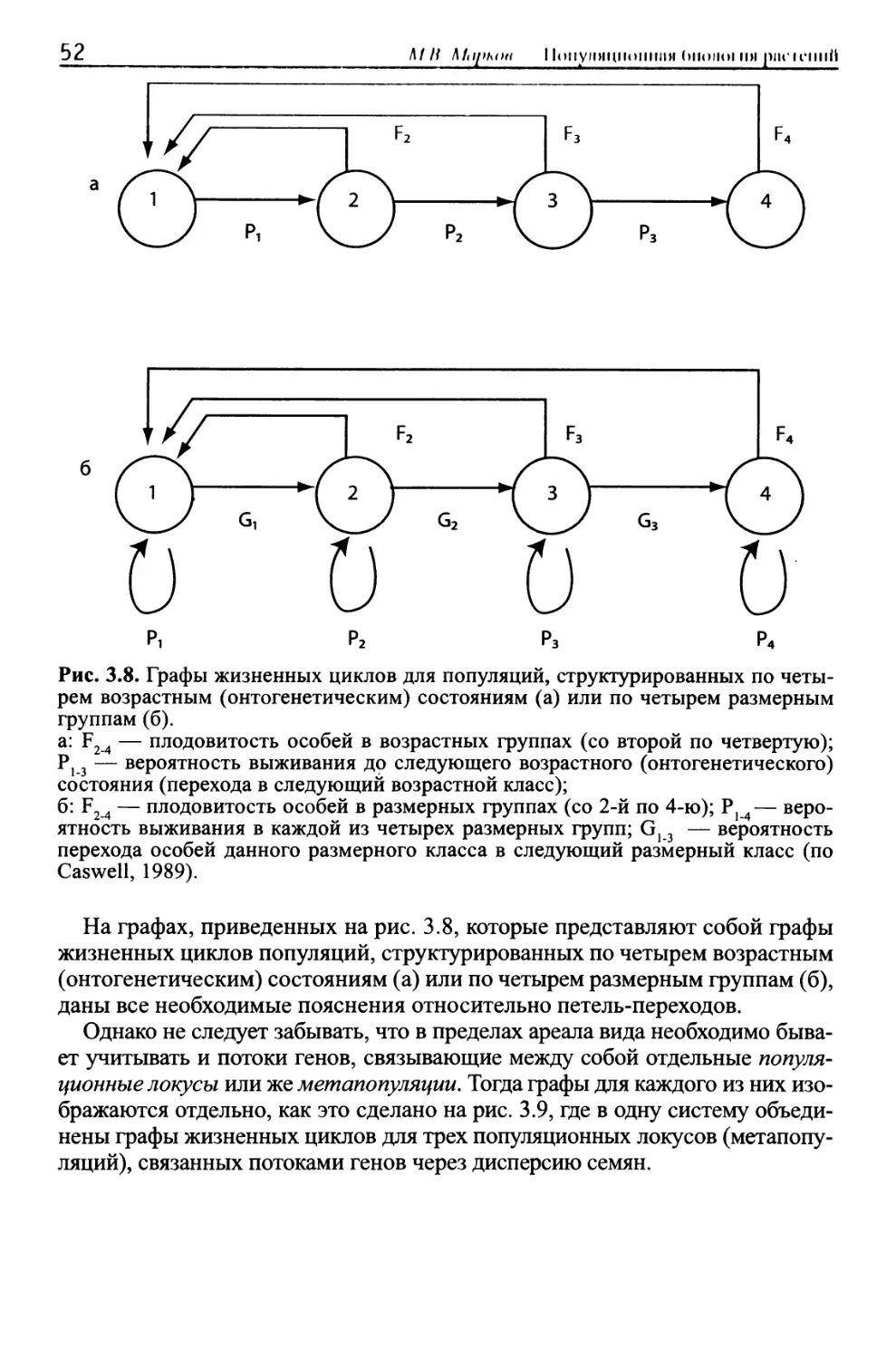
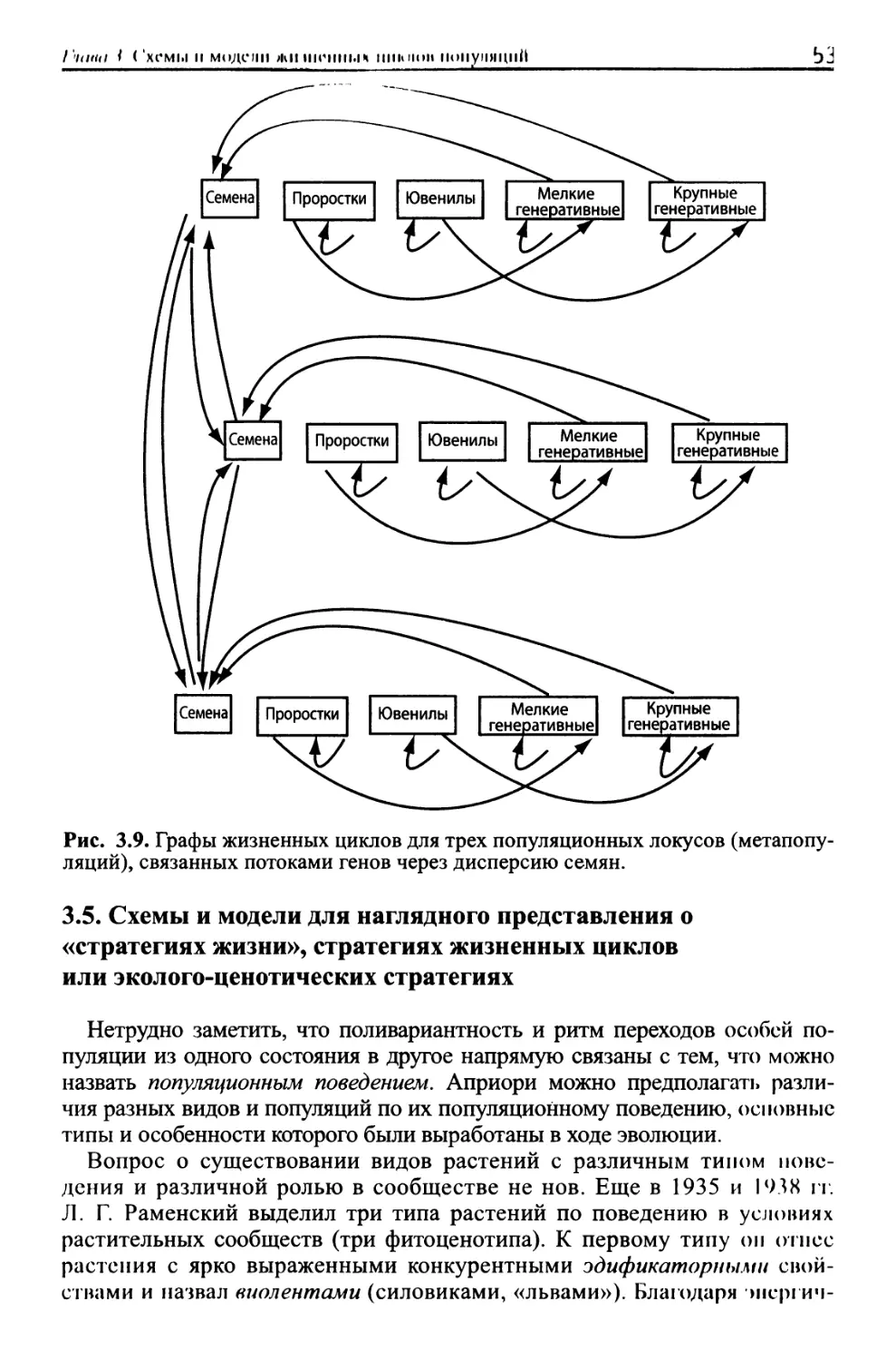
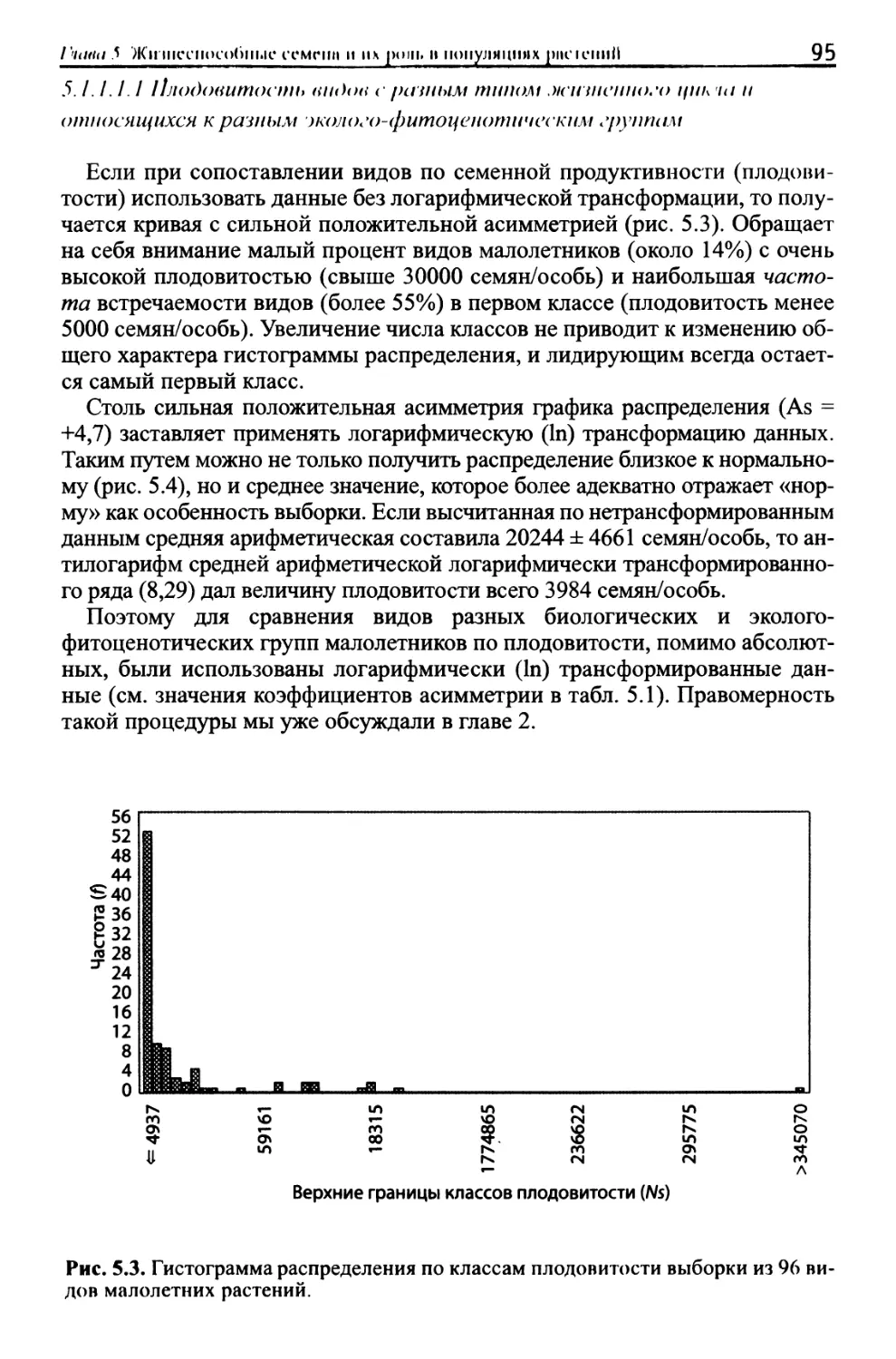
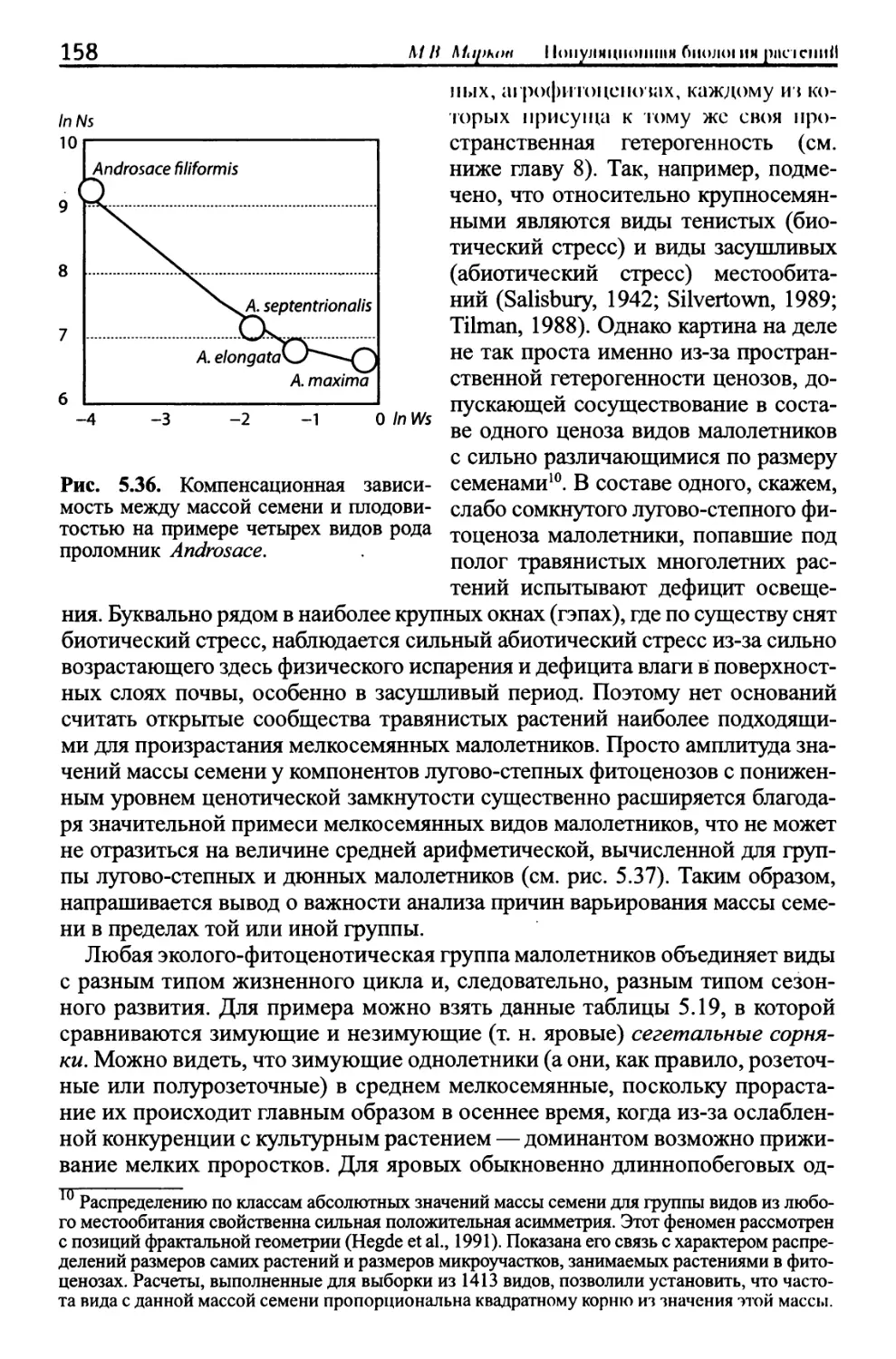
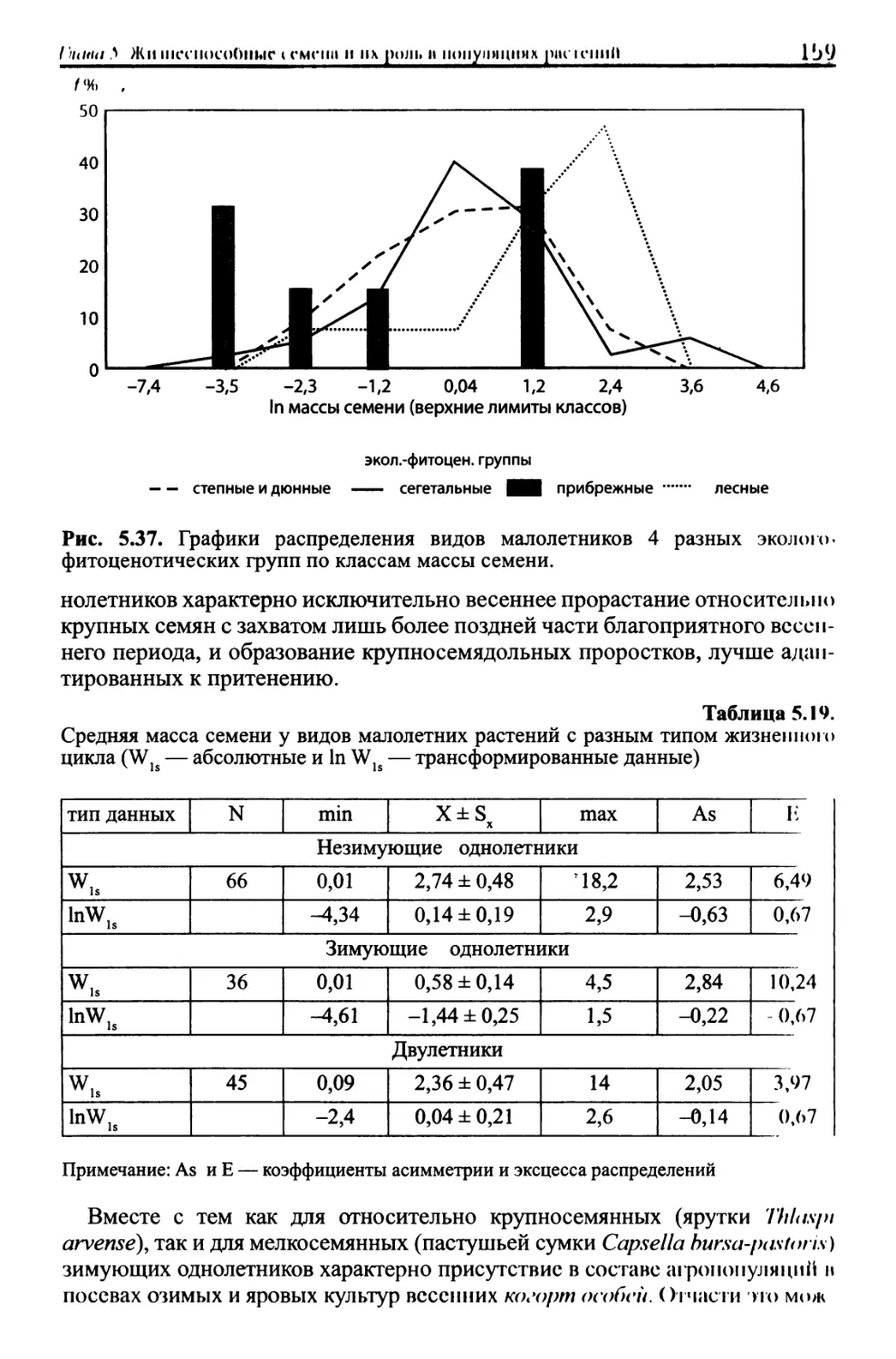
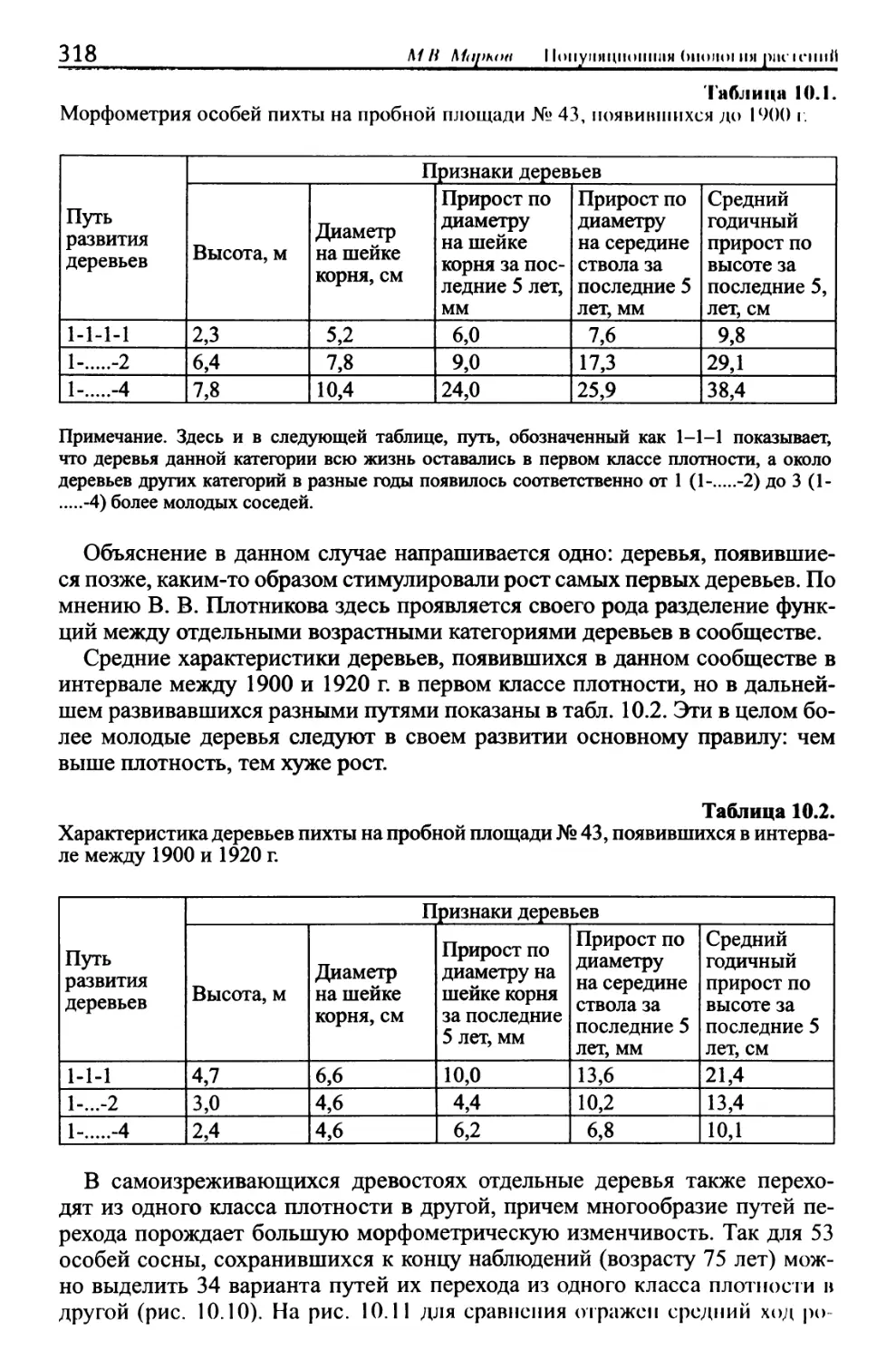
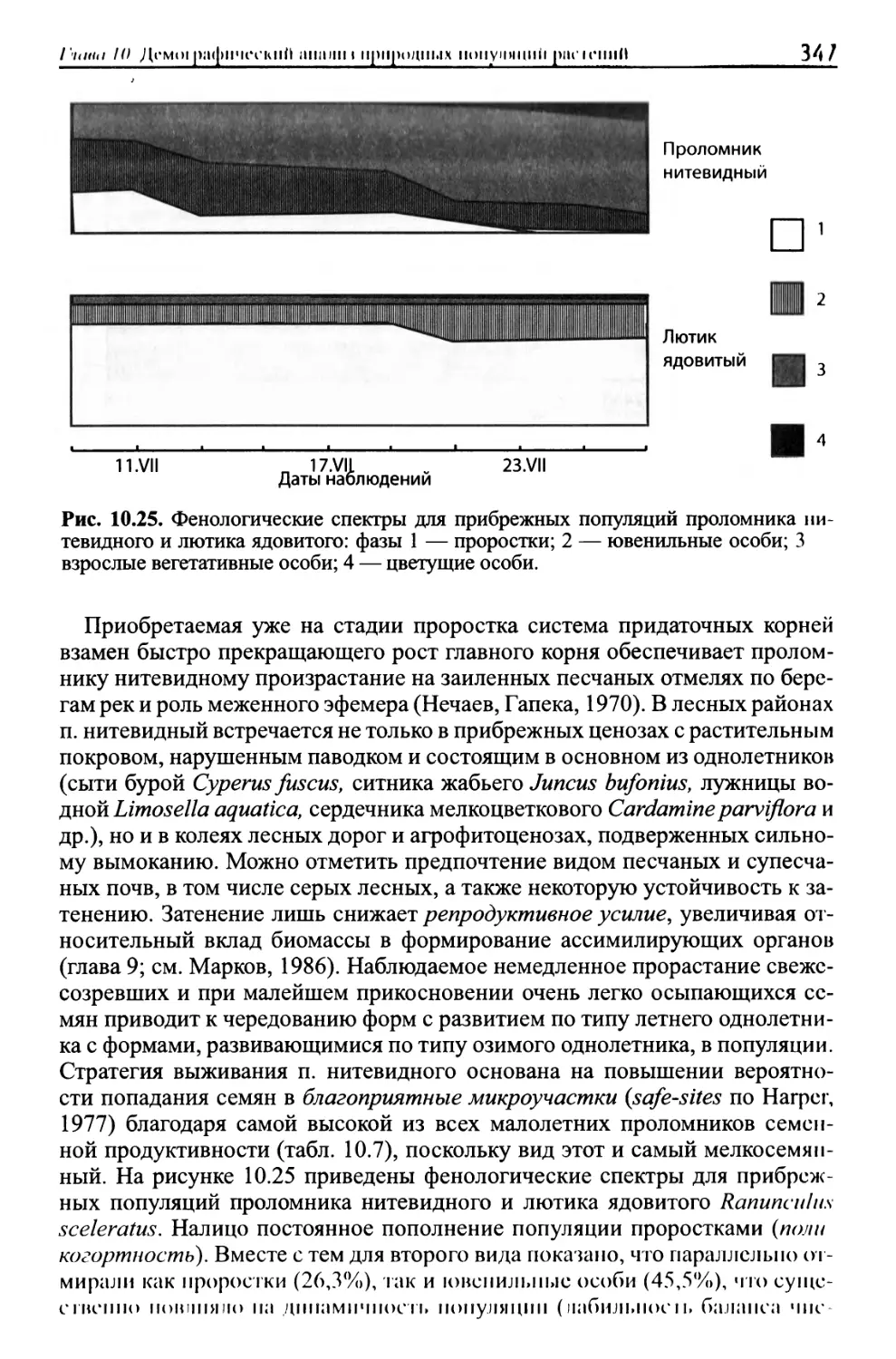
/




































































































































































































































































































































































































Author: Марков М.В.
Tags: общая экология биоценология гидробиология биогеография экология и биогеография охрана живой природы биология фауна
Year: 2012




Text
м.в. Марков Популяционная биология растений
М.В. Марков
Популяционная
биология растений
Учебное пособие
М.В. Марков
Популяционная
биология растений
Учебное пособие
Товарищество научных издании КМК
Москва **’ 2012
УДК 574.3(58)
ББК 28.080(28.5)
М26
Рекомендовано У МО по образованию в области подготовки
педагогических кадров в качестве учебного пособия для студентов
высших учебных заведений, обучающихся по специальности Биология
(направление подготовки магистров — 050100.68 Педагогическое
образование, магистерская программа «Биолого-экологическое
образование»).
Марков М.В. Популяционная биология растений. Москва: Товарищество на-
учных изданий КМК. 2012. — 387 с.; илл.
Данное учебное пособие к университетскому курсу «Популяционная биология
растений», знакомит студентов и читателей иного ранга с современным состоя-
нием популяционной биологии растений, основными понятиями, концепциями и
методами популяционной ботаники. Дана общая картина — назван и подкреплен
примерами весь набор общих положений, выработанных в этой науке к сегодняш-
нему дню. Ознакомление с основными методами получения данных в популяци-
онной биологии растений, а также методами обработки этих данных и методами
их обобщения иллюстрируется, в немалой степени, с привлечением данных, полу-
ченных самим автором. По поводу выделенных и особенно подробно рассмотрен-
ных дискуссионных положений и положений, по которым имеются противоречи-
вые результаты, автор высказывает свою точку зрения. Привлечен обширный ил-
люстративный материал, способствующий усвоению курса. Для студентов и всех
интересующихся популяционной биологией растений.
Рецензенты: доктор биологических наук, профессор В.Г. Онипченко
доктор биологических наук, профессор ВА. Черемушкина
" 1 (< 111 jc< тво । i.i уч 11 ы х и 1Д;| 111 ii i КМК. и <д;1111 tc, 201 2
ПРЕДИСЛОВИЕ АВТОРА
Эта книга — о жизни популяций цветковых растений. Даже если бы она
была названа просто «Биология растений», то избежать разговора о популя-
циях было бы невозможно. Оно и понятно — ведь виды растений в приро-
де живут популяциями - группами, множествами отдельных, но тесно взаи-
модействующих друг с другом особей. Одиночной особи какого-либо вида,
если она растет не в горшке или на грядке под присмотром заботливого са-
довника, а в природе, в условиях интенсивной конкуренции за ресурсы с
другими видами растений, несдобровать, не выжить. Точнее сказать, такое
выживание было бы слишком маловероятным. Существует даже научная
проблема установления для разных видов растений минимальных размеров
(минимальной численности) их популяций, которые гарантировали бы или
значительно повышали шансы выживания и сохранения.
Особи растений в составе популяции (множества) многолики, разноо-
бразны! Это неотъемлемое свойство популяции, которое воспринимается
первоначально (и подчас весьма эмоционально!) на качественном уровне,
и подлежит внимательному изучению путем измерений, подсчетов с при-
менением количественных методов анализа результатов. Жизнь популяции,
слагающих ее особей, как стало понятно из сказанного выше, подвласт-
на лишь не очень строгим законам вероятности. Стало быть, инструмен-
том для познания жизни популяций выступает статистика со своим арсе-
налом. Поэтому приготовьтесь к восприятию статистически детерминиро-
ванных закономерностей популяционной жизни растений. Но не пугайтесь!
Математической статистики в книге минимум — ровно столько, сколько
надо для повышения достоверности наших знаний, и ни формулой боль-
ше! Что же поделать — всякая наука только тогда и доказывает свою значи-
мость, когда дает возможность с достаточной долей вероятности прогнози-
ровать результат.
Но инструмент, как бы он ни был важен, не должен слишком много о
себе понимать! Статистические методы есть только методы, позволяющие
грамотно представить результаты, но отнюдь не самоцель. Главное в кни-
ге — сведения об особенностях жизни растений в популяциях, роста, раз-
множения и, конечно, эволюции, в свете которой нам следует рассматри-
вать все факты, чтобы наши исследования не потеряли основной биологи-
ческий смысл.
Итак, приступаем к изложению, вооружась ранее полученными знания-
ми по ботанике, а также генетике и биометрии (биологической статистике).
М Марков
Глава 1. ВВЕДЕНИЕ
«Только то в биологии имеет
смысл, что рассматривается
в свете эволюции»
Ф. Г. Добржанский
1.1. Что за наука «популяционная биология растений» и как она
соотносится с другими ботаническими науками
Поскольку название науки «популяционная биология» прямым образом
указывает на основной ее объект — популяцию, казалось бы логично дать
«во первых строках» учебника исчерпывающее определение популяции, что-
бы стало ясно, чем эта наука занимается. Увы, сделать это нелегко, а глав-
ное. .. нецелесообразно! Популяция представляет собой настолько сложное и
многоликое явление, что определить ее, равно как и биологический вид, ла-
конично и исчерпывающе, а значит приемлемо для всякого биолога, не пред-
ставляется возможным. Впрочем, надо не усматривать в этом «криминал»,
а, вооружась неким набором рабочих понятий и определений, двигаться от
главы к главе, «обрастая» фактическим материалом и, вместе с ним, правом
определить, что же представляет собой популяция для биолога-ботаника, из-
учающего семенные преимущественно цветковые растения.
Первое рабочее определение уже дано в предисловии: популяция — это
множество особей одного вида, живущих вместе, И не беда, что оно поро-
дило немало вопросов — на них мы будем планомерно отвечать или пытать-
ся находить ответы на протяжении всего дальнейшего изложения.
1,1,1 Общая экология, экология растений и популяционная биология
растений
В общей экологии в период бурного ее развития, т. е. в двадцатых-
сороковых годах XX века, когда сложился понятийный аппарат этой нау-
ки, и были оформлены доминирующие экологические концепции, сформи-
ровались и два основных методологических подхода: экосистемный и по-
пуляционный. Эти два подхода, практически исчерпывающие в своей сово-
купное ги содержание общей экологии сосуществуют именно с этого време-
ни (Гиляров, 1990).
Экосистемный подход, как следует из названия, ставит во главу угла
понятие экосистемы совокупности организмов и неживых компонен-
тов, связанных погонами вещества и энергии. Термин и понятие «экоси-
стема» 1)1>1п введен А Тщспн в 19 И । |< числу нанСюлсс удачных опрс-
llddd / |1|1ГДГ1П1е 5
делений жошитемы можно отнести определение, данное Р. Линдеманом
(Lindeman, 1942, см. также Гиляров, 1990). Он определил ее как систему
фишко-химико-биологических процессов, протекающих в пределах некото-
рой пространственно-временной единицы любого ранга.
С точки зрения ученых, которые работают в рамках экосистемного под-
хода, тяготеющего к целостному (холистическому) описанию, экология
это наука об экосистемах, а любое изучаемое экологами явление представ-
ляет интерес, прежде всего постольку, поскольку оно имеет значение для
экосистемы в целом. В становление экосистемного подхода отечествен-
ной наукой был внесен заметный вклад: достаточно назвать имя академика
В. Н. Сукачева — автора понятия «биогеоценоз», примененного им к лесно-
му сообществу или Г. Г. Винберга, развивавшего «балансовое» направление
в изучение водных экосистем1.
Почему именно лесные и водные сообщества стали объектами ученых,
проводивших исследования в рамках экосистемного подхода? Случайно
это или не случайно? Давайте проанализируем особенности этих объектов.
Лесные сообщества, и вы могли убедиться в этом на практике, — это наи-
более стабильные (т. н. климаксовые) сообщества с хорошо выработанной
структурой и устоявшимися связями, которые можно детально изучать па
стационарах. Человек, входя под полог леса, оказывается как бы внутри со-
общества, может изучать его структуру «изнутри», отчетливо ощущает на
себе влияние растений и способен четко распознать границу этого влияния.
Приборы, которые можно применить в данном случае, могут быть достаточ-
но крупными, удобными в обращении. Лесное сообщество (лесной биогеоце-
ноз) формируется длительное время, не испытывая каких-либо серьезных на-
рушений со стороны внешних факторов, и вырабатывает хорошо развитую
ярусную структуру. Деревья сильно меняют среду под своим пологом, прояв-
ляя тем самым свою так называемую эдификаторную роль и во многом опре-
деляя довольно высокую стабильность потоков вещества и энергии.
Травяные сообщества (заметьте — правильно говорить не «травяни-
стые», а именно «травяные»!), на которые человек смотрит чаще всего не-
сколько «свысока», оказывают относительно меньшее влияние на среду, и
потому границы между ними, как правило, нерезкие. Один фитоценоз плав
но переходит в другой (континуум). Эти сообщества, как правило, не име-
ют четко выраженной ярусной структуры и способны сильно меняться под
воздействием меняющейся внешней среды. Здесь фиксировать потоки ве-
щества и энергии будет намного сложнее в силу лабильности компонентов
и всего сообщества в целом.
Выгодны для изучения потоков веществ и энергии, а также моделирования,
водные биогеоценозы, поскольку вода представляет собой среду физически
гомогенную и создает комплекс условий, который может повторяться в са
мых разных частях земного шара, даже на фоне разного климата. Не случай
1 Коротко о соотношении между понятиями «экосистема» и «биогеоценоз». Экосистема
понятие безранговое, включающее в себя, образно говоря, все объекты «от кочки до обопоч
ки» от болотной кочки до биосферы. Биогеоценоз же теоретически имеет границы, про
ходящие по границам его автотрофного блока, т.е. фитоценоза. Практически, в силу коп in
iiyaabiioci и растительного покрова, границы ни чаек» весьма расплывчаты.
6 _ Маркой 11онуаяцнонния (hiojiqi ня рас юний
но именно среди водных растений гидрофитов очень miioi ис виды космо-
политы. Режимы температуры, влажности и химизма воды относительно лег-
ко задавать как параметры в моделях. Прогностическая сила таких моделей
обычно бывает по этой причине весьма внушительна.
Популяционный подход, концентрирующий основное внимание на попу-
ляциях, т.е. совокупностях особей одного вида, населяющих определенную
территорию (или акваторию), в отличие от экосистемного подхода более
склонен к аналитическому объяснению. Это отражается и в определении
экологии, которое дал с позиций популяционного подхода канадский иссле-
дователь Ч. Кребс: экология — это научное познание взаимодействий, опре-
деляющих распространение и численность организмов. Можно видеть, что
основное содержание экологии сторонники популяционного подхода видят
в познании взаимодействий между популяциями организмов и их окружа-
ющей средой. На поверку популяционный подход в экологии оказался тоже
не однородным. Внутри него не только сложились разные методические
подходы к изучению популяций, но и были выработаны различные вариан-
ты определения того, что представляет собой популяция с акцентом, выда-
ющим специализацию исследователя.
1.1,2. Популяционная биология растений как наука о типах или
«стратегиях» жизненных циклов
По мнению автора вузовского учебника по общей экологии И. А. Шилова
(1997) сложность представления о популяции определяется ее местом в ие-
рархии биологических систем, а точнее — двойственностью ее положения в
рядах иерархически соподчиненных систем, отражающих различные уров-
ни организации живой материи.
С одной стороны, популяция включается в генетико-эволюционный ряд,
который отражает филогенетические связи таксонов разного уровня как ре-
зультат эволюции:
организм - популяция - вид - род -... - царство.2
Главными особенностями популяции как формы существования вида в
соответствии с этим выступают: во-первых, общность и специфичность ге-
нофонда, определяющие известную относительную адаптированность всех
особей и популяции в целом к конкретным условиям среды; во-вторых, ге-
нетическая гетерогенность особей (или иных структурных или счетных
единиц в популяции — М.М.) как источник материала для микроэволюци-
онного процесса в условиях меняющейся среды или возникшей по тем или
иным причинам изоляции.
Генетико-эволюционный подход, как будет показано ниже, обладает при-
оритетом в части изучения популяций, а продуктом его исторического раз-
вития стало широкое определение популяции как «совокупности особей
2 Уже в мой веренице иерархически соподчиненных таксонов может проявляться специфи-
ка подходов бцолоюв разной специализации. Например, таксоны, которые ботаники назы-
ваю! порядками н орденами, зоологи по традиции именуют отрядами и тинами. Это неудоб-
ное рп П1НЧ11Г нмес। не юрпчес кис корни (Рсйвн и др , 1990).
Г’1(1в(1 I Введение
7
определенного вида, в течение достаточно длительного времени (большого
числа поколений) населяющих определенное пространство, внутри которо
го практически осуществляется та или иная степень панмиксии (свобод
ного равновероятного скрещивания между особями — М.М.), и нет замет
пых изоляционных барьеров, которая отделена от соседних таких же со-
вокупностей данного вида той или иной степенью давления тех или иных
форм изоляции» (Тимофеев-Ресовский, Яблоков, Глотов, 1973).
С другой стороны, по совершенно справедливому замечанию И. А. Шилова,
популяция как репрезентативная совокупность особей вида вступает в тро-
фические и иные отношения с популяциями других видов. Именно поэтому,
включаясь в состав конкретного фитоценоза и, вместе с ним, в состав конкрсг-
ного биогеоценоза, она занимает соответствующее место и в функционально-
энергетическом ряду:
организм - популяция - фитоценоз - биоценоз -
биогеоценоз - биосфера.
Здесь популяция выступает в качестве функциональной субсистемы кон-
кретного фитоценоза и биогеоценоза; ее функция — участие в трофических
цепях и сетях — определяется видоспецифическим типом обмена.
Как отражение специфики растительных объектов с характерной для них
оседлостью (седиментарностью) и автотрофностью (способностью к форми-
рованию автономного автотрофного блока биогеоценоза) можно рассматривать
возникновение фитоценологического подхода к определению и изучению попу-
ляций в нашей стране (Работнов, 1945,1950 а, б). По Т. А. Работнову любой вид,
если он встречается в нескольких сообществах, бывает представлен в них осо-
быми популяциями.
Вместе с определением популяции как совокупности особей одного вида
в границах фитоценоза в науку вошел и ряд соответствующих новых тер-
минов. В 1964 г. А. А. Корчагин предложил использовать для обозначе-
ния популяции в узком конкретном смысле термин «ценозная популяция»
(ценотическая популяция) или «ценопопуляция». употреблявшийся ранее
В. В. Петровским (1961). В последние годы жизни Т. А. Работнов отдавал
предпочтение термину «фитоценотическая популяция».
Во всех случаях, когда речь идет о популяции, приуроченной к конкрс г-
ному фитоценозу, следует говорить о ценопопуляции или фитоценотической
популяции. Однако из-за объективной невозможности подчас доказать эту
приуроченность, невозможности провести границы между фитоценозамп и
просто для удобства изложения мы будем пользоваться в настоящем пособии
только термином «популяция», учитывая, что главное свойство любой нону
ляции — наличие разнообразных взаимоотношений между слагающими се
особями. При таком подходе искусственно созданные одновидовые груипи
ровки растений можно назвать модельными популяциями, где действую! ре
гулялорные механизмы, придающие группировке эмерджентные свойс i вл.
Знаменателен и тот факт, что отечественный классик популяционной (ю
таники Г. А. Работнов (см. портрет) первым обратил внимание на важное и,
8
МН Марков 11 < > । iy 11 >i I ц I < > 1111; । и (hioiioi нм рас leiniii
Т.А. Работнов.
исследования популяции рас гений
для познания «стратегии жизни»\
свойственных тому или иному виду.
Используя оригинальный подход
Л. Г. Раменского (1935, 1938), выделив-
шего три фитоценотипа (виоленты, па-
тиенты и эксплеренты) — три типа
растений по их поведению в усло-
виях растительных сообществ, Т.
А. Работнов (1975) предложил рас-
сматривать эти типы как страте-
гии жизни. Здесь же он дал и опре-
деление стратегии жизни как со-
вокупности приспособлений к про-
израстанию данного вида в опре-
деленных условиях среды в сообще-
стве с другими видами растений.
Т. А. Работновым была реализована воз-
можность дальнейшего классифициро-
вания стратегий и подразделения пати-
ентов и эксплерентов на более мелкие
подтипы стратегий с соответствующи-
ми названиями (см. ниже — главу 3).
Именно он акцентировал внимание
ботаников на очень важном тезисе
Л. Г. Раменского о возможности переходов между фитоценотипами и, вме-
сте с ней, о реальности оценки виолентности, патиентности и экспле-
рентности как свойств, поддающихся количественной оценке.
Дальнейшее свое развитие этот тезис получил в работах Д. Грайма
(Grime, 1979), независимо переоткрывшего триаду макростратегий (прав-
да, под другими названиями, что вполне естественно) и обосновавшего не-
обходимость ординации стратегий в специально разработанных для этого
и градуированных соответствующим образом осях (см. подробнее главу 3).
Д. Грайм пытался использовать для характеристики стратегий 19 показате-
лей (свойств), в число которых вошли наиболее важные биоморфологиче-
ские особенности. Путем шкалирования количественных параметров этих
особенностей Д. Грайму удалось создать базу данных по травянистым лу-
говым растениям с указанием для каждого вида варианта стратегии, обо-
значенного комбинацией начальных букв в названиях макростратегий по
Д. Грайму (С — Competitors, S — Stress-tolerators и R—Ruder als). Принцип:
один вид — одна стратегия был некритически воспринят немецкими ис-
следователями, создавшими комбинированные экологические шкалы для
’ Термины «стратегия» и «тактика», заимствованные из области военного искусства, вот уже
более 30 лег успению используются в популяционной биологии растений. Причины успеха,
ио-видимому, кроются в гом, чю названные термины отражают диалектическую взаимосвязь
между возможностью и дснсгви1ены1ос1ыо, а в более конкретном биологическом понимании —
мсж/iy i гигиена коп iipoi paMMoii pa ibiiiiim opiainriMa (iviioiином) п вариантами ее реализации в
<(мив(*н ।вук»|цнч уз повняч окружавнцеп среды (фентинами)
Гпаиа /. Введение
9
Рис. 1.1. Схема нарастания количества информации наподобие «снежного кома»
по мере движения от звена к звену по алгоритму: Архитектурная Модель (AM) —*
Жизненная форма (ЖФ) —> Стратегия Жизненного Цикла (СЖЦ) или Эколого-
Ценотическая Стратегия (ЭЦС).
2002 видов европейской флоры (Frank, Klotz, 1988). В итоге для каждого
вида в этих шкалах, помимо отношения к главным экологическим факто-
рам, указан и тип стратегии все в том же буквенном варианте их обозначе-
ния. Подобная информация вполне может создать иллюзию завершенности
исследования. На самом деле, что же еще изучать, если мы уже знаем тип
стратегии жизненного цикла для столь большого числа видов?!
Наиболее последовательным критиком присвоения виду в целом какого-
либо «стратегического» ярлыка выступил зоолог Ю. Э. Романовский (1989).
Он первым среди зоологов прочувствовал преимущества системы выделя-
емых ботаниками стратегий Раменского-Грайма по сравнению с применяе-
мыми в зоологии r-К стратегиями Р Мак-Артура и Е. Вильсона (McArthur,
10 мн м,1/ >К(>« I lolly 11ЯЦ11О111ШЯ bllOHOI пи рпоснпй
Wilson, 1967; Pianka, 1970), и вполне резонно обосновал необходимость ана-
лиза стратегий на уровне отдельных популяций. Он же предложил говорить,
как это принято за рубежом, не о стратегиях жизни, а о стратегиях жизненно-
го цикла (life-history strategy) популяций и тем самым сделать более строгим
и претендующим на большую точность само понятие стратегии.
Мы не только будем в дальнейшем придерживаться этого подхода, но и
возьмем на себя смелость утверждать, что популяционная ботаника — это на-
ука, главной целью которой выступает научное познание стратегий жизнен-
ных циклов растений. Можно также предложить своеобразный алгоритм на-
копления информации о том или ином виде (см. Марков, 1987). Начиная с
изучения архитектуры и архитектурной модели, свойственной виду, целе-
сообразно переходить к анализу приспособительного значения морфологи-
ческой конструкции, а значит — особенностей жизненной формы (см. гла-
ву 7), чтобы на последнем этапе, обладая данными об особенностях динами-
ки популяций судить о стратегии жизненного цикла (рис. 1.1). Именно та-
кой подход обеспечит нам рассмотрение всех используемых в тексте учеб-
ника фактических материалов в эволюционном аспекте и следование тезису
Ф. Г. Добржанского, взятому в качестве эпиграфа для данной главы и книги
в целом.
1.2. Краткая история формирования популяционной биологии
растений («популяционной ботаники») как науки
и университетского спецкурса
Итак, на основании сказанного выше можно заключить, что в России по-
пуляционная биология растений имеет «фитоценологическое» происхо-
ждение. Такого рода заключения можно встретить и в специальной лите-
ратуре (Миркин, 1985). Но не будем забывать уже упомянутую двойствен-
ность положения популяции в иерархических рядах биологических систем.
А потому необходимо признать, что в развитие отечественной популяци-
онной биологии растений внесли неоценимый вклад исследователи, про-
водившие анализ многих сторон жизни растений в популяциях и, в пер-
вую очередь, внутривидовой изменчивости с применением статистики,
даже не употребляя термина «популяция». Так, например, с популяциями
в ключе популяционной биологии растений работали все русские ученые,
принимавших участие в разработке концепции вида у растений, такие как
С. И. Коржинский, Н. В. Цингер, Н. И. Вавилов и др. (см. Завадский, 1968).
Следует также принять во внимание тот факт, что генетический аспект
изучения популяций заинтересовал русских ботаников-прикладников (се-
лекционеров) с момента выхода работы В. Иоганнсена о популяциях и чи-
стых линиях, то есть с 1903 г. (см. Аверьянова, 1975 и главу 2). В эколо-
гическом отношении (в плане изучения экотипической дифференциации
популяций) русские ботаники (ученицы Н. И. Вавилова М. А. Розанова и
Е. Н. Синская) приступили к углубленному изучению популяций дикора-
стущих растений после публикации работ Г. Турессона (Turesson, 1922а,b,
1925), в которых было развито учение об экотипах. В выполненных ими
монографических работах но генэкопогни (Розанова, 1935; Синская, 1948)
/ / Введение
11
изложены результаты изучения люрфобиологическои изменчивоеi и, нону
ченныс на популяционно-видовом уровне. Были охарак'гернзованы некою
рые нуги приспособления сорных растений к произрастанию в агроценозах
(уподобление и дифференциация) в условиях действия искусственного бес-
сознательного отбора человеком, изучены примеры мимикрии у рас гений
(Н. И. Вавилов, Б. М. Козо-Полянский), обоснованы попытки выявления
границ природных популяций по условиям местообитания.
Изложение основ биологической статистики, столь необходимой для
корректного популяционного анализа, впервые специально для ботаников-
селекционеров осуществленное В. Иоганнсеном (в русском переводе боль-
шая часть книги вышла в 1933 г.) тоже очень часто иллюстрировалось бота-
ническими примерами (см. Филипченко, 1915,1929, а также главу 2). Это не
могло не спровоцировать видных отечественных ботаников на проведение
работ по изучению статистическими методами внутривидовой изменчиво-
сти (см., например, Гордягин, 1907).
Даже такой печальный для отечественной биологии факт как «лысенковщи-
на» в известном смысле стимулировал проведение исследований в ключе попу-
ляционной экологии растений. Так дискуссия с лысенковцами по поводу значе-
ния дарвиновской внутривидовой борьбы за существование побудила к обсуж-
дению и переосмыслению результатов многих агрономических и лесоводствен-
ных опытов, поставленных для выявления оптимальных норм высева и густот
насаждений древесных пород. Кроме этих достаточно старых лесоводственных
опытов было также поставлено немало новых опытов с выращиванием осо-
бей какого-либо вида в условиях разной (и в том числе сильно повышенной)
густоты насаждения (= плотности популяции). Хороший обзор таких опытов
можно найти в книге «Борьба за существование как фактор эволюции» (Галл,
1975). Эта работа уже была по существу фитоценологической, и возглавлял ее
В. Н. Сукачев — в то время признанный лидер русской фитоценологии, автор
первого учебника «Основы фитосоциологии» (^фитоценологии — Л£Л£). В
тексте этого учебника приведены данные всемирно известного опыта с ромаш-
кой Matricaria inodora (^Matricaria perforata), которую выращивали не только
в условиях специальных ее посевов (= модельных популяций) разной густоты,
но и на фоне удобренного и бедного субстратов (см. главу 7). Мировую извест-
ность результатам этого опыта обеспечил авторитетнейший английский бота-
ник Джон Л. Харпер, присвоив выявленной на основе этого опыта закономерно-
сти название «эффект Сукачева».
Ранее в России такое же, в сущности, название («правило Сукачева») по-
лучила иная закономерность (Завадский, 1961), на которую обратил внима-
ние ботаников В. Н. Сукачев. Помимо дифференциации особей по мощно-
сти в насаждениях разной плотности анализировали темпы развития расте-
ний (смену фенологических фаз). В. Н. Сукачев нс только подметил, но и те-
оретически обосновал факты ускоренного развития (перехода к репродук-
ции) однолетних и замедленного развития многолетних растений в переза-
гущенных популяциях. В дальнейшем из-за накопившихся новых данных но
У!ому вопросу неоднократно подвергалась критике излишняя строгость фор-
мулировки этого «русского правила Сукачева» (см. Агаев, 1972 и др.), хотя
В. II. ( укачев и сам отмечал и пытался объяснить неподчинение или неполное
12 МН Маркин 11()!!у1!яц11(>нн;1я (Mfoiioi пи paciciinii
подчинение некоторых видов при некоторых гусгогах «правилу Сукачева»
(Завадский, 1968). Несмотря на немалое число примеров отклонения в по-
ведении растений от этого правила, ускорение развития однолетних расте-
ний при ухудшении условий произрастания вообще и при возрастании на-
пряженности конкуренции за ресурсы из-за повышенной плотности популя-
ции, в частности, безусловно, можно рассматривать как адаптивную, полез-
ную для сохранения популяции реакцию. Даже если момент наступления той
или иной фенологической фазы в условиях загущения задерживается во вре-
мени, то часто сам габитус (= облик) особи однолетника свидетельствует о
неотении (выпадении фаз формирования фитомеров взрослого типа из он-
тогенеза). Поэтому в данном случае, как и в случае с анализом возрастной
структуры популяций многолетних растений, нужно в большей степени по-
лагаться на биологический, нежели хронологический календарный возраст.
И все же бурное формирование популяционной экологии растений как обо-
собляющейся ветви фитоценологии (в России) или экологии растений (за рубе-
жом) по-настоящему началось с появления классических трудов Т. А. Работнова
(1950а, б). Это признано мировой наукой (White, 1985), и признание по праву
включено в учебники зарубежными ботаниками (см. Urbanska, 1992).
Профессор МГУ Т. А. Работнов привлек внимание отечественных бо-
таников к необходимости учета двух аспектов в изучении популяций рас-
тений — численности и состава. Эти аспекты тесно связаны друг с дру-
гом, хотя в зависимости от целей, преследуемых исследователем, то один,
то другой аспект выдвигается на первый план. Один из путей связи — от-
носительная численность входящих в состав популяции внутрипопуляци-
онных элементов. Численность здесь служит для количественной харак-
теристики популяционного состава или спектра. Однако, изучение соста-
ва или спектра популяций отнюдь не предполагает только аналитическое
расчленение популяции на составные элементы с целью их последующе-
го изучения. Сюда входит также и синтетическое определение взаимоза-
висимого функционирования этих элементов в популяции как в известной
мере целостной системе. Примером такого рода исследований и послужи-
ли труды Т. А. Работнова, связанные с установлением возрастных спек-
тров (или спектров онтогенетических состояний) популяций многолет-
них растений и труды его многочисленных последователей. Здесь нагляд-
но выступают обе фазы популяционного исследования. На первом этапе,
изучая онтогенез растения, исследователи аналитически расчленяют его
на возрастные этапы или, онтогенетические состояния, имеющие соответ-
ствующие индикаторные морфологические признаки. Содержание второ-
го этапа — определение роли каждой возрастной группы (группы особей,
находящихся в каждом из онтогенетических состояний) в жизни популя-
ции и численного соотношения групп (спектра). В целом, руководствуясь
данными, накопленными на этих этапах, исследователь составляет сужде-
ние нс только о прошлом и настоящем положении вида в фитоценозе, но
может и прогнозировать будущие его изменения.
(' момента вывода основных трудов Т. А. Работнова развитие отечествен-
ной нонуляциошк^й биологии растений получило мощный толчок и потому
I 'hltid I ВПСЦСПНС
П
А. А. Уранов.
Дж. Л. Харпер.
значительно ускорилось, хотя и сохранило некоторую инерцию в плане под-
бора объектов и методов популяционного исследования.
Известную самобытность отечественная популяционная ботаника по-
лучила благодаря привлечению к анализу особей в составе популяций раз-
ных видов данных по морфогенезу. Использование Т. А. Работновым мор-
фологических критериев для индикации возрастных ^онтогенетиче-
ских) состояний привлекло внимание известного ботаника-биоморфолога
И. Г. Серебрякова, разработавшего известную классификацию жизненных
форм (см. ниже). Большой вклад в развитие популяционной биологии рас-
тений внес А. А. Уранов, выступивший наряду с И. Г. Серебряковым и зоо-
логами Ф. И. Правдиным и С. П. Наумовым в качестве организатора един-
ственной в бывшем СССР научной лаборатории по изучению популяций и
возглавлявший ботанический отдел в этой лаборатории. А. А. Уранову вме-
сте с его учениками (О. В. Смирновой, Л. Б. Заугольновой, Л. А. Жуковой,
Ю. А. Злобиным и др.) удалось существенно уточнить терминологию, про-
двинуться в оценках жизненного состояния особей в популяции, выявле-
нии пространственной неоднородности популяции и асинхронности разви
тия ее локусов. Были разработаны и представления о динамике возрастных
спектров популяций и подвергнута дальнейшей разработке классификация
типов популяций по характеру их возрастных спектров. Не случайно нот
тому представители школы И. Г. Серебрякова и А. А. Уранова продолжи
ли исследование популяций с привлечением в качестве объектов видов еа
мых разных жизненных форм, но (по примеру Т. А. Рабогнова), как нра
вино, многолетних рас гений. Такую инерцию в выборе объектов можно в
1 4 Л/// AAyj/uw I lonyiiMiuioiiiniM (nioiioi ня pacicmib
какой io c i ciici111 01ipaiyja i ь ведь анализ спектра возрастных (онтогенети-
ческих) сое loMiiiiii даст по-пастоящсму хорошие и полезные для фитоцено-
логии результаты только при работе с популяциями многолетних растений.
Да и выделение самих онтогенетических состояний, достаточно долго (не
менее одного сезона вегетации) сохраняющих свои индикаторные призна-
ки, целесообразно только для многолетних (моно- и поликарпических) рас-
тений. Быстрое же развитие однолетних растений, приводящее к одновре-
менному присутствию всех индикаторных признаков, которые диагности-
руют скоротечные возрастные изменения у одной особи, не дает возможно-
сти получать и в этом случае полезную фитоценологическую информацию.
Попытку некоторых отечественных ботаников применять традиционную схе-
му исследования возрастных (онтогенетических) состояний и их спектра к по-
пуляциям малолетних растений (и даже незимующих однолетников!) приходит-
ся квалифицировать не иначе как методическую инерционность или инерцион-
ность мышления по причине слабой информативности получаемых данных.
За рубежом в конце 1950-х гг. профессором университета в Бангоре
(Северный Уэльс, Великобритания) Джоном Л. Харпером было развито де-
мографическое направление в изучении популяций растений. Из большо-
го числа работ Дж. Л. Харпера до недавнего времени лишь одна была напе-
чатана в русском переводе (Харпер, 1964), и поэтому принципы развивае-
мого им направления в популяционной биологии растений очень медленно
приобретали известность в нашей стране (см. Марков, 1976). Ситуация не-
сколько улучшилась после установления в начале 1970-х гг. личной перепи-
ски Дж. Харпера с Т. А. Работновым и совместного участия ряда бывших
стажеров Харпера (J. White, Р. Werner, Р. Cavers, J. Sarukhan) с представите-
лями школ Т. А. Работнова - А. А. Уранова в работе XII Международного
Ботанического Конгресса в Ленинграде (1975 г.).
Через два года была опубликована монография Дж. Харпера ’’Population
Biology of Plants”, вобравшая в себя огромный материал, который был нако-
плен ее автором в совместной работе с коллективом стажеров.
1.3. Задачи, стоящие перед настоящим учебным пособием
В данном учебном пособии будут затронуты различные стороны популяци-
онной экологии и биологии семенных, преимущественно цветковых растений.
Акцент сделан именно на экологии и биологии, а не генетике популяций. Вместе
с тем некоторые (правда, только самые основные) генетические аспекты, и это
вполне закономерно, тоже подлежат обязательному рассмотрению. Если строго
следовать содержанию дисциплин и принять, что популяционная биология рас-
тений в равной степени должна затрагивать популяционную экологию и попу-
ляционную генетику растений, то приходится признать — соотношение, приня-
тое в настоящем учебном пособии, наверное, более всего соответствует дисци-
плине с осторожным названием «популяционная ботаника». Оставив право ре-
шения этого вопроса за читателем, проанализируем ситуацию, которая сложи-
лась к настоящему времени с преподаванием нашего курса.
Опыт многолетнего преподавания подсказывает, что изложение материа-
ла лучше всего проводить по какой-то схеме, позволяющей, хотя и упрощен-
Г'К1П(1 / Введение
15
но, редуцированно, выс i poiri ь звенья
логической цени. Эго может быть, к
примеру, редукционный ряд жизнен-
ных форм растений, иллюстрирую-
щий наши современные представле-
ния об эволюции цветковых растений
безотносительно того, в каком имен-
но направлении мыслятся эволюци-
онные преобразования разными ис-
следователями. Это может быть и
обобщенная схема жизненного цик-
ла популяции, намечающая основные
моменты жизни и точки приложения
регуляторных сил.
Схематизация жизненного цикла
с выделением основных его фаз, ко-
торые удобно рассматривать как от-
дельные главы при изложении ма-
териала, — подход не оригиналь-
ный. Он применен в монографии Дж. Д. Сильвертаун.
Харпера, которая из-за сильной пере-
груженности фактическим материалом так и не стала использоваться в ка-
честве учебного пособия.
Однако материал, представленный в этой монографии, наши зарубежные
коллеги уже неоднократно использовали для учебных целей. В 1981 г. вышло
первое издание учебного пособия Мишеля Бигона и Мартина Мортимера
«Популяционная экология» с подзаголовком «Унифицированное изучение
животных и растений». Эта книга, в которой представленные ботанические
примеры были заимствованы из монографии Дж. Харпера, выдержала к на-
стоящему времени уже три издания. Последнее третье издание было выпу-
щено под тройным авторством — к Бигону и Мортимеру присоединился
Дэвид Томпсон (Begon, Mortimer, Thompson, 1996).
По мнению зарубежных коллег Джонатан Сильвертаун сделал на основе
собственных исследований и извлечения из монографии Дж. Харпера весь-
ма удачное учебное пособие «Введение в популяционную экологию расте-
ний» (Silvertown, 1982, 1987). Оно выдержало к настоящему времени уже
четыре издания. Третье, переработанное и дополненное преимущественно
генетическим материалом, опубликовано в соавторстве с J. Lovett-Doust в
1993 г. под названием «Введение в популяционную биологию растений».
В процессе подготовки четвертого издания у книги появился еще один со-
автор — Дебора Чарлзворт (D. Charles worth). Готовя первое издание свое-
го учебника, Д. Сильвертаун, видимо из скромности или из-за боязни стать
уязвимым для критики за неполноту использования в тексте накопленного
материала, назвал свое пособие лишь введением в популяционную эколо-
гию растений. Однако, судя по очень хорошим отзывам (частью они приве-
дены па задней странице обложки второго издания пособия), книга удалась
именно как учебник. В последующих изданиях слово «введение» из титула
1 б Л/// Марков 11(>!!у1!>|ц||(>11пая ня рас । сннй
нс исчезло, и это свндсгсльс!вуст об осознании автором их конспективно-
го характера рядом с огромной но объему (892 стр.) и глубокой по содержа-
нию хрестоматийной монографией Дж. Харпера.
Конспективный, но хорошо иллюстрированный учебник по популяцион-
ной биологии растений был не так давно издан на немецком языке извест-
ной швейцарской исследовательницей польского происхождения Кристиной
Урбанской (Urbanska, 1992). Книга имеет карманный формат, но в отличие
от учебников, изданных на английском языке, иллюстрирована не только
графиками, но даже фотографиями!
Однако в нашей отечественной литературе до сих пор нет даже такого
конспективного учебного пособия по популяционной экологии растений4,
хотя специфика высших и, особенно, цветковых растений безусловно тре-
бует отдельного рассмотрения экологии и биологии их популяций в допол-
нение к общебиологическим представлениям, изложенным, к примеру, в
книге А. В. Яблокова «Популяционная биология» (1987) или книге И. А.
Шилова (1996) «Экология» и проиллюстрированным данными о популя-
циях животных. Во второй книге популяциям растений уделено несколь-
ко больше внимания, чем в первой (чуть более трех страниц!), но все рав-
но этого явно недостаточно, если учесть наблюдающееся в настоящее вре-
мя стремительное развитие популяционной биологии растений за рубежом.
Единственной современной книгой, которую можно было бы смело ре-
комендовать студентам-ботаникам, можно считать книгу «Экология: особи,
популяции, сообщества» (Бигон, Харпер, Таунсенд, 1989), переведенную на
русский язык и выпущенную издательством «Мир». Правда материал по по-
пуляциям растений там несколько распылен и перемежается примерами из
популяционой биологии животных. Но, в некотором отношении, это можно
рассматривать и как успех книги, демонстрирующей преимущества обще-
биологического популяционного подхода.
Не спасает в данном случае и издание учебного пособия «Популяционная
экология» А. М. Гилярова (1990), под редакцией которого было осуществлено
издание перевода книги М. Бигона, Дж. Харпера и К. Таунсенда. На протяже-
нии трех первых глав автору пособия удавалось обсуждать, и притом весьма
успешно, общие вопросы экологии популяций организмов независимо от их
принадлежности к животному или растительному миру. Однако, в последую-
щих главах сказалась его зоологическая специализация — ботанических при-
меров в книге, по крайней мере для ботаников, явно недостаточно.
Задачу же издания учебника по популяционной экологии и биологии рас-
тений на русском языке для студентов университетов приходится считать,
по сей день нерешенной. Это порождает значительные трудности при вве-
дении этого предмета в число спецкурсов, читаемых студентам, которые
специализируются на ботанических университетских кафедрах.
4 Учебное пособие «Популяционная биология растений», выпущенное автором (Марков,
1986), было, по-видимому, первой попыткой восполнить пробел, но чрезвычайная конспек-
тивность книги, равно как и малый тираж издания, по вполне понятным причинам не позво-
лили существенно изменить ситуацию с преподаванием этой дисциплины в нашей стране.
На популярное ги учебного пособия Ю/А. Злобина (1989), по-видимому, негативно сказался
с шпиком ciuii.in.ni крен в Ьгоропу математических методов и дефицит биологического мате-
рнипа, подаваемо!о inioiaa спишком схема!ично.
I hlUd l Введение
1/
Ниже перечислены задачи, стоявшие перед автором при подготовке на-
стоящего учебника:
1. Познакомить студентов и читателей иного ранга с современным состо-
янием популяционной биологии растений;
2. Дать общую картину — назвать и подкрепить примерами весь набор
общих положений, выработанных в этой науке к сегодняшнему дню;
3. Ознакомить с основными методами получения данных в популяцион-
ной биологии растений, а также методами обработки этих данных и мето-
дами их обобщения;
4. Выделить и особенно подробно рассмотреть дискуссионные положе-
ния или положения, по которым имеются противоречивые результаты, и вы-
сказать свою точку зрения по этим вопросам;
5. Привлечь по возможности обширный иллюстративный материал, спо-
собствующий усвоению курса.
Выполнение этих задач во многом облегчалось благодаря изданию кол-
лективных обобщающих трудов представителями Урановской школы, в
особенности — таких своеобразных хрестоматий по популяционной био-
логии растений, как «Ценопопуляции растений» (1988) с многочисленны-
ми примерами, взятыми преимущественно из отечественных публикаций.
Глава 2. ОСНОВЫ ГЕНЕТИЧЕСКОГО
ПОДХОДА К ОПРЕДЕЛЕНИЮ
И ИЗУЧЕНИЮ ПОПУЛЯЦИЙ
РАСТЕНИЙ. ОБЩИЕ ПРЕДСТАВЛЕНИЯ
ОБ ЭВОЛЮЦИОННОМ ПРОЦЕССЕ
Ни в одной области биологии единство
жизни не выступает с такой ясностью как
во всех вопросах, касающихся наслед-
ственности... можно получить одинаковые
общие результаты путем исследований раз-
личных объектов, причем безразлично, ка-
саются ли эти исследования «венца творе-
ния», или собак и лошадей, птиц и мышей,
моли и тли или, наконец, энотеры и мака,
бобов и гороха, ячменя и овса.
В. Иоганнсен (1935: 25)
Хотя основное содержание настоящей книги будет посвящено экологиче-
ским аспектам жизни популяций, начинать целесообразно именно с рассмо-
трения наиболее общих генетических аспектов. И это будет не просто да-
нью приоритету генетиков в постановке проблемы. Ведь если претендовать
на эволюционную трактовку ботанического материала и говорить о приспо-
собленности — мере относительного преимущества одного генотипа (по-
пуляции, вида) перед другим, и, стало быть, главном количественном попу-
ляционном параметре, то без генетики никак не обойтись. В каждой из по-
следующих глав при рассмотрении различных моментов и явлений в жизни
популяций в той, или иной степени будет затронута и генетическая состав-
ляющая. В этой ясе главе мы рассмотрим только самые общие проблемы и
подходы к изучению того, что под популяцией в настоящее время понима-
10'1 I снстнкн.
/niaa ? Ociioiii.i Iciicni'icckoiо ||(ЩЧ(»/ц|
19
2.1. Роль Ч. Дарвина в создании популяционной биол<н ии
Чрезвычайно важным, хотя и скрытым достижением совершенною
Ч. Дарвином переворота в биологической науке была, согласно Эрнсту Майру
(Маут, 1972), замена типологического мышления в биологии популяционным.
На смену представлениям о виде, формирующимся по «типичному» образцу,
пришло представление об изменчивости вида, подкрепленное и уточненное
средствами статистики. Исследователи стали живо интересоваться именно от-
клонениями любого признака от его средней арифметической, взятой за «нор-
му». Отклонения эти графически должны были отображаться симметричной,
одновершинной кривой т. н. «нормального» распределения (см. вставки 1 и 2).
Ю. В. Чайковский совершенно справедливо считает, что Ч. Дарвин выра-
жал идеи своего века, трактуя биологическое знание как статистику, мир —
как баланс средних, а эволюцию — как сдвиг средних. Например, если усло-
виям среды лучше соответствует более определенная выраженность какого-
либо признака организма или просто увеличенное его значение, то мате-
риал для такого преобразования в популяции этого признака найдется: его
предоставят те особи, которые, отклоняясь по значению данного признака
от средней в нужную сторону, произведут, благодаря этому, больше потом-
ков. Одновершинная симметричная Гауссова кривая нормального распреде-
Рис. 2.1. Действие стабилизирующей (А, С) и ведущей (В, D) форм естественного
отбора по И. И. Шмальгаузену (1946): М — средняя, N — область «нормальных»
особей, Е область элиминируемых уклонений, М’ — новая средняя норма; N’
область уклонений, включаемых в состав новой «нормы», Е’— область элимини-
руемых представиiciieii прежней нормы.
20
Л/Н Mai на hi I loiiyiDiniioiiiiiiM пионыим рас iciiiih
Вставка 1. Основным законом всех явлении индивидуальной изменчи-
вости является закон Адольфа Кетлэ: чем больше уклонение и чем даль-
ше в вариационном ряду отстоит от средней величины данная вариан-
та, тем она попадается реже, и обратно — чем меньше уклонение и чем
ближе варианта к средней величине ряда, тем она представлена в нем
большим числом особей. Приняв более точную математическую форму-
лировку, можно сказать, что распределение особей в каждом вариаци-
онном ряду следует коэффициентам бинома, возведенного в известную
степень. Ф. Гальтоном был сконструирован остроумный аппарат с дро-
бинками и иглами, иллюстрирующий значение множественности, муль-
типликативности причин, ответственных за проявление индивидуально-
сти и наглядно объясняющий закон Кетлэ (А). Позднее подобная «ана-
логовая машина» была предложена как модель Каптейна (Б), объясняю-
щая происхождение столь часто встречающегося в биологическом мате-
риале логнормального распределения, т. е. распределения, которое ста-
новится близким к нормальному только после логарифмической транс-
формации данных.
Кривая и гистограмма нормально-
го распределения с обозначением
частоты (f), на левой вертикальной
оси, и (f %) частости — на правой,
даны здесь для сопоставления с кри-
вой и гистограммой логнормального
распределения (см. внизу).
I ниш ' ( h новы ।еш ।ич< ci.юл ода
21
Вставка 2
1 2 3
D
Характер варьирования коли-
чественных признаков (A-D)
и шкала для логарифмической
трансформации данных (Е):
А — Нормальное распреде-
ление: симметричная кри-
вая со: средней арифметиче-
ской (х), которая совпадает
с модой (нормой); стандарт-
ным отклонением (s) и меди-
аной (Z); В — положительно (
х - D>0) асимметричная кри-
вая логнормального распре-
деления; С — отрицательно
(х - D<0) асимметричная кри-
вая распределения; D — кри-
вые с отрицательным (сле-
ва) и с положительным (спра-
ва) эксцессом рядом с кривой
нормального (без эксцесса)
распределения (в центре)
Шкалы:
Е 1600 6400
50 100 200 400 800 I3200H 2800
50 100 200 400 800160016400125600
1_________ 2_______ 3____4 5 6 7 8 9 10
Логарифическая
01234 56789 10
50 100 150 200 250 300 350 400 450 500 550
50 50 50 50 50 50 50 50 50 50
Арифметическая
См. продолжение вставки 2
22 Л/.//. Маркин I loiiyiiHiutoiiniiH Gifojioi пи растений
Вставка 2. {продолжение)
Трансформация данных
Когда мы сталкиваемся с положительно асимметричным распределени-
ем признака, а это случается достаточно часто при работе с весовыми
и объемными показателями, мы можем откорректировать ситуацию пу-»
тем логарифмической трансформации данных. Это означает, что вме-
сто абсолютных значений признака, определенных (путем измерения,
подсчета, взвешивания) в обычной для нас десятичной шкале, мы берем
их логарифмы. После этого полный анализ вариансы можно проделать
уже для множества трансформированных значений. Логарифмическая
трансформация данных — наиболее часто применяемый тип трансфор-
мации, но наряду с ней можно применять и другие способы, используя,
к примеру, квадратный или кубический корни из значений и т.д.
Коррекция данных путем их преобразования (трансформации) может
вызвать у начинающих исследователей некоторое сомнение в правомер-
ности процедуры. Может сложиться впечатление, что мы искусственно
подгоняем, подправляем данные для получения желаемого результата.
В особенности тогда, когда преобразованные данные действительно по-
зволяют нам обеспечить статистическую значимость тестов, которой мы
не имели до того, как провели трансформацию. Чем же можно оправдать
правомерность преобразования данных? Это требует осознания идеи о
том, что нет научно обоснованной необходимости применять только
линейную или арифметическую шкалу для измерений. Преподавание ма-
тематики в обычных школах привело к наивному представлению о том,
что десятичная система чисел единственно естественная. Потребуется
изрядный опыт в науке и в применении статистических методов, чтобы
осознать тот факт, что линейная шкала, столь знакомая всем нам по на-
шему раннему опыту, занимает положение сходное с положением дру-
гих шкал измерения, подобно тому, как десятичная система чисел сход-
на по своему положению с бинарной (двоичной) или октальной (вось-
меричной) системой или иными системами. Если какое-либо отношение
в линейной шкале принимает мультипликативный характер, то гораздо
больше имеет смысла проанализировать его как аддитивную систему в
логарифмической шкале. Преобразование путем извлечения квадратно-
го корня — другой часто проводимый путь трансформации. Квадратный
корень из величины поверхности организма — это более соответствую-
щая мера фундаментальной биологической переменной, подверженной
влиянию физиологических и эволюционных факторов, чем сама пло-
щадь. Это отражается в нормальности распределения квадратных кор-
ней из значений переменной по сравнению с асимметричным распреде-
лением самих значений площади.
Г'кта 2 Основы генетической» подходи
23
лсния как бы вытянет «голову» и подожмет «хвост», чем и сдвинет среднее
(рис. 2.1 В, D): в следующих поколениях эта смещенная кривая плотности
станет основой для новых сдвигов — вот и элемент эволюции (Чайковский,
1990). Таково было представление о движущей форме отбора. Но всегда
ли у плотности распределения искомого признака есть «хвост», который
может быть поджат, и «голова», которая ползет куда надо, — спрашивает
Ю. В. Чайковский? «Например, признано, что первые организмы не облада-
ли фотосинтезом. Что надо поджимать, чтобы он появился?» (1990: стр. 77).
Действие стабилизирующей формы отбора по И. И. Шмальгаузену (1945)
тоже может быть проиллюстрировано с использованием кривой нормального
распределения (рис. 2.1. А, С), которая в этом случае сохраняет стабильность
«нормы» благодаря отсеканию (элиминации) излишне сильных отклонений
от нее как в области «головы», так и в области «хвоста» (рис. 2.2).
К рассмотрению этого вопроса мы вернемся ниже, а пока обсудим гене-
тические предпосылки эволюции.
Как известно, в теории эволюции путем естественного отбора Ч. Дарвина
есть несколько допущений. Сформулируем их таким образом, чтобы выделен-
ные заглавными буквами слова передавали основную суть того или иного до-
пущения, а остальные — служили необходимым дополнительным пояснением:
1. ИНДИВИДУУМЫ, представляющие какой-либо вид (образующие
популяцию, добавим мы!), не тождественны и ОТЛИЧАЮТСЯ ДРУГ ОТ
ДРУГА, пусть даже незначительно, по размерам, по скорости развития, по
реакциям на температуру и прочие экологические факторы.
2. ОТЛИЧИЯ эти, хотя бы частично, ПЕРЕДАЮТСЯ ПО НАСЛЕДСТВУ
и потому являются материалом для
естественного отбора (иными слова-
ми, внешние свойства особи, предъ-
являемые естественному отбору, в
какой-то мере предопределяются его
генетической конституцией, его гено-
типом — дети наследуют гены родите-
лей и потому, как правило, разделяют
родительские признаки).
3. ПОТЕНЦИАЛЬНО ЛЮБОЙ ВИД
(любая популяция, вновь добавим мы!)
В СОСТОЯНИИ ЗАПОЛНИТЬ ВЕСЬ
ЗЕМНОЙ ШАР. Так бы оно и случи-
лось, если бы каждая особь выжива-
ла и производила максимальное число
потомков. Но такого не случается: мно-
гие особи погибают, не успев произве-
сти потомство, а многие (если не все)
размножаются со скоростью далекой от
максимально возможной.
4. Разные ОСОБИ имеют разную при-
способпеиность (по-английски —fitness)
и поюму ОСТАВЛЯЮТ РАЗЛИЧНО!:
Рис. 2.2. Схема, поясняющая механизм
действия стабилизирующего отбора на
примере обычной вариационной кри-
вой но И.И.Шмальгаузену (1946).
24 МН Марком I loiiyiiHHiioiiiiiiM (hiojioi ия puciений
ЧИСЛО I ЮГОМ КОИ (Здесь подразумеваются не чолько различия в плодови-
тости особей, но и вероятноеть доживания дочерних особей до репродуктивно-
го возраста, и выживание и размножение потомков следующего поколения — и
так до бесконечности).
5. ЧИСЛО ВЫЖИВАЮЩИХ до репродуктивной зрелости ПОТОМКОВ
данного организма ЗАВИСИТ — не всецело, но решающим образом — ОТ
ИСХОДА ВЗАИМОДЕЙСТВИЯ МЕЖДУ ОРГАНИЗМОМ И СРЕДОЙ.
Таким образом, Ч. Дарвин не употреблял в своих работах терминов «гено-
тип» и «популяция», но в его допущениях явно мыслится не типологический,
но популяционный подход к виду с характерной для последнего внутривидо-
вой изменчивостью особей, различающихся по приспособленности.
2.2. Работы В. Иоганнсена
Никто не был столь успешен на поприще внедрения в биологическую на-
уку важнейших и быстро ставших общепринятыми терминов как датский
генетик и селекционер Вильгельм Иоганнсен (см. портрет). Именно ему
принадлежат термины «ген», «фенотип» и «генотип». Есть основания счи-
тать, что термин «популяция» в его современном значении тоже был впер-
вые употреблен им в знаменитой классической работе 1903 года1. В этой ра-
боте, посвященной наследованию признаков в популяциях и чистых лини-
ях, в науку был введен и термин «биотип», который обозначал группу рав-
нонаследственных (=имеющих одинаковый генотип) особей.
«Расу, население (! М.М.), группу какого-нибудь вида, или, как мы будем гово-
рить здесь и дальше, «популяцию» (курсив мой М.М.), нельзя с биологической точ-
ки зрения рассматривать как нечто единое. Это ед иное может и не быть таковым
даже в том случае, когда для каждого из отдельных свойств существует средняя,
якобы «типичная» величина, вокруг которой флюктуируют вариации всех дан-
ных индивидов в виде «идеальной вариационной кривой». Такая популяция мо-
жет содержать... различные самостоятельные, сильно отличающиеся друг от дру-
га типы, которые ни в коем случае не обнаруживаются при непосредственном рас-
смотрении эмпирических кривых или таблиц» (Иоганнсен, 1935:27).
Очень важная роль цитированной выше работы Ю. В. Чайковского состо-
ит в том, что он, подобно В. Иоганнсену, обсудил достаточно частые факты
отклонения эмпирических кривых от нормального распределения для неко-
торых признаков. Несмотря на то, что в таких случаях мы, строго говоря, не
имеем права применять традиционную статистику, оперирующую с абсолют-
ными (=не трансформированными) эмпирическими данными, здесь нет осно-
1 Термин «популяция» встречается уже на первой странице (если не считать самого названия
работы, обозначенного на титульном листе!) и употребляется вскользь, как уже привычный.
Это заставляет считать, что либо термин «популяция» употребляли до В. Иоганнсена, либо
он сам употреблял его где-то раньше. В 48-м томе энциклопедии Брокгауза-Ефрона (вышед-
шем в 1898 г.) приводится наука популяцйонистика — учение о населении, основанное на
статистических данных. Сказано, что термин «популяционистика» введен Христофом Бер-
нулли в его «Руководстве к популяционистике», изданном в 1841 г., но что он не привился в
науке из-за применения более привычных тогда выражений: теория, статистика или полити-
ка населения (в Германии и России) и демография (во Франции).
/ it mt i .* (>ci h >iii.i i cue i и* ir< luu о iKHixirin
Вильгельм Людвиг Иоганнсен (1X57
1927).
BJiiiiii дня пессимизма (см. продолже-
ние вставки 2). К более подробному
рассмотрению этого вопроса мы будем
неоднократно обращаться ниже.
Определяя популяцию как совокуп-
ность (сумму) разных биотипов, т.е.
как генетически неоднородную смесь
особей одного вида. В. Иоганнсен про-
тивопоставил ей генетически одно-
родные множества особей — чистые
линии (^гомозиготные биотипы), на
которые распадается популяция расте-
ний в результате длительного (на про-
тяжении жизни нескольких поколе-
ний) принуждения особей к самоопы-
лению и предотвращения перекрест-
ного опыления. Была показана беспо-
лезность селекционной работы с од-
нородным чистолинейным материа-
лом, из которого отбору (естественно-
му или искусственному) фактически
нечего «отбирать» по причине генетической эквивалентности особей.
Унаследованное от первого определения суммативное представление о
популяции не мешало селекционерам работать до той поры, пока они вели
селекцию растений облигатных самоопылителей. Однако с т. н. «пере-
крестниками» (растениями с той или иной степенью самонесовместимо-
сти. опыляющимися всегда или преимущественно перекрестным путем)
дело оказалось сложнее. Скрещивания интегрировали генофонд популяции,
и популяция демонстрировала генетическую разнородность, которую труд-
но было представить как смесь или результат простого суммирования не-
скольких немногих биотипов.
Вместе с тем, как мы знаем, селекционеры неплохо научились работать
и с перекрестно опыляющимися растениями. Лучший и наиболее впечатля-
ющий тому пример — результат селекционной деятельности человека, г.
е. результат искусственного отбора — разнообразнейшие по облику сорта
капусты — Brassica oleracea (цветная, белокочанная и краснокочанная ка-
пуста, кольраби, брокколи), широко используемые в пищу в разных странах
мира (рис. 2.3). Подобные изменения (сдвиги в генетической структуре по-
пуляций и в наборе фенотипов) аналогичны естественным эволюционным
преобразованиям, только протекают они намного быстрее.
2.3. Закон Харди-Вайнберга как математическая закономерность
и метод генетического анализа панмиктических популяций
Вскоре после выхода классической работы В. Иоганнсена, уже в 1908 1909 и:,
появился и математический «инструмент», пригодный для изучения популяций,
когорые состоят из особей, свободно и на основе голько случайности скрещиваю
26
МН АЛ/у>лп/.- I It )i iy iimiiiioiiii;ih Ituoih)i ни p;ic iciinii
Рис. 2.3. Различные сорта капусты, представляющие собой результат селекционной
работы с одним только видом Brassica oleracea.
щихся между собой, или, иными словами, менделевских или панмиктических по-
пуляций. Это был «закон Харди-Вайнберга» — закономерность, которую выве-
ли независимо друг от друга английский математик Годфрей Харди и немецкий
врач Вильгельм Вайнберг. Именно этот закон вооружил популяционных генети-
ков уравнением, с помощью которого можно высчитать соотношение частот гено-
типов в панмиктической популяции по достижении ею равновесного состояния.
Для любого отдельного локуса с двумя аллелями (А и а), встречающими-
ся в популяции с известными частотами, мы можем констатировать по зако-
ну Харди-Вайнберга состояние равновесия, если соотношение частот этих
двух аллелей составляет (см. также табл. 2.1):
(рА + qa)2= р2АА + 2pq Аа + q2aa = 1
В отечественной литературе закон Харди-Вайнберга иногда трактуется
как имеющий ряд серьезных ограничений. Помимо сомнений в осущест-
влении стопроцентной панмиксии (особенно у растений) указывают, что
постоянное соотношение частот генотипов поддерживается лишь в том слу-
чае, если в популяции нет мутационного процесса, отсутствуют миграции,
и не действует отбор.
Таблица 2.1.
Соотношение генотипов в панмиктической популяции, которая по закону Харди-
Вайнберга находится в состоянии равновесия в отношении двух аллелей (А и а)
Частота аллеля А(р) а(?)
А(р) А А (/г) Аа(рд)
а(</) аА ^2) аа ')
/ Itlfid 2 < >С11С»НЫ I CIICI Il'iri kOl О подходи
2/
Иногда просто niiinyi, чю закон применим только к идеальным менде
невским популяциям. Таким образом, у неспециалиста может сложиться ни
линия ограниченности применения, малой значимости закона и возникнуть
недоумение, почему в этом случае закон остается, по сей день, краеуголь-
ным камнем популяционной генетики. Парадоксальность ситуации состо-
ит в том, что такая трактовка закона в принципе правомерна, просто в ней
неправильно расставлены акценты. Представляется гораздо более коррект-
ным и правильно расставляющим акценты подход к закону как чисто мате-
матическому ожиданию — описанию равновесного состояния популяции.
Отклонения от этого состояния, которые возникают из-за действия разных
и в том числе перечисленных в определении факторов в природных и мо-
дельных реальных популяциях как раз и достойны внимательного изучения.
2.4. Причины отклонений от равновесия Харди-Вайнберга
К настоящему времени известны по меньшей мере семь факторов, спо-
собных смещать «равновесие Харди-Вайнберга» в популяциях растений.
Это: 1.) мутационный процесс; 2.) сцепление генов (linkage); 3.) неслучай-
ное скрещивание; 4.) разделение популяции на генетические соседства, суб-
популяции и т. п.; 5.) поток генов (geneflow); 6.) дрейф генов (genetic drift) и
7.) естественный отбор. Вкратце рассмотрим их по порядку.
2.4,1, Мутационный процесс
Мутации, появляющиеся относительно редко, как правило не оказывают
сколько-нибудь существенного влияния на демографические параметры по-
пуляций, которые обычно изучают относительно недолго.
2,4.2. Сцепление генов
Когда в сочетаниях (комбинациях) друг с другом аллели двух или более
локусов встречаются с более высокой или более низкой частотой, чем это
прогнозируется на основе расчета их индивидуальных частот, говорят о
дисбалансе из-за сцепления генов.
2.4.3. Неслучайное скрещивание
Случайное скрещивание или панмиксия — не более чем удобное теоре-
тическое допущение. Если фенотип растения предопределяет, с какими из
особей той же популяции скрещивание наиболее вероятно, то это называю!
ассортативным или избирательным скрещиванием. Иногда условия благо-
приятствуют скрещиванию сходных особей (положительная избиратель-
ность самосовместимость), а иногда, наоборот, скрещиванию несход-
ных особей (отрицательная избирательность — самонесовместимость).
11оложительная избирательность ведет к самоопылению (инбридингу), а от-
рицательная к перекрестному опылению (аутбридингу). В зарубежной
28 Л/// Марьон I loiiyiinnnoiiiKiM biioiioi ия рас iciinii
литературе относительный вклад в приспособленность через продукцию се-
мязачатков (функцию материнства) и через продукцию пыльцы (функцию
отцовства) называют джендером (gender). У растений существуют три
основных типа джендера или проявления пола (сексуализации).
1. Разделение мужских и женских репродуктивных органов в пространстве
(геркогамия). В дополнение к бесспорно преобладающему гермафродитизму
цветков, в случае которого у растений как механизм разделения нередко вы-
ступает гетеростилия, встречаются однодомность в двух ее вариантах (ан-
дромоноэция и гиномоноэция) и двудомность (чаще в форме гинодиэции).
2. Разделение во времени мужской и женской репродуктивной активно-
сти (дихогамия).
3. Выработка механизмов самонесовместимости.
Варианты джендеров с примерами более подробно рассмотрены в табли-
це 2.2.
Таблица 2.2.
Три основных типа проявления пола (сексуализации) у растений и их варианты. В
процентах определено соотношение типов в выборке из 121492 видов цветковых
растений, что составляет около половины всех известных видов покрытосеменных
(по Yampolsky, Yampolsky, 1922 с изменениями)
Разделение в пространстве мужских
и женских репродуктивных органов
(геркогамия)
Джендер одного класса (все растения
в популяции представляют один и тот
же джендер)
ГЕРМАФРОДИТИЗМ (72% видов)
Представлен только один тип гермафро-
дитного (обоеполого) цветка с органами
обоих полов (большинство видов, вклю-
чая почти все виды семейств Лилейных
и Орхидных)
Гетеростилия: два или три типа гер-
мафродитных особей (морфы) в составе
одной популяции с цветками, в которых
столбик имеет разную длину, а тычин-
ки всегда длиннее или короче столби-
ка, при обычно наблюдающихся допол-
нительных морфологических отличиях
между особями разных типов
Дистилия: два типа особей, из которых
одни несут длинностолбчатые («корот-
котычиночные»), а другие — корот-
костолбчатые («длиннотычиночные»)
цветки (Primulaceae, Linaceae)
Тристилия: три типа особей, которые
несут, соответственно только длинно-,
средне- или короткостолбчатые цвет-
ки и при этом пыльники в каждом слу-
чае находятся в отдалении от рылец
(Lythraceae, Oxalidaceae и др.)
ОДНОДОМНОСТЬ или МОНОЭЦИЯ
g>% видов)
тдельно мужские и женские цвет-
ки формируются на одном растении
(Typha, Zea, Quercus, Cucurbita)
АНДРОМОНОЭЦИЯ (1,7% видов)
Особи несут как гермафродитные, так и
мужские цветки (большинство видов в
сем. Apiaceae)
ГИНОМОНОЭЦИЯ (2,8% видов)
Наряду с гермафродитными цветка-
ми растения формируют женские цвет-
ки (многие виды сем. Asteraceae, в т. ч.
Bellis и Solidago)
Джендер двух классов (наличие двух по-
ловых морф в составе популяции)
ДВУДОМНОСТЬ или ДИЭЦИЯ (4% ви-
дов)
Растения несут только женские или
только мужские цветки, напр. Salix,
Vallisneria, Elodea, Cannabis
ГИНОДИЭЦИЯ (7% видов)
Особи формируют либо гермафродит-
ные, либо женские цветки (многие виды
сем. Lamiaceae, включая Giechoma,
Thymus)
АНДРОДИЭЦИЯ (известна пока толь-
ко у одного вида — Datisca glomerata
(Liston et al., 1990)
/ hlthl 2 Основы I CIICI H'K’ChOI О IIO/14O/UI 29
Разделение во времени мужской и
женской репродуктивной активности
(дико! амия)
ПРОТАНДРИЯ
11ыльники созревают и пыльца высыпа-
ется до того, как рыльца приобретают
способность принимать пыльцу
(например Daucus, Ranunculus acris)
ПРОТОГИНИЯ
Рыльца приобретают способность к
приему пыльцы раньше, чем созреют
пыльники и пыльца высыпается наружу
(например, Magnolia, Brassica)
Наличие или отсутствие самонесов-
местимости
Самонесовместимость
Полиморфизм по одному или более
локусам, определяющим самонесов-
местимость. Оплодотворения семяза-
чатка не происходит, если пыльца или
донор пыльцы несет тот же аллель са-
монесовместимости, который представ-
лен в самом семязачатке. Все подобные
системы срабатывают небезупречно,
из-за чего имеет место некоторое число
иллегитимных случаев оплодотворения
i ami:t()фи гпы1- (।к I г:мы
Тип самонесовмсстимости пыльцевых
зерен, определяемый гаплоидным гено-
типом самих гамет.
Обычны в сем. Fabaceae, Solanaceae,
Poaceae, Rosaceae и Ranunculaceae
СПОРОФИТНЫЕ СИСТЕМЫ
Тип самонесовместимости пыльцевых
зерен, определяемый диплоидным ге-
нотипом родительского растения (спо-
рофита).
Характерен для семейств: Asteraceae,
Brassicaceae, Lythraceae и Primulaceae
Самосовместимость
Растения, не несущие локусов само-
несовместимости. Большинство актов
опыления, включая самоопыление при-
водит к завязыванию плодов.
Примеры можно найти в сем. Apiaceae
2.4.4. Разделение популяции на генетические соседства, субпопуля-
ции (внутрипопуляционная генетическая структурированность).
Эффективная генетическая единица
По причинам, которые особенно очевидны для организмов, ведущих прикре-
пленный образ жизни, вероятность скрещивания между двумя растениями и фи-
зическое расстояние между ними связаны обратной зависимостью. Если доба-
вить к этому ограниченность в пространстве дисперсии зачатков, как составля-
ющей потока генов (gene flow), то вполне понятно, что популяция растения до-
вольно быстро приобретает особую пространственную структуру в связи с фор-
мированием генетических соседств родственных особей. Площадь генетиче-
ского соседства (Ап) определяют как площадь круга, в центре которого находит-
ся данное растение, при условии, что в пределах этой площади можно с веро-
ятностью 86,5% найти его родителей. Например, если вегетативные диаспоры,
семена и пыльцевые зерна распространяются одинаково по всем направлениям
ел родительского растения, то площадь генетического соседства можно опреде-
лить по формуле: Ап = 4п<з2, где а2 — вариансс? расстояния, на которое происхо-
* В отечественных пособиях по биометрии вариансу рассматривают часто как синоним дис-
персии. Однако в контексте данного учебника, где говорится о дисперсии как о процессе и
результате распространения (=рассеивания) диаспор, лучше употреблять во избежание пу i а
iiiiiu.i «чисто биометрический» термин «варианса».
30 М.В. Марков Популяционная биология растений
дит дисперсия генов. Эта формула была впервые приведена С. Райтом (Wright,
1943), который обозначил границы ее применения и для этой цели ввел следу-
ющие допущения: 1) число потомков от одного родителя и расстояния, на кото-
рые происходит дисперсия, должны подчиняться нормальному распределению;
2) соотношение полов при этом должно составлять 1 : 1 и 3) популяция долж-
на находиться в равновесном состоянии. При справедливости этих допущений
появляется возможность произвести теоретическое определение вероятности
(= частоты) перекрестного опыления (ауткроссинга) для какой-либо конкрет-
ной популяции.
Общую вариансу и2 можно для этого разложить на несколько отдель-
ных частных варианс, которые отражают как дисперсию пыльцы и диспер-
сию семян, так и дисперсию клонального разрастания родительских осо-
бей, словом все процессы, составляющие поток генов (см. ниже). Эти пара-
метры можно рассчитывать по эмпирическим данным. Поскольку вариансы
этих компонентов потока генов независимы друг от друга, то их вклад в об-
щую вариансу можно суммировать:
и2 = t/2 а2 пыльцы + и2 клонального разрастания + и2 семян.
где t — частота ауткроссинга.
Другой путь описания генетической структуры популяции лежит через
выявление размера внутрипопуляционной эффективной генетической еди-
ницы3 (ЭГЕ) — Ne , который определяется как эффективное число репро-
дуцирующих растений, расположенных в пределах площади одного гене-
тического соседства: Ne = Ап • N/2 • (1 + /), где N — плотность репроду-
цирующих особей в популяции, a t — частота ауткроссинга. По получен-
ным к настоящему времени данным (см. например, Silvertown, 1993) эффек-
тивная генетическая единица варьирует у травянистых растений несколь-
ко шире (от 3 до 282 особей), чем у древесных (от 11 до 207 особей) при, в
общем-то, сильном перекрывании вычисленных абсолютных ее значений.
В том случае, когда Ne становится равной общему числу взрослых (=репро-
дуцирующих) особей популяции (и), режим скрещиваний приближается к
случайному и популяция не приобретает какой-либо характерной генетиче-
ской структуры. Д. Левин (Levin, 1988) показал, что фактически N — может
сильно варьировать в пределах популяции, но, как правило, бывает намного
меньше ее общей численности. Скорость распространения благоприятного
аллеля в этом случае будет связана с Ne обратной зависимостью, а экологи-
ческие факторы, ведущие к уменьшению Ne относительно п, увеличивают
вероятность того, что разные части одной и той же популяции будут дивер-
гировать по своему генетическому составу.
В популяции, которая состоит из генетически разнородных прерыви-
стых субпопуляций, каждая из которых частично изолирована от других,
степень внутрипопуляционной генетической разнородности (Н) должна
быть выше, чем в гомогенной панмиктической популяции такого же разме-
ра. Аутбредные популяции должны иметь большее разнообразие аллелей * I
' За рубежом се называю! «effective population size», что дословно псреводн1ся как -><|><|>ск
I ши naif ра «мер (и нс псиное и.) i к и i у пя i in н
l idfiu ? Основы I ciicih'icl kih<> подхода
31
па видовом уровне и меньшую мсжнонуляционную дифференциацию, чем
инбрсдные популяции.
Другим индикатором того, что внутри популяции существует некая ге-
нетическая структура, является явное общее снижение частоты гетерози-
гот. Хотя такое снижение и может быть следствием инбридинга, второй его
причиной способно стать присутствие двух или более субпопуляций в со-
ставе той, что мы анализируем. Если каждая из субпопуляций находится в
состоянии равновесия Харди-Вайнберга, но равновесные частоты аллелей
в разных субпопуляциях различны, то выборка генотипов, взятая из популя-
ции в целом, может дать частоты, отклоняющиеся от частот, теоретически
прогнозируемых равновесием Харди-Вайнберга. Такое явление предложе-
но именовать эффектом Валунда (Wahlund effect), а симптомом его приня-
то считать кластеризацию в пространстве (формирование скоплений) осо-
бей со сходными генотипами.
Особый случай, когда в одной популяции соседствуют субпопуляции, раз-
личающиеся по ритму чередования активности и покоя и потому, несмотря
на симпатричность, изолированные друг от друга во времени (см. Марков,
2001). Такая картина свойственна популяциям растений с почвенным бан-
ком семян и/или вегетативных зачатков (клубней, луковиц и т. д.). Подобные
ситуации, наиболее ярко представленные у однолетних растений, а также у
псевдооднолетников, и сопутствующие генетические эффекты лишь совсем
недавно привлекли внимание исследователей (подробнее см. главу 1).
2.4.5. Поток генов (gene flow)
Поступление новых генов в популяцию цветкового растения возмож-
но, как уже было сказано выше, благодаря распространению пыльцы, се-
мян или специализированных вегетативных зачатков. Темпы потока генов
можно определять, отталкиваясь от соответствующего числа мигрирующих
структур, которыми обмениваются популяции за время жизни одного поко-
ления. Благодаря потоку генов все популяции объединены в единую видо-
вую систему.
2.4.6. Дрейф генов
Распределение и концентрация частот аллелей в популяциях может идти
не только под действием панмиксии (неограниченного перемешивания) и
естественного отбора, но и под действием случайных, стохастических фак-
торов, получивших на Западе название «дрейфа генов» (Wright, 1943).
Используя сложный математический аппарат, Рональд Фишер и Сьюэлл
Райт показали, что при отсутствии различий в селективной ценности алле-
лей — т. е. при отсутствии отбора и отсутствии перекрывания поколений
во времени может происходить случайное вытеснение одного аллеля дру-
гим. В конечном счете одни популяции могут со временем оказаться моно-
морфными по аллелю А, а другие — по аллелю а. Это случайное вытесне-
ние одного селекционно нейтрального аллеля другим и было названо дрен
32
MH MtiphlHI I биология pnc ICIHli
фом генов. Модель этого явления, созданная но тину модели («аналоговой
машины») I альтоиа представлена на рис. 2.4.
Ученики и последователи С. С. Четверикова отечественные ученые
Дмитрий Дмитриевич Ромашов и Николай Петрович Дубинин, незави-
симо от Фишера и Райта также обратили внимание на этот важный факт
и в иной форме развили теорию процессов, названных ими генетико-
автоматическими.
Отечественный математик А. Н. Колмогоров в отличие от Фишера и Райта,
исследовавших динамику концентрации аллелей в популяциях при свобод-
ном скрещивании, рассмотрел уклонения от правила Харди-Вайнберга, воз-
никающие в условиях частичной изоляции. Связанный школьной друж-
бой с Колмогоровым Ромашов был первым биологом, объединившим
эволюционно-генетические идеи С. С. Четверикова с математическими иде-
ями знаменитого, а можно сказать и великого, отечественного математика
Частота гена
Рис. 2.4. Модель, иллюстрирующая
дрейф генов. Частота генов ведет себя
аналогично упругому шарику, который,
скатываясь по наклонной доске, отска-
кивает от вбитых в нее штифтиков, пока
не дойдет до одного из краев доски —
пока ген не будет утрачен (частота 0 и
все особи аа) или фиксирован (частота
1.0 и все особи АА).
Андрея Андреевича Маркова, кото-
рый еще в 1907 г. разработал теорию
случайных процессов или «марков-
ских процессов».
Д. Д. Ромашов обратил внимание
на существование инадаптивного
популяционного полиморфизма и на
возможность дивергенции популяций
по инадаптивным признакам за счет
«ошибок выборки», т. е. за счет сто-
хастических, случайных процессов.
В знаменитой статье Н. П.
Дубинина и Д. Д. Ромашова и 1932
г. фактически были описаны та-
кие популяционные феномены, как
«эффект бутылочного горлыш-
ка», и то, что мы теперь вслед за
Эрнстом Майром называем «прин-
ципом основателя», следующие из
четвериковских «волн жизни», из
генетико-автоматических процессов
Дубинина-Ромашова и из «дрейфа ге-
нов» Фишера-Райта-Холдейна.
2.4.7. Естественный отбор
Если упростить ситуацию и не учитывать того, что особи с равными
шансами на выживание могут различаться по числу и оплодотворяющей
способности производимых ими гамет, вероятности успешного переопыле-
ния и т. д., то можно представить такую картину действия естественного от-
бора (см. также Бердников, 1991).
При наличии в модельной популяции локуса, представленного двумя ал-
лелями А и а все особи будут распределяться по трем генотипическим труп-
I hiati J Псионы iciiciii'ivcmmi ikmimhqi J J
нам: гомозиготы A A (nx 'iiiciio обозначим как N ) и aa (их число N ), i c
1срозигогы Aa (их число N ). Общая численность популяции тогда со
ciainn: N + N^
В силу диплоидности особей каждая из них вносит в генофонд популя-
ции два локуса, причем гомозиготы АА вносят 2N^ копий аллеля А. гомо-
зиготы аа вносят 2Naa копий аллеля а, тогда как гетерозиготы Аа внося т N (u
копий аллеля А и столько же — аллеля а. Применяя обозначения частоты
аллелей черезр и q и помня, чтор + q =1, для нашей модельной популяции,
где имеется всего 2N локусов, можно расписать так:
Р = (2N./NJ/2N=(N^ + 0.5NJ/N,
q = (2N +N. )/2N= (N + 0.5N )/N.
1 v aa Aa' v aa Aa'
Примем, что гетерозиготы в нашей гипотетической популяции делятся
поровну на те, которые вносят в генофонд аллель А и те, что вносят аллель
а. Тогда, представляя в составе популяции два класса, вносящих, соответ-
ственно, аллели Айа, запишем:
N = N.. + 0.5N
А АА Аа
N =N + 0.5N..
а аа Аа
Это нам позволит записать частоты аллелей как доли особей, несущих
эти аллели в общей численности:
p = N./N; ? = N/N.
Чтобы представить действие отбора, нужно сопоставить численность
зигот на старте гипотетического поколения (N) с численностью взрослых
особей, доживших до репродуктивного возраста (N'). Сопоставлять будем
с учетом присутствия трех генотипических групп: ; Noa для зигот
и N’^; N'^a ; N'ao—для взрослых особей. Обозначим теперь приспособлен-
ность каждого генотипического класса по выживаемости особей, которые
его представляют:
F • F • F и F =N'/N-
F = N' /N ;F =N'. /N. .
аа аа аа 7 Аа Аа Аа
Логично рассмотреть какую-то среднюю приспособленность как по-
пуляции в целом (F = N’/N), так и особей-носителей каждого аллеля:
F^N’/N^hF^N’/N^.
Теперь осталось выяснить, как под действием отбора изменяется ал-
лельный состав популяции (частоты аллелей) за время жизни поколения.
Отталкиваясь от выше сделанного заключения о долях особей-носителей, и
выражая численности взрослых особей через численности зигот, запишем
Aq как сдвиг частоты аллеля а в результате отбора: (N’/N’ - N/N):
34 MH AftipKO" I loiiyiiHHiKHinihi Giioiioi ия ])ac icniiil
Aq (NJJ/NF N/N q |(F, F)/F|.
Получается, что сдвиг частоты аллеля в генофонде популяции в результа-
те отбора равен частоте этого аллеля до отбора, умноженной на относитель-
ное преимущество в приспособленности его доноров перед средней при-
способленностью популяции в целом.
2.5. Внутривидовая и внутрипопуляционная изменчивость.
Ее природа и наиболее общие моменты проявления
По своей природе внутрипопуляционная изменчивость может быть мо-
дификационной (или фенотипической — Vp)n генетической (или генотипи-
ческой — Vg). Модификационная изменчивость проявляется в формирова-
нии множества различающихся по внешним признакам и потому разноликих
особей. Внешними мы будем называть любые количественные признаки,
которые мы можем определить путем измерений (метрические признаки)
или пересчета (счетные или меристические признаки), а также качествен-
ные признаки, которые мы можем классифицировать по их визуальному вы-
ражению. В любом случае внешний признак является следствием (продук-
том) белкового синтеза, осуществленного по информации, закодированной
в первичной структуре ДНК. Отсюда ясно, что такой признак как морфоло-
гия хромосом (и вообще особенности кариотипа) — признак фенотипиче-
ский, и не только потому, что выявляется визуально путем микроскопиро-
вания, но и потому что форма и величина хромосом зависят от присутствия
структурных белков гистонов.
Характер наследования качественных и количественных признаков суще-
ственно различается. Если качественные признаки часто, как говорят гене-
тики, «менделируют», то количественные признаки зависят от полимерных
генов. Чем большее число неаллельных генов оказывает влияние на количе-
ственный признак, тем более плавные (мелкие) колебания будет проявлять
такой признак в популяции и тем шире будет амплитуда его изменчивости.
Как количественные признаки, которые могут варьировать либо непре-
рывно, либо дискретно, так и качественные признаки, для которых харак-
терен прерывистый характер изменчивости, могут быть исследованы ста-
тистически путем построения спектров. Внутрипопуляционные спектры
для количественных признаков, изменение которых можно представить в
виде вектора, являются ни чем иным как уже рассмотренными выше гисто-
граммами или кривыми распределения. При необходимости возможен пе-
ревод качественного признака в количественный и наоборот (см. вставку
3), но наиболее часто для правомерности использования обычной параме-
трической статистики необходима трансформация данных. Иногда при ра-
боте с таким широко варьирующим количественным признаком как общая
биомасса растения исследователи размерной дифференциации (= размер-
ной иерархии) популяций (см. Solbrig, Weiner, 1984) прибегают для ее срав-
нительной оценки к применению G — коэффициента Джини (Gini\ кото-
рый можно высчитать по формуле (см. вставку 3).
I itidii 2 Основы Icnciii'icckoiо подходи
35
Вставка 3
11омер класса Начало класса, % Конец класса, % Балл
I 0 3.125 1
II 3.126 6.250 2
III 6.260 12.50 3
IV 12.6 25.0 4
V 25.1 50.0 5
VI 50.1 100.0 6
Шестибалльная логарифмическая шкала для перевода количественных при-
знаков в качественные (Расницын, 1965 по Шмидт, 1984)
Примечание: начало и конец класса даны в частостях (процентах
натуральных частот от объема выборки)
Коэффициент Джини (Gini)
Когда необходимо выявить размерную разнородность (неодинако-
вость) особей внутри популяции, а не просто характер распределения
по классам размеров, наиболее адекватной мерой считается коэффи-
циент Джини (G). Для п особей, средний размер которых составляет х,
п п
G = f ij 1------
2n2x
Этот коэффициент, носящий имя своего итальянского автора, ранее при-
менялся только в экономике для определения неравенства состояний и до-
ходов в обществе. Графическим способом для ранжированной по биомассе
популяции коэффициент Джини определяют с помощью кривой Лоренца.
откладывая кумулятивный процент биомассы особей каждого последующе-
го ранга (класса) против кумулятивного процента их числа в популяции.
Отношение площади между диагональю и кривой Лоренца (рис. 2.5) к пло-
щади всего треугольника под диагональю и есть величина коээфициента
Джини (G).
Точкой соприкосновения двух названных типов изменчивости можно
считать норму реакции — совокупность фенотипов, которые потенциаль-
но способен продуцировать один генотип.
Генотип определяет потенциальные возможности особи проявить какой-
либо признак в фенотипе, т. е. внешнем облике, габитусе, жизненных от-
правлениях (интенсивности первичного и вторичного метаболизма, дыха-
ния, регулируемой транспирации). Надо сказать, что полностью все потен-
циальные возможности генотипа реализоваться в фенотипе одной особи нс
могут никогда. Эго и понятно, если учесть, что каждая особь растет в кон
Рис. 2.5. Кривые Лоренца, построен-
ные для трех (1-2-3) последовательно
взятых выборок особей из популяции
пастушьей сумки.
36 Л/ //. Мауков I h>iiyjiMUii(>niia>i (>11(>ло1 ня pacicniiii
кретных условиях среды (микроус-
ловиях), которые в какой-то степени
тоже влияют на выраженность того
или иного признака.
Разноликость особей в составе по-
пуляции, отмеченная выше, вытекает
из того факта, что фенотипы склады-
ваются из многих внешних признаков,
даже если мы из этого множества вы-
деляем некие ключевые признаки, из-
менчивость которых нам особенно ин-
тересна. На рис. 2.6. представлен один
из довольно необычных графических
способов одновременного отображе-
ния изменчивости многих (их могло
быть до двадцати двух!) фенотипиче-
ских признаков в выборке из реальной
популяции декоративного луковичного
эфемероида рябчика шахматовидного
Fritillaria meleagroides (сем. Liliaceae) в
Татарии.
В популяциях рябчика шахматовидного выявлена очень сильная измен-
чивость признаков как вегетативной, так и генеративной сферы.
Большое значение имеет не только изучение изменчивости признаков по
отдельности, но и изучение отношений между количественными признака-
ми и корреляционных связей между ними. Эти связи дают важное представ-
ление о сопряженности в изменчивости разных признаков и потому отража-
ют интегрированный характер фенотипа растения и корреляционную струк-
туру (см. рис. 2.7).
Наши исследования затрагивали сравнительно большие и полночленные
пойменные популяции рябчика, которые изолированы друг от друга рассто-
янием около 300 км. Цель работы состояла в изучении внутрипопуляцион-
ной и межпопуляционной изменчивости признаков не только в статике, но
и в динамике, для чего проводили сравнение выборок, взятых в одной по-
пуляции, но в разные годы. При обработке особей в выборках были учте-
ны 23 признака: общая высота растений, расстояние до первого (нижнего)
листа, расстояние от верхнего листа (узла) до цветка, общее число листьев,
окраска околоцветника, ширина нижнего листа, длина околоцветника, ши-
рина листочков внутреннего и наружного кругов околоцветника, расстоя-
ние от верхушек листочков внутреннего и наружного кругов до максималь-
но широкой их части, общая длина столбика, длина расщепленной части
столбика, длина тычинок, длина пыльников, ширина и длина луковиц, дли-
на нектарника, масса околоцветника, масса тычинок, масса гинецея, общая
масса цветка и др. и ряд отношений между признаками (индексов). Данные
были получены для сезонов разных лет, отличавшихся как по метеороло-
гическим условиям, так и по максимальному уровню воды во время павод-
/ !l(l(i(I 2 ( )c110IIIИ I С11C l 11'ICCK(>I о ПОД XI »Д11
37
Case 1
Case 2
Case 3
Case 4
Case 7
Case 8
Case 13
Case 14
Case 15
Case 10
Case 16
Case 19
Case 20
Case 21
Case 22
Case 11
Case 23
Case 6
Примечание: каждая из 24 особей характеризуется многими признаками, значе-
ния которых фигурируют в отведенной для нее строке статистической таблицы
[Case — строка (англ.)]
С чем вы предпочитаете иметь дело: с
картинками, графиками или с числовы-
ми таблицами? Почти наверное можно
сказать — с картинками. Но если число
компонентов, которые надо показать ве-
лико, то надо решить проблему графиче-
ского отображения многомерной инфор-
мации. В данном случае комплекс при-
знаков отдельных особей популяции ото-
бражается в виде стилизованной челове-
ческой физиономии. Любое изменение
двадцати двух параметров (на деле при-
знаков, обозначенных на этом рисунке X
1-Х 22) вызывает деформацию лица, вы-
ражение которого меняется.
Рис. 2.6. Визуализация изменчивости особей в популяции рябчика шахматовидно-
го с использованием метода икон Чернова (пояснения в тексте).
ка. Последнее было важно при работе с пойменным видом. Обработка дан-
ных состояла в выявлении и сопоставлении корреляционной структуры вы-
борок (и популяций), а также в оценке различий между ними при помо-
щи ступенчатого дискриминантного анализа. Каждый из 23 учтенных при-
знаков хотя бы однажды (в одной популяции или в одну дату) демонстри-
ровал достоверную корреляционную связь с каким-либо другим призна-
ком. Доля достоверно (хотя бы однократно) коррелировавших пар состави-
ла 64,8% от общего теоретически возможного их числа. Из 164 нар призна-
ков, достоверно коррелировавших между собой, 123 были отмечены в обе
их популяциях, 8 н>л1.ко в первой (при общем числе Ml) и И юлг
A 1993
Рис. 2.7. Динамика корреляционной структуры: данные для одной и той же популя-
ции рябчика шахматовидного Fritillaria meleagroides, анализируемой в разные годы
(с 1990 по 1995 г.).
/ itidii ? (>cnoiii.i i снеiii'icchoiо
39
ко во второй (при общем числе 156). От общего числа определенных ко-
эффициентов в первой популяции было 28,3% достоверных (и более вы-
соких), а во второй — 45,0% (при менее высоких их значениях). Только 5
пар признаков (среди них как вегетативные, так и генеративные) всегда (во
всех выборках и во все годы наблюдений) выказывали достоверную корре-
ляцию. Таким образом, изученные популяции отличались по их корреля-
ционной структуре, причем, исследования показали, что последняя может
существенно меняться во времени. Признаки, которые продемонстрирова-
ли наибольшее число корреляций (общая высота растений и масса цветка),
вносили и наиболее значимый вклад не только в различие между популяци-
ями, но и в различие между состояниями одной и той же популяции в раз-
ные годы. Особенно интересным представляется тот факт, что таксономи-
чески важные признаки, картина изменчивости и корреляционная структу-
ра признаков в одной и той же популяции в разные годы могут отличать-
ся очень сильно (Идрисова, Марков, 1997). С учетом наблюдений в специ-
ально созданной интродуцированной популяции рябчика шахматовидного
(Марков, Идрисова, 1996) можно предполагать, что каждый год состав ге-
нотипов, представленных активной частью популяции, различен из-за ухо-
да одних особей в состояние покоя и выхода из этого состояния других осо-
бей (см. следующую главу).
Отразить относительную роль генотипа и среды в проявлении фенотипи-
ческой изменчивости можно при помощи суммации соответствующих ва-
рианс:
V = V + V +2COV ,
р g е g.e’
где 2COVge— ковариация (сопряженная вариация) генотипа и среды, кото-
рая отражает тенденцию растений определенных генотипов к произраста-
нию в определенных местообитаниях. Отношение V /V можно использо-
вать как количественную меру наследования того или иного признака или
его наследуемость (HN) в широком смысле слова. Наследуемость призна-
ков, существенно и прямо влияющих на приспособленность, как правило,
оказывается весьма невысокой.
Глава 3. СХЕМЫ И МОДЕЛИ
ЖИЗНЕННЫХ ЦИКЛОВ
ПОПУЛЯЦИЙ, ТЕОРИЯ И
ПРАКТИКА ДЕМОГРАФИЧЕСКОГО
ПОДХОДА К ИЗУЧЕНИЮ
ПОПУЛЯЦИЙ У РАСТЕНИЙ,
ДЕМОГРАФИЧЕСКИЙ МЕТОД ОЦЕНКИ
ПРИСПОСОБЛЕННОСТИ
«Наглядность — хорошее средство
против неопределенности слов».
Г. В. Лейбниц
Динамика популяции любого вида представляет собой очень сложный
и многоступенчатый процесс, протекающий под влиянием неисчислимого
множества разнообразных факторов. И все же для успешного исследования
популяционной динамики у растений и для рассмотрения ее в свете эволю-
ции, прежде всего, необходима некоторая редукция, схематизация.
3.1. Спектры возрастных или онтогенетических состояний
В отечественной ботанической науке такая схематизация легла в осно-
ву представления о возможности дискретного описания онтогенезов рас-
тений в целях выявления возрастных или онтогенетических состояний
(Работнов, 1950а, б). Введение при этом понятия относительного возрас-
та или «возрастности» несколько смягчило, в общем-то, обескуражива-
ющую ситуацию, связанную с невозможностью определения абсолютно-
го возраста у огромного большинства видов травянистых многолетних рас-
тений. Хотя онтогенез, индивидуальное развитие растения — это процесс
непрерывный, представление его как череды дискретных состояний, по-
добно своеобразной мультипликации оказалось очень полезным и инфор-
мативным для целей фитоценологии и популяционной экологии растений
(см. вставку 4). Для многих видов растений к настоящему времени разра-
ботаны диагнозы онтогенетических состояний. Профессионально выпои-
fhltltl I Схемы II модели ЖИ П1Г11П1.1Ч циклов популяций
41
Вставка 4
Периоды онтогенеза, онтогенетические состояния и их обозначение
в спектре
Периоды онтогенеза Возрастные (онтогенетические) состояния особей Обозначения в спектре
I. Первичного покоя (латентный) Покоящиеся семена se
II. Прегенеративный (виргинильный) Проростки (всходы) Ювенильные Имматурные (прематурные) Виргинильные (молодые вегетатив- ные, взрослые вегетативные) Р j im V
III. Генеративный Молодые (ранние) генератив- ные Средневозрастные (зрелые) гене- ративные Старые (поздние) генеративные OQ OQ OQ
IV. Постгенератив- ный (старческий, сенильный) Субсенильные (старые вегетативные) Сенильные ss s
р — Смешанное питание (за счет веществ семени и собственной ассими-
ляции первых листьев); наличие морфологической связи с семенем (или
семядолей); наличие зародышевых структур: семядолей, первичного (за-
родышевого) корня и побега.
j — Простота организации, несформированность признаков и свойств,
присущих взрослым растениям. Наличие листьев иной формы и располо-
жения, чем у взрослых особей; иной тип нарастания и ветвления (или о i -
сутствие ветвления побегов); возможно изменение типа корневой сис ге-
мы. Сохранение некоторых зародышевых структур (корня, побега); поте-
ря связи с семенем; как правило — отсутствие семядолей.
im — Наличие свойств и признаков, переходных от ювенильных рас ге-
ний к взрослым: развитие листьев и корневой системы переходного (ио-
лувзрослого) типа, появление отдельных взрослых черт в структуре по-
бегов (например, смена типов нарастания, начало ветвления, появление
плагиотропных побегов и др.). Сохранение отдельных элементов первич-
ного побега.
v — Появление основных черт типичной для вида жизненной формы.
Растения имеют характерные для вида взрослые листья, побеги и корне-
вую систему. Генеративные органы пока отсутствуют.
g( - Появление генеративных органов. Преобладание процессов новое >6
разевания над отмиранием, проявляющееся в разных формах. В некою
рых случаях (полурозеточные формы и др.) окончательное формирование
взрослых структур.
g, Уравновешивание процессов новообразования и отмирания
Максимальный ежегодный прирост биомассы, максимальная семенная
продукгнвноегь.
42 Hill MtiphOfl I |()liyil>IUIlOIIIHI)l bliouoi IIH pill 1СППЙ
Вставка 4 (продолжение)
g3— Преобладание процессов отмирания над процессами новообразова-
ния: резкое снижение генеративной функции, ослабление процессов кор-
не - и побегообразования. В некоторых случаях - упрощение жизненной
формы (выражающееся, в частности, в потере способности к образова-
нию побегов разрастания).
ss — Полное отсутствие плодоношения. Резкое преобладание отмирания
над новообразованием. Возможно упрощение жизненной формы, прояв-
ляющееся в смене способа нарастания (или в потере способности к вет-
влению), вторичное появление листьев переходного (имматурного) типа,
s — Накопление отмерших (прекративших рост) частей растения.
Предельное упрощение жизненной формы, вторичное появление некото-
рых ювенильных черт организации (форма листьев, характер побегов и
др.). В некоторых случаях - полное отсутствие почек возобновления и
других новообразований (по Ценопопуляции..., 1976).
ненные рисунки (по определению их автора — «биографические портре-
ты») растений, находящихся на разных стадиях онтогенетического разви-
тия (Нухимовский, 1997) наилучшим образом иллюстрируют используемые
в диагнозах морфологические критерии (рис. 3.1). Большая важная работа
выполнена в этом направлении коллективом авторов ботаников Марийского
государственного университета и их коллег из других вузов, участвующих в
создании «Онтогенетического атласа лекарственных растений». В четырех
томах этого издания иллюстративно представлена череда онтогенетических
состояний для каждого из 144 охарактеризованных видов.
3.2. Базовые спектры онтогенетических состояний
и классификация популяций
Плодотворные идеи Т. А. Работнова о значимости спектра возрастных
или онтогенетических состояний как важнейшей характеристики популя-
ции любого многолетнего растения были подхвачены рядом научных кол-
лективов, до сих пор чрезвычайно популярны и активно эксплуатируются.
Это касается и самого понятия «ценопопуляция», привязывающего популя-
цию к определенному фитоценозу, правда, без снятия проблемы выявления
границ самого фитоценоза, и выявления базовых и теоретических спектров
для ряда видов, для классифицирования популяций по характеру их спектра
(инвазионные, нормальные, регрессивные), и выявления «стратегий жиз-
ни», стратегий жизненных циклов или популяционных стратегий.
На рисунке 3.2 показан принцип выявления базового и теоретического
спектров, как максимально вероятных для популяций того или иного вида и
потому полученных путем усреднения частот в каждом из классов онтоге-
нетического состояния. Особенно важно уловить не просто вариант наибо-
лее часто встречающегося спектра, цо спектр с устойчивыми соотношения-
Ihlflll { ( 'XCMI.I II МОДСНИ Ж1Г1НГIllil.lM llltuuoll
43
Рис. 3.1. Изображения oinoic
нетических состояний у юр
ца живородящего (со схе-
мой) и гравилата речного
(Нухимовский, 1997)
с — семядоли; гп — главный от-
крытый побег; вс — верхушеч-
ное соцветие репродуктивного
побега; св — бульбочка (специ-
ализированная выводковая поч-
ка в соцветии) рп — репродук-
тивный побег; пр---- пророс-
шая бульбочка;
А — проростки; Б — ювениль-
ные; В — имматурные особи.
44
Л / Н Miiphtm I h я ly нищи я и ши (>1101101 пи puci ciinit
2 о--о 3
Рис. 3.2. Принцип выделения базового спектра для популяций некоторых ви-
дов растений (А — бурачка ленского Alyssum lenense, Б — анабазиса безлистно-
го Anabasis aphylla. В — овсяницы бороздчатой Festuca sulcata, Г— о. луговой F
pratensis,)-, 1 — базовый спектр, 2 — зона базового спектра, 3 — теоретический
спектр.
ми во взрослой (наиболее стабильной) части популяции. Ведь вполне есте-
ственно, что популяция меняется во времени, и меняется ее спектр.
Многократное и плохо прогнозируемое повторение пиков семенного раз-
множения и возобновления порождает впечатление инициации волнового
процесса.
Популяции с левосторонним спектром, в которых преобладают особи, нахо-
дящиеся на ранних стадиях онтогенеза, принято называть инвазионными, т. е.
находящимися на стадии внедрения, инвазии в данный фитоценоз или данное
местообитание. Поступление зачатков бывает зачастую неритмичным, случай-
ным, и потому учитывать «молодую» часть популяции при выявлении базового
спектра для многих видов не целесообразно.
I iathi / ( xi’Mi.i и модени ж iriiiciiiiы ч ник нов популяций
45
Успешное приживание проросi ков
н переход растений в следующие он-
тогенетические состояния и, главное,
к репродукции — это атрибуты нор-
мальной популяции, хотя ее полноч-
ленность (наличие в момент анализа
особей всех онтогенетических состо-
яний) не является обязательной.
Регрессивная популяция, наобо-
рот, характеризуется правосторон-
ним спектром и преобладанием в по-
пуляции сенильных или старых гене-
ративных особей.
Развитие популяции от инвази-
онного до регрессивного состояния
предложено рассматривать как осу-
ществление «большой волны раз-
вития» (большого жизненного цик-
ла популяции — по Работнову, 1969).
Продолжительность большой вол-
ны развития у разных видов расте-
ний сильно отличается, завися от био-
логических свойств вида (длительно-
сти жизни особи, жизненной формы
и т. д.) и влияния внешних факторов.
Например, в популяциях монокар-
пических видов есть периоды, когда
Рис. 3.3. Схема жизненных циклов рас-
тений с показом возможной поливари-
антности онтогенеза (по Бигон, Харпер,
Таунсенд, 1999 с изменениями): 3 —
зима; Л — лето; Одн. — однолетник;
Двул. — двулетник; Многол. — много-
летник; штриховка — репродукция.
они представлены только жизнеспо-
собными семенами в почве (рис. 3.3).
Таблица 3.1 дает представление о системе монокарпических растений с уче-
том поливариантности онтогенеза.
Таблица 3.1.
Система монокарпических растений по продолжительности жизни
с учетом поливариантности развития
МОНОКАРПИКИ
I. Малолетние монокарпики
1.1. Малолетние монокарпики с индетерминированным ростом
(т.н. «растянутая монокарпия»)
1.1.1. Незимующие (однолетники)
1.1.2. Зимующие
1.2. Малолетние монокарпики с детерминированным1 ростом
1.2.1. Однолетники
1.2.1.1. Однолетники незимующие (моновариантные)
1 Имес гея в виду детерминация мощности развития репродуктивной сферы предшествующим
во времени развитием сферы вегетативной.
46 Л/ // MaphtHi 11 (>i iy 11 я hi i< н 111 ii и ( hi< > ji< >i пи рас iciinii
Таблица 3.1. {продочжение)
1.2.!.!.!. Одполс! ники весенние (эфемеры)
1.2.1.1.2. Однолетники летние
1.2.1.2. Однолетники зимующие
1.2.1.2.1. Однолетники зимующие поливариантные (с моноцикличе-
ским (эфемерным) и дициклическим вариантами)
1.2.1.2.2. Однолетники зимующие моновариантные (только с дици-
клическим вариантом = озимые)
1.2.2. Двулетники
1.2.2.1. Двулетники моновариантные (только дициклический
вариант) — облигатные двулетники
1.2.2.2. Двулетники поливариантные (моно-, ди- и
трициклический варианты)- факультативные
двулетники
II. Многолетние монокарпики
II. 1. Многолетние монокарпики с детерминированным ростом
II. 1.1. Многолетние монокарпики поливариантные (ди- и полицикличе-
ский варианты)
II. 1.2. Многолетние монокарпики моновариантные (только полицикличе-
ский вариант)
3.3. Схемы-модели пофазного рассмотрения жизненных циклов
популяций
Задача схем и моделей, предложенных зарубежными экологами и, в пер-
вую очередь, лидером зарубежных ботаников популяционистов Джоном
Л. Харпером, была несколько иной. В них визуализируются жизненные ци-
клы растений с обозначением стадий или фаз, которые являются точками
приложения регуляторных сил, определяющих численность всех компонен-
тов популяции, включая семенные пулы или банки. При этом последова-
тельность в жизненном цикле этих фаз удобно использовать как некий план
для подробного рассмотрения связанных с ними событий в жизни популя-
ций разных видов. Именно такие схемы были опубликованы Д. Харпером и
Д. Уайтом (Harper, White, 1971), которые в весьма образной форме охарак-
теризовали содержание каждой из четырех фаз (см. рис. 3.4а, б).
I фаза включает опад семян (дословно — seed «rain»). В ходе дисперсии
семена, сформированные активными особями предыдущего поколения, по-
полняют запас жизнеспособных семян в почве, накопленный популяцией
в предшествующие годы. Этот запас или семенной банк наряду с другими
аспектами биологии и экологии диаспор будет подробнее рассмотрен нами
в главе 5.
II фаза. Часть свежеосыпавшихся, а также часть уже имевшихся ранее в по-
чве семян прорастает и образует проростки под контролем среды, действующей
как «просеивающий», отбирающий фактор (дословно с английского — сито сре-
ды — «environmental sieve»). На схеме ячейки сита показаны жирной пунктир-
ной линией.
I 'hitia 1 ( xcmi.i и модгии Ail iiiriiiii.H iiiiknoii ионуiimiuni
47
III фа за. 11од нснрскращающнмся
пшисипсм среды происходя! рост и
разни гис растений, проявляющиеся в
коррелированных изменениях числа и
размеров особей популяций (влияние
среды показано на схеме в виде двой-
ных жирных линий). Появляющиеся
проростки растут, взаимодействуют и
дают взрослые растения, плодонося-
щие в разной степени.
IV фаза. Продуцирование семян
особями, которые смогли достичь ре-
продуктивной зрелости и которые
формируют вклад в семенной фонд
популяции, реализуя семенную про-
дукцию в процессе диссеминации.
Позднее в своей объемной моно-
графии Д. Харпер (Harper, 1977) ис-
пользовал фазы, выделенные в рас-
смотренной схеме для составления
двуцветных иконок, предваряющих
каждую из глав. Выделенные цветом
или штриховкой фрагменты икон-
ки символизировали содержание гла-
вы, указывая на ту часть жизненного
цикла, которая будет в ней подробно
рассмотрена.
Д. Харпер и Д. Уайт сделали по-
пытку построить на базе этой схемы-
модели жизненного цикла популяции
Рис. 3.4а. Схема Дж. Харпера для одно-
летнего монокарпического растения.
/ // ///-/I/
Рис. 3.46. Схема Дж. Харпера для мно-
голетнего поликарпического растения
(пояснения в тексте).
однолетнего растения модель жизнен-
ного цикла популяции многолетнего
поликарпического растения (см. рис. 3.46).
Можно заметить, что отличительными особенностями здесь выступают:
более сложное, чем в первой схеме «сито среды» из-за присутствия сосед-
ствующей с популяцией модельного вида растительности и опада, а так-
же наблюдающееся из года в год или с перерывами плодоношение. Видно,
что модель пришлось несколько усложнить, исходя из необходимости обо-
значить прорастание семян и появление проростков на поверхности почвы,
уже занятой другими растениями и покрытой слоем отмерших час гей (опа-
да). Некоторым проросткам удается преодолеть этот барьер, но далеко нс
всем. Отразить каким-либо образом наличие вегетативного возобновления
и размножения на схеме-модели авторам не удалось. Отсюда ее весьма огра-
ниченное значение — модель в лучшем случае пригодна для схематизации
жизненного цикла популяций многолетних поликарпических растений, воз
обновляющихся исключительно семенным путем.
48
MH Маркин 11 он у иицшн 1 паи (>1101101 пи рас iciinii
3.4. Графы жизненных циклов популяции
Несколько иную модель для анализа жизненного цикла популяции многолет-
него растения (рис. 3.5) предложил Д. Ф. Грайм (Grime, 1979). Он выделяет в
жизненном цикле популяции любого растения две главных фазы —регенера-
тивную фазу или фазу возобновления (regenerative = immature phase) и фазу
«проявления» или зрелости (establish = mature phase) в смысле проявления
особями наибольшей экологической активности. Каждая из двух главных
потомок (рамета)
Рис. 3.5. Схема жизненного цикла по-
пуляции многолетнего поликарпиче-
ского растения с семенным и со специ-
ализированным вегетативным размно-
жением:
регенеративная фаза
фаза зрелости -----
фаз подразделяется на ряд промежу-
точных стадий. Так регенеративная
фаза состоит из серии стадий (вскры-
вание плодов — освобождение се-
мян, диссеминация, покой, прораста-
ние и приживание проростка), при-
чем каждая отличается по продолжи-
тельности и по действующим на ее
протяжении механизмам, специфич-
ным для видов и популяций. Фаза
зрелости тоже характеризуется набо-
ром взаимосвязанных стадий, вклю-
чающих освоение (capture — дослов-
но — перехват) ресурсов, рост, кон-
куренцию, продуцирование семян.
В схеме Грайма для многолетних
растений фигурируют две «петли»
(по-английски — loops} — учтены
два возможных и параллельно осу-
ществляющихся способа регенера-
ции (= возобновления) — семенным
путем и вегетативным (соответствен-
но, 1 и 2 на рис. 3.5). Поскольку на
схеме предусмотрена дисперсия ве-
гетативных зачатков, то речь идет в
данном случае о специализирован-
ном вегетативном размножении, по-
добном размножению бульбочками
горца живородящего (см. рис. 3.1).
Как видно на схеме, в каждом из двух
вариантов осуществления регенера-
тивной фазы выделяются сходные
стадии. Грайм подчеркивает, что у
большинства многолетних растений
два типа потомков радикально отли-
чаются по свойствам: размерам, про-
дуктивности, эффективности рас-
пространения, степени изреживапия,
конкурентоспособноети и др.
/ hlflil < ( ‘XCMI.I II МОДСИН /КП IIU* 1111 l.l 4 llllk UOll 11 <) 11У! IMIU111
49
Как уже было сказано, nia'icinic рассмотренных схем-моделей cocioin
1лавным образом в выделении основных звеньев в цени поколении п i пан
ных точек приложения регуляторных сил, влияющих на демографические
параметры популяций. Дальнейшая разработка подобных моделей привела
к необходимости включения в них количественных параметров. Так появи-
лись модели, называемые графами жизненных циклов, учитывающие осо-
бенности популяционной структуры как возрастной, так и размерной.
В простейшем варианте они могут быть представлены в виде хронологи-
ческой серии состояний или спектров с учетом активного и, отчасти, пас-
сивного компонентов одной и той же популяции в череде лет. Примером га-
кой презентации данных могут послужить наши результаты изучения мо-
дельной популяции рябчика шахматовидного Fritillaria meleagroides. соз-
данной из собранных в природе и пересаженных на опытный участок 30
луковиц отплодоносивших экземпляров. Размеры луковиц были примерно
одинаковы и в среднем составляли в длину около 1,0 см, а в ширину при-
мерно 1,3 см. В распоряжение каждой луковицы была предоставлена одина-
ковая площадь питания, а место посадки было строго фиксировано. В даль-
нейшем число растений, переходящих к активному росту, и их состояние
отмечали ежегодно. Как и у многих других эфемероидов, каждая активно
растущая особь рябчика ежегодно весной заново формирует надземный ве-
гетативный либо генеративный побег, по состоянию которого можно судить
о процессах, происходящих в подземной сфере.
Главный вопрос, который предстояло выяснить, — возможно ли возник-
новение разногодичной поливариантности поведения особей, взятых на
одной и той же стадии онтогенеза, если они произрастают в искусствен-
но выровненных условиях. Наряду с этим интересовала возможность и ча-
стота перехода особей в состояние вторичного покоя и способность рябчика
шахматовидного к вегетативному размножению в условиях эксперимента.
Несмотря на одинаковый онтогенетический статус особей на старте опы-
та, в ходе семилетних наблюдений была выявлена поливариантность разви-
тия. Поэтому уже на следующий после посадки год оказалось возможным
разделить растения по их онтогенетическому состоянию на 4 группы: юве-
нильные, вегетативные, генеративные и находящиеся в состоянии вторич-
ного покоя2.
На рис. 3.6, где представлена суммарная диаграмма переходов, в которой
наряду с другими онтогенетическими состояниями фигурируют отсутствую-
щие в надземной сфере (=в активной части популяции) особи, можно виден»,
как возникла и развивалась поливариантность онтогенезов особей, находив-
шихся в начале опыта на одной и той же стадии. При этом судьбу некоторых
из них можно было отследить не каждый год, а только в год их активности.
2 Ювенильными считали растения, имевшие один настоящий лист (от 5 до 7 мм шириной)
Вегетативными называли нецветущие особи с числом листьев от 2 до 4-5. Генеративные
(цветущие и плодоносящие) особи могли нести от трех до восьми листьев на монокарпичс
с ком побеге. Если особь ничем себя не проявляла в надземной сфере в какой-либо данный
год, но переходила в активное состояние в другие годы, то в данном году ее считали находи
1ЦСЙСЯ в состоянии вторичного покоя.
50
А/// Маркой I к я iy нм цп< HiiuiM (июнем им рис i сппй
Генеративные
Вегетативные
Отсутствующие
30(100%)
30
13 (43,4%)
7 (23,3%)
10(333%)
В1
14(45,2%)
7 (22,6%)
8 (25,8%)
1993
2 (6,4%)
ВЗ
8 (23,5%)
9 (26,5%)
13 (38,2%)
1994
В4
4(11,8%)
12(30,8%)
13(33,3%)
6(15,4%)
8 (20,5%)
Рис. 3.6. Граф, показывающий поливариантность онтогенезов особей в модельной
популяции рябчика шахматовиднрго — варианты переходов особей из одного он-
тогенетического состояния в другое и связанные с этим погодичные изменения он-
тогенетических (возрастных) спектров.
В — новые особи, появившиеся вследствие вегетативного размножения.
1995
Общее
число
30
31
34
39
На основании семилетних наблюдений (на рис. 3.6 представлены данные
только за 4 года) в опыте удалось выяснить, что переход в состояние вто-
ричного покоя — частое явление в интродуцированной популяции рябчи-
ка. 17 особей демонстрировали такой переход, причем 11 из них пребывали
в состоянии вторичного покоя один год, две особи — два года подряд, одна
особь — два раза по одному году, между которыми был двухлетний активный
Гшни J ( xcmi.i и модели >Kirniriiiii.iк щитов популяций
51
рост, одна особь два раза по одному году, между которыми был пятилетний
активный рост, одна особь три раза по одному году, между которыми па
блюдался ее активный рост в течение одного сезона и одна особь — три года
подряд. Все эти особи не исчезли совсем и были зафиксированы вновь в со-
ставе активной части модельной популяции. Часто выход из состояния по-
коя был сопряжен с сильным омоложением. Например, в популяции было
отмечено появление ювенильных особей, которое можно объяснить толь-
ко вегетативным размножением, обычно (но не всегда!) сопряженным с от-
миранием материнской луковицы. В последнем случае наблюдалось как бы
замещение особи, достигшей генеративного состояния, ее потомком (раме-
той), демонстрирующим статус ювенила. Обычно такая картина наблюда-
лась после ухода особи в состояние покоя, за время которого, по-видимому,
и разрушалась материнская луковица, отделявшая от себя мелкую дочер-
нюю луковицу, дающую начало ювенильной особи.
В конце наблюдений, через 7 лет, вероятно, уже можно считать, что 4 осо-
би погибли, три из них ни разу с момента посадки не были отмечены в ак-
тивных фазах, а одна вегетировала только один раз (на следующий год по-
сле посадки). Еще одна особь ничем не проявила себя в течение четырех по-
следних лет наблюдений. 8 особей проявляли активность каждый год, но из
них только одна ежегодно формировала генеративный побег, на котором од-
нажды развилось два цветка. Другие семь чередовали в разном ритме сезо-
ны, когда формировались генеративные побеги с сезонами, когда наблюда-
лась только активная вегетация.
Влияние, которое такой несинхронный ритм онтогенетического развития,
мог оказать на результаты сравнения активных частей одной и той же популя-
ции по корреляционной структуре признаков в разные годы, мы уже обсуж-
дали выше (см. главу 2).
Рассмотренный нами пример убеждает в том, что в графах жизненного
цикла совершенно необходимо учи-
тывать не только семенной и вегета-
тивный способы возобновления, но и
стадию покоя. Разумеется, для разных
жизненных форм это могут быть раз-
ные формы покоя: у малолетних рас-
тений это будет первичный покой жиз-
неспособных семян, а у прочих — по-
мимо этого, и вторичный покой мно-
голетних подземных органов.
На рис. 3.7 представлен граф жиз-
ненного цикла, который учитывает
как два способа возобновления (ве-
гетативный и семенной), которые мо-
гут происходить с разной периодич-
ностью и интенсивностью, что отра-
жено в размерах соответствующих
«петель», так и вариант («петля») с
уходом в состояние покоя.
Рис. 3.7. Граф жизненного цикла с по-
казом варьирования времени оборо-
та (показано размерами «петель») при
разных способах возобновления и ухо-
де в состояние покоя.
Рис. 3.8. Графы жизненных циклов для популяций, структурированных по четы-
рем возрастным (онтогенетическим) состояниям (а) или по четырем размерным
группам (б).
a: F2 4 — плодовитость особей в возрастных группах (со второй по четвертую);
Р — вероятность выживания до следующего возрастного (онтогенетического)
состояния (перехода в следующий возрастной класс);
б: F2 4 — плодовитость особей в размерных группах (со 2-й по 4-ю); Р — веро-
ятность выживания в каждой из четырех размерных групп; Gj 3 — вероятность
перехода особей данного размерного класса в следующий размерный класс (по
Caswell, 1989).
На графах, приведенных на рис. 3.8, которые представляют собой графы
жизненных циклов популяций, структурированных по четырем возрастным
(онтогенетическим) состояниям (а) или по четырем размерным группам (б),
даны все необходимые пояснения относительно петель-переходов.
Однако не следует забывать, что в пределах ареала вида необходимо быва-
ет учитывать и потоки генов, связывающие между собой отдельные популя-
ционные локусы или же метапопуляции. Тогда графы для каждого из них изо-
бражаются отдельно, как это сделано на рис. 3.9, где в одну систему объеди-
нены графы жизненных циклов для трех популяционных локусов (метапопу-
ляций), связанных потоками генов через дисперсию семян.
Рис. 3.9. Графы жизненных циклов для трех популяционных локусов (метапопу-
ляций), связанных потоками генов через дисперсию семян.
3.5. Схемы и модели для наглядного представления о
«стратегиях жизни», стратегиях жизненных циклов
или эколого-ценотических стратегиях
Нетрудно заметить, что поливариантность и ритм переходов особей по-
пуляции из одного состояния в другое напрямую связаны с тем, что можно
назвать популяционным поведением. Априори можно предполагать разли-
чия разных видов и популяций по их популяционному поведению, основные
типы и особенности которого были выработаны в ходе эволюции.
Вопрос о существовании видов растений с различным типом пове-
дения и различной ролью в сообществе не нов. Еще в 1935 и 1938 гг.
Л. Г. Раменский выделил три типа растений по поведению в условиях
растительных сообществ (три фитоценотипа). К первому типу он отнес
растения с ярко выраженными конкурентными эдификаторными свой-
ствами и назвал виолентами (силовиками, «львами»). Благодаря энергии-
54
М.Н Марков
11 (>i iy 11 я i и к > 1111; i я bitonot пя рас iciini'i
Л. Г. Раменский.
Ф. Грайм.
ному разрастанию вегетативных надземных и подземных органов виоленты,
перехватывают «львиную долю» ресурсов, конкурентно подавляют другие
виды растений и обычно доминируют в сообществах, формирующихся в про-
дуктивных слабо нарушаемых местообитаниях. «Где хочу, там расту — где
хорошо расту, там подавляю» — вот по какому принципу осуществляет-
ся стратегия виолентов. Второй тип поведения — выносливость, а виды
растений, отнесенные к этому типу, были названы Л.Г. Раменским па-
тиентами (выносливцами, «верблюдами»). Для эксплерентов («шака-
лов»), так были названы растения, отнесенные к третьему типу, в отсут-
ствие конкурентов (при временно высокой обеспеченности ресурсами)
характерны быстрый рост и накопление биомассы, которое, однако, бы-
стро реализуется в энергичном семенном размножении. Поэтому харак-
терная для них высокая скорость этого вида размножения лишь отчасти
компенсирует низкую конкурентоспособность, связанную с затратным для
растений процессом формирования семян. Подобно шакалам эксплеренты
исчезают из сообщества в ходе восстановительной сукцессии (демутации).
Разные типы поведения видов в сообществах (ценотипы или фитоцено-
типы) можно с полным правом назвать стратегиями жизни и трактовать
как три линии эволюционного развития. Само понятие о стратегии жизни
у растений вошло в отечественную литературу после появления статьи
Т. А. Работнова «Изучение ценотических популяций в целях выяснения
«стратегии жизни» у растений» в 1975 г. В этой статье было дано опре-
деление стратегии жизни как совокупности приспособлений к произраста-
нию данного вида в определенных условиях среды в сообществах с дру-
гими видами растений. ч
По мнению Т. А. Работнова эволюция растительного мира шла по
гром направлениям: 1) выработка способности к максимально поипому
r>hl(ht < Схемы II МОДСIIII ЖИ IIICIIIII.IH link нов популяций 55
использованию ресурсов мссюобигания, конкуреннюму подавлению дру
। их видов и вытеснению их из местообитаний с благоприятными услови
ями (виоленты)} 2) выработка способности переносить неблагоприятные
условия конкурентного давления со стороны виолентов или избеган» кон
куренции с виолентами, занимая малопродуктивные местообитания (пи-
тиенты)} 3) выработка способности к быстрому захвату временно осво-
бодившихся территорий, быстрому развитию и плодоношению, связанному
с насыщением почвы диаспорами, которые длительное время могут нахо-
диться в состоянии покоя и сохранять жизнеспособность (эксплеренты).
Т. А. Работнов справедливо считал, что для отнесения какого-либо вида
к одному из трех ценотипов, предложенных Л. Г. Раменским, необхо-
дим анализ всего состава популяций этого вида в разных сообществах,
включая изучение запаса жизнеспособных семян в почве. Приведем не-
сколько примеров.
Ель обыкновенная — потенциальный виолент, который сам, обладая зна-
чительной теневыносливостью, сильно ограничивает световое довольствие
видов, произрастающих совместно с ним. Помимо этого, перехваты-
вая воду и элементы минерального питания, ель безраздельно властвует
в образованных ею фитоценозах. Вместе с тем, она, как мощный сре-
дообразователь, создает относительно благоприятную среду для некоторых
видов, свойственных темнохвойным лесам.
Тростник обыкновенный, образующий мощные заросли на мелководьях
или влажных аллювиальных почвах, тоже проявляет себя в этих условиях
как потенциальный виолент. Это травянистое растение конкурентно пода-
вляет растущие рядом другие виды.
Сосну обыкновенную можно отнести к типичным патиентам, посколь-
ку она вытесняется другими видами деревьев (например, елью) из более
продуктивных местообитаний с благоприятными лесорастительными
условиями, где она потенциально способна расти гораздо лучше. Таким
образом, типичными патиентами можно назвать виды, способные в есте-
ственной обстановке успешно произрастать и иметь наибольшую числен-
ность в неоптимальных для них условиях.
3.5.1. Критерии разграничения видов с разными стратегиями жизни
Одним из критериев, позволяющих различать виоленты и патиенты, сле-
дует вслед за Т. А. Работновым признать совпадение и несовпадение аут- и
синэкологического оптимумов. Л. Г. Раменский — основоположник учения
об экологической индивидуальности видов растений ввел в обиход фито-
ценологов амплитудные экологические шкалы, по которым можно опреде-
лить фитоценотический (синэкологический или фактический) оптимум вида.
Сопоставляя эти данные, которые могут быть представлены в форме одно-
вершинных кривых (кривых распределения по градиенту условий, отражаю-
щих количественное участие видов в сложении ценозов), с данными об эко-
логическом или потенциальном оптимуме, можем наблюдать либо совпаде-
ние, либо несовпадение оптимумов. Для виолентов характерно их совпаде-
ние (как уже говорилось, где хочу, там расту!), поскольку виоленты, обладая
ВЫСОКОЙ KOIIKypCII lOCIIOCoGlIOC I 1»1О, ДОМ!111Иру|О1 I! ВЫ1ССНЯ1О1 дру| ИС НИДЫ В
условиях своего jkojioi пиескою оптимум;!. Для 'типичных илтисигов совпа-
дение оитимумов не характерно, г. е. они, как правило, лучше растут вне со-
обществ, в составе которых чаще всего встречаются. Эти растения мирят-
ся с теми условиями, которые предоставляют им эдифицирущие виоленты.
Однако даже такой важный критерий, взятый изолированно от других харак-
теристик сравниваемых видов, не всегда срабатывает. Например, облигатный
патиент белоус торчащий Nardus stricta произрастает, как правило, на бедных
кислых почвах, где часто доминирует. В опыте удалось показать, что мак-
симальный урожай белоус дает уже при 27 мг азота на 1 кг песка, в то время
как другой патиент — полевица тонкая — повышает урожай вплоть до самой
высокой дозы азота, использованной в опыте (243 мг на 1 кг песка). Это озна-
чает, что у белоуса, несмотря на то, что он является патиентом, два типа опти-
мумов совпадают. В данном случае мы сталкиваемся с примером специали-
зированных патиентов, к рассмотрению которых мы еще вернемся ниже.
Если виоленты и патиенты представлены исключительно многолетними
растениями, то среди эксплерентов главенствуют однолетники, хотя вид-
ная роль принадлежит также вегетативно подвижным многолетникам, та-
ким как лютик ползучий, будра плющевидная, иван-чай узколистный и др.
3.5.2. Дальнейшая теоретическая разработка проблемы стратегий
К дальнейшей разработке проблемы стратегий жизни каждый исследова-
тель подходит по-своему. Т. А. Работнов предложил более детальную клас-
сификацию стратегий жизни. Особенно это касается патиентов и эксплерен-
тов. Патиенты логично подразделять на типичные патиенты, не утратившие
способности произрастать лучше при устранении экстремальности условий
среды, и специализированные патиенты, которые могут быть специализиро-
ваны экотопически (например, галофиты) или фитоценотически (например,
ряд видов темно-хвойных лесов — кислица и др.). У специализированных па-
тиентов отмечено полное или почти полное совпадение оптимумов что гово-
рит об утрате ими способности положительно реагировать на ослабление не-
благоприятности условий среды. Среди эксплерентов предложено различать:
1) сезонные эксплеренты, использующие ослабление конкуренции в какой-то
сезон года, например эфемероиды широколиственных лесов; 2) флуктуаци-
онные эксплеренты — растения, способные использовать снижение напря-
женности конкуренции в отдельные годы в связи с флуктуационными изме-
нениями фитоценозов, например, лютик ползучий на лугах в годы вымока-
ния; 3) сукцессионные эксплеренты — виды, разрастающиеся на месте уни-
чтоженных или сильно нарушенных фитоценозов, например, демутационный
эксплерент иван-чай узколистный на гарях и очень многие однолетники.
Популяционный анализ видов, входящих в синузию эфемероидов и
синузию широкотравья, проведенный О. В. Смирновой (1982), еще раз
убедил в необходимости выделения промежуточных типов стратегий. О
наличии переходов между типичными виолентами, патиентами и эксплерен-
тами писал и сам Л. Г. Раменский. Поэтому с полным правом, ориентиру-
ясь на мнение автора концепции, можно говорить о степени выражение-
С III BIIOllCH I ПОС I II, IIIH IICII I HOC I II, ЖСHJiepCII I HOC I II. Наличие Н|)омсжу точ-
ных iiiiioB cipaicruii hoibojimci рассматривал, своеобразный коп।инуум,
oi l рапичеппый с двух сторон двумя крайними с тратегиями: нагиентами, с
одной стороны, и эксплерснтами — с другой.
RGR
Рис. 3.10а. «Триангулярная модель» для ординации стратегий (по Gri^e> 1979 с из-
менениями). RGR — относительная скорость роста (г/г х неделя); М трехкомпо-
нентный морфологический индекс (пояснения в тексте).
Ат — Achillea millefolium, тысячелистник обыкновенный; Apr — Alopecurus pra-
tensis, лисохвост луговой; Ar — Agropyron repens, пырей ползучий; А$е___Arenaria
serpyllifolia, песчанка тимьянолистная; Bt — Bidens tripartita, череда трехраздель-
ная; Cal — Chenopodium album, марь белая; Cpr — Cardamine prat^nsjSf сердеч-
ник луговой; Cv — Clinopodium vulgare, пахучка обыкновенная; De —>.Deschampsia
caespitosa, луговик дернистый; Dg — Dactylis glomerata, ежа сборная; pg_Festuca
gigantea, овсяница гигантская; Fr — Festuca rubra, овсяница красная; Qa_Galium
aparine, подмаренник цепкий; Gv — Galium verum, подмаренник настоящий;
Gu — Geum urbanum, гравилат городской; Kc — Koeleria cristata, тон^оног гребен-
чатый; Lc — Lotus corniculatus, лядвенец рогатый; Mm — Matricaria matricarioi-
des, ромашка душистая; Мп — Melica nutans, перловник поникший; ns_______Nardus
stricta, белоус торчащий; Pa — Роа annua, мятлик однолетний; Pav Polygonum
aviculare, горец птичий; Pl — Plantago lanceolata, подорожник ланцет0ЛИСтный;
Pm — Plantago major, подорожник большой; Ra — Rumex acetosellc^ щавель кис-
лый; Sa — Sedum acre, очиток едкий; Sm — Stellaria media, звездцатка средняя;
Sv — Senecio vulgaris, крестовник обыкновенный; Tm — Trifolium тесцит клевер
средний; Ud — Urtica dioica, крапива двудомная; Va — Veronica arvensjs> верони-
ка полевая.
58 MH Миры nt I loiiyiiHiiiioiiiHiH Giioiioi ii>i piii iciinil
5.5.3. Гриангулярная модель Дж. Ф. Граима
В вышедшей в 1979 г. книге «Стратегии растений и процессы в раститсяь-
ности» Дж. Ф. Грайм (J. Р. Grime) выделяет три стратегии: конкурентную
(competitors — С), стресс-толерантную (stress-tolerators — S) и рудеральную
(Ruderals — R). Нетрудно заметить полное соответствие выделенных им ти-
пов фитоценотипам Л. Г. Раменского, но при этом использованные терми-
ны явно менее удачны. Ведь все без исключения виды растений вступают
в конкурентные отношения и потому являются «конкурентами», под «ру-
деральными» растениями» в ботанике давно понимаются растения мусор-
ных мест, а условия произрастания типичных «стресс-толерантов» для них
не являются стрессовыми. Однако факт появления книги Дж. Грайма за-
ставляет нас более внимательно относиться к разработке положений, выска-
занных ранее нашими соотечественниками. Поставив задачу сопоставления
стратегий по количественным показателям, Грайм не ограничился выделе-
нием дискретных типов и применил очень интересную по замыслу триангу-
лярную (треугольную) ординацию (рис. 3.10). Он наклонил вертикальную
ось координат на 45°, и ограничил (по сути дела исключил), таким образом,
область маловероятных сочетаний параметров, он поместил в углах треу-
гольника три вышеназванных типа основных стратегий. В поле треугольни-
ка каждый вид растений получил свою точку, лежащую на пересечении ко-
ординат, а координаты определяются по двум параметрам. Первый и глав-
ный параметр — это максимально возможная скорость накопления биомас-
сы (RGRmax см. главу 8), которая откладывается на логарифмической шкале
и выражается в единицах биомассы, накопленной в пересчете на единицу
начальной биомассы за неделю (рис. 3.10а). Второй параметр сборный. Это
зависящий от жизненной формы растения морфологический индекс, кото-
рый вычисляется по формуле:
М = (а + Ь + с)/2, [3.1]
где а — максимальная высота растения; b — величина горизонтально-
го разрастания; с — максимально возможная аккумуляция (толщина слоя)
опада в местообитаниях, свойственных данному виду. Величина каждо-
го из трех компонентов морфологического индекса оценивается при помо-
щи шкал: первого — по десятибалльной, а двух других — по пятибалль-
ным. Таким образом максимальное значение морфологического индекса у
какого-либо вида составляет 10 баллов, что соответствует градуировке шка-
лы. На рисунке 3.106 можно видеть, что поле треугольника поделено на
зоны, соответствующие трем основным и четырем промежуточным страте-
гиям: C-R, C-S, S-R и C-S-R.
3.5.4. О г- и К-стпратпегиях
Подобный континуум рассматривается и многими известными зарубеж-
ными исследователями, которые используют для обозначений крайних ва-
риантов константы, фигурирующие в уравнении логистического роста по-
l 1 Схемы и модели жтнгнным пик нов популяций
59
иуляции. '' )кс 11OI ici II |И<1II l»l I ый рост
численности популяции определя-
ется константой г, но продолжается
он сравнительно недолго, пока по-
пуляция не достигнет численно-
сти К — константной при каких-то
определенных условиях. Численность
К — это верхний предел насыще-
ния пространства особями данно-
го вида (см. ниже рис. 3.12, 3.13),
и чем ближе плотность популяции
к своему максимальному значению,
тем медленнее происходит ее уве-
личение из-за обострения конку-
рентных отношений между особями.
Начиная с 1967 г., зарубежные уче-
ные (McArthur, Wilson, 1967) говорят
о К-стратегиях и r-стратегиях и, со-
ответственно, о К-отборе и г-отборе
(К- и r-видах). Для К-видов (видов
с К-стратегией) характерна очень
Рис. 3.106. Сетка, разделяющая орди-
национную карту на фрагменты, со-
ответствующие трем основным и че-
тырем промежуточным стратегиям:
С — компетиторы (конкуренты), S
стресс-толеранты, R — рудералы.
высокая степень насыщения пространства, что сопряжено либо с вы-
сокой конкурентоспособностью и максимально выраженным осуществле-
нием эдификаторной роли, либо способностью довольствоваться остатка-
ми ресурсов под пологом эдификаторов. Можно усмотреть соответствие
К-видов патиентам и виолентам Л. Г. Раменского. Также легко заметить,
что эксплеренты соответствуют r-видам, обладающим высоким репродук-
тивным потенциалом, который позволяет им очень быстро, почти экспо-
ненциально, увеличивать свою численность на первично или вторично сво-
бодных от растительности участках, то есть при отсутствии конкурентов.
«Мир не окрашен только в черное и белое», — так прокомментировал на-
личие переходов между К- и r-стратегиями видный американский эколог
Э. Пианка (1981). Интересен и тот факт, что сукцессии идут обычно в на-
правлении от r-видов, которые благодаря обилию продуцируемых диаспор
первыми поселяются на свободной территории, к К-видам, характерным
для конечных сукцессионных стадий, близких к климаксовым.
Необходимо сказать и еще об одной классификации стратегий, которую
приводит известный американский эколог Р. Уиттекер (1980). К двум уже
названным типам — г и К-видам он добавляет L-виды, которые, произрас-
тая в экстремально неблагоприятных условиях, дают резкие вспышки чис-
ленности, приуроченные к временным улучшениям условий, например, вы-
падению осадков в пустынях и т. п. В неблагоприятные периоды L-виды ни-
чем себя не проявляют, т. к. находятся в состоянии глубокого покоя в виде
семян или многолетних органов в почве.
Сопоставление типов стратегий, выделяемых разными авторами, провел
Б. М. Миркин (1983). Результаты сопоставления представлены в габнице 3.2.
60 МН Миры hi I l<HiyiiMiuioiiiiii>i OiioiioHiH рас 1С1П1Й
Тиблици 3.2.
Соотношение iпион стратегий, выделяемых разными авюрамн
(по Б. М. Миркину, 1983, с изменениями)
Авторы Стратегии
R. McArthur, Е. Wilson г к
J. Р. Grime R с S
R. Whittaker г | L к L
Л. Г. Раменский эксплеренты виоленты патиенты
5.5.5. Типы репродуктивных стратегий
В зарубежной научной литературе часто говорят о репродуктивных стра-
тегиях. Они теснейшим образом связаны с общими стратегиями жизни и
входят в них как главные элементы наряду с темпами накопления общей
биомассы, продолжительностью жизни, размерами тела, экономикой пере-
качивания (аллокации) питательных веществ из отмирающих органов рас-
тений в живые, распределением и перераспределением биомассы в преде-
лах особи. Вопрос о принадлежности любого рассматриваемого вида расте-
ний к одной из групп может решаться только на основе комплексного учета
разных показателей, а не одного, взятого изолированно. Это и понятно, если
учесть существование г—>К континуума.
Дж. Грайм выделяет пять типов репродуктивных стратегий. Это: вегета-
тивная экспансия — V (характеризующаяся замедлением генетических по-
пуляционных процессов, но снижением риска отмирания); сезонная регене-
рация в «окнах» (гэпах — gaps), т. е. локальных нарушениях сообществ, с
подразделением на весеннее и осеннее возобновление — S; возобновление
за счет постоянного банка семян — Bs; возобновление за счет банка (резер-
ва — М.М.) проростков — Bsd; возобновление за счет многочисленных се-
мян и спор, рассеиваемых ветром — W. При этом он допускает возможность
сочетания нескольких репродуктивных стратегий у одного вида.
Из всего сказанного выше следует вывод, что проблема стратегий жизни
у растений, находится в начальной стадии разработки и что динамика рас-
тительных популяций, к рассмотрению которой мы переходим, и все демо-
графические характеристики во многом зависят от репродуктивных стра-
тегий и, в целом, от стратегий жизненных циклов, проявляемых видами.
3.6. Демографический подход к изучению популяций растений
Итак, по нашему рабочему определению популяция — это всегда множе-
ство особей, населяющих определенную территорию. Отсюда вытекает необ-
ходимость применения статистики при изучении биологии популяции как ча-
сти населения вида. Это послужило отправной точкой для исследователей, на-
чавших применять принципы демографии к изучению биологических объектов.
Ведь демография — это «народоописание», раздел статистики, изучающий со-
став и движение (возрастной состав, плотность, рождаемость, сортность и т.
д. - М. М.) населения (см. Словарь иностранных слов, 1954).
I'lKItUI J ( XCMI.I II МОДСНП >IUI niCIIIII.IX HHIUIOII IIOliyJHIHIlil 01
Обратимся к одной из наиболее важных проблем популяционной био
логин проблеме изучения динамики численности и сослана популяции
Называя подобные исследования демографическими, мы вряд ли привносим
большую долю антропоморфизма. Осуществив важный переход от изучения
отдельных особей к изучению их естественных группировок — популяций,
мы сталкиваемся с целесообразностью изучения численности и ее динамики
статистическими методами, а потому заимствуем термин, уже применявший-
ся для обозначения проводимых ранее исследований популяций человека.
Формально демографические исследования, как человека, так и растений
связаны решением основного демографического уравнения. Никакой более
глубокой аналогии здесь не проводится, и проводиться не должно.
3.6.1. Основное демографическое уравнение и его параметры, годич-
ная или конечная скорость роста (X) как мера приспособленности
Основное демографическое уравнение — это простейшая модель роста
(и вообще любого изменения) численности популяции, и интуитивно наи-
более очевидная. Она основана на использовании прямых и относительных
данных о числе рождающихся, отмирающих и мигрирующих (эмигрирую-
щих и иммигрирующих) особей, как о четырех параметрах:
Число особей популя- Число особей попу- ляции в Рождае- мость Смерт- ность Иммигра- ция (Им- Эмигра- ция (Эми- ПТМГН/З
ции в данный момент времени = предше- ствую- щий мо- мент вре- мени + (Родив- шиеся особи) - (Отмер- шие осо- би) + мигриро- вавшие особи) 1рири- вавшие особи)
В буквенном выражении это дифференциальное уравнение выглядит так:
7V/+;=7V4-B-D+I-E
[3.2]
Буквенные символы, взятые для обозначения основных параметров,
представляют собой первые буквы английских эквивалентов (Birth, Death,
Immigration, Emigration). Приходная и расходная части этого уравнения бу-
дут равны, когда популяция пребывает в стабильном состоянии:
B-D + I- E = О или В + I = D + Е
Величины этих популяционных параметров, отнесенные к 7V для вычис-
ления относительной роли (или темпов) каждого из процессов за календар-
ный год, позволяет нам рассчитать (установить) численность популяции
Nnt по истечении года, отталкиваясь от первоначальной численности Nf.
62
М.Н Мирной
I I < >1 ly IIMЦ1К >111 in м Ohqjioi ин рис гений
|3.3|
7V(///V- 1 l-B/A D/ЛМ UN VJN
Скорость, с которой возрастает (или падает) численность за календарный
год, называют годичной скоростью изменения численности популяции. Она
составляет просто отношение конечной численности (по прошествии года)
к первоначальной:
[3-4]
Теперь применим новые обозначения и перепишем уравнение 3.2, обо-
значая в правой его части относительную роль (темпы) основных процес-
сов курсивом (В = B/N'; D = D/N, ;I=l/Nt;E = Е/N,):
Х= 1 +B-D + I -Е
[3.5]
Когда В +1 > D + Е, численность популяции будет экспоненциально ра-
сти со скоростью X, то есть, умножаясь, каждый год в X число раз. Поэтому
X можно использовать в этом случае как постоянный множитель для опре-
деления численности популяции в каждом из последующих годичных от-
резков. Хотя, как уже отмечалось, численность популяции может, не только
расти, но и падать, по сравнению с исходной, для примера рассмотрим си-
туацию с возрастанием численности.
3.7. Теория непрерывного роста численности популяций
в приложении к растениям
Воспользуемся для иллюстрации следующим примером, поясненным схе-
мами. Взяв популяцию некоего однолетника с начальной численностью (Nt) в
3 особи, представим дело так, что она по прошествии последовательных го-
дичных промежутков времени увеличивает свою численность до 9 (Nt+1), 27
(Nt+2) особей и т. д. (рис. 3.11 А). По истечении произвольного числа проме-
жутков времени (х) численность теоретически должна достигнуть значения
Nt+x. Можно написать, что Nt+1 = NtxX = 3x3 = 9 и Nt+2 = Nt+1 хЛ, = 9><3 = 27
или Nt+2 = Nt x Z x Z = Nt x V = 3x3x3 = 27, отметив, что в нашем примере
как универсальный множитель берется X = 3. В общем виде запись будет та-
кой: Nt+x=NtxXx.
Необходимо подчеркнуть также, что здесь мы пока учитываем только
«рождение» новых особей без сохранения старых, поскольку взяли для при-
мера однолетнее монокарпическое растение. При такой ситуации темп ро-
ста численности определяется только плодовитостью особей (в нашем слу-
чае 3 семени/особь). В итоге за два календарных года численность возрос-
ла до 27 особей.
В популяции поликарпического многолетника при той же взятой для при-
мера плодовитости (3 семени/особь), но выживании и вегетативном возоб-
новлении «старых» особей, приступающих к новому плодоношению на сле-
дующий год, динамика изменения численности показана на рисунке 3.11 Б.
Здесь иллюстрируется такой важный момент, как составной характер X как
универсального множителя: X = 1 + 3, учитывающего как плодовитость, так
I iiatia 1 ( xcmi.i и модели *11111011111.1* щитов популяций
OJ
Г. 3 особи 6 семян 6 проростков
4 новых +2 старых особи -► 18 семян
А. 3 особи 9 семян 9 особей -*
27 семян
Б. 3 особи 9 семян -* 9 новых +3
старых особи —► 36 семян
Рис. 3.11. Модели динамики численно-
сти (по Silvertown, 1987 с изменения-
ми):
А — популяции однолетнего растения
из трех особей при условии выжива-
ния всех особей и равной их плодови-
тости (3 семени/особь); Б — популяции
многолетнего поликарпического рас-
тения из трех особей при тех же усло-
виях выживания и равной плодовито-
сти всех (включая старые) особей (3 се-
мени/особь); В — популяции однолет-
него растения из трех особей при усло-
вии выживания 2/3 проростков и равной
плодовитости всех выживших особей (3
семени/особь); Г — популяции много-
летнего поликарпического растения из
трех особей при условии выживания
2/3 проростков и равной плодовитости
всех, доживших до репродукции особей
(3 семени/особь).
Точечной штриховкой обозначены осо-
би семенного происхождения; без то-
чечной штриховки — особи неизвест-
ного происхождения и старые особи,
выживающие до повторного плодоно-
шения, в котором они участвуют вместе
со своими потомками, достигшими ста-
дии репродукции.
В. 3 особи -* 6 семян -► 6 проростков
-► 4 выживших особи 12 семян
06
Л/ Н Miiph'Hi 11< >11 у и >11 ин >11 и ля (>ii( >iu >i ин pin i ch nil
ii веройriюс11» ocymccibiiciiidi нолпкарнии («сырая» особь I l ее потомка).
Итоговая численность за тот же период, что н в нервом случае, состави г 36
особей.
Однако в обоих случаях картина будет гораздо реалистичнее, если учиты-
вать не только «рождаемость», но и смертность особей. Тогда даже при нее-
стественно низкой, взятой лишь для удобства рассмотрения примера, смерт-
ности (-30%) можно вновь сопоставить моно- и поликарпический вариан-
ты жизненных циклов по темпам изменения численности (рис. 3.11 В, Г).
3,7.1. Уравнение экспоненциального роста численности популяций
Чтобы высчитать число особей N после х лет экспоненциального роста
для популяции, начальная численность которой составляла N, мы можем
использовать уравнение:
Рис. 3.12. Кривые биотического потен-
циала (I) и логистического роста (II)
Т — время, N — число особей; заштри-
хованная область соответствует
сопротивлению среды.
Рис. 3.13. Две зоны логистической
кривой, отражающей рост популяции
t — время, N — число особей.
[3.6]
где е — основание натуральных ло-
гарифмов — константа равная 2,718;
г — мгновенная скорость (^истинная
скорость) изменения численности,
результирующая процессы рождения
и отмирания особей.
Для установления связи между X и
г примем, что х = 1, разделим обе ча-
сти уравнения на N и после исполь-
зования натуральных логарифмов по-
лучим:
1пХ = г [3.7]
Если, как в наших выше рассмо-
тренных примерах, X будет иметь
какую-то фиксированную величину и
хотя бы ненамного превышать едини-
цу, то в реальных условиях это будет
возможно только в популяциях низ-
кой плотности (т. е. при малом чис-
ле особей, приходящихся на едини-
цу площади), когда имеется достаточ-
но пространства (=ресурсов) для экс-
поненциального роста численности.
Однако ситуация с длительным уве-
личением численности популяции
по экспоненте явно не реалистична.
В любой популяции помимо роста
численности особей происходит рост
I'hlfld / ( XCMI.I II МОЦСИН ЖН HICHHI.I ч 1111 h > I«»11 IК H lyilHHIlh 65
особен, увеличение их размеров, чю необходимо для приобретения способ
пости к репродукции. Но poc i всех особей в популяции должен бы и» огра
ничеп доступностью некоторых ресурсов, даже если этот ресурс только
общая площадь земной поверхности. Таким образом, в реальности по мерс
роста популяционной плотности величина X должна снижаться (рис. 3.12).
3.7.2. Уравнение логистического роста численности популяции
Уравнение 3.6 можно модифицировать, если принять во внимание огра-
ничение темпов роста популяции, которое начинает действовать по мере
увеличения численности (в природе — чаще и плотности) с момента огра-
ничения доступности ресурсов до полной остановки роста численности.
Необходимо ввести параметр, который будет снижать темпы роста по мерс
приближения численности N к ее верхнему пределу К — т. н. ёмкости ме-
стообитания в отношении данного вида3.
N=Noer [(K-N)/K] t [3.81
Мы получили в интегрированной форме логистическое уравнение ( гак
оно было названо П.-Ф. Ферхюльстом, см. подробнее Гиляров, 1990), а кри-
вая, которая его иллюстрирует, тоже называется логистической (рис. 3.13).
На этой кривой иногда различают две зоны, одна из которых (г) соответству-
ет экспоненциальной, а другая (К) — собственно логистической части кривой.
Символы г и К, как уже говорилось, были использованы для обозначения двух
противоположных стратегий приспособления живых организмов и, соответ-
ственно, двух типов отбора т. н. г- и К-отбора (MacArthur, Wilson, 1967; Пиалка,
1981). Для r-стратегов (r-видов) при благоприятных условиях (наличии в сре-
де обитания свободных ресурсов) характерна способность к быстрому нара-
щиванию численности особей за счет интенсивного размножения, а /Сстратсги
(Х-виды) обладают способностью к произрастанию в сомкнутых сообществах
или бедных ресурсами местообитаниях. И в том и в другом случае им приходит-
ся выживать на фоне изначального (экологически обусловленного) или возник-
шего (ценотически обусловленного) дефицита ресурсов.
Важно подчеркнуть также относительную динамичность ресурсов в ме-
стообитаниях r-видов и относительную стабильность уровня обеспечения
ресурсами в местообитаниях ЛГ-видов. В любом случае речь идет лишь о
потенциальных («стратегических») возможностях видов, и не исключае т-
ся проявление г- и К- свойств одной и той же популяцией в разные фазы ее
становления.
3.8. Чистая скорость размножения
В большинстве случаев бывает полезнее вычислять изменения в N(, взяв
за основу время жизни одного поколения, а не один календарный год. Время
жизни поколения определим как среднее время т (тау), которое проходи i с
’ Подразумевается некий предел насыщения пространства особями данного вида.
momciihi, koi да MaicpniK'koc рас icniic дас i ноюмка до юю, как hoi iioio-
мок сделает то же самое. Скорое и» увеличения численноетп популяции за
время жизни одного поколения называю! чистой скоростью раумножения
Ro. Ее можно высчитать по формуле:
[3.9]
Поскольку Ro вычисляется за время т лет, то годичная скорость увеличе-
ния определяется как корень степени т из Ro:
X = R0Vx [3.10]
Отсюда ясно, что для популяций однолетних растений, у которых время
жизни одного поколения составляет приблизительно один календарный год,
можно констатировать совпадение Ro и К (т.е. Ro = X).
Для поликарпических многолетних растений можно также вычислять R°
по формуле, учитывающей главные параметры приспособленности — вы-
живание и плодовитость в ряду эпизодов репродукции:
R =S1 m , [3.11],
где 1х — вероятность выживания до х лет, а тх— плодовитость, свойствен-
ная особям возраста х. Материал для суммарного вычисления чистой ско-
рости размножения, которая проявилась в ходе нескольких эпизодов репро-
дукции, в этом случае берут из когортных таблиц выживания (survivorship
tables), которые вместе с кривыми выживания будут рассмотрены в главе
10. Если же объектом является популяция монокарпического многолетника,
формирующего семена лишь один раз в жизни после нескольких (х) лет чи-
сто вегетативного роста, то годичная скорость изменения численности по-
пуляции составит:
Wlxmx)1/X [3.12]
3.9. О моделях непрерывного роста численности популяций
Геометрическая (экспоненциальная) и логистическая модели основаны
на допущении, что рост численности популяции непрерывен. Именно по-
этому в них применяется мгновенная скорость роста г. Хотя этот подход и
имеет некоторую описательную ценность в аспекте оценки приспособлен-
ности как скорости размножения генотипов, он не позволяет получить ре-
алистичную картину для большинства растений. Примеры адекватности
этих моделей могут быть, и были получены только в лабораторных опытах
и притом только с водными культурами (некоторых водорослей и видов ря-
ски — Lemnd). когда достижимо максимальное выравнивание условий для
размножающихся организмов.
Возможности подхода ограничены не только редкостью примеров экс-
поненциальное™ роста численности природных популяций, о чем уже го-
ворилось выше. Дело еще и в том, что в природных популяциях репродук-
I'KIthl / ( XCMI.I If модели ЯО1 IIICIIIII.I4 Hill* ПОН IIOliyilHHldi 6 /
цпя обычно приурочена юпько к определенному сезону года п осущест вин
егся юлько особями, досi hi шими какою-io минимального («крт нческо
го») возраста или размера. Учесть это можно, применяя матричные модели.
3.10. Матричный метод анализа динамики численности
и возрастного или размерного спектра популяций
Возможность более глубокого анализа изменения численности, а также из-
менения состава (спектра) популяции дают нам простые матричные модели.
В отличие от моделей непрерывного роста они позволяют смоделирован, из-
менения численности реальной природной популяции, в которой особи, как
правило, принадлежат к разным классам возраста или размера и демоне гри-
руют различные темпы репродукции и отмирания в разном возрасте.
Сначала число особей, представляющих каждый возрастной (или размер-
ный) класс в отдельной популяции, записывают в матрицу-колонку. Число
классов возраста (возрастных групп), взятых для построения матрицы, за-
висит от типа онтогенеза и от того, насколько часто проводится анализ со-
Вставка 5
А
Данный год (t)
oss 0 afs
asr arr afr
о Orf Off
Матрица 3
A2 для многолетника
(Пример 2)
68
Л/ // Маркаи 11(>iiy11>iiиt(>i111nя (mioiioi им piicieiinh
Рис. 3.14. Диаграмма, показывающая
возможные переходы из одного онтоге-
нетического состояния в другое в чере-
де поколений (t —> t + 1 —> t + п) и их
вероятности (а).
c i ana и численноеiи популяции. 1’сли
мы изучаем популяцию одни раз в
год в момент цветения, то можно
для простоты взять только три воз-
растных класса: семена (s), вегетиру-
ющие розетки (г) и цветущие особи
(f). Обозначим число особей в каж-
дом из этих классов и составим ма-
трицу - колонку В (см. вставку 5).
Задача моделирования — прогно-
зировать значения Ns, Nr и Nf в сле-
дующем году. Чтобы сделать это, мы
должны знать, какое число семян даст
не цветущие, вегетирующие особи-
розетки, какое число особей зацветет,
и сколько семян будет образовано в пересчете на каждое цветущее растение.
Подобная информация может быть получена демографическими методами
— достаточно продолжительным наблюдением на постоянных площадках.
В результате демографического изучения нашего модельного однолет-
него растения должны стать известными значения вероятностей перехода
семени в другие возрастные категории. Значения эти, которые можно на-
звать вероятностями (или коэффициентами) переходов принимают значе-
ния от 0 до 1. Они входят в состав матрицы переходов, которую мы обозна-
чим как матрицу А (см. Матрица 7).
Эта матрица представляет собой квадрат со сторонами равными длине
матрицы-колонки. Матрицы переходов размещают в себе коэффициенты,
обозначающие вероятность любого перехода, т. е. перехода из одного воз-
растного класса в любой другой.
В Матрице 1 коэффициент ass— это вероятность того, что семя в этом
году не прорастет и на следующий год так и останется жизнеспособным се-
менем; asr — это вероятность того, что семя превратится на следующий год
в вегетирующую (не цветущую) розетку, a asf — вероятность того, что на
следующий год из этого семени получится цветущая особь и т. д. Первая ко-
лонка говорит о судьбе семян, вторая — о судьбе розеточных вегетирующих
растений, а третья — о судьбе цветущих растений. Для наглядности перехо-
ды показаны стрелками.
Некоторые из коэффициентов, формально обозначенных в матрице пере-
ходов, могут не иметь биологического смысла. Например, по определению,
не зацветшие до конца сезона розетки не могут дать семян. На диаграмме
(рис. 3.14) подобные нереальные переходы просто не обозначают, а при за-
писи в матрице переходов соответствующие им коэффициенты заменяют-
ся нулями. Другие коэффициенты в матрице могут иметь нулевые значе-
ния, когда определенные варианты переходов не наблюдаются из-за особен-
ностей жизненного цикла, присущего виду и, стало быть, его популяциям.
I 'HKKI .1 Схемы II модели >l(lt llirillll.14 щитов Iюиуляций 69
3.10.1. Популяция нешмукицего однолетника (пример 1)
Возьмем в качестве модели однолетнее растение, которое ведет себя как
строгий (к примеру, незимующий) однолетник. Все розетки (точнее ве
гетирующие особи, т. к. у незимующих однолетников розетки обычно вооб-
ще не формируются и заменены удлиненными побегами), которые не пере-
йдут к цветению в первый (и единственный!) год жизни, отомрут, и ни одна
особь не останется в живых после образования семян. В этом случае коэф-
фициенты а^ ars и afr будут равны нулю, а матрица переходов (А{) запишет-
ся так (см. Матрицу 2).
Заметим, что вероятность перехода от одного цветущего к другому цве-
тущему растению aff не равна нулю. Этот переход на деле вероятен, если
идет так: цветущая особь —> семя —> цветущая особь (т. е. за сезон фактиче-
ски успевает «проскочить» еще одно поколение). Поскольку мы условились,
что лишь дважды снимаем информацию и оба раза в период цветения, то тог-
да это необходимый для учета вариант перехода, вероятность которого мы
вынуждены учитывать целиком для всего события: цветущая особь —> цве-
тущая особь.
Еще один промежуточный анализ — в середине года — дал бы больше
информации о популяционной динамике и позволил бы нам построить две
матрицы переходов — одну для оценки вероятности событий в первой по-
ловине года, а другую — во второй его половине. Эти дополнительные воз-
можности метода мы не рассматриваем для простоты.
3.10.2. Популяция многолетнего поликарпического растения (пример 2)
Если же наш объект — многолетник, способный продуцировать розетки
в ходе вегетативного размножения (раметы = дочерние вегетирующие осо-
би), то матрица переходов (А2) при однократном ежегодном освидетельство-
вании будет выглядеть немного иначе (см. Матрицу 3).
Многие семена остаются покоящимися в почве от одного года к другому
и потому ass» 0; очень немногие розетки образуются из семян каждый год,
т. е. asr >0, но ни одно семя не превратится за год в цветущую особь, и поэ-
тому asf =0. При этом большая доля цветущих и вегетирующих особей про-
дуцирует дочерние розетки afr »0 и а^ >0.
Взяв для примера популяцию незимующего однолетника, осуществим
процедуру умножения матриц — матрицы переходов (Матрица 2), и
матрицы-колонки — В(, где t — это время жизни одного поколения (или дру-
гой какой-либо временной интервал), по истечении которого выявляю! воз-
растную структуру популяции. В нашем примере уместно взять за интервал
один календарный год и, соответственно, обозначить колонку-матрицу через
Вг Перемножением матриц А1 и В1 получаем новую матрицу-колонку В „
которая описывает структуру популяции в следующем поколении.
Перемножение производим следующим образом. Начиная с известных
значений коэффициентов в матрице Л7и некоторых известных численно
стсй в ^атрицс Вг мы получаем новые значения N для колонки-матрицы В „
70 Л/// Мауков I loiiyiniiiiioiiiiihi biioiioi пл рас iriiiiii
умножая каждый коэффициент к нервом ряду матрицы /1 па соотвстствую-
щие значения Ns и N в колонке матрице В р а зачем еуммируя произведения.
Новое значение для Nr получаем, умножая каждый коэффициент второй
строчки матрицы А / на соответствующую величину в колонке-матрице В} с
последующим суммированием и т. д.
Может быть полезно для понимания биологической сущности перемно-
жения А] и В1 вспомнить объясняющую матрицу переходов со стрелками.
Когда любую матрицу переходов подвергают итерации, т. е. последова-
тельному неоднократному умножению: А1 х В1 = В2 *В2 = В3 А3* В3 =
В4 и т. д., возрастная структура популяции в конечном итоге стабилизирует-
ся по достижении какого-то константного возрастного спектра, который за-
висит от значений коэффициентов в матрице переходов А 9 но не зависит от
относительных численностей в матрицах-колонках — В14. Следует только
сразу пояснить, что переход к стабильному возрастному спектру представ-
ляет собой математическое свойство матричных моделей, а не биологиче-
ское свойство объекта. При этом, однако, не будем забывать и о достаточно
часто наблюдающейся в природных популяциях многолетних растений из-
вестной стабильности их возрастного спектра.
Если А х Bfe процессе итерации достигло точки, когда устанавлива-
ется стабильный возрастной спектр, то отношение числа особей любого
возрастного класса к числу особей этого же класса в следующем поколении
дает нам Ro или чистую скорость репродукции для изучаемой популяции.
Преднамеренный акцент, который был сделан в этой главе на матрицах
как методе исследования, несколько сместил на второй план биологическое
значение экспериментально получаемых данных о вероятностях переходов
и возрастных спектрах. Однако, великое преимущество метода матриц со-
стоит в том, что он позволяет нам выявлять эффект изменения вероятностей
переходов между онтогенетическими состояниями: к примеру, семя - ро-
зетка или цветущая особь - семя. Метод был рассмотрен очень схематично.
Это было необходимо для выявления его места среди других простых моде-
лей популяционной динамики.
3.10.3. Анализ «чувствительности» (sensitivity) и «эластичности»
(elasticity)
Матричные модели часто используют для изучения популяционной ди-
намики, например в целях выявления наиболее важных факторов при реше-
нии проблемы сохранения редких видов. Матричная презентация полезна в
плане получения суммативных комплексных демографических данных, по-
скольку отдельные аналитические параметры матрицы могут получить при
этом интересные биологические толкования, интерпретации.
Запишем, используя выше введенные обозначения,: N ж = А х Nt , где N —
это вектор, обозначающий число всех особей в популяции в момент времени
t, распределенных по возрастным, либо по размерным категориям, N t+1 —
это вектор для обозначения численности популяции в следующий момент
учета, при том что А — это матрица переходов, которая показывает, какой
вклад особи каждой возрастной (онтогенетической) или размерно!! группы
Г>11ШИ 3 ( ХОМЫ II МОДСИИ ЖН Н1СНП1.1Ч HIIKIIOll популяций /1
вносяг в Каждый из возрасшых или размерных классов за промежуток врс
мели до следующего учета. Как уже было сказано, после мноюкрачно но
в торенного перемножения (итерации) значений вектора и матрицы перехо-
дов возникает «стабильное распределение» по возрастным (онтогенстичс
ским) или размерным классам, после чего вместо матрицы переходов мож-
но фактически умножать вектор числа особей на величину «конечной ско-
рости изменения численности»: А х Nt = X х Nt
Однако, такая суммарная статистика не дает нам информации о влиянии
отдельных элементов на свойства матрицы. Вместе с тем матричные моде-
ли можно использовать для определения «чувствительности» X к неболь-
шим пертурбациям — изменениям величины элементов а., в матрице А.
К изменению каждого элемента а., имеется своя чувствительность sy. Эти
величины могут быть применены для очень важных целей. Во-первых, но
ним мы можем судить об относительной значимости различных переходов
в поддержании темпов популяционного роста. Тогда, если нам нужно уве-
личить или уменьшить X, анализ «чувствительности» выявит наиболее уяз-
вимые точки в популяционной динамике, на которые могут быть направле-
ны, по обстоятельствам, истребительные или защитные воздействия. Во-
вторых, поскольку X может быть использована для подсчета приспособлен-
ности, величины чувствительности s. покажут, как изменения некоторых а(|
отразятся на величинах приспособленности особей в составе популяции.
Анализ чувствительности основан на учете зависимости годичной скоро-
сти роста численности популяции (X) от изменения имеющих возрастную
или стадийную специфику вероятностей переходов. Подсчитывают влия-
ние на X очень малых абсолютных изменений в каком-либо элементе матри-
цы А, к примеру, — a.j5 который отражает какие-то особые переходы на про-
тяжении жизненного цикла.
Значениями «чувствительности» меряют относительное влияние на X не-
больших, но постоянных, изменений в а... Значения а., представляющие ве-
роятности выживания могут изменяться только в пределах от 0 до 1, тог-
да как значение а.., отражающее изменение плодовитости, может прини-
мать любое значение. Это создает сложности при сравнении величин «чув-
ствительности», поскольку фиксированное изменение вероятности выжи-
вания, скажем, от 0,05 до 0,5 намного важнее, чем такое же фиксированное
изменение плодовитости от 10 до 100. Для преодоления этих сложностей
де Крун (De Kroon, van Groenendael, Ehrle, 2000) усовершенствовал анализ
«чувствительности», введя модифицированный индекс, который называю!
«эластичностью» (е„) и высчитывают по формуле: е..= а../ X х s
Значения эластичности могут быть использованы для тех же целей, кото-
рые мы обозначили для sij? но имеют то дополнительное полезное свойство,
что в сумме своей в пределах матрицы дают единицу. Это означает, чю с
представляет собой долю X, связанную с каким-то конкретным переходом а
хотя мы, конечно, должны помнить, что для того, чтобы иметь любое знп
чение X требуется знание всех переходов в матрице, поскольку рас iciihc нс
может обходиться какой-нибудь одной стадией своего жизненною цикла.
Математические свойства и биологическая интерпретация значений чун
ствитслыюсти и эластичности сильно разнятся между собой. 1\сли значс
7 2 Л/// Марков 11 < >i iy ч >i । ц к »i 11 hi я (я ionol пи p;u iciinii
ния чувствительности отражают влияние на X абсолютных значении демо-
графических параметров, измеряемых в самых разных шкалах: от вероят-
ностей переходов, которые не могут превышать единицу до высоких значе-
ний плодовитости, то эластичность любого перехода можно всегда предста-
вить в долях единицы.
Как видно, особенности матричного метода состоят в делении года или
периода жизни одного поколения на подпериоды (от анализа до анализа),
а его цель — получение стабильных возрастных или размерных спектров,
возникающих при достижении вероятностями переходов константных зна-
чений и т. д. При этом особенности метода не следует смешивать со свой-
ствами самих популяций. Ведь наш главный объект — природные популя-
ции — часто испытывают весьма продолжительные периоды разнонаправ-
ленных изменений.
Глава 4. ФУНДАМЕНТАЛЬНЫЕ
ОСОБЕННОСТИ РАСТЕНИЙ КАК
ОБЪЕКТОВ ПОПУЛЯЦИОННОГО
АНАЛИЗА: СООТНОШЕНИЕ ПОНЯТИЙ
«ОСОБЬ» И «ПОПУЛЯЦИЯ»
У РАСТЕНИЙ
«Боковые ветви, отходящие от узлов рас ге-
ния, можно рассматривать как отдельные
дочерние растеньица, которые прикрепле-
ны к телу материнской особи так же, как
она сама прикрепляется к земле»
И. В. Гете «Метаморфозы растений», 17()()
«Растение — это матриархальное племя
побегов»
А. Арбер «Goete's botany», 1946
Специальная глава, посвященная соотношению понятий «особь» и «по-
пуляция» в основном нужна для обсуждения специфики растений как объ-
ектов популяционного анализа. Но вначале целесообразно сказать несколь-
ко слов о соотношении биологии растений, которую изучали со времен К).
Визнера (1892) и А. Кернера, и современной популяционной биологии рас
тений. В отечественной популяционной ботанике исторически сложилась нс
вполне логичная ситуация с определением того, какие исследования право
мерно назвать исследованиями популяционной биологии, а какие «11р<>с i < >
исследованиями биологии растений». Автору приходилось сталкиваться с
мнением некоторых отечественных ботаников, будто отсутствие в какой
либо работе традиционных данных по возрастному (онтогенетическому)
спектру уже означает, что работа посвящена «просто биологии или жопо
гии вида»1. По образному выражению К. М. Завадского (1968) виды жп
вут популяциями. Поэтому любое исследование, затрагивающее вну гриви
1 Применив такую мотивацию, редколлегия авторитетного отечественного журнала прннуди
ла меня однажды изменить название статьи «К популяционной биологии катрана i a i лр< ко
ю», коюрая по ному вышла под названием «К (hiohoi ни мн рапа нилре кою»
74 Л/// I lotiyiiHmioiitiaH (нкмкн пи рас iriniii
довую изменчивость, размерную структуру, иоливариан 1'иоеи» онтогенеза у
какого-либо вида и т. п., вполне нравомерно снизать исследованием популя-
ционной биологии этого вида. Строго говоря, и особенности «просто био-
логии» редко изучались и изучаются на примере одиночных изолированных
особей, так что грань здесь будет заведомо условной. Тем более, как явству-
ет из следующих параграфов, у высших цветковых растений грань между
особью и популяцией подчас тоже весьма условна.
Автотрофные цветковые растения во всем многообразии их жизненных
форм — наши основные объекты — обладают рядом общих специфических
черт, отличающих их от других живых организмов. Поэтому перед изложе-
нием основ популяционно-биологического анализа целесообразно предва-
рительно рассмотреть эти специфические особенности, а также содержа-
ние и взаимоотношения понятий «особь» и «популяция» у растений разных
жизненных форм.
4.1. Наличие банка (=пула) покоящихся диаспор. Понятие
о субпопуляциях и криптопопуляциях
В условиях сезонного климата с чередованием вегетационного и вне-
вегетационного сезонов популяции травянистых цветковых растений, как
правило, имеют почвенные пулы или «банки»2 зачатков (Grime, 1979;
Thompson, 1987; Leek, Parker, Simpson, 1989). Поэтому каждая популяция в
сезон вегетации бывает представлена двумя своими частями или субпопуля-
циями. Активную составляют особи, проявляющие активность в надземной
сфере, а пассивную — покоящиеся в почве зачатки. Это характерно в пер-
вую очередь для терофитов однолетников (зимующих и незимующих) с их
почвенными семенными банками и эфемероидов-геофитов с банком покоя-
щихся луковиц, клубней и т. п. Нетрудно вообразить себе и ситуацию, когда
вся популяция будет представлена только своей пассивной частью, т. е. бу-
дет по существу в этот момент криптопопуляцией3.
Наверное, не будет ошибкой, если рассматривать наличие почвенного
банка зачатков в качестве фундаментальной особенности популяций цвет-
ковых растений наряду с другой их фундаментальной особенностью — ме-
тамерным (модульным) строением особей (рис. 4.1). Мысля теоретически,
можно прогнозировать, что и та и другая особенности могут оказывать су-
щественное влияние на генетическую структуру популяций (см. ниже) и по-
тому играть важную роль в эволюции на фоне окружающей среды, меняю-
щейся случайным образом.
Границы особи у многолетних цветковых растений четко выявляются
только на стадии семени или специализированного вегетативного зачатка
2 Далее в силу общеупотребительности данного термина термин «банк» используется без ка-
вычек
3В пустынных сообществах Т. А. Работнов (1981) вслед за Р. Уиттэкером, выделяющим
L-стратегии (см. табл. 3.2), предлагает даже рассматривать способность к длительному су-
ществованию в форме криптопопуляции как особый вариант стратегии жизненного цикла
(seed surviver).
/ itifiii 7 Фупдимеи i mii.iii.ir «и пьгшнн in pm iciniil
(бульбомkji, как у горца живородяще-
го Polygonum viviparum L.; клубень-
ка, как у чистяка Ficaria verna Dostal.,
рдеста Potamogeton pectinatus L. и
т. д.). У однолетних растений особь
дискретна только на стадии жизне-
способного семени (рис. 4.1), ког-
да представлена всего одним эле-
ментарным метамером (метамером
s. str) с зачатком корневой системы
(зародышевым корешком).
Не следует забывать, что каждое
семя с момента его отделения от ма-
теринского растения действитель-
но есть особь, то есть тот самый
по образному выражению Э. Майра
(1974) недолговечный сосуд, в кото-
ром популяция хранит часть своего
генофонда. Впрочем, не стоит осо-
бенно подчеркивать и недолговеч-
ность. Особь даже у очень многих
однолетних растений проводит боль-
шую часть своей жизни (иногда мно-
го лет!) именно на стадии жизнеспо-
собного семени в составе почвен-
ного семенного банка популяции.
Пусть она на этой стадии предель-
но пассивна или, точнее, малоактив-
на в аспекте взаимодействия с внеш-
ней средой, поскольку так легче пе-
реносить неблагоприятность эколо-
гических условий. Но при этом семя
может действительно сохранять геноп
Рис. 4.1. Общий план строения дву-
дольного растения, указывающий на
его метамерный характер. Слева
стадии развития биполярного зароды-
ша: Н — гипокотиль; Со — семядо-
ли; R — корневой полюс зародыша;
Ra — придаточные корни, отходящие
от узлов фитомеров (по Troll, 1937).
довольно продолжительное время
in как часть генофонда популяции.
4.2. «Неограниченный рост» растений. Увеличение размеров
тела и изменение соотношений между поверхностью и объемом.
Фракталы.
Главная особенность растений вообще и наших объектов автотрофных
цветковых растений заключается в их способности к потенциально неогра-
ниченному росту. Прикрепленный образ жизни связан с извлечением ресур-
сов из окружающего пространства, до которого «дотягиваются», дорастают
органы поглощения. В почве это корни, достигающие в ходе роста все но-
вых частей одного и того же или разных почвенных горизонтов и увеличи-
вающие поверхность всасывания, а в надземной сфере — побеги (стебли с
7 6 MH Миры hi I h >i ly и >i nt к »i 11 m >i (>ik>)ioi ня paciciniil
зелеными листьями), задача которых дорастать до воздушных горизон-
тов, где возможно достаточное поглощение (перехват) световой энергии, и
увеличивать фотосинтетически активную поверхность.
Отсюда понятно, что для нормальной жизнедеятельности растению необ-
ходима достаточно большая поверхность соприкосновения со средой — ис-
точником ресурсов. Вместе с тем нормальный рост влечет за собой увеличе-
ние размеров тела, его объема, который может войти в противоречие с раз-
мерами не столь же быстро увеличивающейся поверхности и породить вну-
тренние проблемы для растения. Образно говоря, может наступить момент,
когда поверхность будет не в силах прокормить объем.
Каждое растение, подчиняясь определенным правилам роста, в итоге за-
полняет какой-то объем пространства. Особи одного вида (и, стало быть,
одной популяции), находясь на одной и той же стадии развития, несмотря
на порой очень разные размеры по понятным причинам самоподобны, что
дает основание для применения к ним понятия о фрактале — нелинейной
структуре, сохраняющей самоподобие при неограниченном уменьшении
(или увеличении) масштаба (см., например, Чайковский, 1993). Во фракталь-
ной геометрии степень заполнения пространства называют размерностью
фрактала. Обычно мы имеем дело с целыми размерностями (у линии 1, у по-
верхности 2, у объема 3, у процесса в пространстве 4 и т. д.).
Если два тела при сохранении их геометрического подобия отличаются
линейным размером, то они закономерно отличаются и отношением поверх-
ности к объему: при увеличении линейного размера поверхность возраста-
ет в квадрате, а объем — в кубе. Помимо внутренних проблем существу-
ют и внешние — в обеих названных сферах растениям, как правило, при-
ходится конкурировать с соседними растениями, принадлежащими к этому
или иному виду или видам. Это означает, что от энергии ростовых процес-
сов во многом зависит приспособленность — статус, который данное рас-
тение сможет приобрести в популяции или сообществе и, в конечном итоге,
то число потомков, которое оно сможет произвести себе на смену, и, вместе
с ним, перспективы в процессе естественного отбора.
4.3. Метамеризация. Проявление метамеризации (модульности)
у цветковых растений
Как в животном, так и в растительном мире проблема частичной (!) ком-
пенсации опережающего увеличения объема по сравнению с поверхностью
решается в принципе одинаково — благодаря усложнению формы органов и
тела в целом за счет расчленения на мелкие взаимосвязанные друг с другом
субъединицы. Более или менее однотипные и в той или иной степени авто-
номные (особенно у растений) субъединицы, которые, дифференцируясь, на-
капливаются в ходе роста, и из которых складывается тело, называют ме-
тамерами4. Благодаря метамерности, многолетнее или однолетнее тело рас-
тения имеет ритмическое строение и по существу представляет пластический
слепок онтогенеза особи. Структурная ритмичность — отражение времен-
4 Общая генетически закрепленная программа роста носит название «архитектурная
модель» см. ниже.
Июни 4 Фундаментальные особеiiiioi i и растений 77
ной ритмичности. Панбоисс полным этот слепок оказывается в случае мио
гих древесных растений, а также однолетних т рав (Юрцев, 1976). Именно иг
тамерность строения растений из-за относительной автономност и мсчамс
ров порождает немало проблем при определении границ особи.
4.3,1. Особь или индивид. Простой и сложный индивиды
Итак, что же такое особь? Вроде бы все просто. Это — структурно обосо-
бленный и физиологически самостоятельный организм, размножающийся
половым и/или бесполым путем. Даже этимологически термин «обособлен-
ный» прямым образом указывает на главную черту особи.
Однако фактически только на самых ранних стадиях онтогенеза границы
особи у цветковых растений установить легко. Как уже говорилось выше,
жизнеспособное семя — вот индивидуум, границы которого четко опреде-
лены. Однако с началом активной жизнедеятельности после прорастания
семени, с началом метамерного роста трудности определения границ могуч
существенно возрасти. Уже каждый элементарный метамер (фитомер). на-
чиная с самого первого (состоящего из
гипокотиля, семядольного узла, семя-
долей с их пазушными почками), об-
ладает, благодаря наличию меристе-
матических очагов (или очага — если
семядоля одна) в пазушных почках,
потенциями самостоятельного авто-
номного организма (рис. 4.2).
Опытами с декапитацией пророст-
ков некоторых двудольных доказана
возможность получения нормальных
растений из почек, находившихся в
пазухах семядолей (Флеров, 1958). А
в ряде случаев даже удавалось пока-
зать более высокую продуктивность
модельной популяции, которая состо-
яла из декапитированных двустебель-
ных растений, по сравнению с попу-
ляцией из нормальных одностебель-
ных особей (Смирнов, 1962).
Сложным приходится признать и
жизненный цикл (онтогенез) особи у
высшего растения, выделяя в его гра-
ницах частные онтогенезы слагающих
его метамерных образований. Иными
словами в пределах большого жизнен-
ного цикла выделять несколько малых
жизненных циклов (Смелов, 1947).
Своеобразие природы (архитекту-
ры чела) высших сосудистых и осо-
Рис. 4.2. Границы I -го, 2-го, 3-го и 4 i о
фитомеров С метамеров s. str.) в чепе
двудольного растения. Черным цветом
обозначены меристематические очш и
№H. Миркин 1 loiiyjiMniioiiiniM Onojioi idi paciciinii
бенно цветковых растений способно породить сомнение даже в точности
самого определения их популяций. Именуя популяцией множество особей
(!) одного вида, по аналогии с другими организмами, мы вынуждены закры-
вать глаза не только на частую невозможность выявления границ особей в
природе, но и на значительную автономность (самостоятельное укоренение,
фотосинтетическую деятельность и минеральное питание, фитоценотиче-
ское воздействие на среду и соседей и прочие проявления приспособленно-
сти) фрагментов особи разного статуса.
Вместе с тем центральное звено в популяционной биологии — это иссле-
дование демографических процессов, связанных с плотностью популяций
(см. главы 6 и 8). И вот снова проблема: что будет приниматься за основу
при выявлении плотности?
Проблема к настоящему моменту была отчасти решена введением поня-
тия «счетная единица» (см. ниже). Однако биологический статус остается в
этом случае совершенно неясным — пересчитать можно, как известно, все
дискретное, но будет ли сопоставима в количественном плане реакция на
плотность действительно дискретных особей с реакцией на плотность по-
бегов, представляющих неизвестное число особей?
4.3.2. Иерархичность метамерных (модульных) субъединиц
Сложность сопряжена еще и с тем, что у цветковых растений метамериза-
ция тела как правило носит характер соподчинения (иерархии). По этой при-
чине в составе особи мы можем найти метамеры, которые представляют раз-
ные иерархические уровни и в разной степени повторяемы в пределах тела
растения и в разной степени автономны. Здесь вновь мыслится применимость
понятия о фракталах, если принять во внимание самоподобие метамеров раз-
ных иерархических уровней и особи в целом. У некоторых жизненных форм
автономизация метамеров (или их омоложенных фрагментов — почек) при-
водит к их полному обособлению, т. е. они становятся особями в полном
смысле этого слова. Отсюда понятно, что в отношении растений приме-
нять термин «индивид» как синоним термина «особь» небесспорно в силу
«делимости» особи на метамеры и в силу того, что рост у цветковых рас-
тений — это не только увеличение объема, но и процесс размножения ме-
ристем — потенциально самостоятельных организмов со своими жизнен-
ными циклами. Меристемы представляют собой своеобразный «ресурс»
растений, определяющий потенциальные возможности роста и развития.
Меристема может быть необратимо «потреблена» на формирование нефо-
тосинтезирующих структур (органов репродуктивной сферы, придаточных
корней) или на формирование новых вегетативных метамеров с новыми ме-
ристемами и новыми возможностями. Вот почему известны попытки ис-
пользования судьбы и типа аллокации меристем особи как показателя типа
стратегии жизненного цикла (Bonser, Aarssen, 1996).
I <I>yii;uiMCiniiJii»iii.ir (>( (>бгн11(>(1 и рис гений 79
4.4. Структурные (архитектурные) тины рас гительных
организмов. Типы взаимодействия надземных и подземных
структур
За рубежом предпочитают простое решение проблемы — выделять два
типа особей: «генету» (genet), которая появляется из семени, т.е. по проис-
хождению своему генетически уникальна и «решету» (ramet), которая по-
является из вегетативной почки или диаспоры, обладая, тем же генотипом,
что и материнская особь, и может находиться на разных стадиях автономи-
зации вплоть до полного отделения (Harper, 1977).
Г. Г. Левин (1961) предложил выделять у высших растений два типа физи-
чески непрерывных образований: простой индивид и сложный индивид (см.
рис. 4.3). Простой индивид — физически непрерывное образование, у кото-
рого морфологическая целостность преобладает над автономностью частей
и отсутствует ясно выраженная морфологическая дезинтеграция.
Сложный индивид — физически непрерывное образование, у которо-
го автономность частей преобладает над морфологической целостностью
(хотя бы во взрослом состоянии) и явно выражена морфологическая дезин-
теграция.
Проблема разграничения простых и сложных индивидов, равно как и
рассмотрение дезинтеграции как способа обособления метамеров (рамет),
должны, как правило, учитывать события, происходящие не только в над-
земной, но и подземной сфере. Лишь по отношению к вторично аризным
(бескорневым) растениям, подобным некоторым эпифитам («исландскому
мху» Tillandsia usneoides из бромелиевых), дезинтеграция фотосинтезиру-
ющих побегов есть одновременно обособление новых особей. В огромном
же большинстве случаев об автономизации мы судим по самостоятельно-
му укоренению фрагментов тела материнской особи. Таким образом, взаи-
мосвязь между сферами, ее тип, приобретает большой вес. На рис. 4.4 мож-
но видеть все теоретически мыслимые, возможные варианты взаимосвязи
между системами побегов и корневыми системами (по Groff, Kaplan, 198К
с изменениями).
К классу 1 относятся «биполярные» растения, у простых индивидов ко-
торых не формируется ни побегообразующих корней, ни корнеобразующих
побегов. Корневая система контактирует с системой побегов в одной един
ственной точке (зоне), то есть сохраняется та же картина, что в биполярном
зародыше. Класс 2 включает растения с корнеобразующими побегами, по
без побегообразующих корней. Одна единственная побеговая система мо-
жет нести при этом множество отдельных (напрямую не связанных между
собой) корневых систем. В класс 3 выделены растения с побегообразующи
ми корнями, но без корнеобразования на побегах. Одна единственная кор
невая система несет множество отдельных (напрямую не связанных меж
ду собой) побеговых систем. Растения 4-го класса способны к формирова
нию как корнеобразующих побегов, так и побегообразующих корней. Одна
и та же особь (генета) формирует множество побеговых и множество корне
вых систем. Рисунок даст только ряд возможных примеров растений, кого
рыс понадают в кп или иной класс. Если приняп», что paciгния I го клас
80
МП Марков Нопуииционппи (»иол<>1 им рис icuni
Рис. 4.3. Физически непрерывные образования у растений. I — простой индивид,
II — сложный индивид, III — колония; границы: 1 физически непрерывного об-
разования, 2 — особи, 3 — условной особи (раметы), 4 — элемента популяции.
са являют пример простых, а растения 4-го класса — безусловно, сложных
индивидов, то два оставшиеся класса (2 и 3) дают примеры индивидов про-
межуточного типа.
Конкретным примером простых индивидов могут служить многие дере-
вья, многолетние стержнекорневые растения и преобладающее большин-
ство малолетних (двулетних и однолетних) растений, а примером слож-
ных — взрослые особи длиннокорневищных и наземноползучих трав, вклю-
чая некоторые малолетники (мшанка Sagina procumbens, лужница Limosella
aquatic а).
Л. А. Жукова и О. В. Смирнова (1988) полагают, что кроме простого и
сложного индивида у растений встречаются физически непрерывные обра-
зования, у которых каждая элементарная структура, возникающая из веге-
тативного зачатка, проходит свой собственный онтогенез, аналогичный он-
тогенезу простого или сложного индивида от рождения до смерти. Этот тип
физически непрерывных образований они предлагают называть колонией
(см. рис. 4.3), в которой каждая элементарная структура может условно рас-
сматриваться как особь (рамета см. выше), а в качестве примера растений,
формирующих колонии, приводят корнеотпрысковые виды. Вместе с тем
по их совершенно справедливому мнению между простыми индивидами на
одном полюсе и колониями физически связанных, но фактически автоном-
ных рамет, — на противоположном — существует континуум. Это сильно
затрудняет общее (подходящее для всех жизненных форм) определение по-
нятия «онтогенез» у растений и заставляет, как уже было показано выше, го-
ворить о «полном» (для особи в целом) и «частном» (для парциальных об-
разований) типах онтогенезов.
С потерей физической непрерывности колония превращается в клон —
группу равнонаследственных потомков одной генеты. В этом случае при-
нято говорить о вегетативном размножении, которое в таком его вариап-
Ihldtl -I Фупдпмсп IIIJII.IIIJC (H'tlGVHIHK III pile ICIIIlii
Hl
Вставка 6
Наиболее известные определения того, что называется размножени-
ем у растений и связанное с ними определение биологической особи у
растений:
Размножение состоит в формировании новой особи из одной един-
ственной клетки. Обычно (но не всегда, если взять, к примеру, апомик-
ты) — это зигота. В этом процессе новый индивидуум воссоздается по
информации, имеющейся в закодированной форме в этой клетке. Рост
же происходит в результате развития организованных меристем. Клоны
формируются благодаря росту, а не размножению (Harper, 1977: 27).
Размножение — это формирование половым или бесполым путем
пропагулы (диаспоры), которая начинает новый цикл развития и дает но-
вый организм (новую особь) (Dawkins, 1982; Tuomi, Vuorisalo, 1989).
Размножение есть биологический процесс распространения роди-
тельских генов во времени и пространстве, результатом которого являет-
ся формирование физиологически независимых потомков, представляю-
щих новое поколение в демографическом смысле этого слова (Urbanska,
1990).
Биологический индивид — это структурно единообразный, физиоло-
гически автономный организм, возникший половым, вегетативным либо
бесполым путем. Поэтому он может представлять собой, во-первых, одну
генету или одну рамету, а во-вторых, - группу взаимосвязанных рамет,
соответствующих либо целому клону, либо части клона.
Биологический индивид может представлять собой часть клона, от-
делившуюся в результате фрагментации или образоваться в результате
размножения из:
семени, образованного половым путем;
семени, образованного бесполым (агамным1) путем;
специализированного вегетативного зачатка.
1 Агамоспермия — бесполое размножение семенами, которое происходит в форме по-
явления потомков, представляющих собой генетические копии материнского организ-
ма. Характерный контраст между физиологической и структурной автономией отдель-
ных особей и общим генетическим единообразием всего поколения потомков было хоро-
шо продемонстрировано на ястребинке Hieracium villosum (2п=27), у которой одна раме-
та может продуцировать до нескольких сотен проростков, обладающих генотипом мате-
ринской особи.
те оказывается прямым следствием роста. Дж. Харпер предпочитает узкую
трактовку термина «размножение» и противопоставляет размножение
(reproduction) росту (growth). Другие зарубежные исследователи склонны
трактовать термин «размножение» более широко и, соответственно, призна-
вать растительные индивиды разного происхождения (см. вставку 6).
иг
м.н MnphtHi
I I(ИiyЛЯЦ1К
НИШИ ()1Н>Л4>1 нм
pile 1СНИЙ
Рис. 4.4. Варианты взаимодействия по-
беговых и корневых систем:
1 — одна корневая система и одна побе-
говая система контактируют в теле рас-
тения в одной точке — так же как кон-
тактировали до того в биполярном заро-
дыше; 2 — растение формирует толь-
ко корнеродные побеги; 3 — растение
формирует только побегородные кор-
ни (корнеотпрысковость); 4 — растение
формирует как побегородные корни,
так и корнеродные побеги (для каждого
класса приведено только по одному из
нескольких возможных примеров).
Рис. 4.5. Побеговые системы у рыхло-
кустового (1), плотнокустовых (2, 3) и
длиннокорневищного (внизу) злаков.
Возникает впечатление, что решение вопроса о границе между сложным
индивидом и колонией зависит от способа связи между побеговой и корне-
вой системами (см. рис. 4.4). Развитие корневого отпрыска представляется
наиболее автономизированным и потому соответствующим полному онто-
генезу элемента колонии.
«Фитоценологическое» происхождение отечественной популяционной
биологии растений заставило рассматривать элементы особи и еще под од-
ним углом зрения — в плане локализации их воздействия на окружающую
среду и соседние растения.
С учетом индивидуального воздействия на среду любого метамерно-
го образования, обладающего известной автономностью (к примеру, само-
стоятельной побеговой и корневой системами), но не обязательно физиче-
ски да и физиологически отделенного, можно признать важной характери-
стикой популяции абсолютное число (численность) и число в пересчете на
единицу площади (плотность) таких образований. Этот вывод подкрепля-
ется чрезвычайной методической сложностью (если не невозможностью!)
подсчета именно особей как таковых в популяциях многих видов, особен-
Г'1(1(К1 4 Фундпмсп i ильные осоАсшнк i и paciciinii
83
Рис. 4.6. Типы биоморф цветковых растений и счетные единицы: I — моноцен-
трические, II — неявнополицентрические, III — полицентрические; счетные еди-
ницы: 1 — особь семенного происхождения (генета), 2 — партикула (рамета),
3 — клон, 4 — парциальный куст или побег (рамета); границы: А — морфологи-
ческой; Б — фитоценотической счетной единицы.
но злаков и прочих граминоидов (рис. 4.5). Именно так возникло представ-
ление о биологических счетных единицах, в роли которых могут выступать
как единицы онтогенетического развития (морфологические), так и едини-
цы воздействия на среду. На рис. 4.6 показаны различные варианты счет-
ных единиц, каждая из которых является центром воздействия на среду, об-
ладая собственным «фитогенным полем» (Уранов, 1965), и выделенные на
основе таких теоретических представлений моноцентрические, неявнопо-
лицентрические и явнополицентрические (= полицентрические — М.М.)
типы биоморф (Смирнова и др., 1976).
К типу моноцентрических биоморф относят главным образом вегетативно
неподвижные и малоподвижные виды: стержнекорневые, луковичные, клуб-
невые травы, большинство деревьев, а также, что более спорно, дерновинные
травы. В качестве счетных единиц могут выступать компактный клон или от-
дельные партикулы (раметы = особи вегетативного происхождения).
Для полицентрических биоморф характерно наличие нескольких четко
выраженных центров сосредоточения корней, побегов и почек возобнов-
ления, соединенных между собой протяженными коммуникациями, кото-
рые могут находиться как под землей (гипогеогенные корневища, подзем-
ные столоны), так и на ее поверхности (столоны, плети). К этому типу био-
морф относятся длиннокорневищные, столонообразующие, наземноползу-
чие травы, а также длиннокорневищные кустарнички, кустарники и деревья
(Жукова, Смирнова, 1988).
За рубежом предпочитают использовать (см. Lovett-Doust, 1981) для дср-
новинного типа разрастания термин «фаланга» {«phalanx»), а для форм со
значительно более протяженными коммуникациями — термин «партизан»
Л/ H Маркин I Ituiyiniциопнни (hioiioi пи растений
(«цнепНа»). 'Термины ин, чго весьма ciwiic гвеппо зарубежной популяцион-
ной биологической литературе, морфопогнчсскн не строгие, образные и на-
мечают не более чем два крайних тина стратегии разрастания.
Промежуточным типом являются неявнополицентрические биоморфы,
имеющие несколько слабо различимых центров сосредоточения корней, по-
бегов и почек возобновления. В качестве счетной единицы только на первых
этапах онтогенеза до начала вегетативного размножения выступает особь, а
впоследствии — партикула. Примеры неявнополицентрического типа био-
морф можно найти среди короткокорневищных и кистекорневых трав, ряда
древесных растений (Жукова, Смирнова, 1988).
Весьма принято рассматривать моноцентрические, неявно- и явнополи-
центрические биоморфы как эволюционный ряд, в котором возрастает сте-
пень автономности структурных элементов особей, дезинтеграция стано-
вится специализированной (Смирнова и др., 1976; Жукова, Смирнова, 1988),
усиливается роль и совершенствуются формы вегетативного размножения
при возрастании вегетативной подвижности. Однако в этом случае прихо-
дится признать существование и другой линии эволюции, которая привела к
полной утрате автономности метамерных структур и «узаконила» облигат-
ную мондцентричностъ — линии эволюции, давшей начало монокарпиче-
ским двулетним и однолетним растениям.
4.5. Иерархическая «лестница» метамерных образований
у цветковых растений
В отечественной литературе, помимо введения общих терминов, иссле-
дователями разработана классификация наименований метамерных струк-
тур разного уровня. Если оставить за термином «метамер» первоначальный,
то есть широкий смысл и понимать под этим термином любую повторяю-
щуюся субъединицу, то в иерархии метамеров каждому рангу потребуется
свое название.
Л. Е. Гатцук (1974) разработала иерархическую классификацию метамер-
ных структур у геммаксилярных (=почкопазушных — М.М.) многолетних
растений, в которой представлено 7 разных уровней:
Элементарный метамер (собственно метамер или метамер sensu stricto) -
Элементарный побег или побег ритма - Моноподиальный побег - Система
моноподиального (скелетного) побега - Система моноподиальных побегов -
Первоначальный куст и куст замещения - Первичный куст (возникший из
проростка) и парциальный куст.
Определение элементарного метамера выглядит так — группа разных
органов — один узел с листом (листьями), пазушной почкой (осью), ниже
расположенным междоузлием и (не всегда!) корнем (корнями). Для кратко-
сти можно обозначить такой элементарный метамер в теле однодольных и
двудольных растений термином фитомер, который с успехом был исполь-
зован Т. И. Серебряковой (1971) при рассмотрении морфологии и архитек-
туры злаков. Нужно только помнить, что в строении фитомеров двудоль-
ных (см. рис. 4.1 и 4.2), и однодольных и голосеменных имеются некото-
рые отличия. Например, пазушная почка у однодольных но своим функ-
! ниш / Фундлмсп iaiii.iii.ic o< oIh iiiuk i и рас icnnh
цноналыйлм связям ошосшся к
верхнему по положению структур-
ному элементу побега, а не к тому,
в пазухе которого она находится
(рис. 4.7).
При всем при этом фитомер про-
должает оставаться повторяющейся
в ходе роста (итеративной) структу-
рой и элементарной единицей сложе-
ния побегов.
По мнению М. М. Магомедмирзаева
(1990) вряд ли целесообразно обо-
значать как следующий за фитоме-
ром ранг метамерной единицы эле-
ментарный побег, включенный
Л. Е. Гатцук в разработанную ею си-
стему метамеров. Он считает, что яв-
ление ритмичности побегообразова-
ния представляет несомненный ин-
терес для интерпретации развития
побегов в аспекте морфологической
эволюции, но добавлять более дроб-
ные подразделения побега, образую-
щегося часто с интервалом промежу-
8‘j
точного покоя, называя их также ва-
риантами побега (элементарными Рис- Особенности строения фито
побегами или побегами ритма sensu меР0В У злака-
Сабинин, 1957) не следует. Лучше,
если побег будет фигурировать как единица первого порядка конструкции
особи, подобно тому, как фитомер фигурирует как единица первого порядка
в конструкции побега. С таким мнением трудно не согласиться.
Система соподчиненных метамерных структурных единиц была созда-
на Л. Е. Гатцук, по ее собственным словам, для того, чтобы в дальнейшем
можно было использовать данный структурный элемент для характеристи-
ки разных жизненных форм (Гатцук, 19746). При этом было рекомендовано
использовать, по крайней мере, три существенные черты жизненных форм:
1) тип внешнего строения метамерных образований, создающих структуру
побегового тела; 2) характер их «сочленения» и 3) положение места «сочле-
нения» по отношению к поверхности почвы. В главе, посвященной анализу
роста особей в популяциях в его структурном аспекте, мы вернемся к рас-
смотрению этих вопросов.
4.5.1. Термины «метамер» и «модуль»
После появления концепции архитектурных моделей (Halle, Oldemann,
1970, 1978) J. White (1979), обсуждая некоторые наиболее интересные во
просы мстамерности строения растений, попытался терминологически обо
86 Л//* Марыш I h»iiyiinum>iimi>i Ощякяии pncicmiii
значить следующую в иерархии за метамером s. str. или элементарным ме-
тамером мстамерную субъединицу и дал определение «модуля».
Модуль — это продукт деятельности одной верхушечной меристемы,
представляющий собой ось, несущую узлы с листьями, пазушными почка-
ми, междоузлиями, (и, возможно, корнями — М. и завершающуюся со-
цветием или вегетативными структурами.
Побеговое тело малолетнего растения (а, в известном смысле, и дерева),
как уже говорилось, являет собой полный «пластический слепок онтогене-
за», отражающий в своей пространственной структуре временную ритми-
ческую последовательность образования метамеров. Иными словами, ис-
ключительно для малолетних растений понятия «архитектурная модель»
и «тип морфологической конструкции» или «архитектура» оказываются
очень близкими по содержанию.
Теперь после рассмотрения непрерывных рядов метамерных образова-
ний и выделенных на их основе биоморф нам предстоит уточнить, что же
с точки зрения теории представляет собой при таком подходе популяция.
Вполне осознавая всю сложность выделения границ особей у растений,
мы, тем не менее, вынуждены давать определение популяции цветкового
растения, используя термин «особь».
Популяция — это группа особей, принадлежащих к одному виду и пото-
му произрастающих в одном местообитании, которая в той или иной сте-
пени изолирована от других групп особей того же вида.
Можно видеть, что в этом определении подчеркивается с одной стороны
таксономическая общность (принадлежность к одному виду), а с другой —
экологическая общность (приуроченность к одному типу условий — одно-
му местообитанию). Помимо связей такого типа, это, безусловно, очень об-
щее определение не указывает на какие-либо другие связи, которые объеди-
няют компоненты популяции, интегрируют ее, отделяя, в то же самое время,
от других групп внутривидового (=популяционного) уровня.
Дальнейшее уточнение в определение, модифицируя его, привносят част-
ные разделы популяционной биологии растений (популяционная генетика,
популяционная экология) и фитоценология.
Как уже говорилось выше (глава 2), с позиций генетики наиболее важ-
ным интегрирующим началом является панмиксия (свободное равноверо-
ятное скрещивание между особями), которая выступает как необходимый
фактор при анализе менделевских популяций с помощью уравнения Харди-
Вайнберга. Однако и здесь мы сталкиваемся с континуумом форм, отли-
чающихся особенностями репродуктивной системы — соотношением двух
форм опыления, унипарентальностью и бипарентальностью и т. д.
К. Урбанска (Urbanska, 1992) приводит схемы, наглядно изображающие
степень генетической общности в панмиктической популяции, популяции,
разделенной на соседства и в популяции унипарентальных видов.
С точки зрения популяционной биологии больший вес, чем внутрипопу-
ляционные скрещивания, для интеграции имеют отношения взаимоблаго-
приятствования и конкурентные отношения, зависящие от плотности попу-
ляции.
/ 7 <1>уПДПМС11 inill.lll.IC 04 (»(>1‘||||<ц III |П11‘1С1111Й И /
Для фнтоцснолога нопупяция это, прежде всею, комнопеш фитоцс
поза, и потому преимуществом при изучении популяции фптоцепологамн
возобладал ценотический принцип выделения популяций г. с. определение
их границ по границам фитоценозов (Работнов, 1950 а, б). Популяция со-
вокупность особей данного вида, произрастающих в данном фитоценозе.
Позднее этот принцип был закреплен терминологически — В. В.
Петровский (1961) употребил впервые, а А. А. Корчагин (1964) ввел в оби-
ход фитоценологов термин «ценотическая популяция» (сокращенно — це-
нопопуляция). Популярность, которую приобрели этот термин и это поня-
тие, не решили проблем выявления границ между популяциями, поскольку
вопрос о границах фитоценозов также в большинстве случаев спорен.
4.5.2. Особи у растений как популяции: метапопуляции
Джеймс Уайт (White, 1979), учитывая «сборный», метамерный характер
особи высшего растения, писал о ней как о своеобразной метапопуляции или
популяции метамеров. Учитывая наибольшую самостоятельность в теле выс-
шего растения модулей, целесообразно говорить именно об особи как о мс-
тапопуляции модулей. Выше уже говорилось, что границы особи у семенных
растений четко выявляются только на стадии жизнеспособного семени.
Позднее из-за метамерного и итеративного характера роста и сохранения
меристематичности отдельных участков тела в структуре особи можно вы-
явить раметы (реально — модули, а потенциально — особи вегетативно-
го происхождения), обладающие определенной степенью физиологической
автономности (автономностью в части ассимиляции углерода, самостоя-
тельным укоренением и т. д.). Последнее может иметь существенное зна-
чение не только для выявления демографических параметров популяций на
основе числа модулей как счетных единиц, но и для приложения сил есте-
ственного отбора. В данном случае концептуальное разграничение индиви-
дуального и группового вариантов отбора, например, отнюдь не будет са-
моочевидным (Tuomi, Vuorisalo, 1989). Ведь по определению групповой от-
бор — это «процесс, при котором могут отбираться признаки, благоприят-
ные для группы, но не для особи» (Майр, 1974). Поскольку группа полуав-
тономных модулей, составляющих коллективную особь (метапопуляцию) у
растений, мало чем отличается от группы особей, как таковых, можно вести
речь об отборе, действующем на уровне групп модулей, как одном из вари-
антов группового отбора.
Однако термин «метапопуляция» сейчас используется в несколько ином
смысле — для обозначения группы связанных между собой какими-либо
специфическими узами популяций, т. е. затрагивает явления межпопуляци-
онного уровня. Для примера можно рассмотреть различные, иерархические
уровни существования вида в форме популяций различного ранга, популя-
ционных локусов (см. рис. 6.2). Однако нетрудно заметить, что изображен-
ный иерархический ряд мог бы быть даже продолжен, если перейти на уро-
вень модулей, составляющих побеговое тело отдельных особей Clematis.
Итак, резюмируя все, что сказано в данной главе, можно заключить, что
биологический индивид — это структурно единообразный, физиологически
88 М // I loiiyihimnuiiiiiM biioiioi нм pin iciihii
автономный организм, который у растений возникас! половым, вегетативным
либо бесполым путем. Поэтому он можег представлять собой, во-первых,
одну генету или одну решету, а во-вторых, — группу взаимосвязанных ра-
мет, соответствующих либо целому клону, либо части клона. Поскольку клон
представляет собой явление уже популяционного уровня, то можно констати-
ровать наличие плавного перехода между особью и популяцией у растений,
или, иными словами, отсутствие не только в теории, но и на практике четких
границ между этими двумя нашими объектами. Важнейшим методическим
следствием такого вывода будет применение одних и тех же подходов коли-
чественного анализа к изучению динамических процессов, происходящих
на уровне особи и на уровне популяции.
Глава 5. ЖИЗНЕСПОСОБНЫЕ
СЕМЕНА И ИХ РОЛЬ
В ПОПУЛЯЦИЯХ РАСТЕНИЙ
Тайная сила лишь дремлет в сухих семенах:
Согнут прообраз растения под кожурою.
Бледные почечку, корень и листья
в бескрасочных снах —
Жизни пружину — таит в себе семя сухое.
И.В. Гёте. Метаморфоз растений
(перевод мой, М.М.)
Роль семени в жизни цветковых растений чрезвычайно велика.
Независимо от преобладающего способа возобновления любой вид этого
отдела высших растений на каком-то этапе своего жизненного цикла не-
пременно образует семена. Если даже травянистые многолетники, возоб-
новляющиеся преимущественно вегетативным способом (сныть обыкно-
венная, лапчатка гусиная, седмичник европейский и др.), время от време-
ни прибегают к семенному размножению, то у малолетних растений и мно-
гих деревьев семя — это незаменимое звено в цепи поколений. Не случайно
Е. Симонидес (Symonides, 1988а, Ь), характеризуя наиболее общие, фунда-
ментальные свойства малолетников, наряду с краткостью жизненного цик-
ла и обязательной монокарпичностью подчеркивает их третье общее свой-
ство — исключительно семенное возобновление.
Семена уже очень давно служат объектом пристального внимания уче-
ных. Однако, по мнению Р. Е. Левиной (1981), в отечественной науке, да
и в зарубежной тоже, активно разрабатывалось лишь одно направление,
имеющее непосредственное отношение к популяционной экологии рас-
тений. Это — лабораторно-физиологическое исследование покоя семян.
Действительно, как отечественными, так и зарубежными исследователя-
ми опубликованы важные обобщающие сводки по этому вопросу (Леман,
Айхеле, 1936; Крокер, Бартон, 1955; Николаева, 1967; Овчаров, 1969; Baskin,
Baskin, 1988). Вместе с тем изучение покоя и прорастания семян именно в
составе природных популяций — это чрезвычайно сложная, но столь же ин-
тересная биологическая проблема. Дело в том, что биология прорастания
затрагивает и экологию, и морфологию, и физиологию этого процесса, а
«лабораторная экофизиология», применяя чашки Петри и фильтровальную
бумагу, зачастую дает лишь приблизительное представление о прорастании
того же вида в естественных условиях. Являясь с одной стороны весьма
У и
М II Миры hi 11<>пулн1 икнншм bnoJioi ия рис Ieniiil
Рис. 5.1. Покой и прорастание семян (процессы, состояния и факторы).
удобным объектом для экспериментального изучения, семена редко обеспе-
чивают повторяемость результатов опытов, и потому продолжают оставать-
ся во многом загадкой для исследователя.
Поведение семян как один из важнейших элементов стратегии жизнен-
ного цикла очень трудно прогнозировать — уж очень много разнообраз-
ных факторов, внешних и внутренних, включая генетические, может быть
задействовано в этом процессе (рис. 5.1). А между тем, ряд важнейших мо-
ментов в динамичной жизни популяции, и особенно ее возобновлении свя-
зан именно с поведением семян.
К сожалению, работы по изучению биологии семян в аспекте эколого-
ценотических стратегий проводятся почти исключительно за рубежом
(Grubb, 1976; Grime et al., 1981; Gross, Werner, 1982; During et al., 1985), хотя
и там отмечены пробелы в познании демографического и эволюционного
аспектов проблемы (Cook, 1980). Поэтому очень ценными представляют-
ся статьи Т. А. Работнова (1981, 1986), ориентирующие отечественных ис-
следователей на изучение этого важного вопроса популяционной экологии
растений.
5.1. Основные биологические функции семени
Дж. Л. Харпер (Harper, 1977) перечисляет пять основных функций се-
мени, которые можно кратко обозначить следующим образом: 1) единица
размножения; 2) единица распространения (вид диаспоры или ее основная
составная часть); 3) структура и средство, предназначенные для пережива-
ния неблагоприятного сезона; 4) зародыш «в упаковке» со стартовым запа-
сом питательных веществ, необходимым для приживания проростка; 5) ге-
Г'1<1«<1 Л Жизнеспособные семени и их роль и популяциях pn< ICIIIlit 91
пегичсский вариант с заложенными в нем последе гпиями рекомбинации
Прокомментируем эти функции в приложении к цветковым растениям
5.1.1. Семя как единица размножения. Плодовитость (семенная
продуктивность) цветковых растений
Наиболее высокой семенной продуктивностью, которая способствует
резкому повышению их биотического потенциала, характеризуются мало-
летние растения. Это, казалось бы, бесспорное утверждение, однако, остро
нуждается в конкретных, а главное, достоверных количественных данных.
Данные о варьировании плодовитости целесообразно рассматривать па
двух уровнях: межвидовом и внутривидовом.
5.1.1.1. Плодовитость растений (межвидовой уровень)
В 1970-1980-х гг. на страницах журнала «Американский Натуралист»
("American Naturalist") развернулась весьма эмоциональная дискуссия
о двулетних растениях. Открыта она была статьей Р. Харта (Hart, 1977)
«Почему двулетники столь редки?» Р. Харт, обратив внимание на относи-
тельно низкий процент двулетних растений во флоре Северной Америки
200 видов (1,4%) при 3155 (21,8%) видах однолетников и общем числе ви-
дов 14500', — пытался объяснить этот факт их меньшей приспособленно-
стью. Мерой приспособленности (fitness) он избрал конечную или годич-
ную скорость размножения X, которая, как мы выяснили (см. Главу 3), свя-
зана зависимостью:
Х = Я |/т
о
с чистой скоростью размножения, определяемой произведением выживае-
мости /, свойственной возрасту х9 на плодовитость (т), свойственную это-
му возрасту: Ro = Z lx тх.
Если обозначить выживаемость в первый и во второй год жизни, соот-
ветственно, через 1Х и 12, то для однолетних растений: Ха = lx та, для дву-
летних монокарпических растений: Хь= /2ть)1/2, а для многолетних по-
ликарпических: Хр = /, m + /2, где та, и тр — плодовитость, соответ-
ственно, однолетних, двулетних и многолетних поликарпических расте-
ний. Нетрудно заметить, что здесь, как в главе 3 и на рисунке 3.11 Б, налицо
составной характер универсального множителя к, отражающего не только
плодовитость, но и вероятность осуществления поликарпии.
Используя как модель и сопоставляя эти уравнения друг с другом,
Р. Харт вывел, что для достижения потенциальных темпов роста попу-
ляций, свойственных многолетникам, двулетние растения должны имен»
вчетверо большую плодовитость, чем у многолетников, т. е. mb = 4m
Однолетние же растения им необходимо превзойти по плодовитости, но
1 Очень сходные соотношения наблюдаются, как показывает рис. 5.2, и но флоре енропей
ской части России («Флоре» П. Ф. Маевского).
92
М Н. Марков
I louyiiMHitc
НП1Ш1 bnonoi ин
рпсюпий
Рис. 5.2. Гистограммы, показывающие соотношение частоты встречаемости раз-
ных жизненных форм у видов флоры П.Ф.Маевского (1964) из класса двудоль-
ных (1а) и однодольных (16), а также кривые распределения по логарифмическим
классам массы семени (II: 1 — 0,01—0,03; 2 — 0,032-0,099; 3 — 0,1-0,315; 4 —
0,316-0,999; 5 — 1,0-3,161; 6 — 3,152-9,999; 7 — 10,0-31,611; 8 — 31,61 -99,99) и
плодовитости (III: 1 — 100-315; 2-317-1000; 3 — 1001-3162; 4 — 3162-10000;
5 — 10001-31 622; 31623-100000; 7 — 100001-316227, 8 — 316228-Ю6) для дву-
летних (пунктир) и всех монокарпических (сплошная линия) видов сорных расте-
ний; 1 — однолетники, 2 — двулетники, 3 — травянистые и 4 — древесные мно-
голетники (принятые во флоре обозначения основных типов жизненных форм).
крайней мере, в два раза (mb = 2ma), чтобы сравняться с ними по величи-
не X. И это при условии, что и 12 будут принимать оптимальные, а имен-
но — какие-то средние значения.
Дж. Сильвертаун (Silvertown, 1983), продолжив обсуждение этой темы
в статье с полемическим названием «Почему двулетники иногда не столь
редки?», усомнился в справедливости такого объяснения относитель-
ной редкости двулетних растений. Он подчеркнул, что если смотреть не
в масштабе целой флоры, а в масштабе отдельных таксонов, то в семей-
стве Apiaceae, например, около 30%, а в семействе Asteraceae — около 20%
дву-многолетних монокарпических видов. По-видимому, в такой форме
была впервые высказана идея о необходимости учета филогенетических
связей видов, когда их сравнивают по каким-либо биологическим свой-
ствам или параметрам жизненных циклов. Впоследствии это сделалось
правилом, сформулированным как методическое предписание выявлять
и анализировать только филогенетически независимые различия — PIC
(Phylogenetically Independent Contrasts).
Факт существования в некоторых семействах цветковых растений мно-
голетних монокарпических видов сам по себе весьма интересен в аспекте
Iiiiuti) Mihiiccikk oIhii.ic ccmciiii и 11 \ рин и uonynMnini.x рш icinni lJJ
дискутируемой проблемы Ведь скорость размножения мио1 one।пего мо-
нокарника, как уже говорилось выше, составляет: X (I in )|z', |де \ чис
ло лет, которые растение проводит до зацветания, a m нлодовиюсн,,
проявленная во время единственного в жизни репродуктивного эпизода.
Иными словами, при задержке репродукции (увеличении х) скорость роста
численности фактически при любой плодовитости резко падает.
Вместе с тем наблюдения Т. А. Работнова (1964) показали, что задерж-
ка репродукции у монокарпических многолетников (потенциально — фа-
культативных двулетников) на срок до 10 и более лет отнюдь не редкос ть.
В ходе наших наблюдений за природной популяцией катрана татарского
Crambe tataria Sebeok. (сем. Brassicaceae) было выявлено, что особи этого
трициклического в культуре монокарпика зацветают не ранее, чем на седь-
мой год жизни (Марков, 1989а, см. также главу 10). Стало быть, если од-
нолетние и многолетние поликарпические растения имеют определенное
преимущество, то надо искать свойства, компенсирующие относительно
низкий потенциальный темп размножения двулетников и многолетних мо-
нокарпических растений, который по теории может существенно повлиять
на их приспособленность.
По мнению Дж. Сильвертауна, преимущества однолетников перед дву-
летниками несколько преувеличены. Ведь если взять наиболее распростра-
ненный тип однолетников, для которых обязательным компонентом (хотя и
довольно пассивной частью) любой популяции выступает постоянный по-
чвенный банк семян, то резкое снижение темпов размножения можно на-
блюдать, если семенное возобновление (успешное прорастание) идет не
каждый год, а только через интервал х лет. Такая ситуация отнюдь не ред-
кость в популяциях пустынных однолетников, возобновление которых за-
висит от редкого и нерегулярного выпадения осадков. Семена долгое вре-
мя остаются в почвенном банке, и тогда даже при смертности равной нулю
для любого однолетнего растения оказывается справедливым выражение X
= (/ mx)1/x. Любопытно, что в этом случае из-за долгого пребывания семе-
ни в почве особь фактически становится многолетней монокарпической,
и дает еще одну хорошую иллюстрацию поливариантности онтогенеза.
свойственной одному виду.
Отсюда ясно, что в данном случае двулетники могут компенсировать
преимущество однолетников меньшим возрастанием плодовитости, чем
это предсказано моделью Р. Харта.
Третий дискутирующий автор Д. Келли (Kelly, 1985а), полагает, что нель-
зя недооценивать и смертность семян в почвенном банке однолетников.
При учете этого параметра вместе с прерывистой доступностью микроу-
частков —регенерационных ниш (Grubb, 1977) или прерывистым возник-
новением регенерационных ситуаций (Работнов, 1986) еще больше снижа-
ются потенциальные преимущества однолетнего онтогенеза в сравнении с
двулетним по приспособленности. Преимущества многолетних поликарпи
ческих растений в расчетах Р. Харта тоже несколько завышены, нисколь
ку долгоживущие (свыше 5 лет) травянистые многолетники крайне редко
в естественных условиях переходят к репродукции на первом году жизни.
li Л1,Ч>к,,а I loll yi|>IIHK>IIIUI>l 1>1П>1Ц)| ПИ put ЦНИИ
Коро I КожинуЩИС MIIOI OJIC I ИКС HOHUKiipnUkU В широком ДШННГЮИС 1ПМСПС-
ний /( и / демонстрирую! темпы размножения, сходные с геми, какие ха-
рактерны для долгоживущих поликарпичееких трав (Bender, Baskin, 19X8).
Таким образом, дискутировавшие исследователи пытались, в основном,
доказать, что у однолетников и многолетников значительно меньше пре-
имуществ в приспособленности перед двулетниками, чем получается по
расчетам Р. Харта. Это не означает, однако, что у двулетников в ходе эво-
люции не вырабатывалось свойств, позволяющих компенсировать отно-
сительно низкую потенциальную скорость размножения. Безусловно, на
одном из первых мест — высокая плодовитость двулетников.
К сожалению, данные по плодовитости разных видов растений встреча-
ются в литературе очень редко. Из зарубежных публикаций одной из са-
мых надежных и часто цитируемых остается работа Э. Солсбери (Salisbury,
1942). Согласно данным, приведенным в этой книге, двулетники действи-
тельно обгоняют по плодовитости сравниваемые с ними другие жизнен-
ные формы растений. Так средняя плодовитость однолетних, двулетних и
многолетних поликарпичееких растений составляет, соответственно, 6368,
28790 и 5869 семян на особь (см. также Fenner, 1985).
Принимая во внимание методические сложности и особый дефицит био-
логических данных о жизненных формах, находящихся в правой части ре-
дукционного ряда, проведем сравнительный анализ плодовитости только
малолетних (короткоживущих) монокарпических растений.
В отечественной литературе вопрос о плодовитости наиболее интен-
сивно исследовался в отношении сорных растений, хотя нельзя не отме-
тить приблизительность оценок и погоню за астрономическими цифра-
ми, которые, по-видимому, должны были служить своего рода устрашени-
ем и призывом к беспощадной борьбе с сорняками. В итоге своеобразно-
го соревнования «кто больше?» справочники изобилуют данными о мак-
симально возможной (и, тем не менее, часто все же завышенной!) семен-
ной продукции сорных растений без учета внутри- и межпопуляционной
изменчивости особей одного вида по плодовитости (см. ниже). Нам пред-
ставляется, что несколько более объективную оценку плодовитости сор-
ных растений дает книга В. Н. Доброхотова (1961), хотя бы потому, что
для многих видов здесь примерно обозначены пределы варьирования это-
го показателя.
Критический отбор данных О. Стевенса (Stevens, 1932, 1957),
Э. Солсбери (Salisbury, 1942,1963), В. Н. Доброхотова (1961), Е. Симонидес
(Symonides, 1988), а также наших собственных данных позволило полу-
чить выборку из 96 видов малолетних растений с известной максималь-
ной плодовитостью. Проанализируем ее целиком, а также с разбивкой на
подвыборки по основным типам жизненного цикла и условным эколого-
фитоценотическим группам.
I ’litfid 5 )KiniiccIioc<)()iii.ic ccMriin и их рош. н популяциях рис юпий 95
5. /. /. /. / Плодовитость ендов с разным типом жизненного цнк ia н
относящихся к разным эколого-фитоценотическим группам
Если при сопоставлении видов по семенной продуктивности (плодови-
тости) использовать данные без логарифмической трансформации, то полу-
чается кривая с сильной положительной асимметрией (рис. 5.3). Обращает
на себя внимание малый процент видов малолетников (около 14%) с очень
высокой плодовитостью (свыше 30000 семян/особь) и наибольшая часто-
та встречаемости видов (более 55%) в первом классе (плодовитость менее
5000 семян/особь). Увеличение числа классов не приводит к изменению об-
щего характера гистограммы распределения, и лидирующим всегда остает-
ся самый первый класс.
Столь сильная положительная асимметрия графика распределения (As =
+4,7) заставляет применять логарифмическую (In) трансформацию данных.
Таким путем можно не только получить распределение близкое к нормально-
му (рис. 5.4), но и среднее значение, которое более адекватно отражает «нор-
му» как особенность выборки. Если высчитанная по нетрансформированным
данным средняя арифметическая составила 20244 ± 4661 семян/особь, то ан-
тилогарифм средней арифметической логарифмически трансформированно-
го ряда (8,29) дал величину плодовитости всего 3984 семян/особь.
Поэтому для сравнения видов разных биологических и эколого-
фитоценотических групп малолетников по плодовитости, помимо абсолют-
ных, были использованы логарифмически (In) трансформированные дан-
ные (см. значения коэффициентов асимметрии в табл. 5.1). Правомерность
такой процедуры мы уже обсуждали в главе 2.
Рис. 5.3. Гистограмма распределения по классам плодовитости выборки из 96 ви-
дов малолетних растений.
96
Л/ //. M(lpKt)t) I lolly 11ИЦ1К>111111)1 (HIO)IOI II» pile ICIIIlO
Верхние границы классов логарифма плодовитости (InNs)
Рис. 5.4. Гистограмма распределения по классам логарифма (In) плодовитости
для выборки из 96 видов малолетних растений (X = 8,29±0,20).
Таблица 5.1.
Статистика данных о плодовитости у двулетников, зимующих и незимующих од-
нолетников
Тип данных Объем выбор- ки Мини- мальное значение X + Sx (антилога- рифм) Макси- мальное зна- чение Асим- метрия As Эксцесс Е
Незимующие однолетники
N S 43 8 16419 + 5154 120000 2,22 3,85
InN S 2,1 7,97 ± 0,33 (2893) 11,7 -0,19 0,2
1. Зимующие однолетники
N S 24 300 26158± 15629 350000 3,9 16
InN S 5,7 8,59 + 0,38 (5378) 12,8 0,3 0,3
2. Двулетники
N S 29 137 24600 + 7283 136000 1,64 1,71
InN S 4,9 8,80 ± 0,34 (6634) 11,8 0,05 -0,72
97
I iiHitt .5 >l< и iiicciiocobin.ic ссмгий и и \ jH>!ii. н популяциях рис I C1111Й
' Таблица 5.2.
( сменная продукция (нлодовн гос i ь), масса диаспор, и i пн ccmchiioi о (kiihgi у неко
юрых видов лесных и стенных двулетников
Виды двулетних растений Тип семен- ного банка no J. Grime (1979) Средняя масса одной ди- аспоры мг Семенная продукция (плодовитость) Источ- ник1*
Alliaria petiolata (II) ТВ’ 2,6(5") 132-440 2,4,9
Lapsana communis (I-II) ТВ 1,3(4) 850-1000 6, 7,9
Geranium robertianum (II) ТВ 1,6(4) 200-550 6, 8,9
Torilis japonica (II) ТВ 2,0(4) 160-500 3, 7, 8,9
Androsace septentrionalis (I-II) ТВ 0,25(2) 952-1300 5,9
Dracocephalum thymiflorum (I-П) ТВ 0,5 (2-3) 48-3212 9
Isatis costata (II) ТВ 2,5 (5) 25-1183 9
Verbascum lychnitis (III?) РВ 0,13(1) 82000-94000 1
Verbascum thapsus (III?) РВ 0,09(1) 19670-1106280 1
* ТВ — временный банк, РВ — постоянный банк.
** Классы размера (массы) диаспор, по Grime et al. (1988).
Источники, на основании которых составлена таблица: 1 — Salisbury, 1942; 2 — Мельникова,
1972; 3 — Baskin, Baskin, 1975; 4 — Lhotska, 1975; 5 — Symonides, 1979; 7 — Grime, 1979;
Grime, Hodgson, Hunt, 1988; 8 — Петров, 1989; 9 — Марков, 19916.
Прежде всего, обратимся к двулетним растениям. Данные таблиц 5.1 и
5.2 заставляют усомниться в обязательной «сверхплодовитости» наших
двулетних растений и поискать другие факторы, влияющие на потенциаль-
ные темпы их размножения и компенсирующие задержку репродукции.
По величине антилогарифмов средних величин, вычисленных для транс-
формированных данных можно заключить, что плодовитость незимую-
щих однолетников несколько ниже, чем у зимующих однолетников и дву-
летников. Две последние группы видов по плодовитости почти не отли-
чаются друг от друга. Что же касается эколого-фитоценотических групп,
то достоверная разница по плодовитости между слагающими их видами
не выявляется. Приходится констатировать совмещение в разных эколого-
фитоценотических ситуациях видов малолетних растений с сильно разли-
чающейся потенциальной семенной продуктивностью.
Отдельно следует остановиться на случаях очень низкой плодовитости
среди малолетних растений, рассмотрев для примера семенную продукцию
лесного незимующего однолетника недотроги обыкновенной Impatiens noli-
tangere. Методическая трудность изучения плодовитости этого растения со-
стоит, как и во многих других случаях, в неодновременном вскрывании пло-
дов на одном растении.
98
Л/ Н Марков 11<>пупмцио!ишя (июлем пя piK iciiiiO
Нели производить одноразовый подсчет, то получаются краппе низкие
значения плодовитости, отражающие на деле только число зрелых пло-
дов и семян, которые находились на растении в момент учета (рис. 5.5а,
б). Близкие величины (от 1,6 ± 0,8 до 4,9 ± 2,4 семян/особь) приводит для
недотроги обыкновенной М. Фаленка (Falencka, 1983), изучавшая этот
вид в условиях Польши. Там же проводила свои исследования Кристина
Фалинска (Falinska, 1986), усреднив данные которой, мы получили такие
величины плодовитости для недотроги обыкновенной в двух различаю-
щихся по условиям местообитаниях: 21,2 ± 3,4 цветков и плодов на особь и
6,0 ± 0,7 плодов на особь.
Путем одновременного учета всех репродуктивных единиц (бутонов вме-
сте с клейстогамными цветками, хазмогамных цветков, плодов и плодоно-
Рис. 5.5а. Гистограмма распределения особей популяции недотроги обыкновен-
ной по классам плодовитости (все особи несли семена только клейстогамного
происхождения).
| | беспл. особи ПТ! популяция 1 популяция 2
Рис. 5.56. Гистограммы распределения особей двух популяций недотроги обык-
новенной по плодовитости (в популяции 1 некоторые особи семян не сформиро-
вали совсем).
Гкта 5 Жизнеспособные семени них рош. в популяциях рисichhB 99
жск, оставшихся после вскрывания или аборгации плодов) у особей в по
пуляциях недотроги обыкновенной под пологом широколис ! венного псе а,
мы вычисляли потенциальную семенную продуктивность. Опа достоверно
(Р<0,001) различалась у особей, имевших хазмогамные цветки наряду с клеи
стогамными, и особей, имевших только клейстогамные цветки. У первых она
составила 68,9 ± 2,8, а у вторых — 33,8 ± 2,4 семян/особь (среднее число се-
мян в плоде было высчитано ранее и принималось равным 2,5 — см. ниже).
После выявления того, что реальная семенная продукция составляла не бо-
лее 48% от потенциальной, стало ясно — в изученных нами популяциях не-
дотроги особи формировали от 14 до ПО семян на особь. При этом сред-
няя плодовитость для особей, сформировавших как клейстогамные (нерас-
крывающиеся, самоопыляющиеся в бутонах), так и хазмогамные цветки,
составила 34,5 ± 1,7, а для особей только с клейстогамными цветками
18,2 ± 1,6 семян/особь. Реальную плодовитость этого вида определяли пу-
тем периодического сбора семян с маркированных особей в природных по-
пуляциях. Она колебалась от 9 до 37 семян/особь при среднем значении
20,9 ± 1,6 семян/особь. Не намного выше, чем у недотроги обыкновенной,
плодовитость особей у недотроги мелкоцветковой — Impatiens parviflora
DC. в популяции из рудерального сообщества. Значение потенциальной
плодовитости этого вида варьировало от 61 до 194 семян/особь, при том,
что реальная плодовитость составляла около 80% от потенциальной.
К числу сравнительно малоплодовитых малолетних растений принадле-
жат также и такие виды, как Raphanus raphanistrum (до 160 семян/особь),
Alliaria petiolata, Torilis japonic a, Geranium rohertianum, Lapsana communis
(см. табл. 5.2) и некоторые другие. Наши данные вместе с данными литера-
туры (табл. 5.3) дают основание заключить, что в естественных популяциях
многих (по-видимому, более 50% от общего числа видов) и в том числе се-
гетальных (редька дикая Raphanus raphanistrum) малолетних растений пло-
довитость особей относительно низкая. Во всяком случае, она может быть
намного ниже среднего значения, высчитанного для выборки из 96 видов,
данные которой мы подвергли трансформации и статистическому анализу.
Таблица 5.3.
Данные о плодовитости некоторых однолетних растений (по Symonides, 1988 с из-
менениями)
Виды Число семян на особь Местообитание Источники информации
Vulpiafasciculata 1,7 Прибрежные дюны Watkinson, Harper, 1978
Floerkea proserpinacoides <23 Листопадный лес Smith, 1983a,b
Anisantha tectorum 250 Злаковник Mack, Pyke, 1983
Erophila verna 35 Злаковник Symonides, 1983
Koenigia islandica 8,2 Высокогорья Reynolds, 1984
Cerastium atrovirens 7,3 Прибрежные дюны Mack, 1976
Sedum smallii 114 Гранитная осыпь Sharitz, McCormick, 1973
100 МН Маркой I loiiy ня цн< >n i hi я (hioiioi им pur iciinh
5.1.1.2. Плодовитость растении ни внутривидовом
(внутрипопуляционном) уровне
Как и многие другие биологические показатели, плодовитость сильно ва-
рьирует даже у особей одной популяции, из-за чего возникают серьезные
методические затруднения при вычислении достоверных средних величин,
которые бы адекватно отражали плодовитость какого-либо вида.
Рисунки 5.6 и 5.7 демонстрируют сильную положительную асимметрию
кривых распределения особей по классам плодовитости (Ns) в популяциях
облигатных двулетников змееголовника тимьяноцветкового Dracocephalum
Рис. 5.6. Гистограммы распределения особей по классам плодовитости для попу-
ляции чесночницы Alliaria petiolata (N=103 — черные столбики) и змееголовни-
ка Dracocephalum thymiflorum (N=50).
Рис. 5.7. Гистограммы распределения особей по классам плодовитости для попу-
ляции рогоглавника Ceratocephala falcata (N=96 — черные столбики) и горца ше-
роховатого Polygonum scabrum (N=94).
Гчини Жи 111СС11<>с<»(»11ыг гсмгпи и их роль и популяциях pin iciiiih
101
thymijloYum (As *2,32) и чесночницы
AHiariu petiolata (As I 1,29), озимого
однолетника рогоглавника серповид-
ного Ceratocephala falcata (As=+1,65)
и незимующего однолетника гор-
ца шероховатого Polygonum scabrum
(As=+1,86). Подобную картину мож-
но наблюдать и у многих других ма-
лолетних растений. Например, на ри-
сунке 5.8 приведены данные для двух
видов клоповника с указанием раз-
махов варьирования признака, и для
двух популяций пастушьей сумки,
произраставших в разных условиях.
Любопытно в этом случае отме-
тить видоспецифичные различия в
архитектуре при равенстве высоты
главного побега и сравнительно не-
больших различиях средних значе-
ний массы крупных особей. Дело в
том, что у клоповника густоцветково-
го L. densiflorum такие особи характе-
ризуются акротонией — ветвлением
в верхней части главного побега, а у
клоповника сорного L. ruderale — ба-
зитонией — ветвлением от основа-
ния, и связать с этим различия в на-
блюдаемой плодовитости (табл. 5.4).
Рис. 5.8 Характер распределения осо-
бей по числу плодов (Nr) в популяци-
ях: клоповников сорного и густоцвет-
кового (вверху) — показаны разы а-
хи варьирования признака у Lepidium
ruderale (L.r.) и у L. densiflorum (L.d.)
и пастушьей сумки (внизу) — гисто-
граммы для популяций с питомника, в
условиях свободного роста (П) и из по-
сева озимой ржи (О.Р.).
Таблица 5.4.
Статистика некоторых биоморфологических показателей, связанных с плодовито-
стью для выборок из популяций Lepidium ruderale и L. densiflorum
Название вида Показатели
Число семян Число побегов Высота главного побега h (см) Высота прикрепле- ния первой снизу ветви h, (см) Биомасса особи мг
Lepidium ruderale 961 ± 114,0 13,1 + 1,0 26,8 + 0,9 2,2 ± 0,6 0,56 ± 0,06
Lepidium densiflorum 597 ± 39,4 8,4 ± 0,6 24,9 + 0,7 12,9 ± 0,5 0,80 ± 0,07
Приведенные примеры указывают на настоятельную необходимость при-
водить в работах, по крайней мере, максимальное и минимальное значе-
ния плодовитости в дополнение к средним. За минимальную плодовитость
можно, по-видимому, условно принять среднее число семян, содержащее
102 МН Марков I loiiyiiMiuioiiiuiH биоло! ия piivicindl
ся в одном плоде, гак как нередки случаи образования мелкой особью мало-
летнего растения одного единственного плода.
Насколько условен этот минимум? Очевидно, в той мере, в какой не фикси-
ровано число семян, образующихся в пересчете на один цветок. Ведь размеры
как односеменного, так и многосеменного плода и число семян в последнем,
могут меняться, отражая уровень жизненного состояния особи. Мелкие гене-
ративные особи с минимальной биомассой, как правило, преобладают в есте-
ственных популяциях малолетних растений, особенно зимующих однолетни-
ков и облигатных двулетников, и обусловливают столь характерное для них
положительно асимметричное распределение по классам биомассы.
Положительная асимметрия распределения по общей биомассе, которое
иногда не удается «нормализовать» даже путем логарифмической транс-
Рис. 5.9а. Линия регрессии числа плодиков (семян) на общую биомассу растения
в популяции рогоглавника: Nf = 4,1 + 0,83W (г = +0,988; N = 90; р<0,001).
Доверительный
уровень 95%
Рис. 5.96. Линия регрессии числа плодов на общую биомассу растения в популя-
ции клоповника густоцветкового: Nfr = 2,10 + 0,83W (г = +0,992; N 95; р< 0,001).
Доверительный
уровень 95%
I iidtiii 5 Жmiicvilocobni.ir семени и их роль it популяциях рИС I Cllllii
103
Рис. 5.9в. Линия регрессии числа плодов (семян) на общую биомассу особи в по-
пуляции горца шероховатого: Nfr = 4,33 + 0,08W; г = +0,901; N = 93; р<0,001).
Доверительный
уровень 95%
Рис. 5.10. Варианты связи с различным числом промежуточных звеньев между
вегетативными органами и отдельным семенем.
формации (Марков, 1980), теснейшим образом связана с асимметрией рас-
пределения по плодовитости, что говорит о глубокой связи характера варьи
104 MH Маркин 11 < >i iy ji >i hi к >i 11 ui >i < >1 к > л( >1 и >i piui ci 11И
ровапия этих показателей. Данные рисунка 5.9а,6,в свидетельствуют о ли-
нейности и высокой положительной корреляции между числом плодов (и
семян) на особь и общей биомассой особи в популяциях рогоглавника, кло-
повника густоцветкового и горца шероховатого.
Если взять пластичность различных структур в пределах особи, то пла-
стическое изменение вегетативных и вспомогательных репродуктивных ор-
ганов, выполняет буферную роль, (см. рис. 5.10) как бы ступенчато осла-
бляя воздействие внешних условий на репродуктивную сферу (Bonaparte,
Brawn, 1975; Марков, 1986, 1992). Рассмотрим варьирование плодовитости
в его связи с варьированием общей биомассы у видов как с не фиксирован-
ным, так и с фиксированным числом элементов цветка.
5.1.1.2.1. Внутрипопуляционное варьирование плодовитости особей у видов
с нефиксированным числом плодолистиков в цветке
При большом нефиксированном числе плодолистиков, например у лю-
тиковых из подсемейства Ranunculoideae, возможно значительное влияние
экологических условий на размеры цветка и его продукта — многоорешка
(Хохряков, 1974). Отсюда можно предполагать и заметное внутрипопуляци-
онное варьирование многоорешков по размерам. Чтобы выяснить степень
внутрипопуляционной пластичности многоорешков, оценить вариабель-
ность их длины, числа орешков в них и корреляцию этих показателей с об-
щей биомассой растений были взяты выборки из естественных популяций
рогоглавника Ceratocephala falcata и мышехвостника Myosurus minimus.
Рогоглавник и мышехвостник озимые однолетники. Морфологически
они довольно сходны и представляют собой розеточные растения с гипоко-
тилем хорошо заметным в течение всей жизни (см. главу 6). После форми-
рования цветка главной осью начинается заложение и развитие параклади-
ев, каждый из которых несет один-два сближенных листа и терминальный
цветок. Конические цветоложа покрыты тесно сидящими спирально распо-
ложенными карпеллами и могут, особенно у мышехвостника, сильно варьи-
ровать по длине. Розеточный рост, короткая осенняя вегетация (октябрь-но-
ябрь) и быстрый переход к цветению и плодоношению весной (май) — вот
основные черты жизненного цикла этих видов.
Выборка, состоящая из 90 плодоносящих особей, была взята весной в по-
пуляции рогоглавника, расположенной на мергелистых холмах. Той же вес-
ной выборку брали в популяции мышехвостника на опушке широколистве-
ного леса. За неделю до этого была сделана фиксация цветков с цветоложа-
ми, удлиненными в разной степени для эмбриологического анализа. При ка-
меральной обработке высушенных до воздушно-сухого состояния особей для
каждой из них определяли общую биомассу (W), число многоорешков (Ng),
длину каждого многоорешка с первого (L — самого крупного) до последне-
го и суммарную длину многоорешков для особи (Lg). Кроме этого, у особей
рогоглавника подсчитывали число орешков в составе каждого многоорешка
(Nfj-Nf6) и суммарное число орешков (плодовитость) на особь (Nf).
Данные таблицы 5.5 указывают на большую размерную дифференциацию в
обеих обследованных популяциях, особенно в популяции рогоглавника, где са-
///<///</Л Жи ineciiocobiii.ic семени ii их р<нн. it популяциях pin iciiidl 105
мая крупная особь в 52 раза превышала самую мелкую но биомассе н в 47,2 раза
но плодовитости (N). 11 популяции мышехвостника все iюказачсип варьировапп
не так сильно, хотя значения коэффициента вариации и здесь часто превышали
30%-ный уровень. В обеих популяциях обращает на себя сильная положи гель
ная асимметрия распределения (As) абсолютных весовых показателей: or I 1, I8
до +1,77 у рогоглавника и от +0,81 до +1,57 — у мышехвостника.
Таблица 5.5.
Статистика некоторых биоморфологических показателей для популяций
рогоглавника и мышехвостника
Показатель Мини- мальное значение X + sx Среднее ± ошибка Макси- мальное значение Асим- метрия As
Популяция рогоглавника (плотность — около 360 особей/кв.м)
Общая биомасса особи (мг) W 6,5 80,3 ± 6,8 338 1,77
Число многоорешков/особь (Ng) 1 2,1 ±0.1 6 1,18
Суммарная длина многоорешков (см)/ особь (L) 0,3 1,7 ±0.1 6 1,53
Суммарное число орешков/особь (Nf) 6 70,5 ± 5,6 283 +1,65
Популяция мышехвостника (плотность — около 3200 особей/кв.м)
Общая биомасса особи (мг) W 6 15,4 ± 1,1 38 1,57
Число многоорешков/особь (Ng) 1 1,6 ±0,1 3 0,81
Суммарная длина многоорешков (см)/ особь (LJ 0,4 0,9±0,1 2,4 1,23
Анализ объединенной корреляционной матрицы (табл. 5.6) указывает на
наличие тесных положительных корреляционных связей между признака-
ми, характеризующими репродуктивную сферу, а также между каждым из
этих признаков и общей биомассой растений у обоих видов. Один из глав-
ных результатов, отраженных в таблице 5.6, — это возрастание тесноты
корреляционной связи с общей биомассой в ряду: Ng - Lg - Nf у рогоглавни-
ка и N -L у мышехвостника.
g gJ
Таблица 5.6.
Матрица коэффициентов корреляции между показателями особей в популяции
рогоглавника (над диагональю) и мышехвостника (под диагональю)
L g Nf w
N g 0 943*** 0 944*** 0,923***
L г 0,64 "s 0,983*** 0,982***
Nf H.O. H.O. 0,988***
W 0,47* 0,53** H.O.
Примечание: но. показатель не определяли; Уровни значимости: *
*** 0,001; n.s недостоверно.
0,05;** 0,01,
106
Л/ // Маркин 11о11уш1Ц11<>||нл>1 bitouoi пи расiciuii
Рис. 5.11а. Зависимость длины перво-
го (Lg2) и второго (Lgl) многоорешков
и числа орешков в них (Nfl и Nf2) от
общей биомассы особи (W) в популя-
ции рогоглавника серповидного.
Рис. 5.116. Стабилизация числа семян
в плоде по мере улучшения жизнен-
ного состояния особей — возрастания
общего числа плодов на растении (Nfr)
у ярутки полевой.
Изменения отдельных многоо-
решков по длине и но числу состав-
ляющих их орешков при увеличе-
нии мощности особей можно проил-
люстрировать графиком (рис. 5.11а).
Налицо поначалу резкое увеличение
длины (Lgl) и числа орешков (Nfl) с
последующим плавным асимптоти-
ческим приближением к максималь-
ным значениям (1,7 см и 67) у пер-
вых, самых крупных, а для 35 осо-
бей и единственных многоорешков.
Динамика тех же двух показателей
(L _ и и максимальные их значе-
ния (1,3 и 67) у вторых многоореш-
ков очень сходны, но кривые берут
свое начало от общего веса в области
28,0 мг — нижнего предела мощно-
сти особей с двумя многоорешками.
Можно видеть, что в момент начала
формирования второго многоореш-
ка первый многоорешек уже состоит
примерно из 38 орешков.
Непропорциональность установ-
ленных в популяциях рогоглавника и
мышехвостника сильного варьирова-
ния общей биомассы особей и очень
слабой изменчивости числа многоо-
решков (соответственно — от 1 до 6,
и от 1 до 3) побуждает к поиску того
уровня, на котором приводятся в со-
ответствие отображаемое общей био-
массой жизненное состояние особей и
их плодовитость (см. рис. 5.9а,б,в).
5.1.1.2.2. Факторы внутрипопуляцион-
ного варьирования плодовитости у ви-
дов с фиксированным числом элементов
в цветке
В качестве примера рассмотрим
варьирование плодовитости у трех представителей семейства крестоцвет-
ных: пастушьей сумки Capsella bursa-pastoris, чесночницы Alliariapetiolata
и ярутки Thlaspi arvense, а также у двулетника из семейства губоцветных
змееголовника тимьяноцветкового Dracocephalum thymiflorum.
Таблица 5.7 дает представление о том, насколько архитектура особей па-
стушьей сумки вносит свой вклад в варьирование плодовитости. По мере
I'hifni Жи ineciiocoGiii.ic сгмгпп и их роль в иоиупяциях р;к iciinii 107
увеличения общей мощпоеги растений в пределах популяций >ioiо вида ча
ею можно наблюдать такую последовательность архитектурных i инов от
сутствие какого-либо ветвления - акротония - базитония (в таблице 5. /,
соответственно: 1-2-3 типы). Можно видеть, что определенное наращива
ние мощности до поры возможно на базе первого типа, но свыше какого- то
предела плодовитость (число плодов) может нарастать лишь за счет пере
хода ко второму типу архитектуры, а максимальных своих значений она до-
стигает только при третьем типе архитектуры. Нельзя не сказать и о имею-
щейся генетической детерминации плодовитости у пастушьей сумки, ко-
торая осуществляется опять-таки через архитектуру. Генетически обуслов-
ленная акротония у биотипа С. b-p. heteris и базитония у биотипа С. Ь-р.
simplex (о биотипах у пастушьей сумки подробнее см. Shull, 1909; Марков,
19766, 1978 и глава 4) явно предопределяют и разницу между ними в сред-
ней плодовитости, которая была отмечена еще Э. Солсбери (1819 ± 232 н
3739 ± 292, соответственно, по Salisbury, 1942).
Таблица 5.7.
Биологическая характеристика разных архитектурных вариантов, представленных
в агропопуляции пастушьей сумки из посева озимой пшеницы
Архитектурные типы Биологические показатели
Общая биомасса (г) Число плодов
min. max. х ±s X min. max. X ±s
1 (N=26) 0,05 0,3 0,12 ±0,01 6 57 18,3± 2,1
2 (N=20) 0,1 1,6 0,42± 0,07 30 160 55,4± 7,7
3 (N=28) 0,15 2,5 0,72 ±0,12 38 284 94,2 ±15,1
4* (N=ll) 0,15 0,95 0,58 ±0,08 66 140 76,2 ±13,5
* растения с поврежденным на стадии розетки апексом.
Однако пластичность плодовитости у пастушьей сумки этим не ограни-
чивается, и нижняя ее граница очевидно может быть установлена только на
уровне числа семян в плоде. При сравнении двух популяций этого вида из
двух контрастных по условиям местообитаний была выявлена существен-
ная разница по числу семян в плоде (рис. 5.12). Неблагоприятные условия
произрастания вызвали резкое уменьшение значений этого показателя.
Реальная плодовитость облигатных двулетников чесночницы Alliaria
petiolata и змееголовника Dracocephalum thymiflorum тоже существенно за-
висит от числа семян в плоде — у первого вида, или от числа нормально
развитых эремов в цветках — во втором случае. Варьирование числа семян
в плоде (стручке) чесночницы показано на рисунке 5.13.
Нормально развитый цветок змееголовника содержит 4 односсменных
эрема, на которые распадается при созревании его завязь. У особей с еще нс
вполне дозревшими семенами, поделенных на четыре класса по высоте как
признаку жизненного состояния, был проведен анализ состояния кремов
и определена частота встречаемости цветков с одним, двумя, тремя и чс
гырьмя полноценными по размеру эрсмами. Распределения частот но ня т и
1U8
Л/// Марков 11<>||у)1мц|1<>н1Н1И (moJioi ни iciiiHI
УСЛОВИЯ ДЛЯ рОС Id р<К 1С‘НИИ
Рис. 5.12. Зависимость числа семян в плоде у особей популяций пастушьей сум-
ки в различных по благоприятности условиях для роста.
Рис. 5.13. Варьирование числа семян в
стручке у особей в популяции чесноч-
ницы черешчатой.
классам (от 0 до 4) оказались различ-
ными для особей разного жизненно-
го состояния (рис. 5.14). Однако про-
слеживается общая закономерность
типа «все или ничего», т. е. при всех
уровнях жизненного состояния ча-
стоты в промежуточных классах (1-
3) явно ниже, чем в нулевом или по-
следнем классе (4). Чем лучше жиз-
ненное состояние особей, тем ниже
частота нулевого класса и, соответ-
ственно, выше частота в последнем
классе (4) при почти одинаково низ-
ких частотах одно-, дву- и трехэрем-
ных цветков.
На примере трех модельных популяций ярутки полевой рассмотрим зави-
симость числа семян в плоде от общей мощности растений, которую мож-
но выразить как, по примеру Э. Солсбери, через один из параметров самой
плодовитости — число плодов на особь, так и через общую биомассу.
Д. Келли (Kelly, 1984) использовал в своей работе первый тип зависи-
мости, изучая специально выбранные популяции шести видов малолетних
растений (Linum catharticum, Gentianella amarella, Erophila verna, Cerastium
semidecandrum, Myosotis ramosissima и Euphrasia pseudokerneri) из относи-
тельно неблагоприятных по условиям местообитаний. Ему удалось пока-
зать, что по мере возрастания числа плодов на особь от одного до 10-25 чис-
ло семян в плоде возрастает. Зависимость была или линейной, или чаще от-
ражалась кривой, плавно выходившей на «плато», что соответствовало ста-
/ hiati *> Жн 1ПСС1КН obni.ic I смени и их рош. и популяциях ]»Ш к null
иг/
Рис. 5.14. Чагины цис ниш с различным
числом нормаiii.no ранимых ipcMon дни
четырех групп особей змееголовники
разного жизненного состояния. Классы
особей по высоте как показателю жиз-
ненного состояния (см): I — 7,3 II ,6,
II — 11,7-14,7; III — 14,8-20,9; IV
21,8-38,8; по оси X — число нормально
развитых эремов в цветке.
билизации отношения число семян/плод у особей, начиная с какого-то ви-
доспецифичного уровня жизненного состояния.
Нами были подвергнуты анализу две чистые модельные популяции
Thlaspi arvense (I и II), произраставшие с весны в хороших условиях, и одна
модельная популяция (III), испытавшая сильное фитоценотическое пода-
вление со стороны специально подсеянной гречихи и потому отличавшая-
ся от двух других резко сниженным размахом варьирования изученных мор-
фобиологических показателей (табл. 5.8).
Таблица 5.8.
Статистика биоморфологических показателей в трех модельных
популяциях ярутки Thlaspi arvense
Популяции показатели X min X ± S X X max As
I (N=100) высота (см) 9,1 27,3 ±0,6 41 -0,79
число:
плодов/особь 1 22,2 ±1,4 72 0,84
семян/особь 2 145,3 ±11,3 489 1,03
семян/плод 1 5,9 ±0,2 12 -0,28
II (N=39) высота (см) 14,5 29,8 ± 0.9 40 -0,55
Общая биомасса (г) 0,05 1,1 ±0,2 4,2 1,16
число:
плодов/особь 8 81,3 ±12,9 303 1,17
семян/особь 18 460,9 ±84.5 2025 1,47
семян/плод 1 5,2 ± 0,4 11 0,39
III (N=170) число:
плодов/особь 1.0 2,4 ±0,08 6 0,75
семян/особь 1.0 10,1 ± 0,53 35 1,16
семян/плод 1.0 . 4,1 ±0,14 10 0,51
С изменчивостью основных показателей жизненного состояния резко
контрастируют слабо меняющиеся от популяции к популяции размах ва-
рьирования и средняя арифметическая числа семян в плоде при некотором
возрастании коэффициента асимметрии распределения в подавленной по-
пуляции (1П). Такое свидетельство возрастания числа плодов с минималь
110 Л/ II Марков 1l< >i lyjini ин ин ши (>1101101 им рис i cuiiil
НЫМ ЧИСЛОМ ССМЯП В популяции ВМСС ГС С очень С IKlOl.lM изменением средне-
го числа семян в плоде но мере увеличения числа плодов па особь убежда-
ет в том, что стресс, испытываемый популяцией, очень сильно сказался на
репродуктивной сфере. На рисунке 5.116 можно видеть, как в форме асим-
птотической кривой достигалась стабилизация числа семян в плоде по мере
улучшения жизненного состояния особей — возрастания общего числа пло-
дов на растении. Не случайно этот рисунок совмещен на одной странице с
рисунком, отражающим стабилизацию числа орешков в многоорешках ро-
гоглавника — эти явления безусловно связаны друг с другом и играют сход-
ную роль в пластической регуляции плодовитости.
Как уже говорилось выше, взаимосвязь структур разных уровней, на ко-
торых осуществляется регуляция, обеспечивающая поэтапное затухание
изменчивости от лабильных вегетативных структур до консервативных ре-
продуктивных структур, кончая массой отдельного семени, можно показать
при помощи схемы (см. рис. 5.10). Видно, что существует, по крайней мере,
пять вариантов взаимодействия между структурами разного уровня, отли-
чающихся неодинаковым числом промежуточных звеньев.
У рогоглавника и мышехвостника наблюдаем вариант взаимоотношений
между вегетативной и репродуктивной сферами, фигурирующий в нашей
схеме под номером 2. Многоорешек, как видно, играет роль многосеменно-
го ценокарпного плода или даже уподобляется соцветию по своей регуля-
торной роли. Таким образом достигается более тонкая пластическая «при-
гонка» плодовитости к общей биомассе, о чем свидетельствуют более вы-
сокие коэффициенты корреляции между L , Nf и W, чем между N и W (см.
табл. 5.6). 8 g
При плавном нарастании общей биомассы соответствующее нарастание
плодовитости происходит вначале за счет увеличения числа орешков в со-
ставе первого многоорешка и, как следствие, увеличения его размеров. На
фотографии, где приводится продольный срез многоорешка мышехвостни-
ка, и сопроводительных рисунках (5.15) видно, что в верхней части он не за-
крыт, здесь продолжается рост — расположенные в самых верхних орешках
семязачатки содержат еще лишь археспориальные клетки, тогда как нижние
орешки уже содержат зрелые зародышевые мешки. Только во вторую оче-
редь плодовитость регулируется изменением числа самих многоорешков.
При константном размере многоорешка и постоянном числе плодиков на
нем такая пластичность плодовитости была бы вряд ли достижима.
Внутри соцветия, внутри плода или в составе апокарпного сборного пло-
да механизм регуляции плодовитости однотипен: изначально закладывает-
ся избыточное число цветков или семян, а позднее часть их недоразвивает-
ся. Однако важен и момент, когда происходит прекращение доступа ассими-
лятов к развивающимся цветкам, плодам и/или семязачаткам. Ведь от него
зависит, насколько жизнеспособными будут репродуктивные структуры, за-
ложившиеся последними. Поскольку, так или иначе, это связано с разме-
рами образующихся диаспор, то рассмотрение этого вопроса продолжим в
разделе 5.1.4.
/ //</«</ Л Ж inure noct >()i ii.iv семени и п.ч рощ, в популяциях pnciciiiill 111
5.1.2. Семя как единица дисперсии
Одним из увлекательнейших разделов ботаники и популяционной эколо-
гии растений можно назвать ту ее часть, которая изучает распространение
зачатков или диаспор.2 Именно здесь нам предоставлена возможность ра-
ботать с приспособлениями (т. н. адаптивными признаками или признака-
ми, которые должны повышать приспособленность), получающими хоро-
шее «морфологическое оформление». Летучки одуванчика, крылатки кле-
на, перистые ости чешуй при зерновках ковылей, прицепки на семянках че-
реды и много, много других. Хорошо известны прекрасные иллюстрации
с изображением этих приспособлений, кочующие из одной ботанической
книги в другую. Однако столь ли уж очевидна необходимость выработки
подобных приспособлений и насколько способствуют они повышению при-
способленности популяции и вида? Какие факторы отбора могли и могут
поддерживать их существование?
Начнем с того, что распространение или рассеивание диаспор — это один
из способов осуществления потока генов (gene flow), процесс, который над-
лежит рассматривать на нескольких уровнях. С учетом этих уровней и сле-
дует оценивать его значимость.
Процесс и результат рассеивания диаспор какой-то конкретной плодо-
носящей особью принято называть дисперсией зачатков.3
Если считать это первым уровнем, то следует уточнить, с какого момен-
та мы вправе говорить о результатах дисперсии. По крайней мере, для неко-
торых видов и характерных для них местообитаний важно подчеркнуть зна-
чимость не только первичной дисперсии как процесса отделения зачатков
от материнской особи и достижения ими поверхности субстрата в ходе
преимущественно горизонтального распространения. Важно отметить ре-
альность и значение вторичной дисперсии — дальнейшего перемещения за-
чатков при участии разных внешних агентов. Она может происходить как в
горизонтальном, так и в вертикальном направлении, оказывая заметное вли-
яние на площадь репродуктивной активности особей (см. ниже). Наконец,
можно рассматривать миграции — распространение, расселение вида и фор-
мирование его ареала в географическом масштабе и историческом времени.
Как будет показано ниже, на этом уровне прямую связь со способом первич-
ной дисперсии нам удастся выявить далеко не всегда.
2 Термин «зачаток», как образование, посредством которого осуществляется размножение
и расселение растений, впервые использовал, по-видимому, В. Н. Хитрово (1912), а очень
близкий (если не эквивалентный) по значению термин «диаспора» в ботаническом контек-
сте впервые был предложен шведским ботаником Р. Сернандером в 1927 г. (см. Semander,
1927).
3 Термин «дисперсия» (от латинского dispersio) употребляется здесь, как и в отечественной
литературе по физике или статистике именно в своем основном значении, то есть как «рас-
средоточение или рассеивание». В статистике он обозначает величину, характеризующую
рассеивание значений признака вокруг средней арифметической. В зарубежной литературе
в биологии и биометрии для обозначения этих понятий применяются разные термины dis-
persal и variance, соответственно.
112
МН Маркой I loily пицпопппи (>1101101 пи piuieniHi
Рис. 5.15. Многоорешек мышехвост-
ника маленького Myosurus minimus в
продольном разрезе. Справа (в соот-
ветствии с их размещением) показаны
стадии развития зародышевых мешков
в семязачатках, расположенных на раз-
ных уровнях многоорешка.
5.1.2.1. Первичная дисперсия
зачатков у цветковых растений.
Горизонтальное распространение
(рассеивание) зачатков и расселение
растений
Уже Карл Линней отмечал, что
плоды и семена снабжены структура-
ми, обеспечивающими их рассеива-
ние. Он и его ученики, по-видимому,
были первыми, кто пытался провести
опыты по изучению дисперсии зачат-
ков. Подобного рода эксперименты
ставил и Ч. Дарвин, который впервые
в науке пытался изучать флору жиз-
неспособных семян, извлеченных из
почвы.
После научно-популярного обзо-
ра морфологических приспособле-
ний для дисперсии, предпринятого Э.
Ульбрихом (Ulbrich, 1928) главным
классическим исследованием дис-
персии за рубежом принято считать
монографию X. Ридли (Ridley, 1930)
«The Dispersal of Plants throughout the
World» (Расселение растений по зем-
ному шару). Из названия книги вид-
но, что X. Ридли интересовался во-
просами биогеографии, миграции ви-
дов и формированием флор. В своей
монографии он пытался выявить вза-
имосвязь между типами расселения видов и наличием у них определенных
механизмов дисперсии.
Изучению дисперсии зачатков у растений много работ посвятила Р. Е.
Левина, которой, предложено и несколько наиболее полных и детальных
классификаций типов распространения диаспор (Левина, 1957,1987), а так-
же ее ученик и последователь В. Ф. Войтенко (1974). Поскольку для попу-
ляционной экологии важны в первую очередь некие общие закономерности
дисперсии, то для начала рассмотрим упрощенную классификацию типов
первичной дисперсии зачатков.
5.1.2.1.1. Классификация типов первичной дисперсии зачатков
Ахория — способы обеспечения минимального перемещения диаспор от
материнской особи:
Геокарпия — созревание плодов в почве, где они формируются из под-
земных цветков, или куда внедряются вместе с цветками после оплодот-
I'hltui Л Ж III11Cl Il( >C( >( >11I.IC < СМ4Ч1И II 114 jlOIII. II нопуиициич pm Hllllll
11 i
Рис. 5.16a. Геокарпия у арахиса.
ворения. Классическим примером ге-
окарпного растения можно считать
арахис Arachis hypogaea (рис. 5.16) с
его зарывающимися в почву и дозре-
вающими в ней бобами из нижних
цветков особи, которые в отличие от
верхних плодовиты.
Амфикарпия — частичная гео-
карпия (от греческого амфо — оба;
амфи — в смысле двоякий), когда в
дополнение к подземным плодам, ко-
торые развиваются из клейстогам-
ных цветков, на растении формиру-
ются и, созревая, дают семена плоды
из обычных надземных хазмогамных
цветков4 [Vicia amphicarpa — горошек
двоякоплодный, Lathyrus amphicarpus — чина двоякоплодная (рис. 5.17),
Ceratocarpus arenarius — рогач песчаный и др.].
Автохория — участие структур самого материнского растения в распро-
странении диаспор:
Автобарохория — опадание диаспор под действием силы тяжести и блаю-
даря формированию отделительной ткани на рубчике (Quercus robur — дуб чс-
решчатый, Polygonum scabrum — горец шероховатый, Cannabis ruderalis — ко-
нопля, Avena fatua — овсюг и множество других видов, если не все семен-
ные растения вообще!);
Автомеханохория — механическое разбрасывание семян благодаря осо-
бым и разнообразным анатомическим приспособлениям, обеспечивающим
взрывное вскрывание плодов (Impatiens noli-tangere — недотрога обыкно-
4 Е. Symonides (1988) рассматривает как амфикарпию формирование двух типов над земных
плодов из клейстогамных и из хазмогамных цветков — у недотроги обыкновенной (1трч
ficus noli-tangere I г. е. вне всякой связи с геокарпией, что вряд ли целесообразно.
114 Л / //Л/<у>л<т I loHyiniiuioiiiiiiH (hioiioi ни pm iciniii
венная, Caragana arboresccns uapaiana древовидная, ()\ahs acetosclla
кислица обыкновенная, Viola arvensis фиалка нолевая);
Автобаллистохория — разбрасывание семян благодаря упругим силь-
но пружинящим плодоножкам, реагирующим на толчки дождевых ка-
пель, прыгающих насекомых и прочие механические воздействия (про-
ломник наибольший Androsace maxima, змееголовник тимьяноцветко-
вый Dracocephalum thymiflorum, Черноголовка обыкновенная Prunella
vulgaris, синяк обыкновенный Echium vulgare, оносма простейшая Onosma
simplicissima и др.).
Аллохория — дисперсия, происходящая благодаря внешним по отноше-
нию к популяции факторам разной природы:
Анемохория — дисперсия зачатков, главным фактором которой выступает
ветер, разносящий либо сами диаспоры, отделившиеся от материнского рас-
тения (Acer platanoides — клен платановидный, Taraxacum officinale — оду-
ванчик лекарственный, Isatis costata — вайда ребристая), либо перемещаю-
щий само оторванное от корня мате-
ринское растение в форме «перекати-
поле» (Ceratocarpus arenarius,
Crambe tataria — катран татарский,
Lepidium ruderale — клоповник сор-
ный, Rheum tataricum — ревень та-
тарский);
Гидрохория — распространение
зачатков водой, к примеру у кокосо-
вой пальмы (рис. 5.166);
Зоохория — растаскивание диа-
спор разной природы от семени до
соплодия животными, переносящи-
ми зачатки либо на себе (эпизоохо-
рия — Ceratocephala falcata — ро-
гоглавник серповидный, Torilis
japonica — торилис японский,
1 Arctium lappa — лопух большой,
Xanthium strumarium — дурнишник
3 обыкновенный, Bidens tripartita —
5 череда трехраздельная, Cynoglossum
officinale — чернокорень лекарствен-
7 ный), либо внутри себя (эндозоохо-
рия — растения с сочными плодами
— ягодами, костянками и т. д.).
Антропохория — распростране-
Рис. 5.17. Амфикарпия у бобовых
а — чины двоякоплодной Lathyrus
amphicarpus', б — горошка двоякоплод-
ного Vicia amphicarpa Lam. (по Ulbrich,
1928) и (в) у рогача песчаного (с — се-
мядоли, п — плоды в их пазухах).
ние диаспор человеком в процессе
его сельскохозяйственной деятель-
ности и потому выделяемое в особый
тип дисперсии.
Явления, когда одному виду рас-
тений свойственно распространение
I 'lhifiii 5 Ж|| |||сс11()со()11|.1с семени и их роль и популяциях pilCICIIIlH 115
одинаковых (или очень сходных) но морфологии зачатков двумя ппп iicckoiii.
кими способами, Р. К Левина предлагает обозначать понятиями «()imuo\<>
рия» и «полихория» (от греч. «диплос» — двойной, «поли» много). 11а наш
взгляд дисперсия зачатков у любого вида семенных растений осуществляем
ся, как правило, не одним, а несколькими способами, если мы будем прини-
мать во внимание возможности и вторичной дисперсии. Иными словами, но
существу все семенные растения по природе своей полихорны. Можно лишь
пытаться выделить преобладающий (или преобладающие) способ дисперсии
у того или иного вида. Сама Р. Е. Левина приводит как пример диплохории
вторичную мирмекохорную дисперсию автомеханохорно разбросанных се-
мян у растений, таких как фиалки, пролесник многолетний и др. Помимо
внезапно и резко вскрывающихся плодов эти виды имеют мясистые придам -
ки — элайосомы на семенах, из-за чего семена активно растаскиваются му-
равьями.
Распространение же различными способами морфологически различ-
ных зачатков одной и той же особи (при гетерокарпии — см. ниже), Р. К
Левина предложила называть гетерохорией. Здесь мы теоретически можем
столкнуться с различиями в способах первичной дисперсии зачатков одной
особи. Однако чаще, наверное, можно наблюдать лишь различия в эффек-
тивности (дальности) рассеивания каким-либо одним агентом (к примеру,
ветром) морфологически разных диаспор, формируемых одним гетерокарп-
ным материнским растением.
В качестве примера гетерохории можно рассмотреть изученную в опы-
те вторичную анемохорную дисперсию гетерокарпных плодов ревеня ком-
пактного (Rheum compactum). Для четырех хорошо различимых морфотипов
у этого вида были получены разные расстояния и, соответственно, скорости
перемещения диаспор под действием турбовентилятора (см. табл. 5.9).
Таблица 5.9.
Результаты экспериментального изучения вторичной анемохорной дисперсии
крылатых односеменных плодов ревеня компактного
Параметры Морфы плодов ревеня компактного
Двухгран- ные Мелкие трех- гранные Крупные трех- граннЫе Четырех- гранные
Среднее расстояние, м 16,43 16,13 44,55 30,8
Среднее квадратическое отклонение 21,55 10,98 31,69 21,52
Скорость полета плодов, м/сек 0,08 0,08 0,22 0,15
К. Урбанска (Urbanska, 1992) после рассмотрения типов первичной диспер-
сии разбирает особые стратегии распространения зачатков, сопоставляя ту
часть диаспор, которая рассеивается на значительные расстояния, с той, ко-
торая остается вблизи материнского растения и иногда даже имеет спсциаль
ныс приспособления для этого. Именно в этом контексте она рассматриваем:
116 Л/.// MdpKoti I h>ilyиИЦП01 иnni <>1101101 ни pncieniili
1. 11олихорные растения, вкладывая в no понятие roi же смысл, что и Р.
Е. Левина, но подчеркивая, как важнейшее их свойство, очень выеокую пло-
довитость и связанный с ней вероятностный (основанный на случайности)
характер их рассеивания и возобновления {«Оппортунистическая страте-
гия» — от английского opportunity — случай);
2. Амфикарпические растения, называя так виды с ярко выраженной
«Пессимистической стратегией», заставляющей их зарывать в почву уже
освоенного ими местообитания рядом с собой значительную часть своих за-
чатков (плодов с семенами);
3. Гетерокарпные растения, обсуждая, на деле, виды, которые демон-
стрируют гетерохорию (sensu Левина, 1957).
Крупный специалист по части карпологии и изучения способов распро-
странения плодов и семян Р. Е. Левина считала, что термин «диссеминация».
как чисто биологическое понятие, предпочтительнее термина «дисперсия».5
По мнению Р. Е. Левиной диссеминация — это сложный экологический про-
цесс биоценотического уровня, который нельзя сводить лишь к биологии пло-
дов. Полностью соглашаясь с Р. Е. Левиной в оценке сложности и важности
процесса диссеминации, нужно лишь подчеркнуть, во-первых, больший объ-
ем данного понятия, которое вбирает в себя и процесс формирования на рас-
тении диаспор или диссеминул, помимо собственно процесса их рассеивания
(дисперсии), и, во-вторых, биоценотический уровень самого акта дисперсии,
в котором бывают задействованы не только посторонние для популяции аби-
отические агенты (вода, ветер), но и организмы, связанные с рассеивающим
семена растением консортивными связями.
5.1.2.1.2. Рассуждения о причинах эволюции приспособлений для первичной
дисперсии зачатков
И все же рассмотрение любой классификации типов дисперсии не может
не вызвать вопроса о том, каким образом и какие силы отбора смогли закре-
пить столь очевидную «тягу» растений к распространению. Ведь если мате-
ринское растение смогло успешно пройти свой онтогенез до генеративной
стадии и сформировать некое число диаспор в условиях данного местоо-
битания, значит это местоообитание и впрямь благоприятно для данного
вида. Зачем же его покидать, а не возобновляться здесь устойчиво, усили-
вая ценотическую позицию?
Отправной точкой для рассуждений должен стать тезис о том, что есте-
ственный отбор должен поддерживать выработку приспособлений для
дальнего переноса диаспор лишь в том случае, если вероятность достиже-
ния мест с более благоприятным комплексом условий выше, чем вероят-
ность отмирания зачатков в процессе диссеминации или чем вероятность
достижения менее благоприятных экотопов (Gadgil, 1971).
Издавна доминирующим взглядом на причину выработки приспособле-
ний для дисперсии было мнение, что рассеивание зачатков необходимо для
5 В опубликованных к настоящему времени словарях биоморфологических терминов между
этими двумя понятиями часто ставят знак равенства.
/'/<«<</> Жн нкч н<иobni.ir t умгни и и\рош. и iiiHiyinmihix put H indi 11/
удаления растений друген друив и ослабления впу i рниндонон конкуренции
(Визпср, 1892; Вуль<|), 1933).
Некоторые авторы (Ridley, 1930; Farghali, 1940) c’iniaioi, чю рассредо
точение или рассеивание диаспор может поощряться отбором, поскольку
именно таким способом можно избежать их массового повреждения фи-
тофагами (Левина, 1957). Признавая логичность данного предположения,
нельзя, вместе с тем, не отметить многочисленные факты задержанной дис-
персии и зимовки диаспор на надземных (живых или отмерших) побегах
материнских особей в составе воздушного банка (см. ниже). В этом случае
семена подвергаются особенно высокому риску из-за фитофагии.
О. Эрикссон и К. Кивиниеми (Eriksson, Kiviniemi 1999) обобщают гипо-
тетические представления о причинах выработки приспособлений к дис-
персии в виде таблицы (табл. 5.10).
Таблица 5.10.
Гипотезы о факторах эволюции дисперсии зачатков у растений (по Eriksson,
Kiviniemi, 1999)
Гипотезы Условия Благоприят- ствует дис- персии: Ссылки
1. Стратегия ком- пенсированно- го риска («bet hedging») Вариабельность во вре- мени Во времени и пространстве Cohen, 1966; Levin et al., 1984
2. Избегание пере- загущения Вариабельность во време- ни и пространстве В простран- стве Howe, Small- wood, 1982; Ven- able, Brown, 1993
3. Относитель- ность преиму- ществ дисперсии в зависимости от внешних условий Вариабельность в про- странстве или во времени Во времени и пространстве McPeek, Holt, 1992
4. Эффект метапо- пуляций Наличие метапопуляцион- ной структуры Во времени и пространстве Olivieri, Gouyon, 1997
5. Избегание кон- куренции с бли- жайшими род- ственниками Генетическая структура* Во времени и пространстве Hamilton, May, 1977; Venable, Lawlor, 1980
6. Избегание есте- ственных врагов (фитофагов) Увеличенный риск нападе- ния со стороны естествен- ных фитофагов около ро- дительского растения В простран- стве Howe, Smallwood, 1982
7. Направленная дисперсия Известны вектора, влияю- щие на направление дис- персии В простран- стве Howe, Small- wood, 1982; Ven- able, Brown, 1993
* Примечание: в данном случае подразумевается тот факт, что в среднем родственность
особей в пределах «ореола первичной дисперсии» любой особи всегда выше, чем между
особями разных ореолов.
Рис. 5.18. Модель ореола рассеяния,
возникающего в процессе первичной
дисперсии диаспор, и кривая, отража-
ющая резкое падение вероятности при-
земления диаспор по мере удаления от
материнской особи (местоположение
ее показано стрелкой 1).
118 А/// Марков I loiiyiiMHiioiiiniH (hioiioi ия рис гений
Главным принципом, по которо-
му происходит дисперсия семян у
большинства видов, является, по-
видимому, сочетание по возможно-
сти дальнего разноса диаспор с mo-
no спорней, т. е. закреплением в уже
освоенном экотопе. Количественное
соотношение дальнего переноса и то-
поспории при рассмотрении в систе-
ме координат расстояние — вероят-
ность достижения отражается хоро-
шо известной асимметричной кривой
(см. например, Howe, Westley, 1986;
Марков, 1986), которая показывает,
что преимущество (самая высокая ве-
роятность) всегда за топоспорией, а
вероятность дальнего переноса семян
резко снижается по мере увеличения расстояния (рис. 5.18). В зарубежной
англоязычной литературе такое графическое изображение дисперсии образ-
но называют «seed shadow»6, а в отечественной — ореолом рассеивания,
определяющим площадь репродуктивной активности особи. При рассмо-
трении в трехмерном пространстве кривая дисперсии вокруг особи всегда
одновершинна, более или менее симметрична и может отличаться лишь экс-
цессом (Salisbury, 1942, 1961; Willson, 1983).
Благоприятность условий местообитания в аспекте дисперсии можно
оценивать по частоте встречаемости микроучастков (safe-sites — по терми-
нологии Дж. Харпера), которые пригодны для приживания проростков.
Все теоретически возможные последствия, связанные с этой закономер-
ностью и конкурентными отношениями, которые возникают между мате-
ринской особью и ее потомством, рассмотрены недавно С. Мак-Канни
(МсСаппу, 1985). Если общая форма кривой по существу универсальна, то
конкретная форма видоспецифична, поскольку зависит от наличия специ-
альных приспособлений, обеспечивающих с одной стороны топоспорию, а
с другой — дальний перенос. Здесь уместно уточнить, что приспособлени-
ями, обеспечивающими эти два процесса, обладают зачастую не сами семе-
на, а плодики, соплодия, мерикарпии, эремы и т. д.
Не все семенные растения, и особенно однолетники, обладают какими-
либо ярко выраженными приспособлениями для рапространения плодов
и семян. Но, как справедливо замечает Р. Е. Левина (1957), видов, лишен-
ных каких бы то ни было приспособлений для распространения диаспор, в
природе не существует. При более внимательном рассмотрении выясняет-
ся, что каждый из видов цветковых растений характеризуется полихорией
и использует не один, а по меньшей мере два способа распространения за-
чатков (табл. 5.11), сочетая, например автобарохорию и баллистохорию или
анемохорию и автобарохорию и т. д. Можно заметить, что один из этих спо-
6 При дословном переводе это — «семенная тень», или «осеняемая» или осыпаемая
распространяющимися семенами поверхность, часть территории попупяции.
Г/hitui > Ж|Г111сс11(>со(>11ис cvMPihi и их р<нн, и популяциях рисICIIIlit 119
собой обеспечивает тотн норию, а другой дисперсию на более дальние
дистанции.
Таблица 5.11.
Способы диссеминации некоторых розеточных и полурозеточных малолетних рас-
тений
Виды растений, сгруппированные по фитоценотической приуроченности Характер диаспор4* Способы распространения (дисперсии)5**
Сегетальные: Capsella bursa-pastoris Arabidopsis thaliana Erysimum cheiranthoides Thlaspi arvense ОС или створка стручочка с семенем НОС ОС нос автобарохор, анемохор, антропохор автобарохор, антропохор автобарохор, антропохор автобарохор, антропохор
Рудеральные: Lepidium ruderale Lepidium densiflorum ОС ОС баллист, анемогеохор (перекати-поле) баллист, антропохор
Степные: Androsace maxima A. elongata A. septentrionalis Myosurus minimus Ceratocephala falcata Dracocephalum thymiflorum Draba nemorosa Isatis costata нос нос нос орешек орешек ОС НОС крылатый односеменной стручочек баллист, автобарохор баллист, автобарохор баллист, автобарохор автобарохор, гидрохор ? эпизоохор, антропохор баллист, антропохор автобарохор, антропохор анемохор, автобарохор
Прибрежные: Rorippa palustris Limosella aquatica Plantago major Androsace filiformis НОС ОС ОС НОС автобарохор, гидрохор гидрохор, автобарохор автобарохор, антропохор баллист, гидрохор
.ОС — ослизняющиеся, НОС — неослизняющиеся или очень слабо ослизняющиеся семена.
Первым значится либо преобладающий способ распространения диаспор, либо
наблюдающийся в начале диссеминации и сменяющийся другим способом на более позднем
ее этапе
В этом аспекте хорошим примером может служить Ceratocarpus arenarius
(рис. 5.17в), у которого первые два плода образуются при самом основании
стебля в пазухах семядолей. Находясь в тесном контакте с поверхностью
почвы, а иногда и будучи втянутыми из-за контрактильности корня и замы-
тыми грунтом, эти плоды обеспечивают топоспорию, тогда как диаспоры,
находящиеся в верхней части обычно растопыренно ветвистого стебля (т. н.
«перекати-поле»), могут распространяться анемогеохорно (Левина, 1987).
120 МН Марком 11о11уляц||(>И1П1я (hiojioi пи piicivniih
Из таблицы явствует, что в качестве естественных факторов, способству-
ющих дальнему разносу диаспор наших объектов, могут выступать ветер и
вода, к которым можно еще добавить миксоспермию — ослизнение семян.
Миксоспермия — универсальный фактор, «убивающий сразу двух зай-
цев». Она обеспечивает надежное закрепление (приклеивание слизью) се-
мян вблизи материнской особи и в то же время прилипание к колесам и но-
гам, разносящим семена на очень большие расстояния. Темпы выработки
слизистого слоя, от которых зависит длительность сохранения диаспорами
способности к перемещению, были изучены нами в опыте со змееголовни-
ком тимьяноцветковым. Семена эремы этого вида, размещенные на влаж-
ной фильтровальной бумаге, через каждые 5 минут пытались сместить па-
дающими каплями воды. Если с какой-либо высоты сдвинуть семена не уда-
валось, то высоту увеличивали, каждый раз отмечая процент сдвинувшихся
с места семян. Результаты опыта приведены в таблице 5.12. Можно видеть,
что уже через 5 минут после начала опыта — смачивания подстилки — 50%
диаспор перемещалось под действием капель, падающих с высоты 5 см, а
через 25 минут после начала — ни с какой доступной высоты нельзя было
сдвинуть ни одного семени.
Таблица 5.12.
Результаты опыта по изучению скорости выработки слизистого слоя семенами зме-
еголовника тимьяноцветкового (Dracocephalum thymiflorum)
Время (минуты) с момента помещения семян на влажную подстилку Минимальная высота, с Которой можно было сдвинуть семена каплями воды % переместившихся семян
5 5 50
10 7 20
15 15 10
20 20 20
25 - 0
К разной степени ослизнения семян способны малолетние растения из се-
мейства крестоцветных. Например, благодаря сильному ослизнению семян па-
стушья сумка стала вторым из «вездесущих» растений в мире (Coquillat, 1951).
В ходе изучения биологии этого вида выявлено существование способа распро-
странения семян на средние дистанции при помощи ветра. Каждая из двух ство-
рок стручочка отлетает, унося с собой одно верхнее семя, находящееся в своео-
бразной камере (см. рис. 5.19), которая отделена от общей полости неглубокой
перетяжкой (Марков, 1978, 1986). Коэффициент парусности такой структуры
равен 190-200 против 49-67 у самих семян, то есть в 3^4 раза выше. При про-
ращивании на фильтровальной бумаге проросток появляется через отверстие
створки. В полевых условиях нам неоднократно попадались особи пастушьей
сумки, корень которых пробил Насквозь полуистлевшую створку.
Androsace filiformis — проломник нитевидный, как и другие три мало-
летних вида этого рода во флоре средней полосы, это баллист, разбрасы-
I 'KItKI Ь Ж» IIICCIIOCoGlIblt* ГСМСНП и нч рош> в понулициих рис ICIIIlil
121
Рис. 5.19. Раскрытый стручочек пасту-
шьей сумки (пояснения в тексте).
вающий свои мелкие семена благода-
ря упругости стрелки, несущей зон-
тик. Однако в отличие от остальных
видов, произрастающих на остепнен-
ных сухих склонах, этот гигрофиль-
ный вид прибегает к гидрохории —
его неослизняющиеся семена раз-
носятся водой. Влажные заиленные
участки береговой линии иногда по-
крываются густыми скоплениями ро-
зеток, а позднее — цветущих и пло-
доносящих растений. Такую картину
наблюдали летом 1984 г., когда уро-
вень воды в реке Волге в межень был
очень низким.
На реке Амур, по свидетельству
А. П. Нечаева (Нечаев, Гапека, 1970),
A. filiformis в числе других так назы-
ваемых меженных эфемеров укла-
дывается со своим жизненным ци-
клом в относительно краткий проме-
жуток времени между двумя паводка-
ми. Во время паводка его семена раз-
носятся водой по всему побережью.
Отмечено значение полых вод и в разносе семян желтушника левкойного
Erysimum cheiranthoides, пастушьей сумки Capsella bursa-pastoris и кло-
повника сорного Lepidium ruderale (Работнов, 1956).
Как уже говорилось, в большинстве случаев результат горизонтальной
дисперсии оценивают по достижении поверхности субстрата диаспора-
ми. Однако, как показали исследования А. Мортимера (по Harper, 1977),
диаспоры подорожника ланцетолистного Plantago lanceolata, райграса
Arrhenatherum elatius и овсяницы красной Festuca rubra в ходе вторичной
дисперсии способны перемещаться по поверхности почвы. Скорость пере-
мещения зависела от способности зачатков к ослизнению и от неровности
поверхностного слоя почвы, а последний фактор специально меняли в опы-
те. Изучать подобные процессы сложно без применения радиоактивных ме-
ток и монитора для обнаружения меченых семян, то есть без метода, приме-
ненного, к примеру, А. Уоткинсоном (Watkinson, 1978) при изучении попу-
ляции дюнного однолетника Vulpia fasciculata.
5.1.2.2. Вертикальное перемещение (распространение) диаспор
в ходе вторичной дисперсии
Под распространением (дисперсией) обычно понимают перемещение се-
мян и вообще диаспор в горизонтальном направлении. Но в жизни некоторых
наших малолетних и многолетних травянистых растений важное место при
надлежит и перемещению диаспор в вертикальном направлении, г. е. загну
122 МН Марков 11(Н|уляц|1оппп>| (nioiioi ня рпс1СН11й
бдению в почву и возвращению на поверхность. В а1рофитоцепозах ни про-
цессы сопровождают каждую вспашку, в ходе которой зачатки малолетних сс-
гетальных сорняков в почве основательно перемешиваются. Именно поэтому
распределение семян по горизонтам в почве агрофитоценозов и естествен-
ных, например, луговых, сообществ различно. Если в луговых и степных фи-
тоценозах число диаспор чаще всего, за редким исключением (см. Мовчан,
Осычнюк, 1985), закономерно убывает от самого верхнего десятисантиме-
трового слоя к нижним (Работнов, 1984), то в полевых растительных сообще-
ствах такая картина наблюдается лишь осенью перед вспашкой, когда на по-
верхности почвы находятся только что осыпавшиеся семена. Вспашка меня-
ет картину, и максимум зачатков может приходиться после нее на более глу-
бокий слой — от 10 до 20 см (Казанцева, Туганаев, 1972).
Влияние вспашки может распространиться и на генетическую структуру
популяций. Сегетальные популяции пастушьей сумки (рис. 5.20а) всегда де-
монстрируют большое генетическое разнообразие (Марков, 1976; Bosbach,
Hurka, Haase, 1982). Однако при отсутствии повторяющихся вспашек под
влиянием межвидовых конкурентных отношений, складывающихся в ходе
демутации, и сукцессионного вытеснения как фактора отбора, популяции
становятся более однородными в генетическом отношении (рис. 5.206).
Можно видеть повышение концентрации одного генотипа (обозначенного
черным кружком) в активной части популяции и самой активной припо-
верхностной части семенного банка.
Вместе с тем, говоря о распределении семян малолетников в почве при-
родных фитоценозов, не следует принижать роли естественных «переме-
шивающих» агентов, в качестве которых выступают выпасаемые и дикие
копытные животные, землерои и особенно дождевые черви. В экспери-
менте, проведенном с семенами пастушьей сумки Capsella bursa-pastoris
Гербертом Хуркой и Р. Хаазе (Hurka, Haase, 1982) получили данные, пред-
ставленные в таблице (табл. 5.13).
Таблица 5.13.
Перемещение семян пастушьей сумки Capsella bursa-pastoris дождевыми червями
в контейнерах, наполненных песком: I — сверху вниз и II — снизу вверх (по Hurka,
Haase, 1982)
Слои почвы (см) 1-2 2-3 4-5 6-7 9-10 13-14 17-18
I Содержа- ние семян (по числу проростков) А Б 10 85 20 10 20 40 20 40 10
II Содержа- ние семян (по числу проростков) А Б - — - 15 15 20 10 75
Примечание: А — контейнеры с червями; Б — контрольные контейнеры.
I hltltl *> >kll IHCt HOC (H >11 Lie ItMi I in II 114 j>* > ll I, Il I lolly II ЯЦНИК pile I <111 Hi 1 / j
Как видно из таблицы дем i слышен» дождевых черней по перемещению
зачатков из верхних споен в нижние и обратно досiaiочно тффек! инна
Однако, количество дождевых червей и их активное и» в рашых почвах не
Рис. 5.20. Два варианта почвенных банков в популяциях пастушьей сумки (рнзны
ми значками в кружках обозначены разные генотипы).
124 МН Мирное I loiiyiiHiuioiiiiiiM biHHioi ini pile iciiidl
одинаковы. Особенно значительно их участие в перемещении семян в лес-
ных, луговых и лугово-стенных биоценозах (Работнов, 1986).
5.1.2.3. Расселение и миграционные способности растений
И. Ф. Удра (1988: 98) считает необходимым «четко и однозначно разли-
чать термины — расселение и миграция». Собственно расселением он пред-
лагает называть возобновление вида в пределах его ареала, связанное с фор-
мированием конкретных ореолов рассеивания, а также внутри и межпопу-
ляционными перемещениями зачатков. «Все же расселения видов за преде-
лы своего ареала (на период его рассмотрения) должны означать миграцию
или же иммиграцию (по отношению к исследуемому региону, в который
внедряется ранее отсутствовавший здесь вид)» (там же). Поскольку и в том,
и в другом случае речь идет о расселении, терминологической «четкости
и однозначности» нам, к сожалению, здесь не обрести, как не обрести на
Рис. 5.21. Инвазия недотроги мелкоц-
ветковой (пояснения в тексте).
каждый конкретный момент времени
точных исчерпывающих сведений об
ареале того или иного вида и его ми-
грационных возможностях.
Например, в цитируемой выше ра-
боте (стр. 23) И. Ф. Удра, кратко рас-
смотрев основные способы диспер-
сии, заключает, что наиболее эффек-
тивный способ распространения пло-
дов и семян для растений — автоба-
рохорный, т. е. путем свободного па-
дения под влиянием силы тяжести.
Можно согласиться с И. Ф. Удрой,
что автобарохорный способ в разных
модификациях — это самый распро-
страненный среди семенных расте-
ний, и что в лесоведении меропри-
ятия по восстановлению дубовых и
буковых лесов рассчитаны именно на
этот способ дисперсии. Однако, нель-
зя не отметить, что наиболее эффек-
тивным для расселения (миграции)
его вряд ли можно считать. На самом
деле, если оценивать эффективность
по тому максимальному расстоянию,
на которое возможен разлет диаспор,
то автобарохория окажется, наоборот,
наименее эффективной. В небольшом
обзоре, посвященном миграцион-
ной способности растений, К. Беннет
(Bennett, 1998) ссылается на работу
Д. Кларка (Clark, 1997) и работу М.
Рниш > Я(|Г111СС11(>с(>(>пыг семени и им рощ, в понупяцнях pnririnih 125
К aiiiia (( ain, Damman, Mun, 1998) в которых копеraiпропаны Сюисс высокие
реализованные темпы миграции некоторых древесных (б>//<v/.v) iripaioiiin
стых (Asarum canadense) растений по сравнению с теоретическим ожидапи
ем. Основными заключениями в соответствии с этими данными были сне
дующие. 1. Именно «хвосты» ореолов рассеивания диаспор имеют большое
значение, а не средние арифметические дистанций разлета; 2. Случайные
(маловероятные) события (торнадо, ураганы), приводящие к максимально
дальнему распространению, должны иметь особое значение для колони
зационных явлений в северных умеренных лесах; 3. Вторичная дисперсия
(особенно зоохорная) может способствовать гораздо более дальнему заносу
диаспор, чем первичная анемохорная, причем в документировании подоб-
ных явлений исключительно важная роль должна принадлежать генетиче-
скому анализу; 4. При рассмотрении в соответствующем (адекватном) мас-
штабе распределение растений предстает чрезвычайно (высоко) мобиль-
ным, и не удивительно, что механизмы, лежащие в основе такой мобильно-
сти, могут быть абсолютно иными, нежели те, что работают во временных
масштабах, при которых распределение кажется статичным. Экстраполяция
данных, полученных о дисперсии зачатков в рамках относительно коротких
отрезков времени, на более длинные временные отрезки оказывается нс
правомерной. Иное дело, когда внедрение вида на новые территории в рс
зультате расширения его ареала, хорошо документировано.
Так наглядное представление об инвазии синантропного вида недотроги
мелкоцветковой Impatiensparviflora на территорию Чехии может дать серия
карт (рис. 5.21), составленных по данным разных лет.
5.1.2.4. Понятие о семенных банках популяций
Задержку дисперсии — сохранение части (иногда большей части!) ди-
аспор на уходящих в зиму живых или отмерших побегах можно тракто-
вать как формирование воздушного семенного банка. В зарубежной англоя-
зычной литературе это очень слабо изученное явление получило название
«serotiny», что означает в дословном переводе просто запаздывание, позд-
нюю активность, позднеспелость, а конкретно — задержку семян в шиш-
ках или плодах (Lamount, LeMaitre, Cowling, Enright, 1991; Hanley, Lamounl,
2001). В условиях сезонного климата и холодных зим подобная задержки
делает возможной эффективную по дальности распространения вторичную
дисперсию по поверхности снежного покрова, насту. Хорошо известна ia-
кая дисперсия у ели, сосны и других хвойных, шишки которых открываю!
ся в зимнее время, а также у многих лиственных деревьев. Однако, зимов-
ка зрелых плодов и семян на растении свойственна и очень многим травя
нистым растениям (см. например, Марков, 1992). Опыты показываю!, что
зимующие прямо на растениях семена вполне жизнеспособны и находятся
в состоянии вынужденного покоя только из-за низких температур воздуха.
Почвенный запас семян, расположенных вблизи поверхности почвы в
условиях, благоприятствующих прорастанию, называют активным (Нагрет.
I960, 1977; Работнов, 1974,1984,1986), а если семена находятся вболссглу
боких слоях, io говорят о потенциальном запасе (Работнов, 1984, 1986) пап
126
М Н Миркон I loiiyiiHHiioiiiHHi (hiojioi ия рис iciinh
АМИИАСОНДЯФМАМИИАСОНДЯФМА
Рис. 5.22. Различные типы почвенных
семенных банков по Grime, 1977 с изме-
нениями: I и II — временные (transient)
III и IV — постоянные (persistent); (чер-
ным цветом обозначена длительность
пребывания семян и их относительное
количество в составе того*или иного по-
чвенного банка — пики соответству-
ют моментам обсеменения растений)
остальные пояснения в тексте.
банке (Grime, 1979) покоящихся се-
мян. Д. Грайм (Grime, 1979) выделя-
ет 4 типа банков семян, два из кото-
рых (I и II) он называет временными
{transient) и два (III и IV) — посто-
янными {persistent). Внутри каждо-
го из этих типов выделены два под-
типа по продолжительности пребы-
вания семян в их составе (рис. 5.22).
Растянутый во времени переход
семян из пассивного запаса в ак-
тивный и растянутое во времени
появление проростков по анало-
гии с дисперсией в пространстве
образно называют дисперсией во
времени (Harper, 1977; Symonides,
1988). Естественные ритмически
возникающие паузы, которые соз-
дают «дисперсию во времени» вы-
званы вклинивающимися неблаго-
приятными периодами. Здесь мы подошли к разговору о покое как о со-
стоянии диаспор, в котором они способны переносить неблагоприятные
условия.
5.1.3. Семя как структура и средство переживания
неблагоприятных периодов
Что такое семя? Потомок материнского растения, на время запрограммиро-
ванно приостановивший свою активную жизнедеятельность, т.е. пребываю-
щий в состоянии покоя. Выход из состояния покоя — важнейший элемент по-
ведения особи в составе популяции. В осуществлении альтернативной линии
поведения: прорастание — непрорастание (покой) важна как роль собственно
потомка (зиготы, зародыша), так и роль материнского растения, снабдивше-
го семя семенной кожурой (формирующейся из интегументов семезачатка).
Семенная кожура, таким образом — это структура, доставшаяся в наследство
от материнского растения, влияние которого на потомка (своеобразная «забо-
та»), таким образом, продлевается вплоть до ответственного события в жиз-
ни этого потомка — процесса прорастания. Изменчивость структуры и состо-
яния семенной кожуры у жизнеспособных семян одной популяции представ-
ляется важным элементом «стартовой» изменчивости, которая может сказать-
ся на более поздних стадиях жизненного цикла популяции.
Термин «покой семян» не всеми исследователями понимается однознач-
но. В относительно редких случаях понятие сужают и трактуют как непро-
растание в условиях влажности температуры и аэрации благоприятных для
вегетативного роста (Amen, 1968). Гораздо чаще термин используют в ши-
роком смысле как синоним непрорастания независимо от причин, но диф-
ференцируют типы покоя, выделяя вынужденный и органический покой
Гита Жи iiiccnocobiii.ic u^Mrihi п их роль в популяциях piu ICIIHll 12/
(Николаева, 1982)либо вынужденный, врожденный и индуцированный но
кой (Harper, 1957, 1977).
Клаесификация типов покоя учитывает то обстоятсльсню, что алыерна
тивная реакция семени (прорастание-непрорастание) зависит как от внеш
них (экзогенных), так и от внутренних причин. Прорастание есть итог со
впадения внутренней готовности семени к активизации жизненных про
цессов с благоприятностью внешних факторов. Несовпадение возможно
по вине либо внешних, либо внутренних факторов, либо по вине и тех и
других, и это лежит в основе выделения разных типов покоя.
В вынужденном (экзогенном) покое пребывают семена внутренне го-
товые к прорастанию, но находящиеся в экологических условиях (низкие
или высокие температуры, недостаток влаги, отсутствие света и др.), пре-
пятствующих прорастанию7. Непрорастание семян в условиях низких тем-
ператур во время зимовки в почве, в условиях дефицита влаги и, наобо-
рот, слишком высокой температуры в разгар лета — вот наиболее типич-
ные примеры вынужденного покоя. В вынужденном покое находились се-
мена проломника Androsace maxima, пролежавшие около 20 лет в гербарии,
после чего проросли на 50% в нашем опыте. Вынужденный покой обуслов-
лен действием экзогенного фактора, например, холода и продолжается толь-
ко в течение времени, пока действует этот фактор, после чего семена про-
растают при обычных условиях и без применения специальных обработок.
Типичный пример вынужденного покоя — непрорастание семян в течение
зимы. Если перенести семена из мерзлой почвы в тепло, то они сразу же
начнут энергично прорастать. Другим примером является вынужденный
покой семян, находящихся на значительной глубине в почве. Роль одного из
ведущих факторов играет повышенная концентрация углекислоты, ане-
стезирующей зародыш.
Врожденный покой обусловлен внутренними причинами, поскольку в этом
случае мы наблюдаем непрорастание семян, находящихся в оптимальных
условиях. Оптимальность условий доказывается хорошим прорастанием по-
сле устранения внутренних причин. Врожденный покой семян может прео-
долеваться по мере дозревания зародыша, однако, у некоторых видов, напри-
мер у ярутки Thlaspi arvense и пастушьей сумки Capsella bursa-pastor is, заро-
дыш дозревает раньше, чем сформируются покровы семени. Зеленые семе-
на, извлеченные из невскрывшихся плодов второго вида, за 16 дней проросли
на 10%. Зеленые и желтые семена, взятые нами из ниже расположенных пло-
дов той же особи, за это время не дали ни одного проростка — по-видимому,
сформировавшиеся покровы полностью блокировали прорастание. По мне-
нию физиологов растений (Максимов, 1958) семенная кожура как бы изол и
рует зародыш от внешней среды, и в результате создается внутренняя сре-
да с повышенной концентрацией продуктов жизнедеятельности, например
углекислоты, обладающих ингибиторными свойствами.
7М. Г. Николаева (1982) причисляет непроницаемость и другие свойства покровов к числу
экзогенных факторов покоя семян. На наш взгляд целесообразно все причины покоя, связан
ныс с устройством самих диаспор, называть эндогенными. Покровы, отделяющие зародыш
от внешней среды, хоть и находятся кнаружи от него, все же являются неотъемлемыми атрн
бутами целой диаспоры. Так же как мы трактуют экзогенный покой D. Zohary (I 965), A S
Sussman, II. О. Halvorsen (1966) и многие другие исслсдовагепи.
128 МН Маркой I Ion ушник шипя (и юшн ня рисiciiiH1
Важными факторами врожденного покоя являются: миксоспермия, фи-
тохромная система и ингибиторы.
Миксоспермия, значение которой для распространения диаспор обсужда-
лось выше, по всей видимости может сильно влиять на прорастание, огра-
ничивая доступ кислорода к зародышу. Группе ученых (Witztum et al., 1969;
Gutterman et al., 1973) удалось показать, что слизь выделяемая клетками ко-
журы семян блефариса Blepharis persica препятствует доступу кислорода
и прорастанию не как простой физический барьер. Прохождение кислоро-
да затруднено из-за деятельности микроорганизмов, которые, развиваясь в
слое слизи, конкурируют с зародышем семени за поступающий извне кис-
лород.
Семена таких наших видов, как пастушья сумка Capsella bursa-pastoris.
желтушник левкойный Erysimum cheiranthoides и крупка дубравная Draba
nemorosa ослизняются, что, по-видимому, еще более затрудняет газообмен
между семенем и внешней средой. Всякая обработка, уменьшающая ослиз-
нение семян этих видов, может играть роль стимула прорастания. Показано,
например, что высушивание набухших семян приводит к «вспышке» про-
растания после нового намачивания (Марков, 1986). В опытах с Capsella
bursa-pastoris (рис. 5.23) и Draba nemorosa такое воздействие приводило
к повышению числа проросших семян, соответственно, до 60 и 84% (про-
тив 27 и 10% в контроле). Замечено, что при новом намачивании степень
ослизнения заметно ниже. Сказанное отнюдь не означает, что стимулирую-
щий эффект высушивания-намачивания связан только с подавлением мик-
соспермии. Эффект этот хорошо известен физиологам (см. например Berrie,
Drennan, 1971), предлагающим некоторые его объяснения. Так Т. Хегарти
(Hegarty, 1977) утверждает, что повторное намачивание приводит к более
быстрому начальному набуханию семян в результате изменения физиче-
ских свойств семенной кожуры и в результате ряда метаболических пре-
вращений, происходящих при первом намачивании и готовящих более бы-
строе прохождение предростовой стадии. По нашим данным стимуляция
проявляется и у семян с неослизня-
ющимися покровами, например у
проломника наибольшего Androsace
maxima. Корешки, появившиеся из
наклюнувшихся семян этого вида,
при высушивании подсохли, но по-
сле нового намачивания продол-
жили рост, и проростки нормально
укоренились в почве. Принимая во
внимание указание Н. В. Обручевой
(1982) на необратимость измене-
ний хроматина, прекращение син-
теза ДНК и потерю зародышем спо-
собности переносить обезвожива-
ние при иссушении семян уже на-
чавших прорастание, можно заклю-
чить, что некоторые виды растений
Рис. 5.23. Повышение всхожести при вы-
сушивании набухших семян пастушьей
сумки с последующим намачиванием.
+ — дата намачивания семян (после вы-
сушивания). В чашках: 1 — без высуши-
вания, 2 — с высушиванием.
L'ldfid Жи tIICCIloroftlll.ir t (’MCIIII II их роль и популяциях piK ICIIIlil
12(J
лучше других адат пропины к перепадам влажное i и поперхное i ного слоя
почвы. В еамом деле, ведь экспериментальное намачивание высуншна
нис — это своеобразная «встряска», моделирующая естественный процесс,
который имеет место в самом поверхностном слое почвы, и приводящая к
«разрыхлению» покровов семени, улучшению газообмена, стимуляции за-
родыша. Эффект повторной процедуры высушивания - намачивания, при-
мененной к не проросшим после первой обработки семенам, заметно сни-
жается по сравнению с первой обработкой (рис. 5.23). В естественных усло-
виях жаркая погода в период созревания и осыпания семян на почву может
вызвать массовое прорастание после первого же дождя свежесозревших се-
мян, которые в обычной ситуации не прорастают. Такая картина наблюда-
лась в годы с жаркой засушливой осенью (1972 и 1975) у пастушьей сумки.
Ослизнение покровов семян во влажном состоянии, судя по всему, лишь
усугубляет действие какого-то более сильного фактора непрорастания, по-
скольку сильно ослизняющиеся семена клоповников Lepidium ruderale.
L. densiflorum, змееголовника Dracocephalum thymiflorum и подорожни-
ка Plantago major хорошо прорастают, даже будучи свежесобранными, но
только на свету.
Чувствительность семян к свету обусловлена наличием в них системы
фитохромов, способных реагировать не только на интенсивность света, по
и на его спектральный состав (в частности, соотношение дальнего красно! о
и красного света в спектре). Различное отношение семян разных видов рас-
тений к свету общеизвестно. У одних растений прорастание возможно толь-
ко на свету, а у других — только в темноте. Семена третьей категории растс-
ний относятся к этому фактору безразлично. Из 964 видов, исследован и ых
Кинцелем (Крокер, 1950), свет способствовал прорастанию семян у 672 ви-
дов, что составляет 70% от числа испытанных видов, но у многих его стиму-
лирующее действие могло быть заменено действием других агентов. В це-
лом проявляется следующая закономерность: наиболее чувствительны и хо-
рошо прорастают на свету мелкие семена. Это и понятно — эволюция при-
вела к выработке фитохромной системы, способствующей прорастанию се-
мян с относительно небольшими запасами питательных веществ на поверх-
ности почвы или близко к ней. Сигнальная роль света в числе других фак-
торов прорастания семян подробно обсуждается в работах Н. М. Ковриго и
3. Г. Дядькиной (Ковриго, Дядькина, 1976; Ковриго, 1977; Дядькина,
Ковриго, 1977).
Наши розеточные и полурозеточные малолетние растения демонстриру-
ют, в основном, положительное отношение к свету как фактору прорас га-
ния за исключением наиболее крупносемянных видов, таких как ярутка но-
левая Thlaspi arvense, которая хуже прорастала на свету (табл. 5.14), и пролом-
ник наибольший Androsace maxima, который к свету относился безразлично.
Однако, не следует забывать, что светочувствительность семян — свойство
довольно изменчивое и зависящее от многих условий вплоть до условий
жизни материнского растения. Она может возрастать со временем или у гра-
чиваться. Так, у Capsella bursa-pastoris первоначально высокая светочув-
ствительность семян со временем ослабевает, и семена становятся безраз-
личными к влиянию света. Свсжесозревшие семена проломника нитевидно-
130 МН. Марков 11(И1уляц11<>ни11>1 (nioiioi ия pm iciiiih
го AndrosaceJiliformis одинаково хороню прорастали в наших опытах в тем-
ноте и на свету при прочих благоприятных условиях, но скоро они станови-
лись настолько светочувствительными, что прорастая на 100% при освеще-
нии, не давали ни одного проростка в темноте.
Таблица 5.14.
Динамика прорастания семян ярутки полевой на свету и в темноте
Дни прорастания с момента закладки опыта Процент проросших семян
без света на свету
утро вечер утро вечер
3-й 0 0 0 0
4-й 3 4 1 1
5-й 7 13 2 2
6-й 23 30 2 2
7-й 38 40 3 3
8-й 41 41 3 3
9-й 41 42 3 3
10-й 42 43 3 3
Чувствительность семян к спектральному составу света интересна не
только с точки зрения физики. Дело в том, что листья растений уже расту-
щих на данном участке, играют роль светофильтров, пропуская под свой
полог свет определенного спектрального состава. Семена, лежащие на по-
верхности почвы, могут воспринимать лучи, проникающие через полог, и
от этого будет зависеть их поведение. Фитохромная система включает две
формы фитохрома. Красный свет (КС — длина волны 660 нм) вызывает об-
разование активной формы фитохрома, которая стимулирует прорастание,
но стимуляция может сменяться ингибированием, если в спектре возрастает
доля дальнего красного света (ДКС — 700-730 нм), вызывающего обратную
инверсию фитохрома. Поэтому значительное снижение отношения красный
свет/дальний красный свет (КС: ДКС или Red : Far Red ratio) — под ли-
стовым пологом выступает как сильный ингибитор прорастания (Taylorson,
Borthwick, 1969; Monteith, 1976), причем влияние этого фактора сказыва-
ется не только в лесных ценозах (Black, 1969), но и в луговых (King, 1975;
Silvertown, 1980; Smith, 1982; Senden et al., 1986). Важно отметить, что че-
рез врожденную светочувствительность и избирательное отношение семян
к свету естественный отбор привел к закреплению еще одной связи между
внутренним акцептором и внешним сигналом.
Третья из названных выше причин врожденного покоя состоит в со-
держании тормозителей прорастания ингибиторов в семенах или плодах.
Рассматривая этот фактор, вновь никак нельзя не подчеркнуть, что диа-
споры (или часть диаспор) у многих видов состоят не только из семян, но
и структур околоплодника. Часто именно эти структуры берут на себя не
только обеспечение дисперсии и механическую защиту семени, но, од-
новременно, механический и биохимический контроль его прорастания.
Семена двулетника вайды ребристой Isatis costata, распространяющиеся,
как уже говорилось выше, при помощи крылатого околоплодника, не про-
/ 7</Г/</ > Жн 11КЧ ll<>< (>(>llhir t ГМ>Ч1П II 114 р(>)1Ь II IIOliyilHHiniX р<1( ICIIIlil 1JI
p.ic'iа ют, судя но всему пока нс разрушится околонподпии It пашем oiii.i
ic семена этого вида дружно прорастали юлько в гом случае, если их hi
вл с кал н из одпоссмсниых стручочков. Примечательно, чю ыким оЬраюм
можно было побудить к прорастанию даже свежесозревшие семена. Очень
сходная картина отмечена у катрана татарского Crambe tat aria, произрас-
тающего в одном местообитании с вайдой. Четырехлетние наблюдения за
единственными полночленными популяциями вайды ребристой и катрана
татарского известными на территории Татарии (Марков, Ситников, 1985),
показали, что численность особей в активной части популяции резко меня-
ется от года к году (см. Главу 10). Вслед за годом массового цветения (1984)
шел год, когда на всей территории, занятой популяцией, с трудом удава
лось отыскать единичные цветущие экземпляры. Периодическое состоя-
ние криптопопуляции и синхронизацию цветения логично связать с поко-
ем семян и ингибирующим действием перикарпа. Разрушение перикарна в
естественных условиях (т. е. процесс, который моделировался в опыте) у
большинства диаспор занимает определенное время, в нашем случае, но-
видимому, одну-две зимовки, после чего наблюдается массовое прорас та-
кие, а на следующий год — пик цветения.
Сходную картину со стопроцентным прорастанием семян после снятия
перикарпа мы наблюдали у рогоглавника Ceratocephala falcata.
Вместе с тем ингибиторы часто содержатся и в самих семенах, и тогда
они могут быть экспериментально удалены, например длительным промы-
ванием. Продолжительность промывания водой семян Thlaspi arvense поло-
жительно коррелировала с энергией и конечным результатом прорастания в
опыте (Марков, 1983): в контроле свежесобранные семена, не подвергавши-
еся промыванию, проросли всего на 7%; промывание в течение 3-6 часов
увеличило всхожесть до 37%, а промывание в течение 9 часов — до 59%.
Подобным образом, очевидно, вымываются ингибиторы из семян пустын-
ных однолетников, которые приобретают всхожесть только после достаточ-
но продолжительных ливней (Went, 1957).
В возникновении третьего типа покоя — индуцированного покоя се-
мян — замешаны и внешние и внутренние факторы. Под действием какого-
либо фактора внешней среды покой индуцируется у поначалу непокоящихся
семян, и они, внутренне изменяясь биохимически и (или) физиологически,
теряют способность прорастать при обычных благоприятных для этого усло-
виях даже после снятия действия индуктора. Индукция покоя может проис-
ходить под действием повышенной концентрации углекислого газа в почвен-
ном воздухе, как было показано известными опытами Ф. Кидда и К. Уэста
(Kidd, West, 1917), под действием низких температур (Baskin, Baskin, 1978).
У некоторых растений повышенная концентрация углекислоты, букваль-
но анестезирующая зародыш, способна переводить семена в состояние
глубокого покоя. Ф. Кидд и К. Уэст помещали семена разных растений на
слой торфа и засыпали почвой слоем 7,5, 15,0 и 22,5 см. В опытном вариан-
те под слоем торфа помещали перегнивающие остатки растений, из-за ко-
торых концентрация СО2 в почвенном воздухе была на 12,4-18,8% выше,
чем в обычной почве. Если в контроле семена прорастали с любой глуби-
ны, то в опыте даже с минимальной глубины проросло всего три семени.
M.H Маркин
I loilyН>1Ц11О1111ПИ (IIIOIIOI ИМ pill Cllllil
Рис. 5. 24. Взаимосвязи между различ-
ными типами покоя семян. Стрелки по-
казывают направления переходов в соот-
ветствии с определениями разных типов
покоя.
Апр. Июнь Авг. Окт. Дек. Февр. Дек. Июнь
Рис. 5.25. Динамический спектр для пас-
сивной части популяции лютика ползу-
чего Ranunculus repens, показывающий
судьбу фракций семян (в %), периоди-
чески извлекаемых из почвенного банка.
G — число проросших семян в пробе;
D — число семян, отмирающих без про-
растания; ID — число семян, находя-
щихся в состоянии первичного (врож-
денного) или индуцированного покоя;
ED — число семян, находящихся в со-
стоянии вынужденного покоя; Р — чис-
ло семян, потребленных фитофагами (по
Sarukhan, 1974).
Исследователи показали, что если у
многих рас гений анестезирующий
эффект углекислоты снимается по-
сле обеспечения нормальной аэра-
ции, то у белой капусты (Brassica
alba) семена переходили в состо-
яние такого глубокого покоя, что
преодолеть его можно было только
применяя специальные обработки:
чередование намачивания и высу-
шивания, прокалывание или снятие
кожуры семени.
Схема на рисунке 5.24 представ-
ляет собой попытку наглядно изо-
бразить три типа покоя семян и по-
казать взаимосвязи между ними. В
популяциях многолетних и малолет-
них растений, имеющих почвенный
запас (банк) жизнеспособных семян,
почти всегда можно наблюдать ком-
плекс различных вариантов покоя.
Прекрасное демографическое иссле-
дование X. Сарукана (Sarukhan, 1974)
дает пример динамического рассмо-
трения фракций семян: а) находя-
щихся в разных типах покоя; б) отми-
рающих и в) потребляемых фитофа-
гами (рис. 5.25). Полученные спек-
тры динамики резервной части попу-
ляций трех видов лютиков представ-
ляют собой наиболее удачный и при-
том наглядный методический прием.
При осеннем анализе семенного за-
паса без мечения свежеосыпавшихся
семян трудно отличить врожденный
покой от индуцированного. Однако,
после дозревания семена, извлечен-
ные из почвы, жизнеспособность ко-
торых можно установить, применив
тест с тетразолиумом, вероятнее все-
го находятся в состоянии индуциро-
ванного покоя, если не прорастают в
оптимальных условиях температуры
и влажности.
Итак, покой семян как способ пе-
ренесения ими неблагоприятных
условий и сохранения определенной
/ //</«</ > >l< II IlirCIIOl oblll.ic СГМУПП II ll\ jH))ll> II популяциях pih ICIIIlil j 5 j
резервной части популяции в патентном cociomiiiih можно pa< смагриваi i. с
двух сторон: «изнутри» анализируя различные вариан i ы покоя, и «снару
жи» выявляя комплекс экологических факторов, удовлс! воряющий спец
ифические требования семян к условиям прорастания. Нс следует забывать,
что эти требования, разнящиеся у разных видов, все же соответствую г той
обычной естественной эколого-ценотической обстановке, в которой оказы-
ваются семена данного вида, тому сочетанию факторов, к которому в тече-
ние длительного времени осуществлялось приспособление в ходе эволю-
ции. Не случайно мы уделили особое внимание таким факторам, как чере-
дование намачивания и высушивания, освещение и темнота, спектральный
состав света, изменяемый листовым пологом, словом тем сигналам, по кото-
рым семена в реальной обстановке распознают регенерационную нишу (тер-
мин П. Грабба — Grubb, 1977) или благоприятную регенерационную ситу-
ацию (Работнов, 1986а,б). Второй термин явно предпочтительнее, посколь-
ку отражает не только пространственный, но и временной характер распо-
знавания в эколого-ценотическом смысле. Резкие колебания влажности и
температуры поверхностных слоев почвы, интенсивное освещение, высо-
кое значение отношения красный/дальний красный свет — вот сигналы для
распознавания в пространстве и во времени «окон», гэпов (gaps) — локаль-
ных регенерационных ситуаций — в которых только и возможно достаточ-
но успешное возобновление малолетних растений.
5.1.3.1. Оценка семени как средства перенесения неблагоприятных
периодов в различных эколого-ценотических условиях
5.1.3.1.1. Динамика жизнеспособных семян как неактивного компонента
в популяциях
Схемы, отражающие динамику жизнеспособных семян, показывают, что
запас их в почве периодически уменьшается за счет прорастания части из
них. Ежегодно некоторое количество семян превращается в проростки, то
есть некоторое количество особей переходит в активную часть популяции.
Несмотря на известную автономность резервной части популяции, позво-
лившую нам рассматривать ее динамику изолированно от других фаз, связь
ее с этими другими фазами существует и притом очень тесная. Пока для
получения целостного представления о цикличности процессов жизнеде-
ятельности популяций мы будем рассматривать лишь односторонние свя-
зи. Однако всегда следует помнить, что фазы в жизненном цикле популя-
ции связаны и многочисленными обратными связями. Это и понятно, по-
скольку никакая регуляция невозможна без обратных связей. Первое, что
нам надо рассмотреть здесь, это зависимость числа ежегодно появляющих-
ся проростков от размеров семенного банка. К настоящему времени мы пс
располагаем обширными данными по этому вопросу. Для некоторых рас-
тений и, в первую очередь, сорных однолетников было показано, что чис-
ло ежегодно появляющихся проростков представляет лишь незначительную
часть того количества жизнеспособных семян, которое содержится в почве,
а иногда встречаются криптопопуляции однолетников.
134 MH I loiiyii>iHii(Hiiiii>i (miohoi ни р<н iciinil
И. Д. Богдановская-Гнсн эф еще в 1926 г. отмечала, что в почве луговых
ценозов содержи гея огромное количество семян видов, которые в момен т
наблюдения не встречаются в составе ценоза или встречаются в ничтожном
количестве. Часто это бывают семена полусорных трав — вероники тимья-
нолистной, ситника развесистого, ситника лягушачьего и др. Вспашка при-
вела бы к массовому прорастанию семян этих ценотически слабых видов.
Выше уже было показано, что при различном режиме культивации процент
семян, ежегодно превращающихся в проростки, сохраняется почти посто-
янным в том случае, когда предотвращено осыпание новых семян. Однако,
судя по данным Р. Нэйлора (1970), у некоторых видов в формировании ак-
тивной части популяции принимают участие главным образом относитель-
но свежие семена. Нэйлор пометил часть семян в популяции лисохвоста
мышехвостниковидного, естественным путем упавших на обследованную
площадь возделываемой пашни. Процедура маркировки включала опры-
скивание семян каплями флуоресцирующей краски, а появлявшиеся про-
ростки могли быть затем распознаны как принадлежащие к маркированно-
му урожаю. В результате было показано, что для этого вида в условиях паш-
ни около 80% проростков появляется из семян, опавших в предшествую-
щем году и только 20% — из более старых семян.
Даже при высокой плотности появившихся проростков довольно трудно
бывает судить о размерах семенного банка. Чанселор, например, показал, что
в плотном травостое ромашки аптечной (1280 особей на кв. м) представлено
только 4% всех жизнеспособных семян этого растения, имевшихся в почве.
Таким образом, мы убеждаемся на многих примерах в существовании очень
сильной зависимости между резервной и активной частями популяции.
По Харперу, число семян, которые прорастают в данный сезон, опреде-
ляется комплексом механизмов «врожденного», индуцированного и вынуж-
денного покоя, которым обладают семена, а также потенциальными воз-
можностями среды, создающей условия, необходимые для прорастания.
Из этого заключаем, что II фаза, как и I фаза в динамике популяций, пред-
ставляет собой точку приложения регуляторных сил — факторов, опреде-
ляющих основные демографические характеристики популяцищ факторов
внешних и внутренних.
При экологических исследованиях целесообразно говорить не о специфи-
ческих требованиях семян к условиям прорастания. Под потенциальными
возможностями среды Харпер понимает частоту микроучастков, в которых
эти требования удовлетворяются (safe-sites — дословно, с английского, «спа-
сательные участки»). Гетерогенность поверхности почвы, чередование на ней
благоприятных и неблагоприятных микроучастков, играет роль отбирающего
фактора «сита», которое обусловливает прорастание одних семян и непрорас-
тание других. При таком подходе необходимо учитывать следующее.
Во-первых, у одного вида растения может быть два или более типов семян,
т. е. иметь место полиморфизм, генетический или соматический8. Уильямс и
8 Соматический полиморфизм связан с размещением разных семян на разных частях мате-
ринского растения и, следовательно, в разных условиях обеспечения питательными веще-
ствами в период их формирования. Генетический полиморфизм наблюдается в том случае,
когда особи одного вида дают различные семена из-за различий в генотипе.
I'fiatta > Жn iiicciK>co(>iii.ic семени и их роль n популяциях piicicniHl 13b
Харнер (Williams, Harper, 1965) изучали соматический полиморфизм семян
мари белой. г)то растение имеет четыре тина семян: коричневые гладкие, ко
ричневые сетчатые, черные гладкие и черные сетчатые, коюрые можно обна
ружить на каждой особи. Семена различной окраски и с различиями в харак-
тере поверхности кожуры прорастают неодинаково. Коричневые семена про-
растают быстро, если им дать влагу, даже при температуре около 0°С; черные
сетчатые семена находятся в состоянии глубокого покоя, который нс удается
нарушить охлаждением, тогда как черные гладкие частично прорастают по-
сле обработки холодом. Очевидно, полиморфные семена могут использовать
два или несколько типов микроучастков, поскольку они предъявляют различ-
ные требования к условиям прорастания.
Во-вторых, одно и то же семя может менять свои требования к условиям
прорастания с течением времени. Так, по данным Попэя и Робертса (1970),
семенам пастушьей сумки для стратификации необходима пониженная тем-
пература (+4°), после чего они лучше прорастают при более высокой тем-
пературе (t ~ +30°).
В-третьих, сами «спасательные» микроучастки могут быть лишь времен-
но благоприятными для прорастания, в связи с чем надо учитывать не толь-
ко пространственный, но и временной аспект потенциальных возможно-
стей среды. Обращаясь к наиболее важному фактору прорастания — фак-
тору влаги, можно привести такой пример. После дождя те микроучастки,
которые были неблагоприятными для прорастания из-за дефицита влаги,
стали временно благоприятными, а затем, по иссушении, вновь перешли в
разряд неблагоприятных участков. Такая динамика влаги, свойственная по-
верхностным слоям почвы, сама по себе налагает отпечаток на прораста-
ние семян. Периодическое намачивание-высушивание повышает всхожесть
семян некоторых видов растений и обусловливает вспышковый или «кван-
товый» характер их прорастания. П. Кэйвере (Cavers, 1975) указывает, что
такой стимул играет большую роль в процессе прорастания семян некото-
рых видов горца. Судя по его наблюдениям, даже подсушенные корешки,
уже успевшие появиться за время намачивания, после высушивания и но-
вого намачивания продолжают нормально расти. Замечена и такая особен-
ность — чем дольше семена набухали перед высушиванием, тем иптснсив-
нее прорастание после него. Сходная закономерность, отмеченная нами у
пастушьей сумки, для которой характерно появление вспышек прорастания
семян в летний период, уже была обсуждена выше, когда обсуждалось зна-
чение миксоспермии.
Различная степень гетерогенности почвенной поверхности может быть
смоделирована в опыте. Так, в опыте Д. Уайта (1968) была установлена и на-
глядно продемонстрирована зависимость прорастания семян от диаметра ча-
стиц субстрата. Для опыта были взяты семена брюквы и редиса, имеющие
округлую форму и прораставшие на фильтровальной бумаге при I 15°С на
96%. В четырех вариантах в качестве субстрата были использованы час i ицы
вермикулита разного диаметра: Т1 — 5,5-9,0 мм; Т2 — 3,2-5,5 мм; 2,1
3,1 мм; Т4 — 2,1 мм, которые получали путем просеивания. Прочие условия
во всех вариантах были одинаковые. Семена высыпали на субстрат с одина-
ковой высоты. Уаигу удалось показан», что даже относи юльно небольшие
136
МН Марков
1 loi iy пи I uioiiiiiiH biioiioi ня рис । ciinii
Рис. 5.26. Прорастание семян брюквы
(I) и редиса (И) на физически различных
субстратах (пояснения в тексте).
отличия в величине семян у двух ви-
дов могут обусловить значительную
разницу в прорастании, зависящую
от соотношения диаметра семян и
диаметра частиц субстрата. По ис-
течении четырех недель со дня за-
кладки опыта наблюдалась следу-
ющая картина (рис. 5.26). Процент
проросших семян брюквы, имею-
щих диаметр около 1,5 мм, снижал-
ся от 90 до 70% по мере уменьшения
диаметра частиц субстрата. Гораздо
более резко (от 60 до 5%) снижал-
ся процент проросших семян реди-
са, имеющего более крупные семена
(2,5 мм в диаметре).
Большую роль субстрата в обеспече-
нии условий прорастания семян де-
монстрирует опыт Харпера и Сагара
(Harper, Sagar, 1960) с тремя видами
подорожников. Проращивание се-
мян подорожников (большого, сред-
него и ланцетолистного) проводили
с использованием трех разных суб-
стратов: фильтровальной бумаги,
вермикулита и почвы. Все образцы семян были подвергнуты предваритель-
ному охлаждению (при +5°) в течение 14 дней, а затем помещены в условия
освещения и температуры t° = +20°. Как видно из таблицы, процент про-
росших семян подорожника ланцетолистного значительно увеличивался на
почве и вермикулите по сравнению с фильтровальной бумагой (табл. 5.15).
Следовательно, фильтровальная бумага — неудовлетворительный субстрат
для экспериментального выявления всхожести семян этого вида. Влияние
же субстрата на прорастание семян других видов было, как видно, невели-
ко. Здесь вновь налицо наиболее ярко выраженная зависимость прораста-
ния от субстрата у самого крупносемянного вида.
Таблица 5.15.
Влияние субстрата на прорастание семян подорожников
большого, среднего и ланцетолистного (в %)
Виды подорожника (Plantago)'. Субстрат
Фильтровальная бумага Вермикулит Почва
П. большой 92 96 98
П. средний 92 91 92
П. ланцетолистный 45 54 72
I 7</Г/</ Ж|| IIICCIKH oOlll.lC I CMCIHI II IIX рощ. II популяциях put 1Г1П1Й
и/
Рис. 5.27. Влияние соотношения разме-
ров семени (С) и частиц субстрата на ве-
личину поверхности контакта (зона кон-
такта обозначена жирной линией).
Рис. 5.28. Схема, показывающая два по-
ложения семени подорожника ланце-
толистного (в поперечном сечении) на
поверхности субстрата, а — «спинкой»
вверх; б — «спинкой» вниз; в — общий
вид семени с двух сторон.
Поскольку благонрня । носи, ми-
кроучастка онрсдслиосм, прежде
всего способностью попавшего в
пего семени поглощать из субстра-
та достаточное количество влаги, то
не последнюю роль в этом процес-
се будет играть величина поверх-
ности контакта семени с частица-
ми субстрата, зависящая от соотно-
шения размеров почвенных частиц
(элементов почвенной микротопо-
графии) и размеров семян. Третьим
важным компонентом является фор-
ма семян. В вышеописанном опы-
те Уайта были взяты семена отно-
сительно простой, округлой формы.
На рис. 5.27 схематично показано,
как соотношение размеров частиц
субстрата и округлых семян может
повлиять на величину поверхности
контакта.
Если же семена имеют более
сложную форму, то поверхность
контакта с субстратом зависит от
положения семени на поверхности
субстрата. Так, в опыте с подорож-
ником ланцетолистным было пока-
зано, что семян, лежавших на филь-
тровальной бумаге плоской сторо-
ной вниз, проросло на 20% больше,
чем семян, лежавших на выпуклой
«спинке» (рис. 5.28).
Опыты по выяснению влияния различных химических агентов на прорас-
тание семян должны обязательно проводиться не только на искусственных
субстратах, например, на фильтровальной бумаге, но и на почве. Как пока-
зали опыты с семенами пастушьей сумки, субстрат способен вносить су-
щественные коррективы в действия различных факторов на прорастание.
Прибавка Ca(NO3)2 — азотного компонента раствора Кнопа к воде, смачи-
вающей субстрат, вызывала на фильтровальной бумаге сильное повыше-
ние энергии прорастания, а на почве привела к обратному эффекту. В обоих
опытах семена помещали на поверхность субстрата.
Для заглубленных в почву семян нитраты иногда способны заменять с ти-
мулирующее действие освещения. Действительно, несмотря на то, что мно-
гие семена прорастают, находясь на поверхности почвы, в естественных
условиях, особенно в агрофитоценозах, семена бывают в той или иной сте-
пени заглублены в почву. Проблема поглощения воды за счет увеличения
контакта с почвенными частицами при этом практически снимается, но в
1J6 MU l\hipK(Hi 1 h и iy 11 и Ц1 ни 11 ni >i (> 11< > 11< >111 >i pm i c n nil
го же самое время семена inoniipyioiся oi света одною на важнейших
стимулов прорастания. В ходе эволюции сорных pacieiinii выработались
приспособления, не допускающие прорастания семян на глубине, с кото-
рой проростки не могут пробиться к поверхности, к свету. Семена сорняков
чувствительны к условиям освещения, температурным условиям и концен-
трации углекислого газа в почвенном воздухе. При определенном сочетании
этих факторов (пониженная температура, при отсутствии освещения и при-
сутствии высокой концентрации СО2) семена переходят в состояние покоя и
не прорастают на губительной для проростка глубине.
Семена разных видов растений предъявляют различные специфические
требования к условиям прорастания, но, как правило, эти требования со-
ответствуют той обычной естественной обстановке, в которой оказывают-
ся семена данного вида, то есть тому сочетанию факторов, к которому в те-
чение длительного времени осуществлялось приспособление в ходе эволю-
ции. В настоящее время наше понимание регуляции прорастания не идет
дальше выявления и изучения отдельных факторов микросреды, которые
играют наибольшую роль при «отборе семян средой». Температура, вла-
га, запас питательных веществ, глубина заделки, интенсивность и спек-
тральный состав света — все эти переменные широко используются семе-
нами для «распознавания» условий, в которые они попали. Изучать эти фак-
торы можно только в экспериментальных системах. В природных условиях
их напряженность и ритмы вряд ли можно регулировать. В заключение сле-
дует сказать, что эволюция привела к возникновению почти универсальных
механизмов, регулирующих прорастание, которые обеспечивают сохране-
ние части семенной продукции и вместе с ней возможность попадания се-
мян в лучшие для вида условия произрастания.
Учет количества семян, продуцируемых растениями на данном участке
территории и имеющихся в почве, является, пожалуй, одним из относитель-
но простых способов получения демографических данных, если не касаться
сложностей технического, методического характера. Можно даже отвлечься
от особенностей самих особей, давших семена, рассматривая особь только
как способ получения из одного семени многих семян. Харпер, употребив-
ший такую модификацию афоризма Спенсера «Курица есть только способ,
которым из одного яйца получается много яиц», считает эту своеобразную
«демографию семян» очень важной, особенно для исследования засоренно-
сти почвы сельскохозяйственных посевов семенами малолетних сорняков
в целях предсказания вспышек засоренности — инфестаций. В агрофито-
ценозах растения сталкиваются с чрезвычайно своеобразными условиями
произрастания, связанными с культивацией, которая оказывает влияние,
прежде всего на распределение в почве зачатков (диаспор). Сильно уско-
ряется процесс заглубления семян и возвращения их на поверхность, кото-
рый в естественных условиях идет довольно медленными темпами и свя-
зан обычно с деятельностью животных агентов — дождевых червей и пр.
Исходя из этого, некоторые исследователи попытались построить модель, в
которой динамика популяции однолетника в агрофитоценозе представле-
на как динамика жизнеспособных семян в почве. Подобная модель была по-
строена и Дж. Харпером (рис. 5.29). В модели отражена зависимость чис-
Рис. 5.29. Модель, отображающая динамику жизнеспособных семян (пассивной
части популяции) в почве: S, G и D — параметры видоизмененного основного де-
мографического уравнения (по Harper, 1977 с изменениями).
ленности популяции семян9 сорного растения на культивируемом участке
от четырех важнейших факторов: 1. накопления семян, опавших с расте-
ний, которые, произрастали на данной территории; 2. заноса семян (инва-
зии) в процессе их распространения с соседних территорий; 3. внесения в
почву семян сорняка, засоряющего посевной материал; 4. отмирания семян.
Рассмотрим эти факторы по порядку.
1. Исследования, проведенные рядом отечественных и зарубежных уче-
ных, показали, какое огромное количество семян может накапливаться в по-
чве. Так, например, в верхнем (0-10 см) слое почвы пойменного луга число
жизнеспособных семян достигало 17 000 на 1 м2 (Работнов, 1956), в почвах
материковых лугов Московской области обнаружено от 13 600 до 38 200
всхожих семян на 1 м2 (Пятин, 1970), в почвах лугов Великобритании от
4 000 до 70 000 (Champness, Morris, 1948). Если же взять данные о содержа-
нии семян сорняков в почве агрофитоценозов, то цифры и вовсе покажутся
астрономическими. Английские исследователи У. Бренчли и К. Уоррипггоп
обнаружили в пахотной почве 75 000 жизнеспособных семян сорняков, а
Л. И. Мальцев — от 50 тыс. до 400 тыс. семян на одном квадратном метре.
Однако, основываясь на данных Бренчли и Уоррингтон о восстановлении
в течение двух-трех лет популяции погребенных в почве семян, по извест-
9 Гермин «популяция семян» часто используется зарубежными исследователями. У пас при
пято считать, что жизнеспособные семена, входят в состав популяции растения в качссте
неактивной ее части. Липп» в том случае, когда временно в популяции предсчавлсна iiiinii. ни
час 11., можно в п ии-c i ном смысле i oBopiri i> о популяции семян
140 Л/ Н ЛЛ/у/лиг/ 11(>11у||яцп<>111111>1 (пинки им рис iciinh
ным причинам снизившей перед этим свою численность до уровня, уста-
новившегося за многие годы сельскохозяйственной обработки ноля, можно
предположить наличие стабилизирующего механизма, который препятству-
ет неограниченному накоплению жизнеспособных семян в пахотной почве.
К сожалению, пока не проведено исследований, результаты которых могли
бы подтвердить или опровергнуть данное предположение. Лишь пользу-
ясь косвенными данными, мы можем предполагать действие регуляторных
механизмов, стабилизирующих популяцию на этом этапе. В качестве свой-
ства, которое позволяет накапливаться в почве огромному количеству жиз-
неспособных семян и способствует формированию резервной части попу-
ляции, выступает покой семян. Это свойство усовершенствовалось под дей-
ствием естественного отбора из-за неоспоримого адаптивного значения для
очень многих и, в первую очередь, сорных растений. Покой семян обеспе-
чивает виду своеобразную буферную систему, позволяющую переживать
катастрофические воздействия среды (например, вспашку) и допускающую
периодическое прорастание, т. е. играющую роль, сходную с ролью диапа-
узы у насекомых. В следующей главе явление покоя семян, его различные
типы будут рассмотрены подробно.
Размеры накопления семян в почве агрофитоценозов в большой степени
зависят от интенсивности ее обработки. В этом нас убеждают данные мно-
голетних опытов, проведенных Е. Робертсом и П. Фиет (Roberts, Feast, 1972,
1973). Опыты заключались в сравнении динамики семян сорных растений в
обрабатываемой и необрабатываемой почве.
Первый опыт. Семена 20 видов сорных растений (ромашки пахучей,
трехреберника непахучего, крестовника обыкновенного, дымянки лекар-
ственной, торицы посевной, ярутки полевой, мари белой и др.) помести-
ли по 300 в заполненные полевой почвой цилиндры и в течение 6 лет реги-
стрировали количество жизнеспособных семян, оставшихся в цилиндрах.
В одном варианте почву перемешивали, имитируя вспашку, а в другом —
оставили необработанной в течение всего опыта. Со временем в обоих ва-
риантах число жизнеспособных семян убывало, причем на обрабатываемой
почве этот процесс шел более быстрыми темпами — в среднем на 32% в год
с колебаниями для разных видов от 20 до 48%. На необрабатываемой почве
уменьшение числа жизнеспособных семян было менее резким и составило
только 12% (в среднем) в год с колебаниями от 6 до 21%. К концу опыта, т.
е. через 6 лет в варианте с обработкой почвы сохранилось 5,9% от первона-
чального числа семян, а в варианте без обработки — 27,5%.
Второй опыт. Смесь заданного числа семян сорняков с суглинистой по-
чвой помещали в цилиндры слоем 2,5, 7,5 и 15 см. Часть почвы оставили не-
тронутой, а другую часть ежегодно перемешивали. Во всех вариантах прово-
дили учет числа проросших сорняков, а с 5 лет подсчитали число оставших-
ся в почве жизнеспособных семян. В итоге в перемешиваемой почве толщине
слоя 2,5,7,5 и 15 см сохранилось 2,3,4,0 и 7,7% жизнеспособных семян соот-
ветственно. В нетронутой почве число сохранившихся жизнеспособных се-
мян было больше и составляло в тех же слоях: 6,8, 16,5 и 31,6%.
Результаты этих опытов не должны привести к неверному заключению,
будто в почве под агрофитоценозами условия не позволяют накапливаться
/ hitia Ь Жишсч носочные сгмгнп и их роль в популяции,ч piiciciiiiB 14 1
большому количеству семян, что вопию бы в противоречие с приведеппы
ми выше цифрами. Надо учитывать, что преобладающие в агрофитоцсио
зах однолетники в естественной обстановке осыпают семена ежегодно и и
огромных количествах, а перепашка в данном случае может служить одним
из факторов, стабилизирующих размеры накопления.
С явлением накопления жизнеспособных семян в почве очень тесно свя-
зана эволюция сорных растений и, в первую очередь, сегетальных. Судя по
всему, многие виды, относимые сейчас к рудеральным и сегетальным сор-
някам, были распространены по краю отступающего ледника, после тая-
ния которого их вытеснили более конкурентоспособные лесные, луговые и
степные растения. В степях, также как и в лесных сообществах, сорные рас-
тения могли сохраниться только в виде накопленных жизнеспособных се-
мян в почве, которые прорастали после вынесения на поверхность земле-
роями. На выбросах землероев в условиях ослабленной конкуренции по-
явившиеся особи проходили весь свой жизненный цикл и завершали его
плодоношением, пополняя и обновляя при этом семенной банк популяций.
Распашка степей как своеобразная имитация деятельности землероев при-
вела, очевидно, к превращению этих растений в засорителей посевов.
2. Семена сорняков могут попасть на данную территорию при их есте-
ственном распространении с других участков или, наоборот, с данной тер-
ритории распространиться на другие участки. Именно распространени-
ем обычно пытаются объяснить вспышки засорения (т.н. инфестации).
Наиболее часто в этом обвиняют сорные растения из семейства сложноц-
ветных, семянки которых снабжены хохолком и распространяются при по-
мощи ветра. Однако, как показано Беккером на примере бодяка полевого,
большинство семянок отлетает совсем недалеко от родительских растений,
а то, что разлетается на большие расстояния, оказывается хохолками, не не-
сущими семянок. Сходные данные о редкости переноса семянок на боль-
шие расстояния получены для крестовника Якова. Исходя из этого, распро-
странение можно рассматривать только как способ инициации вспышки за-
сорения и предпосылку её возникновения, а собственно инфестация проис-
ходит за счет быстрого размножения популяции сорного растения на новом
месте. Если популяция сорняка невелика по размерам, то для нее эмиграция
семян в процессе распространения на большие расстояния может быть при-
чиной ощутимых потерь.
3. При засорении семенами сорняка посевного материала сорный вид
может перемещаться от поля к полю, оставаясь всегда связанным с посе-
вом одного культурного растения и одним типом обработки поля, к кото-
рому он приспособлен. Этим вопросом у нас занимались Н. В. Ципгер и
Е. Н. Синская, которым принадлежат интересные исследования по выявле
нию уподобления семян сорняков семенам культурных растений как пути
их приспособления к произрастанию в посевах, этого яркого примера мп
микрии в растительном царстве.
Вхождение в состав полевого растительного сообщества приводит к оео
бым адаптивным изменениям в популяциях сорных растений. Человек при
подготовке почвы к посеву, при севе, уходе за посевом, сборе урожая п ob
14Z Л/// Л/<//>Л<>6 I loiiyilHHIIOIIIHlH (HIOIIOI П>1 pin ICIIlHI
работке части его на посевной материал бессознательно оiбирал соответ-
ствующие формы сорных растений. Кроме этого, всячески способствуя наи-
лучшему развитию культурного растения, он пытается усилить эдификагор-
ную способность последнего и тем самым косвенно участвует в отборе сор-
ных форм, наиболее приспособленных к фитоценотической обстановке в
посевах. Н. В. Цингер и Е. Н. Синская показали действие бессознательного
отбора при отвеивании в популяциях сорных растений, засоряющих семе-
нами посевной материал и вместе с ним проникающих в посев, а также фор-
мообразующее влияние культурного растения, которое через создание в по-
севе соответствующей фитоценотической среды толкает сорные растения
на путь уподобления. Для основных объектов их исследования — специа-
лизированных засорителей посевов льна, торицы льняной и рыжика льня-
ного — характерна не только соответствующая высота стеблей, обеспечива-
ющая срезание сорняков при уборке, но и форма и коэффициент парусности
семян, близкие по значению к таковым у семян льна.
Существует, однако, и другая большая группа сорных растений, которые
используют иной путь к приспособлению — «дифференциацию» (термин
Е. Н. Синской). Растения из этой группы образуют мелкие семена, засоряю-
щие почву. При анализе динамики популяций таких растений третий из вы-
шеупомянутых факторов, очевидно, не будет иметь существенного значения.
4. Об условиях отмирания семян в почве и сохранении их всхожести из-
вестно относительно немного. В разное время было поставлено несколько
экспериментов по изучению сохранения жизнеспособности семенами, зако-
панными на разную глубину в почву. В одном из опытов семена 107 видов,
помещенные в цветочные горшки, которые были наполнены стерильной по-
чвой и закрыты пористыми глиняными крышками, закопали на глубину 20,
55 и 105 см. Через 1,3,6, 10, 16 и 20 лет извлекали образцы для проращива-
ния. Было установлено, что глубина закапывания семян в почву не оказыва-
ет заметного влияния на продолжительность жизни семян. Большинство се-
мян культурных растений после пребывания в почве в течение 1 года оказа-
лись мертвыми. Семена же дикорастущих растений сохранили в течение 20
лет высокую всхожесть.
Аналогичный опыт был заложен Билом еще в 1879 г. Семена 20 дико-
растущих видов и, в первую очередь, сорняков были смешаны с песком и
помещены в не закупоренные бутылки, повернутые горлышком вниз и за-
копанные в почву. Через 20 лет остались живыми семена 11 видов, через
40 лет — семена 8 видов (щирицы запрокинутой, клоповника виргинского,
ослинника двулетнего, подорожника большого, щавеля курчавого и др.) и
через 60 лет — только семена ослинника и щавеля курчавого.
Известный опыт Дювеля, поставленный по сходной методике, показал,
что семена сорняков, характерных для данной местности, дольше остают-
ся живыми при закапывании в почву, чем семена культурных растений или
семена сорняков, не свойственных данному району. Стало быть, одним из
факторов, определяющих, какой вид в данной местности станет сорным, яв-
ляется способность его семян сопротивляться особым процессам разложе-
ния, характерным для почв местности.
Жизнеспособные семена Цыс./м')
Рис. 5.30. Экспоненциальное сниже-
ние числа жизнеспособных семян в по-
чве при четырех различных вариантах ее
обработки: сверху вниз — без вспашки,
вспашка на глубину 23 см 4 раза в год,
вспашка на глубину 23 см 2 раза в п>д,
интенсивная частая вспашка на глубину
15 см.
Гчава 5 >l<iniiccii()C()6iii»ir семени и их роль в популяциях расicindt 143
Одной из замечательных особен-
постей семян сорняков, способных
длительное время сохранять жиз-
неспособность в почве, является то,
что большинство из них не обладает
водонепроницаемыми покровами —
одной из распространенных причин
длительного покоя семян. Это озна-
чает, что, осыпавшись на землю, эти
семена вскоре достигают полно-
го набухания и остаются в этом со-
стоянии непроросшими до тех пор,
пока не попадут в необходимые для
прорастания условия. Установлено
даже, что семена некоторых сорня-
ков фактически сохраняют жизне-
способность в почве в течение бо-
лее длительных сроков, чем в усло-
виях сухого хранения в лаборатории
, 1940, по: Крокер, Бартон,
Исследования динамики резерв-
ной части популяций некоторых
сорных растений, проведенные X.
Робертсом и Докинзом (Roberts, Dawkins, 1967), показали, что семена сор-
ных растений, находящиеся в почве, постепенно отмирают, причем темпы
отмирания зависят от видовой специфики, но, как правило, остаются посто-
янными во времени, т. е. это процесс равномерный. Поставленный опыт со-
стоял в следующем. Четыре участка отличались друг от друга по способу их
обработки. Один из них перепахивали четыре раза в год, другой — два раза
в год, третий вообще оставался в течение года необработанным, а на четвер-
том устанавливали режим частых культиваций как для пропашных культур.
Ежегодно из почвы (до глубины 23 см) извлекали образцы семян и проверя-
ли их жизнеспособность. Опадение на почву новых семян было предотвра-
щено. Как можно видеть на графике (рис. 5.30), во всех вариантах наблю-
далось экспоненциальное уменьшение числа живых семян. Даже при от-
сутствии вспашки вероятность отмирания от года к году оставалась посто-
янной. Видовая специфичность темпов отмирания семян в почве отражена
на рисунке 5.31. Параллельно в опыте учитывали число ежегодно появля-
ющихся проростков, которое представлено в табл. 5.16 как процент от чис-
ла жизнеспособных семян, содержащихся в момент учета в почве. Можно
видеть, что, несмотря на довольно быстрое снижение числа живых семян и
почве, процент появлявшихся проростков менялся незначительно от года к
году за исключением варианта без вспашки.
144 МН Марков I loiiyiuiniioiiiiini (Я1О11О1 пя pnciciinil
Гибл и ци 5.16.
Число проростков, появляющихся от года к году в процентах от общею числа жиз-
неспособных семян, присутствовавших в верхнем слое почвы (0 23 см) в начале
каждого года
Тип обработки почвы Годы наблюдений
1961 1962 1963 1964 1965 1966
Вспашка четыре раза в год 10,7 6,1 9,4 8,3 8,7 9,6
Вспашка два раза в год 7,8 4,7 6,7 6,1 9,0 7,2
Без вспашки 4,5 1,7 0,6 0,3 0,2 0,3
Если учесть, что в ненарушенной почве семена отмирают, и это было по-
казано выше, также экспоненциально, то здесь мы наблюдаем действие яв-
ного регуляторного механизма, базирующегося на приспособлении сеге-
тальных сорняков к обязательному периодическому нарушению почвы.
5.1.3.1.2. Жизнеспособные семена в почве и стратегии жизни видов растений
По Т. А. Работнову (1981) можно выделить три основных типа популяций
по уровню накопления жизнеспособных семян в почве. Это:
1) популяции, в состав которых жизнеспособные семена входят только
в годы массового обсеменения и лишь в течение непродолжительного вре-
мени;
2) популяции, в составе которых всегда присутствуют в больших количе-
ствах жизнеспособные семена;
3) популяции пирофитов с присутствием большого количества накопив-
шихся в почве плодов с семенами, прорастающими после воздействия огня.
Подробнее охарактеризуем первые два типа популяций.
Популяции первого типа довольно характерны для виолентов и, в первую
очередь, для древесных растений, доминирующих в климаксовых сообще-
ствах лесной зоны — ели, дуба и др. Семена этих растений имеют довольно
большой запас питательных веществ и потому в большом количестве пое-
даются животными. Как эволюционное приспособление для увеличения ве-
роятности семенного размножения названных видов выступает очень энер-
гичное прорастание свежеосыпавшихся семян. По этим двум причинам се-
менной банк в популяциях древесных виолентов обычно не формируется.
Не характерно создание больших семенных банков и для популяций
вегетативно-подвижных лесных травянистых растений. Большая энергия
вегетативного размножения компенсирует редкость образования семян, ко-
торое возможно, как, например, у сныти обыкновенной, только в условиях
«окон», т. е. нетипично для леса высокой освещенности.
Отсутствие жизнеспособных семян в составе популяций так называемых
«кочующих» видов, например, ив, тополей на аллювиальных наносах, иван-
чая на гарях можно объяснить тем, что огромное количество продуцируе-
мых ими семян расходуется на заселение новых территорий, свободных от
растительности, а семенное возобновление под материнским пологом не-
возможно.
I'UIHil Ж Illi ICCI lovobl U.K' ( ('MCIIII II II.ч poll!» в иопуницпях pile I Cllllil
145
Кроме этого нетипично!о случая
виды-эксплеренты и в юм числе вес
дикорастущие однолетники, образу-
ют популяции второго типа.
Ко второму типу относятся и по-
пуляции большинства патиентов,
у которых иногда способность на-
капливать в почве жизнеспособ-
ные семена сочетается со способ-
ностью к вегетативному размноже-
нию. Примером таких видов могут
служить будра плющевидная, кле-
вер ползучий и др.
В связи с тем, что нарушения рас-
тительного покрова происходили за-
долго до появления на земле челове-
ка (пожары, ветровал, массовое раз-
множение фитофагов и землероев),
некоторые виды растений в процес-
се эволюции выработали стратегию,
включающую способность их семян
десятилетиями и столетиями нахо-
диться в почве в состоянии покоя.
После того как происходит новое
IgN
10
5
1
0,5
0,1
0,05
Рис. 5.31. Кривые выживания семян в
почвенных банках пяти видов сорных
растений при вспашках, производимых
четыре раза в год
а) марь белая, б) пастушья сумка, в) мяч-
лик однолетний, г) паслен черный, д)
звездчатка средняя; Т — время (годы).
нарушение сложившегося растительного покрова, эти растения участвую!
в формировании фитоценозов и гарях, вырубках и т. д. В зоне хвойных бо-
реальных лесов к таким растениям относятся малина, луговик дернистый,
полевица тонкая, некоторые виды осок (буреющая и др.). По своему пове-
дению эти виды сходны с пустынными однолетниками с L-стратегией, у ко-
торых стадии роста и размножения чередуются со стадиями «криптопопу-
ляций» (от греческого «криптос» — скрытый), когда все особи находятся в
почве в виде жизнеспособных семян.
В сезонном ритме температуры и влажности, отрастания и отмирания ли-
стового полога, т. е. ритме возникновения и исчезновения регенерационных
ситуаций для малолетников большая роль принадлежит человеку и его хо-
зяйственной деятельности. Выше уже говорилось о слабой предсказуемо-
сти среды в ценозах, предпочитаемых малолетними видами. Когда говоря г
о слабой предсказуемости среды, в которой живут малолетние растения, то
имеют в виду в первую очередь антропогенный фактор, момент воздействия
которого популяции трудно прогнозировать главным образом из-за того, что
он часто не имеет постоянного ритма. В том случае, когда воздействие это-
го фактора имеет довольно четкий ритм (сенокошение), мы сталкиваем-
ся, например, с возникновением сезонных рас, о которых писал Веттппейн
(Wettstein, 1901, по: Завадский, 1968). «Шаблон» пашенного местообитания
диктует жесткий регламент продолжительности жизненного цикла, создает
среду в очень слабой степени предсказуемую для сорняков, поскольку че-
146 МН Мирков I loiiyihiiuuniiiiHi biioiioi ни pnciriiiiH
ловек своими агротехническими приемами вклинивается и во многом нару-
шает сезонный ритм. С одной стороны, создавая агрофитоцснозы, человек
создает одновременно множество регенерационных ситуаций в силу разру-
шения естественного покрова и относительно слабого ценотического влия-
ния культурных растений (Туганаев, 1984). Однако, провоцирование семян
малолетников к прорастанию в любой момент может обернуться большими
потерями для их популяций, так как человек, ухаживая за посевами, приме-
няет истребительные меры по отношению к сорным сегеталам. Возникает
пониженная вероятность того, что появившиеся проростки достигнут ре-
продуктивной зрелости. Наглядно представить и рассчитать последствия
того или иного поведения малолетников в случайно флуктуирующей сре-
де можно только при помощи математических моделей. Наилучшим обра-
зом для моделирования подходят периодические оценки семенного банка,
которые, как известно, являются простейшим (если не учитывать трудности
методического характера) способом получения демографических данных о
популяции малолетнего растения. Математическая модель динамики семян
в почве, учитывающая случайный характер внешних условий (Cohen, 1966),
сводится к уравнению:
Sl+I = Sl-SlG-D(Sl-SlG)+SlGYl,
где St — число семян в почве в момент времени /; S {+] — число семян в по-
чве в следующий момент времени; G — процент прорастающих семян; D —
процент семян, которые отмирают; Y( — число семян, продуцируемое в пе-
ресчете на каждый появившийся проросток — вероятностная переменная,
зависящая от условий среды. Графическое решение с обозначением прогно-
зируемой скорости увеличения семенного банка в зависимости от вероят-
ности выживания проростка до зрелости (Р), а также параметров G, D и Yt
приведено на рисунке 5.32.
Теперь, пользуясь этим уравнением, можно теоретически определять воз-
можные стратегии для различных типов условий среды обитания растений.
Вот некоторые теоретические ситуации и соответствующие им стратегии.
I. Если имеется высокая вероятность успешного обсеменения (Р = 0,8),
то при большой плодовитости растения (Y = 500) оптимальной стратегией
будет увеличение числа ежегодно появляющихся проростков. Необходимо
учесть, что резервная часть популяции увеличивается при уменьшении па-
раметра D и увеличении параметра Y и, стало быть, названная стратегия —
наиболее оптимальный путь снижения параметра D.
II. Для растения, обладающего низкой плодовитостью (Y = 5) при боль-
шой вероятности приживания проростков и обсеменения (Р = 0,8), наибо-
лее благоприятным будет энергичное прорастание свежеосыпавшихся се-
мян (G—>100%). Если же G«10% из-за перехода части семян в состояние
покоя, то значение D возрастает.
III. При низкой вероятности приживания проростков и успешного обсе-
менения (Р = 0,1) независимо от плодовитости растения (Y = 5 или Y =
500) оптимальная стратегия — накопление большого покоящегося семен-
ного банка и прорастание небольшими порциями (G«10%). Если же сс-
1'1/
мена будут ежегодно прорастать в
больших количествах (G ► !()()%),
го это опасно для популяции из-за
сильного возрастания параметра D.
1 и 11 ситуации и соответствую-
щие им стратегии характерны для
виолентов, обладающих, как уже
было сказано выше, большой кон-
курентоспособностью и энергич-
ным ростом. Фактор поедания се-
мян животными, который име-
ет большое значение в популяциях
крупносемянных виолентов, в при-
веденном уравнении не фигури-
рует, поскольку это создало бы из-
лишнюю сложность. Однако II си-
туация вполне подходит для таких
видов, если допустить, что низкая
плодовитость получается на базе
первоначально высокой плодовито-
сти, которая снижается за счет по-
Рис. 5.32. Графическая презентация мо-
дели Коэна (Cohen, 1966), прогнозирую-
щей зависимость скорости роста числен-
ности семян в почвенном банке от доли
семян, прорастающих ежегодно (поясне-
ния в тексте).
едания, а вероятность приживания
проростков благодаря значительному запасу питательных веществ в семе-
ни в общем достаточно высока. III ситуация чрезвычайно характерна для
популяций сорных видов и в особенности — для однолетних эксплерентов.
Можно видеть, что в условиях пониженной вероятности успешной ре-
продукции для поддержания максимально возможных темпов роста попу-
ляции модель прогнозирует оптимальность стратегии, которая основана
на прорастании семян небольшими порциями (например, по причине раз-
ного типа покоя) и накоплении значительного семенного запаса в почве.
Такую стратегию принято именовать стратегией компенсированного ри-
ска {«bet hedging» strategy — Steams, Crandall, 1981). Однако, другая мо-
дель, построенная Д. Коэном (Cohen, 1967) для определения оптимально-
го значения параметра G у пустынных растений с учетом соотношения «хо-
роших» и «плохих» по условиям лет, уже не дала столь однозначного реше-
ния. Работавший с этой моделью Р. МакАртур (MacArthur, 1972), отметил,
что сохранение состояния покоя не обязательно будет наилучшей стратеги-
ей у пустынных растений, если учесть возможность реколонизации терри-
торий — ведь успешная дисперсия на дальние расстояния обычно сопряже-
на с отсутствием эндогенного покоя у диаспор.
Таким образом, меняя исходные посылки, и проигрывая различные ситу-
ации с помощью моделей можно получить диаметрально противоположные
выводы. Это наводит на мысль, что, хотя способность накапливать жизне-
способные семена в почве как непременный атрибут сегетальности прису-
ща огромному большинству малолетников, стойко удерживающихся в агро-
фитоценозах, все же следует особо проанализировать в этом плане малолет-
нис растения естественных местообитаний.
148 М. И, Маркин I loiiyiuiHiioiiiiiui (dipnoi пи рис icnid
Хорошее прорастание свежсосынавшихся семян малолетника, сразу же
помещенных на влажную подстилку, ставит под сомнение наличие у него
большого семенного банка в популяциях. Поэтому для ряда лесных, стен-
ных, прибрежных и рудеральных малолетних растений автор в опыте изу-
чал этот показатель. Из данных таблицы 5.17 видно, что для многих видов
прибрежных и степных малолетних растений характерно прорастание све-
жих семян, не зимовавших в почве. Семена другой большой группы видов
резко повышали всхожесть после высушивания их вслед за первым нама-
чиванием, т. е. уже к концу лета. Стало быть при наличии благоприятных
внешних условий свежеосыпавшиеся семена многих малолетников способ-
ны к немедленному прорастанию и не находятся в этот момент в состоянии
эндогенного покоя. Иное дело, что такие условия далеко не всегда и не во
всех фитоценозах создаются с момента осыпания до зимовки.
Таблица 5.17.
Данные опыта по проращиванию семян розеточных (Р), полурозеточных (П), вре-
меннорозеточных (В) и безрозеточных (Б) малолетних растений
Виды растений: % прорастания Примечание
1. Прорастали без периода покоя (сразу по высыпании)
Р Androsace septentrionalis 100
Р Androsace filiformis 100
В Dracocephalum thymiflorum 100
П Isatis costata 100 Без перикарпа
P Limosella aquatica 100
П Verbascum lychnitis 100/50 На свету/в темноте
В Acinos arvensis 90
П Lepidium ruderale 80
П Berteroa incana 85
P Androsace maxima 76
П Verbascum thapsus 72/59 На свету/в темноте
Б Eremopyrum triticeum 68
П Ranunculus sceleratus 52
Б Trifolium arvense 40
В Veronica vema 22
Б Anizantha tectorum 20
Б Psammophiliella muralis 8
2. Прорастали после сухого хранения в лаборатории в течение 3-х месяцев
В Veronica vema 94
Б Meniocus linifolius 76
В Acinos arvensis 40
П Draba nemorosa 26
Б Alyssum turkestanicum 6
P Myosurus minimus 2
I ldtill У Ж|| IIICCIIQCOClIlhir ГГМ<*1111 II ИХ роЛЬ II IIOliyilllllllHX pilC ICIIIlll 149
Гибл и цп 5.17. (окончание]
3. Прорас 1али после краткого периода эндогенного покоя (после намачивания, высушивания и нового намачивания в конце лета)
Б Anizantha tectorum 100
Р Ceratocephala falcata 100 без перикарпа
В Descurainia sophia 100
Б Eremopyrum triticeum 100
В Veronica verna 100
Б Alyssum turkestanicum 90
P Myosurus minimus 76
В Plantago indica 10
4. He прорастали совсем:
П Alliaria petiolata 0
P Ceratocephala falcata 0 С перикарпом
Б Impatiens noli-tangere 0
П Isatis costata 0 С перикарпом
Б Juncus bufonius 0
Б Thymelaea passerina 0
5.1.3.1.3. Особенности двулетних растений
Тезис Дж. Л. Харпера о том, что у двулетних растений слабую дисперсию
в пространстве компенсирует существенная дисперсия во времени, судя по
всему, можно адресовать лишь небольшому числу мелкосемянных видов
подобных коровякам Verbascum lychnitis, Verbascum thapsus (см. выше, табл.
5.2) или наперстянке Digitalis purpurea (Silvertown, 1982), которым свой-
ственны значительные по размерам постоянные банки покоящихся семян,
обеспечивающие возможность реагирования популяции на всевозможные
нарушения растительного покрова и возникновения регенерационных ниш
и ситуаций.
При этом именно среди мелкосемянных двулетников мы находим при-
меры поразительной долговечности семян в составе почвенного банка (уже
упомянутые опыты Била). Вместе с тем у крупносемянных двулетников мы
наблюдаем другую картину и, прежде всего — отсутствие постоянного се-
менного банка у многих (по крайней мере для 50% — Odum, 1978 по: de
Jong, Klinkhamer, Metz, 1987; см. также табл. 5.2) из них. Семена многих
двулетников и в том числе даже сравнительно мелкосемянных (из наших
объектов — коровяк метельчатый Verbascum lychnitis, змееголовник тимья-
ноцветковый Dracocephalum thymiflorum, проломник северный Androsace
septentrionalis) способны прорастать буквально сразу по осыпании, причем
в лабораторных условиях дают почти стопроцентное прорастание. Можно
полагать, что их непрорастание в природных условиях вызвано внешни-
ми причинами, т. е. это экзогенный или вынужденный покой, который мо-
жет быть связан с необходимостью освещения семян для прорастания (что
показано нами для обоих видов коровяка), неблагоприятной влажностью и
(или) температурой среды. Некоторые трудности возникают в распознана
нии случаев покоя семян, индуцированного внешними условиями.
150 МН. Маркой I loiiyiiHiuioiiiuiH Gitonoi ня piiciciiidi
Хорошее прорастание свежесозревших семян наших объектов нельзя
считать неоспоримым доказательством главенствующей роли вынужденно-
го покоя, поскольку у семян, поначалу непокоящихся, покой может быть ин-
дуцирован внешними факторами, после чего они не прорастают даже в бла-
гоприятных для этого условиях, пока каким-то особым стимулом не будут
выведены из этого состояния. Например, Джерри и Кароль Баскин (Baskin,
Baskin, 1975) отводят значительное место покою семян Torilis japonica, ин-
дуцированному низкими зимними температурами, в определении характера
жизненного цикла у этого вида. По их данным, этот вид ведет себя как зиму-
ющий однолетник из-за непрорастания семян весной. Однако в наших усло-
виях под пологом липового леса оподзоленных почв на карбонатной мате-
ринской породе (на правом крутом берегу р. Волги), Torilis japonica, как и
многие другие двулетники, дает несколько когорт проростков, в том числе и
весеннюю, которая поэтому развивается по типичному двулетнему циклу с
яровизацией в период зимовки и плодоношением вплоть до августа второ-
го года жизни.
Как два варианта эндогенного покоя можно рассматривать непрораста-
ние свежесозревших семян чесночницы Alliariapetiolata и вайды ребристой
Isatis costata, поскольку во втором случае сами семена способны к энер-
гичному прорастанию, но прорастание тормозит крылатый перикарп (см.
выше). У чесночницы покой прерывается уже после первой зимовки, в ходе
которой проходит их стратификация (Мельникова, 1972; Lhotska, 1975), а у
вайды разрушение перикарпа и снятие его ингибирующего действия, оче-
видно, требует одну-две зимовки, о чем свидетельствует ритм возобновле-
ния ее популяции (см. выше).
Если готовность к прорастанию свежесозревших семян Verbascum
lychnitis — дву-многолетника, который переходит к репродукции не ранее,
чем накопит достаточную биомассу, — не вызывает удивления, то этого нель-
зя сказать о других наших объектах. В такой ситуации двулетники, для кото-
рых стимулом перехода к репродукции является зимняя яровизация, должны
иметь приспособление для предотвращения прорастания свежесозревших се-
мян. Иначе вся активная часть популяции будет развиваться по озимому уко-
роченному пути, а это неблагоприятно скажется в дальнейшем на общей се-
менной продукции популяции. Проблема может быть решена либо благода-
ря эндогенному покою, как у чесночницы или вайды, либо путем задержки
диссеминации. Формирование воздушного или надземного банка семян из-за
длительной задержки некоторой части семян в засохших чашечках змееголов-
ника (Dracocephalum thymiflorum), в плодах проломника, икотника и донни-
ка (Androsace septentrionalis, Berteroa incana, Melilotus alba), в обертках кор-
зинок бородавника (Lapsana communis), на столбочках двусемянок пупырни-
ка (Torilis japonica), судя по нашим наблюдениям, представляет собой вполне
нормальное явление. Благодаря этой задержке многие зрелые семена просто
не успевают достичь поверхности почвы до наступления неблагоприятных
для прорастания температурных условий. В ноябре-феврале семена Berteroa
incana, эремы Dracocephalum thymiflorum и семянки Lapsana communis, из-
влеченные, соответственно, из сухих невскрывшихся стручочков, чашечек и
оберток, которые находились на совершенно сухих стеблях, торчащих из-под
I ii/fiti Л Жтнссшкo()iii.ic tсмени n нч рощ, в иопуницпих pm iciuiH 15 1
снега, оказались немедленно всхожими и дали, cooibcici вгппо, 98, 30 и 28°»
прорастания. Наблюдения показали, что воздушный банк вепда формируо
ся и у некоторых факультативных двулетников, например у юх же двух видов
Verbascum, видов Arctium и др. Логично предположить, что таким способом
обеспечивается пополнение весенней когорты проростков, которая, судя но
нашим наблюдениям за популяциями лесных и степных двулетников, всегда
появляется и часто бывает представлена большим числом особей, чем после-
дующие когорты в данном сезоне. Здесь важно отметить, что так большин-
ство семян двулетних растений достаточно быстро вовлекается в популяци-
онный оборот, а это несомненно положительно отражается на темпах размно-
жения и опять-таки отчасти компенсирует задержку с переходом к репродук-
ции (см. выше раздел 5.1.1).
Таким образом, не обладая отчетливо выраженным эндогенным покоем
семян, многие из двулетних растений адаптированы к использованию эк-
зогенного (вынужденного) покоя как средства сохранения резервной ча-
сти семенной продукции несмотря на повышенный риск неблагоприятно-
го воздействия фитофагии со стороны птиц и прочих позвоночных в тече-
ние зимы.
Если приведенные факты еще не говорят напрямую об отсутствии посто-
янного банка семян в почве у некоторых малолетников, то они, по крайней
мере, заставляют отводить более заметную роль в популяционной жизни
малолетних растений внешним факторам прорастания или экзогенного по-
коя и поставить под сомнение общее значение тезиса о том, что среда оби-
тания для всех малолетних растений в одинаковой степени непредсказуема.
Встречающиеся в литературе (Crawley, 1986) высказывания о том, что для
однолетних растений самоубийственно иметь синхронное прорастание, вы-
глядят чересчур категоричными. На примере лесного незимующего одно-
летника недотроги обыкновенной Impatiens noli-tangere нам удалось пока-
зать наличие временного банка II типа (по Grime, 1979), в котором семена
пребывают лишь одну зиму, чтобы почти синхронно на 100% прорасти вес-
ной (Марков, 1991а). Такую же картину удалось выявить даже у сегеталь-
ного сорняка редьки дикой Raphanus raphanistrum, проявляющего в этом
стратегию, которая не свойственна большинству однолетних сегеталов. По
данным литературы такая же картина наблюдается и у некоторых других
относительно крупносемянных видов. Очевидно, у растений такого типа
каким-то образом снижается риск отмирания проростка до зрелости — этот
существеннейший показатель, фигурирующий во всех демографических
моделях. Один из путей такого снижения риска лежит через укрупнение се-
мян — через увеличение в них резерва питательных веществ.
5.2. Семя как зародыш в «упаковке» со стартовым запасом
питательных веществ
Осуществление роли семени в процессе адаптации вида всегда сопряже-
но с определенным компромиссом или компенсациями (trade-off), посколь-
ку некоторые из пяти перечисленных и последовательно рассматриваемых
функций находятся в известном противоречии друг с другом.
152 M.H. Маркин I loiiyjiHiuioiiiiiui Oihuioi ни pin iciniil
Так, быстрое размножение самоопыляющегося однолетника, связанное с
высокой семенной продукцией, происходи! на базе замкнутой рекомбина-
ционной системы (Грант, 1984). Налицо противоречие между первой и пя-
той функциями — множество формируемых семян не обеспечивает соот-
ветствующего множества генетических вариантов. Не надо, однако, забы-
вать, что системы скрещивания, свойственные многим однолетним расте-
ниям, позволяют в большинстве случаев самоопыляться, обеспечивая бы-
строе размножение определенных адаптивно ценных генотипов, но с сохра-
нением способности к скрещиванию и вместе с ней способности давать ге-
нетически разнородное потомство, осваивающее мозаику различных место-
обитаний (Allard, 1975).
Крупное тяжелое семя с относительно большим запасом веществ способ-
ствует успешному приживанию проростка, но из-за большой массы, за ред-
ким исключением, когда дисперсия осуществляется океаническими течени-
ями (рис. 5.166), не может подчас служить эффективной единицей распро-
странения на значительные расстояния. Если учесть, что вклад биомассы в
формирование семян всегда регламентирован (Kawano, 1981; Haig, Westoby,
1988), то, очевидно, такие крупные семена образуются в относительно ма-
лом количестве и потому не вполне отвечают требованиям функции размно-
жения. Прибавим к этому, что большой запас питательных веществ в семе-
ни — это большой соблазн для животных фитофагов, для которых сезон
неблагоприятный для растений (например, зимовка) тоже неблагоприятен.
Отсюда, повышенная вероятность поедания крупных семян снижает эффек-
тивность их как средства для перенесения сурового сезона.
Ради оптимизма, однако, следует заметить, что крупносемянные виды
растений в природе не перевелись, и в ближайшее время быстрого их выми-
рания не ожидается. Компромисс адаптации и состоит в сопряжении «про-
игрыша» по одним биологическим параметрам с «выигрышем» по другим.
Так, например, животные, поедающие семена, оказывают растению ответ-
ную услугу, обеспечивая эффективное распространение — например, тя-
желые диаспоры дуба черешчатого (желуди) эффективно распространяют-
ся сойкой.
В этой связи весьма интересными выглядят попытки некоторых исследо-
вателей (Jain, 1979; Haig, Westoby, 1988; Winn, 1988) обозначать стратегию
вида или популяции как функцию: выгода за вычетом цены (Benefit - Cost),
выраженную в общем виде через уравнение:
F = f(B)-f(C),
где f(C) — это затраты организмов на формирование каких-либо адаптив-
ных структур, a f(B) — это выгода в форме повышенной выживаемости.
Приоритет в теоретическом осмыслении такого соотношения, однако, все
же принадлежит нашему выдающемуся отечественному генетику А. С.
Серебровскому, рассматривавшему значение любого признака как разницу
между двумя параметрами: польза - вред.
Момент прорастания диаспор как переход от латентной фазы (с присущи-
ми ей заторможенностью метаболизма и устойчивостью) к активному росту
Iiddd Л Ж пн icci кнобныс ссмспи и их роль в попунмцпях pin 1СН11Й 153
является критическим и крайне важным для оценки jkoiioi о-цсногичсскои
стратегии растения гак же как и размеры самих зачатков. ')ю явствуец на
пример, из схемы М. Angevine и В. Chabot (1979), которая, в несколько из-
мененном виде, представлена в таблице 5.18. Поведение семян, безуслов-
но, является лишь элементом общей эколого-ценотической стратегии, при-
сущей малолетнему растению, но явно определяющим, базисным для всех
остальных элементов. Рассмотрим схему чуть подробнее. Любой стресс
или комплекс неблагоприятных условий требует затрат на его преодоление.
Можно либо избежать действия неблагоприятных условий (стресса), либо
выдержать это неблагоприятное влияние, понеся соответствующие затраты.
Отсюда понятна первоначальная дихотомия с разделением стратегии избе-
гания и стратегии толерантности. Если избежать неблагоприятного абиоти-
ческого или биотического воздействия, как видно из таблицы 5.17, можно
в основном двумя путями: «разойдясь» с ним либо в пространстве, либо во
времени, то путь повышения толерантности один. Независимо от природы
действующего неблагоприятного фактора повысить устойчивость пророст-
ка можно только путем увеличения резерва питательных веществ или путем
увеличения размеров семени.
Таблица 5.18.
Схема, демонстрирующая варианты поведения на стадии прорастания семян
(Angevine, Chabot, 1979)
Основная стратегия Избегание Толерантность
Характер стресса Абиотический Биотический Абиотический Биотический
Конкретная тактика поведения Избегание холода Избегание засухи Избегание (конкуренции) в пространстве Избегание (конкуренции) во времени Увеличение вклада биомассы в каждое семя
Биологическое значение массы семени и ее вариабельности на разных
уровнях можно рассмотреть в трех аспектах: морфологическом, физиоло-
гическом и генетическом. Выделение третьего аспекта, пожалуй, наиболее
условно, поскольку влияние генотипа сказывается как на морфологических
признаках, так и на физиологических свойствах растений. Краткое обсуж-
дение генетического аспекта вынесено в последний раздел данной главы, а
здесь при помощи схемы (рис. 5.33) представим возможные каналы влия-
ния морфологии репродуцирующих растений на размеры (массу) и измен-
чивость образуемых ими семян, а также зависящие от массы семени физи-
ологические показатели.
154
Л/.// Миры hi
I h)lIVIIНЦИО1ПШН (IIIOHOI IIH pnc ICIIIli
Морфологический аспект
Физиологический аспект
Рис. 5.33. Факторы, связанные с массой семени (морфологический и физиологи-
ческий аспекты).
5,2,1. Межвидовой уровень
Очень важную роль в определении диапазона изменчивости массы семе-
ни на межвидовом уровне, несомненно, играет филогенетический фактор.
Тип гинецея и плода, тип плацентации, число и структура интегументов се-
I'llUthl 5 XirillCCIIQCofillMC СГМГПП II ИХ роль К популициих |ШС1СП11Й 155
мязачатка, тип эмбрио! снеза, наличие или отсутствие эндосперма и про
чие признаки, входящие на правах нсогьсмлсмых атрибута в харакгсрп
стику семейств и родов, накладывает естественные ограничения на размер
единичного зачатка, наблюдаемый у того или иного вида (Hodgson, Mackey,
1986). Дж. Харпер (Harper, 1977) приводит в этой связи данные О. Стевенса
(Stevens, 1932, 1957) о пределах варьирования массы семени у сорных рас-
тений из разных семейств: от 0,00001 до 0,02 г (Gramineae, Сурегасеае,
Ranunculaceae, Chenopodiaceae, Caryophyllaceae, Cruciferae, Rosaccae,
Scrophulariaceae, Compositae); от 0,0003 до 0,01 г (Liliaceae, Polygonaccae);
от 0,001 до 0,02 г (Fabaceae, Umbelliferae).
Вместе с тем на осуществлении эволюционных преобразований в так-
сонах не могут не сказываться общие количественные закономерности и
в первую очередь компенсационные связи между элементами (Хохряков,
1975; Бигон, Харпер, Таунсенд, 1989). В качестве примера для иллюстра-
ции разберем компенсаторную связь между массой одного семени и плодо-
витостью с привлечением массива данных о 151 виде малолетних растений
(см. раздел 5.1). Также как и плодовитость, масса одного семени в абсолют-
ном выражении дает распределение с сильной положительной асимметрией
при почти симметричном (логнормальном) распределении натуральных ло-
гарифмов массы (рис. 5.34). Рисунок 5.35 демонстрирует линейность отри-
цательной корреляционной связи между максимальной плодовитостью (Ns)
и средней массой одного семени (Wls). Статистически достоверный коэф-
фициент корреляции (г = -0,631; Р<0,001) подтверждает основной вывод:
существенное повышение плодовитости у того или иного вида растения мо-
жет происходить преимущественно за счет соответствующего измельчания
семян.
Весьма показательно, что при разбивке массива на подвыборки, соответ-
ствующие основным типам жизненных форм, эта компенсаторная зависи-
мость проявляется и в каждой из отдельно взятых групп.
Меняется лишь наклон линий регрессии (коэффициенты уравнений) и
достоверность корреляционной связи. Последнее обусловлено, главным об-
разом, уменьшением объема выборки.
Уравнение, полученное для всей выборки европейских малолетних видов
в целом: lnWls = 2,3 - 0,33 lnNs (г = -0,365; р<0,001) по своим параметрам
довольно близко к тому, которое получил Ш. Кавано (Kawano, 1981) для вы-
борки из 60 дикорастущих японских и 38 североамериканских видов расте-
ний: logWls = 1,2 - 0,46 log Ns (г = -0,10; р<0,001), причем японский иссле-
дователь совместил в своей выборке многолетние поликарпические и мало-
летние виды.
На примере четырех видов из рода проломник Androsace показано (рис.
5.36), что компенсационную зависимость можно наблюдать и сравнивая
разные виды в пределах одного рода.
В контексте обсуждения теории «г» и «К» отбора был выдвинут тезис,
что многолетние растения имеют обычно мало крупных, а малолетние
много мелких семян. X. Бэйкером (Baker, 1972) на примере калифорний-
ских видов растений показано, что в ряду жизненных форм от однолетни-
ков до деревьев-фанерофитов (двулетники как отдельная жизненная фор-
156
МН Маркин I loiiyii>iniioiiin»i (hiojioi им piicicind
Рис. 5.34. Гистограммы распределения по классам абсолютных значений массы одно-
го семени (вверху) и In массы одного семени (внизу) для выборки из 155 видов мало-
летних растений.
ма не выделялись) средняя масса одного семени возрастает, что позднее на-
шло свое отражение в обосновании «аллометрической гипотезы» — круп-
ные растения дают крупные семена (Thompson, Rabinowitz, 1989). Вместе
с тем в литературе был высказан (Salisbury, 1942) и впоследствии неодно-
кратно цитирован (Silvertown, 1989) тезис о более тесной корреляции меж-
ду массой семени и типом местообитания (фитоценоза), чем между массой
семени и типом (продолжительностью) жизненного цикла. Разберем, на-
сколько сильна связь массы семени с типом жизненного цикла и с эколого-
фитоценотическими условиями местообитания, используя группу из 147
видов малолетних монокарпических растений.
Статистика массы семени для малолетних монокарпических видов растений
с разным типом жизненного цикла приведена в таблице 5.19, а распределение по
классам массы семени для видов каждой из четырех эколого-фитоценотических
групп представлено на рисунке 5.37. Силу связи в каждом случае оценивали при
помощи непараметрического рангового теста Крускала-Уоллиса в дисперсион-
/ ildtid Л
>l< ir nice i iocoOi ii.ir ICMCIIII и их
poJIl. II ПОПУЛЯЦИЯХ j)IK irniiil
15/
Рис. 5.35. Компенсационная зависимость между массой семени (Ws) и плодовито-
стью (Ns) в логарифмических координатах для выборки видов растений разных жиз-
ненных форм: InNs = 8,13 - 0,48 InWs (г = -0,631).
ном анализе. Тип жизненного цикла оказал более сильное влияние на массу се-
мени (Н = 24,961; р<0,0001) нежели тип местообитания (Н = 11,569; р=0,0208),
хотя влияние каждого из этих факторов было статистически достоверным.
Впрочем, здесь вполне могла сказаться большая доля условности в отнесении
видов малолетников к тому или иному эколого-фитоценотическому типу и во-
обще меньшая зависимость малолетних растений — эксплерентов от фитоцено-
тических режимов.
Преобладающее большинство малолетних растений, проявляя эксплерент-
ность, избегают действия неблагоприятных условий абиотического или био-
тического происхождения. Это выражается в осеннем или весеннем прораста-
нии семян. Осень и весна — периоды пониженного конкурентного давления
со стороны многолетних растений, причем менее интенсивной, в силу есте-
ственных сезонных циклов, становится конкуренция и за влагу, и за элемен-
ты минерального питания, и, что особо важно, за свет. От лета к осени наблю-
дается снижение конкуренции за перечисленные ресурсы. Весной же, наобо-
рот, интенсивность конкуренции от минимальной резко возрастает. Поэтому
растениям, демонстрирующим позднее прорастание, приходится уже в фазе
проростков сталкиваться с биотическим стрессом (затенением) или абиоти-
ческим стрессом (дефицитом влаги, высокими температурами).
Означенная динамическая тенденция, конечно же, по-разному проявля-
ется в разных фнтоценозах — лугово-степных, дюнных, прибрежных, лес-
158
М Н Miipkim I h >иул мцноннпн ftiiojioi им рис i спи Л
Рис. 5.36. Компенсационная зависи-
мость между массой семени и плодови-
тостью на примере четырех видов рода
проломник Androsace.
пых, агрофитоцспозах, каждому из ко-
торых присуща к кому же своя про-
странственная гетерогенность (см.
ниже главу 8). Так, например, подме-
чено, что относительно крупносемян-
ными являются виды тенистых (био-
тический стресс) и виды засушливых
(абиотический стресс) местообита-
ний (Salisbury, 1942; Silvertown, 1989;
Tilman, 1988). Однако картина на деле
не так проста именно из-за простран-
ственной гетерогенности ценозов, до-
пускающей сосуществование в соста-
ве одного ценоза видов малолетников
с сильно различающимися по размеру
семенами10. В составе одного, скажем,
слабо сомкнутого лугово-степного фи-
тоценоза малолетники, попавшие под
полог травянистых многолетних рас-
тений испытывают дефицит освеще-
ния. Буквально рядом в наиболее крупных окнах (гэпах), где по существу снят
биотический стресс, наблюдается сильный абиотический стресс из-за сильно
возрастающего здесь физического испарения и дефицита влаги в поверхност-
ных слоях почвы, особенно в засушливый период. Поэтому нет оснований
считать открытые сообщества травянистых растений наиболее подходящи-
ми для произрастания мелкосемянных малолетников. Просто амплитуда зна-
чений массы семени у компонентов лугово-степных фитоценозов с понижен-
ным уровнем ценотической замкнутости существенно расширяется благода-
ря значительной примеси мелкосемянных видов малолетников, что не может
не отразиться на величине средней арифметической, вычисленной для груп-
пы лугово-степных и дюнных малолетников (см. рис. 5.37). Таким образом,
напрашивается вывод о важности анализа причин варьирования массы семе-
ни в пределах той или иной группы.
Любая эколого-фитоценотическая группа малолетников объединяет виды
с разным типом жизненного цикла и, следовательно, разным типом сезон-
ного развития. Для примера можно взять данные таблицы 5.19, в которой
сравниваются зимующие и незимующие (т. н. яровые) сегеталъные сорня-
ки. Можно видеть, что зимующие однолетники (а они, как правило, розеточ-
ные или полурозеточные) в среднем мелкосемянные, поскольку прораста-
ние их происходит главным образом в осеннее время, когда из-за ослаблен-
ной конкуренции с культурным растением — доминантам возможно прижи-
вание мелких проростков. Для яровых обыкновенно длиннопобеговых од-
10 Распределению по классам абсолютных значений массы семени для группы видов из любо-
го местообитания свойственна сильная положительная асимметрия. Этот феномен рассмотрен
с позиций фрактальной геометрии (Hegde et al., 1991). Показана его связь с характером распре-
делений размеров самих растений и размеров микроучастков, занимаемых растениями в фито-
ценозах. Расчеты, выполненные для выборки из 1413 видов, позволили установить, что часто-
та вида с данной массой семени пропорциональна квадратному корню из значения этой массы.
I Utltltl Л
Жи пкч иособпыс с смени и их роль и попуннцннх pm iciinil
159
экол.-фитоцен. группы
---степные и дюнные -- сегетальные НН прибрежные .. лесные
Рис. 5.37. Графики распределения видов малолетников 4 разных эколокь
фитоценотических групп по классам массы семени.
нолетников характерно исключительно весеннее прорастание относительно
крупных семян с захватом лишь более поздней части благоприятного весен-
него периода, и образование крупносемядольных проростков, лучше адап-
тированных к притенению.
Таблица 5.19.
Средняя масса семени у видов малолетних растений с разным типом жизненного
цикла (Wls — абсолютные и In Wls — трансформированные данные)
тип данных N min X±S X max As E
Незимующие однолетники
Wls 66 0,01 2,74 ± 0,48 48,2 2,53 6,49
lnWls -4,34 0,14 ±0,19 2,9 -0,63 0,67
Зимующие однолетники
W„ 36 0,01 0,58 ±0,14 4,5 2,84 10,24
lnWls -4,61 -1,44 ±0,25 1,5 -0,22 0,67
Двулетники
W, 1s 45 0,09 2,36 ± 0,47 14 2,05 3,97
InW. Is -2,4 0,04 ±0,21 2,6 -0,14 0,67
Примечание: As и Е — коэффициенты асимметрии и эксцесса распределений
Вместе с тем как для относительно крупносемянных (ярутки Thlaspi
arvense), так и для мелкосемянных (пастушьей сумки Capsella hursa-pasforis)
зимующих однолетников характерно присутствие в составе агропопуляций в
посевах озимых и яровых культур весенних когорт особен. Отчасти это мож
160
MH. MtipkiHi
I loiiyiiHUiioiiiiiiM (нюлогня pnciciiiiil
Популяции:
Европейская (N = 256) ИИ Дальневосточная (N = 194)
Рис. 5.38. Сравнение европейской и дальневосточной популяций недотроги обык-
новенной по числу семян в плоде (N — объемы выборок плодов).
но объяснить тем, что в яровом, а тем более в озимом посеве весенние ко-
горты захватывают ранневесенний сравнительно благоприятный период, но
краткосрочность этого периода обусловливает в итоге очень существенное
снижение их плодовитости по сравнению с плодовитостью осенних когорт.
В контексте выше начатого разговора о двулетниках нельзя не остано-
виться на факте их явной относительной крупносемянности. Значение сред-
ней арифметической, фигурирующее в таблице 5.18, которое приближается
к максимальному — отмеченному для незимующих однолетников — следу-
ет рассматривать с учетом того, что группа двулетников заведомо неодно-
родна и включает подгруппу мелкосемянных факультативных двулетников
из семейства норичниковых Scrophulariaceae (коровяков Verbascum lychnitis,
V. thapsus, наперстянки Digitalis purpurea). Свойственная норичниковым от-
носительная мелкосемянность, из-за которой это семейство входит в проти-
воречие с уже упомянутой «аллометрической гипотезой» присуща, таким
образом, и двулетним растениям из этого семейства, и это существенно вли-
яет на их демографические показатели. Такие виды могут быть ориентиро-
ваны, во-первых, на формирование массовых скоплений мелких пророст-
ков со своим микроклиматом, положительным эффектом группы (см. главу
8), а впоследствии и интенсивным изреживанием. А во-вторых, — на всту-
пление в симбиоз с грибами и образование микоризы, что по всей вероят-
ности тоже может способствовать выживанию мелких проростков даже в
столь суровых экотопах как песчаные дюны или эродированные остепнен-
ные склоны (Verbascum thapsus, V lychnitis)n.
11 Сравнение 62 видов малолетников, охарактеризованных Д. Граймом (Grime, Hodgson,
Hunt, 1988), и разделенных нами на микоризные (40) и безмикоризные (22), по массе семе-
I 'ltirtii > Жп nicciiocoGiii.ir и мени и их рош. н нонуинцних рас iciiiiil 161
Подгруппа мелкосемянных двулсгников, нссмогря па свою малочислен
носи», несомненно, повлияла па среднюю величину noi<a »aicini дня всей i рун
ны, и представляегся важным, что даже при лом условии двулегннки оказались
в числе наиболее крупносемянных малолетних растений. Можно рассматривать
относительную крупносемянность двулетников как средство повышения толе-
рантности и выживания проростков, иными словами, как один из способов уве-
личения (см. формулы, приведенные в разделе 5.1.1), и через него — темпов
размножения. Можно связать ее и с характером населяемых двулетниками ме-
стообитаний — либо тенистых, либо засушливых (Марков, 19916).
Современными статистическими методами была показана индивидуаль-
ность сукцессионного поведения двулетников как биологической группы на-
ряду с индивидуальностью поведения однолетников. Тот факт, что в ходе
сукцессии пик двулетников наблюдается позже, чем пик однолетников, и
что двулетники удерживаются до более поздних сукцессионных стадий, го-
ворит о меньшей их ценофобности, которая зависит от особенностей био-
логии диссеминации. Толерантность к факторам среды и способность до
определенного момента к нейтрализации конкурентного воздействия со
стороны других, и в первую очередь многолетних, видов (Myster, Pickett,
1988) — выступают у них достаточно отчетливо.
Итак, с учетом всех высказанных оговорок, рассматривая массу семени у
малолетников на межвидовом уровне, можно отметить следующие законо-
мерности (см. табл. 5.18, 5.19 и рис. 5.37). В среднем более крупные семе-
на характерны для: 1) незимующих (длиннопобеговых) однолетников и дву-
летников; 2) лесных и степных малолетников. В среднем более мелкие се-
мена свойственны: 1) зимующим (розеточным, полурозеточным) однолет-
никам; 2) прибрежным малолетникам.
5.2.2. Внутривидовой уровень
Говоря о размере и массе семян как о важном параметре эколого-
ценотической стратегии, нельзя упускать из виду изменчивость семян вну-
три популяции и даже в пределах одной особи.
Несмотря на то, что факт вариабельности массы семени установлен ныне
для многих видов растений (Schaal, 1980; Pitelka, Thayer, Hansen, 1983; Dolan,
1984; Thompson, 1984; Tripathi, Khan, 1990 и др.), все же во многих работах
проглядывает недооценка значимости этого факта для популяционной биоло-
гии. В этой связи уместно вспомнить высказывание Дж. Харпера о том, что
нет противоречия между тем, что с одной стороны мы находим крайне малую
внутривидовую изменчивость размера семян, а с другой — подчеркиваем
огромную важность изменчивости (генетической или фенотипической) этого
показателя (Harper, 1977). Обыкновенно «константность» массы одного семе-
ни противопоставляют сильной внутрипопуляционной изменчивости особен
по числу формируемых семян, т. е. по плодовитости, забывая при этом, что
рассматривают организмы метамерного строения с незавершенным ростом,
ни показало, что первая группа характеризуется более мелкими семенами (1,04 J 0,3 мг), чем
вторая (1,55 । 0,5 мг), хотя разница и не была статистически достоверной.
IbZ МН Марком I loiiyjiHUiioiiihhi (hiojioi ни р<к iciniil
у которых реальнее находить отличия между повторяющимися субъедини-
цами в составе одной особи, нежели в составе популяции (Silvertown, 1989).
Издавна, по-видимому, с конца прошлого века, известны явления гете-
рокарпии (разноплодия) и гетероспермии (Гордягин, 1892; Goebel, 1911),
называемые в зарубежной литературе полиморфизмом или гетеромор-
физмом плодов и семян. В таких обширных «сорных» семействах как
Chenopodiaceae, Brassicaceae, Asteraceae и Роасеае сам характер соцветий
ставит образующиеся плоды и семена в разные условия обеспечения водой
и ассимилятами. Изменчивость размеров, а иногда и морфологии семян, ко-
торую можно наблюдать в этом случае, иногда называют соматическим по-
лиморфизмом (Williams, Harper, 1965; Harper, 1977). В. Ф. Войтенко (1968)
пишет о широко распространенных в семействе крестоцветных гетерокар-
пии и ярусной разнокачественности семян.
Возвращаясь вновь к схеме гипотетического поэтапного регулирования
плодовитости особи и массы одного семени, изображенной на рисунке 5.10,
можно попытаться проверить гипотезу о зависимости степени варьирова-
ния массы семени от числа «буферных уровней», ослабляющих влияние на
нее изменчивых органов вегетативной сферы. Для этого была изучена кар-
тина вариабельности массы семени у 29 видов из 12 семейств.
В качестве показателя вариабельности было использовано отношение
размаха варьирования признака к средней арифметической, которое приме-
няли совместно с коэффициентом вариации и коэффициентом Джини.
Обращает на себя внимание хорошее соответствие между значениями
обоих показателей варьирования — виды в таблице 5.20 были сортированы
в порядке возрастания коэффициента вариации, и выбранный нами допол-
нительный показатель, вместе с коэффициентом Джини также оказались в
значительной степени сортированными. Этот вывод подтверждается и до-
стоверными значениями коэффициента корреляции между показателями.
Особенно тесно коррелируют значения коэффициентов вариации и Джини.
Таблица 5.20.
Средняя масса семени и показатели ее внутривидового варьирования у 29 видов
малолетних растений из 12 семейств
Виды растений: (семейство) Русское название X ± S X CV R/X Gini As
1 2 3 4 5 6 7
Meniocus linifolius (Cru.) Плоскоплодник льнолистный 0,20±0,02 5,0 0,2 0,03 0,48
Chenopodium album (Che.) Марь белая 0,70±0,10 7,1 0,29 0,03 н.о.
Alyssum turkestanicum (Cru.) Бурачок пустынный 0,47±0,10 11,0 0,43 0,08 0,66
Geranium robertianum (Ger.) Герань Роберта 1,19±0,03 11,8 0,42 0,07 н.о.
Lapsana communis (Com.) Бородавник обыкновенный 0,59±0,02 12,0 0,35 0,07 0,01
Гчани 5 Жизнеспособные ссмспн п их роль в нонуимциих piiciciiiiO
163
In блицы 5.20. (Продолжение)
3 I 4 | 5 ' I 6 I / '
Echinochloa crus-galli (Gra.) Ежовник куриное- просо 2,78±0,07 13,4 0,54 0,73 0,71
Eremopyrum triticeum (Gra.) Мортук пшеничный 1,12±0,03 14,6 0,54 0,01 0,81
Spergula arvensis (Car.) Торица полевая 0,34±0.01 14,7 0,59 0,11 0,58
Acinos arvensis (Lab.) Щебрушка полевая 1,08±0,03 15,2 0,65 0,09 0,37
Polygonum aviculare (Pol.) Горец птичий 1,49±0,05 17,1 0,74 0,09 -1,02
Arabis pendula (Cm.) Резуха повислая 0,13±0,01 18,4 0,75 0,10 1,2
Ceratocephala falcata (Ran.) Рогоглавник серповидный 0,91±0,02 18,7 1,5 Н. О. 1,32
Dracocephalum thymiflorum (Lab.) Змееголовник ти- мьяноцветковый 0,38±0,02 19,8 0,81 0,10 -0,61
Thymelaea passerina (Thy.) Тимелея воробьиная 1,12±0,03 20,1 1,07 0,09 -0,93
Alliaria petiolata (Cm.) Чесночница черешчатая 2,12±0,04 20,3 0,66 0,09 0,33
Impatiens noli-tangere (Bal.) Недотрога обыкновенная 3,44±0,15 21,1 0,87 0,12 -0,19
Stachys neglecta (Lab.) Чистец незамечаемый 1,18±0,05 22,0 0,93 0,13 -0,19
Rumex maritimus (Pol.) Щавель приморский 0,13±0,01 22,2 0,68 0,12 -0,39
Androsace maxima (Pri.) Проломник наибольший 0,73±0,02 22,9 0,96 0,13 0,26
Polygonum scabrum (Pol.) Горец шероховатый 1,08±0,05 24,0 0,93 0,13 0,28
Descurainia sophia (Cm.) Дескурайния Софии 0,13±0,01 24,5 0,85 0,13 1,о
Berteroa incana (Cm.) Икотник серый 0,52±0,2 27,0 1,02 0,16 0,09
Fallopia dumetorum (Pol.) Горец заборный 3,60±0,18 27,0 1,11 0,20 -0,53
Secale sylvestre (Gra.) Рожь лесная 5,38±0,40 30,9 0,93 0,17 0,43
Consolida regalis (Ran.) Сокирки полевые 1,33±0,07 32,0 1,2 0,18 0,4
Raphanus raphanistrum (Cm.) Редька дикая 6,79±0,95 33,2 1,15 0,19 II. о.
Arctium lappa (Com.) Лопух обыкновенный 6,29±0,46 39,7 1,38 0,23 0,1
Impatiens glandulifera (Bab) Недотрога железконосная 7,32±0,59 46,9 2,38 0,25 1,35
164
МН Марков 1 loriyjinuiioinuiM Ihioiiohih pnciciinii
Рис. 5.39. Компенсационная зависи-
мость между массой семени (Wls) и
числом семян в плоде (Ns) у недотроги
обыкновенной (N=153; Wls = 3,26 - 0,24
Рис. 5.40. Кривые изменчивости длины
семени (1s мм) в пределах особи у кло-
повников сорного (кружки) и густоцвет-
кового (квадраты).
11ровсрка скоррелированное ти
параметров варьирования и сред-
ней массы семян (г между In Wls
и Джини — +0,50; р=0,007; InWls
и R/X — +0,48; р=0,008; In Wls и
CV — +0,53; р=0,003) подтвердила
опасения насчет влияния заведомо
большей точности взвешивания бо-
лее крупных семян. Мелкие семена
некоторых видов приходилось взве-
шивать по два, три или пять, что
также способствовало получению
искусственно сглаженных оценок
варьирования. И все же следует за-
метить, что среди видов, лидирую-
щих по вариабельности массы семе-
ни (коэффициент вариации превы-
шает 20%), фигурируют представи-
тели разных семейств. Причины по-
падания в лидирующую группу мо-
гут быть разные, но наиболее ин-
тересны, безусловно, биологиче-
ские. Три представителя семейства
Balsaminaceae, к примеру, характе-
ризуются заметно повышенным ва-
рьированием массы семени по срав-
нению с другими видами благода-
ря образованию двух типов пло-
дов: клейстогамных и хазмогамных.
Если клейстогамные плоды как пра-
вило односеменные, то хазмогам-
ные содержат всегда более одно-
го семени и тогда у всех трех видов
можно наблюдать варьирование массы семени внутри одного плода. Форма
коробочек при этом выдает различия в порядке выполнения семян. Если у
Impatiens noli-tangere. раньше всего оформляется семя в проксимальной ча-
сти коробочки, и потому наблюдается характерное для многих плодов по-
степенное сужение к дистальному концу, то у L parviflora и I. glandulifera
все происходит в обратном порядке, а расширена бывает, как правило, дис-
тальная часть коробочек. Сказывается это и на числе семян, содержащихся
в плоде к моменту его созревания.
Компенсационные связи типа число — масса (размеры) у некоторых рас-
тений, по-видимому, могут осуществляться и на уровне отдельных плодов,
что должно быть связано с лимитированностью общей биомассы семян, ко-
торые могут формироваться внутри одного плода. Рассмотрим несколько
примеров, используя крупносемянные виды малолетников, у которых мож-
но с максимальной точностью определить массу семян.
/ //(/«(/ Ж|| 111СЧ11ОСОС)11ЫГ IГМГНИ II ИХ роль II IIOliyilHUIIHX рИС1С11НЙ 16‘j
Потенциальное число семян, способных ра нни вся в одном иноде, мож
но определить по числу закладывающихся семязачатков. Гак, например, в
условиях средней полосы европейской части России в популяциях исдо'ро
ги обыкновенной нам удалось насчитать до 9 семязачатков на одну коро
бочку. Вместе с тем, график распределения плодов по числу семян в тех же
популяциях (рис. 5.38) показывает, что реализуется всего от 11,0 до 66,7%
числа семязачатков, и среднее значение составило 2,1 ± 0,2 семян/плод1
На примере одно-трехсеменных коробочек на фоне существенного варьи
рования массы семени в масштабе популяции можно зафиксировать хон. н
слабую, но статистически достоверную отрицательную корреляцию между
числом семян в плоде и средней массой семени (см. рис. 5.39).
У Raphanus raphanistrum можно было найти в агропопуляции из посе-
ва яровой пшеницы плоды, содержащие от 1 до 10 семян. При возрастании
числа семян от 1 до 5 в стручке наблюдалась очень слабая компенсация, по-
сле чего наклон линии стал намного круче — lnWls= 2,7-0,94 InNs, а коэф-
фициент при независимой переменной близок к единице. По всей вероя i по-
сти эта картина является следствием того, что ресурсы стручка при выпол-
нении от одного до пяти семян явно недоиспользуются, и только, начиная с
6 семян/стручок, достигается предел суммарной биомассы и включается ли-
митирующий фактор.
Однако, компенсационные связи такого рода наблюдаются не у всех
видов. Так, несмотря на то, что средняя масса семени в пределах особен
Alliaria petiolata плавно снижается от нижнего плода кисти к верхнему,
крайне слабая отрицательная корреляция между числом семян в стручке и
средней массой семени оказалась статистически недостоверной.
Хотя внутривидовое и внутрипопуляционное варьирование массы семе-
ни само по себе представляется очень интересным явлением, достойным
внимательного исследования, все же главный интерес в аспекте популяци-
онной биологии имеют его популяционные следствия. Из этих следствий
наиболее важными, на наш взгляд, являются, во-первых, разновременное
прорастание семян и, во-вторых, возможность сохранения первоначаль-
ных преимуществ по величине стартовой биомассы у особей, появившихся
из наиболее крупных семян, в течение всего дальнейшего роста. Как наши
данные, так и данные литературы дают противоречивые результаты при по-
пытке ответить на вопрос, какие семена прорастают раньше и более энер-
гично, крупные или мелкие.
Нами изучено варьирование величины семени в пределах особи у двух
видов родаLepidium*. L. ruderale и L. densiflorum. Кривые изменчивости дли-
ны семени на рисунке 5.40 показывают, что величина семени у L. ruderale
сильнее, чем у L. densiflorum, отражая уменьшение размеров плодов в верх-
них частях соцветий, свойственное первому виду. Отрицательная асимме-
трия кривой (для L. ruderale) наглядно свидетельствует о наличии фрак-
ции более мелких семян, отсутствующей у L. densiflorum. В ходе опытов
выяснилось, что величина семян сказывается на характере их прорас гания:
12 Интересно, что в популяции этого вида на территории Уссурийского заповедника и При
морьс число семян в одном плоде в среднем выше и варьирует в более широких пределах
(рис. 5.38).
1UU
л/ /( Hhiph'Hi I h>ilyii>ihi10111ши oiioiioi ii>i pin iciiihi
Рис. 5.41. Динамика прорастания мел-
ких (м) и крупных (к) семян у клопов-
ника сорного; G — процент прораста-
ния; по оси X — даты наблюдений.
крупные семена (1,1 1,5 мм) про-
растали более энергично, чем мел-
кие (0,7 1,0 мм), и к концу они та их
проросло на 20% больше (рис. 5.41).
Варьирование размеров может
наблюдаться и среди плодиков и се-
мян, образовавшихся в одном цвет-
ке. Это явление мы изучали на при-
мере Myosurus minimus, у которого
односеменные плодики-орешки рас-
полагаются на длинном коническом
цветоложе, образуя многоорешек.
Как уже было показано выше, дли-
на многоорешка сильно варьирует
даже в пределах одной особи, из-за
чего меняется число, и, в меньшей
степени, величина орешков и обра-
зующихся в них семян. На прора-
щивание были поставлены семена, взятые из многоорешков максимальной
(для исследованной популяции этого вида), средней и минимальной дли-
ны: 16-23 мм, 9-14 мм и 2-7 мм, соответственно. Энергия и конечный про-
цент прорастания были связаны с величиной многоорешка прямой зависи-
мостью (табл. 5.21).
Таблица 5.21.
Прорастание семян мышехвостника из многоорешков разной длины (данные двух
опытов)
Категории по длине многоорешков % проросших семян (среднее из трех повторностей с ошибкой)
Опыт I
дни прорастания 1-й 3-й 4-й 9-й
I (16-23 мм) 74,7±8,7 84,0±6,9 90,7±4,8 93,3±3,5
II (9-14 мм) 44,0±16,0 64,0±16,0 68,0±16,0 76,0±12,0
III (2-7 мм) 48,0±12,0 61,3±8,1 68,0±9,2 69,0±9,3
Опыт 2
дни прорастания 1-й 2-й 5-й 6-й
I (16-23 мм) 9,3±4,8 28,0±6,1 73,3±5,8 82,7±3,5
II (9-14 мм) 3,0±2,2 17,3±7,4 57,3±4,8 66,7±7,4
III (2-7 мм) 1,3±1,0 8,0±4,6 28,0±8,3 37,3±5,8
Иногда мелкими могут быть недозревшие и потому недовыполненные
семена, которые, однако, оказываются подчас не менее жизнеспособными,
чем нормальные выполненные. Семена, извлеченные нами из недозрелых
Г’шна .5 Жизнеспособные семени и их роль п нонуиицних piicicinih
167
Рис. 5.42. Независимо от конечного строения семени и степени развития составляю-
щих его компонентов (кожуры, эндосперма, остатков женского гаметофита, зароды-
ша), налицо система из структур, представляющих 4 различных генотипа [2п (п+п), п,
Зп (2п+п’), 2п (п+п’)], которая поэтому обладает составной приспособленностью, т. с.
суммой приспособленностей каждого из этих четырех генотипов.
плодов проломника наибольшего Androsace maxima, были намного мельче
нормальных (масса одного семени составляла 0,30 мг против 0,77 мг). И те
и другие проросли, причем через день нормальные семена дали 100% про-
растания, а невыполненные — 30%, но по прошествии еще трех дней и они
проросли на 100%. И вновь приходится констатировать различную энергию
прорастания семян, отличающихся по размерам.
Для популяций малолетников асинхронность прорастания семян играет
очень важную роль, поскольку обеспечивает поддержание внутрипопуля-
ционного разнообразия. Интересное предположение Р. Е. Левиной (1967)
относительно приспособительного значения гетероспермии у апомиктов
как источника разнообразия, компенсирующего отсутствие скрещиваний,
можно адресовать и нашим преимущественно самоопыляющимся однолет-
ним растениям с их замкнутыми рекомбинационными системами. Здесь мы
затронули вопрос о пятой последней из перечисленных функций семени.
5.3. Семя как генетический вариант с заложенными в нем
последствиями рекомбинации
По данным М. Негшу (1985) уже при первой попытке разделить растения
на две группы по особенностям жизненного цикла (т. е. на два типа стра-
тегии), которая была осуществлена в 1894 г. Мак Леодом, оказались учте-
ны системы скрещивания. Многолетние растения — «капиталисты» по Мак
Леоду являются преимущественно перекрестноопыляющимися с относи-
тельно крупными нектароносными цветками, тогда как однолетние «проле-
тарии» в большинстве своем самоопылители. Основные типы проявления
сексуализации у растений уже были рассмотрены нами выше (табл. 2.2).
168
Л/,// Марков I loiiyiiHUiioiiiiiiH biioiioi пи piiciciiiiil
Рис. 5.43. Участие особей разной мощ-
ности в воспроизводстве популяций
клоповника густоцветкового Lepidium
densiflorum из относительно благо-
приятных (вверху) и неблагоприятных
(внизу) условий.
Nr — суммарное число плодов в клас-
сах, W — общая биомасса (мг) на фоне
кривой распределения по классам об-
щей биомассы.
5.3.1. Особенности
репродуктивных систем
многолетних растений
Почти исключительно среди мно-
голетних растений встречаются апо-
микты — «бесполосеменные» рас-
тения, которые способны продуциро-
вать семена без оплодотворения пу-
тем митоза или модифицированного
мейоза, происходящего без редукции
числа хромосом. Апомикты — это
чаще всего полиплоиды и особенно
характерны они для семейств слож-
ноцветных, злаков и розоцветных,
хотя могут принадлежать и к другим
семействам цветковых, например се-
мейству гречишных. Так около 90%
известных «видов» одуванчиков это
апомикты, за что их принято имено-
вать микровидами. Практически все
апомикты имеют близкородственных
неапомиктов. Хотя у апомиктов сли-
яния гамет не происходит, большин-
ство из них псевдогамно, т. е. нуж-
дается в процессе опыления и росте
пыльцевых трубок для слияния, га-
рантирующего формирование эндо-
сперма и обеспечение зародыша пи-
танием. Немаловажно, что апомик-
сис всегда сочетается с самонесовместимостью и связанным с ней недостатком
опылителей как фактором, лимитирующим продукцию семян. Поскольку апо-
микты — это часто полиплоидные потомки двух или более диплоидных не апо-
миктичных предков, уровень их гетерозиготности достаточно высок, как высок
и уровень их экологической толерантности (общее свойство полиплоидов).
5.3.2. Репродуктивные системы малолетних растений
В генетическом отношении большинство малолетних растений характеризу-
ются высокой степенью гомозиготности популяций из-за самоопыления, проис-
ходящего с большей вероятностью, чем опыление перекрестное. По мнению М.
Г. Агаева (1978:245), автогамия в мире однолетников отнюдь не снижает эволю-
ционной лабильности их популяций. Наоборот, сохраняя возможность гибриди-
зации, рекомбинации, а также все выгоды семенного способа воспроизведения,
она «кладет конец «рабской» зависимости популяционно-видовых систем от из-
менчивых агентов опыления, тем самым сильно повышает их биотический по-
тенциал и при этом сокращает плату за размножение».
Ittifhi >l<iriiiccii()co(>iii>ir icMciHi п пч poui. п попуняцпях paciriinH 169
Говоря о высокой iipiiciioco(niciiiiocrii некоторых ccivi;нн.пых крести
ветных (пастушьей сумки Capsella bursa-pastoris, женiушника нсвкои
пого Erysimum cheiranthoides, ярутки полевой Thlaspi arvense,речушки
Arabidopsis thaliana, жерушника болотного Rorippa palustris и др.) мы де-
лали упор на их системе размножения, связанной с почти облигатным са-
моопылением (Марков, 1983, 1991в). Действительно, эта важная особен-
ность стратегий названных видов, находящая закономерное отражение в
генетической структуре их популяций и, в первую очередь, популяций се-
гетальных, обеспечивает страховку от недостатка опылителей, проникаю-
щих в средний и нижний ярусы агрофитоценозов. Подобные ограниченные
и закрытые рекомбинационные системы позволяют успешно воспроизво-
дить адаптивный(ые) генотип(ы), быстро увеличивая численность в пре-
делах освоенного открытого местообитания, и свести к минимуму «экспе-
риментирование» с новыми типами рекомбинантов, могущими понизить
успех размножения (Грант, 1984). Однако, примечательно, что устойчивым
ложносреднеэдификаторным ассектатором (Туганаев, 1984) является дру-
гое крестоцветное — редька дикая (Raphanus raphanistruni), которая уна-
следовала очень характерное для всей трибы Brassiceae облигатное (!) пере-
крестное опыление.
Очевидно, что один из наших объектов — редька дикая сочетает в себе
черты «капиталиста» (как облигатный перекрестник с крупными цветками)
и «пролетария» (незимующий моновариантный однолетник, терофит). О
помощью простой математической модели Д. Ллойду (Lloyd, 1980) удалось
показать, что флуктуации семенной продуктивности в меньшей степени
снижают адаптивность многолетних, чем однолетних растений. При этом
влияние первичного или задержанного самоопыления на процесс подъе-
Общая биомасса — верхние лимиты классов (мг)
Рис. 5.44. Участие особей разной мощности в воспроизводстве популяции рот
главпика серповидного на фоне кривой распределения но классам общей биомассы
170 Л/// Миркин I loiiyjiHHiioiiiiiiH OiioJioi пн piiciciiiii!
ма флуктуирующей семенной продукции будет менее важно для многолет-
них, нежели для однолетних растений. Следовательно, перекрестно опыля-
ющийся однолетник должен в своих приспособительных свойствах чем-то
компенсировать риск снижения плодовитости особи. У редьки дикой, судя
по всему, эта компенсация связана с увеличением размера и веса единично-
го зачатка (6,79 ± 0,95) и, отсюда, возрастанием толерантности и выживае-
мости проростков.
Р. Накамура (Nakamura, 1988) вслед за Д. Харпером (Harper, 1977) и
О. Солбригом (Solbrig, 1980) призывает рассматривать демографию как
описание динамики популяций от зиготы до зиготы. В этом случае зрелые
семена выступают как популяция зародышей, которые у перекрестников в
отличие от самоопылителей в большей степени генетически индивидуаль-
ны. По О. Солбригу (Solbrig, 1980) при генетической идентичности семян
и материнского растения продуцируется ли много семян с низкой адаптив-
ностью (мелких — М.М.) или мало семян с высокой адаптивностью (круп-
ных — М.М.) — для популяции генетически безразлично. Но когда семя
имеет другой генотип, нежели материнское растение, то конкуренция зи-
гот будет благоприятствовать семени, извлекшему больше ресурсов из ма-
теринской особи.
Каждое жизнеспособное семя с момента его отделения (обособления)
от материнского растения13 действительно есть особь, то есть тот самый,
по образному выражению Э. Майра (1974), «недолговечный сосуд», в ко-
тором популяция хранит часть своего генофонда. Впрочем, не стоит осо-
бенно подчеркивать и недолговечность. Особь даже у очень многих одно-
летних растений проводит большую часть своей жизни именно на стадии
жизнеспособного семени в составе почвенного семенного банка популяции.
Пусть она на этой стадии предельно пассивна или, точнее, малоактивна в
аспекте взаимодействия с внешней средой, поскольку так легче переносить
неблагоприятность экологических условий. Но при этом она действительно
не только хранит часть генофонда, но еще и несет в себе составную приспо-
собленность, которая складывается из приспособленностей четырех разных
генотипов: сформировавшей ее материнской особи (покровы семени — 2п),
женского гаметофита — п, эндосперма — 2п + п’ и зародыша — п + n’ (Haig,
Westoby, 1988).
Теоретически соседствующие в составе семени четыре генетически раз-
личных элемента: покровы (семенная кожура), гаметофит, эндосперм и за-
родыш наглядно представлены на рисунке 5.42. Поскольку от генетической
природы зародыша и эндосперма, выполняющего посредническую функ-
цию (Смирнов, 1982), зависит интенсивность извлечения питательных ве-
ществ из ткани материнского растения (спорофита), то мы вправе ожидать
и большей вариабельности массы единичных зачатков у перекрестников во-
обще и у редьки дикой в частности. Выше уже была показана явно повы-
шенная вариабельность семян редьки дикой по массе, компенсационные яв-
ления, имеющие место в ее плодах, а ниже будет продемонстрировано зна-
13 Строго говоря, особью в известном смысле можно считать уже зиготу и формирующийся
из нее зародыш, тем более, что в многосемянных плодах зиготы и зародыши уже конкуриру-
ют между собой за ресурсы, которыми снабжает их материнское растение, и в этом процессе
проявляют активность, от которой зависят их размеры к моменту отделения.
I hithi У Ж11 iiiccihh obiii.ir c tMriiii и их рощ» и понупицпих рисiciiiHi 1/ j
чепце огих явлений для роста и развития особей, вырасниощих из семян
разной массы.
На базе ограниченного набора генотипов в популяциях самоопыляющих
ся малолетних растений в очень яркой форме выступает фенотипическая
пластичность. Исходя из положительной асимметричности распределения
по массе особей в популяциях, некоторые исследователи (Solbrig, 1981) по-
лагают, что в общий урожай семян популяции немногочисленные крупные
особи вносят непропорционально высокий вклад и что это может имен»
определенные генетические последствия. Не ставя под сомнение выдающу-
юся роль крупных экземпляров, имеющих, кстати, больше шансов опыли гь-
ся перекрестно, в формировании общего урожая семян популяции, нужно
отметить, что преобладающие в популяции мелкие особи иногда в зависи-
мости от условий могут вносить более высокий суммарный вклад в воспро-
изводство популяции. На рисунке 5.43. видно, как в относительно благопри-
ятных условиях крупные, а в неблагоприятных — мелкие особи клоповни-
ка Lepidium densiflorum формируют наиболее весомый суммарный вклад в
общий урожай семян популяции. Такие же данные о большем суммарном
вкладе класса самых мелких в популяции особей в семенную продукцию
приведены на рисунке 5.44 и другого малолетнего вида.
Таким образом, вызванная внешними факторами фенотипическая пла-
стичность, эволюционное значение которой для нас все еще далеко не ясно
(вспомним факты наследования у льна Ыпит, о которых уже говорилось,
см. также Sultan, 1987), по-видимому, может существенно перекрывать ге-
нетические эффекты и обеспечивать выживание малолетних растений даже
при низком уровне генетической изменчивости.
Глава 6. РАЗМЕРЫ И
ГОРИЗОНТАЛЬНАЯ СТРУКТУРА
ПОПУЛЯЦИЙ (ПРОСТРАНСТВЕННЫЙ
АСПЕКТ ПОПУЛЯЦИОННОЙ
БИОЛОГИИ РАСТЕНИЙ)
Plants stand still and wait to be counted.
J. L. Harper1
Уже в самом общем определении популяции присутствует территориаль-
ный и, значит, пространственный аспект. От размеров и размещения попу-
ляции, ее компонентов в пространстве или, иначе говоря, ее пространствен-
ной горизонтальной структуры зависят очень многие важные стороны по-
пуляционной жизни. Каковы могут быть количественные подходы к опреде-
лению размеров популяций? В англоязычной популяционно-экологической
литературе слово «size» (размер) может употребляться и по отношению к чис-
ленности (например, effective population size — эффективная численность по-
пуляции, см. главу 2), и по отношению к объему (например, gap size — объем
гэпа. «окна» в растительном пологе), и по отношению к массе (например,
seed size — под этим, как правило, подразумевается масса семени).
6.1. Размер популяции
Говоря о размерах популяции, отечественный исследователь обычно име-
ет в виду либо размер территории, на которой обитают (произрастают) осо-
би данной популяции, либо общую численность популяции.
6.1,1. Размер территории, которую занимает популяция; вид как
система популяционных единиц; элементарная демографическая
единица
Наиболее впечатляющие примеры популяций, занимающих огромные
территории, сопоставимые с территориями ботанико-географических зон,
конечно же, дают древесные растения, такие как ель (Picea), лиственница
(Larix) и др., хотя космополитные ареалы неизменных спутников человека
1 Растения стоят тихо и ждут, пока их пересчитают. Дж. Л. Харпер
I hidd PiHMcpi.i и lopit loiiuiiii.miH cipykrypii iioiiyiiHunii 1/3
пастушьей сумки (\tpsella biirsa-pastoi is и подорожника (мни.того !4antayo
major., безусловно, южс весьма интересны в указанном oi ношении.
Выше, во введении, уже обсуждалось двойственное положение нонуля
ции в иерархических рядах биологических систем.
Вопрос о виде как системе популяционных единиц и подходах к выделе-
нию популяций наиболее обстоятельно излагает Л. Б. Зауголыюва (1994).
Из двух подходов, наметившихся к настоящему времени и названных ею
фенетико-генетическим и ландшафтно-территориальным, она как бота-
ник и фитоценолог отдает предпочтение второму. Из-за сложности выявле-
ния барьеров для скрещивания и, вместе с ними, границ локальной, мсст-
ной или панмиктической популяции, и нисколько не умаляя важности ее
исследования фенетико-генетическими методами, в ботанике принято вы-
делять в качестве основного элементарного популяционного объекта фито-
ценотическую популяцию. Однако, как совершенно справедливо указывает
Л. Б. Заугольнова, для подавляющего большинства видов растений прихо-
дится признать и учитывать существование ряда соподчиненных популя-
ционных единиц более высокого уровня: экотопическая популяция - ка-
тенная (или катенеарная) популяция - популяция речного бассейна - ре-
гиональная популяция. Для иллюстрации предложена система популяци-
онных единиц у травянистого растения — ковыля перистого Stipa pennata.
(рис. 6.1). Пояснения, которыми автор сопровождает представленную схе-
му, убеждают в том, что каждая из обозначенных ею единиц может быть
вполне конкретно охарактеризована, что они действительно реальны и нс
носят отвлеченного характера.
Изучение пространственной структуры и динамики популяций ряда
степных травянистых растений, а также некоторых древесных видов с раз-
ными типами стратегий жизненного цикла побудило Л. Б. Заугольнову
(Заугольнова и др., 1992) выделить еще одну структурную единицу попу-
ляционного уровня. Эту единицу, наиболее важным демографическим при-
знаком которой является обеспечение непрерывного оборота поколений,
предложено называть элементарной демографической единицей (ЭДЕ). Для
Рис. 6.1. Система соподчиненных популяционных единиц на примере ковыля нерп
стого Stipa pennata. ЦП — ценотические популяции, ЭП — экотопические нонуля
ции, ПБ — популяции бассейна, РП — региональные популяции (по Зауголыювой
1994).
174 Л / /< I к)||уи>|Ц||()||||11>| (hiojioi пи pnciciinii
наращивания пробных площадок до такого размера, при котором большая
их часть (с вероятностью 0,8-1,0) будет содержать практически все онтоге-
нетические группы особей, необходимые для оборота поколений. За мини-
мальный размер, с которого необходимо начинать наращивание, взята пло-
щадь репродуктивной активности особи данного вида. Отсюда понятно, что
размеры ЭДЕ видоспецифичны, и порядок величин связан с размером рас-
тений и дальностью разноса семян, а более тонкие различия — с экологиче-
скими свойствами видов (табл. 6.1).
Таблица 6.1.
Размеры ЭДЕ и продолжительность оборота поколений в популяциях ряда видов
лесных растений (по Заугольновой и др., 1992 с изменениями)
Вид Продолжительность обо- рота поколений Размеры площади, занимаемой ЭДЕ, м2
Alliaria petiolata 2 1 Х1О°
Corydalis solida 10 0,25 х Ю°
Juniperus sibirica 30 5 х Ю1
Picea excelsa 300 1 х ю3
Carpinus betulus 100 12 х Ю3
Fraxinus excelsior 200 18 х Ю3
Quercus robur 500 20 х Ю4
Устойчивое существование ЭДЕ возможно при условии непрерывности
оборота поколений и близкой к нулевой скорости роста численности осо-
бей, а устойчивость популяций от соотношения ее размеров с размерами
ЭДЕ. Чем меньше размеры ЭДЕ по сравнению с популяцией в целом, тем
меньше опасность для популяции утратить стабильность, но если размер
ЭДЕ больше, чем размер конкретной популяции, то есть основания состоя-
ние последней считать критическим.
Другой вариант наглядного представления данных об иерархии популя-
ционных единиц у травянистого многолетника ломоноса Clematis fremontii
var. riehlii (no Erickson, 1945) можно видеть на рис. 6.2. Очевидно, что боль-
шинство видов растений и особенно редких имеют подобное пятнистое или
мозаичное распространение. Отдельные пятна в этой мозаике могут быть
связаны через посредство мигрирующих диаспор. Зарубежные исследова-
тели предпочитают говорить о метапопуляциях или популяциях популяций
(Levins, 1970) как о системе популяций, связанных существенными потока-
ми мигрантов — мигрирующих особей. У растений мигрируют, как прави-
ло, семена в ходе их естественной первичной и вторичной дисперсии (см.
главу 5).
I iiiiiiii f> Pn [меры и i ори к>n iальная сц>ук lypa 1101 lyuiinnii
l/'i
6.1.2. Общая численность
популяции. Типы популяций по
общей численности
О выделении типов популяций по
общей численности не раз упоми-
налось в ботанической научной ли-
тературе (Seybold, 1980; Urbanska,
Landolt, 1990).
3. Зибольд привел результаты ста-
тистической обработки данных по
4451 местообитанию и, подчеркнув
объективные сложности выявления
популяционных границ, предложил
выделять 5 классов популяций по их
размеру (die GroBe). Число особей в
классах составляло: I — 1; II — 2-5;
III — 6-20; IV — 21-100; V — более
100. В материале 3. Зибольда наи-
большее число популяций попало в
III класс, хотя привлечение данных
по широко распространенным ви-
дам тут же давало второй максимум
в V классе.
К. Урбанска и Е. Ландольт пред-
ложили различать тоже 5 классов,
но с несколько иными градациями.
Каждый класс имеет краткую сло-
Рис. 6.2. Иерархия популяционных ед и
ниц у Clematis fremontii var. riehlii (по
Erickson, 1945) a — географический аре-
ал (площадь 1129 км2); b — фрагмет
ареала; с — группа полян с выходом из-
вестняков; d — поляна с общими кончу-
рами локуса; е — популяционный локус
(скопление отдельных особей).
вескую характеристику:
1 — очень маленькие популяции примерно из 15 особей; 2 — маленькие-
популяции из не более, чем 40 особей; 3 — популяции средней величины,
имеющие в составе от 40 до 70 особей; 4 — крупные популяции из 70- 100
особей; 5 — очень большие популяции из неопределенно большого числа
особей, которых всегда больше 100.
Нельзя не отметить расхождение взглядов у авторов двух приведен
ных шкал общей численности популяций по поводу классов популяции
малой численности и, вместе с этим, субъективность в выборе градаций.
Совершенно очевидно, что в суждении о том, какую популяцию называл,
маленькой, а какую — большой, нужны определенные биологические осно-
вания. Тогда одна и та же общая численность популяции будет иметь для
разных видов существенно разное содержание. Таких биологических осно
ваний можно назвать немало, и среди них будут фигурировать и размеры
взрослых особей, и дальность переноса жизнеспособной пыльцы или ди-
аспор и т. д. Не обойтись и без учета естественной плотности популяции.
176 Л/// Марков I loiiyiiMiuioiiiniH 61101101 пи piicieuid!
6.2. Плотность и способы ее определения
Плотность, понимаемая как среднее для данной популяции число осо-
бей на единицу площади, в специальных случаях имеет ряд производных:
например, проективное покрытие, обилие и сомкнутость в фитоценоло-
гии, полнота древостоя и сумма площадей сечения стволов в лесоводстве.
Оценивают плотность обычно выборочными методами.
Несмотря на то, что, по сути, общепринятым методом определения плот-
ности является подсчет особей, побегов, куртин и иных счетных единиц на
единице площади, любому исследователю популяций понятно, что плот-
ность — не простой фактор. Это можно легко уяснить хотя бы из того при-
мера, что даже при одинаковом числе особей, произрастающих на площад-
ке в 1 м2, многое во взаимодействии особей будет зависеть от конкретного
рисунка размещения этих особей, от размера и даже конфигурации площа-
ди, приходящейся на долю каждой особи популяции, от специфической ре-
акции вида на загущение — усиление напряженности внутривидового вза-
имодействия, конкуренции (см. главу 7) и прочих обстоятельств.
6.2.1. Случайное, регулярное и пятнистое размещение особей, семян
в почве или иных счетных единиц на территории популяции или
учетной площадке
Выявление характера (рисунка) размещения особей, который может быть
регулярным или равномерным, случайным и контагиозным или пятни-
стым, обычно сводится к построению графика распределения элементар-
ных площадок по классам числа особей для последующего сопоставления
его частот с частотами теоретического распределения. В качестве послед-
него обыкновенно используют распределение Пуассона, для которого ха-
рактерной и важной в данном контексте особенностью выступает равенство
средней арифметической и вариансы (квадрата стандартного отклонения).
Установленное методом %2 соответствие эмпирического распределения тео-
ретическому распределению свидетельствует о случайном размещении осо-
бей или других счётных единиц по площади популяции. Отношение дис-
персии к средней арифметической при этом равно единице. Когда это от-
ношение принимает значения меньше единицы, мы констатируем возрас-
тание регулярности в размещении особей, а если оно превышает единицу,
то налицо пятнистое или контагиозное размещение. Для растений как орга-
низмов, ведущих прикрепленный образ жизни, контагиозность размещения
особей — это вполне нормальное явление, если вспомнить особенности их
дисперсии и вегетативного разрастания и размножения.
Поэтому в пределах любой природной популяции можно наблюдать раз-
ные по плотности скопления — сгущения и разрежения особей, выявлять
популяционные локусы с различной плотностью и учитывать это при разме-
щении пробных площадок для изучения внутрипопуляционных процессов.
Когда для иллюстрации типа размещения особей в работах приводятся
планы или карты, где обозначено расположение каждой особи в пределах
пробных площадей, можно применить более гонкий и формальный ено-
I 6 PiiiMcpm и lopii loiiiiiiii.iiihi cipyurypn иопуняцпп 177
соб анализа, предложенный для пой цели В. В. Плотниковым (196S; 1979)
г Пот способ оценки it сравнения характера размещения особей на пробных
площадях, основанный на статистическом анализе варьирования плотно-
сти и предназначенный для камеральной, машинной обработки материалов,
сам по себе не нов. Новым является лишь предложение учитывать числен-
ность особей по плану участка на площадках, перекрывающихся между со-
бой (рис. 6.3.). Операция заключается в сканировании плотности, параме-
тры которого задаются формулой:
п = [(А- а)/Р1 + 1 ] х [(в - Ь)/Р2 + 1], [6.11
где п — число наблюдений плотности на учетных площадках, А — шири-
на пробной площади; В — ее длина; а — ширина учетной площадки; b ее
длина; — расстояние (шаг) между центрами смежных учетных площа-
док по ширине пробы; Р2 — шаг по длине пробы. Если пробная площадь и
учетные площадки квадратные, а шаг постоянен, то формула сканирования
упростится до вида:
п = [(А-а)!Р + I]2 [6.2]
Тип размещения особей при этом
оценивается по характеру распреде-
ления частот плотностей (см. рис.
6.3). При регулярном размещении
особей плотность может иметь не-
сколько определенных значений, ча-
стоты которых зависят только от си-
стемы учета и могут быть предска-
заны. При случайном размещении
распределение частот плотностей
подчиняется законам нормального
распределения; при групповом раз-
мещении особей кривая распределе-
ния частот плотностей будет много-
вершинной. Разумеется, на практике
можно встретить множество пере-
ходных состояний между тремя пе-
речисленными типами размещения,
но в любом случае даже ничтожные
различия в размещении можно уло-
вить таким способом весьма тонко
и выразить в количественной фор-
ме, если распределение частот будет
приведено к относительным показа-
телям.
Размеры учетных площадок жела-
тельно выдерживать в пределах пло-
Рис. 6.3. Принцип определения типа раз-
мещения особей на пробных площадях
методом сканирования плотности по В.
В. Плотникову (1979):
I — фрагменты карт пробных площадей
и ход сканирования; II — типы распре
делений частот плотностей при регул яр
ном (1), случайном (2) и контагиозном
(3) размещении особей. По оси X чис
ло особей па учетной площадке (iijioi
iioci к); по оси Y частота.
178 M H. I 1<)||уИЯЦ11О1111ПЯ GltOJIOl IfH pilCICItllh
щади, приходящейся в среднем на одну особь в данном элементе сообщества.
По мнению самого автора метода в целом этот способ весьма трудоемок и мо-
жет быть рекомендован лишь для особых случаев, когда камеральной обра-
ботке подвергается массовый фактический материал картирования.
6.2.2. Плотность как фактор существования особи
При определении плотности популяции в пересчете на какую-то площадь
мы вынуждены усреднять характеристику плотности. Немаловажно, и то, что
усреднение приходится проводить не единожды, поскольку, оценивая выборку
элементарных площадок, мы уже получаем среднюю для выборки, хотя, строго
говоря, для каждой особи плотность даже в пределах элементарной площадки
будет какая-то индивидуальная, своя. Ведь в силу нерегулярности размещения
особей будут различаться и их индивидуальные площади питания.
Поскольку эти методические вопросы наиболее интенсивно разрабатыва-
лись в лесоведении, рассмотрим их в приложении к популяции какого-либо
древесного растения, взятого в качестве примера.
6.2.3. Полигон площади питания
О точной конфигурации площади питания, имеющейся в распоряжении
одного дерева, можно только догадываться. В лесоведении говорят о «по-
лигоне местообитания» (за рубежом их называют «полигонами Тиссена»)
каждого конкретного дерева в популяции и сообществе. Считается, что
только 5-6 ближайших соседей оказывают влияние на величину и конфигу-
рацию такого «полигона местообитания».
Построение самих полигонов для каждого дерева на плане участка — это
чрезвычайно кропотливая операция. Суть ее состоит в том, что от каждо-
Рис. 6.4. Принцип определения «полигона местообитания» (или полигона Тиссена)
особи с учетом пяти соседних растений. Кружки показывают размещение деревьев и
диаметры их стволов. Формула показывает, в какой пропорции делят отрезки, соеди-
няющие центры соседних стволов (Плотников, 1979).
I'KItHI (i PlItMCpi.l Il lopit IOII Г11ЛЫ1ПЯ cipyxiypii популяций
179
Рис. 6.5. А — карта первоначального размещения проростков в модельной популя-
ции бородавника обыкновенного Lapsana communis L. с их полигонами; Б кари»
размещения взрослых особей (выделенных на первой карте кружками), доживших до
репродуктивного состояния, с их полигонами; В — гистограммы распределения ио
лигонов (вверху — проростков; внизу — генеративных особей) по классам их пло-
щади (мм2); G значения коэффициента Джини, показывающие степень варьпрова
пия площади полигонов.
180 МН Марков I hHiyihimioiiiinH (Hioiioi им piu I viniti
го дерева ио направлению к его 5 6 ближайшим соседям проводили лучи.
Каждый луч затем делится перпендикулярной ему линией либо на два рав-
ных по длине отрезка или на два отрезка, пропорциональных по длине ди-
аметрам соединяемых лучом деревьев. Чем больше диаметр дерева, тем
больший отрезок луча считается принадлежащим к его полигону. Далее во-
круг каждого дерева по концам тяготеющих к нему отрезков лучей строится
в виде ломаной линии конкретный полигон (рис. 6.4).
Материал, получаемый исследователем после построения полигонов, мо-
жет быть обработан разными путями.
Во-первых, представляет интерес сама изменчивость размеров поли-
гонов или площадей питания и ее характер. В модельных популяциях, по
крайней мере, на самом первом этапе можно создавать разные схемы раз-
мещения особей. Во-вторых, варьирование размеров полигонов хорошо со-
гласуется с особенностями размерной дифференциации особей внутри по-
пуляции. Даже естественная динамика численности и размерной дифферен-
циации особей находит свое отражение не только в изменении размеров по-
лигонов (результат изреживания), но и в изменении характера варьирования
этих размеров (Mithen et al., 1984). Бывшее поначалу положительно асимме-
тричным распределение по классам размеров полигонов в модельной попу-
ляции бородавника обыкновенного Lapsana communis к концу сезона сме-
нялось на распределение близкое к нормальному при заметном возрастании
общей амплитуды изменчивости (рис. 6.5). Однако, несмотря на то, что ва-
рьированием размера полигонов объяснялось 60% варьирования биомассы
особей, все же взаимосвязь между площадью полигонов и размерами осо-
бей оказались сложнее, чем это можно было предположить априори.
Конечно, очень заманчиво было бы досконально исследовать связь меж-
ду площадью питания или «полигоном местообитания» каждой конкретной
особи и ее морфометрическими характеристиками. На первый взгляд, такой
подход имеет ряд преимуществ перед традиционным «усредняющим» ме-
тодом изучения эффекта плотности (см. главу 8), но рассматриваемый ме-
тод полигонов при всей его трудоемкости, наверное, не поможет нам ре-
шить все проблемы.
Поскольку размеры полигонов, не говоря уж об их конфигурации, отли-
чаются большой и непрерывной изменчивостью, вновь возникает необходи-
мость особи с близкими характеристиками полигонов по каким-то принци-
пам объединять в классы, то есть опять-таки усреднять.
6.2.4. Плотность в точке
Отталкиваясь от материалов картирования особей древесных растений
в популяциях, В. В. Плотников (1979) предлагает несколько видоизменить
содержание традиционного понятия «плотность» (Плотников, 1973, 1974).
Зная густоту древостоя на пробной площади, или плотность в об-
щепринятом смысле, можно рассчитать площадь, приходящуюся в среднем
на одно дерево.
Г’ккш A 1*и 1мсры и Hipu uni hiui.iiiui cipyuiypn imiiyiitiuiiii iai
1'л,Л11*прсд( iaini 11. площадь, приходящуюся в среднем па одно дерево, в
виде кру| а, ю можно рассчитать радиус лого круга но формуле.
R = ^(S/nN),
|6.3|
где S — площадь пробы; N— число деревьев на ней. Далее по карте проб-
ной площади можно подсчитать число деревьев, окружающее каждое из
них в пределах круга, с радиусом R. Это число и устанавливается нами для
каждого дерева как показатель плотности в точке или класс плотности.
У немецких лесоведов (Prodan, 1968, цит. по: Плотников, 1979) круговая
площадка площадью: S= 10000/и, где п
«пробным кругом». Число деревьев
в пробном круге соответствует поня-
тию «плотность в точке». Однако у
В. В. Плотникова это число ставится
в характеристику особи, и количество
наблюдений плотности соответству-
ет численности деревьев на пробной
площади, а немецкие исследователи
ставят эту величину в характеристику
площадки и производят наблюдения
плотности выборочными методами.
Метод В. В. Плотникова и его новое
понимание плотности более инфор-
мативны, поскольку показатель плот-
ности привносится в характеристику
каждого дерева на пробной площади,
а не дается обезличенно для пробы
число деревьев на 1 га, называется
Рис. 6.6.А. Фрагмент карты природной
популяции с приведением формулы для
вычисления радиуса пробной площади
и показом методики определения плот-
ности в точке (от 1 до 5 особей/пробная
площадь) для каждой из особей.
в целом или отдельных, сколь угод-
но мелких ее частей. В смешанных
древостоях показатель плотности для
особи может быть сопровожден не-
которым «весом», в зависимости от
видового или возрастного состава
окружения этой особи.
Принцип определения класса
плотности для отдельных особей в
природной популяции показан на
рис. 6.6А. Показатель плотности
при этом выражается в целых чис-
лах натурального ряда, то есть его
изменчивость дискретна, что выгод-
но отличает его от таких признаков,
как площадь питания или среднее
расстояние до ближайших растений.
Естественно, что для дальнейшей,
конкретной обработки особи с оди-
Рис. 6.6.Б. Определение плотности в
точке для особей, посаженных в усло-
виях опытного участка с соблюдением
единого принципа: соседние 6 особей
были удалены от центральной в груп-
пе на равное расстояние и потому <|)ор
мировали правильный шестиугольник.
Показанные варианты плотности от 7 до
1 возникли в результате выпадения ча
стн особей в ходе роста.
182 Л/// MuphOtt I loiiyJIMIUIOIIIIIIH (HIOJIOI ЛЯ pilCICIIIlh
паковым показа телем плотности группируются в классы плотности и вот
здесь проявляются некоторые «узкие места» метода. Например, сам автор
указывает на то, что природа деревьев 1-го класса плотности в естествен-
ном древостое сложна. Если в среднем на одно дерево здесь приходилась
площадь S/N, а в популяции обычно имеются деревья 2, 3 и 4-го классов
плотности (деревья, на которые соответственно приходились площади S/2N,
S/3N, S/4N), то очевидно, что среди деревьев 1-го класса плотности будут
деревья, на которые приходились площади 2S/N, 3S/N и 4S/N. Иными сло-
вами, деревья 1-го класса плотности по своему фитоценотическому и вну-
трипопуляционному положению могут быть чрезвычайно гетерогенны, что
весьма сложно учесть при популяционном анализе полученных данных.
В модельных популяциях, например в полевых и лесных культурах, где
первоначальное размещение особей приближено к равномерному или ре-
гулярному, содержание показателя плотности в точке несколько видоизме-
няется (Плотников, 1979). Плотность и размещение особей в культурах за-
даются технологической схемой посадки или посева, с учетом биологиче-
ских свойств культивируемых растений. Большинство схем предусматри-
вает создание равных условий для каждого из посаженных или посеянных
растений.
Как и в случае природных популяций, показатель плотности в модельных
популяциях желательно отразить в характеристике каждой особи. При этом
следует учитывать, что часть растений может не прижиться, и потому в каж-
дый конкретный момент наблюдения выявлять число сохранившихся бли-
жайших по схеме посадки соседних данной особи растений. На рис. 6.6Б. по-
казан принцип определения показателя плотности в точке для особи в куль-
турах при различных схемах посадки растений. Распределение численности
особей по классам плотности на том или ином фрагменте модельной популя-
ции будет отражать меру отклонения размещения растений от первоначаль-
ной схемы посева или посадки. Ведь при стопроцентном выживании расте-
ний все они, кроме краевых, должны находиться в одном расчетном классе.
Хотя традиционная методика и предписывает исключение из обработки
крайних, «пограничных» особей и, вместе с ними, так называемого краево-
го эффекта (edge effect — в англоязычной литературе), данная процедура
чревата потерей информации. Отсутствие в природе и сельскохозяйствен-
ной практике «бескрайних» популяций и искомое преимущество рассма-
триваемого метода, связанное с индивидуальным подходом в оценке эффек-
та плотности — два весомых аргумента в пользу учета морфометрии и кра-
евых особей.
Завершая данную главу, отметим, что в последующих главах нам еще
предстоит обращаться к рассмотренным здесь понятиям и методам. Это и
не удивительно, если принять во внимание важность плотности как попу-
ляционного параметра для растений — организмов, ведущих прикреплен-
ный образ жизни.
Глава 7. СТРУКТУРНЫЕ АСПЕКТЫ
РОСТА РАСТЕНИЙ
Рост — процесс новообразования элемен-
тов структуры организма.
Д.А. Сабинин,
Физиология развития растений, 196J
7.1. Архитектура растений
Своеобразие явления роста у высших растений, связанное с метамерно-
стью строения, первым из отечественных ботаников отметил известнейший
физиолог растений Д. А. Сабинин (1940), который считал, что биологиче-
ски обоснованной мерой скорости роста может быть лишь число элемен-
тарных актов роста (число сформированных метамеров, а точнее — фито-
меров), пройденных в единицу времени.
В последнее время, проводя анализ структуры растений, многие ботани-
ки предпочитают говорить об их архитектуре. Понятие «архитектура» тес-
нейшим образом связано с повторением более или менее неизменных (ин-
вариантных) элементов конструкции (т. н. модулей), которые служат сво-
еобразными единицами измерения и придают постройке соразмерность
(Словарь иностранных слов, 1964).
Заимствование термина «архитектура» ботаниками вероятнее всего
было следствием новой «послегетевской» волны интереса к метамерности
строения растений. Если слегка перефразировать определение архитектуры
растения (Devades, Beck, 1971), то можно заключить, что она создается бла-
годаря пространственному упорядочиванию и координации метамеров раз-
ного ранга (фитомеров, модулей и т. д.), как повторяющихся субъединиц,
изменяясь для особых функций и отражая в своих пропорциях влияние усло-
вий среды. Именно пространственное упорядочивание и координация по-
вторяющихся элементов конструкции составляет суть явления, и поэто-
му заимствование терминологии можно, пожалуй, и оправдать.
7.2. Архитектурные модели (или -модели побегообразования)
Если какое-либо здание возводится по плану архитектора, то тело рас ге-
ния создается по программе, заложенной в генотипе и потому реализуемой
в процессе роста автономно, независимо от экологических условий, сово
184 Л/ // Маркой I loiiyiiHHiioiiiiiiH (>1101101 ин рас iciiidi
купнои dcHincnbiiociiibio меристем'. Различные i ины uikiix программ бы ли
названы архитектурнымн моделями (I lallc, ()klcman, I(>7<); Halle ct al., 1978
по Жмылев и др., 2002). Описание той или иной архитектурной модели
предполагает известное абстрагирование от биологии и экологии объекта
и смещение акцента на макроморфологию как таковую, то есть сводится к
выявлению типа роста, посредством которого из более или менее однотип-
ных и одноразмерных субъединиц может создаваться определенная прост-
ранственная конструкция растения, иначе говоря, его архитектура.
В зависимости от конкретных целей исследования концепцию архитек-
турных моделей можно использовать для описания морфологии роста био-
логической либо таксономической группы растений.
Когда ставится задача изучить эколого-географические закономерности
распространения и возможные пути эволюционной трансформации архи-
тектурных моделей, исследователь обращается к анализу таксономической
группы. Например, Т. В. Шулькина (1988) провела такой анализ для семей-
ства Campanulaceae.
Если же исследователя интересуют архитектурные особенности как
средство достижения определенного фитоценотического статуса, то он мо-
жет обратиться к анализу биологической группы растений, в которую вхо-
дят представители различных таксонов, не обязательно связанные узами
близкого родства. Именно в работах такого рода названная концепция ар-
хитектурных моделей нашла себе применение. В начале, например, в этом
ключе было дано описание роста тропических деревьев (Halle, Oldeman,
Tomlinson, 1978), а чуть позже и травянистых многолетних, преимуще-
ственно вегетативно подвижных, растений (Серебрякова, 1977, 1981; Bell,
Tomlinson, 1980; Hutchings, Slade, 1987).
T. И. Серебрякова, отождествляя, в какой-то степени, архитектуру с мо-
нументальностью, предпочитала говорить о моделях побегообразования у
травянистых растений, поскольку к этому располагают гораздо более мел-
кие, чем у деревьев, размеры их вегетативного тела. Однако нельзя не заме-
тить, что тогда полностью выпадают из поля зрения корневые системы и их
архитектура.
Схематическое изображение основных архитектурных моделей много-
летних травянистых растений приведено на рис. 7.1. Корневые системы, как
видно на схеме, только чуть обозначены как не существенный для характе-
ристики моделей атрибут.
Исследование малолетних растений как особой и своеобразной биоло-
гической группы в этом аспекте по сути дела только начинается. И в то же
время можно привести, по крайней мере, четыре довода в пользу того, что-
бы при рассмотрении травянистых растений и в том числе малолетников
говорить именно об архитектуре, а не только о побегообразовании:
1) Применение термина «архитектура» обязывает учитывать мета-
мерность строения и рассматривать процессы роста и репродукции в тес-
ной связи с типом проявления метамерности;
1 Под независимостью в данном случае понимается способность поддерживать констант-
ность видовых морфологических признаков и в Том числе типа роста, меняя лишь пропор-
ции, число и (или) размеры повторяющихся элементов конструкции.
r>idti(i / ( ipyu iypiii.ii* tn nrkii.i j)ocia piiciciiiill
185
2) Выше (liiami -I) уже говори-
лось, что побеювое тело малолет-
них растений (как и у деревьев), по
образному замечанию Б. А. Юрцева
(1976), содержит полный «пласти-
ческий слепок онтогенеза», отра-
жающий в своей пространствен-
ной структуре временную ритми-
ческую последовательность обра-
зования метамеров. Иными слова-
ми, исключительно для малолетних
растений понятия «архитектурная
модель» и «тип морфологической
конструкции» или «архитектура»
оказываются очень близкими по со-
держанию.
3) Совершенно необходимый
учет особенностей строения (архи-
тектуры) корневых систем и спо-
соб их связи с побеговыми система-
ми (см. Главу 4) не позволяет огра-
ничиться моделями побегообразо-
вания Т. И. Серебряковой и, несмо-
тря на более чем скромные размеры
большинства малолетних растений,
заставляет все же говорить об архи-
Рис. 7.1. Схема, показывающая основ-
ные модели побегообразования (архи-
тектурные модели) многолетних трав:
I — «полурозеточная симподиальная»;
II — «длиннопобеговая симподиальная»;
III — «розеточная моноподиальная»;
IV — «длиннопобеговая моноподиаль-
ная».
Треугольниками обозначены соцветия,
стрелками — вегетативные точки роста,
крестиками — отмирающие и недораз-
вивающиеся точки роста, сплошной ли-
нией — живые, пунктиром — отмершие
части растения (Серебрякова, 1981).
тектуре их надземных и подземных органов;
4) Четвертый довод — это просто стремление ограничиться при изучении
в популяционном аспекте архитектуры травянистых растений, включая ма-
лолетники, уже имеющимися общепринятыми терминами.
7.3. Архитектурная модель и жизненная форма
В связи с новой волной отраженного в отечественных и зарубежных пу-
бликациях интереса к метамерности строения растений и появлением кон-
цепции архитектурной модели как процесса и итога реализации генотипа
в соответствующей морфологической конструкции особи вполне законо-
мерно возник вопрос о соотношении понятий «архитектурная модель « и
«жизненная форма». В целом наиболее логичной представляется точка зре-
ния Т. И. Серебряковой, согласно которой во втором понятии определяю-
щим должен быть адаптивный аспект толкования общего габитуса расте-
ний, и что на базе одной и той же архитектурной модели могут возникну! ь
разные жизненные формы (Серебрякова, 1981).
Последний тезис не выглядит столь бесспорным как первый главным об-
разом из-за большой запутанности к настоящему времени вопроса о жиз-
ненных формах (см. Алеев, 1986), критериях их выделения, классификации.
186 A/// MtipKtHi 11 (>ну и я i (in hi i hi и (hioiiqi ня piicicindl
Обосновывая необходимое н> оформления специальной науки о жиз-
ненных формах, которую он называет экоморфологией7К). Г. Алеев счи-
тает, что не следует менять исторически сложившееся в биологии поня-
тие об экоморфе. Во-первых, как о комплексе габитуалъно проявляющихся
свойств организма, и притом не только морфологических, а во-вторых, как
о комплексе, представляющем определенную фазу онтогенеза, который за-
кономерно сменяется другим комплексом с переходом к следующей фазе.
Однако вся беда в том, что границы комплекса отнюдь не ясны. Сколько
и какие свойства и черты подлежат учету, не ясно. А потому всегда есть
риск, что в погоне за охватом возможно большего набора признаков низ-
шим звеном классификации окажется популяция, если не особь! То есть
случится, как раз то, против чего очень категорично выступает Ю. Г. Алеев.
Критикуя ботаников за односторонность подхода и переоценку морфологи-
ческих критериев, он, как и многие другие зоологи, недооценивает откры-
тый характер роста растений и автономность субъединиц, слагающих тело
растения. А между тем именно эта черта высших растений не дает проя-
виться той степени дискретности организма, которая необходима по Ю. Г.
Алееву для описания экоморфы.
Т. И. Серебрякова пишет: «Представление о моделях как будто возвра-
щает нас к отнюдь не новым идеям, лежавшим в основе учений о «пла-
нах строения» и «типах». Однако на современном этапе в сочетании с нако-
плением совершенно новых данных об онтогенетическом развитии струк-
тур, об их взаимоотношениях друг с другом в многоступенчатой системе
соподчиненных единиц и с внешней средой, а также в связи с достижения-
ми и перспективами генетики эти идеи получают новое звучание. Именно
моделирование структур и процессов становится в биологии одним из са-
мых плодотворных путей познания и прогнозирования. Не случайно, по-
этому представители «типологической школы» В. Тролля, издавна расце-
нивавшейся как «идеалистическое» направление, сейчас усиленно развива-
ют его позитивные стороны, находя в нем инструмент для эволюционных
построений и филогенетических реконструкций» (Серебрякова, 1977: 113).
Знакомство с уже упомянутым обзором Д. Каплана (Kaplan, 2001) даст пре-
красное иллюстрированное подтверждение этим словам.
7.3.1. Место архитектурных моделей в алгоритме изучения
стратегий жизненного цикла
Несмотря на то, что исследование архитектуры и архитектурных моде-
лей, безусловно, само по себе представляет интерес и рассматривается мор-
фологами растений как вполне самостоятельная проблема, все же говорить
об архитектурных моделях «в чистом виде», не касаясь совершенно приспо-
собительного значения той или иной морфологической конструкции, до-
вольно сложно. Не случайно А. Бэлл и П. Томлинсон (Bell, Tomlinson, 1980),
применившие концепцию архитектурных моделей к вегетативно подвиж-
ным травам, предпочитают говорить об адаптивной архитектуре. Вместе с
2 Исключение из термина греческого корня био- (bios — жизнь) вряд ли целесообразно.
I 'lllltui / ( 1румурныг Ш ПГ1О1.1 рос III piuicinih 187
тем емкое hoiimiiic «стратегия жити» пии «нсоцого цснотичсская етра
тегия», подразумевает совокупное и» свойств, среди которых <|)iнурпруеi
жизненная форма (архитектурная модель и архитектура с известной долен
их адаптивного значения), обусловливающая экологически и фитоценогн
чески целесообразный, способствующий выживанию популяции, тин по-
ведения в соответствующих эколого-ценотических условиях. А ведь имен-
но рост и представляет собой одно из главных слагаемых поведения у рас-
тений. Таким образом, через алгоритм, три звена которого в порядке воз-
растания информативности можно расположить следующим образом: архи
тектурная модель (и архитектура per se) - жизненная форма - стратегия
жизни, мыслится связь концепции архитектурных моделей с популяцион-
ной экологией и биологией растений (см. рис. 1.1).
7.4. Основные аспекты в изучении архитектуры растений
Анализ архитектуры травянистых растений может иметь своей целью
выяснение: характера размещения ассимиляционной поверхности вдоль но
главному побегу при оценке структуры главного побега (модуля); направле-
ния разрастания особи по мере увеличения ее мощности; «запечатленного»
в архитектуре (характере размещения на растении репродуктивных струк-
тур) способа переключения побегов с чисто вегетативного роста на репро-
дукцию. Третья задача особенно важна при работе с малолетними растени-
ями.
Рассмотрим эти аспекты на примере малолетних растений, причем сде-
лаем это на двух уровнях: межвидовом и внутривидовом. Так можно проде-
монстрировать, с одной стороны, архитектурное разнообразие, вытекающее
из таксономической принадлежности сравниваемых малолетних растений,
а с другой — разнообразие архитектурных «решений» у особей, принад-
лежащих к одному виду, но различающихся по жизненному состоянию и за-
нимающих разное положение во внутривидовой и внутрипопуляционной
размерной иерархии. Следует заметить, только, что если в первом случае
различия носят качественный характер, то во втором случае мы будем на-
блюдать широкий, созвучный чрезвычайной пластичности растений, спектр
вариантов, различающихся, как правило, количественно.
7.4.1. Размещение ассимиляционной поверхности вдоль по главному
модулю (структура модуля)
Некоторые исследователи, имея в виду главным образом размещение ор-
ганов ассимиляции, предпочитают говорить о «геометрии» растений и де-
монстрируют ее влияние на эффективность создания биомассы в условиях
конкуренции (Lonsdale, Watkinson, 1983) или на световое довольствие рас-
тений под пологом леса (Menges, 1987). Для примера рассмотрим архитек-
туру ряда малолетних растений, у которых характер нарастания листовой
поверхности, от первых до последних фитомеров главного модуля, можно
трактовать как некую стратегию роста.
188
MH MupKiHi
I l<>1 lyJIMHIK>111111»! (HIOIIOI ИМ рис 1СИНЙ
S (Площадь листовой поверх»юс ти в % oi общей всего побсча)
Рис. 7.2. Кумуляты нарастания площади листовой поверхности (S) с ярусом при дви-
жении вверх по главному побегу, имеющему разную структуру: 1— с нижней розет-
кой (проломник северный — Androsace septentrionalis), 2 — без розетки (тимелея во-
робьиная — Thymelaea passerind) и 3 — с верхней розеткой (недотрога обыкновен-
ная — Impatiens noli-tangere).
а) межвидовой уровень. Архитектурные особенности розеточных ма-
лолетних растений в сравнении с растениями полурозеточными и длин-
нопобеговыми хорошо проявляются при анализе нарастания асси-
Высота узла на главном побеге
(% от общей)
Рис. 7.3. Кумуляты нарастания площади
листовой поверхности с ярусом при дви-
жении вверх по главному побегу у гера-
ни Роберта (Geranium robertianum).
миляционнои поверхности по мере
движения снизу вверх по стеблю.
На рисунке 7.2 приведены кумуля-
ты, полученные в результате отк-
ладывания суммируемой листовой
поверхности в процентах от об-
щей (образованной за сезон веге-
тации) против высоты расположе-
ния соответствующего узла в про-
центах от общей высоты растения.
Можно видеть, что розеточностъ
(на примере проломника северно-
го Androsace septentrionalis) — это
способ быстрого наращивания ас-
симиляционной поверхности как
бы за счет экономии на росте меж-
доузлий. График, наряду с этим,
показывает, как равномерно, по во-
гнутой кривой, идет «накопление»
листовой поверхности у длинно-
Гиа(<(1 / ( ipykiypiihir ncnckii.i pocia pncieiiiih
189
Рис. 7.4. Пример архитектуры однолег
них растений только с присоцветной (мо
лочай солнцегляд Euphorbia helioscopia)
и как с прикорневой, так и присоцветной
(проломник наибольший) розетками ли-
стьев.
набегового cieiiiioro незимующе!о
однолетника шмелей воробьиной
Thymelaea passerina и как сосредо-
точивается в верхней части стебля
основная ассимиляционная поверх-
ность у незимующего лесного од-
нолетника недотроги обыкновенной
Impatiens noli-tangere.
У лесного полурозеточного дву-
летника герани Роберта Geranium
robertianum генеративный побег, об-
разуемый на второй год жизни, оли-
ствен неравномерно — первое длин-
ное междоузлие выносит наверх
своеобразную «розетку» (несколько
сильно сближенных листьев) в усло-
вия лучшего освещения. На графи-
ке это отражено как резкое (в виде
ступеньки) повышение концентра-
ции листовой поверхности ближе
к верхней части стебля (рис. 7.3).
Аналогичную картину мы наблюда-
ем у степного озимого однолетника
проломника наибольшего Androsace
maxima (рис. 7.4). Безлистная стрел-
ка отделяет прикорневую розетку от
«розетки» прицветников (брактей),
приподнимая последнюю над по-
чвой. Логично предположить, что
такая «розетка» может снабжать цветки и плоды растения дотацией ассими-
лятов после засыхания главной розетки, подобно тому, как это делает верх-
ний, «флаговый» лист у злаков. У сегетального незимующего однолетни-
ка Euphorbia helioscopia на верхушке равномерно олиственного побега рас-
полагается розетка прицветных (точнее присоцветных) листьев (рис. 7.4),
что можно рассматривать как полезное свойство при недостатке освещения
под пологом культурного растения. Таким образом, как и у розеточного, и
у полурозеточного, и у длиннопобегового растений вследствие особого рит-
ма чередования брахиморфных и долихоморфных структур возникает сход-
ная архитектурная деталь, которая, вероятно, может служить решению ряда
эколого-ценотических проблем.
Среди граминоидов (и в том числе малолетних) тоже наблюдается раз
нообразие по обсуждаемому признаку. Кроме афилльных модулей (напри-
мер, у болотницы яйцевидной Eleocharis ovatd), здесь лишь изредка мож-
но встретить почти равномерное олиствение. Гораздо чаще можно ветре
тить некоторую концентрацию узлов (и листьев) в нижней, а иногда и верх-
ней нрисоцветной части модуля в дополнение к узлам (и листьям), отделен
ним нормально развитыми междоузлиями (например, у сычи бурой Cypcrus
190
Л/// Марков I loiiyiiMunoiiiinn bifoiioi ия piiciciniit
Рис. 7.5. Нормальная форма роста луж-
ницы водной Limosella aquatica (слева)
и полученная путем притенения длинно-
побеговая форма этого растения (спра-
ва).
juscus). Особенности строения ли-
стьев у злаков я, в частности, нали-
чие у листьев влагалищ разной дли-
ны, принимающих очень активное
участие в фотосинтетической дея-
тельности модулей, не позволяет на-
прямую связывать высоту располо-
жения узла с уровнем размещения
ассимиляционной поверхности ли-
стовых пластинок.
б) внутривидовой уровень.
Известны случаи изменения струк-
туры побега при неблагоприятных
условиях. Например, у пастушьей
сумки Capsella bursa-pastoris в загущенных популяциях или популяциях,
произрастающих при дефиците элементов минерального питания, особи
утрачивают розетку и по существу становятся длиннопобеговыми (безрозе-
точными). Подобная вынужденная длиннопобеговость может быть вызвана
в условиях опыта у розеточных растений типа лужницы водной Limosella
aquatica с целью изучения размещения на укороченном в норме побеге па-
зушных репродуктивных структур (рис. 7.5). Лишь относительно редко
даже под действием экстремальных условий растения не теряют розеточ-
ности. Так, деятельность плоского или вогнутого апекса у некоторых ви-
дов Plantago (например, подорожника большого Plantago major) не меня-
ется даже после его заглубления в почву — наблюдается лишь удлинение
черешков, но междоузлия не удлиняются никогда.
Поскольку в розеточной стадии розеточные и полурозеточные ма-
лолетники пребывают вплоть до перехода в генеративное состояние, то
все основные возрастные изменения у них приурочены именно к этой ста-
дии. Последовательно появляющие-
ся друг за другом розеточные сбли-
женные листья своей формой, ко-
нечными размерами, весом и анато-
мической структурой диагностиру-
ют наступление тех или иных воз-
растных этапов (Кренке, 1933-1935;
Сабинин, 1963). Течение возраст-
ных изменений в деятельности апек-
са наиболее ярко проявляется у ро-
зеточных и полурозеточных видов с
хорошо выраженной гетерофилли-
ей или гетеробластным развитием
(Goebel, 1928-1933, по: Серебряков,
1952). Длиннопобеговым малолет-
никам она свойственна не в такой
яркой форме. Однако даже в соста-
ве одного вида иногда можно най-
Рис. 7.6. «Конвергенция» четырех раз-
ных биотипов пастушьей сумки в на-
правлении очень сходного фенотипа в
условиях неблагоприятных для розе-
точного роста (биотипы слева направо:
Capsella b.-p. simplex, rhomboidea, tenuis,
het er is).
Гтм<1 / ( ipyn iypiti.iv ih iicmi.i рос mi pnciгннй
191
ти примеры как гетеро-, гак и гомо
бластного разниi им.
Рассмотрим )то на примере пасту-
шьей сумки, которая, являясь почти
облигатным самоопылителем, об-
разует популяции, распадающиеся
на 4 гомозиготных биотипа (Shull,
1909; Марков, 1976, 1978). Биотипы
можно уверенно различить лишь в
том случае, когда растения растут
в благоприятных дифференцирую-
щих условиях. При неблагоприят-
ных для розеточного роста услови-
ях из-за выпадения фазы взрослых
листьев из онтогенеза имеет место
«конвергенция» к очень сходному
фенотипу (см. рис. 7.6), и биотипы
практически неразличимы (Марков,
1976). Три биотипа {heteris, tenuis
и rhomboidea см. рис. 7.6 — спра-
ва налево) характеризуются гетеро-
бластным, а один {simplex — край-
ний слева) — гомобластным разви-
тием. Рисунок 7.7 показывает, что
Рис. 7.7. Изменение значений листовоп)
индекса (отношения периметра листа к
его площади — Р/S) с ярусом у 4 биоти-
пов пастушьей сумки (S, R, Т, Н — био-
типы).
онтогенетическая динамика индекса
расчлененности (отношения периметра листа к его площади) для розеточных
листовых серий разных биотипов только вначале однотипна. С переходом к
Рис. 7.8. Листовые серии главных побегов для особей разных биотипов пастушьей
сумки (а — Carsella b.-p. simplex', б — С. h.-p. tenuis}.
192 Л/// Марков I loiiyiuiniioiiiiiui (hioiioi ия рис icnnh
листьям взрослого типа главным индикаторам генетической природы осо-
би — ход кривых существенно различается. Любопытно отметить, что макси-
мальное значение индекса наблюдается у доминантно-гомозиготного биотипа
heteris (ААВВ), минимальное — у рецессивно-гомозиготного simplex (aabb),
а промежуточное (и притом примерно равное) — у двух доминантно-рецес-
сивных гомозигот tenuis и rhomboidea (AAbb и ааВВ). Распознавание биоти-
пов пастушьей сумки в природе возможно только в благоприятных диффе-
ренцирующих условиях, когда каждая особь может развить соответствую-
щую потенциальным возможностям ее генотипа серию фитомеров взрос-
лого типа. На рисунке 7.8 показаны листовые серии (вместе с пазушными
побегами) особей двух биотипов (simplex и tenuis), выращенных в услови-
ях оптимальных для роста. Хорошо заметна потенциальная скороспелость
биотипа simplex (а) — при более короткой листовой серии главного побега в
пазухах верхних розеточных листьев уже сформированы цветоносные побе-
ги. Более сильное развитие всех пазушных побегов у особи биотипа simplex
обеспечивает ей более высокую плодовитость (см. главу 3) по сравнению с
особью биотипа tenuis и говорит о большей роли листового аппарата этих
побегов в создании всей ассимиляционной поверхности особи. Таким обра-
зом, мы наблюдаем здесь два разных генетически обусловленных направле-
ния в период розеточного роста. Одно из них — увеличение числа фитоме-
ров на главной оси, сопряженное с замедлением генеративного развития, а
второе — на фоне ускоренного генеративного развития главной оси и сня-
тия апикального доминирования быстрое формирование пазушных побе-
гов паракладиев со «вторичнымирозетками» (Марков, 1978), существенно
ускоряющих образование семян и повышающих плодовитость особи.
Ухудшение условий освещения или обеспеченности влагой приводит к
сильному укорочению листовых серий за счет неотенического3 выпадения
фаз продуцирования листьев взрослого типа, т. е. за счет утери индикатор-
ных фенотипических признаков, выявляющих генотип. Это явление уже не
раз было отмечено в литературе у других видов крестоцветных с гетеро-
бластным развитием [завитка пустынного Streptoloma desertorum (Нечаева,
1958) Hymenolobus procumbens (Келлер, 1948)]. Поскольку при произраста-
нии в фитоценозах всегда имеют место некоторое притенение и конкуренция
за влагу, то выявить биотипическую принадлежность особей в естественных
популяциях пастушьей сумки бывает затруднительно. Даже осенние условия,
благоприятствующие розеточному росту, далеко не всегда обеспечивают осо-
бям в фитоценозе развитие индикаторных признаков. Но это не означает, что
в таких условиях вообще нет возможности для гетеробластного развития. В
агрофитоценозах озимых культур в период осеннего розеточного роста мож-
но выделить три типа хода возрастных изменений, два из которых представ-
ляют варианты гетеробластного развития. На рисунке 7.9 приведены соот-
ветствующие три кривые изменения длины розеточного листа в зависимости
от яруса с условным обозначением степени расчленения листовых пластинок
3 Неотенией называют переход в репродуктивное состояние особей, еще не утративших
ювенильного габитуса, по причине выпадения из онтогенеза каких-то промежуточных
(взрослых) этапов (и органов) вместе с характерными для них морфологическими
особенностями
/ hifiti / ( ipyiuypin.ic ih иск ii.i pocia paciciniil 193
(а) и три кривые, oi ражающис изменение индекса рассеченное th (oiiioine
ине nepiiMcipa лис la к его длине) с ярусом (б). Два вариант rciepoGuaciiio
го развития отличаются количеством слаборассеченных переходных листьев
иммагурного типа в листовой серии главной розетки и, отсюда, плавностью
перехода во взрослое вегетативное состояние. Заметно, также, что для вари-
анта с одним листом иммагурного типа характерен переход во взрослое со-
стояние на восходящей части кривой, а для варианта с серией листьев имма-
турного типа этот переход приурочен к нисходящей части кривой.
Длина листа, см
Рис. 7.9а. Основные варианты изменения
длины и формы листовых пластинок у ро-
зеточных листьев пастушьей сумки с яру-
сом: ЯЛ — ярус листа; 1 — цельные ли-
стья ювенильного типа; 2 — переходные
листья иммапурного типа со слабо разре-
занными (лопастными или раздельными)
пластинками; 3 - сильно рассеченные ли-
стья взрослою типа; 4 — листья па удли-
ненной части полурозеточного побега (ею
блевые); б. Кривые, отражающие изме-
нение индекса рассечен пости (отношение
ncpiiMei ра лис ia к ci о длине) с ярусом
194 MH Марков I lonyihiniioiiiiiiH (hioiioi пя pnciciiiiit
7.4.2. Фотосинтетическая эффективность листьев одного модуля
Оптимизация фотосинтетической функции на морфологическом уровне
достигается максимизацией поверхности перехвата физиологически актив-
ной радиации и минимизацией затрат строительного материала на образо-
вание этой поверхности (Корона, 1987). На межвидовом уровне сравнению
эффективности листьев может служить поверхностная масса (масса одно-
го квадратного сантиметра листовой пластинки), определенная для листьев
сходного яруса.
Если взять длину листа как линейный показатель его размера и, в извест-
ной степени, эффективности его функции, то можно сопоставить ее онто-
генетическое изменение в розетках с изменением массы листа, как меры
затрат строительного материала, для короткой и длинной листовых серий
у пастушьей сумки. Рисунок 7.10 показывает, как медленно возрастает от-
ношение масса листа/длина листа в ходе розеточной фазы онтогенеза, де-
монстрируя очень плавное и, в целом, незначительное снижение эффектив-
ности листьев. Одна из причин этого — увеличение затрат материала на
формирование жилок с их механическими (непроизводительными) элемен-
тами, общая длина которых закономерно {закон Заленского) возрастает от
нижних к верхним листьям розетки. Сходную картину можно наблюдать и
сравнивая листья разных ярусов у безрозеточного растения.
ДЛ., мм
100
50
10
ЯЛ
Рис. 7.10. Изменение отношения масса листа/длина листа с ярусом в двух розетках
пастушьей сумки с короткой и длинной листовыми сериями: Я.Л. — ярус листа сни-
зу вверх по побегу; М.Л. — масса листа; цифры на графиках — величины отношения
для конкретных листьев в сериях.
Гчавч 7 ('ipyhiypiii.ir nt nckii.i poem pnciciiiiH 195
7.5. Архитектурные варианты разрастания особи
Направления, по которым идет разрастание особей по мерс увеличения
их мощности, могут быть рассмотрены отдельно на межвидовом и внутри
видовом уровнях.
7.5.1. Межвидовой уровень
Характер олиствения главного модуля определяет пространственное раз-
мещение пазушных меристем, которые при наличии соответствующих бла-
гоприятных условий формируют боковые модули с их собственными набо-
рами меристематических зон — пазушных точек роста. Происходит форми-
рование органогенного потенциала (Седова, 1989), временная и простран-
ственная реализация которого различается у разных видов. Один из ярких
примеров — отличия между малолетними граминоидами с их «дернистым»
характером разрастания, обусловленным активизацией пазушных сближен-
ных меристем в нижней части главного модуля, и малолетними двудоль-
ными. Ветвление в базальной части стебля (или так называемое кущение)
свойственно и некоторым двудольным малолетникам особенно розеточным.
Побеги кущения при этом соответствуют побегам возобновления, которые
мы находим у многолетних травянистых растений, и повторяют в той или
иной степени структуру главного побега. Если такие побеги кущения мы
без колебаний можем называть модулями, то с паракладиямщ имеющими
сильно укороченные листовые серии или не имеющими их совсем, и с побе-
гами ветвления у злаков (Суворова, 1961) дело обстоит несколько сложнее.
Сложность с интерпретацией паракладиев и побегов ветвления злаков на-
ряду с побегами кущения при наличии или, чаще, отсутствии зоны тормо -
жения как модулей возникает из-за плавности перехода от побегов кущения
к паракладиям и далее, подчас, к цветкам (плодам) на цветоножках (плодо-
ножках) в составе соцветия (как у эбрактеозных соцветий крестоцветных).
Однако, при анализе тела малолетника как системы, устремленной к макси-
мально мощной репродукции, и с учетом того, что побеги кущения как пра-
вило не могут служить у малолетников целям вегетативного возобновления
даже в случае их самостоятельного укоренения (образования придаточных
корней), модулями, правомерно называть любые побеги, включая паракла-
дии и побеги ветвления злаков. Подобный тип разрастания особенно харак-
терен для однолетних граминоидов подобных однолетнику камчатских ги-
дротерм фимбристилису охотскому [Fimbristylis ochotensis (Mensh.) Koni.|.
Дерновинка этого однолетника по густоте очень близка дерновинкам мохо-
видных, хотя в подземной сфере ей свойственна повышенная плотность на-
стоящих корневых систем, а не ризоидов, как у мхов.
Вместе с тем, чтобы как-то отделить два существенно различающихся ха-
рактера разрастания и репродукции у травянистых растений целесообразно
условно выделять изомодульный и гетеромодулъный типы роста и репро-
дукции (см. схему на рис. 7.11).
Гетеромодул ьн ые
Изомодульные
Рис. 7.11. Схематичное изображение ге-
теромодульных (вверху) и изомодуль-
ных (внизу) растений и путей разраста-
ния особей в процессе увеличения мощ-
ности (жизненного состояния).
196 Л/// Марков I loiiyihiHiKHiiuui (nioiioi им рис iciiiiii
/ 'етсромодуньная структура воз-
никает в случае слабой выражен-
ности зоны торможения или се
полного отсутствия, когда на раз-
ных уровнях хорошо выделяюще-
гося главного побега формируются
пазушные модули разных размеров
с различными листовыми сериями и
обязательным присутствием репро-
дуктивных структур. Графики рас-
пределения модулей в составе осо-
би по классам их общей биомассы,
длины и числа плодов (рис. 7.12)
показывают, насколько разными мо-
гут быть плодоносные модули, раз-
вившиеся на одном растении у дву-
дольных. У клоповника густоцвет-
кового Lepidium densiflorum, напри-
мер, максимальный по размерам мо-
дуль имел биомассу в 107 раз боль-
шую, чем минимальный, притом,
что на обоих были нормальные пло-
ды.
Как показывает схема, в пределах
гетеромодулъного типа роста и ре-
продукции выделены два подтипа,
один из которых характеризуется за-
крытыми (с терминальными цветка-
ми или соцветиями), а второй — открытыми модулями. При втором варианте
гетеромодульной архитектуры может возникать система разных по длине от-
крытых модулей с фитомерами, состоящими из вегетативных и генеративных
структур. Каждый фитомер состоит из вегетативной части (листа, узла и меж-
доузлия) и репродуктивной части (пазушного цветка или соцветия). Таким
образом, обе сферы развиваются параллельно, а взаимная детерминация меж-
ду ними если и проявляется, то только на уровне фитомера. Вот почему рас-
тения, имеющие такую архитектуру, чаще всего характеризуются индетер-
минированным ростом. Помимо очень раннего перехода к репродукции (пер-
вый цветок может закладываться в пазухе уже 3-5-го настоящего листа), у
них можно наблюдать либо ритмичное чередование брахиморфных и доли-
хоморфных структур (лужница Limosella), либо длиннопобеговость (очный
цвет Anagallis, низмянка Centunculus) (рис. 7.13).
В средней полосе относительно не часты случаи гетеромодульности у ма-
лолетних злаков. Например, у скрытницы Crypsis shoenoides разные по раз-
мерам модули в составе куста в пазухах всех стеблевых листьев несут ко-
роткие модули (побеги ветвления), увенчанные султанами. Число султанов
/ /( ipykiypiihir испекii.i pocni pnciciiiill 197
в пересчете па один побег кущения колеблется oi одного до шести, чю и
создает iGipinny гетеромодул ыюй структуры.
Изомодульный тип разрастания присущ растениям, у которых очень oi
четливо выражена зона торможения, и не развиваются паракладии или но
беги ветвления. Подобно тому, как гетеромодульные растения с открыты-
Рис. 7.12а. График распределения побегов в составе двух особей клоповника густоц-
веткового Lepidium densiflorum по классам биомассы.
Рис. 7.126. График распределения побегов в составе особи песчанки тимьянолист-
ной Ап’папа serpvllifolia по классам числа плодов.
198
Л/.// Марков I loiiyпицнонппи bitonoi пя рпсicniiil
Рис. 7.13. Однолетние растения с откры-
тыми осями (они показаны стрелками):
Anagallis arvensis (вверху) и Limosella
aquatica (внизу).
ми модулями в ходе роста нака-
пливают очень сходные фитомсры,
малолетники с изомодульным ти-
пом структуры накапливают очень
сходные модули. Очень часто это
наблюдается у злаков и других гра-
миноидов (представителей семейств
Cyperaceae, Juncaceae).
Для примера рассмотрим дан-
ные таблиц 7.1 и 7.2, где по не-
скольким биоморфологическим па-
раметрам сравниваются модули из
состава одно-четырехстебельных
особей у ситника жабьего Juncus
bufonius и пшеницы Triticum vulgare.
Случайную выборку особей яровой
пшеницы обрабатывали, анализи-
руя размеры и структуру побегов-
модулей в масштабе популяции и
кодируя модули, находившиеся в со-
ставе особей из разного числа про-
дуктивных побегов. Сходным образом была обработана и случайная выбор-
ка из популяции ситника.
Таблица 7.1.
Сравнение модулей из состава одно-, двух-, трех-, и четырехпобеговых особей в по-
пуляции ситника жабьего Juncus bufonius по ряду морфобиологических признаков
Признаки Средние значения по группам: Уровни значимости различий между:
I- 1 2 | 1 з 4 1 и2 | 2иЗ | Зи4
Метрические и меристические:
Высота (см) 17,3 12,3 16,0 16,8 0,001 0,070 0,804
Число листьев 3,5 3,1 3,0 3,3 0,388 0,631 0,540
Масса (мг): Листьев 2,9 2,3 2,2 3,0 0,303 0,906 0,390
Стебля 4,5 3,2 3,4 5,2 0,088 0,816 0,193
Репродуктивных структур 10,9 7,9 12,4 10,0 0,086 0,101 0,660
Аллометрические:
Относительный вклад (%) биомассы в:
Листья 15,1 17,8 13,4 17,4 0,325 0,258 0,312
Стебель 26,6 25,1 25,1 28,7 0,711 0,988 0,654
Репродуктивные структуры 58,3 57,1 61,5 54,0 0,824 0,557 0,495
* Примечание: номера групп соответствуют числу модулей в составе особи.
Рима/ ( 'ipyк iурныг m iick i ы рос in piiciсииЙ 199
' I nftjiinui 7.2.
( равнение модуисн из состава одно-, двух- н i pcxuoGciоных ocoOcii
। iiiieiini u>i ТпПснт vulture но ряду признаков
Признаки Средние значения величин для групп: Уровни значимости различий между:
1 2 3 1 и2 2иЗ 1 и 3
Высота (см) 65,6 60,9 62,0 0,29 0,72 0,14
Длина колоса (см) 7,5 6,7 7,0 0,40 0,35 _ 0,09
Масса колоса (г) 0,8 0,6 0,7 0,52 0,29 0,07
Число колосков 11,4 11,0 11,8 0,62 0,16 __0153
Число зерен 20,4 15,5 18,6 0,49 0,14 0,04
Масса зерен в одном колосе(г) 0,5 0,3 0,5 0,73 0,07 0,04
Число листьев 3,9 3,9 3,8 0,95 0,75 0,81
Масса листьев (г) 0,2 0,2 0,2 0,09 0,72 0,20
Масса стебля (г) 0,7 0,6 0,6 0,38 0,55 0,14
Масса надземных органов (г) 1,7 1,4 1,5 0,35 0,37 0,06
Общая масса (г) 2,0 2,1 2,5 0,09 0,19 0,61
Масса одного зерна (г) 0,025 0,023 0,026 0,53 0,24 0,58
Как показывают данные таблиц, почти не нашлось абсолютных или ал-
лометрических параметров, по которым модули из состава особей разной
мощности статистически достоверно отличались бы друг от друга. Оба
вида, выбранные для иллюстрации морфологической изомодульности дали
основание для такого вывода.
При этом важно не только то, что формирующиеся в составе куста модули
бывают очень однотипными и то, что они, благодаря высокой степени авто-
номности, выступают в качестве демографических счетных единиц как на
уровне особи, так и на уровне популяции. Очень значимым представляется
тот факт, что в отличие от двудольных, обычно теряющих в ходе роста только
отдельные части фитомеров (например, розеточные листья с реутилизацией
накопленных в них ассимилятов), у изомодульных граминоидов у грачивасг-
ся часть модулей, а находившиеся в них ассимиляты также реутилизируются.
Сходство модулей (рамет\ формирующихся в составе одного куста (од-
ной генеты), может проявляться в одинаковом или близком числе слага-
ющих их фитомеров, одинаковых размерах модулей — это можно назван»
морфологическим аспектом изомодульности, — а также, у некоторых ма -
лолетних граминоидов — в одинаковом поведении. Анализ изомодул ыю-
сти в поведенческом аспекте позволил наметить три направления измене-
ний структуры особи в онтогенезе малолетних граминоидов.
Первый путь связан с формированием избыточного числа модулей в фазу
кущения, когда морфологическая изомодульность проявляется особенно
ярко (рис. 7.14а). В ходе начинающегося вслед за этим стеблеогбора (см.
Муравьев, 1973) часть модулей выбраковывается. Опп остаются ненро
дукгивными и отмирают, отдавая пластические вещества остающимся про
200
МН. Марков I loi lyjiM Ц1К>1 и in>i (hiojioi пн pnci cinif
дуктивным модулям. Число образу-
ющихся и отмирающих в ходе сгс-
блеотбора модулей может быть на-
столько значительным, что у неко-
торых малолетних злаков (ковыль
капский Stipa capensis) образует-
ся небольшая дерновинка, которая,
вероятно, играет роль аналогичную
роли розетки листьев у многих ма-
лолетних двудольных.
У малолетних граминоидов, де-
монстрирующих второй путь,
модули последовательно друг за
другом дорастают до какого-то
Рис. 7.14. Мыслимые пути стеблеотбо- определенного уровня (как бы под-
ра у злаков. равниваются) и переходят в репро-
дуктивное состояние (рис. 7.146).
Третий путь лежит через сохра-
нение всех модулей, развившихся в фазу кущения, которые все более или
менее синхронно переходят к репродукции, и потому морфологическую
изомодульность в этом случае можно наблюдать как в конце фазы кущения,
так и на стадии репродукции (рис. 7.14в).
Вряд ли какой-либо из этих трех путей видоспецифичен. Вполне можно
допустить, что в зависимости от внешних условий изомодульность на ста-
дии репродукции может быть достигнута разными путями у одного и того
же вида.
Подробнее остановимся на характеристике первого пути, который, по-
видимому, встречается наиболее часто у малолетних злаков средней поло-
сы. Рассмотрим вновь наиболее удобный для опыта объект — культурное
растение — яровую пшеницу.
На засеянных делянках в пределах постоянных квадратов вели наблюде-
ния за появлением и судьбой побегов у каждой из 103 маркированных осо-
бей. Три раза на протяжении жизненного цикла растений было учтено число
побегов, и таким путем выявлено 84 варианта переходов особей от началь-
ного (в фазе кущения) до конечного (в фазе зрелости) состояния. Используя
число побегов в каждый из сроков учета, переходы (пути стеблеотбора) обо-
значали следующим образом: 1-2-1; 3-2-1 и т.д. Теоретически (комбинатор-
но) таких переходов могло быть 280.
Частота встречаемости разных вариантов переходов (из 84) сильно раз-
личалась и в максимуме составляла 47%. На рисунке 7.15. можно видеть
и сравнить частоту тех или иных вариантов переходов для изначально (в
момент первого учета) одно-, двух-, трех-, и четырехпобеговых особей. Во
всех случаях (вариантах начала) наиболее высокой была вероятность отми-
рания уже ко второй дате всех побегов кроме одного, который и становил-
ся репродуктивным.
Вероятность отмирания уже в первом интервале времени была высока
только у изначально однопобеговых особей (18,0%) и резко снижалась у осо-
I 'Kiad / ( ipyn lypni.ic iu ИСК1Ы pocni pnciriiiiH ~20 1
бей c изначально Волыним числом модулей (у двухпобеговых 6,ХП<>; i рех
нобеговых М%; четырех-; пяти-; iiicci ппобеговых 0%). Нерол iпосiг
отмирания во втором интервале времени была значительна (12,2%) юже
только у изначально однопобеговых растений.
Из данных таблицы 7.3 видно, что с возрастанием изначально развито-
го числа модулей снижалась вероятность «константных переходов» (пере-
ходов без изменения модульной структуры), причем во втором интервале
времени это снижение было явно более плавным. Можно также отметить
Пути стеблеотбора
В начале наблюдений:
|1 ‘ I — 3 модуля
— 4 модуля
Рис. 7.15. Пути стеблеотбора у особей в популяции пшеницы, которые имели в нала
ле наблюдений 1,2, 3 или 4 модуля (f частота встречаемости варианта стебпео|бо
pa; X отмирание всей особи).
202 Л/ Н Мирков I loilyШ1Ц11О111111И (hhhioi ii>i pin iciinh
закономерноеть такого рода: чем больше побегов имела особь в (разе ку-
щения, тем ниже была вероятность появления новых модулей в нервом вре-
менном интервале при практически нулевой вероятности этого события во
втором интервале для любого класса начального состояния. Наоборот, веро-
ятность отмирания оказалась связанной с первоначальным числом модулей
положительной связью, что проявилось в течение обоих интервалов време-
ни, и особенно первого.
Таблица 7.3.
Динамика модулей у особей пшеницы с изначально разной кустистостью
Вероятность (%): Однопобе- говые Двухпобе- говые Трехпобе- говые Четырех- побеговые
I II I II I II I II
Увеличения числа модулей 21,0 0 23,5 0,5 14,4 0 10,6 0
Постоянства числа модулей 60,2 61,0 26,6 54,5 16,6 43,8 15,3 45,9
Отмирания модулей 18,8 39,0 49,9 45,0 68,7 56,2 74,1 54,1
Примечание: I — первый временной интервал; II — второй временной интервал,
Основываясь на доказанной однотипности модулей в составе особей раз-
ной архитектуры (морфологической изомодульности) была сделана попыт-
ка теоретически прогнозировать продуктивность и количественную морфо-
логию двух-, трех-, четырехстебельных растений путем простого умножения
средних значений, вычисленных для всей совокупности модулей в популя-
ции, на два, три, четыре, соответственно. Затем была получена разница меж-
ду эмпирическими и модельными значениями для зрелых особей, которую
выразили в процентах от модельных значений с соответствующим знаком.
Разные признаки вели себя очень сходно. Эмпирические значения характе-
ристик у двумодульных особей как правило были выше теоретических, что
можно трактовать как наиболее высокую эффективность растений, имевших
в итоге два продуктивных модуля. Обратную картину можно было наблю-
дать у трехмодульных особей. Оценка поведения четырехмодульных особей
не могла быть статистически значимой из-за малого их числа в популяции.
Говоря об относительной стабильности структуры и мощности модулей,
не следует забывать, что данный анализ захватывал только конечный итог
развития особей, который мог быть достигнут разными путями в ходе сте-
блеотбора. Наибольшее разнообразие путей, приводящих в конечном ито-
ге к одностебельным репродуктивным особям, может служить объясне-
нием наиболее высоких значений коэффициента вариации, которые были
высчитаны для всех взятых морфобиологических показателей таких осо-
бей. Хорошо прослеживаемая тенденция к снижению этого коэффициента в
ряду: одно- дву- трех- четырехстебельные растения вполне может отражать
снижение частот встречаемости путей стеблеотбора, посредством которых
были получены эти архитектурные типы особей.
Гинеи/ ( 1рум YjHii.ir iu iiriai.i pocin piu iriiiiH 203
I Iccmoi pH ihi io, что у большинства двудольных главный nobci выдсняс1см
средн ос । аныii.ix очень четко, все же в ряде случаев рано начинающееся оа
зитонное ветвление и выраженная зона торможения приводят к морфоло! и
ческой (но не поведенческой) изомодульности у некоторых малолетних рас
тений из этого класса. При этом возможна ситуация, когда боковые моду-
ли начинают развиваться в пазухах семядольных листьев еще до видимого
перехода главного побега к репродукции как у бурачка пустынного Alyssum
turkestanicum, плоскоплодника Meniocus linifolius, молочая-солнцсгляда
Euphorbia helioscopia. Развитие побегов второго порядка тоже может проис-
ходить в самых нижних частях модулей первого порядка, и тогда на стадии
репродукции мы наблюдаем куст из однотипных модулей, т. е. морфологи-
ческую изомодульность. Интересно отметить, что подобная же структура
может развиваться в том случае, когда главный побег с немногочисленны-
ми удлиненными междоузлиями, развивающийся на первом году жизни, о г
мирает за время зимовки, а репродуктивные побеги развиваются «кустом»
на втором году жизни. Такая картина закономерно возникает у щебрушки
Acinos arvensis.
Внешняя картина изомодульности (без явления, соответствующего сте-
блеотбору) может возникать у травянистых растений в результате воздей-
ствия фитофагов и даже у древесных. Судя по нашим данным (Марков,
1985), в липняке порослевого происхождения за счет многоствольное™ до-
стигается существенное повышение суммарного диаметра стволов в пере-
счете на единицу площади.
7.5.2. Внутривидовой уровень
При рассмотрении направлений разрастания особей на внутривидовом
уровне возникают следующие вопросы:
1) Являются ли мелкие особи, преобладающие в популяции (положи-
тельная асимметрия распределения по классам биомассы особей — обыч-
ная картина в популяциях малолетников), всего-навсего уменьшенными ко-
пиями крупных особей при сходстве основной архитектуры?
2) До какой степени возможно увеличение общей мощности и плодо-
витости особи на базе одной и той же архитектуры, например однос-
тебельного неветвящегося растения, растения всего с одной ветвью и т. д.
Иными словами, насколько существенными должны быть различия в общей
мощности растений, чтобы проявились заметные количественные архитек-
турные особенности?
Следует заметить, что отвечая на эти вопросы, мы совершенно опреде-
ленно выйдем за рамки чисто макроморфологического анализа и сделаем
шаг в направлении исследования биологических последствий в изменении
архитектуры. Поиски архитектурного аспекта в обеспечении подобия меж-
ду особями, занимающими разное положение во внутрипопуляционной раз-
мерной иерархии у малолетних растений, имеют биологический интерес,
поскольку на этой основе можно будет оценивать биологическую эффек-
тивность особей разного жизненного состояния у разных видов. Здесь мы
пока ограничимся рассмотрением теоретически мыслимых путей разраста-
204 МН Марков I loiiyiiMUiioiiiiiiM Г>иоп<>1 ня рис I ciiiiif
ния особей на основе то- или сетеромодульного строения, чтобы в даль-
нейшем вновь вернуться к этому вопросу при аллометрическом анализе ре-
продукции.
7.6. Способ переключения на репродукцию
На межвидовом уровне анализ архитектуры генеративных особей мало-
летних растений, как правило, позволяет по размещению и состоянию (на-
пример, степени зрелости) репродуктивных структур составить суждение о
типе взаимоотношений между органами вегетативной и репродуктивной сфе-
ры. По-видимому, существует три основных типа взаимоотношений между
сферами (рис. 7.16). Два из них связаны с симподиальным и один — с мо-
ноподиальным нарастанием побегов. Первый тип, наиболее часто встречаю-
щийся, связан с образованием достаточно большого числа чисто вегетатив-
ных фитомеров перед тем как апекс будет «израсходован» на формирование
соцветия — системы репродуктивного побега, мощность которого детерми-
нируется мощностью вегетативного элементарного побега. Второй тип осно-
ван на образовании сравнительно небольшого числа вегетативных фитоме-
ров с последующим формированием цветка, завершающего рост главной оси.
Дальнейшее разрастание идет на базе параллельного развития обеих сфер —
каждый формирующийся в ходе симподиального нарастания фитомер, поми-
мо листа, связан с цветком, завершающим его ось. Третий тип характеризу-
ется открытыми моноподиально нарастающими осями и пазушными одиноч-
ными цветками или небольшими соцветиями.
Порядок рассмотрения типов соответствует трем этапам на пути ослабле-
ния детерминирующего влияния вегетативной сферы, с формирования ко-
торой начинается онтогенез особи малолетнего растения, на репродукцию
и на пути к максимальному ускорению темпов развития.
Насколько нам известно, о воз-
можности появления различных ва-
риантов перехода к репродукции из
отмеченных выше у особей одно-
го вида сведений в литературе нет.
Однако именно в этом разделе надо
хотя бы вкратце затронуть факты аль-
тернативного поведения пазушных
почек у ряда малолетних растений.
На примере однолетника Floerkea
proserpinacoides Б. Смит (Smith,
1984) продемонстрировал альтерна-
тивные варианты развития из пазуш-
ной почки либо одного цветка, либо
побега с листовой серией, причем в
зависимости от условий года наблю-
далась разная частота особей с тем
или иным вариантом архитектуры.
Любопытным было заключение, что
Рис. 7.16. Три основных типа взаимоот-
ношений между вегетативной и генера-
тивной сферами: два (1 и 2) — с симпо-
диальным и один (3) — с моноподиаль-
ным нарастанием побегов (пояснения в
тексте).
Г'Кши / ( 1 ipyu i ypni.ic in nrkii.i рос iи pnc i rinill 201
pca/iiкуемi.iи iiapiiani архитектуры (no ic|>miiiiojk>ri111 ашора дизайн) iuei
да обеспечивал более высокую плодов!i юсть (и т. о. уровень прнспособиенно
сти), чем альтернативный вариант, просчитанный теоретически. Аналогичпук
картину мы наблюдали в популяциях лужницы Limosella aquatica, у Koiopoi
альтернативой одиночного пазушного цветка выступает столон.
7.7. Признаки, положенные в основу классификации
архитектурных типов малолетних растений
Во многих систематических группах цветковых растений намечается ре
дукционный ряд жизненных форм от деревьев до однолетников (Попои
1948; Хохряков, 1975 и др.). На рис. 7.17 представлена детализирован на»
правая часть такого редукционного ряда с показом поливариантносгп он
тогенеза, а вернее, связанного с ней значительного перекрывания звеньев.
Параллельная эволюция разных таксонов имела по существу биологичс
ски близкий итог, но выраженный средствами каждого отдельного таксон;
и потому характеризующийся известной уникальностью макроморфоло! и
ческой конструкции. Меньше других разработке подверглась в интересую
щем нас аспекте именно группа малолетников, короткоживущих — одно
двулетних растений. В наборе морфологических признаков любого вида ма
лолетнего растения можно усмотреть компромисс между наследственные
консерватизмом конституции, связанным с систематическим положением, i
адаптацией к комплексу эколого-ценотических условий местообитания, ко
торое этот вид делит с другими видами малолетников.
Список сообществ средней полосы, в состав которых входят малолетние
растения, достаточно велик, и это создает представление об очень шири
кой эколого-фитоценотической амплитуде малолетних растений. Эта шире
та амплитуды, по-видимому, должна как-то коррелировать с разнообрази
ем морфологических конструкций, архитектуры малолетников. Однако еле
дует помнить о том, что и в пределах одного фитоценоза можно наблюди11
значительную пространственную и временную гетерогенность эколого
ценотических условий. В особенности это характерно именно для фи к ни-
возов в той или иной степени нарушенных, в которых играют заметную рол ь
малолетние растения. Поэтому совмещение малолетних видов с разными
архитектурными моделями, разной архитектурой в составе одного фитоцс
ноза вполне закономерно. Наконец, нельзя не учитывать и связанную с но
ливариантностью онтогенеза внутривидовую изменчивость архитектуры
Рис. 7.17. И| xiiuim детализированная часть редукционного ряда жизненных <|><>рм
цветковых pacieiiiiii с показом i iojii тарпан и юс ги онтогенеза, а вернее, связанною <
iieii nia'iii leui.iioi о перекрывания звеньев.
206 Л/ Н Марков I loiiyiiHHHoiimui (hiojioi idi pnc iciinh
Определив «противоборствующие» моменты, вызывающие, с одной сторо-
ны, дифференциацию, а с другой — конвергенцию параметров архитектуры
малолетних растений, перейдем к обсуждению этих параметров.
При всякой попытке классифицировать архитектурные типы и жизненные
формы исследователь неминуемо сталкивается с необходимостью выбо-
ра признаков, обоснования их числа, ранжирования их по значению и т.д.
В основу выделения архитектурных моделей (моделей побегообразования)
Т. И. Серебряковой (1977, 1981) положено всего два признака — тип нарас-
тания осей (2 варианта: симподиальный и моноподиальный) и структура по-
бега (3 варианта: розеточность, полурозеточность и длиннопобеговость, или
безрозеточность). Из шести возможных комбинаций между вариантами при-
ведены примеры для четырех (моноподиальные розеточные, моноподиаль-
ные длиннопобеговые, симподиальные полурозеточные и симподиальные
длиннопобеговые), которые и были зафиксированы как архитектурные моде-
ли. Вместе с тем, по признанию самой Т. И. Серебряковой, разнообразие ар-
хитектурных моделей трав этим не исчерпывается. В. Н. Голубевым (1965),
который понимает полурозеточность более широко, чем И. Г. Серебряков
(1952), только из числа травянистых поликарпических многолетников выде-
лены полурозеточные моноподиальные растения, а для много- и малолетних
растений — розеточные симподиальные. Таким образом, матрица вариантов,
учитываемых Т. И. Серебряковой, оказывается заполненной до конца.
В основу нашей классификации архитектурных типов и архитектурных
моделей малолетних растений средней полосы России положено 6 призна-
ков (16 вариантов):
1. Ранг повторяемых метамеров (2 варианта). Например, итерация од-
нотипных модулей как элементов латерального разрастания особи (у изомо-
дульных однодольных) или итерация фитомеров (=фитонов) или разнораз-
мерных модулей (у гетеромодульных, преимущественно двудольных мало-
летников).
2. Характер корневой системы (2 варианта). Этот, по определению В. Н.
Голубева (1971), стенотопный признак необходимо учитывать, как он учи-
тывался Т. В. Шулькиной (1988) при анализе ею архитектурных моделей в
семействе Campanulaceae. Единственная известная нам попытка классифи-
цировать корневые системы малолетних растений, была предпринята В. Н.
Голубевым (1965). Однако его классификацией оказались затронуты лишь
аллоризные виды степных малолетников.
Особого внимания, на наш взгляд, заслуживает явление вторичной гомо-
ризии у двудольных, т.е. очень раннее (обычно у всходов в фазе двух семя-
долей — рис. 7.18) прекращение роста главного корня и замена его прида-
точными. Такую картину можно наблюдать у Ceratocephala falcata, Myosurus
minimus, Limosella aquatica, Androsacefiliformis. Полное отсутствие выражен-
ного главного корня у Impatiens noli-tangere вообще представляется для дву-
дольных уникальным явлением. На стадии проростка у Impatiens parviflora
главный корень можно выделить только в самом начале — еще до развора-
чивания семядолей. Позднее его догоняют по размеру придаточные корни,
возникающие из нижней расширенной части гипокотиля, и он теряется среди
них (фото 7.1. и рис. 7.18).
/ / ( Ipy It I ypHLIC lit ИСК I 1,1 рос III pile I r 111111
20/
3. ('fhpyemvpu . пивного и боковых
побегов ( \ варнашa). 11редс'гавляс1
особый интерес ритм чередования
в составе побегов брахиморфных и
долихоморфных структур (Голубев,
1965), т. е. серий а- и Р-фитомеров.
Этот признак имеет большое значе-
ние в оценке размещения ассими-
ляционной поверхности вдоль по
телу растения и формирования со-
ответствующих архитектурных ти-
пов (см. выше 7.2). К основным ти-
пам — розеточным, полурозеточ-
ным и длиннопобеговым — следует
добавить, как уже говорилось выше,
временнорозеточный, который целе-
сообразно рассматривать в качестве
подтипа в типе длиннопобеговых.
4. Листорасположение (3 вари-
анта). Признак, тесно связанный с
предыдущим признаком, и теоре-
тически может быть к нему сведен,
если рассматривать супротивное и
мутовчатое листорасположение как
равномерно-ритмическое чередова-
ние укороченных (-ого) и удлинен-
ных (-ого) междоузлий. Именно в
случаях супротивного и мутовчатого
листорасположения чаще всего воз-
Фото 7.1. Стадии прорастания подотри
ги мелкоцветковой.
никает временная розеточность.
5. Характер нарастания и вет-
вления побегов (4 варианта). Целе-
сообразно учесть не только моно-
подиальность или симподиальность
нарастания и не только мощность
ветвления, но и уровень отхождения
боковых модулей и степень выра-
женности главного модуля при фор-
мировании серии модулей в соста-
ве одной особи. Заслуживает особо-
го внимания способность некоторых
малолетних растений образовывать
боковые модули уже в пазухах семя-
Рис. 7.18. Вторично гоморизные корне-
вые системы у ряда однолетних расте-
ний (схема): I — мышехвостник малень-
кий Myosurus minimus’, II — недотрога
обыкновенная Impatiens noli-tangere (см.
также фото внизу); III — н. мелкоцвет-
ковая Impatiens parviflora (1 — главный
корень; 2 — придаточный корень; 3
основание розетки).
дольных листьев при неповрежденной главной оси.
6. Морфология фитомеров (=фитонов) в пределах модуля (2 вариан-
та). Здесь имеется в виду гомо- и гетеробластное развитие по К. I ебслю.
Иногда оба варианта развития встречаю'! ся у одного н юго же вида мало
208
Л/ // I hnlyи>iiuкn11m)i (hioiioi ibi pncicinih
летнего рас гения (Марков, 1976). ')тот признак важно учитывал, и пото-
му, что основной путь пластического изменения oirroi снеза у очень мно-
гих малолетников лежит через образование неотенических модификаций
(Васильченко, 1948; Колев, 1960; Маркова, 1971; Марков, 1976, 1978, 1986).
Для таких модификаций характерна упрощенная ювенильная архитектура
особи и гомобластное развитие из-за выпадения фазы продуцирования фи-
томеров взрослого типа из онтогенеза.
7.8. Классификация архитектурных типов малолетних растений
В хорошо известной работе, характеризуя достоинства «линеинои
классификации жизненных форм», В
Рис. 7.19. Основные архитектурные типы
малолетних растений (на примере ряда
модельных видов): I — граминоиды (сле-
ва направо: мятлик однолетний, сыть бу-
рая, мортук пшеничный); II — розеточ-
ные (слева направо: проломник север-
ный, п. нитевидный, мышехвостник ма-
ленький, лужница водная, мшанка леачая;
III — полурозеточные (слева направо:
вайда ребристая, пастушья сумка биотип
simplex, п. с. биотип heteris; IV — безро-
зеточные (слева направо: недотрога обык-
новенная, тимелея воробьиная, горец ше-
роховатый, бурачок пустынный, подо-
рожник индийский, звездчатка средняя.
Остальные пояснения содержатся в дихо-
томическом ключе.
Н. Голубев (1968) подчеркивает два
важных пункта: 1) из многообразия
признаков наиболее существенны
те, которые обусловливают фитоце-
нотическую роль видов, а она зави-
сит от строения подземных и над-
земных органов, физиологических
особенностей растения, ритма раз-
вития и т. д.; 2) нужно выявлять и
учитывать динамику признаков.
Предлагая свою «линейную клас-
сификацию жизненных форм» В. Н.
Голубев (1968) пишет, что каждый
вид представляет неповторимую
систему эколого-биологических
приспособлений и может рассма-
триваться как отдельная жизненная
форма. Аналогичная картина возни-
кает и при попытке детального опи-
сания типов морфологической кон-
струкции или архитектурных ти-
пов. При этом, чем больше призна-
ков мы хотим учесть, тем более бу-
дет ощутим уход от широкого обоб-
щения в сторону мелких единиц
вплоть до отдельных особей вну-
три вида. Поскольку мы искусствен-
но акцентируем внимание только на
шести параметрах архитектуры (В.
Н. Голубев перечисляет 36 биоэко-
логических признаков жизненных
форм), на данном этапе нам уда-
лось свести все многообразие ма-
лолетних растений средней поло-
сы России к пятнадцати архитектур-
ным типам. Место архитектурных
Г>и1пч / ( ipyhiypiii.iv iininui.i pocin piu iciiiHI 209
Г1ПЮ1Ы1 cooi nr i г i нующих архитектурных моделей в характеристике жи i
ценных форм малолетних растении показано в таблице 7.4. Основные ihih.i
с соотвстс жующими модельными видами представлены на рис. 7.19.
Таблица 7.4.
Место архитектурных моделей в характеристике жизненных форм
малолетних растений
ЖИЗНЕННАЯ ФОРМА
АРХИТЕКТУРНАЯ МОДЕЛЬ | ТИП ЖИЗНЕННОГО ЦИКЛА
вторично го м о р и з н ы е
1. Растения с однотипными по размерам и строению модулями («изомодульные») Поли- и моновариантные малолег ние граминоиды иногда с возможно стью индетерминированного роста (за счет итерации однотипных мод ул ci i) и иногда даже поликарпии
2. Длиннопобеговые симподиальные Незимующие (моновариантные) мо- нокарпики
3. Полурозеточные симподиальные Дву-однолетние (поливарианп i ы е) монокарпики
4. Розеточные симподиальные с сохраня- ющимся в течение всей жизни гипокоти- лем Зимующие (поливариантные) моно карпики
5. Розеточные симподиальные без замет- ного на стадии репродукции гипокотиля Дву-однолетние (поливарианп i ы с) монокарпики
6. Розеточные с сочетанием моно- и сим- подиального роста и способностью к сто- лонообразованию Незимующие однолетние монокарнп ки с возможностью индетерминиро ванного роста
аллоризные
7. Длиннопобеговые или временнорозе- точные моноподиальные Незимующие однолетние монокарпп- ки
8. Длиннопобеговые или временнорозе- точные симподиальные с супротивным листорасположением Дву-однолетние (поливарианп i ыс) монокарпики
9. Длиннопобеговые или временнорозе- точные симподиальные с очередным ли- сторасположением и базитонным ветвле- нием Незимующие или реже (поливариан! ные) зимующие однолетние монокар- пики
10. Длиннопобеговые симподиальные с акротонным ветвлением Незимующие (моновариантные) мо нокарпики
11. Полурозеточные симподиальные с го- мобластным развитием Дву-однолетние (поливариа! иные) монокарпики
12. Полурозеточные симподиальные с ге- теробластным развитием Дву-однолетние (поливариантные) монокарпики
13. Розеточные с сочетанием моно- и сим- подиального роста и способностью к сто- лонообразованию Много-дву-однолетпие (поливарианi ные) монокарпики с возможное и.ю индстермипироваш юго роста
14. Розеточные симподиальные Двулетние или зимующие ()Д11О11С 1 1IIIC (иол и вариантные) монокарпики
15. Розеточные мопоподпальпые Зимующие одполе гипс (поппварпап i ные) мопокарппкп |
210 Л/ Н Миры hi I loi iy и hi цк >111П1И biionoi ни рис । спи<!
Каждый архитектурный тип обязательно характеризуется сочетанием не-
скольких параметров (точнее их вариантов). Относительно большое число
выбранных нами параметров не позволяло применить матричный принцип
построения классификации, распространенный у морфологов. Поэтому в
целях некоторого упорядочивания выделенных типов здесь приведем дихо-
томический ключ.
1. Разрастание на основе итерации более или менее инвариантных (изо-
модульность) или, реже, существенно различающихся по размерам и
структуре (гетеромодульность) модулей как элементов горизонтальной
структуры; системы придаточных корней.
Модельные виды: мятлик однолетний Роа annua, мортук пшеничный
Eremopyrum triticeum. сыть бурая Cyperus fuscus. ситник жабий Juncus
bufonius и другие малолетние граминоиды из класса Однодольных
(представители семейств Gramineae, Cyperaceae, Juncaeae).
+ Разрастание на основе итерации фитомеров и модулей как элементов вер-
тикальной структуры; системы стержневого и, гораздо реже, придаточ-
ных корней............................................... 2
2. Системы придаточных корней (вторичная гоморизия)............3
+ Системы стержневого корня....................................6
3. Длиннопобеговые (безрозеточные) или верхнерозеточные растения, при-
даточные корни развиваются на хорошо выраженном в течение всей
жизни гипокотиле.
Модельные виды: недотроги Impatiens noli-tangere. I. parviflora (Balsaminaceae)
+ Розеточные растения..............................................4
4. Розеточные растения с сохраняющимся в течение всей жизни ги-
покотилем, от которого отходят придаточные корни.
Модельные виды: мышехвостник Myosurus minimus, рогоглавник Ceratocephala
falcata (Ranunculaceae).
+ Гипокотиль, хорошо заметный у проростков, довольно быстро исчезает за
счет контрактильности придаточных корней..........................5
5. Вертикальное (ортотропное) ритмическое чередование брахиморфных и
долихоморфных структур.
Модельный вид: проломник нитевидный Androsacefiliformis (Primulaceae).
+ Горизонтальное (плагиотропное) чередование брахиморфных и до-
лихоморфных структур, образование столонов.
Модельный вид: лужница Limosella aquatica (Scrophulariaceae).
6. Безрозеточные (иногда временнорозеточные) растения..............7
+ Растения образуют в начальной стадии онтогенеза розетку листьев, кото-
рая в дальнейшем не растягивается и сохраняется по крайней мере до
цветения......................................................10
7. Листорасположение супротивное...................................8
+ Листорасположение очередное .....................................9
8. Моноподиально нарастающие растения с пазушными безлистными побе-
гами, несущими терминальные соцветия.
Модельный вид: подорожник индийский Plantago indica (Plantaginaceae).
Г/ititui / ( ipykiypiii.ir n< nrkii.i pocin piicirimB 211
< PaciCiiiiM с симнодпальным характером разрастания, у которых главная
ось заканчивается терминальным цветком или соцветием.
Модельные виды: крапива жгучая Urtica urens (Urticaccac), конопля сорная
Cannabis ruder alts (Cannabaceae), звездчатка средняя St el I aria media,
песчанка тимьянолистная Arenaria serpyllifolia (Caryophyllaceae), веро-
ника весенняя Veronica verna (Scrophulariaceae), змееголовник тимья-
ноцветковый Dracocephalum thymiflorum (Labiatae).
9. Ветвление главной оси начинается с самого низа (базитония) — первые
боковые модули закладываются в пазухах семядольных листьев иногда
уже в фазе всхода — ювенильного растения.
Модельные виды: бурачок Alyssum turkestanicum, плоскоплодник Meniocus
linifolius (Cruciferae).
+ Главная ось или не ветвится, или ветвится обыкновенно в верхней час ти
(акротония).
Модельные виды: желтушник левкойный Erysimum cheiranthoides
(Cruciferae), горцы Polygonum scabrum, P minus, щавель примор-
ский Rumex maritimus (Polygonaceae), тимелея Thymelaea passerina
(Thymelaeaceae), дурнишник Xanthium strumarium (Compositae)
10. Полурозеточные растения...................................II
+ Розеточные растения .......................................12
11. Растения с гомобластным развитием
Модельные виды: вайда Isatis costata, пастушья сумка Capsella bursa
pastoris (биотип simplex), Arabidopsis thaliana, крупка дубравная
Draba nemorosa (Cruciferae), герань Роберта Geranium robertianum
(Geraniaceae).
+ Растения с гетеробластным развитием.
Модельные виды: клоповники Lepidium ruderale, L. densiflorum, пастушья
сумка Capsella bursa-pastoris (биотипы: heteris, tenuis, rhomboidea), гу-
лявник Лезеля Sisymbrium loeseli (Cruciferae).
12. Розеточные симподиальные растения..............................13
+ Розеточные моноподиальные растения.
Модельный вид: подорожник Plantago intermedia (Plantago major sbsp.
pleiosperma) (Plantaginaceae).
13. Вертикальное (ортотропное) чередование брахиморфных и доли-
хоморфных структур.
Модельные виды: проломники Androsace maxima, A. elongata,A. septentrio-
nalis (Primulaceae).
+ Горизонтальное (плагиотропное) чередование брахиморфных и до-
лихоморфных структур.
Модельный вид: мшанка лежачая Sagina procumbens (Caryophyllaceae).
Все сказанное в данной главе приводит к убеждению, что анализ архитек-
туры малолетних растений может и должен предварять углубленное изуче-
ние биологии их популяций, поскольку позволяет наметить ответы па важ-
нейшие вопросы, касающиеся стратегии жизненного цикла. В самом деле,
структура малолетних растений сама но себе дает ценную информацию об
212 М.Н Марков I loiiyihiunoiiiiihi (>110)101 пя рпсюппй
особенностях роста, развития и внутринонуляционной дифференциации но
этим показателям. Изучив архитектурные модели и архитектуру, можно вы-
явить пути, по которым: 1) осуществим максимально быстрый переход к
репродукции; 2) возможно при наличии благоприятных условий наращи-
вание мощности репродукции; 3) обеспечивается максимальная эффектив-
ность репродукции.
Глава 8. КОЛИЧЕСТВЕННЫЕ АСПЕКТЫ
РОСТА РАСТЕНИЙ В ПОПУЛЯЦИЯХ
«...самой важной стороной роста является
непрерывное увеличение объема... орга-
низма»
Э. Синнот, Морфогенез растений, / 963
По определению Д. А. Сабинина (1940, 1963) рост — это необратимое
увеличение размеров и массы растения, связанное с новообразованием эле-
ментов структуры. На первое место, как видим, поставлено увеличение
массы, которое мы условно и будем здесь рассматривать как количествен-
ный аспект роста.
Изучение роста растений — это важнейшее звено, связывающее физио-
логическую экологию с популяционной экологией растений. Ведь физиоло-
гические реакции у растений, как правило, можно объяснять именно росто-
выми процессами. Сами размеры (биомасса) растения, приобретенные им
за какое-то известное время, можно использовать при этом как важный по-
казатель для описания варианта его онтогенеза. При этом отдельные струк-
турные элементы (метамеры sensu lato) особи будут проявлять характерный
для данного варианта онтогенеза тип сопряжения выживаемости и плодови-
тости как двух альтернативных параметров адаптации. Для физиологов рас-
тений рост — это следствие успешного сочетания двух важнейших процес-
сов: фотосинтетической деятельности (углеродного питания) и минерально-
го питания, в ходе которых накапливается биомасса тела.
8.1. Относительная скорость роста
Д. А. Сабинин первым из отечественных ученых обратил внимание на
уравнение экспоненциального роста особи, с которым работал В. Блэкман
(Blackman, 1919):
W= WoeRGRf, [8.11
где t — промежуток времени; Wo — начальная биомасса, a FT — биомасса но
истечении времени t. Нетрудно заметить соответствие между этим уравне-
нием и уравнением экспоненциального роста численности популяции [3.61,
214 Л/./< Миркой I 1()||уЛИЦ11<)111111>1 (ШОЛОГПИ pilCICHIlil
рассмотренным в главе 3. Обозначив начальный и конечный моменты вре-
мени, соответственно, через / и /, можно рассчитать величину RGR:
RGR = (In W-In Wo)/( t - to) [8.2]
Показатель RGR в современной литературе называют относительной ско-
ростью роста (Relative Growth Rate — общепринятое сокращение RGR). Ее
вычисляют как приращение биомассы в пересчете на единицу уже имев-
шейся в начале наблюдений какой-то «стартовой» биомассы и потому выра-
жают в единицах: мг/мгхеутки, мг/мг^неделя или мг/мг*единица времени.
Определяя биомассу растения взвешиванием, мы в огромном большин-
стве случаев вынуждены убивать растения выкапыванием и сушкой их до
приобретения постоянного веса. Только рост биомассы у водных плаваю-
щих неприкрепленных растений, таких как ряски Lemna, телорез Stratiotes
aloides и др. можно пытаться изучать во влажном состоянии, не убивая и ис-
пользуя для взвешивания одни и те же особи несколько раз.
С учетом метамерности строения растений и ритмичного формирования
элементарных метамеров апексом побега в ходе роста правомерно исполь-
зовать для оценки интенсивности ростовых процессов особи темпы измене-
ния числа ее метамеров. Это будет своеобразным «взвешиванием» растений
в единицах числа элементарных или иных метамеров и сочетанием струк-
турного и количественного подходов к изучению роста. При работе, к при-
меру, с редкими охраняемыми растениями такой «неповреждающий» инде-
структивный способ определения темпов роста обладает рядом серьезных
преимуществ перед традиционным весовым методом.
Действительно, ведь прежде чем рассматривать накопление собственно
биомассы, можно, отталкиваясь от относительной однотипности (и равно-
великости) метамеров, слагающих тело растения, регистрировать увеличе-
ние в ходе роста числа метамеров (р) как своеобразных единиц биомассы и
тогда работать с формулой:
RGR’= (In - In >?о)/(Q. [8.3]
Нужно только определиться с минимальным размером растущего метаме-
ра, с момента достижения которого он будет сосчитан как сформированный.
Первая попытка использовать темпы относительного роста (RGR) как
наиболее существенный параметр для выявления эколого-ценотической
стратегии у того или иного вида принадлежит Д. Ф. Грайму (Grime, Hunt,
1975; Grime, 1979). Напомним, что в единицах этого количественного пока-
зателя проградуирована горизонтальная ось в его треугольной диаграмме
для ординации стратегий (см. главу 3). Им же было начато накопление базы
данных об RGR, определяемой в условиях опыта у разных видов. А с уче-
том того, что чисто экспоненциальный рост наиболее характерен для ран-
них стадий онтогенеза, Д. Грайм и R Хант определяли показатель и его из-
менчивость у десятидневных проростков, выращенных в песчаной культуре
гидропонным способом в стандартных условиях опытной установки.
I 'latia S Koinrirt i nciini.ir испекiы росin pm iriiiiB п попуияцпях 215
8.2.0 lieicoiорых методических вопросах выявлении RGR
в популяционных исследованиях
Наблюдаемые отклонения эмпирических данных о росте особей в популя-
циях от экспоненциальной кривой чаще всего связаны с неравномерностью
темпов роста, которая наиболее заметна у растений, растущих в естествен-
ных условиях. Даже если проростки появляются одновременно, а мы и рас-
смотрим вначале именно такие случаи, скорость роста в разные моменты
времени будет разной. Многое зависит от плотности размещения и, отсюда,
от времени возникновения и интенсивности контактов между растениями в
популяциях. Для получения большей информации в этом случае весь пери-
од роста необходимо подразделять на отдельные отрезки и для каждого о г-
резка вычислять свое значение RGR.
Вычисление RGR выгодно отличается от определения абсолютных при-
ростов биомассы. Во-первых, потому что приросты эти зависят от первона-
чальной (уже накопленной до начала наблюдений) биомассы. Во-вторых,
потому, что по получаемым цифрам правомерно сопоставлять разные виды
или популяции одного вида из разных по экологическим условиям местоо-
битаний.
Зависимость была бы наиболее строгой в идеальной ситуации, когда мас-
са растения складывалась бы только из фотосинтезирующих тканей со спо-
собностью всех клеток к фотосинтезу и делению. Очевидно розетка, состо-
ящая из сближенных листьев при недоразвитии междоузлий, более всего
удовлетворяет этому условию.
8.2.1. Особенности розеточного роста
Методически важными представляются два момента. То, что одна из при-
чин отклонений от роста биомассы по экспоненте — образование струк-
турных веществ типа целлюлозы, не вовлекаемых больше в обмен и то,
что розеточное растение с позиций концепции source-sink, т. е. донорно-
акцепторных отношений — это система с непосредственным контактом до-
нора (розеточных листьев) и акцептора (корневой системы).
Первый момент важно учитывать при сравнении роста в разной степени
ксерофилизованных видов. Мезоморфный лист, а розеточный лист в боль-
шинстве случаев именно мезоморфный, метаболически дешевле и потому
выгоднее для растения, чем ксероморфный, содержащий большее относи-
тельное количество структурных веществ. Поэтому даже в аридных усло-
виях многие виды растений формируют сравнительно недолговечные ли-
стья мезоморфной структуры (Orians, Solbrig, 1977; Mooney, Gulmon, 1979).
На недолговечность и мезоморфность розеточных листьев указывали еще
Е. Варминг и К. Раункьер (по Серебрякову, 1952). П. А. Генкель и Р. Г.
Абдиева (1971) относят пустынные эфемеры, среди которых преобладаю!
розеточные и полурозеточные растения, к псевдоксерофитам, подчеркивая
своеобразие их борьбы с засухой путем ухода от нее — избегания во време-
ни. Однако думать, что все розеточные и полурозеточные растения в одина-
ковой степени мезоморфны, конечно же, ошибочно.
216 А/ Н Марк(к, 11< >i iy ня щи >i 11 him (Hioiioi ня рас iciinh
Поскольку, как уже было отмечено выше, приблизительное представле-
ние о степени ксерофилизации даст сырой или сухой вес единицы площа-
ди листа (т. н. поверхностная масса), то можно, используя этот показатель,
сравнить некоторые экологические группы растений (табл. 8.1).
Таблица 8.1.
Поверхностная масса сырых листьев (мг/см2) в разных экологических группах рас-
тений (по Шульгину, 1963 с изменениями)
Экологические группы растений Поверхностная масса
минимальная средняя максимальная
Гигрофиты и гидрофиты 11,8 30,8 46,6
Мезофиты 6,8 22,4 66,5
Ксерофиты (склерофиты) 18,1 45,5 180
Суккуленты 26,8 223,5 610
Еще более интересно сопоставлять по поверхностной массе листья ви-
дов растений, относящихся к одному роду. Так, к примеру, розеточные ли-
стья двух видов проломника: наибольшего Androsace maxima и северного А.
septentrionalis существенно отличаются по значению поверхностной мас-
сы — у A. maxima она в 2,5 раза больше. Как будет более подробно показа-
но ниже, оба вида растут в степных фитоценозах, но занимают разные, хотя
и подчас контактирующие местообитания. Проломник наибольший — зи-
мующий однолетник, розетка которого продолжает некоторое время функ-
ционировать весной, захватывая засушливый период. Именно в этот пери-
од, когда открытые участки субстрата, занятые проломником наибольшим,
сильно (до 45°) нагреваются, более высокая относительная ксероморфность
его розеточных листьев может быть полезной. Второй вид — двулетник
проломник северный растет на участках с гораздо более густым травяным
покровом и предпочитает склоны не южных экспозиций. Розетка его содер-
жит, обычно, больше листьев (до 50), чем у Л. maxima (см. рис. 8.1) и может
мириться с некоторым притенением. Многие розеточные листья, прижатые
в норме к поверхности почвы, в условиях притенения меняют ориентацию
в пространстве и располагаются под острым, иногда почти прямым углом к
линии горизонта.
Второй момент связан с тем, что растения с розеткой листьев относят-
ся по классификации А. Т. Мокроносова (1983) к системам I типа, у кото-
рых ассимиляты передаются сразу и непосредственно от донора (source) к
акцептору (sink). Действительно, в период розеточного роста главным ак-
цептором и аттрагирующим центром является энергично растущая кор-
невая система (а иногда, как у редиса, и гипокотиль). Однако необходимо
подчеркнуть лабильность этих отношений, которые у ряда видов (пасту-
шья сумка Capsella bursa-pastor is, резуховидка Таля Arabidopsis thaliana и
др.) легко могут меняться на обратные под влиянием внешних условий, на-
пример затенения, или дефицита влаги. В этом случае вновь появляющийся
аттрагирующий центр — зачаточное соцветие — вызывает противополож-
ное (снизу вверх) перемещение ассимилятов, а корневая система нрекра-
X
217
I hlfitl <S' Koilll'irt 111С11ПЫГ ЛСНСК1Ы
poem pm
‘ I СНИП и популяция
щаст расти н начинает отдавать ас-
симиляты, накопленные ею в пред-
шествующий период времени.
Второе звено упомянутого выше
алгоритма (архитектурная модель -
жизненная форма - стратегия жиз-
ненного цикла) предусматривает
анализ адаптивности выделенных
архитектурных типов растений или,
иными словами, выяснение приспо-
собительного значения той или иной
совокупности параметров архитек-
туры для жизни в соответствующих
эколого-ценотических условиях.
Рис. 8.1. Сопоставление двух видов про-
ломника (Androsace) по числу розеточ-
ных листьев (пояснения в тексте).
8.2.2. Приспособительное
значение розеточного роста
Поговорим о приспособительном значении розетки. В жизненном ци-
кле многих многолетних, но, особенно малолетних, растений розеточный
рост выступает как хорошо обособленная фаза. По существу вся вегетатив-
ная сфера розеточных малолетников формируется в ходе розеточного ро-
ста, когда апекс главного побега продуцирует сильно сближенные узлы с
листьями, отражающими в своих размерах и форме скоротечные возраст-
ные изменения. Перемена деятельности апекса — переход к формированию
одного или нескольких удлиненных междоузлий означает переход в генера-
тивное состояние. Поэтому розетка может рассматриваться как элементар-
ный побег и одновременно как способ быстрого наращивания ассимиля-
ционной поверхности благодаря экономии на росте междоузлий (см. выше
рис. 7.2).
Из 15 архитектурных типов малолетних растений, описанных в главе 7,
для девяти розетка является обязательным компонентом архитектуры особи,
а еще для трех обычным явлением выступает временная розеточность (см.
табл. 7.5). Таким образом, в сочетании с системой придаточных корней или,
что гораздо чаще, с системой стержневого корня розетка встречается у боль-
шинства малолетних растений средней полосы России. Продолжительность
розеточной фазы у разных видов малолетников различна.
Часть монокарпических многолетников (катрана татарского Crambe tataria),
факультативных двулетников (репейников Arctium minus, A. tomentosum) и по-
ликарпических многолетников (виды ревеня Rheum, подорожника Plantago)
сбрасывает розеточные листья на зиму и восстанавливает розетку весной.
Другой части (герань Роберта Geranium robertianum, коровяки Verbascum
thapsus, V. lychnitis и др.) присущи зимнезеленые розетки, причем розеточная
фаза может даже растягиваться на несколько лет. У облигатных двулетников
(чесночница AUiariapetiolata, проломник северный Androsace septentrionalis,
змееголовник тнмьяноцветковый Dracoccphalum thymiflorum и др.) в розсточ
нон фазе ирохо/цн весь первым год жизни (сезон вегетации). За зиму пни по
218 М Н. Марыш I loiiyiiHiuioiiiiihi (Hioiioi ия pnciciiiih
СЛС перезимовки ЛИСТЬЯ розетки полное ГЫО ИЛИ 110'1111 полное гыо отмирают,
и она заменяется по весне либо равномерно олпственнон частью побега с уд-
линенными междоузлиями, либо безлистной стрелкой, выносящей наверх со-
цветие. В связи с этим характерные особенности архитектуры главного дици-
клического побега для двулетних растений можно только воссоздавать, учи-
тывая рост в первый и второй год жизни.
Озимым и зимующим однолетникам, семена которых прорастают в кон-
це лета или начале осени, свойственны осенний розеточный рост и зимовка
в стадии розетки. Весной розетка отмирает из-за оттока ассимилятов, запа-
сенных ею, в быстро растущие органы генеративной сферы. Этот процесс
можно графически отобразить, зафиксировав перераспределение биомассы
по вертикальным слоям или по структурам растения (см. главу 9).
По вопросу о приспособительной биоэкологической роли розеточности в
научной литературе имеется относительно мало данных. И. Г. Серебряков
(1964), создавая классификацию жизненных форм, решил не использовать
этот признак для выделения крупных таксономических единиц в своей
классификации. Он полагал, что экологическое значение розеточности еще
недостаточно выяснено. Нельзя не согласиться с В. Н. Голубевым (1965),
который считает, что биоэкологическая роль розетки должна оцениваться
по-разному в зависимости от особенностей экологии, ввиду необычайной
широты географического распространения розеточных растений. В услови-
ях средней полосы главное биоэкологическое значение розеточности у ма-
лолетних растений, несомненно, состоит в обеспечении успешной перези-
мовки. Об этом свидетельствует розеточность или временная розеточность
всех озимых и зимующих однолетников, а также розеточность двулетников.
Однако другое значение розеточности состоит в обеспечении «стратегии
избегания» (avoidance strategy см. главу 5).
Розетка способствует быстрому накоплению ассимилятов в осенний и ве-
сенний периоды временно повышенной влажности, временно повышенного
содержания нитратов в почве, а также ослабленной конкуренции за назван-
ные ресурсы и свет с многолетними (или культурными озимыми) растени-
ями. Резерв пластических веществ, накопленный в системе:
розетка <----> корень,
как бы высвобождает, делает менее зависимым дальнейший рост органов
генеративной сферы от быстрого нарастания дефицитов влаги, нитратов,
света (Марков, 1980). Безрозеточная, длиннопобеговая архитектура у мало-
летних растений, наоборот, часто сопряжена со стратегией толерантности
или выносливости к дефициту влаги и нагреву субстрата (тимелея воро-
бьиная Thymelaea passerina), дефициту света (недотрога обыкновенная
Impatiens noli-tangere).
Все сказанное в данном разделе свидетельствует о значительном морфо-
биологическом разнообразии малолетних растений, что не может не отраз-
иться на темпах накопления биомассы или относительной скорости роста,
которые характерны для того или иного вида малолетника.
I 'latta 5 Ионичи i ионные испек гы pocin pm iciiiHI it нонуняцимх 219
8.3. 11опу.111ци<)11111>1Й подход к аппли iy относительной скорости
роста растений
Средние значения RGR, которые могут быть получены для некоторых ни
дов растений в природных популяциях, недостаточно информативны. Дело
в том, что к изучению темпов роста розеточных и полурозеточных малоле i -
них растений целесообразно подходить с позиций популяционного подхо-
да. А это значит, что заранее следует предполагать: 1. изначальную («стар-
товую») вариабельность особей по массе (внутрипопуляционную вариа-
бельность массы одного семени); 2. различия в темпах роста разных особей
(различия индивидуальной RGR) в составе природной или модельной по-
пуляции и влияние плотности популяции на накопление биомассы отдель-
ными особями и урожай; 3. возникновение и динамику «размерной иерар
хии особей» (size chierarchy) в популяциях разной плотности (White, Harper,
1970; Harper, White, 1971; Burdon, Harper, 1980; Weiner, Solbrig, 1984; Weiner,
1988). Рассмотрим эти допущения подробнее.
8.3.1. Влияние различий в стартовой биомассе особей (массе семян)
на рост и конечные размеры особей
В главе 5 была наглядно продемонстрирована внутривидовая (и внутри-
популяционная) изменчивость массы одного семени для некоторых расте-
ний.
Чтобы изучить влияние различий в стартовой биомассе на рост и разви-
тие особей, можно проводить как опыты с созданием модельных популя-
ций, так и наблюдения в природных популяциях растений. Рассмотрим объ-
екты, методики и результаты ряда подобных опытов.
Очевидно, что объекты для такого исследования должны удовлетво-
рять определенным условиям. Хорошо, если они: имеют предельно корот-
кий жизненный цикл (тогда есть надежда получить результаты побыстрее);
проявляют монокогортностъ (проростки их появляются достаточно синх-
ронно); относительно крупносемянны (что сделает возможным достаточ-
но точное взвешивание отдельных семян) имеют обусловленное биологи-
ческими причинами заметное внутрипопуляционное варьирование массы
(Wls) жизнеспособных семян.
На экспериментальном участке биологической станции был проведен
следующий опыт. Семена временнорозеточного незимующего однолетни-
ка редьки дикой Raphanus raphanistrum были разделены на две группы но
диаметру (и, соответственно, массе): 1-я группа — 4,0-4,6 мг и 2-я груп-
па — 6,9-8,4 мг.
Предварительно проведенные проверки влияния размеров семян у редьки
дикой на время прорастания их на фильтровальной бумаге в чашках Петри
дали отрицательные результаты — масса семян не оказывала сколько-нибудь
заметного влияния на время прорастания семени. На это же указывала н ди-
гги
МН Марьин 11<>iiyiiiminшипи (hiojioi ни pm iciinli
Рис. 8.2. Динамика роста семядолей у
проростков редьки дикой, появившихся
из крупных (I) и мелких (II) семян: на-
растание длины (Д) — вверху; нараста-
ние ширины (Ш) внизу; Т — время (дни
от начала наблюдений).
иамика появления проростков в дан-
ном мслкодсляночном опыте в усло-
виях открытого груша.
Как показывает график (рис. 8.2),
и длина и ширина семядолей у рас-
тений, развившихся из более круп-
ных семян, на протяжении всего пе-
риода наблюдений (около 20 дней)
превышали по значениям длину и
ширину семядолей у растений, поя-
вившихся из фракции более мелких
семян. Детализированные феноло-
гические спектры, построенные для
этих двух групп растений (рис. 8.3),
наглядно свидетельствуют о том,
что при свободном стоянии (пло-
щади питания 256 см2/особь) осо-
бей, исключающем ощутимые кон-
курентные взаимодействия, расте-
ния из крупных семян росли более
энергично и опережали по темпам
разворачивания листьев на главной
оси и длине первого настоящего ли-
ста (табл. 8.2.) те экземпляры, кото-
рые выросли из более мелких семян.
Таблица 8.2.
Динамика роста в длину первого настоящего листа у особей редьки дикой, выра-
щенных из крупных (I) и мелких (II) семян
Даты периодических промеров Варианты размера семян Середины классов (мм)
5 10 25 35 45 55
12. VII I 100 0
II 0
14.VII I 76 24 0
II 100 0
16. VII I 22 66 12 0
II 84 16 0
18.VII I 4 52 42 2 0
II 36 52 12 0
20.VII I 0 8 48 36 8 0
II 0 44 52 4 0
22.VII I 0 0 22 54 24 0
II 0 16 60 20 4 0
Примечание: Все числа в таблице — частоты в % для классов, по которым строилось
распределение, причем модальные выделены жирным шрифтом.
/ Л’ Коипчгс । ц<ч11н.п* пенек।и
рост pnciriiiiB н популяциях
22
Изнача ныюс нрсимущсс i но
сохранялось внноть до цветения, ко
торос наступило, по существу, син-
хронно в обеих группах растений.
Таким образом, повлияв на темпы ро-
ста, стартовый запас биомассы почти
не оказал влияния на скорость разви-
тия, т. е. на фенологию как таковую.
Выявление различий между дву-
мя группами, на которые разбит, в
общем-то, непрерывный ряд варьи-
рования семян по массе, может дать
не очень надежные результаты по-
добно линии регрессии, построен-
ной только по двум точкам.
Приняв это во внимание, в после-
дующих опытах с редькой дикой, а
также культурным ее сородичем ре-
дисом R. sativus, высевали семена с
известной массой в центры квадра-
тов (16x16 см), образованных натя-
нутыми капроновыми нитями, на
одинаковую глубину (1 см) при со-
блюдении случайности размещения
8 12 16 20 24 28.VII. /
Рис. 8.3. Детализированные фенологи
ческие спектры для групп особей редь
ки дикой, выращенных из крупных (I) г
мелких (II) семян: С — присутствие се
мядолей; 1-6 — наличие одного iiiecin
настоящих листьев; фазы: б — бутон и
зации; ц — цветения; цп — цветения
плодоношения; Т — время (даты в ваблю
дений).
(в отношении массы).
В таблице 8.3 приведена статистическая характеристика изменчивости
массы семени для обоих видов. При этом можно сравнить по таким пара-
метрам как максимальное (Lim ) и минимальное (Lim ) значения, коэф-
фициенту вариации (CV), коэффициенту асимметрии (As) и коэффициенту
Джини (Gini) образцы семян, взятые для опыта, с фракциями, которые про
росли.
Таблица 8.3.
Статистические параметры для оценки изменчивости массы семени у редьки дикой
Raphanus raphanistrum и редиса R. sativus в образцах, использованных для опытов
Параметры Raphanus raphanistrum R. sativus
N 102 (39) 113(76)
Ыт . тт 1,0 (2,0) 5,0 (6,0)
Ыт max 8,0 (8,0) 19,5 (19,5)
X±Sx 3,2 ± 0,2(4,4 ± 0,2) 11,6 + 0,3 (11,7 + 0,3)
CV (коэф, вариации) 57,0 (33,2) 24,2 (23,3)
As (коэф, асимметрии) +1,2 (+0,75) +0,04 (+0,01)
Gini (коэф. Джини) 0,245 (0,198) 0,135 (0,131)
Примечание: В скобках указаны величины, высчитанные отдельно голь ко для семян,
коюрые иноеiic ii iипп проросли в опыте.
222
Л/.// Марков
I loi|улЯЦПО1 шин (hiojioi ия pnc iспий
UVs — масса семени, мг
Рис. 8.4. Кривые распределения проросших (А) и не проросших (В) в опыте семян
редьки дикой (а) и редиса (б) по классам массы.
Более наглядную картину для такого сравнения дают рисунки 8.4а, б.
Можно видеть, что кривые распределения для всего образца и для фракции
проросших семян у редиса отличаются очень мало, а вероятность прораста-
ния весьма слабо зависит от массы семени. Логично связать такую картину
с тем, что семена редиса как культурного растения подвергались какой-то
предварительной сортировке, прежде чем поступили в продажу. У редьки
дикой асимметрия полигона для фракции проросших семян, равно как и все
показатели изменчивости, существенно ниже, чем для всего образца, а ве-
роятность прорастания при увеличении массы семени резко повышается.
223
I 'fitifiti Kojnrici Ittriiiii.ic ih iickii.i pociii pm iriiiill и попупяцимх
В обеих МОДСНЫ1ЫХ популяциях проростки ПОЯВИЛИСЬ 1104 1 и синхронно
Таким образом, каждая популяция бы на по существу предс тавлена одной
когортой особей. Данные таблиц 8.4 и 8.5, в которых приведены средние
значения конечной биомассы особей для каждого из четырех логарифми-
ческих (In) классов размера (массы) семян убеждают в том, что конечная
биомасса особей у обоих видов во многом предопределялась массой семян,
давших им начало. Данная закономерность проявляется на фоне заметного
возрастания изменчивости общей биомассы, о чем свидетельствуют приве-
денные в тех же таблицах значения коэффициента Джини.
Таблица 8.4.
Зависимость некоторых биоморфологических показателей у особей редьки дикой
Raphanus raphanistrum от массы семян, из которых они появились
Классы по массе семян (мг) Ширина семядолей у проростка (мм) Среднее число листьев на главном побеге Средняя для класса высота особи (см) Средняя для класса общая биомасса особи (мг)
1,8-2,7 15,3 ± 1,7 4,0 ± 0,6 6,8 ± 2,8 322,0 ± 153,7
2,8-4,1 16,5 ±1,2 5,8 ± 0,5 11,0 ± 1,4 619,8 ± 79,6
4,2-6,1 22,2 ± 0,6 6,9 ± 0,7 18,2 ±2,3 930,1 ±93,8
6,2-9,0 23,2 ± 0,8 7,8 ± 1,0 19,0 ± 3,9 1193,3 ±84,5
Коэффициент Gini=0,198 Коэффициент Gini=0,290
Таблица 8.5.
Зависимость конечной общей биомассы особей редиса от массы семян,
из которых они появились
Логарифмические (In) классы по мас- се семени f частота f % Среднее (для класса) значение ширины семядолей (мм) Среднее значение общей биомассы (г) для данного класса
1,5-7,0 7 8,2 26,0 ± 0,9 7,6 ±2,1
7,1-9,9 13 15,3 31,8 ± 1,2 15,0 ±2,6
10,0-14,9 53 62,4 32,3 ± 0,8 23,7 ± 2,0
15,0-22,2 12 14,1 34,4 ±1,1 20,2 ± 3,4
Коэффициент Gini = 0,131 Коэффициент Gini = 0,299
Заслуживает особого внимания и вновь наметившаяся связь между мас-
сой семени (PF) и размерами семядолей у проростка (Ds). К этому можно
дать следующие два комментария.
Первый комментарий. Тесная положительная корреляционная связь, ко-
торую можно зафиксировать между Ds и конечной биомассой особей (W и
InFT) часто оказывается более сильной, чем связь Wx и W(табл. 8.6, см. также
рис. 8.5, показывающий результаты path-анализа — анализа путей корреля-
ционных связей). Судя по всему, именно на этапе формирования проростка
может происходить некоторое «расшатывание» корреляционной связи, но-
224
Л/.//. Марьин
I 1<>1 ly ll>ll Ц К >1111И>1 (>1К> l|( >1 II >1 ]>ПС IVIIIlH
Рис. 8.5. Анализ путей корреляционных связей (path analysis) для модельных попу-
ляций редьки дикой (вверху) и редиса (внизу): жирным шрифтом выделены коэф-
фициенты корреляции между логарифмически (In) трансформированными данными;
уровни значимости — * р<0,05, *** р<0,001, n.s. — недостоверно.
еле чего, по крайней мере, на протяжении относительно короткого перио-
да времени (продолжительность жизни незимующих летних однолетников),
наблюдается ее «восстановление» — связь между стартовой и конечной
биомассой оказывается в итоге статистически высоко достоверной.
Таблица 8.6.
Матрица коэффициентов корреляции между параметрами в модельных популяциях
Raphanus raphanistrum (над диагональю) и R. sativus (под диагональю)
W S 1пЖ D W InlF RGR
W 0,98*** 0,53*** 0,63*** 0,61*** 0,053 n.s.
W 0,61*** 0,66*** 0,67***
D 0,25* 0,57*** 0,60*** 0,34**
W 0,27** о 51*** q 94***
\nW 0,38*** 0,61***
I'flddll N КоЛИЧСС I ВСНН1.1Г 11С11СК1Ы pocill piicirilllil II популяциях
225
Таблица 8.6. (окончание)
RGR 0,52***
In Ий A 0,43***
ln»W 0,32***
Примечание: Уровни значимости: * р<0,05; ** р<0,01; *** р<0,001;
названия параметров см. в тексте
Второй комментарий. Нужно признать, что в полевых условиях прямое
исследование варьирования массы зачатков, из которых появились пророст-
ки, в большинстве случаев провести невозможно.
Косвенным показателем, позволяющим оценить изменчивость веса семян,
т. е. стартового запаса биомассы, является размер семядолей. Наши данные
по двум видам Raphanus хорошо согласуются с данными М. Стантон (Stanton,
1984), которая также обнаружила высокую положительную корреляцию меж-
ду размером семени (массой) и шириной семядолей у редьки дикой.
Редька дикая — очень удобный объект для изучения интересующей нас за-
висимости стартовая биомасса — конечная биомасса еще и потому, что
семядоли у ее особей не только в модельных, но и в природных популяциях
сохраняются часто вплоть до плодоношения. Это позволило однажды проа-
нализировать корреляцию (по величине коэффициента линейной корреляции
Пирсона — г) между массой семядолей (Wsd) и общей массой растения (IV).
используя данные трех последовательно взятых выборок из агропопуляции в
посеве яровой пшеницы. Получены следующие уравнения регрессии:
для абсолютных данных
20 июня №= 3,6 + 0,06 Wd (N = 43; г = +0,497; р<0.001)
4 июня W= 10,01+0,03 ^(N = 38; г = +0,691; р<0.001)
18 июля W= 12,22 + 0,01 ^(N = 27; г = +0,477; р=0,012)
для логарифмически трансформированных данных
20 июня In W= 0,23 + 0,40 1пИ^(г = +0,611; р<0,001)
4 июня In W= 0,25 + 0,50 ln^/(r = +0,610; р<0,001)
18 июля In PF= 0,37 + 0,38 1п^ (г = +0,684; p<0,001)
Как видно из этих уравнений, на протяжении всего периода наблюдений
за агропопуляцией редьки дикой наблюдалась статистически достоверная
корреляция между начальной биомассой (по массе семядолей) и конечной
биомассой (биомассой плодоносящих особей).
В течение нескольких лет связь между размерами семядолей у пророст-
ка и конечной биомассой особи изучали на примере лесных1 незимующих
однолетников — недотрог обыкновенной и мелкоцветковой Impatiens noli-
tangere и /. parviflora.
В 1987 г., проводя наблюдения на постоянных квадратах в двух популя-
циях недотроги обыкновенной (подробнее см. главу 10), в числе других па-
раметров мы отмечали у особей величину семядольного индекса (произве-
дение длины наибольшей семядоли па ее ширину). Это позволило в конце
1 Имеется и виду голераптность к чат ci icii н io и приуроченность к затененным Mecioobiiin
пням.
Даты наблюдений
Рис. 8.6. Рост недотроги (динамика крайних и средних значений общей биомассы) в
двух местообитаниях (пояснения в тексте).
наблюдений определить зависимость выживания и роста особей от старто-
вых размеров (по семядольному индексу) и сравнить эти показатели в двух
местообитаниях, отличающихся по влажности и интенсивности нарушений
со стороны человека. Выжившие особи в «сухом» местообитании достовер-
но отличались от отмерших особей более высокими значениями семядоль-
ного индекса. Во влажном местообитании, вероятно из-за более интен-
сивных антропогенных нагрузок, приводивших к неизбирательной гибели
особей от случайных причин, наблюдалось то же различие, но статистиче-
ски недостоверное.
Для недотроги обыкновенной, как растения затененных местообитаний,
чрезвычайно важен рост особей в высоту, который обеспечивает особям,
опередившим остальных, существенное одностороннее преимущество.
Поэтому представляется весьма показательной высокая и статистически
достоверная положительная корреляция (+0,71 - 0,82, р<0,01) между семя-
дольным индексом, определенным в начале наблюдений, и конечной высо-
той растений. Интересно и то, что корреляция в ходе роста особей почти не
снижалась.
Постоянные наблюдения за ростом особей в высоту дали возможность
сравнить особи, относящиеся к разным классам семядольного индекса, не
только по высоте, но и по биомассе. Биомассу вычисляли по уравнению:
W = 77,4 — 8,4 h + 0,28 h2, где W — общая биомасса (мг); h — высота (см).
Использование уравнения было возможно благодаря высокой положитель-
ной корреляции, которая наблюдалась между высотой и общей биомассой в
течение всего периода наблюдений.
Прямое наблюдение за ростом биомассы проводили с помощью пе-
риодических выборок из обеих популяций. График роста, построенный в
полулогарифмических (In) координатах (рис. 8.6), показывает, что первона-
чальные относительные различия по биомассе между особями сохраняются
I’hifia S Koinpiri niriiiii.ir m iickii»i pocin pm iriniB и популяциях 2?/
на ii|A)тяжспии всего периода наблюдении. 1сли продолжи i ь влево выпрям
ленную кривую роста до пересечения с дач он прорастания, то тсорепгвч г и
можно оценить варьирование веса семян. По нашим данным масса одною
семени колебалась от 1,8 до 6,3 мг. Сравнение с массой семян, извлеченных
из почвы, варьировавшей от 2,4 до 6,5 мг показало хорошее совпадение.
Для изучения динамики семядольного индекса и выяснения момента
стабилизации его значений у особей на самых ранних этапах становления
проростков из-под снега 19 марта 1988 г. были извлечены семена недотро
ги обыкновенной и поставлены вместе с почвой на окно в лаборатории.
Значения индекса и обоих его компонентов стабилизировались примерно
через неделю после начала прорастания.
Данные в целом подтверждают значительную роль различий стартовой
биомассы в обеспечении конечной размерной дифференциации в природ
ных популяциях недотроги обыкновенной Impatiens noli-tangere (см. табл.
8.7). Помимо значений высоты и биомассы (высчитанной опять-таки по
высоте), а также уравнений регрессии и коэффициентов корреляции меж-
ду семядольным индексом и конечной высотой, доказывающих правомер-
ность нашего вывода, в таблице можно видеть различия по длине гипоко-
тиля. Этот показатель представляется очень важным, поскольку от него в
большой степени зависит статус проростка и его дальнейшая судьба в кон-
курентной борьбе за свет.
Таблица 8.7.
Семядольный индекс, длина гипокотиля, конечная высота и конечная биомасса у
особей недотроги обыкновенной на четырех постоянных пробных площадях во
«влажном» (I) и «сухом» (И) местообитаниях
Популя- ция и номер пробной площади Средняя величина семядоль- ного индекса Средняя длина гипоко- тиля (см) 2.VI.89 Конечная высота растений (средние значения) Конечная средняя биомасса особи (мг) Коэффициент корре- ляции между семядольным индексом и высотой (г)
I(N1) 0,95 ±0,1 4,4 ±0,2 32,2 ± 0,6 99,0 +0,80
II(N2) 1,16 ± 0,1 6,9 ± 0,4 34,7 ± 1,3 124,2 +0,69
I(N2) 1,27 ±0,1 8,2 ±0,4 42,9 ± 1,4 233,0 +0,84
II(N1) 2,03 ±0,1 9,1 ±0,3 46,9 ±2,1 302,0 +0.71
Осенью 1989 г. из почвы под популяциями недотроги обыкновенной
были извлечены семена, которые вместе с семенами, собранными в том же
году из зрелых раскрывающихся коробочек, были взвешены. Данные с та ги-
стической обработки приведены в таблице 8.8. Эта работа была проделана,
чтобы получить представление об изменчивости массы семени в двух по-
пуляциях, где в 1990 г. планировалось провести эксперимент с маркиров-
кой проростков.
228
А/ // Марков 11<>nyiihiuu>iищи (iiiojioi ня piiciciinh
Таблица 8.8.
Статистическая характеристика изменчивости массы семени в двух популяциях не-
дотроги обыкновенной (август 1989 г.)
Номер популяции N Масса семени (мг) Коэффициенты:
min X ± Sx max вариации Джини
I 78 1,5 2,8 ± 0,07 4,1 23,2 0,133
II 59 2,0 3,4 ±0,15 5,0 21,1 0,126
В популяции 1(15 мая 1990 г.) и в популяции II (23 мая 1990 г.) все про-
ростки, попавшие в пределы четырех площадок 50 х 50 см, были маркиро-
ваны проволочными кольцами разного цвета в зависимости от значения их
семядольного индекса. Всего было выделено 5 классов индекса, из кото-
рых в популяции I были представлены лишь первые четыре (см. табл. 8.9.).
Частоты особей, представляющих тот или иной класс семядольного индек-
са (они были высчитаны как в момент маркировки, так и в конце опыта, ког-
да растения были выкопаны и высушены) приведены для обеих популяций
в таблице 8.9.
Таблица 8.9.
Частоты распределения особей недотроги обыкновенной по классам
величины семядольного индекса (/) и его статистика в момент маркировки про-
ростков (начало опыта) и в момент выкапывания особей (конец опыта)
Середины классов семя- дольного индекса Популяция I 15.05.90 Начало опыта Популяция II 23.05.90 Начало опыта Популяция I 3.07.90 Конец опыта
f f% f f f%
0,5 21 18,1 6 11,0 14 17,9
1,5 48 42,4 23 41,2 30 38,5
2,5 35 30,7 22 39,4 26 33,3
3,5 11 9,0 3 5,2 8 10,3
4,5 0 1 1,6
N=115 X=l,81±0,08 CV=48,7 Коэффициент Джини 0,206 N=55 X=l,96±0,ll CV=42,7 Коэффициент Джини 0,176 N=78 X=l,86±0,10 CV=48,3 Коэффициент Джини 0,464
Можно видеть, что в популяции I, где в ходе роста растений наблюдалось
существенное изреживание, частоты к концу опыта несколько изменились.
В популяции II все особи сохранились, и потому частоты для них в таблице
8.9 не приводятся.
В популяции I характер распределения по классам семядольного индекса
за время роста почти не изменился.
Гнана S Koiiipici Micinibir m иск I i>i рос i it piu iciinil и иоиуняцпях 2?9
Из данных iа()ницы 8.9 можно закпючить, что выживаемоеiь особей в
популяции I мапо зависела от величины семядольного индекса, чею нспыя
сказать о высоте и биомассе особей на момент взятия выборки. В обеих по
пуляциях тесную зависимость конечных размеров от величины семядоль-
ного индекса демонстрируют уравнения регрессии:
Популяция I Популяция II
W= ~ 56,7 + 160,8 / (г=+0,67*) №= - 48,3 + 68,1 / (г=+0,72*)
In 17= 3,3 + 0,92 /(r=0,78***) lnW= 2,8 + 0,62 /(г=+0,71**).
Зависимость конечного размера (биомассы) особей от размеров семядо-
лей была прослежена и у недотроги мелкоцветковой Impatiens parvijlora,
причем как в модельных популяциях разной плотности (эти результаты см.
ниже), так и в природных.
13 июня 1990 г. из природной популяции, расположенной в парковой
зоне г. Казани под пологом дубово-
липовой рощи, была взята выборка
особей недотроги мелкоцветковой в
стадии цветения и начала плодоно-
шения с сохранившимися зелеными
семядолями. Особи были высушены
до воздушно-сухого состояния, а за-
тем проанализированы по ряду по-
казателей. В частности был опреде-
лен семядольный индекс, измерена
высота, а весовые показатели опре-
деляли после расчленения каждой
особи на части, соответствующие
основным сферам. Весовым мето-
дом у каждой особи была определе-
на площадь листьев.
Как и ожидалось, семядольный
индекс достоверно положительно
коррелировал с массой семядолей,
и от обоих этих показателей силь-
но зависели все остальные абсолют-
ные морфо-биологические показа-
тели, включая массу органов репро-
дуктивной сферы. Ниже на приме-
ре той же недотроги мелкоцветковой
будет показано, что величина семя-
дольного индекса (как коррелят мас-
сы семени) дает себя знать и при си-
туации обострения внутривидовой
конкуренции, связанной с загущени-
ем, обеспечивая особям, появившим-
Рис. 8.7. Гистограмма распределения
проростков по длине семядолей (Isd) и
изменение суммарной длины листьев
(£1) в ходе роста розеток особей про-
ломника нитевидного, имевших на ста-
дии проростков разные по величине се-
мядоли и поделенных по этому при зна-
ку на пять классов: 1 <1,4; 2 1,4 1,6;
3 — 1,7-1,9; 4 — 2,0 2,2; 5 -2,2 мм.
Т время (даты наблюдений).
230 MH MtiphOti I lony IIHUIIOIIIIIDI (>1101101 им рис I Cllllil
ся из наиболее крупных семян, возможность заиметь важное нрснмущсс1 во
в конкурентной борьбе.
Примененный в опытах с двумя видами недотроги метод оценки вариа-
бельности массы семени в популяции по варьированию величины семядо-
лей можно использовать при работе с чрезвычайно мелкосемянными ви-
дами. Например, у проломника нитевидного Androsace filiformis такая ин-
формация была получена путем периодических промеров семядолей. После
прекращения роста семядолей особи из выборки, взятой из агропопуляции
этого вида и пересаженной в вегетационный сосуд, были распределены по
длине семядолей на пять классов. Периодические промеры длины всех ро-
зеточных листьев позволили по суммарной длине получить представ-
ление о темпах роста особей, имевших разные по длине семядоли. Рисунок
8.7 показывает, как сильно сказались на темпах розеточного роста перво-
начальные, казалось бы слабые, различия в размерах семядолей. При этом
почти параллельный ход кривых (отсутствие пересечений) показывает, что
первоначальный размерный статус особей, в ходе роста не существенно не
менялся.
Судя по данным анализа выборки особей облигатного двулетника чесноч-
ницы Alliariapetiolata в конце первого весеннего периода интенсивного ве-
гетативного роста, длина семядолей, как коррелят массы семени, также ока-
зала существенное влияние (г = +0,77***) на темпы накопления биомассы в
этот столь важный для растения интервал времени.
Итак, результаты опытов и наблюдений в природных популяциях при-
водят к выводу о важной роли варьирования начальной биомассы (Wo) в
предопределении размерной дифференциации особей к концу жизненного
цикла, причем складывается впечатление, что условия конкуренции за свет
способствуют не только сохранению, но даже усугублению первоначаль-
ных различий. При этом вероятность повышения статуса особи, занимав-
шей подчиненное положение в начале роста, крайне низка.
Теперь, доказав значение параметра Wo в уравнении экспоненциального
роста: Wt = We RGR 4, обратимся к анализу параметра, который, как уже гово-
рилось, представляет собой относительную скорость роста (RGR).
8.4. Внутрипопуляционные различия RGR и размерная
дифференциация особей
Уже говорилось, что прямое определение темпов нарастания массы всег-
да сопряжено с уничтожением изучаемых особей и невозможностью их но-
вого анализа через какой-то промежуток времени. Для количественного из-
учения роста приходится применять периодическое взятие случайных вы-
борок из популяции, которое позволяет непосредственно определять воз-
душно-сухую (а при необходимости и сырую) массу каждой особи в вы-
борке. Отталкиваясь от допущения, что в исследуемых популяциях все осо-
би появились одновременно и растут параллельно (а это, как будет показано
ниже, далеко не всегда соответствует истине), при достаточном объеме вы-
борок можно получить адекватное представление о динамике среднего зна-
чения массы и его крайних значений.
I uithi S коцц>нч iih iiiii.h in iick i i.i рос iit ])ih iriinh и иопуиицних
Рис. 8.8. Динамика средних и крайних ве-
личин биомассы в абсолютном (а) и полу-
логарифмическом (б) масштабе (по оси X
даты взятия выборок) и динамика логнор-
мальных распределений (в) по классам
биомассы надземных органов (1, 2, 3
последовательные даты взятия выборок) в
агропопуляциях пастушьей сумки из посе-
вов гороха (ромбы) и овса (круги).
По такой методике изучены агропопуляции пастушьей сумки Capsella
bursa-pastoris в посевах овса и гороха (рис. 8.8а). График даст представ-
ление о темпах нарастания среднего и крайних (Lim mm и Lim nix) значений
в абсолютных шкалах, иллюстрирует типичнейшую картину увслпче-
232
Л/ Н Марков 11(>11уляц||он11ия finojioi пя puciciiiih
Рис. 8.9. Ход теоретических кривых роста восьми гипотетических особей в популя-
циях малолетников.
ние размаха варьирования из-за отрыва немногочисленных крупных осо-
бей от основной массы особей, подавленных в росте и обусловивших из-
вестную «инерцию» среднего арифметического и возникновение положи-
тельной асимметрии распределения. Подобные графики роста были полу-
чены нами для многих других видов, включая двулетники, зимующие и не-
зимующие однолетники.
На основании многочисленных эмпирических данных можно теоретиче-
ски представить наблюдаемую картину в виде графической модели, которая
отображает траектории роста всех особей популяции некоего малолетника
(рис. 8.9). Как явствует из этой модели, экспоненциальный рост демонстри-
руют в популяции только самые крупные и средние особи, тогда как основ-
ная масса растений растет в лучшем случае линейно. Размерная дифферен-
циация, скорее всего, возникает очень рано (если уже не на стадии прорас-
тания семян), и особи редко способны в ходе роста существенно изменить
свой статус в сложившейся размерной иерархии.
Следует отметить, что период роста биомассы может захватывать не
только вегетативный рост, но и время генеративного развития, когда перво-
начально полученные преимущества некоторых особей усиливаются.
Переход к полулогарифмическим (In) шкалам (рис. 8.86) позволил вычис-
лить RGR в выше приведенном примере с пастушьей сумкой и установить,
что относительная скорость роста средних особей (в посеве гороха — 0,42
мг/мг><неделя; овса — 0,32 мг/мгхнеделя) меньше, чем темпы роста са-
мых крупных особей (в посеве гороха — 0,56; овса 0,40 мг/мгхнсдсля).
Г/шиа Конину* iiiriiin.ic испек I i.i рос hi ptii iritiiH и нонупяцинх ?JJ
Даже гакам, ка laiioci» бы, небольшая разница в о и ioci и с и ы к и i скорое 1 и ро
era нрнвеиа к юму, что но абсолюнюму значению воздушно сухою веса
надземных органов наиболее крупные особи превосходили средние в 8 рас
в посеве гороха и в 5 раз — в посеве овса (см. рис. 8.8а). При этом нс сто
ит забывать, что примененный графический способ предоставляет нс нои
ную информацию о внутрипопуляционной изменчивости в каждую дату на-
блюдений, поскольку дает картину как бы при взгляде на кривую распреде-
ления сверху. Реальные же логнормальные распределения (рис. 8.8в) в этом
смысле предпочтительнее.
Приведенные данные получены выборочным методом, который при всех
преимуществах прямой оценки имеет, однако, и существенные недостатки.
Во-первых, каждая выборка особей все же уникальна, а увеличение сс
объема в какой-то мере влияет на объективную «случайность» взятия но
следующих выборок.
Во-вторых, изъятие особей может достаточно безболезненно проходить
в опыте, где все можно учесть, но в естественных популяциях такое изъя-
тие влечет за собой последствия эколого-ценотического плана и возраста-
ние роли артефакта.
В-третьих, мы не можем получить полную картину внутрипопуляцион-
ной вариации RGR, изучить характер распределения особей по этому пока-
зателю, а ориентируемся в лучшем случае на сравнение крайних и средних
величин. А между тем чрезвычайно важным представляется мониторит
каждой особи из репрезентативной их совокупности в популяции.
Все это обосновывает целесообразность применения неповреждающего
индеструктивного метода — проведения косвенных оценок роста особей но
линейным, меристическим признакам или их комплексам, позволяющих пе-
риодически наблюдать за одними и теми же маркированными или же кар ти-
рованными экземплярами.
Как у розеточных, полурозеточных, так и у длиннопобеговых видов в пе-
риод вегетативного роста, с учетом представлений о росте как процессе на-
копления фитомеров, такими признаками могут быть число листьев, их раз-
меры (например, максимальный радиус или диаметр розетки), высота рас-
тений. Каждый из этих показателей можно использовать по отдельноеги,
поскольку корреляция их с общей биомассой, как правило, бывает достаточ-
но тесной. А установить это можно с помощью одной, лучше двух, выборок
для вычисления коэффициентов корреляции. Вот один пример. 16 октября
была взята выборка из популяции крупки дубравной Draba nemorosa, про-
израставшей на участках с нарушенной дерниной на железнодорожной на-
сыпи. Все особи вегетировали и находились в розеточной фазе. Через две
недели была взята еще одна выборка. В воздушно-сухом состоянии опре-
деляли массу растений, измеряли длину наибольшего листа (радиус розе г-
ки) и подсчитывали число розеточных листьев. Данные таблицы 8.12 сви-
детельствуют о высокой достоверной положительной корреляции ради
уса розетки и числа розеточных листьев с общим весом растения, причем
корреляция не ослабевала в процессе розеточного роста.
40
1 2
11 Т
Рис. 8.10. Резкое ускорение роста особей — ускорение развертывания розеточных
листьев после искусственного ослабления конкуренции.
Таблица 8.10.
Показатели розеточного роста крупки дубравной Draba nemorosa и корреляция
между ними
Дата взятия выборки Число розеточных листьев Длина наибольшего листа (радиус розетки) см Масса особи мг Коэффициенты корреляции между показателями:
1-м и 2-м 1-м и 3-м
16 окт. 8,3 ± 0,74 1,08 ±0,05 11,9± 1,1 +0,81*** +0,73***
28 окт. 11,6 ±0,70 1,33 ±0,09 23,8 ± 2,8 +0,75*** +0,81***
Число розеточных листьев было использовано для сравнительного изуче-
ния роста вегетирующих особей первого года жизни проломника северного
Androsace septentrionalis в условиях конкуренции с луговыми злаками и при
ее искусственном ослаблении. Ослабление конкуренции достигалось пери-
одическим подрезанием надземных органов злаков на уровне поверхности
Г’1(Н<(1 «V KoHII'ICt I ItCIIIII.IC НСНСК1Ы рос HI put irniiB it нонунчцинх 235
почвы. Для сравнения брались осо-
би пролом! шка северного, у которых
в начальный момент времени было
одинаковое с опытными растения-
ми число розеточных листьев. На
рисунке 8.10. хорошо видно резкое
ускорение роста особей — ускоре-
ние развертывания розеточных ли-
стьев после искусственного осла-
бления конкуренции.
Следует, однако, учесть, что ро-
зеточный рост сопряжен не только
с постоянным разворачиванием но-
вых листьев розетки, но и, с неко-
торого момента, с параллельно иду-
щим процессом отмирания ниж-
них старых листьев1 2. Поэтому важ-
но знать баланс между этими двумя
процессами. Особенно резко усили-
вается отмирание розеточных ли-
стьев при переходе особей к ре-
продукции, хотя в немалой степе-
ни зависит оно и от фитоценотиче-
ских влияний. На рисунке 8.11 мож-
но наблюдать динамику моды рас-
пределений по числу розеточных
листьев в агропопуляциях пасту-
шьей сумки из посевов гороха, овса
и гречихи. Можно видеть, что все
три траектории имеют восходящую
и нисходящую части, хотя ценоти-
ческие условия различны (наибо-
лее благоприятны они были в посе-
ве гороха, наименее благоприятны
— в посеве овса), и потому каждая
из траекторий имеет свои неповто-
римые особенности.
Диаметр розетки может быть ис-
пользован для прогноза будуще-
го роста и развития особи, как это
Рис. 8.11. Динамика моды распределе-
ний по числу розеточных листьев в аг-
ропопуляциях пастушьей сумки из по-
севов гороха (1), овса (2) и гречихи (3):
N1 — число розеточных листьев; Т
время (даты наблюдений); стрелки пока-
зывают, в какую сторону от моды распо-
ложен класс со второй по величине ча-
стотой.
Рис. 8.12. Линия регрессии массы над-
земных органов на диаметр розетки для
особей пастушьей сумки, перезимовав-
ших в посеве озимой ржи: 1 — диаметр
розетки; Wa — масса надземных орга-
нов (мг).
было сделано П. Вернер (Werner, 1975) для ворсянки Dipsacus fullonum. 1 lo
параболической линии регрессии массы надземных органов в период пло-
доношения на диаметр розетки только что вышедших из-под снега зимовав-
ших особей пастушьей сумки Capsella bursa-pastoris можно было прогиозп
1 Вновь следует подчеркнуть, что в отличие от растений с типичной изомодулыюй opiaiiii
записи мы наблюдаем в данном случае не отмирание фитомеров, а отмирание шниь О1дсш>
них 'lacieii фнюмсрон.
236
Л/ Н Марков I loiiyihiHiioiiiiiiH (>1101101 ня pin1 iciinii
Рис. 8.13. Рост розеточных особей (ди-
намика крайних и средних величин био-
массы W) в популяциях герани Роберта
(вверху, RGR=0,162 мг/мг.нед) и чесноч-
ницы черешчатой (внизу, RGR=0,056
мг/мг.нед). Показан способ измерений
параметров листьев, которые были ис-
пользованы для вычисления индекса
ndl — коррелята общей биомассы.
ровать развитие этого растения в по-
севе озимой ржи (рис. 8.12).
Хотя взятые по отдельности при-
знаки могут служить для косвенных
оценок процесса роста, в ряде случа-
ев, по всей вероятности, целесообраз-
но учитывать их в комплексе. Этого
можно добиться, вычисляя индексы:
1) суммарной длины всех розеточ-
ных листьев
п
I,-
i-i
2) произведения числа розеточных
листьев на радиус розетки
I2 = (n X 1);
3) суммы произведений двух пара-
метров (длины и ширины) каждого
листа
п
13= 2(^x1);
i=l
или
4) произведения всех трех параме-
тров розетки
14 = (п X d X 1).
Выше уже был приведен пример
использования первого индекса для
анализа розеточного роста пролом-
ника нитевидного. Второй индекс
был применен при изучении ро-
ста особей проломника северного
Androsace septentrionalis на постоян-
ных квадратах, заложенных в лугово-
степном фитоценозе (см. ниже), тре-
тий — при анализе роста розеток пер-
вого года жизни лесных двулетников
чесночницы Alliaria petiolata и гера-
ни Роберта Geranium robertianum. На
рисунке 8.13 показано, как снимались
промеры с каждого розеточного ли-
ста у этих видов. По модельным гер-
барным экземплярам определяли па-
раметры линии регрессии на величину индекса (13) и получили два урав-
нения: W = 1,52 I3 -I- 0,75 для чесночницы и W = 0,64 1 1,70 для герани.
I 'lllfid Л КоШРНЧ 111Г1ПИ.1С in иск гы рос III pilC I CIIIlll II нонупициич 23/
11угсм би рс де пенни индекса (IJ у особен на ра и них с ыднях, начиная с <|>азы
всходов, cociaBiiena картина pocia н вычислена величина R(iR.
8.5. Влияние плотности популяции на рост растений
В главе 6 уже были достаточно подробно рассмотрены понятия плот-
ность популяции, площадь питания, а также разные методы и подходы к их
определению. Рассматриваемый в данной главе рост растений в его коли-
чественном аспекте как увеличение их размеров делает фактор плотности
весьма динамичным. Этот фактор включается в действие тогда, когда расту-
щие особи, «дорастая» друг до друга, входят в контакт, проявляют результат
взаимодействия. Знак этого взаимодействия тоже динамичен и меняется во
времени. Рассмотрим данные, которые заставляют предполагать закономер-
ной смену знака от 0 (нейтральности) через + (плюс) к - (минусу).
8.5.1. Критерии оптимальной плотности популяции
Исследователю популяционной биологии растений приходится решать
вопрос о том, какая плотность оптимальна для изучаемой популяции и ка-
кие критерии следует применять при определении оптимальной плотности.
Казалось бы, определение оптимальной плотности может быть основано на
состоянии средней особи с учетом ее биомассы и плодовитости. При этом за
оптимальную густоту можно было бы принять ту, при которой масса и пло-
довитость среднего растения будут самыми большими. Однако к определе-
нию оптимальной плотности популяции необходимо подходить, основыва-
ясь на характеристике насаждения в целом.
Дело в том, что, как справедливо указывает К. М. Завадский, густота сто-
яния — это явление популяционной жизни, т. е. явление более высокого
уровня, чем моменты, связанные с жизнью одной особи. Поэтому главны-
ми критериями для определения оптимальной плотности должны служить
выживаемость, урожай биомассы и урожай семян (численность биологиче-
ски полноценных семян с единицы площади), которые объективно отража-
ют процесс осуществления видовой функции. Наиболее целесообразно
использовать эти показатели в комплексе с массой средней особи, не абсо-
лютизируя значение какого-либо одного из них.
8.5.2. Методы исследования влияния популяционной плотности
Изучая влияние повышающейся плотности на рост и развитие растении,
мы решаем важный вопрос о фитоценотической реакции вида. Наиболее
детально это можно выявить в опытах по изучению внутривидовых конку-
рентных и иных (!) взаимоотношений, возникающих при разных степенях
сближения особей одной жизненной формы и качественно одной архнтск
туры. Эксперименты такого рода были широко использованы представлю
лями школ В. Н. Сукачева и Дж. Л. Харпера, а также японскими исследова
гелями (Yoda е( al., 1963 и др.).
238 Л/. //. Марков I 1(>11уЛЯЦ1К>11Н11Я (HIOJIOI НЯ pil(‘ I Cllllil
Методически эти опыты очень сходны: в каких-либо вегетационных со-
судах или на микроделянках одновременно выращивают модельные попу-
ляции разной плотности, чтобы потом иметь возможность рассмотреть и
графически изобразить меняющуюся плотность как ось, вектор. На деле ис-
следователю приходится соединять точки, каждая из которых характеризу-
ет отдельную модельную популяцию через общую биомассу (=урожай био-
массы) и (или) состояние средней особи, ее массу. Естественно стремление
привнести в опыты и истинно динамический аспект путем анализа модель-
ных популяций в разные моменты их роста и развития и вычерчивания тра-
екторий моментального сравнения средней массы особи в модельных попу-
ляциях разной плотности.
8.5.3. Зависимость общего урожая и его компонентов от
плотности
В характере зависимости общего урожая биомассы популяции от плотно-
сти действие лимитирующих факторов проявляется столь же отчетливо, как
в рассмотренной выше логистической модели роста численности (глава 3).
Только по оси X вместо времени фигурирует возрастающая плотность как
мера снижения доступности ресурсов каждой особи, а по оси Y — степень
заполнения одинакового объема пространства (ограниченного взятыми для
опыта сосудами) биомассой.
По мере роста растения занимают все больше и больше пространства, и по-
тому рано или поздно заполняются свободные промежутки между растения-
ми (гэпы) и растения начинают взаимодействовать друг с другом, конкурируя
за ресурсы. Общий урожай (биомасса на единицу площади) при этом при-
Рис. 8.14. Влияние плотности попу-
ляции (число особей на м2) на урожай
надземной биомассы (г/м2) при низких
(1) и высоких (2) дозах удобрений на
примере риса (Алешин и др., 1976, по
Заугольнова, 1988).
ближается к определенному уровню,
который, завися, в общем, от вида
растения и условий местообита-
ния (условий для роста), становит-
ся независимым от плотности выше
ее какого-то порогового значения.
Если на графике величины общего
конечного урожая (сформированного
к концу сезона вегетации или концу
жизненного цикла растений) откла-
дывать против соответствующих зна-
чений плотности, получается харак-
терная кривая с отчетливо выражен-
ной зоной плато. За это названную
закономерную связь между двумя по-
казателями именуют «законом посто-
янства конечного урожая»(соп81аЩ
final yield), и таким образом, уже в са-
мом наименовании зависимости про-
является статичность. Помня о том,
что для роста растениям необходимо
I Л* Копингеiигпныс аспект росni piiciciiiilt к нонуияцнях
234
Рис» 8.15a. Влияние плотности стояния растений на общий урожай и его структуру у
куколя: I — общее число семян на 1 м2; II — средняя масса семян; III — число семян
на коробочку; IV — число коробочек на растение.
время, мы должны учесть — полное описание изменения урожая в зависимо-
сти от плотности популяции должно включать фактор времени. Если модель-
ные популяции (посевы) при каждой плотности посева находятся под наблю-
дением в течение достаточного промежутка времени, то в каждом варианте
плотности кривая хода изменения урожая во времени по характеру прибли-
жается к кривой в координатах урожай — плотность.
Итак, при увеличении плотности популяции от каких-то низких ее зна-
чений общий урожай (сухое вещество, произведенное на единицу площа-
ди) растет сначала, в основном, пропорционально плотности3. Затем ско-
рость роста снижается, и кривая часто выходит на плато, характеризующее
более или менее постоянную урожайность, так называемый постоянный ко-
нечный урожай. Каковы же причины возникновения специфической формы
кривой, отражающей зависимость урожая биомассы от плотности.
Сначала о восходящей части кривой. Первоначальное увеличение уро-
жая при повышении плотности связано с недоиспользованием наличных
ресурсов из-за слишком редкого стояния и вследствие недостаточной на-
сыщенности пространства активными органами растений. Более интенсив-
ная работа некоторых органов растений, наблюдаемая в разреженных по-
севах, не может компенсировать дефицит этих органов в масштабе нонуля-
1 Теорешческн |дсс1,, конечно же, можно ожнд;нь нрояниеиня элементов inaiiMoCuiaiонрпяi
с । кования, о koiopi.ix речь нондс! ниже
240
МН. Мирков
I lonyпнiuioiiiши (hiojioi ihi рас i ciiий
Рис. 8.156. Влияние плотности стояния растений на общий урожай и его структуру у
мака опийного: I — урожай семян; II — масса семян на одну коробочку (г); III — чис-
ло коробочек на растение.
ции в целом. Зона «плато» соответствует «емкости местообитания» и мак-
симальному использованию ресурсов, которые играют роль лимитирующе-
го фактора, не допускающего дальнейшее увеличение урожая биомассы.
Протяженность зоны «плато» у разных видов растений различна (иногда
эта зона превращается в точку) и зависит от индивидуальных свойств вида,
его пластичности (рис. 8.14, 8.15, 8.16).
Возвращаясь к вопросу об оптимальной плотности, можно сделать вы-
вод, что она соответствует зоне «плато». На двух из трех приведенных ри-
сунков (8.15 и 8.16) можно видеть кривые, отражающие снижение массы
одной особи при возрастании плотности. Оказывается, точка максимума
урожая может соответствовать биомассе средней особи, составляющей все-
го около 5-10% от средней биомассы одиночно растущих растений. Это еще
раз убеждает нас в том, что исходить только из состояния средней особи
при определении оптимальной плотности популяции нельзя.
1'ihitui Л Ионичем I iwiiiii.ir m ucKibi
рос hi pnciciiiill и нонуияциях
241
8.5.4: Положительный эффект
группы и отрицательный
эффект плотности или C-D
эффект
В течение нескольких дней после
появления (прорастания) размеры
новых проростков могут стать за-
висимыми от плотности. Чем выше
плотность, тем раньше скажется ее
влияние на накоплении биомассы
взаимодействующими проростка-
ми, и на первых порах это влияние
может быть положительным.
Рис. 8.16. Влияние плотности стояния
растений подсолнечника (числа семя-
нок, высеянных на 1 м2) на общий уро-
жай и его структуру: I — урожай се-
мянок; II — среднюю массу семянки и
III — число семян на корзинку (Clemcnls
et al., 1929, по: Харпер, 1964).
Так, согласно исследованиям П. Г.
Вдовина (1959), при посеве желудей
кучками в лунках происходит хо-
рошо заметное изменение теплово-
го режима почвы в положительную
сторону, одновременно возрастает
разрушающая сила давления про-
ростков на почвенную корку. В ре-
зультате появление всходов ускоря-
ется на 1-7 дней. Повышение температуры почвы в сфере влияния на нес
прорастающих желудей обусловлено усилением их дыхания. Тепло, выде-
ляемое при дыхании одним желудем, почти незаметно, тогда как тепло, вы-
деляемое кучкой желудей, может в известной степени оказать влияние на
тепловой режим почвы.
Подобное же явление установил В. О. Гулканян (1959) в своих опы-
тах с проращиванием семян хлопчатника. Полученные данные позволили
Гулканяну заключить, что при проращивании семян в группе взаимоотно-
шения, возникающие между ними, имеют, видимо, химическую и физиче-
скую природу, причем последняя связана с температурными микроуслови-
ями в гнездах, а первая — с какими-то тонкими химическими процессами.
Интересный тип взаимодействия между прорастающими семенами был
обнаружен А. А. Часовенной (1951). Изучая взаимодействие корней одного
растения, а также корневых систем разных особей на начальных этапах раз-
вития у озимой ржи, овса, ячменя, озимой пшеницы, полевой горчицы, ти-
мофеевки и клевера, Часовенная установила, что с начала прорастания се-
мян до фазы второго-третьего листа наблюдается явление хеморизотакси-
са — стремления зародышевых корней друг к другу. Групповое произраста-
ние корней в указанный период, когда корневые системы еще слабо разви-
ты, благоприятно для роста растений, так как при этом повышается насы-
щенность ризосферы микробным населением и продуктами, его жизнедея-
тельности, усвояемыми растениями. Смыкание корней и корневых систем
на начальных этапах развития растений не означает начала конкурентных
242 М Н. Марков 11(>iiyлмЦ11<>|iiian Giiojioi ня рас I cniiit
отношений, а является моментом взаимоблаатриятствования. что и под-
тверждается более интенсивным ростом корней у растений в группах.
Стадия проростков, совершающих переход от гетеротрофного питания за
счет запасов питательных веществ семени к автотрофному является крити-
ческой. Именно в этот период всходы и молодые растения наиболее беспо-
мощны и в массе могут отмирать. Поэтому проявление элементов взаимо-
благоприятствования в данный момент развития популяции особенно важ-
но. Часто гибель всходов связана с действием внешних факторов, губитель-
ных воздействий среды. Если всходы находятся далеко друг от друга (плот-
ность их стояния низкая), то они очень часто страдают, например, от ударов
капель дождя или забрызгивания грязью. При более плотном стоянии про-
ростков они лучше сопротивляются таким воздействиям, конечно, до опре-
деленного момента загущения.
На вышеописанных закономерностях зиждется применение в сельско-
хозяйственной практике гнездового способа посева. В гнезде, содержащем
группу семян, благодаря временному взаимоблагоприятствованию, происхо-
дит более продуктивная интегральная «ассимиляция условий», а затем в силу
вступают внутривидовые конкурентные отношения, известный фактор есте-
ственного отбора. Наиболее энергично растущее растение одерживает верх и
обеспечивает получение продукции. Особо отметим, что интенсивный рост
его в условиях кратковременного первоначального благоприятствования мо-
жет сказываться вплоть до самых конечных стадий развития.
Вся совокупность элементов взаимоблагоприятствования, проявляю-
щихся на ранних стадиях развития организмов, названа зоологами эффек-
том группы. Ю. В. Титов (1978), предложивший использовать этот термин
в ботанике, полагает, что в ходе развития популяции эффект группы посте-
пенно сменяется эффектом плотности, т. е. происходит смена знака во вза-
имоотношениях между особями одного вида. «Эффект группы,— пишет
Ю. Н. Титов,— возникает уже в случае двух взаимодействующих особей или
во всяком случае в небольших группах растений и проявляется в устойчиво-
сти, стабилизации и активном функционировании группы в целом при всех
возможных частных неблагоприятных отклонениях у отдельных особей. В
большинстве случаев эффект группы является следствием взаимопомощи,
кооперации организмов... В ходе развития естественных или искусственных
сообществ от проростков до зрелого возраста взаимодействия между рас-
тениями могут проявляться как в эффекте группы, так и в эффекте плотно-
сти. На ювенильных стадиях растений проявляются положительные эффекты
группирования особей. При увеличении размера растений с возрастом в со-
обществе часто возникает пороговая плотность особей, при которой начинает
проявляться эффект плотности» (Титов, 1978: 18). Представляется логичным
некоторое уточнение терминологии и отражение в двух названных явлени-
ях знака взаимодействия. Хоть ради этого и придется слегка «удлинить» тер-
мины, но в названиях «положительный эффект группы» и «отрицательный
эффект плотности» гораздо больше четкости и определенности.
В зарубежной литературе кратковременное взаимоблагоприятствованис
между проростками никак не комментируется, хотя сам термин «эффект
Рис. 8.17. Влияние плотности популя-
ции клевера подземного (D) на среднюю
массу особи (W), учтенное в разные сро-
ки после посева и представленное в ло-
гарифмическом масштабе (Donald, 1951,
по: Харпер, 1964).
Iiathi <S' Kouiriri i пенные m пгки.1 рос in pin irnnil н популяциях 243
группы», I ipaivui проиллюсгрпро
ванный на ioojioi пчсском материа-
ле, пришел к нам именно из-за ру-
бежа. Даже когда на графиках в ста-
тьях, посвященных влиянию плот-
ности на рост растений, проявляется
этот эффект, авторы воздерживают-
ся от комментариев, возможно пола-
гая его находящимся на грани стати-
стической достоверности. Однако,
и весьма слабые эффекты представ-
ляют для популяционной биологии
изрядный интерес, а завидная по-
вторяемость, с которой проявляет-
ся в опытах положительный эффект
группы на ранних этапах формиро-
вания активной части популяций у
растений, уже сама по себе набира-
ет искомую поддержку статистиче-
скими методами.
Рассмотрение полученных нами
графических свидетельств проявле-
ния положительного эффекта груп-
пы предварим анализом данных С. Дональда, которые были получены в его
классических опытах с загущенными в разной степени популяциями клеве-
ра подземного (рис. 8.17), а также анализом данных японских ученых (Yoda
et al., 1963, по: Harper, 1977) и Д. Харпера.
Обсуждая график Дональда, отметим несколько важных моментов.
Можно видеть, что в ряду (градиенте) испытанных плотностей на протя-
жении 182 дней опыта отмирания растений не наблюдалось. Именно поэ-
тому в каждом варианте плотности можно, мысленно соединив точки, по-
лучить строго вертикальную линию — траекторию роста средней массы
особи (W), а относительная длина составляющих линию отрезков отражает
приращение показателя за время между освидетельствованиями, иначе го-
воря, темпы роста.
Протяженность этих вертикальных линий с возрастанием плотности убы-
вает — рост массы особей в разных вариантах останавливается на разном
уровне. Поэтому сопоставление вариантов по горизонтали (в каждый из вы-
бранных моментов времени) дает информацию о влиянии разной плотности.
Начало опыта (0 дней) задается горизонтальной траекторией, иллюстри-
рующей равенство средней массы семени во всех комплектах, взятых для
создания модельных популяций, и отсутствие какого-либо влияния плоз по-
сти на эту массу. Однако в дальнейшем (на 61-й, 131-й и 182-й день опы-
та) взаимодействие между особями (внутривидовая конкуренция) привело
к пропорциональному снижению средней массы особи, особенно заме гно-
му при самых высоких плотностях. Если записать зависимость между ин-
244 М Н. Маркин I loiiyihiniioiniihi 61101101 ия paciciiiiri
тересующими нас параметрами как W - Cd11 (где а показатель изменений
в степени использования пространства в процессе роста, и С констан-
та пропорциональности), то с течением времени наклон прямой становил-
ся все круче, выявляясь при все более низких плотностях, пока, наконец, не
достиг угла 45° для любого значения плотности. Показатель «а», следова-
тельно, стал равен единице, поскольку tg45°=l. Зависимость средней массы
особи от плотности приобрела вид W = Cd1.
Нетрудно убедиться, что эта зависимость соответствует зоне плато кри-
вой постоянного конечного урожая. Достаточно умножить обе части урав-
нения на d, чтобы получить в левой части урожай биомассы (Wxd=Y) и за-
писать У = С, то есть урожай оказывается равным константе.
Прокомментируем, однако, сущность и роль константы С. Очевидно, что
ее значение не сводится только к обеспечению пропорциональности. В тео-
рии — это точка пересечения линией регрессии оси У, соответствующая ми-
нимальной возможной плотности (lgO=l), а, стало быть, величина ее как не-
коего коэффициента свободного развития должна отражать массу особи, ра-
стущей свободно, без всяких помех со стороны соседей. Заметим, без всяких
помех, но и без всяких благоприятных влияний! Не потому ли в самой верх-
ней левой части траектории, полученной в конце опыта, мы видим «ступень-
ку» — растущие «на свободе» особи как бы не добирают биомассы.
Виды растений отличаются по значению этой константы, а эмпириче-
ски определенная амплитуда ее изменения находится в области между 3,5 и
5,0 (White, 1985). Нетрудно заметить, что этот размах варьирования на деле
очень внушителен: если в логарифмическом масштабе он и кажется неболь-
шим, то при переводе в линейные шкалы он составляет около 105.
Последний комментарий к рис. 8.17 адресуем времени проведения пер-
вого освидетельствования. Оно в опыте Дональда было осуществлено толь-
ко на 61-й день опыта, а между тем за это время как раз и могла произойти
смена знака взаимоотношений.
В модели Харпера-Уайта (см. главу 3) третья фаза включает коррелиро-
ванные изменения в числе и размерах особей. Таково концентрированное
выражение сущности этой фазы. Харпер считает, что на увеличение плот-
ности популяции растения реагируют двояко. Одни виды проявляют боль-
шую пластичность, изменяя размеры особей без потери способности к раз-
множению, а другие довольно быстро изреживаются. И та, и другая реак-
ции являются регуляторными, причем, если во втором случае численность
и плотность популяции уменьшаются за счет выпадения части особей, т. е.
в ходе изреживания, то в первом случае плотность остается постоянной. У
одних видов растений сильнее развита первая, а у других — вторая реак-
ция, но, как правило, регуляция плотности обеспечивается неким сочетани-
ем этих адаптивных регуляторных реакций.
Наиболее широкой зоной плато кривой постоянного конечного урожая
отличаются виды растений, отвечающие на загущение их популяций в пер-
вую очередь пластической реакцией (Харпер, 1964) или, иными словами,
способные к сильной аккомодации роста (Завадский, 1954, 1968). Под ак-
комодацией роста Завадский понимал способность растений при измене-
Г'ПНШ S КоЛИЧСЧ I ВГНН1.1С aCIICKI l.l рос III pill irniih и нонуияцпих ?/|*
inii! iijioi hoc i и популяции ада111 innio immciimti» уровень рос юных пронес
сов, сохраняя соответствующую иому уровню плодовн ion ь. Пскоюрыс
виды, особенно однолетние сорняки способны к ростовым аккомодациям и
удивительно широких пределах. В опытах Завадского (1957) с марью (>сион
были получены следующие данные: при увеличении плотности популяции
в 2852 раза (с 2,3 до 6552 особей/м2) средняя плодовитость растений сокра
тилась в 2987 раз (с 170260 до 57 семян на особь) т. е. снизилась почти во
столько же раз, во сколько возросла плотность. Урожайность же при ном
осталась почти стабильной, варьируя в пределах между 329 и 360 тысячами
семян на 1 м2. Таким образом, повышение плотности популяций таких рас
тений влечет за собой лишь пропорциональное снижение массы отдельных
особей без изменения суммарной биомассы с единицы площади, причем
даже самые мелкие особи способны переходить в генеративное состояние.
Однако и у столь пластичных видов может наступать момент загущения
популяций, когда начинает в результате внутривидовой конкуренции про
являться самоизреживание, которое усиливается по мере дальнейшего ро
ста плотности.
Большинство экологов понимает под конкуренцией преуспевание одного
организма за счет другого, причем подразумевается, что организмы оказы
вают друг на друга взаимонеблагоприятное влияние.4
Л. Г. Раменский подчеркивал, что само по себе слово «конкуренция» не
сет очень мало информации. Нельзя говорить о конкуренции только в общем
смысле, не указывая, за что происходит конкурентная борьба. Конкуренция
возникает там, где в распоряжении нескольких особей имеются ограничен
ные ресурсы, т. е. конкурируют растения, перехватывая друг у друга что-то,
находящееся в дефиците.
Не раз исследователи пытались заменить термин «конкуренция», усма-
тривая в нем некоторую долю антропоморфизма. Джон Харпер, например,
предлагал заменить этот термин термином «интерференция». Однако згу
замену нельзя признать удачной. Замена английского слова competition, что
означает конкуренцию, на interference, что можно перевести как взаимную
помеху (от глагола to interfere — мешать), еще может быть оправдана. 11о
в русском языке термин «интерференция» выглядит еще более расплывча
тым, чем более привычный термин «конкуренция».
Изучение конкуренции в популяциях растений можно разделить на два
этапа. На первом этапе можно анализировать проявления конкуренции, г.
е. результаты ее действия на популяцию в целом и на отдельные особи.
Второй этап включает анализ причин, наблюдаемых явлений, т. с. ноны г
ки связать внешние проявления конкуренции, и в том числе изреживание, с
изменениями условий внешней среды, которые возникают при увеличении
плотности популяции.
В России изучение причин и закономерностей изреживания началось
раньше, чем этим занялись зарубежные исследователи. Первоначально по
4 В системе взаимоотношений, где характер воздействия одного вида на другой или
воздействие одной особи на другую обозначены гремя значками (I положи rcin.iior
воздействие; о|рицател1.ное воздействие; 0 безразличное отношение), конкуренция
oiмечена двумя минусами
246 Л / // Марков I h >i i у 11 и ц i к я 11 him (hiojioi ня рас i riniii
явление заинтересовало лесоводов, гак как в лесных насаждениях оно на-
блюдалось в очень яркой форме. Но о древесных насаждениях речь пой-
дет позже, а сейчас рассмотрим процесс изреживания и его закономер-
ности в популяциях однолетников. Зависимость числа отмирающих осо-
бей популяции от плотности была наглядно показана В. Кушниренко, Ю.
Хоментовским и Н. Куликом (1960) в опытах с бобами. Как показывают дан-
ные таблицы 8.11, выживаемость увеличивалась с уменьшением плотности,
но в самом редком посеве она опять становилась несколько ниже. Из пред-
ыдущего материала нам становится ясно, что последнее может быть связа-
но с отсутствием взаимоблагоприятных влияний на ранних этапах развития
растений из-за слишком редкого стояния и гибелью особей от случайных
причин.
Таблица 8.11.
Влияние плотности популяции на выживаемость особей у бобов
Площадь питания см2 (величина обратная плотности) 4 16 64 256 1024 4096
Процент отмирающих особей 88,0 59,6 18,8 13,7 10,7 13,9
Регуляторное значение отмирания части особей в популяциях показано в
работе Харпера и Мак Навтона (Harper, Me Naughton, 1962). Авторам уда-
лось показать, что количество зрелых плодоносящих растений в популяци-
ях различных видов мака становится независимым от количества семян,
посеянных свыше определенной плотности, т. е. плотность регулируется и
остается на каком-то постоянном уровне за счет отмирания соответствую-
щей части проростков и молодых растений. Для сравнения исследователи
привели данные о реакции на плотность куколя. Куколь нейтрализует влия-
ние плотности за счет индивидуальной пластичности, и отмирание в его по-
пуляциях стабилизирует плотность только на уровне 2300 особей на 1 м2, а
у маков эта граница находится на уровне 1000 особей на 1 м2.
В нашей стране одним из первых экспериментально изучать изрежива-
ние у травянистых растений начал В. Н. Сукачев. В качестве еще одной пе-
ременной в его классических опытах брались варианты с различным уров-
нем плодородия почвы, которое, как было показано, существенно влияет на
исход фитоценотических взаимодействий.
Результаты его опыта с ромашкой непахучей Matricaria inodora, одни из
немногих, ставших известными за рубежом, показали, что в посевах этого
растения на удобренной и неудобренной почвах происходит изреживание,
зависящее от плотности (табл. 8.12). Тот вывод, что в посеве максимальной
плотности на удобренной почве убыль особей была наибольшей, позднее
назвали «эффектом Сукачева» (Harper, 1977: 182).
Вполне закономерно, что многие особи погибли при самой высокой плот-
ности — мы уже рассматривали примеры, когда изреживание возрастало
при увеличении плотности — но почему процент выпавших растений был
выше на удобренной почве?
Г'нкш <4 Kojiiimci iHciiiii.ic nciiriai.i рос hi piiciriiiHt и нопуняцняч 24 7
Гибл и цн 8.1 2.
I Проживание модсиьных популяции ромашки псиахучсй па улоОрспион и псуш»
бренной почвах в зависимости от плотности
Густота посева (плотность популяции) Почва Убыль особей в % от первоначального числа
Густой посев Неудобренная 5,8
Удобренная 25,1
Посев средней густоты Неудобренная 0
Удобренная 3,1
Редкий посев Неудобренная 0
Удобренная 0
Сам В. Н. Сукачев дал этому следующее объяснение: «Более благо-
приятные почвенные условия влекут за собой более энергичное взаим-
ное состязание, а следовательно, и большую убыль особей». Это объясне-
ние выглядит вроде бы убедительным, но если вдуматься, то оно входит в
противоречие с определением, которое мы даем конкуренции, как борьбе за
что-то, находящееся в дефиците. И вот тогда, когда мы даем растениям удо-
брение, т. е. фактически снимаем конкуренцию за питательные вещества,
среди них усиливается отмирание. В чем же дело? Рассмотрим несколько
предположительных ответов.
Во-первых, можно предположить, что в дефиците находится простран-
ство, за которое и борются интенсивно растущие особи. Примером пря-
мой борьбы за пространство служит физическое выталкивание растущи-
ми корнеплодами, например, морковью, слабых отставших в росте осо-
бей, которое наблюдал в своих опытах с загущенными посевами моркови
К. М. Завадский. Однако этот пример достаточно уникален, и вряд ли ему сле-
дует отводить решающую роль.
Еще один вариант ответа основан на учете динамики обеспеченности рас-
тений питательными веществами. Ф. Милторп (1964: 435) пишет по это-
му поводу: «Часто считают, что наличие среды с первоначально высоким
содержанием питательных веществ снимает значение запаса этих веществ
как фактора конкуренции. Однако это неверное допущение; этот фактор не
будет играть роли лишь в том случае, если почвенный раствор непрерывно
обновляют, доводя его до первоначального состава на протяжении всей кор-
невой зоны»
Реальнее предположить, что, несмотря на первоначально благоприятный ре-
жим питания, растениям все же чего-то не хватает, и этот дефицит приводит
часть из них к гибели. Например, особям, растущим в густом посеве на удобрен-
ной почве и развивающим довольно большую листовую поверхность, конечно
же, может не хватать света. А пластическая реакция, которая могла привести к со-
ответствию величины листовой поверхности степени загущения популяции, нс
срабатывает из-за «асимметричности» конкуренции за свет в отличие от «симме-
тричной» конкуренции за элементы минерального питания, и потому отпад осо-
С)сй возрасгаег.
248
МН Марков 11< >i ly и >i ш к >i 11 in я (>1101101 n>i pnc icinii
Рис. 8.18. Траектории роста средней
массы особи (W) в четырех самоизре-
живающихся модельных популяциях
плевела многолетнего Lolium регеппе с
разной начальной плотностью (d): Н1-
Н5 — последовательные моменты ана-
лиза (Kays, Harper, 1974).
I (ринимая во внимание естествен-
ный процесс изреживания природ-
ных и модельных популяций, отме-
тим, что W (масса растений) во вре-
мени растет, a d (плотность) в опре-
деленный момент начинает сни-
жаться. Это снижение d в наиболее
загущенных популяциях может на-
чинаться, скорее всего, уже и при са-
мых минимальных размерах W.
За рубежом зависимость W от
плотности называется эффектом
конкуренции-плотности (competition-
density) или С-D эффектом.
Хотя часто С-D эффект и сме-
шивают с самоизреживанием (self-
thining), это различные процессы.
Самоизреживание отражает как
плотность и урожай изменяются во
времени в пределах одной и той же
модельной популяции в случае про-
явления зависящей от плотности
смертности особей. Его траектория
формирует верхнюю границу для
траекторий С-D эффекта, описыва-
ющих в какие-то отдельные момен-
ты времени зависимости средней
массы одной особи от плотности в
серии все более загущенных модельных популяций.
На рис. 8.18 представлены результаты опыта Кейса и Харпера (Kays,
Harper, 1974) с плевелом многолетним или райграсом. В отличие от рассмо-
тренного выше графика Дональда на основе пятикратного анализа реаль-
ных плотностей и средней массы генет (а в опыте учитывались также и ра-
меты) вычерчены вертикальные траектории для каждого из четырех вариан-
тов первоначальной плотности. Можно видеть, как все они загибаются вле-
во, отражая снижение плотности в результате самоизреживания.
Пока продолжаются рост (увеличение W) и параллельное отмирание ча-
сти особей, модельные популяции разной плотности дают на графике, от-
ражающем самоизреживание, траектории, которые приближаются к линии
с постоянным наклоном в 56° (tg 56° = 3/2). Осуществляется эмпирическая
связь между средней массой особи (W) и плотностью выживших особей (d),
которая описывается уравнением:
W = Cd'3/2.
I'lllltui S KoilllMVt IIH4IIII.IC IICIICKIbl pOCIlipiK irilllh и попуиициях 249
Значение переменных данного уравнения nci чс уяснить, если иснользо
вать логарифмический масштаб, приводящий зависимость к линейному виду
log W = log С -1.5 log d.
Рассмотренную зависимость (закономерность) стали именовать «зако-
ном степени минус трех вторых» или проще «законом минус трех вторых».
Столь же часто, учитывая явно излишнюю в данном случае торжествен-
ность слова закон, говорят о «правиле минус трех вторых».
Помимо, а иногда и вместо средней массы особи в опытах по провер-
ке универсальности данного правила анализируют популяционный показа-
тель — урожай биомассы (продуктивность), который в конечном итоге ока-
зывается связанным с плотностью зависимостью Y = С d1/2.
На рисунке 8.19 приведены данные, полученные японскими учеными,
впервые выявившими закономерность, Йодой, Кирой и др. (Yoda et al., 1963)
при исследовании природных и модельных популяций мелколепестника ка-
надского. Мы вновь можем видеть на графике, что плотность модельных
популяций с течением времени уменьшалась за счет изреживания. Точки,
каждая из которых обозначает сред-
нюю массу особи при соответству-
ющей плотности и соответствую-
щем уровне минерального питания,
проявляют отчетливую тенденцию
к группированию вдоль погранич-
ной линии с наклоном 56° (tg56° =
3/2). Важно уяснить, что внешние
условия, в частности почвенное пло-
дородие, не вносят существенных
корректив в указанную закономер-
ность. При увеличении скорости ро-
ста в связи с повышенным плодо-
родием почвы число отмирающих
особей в соответствии с «эффектом
Сукачева» возрастает — плотность
уменьшается, но оставшиеся особи
развиваются лучше за счет освобо-
дившегося пространства, а точка все
равно располагается вблизи линии,
графически иллюстрирующей закон
трех вторых.
Одним из главных доводов того,
что траектории не могут пересечь
пограничную линию, выступает гео-
метрическая закономерность, на ко-
торую ссылался Йода с соавторами
(Yoda et al., 1963), объясняя выдви-
гаемое ими нравипо.
Рис. 8.19. Изменение зависимости сред-
ней массы особи от плотности осо-
бей, доживших от начала опыта (7.XI)
до каждого из пяти моментов анализа
(16.XI, 2.IV, 13.V, 25.VI) на фоне в раз-
ной степени удобренной почвы (1-5
участки, на которые вносили удобрения
(N, Р, К, Mg) с постепенным снижением
их дозы в соотношении 5: 4: 3: 2: 1 (по
Yoda et al., 1963).
log W
10'
103
105
III
I — деревья
II — клоно-
образуюгцие
многолетние травы
III — травянистые
многолетники
и однолетники
10‘3
10-
102 1 04 1 06 1 0е logD
10’2 1
Рис. 8.20. Обобщение результатов экспе-
риментальной проверки универсально-
сти закона — 3/2 в чистых насаждениях
древесных видов и модельных популя-
циях травянистых растений разных жиз-
ненных форм (пояснения в тексте) и раз-
ных экологических групп.
250 М.Н. Майкоп I loiiyiiMiuioiiiiiiM (hiojioi пи piiciriiiiil
Действительно, с учетом геоме-
трии формула закона (или правила)
может быть выведена логическим
путем. Если взять какой-то линей-
ный параметр растений, например
высоту, и обозначить его через L, то
масса растения, пропорциональная
объему, который оно занимает в про-
странстве, будет одновременно про-
порциональна кубу линейного пара-
метра L3, а площадь S, занимаемая
этим растением (величина обратная
плотности, т. е. S=l/d), будет про-
порциональна квадрату этого линей-
ного параметра: L2. Если растение,
располагаясь на этой площади, за-
полняет весь доступный ему объем,
как это и происходит в условиях за-
гущения, когда проявляется самоиз-
реживание, отношение объема (или
массы) к площади не должно превы-
шать 3:2. Выразив в обеих формулах
L через плотность d и W, получим 1/
d1/2 пропорционально W1/3, а возве-
дя обе части в куб и вводя констан-
ту получим уравнение закона минус
трех вторых: W = Cd'3/2.
Многочисленные попытки экспериментально проверить универсаль-
ность закона дали результат, приведенный на рис. 8.20. Для множества ви-
дов разных жизненных форм от самых крупных деревьев, например сек-
войи (верхний черный кружок) до рясок (три светлых кружка внизу) пока-
зано хорошее совпадение множества траекторий (показаны в виде заштри-
хованных областей) с теми, которые декларирует закон. Там где популяции
самоизреживаются закономерно, например у деревьев, там мы находим осо-
бенно хорошее соответствие (см. рис. 8.21).
Поскольку закон — 3/2 обязательно предполагает самоизреживание и
снижение общего урожая биомассы с увеличением плотности, формула его
отличается от формулы: W = Cd* 1. Можно предположить в связи с этим, что
область применения закона сдвигается для пластичных видов в сторону
очень высоких, почти нереальных плотностей.
После такого теоретического вступления обратимся к рассмотрению ре-
зультатов ряда наших опытов с модельными популяциями некоторых одно-
летних растений, которым как никаким другим свойственна пластичность.
При анализе роста особей в загущенных популяциях перед иссле-
дователем встает все та же дилемма — применять выборочный метод со
всеми его недостатками или же периодически наблюдать за группой модсль-
S k(>l||l4(,l | lUIIIII.IC H( ИСК I l.l po< III pIH'IfllllB и нонуняциях
log
( родней
MdCCbl
особи W
fo
fo
4L-
Низкая плотность
Высокая плотность
log Плотности популяции (D)
Рис. 8.21. Динамика популяций (от начального момента времени t0 до момента t|0)
древесного растения низкой и высокой плотности, показывающая основные свойства
процесса самоизреживания (по Silvertown, 1987, с изменениями).
252 M.HMapkoti I h>iiyii>ii(ii<>iiii;i>i buoiioi ин рис’iciinii
ных особей и оценивать прирост биомассы косвенными индссгрукгивными
методами. В работе мы пользовались обоими методами.
8.5.5. Опыты с пастушьей сумкой (Capsella bursa-pastoris)
Способностью пастушьей сумки давать в неблагоприятных условиях не-
отенические модификации (Марков, 1976) объясняется то, что популяции
этого растения обладают довольно совершенным механизмом пластичности
или аккомодации роста, в чем убеждает следующий опыт. Прямоугольные
деревянные ящики глубиной 5 см с дном из оцинкованного железа были
разделены перегородками на 10 равных отсеков. Площадь каждого отсека
составляла 200 см2. В отсеки, заполненные светло серой лесной почвой из
пахотного слоя, было посеяно по 1, 8, 50, 100 и 200 семян пастушьей сумки
в трехкратной повторности. Семена высевались прямо на поверхность на
равном расстоянии друг от друга и от края отсека. Опыт проводили в веге-
тационном домике в июле-августе 1974 г.
Наблюдения за прорастанием семян позволили установить небольшое по-
вышение всхожести по мере увеличения плотности посева. Закономерность,
однако, была выражена слабо — разница между крайними вариантами со-
ставила 27%. Образовавшиеся модельные популяции пастушьей сумки име-
ли плотности несколько ниже расчетных, но ряд посевов возрастающей
плотности все же был получен. Фенологические наблюдения выявили не-
которое ускорение развития особей с повышением плотности модельных
популяций (Марков, 1977). Опыт длился около 50 дней. За это время в мо-
дельных популяциях отмирания особей не отмечалось. Только в самом за-
гущенном варианте было несколько растений, сильно отставших в развитии
и находившихся в состоянии крайне сильного угнетения, не оставлявшего
надежды на выживание.
После окончания опыта надземные органы растений были высушены до
воздушно-сухого состояния и взвешены в каждом варианте. На рисунке 8.22
приведена кривая (I), отражающая зависимость между плотностью популя-
ций и средней массой одного растения в абсолютных шкалах. Она имеет вид
гиперболы и в математически выравненном виде (II) очень сходна с кривой,
полученной Р. Пальмбладом (Palmblad, 1968), исследовавшим зависимость
между плотностью и разными биологическими показателями в искусствен-
ных популяциях пастушьей сумки и других растений. Резкое поначалу па-
дение массы особи сменяется затем плавным, асимптотическим снижением
в модельных популяциях увеличивающейся плотности. Зависимость меж-
ду плотностью и урожаем надземной биомассы иллюстрируется кривой III
на рис. 8.22, являющейся наглядным подтверждением компенсации слабого
развития отдельных особей большим их числом. Максимальное загущение
в опыте было, по всей вероятности, допороговым, поскольку у кривой нет
хорошо выраженного «плато».
Р. Пальмблад получил в своих опытах «плато», но площадь вегетаци-
онного сосуда была у него несколько меньше — 182 см2 при одинаковых ко-
личествах высеваемых семян (от 1 до 200).
Рис. 8.22. Сглаженные линии регрессии
средней массы (W) особи (I) и урожая
биомассы (И) на плотность модельной
популяции пастушьей сумки (D): макси-
мальная плотность, (-5850 особей на I
м2) была, очевидно, допороговой, и кри-
вая не вышла на плато.
за: 6 и 24 августа и 1 сентября
года.
I S Kouirirt i hciiiii.ic ;к иск i i.i рос in piu iriiini п нинуняцнях 253
Для aiHimru динамики нараста-
ния биомассы в посевах пас'гушьсй
сумки разной плотности был зало-
жен другой опыт. В нем, как и в пер-
вом опыте, были применены ящики
с отсеками. Было взято две плотно-
сти высева — 25 и 50 растений на
отсек в трехкратной повторности.
Вследствие того, что часть се-
мян не взошла, реальные плотно-
сти оказались иными, но соотноше-
ние их осталось близким к расчет-
ному (1:2). Число растений на отсек
по повторностям распределялось
следующим образом: I вариант —
12, 15, 10; II вариант — 32, 34, 35.
Порядок повторностей в вариантах
соответствует очередности, в кото-
рой они попарно (по одной из каж-
дого варианта) брались для анали-
1974
Первая проба, взятая 6 августа,
показала, что средняя масса одного
растения была почти одинаковой в
обоих вариантах. В более густом по-
севе она была даже несколько выше,
чем в редком. Возможно, это было связано с временным взаимоблагоприя г-
ным влиянием ювенильных особей друг на друга на первых этапах развития
модельных популяций или положительным эффект группы. В дальнейшем
ход увеличения среднего веса был различным в посевах разной плотности,
и благодаря этому разница между вариантами сменилась на обратную и со
временем возросла. 1 сентября, по данным последней пробы, вес одного
растения в первом варианте был в три раза выше, чем во втором (рис. 8.23).
В обоих вариантах второго опыта было отмечено нарастание по-
ложительной асимметрии распределений по классам массы надземных орга-
нов. Только в самой первой пробе (6.VIII), где все растения были на началь-
ных этапах роста и слабо взаимодействовали, кривые распределения в обоих
вариантах были почти симметричными. Дальнейшее увеличение проходи ио
быстрее в загущенном варианте, отражая более быстрое нарастание ос i poi ы
конкурентных взаимодействий между особями. Загущение отразилось и па
динамике развертывания розеточных листьев. Синхронность этого процес-
са нарушалась у особей, слагавших загущенные модельные популяции, и ио
явилось одной из причин возникновения положительно асимметричных кри-
вых распределения по массе.
254
Л/ Н Маркин 11 Qi । у и я । цк>1 пшя luiouoi пи рис icniih
Рис. 8.23. Проявление положительного
эффекта группы в модельных популяци-
ях пастушьей сумки разной плотности.
Процесс пей|раличации повы-
шенной популяционной плотности
за счет компенсационных явлений и
связанный с ним процесс возникно-
вения положительной асимметрии
распределений можно считать ре-
гуляторными. Заслуживает внима-
ния тот факт, что в данном случае ре-
гулируется не численность особей,
а насыщение доступного простран-
ства ассимилирующими органами,
т.е. полнота использования ресурсов
среды популяцией. Степень такого
насыщения достигает у пастушьей
сумки внушительных величин.
8.5.6. Опыты с модельными
популяциями клоповников
сорного (Lepidium ruderale) и
густоцветкового (L. densiflorum)
Розеточный рост двух видов кло-
повника в июне-ноябре 1982 г. изу-
чали в пластмассовых кубических вегетационных сосудах объемом 7 х 7 х
7 см, заполненных серой лесной почвой. В двух опытах плотность модель-
ных популяций менялась от 1 до 100 особей на сосуд, что соответствовало
200-20000 экз./м2.
В первом опыте, где набор плотностей был 1-50-100 особей на сосуд,
создали два варианта плодородия. Более бедную почву получали путем до-
бавления к серой лесной почве речного песка в соотношении 5:3. При об-
суждении результатов для краткости почву будем условно именовать «бед-
ной» и «нормальной». Семена клоповника густоцветкового были высеяны 7
июня, а сорного — 16 июня прямо на поверхность. Размещение регулярное.
Опыт проводили в вегетационном домике. Прорастание у кл. густоцветко-
вого началось 13, а у сорного — 18 июня, было дружным, и за 4 дня в обо-
их случаях оно завершилось. Судя по графикам (рис. 8.24), никакой отчет-
ливой зависимости между плотностью и прорастанием не прослеживалось.
На бедной почве энергия прорастания была чуть ниже.
В каждом варианте плотности и плодородия было выделено по 5 модель-
ных растений, у которых 4 раза за время опыта (5,10,17 и 22 июля у клопов-
ника сорного и 6, 11, 18 и 22 июля — у кл. густоцветкового) подсчитывали
число розеточных листьев и измеряли длину наибольшего из них. Помимо
этого, 5 и 19 июля (у клоповника густоцветкового), 9 и 25 июля (у кл. сорно-
го), были взяты пробы по одной повторности из каждого варианта для анали-
за веса растений и прочих показателей, а 5 ноября опыт был завершен выкоп-
I Конпчгс I IICIIIH.ir IICIICKII.I рос I II piu ICIlIlh и нонупяцпях
кой всех рас iciinii, высушиванием до воздушпосухого веса, измерением дин
цы наиболынс!о розеточного листа (радиуса розеток), нодсчеюм числа рои*
точных листьев, определением биомассы надземных и подземных органов.
Во всех вариантах плотности наблюдался переход некоторых особей к л
густоцветкового к репродукции (в первый год жизни!). Плодоносящих осо
бей было больше на бедной почве при плотности 50 (12-24%), а при нлоз
ности 100 на бедной почве к репродукции перешло лишь 4% особей, тогд;
как на нормальной почве — 8%. У кл. сорного к репродукции перешла вес
го одна особь, росшая изолированно (плотность 1) на нормальной почве.
В таблице 5.13 приведены данные о росте модельных особей в виде ди
намики средних (из пяти значений) величин радиуса розетки и числа розе
точных листьев. Кривые распределения по числу розеточных листьев и ра
диусу розетки (рис. 8.25) показывают, что оба эти показателя могут быть ис
пользованы для анализа розеточного роста в условиях различной плотное и
и различного плодородия почвы. Трудно сказать, который из этих параме-
тров является более чутким. То один, то другой позволяют улавливать ела
бые различия в условиях роста. Хорошо видно, что к концу опыта (5 ноя
бря) в модельных популяциях клоповника густоцветкового резко возросл.
дифференциация особей по числу розеточных листьев. На фоне этого усн
ления высокая плотность популяций (100) практически нивелировала раз
личия в плодородии почвы, тогда как при меньшей расчетной плотное и
(50) проявился не только более интенсивный рост, но и разница в условия:
минерального питания.
Рис. 8.24. Динамика прорастания семян в опытах с модельными популяциями kjioi к>i
ников густоцветкового (слева) и сорного (справа) разной плотности. Выращенным
при двух уровнях плодородия: Т — время (даты наблюдений); G — процент нр<
росших семян; плотность популяций число экз./сосуд: на нормальной почве (I 5(
? 100) па бедной почве (3 50; 4 100).
256
МП. Мирков
I loiiyцяцпопнпя biioiioi ня piicieiini
Рис. 8.25. Кривые распределения по числу розеточных листьев (А) и радиусу розет-
ки (Б) особей модельных популяций клоповника густоцветкового (L.d.) (L.r.) на нор-
мальной (темные значки) и обедненной (светлые значки) почвах в разные моменты
времени: левые графики для расчетной плотности 100 экз./сосуд, правые — для рас-
четной плотности 50 экз./сосуд.
Гики! S Kouipiri iiu’iiiii.ic аснскi ы рос hi pm iriinii нпопуляциях 257
У клоповника сорного кривые распределения но числу ihici ьев 25 июня
отличались мало: и для двух вариантов плодородия, н для двух высоких
плотностей можно отметить совпадение модальных классов. Длина листа,
наоборот, хорошо диагностировала и различия в плодородии и разницу в
плотности модельных популяций.
Результаты графического анализа зависимости средней массы надземных
органов особи от плотности в логарифмических координатах приведены на
рисунке 8.26. Видно, что наклон всех линий регрессии приближается к 45°,
т. е. это линии компенсации слабого роста особи при повышенной плот-
ности большим числом особей при слабо меняющемся урожае биомассы.
Графики также указывают на большую отзывчивость кл. сорного на обедне-
ние субстрата по сравнению с кл. густоцветкового — в каждую из дат взя-
тия проб, кроме первой, варианты по плодородию значительно отличаются
друг от друга, и линии регрессии рассредоточены. Иная картина наблюдас г-
ся у кл. густоцветкового — линии регрессии, соответствующие двум вари-
антам плодородия, почти сливаются друг с другом в каждый из сроков ана-
лиза.
Во втором опыте, заложенном 30 июня 1982 г., был взят следующий на-
бор плотностей в пятикратной повторности: 1, 4, 9, 16, 25 особей на сосуд,
что соответствовало 200-5000 особей/м2. В качестве субстрата использова-
ли серую лесную почву. Так же как и в первом опыте, семена раскладыва-
ли на поверхности почвы с тем же расчетом, чтобы они находились на рав-
ных расстояниях друг от друга. Прорастание обоих видов началось 2 июля и
Рис. 8.26. Зависимость средней массы особи (W) от плотности (D) в модельных по-
пуляциях клоповников сорного (L.r.) и густоцветкового (Ld.) в логарифмических (In)
коорди птах
258
М Н Маркин I h >i iy л я цп< >i 11 тя biioiioi пн p;ic iciuiN
завершилось в течение трех дней. Реальные плотности мало отличались от
расчетных, но в варианте с одной особью на сосуд у кл. густоцветкового две
повторности выпали из-за непрорастания. 21 и 30 июля по одной повторно-
сти из каждого варианта (кроме варианта плотности 1 у кл. густоцветкового)
и 6 ноября по три повторности из каждого варианта были взяты для анализа
Рис. 8.27а. Проявление положительного эффекта группы в опыте с модельными по-
пуляциями разной плотности (варианты от 1 до 25 особей на единицу площади) кло-
повника густоцветкового — Lepidium densiflorum (вверху) и клоповника сорного —
Lepidium ruderale (внизу). Слева: ход изменения средней биомассы особи во време-
ни (вертикальными пунктирными линиями выделен период перестройки отношений
и, соответственно, статуса варианта по значению средней биомассы); справа: пере-
ход от взаимоблагоприятных отношений к конкурентным, который заметен по изме-
нению траекторий связи логарифма средней биомассы особи (InW) с логарифмом
плотности (In d) в модельных популяциях, проанализированных в разные моменты
их развития: 21 .VII - 5.XI.
/ /игб/5 К()Л||*|<ч ihriiiii.ic аспект pen in pm iciiiiii n iн>ily11яiиiих 259
воздушно cyxoii массы наряду с дру-
гими парамо i рами, как в нервом опы-
те.
Раннее взятие первой пробы по-
зволило более ярко, чем в опыте с
пастушьей сумкой, выделить поло-
жительный эффект группы. Графики
(рис. 8.27а) показывают, что для кл.
сорного удалось получить траекто-
рию слабого возрастания средне-
го веса особи по мере увеличения
плотности (InW = 0,09 InD + 2,3),
а первоначальное отставание в ро-
сте одиночных особей сказывалось
даже на более поздних пробах. Для
кл. густоцветкового в первую дату
зафиксирована менее определен-
ная картина, а линейная регрессия с
учетом плотности 4 особи на сосуд
имела вид InW = 1,95, т. е. указыва-
ла на константность биомассы при
разных плотностях. При графиче-
ском анализе темпов относительно-
го роста (рис. 8.27а) для обоих видов
были получены S-образные кривые,
которые условно можно было поде-
лить на три отрезка. Хорошо вид-
но, как первоначальный порядок то-
чек на первой вертикальной линии (дата первого взятия и условная граница
первого этапа роста) менялся в промежутке между первой и второй датами
взятия проб, после чего оставался почти неизменным до окончания опыта.
Перестановку точек в данном случае можно рассматривать как иллюстра-
цию перехода от эффекта группы к эффекту плотности.
А. Д. Рогаченко (1988) в своих опытах с кукурузой получила данные,
представленные на рис. 8.276, которые также хорошо иллюстрируют этот
переход. По сравнению с посевами традиционной плотности (4-5 особей/
м2) особи кукурузы в условиях повышенной плотности (16 особей/м2) со-
храняют преимущество по высоте в течение всего онтогенеза, а при сверх-
загущении — до 40-го дня после появления проростков.
В течение 30-40 дней после появления проростков масса одного расте-
ния кукурузы выше в популяциях высокой плотности.
Подсчитанные значения RGR для трех отрезков времени приведены в
таблице 8.13. Они подтверждают данные графического анализа.
Рис. 8.276. Проявление положительно-
го эффекта группы в посевах кукурузы:
соотношение масс одной особи (mn), вы-
ращенной при высокой плотности (п)
и плотности 4 особи/м2 (т4) превыша-
ет единицу на протяжении 30 (1) - 40
(2) дней после появления проростков и
лишь на 50-й день (3) стабильно ниже
единицы (по: Рогаченко, 1988).
260
Л/ Н Маркон I loi iy и я щи >i 111 п я biionoi им pnc iciiiiii
Таблиц» 8.13.
Значения относительной скорости роста (RGR) для 'грех отрезков времени в опыте
с разными плотностями модельных популяций клоповников (мг/мг.неделя)
Плотность модельных популяций L. ruderale L. densiflorum
I II III I II III
1 0,86 2,76 0,10
4 0,97 1,67 0,06 0,84 1,70 0,06
9 1,08 0,90 0,06 0,68 1,20 0,08
16 1,08 0,40 0,06 0,76 1,20 0,04
25 0,88 0,70 0,06 0,84 0,60 0,06
Примечание: I, И, III — временные интервалы (см. рис. 8.27).
Если вновь обратиться к двум важным параметрам розеточного роста — чис-
лу розеточных листьев и радиусу розетки, то линии регрессии их на плот-
ность в полулогарифмическом масштабе в целом дают такую же картину,
как и биомасса надземных органов (рис. 8.28), причем поведение перво-
го параметра позволяет говорить об эффекте группы у обоих видов с боль-
шим основанием. Поведение второго параметра может быть менее опреде-
ли
• 21.VII
О 30.VII
5.XI
гг
4 21.VII
A 30.VII
□ 5.XI
Рис. 8.28. Проявление эффекта группы в модельных популяциях клоповника сорно-
го (справа) и густоцветкового (слева).
Г'пкш N Konirict iMriiin.tr nciicia i.i р<н 111 piu iriiiih it популяциях 261
Леино в силу ioio, что при загущении иногда наблюдается известная и ио
ляция, проявляющаяся в удлинении листьев за счет удлинения их черешков
Возвращаясь вновь к рисунку для рассмотрения линий регрессии, соотвст-
ствующих двум последним датам взятия проб, можно сделать вывод, что за-
висимость между биомассой и плотностью приобрела следующий характер:
lnW= 4,9 - 0,641nD; lnW= 5,8 - 0,66 InD (у кл. сорного) и
lnW= 4,4 - 0,531nD; InW = 5,9 - 0,81 InD (у кл. густоцветкового),
т.е. приблизилась лишь к W = С D1, но показатель степени так и не достиг
значения - 3/2. Сходный вид имела и зависимость между массой корня и
плотностью.
Теперь обратимся к зависимости урожая биомассы от плотности (рис.
8.29). Более или менее типичную кривую постоянного конечного урожая мы
видим только в опыте с плотностями 1, 50, 100 и только в первые две даты
взятия проб. В последнюю дату взятия выборок одиночно росшие особи до-
гоняют (или почти догоняют) по продуктивности загущенные модельные
популяции. Еще более ярко выступает чувствительность клоповника сорно-
го к обеднению субстрата, которое приводит к резкому снижению продук-
тивности у этого вида.
Несколько иной облик имели кривые в опыте с плотностями 1, 4, 9, 16,
25. Сказывается меньшая продолжительность опыта и допороговые вели-
чины плотности — с увеличением плотности модельных популяций уро-
жай биомассы (общей и надземной) растет, а «плато» заметно лишь в пер-
вой пробе. Раздельный учет общей и надземной биомассы в данном случае
Рис. 8.29. Зависимость урожая общей биомассы (Y) от плотности модельных попу-
ляций (D) клоповников густоцветкового (А) и сорного (Б), выращенных на нормаль-
ной (сплошные* IIIIIHHI) и бедной (пунктир) почвах.
262
М/1. Марков I loilyлящи>iiiinn (hioiiqi ия рис iciinii
Рис. 8.30. Зависимость урожая общей и
надземной биомассы от плотности мо-
дельных популяций клоповника сорно-
го и ее динамика: Y — урожай общей
биомассы;Yh — урожай надземной био-
массы; D — популяционная плотность
(экз./сосуд).
служил выявлению все большего
расхождения между этими показате-
лями (рис. 8.30) и демонстрации, та-
ким образом, постоянного возраста-
ния относительной роли подземных
органов в создании общей продук-
ции биомассы.
8.5.7. Опыты с модельными
популяциями подорожника
большого (Plantago major)
и проломника нитевидного
(Androsace filiformis)
Поскольку в реакции вида на
популяционную плотность нема-
лая роль принадлежит особенно-
стям его архитектуры (Lonsdale,
Watkinson, 1983), по сходной мето-
дике были проведены еще два опы-
та с модельными популяциями двух
видов с другой, нежели у пастушьей
сумки и видов клоповника, архитек-
турой. Были взяты розеточные рас-
тения с системой придаточных кор-
ней подорожник большой и проломник нитевидный.
Подорожник большой Plantago major — это потенциальный поликарпи-
ческий многолетник, который был использован в качестве объекта в уже
упомянутом опыте Р. Пальмблада. Р. Пальмблад, испытавший подорожник
большой на отношение к повышенной плотности, отметил, что этот вид
способен дать семена в первый год жизни «любой ценой». Это свойство и
позволяет п. большому выживать в условиях агрофитоценоза и в прибреж-
ных ценозах фактически путем перехода к однолетнему образу жизни.
В опыте, проведенном в лаборатории с 17 апреля по 23 июня 1983 г., соз-
давали 4 плотности — 1, 9, 25 и 49 особей на сосуд, что соответствовало
200-10 000 особей/м2. Эти плотности были выбраны с таким расчетом, что-
бы захватить некоторый средний диапазон загущения, но исключить очень
высокие плотности, иногда встречающиеся в природе. Дело в том, что на
первом этапе наблюдения вели за каждым семенем и появляющимся про-
ростком с помощью картирования (в целях точной регистрации момента по-
явления особи), и потому использование более высоких плотностей сдела-
ло бы процесс наблюдений излишне трудоемким. На графике (рис. 8.31.),
где приведены линии регрессии общего веса и веса подземных органов на
плотность в логарифмических (In) координатах, отчетливо прослеживается
отставание в росте одиночных особей, заметное в оба срока взятия проб
так проявлялся в опыте эффект группы. Линии регрессии для обеих дат по-
Г'ШК<1 N КоПИЧСГ I НСНПЫС IICIICK’l 1.1 рос I'll рис icimit n популяциях ?63
строены без учета массы одиночных
растений н описываются следующи-
ми уравнениями:
lnW= 5,5 - 0,591nD; InW = 4,8 -
0,63 InD (17.V) и
lnW= 8,2 - l,001nD; InW =7,1-
1,00 InD (23.VI),
где Wr — биомасса корневой систе-
мы, a W — общая масса растения.
Продолжение каждой из линий до
пересечения с осью ординат показы-
вает разницу с реальным весом оди-
ночной особи или ее корневой систе-
мы. Эта разница, уменьшавшаяся со
временем (от 17 мая до 23 июня), по-
видимому, может рассматриваться
как своеобразная количественная
оценка эффекта группы, а точнее его
последействия.
В опыте с проломником нитевид-
ным A. filiformis, выращиваемым
при реальных плотностях 1, 4, 6, 9,
11 экз./сосуд, проба была взята на-
столько поздно, что масса одиноч-
но росших растений оказался выше,
чем средняя масса особи при плот-
ности 4. Но и здесь можно усмотреть
(рис. 8.32) известную инерцию —
свидетельство первоначального от-
ставания в росте одиночных особей.
Данные (они приведены для общей
биомассы, и для массы подземных
органов) по четырем другим плотно-
стям (от 4 до 11) строго выстроились
в линию в логарифмических (In) ко-
ординатах, причем для общей массы
и массы подземных органов линии
оказались параллельными. Их урав-
нения, соответственно InW = 10,9 -
1,23 InD и lnWr = 9,2 - 1,22 InD, пока-
зали, что даже без отмирания особей
(без самоизреживания) наклон линии
W-I) может быть выше 45°.
Рис. 8.31. Динамика зависимости общей
массы (W) и массы подземных органов
(Wr) от плотности (D) модельных попу-
ляций подорожника большого.
Рис. 8.32. Динамика зависимости общей
массы (W) и массы подземных органов
(W ) от iiJKiriiocin (I)) модельных нону
пяцнй нропомника пи 1СВ11ДНОГО.
264 Л/// Маркин 1 l<Hiymiiuioiiiin>i biioiioi пя расiciiiih
При учете минимальной плотности наклон заменю меняется в сторону
уменьшения угла:
InW = 8,9 - 0,9 InD и lnWr = 7,4 - 0,8 InD
Масса систем придаточных корней при разной плотности, как мы ви-
дим, подчиняется общей закономерности. Урожай биомассы проломника А.
filiformis (табл. 8.16) оказался очень сходным при всех плотностях.
Таблица 8.14.
Урожай биомассы в мг на сосуд (49 см2) при разной плотности модельных
популяций проломника нитевидного
Плотность модельных, популяций 1 4 6 9 11
Урожай 1102,0 1333,6 1070,4 978,0 1133,0
8.5.8. Опыты с недотрогой мелкоцветковой Impatiens parviflora
В качестве длиннопобегового вида для изучения влияния плотности на
рост особей выбрали недотрогу мелкоцветковую Impatiens parviflora, при-
чем наблюдения вели как в модельных, так и природной популяциях.
Модельные популяции были созданы в кубических пластмассовых веге-
тационных сосудах объемом 7x7x7 см, куда вместе с почвой пересадили
проростки в фазе двух семядолей из естественной популяции. Популяция,
из которой были взяты проростки, занимала плоскую горизонтальную тер-
расу, расположенную на склоне оврага в черте г. Казани и покрытую, по-
мимо зарослей недотроги, крапивой двудомной и кое-где редко разбросан-
ными проростками клена американского Acer negundo. Плотности, выбран-
ные для опыта, составили в пересчете на 1 м2: 200,400, 600, 800,1400,1600,
2000 особей. Повторность в каждом варианте плотности была пятикратная.
Все вегетационные сосуды были составлены плотно друг к другу, окруже-
ны защитной полосой из одного ряда сосудов также с пересаженными рас-
тениями и каждый день (за исключением воскресных) порядок расстановки
опытных сосудов меняли по принципу случайности.
Растения хорошо перенесли пересадку — ни одно растение после пере-
садки не погибло, а отмирание единичных особей наблюдалось значитель-
но позже из-за поражения средней части гипокотиля патогенным грибом.
В природной популяции была намечена сеть квадратных площадок 7x7
см и подобраны площадки с разными плотностями. Таким путем был полу-
чен ряд возрастающих плотностей, в основном соответствующий ряду, ко-
торый фигурировал в эксперименте (200, 600, 1000, 1400, и 2000 особей/м2.
И в модельных, и в природной популяциях недотроги мелкоцветковой с 22
мая до 26 июня 1989 г. вели периодические наблюдения: измеряли длину ги-
покотиля и каждого междоузлия, а также общую высоту растений. У особей
модельных популяций, кроме этого, для определения семядольного индекса
(см. выше) измерили размеры (длину и ширину) семядолей i юс нс достижс-
/'///(/</ 5 К(>1|||»|(4 incillll.ir ЛСПСКН»! pocill pm irilllit И 11ОИУИИЦ1И1Х
пня ими koi lei i к»i i величины. В конце oiiuia ii в конце наблюдении (2(> 2/
июня) все особи были выкопаны bmccic с корневыми сис1смамн, высушены
до воздушно сухого состояния, после чего подвергнуты взвешиванию но ча
стям. Отдельно были взвешены репродуктивные структуры, листья, стебпи
и корневые системы, определена площадь листьев.
Наклон линий регрессии общей массы особей на плотность модельных
и природных популяций был ниже и только приближался к 45°. Это сви-
детельствует, во-первых, о том, что объем местообитания почти заполнен
биомассой недотроги мелкоцветковой, но не настолько, чтобы у нес рас
тения теневыносливого — проявилось изреживание. В природных условиях
действие повышенной плотности было явно смягчено отсутствием псрего
родок, отграничивающих друг от друга площади питания растений, как в
опыте.
Вместе с тем в условиях опыта плотность явно повлияла на рост рас ге-
ний в высоту, длину и число междоузлий, заложение органов репродукции
и его динамику. Последнюю хорошо представляет рисунок 8.33, который
Номер у (ни iy пигрх по главному побегу
Номер узла снизу вверх по главному поСмч у
со
>сгв
ДИ
по-
огн
266 МН Мауков I loiiyiiMnnoiiiniM (hiohoi ин ])iu iciiidi
позволяет сделать следующие выводы: 1. Заложение генеративных побегов
идет снизу вверх — от третьего узла и выше; 2. Только у одиночно растущих
особей со 100%-ной частотой возможно заложение генеративных побегов в
третьем снизу узле. При более высоких плотностях модельных популяций
доля особей, сформировавших генеративные побеги в третьем узле снизи-
лась до 72-42%; 3. При минимальной плотности к 26 июня соцветия зало-
жились во всех узлах, начиная с третьего и выше. При средней плотности
(5 экз./сосуд) процесс заложения соцветий продолжался вплоть до 26 июня,
но только в четвертом узле соцветия заложились у всех без исключения осо-
бей, а частота заложения соцветий во всех выше расположенных узлах рез-
ко снижалась.
Таблица 8.15 содержит данные о взаимосвязи семядольного индекса со
средними значениями конечной биомассы особей, выросших в условиях
разной плотности модельных популяций. Налицо статистически достовер-
ная детерминация конечного размера особей размерами семядолей как кор-
релята массы семени, причем в эту детерминацию плотность популяций
вносит свои коррективы, не маскируя, однако, указанную взаимосвязь.
Таблица 8.15.
Взаимосвязь конечной биомассы особей недотроги мелкоцветковой Impatiens
parviflora с величиной семядольного индекса в разных вариантах плотности мо-
дельных популяций
Классы семядольного индекса плотность модельных популяций (экз./сосуд)
1 5 10
1,0-1,5 245,3 ±75,1* 127,6 ±25,7***
1,6-2,1 1078,0 ±212,2 412,3 ± 140,7* 222,2 ± 22,6***
2,2-2,7 563,8 ± 131,8* 235,0 ±38,8**
Примечание: Уровни значимости для средних: * р<0,05; ** р<0,01; *** р<0,001.
8.6. Динамика корне-побеговых и корне-листовых отношений в
ходе розеточного роста
Биомасса, накапливаемая в ходе розеточного роста, распределяется меж-
ду компонентами системы source-sink, т. е. между розеткой и корневой си-
стемой. Обычно тесная корреляция между побегом и корневой системой
дает основание оперировать биомассой (весом) надземных органов, кото-
рые легче взять для анализа неповрежденными, как «полпреда» растения
в целом. При работе с многолетними растениями из-за трудностей мето-
дического характера редко учитывают биомассу подземных органов, обе-
спечивающих многодетность (переннизацию). Малолетние растения в этом
аспекте изучать много легче, и непростительно игнорировать данные о ро-
сте корневых систем, хотя бы скелетных корней, тем более что раннее пре-
кращение их формирования — одна из главных черт малолстпости. Вот по-
nh и ।к>11уияц||>1 х 267
I lang S К()1|нчс> i ценные acnckibi pocin pin ici
чему, ila111)11мсp, первый вариаш мо
дели размещения и динамики аеси
милятов, разработанный Д. Коэном
(Cohen, 1966), был заменен другим,
в котором в качестве посредника в
перемещении продуктов ассими-
ляции выступает корневая система
(Schaffer et al., 1982). Каковы пути
изучения взаимоотношений надзем-
ных вегетативных органов и корне-
вой системы? Как влияют внешние
условия на динамику этих взаимо-
отношений? В чем состоят особен-
ности системы придаточных кор-
ней? Последовательно рассмотрим
эти вопросы.
Один из методических приемов
изучения раннего роста стержневой
корневой системы основан на выра-
Рис. 8.34. Темпы роста корня в длину
у проростков катрана татарского: Lr.
длина корня; Т — время (дни от начала
опыта).
щивании всходов на узких почвен-
ных монолитах, заключенных между тонкими стеклами (т. н. метод стек-
ла). Так были проведены наблюдения и визуально отмечены линейные при-
росты в надземной и подземной сфере у проростков полурозеточного мо-
нокарпика катрана татарского Crambe tataria, который, как уже говорилось
выше, в условиях культуры способен стать трицикликом или даже зацвести
на второй год жизни. Показано, что средний прирост главного корня по дли-
не за сутки может составлять до 2,5 см (рис. 8.34) при очень малых, прак-
тически незаметных изменениях в надземной сфере. Адаптивная ценность
возникновения мощного аттрагирующего центра в корне, куда интенсив-
но транспортируются ассимиляты из крупных семядолей (как запасенные в
семени, так и вновь образуемые в ходе фотосинтетической деятельности се-
мядолей) вполне понятна, если принять во внимание большое значение бы-
строго достижения глубоких влажных слоев субстрата, который на поверх-
ности сильно нагрет и пересушен. В дальнейшем у катрана формируется ро-
зетка листьев, которая в естественных условиях обитания этого вида — на
крутых известняково-мергелистых сильно инсолируемых склонах — к осе-
ни сбрасывается, и взамен ее весной отрастает новая розетка. Таким обра-
зом, в ходе вегетации наибольшая порция ассимилятов ежегодно вкладыва-
ется в формирование розетки, которая фотосинтезирует, транспортируя пла-
стические вещества в корневую систему (рис. 8.35). Процесс вегетации про-
должается до тех пор, пока корень не достигнет определенного диаметра (у
катрана в условиях Татарии — это 2,0-2,5 см) и соответствующей «крити-
чсской массы». Перестройка деятельности верхушечной меристемы побе-
га приводит к заложению мощного соцветия, потребляющего все ассимиля-
ты, запасенные в корне за годы розеточного роста и подводящего черту жиз-
ненному циклу особи катрана (рис. 8.35). Там же на рисунке 8.35 показано,
как можно изобразить схему двулетнего цикла, который наблюдается у еди-
268
МН Марков 11 oi i у 11 и i tno 111 1.1 я bnouoi им p;ic iciinii
Рис. 8.35. Схема взаимоотношений над-
земных и подземных органов: А — у
многолетнего (катран), Б — двулетне-
го (редис) и В — однолетнего (редька
дикая) монокарпиков в ходе онтогене-
за. I-VI — годы жизни. Вклад биомассы:
1 — в надземные вегетативные органы;
2 — подземные органы; 3 — в соцветие.
АЗ Б
В
Рис. 8.36. Динамика отношения масса
корня/общая масса (RWR) в модельных
популяциях клоповников густоцветково-
го (L.d.) и сорного (L.r.) разной плотно-
сти (разные плотности обозначены как
на рис. 8.27).
пичных особен катрана на ннтомпп-
кс и у всех двулетников в нормаль-
ных естественных условиях. Если
взять для примера растения из той
же трибы (Brassiceae), что и катран,
двулетнее растение редис Raphanus
sativus и незимующий временноро-
зеточный однолетник редька дикая
Raphanus raphanistrum, то по схемам
А-Б-В можно хорошо проследить
один из возможных эволюционных
путей возникновения однолетности,
лежащий через укорочение вегета-
тивной розеточной фазы и време-
ни транспортировки ассимилятов на
формирование корневой системы.
Здесь опять нельзя не вспомнить о
цветушных экземплярах двулетни-
ков, которые вырастают при небла-
гоприятных внешних условиях, су-
щественно влияющих на процесс
распределения и перераспределения
ассимилятов. Подчеркнув, что вто-
рой путь исследования взаимоотно-
шений в системе корень — розетка
— периодическая оценка биомассы
надземных и подземных органов с
вычислением аллометрических по-
казателей типа: вес корней/общий
вес растения (root weight ratio —
RWR) или вес корня/вес побега (root
shoot ratio — RSR), перейдем к рас-
смотрению роли среды.
Любопытный материал для ил-
люстрации внутривидовых влияний
на взаимодействие розетки и корне-
вой системы особей дают уже опи-
санные опыты с различной плот-
ностью модельных популяций. У
растений, развивающихся наибо-
лее часто по двулетнему циклу, кло-
повников сорного и густоцветкового
на протяжении розеточного роста в
первый год жизни наблюдалось по-
вышение RWR, т. е. относительная
масса корня постепенно возрастала
(рис. 8.36). В опыте с плотностями
IldfitlS Кошгнч 111Г111И.1Г IICIICKII.I pot HI put irillHt п ноиуиицпих 26*J
oi I до 25 ii и пи пне плотности на hoi процесс было мало ищу i пмым, по при
наиболее высоких плотностях нее же были отмечены несколько меньшие
значения RWK. В более продолжительном опыте с высокими плотностями,
когда все одиночно растущие и часть особей из модельных популяций вы
сокой плотности (50 и 100) перешли в генеративное состояние, отношение
вес корней/общий вес у них резко уменьшилось — от 0,47 до 0,12 (плот-
ность 1), от 0,40 до 0,12 (плотность 50) и от 0,32 до 0,20 (плотность 100).
Потенциально многолетний подорожник большой Р. major только на пер-
вых этапах роста реагировал на плотность изменением величины RWR, а
на более позднем этапе значение RWR при всех плотностях стало одинако-
вым. Стабилизация RWR, по-видимому, каким-то образом связана с потен-
циальной многолетностью подорожника большого, обладающего способно-
стью переннировать при разных плотностях популяции. Очень интересным
представляется и то, что переход некоторых особей в генеративное состо-
яние к концу опыта (23-24 июня 1983 г.) практически не повлияло на ста-
бильность RWR.
Межвидовые (фитоценотические) влияния продемонстрируем на приме-
ре агропопуляций зимующих однолетников из семейства Cruciferae: жеруш-
ник болотный Rorippa palustris, резуховидка Arabidopsis thaliana и пасту-
шья сумка Capsella bursa-pastoris. Все названные растения зимуют в посе-
вах или на паровых участках в виде зеленых розеток, способных к фотосин-
тезу сразу после схода снега.
w
Рис. 8.37. Динамика: А — распределений по общей биомассе (W); Б отношения
масса корня/общая масса (RWR); максимальной (RGRmax) и средней (RGR) отно-
сительной скорости роста в популяции жерушника болотного. t( 24.IV; t, 4.V;
t, ll.V. 19831.
270
Л/// MupKOtt I 1о11уНИЦ11О1111ПИ OllOJIOl НИ рис ICIIIlil
Рис. 8.38. Регрессия массы корня (Wr)
на общую массу (W) для популяции же-
рушника болотного по данным трех вы-
борок, взятых: 28.IV, 4.V и 11.V 1983 г.
Три последовательно взятые вы-
борки из популяции жерушника па
паровом, осенью вспаханном участ-
ке свидетельствовали об энергичном
росте биомассы, который у каж-
дого экземпляра зависел от разме-
ров розетки, вышедшей из-под сне-
га. Динамика гистограмм распреде-
ления (рис. 8.37А) по общему весу
хорошо иллюстрирует эту зависи-
мость (наиболее энергичный рост у
самых крупных особей — в правой
части гистограммы распределения).
Более точные характеристики мож-
но получить при анализе графика
«Б» на рисунке 8.37 — относитель-
ная скорость роста у самых крупных
особей составляла 0,3 мг/мг неделя,
а у средних — 0,19. На этом же гра-
фике приведена кривая изменения
среднего значения RWR от 29 апре-
ля до 11 мая 1983 г. Названный па-
раметр возрастал, причем на первом
отрезке времени было отмечено более резкое, а на втором — более плавное
нарастание.
Памятуя о необходимости оперировать не только средними данными при
характеристике популяции, рассмотрим связь между массой корневой си-
стемы (Wr) и общей массой (W) в логарифмических (In) координатах с ис-
пользованием всех присутствовавших в трех выборках особей (рис. 8.38).
Явная линейность связи позволяет перейти к следующим уравнениям:
In W = 0,96 In W - 0,94 (28.IV)
In W = 1,06 In W-1,07 (4.V)
In W, = 0,89 In W - 1,09 (11 .V).
Все значения аллометрического коэффициента близки к единице, но по-
следнее значение уже свидетельствует о намечающемся сдвиге, кото-
рый, без сомнения, связан с переходом растений в репродуктивное состоя-
ние, когда общий вес нарастает преимущественно за счет надземных орга-
нов. Ранневесенний рост жерушника R. palustris происходил по существу без
какого-либо значительного фитоценотического влияния из-за отсутствия на
паровом участке культурных растений. Поэтому его можно рассматривать как
своеобразный контроль, как пример быстрого, но кратковременного запаса-
ния биомассы, иными словами, как элемент стратегии избегания. В посе-
ве вики с овсом в том же году максимальное значение RWR у жерушника
составило всего 0,23 (против 0,53 на паровом участке) и в начале перехода по-
пуляции в генеративное состояние (от 14 до 19 июня) снизилось до 0,09 0,10.
271
Г’1(1«и Л Коничгч lliriflll.ic ИСПСКИ»! рос I II pill irilllil и попупяциях
Рис. 8.39. Кривые распределения логарифмически трансформированных значений
общей биомассы (W) для периодических выборок из агропопуляций резуховидкн
Галя в посевах озимой и яровой пшеницы; внизу для удобства сравнения кривые
для особей, находившихся в фазе плодоношения из обеих популяций, совмещены на
одном графике
272
MH MtinhtHi I h >i i у ни i un >111 him (>1101101 им рас iciinii
В присутствии па поле озимого культурного растения, например, озимой
ржи или пшеницы, использование сегетальпым сорным зимующим одно-
летником ранневесеннего периода сильно осложняется. Сравнение агропо-
пуляций резуховидки Arabidopsis thaliana в посевах озимой и яровой пше-
ницы показало, что в первом случае RWR уже 29 апреля было очень низ-
ким (0,07) и мало менялось с течением времени. Во втором случае наблю-
дался заметный перепад в динамике параметра от 0,13 в первую неделю
наблюдений (12-15 июня) до 0,06-0,04 к концу (с 22.VI по 8.VI). В посе-
ве яровой культуры динамика значений RWR диагностирует наличие ко-
роткого весеннего благоприятного периода, который может и не использо-
ваться агропопуляцией из озимого посева из-за того, что перезимовавшее
культурное растение быстрее набирает силу, чем яровое, посеянное весной,
и из-за быстрого перехода вышедших из-под снега особей резуховидки А.
thaliana в генеративное состояние и прекращение роста. Вместе с тем полу-
ченные выборочным методом данные на рис. 8.39 свидетельствуют о том,
что общая мощность особей A. thaliana в озимом посеве существенно пре-
восходит таковую в яровом посеве. Так подчеркивается наибольшее значе-
ние осеннего розеточного роста в осуществлении стратегии избегания. Мы
приходим к выводу, что динамику аллометрических показателей RWR и
RSR можно использовать как своеобразный критерий благоприятности фи-
тоценотических условий для роста полурозеточного малолетнего растения.
Действительно, анализ траекторий RSR в агропопуляциях пастушьей сумки
(рис. 8.40) показал, что самые благоприятные условия для роста этого сеге-
тала складывались в посеве гороха, а в двух других посевах они были замет-
но хуже. Это подтвердил прямой анализ мощности растений. Равномерно
низкое значение RSR в посеве гречихи связано, по всей видимости, с фор-
Рис. 8.40. Динамика отношения масса
корня/масса побега (RSR) в трех агропо-
пуляциях пастушьей сумки из посевов:
I — гороха, II — гречихи и III — овса;
Т — время (даты наблюдений).
мированием затеняющего полога
культурным растением. Затенение
и обусловило слабое относительное
развитие корневой системы.
Данные литературы о том, что
система придаточных корней обхо-
дится растениям дороже, чем
стержневая (Caldwell, 1979), подт-
верждаются нашими данными на
примере розеточных малолетних
растений. У трех видов, представ-
ляющих разные семейства, мы на-
блюдаем даже на стадии репродук-
ции высокие значения RWR: луж-
ница водная Limosella aquatica
(Scrophulariaceae) — 0,28 ± 0,05,
мышехвостник маленький Myosurus
minimus (Ranunculaceae) —
0,27 ± 0,02, проломник нитевидный
Ansdrosacefiliformis (Primulaccac)
0,19 0,01.
I'hifhi S Конп'нч i иен it i.ic iicnckii.i pot* in pm iciniil и попуимциих
?7
Для сравнения можно напомни i в,
что у гсисра। пвпых особей изучи
емых нами стержнекорневых мало-
летников RWR не превышает 0,1.
В опыте с разными плотностя-
ми вегетирующие особи проломни-
ка нитевидного обладали довольно
стабильным и высоким отношени-
ем RWR — 0,38 ± 0,08 (плотность 1);
0,32 ± 0,06 (плотность 4); 0,38 ± 0,04
(плотность 6); 0,31 ± 0,03 (плот-
ность 9); 0,29 ± 0,03 (плотность 11
экз./сосуд).
Отсутствие явной (как в надзем-
ной сфере) метамерности корневой
системы в большинстве случаев не
позволяет получать количественные
соотношения между числом мета-
меров в надземной сфере и числом
элементов корневых систем. Однако
такие связи, очевидно, поддержива-
ются и у растений с придаточными
Рис. 8.41. Регрессия числа розеточиы
листьев (N1) на число придаточных кор
ней (Nr) для популяции мышехвостник
(17.V. 1986).
корнями, которые можно рассматри-
вать как рассредоточение несколь-
ких корневых систем (Хохряков, 1982), по крайней мере, на ранних этанах
роста их можно выявить. На рис. 8.41. приведена линия регрессии числа ро
зеточных листьев на число придаточных корней у молодых экземпляров г
популяции мышехвостника Myosurus minimus ( пойма р. Ик, 17 мая 1986 г.)
Налицо четкая линейная связь, указывающая на «окорененность» каждою
из фитомеров в составе розеточной особи мышехвостника.
8.7. Размерная дифференциация особей, возникающая в ходе
роста в популяциях растений
Изначальное определение популяции как системы, включающей разня
щиеся по многим признакам особи, предполагает дифференциацию. Потому
в каждой главе вопрос о дифференциации особей в популяциях растений за
тронут в той или иной степени как имеющий самостоятельное общебиоио
гическое значение (Злобин, 1980).
Особи в популяции любого вида растения различаются, прежде всего
по биомассе, которая выступает как интегрированный показатель жизнен
ного состояния. Общая биомасса (или общий вес) складывается из многих
других параметров, в той или иной степени коррелятивно связанных с licit
но при характеристике роста и оценке жизненного состояния малолетних
растений главенствующее место правомерно оставить именно за биомаг
сой. В этой связи, на наш взгляд, нет противоречия между гем, что К). А
Злобин (1989) призывает использовать весь комплекс морфометрических
274 МН Марьин I louyiihhiioiiihui (hioiioi пи pnciспнй
параметров, не замыкаясь па биомассе, и гем, что зарубежные исследова-
тели, изучающие внутрипопуляциоппую дифференциацию как i аковую, бе-
рут за основу именно гетерогенность популяции по биомассе. Именуют се
при этом иерархией использования ресурсов (White, Harper, 1970), иерар-
хией доминантности или просто размерной иерархией (Ross, Harper, 1972;
Weiner, Solbrig, 1984). Ю. А. Злобин полагает, что биомасса как показатель
перегружена неживыми инертными структурами и в большей мере отража-
ет прошлую историю особи, чем ее современный статус, но при этом ого-
варивается, что в первую очередь это относится к многолетним растени-
ям. Здесь уместно вновь напомнить уже цитированное выше высказывание
Б. А. Юрцева о том, что у однолетних растений побеговое тело представ-
ляет собой «наиболее полный пластический слепок онтогенеза». Биомасса
этого тела наилучшим образом информирует нас о жизненном состоянии
и на разных промежуточных этапах роста, и в конце как итог онтогенеза.
Поэтому, не отвергая применимость анализа биомассы при работе с популя-
циями многолетних растений, рассмотрим примеры возникновения и разви-
тия размерной дифференциации в популяциях ряда малолетних растений.
Очень важно, что при таком понимании не усматривается противоречий
между понятием «поливариантность развития» с ощущающимся в нем ди-
намизмом и понятием «размерная иерархия» с его статичностью. Каждый
вариант онтогенеза подразумевает определенный итог, овеществленный в
архитектуре и биомассе, выступающей как индикатор варианта развития
без выяснения того, имеет ли он генетическую природу или обусловлен
влиянием внешней среды. Сложные и далеко еще не выясненные отноше-
ния между генотипом и фенотипом (Sultan, 1987) не позволяют связывать
напрямую генетическую и фенотипическую дифференциацию. Мы уделим
основное внимание последней как имеющей большое значение в популя-
циях растений.
8.8. Спектры жизненных состояний и их характеристика
Главным объектом анализа внутрипопуляционной дифференциации была
и остается кривая (или гистограмма) изменчивости веса (биомассы). Она
является по сути дела спектром жизненных состояний (виталитетным спек-
тром — по Ю. А. Злобину) популяции. По ней мы судим об амплитуде ва-
риабельности массы и характере распределения, который совершенно не-
обходимо учитывать при любой статистической обработке. Обширный ма-
териал, собранный исследователями популяционной биологии, показывает,
что в популяции растения первоначально нормальное (или близкое к нор-
мальному) симметричное распределение по массе всегда приобретает по-
ложительную асимметрию со сдвигом моды в первые (если не в самый пер-
вый) классы. Приобретение и увеличение асимметрии начинается тем рань-
ше и происходит тем резче, чем более загущена популяция или чем сильнее
на нее стрессовое влияние соседствующих видов. Дж. Уайт (White, 1981)
рассматривает это явление как одно из следствий «закона — 3/2». С момен-
та приобретения положительной асимметрии применение обычных стати-
стических параметров, основанных на нормальности распределения, era-
Гчаад N Коиичсс 1 псиные испек 1 ы pm iи piu leniiil n популяциях ? 7S
новптся неправомерным. В часiпосiн, iai<on показалсиь дня оценки hi
мспчнвос।н, как коэффициент вариации, н эго отмечал еще в l()ISi В II
С укачев, перестает быть адекватным мерилом изменчивости, поскольку за
висит от основных статистических параметров (стандартного отклонении,
среднего арифметического) и является параметрическим. Поэтому в пас юя
щее время практикуется применение непараметрических или «малопарамс
трических» критериев, один из которых, заимствованный из экономики (см.
вставку 3 в главе 2), называют по имени предложившего его итальянского
ученого коэффициентом Джини (Gini). Коэффициент Джини (G) равен по
ловине относительных отклонений или средней арифметической различий
всех пар особей выборки:
п п
S S [х - х ]
r_*=v=i
Cj---------------
2п2х
В ряде работ (Weiner, Solbrig, 1984; Weiner, 1985, 1986, 1988), где апроби
ровано применение коэффициента, предложено умножать его значение па н/
(п-1) для снижения искажений из-за разных объемов выборок (п), и ему о гда
ется предпочтение в отражении размерной иерархии и оценке эффекта копку
ренции при сопоставлении, скажем, с коэффициентом асимметрии. По мне
нию Д. Вейнера и О. Т. Солбрига (Weiner, Solbrig, 1984), можно выделил» три
аспекта размерной иерархии: 1) популяция демонстрирует большую измен
чивость особей по весу; 2) в ней много мелких и мало крупных особей; 3) эти
немногие крупные особи вносят наибольший вклад в урожай биомассы по
пуляции. Исследователи доказывают, что асимметрия отражает только вто-
рой из этих аспектов, приводя графические примеры одинаковой асимметрии
распределений при сильно различающейся гетерогенности по весу.
Научный интерес биологов к коэффициенту Джини может быть обуслов-
лен, на наш взгляд, двумя причинами: во-первых, в методическом плане ре-
шается проблема оценки гетерогенности или размерной дифференциации
«малопараметрическим» критерием; во-вторых, появляется возможность
использовать адекватный показатель изменчивости биомассы в теории кон-
куренции для проверки двух существующих гипотез «симметричной» и
«асимметричной» конкуренции. Полагая, что по поводу первой причины
уже достаточно сказано выше, прокомментируем вторую.
Если конкуренция «симметрично» приводит к общему подавлению
особей, то размерная дифференциация по весу, или размерная иерар-
хия, должна уменьшаться по мере усиления напряженности конкуренции.
«Асимметричность» конкуренции должна состоять в выделении группы
резко доминирующих растений, односторонне подавляющих соседние мел-
кие экземпляры и увеличивающие свой отрыв в ходе роста.
Об увеличении этого отрыва и будет свидетельствовать возрастающая
размерная дифференциация. Данные литературы показывают, что корневая
конкуренция, обыкновенно, «симметрична», тогда как конкуренция за свет
часто принимает форму одностороннего (асимметричного) подавления. В
интересном опыте Д. Вейнера (Weiner, 1986), где удалось разделить вли
276
М Н Маркие I It >i iy и >1 щк >i и t л я biioiioi ii>i pn< iciiid1
h высота, cm
Рис. 8.42. Регрессия логарифма (In) об-
щей массы на высоту в популяции недо-
троги мелкоцветковой (по семи последо-
вательным выборкам): траектории для
самых мелких (Ряд 1) и самых крупных
(Ряд 2) особей в выборках.
япис корневой конкуренции и кон-
куренции за снег у однолетней ли-
аны Ipomoea tricolor, были получе-
ны результаты, поддерживающие
гипотезу об «асимметричности»
конкуренции за свет и демонстри-
рующие возрастание вариабельно-
сти размеров при повышении плот-
ности популяций. Т. Митчелл-Олдз
(Mitchell-Olds, 1987) на примере не-
дотроги Impatiens capensis также по-
казал, что одностороннее подавле-
ние может иметь место при конку-
ренции за свет, а при конкуренции за
воду и элементы минерального пи-
тания такой односторонности не на-
блюдалось.
Архитектура немногих видов ма-
лолетников (например видов рода
недотрога Impatiens — см. главу 4)
располагает к возникновению асим-
метричной конкуренции. По нашим наблюдениям за траекториями роста
самых мелких и самых крупных особей в последовательных выборках из
популяций недотроги мелкоцветковой мелкие росли вначале даже более
энергично, чем вторые (т. н. «подгон»). Впоследствии, однако, рост пер-
вых фактически прекращался, а темп дифференциации с этого момента
резко ускорялся. Весьма характерно, что указанную закономерность мож-
но было наблюдать только при использовании для изображения динамики
средней и крайних величин полулогарифмического масштаба (рис. 8.42).
Количественно оценить динамику дифференциации можно было как по
значениям коэффициента асимметрии, так и по значениям коэффициента
Джини, хотя последний отличался более устойчивой динамикой. Итогом
роста была достаточно хорошо заметная бимодальность (двувершинность)
кривой изменчивости по высоте в популяции (рис. 8.43).
Для экспериментов по изучению конкуренции за свет, как правило, исполь-
зуются безрозеточные, длиннопобеговые растения. Реже берутся полурозе-
точные виды, и тогда гипотезы приходится корректировать. Например, при
работе с редисом (Schmitt et al., 1986), когда в опыте испытывались не толь-
ко разные плотности, но и три разных уровня освещения, был сделан вывод,
что более сильная размерная иерархичность возникает при высокой плотно-
сти, но только при минимальной освещенности (25% от полного света). На
полном свету возникновение гетерогенности по весу можно было объяснить
только разными темпами роста у разных особей, т.е. различиями индивиду-
альных RGR. Однако тут же возникает вопрос, чем объясняются эти отличия
в относительной скорости роста, и чем обусловлен отрыв отдельных особей
от большинства.
Г'шна Л Коннчгс I ценные aciicKi 1.1 poem рисiгний н поиуиицпях
277
А %
h (середины классов, см)
----30.V -н- 4.VI
11.VI
18.VI
-и-12.VII —20.VII 27.VII
Рис. 8.43. Динамика роста в высоту особей популяции недотроги мелкоцветковой ~
появление бимодальности распределения.
Можно назвать следующие причины: 1) различие стартовых запасов био-
массы; 2) генетические причины; 3) более раннее появление некоторых
особей (или их когорт) — упреждение по моменту начала роста; 4) локаль-
ная благоприятность микроучастков.
В популяциях почти облигатно самоопыляющихся малолетников типа па-
стушьей сумки Capsella bursa-pastoris и пр. на фоне большинства особей ин-
бредного происхождения могут выделяться сравнительно малочисленные
аутбредные экземпляры с признаками гетерозиса и в их числе способностью
к более быстрому росту. Для проверки этого предположения нужны специ-
альные опыты. Косвенные же данные мы можем получить путем анализа
видов, у которых генетическая природа семян проявляется в их размерах.
Семена хазмогамного происхождения у недотроги обыкновенной Impatiens
noli-tangere крупнее по размерам, чем семена клейстогамного происхожде-
ния, что, как мы видели, отражается не только на размерах семядолей у про-
ростков, но и на темпах роста, конечной мощности и плодовитости особей.
Во всех упомянутых выше опытах по изучению размерной иерархии обя-
зательным условием была одновозрастность особей, базирующаяся на од-
новременном прорастании семян. В естественных популяциях малолетни-
ков более обычным явлением бывает поликогортностъ — растянутое во
времени прорастание семян, которое можно зарегистрировать, только про-
водя демографические наблюдения на постоянных площадках (см. напри-
мер, Марков, 1976). Проростки, появившиеся первыми, оказываются в луч-
ших, а появляющиеся позднее — в худших условиях.
Один из вариантов разновременности прорастания мы наблюдали в агро-
популяциях резуховидки Arahidopsis thaliana и пастушьей сумки С. bursa-
pastoris в посевах озимых культур, где наряду с когоргой особей, появпв-
278 МН Маркин 11(>11у||яц||(>1111пя (hio)ioi пя рис 1СПИЙ
шихся осенью, появляются весенняя н иногда летняя когорты. Особи из ве-
сенней когорты, как явствует из рисунка К.44, существенно оз стают в росте
от экземпляров, появившихся осенью, и их присутствие в составе популя-
ции усиливает размерную гетерогенность. Однако, на рис. 8.44 для каждой
из когорт дана отдельная кривая распределения.
Когда же рассматривали взятую ранней весной общую выборку из популя-
ции, то кривая распределения абсолютных значений массы оказалась силь-
но положительно асимметричной, но без всяких признаков бимодальности.
Лишь при логарифмировании данных в один из ранних сроков (14.V.1975)
была выявлена бимодальность как свидетельство заведомой размерной гете-
рогенности изучаемой агропопуляции, но со временем она стала почти неза-
метной (рис. 8.44). Наиболее быстро растущие особи весенней группы бы-
стро достигали уровня самых слабых особей из осенней группы, что привело
к получению непрерывного (без деформаций — по В. С. Ипатову, 1969) асим-
метричного графика распределения вместо изначально бимодального.
Некоторые исследователи считают, что односторонность, «асимметрич-
ность» конкуренции должна приводить к возникновению бимодальности
распределения по массе даже в одновозрастной популяции.
Э. Форд (Ford, 1975) пишет, что многие мельчайшие особи имеют RGR близ-
кое к нулю, и что наличие резкого перехода к высоким значениям RGR у круп-
ных особей порождает бимодальность. Имитационная модель роста популя-
ции, предусматривающая различия RGR и наличие односторонней конкурен-
ции, дала бимодальное распределение (Aikman, Watkinson, 1980). Однако, зави-
симость поли- и биомодальности от числа взятых классов и отсутствие прямых
статистических тестов на бимодальность в значительной мере затрудняют осо-
знание ее значения в теории конкуренции (Hutchings, 1986).
Последовательное появление групп проростков (т. н. когорт) связанное
с «квантовым», вспышковым характером прорастания семян, наблюдалось
нами и в агропопуляциях пастушьей сумки из посевов яровых культур: горо-
ха, гречихи, овса. Наблюдения на постоянных квадратах с маркировкой всех
Рис. 8.44. Полигоны распределения логарифмов массы (W) надземных органов для
двух выборок из агропопуляции пастушьей сумки, взятых в разные даты: 14.V и
30.V.1975 г. I — осенняя и II — весенняя когорты, границы которых на фоне полиго-
нов показаны пунктиром; f — частость.
Рис. 8.45. Влияние разных сроков поя в
ления когорт проростков (Т) на конеч-
ную среднюю массу особи в когорте (W)
в агропопуляциях пастушьей сумки из
посевов гороха (I), овса (II) и гречихи
(III).
Г'кшиН Kouirit’t IHt’inii.ic аспекIы pot'In piH H'iiiih н in>нуимцннх 279
появляющихся особей позволили за
регистрирован» группы пророс гков
{когорты), появлявшиеся 24, 29 мая,
1,7 июня 1974 г. Ценотическая обста-
новка в посеве, эдификаторное влия-
ние культурных растений сказались
на процессе появления всходов — в
более благоприятных условиях, в по-
севе гороха, оно было более растяну-
тым во времени, чем в худших усло-
виях, в посеве овса. Относительную
благоприятность режимов в данных
агрофитоценозах можно оценить по
параметрам агропопуляций (табл.
8.16). Рис. 8.45 показывает, насколь-
ко резко, гораздо резче, чем теоре-
тическая линия снижалась средняя
масса особи в последовательно появлявшихся когортах, а на рис. 8.46 видно,
как это сказалось на репродуктивной мощности особей в группах, их суммар-
ном вкладе (числе сформированных плодов) в воспроизводство популяции.
Таблица 8.18 показывает также резкую положительную асимметрию кривых
распределения по биомассе при близких к нормальным, мало асимметрич-
ных распределениях по высоте. Особи из поздно появлявшихся когорт несо-
мненно внесли общую лепту в размерную дифференциацию популяции, уси-
лив асимметрию.
Рис. 8.46. Участие в воспроизводстве популяций особей из последовательно ноя в
лившихся когорт в агропопуляциях пастушьей сумки из посевов гороха (а), гречихи
(б) и овса (в); Nfr — среднее число плодов в когорте; по горизонтальной оси да i ы
появления когорт.
280 МН Марко" I loiiyininiioiuiaii (iiioiioi ня рас leiinii
Таблица 8.16.
Сравнение агропопуляций пастушьей сумки из посевов 'грех яровых культур по
массе надземных органов и высоте
В посеве Масса надземных органов (г) Высота (см)
X ± S X As X ± S X As
гороха 0,240 ±0,017 + 2,0 26,4 ±0,9 ±0,07
гречихи 0,132 ±0,012 ±4,5 22,2 ±1,0 ±0,52
овса 0,043 ±0,003 ±13,9 14,5 ±0,6 ±0,57
М. Росс (Ross, Harper, 1972) в эксперименте изучал процесс появления
проростков в популяции ежи сборной, влияние размеров пространства, пре-
доставленного особи, и положения, которое она в нем занимала, на ее рост. С
помощь регистрации даты появления каждого проростка Россу удалось по-
казать, что преимущества, которые проросток получает благодаря более ран-
Рис. 8.47. Вверху: Влияние продолжи-
тельности периода от момента появле-
ния особей до уборки на среднюю мас-
су растения у ежы сборной. Т — вре-
мя (дни) от появления до уборки; W —
средняя масса особи (мг).
Внизу: Схема опыта М. Росса с ежой
сборной, отдельные особи которой вы-
севались на свободных круглых пло-
щадках среди плотного травостоя из-
учаемого вида: I — особи высевали в
центр свободных площадок разной ве-
личины; II — особи занимали разное
положение в пределах площадок одина-
кового размера.
нему появлению, не ограничиваются
тем, что при таких условиях он про-
сто имеет больше времени для роста.
На рис. 8.47 приведена теоретически
высчитанная линия, которая отра-
жает снижение массы особей толь-
ко как результат сокращения про-
межутка времени, отпущенного им
для накопления биомассы. Однако
на деле даже небольшая задержка в
прорастании обходится особи гораз-
до дороже. На том же рис. 8.47 мож-
но видеть, насколько круче снижа-
лась реальная масса особей ежи при
более позднем их появлении. В опы-
те Росса особям предоставляли так-
же разное по размерам простран-
ство, или же в одинаковом простран-
стве особь помещали по-разному.
Оказалось, что размер простран-
ства, доступного особи, имеет боль-
шее значение для ее роста, чем по-
ложение особи в этом простран-
стве. Малая вероятность вычлене-
ния большого пространства, оказы-
вающегося в распоряжении особи,
приводит к относительной редкости
появления крупных особей в попу-
ляциях многолетних растений и пре-
обладанию более мелких особей, то
есть к асимметричности распределе-
ния по размерам.
Г’кши Л' кошгкч । ценные ас иск i и pot hi pm iciiuh и n< hividiuiihx
281
Задавшись целью охарактеризо-
вать динамику развития размерной
иерархии и принимая во внимание
очень раннее появление положи-
тельной асимметрии распределе-
ний, мы использовали коэффици-
ент Джини для характеристики агро-
популяции пастушьей сумки в посе-
ве гороха. 22, 27 мая и 4 июня 1975
г. получены значения коэффициента,
соответственно: 0,375, 0,413 и 0,460.
Налицо медленное нарастание раз-
мерной дифференциации, несмотря
на то, что в 1975 г. все особи пасту-
шьей сумки появились в посевах од-
Рис. 8.48. Динамика «логнормальных»
распределений по общей биомассе (W)
для дюнной популяции подорожника
индийского: 1 — 6.VII; 2 — 18. VII; 3
16.VIII; 4 — 30.VIII; f % — частость.
повременно, составив одну когорту.
Значит, в данном случае в осно-
ве возрастающей гетерогенности лежат различия в RGR у одновозрастных
особей, растущих в условиях конкурентного подавления, в частности, при-
тенения культурными растениями.
В популяции дюнного незимующего временнорозеточного однолетни-
ка подорожника индийского Plantago indica коэффициент Джини позво-
лил выявить обратную тенденцию — снижение гетерогенности. Популяция
подорожника индийского, расположенная на песчаных дюнах в условиях
полного освещения, имела невысокую плотность (40-104 экз./м2), так что
сколько-нибудь заметной конкуренции между редко стоящими синхронно
появившимися особями не наблюдалось. Как показывают логнормальные
распределения (рис. 8.48), заметный рост наблюдался только между дата-
ми взятия первой и второй пробы, после чего он резко замедлился — мак-
симум (мода) кривых распределения перестал смещаться вправо. Была за-
метна лишь перегруппировка — изменение частот в классах, отражающая,
судя по всему преимущественный рост отставших по размерам особей. То,
что эти особи подтянулись в росте, и отразилось на значениях коэффициен-
та, которые для 16 июля, 16 и 30 августа 1984 г. составили, соответствен-
но, 0,465, 0,375 и 0,373. Возможно, в данном случае сказывается полное от-
сутствие эффекта фитоценотического взаимодействия между растениями.
Фитоценотическая среда может сильно влиять на степень дифференциро-
ванности популяций. В этом убеждает следующий пример. Доминирующее
в агрофитоценозе культурное растение отражает своим ростом и развити-
ем те экологические режимы, которые были оставлены предшественниками
и подготовлены человеком. На всякое улучшение этих режимов доминант
реагирует положительно; его поведение по терминологии JI. Г. Раменского
(1924, см. сборник избранных работ) следует назвать «экологичным» (по
К. А. Куркину, 1973 — ортоэкотопичным), т. е. в известной степени неза-
висимым от ценотических отношений, складывающихся в агрофитоцепозе.
Ссгстальные сорные растения — ассектаторы (Сукачев, 1928), рост и раз-
витие ко юры ч сильно зависят от влияния культурного растения, оиределяе-
282
МН Маркин
I loi । у и я цио! 11 nix biionoi ия рас iciniii
Рис. 8.49. Сужение размаха варьирова-
ния общей биомассы (W) и высоты (h)
в трех агропопуляциях пастушьей сум-
ки (уменьшение разницы между когор-
тами) по мере ухудшения условий про-
израстания в агроценозах. I — осенняя,
II и III весенне-летние когорты в посевах
озимой ржи по: Ч.П. — чистому; Г.П. —
гороховому и В-О.П — вико-овсяному
парам.
мою в свок) очередь его конкурент-
ной мощностью, отрицательно ре-
агируют на воздействие, усиливаю-
щее положение доминанта; их по-
ведение поэтому является «анэко-
логичным» (Раменский, 1924), или
«анэкотопичным» (Куркин, 1973).
В сегетальных популяциях па-
стушьей сумки сезонные когор-
ты особей отличаются не только по
морфо-биологическим показате-
лям, но и по «экологичности» свое-
го поведения. В посевах озимой ржи
осенняя когорта развивается в соот-
ветствии с экологическими услови-
ями, которые рожь не успевает еще
изменить в достаточно сильной сте-
пени. Данные 1971 г. показали, что
растения осенней когорты (I — на
рис. 8.49) росли в соответствии с
теми режимами, которые были под-
готовлены предшественниками, т.
е. выказывали «экологичное» пове-
дение. Другие (весенняя и летняя)
когорты (на рис. 8.49 — II и III) в
своем росте сильно зависели от со-
стояния озимой ржи и были связа-
ны с ним обратной зависимостью,
т. е. вели себя «анэкологично». Разница между агропопуляциями пастушьей
сумки из посевов озимой ржи по разным предшественникам, установлен-
ная по состоянию осенней когорты, сменилась на обратную, когда для срав-
нения были взяты весенние и летние когорты особей. По мере ухудше-
ния экологического фона (тенденция установлена по состоянию культурно-
го растения — доминанта агроценоза) сужался размах варьирования между
средними размерами (средней биомассой и средней высотой) особей трех
групп. В полной мере, таким образом, стратегию избегания проявляла толь-
ко осенняя когорта, наименее зависящая от усиливающегося фитоценоти-
ческого стресса благодаря формированию розетки на ранних этапах онтоге-
неза. Всякое изменение, ставящее и осеннюю когорту в тесную зависимость
от развития озимой ржи, приводило к «анэкологичному» ответу всей попу-
ляции в целом на режим в посеве. Позднее появление осенью 1972 г. всхо-
дов осенней когорты (из-за летне-осенней засухи) обусловило уход расте-
ний под снег в малоразвитом состоянии, в связи с чем усилилась их зави-
симость от состояния доминанта агрофитоценоза: чем лучше была развита
озимая рожь в 1973 г. (завися опять-таки от предшественников), тем хуже в
ее посевах было жизненное состояние особей агропопуляций пастушьей
сумки.
l i(Hi(iS Konirict iпенные аспект pot mi piu iriinh и попуиицпих
283
Значение ннутрипоиуляциоиных
конкурентных отношений для раз-
мерной гетерогенности можно про-
иллюстрировать результатами на-
шего опыта с редисом (Raphanus
sativus), который выращивали на
почве из пахотного горизонта без
удобрений при низкой плотности
(площадь питания 100 см2/особь).
Появление когорт проростков отме-
чено 30-31 июля, 1, 2-3 и 4-5 авгу-
ста 1986 г. Из всех когорт периоди-
чески (через 3-4 дня) брали выбор-
ки особей, подсчитывали число ро-
зеточных листьев и вычисляли RGR.
Рисунок 8.50 свидетельствует о том,
что время появления проростка поч-
ти не сказалось в условиях свобод-
ного роста (без конкуренции) на ве-
личине RGR и скорости разворачи-
вания розеточных листьев. Все эк-
Рис. 8.50. Темпы роста особей редиса из
разных когорт в условиях низкой плот-
ности модельной популяции (ослаблен-
ной конкуренции): I — динамика изме-
нения общей биомассы (W); II — ди-
намика числа розеточных листьев (N1);
Т — время (дни с начала опыта); разны-
ми значками помечены кривые для раз-
ных когорт.
земпляры росли примерно с одинаковой скоростью, и первоначальная диф-
ференциация, связанная с задержкой на 1-6 дней прорастания некоторых
семян, не усугублялась, как это имело место в уже охарактеризованных аг-
ропопуляциях пастушьей сумки из посевов яровых культур.
Завершая эту главу, приходится констатировать, что множественность
факторов, влияющих на процесс внутрипопуляционной размерной диффе-
ренциации, затрудняет решение вопроса о том, какой именно фактор или
группа факторов обусловливает наблюдаемое повышение или понижение
уровня вариабельности популяции по биомассе особей с течением време-
ни (в ходе онтогенеза) в конкретной эколого-ценотической ситуации. Ясно
одно, что степень иерархичности размеров, биомассы в популяциях мало-
летних растений всегда достаточно велика и даже в том случае, когда не г
условий для «асимметричной» конкуренции. При этом различия в относи-
тельной скорости роста выступают скорее как следствие, нежели как при-
чина, поскольку они-то и проявляются под действием многочисленных фак-
торов. Фитоценотическая среда, в основном, усугубляет различия в темпах
роста малолетних растений и потому часто усиливает дифференциацию.
Однако постепенное возрастание фитоценотического влияния, из-под ко-
торого часть популяции почти полностью, а другая лишь в небольшой сте-
пени уходят благодаря стратегии избегания, осуществляемой при содей-
ствии розеточного роста, может приводить и к сглаживанию различий.
В главе 10 будет показано, что дифференциация затрагивает нс только
темпы роста, но и фенологию — темпы развития растений.
Глава 9. АЛЛОКАЦИЯ БИОМАССЫ,
ЕЕ СВЯЗЬ С РАЗМЕРНЫМ
СПЕКТРОМ ПОПУЛЯЦИИ
И ДИНАМИКА
«Наиболее изощренную часть теории жиз-
ненных циклов (онтогенезов) представля-
ет принцип аллокации, согласно которо-
му организмы с ограниченными ресурса-
ми должны делить эти ресурсы между не-
сколькими конкурирующими сферами»
Cody, 1966
“Resource allocation is a tenuous compromise
between fecundity, survival and growth”
«Размещение (аллокация) ресурсов
есть тонкий компромисс между
плодовитостью, выживанием и ростом»
Silvertown & Dodd, 1999
Помимо того, что общая биомасса растения структурирована в плане ме-
тамерности, из нее сформированы как фотосинтезирующие (фототрофные)
структуры, так и структуры, потребляющие ассимиляты (продукты фото-
синтеза). Специфика роста особи какого-либо вида растения состоит в кор-
релированных определенным образом изменениях биомассы этих двух ти-
пов структур. Продукты фотосинтеза (ассимиляты) распределяются в теле
особи, обеспечивая три альтернативных процесса: рост, самоподдержание
и репродукцию (рис. 9.1). Увеличение вклада в один из названных процессов
неминуемо влечет за собой соответствующее недовложение в остальные. У
растений, и только у них, между ростом и одним из типов репродукции (т.
н. вегетативным размножением, особенно неспециализированным) грань
провести очень сложно.
Важнейшая заслуга Д. А. Сабинина состоит еще и в том, что он впервые
обратил внимание ботаников на проблемы аллометрии или оценки относи-
тельного роста, изложенные в работе Дж. Гексли (Huxley, 1932).
I hl«ii V AllllOIHIHIIII (HIOMJICCIJ 28b
p . , , .... , ,
MbicjnricH прямая неразрывная связь icx рабоч с современными оцеп
ками размещения биомассы (аллокации) в целях определения чао и, taipa
ченной на выживаемость (вегетативный рост) и части, затраченной на пло-
довитость (репродуктивный рост). Описывая вслед за Дж. Гексли процесс
относительного роста при помощи линий регрессии в логарифмических
координатах и уравнений типа:
Y=bXan
log Y - log b + a log X,
[9.11
где Y — вес части растения; X — вес целого растения или вес остальной
части растения; а — константа (т. н. константа относительного роста или
аллометрический коэффициент) и b — константа, Д. А. Сабинин отмечал,
что цветение является столь резким переломным моментом в жизнедеятель-
ности растений, что не приходится ожидать сохранения константы относи-
тельного роста а при переходе к репродуктивной фазе. «Мы уверены, — пи-
шет он, — что применение уравнения относительного роста к изучению со-
отношения веса частей организма при росте позволит вскрыть ряд интерес-
ных закономерностей, характеризующих продуктивность растения. До сих
пор (к 1940 г. — М.М.) по этому важнейшему для практики вопросу мы, к
сожалению, не имеем обоснованных количественных данных» (Сабинин,
1940: 277). Насколько нам известно, и по сей день эти уравнения нашли в
отечественной ботанике лишь очень ограниченное применение, по крайней
мере, в популяционной биологии растений.
Процесс и результат размещения биомассы в пределах растения за рубе-
жом называют аллокацией (allocation). Будем и мы так называть, не перево-
дя, относительный вклад биомассы в ту или иную сферу в процентах или
долях единицы, который служит хорошим показателем стратегии жизнен-
ного цикла (Steams, 1976; Solbrig,
1980) или так называемой репро-
дуктивной стратегии (Harper, Ogden,
1970). Благодаря этому в 1980-х гг.
появилось много статей зарубеж-
ных ученых, затронувших методи-
ческие и теоретические стороны из-
учения аллокации (Hickman, 1975;
1977; Law, 1979; Bazzaz, Carlson,
Harper, 1979; Lee, Cavers, 1981;
Thompson, Stewart, 1981; Zeide, 1978;
Abrahamson, Caswell, 1982; Trivedi,
Tripathi, 1982; Stewart, Thompson,
1982; Reekie, Bazzaz, 1987a, б, в и
др.). Центральным звеном теории
аллокации в общей теории страте-
гий жизненного цикла, несомненно,
выступает репродуктивное усилие
(Марков, 11 лсщинская, 1987).
Рис. 9.1. Триада альтернативных про-
цессов, идущих с затратами ассимиля-
тов (А), которые берутся из общей «ко-
пилки» и потому возрастание вложения
в один из процессов влечет за собой ком-
пенсационное уменьшение вложения в
остальные.
286 Л/// Маркой Популяционная Ghojioi ия piicicniih
9.1. Некоторые методические вопросы изучения
репродуктивного усилия и аллокации в целом
Виды растений отличаются друг от друга по относительному вкладу био-
массы в формирование генеративных органов и образование семян, кото-
рый называют репродуктивным усилием (reproductive effort — RE).
9.1.1. Как определять?
Существует два основных способа определения репродуктивного усилия.
Первый способ основан на вычислении отношения количества энер-
гии, заключенной в генеративных органах, к общему количеству энергии,
заключенной в теле растения (REj = Eg/Ew). Количество энергии при этом
определяют методом калориметрирования. Более простой способ заклю-
чается в оценке доли веса генеративных органов в общем весе растения
(RE2= Wg/W). Здесь необходимо уточнить несколько моментов.
Определение количества энергии, заключенной в разных органах рас-
тения, несравненно более трудоемко, чем простое взвешивание. Поэтому
в большинстве случаев исследователи прибегают ко второму способу из-
учения репродуктивного усилия. Сравнительное исследование двух спо-
собов привело к выводу о том, что для растений, семена которых содер-
жат много углеводов, весовой способ позволяет получить вполне адекват-
ную информацию о величине репродуктивного усилия. Ошибка увеличива-
ется в том случае, когда исследуемые растения образуют семена с высоким
содержанием жиров, то есть органических веществ с повышенной энерго-
емкостью. Однако и в этом случае ошибка редко серьезно грозит достовер-
ности вычисленного показателя.
9.1.2. Когда определять ?
Определению репродуктивного усилия как биологического параметра
какого-либо вида должно предшествовать выявление момента времени, ког-
да оба типа структур (и вегетативные, и генеративные) близки к своему мак-
симальному развитию. Здесь мы сталкиваемся со значительными трудно-
стями, которые связаны с известной альтернативностью в развитии этих ти-
пов структур у всех монокарпических видов и особенностями «перекачива-
ния» ассимилятов из отмирающих органов в живые. Наиболее типична си-
туация, когда растение сначала растет и формирует свою вегетативную сфе-
ру, а потом, переходя к репродукции, резко замедляет ростовые процессы.
Для пояснения можно привести рисунок 9.2, иллюстрирующий «перекачи-
вание» биомассы из розетки в органы генеративной сферы у полурозеточ-
ного (пастушья сумка) и розеточного (проломник северный) малолетников.
Еще больше проблем возникает при попытке определить репродуктивное
усилие у растений с индетерминированным ростом, поскольку период фор-
мирования пазушных генеративных структур у них бывает очень продолжи-
тельным, и вероятность потерь как вегетативных, так и генеративных струк-
тур за это время чрезвычайно велика и вполне закономерна. Теорем пческн
/ i(id(i V Aiiiiohitmni ыюмассы
28/
Рис. 9.2. Перераспределение (аллока-
ция) биомассы из розетки в органы гене-
ративной сферы: а — у проломника се-
верного процент биомассы 1 — в орга-
нах генеративной сферы, 2 — в листьях,
3 — в корнях; б — у пастушьей сумки.
Т — даты анализа; 1 — слои (см).
оценить репродуктивное усилие у таких видов можно путем подсчета обще-
го числа фитомеров, за вычетом первых «чисто вегетативных», и определе-
ния репродуктивного усилия одного фитомера.
9.1.3. Как провести границу между сферами ?
Значительные трудности встают на пути исследователя, пытающегося
определить репродуктивное усилие у растений, для которых четкое разде-
ление вегетативной и генеративной сфер не характерно, а таких растений
немало (см. рисунки архитектурных типов розеточных и прочих малолетни-
ков — рис. 7.19). К их числу, например, принадлежат многочисленные полу-
розеточные виды семейства Крестоцветных (Cruciferae). При работе с кре-
стоцветными, помимо отсутствия четкой границы между вегетативными и
генеративными структурами, затруднения связаны также с альтернативно-
стью развития двух сфер и высоким содержанием жира в семенах. Чтобы
как-то оценить репродуктивное усилие и репродуктивные стратегии у пред-
ставителей этой интересной группы растений, можно в качестве показателя
использовать отношение числа плодов (семян) к общему весу и считать это
третьим способом оценки репродуктивного усилия (RE3 = Nfr/W). Однако
следует отметить, что RE3 напрямую несопоставимо с REj и RE2.
9.1.4. Как графически представлять данные?
При вычислении репродуктивного усилия необходимо учитывать мас-
су (или энергию) всех генеративных структур, включая вспомогательные, то
есть цветоносы, прицветники, оси соцветий. Иногда целесообразно приво-
дить на аллокационной диаграмме относительные вклады в каждую из этих
структур в отдельности, а в других случаях ограничиваться определением по-
казателя, суммирующего эти вклады. Аллокационные диаграммы непременно
должны содержать данные о вкладе в вегетативную (отдельно надземную и
подземную) сферу. Методика построения аллокационных диаграмм в каждом
случае обяза1еиыю должна быть специально оговорена в тексте.
288 Л/ Н Марков 11 < >i iy 11 >1 ц| । < я 11 hi и bnonoi пи рис iciinii
Как и всякий другой биологический показатель, репродуктивное усилие
особи в популяции может варьировать по ряду причин. Одна из них пе-
синхронность формирования вкладов биомассы в размножение у разных
особей в составе популяции. Это явление, назовем его фенологической раз-
нородностью особей, можно исследовать путем взятия периодических вы-
борок из популяции и построения динамических аллокационных спектров.
Вторая причина — биологическая неравноценность особей популяции.
Имеется в виду вариабельность репродуктивного усилия как следствие вну-
трипопуляционной дифференциации особей по жизненному состоянию.
Такую вариабельность, как было выяснено выше, можно отобразить при
помощи спектра жизненных состояний, а на нем базируется статический
аллокационный спектр. В отличие от общей биомассы особи и некоторых
других «объемных» показателей, зачастую подчиняющихся, как было пока-
зано, положительно асимметричному распределению, репродуктивное уси-
лие дает распределение близкое к нормальному. Это означает, что вычисле-
ние среднего арифметического и прочих статистических параметров даже
для нетрансформированных данных вполне правомерно. Однако некоторые
исследователи (Grace, Wetzel, 1982) сетуют на то, что данные о средних ве-
личинах лежат в основе выделения стратегий у растений. Поэтому средни-
ми величинами репродуктивного усилия целесообразно пользоваться лишь
в дополнение к анализу варьирования этого показателя и в тех случаях, ког-
да необходимо графически представить тип аллокации особи.
9.2. Аллокация и спектр жизненных состояний (размерный
спектр) в популяциях цветковых растений; статические
аллокационные спектры
Поскольку аллокацию можно рассматривать как процесс и одновремен-
но как результат, иными словами, как перераспределение и распределение
(размещение) биомассы в пределах особи, то иллюстрацией ее могут слу-
жить два вида спектров: статический и динамический.
Статический аллокационный спектр строится для какого-то момента
времени и демонстрирует аллокацию у особей разного жизненного состоя-
ния, устанавливаемого по их общей биомассе. По общей биомассе их и ран-
жируют слева направо от минимальной до максимальной. Вычислив для
особей каждого ранга средний относительный вклад биомассы в три сфе-
ры: подземную, надземную вегетативную и репродуктивную (при необхо-
димости можно применять и еще более дробные характеристики аллока-
ции) и построив серию соединенных аллокационных диаграмм, мы получа-
ем статический аллокационный спектр. Такой спектр, несмотря на его ста-
тичность, в известной мере может отражать и динамику аллокации, если
принять во внимание, что по ходу роста особи могут переходить из ранга в
ранг и перестраивать аллокацию сообразно следующему рангу. Графически
можно отразить и зависимость относительного вклада только в одну какую-
либо сферу, например, репродуктивного усилия от общей биомассы. В связи
с этим возникают два вопроса: 1) как нейтрализовать положительную асим-
метрию распределения по общей биомассе, которая приводи! к различиям
/ id«(i V AJuioKiniiiM (hiomiicci.i 289
no числу особей, представляющих каждый класс или ранг и влияет, ааким
обратом, на репрезентативность средних величин аллокации? 2) каков ха
рактер варьирования аллокационных параметров в разных классах (внут ри
каждого класса)?
Положительная асимметрия может быть преодолена применением лога-
рифмической трансформации и логарифмических классов. Опыт показыва-
ет, что на фоне возрастающей репрезентативности средних в каждом ран-
ге (классе) общая закономерность связи аллокации и жизненного состояния
остается неизменной (см. раздел 9.3).
Статический аллокационный спектр, намечающий общую тенденцию ва-
рьирования аллокации по ходу улучшения жизненного состояния особи,
базируется на средних величинах для каждого ранга. Благодаря тому, что
внутри каждого класса особи распределяются по нормальному закону, эти
средние получаются достаточно корректными, но сама степень варьирова-
ния внутри каждого класса остается вне поля зрения. Для того чтобы полу-
чить картину этого варьирования, которая, вне всякого сомнения, представ-
ляет чисто биологический интерес, можно, взяв за основу микромассивы по
три особи и двигаясь вдоль по ранжированному ряду методом скользящей
средней, вычислять для каждого микромассива и откладывать графически
дисперсию репродуктивного усилия.
9.2.1. Аллометрический метод анализа аллокации
Наиболее строгое описание зависимости аллокационных параметров от
общей массы растений получим в том случае, если воспользуемся регресси-
онными уравнениями внутрипопуляционной аллометрии и перейдем к ло-
гарифмическим координатам и трансформированным данным, обеспечива-
ющим достаточно строгую линейность. Аллометрический характер связи
между разными сферами растения позволяет надеяться на получение тео-
ретически, регрессионным путем высчитанных значений порогового раз-
мера особи, допускающего переход к репродукции для разных видов и по-
пуляций.
9.2.1.1. Минимальный (пороговый) размер особи, допускающий ее
переход к репродукции
Среди зарубежных исследователей нет единства в оценке возможностей
методов регрессионного анализа для решения подобных задач. Одни сету-
ют на возрастание автокорреляции при использовании общей биомассы как
независимой переменной (Samson, Werk, 1986), другие считают не вполне
корректным использование натуральных логарифмов для трансформации
данных, получения логнормальных распределений и приведения графи-
ка показательной функции к линейному виду (Thompson, Weiner, Warwick,
1991), а третьи, полагая, что аллометрическая (In - In) регрессия представ-
ляет данные лучше, чем линейная, все же для построения линии регрессии
не используют общую биомассу растений (Sugiyama, Bazzaz, 1998). Нам
прсдстаиияе1ся, что, используя репрезентативные выборки репродукции
290 Л/.// Марков I loiiyiiMniioiinaM biiojioi ия ])асiciinii
ных особей, взятых вместе с корневыми системами (системами скелетных
корней) из популяций малолетних растений, можно сопоставить результа-
ты теоретических расчетов, произведенных разными способами, с эмпири-
ческими величинами, реально представленными в изученных популяци-
ях. Сложности, связанные с возрастанием автокорреляции из-за высокой
доли массы репродуктивных структур в общей массе (Samson, Werk, 1986),
не следует преувеличивать. При использовании же для анализа регрессии
массы репродуктивных структур на массу вегетативных не осуществляет-
ся сама идея количественного определения подобия особей по пропорци-
ям части и целого.
Данные таблицы 1 (величина отношения Wmin/Wo) показывают, что эмпи-
рически определенная минимальная пороговая масса генеративной особи (в
воздушно сухом состоянии) у многих малолетних растений ненамного пре-
вышает массу жизнеспособного семени и, в основном, положительно кор-
релирует с ней (г = +0,643; р<0,001; N=23). Исключение составляет факуль-
тативный двулетник коровяк метельчатый, особи которого переходят к ре-
продукции не ранее, чем накопят значительную «критическую» массу, спо-
собную воспринять яровизационный стимул во время зимовки.
Таблица 9.1.
Сопоставление массы семени (Wo) с эмпирическими величинами минимальной
биомассы необходимой для перехода к репродукции (Wmin) и величинами макси-
мальной биомассы (Wmax*) у ряда видов малолетних растений
Виды растений W (мг) Эмпи- рии. W . (мг) Высчи- танное W . (мг) min v 7 Эмпирич. W (мг) Отноше- ние W . / w m,n О
Чесночница черешчатая 2,60 45,5 5,3 2500,0 17,5
Вайда ребристая 2,50 50,0 24800,0 20,0
Коровяк метельчатый 0,13 7200,0 244000,0 55384,6
Клоповник густоцветковый 0,25 1,9 1,8 1126,8 7,6
Проломник нитевидный 0,02 2,5 2,9 1800,0 125,0
П. удлиненный 0,40 1,5 1,5 117,0 3,8
П. северный 0,25 2,2 1,4 827,0 8,8
П. наибольший 1,00 2,0 2,3 4444,0 2,0
Крупка дубравная 0,02 2,9 1580,0 145,0
Веснянка обыкновенная 0,03 1,2 21,0 40,0
Липучка обыкновенная 1,35 37,2 1410,0 27,6
Горец малый 2,20 12,5 22,6 343,5 5,7
Г. шероховатый 3,50 14,0 7,6 826,0 4,0
Клевер пашенный 0,39 14,0 7,8 354,0 35,9
Щавель приморский 0,22 3,5 3,0 469,9 15,9
Iiatai V А н л< л I lit »t (нюмассы
291
Таблица 9.1. (окончание)
Виды растений W (мг) Эмпи- рии. W . (мг) Высчи- танное W . (мг) Эмпирии. W (мг) max v 7 Отношс нис W / пип W о
Редька дикая 6,60 48,0 7,0 3254,0 7,2
Недотрога мелкоцветковая 6,91 29,4 50,9 3104,0 4,3
Н. обыкновенная 2,80 10,5 17,7 1156,0 3,5
Щирица запрокинутая 0,38 72,0 12,8 3944,0 189,5
Лужница водная 0,009 0,8 15,5 268,0 88,9
Пикульник ладанниковый 2,00 26,0 4,07 1666,5 13,0
Чистец забытый 1,18 24,0 8607,0 20,3
Подорожник индийский 1,75 18,6 3,3 781,4 10,6
Череда поникшая 1,08 7,0 8,8 27075,0 6,5
Ч. трехраздельная 3,07 9,0 6,1 35576,0 2,9
Бурачок пустынный 0,47 4,7 3,1 16500,0 10,0
Лютик ядовитый 0,16 12,0 11,0 822,0 75,0
Змееголовник тимьяноцветковый 0,38 21,0 10,1 969,0 55,3
Фиалка полевая 0,49 4,5 6,0 8400,0 9,2
Рогоглавник серповидный 0,90 6,5 1,5 338,0 7,2
* Биомасса самых крупных особей, которые удалось найти в природных или модельных
популяциях, и потому многие значения, скорее всего, несколько занижены.
Такая масса может быть накоплена в природных условиях не за один год,
а за два и более лет. Следует также отметить, что коровяк метельчатый —
представитель семейства Норичниковых, которое, благодаря своей биоло-
гической специфике, не вписалось в уже упомянутую выше «аллометриче-
скую гипотезу», предполагавшую проявление на межвидовом уровне до-
стоверной положительной корреляции между размерами (массой) семени
и конечными размерами взрослых растений (Thompson, Rabinowitz, 1989).
Здесь можно отметить, что на внутрипопуляционном уровне подобная ал-
лометрия проявляется очень хорошо и выражается как в вырастании более
крупных особей из более крупных семян, так и в формировании более круп-
ными особями более крупных семян.
Константа h аллометрического уравнения связи массы репродуктивных
структур (W^ с общей биомассой (W): In W In b ♦ a In W теоретически
292
МН. Маркин I к >i ly л >i hi к >i 11 ia>i (hiojioi ii>i рас iciniii
должна соответствовать минимальному размеру особи, который достаточен
для перехода к репродукции. Графически пороговый размер выявляется в
области пересечения линией регрессии оси абсцисс. В таблице для боль-
шинства видов (для всех видов, по которым имелись достаточно большие,
репрезентативные выборки) представлены результаты его вычисления по
константам аллометрического уравнения. Теоретические значения порого-
вого размера не дали статистически достоверной корреляции с эмпириче-
скими (г = + 0,267; р = 0,225; N = 24), хотя, подобно им, положительно кор-
релировали с массой семени (г = +0,575; р = 0,003; N = 24).
Значения, высчитанные по линии регрессии Wg на W без логарифмической
трансформации, плохо соответствовали как эмпирическим, так и получен-
ным путем вычислений по аллометрическому уравнению, а некоторые из них
даже были представлены отрицательными числами. Линия регрессии, как из-
вестно, строится на основе множества («облака») эмпирических точек по ме-
тоду наименьших квадратов. На каждом из двух концов мы имеем снижение
точности, которое отражается на графиках расхождением вспомогательных
кривых, обозначающих доверительные границы, и которое весьма затрудня-
ет экстраполяцию. Хотя ждать большой точности от теоретически высчитан-
ных по уравнению значений пороговой биомассы и не следует, все же проце-
дура вычислений не лишена смысла. Заманчиво иметь инструмент для про-
гнозирования репродуктивных возможностей растений. Нужно только с пре-
дельной корректностью собирать эмпирический материал и совершенство-
вать методы статистического анализа. К примеру, для обеспечения большего
соответствия теоретических величин эмпирическим можно применять метод
Рис. 9.3. Схематизированные статиче-
ские аллокационные спектры для популя-
ций трех видов проломника: верху — наи-
большего (7.VI.1983); внизу слева — се-
верного (7. VI. 1983); внизу справа — удли-
ненного (24.V.1984). W — общая биомас-
са особи в мг (по горизонтальной оси), по
вертикальной оси процентный вклад био-
массы в: 1 — генеративную сферу, 2 —
прицветные листья, 3 — надземные веге-
тативные органы, 4 — корневую систему
(далее на всех аллокационных спектрах
такие же обозначения.
jack-knife (поочередное исключение
максимальных и/или минимальных
по размерам особей из выборки), что-
бы иметь возможность подвергнуть
полученную при этой процедуре се-
рию значений искомого порогового
размера (для одной выборки из попу-
ляции!) статистической обработке.
9.3. Закономерности аллокации
у малолетних растений
Изучение статических аллокаци-
онных спектров, построенных для
трех лугово-степных видов пролом-
ника (рис. 9.3) дает основание для
вывода, что при любом жизненном
состоянии относительный вклад
биомассы в репродуктивную сфе-
ру (RE2) более или менее одинаков
у всех генеративных особей каждого
из видов. Обращает па себя внима-
ние и конеlain noc 11, oi носи iciiliioi о
I 'Itltld 9 Л II in »k n 11IIII bllOMIICCI.1
293
Рис. 9.4. Статические аллокационные
спектры для подорожников: индийско-
го (вверху) и большого (внизу). W — об-
щая биомасса в мг. Прочие обозначения
как на рис. 9.3.
вклада в корневую систему. Ila an
локационном спектре для пшмпш
отдельно показан вклад в «розетку»
бракгей, о которой говорилось выше
(см. главу 4).
Сходную картину дает статиче-
ский аллокационный спектр для по-
пуляции подорожника индийско-
го (рис. 9.4). Этот спектр постро-
ен в отличие от предыдущих, с при-
менением логарифмических клас-
сов, но кроме большей достоверно-
сти средних для каждого класса, ни-
каких отличий от первоначально по-
строенного спектра по абсолютным
рангам не наблюдалось. Для срав-
нения на том же рис. 9.4. приводит-
ся спектр для подорожника большо-
го, построенный по данным опыта
с разной плотностью (см. главу 8).
При постоянстве вклада в корневую
систему репродуктивное усилие
растет вместе с мощностью особей.
При анализе особей подорожника
большого из естественной популя-
ции репродуктивное усилие выказа-
ло сходную тенденцию. Например,
в популяции этого вида, произраста-
ющей на песчаной дамбе в условиях
полной освещенности, особи были разделены на три группы по мощности:
мелкие, средние и крупные (средние для групп значения биомассы были,
соответственно, 112,5 ± 7,8; 1549,8 ± 94,6; 12558,3 ± 145,3 мг). Диаграммы
аллокации (рис. 9.5) показывают, что по мере увеличения мощности особей
репродуктивное усилие возрастало за счет уменьшения вклада биомассы в
корневую систему при почти неизменном вкладе в ассимилирующие орга-
ны. Таким образом, стабильный вклад в корневую систему у подорожни-
ка большого, который наблюдался в опыте, по-видимому, поддерживается
только при низкой интенсивности репродукции, а резкое увеличение мощ-
ности плодоношения может повлечь за собой пропорциональное снижение
мощности подземных органов, обеспечивающих переннизсщию, и ставит
под сомнение саму возможность последней.
Найденный поздней осенью экземпляр подорожника большого, демон-
стрирующий мощную репродукцию (9 крупных колосьев), к моменту на-
ходки почти уже не имел живых тканей, и таким образом у него практиче-
ски не было никаких шансов для выживания в течение зимы. Но совокуп-
ности при знаков эта особь представляла подвид Р. major L. ssp. pleiosperma
Pilger, Koiopi.iii раньше называли Plantago intermedia (п. промежуточный)
294
МН Марком I loiiyihinnoiiiiaM Giioiioi ini рас iciiiih
100%
50
Рис. 9.5. Диаграммы аллокации для че-
тырех типов особей подорожника боль-
шого: а — мелкие, б — средние, в —
крупные (а, б, в, — особи с открыто-
го участка); г — с признаками формы
Р . major ssp: pleiosperma из агроценоза
озимой ржи (обозначения те же, что и на
рис. 9.3 и 9.4).
1 2
Рис. 9.6. Зависимость репродуктивно-
го усилия от эколого-фитоценотических
факторов у проломника нитевидного:
1 — под пологом леса; 2 — на опушке.
Особи этого подвида часто высту-
пают как сегеталы и встречаются в
посевах озимых и даже яровых куль-
тур, причем обычно плодоносят, со-
ответственно, на втором и даже на
первом году жизни. На рис. 9.5 при-
ведена для сравнения диаграмма ал-
локации биомассы у однолетних осо-
бей, выросших в посеве пшеницы в
условиях значительного затенения.
Форма листьев, опушение, тон-
кий редкоцветный колос у этих рас-
тений указывали на принадлеж-
ность их к выше названному подви-
ду. Налицо увеличение вклада био-
массы в формирование ассимиля-
ционного аппарата и снижение ре-
продуктивного усилия. Если, од-
нако, учесть, что здесь мы сталки-
ваемся с плодоношением потенци-
ального многолетника в первый год
жизни, то приходится признать, что
в данном случае фитоценотическая
среда влияет на величину репродук-
тивного усилия и вызывает прояв-
ление иной заложенной в генотипе
эколого-ценотической стратегии.
Аллокационные диаграммы на
рис. 9.6 показывают, как влияет зате-
нение на величину репродуктивного
и фотосинтетического усилия1 у про-
ломника нитевидного. Несмотря на
то, что это растение часто растет на
лесных дорогах и устойчиво к прите-
нению, более высокое RE2 характер-
но для популяции, произрастающей
на осветленном месте, где меньше за-
траты на формирование ассимиляци-
онной поверхности.
Слабое варьирование репродук-
тивного усилия у особей трех лугово-
степных видов проломника и подо-
рожника индийского, установленные
по статическим аллокационным спектрам, подтверждается анализом алломе-
трической связи массы репродуктивных структур с общей массой.
’Термин «фотосинтетическое усилие» применяется Ю. А. Злобиным.
Гнша V A u n< ши i nt к (шомассы
295
I рафики (рис 9.7.) отчетливо демонеiрируют: во-первых, oicy ic 1 вне oi
KJioiieiniii oi нипейной зависимости в noi арпфмических (In) коордииачах. a
во-вторых, независимость репродуктивного усилия (RE2) от общей биомас-
сы. Аллометрические коэффициенты всех трех уравнений регрессии, попу
ченных для трех видом проломника, близки к единице:
InWg = 1,08 InW — 0,47 (П. наибольший),
InWg =1,01 InW — 0,32 (П. северный),
InWg = 1,04 InW — 0,41 (П. удлиненный).
Похожие уравнения получены и
для розеточных малолетников с си-
стемами придаточных корней: для
рогоглавника (InWg = 0,95 InW —
0,06) и для мышехвостника (InWg =
0,93 InW— 1,01).
На тех же графиках приведены
данные о связи вклада биомассы в
корневые системы с общей массой.
Несколько больший разброс значе-
ний отчасти может быть связан с по-
вышенной вероятностью поврежде-
ния корневых систем при выкапыва-
нии, но крайне слабая тенденция к
снижению относительного вклада в
корни все же намечается для некото-
рых видов.
Это подтверждают и уравнения
регрессии:
Рис. 9.7. Аллометрические зависимо-
сти биомассы репродуктивных струк-
тур (Wg) и корневых систем (г) от об-
щей биомассы у трех видов проломни-
ка: А.ш. — наибольшего, А.е. — удли-
ненного и A.s. — северного.
296 МН Марков I loi iy л>1Ц1к>1 и uni бпоногия расicnnil
InWr 0,81 in W 1,54 (Нроломпик наибольший)
InWr = 1,00 In W — 3,20 (I I. северный)
InWr = 0,79 In W — 2,08 (П. удлиненный)
InWr = 0,86 In W — 1,14 (Подорожник индийский)
InWr = 0,70 In W — 1,90 (Рогоглавник серповидный)
InWr = 1,01 In W — 1,33 (Мышехвостник маленький).
На примере проломников наибольшего и северного методом, описанным
в разделе 9.2, продемонстрирована более высокая вариабельность репродук-
тивного усилия у наиболее мелких особей — в левой части ряда. Величина
дисперсии при сопоставлении средних и крупных особей в микромассивах
заметно снижалась (рис. 9.8 вверху).
Заманчиво было бы предположить, что внутрипопуляционная констант-
ность репродуктивного усилия и изометрическое (или близкое к нему) со-
отношение между биомассой репродуктивных структур и общей биомас-
сой — отличительная черта многих, если не всех малолетних растений
(табл. 9.2).
Таблица 9.2.
Аллометрические коэффициенты в уравнениях связи вклада биомассы в репро-
дукцию (Wg) с общей биомассой (W) у ряда малолетних растений
Названия растений Уравнения r P
Подорожник индийский InWg =-0,69 + 1,00 InW +0,973 <0,001
Щавель приморский lnWg =-1,09 +0,98 InW +0,989 <0,001
Щирица запрокинутая lnWg =-2,58+ 1,01 InW +0,803 <0,001
Бурачок пустынный InWg = -1,05 + 0,94 InW +0,971 <0,001
Лютик ядовитый InWg = -2,42 + 1,01 InW +0,855 <0,001
Пшеница (модули) InWg = -0,92+ 1,19 InW +0,894 <0,001
Пшеница (особи) InWg = -1,03 +1,05 InW +0,923 <0,001
Клоповник густоцветковый InWg = -0,59 +1,06 InW +0,992 <0,001
Лужница водная (центральные особи в клонах) InWg = -2,13 + 0,94 InW +0,570 <0,001
Чесночница черешчатая InWg = -1,86 +1,12 InW +0,979 <0,001
Змееголовник тимьяноцветков. InWg = -1,14+ 1,12 InW +0,989 <0,001
Редька дикая InWg = -2,52 + 1,30 InW +0,920 <0,001
Лужница водн. (одиночные особи) InWg = -3,05 +1,21 InW +0,490 <0,003
Лужница водная (клоны) InWg = -1,86+ 1,27 InW +0,717 <0,001
Горец малый InWg = -2,17+ 0,87 InW +0,894 <0,001
Горец шероховатый InWg = -1,66 + 0,89 InW +0,880 <0,001
Примечание: в уравнениях выделены аллометрические коэффициенты.
Однако есть исключения. Д. Хикманом (Hickman, 1975) для незимующе-
го безрозеточного однолетника горца каскадного Polygonum cascadense и
Ю. А. Злобиным для горца шероховатого Р scabrum со сходной архитекту-
рой и жизненным циклом показано снижение репродуктивного усилия по
мере увеличения мощности особей.
I'KIthl u A II ll( th ill II I M (IIIOMIK'CI.I
29/
Рис. 9.8. А: изменение дисперсии репродуктивного усилия вдоль по ранжированному
ряду общей биомассы у проломника наибольшего (слева) и северного (справа); Ь: ста-
тические аллокационные спектры для двух популяционных локусов змееголовника ги
мьяноцветкового. W — общая биомасса (мг); остальные обозначения как на рис. 9.4.
Наши данные подтвердили эти наблюдения. Особи горца шероховатого
были разделены нами на две группы: средняя общая биомасса особи в пер-
вой группе составляла 51,4 ± 10,9, а во второй — 967,2 ± 245,8 мг. Средние
значения репродуктивного усилия в группах составили соответственно
38,3 ±1,7 и 6,2 ±0,6%.
Есть среди малолетних растений примеры и положительной корреляции
репродуктивного усилия с общей мощностью. У облигатных двулетников:
временнорозеточного змееголовника тимьяноцветкового (рис. 9.8. внизу) и
полурозеточного чесночницы для наиболее крупных особей характерно н
наибольшее в популяции репродуктивное усилие.
Вероятнее всего причина появления этих исключений состоит в особен-
ностях архитектуры уклоняющихся видов. Плодоносящие особи змееголов-
ника тимьяноцветкового, например,
не имеют розеточного строения и
достаточно четкой границы меж-
ду генеративными и вегетативными
структурами. На схеме (рис. 9.9) по-
казано, что позднее формирующие-
ся ложные мутовки цим в составе
тирса спускаются у мощных особей
почти до нижней пары стеблевых
листьев, из-за чего перерезать рас-
тение, отделяя его репродуктивную
сферу, приходится все ниже и ниже.
Ветвление мощных особей, судя по
всему, тоже усугубляет проявление
положительной корреляции — во
второй популяции как раз присут-
ствовали крупные сильно ветвящи-
еся особи. Аллометрический коэф-
Рис. 9.9. Изменения архитектуры особи
у змееголовника тимьяноцветкового при
увеличении мощности.
298 МН Маркой I loiiyiiMHiioiiiiiui (hioiioi ня рис iciihB
фициенг и для первой и гем более для второй популяции существенно пре-
вышает единицу:
InWg =1,16 InW — 1,53 (I популяция)
InWg = 1,28 InW — 2,40 (II популяция)
По существу те же проблемы возникают и при анализе репродуктивного
усилия в популяциях чесночницы, поскольку, как и у других крестоцветных
границу между репродуктивной и вегетативной сферами приходится прово-
дить по уровню самого нижнего плода в кисти.
Полученные графики и уравнения также указывают не только на прямую
зависимость между размерами (общей биомассой) особи и ее относитель-
Рис. 9.10. Зависимость репродуктивного
усилия (RE3) от общей биомассы (W) в
популяциях клоповника густоцветково-
го (а) и ярутки (б).
Рис. 9.П. Кривые распределения по
классам репродуктивного усилия (RE3)
для особей клоповника густоцветкового
трех уровней мощности. Общая биомас-
са особей: 1 — 0,1-6,0 мг; 2 — 6,1-12,0
мг; 3 — 12,1-18,0 мг.
ным вкладом биомассы в репродук-
тивную сферу, но и наиболее яркое
проявление этой зависимости при
наличии в популяции ветвящихся
особей.
Закономерен вопрос: не являют-
ся ли наблюдаемые отклонения от
явно преобладающей в популяци-
ях малолетних растений изометрии
результатом методической ошиб-
ки при проведении границы между
сферами? Один из возможных пу-
тей проверки данной гипотезы ле-
жит через использование для вы-
числения RE2 только чисто репро-
дуктивных структур с исключением
структур вспомогательных.
Если в первом нашем примере с
горцем шероховатым репродуктив-
ное усилие вычисляли по отноше-
нию массы всех соцветий к общей
массе (RE2), то, используя выборку
плодоносящих особей из агропопу-
ляции, помимо общей массы соцве-
тия, при вычислении репродуктив-
ного усилия мы брали массу одних
плодов. Полученные графики алло-
метрической зависимости и уравне-
ния (табл. 9.2) подтвердили перво-
начальный вывод.
Использование в расчетах RE2 от-
дельно цветков и чашечек с эрема-
ми, у змееголовника, и только пло-
дов, у чесночницы, привело опять-
таки к прежним выводам, хотя вели-
/ 9 Д и и* ж к 1111 и биомассы
299
чипа annoMciрпчсского коэффициента и оказалась несколько ниже, чем и
нервом случае.
Для некоторых других видов из семейства крестоцветных был примене-
нен другой способ вычисления репродуктивного усилия RE3.
Графики показывают (рис. 9.10а,б), что при увеличении размеров (веса)
растения число плодов, произведенных на единицу биомассы, у клоповни-
ка густоцветкового и особенно ярутки полевой резко снижается. В случае
с к. густоцветковым разбивка самого первого класса, характеризовавшего-
ся наибольшей частотой, на микромассивы: 0,1-6,0; 6,1-12,0 и 12,1-18,0 мг
позволила еще раз (теперь на примере RE3) показать уменьшение ампли-
туды варьирования репродуктивного усилия по мере увеличения размеров
особи (рис. 9.11).
На основании наблюдаемых зависимостей можно высказать предполо-
жение о меньшей эффективности очень крупных особей и меньшем соот-
ветствии их «интересам» популяции некоторых малолетних растений. Нс
случайно при неблагоприятных условиях, с которыми ей чаще приходится
сталкиваться, популяция возобновляется преимущественно за счет преоб-
ладающих в популяции мелких, но плодоносящих в силу пластичности осо-
бей (см. главу 5.5).
9.4. Изменение репродуктивного усилия и аллокации в целом
во времени; аллокационные динамические спектры
Динамический аллокационный
спектр объединяет усредненные ал-
локационные диаграммы, построен-
ные для разных моментов развития
популяции. Из предыдущего раздела
становится ясно, что для разных ви-
дов достоверность усредненной ди-
аграммы аллокации несколько отли-
чается. Например, на диаграммах,
приведенных в методическом разде-
ле данной главы для иллюстрации
перераспределения биомассы, ре-
продуктивное усилие возрастает от
47 до 67%. Если же эту картину дать
в динамике кривых распределения,
то для каждой даты репродуктивное
усилие дает существенный разброс
значений (рис. 9.12).
Вместе с тем сдвиг моды одновер-
шинных кривых, происходящий на
фоне отсутствия каких-либо суще-
ственных изменений в общей био-
массе (см. графики распределения по
общей биомассе на том же рисунке),
Рис. 9.12. Динамика распределений ио
классам общей биомассы W (вверху) и
репродуктивному усилию RE2 (внизу)
для популяции проломника северной):
18.V сплошная линия; 4.VI пун-
ктир.
300
МП Мирков I loiiyiiHHiioiiiiaH 61101101 ни pur icinih
InW
Рис. 9.13. Относительная скорость ро-
ста (вверху), динамический аллокацион-
ный спектр и линии связи репродуктив-
ного усилия (RE2%) с общей биомассой
W (внизу) для дюнной популяции подо-
рожника индийского: Т1 — 6.VII; Т2 —
18. VII; ТЗ — 16. VIII; Т4 — 30.VIII.1984.
Обозначения на аллокационном спектре
те же, что и на рис. 9.4: 1 — 18.VII; 2 —
16.VIII; 3 — 30.VIII. 1984.
конечно же более ярко демонстри-
руется аллокационными диаграм-
мами, показывающими к тому же за
счет какой из сфер происходит пе-
рераспределение биомассы в репро-
дуктивную сферу. Так что динами-
ческий спектр наряду с параллель-
ной демонстрацией динамики общей
биомассы и со статическими спек-
трами и другими средствами ото-
бражения варьирования аллокацион-
ных параметров применять целесо-
образно. Тем более, что сам общий
характер зависимости между общей
мощностью и репродуктивным уси-
лием во времени может не менять-
ся. Для подорожника индийского это
показано на рис. 9.13а, б, где приве-
дены: кривая относительной скоро-
сти роста, динамический аллокаци-
онный спектр и линии связи репро-
дуктивного усилия с общей массой.
Обращает на себя внимание несколь-
ко повышенный по сравнению с дру-
гими стержнекорневыми малолетни-
ками вклад в корневую систему, оста-
ющийся на уровне 12% даже в пери-
од репродукции. Это можно связать
с дополнительными затратами на то-
лерантность к напряженности водно-
го режима на песчаных дюнах — в
местообитании п. индийского.
Выносливость к затенению также
требует затрат и прежде всего, как мы
уже видели на примере проломника
нитевидного и увидим ниже на при-
мере недотроги обыкновенной, повышения фотосинтетического и соответ-
ственного снижения репродуктивного усилия. Вообще для видов, имеющих
длиннопобеговую архитектуру (недотроги обыкновенная и мелкоцветковая,
щирица запрокинутая и др.), наблюдается следующая закономерность: отно-
сительный вклад биомассы в стебель коррелирует с общей биомассой поло-
жительно, а относительный вклад в листья — отрицательно. Объяснение, ве-
роятнее всего в том, что особо крупным особям, возвышающимся над осталь-
ными благодаря мощно развитому стеблю, можно обеспечить достаточный
перехват солнечной энергии посредством меньшей листовой поверхности.
У проломника наибольшего постоянство характера связи между репро-
дуктивным усилием и общим весом доказывается близкими значениями ал-
Гита V A innihniiiiH (iiioMiicci.i
301
14.VI 27.VI 11.VII 25.VII
Рис. 9.14. Динамические аллокацион-
ные спектры для популяций жерушни-
ка болотного (а) и резуховидки Таля (б).
Популяции жерушника: I — на песча-
ном заиленном пляже; II — в агроцено-
зе вики с овсом.
б
Рис. 9.15. Динамический аллокацией
ный спектр популяции змееголовни-
ка, состоящей из вегетирующих особей.
Относительный вклад биомассы: I и
листья; 2 — в стебли; 3 — в корневую
систему.
лометрического коэффициента для популяции на разных стадиях. На ста-
дии цветения уравнение выглядело так: InWg = 1,03 InW — 0,81, а на ста
дии плодоношения — так: InWg = 1,08 InW — 0,50.
Динамический аллокационный спектр может начинаться еще со стадии ве-
гетации, когда он иллюстрирует динамику соотношения массы корневой сн
стемы с общей массой (RWR). Сравнение двух популяций жерушника боло i -
ного, одна из которых произрастала на заиленном песчаном пляже в услови-
ях почти полного освещения (I), а другая — в посеве вики с овсом в услови-
ях значительного затенения (II), по параллельным динамическим аллокаци-
онным спектрам показало (рис. 9.14а): 1) Условия освещения сказываю гея па
относительном развитии корневых систем — во II популяции вклад в корне-
вую систему был постоянно ниже, чем в I; 2). Более высокое фотосинтетиче-
ское усилие у особей из посева было сопряжено с менее высоким репродук-
тивным усилием (4 июля —16% против 30%); 3) После уборки посева в I ре-
продуктивное усилие некоторое время продолжало увеличиваться главным
образом за счет вегетативной надземной сферы, после чего уменьшилось, от
ражая отсыхание и осыпание плодов и семян.
Для агропопуляции резуховидки в посеве яровой пшеницы динамический
аллокационный спектр позволил уловить кратковременное накопление ас-
симилятов в корневой системе, о котором шла речь в главе 8, после чего пе-
реход к репродукции повлек за собой снижение относительного вклада бно
массы в корневую систему (рис. 9.146). У незимующего однолетнего core
тала редьки дикой это снижение, несмотря на резкое возрастание ре ироду к
тивного усилия, было почти незаметно.
У растений змееголовника первого года жизни динамика аллокации хоро
шо показывает первоначальное преобладание в спектре вклада в надземные ас
симилирующие органы, который возрастал со временем. Однако в последнюю
дату наблюдений (10.Х.1987) соотношение несколько изменилось в пользу под
земной сферы, вклад в которую, отражая подготовку растений к зимовке и не
пользование благоприятной) осеннего периода, несколько возрос (рис. 9.1 S).
302 МН Марков I loiiyiiHHiioiiiiaH (nionoi ня рас iciniH
9.5. Репродуктивное усилие у малолетних растений с разными
вариантами эколого-ценотической стратегии
Резюмируя все, что сказано в этой главе, мы вынуждены признать труд-
ность сопоставления репродуктивного усилия, вычисленного одним каким-
то способом у разных видов малолетних растений из-за различий в архитек-
туре особей. В случае сопоставления видов одного рода со сходной архитек-
турой задача несколько упрощается, хотя даже внутри вида, как было пока-
зано выше, возможны количественно, а в случае тератов (рис. 9.16) даже
качественно различные архитектурные варианты.
И все же есть основания для общего вывода о наиболее высоких значе-
ниях репродуктивного усилия у видов, проявляющих стратегию избегания,
особенно розеточных. Но виды, в генофонде которых запрограммированы
элементы толерантности к затенению или недостатку влаги в почве, отра-
жаемые иногда и в особенностях архитектуры, репродуктивное усилие, как
правило, снижается за счет возрастания фотосинтетического усилия или от-
носительного вклада в корневую систему (табл. 9.3).
Таблица 9.3.
Репродуктивное усилие (RE2%) в популяциях некоторых малолетних растений
Виды растений Lim min х ± s X Lim max
Недотрога обыкновенная 0,8 1,8 ±0,1 5,1
Н. мелкоцветковая 0,8 5,5 ± 0,5 10,9
Горец малый 1,6 6,5 ± 0,5 16,5
Щирица запрокинутая 1,5 10,2 ± 0,7 32,9
Лютик ядовитый 2,8 10,7 ±1,0 22,1
Лужница водная (клоны) 0,9 11,3 ±0,9 30,0
Л. водная (центральные особи в клонах) 1,3 12,2 ± 0,9 36,4
Л. водная одиночные особи без столонов (аклонисты) 0,9 12,3 ± 1,5 35,7
Подорожник промежуточный 5,4 14,6 ± 0,8 19,1
Пикульник ладанниковый 14,2 17,3 ± 0,5 18,9
Тимелея воробьиная 2,1 17,3 ± 1,1 45,0
Бурачок пустынный 21,9 30,1 ±2,1 42,9
Чесночница черешчатая 10,3 30,2 ± 0,6 41,0
Мышехвостник маленький 14,3 30,7 ± 1,3 43,8
Щавель приморский 20,7 31,5 ±2,9 40,6
I 'IHflil 9 A IHKHUIHini (HIOMIICC'I.I
303
Tnftjiiiuii 9.3. {окончание)
Виды рас iciinii 1 лт mm X 1 S ч 1 ,1111 ПН1Ч
Подорожник индийский 14,7 36,5 ± 0,4 54,5
Змееголовник тимьяноцветковый 17,4 36,8 ±1,0 59,8
Вайда ребристая 29,5 43,9 ± 1,1 52,5
Редька дикая 17,7 44,2 ± 2,3 51,0
Пастушья сумка 40,2 48,2 ± 3,5 73,1
Ярутка полевая 38,0 58,4 ±3,1 70,0
Клоповник густоцветковый 37,2 65,9 ± 0,9 81,3
Проломник северный 60,6 75,6 ± 0,9 83,7
П. удлиненный 64,7 76,0 ± 0,7 84,3
Рогоглавник серповидный 57,5 80,9 ± 0,6 89,6
П. наибольший 65,1 85,6 ± 1,1 91,4
Количественно оценивать эффек-
тивность тела малолетнего растения
в плане достижения максимального
репродуктивного усилия, т. е. отно-
сительного вклада биомассы в раз-
множение, как мы выяснили, мож-
но применяя принципы популяци-
онной аллометрии. Основной во-
прос при этом можно сформулиро-
вать примерно так: можно ли в пре-
делах популяции выделить особи та-
кого размера, который бы обеспечи-
вал максимальную эффективность
репродукции? Анализ показал, что
возможны три варианта ответа: 1)
выделить такую группу особей не
представляется возможным, ибо все
особи («от мала до велика») имеют
примерно одинаковое репродуктив-
ное усилие; 2) чем крупнее особи,
тем эффективнее их репродукция;
3) оптимален минимальный (разуме-
ется, вообще возможный для репро-
дукции) размер особей. Каждому из
ответов соответствует группа видов
малолетних растений.
Графически решать поставленную
задачу можно двумя способами. Во-
Рис. 9.16. Особь- герат проломника пию
ВИДНОГО С МПОГОЯруСНЫМ IOII IHKOM
304
Л/.Л Л/и/
I 1<>1 iy 11>1ЦП<>11ИИ>1 ()||<>И<>| ||>| рис ICIIIfil
25.1V 30.1V 4.VI 11.VI 18.VI 12.VII 20.VII 27.VII
Даты наблюдений
Относительный вклад биомассы (%)
| корнев. сист. | | стебли | | листья ИВ РепР стРУкт-
Рис. 9.17. Статический (вверху) и динамический (внизу) аллокационные спектры для
популяции недотроги мелкоцветковой.
первых, можно рассматривать связь между репродуктивным усилием и лога-
рифмом (In) общей биомассы растений, избирая полулогарифмический мас-
штаб для компенсации положительной асимметрии распределения особей по-
пуляции по классам общей биомассы. Второй, более строгий способ связан с по-
лучением аллометрической зависимости и аллометрического коэффициента.
1. Критерием для отнесения видов к первой группе является наиболее
близкое к единице значение аллометрического коэффициента или наклон
линии регрессии биомассы репродуктивных структур на общую биомассу в
логарифмических (In) координатах наиболее близкий к 45°. Это тот случай,
когда репрезентативная выборка особей, сортированная в порядке возраста-
ния общей биомассы, помимо своей статичности, как моментального сним-
ка размерной структуры популяции, содержит явный динамический аспект.
Она представляет как бы процесс увеличения размеров при сохранении но-
I ’lana V A IIinЖП1Ц1>i (нюмассы 30b
стоянных пропорций. Хорошиii пример д;к-1 к ном ключе сопосiлвлеппе
етатическо! о и динамического аллокационных енеклров для популяции не
дотроги мелкоцветковой (рис. 9.17). Здесь в известном смысле можно ю
корить о внутрипопуляционной изометрии особей. М. Reiss (1989) выека
зал критические замечания по поводу все более широкого использования
ботаниками такого показателя как репродуктивное усилие и вместе с гем
редкого использования аллометрического анализа, поскольку по его мне-
нию (мнению зоолога) репродуктивное усилие будет почти наверняка зако
номерно варьировать, завися от размеров растений.
Соглашаясь с М. Reiss в том, что аллометрический анализ совершенно необ-
ходим и в ботанических исследованиях, все же нельзя не отметить наиболее ча-
сто встречаемую изометрию малолетних, особенно зимующих однолетних, рас-
тений (Harper, Ogden, 1970; van Andel, Vera, 1977; Harris, Lovell, 1980; Марков,
1990). В изометрии особей популяции малолетнего растения, в общем-го, nci
ничего удивительного. Представления о росте с позиций учения о метамер! юс п1
как о процессе накопления инвариантных субъединиц как нельзя более соогвс i -
ствуют картине сохранения подобия и пропорциональности по достижении осо-
бями одной и той же биологической стадии развития при существенно различ-
ных размерах. Вместе с тем наблюдаемые отклонения от изометрии, о кагоры х
речь пойдет ниже, несомненно достойны особого внимания.
2. Известно, что задержка репродукции до второго сезона вегетации (а ино-
гда и на более долгий срок), которую мы наблюдаем у двулетних монокарпи-
ческих растений, должна отчасти компенсироваться усиленной репродукци-
ей благодаря накоплению за первый вегетационный сезон значительной ре-
зервной биомассы. Известно и то, что большой группе двулетних растений
свойственна относительная крупносемянность (Марков, 19916). По крайней
мере для некоторых двулетников с присущей им гетеромодульной архитскту-
рой (чесночница, змееголовник) нами отмечено существенное превышение
единицы аллометрическим коэффициентом, что свидетельствует о возрас i а-
нии репродуктивного усилия по мере увеличения общей мощности особей и
популяции. Указанная закономерность наблюдалась независимо от того, что
было включено при обработке в состав репродуктивных структур, одни ли се-
мена и цветки или все соцветие вместе с осями и прочими вспомогатель.
ми структурами, отделенное на уровне самого нижнего цветка. Возрастание
эффективности репродукции вместе с общими размерами особей — обычкам
картина у многолетних растений (Марков, Плещинская, 1987).
Интересным, на наш взгляд, фактом выступает изометрия централ .....
особей в клонах лужницы водной при явном аллометрическом othohiciiiiii у
одиночных (аклоналъных) особей и клонов в целом (табл. 9.2). Очевидно, и
здесь сказывается своеобразие архитектуры с максимально возможной для
малолетников автономностью фитомеров. Достаточно сказать, что ночи!
каждый фитомер в составе одного брахиморфного модуля имеет свой соб
ственный придаточный корень, и потому можно даже вычислить репродук-
тивное усилие отдельного окорененного фитомера. Оно составляло в иссле-
дованной популяции от 23,1 до 65,4%. Значения RE2 для особей и клонов
лужницы приведено в таблице 9.3.
306 МН Маркин I loiiyiiHUHoiiiuui (>по1К>И1>1 piHicmth
У изомодульных малолетних растений, судя но данным, полученным
нами на яровой пшенице, может происходить внугримодульная детерми-
нация развития репродуктивной сферы предшествующим развитием сфе-
ры вегетативной. Это отразилось на величине аллометрического коэффици-
ента в уравнениях (табл. 9.2) высчитанных для выборки модулей (N=106).
Однако, на уровне особей (N=105), судя по аллометрическому коэффици-
енту, вновь можно наблюдать ситуацию близкую к изометрии, что по всей
вероятности связано с повышенной автономностью модулей в составе изо-
модульных особей пшеницы. Значения репродуктивного усилия, высчитан-
ные на основе выборок модулей для других малолетних граминоидов при-
ведены в таблице 9.4.
Таблица 9.4.
А. Значения репродуктивного усилия, высчитанные для некоторых
малолетних граминоидов на основе модуля
Вид растения Репродуктивное усилие (RE7) модуля (%)
Limrnin X ± sx Lim max
Анизанта кровельная • 23,1 54,3±1,9 75,6
Скрытница 54,0 60,2±1,4 66,4
Сыть бурая 16,7 44,5±2,7 62,2
Ситник жабий 15,4 58,9±2,1 82,6
Мятлик однолетний 15,8 29,6±1,5 58,3
Щетинник зеленый 27,4 44,5±4,1 60,2
Б. Аллометрические уравнения связи биомассы репродуктивных структур
с биомассой модулей Wab:
Вид растения Уравнения r P
Анизанта кровельная InWg = -1,02 + 1,07 In Wab +0,978 <0,001
Скрытница InWg = -0,54 + 0,99 In Wab +0,997 <0,001
Сыть бурая InWg = -1,29+ 1,22 In Wab +0,962 <0,001
Ситник жабий InWg = -1,06 + 1,18 In Wab +0,914 <0,001
Мятлик однолетний InWg = -1,71 +1,11 In Wab +0,803 <0,001
Среди гетеромодульных малолетников известны и виды, у которых эф-
фективность репродукции падает с увеличением общей мощности особи.
Для популяций горцев Polygonum minus и Р. scabrum аллометрический ко-
эффициент составил, соответственно, 0,87 и 0,89.
Подводя итоги, можно предположить, что виды со вторым типом зависи-
мости ориентированы на низкую, а виды с третьим типом зависимости — на
высокую плотность популяции. Возникает мало сомнений в том, что рас-
смотренные аллометрические зависимости, играющие важнейшую роль в
формировании стратегий жизненного цикла малолетних растений, напря-
мую связаны с особенностями их архитектуры, которая и подлежит даль-
нейшему сравнительному изучению в этом аспекте.
Глава 10. ДЕМОГРАФИЧЕСКИМ
АНАЛИЗ ПРИРОДНЫХ ПОПУЛЯЦИЙ
РАСТЕНИЙ РАЗНЫХ ЖИЗНЕН-
НЫХ ФОРМ В РАЗНЫХ ЭКОЛОГО-
ФИТОЦЕНОТИЧЕСКИХ УСЛОВИЯХ
Уланы с пестрыми значками,
Драгуны с конскими хвостами,
Все промелькнули перед нами,
Все побывали тут...
М. Ю. Лермонтов, «Бородино»
Во введении демографические параметры популяций названы главным
объектом популяционной биологии растений. В этой связи может показать-
ся странным, что рассмотрению демографических характеристик посвяще-
на самая последняя глава учебного пособия. Дело в том, что многие важ-
ные вопросы демографии: «плата» за размножение, особенности диссеми-
нации, реакция видов на популяционную плотность, одним словом, мно-
гие биологические предпосылки регуляции численности уже рассмотрены
в соответствующих главах. В последнюю же вынесены данные о наблюде-
ниях за темпами развития особей в естественных популяциях и собственно
динамика численности некоторых розеточных, полурозеточных, временно-
розеточных и длиннопобеговых малолетников, т. е. данные, которые по об-
разному выражению Дж. Уайта (White, 1987), составляют «ботаническую
арифметику». Впрочем, показанная в предыдущих главах тесная связь меж-
ду численностью (плотностью) популяции и биоморфологическими харак-
теристиками слагающих ее особей не может быть проигнорирована, по-
скольку роль численности в ее динамике именно через эти-то характерно ги-
ки и проглядывает наиболее рельефно.
10.1. Методические аспекты изучения демографии популяций
растений разных жизненных форм
Краткое описание некоторых методов популяционной ботаники приводе
но здесь для того, чтобы различие между биологией растений н нонуляцн
онной oiKHioi иен выс1унало наиболее рельефно. (huieaiiiie биоло! ни кно
308 Л//< Мирков Популяционная bnojionni рас leniiii
или иного вида идет как бы в статике и, как правило, по среднему значении)
любого показателя, без учета спектра его значений в реальности. Главные
отличительные черты популяционной биологии растений состоят в анализе
количественными методами популяций:
1) как динамических систем;
2) как систем структурированных, гетерогенных по разным биологиче-
ским показателям с показом всего спектра возможных количественных зна-
чений.
Суть анализа динамики численности популяций состоит в статистиче-
ской регистрации убыли-прибыли особей и выявлении соотношения этих
двух процессов. Учет ростовых процессов, на фоне которых осуществляет-
ся динамика численности, и которые могут предопределять названное соот-
ношение, тоже чрезвычайно желателен.
Пожалуй, единственным методом получения достоверной количест-
венной информации о фенологии и динамике численности популяций лю-
бых видов растений со времен проведения классических исследований по-
пуляций субальпийских травянистых многолетников Т. А. Работновым был
и остается метод постоянных квадратов (см., например, Понятовская, 1951;
Markov, 1985). Короткоживущие виды растений — не исключение. Мало
того, повышенная лабильность популяций малолетних растений в сравне-
нии с многолетниками делает этот метод незаменимым, несмотря на его ис-
ключительную трудоемкость.
Д. Сильвертаун (Silvertown, 1982) метко сравнивает популяцию с вокзалом,
на котором постоянно меняется публика — одни приезжают, другие уезжа-
ют. Человеку, дважды зашедшему на вокзал в течение дня, может показать-
ся, что ничего не изменилось — общее число людей осталось вроде бы преж-
ним, но, если приглядеться, люди-то уже в основном не те. Такая же по анало-
гии картина предстает и перед человеком, изредка наблюдающим за популя-
цией какого-либо растения, в которой за время отсутствия наблюдателя могут
произойти существенные смены почти без изменения общего числа особей.
Уловить не только изменение численности, но и количество вновь появив-
шихся и отмерших особей растений можно, лишь применяя маркировку осо-
бей или картирование. Даже при этом, к сожалению, нет гарантии, что учи-
тываются все без исключения особи. Наблюдения заставляют предположить,
например, что многие особи в популяциях малолетних растений, и в первую
очередь мелкосемянных, отмирают уже в момент появления проростка из се-
мени, те. до момента, когда можно распознать их видовую принадлежность.
Эти особи остаются неучтенными, и подозревать об их кратковременном су-
ществовании заставляет лишь несоответствие между численностью семян в
семенном банке и численностью активной части популяции в динамике. Но в
том-то и состоит специфика демографических процессов в популяциях расте-
ний, что исследованию подлежат и скорость оборота поколений, и соотноше-
ние длительности пребывания особей в виде зачатков и в виде сомы, и плодо-
витость, а также размерные спектры потомков.
Маркировка особей этикетками дает удовлетворительные результаты при
работе на огороженной территории стационара. В условиях же постоянно-
го риска вмешательства посторонних людей более пригодным приходил
I'laea 10 Дсм<>1 рифпчсскпй анализ природных п<>пупяnull pacieiinh 309
ся признан» картирование. В зависимости oi обстановки следуем примени!ь
оба метода, причем картирование проводин» пли па бумаге, применяя попе
вой пантограф, или путем зарисовки особей (с отражением размеров рою
ток) на прозрачных пленках для графопроектора по методу В. Г. Кияка (1984).
Достаточную точность может обеспечить и периодическое, в нужном ритме,
фотографирование цифровой камерой, которую фиксируют на штативе в но
стоянных точках над площадками. Сопоставление с картой или ранее сделан
ными снимками позволяет не только следить за ростом, но и учитывать появ
ление новых и отмирание уже отмечавшихся ранее растений.
Дополнительная полезная информация, получаемая на постоянных ква
дратах, касается ростовых процессов и переходов особей из одного фено
логического или онтогенетического (возрастного) состояния в другое. При
этом основным объектом наблюдения становится когорта (или когорты)
особей, а иногда и метамеров разного ранга или даже их частей (листьев и
т. д.). Динамичность же популяции будет зависеть от моно- или поликогорт
ности, процессов внутрикогортного изреживания на фоне каких-то внеш
них и внутрипопуляционных условий, сопряженного с соответствующими
ритмами процессов плодоношения и семенного возобновления.
Привязка исследуемых популяций растений к определенным фитоценозам
делает необходимой достаточно подробную характеристику последних с ука
занием видового состава, продуктивности и пространственной структуры.
Темпы развития особей в популяциях изучают путем ритмичной реги
страции фенофаз {фенологических фаз) с построением фенологических
спектров, которые, как уже отмечалось (Марков, 1980, 1987), для аналп
за популяций малолетних растений являются более информативным сред
ством отображения динамики, чем возрастные спектры. По фенологиче
ским спектрам, построенным для всей популяции и отдельных внутрипо
пуляционных элементов (например, для разных когорт, т. е. групп особей,
появившихся неодновременно), можно судить о темпах развития как одном
из показателей жизненного состояния и о синхронности или растянутой п
переходов из одной фенологической фазы в другую. Параллельно, когда по
возможно, фенологические спектры детализируют, отмечая на них, помимо
фенофаз. также фазы роста, например — число фитомеров на главной осн,
наличие или отсутствие семядолей и т. д.
Темпы изреживания можно оценивать также по кривым выживании
{survivorship curve), построенным для одновозрастных групп {когорт\)у\с
тений, или по кривым исчерпания {depletion curve), когда когортный состав
особей наблюдателю неизвестен. Из числа других демографических пока
зателей целесообразно использовать периоды полуотпада или полуизрежи
ваниях (Марков, 1982, 1986; Заугольнова,1985, 1986), которые определяю!
по формуле:
H1/2=tln2/(lnN -lnN+t), [10.11
1 Иногда употребляют термин «период полужизни», пытаясь дословно перевести с aHinnii
ского half-life. Перевод этот как лишь приблизительно передающий смысл показа геля при
ходи । си при hi л । ь крайне неудачным
310 М.Н Марков 11(>11у1!ящ1()1111п>| (HioJioi ня рпс 1С1П1Й
где HJ/2— период {время) полуотпада (half-life), Nx начальная числен-
ность, Nx+t— численность популяции по прошествии времени t, а также
лабильность баланса численности (Заугольнова, 1985), которую определя-
ют по формуле:
В = 1 - Np/SN, [10.2]
где В — лабильность баланса численности, Np — число особей, присут-
ствовавших на постоянном квадрате в течение всего периода наблюдений, а
S N — общее число особей, зарегистрированных за все время наблюдений,
включая вновь появившиеся и выпавшие.
В ряде случаев, помимо величины общей лабильности баланса числен-
ности, можно отдельно вычислять частные значения этого показателя, фик-
сируя изменение численности либо только из-за появления, либо — только
из-за отмирания особей. Темпы изреживания оценивали также по кривым
выживания, построенным для одновозрастных групп (когорт) растений.
Каждая постоянная площадка представляет какой-то определенный попу-
ляционный локус (Уранов, Г977), характеризующийся, как правило, извест-
ным скоплением особей изучаемого вида и фрагмент фитоценоза, в соста-
ве которого наблюдают популяцию. Локусы для постоянных наблюдений и
сравнения следует выбирать с таким расчетом, чтобы были представлены
разные фитоценозы или разные фрагменты одного и того же фитоценоза (к
примеру, на склонах разных экспозиций и разной крутизны).
В дополнение к наблюдениям на постоянных квадратах периодически
брались выборки особей с временных площадок. Размеры постоянных пло-
щадок были в основном 25 х 25 см, 33 х 33 см, реже — 50 х 50 см, а размер
временной площадки зависел от числа экземпляров, которых должно было
хватить для полноценной статистической обработки, но не менее 50.
Рассмотрим в качестве примеров некоторые результаты исследований,
которыми были охвачены виды древесных растений и виды малолетников,
относящихся к разным условным эколого-фитоценотическим группам. Это
в первую очередь рудеральные виды и виды агрофитоценозов, фитоценозов
эродированных остепненных склонов, фитоценозов береговой полосы во-
доемов, лесных и лесопарковых фитоценозов.
Продемонстрируем возможности перечисленных методов, используя в
качестве примеров данные литературы, а также некоторые результаты на-
ших исследований ряда древесных, а также травянистых многолетних и ма-
лолетних монокарпических видов растений, относящихся к разным услов-
ным эколого-фитоценотическим группам, — приуроченных к агрофитоце-
нозам, фитоценозам эродированных остепненных склонов, фитоценозам
береговой полосы водоемов, лесным фитоценозам. Примеры подобраны с
таким расчетом, чтобы были охвачены популяции с различными типами ди-
намики и различной степенью лабильности численности. Хотя сообщества,
в составе которых были изучены популяции травянистых растений, были,
как правило, слабо сомкнутые, их структура в целом тоже была учтена, по-
скольку заметно влияла на динамические популяционные процессы.
Г'икш 10 Дсм<>1 рифпчсскпй апаши природных попуниций р;ц iriiiih
311
10.2. Изучение динамики популяций древесных растений
10.2.1. Изреживание древостоев
Лесоводами очень давно замечена та удивительная равномерность про-
цесса изреживания, которая свойственна популяциям древесных растений.
Убыль деревьев происходит в строгом соответствии с процессами роста (см.
рис. 8.21). Средний диаметр ствола на уровне груди — наиболее распростра-
ненный показатель жизненного состояния деревьев — все время остается
связанным с плотностью (числом стволов на единице площади) соотноше-
нием: d = С D1/2. Эта формула может быть подобно формуле «закона - 3/2»
выведена геметрически и подкрепляется эмпирическими данными, получен-
ными при анализе древостоев сосны разного возраста (рис. 10.1). Отмирание
дерева — процесс постепенный и протекает он параллельно разрастанию со-
седних деревьев, т. е. в форме пространственного исключения, так что к мо-
менту полной остановки жизненных процессов пространство, некогда зани-
маемое отмершей особью, в сущности уже может быть поделено соседними
растениями. Этим, по-видимому, и объясняется постоянство указанного со-
отношения между диаметром ствола и плотностью.
Определение массы отдельных деревьев проводятся редко и не в массовых
количествах, допускающих статистическую обработку. Поэтому для анали-
за динамики распределения как показателя изменчивости размера деревьев
приходится использовать тот же диаметр ствола как коррелят его объема и
массы. Дж. Уайтом показано, что положительно асимметричное распределе-
ние по диаметру ствола с увеличением возраста древостоя сосны постепенно
становится симметричным, близким к нормальному (рис. 10.2).
На первый взгляд это противоречит закону — 3/2, но надо принимать во
внимание следующее. Уайт использовал для анализа динамики распределе-
ния данные, полученные лесоводами, а лесоводы обычно начинают интере-
соваться древостоем не раньше, чем
с десяти- двадцатилетнего возраста
деревьев. Процессы, протекавшие в
популяциях до этого возраста, оста-
ются без внимания. Вместе с тем,
логично представить динамику по-
пуляции древесного растения, начи-
ная с семян, пополняющих актив-
ную часть популяции всходами, сле-
дующим образом. Сами семена, как
и появившиеся из них всходы, бу-
дут подчиняться нормальному рас-
пределению (рис. 10.3а), поскольку
здесь преобладают случайные про-
цессы, а внутривидовые взаимодей-
ствия из-за малой величины особей
и относительно больших расстоя-
ний между ними, осин и нроянпяют-
Рис. 10.1. Эмпирическая линия регрес-
сии среднего диаметра ствола на плот-
ность в древостоях (популяциях) сосны
разного возраста. D плотность; d
диаметр ствола.
312
Л/Л Марков I loiiyiiHniioiiiiiiH (>1101101 пи piiciciinh
Рис. 10.2. Динамика распределений по
классам диаметра ствола в древостоях
сосны разного возраста. 20 — 160 —
возраст древостоев; f — частота; d —
классы диаметра ствола на высоте груди.
ся, го лишь в формс взаимного бла-
гоприятствования. По мерс роста
растений эти взаимоотношения сме-
няются конкурентными, причем на-
пряженность конкуренции со време-
нем возрастает, что и находит свое
отражение в возникновении поло-
жительной асимметрии распреде-
ления. Наибольшего значения кон-
курентные отношения достигают в
фазе жердняка, т. е. как раз к двад-
цатилетнему возрасту. С этого вре-
мени деревья начинают изреживать-
ся. Изреживание приводит к тому,
что вызванный повышенной плот-
ностью стресс постепенно снима-
ется, благодаря чему распределение
опять становится близким к нор-
мальному.
Изучение динамики численно-
сти древесных растений чаще всего
проводят путем сопоставления дре-
востоев разного возраста на разных
площадках, которые стараются за-
кладывать в однотипных экотопах.
В этом случае о характере динами-
ческих процессов, происходящих в
Рис. 10.3. Теоретическая схема: динамика популяций древесного растения (вверху) и
однолетника (внизу), представленная как динамика кривых распределения
Г'1(1Н(1 10 Дгмо| |)1н|)ИЧССКП11 ЛИНИИ! Iipiipo/Ull.l
популяциях деревьев, судят ПО КрН
вым выживания или кривым исчср
пания и ряду выше приведенных ко-
личественных показателей, которые
используются для анализа этих кри-
вых.
Шанс перейти в более высокий
класс жизненного состояния может
сохраняться несколько лет. Отсюда
понятно, что при многолетнем об-
разе жизни особи имеют больше
шансов воспользоваться простран-
ством, освободившимся после от-
мирания других особей. Поэтому
отмирание как процесс регуля-
ции плотности играет в популяци-
ях многолетников, и особенно дере-
вьев, ведущую роль. Проявлением
этого можно считать довольно стро-
гое подчинение популяций древес-
ных растений закону -3/2. Правда,
вместо массы особи приходится ис-
пользовать пропорциональный мас-
се объем. На рисунке 10.4, где при-
ведены данные Г. Ф. Хильми (1957)
для дуба, сосны и ели, хорошо вид-
но, что все три линии имеют почти
одинаковый угол наклона, близкий
к 56°. Довольно просто перейти от
обсуждаемого выше соотношения
между диаметром ствола и плотно-
стью к формуле закона. Объем про-
порционален кубу линейного разме-
ра, т. е. диаметра. Поэтому, возведя
в куб обе части формулы d = С D1/2,
получим d3 = С3 D’3/2, а заменив кон-
станту для перехода к массе, — W =
С’ D’3/2.
Почти во всех руководствах по фи-
тоценологии приводятся кривые вы-
живания для «насаждений разных
бонитетов и древесных пород, от-
личающихся по теневыносливости.
Здесь уместно уточнить, что по-
нимается под кривой выживания и
остановиться на рассмотрении раз-
ных типов кривых выживания, свой-
Рис. 10.4. Зависимость среднего объема
одного дерева от плотности в древостоях
сосны (1), дуба (2) и ели (3). D — плот-
ность древостоя (число деревьев на га);
V — средний объем одного дерева (м3).
Рис. 10.5. Основные типы кривых выжи-
вания (Pearl, 1928). N — численность ко-
горты; Т возраст (время); I 111
тины кривых.
314
МВ. Марков I к>п у/1ЯЦИО1ИШЯ Oiiojioi ня рис icinih
Рис. 10.6. Однотипность хода изреживания
в насаждениях сосны разных бонитетов.
Рис. 10.7. Ход изреживания насаждений
пихты, ели и сосны: Т — время (возраст
насаждения).
ствснных разным видам растений.
Кривая выживания иллюстрирует
динамику отмирания особей одного
поколения (=одной когорты) во вре-
мени. Выделяют (Pearl, 1928) три
основных типа кривых выживания
(рис. 10.5).
Кривая I типа, показывающая
резкое увеличение смертности в
относительно старом возрасте, свой-
ственна некоторым животным (на-
пример, дрозофиле) и человеку.
Для растений наиболее характерны
другие два типа кривых. Особенно
часто встречаются кривые, кото-
рые выпрямляются при использо-
вании не абсолютной численности,
а ее логарифма (тип II на рис. 10.5)
Линейность зависимости в этом слу-
чае свидетельствует о постоянной
скорости изреживания и одинако-
вой вероятности отмирания особей
любого возрастного класса.
Если при иллюстрации процес-
са изреживания древесных по-
род использовать не абсолютную
численность, а ее логарифм, ста-
новится очевидной однотип-
ность хода изреживания в на-
саждениях разных бонитетов (рис.
10.6), притом, что различия в «пер-
воначальной» (в возрасте 20 лет)
плотности популяций сохраняются
в течение всего времени. Для срав-
нительной оценки скорости изре-
живания в насаждениях можно ис-
пользовать «время полуотпада» —
время, за которое отмирает полови-
на особей популяции. Этот показа-
тель по своей сущности аналогичен
«времени полураспада» радиоак-
тивных элементов, поскольку тоже
отражает логарифмическую зави-
симость. Сравнение по времени по-
луотпада не подтверждает вывода о
более быстром переживании свето-
любивых пород деревьев Чем коро-
Гаана 10 Дгм<>|р»ф||чсск11Й инаниз природных нопуииций pacieiniB 315
че время полуотпада, чем быстрее пзрсжинается популяция. Время iюлуо1
пада (с 20-летнего возраста) для насаждений пихты, ели и сосны сосзавпя
ет, соответственно, 10, 15 и 17 лет (рис. 10.7).
Рис. 10.8. Ход изреживания нормальных древостосв по Н. В. Третьякову,
П. В. Горскому, Г. Г. Самойловичу (1952). 1 —дуб семенной, 2 дуб порослевой,
3 бук, 4—осина, 5 — липа, 6 — береза, 7 ель, 8 сосна, 9 пихта. По вер-
тикальной оси численность особей на га, по горизонтальной возраст древосто-
ев (лет).
316 А/// Марков I loiiyiiHUiioiinihi ()11<>и<>1 пи рдс 1СИПЙ
Приведенные на рис. 10.8 данные о динамике изреживания особей в нор-
мальных древостоях по Н.В. Третьякову, П.В. Горскому, Г.Г. Самойловичу
(1952) указывают на однотипность кривых выживания, построенных для
популяций многих видов деревьев, включая хвойные и лиственные.
10.2.2. Возрастание численности особей в древостоях
Динамика численности особей в сообществах древесных растений отне-
сена В. В. Плотниковым (1979) к числу «наиболее сложных и «актуальных»
моментов» в лесоведении. Он сожалеет о том, что возрастная динамика
численности древесных растений в сообществе трактуется зачастую од-
носторонне — как «самоизреживание» (Эйтинген, 1944; Хильми, 1957).
«Достаточно заглянуть в любые таблицы хода роста древостоев, чтобы убе-
диться, что в лесной таксации, признается практически только один тип
возрастной динамики численности — динамика, основанная на убыли осо-
бей», а между тем возрастная динамика сообщества может сопровождаться
и увеличением численности особей (Плотников, 1979: 177). Выше (главы 6,
8) мы уже обсуждали влияние взаимного расположения особей в популяци-
ях на их рост. Именно у таких долгоживущих растений как деревья всякие
изменения в расположении особей, связанные с динамикой их численности,
непременно должны отражаться на их росте и развитии.
В природе после ветровала, рубок или в ходе зарастания заброшенных
сельскохозяйственных угодий процесс лесообразования растянут во време-
ни и сопровождается постепенным и непрерывным увеличением числен-
ности особей. Воспользуемся данными В. В. Плотникова (1979) для иллю-
страции динамики условий роста отдельных деревьев в насаждениях ели и
пихты.
Понятно, что возрастание численности особей в древостое должно со-
провождаться перераспределением их по классам плотности. При этом чис-
ло деревьев, находившихся в том или ином классе плотности, увеличивает-
ся к моменту очередного наблюдения не только за счет абсолютного прира-
щения численности новых особей, но и за счет перехода части деревьев из
одного класса плотности в другой. Даже одновозрастные деревья, оказав-
шиеся к определенному моменту в каком-то классе плотности, гетероген-
ны по признаку перехода их из класса в класс в период, предшествовавший
последнему наблюдению.
Если проследить детально судьбу любого конкретного дерева на выбран-
ной пробной площади, то можно установить, что его окружение со време-
нем менялось. Когда вблизи него появлялись новые, более молодые осо-
би класс плотности для этого дерева менялся. Зная возраст деревьев и их
взаимное расположение, для каждого дерева можно восстановить его «путь
развития» — порядок перехода его из одного класса плотности в другой
класс из-за динамики его окружения. На рис. 10.9 для рассматриваемой
пробной площади показана динамика численности особей и способы или
варианты перехода их по плотности из класса в класс в разрезе 20-30-лет-
них интервалов. На этой диаграмме хорошо видно, что как у слн, гак и у
/ hiaa 10 Дгмо|]>л([>11ческ11Й aiiainrt прпроцпых нопунициЙ рпси-ппЙ 3 1 /
пихты судьбы отдельных растений в данных сообществах чравычанно
разнообразны При этом существенно различат вся между собой могу i даже*
деревья одинакового возраста; одно, например, может всю жизнь осгаван.
ся в первом классе плотности, а другое — совершить несколько переходов
из класса в класс и оказаться в итоге в условиях повышенной локальной
плотности. Новые растения, в свою очередь, могли появиться либо вбли-
зи от имевшихся (во втором и выше классах плотности), либо на некото-
ром удалении от них, т. е. в первом классе плотности. Строить традицион-
ными способами какой-то единый «ход роста», средний или основной вари-
ант развития для всего древостоя пробной площади как совокупности осо-
бей чрезвычайно трудно, если не невозможно. Но при этом мы вправе ожи-
дать, что на фоне прочих равных условий рост каждого дерева как-то зави-
сит от динамики его окружения.
На рис. 10.9 можно видеть, что из шести экземпляров пихты, появивших-
ся в данном сообществе до 1900 г. в первом классе плотности, два оста-
лись в нем до 1970 г., два в разные годы перешли во второй класс и два — в
четвертый. В табл. 10.1 даны некоторые морфометрические показатели этих
деревьев.
Данные табл. 10.1 свидетельствуют о том, что деревья, перешедшие из
первого класса плотности во второй, растут значительно лучше деревьев,
остающихся в первом классе, но хуже деревьев, перешедших из первого
класса плотности в четвертый.
Год
1970
1940
1920
1900
1970
1940
1920
1900
Классы плотности
Рис. 10.9. Пути развития деревьев пихты (вверху) и ели (внизу) в связи с динамикой
их численности. Цифры в прямоугольниках численность особей (по Плотникову,
1979)
318
М Н Марков I lol I у я и циои и л м (>1101101 ня рис 1гннй
Таблица 10.1.
Морфометрия особей пихты на пробной площади № 43, появившихся до I960 г.
Путь развития деревьев Признаки деревьев
Высота, м Диаметр на шейке корня, см Прирост по диаметру на шейке корня за пос- ледние 5 лет, мм Прирост по диаметру на середине ствола за последние 5 лет, мм Средний годичный прирост по высоте за последние 5, лет, см
2,3 5,2 6,0 7,6 9,8
1- -2 6,4 7,8 9,0 17,3 29,1
1- -4 7,8 10,4 24,0 25,9 38,4
Примечание. Здесь и в следующей таблице, путь, обозначенный как 1-1-1 показывает,
что деревья данной категории всю жизнь оставались в первом классе плотности, а около
деревьев других категорий в разные годы появилось соответственно от 1 (1-.-2) до 3 (1-
..-4) более молодых соседей.
Объяснение в данном случае напрашивается одно: деревья, появившие-
ся позже, каким-то образом стимулировали рост самых первых деревьев. По
мнению В. В. Плотникова здесь проявляется своего рода разделение функ-
ций между отдельными возрастными категориями деревьев в сообществе.
Средние характеристики деревьев, появившихся в данном сообществе в
интервале между 1900 и 1920 г. в первом классе плотности, но в дальней-
шем развивавшихся разными путями показаны в табл. 10.2. Эти в целом бо-
лее молодые деревья следуют в своем развитии основному правилу: чем
выше плотность, тем хуже рост.
Таблица 10.2.
Характеристика деревьев пихты на пробной площади № 43, появившихся в интерва-
ле между 1900 и 1920 г.
Путь развития деревьев Признаки деревьев
Высота, м Диаметр на шейке корня, см Прирост по диаметру на шейке корня за последние 5 лет, мм Прирост по диаметру на середине ствола за последние 5 лет, мм Средний годичный прирост по высоте за последние 5 лет, см
1-1-1 4,7 6,6 10,0 13,6 21,4
1-...-2 3,0 4,6 4,4 10,2 13,4
1- -4 2,4 4,6 6,2 6,8 10,1
В самоизреживающихся древостоях отдельные деревья также перехо-
дят из одного класса плотности в другой, причем многообразие путей пе-
рехода порождает большую морфометрическую изменчивость. Так для 53
особей сосны, сохранившихся к концу наблюдений (возрасту 75 лет) мож-
но выделить 34 варианта путей их перехода из одного класса плотности в
другой (рис. 10.10). На рис. 10.11 для сравнения отражен средний ход ро-
Класс плотности
Численность особей, экз
Рее 10.10. Пуги сосен из одного класса плотности в другой в ходе самоизреживания искусственного древостоя (обозначения те
же. что и на рис. 10.9).
Г>ннт 10 Дсм(ирпфичсский uiiiiiiin природных популяций рапсниЙ 319
J^U MH I hniyiinuiioiniiiM (hioiioi ни pncicuiH
Рис. 10.11. Ход роста по диаметру на
высоте груди деревьев, переходивших
из одного класса плотности в другой по
схеме 5-3-1 -1 -1 (кривая 1) и по схеме 5-4-
3-2-1 (кривая 2) на пробной площади. D
— средний диаметр ствола, Т — время
(возраст древостоев).
с га по диаметру для двух категорий
деревьев па данной пробной площа-
ди; для деревьев, проделавших путь
по схеме 5-3-1-1-1 и переходивших
из одного класса плотности в дру-
гой по схеме 5-4-3-2-1. Графики от-
четливо показывают, что наилуч-
шим ростом отличались не просто
деревья, сгруппировавшиеся в пер-
вом классе плотности в результате
самоизреживания, не деревья, кото-
рые всю жизнь росли в каком-либо
одном классе, а деревья, переходив-
шие из класса в класс определенны-
ми оптимальными путями.
10.3. Виды агрофитоценозов
Из числа видов типичных сеге-
тальных малолетников выбраны па-
стушья сумка, резуховидка Таля и
редька дикая.
У резуховидки Таля, рост и разви-
тие которой наблюдали в посевах озимой и яровой пшеницы, каждая агро-
популяция была представлена по существу одной когортой2. В озимом посе-
ве особи агропопуляции появились осенью, зимовали в фазе розеток разно-
го размера и массы, которая во многом зависела от благоприятности микроу-
частка, где появился проросток. Кривые распределения показывали, что био-
масса розеток, готовящихся к зимовке, варьировала довольно сильно (от 15 до
105 мг). Весной перезимовавшие особи дружно перешли в генеративное со-
стояние, демонстрируя рост, соответствующий резерву биомассы, накоплен-
ной осенью. Их мощность и плодовитость оказались существенно выше, чем
у не зимовавших особей из агропопуляции ярового посева (см. рис. 8.39).
Фенологические спектры (рис. 10.12) показывают почти синхронный пере-
ход осенней когорты к бутонизации и очень низкий процент несколько задер-
жавшихся в развитии особей, среди которых преобладали немногочисленные
особи весенней когорты.
В посеве яровой пшеницы фенологический спектр демонстрирует боль-
шую растянутость фенофаз. При этом характерно полное отсутствие пе-
рекрывания фаз цветения в агропопуляциях из озимых и яровых посевов,
из чего можно заключить, что обмена генетической информацией между
популяциями, территориально близкими, но развивающимися одна в ози-
мом, а другая — в яровом посевах, не происходит. Несинхронный, растяну-
тый переход особей резуховидки в генеративное состояние в посеве яровой
2 Весной на постоянных площадках в посеве озимой пшеницы появилось всего 9 особей ре-
зуховидки.
321
Рис. 10.12. Фенологические спектры для
агропопуляций пастушьей сумки из по-
сева гороха (I) и овса (II) и для резухо-
видки Таля из посева озимой (III) и яро-
вой (IV) пшеницы. Фенологические
фазы: 1 — вегетация; 2 — бутонизация;
3 — цветение; 4 — плодоношение.
l /uithi 10 ДсмтрпфичсскнЙ шпини природных iioiiyiiHiiiiii рис icintil
кулыуры, по видимому, представля-
ет собой вполне нормальное явле-
ние, если учесть почти облигатное
самоопыление в природных популя-
циях этого вида.
В течение нескольких лет (с 1972
по 1976 г.) нами проводились на-
блюдения в агропопуляциях пасту-
шьей сумки посевов озимых и яро-
вых культур. Как и у резуховидки,
жизненный цикл пастушьей сумки
может идти по двум путям: с зимов-
кой (в посевах озимых) и без нее (в
посевах яровых культур). Стратегия
избегания наилучшим образом осу-
ществляется группой зимующих
растений, использующих благопри-
ятные условия и осеннего, и весен-
него периодов. Весенний период ис-
пользуется с меньшей эффективно-
стью, ибо зимовка резко переводит
все без исключения особи в репро-
дуктивное состояние. Осеннее появ-
ление всходов, как и у резуховидки,
очень дружное и несколько растяги-
вается только в случае сухой осени.
В посевах яровых культур у попу-
ляций пастушьей сумки имеется воз-
можность использовать только весен-
ний, относительно благоприятный
период, притом его более позднюю
часть, так как сев яровых производит-
ся обычно не сразу вслед за сходом снега. Параллельное развитие пастушьей
сумки и культурного растения на первых этапах обеспечивает ее особям осла-
бление помех из-за конкурентных взаимодействий. Довольно резкое ухудшение
режимов в яровых посевах сказывается неблагоприятно на особях пастушьей
сумки, появившихся из семян более поздно. Последовательно появлявшиеся
группы особей в посевах гороха, овса и гречихи (о них уже говорилось в главе 6)
представляли иерархию по степени подавления. Более позднее появление рас-
тений приводило к более слабому их росту, развитию, плодоношению (Марков,
1976). В 1974 г. плотность агропопуляций в посевах гороха, гречихи, овса соста-
вила соответственно: 8,3 ± 1,02, 5,9 ± 0,44 и 10,3 ± 1,6 на 0,1 м2). Благодаря по-
явлению вышеописанных групп лабильность баланса численности приобрела в
агропопуляциях пастушьей сумки из посевов гороха и гречихи довольно высо-
кие значения (табл. 10.3).
322 МН Марков 11<>11уници()||||||я бионогии рисюпиЙ
Таблица 10.3.
Лабильность баланса численности в агропопуляциях пастушьей сумки из
посевов разных яровых культур в 1974 и 1975 гг.
Агропопуляции из посевов: 1974 1975
гороха 0,69 0,43
гречихи 0,72 0,21
овса 0,23 0,36
Наибольшая синхронность появления всходов пастушьей сумки в посе-
ве овса проявилась в самом низком значении лабильности баланса числен-
ности.
Условия жизни пастушьей сумки и других сегеталов в посевах разных яро-
вых культур различны. Наименее благоприятны они в посеве овса — сильно-
го эдификатора с аллелопатическим эффектом, развивающего большое чис-
ло побегов на единицу площади. Лучшие условия для роста создаются в по-
севе гороха, поскольку последний не развивает большого числа побегов, мед-
ленно формирует затеняющий полог и довольно быстро полегает, улучшая
тем самым условия освещения для сорных растений. Об условиях в посевах
можно судить по количественным данным об урожае общей биомассы всех
сорных растений. В 1974 г., например, этот показатель составил в посеве го-
роха 9,28 ±1,3; гречихи — 4,51 ± 0,7 и овса — 2,63 ± 0,3 г/0,1 м2.
И в озимом, и в яровом посевах немногочисленные группы особей (ко-
горты) пастушьей сумки могут появляться после летних дождей и исполь-
зовать чрезвычайно непродолжительные периоды повышения влажности.
Такие группы появлялись в 1972 и 1975 гг. в посевах яровых культур. В 1972
г. поздно появившиеся особи прошли в ускоренном темпе цикл развития и
завершили его плодоношением. В засушливом 1975 г. растения, не дав се-
мян, отмерли. Наибольшая скорость отмирания отмечена в посеве гороха
(Н1/2= 0,90), чуть ниже — в посеве овса (Н1/2= 1,35) и самая низкая — в посе-
ве гречихи (Н1/2= 2,70). Таким образом, оптимальные для роста условия мо-
гут быть отнюдь не оптимальными для выживания. В засушливом году, оче-
видно, затеняющий полог играл отчасти положительную роль, улучшая ми-
кроклимат приземного слоя воздуха и способствуя поддержанию чуть бо-
лее высокого содержания влаги в самых поверхностных слоях почвы. Более
резко дифференцированные условия, наблюдаемые в посеве гороха, уси-
ливали дифференциацию особей пастушьей сумки (см. главу 8), что сказа-
лось на демографической стабильности популяций — лабильность баланса
численности агропопуляции в посеве гороха была наивысшей (табл. 10.3).
Интересно отметить, что изменение лабильности баланса численности в
разные годы зависело от разных причин. В одном случае от появления но-
вых особей при стопроцентном выживании, а в другом — от отмирания осо-
бей, появившихся в одно и то же время.
Растянутое появление всходов, наблюдавшееся в 1974 г. в яровых посевах,
было связано не только с разновременным прорастанием, но, по-видимому, и
прорастанием с различной глубины. Фенологические спектры, построенные
для каждой из этих групп, демонстрируют ускорение развития сокращение
природных llOliyilHIIIIII |><К iciiiiii 1
l'l(l(<(l 10 Дгм<>| рпфпЧССКПВ niHIIIII I
продолжи।слыюсти пребыва-
ния в внргинилыюм СОСТО-
янии особей, появлявших-
ся во все более поздние сро-
ки (рис. 10.13.), т.е. встречав-
шихся со все более суровы-
ми условиями. Здесь мы на-
блюдаем одно из проявлений
«правила Сукачева». Вместе
с тем, по нашим наблюдени-
ям, более или менее друж-
ный переход из одной фазы
в другую наблюдается толь-
ко при благоприятных для
роста условиях, когда одно-
временно появившиеся осо-
би растут изолированно друг
от друга и не испытывают
конкурентного давления со
стороны какого-либо эдифи-
катора. Произрастание в су-
ровых ценотических услови-
ях отличается растянутыми
переходами из одной фазы
в другую и растянутостью
фенологических спектров.
Последнее типично для есте-
ственных популяций пасту-
шьей сумки, что заставляет
Рис. 10.13. Фенологические спектры для когорт
особей пастушьей сумки. НП — не плодоносив-
шие особи.
предположить существова-
ние в популяциях этого вида особей двух типов. Особи первого типа ускоряют
развитие при произрастании в неблагоприятных условиях. Наличие в популя-
ции таких особей обусловливает более ранний переход к репродукции в по-
севах с наиболее суровым режимом. Особи второго типа замедляют развитие
при плохих для роста условиях, чем и обусловливают растянутость феноло-
гических спектров. Крайнее выражение этого явления можно наблюдать у от-
ставших особей, настолько замедливших свое развитие, что некоторые из них
так и остались в вегетирующем состоянии, не дав семян (НП на рис. 10.13).
10.4. Виды степных эродированных склонов
Исследование роста и репродукции особей в природных популяциях не-
скольких потенциально многолетних монокарпических видов: катрана та-
тарского, коровяка метельчатого и коровяка медвежье ухо было предприня-
то с целью эмпирического выявления «критического» размера особи, кото-
рый позволяет ей зацвести, а также с цепью анализа впутринопуляциопой
дифференциации как одною из крик-рнсн при выяснении iина жизненною
324 М Н Мауков 11(>11уняц11(>111шя Giioiioi пя piiciciiiiii
цикла. Основные вопросы, которые предполагалось решить в ходе наблюде-
ний за тем или иным способом маркированными особями, были:
1) Какое время требуется для достижения критических размеров при на-
блюдаемой в природной обстановке скорости роста?
2) Может ли быть добрана особью критическая биомасса в начале сезо-
на вегетации с соответствующим сдвигом цветения на более позднюю его
часть?
3) Могут ли какие-либо отдельные признаки служить надежным критери-
ем для суждения о том, достигла ли особь в процессе вегетации своих кри-
тических размеров?
4) Существуют ли определенные ритмы цветения популяций моно-
карпических растений в условиях средней полосы?
5) Насколько вероятна и при каких условиях возможна поликарпия у на-
званных, в норме монокарпических, видов?
Наблюдения за ростом и развитием особей в природных популяциях
должны удовлетворять двум условиям. Во-первых, должна быть обеспе-
чена возможность проследить за ростом репрезентативной группы особей,
чтобы получить именно популяционные характеристики и, вместе с ними,
представление о показателях роста и их варьирования в масштабе популя-
ции. Во-вторых, должен соблюдаться принцип минимального вмешатель-
ства в жизнь популяции, чтобы в процессе наблюдений не привнести су-
щественную долю искусственности в среду обитания растений и не пло-
дить артефакты. Как уже говорилось, этим требованиям в наибольшей сте-
пени удовлетворяет индеструктивный метод наблюдений за особями на
постоянных квадратах или за маркированными особями, который связан
с периодическим измерением высоты особей, измерением листьев и под-
счетом их числа. Однако на первой стадии изучения необходимо получить
уравнения аллометрической связи между измерениями, подсчетами и об-
щей массой, для чего бралась одна (или две) выборка особей из той же по-
пуляции (отражающая присущую изучаемой популяции размерную диффе-
ренциацию), но за пределами пробных площадей. Опыт показывает, что для
какой-либо данной архитектуры, например свойственной изучаемому виду,
это уравнение оказывается в достаточной степени стабильным.
10,4.1 . Результаты изучения популяции катрана Crambe tataria
Катран татарский Crambe tataria Sebeok — стержнекорневое многолет-
нее монокарпическое растение, представляющее очень интересную и мало-
изученную жизненную форму «перекати-поле». Это редкое растение про-
израстает в степях европейской части России, восточная граница его ареа-
ла проходит по Южному Уралу (Павлов, 1948). Редкость катрана татарского
обусловлена распашкой степей, из-за которой он вынужден занимать непри-
годные для пахоты крутые склоны. Отмечена приуроченность этого вида к
каменистым степям (Мальцев, 1923). Катран относится к числу пищевых и
кормовых растений перспективных для введения в культуру. В Башкирии
уже ведутся эксперименты по возделыванию катрана, и изучается рост и
развитие его в условиях культуры (Кучеров, 1979).
Iiiihu 10 Дгм<>|ра([)11чеу|сий шпини природных нонуницнЙ расiriinh 3?5
В Гагарий, где были проведены наши исследования, с I950-х м Ьыпо
извссгно лишь одно местонахождение данною вида окреп пои и л
Хансверкино (Бавлинский район), г. е. населенного пункта, находящеюся
на крайнем юго-востоке Татарского Заволжья. В 1983 г. в указанном ну и
кте после долгих поисков была найдена одна единственная вегетирующая
особь катрана, но несколько севернее, недалеко от поселка Бавлинский уда-
лось обнаружить относительно большую и полночленную популяцию этого
вида. Следует подчеркнуть, что в заволжских районах Татарии на дневную
поверхность выходят известняки и мергели татарского яруса пермской си-
стемы (Спрыгин, 1934; Сементовский, 1951). Отчетливо выраженная асим-
метричность речных долин обусловливает расположение длинных гряд
известняково-мергелистых склонов вдоль ныне текущих, а порой уже пе-
ресохших речек. Именно такую гряду вдоль сухого русла занимает найден-
ная нами популяция, входящая в состав фитоценозов каменистой степи, ко-
торые покрывают узкую полосу плакорного водораздела, непосредственно
примыкающую к склонам, и сами крутые склоны (Марков, 1989).
Особенно крупные фрагменты (локусы) популяции приурочены к наибо-
лее крутым (30-45°) частям склонов, ориентированным на юг и юго-запад
(на склонах балок, имеющих юго-восточную экспозицию, катран не встре-
чается). Один такой фрагмент в пределах урочища, отделенного от сосед-
них глубокими балками, был наиболее подробно изучен. Здесь был зало-
жен профиль поперек крутого склона чисто южной экспозиции (на трансск-
те шириной 2 м, протянувшейся от сухого русла временного потока у под-
ножья склона до плоской его вершины) и зарегистрированы все виды рас ге-
ний в том порядке, в каком они были встречены при движении снизу вверх
по профилю. В таблице 10.4, дающей представление о видовом составе со-
общества с участием катрана, с помощью балльных оценок обилия по ком-
плексной шкале Браун-Бланке показано размещение каждого вида. Для это-
го весь профиль условно разделен на отрезки: сухое русло (I); бровка об-
рыва (II); бровка обрыва — Юм вверх по профилю (III); 10-20 м (IV), 20-30
м (V); 30-40 м (VI); 40 м — верхняя кромка склона (VII), верхняя кромка
склона и плоская вершина (VIII). Можно видеть, что особи катрана встреча-
лись почти по всему профилю, за исключением самых нижних его отрезков.
Все встреченные в границах данного урочища особи этого вида были за-
картированы. Около 100 особей, находящихся на разных этапах роста было
маркировано этикетками. Их состояние (радиус розетки, число листьев и
диаметр корня — у вегетирующих; диаметр корня, высоту, ширину и длину
проекции растопырено ветвящегося стебля — у цветущих экземпляров) о г-
мечали в разные по условиям годы (1983-1986) и в разные периоды сезона.
Параллельно изучали процесс семенного возобновления, проводя учет по-
являющихся проростков, а также процесс дисперсии. В лабораторных опы-
тах изучали прорастание семян и влияние внешних условий на этот про-
цесс, наблюдали за ростом корня и надземных органов у проростков, выра-
щиваемых на тонких (1 см) черноземно-известняковых монолитах, заклю-
ченных между двумя тонкими стеклами (см. выше рис. 8.35).
326 М Н. Мирков I loiiyihiuiioiiiiiBi Giiojioi ня pnciciiiiil
Таблица 10.4.
Размещение видов растений по профилю склона южной экспозиции
Виды растений I II III IV V VI VII VIII
Verbascum lychnitis 1 + 1 + + 1 1 +
Tussilago farfara 1 +
Dracocephalum thymiflorum 1 1 1 +
Euphorbia virgata 1 +
Convolvulus arvensis 1 1 +
Achillea millefolium + 1 +
Festuca valesiaca + 1 2 1 +
Sisymbrium loeselii + 1 +
Salvia stepposa 1 2 1 2 1 + 1
Caragana frutex 1 2 1 • 2 1 +
Amygdalus папа + 1 +
Camelina sylvestris 1 +
Artemisia campestris 1 +
Artemisia absinthium 1 + + 1 +
Talictrum minus 1 1 1 1 1 +
Poa compressa 1 +
Pimpinella saxifraga 1 +
Meniocus linifolius 1 1 +
Androsace maxima 1 1 +
Bertheroa incana 1 + + 1 +
Lactuca tatarica + 1 1 +
Koeleria cristata 1 2 1 +
Achillea nobilis 1 1 1 1 +
Crambe tataria 1 1 1 1 1 1
Sisymbrium polymorphum 1 2 1 1 2 1
Nonea pulla 1 + 1 1 1 +
Astragalus wolgensis 1 1 1 +
Hypericum perforatum 1 +
Galium ruthenicum 1 1 2 1 +
Asparagus officinalis 1 1 1 ' +
Thesium arvense 1 + 1 + 1 +
Isatis costata + 1 2 1 +
Stipa pulcherrima + 1 1 2 1 2
Falcaria vulgaris + 1 2 1 +
Erysimum hieracifolium + 1 +
Linum flavum + 1 2 3 2 1
Centaurea ruthenica + 1 2 1 2 +
Thymus serpyllum s. 1. + 1 2 1 2 +
Reseda lutea + 1 1 +
Thymelaea passerina. + 1 1 4-
I'ltiKa 10 Дсмо»рифичсскиП жииин природных попупицпй рисicimi! 32
Таблица 10.4. (ироОопжгти'
Виды растений 1 II III IV V VI VII VIII
Erucastrum armoracioides + 1 1 1 1
Euphorbia seguieriana + 1 1 1 1
Artemisia austriaca + 1 +
Atraphaxis frutescens + 1 +
Hedysarum grandiflorum + 1 1 1 _ 1
Astragalus macropus + 1 +
Centaurea sumensis + 1 + 1
Peucedanum alsaticum + 1 +
Cytisus ruthenicus 1 +
Anlhemis subtinctoria 1 +
Gypsophila altissima 1 1 +
Onosma simplicissima + + 1 +
Alyssum lenense + 1
Arenaria ucranica + 1 _2
Jurinea arachnoidea + _l
Aster alpinus + _1
Ephedra distachya I
Linosyris villosa
Potentilla arenaria _1
Trinia muricata I
Clausia aprica I
Astragalus austriacus 1
В результате картирования фрагмента популяции катрана была получе
на карта размещения 182 вегетирующих и 113 цветущих особей, на которо!
была хорошо заметна контагиозность. Сравнение эмпирического и теорстн
ческого распределений методом %-квадрат подтвердило визуальную оцеп
ку. Причины контагиозности следует, по-видимому, искать, прежде всего, i
особенностях анемогеохорной дисперсии катрана. Поэтому с процесса дис-
персии мы и начнем пофазное рассмотрение жизненного цикла этого рас
тения. Шатрообразная, по определению А. И. Мальцева (1923), форма ку
ста перекати-поле, как известно, призвана обеспечивать дальний перенос
диаспор. Однако, судя по нашим наблюдениям, у катрана, как и огромногс
большинства малолетних растений (см. главу 5), преобладает топоспория i
происходит она еще до того, как стебель отломится в основании. На одно\
уже отломившемся, но еще не перемещавшемся кусте оказалось 1860 пло
дов. Многочисленные плоды лежали рядом на почве. Если учесть, что се-
менная продукция особи катрана приближается к 3-5 тыс., то логично за
ключить, что к путешествию на более далекие расстояния приготовилаа
лишь меньшая часть урожая особи, и что наблюдаемая контагиозность раз
мещения особей в популяции может быть обусловлена преимущественно!
топоспорией.
328 Л/ /? Марков I l()iiyii>iniioiiiia>i (nioiioi ии pacicindl
Стручочки катрана татарского, как и у всех других видов этого рода, сосю-
ят из двух члеников, нижний из которых недоразвит, а верхний плодущий.
Сочленение между члениками стручочка гораздо более хрупкое, чем между
нижним члеником и плодоножкой. Поэтому верхний членик, содержащий
семя, которое заключено в твердый по высыхании перикарпий, всегда отла-
мывается от нижнего и в силу упругости подсохшей плодоножки «отстре-
ливается» на некоторое расстояние (баллистохория). Снаружи сухой пери-
карпий верхнего членика имеет бугорчатую скульптуру и хорошо заметные
ребра, делающие членик четырехгранным. Основные параметры диаспор
(длина и масса плодиков-члеников, диаметр собственно семян), имеющие,
как и их форма, диагностическое значение, дают близкое к нормальному
одновершинное распределение. Необходимо отметить, что длина верхнего
членика в 87% случаев колеблется от 3,7 до 4,5 мм со средним значением
4,1 ± 0,04, лежащим на нижнем пределе этого признака, который фигури-
рует во «Флоре европейской части СССР» для С. tataria. Внутри плодика-
членика семя прикрепляется на тонком фуникулусе, длина которого пре-
вышает длину семени. От фуникулуса семя отделяется часто со значитель-
ным повреждением семенной кожуры, которая сама по себе слабо защища-
ет семя от неблагоприятных воздействий. Прочный бугорчатый перикарпий
служит не только защитой от повреждения зародыша семени, но и механи-
ческим и биохимическим препятствием для прорастания. По данным Г. А.
Кузнецовой (1959), даже выдерживание под снегом в течение 60 дней не
оказало существенного влияния на прорастание семян катрана татарского.
При этом известно, что холодная стратификация положительно сказывает-
ся на всхожести катранов благодаря инактивации фенольных ингибиторов,
содержащихся в перикарпии и семенной кожуре, и нарушению целостности
покровов (Коломиец и др., 1968). Посеяв плодики-членики катрана татар-
ского в начале зимы под снег, весной мы наблюдали всхожесть более 50%.
В лабораторных условиях, даже при очень длительном выдерживании се-
мян с перикарпием в холодильнике, прорастания не наблюдалось — пре-
доставленные условия не обеспечивали разрушения природных тормози-
телей в перикарпии. Сильно растянутое во времени прорастание началось
прямо в холодильнике (при температуре +4°), когда семена были извлечены
из околоплодника. Это подтверждает отмеченную Н. Амирхановым и Ш.
Хамракуловым (1986) оптимальность пониженных температур для прорас-
тания семян катрана татарского.
В природных условиях появление проростков, отмеченное только в
1985 г., тоже было сильно растянуто во времени, и продолжалось с нача-
ла мая до начала июня. Осеннего появления всходов зарегистрировано не
было. Проростки катрана располагались группами. Такие группы были об-
наружены в мае и июле 1985 г. на щебнистых известняковых осыпях юго-
западной экспозиции в верхней части исследуемого урочища. Плотность в
одной из групп достигала 75 особей на 1 м2, а на квадрате 25 х 25 см2 мож-
но было насчитать до 15 проростков.
Семядоли у проростков катрана начали засыхать в середине первого се-
зона вегетации, т.е. почти до конца июля это растение пребывает па нер-
вом году жизни в фазе проростков. В условиях известняковом щебнистом
I idtid 10 Дсмо!рпфичсч кий апаши природных понуиицнЙ рас icindl 329
осыпи па первом году жизни особи катран образует всею дна мелких (? 1
см длиной) настоящих листа с почти нс расчлененной micioiioii ниаенш
кой. В. М. Покровская (I960), изучавшая проростки катрана в Центрально-
черноземном заповеднике, отмечала засыхание семядолей только после по-
явления 7-8-го настоящего листа. Для нас этот факт совершенно непоня-
тен, поскольку в изучаемой нами популяции 7-8 листьев в розетке развива-
ли только взрослые многолетние особи, давно лишившиеся семядолей. Из
отмеченных весной 1985 г. проростков уже к середине лета 13% отмерло.
На стадии проростка и однолетнего ювенильного растения при крайне
слабо заметных ростовых процессах в надземной сфере происходит очень
интенсивное нарастание главного стержневого корня в длину (см. главу 8,
рис. 8.35). Приспособительное значение этого явления очевидно и стано-
вится еще более понятным в условиях щебнистых осыпей на крутых скло-
нах южных экспозиций. Достаточно проследить, как в жаркий июльский
день резко меняется температура (от +45°C) субстрата, на котором растут в
природе молодые особи, с глубиной, в какой-то степени отражая и измене-
ние влажности. Чрезвычайно глубоко идущая стержневая корневая система
катрана татарского, описанная М. С. Шалытом (1950) в условиях степных
фитоценозов, делает это растение мало зависимым от метеорологических
условий года, на что указывает Г. А. Кузнецова (1959).
Интенсивный рост корневой системы может осуществляться только бла-
годаря энергичной транспортировке ассимилятов из листьев в корень, т. е. за
счет медленного роста наземных вегетативных органов. Этот процесс ове-
ществляется в биомассе всходов, у которых в естественных условиях интен-
сивного освещения вес подземных органов равен или больше веса надзем-
ных (RWR = 0,88-1,10). В дальнейшем, учитывая листопадность розеток ка-
трана (розеточные листья засыхают и опадают осенью, примерно к началу
октября), этот процесс приобретает ритмический характер, о чем уже шла
речь выше (см. главу 8, рис. 8.36). Поскольку взрослые особи катрана по по-
нятным соображениям без крайней на то необходимости не выкапывали,
проследить этот ритм хотя бы частично можно было путем периодических
замеров диаметра корня у корневой шейки.
Из четырех признаков, отражающих накопление биомассы и возрастные
изменения особей в популяции катрана, — радиус розетки, число розеточ-
ных листьев, увеличивающаяся с возрастом расчлененность их листовых
пластинок, диаметр корня у корневой шейки — последний наиболее удо-
бен для измерений и, по-видимому, наиболее тесно коррелирует с абсолют-
ным возрастом. Отчасти в этом убеждает анатомическое изучение попереч-
ных срезов корня, на которых, правда, годичные приросты не всегда хорошо
заметны. Периодические замеры диаметра корня у маркированных особей
показали, что ежегодный прирост у ювенильных и взрослых вегетативных
экземпляров составляет всего 2-3 мм в год, но у особей, готовящихся к за-
цветанию на следующий год, он увеличивается до 8-9 мм. Усиленный отток
ассимилятов, вызывающий это резкое увеличение, сопровождается более
ранним, начинающимся уже в июле засыханием и опадением листьев ро-
зетки и переходом особи в состояние покоя, который продолжается до сле-
дующей весны, до развития системы цветоносного элементарного побега.
330
М.Н Мариин I loiiyiiHiUHUiiniH Itiioinn пн pncicimh
Рис. 10.14. Регрессия индекса Q на диа-
метр корня (Dr) как возрастной показа-
тель.
Рост особей катрана количествен-
но оценивали но изменению величи-
ны произведения числа розеточных
листьев на вес наибольшего из них,
а вес листа определяли по его дли-
не с помощью специально высчи-
танной линии регрессии. Поскольку
связь между длиной листа и его мас-
сой линейна лишь в логарифмиче-
ских координатах, в расчетах ис-
пользовали логарифм массы, т.е. вы-
числяли индекс: Q = InWl х N1, где
W1 — масса листа; N1 — число ро-
зеточных листьев. Рисунок 10.14 по-
казывает, как медленно увеличива-
ется биомасса (по индексу Q) вплоть
до достижения «критического» диа-
метра корня (=2,0 см), после чего наблюдается очень резкий подъем. Этот
подъем соответствует бурному накоплению биомассы в год, предшествую-
щий году цветения.
Апекс побега, сформировавший до того момента розетку весь «потребля-
ется» на образование мощного соцветия с диаметром проекции (в изучае-
мой популяции) от 44 до 90 см (X = 64,0 ± 2,9 см). N. Simmonds (1980) ука-
зывает, что для монокарпиков этого типа нет доказательств существования
точного эндогенного «календаря» цветения, но некоторые механизмы от-
счета времени (например, по числу образованных апексом листьев) можно
предполагать. На рис. 10.15 приведены данные, которыми мы располагаем
в отношении изучаемой популяции. Гистограммы распределения особей по
диаметру корня (отдельно для вегетирующих и для цветущих экземпляров)
приведены в дополнение к диаграммам цветения катрана. В 1983 г., когда
была обнаружена изучаемая популяция, в ее составе насчитывалось очень
немного цветущих особей, однако точного учета их доли в составе попу-
ляции не проводилось. Весной следующего года в данном урочище (как и
во всей популяции) наблюдалось массовое цветение катрана — цвело око-
ло 38% особей. Это обильное цветение и благоприятные условия влажно-
го лета 1985 г. создали благоприятную регенерационную ситуацию, в ре-
зультате чего популяция существенно пополнилась всходами в соответству-
ющих регенерационных нишах (термин Р. J. Grubb, 1977). Весной 1985 г.
цветение наблюдалось лишь у 13% особей, но часть экземпляров в популя-
ции, как это видно на гистограммах, перешла к бурному росту, т.е. пригото-
вилась цвести в следующем году. Все эти особи, доля которых была очень
невелика (4%), действительно зацвели в 1986 г. По данным 1985-1986 гг.
построены кривые распределения цветущих особей по диаметру корня, и
выяснилось, что средние значения этого признака в разные годы (2,74 ± 0,11
см в 1985 г. и 2,65 ±0,18 — в 1986 г.) весьма близки. Линия регрессии чис-
ла розеточных листьев на диаметр корня показала, что вегетирующие особи
с таким диаметром корня имеют, как правило, 8 листьев.
I iatid 10 Дсм<>1 рпфичсскиЙ шпини природных популяций рис iгннЙ
331
II
]lll
20.VI11984
31.V.1985
21.VI11985
Рис. 10.15. Динамика изменчивости вегетирующих (гистограммы) и цветущих (кри-
вые) особей по диаметру корня и процент цветущих особей в природной популяции
катрана татарского
Dr — диаметр корня (см); f — частость; I — пополнение популяции проростками
(показано вне масштаба); II — процент цветущих и III — процент вегетирующих
особей в популяции на диаграммах цветения.
По совокупности данных (с привлечением сведений Е. В. Кучерова, 1976)
была составлена примерная таблица хода роста особей катрана татарско-
го в изучаемой популяции в природе (А) и в питомнике (Б). Из нее явству-
ет, что в природной популяции катран зацветает не раньше, чем на седьмой
год жизни.
332 Л/.// Марков I loiiyiiHHiioiiiiihi (>1101101 пи рис iciniil
По данным Е. В. Кучерова (1976), в условиях питомника небольшая
часть особей катрана татарского (2,2 8,0%) зацветает уже на второй год
жизни. На третий год цветение усиливается, что согласуется с данными
Л. И. Казакевича (1922) о катране как о потенциальном трициклике. Мы ис-
пользовали приведенные Е. В. Кучеровым данные о размерах и числе розе-
точных листьев в первый и во второй годы жизни в питомнике и вычисли-
ли индекс Q. Величина индекса свидетельствует о том, что условия питом-
ника способствуют резкому ускорению накопления биомассы катраном и за
один-два года достижению уровня, которого особи природной популяции
достигают за семь или более лет (табл. 10.5).
Таблица 10.5.
Модель хода роста особей катрана татарского в природных условиях (А)
и на питомнике (Б)
Показатели роста Годы жизни
А
1 2 3 4 5 6 7
Диаметр корня (см) 1 0,5 0,8 1,1 1,4 1,7 2,6
Число листьев (N1) 0.2 2-3 2-3 3^4 4-5 6-7 8
длина листа в см (1) 2 4-10 10-15 20-35 35—45 45-50 50
индекс Q пропорцио- нальный биомассе 2-3 11 18 24 29 35 80
Б
индекс Q пропорцио- нальный биомассе 80 86
Судя по нашим наблюдениям, не все особи катрана отмирают на следу-
ющий же год после плодоношения. Лишь единичные особи демонстрирует
постгенеративную вегетацию, т. е. сенильность — образуют нескольких
пучков относительно мелких почти цельных листьев похожих на листья
ювенильного типа, а особи, у которых был поврежден главный цветонос-
ный побег, могут образовать и относительно мелкий репродуктивный по-
бег. Однако в последнем случае нет никаких оснований говорить о насто-
ящей подикарпии, поскольку налицо ослабленная репродукция по причи-
не недореализации репродуктивного потенциала особи в предшествующем
году из-за травмы.
10.4.2 . Результаты изучения природных популяций коровяков
Verbascum lychnitis и V. thapsus
Популяцию коровяка метельчатого в течение сезонов вегетации 1987-
1989 гг. изучали в условиях лугово-степного фитоценоза класса Festuco-
Brometea Br.-Bl. et Tx. 1943. Фрагменты данного сообщества расположены
на дерново-карбонатных почвах на склонах холмов южной и юго-западной
экспозиций близ д. Петропавловка Верхнеуслонского района Татарии.
I 'Kirtii 10 ^cmoi рифичсскиЙ iiiiiuim природных понуиицнй рисleiiidl
333
Анализ фитоценоза включал выявление фнорисihmcckoiо состава, о(>
щей продую ивносги, участия в создании поп и роду к i ивиос i и отделы i ы х
видов, а также горизонтальной неоднородное!и размещения фитомассы в
пространстве.
Для оценки продуктивности с площадок 50 х 50 см;, разделенных на
100 элементарных квадратов (5><5 см2), выкапывали все растения с по-
следующим разделением на надземные и подземные органы, взвешивани-
ем в воздушно-сухом состоянии. Горизонтальная неоднородность в сложе-
нии фитоценоза выявлена путем получения данных о продуктивности для
каждого элементарного квадрата (рис. 10.16). На склоне южной (А) и юго-
западной (Б) экспозиций фитоценозы отличались не только общей продук-
тивностью (ряд III), но и разным участием в создании фитомассы злаков
(ряд I) и разнотравья (ряд II). Границы одиннадцати логарифмических (In)
классов значений фитомассы (мг) таковы: 1 — 0; 2 — 0,9—1,1; 3 — 1,2-1,4;
4 — 1,5-1,7; 5 — 1,8-2,0; 6 — 2,1-2,3; 7 — 2,4-2,6; 8 — 2,7-2,9; 9 — 3,0-3,2;
10-3,3-3,5; 11 — >3,6.
Взрослые особи коровяка метельчатого были закартированы на двух проб-
ных площадях по 16 м2 (8 х 8 м2) каж-
дая и маркированы деревянными ко-
лышками с номерами.
Помимо этого специально для на-
блюдений за ростом особей первого
года жизни 16 мая 1989 г. были за-
ложены дополнительные проб-
ные площадки, в пределах которых
особи коровяка метельчатого про-
сто картировали. С интервалом в 5
дней проводили учет числа листьев
у каждой особи, а также измеряли
длину и ширину максимального по
размерам листа. Если особь перехо-
дила к репродукции, то у нее перио-
дически измеряли высоту цветонос-
ного стебля.
Аналогичные наблюдения про-
водили в течение вегетационно-
го сезона 1990 г. в популяции ко-
ровяка медвежье ухо, которая в
составе сообщества класса Sedo-
Scleranthetea Вг.-В1. 1955 произ-
растала на песчаных дюнах, при-
легающих к опушке смешанного
березово-соснового леса.
Аллометрический анализ выборок
из популяций обоих видов (см. табл.
10.6) приводит к выводу, что любой
из снятых показателей и длина, и
1 2 3 456789 10 11
Рис. 10.16. Гетерогенность растительно-
го покрова в местообитаниях коровяка
(пояснения в тексте).
334
М Н Маркой I lonyпяти>нпни (hiojioi им рис icinih
ширина листа, а также число розеточных листьев может быть использован
для изучения динамики роста благодаря высокой и статистически достовер-
ной положительной корреляции с биомассой. И все же предпочтение, оче-
видно, следует отдать двух или трехкомпонентным индексам, которые свя-
заны линейной зависимостью с общей биомассой при наиболее высоких
значениях коэффициента корреляции между этими двумя показателями.
Особи коровяка метельчатого переходили в репродуктивное состояние не
ранее, чем накапливали биомассу 4148,8 мг при следующих параметрах ро-
зеток: N1 = 10,2 ± 0,7; D1 = 6,8 ± 0,3; L1 = 16,5 ± 0,7.
Таблица 10.6.
Матрица корреляции показателей для особей коровяка метельчатого (над диагона-
лью) и к. медвежье ухо (под диагональю)
По- каза- тели N. L, max Di max W, w г w InW InW, InW Г NDL
N1 0,77 0,77 0,84 0,77 0,83 0,80 0,60 0,77 0,82
L1 max 0,80 0,94 0,88 0,80 0,86 0,89 0,88 0,86 0,87
D1 max 0,78 0,97 0,84 0,76 0,83 0,93 0,93 0,90 0,81
W1 0,86 0,87 0,85 0,96 0,99 0,78 0,77 0,77 0,99
Wr 0,27 0,64 0,54 0,85 0,98 0,69 0,67 0,73 0,96
W 0,37 0,76 0,73 0,97 0,96 0,76 0,74 0,77 0,99
InW 0,49 0,76 0,73 0,89 0,76 0,86 0,99 0,96 0,92
InWl 0,84 0,94 0,95 0,81 0,69 0,83 0,98 0,93 0,71
InWr 0,43 0,65 0,64 0,80 0,81 0,84 0,95 0,87 0,73
NDL 0,87 0,89 0,86 0,97 0,71 0,86 0,83 0,81 0,71
Уравнения регрессии приведены ниже:
Коровяк метельчатый: W = 6,1 + 3,62 ND1 maxLl max (N=68; r=+0,975***)
К. медвежье ухо: W = 118,3 + 3,22 ND1 maxLl max (N=54; r=+0,988***),
где W — общая биомасса (мг); N1 — число розеточных листьев, DI max —
ширина максимального (по размеру) листа, LI max — длина максимального
листа. Выборки одновременно продемонстрировали существенную размер-
ную гетерогенность особей в популяциях обоих видов коровяка.
Из изучаемой популяции были взяты две группы генеративных особей —
самые крупные и самые мелкие. Вычисление индекса Левинса дало следую-
щие результаты: при сравнении средних для групп его величина составила
26,3, а при сравнении крайних особей в группах — 33,9. Повреждение глав-
ной точки роста у особей, переходящих к репродукции приводило к замет-
ному их отставанию по росту в высоту, хотя они развивали от двух до пяти
цветоносных побегов из пазух розеточных листьев.
Применение индеструктивного метода позволило получить в популяции
коровяка медвежье ухо кривые распределения особей по значениям RGR и
проследить за динамикой этих кривых и средних величин. Среднее значение
Ю Д(-М()1 рпфичсч кий JIIUIJIII I природных НОНуПИЦИЙ pilC 1Г11ПЙ 3 V)
RGR в интервале, соответствующем самому засушливому периоду сезона (с
26.06 12.07.1990), опускалось в область оrpiпепельных величин расiсиня
теряли биомассу. После этого (в интервале от 12.07 до 11.10) наблюдался не-
который подъем показателя и переход в область положительных значений.
Из 79 особей к. медвежье ухо, отмеченных 1 июня 1990 г., 16 особей пе-
решли к репродукции (на момент учета они имели диаметр розетки 27,0 I
9,4 см) и 89% их дали семена, а остальные погибли до обсеменения; 24
особи, имевшие средний диаметр розетки 6,8 ± 0,3 см, отмерли в вегета-
тивном состоянии и 36 особей (средний диаметр розетки которых составил
8,6 ± 0,4 см) остались в вегетативном состоянии.
Поскольку наблюдение за особями вели не с момента их появления, т. е.
под наблюдением находилась не одна когорта, то было возможно проанали-
зировать лишь кривые исчерпания и по ним вычислить период полуизрежи-
вания. Он составил для первого интервала 124,2 дня, для второго — 368,0
дней, а для третьего — 142,0 дня. Таким образом, можно констатировать
картину, уже отмеченную для других видов — максимальные темпы отми-
рания приходились на периоды наиболее быстрого роста и, наоборот, де-
прессия ростовых процессов соответствовала минимальным темпам изре-
живания популяции.
10.4.3 . Результаты наблюдений в популяциях проломников наиболь-
шего, удлинённого, северного и змееголовника тимьяноцветкового
На базе даже очень сходной архитектуры (и архитектурной модели) особи,
обусловленной известной морфологической консервативностью таксона, воз-
можна адаптация к произрастанию в различных экотопах и фитоценозах либо за
счет частичного видоизменения мор-
фологических признаков в ходе онто-
генеза, либо путем модифицирования
некоторых демографических параме-
тров популяций.
Отпечаток обоих названных пу-
тей, преимущественно первого, не-
сет популяционная биология гигро-
фильного зимующего однолетника
проломника нитевидного, лишь не-
давно выделенного в особую сек-
цию. Этот вид будет рассмотрен в
следующем разделе, а здесь озна-
комимся с приспособлениями трех
других видов проломника, произ-
растающих на остепненных эроди-
рованных склонах. Дадим краткую
характеристику всех трех видов, по-
сле чего подробнее ознакомимся с
демографическими характсрисгика-
ми пропомника северного.
Рис. 10.17. Архитектура и аллокацион-
ные диаграммы особей у четырех видов
проломника: 1 — П. наибольший; 2
П. удлиненный; 3 — П. северный; 4
П. нитевидный. Относительный вклад
биомассы: а в репродуктивную севе-
ру, b в надземные веге i азивные opia
ны (розезку), с в корневую спс1сму
336 М-Н Марков I loiiyininiioiiiinH (nioiioi пн рисiciiiHi
Зимующий однолетник проломник удлиненный Л. dongata (рис. 10.17)
способен произрастать в условиях нарушенных лугово-стенных фитоце-
нозов с примесью типчака Festuca valesiaca, мятлика узколистного Роа
angustifolia и др. Относительная крупносемянность компенсируется для
вида некоторым снижением семенной продукции (табл. 10.5) и потенци-
альной скорости роста численности популяции (биотического потенциа-
ла). Главными характерными особенностями стратегии выживания пролом-
ника удлиненного A. elongata являются дружное появление всходов осе-
нью, непродолжительный осенний рост в условиях достаточной влажно-
сти и сезонно ослабленной конкуренции со злаками и другими растения-
ми, быстрое весеннее развитие с зацветанием в конце апреля и полное ис-
чезновение из травостоя к середине июня. Сходное сезонное развитие отме-
чено для озимого однолетника проломника наибольшего, который, однако,
весной несколько дольше сохраняет розетку, а в виде сухих стрелок с зон-
тиками заметен вплоть до октября. Сдвиг вегетации проломника наиболь-
шего в сторону обычно засушливого конца весны возможен благодаря его
относительной ксероморфности (табл. 10.7; см. также главу 4).
Таблица 10.7.
Некоторые биоэкологические свойства видов рода проломник Androsace.
Виды: Экол. тип Высота (см) Семенная продукция средн.(тах) Средняя масса семени(мг) Среднее ре- продук- тивн._уси- лие (%)*
77. нитевидный м 3-15 4800(9600) 0,02 63
П. северный км 5-25 750(1650) 0,15 72
П. удлиненный м 2-10 588(1008) 0,30 76
77. наибольший км 3-15 540( 756) 0,77 86
Примечание: М — мезофит; КМ — ксеромезофит.
* Биомасса репродуктивных структур, включая стрелки/общая биомасса
О предполагаемом адаптивном значении «розетки» брактей уже гово-
рилось выше. Режим конкуренции и ухудшенной аэрации, возникающий
в сомкнутых степных ценозах на черноземе, крайне неблагоприятно ска-
зывается на приживании проростков, розеточном росте и последующей
репродукции у этого вида.
Поскольку у проломника наибольшего, как и у других малолетних ви-
дов рода Androsace, зимовка делает переход к репродукции обязательным
для всех особей независимо от их жизненного состояния, т. е. мощности ро-
зетки, то условия, в которых протекает осенний розеточный рост, в конеч-
ном итоге определяют репродуктивную мощность особей. При мало меня-
ющемся репродуктивном усилии (см. главу 7) семенная продукция вместе
с размерами особей снижается очень резко — в популяции начинают пре-
обладать особи с одним-двумя плодами, которые не обеспечивают ее нор-
мальное воспроизводство, отчасти из-за снижения вероятности попадания
семян на рыхлый свободный субстрат. Проломник наибольший положи-
IhltUI 10 ДгМО! |Шф|1ЧСЧКПИ IIIUIHII I природных НОНУИИЦПЙ plU 1СНПЙ 33/
icjii.iio омывается па всякое рыхление субсipaiа, даже происходящее в pc
зульгаю эрозии и выхода на повсрхиос11. бедной материнской породы
мергеля с известняковыми плитками (петрофидьность). Физическая исола
гоприятность такого субстрата (незакономерные колебания содержания эле-
ментов минерального питания, резкие колебания температуры и влажно-
сти), очевидно, компенсируются крупносемянностью и связанной с ней эко-
топической толерантностью. К переувлажнению, наоборот, проломпик наи-
больший относится по нашим наблюдениям крайне отрицательно. Хорошо
развитые мощные розетки, выращенные в условиях опыта на ровном участ-
ке почти все погибли за зиму, не выдержав переувлажнения, возникшего из-
за отсутствия наклона поверхности и стока. Те же особи, которые остались
живыми (2,3%), вышли из-под снега в крайне угнетенном состоянии и дали
по одному жалкому соцветию. Семенная продукция оказалась недостаточ-
ной для возобновления модельной популяции, и растение в итоге не при-
жилось.
Ксеромезофит проломник северный демонстрирует черты облигатного
двулетника. Весной, когда перезимовавшие особи дружно зацветают и пло-
доносят, сразу же высыпая многочисленные мелкие семена, способные к
немедленному прорастанию, популяция пополняется новыми всходами, и
они могут продолжать появляться вплоть до осени, если позволяют мете-
орологические условия лета. Разная продолжительность розеточного ро-
ста, накладывающаяся на пространственную гетерогенность довольно сом-
кнутого (покрытие 85%) лугово-степного фитоценоза с участием типчака
Festuca valesiaca, тонконога Koeleria cristata и часто мха (Abietinella abietina
(Hedw.) Fleisch.), обусловливает значительную дифференциацию особей по
жизненному состоянию. Выживание относительно мелкосемянного про-
ломника северного основано на более стабильной и чуть повышенной влаж-
ности самого поверхностного слоя почвы под сомкнутыми фитоценозами и
физиологической устойчивости к некоторому притенению благодаря более
высокому, чем у п. наибольшего, числу розеточных листьев (см. главу 4).
Эрозия почв как фактор нарушения фитоценозов на склонах разной экс-
позиции и крутизны выражена неодинаково — более сильно эродированы
крутые склоны, а они в Татарии чаще всего, хотя и не всегда, имеют южные
экспозиции. Из-за смыва опада и верхнего плодородного слоя почвы на бо-
лее крутых склонах развиваются обедненные варианты тех сообществ, ко-
торые занимают соседние менее крутые склоны.
Сравнение фитоценозов, предпочитаемых нашими видами, которое было
проведено в ходе маршрутных обследований Бугульминского и Бавлинского
районов, интересно, на наш взгляд, потому, что эти фитоценозы непосред-
ственно и плавно переходили друг в друга, поскольку занимали разные
экспозиции одного и того же склона и на одном и том же уровне высоты.
Популяции проломников северного и наибольшего были разделены бук-
вально несколькими метрами, т. е. почти примыкали друг к другу. При этом
на склоне южной и юго-западной экспозиций видов, которые не встреча-
лись на склонах северной и северо-западной экспозиций, было отмечено со-
всем мало. Отсюда есть все основания считать типчаково-разнотравный фи-
тоценоз на склоне южной экспозиции обедненным вариантом с существен-
338
МН Мирыш 11<иiyимщк>i111пи biioiioi ни рш iciinil
но более низкой продуктивноетыо,
а диаграмма на рие. 10.18 наглядно
показывает не только обеднение ви-
дового состава и снижение продук-
тивности, но и более низкое значе-
ние отношения надземная фитомас-
са/подземная фитомасса.
В Верхнеуслонском районе, где
проводились стационарные наблю-
дения на постоянных квадратах за
проломником северным, фрагменты
фитоценозов на склонах разной кру-
тизны и экспозиции тоже сущест-
венно отличались по видовому со-
ставу и продуктивности. Наиболее
обедненные и малопродуктивные
фрагменты были приурочены здесь
к склонам северо-западной экспо-
Рис. 10.18. Диаграмма продуктив-
ности наименее нарушенных фраг-
ментов типчаково-разнотравного (в
Бугульминском районе) и кострецово-
тысячелистникового (в Верхне-
Услонском районе) фитоценозов.
зиции. Таким образом, эрозия, сопряженная со смывом, делает крутизну
склона решающим фактором, пересиливающим влияние экспозиции. Ди-
аграмма (рис. 10.18) свидетельствует о сходной продуктивности наименее
нарушенных фрагментов типчаково-разнотравного (в Бугульминском райо-
не) и кострецово-тысячелистникового (в Верхне-Услонском районе) фито-
ценозов при некотором смещении равновесия в сторону подземной фито-
массы в первом случае. Обедненный вариант в Верхне-Услонском районе
по продуктивности был почти вдвое выше, чем в Бугульминском.
Поскольку регенерационной нишей для обоих видов проломника можно счи-
тать только микроучастки, свободные от какой-либо растительности, то необ-
ходимо было оценить размещение и величину таких свободных микроучастков
в различных фрагментах фитоценозов. Размещение микроучастков (5><5 см) с
различным количеством общей (суммарной для всех видов) наземной фитомас-
сы в нескольких фрагментах фитоценозов с участием проломников наибольше-
го и северного показано на рис. 10.19 (I). Сильное варьирование абсолютных
значений фитомассы послужило причиной выбора логарифмических (In) клас-
сов для графического изображения. Свободные (в надземной сфере) от расти-
тельности микроучастки учитывали особо, а занятые подразделяли по натураль-
ному логарифму фитомассы на следующие классы: 0-3; 3-4; 5-6; 6<. Для оцен-
ки роли мха Abietinella abietina в создании пространственной горизонтальной
структуры сравниваемых сообществ фрагменты с развитым моховым покровом
изображены с учетом и без учета фитомассы мха.
Выделение элементарных квадратов хорошо иллюстрирует гетеро-
генность фитоценозов, но нельзя не отметить произвольность выбора ве-
личины этих квадратов, которая не позволяет получить представление о ре-
альных размерах свободных и занятых микроучастков. Поэтому в дополне-
ние к другим оценкам были проделаны линейные промеры (в см) трех ти-
пов микроучастков: 1) свободных (в наземной сфере); 2) занятых цветковы-
ми растениями; 3) занятых мхом с вычислением процента от общей нротя-
lianu 10 Деменрафичсскип апаши природных нопушщип рае icnidt
119
жепностп промеренных линий. Множсс!венные промеры делали е помо
щью металлического складного метра, коюрый был орпснiпропан каждым
раз случайным образом.
Для Бугульминского района данные показывают (рис. 10.19, II), что
средние размеры свободных микроучастков в фитоценозе с проломником
наибольшим A. maxima, где их было 80%, примерно в 6 раз (8,8 ± 1,6 про-
тив 1,3 ± 0,1) превышают размеры таковых в фитоценозах с участием про-
ломника северного, где их протяженность составляла менее 20%.
Средняя линейная протяженность микроучастков, занятых мхом и цвет-
ковыми растениями в фитоценозах с участием проломника северного, тоже
невелика (2,3 ± 0,4 и 1,5 ± 0,2 соответственно). Для Верхне-Услонского рай-
она, где сравнивались два фрагмента, размеры и процент свободных ми-
кроучастков варьировали от 1,3 ± 0,2 — 15,3% (на пологом склоне север-
ной экспозиции) до 3,8 ± 0,3 — 65,0% (на крутом склоне северо-западной
экспозиции).
Средние размеры и процент микроучастков, занятых мхом и цветковыми,
на одном фрагменте составили 2,6 ± 0,3 см — 31,0%; 2,3 ± 0,4 см — 54,2%, а
на другом фрагменте, где не
было мха, — 1,9 ± 0,2 см —
35%.
Приведенные результаты
наглядно убеждают в том,
что фитоценотические аре-
алы проломников северного
и наибольшего практически
не перекрываются.
Наблюдения, проведенные
на постоянных квадратах за
динамикой численности п.
северного, позволили уста-
новить следующее. На фраг-
менте с более высокой про-
дуктивностью травостоя по-
явление проростков было от-
мечено неоднократно в тече-
ние периода наблюдений; а
на обедненном фрагменте с
наиболее изреженным пок-
ровом, в котором отсутство-
А.т. Бугульминский р-н As.
Рис. 10.19. I. Горизонтальное размещение надзем-
ной фитомассы во фрагментах фитоценозов с уча-
стием проломника наибольшего (А.т.) и пролом-
ника северного (A.s.) в Бугульминском и Всрхне-
Услонском районах Татарии: Wt - надземная ф|I-
томасса; а — без учета мха, б - с учетом мха, в
при отсугствии мха. II. Линейные средние размеры
микроучастков (см), занятых мхом (з.м ), цветковы-
ми рас I синями (з.ц.) и свободных (св )
вали мхи, проростки появи-
лись однократно — в июле
только после резкого повы-
шения влажности почвы из-за
выпадения дождей с некото-
рым понижением температу-
ры. Па площадках с наименее
нарушенным покровом отмн-
340 МВ Марков 11< я iy и >i 111 к я 111.> я Uiioiioi нм р;к iciniii
рание особей наблюдалось в очень небольших маснпабах в течение бездождно-
го периода, хотя смертность превалировала в динамике и определяла неуклон-
ное снижение общей численности несмотря на постоянное параллельное попол-
нение популяции проростками (рис. 10.20). Несколько выше была смертность в
фитоценозах, где сомкнутый полог отсутствовал. Периоды полуотпада соста-
вили соответственно от 26,5 до 116,8 дней и 13,8 дня (табл. 10.8). Высчитанная
для площадки № 8 (фрагмент П1) лабильность баланса численности была наи-
высшей (0,76), что вполне понятно с учетом влияния более энергично протекав-
ших здесь процессов возобновления и отмирания особей. Для сравнения мож-
но взять данные о популяционных локусах с площадок № 3 и 4 (фрагмент I), на
которых лабильность баланса численности составила 0,63 и 0,22 соответствен-
но (табл. 10.8).
Таблица 10.8.
Демографические параметры нескольких популяционных локусов проломника
северного
Фрагмент № площадки Общее число появившихся проростков н1/2 (дни) В
до начала дождей после начала дождей
I 2 — 2 39,2 0,57
3 4 4 26,5 0,63
4 1 1 116,8 0,22
II 5 — — 0
6 — — 48,9 0,27
7 — — 62,5 0,27
III 8 - 6 13,8 0,76
Для определения скорости роста особей проломника северного в разных
локусах мы воспользовались косвенным, индеструктивным методом, ис-
пользуя индекс — произведение числа розеточных листьев на диаметр ро-
зетки (I2=DxN1). По растениям из выборок была оценена связь между нату-
ральным логарифмом индекса (In I2 = X) и общей биомассой (W). Она ока-
залась параболической:
W = 36,7 — 20,0Х + 2,8Х2.
Графики на рисунке 10.21 показывают, насколько энергично росли рас-
тения на площадке № 8 по сравнению с растениями на площадках № 3 и 4.
Судя по этим данным, масса особей на площадках № 3 и 4 очень мало из-
менилась за время наблюдений. В засушливый период растения прекраща-
ли свой рост, а некоторые (чаще всего наиболее мелкие по размеру) теря-
ли массу и отмирали. Потеря массы происходила за счет отмирания розе-
точных листьев. Сухой период сменился очень резким повышением влаж-
ности из-за выпадения дождей, что существенно нс повлияло па размеры
I'liathi 10 Дгмо!рафпчсский iiiiiuiii t природных попуиицпй рисirniill 341
особей с площадок №3 и 4, по отчет-
ливо сказалось на росте особей пло-
щадки №8.
Известно, что малолетники из-
за слабой конкурентоспособности
приурочены в той или иной степе-
ни к открытому субстрату. Они спо-
собны к быстрому захвату времен-
но освободившихся микроучастков,
когда ослабляется конкуренция.
Однако в некоторых случаях одно-
летние растения способны произ-
растать и на участках с некоторым
задернением. В связи с этим разли-
чают совместимые и несовместимые
с задернением (turf compatible и turf
incompatible) виды малолетних рас-
тений. При этом постулируется от-
рицательное влияние полога из мно-
голетних ценозообразующих и эди-
фицирующих растений для роста
малолетников. Однако в присут-
ствии полога существенно снижает-
ся физическое испарение с поверх-
ности почвы и создается чуть бо-
лее благоприятный режим влажно-
сти в самых поверхностных слоях.
На это указывают также X. Веркаар
и др. (Verkaar, Schenkeveld, Brand,
1984), которые отмечают оптималь-
Рис. 10.20. Динамика общей численно-
сти (светлые круги), кумуляты пополне-
ния (квадраты) и потерь (темные круги)
в популяционном локусе проломника се-
верного.
ность светового режима, но экстре-
мальность температурного и водного режимов в верхних (до 4 см) слоях по-
чвы под сильно разреженными фитоценозами. Л. Павон и Р. Ридер (Pavone,
Reader, 1985) на примере люцерны хмелевой Medicago lupulina наглядно
продемонстрировали худшее выживание растений в ценозах без мохового
покрова, объясняя это влиянием стресса засухи, и вместе с тем лучшее жиз-
ненное состояние и более высокую плодовитость выживших экземпляров в
этих условиях.
Как показали наши результаты, проломник северный способен давать
проростки даже в относительно засушливый период в фрагментах с более
развитым пологом из сопутствующих растений и при соседстве мха. Если
считать лабильность баланса численности показателем, по которому мож-
но судить о стабильности популяционного локуса, то наиболее стабильны
локусы популяции п. северного под пологом сомкнутого и продуктивно-
го фитоценоза. За время засухи относительно высокая смертность (выше
60%) вызвала сильное изреживание популяционного локуса на фрагмен-
те III. Однако выжившие особи, которые в момент повышения влажности
342
МН Марков IloiiyiBimioiiiiini 61101101 пи pnciciuiil
Рис. 10.21. Сравнение по темпам роста
особей проломника северного, произрас-
тавших в условиях разреженного (пло-
щадка № 8) и более сомкнутого (пло-
щадки № 3 и № 4) травостоя: W — об-
щая биомасса (мг); Т — время (дни с на-
чала наблюдений).
ный цикл озимого однолетника.
лишь слегка превосходили по мощ-
ности экземпляры с 1 и II фрагмен-
тов, продемонстрировали очень бы-
стрый рост (рис. 10.21), и к концу
наблюдений их преимущество было
усилено многократно (20,0 против
5,0 мг). Снятие стресса засухи мож-
но расценивать как более или менее
случайное событие, если учесть, что
июль, по средним многолетним дан-
ным, более сухой месяц, чем июнь.
Все это свидетельствует о снижении
вероятности выживания проломни-
ка северного в излишне разрежен-
ных фитоценозах, где он вследствие
отмирания весенней когорты особей
и появления летне-осенних когорт
фактически демонстрирует жизнен-
Однако именно такие фитоценозы минимальной продуктивности и сомкну-
тости предпочитает проломник наибольший. Стратегия выживания у этих ви-
дов, судя по всему, различна, поскольку они довольно сильно отличаются по се-
менной продуктивности (табл. 10.7). Более высокая продуктивность п. северно-
го, по всей видимости, повышает вероятность попадания диаспор в относитель-
но мелкие свободные микроучастки. Стратегия п. наибольшего, по-видимому,
направлена, в основном, не на «отыскание» мелких микроучастков, а на повы-
шение устойчивости, толерантности за счет более высокого вклада биомассы в
каждое семя. Как следствие этого семенная продукция у него заметно ниже, чем
у проломника северного, а мы вновь наблюдаем пример компенсационной зави-
симости.
Очень сходная картина демографических параметров в тех же фрагмен-
тах фитоценозах в 1987 г. получена для облигатного двулетника змееголов-
ника тимьяноцветкового (табл. 10.9).
Таблица 10.9.
Периоды полуотпада для популяционных локусов змееголовника тимьяноцветко-
вого на двух фрагментах кострецово-тысячелистникового фитоценоза
Фрагмент фитоценоза Номер постоянного квадрата Н./2 периоды полуот- пада (дни)
На крутом склоне северо- западной экспозиции I 20,7
II 9,2
III 25,9
На пологом склоне южной экспозиции I >100
II >1(Ю
ш 41,4
Г'1(1Н(1 IО Дсм<)1 рпфичсскпй HIHIJI1I I природных попуимцпй pill IC11ий
343
Рис. 10.22. Динамики
общей ’ihcjicihioc । и
(I), кумуляты ионол
нения (2) и потерь (3)
в двух популяцион-
ных локусах змеего-
ловника тимьяноцвет-
кового (вверху) и лю-
тика ядовитого (вни-
зу).
В 1987 г. засушливым был июнь, а с начала июля пошли дожди, и потому
в обоих популяционых локусах (табл. 10.9) процесс отмирания особей про-
должался параллельное процессом появления проростков (рис. 10.22). При
внешнем сходстве кумулят необходимо отметить разный масштаб графиков
344 М.Н Марков I louyihimioiiiHin (hioiioi ия ])!ic
(шкалы численности), и тогда станет понятна более высокая динамичность
популяционного локуса на I фрагменте, чем на II фрагменте (лабильное и>
баланса численности: 0,66 ± 0,03 против 0,53 ± 0,04) за одно и то же время
в популяционном потоке (population flux) участвовали около 150 особей на
первом (I) и около 60 особей — на втором (II) фрагментах. Помимо общей
лабильности баланса численности (с учетом любых изменений численно-
сти в локусе) определяли частные значения этого показателя, вычисленные
либо с учетом только смертности (Вт), либо только пополнения всходами
Рис. 10.23. Динамика численности особей змееголовника на двух пробных площа-
дях: А — летняя когорта, В — плодоносившие особи и С осенняя uoiopia.
365
Рис. 10.24. Рост особей весенней (в) и летней (л) когорт в двух популяционных локу-
сах змееголовника, представленный как динамика значений индекса Nxl. Крестиками
помечены значения индекса для вегетировавших, но отмерших особей.
(Bg) (табл. 10.10). Раскладывая таким образом общую лабильность балан-
са численности на составляющие, мы получаем возможность оценить отно-
сительную роль двух процессов по их роли в разных по продуктивности и
сомкнутости фрагментах фитоценозов.
346 А/ // Марков I h>iiyпицце>1 шан (як>и<и пи pin iciiiifi
Таблица 10.10.
Частные значения лабильности баланса численности для двух популяционных ло-
кусов змееголовника тимьяноцветкового в сезоны вегетации 1987 и 1988 гг.
№ постоянного 1987 г. 1988 г.
квадрата I локус II локус I локус II локус
I Вт = 0,14 Bg = 0,50 Вт = 0,05 Bg =0,39 Вт = 0,59 Bg =0,45 Вт = 0,72 Bg = 0,50
II Вт = 0,34 Bg =0,39 Вт = 0,02 Bg =0,54 Вт = 0,45 Bg =0,55 Вт = 0,30 Bg = 0,40
III Вт = 0,20 Bg =0,55 Вт = 0,21 Bg = 0,40 Вт = 0,42 Bg =0,65 Вт = 0,61 Bg =0,28
Роль различий в динамичности локусов особенно ярко проявилась в 1988 г.,
когда из-за большей продолжительности засушливого периода, охватившего
конец июня и весь июль, весенние когорты особей змееголовника вообще по-
гибли в менее сомкнутом и менее продуктивном фрагменте I. Под разнотрав-
ным пологом фрагмента II некоторые особи этой когорты пережили засушли-
вый период и, развиваясь по типично двулетнему циклу, намного превзошли
по мощности особей позднелетней когорты, которые появились после того,
как пошли дожди. На рисунке 10.23 можно видеть общую картину динамики
численности на постоянных квадратах в двух популяционных локусах зме-
еголовника тимьяноцветкового с учетом генеративных особей, демонстри-
рующих закономерное отмирание после плодоношения (кривые 2). Здесь же
показана динамика численности вновь появлявшихся когорт (летней — 1 и
осенней — 3), судьба которых складывалась неодинаково.
Рисунок 10.24 демонстрирует при помощи индекса Nxl, как росли особи
разных когорт в разных фрагментах, причем отдельно (крестиками) выделе-
ны особи, которые не дожили до очередного срока наблюдений.
Яровизация перевела все перезимовавшие растения в генеративное со-
стояние, а мощность репродукции прямо зависела от роста вегетативной
сферы в предыдущий сезон вегетации. Приходится констатировать не толь-
ко разную динамичность локусов одной популяции в зависимости от разви-
тия полога и метеорологических условий, но и выживания популяций дву-
летников за счет перехода отдельных популяционных локусов от двулетнего
жизненного цикла к циклу озимых однолетников или возникновения в раз-
ных локусах неодинакового соотношения когорт, развивающихся по тому
или иному циклу.
10.5. Виды прибрежной зоны водоемов с переменным уровнем
воды
Характерными особенностями среды прибрежных сообществ являются:
1) высокая обеспеченность растений элементами минерального питания,
2) частые, а главное непредсказуемые для популяции нарушения, вызван-
ные зарегулированностью стока, 3) высокая влажность почвы и воздуха и 4)
плохая аэрация почвенного горизонта из-за переувлажнения.
/ 'itifhi 10 Демо! рафнчсскпй aiiiuim природных попуинций pin1 iriniB
34 7
Проломник
нитевидный
I....Ill.....Illi....................II..IIIIHII......................I..........Hill..........IM
11.VII n 17.VIL „ 23.VII
Даты наблюдении
Лютик
ядовитый
Рис. 10.25. Фенологические спектры для прибрежных популяций проломника ни-
тевидного и лютика ядовитого: фазы 1 — проростки; 2 — ювенильные особи; 3
взрослые вегетативные особи; 4 — цветущие особи.
Приобретаемая уже на стадии проростка система придаточных корней
взамен быстро прекращающего рост главного корня обеспечивает пролом-
нику нитевидному произрастание на заиленных песчаных отмелях по бере-
гам рек и роль меженного эфемера (Нечаев, Гапека, 1970). В лесных районах
п. нитевидный встречается не только в прибрежных ценозах с растительным
покровом, нарушенным паводком и состоящим в основном из однолетников
(сыти бурой Cyperus fuscus, ситника жабьего Juncus bufonius, лужницы во-
дной Limosella aquatica, сердечника мелкоцветкового Cardamineparviflora и
др.), но и в колеях лесных дорог и агрофитоценозах, подверженных сильно-
му вымоканию. Можно отметить предпочтение видом песчаных и супесча-
ных почв, в том числе серых лесных, а также некоторую устойчивость к за-
тенению. Затенение лишь снижает репродуктивное усилие, увеличивая от-
носительный вклад биомассы в формирование ассимилирующих органов
(глава 9; см. Марков, 1986). Наблюдаемое немедленное прорастание свеже-
созревших и при малейшем прикосновении очень легко осыпающихся се-
мян приводит к чередованию форм с развитием по типу летнего однолетни-
ка с формами, развивающимися по типу озимого однолетника, в популяции.
Стратегия выживания п. нитевидного основана на повышении вероятно-
сти попадания семян в благоприятные микроучастки (safe-sites по Harper,
1977) благодаря самой высокой из всех малолетних проломников семен-
ной продуктивности (табл. 10.7), поскольку вид этот и самый мелкосемяп-
ный. На рисунке 10.25 приведены фенологические спектры для прибреж-
ных популяций проломника нитевидного и лютика ядовитого Ranunculus
sceleratus. Налицо постоянное пополнение популяции проростками (поли
когортностъ). Вместе с тем для второго вида показано, что параллельно от-
мирали как проростки (26,3%), гак и ювенильные особи (45,5%), что суще-
ственно 1ЮВ1П1Яио на динамичность популяции (иабилыюсiь баланса чис
348
Л/ Н М(ЦЖОв I |( > 11УII >1 Ц| К >1111Ш1 bllOIIOI II >1 |)IH ICIIlH
11.V 14.VI 24.VI30.VI 10.VII18.VII25.VII даТы наблюдений 21.IX
EZZJ Вегетация
1 Бутонизация
ИИ Цветение
В Плодоношение
Рис. 10.26. Фенологические спектры для двух популяционных локусов пупырника.
ленности — 0,75-0,79) при сравнительно небольших колебаниях от срока к
сроку общей численности. Кумуляты пополнения и убыли особей в двух по-
пуляционных локусах лютика ядовитого приведены на рисунке 10.22 вме-
сте с таковыми для змееголовника, чтобы показать сходство проявления по-
ликогортности у видов с разной экологией.
Наиболее стабильную численность демонстрировала популяция лу-
жницы водной Limosella aquatica — вида с крайне нетипичной для мало-
летников архитектурой (лабильность баланса численности всего 0,26).
Укоренение дочерних розеток, отодвинутых от материнской на длину сто-
лонов, по всей вероятности существенно повышает устойчивость такой си-
стемы за счет внутриклональных коммуникаций.
l iiatiti 10 Дсмо!рп([)11чсский iiimiiiii природных иопуцицпй piicirniill
349
14.VI 24.VI 13.VII 25.VII
I | вегетация
IMIBI бутонизация
цветение
плодоношение
Рис. 10.27. Фенологический спектр для популяции бородавника обыкновенного: по
горизонтальной оси даты наблюдений (1989 г.).
10.6. Лесные виды
10.6.1. Наблюдения в популяциях лесных двулетников бородавни-
ка обыкновенного Lapsana communis и пупырника японского Torilis
japonica
Выборочный метод был использован при изучении популяций лесных
двулетников только на первых порах для получения достоверных алломе-
трических зависимостей, после чего основным методом получения данных
был индеструктивный метод.
Наблюдения за ростом вегетирующих особей первого года жизни показа-
ли, что в летнее время ростовые процессы сильно заторможены. Так в 1988
г. у пупырника (с 11.07 по 26.07) и у бородавника (с 20.07 по 26.07) сред-
ние значения RGR составили, соответственно, 0,033 и 0,09 мг/мг. нед. К осе-
ни рост значительно активизировался, и RGR составила 0.227 у первого и
0,150 — у второго вида. В 1989 году отмечен интенсивный весенний рост
Т. japonica (RGR = 0,242 мг/мг. нед) и летняя депрессия (RGR = 0,131 мг/
мг.нед). Сходная картина получена для обоих видов и в 1990 году.
Индеструктивным методом выявлена широкая вариабельность отно-
сительной скорости роста в пределах популяций обоих видов, причем, как и
в случае со степными двулетниками, многие особи в период летней депрес-
сии теряли биомассу и демонстрировали отрицательные значения RGR.
Однако, даже несмотря на летнюю депрессию ростовых процессов, на-
блюдавшуюся в широколиственных лесах из-за недостатка света, исполь-
зование двух благоприятных периодов для вегетативного роста приводит к
значительному возрастанию вегетативной мощности особей лесных обли-
гатных двулетников, а это в свою очередь служит залогом более мощной ре-
продукции на втором году жизни для особей весенних когорт.
350 Л/// Маркин I loiiyihimioiiiiiui (dipnoi пи piiciciiiiit
Фенологические спектры (рис. 10.26 и 10.27) указываю! па более растяну-
тую во времени репродукцию у пупырпика, чем у бородавника.
10.6.1.1. Результаты наблюдений в популяциях недотрог обыкновенной Impatiens
noli-tangere и мелкоцветковой Impatiens parviflora
Оба вида имеют семена с прямым зародышем, дифференцированным на
семядоли, которые у жизнеспособного зародыша окрашены в сине-зеленый
цвет, гипокотиль, несущий зачатки придаточных корней в виде бугорков.
Покровы непрочные с характерной бороздчато-бугристой скульптурой, и
потому легко повреждаются при механическом воздействии. Семена, ко-
торые, как уже говорилось, существенно варьируют по массе, находятся
в глубоком врожденном (эндогенном) покое, из которого их можно выве-
сти, подвергнув длительной (не менее трех месяцев) холодной стратифи-
кации во влажных условиях. Однако, после такой стратификации, проис-
ходящей во время зимовки в почве, фактически все жизнеспособные семе-
на приобретают способность к прорастанию дружно весной в конце апреля
- начале мая. Дружность прорастания и появления проростков нарушается
в том случае, если весной наблюдается засушливый период. Тогда первая
когорта всходов может отделяться от последующих единичных проростков
некоторым промежутком времени (1-2 недели), и это сильно сказывается на
степени дифференцированности особей в составе популяции. Прорастание
надземное — проросток выносит наверх две широкояйцевидных (н. обык-
новенная) или овальных обратнояйцевидных (н. мелкоцветковая) семядоли
с небольшой выемкой на верхушке. Семядоли недотроги обыкновенной яв-
ственно сизые и очень часто (при достаточно сильном весеннем освещении)
направлены вниз под острым углом к гипокотилю благодаря характерно-
му изгибу черешков. Нижняя якореобразно вздутая часть гипокотиля сразу
одновременно (н. обыкновенная) или сразу вслед за формированием слабо
выделяющегося главного корня (н. мелкоцветковая) дает 8-9 придаточных
корней (см. рис. 7.18). Гипокотиль проростка способен сильно удлиняться, а
степень его удлинения во многом зависит от глубины прорастания. Позднее
уже у взрослых особей гипокотиль может, изгибаясь в нижней части, как бы
частично полегать, и на всем протяжении зоны соприкосновения с влажной
почвой и даже несколько выше ее образуются придаточные корни.
У проростка главная точка роста формирует два близко друг к другу рас-
положенных настоящих листа, и сразу же удлиняющееся подсемядольное
колено выносит их на значительную высоту (8 см и выше). Семядоли мо-
гут сохраняться почти до середины июня, и время их опадения примерно
приходится на момент перехода всех особей к репродукции — к заложению
клейстогамных цветков, с которого она начинается.
В 1987 г. наблюдения на постоянных площадках проводили в двух популя-
циях недотроги обыкновенной под пологом липового леса. Местообитания
популяций различались по условиям увлажнения: первая популяция распола-
галась на делювиальном шлейфе лесного оврага, периодически омываемом
водой, стекающей по оврагу с водораздела; вторая на бугре в mouieii части
второй надпойменной террасы долины р. Волги, где увлажнение <н \ nicci нам
/ 7fW</ 10 Деменрафичсский пинии 1 природных iioiiyuHiuiii рн 1гппй 351
лось только атмосферными осадками. В даиыiciinicM дня краi кос i и нсрнос
мес гооби гаинс будем условно именован, влажным, а втрое сучим.
В течение всего периода наблюдений в обоих местообитаниях проводили из-
мерения освещенности (в люксах) и температуры почвы па поверхнос ! и и uiy-
бинах 5 и 10 см. С момента первого измерения (12.V) освещенность па уровне
исследуемых растений резко снижалась вплоть до первых чисел июня (с 20 до
1,5 тыс. люксов), отражая быстрое формирование листового полога деревьев и
кустарников. Существенной разницы в интенсивности и ходе изменений осве-
щенности, а также в температурных режимах почвы на глубинах 5 и 10 см меж-
ду двумя местообитаниями не наблюдалось. Однако в сухом местообитании в
период бездождья (июнь) колебания температуры поверхности почвы были бо-
лее резкими, чем в более тенистом влажном местообитании. Анализ суточною
хода изменений влажности воздуха при помощи гигрографа выявил почти пол-
ное отсутствие колебаний в обоих местообитаниях.
В каждой популяции имелись локусы, представлявшие собой почти чи-
стые заросли недотроги. Заложенные для периодического наблюдения посто-
янные квадраты (40 х 40 см2) с закартированными со стадии всходов особями
были приурочены к таким локусам.
Присутствие таких видов, как чистотел Chelidonium majus, крапива дву-
домная Urtica dioica, и лопух Arctium lappa, встреченных рядом, но не по-
павших в пределы квадратов (табл. 10.11), указывало на несколько боль-
шее богатство почвы в сухом местообитании. Во влажном местообитании к
обычному режиму нарушений растительного покрова (из-за вытаптывания)
присоединялось замывание песком, к которому недотрога обыкновенная
достаточно хорошо адаптирована, а в сухом — нарушение было обусловле-
но только антропогенным воздействием, причем интенсивность последнего
была здесь существенно ниже, чем во влажном местообитании.
Таблица 10.11.
Виды растений в пределах постоянных площадей или встреченные
по соседству с ними (в скобках)
Влажное местообитание Сухое местообитание
Сердечник луговой Cardamine pratensis Ветреница лютичная Anemone ranunculoides
Селезеночник очереднолистный Chrisosplenium altemifolium Чистяк весенний Ficaria verna
Хвощ луговой Equisetum pratense Будра плющевидная Glechoma hederacea
Чистяк весенний Ficaria verna Яснотка крапчатая Lamium maculatum
Вербейник монетчатый Lysimachia nummularia Крапива двудомная Urtica dioica
Мятлик дубравный Роа nemoralis
Звездчатка жестколистная Stellaria holostea
352
(Tilia cordata, Aconitum excelsum, Gagea lutea,
Geum urbanum, Matteuccia struthiopteris,
Mercurialis perennis, Paris quadrifolia, Euonimus
verrucosa)
Л / // Мауьон 11 < > 11 у и и । u । < н 111 и я ()ПО1К>1 ин pm iciinh
Табл и цн 10.11. (продопжснис)
(Corylus avelana, Anthriscus
sy Ives tris, Aegopodium
podagraria, Arctium lappa, Geum
urbanum, Chelidonium majus,
Tilia cordata, Acer platanoides)
h (cm)
30
Периодические замеры высоты особей, подсчет числа листьев, измере-
ния ширины и длины семядолей и настоящих листьев, длины гипокотиля и
междоузлий, анализ изменения численности особей и фенологические на-
блюдения с 12 мая по 20 июля 1987 г. проводили на трех постоянных квадра-
тах в сухом и на четырех — во влажном местообитаниях. Параллельно при-
мерно через неделю из каждой популяции брали в среднем по 16-20 расте-
ний недотроги обыкновенной для морфобиологического анализа. У таких
растений измеряли высоту, длину нижних четырех междоузлий и гипокоти-
ля, длину и ширину семядолей, подсчитывали число листьев и определяли
их массу (отдельно на главном и пазушных репродуктивных побегах), опре-
деляли массу подземных органов, стеблей и репродуктивных структур. Как
уже говорилось выше, помимо более высокой влажности, во влажном ме-
стообитании была более низкая освещенность. В связи с этим, а также из-за
того, что крайне слабое плодоношение недотроги обыкновенной во влаж-
ном местообитании привело в последующие годы к резкому снижению чис-
ленности этой популяции, в 1989 г., когда было продолжено детальное изу-
чение этого вида, была изучена другая популяция. Эта популяция располо-
жена вдоль русла небольшого пересыхающего летом ручья в более широком
овраге, и потому по условиям осве-
щения данное местообитание было
близко к сухому, в котором изучение
недотроги продолжали и в 1989 г.
Содержание жизнеспособных
семян недотроги в почве оценива-
ли дважды: вскоре после появле-
ния всходов в популяциях (20 мая)
и после окончания диссеминации и
естественного отмирания растений
(7 октября). В первом случае был
применен метод проращивания, а во
втором — метод промывания на си-
тах с размерами ячеи 1x1 мм2, по-
скольку свежие семена недотроги
не прорастают без стратификации
(Jouret, 1976, 1977).
Динамика роста гипокоти-
ля и междоузлий у недотроги
обыкновенной в 1987 г. (рис. 10.28)
показывает, что в основе характер-
ной архитектуры особи (см. главу 4,
ю
20.V 1.VI 9 .VI
Даты наблюдений
Рис. 10.28. Динамика роста гипокотиля
и первых четырех междоузлий у недо-
троги обыкновенной.
I ’Kifui 10 Демен рпфическпй niiiuini природных iюпунищiЙ рис iспий
353
-fr— Wmin
—О- IV max
-о- X
стат, параметры
Wmin
Wmax
Рис. 10.29. Динамика роста особей в популяции недотроги обыкновенной (1989 г.)
как динамика средних ( х ± SJ и крайних (Wmin и Wmax) величин общей биомас-
сы в абсолютном (вверху) и логарифмическом (In) выражении.
рис. 4.1) лежи г соответствующий рн гм деятельноеi и анекеа и юпько во вто-
рую очередь неравномерное рас1яженнс уже обраюнанныч междоузлий.
Рис. 10.30. Детализированные фенологические спектры для популяций недотроги
обыкновенной из сухого (I) и влажного (II) местообитаний:
1с, 2с — наличие одной или двух семядолей; в — вегетация; г — генеративное состо-
яние; 2-18 — фазы двух-восемнадцати листьев.
В образцах почвы, взятых весной сразу за дружным появлением всходов,
семян недотроги не обнаружено. После обсеменения и полного отмирания
растений в изучаемых популяциях новый анализ выявил присутствие очень
небольшого числа семян этого вида (= 24 семени/м2) в самых верхних (0-5
см) слоях почвы.
На фоне колебаний относительной скорости роста намечалась стойкая
тенденция к снижению этого показателя с течением времени. Максимальные
значения в обоих местообитаниях были зарегистрированы в первую неде-
лю роста. На графике (рис. 10.29) можно наблюдать динамику среднего и
крайних значений общей биомассы у недотроги обыкновенной. Хорошо
видно, что в популяции в течение всего периода наблюдений наблюдался
высокий процент очень мелких особей, и среди них были и такие, которые
еле достигли критических размеров (=20 мг), допускающих репродукцию.
Условия лучшей освещенности в сухом местообитании положительно ска-
зались на темпах роста особей и биомассе, накопленной ими к концу жиз-
ненного цикла, но и здесь также было немало мелких угнетенных особей с
минимальной плодовитостью. В главе 8 уже приводились данные о возрас-
10 Дсмо|piKpiiueckiiii iiimiiin природ! 11.1ч noiiyiniiiiiit pm iriiiH!
35‘)
12.V 20.V 26.V 1.VI 9 .VI 18.VI 2.VII 14.VII
Даты наблюдений
11 □ I
%
12.V 20.V 26.V 1.VI 9.VI 18.VI 2.VII 14.VII
Даты наблюдений
Рис. 10.31. Динамические аллокационные спектры для двух популяций недотроги
обыкновенной из влажного (вверху) и сухого (внизу) местообитаний: относится!»
ный вклад биомассы в корпи (к); семядоли (с); стебли (ст); лисп.я (и) и репродукции
iii.ii' opi апы (р.о )
356 Л/// MiipKOtl I lonyilMUIIOIIIIini (MIOIIOI НН piH ICIIIlil
тан и и значения коэффициента Джинн в ходе pocia, н указано, «но данная
тенденция была более заметной во влажном местообитании. Интересно oi-
метить, что отдельно высчитанные значения коэффициента для маееы се-
мядолей — коррелята массы семени — диагностируют большую изменчи-
вость в сухом местообитании (0,315 против 0,208). Можно лишь предпола-
гать, что это отражает наличие примеси более крупных семян хазмогамного
происхождения, присутствовавших во временном семенном банке более
благоприятного по условиям (что явствует из итогов роста) сухого место-
обитания.
На рисунке 10.30. приведены два вида фенологических спектров. Одни из
них (нижние) построены с учетом наличия или отсутствия семядолей и пе-
рехода к репродукции, а другие (вверху) — с учетом стадий роста, т. е. ста-
дий двух-, четырех, шести и т. д. настоящих листьев. Обращает на себя вни-
мание более длительное сохранение семядолей и более раннее наступление
всех выделенных фенофаз и стадий у растений в сухом местообитании, т. е.
в относительно более благоприятных для роста условиях.
На статическом аллокационом спектре выявляется различное раз-
мещение биомассы в пределах особи (т. е. различный тип аллокации) у рас-
тений, принадлежащих к разным классам по общей биомассе как показате-
лю жизненного состояния. Так у особей, имеющих малую биомассу, наи-
большая часть биомассы (и энергии) вкладывается в создание листовой по-
верхности, т. е. в вегетативные органы, а репродуктивное усилие остается
очень низким. С возрастанием общей биомассы (улучшением жизненного
состояния особи) увеличивается вклад в репродуктивные структуры за счет
уменьшения относительного веса листьев или фотосинтетического усилия.
Динамические аллокационные спектры для обеих популяций показывают,
что у растений во влажном местообитании относительный вклад биомас-
сы в корневую систему был в среднем на 5% выше, чем у растений сухого
местообитания (рис. 10.31). Фотосинтетическое усилие лишь в начале на-
блюдений, т. е. до формирования затеняющего полога, было на 5-7% выше
у растений сухого местообитания, после чего разница сменилась на об-
ратную, достигнув к концу наблюдений 12%. Более высокие темпы разви-
тия особей в сухом местообитании и дружный переход к репродукции, уже
установленные по фенологическим спектрам, хорошо прослеживаются и в
динамике аллокации.
Численность особей в обеих популяциях изменялась только по причи-
не отмирания, так как появление новых всходов после первой когорты не
наблюдалось. Процессы естественного выпадения особей были замедлены,
и ускорение их всегда было связано с действием посторонних нарушений.
Именно поэтому во влажном местообитании, подвергавшемся более силь-
ным нарушающим воздействиям, период полуотпада был в 3-6 раз короче,
чем в сухом местообитании (25-28 против 68-186 дней). Кривые выжива-
ния в первом случае можно было отнести ко II типу (Pearl, 1928), тогда как
во втором случае наблюдались гораздо более типичные для таких крупносе-
мянных видов кривые I типа.
35/
I'hiiki 10 Дгмо! рпфичссипй пинит природных iioiiyiiMiuili pat irimh
10.6. / 2 Ре п'чьтаты опыта с пересадкой ocoocii под почос пере юной рощи
Под пологом березовой рощи в местообитании, куда осуществляли не
ресадка особей недотроги обыкновенной в фазе двух семядолей, освещен
ность снизилась в период с 28 мая по 15 июля 1987 г. от 930 до 280 пк.
Пересаженные как с сухого (20 экз.), так и с влажного местообитания (20
экз.) особи хорошо (на 100%) прижились, хотя в дальнейшем и у тех и дру-
гих наблюдалось изреживание. Более интенсивно изреживались особи, пе-
ресаженные из сухого местообитания, и изреживание их началось раньше
(12-13.VI) в момент перехода в генеративное состояние. Бутонизировать
все особи начали почти одновременно и приступили к формированию
клейстогамных плодов.
28 мая особи, пересаженные из влажного местообитания, несколько усгу
пали по высоте (4-17 против 3-20) при одинаковом числе листьев на глав-
ной оси (3-6), но если у части особей, пересаженных из сухого местооби га-
ния, еще оставались семядоли, то у другой группы семядолей уже нс было.
В итоге к 15 июля численность выровнялась, но сохранились некоторые
преимущества у особей из сухого местообитания по высоте (10-25 прогни
14-29 см) и по числу листьев на главной оси (9-12 против 11-14).
Таким образом, можно еще раз заключить важность приобретения пре-
имуществ, и в первую
очередь, по высоте, на
самых ранних этапах,
после чего даже та-
кая «встряска» как пе-
ресадка в иное мес-
тообитание не способна
изменить статус особей,
демонстрируемый ими
на момент их переноса
в одинаковые условия.
В 1989 г. работа по
наблюдению за популя-
циями недотроги обык-
новенной велась по той
же схеме, что и в 1987
г., но, как уже было от-
мечено, из-за резко-
го снижения численно-
сти во влажном место-
обитании, была взята
другая популяция так-
же из влажного, но бо-
лее светлого местооби-
тания.
Поскольку в преде-
лах каждой из । шпуля-
Рис. 10.32. Динамика средней арифметической и но
явления бимодальности (выделена черным цветом) на
гистограммах распределения по классам bncoim (//) в
популяции недотроги обыкновенной: /' час гос и»; но
горизонтальной оси даты наблюдений.
358 Л/// Марков I l(>iiyiiMHii(>iiiin>i bnoiioi п>| рис 1СППЙ
ций недотроги обыкновенной наблюдения велись за особями двух локусов
— на двух пробных площадях, то сравнение было проведено не только меж-
ду популяциями в целом, но и между отдельными локусами. На рисунке
10.32 приведены гистограммы распределения по классам высоты растений,
построенные для популяций в целом в каждую дату наблюдений. Обращает
на себя внимание тот факт, что при очень близкой динамике среднего ариф-
метического высоты особей в двух популяциях характер варьирования по-
казателя довольно сильно отличается. Если во влажном местообитании, на-
чиная с 10 июня, кривые имели четкий максимум (модальный класс), то в
сухом местообитании уже со 2 июня (кривая выделена на рисунке) у кривых
появлялись признаки бимодальности на фоне более широкого (чем в первом
случае) размаха варьирования показателя.
Хотя ни первая (во влажном местообитании), ни вторая (в сухом местоо-
битании) популяции не были перезагущены (60-84 и 74-80 экз./м2, соответ-
ственно), налицо проявление асимметричности конкуренции (см. главу 8)
между особями за свет. Во влажном местообитании все особи находились
в условиях «симметрично» пониженной интенсивности света, и это мог-
ло снизить значение соревнования между ними. Вероятно если бы наибо-
лее мелкие особи в популяциях не могли осуществлять «подгон» (см. главу
8 и рис. 8.42), то эффект «асимметричности» конкуренции за свет выявился
бы еще ярче. Еще одним примером огромной значимости размеров семени,
семядольного индекса и сообразного этим показателям изначального стату-
са может служить динамика роста особей в высоту на четырех постоянных
квадратах — средние для квадратов величины дали траектории, которые ни
разу между собой не пересеклись, что говорит о поддержании изначально-
го status quo.
Отмирания особей на постоянных квадратах ни в одном из двух местооби-
таний не наблюдалось, что по-видимому, свидетельствовало о большей бла-
гоприятности метеорологических условий вегетационного сезона 1989 г.
Темпы роста особей недотроги обыкновенной летом 1989 г. были не-
сколько выше, чем в 1987 г. (RGR = 0,47-0,52 мг/мг.неделя), причем суще-
ственных различий между двумя популяциями зарегистрировать не удалось
ни по числу фитомеров главной оси, ни по высоте, ни по биомассе. Это при-
водит к выводу, что для недотроги обыкновенной при достаточной влаж-
ности почвы и воздуха основным фактором, определяющим темпы роста,
накопление биомассы и в итоге мощность репродукции, является свет.
Исследуемый нами вид, занимающий крайне узкую и специфическую
экологическую нишу в лесных ценозах, демонстрирует своеобразный ком-
плекс свойств. Наблюдения показали, что недотрога обыкновенная растет
только там, где присутствуют хотя бы незначительные нарушения сложив-
шегося под пологом леса травяного покрова. Режим этих нарушений может
зависеть от деятельности человека и некоторых вполне естественных факто-
ров, таких как временные потоки воды. Узость экологической ниши недотро-
ги (стенотопность) в большой степени обусловлена гигрофильностью этого
вида — в предпочитаемых ею сообществах увлажнение колеблется от влаж-
нолугового до болотного (от 68-й до 100-й ступени по шкале Л. Г. Раменского
(Раменский и др., 1956). Высокая влажность воздуха это фак'юр, лимитп-
Гтша 10 Д(-мо1 рафичсскнй пинит природных попуимипй рисiriinU 349
рующнй распространение недотроги обыкновенной Адвепiпвпый вид не-
дотрога мелкоцветковая отличается мспыисй i ш рофнныюс i ыо п, как cnci
ствие, более широкой экологической амплитудой, что позволяет ему успеш-
но конкурировать с недотрогой обыкновенной в местах контакта этих ви-
дов (Weise, 1968).
В 1989 г. проводились наблюдения и в популяции н. мелкоцветковой, рас-
положенной на краю оврага под пологом паркового липового леса в черте
г. Казани. Сочетая метод постоянных и временных площадок, удалось про-
следить за динамикой численности особей, ростом особей в высоту и нако-
плением биомассы у вида, который как по особенностям архитектуры осо-
бей, так и по многим другим важным биологическим показателям довольно
сходен с н. обыкновенной.
Для недотроги мелкоцветковой во всех исследованных популяционных
локусах зарегистрированы кривые выживания I типа со смертностью рез-
ко сдвинутой к концу сезона вегетации. Были в популяции и особи, кото-
рые не смогли участвовать в воспроизводстве популяции по причине силь-
ного угнетения.
Динамика средней и крайних величин общей биомассы как в по-
лулогарифмическом (In), так и в абсолютном масштабе дает картину очень
сходную не только с недотрогой обыкновенной, но и со многими другими
малолетними видами. Для каждой из семи выборок по данным об общей
биомассе особей высчитано значение коэффициента асимметрии распреде-
ления и коэффициента Джини. Оба коэффициента обнаруживают тенден-
цию к нарастанию во времени, но, в отличие от коэффициента асимметрии
распределения, траектория коэффициента Джини гораздо стабильнее и по-
тому ему следует отдать предпочтение в характеристике степени размерной
дифференциации.
Данные сравнения особей по высоте на самом раннем и на самом позд-
нем этапах убеждают еще раз в крайне важном значении первоначального
статуса особи в размерной иерархии для ее дальнейшего роста и развития.
Статус особей по высоте, этому важнейшему в условиях обостренной кон-
куренции за свет показателю, очень редко менялся в ходе роста, и пото-
му итоговая картина предъявляла усугубленные различия, характерные для
случаев асимметричной конкуренции.
Интересным представляется уже обсуждаемое выше большое сходство
статического (по данным 27 июля 1989 г.) и динамического аллокационных
спектров (рис. 9.17), поскольку это наглядный пример осуществления ал-
лометрической зависимости при слабых изменениях архитектуры особей в
ходе роста. Подобные примеры дают ценную возможность получения ди-
намической популяционной информации всего лишь по одной выборке из
популяции. Однако следует подчеркнуть, что речь в данном случае идсг о
последней, т. е. достаточно поздно взятой выборке, которая отражает ко-
нечный итог развития размерной внутрипопуляционной дифференциации.
Промежуточные же выборки тоже несут ценную информацию биологиче-
ского плана. Например, наблюдаемое снижение репродуктивного усилия но
мере возрастания общей биомассы особей, заметное на статическом алло-
кационном спектре для первой выборки (30.V. 1989) могло имен», по всей
360
MH Maphtxi I Ion у и ицноп i in и bnoiioi ihi рис i ci i nil
вероятности, только одно объяснение роль внешних условий в переходе
особей недотроги мел ко цветковой к репродукции очень велика. Иными сло-
вами, в какой-то определенный момент времени все особи популяции, нахо-
дящиеся выше известного предела мощности (общей биомассы) претерпе-
вают заложение генеративных структур. Именно поэтому в тот момент при-
мерно одинаковая и относительно небольшая биомасса заложившихся ор-
ганов репродукции имеет разный удельный вес у разнящихся по мощности
особей. В дальнейшем зависимость менялась на обратную, и к концу жиз-
ненного цикла мы уже наблюдали статистически достоверный положитель-
ный коэффициент корреляции между репродуктивным усилием и общей
биомассой или биомассой вегетативных органов. Это отступление от гораз-
до более типичной изометрии, вероятно, следует приписать большому эко-
логическому своеобразию малолетника, способного переносить затенение.
ЗАКЛЮЧЕНИЕ
В заключение можно еще раз резюмировать особенности растений как
объектов популяционной биологии (популяционной ботаники).
Модульность (метамерность) — одна из фундаментальных особенностей
растений, отличающая их от унитарных организмов — вводит уровень, на
котором, в дополнение к организменному, необходимо рассматривать про-
явление популяционных закономерностей. Без учета этого свойства, за-
дающего особую структуру и квантированность непрерывных, на первый
взгляд, процессов, представление о росте и развитии растений в составе по-
пуляций будет далеко не полным.
Прикрепленный образ жизни делает очертания, границы популяций рас-
тений более значимыми, влияя на все типы взаимодействий между особя-
ми, начиная с генетических (поток генов) и кончая взаимоблагоприятными
и конкурентными, обусловленными популяционной плотностью. Да и сама
плотность в популяциях растений как важнейший демографический фак-
тор, проявляется опять-таки на двух уровнях (модулей и организмов).
Несмотря на длительную историю изучения популяций в науке явственно
ощущается дефицит репрезентативных данных, полученных путем длитель-
ных скрупулезных количественных наблюдений, проверяемых или углубля-
емых экспериментами. Свидетельство тому — переходящие из одного учеб-
ного пособия в другое удачно уловленные и проиллюстрированные законо-
мерности, среди которых есть и такие, которые остро нуждаются в даль-
нейшей разработке. Помимо таких все еще далеких от окончательной раз-
работки и недоосмысленных концепций как концепция архитектурных ти-
пов и моделей, концепция жизненных форм и эколого-ценотических страте-
гий, можно назвать и обсужденную в пособии концепцию положительного
эффекта группы. Лишь чрезвычайная тонкость таких взаимовлияний между
растениями, ставящая результат на грань статистической достоверности, но
повторяющаяся — наблюдающаяся в популяциях разных жизненных форм
и в разных природных и искусственных ситуациях мешала до сих пор про-
должению исследований в этом направлении. И вновь специфика модульно-
го растительного организма требует чрезвычайной кропотливости и скрупу-
лезности от исследователя, суля взамен перспективы прогноза, делающею
занятие популяционной биологией растений теоретически и практически
значимой современной областью ботанической науки.
Кто-то по прочтении этого учебно-методического пособия наверняка
спросит, а где же здесь разговор об эволюции, вроде бы обещанный в глав-
ном эпиграфе. Действительно, в явной форме об эволюции, за исключением
одной только главы, в книге речи не было. Кто ж виноват, что все наше про
362
Л/.// Мирков I loiiyинцн<»i11ши G1101101 ин рис iciiiiM
движение биологической пауки в этом направлении было па протяжении
последних десятилетий более чем скромным, а некоторые известные иссле-
дователи за это время от одной своей книги к другой даже вынуждены были
коренным образом поменять свои воззрения на эволюцию (см. Чайковский,
2007,2008). Однако некоторые принципиальные моменты, связанные с раз-
личными формами изменчивости, а также признанием популяций, которы-
ми живут виды, ареной микроэволюционных процессов, никто пока не пе-
ресматривал. Потому, материалы, приведенные в этой книге, можно пола-
гать небесполезными для размышлений об эволюции, а испытанные мето-
ды — подлежащими усовершенствованию в аспекте статистической стро-
гости и применению к популяциям других не затронутых здесь видов рас-
тений.
СЛОВАРЬ
специальных терминов, употребляемых в популяционной
биологии растений (зарубежной и отечественной)
Акротония — ветвление в верхней части главного побега.
Аллокация — размещение биомассы или каких-либо ресурсов в преде-
лах тела растения. Распределение биомассы (и) или энергии между разны-
ми сферами растения.
Аллокация меристем — соотношение в пределах тела растения мери-
стем трех условно выделяемых типов: 1) покоящихся (постоянно в тече-
ние всей жизни растения или временно, пережидая стрессовую ситуацию);
2) формирующих вегетативные побеги («потраченных» на формирование
структур, которые создают ассимиляты и обеспечивают рост) и 3) формиру-
ющих репродуктивные структуры («потраченных» на формирование струк-
тур, которые потребляют ассимиляты).
Аллокационный спектр динамический — спектр, отражающий дина-
мику размещения и перераспределения биомассы между органами разных
сфер во времени.
Аллокационный спектр статический — спектр, отражающий своео-
бразие аллокации (размещения биомассы между органами разных сфер) у
особей разных размерных классов.
Аллометрия — изменение пропорций между разными структурами осо-
би при изменении (увеличении или уменьшении) общих размеров ее тела.
Аллометрический коэффициент — коэффициент при независимой
переменной в уравнении регрессии, описывающего зависимость изме-
нения какой-либо части (структуры) от целого. Может быть равен едини-
це (Изометрия), принимать значения больше единицы (положительная
Аллометрия) или меньше единицы (отрицательная Аллометрия).
Анемогеохорная дисперсия — дисперсия диаспор катящимся шароо-
бразным кустом — перекати-поле.
Аутбридинг (=ауткроссинг) — перекрестное опыление — опыление
между цветками разных особей.
Базитония — ветвление от основания главного побега.
Баллистохория — плод, плодик или семя в силу упругости подсох шеи
плодоножки «отстреливается» на некоторое расстояние.
Бипарентальность — участие в продукции семян двух родительских
растений.
Виоленты, патиенты и эксплеренты — три типа (фитоценотипа) рас-
тений по их поведению в условиях растительных сообществ (Раменский,
1935,1938), в соответствии Дж. Ф. Праймом (J. Р. Grime) выделены три стра-
364 МП Марков I loiiyiimiHoiiniui (>ионо1 нм pnc iciniit
тегии: конкурентную (competitors С), стресс-толерантную (strcss-tolera-
tors — S) и рудеральную (Ruderals R).
Генета — генетический индивид, особь семенного происхождения.
Геркогамия — разделение мужских и женских репродуктивных органов
в пространстве. В дополнение к бесспорно преобладающему гермафроди-
тизму цветков, в случае которого у растений как механизм разделения не-
редко выступает гетеростилия, встречаются однодомность в двух ее вари-
антах (андромоноэция и гиномоноэция) и двудомность (чаще в форме гино-
диэции).
Графы жизненных циклов — модели в виде петель (loopes) с обозначе-
нием вероятностей переходов особей из одного состояния (возрастного или
онтогенетического, жизненного) в другое.
Групповой отбор — это «процесс, при котором могут отбираться призна-
ки, благоприятные для группы, но не для особи».
Движущая форма отбора — форма отбора с направленным изменени-
ем значения показателя (признака), проявляющимся в смещении моды рас-
пределения.
Джендер или гендер (gender) — относительный вклад в приспособлен-
ность через продукцию семязачатков (функцию материнства) и через про-
дукцию пыльцы (функцию отцовства).
Дихогамия — разделение во времени мужской и женской репродуктив-
ной активности растения.
Дрейф генов — случайное вытеснение одного селекционное нейтраль-
ного аллеля другим (другое название — генетико-автоматические про-
цессы).
Емкость местообитания (carrying capacity) — способность местооби-
тания поддерживать определенную максимальную численность популяции
(К) данного организма, по достижении которой прекращается ее дальней-
ший рост.
Жизненная форма — внешний облик (габитус, совокупность морфоло-
гических черт) растения, формирующийся в онтогенезе и отражающий ре-
зультат приспособления к определенному комплексу экологических усло-
вий.
Изометрия — сохранение постоянных пропорций между разными струк-
турами особи [или между частью (специализированной структурой) и це-
лым] при изменении (увеличении или уменьшении) общих размеров тела
— фактически — альтернатива Аллометрии (см.) или частный ее случай,
когда Аллометрический коэффициент равен единице.
Климакс — завершающая и наиболее устойчивая из конечных сукцесси-
онных стадий развития растительного покрова на какой-либо территории.
Контагиозность — пятнистость в размещении особей на площади
Корреляционная структура — совокупность корреляционных связей
между количественными признаками особей в популяции, которую чаще
всего графически представляют в виде корреляционного цилиндра.
Менделевские или панмиктические популяции — популяции, между
особями которых наблюдается свободное равновероятное скрещивание.
Метамер — повторяющийся однотипный элемент в геле рас i сипя.
( i(Hhiph t Kh'pyffiHtHt JQ5
Mei имериЗинии гели нодр<1здслс11пс icna p;icicnii>i н-i меымсры pa i
пого ранга, носящее, как правило, характер соподчинении (иерархии).
Модуль — мегамер определенного ража, продут дем leniaiociii одном
верхушечной меристемы, представляющий собой ось, несущую узлы с ли-
стьями, пазушными почками, междоузлиями, (и, возможно, корнями
М. М.) и увенчанную завершающуюся соцветием или вегетативными
структурами.
Моноцентрические виды — вегетативно неподвижные и малоподвиж-
ные виды: стержнекорневые, луковичные, клубневые травы, большинство
деревьев
Морфобиологическая внутривидовая изменчивость — разнообразие
особей одного вида по ряду морфологических и/или биологических при-
знаков, свойств.
Неявнополицентрические виды — виды, занимающие промежуточное
положение между моно- и явнополицентрическими видами, поскольку име-
ют несколько слабо различимых центров сосредоточения корней, побегов и
почек возобновления. В качестве счетной единицы только на первых этапах
онтогенеза до начала вегетативного размножения выступает особь, а впо-
следствии — партикула. Примеры неявнополицентрического типа биоморф
можно найти среди короткокорневищных и кистекорневых трав, ряда дре-
весных растений
Популяционный подход в экологии — концентрирует основное вни-
мание на популяциях, населяющих определенную территорию (или аквато-
рию), и в отличие от экосистемного подхода (см.) более склонен к анали-
тическому объяснению, что отражается и в определении экологии, которое
дал с позиций популяционного подхода канадский исследователь Ч. Кребс:
экология — это научное познание взаимодействий, определяющих распро-
странение и численность организмов.
Оседлость (седиментарность) — прикрепленный образ жизни, свой-
ственный огромному большинству растений, связанный с извлечением ре-
сурсов из окружающего пространства, до которого «дотягиваются», дорас-
тают органы поглощения.
«Партизан» — (guerilla) — образное название экспансивного характера
роста (разрастания) особи, связанное с формированием протяженных ком-
муникаций, отдаляющих друг от друга дочерние структуры, в зарубежной
литературе (см. Lovett-Doust, 1981).
«Перекрестники» — растения с той или иной степенью самонесовме-
спгимости (см), опыляющимися всегда или преимущественно перекрест-
ным путем)
Петрофильность — приуроченность (связанная с предпочтением) видов
растений к каменистым субстратам, например, мергелям с известняковы-
ми плитками
Поливариантность развития (или поведения) — реализация разных
вариантов онтогенеза особями популяции (популяций) одного вида.
Популяционный локус — фрагмент популяции с более или менее отчет-
ливым контуром и какой-то характерной плотностью.
366 МН Мч[ >К(М1 I I <) 11УI HI 11 Н< >1II III >1 (HI0H0I 114 рис 1СННЙ
«Правило Сукачева» (К.М.Завадский, 1947) ускорение развития од-
нолетними и замедление — многолетними растениями в неблагоприятных
экологических условиях.
Приспособленность — количественная мера относительного превосход-
ства одного генотипа перед другим или одного вида перед другим.
Простой индивид — физически непрерывное образование, у которого
морфологическая целостность преобладает над автономностью частей и от-
сутствует ясно выраженная морфологическая дезинтеграция.
Псевдооднолетники — многолетние растения, активно вегетирующие
только летом и переносящие зимовку в виде мелких клубеньков как вегета-
тивныхетруктур обеспечивающих многолетность (переннизацию).
Рамета — потомок вегетативного происхождения на разных (и иногда
так и не реализуемых) стадиях отделения от материнского растения.
Регенерационная ниша — (термин Р. J. Grubb, 1977) локализованный в
пространстве комплекс условий, который наиболее подходит для прораста-
ния семян и приживания проростков данного вида растения.
Репродуктивное усилие — доля от общей энергии или биомассы расте-
ния, направляемая на формирование репродуктивных структур.
Симпатричность — формирование нового вида «в недрах» популяции
материнского распадающегося вида.
Сложный индивид — физически непрерывное образование, у которо-
го автономность частей преобладает над морфологической целостностью
(хотя бы во взрослом состоянии) и явно выражена морфологическая дезин-
теграция.
Стабилизирующая форма отбора — по И. И. Шмальгаузену (1945) —
сохранение стабильности «нормы» благодаря отсеканию (элиминации) из-
лишне сильных отклонений от нее как в области «головы», так и в области
«хвоста» кривой распределения по классам признака
Стратегия жизни (=эколого-фитоценотическая стратегия, страте-
гия жизненного цикла) — совокупность приспособлений к произраста-
нию данного вида в определенных условиях среды в сообществе с другими
видами растений
Счетная единица — в большей или меньшей степени самостоятельный
и укорененный фрагмент особи, который учитывается при оценке воздей-
ствия на среду обитания и соседние особи.
Топоспория — преимущественное осыпание (и, иногда, с механизмом
закрепления на субстрате) диаспор в ходе первичной дисперсии вблизи ма-
теринской особи
Унипарентальность — участие в продукции семян одного родительско-
го растения (например, при самоопылении или апомиксисе).
Уподобление и дифференциация — два альтернативных пути приспо-
собления к произрастанию в агроценозах
«Фаланга» — (phalanx) — образное название дерновинного характера
роста (разрастания) особи, связанное с формированием плотного куста.
«Ценозная популяция» (ценотическая популяция) или «ценопопуля-
ция» — термин употреблявшийся ранее В. В. Петровским (1961). В послед-
ние годы жизни Т. А. Работнов отдавал предпочтение термину «фитоцепо-
J()/
< 'чот/рь ( терминов
тическай популяция» совокупное*!!» особей одного вида в i рапидах фи
тоцепоза.
Чистая скорость размножения — скорость увеличения численноеiи
популяции за время жизни одного поколения называют
Экосистемный подход в экологии — ставит во главу угла ионятис эко-
системы — совокупности организмов и неживых компонентов, связанных
потоками вещества и энергии.
Экотипическая дифференциация популяций — формирование раз-
ными популяциями одного вида в экологически существенно различаю-
щихся местообитаниях экологических рас (экотипов — по терминологии
Турессона).
Элементарная демографическая единица (ЭДЕ) — структурная еди-
ница (и одновременно система) популяционного уровня, наиболее важным
демографическим признаком которой является обеспечение непрерывного
оборота поколений. Размер ЭДЕ определяется площадью, в пределах кото-
рой с вероятностью не ниже 0,8-1,0 будут представлены практически все
онтогенетические группы особей, необходимые для оборота поколений.
Эмерджентные свойства — свойства целого, не сводимые к сумме
свойств его отдельных частей.
«Эффект Джека Спрата» — дифференциация экологических ниш, свой-
ственных особям разного пола, у двудомных (=диэцичных) растений.
Jack Sprat could eat no fat,
His wife could eat no lean,
And so between the two of them,
They licked the platter clean!
Джек Спрат не ел жиров,
Его жена — бобов,
Так что вдвоем они всегда
Тарелку чистили до дна.
«Эффект бутылочного горлышка», и то, что мы теперь вслед за Эрнстом
Майром называем «принципом основателя», вытекающие из четвери-
ковских «волн жизни», генетико-автоматических процессов Дубинина-
Ромашова и из «дрейфа генов» Фишера-Райта-Холдейна отклонения в гене-
тической структуре популяции..
Эффект Сукачева (по Harper, 1977) — более высокая (более низкая)
смертность особей в популяции из-за внутривидовых конкурентных отно-
шений, складывающихся на фоне повышенного (пониженного) плодородия
субстрата.
Эффективная генетическая единица — эффективное число репроду-
цирующих растений, расположенных в пределах площади одного генетиче-
ского соседства
Явнополицентрические виды — для особей характерно наличие не-
скольких четко выраженных центров сосредоточения корней, побегов и по-
чек возобновления, соединенных между собой протяженными коммуника-
циями, которые могут находиться как под землей (гипогсогенные корневи-
ща, подземные столоны), так и на ее поверхности (столоны, плети).
ЛИТЕРАТУРА
Аверьянова Т.М. 1975. Популяционные исследования в прикладной ботанике. Л.:
Наука. 140 с.
Агаев М.Г. 1972. Реагирование однолетних растений на повышение плотности по-
пуляций // Бот. журн. Т.57. № 5. С.434-445.
Агаев М.Г. 1978. Онтогенетическое реагирование однолетних растений на популя-
ционную плотность // Бот. журн. Т.63. № 2. С.244-249.
Агаев М.Г. 1980. Популяционно-онтогенетическая двойственность однолетних
растений //Докл. АН СССР. Т.250. № 6. С.1508-1512.
Алеев Ю.Г. 1986. Экоморфология. Киев: Паукова думка. 423 с.
Амирханов Н., Хамракулов Ш. 1986. Биология видов рода катран и их хозяйствен-
ное использование в Узбекистане. Ташкент. 115 с.
Барыкина Р.П., Туленкова М.А. 1983. Элементарный метамер побега цветкового
растения // Бюл. Моск, о-ва испыт. прир. Отд. биол. Т.88. Вып. 4. С. И 4-124.
Бердников В.А. 1991. Эволюция и прогресс. Новосибирск: Наука. 192 с.
Бигон М., Харпер Дж., Таунсенд К. 1989. Экология: особи, популяции и сообще-
ства. М.: Мир. Т.1. 667 с.
Васильченко И.Т. 1948. Неотения у цветковых растений // Тр. Бот. ин-та им. В.Л.
Комарова АН СССР. Флора и систематика высших растений. Сер.1. Вып.7.
С. 149-162.
Визнер Ю. 1892. Биология растений. СПб.: Издание А.Ф. Девриена. 293 с.
Войтенко В.Ф. 1968. Формы гетерокарпии в семействе Brassicaceae Bum. и их эво-
люционная оценка // Бот. журн. Т.53. № 10. С.1428-1439.
Войтенко В.Ф. 1974. Основные черты дисперсии гетерокарпных крестоцветных //
Вопросы биологии семенного размножения. Ульяновск: Изд-во Ульяновск, пед.
ин-та. С.67- 90.
Воронцова Л.И., Заугольнова Л.Б. 1978. Мультивариантность развития особей в те-
чение онтогенеза и ее значение в регуляции численности и состава ценопопуля-
ций растений // Журн. общ. биол. Т.39. № 4. С.555-562.
Галл Я.М. 1976. Борьба за существование как фактор эволюции. Л.: Наука. 155 с.
Гатцук Л.Е. 1974а. Геммаксиллярные растения и система соподчиненных единиц
их побегового тела // Бюл. Моск, о-ва испыт. прир. Отд. биол. Т.79. Вып. 1. С. 100-
113.
Гатцук Л.Е. 19746. К методам описания и определения жизненных форм в сезон-
ном климате // Бюл. Моск, о-ва испыт. прир. Отд. биол. Т.79. Вып.З. С.84-100.
Генкель П.А., Абдиева Р.Г. 1971.0 жароустойчивости гемиксерофитов // Физиология
засухоустойчивости растений. М. С.228-246.
Гиляров А.М. 1990. Популяционная экология. М: Изд-во МГУ. 191с.
Голубев В.Н. 1962. Основы биоморфологии травянистых растений центральной ле-
состепи // Тр. Центр. Черноземного гос. заповедника им В.В. Алехина. Воронеж.
Вып.7. 511с.
Голубев В.Н. 1965. Эколого-биологические особенности травянистых растений и
растительных сообществ лесостепи. М. 286 с.
Голубев В.Н. 1968. Об изучении жизненных форм растений дня целей фитоценоло-
гии // Бот. журн. Т.53. X. С. 10X5 1093.
Литература
Голубей В.11. 1971. Морфологические при знаки paciciiiin как ипдикаюры И hop
вопросы (|)и гоиндикацип. Л.: Наука. (’. В7 М2.
Гордягин А.Я. 1892. Несколько ботанических данных о семенах лебеды И Дпсви
о-ва врачей при Казанск. университете. Казань. Вып.2. С.81 96.
Гордягин А.Я. 1907. Биометрические исследования над Chrysanthemum si hi пент
(DC.) // Тр. о-ва естествоиспыт. при Казанск. ун-те. Казань. Т.40. Вын.5. С. I 41.
Гордягин А.Я. 1920. Наблюдения над изменчивостью Anemone patens L. II Тр. о-ва
естествоиспыт. при Казанск. университете. Казань. Т.49. Вып.5. С. 1 88.
Грант В. 1984. Видообразование у растений. М. 528 с.
Доброхотов В.Н. 1961. Семена сорных растений. М. 414 с.
Дубинин Н.П., Ромашов Д.Д. 1932. Генетическое строение вида и его эволюция И
Биол. журн. Т.1. Вып. 1-6. С.52-95.
Евстигнеев О.Е. 2004. Популяционные стратегии видов деревьев // Восточно-
европейские леса. С. 176-205.
Евстигнеев О.Е., Диденко Н.Г. 2004. Популяционные стратегии видов кустарников
// Восточно-европейские леса. С.205-224.
Жмылев П.Ю., Алексеев Ю.Е., Карпухина Е.А., Баландин С.А. Биоморфология рас-
тений: иллюстрированный словарь. М.: 2002. 240 с.
Жукова Л.А., Смирнова О.В. 1988. Элементы популяций и их дифференциация //
Ценопопуляции растений. М.: Наука. С. 13-33.
Жукова Л.А., Заугольнова Л.Б., Смирнова О.В. 1991. Развитие концепции А.А.
Уранова в популяционной экологии растений // Популяции растений: принципы
организации и проблемы охраны природы. Йошкар-Ола. С.3-6.
Завадский К.М. 1954. О причинах выпадения растений в гнездовых посевах // Бог.
журн. Т.39. № 4. С.515-544.
Завадский К.М. 1957. Перенаселение и его роль в эволюции // Бот. журн. Т.42. №3.
С.426-449.
Завадский К.М. 1968. Вид и видообразование. Л.: Наука. 404 с.
Заугольнова Л.Б. 1985. Оценка степени динамичности ценопопуляций растений в
пределах одного фитоценоза // Динамика ценопопуляций растений. М. С.45-62.
Заугольнова Л.Б. 1986. Сбор материала в слабосомкнутых травяных сообществах.
Методика обработки материала // Изучение структуры и взаимоотношений по-
пуляций (Методические разработки для студентов биологических специально-
стей). М. С.22-27.
Заугольнова Л.Б. 1988. Функциональная структура. Внутрипопуляционные взаи-
модействия // Ценопопуляции растений (очерки популяционной биологии). М.:
Наука. С.90-102.
Заугольнова Л.Б., Жукова Л.А., Попадюк Р.В., Смирнова О.В. 1992. Критическое
состояние ценопопуляций растений // Проблемы устойчивости биологических
систем. М. Наука. С.51-59.
Злобин Ю.А. 1980. О неравноценности особей в ценопопуляциях растений И Бот.
журн. Т.65. №3. С.311-322.
Злобин Ю.А. 1989. Принципы и методы изучения ценотических популяций расте-
ний. Казань. 146 с.
Идрисова Г.И., Марков М.В. 1999. Сопоставление двух географически изолиро-
ванных природных популяций рябчика шахматовидного методами биосистема-
тики // Уч. зап. Тверского гос. университета. Т.5. Тверь. С.185-191.
Иоганнсен В.Л. 1935. О наследовании в популяциях и чистых линиях. М.-Л.: Огиз-
Сельхозгиз. 77 с.
Ипатов В.С. 1969. Дифференциация древостоя. 2. Выявление деформаций у кривых
распределения деревьев по толщине // Вести. Л1 У. Сер. биол. № 15. С.43 53.
370 МН Мары hi 11»>i t у 11 >i i ii н >i 11111 >t (inoiioi ин piu iciinh
Казакевич Л.И. 1922. Материалы к биологии растений К )ю Восюка России // Бюл.
отд. прикл. ботан. Саратовск. обл. с-х. оиытн. стаиции. 1К. I 21.
Келлер Б.А. 1948. Основы эволюции растений. М.-Л. 207 с.
Кияк В.Г. 1984. К методике картирования особей ценопопуляций растений в био-
геоценологических исследованиях // Биогеоценологические исследования на
Украине. Львов. С. 125-126.
Колев И.Д. 1960. Неотения у сорных растений // Вестник с-х. науки. №9. С. ИО-
152.
Корона В.В. 1987. Основы структурного анализа в морфологии растений.
Свердловск. 269 с.
Корчагин А.А. 1964. Внутривидовой (популяционный) состав растительных сооб-
ществ // Полевая геоботаника. Т.З. С.63-131.
Кренке Н.П. 1933-1935. Феногенетическая изменчивость. Сборник работ. Т.1. Изд.
Биол. ин-та им. Тимирязева. М. 860 с.
Кренке Н.П. 1940. Теория циклического старения и омоложения растений. М. 136 с.
Крокер В. 1950. Рост растений. М. 359 с.
Крокер В., Бартон Л. 1955. Физиология семян. М. 399 с.
Куркин К.А. 1973. Эколого-ценотический режим замкнутости луговых биогеоцено-
зов // Проблемы биогеоценологии. М.: Наука. С. 137-147.
Кучеров Е. В. 1979. Ресурсы и интродукция полезных растений в Башкирии. М.:
Наука. 264 с.
Левин Г.Г. 1961. Проблемы индивидуальности у растений // Бот. журн. Т.46. № 3,
С.432^147.
Леман Е. Айхеле Ф. 1936. Физиология прорастания семян злаков. М.-Л.: Сельхозгиз.
483 с.
Левина РЕ. 1981. Репродуктивная биология семенных растений. М. 96 с.
Левина РЕ. 1987. Морфология и экология плодов. Л.: Наука. 160 с.
Любич Ф.П. 1950. Гетерокарпия некоторых растений, ее биологическое и практи-
ческое значение // Сборник материалов по флоре и растительности юго-востока.
Саратов. С.3-35.
Магомедмирзаев М.М. 1990. Введение в количественную морфогенетику. М.:
Наука. 229 с.
Мазинг В .В. 1973. Что такое структура биогеоценоза? // Проблемы биогеоценологии.
М.: Наука. С. 148-157.
Майр Э. 1974. Популяции, виды и эволюция. М.: Мир. 460 с.
Макрушин Н.М. 1989. Основы гетеросперматологии. М.: Агропромиздат. 287 с.
Максимов Н.А. 1958. Краткий курс физиологии растений. М. 559 с.
Мальцев А.И. 1923. Несколько слов о катране Crambe tatarica Jacq. // Тр. по прикл.
бот., генет. и селекции. Т.13. Вып.З.
Марков М.В. 1976а. О направлении, развиваемом Джоном Л. Харпером в популяци-
онной экологии растений // Бот. журн. Т.61. № 7. С.999-1004.
Марков М.В. 19766. Проявление пластичности и состав популяций пастушьей сум-
ки в посевах разных полевых культур // Бюл. Моск, о-ва испыт. прир. Отд. биол.
Т.81.Вып.4. С. 118-123.
Марков М.В. 1978. Пастушья сумка обыкновенная // Биологическая флора
Московской области. М. Вып.4. С.86-95.
Марков М.В. 1980. Популяционный состав зимующих однолетников и его динами-
ка в разных агрофитоценозах // Проблемы агрогеоботаники. Ижевск. С.57-67.
Марков М.В. 1986. Популяционная биология растений (учебно-методическое посо-
бие). Казань. 108 с.
Литература J/l
Марков М.'В. 19X7. Популяционная биология uopoi коживущих монокарпических
растений // Биол. науки. №» X. ( '. 39 46.
Марков М.В. 1989. К биологии катрана пиарского Crambe tatami Sebeok.//Inon.
Моск, о-ва испыт. прир. Отд. биол. Т.94. Вын.З. С.65 74.
Марков М.В. 1990. Популяционная биология розеточных и иол у розеточных мало-
летних растений. Казань: Изд-во Казанского ун-та. 178 с.
Марков М.В. 1991а. Популяционная биология недотроги обыкновенной Impatiens
noli-tangere L. II Экология. № 1. С. 17-26.
Марков М.В. 19916. Жизненный цикл двулетних растений с общебиологических
позиций // Бюл. Моск, о-ва испыт. прир. Отд. биол. Т.96. Вып.4. С.51-67.
Марков М.В. 1992. Роль многоорешков в обеспечении пластичности репродуктив-
ной сферы у Ceratocephala falcata (L.)Pers. и Myosurus minimus L. И Бюл. Моск,
о-ва испыт. прир. Отд. биол. Т.97. Вып.1. С.81-88.
Марков М.В. 2001. Особенности взаимодействия активной и пассивной частей по-
пуляции у некоторых жизненных форм цветковых растений И Экология. № 3.
С.338-345.
Марков М.В., Идрисова Г.И. 1996. Поливариантность развития и разногодичного
ритма активности особей Fritillaria meleagroides в условиях интродукции // Флора
и растительность Тверской области. Тверь: Изд-во Тверского ун-та. С.55-61.
Марков М.В., Идрисова Г.И. 1998. Сравнение при помощи корреляционного и дис-
криминантного анализа двух изолированных популяций Fritillaria meleagroides
Patrin ex Schult, et Schult. Fil И Жизнь популяций в гетерогенной среде. Часть 1.
Йошкар-Ола: Периодика Марий Эл, С. 141-143.
Марков М.В., Плещинская Е.Н. 1987. Репродуктивное усилие у растений И Журн.
общ. биол. Т.68. № 1. С.77-83.
Маркова С.А. 1971. Влияние фитоценотической среды на рост и развитие желтуш-
ника левкойного Erysimum cheiranthoides L. И Бюл. Моск, о-ва испыт. прир. Отд.
биол. Т.76. № 5. С.86-95.
Мельникова Т.М. 1972. Биологические особенности прорастания семян некоторых
травянистых растений И Бюл. Главн. Бот. сада. Вып.85.
Милторп Ф. 1964. Природа и анализ конкуренции между растениями разных видов
// Механизмы биологической конкуренции. М.: Мир. С.413-443.
Миркин Б.М. 1981. Рецензия на книгу: Грайм Дж. Стратегии растений и процессы
в растительности (1979) И Журн. Общ. биол. Т.42. № 4. С.628-631.
Миркин Б.М. 1983. О типах эколого-ценотических стратегий у растений //Журн.
Общ. биол. Т.44. № 5. С.603-613.
Миркин Б.М. 1985. Теоретические основы современной фитоценологии. М. 136 с.
Муравьев С.А. 1973. Стеблеотбор в злаковом фитоценозе. Рига. 73 с.
Нечаев А.П., Гапека З.И. 1970. Эфемеры меженной полосы берегов нижнего Амура
И Бот. журн. Т.55. № 8. С.1127-1137.
Нечаева Н.Т. 1958. Динамика растительности Кара-Кумов под влиянием метеороло-
гических условий. Ашхабад. 214 с.
Нечаева Н.Т., Василевская В.К., Антонова К.Г. 1969. Экологическая классификация
однолетних растений Каракумов И Бот. журн. Т.54. №11. С.1689-1704.
Николаева М.Г. 1967. Физиология глубокого покоя семян. JL: Наука. 203 с.
Николаева М.Г. 1982. Покой семян и фак горы его контрол и рующие//Физиология и
биохимия покоя и прорастания семян. М. С.72 96.
Нухимовский Е.Л. 1997. Основы биоморфологии семенных растений. М. 630 с.
Овчаров К.Е. 1969. Физиологические основы всхожее ш семян. М.: Наука. 279 с.
Павлов Н.В. 1948. Ботаническая география (’(’( Р. Алма-Ain.
372 МН Маркой I loiiyininnoiiiinH (nioiioi ия рас 1ГННЙ
Петров В.В. 1989. Банк семян в почвах лесных фичоценолов европейской части
СССР. М. 176 с.
Петровский В.В. 1961. Синузия как форма сосуществования растений // Бот. журн.
Т.46. № 11. С. 1615-1626.
Пианка Э. 1981. Эволюционная экология. М. 399 с.
Плотников В.В. 1979. Эволюция структуры растительных сообществ. М.: Наука.
275 с.
Понятовская В.М. 1951. К вопросу о семенном возобновлении растений в степных
фитоценозах // Тр. Бот. ин-та АН СССР. Сер.З. Геоботаника. № 7. С.7-22.
Попов М.Г. 1948. Род Erigeron в горах Средней Азии И Труды Бот. ин-та АН СССР.
Сер.1. Вып.7. С.7-44.
Работнов ТА. 1950а. Вопросы изучения состава популяций для целей фитоценоло-
гии // Проблемы ботаники. Вып.1. С. 465-483.
Работнов ТА. 19506. Жизненный цикл многолетних травянистых растений в лу-
говых ценозах // Тр. Бот. ин-та АН СССР. Л. Сер.З. Геоботаника. № 6. С.7-204.
Работнов ТА. 1951. К методике наблюдения над травянистыми растениями на по-
стоянных площадках // Бот. жур. Т.36. № 6. С.643-645.
Работнов ТА. 1964. Некоторые проблемы изучения ценотических популяций //Бюл.
Моск, о-ва испыт. прир. Отд. Биол. Т.69. Вып.6. С.47-55.
Работнов ТА. 1969. О биологии монокарпических многолетних растений И Бюл.
Моск, о-ваиспыт. прир. Отд. биол. Т.74. Вып.1. С. 141-149.
Работнов ТА. 1975. Изучение ценотических популяций в целях выяснения «стра-
тегии жизни» видов растений // Бюл. Моск, о-ва испыт. прир. Отд. биол. Т.80. №
2. С.5-17.
Работнов ТА. 1981. Жизнеспособные семена в составе ценотических популяций
как показатель стратегии жизни видов растений // Бюл. Моск, о-ва испыт. прир.
Отд. биол. Т.86. № 3. С.68-78.
Работнов Т.А.1984. Луговедение. М. 319 с.
Работнов Т.А. 1986. Рец.: Нахуцришвили Г.Ш., Гамцемлидзе З.Г. Жизнь растений в
экстремальных условиях высокогорий И Экология. № 4. С.91-92.
Раменский Л.Г. 1935. О принципиальных установках, основных понятиях и терми-
нах производственной типологии земель, геоботаники и экологии И Сов. ботани-
ка. № 4. С.25-42.
Раменский Л.Г. 1938. Введение в комплексное почвенно-геоботаническое исследо-
вание земель. М. 620 с.
Раменский Л.Г, Цаценкин И.А., Чижиков О.Н., Антипин Н.А. 1956. Экологическая
оценка кормовых угодий по растительному покрову. М.: Сельхозгиз. 472 с.
Рогаченко А.Д. 1988. Ценотическое взаимодействие растений Zea mays И
Перспективы теории фитоценологии. Тарту. С. 98-104.
Розанова М.А. 1946. Экспериментальные основы систематики растений. М.; Л.
Изд-во АН СССР. 254 с.
Романовский Ю.Э. Современное состояние концепции стратегии жизненного цик-
ла // Биол. науки. 1989. № 11. С. 18-31.
Рытова Н.Г. 1967. Некоторые закономерности роста листьев и вегетативных по-
бегов у злаков // Бот. журнал. Т.52. № 2. С.249-256.
Сабинин Д.А. 1940. Минеральное питание растений. М. 307 с.
Сабинин Д.А. 1957. О ритмичности строения и роста растений И Бот. журн. Т.42.
№7. С.991-1010.
Сабинин Д.А. 1963. Физиология развития растений. М. 196 с.
Седова Е.А. 1989. Органогенный потенциал апикальных меристем цветковых рас-
тений и продуктивность // Вести. Моск, ун-та. Сер. 16, биология. 4. С.25 33.
Литература J / J
Семсптовскнй В.11. 1951. Тектоника Ta lapilli в связи с рсиьсфом // 1\и1.сф laiapmi
Казань. С.23 50.
Серебряков И.Г. 1952. Морфология вегсгагивных органов высших расiciinii. М. 391 с
Серебряков И. Г. 1964. Жизненные формы высших растений и их изучение //
Полевая геоботаника. Вып.З. М.- Л. С. 146-205.
Серебрякова Т.Н. 1971. Морфогенез побегов и эволюция жизненных форм у злаков.
М.: Наука. 359 с.
Серебрякова Т.Н. 1977. Об основных «архитектурных моделях» травянистых
многолетников и модусах их преобразования И Бюл. Моск, о-ва испыт. прир.
Отд. биол. Т.82. № 5. С. 112-128.
Серебрякова Т.Н. 1981. Жизненные формы и модели побегообразования наземно-
ползучих многолетних трав И Жизненные формы: структура, спектры и эволю-
ция. М. С.161-179.
Синская Е.Н. 1948. Динамика вида. М.; Л.: ОГИЗ-Сельхозгиз. 525 с.
Смелов С.П. 1947. Биологические основы луговодства. М.: Сельхозгиз. 230 с.
Смирнов А.Г. 1982. Женский гаметофит покрытосеменных и его эволюция. Казань.
120 с.
Смирнова О.В. 1985. Динамика ценопопуляций травянистых растений ши-
роколиственных лесов европейской части СССР // Динамика ценопопуляций
растений. М. С.23-36.
Смирнова О.В., Заугольнова Л.Б., Торопова Н.А., Фаликов Л.Д. 1976. Критерии вы-
деления возрастных состояний и особенности хода онтогенеза у растений раз-
ных биоморф // Ценопопуляций растений (основные понятия и структура). М.:
Наука. С. 14-44.
Солбриг О.Т., Солбриг Д. 1982. Популяционная биология и эволюция. М.:Мир. 488 с.
Спрыгин И.И. 1934. Выходы пород татарского яруса пермской системы в Заволжье
как один из центров видообразования в группе калькофильных растений // Сов.
ботаника. № 4. С.61-74.
Суворова Т.Н. 1961. Типы побегов у злаков И Бот. журн. Т.46. № 2. С.208-211.
Сукачев В.Н. 1928. Растительные сообщества (Введение в фитосоциологию). М.-Л.
(Избр. Труды. 1975. ТЗ. С.42-93.)
Сукачев В.Н. 1935. Опыт экспериментального изучения межбиотипной борьбы за
существование у растений И Труды Петергофск. биол. ин-та. № 15. С.69-86.
Сукачев В.Н. 1941. О влиянии интенсивности борьбы за существование между рас-
тениями на их развитие. (Избр. Труды. 1975. Т.З. С.334-337.)
Тимофеев-Ресовский Н.В., Яблоков А.В., Глотов Н.В. 1973. Очерк учения о попу-
ляции. М.: Наука. 277 с.
Титов Ю.В. 1978. Эффект группы у растений. Л. 151 с.
Третьяков Н.В., Горский А.В., Самойлович ГГ. 1952. Справочник таксатора. М.; Л.:
Гослесбумиздат. 853 с.
Удра И.Ф. 1988 Расселение растений и вопросы палео- и биогеографии. Киев: Наук.
Думка. 1988.195 с.
Уиттекер Р. 1980. Сообщества и экосистемы. М. 327 с.
Уранов А.А. 1965. Фитогенное поле И Проблемы современной ботаники. Т.1.
С.251-254.
Уранов А.А. 1977. Вопросы изучения структуры фитоценоза и видовых ценопопу-
ляций // Ценопопуляций растений. Развитие и взаимоотношения. М.
Филипченко Ю.А. 1929. Изменчивость и методы ее изучения. 4-е изд. М.-Л.:
Госиздат. 275 с.
Флеров В.А. 1958. Возобновление органов у двусеменодольных растений // Уч. зап.
Ростовского-на-Дону гос. ун-та. Т.38. Вып.З. С.71 I К).
374 МН Маркин I lonyum пк ниши (hioiioi пн pm icmiii
Харпер Дж. 1964. Некоторые подходы к изучению конкуренции у paciciinii //
Механизмы биологической конкуренции. М.: Мир. (.6 24.
Хильми Г.Ф. 1957. Теоретическая биогеофизика леса. М.: Изд. АН СССР. 266 с.
Хитрово В.Н. 1912. О парусности зачатков полевых сорняков различных горизон-
тов // Тр. Бюро по прикл. бот. СПб. Т.5.
Хохряков А.П. 1974. О некоторых закономерностях эволюции цветка // Проблемы
филогении высших растений. М. С. 163-175.
Хохряков А.П. 1975. Закономерности эволюции растений. Новосибирск: Наука. 202
с.
Хохряков А.П. 1982. Уровни полимеризации в эволюции растений // Докл. АН
СССР. Сер. биол. № 5. С.722-737.
Цвелев Н.Н. 2000. Зародыши семян покрытосеменных растений (Magnoliophyta)
как первичные фитомеры И Бюл. Моск, о-ва испыт. прир. Отд. биол. Т.5. Вып.4.
С.61-65.
Ценопопуляции растений (основные понятия и структура). 1976. М.: Наука. 217 с.
Цингер Н.В. 1928. О подвидах большого погремка (Alectorolophus major). Вологда.
97 с.
Чайковский Ю.В. 1990. Элементы эволюционной диатропики. М.: Наука. 272 с.
Чайковский Ю.В. 1993. К общей теории эволюции // Путь. № 4. С. 101-114.
Шафранова Л.М. 1980. О метамерности и метамерах у растений // Журн. общ. биол.
Т.41. №3. С.437-447.
Шафранова Л.М. 1981. Ветвление растений: процесс и результат И Жизненные фор-
мы: структура, спектры и эволюция. М. С. 179-212.
Шилов И.А. Экология. 1996. 432 с.
Шульгин И.А. 1963. Морфофизиологические приспособления растений к свету
(оптические свойства листьев). М.: Изд-во МГУ. 74 с.
Шулькина ТВ. 1988. Архитектурные модели в семействе Campanulaceae s. str., их
география и возможные пути преобразования // Бот. журн. Т73. № 1. С.3-15.
Юрцев Б.А. 1976. Жизненные формы: один из узловых объектов ботаники //
Проблемы экологической морфологии растений. М. С.9-54.
Яблоков А.В. 1987. Популяционная биология. М.: Высшая школа. 303 с.
Abrahamson W.G., Caswell Н. 1982. Comparative allocation of biomass, energy and
nutrients in plants 11 Ecology. VoL63. No.4. P.982-991.
Aikman D.P., Watkinson A.R. 1980. A model of growth and self-thinning in even-aged
monocultures of plants //Ann. Bot. Vol.45. No.3. P.419-427.
Angevine M.W. Chabot B.F. 1979. Seed germination syndromes in higher plants И O.T.
Solbrig et al. (eds.) Topics in plant population biology. N.Y. P. 188-206.
Baker H.G. 1972. Seed weight in relation to environmental conditions in California//
Ecology. Vol.53.No.5. P.997-1010.
Baskin C., Baskin J. 1988. Germination ecophysiology of herbaceous plant species in a
temperate region //Amer. J. Bot. Vol.75. No.2. P.286-305.
Bazzaz F.A. Carlson R.W. Harper J.L. 1979. Contribution to reproductive effort by pho-
tosynthesis of flowers and fruits 11 Nature. Vol.279. P.554-559.
Begon M.E., Mortimer M., Thompson D.J. 1996. Population ecology (A unified study of
animals and plants). Blasckwell Science. 247 p.
Bell A.D. Tomlinson P.B. 1980. Adaptive architecture in rhizomatous plants // Bot. J.
Linnean Soc. Vol.80. No.2. P. 125-160.
Bender B., Baskin J. 1988. Theoretical rates of increase in annual, biennial and perennial
plants: effect of delayed reproduction in perennials // Ibid. Vol.75. No.6, Pari 2. P.69.
JluillV[himy[hl j/t)
Blackman V.ll. 1919. The compound inlciesl law and plant giowlh // Annals ol Hol
Vol.33. No. 13 I. P.353 360.
Bonaparte E.E., Brawn R.L 1975. The effect of ml inspectin' competition on the plastic ii \
of morphological and agronomic characters of four maize hybrids // Annals of Bot.
Vol.39. P.863-869.
Bonser S.P., Aarssen L.W. 1996. Meristem allocation: a new classification theory for
adaptive strategies in herbaceous plants H Oikos. Vol.77. P.347-352.
Bosbach К., Hurka H., Haase R. 1982. The soil seed bank of Capsella bursa-pastoris
(Cruciferae): its influence on population variability // Flora. Vol. 172. No.l. P.47-56.
Burdon J.J., Harper J.L. 1980. Relative growth rates of individual memebers of plant
population 11 J. of Ecol. Vol.68. No.3. P.953-957.
Caldwell M.M. 1979. Root structure: the considerable cost of belowground function//
O.T. Solbrig et al. (eds.). Topics in plant population biology. N.Y. P.408-427.
Cohen D. 1966. Optimising reproduction in randomly varying environment И J. Thcor.
Biol. Vol. 12. No.l.P. 119-129.
Cohen D. 1967. Optimising reproduction in a randomly varying environement when a
correlation may exist between conditions at the time a choice has to be made and the
subsequent outcome 11 J. Theor. Biol. Vol. 16. No.l. P.1-14.
Cook R. 1980. The biology of seeds in the soil // O.T. Solbrig et al. (eds.). Demography
and evolution of plant populations. Oxford. P. 107-129.
Crawley M.J. 1986. Life history and environment И M.J. Crawley (ed.). Plant ecology.
London. P.253-290.
Devades C., Beck C.B. 1971. Development and morphology of stelar components in the
stems of some members of the Leguminosae and Rosaceae // Amer. J. Bot. Vol.58.
No.2. P.432-446.
During H.J., Schenkeveld A.J., Verkaar H.J., Willems J.H. 1985. Demography of short-
lived forbs in chalk grassland in relation to vegetation structure // J. White (ed.). The
population structure of vegetation. Handbook of vegetation science. Vol.3. P.341-370.
Eriksson O., Kiviniemi K. 1999. Evolution of plant dispersal И Life history evolution in
plants. Vol.7. P.215-238.
Falencka M. 1983. The effect of the size of cotyledons on the development of individuals
in Impatiens noli-tangere L. population // Ecol. Polsk. Vol.31. No.l. P. 123-143.
Falinska K. 1986. Pojecie osobnika w demografii roslin И Wiadomosci Ecologiczne.
Vol.32. No.4. P.361-380.
Fenner M. 1985. Seed ecology. 151 p.
Fenner M. Thompson K. 2005. The ecology of seeds. 250 p.
Ford E.D. 1975. Competition and stand structure in some even-aged monocultures 11 J. of
Ecol. Vol.63. No.l. P.311-333.
Frank D., Klotz S. 1988. Biologisch-dkologische Daten zur Flora der DDR. Halle
(Saale). 103 S.
Gadgil M.D. 1971. Dispersal: population consequences and evolution // Ecology. Vol.52.
No.l. P.233-261.
Grime J.P. 1979. Plant strategies and vegetation processes. 222 p.
Grime J.P., Hunt R. 1975. Relative growth rate: its range and adaptive significance in a
local flora П J. of Ecol. Vol.63. No.2. P. 393-422.
Grime J.P., Mason G., Curtis A.V. et al. 1981. A comparative study of germination
characteristics in a local flora// J. Ecol. VoL69. No.3. P. 1017-1059.
Groff P. A., Kaplan D.R. 1988. The relation of root systems to shoot systems in vascular
plants //The Botanical Review. Vol.54. P.387-422.
376 MH Мары hi Нопул ИЩИ HIIIIIM ()IH)IH>I ни pm 1СНИЙ
Gross K.L. Werner P.A. 19X2. Colonizing abilities ol “biennial” plant species in relation
to ground cover: implications lor their distributions in a sueeessional sere // Ecology.
Vol.63.No.4. P.921-931.
Grubb P.J. 1976. A theoretical background to the conservation of ecologically distinct
groups of annuals and biennials in the chalk grassland ecosystem // Biol. Conserv.
Vol. 10. No. 1. P.53-76.
Haig D., Westoby M. 1988. Inclusive fitness, seed resources and maternal care I I J. Lovett
Doust. L. Lovett-Doust (eds.). Plant reproductive ecology: patterns and strategies.
N.Y. P.60-79.
Halle F., Oldeman R.A. 1970. Essai sur 1’architecture et la dynamique de croissance des
arbres tropicaux. Paris. 178 p.
Halle F., Oldeman A.A., Tomlinson P.B. 1978. Tropical trees and forests: an architectural
analysis. N.Y. 441 p.
Hanley M.E., Lamont B.B. 2001. Herbivory, serotiny and seedling defence in Western
Australian Proteaceae И Oecologia. Vol. 126. P.409-417.
Harper J.L. 1977. Population biology of plants. London. 892 p.
Harper J.L., Lovell P.H., Moore K.G. 1970. The shapes and sizes of seeds //Ann. Rev. of
Ecol. and Syst. No.l. P.327-356.
Harper J.L., Me Naughton I.H. 1962. The comparative biology of closely related species
living in the same area. VII. Interference between individuals in pure and mixed
populations of Papaver species // New Phytol. Vol.61. Pl75-88.
Harper J.L., Ogden J. 1970. The reproductive strategy of higher plants. I. The concept of
strategy with special reference to Senecio vulgaris // J. of Ecol. Vol.58. No.3. P.681-
698.
Harper J.L., White J. 1971. The dynamics of plant populations П P.J. den Boer, G.R.
Gradwell (eds.). Dynamics of populations. Wageningen. P.44-63.
Harper J.L., White J. 1974. The demography of plants // Ann. Rev. of Ecol. and Syst.
Vol.5. P.419-463.
Hart R. 1977. Why the biennials are so few? П Am. Natur. Vol.111. No.9. P. 792-799.
Harris G.R., Lovell P. H. 1980. Adventitious root formation in Veronica species // Annals
of Botany. Vol.45. No.3. P. 459-468
Hegde S.G., Lokesha R. et al. 1991. Seed size distribution in plants: an explanation based
on fractal geometry П Oikos. Vol.62. No.l. P. 100-101.
Hickman J. 1975. Environmental unpredictability and plastic energy allocation strategies
in the annual Polygonum cascadense (Polygonaceae) 11 J. Ecol. Vol.69. P.689-701.
Hickman J. 1977. Energy allocation and niche differentiation in four co-existing annual
species of Polygonum in Western North America // J. Ecol. VoL65. No.l. P.317-326.
Howe H.F., Westley L.C. 1986. Ecology of pollination and seed dispersal H M. Crowley
(ed.). Plant Ecology. P. 185-215.
Hurka H., Haase R. 1982. Seed ecology of Capsella bursa-pastoris (Cruciferae): dispersal
mechanism and the soil the seed bank И Flora. Vol. 172. No.l. P.35-46.
Hutchings M.J. 1986. The structure of plant populations 11 M. Crawley (ed.). Plant
ecology. P.97-136.
Hutchings M.J., Slade A. J. 1987. Modular structure and plant architecture 11 Bull, of
British Ecol. Soc. Vol. 18. No.l. P.31.
Huxley J. 1932. Problems of relative growth. N.Y.
lohannsen W. 1903. Uber Erblichkeit in Populationen und in reinen Linien. Jena.
Jain S. 1979. Adaptive strategies: polymorphism, plasticity, and homeostasis // O.T
Solbrig et al. (eds.). Topics in plant population biology. N.Y. P. 160-187.
Jong de T.J., Klinkhamer P.G., Metz A.J. 1987. Selection for biennial life histories in
plants //Vegetatio. Vol.70. No.3. Pl49-156.
Литература 5 / /
Jourct M.-I .>1976. Ideologic de hi dorinancc sciniiialc cl de hi geiinitialк»n dies divcisrs
especes du genre Impatiens L. //Bull. Sec. Roy. Hol. Belg Vol !()*> No .’ I* ?H ??s
Jouret M.-F. 1977. Relation entre la doiniancc sciniiialc cl la dioiologic de divcises
especes du genre Impatiens L. // Bull. Soc. Roy. Bot. Belg. Vol. 110. No. I 2. PI19
128.
Kaplan D.R. 2001. The science of plant morphology: definition, history, and role in
modem biology//Am. J. Bot. Vol. 88. No. 10. P. 1711-1741.
Kawano S. 1981. Trade-off relationships between some reproductive characteristics
in plants with special reference to life history strategy // Bot. Mag. Tokyo. Vol.94.
No. 1035. P.285-294.
Kays S., Harper J.L. 1974. The regulation of plant and tiller density in a grass sward // .1.
Ecol. Vol.62. P.97-105.
Kelly D. 1984. Seeds per fruit as a function of fruits per plant in “depauperate” annuals
and biennials//New Phytol. Vol.96. No.l. P. 103-114.
Kelly D. 1985. Why are biennials so maligned? //Am. Naturalist. Vol. 125. No.3. P.473
479.
Kidd F., West C. 1917. The controlling influence of carbon dioxide. IV. On the produc-
tion of secondary dormancy in seeds of Brassica alba following treatment with carbon
dioxide and the relation of this phenomenon to the question of stimuli in growth pro-
cesses //Ann. Bot. Vol.31. No.3. P.457-487.
Law R. 1979. The cost of reproduction in annual meadow grass H Am. Naturalist. Vol. 113.
No.l. P.3-16.
Leek M.A., Parker V.T., Simpson R.L. (eds.) 1989. Ecology of soil seed banks. London:
Acad. Press. 382 p.
Lee S.M. Cavers P.B. 1981. The effects of shade on growth, development and resource
allocation patterns of three species of foxtail (Setaria) 11 Can. J. Bot. Vol. 59. No. 9.
P.1776-1786.
Levin D.A. 1988. Local differentiation and the breeding structure of plant populations //
L.D. Gottlieb, S.K. Jain (eds). Plant Evolutionary Biology. P.305-329.
Levins R. 1970. Lectures on mathematics in the life science. Vol.2. P. 77-107.
Lhotska M. 1975. Notes on the ecology of germination of Alliaria petiolata И Folia
geobot. et phytotaxon. Vol.10. No.2. P.179-183.
Liston A., Riesberg L.H., Elias T.S. 1990. Functional androdioecy in the flowering plant
Datisca glomerata H Nature. Vol.343. P.641-642.
Lloyd D. 1980. Demographic factors and mating patterns in Angiosperms И O.T. Solbrig
(ed.). Demography and evolution in plant populations. Oxford. P.67-88.
Lonsdale W.M., Watkinson A.R. 1983. Plant geometry and self-thinning П J. Ecol. Vol.71.
No.l. P.285-297.
Lovett-Doust L. 1981. Population dynamics and local specialization in a clonal perennial
(Ranunculus repens) 11 J. Ecol. Vol.69. No.3. P.743-755.
Mac Arthur R.H. 1972. Geographical ecology. N.Y. 269 p.
Mac Arthur R.H., Wilson E.O. 1967. The theory of island biogeography. Princeton:
Princeton Univ. Press. 203 p.
Mack R.N. 1976. Survivorship of Cerastium atrovirens at Aberffraw, Anglesey // J. Ecol.
Vol.64. No.l. P.309-312.
Mack R.N., Pyke D.A. 1983. The demography of Bromus tectorum\ variation in time and
space 11 J. Ecol. Vol.71. No.l. P.69-93.
Markov M.Vit. 1985. Research on permanent quadrats in the USSR // J. White (ed.). rhe
population structure of vegetation, Handbook of vegetation science, Vol.3. Pill 119.
Mayr E. 1972. The nature of the Darwinian revolution // Science. Vol. 176. P.981 989.
378 A/.// MiiphiHt I h >i i yii ищк>1 и him (>ih>ih>i ни pin icinii
Me Canny S.J. 19X5. Alternatives in parent-offspring lelationships in plants // Oikos.
Vol.45. No.l. P. 148 149.
McGraw J.B., Wulff R.D. 1983. The study ofplanl growth: a link between the physiological
ecology and population biology of plants // J. Theor. Biol. Vol. 103. No. I. P. 21 -28.
Menges E.S. 1987. Biomass allocation and geometry of the clonal forest herb Laportea
canadensis', adaptive responses to the environment or allometric constraints? //Am. J.
Bot. Vol.74. No.4. P.551-563.
Mitchell-Olds T. 1987. Analysis of local variation in plant size 11 Ecology. Vol.68. No.l.
P.82-87.
Mithen R., Harper J.L., Weiner J.l 984. Growth and mortality of individual plants as a
function of “available area” 11 Oecologia. Vol.57. No.l. P.57- 60.
Mooney H.A., Gulmon S.L. 1979. Environmental and evolutionary constraints on the
photosynthetic characteristics of higher plants 11 O.T. Solbrig et al. (eds.). Topics in
plant population biology. N.Y. P.316-337.
Mountford M.D. 1971. Population survival in a variable environment 11 J. Theor. Biol.
Vol.32. No.l. P.75-79.
Myster R.W., Pickett S.T.A. 1988. Individualistic patterns of annuals and biennials in
early successional oldfields // Vegetatio. VoL78. No.l. P.53-60.
Nakamura R.R.1986. Maternal investment and fruit abortion in Phaseolus vulgaris!I
Amer. J. Bot. Vol.73. P. 1049-1057.
Nakamura R.R. 1988. Seed abortion and seed size variation within fruits of Phaseolus
vulgaris', pollen donor and resource limitation effects // Amer. J. Bot. Vol.75. No.7.
P.1003-1010.
Newell S.J., Tramer E.J.1978. Reproductive strategies in herbaceous plant communities
during succession // Ecology. VoL59. No.2. P.228-234.
Orians G.H., Solbrig O.T. 1977. A cost-income model of leaves and roots with special
reference to arid and semiarid areas П Am. Naturalist. Vol. 111. No.980. P.677-690.
Palmblad LG. 1968. Competition in experimental populations of weeds with emphasis on
the regulation of population size // Ecology. Vol.49. No.l. P.26-34.
Pavone L.V., Reader R.J. 1985. Reproductive schedule of Medicago lupulina
(Leguminosae) in a patchy environment // Can. J. Bot. VoL63. No. 10. P.2044-2048.
Pearl R. 1928. The Rate of Living. N.Y: Knopf. 185 p.
PiankaE.R. On r- and К-selection //Am.Natur. 1970. Vol. 104. No.5. P.592-597.
Pitelka L.F., Thayer M.E., Hansen S.B. 1983. Variation in achene weight in Aster
acuminatus И Can. J. Bot. Vol.61. No.6. P.1415-1420.
Popay A. I., Roberts H.A. 1970. Factors involved in the dormancy and germination of
Capsella bursa-pastoris (L.) Medik. and Senecio vulgaris L. // J. Ecol. Vol. 58, No.
103. P.87-90.
Primack R. 1979. Reproductive effort in annual and perennial species of Plantago
(Plantaginaceae) //Am. Naturalist. Vol. 114. No.l. P.51-62.
Primack R.B. 1987. Relationships among flowers, fruits, and seeds // Ann. Rev. Ecol.
Syst. Vol. 18. P.409-430.
Raunkiaer C. 1934. The life-forms of plants and their bearing on geography H The
life-forms of plants and statistical plant geography being the collected papers of C.
Raunkiaer. P.2-104.
Reekie E.G., Bazzaz F.A. 1987. Reproductive effort in plants. 1. Carbon allocation to
reproduction //Am. Naturalist. Vol. 129. No.6. P.876-896.
Reekie E.G., Bazzaz F.A. 1987. Reproductive effort in plants. 3. Effect of reproduction
on vegetative activity //Am.Naturalist. Vol. 129. No.6. P.907-919.
Reinartz J. 1984. Life history variation of common mullein (Verbascum thapsus): 3.
Differences among sequential cohorts // J. Ecol. Vol.72. No.3. P.927 936.
Литература 379
Reiss M. 1989. Plant resource allocation // Trends of I colop.v and I vohilion. Vol I
No. 12. P.233.
Ridley H.N. 1930. The dispersal of plants throughout the world. I... Kent. 744 p.
Roberts H.A., Dawkins P.A. 1967. Effect of cultivation on the numbers of viable weed
seeds in soil 11 Weed Research. No.7. P.290-301.
Ross M.A., Harper J.L. 1972. Occupation of biological space during seedling establishment
// J. Ecol. Vol.60. No.l. P.77-88.
Sagar G.R., Mortimer A.M. 1976. An approach to the study of the population dynamics of
plants with special reference to weeds //Applied Biology. Vol.l. P.1-43.
Salisbury E.J. 1942. The reproductive capacity of plants: studies in quantitative biology.
London. 244 p.
Salisbury E.J. 1961. Weeds and aliens. London. 384 p.
Salisbury E.J. 1974. Seed size and mass in relation to environment 11 Proc. Roy. Soc. Ser.
B186. No. 1083. P.83-88.
Samson D.A., Werk K.S. 1986. Size-dependent effects in the analysis of reproductive
effort in plants //Am. Naturalist. Vol. 127. P.667-680
Sarukhan J. 1974. Studies on plant demography: Ranunculus repens L., R. bulbosus L.
and R. acris L. II. Reproductive strategies and seed population dynamics H J. of Ecol.
Vol.62. No.l.P. 151-177.
Schaal B. A. 1980. Reproductive capacity and seed size in Lupinus texensis И Am. J. Bot.
Vol.67. No.5. P.703-709.
Schaffer W., Lnovyc R., Whittam T. 1982. Energy allocation by annual plant where
the effects of seasonality on growth and reproduction decoupled П Am. Naturalist.
Vol.120. No.6. P.787-815.
Schmitt J., Ehrhardt D.W., Cheo M. 1986. Light-dependent dominance and suppression
in experimental radish populations // Ecology. Vol.67. No.6. P. 1502-1507.
Schenkeveld B., Verkaar D. 1983. On the ecology of short-lived forbs in chalk grassland.
Thesis. Utrecht. 64 p.
Senden J.W., Schenkeveld A. J., Verkaar H. 1986. The combined effect of temperature and
red:far red ratio on the germination of some short-lived chalk grassland species //Acta
Oecologica, Oecol. Plant. Vol.7(21). No.3. P.251-259.
Semander R. 1927. Zur Morphologic und Biologie der Diasporen. Nova Acta Regiae
Societatis Scientarum Uppsaliensis. Uppsala. 104 p.
Sharitz R.R., McCormick J.F. 1973. Population dynamics of two competing annual plant
species И Ecology. Vol.54. No.3. P.723-740.
Shull G.H. 1909. Bursa bursa-pastoris and Bursa heegeri\ biotypes and hybrids H
Carnegie Inst. Publ. P.3-57.
Silvertown J. 1980. Leaf-canopy-induced seed dormancy in grassland flora // New Phytol.
Vol.85. No.l. P.109-118.
Silvertown J. 1982. Introduction to plant population ecology. London.
Silvertown J. 1983. Why are biennials sometimes not so few? // Am. Natur. Vol. 121.
No.3. P.448-453.
Silvertown J. 1989. The paradox of seed size and adaptation 11 Trends in Ecology and
Evolution. Vol.4. P.24-26.
Simmonds N.W. 1980. Monocarpy, calendars and flowering cycles in angiosperms // Kcw
Bull. Vol.35. No.2. P.235-245.
Silvertown J., Lovett Doust J. 1993. Introduction to plant population biology. 210 p.
Slavik B. 1996. Rod Impatiens v Ceske republice // Preslia. Praha. Vol.67. No.2. P. 193
211.
Smith H.B. 1927. Annual versus biennial growth habit and its inheritance in Melilotus
alba// Am. .1. Bot. Vol. 14. No.l. P.I29 146.
380 Л/// МарКОН I joliyilHUIIOIIIIlIH ИЯ pill ICIIIlil
Smith Н. 1982. Light quality, photoperception and plant strategy // Ann. Rev. Plant
Physiol. Vol.33. No.3. P.481518.
Smith B.H. 1983a. Demography of Floerkea proserpinacoides, a forest-lloor annual. I.
Density dependent growth and mortality 11 J. Ecol. Vol.71. No. 2. P.391-404.
Smith B.H. 1983b. Demography of Floerkea proserpinacoides, a forest-floor annual. II.
Density dependent reproduction 11 J. Ecol. Vol.71. No.2. P.405-412.
Smith B.H. 1984. The optimal design of herbaceous body H Am. Naturalist. Vol. 123.
No.2. P. 197-211.
Sokal R.R., Rohlf F. J. 1995. Biometry: the principles and practice of statistics in biological
research. New York. 887 p.
Solbrig O.T. 1980. Demography and evolution in plant populations H Bot. Monographs.
Vol. 15. P. 1-20.
Solbrig O.T. 1981. Studies on the population biology of the genus Viola II.The effect of
plant size on fitness in Viola sororia 11 Evolution. VoL35. P. 1080-1093.
Southwood T.R.E. 1977. Habitat, the templet for ecological strategies? 11 J. of Animal
Ecology. Vol.46. No.2. P.337-365.
SouthwoodT.R.E. 1988. Tactics, strategies and templets//Oikos. Vol.52. No.l. P.3-18.
Stanton M. 1984. Seed variation in wild radish: effect of seed size on components of
seedling and adult fitness // Ecology. Vol.65. No.4. P. 1105-1112.
Stanton M.L. 1985. Seed size and emergence time within a stand of wild radish (Raphanus
raphanistrum L.): the establishment of a fitness hierarchy H Oecologia. Vol.67. No.3.
P.524-531.
Steams S.C. 1976. Life-history tactics: a review of the ideas // Quart. Rev. Biol. VoL51.
No.l. P.3-47.
Steams S.C. 1977. The evolution of life history traits: a critique of the theory and a review
of the data//Ann. Rev. Ecol. Syst. VoL8. P.145-171.
Stevens O.A. 1932. The number and weight of seeds produced by weeds H Amer. J. Bot.
Vol. 19. No.6. P.784-794.
Stewart A.J., Thompson K. 1982. Reproductive strategies of six herbaceous perennial
species in relation to a successional sequence H Oecologia. VoL52. No.2. P.269-272.
Sugiyama S., Bazzaz F. A. 1998. Size dependence of reproductive allocation: the influence
of resource availability, competition and genetic identity H Functional Ecology. Vol. 12.
No.2. P.280-288.
Sultan S.E. 1987. Evolutionary implications of phenotypic plasticity in plants //
Evolutionary Biology. Vol.21. No.l. P.127-178.
Sussman A.S., Halvorsen H.O. 1966. Spores: their dormancy and germination. N.Y. 354 p.
Symonides E. 1979. The structure and population dynamics of psammophytes on inland
dunes II. Loose-sod populations 11 Ecologia Polska. Vol.27. No.2. P. 191-234.
Symonides E. 1983. Population size regulation as a result of intra-population interactions.
I. Effect of density on survival and development of individuals of Erophila verna (L.)
C.A.M. 11 Ecologia Polska. Vol.31. 2. P.839 - 882.
Symonides E. 1988a. On the ecology and evolution of annual plants in disturbed
environments 11 Vegetatio. VoL77. No.l. P. 21-31.
Symonides E. 1988b. Population dynamics of annual plants H A. J. Davy, M.J. Hutchings,
A.R. Watkinson (eds.). Plant Population Ecology. London. P.221-248.
Taylorson R.B., Borthwick H.A. 1969. Light filtration by foliar canopies: significance for
light-controled weed seed germination // Weed Sci. Vol. 17. No. 1. P.48-51.
Thompson J.N. 1984. Variation among individual seed masses in Lomatium grayi
(Umbelliferae) under controlled conditions: magnitude and partitioning of the vari-
ance 11 Ecology. Vol.65. P.626-631.
Thompson K. 1987. Seeds and seed banks//New Phytol. Vol. 106 (Suppl.). P.23 34
Литература 381
'Thompson К., Grime J. 1979. Seasonal variation in Ihc seed hanks ol hcibiii eons species
in ten contrasting habitats. // .1. T’col. Vol.67. P. 89? 89 t
Thompson J.N., Rabiniwitz I). 1989. Do big plants have big seeds*? // Am Natuiahst
Vol.l33.No.5. P.722-728.
Thompson K., Stewart A.J. 1981. The measurement and meaning of the reproductive ef-
fort in plants // Am. Naturalist. Vol. 117. P.205-211.
Thompson B.K., Weiner J., Warwick S. I. 1991. Size-dependent reproductive output in
agricultural weeds // Can. J. Bot. Vol.69. No.3. P.442^446.
Tilman D. 1988. Plant strategies and the dynamics and structure of plant communities.
Princeton. 360 p.
Trivedi S., Tripathi R.S. 1982. Growth and reproductive strategies of two annual weeds
as affected by soil nitrogen and density level // New Phytol. Vol.91. No.3. P.489-500.
Tripathi R.S., Khan M.L. 1990. Effects of seed weight and microsite characteristics on
germination and seedling fitness in two species of Quercus in a subtropical wet hill
forest // Oikos. Vol.57. No.3. P.289-296.
Troll W. 1937. Vergleichende Morphologic der hoheren Pflanzen. Berlin: Bomtraeger. .
S. 288-304, 499-511.
Tuomi J., Vuorisalo T. 1989. Hierarchical selection in modular organisms // Trends in
Ecology & Evolution. Vol.4. No.7. P.209-213.
Turesson G. 1922a. The species and variety as ecological units // Hereditas. Vol.3. P.100-
113.
Turesson G. 1922b. The genotypical rersponse of the plant species to the habitat //
Hereditas. Vol.3. P.211-350.
Turesson G. 1925. The plant species in relation to habitat and climat // Hereditas. Vol.6.
P. 147-236.
Turner M.D., Rabinowitz D. 1983. Factors affecting frequency distributions of plant
mass: the absence of dominance and suppression in competing monocultures of
Festucaparadoxa // Ecology. Vol.64. No.3. P. 469^475.
Ulbrich E. 1928. Biologie der Fruchte and Samen (Karpobiologie). Berlin: Springer Verl.
128 S.
Urbanska K. 1990. Biology of asexually reproducing plants // S. Kawano (ed.) Biological
Approaches and Evolutionary trends in Plants. Academic Press. P. 273-292.
Urbanska К. 1992. Populationsbiologie der Pflanzen. Grundlagen, Probleme, Perspectiven.
Gustav Fischer Verlag. Stuttgart. 374 S.
Urbanska K., Landolt E. 1990. Biologische Kenn wer von Pflanzenarten // Ber. Geobot.
Institute ETH, Stiftung Rubel, Zurich. Bd.56. S.61-77.
Van Andel J. Vera F. 1977. Reproductive allocation in Senecio sylvaticus and Chamaenerion
angustifolium in relation to mineral nutrition // J. Ecol. Vol.65. No.3. P.747-758.
Watkinson A.R. 1978. The demography of a sand dune annual Vulpia fasciculata. II. The
dynamics of seed populations // J. Ecol. Vol. 66. No.l. P.35-44.
Watkinson A.R., Harper J.L. 1978. The demography of a sand dune annual: Vulpia
fasciculata. I. The natural regulation of populations // J. Ecol. Vol. 66. No.l. P. 15-33.
Watkinson A.R., White J. 1985. Some life-history consequences of modular construction
in plants // Phil. Trans. R. Soc. London. Vol.B313. P.31-51.
Weiner J. 1985. Size hierarchies in experimental populations of annual plants // Ecology.
Vol.66. No.3. P.743-752.
Weiner J. 1986. How competition for light and nutrients affects size variability in Ipomoea
tricolor populations // Ecology. Vol.67. No.5. P. 1425-1427.
Weiner J. 1987. Size variation in plant populations // Bull, of British Ecol. Soc. Vol. 18.
No.l. P.32.
382 Л/// I lonyiiMUnoiiiiiMi (>1101101 пя pin 1С1ШЙ
Weiner J. 1988. The influence of competition on plant reproduction // .1. I .ovetl-Doust,
К Lovett-Doust (eds.). Plant Reproductive Ecology: patterns and strategies. N.Y.
P.228-245.
Weiner J. 1988. Variation in the performance of individuals in plant populations // A.J.
Davy, M.J. Hutchings, A.R. Watkinson (eds.). Plant Population Ecology. P.59-81.
Weiner J., Solbrig O.T. 1984. The meaning and measurement of size hierarchies in plant
populations // Oecologia. Vol.61. No.3. P.334 -336.
Went F.W. 1955. The ecology of desert plants // Scientific American. Vol. 192. P.68-75.
Werner P. A. 1975. Predictions of fate from rosette size in teasel (Dipsacus fullonum L.) //
Oecologia. Vol.20. P. 197-201.
White J. 1979. The plant as a metapopulation //Ann. Rev. Ecol. Syst. Vol. 10. No.l. P.
109-145.
White J. 1980. Demographic factors in populations of plants // O.T. Solbrig (ed.).
Demography and evolution of plant populations. Oxford. P.21—48.
White J. 1981. The allometric interpretation of the self-thinning rule // J. Theor. Biol.
Vol.89. P.475-500.
White J. 1984. Plant metamerism // R. Dirzo, J. Sarukhan (eds.). Perspectives on plant
population ecology. P.15—47.
White J. 1985. The census of plants in vegetation // The population structure of vegetation.
P.33-88
White J., 1987. Demography of plants // Abstracts of XIV Intern. Bot.Congress. Berl.
(West.)Germany. P.8.
White J., Harper J.L. 1970. Correlated changes in plant size and number in plant
populations // J. Ecol. Vol.58. No.2. P.467-485.
Wikberg S. 1985. Fitness in clonal plants // Oikos. Vbl.72. No.2. P.293-297.
Williams J.T., Harper J.L. 1965. Seed polymorphism and germination. Part 1. The
influence of nitrates and low temperatures on the germination of Chenopodium album
//Weed Res. Vol.5. P. 141-150.
Willson M.F. 1983. Plant reproductive ecology. N.Y. 282 p.
Winn A. 1988. Ecological and evolutionary consequences of seed size in Prunella
vulgaris // Ecology. Vbl.69. No.5. P. 1537-1544.
Witztum A., Gutterman Y, Evenary M. 1969. Integumentary mucilage as an oxygen
barrier during germination of Blepharis persica (Burm) Kuntze // Bot. Gaz. Vol. 130.
No.4. P.238-241.
Wright S. 1943. Isolation by distance // Genetics. Vbl.28. P. 114-138.
Yampolsky E., Yampolsky H. 1922. Distribution of sex forms in the phanerogamic flora
// Bibliotheka Genetica. Vol.3. P.1-62.
Yoda K. Kira T., Ogawa H., Hozumi K. 1963. Self-thinning in overcrowded pure stands
under cultivated and natural conditions // J. Biol. Osaka City Univ. Vol. 14. No.4.
P.107-129.
Zeide B. 1978. Reproductive behaviour of plants in time // Am. Naturalist. Vol. 112.
No.985. P.636-639.
Zohary D. 1965. Colonizer species in the wheat group // H. Baker, G.L. Stebbins (eds.).
The genetics of colonizing species. N.Y. Academic Press. P.403—419.
ОГЛАВЛЕНИЕ
ПРЕДИСЛОВИЕ АВТОРА.........................................3
Глава 1. ВВЕДЕНИЕ..........................................4
1.1. Что за наука «популяционная биология растений» и как она
соотносится с другими ботаническими науками...............4
1.1.1. Общая экология, экология растений и популяционная
биология растений..................................... 4
1.1.2. Популяционная биология растений как наука
о типах или «стратегиях» жизненных циклов...........6
1.2. Краткая история формирования популяционной
биологии растений («популяционной ботаники»)
как науки и университетского спецкурса...................10
1.3. Задачи, стоящие перед настоящим учебным пособием....14
Глава 2 ОСНОВЫ ГЕНЕТИЧЕСКОГО ПОДХОДА
К ОПРЕДЕЛЕНИЮ И ИЗУЧЕНИЮ ПОПУЛЯЦИЙ РАСТЕНИЙ.
ОБЩИЕ ПРЕДСТАВЛЕНИЯ ОБ ЭВОЛЮЦИОННОМ ПРОЦЕССЕ...........18
2.1. Роль Ч. Дарвина в создании популяционной биологии...19
2.2. Работы В. Иоганнсена................................24
2.3. Закон Харди-Вайнберга как математическая закономерность
и метод генетического анализа панмиктических популяций...25
2.4. Причины отклонений от равновесия Харди-Вайнберга....27
2.4.1. Мутационный процесс.............................27
2.4.2. Сцепление генов..............................
2.4.3. Неслучайное скрещивание.........................27
2.4.4. Разделение популяции на генетические соседства,
субпопуляции (внутрипопуляционная генетическая
структурированность). Эффективная генетическая единица.29
2.4.5. Поток генов.....................................31
2.4.6. Дрейф генов.....................................31
2.4.7 Естественный отбор...............................32
2.5. Внутривидовая и внутрипопуляционная изменчивость.
Ее природа и наиболее общие моменты проявления...........34
Глава 3. СХЕМЫ И МОДЕЛИ ЖИЗНЕННЫХ ЦИКЛОВ
ПОПУЛЯЦИЙ, ТЕОРИЯ И ПРАКТИКА ДЕМОГРАФИЧЕСКОГО
ПОДХОДА К ИЗУЧЕНИЮ ПОПУЛЯЦИЙ У РАСТЕНИЙ,
ДЕМОГРАФИЧЕСКИЙ МЕТОД ОЦЕНКИ
ПРИСПОСОБЛЕННОСТИ.........................................40
384 Л/ 11<>11упяц1к>1111Ш1 bitoiioi и>1 paciriinh
3.1. Спектры возрастных или онтогенетических состояний ....40
3.2. Базовые спектры онтогенетических состояний и
классификация популяций....................................42
3.3. Схемы-модели пофазного рассмотрения жизненных циклов
популяций..................................................46
3.4. Графы жизненных циклов популяций......................48
3.5. Схемы и модели для наглядного представления
о «стратегиях жизни», стратегиях жизненных циклов
или эколого-ценотических стратегиях........................53
3.5.1. Критерии разграничения видов с разными
стратегиями жизни........................................55
3.5.2. Дальнейшая теоретическая разработка
проблемы стратегий.......................................56
3.5.3. Триангулярная модель Дж.Ф.Грайма..................58
3.5.4. О г- и К-стратегиях...............................58
3.5.5. Типы репродуктивных стратегий.....................60
3.6. Демографический подход к изучению популяций растений..60
3.6.1. Основное демографическое уравнение и его параметры.
Годичная или конечная скорость роста (X) как мера
приспособленности........................................61
3.7. Теория непрерывного роста численности популяций
в приложении к растениям...................................62
3.7.1. Уравнение экспоненциального роста численности
популяций................................................64
3.7.2. Уравнение логистического роста численности популяции.65
3.8. Чистая скорость размножения...........................65
3.9. О моделях непрерывного роста численности популяций....66
3.10. Матричный метод анализа динамики численности
и возрастного или размерного спектра популяций.............67
3.10.1. Популяция незимующего однолетника (пример 1).....69
3.10.2. Популяция многолетнего поликарпического
растения (Пример 2).........................................69
3.10.3. Анализ «чувствительности» (sensitivity)
и «эластичности» (elasticity)...............................70
Глава 4. ФУНДАМЕНТАЛЬНЫЕ ОСОБЕННОСТИ РАСТЕНИЙ
КАК ОБЪЕКТОВ ПОПУЛЯЦИОННОГО АНАЛИЗА:
СООТНОШЕНИЕ ПОНЯТИЙ «ОСОБЬ» И «ПОПУЛЯЦИЯ»
У РАСТЕНИЙ..................................................73
4.1. Наличие банка (=пула) покоящихся диаспор.
Понятие о субпопуляциях и криптопопуляциях....................74
4.2. «Неограниченный» рост растений. Увеличение размеров
тела и изменение соотношений между поверхностью
и объемом. Фракталы........................................75
4.3. Метамеризация. Проявление метамеризации (модульности)
у цветковых растений..........................................76
4.3.1. Особь или индивид. Простой и сложный индивиды.....77
4.3.2. Иерархичность метамерных (модульных) субъединиц......78
/ 'land 10 Деменр;|фи'1гск11Й niiiuiii i природных iioiiy’ImiiiHi pn< iriniil 385
4.4. ( грунту pi iue (архiriск'гурныс) тины p;ic i incni.iii.ix opi aiiiriMoii.
Тины взаимодействия надземных и подземных cipyniyp . 79
4.5. Иерархическая «лестница» мстамсрных образований
у цветковых растений.......................................84
4.5.1. Термины «метамер» и «модуль».......................85
4.5.2. Особи у растений как популяции: мстапонуляции......87
Глава 5. ЖИЗНЕСПОСОБНЫЕ СЕМЕНА В СОСТАВЕ
ПОПУЛЯЦИЙ ЦВЕТКОВЫХ РАСТЕНИЙ.................................89
5.1. Основные биологические функции семени.................90
5.1.1. Семя как единица размножения. Плодовитость (семенная
продуктивность) цветковых растений........................91
5.1.1.1. Плодовитость растений (межвидовой уровень).....91
5.1.1.1.1 Плодовитость видов с разным типом жизненного
цикла и относящихся к разным эколого-фитоценотическим
группам................................................95
5.1.1.2. Плодовитость растений на внутривидовом
(внутрипопуляционном) уровне............................100
5.1.1.2.1. Внутрипопуляционное варьирование
плодовитости особей у видов с нефиксированным
числом плодолистиков в цветке.........................104
5.1.1.2.2. Факторы внутрипопуляционного варьирования
плодовитости у видов с фиксированным числом
элементов в цветке................................... 106
5.1.2. Семя как единица дисперсии.........................111
5.1.2.1. Первичная дисперсия зачатков у цветковых растений.
Горизонтальное распространение (рассеивание) зачатков
и расселение растений...................................112
5.1.2.1.1. Классификация типов первичной дисперсии зачатков ..112
5.1.2.1.2. Рассуждения о причинах эволюции приспособлений
для первичной дисперсии зачатков.......................116
5.1.2.2 Вертикальное перемещение (распространение)
диаспор в ходе вторичной дисперсии ....................121
5.1.2.3. Расселение и миграционные способности растений. 124
5.1.2.4. Понятие о семенных банках популяций............ 125
5.1.3. Семя как структура и средство переживания
неблагоприятных периодов..................................126
5.1.3.1. Оценка семени как средства перенесения
неблагоприятных периодов в различных эколого-ценотических
условиях................................................ 133
5.1.3.1.1. Динамика жизнеспособных семян как неактивного
компонента в популяциях................................13 3
5.1.3.1.2. Жизнеспособные семена в почве и стратегии
жизни видов растений................................. 144
5.1.3.1.3. Особенности двулетних растений.............149
5.2. Семя как зародыш в «упаковке» со стартовым запасом
питательных веществ..................................... 151
5.2.1. Межвидовой уровень............................... 154
5.2.2. Внутривидовой уровень.......................... 161
386 МП Мчпкоч I loiiyiiHUiioiniaM bnonoi ня pm iciiiiii
5.3. Семя как генетический вариант с заложенными в нем
последствиями рекомбинации................................167
5.3.1. Особенности репродуктивных систем многолетних
растений................................................168
5.3.2. Репродуктивные системы малолетних растений.......168
Глава 6. РАЗМЕРЫ И ГОРИЗОНТАЛЬНАЯ СТРУКТУРА
ПОПУЛЯЦИЙ (ПРОСТРАНСТВЕННЫЙ АСПЕКТ
ПОПУЛЯЦИОННОЙ БИОЛОГИИ РАСТЕНИЙ)............................172
6.1. Размер популяции.....................................172
6.1.1. Размер территории, которую занимает популяция;
вид как система популяционных единиц; элементарная
демографическая единица.................................. 172
6.1.2. Общая численность популяции. Типы популяций
по общей численности......................................175
6.2. Плотность и способы ее определения....................176
6.2.1. Случайное, регулярное и пятнистое размещение
особей, семян в почве или иных счетных единиц
на территории популяции или учетной площадке..............176
6.2.2. Плотность как фактор существования особи...........178
6.2.3. Полигон площади питания...........................178
6.2.4. Плотность в точке.................................180
Глава 7. СТРУКТУРНЫЕ АСПЕКТЫ РОСТА РАСТЕНИЙ..................183
7.1. Архитектура растений..................................183
7.2. Архитектурные модели (или -модели побегообразования).. 183
7.3. Архитектурная модель и жизненная форма................185
7.3.1. Место архитектурных моделей в алгоритме изучения
стратегий жизненного цикла.............................. 186
7.4. Основные аспекты в изучении архитектуры растений......187
7.4.1. Размещение ассимиляционной поверхности вдоль
по главному модулю (структура модуля).....................187
7.4.2. Фотосинтетическая эффективность листьев одного модуля.... 194
7.5. Архитектурные варианты разрастания особи. Направления
разрастания особи по мере увеличения ее мощности.
Изомодульный и гетеромодульный типы архитектуры и роста...195
7.5.1. Межвидовой уровень............................... 195
7.5.2. Внутривидовой уровень..............................203
7.6. Способ переключения на репродукцию....................204
7.7. Признаки, положенные в основу классификации
архитектурных типов малолетних растений....................205
7.8. Классификация архитектурных типов малолетних растений.208
Глава 8. КОЛИЧЕСТВЕННЫЕ АСПЕКТЫ РОСТА РАСТЕНИЙ...............213
8.1. Относительная скорость роста..........................213
8.2. О некоторых методических вопросах выявления RGR
в популяционных исследованиях..............................215
8.2.1. Особенности розеточного роста......................215
Глава 10 Дсм<>|рафнчсский шнинн прпр<у un.ix попу i ni 1111 hnacicnnii 387
8.2.2. 11риспособительное значении розеточного рос ia.. .217
8.3. Популяционный подход к анализу оапостельной
скорости роста растений....................................219
8.3.1. Влияние различий в стартовой биомассе особей
(массе семян) на рост и конечные размеры особей...........219
8.4. Внутрипопуляционные различия RGR и размерная
дифференциация особей.......................................230
8.5. Влияние плотности популяции на рост растений.......... 237
8.5.1. Критерии оптимальной плотности популяции............237
8.5.2. Методы исследования влияния популяционной плотности.237
8.5.3. Зависимость общего урожая и его компонентов
от плотности............................................. 238
8.5.4. Положительный эффект группы и отрицательный
эффект плотности или С-D эффект............................241
8.5.5. Опыты с пастушьей сумкой (Capsella bursa-pastoris)..252
8.5.6. Опыты с модельными популяциями клоповников сорного
(Lepidium ruderale) и густоцветкового (L. densiflorum)....254
8.5.7. Опыты с модельными популяциями подорожника
большого (Plantago major) и проломника нитевидного
(Androsace filiformis)....................................262
8.5.8. Опыты с недотрогой мелкоцветковой
Impatiens parviflora.......................................264
8.6. Динамика корне-побеговых и корне-листовых
отношений в ходе розеточного роста..........................266
8.7. Размерная дифференциация особей, возникающая в ходе
роста в популяциях растений.................................273
8.8. Спектры жизненных состояний и их характеристика.......274
Глава 9. АЛЛОКАЦИЯ БИОМАССЫ, ЕЕ СВЯЗЬ С РАЗМЕРНЫМ
СПЕКТРОМ ПОПУЛЯЦИИ И ДИНАМИКА................................284
9.1. Некоторые методические вопросы изучения
репродуктивного усилия и аллокации в целом.................286
9.1.1. Как определять?....................................286
9.1.2. Когда определять?..................................286
9.1.3. Как провести границу между сферами?................287
9.1.4. Как графически представлять данные?................287
9.2. Аллокация и спектр жизненных состояний (размерный
спектр) в популяциях цветковых растений; статические
аллокационные спектры......................................288
9.2.1 Аллометрический метод анализа аллокации.............289
9.2.1.1. Минимальный (пороговый) размер особи,
допускающий ее переход к репродукции......................289
9.3. Закономерности аллокации у малолетних растений........292
9.4. Изменение репродуктивного усилия и аллокации в целом
во времени: аллокационные динамические спектры..............299
9.5. Репродуктивное усилие у малолетних растений с разными
вариантами эколого-ценотической стратегии...................302
388 Л/ Н Mii/ihnn I It >i iy и >i i и к и 11 in >i (iikiiioi ini pm leiinii
Глава 10. ДЕМОГРАФИЧЕСКИЙ ДИАЛИЗ ПРИРОДНЫХ
ПОПУЛЯЦИЙ РАСТЕНИЙ РАЗНЫХ ЖИЗНЕННЫХ <1>()РМ
В РАЗНЫХ ЭКОЛОГО-ФИТОЦЕНОТИЧЕСКИХ УСЛОВИЯХ......................307
10.1. Методические аспекты изучения демографии популяций
растений разных жизненных форм.......................307
10.2. Изучение динамики популяций древесных растений.311
10.2.1. Изреживание древостоев......................311
10.2.2. Возрастание численности особей в древостоях.316
10.3. Виды агрофитоценозов...........................320
10.4. Виды степных эродированных склонов.............323
10.4.1. Результаты изучения популяции катрана Crambe tataria.324
10.4.2. Результаты изучения природных популяций коровяков
Verbascum lychnitis и V. thapsus....................332
10.4.3. Результаты наблюдений в популяциях проломников
наибольшего, удлинённого, северного и змееголовника
тимьяноцветкового ..................................335
10.5. Виды прибрежной зоны водоемов с переменным
уровнем воды.........................................346
10.6. Лесные виды....................................349
10.6.1.. Наблюдения в популяциях лесных двулетников
бородавника обыкновенного Lapsana communis и пупырника
японского Torilis japonica..........................349
10.6.1.1. Результаты наблюдений в популяциях недотрог
обыкновенной Impatiens noli-tangere и мелкоцветковой
I. parviflora.....................................350
10.6.1.2. Результаты опыта с пересадкой особей
под полог березовой рощи..........................357
ЗАКЛЮЧЕНИЕ.............................................361
СЛОВАРЬ СПЕЦИАЛЬНЫХ ТЕРМИНОВ...........................363
СПИСОК ЛИТЕРАТУРЫ......................................368
Книги Товарищества научных изданий КМК
БИОЛОГИЯ
СЕРИЯ «ОПРЕДЕЛИТЕЛИ ПО ФЛОРЕ И ФАУНЕ РОССИИ»
Деревья и кустарники зимой. 2-е изд. [Вып.9]. / /. Валягина-Малютина. 200/. 268 с
Формат 170 х 240 мм. Те. перепл. — Цена 220 руб. Булавоусые чешуекрылые Вос-
точной Европы. [Вып.8]. А.Л. Львовский, Д.В. Моргун. 2007. 443 с., 8 цв.вкл. Формат 170
х 240 мм. Те. перепл. — Цена 400 руб. — Флора Нижнего Поволжья. Том 1 (споровые,
голосеменные, однодольные). [Вып.6]. А.К. Скворцов (ред.). 2006. 435 с. Формат 170 х 240
мм. Те. перепл. — Цена 400 руб.
Планируется: Е.Т. Валягина-Малютина. Деревья и кустарники средней полосы европейской
части России (иллюстрированный определитель).
ПРОЧИЕ ОПРЕДЕЛИТЕЛИ ПО ФЛОРЕ И ФАУНЕ
Краткий определитель беспозвоночных пресных вод центра Европейской России.
М.В. Чертопруд, Е.С. Чертопруд. 4-е изд. 2011. 219 с., ил. Формат 145 х 217 мм. В обл. -
Цена 180 руб. — Иллюстрированные определители свободноживущих беспозвоноч-
ных евразийских морей и прилегающих глубоководных частей Арктики. Том 2. Не-
мертины, головохоботные, малощетинковые черви, пиявки, погонофоры, эхиуры,
сипункулы, форониды, плеченогие. Б.И. Сиренко (ред.). 2010. 186 с., ил. Формат 170 х
240 мм. Те. перепл. — Цена 300 руб. — Определитель зоопланктона и зообентоса прес-
ных вод Европейской России. Том 1. Зоопланктон. В.Р. Алексеев (ред.). 2010. 495 с., ил.
Формат 170 х 240 мм. Те. перепл. — Цена 600 руб. — Флора и фауна Белого моря.
Иллюстрированный атлас. А.Б. Цетлин и др. (ред.). 2010. 471 с., цв. фотоатлас. Фор
мат 175 х 245 мм. Те. перепл. — Цена 1500 руб. — Растения Российского Западного
Кавказа. Полевой атлас. А.С. Зернов. 2010. 448 с., цв. фотоатлас. Формат 155 х 220
мм. Те. перепл. — Цена 600руб. — Стрекозы Восточной Европы и Кавказа: Атлас-опре-
делитель. В.Э. Скворцов. 2010. 623 с., ил. Формат 205 х 145 мм. Те. перепл. — Цена 400
руб. — Определитель наземных моллюсков лесостепи Правобережного Поволжья.
Т.П Стойко, О.В. Булавкина. 2010. 96 с., ил., 17 цв.табл. Формат 165 х 235 мм. В обл.
Цена 220 руб. — Иллюстрированные определители свободноживущих беспозвоноч-
ных евразийских морей и прилегающих глубоководных частей Арктики. Том 1. Ко-
ловратки, морские пауки, ракообразные: усоногие, тонкопанцирные, эуфаузииды,
неполнохвостные, мизиды, капреллиды. Б.И. Сиренко (ред.). 2009. 189 с., ил. Формат
170 х 240 мм. Те. перепл. — Цена 300 руб. — Атлас дикорастущих растений Ленинград-
ской области. 2010. 664 с., цв. фотоатлас. Бум. мелов. Формат 130х 170 мм. Те. перепл.
— Цена 600 руб. — Определитель сосудистых растений Оренбургской области. З.Н.
Рябинина, М.С. Князев. 2009. 758 с., ил. Формат 170 x240 мм. Те. перепл. — Цена 600 руб.
— Растения средней полосы Европейской России. Полевой атлас. И.А. Шанцер. 3-е
изд. 2009. 470 с., цв. фотоатлас. Бум. мелов. Формат 130 х 170 мм. Те. перепл. — Цена
500 руб. — Флора водоемов Волжского бассейна. Определитель сосудистых расте-
ний. Л.И. Лисицына и др. 2009. 219 с., ил. Формат 143 х 210 мм. В обл. — Цена 150 руб. —
Определитель грибов России. Порядок Афиллофоровые. Вып.З. И.В. Змитрович. 2008.
278 с. Формат 145 х 218 мм. Те. перепл. — Цена 300 руб. — Определитель сосудистых
растений Соловецкого архипелага. К.В. Киселёва и др. 2004. 175 с., цв. фото. Формат
170 х 240 мм. Те. перепл. — Цена 220 руб. — Иллюстрированное руководство для бота-
нических практик и экскурсий в Средней России. В.Э. Скворцов. 2004. 506 с. Формат
170 х 240 мм. Те. перепл. — Цена 300 руб. — Иллюстрированный определитель расте-
ний Средней России. Том 3. И.А. Губанов и др. 2004. 520 с. Формат 210 х 295 мм. Те.
перепл. — Цена 350 руб. Том 2 (распродан; готовится 2-е издание) — Том 1. 2002. 526 с. с
портр. — Цена 350руб. — Флора мхов средней части Европейской России. Том 2. М С.
Игнатов, Е.А. Игнатова. 2004. С. 609-944. Бум. мелов. Формат 195 х 270 мм. Те. перепл.
— Цена 300 руб. Том 1. 2003. С.1-608, илл. — Цена 500 руб. — Определитель грибов
России. Дискомицеты. Вып.1. Копротрофные виды. В.П. Прохоров. 2004. 255 с. Фор-
мат 145 х 218 мм. Те. перепл. — Цена 200 руб.
Планируется: Ан.А. Шилейко, Т.С. Рымжанов. Фауна наземных моллюсков Казахстана и сопре-
дельных территорий.
СЕРИЯ «РАЗНООБРАЗИЕ ЖИВОТНЫХ»
Человек и подводный мир [Вып 6] А Н Островский 2011 231 с , 16 с цв вкл Формат
145 х 205 мм Цена 220 руб Повелители бездны |Вып !>| АН Островский 2004
216 с., 16 с. цв. вкл. Формат 146 х 206 мм. Цини 220 руб. Мамонт |Вып,3|. АН 1ихо
нов. 2005. 90 с., цв. вкл. Формат 145 х 205 мм. Цена 100 руб. Гидра: от Абраама
Трамбле до наших дней [Вып.1]. С.Д. Степаньянц и др. 2003. 101 с. + цв.вкл. Формат 145
х 205 мм. — Цена 100 руб.
УЧЕБНИКИ ДЛЯ ВУЗОВ
Общая арахнология. Краткий курс. Часть 1. Введение. Малые отряды. КГ. Михайлов.
2011. 65 с. Формат 145 х 203 мм. В обл. — Цена 60 руб. — Протистология: Руководство.
К. Хаусман и др. Пер. с англ. 2010. 495 с., ил. Формат 170 х 240 мм. Те. перепл. — Цена
600 руб. — Жизнь на дне. Био-экология и био-география бентоса. И.А. Жирков. 2010.
453 с., ил. Формат 170 х 240 мм. Те. перепл. — Цена 600 руб. — Морская биогеоценоло-
гия. И. В. Бурковский. 2006. 285 с. Формат 170 х 240 мм. Те. перепл. — Цена 220 руб. —
Основы микологии (морфология и систематика грибов и грибоподобных организмов).
Л.В. Гарибова, С.Н. Лекомцева. 2005. 220 с. Формат 170 х 240 мм. Те. перепл. — Цена 220
руб. — Малый практикум по зоологии беспозвоночных. Часть 1. 2-е изд. И.А. Тихоми-
ров и др. 2008. 304 с., 14 ч/б вкл. Формат 170 х 240 мм. Те. перепл. — Цена 250 руб. —
Основы биогеографии. В.Г. Мордкович. 2005. 236 с., 1 цв. вкл. Формат 170 х 240 мм. Те.
перепл. — Цена 220 руб.
Планируется'. Малый практикум по зоологии беспозвоночных (часть 2).
СЕРИЯ «СОВРЕМЕННАЯ ОТЕЧЕСТВЕННАЯ БИОЛОГИЯ»
Избранные труды. Н.В. Кокшайский. 2008. 410 с. с портр. Формат 170 х 240 мм. Те.
перепл. — Цена 400 руб. — Избранные труды. В.В. Кучерук. 2006. 523 с. с портр. Фор-
мат 170 х 240 мм. Те. перепл. — Цена 300 руб. — Избранные труды. Е.Н. Матюшкин.
2005. 658 с. с портр. Формат 170 х 240 мм. Те. перепл. — Цена 400 руб. — Избранные
труды по эволюционной биологии. А.П. Расницын. 2005. iv + 347 с. с портр., 16 фото-
таблиц. Формат 170 х 240 мм. Те. перепл. — Цена 250 руб. — Избранные труды. Орга-
низм, геном, язык. Б.М. Медников. 2005. 452 с. с портр. Формат 170 x240 мм. Те. перепл.
— Цена 300 руб.
Планируется'. С.М. Разумовский. Избранные труды.
НАУЧНО-ПОПУЛЯРНЫЕ ИЗДАНИЯ
Цокотуха ли муха? В.Н. Танасийчук. 2011. 410 с., ил., 8 цв. вкл. Те. перепл. Формат 150х
220 мм. — Цена 400 руб. — Невероятная зоология. В.Н. Танасийчук. 2-е изд. 2011. 372 с.,
в обл. Формат 140 х213 мм. — Цена 220 руб. — Моя биосфера: научно-художественное
изложение биосферных знаний. Г.И. Флёрова. 2010. 247 с., ил. Формат 170 x240 мм. Те.
перепл. — Цена 300 руб. — Киты и история китобойного промысла: взгляд из Японии.
М. Комацу, С. Мисаки. 2005. 142 с., в те. перепл., цв. вкл. Формат 145 х 215 мм. — Цена
200 руб.
СПРАВОЧНЫЕ ИЗДАНИЯ, ИСТОРИЯ БИОЛОГИИ
Научное наследие Джаныбекского стационара. М.К. Сапанов (отв. ред.). 2012. 97 с., 16
с. цв. вкл., в обл. Формат 140 х 200 мм. — Цена 100 руб. — Очерки о биологах второй
половины XX века. Ю.Ф. Богданов. 2012. 508 с., ил. Формат 150 х 220 мм. — Цена 500
руб. — Растительные ресурсы России. Дикорастущие цветковые растения, их компо-
нентный состав и биологическая активность. Том 4. А.П. Буданцев (отв. ред.). 2011.
630 с., в те. перепл. Формат 170 х 240 мм. — Цена 600 руб. — Зоотомический кабинет
(кафедра зоологии беспозвоночных) Санкт-Петербургского университета. К 140-ле-
тию основания. С.И. Фокин (ред.). 2011. 285 с., 36 ч/б вкл., в обл. Формат 145 х 220 мм. —
Цена 300 руб. — Новоселье биологов МГУ на Ленгорах. Л.И. Лебедева (сост.). 2011. 736
с., 1 цв. вкл., те. перепл. Формат 145 х 220 мм. — Цена 700 руб. — Пауки Cn6nplSBN 978-
5-87317-772-1. 2011. 344 с., цв. фотоатлас. Бум. мелов. В те. перепл. Формат 170 х 240
мм. — Цена 600 руб. — Столовые корнеплоды в России. В.И. Леунов. 2011. 272 с., ил.,
цв. книга. В те. перепл. Формат 210 х 290 мм. — Цена 900 руб. — Промысловые, перс-
пективные для промысла и кормовые беспозвоночные Российских морей. Э.Н. Его-
рова, Б.И. Сиренко. 2010. 285 с., 8 вкл., 16 цв. вкл. В те. перепл. Формат 170х 240 мм. —
Цена 350 руб. — Методы исследования сообществ микроартропод. М.Б. Потапов, Н.А.
Кузнецова. 2011. 84 с., ил., 4 цв. вкл., в обл. Формат 145 х 215 мм. — Цена 60 руб. —
Методические подходы к экологической оценке лесного покрова в бассейне малой
реки. Л.Б. Заугольнова, Т.Ю. Браславская (отв. ред.). 2010. 383 с., ил., 14 цв. вкл., в те.
перепл. Формат 170 х 240 мм. — Цена 500 руб. Биологическая систематика: эволю-
ЦИИ ИДО И. И >1 I ItltUHIIKHI, I HI I unuHfH г\чч (|..г.......
то iwptmii Формат 170 x 240 мм Цтт ООО руО Клк иомо(можно жили мы /1/1
Меншуткин (пост ). 2011 2/9 с, //<; вкп , воОп Формат 140 * 200 мм I (они 2/9 руО
— Каталог типовых образцов сосудистых растении Восточной Л 1ии, хранящихся и
Гербарии Ботанического института им. В.П. Комарова (I I ) часть 2 (Китаи). А / /ра
бовская-Бородина (отв. ред.). 2010. 517 с., в те. поронп Формат 150 х 210 мм Цонп
500 руб. — Культурные растения Главного ботанического сада им. Н.В. Цицина Рос-
сийской академии наук: 60 лет интродукции. А.С. Демидов (отв. род.). 2011. 511 с.. 50
цв. вкл., в те. перепл., суперобл. Формат 170х 240 мм. - Цена 700 руб. — Растительные
ресурсы России. Дикорастущие цветковые растения, их компонентный состав и био-
логическая активность. Том 3. А.Л. Буданцев (отв. ред.). 2010. 601 с., в те. перепл. Фор-
мат 170 х 240 мм. — Цена 600 руб. — Большая энциклопедия цветочных многолетни-
ков. М.М. Диев. 2011. 515 с., цв. фотоатлас. Формат 210 х 295 мм. — Цена 1500 руб. -
«Философия зоологии» Жана-Батиста Ламарка: взгляд из XXI века. А.И. Шаталкин.
2009. 606 с., ил., в те. перепл. Формат 150 х 220 мм. — Цена 500 руб. — Справочник по
семеноведению. В. Броувер, А. Штелин. Пер. с. нем. 2010. 694 с. в те. перепл. Формат
210 х 290 мм. — Цена 1100 руб. — Эволюция и систематика: Ламарк и Дарвин в совре-
менных исследованиях. Сб. тр. Зоол. музея МГУ. Том 50. А.В. Свиридов, А.И. Шаталкин
(отв. ред.). 2009. 366 с., 40 с. вкл., в те. перепл. Формат 150x220 мм. — Цена 300 руб. -
Растительные ресурсы России. Дикорастущие цветковые растения, их компонент-
ный состав и биологическая активность. Том 2. А.Л. Буданцев (отв. ред.). 2009. 513 с.,
в те. перепл. Формат 170 х 240 мм. — Цена 600 руб. — История Зоологического музея
МГУ: Идеи, люди, структуры. ГЮ. Любарский. 2009. 744 с., ил., в те. перепл. Формат
210 х 290 мм. — Цена 900 руб. — Вид и видообразование. Анализ новых взглядов и
тенденций. (Тр. Зоол. ин-та РАН, Прил. № 1). 2009. 298 с., в те. перепл. Формат 210 х 267
мм. — Цена 500 руб. — Международный кодекс ботанической номенклатуры. 2009. 282
с., в те. перепл. Формат 145 х 220 мм. — Цена 300 руб. — Виды и формы хвойных,
культивируемые в России. Часть 2. Picea, Thuja. Д.Л. Матюхин и др. 2009. 288 с., бум.
мелов., цв. фото. Те. пер. Формат 170 х 240 мм. — Цена 700 руб. — Виды и формы
хвойных, культивируемые в России. Часть 1. Juniperus, Cephalotaxus, Taxus, Torreya.
2-е изд. Д.Л. Матюхин и др. 2009. 259 с., бум. мелов., цв. фото. Те. пер. Формат 170 х 240
мм. — Цена 700 руб. — Учёные в вихре времени. Воспоминания об орнитологах, за-
щитниках природы и других натуралистах. Е. Новак. Пер с нем. 2009. 473 с., в те. пе-
репл. Формат 150x207мм. — Цена 500руб. — Страна ББС. Е. Каликинская. 2008. 534 с.,
в те. перепл. Формат 170 х 245 мм. — Цена 500 руб. — Каталог чешуекрылых России.
С.Ю. Синев (ред.). 2008. 424 с., в те. перепл. Формат 220 х 290 мм. — Цена 620 руб. —
Растительные ресурсы России. Дикорастущие цветковые растения, их компонент-
ный состав и биологическая активность. Том 1. А.Л. Буданцев (отв. ред.). 2008. 421 с.,
в те. перепл. Формат 170 х 240 мм. — Цена 500 руб. — Введение в палеоэнтомологию.
В.В. Жерихин и др. 2008. 371 с., в те. перепл. Формат 170 х 240 мм. — Цена 300 руб. —
Мозаика судеб биофаковцев МГУ 1930-1960 годов поступления. Том II. 1951-1960-е годы.
Л.И. Лебедева (сост.). 2007. 640 с., те. перепл. Формат 145 х 220 мм. — Цена 300 руб. —
Том 1.1930-1950-е годы. 2007. 479 с., те. перепл. Формат 145 х 220 мм. — Цена 200 руб. —
Ваш любящий Валя. Валентин Александрович Догель. Письма домой. С.И. Фокин (ред.).
2007. 266 с., + 40 с., ил., в обл. Формат 145 х 210 мм. — Цена 150 руб. — Эрнст Майр и
современный эволюционный синтез. Э.И. Колчинский. 2006. 149 с., ч/б вкл., в обл. Фор-
мат 145 х215 мм. — Цена 100 руб. — Александр Николаевич Формозов: Жизнь русского
натуралиста. А.А. Формозов. 2006. 208 с., в обл. Формат 135 х 203 мм. — Цена 100 руб. —
Пока горит свеча... Очерки по истории кафедры зоологии беспозвоночных МГУ. 2-е изд.
В.В. Малахов. 2006. 153 с., бум. мелов., в обл. Формат 145 х 210 мм. — Цена 120 руб.
Заказать эти и другие издания изд-ва КМК (биология, география, история, медицина) мож-
но по адресу: 123100 Москва, а/я 16 изд-во КМК, Михайлову Кириллу Глебовичу
Комп, почта: mikhailov2000@gmail.com
Интернет: http://avtor-kmk.ru (аннотации изданных книг)
Факс: (495) 629-4825
МАРКОВ Михаил Витальевич
Популяционная биология растений
Москва: Т-во научных изданий КМК. 2012. 387 с.
при участии ИП Михайлова К.Г.
Редактор издательства К.Г. Михайлов
Оригинал-макет: М.В. Скороходова
Для заявок:
123100, Москва, а/я 16, Издательство КМК
электронный адрес mikhailov2000@gmail.com
http ://avtor-kmk.ru
Подписано в печать 29.02.2012. Формат 70x100/16. Объём 24,5 п.л.
Бумага офсеты. Тираж 1000 экз.
Отпечатано в ООО «Галлея-Принт», Москва, 5-я Кабельная ул., 56
Данное учебное пособие к университетскому курсу «Популя-
ционная биология растений», знакомит студентов и читателей
иного ранга с современным состоянием популяционной био-
логии растений, основными понятиями, концепциями и ме-
тодами популяционной ботаники. Дана общая картина — на-
зван и подкреплен примерами весь набор общих положений,
выработанных в этой науке к сегодняшнему дню. Ознаком-
ление с основными методами получения данных в популяци-
онной биологии растений, а также методами обработки этих
данных и методами их обобщения иллюстрируется, в немалой
степени, с привлечением данных, полученных самим автором.
По поводу выделенных и особенно подробно рассмотренных
дискуссионных положений и положений, по которым имеют-
ся противоречивые результаты, автор высказывает свою точ-
ку зрения. Привлечен обширный иллюстративный материал,
способствующий усвоению курса. Для студентов и всех инте-
ресующихся популяционной биологией растений.
’txVKJ*