As. Ill, Petropoli (Крым). 1834 — Lacerta sylvicola Eversmann, Nouv. Mem. Soc. Natural. Moscou, (2)3: 344 (между Симбирском и Сызранью).
1856 — Lacerta dilepis Lichtenstein, Nomenci. Rept. Amph. Mus. zool. berolin.: 14 (киргизские степи).
1874 — Lacerta doniensis Bedriaga, Entst. Farb. Eidechs.: 14 (Дон у Воронежа и Волга у Самары).
1878 — Lacerta agilis var. orientalis Kessler, Tp. СПб. о-ва еств. 8,2: 151 (юго-восток Европейской России).
1887 — Lacerta agilis var. exigua, Boulenger, Cat. Liz. Brit. Mus., 3:21.
1899 — Lacerta agilis var. altaica Kaschenko (syn. fide Boulenger 1920). Pe-зульт. алтайск. экспед.: 116 (Алтай).
1912 — Lacerta agilis var. concolor Schreiber, Herpetol. europ., Ed. 2:482, (Крым).
1912 — Lacerta agilis var. eremioides Schreiber, Herpetol. europ. Ed. 2:482, (Крым).
1926 — Lacerta agilis tauridica Suchow, 36. нраць зоол. муз. АН УРСР, 2 : 327 (Крым).
1960—Lacerta agilis exigua Mertens u. Wermuth, Die Amph. u. Rept. Europas.: 101.
Описание. L. mtn_max 30,0—114,0 (данные из разных мест ареала представлены в табл. Г.); L.cd.mln_max 36,0—190,5; —?—сМ (по данным из Крыма) de? 1,58—2,03 (А1= 1,74±0,02; п=27); 99 1,38—1,68 (М = 1,50 ±0,02; n=22). Sq. т1п-ти1 38—54 (данные из разных мест ареала представлены в табл. 2, кроме того: 42,10 ± 0,35 (для особей из Томска), 42,50 ± 0,46 (Горноалтайск), 44,93 ± 0,57 (ст. Иланская, восточнее Красноярска). Ventr. mln_max 3V 23—32 (см. табл. 2) 9$ 25—34 (видовой максимум). Р. f.min-maj, 12—18 (средние показатели см. табл. 2). G-min-max 15—23 (средние показатели — табл. 2).
Зернышки между верхнересничными и надглазничными щитками обычно отсутствуют (их количество несколько возрастает У западных популяций: так, у крымских особей они встречаются У 30,6 ± 3,8%, в Западном Предкавказье—только у 1,0 ± 0,5%,
77
Рис. 30
Различные типы рисунка верхней стороны тела у самок L. a. brevicaudata (а—в), •по сравнению с типичным рисунком самки L. a. exigua (г), в— зеленая самка , окрашенная по типу L. a. grusirrca (Peters, 1958)
в Горьковской обл. у 328 экз. ящериц зернышки не найдены). Задненосовых щитков обычно 2 (см. рис. 24), скуловых также 2, следовательно, в отличие от западной и южной прыткой ящерицы — у восточной наиболее обычная комбинация в задненосовой области 2/2. Количество скуловых, как правило, возрастает у восточных популяций. Так, у особей из Крыма комбинация 2/2 найдена у 33,4% особей, а 2Д у 25,8%- В Предкавказье максимум для комбинации 2/2 доходит до 97,0±3,1%. Преанальные щитки обычно расположены двумя полукругами (в отличие от номинативной формы), пара средних щитков внутреннего круга, как правило, увеличены. Анальный щиток у самцов почти всегда относительно шире, чем у самок; отношение его ширины к длине — 2,00—2,65. Широкая темная спинная полоса, разделенная светлой хребтовой линией занята 13—17 чешуями (у западных форм
78
прыткой' ящерицы здесь в поперечном направлении 8—14 чешуи). Эта полоса кроме того, в отличие от L. a. agilis, L. a. bos-nica, L. a. euxinica и L. a. chersonensis, не выделяется ребристостью чешуи.
Рисунок и окраска. Пилеус и спина половозрелых самцов преимущественно (до 75%) зеленая, самки бурые (иногда зеленый цвет бывает и у самок). Бурые самцы чаще (до 90%) встречаются в междуречье Волги и Урала. В основе рисунка спины восточных прытких ящериц три непрерывные светлые линии (две теменные и одна хребтовая). Как было показано выше, у особей западных форм в основе рисунка чаще выступают две светлые теменные линии. В зонах интерградации и некоторых других местах возможны находки особей, уклоняющихся от этой схемы рисунка спины. Так, например, теменные полосы могут прерываться (у особей из крайне восточных популяций). Наибольшее число отклонений окраски и рисунка отмечено у особей, из Восточного Казахстана и Алтая. Не исключено, что в результате специальных исследований там будут описаны новые формы или восстановлена форма «altaica», описанная еще в. 1899 г. Н. Ф. Кащенко.
79
Таблица 1
Размеры тела L. a. exigua из разных мест ареала (по данным В. К. Булахова^ Н. Ф. Константиновой, Н, Н. Щербака, В. К. Жарковой, Л. В. Турутиной, Н. М. Окуловой
А. С. Баранова)
п Пол L L. cd.
М±т М±т
240 d+9 Юго-Восток Украины 68,95 +0,23 159,3+1,48
253 <5 Крым 30—108; 84,7+1,20 36,3-190,5
9 30—114; 82+1,20 36,0—165,0
Рязанская обл., лесостепь
33 1 d I 73,33±1,03 1 114,0+2,51
1 9 | 71,9±1,64 116,6±3,5
Лес
54 | 71,44^3,67 1 122,0±4,45
9 1 75,42±1,23 116,6+3,5
Горьковская обл., популяция I
30 1 d 1 75,2+0,74 1 121,58+2,4
63 1 9 I 74,48±0,76 1 115,98±1,2
Популяция II
32 1 1 d 1 I 73,31+1,31 1 | 126,36+2,96
87 1 9 1 80,55+0,84 120,71+1,6
Популяция III
36 1 J 1 73,92+1,43 1 119,6+2,72
80 1 9 1 79,59+1,04 1 122+1,8
Междуречье Волга—Урал
26 <5 80,3+3,8 135,1+8,6
19 9 76,8+3,7 126,9+5,7
Окрестности г. Кургана
8 М=80,56 М=121,25
16 9 М=83,91 М=128,75
Рисунок верхней поверхности туловища как самцов, так и самок, дополняется яркими светлыми «глазками», которые в два—три ряда расположены на боках. Горло самцов желто-зеленое у 50—75% особей. В окраске брюха как самцов, так и самок часто присутствуют зеленые тона (в Восточном Казахста-
80
Скуловые щитки, М или % СО o' -н XF Q 2 у 51,2% о 11 1 ед кО । i5 1 т-1 +1 со СО 1,59+0,05 1,76+0,05 го со
Задненосовые щитки, М или % СО О, o' -Н о eq со ед ЧДЭ о+ Ю eq 1 1 1 1 1 О Н 1 о ej 1 1 xF О +j 1 со О СО 1 О 4-1 со 1 СО 44 03 со । 5 со ттч т-1
а Ьл И о rf ГО ЕЕ 1 +1 г 6 хН o' 4-1 СО_ оГ т-1 1 1 1 1 1 1 1 о o' й 1 ед ед О -н eq о xF г- о +1 о eq eq со о" о" +1 +1 eq со Ю 05 05 20,73±0,27 20,52+0,24 ко eq сГ +1 со со С5 20,1+0,16 20,56+0,19 o' +1 1 о со о’ ед со СО ю eq eq
о О. га 3 а м и» X го 1 Ё с+] а О о' +1 т-1 12-18 14,7=1=0,06 1 1 1 1 00 1 ед Т-< со Л 1 о КО nf XF +1 eq xF со eq F 00 xf со •ч-i eq к-со КО о" о +1 -н о S 14,77±0,22 14,32+0,14 СО ? Ю 00 СО o' 4-1 со кО СО о” +1 1 со SF, 3 coco
л и ц а 2 и фолидоза из р X го Е is ~+1 J? eq тН о" и т-* ко о eq 1 1 о eq 1 со eq о о +1 со со eq T-i со 1 Ю eq eq Т“< 1 СО eq 1 1 eq о -н КО СО ед eq +i со 28,21+0,65 30,01±0,75 СО чн eq -н eq о о" о +1 +1 +1 СО «4-1 ' со *<F со ei о СО со NF ЧтН o' о е5 eq со eq О -н о Й О +1 1 СО С5 eq со 1 1
Таб торым признака! ХЕШ— ЩШфд ei о* й ед NF 38-50 43,73+0,14 1 1 *<F КО •nF 1 1 СО vr 45.66+0.39 44.48+0.59 xF eq хн 00 +i +1 ei о СО xF xF xF со со ко eq со *<F со eq о o' о о +1 +1 +1 +1 чгИ хГ О со - СО со СО SF - - ST nF nF 'F со л ОЭ СО СО eq о" 41 со со ^F о г? KO CO sF *<F
1 | О О К О + + *0 *о от О+- ’О оь ’О оь оь а *О оь ’OOb
S К а Kt ьг с eq СО КО eq S со со *cF ко О СО СО СО ei со СО со со О ОО co co T“1
Сравнение о аз го К о го Юго-Вост. УССР (Булахов и др.) Крым (Щербак, 1966) Западное Предкавказье (Лукина) га m S о га * ч с tK га * а Д га г а из с н о и ф Ф га н Н to ХО о к га р: С tc и: С и 2 и: с, С ь с Сухобезводное га X S д га 1 е и ► S' 0 Д ra ra 0^ к га 4 д я й га я
не встречено 10—45% самок с зеленым брюхом, напротив, зеленые тона отсутствуют в окраске брюха особей из междуречья Волги и Урала), Встречаются и полные меланисты.
Распространение. Западная граница ареала подвида проходит приблизительно (широкая зона интерградации) через 'Ленинград—Новгород—Калинин—Москву—Курск—Днепропетровск— Крым. На юге ее распространение ограничено северными склонами Большого Кавказа, на востоке—граница видового ареала, доходящая на востоке до Байкала, Тувы и Иссык-Куля (см. рис. 28).
Прыткая ящерица боснийская — Lacerta agilis bosnica Schreiber, 1912
Terra typica: Босния (Западная Болгария).
1897 — Lacerta agilis var. spinalis Werner (Part) Rept. Amph. Oesterr.—Ung.; 39.
1912 — Lacerta agilis var. bosnica Schreiber, Herpet. Europ. ed 2:483, 944 (Босния).
1940 — Lacerta agilis bosnica, Mertens und L. Muller, Abh. senckenberg, naturf. Ges. Frankfurt am Main, 451 : 43.
Описание. Наименее изученная из всех подвидовых форм прыткой ящерицы. L.m)n_max dd+$? 72—87; Sq.mln_mai dd-)-?? (n=15) 30—39, в среднем 36; P.f. 10—14, в среднем 12; G.mln_max 13—17, в среднем 15.
Наиболее обычное расположение щитков задненосовой области (около 60%)—’/1- Реже встречаются соотношения */2 и 2/2 и 2/с- Спинная чешуя несет на себе сравнительно слабо выраженные ребрышки и не резко отличается от боковой. Анальный щиток окружен спереди обычно одним рядом преанальных.
Окраска и рисунок. Ограниченная светлыми теменными линиями спинная полоса типа «exigua» разделена посередине сплошной затылочной линией. Тело самцов полностью или только на боках зеленое; в последнем случае верхняя сторона головы и спина бурая или буровато-коричневая. Самки серовато-коричневые или бурые.
Распространение. Западная и, вероятно, северо-восточная Болгария, Югославия (см. рис. 28).
Прыткая ящерица западная — Lacerta dgilis agilis Linnaeus, 1758
Terra typica restricta: Южная Швеция.
1804 — Lacerta agilis grisea Hermann, Observ. zool., 1 : 258 (Штрассбург). 1826 — Lacerta agilis var. erythronotus Fitzinger, Neue Classif. Rept.: 51.
1874 — Lacerta agilis var. ischliensis Bedriaga, Entst. Farb. Eidechs.: 18 (Him.) 1897 — Lacerta agilis var. albolineata Diirigen, Deutschl. Amph. Rept.: 151 (Германия).
1897 — Lacerta agilis var. imrnaculata Diirigen, Deutschl. Amph. Rept. : 153 (Германия).
1897 — Lacerta agilis var. melanota Diirigen, Deutschl. Amph. Rept.: 153 (Германия).
82
1897 — Lacerta agilis var. nigricans Diirigen, Deutschl. Amph. Rept.: 153 (Германия).
1897—Lacerta agilis var. annulata Werner, Rept. Amph. Osterr.— Ungarn.: 30 (Австро-Венгрия).
1897 — Lacerta agilis var. dorsalis Werner, Rept. Amph. Osterr.— Ungarn.: 30 (Австро-Венгрия).
1897 _ Lacerta agilis var. spinalis Werner, Rept. Amph. Osterr.— Ungarn.: 30 (Австро-Венгрия).
1915 — Lacerta agilis agilis, Никольский, Фауна России, Рептилии, 1:292.
Описание1. L.mln_max ad. 50—98, L. de? (n=68) 50—86 (69,2± ±1,13); L. 99 (n=73) 65—98 (78,1 ±0,73); L. cd.mln_max dd 102-130 (M=119); 99 80—138 (M= 115,41). dd (n=33) 1,24—1,86; 99 (n=36) 1,18—1,81. Sq.mln_max 33—49, dd (n=53) 35—46 (39±0,36); 9 9 (n=73) 33—45 (38±0,23). Sq. d d + 9 9 2 (n=15) 40,26. Ventr.mln-max 25—32, dd (n=56) 25—30 (28±0,2); 99 (n=68) 28—32 (30±0,12); P.f.mln-max 9—16, dd (n=53) 11—15 (12±0,15; 13±0,16); 99 (n=72) 9—15 (13±0,21); dd+992 (n=l5) 12, 73; G.mln_mai 14—22, dd (n=52) 14—22 (18±0,2); 99 (n=69) 14—22 (18±0,18). G. dd+99 2 (n=15) 19,53.
Зернышки между верхнересничными и надглазничными щитками имеются у 36% (п=152). Характер щиткования задненосовой области (задненосовые/скуловые) =‘/2. У особей из Румынии (п=252) 1/з==36,5%; 1/1=34%; ‘Л—‘/2=10,3%, остальные комбинации не превышают 3,5 %. У особей из Англии (и = 15) задненосовых 1—63,4%, 2—36,6%, скуловых 0—3,3%, 1—9,9%, 2—83,5%, 3—3,3%. Преанальные щитки расположены в один ряд (.44=7,46 2), два ряда встречаются редко, и только у особей из восточной части ареала подвида. Анальный щиток у dd всегда шире, чем у 99.
Окраска и рисунок. У самцов пилеус и спина бурые (до 100%), бока зеленые, самки— бурые.. Спинные линии — белые, как правило две; иногда выражены 3 линии: две теменные и одна хребтовая (сплошная отсутствует не менее, чем у 75%), у самок сплошные полосы на боках отсутствуют у 100%. Темная спинная полоса от темно-коричневого до красно-коричневого цвета, ее ширина соответствует размерам двух внутренних половин теменных щитков или 8—14 килеватых чешуй поперек, посередине более светлая (серая или светло-коричневая). На этом фоне может быть заметен пунктир более светлых пятен (остатки хребтовой линии). На темной спинной полосе часто выделяются разнообразные по форме темно-коричневые или черные пятна. Бока туловища с одним-двумя рядами темных пятен и «глазков».. Брюхо — зеленое, у самок — беловатое, иногда с темными пятнышками.
По Фун, Ванци (Fuhn, Vancea, 1964), материалы из Румынии.
По материалам из Англии (колл. Брит, музея).
83
Распространение: Франция, Бельгия, Голландия, Дания, Англия, Швеция, ФРГ, ГДР, Польша, Швейцария, Австрия, Венгрия, Чехословакия, Северо-Западная Югославия, Западная Румыния на юг и восток до границы предгорий Карпат и равнинных степей, в Молдове до р. Серет). В СССР особи, относимые к номинальной форме, найдены в Закарпатской и частично во Львовской и Ивано-Франковской областях УССР, а также на западе Молдавии. Восточнее начинается зона гибридизации с L. a. chersonensis (рис. 28).
Прыткая ящерица южная — Lacerta agilis chersonensis Andrzejowski, 1832
Terra typica restricta: Херсон; Волынь (УССР).
1832 — Lacerta chersonensis Andrzejowski, Nouv. Mem. Soc. Natural. Moscou, (2), 2 : 327.
1916 — Lacerta agilis var. chersonensis, Boulenger, Transact, zool. Soc. London, 21 :27.
1949 — Lacerta agilis agilis (part.) Терентьев, Чернов. Определитель пресмык, и земн. М.: 181.
1960 — Lacerta agilis chersonensis Mertens u. Wermuth, Die Amphibien und Reptilien Europas: 101.
Описание1 L. mln_roai, ad 64 — 94; L. dd (n—23, Валахия) 64—82 (72,9+1,05); («=47, Молдова) 68—91 (78,7±0,75); L. 22 («=25, Валахия) 64—84 (71,1 + 1,02); («=31, Молдова) 70—94 (83,8 + 1,05). L. cd. dd 98—156 (115); L. cd. 2£ 84—117 L. cd.
(103). dd (n=6, Валахия) 1,58—1,90 (l,73±0,04);
(n=13, Молдова) 1,60—1,75 (1,69+0,008); ?? («=15,
Валахия) 1,30—1,64 (l,49±0,02); (n=13, Молдова) 1,36—1,54 (l,46±0,13). Sq.mln_max dd-j- 9 2 («=65, Валахия) 37—49 (41,5± +2,13); (n=79, Молдова) 36—47 (40,2+0,25). Ventr.^m-^, dd («=34, Валахия) 25—29 (27,1+0,2); («=49, Молдова) 26—29 (27,7±0,14); Ventr. 22 («=31, Валахия) 26—32 (29,2+0,2); (« = 31, Молдова) 28—34 (29,3±0,2). P. f. mln_max dd + 22 (« = = 65, Валахия) 11—18 (14,5+0,16); P. f. dd («=49, Молдова) 12—19 (14,5+0,15); P. f. 29 (n=31, Молдова) 13—17 (14,8+ +0,04). G.mln_max dd+2 2 («=65, Валахия) 15—23 (18,8+0,2), («=80, Молдова) 14—25 (18,4±0,19).
Зернышки между верхнересничными и надглазничными (« = = 172) найдены у 4% особей. Задненосовых щитков 1—2, чаще один. Скуловых 0—2, чаще 1. Наиболее частая комбинация щитков в задненосовой области */i (59%), 11г (10,4%) и 2/, (10%). Остальные комбинации занимают незначительное ме
1 По Фун, Ванци (Fuhn, Vancea, 1964), материалы (п=72) из юго-восточной Румынии.
84
сто (от 0,6 до 7%). Преанальные щитки расположены в два ряда, редко в один. Почти у всех ящериц пара средних преанальных щитков заметно увеличена (признак, сближающий с L. a. exigua).
Рисунок и окраска. Пилеус и спина половозрелых самцов — зеленые (до 100%), молодые и самки — бурые. Темная непрерывная спинная полоса по бокам окаймлена узкими светлыми (теменными) линиями, которые у 55—90% особей прерывистые. Центральная (хребтовая), как правило, отсутствует (не менее, чем у 75% особей). На темной спинной полосе выделяются более темные (черные) пятна средней величины или точки. На боках туловища заметны (до трех) ряды белых глазков. Изредка встречаются у этих ящериц цветовые аберрации: imma-culata Dur. (спина зеленая, без пятен) и erythronotus Fitz, (спина серая или красно-бурая, без пятен). Брюхо самцов зеленовато-голубоватое у 100%, самок — зеленое до 75% особей.
Распространение. Румыния южнее и восточнее Карпат (кроме Добруджи и прилегающего побережья Черного моря). В СССР — Молдавия и правобережная Украина (на север до Черкасс), а также Голопристанский район Херсонской области. Широкая зона интерградации с восточным подвидом прослеживается на север в Белоруссии, восточной Литве, до восточной Фенно-Скандии, а также на восток до Курской и Белгородской областей (см. рис. 28).
Прыткая ящерица добруджинская — Lacerta agilis euxinica Fuhn et Vancea, 1964
Terra typica — Караорман, дельта Дуная, Румыния.
1961 — Lacerta agilis chersonensis Fuhn et Vancea, Fauna RPR, Reptilia, XIV, 2 : 197.
1964 — Lacerta agilis euxinica Fuhn et Vancea, Senck. biol., 45, 3/5 : 481.
Описание1. L.min_max, ad. 44—91; L. dd 44—91 (73±3,08);
L. 99 54—90 (68+0,86) dd 1,65—1,92 (l,82±0,02), 9?
1,43—1,75 (1,68+0,02) (сравнительно c L. a. chersonensis более длиннохвостые особи). Sq.mm-max 35—46 (39,1+0,39); d <d 38—46 (40,7+0,54), $2 39—45 (41,2+0,6). Ventr.mIn_mM 23—32; dd 23—29 (26,4±0,5), 29 27—32 (29,5+0,39). P.f.mln_max 12—16; dd 13—16 (14,3+0,02), 92 12—15,5 (14+0,26). G.mln_max 15—22 (18,8±0,4). Задненосовых щитков 1—2, чаще один. Скуловых щитков 0-—2, чаще один. Наиболее частая комбинация Щитков в задненосовой области */, (60%), в отличие от L. a. agi-lis, у которой на втором месте стоит комбинация ’/2, здесь 71
1 По Фун, Ванци (Fuhn, Vancea, 1964) материалы (л = 57) из Добруджи (Румыния).
85
(10,5%). Преанальные состоят из двух рядов, среди которых пара средних, как и у L. a. exigua, увеличены.
Рисунок и окраска. Пилеус у самцов светло-коричневатый, спинная полоса от песочного до шоколадно-коричневого цвета доходит до конца хвоста. Снаружи она окаймлена ярко белыми теменными линиями, которые не прерываются (так же, как и у самок). Форма, наиболее близкая к L. a. chersonensis.
Распространение. Обитает только в Добрудже, вблизи морского побережья, на песчаных пляжах и дюнах (см. рис. 28).
КРАТКИЕ СРАВНИТЕЛЬНЫЕ ХАРАКТЕРИСТИКИ ПОДВИДОВЫХ ФОРМ ПРЫТКОЙ ЯЩЕРИЦЫ
Наиболее частое (40%) расположение щитков задненосовой области 70; реже наблюдаются комбинации */i и */г; более чем у половины (60%) особей в популяции скуловой щиток не выражен. Зернышки между верхнересничными и надглазничными щитками в числе 1—2 имеются у 7% особей. Sq. максимальное для вида (44—54; М—49,16). Р. L 12—18; 7И=15,03. Средний индекс анального щитка 1,98 у самцов, 1,79 у самок. Отношение длины хвоста к длине тела в среднем 1,79 у самцов и 1,61 у самок. Верхняя сторона тела самцов и самок зеленая, оливковая или коричневая, одноцветная без рисунка или со слабо проступающей сдвоенной спинной полосой типа «exigua» без окантовки из светлых линий. Бока тела с мелкими пятнышками, светлые глазки, если они имеются, расположены лишь в области шеи...............................L. a. grusinica
Самое частое (до 65%) расположение щитков задненосовой области 2/2. Верхний скуловой обычно выступает на верхнюю сторону головы, где соприкасается с заднескуловыми, предлобным, лобоносовым и носовыми щитками. Зернышки между надглазничными и верхнересничными щитками в числе 1—3 имеются у 5% особей. Sq. 40—54, 714=45,7; Р. f. 11—19, М= 14,2. Средний анальный индекс 2,36 у самцов и 1,77 у самок (абсолютный максимум для (вида). Отношение длины хвоста к длине тела 1,48—1,54 у самцов, 1,32—1,45 у самок (одна из наиболее короткохвостых форм). Верхняя сторона тела самцов зеленая или салатовая, самок — коричневая, коричнево-бурая или, как у самцов, зеленая. Спинная полоса типа «exigua» слагается из сдвоенного ряда темных пятен; такого же типа пятна особенно крупные у самок, имеются обычно на боках тела, где они бывают окантованы белым (рис. 30). Встречаются особи с окраской и рисунком, как у L. a. grusinica, а также одноцветные, без всякого рисунка...........................L. a. brevicaudata
В задненосовой области резко преобладает (до 92% особей) комбинация 2/ъ в остальных случаях чаще всего наблюдается 2/г- Между верхнересничными и надглазничными щитками у 97% особей располагается до 12 зерньцпек, образующих пре-
86
рванный или сплошной ряд. Sq. 38—49, Л4=49,2; Р. f. 13—19, 714:= 15,9 (абсолютный максимум для вада). Средний индекс анального щитка у самцов 2,0, у самок—1,77 (у самок резко снижен). Центральные щитки внутреннего ряда преанальных увеличены примерно у 60% особей; нередко впереди анального расположен один крупный преанальный. Отношение длины хвоста к длине тела в среднем 1,74 у самцов и 1,67 у самок, половой диморфизм, в отличие от других подвидов, по этому признаку не выражен. Окраска верхней стороны тела самцов зеленая или оливковая, у самок коричневая, коричневато-бурая, реже — зеленая. Спинная полоса типа «exigua» обычно хорошо выражена, встречаются особи лишенные рисунка вовсе. В период размножения и осенью горло, бока головы и шеи, а также грудь приобретают яркую сине-голубую окраску; синие, голубые или фиолетовые пятна появляются иногда и на брюхе.......................................L. a. boemica
В задненосовой области резко (до 92%) превалируют 2/i, в остальных случаях наблюдаются г/2 или */2. Между верхнересничными и надглазничными щитками у 64% особей лежит от 1 до 4 зернышек Sq. 38—46, 7И=41,5; Р. f. 13—18, 714=15,8. Средний индекс анального щитка 1,9 у самцов и 1,5 у самок. Отношение длины хвоста к длине тела 1,42—1,43 у самцов и 1,28— 1,29 у самок (самый короткохвостый подвид). Окраска зеленая или оливковая у самцов, коричневая или коричневато-бурая у самок, зеленые самки видимо отсутствуют. Пятна на спинной полосе типа «exigua» четкие или слабо заметны, в последнем случае ограничивающие их светлые линии почти не выражены. ....................................... . . . L. a. iorensis
Наиболее обычная (обычно более 50%) комбинация щитков задненосовой области г/2, реже наблюдаются 2/ъ 2/о или 3/0. Зернышки между верхнересничными и надглазничными щитками на большей части ареала обычно отсутствуют, однако в Крыму они в числе 1—3 имеются у 30% особей. Анальный индекс 1,83—2,65 у самцов и 1,51—1,73 у самок. Анальный щиток обычно окружен двумя рядами преанальных, два средних внутреннего ряда как правило увеличены. Sq. 38—54, 7Vl=45,0; Р. fm. 12—18, М=14. Отношение длины хвоста к длине тела 1,58— 2,4 у самцов и 1,38—1,69 у самок. Верхняя сторона тела самцов преимущественно зеленая, реже бурая. Самки бурые, гораздо реже зеленые. Сдвоенная спинная полоса обычно хорошо выражена с четкими светлыми краевыми и срединной линиями. Встречаются и одноцветные, лишенные всякого рисунка особи. На боках тела самцов и самок в 2—3 продольных ряда обычно располагаются светлые, иногда в темной окантовке, глазки.
•...................................... . . . L. a. exigua
Наиболее обычная (более 60%) комбинация щитков задненосовой области */,, реже наблюдаются . ‘/г или г/2. Зернышки между верхнересничными и надглазничными щитками обычно
87
'отсутствуют. Sq. 30—39, Л4=36; Р. f. 10—14, Л4=12. Верхняя сторона тела зеленая или салатовая, у самцов, и коричневая или буровато-коричневая у самок. Сдвоенная спинная полоса типа «exigua» хорошо выражена и разделена сплошной светлой затылочной линией................................L. a. bosnica
Наиболее обычная комбинация щитков задненосовой области (до 60%) —‘/б реже наблюдается 1/2 или 71- Зернышки между верхнересничными и надглазничными щитками крайне редки, встречаются не более чем у 4—5% особей. Преанальные расположены обычно в два ряда, пара средних преанальных, как правило, увеличена Sq. 36—49, 7И=40,85, Р. f. 11—19, Л4=14, 5. Отношение длины хвоста к длине тела 1,58—1,90, Af = l,73 у самцов и 1,30—1,64, М—1,49 у самок. Верхняя сторона тела самцов всегда зеленая, самок — бурая. Темная спинная полоса «agilis», окаймленная по краям светлыми, сплошными или прерывистыми линиями. Светлая срединная линия примерно у 75% особей отсутствует. На боках туловища до 3 продольных рядов светлых глазков. Встречаются особи, у которых рисунок полностью или частично (на спине) отсутствует...................
........................................L. a. chersonensis
Наиболее обычное расположение щитков задненосовой области (60%) —71, реже наблюдается комбинация z/i- Зернышки между верхнересничными и надглазничными щитками, как правило, отсутствуют. Анальный щиток окружен спереди двумя рядами преанальных, два средние щитка внутреннего ряда увеличены. Sq. 35—46, 714=40,1; Р. f. 12—16, Л4= 14,1. Отношение длины хвоста к длине тела 1,65—1,92, М= 1,73, у самцов и 1,43—1,75, 7И= 1,68 у самок.
Верхняя сторона тела самцов, как правило, зеленая, самок -— коричневая или бурая. Спинная полоса типа «agilis» песочного или шоколадно-коричневого цвета, окаймлена сплошными белыми линиями. В редких случаях имеется пунктирная срединная линия................................L. a. euxinica
Самая обычная комбинация щитков задненосовой области 7г, редко 71 или другие комбинации. Единичные зернышки между верхнересничными и надглазничными щитками в разных местах ареала имеются у 5—35% особей. Анальный щиток окружен спереди обычно одним рядом преанальных; особи с двумя рядами наблюдаются в восточных районах ареала. У самцов анальный всегда шире, чем у самок. Sq. 33—49, Л4=38,5; Р. f. 9—16, М=Г2,5. У самцов спина и верхняя сторона головы бурые, бока — зеленые. Самки сплошь бурые. Спинная полоса типа «agilis» темно-коричневого или красно-коричневого цвета с окаймляющими ее прерывистыми или сплошными белыми линиями. Срединная линия, если она имеется (примерно у 25% особей), всегда прерывистая. Бока тела с 1—2 рядами темных пятен и глазков..................................L. a. agilis
88
ЗОНЫ ИНТЕРГРАДАЦИИ ПРИЗНАКОВ И ГИБРИДИЗАЦИЯ
Наличие более или менее выраженных зон интерградации между подвидами — явление, широко распространенное у пресмыкающихся, и прыткая ящерица не представляет в этом отношении исключения.
С. Д. Перелешин (1928) был, вероятно, одним из первых, кто обнаружил на юге Московской области зону соприкосновения подвидов L. a. chersonensis и L. a. exigua. К сожалению, при биометрической обработке особей из смешанных популяций он сознательно отказался от анализа признаков окраски и рисунка,, как не поддающихся, по его мнению, точному математическому выражению. Тем не менее, вычислив так называемое «среднее квадратичное расстояние» (т. е. величину, характеризующую отклонение признаков отдельных особей от принятого стандарта), Перелешин обнаружил своеобразное нарастание различий между сравниваемыми формами в зоне контакта и высказал предположение о связи этого явления с вторичной интерградацией. Аналогичная зона взаимного проникновения названных подвидов была обнаружена А. К. Крень (1939) в Белгородской области в районе «Лес на Ворскле». Позднее Г. Петерс (.в печати) показал, что обитающие здесь прыткие ящерицы характеризуются повышенной степенью изменчивости ряда признаков, обусловленной вторичной гибридизацией. Что касается в частности рисунка, то было установлено что в зоне контакта встречаются самые различные варианты, включая крайние его типы, характеризующие тип «exigua», с одной стороны, и тип «agilis», с другой (рис. 31). Такого же рода зона интерградации признаков между этими подвидами обнаружена и в Калужской области (Стрельцов, Воронин, 1973). В настоящее время считается установленным, что зона вторичного контакта между восточным и западным подвидами прыткой ящерицы занимает обширную территорию в пределах северо-восточной Украины, Белоруссии и восточной Литвы на западе до Белгородской, Курской, Калужской, юга Московской, Калининской и части Ленинградской областей на востоке (см. рис. 28).
В литературе имеются данные о существовании промежуточных популяций также между L. a. agilis и L. a. chersonensis в северо-западной Румынии (Fuhn, Vancea, 1964) и в западной Украине (Щербак, 1966). Некоторые факты позволяют предполагать переходную зону и между южным подвидом L. a. grusini-са и восточным — L. a. exigua, ца что указывает, в частности, существование зеленых самок последнего на крайнем юго-западе Краснодарского края. Наконец, Шебзухова (1969) привела известные доказательства совмещения ареалов L. a. boemica и A. a. exigua в Кабардино-Балкарии. Как и следовало ожидать, промежуточные популяции не поддаются точному определению и не могут быть альтернативно целиком отнесены ни к одному
89
Рис. 31
Вариации спинного рисунка в зовах вторичной интерградации Л. a. exigua и L. a. cherso-nensls
Верхний ряд — «Лес на Ворскле», Белгородская обл., средний ряд — окр. г. Курска, нижний ряд — д. Куриловка, Днепропетровской обл. (виден постепенный переход от рисунка с полосой типа «agilis» (крайние слева) к типу рисунка «exigua» (крайние справа) (рис. Петерса)
из пограничных подвидов. Рассуждая теоретически, в зонах контакта должны обнаруживаться также и «чистые» особи, однако учитывая, что обмен генами происходит здесь очень длительное время, чистоту их следует признать лишь относительной. Во всяком случае, проведение специального гибридологического анализа крайне желательно, причем особенно перспективным представляется в этом плане применение метода электрофореза.
Остановимся также на вопросе о 'возможной гибридизации прыткой ящерицы с другими представителями рода Lacerta. Известно, что симпатрические виды этого рода сравнительно легко гибридизируют в природе, следствием чего является появление гибридных особей (Mertens, 1950, 1956, 1964, Даревский 1967).
Что касается, в частности, прыткой ящерицы, то в разных местах своего ареала она контактирует с видами Lacerta viridis, L. trilineata и L. strigata, причем местами, например на Кавказе, зоны симпатрии имеют значительное протяжение. Детальное изучение особей из смешанных популяций L. agilis и L. strigata в Северной Армении действительно свидетельствует, по-види: мому, о существовании здесь промежуточных особей, хотя воп
90
рос этот нуждается еще в изучении (А. А. Симонян, уст. сообщ.) Отсутствие значительной интерградации признаков между этими видами косвенно свидетельствует о действенности докопуляцион-ных изолирующих механизмов, заключающихся, возможно, в различиях сигнальной окраски. В данной связи интересно отметить, что в Дагестане в зонах контакта указанных видов наблю-даются особи L. agilis, у которых глазки на боках тела сливаются в светлые продольные линии, что в норме характерно для L. strigata (Кутузова, уст. сообщ.). Не исключено, что в данном случае это объясняется интрогрессией генов в результате гибридизации этих видов.
Возможность гибридизации полосатой и прыткой ящериц, (а также близких видов этого рода — средней и зеленой) в последнее время была доказана экспериментально (Bischoff, 1969* 1973; Nettmann, Silke, 1974).
ПРОИСХОЖДЕНИЕ И ФИЛОГЕНИЯ ВИДА LACERTA AGILIS
Едва ли можно сомневаться в том, что описанная выше картина географического распространения и 'внутривидовая структура прыткой ящерицы в историческом плане сложились относительно недавно, только в послеледниковое время. Однако в целом как вид Lacerta agilis, судя по ряду косвенных доказательств, возникла в пределах современного Кавказского перешейка значительно раньше, скорее всего уже на границе миоцена и плиоцена.
Мы уже говорили, что многие важные морфологические признаки рассматриваемого вида обнаруживают четкую радиальную изменчивость с центром кульминации на Кавказе. К их числу относится количество чешуй вокруг середины тела (максимум у L. a. grusinica в Западной Грузии), число бедренных пор (максимум у L. a. boemica в Кабардино-Балкарии, Северной Осетии и Дагестане), величина анального индекса (максимум у L. а. Ьге-vicaudata в Армении), комбинация 2Л в щитковании задненосовой области (максимум у L. a. boemica) и максимальное число «зернышек» между надглазничными и верхнересничными щитками (£. a. boemica и L. a. iorensis в восточном Предкавказье и Восточной Грузии). По ряду признаков, включая окраску, именно на Кавказе прыткая ящерица обнаруживает наибольшее сходство с встречающимися здесь родственными видами L. strigata и L. trilineata (наличие синих и голубых тонов в окраске самцов, большее число бедренных пор и «зернышек» и некоторые другие признаки). Рисунок L. a. grusinica, связанный у этого подвида с максимальным числом чешуй вокруг середины туловища, также возможно наиболее близок к исходному, что доказывается в частности отсутствием в онтогенезе пятен на темных спинных полосах у молодых особей.
91
Рис. 32
Схема вероятных филогенетических связей подвидовых форм прыткой ящерицы
но и образование L. a. agilis, вое время от L. a. chersonensis.
в Балканском ледниковом рефугиуме, то, по-видимому, она дала, с одной стороны, начало болгарско-югославскому подвиду L. a. bosnica (до настоящего времени сохранившего сдвоенную спинную полосу типа «exigua»), а с другой, ди-вергировала в подвиды L. а. chersonensis и L. a. euxinica, которые приобрели спинную полосу типа «agilis». Начавшие расселяться в послеледниковое время по Евро-Азиатской равнине восточная (L. a. exigua) и западная (L. a. chersonensis) популяции прыткой ящерицы встретились затем в районе Днепра и образовали здесь со временем обширную зону вторичной интерградации, подробно о которой мы уже говорили выше. По-видимому, с этим процессом расселения связа-отделявшейся в послеледнико-
На молодость номинативного
подвида указывает его сравнительно слабая изменчивость в рисунке и признаках чешуйчатого покрова.
Изложенные выше соображения о филогенетических отношениях внутри вида Lacerta agilis наглядно представлены схематически (рис. 32). В заключение следует подчеркнуть, что процесс внутривидовой дивергенции несомненно продолжается у прыткой ящерицы и в настоящее время, что относится прежде всего к обладающему очень широким ареалом восточному подвиду L. a. exigua. Примером такой дивергенции, не достигшей .подви-
дового уровня, служит в частности изолированная таежная популяция прыткой ящерицы в окрестностях г. Иланского в северных предгорьях Восточных Саян. Как свидетельствуют данные Чугунова (1911) и позднейшие исследования Петерса, особи из этой популяции по ряду признаков (количеству чешуй вокруг середины тела, анальному индексу, абсолютному максимуму комбинации 2Д в задненосовой области, повышенному числу брюшных щитков, отношению длины хвоста к длине тела) значительно уклоняются от соседних популяций в Сибири и на Алтае. Все это вместе взятое позволило в свое время С. М. Чугунову отнести иланских ящериц к особой вариации L. a. var. altaica Kas-tschenko. Вопрос о подвидовой самостоятельности алтайских ящериц нуждается, однако, в дальнейшем изучении. Можно также
94
упомянуть находящуюся в изоляции на берегах оз. Иссык-Куль киргизскую популяцию L. a. exigua, несколько уклоняющуюся от подвидовой нормы и находящуюся, по мнению Петерса (Peters, 1958), на пути превращения в горную ящерицу, т. е. повторяющую путь, пройденный в свое время на Кавказе L. a. brevi-caudata. Перспективна в эволюционном плане и описанная этим автором (Peters, 1959) изолированная популяция L. a. cherso-tiensis в долине р. Свирь на северо-востоке Ленинградской области. Несомненно, что примеры такого рода в пределах видового ареала Lacerta agilis в целом могут быть умножены.
Изучение географической изменчивости указывает также на существование в различных частях видового ареала так называемых сгущений частот встречаемости отдельных признаков (фенов). Это обстоятельство при изучении внутривидовой изменчивости прыткой ящерицы еще не получило необходимого освещения, и здесь открывается широкое поле для дальнейших исследований.
ГЛАВА ТРЕТЬЯ
МОРФОЛОГИЯ 1
При всяком монографическом исследовании вида морфологические данные являются одними из исходных и важнейших для понимания места вида в биогеоценозах (через определение массы тела отдельных возрастно-половых групп) и для понимания особенностей современного этапа микроэволюционного процесса в элементарных эволюционных единицах — популяциях— данного вида и в их группах. Данные морфологии оказываются важными для систематики, экологии и этологии. В то же время, несомненно, что достаточно подробное морфологическое описание даже одного вида само по себе требует отдельной монографической работы. Поэтому в настоящей главе делается попытка дать лишь основные черты морфологии в расчете на то, что лица, заинтересованные в специальном морфологическом исследовании, смогут использовать материал настоящей главы как своего рода «стартовую площадку» для углубленного изучения в любой из основных областей современной морфологии— функциональной, сравнительной или попу-ляционной. При этом описание общей морфологии в настоящей работе облегчено тем обстоятельством, что прыткая ящерица как удобный объект для практического знакомства с анатомией рептилий издавна привлекала внимание анатомов, и имеется целый ряд весьма подробных общих руководств, части которых посвящены разным сторонам морфологии этого вида (Wiedersheim, 1892; Bolk, 1932—1938; Osborn, 1903; Паркер, Гасвель, 1899—1914; Беляев, 1947 и др.). Приводимые ниже описания морфологии скелета и ряда других внутренних органов обычно даются по М. М. Беляеву (1947).
Некоторые морфологические данные, в основном касающиеся внешнего вида (особенности окраски, фолидоза, размеров), приводятся в главе «Систематика», а другие — в главах, посвященных физиологии (гл. IV), размножению (гл. VII), развитию и росту (гл. VIII).
Авторы: А. С. Баранов, А. В. Валецкий, А. В. Яблоков, при участии Г. П. Лукиной, М. Ф. Тертышникова, Н. М. Окуловой, Л. В. Ту рутиной, В. А. Кутузовой, А. А. Симоняна, А. Б. Стрельцова.
96
ВНЕШНИЙ ВИД И РАЗМЕРЫ ТЕЛА
Внешний вид прыткой ящерицы (рис. 33) определяется несколько суженной кпереди плоской головой, длинным и гибким туловищем со сравнительно небольшими, симметрично расположенными передними и задними конечностями и длинным сужающимся хвостом.
Голова ящерицы имеет округленно пирамидальную форму. На вершине головы, в центре межтеменного щитка, располагается теменной (третий) глаз—(париетальный орган), значение которого до сих пор не вполне ясно. Глаза располагаются по бокам в средней части головы. Характерной чертой глаз прыткой ящерицы, как и всех рептилий, является хорошее развитие третьего века (мигательной перепонки). Верхние и нижние веки ящерицы подвижны. На конце морды располагаются парные ноздри, а по бокам в задней части головы — вертикальные, овальной формы слуховые отверстия, затянутые в глубине барабанной перепонкой.
Туловище ящерицы удлиненное и гибкое, отделено от головы выраженным шейным перехватом. Границей между туловищем и хвостом является клоака, открывающаяся наружу поперечным отверстием на вентральной стороне тела (см. рис. 33). Хвост в поперечном сечении округлый, постепенно сужающийся к концу. Основание хвоста несколько более массивное у самцов (это связано с расположением крупных парных копулятивных органов позади клоаки).
Хвост ящерицы часто отсутствует в результате аутотомии (отбрасывания) — приспособления, помогающего ящерице избежать преследования хищника. Поэтому по числу особей с аутотомированными животными в популяциях можно косвенно судить о давлении хищников (см. гл. X). При регенерации остающейся части хвоста (или как врожденное уродство) изредка возникает двойной хвост.
Конечности сравнительно небольшие, причем задние развиты заметно сильнее передних. Кисть и стопа оканчиваются пятью тонкими пальцами, вооруженными острыми коготками.
Как линейные размеры, так и вес тела являются признаками комплексными, характеризующимися сравнительно низкими величинами наследуемости; это значит, что они могут рассматриваться в первую очередь в качестве эколого-биогеоценотиче-ских показателей. При этом более удобным и точным показателем является общий вес тела, который более объективно, чем любые линейные промеры, может характеризовать место как отдельной особи, так и разных возрастно-половых групп в потоках вещества и энергии, проходящих через населяемые данным видом биогеоценозы.
4 Прыткая
ящерица
97
Рис. 33
Общий вид взрослых прытких ящериц со спины (4 ) и с брюшной стороны ( Ь)
Видны основные особенности распределения групп щитков на голове, шее, туловище и основании хвоста (фото А. В. Валецкого)
Вес тела
Данных об изменении веса тела по всему ареалу того или иного вида рептилий мало, и они основаны, как правило, на случайном и песпециально собранном материале (Терентьев, 1946, 1947; Шварц и др., 1968; Lindsey, 1966). Имеющиеся к настоящему времени в нашем распоряжении сведения, включающие Данные по 5423 экз., позволяют проанализировать географические изменения веса тела прыткой ящерицы на большей части ее ареала.
4*
99
При сопоставлении веса тела ящериц из разных популяций в качестве основного исходного показателя нами был взят максимальный вес самцов. Выбор самцов определен тем, что вес взрослых самок сильно колеблется по месяцам из-за развития яиц и их откладки. Выбор максимального, а не среднего веса определен, во-первых, тем, что, как показал специальный анализ (Смирина, 1974), на средний вес ящериц может оказывать большое влияние количественное соотношение в популяционной выборке разных недифференцируемых при возрасте свыше 2 лет возрастных групп, во-вторых, тем, что не всегда возможно по внешнему виду отличить крупных неполовозрелых особей от мелких половозрелых.
Для удобства анализа полученные данные по максимальному весу самцов были сгруппированы нами в три класса: 8—14, 15— 21, 22 и более грамм. Такое определение данных по весу, безусловно, несколько «огрубляет» их, но вместе с тем дает более четкую картину для рассмотрения изменений этого показателя в масштабах ареала.
При рассмотрении общей картины изменения веса тела прыткой ящерицы (рис. 34) можно проследить определенную тенденцию к увеличению максимального веса особей по мере продвижения от северной границы ареала к южной. Исключением являются районы рек Урала, Дона и северного берега Азовского моря (здесь встречаются ящерицы максимального веса), а также высокогорные районы Закавказья и Заилийского Алатау (где встречаются ящерицы среднего, а не максимального веса).
Причины, обусловливающие различия по весу тела ящериц названных районов от географически близких, можно понять, проведя сравнение близко расположенных одна от другой, но занимающих разные биотопы популяций. Так, выборки из популяций на правом берегу р. Дон в окрестностях г. Верхний Мамон находились на расстоянии 300—400 м одна от другой и были отловлены 31 июля — 1 августа 1971 г. Первая выборка взята из прибрежной балки с невысоким полусухим травостоем, редким невысоким кустарником и обилием мест с оголенной почвой. Вторая — в пойменном лесу с густой высокой травой, обилием лещины, ежевики и шиповника. При сравнении весовых характеристик этих выборок оказалось, что взрослые самцы из балки в среднем заметно мельче представителей поймы (5,1 — 15,0 г, а в пойменном лесу 7,1—21,0 г). Другой пример касается трех популяций ящериц из района Нарынского залива Бухтар-минского водохранилища (Алтайский край). Одна из популяций была отловлена на острове в 600—700 м от следующей береговой популяции, третья находилась в 14 км по берегу водохранилища от второй. К моменту нашего исследования период изоляции островной популяции, связанный с затоплением Бухтарминского водохранилища, длился 11—12 лет. При сравнении этих трех популяций видно, что животные с острова значительно мельче пред
100
ставителей береговых популяций. Оба примера подтверждают, что популяции, расположенные географически очень близко одна от другой, могут различаться между собой, и эти различия, видимо, связаны с микробиогеографическими (биоценотическими) различиями.
В свете только что рассмотренных примеров можно в какой-то степени объяснить и некоторые исключения из общей тенденции изменения веса тела ящериц по ареалу (см. рис. 34), в большинстве случаев относящиеся к горным районам. Последние,
Таблица 3
Вес (в г) прытких ящериц разных возрастных групп (сводные данные по всему ареалу)
Возрастные группы lim X С V*
Сеголетки (0—3 мес.) 0,50—3,5 1,5 11
Неполовозрелые (0,5—1,5 года) 2,5—6,0 4,5 22,4
Половозрелые (2 года и старше) 5,0—40,0 12,0 11
* ПО данным С. С. Шварца и др. (1968) для Южного Урала.
как известно, отличаются исключительной пестротой микробио-географических условий. В подавляющем же большинстве популяций, расположенных на равнинных участках ареала, общее клинальное изменение максимального веса проявляется достаточно отчетливо.
В целом полученные данные показывают, что у прыткой ящерицы существуют четко выраженные клинальные изменения максимального веса, прямо противоположные «правилу Бергмана» (Bergman, 1847; Терентьев, 1946). Четко выраженная географическая изменчивость веса указывает на изменение места и значения ящериц в биоценозах разных ландшафтов в пределах всего их огромного ареала.
Как уже говорилось, в отношении самок и молодых животных подобный анализ географической изменчивости веса тела значительно затруднен из-за невозможности точного определения различных возрастных групп при обработке массового материала, высокой степени онтогенетических и сезонных изменений веса. Так, например, недельное голодание приводит к потере до 22, а двухнедельное голодание — до 34% исходного веса взрослыми ящерицами (Шварц и др., 1968). Поэтому мы вынуждены ограничиться здесь приведением лишь общих сводных данных по весу тела основных возрастных и половых групп (см. табл. 3); более подробно данные по весовой динамике роста яШериц'рассматриваются в главе VIII (в приложении к биомас-се вида и отдельных популяций в гл. XV).
101
Рис. 34
Географическая изменчивость веса взрослых самцов прыткой ящерицы в центральной и восточной частях ареала Вес взрослых самцов, в г: I - 8-14; 2-15-21; 3 - больше 21; 4 - граница ареала
\1Ло А. с, Ъэранова м А.. В. В алецкого) в
Рис. 35
Географическая изменчивость особенностей полового диморфизма по весу тела прытких ящериц центральной и восточной частей ареала 1 — самки больше самцов; 2 — половой диморфизм по весу отсутствует; 3 — самки меньше самцов (по данным А. С. Баранова
и А. В. Вал едкого)
Рис. 36
Распределение самцов и самок (взрослых) по длине тела в трех популяциях
с —Крым (Н. Н. Щербак, 1966): б —Предкавказье (Г. П. Лукина, 19666); в — Рязанская обл. (В. к. Жаркова, 19736)
Данные по резкому увеличению коэффициентов вариации веса тела перезимовавших ящериц указывают на несколько интересных биологических моментов (Шварц и др. 1968). Во-первых, сравнительно невысокая изменчивость молодых перед спячкой свидетельствует об их относительно однородном физиологическом состоянии; резкое увеличение изменчивости вышедших после зимовки ящериц — о разном, видимо, расходе питательных веществ (и энергии) отдельными особями; наконец, в третьих, снижение изменчивости взрослых вновь свидетельствует о стабилизации основных физиологических отправлений. К сожалению, приведенные данные не позволяют проанализировать изменчивость веса на всем протяжении ареала ящерицы.
По имеющимся данным, не существует выраженного полового диморфизма по весу тела для вида в целом (рис. 35), поскольку то в одной, то в другой популяции взрослые самки могут быть и больше и значительно меньше, чем самцы.
Длина и другие промеры тела
Данные по ряду стандартных промеров тела, входящих
ких ящериц разных географических районов, приведены в табл. 4—12.
Здесь нам кажется целесообразным обратить особое внимание на недостаточную сравнимость приводимых размеров. Это обстоятельство связано с тем, что прыткая ящерица интенсивно растет на протяжении первых лет жизни (см. гл. VIII), и при
Таблица 4
Длина головы взрослых прытких ящериц в разных районах ареала (по В. А. Кутузовой, В. К. Жарковой, Л. В. Турутиной, А. А. Симоняну)
Популяция Пол X±Sx Llm cv±sft, n
Дагестан
Окрестности г. Махачкала d 19,92+0,26 17,0—23,7 8,0±0,92 38
5 15,63±0,24 14,3—18,1 7,5+1,08 24
Сергокалинский район о 17,23±0,30 13,9—20,4 8,1±1,23 22
9 14,19+0,22 12,6—17,0 7,8±i,08 26
Буйнакский район d 17,64+0,30 15,5—20,0 8,8+1,17 28
9 15,35+0,25 13,6—20,4 8,6+1,16 27
Рязанская обл.
«Лесостепная популяция» d 15,50+0,31 — 11,2±1,44 33
9 14,03±0,29 — . 12,81+1,57 33
«Лесная популяция» d 15,36+0,39 — 14,7+1,81 54
9 14,00+0,42 — 17,4+2,14 54
Горьковская обл.
Окрестности с. Старая <3 14,70+0,19 — 7,10+0,92 30
Пустынь 13,59+0,14 •— 8,0+0,71 63
Окрестности ст. Сухобез- d 15,03+0,25 — 9,3+1,17 32
водное 9 14,16+0,12 — 8,l±0,61 87
Армения
Севан, островная популя- d 15,90+0,30 —— 12,7+1,69 28
ция 9 13,80+0,44 —• 14,9±2,30 23
Окрестности с. Семеновки d 13,99+0,40 — 9,01+2,02 10
(Севанский район) 9 12,99+0,38 — 13,0+2,1 21
в таксономическую характеристику данного вида, уже приводились в главе «Систематика». Эти данные свидетельствовали о существовании заметной географической изменчивости многих размерных признаков. Другие промеры тела, характеризующие прыт-
этом оказывается, что материал, полученный даже в одной и той же популяции в разные месяцы вегетационного сезона, показывает различия. Это относится особенно к младшим и средним возрастным группам. Обычно же авторы суммировали материал, получаемый за все месяцы и за все годы сбора наблюдений. Такие суммарные данные сравнимы только между собой, но не могут дать истинного представления о сезонной и многолетней Динамике размеров тела в тех или иных популяциях, позволяя
105
Таблица 5
Ширина головы непосредственно за ушами взрослых прытких ящериц в разных районах ареала (по В. А. Кутузовой, В. К. Жарковой^ Л. В. Турутиной, А. А. Симоняну)
Популяция Пол X±sx Lim cv±sro';; п
Окрестности г. Махачкала <3 Дагестан 10,18+0,20 8,4—13,0 12,3+1,41 38.
9 7,83+0,17 7,0—10,0 10,9+1,57 24
Сергокалинский район d 8,45+0,18 7,0—10,6 9,9±1,50 22
9 7,04+0,13 6,0-8,8 9,2+1,28 26
Буйнакский район <3 8,93 ±0,24 7,0—12,3 14,3±1,91 28
9 7,44+0,14 6,8-10,0 9,8+1,32 27
Рязанская обл.
«Лесостепная популяция» d 9 10,0±0,25 9,4+0,28 — 13,9±1,79 19,1+2,35 33 33
«Лесная популяция» d 10,06+0,30 — 17,6+2,16 54
9 9,41+0,30 — 18,4±2,27 54
Горьковская обл.
Окрестности с. Старая Пустынь d 9 9,77+0,13 8,78+0,10 — 7,4+0,95 8,7+0,78 30 63
Окрестности ст. Сухобез- d 12,69+0,30 — 13,4+1,67 32
водное 9 11,51+0,14 Армения — 11,2+0,85 87
Севан, островная популя- d 10,20+0,20 — 12,4+1,65 28
ция 9 9,52±0,22 — 11,0+1,70 23
Окрестности с. Семеновки d 10,20+0,41 — 13,0±2,91 10
(Севанский район) 9 9,00±0,26 — 13,l±2,07 21
улавливать лишь самые общие тенденции. В качестве примера анализа таких общих тенденций в размерах тела на рис. 36 приведена длина тела взрослых ящериц из трех разных районов: Крымского полуострова, Предкавказья и Рязанской области. Единственным общим выводом из сопоставления таких данных может быть вывод о существовании во всех этих районах известного полиморфизма по длине тела: более или менее четко выделяются три крупные размерные группы (абсолютные величины, характеризующие каждую из групп особей, везде несколько различны). Как показано ниже (глава «Рост и возраст»), две первые из этих размерных групп более или менее соответствуют ящерицам второго и третьего года жизни, тогда как третья группа, видимо, включает ящериц нескольких старших возрастов. Несомненно, для получения точных сравнимых данных по раз-
106
Т а б,л и ц а 6
Расстояние от переднего’края теменного глаза до переднего края верхней челюсти взрослых прытких ящериц в разных районах ареала (по В. А. Кутузовой, В. К. Жарковой, Л. В. Турутиной, А. А. Симоняну)
Популяция Пол X±sx Lim cv±sm n
Дагестан
Окрестности г. Махачкала 6 15,82±0,21 13,5—18,6 8,3±0,95 38
9 12,71+0,22 11,0—15,5 8,3±l,20 24
Сергокалинский район d 13,82±0,23 11,4—16,0 7,7±1,17 22
? ll,58±0,19 9,7—13,7 8,4±1,17 26
Буйнакский район d 14,14±0,27 12,0-17,0 9,9±1,32 28
? 12,44+0,20 11,6—16,6 8,5±1,15 27
Рязанская область
«Лесостепная популяция» d 13,26+0,32 •— 13,3±1,84 33
9 11,66+0,46 — 22,8±2,80 33
«Лесная популяция» d 12,78±0,23 — 10,4±l,28 54
9 11,34+0,36 — 18,3±2,24 54
Горьковская область
Окрестности с. Старая d 12,43+0,14 — 6,l±0,79 30
Пустынь Q 11,46+0,10 — 7,2±0,64 63
Окрестности ст. Сухобез- d 12,06+0,18 — 7,5±0,94 32
водное 9 11,44+0,12 — 9,4±0,7i 87
Армения
Севан, островная популя- d 13,20±0,20 — 8,l±l,08 28
ция 9 ll,42±0,33 — 13,4±2,06 23
Окрестности с. Семеновки d 10,70+0,44 — 13,2±2,96 10
(Севанский район) 9 10,8±0,35 — 14,8±2,33 21
мерам тела ящериц из разных частей ареала необходимо сравнение ящериц обязательно одного возраста (или, во всяком случае, пойманных в одни и те же достаточно сжатые сроки).
КОЖНЫЙ ПОКРОВ
Кожа прыткой ящерицы, так же как и других рептилий, сухая, лишена желез и покрыта роговыми образованиями в виде чешуй. Такое строение кожи предохраняет тело от высыхания, а также от механических повреждений в связи с жизнью на твердом субстрате.
У прыткой ящерицы, как и у других позвоночных животных, кожа состоит из двух слоев: верхнего (эктодермального) —эпи-
107
Таблица 7
Длина тела без хвоста взрослых прытких ящериц в разных районах ареала (по В. А. Кутузовой, В. К. Жарковой, Л. В. Турутиной, А. А. Симоняну)
Популяция Пол X±sx Lim CV+Sm п
Дагестан
Окрестности г. Махачкала d 93,84±1,01 81,0—107,8 6,6+0,76 38
? 85,08+1,21 70,0—97,0 7,1=Ы,02 24
Сергокалинский район d 84,73+1,35 70,5—97,0 7,5+1,14 22
? 77,38+1,27 60,0-87,6 8,4±1,17 26
Буйнакский район d 86,28+1,52 70,2-106,6 9,3+1,24 28
¥ 82,18+1,10 75,4—97,0 6,9+0,93 27
Рязанская область
«Лесостепная популяция» d 73,33±1,03 — 7,7+0,99 33
¥ 71,90+1,64 —• 13,4+1,64 33
«Лесная популяция» d 71,44+3,67 — 29,5+3,63 54
¥ 75,42+1,23 — 9,4+1,15 54
Горьковская область
Окрестности с. Старая Пустынь d 75,29+0,74 74,48+0,76 — 5,4+0,70 8,l±0,72 30 63
Окрестности ст. Сухобез- d 78,31+1,31 — 9,4+1,18 32
водное ? 80,55+0,84 Армения — 9,7+0,73 87
Севан, островная популя- d 77,00+0,76 —. 8,0+1,07 28
ция ? 73,71+1,52 — 9,4+1,45 23 ;
Окрестности с. Семеновки d 69.00+2,20 — 10,2+2,30 10
(Севанский район) ¥ 69,10+2,40 — 15,8+2,50 21
дермиса и нижнего (мезодермального)—кориума (рис. 37). В свою очередь эпидермис дифференцируется на роговой и мальпигиев слои. Клетки рогового слоя в процессе жизни животного претерпевают особого рода перерождение: они переполняются капельками жироподобного вещества элеидина, которое постепенно уплотняется, в то время как сами клетки становятся все более плоскими. Ядро перестает быть различимым, и клетка превращается в твердую роговую чешуйку. По мере того как поверхностные слои ороговевших клеток изнашиваются и сбрасываются, роговой слой непрерывно возобновляется снизу за счет нижнего слоя клеток с ядрами, а также за счет 5—10 рядов клеток мальпигиева слоя. Сбрасывание износившихся поверхностных клеток совершается путем линьки. Иногда в более глубоких
108
Таблица 8
Высота головы взрослых прытких ящериц в разных районах ареала (по В. А. Кутузовой, В. К- Жарковой, Л. В. Турутиной,- А. А. Симоняну)
—_ Популяция Пол X±sx Lim cv±sm n
Дагестан
Окрестности г. Махачкала <5 13,11+0,27 10,0—16,4 12,4+1,42 38
9 9,04+0,22 7,9—11,5 11,7+1,69 24
Сергокалннский район d 10,18±0,29 7,4—13,6 13,2+2,00 22
$ 7,85 ±0,20 6,7—10,0 13,1±1,82 26
Буйнакский район d 10,36±0,28 8,0—14,3 14,5±1,93 28
9 8,33±0,18 7,0—12,5 11,3±1,53 27
Рязанская область
«Лесостепная популяция» <5 10,16±0,33 — 18,1+2,33 33
9 8,54±0,22 — 15,0±l,84 33
«Лесная популяция» d 10,00±0,35 —. 20,0±2,46 54
9 8,65±0,38 — 26,5±l,06 54
Горьковская область
Окрестности с. Старая d 10,77±0,16 — 8,2+1,06 30
Пустынь 9 9,70±0,10 — 7,9+0,71 63
Окрестности ст. Сухобез- d ll,44±0,23 — 11,5±1,44 32
водное 9 10,63±0,12 — 10,2±0,77 87
Армения
Севан, островная популя- d 10,40±0,20 —- 11,5+1,53 28
ЦИЯ 9 8,C0±0,32 — 17,0±2,61 23
Окрестности с. Семеновки d 8,79±0,23 — 8,5+1,90 10
(Севанский район) 9 7,89±0,29 — 16,9+2,67 21
слоях эпидермиса обнаруживается красящее вещество — пигмент, обуславливающий окраску кожи животного, но основная часть пигмента находится в кориуме (Полякова, 1962).
Кориум состоит из двух слоев: наружного рыхлого слоя, образующего сосочки роговых чешуй, и внутреннего, более толстого слоя из плотной волокнистой соединительной ткани. Глубже следует слой рыхлой подкожной клетчатки. Наиболее глубоко в плотной соединительной ткани, особенно там, где она переходит в рыхлую ткань, лежат меланофоры, выше располагаются различно окрашенные хроматофоры (у прыткой ящерицы среди них преобладают желтые ксантофоры и красные эритрофоры).
Из роговых образований прыткой ящерицы необходимо упомянуть когти, которые отрастают так же, как и у млекопитаю-
109
Рис. 37 '
Разрез через чешую прыткой ящерицы
1 — эпителиальный слой; 2 — роговые клетки; 3 — пигментная клетка; 4— клетки рогового слоя вне чешуи; 5 — мальпигиев слои; 6 — дерма (ло'А. Э. Шиплею, Э. В. Мак-Брайту, 1911)
щих, с основания. В зависимости от образа жизни и условий существования в разных районах (твердый или мягкий грунт, наличие или отсутствие естественных убежищ и т. д.) когти прыткой ящерицы могут быть по-разному изогнуты и стерты (Кашкаров, Станчинский, 1940; Булахов, Константинова, 1972).
ФОЛИДОЗ
Все тело прыткой ящерицы покрыто чешуями (рис. 38, см. рис. 33, 37), которые в зависимости от расположения на теле сильно различаются по форме, строению, размеру и имеют большое систематическое и диагностическое значение. Роговой слой между чешуями остается тонким, что не препятствует движению тела. На частях тела, не требующих подвижности при движении, развиваются плоские роговые щитки (scutae). Места, требующие большой подвижности, покрыты мелкими чешуйками-зернышками (granulae). Особого развития щитки достигают на голове. Часть верхней поверхности головы, покрытая ими, называется пилеусом (pileus) (Терентьев, Чернов, 1949).
При рассмотрении головы сверху, по оси черепа различают пять непарных щитков: межчелюстной, лобноносовой, лобный, межтеменной и затылочный. Посредине межтеменного щитка находится светлое пятно — непарный рудиментарный глаз. Остальные щитки — парные: передний и задний носовые, передний и задний скуловые, предлобные, верхнересничные, надглазничные, I лобнотеменные, теменные.
При рассмотрении головы сбоку различают передний и задний скуловые щитки, подглазничный, предглазничные и заднеглазничные, а также центральновисочный, височные, верхневи
110
сочные и барабанный щитки. Нижний край верхней челюсти обрамлен верхнегубными щитками, а верхний край нижней челю-стМ — нижнегубными. В месте соединения нижнегубных щитков находится подбородочный щиток.
Шея отделяется от туловища рядом увеличенных чешуй — воротником (collare) (см. рис. 38), между этим рядом и перед-
Таблица 9
длина туловища от шейной склтдки до клоакальной щели взрослых прытких ящериц ] в разных районах ареала (по В. А. Кутузовой^ В. К. Жарковой, Л. В. Турутиной,
А. А. Симоняну) /
Популяция Пол X±SJ Lim cv±sra n
Окрестности г. Махачкала d Дагестан 72,24±0,89 64,0—86,0 7,4±0,85 3J
$ 69,50+1,18 57,8—80,0 8,3±l,20 24
Сергокалинский район d 67,91±1,27 56,0—78,0 8,8±1,33 22
9 62,77+1,27 48,0—72,0 10,3+1,43 26
Буйнакский район d 68,42±l,06 55,0—79,6 8,2±0,97 27
9 68,11+1,16 — 10,4+1,34 33
Рязанская область
«Лесостепная популяция» d 61,16±1,16 — 10,4±l,34 33
9 60,00+1,56 — 15,Oil ,84 33
«Лесная популяция» d 58,62±i,79 — 17,6±2,16 54
9 62,78+1,41 — 12,8±l,60 54
Горьковская область
Окрестности с. Старая Пустынь d 9 60,80+0,63 61,48+0,69 — 5,6i0,73 8,9+0,79 30 63
Окрестности ст. Сухобез- d 62,28+1,10 — 10,0+1,25 32
водное 9 65,45±0,70 Армения — 10,0+0,76 87
Севан, островная популя- d 61,00+1,10 — 8,4+1,12 28
Ния 9 60,57±l,54 — 11,7±1,80 23
Окрестности с. Семеновки d 55,19±1,71 — 9,8+2,18 10
(Севанский район) 9 55,80+2,09 — 16,7+2,60 21
ней частью головы находится горловая складка (plica gularis или sulcus gularis) (Терентьев, Чернов, 1949).
Чешуи спины и боков мелкие, узкие, с хорошо выраженными продольными ребрышками. Брюшные щитки значительно крупнее чешуй спины и боков и располагаются в шесть продольных Рядов. Крупный анальный щиток спереди окружен I—3 рядами прианальных, форма и количество которых в каждом варьирует.
111
Таблица 10
Ширина основания хвоста взрослых прытких ящериц в разных районах ареала (по В. А. Кутузовой, В. К. Жарковой, Л. В. Турутиной, А. А. Симоняну)
Популяция Пол 1 * z +1 I* Lim n
Дагестан
Окрестности г. Махачкала d 9,29+0,16 7,7—11,0 10,8+1,24 38
9 7,80+0,19 6,8—10,4 11,7±1,69 24
Сергокалинский район d 7,91+0,17 6,0—9,3 10,0+1,51 22
9 6,73+0,14 5,42—8,8 10,5+1,46 26
Буйнакский район d 8,32+0,19 7,4—10,8 12,0+1,60 28
9 7,33+0,15 6,4—9,6 10,5+1,42 27
Рязанская область
«Лесостепная популяция» d 7,70+0,20 — 14,9+1,92 33
$ 6,42+0,49 — 25,3+3,10 33
«Лесная популяция» d 7,58+0,20 .— 15,0+1,84 54
¥ 6,67+0,12 — 10,9+1,34 54
Горьковская область
Окрестности с. Старая d 8,40+0,10 —. 6,5±0,84 30
Пустынь 9 7,81+0,11 .—. 11,1+1,99 63
Окрестности ст. Сухобез- d 8,75+0,18 .—. 11,4+1,43 32
водное 9 8,86+0,12 — 12,5+0,94 87
Лрмения
Севан, островная популя- d 7,60+0,25 —. 13,7+1,83 28
ция 9 6,66+0,34 — 23,3±3,59 23
Окрестности с. Семеновки d 5,50±0,25 14,7+3,27 10
(Севанский район) 9 5,30+0,32 — 27,4+4,33 21
На внутренней поверхности бедер располагается ряд чешуек, имеющих в середине отверстие, из которого выделяется столбик ороговевших клеток. Эти чешуйки называются бедренными порами (pori femorale). Назначение бедренных пор пока еще точно не выяснено, но число их специфично для вида и пола.
Хвост покрыт прямоугольной формы чешуйками, постепенно уменьшающимися к кончику хвоста. В случае аутотомии восстановленный (регенерированный) хвост можно отличить по цвету и по более примитивным чешуйкам (Меландер, 1935; Терентьев, Чернов, 1949).
В целом расположение чешуй и щитков на теле прыткой ящерицы закономерно, но подвержено географической и индивидуальной изменчивости.
112
Таблица II
Длина задней ноги взрослых прытких ящериц в разных районах ареала (по В. А. Кутузовой, В. К. Жарковой, Л. В. Турутиной)
Популяция Пол X±S~ Lim cv±sra п
Окрестности г. Махачкала 3 Дагестан 37,89+0,72 33,8—47,0 11,7+1,34 38
Q 34,78+0,50 27,0—39,7 6,8+0,98 24
Сергокалинский район 3 37,68+0,67 31,3—41,0 8,3+1,26 22
9 32,94+0,40 28,0—38,8 6,3+0,88 26
Буйнакский район d 38,07+0,41 33,0—43,0 5,6+0,75 28
9 33,78+0,48 28,0—40,0 7,3+0,99 27
Рязанская область
«Лесостепная популяция» 3 31,26+1,47 — 25,9+3,33 33
9 29,67+0,64 —• 12,5+1,54 33
«Лесная популяция» 3 30,34+1,54 — 28,7+3,53 54
9 29,67+0,64 — 12,5+1,54 54
Горьковская область
Окрестности с. Старая 3 32,07+0,34 — 5,7+0,74 30
Пустынь 9 30,24+0,22 — . 5,7+0,51 63
Окрестности ст. Сухобез- 3 34,92+0,56 — 8,1+1,14 32
водное 9 33,38+0,29 — 7,7+0,60 87
Таблица 12
Длина IV пальца задней лапы взрослых прытких ящериц в разных районах ареала (по В. А. Кутузовой, Л. В. Турутиной)
Популяция Пол Lim п
Дагестан
Окрестности г. Махачкала 3 16,89+0,20 14,5—19,5 6,9+0,79 38
9 14,42+0,21 11,0—16,4 7,2+1,06 23
Сергокалинский район 3 15,68+0,17 14,2-17,4 5,2+0,79 22
9 13,73+0,17 12,8—16,0 6,3+0,88 26
Буйнакский район 3 16,19+0,16 14,8—18,0 5,3+0,71 28
О т 14,67+0,19 13,0—16,6 6,5+0,88 27
Горьковская обл.
Окрестности с. Старая Пустынь 3 12,47+0,13 —- 5,8+0,75 30
9 11,70+0,11 — 7,5+0,67 63
Окрестности ст. Сухобез- 3 14,00+0,28 — 11,4+1,60 32
водное 9 13,05+0,08 — 5,7+0,44 87
113
Рис. 38
Некоторые элементы чешуйчатостм и щиткования прыткой ящерицы
А — голова сверху; Б — голова сбоку; В — голова снизу; Г — вид с брюшнбй стороны заднего отдела туловища (Фото А. В. Валецкого)
ОКРАСКА
Окраска прыткой ящерицы определяется двумя главными компонентами: цветом и рисунком. Сочетание различных цветовых вариантов с многочисленными вариантами рисунка характеризует окраску как весьма комплексный морфологический признак. На такую сложность окраски неоднократно указыва-
114
лось в литературе (Никольский, 1915; Иоганзеп, 1923; Чернов, 1937; и др.). Опыт показывает, что только при дифференцированном исследовании составляющих окраску элементов цвета и рисунка возможно выделение определенных типов окраски ящериц (Сухов, 1948; Терентьев, Чернов, 1949; Яблоков, Баранов, 1971; Баранов, 1973). Сложность выделения разных типов окраски усугубляется четко выраженными возрастными изменениями окраски и различной степенью полового диморфизма (обычно более четко выраженного по цвету, чем по рисунку). Наконец, в настоящее время становится все более ясным и существование значительной географической изменчивости окраски (Баранов, 1973).
115
Особенности цвета
Молодые особи обоих полов окрашены одинаково. Спина буровато-серая или коричневая, такого же цвета голова и конечности. Брюхо и горло светлые (от бело-голубоватого до пастельно-бежевого). При этом у самцов довольно часто брюхо и горло зеленого (желто-зеленого) цвета (по этому признаку в первом приближении можно без вскрытия определить пол молодых ящериц).
Самцы. Пожалуй, наибольшего разнообразия цветовая гамма окраски достигает у самцов. Цвет спины даже одной особи на протяжении одного сезона сильно колеблется. Так, во время брачного периода в южных частях ареала в окраске самцов проявляется ярко-зеленый цвет. В северных районах ареала процент самцов с ярко-зеленой окраской меньше (а в некоторых популяциях зеленых самцов вообще не встречается). По мнению некоторых авторов, цвет самцов может резко меняться от зеленого в брачный период до коричневого вне сезона размножения (Олигер, 1966; Кузнецов, Макковлева, 1959 и др.). Специальное изучение этого вопроса показывает, что происходит лишь изменение тональности и интенсивности: коричнево-желтые тона в окраске самцов присутствуют всегда (даже в южных популяциях вида), только в брачный период они маскируются более ярким зеленым цветом (см. также гл. II). Во время зимней спячки цвет покровов становится значительно темнее.
Голова самцов обычно такого же цвета, как и спина, но в некоторых случаях черные и коричневые пятна, покрывающие головные щитки, сливаются. Горло у взрослых самцов обычно зеленовато-желтоватое (на Кавказе у некоторых ящериц горло голубого цвета, что напоминает окраску горла у зеленой ящерицы). В некоторых популяциях встречаются самцы с белым горлом.
Брюхо самцов окрашено обычно в зеленый цвет (цвет брюха может не совпадать с цветом спины). В разных географических районах цвет брюха может быть белым, голубоватым, коричневатым, желто-зеленым. По сравнению с окраской спины цвет брюха более постоянен на протяжении сезона и при полевом определении зеленоватыми тонами брюха можно руководствоваться для выделения самцов.
Конечности самцов обычно окрашены в такой же цвет, как и спина, однако в некоторых географических районах конечности коричневого цвета, в то время как спина и бока зеленые (интересно, что коричневая окраска конечностей характерна для Lacerta strigata).
Самки. Общий тон окраски взрослых самок сходен с таковым молодых особей. На большей части ареала вариации цвета спины самок незначительны и включают гамму серо-коричневых тонов, но в некоторых районах (в основном южных) можно
116
встретить самок с зеленой спиной; самый большой процент таких самок характерен для ряда Кавказских популяций.
Голова самок такого же цвета, как спина. Горло обычно беловатое, голубоватое, редко с коричневатым оттенком. У самок с зеленой окраской спицы цвет горла также зеленый. Окраска брюха обычно такая же, как и горла, но несколько более темная; у животных с зеленой окраской цвет брюха светло-зеленый. Конечности такого же цвета как и бока туловища. Однако как и у самцов конечности некоторых самок с зеленым цветом спины могут быть коричневыми.
Анализ литературного материала по окраске (цвету) прыткой ящерицы позволяет высказать предположение об определенной хронографической изменчивости этого .признака. Все авторы, изучавшие окраску ящериц в первой трети XX в., отмечают, «что основной цвет у большинства ящериц — серый, или серовато-бурый» (Кащенко, 1900, 1909; Браунер, 1904; Никольский, 1905, 1907, 1915; Рузский, 1916; Платонов, 1926; Шестаков, 1926; Сухов, 1928; Соболевский, 1929 и др.). Однако большой современный материал показывает, что в настоящее время основным цветом прыткой ящерицы является коричневый. Особи с серой окраской встречаются в популяциях как редкое исключение (в основном в районах северо-востока).
Особенности рисунка
Можно выделить следующие элементы рисунка: полосы спины (центральная и боковые), пятна спины, располагающиеся между спинными полосами, ряды глазков по бокам тела, рисунок головы, пятна на брюшных щитках, рисунок конечностей. Географический анализ изменчивости некоторых элементов рисунка рассмотрен 'в гл. II, ниже рассматриваются основные встречающиеся варианты элементов рисунка.
Полосы спины. Основные варианты боковых полос спины у самцов и самок приведены на рис. 39. Наибольшего разнообразия достигает рисунок центральной полосы (см. рис. 33, 39, г).
Пятна спины. Между полосами на спине располагаются темные пятна, среди которых можно выделить четыре группы (рис. 33, 39, в). К первой группе относятся точечные пятна (размером в 1—2 спинные чешуи); ко второй группе относятся все пятна, превышающие по своим размерам первые; к третьей —«пятна», которые полностью слились и образовали один сплошной «ремень», и, наконец, четвертую группу составляют особи, у которых пятна на спине отсутствуют.
По форме пятна обычно округлые, но встречаются и угловатые, причем и те и другие могут встречаться у одной особи. Цвет пятен разнообразен: от черно-коричневых до желтовато-зеленоватых. В некоторых популяциях встречаются особи, пятна спины которых заключены в своеобразные «оторочки» несколько
117
Рис. 39
Основные элементы рисунка спинной поверхности прыткой ящерицы
a — характер боковых полос:
1 — сплошные;
2 — прерывистые;
3 — полосы отсутствуют;
б — варианты захождения боковых полос на головные щитки:
1 — прямое;
2 — с ответвлением;
в — характер темных пятен:
1 — слились в один сплошной «ременьж;
2 — крупные;
3 — мелкие (каждое пятно занимает не более 3 спинных чешуй);
4 — пятна отсутствуют;
г — характер центральной полосы:
1 — темная;
2 — светлая;
3 — прерывистая;
4 — петлевидная;
5 — полоса отсутствует (рис. А. С. Баранова)
более светлых тонов (особенность, характерная только для крупных пятен спины).
«Глазки» по боку тела. Практически все прыткие ящерицы имеют «глазки» по боку тела. Глазки представляют собой светлые пятнышки, отороченные по краям темными ободками. Число их в одном ряду различно и достигает 18. Иногда глазки сливаются в одну сплошную светлую полоску, причем такое их слияние наиболее часто в передней части туловища. Если такое слияние наблюдается в 2 и 3 ряду глазков, то прыткая ящерица делается похожей на полосатую ящерицу (Кутузова, 1971; см. также гл. II).
Рисунок головы. У молодых особей (как самцов, так и самок) рисунок на голове отсутствует. Для взрослых самок характерно наличие округлых черных и темиощоричневых пятен на теменных щитках (у некоторых особей заходят на спинную чешую). В некоторых популяциях встречаются особи, у которых боковые полосы спины заходят на шов между теменными щитками и группой височных (см. рис. 33, 39, б). Встречаются и другие варианты в рисунке дорсальной поверхности головы (рис. 33).‘На боковых поверхностях головы самок имеются темные и светлые пятна, которые в основном сосредоточены в центральновисочной области и на щитках по краю верхней челюсти (см. рис. 38, б). Горло у самок без каких-либо пятен. Встречаются самки, у которых рисунок на голове вообще отсутствует.
У взрослых самцов голова сверху обычно покрыта червеобразными черными пятнами, которые у некоторых особей сливаются. В разных популяциях число таких пятен различно (у некоторых особей они вообще могут отсутствовать). Боковая поверхность головы самцов изобилует пятнами. Пятна в основном располагаются по краям крупных щитков. У некоторых самцов рисунок боковой поверхности может вообще отсутствовать. Горло самцов, как правило, всегда покрыто мелкими черными пятнышками. Они приурочены, по мере продвижения к воротничку, к окончанию чешуек и у некоторых экземпляров образуют четкую оторочку. Впрочем, иногда встречаются особи, у которых горло лишено пятнистости.
Рисунок брюха. Молодые особи обоих полов не имеют пятен на брюшных щитках. Только после первой зимовки начинают встречаться особи с небольшими черными пятнышками на внешних брюшных щитках, в основном самцы.
.Для взрослых самок характерно отсутствие пятен на брюшной стороне тела. Лишь в некоторых популяциях встречаются особи, у которых пятна приурочены к двум первым наружным рядам брюшных щитков (см. рис. 33). Такие пятна расположены не на всех щитках одного ряда и могут сосредотачиваться в области задних конечностей или же ближе к голове.
У большей части взрослых самцов вся брюшная поверхность покрыта -мелкими черными пятнами (т. е. пятнистость брюха
119
является вторичным половым признаком). Характер пятен, покрывающих брюшные щитки самцов, несколько отличен от таковых у самок. Пятна самцов не округлой формы, а несколько эллипсоидного типа, и у некоторых особей имеют такую же оторочку, как и на горловых чешуях. Эта оторочка может занимать до половины каждого брюшного щитка. Наряду с животными, у которых каждый брюшной щиток несет пятно, в популяциях встречаются особи, брюшные щитки которых вообще без пятен или же пятна очень мелкие и расположены лишь на наружных рядах чешуй.
Рисунок на конечностях. Передние конечности молодых особей без рисунка. У взрослых самцов и самок они покрыты хаотично разбросанными темными пятнами. Наибольшее число пятен и пятнышек наблюдается, как правило, в области 'предплечья. У некоторых животных пятна сливаются в своеобразные образования звездчатого характера. Наряду с темными пятнами на конечностях бывают и белые пятнышки, чаще у самок, чем у самцов.
На задних конечностях молодых прытких ящериц имеется рисунок в виде двух светлых пятен, обрамленных темной окантовкой. С возрастом этот признак исчезает у самцов, окраска конечностей которых приобретает зеленый цвет. У самцов с коричневой окраской задних конечностей этот признак остается, но интенсивность окраски «глазков» слабеет. Наряду с этим признаком наблюдаются черные или коричневые пятна, в беспорядке разбросанные на щитках и чешуях, покрывающих заднюю конечность.
У самок «глазчатость» задней конечности с возрастом не исчезает, а в некоторых популяциях их «резкость» становится еще большей. Как и у самцов, помимо «глазков» имеются темные пятна.
Некоторые типы окраски
Наряду с типичными формами окраски, определенными для каждого из подвидов (см. гл. II), в популяциях встречаются особые цветовые аберрации (рис. 40), обычно локализованные определенным распространением. В основном все аберрации связаны с исчезновением рисунка (отсутствие линий спины, в некоторых случаях пятен, а иногда и глазков). Надо отметить, что все такие аберрации четко отличаются от тех форм окраски, которые встречаются наряду с ними в популяции (Самош, 1953; Сухов, 1948 и др.).
I. Аберрация erythronotus {rubra).
Вся середина спины до верхнего ряда глазков, одноцветна, ржаво-коричневого, красновато-коричневого или кофейного цвета, без пятен и полос. Такая окраска присуща и самцам и самкам. Бока, как у типичной формы. В своих исследованиях
120
1‘не. 40. Некоторые варианты окраски прыткой ящерицы
Верхний ряд — самцы; нижний ряд—самки; слева направо: abb. iipica, abb. «rytTonothus (rubra) abb. immaculata. (рис. А. С. Баранова)
С. Држевицкий (Drzewicki, 1926) показал, что эта вариация хорошо наследуется и является доминантной.
2. Аберрация immaculata.
Самки одноцветного серо-мышиного цвета, без каких-либо пятен и полос. У молодых самцов окраска такая же, как и у самок, взрослые самцы ярко-зеленого цвета, без рисунка.
3. Аберрация punctatta.
Отличается от аберрации erythronotus (rubra) присутствием мелких крапинок по ржаво-коричневому фону спины. Г.,Ф. Сухов (1948) считает, что эта аберрация не является самостоятельной и образуется в результате скрещивания особей ab. erythronotus с ab. tipica.
4. Аберрация nigra.
Спина, бока, конечности и брюхо — черного цвета без пятен и полос. Известен единственный экземпляр такой особи, отловленный на Западном Алтае экспедицией ИБР АН СССР.
СКЕЛЕТ
Строение скелета как главной опорной системы тела в наибольшей степени отражает эволюционно сложившийся тип строения тела у разных групп позвоночных. Рассмотрим основные черты строения скелета прыткой ящерицы.
Череп
В черепе рептилий со сравнительно-анатомической точки зрения выделяют два отдела, различающихся по своему происхождению: 1) черепная коробка и 2) висцеральный аппарат. Последний состоит из челюстной дуги (нёбно-квадратная кость, квадратная кость и меккелев хрящ) и подъязычной дуги (слуховая косточка — подъязычная кость). Череп образуют как первичные (замещающие), так и многочисленные вторичные (покровные) кости. Одной из важнейших функциональных харак теристик черепа ящериц является кинетизм — развитие подвижности затылочного сегмента черепа относительно максил-лярного (Иорданский,1971).
Череп прыткой ящерицы (рис. 41)—тропибазальный (с узким основанием и сближенными глазницами), окостеневающий; хрящ сохраняется лишь в обонятельной и слуховой областях. Сзади черепа располагается большое затылочное отверстие,, окруженное четырьмя затылочными костями, из них основная затылочная несет один мыщелок для сочленения с первым шейным позвонком (см. рис. 41, 15—16).
При рассмотрении черепа ящерицы сверху (см. рис. 41, А) прежде всего бросается в глаза, что крыша черепа покрыта отпечатками контуров роговых головных щитков (остеодермы),, затрудняющих в некоторых случаях выявление истинной карти-
121
Рис. 41
Схема строения черепа прыткой ящерицы
А — вид сверху; Б — вид снизу; В — вид сбоку;
1 — parietale;
la — foramen parietale;
2 — frontale;
3 — praefrontale;
4 — supraorbitale;
5 — postorbitale;
6 — nasale;
7 — maxillae;
8 — praemaxillare;
9 — vomer;
10 — palatinum;
11 — internasalia;
12 — pterygoideum;
13 — quadratum;
14 — transversum;
15 — basioccipitale;
16 — condilus;
17 — basisphenoideum;
18 — praesphenoideum;
19 — jugale;
20 — lacrimale;
21 — epipterygoideum;
22 — squamosum;
23 — supratemporale;
24 — dentale;
25 — articulare;
26 — angulare;
27 — supraeangulare;
28 — proc. coronarium (Зограф, 1900)
Рис. 42
Схема строения подъязычного аппарата (А ) и опорной части переднего плечевого пояса*. £) прыткой ящерицы
А: 1 — cornua hyalia; 2 — с. branchialia; 3 — proc, entoglossus (.no Wiedersheim, 1892; Gegenbaur, 1864—1865); Б: / — sternum; 2 — episternum; 3 — clavicula; 4 — scapula;. 5— coracoid (no Gegenbaur, 1864—4865)
ны расположения черепных костей. В крыше черепа, кроме те-, менных сливающихся вместе и прободенных отверстием для теменного глаза парных и носовых костей, имеются также маленькие предлобные, надглазничные (несколько пар), заглазничные кости.
При рассмотрении черепа снизу видно, что контуры передней половины черепа образованы парными верхне-челюстными и предчелюстными костями (см. рис. 41, Б). Обе пары костей; имеют зубы и образуют так называемую вторичную челюсть. К парному сошнику сзади прилегают нёбные кости. Между их переднебоковыми концами и небольшими нёбными отростками верхнечелюстных костей находятся хоаны (внутренние ноздри) . Упираясь концами в нёбные кости, назад тянутся крыловидные кости, задние концы которых в свою очередь упираются в квадратные кости. Нёбные, крыловидные и квадратные кости являются окостенениями нёбноквадратного хряща. Квадратная кость, прыткой ящерицы подвижна относительно черепа и на нижней поверхности ее есть сочленованная поверхность, соединяющая ее с нижней челюстью.
Поперечные кости идут в виде распорок между передними концами крыловидных костей и верхнечелюстными. Дно собственно осевого черепа образовано основной затылочной костью, с которой спереди сливается массивная основная клиновидная, дающая кпереди отростки, к которым присоединяются крыловидные кости.
123
При осмотре черепа сбоку (см. рис. 41, В) видна хрящевая обонятельная капсула, окруженная покровными костями: предчелюстной, носовой и верхнересничной. Орбита образована рядом надглазочных костей, а спереди и отчасти снизу — верхнечелюстной и скуловой. В переднем углу глазницы расположена слезная кость. В глубине височной области находится столбчатая (надкрыловидная) кость. Сзади височная область ограничивается квадратной костью. Немного позади последней обнаруживается узкая крючковато изогнутая чешуйчатая кость, которая прикрывает дистальный отдел слуховой капсулы.
Основу нижней челюсти составляет меккелев хрящ (см. рис. 41, В). На нем образуется целый ряд покровных костей. Несущая зубы зубная кость составляет всю переднюю половину нижней челюсти; присутствуют также угловая, надугловая и венечная кости. Окостеневший проксимальный отдел меккелева хряща образует сочленованную кость, сочленяющуюся с квадратной костью.
Подъязычный аппарат налегает с вентральной стороны на гортань и лежит между ветвями нижней челюсти. Он состоит из переднего удлиненного выроста и трех пар рожков (рис. 42, А). Передний удлиненный вырост служит опорой для языка.
Позвоночник
В позвоночнике прыткой ящерицы хорошо дифференцированы все 5 отделов, характерных для высших позвоночных.
Шейный отдел (parscervicalis) представлен обычно8позвонками. На первом из них (atlas) находится суставная поверхность для затылочного мыщелка черепа; 4—8-й позвонки, как правило, несут на себе ребра, не доходящие до грудины (так называемые шейные ребра).
Грудной и поясничный отделы не вполне ясно обособлены друг от друга и образуют единый комплекс (pars thoraco-lumba-1 is). Первым грудным позвонком принято считать тот, присоединяющиеся к которому ребра доходят до грудины. Всего насчитывается обычно 5 грудных позвонков (см. рис. 43). Следующие пары ребер, отходящие от позвонков грудного и поясничного отдела, по мере продвижения к хвосту постепенно уменьшаются.
Крестцовый отдел позвоночника (pars sacralis) представлен двумя позвонками, обладающими мощно развитыми поперечными отростками, с которыми соединяются подвздошные кости пояса задних конечностей. Этим обеспечивается большая прочность соединения пояса задних конечностей с позвоночником.
Хвостовой отдел (pars caudalis) состоит из многочисленных позвонков, строение которых по направлению назад упрощается. Тело каждого хвостового позвонка разделено хрящевой прослойкой на переднюю и заднюю половины (в месте этой прослойки тело позвонка не окостеневает и отрыв хвоста при аутотомии
124
происходит именно в таких местах). Отрыв хвоста прыткой ящерицы при аутотомии происходит только в области определенных хвостовых позвонков (Меландер, 1935). Общее число хвостовых позвонков сильно варьирует внутри вида и, видимо, в разных участках ареала, в общем достигая нескольких десятков. Точных данных по числу хвостовых позвонков в разных участках ареала пока нет, но по аналогии с тем, что известно для других видов позвоночных, можно предположить, что этому признаку должна быть присуща значительная географическая изменчивость. Косвенным подтверждением этого служит разная относительная длина хвоста в разных группах популяций прыткой ящерицы.
Позвонки прыткой ящерицы процельные — вогнутые спереди и выпуклые сзади. Кверху от тела позвонка отходит нервная дуга, завершающаяся остистым отростком. На большинстве позвонков хорошо развиты поперечные отростки, а также две пары сочлененных отростков (передние и задние). В месте сочленения нервной дуги с телом позвонка находится небольшая поверхность для подвижного причленения головки ребра.
Грудина (sternum) представлена в виде ромбической полу-хрящевой пластинки, к которой причленяются с каждой стороны по пять ребер, два из которых обычно соединены с ее задним концом одним общим основанием. Грудина, ребра и грудные позвонки образуют грудную клетку, тесно связанную также с плечевым поясом (см. рис. 42).
Грудная клетка ящерицы подвижна, что достигается как за счет подвижного соединения ребер с позвонками, так и за счет развития легких, не окостеневающих дистальных отделов ребер, причленяющихся к грудине.
Скелет конечностей
Как и у типичных наземных позвоночных, в скелете конечностей прыткой ящерицы выделяются собственно свободные конечности и пояса конечностей, лежащие на туловище и служащие опорами для свободных конечностей (рис. 43). Пояса конечностей имеют у ящерицы вид дуг, охватывающих тело снизу и с боков.
Основу плечевого пояса составляют лопатки и каракоид, которые в месте присоединения плечевой кости срастаются в единое образование (см. рис. 42). Своеобразными элементами, характерными для плечевого пояса всех ящериц, являются надлопатка и прокаракоид. Надлопатка соединяется с хорошо развитой ключицей; правая и левая ключицы внутренними концами соединяются с покровной костью грудины — надгрудинником. Часлю в этой весьма сложной системе первичных и вторичных костей плечевого пояса современных ящериц видят лишь указания на пути филогенеза плечевого пояса конечностей; возможно, однако, что мы просто недостаточно хорошо пока знаем функцио-
125
Рис. 43
Общий вид скелета прыткой ящерицы
Просветленный в глицерине препарат из Британского музея (Ns 1970); Л—вид со спины; Б — вид с брюха; фото А. В. Яблокова
нальное значение отдельных элементов плечевого пояса рептилий и прыткой ящерицы в частности.
Строение заднего, тазового, пояса несколько более просто: он состоит лишь из трех парных костных элементов (рис. 44). Подвздошная кость палочковидной формы располагается параллельно позвоночному столбу в области крестцового отдела и прочно соединена с поперечными отростками двух крестцовых позвонков. В области хорошо развитой суставной впадины эта кость соединяется с лобковой и седалищной костями. Тазовый пояс в целом более прочен, чем передний, и образует более жесткую опору для заметно более развитых у прыткой ящерицы задних конечностей.
126
Рис. 44
Строение тазового пояса прыткой ящерицы / — acetabulum; 2—ileum; 3 — pubis; 4 — ishium; 5 — fenestra ishiopubica; 6 — ca-nalis obturatorius; 7 — proc, hypoishiadi-cus; 3— proc, epipubis (no Wied ershei urn, 1892)
Рис. 45
Схема строения кисти (Л ) н стопы (Б) прыткой ящерицы
А: 1—5 — саграИа distalia; 6 — centrale; 7 — radiale; 8 — ulnare; 9 — intermedium;
10 — pisiforme; 11 — ulna; 12 — radius; I — V — metacar pa li а; Б-. 1 — tarsus; 2 — tarsa-lia distalia; 3 — tibia; 4 — fibula; /— V --metatarsalia (из M. M. Беляева, 1947)
Передняя конечность прыткой ящерицы имеет типичное дл** конечности наземных позвоночных строение и включает плечо, предплечье и кисть. Скелет кости довольно прост (рис. 45, Л).
Типичная формула фаланг пальцев вида (счет начинается с 1-го внутреннего пальца) —2, 3, 4, 5, 3.
Задняя конечность также состоит из трех обычных отделов: бедра, голени и стопы. В результате слияния проксимального ряда плюсневых косточек (см. рис. 45, Б) образуется характерное для всех рептилий интертарзальное сочленение.
Обычная формула числа фаланг пальцев этого вида (начиная с первого внутреннего пальца) — 2, 3, 4, 5, 3.
МУСКУЛАТУРА
Мускулатура рептилий представляет собой дальнейшую ступень дифференциации и усложнения по сравнению с амфибиями. Ниже будет рассмотрено строение мускулатуры тела и конечностей по отделам.
Голова. Из мускулатуры геоидной дуги у прыткой ящерицы имеется две мышцы: сжиматель шеи (m. sphincter colli) и мускул, опускающий челюсть (m. depressor mand.). Мускулами подъязычной дуги являются челюстно-подъязычная (m. mylohyoide-
127
'Ш.
Рис. 46
Схема расположения основных мышц прыткой ящерицы (вид с брюшной стороны) 1 —- m. rectus abdominis;
2 — т. obliquus externus;
3 — т. pectoralis;
4 — т. mylohyoideus;
5 — mm. intercostales;
6 — m. odliquus internus (из M. M. Беляева, 1947)
us) и височная (m. temporalis), а также крыловидная мышцы (m. pterygoideus). В связи с тем, что у прыткой ящерицы еще сохранилась подвижность крыловидной и квадратной костей, у нее еще сохранились мускулы, такие же, как у примитивных амфибий — т. т. protractores levatores pterig., приводящие в движение крыло-видную кость.
Шейный отдел. На шее в связи с ее обособлением дифференцируется мускулатура, служащая для поднимания головы, боковых движений шеи и т. д.
Туловище. Мускулы туловища дифференцируются на три ряда мышц (рис. 46). Вдоль всего позвоночника тянется система поперечно-остистых мышц (m. т. transversospinal), снаружи от них — система длинных мышц (т. longissimus) и вдоль ребер — система подвздошно-реберных мышц (т. ileocostalis).
В связи с активной дыхательной деятельностью с помощью ребер развивается их мускулатура. Всего имеется шесть слоев мышц. Два наружных (m. obliquus externus) — наружная косая мышца (парная). От основания передней конечности идет грудная мышца (m. pectoralis). Бока тела прикрывает уже названная наружная косая мышца. Далее идут межреберные внешние и внутренние мышцы (m. m. intercostalis externus internus). Внутренняя косая мышца (m. obliquus internus) располагается между прямой брюшной и межреберными мышцами.
128
Конечности. В связи с усложнением движения рептилий мускулатура конечностей усложняется, значительно дифференцируется и сильно отличается у различных представителей рептилий в зависимости от их биологии и экологии (Суханов, 1961).
Хвост. Примитивное метамерное расположение мышц с хорошо выраженными миокомами. Жир и мышцы разделены системой септ. Такое положение элементов хвоста способствует аутотомии. Изучая механизм аутотомии, Ш. Леслей и А. Беллайре (Lesley, Bellairs, 1972) показали, что мышцы и отложения жира в хвосте разделены сагитальной и фронтальной септами на 2 эпаксиальных и 2 глипаксиальных ленты. Каждая из мышц расщеплена спереди и сзади на 3 отростка, которые вклиниваются между отростками соответствующих мышц прилежащих аутотомических сегментов. При аутотомии происходит сокращение мышц как позади, так и спереди от схваченного 1участка хвоста.
КРОВЕНОСНАЯ СИСТЕМА
Сердце прыткой ящерицы состоит как у большинства рептилий из двух предсердий и одного желудочка. Желудочек поделен неполной перегородкой на две полости.
В среднем сердце весит у взрослых особей 0,04 г, что составляет около 3,4—3,9% от общего веса тела (Шварц и др., 1968; Антипчук, 1966). Половой диморфизм по весу сердца отсутствует. Интересна динамика изменения относительного веса сердца; сердце сравнительно очень крупное у сеголетков (5,1%) и значительно меньше у взрослых особей (3,4%). При голодании индекс сердца прыткой ящерицы может уменьшиться до 1,4—1,6% общего веса тела (Шварц и др., 1968). Правая аорта дает вперед пару сонных артерий и затем, загибаясь, сливается с левой аортой в общий непарный ствол спинной аорты (рис. 47). От правой дуги аорты, перед слиянием ее с левой, отходят подключичные артерии. Спинная аорта на всем своем протяжении дает ответвления артерий к различным органам тела.
В правое предсердие впадает правая передняя полая вена (см. рис. 47). Слияние яремной и подключной вен образует левую верхнюю полую вену, которая также впадает в правое предсердие. Подобно же образована и правая передняя полая вена, но в нее еще впадает непарная головная вена. Особенности кровоснабжения кожи ящерицы описаны Ст. Кубиаком (Kubiak, 1963). В заднюю полую вену впадают печеночная вена, воротная вена печени, брюшная вена, образующаяся за счет слияния тазовых вен. Венозная кровь также попадает из задних отделов тела и другим путем, так как вены, несущие кровь от задних конечностей, образуют воротные вены почек, к которым присоединяется хвостовая вена. Из воротной системы почек кровь собирается в парные почечные вены, которые, слившись, образуют заднюю полую вену.
5 Прыткая ящерица i oq
Рис. 47
Схема расположения основных кровеносных сосудов прыткой ящерицы
1 — V. cranialis;
2 — a. carotides;
3 — v. jugulares;
4 — ductus caroticus;
5 — arcus aortae;
6 — a. pulmonalis;
7 — легкие;
8—15 — v. cava posterior;
9 — a. dorsalis;
10 — a. vertebralis;
11 — печень;
12—a. coeliaca;
13 — v. porta hepatica;
14 — кишечник;
16 — v. abdominalis;
17 — a. mesenterica caudalis;
18 — a. abdominalis;
19 — v. renalis;
20 — v. lateralis;
21 — a. lateralis;
22 — a. renalis;
23 — a. iliacae;
24 — v. iliacae;
25 — a. caudalis;
26 — v. caudalis
ДЫХАТЕЛЬНАЯ СИСТЕМА
Дыхательные пути начинаются наружными носовыми отверстиями— ноздрями, лежащими на конце морды (см. рис. 38). Далее воздух через носовой проход, обонятельный мешок и внутренние ноздри (хоаны) попадает в ротовую полость (рис. 48), в глубине которой располагается гортань.
Таким образом, в результате слияния дыхательного и пищеводного путей в ротовой полости во время акта проглатывания пищи дыхание должно быть затруднено.
Гортань прыткой ящерицы состоит из перстневидного хряща (cartilago cricoidea) и пары черпаловидных хрящей (с. arytae-noideae). Голосовые связки у ящерицы отсутствуют. Отмеченная способность ящериц издавать звуки (Баранов и др., 1972) объясняется, видимо, вибрацией слизистой оболочки, покрывающей черпаловидные хрящи. Вход в гортань располагается позади основания языка. Гортань ящерицы тесно связана с подъязычным аппаратом. Отходящая от гортани дыхательная трахея образована подвижно соединенными хрящевыми полукольцами. Число колец в трахее у разных животных даже в пределах одной популяции варьирует (в среднем около 50). У легких трахея разделяется на два бронха. Легкие представляют собой тонко
130
стенные полые мешки. Внутренние их стенки, в которых ветвятся кровеносные капилляры, имеют губчатое строение, что увеличивает общую дыхательную поверхность легких.
ПИЩЕВАРИТЕЛЬНАЯ СИСТЕМА
Раскрыв ротовую полость, легко обнаружить на краях верхней и нижней челюстей острые, слегка загнутые и в основном одновершинные зубы. Зубы лишены корней и сидят в одном общем желобке (плевральный тип зубной системы). Зубы подобного строения пригодны лишь для схватывания и удерживания добычи, но не для разжевывания пищи. Раскрыв глубже рот ящерицы, можно видеть две группы мелких зубов, находящихся также и на крыше ротовой полости. В связи с тем, что как на
Таблица 13
Распределение зубов в верхней и нижней челюстях прыткой ящерицы (Закарпатье, сборы М. И. Щербаня, 10 экз., 1970—1972 гг., просчет Г. И. Рузавиной)
Длина тела, мм Пол Верхняя челюсть Нижняя челюсть
справа слева справа слева
74,8(62—82) dd 17,6(13-20) 17,6(14-21) 17,1(11-21) 18,0(13-21)
76,4(64—89) ₽₽ 17,3(13—21) 17,3 (12-21) 17,2(14—21) 17,3(13—21)
верхней, так и на нижней челюстях встречаются двувершинные зубы, действительное их число определить довольно трудно, но возможно (табл. 13).
Как видно из табл. 13, число зубов практически одинаково и с правой и с левой стороны в верхней и нижней челюстях. Пока нет точных данных о числе зубов в разных участках ареала, но несомненно, эта величина несколько колеблется.
Язык-—мускулистый, плоский, суживающийся впереди, с глубокой вырезкой на конце. Он весьма подвижен, может довольно далеко высовываться из ротовой полости и выполняет функцию осязания.
В задней части ротовой полости находится отверстие пищевода. Пищевод в виде мускулистой растяжимой трубки тянется снизу от трахеи и в передней части брюшной полости впадает в объемистый мешкообразный желудок (рис. 49). Полость желудка неполностью перегораживается целой системой более крупных и мелких складок, но настоящего раздела на отделы еще не обнаруживается. От 12-перстной кишки желудок отделен хо-‘ рошо развитой кольцевой складкой (пилорус). Двенадцатиперстная кишка постепенно, без резкой границы переходит в тонкую кишку, которая после нескольких петель переходит в тол-
131
5*
Рис. 48
Общий вид ротовой полости ( Д)-и схема строения начала дыхательных путей ( Б) прыткой ящери цы
1 — носовые отверстия;
2 — хоаны;
3 — челюстная раковина;
4 — обонятельный отдел обонятельного мешка;
5 — дыхательный отдел;
6 — якобсонов орган (Д — рис. Г. И. Рузавипой, Б — из М. М. Беляева, 1947)
стую (заднюю) кишку. Задний отдел толстой кишки представляет собой прямую кишку, которая открывается в клоаку.
Относительная длина кишечника без пищевода и желудка по отношению к длине полости тела составляет у прыткой ящерицы от 76% (у сеголетков) до 147% у взрослых особей (Шварц и др., 1968). Такая длина кишечника обычно характерна для питающихся насекомыми рептилий и амфибий. Размеры отделов кишечного тракта, характеризующие одну из популяций прыткой ящерицы в Закарпатье, приведены в табл. 14.
Таблица 14
Абсолютные и относительные размеры (по 10 с? и 10 Q) пищеварительного тракта и его отделов у прыткой ящерицы (район Ужгорода, сборы М. И. Щербаня, 1970—1972 гг.у промеры по фиксированному материалу)
Показатель Длина
мм % ММ %
Самцы Самки
Длина тела (от кончика морды до анального отверстия) 74,8(62—82) 100 76,4(64,89) 100
Пищевод 25,2(18—30) 33,7 23,4(17-27) 30,6
Тонкий кишечник 63,7(46—91) 85,2 65,2(55—76) 85,3
Толстый кишечник 20,2(15—26) 27,0 19,6(14-27) 25,7
Весь пищеварительный тракт, от глотки до анального отверстия 132,8(118—171) 177,5 129,5(112—146) 169,5
Рядом с желудком и двенадцатиперстной кишкой располагается поджелудочная железа. На брыжейке вместе с желудком располагается небольшая шаровидная селезенка. Вся передняя часть брюшной полости занята крупной 3-лопастной печенью, имеющей форму треугольника, направленного вершиной вперед. На внешней ее стороне, в месте наложения лопастей, находится желчный пузырь. Отходящий от него желчный проток впадает в начало двенадцатиперстной кишки (см. рис. 49).
132
1 — пищевод;
2 — желудок;
3 .— тонкие кишки;
4 — толстая кишка;
.5 — клоака;
6 — печеиь;
у — желчный пузырь;
д — поджелудочная железа;
9 — трахея;
10 — левое легкое;
// — желудочек сердца;
12 — правое предсердие;
13 — правая дуга аорты;
14 _ правая сонная артерия;
15 — левая сонная артерия;
16 — левая дуга аорты;
77 — спинная аорта;
jg __ правая легочная артерия;
19 — задняя полая вена;
20 — легочная веиа;
21 — левый семенник;
22 эпидидимис;
23 —- семяпровод;
24 — левая почка;
25 —впадение семяпроводов в клоаку;
26 — мочевой пузырь;
27 — селезенка;
28 — надпочечник;
29 — копулятивные органы (из М. М. Беляева, 1947, с дополнениями и изменениями)
Рис. 49
Схема расположения внутренних органов самца ящерицы рода Lacerta
Несмотря на то, что печень, несомненно является важной железой пищеварительного тракта, основное се значение связано с общим изменением метаболизма организма ящерицы (Шварц и др., 1968). Об этом свидетельствуют резкие изменения относительного веса печени у самок в разном физиологическом состоянии: индекс печени у самок с неразвитыми фолликулами 45,5%, у самок с яйцами диаметром до 5 мм —71,5%, а у самок с вполне сформировавшимися яйцами индекс печени вновь падает до 39,5% (Шварц и др., 1968). При этом коэффициент вариации индекса печени вне периода размножения достигает 30%, а в период наибольшего развития печени падает до 16% (что косвенно свидетельствует о функциональной важности органа именно в этот период).
Онтогенетические изменения абсолютного и относительного веса печени на большой серии самцов прыткой ящерицы показаны на рис. 50. Видно, что, несмотря на постоянный и значи-
133
700
Рис. 50
Изменение относительного н абсолютного веса печени самцов прыткой ящерицы с возрастом (на основании данных по 76 особям) (Шварц и др., 1968)
тельный прирост веса печени на протяжении всей жизни ящериц, относительный вес печени, достигая максимума у средних по размерам особей, у самых крупных животных, падает.
Печень — самая крупная железа в теле прыткой ящерицы; ее абсолютный вес достигает у крупных самцов более 0,5 г (относительный вес печени 42,6+3,6%) в среднем для 90 изученных особей (Шварц и др., 1968), что значительно превосходит индексы практически всех остальных внутренних органов.
МОЧЕПОЛОВАЯ СИСТЕМА
При определении половой принадлежности той или иной особи уже некоторые внешние признаки (такие, как диморфизм в окраске, форма головы, различие в степени развития бедренных пор и форма основания хвоста), как правило, позволяют определить пол. Но для более точной характеристики степени половой активности и установления достижения половой зрелости следует прибегнуть к рассмотрению половой системы отловленных животных. С половыми органами топографически и филогенетически связана система органов мочеотделения. Рассмотрим эти системы более подробно.
Половая система самцов. Овальные семенники расположены в верхней части брюшной полости несколько на разных уровнях (левый — каудальнее) и тесно связаны посредством брыжейки с развитыми придатками семенника, которые продолжаются в тазовую полость тонкими извитыми семяпроводами (см. рис. 49).
1 Последние открываются маленькими отверстиями в клоаку с дорзальной ее стороны. Вблизи верхних концов семенников лежат небольшие надпочечники, относящиеся к железам внутренней секреции.
Оплодотворение у прыткой ящерицы внутреннее, оно проис-ходит с помощью парных копулятивных органов, которые отхо-1 дят от середины задней стенки до клоаки и залегают параллель-1 но друг другу в особых карманах под кожным покровом. По форме они напоминают удлиненные мешки, плотные на ощупь, так как состоят из плотной фиброзной ткани. Наполняясь кровью
134
в состоянии эрекции, эти копулятивные органы выворачиваются наружу и вводятся при копуляции в клоаку самки. На внутренней стороне каждый из копулятивных органов снабжен бороздкой для стекания семенной жидкости, которая попадает туда из семяпроводов.
Половая система самок. У небеременной самки под позвоночником в поясничной области хорошо заметны яичники (рис. 51), имеющие форму удлиненных овальных образований желтоватого цвета; они подвешены на особой брыжейке. От присутствия находящихся в разных стадиях созревания яиц (обычно, у взрослой ящерицы около 10 в каждом яичнике) яичники имеют гроздевидное строение. К поверхности яичников со стороны спины плотно прилегают надпочечники. Яйцеводы, имеющие вид сильно извитых и сжатых трубок, своим широким концом — воронкой — открываются в переднюю часть брюшной полости, а другим — открываются в дорсальный отдел клоаки. Таким образом, яйцеводы не сообщаются непосредственно с яичниками. Зрелое яйцо выпадает через разрыв стенки яичника в полость тела и, подхваченноерасширен-ной воронкой яйцевода, опускается по нему в результате перистальтических движений. На этом пути происходит оплодотворение яйца сперматозоидами, активно поднимающимися вверх по яйцеводам после успешной копуляции. После оплодотворения в задних, рас-
Рис. 51
Схема строения мочеполовых органов самки прыткой ящерицы
/ — яичники; 2 — правый яйцевод; 3 — воронка яйцевода; 4 — выходное отверстие яйцевода в клоаке; 5 — почка; 6 — отверстие мочеточников в клоаке; 7 — мочевой пузырь; 8 — прямая кишка; 9 — впадение прямой кишки в клоаку (по Wiedersheim, 1892)
135
ширенных у беременных самок, частях яйцеводов, происходит окончательное формирование яиц. Вероятно, степень развития яиц перед откладкой их несколько различается в разных частях ареала; по аналогии с тем, что наблюдается у живородящей ящерицы, можно предположить, что яйца у северных популяций прыткой ящерицы могут задерживаться в задних частях яйцевода на более продолжительный период и откладываются более зрелыми (см. также гл. VII).
Система органов мочеотделения. Система органов мочеотделения у рептилий включает почки, мочеточники и мочевой пузырь. Почки располагаются в тазовой области (см. рис. 51) и являются по своему происхождению вторичными почками (metanephros), имеют продолговатую форму и покрыты париетальным листком брюшины (т. е. расположены, как и обычно у позвоночных, в забрюшинном пространстве). От их задних суживающихся концов отходят короткие мочеточники, впадающие дорсально в клоаку общими отверстиями с семяпроводом. Моча из этих отверстий стекает по стенке клоаки в объемистый мочевой пузырь. По мере наполнения мочевого пузыря, а также в ряде поведенческих актов, имеющих, несомненно, сигнальное значение (например, маркировка территории), мочевой пузырь опорожняется.
ЦЕНТРАЛЬНАЯ НЕРВНАЯ СИСТЕМА
Мозг прыткой ящерицы подразделяется на пять отделов, характерных для мозга всех позвоночных.
Передний мозг прекрасно выражен, имеет колбообразную суживающуюся кпереди, приближающуюся к треугольной, форму (рис. 52). Полушария широкие и слегка выпуклые с дорсальной
Таблица 15
Абсолютный и относительный вес головного мозга н его отделов у трех видов ящериц рода Lacerta и других позвоночных (по Никитенко, 1969)
Вид Вес тела, г Головной мозг Коэффициент стриатализации Средний мозг Мозжечок । Продолговатый мозг Обонятельные луковицы
о са в процентах к весу тела квадратный указатель по Я. Я. Рогинскому
в % к ВС су все о мозга
L. vivipara 8,2 0,043 0,28 0,00060 43,4 12,7 4,3 28,3 11,3 60
L. viridis 51,2 0,130 0,24 0,00030 44,1 14,3 4,2 25,2 11,2 10
L. agilis 12,3 0,048 0,38 0,00020 45,3 13,8 5,6 27,1 8,2 50
Natrix natrix 59,3 0,062 0,11 0,00006 48,6 8,4 9,8 30,4 3,8 22
Vipera berus 60,5 0,091 0,15 0,00014 51,6 8,1 9,6 25,6 4,2 20
Emys orbicularis 648,0 0,207 0,03 0,00007 51,4 10,8 10,2 20,4 7,2 10
Passer montanus 13,0 0,39 3,12 0,01200 56,4 0,4 7,6 20,5 10,2 16
136
Рис. 52
Схема строения головного мозга прыткой ящерицы а — сверху; б — снизу; в — сбоку; 1 — обонятельные доли; 2— передний мозг; 3 — эпифиз; 4 — Средний мозг; 5 — мозжечок; 6 — IV желудочек; 7 — продолговатый мозг; 8 — зрительный нерв; 9 — хиазма; 10 — «воронка; 11 — гипофиз (рис. Л. И. Суховской н Г. И. Рузавиной)
стороны. У прыткой ящерицы, как и у большинства рептилий, имеется мозговая кора. При рассмотрении мозга обращают внимание массивные образования переднего мозга — обонятельные доли, длина которых составляет у этого вида до 22% общей длины головного мозга (Никитенко, 1969).
Передний мозг покрывает промежуточный. От крыши промежуточного мозга, кроме эпифиза, отходит также нерв к теменному глазу, хорошо развитому у прыткой ящерицы.
Средний мозг представлен двухолмием и хорошо развит, хотя по абсолютным размерам уступает переднему мозгу. Средний мозг несколько приподнят и так же, как и передний мозг, налегает на промежуточный (см. рис. 52). Мозжечок представляет собой язычкообразную пластинку и немного прилегает к среднему мозгу, в заднем своем направлении нависает над продолговатым мозгом.
По уровню развития мозга в целом и некоторым энцефало-метрическим показателям среди исследованных видов настоящих ящериц прыткая ящерица стоит на первом месте (Никитенко, 1969). Так, в частности, как видно из табл. 15, прыткая ящерица заметно отличается от других видов рептилий более высоким относительным весом головного мозга.
СТРОЕНИЕ ОРГАНОВ ЧУВСТВ
В настоящее время, видимо, нет достаточно полных данных о функционировании и развитии всех органов чувств у прыткой ящерицы. Поэтому рассмотрим характеристики лишь основных органов чувств.
137
Рис. 53
Взрослая самка прыткой ящерицы в террариуме ИБР АН СССР, январь 1975 г.
Обратить внимание на расположение носового отверстия, форму и величину глаза (фото А. В. Яблокова и А. В. Валецкого)
Обоняние является чувством дистантного действия и развито у прыткой ящерицы очень хорошо, о чем свидетельствуют относительно крупные размеры обонятельных луковиц обонятельной полости (см. рис. 48) и других формаций обонятельного анализатора. Однако отсутствие экспериментальных данных не позволяет пока дать точную оценку экологического значения и степени важности обонятельного анализатора у прыткой ящерицы. К этому следует добавить, что в настоящее время недостаточно известны функциональные возможности якобсонова органа (см. рис. 48), помещающегося на дорсальной поверхности ротовой полости, который также может принимать участие в обонятельной рецепции (хотя, видимо, больше связан с вкусовой рецепцией).
Значительное развитие обонятельных луковиц головного мозга и обонятельной полости в носовом проходе у ящериц находится в противоречии с предположением о сравнительно небольшом значении обонятельного анализатора в жизни ящериц.
138
Только дальнейшие специальные исследования позволят разрешить это противоречие. При этом кажется важным принять в0 внимание одну принципиальную особенность обонятельной рецепции — она позволяет анализировать сигналы через очень длительный период времени. Эта особенность делает обонятельный анализатор чрезвычайно важным органом чувств.
Считается, что у всех пресмыкающихся акустическая сигнализация развита незначительно и играет значительно меньшую роль, чем у птиц, земноводных и млекопитающих. В то же время специальных широких исследований по значению слухового анализатора у большинства рептилий (и прыткой ящерицы, в частности) практически не проводилось (подробности см. в главе IV).
Зрение. Несмотря на сравнительно небольшие размеры, орган зрения у прыткой ящерицы (рис. 53) является хорошо развитым органом чувств. Веки подвижные (особенно нижнее, которое в основном и покрывает все глазное отверстие). Зрачок овальный, его длинная ось расположена поперек глазного отверстия. Хрусталик слегка уплощен. Характер рефлекторных ответов зрачка на изменение интенсивности света (изменение размеров зрачка) у Lacertidae происходит медленно и, по-ви-дпмому, не влияет на их зрение (Prince, 1956).
Глаза ящерицы камерного типа, с большим расстоянием между хрусталиком и сетчаткой, что свойственно дневным животным. У них имеются внутриглазные цветные фильтры, поглощающие свет в синем конце спектра, что уменьшает хроматическую аберрацию на поверхности сетчатки и, следовательно, увеличивает остроту зрения.
Аккомодация глаза осуществляется за счет изменения кривизны хрусталика. Интересно отметить, что у L. agilis, как у большинства других видов ящериц, движения обоих глаз бывают независимы (Welsh, Osborn, 1937; Walls, 1942; Ali, 1959). Сетчатка глаза прыткой ящерицы по своему строению напоминает сетчатку глаза птиц, приматов и человека. В ней встречаются два типа колбочек и сравнительно мало палочек (это указывает на развитие у этого вида цветного зрения (Васильев Б. Д., in litt, см. также гл. IV). Внутренний ядерный слой сетчатки в 2—3 раза мощнее наружного. Система зрительных пигментов имеет типичные для наземных позвоночных черты: в ней встречаются как палочковые (родопсин), так и колбочковые (иодопсин, цианопсин) пигменты, что свидетельствует о развитии цветного зрения. Это подтверждено и экспериментально (Wiand, 1960). Кроме того, показано, что нейроны, формирующие волокна зрительного нерва у ящериц, построены таким образом, что могут обеспечивать цветное зрение (Granit, 1947).
Осязание. Основным органом, выполняющим функцию осязания, является язык. Ящерица часто касается языком находящихся впереди нее предметов. Когда язык прячется в рот, кон-
139
чики его попадают в особое углубление, снабженное нервными чувствующими окончаниями,— якобсонов орган (см. рис. 48), воспринимающий химические раздражения. Тактильная чувствительность кожного покрова значительно понижена в связи с его ороговением и развитием чешуйчатости. В то же время отдельные участки тела, и в частности, нижняя поверхность конечностей, должна обладать достаточно высокой чувствительностью.
Другие органы чувств. Из других органов чувств остается упомянуть о существовании так называемого теменного глаза (см. рис. 38). Считается, что теменной глаз несет определенную фоторецепторную функцию. Возможно также, что назначение теменного органа связано с функцией терморегуляции. Этот орган, несомненно чувствительный к свету, не способен к предметному зрению. Наконец, возможно, что этот орган сохраняется у ныне живущих форм лишь в качестве рудиментарного.
ГЛАВА ЧЕТВЕРТАЯ
ФИЗИОЛОГИЯ и БИОХИМИЯ1
Изучение жизненных процессов и функций — т. е. собственно физиологические исследования, до сих пор в отношении прыткой ящерицы проводились нерегулярно. Имеются лишь немногие отдельные работы, посвященные терморегуляции и газообмену (Либерман, Покровская, 1943; Стрельников, 1944; Тер-тышников, 1973). Сказанное определяет то обстоятельство, что в настоящей главе при описании отдельных функций более широко, чем это сделано в других главах, используются данные, касающиеся других видов рептилий. Это же замечание относится и к данным по биохимии прыткой ящерицы. Более или менее подробные оригинальные данные по составу микроэлементов в теле прыткой ящерицы, по существу, представляют собой основу этой части главы. Немногочисленные опубликованные работы (Vladescu et all., 1971; Hadley, Christie, 1974; Hahn, Tinkle, 1965) дополняют краткий биохимический очерк.
ОСНОВНЫЕ ЧЕРТЫ ФИЗИОЛОГИИ ПРЫТКОЙ ЯЩЕРИЦЫ И ДРУГИХ РЕПТИЛИЙ
Имеющийся материал позволяет с той или иной степенью подробности рассмотреть следующие жизненные процессы и функции прыткой ящерицы и других рептилий: дыхание и газообмен, терморегуляцию, кровообращение, пищеварение, выделение и некоторые особейности обмена веществ.
Дыхание и газообмен
Прыткой ящерице присуще как кожное, так и легочное дыхание. Точных данных по кожному дыханию у этого вида нет, но по аналогии с другими видами ящериц (Fitzgerald, 1957) можно предположить, что значительная часть углекислоты может выделяться через кожу (до 85% выделения ее через легкие).
1 Авторы: Ю. П. Антппчук (физиология) и Л. М. Смеловский (микроэлементы), при участии М. Ф. Тертышникова, В. Е. Боченко, А. В. Яблокова.
141
Основное значение при вдохе — выдохе имеет работа мышц боковых стенок грудной полости и шейного отдела. У спокойно сидящей ящерицы дно ротовой полости опускается и поднимается с частотой 2—4 раза в 1 сек. Эти движения имеют определенное значение для вентиляции передних частей легких, равно как и для постоянного восприятия запахов при помощи обонятельного мешка, располагающегося по ходу проводящих воздух путей. Значительно более редки глубокие дыхательные движения, совершающиеся за счет сокращения мышц стенок грудной полости. В зависимости от активности животного эти движения совершаются с частотой 1—2 в 1 сек., но временами между серией глубоких вздохов может наступать пауза в несколько секунд. Число дыханий резко уменьшается в периоды пониженной активности (ночью, в течение летней и зимней спячки). Вероятно, в такие периоды увеличивается относительное значение кожного дыхания.
Как показывают наши эксперименты, у прыткой ящерицы увеличиваются частота и амплитуда дыхания с повышением концентрации СО2 во вдыхаемом воздухе (подобное явление отмечается и у других видов пресмыкающихся). Таким образом, дыхательный центр ящериц можно рассматривать как прототип дыхательного центра высших позвоночных (Smith, 1939).
Проведенные М. Ф. Тертышниковым (1973) эксперименты-показывают, что с повышением температуры тела потребление
Таблица 16
Потребление кислорода прыткой ящерицей и разноцветной ящуркой при разной температуре тела (В сма на 100 г веса в течение 1 часа при 0° и 760 мм рт. ст.) (по материалам
М. Ф. Тертышникова* 1968—1971 гг., сводные данные по животным разного пола и возраста)
Температура, °C Л М±т п М±т
Прыткая ящерица Разноцветная ящурка
0 8 3,0+1,0 5 14,4±5,8
5 9 47,8±4,6 23 72,2±12,0
10 15 66,0±28,94 •14 98,5+11,3
15 20 84,2+10,43 14 124,7±11,3
20 37 128,5±17,0 18 195,0±14,1
25 18 242,9zbll,2 14 347,0±23,4
30 16 532,0±91,97 11 1102,0±132,7
31 4 169,0±27,75 5 264,0± 73,63
. 32 8 260,0 +27,0 4 256,0±98,65
33 8 236,0+ 20,10 3 340,0± 140,6
34 7 113,0+40,0 4 243,0+ 61,0
35 13 127,0+46,0 26 273,0±13,6
40 3 24,3+2,00 3 138,0±15,18
.47 — 0,00 4 0,00
142
кислорода увеличивается, а с понижением уменьшается (табл. 16, рис. 54), что характерно для всех пойкилотерм-ных животных, к которым ОТНОСЯТСЯ и прыткие ящерицы. Аналогичные данные известны и для других рептилий (Dawson, Bartholomeu, 1958; Dawson, Templeton, 1963; Калашников, 1964; Benedict, 1932, 1938 и др.).
Наиболее низкое потребление кислорода прыткими ящерицами было отмечено нами при 0е. При такой температуре энергетический обмен очень низок, что. несомненно, биологически оправдано, так как удлиняет продолжительность жизни этих животных в отсутствие корма. Повышение количества потребляемого кислорода происходит при подъеме температуры тела до 30°. При ее увеличении с 30 до 35° количество потребляемого кислорода резко падает и находится на более или менее постоянном уровне. Дальнейшее повышение температуры тела приводит к тому, что газооб-
Рис. 54
Мен прЫТКОЙ ЯЩерИЦЫ прекра- Характер потребления кислорода прыткими щается при 41,6—42°, а у раз- ящерицами (() и разноцветными ящурка-ми (2) в зависимости от температуры тела (no М. Ф. Тертышникову)
ящерицы погибают).
Удержание уровня потреб-
ления кислорода в узкой зоне температур от 30 до 35° на более
ноцветнои ящурки — при 47—
47,2° (при этих температурах
или менее постоянном уровне свидетельствует о наличии у прыткой ящерицы химической терморегуляции. Однако интенсивность этих процессов невысока, что подтверждается их неспособностью удерживать температуру тела на оптимальном уровне (около 30°) при ее снижении или повышении.
Изменение температуры среды на протяжении суток и сезонов сказывается на уровне газообмена. Весной, летом и осенью прослеживается установленная зависимость между количеством потребляемого ящерицами кислорода и температурой. Кроме то
143
го, следует отметить, что у прыткой ящерицы уровень газообмена осенью выше, чем летом, а летом выше, чем весной.
Проведенные эксперименты не обнаружили разницы между количеством потребляемого кислорода днем и 'ночью (при условии, что температурные условия среды не изменялись на протяжении суток).
Между ящерицами разного пола и возраста существуют довольно заметные отличия в уровне газообмена (табл. 17).
Таблица 17
Суточное потребление кислорода прыткими ящерицами разных половых и возрастных групп (данные М. Ф. Терты пникова, 1938—1971 гг., в см3 иа 100 г веса при 0° и 760 мм рт. ст.)
Группа особей по 5 экз. Количество кислорода, см3 Температура тела, °C Вес животного, г
Взрослые самцы 1410 11,5-25 13,0—25,0
Взрослые самки 1053 14,0—19,5 13,0—20,0
Полувзрослые самцы и самки 3330 17,0—24,0 1,4-2,8
Возрастные отличия в количестве потребляемого кислорода отмечены и для других групп рептилий (Калашников, 1964). Причину данного явления следует искать, видимо, во взаимосвязи между уровнем окислительных процессов в организме и размерами тела. Более интенсивное поглощение кислорода молодыми животными объясняется относительно большей поверхностью их тела (по отношению к весу), чем у взрослых животных (Rub-’ пег, 1883).
Терморегуляция
Обычно наибольшая активность прытких ящериц наблюдается при размахе предпочитаемых температур тела 18,0—36,0° (измерения в клоаке), причем в разных географических районах температура наибольшей активности несколько различна и, видимо, является локальной адаптацией к конкретным условиям существования (Тертышников, 1973а; Боченко, уст. сообщ. и др.). В среднем максимальная активность наблюдается при температуре тела, близкой к 30° (рис. 55). При температуре тела ниже 7,5° прыткая ящерица впадает в оцепенение, при температуре тела ниже —4,9° и выше 42° погибает (Либерман, Покровская, 1943; Стрельников, 1944).
Для прыткой ящерицы характерны значительные суточные и сезонные колебания температуры тела. Данные о топографии температуры тела отсутствуют, однако прямые наблюдения в природе и террариуме показывают, что температура конечностей и хвоста обычно заметно ниже температуры брюха и горла.
144
зо
25
гП
50 25 30 35 40 45 50
Температура
Рис. 55
Основные температурные параметры прыткой ящерицы
1 — температура активных ящериц в природе; 2 — температура, избираемая ящерицами в эксперименте в аппарате Гертера;
3— температура тела ящериц при температуре воздуха 32,8° С (оптимальной); 4 — температура теплового окоченения прыткой ящерицы; 5 — летальная температура. Число исследованных особей соответственно: 37, 65, 79, 21 и 14 экз. (по данным С. С. Либермана, Н. В. Покровской, 1943)
10
5
Итак, несомненно, что температура тела ящерицы может значительно отличаться от температуры окружающей среды. По наблюдениям М. Ф. Тер-тышникова, эти различия достигают 18°. А. В. Яблоков и А. В. Валецкий в апреле 1973 г. в районе г. Караганды наблюдали прытких ящериц, активно бегающих по снегу в солнечную ветреную погоду при температуре воздуха до —10е; в этом случае разница между температурой среды и температурой тела должна была быть не менее 25—30° (!). Поэтому, строго говоря, отнесение этого вида к пойкилотермным животным, у которых отсутствует терморегуляция, может быть сделано лишь с большими оговорками. Видимо, правильнее считать этот вид ящериц (как видимо и большинство других рептилий) гетеро-термным. Пока мало известно о механизмах химической терморегуляции данного вида, хотя сам факт такой терморегуляции несомненно имеет место (М. В. Тертышников). Значительно больше данных имеется по физической терморегуляции, которая у прыткой ящерицы осуществляется всеми тремя известными способами — проведением, излучением и испарением —-(Либерман, Покровская, 1943; Стрельников, 1944; Тертышников, 1973 и др.) и тесно связана с поведенческой активностью (см. гл. X).
Поскольку прыткая ящерица покрыта прочной роговой чешуей, до последнего времени считалось, что животные теряют мало воды путем испарения. В связи с этим предполагалось, что температура тела прыткой ящерицы будет мало зависеть от влажности. В пользу этого суждения свидетельствует тот факт, что у жабровидной ящерицы снижение влажности со 100 до 7% при 20° вызывало понижение температуры тела всего на 1° (Hall, Root, 1930).
Однако в последние десятилетия накопилось немало данных, показывающих, что чешуйчатый покров ящериц не является препятствием для испарения влаги организмом. Прямые экспе
145
рименты показали, что через кожу ящерицы могут испарять от 25 до 50% всей теряемой организмом влаги, и кожа является таким образом важным органом активной физической терморегуляции (Claussen, 1967; Kerfoot, 1970 и Др-)- Об этом же косвенно свидетельствует высокая положительная корреляция числа чешуи у некоторых видов североамериканских ящериц с минимальными зимними температурами, с одной стороны, и величиной осадков в летние месяцы с другой (Soule, Kerfoot, 1972). К этому остается добавить, что существует клинальная изменчивость числа чешуй у некоторых вйдов рептилий, что также свидетельствует о возможной роли чешуйчатого покрова в терморегуляции.
Итак, температура тела ящериц во многом зависит от температуры внешней среды, однако прыткая ящерица имеет некоторые механизмы, регулирующие температуру. Эта регуляция у прыткой ящерицы достигается изменением как поведения (см. гл. X), так и метаболизма.
Характеристика крови
У прыткой ящерицы содержание гемоглобина составляет 7,8 г на 100 мл крови (Тертышников, 1972). О продолжительности жизни эритроцитов крови у прыткой ящерицы сведения отсутствуют, но можно предположить, что она равна 11 месяцам, как у большинства других пресмыкающихся (Berlin, 1959).
По нашим данным, количество эритроцитов у прыткой ящерицы составляет 11 670 в мм3 крови, РОЭ —4 мм в час, гемоглобин—11,5%. У близких видов ящериц полосы поглощения различных форм пигмента крови характеризуются такими величинами: НЬО2—576,2 ммк, НЬСО—571,5, расстояние между a-полосами НЬО2 и НЬСО—4,7 ммк (Prosser, Brown, 1962). Нет данных по прыткой ящерице о видах гемоглобина, которые у пресмыкающихся отличаются по молекулярному весу, изоэлектрической точке и времени денатурации (Ramirez, Dessauer, 1957). Константа седиментации гемоглобина у L. vivipa'ra равна 4,6 S. Поскольку эта константа свойственна рыбам, пресмыкающимся, птицам и млекопитающим (Svedberg, Pedersen, 1959), то и прыткая ящерица, видимо, не должна составлять исключения.
Дыхательные свойства крови у настоящих ящериц, к которым относится и L. agilis, характеризуются следующими величинами: кислородная емкость крови составляет 10—12,5 об.%, напряжение О2 при 50% насыщения крови (Р50) равно 19—31 мм рт. ст. (Edwards, Dill, 1935; Dawson, Bartholomew, 1960).
Касаясь взаимосвязи дыхательных пигментов у ящериц с экологическими условиями следует сказать, что, чем доступнее кислород для организма, тем выше давление, при котором происходит насыщение гемоглобина кислородом. У прыткой ящерицы, по нашим данным, не отмечено усиления эритропоэза при
146
Таблица 18
процентное соотношение белковых фракций крови двух популяций прыткой ящерицы (Страутман и др., 1965)
Популяция (УССР) Альбумины Глобулины
I 2 «1 Т1
Равнинная (Подолия) Субальпийская (Карпаты) 36,699 39,062 33,854 43,615 12,500 4,265 3,645 15,417 10,937
низком парциальном давлении кислорода. При этом у прыткой ящерицы имеет место лишь усиление дыхания. О переносе углекислого газа у ящериц известно лишь то, что его содержание в крови колеблется от 38,6 до 44 об.% (Hopping, 1923).
Рис. 56
Денситограмма сыворотки крови прыткой ящерицы
а —равнинная популяция в Подо-лии; б—горная популяция на альпийском лугу в Карпатах (Страутман и др., 1965)
У ящериц сывороточные белки крови имеют хорошо выраженные буферные свойства (Redfield, 1933, 1934). У прыткой ящерицы обнаружено несколько фракций крови. Данные о составе белковых фракций сыворотки крови для двух западных популяций прыткой ящерицы приведены в табл. 18, а денсито-метрическая кривая — на рис. 56.
Из приведенных данных видно, что характер фракций крови у прыткой ящерицы значительно колеблется в разных популяциях (Черкащенко, Кушнирук, 1973).
Относительный объем крови у пресмыкающихся сильно варьирует (Coulson, 1950; Semple, I960). По нашим наблюдениям, у прыткой ящерицы объем крови составляет 6,3% веса тела.
Кровообращение
Как отмечено в главе III, вес сердца у прыткой ящерицы составляет 3,4—3,9% веса тела, что очень близко к показателям у других представителей этого рода (Quiring, 1950). Наши наблюдения показывают, что число сокращений сердца при 20° у
147
взрослых ящериц составляет 52+2 ударов/мин. Другие авторы (Laurens, 1920) приводят несколько меньшие величины, которые, однако, даже еще меньше, чем у других представителей этого рода (Buchanan, 1909). При этом надо отметить, что изменение температуры среды на 1° в диапазоне 18—32° вызывает у прыткой ящерицы изменение частоты сокращения сердца в среднем на 1 удар в минуту.
Данных по кровяному давлению в малом и большом кругах кровообращения у прыткой ящерицы нет, однако, по всей вероятности, они должны быть близки к этим показателям у других видов ящериц. У серого варана (Varanus griseus), например, по нашим данным, кровяное давление в левом «желудочке» сердца равно 65—75/0, а в дуге аорты — 65—75/30 мм рт. ст. В малом круге кровообращения уровни кровяного давления характеризуются такими величинами: в правом «желудочке» сердца — 42± ±1/0, а в легочной артерии — 41 +1/22+1 мм рт. ст. Эти показатели близки к таковым у других видов ящериц (Harrison, 1965; White, 1968). Как показывают наши эксперименты, минутный объем сердца на килограмм живого веса у серого варана равен 112 мл, общее сосудистое сопротивление большого круга кровообращения составляет 19 980, а малого круга кровообращения — 8880 дин/сек/см~5.
Известно, что у пресмыкающихся повышение температуры тела ведет к повышению кровяного давления. Таким образом, у ящериц, вероятно, существуют вазомоторные механизмы экзотермической терморегуляции, когда расширение сосудов способствует поглощению тепла извне, а сужение их препятствует этому явлению (Prosser, Brown, 1962).
Несмотря на то, что у ящериц межжелудочковая перегородка неполная (Foxon, 1955; Татаринов, 1960; Жеденов, 1961; Ludicke, 1964; Антипчук, 1966, 1967; Антипчук, Гибрадзе, 1973), существ вует мнение, что потоки крови в трехкамерном сердце ящериц в какой-то мере разделены (Hoffman, 1879—1890; Соре, 1900; Goa odrich, 1930; Шмальгаузен, 1938; Meinertz, 1952, 1966; Foxon, 1956; Татаринов, 1960; Жеденов, 1961; Ludicke, 1964). С одной стороны, физиологические данные свидетельствуют о том, что у ящериц существует незначительное смешивание крови (Prakash, 1952; Foxon et all., 1953; Foxon, 1955; Khalil, Zaki, 1964; Grasse, Devillers, 1965) в трехкамерном сердце. С другой стороны, в противоположность приведенным данным,, существует мнение, что при оптимальной температуре тела у ящериц имеет место полное разделение артериальной и венозной крови в трехкамерном сердце (White, 1955, 1956, 1959, 1968; Антипчук, 1966, 1967). Объяснение подобного противоречия приводится в книге А. П. Антипчука, Т. А. Гибрадзе (1973). Поскольку у прыткой ящерицы сердце трехкамерное, то и у нее при оптимальных температурах должно иметь место полное разделение потоков артериальной и венозной крови.
148
Пищеварение
Известно, что способ питания данного организма определяется главным образом средой обитания и характером доступной пищи. Гистохимическое исследование двенадцатиперстной кишки у 33 видов ящериц (в том числе и у прыткой ящерицы) показало, -что строгая зависимость между характером питания и гистоструктурой отсутствует (Gabe, Girons, 1972).
Касаясь переваривания углеводов, можно сказать, что у ящериц обнаружена в кишечнике активная мальтоза.' Поскольку мальтоза и сахароза, как правило, встречаются вместе и оптиму-мы их pH близки, можно предположить, что мальтоза и сахароза перевариваются при помощи одного и того же фермента (Yon-ge, 1928; Manseur-Bek, 1932, 1933, 1948). Кроме того, имеются данные, показывающие, что у прыткой ящерицы лактоза в кишечнике не переваривается. По нашим данным, у взрослых прытких ящериц химус желудка имеет кислое pH, и в нем обнаружен пепсин.
О переваривании жиров у прыткой ящерицы прямых данных нет, однако известно, что у большинства рептилий в кишечнике имеется триоксикопростановая и питохолевая кислоты (Bergmann, Расе, 1943). Эти кислоты также должны быть присущи и прыткой ящерице, у которой, как и у прочих рептилий, активность пищеварительного сока связана с панкреатической липазой. Однако у ящериц не установлено, какой фермент принимает участие в переваривании жиров — липаза или эстераза. Состав потребляемого жира у ящериц до некоторой степени определяет состав депонированного жира, в который входит значительное количество ненасыщенных жиров. Депонированный жир у ящериц содержит много олеиновой кислоты и меньше пальмитиновой и стеариновой кислот (Klenk, 1933, 1935; Крепе, 1964).
Данные по скорости переваривания пищи, количественному и качественному составу пищи в разных частях ареала приведены в главе VI.
Выделение
У ящериц, как и у других пресмыкающихся, конечные продукты обмена веществ удаляются из организма путем диффузии гг активных процессов выделения. Аминокислоты подвергаются окислительному или гидролитическому дезаминированию с выделением свободного аммиака. По нашим данным, у прыткой ящерицы концентрация аммиака в крови достигает 0,12 мг/100 мл, т. е. примерно такая же, как и у других видов пресмыкающихся. Ящерицы способны превращать аммиак в мочевую кислоту. Последняя мало токсична и плохо растворима; она может откладываться или выделяться в кристаллической форме. Точка замерзания мочи у прыткой ящерицы, по нашим наблюдениям, 0,2, а;
149
крови 0,62, т. е. находится в пределах, свойственных другим пресмыкающимся (Smith, 1932; Bentley, 1959).
Непосредственным источником аммиака у ящериц является глютаминовая кислота, а источником выделяемого азота — продукты обмена нуклеиновых кислот — пурины. Азот пиримидинов выделяется у ящериц в виде мочевины и аммиака, причем мочевая кислота образуется как конечный продукт окислительного дезаминирования пуринов. Птерины насекомых, которыми питаются ящерицы, входят в состав фолиевой кислоты — птеро-ил-Ь-глютаминовой кислоты. Биосинтез птеринов во многих чертах напоминает биосинтез мочевины. Кроме того, существует связь между биосинтезом птеринов и биосинтезом рибофлавина. -Эти вещества встречаются в хроматофорах пресмыкающихся .(Forrest, et al., 1962).
У большинства рептилий процентное соотношение конечных продуктов азотистого обмена (по азоту) составляет: аммиака — 8,7, аминного азота — 2,3 и мочевой кислоты — 89% (Bacon, 1909). Следовательно, у ящериц большая часть азота выделяется в виде мочевой кислоты (Khalil, Haggag, 1955) и, вероятно, прыткая ящерица не составляет исключения; поэтому ее можно отнести к уриколитическим животным. Различие между уриколити-ческими иуролитическими организмами является чисто количественным, и наличие мочевой кислоты в крови еще совсем не означает, что мочевая кислота является основным продуктом азотистого обмена.
Известно, что у чешуйчатых рептилий катаболизм белков из-•меняется на различных стадиях онтогенеза. На ранних стадиях развития конечные продукты белкового обмена представлены аммиаком: далее возрастает удельный вес мочевины и затем •большая часть азота выделяется в виде мочевой кислоты, скапливающейся в аллантоисе. Кроме того, у чешуйчатых в период уменьшения выделения мочевины в аллантоисе во время развития зародыша обнаруживалась уреаза в печени и почках (Clark, 1953). Подобное изменение в составе конечных продуктов азотистого обмена в раннем онтогенезе у ящериц биологически оправдано, так как токсичность азотистых продуктов обмена снижается в направлении от аммиака к мочевой кислоте.
Для развития яиц ящерицы большое значение имеет процесс задержки расходования воды. В связи с этим в развивающихся яйцах ящериц продукты азотистого обмена сохраняются до вы-.лупления и, как показывают наши данные, эти продукты у прыткой ящерицы представлены в основном мочевой кислотой.
Известно, что у пресмыкающихся отщепление аммиака от аминокислот катализируется дезаминазами. Мочевина образуется в орнитиновом цикле при действии аргиназы на аргинин и аллантоиказы на аллантоиновую кислоту. В пользу этого суждения свидетельствует тот факт, что у чешуйчатых в почках обнаружена аргиназа, расщепляющая экзогенный аргинин (Clark,
150
1953; Clark, Sisken, 1956; Goldie, 1959). Возможно, что у чешуйчатых мочевая кислота синтезируется при участии карбамилфосфата, а источниками аммиака являются глютаминовая и аспар-гиновая кислоты. При этом требуется участие кислот трикарбонового цикла и энергия АТФ. Кроме того, синтез мочевины возможен путем расщепления пуринов ксантинокиназой (Prosser, Brown, 1962).
Поскольку у ящериц из ферментов, расщепляющих мочевую-кислоту, обнаружена лишь ксантиноксидаза, у них пурины могут дезаминироваться до ксантина или гипоксантина, которые затем превращаются в мочевую кислоту (Florkin, Duchateau, 1943; Laskowski, 1951; Albritton, 1954). По всей вероятности, прыткая ящерица в этом отношении не является исключением.
Данные о функции почек у ящериц не многочисленны. Можно отметить в этой связи, что почки бедны кровеносными сосудами. Нет сведений о резорбции воды в мочевом пузыре. Считается, что почки лишь частично регулируют экскрецию-воды и электролитов. По современным представлениям, у ящериц резорбция большей части воды, имеющейся в моче, происходит в клоаке (Green, 1972). Наши наблюдения показывают, что клоакальная моча прыткой ящерицы может быть вязкой вследствие наличия кристаллов мочевой кислоты. Количество мочи, выделяемой прыткой ящерицей, составляет 5,1 мл/час на-кг веса тела, т. е. примерно так же, как и у прочих видов ящериц (Bentley, 1959).
Органы чувств
В последнее время появились специальные исследования органов чувств у прыткой ящерицы (Swiezawska, 1949; Dudziak, 1955; Viand, 1960; Васильев, 1973). Однако работа органов чувств-данного вида -все же остается далеко не полно изученной.
Слух. Имеются данные (без указания вида), что ящерицы способны воспринимать звуки в диапазоне всего лишь 7,4— 8,2 кгц (Buddenbrock, 1952; по Никитенко, 1969); эти данные нуждаются в проверке на более совершенном современном оборудовании. Известно, что прыткая ящерица иногда шипит, с силой выпуская струю воздуха сквозь суженный просвет глотки.. Этот звук, несомненно, воспринимается другими особями как сигнал угрозы. Недавно описаны (Баранов и др., 1972) звуки прытких ящериц, напоминающие пение, однако биологическое значение их остается неясным (звуковая активность в условиях этих наблюдений была, по всей видимости, связана с половой активностью). Не исключена возможность того, что при тщательном специальном исследовании окажется, что ящерицы способны к восприятию ультразвуковой и инфразвуковой сигнализаций. Инфразвуки, хорошо передающиеся через твердый грунт на-значительное расстояние, несомненно должны улавливаться через участки тела, прилегающие к грунту.
151
Известно, что многие виды ящериц реагируют на звуки в диапазоне частот от 8000 до 10 000 гц, однако нет никаких данных о том, что они способны различать звуки различной частоты (Adrian, 1938, 1943; Wever, Vernon, 1956).
Исследование чувствительности слухового анализатора у прыткой ящерицы путем отведения биопотенциалов от слуховых ядер продолговатого мозга показало, что они обладают хорошей чувствительностью к высоким звукам. Низкие звуки воспринимаются прыткими ящерицами в диапазоне частот 200— 6000 гц. При этом ящерицы воспринимают разнокачественные сигналы с четкой дифференцировкой и изменение амплитуды ответной реакции на щелчок происходит скачкообразно (с быстрым переходом к насыщению), а изменение амплитуды ответов на тоны меняющейся интенсивности носит почти линейный характер (Васильев, 1973). Способность медуллярных ядер продолговатого мозга воспринимать повторные раздражения на 99% восстанавливается у прыткой ящерицы уже через 60 мсек. Увеличение фронта нарастания амплитуды стимулирующего сигнала у прыткой ящерицы приводит к падению амплитуды ответа, хотя и при различных значениях экспоненты (Васильев, 1973).
Зрение. Строение зрительного анализатора (см. главу III) свидетельствует о том, что для прыткой ящерицы должна быть характерна чувствительность к красному, оранжевому и желто-зеленому цветам (Forbes a. oth., 1959). Экспериментально показано, что прыткая ящерица может различать от 5 до 8 цветов (Viand, 1960). Вывод о развитии цветового восприятия у прыткой ящерицы подтверждается косвенно и яркой окраской наружных покровов. Язык зрительных сигналов у многих пресмыкающихся основан на смене и показе в определенной последовательности специфически окрашенных участков тела, поз и движений. Если основываться на наблюдениях Г. Китцлера (Kitzler, 1941), который показал, что закрашивание в синий цвет горла у самки зеленой ящерицы (Lacerta viridis) приводит к тому, что на нее нападают самцы (сине-голубое горло является вторичным половым признаком самца), то вполне понятным становится такое же поведение самцов прыткой ящерицы, нападающих на самок, тело или горло которых окрашены в зеленый цвет. Острота зрения прыткой ящерицы была определена в эксперименте, проведенном я. Дудчеком (Dudziak, 1955). Ящерицы различали расстояние между предъявляемыми им полосами в 1,8 мм с дистанции 15 см.
Другие органы чувств. Данных по функционированию других органов чувств у прыткой ящерицы практически нет. Некоторые заключения, основанные на наблюдениях в природе и анализе строения специальных структур, касающиеся работы органа юсязания, обоняния и теменного глаза, приведены выше (см. главу III).
152
О физиологии нервно-мышечного аппарата
Прямых данных о физиологии нервно-мышечного аппарата у прыткой ящерицы не имеется, однако процесс нервно-мышечного проведения изучен на ящерицах других видов (Eyzdguirre, 1957). У некоторых пресмыкающихся установлена зависимость между скоростью проведения нервных импульсов и диаметром нервного волокна (Tasaki, 1959). Как показывают наши наблюдения, мышцы легких прыткой ящерицы способны к длительному спонтанному сокращению, что свойственно также ящерицам других видов (Serfaty, Peyrand, 1960). У некоторых пресмыкающихся показано существование центров вдоха и выдоха, но не установлено наличия пневмотоксического (тонического) центра. У ящериц с удалением мозжечка уменьшается спонтанность движения и изменяется походка, что свидетельствует об участии мозжечка в процессе координации движений (Prosser, Brown, 1962).
У ящериц локальное раздражение переднего мозга может вызывать моторные реакции (Prechtl, 1949), а раздражение среднего мозга усиливает электрическую активность переднего мозга и вызывает судорожные реакции (Winkel, Caspers, 1953).
Физиология желез внутренней секреции
Наши знания об эндокринных регуляциях пресмыкающихся пока еще скудны. Ниже приводятся отрывочные данные по этому разделу физиологии рептилий, которые позволяют в общей форме представить основные черты этих процессов и у прыткой ящерицы.
Известно, что у чешуйчатых рептилий длительное кормление щитовидной железой ведет к повышению интенсивности обмена веществ, повышает возбудимость и ведет к значительной потере веса ( Krockert, 1941).
Чувствительность тканей пресмыкающихся к инсулину очень низкая (Sturkie, 1954; Miller, Wurster, 1956), а удаление поджелудочной железы ведет у них к развитию гипергликемии (Fog-lia, et all'., 1955). Вместе с тем пресмыкающиеся весьма чувствительны к глюкагону, их поджелудочная железа особенно богата а-клетками, которые и продуцируют глюкагон (Houssay, 1959). У прыткой ящерицы кормление в эксперименте вызывало гликемию и увеличение содержания глюкогена в печени (Vladescu, et all., 1971). Введение инсулина при этом приводило к развитию гипогликемии.
У пресмыкающихся, как у прочих пойкилотермных животных, нейрогипофизарные гормоны характеризуются тем, что их действующим началом является вещество, близко стоящее к вазопрессину, а именно — аргинин-вазопрессин (Heller, 1942;
153
Ewer, 1952; Sawyer, 1955, 1960; Sawyer, Schisgall, 1956; Sawyer, et all., 1959, I960).
Как и у прочих пойкилотермных животных, очевидно, у них имеется меланоцитостимулирующий гормон, который продуцируется аденогипофизом. Этот гормон вызывает рассеивание пигмента в меланофорах животных, что ведет к сезонному изменению тональности их окраски (Leloup, Berg 1954; Geschwind et ^all., 1956, 1957; Lee, Lerner, 1957; Harris, Roos, 1956; Harris, Lerner, 1957; Gilbert, Schneiderman, 1958). При этом на изменение окраски оказывает влияние кроме гормональной также и нервная системе (Redfield, 1918; Slome, Hogben, 1928; Parker, 1940; .Matsumoto, 1954). Удаление гипофиза у ящериц приводит к по-светлению кожных покровов. Хроматофоры ящериц могут принимать участие в терморегуляции. Кроме того, существуют данные о регуляции изменения формы меланофоров при участии вегетативной нервной системы (Parker, 1906; Bauer, 1914; Kruger, Kern, 1924). У пресмыкающихся при повышении температуры среды происходит концентрация меланина в меланоцитах, а при понижении — его дисперсия. По всей вероятности, это служит регулятором света, поглощаемого меланином (Parker, 1940). Все изложенное должно быть справедливо и для прыткой ящерицы.
Существует мнение, что пресмыкающимся свойственны гонадотропные, фолликулостимулирующие, лютеинизирующие и лю-теотропные гормоны (Hisaw, 1959). Доказано, что функция половых желез и развитие вторичных половых признаков у пресмыкающихся регулируется гормонами гипофиза (Combescot, 1955). У ящериц при введении фолликулостимулирующего и лю-теотропного гормонов наблюдается увеличение яичников (Ме1-lish, Meyer, 1937). Андрогены и эстрогены обнаружены в крови ящериц (Bragdon et all., 1954; Amoroso, 1955; Matthews, 1955; Panidel, 1956). Все сказанное присуще, очевидно, и прыткой ящерице.
В регуляции основных физиологических процессов в организме ящериц важную роль играют жировые тела (Hann, Tinkle, 1965). Во время спячки и размножения биохимический состав последних заметно изменяется и, в частности, резко колеблется содержание жирных кислот, холестерола, фосфолипидов, гликогедена и каротиноидов (Hadley, Christie, 1974). Динамика содержания липидов в развивающихся яйцах прыткой ящерицы описана в работе 3. Гродзинского (Grodzinski, 1949).
Из всех известных внешних раздражителей, влияющих на репродукцию, наиболее действенным, очевидно, является свет (Rowan, 1938; Bartholomew, 1949, 1956; Jenner, Engels, 1959; Earner, 1955, 1959; Wolfson, 1959). Прыткой ящерице, как и всем остальным видам ящериц, присущи резкие фотопериодические изменения полового цикла. Об этом свидетельствуют как наблюдения в природе, так и экспериментальные данные. По наблюдениям А. Б. Стрельцова, отловленные в июне—августе
154
и содержащиеся в террариуме ящерицы к ноябрю стали вялыми, перестали питаться, по бокам появились складки кожи, свидетельствующие об истощении. Ящерицы начали прятаться под камни, закапываться в землю, хотя температура под электролампой в террариуме достигала 6'0°.
После установления ультрафиолетовой лампы (ПРК-8 на тлеющем разряде ежедневно 5 час. облучения) через две недели активность ящериц резко возросла. Они интенсивно передвигались по террариуму, нормально питались, началась массовая линька. Самцы, окраска которых после отлова постепенно бледнела, становясь все более желто-коричневой, после облучения стали зеленеть и приобрели ярко-зеленую окраску, типичную» для многих самцов в разгар половой активности. Наконец, 27 декабря наблюдалось первое спаривание.
У некоторых ящериц гормон щитовидной железы стимулировал в экспериментах линьку (Maderson and oth., 1970). Интересно, что у змей тиреоидные гормоны не оказывали такого действия.
СОДЕРЖАНИЕ МИКРОЭЛЕМЕНТОВ В ОРГАНИЗМЕ ПРЫТКОЙ ЯЩЕРИЦЫ
Элементы, содержание которых в организме составляют тысячные, десяти- и стотысячные доли процента, носят название микроэлементов. Они играют в организме, как становится все более ясным, важную роль.
Большинство микроэлементов входит в состав ферментов,, дыхательных пигментов, гормонов и витаминов. Исследованию-связи микроэлементов с функциями ферментов и гормонов посвящены работы многих авторов (Галахова, 1958; Беренштейн,. 1947; Логинов, 1962; Нейве, 1960; Школьник, 1964; Эюбов,. 1967; и др.). При изучении биологической роли микроэлементов в организме животных исследователями рассматриваются такие вопросы, как специфичность действия отдельных микроэлементов, содержание и возрастная их динамика, взаимосвязь-между микроэлементами и многие другие (Белехов, Чубинская, 1965; Беренштейн, 1958, 1966; Войнар, 1952, 1960; Дьяков, Голубенцова, 1947; Ковалевский, 1963; Михлин, 1950; Одынец, 1966; Рудин, 1959; Савронь, 1966; и др.). Такие элементы, как марганец, медь, цинк, кобальт, йод, являются жизненно необходимыми.
До сих пор сведения о содержании микроэлементов в организме различных групп животных, обитающих в естественных условиях, весьма ограничены. В литературе до последнего времени не было сведений о микроэлементах в организме пресмыкающихся. Поэтому в качестве первого этапа необходимо было^ выяснить общие черты микроэлементарного состава организма прыткой ящерицы. С этой целью изучалось распределение, половые, возрастные различия и некоторые другие особенности;
155
•содержания в организме прыткой ящерицы четырех жизненно необходимых микроэлементов — марганца, меди, цинка и кобальта. Методом определения микроэлементов был избран эмиссионный спектральный анализ, обладающий достаточной чувствительностью (Бураков, Яновский, 1960; Кустанович, 1962; Гордон, 1963; Зайдель и др., 1964; Зайдель, 1965; и др.). При работе использовалось стандартное оборудование: генератор ДГ-2, спектрограф ИСП-22, спектропроектор ПС-18 и микрофотометр МФ-2.
Содер?кание микроэлементов в организме ящериц выражено в мкг % на сырое вещество. Приводимые далее табличные данные являются средними из пяти анализов. Общее представление о содержании микроэлементов в целом в организме ящериц можно составить из приведенных данных (табл. 19).
Таблица 19
Содержание микроэлементов (в мкг%) в организме прыткой ящерицы разного возраста и пола (Волгоградская обл., 1973 г., данные Л. М. Смеловского)
Возраст *, годы Zn Со Си Мп Zn Со Си Мп
Самки Самцы
Сеголетки 12 050,0 872,8 325,2 179,9 17 830,0 154,0 319,4 193,3
1 27 720,0 446,1 821,5 311,6 33 333,0 957.9 842,9 619,6
2 26 920,0 443,1 511,2 443,1 35 009,0 739,8 500,0 232,3
3 15 580,0 637,4 265,6 336,4 18 730,0 550,5 356,2 307,6
* Определен приблизительно по размерам и окраске тела. Подробнее см. главу VIII.
Из рассматриваемых микроэлементов в организме прыткой ящерицы цинк содержится в наибольших количествах, а марганец в наименьших. Как правило, в большинстве возрастных групп наблюдаются половые различия в содержании микроэлементов. Так, значительны половые различия в содержании кобальта, а у двухлеток—марганца. Интересно, что практически отсутствуют половые различия в содержании меди.
Изменение содержания микроэлементов в организме ящериц с возрастом имеет определенную направленность. Количество цинка, меди и марганца в организме сеголеток минимальное. •Оно значительно возрастает у особей одно-, двухлетнего возраста и резко уменьшается в организме трехлеток. В отличие от описываемых микроэлементов кобальт в наибольших количествах содержится в организме сеголеток. С возрастом уровень его в организме снижается. Минимальные количества этого элемента обнаружены у трехлетних особей. Изложенное выше свидетельствует о специфичности депонирования организмом каждого из указанных микроэлементов.
156
Таблица 20
Содержание некоторых микроэлементов (в мкг%) в разных органах прыткой ящерицы (Волгоградская обл.? 1973 г., данные Л. М. Смеловского)
—— ' Орган Zn Со Си Мп Zn Со Си Мп
Самки Самцы
Кожа 8 007,0 298,9 624,8 193,9 17 693,0 315,5 617,2 150,9
Мышцы 2 937,2 640,8 106,8 64,0 6 966,8 361,7 134,0 60,3
Почки 9 234,8 445,3 230,3 214,4 6 873,8 329,4 286,4 128,9
Печень 10 300,8 947,7 947,7 350,2 17893,9 332,5 395,9 269,2
При изучении распределения цинка, кобальта, меди и марганца в организме ящериц мы исследовали содержание их в коже, мышцах, почках и печени (табл. 20).
Как видно из табл. 20, распределение в организме прыткой ящерицы цинка, меди и марганца имеет некоторые сходные черты. Сходство заключается в том, что небольшое количество указанных элементов находится в печени. Далее в порядке снижения содержания микроэлементов следуют: кожа, почки и мышцы. Уровни кобальта в тканях и органах также различны. Но характер распределения в организме этого микроэлемента несколько иной. Минимальное количество его содержится в коже, тогда как мышцы богаты этим микроэлементом.
Не вызывает сомнения тот факт, что мнкроэлементный фон района сбора влияет на содержание в организме ящериц тех или иных элементов. С целью выяснения такого влияния были отловлены ящерицы в различных точках ареала в пределах Волгоградской области. Эти места, изолированные географически, различаются экологическими условиями и населены, несомненно, разными популяциями ящерицы (рис. 57). Пунктами сбора были правый берег Волги, окрестности пос. Светлый Яр, место сбора — степь; левый берег Волги, окрестности хутора
Та блица 21'
Содержание микроэлементов (в мкг%) в организме прытких ящериц из разных популяций Волгоградской обл. (данные Л. М. Смеловского, сборы 1972—1973 гг.)
Популяция Zn Со Си Мп Zn Со Си Мп
Самки Салщы
Пос. Светлый Яр 21 630,0 262,2] 180,0 72,1 35700,0 892,5 501,2 143,9
Хут. Громки 19150,0 496,0 294,4 177,4 24 860,0 637,4 253,4 184,6
о. Сарпинский 26 538,0 500,0 423,1 215,4 27 540,0 528,0 377,4 196,2
г. Калач 22 970,0 1108,9 514,9 301,0 11 630,0 332,3 235,4 218,8
157
Громки, место сбора — пойменный лес; остров Сарпинский на Волге, против г. Волгограда, место сбора — пойменный лес; окрестности г. Калача на Дону, место сбора — степь. В организме прытких ящериц, отловленных в- указанных местах, микроэлементы содержались в различном количестве (табл. 21). Микроэлементный состав указанных ящериц разнороден и отличия, обусловленные местом отлова, отмечены для всех рассматриваемых микроэлементов. Ни один элемент не имеет определенной концентрации в организме ящериц, отловленных в разных местах (см. рис. 57). Уровни их различны для цинка, меди, марганца. Лишь кобальт в организме ящериц из трех различных пунктов сбора содержится примерно в одинаковом ко-
Рис. 57
Содержание некоторых микроэлементов в организме прытких ящериц из нескольких популяций Среднего Поволжья и междуречья Дона и Волги
158
личестве. Однако указанная разнородность микроэлементного состава ящериц из разных мест не нарушает закономерности соотношения в организме рассматриваемых микроэлементов. В организме прытких ящериц, добытых в четырех различных точках ареала, содержание микроэлементов в порядке их убывания вполне определенное — цинк, кобальт, медь, марганец (см. рис. 57).
Можно считать, что содержание микроэлементов в органах и тканях ящериц отличается большим разнообразием, обусловленным как местом сбора, так и половыми различиями (табл. 22).
о—самки, б — самцы. 1 — окрестности г. Калач; 2 — о. Сарпинский; 3 — окрестност*! хутора Громки; 4— окрестности пос. Светлый Яр. (по данным Л. М. Смеловскоги)
159
Таблица 22
Содержание некоторых микроэлементов (в мкг%) в органах прытких ящериц из разных популяций Волгоградской обл. (данные Л. М. Смеловского, сборы 1972—1974 гг.)
Орган . х Zn Со Си Мп Zn Со Си Мп
Сарпинская популяция Самки Калачская популяция Самки
Кожа 5333,7 216,0 317,7 85,1 5 654,1 154,2 321,3 179,9
Мышцы 1 849,0 336,1 100,8 16,8 3148,6 94,5 42,0 9,4
Почки 5060,0 480,0 512,0 204,1 3 454,2 252,4 345,4 63,7
Печень 7 979,9 Са 433,2 чцы 752,4 296,4 10933,9 168,7 Са 578,3 мцы 100,0
Кожа 5 840,5 177,8 304,7 77,5 6 934,1 101,6 165,1 73,7
Мышцы 1 580,3 474,1 98,3 40,3 5 905,5 348,3 54,5 15,1
Почки 3 599,4 22,3 111,7 48,6 1827,1 351,3 238,9 98,3
Печень И 212,8 4’1,1 1027,8 48,5 12 706,7 352,1 643,0 199,0
Помимо изучения микроэлементного состава организма прыткой ящерицы в условиях естественной свободы были проведены эксперименты по выяснению динамики микроэлементов ящериц при действии разных факторов, в частности, голода. С этой целью исследовано содержание микроэлементов в организме ящериц, голодавших в течение восьми и шестнадцати дней. Ящерицы содержались в террариумах при температуре 26—28° и относительной влажности 41—43% (табл. 23). Голодание ящериц привело к изменению содержания микроэлементов в органах и тканях и некоторому перераспределению их внутри организма. По содержанию и распределению микроэлементов цинка, меди и марганца органы и ткани ящериц, голодавших 8 дней, мало отличаются от контроля. Изменения же уровня кобальта при голодании ящериц значительны только в печени. У ящериц в опыте резко снижаются его запасы.
Более продолжительное голодание в 16 дней усиливает изменение содержания микроэлементов в органах и тканях ящериц по сравнению с контролем. Содержание цинка резко уменьшается в коже, мышцах и почках голодавших ящериц, но при этом наблюдается значительное увеличение его количества в печени. Уровень кобальта у голодавших ящериц по сравнению с контролем ниже лишь в печени. Кожа, почки и еще в большей степени печень у голодавших ящериц обогащены медью. Сходная картина наблюдается в отношении марганца.
Таким образом, при длительном голодании ящериц все рассматриваемые микроэлементы (за исключением кобальта) кон-
160
Т а б л и ц a 23
Содержание некоторых микроэлементов (в iv.fi %) в органах и тканях прытких ящериц при двух различных сроках голодания (данные Л. М. Смоленского)
Орган Zn Со Си Мп Zn Со Си Мп
г Кожа олодание Сам 26128,6 8 дней ки 290,3 653,2 174,2 5 649,7 Г олода ( 295,9 ние 16 дн Замки 1076,1 й 242,1
Мышцы 5 952,4 739,5 88,4 37,9 1800,0 240,0 190,0 80,0
Почки 7 568,8 333,0 287,4 166,5 4096,0 436,9 1092,2- 150,1
Печень 16 646,5 241,6 698,0 174,5 17 531,8 397,2 6848,3 493,0
Кожа Сам 7818,4 цы 300,7 481,1 118,8 3 089,1 397,1 Замцы 2353,6 426,5
Мышцы 4692,1 293,3 98,2 30,8 2 456,4 859,7 204,7 112,5
Почки 9 716,3 391,7 376,1 188,0 5900,0 300,0 1100,0 240,0
Печень 12241,5 379,9 802,0 379,9 19 981,5 394,3 6572,8 315,4
центрируются в печени. Вполне возможно, что такой процесс служит адаптацией организма к перенесению неблагоприятных условий. При депонировании печенью микроэлементов в других органах и тканях возможно поддержание минимально необходимого уровня их более длительное время за счет запасов печени. Появление большого количества микроэлементов меди и марганца в коже при длительном голодании ящериц может быть связано с процессами, обеспечивающими усиленную линьку ящериц, находящихся в неблагоприятных условиях. Предположение об адаптивном характере перераспределения микроэлементов в организме длительно, голодавших подтверждается, тем, что менее продолжительное голодание (8 дней) не вызывает в организме подобных процессов. По-видимому, у прытких ящериц реакция на неблагоприятные условия такой продолжительности обеспечивается каким-либо другим компенсаторным механизмом, не влекущим за собой изменения микроэлементного состава органов и тканей. Приведенные в этом разделе данные показывают, что в организме прыткой ящерицы наблюдается определенная возрастная динамика, и существуют различные уровни концентрации микроэлементов в различных органах и тканях. Таким образом, микроэлементарный состав организма прыткой ящерицы может служить в некоторых случаях своеобразным индикатором условий внешней среды.
Приведенные в этой главе данные по некоторым особенностям физиологии и биохимии прыткой ящерицы показывают, насколько ограничены имеющиеся данные в этой области и насколько необходима интенсивная дальнейшая работа в этом направлении.
6 Прыткая ящерица
ГЛАВА ПЯТАЯ
СРЕДА ОБИТАНИЯ 1
Для понимания места вида в природе, его значения в биосферном круговороте недостаточно знать общий характер его распространения (глава I). Прыткая ящерица, населяя несколько природных зон — от подзоны тайги до зоны полупустынь, в каждой из этих зон занимает определенные .местообитания. В настоящей главе рассматривается конкретная среда обитания прыткой ящерицы. Это рассмотрение ведется как бы в двух масштабах. Во-первых, описываются биотопы, населяемые данным видом в каждой из природных зон, и дается их систематизация и классификация, что составляет как бы популяционный, групповой уровень в проблеме среды обитания. Во-вторых, анализируется среда обитания, характерная для отдельных индивидуумов. Приводимые в этой главе данные являются основой для рассмотрения проблем, связанных с перемещениями и миграциями (глава IX), активностью (глава X), структурой популяции (глава XI) и всех других, определяющих место вида в биогеоценозах (глава XIII—XV).
ЗОНЫ ОБИТАНИЯ
Ареал прыткой ящерицы охватывает несколько природных зон: лесную, лесостепную, степную, полупустынную. Кроме того, этот вид обитает и в горных ландшафтах (рис. 58).
В таежной лесной подзоне прыткая ящерица встречается в основном в районах сосновых боров, на окраинах, вырубках, зарастающих гарях (Словцов, 1892; Дрягин, 1926; Кащенко, 1902; Иоганзен, 1923 и др.), в кустарниковых зарослях на возвышенностях (Румянцев, 1926; Верещагин, Громов, 1947), на склонах шоссейных дорог и железнодорожных насыпей (Чащин, Литвинов, сообщ.), по хорошо прогреваемым участкам.' речных долин. Речные долины, а также антропогенные «коридоры» (насыпи, кюветы, искусственные насаждения) в сплош-
4 Автор: М. Ф. Тертышников, при участии Н. В. Щепотьева, В. Л. Булахова, Н. Ф. Константиновой, И. С. Даревского, Г. П. Лукиной, Н. А. Рашкевича, Н. М. Окуловой, 3. П. Хонякиной, А. Б. Стрельцова, М. И. Щербаня, Л. М. Смеловского, С. П. Чащина, Н. А. Литвинова, В. К- Жарковой, А. С. Баранова, Г. Добровольской и К- Шафраньской.
162
ных лесных районах служат основными путями распространения прыткой ящерицы на север и восток в этой зоне. В Западной Европе такие антропогенные биотопы, не несущие рекреационной нагрузки, служат, напротив, местами сохранения популяций этого вида. Основным требованием, обеспечивающим пригодность биотопа, в данном случае является его достаточно большая протяженность, позволяющая время от времени отдельным популяционным «островкам» обмениваться особями и поддерживать достаточно высокую и стабильную численность.
В северной части подзоны смешанных лесов прыткая ящерица поселяется на песчаных буграх и южных склонах холмов, покрытых сосновыми борами, на водораздельных лугах с редким кустарником (Формозов, 1935; Красавцев, 1936; Пузанов, 1942; Либерман, Покровская, 1943), на зарастающих просеках сосновых боров, по обочинам дорог и на склонах железнодорожных насыпей (Турутина, 1972). В более южных частях данной зоны прыткая ящерица заселяет солнечные поляны и опушки лиственных лесов, южные склоны холмов с редким кустарником, березовые колки, лесистые дюны, перелески, овраги, уцелевшие степные участки среди пахоты, межи на полях (Шарлемань, 1917, 1936; Меландер, 1935; Красавцев, 1936), встречается в молодых сосновых посадках, по возвышенным местам речных долин.
В лесостепной зоне ящерица обитает на лесных опушках и полянах, в светлых кустарниковых зарослях, по склонам оврагов и болот, на участках ковыльных степей, на паре и межах полей, на заливных лугах, берегах рек и окраинах болот (Силантьев, 1894; Браунер, 1907; Огнев, 1908; Огнев, Воробьев, 1924; Горбачев, 1925; Аммон, 1928; Сухов, 1928; Гумилевский, 1931; Птушенко, 1934, 1938а), в борах и на разных склонах, достаточно освещаемых солнцем (Положенцев, 1935, 1937), на лесных просеках и вырубках, на участках спелого леса с редким древостоем, в лесных островках среди пашен и степи, в полезащитных и снегосборных полосах, по обочинам дорог, в кюветах, в различных понижениях с богатой растительностью среди степей, среди пятен ковыля в разнотравной степи и по склонам железнодорожных насыпей (Щепотьев, 1950, 1952).
В степной зоне ареала прыткая ящерица заселяет залежные земли, поля с зерновыми культурами и кормовыми травами, кустарниковые заросли (Браунер, 1905, 1923; Орлов, 1929; Иваненко, 1938), встречается на глинистых равнинах п песках (Зарудный, 1895; Дидусенко, 1964), поселяется в окрестностях или даже в черте населенных пунктов—на пустырях, кладбищах, бахчах, в садах, виноградниках, парках (Лукина, 1966; Тертышников, 19726), в лесопарках (Боченко, сообщ.) и широколиственных лесах по берегам рек (Беме, 1929;
163
6*
Рис. 58
Некоторые типичные местообитания прыткой ящерицы
А — горный склон у реки (Груз. ССР, Гегечкорский p-и, долина р. Техури, 4 км вверх .от пос. Курзу); В—склоны и долины балки (Каз. ССР, Кокчетавская обл., вблизи левого .берега р. Селеты в б км. ниже гидроузла Селетинского водохранилища); В—каменистые
164
россыпи (Груз. ССР, Цалкский р-н, окоестности пос. Цалка); Г—поляна в сосковом лесу (Каз. ССР, Цел иноградская обл., окрестности пос. Кейес) (Фото А. В. Валецкого и А. В. Яблокова)
165
Красовский, 1929), в линейных древесно-кустарниковых насаждениях вдоль дорог, по склонам шоссейных и железнодорожных насыпей, в лесных полосах, по берегам рек, прудов, озер, водохранилищ и оросительных каналов, на окраине, полянах и под пологом искусственных лесов, на лесных полянах, вырубках, просеках и лесных дорогах, в целинных степях с различными растительными ассоциациями, на остаточных участках целинной степи среди пашен, па толбках и черном паре (Богачев, 1938; Утробина, 1956; Лукина, 1966; Булахов, Константинова, 1972; Тертышников, 19726; Рашкевич, сообщ.).
В зоне полупустынь прыткая ящерица поселяется на пойменных лугах, в бугристых степях с высоким горизонтом грунтовых вод (Орлов, Фенюк, 1927; Смеловский, сообщ.), на зеленых луговинах среди песков и пустынных биотопов (Орлов, 1928; Баранов и др., сообщ.), в кустарниковых зарослях на участках степной растительности лугового и лугово-степного типа, по берегам водоемов, на возвышенностях среди заливаемых при половодье участков, в заросших карьерах и окопах, в населенных пунктах (вдоль заборов, в садах), в лесопитомниках, в лесных посадках, на опушках пойменных лесов (Кутузова, 1971; Пархоменко, 1973; Смеловский, сообщ.). В полупустынях она также встречается на низменных участках — в камышах по берегам рек и озер, на мокрых луговых участках и болотах (Шнитников, 1928, 1935; Бобринский, 1929; Кашкаров, 1932), в понижениях среди кустарников у ключей и колодцев (Параскив, 1956), на полынных, злаково-полынных, глинисто-полынных степных участках, а также на чернополынных (Окулова, сообщ.). Изредка ящерица в этой зоне, как и в других зонах, встречается в каменистых россыпях.
Наконец, в горных ландшафтах этот вид встречается также, как и в других природных зонах, на участках редкого леса, на лесных полянах и в кустарниковых зарослях (Банников, 1954), на черном паре, межах полей, по обочинам дорог и берегам водоемов (Мусхелишвили, 1970; Щербань, сообщ.), по краям карстовых воронок, на каменистых гребнях холмов с древесной растительностью (Щербак, 1966), в горных степях, на южных склонах альпийских и субальпийских лугов (Рашкевич, сообщ.). В горы прыткая ящерица поднимается на высоту до 2200 м над уровнем моря (Кавказ) (Стрельников, 1944).
Огромный ареал должен был бы, казалось, определять большое разнообразие занимаемых ящерицей биотопов. Однако этого не происходит. Те биотопы, которые она заселяет, в разных зонах во многом сходны. В результате анализа населенных ящерицами биотопов в разных зонах можно предложить следующую типологию биотопов.
166
Естественные биотопы
Лесные биотопы. Связаны с широколиственными, смешанными (всегда с сосной) лесами.
1) лесные опушки; 2) гари зарастающие; 3) поляны (см. рис. 58, Г); 4) островки леса; 5) редколесье; 6) кустарниковые заросли; 7) пойменный лес; 8) покрытые редким лесом дюны.
Биотопы открытых пространств.
1) луг пойменный; 2) луг суходольный (водораздельный); 3) луг горный (субальпийский, альпийский); 4) разнотравная степь; 5) полынно-злаковая степь; 6) полынная степь; 7) ковыльная степь; 8) чернополынные солонцы; 9) типчаково-полынная степь; 10) окраины болот и заболоченных участков; 11) склоны оврагов, балок, карстовых воронок и других естественных понижений, береговые склоны (см. рис. 58, Б) ; 12) горные склоны (см. рис. 58, А); 13) каменистые россыпи разного типа (см. рис. 58, В).
Антропогенные биотопы
Сельскохозяйственные биотопы.
1) посевы зерновых; 2) посевы кормовых трав; 3) бахча; 4) межа; 5) пар; 6) залежи; 7) сады; 8) виноградники; 9) огороды; 10) толока (скотобойные участки).
Другие биотопы, связанные с деятельностью человека.
1) парки; 2) вырубки и просеки; 3) обочины дорог; 4) железнодорожные и другие насыпи; 5) лесные полосы разных типов; 6) лесовосстановительные посадки и лесопитомники; 7) кладбища; 8) пустыри и промышленные пустоши; 9) склоны прудов, водохранилищ, оросительных каналов; 10) заброшенные карьеры, рвы и окопы.
При сравнении различных природных зон по количеству биотопов, занимаемых прыткой ящерицей, можно видеть, что наибольшее количество их приходится на степную зону, которую, по всей видимости, можно считать зоной оптимума в ареале данного животного (табл. 24).
ФАКТОРЫ, ОПРЕДЕЛЯЮЩИЕ ВЫБОР ЯЩЕРИЦЕЙ БИОТОПА И СТАЦИИ
Из приведенных выше данных следует, что у прыткой ящерицы нет «типичного» биотопа. Более того, даже в пределах одного биотопа распределение ящериц крайне неравномерно. Выбор ими того или иного биотопа, тех или иных участков в нем определяется, видимо, многими факторами. Выбор ящерицей места обитания во многом зависит от освещенности, влажности, высоты снегового покрова (определяющего промерзание почвы), температурных условий, наличия корма и убежищ, величины покрытия и высоты травостоя, характера почвы биотопа (Щепотьев,
167
Таблица 24
Биот.оническое распределение прыткой ящерицы в разных природных зонах
Биотоп Основная природная зона
лесная лесостепная степная полупустынная горные районы
Естественные биотопы
Лесные бистопы
Лесные опушки + + + — —
Гари зарастающие + 4“ + — —
Поляны + + — — —
Островки леса + + —.
Редколесье + + — — —
Кустарниковые заросли + + + + +
Пойменный лес — + + +
Покрытые редким лесом дюны — — — — —
Биотопы открытых пространств
Луг пойменный + + + — —
Луг суходольный (водораздельный) — + + — —
Луг горный (субальпийский* альпийский) — — + — +
Разнотравная степь — + + —
Полынно-злаковая степь +
Полынная степь .—. + ,—
Ковыльная степь — —
Чернополынные солонцы — — —. + —.
Типчаково-полынная степь — .— + + .
Окраины болот и заболоченных участков + + + + .
Склоны оврагов, балок, карстовых воронок
и других естественных понижений, береговые
СКЛОНЫ + + + + +
Горные склоны —. — — — +
Каменистые россыпи разного типа — — + + +
Антропогенные биотопы
Сельскохозяйственные биотопы
Посевы зерновых +* +* < —
Посевы кормовых трав +* +* +* — .
Бахча +* + + —.
Межа + + + —
Пар — + + — —.
Залежи + + + — —
Сады .—. + + + +
Виноградники — -—. + + +
Огороды +* +* +* +
Толока (скотобойные участки) —. — +
Парки — + + + —
168
Таблица 24 (окончание)
Биотоп Основная природная зона
лесная лесостепная степная полупустынная горные районы
Выручки и просеки + + — — —
Обочины дорог + + + + +
Железнодорожные и другие насыпи + + + —• —
Лесные полосы разных типов — + + + —
Лесовосстановительные посадки и лесопитомники + + + + +
Кладбища — + + + —
Пустыри и промышленные пустоши + + + •— —
Склоны прудов, водохранилищ, оросительных каналов + + + + —
Заброшенные карьеры, рвы и окопы + + + — —
Общее число биоценозов, занимаемых прыткой ящерицей в каждой зоне 20 29 33 18 9
• Пребывание временное.
1950, 1952; Лукина, 19666; Булахов, Константинова, 1972; Тер-тышников, 19726). Рассмотрим некоторые из этих факторов (табл. 24а).
Пища
Прыткая ящерица поедает различных беспозвоночных (см. главу VI), поэтому она поселяется в местах с развитым и достаточно разнообразным растительным покровом, который обеспечивает обилие беспозвоночных на протяжении всего сезона активности (Щепотьев, 1950) на сухих солнечных склонах холмов в борах северной части ареала, на опушках и лесных полянах в лесной зоне, в кустарниковых зарослях, в молодых лесных насаждениях, в понижениях степной и лесостепной зон, а у южных границ ареала — на более увлажненных участках.
Убежища и условия зимовки
В качестве убежищ прыткая ящерица использует норы различных животных, многие случайные укрытия и сама роет норы.
По данным С. С. Либермана, Н. В. Покровского (1943), прыткая ящерица погибает при —4,9°. Поэтому глубина ее нор должна быть такой, чтобы исключалась гибель ящерицы при действии низкой температуры зимой. В связи с этим определенное значение приобретает фактор снежности (Формозов, 1946). Выбор места обитания во многом определяется распределением сне
169
гового покрова: ящерицы заселяют преимущественно кустарниковые заросли, овражки и прочие понижения, где зимой накапливается достаточно снега.
Влажность
Н. В. Щепотьев (1950) относит прыткую ящерицу к влаголюбивым животным. В подтверждении этого можно сослаться на то, что ящерицы предпочитают влажные участки при выборе места для откладки яиц (Либерман, Покровская, 1943). В Этой же связи значительный интерес представляет отношение прыткой ящерицы к биотопам с неодинаковым увлажнением в той или иной природной зоне. Так, например, в зоне тайги она поселяется на сухих и хорошо инсолируемых склонах холмов в сосновых лесах. В зоне смешанных лесов ящерица придерживается биотопов с пониженной влажностью в сосновых и широколиственных лесах и избегает лугов, болот, ольховых лесов и зарослей кустарника по берегам водоемов (Шарлемань, 1936). В лесостепной зоне это животное избегает не только сырые луга и леса (Гааке, 1901; Шарлемань, 1917; Roszkowski, 1924), но и глинистые и солонцеватые участки (Щепотьев, 1952). Однако на границе лесостепи и степи ящерица уже начинает заселять места с повышенной влажностью — сырые луга, долины рек, болота (Силантьев, 1894; Сухов, 1928). В степной зоне она обитает в различных понижениях в овражках с более влажными по сравнению с окружающими биотопами условиями обитания. В условиях степей Западного Предкавказья прыткая ящерица не избегает влажных мест обитания и в ее распространении, что характерно и для остальных районов степной зоны, намечается явное смещение в сторону мезофитных лугов (Лукина, 1961, 19666). В лесах степной зоны эта ящерица ведет себя несколько иначе. Так, по данным Л. В. Булахова, Н. Ф. Константиновой (1972), в лесах юго-восточной части УССР наиболее оптимальные условия для ящерицы складываются в суховатых биотопах с постепенным снижением в сторону сухих и резким снижением в сторону влажных и сырых биотопов. И, наконец, в полупустынной и пустынной зонах данное животное явно предпочитает биотопы с повышенной влажностью.
В сухих районах ареала водоемы играют положительную роль в биотопическом размещении прыткой ящерицы. В поисках пищи она часто мигрирует к воде, где иногда концентрируется в большом количестве. Так, Г. П. Лукина в 1960 г. на 0,75 м2 прибрежного ивняка насчитала 13 животных. По ее же данным (Лукина, 19666), на берегах оросительных каналов у станицы Ново-Троицкой Ставропольского края численность прыткой ящерицы в период миграции к воде летом достигала 1000 экз./га, в то время как на отдаленных от каналов степных участках численность ящериц была в несколько раз ниже.
170
В. Л. Булахов и Н. Ф. Константинова также отметили повышение плотности населения ящерицы на склонах берегов различных водоемов и особенно там, где в непосредственной близости от водоема расположены различные лесные участки и кустарниковые заросли. По их мнению, повышенная концентрация ящериц по берегам водоемов может быть объяснена мозаичностью микроландшафта, сочетающего в себе открытые поляны, настоящие микростепные участки, овраги, кустарниковые заросли, разнотравье, обеспечивающие достаточную освещенность, кормность и защитные условия биотопов. Значительное удаление от водоема влияет на количественное соотношение ящериц, но" даже в безводных биотопах они поселяются все же приближаясь к различным лесным участкам.
Растительный покров
Данный фактор часто играет значительную роль при распределении ящериц в степных и лесных участках. Голый, без травянистого покрова грунт, толока, посевы зерновых культур и кормовых трав ящерицы обычно избегают и они практически выпадают как постоянные места обитания. В них ящерицы встречаются только по окраинам, примыкающим к лесным массивам, берегам водоемов, участкам целинных степей. Очень мало ящериц на черном паре, на бахчах, виноградниках, залежных землях, в садах, лесопитомниках и огородах.
Установлено, что существует определенный минимум проективного покрытия травянистой растительностью, при котором ящерицы поселяются в том или ином биотопе. Так, например, в лесах юго-востока Украины — это 20—40% (Булахов, Константинова, 1972), на Ставропольской возвышенности — 50% (Терты-шников, 19726), в Армении и Мещерской низменности — 60— 65% (Даревский, Жаркова, сообщ.). Точно также существует и максимум проективного покрытия травянистой растительностью, превышение которого приводит к уменьшению и даже к полному исчезновению ящериц из биотопа. В лесах юго-востока Украины—это 80% (Булахов, Константинова, 1972), на Ставропольской возвышенности — 95% (Тертышников, 19726).
По мнению И. С. Даревского, выбор ящерицами мест обитания определяется не только степенью проективного покрытия, но и ярусностью травяного покрова. По данным В. Л. Булахова, Н. Ф. Константиновой (табл. 24а), в лесах юго-востока Украины наиболее оптимальная высота травостоя равна 20—30 см. По-видимому, в связк е этим ящерица в открытых ландшафтах предпочитает целинные участки с более или менее высокой травянистой растительностью. На открытых участках кустарниковая растительность в некоторой степени может замещать высокий травостой и играть определяющую роль в образовании защитных условий.
171
Таблица 24а
Влияние различных типологических особенностей мест обитания в лесостепной зоне на биотопичсское распределение прыткой ящерицы (в % от общего количества учтенных ящериц, по данным В. Л. Булахова, Н. Ф. Константиновой)
Освещенность местообитания Осветленные Полуосветлен-иые Полутеневые Теневые
40,2 32,9 26,1 0,8
Процент сомкнутости кроны леса 0 10—20 30—40 50-60 70—80 90—100
23,3 28,8 24,7 18,4 4,8 0
Природный состав насаждений Гледичия Акация белая Акация + 4- ясень Береза 4-4- осина Сосна Дуб4-ясень Дуб 4-4-клен Ольха
25,6 22,1 21,2 9,0 16,7 4,6 0,8 0
Почва Суглинки Супеси Песок Глнна
тяжелые средние рыхлые тяжелые легкие уплотненные сыпучие
3,3 14,1 11,3 17,5 21,5 20,8 3,3 8,1
Степень увлажнения почвы, в баллах, по А. Л. Бель-гарду'(I960) 0 0-1 I 1—2 2 3 4 5
очень сухая сухая суховатая свежеватая свежая влажная сырая мокрая
11,6 24,8 38,3 19,2 4,1 1,6 0,4 0
Травяной покров, в % покрытия 0—20 20—40 40—60 60—80 80—100
Высота травяного покрова, в см 1,8 17,4 26,7 39,3 14,8
10 10—20 20—30 30-40 40—50
9,9 28,6 33,8 15,1 12,6
172
Освещенность
Особенности биотопического распределения прыткой ящерицы в биоценозах с древесной растительностью обусловлены также и освещенностью. По этой причине определяющее значение в количественном распределении этого животного в подобных биотопах зависит от экологической структуры древесной растительности, общей сомкнутости кроны, типа древостоя и его возраста. Наиболее благоприятные условия для обитания ящериц складываются в осветленных и полуосветленных структурах, состоящих из акациевых, акациево-ясеневых и сосновых насаждений. В лесополосах этим условиям соответствуют гледичие-вые и акациевые насаждения (см. табл. 24а). Указанные насаждения состоят в основном из ажурнокронных, полуажурнокрон-иых и полуплотнокронных деревьев, различные сочетания пород которых обуславливают степень осветленности насаждений. Если в Саратовской области отдают предпочтение посадкам из дуба, липы и клена, опавшая листва которых образует толстую подстилку, в которую они могут прятаться (Щепотьев, 1952), то на юго-востоке Украины в дубовых и кленовых насаждениях складываются не совсем благоприятные условия для обитания этих животных (Булахов, Константинова, 1972). В таких насаждениях они поселяются только при очень малой сомкнутости кроны или же при условии, что высота древостоя не превышает 1 м.
Сила освещенности обуславливается сомкнутостью кроны. В молодых посадках всегда отмечается большое количество ящериц. Наибольшее их количество зарегистрировано при сомкнутости 0,1—0,2. Значительное количество ящериц встречается и в насаждениях с сомкнутостью кроны от 0,3 до 0,6. С увеличением возраста деревьев, их высоты и степени смыкания кроны идет постепенное вытеснение этих животных на краевые участки — на опушки и поляны, на просеки, вырубки и в более разреженные посадки. При сомкнутости кроны от 0,7—0,8 количество ящериц резко снижается (Булахов и др., 1970), а при сомкнутости от 0,9 до 1,0 эти животные практически отсутствуют.
Механический состав почвы
В. Л. Булахов, Н. Ф. Константинова (1972) установили, что на юго-востоке Украины наиболее благоприятными для ящериц являются почвы средней плотности, суглинки и уплотненный песок. Значительно меньшее количество их поселяется на глинистых почвах. В других частях ареала ящерицы предпочитают почвы средней плотности, мягкие суглинки и песок, так как на них этим животным легче строить постоянные и временные норы.
173
ТЕРРИТОРИАЛИЗМ
Прыткая ящерица ведет одиночный образ жизни (см. главы IX и X). Нахождение меченых ящериц в течение продолжительного времени на одних и тех же участках указывает на существование у них индивидуальной территории, которая удовлетворяет потребность в пище и убежищах и на которую приходятся суточные и сезонные перемещения этих животных.
Данные о территориализме прыткой ящерицы незначительны и противоречивы. В. К. Жаркова (19736), например, считает, что в условиях Рязанской области индивидуальная территория имеется у тех особей, которые равномерно размещены на территории биотопа, а у других животных, обитающих в условиях значительной плотности на ограниченной территории, имеются только индивидуальные дорожки, по которым они 3—5 раз в день перемещаются в поисках корма и особей другого пола па расстояние, не превышающее 12 м от норы.
М. И. Щербань установил, что в условиях Закарпатской области прыткая ящерица имеет небольшую индивидуальную территорию, размер которой на равнине варьирует от 75 до 230 м2 (п=Ю), в предгорной зоне — от 50 до 180 м2 (п= 10) и в горных районах — от 25 до 100 м2 (п—10).
Н. А. Рашкевич сообщает, что в степных биоценозах возле г. Грозного размеры индивидуальной территории взрослой прыткой ящерицы составляли 190—250 м2.
Г. Добровольска и К. Шафраньска, исследуя подвижность и территориализм прыткой ящерицы на участке соснового леса в окрестностях населенного пункта Новый Дунинов (Польша), установили, что 30% исследованных ящериц (/?—303) не имеют индивидуальной территории, причем самки меньше связаны с определенной территорией, чем самцы. 70% других особей имеют индивидуальную территорию, которую используют в течение разного числа дней. Размеры индивидуальной территории этих । животных колеблются от 3,75 до 136,5 м2.
Шестилетние наблюдения М. Ф. Тертыщникова показали, что | на Ставропольской возвышенности прыткая ящерица имеет индивидуальную территорию, размеры которой варьируют в разных биотопах от 69 до 274 м2 (135,0+25,4 м2, при м=9) (Тер-тышников, 1967, 1970, 19726). Самая большая индивидуальная территория отмечена у ящериц, обитающих в открытых биотопах; в типчаково-полынной степи — 274,0 м2 и разнотравно-злаковой степи — 229 м2, а наименьшая — в закрытом биотопе (лесная полоса)—71—170 м2 (108,3±31,1 м2). По мнению этого исследователя, размеры индивидуальной территории находятся в зависимости от особенностей растительного покрова и обилия корма.
В отличие от Л. Г. Динесмана и М. Л. Калецкой (1952) М. Ф. Тертышников (1970) обнаружил, что не только самки, но
174
1709
Рис. 59
Дневные участки прытких ящериц разного пола и возраста в разных биотопах Предкавказья
I— типчаково-полынная степь; II— разнотравно-злаковая степь; III — лесная полоса; 1 — 5 — дни наблюдения; 6 — нора ящерицы (по М. Ф. Тертышникову, 1970)
и самцы имеют индивидуальную территорию. Последние в период размножения более активны, нежели самки, и их часто можно видеть в местах перекрытия их индивидуальных территорий
bo с таковыми особей другого пола. По данным Н. А. Рашкевича, индивидуальная территория взрослых самцов варьирует от 190 до 250 м2, взрослых самок — от 190 до 220 м2, а годовалых ящериц — от 180 до 300 м2.
Установлено, что форма индивидуальной территории весьма разнообразна (рис. 59) и зависит от микрорельефа, характера распределения корма, убежищ, растительности (Тертышников, 1970, 19726; Баранов, Яблоков и др., сообщ.).
Как показал М. Ф. Тертышников (1970, 19726), ящерица в поисках корма редко использует полностью индивидуальную территорию в течение дня. Выедая корм на одних участках, она обычно переходит на другой и только через некоторое время (1 и более дней) возвращается на прежнее место охоты (рис. 60).
175
По-видимому, именно неравномерность распределения корма на индивидуальной территории приводит к последовательном, ее использованию. Размеры таких участков (дневных) неодин аковы в разные дни и варьируют от 14 до 181 м2 в типчаковс ' лынной степи, от 44 до 114 м2 в разнотравно-злаковой степи . от 28 до 127 м2 в лесной полосе. В хорошую погоду при кормовых миграциях ящерицы используют большие площади, чем в плохую погоду, причем в типчаково-полынной степи (137,0±24,7 м2) больше, чем в разнотравно-злаковой степи (83,3 ± 8,7 м2) и в лесной полосе (71,6 ± 16,8 м2}.
знз.
f И 30 45 00 70 90 Shu
Рис. 60
Период пребывания ящерицы на исследуемой территории
1 — разовые встречи меченых особей; 2 — меченые особи, встреченные повторно (по М.. Ф. Тертыш-ннкову)
Анализируя подвижность ящериц разных половых и возрастных групп при кормовых миграциях, М. Ф. Тертышников (1970, 19726) установил, что взрослые самцы и самки в типчаково-полынной степи приблизительно в равной мере используют индивидуальную территорию. В разнотравно-злаковой степи и лесной полосе взрослые самцы «обрабатывают» большие участки, чем самки. При сравнении дневных участков взрослых и молодых особей оказалось, что только в типчаково-полынной степи дневные участки взрослых животных больше, чем молодых, а в других биотопах (с иной растительностью) сравниваемые возрастные группы животных используют индивидуальную территорию приблизительно одинаково. Следует полагать, что по мере приближения спячки (см. главу X) и падения активности ящериц размеры их дневных участков уменьшаются.
УБЕЖИЩА
В качестве убежищ прыткая ящерица использует щели и пустоты в почве, камнях, прячется под кучами камней, хвороста, в опавших листьях, мусорных ямах и гнилых пнях, среди густой травы и веток кустарника, под корнями деревьев. А. С. Баранов наблюдал в районе г. Первомайска (УССР), как это животное пряталось в норы земляных ос, в дупла деревьев, в бутылки из-под молока и пива, под брошенную бумагу. Пожалуй, наибольшее значение в жизни прыткой ящерицы имеет постоянный и обычный тип убежищ, которым являются норы. В степных райо
176
нах она редко сама роет норы. Значительно чаще ящерица поселяет”; в пустующих норах различных грызунов и птиц-норпи-ков ^ Лукина, 19666; Тертышников, 19726; Окулова, сообщ.). Моле.': животные используют норы крупных навозников и чернотелок (Щепотьев, 1950).
По мнению Н. Н. Щербака (1966) и В. А. Кутузовой (1971), у прыткой ящерицы существуют зимние и летние норы. Последние из них характеризуются незначительной протяженностью ходов и глубиной их расположения. Так, по данным В. А. Кутузовой (1971), длина их колеблется от 12 до 15, а глубина — от
Рис. 61
Схема строения основных типов нор прыткой ящерицы (рис. М. Ф. Тертышникова)
3,5 до 32 см. Однако М. Ф. Тертышников (1972а) считает, что постоянные норы прыткой ящерицы в одно и то же время являются как летними, так и зимними. Изучая строение нор, в которых длительное время обитали прыткие ящерицы, указанный исследователь пришел к выводу о том, что в условиях Ставропольской возвышенности все многообразие нор можно свести к 4 типам (рис. 61).
Норы первого типа наиболее просты по строению. Это одноколенчатые ходы, длина которых не превышает 40, а глубина расположения—-13—16 см (рис. 61, а). По-видимому, их истинными хозяевами (изготовителями) являются сами ящерицы. Норы второго типа (рис. 61, б) имеют вид двухколенчатых ходов с несколькими отнорками. Длина их достигает 44, а глубина расположения—39 см. К третьему типу (рис. 61, в) относятся более сложно устроенные норы с двумя-тремя входными отверстиями и несколькими отнорками, общая длина которых равна 30—70, а глубина расположения 13—22 см. Норы четвертого типа (рис. 61, г) наиболее сложны по строению. Количество входных отверстий еще больше. Общая протяженность ходов достигает 170—200, а глубина их расположения—12—47 см. Вероятнее всего, норы второго, третьего и четвертого типов принадлежали раньше различным грызунам.
177
Длина собственных нор прыткой ящерицы варьирует в значительных пределах. 3. П. Хонякина (1964) предполагает, что длина их зависит от сезона, возраста ящерицы и продолжительности использования убежища. По одним данным (Тертышников, сообщ.), длина собственных нор прыткой ящерицы не превышает 40 см (см. выше), по другим данным (Щепотьев, 1952; Тара-щук, 1959 и др.)—60—70 см, а по третьим данным — 100—150 см. Интересно, что норы ящериц в степных и лесных биоценозах отличаются по строению (Кутузова, 1971). Так, в лесу они имеют вид длинных коридоров сложной конструкции, достигая длины 200 см, а в степи их строение проще. В болотистых местах норы также не сложны и расположены обычно на возвышенных местах, поросших травой.
В. Л. Булахов и Н. Ф. Константинова (1972) отметили определенную зависимость экспозиции входа в убежище от места поселения ящерицы. Большинство входов направлено на запад и юго-запад (62,5%), а остальные — 37,5%—на восток и юго-восток. На открытых участках и в осветленных насаждениях норы имеют выход на запад, а в полуосветленных насаждениях, полутеневых и теневых — на восток. Как отметил М. Ф. Тертышников (19726), в степи отверстия нор расположены под пучками травы, а в лесу и лесной полосе — у корней кустарников и деревьев.
В норах прыткой ящерицы нередко находят «гнездовую камеру», имеющую круглую или эллиптическую форму (Кутузова, 1971). Стенки ее хорошо уплотнены и выстланы сухой травой, листьями, мхом. Следует полагать, что норы с подобными сооружениями могли принадлежать ранее грызунам.
Ареал прыткой ящерицы охватывает несколько природных зон, наиболее оптимальной из которых по числу населяемых биотопов является зона степей. Являясь эвритопным видом, это животное заселяет многие биотопы, выбирая в них те участки, в которых имеется достаточное количество корма, убежища, необходимая влажность, освещенность и пр. На этих участках ящерица занимает территорию, размеры которой не превышают 300 м2 и варьируют в разных биотопах.
В пределах индивидуальной территории у ящериц имеются временные и постоянные убежища разного типа. Наиболее обычными являются норы. Ящерицы чаще поселяются в норах разных животных, но роют и сами.
До настоящего времени остаются недостаточно решенными вопросы, связанные с причиной выбора ящерицами тех или иных биотопов, территориализмом ящериц разных возрастных и половых групп, связь его с разными биотопами и сезонами.
ГЛАВА ШЕСТАЯ
ПИТАНИЕ1
Имеющиеся в литературе сведения по питанию прыткой ящерицы не многочисленны, а если учесть величину ареала этого вида, то совсем недостаточны. Они охватывают Центральночерноземную полосу Европейской части СССР (Изосов, 1951; Жаркова, 1969; Утробина, 1952; Щепотьев, 1952), юг материковой Украины и Крым (Щербак, 1966; Пащенко, 1954; Булахов, Константинова, 1972). К сожалению, большинство авторов обычно устанавливает только состав пищи ящериц вне какой-либо связи с их численностью и численностью беспозвоночных в природе. Кроме того, только изредка приводятся сведения о весе поедаемой ящерицами добычи и о продолжительности ее переваривания (Пащенко, 1954; и др.). Чаще в работах содержатся лишь отрывочные сведения о питании прыткой ящерицы (например, Мусхелишвили, 1970; и др.).
В настоящей главе сделана попытка обобщить уже имеющиеся сведения о питании прыткой ящерицы, дополнив их новыми обширными данными.
Изучение питания прыткой ящерицы имеет большое значение для определения ее значения в биоценозах. Для характеристики состава пищи прыткой ящерицы нами обработан материал из следующих 15 районов ареала: Калужская обл. (50 желудков, материал А. Б. Стрельцова), Рязанская обл. (1025 желудков, материал В. К. Жарковой), Ивановская обл. (121 желудок, материал Б. А. Красавцева), Саратовская обл. (485 желудков, материал Н. В. Щепотьева), Западный Казахстан (62 желудка, материал Н. М. Окуловой), Чувашская АССР (43 желудка, материал В. И. Козлова), Татарская АССР (204 желудка, материал Н. М. Утробиной). Кроме того, исследованиями охвачены южная часть ареала: Закарпатская обл. (62 желудка, материал М. И. Щербаня), Кировоградская обл. (16 желудков, материал В. Е. Боченко), Днепропетровская обл. (263 желудка, материал В. Л. Булахова, Н. Ф. Константиновой), области Крыма
1 Автор: Г. П. Лукина, при участии В. К- Жарковой, Н. В. Щепотьева, В. Л. Булахова, Н. Ф. Константиновой, Н. Н, Щербака, М. Ф. Т ертышникэ-ва, Н. А. Рашкевича, 3. П. Хонякиной, В. А. Кутузовой, М. И. Щербаня, В. Е. Боченко, А. Б. Стрельцова, Н. М. Окуловой, В. И. Козлова, Н. М. Утробиной.
179
Таблица 25
Состав пищи прыткой ящерицы L. agills
Вид пищи Встречаемость Число съеденных объектов
абсолютная % абсолютное %
K«i. Oligocheta 104 2,64 167 0,89
Сем. Lumbricidae 82 2,08 103 0,55
Lumbricus terrestris L. 18 0,46 22 0,12
Отр. Gastropoda 86 2,18 108 0,58
Сем. Pulmonata 63 1,60 77 0,41
Planorbis corneus L. 5 0,13 5 0,03
Helix sp. 9 0,23 12 0,06
Clausilia sp. 2 0,05 2 0,01
Galba t runcat ala L. 4 0,10 4 0,02
Отр. Stylommatophora 3 0,08 4 0,02
Кл. Crustacea 176 4,46 143 0,77
Отр. Isopoda 172 4,36 139 0,74
Сем. Oniscoidea 146 3,70 97 0,52
Отр. Amphipoda 4 0,10 4 0,02
Кл. Aracb noidea — — 63 0,34
Отр. Phalangidae 1 0,03 1 0,01
Отр. Araneinae 49 1,24 58 0,31
Сем. Lycosidae 15 0,38 19 0,10
Сем. Gnaphosidae 5 0,13 5 0,03
Сем. Thomisidae 17 0,43 19 к0,10
Сем. Salticidae 4 0,10 5 0,03
Сем. Amaurobiidae 2 0,05 2 0,01
Сем. Clubionidae 3 0,08 3 0,02
Сем. Theridiidae 1 0,03 1 0,01
Сем. Araneidae 1 0,03 1 0,01
Сем. Tetragnathidae 1 0,03 3 0,02
Отр. Acarina 3 0,08 3 0,02
Сем. Ixodidae 1 0,03 1 0,01
Кл. Miryapoda — — 70 0,38
Отр. Diplopoda 55 1,39 46 0,25
Сем. lulidae 15 0,38 19 0,10
Сем. Geophylidae 8 0,20 8 0,04
Geophylus sp. 1 0,03 1 0,01
Scolopendra sp. 4 0,10 4 0,02
Отр. Chilopoda 18 0,46 24 0,13
Сем. Lithobiidae 2 0,05 2 0,01
Сем. Oniscidea 7 0,18 10 0,05
Кл. Insecta 14 354 76,9
180
Таблица 25 (продолжение)
Вид пищн Встречаемость Число съеденных объектов
абсолютная % абсолютное! %
Отр. Odon ata 10 0,05
Odonata (larvae) 1 0,03 4 0,02
Agrion sp. 1 0,03 1 0,01
Leptetrum sp. 3 0,08 3 0,02
Libellula sp. 2 0,05 2- 0,01
Отр. Blattodea 4 0,10 6 0,03
Сем. Ectobiidae 1 0,03 1 0,01
Отр. Mantodea 9 0,23 9 0,05
Сем. Mantidae (larvae) 2 0,05 2 0,01
Отр. Orthoptera 1193 6,39
Сем. Scaphuridae 18 0,46 20 0,11
Barbitistes sp. 1 0,03 2 0,01
Poecilimcn sp. 17 0,43 18 0,10
Сем. Tettigoniidae 126 3,19 246 1,32
Tettigonia caudata Ch. 9 0,23 13 0,07
Сем. Gryllidae 87 2,21 128 0,69
Gryllus desertus Pall. 55 1,39 84 0,45
Сем. Gryl lotalp id ae 46 1,17 53 0,28
Сем. Acrididae 455 11,53 746 4,00
Acrididae (larvae) 32 0,81 77 0,41
Acridium subulatum L. 2 0,05 2 0,01
Chorthippus mollis Charp. 4 0,10 4 0,02
Ch. biguttulus L. 3 0,08 3 0,02
Stenobothrus sp. 26 0,66 38 0,20
Dociostaurus brevlcollis Ev. 1 0,03 2 0,01
Gemphoceras sp. 7 0,18 7 0,04
П/сем. Locustinae 7 0,18 10 0,05
Ocdipoda coerulescens L. 3 0,08 3 0,02
Отр. Plecoptera 2 0,05 2 0,01
Perla sp. 2 0,05 2 0,01
Отр. Dermaptera 31 0,79 37 0,20
Forficula auricularia L. 3 0,08 5 0,03
Отр. Homoptera 100 2,53 200 1,07
Сем. Diaspididae 5 0,13 5 0,03
Сем. Jassidae 37 0,94 54 0,29
Сем. Membracidae 9 0,23 20 0,11
Сем. Cercopidae 15 0,38 . 24 0,13
Сем. Cicadidae 30 0,76 58 0,31
Cicada sexnotata FLL. 2 0,05 2 0,01
Melampsalta montana Scop. 2 0,05 3 0,02
Сем. Aphididae 4 0,10 39 0,21
Отр. Elemi ptera 272 6,89 452 2,42
181
Таблица 25 (продолжение)
Таблица 25 (продолжение)
Вид пищи Встречаемость Число съеденных объектов Вид пищи Встречаемость Число съеденных объектов
абсолютна f 1 % абсолютное 1 %
абсолютная] % абсолютное] %
Hemiptera (larvae) Сем. Miridae 1 15 0,03 0,38 1 37 0,01 0,20
Pterostichus sp. 13 0,33 17 0,09
Lygus sp. 1 0,03 1 0,01 Pt. melanarius L. 2 0,05 2 0,01
Noto stir a sexnotata L. 4 0,10 4 0,02 Pt. lepidus Leske 7 0>18 10 0,05
Сем. Nabiae 11 0,28 13 0,07 Pt. cu preus L. 8 0,20 14" 0,08
Reduviolis sp. 9 0,23 И 0,06 Pt. punctulatus Schall 1 0,03 1 0,01
Сем. Nepidae 7 0,18 24 0,13 Harpalus sp. 128 3,24 216 1,16
Сем. Pyrrhocoridae 15 0,38 65 0,35 M. psittaceus Geoffr 11 0,28 13 0,07
Pyrrhocoris apterus L. 20 0,51 40 0,21 Mycrolestes sp. 1 0,03 1 0,01
Сем. Coreidae 4 0,10 8 0,04 Сем. Gyrinidae 2 0,05 3 0,02
Mesocerus morginatus L. 1 0,03 1 0,01 - Gyrus sp. 2 0,05 3 0,02
Syromastus sp. 1 0,03 1 0,01 Сем. Hydrophilidae 2 0,05 2 0,01
Сем. Pentatomidae ИЗ 2,86 162 0,87 Сем. Histeridae 13 0,33 19 о,ю
Eurygaster sp. 2 0,05 2 0,01 Hister sp. 4 0,10 6 0,03
E. integriceps Put. 29 0,74 49 0,26 Сем. Searabeidae 276 7 484 2,59
Palomena prasina L. 1 0,03 1 0,01 П/сем. Coprinae 12 0,30 21 о,н
Eurydetna sp. 1 0,03 1 0,01 Aphodius sp. 44 1,12 78 0,42
E. oleracea L. 1 0,03 1 0,01 Geotrupes stercorarius L. 7 0,18 15 0,08
E. festiva L. 4 0,10 4 0,02 Onthophagus sp. 79 ' 2,00 128 0,69
Отр. Coleoptera 1348 34,17 5443 29,16 O. amyntas Ol. 2 0,05 6 0,03
Coleoptera (larvae) 81 2,05 126 0,68 O. taurus L. 4 о.ю 4 0,02
Сем. Carabidae 603 15,29 977 5,23 O. austriacus Panz. 26 0,66 38 0,20
Cicindela silvatica L. 1 0,03 1 0,01 O. nuchicornis L. 2 0,05 4 0,02
C. germanica L. 1 0,03 1 0,01 Coccobius schreberi L. 4 о,ю 10 0,05
C. campestris L. 1 0,03 1 0,01 Coccobius sp. 4 о,ю 6 0,03
Carabas cancellatus Ill. 35 0,89 65 0,35 Oniticellus sp. 2 0,05 2 0,01
C. arcensis Hbst. 28 0,71 47 0,25 П/сем. Melolonthinae 194 4,92 411 2,20
C. nit ens L. 14 0,35 27 0,14 Pentodon idiota Hbst. 4 о,ю 4 0,02
C. granulatus L. 7 0,18 18 0,10 Arnphimallon solstitialis L. 53 1,34 100 0,54
C. netnoralis Muel. 8 0,20 15 0,08 Rhizotrogus sp. 23 0,58 23 0,12
Bembidion sp. 27 0,68 39 0,21 Melolontha hippocastani F. 33 0,84 53 0,28
B. lampros L. 1 0,03 2 0,01 Anomala bubia Scop. 32 0,81 80 0,43
Dolichus sp. 1 0,03 1 0,01 Phyllopertha horticola L. 41 1,04 111 0,59
Dolichus halensis 1 0,03 1 0,01 Ph. acstivus Oliv. 22 0,56 37 0,20
Dromius sp. 5 0,13 5 0,03 Anisoplia segetum Hbst. 2 0,05 5 0,03
Ago num sp. 3 0,08 3 0,02 A. anstriaca L. 15 0,38 17 0,09
A. septempunctatum L. 7 0,18 И 0,06 A. agricola Pod a. . 1 0,03 1 0,01
Amara sp. 25 0,63 29 0,16 Hoplia parvula Kry. 1 0,03 1 0,01
Zabrus tenebrioides Goeze 34 0,86 6t 0,33 Epicometis hirta Poda. 1 0,03 2 0,01
Panageus cruxmajor L. 19 0,48 30 0,16 ! Cetoma aurata L. 22 0,56 41 0,22
Ophonuscalceatus Duft 16 0,41 31 0,17 C. hirta Poda. 8 0,20 3 0,02
Oph. pubescens Miill 6 0,15 6 0,03 Potonia metallica Hbst. 13 0,33 32 0,17
—— — — Сем. Sylphidae 18 0,46 35 0,19
182
183
Таблица 25 (продолжение)
Вид пищи Встречаемость Число съеденных объектов
абсолютная] % абсолютное. %
Xylodepa i punctata L. 4 0,Ю 4 0,02
Сем. Staphylinidae 26 0,66 47 0,25
Stenus sp. 1 0,33 1 0,01
Сем. Elateridae 319 8,09 683 3,66
Elateridae (larvae) 4 0,10 5 0,03
Elater sp. 4 0,10 4 0,02
Athous sp. 2 0,05 2 0,01
Ludius sp. 1 0,03 1 0,01
Selatosomus aeneus L. 30 0,76 18 0,10
S. latus F. 143 3,75 526 2,82
Corymbites sp. 6 0,15 6 0,03
Agrioles sp. 3 0,08 5 0,03
A. sputator L. 25 0,63 26 0,14
A. llneatus L. 22 0,56 25 0,13
A. obscuras L. 1 0,03 1 0,01
Drasterlus bimaculatus Rossi. 2 0,05 4 0,02
Сем. Cantharididae 37 0,94 53 0,28
Cantharis sp. 2 0,05 2 0,01
C. rustica Fall. 7 0,18 7 0,04
C. fusca L. 1 0,03 1 0,01
Lampyris sp. 2 0,05 2 0,01
Сем. Dermestidae 8 0,20 14 0,08
Сем. Byrrhidae 4 о,ю 4 0,02
Byrrhus fasciatus Forst 2 0,05 2 0,01
Byrr. pibula L. 2 0,05 2 0,01
Сем. Melyridae 1 0,03 1 0,01
Malachius sp. 1 0,03 1 0,01
Сем. Coccinellidae 36 0,91 137 0,73
Coccinella sp. 5 0,13 10 0,05
C. quinque punctata L. 1 0,03 1 0,01
C. septempunctata L. 18 0,46 27 0,14
C. quatuordecimpunctata L. 1 0,03 1 0,01
Propylaea qtialuordecimpuno-
tata L. 2 0,05 2 0,01
Halyzia sp. 1 0,05 2 0,01
H. quatuordecimpunctata L. 2 0,05 2 0,01
Micraspis sp. 2 0,05 2 0,01
Hippodamia tredecimpune-
tata L. 3 0,08 5 0,03
Сем. Meloidae 24 0,61 30 0,16
Meloe violaceus Marsh. 1 0,03 1 0,01
Сем. Alleculidae 2 0,05 4 0,02
184
Таблица 25 (продолжение)
Вид пищи Встречаемость Число съеденных объектов
абсолютная % абсолютное %
Сем. Lagriidae (L hirta L.) 2 0,05 2 0,01
Сем. Tenebrionidae 67 1,70 90 0,48
Blaps sp. 1 0,03 1 0,01
В. halophila Fisch. 4 0,10 4 0,02
Opatrum subulosum L. 11 0,28 15" 0,08
Pedinus fetnoralis L. 2 0,05 2 0,01
Platyscelis sp. 1 0,03 1 0,01
Сем. Cerambycidae 48 1,22 65 0,35
Dorcadion sp. 12 0,30 20 0,11
D. Caucasians F. 24 0,61 24 0,13
D. elegaus Kr. 7 0,18 17 0,09
D. aethups Scop. 1 0,03 1 0,01
D. striatus Dalm. 1 0,03 2 0,01
Acauthocinus aedilis L. 1 0,03 1 0,01
Leptura rubra L. 1 0,03 1 0,01
Monochamus sp. 1 0,03 1 0,01
Oberea sp. 7 0,18 13 0,07
Сем. Chrysomelidae 282 7,15 700 3,75
Chrysomelidae (larvae) 2 0,05 4 0,02
Cryptocephalus sp. 2 0,05 2 0,01
Ghrysomela sp. 3 0,08 5 0,03
Gastroidea polygoni L. 10 0,25 39 0,21
Metasoma populi L. 2 0,05 6 0,03
Haltica oleracea L. 1 0,03 1 0,01
Donacia crassipes F. 9 0,23 15 0,08
Pilemostoma fastuosa Schall. 1 0,03 1 0,01
Entomoscells adonidis Pall. 5 0,13 13 0,07
Phyllodeda rufipes Deg. 3 0,08 4 0,02
Ph. vulgaris L. 3 0,08 4 0,02
Agelastica alni L. 5 0,13 16 0,09
Cassida sp. 29 0,74 35 0,19
C. viridis L. 2 0,05 2 0,01
C. Nebulesa L. 56 1,42 111 0,59
C. nobilis 1 0,03 1 0,01
П/сем. Halticinae 12 0,30 16 0,09
Сем. Bruchidae 5 0,13 8 0,04
Сем. Anthribidae 2 0,05 2 0,01
Сем. Curculionidae 668 16,93 1657 8,88
Otiorrhynchus sp. 24 0,61 44 0,24
Of. tristis Scop. 5 0,13 5 0,03
Of. ligustici L. 75 1,90 108 0,58
Phyllobius sp. 12 0,30 46 0,25
185
Таблица 25 (продолжение)
Вид пищи Встречаемое ть Число съеденных объектов
абсолютная % абсолютное] %
Ph. urticea Deg. 18 0,46 30 0,16
Ph. argentatus L. 12 0,30 35 0,19
Eusomus ovulum Gern. 1 0,03 1 0,01
Sitona sp. 53 1,34 212 1,14
Chromoderus fasciatus Miill- 1 0,03 1 0,01
Tanymecus palliatus F. ИЗ 2,86 324 1,74
Cleonus sp. 3 0,08 5 0,03
C. glaucus turbatus Fah. 4 0,10 4 0,02
Lixus sp. 1 0,03 1 0,01
Hylobius abletis L. 24 0,61 53 0,28
H. pinastris Gyll. 19 0,48 43 0,23
Phytonomus arator L. 15 0,38 40 0,21
Ceuthorrhynchus sp. 3 0,08 3 0,02
Bams sp. 2 0,05 2 0,01
Anthonomus sp. 1 0,03 1 0,01
Apion sp. 6 0,15 17 0,09
A. apricans Hbst. 2 0,05 5 0,03
Сем. Ipidae 1 0,03 1 0,01
Отр. Raphidioptera 2 0,05 2 0,01
Raphidia sp. 2 0,05 2 0,01
Отр. Neuroptera 6 0,15 8 0,04
Chrysopa sp. 1 0,03 1 0,01
Myrmeleon fortnicarius L. 2 0,05 2 0,01
Отр. Hymenoptera 703 17,82 3900 20,90
Hymenoptera larvae 1 0,03 1 0,01
Hymenoptera pupa 1 0,03 1 0,01
Сем. Te'nthredinidae 15 0,38 22 0,12
Dolerus sp. 4 о,ю 4 0,02
Сем. Siricidae 6 0,15 9 0,05
Siricinae gigas L. 5 0,13 8 0,04
Сем. Cephidae 33 0,84 52 0,28
Cephus pygmacus L. 9 0,23 16 0,09
Rhogogaster viridis L. 7 0,18 13 0,07
Athalia colibri Christ. 7 0,18 13 0,07
Di prion pini L. 14 0,35 18 0,10
Сем. Ichneumonidae 58 1,47 93 0,50
Trogus lutorius F. 4 0,Ю 6 0,03
Try phon signata Graw. 6 0,15 10 0,05
Ichneumon pisorius L. 6 0,15 8 0,04
Сем. Proctotrupoidea 4 0,10 4 0,02
Сем. Braconidae 19 0,48 30 0,16
Сем. Chalcididae 1 0,03 1 0,01
186
Таблица 25 (продолжение)
Вид пищи Встречаемость Число съеденных объектов
абсолютная % абсолютное %
Сем. Mutillidae 1 0,03 1 0,01
Сем. Pompilidae 2 0,05 2 0,01
Сем. Chrysididae 10 0,25 15 0,08
Сем. Vespidae 15 0,38 19 0,10
Vespa sp. 1 0,03 1 - 0,01
Vespula media De Geer. 5 0,13 5 0,03
Сем. Sphecidae 12 0,30 18 0,10
Sphex maxillcsus F. 5 0,13 3 0,02
Ammo phi la sabulosus 7 0,18 10 0,05
Сем. Apjdae 91 2,31 123 0,66
Apis mellifera L. 50 1,27 61 0,33
Anthophora sp. 14 0,35 21 0,11
B. terrestris L. 3 0,08 3 0,02
Bombas sp. 6 0,15 7 0,04
Halidas sexcinctus L. 5 0,13 8 0,04
Psithyrus vestalis Geffr. 1 0,03 1 0,01
Сем. Formicidae 290 7,35 1028 5,51
Formica rufa L. 19 0,48 ИЗ 0,61
Camponotus sp. 23 0,58 129 0,69
Last us sp. 10 0,25 39 0,21
Отр. Mecoptera 8 0,20 8 0,04
Сем Panorpidae 7 0,18 7 0,04
Отр. Diptera 567 14,37 900 206
Diptera (larvae) 7 0,18 7 0,04
Diptera pupa 1 0,03 1 0,01
Сем. Tipulidae 22 0,56 33 0,18
Сем. Culicidae 40 1,01 69 0,37
Сем. Bibionidae 12 0,30 13 0,07
Bibion sp. 1 0,03 1 0,01
Brachycera sp. 10 0,25 И 0,06
Сем. Mycetophilidae 2 0,05 2 0,01
Сем. Stratjomyidae 6 0,15 12 0,06
StraHomys sp. 2 0,05 2 0,01
Сем. Tabanidae 29 0,74 44 0,24
Tabanus bovinus W. 14 0,35 20 0,11
Chrysops sp. 1 0,03 1 0,01
Ch. caecutiens L. 5 0,13 7 0,04
Hematopota pluvialis L. 9 0,23 16 0,09
Сем. Rhagionidae 13 0,33 20 0,11
Сем. Asilidae 38 0,96 57 0,31
Asilus sp. 11 0,28 12 0,06
Laptogaster cylindrica Deg. 18 0,46 25 0,13
187
Таблица 25 (продолжение)
Вид НИЩ.Н Встречаемость Число съеденных объектов
абсолютная % абсолютное %
Laphria flava L. 6 0,15 15 0,08
Сем. Syrphidae 44 1,12 61 0,33
Sphaerophoria scripta L. 6 0,15 8 0,04
Baecha elongata L. 9 0,23 12 0,06
Syrphus ribesli L. .8 0,20 11 0,06
Velucella pellucens L. 4 0,10 7 0,04
Chrysotoxum bicinciwn L. 3 0,08 6 0,03
Ch. festivum L. 7 0,18 10 0,05
Сем. Larvivoridae 13 0,33 30 0,16
Tachina sp. 2 0,05 2 0,01
Callifora sp. 1 0,03 1 0,01
Sarcophaga sp. 40 1,01 27 0,14
Сем. Muscidae НО 2,79 204 1,09
Musca sp. 5 0,13 7 0,04
Stomoxys caldtrans L. 9 0,23 16 0,09
Pseudophyrellia cornicina F. 1 0,03 1 0,01
Сем. Trichoceridae 5 0,13 10 0,05
Сем. Chloropidae 8 0,20 18 0,10
Сем. Anthomylidae 26 0,66 34 0,18
Delia brassicea Bouche. 10 0,25 18 0,10
D. floratis FH. 16 0,41 16 0,09
Отр. Triclioptera 2 0,05 16 0,09
Отр. Lepidoptera 1377 34,90 2168 11,62
Lepidoptera (larvae) 280 7,10 438 2,35
Lepidoptera pupa 3 0,08 3 0,02
Сем. Zygaenidae 5 0,13 5 0,03
Сем. Tortricidae 5 0,13 6 0,03
Сем. Lycaenidae 22 0,56 24 0,13
Сем. Pieridae 2 0,05 3 0,02
Сем. Nymphalidae 16 0,41 28 0,15
Vanessa polichloros L. 10 0,25 18 0,10
Melitaea cinxia L. 6 0,15 10 0,05
Сем. Satyridae 25 0,63 42 0,23
Сем. Sphingidae 58 1,47 122 0,65
Sphinx pinastri L. 20 0,51 37 0,20
Marumba guercus Schiff. 18 0,46 35 0,19
Amorpha populi L. 9 0,23 25 0,13
Сем. Geometridae 242 6,13 387 2,07
Cosymbia annulata Schultze 36 0,91 51 0,27
Phigalia guercimontaria 30 0,76 42 0,23
Eranuis bajaria Schiff 22 0,56 34 0,18
Chloroclystis rectangulataHb. 13 0,33 31 0,17
188
Таблица 25 (окончание)
Вид ПИЩИ Встречаемость Число съеденных объектов
абсолютная % абсолютное %
Lomographa cararia Hb. 12 0,30 30 0,16
Сем. Lasiocampidae 27 0,63 53 0,28
Сем. Liparidae 5 0,13 7 0,04
Сем. Noctuidae 402 10,19 457 2,45
Agrotis segetum Schiff. 20 0,51 38 0,20
Catocasia coryli L. 25 0,63 40 X 0,21
Ochpteura praecox L. 22 0,56 35 0,19
Catlierges rotnosa Esp. 10 0,25 25 0,13
Casmia lutea Strom 18 0,46 22 0,12
Meristis trigrammica 18 0,46 22 0,12
Cerapteryx graminis L. 14 0,35 20 0,11
Mythimna L—alba L. 15 0,38 17 0,09
Triphaena pronula L. 8 0,20 15 0,08
Сем. Arctiidae 16 0,41 30 0,16
Сем. Tineidae 1 0,03 1 0,01
Сем. Hypononentidae 2 0,05 3 0,02.
Сем. Pyralididae 3 0,08 6 0,03
Сем. Derpanidae 1 0,03 1 0,01
Microlepidoptera 13 0,33 14 0,08
Кл. Reptilia 12 0,30 12 0,06
Сем. Lacertidae. 9 0,23 9 0,05
Lacerta vivipara Jaeg 2 0,05 2 0,01
L. agilis exigua Eichn. 4 0,10 4 0,02
Eremias ar gut a Pall 2 0,05 2 0,01
Сем. Psilidae 1 0,03 2 0,01
Растительные остатки 61 1,5 — —
Камни 64 1,6 —. —
Шерсть 1 0,03 — —
(100 желудков, материал Н. Н. Щербака), Западного Предкавказья (485 желудков, материал Т. П. Лукиной), Ставропольского края (462 желудка, материал М. Ф. Тертышникова), Чечено-Ингушской АССР (140 желудков, материал Н. А. Рашкеви-ча) и Дагестанской АССР (427 желудков, материал 3. П. Хоня-киной и В. А. Кутузовой). Всего проанализировано 3945 желудков, в которых определено 18 663 экз. животных (табл. 25).
ОБЩАЯ ХАРАКТЕРИСТИКА ПИТАНИЯ
Судя по результатам анализа, основу питания ящериц составляют насекомые. Представители других классов содержатся в пище в сравнительно небольшом количестве. Так, малощетин
189
ковые черви составляют 0,89%, брюхоногие моллюски — 0,58%, ракообразные — 0,77%, паукообразные — 0,34%, многоножки — 0,38%. В редких случаях ящерицы поедают позвоночных, в основном пресмыкающихся (0,06%). Поедаются особи своего и других видов. Например, в желудке прыткой ящерицы из Крыма была найдена разноцветная ящурка—Eremias arguta Pall, длиной 43 мм. Чаще в желудках встречаются лишь хвосты ящериц, возможно, они поедают подвижные автотомированные хвосты.
Известны факты поедания самцами прытких ящериц яиц, отложенных самками (Даревский, 1946). Например, для Рязанской обл. встречаемость яиц в желудках составляет 0,8% (Жаркова, 1969).
Несмотря на трудности при таксономическом определении животных, содержащихся в желудках, было определено до вида 125 животных, до рода 232.
Преобладающая часть насекомых принадлежала к следующим отрядам: жуков (29,16%), перепончатокрылых (20,9%) и чешуекрылых (11,62%); кроме того, встречаются прямокрылые (6,39%), двукрылые (4,82%), клопы (2,42%) и равнокрылые хоботные (1,07%). Остальные 8 отрядов в пище ящериц очень малочисленны и составляют от 0,2 до 0,1 %.
По частоте встречаемости в желудках прытких ящериц на первом месте стоят представители класса насекомых (100%), затем паукообразных (21,3%) и ракообразных (4,4%). Среди насекомых наиболее часто встречались жуки (94,0%), затем чешуекрылые (39,4%), прямокрылые (30,4%), перепончатокрылые (18,9%), двукрылые и клопы (10,8%).
Среди объектов питания прытких ящериц встречаются как медленно передвигающиеся (моллюски, некоторые виды жуков, гусеницы), так и быстро передвигающиеся формы (жужелицы, пчелы, осы, бабочки и даже молодые ящерицы). В пище ящериц присутствует большое количество криптически или апосематиче-. ски окрашенных животных, что было отмечено С. С. Шварцем (1946) для некоторых пойкилотермных животных. В их пище можно встретить ядовитых, жалящих или неприятно пахнущих ) животных. |
В пищеварительном тракте ящериц встречаются раститель- j ные остатки (1,5%). Чаще всего это случайно схваченные вме- ! сте с животными кусочки листьев или части цветка; однажды было найдено семя Geranium sp. В некоторых районах, например, в окрестностях с. Красногвардейского (Ставропольский край) и в окрестностях г. Грозного, мы наблюдали, как прыткие ящерицы поедали упавшие на землю ягоды шелковицы. Эту же пищу мы обнаружили и в просматриваемых желудках, причем в одном желудке находили до 4 ягод. По наблюдениям А. И. Рыжикова (устное сообщение), прыткая ящерица поедает землянику (Fragaria vessa), обрывая ее прямо с ветки.
Потребность в воде у ящериц незначительна, так как основное
190
ее количество ящерицы поглощают, поедая различных беспозвоночных. Тем не менее нам неоднократно приходилось наблюдать этих животных, слизывающих капли воды в жаркие дни с различных предметов.
Довольно редко в содержимом желудков ящериц встречаются небольшие камешки, которые, возможно, играют роль гастролитов (Коштоянц, 1950). По-видимому, они имеются у всех ящериц, но время от времени выводятся из желудка вместе с перевариваемой пищей.
ИЗБИРАТЕЛЬНАЯ СПОСОБНОСТЬ К ОПРЕДЕЛЕННЫМ КОРМАМ
Список кормовых объектов прыткой ящерицы так обширен, что казалось бы приводит к выводу, что избирательность в объектах питания у прыткой ящерицы может, видимо, вообще отсутствовать. Об этом свидетельствует и то, что по мере расширения района исследований список кормовых объектов постоянно увеличивается. Различия в составе корма зависят главным образом от географической точки, наличия корма в природе и легкости его добывания.
Об отсутствии избирательной способности к пищевым объектам у прытких ящериц упоминали Б. А. Красавцев (1936, 1939), А. М. Андрушко (1954), А. А. Изосов (1951), А. В. Бочарникова (1964), Н. Н. Щербак (1966), Г. П. Лукина (1966а) и М. Ф. Тертышников (1972а). Но в этих работах не приводилось сравнения частоты встречаемости различных животных в желудках ящериц и в природе. Однако Н. М. Утробина (1956), проводя параллельно с отловом ящериц и сбор насекомых в лесозащитных насаждениях Татарской АССР, указывает на то, что у ящериц наблюдается как бы избирательная способность к определенным пищевым объектам. Далее Н. М. Утробина пишет, что большое количество щелкунов в пище ящериц совпадает с периодом их активности. Объясняя увеличение встречаемости хрущей наступлением у них периода яйцекладки, она противоречит себе, так как из ее же слов явствует, что ящерицы в этом случае питались наиболее часто встречающимися в природе видами.
Для выявления .избирательной способности ящериц было проведено количественное и качественное сравнение найденных в желудках видов насекомых с числом тех же видов, собранных в районе охоты «хищника» (Шварц, 1948).
Результаты учета численности беспозвоночных в молодых сосновых посадках и анализ содержимого пищеварительных трактов ящериц, отловленных в этом биотопе, по отрядам представлен в табл. 26.
Из табл. 26 видно, что в пище ящериц частота встречаемости жесткокрылых (38,0%) и прямокрылых (16%) близка к встречаемости их в природе. Встречаемость чешуекрылых и пауков
191
в пище ящериц в 4,5—5 раз выше, чем в природе, и, наоборот, двукрылых и перепончатокрылых в 5,7—2,3 раза меньше, чем в природе. Говоря о двух основных кормах, следует отметить, что жуки встречаются в пище ящериц с той же частотой, что и в природе, а чешуекрылые явно избираются, что находится в соответствии с результатами кормления в террариумах. Интересные результаты получены по группам кормов. Из отряда жесткокрылых в пище ящериц найдено божьих коровок в 8 раз меньше (2,2%) от числа их встреч в природе (17%), долгоно-
Таблица 26
Частота встречаемости пищевых объектов (в %) в природе и в пище прытких ящериц в Шацком районе Рязанской обл. (по Жарковой, I960 г.)
В стречаемость Жуки Чешуекрылые Прямокрылые Двукрылые . Пауки Перепончатокрылые
В природе 39,2 4,8 18,6 25,1 4,5 23,0
В желудках, «=187 38,0 20,2 16,6 4,8 22,1 8,1
сиков и хрущей в 3 раза больше (29,8 и 30,3%), чем в природе (11 и 10%). В пище прыткой ящерицы был обнаружен крапивный слоник (Phyllobius urticae Dej.), который не был найден в природе и, напротив, во время учета часто встречающаяся синяя муха {Musca sinicus — 5,6%) в пище ящериц не была обнаружена.
В Днепропетровской области количественное соотношение объектов питания в составе пищи ящериц в природе при сопоставлении с данными наблюдений в террариуме показало, что как самцы, так и самки в одинаковой мере в первую очередь выбирают наиболее ярко окрашенных, подвижных и средней величины насекомых (жуков длиной до 6—8 мм, двукрылых 8—10 мм, прямокрылых 10—15 мм). Более крупных беспозвоночных, таких, как хрущ майский или навозники, ящерицы берут неохотно. В террариумах ящерицы совершенно не ели божьих коровок, хотя в природе они встречаются в их корме. Из смешанного корма, предлагаемого террариумным ящерицам, в первую очередь поедались гусеницы, прямокрылые и двукрылые, а уже в последнюю очередь жесткокрылые. Последние совсем не поедались, если гусениц, прямокрылых и двукрылых было достаточно для насыщения. В этой связи можно думать, что жуки как наиболее массовая пища в природе имеют большее значение в питании не потому, что ящерицы их избирают, а потому, что это наиболее массовый объект. Все перечисленные выше данные свидетельствуют о том, что прыткая ящерица (по крайней мере в данном эксперименте) различает пищу по размеру, цвету и, очевидно, по каким-то другим качествам. Наличие избиратель
192
ной способности к объектам пищи существует, но эта избирательность подвержена большим изменениям и зависит от набора пищевых компонентов (Даревский, 1963; Хонякина, 1965).
СПОСОБЫ ДОБЫВАНИЯ ПИЩИ.
ВЕЛИЧИНА И ОСОБЕННОСТИ КОРМОВЫХ ОБЪЕКТОВ
Большое разнообразие объектов пищи ящериц характеризует их как активных энтомофагов, поедающих всех двигающихся беспозвоночных животных. Однако ящерицы иногда все же поедают не только неподвижных, но и мертвых беспозвоночных. Еще не видя добычи, они реагируют на шум, производимый ею, и продвигаются по направлению к ней. Не исключено, что ящерицы могут при помощи слухового анализатора безошибочно отыскивать зарывшихся в почву личинок насекомых, как это было установлено для быстрой и сетчатой ящурок, а также и для ящурки Штрауха (Васильев, 1969). Не меньшее значение имеет зрение. Заметив сидящее насекомое, ящерица продвигается в ее сторону и старается схватить.
Пищевые объекты ящериц невелики. Однако в желудках нередко находили довольно крупную добычу. Так, при вольерном содержании ящериц было замечено, что они могут проглотить даже таких крупных беспозвоночных, как стрекозы или дождевые черви. В окрестностях г. Грозного у ящерицы длиной 10,2 см в желудке наряду с другой пищей был обнаружен дождевой червь длиной 10,5 и толщиной 0,5 см.
Различную по размерам добычу ящерицы съедают не одинаково по времени: меньших животных они глотают быстро, а для того, чтобы проглотить крупную добычу, им приходится затрачивать до 3 мин.
Наблюдения за питанием прытких ящериц в террариумах' показали, что ящерицы, увидев добычу, хватают ее поперек тела, сильно сжимают челюсти, начинают энергично трясти головой из стороны в сторону, бьют о пол террариума, иногда отпуская добычу, затем снова хватают ее, широко раскрывая рот и, наконец, раздавливают и глотают ее. Затем, медленно извивая тело, помогают продвижению пищи по пищеводу. Это наблюдение проведено в момент пожирания ящерицей крупной бабочки бражника, имеющей длину туловища 4 и толщину 0,8 см. Ящерица справилась с бабочкой в течение 2 мин. При этом ящерица с помощью языка «поправляет» положение добычи в ротовой полости таким образом, чтобы она располагалась параллельно оси тела и могла быстрее пройти в желудок.
Любопытны методы лова ящерицами добычи. Можно часто видеть, как они прыгают и ловят пролетающих над ними мух и бабочек. В других случаях ящерицы располагаются на небольшом возвышении, в 20—30 см от которого находится какое-либо невысокое растение. Заметив появившихся на нем мух, муравьев
7 Прыткая ящерица 193
или бабочек, ящерица, сделав 2—3 шага, прыгала и хватала насекомое. Обладая способностью лазить по деревьям, кустарникам и травянистым растениям, имеющим прочные стебли, ящерицы и там нередко добывают себе пищу.
КОЛИЧЕСТВЕННОЕ СООТНОШЕНИЕ КОМПОНЕНТОВ ПИЩИ ПРЫТКОЙ ЯЩЕРИЦЫ В РАЗЛИЧНЫХ ЧАСТЯХ АРЕАЛА
Как уже говорилось, список видов, входящих в состав рациона прыткой ящерицы, очень обширен. Для того, чтобы представить себе качественное и количественное соотношение отдельных компонентов в пище ящерицы в различных частях ее ареала, проанализирован большой фактический материал (рис. 62). Что же представляют собой спектры питания ящерицы в различных пун-ктах?
; Пищу прыткой ящерицы в Калужской области и по количеству экземпляров и по частоте встречаемости составляют в основном жуки (соответственно 86,0 и 32,82%), прямокрылые (54,0 и 23,0%), чешуекрылые (46,0 и 18,5%), перепончатокрылые (24,0 и 8%) и пауки (22,0 и 5%). Менее часто попадаются в желудках равнокрылые хоботные (12,0 и 3,0%), двукрылые и клопы (10,0 и 2,5%) и еще реже моллюски (6,0 и 2,5%). Из жуков наиболее многочисленны слоники (40,0 и 19,0%), божьи коровки (44,0 и 4,5%) и щитовки (10,0 и 2,5 %).
В Рязанской области в желудках прытких ящериц были обнаружены: насекомые — у 80% особей, пауки — у 17,3%, олигохеты—у 1,1 % и легочные моллюски — у 0,8%. Кроме того у 0,8% особей обнаружены отложенные яйца этого вида. Из класса насекомых в пище прытких ящериц встречены жуки (61,5%), чешуекрылые (36,3%), прямокрылые (21,3%), двукрылые (15,7%). Из отряда жуков обнаружены представители 10 семейств. Наиболее часто встречаются хрущи (Melolonthinae) —28,5%, жужелицы (Carabidae)—26,1%; долгоносики (Curculionidae) — 23,7%; листоеды (Chrysomelidae) —3,9% и божьи коровки (Сос-cinellidae)—2,5%. Остальные 5 семейств — дровосеки (Сегаш-bycidae), щелкуны (Elateridae), мягкотелки (Cantharidae), вер-течки (Gyrinidae) и нарывники (Meloidae) встречаются значительно реже — у 0,3— 1,1% особей. Из семейства хрущей чаще всего поедаются кузька зеленый (Anomoladubia Scop.) —5,2%’, кузька садовый (Phyllopertha horticola L.) —4,8%; хрущ майский (Melolontha hoppocastani F.) —3,6%; бронзовка золотистая (Cetonia aurata L.) —3,5% и нехрущ весенний (Phyllopertha ас-stivus Oliv.) —3,3%. Из семейства жужелиц наиболее часто нами обнаружены жужелица красноногая (Carabas cancellatus Ill.)—5,6%; полевая (С. arcensis Hbst.)—4,5%; хлебная (Са-brus tenebrioides Goeze.) —3,6% и большой крестоносец (Рапа-gaeus crux-major L.) —3,0%. Из семейства долгоносиков чаще всего встречены долгоносик серый многоядный (Tanymecus palli-
194
Рис. 62
Качественное соотношение основных компонентов пищи прыткой ящерицы в восточноевропейской части ареала
/— Днепропетровская обл.; 2— Кировоградская обл.; 3—-Крым; 4— Западное Предкавказье; 5 — Ставропольский край; 6 — Чечено-Ингушская АССР; 7 — Дагестан; 8 — Калужская обл.; 9—Рязанская обл.; 10— Ивановская обл.; // — Чувашская АССР; 12 — Саратовская обл.; /3 —Татарская АССР; /4 — Закарпатская обл.; условные обозначения компонентов пищи см. на рис. 63
atus F.)—5,4%; большой сосновый слоник (Hylobius abietis L.) —3,8%; хвойный слоник (Н. pihastri Gill.) —3,0% и грубко-верт дубовый (Лttelabus nitens Scop.)—2,9%. Встречаемость остальных видов долгоносиков (см. табл. 25) колеблется от 0,8 До 2,4%.
Из отряда чешуекрылых в пище прыткой ящерицы обнаружены представители 8 семейств: чешуекрылые в стадии гусениц — у 28,1% особей, а в стадии имаго —у 8,2%. Чаще других встречены совки (Noctuidae) — у 40,1%, в том числе 33,6% гусениц, 6,5% имаго; пяденицы (Geometridae) —33,6%; из них гусениц — 29,8%, имаго—3,8%; бражники (Sphingidae) —12,6%, в том числе 11,2% гусениц и 0,6% имаго. Гусеницы в первых двух семейст
195
7*
вах составляют 7s всех особей, а семейство бражников полностью состоит из гусениц, исключая двух особей.
Из семейства совок, встреченных в желудках прыткой ящерицы, нами определено 9 видов. Наиболее часто встречены совка шелкопряд (Catocasia coryli L.) —6,7%; совка земляная (Ocho-pleura praecox L.)—5,9%, озимая (Agrotis segetum Schiff.) — 5,3%; желтая и тупокрылая (Merististrigrammica. Hyfn.) no 4,0; травяная (Cerapteryx grammi L.)—3,7%; гладконогая (Callier-des ramosa Esp.) —2,7% и большая ленточница (Triphsena pro-nula L.)—2,1%. Из семейства пядениц найдено в желудках прыткой ящерицы 6 видов. Наиболее часто встречены — пяденица кленовая-кольчатая (Coaymbia annulata Sshultze)—9,6% и волосистая (PhigaliapedariaE.) — 8,0%.Несколько реже — пяденица щавелевая (Cosymbia quercimontaria Basteb.) —5,9%; пятнистая (Chlerechustia reetanqulata L.) — 3,5% и Eranuis bajaria Schiff. — 3,5%, Lanographa cararia Hb. — 3,2%.
В желудках прыткой ящерицы обнаружено три вида бражников, частота встречаемости которых колеблется от 2,4 до 5,3%.
Из отряда прямокрылых в желудках прыткой ящерицы встречены представители двух семейств — саранчевых (Acridi-dae) 67,2% и кузнечиковых (Tettigonidae) —38,2%, род и виды которых нами не были определены.
Отряд двукрылых представлен в пище прытких ящериц 9 семействами. Из них наибольшее число встреч приходится на семейства: журчалки (Syrphidae)—23,0%, настоящие мухи (Musci-dae) —21,7%, слепни (Tabanidae) — 17,4% и ктыри (Asilidae) — 14,9%. Из семейства журчалок одинаково часто встречены журчалка продолговатая (Baecha elongata F.) — 5,6 и сирф перевязанный (Syrphus ribesii L.) —5,0%.
В Ивановской области в пище ящериц также преобладают жуки 36,9%, чешуекрылые (29,7%), пауки (21,06%), прямокрылые (11,61%) и двукрылые (11,6%).
В Чувашской АССР пищевой спектр несколько отличается от других мест, но, по-видимому, здесь нужно учесть, что материал собран в конце лета. По численности животных в желудках ящериц на первом месте перепончатокрылые (57,44%), далее идут жуки (10,7%), прямокрылые (9,3%) и пауки (4,5%).
В Татарской АССР в пище прыткой ящерицы преобладают жуки (64,01%), двукрылые (5,71%), чешуекрылые (5,86%), а пауков (3,67%) и перепончатокрылых (3,03%) попадалось значительно меньше.
В Западном Казахстане большое место в питании ящериц занимают гусеницы. Они были встречены в 65% желудков. Из них часто встречаются гусеницы озимой совки (Agrotis segetum) — важного вредителя сельского хозяйства. Эта гусеница встречена в 35% желудков. Кроме того, находили гусениц различных пядениц, а также мохнатых гусениц (до 5 гусениц совок в одном желудке).
196
Саранчовые были найдены в 57% желудков. Среди них — личинки и взрослые формы итальянского пруса (Calliptamus itali-cus), а также представители сем. Tettigonidae. В желудках находили до 4—5 экз. одновременно.
Жуки отмечены более чем для трети желудков (36,5%). Также были встречены бронзовики (Potosia sp.), жужелицы (Наг-palus sp.), чернотелки (Tenebrionidae). Чаще встречаются более мелкие жуки, например, щелкуны (Salatosomus sp.) (в одном желудке обнаружено до 12 экз.). В одном из желудков был обнаружен наездник (Ichneumonidae), а у другого самца прыткой ящерицы была обнаружена молодая разноцветная ящурка, т. е. и в данном случае, как и в случае совместного обитания с живородящей ящерицей в других частях ареала (Параскив, 1956), прыткая ящерица прибегает к хищничеству.
В южной и западной частях ареала на территории СССР состав пищи ящериц практически не меняется. Так, в Закарпатской обл. также преобладали жуки (33,05%), затем чешуекрылые (18,77%). В Кировоградской обл. пища ящериц также состояла в основном из жуков (32,0%), но здесь попадалось несколько больше прямокрылых (12,0%), и равные количества (10,0) чешуекрылых и перепончатокрылых.
В Днепропетровской области основу питания составляют насекомые (88,49% всего количественного состава пищи и 91,11 % — весового), среди которых превалируют жуки (соответственно 46,7 и 30,26%), чешуекрылые (14,3 и 37,54%) и перепончатокрылые (14,48 и 10,94%). Среди жуков в наибольшей степени потребляются различные долгоносики (13,28% по количеству; 7,69%—по весу; 10,48% по встречаемости *). Это Phyllobius ablon-gus (соответственно 3,87; 2,1; 3,8), Otiorrynchus sp. (3,5; 2,5; 3%), О. Caucasians, О. ligustici и ряд других (в пределах 0,14—-1,1; 0,1—0,4; 0,4—3,8%). Затем идут различные жужелицы 11,3; 5,3; 8,8% —Harpalus smaragdinus, А тага similata, Carabas sp. и другие виды (в пределах 0,3—2,9; 0,3—2,1 и 0,8—3,0%), затем почти равное значение в питании имеют божьи коровки, чернотелки, хрущи, стафилины и листоеды (в пределах 2,5—3,1% всего количественного состава). Это Coccinella septempunctata (0,3—1,0; 0,3—0,7 и 0,8—1,1%); Gnaptor spinimantts, Platyscelis gages, Pedinas femoralis, Cetonia aurata, Aphodius acureus, Anisoplia segetum, A. austriaaca и другие хрущи (в пределах 0,1—1,2%, 0,2— 1,0%; 0,4—0,8%); Labidostonis humeralis, Criptocephalus virens, Gastroidae poligoni, Chrysometa limbata, Ch. fastuosa, Galeruca tanaceti (0,1—0,6; 0,1—0,4; 0,4—0,8%).
Несколько меньшее значение имеют щелкуны, кожееды, зерновки, карапузики (в пределах 0,8—1,8% количественного состава пищи, 0,7—2,0%—по весу и 1,5—3,5%—по встречаемости).
1 Во всех последующих первый процент означает количественное соотношение объектов питания, второй — весовой, третий — встречаемость.
197
Из них в количественном отношении разнообразны следующие виды: щелкуны (Athas hacmorroidalis, Elater sanguineus, E. sanquinolentus, и усачи (Stenocorus apterus, Strangalia attenuate). Незначительны мягкотелки и мертвоеды (0,3—по количест-. ву, 0,2—0,4%— по весу и 0,4—<по встречаемости). Все жуки представлены в основном имагинальными стадиями развития. Личинки составляют 4,7% от всего количества (усачи, щелкуны, листоеды, кожееды, божьи коровки). Среди чешуекрылых в наибольшем количестве потребляются различные совки —7,0% по количественному составу, 29,1% по весовому и 17,25% по встречаемости, основу которых составляют гусеницы (5,7% весового состава). Это в основном Atnphipyra pyraniidea, Taeniacampasp., Agrotis segetum-, затем медведицы (Arcticavilica — 2,3; 2,1; 3,5%), пядиницы (Operophthera brumata, Boarmia sp.—1,8; 3,2; 3,8%), листовертки (Argyroploce variegana, Tortrix viridana 0,8, 0,6, 1,9%). Остальные объекты чешуекрылых представлены 0,1—0,4% всего количественного состава. Это волнянки (Euprectis chrisor-roea, Lymanthria dispar), моли (Hyponomeuta padellum), серпокрылые моли (Drepana sp.), белянки (Pieris rapae), нимфелины (Melitaea didyma), пестрянки (Procrus statices).
Чешуекрылые представлены преимущественно (27,7% всего весового состава) гусеницами. Среди перепончатокрылых в питании ящерицы преобладают муравьи (18,6% количественного состава). Из муравьев наибольший удельный вес занимают Formica cinerea (3,2%), Е. fusca (1,4%), Lasius niger (0,7%). Остальные объекты — F. polyctena, F. cunicularia,Tetranioriumcaespitum встречаются в пределах 0,1—0,4%.
Последующие места среди насекомых занимают различные двукрылые (4,2% всего количественного состава, в основном за счет мух); равнокрылые (2,9%, в том числе Aphrophora spumaria, Oncopsis lanio, Philaenus spumaria и др.); клопы (2,5, в том числе щитники Palomena prasina, Р. viridissima и краевики); прямокрылые (2,2%, в том числе личинки саранчи и кузнечиков, медведка обыкновенная); ручейники (0,6%), стрекозы и личинки богомолов (по 0,3%).
В основе все насекомые представлены имагинальной стадией развития (71,0% по количеству и 52,7% по весу), затем — личиночной (17,5 и 38,4%) и незначительно — куколками (0,4—0,6%). Остальная часть пищи прыткой ящерицы приходится на паукообразных (7,95% по количеству и 6,11% по весу); многоножек— (соответственно 3,01 и 2,33%); брюхоногих моллюсков (0,55 и 0,45%).
Среди паукообразных очень редко встречаются клещ Ixodes ricinus (0,14), а основную массу составляют различные пауки. Наиболее часто отмечены бокоходы (2,6% по количеству) —Ху-cticus cristatus, X. stratipes, X. kochi, X. cambridgei, Philodromus aereolus, Thanaius arenarius, Th. sabulosus и др.; пауки-волки (2,5%) —Pardosa arcuata, Alopeco sp., остальные пауки (Evarcha
198
arcuata, Heliophamus sp., Haplodrascus signifer, Gnaphosa sp.) составляют 0,14—0,7%. Из многоножек встречаются Oniscidae (1,3%), Lithobiidae (0,8%), Julidae (1,37%).
’ Перечисленные различные виды и группы беспозвоночных животных представлены в основном обитателями подстилки, травяного покрова и падающими с кроны деревьев в незначительном количестве дендрофильными насекомыми.
В Крыму рацион питания ящериц такой же, как и на юге материковой Украины, но процент попадания в желудках перепончатокрылых несколько выше и составляет 25,43% против 10% в Днепропетровской области.
В степных участках Предкавказья количество жуков -в составе пищи ящериц значительно выше, чем в горных. Так, в Ростовской области, Краснодарском и Ставропольском краях жуки составляют соответственно 53,55 и 61,7%, а в Дагестанской и Чечено-Ингушской АССР — лишь 17,21 и 28,5%. Что касается других групп животных, то их количество различно. Например, в Западном Предкавказье и Чечено-Ингушской АССР преобладают чешуекрылые (15,56, 13,7%), в Ставрополье — перепончатокрылые (16,06%), а в Дагестане — пауки (16,26%).
Таким образом, пищевой рацион прыткой ящерицы на большей части ее ареала сравнительно мало изменяется. Основу ее пищи во всех пунктах составляют пауки, далее перепончатокрылые и чешуекрылые, а затем уже пауки и прямокрылые. По частоте встречаемости отдельных компонентов пищи наблюдается подобное соотношение.
ПОЛОВОЙ ДИМОРФИЗМ В ПИТАНИИ
По-видимому, по составу пищи у самцов и самок прытких ящериц существенных различий не наблюдается, хотя О. П. Богданов (1960) указывает, что в период беременности состав кормов самок отличается от такового у самцов. Наблюдения в Ставропольском крае, Чечено-Ингушской АССР и Рязанской области (рис. 63) не подтверждают этих результатов. По нашему мнению, пища самцов и самок состоит из однородных компонентов, хотя в разном соотношении.
Таблица 27
Состав кормов прыткой ящерицы Рязанской области в зависимости от пола (Жаркова, 1973а)
Пол и число особей Жесткокрылые Чешуе -, крылые S ф «2 Прямокрылые Пауки Перепончатые Стрекозы Полужесткокрылые Земляные черви Легочные МОЛЛЮСКИ Позвоночные
Самцы, п-331 Самки, п-348 58,6 40,9 37,2 39,1 25,0 16,7 20,6 20,9 19,3 19,6 18,4 7,3 4,4 0,2 2,8 2,9 0,9 1,3 0,5 0,9 4,1 0,0
199
в
Рис. 63
Качественная и количественная характеристика питания различных возрастно-половых групп прытких ящериц в разных участках ареала
/ — Ставропольский край; 2— Чечено-Ингушская АССР; 3— Дагестанская АССР; 4 — Рязанская обл. Компоненты пищи: 1—жуки; 2— перепончатокрылые; 3— двукрылые; 4 — кивсяки; 5 — мокрицы; 6 — чешуекрылые; 7 — пауки; 8 — прямокрылые; 9 — другие компоненты. А— молодые; Б—самки; J3 — самцы; I—IV — возрастные группы, ориентировочно определенные по размерам тела (рис. Г. П. Лукиной по сводным данным разных авторов)
М. Ф. Тертышников (1972а) указывает на то, что беременность снижает подвижность самок и ведет к увеличению в рационе малоподвижных перепончатокрылых и двукрылых. В табл. 27 представлен состав пищи самцов и самок из Рязанской области.
Как видно, в пище самцов и самок преобладают жуки (40,9, 58,6%) и чешуекрылые (37,2, 39,1%). Однако самцы значительно чаще поедают жуков, а самки — чешуекрылых. Почти в одинаковой степени поедаются особями обоего пола прямокрылые (20,6, 20,9%), пауки (19,3, 19,6%) и клопы (2,8, 2,9%). Кроме того самцы на 9% более поедают двукрылых, в 2 раза-—перепончатокрылых и в 26 раз больше стрекоз. Следует-отметить также, что самцы поедают яйца, отложенные самками. По весу у самок прыткой ящерицы из Чечено-Ингушской АССР (окрестности г. Грозного), наоборот, преобладали жуки (66,6, 44,4%), у самцов перепончатокрылые (13,8, 9,0%), пауки (8,3, 6,0%) и гусеницы (19,7, 9,0%) (см. рис. 63). Прямокрылые были найдены только в желудках самок, но, вероятно, это — случайное явление, не зависящее от пола ящериц, поскольку в одних точках ареала в рационе питания самцов преобладают жуки, а в других жуки преобладают в рационе самок. Та же картина наблюдается в отношении других компонентов питания. Различий в питании самцов и самок прыткой ящерицы нами не отмечено ни по составу пищи, ни по частоте ее встречаемости в желудках. Исключение могут составлять беременные самки, в рацион питания которых входят малоподвижные формы насекомых.
ВОЗРАСТНЫЕ ИЗМЕНЕНИЯ В ПИТАНИИ
Состав кормовых объектов прыткой ящерицы с возрастом несколько изменяется. О. П. Богданов (1965) отмечает как общую закономерность для всех ящериц — преобладание мелких объектов в пище молодых и наличие большого числа муравьев, которых взрослые особи поедают значительно реже. Существенные отличия в питании различных возрастных групп отмечает Б. А. Красавцев (1939).
Наши данные из различных точек ареала прыткой ящерицы (см. рис. 63) указывают на отсутствие качественного отличия в рационе взрослых и молодых особей. Но если в Ставропольском крае в пище взрослых ящериц жуки играют основную роль, то в пище молодых, кроме жуков, не меньшее значение приобретают перепончатокрылые, чешуекрылые, а в Дагестанской АССР пауки. Понятно, что молодые ящерицы в силу своей меньшей величины и подвижности питаются более мелкими и более доступными животными.
Сравнительный анализ трех размерных групп ящериц из Днепропетровской области (35—50, 51—70 и свыше 70 мм) показал,
201
что общий коэффициент сходности питания их составляет по качеству 3,5% и по количеству —13,04%.
Общими объектами для всех возрастов являются различные пауки-волки, мелкие жужелицы, Stenocopus apterus из усачей, божьи коровки, мелкие долгоносики (Phyllobius ablongus), гусеницы зимней пяденицы, мелкие мухи. Для первой возрастной группы характерны мелкие беспозвоночные, лишенные твердых оболочек: в основном молодь сухопутных брюхоногих моллюсков с крупными раковинами, различные пауки (22,2% всего количества пищи), затем чешуекрылые (33,3%) и мелкие жуки (27,7%). У второй размерной группы пищевой рацион более разнообразен. При сохранении тех же объектов питания ассортимент пищи увеличивается за счет включения большого количества крупных пауков, жужелиц, хрущей, усачей, крупных долгоносиков, листоедов, кузнечиков, цикад, гусениц-совок, листоверток, различных муравьев. За счет прибавления новых объектов питания качественное и количественное сходство в питании первой и второй размерных групп ящериц очень незначительное (12,8 и 17,4%).
У наиболее старшей возрастной группы (третья размерная) увеличивается количество жуков, стрекоз, но характер питания изменяется значительно меньше. Качественное сходство состава пищи 35,1, количественное 45,8%. Почти полное сходство наблюдается по таким объектам, как пауки, жужелицы, усачи, долгоносики, саранча, зеленые кузнечики, цикады, гусеницы совок, зеленой листовертки, зимней пяденицы, различные мухи, песчаный муравей и др.
Анализ содержимого пищеварительных трактов был проведен также в Рязанской области по четырем возрастным группам (см. рис. 63, 2). Как видно, ящерицы на первом году жизни наиболее часто поедают чешуекрылых (38,9%), жуков (33,2%) и пауков (24,7%), несколько меньше прямокрылых (20,8%) и почти с одинаковой частотой употребляют двукрылых (10,6%) и перепончатокрылых (10,1%). Встречались также дождевые черви и моллюски.
На втором году жизни в пище прытких ящериц в 2 раза увеличивается встречаемость жуков (68,1%), двукрылых (20,0%) и в 4 раза — дождевых червей (1,6%). Кроме того на %4 часть увеличивается поедание прямокрылых (25,2%) и совсем не найдены моллюски.
На 3-м году жизни частота встречаемости жуков увеличивается в 1,5 раза, полужесткокрылых в 3 раза (7,1%) и отмечается максимальная частота встреч легочных моллюсков; несколько чаще поедаются чешуекрылые, пауки и стрекозы.
В четвертой возрастной группе уменьшается частота поедания двукрылых, пауков и жуков и совсем не обнаружены дождевые черви и моллюски. Таким образом, наиболее существенно изменяется по возрастам частота поедания жуков (от 33,2 до
202
97,1%), чешуекрылых (от 31,1 до 42,5%), двукрылых (от 20 до 7,5%), пауков (от 24,7 до 7,5%), стрекоз (от 4,0 до 7,5%) и полужесткокрылых (от 2,6 до 7,1%).
Н. В. Щепотьев (1952) указывает, что пища молоди ящериц из Саратовской области состоит в основном из чешуекрылых (23,6%), прямокрылых (16%), жуков (14,1%) (см. рис. 63, 1). При этом у молодых ящериц в рационе' не отмечено резкого преобладания каких-либо групп животных. Пища взрослых представляется более однообразной из-за резкого преобладания в ней жуков (46,4%) и чешуекрылых (29%), причем разница в питании молоди и взрослых нивелируется в зависимости от условий года. Как видно, значительной вариации в разнообразии кормов у всех возрастных групп не наблюдается. Сходный качественный состав пищи взрослых и молодых свидетельствует о быстроте выработки реакции на внешний вид корма, которую отмечает В. В. Черномордиков (1944).
Интересно отметить, что процент пустых желудков увеличивается ежегодно, до 3-го года жизни включительно, и это, по-видимому, отражает частоту принятия корма ящерицами. Некоторое уменьшение процента пустых желудков в старшей возрастной группе, возможно, связано со старением особей.
ИЗМЕНЕНИЕ СОСТАВА КОРМОВ ПО БИОТОПАМ
Исследования С. С. Шварца (1948) показали, что кормовой рацион вида можно понять только при сравнении частоты встречаемости определенной добычи в пище животного и в местах его охоты. При таком сравнении спектр питания ящериц будет или отражать численное соотношение объектов добычи в биоценозах или указывать на предпочтение ящерицами определенного вида корма.
Если проанализировать состав пищи у ящериц в биотопах центральных районов Европейской части СССР, то окажется, что в Рязанской области наиболее часто встречающимся видом пищи во всех биотопах являются жуки и чешуекрылые (табл. 28).
Биотоп «лиственный лес» характеризуется тем, что частота встречаемости прямокрылых (8,5%), пауков (16,5%), двукрылых (15,5%) в пище ящериц меньше, чем в других биотопах, а стрекоз, наоборот, больше (12,7%). В рационе ящериц, обитающих в лиственном лесу, отсутствовали дождевые черви, которые встречаются во всех других биотопах.
Смешанный лес характеризуется большой встречаемостью в желудках ящериц, прямокрылых (32,5%) и наименьшим числом жуков (43,2%) и двукрылых (7,7%).
В сосновом молодняке чаще всего в пище ящериц встречаются кроме жуков и чешуекрылых прямокрылые (32,5%)•
203
Суходольный луг отличается тем, что по частоте встречаемости на третьем месте после жуков и чешуекрылых стоят пауки (17,5%), Количество прямокрылых (9,8%) и двукрылых (12,4%) здесь меньше, чем в других биотопах, зато частота встречаемости клопов наибольшая (4,7%).
В биотопе «разнотравный луг» по сравнению с другими биотопами встречается наибольшее количество жуков (68,3%), чешуекрылых (39,2%), двукрылых (26,5%) и пауков (22,1%), а
Таблица 28
Состав пищи прыткой ящерицы (в %) в биотопах Шацкого района Рязанской области (Жаркова, 1973а)
Стация и число желудков (л)
Суходольный луг с редким злаковым покровом и кучами известковых камней (НО) Разнотравный луг в пойме реки с кустарником вокруг стариц (40)
Лиственный лес (опушка) с нормальным травостоем (55) Смешанный лес с преобладанием лиственных пород и редким травостоем (115) Сосновые молодые посадки в пойме реки (87)
62,3
68,3
50,0
43,2
59,9
38,4
39,2
29,5
27,7
37,8
12,4
26,5
15,5
7,7
18,0
9,8 17,5
8,3 22,1
8,5 16,5
32,5 15,0
32,5 18,0
7,6
6,3
2,0
10,6
10,6
6,8 4,7
3,8 1,9
12,7 1,5
7,0 1,9
2,6 1,6
1,2
0,6
0,0
0,8
0,8
0,0
1,2
0,0
0,8
0,0
прямокрылые (8,3%), стрекозы (3,8%) и перепончатокрылые (6,3%) встречаются здесь реже. В пище ящериц этого биотопа имеются моллюски (1,2%), которые встречаются также в смешанном лесу (10,8%).
По наблюдениям в Саратовской области, состав пищи прытких ящериц также зависит от занимаемого ящерицами биотопа. У ящериц из лесных стаций чаще встречались листогрызы, а в степных — долгоносики. Близ посевов пшеницы в пище ящериц найден хлебный жук кузька, а по обочинам лесных дорог попадались крупные цикады.
В южной части ареала прыткой ящерицы, например, в Дагестане, близ г. Буйнакска и с. Сергокала, видовой состав пищевых объектов ящериц в лесу более разнообразен, чем на кустарниково-степных участках (табл. 29).
204
Таблица 29
Состав пищи прытких ящериц (в %) в различных биотопах Дагестанской АССР (Кутузова, 1971)
Степь
Лес
I 74,41 13,9 I 30,2
I63,9I 8,3 I 25,0
с. Сергокаса
27,0 23,2 I 34,8
27,8 30,6 | 41,7
8,6 I 9,3
3,3 | -
7,0 I 30,2
2,3 I 38,9
Степь Лес
82,6 21,7 26,1
95,1 24,4 14,6
г. Буйнакск
43,5 I 8,7 26,1
19,5 26,8 43,9
4,3 26,1 | — 8,7 21,7
2,4 17,1 I - 9,8 19,5
В лесах г. Буйнакска по встречаемости первое место принадлежит жукам (95,1%), второе — перепончатокрылым (43,9%), далее паукам (26,8%) и чешуекрылым (24,4%), тогда как в кустарниково-степных участках чаще встречались прямокрылые (43,5%), жуки (99,1%), клопы и перепончатокрылые (26,1%).
Имеется определенная зависимость состава пищи ящериц от биотопа и в Чечено-Ингушской АССР. Как видно из табл. 30, пища ящериц, живущих на опушке пойменного леса, более разнообразна, чем у ящериц в степном биотопе.
Таблица 30
Состав пищи прытких ящериц (в %) в разных биотопах окрестностей г. Грозного (Рашкевич^ 1970)
Биотоп Жуки Чешуекрылые Прямокрылые Пауки Пере пон-чатокры-лые Моллюски Мокрицы Дождевые черви Ягоды шелковицы
Опушка пойменного леса Участок залежи в степи 36,4 45,1 4,7 18,3 5,2 14,1 7,8 14,1 11,7 8,4 2,6 6,5 1,3 3,9
В Ставропольском крае кроме общих компонентов питания Для отдельных ящериц существуют свои объекты добычи, определяемые спецификой биотопа. Так, для рациона ящериц, обитающих в типчаково-полынной степи — это чешуекрылые, прямокрылые и пауки; в разнотравно-злаковой степи — чешуекрылые и прямокрылые; в полынной степи — прямокрылые; в лесополосах прямокрылые, чешуекрылые, пауки; а на посевах — моллюски.
205
Для выявления сходных черт в составе пищи ящериц различных биотопов Днепропетровской области был определен коэффициент пищевого сходства. Сходство в питании прытких ящериц во всех биотопах равно нулю. Характеристика этого сходства в питании- прытких ящериц различных биоценозов степной зоны УССР представлена в табл. 31. Как видно, наибольшее качественное сходство отмечается в пристенных лесах и су-дубравах (23,5 % )> а также в пойменных лесах и сосновом бору (17,6%), наименьшее сходство — в акациевых насаждениях— судубравах (3,8%). Полностью отсутствует сходство в питании
Таблица 31
Характеристика сходства питания прыткой ящерицы (качество/количество в %) в различных биогеоценозах степной зоны УССР (Булахов, Константинова,? 1972)
Биогеоценоз Целинные участки Байраки Акациевые насаждения Пристенные леса Пойменные леса Судуб-рава Сосновый бор
Целинные участки — 9,1 6,7 7,6 0 4,3 0
Байраки 19,1 —• 7,1 10,9 11,5 12,7 11,5
Акациевые насаждения 17,3 5,7 —- 8,62 5,4 3,8 5,0
Пристенные леса 10,4 14,8 9,0 —. 10,3 23,4 10,3
Пойменные леса 0 15,1 3,4 12,6 .— 10,4 17,7
Судубрава 3,2 18,0 3,2 31,9 14,8 — 12,8
Сосновый бор 0 11,3 21,4 12,6 24,8 19,9 —
на целинных участках и пойменных лесах; целинных участках и сосновом лесу.
В остальных попарных сравнениях сходство питания равно 6,7—12,8%. Наибольшее количественное сходство наблюдается также в тех же самых биогеоценозах (соответственно 31,9 и 24,8%). Высокий коэффициент сходства питания в этих биогеоценозах складывается за счет кивсяков, пауков-волков, божьих коровок, жужелиц, долгоносиков, пилильщиков, зерновок, совок, пядениц, наездников, мух, песчаных муравьев, саранчи. Кроме того, большое сходство по количественному составу пищи отмечается в сосновых борах судубравах (19,9%), особенно за счет пауков-волков (5,3%), совок (6,4%), зерновок (2,1%) и др., затем в байраках и целинных участках (19,1%) за счет жужелиц и цикадки; в судубравах и байраках (18%) за счет совок, кивсяков, цикад, пауков, муравьев. Незначительное количественное сходство, как и качественное, в пойменных лесах — акациевых насаждениях, судубравах — целинных участках (3,4—3,2%). В остальных биогеоценозах количественное сходство равно 5,7—17,3%.
206
СЕЗОННЫЕ ИЗМЕНЕНИЯ СОСТАВА КОРМОВ
В связи с тем, что соотношение численности беспозвоночных Б биоценозах изменяется по сезонам, пищевой рацион ящериц различен в разное время года. При сравнении сезонных изменений состава кормов в центральной полосе Европейской части СССР (Рязанская, Саратовская области) и в южной части ареала (Ставропольский край, Чечено-Ингушская и Дагестанская АССР) оказалось, что весной пищевой рацион прыткой ящерицы в Ставропольском крае содержит в основном жуков (38,44%),
Таблица 32
Частота встречаемости кормовых объектов (в %) по месяцам в Рязанской обл.
(Жаркова ? 1973а)
Месяц и число желудков (п) Жуки i £ «я 9 S гг й О) 3 Ц& Перепончатокрылые S С Прямокрылые Стрекозы Полужесткокрылые Земляные черви МОЛЛЮСК! Ящерицы
Май (123) 72,0 36,0 12,5 10,5 18,2 5,5 8,5 5,0 0,5 0,0 0,0
Июнь (223) 69,2 39,6 16,5 10,0 18,4 15,7 5,2 0,0 0,7 1,2 0,0
Июль (108) 58,8 40,1 17,7 10,4 20,8 39,0 4,6 3,3 2,2 1,1 4,1
Август (105) 49,0 47,4 22,6 10,1 18,6 44,0 3,3 3,3 0,0 1,1 .0,0
чешуекрылых (25,46%) и перепончатокрылых (13,89%). В меньшей степени встречаются двукрылые (6,01%), пауки (5,56%), равноногие ракообразные (5,1%) и прямокрылые (0,46%). В весеннем рационе ящериц Ивановской области Б. А. Красавцев (1936, 1939) наряду с отмеченными выше животными отмечает большое количество прямокрылых (30,34%). Кроме того, В. К. Жаркова (1969, 1970, 1973а) отмечает в Рязанской области и И. М. Утробина (1956) в Татарской АССР большое количество пауков (18,4%), а И. В. Щепотьев (1952) пауков и муравьев. Только весной в Крыму (Щербак, 1965) встречаются крылатые формы муравьев и личинки двукрылых.
В летнем рационе ящериц основное значение имеют жуки. Соотношение других видов пищи, перепончатокрылых, чешуекрылых, двукрылых, пауков и прямокрылых изменяется в зависимости от биоценоза и, по-видимому, испытывает влияние изменений в составе энтомофауны по годам.
Так, в Ставропольском крае и Татарской АССР первое место по частоте встречаемости принадлежит перепончатокрылым (15,56%), затем чешуекрылым (4,33%) и двукрылым (4,29%); пауки составляли лишь 1,65%. Аналогичны данные для Саратовской области и Чечено-Ингушской АССР, где в пище ящериц второе место после жуков также занимали чешуекрылые (соответственно 16,6 и 60,0%), а в конце лета они даже превышают жуков (жуков —16,6 и 50%; чешуекрылых — 22,4 и °4,0%), затем идут пауки (15,5 и 27,2%).
207
В Рязанской области (табл. 32) третье место в пище лщериц по частоте встречаемости занимают не пауки, а прямокрылые (39,0%), а основу пищи также составляют жуки (58,8%) и чешуекрылые (40,1 %).
В Дагестанской АССР в июне-июле преобладают жуки (71,9 и 84,4%), затем перепончатокрылые (34,4 и 40,6%), на третьем месте в июне были пауки (25,0%), а в июле прямокрылые (34,4%) и чешуекрылые (21,9% и 25,0%). цам.
Что касается Б. А. Красавцева (1936, 1939), то он в летнем рационе прытких ящериц отводит главную роль гусеницам.
Основу осеннего рациона в Ставропольском крае о оставляют перепончатокрылые (38,7%), чешуекрылые (12,0%), жуки (9,0%), двукрылые (8,9%) и равноногие ракообразные (8,9%)- Пауки составляют лишь 6% встреч. Характеризуя осенний рацион, Ю. И. Пащенко указал, кроме того, на увеличение на Украине в пище ящериц количества пауков и чешуекрылых. Видовой состав поедаемых ящерицами животных также изменяется по месяцам. Практически нет почти ни одного вида, встречающегося в пище ящериц на протяжении всего сезона активности.
Таким образом, в изменении состава кормов по сезонам характерным является уменьшение частоты поедания жуков от мая к августу и, наоборот, увеличение встречаемости в пище ящериц прямокрылых, двукрылых, чешуекрылых и пауков, причем в зависимости от местных условий соотношения этих отрядов в пище ящериц могут меняться. Общим правилом является то, что при уменьшении частоты встречаемости жуков в желудках пища ящериц становится более разнообразной. Особенности изменений зависят от местных погодных условий.
ИЗМЕНЕНИЕ СОСТАВА ПИЩИ ПО ГОДАМ
Многолетние наблюдения показали, что рацион ящерицы не остается неизменным на протяжении ряда лет. Климатические условия года оказывают влияние на состав энтомофауны и размещение ящериц на занимаемой ими территории. Поскольку условия года непостоянны, состав корма также изменяется по годам.
Например, засушливый 1946 год в Саратовской области в сравнении с нормально влажными 1940 и 1941 гг. внес существенные изменения в количественный состав поедаемой ящерицами энтомофауны. В годы с нормальными погодными условиями прямокрылые (саранчевые) имеют большой вес в питании ящериц, а перепончатокрылые (муравьи) взрослыми особями почти не поедаются. Сосредоточение ящериц в степных балках с болотцами в период засухи (июнь) ограничило выбор пищи и повлекло резкое увеличение количества муравьев в желудках
208
ящериц падение числа саранчевых и появление водных моллюсков.
Встречаемость отдельных групп беспозвоночных колеблется по годам. Каждый год имеет наибольшую встречаемость каких-либо определенных групп, которые в последующие годы не достигают максимума. Так, в 1965 г. в Рязанской области наиболее часто в пище ящериц встречались пауки (23,8%) и чешуекрылые (43,3%), в 1966 г.— стрекозы (14,69), в 1967 г.— прямокрылые (27,1 %) и в 1969 г. перепончатокрылые (14,7%). В Дагестане (окрестности г. Махачкалы) в 1968 г. ящерицы чаще поедали пауков, а в 1970 г.— мокриц, дождевых червей, перепонча-токры,- ix и чешуекрылых.
Рис. 64
Сезонные изменения интенсивности питания прытких ящериц (в г) в Ставропольском крае 1 — взрослые самцы;
2 — взрослые самки;
3 — молодые самцы и самки (Лукина, 19666)
Таким образом, изменение в составе корма прыткой ящерицы зависит по годам от наличия и обилия данного корма в природе.
Наблюдения показали, что интенсивность питания ящериц зависит от погодных условий. В дождливую погоду при низкой или при очень высокой температуре среды ящерицы только на короткое время выходят из укрытий. Понятно, что в таких случаях вес их пищи будет минимальным.
Можно предположить, что интенсивность питания в какой-то степени отражает готовность организма к размножению, а в дальнейшем и к спячке. Как видно из рис. 64, интенсивность питания у прытких ящериц в Ставропольском крае увеличивается сразу же после пробуждения в апреле и достигает максимума у самцов во второй половине мая, а у самок — в первой половине июля. Затем интенсивность питания быстро уменьшается и достигает минимума к началу третьей декады июля — времени ухода большинства взрослых и полувзрослых особей в спячку. Вероятно, они успевают к этому времени восстановить затраченную при размножении энергию и накопить необходимые для зимовки запасы питательных веществ.
Молодые особи появляются в промежутке между второй половиной июля и второй половиной августа. В первые дни они поедают мало корма, но уже через 7—11 дней его объем увеличивается и достигает максимума в конце июля —начале августа. По мере накопления энергетических запасов интенсивность питания уменьшается вплоть до времени их ухода в спячку (первая-вторая декады октября).
209
КОЛИЧЕСТВО ПОЕДАЕМОГО КОРМА
Прыткие ящерицы достаточно прожорливы, за время активности они съедают большое количество пищи, хотя, подобно другим рептилиям, способны длительное время голодать. Энергическая сторона питания прытких ящериц практически не изучена.
По данным Н. В. Щепотьева (1952), в Саратовской области взрослая ящерица съедает за день около 1 г пищи. Это количество пищи соответствует 10—12 жукам среднего размера типа щелкунов или 40—60 экз. мелких листогрызов или долгоносиков. Вес гусениц, которых может проглотить ящерица, достигает 3—5 г.
Взвешивание содержимого желудков 37 ящериц из Ставропольского края в различные сезоны показало, что молодые самки и самцы съедают за один день до 0,4 г пищи. Взрослые самки съедают до 3,6, а самцы до 4,7 г.
Некоторые ящерицы могут поглощать большее количество пищи. Так, у одной самки, добытой 12 июня в окрестностях г. Грозного на опушке лесополосы, в желудке было обнаружено: жуков 24 экз. (16 листоедов, 2 божьих коровки, 1 нарывник и 5 экз. не определенных жуков), перепончатокрылых — 4 экз., гусениц— 1 экз., клопов 1 экз. Всего при весе ящерицы 32 г вес пищи составил 12 г. У самца, добытого там же, в желудке обнаружены: дождевой червь длиной 10,5 см (длина тела ящерицы— 10,2 см) и толщиной 0,5 см, уховертки — 2 экз., паук — 1 экз., гусеница — 4 экз., муравьи —4 экз., кобылки — 3 экз., хрущ июньский—1, бабочка-белянка—1 экз., не определенные остатки — 2 г. Всего при весе самца 38 г пища весила 16 г.
Желудок ящерицы наполняется постепенно в течение всего суточного периода активности. Наибольший вес и наименьшее число пустых желудков наблюдается в вечерние часы (рис. 65).
В период спаривания индекс наполнения желудков у взрослых самцов обычно несколько ниже, чем у самок, а у последних он снижается в период кладки яиц. После окончания периода спаривания у самцов, а у самок после откладки яиц, интенсивность питания заметно возрастает. В это время они быстро восполняют все затраты организма в период размножения. Происходит накопление питательных веществ, необходимых для зимовки. К осени интенсивность питания снижается. Немалую роль здесь играет и понижение температуры окружающей среды, снижение активности пищеварительных соков, уменьшение количества пищи и продолжительности суточной активности. У неполовозрелых особей средний индекс наполнения желудков выше, чем у взрослых ящериц, что, по-видимому, связано с относительно быстрым темпом роста молодых животных и, следовательно, большей потребностью в пище.
210
Та же закономерность отмечена 3. П. Хонякиной (1964) для других видов пресмыкающихся.
Б период начала откладки яиц (конец мая) индекс наполнения желудков у взрослых самок не отличается от такового у самцов. Другая картина наблюдается во время массовой откладки яиц (середина июня), когда у самок интенсивность пи-
Рис. 65
Зависимость объема содержимого желудка прыткой ящерицы от времени суток
В каждой часовой пробе средние данные по 10 желудкам;
1 — 1968 г.,
2 — 1969 г.
(Лукина, 19666)
10 12 14 16'
Время сутану
тания несколько понижена. После окончания откладки яиц потребность в корме у самок повышается.
Таким образом, интенсивность обмена у прыткой ящерицы зависит от пола, возраста особи и физиологического состояния организма. Кроме того, она испытывает влияние погоды и изменяется по сезонам.
ПРОДОЛЖИТЕЛЬНОСТЬ ПЕРЕВАРИВАНИЯ ПИЩИ
По данным Н. В. Щепотьева (1952), пища в желудке прыткой ящерицы находится 12—15 час. И. С. Даревский (1967) отмечал, что пища скальных ящериц, состоящая из мучных червей и кузнечиков, полностью переваривается в течение суток. При террариумном и вольерном содержании прытких ящериц было установлено, что на продолжительность переваривания пищи влияет ее состав. Так, при кормлении прыткой ящерицы жуками первая фекация наблюдается через 10—12 час., а жесткокрылые, имеющие твердый хитиновый покров, перевариваются за 18—29 час. При кормлении прямокрылыми и смешанным кормом скорость переваривания равна 6—9 час. Мухи и гусеницы перевариваются за 5—12 час.
Чтобы выяснить быстроту переваривания кормовых объектов, после окончания опыта ящерицы были вскрыты. Первых двух вскрыли спустя час после кормления, следующих двух — спустя два часа и еще двух — через три часа. Через час после кормления состояние содержимого желудков ящериц позволяет легко определить съеденный корм до вида. После двухчасового переваривания сохраняется хитиновый покров, после трехчасового переваривания хитин измельчается в экскрементах, превращаясь в коричневую массу. Элитры жуков даже в экск
211
рементах сохраняют свою форму, изменяя только первоначальный цвет на черный.
По данным С. С. Шварца (1948), переваривающая сила ферментов пойкилотермных животных, в том числе и рептилий, зависит от температуры тела, и наибольший эффект их действия 1 проявляется при оптимальных температурах тела. Резкие колебания температуры действуют отрицательно на переваривающую силу ферментов. Возможно, что в условиях низких ночных ' или дневных температур скорость переваривания съеденной добычи замедляется.
Для выяснений скорости переваривания пищи Н. А. Рашкевич насильно кормил ящериц окрашенным кормом (фуксины) и подсчитывал время, прошедшее от начала кормления до по-’ явления окрашенного кала.
22 июля температура воздуха 22°, пасмурно. Скорость переваривания 22 часа. 25 июля температура воздуха 30°. Скорость передаривания 19 час. 7 августа температура воздуха 18°, дождь. Скорость переваривания 48 час.
Кроме того, существуют и литературные данные, показывающие, что у других пойкилотермных животных оптимальная температура действия пищеварительных ферментов 35—45° (Коржуев, Круглова, 1957; Пятницкий, 1936; Пегель, Попов, 1937; Шварц, 1948). Тем не менее X. С. Коштоянц (1950) считает, что данные, полученные в лабораторном опыте, не могут быть приняты как вполне соответствующие условиям действия на пищу пищеварительных ферментов в организме. Если при выяснении оптимальной температуры действия ферментов учитывать естественные условия их деятельности, то станет ясно, что они будут находиться в пределах от 14 до 36°. В этой связи интересно указать на данные по влиянию температуры на скорость пищеварения у обыкновенного ужа, которые приводит Скасцилас (Skascilas, 1970). Из них следует, что у ужа при 5° пищеварение не отмечено, при 15° проглоченная добыча переваривается очень медленно, а 25° является оптимальной температурой для пищеварения. Дальнейшее повышение температуры замедляет действие пищеварительных ферментов. По-видимому, оптимальные условия действия пищеварительных ферментов уже должны быть близки (если не такие же) к оптимальной температуре тела, что вполне согласуется с данными, которые приводит X. С. Коштоянц (1950) для рептилий.
В связи с изложенным выше можно допустить, что оптимальные условия действия пищеварительных ферментов у прыткой ящерицы будут находиться в области оптимальной температуры тела — 30°.
Мы понимаем, что лучшим заключением данного раздела была бы оценка питания с точки зрения энергетики, но мы не располагаем достаточным количеством данных по этому вопросу. Методика подобных работ применительно к рептилиям отсут
212
ствует. По этой причине литературные данные о энергетике питания ящериц скудны. Анализ пищи ящериц, состоящей из беспозвоночных основных групп, проведенный по нашей просьбе в Лаборатории физиологии Ставропольского научно-исследовательского сельскохозяйственного института, показывает, что дневной рацион ящерицы, вес которого равен 1 г, содержит 3964 кал. (3964+53; п=3).
Часть поглощенного корма выводится в форме экскрементов, но заключенная в них энергия нами не была учтена. Кроме того, при расчете калорийности суточного рациона ящериц мы допускали, что он не содержит воды.
Судя по этим данным, ящерицы способны сохранить жизнь без экзогенного питания в течение длительного времени. Запасы энергии, накопленной животными за сезон активности и сконцентрированной в организме в виде жировых и других веществ, дают возможность ящерицам в условиях зимней спячки существовать за счет эндогенного питания. Большая часть этих веществ после спячки затрачивается на развитие половых продуктов.
ГЛАВА СЕДЬМАЯ
РАЗМНОЖЕНИЕ 1
В настоящей главе мы рассматриваем проблему размножения прыткой ящерицы. Многие вопросы биологии ящериц, связанные с размножением, рассмотрены в главах «Морфология» (гл. III), «Физиология и биохимия» (гл. IV), «Питание» (гл. VI) и особенно в главах «Развитие и рост» (гл. VIII) и «Поведение» (гл. X).
НАСТУПЛЕНИЕ ПОЛОВОЙ ЗРЕЛОСТИ
Наступление половой зрелости можно объективно определить по развитию гонад, состоянию наружных половых органов и по поведению животного (спариванию). Эти методы пригодны только в период размножения. Поэтому более общими, хотя, несомненно, в известной степени основанными на косвенных данных методами являются определение половозрелости по возрасту и размерным признакам (весу, длине) тела и появлению вторичных половых признаков (развитие полового диморфизма по окраске).
У самцов прыткой ящерицы сразу же после выхода из спячки отмечается весенняя активация семенников, что приводит к сильному увеличению последних. Своего максимального размера они достигают в апреле-мае в зависимости от географического района исследования. В этот период семенники принимают цвет яичного желтка (Щепотьев, 1950). Изменение гонад в процессе размножения тесно связано с изменением жировых тел (Лукина, 19666; Тертышников, 1972а, б; Симонян, 1973, и ДР-)-
Жировые тела располагаются у нижнего края брюшной полости ящериц и имеют вид двулопастного образования беловато-желтого или ярко-желтого цвета. Известно, что у некоторых рептилий они состоят на 80—-90% из различных липоидов (Des-sauer, 1955) и в основном из арахидоновой и линолевой кислот
1 Авторы: А. С. Баранов, А. Б. Стрельцов, М. Ф. Тертышников, при участии В. К. Жарковой, А. М. Андрушко, И. М. Окуловой, Г. Добровольской, М. И. Щербаня, К. Шафраньской, В. А. Кутузовой, А. А. Симоняна, Ф. Д. Даниеляна, В. И. Подмарева, 3. П, Хонякиной, В. Е. Боченко.
214
Рис. 66
Сезонные изменения веса жировых тел (1.1), максимальной длины семенников (II. 2 , нахождение зрелых сперматозоидов (3)и максимальные размеры овоцитов (Па) у прытких ящериц Западного Предкавказья
7, II, IIа по Г. П. Лукиной (19666); 270 самцов и 124 самки; 2, 3 — по М. Ф. Тертышни-кову (19726)
(Чеченкин, 1954). В период максимального развития жировые тела имеют большие размеры, а при полном истощении они едва заметны.
Сезонная динамика жировых тел и гонад характеризует зависимость между питанием и размножением ящериц, что хорошо было прослежено Даревским (I960) в Армении. Для прытких ящериц Западного Предкавказья Г. П. Лукиной (1966) установлена взаимная зависимость изменения жировых тел и максимальных размеров гонад (рис. 66). При рассмотрении кривой видно, что у самцов после выхода из спячки длина семенников достигает значительной величины (до 10 мм), в то время как жировые тела расходуются целиком. С середины апреля до конца июня размер гонад постепенно уменьшается (до 4 мм), одновременно происходит быстрое нарастание веса жировых тел. За первую половину июля вес жировых тел увеличивается с 0 до 720 мг. К концу лета семенники снова несколько увеличиваются, что объясняется подготовкой их новому половому циклу. Применительно к данному виду ящериц летнее увеличение семенников было отмечено еще Н. Н. Щербаком (1962). В своей работе Н. Н. Щербак предполагает, что этот процесс связан, возможно, с понижением температуры и сокращением светового дня.
Аналогичные данные о динамике развития гонад самцов прыткой ящерицы приводятся и в других работах (Щепотьев, 1948; Краснова, 1961; Щербак, 1966 и др.).
Интересный факт приводит М. Ф. Тертышников (19726) для прытких ящериц Ставрополья. Несмотря на то, что происхо-
215
дит прекращение сперматогенеза в летний период, зрелые спермин сохраняются на протяжении мая, июня и начала июля (см. рис. 66). По мнению И. С. Даревского (1967), подобная длительная половая активность самцов приводит к тому, что почти все половозрелые самки в популяции оказываются оплодотворенными; при соответствующих условиях в южных районах ареала возможна вторая кладка. При этом, по-видимому, не
Рис. 67
Размеры ооцитов прытких ящериц в мае
Западное Предкавказье, 162 экз-(Лукина, 19666) ?
меньшее значение имеет установленный Д. И. Гениным (1955) факт длительного сохранения зрелых спермиев в половых путях самок.
Весной у пробудившихся самок в яичниках уже имеются несколько желтых и крупных (3—4 мм), а также матовых и мелких (1—2 мм) овоцитов.
Процесс развития яиц в яичниках половозрелых самок начинается с конца апреля. При этом у каждой самки развиваются 15—20 ооцитов, которые постепенно приобретают желтую окраску. Остальные, ооциты отстают в росте и сохраняют белую окраску. Перед переходом в яйцеводы ооциты в яичнике представлены хорошо отличимыми по размерам четырьмя генерациями (рис. 67). Количество ооцитов в третьей генерации всегда мало (не более 4), а в четвертой оно, как правило, равно возможному количеству откладываемых яиц для данного вида.
Таким образом, четвертая генерация наиболее крупных ооцитов (8,5 мм) или ее какая-то часть развиваются дальше в яйцеводах. Ооциты третьей генерации редуцируются и в следующем сезоне, по-видимому в яйцеводах развивается вторая из отмеченных генераций.
Достигнув 8 мм в диаметре, яйца выпадают в полость тела, откуда переходят в яйцеводы (рис. 68). В это время и происходит оплодотворение. Развитие мелких ооцитов задерживается. В мае-июне готовые для откладки яйца, вес которых достигает 0,6 г, встречаются в яйцеводах половозрелых самок. У самок, окончивших размножение, перед уходом на зимовку в каждом яичнике уже имеется группа желтоватых ооцитов размерами до 3 мм.
216
Рис. 68
Расположение зрелых яиц в яйцеводах прыткой ящерицы
Фото В. И. Подмарева
У самок, так же как и у самцов, накопление и расходование жировых тел находится в тесной связи с динамикой развития половых продуктов (см. рис. 66).
Весной у самок при быстром созревании половых продуктов происходит истощение жировых тел. Однако их энергетических
217
запасов видимо хватает на развитие яиц. По окончании размножения самки усиленно питаются и вес жировых тел быстро увеличивается, достигая максимума в конце июля. Часть вновь накапливаемых энергетических запасов расходуется на подготовку к очередному половому циклу.
Как видно (см. рис. 66), пробудившиеся от зимней спячки самки (апрель) содержат в яичниках 10—12 крупных ооцитов диаметром 3—4 мм и большое количество мелких, не превышающих 0,5—1,5 мм ооцитов. Размеры крупных ооцитов быстро увеличиваются, в связи с чем происходит столь же быстрое истощение жировых тел, как и у самцов. К моменту откладки яиц жировые тела оказываются полностью израсходованными, а затем (к концу июля) их вес снова начинает нарастать.
Помимо изменения жировых тел происходят и связанные с половым циклом изменения в работе гипофиза, гипоталамуса и щитовидной железы (повышенная активность и нейросекреторная деятельность в процессе размножения) (Фел1кс, 1970).
Таким образом, приведенные данные показывают, что развитие половых продуктов у прыткой ящерицы происходит не только за счет вещества жировых тел, но и изменения секреторной деятельности щитовидной железы и гормональной работы гипоталамуса и гипофиза.
Возраст
Точные данные о возрасте наступления половой зрелости у самцов и самок прыткой ящерицы в литературе отсутствуют.
Некоторые авторы (Щепотьев, 1948; Андрушко, 1954; Хоия-кина, 1964, и др.) предполагают, что половозрелость у прыткой ящерицы наступает в возрасте около двух лет. М. Ф. Тертышников (19726) определяет наступление половой зрелости для ящериц Ставрополья в 23 месяца. Существует также предположение (Чернов, 1954), что половозрелыми прыткие ящерицы становятся в конце третьего года жизни. Г. П. Лукина (19666) определяет наступление половой зрелости у самцов и самок в начале третьего года жизни.
Размеры животных
При достижении половой зрелости ящерицы (самцы и самки) имеют вполне определенные размеры (вес, длина). Если о длине тела этой группы можно говорить вполне определенно, то о весовой характеристике половозрелых особей (особенно самок), впервые вступающих в размножение, сказать затруднительно (см. гл. III). Это вызвано тем, что развивающиеся к моменту спаривания яйца искажают действительную весовую характеристику этой группы.
Длина тела. К сожалению литературных данных такого рода очень мало и в некоторых случаях довольно трудно определить.
218
какого пола они касаются. Материалы табл. 33 свидетельствуют о том, что размеры самцов в лесостепной зоне несколько больше, чем самок (Жаркова, 1973а). Сведения по степной зоне скудны, а имеющиеся данные не позволяют говорить о превосходстве в размерах одной половой группы над другой. По данным Г. П. Лукиной (19666), самцы значительно меньше самок, достигших половой зрелости. Самки степной зоны тоже отлича-
Таблица 83
Длина тела (в мм) прытких ящериц при наступлении половой зрелости (Щепотьев? 1950; Лукина, 19666; Кутузова^ 1971, Жаркова, 1973а; Тертышников, 19726)
Природная зона Самцы Самки
Lim М±т п Lim X±sx п
Лесостепная 61—80 70,0—0,9 190 55-76 66,l±o,13 169
Степная 60—75 — 298 76—90 81,0+0,08 23
— — —- 60—80 71,0±0,06 65
— —. — 78-92 85,0±0,0i 60
Горные районы — — — 70—85 — 15
ются по своим размерам в разных районах. Они несколько мельче в волжских степях (71,0 + 0,06, Щепотьев, 1950), чем в более южных районах (Ставрополье и Ростовская обл.), где их длина составляет в среднем 81—85 мм (Лукина, 19666; Тертышников, 19726). В горных районах (Кутузова, 1971) самки чуть мельче своих сородичей из степной зоны (см. табл. 33). Больше сведений и конкретных данных по другим выделяемым природным зонам (см. гл. V) в литературных источниках нами не обнаружено.
В целом можно заключить, что ящерицы, вступающие в размножение, по мере продвижения от северных границ ареала к южным отличаются по своей длине, и южнее они становятся гораздо крупнее. Видимо исключение составляют горные популяции, которые дают пеструю картину по аналогии с изменением веса в этих районах (см. гл. III).
Вес животных. В литературе не существует данных, которые бы позволили определить весовую категорию, при которой ящерицы достигают половой зрелости. Об этом можно судить лишь по косвенным данным. В главе III были рассмотрены весовая и возрастная структуры популяции, позволившие выделить у самцов весовые классы. Особи, достигшие половой зрелости (возраст был определен по слоистой структуре кости) и впервые вступившие в размножение, распределились в диапазоне от 7 до 11 г. Но и эти данные, по-видимому, не точны, так как здесь играет роль участие в размножении, требующее определенной затраты общей энергии (Щепотьев, 1950). На весовые данные влияют также и климатические условия данного года и связанные с этим условия кормности.
219
ОСОБЕННОСТИ ПОЛОВОГО ЦИКЛА
После рассмотрения процессов, связанных с наступлением половой зрелости у самцов и самок прыткой ящерицы, можно перейти к рассмотрению особенностей ежегодного полового цикла, включающего спаривание, формирование яиц в теле самки, кладку яиц.
Сроки спаривания (начало и конец)
Спаривание прытких ящериц начинается вскоре после пробуждения. Наиболее ранние сроки спаривания отмечаются для Предкавказья, Ставропольской возвышенности, Крыма, междуречья Волги и Урала, равнинных районов Закарпатья, южных районов Украины и приходятся на вторую (в основном) половину апреля; последние особи, которые спаривались, в природе были отмечены для этих районов в первой половине июня (Красавцев, 1939; Сухов, 1948; Косарева, 1950; Щербак, 1966; Дарев-ский, 1946; Лукина, 19666; Тертышников, 19726, и др.). В Казахстане, Нижнем Поволжье, предгорьях Закарпатья, среднем Поволжье и многих других областях Европейской части СССР прыткие ящерицы вступают в спаривание обычно в первой половине мая и заканчивают его в первой половине июня (Положен-цев, 1941; Даревский, 1946; Федосеев, Никитин, 1951; Кащенко, 1909; Пузанов, 1955; Параскив, 1956; Шапошников, 1959; Жаркова, 1973а, и др.). Наряду с совпадающими сведениями о начале спаривания по этим географическим районам встречаются и несколько противоречивые данные. Так, Е. С. Птушенко (1946, 1947) отмечает начало спаривания в Окском заповеднике во второй половине апреля, в то время как В. К- Жаркова (1973а) для этого же заповедника отмечает начало спаривания в начале мая. Скорее всего, разные даты начала спаривания для одной и той же местности объясняются тем, что исследования проводились на разных популяциях, микроклиматические особенности обитания которых несколько различны.
В низменных зонах Дагестана, северо-западе Армении и на Урале отмечаются самые поздние сроки начала спаривания — вторая половина мая (Шварц, 1951, Кутузова, 1971; Симонян, 1973). Соответственно для этих же районов отмечаются и наиболее поздние сроки окончания спаривания: Урал, Армения — вторая половина июня, Дагестан — начало июля.
На основании изложенного выше можно заключить, что на юге ареала спаривание начинается раньше, чем на севере. Исключение составляют горные районы, в которых начало спаривания приходится на вторую половину июня. Интересен тот факт, что несмотря на то, что сроки начала спаривания на юге и севере ареала различны, конец периода спаривания приходится на июнь, с незначительными колебаниями в пределах этого месяца.
220
Общая продолжительность спаривания
Общая продолжительность спаривания может быть определена по длительности периода между началом спаривания и его окончанием. Оказывается, что в южных районах этот период растянут со второй половины апреля до первой половины июня, т. е. на 2 месяца (Лукина, 19666; Щербак, 1966; Тертышников, 19726; и др.). В северных же районах ареала протяженность этого периода 1—1,5 месяца (с первой половины мая по первую половину июня), т. е. спаривание происходит в более сжатые сроки (Граве, 1951; Федосеев, Никитин, 1951; Пузанов, 1955; Жаркова, 1973а; и др.).
Популяции ящериц (половозрелые особи), обитающие в горных районах, различаются между собой различной продолжительностью спаривания. Так, в горных районах Дагестана этот период длится со второй половины мая до начала июля, т. е. 1,5 месяца (Хонякина, 1964; Кутузова, 1971). Для горных районов Армении и Урала этот период несколько меньше — месяц с небольшим (Шварц, 1951; Симонян, 1973).
Таким образом, период спаривания в северных районах обитания прыткой ящерицы характеризуется более сжатыми сроками, чем в южных. Такое же положение характерно и для других видов животных, имеющих большой ареал. Это вызвано тем, что благоприятный климатический период, обеспечивающий оптимальные условия для развития яиц ящериц, в северных районах гораздо меньше, чем в южных областях ареала.
Вторичное спаривание
Вторичное спаривание отмечается для популяций ящериц некоторых южных районов (Хонякина, 1964; Щербак, 1966;-Шеб-зухова, 1969; Симонян, 1973; Кутузова, 1971). Прямых наблюдений в природе второго спаривания нет. Так, о наличии такого спаривания можно судить по данным Н. Н. Щербака (1966) для Крыма. Прыткая ящерица заканчивает спаривание в этих местах в июне, а в середине июля уже появляются молодые ящерицы. В то же время при обследовании этих же районов в августе встречались самки с хорошо развитыми яйцами и обнаружены кладки яиц, количество которых в ней было значительно меньше, чем обычно для этих мест (почти в два раза). Принимая во внимание, что беременность ящериц длится один месяц, нетрудно подсчитать, что повторное спаривание происходит в первой половине июля, когда окончена первая кладка. Аналогичным путем было сделано предположение о наличии вторичного спаривания и для других районов.
В литературе существует и противоположное мнение по поводу вторичного спаривания. Некоторые авторы считают, что оно Пе происходит. Наличие самок со столь поздними сроками
221
откладки яиц вызвано тем, что они слишком поздно произвели спаривание (Лукина, 1966, Тертышников, 19726, и др.).
Таким образом, вопрос о наличии вторичного спаривания и откладки яиц остается открытым.
Продолжительность беременности
Большинство авторов (Щепотьев, 1950; Лукина, 1966; Щербак, 1966; Тертышников, 19726, и мн. др.) считает, что беременность прытких ящериц длится 1 месяц. Но непосредственных наблюдений в природе практически нет. В основном вывод о продолжительности беременности делается по времени между началом спаривания и временем откладки яиц. О продолжительности беременности можно судить по наблюдениям за ящерицами в террариуме.
Особенности кладки яиц
В этом разделе рассматриваются данные, относящиеся к срокам и числу яиц в кладках и факторах, влияющих на это число, вопрос о наличии порционных (вторичных) кладок и, наконец, характеристика биотопов, в которых обычно происходит откладка яиц.
Сроки откладки яиц и наличие второй кладки
Сроки откладки яиц на протяжении ареала варьируют так же, как и сроки спаривания и, безусловно, зависят от последних. Наиболее ранние кладки встречаются в мае (Крым, Ставропольская возвышенность, Предкавказье, Казахстан, окрестности г. Ворошиловограда и Волгограда, низменные зоны Дагестана, Смоленская и Рязанская области). В остальных частях ареала откладка яиц начинается в июне и даже в июле (на Урале, в Ленинградской обл.). Время откладки яиц может быть растянуто на 1—-2 месяца. Ряд авторов объясняет это наличием второй кладки.
Данные о количестве кладок у прыткой ящерицы весьма противоречивы. Так, Н. В. Щепотьев (1948) в условиях Нижнего Поволжья, М. Ф. Тертышников (19726) для Ставропольского края считают, что прыткие ящерицы в течение сезона откладывают яйца только один раз, а на Северном Кавказе, по данным Э. А. Шебзуховой (1969), прыткие ящерицы за сезон откладывают яйца 2 раза. В Армении у прыткой ящерицы обычно наблюдается одна кладка, но у единичных особей можно обнаружить иногда и вторую кладку. При этом нам неоднократно попадались самки L. apilis, у которых в яичниках имелись крупные овоциты второй генерации размером 7—8 мм. Хорошо развитые в этих случаях желтые тела свидетельствовали о том, что первая кладка накануне уже произошла. Мы располагали также тремя самками L. agilis из окрестностей г. Раздана в Армении,
222
которые отложили яйца в лабораторных условиях, тем не менее, спустя месяц, при вскрытии этих самок в их яичниках были обнаружены крупные желтые овоциты, уже готовые к выходу в яйцевод (Симонян, 1973). Вторая кладка на протяжении сезона встречается у ящериц Крыма (Щербак, 1966), низменного Дагестана (у молодых здесь бывает, по-видимому, только одна кладка (Кутузова, 1972), Закарпатья (Щербак, 1966), Рязанской области (Жаркова, 1973а), Нижнего Поволжья (Щепотьев, 1948). Наблюдалась повторная кладка и в террариуме (Щепотьев, 1950; Стрельцов, сообщ.). Во всех других исследованных частях ареала обычно наблюдается только одна кладка (Тертышников, 19726; Лукина, 1966; Мусхелишвили, 1970; Тертышников, 19726; Либерман, Покровская, 1973; Симонян, 1973, и др.).
Число яиц в кладке
Число яиц в кладках изменяется от 1 (Щепотьев, 1948) до 16—18 (Мусхелишвили, 1970; Стрельцов, сообщ.). Наиболее
Таблица 34 Число яиц в кладке и их характеристика в разных географических зонах
Зона Число яиц в кладке Длина яйца, мм Ширина яйца, мм Средний вес яйца, г Автор
Лесная 6-9 — — 0,6—0,7 Стрельцов, уст. сообщ.
3—15 — — — Шапошников, 1959
6-8 — — — Андрушко, уст. сообщ. .
8—10 — — — Олигер и др., 1966
Лесостепная 6—10 12,0—17,0 7,0—9,0 — Андрушко, уст. сообщ.
2—11 12,0—17,0 —. Щепотьев, 1948
4-12 7,0—16,3 4,4—10,7 —. Жаркова, 1973
11—12 — — — Красавцев, 1936
Степная 3—15 11,5—16,8 7,0—10,0 — Щербак, 1966
2—12 17,2—18,8 7,0—8,0 — Лукина, 19666
1—12 14,0 8,0 0,55—0,62 Тертышников, 19726
4—12 14,0-20,0 7,0—10,0 — Андрушко, уст. сообщ.
Полупустынная 6—11 6—10 14,0—17,0 13,0—16,0 7,0—9,0 — Захидов, 1Э71
— 16,0—17,0 — — Окулова, уст. сообщ.
Горные районы 6-8 12,9—15,2 6,8—8,5 — Симонян, 1973 Даниелян, уст. сообщ.
9—11 11,5 5,5 0,42 Мусхелишвили, 1970
4-9 13,0—16,0 —- Параскив, 1956
3—15 13—18 7,5-9,5 0,65 Хон якина, уст. сообщ. Кутузова, 1971
223
Таблица 35
Сводные данные по числу яиц в кладках прыткой ящерицы разных размерно-возрастных групп
Показатель Число яиц в кладке Место кладки яиц
Lim X±sx
Размер самок, мм
63—76 4-9 6,3 Рязанская сбл. (Жаркова, 1973а)
78—92 6-9 7,9
93-102 8—12 9,0
60—75 2—7 4,0±0,3 Краснодарский край (Лукина»
76—90 9—12 10,0±0,1 1966), данные по 54 кладкам
91—102 6—12 9,0+0,6
Возраст самок, годы
2 1—9 . Ставропольская возвышенность
3 и более 3-12 — (Тертышников, 19726)
2 1—9 5,6+0,4 Армения (Даниелян, уст. сообщ.).
3 и более 3—12 8,1+0,6 данные по 279 кладкам
Взрослые 7—15* Обычно 7—10 Казахский мелкосопочвик (Ан-
Полувзрослые 5—10 Обычно 5—8 друшко,_уст. сообщ.)
Взрослые 10—20* Обычно 12—15 Сев. Кавказ (Андрушко, уст.
Полувзрослые 7—12 Обычно 8—10 сообщ.)
В среднем 1—20 6,5+0,80
* Число фолликулов в одном яичнике.
Таблица 36
Среднее число яиц в кладках прытких ящериц разных размерных групп в трех популяциях Дагестана (Кутузова, 1971)
Размерные группы самок, мм Популяция
Махачкала-1 Аргокала Буйнакск
60—70 — — 6,0
71—80 10,7 7,0 6,8
81—90 14,1 7,3 8,7
91—100 13,4 10,7 —
часто встречаются кладки с 6—11 яйцами. С. А. Чернов, П. В. Терентьев (1949) предполагают, что число яиц в кладках зависит от климата: к северу и востоку их число в кладке становится меньше. По сводным материалам такой четко выраженной закономерности не наблюдается (табл. 34).
Число яиц в кладке в основном, видимо, зависит от индивидуальных размеров и возраста самки. Такая закономерность для
224
прыткой ящерицы в Крыму была обнаружена Н. Н. Щербаком (1966) и другими исследователями (Красавцев, 1936, 1939; Либерман, Покровская, 1943; Пащенко, 1954); об этом же свидетельствуют и новейшие данные (табл. 35—36).
Наконец, есть данные, указывающие на существование определенных различий в числе откладываемых яиц в пределах одной и той же популяции в зависимости от стации (табл. 37).
Таблица 37
Среднее число яиц в кладках прытких ящериц в разных стациях Дагестана (Кутузова, 1971)
Популяция Среднее число яиц в кладке Лесная стация Кустарниковостепная стация
Сергокала 7,03 9,0 6,80
Буйнакск 7,60 8,30 6,50
В среднем 7,31 8,65 6,65
Размеры второй кладки обычно меньше, чем первой (Щербак, 1966; Хонякина, 1964; Кутузова, 1971, и др.). Так, например, в 1970 г. в окрестностях г. Махачкалы в первой кладке в среднем было 7,3 яйца, а во второй только 5,8.
Место и длительность кладки
Развитие яиц прытких ящериц требует определенного комплекса условий, ведущими среди них являются температура и влажность. Поэтому в природе яйца откладываются не только в определенные сроки, но и в определенных местах. Обычно ящерицы откладывают яйца в специально вырытую ямку, на солнечных, хорошо прогреваемых местах. Встречаются кладки в норах, в песке, под опавшими листьями, на южных и восточных склонах (Кутузова, 1971; Стрельцов, сообщ.; Тертышников, 19726; Щербань, сообщ.). Кладка располагается обычно неглубоко, в нескольких сантиметрах от поверхности, что, видимо, обеспечивает оптимальный температурный режим. При откладке яиц в террариуме яйца также располагаются на глубине 2—3 см от поверхности, часто под различного рода укрытиями.
Существует мнение, что в лабораторных условиях в сухой песок яйца ящерицами не откладываются (Либерман, Покровская, 1943). Но по наблюдениям Н. В. Щепотьева (1950) и В. И. Подмарева, хотя стремление откладывать яйца во влажную среду безусловно существует, в террариуме возможна откладка яиц в воздушно-сухой субстрат (яйца конечно гибнут). Оптимальной влажностью субстрата считается 16,2% (Либерман, Покровская, 1943) (более подробно см. гл. VIII).
8 Прыткая ящерица 225
Процесс откладки яиц длится от 1—2 до 4—7 час. (Либерман, Покровская, 1943). Аналогичные результаты получены А. Б. Стрельцовым. По другим сведениям (Хонякина, 1964; Кутузова, 1971) процесс кладки яиц растянут до 5 суток. Но в связи с тем, что в природе яйца в кладке обычно расположены в одном месте (а не разбросаны мелкими порциями), видимо более правильным надо считать мнение о более коротких сроках откладки яиц порядка не более нескольких часов. К сожалению, прямых наблюдений в природе пока нет и все указанные выше наблюдения проводились в террариуме.
Длительность инкубации и выход молодых особей
О сроках инкубации яиц в природе имеются только косвенные данные, вычисленные по срокам откладки яиц и появления первых новорожденных ящериц. Большинство исследователей придерживается мнения, что длительность инкубации в среднем около 2 месяцев (Щербак, 1966; Тертышников, 19726: •Жаркова, 1973а). Однако П. А. Положенцев (1941) утверждает, что молодые выходят спустя уже 3—4 недели после откладки яиц. Очевидно, что утверждение следует считать неправильным, так как в террариумах инкубация яиц занимает от 43—45 (Стрельцов, сообщ.) до 50 (Симонян, 1973; Подмарев, сообщ.) дней (см. также гл. VIII).
Таблица 38
Сроки выхода молодых прытких ящериц из яиц в разных природных зонах (по сводным данным)
Зона Начало Конец
Лесная Лесостепная Степная Полупустынная Горные районы Конец июля Вторая половина июля Середина июля Вторая половина июля Июль Конец августа Начало августа * Начало октября Сентябрь Сентябрь
Сроки выхода молодых, как видно из табл. 38, значительно .варьируют в зависимости от зоны.
В целом вопрос о сроках выхода молодых несомненно прямо связан с вопросом о порционности кладок: естественно, что в районах, где кладка происходит дважды на протяжении одного сезона размножения, сроки выхода молодых будут более растянутыми. При этом надо учесть, что момент выхода молодых из яиц обычно более точно может быть зафиксирован при стационарных наблюдениях в природе, чем факт вторичной кладки. Поэтому очень растянутый срок выхода молодых может косвенно свидетельствовать о значительной растянутости (или наличии второй) кладки.
ГЛАВА ВОСЬМАЯ
РАЗВИТИЕ И РОСТ1
ЭМБРИОНАЛЬНОЕ РАЗВИТИЕ
Эмбриогенезу рептилий посвящено большое число работ, в которых в основном содержатся данные по развитию отдельных органов. Формирование провизорных органов некоторых видов ящериц рода Lacerta изучены Шауинсландом (Schauinsland, 1906). А. М. Сергеев (1940—1943) исследовал функциональное значение этих органов рептилий в связи с развитием на суше. Эмбриональному развитию ящериц посвящены работы Дж. Па-стилса (Pasteels, 1957), который описал процесс гаструляции и некоторые стадии развития, и Г. Луппа (Luppa, 1961), исследовавшего формирование и развитие пищеварительного тракта. Темпам развития органов пищеварения и особенностям питания плодов в эмбриогенезе, а также различным температурным адаптациям на ранних стадиях развития зародышей пресмыкающихся посвящен цикл работ И. Н. Зусмана (1964—1974).
Особый интерес для изучения развития ящериц представляют работы Дж. Дюфора, Дж. Губерта (Dufaure, Hubert, 1961), в которых выделено 40 стадий развития эмбрионов живородящей ящерицы (L. vivipara), описаны гаструляция, ранние этапы развития амниона и аллантоиса, а также подробно изучено эмбриональное развитие гонад этих ящериц (Dufaure, Hubert, 1966). В дальнейшем указанные авторы в таком же плане провели изучение эмбрионального развития Vipera aspis, для которой выделили 13 стадий эмбриогенеза. Эти стадии теперь часто принимаются за основу при описании эмбрионального развития других видов пресмыкающихся.
Среди многочисленных работ, посвященных эмбриональному развитию разных видов рептилий (Даниелян, 1970—1974; Корнева, 1967, 1969; Lehr, 1964; Deraniyagala, 1958; Miller, 1963; Nathukkaruppan et all., 1970, и др.), эмбриональное развитие прыткой ящерицы рассматривается только в работе К. Петера (Peter, 1904).
Ниже будут рассмотрены некоторые вопросы эмбрионального развития и постэмбрионального роста прыткой ящерицы.
1 Авторы: Ф. Л. Даниелян и А. А. Симонян (эмбриональное развитие), А. В. Яблоков (постэмбриональное развитие), Э. М. Смирина (определение возраста).
227
8*
Эксперименты по инкубации яиц проводили в террариумах Кафедры зоологии Ереванского университета Ф. Д. Даниелян, в Калужском областном краеведческом музее А. Б. Стрельцов и в Институте биологии развития АН СССР А. С. Баранов и В. И. Подмарев. Последующие данные по эмбриональному развитию приводятся по наблюдениям Ф. Д. Даниеляна.
Условия инкубации яиц
Для изучения развития прыткой ящерицы самок после спаривания вылавливали за две-три недели до начала кладки. Всех пойманных ящериц размещали в отдельных банках с песком, где они и откладывали яйца. В песке делали углубление, куда помещали отложенные яйца и прикрывали картонным кружком. В каждой кладке учитывали число яиц, начальные размеры и их вес, в дальнейшем следили за изменением размеров и веса яиц во время инкубации. В общем было получено 550 яиц, которые были инкубированы в одинаковых условиях при определенной температуре. Как показали в своей работе Либерман и Покровская (1943), для развития яиц прыткой ящерицы нормальная влажность лежит в пределах 2,5—17,5%, т. е. для развития яйца используется лишь капиллярная вода. Яйца прыткой ящерицы инкубировали при 25° и влажности— 16,5%.
При проведении инкубации яйца вскрывали каждый день, начиная со дня откладки, а с пятого дня развития — каждые пять дней. В каждой пробе брали в среднем по 10 эмбрионов.
Изменение размеров яиц в процессе развития
Размеры яиц прыткой ящерицы варьируют от 12,1 до 15,2 мм в длину и от 7,2 до 8,5 мм в ширину. Не менее значительно варьируют яйца и по весу. Вес целого недавно отложенного яйца колеблется от 550 до 940 мг (средний вес 600,0+0,05 мг).
Скорлупа яиц прыткой ящерицы весит около 0,30 мг, что составляет около 5,6% от общего веса яйца (Либерман, Покров-
Таблица 39
Динамика увеличения размеров и веса яиц прыткой ящерицы (экспериментальные данные Ф. Д. Даниеляна; 95 кладок, 240 яиц)
Размер яйца, мм - День развития
1-й 8-й 10-й 15-й 20-й 25-й 30-й 35-й | 40-й 45-й 50-й
Длина 14,5 15 15 15 16 16,2 16,4 16,5 17,4 20,2 20
Ширина 8,9 10,3 10,3 11 11,3 11,4 12 13 14 15 15,3
Средний вес, мг 850 930 1020 1075 1430 1560 1620 1640 1900 2380 2420j
228
ская, 1943). Свежеотложенные яйца имеют мягкую кожистую скорлупу, которая, как показал А. М. Сергеев (1943), состоит из нескольких слоев. В ней содержится некоторое количество извести, иногда выделяющейся на поверхность в виде кристаллов.
В процессе развития яиц их размеры и вес постепенно увели-
Рис. 69
Увеличение длины тела эмбрионов прыткой ящерицы в разные дни развития
По данным Ф. Д. Даниеляна, А. А. Симоняна
чиваются (табл. 39). Однако этот рост нельзя использовать для определения степени развития, так как, несмотря на различие в размерах, в яйцах одного дня откладки, как выяснилось, содержатся эмбрионы, находящиеся на одинаковой стадии развития. Это хорошо объяснимо: для некоторых Squamata, откладывающих яйца с мягкими оболочками, уже давно описано явление
Таблица 40
Размеры эмбрионов прыткой ящерицы на разных стадиях развития (средние данные по 300 эмбрионам, материал Ф. Д. Даниеляна)
Возраст эмбриона, дни Стадия по Дюфору Промер эмбриона
вес тела, мг Длина тела с хвостом, мм Длина передних конечностей, мм
В момент откладки 25 — —
2 26 — — —
4 27 — —• —
5 27 — — —•
8 27—23 9 8 1
10 29-30 15 11,2 1,1
15 30 27 14,3 1,6
20 31 65 18,7 2,1
25 32—33 112 23,3 2,9
30 34-35 169 34,5 3,5
35 36 234 38,8 4,2
40 37—38 296 41,3 4,5
45 39—40 353 42,7 4,9
50 40 419 43,1 5,2
229
Рис. 70
Общий вид эмбрионов прыткой ящерицы на разных стадиях развития
1 — 2-й день развития; 2 — 5-й день развития; 3— 8-й день развития; -/—10-й день развития; 5— 15-день развития; 6—20-й день развития; 7—5—25-й день развития; 9—30-й день развития; 70 —35-й день развития; //—40-й день развития; /2 — 45-й день развития. Фото Ф. Д. Даниеляна, А, А. Симоняна
увеличения размеров яиц по мере развития эмбриона в силу поступления в яйцо воды извне через оболочку, приводящего как бы к набуханию яйца (Brimley, 1903; Mulherkar, 1962; Сергеев, 1943; и др.). Увеличение длины тела эмбриона в яйцах прыткой, ящерицы представлено на рис. 69.
Развитие эмбрионов по стадиям
Нами изучено более 300 эмбрионов прыткой ящерицы ла разных стадиях развития.
В табл. 40 приведены основные промеры эмбрионов, начиная с момента откладки яиц.
В момент откладки яиц амнион покрывает весь эмбрион. Зародыш лежит на боку. Аллантоис в виде небольшого бугорка располагается в нижней части тела.
230
Эмбрион 2-дневный. Начинают выпячиваться глазные пузырьки и формироваться хрусталик. Аллантоис хорошо различим в виде округлого выпячивания. Начинают появляться 1—2 жаберных отверстия, присутствует 25 пар сомитов (рис. 70, )).
Эмбрион 4-дневный. Аллантоис занимает */4 часть тела. Глазной бокал продолжает углубляться (рис. 70, 2).
Эмбрион 5-дневный. Зачаток глаз приобретает форму подковы. Отверстие сосудистой оболочки глаза открыто. Появляются зачатки передних и задних конечностей. Видно третье жаберное отверстие (см. рис. 70, 2; 71, /).
Эмбрион 8-дневный. Появляются очертания глазной чаши (см. рнс. 70, 3).
Эмбрион 10-дневный. Формируется головной мозг в виде бугорка. Начинается развитие нижней челюсти, верхнечелюстные отростки широко раздвинуты. Конечности видны в виде почкообразпых выростов. Открывается четвертое жаберное отверстие (см. рис. 70, 4\ 71, 2).
Эмбрион 15-дневный. Нижняя челюсть не доходит до верхней челюсти. Конечности уплощены на краях, лопастевидные. Глаза слабо пигментированные и выпуклые. Вокруг глазной чаши образовалось веко, внутренний край которого имеет круглую форму. Видна печень. Копулятивные выросты в виде двух бугорков (см. рис. 70, 5; 71,3, 4).
Эмбрион 20-дневный. Глаза хорошо пигментированные. Веко стало более широким. Конечности лопастевидные, уплощены на краях, формируются зачатки пальцев. Копулятивные выросты хорошо выражены в виде двух выпячиваний (см. рис. 70, 6; 71, 5).
Эмбрион 25-дневный. Видны полушария переднего мозга и теменной глаз. Веко имеет более или менее овальную форму. Нижняя челюсть еще не доходит до края верхней. Пальцы передних и задних конечностей зазубренные, между пальцами имеется перепонка. На хвосте проступают очертания чешуй (см. рис. 70, 7, 3; 71, 6).
Эмбрион 30-дневный. Появляется пигментация на голове. Начинает закладываться слуховая перепонка. Нижняя челюсть
231
Рис. 71
Передние конечности эмбриона прыткой ящерицы на разных стадиях развития
/ — эмбрион в возрасте 5—8 дней; 2—10 дней; 3 — 4—15 дней; 5 — 20 дней; 6 — 25 дней; 7 — 30 дней; 5 — 35 дней; 9—40 дней; 10 — 45 дней (фото Ф. Д. Даниеляна, А. А. Симоняна)
окончательно сформировалась. Мозг закрывается тонкой перепонкой.
Появляются очертания щитков на брюхе. Хвостовые чешуйки дифференцированы в виде колец. В лапке замечается дифференциация 5 пальцев. На кончиках пальцев видны густо пигментированные участки — места образования когтей. Копулятивные выросты у самцов начинают раздваиваться (у самок они ввернуты вовнутрь) (см. рис. 70, 9; 71, 7).
Эмбрион 35-дневный. Внутренний край века доходит почти до зрачка. Отчетливо видна слуховая перепонка. На голове заметны очертания появившихся щитков. На кончиках пальцев закладываются чешуи. Видны прианальные ,и анальные щитки. Появляется пигментация на спине. У самцов вершины копулятивных выростов раздвоились на две лопасти (см. рис. 70, 10;. 71, 8).
232
Эмбрион 10-дневный. Веки хорошо развиты. Очерчиваются нижнечелюстные и нижнегубные щитки. На горле видны чешуи, но они непигментированные. Хорошо выражены бедренные поры. Когти полностью сформировались (см. рис. 71, 9).
Конечности пигментированные. Копулятивные выросты вросли вовнутрь. Все тело слабо пигментировано, на спине появляются темноватые продольные полоски (см. рис. 70, 11).
Эмбрион 45-дневный. На голове дифференцированы предлобные, височные, носовые, надглазничные, межчелюстные, лобные щитки. Теменная часть сильно пигментирована. Ушной щиток хорошо выражен. Окраска еще слабая, но рисунок (полосы, пятна, глазки и т. д.) вполне различим (см. рис. 70, 12\ 71, 10).
Эмбрион 50-дневный. На голове и теле как щитки, так и чешуи вполне развиты. Эмбрион приобретает пигментацию, которая имеется при рождении. Хорошо виден рисунок спины. На верхней челюсти различим слегка возвышающийся яйцевой зуб, используемый эмбрионом при разрезании яйцевой оболочки (см. рис. 70, 12).
В этой стадии эмбрион вполне готов к выходу из яйца. При выходе молодые надрезают изнутри оболочку яйца яйцевым зубом, 'И оболочка сразу отпадает. Иногда на яйце видно несколько разрезов, но используется лишь один. Через разрез можно видеть голову и кончик хвоста животного (рис. 72). Время от времени
ящерята делают конвульсивные движения всем телом и через несколько часов, а некоторые даже через сутки покидают оболочку яйца.
Продолжительность инкубации яиц, по имеющимся данным, несколько колеблется и составляет 43—50 дней. Выход молодых из яиц даже в одной кладке может быть растянут на 2 дня (наблюдения Г. Рузавиной и В. Подмарева в террариумах Института биологии развития, 1974 г.).
ПОСТЭМБРИОНАЛЬНОЕ РАЗВИТИЕ
Постэмбриональный рост рептилий изучен недостаточно. В основном в литературе имеются лишь отрывочные данные, полученные в результате измерений, проведенных на отдельных лабораторных экземплярах или в природе на меченых особях при
233
рис. 72
Момент выхода прыткой ящерицы из яйцевых оболочек в террариуме ИБР АН СССР
Фото А. В. Яблокова, Г. И. Рузавиной, Г. Т. Шваяковой
РИС. 73
Ход развития (вес тела) новорожденных прытких ящериц в первые недели жизни в террариуме ИБР АН СССР
По данным В. И. Подмарева, Г. И. Рузавиной
Рис. 74
Сравнение весового состава популяций прыткой ящерицы Кавказа и Закавказья на протяжении августа-сентября
/—24 августа, берег.оз. Севан; 2—31 августа—2 сентября, окр. г. Ленинакана; 3 — 9— 12 сентября, берег р. Техурн (Зап. Кавказ); 4— 20—21 сентября, окр. пос. Красная Поляка (данные А. С‘ Баранова и А. В. Валецкого)
их повторных выловах (Сергеев, 1937, 1939а, 19396; Андрушко, 1954; Параскив, 1956; Чернов, 1959; Богданов, I960, 1965; Даревский, 1960а, 1967; Хонякина, 1964; Зинякова, 1964; Щербак, 1966; Мусхелишвили, 1970; Babcock, 1939; Cagle, 1946, 1948; Cruce, 1971; Davis, 1967; Jackson, 1970; Dobie, 1971; Michael, 1972; Ruibal et all., 1972; и др.).
235
Б результате специальных исследований, проведенных в ИБР АН СССР в 1970—1974 гг., удалось получить некоторые данные о темпах роста прыткой ящерицы.
При выходе из яйца вес новорожденных ящериц колеблется от 540 до 870 мг (данные по двум кладкам в террариуме ИБР АН СССР). Динамика дальнейшего развития показана на рис. 73, где приведен вес молодых в первые недели жизни в террариуме (при кормлении дрозофилами). К сожалению все ящерицы через 2 месяца погибли и динамика изменения веса, полученная в эксперименте, не вполне правильно отражает изменения, происходящие в природе. Видимо, лишь на протяжении первых двух недель жизни полученные данные могут дать представление о нормальном темпе роста. Видимо, не будет ошибкой считать, что за первые две недели жизни вес молодых может возрасти на 100—150 мг, что составляет около 20% первоначального веса.
Некоторые из полученных в природе данных позволяют более точно определить средний темп роста на протяжении первых месяцев жизни особей. Так, зная диапазон изменчивости веса новорожденных и сроки выхода молодых, можно рассчитать ежемесячные приросты, анализируя диапазон изменчивости сеголетков поздней осенью. Например, в популяции ящериц в долине р. Те-хури (Западный Кавказ), собранной 9—12 сентября, находили сеголетков размером от 1 до 2,5 г. Принимая, что выход молодых начинается с конца июля и новорожденные имеют вес от 0,4 до 0,9 г, легко рассчитать, что к середине сентября ежемесячные приросты составили здесь несколько больше 1 г/мес. Больший темп прироста определяется при сравнении среднего веса сеголетков в близко расположенных популяциях, исследованных последовательно на протяжении нескольких месяцев. Из приведенных на рис. 74 данных видно, что средний прирост за сентябрь для 4 кавказских популяций составил около 2 г.
Б период зимней спячки темп роста резко замедляется, и при выходе весной ящерицы имеют вес, близкий к тому, которого они достигли при залегании в спячку. Сразу же после выхода из спячки ящерицы начинают резко прибавлять в весе и средний темп их роста составляет в это время не менее 2,0—2,6 г/мес. (рис. 75). Ориентировочные расчеты показывают, что после второй зимовки темп прироста веса несколько снижается и составляет, видимо, около 1—1,5 г/мес.
При обработке собранного материала по длине тела ящериц разного возраста выяснилось, что сеголетки после вылупления имеют длину тела около 30—35 мм. К моменту ухода в спячку длина тела увеличивается до 35—55 мм. Средняя скорость роста при этом составляет 5—7 мм/месяц, достигая 8—9 мм/мес. (Андрушко, сообщ.).
Во время зимней спячки темпы роста настолько незначительны, что размер ящериц практически не увеличивается. Но после
236
Рис. 75
Сравнение весового состава одних и тех же популяций прытких ящериц в начале и конце лета
1 — юго-западная Белоруссия (окр. с. Голобы); 2—берег оз. Севан (Армения); 3 — берег р. Техури (Зап, Грузия); Средний прирост ящерицы после первой зимовки 2—2,6 г (данные А. С. Баранова и А. В. Валецкого)
Число особой
Лпчиа тела, мм
Рис. 76
Распределение особей по весу («) и длине тела ( б) в соответствии с возрастом, определенным по числу слоев в костях
Выборка из популяции окр. г. Умани, УССР 19 мая 1970 г.
выхода из спячки процесс роста возобновляется и линейный прирост составляет до 2—3 мм/месяц. Интенсивный рост продолжается до наступления половой зрелости. После ее наступления темп роста резко снижается и у самых старых животных зафиксировать его измерением ящерицы невозможно.
Таким образом, весной, по выходе ящериц из спячки, в популяции существуют три размерные группы. В первую группу входят ящерицы, перезимовавшие лишь одну зиму. Вторую группу составляют дважды перезимовавшие, но еще не размножавшиеся особи. В третью входят в основном половозрелые особи, но разделение двух последних групп не четкое (рис. 76).
238
ОПРЕДЕЛЕНИЕ ВОЗРАСТА
При исследовании большой единовременно взятой выборки можно определить относительный возраст особи по ее положению в ряду размерного состава. Однако эта методика дает тем более надежный результат, чем более сжаты сроки размножения. Учитывая, что у прыткой ящерицы сроки размножения достаточно растянуты, а также и то, что индивидуальная изменчивость темпа роста может быть велика, следует с осторожностью интерпретировать выделяемые группы как годовые классы. Ниже будет показано, что четко выделяется только первая возрастная группа (сеголетки — летом и осенью и перезимовавшие один раз — весной). Особей старших возрастных групп нельзя наверняка отнести к определенной возрастной группе. Поэтому проблема разработки методов точного определения абсолютного возраста ящериц становится актуальной.
Четкая методика определения абсолютного возраста рептилий до сих пор отсутствует. Обычно применяемое разделение на половозрелых и неполовозрелых по состоянию половых желез по сути не является определением возраста. Выделение возрастных классов по размерам тела (Залежский, 1938; Банников, 1940; Щепотьев, 1948; Терентьев, Чернов, 1949; Банников, Денисова, 1956; Богданов, 1965; Blanchard, Blanchard, 1931; Hamilton, 1934; Michael, 1972; Cruce, 1971; Peters, 1964; Gibbons, 1972; и др.) нельзя считать удовлетворительным прежде всего потому, что по достижении половой зрелости темп роста животных сильно снижается и чем дальше, тем медленнее увеличиваются размеры тела. Это ведет к тому, что в последнюю размерную группу из тех, которые можно выделить по длине тела, входят взрослые животные разного возраста. Кроме того эта методика не учитывает индивидуальных различий скорости роста. Было предпринято довольно много попыток определения возраста рептилий по слоям в кости. То, что в костях рептилий, так же как и остальных позвоночных, имеются слои, отражающие периодичность роста животных, известно давно. Слоистая структура кости видна на тонких плоских костях или на тонких шлифах костей (просветленных в глицерине или других просветляющих жидкостях), а также на срезах кости, окрашенных гематоксилином. Узкие линии, образующиеся во время задержки роста и разграничивающие широкие слои костной ткани, называют линиями задержки роста, или линиями склеивания и по их числу определяют возраст животного (Клевезаль, Клейненберг, 1967). Еще в 1907 г. была описана слоистость периостальной кости в диафизе у нескольких видов вымерших и современных рептилий (Seitz, 1907). Тогда же было высказано предположение, что широкие зоны костной ткани отлагаются летом, а узкие линии в холодное время года. Затем слои в кости рептилий были обнаружены целым рядом исследователей у разных
239
видов рептилий (Wallis, 1928; Емельянов, 1929; Mattox, 1935; Брюзгин, 1939; Peter-Rousseaux, 1953; Saint-Girons, 1957, 1965; Peabody, 1958, 1961; Thireau, 1967; Castanet, 1974; и др.).
Сопоставление размеров тела животного с числом слоев в кости дало исследователям основание полагать, что число слоев в кости соответствует их возрасту. Однако никто из исследователей не приводил достаточно веских доказательств того, что видимые слои действительно годовые.
Рис. 77
Соотношение длины тела и числа «зимних» линий склеивания в кости прыткой ящерицы
Популяция окр. г. Умани, УССР
Некоторые авторы выступили с критикой методики определения возраста рептилий по слоям в кости (Dobie, 1971; Griffiths, 1961). Дж. 3. Доби (Dobie, 1971), сравнивая картину слоистости на щитках карапакса черепах (подсчет числа слоев на щитках карапакса — общепринятая методика определения возраста черепах) со слоистостью, видимой на срезах ее позвонков и нижней челюсти, обнаружил лишь очень слабую положительную корреляцию числа слоев на щитках и срезах костей. Гриффитс (Griffiths, 1961), изучавший слои в костях змей, возраст которых был известен, так как их инкубировали и выращивали в неволе в условиях, близких к естественным, сделал вывод о ненадежности этой методики. Одним из его возражений было то, что в длинных костях кроме отложения новых слоев костной ткани со стороны периоста, в процессе роста со стороны эндоста идет разорбция ранее отложившейся кости, и первые слои не сохраняются.
Для решения вопроса о возможности определения возраста рептилий по слоям в кости идеальным было бы провести исследование животных точно известного возраста, взятых из естественных условий. Второй путь—-исследование большой, единовременно взятой из естественных условий выборки. Поскольку у нас не было животных известного возраста, мы выбрали второй путь.
240
В нашем распоряжении была выборка прыткой ящерицы {93 экз.), добытых в середине мая 1971 г. в районе г. Умани, и 33 экз. сеголетков, добытых в августе того же года в Новгородской области.
Для изучения закономерности образования слоев в костях прытких ящериц были изготовлены поперечные срезы из середины диафизов плечевой и бедренной костей. Кости декальцинировали и срезы окрашивали гематоксилином Эрлиха, пользуясь методикой, которая была применена при определении возраста амфибий (Клейненберг, Смирина, 1969).
В уманской выборке по длине тела нами были выделены только 2 группы: годовики и взрослые ящерицы (рис. 77.). Первая возрастная группа, которую можно выделить и по длине тела и по картине на поперечных срезах кости — это сеголетки. Длина тела сеголетков, имевшихся в нашем распоряжении, колебалась от 29 до 42 мм. Картина, видимая на срезах, была неодинаковой. У особей длиной тела от 29 до 32 мм в кости не было видно ни одной линии склеивания, а у особей длиной тела от 32 до 42 мм была видна линия склеивания.
У особей второй возрастной группы, которую мы выделяем из уманьской выборки, длиной тела от 31 до 47 мм, видны две линии склеивания (рис. 78, I). Этих ящериц можно считать зимовавшими один раз, или годовиками.
Очевидно, у сеголетков в первое лето жизни рост идет неравномерно; как правило, у них бывает одна задержка роста, которая и характеризуется первой линией склеивания. Вторая линия образуется в результате зимней задержки роста.
Следующая возрастная группа, которую можно выделить по картине на поперечных срезах костей, это ящерицы с двумя группами линий склеивания (см. рис. 78, //). У них около костномозговой полости обычно хорошо видны две частично резорбированные линии склеивания, сближенные одна с другой, которые мы считаем первой группой линий. Это, очевидно, следы от линии, которая имелась еще у сеголетков и от первой зимовки. Далее виден большой прирост кости и ближе к наружному краю еще одна, часто двойная или даже тройная линия склеивания (вторая группа линий). Ящериц с двумя группами линий склеивания можно считать зимовавшими два раза, т. е. двухгодовиками.
В исследованной выборке из 28 двухгодовиков у 20 видны линии склеивания от двух зимовок, а у 8 экз. линия склеивания второй зимовки не видна. На срезах костей этих экземпляров видны только обе линии склеивания первого года жизни и затем большой прирост костной ткани. Трудно сказать, является ли это особенностью исследованного среза или это нарушение ритма роста и его записи в костях. Однако наша уверенность в том, что эти ящерицы зимовали два раза, а не один, основана на том, что прирост кости от последней линии склеивания до наружного края кости слишком широк, чтобы считать его сформированным
241
Рис. 78
Поперечные срезы плечевой и бедренной костей прыткой ящерицы
I — первая зимовка; II — вторая зимовка; III — третья зимовка; ZV — четвертая зимовка; V — случай образования пяти линий склеивания в течение одного года жизни;, линия склеивания: а — у сеголетков; б — первая зимовка (IV, о); в— вторая зимовка (IV, б); г — третья зимовка (IV, в); д — четвертая зимовка (IV, г) (поперечный срез, декальцинированный препарат, окраска гематоксилином, об. 9Хок. 15 (препараты и фото Э. М.. Смнрнной)
со времени пробуждения из последней спячки до середины мая. возможно, что эти ящерицы еще не начали активно расти и последняя линия склеивания еще не отделилась от наружного края кости. Об этом свидетельствует тот факт, что у некоторых двух-годовиков с линиями склеивания от обеих зимовок, след от второй зимовки проходит около самого 'наружного края кости, едва отделяясь от него. Длина тела этих ящериц (65—76 мм) также дает основание считать их двухгодовиками {см. рис. 77). Вероятно, несоответствие числа слоев возрасту в данном случае можно считать следствием индивидуальных особенностей ритма роста и его записи в костях.
Далее можно выделить группу ящериц, которых нужно считать зимовавшими три раза. От линии склеивания, которая видна у сеголетков, у них обычно остается лишь небольшой участок, и видны.еще три линии (одинарные или расслоенные), которые можно считать соответствующими первой, второй и третьей зимовкам (см. рис. 78, III).
. И, наконец, встречались ящерицы, которых можно считать зимовавшими 4 раза. У них линия склеивания, которая видна у сеголетков, резорбируется и лишь иногда может оставаться совсем маленький ее участок. Линия склеивания, соответствующая первой зимовке, у них также частично резорбирована, а далее следуют еще три линии (см. рис. 78, IV).
Так как в нашем распоряжении не было ящериц точно известного возраста, для того, чтобы выяснить, сколько линий склеивания успевает резорбироваться со стороны эндоста при активном росте молодого животного, сделали следующие промеры на поперечном срезе кости: у годовиков измеряли диаметр круга, ограниченного первой линией склеивания (которая видна у сеголетков), и диаметр круга, ограниченного второй линией склеивания (которая появляется после первой зимовки). У двух-годовиков и старших особей измеряли диаметр костномозговой полости. Промеры брали у 10 экз. каждой возрастной группы (по три среза от каждой особи). Результаты приведены в табл. 41.
Таблица 41
Измерение диаметра поперечных срезов из середины диафиза бедренной кости прыткой ящерицы (в делениях окулярмикрометра при увеличении 10X10, данные Э. М. Смириной)
Г оДовики ДвухгодовИКИ Трехгодов и ки и старше
Диаметр * кости до первой линии склеивания Диаметр кости до второй, линии склеивания Диаметр костномозговой полости
33,4X31,2 41,2X39,4 36,6X34,5 42,6X38,5
Измерялся наибольший н наименьший диаметры, поскольку срез представляет собой скорее эллипс, а не круг. -______•___' ________ ' ________________
243 ' '
V.
> J ж
Plflf .o
’ •W’’ I
.. *r
''"*> % J%L *ъфл ’
^Лхяо
'• ^’V- -~c* j Ж» >
Рис. 78
Поперечные срезы плечевой и бедренной костей прыткой ящерицы
7 — первая зимовка; Н — вторая зимовка; 777 —третья зимовка;
7V — четвертая зимовка; V —случай образования пяти линий склеивания в течение одного года жизни; линия склеивания: а - -у сеголетков; б — первая зимовка (IV, я); s — вторая зимовка (IV, б); г — третья зимовка (IV, в); д — четвертая зимовка (IV, г} (поперечный срез, декальцинированный препарат, окраска гематоксилином, об. 9Хок. 15. (препараты и фото Э. М., См ириной)
Исходя из этих промеров, можно было бы ожидать, что у двухгодовиков полностью резорбируется первая линия склеивания, а вторая, соответствующая первой зимовке, остается. На самом деле, и с первой этого не происходит, так как резорбция идет не равномерно по кругу, а сильнее захватывает одну сторону кости: поэтому и от первой линии у двухгодовиков виден остающийся участок, который может сохраняться даже у особей, зимовавших три раза. Остаток линии склеивания, соответствующий первой зимовке, сохраняется, очевидно, и у четырех-, летних, а, возможно, и более старших особей. Иногда эта линия сохраняется даже целиком.
Таким образом, в выборке из Умани из 93 экз., по нашим предварительным данным, оказалось 49 экз., зимовавших один раз, 28 экз. зимовавших два раза, 10 экз,—три раза и 4—четыре раза (число зимовок 2 экз. трудно определить).
Следует отметить, что на поперечных срезах костей самых старых из пойманных ящериц ширина каждого из слоев, отложенных после первой, второй, третьей и т. д. спячек, уменьшается незначительно, тогда как у большинства более молодых особей слой, отложенный после первой спячки, намного шире, чем слой, отложенный после второй и последующих спячек. Это может быть следствием одной из двух причин: либо линия склеивания, соответствующая первой зимовке, резорбировалась целиком, и первая линия, которую мы видим, соответствует второй зимовке, либо темп роста этих ящериц был медленным и более равномерным в течение всей жизни и вследствие этого ширина слоя, отложенного после первой зимовки уже, чем обычно. Первое вряд ли вероятно, как видно из приведенных выше промеров костномозговой полости. Второе объяснение кажется более вероятным, и, если это так, то относительное количество экземпляров с равномерно расположенными слоями должно возрастать в каждой последующей возрастной группе. Эта тенденция прослеживается даже на нашем небольшом материале. У всех четырех ящериц, которых мы считаем четырехлетними, ширина слоев на срезах в кости различалась незначительно. Из десяти трехлетних у 6 экз. между линиями склеивания, соответствующими первой и второй зимовкам, был большой разрыв, а у трех линий склеивания располагались почти на одинаковом расстоянии друг от друга (один экземпляр оказался сомнительным). Из 14 экз. двухлетних ящериц лишь у двух линии склеивания разделены небольшими промежутками.
В ихтиологической литературе имеются данные о том, что более быстрорастущие рыбы гибнут раньше, чем медленнорастущие. Поэтому у рыб самая старшая возрастная группа иногда представлена только самыми медленнорастущими особями (Richer, 1969). Может быть, что и у прыткой ящерицы до самых старших возрастов доживают лишь особи с более медленным и равномерным темпом роста. Это положение подтверждается и
244
тем, что размеры тела самых старых из определенных нами: ящериц были не самые крупные (см. рис. 76, 77).
Кроме линий склеивания, разграничивающих годовые слои,, на срезах часто видны дополнительные линии, которые, как правило, окрашиваются менее ярко, чем основные линии склеивания, соответствующие зимовкам, но иногда малоотличимы от последних и затрудняют определение возраста. Причиной образования таких линий могут быть, очевидно, влияния любых экзогенных или эндогенных факторов: сильное похолодание летом,, засуха, болезнь и т. д., т. е. любой фактор, вызывающий остановку роста. У ящериц на 1—2-м годах жизни в костях может образовываться до 4—5 близко расположенных полос (см. рис. 78, V). Это можно объяснить тем, что зимой период спячки у молодых ящериц иногда прерывается короткими периодами активности, вследствие чего у них, хотя и незначительно, могут идти процессы роста (Сергеев, 1937; Davis, 1967), а в летний период и остановки роста (например, в течение летней спячки).
Полученный в данном исследовании материал позволяет сделать вывод о том, что у прытких ящериц ежегодно в период активного весенне-летнего роста в трубчатых костях образуется широкий слой костной ткани, отграниченный от последующего слоя одиночной или расслоенной линией склеивания, образующейся в зимний период. Кроме «зимних» линий склеивания, отграничивающих ежегодные приросты кости, имеется еще одна самая первая линия склеивания, образующаяся у сеголеток вскоре после вылупления. Процессы резорбции костной ткани СО' стороны эндоста идут у прыткой ящерицы медленно. Первая линия склеивания, образующаяся у сеголетков, целиком резорбируется лишь к 3—4 годам. Линия, соответствующая первой зимовке, даже у четырехлетних ящериц целиком может не резорбироваться и, возможно, сохраняется всю жизнь.
Исходя из изложенных выше данных, определение возраста прытких ящериц по слоям в кости можно считать вполне возможным.
ГЛАВА ДЕВЯТАЯ
ПЕРЕМЕЩЕНИЯ И МИГРАЦИИ1
Случаи перемещений и миграций прыткой ящерицы изучены мало. Имеющиеся данные (Щепотьев, 1950, 1952) свидетельствуют только о наличии местных миграций, обусловленных временными изменениями фактора влажности.
МЕТОДЫ ИЗУЧЕНИЯ ПЕРЕМЕЩЕНИЙ И МИГРАЦИЙ
Основным методом служат визуальные наблюдения за видимыми перемещениями большого количества меченных ящериц в течение одного или нескольких сезонов активности. При изучении суточных перемещений можно использовать следующую методику. В различных биотопах находят обитаемые норы этих животных, вокруг которых намечают участок площадью 20X20 м и который затем разбивают на квадраты 1X1 м. Эти квадраты размечают на местности. Перед началом наблюдений ящериц отлавливают, определяют их пол и возраст и метят путем сечения фаланг пальцев в различных комбинациях, а для удобства наблюдений у меченных животных окрашивают спину белой или красной быстро высыхающей краской. За перемещением животных наблюдают в течение всего периода их суточной активности (с 5—7 до 17—20 час.) с расстояния не менее 15 м. Дневные перемещения ящериц наносят на схему участка, а время их движения хронометрируют. Во всех случаях следует регистрировать влажность и температуру приземного слоя воздуха, температуру почвы, скорость ветра и осадки, чтобы в дальнейшем можно было бы выяснить влияние того или иного климатического фактора на особенности перемещения ящериц.
При изучении более дальних перемещений в различных биотопах отлавливают и метят большое количество ящериц. Два — три раза в неделю на протяжении одного—двух сезонов активности проводят повторные отловы этих животных. На имеющиеся схемы участков наносят точки первоначального и всех последующих поимок меченных ящериц.
л Автор: М. Ф. Тертышников, при участии Н. В. Щепотьева, Н. А. Рашкевича, А. Б. Стрельцова, В. Д. Булахова, Н. Ф. Константиновой, В. К. Жарковой.
246
ПЕРЕМЕЩЕНИЯ
Под перемещениями имеются в виду обычные передвижения ящериц на сравнительно небольшой территории.
Суточные перемещения
Прыткая ящерица — животное со строго дневным циклом активности (см. гл. X). В течение дня она непрерывно перемещается в поисках корма, особей другого пола в период размножения, .при расселении и по другим причинам. Длина дневных перемещений ящерицы может варьировать в широких пределах и определяется влиянием многих факторов, в частности таких, как особенности биотопа и погоды, обилием корма и характером его распределения, присутствием врагов и др.
Данные о длине суточных перемещений прыткой ящерицы незначительны. Б. А. Красавцев (1936) пишет, что каждая ящерица имеет свою нору, от которой далеко не уходит. Другие исследователи находили ее на расстоянии 3—20 м от норы (Щепотьев, 1952; Костин, 1956; Щербак, 1966; Булахов, Константинова, 1972; Стрельцов сообщ.). В биоценозах Ставропольской возвышенности длина дневных перемещений прыткой ящерицы не превышает 225 м (Тертышников, 1970, 19726), а в условиях Чечено-Ингушской АССР — до 290 м (Рашкевич, сообщ.). Как установил М. Ф. Тертышников (1970), в нормальных условиях средняя скорость дневных перемещений ящерицы варьирует от 0,17 до 0,68 м/мин.
Перемещения ящериц разного пола и возраста
Л. Г. Динесман и М. Л. Калецкая (1952) считают, что в отличие от самок самцы прыткой ящерицы очень подвижны, широко кочуют и за короткий срок перемещаются на десятки метров (рис. 79). Исследование, проведенное М. Ф. Тертышниковым (1970, 19726), показало, что длина дневных перемещений самцов и самок в биоценозах Ставропольской возвышенности зависит от особенности биотопа. Так, в типчаково-полынной степи средние размеры дневных перемещений взрослых самцов и самок (соответственно 158+37,0 и 174 + 51,1 м) приблизительноодинаковы. Подобная закономерность установлена для взрослых самцов и самок, обитающих в разнотравно-злаковой степи (94,6 + 24,5 и 94,0+15,0 м). В лесных полосах взрослые самцы-перемещаются на более значительные расстояния (83,7+18,3 м), чем взрослые самки (40,0+11,7 м). Средние расстояния перемещений молодых особей в типчаково-полынной степи (91,0 + ±47,1 м), в разнотравно-злаковой степи (129,5 + 32,5 м) и лесной полосе (79,0+21,0 м) незначительно отличаются от средних размеров дневных перемещений взрослых особей в соответствующих биотопах (168,1 + 21,4; 94,4+17,2 и 82,0+13,0 м)..
247
Рис. 79.
Повторные встречи окольцованных прытких ящериц в лесной посадке около пос. Джаны-бек 28 апреля — 8 мая 1950 г. -
1—места встреч самцов; 2 — места встреч самок; 3 — границы индивидуальных участков самок; 4—линии, соединяющие места повторных поимок самцов; 5 — посадки клена, тополя, лоха (по Дннесману, Калецкой, 1952)
Перемещение ящериц в разных биотопах
По данным М. Ф. Тертышникова (1970, 19726),, длина дневных перемещений ящериц неодинакова в различных биотопах и ^варьирует от 44 до 225 (147,6±21,6 м) в типчаково-полынной «степи, от 58 до 191 (102,2± 15,2 м) в разнотравно-злаковой сте-
248
,пи и от 16 до 119 (65,3+11,0 м) в лесной полосе. По-видимому,. Б открытых биотопах ящерицы передвигаются на более значительные расстояния, чем в закрытых. Подобную закономерность отмечает и Н. А. Рашкевич, по данным которого длина передвижения прыткой ящерицы на лугах и в степи больше (80 м), чем Б лесной полосе или на опушке пойменного леса (20—50 м).
Влияние погоды на длину дневных перемещений
О перемещениях ящерицы, зависящих от погодных условий,, подробно сказано в главе X. В этом же разделе мы отмечаем, что ящерицы при хорошей погоде перемещаются дальше, чем при плохой погоде. Исследования М. Ф. Тертышникова (1970, 19726) показывают, что в типчаково-полынной, разнотравнозлаковой степях и в лесной полосе ящерицы в хорошую погоду перемещаются больше, чем в плохую. Интересно отметить, что даже при плохой погоде ящерицы в типчаково-полынной степи перемещаются больше, чем в лесной полосе.
Изменение суточных перемещений ящериц в разные сезоны
Как установил Н. А. Рашкевич, в Чечено-Ингушской АССР длина дневных перемещений ящериц весной на 20—50% больше, чем, например, летом и осенью. Такая разница в особенностях перемещений может быть объяснена не только усиленными: поисками корма, но и проходящим весной периодом размножения.
С середины лета, по окончании размножения, и вплоть до ухода в зимнюю спячку продолжительность дневной активности прыткой ящерицы постепенно уменьшается (см. гл. X).
Перемещения, связанные ;с изменением освещенности
По наблюдениям В. Л. Булахова и Н. Ф. Константиновой,, ящерицы обычно не покидают своего участка, но их постепенное смещение по территории биотопа наблюдается при увеличении возраста насаждений и связано с уменьшением освещенности..
МИГРАЦИИ
Под миграцией понимают длительные и на сравнительно значительные расстояния перемещения ящериц. ,
Сезонные миграции
Для прыткой ящерицы сезонные миграции также известны. Они выработались в результате приспособления вида к ритму
249
окружающей среды. Причины их сложны и объясняются в значительной мере влиянием температуры, влажности, наличием корма и др.
Миграции, связанные с засухой
Данные некоторых исследователей (Птушенко, 19386; Гумилевский, 1941; Щепотьев, 19'50, 1952; Хонякина, 1964; Кутузова, 1971) позволяют говорить о наличии у прыткой ящерицы местных миграций, связанных с временным понижением влажности среды. Указанными исследователями замечено, что в засушливые годы ящерицы перемещаются в места с повышенной влажностью: впадины, ложбины с более густой и свежей растительностью, к берегам ручьев, на болотца с кочками и дерновинами, поросшими камышом и кустарником, и др. В сырые же годы миграции идут в направлении сухих биотопов. В. К. Жаркова (1973а), например, указывает, что повышение влажности воздуха до 76% в результате поднятия уровня воды в водохранилище Марьинской ГЭС привело к увеличению численности ящериц на пойменном лугу в 4 раза, а в лесных посадках — в 2,5 раза.
Сведения о дальности перемещений такого рода отсутствуют, но можно предположить, что их протяженность по прямой не превышает 125 м (Тертышников, 19726).
J
Миграции, связанные |с изменением питания по сезонам
Н. В. Щепотьев (1950) полагает, что засуха играет определенную роль в биотопическом распределении запасов пищи ящериц. В связи со сказанным ящерица может совершать и пищевые миграции, которые могут быть довольно значительными по протяженности. По наблюдениям Г. П. Лукиной (19666), после уборки урожая раздельным способом большое количество ящериц переселяется с «неудобных» земель на стерню, где в случае опасности могут прятаться в норы грызунов и под валками еще не обмолоченного хлеба. Не исключено, что сюда их привлекает скопление большого количества насекомых.
Миграции, связанные со спячками
Исчезновение прытких ящериц в июле — августе из обычных для нее биотопов было отмечено в литературе для Воронежской (Северцов, 1950) и Харьковской (Чернай, 1852/1853) областей. Существование летней спячки у прыткой ящерицы было установлено М. Ф. Тертышниковым (19726) и на территории Ставропольской возвышенности. В данном регионе спячка у большей части взрослых и полувзрослых ящериц начинается также в июле — августе и переходит в зимнюю спячку. Причиной ее является, по всей видимости, наступление длительного понижения влажности воздуха (до 30% и ниже) и высокие температу
250
ры среды летом. Вероятно, миграции, связанные со спячкой, незначительны по протяженности, так как ящерицы залегают в спячку преимущественно в тех убежищах, которые расположены в пределах их индивидуальных территорий.
Миграции, связанные с половодьем
Летнее половодье оказывает значительное влияние на образ жизни прыткой ящерицы. По наблюдениям В. К- Марковой (1973а), до июльских дождей 1967 г. на 1000-метровом участке железнодорожного полотна обитало 15 ящериц. После пятидневного наводнения на этом участке была обнаружена только одна особь, через месяц —еще три, но только через год численность этого животного восстановилась. Можно предположить, что в данном случае произошла миграция ящериц на железнодорожное полотно из близлежащих биотопов.
Миграции, связанные с размножением
В первые дни после вылупления сеголетки задерживаются на некоторое время в пределах индивидуальной территории «матери», т. е. там, где были отложены яйца, или же начинают мигрировать на свободные участки, пригодные для обитания, и располагающиеся поблизости. По-видимому, только в редком случае сеголетки мигрируют на значительные расстояния. На один из таких случаев указывает Н. А. Рашкевнч. По его данным, одна молодая ящерица прошла за месяц около 1200 м от места первоначальной поимки. Обычно миграции происходят весной будущего года, когда после спячки появляются взрослые и полувзрослые особи и которые «изгоняют» молодых конкурентов из пределов индивидуальной территории. Следует полагать, что миграции сеголетков идут в том направлении, где имеются участки, более или менее свободные от посещения взрослых ящериц, и пригодные для обитания.
Миграции ящериц в разные годы
Сведения такого рода ограничены. Как установил Н. А. Раш-кевич, в засушливый 1972 год перемещения ящериц в степных биотопах Чечено-Ингушской АССР были больше, чем в другие годы с нормальными условиями увлажнения.
Таким образом, у прыткой ящерицы существуют миграции, которые имеют различную протяженность и обусловлены такими причинами, как климатические условия, поиск пищи и особей Другого пола в период размножения, переселение после вылупления и др.
До настоящего времени остаются недостаточно выясненными вопросы дальности перемещения сеголетков и молодых ящериц от места вылупления, вопросы дальности перемещения этих жи-' вотных по годам,.сезонам, в разных биотопах и др.
ГЛАВА ДЕСЯТАЯ
ПОВЕДЕНИЕ И АКТИВНОСТЬ1
Говоря о поведении ящерицы, представляется целесообразным разграничение собственно поведения и явления активности.
ПОВЕДЕНИЕ
Поведение прыткой ящерицы базируется на безусловных рефлексах, которые по сравнению с более высокоорганизованными животными еще просты и не достигают большой степени сложности. Эволюция рептилий, в том числе и прыткой ящерицы, шла, по всей видимости, по линии выработки морфологических адаптаций, средств адаптивной защиты и нападения на добычу (Кашкаров, Станчинский, 1940). Многие двигательные приспособления и моторные механизмы поведения имеют важное значение во внутривидовых и межвидовых контактах и обеспечивают целостность вида. В этой связи становится ясным важность изучения поведения данного животного.
•Отношения с животными других видов
Поведение при преследовании
Активные и пассивные способы защиты от врагов. Одним из средств защиты является аутотомия, когда животное в ответ на болевое раздражение энергичным боковым движением обламывает хвост, схваченный врагом. Отброшенный хвост извивается и отвлекает внимание врага от туловища, а ящерица нередко благополучно скрывается. Имеющиеся данные указывают на то, что многие ящерицы подвергаются нападению врагов. Так, в Горьковской и Ивановской обалстях количество аутотомирован-ных ящериц достигает 12,6% (Красавцев, 1936), в Саратовской области—17,3% (Щепотьев, 1948, 1950), в Крыму — 39,0% (Щербак, 1966), возле г. Ставрополя—31,1% (Красавцев, 1939), а в Александровском районе Ставропольского края —
1 Автор: М. Ф. Тертышников, при участии А. С. Баранова, А. В. Яблокова, В. И. Борисова, Н. А. Рашкевича, В. А. Кутузовой, Г. П. Лукиной, С. П. Чащина, Н. А. Литвинова, Е. Н. Инщиковой, А. Б. Стрельцова.
252
4 5% (Тертышников, 19726). Как правило, в лесных биоценозах количество аутотомиров энных ящериц ниже, чем в степных биоценозах.
Другим защитным приспособлением является выделение из клоаки экскрементов, что часто можно наблюдать при поимке ящерицы. Животное при этом всегда старается вывернуться из рук; оно извивается всем телом, шипит и делает попытку укусить схватившего его человека. Как заметили А. С. Баранов, А. В. Яблоков и В. И. Борисов, самцы в подобных ситуациях намного агрессивнее самок.
Наиболее редко встречается такой метод защиты, как устрашение. М. Ф. Тертышников (19726) наблюдал, как самец прыткой ящерицы шипел и прыгал с разинутой пастью навстречу нападавшим на него кошке и человеку.
От своевременно замеченного врага ящерица прячется в различных убежищах. Она, например, может взбираться на деревья и, как пишет И. С. Даревский (1946), подобно белке, проворно и быстро поднимается по спирали каждый раз в сторону, обратную той, откуда пытаешься обойти. По его данным, прыткая ящерица может подниматься по стволу дерева на высоту до 25 м. Другие исследователи наблюдали, как ящерица поднималась по стволам деревьев на высоту 3—4 м (Тертышников 19726) и 6—8 м (Кутузова, 1971). Находясь в безвыходном положении при приближении врага, ящерица прыгает вниз и пытается скрыться в ближайшем убежище. А. С. Баранов, А. В. Яблоков и В. И. Борисов указывают, что самцы чаще самок влезают на стволы деревьев три опасности и что молодые особи обычно не используют этого метода защиты. По их мнению, «равнодушие» самок к защите такого рода связано с беременностью. Что же касается молодых, то они еще слабы для того, чтобы подняться на дерево.
Не менее действенным средством защиты является бег. Завидев врага, ящерица настораживается, приподнимает тело, а при его приближении — приподнимает над землей хвост и быстро бежит. Экспериментальным путем установлено (Тертышников, 19726), что скорость бега взрослого самца достигает 8 м/сек. Однако долго бежать с такой скоростью животное не может. Через 10—20 м оно устает и становится малоподвижным. Если хвост аутотомирован, то скорость передвижения ящерицы снижается и она менее ловко лавирует между различными предметами, встречающимися на ее пути.
Убегая от врага, ящерица часто вводит в заблуждение преследователя, так как перемещается обычно не по прямой, а «зигзагом». Если, например, человек находится возле норы, то она вертится у него под ногами, резко меняя направление бега, и пытается проникнуть в нору. А. С. Баранов, А. В. Яблоков и В. И. Борисов наблюдали, как прыткая ящерица забралась на погу преследователя и скребла ее, стараясь освободить себе
253
проход к норе. По их же сведениям, самец бросался преследователям под ноги, возможно отвлекая их внимание от самки, которая в тот момент находилась рядом. Кроме того, эти же исследователи указывали на существование некоторых отличий в поведении между взрослыми самцами, самками и молодыми особями при опасности. Так, самки убегают от преследования как-то «элегантно», самцы более резковаты в движениях, ну а молодые при беге как бы семенят и «стелятся» по земле.
Нередко при преследовании ящерицы совершают прыжки с холмиков и обрывов в густую траву, кустарник, пролетая по воздуху 20—60 см. Иногда они переплывают и водную преграду. По данным Б. А. Красавцева (1936), загнанная в воду прыткая ящерица проплывает 5—10 м, а по наблюдениям В. А. Кутузовой (1971), только 3—4 м. Н. Н. Щербак (1966) также отмечал у прыткой ящерицы способность плавать. Тело у плавающей ящерицы извивается как у змеи, а голова находится над поверхностью воды. По наблюдениям А. С. Баранова, только взрослые особи с целыми хвостами могут переплывать более или менее широкий водоем (по-видимому, не более 8—10 м). Молодые животные и те из взрослых, у которых хвост аутотомирован, тонут. По мнению А. С. Баранова, хвост служит не только рулем при передвижении по суше, но и локомоторным органом при плавании.
Характер использования убежищ. Наиболее часто ящерица уходит от врагов в норы. Если ее не тревожить, то она через некоторое время успокаивается, подползает к выходу и, осторожно осматриваясь, постепенно начинает выползать наружу. Когда раскапывают нору в поисках ящерицы, то она или выскакивает из запасного выхода, или же оказывается в каком-либо отнорке. Забежав в чужую нору, она далеко никогда не заходит. Как правило, ящерица всегда располагается у входа и независимо от вида, пола и возраста «хозяина» норы покидает его убежище и передвигается в направлении своей норы (Тертышников, 19726; Жаркова, 1973а; Баранов, Яблоков, Борисов, сообщ.). Нередко в одно убежище могут скрыться несколько особей.
Поведение во время питания (способы добывания корма и его поедания)
Уже в конце первой недели самостоятельной жизни у сеголетков прыткой ящерицы вырабатывается реакция на внешний вид объекта добычи (Черномордиков, 1944). Не исключено, что в этот же период ее жизни происходит становление инстинктов добывания и поедания корма.
Способы добывания пищи. Утром, выйдя из норы, ящерица некоторое время греется в лучах солнца. Свою добычу она либо подкарауливает, сидя в засаде, либо активно разыскивает. Заметив добычу, ящерица настораживается, следит за ее передви-
254
гениями, а затем стремительно срывается с места и схватывает €е в точном броске. По Г. П. Лукиной (19666), если в момент нападения ящерицы насекомое улетало, а рядом с ним шевелилось другое насекомое, ящерица мгновенно изменяла направление броска и хватало его. Нередко можно видеть, как ящерица подпрыгивает и ловит пролетающих над ней мух и бабочек. Д. С. Баранову удалось наблюдать в Молдавии охоту прыткой ящерицы, которая совершала описываемые выше прыжки на высоту до 40 см. В других случаях (Тертышников, 1972а, 19726; Баранов, Яблоков и Борисов, сообщ.) ящерица подкарауливает добычу, притаившись на каком-либо возвышении, в 20—30 см пт которого находится невысокое цветущее растение. Тело ящерицы в такой момент прижато к субстрату, а голова несколько приподнята. Заметив появившихся на растении мух, муравьев, бабочек и других беспозвоночных, ящерица, сделав 2—3 шага, прыгает и хватает добычу.
Обладая способностью лазать по стволам деревьев, ветвям кустарников и прочным стеблям травянистых растений, ящерица часто добывает себе пищу и там. Своеобразный метод охоты отметил А. С. Баранов в окрестностях г. Первомайска (УССР). По его мнению, данный метод охоты характерен для местной популяции прыткой ящерицы. Ящерица влезала на дерево и затаивалась в его ветвях. Когда под ней пролетала муха или другое летающее беспозвоночное, ящерица прыгала на него сверху с широко разинутым ртом, действуя им наподобие сачка, причем прыжки ящериц очень точны, хотя самки менее прыгучи и проворны, чем самцы.
М. Ф. Тертышников (1972а, 19726) установил, что разную по размерам добычу прыткая ящерица поедает далеко не одинаково. ДАелких животных она глотает быстро, а крупных — медленно, в течение 2—3 минут. Крупных кузнечиков и жуков она долго треплет, выпуская иногда на землю, а потом вновь подхватывает, время от времени сдавливая челюстями и отрывая у них конечности и надкрылья. На подобную «обработку» ящерицей объектов добычи указывает и Н. В. Щепотьев (1950). Затем ящерица «поправляет» положение добычи таким образом, чтобы она располагалась параллельно оси тела и могла быстрее пройти через пищевод в желудок. Добычу, принявшую «нужное положение», ящерица постепенно заглатывает, а проглотив облизывается и, приподнявшись на передних конечностях, медленно изгибается передней частью тела, помогая скорейшему прохождению пищи.
В естественных условиях очень редко, а в вольерных условиях довольно часто прыткая ящерица пьет воду, лакая подобно кошке или собаке.
255
Непищевые взаимоотношения \
Территориальное поведение
Выше (см. гл. V) было отмечено, что прыткая ящерица обычно оседла, хорошо знает индивидуальную территорию и ведет одиночный образ жизни. По данным М. Ф. Тертышникова (1970, 19726), прыткая ящерица посещает одни участки своей территории чаще других (см. рис. 60). Это могут быть освещенные солнцем участки, небольшие холмики, отдельно расположенные пучки травы, кучи камней и т. д. В разнотравно-злаковой степи были найдены даже своеобразные «тропинки»; проложенные ящерицей к наиболее часто посещаемым участкам (предпочитаемым местам кормления). По-видимому, на таких участках они находят достаточное количество корма и имеют возможность быстро скрыться при нападении врагов. Участки с низкой травой ящерицы обычно пробегают быстро, а в высокой и густой траве они ползут медленно, также медленно переползая через встречающиеся препятствия. Свои убежища и предпочитаемые места кормления ящерицы активно защищают при вторжении других особей (претендентов и случайно зашедших соседей). Если вдали от нор и мест кормления ящерицы более или менее терпимо относятся друг к другу, то вблизи них хозяева становятся очень агрессивными. Замечено, что более четко выражено агрессивное поведение во взаимоотношениях крупных особей с крупными, самцов с самками, мелких с мелкими. Между крупными и мелкими особями подобные взаимоотношения проявляются в сглаженной форме.
Агрессивность проявляется не только по отношению к животным своего вида. М. Ф. Тертышников неоднократно наблюдал столкновения прыткой ящерицы и с разноцветной ящуркой и с жабой зеленой, которую ящерица вытаскивала из норы, ухватив пастью за ее бок.
По наблюдениям В. К. Жарковой (1973а), ящерицы обычно сталкиваются на пересечении «дорожек» (см. гл. V), по которым они перемещаются, причем на таких дорожках агрессивное поведение проявляется у особей одинакового- пола. На других же’участках ящерицы спокойно греются на солнце в 30 см друг от друга. При содержании в террариуме ящерицы быстро привыкают к совместной жизни на ограниченной территории, между ними не наблюдается драк, и часто вблизи источника тепла и света они лежат, тесно прижавшись друг к другу, иногда в два — три слоя (рис. 80).
256
Рис. 80
Группа взрослых прытких ящериц в террариуме ИБР АН СССР, январь 1975 г.
Фото А. В. Яблокова, А. В. Валецкого
Иерархия
По наблюдениям А. Б. Стрельцова и М. Ф. Тертышникова, среди ящериц, индивидуальная территория которых значительно перекрывается, устанавливается иерархия. Самый крупный и сильный самец занимает главенствующее положение. Прочие самцы в большинстве случаев быстро убегают при приближении доминантной особи или же принимают определенную сигнальную позу (охранную). В противном случае между ними происходит драка. Доминирующий самец самок не трогает, но иногда в момент сильного возбуждения после драки он кусает и находящихся рядом самок. Среди самок иерархические взаимоотношения не отмечены. Кроме того, и сами самки не замечены в роли доминанта.
9 Прыткая ящерица
257
Поведение во время размножения
В брачный период самцы часто дерутся между собой за овладение самкой. Подробную картину таких сражений описывают И. С. Даревский (1946), Н. В. Щепотьев (1948) и М. Ф. Тертышников (19726). Из их описаний следует, что при встрече самцы принимают угрожающую позу-—поперечную ориентацию по отношению один к другому, приподнимаются на вытянутых передних конечностях, сжимают с боков переднюю часть тела, рассматривают друг друга и постепенно сближаются. Более слабый из них нередко убегает. Чаще все же происходят бои. Каждый самец стремится схватить противника за шею или голову и опрокинуть на спину. Упавший лежит несколько секунд, а затем стремительно вскакивает и убегает. Победитель гонится за убегающим врагом всего несколько метров. Иногда сражения между самцами кончаются тем, что один из них хватает за челюсть другого и оба они в полном исступлении катаются по земле с закрытыми глазами до тех пор, пока кто-либо не вырвется и не убежит.
Ухаживание
Данные В. А. Кутузовой (1971) свидетельствуют о том, что в брачный период самцы очень возбуждены, неосторожны и оживленно бегают в поисках самки. Будучи пойманными, они усиленно стараются вырваться, хлеща из стороны в сторону хвостом, широко раскрывают рот, пытаясь укусить и выворачивают гениталии. После линьки, проходящей перед размножением, окраска ящериц становится более яркой. В этот же период у самцов появляются характерные сигнальные телодвижения и позы, предшествующие брачным играм. По мнению Е. Н. Инщи-ковой, они заключены в демонстрации самцами яркого брюшка, что достигается в результате приподнимания передней части тела и частого поглядывания вокруг. Самка, обратившая внимание на самца, отвечает своеобразными ритуальными движениями. Заметив их, самец начинает преследовать самку, а настигнув, хватает зубами за различные части тела. Самка останавливается, движения ее замедленны, а хвост при этом быстро извивается.
Спаривание
А. Б. Стрельцов отметил, что перед спариванием самец приподнимается на передних лапах, раздувает горловой мешок и поворачивается к самке боком. Оказавшись несколько позади самки, он хватает ее за хвост и, не выпуская его изо рта, начинает постепенно продвигать голову к основанию хвоста, пока не схватит самку за бок. Затем самец резким движением изгибает свое тело так, что его клоака оказывается напротив клоаки самки. Происходит спаривание (рис. 81). По синим подковообраз-
258
Рис. 81
Последовательные этапы спаривания прытких ящериц в террариуме ИБР АН СССР
Фото А. В. Яблокова, А. В. Валецкого
9*
ним следам, длительное время остающимся на теле самок, можно определить, происходило ли спаривание, когда оно началось и как часто животное спаривалось. И. С. Даревский (1967) отметил, что указанный выше тип фиксации вырывающихся самок при спаривании свойствен многим ящерицам сем. Lacertidae. При спаривании самки тоже покусывают самца, но осторожно, как бы «лаская». Однако нередко самки ведут себя при спаривании агрессивно. Они пытаются укусить самца, вырываются и обращаются в бегство. По-видимому, кивки головой в вертикальной плоскости имеют сигнальное значение не только при испуге, но и при ухаживании. Часто молодые самки при виде приближающегося самца предупреждают его о «нежелании» контакта, открывая пасть и производя движения головой сверху вниз. По предположению А. Б. Стрельцова, на них действует окраска самца, так как они не обращают внимания на приближение взрослых самок. Самец ведет себя демонстративно по отношению ко всем особям вида, независимо от пола, а также готовности самки к спариванию. Его стремление к спариванию с той или иной особью затухает, если он получает отпор или не получает соответствующего ответа. По-видимому, самец может определять пол ящерицы по ее поведению, а возможно, и по окраске.
Е. Н. Инщикова считает, что после спаривания животные поселяются парами в одной норе. А. С. Баранов, А. В. Яблоков и В. И. Борисов полагают, что самец и самка образуют брачное сожительство, которое длится в течение всего периода спаривания. По данным В. А. Кутузовой (1971), подобного рода сожительство длится всю весну и лето, а по мнению И. С. Дарев-ского (1946), даже 2—3 года. М. И. Щербань в своих предположениях идет еще дальше. Он считает, что сеголетки в первый период находятся все вместе и лишь перед зимовкой расходятся, отыскивая себе убежище. Весной будущего года они нередко мигрируют в поисках пары, что особенно характерно для самцов. В большинстве случаев пары не расстаются без причины, каковой может быть гибель одного из партнеров. Есть и противоположное мнение по вопросу о наличии брачных пар. Так, Н. В. Щепотьев (1950) и М. Ф. Тертышников считают, что самцы и самки встречаются только в короткий период спаривания, когда они действительно при опасности могут прятаться в одну нору.
Откладка яиц и выход сеголетков
Ящерица откладывает яйца в специально вырытую ямку как в собственных норах, так и в других местах, расположенных в достаточно увлажненных и освещенных солнцем местах. По наблюдениям 3. П. Хонякиной (1964), песок, завезенный для строительства, привлекает ящериц в период откладывания яиц, по-видимо.му, рыхлостью, хорошим прогревом и влажностью.
260
М- Ф- Тертышников находил в степных биотопах Ставрополья кладки яиц прыткой ящерицы, отложенные в выбросах земли из нор слепышей.
Процесс откладки яиц прыткой ящерицы наблюдал А. Б. Стрельцов. Самка располагается таким образом, что клоака ее оказывается над серединой склона ямки. По телу животного проходит волнообразное движение, и из клоаки выталкивается яйцо, которое покрыто небольшим слоем слизи. Между откладыванием каждого яйца проходит около двух минут. Яйца в ямке располагаются обычно в горизонтальном положении. По окончании откладки яиц ящерица засыпает их землей, и они оказываются на глубине 4—6 см.
Сеголеток перед вылуплением прорывает оболочку яйца, а после выхода из него отползает в сторону.
Поведение при рытье нор
И. С. Даревский (1946) описывает, что перед рытьем норы ящерица расчищает головой выбранное место. После этого она копает грунт несколько наклонно вниз поочередно то правой, то левой передними конечностями, помогая бурящими движениями головы с закрытыми глазами.
Поведение в неволе
Ящерица, помещенная в вольер, непрерывно бегает, притрагивается языком к различным предметам и ищет из него выход. Убедившись в отсутствии такового, она пытается выкопать нору и спрятаться в нее или под какой-либо предмет. Уже через несколько дней она начинает привыкать к обстановке, меньше боится человека, а на других ящериц почти не обращает внимания (см. рис. 80). Некоторые ящерицы после нескольких дней содержания в террариуме настолько привыкают к человеку, что не пугаются его и легко дают взять себя в руки, погладить и пр.
Реакция содержащихся в неволе ящериц на погоду та же, что и в естественных условиях: в пасмурные дни активность резко понижена, в солнечные дни, даже в середине зимы, ящерицы значительно более активны. В то же время в условиях террариума общая продолжительность суточной активности видимо меняется. Так, при постоянной комнатной температуре ящерицы не уходят в спячку, а период спаривания и откладки яиц наступает (как показывают наблюдения на протяжении нескольких лет) в декабре—феврале.
261
АКТИВНОСТЬ
Методы учета активности
Наибольшее распространение в герпетологических исследованиях получил учет численности на ленточных пробах (трапсек-тах). Эта методика достаточно подробно изложена .в работах Л. М. Андрушко (1936), Л. Г. Динеомана, М. Л. Калецкой (1952) и М. Ф. Тертышникова (19726; 1973в) и заключается в следующем.
Прыткую ящерицу учитывают па ленте, ширина которой не превышает 2 м, а длина — от нескольких десятков до 1000 м. Каждая учетная лента должна находиться .в пределах одного биотопа. В связи с неодинаковой плотностью поселения ящериц в разных частях биотопа рекомендуется многократная прокладка пробных лент в пределах изучаемого биотопа на расстоянии 5—10 м, причем маршрутный учет необходимо проводить при оптимальных условиях активности ящериц. В каждой географической зоне следует опытным путем определить эти условия. Так, для Ставропольской возвышенности установлено, что прыткая ящерица наиболее активна в период размножения — с середины мая до середины июня в первой половине дня при температуре воздуха 25—34°, почвы — 24—32°, относительной влажности воздуха 50—80% и скорости ветра от 0 до 12 м/сек.
Учет ящериц на ленточных пробах не позволяет определить их абсолютную численность, но остается незаменимым для получения ориентировочных данных о численности и биотопическом распределении этих животных и позволяет решать некоторые вопросы, связанные с их активностью и пр.
Для установления абсолютной численности следует закладывать изолированные площадки, размер которых должен быть равен 300 м2. Понятно, что изолированную площадку необходимо закладывать в пределах одного биотопа и отлавливать на ней ящериц в указанные выше сроки максимальной суточной и сезонной активности при наиболее благоприятных погодных условиях. Несмотря на трудности, связанные с оборудованием площадки (на ее закладку, включая изоляцию заборчиком из рубероида или полиэтиленовой пленки, требуется 6—8 человекочасов), данный метод позволяет за первые три дня работы определить абсолютную численность ящериц.
Естественно, что для определения оптимальных погодных условий для ящерицы необходимо в процессе предварительной работы измерять влажность приземного слоя воздуха, температуру почвы, скорость ветра, температуру тела ящериц, фиксировать наличие осадков.
262
Суточная активность
Общий ход суточной активности
Известно, что прыткая ящерица имеет дневной цикл активности (Hoffmann, 1955, 1957; Тертышников, 19726, и др.). По литературным данным, самый ранний .выход ящерицы из убежища отмечен в 5 час. 45 мин. (Кутузова, 1971; Дагестан), а наиболее поздний уход —в 20 час. (Тертышников, 19726; Ставропольская возвышенность). Летом это животное появляется из убежищ раньше, а уходит позже, чем осенью и весной. В зависимости от нарастания дневных температур у прыткой ящерицы может быть один — два пика активности, причем, весной и осенью их активность имеет один пик, а летом—два: утренний и вечерний с перерывом в самое жаркое время дня (Браунер, 1923; Красавцев, 1939; Либерман, Покровская, 1943; Даревский, 1946; Киселев, 1950; Хонякина, 1964; Щербак, 1966; Кутузова, 1971; Тертышников, 19726; Боченко, Стрельцов, сообщ.). Основными факторами, влияющими на время выхода ящериц из убежищ, являются температура и осадки. Исследования М. Ф. Тертыш-никова (19726) показали, что на Ставропольской возвышенности в хорошую погоду прыткая ящерица выходит из убежищ при интенсивности освещения около 200 лк. Из убежищ она появляется не сразу, а некоторое время лежит у входа, выставив голову наружу. В дальнейшем она полностью выползает из норы и греется в лучах солнца. При разной по температуре погоде прыткая ящерица затрачивает на обогрев от 0 до 160 мин. Достаточно прогревшись, ящерица начинает перемещаться в пределах индивидуальной территории. Интересно привести некоторые наблюдения М. Ф. Тертышникова за перемещениями взрослого самца прыткой ящерицы в разнотравно-злаковой степи на Ставропольской возвышенности. Выход животного из норы был отмечен в 7 час. 45 мин. В течение дня он прошел 145 м, сделав 10 остановок разной продолжительности, на которые затратил всего 30 мин. Несколько раз самец возвращался в нору и провел в ней 6 час. Окончательный уход в нору был отмечен в 18 час. 30 мин. Таким образом, на поверхности это животное провело всего 4 час. 15 мин.
В плохую погоду выход ящерицы из убежища затягивается и может даже и не произойти. Перед заходом солнца ящерица тоже греется в лучах солнца у входа в нору. На ночь она прячется в убежище при интенсивности освещения от 21 000 до 280 лк. Ниже мы приводим рис. 82 *, показывающий характер сезонной и суточной активности данной ящерицы в условиях Ставропольской возвышенности.
Для выяснения активности Lacerta agitia в лабораторных условиях был поставлен следующий эксперимент (Подмарев).
Форма обработки материала заимствована у Н. М. Окуловой (1969).
263
Месяц и декада. Часы суток
7 8 I 9 70 /7 12 13 14 15 16 17 18 19 20
ш 3
/ . ':-Ж
2
3
Т 1
г 3 —
ж 1
2 —-
3
тш / 2 III—
3 у Ж
тли 1 Цж Ж
г in и 1 я ж жж
3
Жж '//// Ц ш ж
п / г 3
///^ Ж
W ->Ж/ У/// У///, У///
щ Ц ж ж ж
Z 1
\УУ/У ЖЖ ж ж /ууу, ж У/у
г ж 1 ж
/ Ик И-?
Рис. 82
Суточная и сезонная активность прыткой ящерицы на Ставропольской возвышенности 1 —• вся популяция, кроме сеголетков; 2 — единичные встречи взрослых и полувзрослых особей; 3 — сеголетки (по Тертышникову, 19726)
Ящериц разделили на три группы. Первая группа обогревалась лампами накаливания круглые сутки. Вторая группа обогревалась с 8 час. утра до 18 час. вечера. Третья — контрольная группа — содержалась при комнатной температуре с естественным освещением и общим освещением лаборатории, но без дополнительного обогрева. Опыт проводится в течение года.
При сравнении активности ящериц первой и второй групп различия в продолжительности суточной активности не были обнаружены. Наблюдения показали, что максимальная активность этих групп приходится на период с 11 до 14 час. дня. К 17—
264
18 час. все ящерицы постепенно уходят в укрытия. С 8—9 час. утра ящерицы обеих групп выходят из укрытия и скапливаются у источника тепла, и к 11 час. все находятся в активном состоянии. В это же время замечены активное кормление и поиск пищи.
В контрольной группе, содержащейся без дополнительного обогрева, суточная активность выражена не так четко по времени, как у первых двух групп. Ящерицы контрольной группы редко выходят из укрытия на свет, плохо поедают корм в сравнении с первыми группами. Ящерицы контрольной группы подвержены различным заболеваниям (кожным, заболеваниям глаз и др.) и гибели. Следует отметить, что в первых двух группах этого эксперимента случаев заболевания ящериц не было.
За осенне-зимний период эксперимента существенных сезонных колебаний активности замечено не было. Лишь к весне активность выживших ящериц из контрольной группы стала приближаться к активности первых двух групп. В летний период все три группы по активности были одинаковыми.
Влияние погоды на суточную активность ящерицы. Температура тела и активность
По данным С. С. Либермана, Н. В. Покровской (1943; Московская область), температура тела активной прыткой ящерицы находится в пределах от 7 до 41° (рис. 83, см. рис. 57). В Крыму температура тела активной прыткой ящерицы варьирует от 9 до 40° (Щербак, 1962, 1966), а на Ставропольской возвышенности— от 14 до 36° (Тертышников, 19726).
Наблюдения показывают, что прыткая ящерица активна при температуре воздуха от 4—7 (Стрельников, 1944) — 10,0 (Тертышников, 19726) до 28,0 (Щербак, 1966) —32,5° (Тертышников, 19726). Изучая действие температуры среды1 на прыткую ящерицу в условиях Ставропольской возвышенности, М. Ф. Тертышников (19726) установил, что температура ее тела до 14° совпадает с температурой среды (рис. 84). При повышении температуры среды до 32—35° температура тела ящерицы оказывается выше на 0—7,5° или ниже на 0—14,5°. Однако при дальнейшем повышении температуры среды до 45° температура тела этого животного оказывается ниже ее на 0—7 или выше на 0—8°. Здесь же следует указать и на то, что молодые особи более пластичны к низкой температуре, но хуже переносят высокую температуру.
В различных регионах наибольшая активность ящериц проявляется при разной температуре. Так, например, в Московской области наибольшая активность ящериц зарегистрирована при температуре тела от 28 до 35° (Либерман, Покровская, 1943) (см. рис. 57, 83), а в Крыму — от 18 до 26° (Боченко, сообщ.).
В данном случае и далее в тексте этой главы под «средой» понимаются почва и воздух.
265
Избираемая оптимальная i, °C
t активных ящериц В прироое
i тела в оптимальных условиях
Смерть
Оцепенение
___I____\___
Неполная активность
Полная активность
Смерть
5137 11 15 13 53 27 __31 35 33 43 45
Температура, °C
Рис. 83
Сводная схема температурных условий существования прыткой ящерицы в лесной и лесостепной зонах (по Либерман, Покровской, 1943)
В Ставропольском крае это животное наиболее активно при температуре тела, равной 29,8, почвы—29,0 и воздуха — 26,5—28,2° (рис. 84, А, Б). В этой связи интересно отметить, что максимальное количество кислорода прыткая ящерица в условиях данного региона поглощает при 30° (Тертышников, 19726, 1973а).
При температуре среды и тела ниже 4—10° прыткая ящерица малоподвижна и в сезон активности обычно находится в убежище. По мере дальнейшего охлаждения почвы и воздуха ящерица теряет способность к активным перемещениям и впадает в оцепенение. С. С. Либерман и Н. В. Покровская (1943) установили, что в оцепеневшее состояние прыткая ящерица впадает при 7,5° (см. рис. 83). Использование микроклимата — одно из важнейших приспособлений ящерицы. В прохладные, но ясные дни она обогревается в лучах солнца и использует тепло нагретого грунта, прижимаясь к нему телом. Подобное поведение животного указывает на то, что солнечная радиация и тепловое излучение субстрата являются основными факторами, определяющими температуру его тела, и что температура воздуха имеет при этом меньшее значение. В подтверждение этого можно привести данные И. Д. Стрельникова (1944), из которых следует, что при температуре воздуха 4—7° температура тела прыткой ящерицы была равна 29,4—33,0°. Быстрое перемещение в холодную погоду также поднимает температуру тела ящерицы. И. Д. Стрельников (1934, 1944, 1959) отмечает, что после короткой пробежки температура тела прыткой ящерицы поднялась на 10°. Получив заряд тепла в условиях предварительного обогрева или после бега, ящерицы используют его, на некоторое время выходя из зависимости от низкой температуры окружающей среды.
266
Рис. 84
Зависимость температуры тела прытких ящериц разных возрастных и половых групп от температуры почвы (Л) и температуры воздуха (J5)
/ — взрослые самцы; 2 — взрослые самки; 3 — сеголетки (по Тертышникову, 19726)
А. Д. Слоним указывает, что у рептилий не удается обнаружить адаптаций к холоду. Но, по всей видимости, именно наличие таких адаптаций обеспечивает активность прыткой ящерицы при низкой температуре среды и определяет возможность ее обитания высоко в горах на южной границе ареала.
Выше было сказано, что, выходя из убежища, прыткая ящерица в прохладную погоду обогревается в лучах солнца. Обогревается она и после приема пищи. Когда ее тело сильно нагреется, ящерица ложится на брюхо, приподнимает конечности и хвост, покачивает головой и быстро моргает. В таком состоянии она остается на несколько минут, а затем быстро вскакивает и убегает в тень или же перебегает с места на место по освещенному солнцем участку. В таком же возбужденном состоянии ящерица бывает и при высокой температуре среды. Она быстро перемещается по субстрату, избегая прикасаться к нему телом, и вскоре во избежание перегрева уходит в тень от различных предметов, где температура среды ниже, чем на освещенных солнцем участках. В таких местах ящерица избегает также и прямого действия солнечных лучей, а температура ее тела снижается и определяется преимущественно температурой почвы. Кроме того, во время дневной жары прыткая ящерица способна влезать на ветки кустарников и стволы деревьев, где она избегает контакта с нагретым субстратом и где ветер снижает температуру ее тела. Весьма сходную реакцию прыткой ящерицы на действие высокой температуры описывают С. С. Либерман, Н. В. Покровская (1943) и В. В. Черномордиков
267
Изложенный выше материал показывает, что при высокой температуре среды для прыткой ящерицы существуют определенные температурные пределы, ограничивающие пребывание этого животного на поверхности. По-видимому, для каждой географической зоны характерны определенные пределы. Для Ставропольской возвышенности, например, такими пределами, ограничивающими пребывание прыткой ящерицы на поверхности, являются температура почвы 32, воздуха —30,6 и тела 36° (рис. 85).
Температура тела, °C
Активность прыткой ящерицы в зависимости от температуры среды (а ) и тела (б) по материалам наблюдения на стационаре в Ставропольском крае
1 — температура почвы; 2—температура воздуха (по Тертышникову, 19726)
Рис. 85
Температурные пределы, при которых способна жить прыткая ящерица, довольно значительны. Нижняя летальная температура равна —4,9° (Либерман, Покровская, 1943). Верхняя же летальная граница варьирует в значительной мере в разных регионах.
В Московской области, например, она равна 44,7° (Либерман, Покровская, 1943) (см. рис. 83), на Ставропольской возвышенности— 41,0—41,6° (Тертышников, 19726), а в горах Кавказа—43—47° (Стрельников, 1944). Различия такого рода можно объяснить, по-видимому, наличием географической изменчивости терморегуляции ящерицы, которая, по мнению Л. Г. Динесмана (1949), имеет свои особенности в каждой географической зоне. В естественных условиях температура тела данной ящерицы не поднимается до указанных величин: у нее имеются элементы регулирования тепла, благодаря которым усиливаются или ослабляются действия того или иного фактора в каждой конкретной ситуации, а температура тела поддерживается на определенном уровне жизнедеятельности в течение дневной активности для обеспечения нормального протекания жизненных процессов. По И. А. Шилову (1962), эти элементы регулирования тепла не являются терморегуляцией: он называет их температурными адаптациями. Помимо перечисленных
268
выше различного рода термотактических перемещений, к ним можно отнести еще испарение воды с поверхности тела и дыхательных путей.
Влажность, осадки и активность
О влиянии влажности среды на поведение прыткой ящерицы известно сравнительно немного. Так, по наблюдениям Н. Н. Щерг бака (1966), прыткая ящерица в Крыму активна в условиях относительной влажности воздуха от 56 до 100% при оптимальной ее величине, равной 56%. М. Ф. Тертышников (19726) установил, что на Ставропольской возвышенности прыткая ящерица активна при 40—100%-ной относительной влажности воздуха, причем, наибольшая ее активность проявляется при
го 40 60 во т г 4 6 в ю iz 14 № № го гг
Относительная влажность Снорость ветра, м/сен
воздуха, 7О
Рис. 86
Активность прыткой ящерицы в зависимости от влажности воздуха (а ) и скорости ветра (б )
По Тертышникову (19726)
62—90%-ной влажности (рис. 86, Л). По его же мнению, именно низкая влажность воздуха во второй половине лета вынуждает большую часть взрослых и полувзрослых ящериц впадать в летнюю спячку. В сопоставлении с температурой среды влажность имеет подчиненное значение, и ее высокий показатель не ограничивает выхода ящериц из убежищ.
Влажность среды в значительной мере формирует микроклимат и имеет большое значение в биотоппческом размещении прыткой ящерицы: определяет наличие миграций (см. гл. V и IX) и выбор места для откладки яиц; установлено, что яйца могут нормально развиваться только при наличии 2,5—17,5% влаги в почве (Либерман, Покровская, 1943).
Непосредственное влияние на жизнедеятельность ящериц оказывают выпадающие осадки и стихийные бедствия, связанные с ними. Так, Л. Г. Динесман (1950) показал, что отсутствие ящерицы в нижней зоне дельты Волги вызвано ее неспособностью переносить паводки. По данным М. Ф. Тертышникова (19726), периодические ливни в районе Прикалаусских высот на Ставропольской возвышенности весной и летом 1968—1969 гг. привели
269
к гибели части популяций прыткой ящерицы, а отсутствие снегового покрова на некоторых участках не могло предохранить этих животных от гибели при действии низких зимних температур, когда в холодную зиму 1968 г. почва на таких участках промерзла на глубине до 100 см.
Сильный ветер снижает температуру тела ящериц и подавляет их активность при низкой температуре среды, но позволяет им быть активными при высокой температуре среды. По данным М. Ф. Тертышникова (19726), прыткая ящерица на Ставропольской возвышенности активна при скорости ветра от 0 до 22 м/сек. Максимум активности ее отмечен при ветре, скорость которого равна 5 м/сек (см. рис. 86, Б).
СЕЗОННАЯ АКТИВНОСТЬ
Зимняя спячка
По данным различных исследователей, время ухода прыткой ящерицы неодинаково в .разных частях ареала. Так, возле Перми они уходят в спячку в начале сентября (Чащин, Литвинов, сообщ.), на Среднем Поволжье — с конца августа до 20 сентября (Щепотьев, 1948, 1950, 1952), в Ставропольском крае — с июля-августа (Тертышников, 19726) —конца сентября до середины октября (Красавцев, 1939; Тертышников, 19726), в Дагестане— в середине октября —середине ноября (Хонякина, 1964; Кутузова, 1971), в Ростовской области и Краснодарском крае — в начале октября—ноябре (Лукина, 19666), в Кабардино-Балкарии — в октябре — середине ноября (Неемченко, Темботов, 1959), в Казахстане — в середине ноября (Параскив, 1956), а в Крыму —в сентябре — октябре (Щербак, 1966) и даже в ноябре (Браунер, 1923; Киселев, 1950).
Первыми в спячку уходят взрослые и полувзрослые особи, а затем и сеголетки. Последние встречаются только в самое теплое время, которое приходится на середину дня, но продолжительность их дневной активности уменьшается с каждым днем вплоть до ухода в спячку. Столь длительная задержка сеголетков на поверхности осенью, по мнению Н. Н. Щербака (1966), объясняется большей их, чем взрослых особей, пластичностью к температуре среды. И. С. Даревский (1967) видит причину позднего ухода сеголетков в спячку не только в этом, но и в характере накопления жировых запасов, необходимых для успешного проведения зимовки.
Неодинаково и время выхода ящериц после спячки в разных регионах. Так, в Пермской (Чащин, Литвинов, сообщ.), Горьковской, Ивановской (Красавцев, 1936) и Курской (Птушко, 1934) областях и в Среднем Поволжье (Щепотьев, 1948, 1950) ящерицы выходят из спячки во второй половине апре
270
ля — мае; в Калининской (Шапошников, 1959), Брянской (Федосеев, Никитин, 1951), Смоленской (Граве и др., 1951) и Луганской (Таращук, 1959) областях, а также в северной части Казахстана (Окулова, сообщ.)—в апреле; в Дагестане (Хонякина, 1964, 1970; Кутузова, 1971)—с середины и до конца марта, на Ставропольской возвышенности (Тертышников, 19726), в Краснодарском крае и Ростовской области (Лукина, 19726), а также в Крыму (Щербак, 1966)—в конце марта — апреле, в Кабардино-Балкарии (Парфеник, Нодъяпольский, 1951; Шеб-зухова, 1969)—во второй половине марта, на равнинах Чечено-Ингушетии — в первой декаде марта, а на альпийских лугах того же региона — в конце марта — во второй половине апреля (Рашкевич, сообщ.).
Даже в пределах одного региона сроки пробуждения ящериц неодинаковы в разные годы. М. Ф. Тертышников (19726), например, установил, что на Ставропольской возвышенности в 1966 г. прыткая ящерица вышла из спячки 1 апреля, в 1967— 1968 гг.—20—25 апреля, в 1969 г.—27 апреля, в 1970 г.—23 марта, в 1971 и 1973 гг.—26 марта. Как правило, массовое появление ящериц происходит несколько позже. По данным того же исследователя, температура среды в момент первого появления ящериц относительно низка. Например, в 1970 г. температура воздуха и почвы была 12,0°, а в 1971 г.—15,0°. Однако в течение нескольких предыдущих дней почва прогревалась до 30° и выше. Понятно, что в месте нахождения спящей ящерицы почва прогрелась, и ее температура удерживалась в диапазоне температур активности животного, что вывело его из спячки и содействовало выходу на поверхность. Пробудившиеся ящерицы показываются на поверхности только в теплую погоду, а в холодную находятся в убежищах.
Таким образом, несовпадение срока выхода ящериц из спячки в разные годы определяется преимущественно температурным режимом среды, а сроки пробуждения их после зимней спячки зависят от установления такой температуры среды, при которой возможна их активная деятельность.
Весьма разноречивы сведения о половых и возрастных особенностях начала сезонной активности. По данным Б. А. Кра' савцева (1936) и Н. В. Щепотьева (1948, 1950), в Горьковской и Ивановской областях, а также в Среднем Поволжье первыми после спячки появляются взрослые, а потом молодые и полувзрослые ящерицы. В Пермской области первыми появляются сеголетки (Чащин, Литвинов). М. Ф. Тертышников (19726) установил, что на Ставропольской возвышенности появление ящериц разного пола и возраста неодинаково в разные годы. В 1966 и 1969 гг. он первыми обнаружил взрослых самцов и самок, в 1967 и 1969 гг.-—взрослых самцов, а в 1970, 1971 и 1973 гг.— молодых особей и взрослых самцов. По его предположению, сроки пробуждения зависят от глубины расположения
271
спящей ящерицы: чем глубже она находится в убежище, тем позже выйдет на поверхность после спячки.
Прыткая ящерица зимует в различного рода убежищах. Обитая в лесу, она проводит зиму в норах, под корнями деревьев и кустарников, но иногда зарывается в мох, опавшие листья и хвою. Б. А. Красавцев (1936) находил сеголетков под отставшей корой старых пней. В степных же биотопах ящерица, как правило, зимует в норах.
По данным М. Ф. Тертышнпкова (19726), прыткая ящерица на Ставропольской возвышенности зимует одиночно, располагаясь в норах на глубине 26—35 см.
Летняя спячка
Как было отмечено в главе IX, летняя спячка прыткой ящерицы зарегистрирована в Воронежской (Северцов, 1950) и Харьковской областях (Чернай, 1852—1853), а также и на Ставропольской возвышенности (Тертышников, 19726). По-видимому, она может быть и у ящериц в Среднем Поволжье.
Основной причиной, вызывающей спячку летом у прыткой ящерицы, следует считать, по-видимому, повышение температуры среды с одновременным резким понижением относительной влажности среды.
Таким образом, уже накопилось много сведений, касающихся отношения ящериц к климатическим факторам и их поведения в разные периоды жизни. Тем не менее до сих пор остаются невыясненными особенности поведения ящериц в разных биотопах, влияние условий биотопа на суточную и сезонную активность, вопросы, касающиеся особенностей зимней и летней спячки, взаимоотношения ящериц разных полов и возрастов и др.
Общие особенности сезонной активности
Продолжительность дневной активности прыткой ящерицы неодинакова в разные сезоны и отражает отношение ее к сезонным условиям существования. Начиная со дня выхода из спячки, ящерицы еще малоактивны, появляются на поверхности в наиболее теплое время суток. С установлением постоянно теплой погоды со второй половины весны их активность достигает максимума и носит уже двухвершинный характер. С конца июня продолжительность дневной активности начинает постепенно сокращаться вплоть до ухода ящериц в спячку.
Неодинаковые физико-географические условия разных частей ареала определяют различную продолжительность сезонной активности прыткой. ящерицы. Так, в Среднем Поволжье продолжительность сезонной активности ящерицы равна 135—150 дням, в Крыму—165—180 дням, в Ростовской области и Краснодарском крае —180—210 дням, на Ставропольской возвышенности —195—210 дням, а в Дагестане —210—225 дням.
ГЛАВА ОДИННАДЦАТАЯ
СТРУКТУРА ПОПУЛЯЦИИ1
Говоря о структуре популяций высшего позвоночного с длительным периодом онтогенеза, мы имеем в виду возрастной и половой составы популяции, а также характер пространственного распределения популяции (а, соответственно, и всего вида), что необходимо при любом определении места вида в природе и его значения в конкретных биогеоценозах. Рассмотрение вопроса о структуре популяций для прыткой ящерицы особенно интересно в связи с тем, что она занимает огромный географический ареал с самыми разнообразными условиями существования, и основные параметры популяционной структуры могут быть в связи с этим весьма изменчивы.
ПОЛОВАЯ СТРУКТУРА ПОПУЛЯЦИИ
По имеющимся полевым наблюдениям, охватывающим период со второй половины апреля до второй половины сентября, соотношение полов в популяции прыткой ящепицы оказывается
Таблица 42
Среднее соотношение самцов и самок по месяцам (сводные данные по всему ареалу, Яблоков, Баранов, Валец кий, 1970—1973 гг.)
Соотношение Апрель Май Июнь Июль Август Сентябрь
Взрослые самцы/Взрослые самки 2,22 1,05 0,88 0,81 0,77 0,59
Все самцы/Все самки 1,94 1,05 0,80 0,78 0,70 0,65
Число изученных популяций 6 33 21 18 4 2
несколько различным в разные сезоны (табл. 42). В апреле обычно встречается больше самцов, чем самок (Лукина, 19666). За-тем, в мае, относительная численность активных самок резко
1 Автор: А. В. Яблоков, при участии А. С. Баранова, А. В. Валецкого, А. М. Андрушко, М. Ф. Тертыимикова, Г. П. Лукиной, В. К. Жарковой, п Добровольской, К. Шафраньской, А. Б. Стрельцова, В: 1\. Щепотьев.г, Л. В. Турутиной, Н М. Окуловой.
273
возрастает и, начиная с июня, становится несколько выше численности самцов. В сентябре активных самок встречается вдвое больше, чем самцов.
Конечно, приведенные данные не могут отражать истинного соотношения особей разного пола в популяциях, поскольку отражают скорее относительную активность особей того или иного пола (Лукина, 19666). В данном случае можно говорить об «экологической» структуре популяций. Эта структура может изменяться из года в год в одной и той же популяции (Окулова, сообщ.) .
Истинное соотношение полов в популяции ящериц, видимо, во все периоды близко 1:1 (Красавцев, 1936; Даревский, 1946; Щепотьев, 1948; Лукина, 19666). Именно такое соотношение чаще всего наблюдается среди многочисленных выборок мелких неполовозрелых животных (как весной, после зимовки, так и среди сеголетков), а также среди особенно многочисленных выборок, собранных на протяжении нескольких дней в одной популяции (Яблоков, Баранов, Валецкий, сообщ.). В то же время в некоторых случаях достоверно и резко различается встречаемость самцов и самок. Так, например, на р. Техури (Западная Грузия) и в мае и в сентябре при тщательном многодневном сборе самки (и среди молодых и среди взрослых) встречались в 2— 3 раза чаще, чем самцы. Эти наблюдения кажутся особенно интересными в связи с тем обстоятельством, что популяция ящериц по р. Техури состоит из особей с максимально развитым половым диморфизмом по размерам тела (здесь обитают самые крупные из встреченных нами на протяжении всего ареала самцы, тогда как размеры самок сохраняются средними для вида).
Преобладание численности самок над самцами среди взрослых животных известно и для других районов ареала: Калужской области — 67,5% самок (Стрельцов, в печати); Средняя Польша—58,7 £ $ (Добровольска, Шафраньска, в печати); Ставропольский край — 60,5% 2? (Лукина, 19666), 56,4% 99 (Тертышников, 19726); равнинного Дагестана — 61,5% (Хонякина, 1970); окрестностей Киева — 60,5% (Даревский, 1946); Рязанской обл.—64,6% (Жаркова, 19736) и др. Подобные наблюдения позволяют высказать предположение о достаточно сложной половой структуре популяции у прыткой ящерицы, возможно различающейся в разных популяциях и на разных территориях. Данные по резкому половому диморфизму в размерах тела (самки по длине несколько меньше самцов) в некоторых популяциях этого вида (см. рис. 35) позволяют предположить в таких случаях существование усложненной половой структуры популяций, включающей нечто вроде «гаремов» и резкой пространственной дифференцировки особей разного пола и возраста, как это часто наблюдается у полигамных высших позвоночных. Добавим, однако, что такие наблюдения пока единичны и нет достаточных данных, свидетельствующих о возможности
274
Таблица 43
Динамика соотношения самцов и самок на протяжении летнего периода в двух популяциях [по А. М. Андрушко для района Баян-Аула (Казахская складчатая страна), 1936—1938 гг. и для Верхнего парка г. Кисловодска,' 1949 г,]
Месяц Неполовозрелые Половозрелые По всей популяции
1 2 3 4 5 6
Казахстан
Июнь — 1:1,2 1:1,5 1:1,5 1:1,6 —
Июль 1:1 1:1,5 1:2 1:1,6 1:1,6 0:1 1:1,6
Август 1,1:1 1:1,5 1:2 1:2 1:2 —
Сентябрь 1,1:1 1:1,4 1:1,8 1:1,7 1:2 —
Кисловодск
Июль 1,2:1 1:1 1:1,3 1:1,3 1:2,5 1:2 Д • А с
Август 1,2:1 1:0,7 1:1 1:2,8 1:2 1:2 1.1,3
Примечание. Абсолютный возраст разных возрастных групп в разные месяцы следующий: 1 — сеголетки: 2— в июне, 9—10 мес.; 3 — в июне, 1 год) мес.—1 год 10 мес.; 4 —в июне, 2 года 9 мес. — 2 года 10 мес.; 5 — в июне, 3 года 9 мес. — 3 года 10 мес.; 6 — старше 4 лет.
разделения всего ареала вида на зоны с преобладанием того или иного типа соотношений самцов и самок в популяциях (табл. 43).
Решение этого вопроса осложняется и тем обстоятельством, что имеются некоторые данные, косвенно свидетельствующие о большем числе самок самых старших возрастных групп по сравнению с таковыми для самцов, в частности, для популяций из Средней Польши (Добровольска, Шафраньска, сообщ.) и Рязанской области (Жаркова, 19736), и, соответственно, о существовании различного уровня смертности среди старших возрастных групп самцов и самок (по данным Жарковой, 19736). Смертность среди самцов во всех возрастных группах значительно выше, чем среди самок.
ВОЗРАСТНОЙ СОСТАВ ПОПУЛЯЦИЙ
В главе VIII содержится описание точных методов определения возраста ящериц по слоистым структурам скелета. Проведенные исследования позволили установить достаточно надежную корреляцию между числом лет, прожитых ящерицей, и размерами тела (Смирина, 1974). Для общего описания возрастной структуры популяции можно теперь обоснованно пользоваться обычным разделением животных по весу тела, выделяя особей данного года рождения (сеголетки), особей, перезимовавших один раз (годовики), и более старшую возрастную группу, включаю-
275
Рис. 87
Модель динамики возрастного состава популяции прыткой ящерицы
Исходные параметры: / — в августе состав популяции по возрастам (в %) следующий: взрослые — 14, полувзрослые — 28,6, молодые этого года рождения — 57,1; 2— среднее число яиц у каждой взрослой пары — 8 штук; 8 — продолжительность жизни достигает 5 лет; 4 — половая зрелость наступает в возрасте 1,5—2 лет. По оси абсцисс — месяцы и годы, По оси ординат — относительная численность каждой возрастной группы. Сезон выхода из яиц — с середины июля до конца августа; сезон спаривания — с середины апреля до конца мая
щую, как правило, взрослых животных (Красавцев, 1936; Щепотьев, 1948; и мн. др.). В главе III были рассмотрены основные весовые характеристики ящериц на всех этапах их постнаталь-ного онтогенёза и показано существование значительной географической изменчивости размеров тела животных на пространстве всего ареала. При этом известно, что носящая в общих чертах клинальный характер эта изменчивость размеров оказывается весьма сложной по всему ареалу и неполовозрелые ящерицы при данном весе в одном случае будут вполне взрослыми в другом. Все это заставляет при сравнительном описании возрастного состава разных популяций отказаться от указания точных весовых характеристик тех или иных групп животных.
Во второй половине лета с выходом молодых ящериц из яиц (конец июля — август) в популяции присутствуют все три главные возрастные классы: сеголетки, «полувзрослые» (прошлого года рождения) п половозрелые особи в возрасте от двух до пяти лет. Примерное численное соотношение этих групп показано на рис. 87. Ровно через год все особи рождения прошлого года перейдут в разряд «полувзрослых». Этот процесс завершится для одних особей раньше, для других — позже. Аналогичным образом особи из раздела «полувзрослых» постепенно перейдут в разряд взрослых. Этот переход также, видимо, будет дифференциальным для разных особей: возможен случай, когда половая зрелость для одной особи наступит в полтора года, а для другой — в 2,5 года. По данной схеме можно определять возрастную структуру популяций в любое время года.
276
Все сказанное выше относится ко всей популяции в целом. При исследовании лишь части популяционного населения (например, при изучении лишь части занимаемой популяцией территории, либо при изучении популяции в краткий период времени, не охватывающий несколько разной суточной и сезонной активности разных возрастных групп), возможна преимущественная встреча особей только некоторых возрастных групп. Так, например, при изучении популяции в сентябре — октябре встречаются в активном состоянии в основном молодые особи (этого и прошлого года рождения), а весной наблюдаютсящрезвычайно активные взрослые самцы. К этому необходимо добавить, что принятые за исходные на рис. 87 численности разных возрастных групп в популяции, окажутся, видимо, несколько иными в разных районах ареала вида. Имеются данные (см. гл. VII), показывающие, что среднее число яиц на одну самку различно в разных районах [в полупустынной зоне составляет около 6 (Турутина, Окулова, сообщ.), а на севере ареала может снижаться и до 3—4]. Соответственно, различным должно быть и общее число выходящих из яиц молодых (если только темп эмбриональной смертности не «выравнивает» в какой-то степени географическую изменчивость по числу откладываемых яиц). В приводимой выше схеме (рис. 87) за исходную величину был взят темп эмбриональной смертности в 50% (выживает только 50% от общего числа отложенных яиц).
Возрастной состав популяции может быть усложнен и другими факторами. В некоторых популяциях могут развиваться либо две самостоятельные кладки (Красавцев, 1936; Щепотьев 1948; Щербак, 1966; Симонян, 1973, и др.), либо кладка может быть порционной и растянута на очень большой промежуток времени. В случае двух кладок за летний сезон (такое положение для центральных и южных, но не горных (Хонякина, 1970) районов ареала, с длительным вегетационным сезонным и более продолжительным периодом активности], каждое новое поколение может состоять не из одного, а двух приплодов. Каждая старшая возрастная группа включает в себя особей нескольких поколений. Кроме того, имеются данные (Лукина, 19666), показывающие, что темп размножения впервые вступающих в размножение самок значительно ниже, чем самок среднего возраста; в самых старших возрастных группах темп размножения снова несколько снижается. Таким образом, в целом возрастная структура популяций прыткой ящерицы оказывается достаточно сложной (табл. 44).
Последним вопросом этого раздела, требующим рассмотрения в плане настоящей монографии, является вопрос о возможном существовании различий возрастной структуры внутри ареала вида. Пока нет точных данных по этому вопросу. В общей форме утверждение о наличии трех главных возрастных групп (сеголетки; «полувзрослые», в возрасте 1—2 года, и стар-
277
Таблица 44
Относительная численность разных возрастных групп в разные месяцы в трех популяциях (данные А. М. Андрушко по популяции Баян-Аул, 1936—1938 гг,. Верхний парк
г. Кисловодска, 1949 г,, по заповеднику «Лес на Ворскле»^ 1949 г.)
Месяц Возрастная группа
1-я 2-я З-я 4-я 5-я 6-я
Популяция Баян-Аула, Казахская складчатая страна
Июнь — 23 11 43,5 22,5 —
Июль 20,7 11,5 6,9 36,8 23,1 1
Август 35,4 19 11,4 22,8 11,4 —.
Сентябрь 47 13,2 9,9 19 11 —
Популяция Верхнего парка в г. Кисловодске
Июль 1 I Ю,2 1 24,8 1 21,7 21,7 I 16,6 I 1 3
Август 24,4 1 18,5 I 14,8 18,5 I 14,8 9
Июль | 1 - Популяция 1 36 1 «Лес на Ворскле» 18 [ 28,8 | 17,2 | —
Примечание. Абсолютный возраст разных возрастных групп в разные месяцы см. табл. 43.
шая возрастная группа, свыше 2 лет) справедливо для всех изученных популяций вида. Однако, несомненно, что разница между полувзрослыми и взрослыми особями оказывается существенно различной в разных популяциях; не исключено, что последующие исследования обнаружат несколько более раннее наступление половозрелостп в одних популяциях по сравнению с другими. Наконец, возможно, что максимальная продолжительность жизни также окажется несколько различной на разных участках ареала (4—6 лет).
ПРОСТРАНСТВЕННАЯ СТРУКТУРА ПОПУЛЯЦИИ
Точные данные о пространственной структуре значительного числа популяций отсутствуют, поскольку для прыткой ящерицы, как и для всех позвоночных, практически нет сведений о границах популяций (Тимофеев-Ресовский и др., 1973). Будучи достаточно тесно связанными с определенными пригодными для жизни местообитаниями и обладая сравнительно незначительным радиусом индивидуальной активности (порядка 5—15 м), группы особей прыткой ящерицы внутри популяции оказываются распределенными достаточно неравномерно. Эта неравномерность населения определяется неравноценностью- Отдельных участков как большой территории, так и внутри даже одного биотопа. Сплошной тип поселений свойствен лишь степным био-
278
Рис. 88
L -1
Д -2
М ~3^
Ъ -7
(у) ~8
-9
1 — самец;
2 — самки:
3, 4 — молодые самки и самцы;
5 — индивидуальный участок;
6 — иора;
7 — дерево;
8 — граница нескоше-ной травы;
9 — кустарник; 0 — май 1969 г.;
I — июль — август 1969 г.;
II — май 1971 г.;
III — до первого покоса (май — начало июня) 1973 г.;
IV — после покоса (конец июня) 19/3 г.
Распределение особей прыткой ящерицы внутри одного поселения в разные сезоны в годы (данные и рисунок М. В. Тертышникова)
Рис. 89
Распределение групп особей прытких ящериц (семей, а может быть демов) в разных биотопах
Калужская обл.» 1972—1974 гг. (данные А. Б. Стрельцова, рис. А. С. Баранова)
топам. Часто внутрипопуляционные объединения поселения ящериц оказываются «мозаичными», образуя как бы кружевной узор (рис. 88). Однако наиболее типичными оказываются поселения третьего типа — «ленточные», или «диффузные» (Тертышников, 19726), как бы вкрапленные в относительно более обширные пространства, не имеющие постоянного населения. Примером таких поселений может служить распределение группировок прыткой ящерицы по разным биотопам Калужской области (рис. 89).
Следует учитывать, что границы между выделенным!! типами поселений оказываются подвижными: ежегодно, после выхода молодых, территория, занимаемая поселениями ящериц, резко увеличивается за счет распространения молодых особей по соседству с индивидуальной территорией самки (Тертышников, 1972). В годы высокой численности миграции ящериц, направленные обычно со стороны сплошных (т. е. более мощных) поселений в сторону мозаичных и диффузных поселений, также изменяют эти границы.
Внутри популяции взрослые ящерицы распределены неравномерно, обитая вблизи надежных укрытий (подробнее см. гл. V). Видимо, от наличия укрытий зависит и величина внутрипо-пуляционных группировок ящериц. Самые мелкие группировки состоят из нескольких (4—6 взрослых) особей, тогда как самые крупные из отмеченных группировок ящериц (временные скопления особей, например у воды, могут быть очень большими) могут достигать нескольких десятков особей (Щепотьев, 1948; Лукина, 19686; и др.). Являются ли эти группировки особей семейными демами, надо решать в каждом конкретном случае специально, но такая вероятность не исключена. Во всяком случае существуют многочисленные наблюдения, указывающие как на длительность (до 2—3 лет) обитания определенных особей в конкретных норах и на длительные контакты между одними и теми же самками и самцами (Даревский, 1946; Лукина, 1966; Жаркова, 19736; Тертышников, 1972; Щербань и др., сообщ.), так и на встречу самцов и самок на зимовке в одной норе (Яблоков и др., сообщ., район оз. Иссык-Куль), равно как и более частые встречи летом самцов и самок в одной норе. Интересным являются случаи встреч в одной норе нескольких взрослых ящериц (Стрельцов и др., сообщ.), большая часть которых оказалась взрослыми самками и только один — взрослым самцом (см. также рис. 88—89). Максимально тесные группы, состоящие из нескольких десятков особей, обнаружены в Западной Белоруссии (район г. Ковеля). Они обитали в единой колонии: отдельные норы были соединены между собой сетью сообщающихся ходов. Ориентировочная численность особей в такой колонии составляла не менее 25—30 взрослых животных (Баранов, Яблоков, сообщ.). Мелкие группировки типа демов обычно объединяются в более крупные в том случае, если подходящий для
281
обитания ящериц биотоп занимает достаточно обширное пространство, например травянистый склон холма, длинный овраг, мощные заросли кустарников, лесополоса, просека в сосновом лесу и т. п. Численность таких вторичных группировок может достигать весьма значительных величин, порядка нескольких сотен взрослых особей, хотя чаще бывает меньше — несколько десятков особей. Так, например, на склоне холма площадью 300 м2 обитало несколько мелких групп общей численностью (за последовательные три года) 23 самца+33 самки; 19 сам-цов+31 самка; 16 самцов+25 самок (Лукина, 19666).
Итак, имеющиеся данные показывают, что внутри популяции ящерицы распределены неравномерно, небольшими (от нескольких до нескольких десятков взрослых особей) группами, приуроченными к благоприятным микробиотопам. Эксперименты показывают, что такие группы особей (вероятно, демы) могут существовать в относительно изолированном состоянии на протяжении жизни ряда поколений (Даревский, 1946; Боченко, сообщ.). Об этом же свидетельствуете значительная изолированность некоторых из таких демов: например, группа ящериц из 6 взрослых самцов и 6 самок, обитающая на склоне оврага на поляне в сосновом бору в Калужской обл1., была удалена от ближайшего соседнего поселения ящериц на 2000 м, расстояние, многократно превышающее радиус индивидуальной активности в обычных условиях (см. рис. 89, Б). Лишь в периоды резкого подъема численности или миграций, связанных с неблагоприятными условиями, возможно преодоление этого пространственного барьера (Стрельцов, сообщ.).
Сказанное о внутрипопуляционной пространственной структуре относится в общей форме и к разным популяциям внутри ареала всего вида. Наблюдения в природе показывают, что у прыткой ящерицы существуют популяции, лишь незначительными изоляционными барьерами отделенные друг от друга и в совокупности образующие как бы сплошной тип поселения; такое тесное расположение популяций характерно для зоны оптимума, которая располагается в основном в степной зоне и частично в соседних (полупустынной и лесостепной) зонах. В других ландшафтах популяции ящериц оказываются распределенными мозаично, часто — ленточно (например, по поймам крупных рек или предгорным районам). Наконец, на севере и крайнем юге ареала популяции распределены диффузно. Так, например, южная граница ареала вида в районе 54—60° в. д. проходит по р. Эмбе (см. рис. 16). В этом районе в настоящее время происходит тесное соприкосновение полупустынного (левый берег) и сухостепного (правый берег) ландшафтов. Тщательное обследование показало, что единственная популяция L. agilis на протяжении нескольких десятков километров правобережья расположена неподалеку от пос. Жаркомыс, в зеленой долине размером около 1000X250 м, окруженной со всех сторон значи
282
тельными песчаными дюнами. Но уже в 20—30 км севернее популяции ящериц образуют типичный кружевной узор, встречаясь недалеко друг от друга. Сходно и распределение ящериц Б горах Заилийского Алатау, в долине оз. Иссык-Куль. Отдельные группы ящериц (без специального стационарного исследования нельзя сказать, являются ли они малочисленными популяциями или представляют собой остатки каких-то миграционных волн, лишь временно обитающих на данной территории) встречаются по северному берегу озера, начиная от пос. Ананьеве. Но первая настоящая популяция, с высокой численностью и постоянно обитающая на данном месте, встречается лишь в 35—57 км восточнее, по широкой балке и прилежащим окраинам полей у Тюбского залива. Здесь сразу же в непосредственной близости обитает несколько популяций (в том числе и интересные мелкие островные популяции), и эта большая группа популяций (видимо включающая в целом десятки популяций) совершенно изолирована от всего остального ареала прыткой ящерицы в Средней Азии: ни в восточной части Тюбской долины, ни в Кегенской долине ящериц нет. Ближайшие популяции обнаружены лишь на правобережье р. Или, за много десятков километров от оз. Иссык-Куль. В то же время прежние наблюдения показывают, что раньше ящерицы встречались и по левому берегу р. Или, т. е. можно предполагать, что изоляция группы популяций на Восточном Иссык-Куле не постоянная, а временная (Яблоков и др., сообщ.).
* * *
Приведенные в настоящей главе материалы показывают, что структура популяций прыткой ящерицы оказывается достаточно сложной (и в половом, и в возрастном, и в пространственном отношениях). Эта сложность многократно увеличивается в связи с подвижной «экологической» структурой популяции — изменениями активности отдельных возрастно-половых групп в разные сезоны года, и даже в разное время суток. Постоянно происходящие в природе микроэволюционные процессы, определяемые действием таких факторов, как естественный отбор, мутационный процесс, волны численности и развитие разных форм изоляции, постоянно нарушают казалось установившуюся структуру популяций, внося в нее элементы подвижности, текучести, динамики.
Структура популяции прыткой ящерицы оказывается несколько различной в разных участках ареала. Несомненно, существование географических различий в пространственной структуре популяций (преобладание сплошного типа поселений в оптимуме ареала вида и разного рода мозаичных и диффузных поселений на периферии ареала). Наконец, есть основания предполагать существование внутриареальной изменчивости половой и возрастной структуры популяции.
ГЛАВА ДВЕНАДЦАТАЯ
ПАРАЗИТЫ1
Прыткая ящерица — один из наиболее полно и всесторонне изученных в паразитологическом отношении видов пресмыкающихся. Однако широкие исследования этой ящерицы осуществлены в основном в пределах ареала, охватывающего лишь территорию Советского Союза и прежде всего Европейскую часть страны. Сведения о паразитах в других частях ареала в основном отрывочны. В общем к настоящему времени имеются данные различной степени полноты о паразитах прытких ящериц, обитающих на территории СССР, а также Англии, Франции, ФРГ, Голландии, Польши, Чехословакии, Венгрии, Румынии и Югославии.
Всего у прыткой ящерицы зарегистрировано 44 вида паразитов, среди которых — простейшие, гельминты и клещи. Несмотря на большое число обнаруженных паразитов, болезни, вызываемые этими организмами, равно как и непаразитарные заболевания, у прытких ящериц не описаны. Поэтому судить о влиянии паразитарного фактора и непаразитарных болезней на различные стороны жизнедеятельности этих ящериц сейчас еще не представляется возможным.
Ниже в систематическом порядке приводятся сведения о паразитах, зарегистрированных у прыткой ящерицы в пределах ее ареала.
ПРОСТЕЙШИЕ
Среди этой группы паразитических организмов у прыткой ящерицы зарегистрирован 1 вид жгутиконосцев.
Класс Жгутиконосцы (Flagellata)
Karyolysus lacertarum (Dan., 1886)
Паразит эритроцитов. Отмечен на территории СССР у прытких ящериц в Харьковской обл. (Глущенко, 1961).
1 Авторы раздела, посвященного простейшим и гельминтам,— В. П. Шарпило, клещам — Л. Е. Шур.
284
ГЕЛЬМИНТЫ
Эта группа паразитов включает 22 вида, в том числе трематод 9, цестод 3, скребней 1 и нематод 9. Среди зарегистрированных видов представлены облигатные и факультативные паразиты. Значительное место принадлежит также личиночным формам гельминтов, в жизненных циклах которых прыткая ящерица выступает в роли резервуарного или промежуточного хозяина. Дефинитивными хозяевами таких гельминтов являются^ птицы и млекопитающие.
Накопленные к настоящему времени данные позволяют констатировать отличия в видовом составе и степени зараженности гельминтами прыткой ящерицы в различных частях ее ареала. Такие виды паразитических червей, как A. abbreviata, S. lacertae и др., приурочены исключительно к южным частям ее ареала. В Европейской части СССР, например, эти паразиты отмечены только на юге Украины, в Краснодарском и Ставропольском краях. М. molini достоверно известен у прыткой ящерицы только в западной части ее ареала. Несколько видов (О. goezei, Р. elegctns и др.) встречено на большей части ареала этой ящерицы, однако распространены они далеко не равномерно. Так, в северо-западных областях Украины О. goezei — обычный вид, зараженность которым достигает 78—100%. На большей части степной зоны он исключительно редок. В Дагестане зараженность прыткой ящерицы этим видом вновь возрастает до 80—100%. Отмеченные закономерности в распространении перечисленных видов паразитических червей обусловливаются такими факторами, как влажность, температура, наличие или отсутствие соответствующих промежуточных и дополнительных хозяев и др.
При анализе видового состава гельминтов прытких ящериц обращает на себя внимание тот факт, что в их гельминтофау-не нет специфичных видов. Их облигатные паразиты одновременно встречаются и у других ящериц (зеленой, полосатой и др.) Среди факультативных паразитов большинство видов является паразитами амфибий и приурочено в основном к ящерицам, обитающим во влажных биотопах или непосредственно близ водоемов.
В целом гельминтофауна прыткой ящерицы богата и разнообразна. По разнообразию видов гельминтов она значительно превосходит другие палеарктические виды ящериц. В основе этого лежит, несомненно, широкая экологическая пластичность прыткой ящерицы, позволяющая ей обитать в различных, нередко резко отличающихся условиях.
285
Класс Трематоды (Trematoda)
С andldotrema loossi (Afrika, 1930)
Паразит кишечника. Зарегистрирован на территории СССР у прыткой ящерицы в качестве факультативного паразита в Запорожской и Херсонской обл. (Шарпило, Магуза, 1972).
Pleurogeties clavlger (Rud., 1819)
Паразит кишечника. Найден на территории СССР у прыткой ящерицы в качестве факультативного паразита в Астраханской, Киевской и Херсонской обл. (Иванов, 1952; наши данные).
Pleura genoldes medians (Olsson, 1876)
Паразит кишечника. Как факультативный паразит отмечен у прыткой ящерицы на территории СССР в Запорожской, Киевской и Херсонской обл. (Шарпило, 1964), а также в Польше Grabda-Kazubska, 1972) и Чехословакии (Moravec, 1963).
Prosotoc us confusus Looss, 1899
Паразит кишечника. Зарегистрирован у прыткой ящерицы в СССР как факультативный паразит в Ворошиловоградской, Черкасской и Харьковской обл. (Шарпило, 1960, 1961, 1964; Шевченко, 1963; Шевченко, Барбашова, 1958).
Leucochlortdium sp.
Транзитный паразит кишечника прытких ящериц. Молодые, неполовозрелые черви найдены на территории СССР в Черниговской обл. (Шарпило, 1964).
PlaglorMs elegans (Rud., 1802)
Паразит кишечника. Многократно регистрировался под названием Р. mentulatus у прыткой ящерицы на территории СССР в Волгоградской, Калининградской, Московской, Омской и Ростовской обл., на Украине, в дельте Волги, Краснодарском крае и в Дагестане (Власенко, 1930; Иванов и др., 1960; Марков и др., 1962, 1964, 1969; Харченко, Сова, 1972; Шарпило, 1959, 1960, 1961, 1962; Шевченко, 1963; Шевченко, Барбашова, 1958; Muhling, 1898а, Ь). Найден также у прытких ящериц в Англии, Франции, Польше, Венгрии и Чехословакии (Dawes, 1946; Luhe, 1909; Moravec, 1963; Edelenyi, 1962; Dollfus, 1961).
286
Plagiorchis maculosus (Rud., 1802)
Паразит кишечника. Как факультативный паразит отмечен нами у прыткой ящерицы на территории СССР в Киевской обл.
Metaplagi orchis nwlini (Lent et Freitas, 1940)
Паразит кишечника. Выявлен у прыткой ящерицы на территории СССР в Закарпатской, Житомирской, Ивано-Франков-ской, Киевской, Черниговской обл. (Шарпило, 1959, 1960, 1964), а также в Италии, Румынии и ФРГ (Capuse, 1971; Lent Freitas, 1946).
Paradistomum mutabile (Molin, 1858)
Паразит желчного пузыря. Зарегистрирован у прыткой ящерицы в Румынии и Италии (Capuse, 1971; Rizzo, 1902).
Класс Цестоды (Cestoda)
Nematotaenia tarentolae Lopez-Neira, 1944
Паразит кишечника. Отмечен у прыткой ящерицы на территории СССР в Ворошиловоградской и Киевской обл. (Шарпило, 1973).
Oochoristica tub er cut at a (Rud., 1819)
Паразит кишечника. Обнаружен у прыткой ящерицы на территории СССР в Днепропетровской, Донецкой, Запорожской, Крымской, Николаевской, Одесской, Саратовской, Харьковской, Херсонской обл., а также в Краснодарском и Ставропольском краях и Южном Казахстане (Ваккер, 1970; Иваницкий, 1927, 1940; Козловский, 1951; Марков и др., 1964; Спасский, 1948; Шарпило, 1959, 1960, 1961, 1963, 1964; Шевченко, Барбашова, 1958; Шевченко, Белинисова, 1964).
Mesocestoides lineatus (Goecei, 1782) — larvae
Паразит полости тела, печени и других внутренних органов прыткой ящерицы. Обнаружен на территории СССР в Донецкой, Запорожской и Херсонской обл. (Шарпило, 1960, 1961, 1964). Прыткая ящерица — резервуарный хозяин этого скребня.
Класс Нематоды (Nematoda)
Spauligodon lacertae Sharpilo, 1966
Паразит прямой кишки. Зарегистрирован у прыткой ящерицы на территории СССР в Ворошиловградской и Херсонской обл. и Южном Казахстане (Ваккер, 1970; Шарпило, 1966).
287
Oswaldocruzia goecei Skrjabin et Schulz, 1952
Паразит тонкого кишечника. Отличен у прыткой ящерицы в СССР в Волгоградской, Волынской, Житомирской, Закарпатской, Ивано-Франковской, Калининградской, Киевской, Львовской, Черкасской, Черниговской обл. (Марков и др., 1969; Шар-пило, 1960, 1964; Шевченко, 1963; Muhling, 1898). Под названиями О. filiforinis, О. subauricularis, О. bialata, по-видимому, именно этот вид неоднократно зарегистрирован в некоторых странах Западной Европы.
Abbreviate, abbreviate (Rud., 1819)
Паразит желудка. Обнаружен у прыткой ящерицы на территории СССР в Волгоградской, Крымской и Херсонской обл., в Краснодарском и Ставропольском краях (Андрушко, Марков, 1960; Ваккер, 1970; Марков и др., 1962, 1964; Шарпило, 1960).
Тhubutiaea smogorzevskii Sharpilo, 1966
Паразит желудка. Зарегистрирован в СССР у прыткой ящерицы в Херсонской обл. (Шарпило, 1966).
Ascarops strongylinae (Rud., 1819)—larvae
Паразит различных внутренних органов прыткой ящерицы. Выявлен на территории СССР в Донецкой, Житомирской, Киевской, Харьковской, Херсонской и Черкасской обл. (Шарпило, 1961, 1964; Шевченко, 1963; Шевченко, Белинисова, 1964). Прыткая ящерица — резервуарный хозяин этих нематод.
Physocephalus sexalatus (Molin, I860)—larvae
Паразит различных внутренних органов прыткой ящерицы. Зарегистрирован на территории СССР в Ворошиловоградской, Житомирской, Закарпатской, Киевской, Одесской и Херсонской обл. (Шарпило, 1960, 1961, 1964). Прыткая ящерица — резервуарный хозяин этих нематод.
Spirocerca lupi (Rud., 1809)—larvae
Паразит различных внутренних органов и полости тела прыткой ящерицы. Зарегистрирован на территории СССР в Житомирской, Киевской, Крымской, Одесской обл. и Краснодарском крае (Марков и др., 1964; Назарова, 1961; Шарпило, 1960, 1961, 1964). Прыткая ящерица — резервуарный хозяин этих нематод.
288
Physaloptera clausa Rud., 1819—larvae
Паразит слизистой желудка прыткой ящерицы. Обнаружен на территории СССР в Волынской и Херсонской обл. В. и Л. Шарпило, (1969). Прыткая ящерица — резервуарный хозяин этих нематод.
Agamosplrura rnagna Sharpilo, 1964
Паразит различных внутренних органов прытких ящериц. Обнаружен на территории СССР в Ивано-Франковской, Ворошиловоградской, Львовской, Херсонской и Черкасской обл. (Шарпило, 1964). Прыткая ящерица — резервуарный хозяин'этих нематод.
КЛЕЩИ (ACARINA)
На прыткой ящерице в качестве паразитов зарегистрировано 20 видов клещей. Наиболее разнообразны иксодовые клещи — 15 видов (на ящерицах паразитируют только личинки и нимфы). Краснотелковых клещей обнаружено 4 вида. Гамазовые клещи представлены 1 видом — специфичным паразитом ящериц.
Т rombicula pilosa Feider, 1948
Отмечен на прытких ящерицах в Румынии (Feider, 1953).
Тrombicula tragardhlana Feider, 1953
Отмечен на прытких ящерицах в Румынии (Fuhn, Vancea S., 1961; Feider, 1953).
Trombicula automnali s Feider, 1953
Отмечен на прытких ящерицах в Румынии (Feider, 1953).
Trombicula pulctira Schluger, 1955
Отмечен на прытких ящерицах на территории СССР в Ставропольском крае и на западном побережье Аральского моря (Шлугер и др., 1955).
Sauronyssus saurarum (Oudeinans, 1902)
Широкораспространенный паразит прытких ящериц. В СССР найден в Волгоградской, Калининской, Крымской, Ленинградской, Московской, Саратовской обл., а также в Ставропольском крае, Закавказье и Западной Сибири (Брегетова, 1956; Земская, 1951; Марков и др., 1964; Федоров, 1970). Единичные находки этих клещей известны на прытких ящерицах в Голландии и Румынии (Fuhn, Vonceas, 1961; Fronseca, 1948).
10 Прыткая ящерица
289
Ixodes ricinus (L., 1758)
I _• Обычный и наиболее распространенный паразит прытких ящериц. Обнаружен на территории СССР в Закарпатской, Киевской, Крымской, Ровненской, Ростовской обл., в Прикарпатье, а также в Белоруссии, Таджикистане и Дагестане (Арзамасов, 1961; Емчук, I960; Марков и др., 1964; Поспелова-Штром, 1935; Сердюкова, 1956; Тертышников, 19726. Наши неопубликованные данные), а также в Югославии и Румынии (Fuhn, Vonceas, 1961; Oswald, 1939).
Ixodes persulcatus P. Sch., 1930
Отмечен на прытких ящерицах на территории СССР в Ново-| сибирской, Омской, Тюменской обл., а также в Алтайском крае (Федоров 1970).
Ixodes redikorzevi Ol., 1927
1 Отмечен на прытких ящерицах на территории СССР в Крымской и Одесской обл. (Емчук, 1960).
Н ае тор fly salts Inermls Bir., 1895
Имеются данные о находке этого вида на прытких ящерицах в СССР без точного указания места (Сердюкова, 1956).
Н aetnophysalls punctata Сап. et Fanz., 1877
Отмечен на прытких ящерицах на территории СССР в Полесье, лесостепной зоне, в предгорьях Крыма и Ставропольском крае (Емчук, 1960; Сердюкова, 1956; Тертышников, 1972а).
IIaetnophysalls sulcata Can. et Fanz., 1878
Отмечен на прытких ящерицах на территории СССР в Крым-I ской и Ростовской обл. (Емчук, 1960; Сердюкова 1956).
IIaetnopfiysalis otophila Р. Sch., 1918
Отмечен на прытких ящерицах на территории СССР в Крымской обл. и Ставропольском крае (Емчук, 1960; Сердюкова, Д956; Тертышников, 19726).
| .
Иaetnophysalls caucaslca Ol., 1928
Имеются данные о находке этого вида на прытких ящерицах в СССР без точного указания места (Сердюкова, 1956).
290
H aetnophysalls concinna Koch, 1844
Имеются данные о находке этого вида на прытких ящерицах в СССР без точного указания места (Сердюкова, 1956).
Boophilus calcaratus Bir., 1895
Имеются данные о находке этого вида на прытких ящерицах в СССР без точного указания места (Сердюкова, 1956).
Dermacentor pictus Herm., 1804
Отмечен на прытких ящерицах в СССР на Украине и в Западной Сибири (Емчук, 1960; Сердюкова, 1956; Федоров, 1970).
Dermacentor marginal us Sulz., 1770
Отмечен на прытких ящерицах в СССР на Украине и в Алтайском крае (Емчук, 1960; Сердюкова, 1956; Федоров, 1970).
Rhipicephalus bursa Сап. et Fanz., 1877
Имеются данные о находке этого вида на прытких ящерицах в СССР без точного указания места (Сердюкова, 1956).
Rhipicephalus turanicus В. Pom., 1940
Имеются данные о находке этого вида на прытких ящерицах в СССР без точного указания места (Сердюкова, 1956).
Н yalomma plumbeum Panz., 1795
Отмечен на прытких ящерицах на территории СССР в Ставропольском крае (Сердюкова, 1956; Тертышников, 19726).
10*
ГЛАВА ТРИНАДЦАТАЯ
ПРЫТКАЯ ЯЩЕРИЦА
КАК КОНСУМЕНТ И ПРОДУЦЕНТ1
Прыткая ящерица входит в состав многих биоценозов. По этой причине становится понятна важность определения ее места в потоке вещества и энергии, переносимой через популяции данного животного. Решение данной проблемы представляет большую сложность по той причине, что в различных частях ареала отдельные популяции данной ящерицы занимают различное место в биоценозах. Кроме того, задача осложняется еще и тем, что в настоящее время практически отсутствуют сведения о количестве потребляемых ящерицами веществ, калорийности пищи и пр. Поэтому настоящая глава основана на выяснении роли прыткой ящерицы как консумента и продуцента на материалах, полученных на территории Ставропольской возвышенности каи модельном, ключевом участке для всего ареала.
МЕСТО ПРЫТКОЙ ЯЩЕРИЦЫ В БИОЦЕНОЗЕ
Биоценозы, в состав которых входит прыткая ящерица, охватывают большое количество разнообразных организмов, связь с которыми у нее сложна и формы которой весьма разнообразны. Понятно, что в плане данной работы более интересны трофические связи ящерицы, ее место в целях питания (рис. 90).
Взаимоотношения с низшими звеньями в пищевых целях уже рассмотрены в гл. VI («Питание»), Здесь же рассматриваются взаимоотношения прыткой ящерицы с видами «соседями» в цепях питания (конкурентами за пищу) и высшими ’звеньями в цепях питания (враги).
Трофические связи: конкуренты за пищу
Как было показано в гл. VI, основу пищи прыткой ящерицы составляют различные беспозвоночные. По типу питания ящерицы— мелкие хищники-зоофаги первого порядка (Darevskij, Те-
1 Автор: М. Ф. Тертышников, при участии А. В. Яблокова, А. С. Баранова, А. В. Валецкого, Н. М. Окуловой, Г. П. Лукиной, В. Е. Боченко, Н. А. Раш-кевич, В. К. Жарковой, М. И. Щербаня, И. Н. Щербака.
292
| Кокцинеллиды Ц
{Прямокрылые р {Жужелицы р | Пауки |-
{Мелкие млекопитающие
{Насекомоядные птицы
{Дневные хищные птицы
{Паукообразные [ | Многоножки
{Ящерицы~\
\3меи [
{Малощетинковые черви \Брюхоногие моллюски
^Лягушки \Жлды [-притоны
{Мелкиесочные плодырас/пенй^. ^Ракоовразные
Яйца и молодые ящерицы^
Жгутиконосцы^//[
Враги
П и щ S ы
Еж овыкновеннь7й{ ''\Дроят{
Хомяк одынновенный |
Краснотелковые клещиЖ(\
ИксодоЙые клещи (15) |
Гамазовые клещи(1) |
Трематоды(З) | Цестоды(З') |
С к редки (1) |
Нематоды (9) |
{Крупные ящерицы (Ч)к \~Полоза (Ч)\——к‘ | Гадюки(2){--------к
^Медянка |----------2
| Цапли (9)
| Аисты(2)У
| Куриные \к
| Дрофы И
| Крачки(2) {к
| Чайки (2)
| СоВьГ(й)У
{ВраноВые (5)
| Сорокопуты (3)
{Дневные хищные пшицы(15)
| Озера а я лягушка
Перепотатокрылыё~\ Прямокрылые\ Чешукрылые | Равнокрылые | ДВукрылые\ Клопы\ Жуки |
Кошки (2)\
Псовые (Ч){
Рис. 90
Связь прыткой ящерицы с другими живыми компонентами в биогеоценозе Схема составлена по сводным данным разных авторов А. С. Барановым Прыткая ящерица
rentjev, 1967), которые хватают всех движущихся беспозвоночных среднего и мелкого размеров. Благодаря низкой избирательности в питании они способны поедать те виды, которые в данный момент в изобилии находятся в биоценозе. Кроме того, известно, что интенсивность питания этого животного резко колеблется в разное время года.
На базе общности питания у прыткой ящерицы имеются мно-гочисленые конкуренты, к которым относятся различные хищники-зоофаги первого и второго порядков, постоянно или временно обитающие в тех же биоценозах, что и ящерица.
Беспозвоночные конкуренты ящерицы представлены пауками, кокцинеллидами, жужелицами, перепончатокрылыми, некоторыми прямокрылыми и др. По численности и многообразию эти беспозвоночные могут быть наиболее важными конкурентами прыткой ящерицы, но судить об этом можно только весьма ориентировочно, так как сведения о поедаемой этими конкурентами ящерицы добыче, ее количестве и численности данных животных в биоценозах пока практически отсутствуют.
Другую группу конкурентов прыткой ящерицы составляют амфибии и прочие рептилии. У всех из них, кроме медянки, беспозвоночные играют большую роль в питании. Непосредственными конкурентами же ящерицы могут быть только те из них, которые обитают в биотопах совместно с прыткой ящерицей (тритоны, лягушки окрестных водоемов; жаба живородящая, зеленая, средняя, полосатая ящерица — и ряд змей).
Сравнивая состав пищи прыткой ящерицы и насекомоядных птиц, обитающих в общих биотопах, некоторые исследователи (Щепотьев, 1950; 1952; Тертышников, 19726; Жаркова, 1973а) полагают, что за редким исключением и у тех и у других основную роль в питании играют сходные группы беспозвоночных— жуки, перепончатокрылые, двукрылые и др. Исходя из этого можно считать насекомоядных птиц серьезными пищевыми конкурентами прыткой ящерицы. Кроме того, беспозвоночные входят в состав пищи и некоторых хищных птиц (пустельга, ястреб-перепелятник, кобчик, сыч, совки и др.), но процентное содержание беспозвоночных в их пище незначительно (Тертышников, 19726; Рашкевич, сообщ.). Однако эта группа птиц скорее является прямыми врагами ящерицы, охотясь на них (см. далее).
Многие из млекопитающих также поедают беспозвоночных, но только у некоторых из них беспозвоночные являются основой рациона, в те или иные сезоны года (например, у землероек). В целом млекопитающие не являются серьезными пищевыми конкурентами прыткой ящерицы.
293
Трофические связи: враги
Прыткая ящерица служит пищей для многих животных. Среди беспозвоночных, по всей видимости, у нее мало врагов: известен лишь один случай нападения на нее богомола (Mantis religiose) (Кострин, 1959). Однако среди наземных позвоночных у прыткой ящерицы врагов значительно больше.
Амфибии очень редко поедают прыткую ящерицу. Истребительную в отношении ящериц деятельность амфибий можно охарактеризовать как случайную; достаточно крупные размеры ящерицы не позволяет амфибиям нападать на них. Однако есть наблюдения (М. Ф. Тертышников, 19726), из которых следует, что озерная лягушка (Rana ridibunda) иногда нападает и поедает сеголетков прыткой ящерицы.
Многие из рептилий являются прямыми врагами прыткой ящерицы: ее поедают ящерица зеленая (L. viridis), желтобрюхий (Coluber jugular is}, четырехполосый (Elaphe quatocorlinea-ta), узорчатый (E. dione) и эскулапов (E. longissima) полозы, степная гадюка (Vipera ursini) и, особенно, медянка (Coronel-la austriaca), пища которой почти целиком состоит из ящериц (Красавцев, 1936, 1939; Щепотьев, 1950, 1952; Андрушко, 1954; Щербак, 1966; Тертышников, 19726; Окулова, Хонякина, сообщ).
Очень много врагов у прыткой ящерицы среди птиц и млекопитающих (Шершевский, 1931; Жарков, Теплов, 1932а; Жарков, Теплов, 19326; Жарков и др., 1932; Колосов, 1935; Красавцев, 1936; 1939; Озоль, 1941; Чиркова, 1948; Бородулина, 1949; Щепотьев, 1950, 1952; Киселев, 1951; Корнеев, 1954; Обтемперан-ский, 1956; Гизенко, 1957; Смогоржевскпй, 1959; Таращук, 1959; Козлов, 1960; Павлов, Кирис, 1963; Щербак, 1966; Тертышников, 19726; Хонякина, Рашкевич, Боченко, Щербань, сообщ.). Прыткая ящерица была обнаружена в содержимом желудков у цапель серой, рыжей и желтой, кваквы, выпи, аистов белого и черного, осоеда, коршунов черного и красного, ястребов перепелятника и тетеревятника, сарычей рыжего, мохноногого и обыкновенного, канюка, подорлика малого, змееяда, луней полевого, лугового и болотного, кобчика, пустельги обыкновенной и степной, глухаря, фазана, дрофы, крачки чайконосой, чаек хохотуньи и серебристой, сов ушастой и болотной, сыча домового, неясыти серой, сорокопупов большого, чернолобого и жулана, ворона, вороны серой, галки, сойки, грача. Правда, здесь следует заметить, что в годы малой численности грызунов многие пернатые хищники переходят на питание ящерицами, тогда как в другое время ящерицы редко входят в состав их рациона. Кроме того, в определенных местах, в основном вблизи крупных колоний птиц, ящерицы подвергаются значительно более частым нападениям с их стороны. Так, наблюдения показывают, что на участке степи радиусом до 1800 м, прилежащей к грачевникам,
294
ящерицы полностью отсутствовали (Херсонская обл., 1955— 1958 гг., БочеПко, личн. сооб.). Прямые наблюдения на колониях серебристых чаек (Larus argentatus) показывают, что птицы приносят своим птенцам ящериц, пойманных в окрестностях урочищ (оз. Тенги, Центральный Казахстан; 1968, 1973, Валец-кий, Яблоков).
Из млекопитающих ящерицу поедают еж обыкновенный, крот, хомяк обыкновенный, ласка, горностай, хорь черный, перевязка, куница каменная, барсук, лисица, корсак, енотовидная собака, лесной кот, домашние собака и кошка.
Наконец, нельзя не отметить, что значительную опасность для ящериц представляет человек, который зачастую уничтожает их без всякой цели. Однако особенно гибельным для населения ящериц является применение химических препаратов в сельскохозяйственных и лесных угодьях и прилегающих к ним участках; наблюдения и учеты показывают, что в таких местах численность этого вида резко сокращается, а иногда популяции полностью уничтожаются (Жаркова, 1973а).
Интересные данные о «давлении» хищников на популяции прыткой ящерицы дает анализ (по полу и возрасту) числа особей в разных популяциях с аутотомированными хвостами. Этот прием, основанный на способности ящерицы «отбрасывать» хвост при нападении преследователя и затем регенерировать его, позволяет точно учитывать число особей в каждой популяционной выборке с такими «следами нападений врагов» (Красавцев, 1939; Жаркова, 1973а). Следует отметить, что хвост отбрасывается действительно только при нападении врага. Как показывают длительные наблюдения, при драках между самцами или при серьезных повреждениях самцами самок (при захвате их во время акта спаривания) хвост не отбрасывается. Не отбрасывают хвост и прирученные ящерицы, хотя бы несколько дней находящиеся в лаборатории и освоившиеся с новой обстановкой (см. гл. X). Конечно, в случае регенерировавших хвостов невозможно определить конкретных врагов ящериц, но несомненно, процент особей с регенерировавшими хвостами способен косвенно указывать на интенсивность преследования со стороны хищников.
Сводные данные по учету этого признака в 75 популяционных выборках, приведенные в табл. 45, показывают, что средн молодых число особей с регенерировавшими хвостами сравнительно меньше, чем среди старших возрастных групп; это и понятно, поскольку в старших группах происходит как бы «накопление» числа встреч с врагами. Интересны данные по различному числу пострадавших от врагов самцов и самок. Среди взрослых животных (где различия статистически достоверны) соотношение между пострадавшими самцами и самками сдвинуто в сторону самок. Пока трудно определенно сказать, связано ли это с более успешным избеганием нападения самцами по сравнению с
295
Рнс. 91
Частота встречаемости прытких ящериц разных возрастных групп с аутотомнрованными хвостами в Рязанской обл. (1 — самцы, п =а —657 экз.; 2 — самки, п «=547 экз.) и Средней Польше (3— самцы+сам-ки, п=303 экз.). Возрастные группы выделены ориентировочно по размерам тела (Жаркова, 1973а; Добро-вольска, Шафраньска)
самками или несколько разной способностью особей разного пола к аутотомии. Заметим при этом, что в разных популяциях (даже близко расположенных) это соотношение сдвинуто не только в сторону преобладания аутотомировавших самок, но и в сторону самцов (рис. 91). На этом же рисунке видно, что в Мещере
Таблица 45
Среднее число особей прытких ящериц разных возрастно-половых групп с регенерировавшими хвостами [Красавцев (1939), а также по данным А. В. Яблокова, А. С. Баранова, А. В. Валецкого, В. И. Борисова, Н. М. Окуловой]
Группа особей X Число изученных популяций Группа особей X Число изученных популяций
Сеголетки 6,2 34 Взрослые самцы 22,2 63
Полувзрослые самцы 10,0 11 Взрослые самки 26,3 64
Полувзрослые самки 9,3 11 Среднее по всем группам 14,8 75
процент ящериц с аутотомированными хвостами равномерно растет с первого до третьего года жизни (около 5%), а затем резко увеличивается. Такой ход возрастного изменения этого тератологического признака позволяет предполагать, что основную массу ящериц старшего возраста уничтожают хищники, тогда как на первом году жизни основное значение в смертности особей имеют другие причины (Жаркова, 19736).
Анализ всего материала по ареалу не позволяет подтвердить точку зрения о том, что процент ящериц с регенерировавшими хвостами прямо коррелирован со степенью открытости местообитания; в ряде максимально «открытых» популяций степной и полупустынной зон процент достигал 50. Сравнение двух групп
296
Таблица 46
Число особей с регенерированными хвостами в двух соседних биотопах в популяции' по среднему течению р. Дон (А. С. Баранов, А. В. Яблоков, А. В. Валецкий,
В. И. Борисов, июль 1973 г.)
Возрастная группа особей Число особей с регенерированными хвостами % Число особей с регенерированными хвостами %
Безлесая балка Пойменный лес
Сеголетки 2/0 0 7/0 0
Взрослые самцы 35/9 35,7 29/11 37,9
Взрослые самки 35/8 22,9 36/16 44,4
ящериц, населяющих соседние биотопы в окрестностях г. В. Мамон, также опровергает это мнение (табл. 46).
Видимо, более правильным будет общее заключение о том, что число особей с регенерировавшими хвостами в популяциях определяется не открытостью биотопа, а степенью давления хищников и активностью особей.
ЯЩЕРИЦА КАК КОНСУМЕНТ
В гл. VI было отмечено, что прыткая ящерица поедает беспозвоночных, обитающих совместно с ней в одних биотопах. Дать точную количественную энергетическую оценку месту ящериц в цепи питания пока еще сложно. В этой связи весьма интересны данные, полученные М. Ф. Тертышниковым (19726) на Ставропольской возвышенности, в зоне оптимума данного вида.
Учет показал, что прыткая ящерица на стационарном участке обитает в разнотравно-злаковой, типчаково-полынной и полынной степях, в лесной полосе, на просеках и вырубках, в лесу, на посевах и толоках. Общее количество ящериц, определенное по методу «объединенного гектара» (Ралль, 1936), равно 5717 экз. Принимая средний вес пищи, потребляемый прыткой ящерицей в сутки, равным 1,0 г (л=269), можно подсчитать, что 5717 ящериц в сутки съедают 5,7 кг беспозвоночных, а 72 млн. ящериц, обитающих на Ставропольской возвышенности, 72 т в сутки.
Прыткая ящерица относится к категории вторичных консументов. Основной обмен веществ у нее ниже, чем у первичных консументов, и эффективность цепи питания при переходе на данный трофический уровень достигает порядка 10% (Slobodkin, 1960). Таким образом, на этом уровне восстанавливается весьма незначительное количество первоначально затраченной энергии. Рассчитано, что суточная норма питания прыткой ящерицы,
297
имеющей вес 15,0 г, равна 33,3 ккал (Черномордиков, Ковалева; 1964). Средний вес взрослой прыткой ящерицы из популяций Ставропольской возвышенности равен 12,0 г (п=269). Для одного животного такого веса требуется в сутки 26,6 ккал, а всем ящерицам со Ставропольской возвышенности—1915 млн. ккал/сутки.
В этой связи интересно установить степень воздействия населения прытких ящериц на тот или иной объект добычи. Для выяснения этого вопроса была использована методика А. А. Иноземцева (1969). На основании данных о численности ящериц на 1 га биотопов, численности беспозвоночных на 1 м2 в тех же биотопах и количестве беспозвоночных, приходящихся на содержимое одного желудка ящерицы (от 4 до 20 в среднем 7,0+ 1,1 экз. при /7=269), была определена предполагаемая численность основных групп беспозвоночных на 1 га в некоторых биотопах и та их часть, которую уничтожили ящерицы на этой площади в течение месяца (см. табл. 47—51).
Оказалось, что суммарное воздействие прыткой ящерицы на доступную ей фауну беспозвоночных в некоторых биоценозах стационарного участка на Ставропольской возвышенности варьирует от 0,08 до 10,6% (4,48±0,2%). При этом в период максимальной активности прыткая ящерица оказывает наибольшее «давление» на жуков (0,7—12,0%), перепончатокрылых (0,09— 20,3%), пауков (0,7—12,0%) и прямокрылых (0,07—28,8%). .
Таким образом, роль прыткой ящерицы, как консумента может быть довольно значительной в биоценозах целинных степей (разнотравно-злаковая и типчаково-полынная степи) и лесной полосе, но может быть и очень малой, как это установлено для лесных биоценозов и агроценоза. К тому же, как это следует из особенностей питания, роль прыткой ящерицы как консумента, может быть особенно значительной в отношении наиболее массовых видов пищи.
В табл. 52 приведены данные о величине суточного рациона и биомассе фоновых позвоночных животных стационарного участка на Ставропольской возвышенности. Эти данные являются приближенными, так как построены на ряде допущений (вес животного и его суточный рацион считался неизменным в течение всего сезона активности; малочисленные позвоночные не принимались во внимание при расчетах и пр.), но позволяют хотя бы ориентировочно оценить роль прыткой ящерицы, как консумента.
Из табл. 52 следует, что биомасса данного вида ящериц в изученном районе превосходит биомассу других позвоночных животных. Точно также и биомасса съеденных прыткой ящерицей беспозвоночных заметно превосходит биомассу беспозвоночных, съеденных остальными позвоночными животными.
298
Таблица 47
Воздействие прытких ящериц иа их жертвы в разнотравно-злаковой степи (по Тертышникову, 19726)
— Абсолютное число беспозвоночных =я Соотношение, % Sis
Поедаемая группа о * 32 « Ф\О к 5 яще- ia <a 1 га в оз де 4 ельнос по ка: те ви;
видов ЕЛ а о Й о; В я S а-gsg л ж « Ф к £ ф ф S
a 2 R £ ® ф д' £ О Ф ш я
'g са я—« ь С5 са § и S с «
Моллюски 1 1000 1 157 15,7 1,о Г,з +0,1
Пауки 3 3000 1 157 5,0 3,0 1,3 -0,4
Многоножки 3 3 000 1 157 5,0 3,0 3,0 —0,4
Прямокрылые 11 11000 9 1409 12j8 10,0 12,0 +0,1
Полужесткокрылые 4 4000 5 783 19,5 3,6 6,7 +0,3
ТКуки 25 25000 19 2 975 11,9 22,4 25,3 +0,1
Двукрылые 38 38000 9 1409 3,7 34,0 12,0 -0,4
Перепончатокрылые 20 20000 26 4072 20,3 18,0 34,7 +0,3
Чешуекрылые (имаго | гусеницы) 3 3 000 2 213 10,4 3,0 2,7 -0,1
Прочие виды 2 2000 2 213 15,6 2,0 2,7 +0,1
Всего . . . НО НО 000 75 11 745 10,6 100,0 100,0 —
Таблица 48
Воздействие прытких ящериц иа их жертвы в типчако во -полынной степи (по Тертышникову, 19726)
Поедаемая группа видов Абсолютное число беспозвоночных Степень воздействия, % Соотношение, % Избирательность питания по каждой группе видов
обнаружено в 3 пробах на площади 1 га (рассчитано) съедено ящерицами за 1 месяц на 1 га обнаружено в 5 желудках ящериц в степи в пище
Ракообразные — 240 1 — —. 2,9 —
Пауки 1 3 300 720 3 21,8 2,9 8,6 +0,5
Прямокрылые 3 10 000 2880 12 28,8 8,6 34,2 +0,6
Полужесткокрылые 4 13 300 720 3 5,4 11,4 8,6 -0,1
Жуки 6 20 000 2400 10 12,0 17,1 28,5 —0,2
Двукрылые 1 3 300 -— — — 2,9 — —
Перепончатокрылые 16 53 300 480 2 0,9 45,6 5,7 -0,7
Чешуекрылые (имаго-j-гусеницы) 3 10 000 720 3 7,2 8,6 8,6 +1,0
Прочие виды 1 3 300 240 1 7,2 2,9 2,9 +1,0
Всего . . . 35 116 500 8490 35 7,2 100,0 100,0
299
Таблица 49
Воздействие прытких ящериц на их жертвы в лесной полосе (по Тертышниковуу 19726)
Абсолютное число беспозвоночных ед Соотношение, % Я л • о ь £ «
Поедаемая группа видов обнаружено в 3 пробах на площади 1 га (рассчитано) обнаружено в 7 желудках ящериц съедено ящерицами за 1 месяц на 1 га Степень возде ствия, % в лесной полосе й> а-в к я Избирательное питания по ка: дой группе ви
Пауки 2 6 700 1 118 1,7 5,0 2,0 —0,7
Многоножки 4 13 300 7 823 6,2 10,0 14,0 +0,1
Прямокрылые 4 13 300 1 118 0,8 10,0 2,0 —0,6
Полужесткокрылые 5 16 700 7 823 4,9 12,5 14,0 +0,05
Жуки 5 16 700 4 470 2,9 12,5 8,0 —0,1
Двукрылые 5 16 700 6 706 4,2 12,5 12,0 —0,02
Перепончатокрылые Чешуекрылые 10 33300 4 470 1,9 25,0 8,0 —0,5
(имаго-] гусеницы) 5 16 700 20 2352 14,1 12,5 40,0 +0,5
Всего. . . . 49 133 400 50 5880 4,4 100,0 100,0 —
Таблица 50
Воздействие прытких ящериц на их жертвы на посевах (по Тертышникову, 19726)
Абсолютное число беспозвоночных ед Соотношение, %
Поедаемая группа видов обнаружено в 3 пробах ьа площади 1 га (рассчитано) обнаружено в желудках 4 ящериц съедено ящерицами за 1 месяц на 1 га Степень возде ствия, % на посевах ф S’ S к я Избирательное питания по ка: дой группе ви
Моллюски 1 3 300 2 15,0 0,4 2,5 7,1 +0,4
Пауки 1 3 300 3 22,5 0,7 2,5 10,8 +0,5
Полужесткокрылые 4 13 200 1 7,5 0,1 10,0 3,6 -0,4
Жесткокрылые 12 40000 15 112,5 0,2 30,0 53,6 +0,2
Двукрылые 17 56 600 2 15,0 0,02 42,5 7,1 —0,7
Перепончатокрылые 1 3 300 2 15,0 0,4 2,5 7,1 +0,4
Чешуекрылые (гусеницы+имаго) 1 3300 2 15,0 0,4 2,5 7,1 +0,4
Прочие виды 3 10000 1 7,5 0,1 7,5 3,6 -0,3
Всего. . . . 40 133000 28 210 0,15 100,0 100,0 —
300
Таблица 51
Воздействие прытких ящериц на их жертвы на лесной просеке (по Тертышникову? 19726)
— Абсолют ное число беспозвоночных Соотношение, %
Поедаемая группа видов Щаружено 3 пробах 1 площади га (рассчи-iho) жаружено желудках ящериц □едено яще-Щами за месяц на 1 га S о к . к С S ф Й 1 просеке пище збирательнос атания по ка: >й группе пи,
о и к — и О Дсп U о к cq Кеч
Моллюски 5 16 700 4 13 0,08 6,7 JB.5 —0,01
Прямокрылые 8 26 700 6 20 0,07 10,7 9,7 —0,04
Полужесткокрылые 6 20 000 5 17 0,08 8,0 8,1 0,00
Жесткокрылые 22 73 300 17 56 0,07 29,3 27,4 —0,03
Двукрылые 12 40000 10 33 0,08 16,0 16,1 0,00
Перепончатокрылые 22 73 300 20 66 0,09 29,3 32,2 +0,04
Всего. . . . 75 250000 62 205 0,08 100,0 100,0 —
Таблица 52
Сравнительные данные по биомассе фоновых позвоночных (пищевые конкуренты н враги прыткой ящерицы) на стационарном участке площадью 170 га в Ставропольском крае (по Тертышникову, 19726, с изменениями)
Вид животного Биомасса всех животных данного вида, кг Общее количество, % Биомасса съеденных прытких ящериц за день, кг Общее количество, % Биомасса съеденных беспозвоночных за День, кг Общее количество, %
АМФИБИИ: Жаба зеленая 5,1 2,7 0,41 . 3,9
РЕПТИЛИИ: Прыткая ящерица 68,5 36,2 — 5,7 54,3
Другие ящерицы (веретеница и разноцветная ящурка) 12,8 6,7 . — 0,65 6,2
Медянка 5,0 2,6 0,064 50,8 — —
Гадюка 5,2 2,8 0,002 1,6 0,2 1,9
ПТИЦЫ: насекомоядные (дрозды черный и певчий, жаворонок полевой, зяблик, каменка обыкновенная, овсянка обыкновенная, пеночка, синица большая, славка серая) 15,82 8,3 3,114 29,6
ПТИЦЫ: хищные (канюк обыкновенный, коршун черный, подорлик, подорлик малый) 10,8 5,7 0,054 42,84 0,1 1,0
МЛЕКОПИТАЮЩИЕ: лисица, барсук, енотовидная собака 66,0 35,0 0,006 4,76 0,32 3,1
Итого 189,2 100,1 0,126 100,0 10,50 100,0
301
ЯЩЕРИЦА КАК ПРОДУЦЕНТ
В. А. Ковда и И. В. Якушевская (1967) указывают, что на суще доля позвоночных составляет 1—3% всей зоомассы, основная часть которой приходится на млекопитающих. Сведения о биомассе прыткой ящерицы немногочисленны и не позволяют составить даже примерное представление о ее значении в круговороте веществ.
Исходя из данных по Ставропольской возвышенности, попытаемся хотя бы ориентировочно выяснить роль прыткой ящерицы как продуцента. Расчеты показывают, что на долю прыткой ящерицы приходится 71,3% всей герпетофауны стационарного участка. Из табл. 52 следует, что наибольшую биомассу ящериц из биоценозов извлекает медянка (50,8%), а остальную — гадюка, канюк, коршун, подорлик, лисица и енотовидная собака (49,2%), причем необходимо учитывать, что настоящим герпетофагом из всех приведенных животных является только медянка. Так, за сезон активности (составляющий 210 дней) прыткой ящерицы хищники извлекают из ее популяции 37,2% или 26,4 кг общей биомассы. Не исключено, что часть ящериц погибает за счет неблагоприятных климатических условий, болезней, не учтенных нами врагов, погибают отложенные ими яйца, неучтенные особи и пр.
Так, (по Тертышникову, 19726, с изменениями) при фактической численности популяции 5717 экз. доля самок 2710 экз. и предполагаемой численности приплода 17 615 экз. •—фактичес-ская биомасса ящериц (в кг)—44,6 или 62,8%, идеальная же (в кг) —71* или 100%.
Как источник энергии прыткая ящерица, подобно прочим плотоядным животным, менее эффективна, чем, например, консументы первого порядка, так как она затрачивает несравненно больше энергии на поиски и поимку добычи.
❖ * *
Прыткая ящерица имеет, по-видимому, определенное значение как консумент и продуцент. В тех биоценозах, где численность этого животного довольно велика, значение ее как консумента может превышать аналогичное значение птиц и млекопитающих. Во многих степных биоценозах биомасса прыткой ящерицы превышает биомассу других позвоночных животных. Учитывая это и зная, что в годы пониженной численности грызунов она является пищей многих хищных птиц и млекопитающих, можно предположить, что ее роль, как продуцента также может быть значительной.
* Вес сеголетков принимали равным 1,5, молодых —4,5, а взрослых —12 г (см. гл. XV).
ГЛАВА ЧЕТЫРНАДЦАТАЯ
ЧИСЛЕННОСТЬ И ЕЕ ДИНАМИКА *
Данных по численности прыткой ящерицы как в популяциях, так и по биотопам и зонам до последнего времени было немного. В то же время изучение численности является ключевым моментом и для определения биомассы как в пределах одного биотопа, так и в разных географических зонах, а также и для определения 'места вида в биогеоценозах. Остается добавить, что изучение численности и ее колебания в разных биотопах прямо связано и с познанием микроэволюционных процессов внутри каждого вида.
В настоящей главе на основании имеющихся пока отрывочных и немногочисленных данных рассматривается численность отдельных популяций в пределах одного географического региона и колебания численности этого вида. Несмотря на то, что многие из приводимых данных будут неизбежно приближенными, мы надеемся, что в целом они отражают основные черты численности данного вида и ее динамику. В заключение делается попытка вскрыть закономерности колебания численности в популяциях прыткой ящерицы.
МЕТОДЫ УЧЕТА ЧИСЛЕННОСТИ
К настоящему времени разработано и применяется несколько методов учета численности рептилий. Наиболее обычным является широко распространенный в экологии метод ленточных проб, который в исследованиях отечественных авторов обычно используется в модификации Л. Г. Динесмана и М. Л. Калец-кой (1952).
Этот метод сводится к следующему.
1. Учет проводится па ленте (трансекте), ширина которой равна 3 м. Длина такой ленты при учете должна составлять обычно не менее 1—1,5 км.
2. Каждая подобная лента должна находиться в пределах биотопов одного типа.
1 Авторы: А. С. Баранов, А. В. Яблоков, при участии А. В. Валецкого, М. А. Бакрадзе, В. К- Жарковой, Г. П. Лукиной, М. Ф. Тертышникова, В. И. Гаранина.
303
3. Учеты следует проводить при максимальной активности животных (сезонной, суточной).
Этот метод количественного учета применим во всех природных зонах и во всех биотопах.
Другим часто используемым для определения численности пресмыкающихся методом является метод пробных площадок. Этот метод заключается в том, что на точно измеренных площадках путем вылова пересчитывают всех животных и полученные результаты с соответствующими поправками переносят на всю изучаемую территорию. В связи с тем, что у прыткой ящерицы распределение особей внутри биотопа, занимаемого популяцией, неравномерно, оптимальный размер репрезентативных площадок в каждом случае должен определяться опытным путем (Тертышников, 1970, 19726). Если одна популяция занимает несколько различные биотопы, то следует закладывать несколько таких площадок. Подсчет животных на площадках дает возможность определять среднюю плотность. Абсолютное число прытких ящериц, обитающих в исследуемой популяции, будет равно сумме произведений средней плотности животных каждо* го из таких участков на его площадь. Метод площадок применим только в том случае, когда мы можем точно определить границы, занимаемые исследуемой группой особей (остров, небольшая зеленая лощинка между песками и т.п.). Во всех других случаях полученные данные не будут отражать действительную численность в популяции.
Интересным методом для определения численности популяций прыткой ящерицы служит метод кольцевания (Динесман, Калецкая, 1952; Жаркова, 19736). В основу описываемого способа положено определение численности взрослых самцов; численность самок и неполовозрелых ящериц устанавливаются путем дополнительных расчетов, при использовании полученных данных о соотношении половых и возрастных групп в популяции. Численность половозрелых самцов определяется путем повторного их вылова и кольцевания.
Наконец, при определении численности используется метод «площадей вида» (Лаптев, 1930), подсчет численности особей по которому производится также при наивысшей активности животных. Подсчет ведется по следующей формуле:
v х t X w'
где П — плотность вида, п — число встреченных особей, v — скорость движения счетчика, t — продолжительность подсчета, w — ширина обзора.
Для унификации данных, приводимых разными исследователями при определении численности прыткой ящерицы в разных популяциях, нами применялся следующий вариант методики. Маршрут для учета численности ящериц прокладывался в
304
периоды максимальной суточной активности (обычно первая половина дня); приблизительно (в шагах или метрах) вычислялась площадь территории, на которой проходил маршрут; подсчитывалось суммарное число выловленных особей, к которым приплюсовывались особи замеченные, но не пойманные. Опыт показал, что численность особей целесообразно определять для территорий порядка нескольких десятков гектаров. Это вызвано тем, что расчетные 'цифры для 1 га не отражают действительной картины территориального распределения ящериц в популяции (см. гл. II).
Например, наблюдатели двигались в одном направлении вдоль по железнодорожной насыпи. Число выловленных особей составило 55 экз; на одну пойманную ящерицу приходилось в среднем 2 убегавшие. Длина исследованного участка составила 350 м, ширина насыпи—5,5 м. Таким образом, на площади 1925 м2 было обнаружено 55+110=165 экз. ящериц. Средняя плотность населения Данного биотопа составляет в данном случае 8,6 экз/1000 м2. Естественно, что какая-то часть особей уходит от наблюдения незамеченной, другие ящерицы в момент наблюдения оказываются в укрытии, наконец, определенная часть животных на время наблюдения может быть вне пределов данного биотопа. Все это осложняет точный учет населения данной территории, а получаемые таким методом цифры будут несколько заниженными.
Соответственно этому, материалы, взятые для данной главы из литературных источников, были пересчитаны в экземплярах на 1000 м2. Так, например, В. К. Жаркова (1973а) производила учет численности прытких ящериц в северной лесостепи Европейской части СССР по методу «ленточных проб». Длина учетной линии у нее составляла обычно 2000 м при ширине 2 м. Плотность населения характеризовалась средним числом особей на маршрут 1000 м, а абсолютная численность — числом , особей на гектар.
В этом случае обследованная площадь одного маршрута равна 1000X2=2000 м2. Если на этой площади обитает 50 ящериц, то на 1000 м2 число обитающих ящериц составит 25 особей.
ЧИСЛЕННОСТЬ ЯЩЕРИЦ В ОТДЕЛЬНЫХ ПОПУЛЯЦИЯХ
Знание плотности населения ящериц в отдельных биотопах еще не позволяет установить общую численность ящериц в отдельных популяциях. Популяции ящериц могут, несомненно, включать в себя население нескольких биотопов в том случае, если они располагаются рядом и не изолированы друг от друга непреодолимыми для ящериц преградами. Например, в уже приводившемся выше случае, население пойменного леса и окрестных балок в окрестностях г. Верхний Мамон (среднее течение р. Дон, левый берег), без всякого сомнения, составляет единую
305
Таблица 53
Характеристика изолированных совокупностей особей прыткой ящерицы, которые могут рассматриваться как отдельные популяции
Местоположение Биотоп Площадь, 'в скобках, га) Приблизительная общая численность ящериц (без сеголетков) Авторы сообщения
Западный Алтай Западный Алтай Окрестности с. Головы, Северо-западная Украина, на границе с Белоруссией Южный склон большого Кавказского хребта, долина р. Иори, около 1700 м над уровнем моря Урочище Сомадло, 30 км к югу от г. Тбилиси. Мал. Кавказ Окрестности пос. Цал-ка, Мал. Кавказ. Около 2000 м над уровнем моря Северо-западное побережье оз. Севан, высота 2000 м над уровнем моря Окрестности пос.Жар-комыс, Актюбинская обл., левый берег р. Эмбы (кладбище) 10 км к северо-западу от г. Целинограда, по левой Стороне шоссе на Кургальджино С мелкими кустами остров на Бухтарминском водохранилище на стрелке Иртыша и р. Нарыма Травянистый холм с камнями напротив острова на берегу; 300 м пашни отделен от аналогичных биотопов на материке Широкая (для газопровода) просека в чистом сосновом лесу, где ящерицы очень редки Редкие посадки вяза в пойме реки и глубокие борозды, негустая трава, открытые травянистые овражки. Нигде кругом на протяжении многих километров ящериц нет Молодые сосновые посадки, около шоссе и луговины. Во всех окрестных лесах прытких ящериц не найдено Молодые сосновые посадки, открытые травянистые склоны, светлый лиственный лес по дороге на Ингу ри. Изолированы полями, поселком, глубоким ущельем Светлое лиственное мелколесье, местами камыши, высокая трава. Изолировано озером, шоссе, поселком, горами Зеленая долиика, выходящая к реке, выше поселка; изолирована песчаными дюнами Насыпь и кюветы шоссе, берега неглубоких карьеров, луг с густой травянистой растительностью. Изолирован кругом широким болотом 300x150 (4,5) 600 x 300 (18,0) 20X1500 (3,0) 120x600 1000x 300 2000x 500 (100,0) 1000X500 (50) 1000x 250 (25,0) 400x 300 (1,2) 800—1000 600—100 300—600 2000—3000 1500—3000 2500—4000 2500—3500 500—900 700—1400 Яблоков, Ва-лецкий Баранов, Ва-лецкий, Яблоков Баранов, Яблоков Баранов, Яблоков, Валецкий Бакрадзе, Баранов, Яблоков, Валецкий, Баранов, Валецкий, Яблоков Даниелян, Яблоков, Баранов Валецкий Валецкий Баранов, Яблоков Мина, Клеве-заль, Яблоков, Валецкий
популяцию, поскольку лес отстоит от ближайших частей балки на несколько десятков метров, что не превышает радиуса индивидуальной активности взрослой ящерицы (известно, что при расселении молодых и сезонных миграций ящерицы могут преодолевать пространства в несколько сотен метров, см. гл. IX).
Определение численности отдельных популяций прежде всего оказывается связанным с установлением границ популяций. Эта проблема определения популяционных границ является од-
306
ной из злободневых для популяционной биологии большинства видов животных (Тимофеев-Ресовский и др., 1973). Определение границ популяции может быть проведено в каждом конкретном случае лишь в результате специального стационарного исследования, включающего длительные наблюдения за природными концентрациями особей на протяжении ряда поколений. Практически таких данных для прыткой ящерицы в настоящее время нет. Однако с меньшей степенью достоверности, но достаточно надежно в первом приближении можно говорить о популяционных границах в случае исследования заведомо изолированных группировок особей.
Несколько таких группировок нам удалось обнаружить во время полевых исследований в 1970—1974 тг. (табл. 53).
Как видно из табл. 53, ориентировочная численность взрослых особей в девяти описанных популяциях ящериц, выделенных по наличию достаточно заметных при разовом исследовании изоляционных барьеров, колеблется от 300—600 до 3000— 4000 особей (в среднем около 1300 экз.). Надо учесть, впрочем, что эти определения численности очень приблизительны. Они основаны на экстраполяции числа пойманных и замеченных ящериц на населенную животными площадь. Однако, все приведенные цифры относятся к популяциям довольно ограниченным по общей площади и с достаточно высокой средней плотностью животных (только 'в таких популяциях был возможен быстрый отлов нескольких десятков особей). Большинство исследованных популяций занимает большую территорию, которая населена ящерицами не равномерно, а по сетчатому, ленточному или мозаичному типу (см. гл. II). Пока трудно привести даже приблизительные данные, характеризующие численность ящериц в таких пространственно разбросанных популяциях; не исключено, что численность в них может быть и больше, чем в описанных в табл. 53 популяциях. Имеются наблюдения, показывающие, что численность отдельных демов, входящих в популяцию и заселяющих разные биотопы, в отдельные годы может значительно колебаться, причем эти колебания происходят в разных направлениях: во влажный год преимущества в размножении получат демы, обитающие на сухих биотопах, а в сухой год — демы, связанные с биотопами, находящимися около водоемов. Такая различная' численность в отдельных биотопах является одним из обычных проявлений экологической дифференциации в пределах одной популяции. Своеобразная «асимметрия» численности в отдельных биотопах внутри популяции дает возможность поддерживатй общую численность на каком-то оптимальном уровне.
Определение абсолютной численности особей в отдельных популяциях интересно и важно для целого ряда популяционно-генетических и популяционно-экологических исследований. Очевидно, что без определения числа популяций, населяющих ту или
307
иную большую территорию, нет основания для определения общей численности особен даже в ограниченном ландшафте. Уровень же изученности прыткой ящерицы в настоящее время недостаточен, чтобы даже ориентировочно определить число популяций вида, входящих в отдельные ландшафты и географические регионы (что отражает общую чрезвычайно слабую популяционную изученность любого вида животных). Поэтому для определения численности ящериц более перспективным на данном этапе исследования оказывается определение плотности населения в различных биотопах и экстраполяция этих величин на отдельные природные зоны.
ПЛОТНОСТЬ НАСЕЛЕНИЯ ЯЩЕРИЦ В РАЗНЫХ БИОТОПАХ И ГЕОГРАФИЧЕСКИХ ЗОНАХ
Несмотря на то, что прыткая ящерица является в целом эври-топным видом, ее бпотопическая приуроченность заметно отличается в разных частях ареала (см. гл. V). Говоря о сравнении численности ящериц в разных биотопах и разных географических зонах, в качестве основного критерия оказывается целесообразным использовать плотность населения, т. е. численность, отнесенную к определенной территории. Эта плотность, выражаемая в экземплярах взрослых ящериц, встречающихся на площади 1000 м2, позволяет проводить как сравнение разных биотопов и популяций внутри одной зоны, так и сравнение одних и тех же биотопов в разных зонах, и сравнение целых зон между собой (см. табл. 54). В лесной зоне типичные биотопы — в основном лесные опушки, поляны, пойменные леса, пойменные луга, водораздельные луга и береговые склоны и лишь в незначительной степени антропогенные биотопы. Численность ящериц в этой зоне сравнительно незначительна в разных населяемых ими биотопах (до 8,3 экз./ЮОО м2). Максимальная численность вида в этой зоне обнаружена на склонах. Видимо, береговые склоны создают наиболее благоприятные условия обитания — мягкие песчаные почвы, удобные для постройки нор, более постоянную влажность в связи с близостью воды, хорошую освещенность (прогреваемость).
Следующими по плотности населения ящериц в естественных биотопах лесной зоны оказываются опушки леса (4,1 экз./ 1000 м2) и светлые пойменные леса (3,3 экз.). Как видно из табл. 53, численность ящериц в антропогенных биотопах этой зоны заметно превышает численность ее в природных биотопах. Средняя численность ящериц в лесной зоне, в подходящих для нее биотопах, оказывается в целом весьма невысокой —4,4 экз./ 1000 м2.
Численность ящериц в лесостепной зоне оказывается заметно более высокой, чем в лесной. Это подтверждается максимальными величинами численностей в подходящих биотопах (до
308
Таблица S4
Численность прыткой ящерицы в различных биотопах и разных природных зонах (число особей на 1000 м2, в скобках — число изученных биотопов данного типа)*
Биотоп Основная природная зона
лесная лесостепная степная полупустыни горные районы
Лесные опушки 4,1(5) — 3(1) — —
Поляны 1,5(1) — 3(1) — —
Островки леса -— 1,5(2) — / —
Пойменный лес 3,3(1) 6,3(2) — •—•
Луг пойменный 1,0(1) — — 14,4(4) 18(1)
Луг суходольный (водораздельный) 2,5(1) — — — —
Луг горный (субальпийский-альпийский) — — — — 40,9(1)
Разнотравная степь — —- 5,8(1) — —
Полынная степь — — 30(1) — —
Ковыльная степь -— —• 0,5(1) — —
Склоны оврагов, балок, карстовых воронок и других естественных понижений, береговые склоны 5,5(5) 0,6(3) 44,4 (6) 20,1(2) 25,3(1)
Горные склоны — — — — 12,7(1)
Каменистые россыпи разного типа -— —— — 7,1(1) —
Пар — 2,6(1) — — —
Сады — — 44,0(2) — —
Вырубки и просеки 3,3(2) — 0,1(1) — —
Обочины дорог — 38,6(2) 02,5(2) — —
Железнодорожные и другие насыпи — 41,0(1) 92,5(2) — —
Лесовосстановительные посадки и лесопитомники — 38,6(2) 36,0(1) — —
Кладбища — 4,4(1) Ю,1(2) — —
Пустыри и промышленные пустоши -— — 92,5(2) — —
Склоны прудов, водохранилищ, оросительных каналов 5,3(1) — 72,8(3) 26,9(1) —
Заброшенные карьеры, рвы и окопы 1,5(1) — 41,0(1) — —
Все биотопы 4,4 17,3 40,4 25,6 27,6
• По данным Л. Г. Динесмана и М. Л. Калецкой (1952), В. Л. Булахова и Н. Ф. Константиновой (1972), В. К. Жарковой (1973а), Г. П. Лукиной (19666), М. Ф. Тертышникова (19726), 3. П. Хонякиной, В. А. Кутузовой, А. С. Баранова, А. В. Балецкого, А. В. Яблокова, В. И. Борисова.
309
38,6 экз./ЮОО м2) и средней численностью по всей зоне (17,3 экз./1000 м2). Особенности населяемых ящерицей биотопов в этой зоне характерны и для лесной .зоны. В этой зоне численность животных в антропогенных биотопах (обочины дорог, насыпи, лесополосы, посадки, межи на полях и т. п.) значительно выше, чем в лесной зоне.
В степной зоне прыткая ящерица наиболее многочисленна. При этом надо учесть, что и по разнообразию занимаемых биотопов эта зона превосходит остальные (см. гл. V).
Естественно, что среди разнообразных биотопов, населяемых ящерицами, в этой зоне встречаются и биотопы с весьма низкой численностью, не превосходящей численность в большинстве биотопов лесной и лесостепной зон, но в целом и по максимальной плотности населения отдельных природных биотопов (до 92,5 экз./ЮОО м2) и по средней плотности для всей зоны (40,4 экз./ЮОО м2) она, безусловно, превосходит все остальные. В этой зоне прыткая ящерица окончательно сменяет биотопы, связанные с лесом, на биотопы открытых пространств. Однако по-прежнему наиболее предпочитаемыми природными биотопами являются склоны оврагов, балок, береговые склоны и другие естественные понижения, где прыткая ящерица достигает наибольшей численности. Даже в случае обитания в лесных биотопах (обычным для этой зоны лесным биотопом является лишь пойменный лес) численность ящериц в них заметно меньше, чем в непосредственной близости расположенных открытых биотопах. Примером может служить популяция ящериц в окрестностях г. Верхний Мамон (среднее течение р. Дон). Здесь в пойменном лесу плотность ящериц составила 9,6 экз./ЮОО м2, тогда как на склонах балок, покрытых редким кустарником, плотность достигала 44,4 экз./ЮОО м2.
В этой зоне наверное, большая часть популяций обитает в антропогенных биотопах. Это связано со значительной хозяйственной (в основном сельскохозяйственной) освоенностью этой зоны. Могучая сеть шоссейных и грунтовых дорог и железнодорожных путей, с малопосещаемыми человеком кюветами, насыпями, снегозащитными посадками, большая протяженность полезащитных лесных полос, межи между полями — все это создает биотопы, чрезвычайно удобные для прыткой ящерицы. Именно здесь плотность населения вида достигает максимальных величин для всего ареала. Так, в окрестностях г. Крымска (Краснодарский край) Г. П. Лукина в 1966 г. обнаружила плотность населения 100 экз./ЮОО м2, в 1973 г. в окрестностях г. Ро-стова-на-Дону, в овражках, возле небольшого пруда, окруженного со всех сторон полями, плотность населения прыткой ящерицы составила 77,5 экз./ЮОО м2 (Баранов; Валецкий, личное сообщ.). В среднем плотность ящерицы в антропогенных биотопах в этой зоне, как и в предыдущих, описанных зонах, несомненно, много выше, чем в природных биотопах.
310
В полупустынной зоне плотность населения ящериц не очень велика. Предпочитаемые здесь биотопы относятся, как и в степной зоне, к биотопам открытых пространств: пойменные луга, склоны и другие естественные понижения. Плотность населения в такого рода биотопах достигает 32,7 экз./ЮОО м2. В среднем по населяемым ящерицей естественным биотопам полупустынной зоны плотность составляет 13,3 экз./ЮОО м2.
В этой зоне подавляющая часть популяций ящериц обитает в природных, а не в антропогенных биотопах. При этом интересно отметить, что, несмотря на редкость встречаемости ящериц в антропогенных биотопах, плотность ее в таких биотдпах достаточно велика и превышает среднюю плотность населения по всей зоне (26,9 экз./ЮОО м2). Эта редкость встреч ящериц в антропогенных биотопах скорее всего вызвана тем, что полупустынная зона еще слабо освоена человеком и антропогенных биотопов в ней сравнительно мало. Напомним, что в этой зоне, согласно правилу смены стаций, прыткая ящерица старается придерживаться увлажненных мест, столь необходимых для откладки и развития яиц (см. гл. VII). В среднем плотность населения ящериц в подходящих биотопах этой зоны довольно велика (25,6 экз./ЮОО м2) и превосходит плотность населения ящериц в лесной и лесостепной зонах.
В горных районах основными населяемыми биотопами являются пойменные и горные луга, горные склоны, береговые склоны и в некоторых случаях каменистые россыпи разного типа. Плотность населения в такого рода биотопах довольно велика и колеблется сравнительно незначительно: от 10,7 до 40,9 экз./ЮОО м2 *. Средняя плотность населения ящериц в подходящих биотопах в горной зоне сравнительно высока — 27,6 экз./ЮОО м2, что значительно выше, чем в среднем по лесной и лесостепной зонам, и примерно столько же, сколько в зоне полупустынь.
Интересными оказываются определения плотности населения прыткой ящерицы в девяти популяциях, 'выделенных в предыдущем разделе главы. Оказывается, размах колебаний плотности населения ® разных популяциях весьма значителен — от 2,5 до 116,7 экз./ЮОО м2. В среднем плотность населения по популяции близка к средней плотности населения ящериц в биотопах степной зоны и составляет 50,3 экз./ЮОО м2.
В заключение этого раздела рассмотрим некоторые общие тенденции в распределении плотности населения ящериц по всему пространству ареала вида. Наибольшая плотность характерна для степной зоны, наименьшая — для лесной. Горные районы, полупустыни и лесостепная зона в отношении плотности
* По последним данным, в долине ручья на Зап. Алтае (левый берег р. Кульд-жир, высота около 1000 м над ур. м.) максимальная численность ящериц составляла 58 экз./ЮОО м2, а средняя (для всего участка в 9 га) — 27,6 экз./ЮОО м2 (А. С. Розанов, сообщ.).
311
ящериц в населяемых ею биотопах различаются незначительно. Во всех зонах плотность оказывается выше в открытых биотопах. Во всех зонах средняя плотность населения оказывается заметно выше также и в антропогенных биотопах. Это обстоятельство свидетельствует о важном качестве прыткой ящерицы как вида: она легко «уживается» с человеком, и хозяйственное освоение территорий, как правило, не ведет к ее уничтожению, как это происходит часто с другими видами наземных позвоночных. Это качество вида делает его не только важным компонентом атроценозов и антропогенных ландшафтов, но и определяет интерес его исследования как в известной степени модельного вида, за процессом микроэволюции которого удобно вести наблюдения и производить эксперименты.
ОБЩАЯ ЧИСЛЕННОСТЬ ЯЩЕРИЦ В РАЗНЫХ ПРИРОДНЫХ ЗОНАХ
Зная среднюю плотность населения ящериц в подходящих биотопах и общую площадь таких биотопов в каждой природной зоне, можно было бы легко определить численность ящериц в каждой природной зоне. Однако в нашем распоряжении отсутствуют данные по площади отдельных биотопов, населенных ящерицами в разных зонах. Поэтому при расчетах общей численности приходится делать ряд допущений, касающихся средней плотности населения ящериц в каждой зоне. Вероятно, отношение населяемых ящерицами биотопов к ненаселенным будет минимальное в степной зоне, где плотность биотопов, населенных ящерицами, в общем ландшафте сравнительно велика. Так, по результатам анализа территории в 170 га в типичной степной зоне Ставропольского края, из 10 биотопов только два были совершенно лишены ящериц и четыре были на 100% заселены этим видом (Тертышников, 19726). Средняя площадь, занятая ящерицами на довольно значительной исследованной территории, составила в среднем по всем биотопам 68,0%- Судя по этим данным, допущение о средней плотности населения ящериц по всей зоне в */5—*/« по средней плотности населения ею подходящих биотопов не будет преувеличением.
Для зон полупустынь, лесостепной и горной можно принять допущение о средней плотности населения по всей зоне в 7ю— Vso от средней плотности населения обитаемых биотопов. Наблюдения показывают, что биотопы, населенные ящерицами в лесной зоне, значительно более дисперсно распределены среди других биотопов, чем в других зонах; соотношение населенных и ненаселенных биотопов составляет в этой зоне, видимо, величины порядка 76о—‘Лоо-
Следующая неизбежная группа допущений связана с определением точной площади той или иной зоны в пределах ареала прыткой ящерицы. Приводимые в табл. 55 данные получены в
312
Таблица 55
Расчетная численность прытких ящериц по природным зонам СССР (входящим -в границы ареала вида)
Зона Средняя плотность экз./1000 м2 Площадь зоны, км2 Численность, тыс. экз.
в населяемых ящерицей биотопах в среднем по всей зоне
Лесная 4,4 0,04—0,09 1 370 100 41103—123 309
Лесостепная 17,3 0,35-1,73 205 200 73 872—354 996
Степная 40,4 4,04-8,08 1 426 500 5 763060—11 526120
Полупустынная 25,6 0,51—2,56 339 500 173 145—869 120
Горная 27,6 0,55-2,76 280 000 154000—772 800
По всем зонам — — — 6 205180—13 646345
результате выкопировки соответствующих зон по карте расти-тельности СССР *.
Возможен несколько другой путь расчетов, связанный с исключением из общей площади природных зон территорий, занятых городами, промышленными предприятиями, коммуникациями, селами и фермами. Статистические данные показывают, что эти территории .занимают >в Европе 37% общей территории. Экстраполируя эти данные на площадь части ареала, занимаемого прыткой ящерицей в СССР, можно сделать следующие допущения: в степной зоне эти цифры могут быть несколько меньшими (условно примем их 25—30%), в полупустынной и горной зонах эти цифры должны быть заметно меньшими (порядка 10—15%). Исключив соответствующие площади, как абсолютно лишенные прытких ящериц, можно представить, что
Таблица 56
Вариант расчета численности прытких ящериц по природным зонам СССР
Зона Площадь (км2) без индустриальных территорий, городов и'поселков Средняя плотность населения ящериц, экз./км2 Расчетная численность (общая), тыс. экз.
Лесная Лесостепная Степная Полупустынная Горная Всего 864163 149 796 1 069 875—998 550 288 575—305 550 238000—262 000 440—880 1730—3460 4940—8080 2560—5120 2760—5520 380 231—760 463 259 147—518 294 4034142—8 644 590 1 738 752—3 477 594 656 880—1 313 760 7 069 152—14714611
1 Физико-географический атлас мира. М., 1964, карта 241, составленная А. Н. Лукичевой.
313
на остальных территориях средняя плотность населения ящериц может быть в 5—10 раз ниже, чем в населяемых ею биотопах. Соответствующие расчеты приведены в табл. 56.
Как видно из сравнения двух таблиц (табл. 55 и 56), полученные данные весьма близки между собой. Совпадение может быть косвенным доказательством справедливости приведенных расчетов. Полученные данные по численности, естественно, должны рассматриваться лишь как ориентировочные, поскольку основаны на произвольном допущении о средней плотности населения ящериц по каждой зоне. В то же время, эти цифры являются отражением статичного состояния, так как численность ящериц, как и численность всех живых существ, подвержена весьма заметным колебаниям по сезонам и годам.
КОЛЕБАНИЯ ЧИСЛЕННОСТИ И ИХ ПРИЧИНЫ
Изучение и вскрытие причин, определяющих колебания численности животных в природе, является одной из важнейших задач современной зоологии. Эти изменения определяют динамику биомассы вида, а следовательно, и степени участия вида в работе биогеоценозов. Первично количественные изменения числа особей в популяциях представляют собой итог воздействия., факторов биотического и абиотического характера. При этом надо различать истинные изменения численности и временные, которые могут быть вызваны миграциями животных из данных,: биотопов в связи с неблагоприятными условиями (засуха, на- , воднение) или различием периодов активности разных возрастных и половых групп. Наблюдения показывают, что как биоти- г ческие, так и абиотические факторы оказывают особенно массированное воздействие на численность наземных позвоночных, , если они связаны с антропогенным воздействием.
Периодические колебания численности
Периодические колебания численности населения ящериц в основном связаны с размножением и возрастной смертностью, т. е. являются сезонными. В ряде случаев количественные флуктуации численности в популяциях могут быть вызваны периодичностью вспышек кормовых видов и периодических изменений «давления» хищников.
Известно, что всякая популяция имеет свою специфическую ритмику численности (как по полу, так и по возрасту). К сожалению, имеются лишь отрывочные данные по ритмике численности особей разного пола прыткой ящерицы по сезонам. В. К. Жаркова (1973а) исходя из факта, что количественные соотношения разных возрастных групп в популяции иные, на основе имеющегося у нее материала определяет различие в возрастной смертности самцов и самок (табл. 57).
314
Как видно из табл. 57, за первый и второй годы жизни среди ящериц Мещерской низменности самцов погибает почти в два раза больше, чем самок. Лишь на третьем году жизни резко увеличивается процент гибели самок. Такой ход динамики численности приводит к преобладанию в популяции молодых самок. В более старших возрастных группах, наоборот, происходит интенсивная смертность самой, и соотношение полов выравнивается. Возможно, именно такая дифференцированная смерт-
Рис. 93
Обобщенная схема динамики численности модельной популяции прыткой ящерицы
Исходная численность — 156 особей — определена путем суммирования разных возрастных групп трех поколений (см. рис. 87)
ность объясняет несколько большую численность самок в некоторых природных популяциях.
Ритмика колебаний численности особей разного пола подчиняется тем же общим закономерностям, которые действуют при возрастных колебаниях, с некоторыми поправками для разных полов. В настоящее время можно построить модель динамики численности населения прыткой ящерицы, определяемую возрастной смертностью (рис. 92). Как показано в гл. XV, темп
Таблица 57
Возрастная смертность самцов и самок (разница в числе особей предыдущей и последующей генерации *) прыткой ящерицы в Мещерской низменности
[по В. К. Жарковой (1973а) за 1965—1969 гг.]
Генерация Самцы (исходная численность сеголетков 1-й генерации 272 экз.) Самки (исходная численность сеголетков 1-й геиерации 203 экз.)
число особей % число особей %
1-я 82 30,0 34 16,7
2-я 118 67,3 45 26,6 f
3-я 49 68,0 73 58,8
* Под генерацией в данном случае имеется в виду потомство, появившееся на свет в один сезон размножения.
316
смертности животных на протяжении их жизни оказывается различным. Проследим динамику численности одного поколения. Принимая данные табл. 62 (гл. XV) за исходные, получаем, что наибольшая численность в популяции достигается в момент откладки яиц (отложенное оплодотворенное яйцо — это уже особь) и выхода молодых. Учитывая большую смертность среди новорожденных (50%), которые гибнут до зимовки и после нее, к следующей весне в живых останется всего 25% от родившихся этого поколения. На второй год погибнет около 33% от достигших возраста в один год. На третий год жизни смертность, видимо, несколько снижается, но на четвертом и пятом годах жизни достигает соответственно 50 и 100% от числа доживших до этого возраста особей.
Суммарная картина сезонных колебаний численности в популяции прыткой ящерицы будет намного сложнее, так как в каждый момент времени в популяции' будут находиться особи пяти поколений, каждое из которых имеет свой темп смертности. В результате модель численности популяции прыткой ящерицы будет выглядеть следующим образом (рис. 93). Необходимо подчеркнуть, что в этой модели учтены только сезонная циклика; на нее в природе практически всегда накладывается апериодическая цикличность.
Безусловно, что в разных частях ареала динамика изменений сезонной численности популяции должна быть различной. Многолетняя динамика численности представляет собой итог сезонных динамик. Но это не простая сумма, а как бы наложение друг на друга различных часто противоположно направленных явлений, обусловленных различными факторами. Говоря о сочетании факторов, имеются в виду как биотические, так и абиотические факторы, действующие на природные популяции. Проиллюстрируем колебания численности в разные годы на примере пяти групп ящериц в северной лесостепи Европейской части СССР (табл. 58).
Как видно, каждому биотопу присуща определенная динамика колебаний численности. Так, на склонах реки численность
317
ящериц за эти четыре года постоянно увеличивается, в то время как в сосновых посадках ящерица отсутствует, затем резкий «всплеск» численности с последующим сокращением числен-ности населения. В смешанном лесу, на разнотравном лугу происходят изменения численности другого порядка.
Несомненно, колебания численности в популяциях прыткой ящерицы, связанные с действием биотических факторов, могут быть вызваны либо периодическими вспышками численности основных кормов (см. гл. VI), либо периодичностью давления пресса хищников, либо, наконец, резкими вспышками численности конкурирующих видов.
Увеличение численности кормовых видов, безусловно, приводит к некоторому увеличению численности популяции (при условии отсутствия пресса хищников и конкурирующих видов), падение же численности пищевых видов в некоторых случаях приводит к тому, что взрослые особи начинают пожирать молодых (каннибализм), тем самым сокращая численность популяции. В некоторых случаях происходит ложное сокращение численности популяции в результате миграции. Такие миграции могут быть вызваны увеличением численности популяций конкурирующих видов или резким спадом численности кормовых видов. Иногда популяции мигрируют полностью и переходят в биотопы, совершенно ей не свойственные, в стадии переживания (см. гл. IX). Важно добавить, что кормовые виды и виды-конкуренты, видимо, не имеют столь же существенного значения в регуляции численного состава популяций прыткой ящерицы (Лукина, 1966; Тертышников, 1972а, б; и др.)
Немаловажную роль в колебаниях численности популяций играют хищники. Резкое увеличение или падение численности хищников неизбежно приводит к сокращениям или увеличениям численности популяций прыткой ящерицы. В то же время значение хищников как регуляторов численности популяций прыткой ящерицы крайне недостаточно изучено. По расчетам М. Ф. Тер-тышникова, в Ставропольском крае, на стационарном участке, расположенном в верховьях р. Томузловки, за сезон гибнет 37,2% общей биомассы популяции ящерицы в результате воздействия на них учтенных фоновых врагов из позвоночных. В этом районе пресс хищников является дополнительной причиной, задерживающей рост численности в популяции. «Давление» хищников, несомненно, различно и в разных популяциях. Об этом косвенно свидетельствуют данные в гл. XIII о различной доле особей с регенерированными хвостами (т. е. особей, подвергавшихся нападению хищников и удачно их избежавших). Напомним лишь, что в некоторых популяциях больше половины взрослых особей несут на себе следы таких нападений.
В целом можно сказать, что периодические колебания, видимо, не вызывают изменений численности популяций прыткой ящерицы более чем на один порядок.
318
Апериодические колебания численности
Факторы, способствующие массовому размножению ящериц в популяции, и факторы, определяющие наступление следующего за ним периода депрессии численности, могут быть вызваны апериодическими явлениями. Такими явлениями могут быть катастрофические изменения биогеоценозов в результате пожаров, наводнений, засухи, сильных морозов в малоснежные зимы и прочие явления, прямо или косвенно благоприятствующие или, наоборот, препятствующие росту численности. Колебания численности могут быть связаны и с длительными изменениями природных условий, вызванными деятельностью человека (опустынивание, засоление больших территорий и т. п., или длительными изменениями климата типа ледниковых периодов. Иногда такого рода причины могут приводить к массовой гибели животных, тем более что прыткая ящерица, являясь пойкилотермным животным, весьма зависит от климатических условий.
.По мнению многих авторов (Терентьев, 1946; Лукина, 19666; Гаранин, 1971, Тертышников, 19726; Жаркова, 1973а; и др.), основное сокращение численности ящериц происходит в основном за счет гибели яиц. Если принять все отложенные яйца в популяции за 100%, то к моменту выхода молодых гибнет от 40 до 60% • Этот процент резко увеличивается особенно в дождливое и холодное лето (Гаранин, 1971). Другой критический момент, связанный с физическими факторами среды, в жизни прыткой ящерицы — осень с ранними заморозками. Именно в этот период времени, когда животные собираются уходить на зимнюю спячку, такого рода явления наиболее опасны (особенно для молодых этого года рождения, которые позднее уходят на зимовку).
Наконец, последним критическим моментом, влияющим на численность популяций, является зимовка. Численность особенно резко падает в морозные малоснежные зимы (Гаранин, 1971; Тертышников, 19726). Обычно взрослые ящерицы зимуют в своих норах или в старых норах грызунов (см. гл. V). Если в открытых биотопах норы большинства грызунов глубокие и как зимние убежища являются надежными (так как температура в них не опускается ниже 0°), то этого нельзя обычно сказать о норах, вырытых самими ящерицами, обычно неглубоких, а также о трещинах и щелях, где часто в почве зимуют молодые особи. В малоснежные холодные зимы, когда почва недостаточно покрыта снегом, а следовательно нарушается температурный режим в зимних норах, многие животные, замующие в таких норах, погибают. Поэтому значительно более надежными для выживаемости ящериц зимой являются лесные биотопы, лесополосы, придорожные канавы, овраги и т. п., так как именно в этих местах скапливается значительное количество снега, утепляющего почву. Здесь массовая гибель животных в морозные зимы может наступить лишь в исключительных случаях. В. И. Гаранин (1971)
319
приводит такой пример. Малоснежная суровая зима 1968 г. в Волжко-Камско.м заповеднике привела к резкому сокращению численности ящериц. Именно во время зимовки происходит резкое сокращение численности среди молодых животных в популяции. Так, М. Ф. Тертышников показал, что отход яиц и смертность сеголетков в первую зимовку в Ставропольской возвышенности составляет 25,7%.
Наиболее существенные изменения среды обитания, носящие катастрофический характер, приводят к резкому снижению ее численности. К типичным явлениям такого порядка можно отнести пожары, наводнения и засухи. По сведениям В. К. Жарковой (1973а), в 1967 г. после сильных дождей в районе Окского заповедника реки вышли из берегов и затопили биотопы, занимаемые прыткой ящерицей. Так, в пойменном лугу и в пойменных посадках сосны на р. Выше в этот год наблюдалось резкое сокращение численности прыткой ящерицы: на пойменном лугу численность ящериц в среднем сократилась в 4 раза по сравнению с 1966 г., а в пойменных посадках в 2,5 раза. Надо заметить, что при этом могут совершенно исчезнуть отдельные демы или целые их группы. Но популяции в большинстве случаев сохраняются, хотя и могут при этом достигать минимальных численностей, достаточных все же для дальнейшего существования.
Наиболее существенные изменения среды, связанные с длительным изменением природных условий, в настоящее время чаще бывают связаны с антропогенными воздействиями. Например, одной из причин гибели прытких ящериц является создание новых водохранилищ на пойменных землях. На берегах Куйбышевского водохранилища Гаранин (1971) отмечает резкое сокращение численности рептилий и в том числе прыткой ящерицы. В то же время на некоторых островах водохранилища сократившиеся сначала численно популяции затем заметно увеличились.
Более серьезным и распространенным фактором, отрицательно влияющим на численность ящериц, является неумеренное использование ядохимикатов в сельском и лесном хозяйстве. Существует строгая корреляция между обработкой ядохимикатами участков Мещеры и населенностью их ящерицами (Жаркова, 19736). В этом районе не заселенные прыткой ящерицей, но подходящие для них биотопы составляют от 32 до 49% обследованной территории. Есть все основания согласиться с предостережением Е. Рене (Rene, 1969) и К. Корбетта (Corbett, 1969) об опасности вымирания прыткой ящерицы в развитых промышленных районах в связи с полным антропогенным разрушением естественных биотопов.
Есть и другая сторона антропогенного влияния на численность прыткой ящерицы. Наблюдения, проведенные в 1970— 1974 гг. в разных частях ареала прыткой ящерицы, показывают, что она в некоторых районах становится «антропогенным видом». Широкая способность этого вида приспосабливаться к антропо
320
генным биотопам (см. табл. 5) несомненно позволит этому виду не только не сокращать свою численность при контактах с цивилизацией, но, возможно, в некоторых частях своего ареала даже увеличивать ее. Пока же в результате активного антропогенного воздействия численность прытких ящериц резко сокращается на Ставропольской возвышенности (Тертышников, 1972в), в Калужской обл. (Стрельцов), во многих районах Сибири (Баранов и др., сообщ.), Латвийской (Бахарев, 1971) и Эстонской ССР (Вельдре, личное сообщ.), а в окрестностях г. Махачкала (Хоня-кина, Кутузова, личн. сообщ.) прыткая ящерица исчезла вообще.
Таким образом, в настоящее время лишь при очень резких изменениях среды, обычно, связанных с разрушением биоценозов под воздействием человека, численность популяций сокращается ниже критического уровня, за которым следует вымирание популяции.
Периодические (сезонные и годовые) колебания численности прыткой ящерицы, видимо, ке превышают одного порядка величин,'тогда как апериодические колебания оказываются часто более значительными.
11 Прыткая ящерица
ГЛАВА ПЯТНАДЦАТАЯ
БИОМАССА ВИДА
И ОТДЕЛЬНЫХ ЕГО ПОПУЛЯЦИИ1
Биомасса популяции и вида в целом, определяемая как количество вещества, накопленное на данный момент времени, может быть определена, исходя из известной численности особей различных возрастных классов и массы отдельной особи определенного возраста. Численность как отдельных популяций, так и по природным зонам, была рассмотрена в предыдущей главе, данные по размерам (весу) отдельных особей различных возрастных классов приведены в главах III и VIII. Объединение этих данных позволяет определить как биомассу ящериц в отдельных популяциях и на небольших территориях (в биозонах), так и общую биомассу вида по зонам. Полученные данные, конечно, будут лишь некоторым приближением к той реальной биомассе, которая характеризует исследуемый вид в природе, поскольку не отражают реальной постоянно наблюдающейся в природе динамики биомассы. Определению основных черт этой динамики биомассы посвящается вторая часть настоящей главы.
БИОМАССА ОТДЕЛЬНЫХ ПОПУЛЯЦИЙ
Исходными данными для определения биомассы отдельных популяций служат величины абсолютной численности разных возрастных групп в конкретных популяциях и средние величины, характеризующие массу особи в каждом возрастном классе.
Средняя масса отдельных особей в разных участках ареала
Как было показано ранее (гл. Ill, VIII), масса взрослых особей в разных участках ареала оказывается несколько различной (см. рис. 34). Хотя, строго говоря, эти данные относятся к максимальному весу самцов, нет оснований для отрицания возможности их экстраполяции на средние величины биомассы отдельной особи (как для самцов, так и для самок). Исходя из имеющегося в нашем распоряжении фактического материала по весу (массе) отдельных особей из разных популяций (5423 отдельных провесов), средняя масса отдельных особей в каждом возрастном классе оказывается следующей (в г):
1 Авторы: А. В. Яблоков и М. Ф. Тертышников:
322
Новорожденные (сеголетки данного года рождения); возраст 0—3 месяца)
Особи прошлого года рождения (обычно, неполовозрелые; возраст 8—12 месяцев)
Взрослые особи (возраст 2—4 года)
—1,5 (0,5—3,5)
—4,5 (2,5—6,0) —12,0(5—40)
Приведенные средние цифры могут несколько изменяться в разных регионах ареала, что приходится учитывать при расчете биомассы отдельных популяций или населения определенных территорий (биотопов, зон). Так, например, при учете численности обычно не делается различий между встречамц взрослых и полувзрослых ящериц, а при учетах, проведенных во второй половине лета, и молодых этого года рождения. Из этого ясно, что использовать данные по численности в этом случае приходится дифференцированно. В дальнейших расчетах принимается, что средняя масса ящериц в популяциях лесной и лесостепной зон равна 8,0 г (среднее взрослых и полувзрослых), а в остальных зонах — 9,0 г.
Средняя биомасса модельных популяций
Как было показано в предыдущей главе, в настоящее время невозможно с большой точностью определить абсолютную численность не только ящериц в большинстве популяций на всем пространстве ареала, но даже точно определить общее число популяций вида. Такое положение сказывается и на определении биомассы вида: пока можно с удовлетворительным приближением определить биомассу лишь некоторых, весьма немногочисленных популяций. При определении биомассы ящериц для больших территорий (ландшафтов и целых природных зон) приходится экстраполировать средние данные по плотности ящериц в отдельных биотопах на площадь таких биотопов в том или ином регионе. К сожалению, достаточно точные данные по площади разных биотопов в разных природных зонах практически отсутствуют и приходится еще раз прибегать к весьма приближенной экстраполяции (см. гл. XIV). Полученные таким образом данные по численности ящериц в разных зонах оказываются несомненным шагом вперед в изучении данного вида и его места в биосфере.
Как было показано в гл. XI, любая популяция состоит из нескольких в разной степени изолированных демов — групп от нескольких экземпляров до нескольких десятков взрослых особей и их потомков, связанных между собой более близким родством. Эти демы являются сравнительно неустойчивыми во времени образования и могут распадаться через 1—2 поколения. Объединение многих демов составляет популяцию. В гл. XIV было показано, что общая численность взрослых особей, входящих в одну популяцию прыткой ящерицы, видимо, не меньше нескольких сотен экземпляров (см. табл. 53). Именно такой оказывает
323
11»
ся численность популяции на южной границе ареала (среднее течение р. Эмбы) и целого ряда других популяций. Численность популяции в урочище Сомадло в Восточной Грузии (Малый Кавказ) превышает 1000 особей, так же, как и численность популяции в долине р. Йори (южные склоны Большого Кавказа) и других популяций.
Итак, имеющийся сейчас материал показывает, что численность отдельной популяции прытких ящериц вряд ли в среднем превышает 1500 экз. взрослых особей. Видимо, не будет большой ошибкой считать (в первом приближении) среднюю численность многих популяций, близкой к 1000 экз. взрослых особей, а мелких популяций — около 500 взрослых особей.
Исходя из средних величин, характеризующих возрастной состав популяции, можно считать, что на 1000 взрослых особей (500 пар) должно приходиться в среднем 2000 полувзрослых (прошлого года рождения) и во второй половине лета — 4000 сеголетков. В популяции, состоящей из 250 пар взрослых особей, должно находиться около 1000 экз. полувзрослых и 200 экз. сеголетков.
Расчет биомассы популяции целесообразно начинать с сеголетков, поскольку их численность более или менее точно фиксирована определенными временными рамками — периодом выхода из яиц, со второй половины июля то середину августа. Принимая средний вес сеголетка в первый месяц жизни за 1 г (при размахе колебаний в первый месяц жизни от 0,5 до 1,5 г), получаем для нашей крупной популяции биомассу этой группы животных 4000 г., а для мелкой популяции — 2000 г. Соответствующий расчет для полувзрослых особей (средний вес 4,5 г при размахе колебаний от 2,5 до 6,0 г) дает биомассу для крупной популяции 9 кг и для небольшой популяции — 4,5 кг. Аналогичный расчет для взрослых, при среднем весе взрослой особи 12,0 г (при колебаниях от 5 до 40 г) дает соответственно 12 и 6 кг.
Таким образом, во второй половине лета (июль-август) суммарная биомасса крупной популяции прытких ящериц составляет около 25 кг, а для небольшой популяции — около 12,5 кг. Конечно, эти данные статичны и не отражают реально наблюдающейся динамики изменения биомассы, связанной с изменением численности отдельных возрастно-половых групп. Однако эту динамику мы рассмотрим после определения общей условной биомассы вида.
Чтобы связать полученные данные по биомассе с биогеоце-нотическим подходом, напомним, что популяция может занимать территорию нескольких биотопов, что определяет обитание такой популяции в нескольких биогеоценозах. В то же время не так уж редки случаи, когда одна популяция ящериц населяет один биогеоценоз (небольшие островки, долинки и т. п.). Учитывая, что средняя площадь каждой из изученных нами популяций составляет около 30 га и средняя плотность населения в изу-
324
ченных популяциях около 50 экз/1000 м2, биомасса ящериц в этих условиях составит около 4,1 кг/га
42,0 г + 4,5
, 2
X 50x10.
БИОМАССА ЯЩЕРИЦ В РАЗЛИЧНЫХ БИОТОПАХ
На основании полученных данных по плотности ящериц в разных биоценозах можно определить биомассу этого вида в населяемых ящерицами биотопах разного типа (табл. 59).
Таблица 59
Средняя биомасса прытких ящериц в населенных ею биотопах разного типа (в кг/га) *
Группа биотопов Природная зона
лесная лесостепная степная полупустыни горные районы
Природные лесные 0,30 (7) 0,12 (2) 0,42 (4) — —
Природные биотопы открытых пространств 0,35(8) 0,05 (1) 3,50(9) 1,26 (8) 2,48(5).
Сельскохозяйственные — 0,21 (1) 3,96 (2) — —
Другие антропогенные 0,47(4) 2,66(6) 4,85(11) 2,85(1) —
* Средние плотности по биотопам разных типов определены на основании данных табл. 53; средний вес одной ящерицы принимается за 8 г в лесной и лесостепной зонах и за 9 г — в других зонах. В скобках—число изученных биотопов данного типа.
Данные по группам биотопов для лесостепной зоны надо, видимо, исключить из сравнения как недостаточно репрезентативные (основаны на случайных единичных определениях). В целом биомасса ящериц в разного типа населяемых ею биотопах колеблется от 0,05 до 4,85 кг/га, достигая максимальных величин в степной зоне, в антропогенных биотопах. Учитывая, что рептилии находятся на вершине трофических цепей, полученные величины биомассы ясно отражают важную роль, которую играет данный вид в населяемых им биогеоценозах. Приведенные данные касаются биотопов, населяемых ящерицами. Как известно, эти биотопы мозаично или сетчато распределены среди биотопов, где ящерицы не встречаются. Подробный анализ распределения биомассы ящериц в различных биотопах, включающий реальное распределение ящериц на больших территориях, при-веден на площади 170 га на территории стационара на Прика-лаусских высотах (верховье р. Томузловки, Александровский район, Ставропольский край) А1. Ф. Тертышниковым (табл. 60).
Из приведенных в табл. 60 данных видно, что при расчете на значительные территории даже в степной зоне-оптимуме обитания вида биомасса оказывается значительно уменьшенной, чем при расчете на обитаемые биотопы. Этот факт свидетельствует
325
Таблица 60
Плотность населения, биотопическое распределение, биомасса и численность прыткой ящерицы на Стационарном участке, расположенном на Ставропольской возвышенности (По Тертышникову, 19726)
Биотоп Площадь биотопа, га Площадь, занятая видом, % Плотность населения, экз/га Биомасса, кг/га Численность, экз.
колебание средняя
Разнотравно-злаковая степь 80,3 39,4 5—110 58 0,70 4657
Типчаково-полынная степь 4,5 100,0 10—70 40 0,48 180
Полынлая степь 4,5 100,0 10—30 20 0,24 90
Полезащитные лесные полосы 27,0 50,0 0—110 28 0,34 743
Посевы 12,5 20,0 0—30 3 0,04 30
Толока 0,6 50,0 0—10 7 0,08 4
Лесные опушки 0,4 100,0 5—40 30 0,36 12
Вырубки и просеки в лесу 0,2 100,0 0-2 1 0,01 1
Опустыненная степь 31,8 — — — — —
Лес 8,2 — —. — — —
Итого 170,0 68,0 0—110 34 0,40 5717
о том, что такое усреднение данных по территориям, заселен-ним ящерицами, не дает достаточно показательных абсолютных величин, позволяя лишь сравнивать разные биотопы по относительной численности и биомассе ящериц.
БИОМАССА ЯЩЕРИЦ В РАЗЛИЧНЫХ ПРИРОДНЫХ ЗОНАХ
Данные по численности ящериц в разных природных зонах, приведенные в предыдущей главе, дают основание для определения биомассы этого вида в целом по крупным природным зонам (табл. 61).
Таблица 61
Расчетная биомасса прытких ящериц по природным зонам СССР (в пределах ареала вида)
Зона Средняя численность ящериц тыс. экз. Средняя масса особи ** Общая биомасса
в кг в % от общей по всем зонам
Лесная 326 026 8,0 2 608 203 2,8
Лесостепная 301 577 8,0 2 412 616 2,6
Степная 7 491 978 9,0 67 427 802 72,5
Полуп устынная 1 564630 9,0 14081 670 15,1
Горная 724 360 9,0 6 519 240 7,0
По всем зонам 10 408 571 8,9 93 049 536 100
* Среднее значение из табл. 55 и табл. 56 (гл. XIV) для соответствующих зон.
** На первую половину лета, без учета сеголетков.
326
Полученные в табл. 61 результаты свидетельствуют о том, что общая биомасса ящериц в пределах части ареала, входящей в территорию СССР, распределяется по природным зонам крайне неравномерно: подавляющая часть биомассы (почти 7s) приходится на степную зону; на степную зону вместе с полупустынной приходится почти 90% биомассы вида на рассматриваемой территории. Эти величины не могут не отражать относительной роли прыткой ящерицы в биогеоценозах разных зон. В то же время эти сведения хорошо дополняют данные гл. XIII о ведущем значении прытких ящериц как консументов во многих степных биоценозах. Напомним, что там она может превосходить роль всех остальных позвоночных вместе взятых (табл. 52, гл. XIII).
ДИНАМИКА БИОМАССЫ
Выше подчеркивалось, что приводимые выше Данные по биомассе вида являются статичными: они отражают лишь какой-то момент в жизни конкретных популяций населения определенных природных зон. Такой условный момент относится к первой половине сезона активности, к периоду, когда проводится большинство маршрутных учетов численности, — маю — Июню. Биомасса вида в целом и отдельных популяций прыткой ящерицы не сохраняется постоянной по сезонам, испытывая достаточно заметные колебания, связанные с динамикой численности. Кроме таких периодических колебаний биомассы, на больших территориях ареала наблюдаются и порой даже более значительные апериодические колебания. К рассмотрению этих двух типов колебаний биомассы мы и переходим.
Сезонные колебания биомассы
Как было показано в разных главах настоящей монографии темп смертности животных на протяжении их жизни оказывается различным. Принимаемые за исходные данные по естественной смертности приведены в табл. 62.
Таким образом, принимается, что общая смертность в первый год жизни составит около 50% от числа родившихся, во второй год — около 33% от числа достигших возраста в 1 год, в третий и четвертый годы — соответственно 50 и 100% от числа особей, достигших предыдущего возраста.
Суммарная картина сезонных колебаний численности в популяции будет значительно более сложной в связи с одновременным пополнением популяции за счет вновь родившихся особей и разных темпов смертности одновременно живущих в популяции поколений (см. рис. 87). «Всплеск» численности, характерный для периодов выхода молодых из яиц, не дает заметного увеличения биомассы, поскольку масса каждой отдельной молодой особи весьма невелика (эквивалентна биомас-
327
Таблица 62
Динамика естественной смертности прыткой ящерицы (в среднем по всем зонам)
Период Отложено 100 яиц
осталось, экз. (в скобках — процент) погибло, экз. (в скобках — процент)
Осень до зимовки 38 (75) 12 (25)
Первая зимовка 28 (75) 10 (25)
Следующий год (до наступления половой зрелости и спаривания) 21 (75) 7(25)
Третий год (1-й год зрелости) 14 (67) 7(33)
4-й год (2-й год половой зрелости 7(50) 7(50)
5-й год (3-й год половой зрелости) 0(0) 7 (100)
се примерно 8 взрослых особей). Следует учесть при этом, что в середине лета гибнет сравнительно большое количество взрослых ящериц после периода размножения, что должно отразиться и на сокращении общей биомассы в середине летнего сезона.
Кроме сезонной динамики биомассы, характерной для любой популяции, существуют еще более четко выраженные апериодические колебания биомассы, связанные с апериодическими колебаниями численности населения как популяций отдельных биотопов, так и целых природных зон. Причины таких колебаний разнообразны и частично рассмотрены в предыдущей главе. Эти колебания численности прямо влияют на колебания биомассы прытких ящериц, отражая общие процессы колебания биомассы вторичной продукции как в отдельных биогеоценозах, так и по ландшафтам и целым природным зонам.
Несомненно, амплитуды колебаний биомассы отдельных популяций могут быть не особенно тесно связанными даже в соседних популяциях, не говоря уже о популяциях разных ландшафтов и природных зон. К сожалению, пока нет точных количественных данных этого рода. Получение таких данных было бы весьма желательным особенно для степной зоны, учитывая ключевую роль прыткой ящерицы как вида во многих типах биотопов этой природной зоны.
ГЛАВА ШЕСТНАДЦАТАЯ
ПРАКТИЧЕСКОЕ ЗНАЧЕНИЕ 1
Проблема оценки практического значения прыткой ящерицы ставилась не однажды и решалась далеко не однозначно. Это не удивительно, поскольку характер .полезной или вредной деятельности животного определяется путем выяснения его сложнейших и разносторонних отношений со всеми многообразными элементами биоценоза, которые в подобного рода исследованиях сами должны получить объективную оценку с точки зрения их значения в хозяйственной деятельности человека и биологии изучаемого вида.
Недостаточность биоценологических сведений и данных, получаемых по заранее и целенаправленно составленным планам, разнообразие используемых методик и относительность самих понятий «польза» и «вред» в применении к живым объектам неминуемо приводят к различному толкованию практического значения прыткой ящерицы. Так, например, Б. А. Гумилевский (1931) и М. С. Ефименко (1939) полагали, что оно невелико. Напротив, большинство других исследователей (Красавцев, 1936, 1939; Аверин, 1951; Изосов, 1951; Щепотьев, 1952; Андрушко, 1954; Хонякина, 1964; 1965; Лукина, 19666; Щербак, 1966; Тертышников, 1972а, 19726; Тертышников, 1973в; Тертышников, Щербак, 1973; и др.) пришли к выводу о значительной хозяйственной ценности данного вида.
В главе VI было показано, что прыткая ящерица активна в среднем в течение нескольких месяцев. В этот период она усиленно питается. К тому же, специфика физиологии и периодов активности, свойственная данному эволюционному уровню компенсируется у этих животных большой прожорливостью (см. гл. VI). В результате прыткая ящерица уничтожает в различных биоценозах от 0,08 до 10,6% членистоногих (Тертышников, 19726; Тертышников, Щербак, 1973). При этом наибольшее «давление» ящерицы оказывают на популяции жуков, перепончатокрылых, пауков и прямокрылых (см. гл. VI).
1 Авторы: Н. Н. Щербак, М. Ф. Тертышников, В. А. Котляревская, В. П. Шар-пило, А. М. Андрушко.
12 Прыткая ящерица
329
Поскольку, как правило, избирательность в питании прыткой ящерицы не выражена, беспозвоночные встречаются в ее рационе в зависимости от их доступности в данном биотопе и пропорционально их количеству в биоценозе. Думается, однако, что при оценке этого факта не стоит сбрасывать со счета свойственную всем низшим наземным позвоночным тенденцию к периодической смене преобладающих кормов (Котляревская, 1972; Кот-ляревская, сообщ.). Подтвержденная экспериментально, эта осо-
Рис. 94 Соотношение основных и побочных кормов (в %) в питании прытких ящериц в различных стациях Кулундинской степи
А — открытая степь;
Б — обочины дорог и огороды;
В—понижения (овраги, балки);
1 — «основные» корма;
2 — «побочные» корма (рис. А. М. Андрушко)
бенность обеспечивает животным поддержание необходимого энергетического уровня и повышение жизнеспособности организма при нарушениях оптимальных условий и возрастании подвижности популяций.
В то же время указанное явление расширяет сферы влияния прыткой ящерицы в пределах ее пищевых связей, корректирует ее «давление» на наиболее распространенные группы пищевых объектов и при вспышках массового размножения позволяет до известной степени осуществлять контроль как над основными, так и над побочными группами беспозвоночных в данном биотопе (рис. 94).
В целом, по данным большинства из перечисленных выше авторов, в пищевом рационе прыткой ящерицы количество вредных беспозвоночных варьирует от 34,4 до 98,0, полезных — от 2,0 до 63,3, а нейтральных — от 2,0 до 33,0%.
Для получения наиболее объективной оценки хозяйственной значимости данного вида необходимо учитывать, по меньшей мере, четыре фактора: численность ящериц в биоценозе, соотношение в нем вредных и полезных беспозвоночных и их абсолютную численность, характер хозяйственного использования данного биоценоза. В связи с этим, используя формулу Б. А. Красавцева (1936), предложенную им для оценки питания амфибий V— п U, где V — коэффициент полезности; д —число
съеденных экземпляров вредных животных; U — число съеденных экземпляров полезных животных; t — число всех съеденных
330
животных, можно предложить ввести в нее величину С, равную численности ящериц в экземплярах на га. Тогда новый коэффициент окажется следующим:
Используя данные некоторых авторов (Пащенко, 1954; Ма-рисова, 1960; Яковлева, 1964; Щербак, 1966; Тертышников,. 19726; Котляревская, неопубликованные данные), оказывается возможным дать небезынтересную картину соотношений двух указанных коэффициентов, их распределения в популяциях, обитающих в различных биотопах и природных зонах Советского Союза (рис. 95). Первое становится ясным из рис. 96. Коэффициент V, зависящий только от соотношения объектов различных категорий в рационе ящериц, дает наибольшие значения в степях Краснодарского края и Западной Сибири, а также в лесах Западной Украины, резко снижаясь в районе степного Крыма, лесостепного Ставрополья и, особенно,— в горном Крыму. Причины подобного явления следует искать прежде всего в особенностях распределения различных групп беспозвоночных. В качестве: предварительных итогов следует отметить возможность наиболее.' «стандартного» соотношения вредных, полезных и нейтральных; беспозвоночных в наиболее типичных биотопах. Соответствующие зональные изменения коэффициента красноречиво свидетельствуют о необходимости введения в формулу оценки полезности исследуемого вида фактора численности. Этот последний является гораздо более лабильным и, по-видимому, эволюционно менее закрепленным фактором, чем естественное соотношение вредных и полезных форм, представляя собою наиболее чувствительное к давлению отбора звено в механизме популяционных адаптаций. В результате становится понятной сущность резкого возрастания коэффициента полезности k прыткой ящерицы в лесостепных биоценозах (k = 52 и 33 по сравнению со значениями 4,4 для лесной или 19—26 для степной зоны, при V = 0,5 и 0,6, соответственно). Более детальный анализ обоих коэффициентов для популяций, обитающих в различных биотопах, может быть дан на основе рис. 97. При рассмотрении значений Vuk для разных стаций, оказывается, что их изменения идут почти синхронно. По-видимому, стации переживания, а таковыми, в сущности, являются почти все места обитания, приведенные на рис. 97 и роль их биоценологических значений оказываются одинаковыми как для прыткой ящерицы, так и для комплекса ее пищевых объектов, который сохраняет в своих пределах типичные Для него соотношения вредных, полезных и нейтральных компонентов. Таким образом, взаимоотношения коэффициентов V и k, иными словами, взаимоотношения между количественной динамикой хищника и качественной его пищевых объектов, принципиально отличны от разных уровней их биоценологических
1'2*
331
S 8
Рис. 95
Распределение «коэффициентов полезности» прыткой ящерицы в разных природных зонах
/.— лесная; // — лесостепная; /// — степная; /V — горные районы; I—V, 2-К (рис. А. М. Андрушко)
связей. Отсюда следует, что при изучении практического значения исследуемого вида мы можем для получения объективной оценки полезности или вредности географических популяций исходить из данных, относящихся к различным стациям переживания и экологическим нишам в их пределах. Для более конкретной оценки возможного значения ящериц как фактора, регулирующего численность вредных беспозвоночных, небезынтересно привести некоторые данные (табл. 63).
ТаблицабЗ х
Оценка практического значения прыткой ящерицы в разных биотопах южных районов СССР
Биотоп (южные районы) Численность прыткой ящерицы, экз./га К Полные затраты на химическую обработку (РУб-/га)* Реальная доля L. agilis в компенсации затрат (руб./га)
Пастбища До 250** 0,5 До 2,12 До 3,04 АВИА 0,20 0,30
Лесополосы До по*** 0,7 До 11,11 1,11
Посевы зерновых До 30*** 0,2 До 4,64 До 5,80 АВИА 0,46 0,58
Овощные и пропашные культуры До 350** 0,5 До 3,37 0,37
Плодовые сады До 300** 0,5 До 7,02 До 7,94 АВИА 0,70 0,79
Виноградники До 300** 0,5 До 6,66 До 7,94 АВИА 0,67 0,79
* По данным Ин-та защиты растений МСХ УССР, здесь не учитываются расходы по доставке ядохимикатов (11,6% общей стоимости).
** По Ю. И. Пащенко (1954).
*** По М. Ф. Тертышникову (19726).
В целом на обработку каждого гектара комплексных сельскохозяйственных угодий затрачивается 5,68—5,77 рубля, в зависимости от применяемого транспорта и метода обработки. Учитывая среднее значение коэффициента полезности для изучаемого вида (0,4) и силу воздействия вида на популяции беспозвоночных (изымание 0,08—10,6% их числа), можно дать достаточно высокую оценку хозяйственного значения прыткой ящерицы. Кроме того, при более детальном рассмотрении вопроса видно, что прыткая ящерица в ряде случаев является пионером в освоении некоторых биоценозов, например, распаханных степных участков, куда она проникает по полезащитным лесополосам, обочинам дорог и канавам. Именно эти участки (в частности, лесополосы) являются наиболее дорогостоящими при химической обработке (11,11 руб/га) и именно там ящерицы до-
333
Рис. 96
Соотношение коэффициентов полезности (К и V) в различных зонах
Степь:
А — Крым;
2 — Ставрополье:
«3 — Краснодарский край;
•4 — Кулундннская
степь;
Лесостепь:
5 — Приднепровье;
6 — Ставрополье; лес:
7 — средняя полоса РСФСР, Европейская часть;
8 — Западная Украина; горные районы:
9 — Крым,
10 — Средняя Азия.
К,
2— V (по сводным данным разных авторов, рнс. А. М. Андрушко)
Рис. 97
Соотношение коэффициентов полезности прыткой ящерицы в разных био* топах
1 — открытая степь;
2 — полезащитная поло-
са;
3 — обочины и огороды;
4 — сады и виноградники;
5 — луга;
€ — опушки;
7 — леса;
8 — посевы;
I — К, II — V (по сводным данным разных авторов, рис. А. М. Андрушко)
и
стирают высокой численности (до ПО экз./га) и дают наиболее высокий коэффициент полезности (0,7). Следующая по стоимости обработки биотопов (плодовые сады и виноградники) также характеризуется высокой численностью прыткой ящерицы (300 экз./га) и коэффициентом полезности (0,5) при затратах на химобработку 6,66—7,94 руб./га. Посадки овощей, пастбища и посевы зерновых требуют меньших затрат на химическую обработку (2,12—5,80 руб./га) и в среднем характеризуются более низким коэффициентом полезности и численностью обитающих в этих угодьях ящериц (0,4 и 163 экз./га). Указанные соотношения весьма интересны и требуют дальнейших исследований в плане биоценологических взаимоотношений.
В целом подобная ситуация еще раз свидетельствует в пользу охраны и привлечения прыткой ящерицы, которая уничтожением вредителей компенсирует затраты от 2 до 11 рублей на каждые 10 га хозяйственно-полезных угодий. Нетрудно представить, что в пересчете на площадь угодий, занятых ареалом прыткой ящерицы, ее полезная деятельность может выражаться многими миллионами рублей.
Ко всему сказанному следует добавить, что среди вредных беспозвоночных, уничтожаемых прыткой ящерицей, имеются не только вредители сельского и лесного хозяйства, но и переносчики и носители некоторых инфекционных заболеваний. Полезная деятельность данного животного увеличивается еще и тем, что оно хорошо приживается в некоторых угодьях, которые находятся в сфере деятельности человека, а потому характеризуются, с одной стороны, специфическими нарушениями экологического равновесия, нередко ведущими к усилению вредных компонентов фауны беспозвоночных, с другой,— обедненностью их естественными врагами. Нет сомнения в том, что снижение обоих коэффициентов полезности в некоторых окультуренных ландшафтах (0,2 и 6 для посевных площадей) компенсируется их высокими значениями для других (0,8 и 88 на огородах; 0,5 и 150—в садах и виноградниках). Наконец, польза прыткой ящерицы заключается еще в том, что она служит кормом (чаще дополнительным) для многих хищных птиц и млекопитающих, среди которых имеются и ценные в хозяйственном отношении формы (см. гл. VI).
При оценке практического значения прыткой ящерицы необходимо учитывать также и вред, который она может приносить. Одна из нежелательных сторон деятельности данного вида заключается в ее хищническом отношении к полезным животным— сеголеткам своего вида, другим ящерицам (см. гл. VI) и полезным беспозвоночным. По наблюдениям И. С. Даревского (1955) в Армении, одна ящерица поедает к концу дня до 9 пчел. Естественно, что популяция животных, обитающая на пасеке, может принести ощутимый вред. Поскольку исследования некоторых авторов свидетельствуют о том, что прыткие ящерицы
335
способны возвращаться лишь со знакомой части территории, с расстояния, не превышающего четверть, а то и восьмую часть километра, И. С. Даревский рекомендует в аналогичных приведенному примеру случаях попросту отлавливать ящериц и уносить их подальше от нежелательного для нас места обитания.
Говоря о практическом значении прыткой ящерицы, необходимо упомянуть и ее значение в прокормлении личинок и нимф различного вида клещей, в том числе и переносчиков заболеваний, а также — участие в циклах развития некоторых паразитических червей. В различных частях ареала прытких ящериц отмечено 20 видов иксодовых, гамазовых и краснотелковых клещей (см. гл. XII), большинство которых могут питаться и на других позвоночных — млекопитающих и птицах, включая домашних. Зараженность ящериц паразитами этой группы колеблется в широких пределах в различные сезоны, годы и в разных частях ареала. Нередко значение прыткой ящерицы в прокормлении этих животных может быть весьма значительным, как это отмечено для Ставропольской возвышенности, Краснодарского края и Дагестана (Хонякина, 1964; Лукина, 19666; Тертышни-ков, 19726; Тертышников, Щербак, 1973; и др.). Роль прытких ящериц (и пресмыкающихся в целом), вероятно, заметно возрастает в годы депрессии численности мышевидных грызунов и других животных, в большей, чем пресмыкающиеся, степени подверженных колебаниям данного фактора.
Как один из компонентов биоценозов прыткая ящерица оказывается вовлеченной в кругооборот паразитических червей. В настоящее время в различных частях ареала у нее зарегистрировано 22 вида гельминтов (см. гл. XII). Семь видов из них представлены личиночными формами, окончательными хозяевами которых являются птицы и млекопитающие. Для большинства этих паразитов ящерица служит резервуарным хозяином. В их числе можно назвать Mesocestoides lineatus и Spirocer-са lupi — паразитов хищных млекопитающих (лисица, волк и др.), Shaerirostris teres — паразита врановых птиц и др. Поскольку прыткая ящерица обнаружена среди пищевых объектов этих животных, ее роль в их заражении несомненна. Во многих случаях прыткая ящерица может оказаться конечным этапом в цикле передачи гельминтов и других патогенных организмов и способствовать их изъятию из биоценоза. Этот аспект биологии ящерицы нуждается в специальных исследованиях. К сказанному выше можно добавить еще следующее. Проникая, в частности, в степную и лесостепную зоны — по лесополосам, канавам, обочинам и оврагам как в открытую степь, так и в культурные стации, прыткая ящерица одновременно является и изолированным от человека животным и в определенном смысле синантропом. К тому же, подвижность степных популяций, находящихся в наименее благоприятных для животных условиях, ввиду чрезмерного хозяйственного использования биотопов, резко повыше
336
на. Таким образом, ящерицы могут осуществлять обмен паразитами между представителями диких и окультуренных ландшафтов, тем самым расширяя сферу влияния паразитов.
В результате развития промышленности и сельского хозяйст ва, промышленного и гражданского строительства происходит изменение ландшафтов, приводящее к сокращению ареала и падению численности прыткой ящерицы. Принимая во внимание большую положительную роль данного вида в регулировании численности вредных беспозвоночных, не дающей, как и другие формы, биологической борьбы с вредителями, никакого отрицательного побочного эффекта, следует использовать все возможности, чтобы сохранить прыткую ящерицу на значительной территории, не нарушая интересов промышленности и сельского хозяйства. Наиболее рациональным методом является искусственное заселение ящерицами садов, виноградников, лесных полос и непригодных для сельскохозяйственного использования земель с целенаправленным подбором оптимальных для вида биотопов в различных природных зонах. Эта работа прежде всего должна проводиться в лесостепных районах страны. Необходимо, наконец, обратить внимание на проблемы охраны прыткой ящерицы как части общегосударственной задачи охраны природы.
ЛИТЕРАТУРА
Аверин Ю. В. 1953. Вредные и полезные позвоночные животные древеснокустарниковых насаждений степного Крыма.— Труды Крымского филиала АН СССР, 3, вып. 2. .
Алисов Б. П. 1956. Климат СССР. Изд-во МГУ.
Алисов Б. П., Дроздов О. А., Рубинштейн Е. С. 1952. Курс климатологии. Ч. I, II. Л., Гидрометеоиздат.
Алисов Б. П., Берлин И. А., Михель В. М. 1954. Курс климатологии. Ч. I, II, III. Л., Гидрометеоиздат.
Аммон П. 1928. Список амфибий и рептилий Тульской губернии. Тула, «Тульский край».
Андреев И. Ф. 1953. Амфибии и рептилии Прикарпатья.— Уч. зап. Кишиневского ун-та, 8.
Андрушко А. М. 1936. Методика и техника количественного учета пресмыкающихся. В сб. «Вопросы экологии и биоценоза», вып. 3.
Андрушко А. М. 1954. Пресмыкающиеся Казахского нагорья и их хозяйственное значение.— Уч. зап. Ленингр. ун-та, № 181, серия биол. наук, вып. 38.
Андрушко А. М., Марков Г. С. 1960. Материалы о гельминтах ящериц Кавказа и некоторые экологические особенности гельминтофауны представителей семейства Lacertidae.—Вести. ЛГУ, 3, серия биол. наук, 1. Изд-во ЛГУ.
Антипчук Ю. П. 1966. О строении и функции сердца рептилий.— Докл. АН СССР, 169, № 6.
Антипчук Ю. П. 1967. Функциональная морфология малого круга кровообращения некоторых наземных позвоночных.— Вести, зоол., № 3.
Антипчук Ю. П„ Гибрадзе Т. А. 1973. К сравнительной морфологии кровеносных сосудов легких. Тбилиси, «Мецниереба».
Арзамасов И. Т. 1961. Иксодовые клещи. Минск, Изд-во АН БССР.
Атлас СССР. 1962. М„ Главное упр. геодезии и картографии Мин. геологии и охраны недр СССР.
Атлас мира. 1964. М., Главн. Упр. геодезии и картографии МВД СССР.
Банников А. Г. 1940. Экологические условия зимовок травяной лягушки в Московской области. В сб. научных студ. работ Моск, ун-та, зоология, 16.
Банников А. Г. 1954. Материалы по биологии земноводных и пресмыкающихся Южного Дагестана.— Уч. зап. Моск. гос. пед. ин-та, 28, 2.
Банников А. Г. 1958. Материалы по фауне и биологии амфибий и рептилий Монголии.— Бюлл. МОЙП, отд. биол., 63, 2.
Банников А. Г., Даревский И. С., Рустамов А. М. 1971. Земноводные и пресмыкающиеся СССР. М., «Мысль».
Банников А. Г., Денисова М. Н. 1956. Очерки по биологии земноводных. М., Учпедгиз.
Баранов А. С. и др. 1972. «Пение» ящерицы.— Природа, № 9.
Баранов А. С. 1973. Особенности окраски прыткой ящерицы Европейской части СССР и Закавказья.—Вопросы герпетологии. III Всес. герпетол. конф. Л., «Наука».
Баранов А. С., Валецкий А. В. 1975. Географическая изменчивость веса тела прыткой ящерицы (Lacerta agilis) на территории СССР.— Зоол. ж., 54, вып. 6.
338
Бахарев В. А. 1971. Популяционно-морфологические особенности популяций прыткой ящерицы (Lacerta agilis). Дипломная работа. Даугавпилский пединститут Латв. ССР.
Башкиров И. С., Попов В. А. 1934. Материалы по биологии ежа. В сб. «Работы Волжско-Камской зональной биол. станции», вып. 3. Казань.
Бей-Биенко Г. А. 1966. Общая энтомология.
Белехов Г. П., Чубинская А. А. 1965. Минеральное и витаминное питание сельскохозяйственных животных. Л.
Бельгард А. Л. 1960. Введение в типологию искусственных лесов в степной зоне. В сб. «Искусственные леса степной зоны Украины». Изд-во Харь-ковск. гос. ун-та.
Беляев М. М. 1947. Зоотомия позвоночных. М.
Беме Л. 1929. Результаты обследования охотничьего хозяйства Парабочевско-го и Самурского заказников И. К. 3. Дагестанской ССР и краткий обзор фауны наземных позвоночных, их населяющей.— Изв. Горьк. пед. ин-та, 6.
Беренштейн Ф. Я. 1947. К вопросу о влиянии микроэлементов на активность гормонов.— Фпзиол. ж., 2.
Беренштейн Ф. Я. 1958. Микроэлементы, их биологическая роль и значение для животноводства. Минск.
Беренштейн Ф. Я. 1966. Микроэлементы в физиологии и патологии животных. Минск.
Берзин А. А. 1971. Кашалот. М., «Пищевая промышленность».
Бескровный М. А. 1953. Практикум по экологии животных. Ч. I. Харьков.
Бобринский Н. 1929. Обзор и очередные задачи исследования фауны позвоночных Туркестана,—Труды научно-исслед. ин-та зоологии, 3, 2.
Богачев А. В. 1938. Зоологические наблюдения над пресмыкающимися и земноводными в Мильской степи. — Изв. Азерб. филиала АН СССР, 4—5.
Богданов О. П. 1949. Возрастные изменения водяного ужа (Natrix tesselata).— Докл. АН Уз. ССР, 8.
Богданов О. П. 1960. Земноводные и пресмыкающиеся.— Фауна Узбекской ССР, 1. Ташкент, Изд-во АН Уз. ССР.
Богданов О. П. 1962. Пресмыкающиеся Туркмении. Ашхабад, Изд-во АН Туркм. СССР.
Богданов О. П. 1965. Экология пресмыкающихся Средней Азии. Ташкент, ФАН.
Болотников А. М., Хазиева С. М., Литвинов Н. А., Чащина С. П. 1973. Распространение и сезонная активность амфибий и рептилий Пермской области. В сб. «Вопросы герпетологии». Л., «Наука».
Большой всемирный атлас Маркса 1910. СПб., Изд. т-ва А. Ф. Марксы Бородулина Т. А. 1949. В кн.: «Охрана природы», 7. М.
Бочарникова А. В. 1964. Значение ящериц Краснодарского края для лесного и сельского хозяйства. В сб. «Вторая научная конф. пед. ин-тов РСФСР» Краснодар.
Браунер А. 1904. Краткий определитель пресмыкающихся и земноводных Крыма и степной полосы Европейской России.— Из записок Крымского горного клуба, № 3—9.
Браунер А. 1905. Третье предварительное сообщение о пресмыкающихся и земноводных губерний Сувалкской, Минской, Подольской, Черниговской, Бессарабской, Херсонской, Екатеринославской и Днепропетровского уезда Таврической губернии. — Зап. Новорос. об-ва естествоиспыт., 28.
Браунер А. 1907. Гады Бессарабии.— Труды Бессарабск. об-ва естествоиспыт., 1. Ч. 2.
Браунер А. 1923. Сельскохозяйственная зоология. Харьков, Гос. из-во Украины.
Брегетова Н. Г. 1956. Гамазовые клещи. М.— Л., Изд-во АН СССР.
Брюзгин В. Д. 1939. К методике исследования возраста и роста рептилии.— Докл. АН СССР, 23, № 4.
Булахов В. Л. 1968. К методике изучения пластических экстерьерных признаков у наземных позвоночных. В сб. «Вопросы степного лесоведения», вып. 1. Днепропетровск.
Булахов В. Л., Губкин А. А., Константинова Н. Ф., Черныш В. П. 1970. Становление комплекса фауны позвоночных животных в процессе сильвати-
339
зации степной зоны Украины. В сб. «Пятая межвузовская зоогеограф, конф. («Влияние антропогенных факторов на формирование зоогеографических комплексов»). Ч. I, Казань.
Булахов В. Л., Константинова Н. Ф. 1972. К морфологической характеристике Lacerta agilis в различных типах лесных биогеоценозов Присамарья. В сб. «Вопросы степного лесоведения», вып. 2. Днепропетровск.
Бураков В. С., Яновский А. А. I960. Практическое руководство по спектральному анализу. Минск, Изд-во АН БССР.
Буреш И., Цонков' Ft. 1933. Изучвания вверху разпространеиието на влечугите, и земноводните в България и по Балканский полуостров.— Изв. на Царе-ките природнонаучни института в София, кн. VI.
Ваккер В. Г. 1970. Паразитофауна рептилий юга Казахстана и их роль в циркуляции некоторых гельминтов человека и животных. Автореф. канд. дисс. Алма-Ата.
Васильев Б. Д. 1969. Морфо-функциональная характеристика амфибий и рептилий. Канд. дисс. МГУ.
Васильев Б. Д. 1973. Электрофизиологическая характеристика слуха некоторых рептилий. В сб. «Вопросы герпетологии». Рефераты докладов III Всес. герпет. конф. Л., «Наука».
Верещагин Н. К., Громов И. М. 1947. Заметки по биологии рептилий в бассейне Шексны.— Природа, № 1.
Власенко П. В. 1930. До фауни трематод амф!б!й та рептилш околиць м. Хар-кова.— Труди Харшвського т-ва досл!дн. природи. Харк1в, 53, 1.
Войнар А. И. 1952. Биологическая роль микроэлементов в организме животных. В сб. «Микроэлементы в жизни растений и животных». М.
Войнар А. И. 1960. Биологическая роль микроэлементов в организме животных и человека. М.
Воронов Г. А., Демидов В. В. 1973. К фауне и экологии рептилий и амфибий Верхнеленья. В сб. «Вопросы герпетологии». Л., «Наука».
Гааке В. 1901. Звери, птицы и насекомые среднеевропейских лесов. СПб.
Гагина Т. В., Скалой В. Н. 1965. Пресмыкающиеся Восточной Сибири. В сб. «Герпетология». Ташкент.
Галахова В. И. 1958. Влияние меди и кобальта на деятельность щитовидной железы и надпочечников. Третья Всес. конф, по микроэлементам. Баку.
Гаранин В. И. 1971. К вопросу о динамике и численности земноводных и пресмыкающихся. В сб. «Природные ресурсы Волжско-Камского края». Казань, Изд-во Казанск. гос. ун-та.
Гексли Т. Г. 1900. Рак. Введение в изучение зоологии. М., Изд-во Собашни-ковых.
Генин Д. И. 1955. Новые данные об оплодотворении у Lacerta agilis.— Докл. АН СССР, 101, 2.
Гизенко А. И. 1957. Материалы по экологии и численности птиц Лебяжьих островов,-— Труды Крымского гос. заповедника, 4.
Глущенко В. В. 1961. Паразитофауна крови амфибий и рептилий в районе среднего течения Северного Донца. В сб. «Проблемы паразитологии,— Труды Укр. республ. об-ва паразитологов. 1. Киев. Изд-во АН УССР.
Горбачев С. Й. 1925. Позвоночные животные. В сб. «Природа Орловского края», 7. Сб. Орловской губ. план, комиссии.
Гордон Б. Е. 1963. Спектральный эмиссионный анализ в криминалистике, судебной химии и судебной медицине. Киев.
Граве Л. Г. и др. 1951. Животный мир Смоленской области. Смоленск, Смол-гиз.
Гримм О. А. 1876. Каспийское море и его фауна.— Труды Арало-Каспийской эксп., вып. 2, тетрадь 1. Приложение к Трудам СПб об-ва естествоиспытателей.
Гумилевский Б. А. 1931. К вопросу о сельскохозяйственном значении гадов в окрестностях Новосибирска —Труды по защите, растений Сибири, I. Новосибирск.
Гумилевский Б. А. 1941. О некоторых эколого-фаунистических исследованиях на Валдайской возвышенности.— Изв. Всес. геогр. об-ва, 73, 1.
340
Паль С. К. 1936. К изучению фауны наземных позвоночных систем Зеравшан-ского и Туркестанского хребтов.—Труды Узб. гос. ун-та, 7. Самарканд. Даниелян. Ф. Д. 1968. Эколого-фаунистическое исследование бисексуальных и партеногенетических видов скальных ящериц Армении. Канд. дисс.
Даниелян Ф. Д. 1970. Нарушение эмбрионального развития и появление уродств у скальных ящериц Армении.— Зоол. ж., 49, 7.
Даниелян Ф. Д. 1971. Некоторые данные по сравнительному эмбриогенезу бисексуальных и партеногенетических форм скальных ящериц Армении.— Биол. журн. Армении, 24, 10.
Даниелян Ф. Д., Айказян А. К- 1974а. Новый тип эмбрионального уродства у партеногенетического вида скальных ящериц.— Биол. журнал Армении, 27, 2.
Даниелян Ф. Д., Айказян А. К- 19746. Данные сравнительного развития пар-теоногенетического и бисексуального видов скальных ящериц Армении.— Биол. журн. Армении, 27, 5.
Даревский И. С. 1946. К биологии прыткой ящерицы.— Природа, Ns 9.
Даревский И. С. 1953. О полезной роли живородящей ящерицы в связи с вопросом об оценке хозяйственного значения ящериц нашей фауны.— Бюлл. МОИП., отд. биол., 58, 4.
Даревский И. С. 1955. Ящерицы — вредители пчеловедства.— Природа, Ns 9. Даревский И. С. 1960а. Динамика популяций, перемещения и роста такырной круглоголовки в полупустыне долины реки Аракса в Армении.-— Бюлл. МОИП, отд. биол., 15 (6).
Даревский И. С. 19606. Сезонные изменения жировых тел и гонад у некоторых ящериц долины реки Араке в Армении.— Зоол. ж., 39, 8.
Даревский И. С. 1967. Скальные ящерицы Кавказа. Л., «Наука».
Даревский И. С., Даниелян Ф. Д. 1967. Сравнительное изучение скальной ящерицы (Lacerta saxicola Eichw.) в условиях Армении.— Уч. зап. Ереванск. ун-та, 1.
Джафаров Р. Д. 1949. Пресмыкающиеся Азербайджанской ССР. Труды Естественноисторического музея АН Аз. ССР им. Г. Зардаби, вып. III. Баку, Изд-во АН Аз. ССР.
Дидусенко Л. М. 1964. Ящерицы Молдавии. В сб. «Вопросы герпетологии». Изд-во ЛГУ.
Динесман Л. Г. 1949. О распределении и экологии рептилий в связи с зонами солнечной радиации. В сб. «Проблемы физической географии», 14. М, Изд-во АН СССР.
Динесман Л. Г. 1950. Роль паводка в экологии амфибий и рептилий дельты Волги.— Бюлл. МОИП, отд. биол., 55, 5.
Динесман Л. Г. 1953. Амфибии и рептилии юго-востока Тургайской столовой страны.— Труды Ин-та географии АН СССР, 54.
Динесман Л. Г., Далецкая М Л. 1952. Методы количественного учета амфибий и рептилий. В сб. «Материалы учета численности и географического распределения наземных позвоночных». М., Изд-во АН СССР.
Добрынин Б. Ф. 1948. Физическая география Западной Европы. М., Учпедгиз.
Дрягин П. 1926. Рептилии и амфибии Вятского края.—-Труды Вятск. пед. ин-та им. Ленина, 1.
Дубинин Б. Б. 1954. Эколого-фаунистический очерк земноводных и пресмыкающихся Хавастского района Ташкентской обл. Уз. ССР.— Труды Ин-та зоологии и паразитологии, 3. Ташкент. Изд-во АН Уз. ССР.
Дьяков М. И., Голубенцова Л. И. 1947. Минеральное питание сельскохозяйственных животных. М., Сельхозгиз.
Емельянов А. А. 1929. Змеи Дальнего Востока.— Зап. Владивостокск. отд. гос. русск. географ, об-ва, 3, 1.
Емчук Е. М. 1960. Фауна Украши. 1ксодов1 клпц1, 25, 1. Вид-во АН УССР. Ефименко М. С. 1931. Про !жу жаб, ягщрок, як! селяться в канавках б!ля бу-рядових пол1в. — 36. пр. Зоол. музею, 23. Киев.
Жарков И. В., Теплов В. П. 1932а. Материалы по питанию барсука (Meles metes L.) в татарской республике. В сб. «Работы Волжско-Камской зональной охотничье-промысловой биол. станции», 7—8, 2. Казань.
341
Жарков И. В., Теплое В. И. 19326. Материалы по питанию хищных птиц Татарии. В сб. «Работы Волжско-Камской зональной охотничье-промысло-вой биол. станции», 7—8, вып. 2. Казань.
Жарков И. В., Теплое В. П., Тихвинский В. И. 1935. Материалы по питанию лисицы (Vulpes vulpes L.) в Татарской республике. В сб. «Работы Волжско-Камской зональной охотничье-промысловой биол. станции», 7—8, вып. 2. Казань.
Жаркова В. К.. 1969. О выборе пищевых объектов прыткой ящерицей.— Труды МОИП, т. 3.
Жаркова В. К.. 1970. Некоторые интерьерные особенности прыткой ящерицы. Материалы IV научн. конф, зоологов пединститутов. Горький.
Жаркова В. К- 1973а. Экология популяций прыткой ящерицы (Lacerta agilis exigua L.) в северной лесостепи Европейской части СССР. Канд. дисс. Свердловск.
Жаркова В. К. 19736. Специфика лесной и лесостепной популяции прыткой ящерицы в Рязанской области. В сб. «Вопросы герпетологии». Л., «Наука».
Жеденов В. В. 1961. Легкие и сердце животных и человека. М., «Высшая школа».
Зайдель А. И. 1965. Основы спектрального анализа. М.
Зайдель А. ВМалышев Г. М., Шрейдер Е. А. 1964. О чувствительности спектрального анализа.— Оптика и спектроскопия, № 17.
Залежский Г. 1938. К динамике численности некоторых видов амфибий.— Сб. научных студенческих кружков Моск, ун-та, 2.
Зарудный И. А. 1895. Материалы для фауны амфибий и рептилий Оренбург- , ского края.— Бюлл. МОИП, 9, 3.
Захидов Т. 3. (Ред.) >1971. Природа и животный мир Средней Азии. Позвоночные животные, 2. Ташкент.
Земская А. А. 1951. Биология и развитие клещей сем. Dermanysidae, паразитирующих на рептилиях, в связи с проблемой возникновения пастбищного паразитизма.— Бюлл. МОИП, отд. биол., 56, 3.
Зинякова М. П. 1964. Размножение и темпы роста среднеазиатской гюрзы в неволе.— Докл. АН Уз. ССР, 7.
Зограф В. 1900. Курс зоологии. М., Типо-литография Русского Товарищества печатного и издательского дела.
Зусман И. В. .1964. О времени установления связи между трахеей и глоткой в эмбриогенезе рептилий и птиц.— Докл. АН СССР, 157, 2.
Зусман И. И. 1965. О сроках наступления морфологической и функциональной зрелости органов пищеварения в эмбриогенезе наземных черепах (Testu-do) — Докл. АН СССР, 163, 1.
Зусман И. В. 19656. О темпах развития органов в эмбриогенезе рептилий и птиц в связи с особенностями питания их плодов. Автореф. канд. дисс.
Зусман И. В. 1970. Чувствительность ранних зародышей (Sauropsido) к действию низких температур. — Ж- Общей биол., 31, 3.
Зусман И. В. 1973. Некоторые вопросы эволюции эмбриогенеза Sauropsida.— Ж. общей биол., 34, 3.
Зусман В. В., Вскандеров Т. М. 1974. Температурные адаптации в раннем эмбриогенезе рептилий. 1. О приспособительном значении перенашиваемо-сти яиц у черепах.— Зоол. ж., 53, 3.
Иваненко И. Д. 1938. К вопросу об изменениях в животном мире населения степи под влиянием агрокультуры. — Зоол. ж. 17, 5.
Иваницкий С. В. 1927. К фауне трематод позвоночных Украины (по материалам 26 СГЭ) — Вет. д!ло, 8. Харшв.
Иваницкий С. В. 1940. Материалы к гельмитофауне позвоночных Украины (фауна цестод, нематод и колючеголовых).— Труды Харьковского вет. ин-та, 19, 1. Харьков.
Иванов А- С. 1952. Паразитические черви рептилий дельты Волги (сосальщики)..— Труды Астраханского гос. мед. ин-та, 10. Астрахань.
Иванов В. И., Марков Г. С., Викулин В. П., Чернобай В. Ф. 1960. Гельминто-фауна пресмыкающихся Северного Прикаспия. Тезисы докл. научн. конф. ВОТ. М., Изд-во АН СССР.
342
Иголкин И. И. 1967. Нора как микробиоценотоп. В сб. «Проблемы экологи», 1. Томск.
И засов А. А. 1951. Сельскохозяйственное значение прыткой ящерицы (Lacerta agilis exigua) в условиях лесостепной зоны.— Бюлл. Об-ва естественен, при Воронежском ун-те, 7. Воронеж.
Иноземцев А. А. 1969. Трофические связи бурых лягушек в хвойных лесах Подмосковья.— Зоол. ж., 48, II.
Иоганзен Г. 1923. По Чулыму.— Изв. Томского гос. ун-та, 72.
Иорданский И. И. 1971. К функциональному анализу черепа ящериц (Lacer-tilia). Особенности строения, связанные с кинетизмом.— Зоол. ж., 50,
Калабухов И. И. 1951. Методы экспериментальных исследований по экологии наземных позвоночных. М., «Сов. наука».
Калашников В. И. 1964. Физиология газообмена у ядовитых змей Средней Азии. В сб. «Вопросы герпетологии».
Кашкаров Д. И. 1932. Животные Туркестана. Изд. 2. Ташкент.
Кашкаров Д. И., Станчинский В. В. 1940. Курс зоологии позвоночных животных. М.—Л., Изд. АН СССР.
Кащенко И. Ф. 1900 Результаты алтайской зоологической экспедиции 1898 г.— Изв. Имп. Томского ун-та, 16.
Кащенко И. Ф. 1902. Обзор гадов Томского края.— И<зв. Томского ун-та, 22.
Кащенко И. Ф. 1909. Гады, собранные среднеазиатскими экспедициями проф. Сапожникова в 1902 и 1908 гг.— Ежегодник Зоол. музея Акад, наук, 14, 1—2.
Кириченко А. И. 1910. Отчет по командировке на Талыш в 1909 году для собирания коллекций для Зоологического музея Импер. Акад, наук, 15.
Киселев Ф. А. 1951. Сельскохозяйственное значение чайки-хохотуньи. Труды Крымского филиала АН СССР, 2.
Клевезаль Г. А., Клейненберг С. Е. 1967. Определение возраста млекопитающих. М., «Наука».
Клененберг С. Е., Яблоков А. В., Белькович В. М., Тарасевич М. И. 1964. Белуха. М., «Наука».
Клейненберг С. Е., Смирина Э. М. 1969. К методике определения возраста амфибий.— Зоол. ж., 48, 7.
Ковальский В. В. 1963. Роль микроэлементов в животноводстве и неотложные задачи изучения их.— Журн. Всес. хим. об-ва им. Д. И. Менделеева, 8.
Ковачев В. Т. 1912. Херпетологичната фауна на Българпя (влечуги м зем-новодни). Пловдив, Печ. Хр. Данов.
Ковда В. А., Якушевская И. В. 1967. Опыт оценки биомассы суши.— Изв. АН СССР, сер. биол., вып. 4.
Козлов И. П. 1960. Питание и численность степных птиц в засушливых районах Ставрополья. В сб. «Орнитология», вып. 3, Изд-во МГУ.
Козловский П. И. 1951. К вопросу изучения паразитофауны рептилий. Саратовской обл.—Уч. зап. Саратовск. гос. пед. ин-та, 16. Саратов.
Коломиец Л. Г. 1972. Рыжий сосновый пилильщик, Новосибирск, «Наука»..
Колосов А. М. 1935. К биологии корсака и степной лисицы.— Бюлл. МОИП, 44, отд. биол., 4.
Коржуев И. А., Круглова И. А. 1957. Некоторые эколого-физиологические особенности рептилий.— Зоол. ж., 36, 2.
Корнева Л. Г. 1967. О способе потребления желтка в эмбриогенезе змей.—• Докл. АН СССР, 174, 6.
Корнева Л. Г. 1969. Эмбриональное развитие водяного ужа (N. tessellata).—-Зоол. ж., 10.
Корнеев А. П. 1954. Енотовидная собака (Nyctereutes procyonoides) на Украине.— Наук. зап. Киевск. ун-та, 13, вып. 12.—Труды Зоомузея, 4. Киев.
Косарева И. А. 1950. Рептилии юга Сталинградской области.— Уч. зап. Ста-лингр. пед. ин-та, вып. 2.
Костин В. П. 1956. Заметки по распространению и экологии земноводных и пресмыкающихся древней дельты Амударьи и Кара-Калпакского Устюрта.— Труды Ин-та зоол. и наразитол. АН Узб. ССР, 5. Ташкент.
Кострин К- В. 1959. Богомол и ящерица.— Природа, № 7.
343
Котляревская В. А. 1972. Некоторые принципиальные вопросы, связанные с характером и особенностями питания низших наземных позвоночных. Мате риалы IV Всес. конф, по экол. физиол. Краснодар.
Коштоянц X. С. 1950. Основы сравнительной физиологии, т. 1. М.—Л., Изд-во АН СССР.
Красавцев Б. А. 1935. О полезной роли лягушки озерной (Rana ridibunda Pall) в пойменных лугах.— Труды Об-ва естествоиспыт. при Казанском ун-те, 52.
Красавцев Б. А. 1936. Биологические наблюдения над прыткой ящерицей (Lacerta agilis exigua Eichw.). В сб. «Вопросы экологии и биоценологии», вып. 3. Л.
Красавцев Б. А. 1939. Еще о экологии прыткой ящерицы.— Природа, № 9.
Красавцев Б. А. 1941. Герпетологические наблюдения в Предкавказье.— Труды Ворошиловского гос. пед. ин-та, 3, 2.
Краснова М. В. 1961. К морфологии возрастных и сезонных изменений семен ников некоторых рептилий и птиц.— Уч. зап. Казахского гос. ун-та, 31 Алма-Ата.
Красовский Д. В. 1929. Материалы к познанию фауны Reptilia и Amphibia Хасав-Юртовского округа Дагестанской ССР.— Изв. Горек, пед. ин-та, 6.
Крень А. К- 1939. Позвоночные животные заповедника «Лес на Ворскле».— Уч. зап. ЛГУ, № 28, вып. 7.
Крепе Е. М., Манукян К. Г., Смирнов А. А., Чарковская Е. В. 1964. Фосфолипиды и эволюция нервной системы. В сб. «Эволюция функций». М.—Л., «Наука».
Кубанцев Б. С. 1962. Животный мир Волгоградской области. Волгоград.
Кузнецов Н. В., Макковлева И. И. 1959. Животный мир Ярославской области. Ярославль.
Кустанович И. М. 1962. Спектральный анализ. М.
Кутузова В. А. 1971. Прыткая ящерица Дагестана. Дипломная работа. Махачкала.
Лаптев М. К- 1930. Учет наземной фауны позвоночных методом маршрутного подсчета (метод площадей видов).— Труды Среднеазиатского ГУ, сер. Villa, зоол. вып. 11, Ташкент.
Либерман С. С., Покровская Н. В. 1943. Материалы по экологии прыткой ящерицы.— Зоол. ж., 22, 4.
Ликвентов А. В. 1948. Длительные концентрации вредителей в лесных полезащитных полосах.— Лесное хоз-во, № 7.
Лобачев В. С., Чугунов Ю. Д., Чуканина И. Н. 1973. Особенности герпетофауны Северного прпаралья. В сб. «Вопросы герпетологии». Л., «Наука».
Логинов В. П. 1962. О патологии щитовидной железы у молодняка животных в районах йодной недостаточности.— Ветеринария, № 10.
Лукина Г. П. 1961. Материалы по биологии прыткой ящерицы в условиях Западного Предкавказья. В сб. «Материалы III научн. конф, аспирантов». Изд-во Ростовского ун-та.
Лукина Г. П. 1966а. Пресмыкающиеся Западного Предкавказья. Автореф. канд. дисс. Ростов-на-Дону.
Лукина Г. П. 19666. Пресмыкающиеся Западного Предкавказья. Канд. дисс. Ростов-на-Дону.
Марисова I. В. 1960. До вивчення живлення прудко! та живородящо! (широк.— 36. праць Зоол. музею, 29.
Марков Г. С., Иванов В. П., Никулин В. П., Чернобай В. Ф. 1962. Гельминто-фауна пресмыкающихся дельты Волги и прикаспийских степей.— Гельмин-тол. сб. Астраханск. заповеди., 6. Астрахань.
Марков Г. С., Лукина Г. П., Маркова Л. И., Мозгина А. А. 1964. К паразито-фауне пресмыкающихся Северного Кавказа. В сб. «Вопросы морфол., экол. и паразитол. животных.—Уч. зап. Волгоград, гос. пед. ин-та, 16. Волгоград.
Марков Г. С., Косарева Н. А., Кубанцев Б. С. 1969. Материалы по экологии и паразитологии ящериц и змей Волгоградской обл. В сб. «Паразитические животные Волгоградской сбл.» Волгоград.
344
Меландер В. А. 1935. Животный мир Западной области. Смоленск.
Мельников А. П. 1936. Уничтожение пчел ящерицами.— Пчеловодство, № 4. Мельниченко А. Н. ,1937. Полезащитные лесные полосы и размножение вредителей полеводства.— Итоги научн. исслед. работ ВИЗРа за 4936 г. Ч. I. Мельниченко А. Н. 1949. Полезащитные лесные полосы степного Заволжья и воздействие их на размножение животных полезных и вредных для с/х. М„ Изд. МОИП.
Мешков М. М. 1958. Земноводные и пресмыкающиеся Псковской области — Уч. зап. Псковск. гос. пед. ин-та им. С. М. Кирова, 5.
Миночкина Т. П. 1972. Иннервация толстой кишки ящерицы. В сб. «Вопр. морфол., экол. и гематол. позвоночных». Саратов.
Михлин С. Я. 1950. Распределение по органам животного марганца в зависимости от путей его введения и от характера вводимых соединений.— Архив патологии, 6.
Мусхелшивили Т. А. 1970. Пресмыкающиеся Восточной Грузии. Тбилиси, «Мец-ниереба».
Назарова И. С. 1961. К вопросу резервуарного паразитизма Spirocersa Itipi (Rudolphi, 1809). В сб. «Вопросы паразитол. в прибалтийских республ.» Рига, Изд. АН Латв. ССР.
Наумов Н. П. 1954. Типы поселений грызунов и их экологическое значение.— Зоол. ж., 33, 2.
Наумов Н. П. 1963. Экология животных. М.
Неемченко М. Г., Темботов А. К- 1959. К герпетофауне Кабардино-Балкарской АССР.— Уч. зап. Кабардино-Балкарского ун-та вып. 6, сер. сельхоз. и биол. наук.
Никитенко М. Ф. 1969. Эволюция и мозг. Минск, «Наука и техника».
Никольский А. М. 1902. Гады и рыбы по Брэму и другим источникам. СПб. Никольский А. М. 1905. Пресмыкающиеся и земноводные Российской империи (Herpetologia Rossia) —Зап. Акад, наук, 8, сер. физ.-мат. отд., 17.
Никольский А. М. 1907. Определитель пресмыкающихся и земноводных Российской империи. Харьков.
Никольский А. М. 1913. Пресмыкающиеся и земноводные Кавказа. Тифлис, Изд. Кавк, музея.
Никольский А. М. 1915. Reptilia т. I. Sauria. В сб. «Фауна Российской Империи и сопредельных стран». Петроград, Изд. Рос. АН.
Оболенский С. И. 1933. Биологический метод борьбы с вредителями в сельском хозяйстве. Сельхозгиз.
Обтемперанский С. И. 1956. Сравнительный анализ питания енотовидной собаки, лисицы и барсука в Воронежской области.— Бюлл. об-ва естествоиспыт. при Воронежском ун-те, 10. Воронеж.
Огнев С. И. 1908. Материалы для фауны зверей, птиц и гадов юго-восточной части Орловской губернии — Изв. Имп. об-ва естеств. антропол. и этнограф., 98. Дневник Зоол. отд., III, 9.
Огнев С. И., Воробьев К- А. 1924. Фауна позвоночных Воронежской губернии. М., «Новая деревня».
Одынец Р. Н. 1966. Распределение молибдена в организме овец, его обмен и взаимоотношения с другими элементами в обмене веществ. В сб. «Микро-элементы в сельском хозяйстве и медицине». Киев.
Озоль М. К- 1941. Материалы к биологии степной гадюки.— Труды Ворошиловского гос. пед. ин-та, 3, 2. Ворошиловск.
Окулова Н. М 1963. Ландшафтные особенности фауны позвоночных животных на северной окраине Волго-Уральских песков.— Зоол. ж., 42, 6.
Окулова Н. М. 1969. Влияние внешних условий на активность, поведение и температуру тела разноцветной ящурки (Eremias arguta).—Зоол. ж., 48, 10.
Олигер И. М. 1966. Животный мир Чувашии. Чебоксары.
Орлов Е. И. 1928. Материалы к познанию фауны наземных позвоночных Калмыцкой области. В сб. «Материалы к познанию фауны Нижнего Поволжья», вып. 2. Саратов.
Орлов Е. И. 1929. Желтый суслик (Citellus fulvus Licht). В сб. «Материалы к познанию фауны Нижнего Поволжья», вып. 4. Саратов.
13 Прыткая ящерица
345
Орлов Е. И., Фенюк Б. К. 1927. Материалы к познанию фауны наземных позвоночных приморской полосы Калмыцкой области. В сб. «Материалы к познанию фауны Нижнего Поволжья», вып. 1. Саратов.
Орлова В. Ф. 1975. Систематика и некоторые эколого-морфологические особенности лесных ящериц рода Lacerta. Автореферат канд. дисс.
Орлова В. Ф., Орлов В. Н. 1969. Хромосомные наборы и некоторые вопросы систематики ящериц рода Lacerta.— Зоол. ж., 48, 7.
Павлов М. П., Карис И. Б. 1963. Питание енотовидной собаки в темрюкских плавнях.— Труды Всес. научно-исслед. ин-та животного сырья и пушнины, 20. М.
Параскив К. П. 1956. Пресмыкающиеся Казахстана. Алма-Ата., Изд. АН Каз.ССР.
Параскив К. П., Бутовский П. М. 1960. О фауне земноводных и пресмыкающихся Западного Казахстана. Материалы по фауне и экологии наземных позвоночных Казахстана. Алма-Ата.
Паркер Г. Дж., Гасвель В. А. 1899—1914. Руководство к зоологии. Т. 1, 2. М.
Парфеник А., Подъяпольский Г. 1951. Животный мир Кабарды. Нальчик. Пархоменко Л. А. 1973. Пресмыкающиеся Волго-Уральского междуречья. Дипломная работа. Элиста.
Пащенко Ю. И. 1954. Экология и хозяйственное значение прыткой ящерицы (Lacerta agilis) в условиях лесостепного Приднепровья.— Наук. зап. Киевского ун-та, 13, 12.— Труды Зоол. музею, 4. Киев.
Пегель В. А., Попов Ф. Г. 1937. Влияние температуры на пищеварение хладнокровных животных. —Труды Биол. НИИ Томского ун-та, 4.
Лейве Я- М. 1960. Микроэлементы и ферменты. Рига.
Перелешин С. 1928. Опыт биометрического анализа понятия «подвид» (subspecies). К вопросу о взаимоотношениях Lacerta agilis и L. a. exigua Eichw.—-Русек. Зоол. ж., VIII.
Платонов К- (молодший). 1926. Короткий визначник'амф!б!й та рептилш Украши. КиТв, Державне Видавництво Украши.
Положенцев П. А. 1935. К фауне млекопитающих и гадов Бузулукского бора.
В сб. «Материалы по изучению природы Среднего Поволжья», вып. 1.
Положенцев П. Л. 1937. Пресмыкающиеся и земноводные. В сб. «Животный мир Среднего Поволжья». Куйбышев.
Положенцев П. А. 1941. Животный мир Среднего Поволжья. Куйбышев.
Полякова Т. И. 1962. Изучение ороговения в кожном эпителии ящериц (Lacerta agilis, Lacerta vivipara) с помощью S-35 метионина.—Докл. АН СССР, 142, 6.
Померанцев Б. И. 1950. Фауна СССР. Паукообразные. М.— Л., Изд-во АН СССР.
Поспелова-Штром М. В. 1935. О клещах диких животных Таджикской ССР. Таджикская комплексная экспедиция 1932 г.— Труды экспедиции, вып. X.
Птушенко Е. С. 1934. Наземные позвоночные Курского края. Амфибии и рептилии.— Бюлл. МОИП, отд. биол., 43 (новая серия), il.
Птушенко Е. С. 1938а. Некоторые данные по амфибиям и рептилиям Мордовского заповедника. В сб. «Фауна Мордовск. гос. зап. им. Смидовича». М.
Птушенко Е. С. 19386. Засуха 1936 года в условиях Мордовского заповедника. В сб. «Фауна Мордовского гос. зап. им. П. Г. Смидовича». М.
Птушенко Е. С. 1946/1947. Материалы к познанию фауны земноводных и прес-смыкающихся Окского гос. заповедника. Рукописный отчет ОГЗ.
Пузанов И. И. 1942. Звери, птицы, гады и рыбы Горьковской области. Горький.
Пузанов И. И. 1955. Животный мир Горьковской области. Горький.
Пятницкий Н. П. 1936. Сравнительное исследование свойств пепсина хладнокровных и теплокровных животных.— Труды Кузбасского мед. ин-та, 4.
Ралль Ю. М. 1936. Некоторые методы экологического учета грызунов. В сб. «Вопросы экологии и биоценологии», вып. 3. Л.
Родионов В. М. 1938. Некоторые данные по газообмену у рептилий в состоянии переохлаждения.— Бюлл. МОИП, 47, отд. биол., 2.
Рокицкий П. Ф. 1967. Биологическая статистика. Минск, «Вышэйшая школа». Рудин В. Д. 1959. Химические элементы в сельском хозяйстве. Ставрополь.
346
Рузский М. 1916. О зоологических исследованиях в Енисейской губернии, проведенных летом 1915 г.— Изв. Томского ун-та, 4.
Румянцев Б. 1926. Материалы по изучению фауны пресмыкающихся и земноводных Костромской губернии.— Труды Костр. научн. об-ва по изучен, места, края, 37.
Рюмин А. 1939. Температурная чувствительность позвоночных и биологический путь происхождения теплокровных форм.— Сб. студенческих научн. работ, биол., 6. Изд. МГУ.
Савронь Е. С. 1966. Биохимия животных. М.
Самош В. М. 1953. К познанию герпетрофауны Закарпатья.— Научн. зап. Ужг. гос. ун-та, 8.
Сатунин К- А. 1912. Животный мир Мугани. Муганская степь. Природа и население, вып. 3. Тифлис.
Сатунин К. А. 1913. Муганская степь. Тифлис.
Северцов А. Н. 1912. Строение языка рептилий. М., Изд. Ассоц. научн. исслед. ин-тов при физ.-мат. фак. <1 МГУ.
Северцов А. И. 1950. Периодические явления в жизни зверей, птиц и гадов Воронежской губернии. М., Изд. АН СССР.
Сергеев А. М. 1937. Материалы к вопросу о постэмбриональном росте рептилий.— Зоол. ж., 16, 4.
Сергеев А. М. 1939а. Материалы к познанию динамики популяций у рептилий. В сб. «Вопросы экологии и биоценологии», вып. 4. Л.
Сергеев А. М. 19396. Материалы по постэмбриональному росту рептилий.— Зоол. ж., 18, 5.
Сергеев А. М. 1940. О филогении некоторых эмбриональных приспособлений рептилий.— Изв. АН СССР, отд. биол. наук.
Сергеев А. М. 1941. Материалы по биологии размножения степной черепахи (Г. horsfieldi).— Зоол. ж., 20, 1.
Сергеев А. М. 1943. Эволюция эмбриональных приспособлений рептилий. М., «Сов. наука».
Сердюкова Г. В. 1956. Искодовые клещи фауны СССР.— М.—Л., Изд. АН СССР.
Силантьев А. А. 1894. Фауна «Падов» имения В. А. Нарышкина, Балашов-ского уезда, Саратовской губ. В сб. «Пады». Спб.
Симонян А. А. 1973. Морфо-экологическая характеристика некоторых популяций Lacerta agilis и Lacerta strigata. Дипломная работа, Ереванск. ун-т.
Словцов И. Я. 1892. Позвоночные Тюменского округа и их распространение в Тобольской губернии. Материалы к познанию фауны и флоры Росс, имп., 1.
Слоним А. Д. 1971. Экологическая физиология животных. М., «Высшая школа».
Смирина Э. М. 1974. Перспектива определения возраста рептилий по слоям кости.— Зоол. ж. 53, 1.
Смогоржевский Л. О. 1959. РибоТдш птахи УкраЧни. КиЧв, Вид-во КДУ.
Соболевский Н. И. 1929а. Материалы к познанию герпетофауны Южного Алтая.— Изв. ассоциации научн.-исслед. ин-тов при физ.-мат. фак. МГУ, 2, 1.
Соболевский Н. 19296. Герпетофауна Талыша и Ленкораньской низменности (опыт зоогеографической монографии).— Мемуары Зоол. отд. об-ва любит, естест., антропол. и этнографии, вып. 5. М.
Спасский А. А. 1948. Новое семейство цестод Skrjabinochoridae fam. nov., характеризующееся полным отсутствием матки,—Докл. АН СССР, 59, 2.
Страутман Ф. И., Сухомлинов Б. Ф., Кушнирук В. А. 1965. Применение биохимических методов для выяснения формообразующей роли среды. В кн. «Внутривидовая изменчивость наземных позвоночных животных и микроэволюция».
Стрелков Е. И. 1962. К экологии земноводных и пресмыкающихся равнинной тайги Зап. Сибири. В сб. «Вопросы охраны природы Зап. Сибири», вып. 3. Новосибирск.
Стрельников И. Д. 1934. Свет как фактор в экологии животных. Действие
347 13*
солнечной радиации на температуру тела пойкилотермитных животных,— Изв. Естественно-научн. ин-та, им. Лесгафта, 17—-18. Л.
Стрельников И. Д. 1944. Значение солнечной радиации в экологии высокогорных рептилий.— Зоол. ж., 23, 5.
Стрельников И. Д. 1959. О терморегуляции у современных и вероятном тепловом режиме мезозойских рептилий. В сб. «Вопросы палеобиологии и биостратиграфии». М., Госгеолтехиздат.
Стрельцов А. Б., Воронин А. А. 1973. К биологии и систематике прытких ящериц в калужской области. В сб. «Вопросы герпетологии». Л., «Наука».
Суханов В. Б. 1961. Некоторые вопросы филогении и системы Lacertilia (Sauna).— Зоол. ж., 40, 1.
Сухов Г. Ф. 1928. Матер1али до вивчення герпетофавни Полтавщини. 36. Пол-тавського Держ. Музею, 1. Полтава.
Сухов Г. Ф. 1929. Description of a new species of lizards from the environs of Vladicaucasus (L. boemica sp. nov.). Труди физ.-мат. вщд. Укр. АН, 13, 1.
Сухов Г. Ф. 1948. Обзор ящериц подрода Lacerta (Sauria) встречающихся в СССР.— Труды Зоол. ин-та АН СССР, VII.
Сыроечковский Е. Е. 1958. Эколого-географический очерк фауны рептилий западной части пустыни Кызыл-Кум (о биологических группах и типах поселений рептилий пустыни).— Зоол. ж., 37, 2.
Таращук В. I. 1959. Фауна УкраТны, 7. Плазуни. Ки1в, Вид-во АН УРСР.
Татаринов Л. Л. 1960. Эволюция аппарата разделения токов крови в сердце позвоночных.— Зоол. ж., 39, вып. 8.
Терентьев [I. В. 1935. К познанию пресмыкающихся и земноводных Чувашской АССР.— Труды Об-ва естествоисп. при Казанском ун-те, 52, 6.
Терентьев II. В. 1946. Опыт применения математической статистики в зоогеографии,— Вести. ЛГУ, 2.
Терентьев П. В. 1947. О применении правила Бергмана к животным с постоянной температурой тела.— Вести. ЛГУ, 12.
Терентьев П. В., Чернов С. А. 1949. Определитель пресмыкающихся и земноводных. М., «Сов. наука».
Тертышников М. Ф. 1967. Материалы пр экологии двух видов рептилий в Ставрополье. В сб. «Материалы Ш зоол. конф. пед. ин-тов РСФСР». Волгоград.
Тертышников М. Ф. 1970. Индивидуальная территория прыткой ящерицы (Lacerta agilis) и разноцветной ящурки (Eremias arguta) и особенности ее использования.— Зоол. ж., 49, 9.
Тертышников М. Ф. 1972а. Состав пищи у прыткой ящерицы и ее влияние на зооценоз в Предкавказье. В сб. Ставропольского пед. ин-та, биол. науки.
Тертышников М. Ф. 19726. Экологический анализ и биоценологическое значение популяций прыткой ящерицы (Lacerta agilis exigua Eichw, '1831) и разноцветной ящурки (Eremias arguta deserti Gm., 1789) в условиях Ставропольской возвышенности. Канд. дисс. Киев.
Тертышников М. Ф. 1973а. К вопросу об особенностях газообмена прыткой ящерицы и разноцветной ящурки. В сб. «Вопросы герпетологии». Л.
Тертышников М. Ф. 19736. К вопросу о методике учета прыткой ящерицы и разноцветной ящурки. В сб. «Материалы научн. совещ. зоологов пед. ин-тов». Владимир.
Тертышников М. Ф. 1973в. Значение амфибий и рептилий в биоценозах Северного Кавказа. В сб. «Геогр. пробл. изуч. охраны и рац. использов. условий и ресурсов Сев. Кавказа». Ставрополь.
Тертышников М. Ф., Щербак И. Н. 1973. Роль прыткой ящерицы и разноцветной ящурки в биоценозах Ставропольской возвышенности. В сб. «Вопросы герпетологии». Л., «Наука».
Тимофеев-Ресовский Н. В., Яблоков А. В., Глотов Н. В. 1973. Очерк учения о популяции. М., «Наука».
Тофан В. Е. 1964. К изучению популяций прыткой и зеленой ящериц Молдавии.— Тезисы докл. Научн. конф. проф.-препод, состава по итогам науч-но-исслед. работ за 1962 г. Кишинев. «Катря Молдовеняска».
348
Тофан В. Е. 1967. О популяциях прыткой и зеленой ящерицы Молдавии.-. Вести. ЛГУ, биол., 3, 1.
Турутина Л. В. 1972. К изучению популяционной изменчивости прыткой ящерицы. Дипломная работа. Горьковский ун-т.
Утробина Н. М. 1956. Питание ящерицы прыткой в полезащитных насажде-ниях Тат. АССР,— Изв. Казанского фил. АН СССР, сер. биол. и сельско-хоз. наук, 3.
Фаткулова А. В. 1962. К биологии прыткой ящерицы в окрестностях Караганды.— Уч. зап. Карагандинск. пед. ин-та, 3, 2.
Федосеев А. В., Никитин К. Н. 1951. Животный мир Брянской области. Брянск, «Брянский рабочий».
Федюшин А. В. 1935. Речной бобр, его история, жизнь и опыт по размножению. М.
Федоров В. Г. 1970. .О кровососущих клещах на амфибиях и рептилиях в Западной Сибири. В сб. «Второе акаролог. совещ.», 2. Киев, «Наукова думка».
Фел1кс Л. С. 1970. Цитологические изменения гипоталамуса и агеногипофиза ящериц (Lacerta agilis) в период размножения. Вкник КиТв. ун-ту сер. бюл., 12.
Филимонов И. Н. 1963. Сравнительная анатомия большого мозга рептилий. М.
Формозов А. Н. 1928. О пустынном элементе в фауне Южной части Восточной Европы.— Докл. АН СССР, 14—15.
Формозов А. Н. 1935. Очерк фауны наземных позвоночных Горьковского края. В сб. «Природа Горьковского и Кировского краев». Горький.
Формозов А. Н. 1946. Снежный покров в жизни млекопитающих и птиц СССР. Изд. МОИП.
Харченко В. И., Сова Ю. Ф. 1972. К гельминтофауне пресмыкающихся приазовских степей. В сб. «Проблемы паразитологии», 2. Киев, «Наукова думка».
Хонякина 3. П. 1964. Ящерицы Дагестана. Автореф. канд. дисс. Махачкала.
Хонякина 3. П. 1965. Ящерицы Дагестана и их хозяйственное значение. В сб.
«II научн. сессия Сев.-Кавк, совета по координац. научно-исслед. работ по техн, и естеств. наукам». Ростов-на-Дону.
Хонякина 3. П. 1970. Материалы по биологии прыткой ящерицы Дагестана. В сб. «Исслед. по зоол. и паразитол. в Дагестане». Махачкала.
Царевский С. 1914. Пресмыкающиеся (Reptilia). Каталог музея при Зоологич. кабинете Имп. Петроградского университета, 2.— Труды Имп. Петрогр. об-ва естествопспыт., 43, 4.
Цемш /. 1939. До систематики та географ!чного пошпрення амф1б1й та реп-тилш на Укра Tiri. — СтудентсыН наук, пращ КиТвск. Держав Ушвер. 1м. Т. Г. Шевченка, 4.
Чан Кьен. 1967. Систематика и экология обыкновенной гадюки. Автореф. канд. дисс.
Чельцов-Бебутов А. М. 1953. Наблюдения над пресмыкающимися Центрального Казахстана на маршруте пос. Джулек — г. Атбаяр. Труды ин-та географии АН СССР, 54.
Черкащенко И. И., Кушнирук В. А. 1973. Состав белков сыворотки крови некоторых видов земноводных и пресмыкающихся. В сб. «Вопросы герпетологии», рефераты докл. III Всесоюзного герпет. конф.» Л., «Наука».
Чернай Л. 1852—1853. Фауна Харьковской губернии и прилежащих к ней мест. Харьков.
Чернов С. А. 1929. Материалы к познанию фауны Amphibia и Reptilia Горной Ингушетии. В сб. «Мат. к позн. фауны позвоночных животных Ингушской автон. обл.».— Изв. Ингуш, научно-псслед. ин-та краевед., 2.
Чернов С. А. 1937. Определитель змей, ящериц и черепах Армении. М., Изд-во АН СССР.
Чернов С. А. 1947. Материалы к герпетофауне Казахского нагорья, северного побережья Балхаша и гор Кан-Тау.— Изв. АН СССР, 6, сер. зоол.
Чернов С. А. 1954. Эколого-фаунистический обзор пресмыкающихся юга междуречья Волга — Урал.— Труды Обл. ин-та АН СССР, 16.
349
Чернов С. А. 1959. Фауна Таджикской ССР. Пресмыкающиеся, 18. Изд-во АН Тадж. ССР.
Черномордиков В. В. 1943. О температурных реакциях рептилий.— Зоол. ж., 22, 5.
Черномордиков В. В. 1944. О врожденных и приобретенных пищевых реакциях пресмыкающихся.— Докл. АН СССР, 43, 4.
Черномордиков В. В. 1947. Суточный цикл активности некоторых пресмыкающихся.— Докл. АН СССР, 57, 5.
Черномордиков В. В., Ковалева 3. Н. 1964. К вопросу о причинах гигантизма мезозойских пресмыкающихся. В сб. «Вопросы герпетологии». Л.
Чернявская С. И. 1965. Природа Кызыл-Агачского заповедника». — Труды Зап. Азербайджана, 1. М., «Лесная промышленность».
Чеченкин М. Н. 1954. Химия жиров рептилий. Уч. зап. Псковского пед. ин-та, вып. 2. Псков.
Чиркова А. Ф. 1948. Материалы по экологии лисицы. Кормовой режим и зараженность лисиц Ставропольского края,—Труды Всес. научно-исслед. ин-та охотн. промысла, 8.
Чугунов С. М. 1911. Гады, собранные, в окрестностях станции Иланской Сибирской железной дороги в 1911 г.— Ежегодн. Зоол. муз. Имп. Акад, наук XVI.
Шапиро Д. С. 1940. Энтомофауна травяного покрова опушек полезащитных полос. Пращ наук.-доел, зоол.-бюлог. ш-ту. Харк. Держ. ун-ту, сектор Екологп.
Шапошников Я. 1959. Животный мир Калининской области. Калинин.
Шарлемань И. 1917. Заметка о фауне пресмыкающихся и земноводных окрестностей Киева.— Труды Киевск. орнитол. об-ва им. К. Кесслера, II.
Шарлемань М. В. 1936. Зоогеограф1я УРСР. Кшв, Вид-во УАН.
Шарпило В. П. 1959. К познанию гельминтофауны некоторых рептилий УССР.— Труды ГЕЛАН СССР, № 9. М„ Изд. АН СССР.
Шарпило В. П. 1960. К изучению гельминтофауны прыткой ящерицы (Lacerta agilis) на территории УССР. В сб. «Проблемы паразитологии». Труды III научн. конф, паразитол. УССР. Киев, «Наукова думка».
Шарпило В. П. 1961. До вивчення гельмштофауни прудко! яшдрки (Lacerta agilis) на Укра’пн. В сб. «36. праць Зоол. музею АН УРСР. КиТв, Вид-во АН УРСР.
Шарпило В. П. 1963. Рептилии фауны Украинской ССР как резервуарные и дополнительные хозяева. В сб. «Проблемы паразитологии». Труды IV научн. конф, паразитол. УССР. Киев, Изд-во АН УССР.
Шарпило В. П. 4964. Гельминты рептилий фауны Украинской ССР. Автореф. канд. дисс. Киев.
Шарпило В. П. 1966. Новый представитель рода Thubunaea (Nematoda. Phy-salopteridae) — паразит ящериц. В сб. «Паразиты, промежуточные хозяева и переносчики». Киев. «Наукова думка».
Шарпило В. П., Шарпило Л. В. 1969. Експериментальна щентифшащя трьох личинковых форм гельм1нт1в—паразипв плазушв з ix статевозр!лими формами. 36. праць Зоол. музею АН УРСР, 33. Кшв, «Наукова думка».
Шарпило В. П„ Магуза В. С. 1972. Про поширення Candidotrema loossi (Africa, 1930, Dollfus, 1951) Trematoda, Pleurogenidae на территорн CPCP у зв’язку з новими знахщками цього паразита. В зб. «Паразита, паразитов и та шляхи ix лшвщацП, 1. КиТв, «Наукова думка».
Шарпило В. П. 1973. Новий у фаун! СРСР вид циклофипдних цестод Nema-totaenia tarentolae Lopez-Neyra, 1944 (Cestoda, Nematotaeniidea).— 36. праць Зоол. музею АН УРСР, 35, Кшв, «Наукова думка».
Шварц С. С. 1946. Эффективность криптической окраски. Вестник ЛГУ, 4—5.
Шварц С. С. 1948. О значении постоянной температуры тела пойкилотермных животных для скорости пищеварения.— Природа, № 12.
Шварц С. С. 1948. О специфической роли амфибий в лесных биоценозах в связи с вопросом об оценке животных с точки зрения их значения для человека.— Зоол. ж., 27, 5.
Шварц С. С. 1951. Животный мир Урала. Свердловск.
350
Щварц С. С. 1960. Некоторые закономерности экологической обусловленности интерьерных особенностей наземных позвоночных животных. В сб. «Проблемы флоры и фауны Урала».—Труды Ин-та биологии УФАН СССР, вып. 14.
Шварц С. С., Смирнов В. С., Добринский Л. Н. 1968. Метод морфофизиологических индикаторов в экологии наземных позвоночных. —Труды Ин-та экологии растений и животных, вып. 58. Свердловск.
Шебзухова Э. А. 1969. Пресмыкающиеся центральной части Северного Кавказа. Автореф. канд. дисс. Нальчик.
Шевченко Н. Н„ Барабашова В. Н. 1958. К гельминтофауне ящерицы прыткой (Lacerta agilis) и гадюки обыкновенной (Vipera berus) Харьковской области. В сб. «Работы по гельминтол. К 80-летию акад. К. И. Скрябина. М., Изд-во АН СССР.
Шевченко Н. Н. 1963. Гельминты рептилий долины Северного Донца. В сб. «Гельминты человека, животных и растений и борьба с ними. М., Изд-во АН СССР.
Шевченко Н. Н., Белинисова Л. К. 1964. О некоторых новых формах гельминтов рептилий долины Северного Донца в Харьковской области. В сб. «Вопр. генетики и зоол.» Изд-во Харьк. гос. ун-та.
Шершевский Э. И. 1931. Сб. работ по изучению фауны Крымского гос. заповедника. М., Медгиз.
Шестаков А. 1926. Фауна Ярославской губернии.— Труды Яросл. естеств. историч. и краевед, об-ва, 5, 3.
Шилов И. А. 1961. Практикум по экологии наземных позвоночных животных. М., «Высшая школа».
Шилов И. А. 1962. О некоторых понятиях, связанных с изучением проблемы терморегуляции.— Зоол. ж., 41,4.
Шилов И. А. 1968. Регуляция теплообмена у птиц. Изд-во МГУ.
Шилов М. П. 1961. Заметки о некоторых рептилиях Северного Приаралья.— Труды Ин-та зоол. АН Каз. ССР, 15.
Шиплей А. Э„ Мак-Брайдт Э. В. 1911. Курс зоологии. М., Изд-во М. и С. Сабашниковых.
Школьник М. И. 1964. Влияние различных солей кобальта на активность некоторых пищеварительных ферментов.— Уч. зап. Петрозаводского ун-та, 12, 3.
Шлугер Е. Г., Пионтковская С. П., Симонович Е. И. Айзенштадт Д. С. 1955. Надсемейство Tombea Turk. В сб. «Клещи грызунов фауны СССР». М.— Л., Изд-во АН СССР.
Шмальгаузен И. И. 1947. Основы сравнительной анатомии. М., «Сов. наука».
Шнитников В. И. 1928. Пресмыкающиеся Семиречья.— Труды Об-ва изуч. Казахстана, 8, 3.
Шнитников В. Н. 1935. Животный мир Казахстана. Ч. 2.
Щепотьев Н. В. 1948. К изучению популяций прыткой ящерицы в условиях лесостепи Нижнего Поволжья.—Зоол. ж., 27, 4.
Щепотьев Н. В. 1950. Экология и хозяйственное значение Lacerta agilis exigua Eichw. Канд. дисс. Саратов.
Щепотьев Н. В. 1952. К вопросу о хозяйственном значении прыткой ящерицы в полезащитных лесополосах.— Зоол. ж., 31, 4.
Щербак И. И. 1962. Герпетофауна Крыма и ее зоогеографический анализ (Herpetologia Taurica). Автореф. канд. дисс. Л.
Щербак Н. И. 1965. Герпетофауна Украши та охорона природи.— В зб. «Охо-роняйте рщну природу», 4.
Щербак Н. Н. 1966. Земноводные и пресмыкающиеся Крыма. Киев, «Наукова думка».
Эюбов И. 3. 1967. Энзоотический зоб у овец.— Труды НИВИ, 21.
Яблоков А. В., Баранов А. С. 1971. Некоторые итоги и перспективы изучения прыткой ящерицы в пределах ареала.— Матер, засед. межсекционной группы по проблеме «Исслед. продук. вида в пределах ареала», 2. Вильнюс.
Яковлева И. Д. 1964. Пресмыкающиеся Киргизии. Фрунзе, Ин-т биологии АН Кирг. ССР.
351
Adrian E. D. 1938. Effect of socend on the ear in Reptiles.— J. Physiol., v. 93, N 1—2.
Adrian E. D. 1943. Discharges from vestibular receptors in the cat.— J. Physiol., v. 101.
Albritton E. C. 1952. Standard values in blood.— Handbook of biological data. Philadelphia, W. B. Saunders Co.
Albritton E. C. 1954. Standart values in nutrition and metabolism. Philadelphia, W. B. Saunders Co.
Alexander С. E., Whitford W. G. 1968. Energy requirements of Uta stansburia-na.— Copeia, 4.
AH M. A. 1959. The ocular structure retinomotore, and photobehavioral responses of juvenile Pacific salmon.— Canad. J. Zool., v. 37, N 6.
Amoroso E. C. 1955. Ann. Endocrinol., 16.
Arnold E. N. 1973. Relationships of the Palaearctic lizards assigned to the genera Lacerta, Algyroides and Psammodromus (Reptilia: Lacertidae).— Bull. Brit. Mus. (Nature, Hist.), v. 25, N 8.
Babcock H. L. 1939. Growth of an individual Box turtle.— Copeia, 3, 175.
Bacon R. F. 1909.— Philippine J. Sci., 4, 165.
Baldwin E. 1959. Dynamic aspects of biochemistry. 3rd ed. N. Y., Macmillan Co., Chap. 12—44.
Bartholomew G. A. 1949. Bull. Mus. Compar. Zool. Harvard, 101.
Bartholomew G. A. 1956. Photoperiodism and related phenomena in plants and animals. A. A. A. S. Press. N. W.
Bauer V. 1914. Z. allg. Physiol., 16.
Benedict F. G. 1932. The physiology of large Reptilies, Washington.
Benedict F. G., Lee R. S. 1938. Lipogenesis in the animal body, with special reference to the physiology of the goose, Washington.
Bentley P. J. 1959. Stadies on the water and electrolyte metabolism of the lizard Trachysaurus rugosus.— J. of Physiol., v. 145, N 1.
Berger L., Jaskowska J., Mlynarski M. 1969. Katalog fauny Polski, Cz. 4. Plazy i Gady, Warszawa.
Bergmann C. 1847. Uber die Verhaltnisse der Warmeokonomie der Tiere zu ihrer Grosse.— Gottinger Studien, I.
Bergmann M., Pace W. T. 1943.— J. Amer. Chem. Soc., 65.
Berlin N. J., Waldmann T. A., Weissmann S. M. 1959. Life of red blood cell.—• Physiol. Rev., v. 39.
Beskov V., Beron P. 1964. Catalogue et bibliographie des amphibiens et des reptiles en Bulgarie.— Sofia, Acad. Bulg. Sci.
Bischoff W. 1969. Lacertenbastarde. I. Aquar. Terr. Jahrg. 16, H. 8.
Bischoff W. 1973. Lacertenbastarde. II. Zool. Garten N. F. Bd. 43, N 5.
Blanchard F. N., Blanchard F. C. 1931. Size groups and their characteristics in the salamander Memidactylium scutatum (Schlegeb).— Amer. Naturalist, 65, N 697.
Boettger O. 1886. Die Reptilien und Amphibien des Talysch—Gebietes.— In: Die Fauna und Flora des Sudwestlishen Caspi—Gebietes. Leipzig.
Bolk L. 1931—1938. Handbuch der vergleichenden Anatomie der Wirbeltiere, Bd. 1—6. Berlin, Wien, Urban Schwarzeiberg.
Boulenger G. A. 1887. Catalogue of the lizards in the British Museum Natural History, v. 3, 2nd ed. London.
Boulenger A. G. 1920. Monograph of the Lacertidae I. London.
Bragdon D. E., Lazo-Wasem E. A., Zarrow M. X., Hisaw F. L. 1954. Hormone progesterone activity snake.—Proc, of the Soc. for Exp. Biol, and Medic., v. 86.
Brattstrom В. M. 1965. Body temperature of reptiles.— Amer. Midland Naturalist, 73, 2.
Brimley J. N. 1903. Note on the reproduction of certain reptiles.— Amer. Naturalist, 37.
Buchanan F. 1909.— J. Physiol., 39.
Buddenbrock W. 1952. Vergleichende Physiologie. I. Basel.
Cagle F. R. 1946. The growth of the slider turtle, Pseudemys scripta elegans.
Amer. Midland Naturalist, 36.
352
Cagle F. R. 1948. The growth of Turtles in lake glandale, Illinois,— Copeia, 3.
Capuse I. 4971. Contribution e I’etudes des Trematodes parasites chez les reptiles de Roumanie.— Travaux Mus. hist, natur., XI.
Castanet J. 1974. Etude histologique des marques squelettiques de croissance chez Vipera aspis (L) (Ophidia, Viperidae), Zoologica scripta, 3.
Clark H. 1953. Metabolism of the Black snake embrio: I—II.— J. Exp. Biol., v. 30.
Clark H., Sisken B. F. 1956. Nitrogenous excretion by embyos of the viviparous snake.— J. of Exp. Bipl., v. 33.
Combescot C. 1955. Action de 1’hyophysectome stir les elements testicularies d’une tortue terestre Testudo iberia.— Bull. Soc. Hist. Afrique Nord, v. 45.
Cope E. D. 1900. The crocodilians, lizards and snakes of North America. Washington, Rept U. S. Natur. Mus., 2.
Corbett K. 1969. Red light for the sand lizard in Britain.— Oryx, 10, 2.,
Coulson R. A., Hernandez T„ Brazda F. G. 1'950. Biochemical studies on the Alligator.— Proc. Soc. Exp. Biol, and Med. v. 73.
Crescitelli F. 1956. The nature of the lamprey visual pigment.—J. of Gen. Physiol., v. 39, N 3.
Cruce Л1. 1971. Cresterea si structure unei populatii de Lacerta taurica taurica Pall.— Studii si cercetari biol. Ser. zool., 23, 5.
Cuellar O. 1966. Delayed fertilization in the lizard Uta sunsburiana.— Copeia, 3
Cyren O. 1933. Lacertiden der SQdostlichen Balkanhalbinsel. Mitt. Kgl. Natur-wiss. Inst. Sofia, Bd. VI.
Darevskij I. S., Terentev P. V. 1967. Estimation of energy flow amphibian and reptile populations. Secondary productivity of terrestrial ecosystems. Warsa wa — Krakow.
Davis J. 1967. Growth and size of the western fence lizard (Sceloporus occidentalism .— Copeia, 4.
Dawes B. 1946. The trematoda with special refrence to British and other European forms. Cambridge Univ. Press.
Dawson W. R. 1960. Physiological responses to temperature in the Lizard.— Physiol, of Zool., v. 33, N 2.
Dawson W. R., Bartholomew G. A. 1956. Relation of consumption to body weight, temperature and temperature acclimation in Lizards.— Physiol, of Zool., v. 29.
Dawson W. R., Bartholomew G. A. 1958. Metabolic and cardiac responses to temperature in the lizard Dipsosaurus dorsalis.— Physiol. Zool., 31, 2.
Dawson W. R., Templeton J. R. 1963. Physiological responses to temperature in the lizard Crototaphytus collaris.— Physiol. Zool., 36, 3.
Deane H. W. 1959. J. Neurophysiol., 21.
Deraniyagala P. E. P. 1058. Reproduction in the monitor lizard Varanus benga-lensis (Daudin).—Spolia Zeylanica, 28, pt II.
Dessauer H. C. 1955. Seasonal changes in the gross organ composition of the lizard Anolis carolinensis.— J. Exper. Zool., 128.
Dobie J. L. 1974. Reproduction and growth in the alligator spapping turtel. Mac-roclemys temmincki (Troost).— Copeia, 4.
Dollfus R. 1961. Sur Distoma mentulatum Rudolphi, Distoma arrectum F. Dujardin et «Distoma clavigerum» sensu F. Dujardin non Rudolphi.—Ann. para-sitol. humaine et comparee, 26, 3.
Drzewicki S. 1926. Sur le croisement entre les lezards Lacerta agilis forma tipi-ca et Lacerta agilis var. erythronota.-— Compt. rend. Soc. biol., 93.
Dudziak J. 1955. Visual acuity in the Etnys orbicularis tortoise in air and water. Folia biol., 3, 3.
Dufaure J. P., Hubert J. 196Г. Table de developpement du lezarl vivipara Lacerta (Zootoca) vivipare jacquin.— Arch. anat. microsc. et morphol. exper., 50, 3.
Dufaure J. P., Hubert J. 1966. Recherches descriptives et experimentales sur les modalites, et tacteurs du developpement de 1’appareil gemital chez le lezard vivipare (Lacerta vivipara Jacquin).-—Arch, anat, microsc. et morphol. exper., 55, 3.
Edelenyi B. 1962. A hazai hiillok nehany belsoeloskodo forge. Az egri pedagogiai foiskola fiizetei szerkeszti Dr. B. Sandor, 272.
353
Edelenyi В. 1963. Harazi hfillok nehany ujabb belsoeloskodo ferge. Az egri pe-dagogial foiskola fuzetei szerkeszti Dr. B. Sandor, 298.
Edwards H. T., Dill D. B. 1935. Properties of Reptilian blood.—J. of Cell. Comp. Physiol., v. 6.
Ewer R. F. 1952. The effects of posterior pituitary extracts on water balance in Bufo carens and Xenopus laevis, together with some general considerations of anuran water economy.— J. of Exp. Biol., v. 29.
Eyzdguirre C. 1957. Functional organization of neuromuscular spindle in Toad.— Jorn. of Neurophysiol., v. 20.
Farner D. S. 1955. In: Recent studies in avian biology. (Ed.). Illinois Press.
Farner D. S. 1959. Photoperiodism and related phenomena in plants and animals. Washington, A. A. A. S.— Press.
Feider Z. 1953. Citeva larva ale genului trombicula (acarieni) si descrierea unui caz de trombidioza la sopirla Lacerta agilis.— Bui. stiint. sect, stiint, biol., agron., geol. si geogr., 5, 4.
Fitzgerald L. R. 1957. Cutaneous respiration in man.—-Physiol. Rev., v. 37.
Florkin M. 1944. L’evolution biochimique. Paris, Masson et Cie.
Florkin M. 1949. Actualites biochimistry. N-W., Academic Press.
Flor kin M., Duchateau G. 1943. Les formes du systeme enzymatique de 1’uricoly-se et revolution du catabolisme purique chez les animaun,— Arch. Internat. Physiol., v. 53.
Florkin M. F., Mason H. S. 1962. Compar. Biochem., 4.
Foglia V. C., Wagner E. M., De Barros M., Marques M. 1955. La diabete par pancreatectomie chez le tortue normal ou hypophysoprive.— C, R. Soc. Biol., v. 149.
Forbes A., Burleigh S., Neyland M. 1955. Electric responses to color shift in frog and turtle retina.— J. of Neurophysiol., v. 18, N 6.
Forbes A., Milbunn N., Fox S. I960. Electroretinogram of freshwater turtle: selective adaptation to coloured light.— J. Neurophysiol., 22, 6.
Forrest H. S. 1962.— J. Amer. Chem. Soc., 79 [цит. no Florkin M. F., Mason H. $.].
Foxon G. E., Griffith J., Price M. 1953. Circulation in Lacerta viridis.— Nature., v. 172.
Foxon G. E. 1955. Problems of the double circulation in vertebrates.— Biol. Rev., v. 30.
Foxon G. E. FL 1956. Proc. Zool. Soc. London, 126.
Fronseca F. 1948. A monograph of the genera and species of Macronyssidae Oudemans, 1936 (syn. Liponyssidae vitzthum, 4931). London, 118, 2.
Fuhn I. E., Vancea S. 1961. Fauna Republicii Populare Romine Reptilia. Bucures-ti, v. 14, N2.
Fuhn J. et Vancea S. 1964. Die innerartliche Gliederung der Zauneidechse (Lacerta agilis) in Rumanien (Reptilia, Lasertidae).— Senckenberg. biol. Bd. 45.
Gabe M., Saint Girons H. 1972. Rapport entre la position systematique des Sau-riens.— Bull. Biol. France et Belgique, v. 106.
Gaupp E. 1905. Das Hyobrachiolskelett der Wirbeltiere.— Ergebn. Anat. und Entwicklungsgesh., 14.
Gegenbaur C. 1864, 1865. Untersuchungen zur vergleichenden Anatomie der Wirbeltiere. 1. Carpus und Tarsus. 2. Schultergurtel d. Wirbeltiere, Brustflosse der Fische, Leipzig.
Gelineo A., Gelineo S. 1955. La concentration de 1’hemoglobine dans le sany chez des poissons d’eau douce.— C. R. Soc. de Biol., v. 149.
Geschwind I. L, Li С. H.. Burnafi L. 1956. Isolation and structure of melanocytestimulating hormone from porcine pituitary glands.— J. of Amer. Chem. Soc., v. 78.
Ceschwind I. L, Li C. H., Burnafi L. 1957. A comparison between porcine and bovine melanocyte — stimulating hormones.— Fed. Proc., v. 16.
Gibbons J. W. 1972. Reproduction growth and sexual dimorphism in the cane-brake rattlesnake (Crolalus horridus africaudafus).—-Copeia, 2.
Gilbert L. J., Schneiderman H. A. 1958. Occurrence of substances with juvenile hormone activity in adrenal cortex of Vertebrates.—Science, v. 128.
354
Gislen T„ Kauri H. 1959. Zoogeography of the Swedish amphibians and reptiles with notes on their growth and ecology.— Acta Vertebratica nordiska museet and scansen. Stockholm, 1, 3.
Goldie M. 1959. Arginase activity in the developing chick embrio in relation to nitrogen excretion.— Physiol. Zool., v. 32.
Goodrich E. S. 1930. Studies on the structure and development of vertebrates. London, MacMillan and Co.
Grabda-Kazubsky B. 1972. Katalog fauny pasozytniczey Polski. Pasozyty plazow i gadov. Pantwowe wydaunictwo naukowe.
Oranit R. 1941. A relation between rod and cone substances.—Acta Physiol.
Scand., v. 2.
Grand R. 1943. «Red» and «Green» receptors in the retina of Tropidonotas. -Acta Physiol. Scand., v. 5.
Grand R. 1947. Sensory mechanisms of the retina. Oxford, Univ. Press.
Grasse P., Devillers C. 1965. En: Traite de Zoologie, v. 11. Paris.
Green B. 1972. Aspects of renal function in the lizard Varanus gouldii.— Comp.
Biochem. Physiol., v. 43A.
Griffiths J. 1961. Skeletal lamellae as an index of age in heterothermous tetrapods.— Ann. Med. Natur. Hist., 13. 4.
Grodzinski Z. 1949. The fat in the yolk of the sand-lizard Lacerta agilis L.— Bull. Acad, polon. sci. Ser. B, 11.
Hadley N. F., Christie W. W. 1974. The lipid composition and triglyceride structure of eggs and fat bodies of the lizard Sceloporus jarrovi.—Compar. Biochem. and Physiol., 48B.
Hahn W. E., Trinkle D. IF. 1965. Fat body cycling and experimental evidence for its adaptive significance to ovarian follicle development in the lizard Uta stansburiana.— J. Exper. Zool., 158.
Hall F. G., Root R. IF. 1930. The influence of humidity on the body temperature of certain poikilotherny.— Biol. Bull., v. 58.
Hamilton IF. /. 1934. The rate of the toad (Bufo americanus americanus Hol.) under natural conditions.— Copeia, 2.
Harris J. I., Roos P. 1956. Amino-acid sequence of a melanophore-stimulating peptide.— Nature, v. 178.
Harris J. L, Lerner A. B. 1957. Amino-acid sequence of the L-melanocyte-stimu-lating hormone.— Nature, v. 179.
Harrison IF. 1965. Цит. no White F. N., 1968.
Heidermanns C. 1937. Tabul. Biol., 14.
Heller H. 1942. The posterior pituitary principles of a species of reptile with some remarks on the comporeative physiology of the posterior petieitary gland generally.— J. of Physiol., v. 101.
Hisaw E. L. 1959. Comparative endocrinology. N. Y., John Wiley and Sons Press. Hoffmann С. К 1879—1890. Bronn’s Klassen und Ordnungen des Tierreichs, Bd. 3. Hoffmann K. 1955. Aktivitatsregistrierungen bei frisch geschlupften Eidechsen.— Zool. vergl. Physiol., 37, 4.
Hoffmann K. 1957. Angeborene Tagesperiodik bei Eidechsen.— Naturwissenschaf-ten. 44, 12.
Hohenacher R. F. 4837. Enumeratio animalium que in provinciis transcaucasicis. Karabach, Schirwan et Talysch nec non in territorio Elisabethopolensi.—Bull. Soc. imp. Nahuralites Moscou, N 7.
Hopping A. 1923. Seasonal changes in the gases and sugar of the blood and the nitrogen distribution in the blood and urine of the Alligator.— Amer. J. of Physiol., v. 66.
Houssay B. A. 1959. Gomparative endocrinology. N. Y., John Wiley and Sons Press.
Huene F. 1956. Palaeontologie und Phylogenie der Niederen Tetrapoden. Jena.
Jackson C. G. 1970. A biometrical study of growth in Pseudemys concinna su-wanniensis I.— Copeia, 3.
Jenner С. E., Engels IF. L. 1959. Biol. Bull., 103.
Kaisila J. 1948. Herpetologische Notizen I. Die Zauneidechse, Lacerta agilis L., in Ostfennoskandien.— Arch. Soc. Zool. Bot. Fenn. «Vanamo», 3.
J^erfoot W. C. 1970. The effect of functional changes upon the variability of lizard and snake body scales.— Copeia.
355
Khalil F., Uaggag G. 1955. Ureotelism and uricotelism in tortoises.— J. of Exp. Zool., v. 130.
Khalil F„ Zaki K. 1964. Z. vergl. Physiol., 6, 48.
Kimura E„ Hosoya V. 1956. Japan. J. Physiol., 6, 1—11.
Kitzler G. 1941. Z. Tierpsychol., 4.
Klenk E. .1933. Hoppe-Seyler’s Z. Physiol. Chem., 221.
Klenk E. 1935, Hoppe-Seyler’s Z. Physipl. Chem., 232.
Korybska Z. 1971. Morfohistologiczne zmiany tarczycy Lacerta agilis w poszc-zegolnych porach roku.— Ann. UMCS, 97—103.
Krockert G. 1941. Kontinuierliche hyperthyreoidisierung und — epiphysierrung an Pylon bivittatus.— Vitam. und Horm., v. 1.
Kruger P„ Kern H. 1924. Arch. Physiol., 202.
Kruta V. 1957. Acta physiol, et pharmacol neerl., 6.
Kubiak St. 1963. Vascularization of the skin in the lizard (Lacerta agilis L.).— Studia Soc. Sci. Torunensis, sect. E., VII, 2.
Laskowski M. 1951. The Enzymes, v. I. N. Y., Acad. Press.
Laurens H. 1920. Pflugers Arch. ges. Physiol., 182, 50.
Lee T. H„ Lerner A. B. 1957. Nature, 179.
Leloup J., Berg O. 1954. C. r. Acad. sci. Paris, 238.
Lent FL, Freitas T. 1946. Sur la position systematique de Distoma arrectum (Molin, 1859).—Ann. parasitol. humaine et comparee, t. XX, N 5—6.
Lesley S„ Bellairs A. 1972. The mechanisms of autotomy in Lacerta.— Brit. J. Herpetol., 4, 11.
Licht P. 1965. Effects of temperature on heart rates of lizards during rest and activity.— Physiol. Zool., v. 38.
Licht P., Bennett A. 1972. A scaleless snake: test of the role of reptilian scales in water loss and heat transfer.— Copeia, 4.
Lindsey C. 1966. Body sizes of poikilotaerm vertebrates at different latitudes.— Evolution, 20, 4.
Lildicke M. 1964. Handbuch der Zoologie, Bd. 7.
Lithe M. 1909. Parasitische Plattwurmer. I. Trematodes. In: Susswasserfauna Deutschlands, Bd. 17.
Luppa H. 1961. Histologie, Histogeneze und Topochemie der Druzen des saurop-sidenmagens I. Reptilie.—Acta histochem., 12.
Mederson P. F. A., Chiu K. W., Phillips J. C. '1970. Endocrine-epidermal relationships in squamate reptiles. Hormones and Environments Proc. Sympos. Sheffield, 1969., 259—280; Discuss. 281—284.
Manseur-Bek J. J. 1932. Die proteolytischen enzyme von Maja squinado.— Ztschr. Vergl. Physiol., v. 17.
Manseur-Bek J. J. 1933. Uber die proteolytischen enzyme von Murex anquilife-rus.— Ztschr. Vergl. Physiol., v. 20.
Manseur-Bek J. J. 1948. Enzymologia, 12.
Matsumoto K- 1954. Biol. J. Okayama Univ., 1.
Matthews L. H. 1955. Mem. Soc. Endocrinol., 4.
Mattox N. T. 1935. Annular rings in the long bones of turtler and their correlation with size.— Trans. Illinois. St. Acad. Sci., 28.
Meinertz T. 1952. Z. Anat. und Entwicklungsgesch., 116.
Meinertz T. 1966. Eine untersuchung uber den sinus coronarius cordis, die v. cordis media and den arcus aortae sowie den ductus botall.— Morph. Jahrb., v. 109, p. 4.
Mellish С. FL, Meyer R. K- 1937. The effects of various gonadotropic substances and thyroxine on the ovaries of horned lizards (Phrynosoma cornutum).— Anat. Rec., v. 69.
Menetries E. 1832. Catalogue raisonne des objets de Zoology recueillis un fron-tieres actuelles de la Perse. St. Peterbourg.
Mertens R. 1950. Uber Reptilienbastarde. I. Senckenberg. biol., Bd. 3.
Mertens R. 1956. Uber Reptilienbastarde. II. Senckenberg. biol., Bd. 37.
Mertens R. 1964. Uber Reptilienbastarde. III. Senckenberg. biol., Bd. 45.
Mertens R., Wemuth H. ‘I960. Die Amphibien und Reptilien Europas. Frankfurt am Main, Verl. Waldemar Kamer.
356
Michael E. D. 1972. Growth rates in the Anolis carolinensis.— Copeia, 3.
Miller M. R., Wurster D. H. 1956. Endocrinology 58, 114 120.
Miller M. R. 1963. The histogenesis of the endocrine organs of the viviparous lizard Xantusia vigiles. Gen. and Compar. Endocrinol., 1.
Moravec F. 1963. Prispevek к poznani helmintofauna nasich plazu. Spisy priro-dovSd. fak. univ. J. E. Pyrkyne v Brne, 19, 356—359, 363—364, 370, 383.
Milhling P. 1898a. Die Helminthen-Fauna der Wirbeltiere Ostpreussens.—Arch. Naturgesch., Bd. I.
Milhling P„ 1898b. Studien aus Ostpreussens Helminthenfauna. Zool. Anz., Bd. XXI, N 549.
Mulherkop L. 1962. Studies of the absorption of water by the eggs of the garden lizard Colotes versicolor (Daud) using Trypan B1W.=—Proc. Nat. Inst, Sci. India, 28.
Nettmann FL K„ Rykena Silke, 1974. Eine gelungene Kreuzung von Lacerta t. trilineata mit Lacerta v viridis. Salamandra, 10, N 2.
Nutnukkaruppan Vr., Ranakambika P., Manickavel V. and Veerarghavan K. 1970. Analysis of the development of the lizard Calotes versicolor.—J. Morphol., 130, 4.
Osborn FL F. 1903. The Reptilian subsclasses Diasida and Synapsida.— Mem. Amer. Museum Natur. Hist., 1.
Oswald B. d939. On Yugoslavian (Balkan) ticks (Ixodoidea).—Parasitology, 31, 3.
Fanigel M. 1956. Ann. sci. natur. Zool., 48.
Parker W. K. 1879. On the structure and development of the skull in the Lacer-tilia.— Philos. Trans. Roy. Soc. London, A 169.
Parker G. FL 1906. The influence of light and heat on the movement of the me-lanophore pigment, specially in lizards.— J. of Exp. Zool., v. 3.
Parker G. FL 1940. Proc. Amer. Philos. Soc., 83.
Pasteels J. 1957. Une table analytique du developpement des Reptiles. I. Stades de gastrulation chez les Cheloniens et les Lacertiliens.— Ann. Soc. roy. zool. Belgique, 87.
Peabody F. E. 1958. A Kansas drowth recorded in growth zones of a fullsnake.— Copeia, 2.
Peabody F. E. 1961. Annual growth zones in living and fossil vertebrates.— J. Morphol., 108, 1.
Peter K. 1904. Normentafeln zur Entwickluggeschichte der Zauneidechse (Lacerta agilis)Keibel’s Normentafeln, 4.
Peter-Rousseaux A. 1953. Recherches sur la croissance et le cycle d’activite testi-culaire de Natrix natrix Helvetica (Lacepede), Terre et vie, 100, 4.
Peters G. 1958. Die Zauneidechse des Kleinen Kaukasus als besondere Unterart L. agilis brevicaudata.— Zool. Jahrb. 86, 1/2.
Peters G. 1959. Zur Taxonomie and Okologie der Zauneidechsen zwischen Peipus- und Onega-See.— Zool. Beitr. (N. F.), 4, 2.
Peters G. 1960. Die Grusinische Zauneidechse Lacerta agilis grusinica nomen novum, Zool. Anz., 165, 7/8.
Peters G. 1962. Erganzende Bemerkungen zur Grusinischen Zauneidechse (Lacerta agilis grusinica).— Zool. Anz., Bd. 169, H. 5/6.
Peters G. 1964. Sekundare geschlechtsmerkmale Wachstum und Fortpflanzung bei einiger transkaukasichen Eremias formen (Reptilia, Lacertidae).— Senckenberg, biol., 45, 3—‘5.
Peters G., Muskhelischwili T. A. 1968. Lacerta agilis iorensis neue Subspecies der Kaukasischen Zauneidechsen.—Zool. Jahrb. Syst., Bd. 95.
Pettus D. 1958. Copeia.
Prakash R. 1952. Indian J. Radiol., 6.
Prechtl FL 1949. Die bedeutung des vorderhirns fur das verhalten von eidech-sen.— Experimentia, v. 5.
Prince S. 1956. Comparative Anatomy of the eys.
Prosser C. L., Brown F. A. 1962. Comparative animal physiology, Philadelphia— London.
357
Quaranta J. V. 1952. An experimental study of color vision of giant tortoise.— Zoologica, v. 37.
Quiring D. P. 1950. Functional anatomy of the vertebrates. N. Y., Me Graw-Hill.
Radford Ch. D. 1950. The mites (Acarina) parasitic on mammals birds and rep-tilies.— Parasitology, 15, 3/4.
Ramirez J. R., Dessauer H. C. 1957. Isolation and characterization of two hemoglobins found in the turtle.— Proc. Soc. Exp. Biol, and Med., v. 96.
Redfield A. C. 1918. The physiology of the melanophores of the horned toad Phrynosoma.—J. Exp. Zool., v. 26, N 2.
Redfield A. C. 1933. The evolution of the respiratory function of the blood.— Quart. Rev. Biol., v. 8.
Redfield A. C. 1934. The haemocyanins.— Biol. Rev., v. 9.
Regal P. L. 1966. Thermofilic response following feeding in certain reptiles.— Copeia, 3.
Rene E. 1969. De legenwoordinge bedreiging von amfibien an reptielen.—Lacerta, 27, 7.
Richer IF. E. 1969. Effect of size-selective mortality and sampling biar on estimates of growth mortality, production and yield.— J. Fisch. Res. Board Canada. 26, 3.
Richet Ch. 1889. Chaleur animal. Paris.
Rizzo A. 1902. La fauna elmintologica dei rettili nella provincia di Catania.— Arch. parasMoL, 6, 1.
Rose F. 1956. Allometrische Anderungen bei der Ontogenese und Phylogenese des Eidechsengehirns.— Zool. Jahrb. Abt. Anat., 75.
Roszkowski W. 1924. Reptilia-Gady. In Podrecznik do zbierania ikonserwowania zwierrat nalezcych do fauny Polskiey, Warszawa.
Rowan W. 1938. Light and seasonal reproduction in animals.—Biol. Rev., v. 13.
Rubner M. 1883. Uber den Einfluss der Korpergrosse auf Stoff und Kraftwech-sel.— Z. Biol., 19.
Ruibal R., Phillbosian R., Adkins J. L. 1972. Reproductive cycle and growth in the lizard Anolis acutus.— Copeia, 3.
Saint-Girons M. 1957. Croissance et fecondite de Vipera aspis L.— Vie et Milieu, 8, 3.
Saint-Girons M. 1965. Les criteres d’age chez les reptiles et leurs applications a 1’etude de la structure des populations sauvages.— Terre et Vie, 112, 4
Sawyer IF. H. 1955. Oxytocic, antidiuretic and vasopressor activites in the neu-rophypophysis of the sea lamprey Petromyzon narinus.— Fed. Proc., v. 14.
Sawyer IF. H. 1960. Increased water permeability of the bullfrog bladder in vitro in respons to synthetic oxytocin and arginine vasotocin and to neurhy-pophysial extracts from non-mammalian vertebrates.— Endocrinol., v. 66.
Sfcwyer IF. H., Munsick R. A., van Dyke H. B. 1959. Pharmacological evidence for the presence of arginine vasotocin and oxytocin in neurohypophysal extracts from coldblooded vertebrates.— Nature, v. 184.
Sawyer IF. H., Munsick R. A., van Dyke H. B. 1960. Circulation, 101, 21.
Sawyer IF. H., Schisgall R. M. 1956. Increased permeability of the frog bladder to water in response to dehydration and neurophypophysial extracts.— Amer. J. of Physiol., v. 187.
Seitz T. 1907. Vergleichende Studien uber den mikroskopischen Knochenbau fos-siler und rezenter Reptilien und dessen Bedeutung fur das Washstum und Umbildung der Rnochengeweber im allgemeinen.— Nova acta Abh. Kaiserl. Leopold. Carol. Dtsch. Akad. Naturforscher, 87, 2.
Semple R. E. 1960. Exposure to cold and calecholamus, Fed. Proc., v. 19.
Serfaty A., Peyrand C. 1960. Bull. Soc. jis. natur. Toulouse, 95, 145.
Schauinsland S. P. 1906. Entwicklung der Eihaute der Reptilien and der Vogel, Heiberg, Vergl. exper. Entwick. Wirbeltiere.
Skalon IF. N. 1926. Herpetologische Untersuchungen aus der Kusnezker ,Steppe. Arch. Naturgesch., 2.
Skascilas P. 1970. Influens of temperature on ganstreek in the grass snake.— Compar. Biol, and Physiol., 33, 4.
Slobodkin L. B. 1960. Ecological relationships at the population level.— Amer. Naturalist, 94, 876.
358
Slome D., Hogben L. T. 1928. South Afric. J. Sci., 25.
Smith H. W. 1932. Water regulation and its evolution in the fishes.— Quart. Rev.
Biol., v. 7, n. 1.
Smyth D. H. 1939. Control of respiration in the frog.— J. of Physiol., v. 95.
Soule M., Kerfoot W. 1972. On the climatic determination of scale size in a lizard.— Systematic Zoology, v. 21, N 1.
Stebbins and Robinson. 1946. Further analysis of a lizard population (Scele-ropus gracious gracilis').— Univ. Calif. Pubis Zoology, 48, 3.
Stossich M. 1895. Distomi dei Rettili Lavoro monografico, Bolletino della Sosie-ta Adriatica di Scimze Naturrali in Trieste, 16.
Sturkie P. D. 1954. Avian Physiology. N. Y., Camstock Press.
Suchow G. F. 1927. Untersuchung fiber die Anzahl der Schenkelporen bei Lacerta agilis Wolf.— Zool. Anz., Bd. LXXI, H. 3/4.
Svedberg T„ Pedersen К. O. 1959. The Ultracentrifuge. Oxford, Washington, A. A. A. S. Univ. Press, Johnson Reprint Corp.
Swiezawska K. 1949. Colour — discrimination of the sand lizard Lacerta agilis L.— Bull. Acad. Polonaise. Sci, Ser. В. (II).
Tasaki I. 1959. Handbook of Physiology, v. I. Amer. Physiol. Soc. Press.
Taylor R. H. R. 1963. The distribution of amphibians and reptiles in England and Wales, Scotland and Ireland and the Channel Isles: a Revised Survey. • Brit. J. Негр., 3, N 5.
Thireau M. 1967. Analyse descriptive et biometrique de la colonne vertebrale du Serpent marin Enhydrina Schistosa Daudin (Hydzozophiines). Caracteres vertebraux des Hydrophiides.— Bull. Mus. Nat. hist, natur., 39, 6.
Torsten G., Kauri H. 1959. Zoogeography of the Swedish amphibians and reptiles with notes on their growth and ecology. Acta Vertebratica nordiska museet and scansen. Stockholm, 1, 3.
Vladescu C., Baltac M., Trandaburu T., Schmidt D. 1971. Cercetari asupra gli-coreglarii la Lacerta agilis chersonensis. Stud, si cere. biol. Zer. zool., 23, 3.
Viand D. 1960. La vision chromatique chez les animaux (saubles insectes), Iir Meeh, color discrimination, 41—57.
Wald G. 1935. Proc. Nat. Acad. Sci., USA, 41.
Wald G. 1955. Visual pigments and vitamins—A of the clawed toad.— Nature v. 175.
Wallis K. 1928. Zur Knochenhistologie und Kallusbildung beim Reptil (Chemys leprosa Schweigg).— Z. Zeblforsch. und mikrosk. Anat., 6.
Walls G. L. 1942. The vertebrate eye and its adaptive, radiation. Cranbrook Inst. Sci. Bull., 19.
Welsh J. H., Osborn С. M. 1937. J. Compar. Neurol., 66.
Wettstein O. 1953. Herpetologia aegaea.— Sitzungsber. Osterr. Akad. Wiss. Math.-naturwiss. KL, Abt. I, Bd. 162, N 9—10.
Wever E. G., Vernon J. A. 4956. Proc. Nat. Acad. ,Sci. USA, 42.
White F. N. 1955. Dissert. Abstr., 15, 11.
White F. N. 1956. Circulation in the reptilian heart.— Anat. Rec., v. 125.
White F. N. 1959. Circulation in the reptilian heart (Squamata).— Anat. Rev., v. 135.
White F. N. 1968. Functional anatomy of the heart of Reptilia.— Amer. Zoologist, v. 8.
Winkel K-, Caspers H. 1953. Untersuchungen an reptilien uber die beeinflussung der grobhirnrindenrhythmik durch zwischenhirnreizungen mit besonderer be-rucksichtigung des thalamus, Pflugers. Arch Physiol., v. 258.
Wolfson A. 1959. Photoperiodism and related phenomena.— in Plants and Animals. Lond.
Yonge С. M. 1928. Biol. Rev., 3.
Zeuthen E. C. R. 1947. Trav. Lab. Carlsberg, Ser. chim., 26.
359