/



















































































































































































































































































































































































Text
н-н-КАРТАШЕВ
СИСТЕМАТИКА ПТИЦ
Допущено Министерством высшего и среднего специального образования СССР в качестве учебного пособия для студентов универси'гетов и педагогических институтов
МОСКВА «ВЫСШАЯ ШКОЛА» 1974
596.4 К 27
Карташев Н. Н.
К 27 Систематика птиц. Учеб, пособие для ун-тов. М.» «Высш, школа», 1974.
362 с. с ил. Библиогр.: с. 342
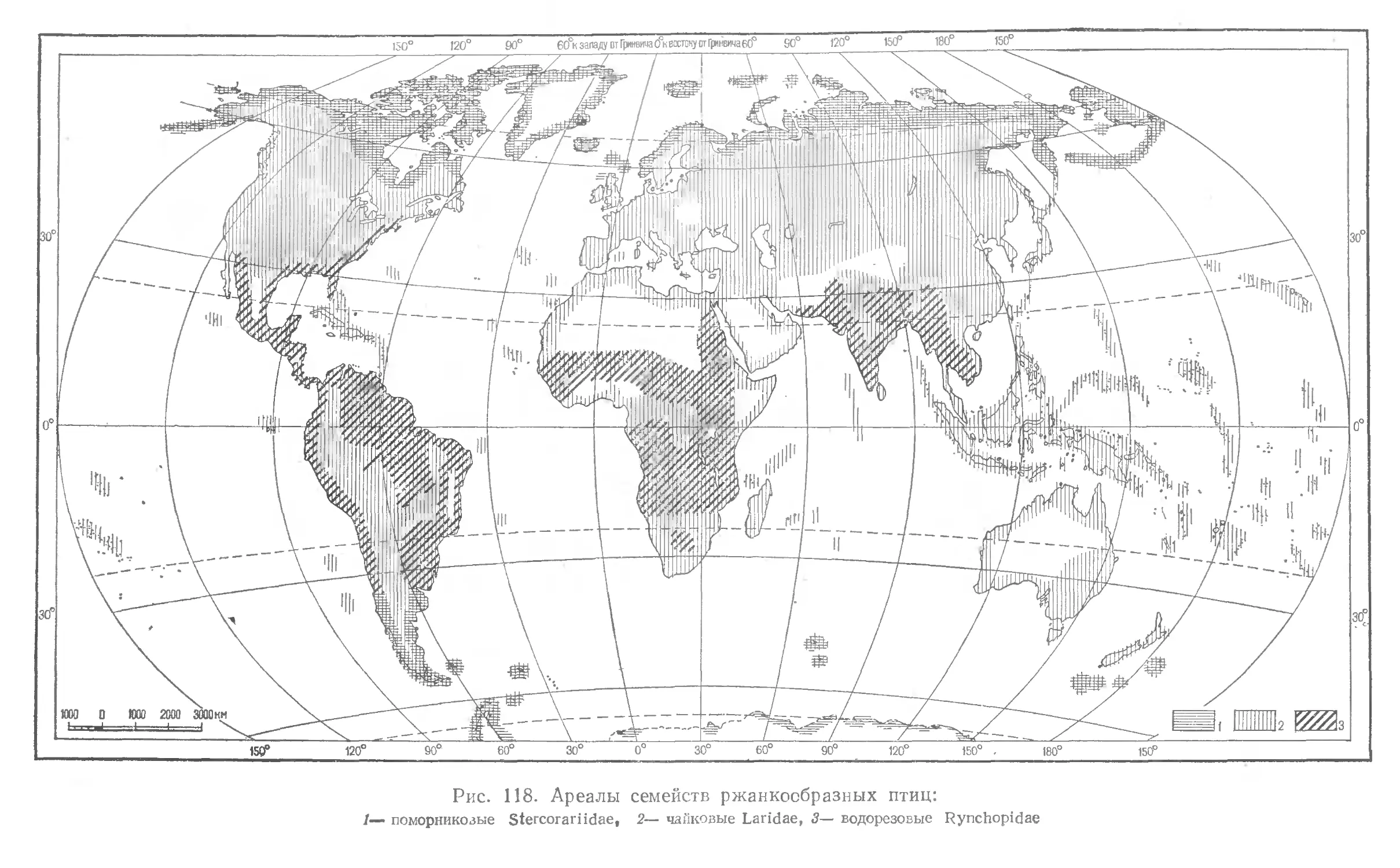
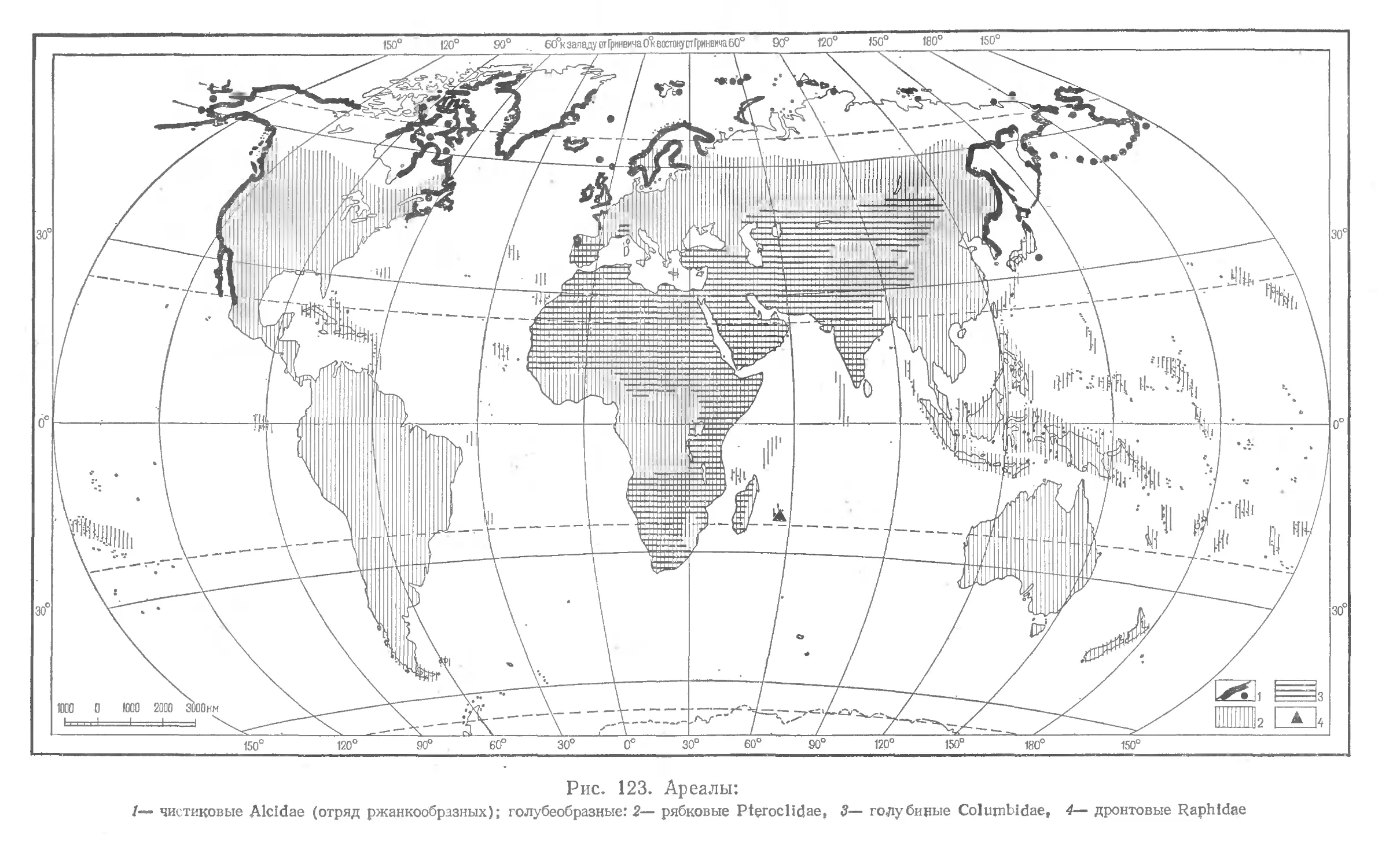
В книге кратко излагаются основные задачи и принципы ^систематики, ха* растеризуются основные систематические категории, приводятся, некоторые правила зоологической номенклатуры. Дается краткий очерк становления системы птиц и описываются более распространенные новые системы класса. Систематический обзор класса птиц составлен по системе Уэтмора (1960); в нём приведены краткие характеристики всех подразделений класса, включая семейства (морфологические особенности, вкология, распространение, числю видов). При описании каждого семейства дается перечень входящих в него рбдов. Книгу иллюстрирует 246 рисуикоц: изображения птиц ижарты ареалов семейств.
К
21008—625 _
-----------89 — 74 001(01)—74
596.4
Издательство «Высшая школа>, 1974 г.
ПРЕДИСЛОВИЕ
«Систематика птиц» входит в учебные планы кафедр зоологии позвоночных биологических и биолого-почвенных факультетов всех университетов либо как самостоятельный курс, либо как один из разделов систематики, составляющий часть большого практикума. Осуществление этого курса крайне затруднено отсутствием доступных современных пособий. Преподаватель вынужден значительную часть материалов излагать на лекциях, что очень сокращает время, отводимое на непосредственную работу студентов с коллекциями. Задача нашего пособия — облегчить усвоение этого курса.
Изложение основных задач и принципов систематики дает тот минимум сведений, который необходим всем студентам, специализирующимся в любой области зоологии позвоночных. Особенно интересующиеся вопросами систематики могут углубить свои знания, используя дополнительную литературу-
Очерк истории становления системы класса птиц важен в методологическом отношении: он показывает, как сказывалось на построении системы накопление фактических знаний и изменение представлений о возникновении и развитии органического мира. Описание наиболее распространенных но
3
вых систем класса не только характеризует современные взгляды на таксономию птиц, но и облегчает ориентировку в специальной орнитологической литературе.
Систематический обзор дает представление о классе птиц в целом и о его подразделениях вплоть до семейств (в некоторых случаях — до подсемейств). Перечень родов характеризует объем каждого семейства и, помимо этого, позволяет использовать это пособие как справочник, облегчающий знакомство с мировой фауной птиц.
Рисунки птиц, иллюстрирующие книгу, выполнены зоологами-художниками В. М. Сми-риным и Ю. М. Смириным. Приношу им мою глубокую благодарность. За ценные советы автор признателен профессору В. Г. Гептнеру и профессору И. А. Шилову. Я крайне благодарен моим коллегам Р. Л. Беме и С. М. Кудрявцеву, взявшим на себя труд просмотреть рукопись. За замечания, способствовавшие улучшению рукописи, автор глубоко благодарен рецензентам — профессору А. С. Мальчевскому и профессору А. К- Сагитову.
Критические замечания просьба сообщать по адресу: 117234, Москва, В-234, Московский государственный университет, биологический факультет, кафедра зоологии позвоночных.
ОСНОВНЫЕ ЗАДАЧИ И ПРИНЦИПЫ СИСТЕМАТИКИ.
СИСТЕМАТИЧЕСКИЕ КАТЕГОРИИ. ПРАВИЛА ЗООЛОГИЧЕСКОЙ
НОМЕНКЛАТУРЫ
ЦЕЛИ И ЗАДАЧИ СИСТЕМАТИКИ
Систематика, или таксономия, — один из наиболее давно сложившихся разделов комплекса биологических наук. Основная цель систематики — выявление многообразия организмов, разработка теории и практики их классификации, выяснение родственных отношений между отдельными видами и группами видов. Без надлежащей классификации совершенно невозможно ориентироваться в громадном многообразии окружающего нас органического мира. К настоящему времени описано более 500 тысяч видов растений и свыше, 1 миллиона видов животных. Класс насекомых, например, объединяет более 750 тысяч видов, тип хордовых — около 43 тысяч видов и т. п. Выявление и описание новых видов продолжается и сейчас (особенно в типе членистоногих).
Первоначально задачи систематики сводились лишь к описанию видов (с целью их различения) и довольно формальной их классификации, т. е. объединению видов в группы разного ранга по сходству. Такая классификация помогала ориентироваться в многообразии уже известных видов. Значение классификации возрастало по мере увеличения числа описанных видов. Это первый период формирования систематики, который начинается трудами Аристотеля (384—322 гг. до нашей эры) и завершается работами Карла Линнея (1707—1778) и его последователей.
Уже с конца XVIII века начинают развиваться эволюционные воззрения, позже четко изложенные Чарльзом Дарвином в его фундаментальном труде «Происхождение видов» (1859). С этого времени начинается второй период развития систематики, когда основное внимание исследователей было обращено на выяснение филогенетических взаимоотношений между различными группами организмов, на поиски «недостающих звеньев» и «примитивных предков». Исследовались преимущественно крупные таксономические группы (семейства, отряды, классы, типы). Одновременно накапливались сравнительные материалы по особенностям строения уже известных видов, продолжалось выявление и описание новых видов и родов. Эти исследования завершились построением первых эволюционных систем всего органического мира и его отдельных групп (типов, классов). В результате в конце
5
XIX века были созданы «вчерне» системы, которые, иногда со многими исправлениями, доработками и дополнениями, существуют и сейчас.
Уже в конце XIX века начинается третий период развития систематики, когда главное внимание исследователей обращается на изучение эволюции внутри вида и на. выяснение путей эволюции видов. Основным объектом изучения становится популяция. Этот период развития систематики продолжается и сейчас, причем в последние десятилетия систематические исследования заметно расширяются и углубляются. Естественно, что наряду с детальным изучением видов и слагающих их популяций продолжается работа по совершенствованию системы всего органического мира и входящих в него таксономических групп разного ранга.
Задачи, которые решает современная систематика, весьма обширны и значимы. Продолжается выявление, описание и обозначение новых видов, как ныне живущих, так и ископаемых. Устанавливаются и уточняются родственные отношения и связи между видами (что выражается объединением их в роды) и между группами видов (объединение родов в семейства и более высокие таксономические группы). Пристальное внимание обращено на выяснение причин и факторов видообразования; основной путь подобных исследований — детальное многообразное сравнительное изучение популяций одного вида, сопоставление материалов по разным видам и т. д. Изучение факторов видообразования дает необходимые обоснования для выявления путей эволюции (исторического развития) разных групп организмов и органического мира в целом. Решая эти сложные и разнообразные задачи, систематика создает все более и более естественную систему (классификацию) органического мира, т. е. размещает все виды в такую систему соподчиненных систематических (таксономических) категорий или групп, которая не только позволяла бы наиболее удобно и быстро ориентироваться во всем многообразии живых форм, но и отражала бы пути эволюции отдельных групп, их родственные связи и отношения. Составление определительных таблиц и определителей — одна из важных практических задач систематики.
При выполнении своих задач систематика, естественно, пользуется специальными систематическими (таксономическими) методиками и приемами: определенными принципами описания и обозначения видов и более крупных таксономических групп, специальными приемами изучения их изменчивости и т. п. Одновременно в систематике сейчас все шире и шире используются методики и приемы других наук: генетический анализ видов и входящих в него популяций, иммунологические и серологические исследования, разнообразные методы вариационной статистики и многие другие. В своей текущей работе систематик широко использует и синтезирует данные многих биологических наук: морфологии, экологии, этологии, биогеографии, генетики, физиологии, палеонтологии, а в последнее время — гистологии, биохимии, биофизики. С другой стороны, результаты систематических исследований представляют необходимую основу для всех остальных биологических наук: дают возможность распознавать виды, характеризуют их родственные связи, показывают направления эволюции отдельных групп и т. п.
Систематика, как один из разделов биологии, естественно подразделяется йа систематику животных, систематику растений и систематику микроорганизмов и вирусов. Систематика животных в свою очередь делится на отделы, занимающиеся построением систем отдельных групп: систематика рыб, систематика птиц и т. п. Общие задачи всех разделов систематики сходны, но частные задачи, применяемые Методики и принципы могут заметно отличаться в связи со спецификой отдельных групп (классов, подтипов, типов) и очень различной степенью их изученности.
ПОНЯТИЕ О ВИДЕ
Конкретная реальная форма существования органического мира — вид. Вид — основная единица жизни, вместе с тем представляющая собой очень сложную биологическую систему. Организмов вне вида нет. В то же время вид — основная систематическая (таксономическая) категория.
Представление о виде существенно менялось по мере накопления знаний об окружающем мире. Впервые четко термин «вид» был введен в науку Рэем (1686). Виды в природе отличаются более или менее ясно выраженными особенностями — видовыми признаками. Поэтому особей, включаемых в данный вид, обычно определяли по их внешнему морфологическому сходству, а один вид от другого отличали по внешним морфологическим различиям. Однако довольно скоро выяснилось, что такой способ распознавания видов нуждается в существенном улучшении и изменении. Оказалось, что у части видов самцы внешне резко отличаются от самок, молодые — от взрослых и т. п. Такие формы иногда описывались как самостоятельные виды. Например, молодых ястребов тетеревятников, отличающихся продольной полосатой исчерченностью груди и брюха, К- Линней относил к виду Falco gentilis, а взрослых птиц с мелкой поперечной исчерченностьюоперения нижней стороны тела — к виду Falco palumbarius. Когда это выяснялось, подобные формы, несмотря, на достаточно резкие внешние морфологические отличия, соединяли в один вид. Многочисленные подобные случаи вызвали необходимость введения дополнительных видовых критериев. Таким основным критерием стала признаваться способность скрещиваться и давать плодовитое потомство. Естественно, что использование этого критерия резко повысило роль полевых наблюдений в систематике. Начиная с середины XIX века, стали накапливаться сведения о том, что у значительного числа широко распространенных видов в разных частях ареала встречаются более или менее четко отличающиеся морфологически географические формы, обычно более или менее постепенно переходящие друг в друга и не обладающие половой изоляцией, т. е. свободно скрещивающиеся и дающие плодовитое потомство. Выявление широко распространенной географической изменчивости вынудило отказаться от чисто морфологической (типологической) концепции вида. Постепенно стала складываться концепция широкого биологического вида, которая сейчас, в общем, принимается большинством систематиков.
Эта биологическая концепция вида основывается на том, что вид определяется не различиями, а обособленностью, состоит не из отдельных не-зависимыхд5собей, а из популяций, репродуктивно изолированных от популяций других видов. По Майру (1940, 1947), вид определяется следующей формулой: «Вид состоит из группы популяций, которые замещают друг друга географически или экологически и из которых соседние интерградируют (или скрещиваются) всюду, где они соприкасаются, или потенциально способны к этому (с одной или несколькими популяциями) в тех случаях, при которых географические или экологические преграды препятствуют контакту». Применяется и более сжатое, краткое определение: «Виды — это группы фактически или потенциально скрещивающихся естественных популяций, которые физиологически изолированы от других таких групп» (Майр, 1940). Предлагались и другие сходные формулировки. Так, по .В. Г. Гепт-неру (1956), «Вид представляет собой ограниченную группу особей — популяцию или группу популяций; особи одного вида характеризуются комплексом определенных, только им свойственных особенностей и свойств (признаков), свободно скрещиваются, дают плодовитое потомство и занимают определенную территорию (ареал). Каждый вид по своим морфологическим и физиологическим признакам отделен от всех других, в том числе и наиболее сходных с ним видов/определенным разрывом (hiatus)».
Все употребляемые ныне определения широкого биологического вида исходят из того, что вид представляет собой генетическое единство, находя
7
щее свое выражение в том, что для всех особей популяций, образующих вид, свойствен определенный набор наследственно обусловленных признаков (свойств) —морфологических, экологических, чисто поведенческих, физиологических, биохимических и т. п., — более или менее четко отграничивающих данный вид от других, даже близких видов. Вид представляет собой .репродуктивное сообщество (особи которого воспринимают друг друга как потенциальных партнеров в размножении), отделенное теми или другими изолирующими механизмами (морфологическими, физиологическими, чисто поведенческими и др.) от других репродуктивных сообществ, т. е. от других видов.
Выявление сложной структуры многомерных политипических видов значительно расширило и углубило наши знания о виде и видообразовании. Последовательное применение политипической концепции вида во многом помогло при выяснении сложных таксономических ситуаций, в ряде случаев позволило отчетливее выявить родственные отношения и приблизиться к лучшему пониманию путей и факторов видообразования (примеры, иллюстрирующие это, будут приведены ниже). Значительно упростилась и классификация класса птиц. Перечень монотипических видов птиц, составленный Шарпом (1909), включал около 19 000 видов. Объединение части этих типологических видов — и нескольких сотен, описанных позже, — в политипи-ческие уменьшило общее число видов птиц примерно до 8600. Соответственно с 6000—7000 до 1700—3000 сократилось число родов. Это, несомненно, облегчило ориентировку в системе класса.
Концепция широкого биологического вида позволяет довольно легко различить (разграничить) большинство видов. Однако иногда возникают определенные трудности, в первую очередь обусловленные тем, что виды в природе находятся на разных этапах эволюционного развития и в ряде случаев, при незавершенном видообразовании, разрывы между группами популяций (морфологические различия, механизмы репродуктивной изоляции ит. п.) могут быть выражены очень неясно. Подобные случаи требуют тщательного анализа и иногда принять однозначное решение бывает трудно (подробности см. Майр, 1968, гл. II).
виды-двойники
Довольно значительные практические трудности представляет выявление так называемых видов-двойников. Это группы популяций, обычно очень нечетко, или практически совсем не различимые морфологически, иногда более явно разграниченные экологически, встречающиеся на одной территории (т. е. имеющие симпатрическое распространение, на более или менее значительной части ареала), но явно и четко изолированные репродуктивно. Виды-двойники довольно часто встречаются среди насекомых (дрозофилы, комары и др.)/ описаны у моллюсков, рыб, грызунов. У птиц к вИдам-двойникам относится менее 5% всех видов (Майр, 1968). Виды-двойники описаны в семействе тиранн Tyrannidae и среди стрижей саланганов рода Collocalia (у последних наилучшим диагностическим признаком служит структура гнезда). Примером видов-двойников нашей фауны может, служить обыкновенный (Luscinia luscinia) и южный (Luscinia megarhyn-chos) соловьи: из используемых для их характеристики 12 диагностических признаков эти виды очень нечетко отличаются только по трем. Видимо, к видам-двойникам должны быть отнесены белоголовый сип Gyps fulvus и гималайский гриф (или кумай) G. himalayensis, большой Aquila clanga и малый A. pomarina подорлики, обыкновенная Cuculus canorus и глухая С. saturatus кукушки, обыкновенный полевой Alauda arvensis и малый полевой A. gulgula жаворонки, малый Sitta пеитауег и большой S. tephronota скалистые поползни, обыкновенная Certhia familiaris и короткопалая С. brachydactyla пищухи, бледная пересмешка Hippolais pallida и бормо-
8
тушка Н. caligata, жемчужный Leucosticte brandti и сибирский L. arctoa горные вьюрки, монгольский Bucanetes mongolicus и толстоклювый В. gi-thagineus пустынные снегири.
Выявление видов-двойников требует всестороннего тщательного анализа: выяснение мелких морфологических различий, сопоставление деталей экологии и поведения (структура гнезда, голос и песня, позы тока и т. п.), выяснение хромосомных, биохимических и физиологических отличий и т. д. При этом удается выявить более или менее явные, хотя и малозаметные разрывы, разобщающие виды-двойники. Очень интересно, что в процессе эволюции при образовании видов-двойников перестройка генотипа, приводящая к взаимной репродуктивной изоляции, не сопровождалась, как это бывает при обычном видообразовании, резкими морфологическими изменениями фенотипа.
Четкая репродуктивная изоляция видов-двойников, устанавливаемая наблюдениями в природе, обычно подтверждается и в лабораторных условиях: от них вообще не удается получить потомства или оно оказывается стерильным. Пожалуй, тут же следует отметить, что наличие репродуктивной изоляции между видами можно установить только наблюдениями в естественных условиях, в природе. При содержании в неволе, видимо из-за нарушения видового стереотипа поведения и из-за отсутствия партнера своего вида, гибридизация, часто с получением плодовитого потомства, довольно свободно идет не только между многими видами птиц одного рода, но и, реже, между видами разных родов. В естественных же условиях такие гибриды встречаются в общем крайне редко. Гибридизация в условиях неволи широко известна для уток, куриных, овсянок, вьюрков, синиц, райских птрц и др.
ИНДИВИДУАЛЬНАЯ ВНУТРИВИДОВАЯ ИЗМЕНЧИВОСТЬ
Большинство видов птиц, как и видов других животных, неоднородно: разные особи более или менее отчетливо отличаются друг от друга. Эти отличия определяются как индивидуальной, так и проявлением возрастной, половой и сезонной изменчивости.
Настоящая' индивидуальная- изменчивость обусловлена специфическими индивидуальными генотипическими и фенотипическими особенностями каждой особи. Эти отличия выражаются в размерах, мелких отличиях окраски, деталях песни и т. п. и обычно не улавливав ются или с очень большим трудом улавливаются человеческим глазом. Однако каждая особь птиц, как и других животных, имеет индивидуальные черты.
Возрастная изменчивость. У всех видов вылупившийся из яйца птенец резко отличается от взрослых особей по размерам, пропорциям, строению и окраске покрова. Однако в пределах вида птенцы одного возраста обладают большим сходством: морфологическим, поведенческим и т. п. У части видов, достигнув размеров взрослых, молодые одевают перьевой наряд, по окраске похожий на оперение взрослых птиц. Если у взрослых особей данного вида выражен половой или сезонный морфизм окраски, то молодые птицы по окраске обычно похожи на самок или на взрослых птиц во внебрачном (годовом) наряде (часть куриных, гусеобразных, голенастых, Многих воробьиных). У других птиц молодые в первом перьевом наряде резко отличаются по окраске от взрослых (часть голенастых, хищников, чайки, мухоловки, скворцы и др.). Наряд взрослых они одевают позже, обычно лишь достигнув половой зрелости (у некоторых крупных чаек на 3—5-е лето жизни).
9
Половой диморфизм. У части видов птиц половой диморфизм не выражен: самцы и самки окрашены сходно. При этом у многих видов самцы обычно чуть крупнее самок, реже самки более или менее крупнее самцов (дневные хищники, совы). У многих видов птиц половой диморфизм выражен более или менее отчетливо. Чаще самцы окрашены ярче самок и заметно превосходят их по размерам (иногда в 1,5—2 раза; куриные, многие гусеобразные, некоторые кулики, многие воробьиные и др.). У трехперсток, куликов плавунчиков, тинаму и некоторых других самки несколько крупнее самцов и ярче окрашены.
Сезонная изменчивость окраски. У довольно большого числа видов птиц самых разных отрядов более или менее четко выражен сезонный морфизм окраски. У этих видов есть более яркий брачный наряд и обычно более тусклый и скромный межбрачный (или годовой) наряд. Межбрачный (годовой) наряд образуется после периода размножения в результате обычно полной послегнездовой линьки, брачный наряд — после частичной или, реже, полной предбрачной линьки (она может идти осенью, на зимовке или ранней весной). У некоторых видов (скворец, чечевица, коноплянка и др.) брачный наряд образуется без линьки в результате снашивания тускло окрашенных кромок опахал контурного пера.
Описанные типы внутривидовой изменчивости свойственны всем особям каждого данного вида, хотя степень их проявления у разных популяций может варьировать в некоторых пределах.
Пол иморфизм. У ряда видов птиц наблюдается изменчивость, не связанная с полом, возрастом, сезоном и проявляющаяся лишь у отдельных особей или части популяции. Такой тип изменчивости обычно называют полиморфизмом. Видимо, у всех видов он может прежде всего проявляться как очень редкие случаи частичного или полного альбинизма (частичное или полное исчезновение пигментов — белая окраска) или меланизма (развитие черной окраски). В мировой фауне примерно у ста видов встречается полиморфизм, охватывающий более или менее значительную часть отдельных популяций и выражающийся в довольно резких отличиях окраски, характере песни, реже — в размерах клюва и т. п. В качестве примера полиморфизма можно привести две цветовые фазы окраски оперения у обыкновенного глупыша Fulmarus glacialis: темная фаза — однотонные дымчато-бурые птицы, светлая — белые с дымчатым налетом на спине и кроющих крыла. Светлая и темная фазы окраски встречаются у орла карлика Aquila pennatus и некоторых других хищников, у поморников р. Stercorarius, голубая и белая фазы — у белого гуся Chen caerulescens, серая и рыжая — у многих сов (у ушастой Asia otus и болотной A. flammeus, обыкновенной неясыти Strix aluco и других), у самок обыкновенной Cuculus canorus и глухой С. saturates кукушек. У обыкновенного глухаря Tetrao urogallus есть белобрюхая и темнобрюхая фазы (первая более обычна на Южном Урале). У самок тетерева Lyrurus tetrix в Зауралье и Западной Сибири встречаются буроватая и сероватая фазы окраски. Варьирует степень развития белого цвета у горихвостки лысушки Phoenicurus phoenicurus на Кавказе, у ласточки касатки Hirundo'rustica в Восточной Сибири, у оляпки Cinclus cinc-lus. Примеры подобного рода можно было бы умножить. У тонкоклювой кайры Uria aalge есть «очковая» фаза: часть птиц имеет узкое белое кольцо вокруг глаза, от которого к уху идет узкая белая полоска.
У части видов фазы окраски контрастны и не имеют переходов (белый гусь, многие хищники и совы), у других наряду с крайними типами окраски встречаются все возможные промежуточные типы (канюки, глупыши, поморники, глухарь, оляпка, горихвостка лысушка, ласточка касатка и др.). Многие из этих цветовых фаз ранее описывались как самостоятельные виды. Но это именно внутрипопуляционная индивидуальная изменчивость, и в естественных условиях особи разных фаз образуют смешанные пары и дают полноценное потомство. В ряде случаев полиморфизм проявляется относительно одинаково на значительной части ареала, в других случаях
10
Рис. 1. Встречаемость «очковой» фазы в атлантических популяциях тонкоклювой кайры в разных частях ареала
он отчетливо выражен лишь у некоторых, географически локализованных популяций. У глупышей встречаются обе фазы, но их соотношение варьирует в разных районах; на Командорских островах (Тихийокеан) белая фаза встречается очень редко, единичными особями, а севернее, в Чукотском море, ойа преобладает. «Очковая» фаза тонкоклювой кайры встречается только в Атлантике, на Тихом океане ее нет; в северных районах Атлантики процент «очковой» фазы в популяции выше, чем в южных (рис. 1). Полиморфизм, видимо, вызывается плейотропией (множественным действием) генов. Его экологическое значение и связь с другими типами изменчивости пока не вполне ясны; здесь необходимы новые исследования.
Описанные выше разные случаи внутривидовой индивидуальной изменчивости не имеют самостоятельного систематического статуса и не обозначаются номенклатурно, т. е. им не присваивается научное название.
ГЕОГРАФИЧЕСКАЯ ИЗМЕНЧИВОСТЬ
У многих видов выявляется популяционная изменчивость, которая выражается в том, что популяции из разных частей ареала более или менее явно отличаются по размерам, пропорциям, деталям окраски (рис. 2), срокам периодических явлений, иногда по степени оседлости, плодовитости, биотопической приуроченности и т. п. Эту изменчивость называют географической.
Географическая изменчивость внутри вида — следствие приспособления его популяций к географически меняющимся внешним условиям. Как чисто эмпирические обобщения, показывающие параллельность морфологической изменчивости внутри вида с некоторыми факторами внешней среды, были сформулированы несколько правил. Степень их достоверности различна для разных районов и для различных групп видов; изменчивость некоторых видов этим правилам не следует, а иногда может и прямо им противоречить.
11
Рис. 2. Различия в окраске некоторых подвидов белой трясогузки Motacilla alba: /—западная М. a. alba L., //—маскированная (туркестанская) М. a. personata Gould., Ill— очковая (якутская) М. a. ocularis Swinh., IV—забайкальская М. a. baicalensis Swinh., V—камчатская М. a. lugens Kit. VI — китайская М. a. leucopsis Gould.
Правило Бергмана (1847): у географически изменчивых видов особи популяций, живущих в более холодных частях ареала, в среднем имеют более крупные размеры тела. По подсчетам Рента (1936), этому правилу соответствует изменчивость около 90% политипических видов палеарктических птиц. В ряде случаев это правило применимо и для популяций, населяющих местности разной высоты над уровнем моря: особи более высокогорных популяций часто крупнее. Обычно это правило объясняется так: поверхность тела возрастает как квадрат, а масса тела — как куб линейных размеров; при увеличении размеров (и массы) тела относительная поверхность уменьшается, что облегчает терморегуляцию.
Правило Аллена (1877) представляет модификацию правила Бергмана: выступающие части тела — клювы, ноги, уши, хвост — в более холодных районах обычно бывают короче, чем в более теплых. Это правило хорошо подкрепляется изменчивостью многих млекопитающих. Изменчивость птиц этому правилу, видимо, подчиняется далеко не всегда. Величина клюва тесно связана с пищевой специализацией, обычно оказывающей на его форму значительно большее влияние, чем климатические условия. Перья крыла и хвоста не участвуют в терморегуляции и их изменчивость определяется особенностями движения. Однако установленная Мэрфи (1938) закономерность, выраженная в том, что островные птицы часто имеют более длинные клювы, чем близкие к ним континентальные подвиды, вероятно, может рассматриваться как частный случай правила Аллена.
Правило Глогера (1833): популяции из теплых или влажных областей обычно пигментированы интенсивнее, чем популяции из более холодных или сухих районов. Это правило более или менее хорошо согласуется с особенностями географической изменчивости многих видов птиц. Физиологи
12
ческий смысл и факторы отбора, вызывающие проявление правила Глогера, пока остаются неясными.
Предлагались и другие правила. Подвиды, совершающие дальние перелеты, имеют более длинное и острое крыло, чем подвиды оседлые или с коротким пролетным путем (Сибом, 1886). Кишечник птиц, питающихся смешанной пищей, относительно короче у тропических подвидов по сравнению с подвидами умеренных широт (Ренш, 1931). Среднее число яиц в кладке может возрастать с увеличением широты (Ренш, 1936; Лэк, 1947). В северном полушарии амплитуда изменчивости возрастает с севера к югу; северные популяции обычно более однообразны (Дементьев, 1941). Действенность этих правил нуждается в дополнительных обоснованиях. Все эти правила отражают экотипические, а не филогенетические явления, и поэтому их можно применять только для характеристики внутривидовой географической изменчивости. Их нельзя использовать при сравнении даже близких видов (в некоторых работах такие попытки бывают).
Сравнительно редкие случаи полного отсутствия географической изменчивости или более частые случаи слабой географической изменчивости, вероятно, объясняются следующими обстоятельствами или их сочетанием (Майр, 1968):
1. Общий ареал вида столь мал, что географическая изменчивость невозможна из-за однородности условий по всему ареалу (овсянка Янковского Emberiza jankowskii населяет лишь Приморье и прилегающие районы Корейского полуострова и Китая, весь ареал желтобровой овсянки Emberiza chryzophrys ограничен средними частями Восточной Сибири и т. п.).
2. Способы расселения столь эффективны, что между всеми популяциями, несмотря на большие размеры ареала, в широких размерах идет перемешивание и свободное скрещивание (панмиктические виды). (Видимо, многие виды уток, некоторые воробьиные и другие группы).
3. Мощные гомеостатические механизмы, сущность которых пока не выяснена, обеспечивают стабильность фенотипа, не давая проявиться значительной части наследственной генетической изменчивости.
4. Иногда, вероятно, можно допустить слабую наследственную изменчивость — генетический гомеостаз.
Анализируя обширные материалы по географической изменчивости птиц, Майр (1968) указывает, что при использовании достаточно чувствительных методик удается выявить разнообразные отличия (генетические, физиологические, биометрические, в голосе, поведении ит. п.) одной популяции от всех других популяций данного вида. У разных видов степень различий между популяциями может варьировать в очень широких пределах: от почти полной идентичности до различий почти видового уровня. Различные признаки часто варьируют независимо. Поэтому соседние популяции могут быть сходны по одним и различны по другим признакам (свойствам). Признаки каждой данной популяции если не полностью, то хотя бы отчасти имеют наследственную основу и в большинстве случаев остаются неизменными относительно длительное время. В целом географическая изменчивость адаптивна и обеспечивает приспособленность каждой популяции к условиям соответствующей местности. Однако не все внешние проявления данной генотипической адаптации обязательно имеют приспособительный характер; они могут быть лишь внешним проявлением генотипов, контролирующих поддерживаемые отбором скрытые физиологические признаки, не улавливаемые применяющимися методиками исследования. Адаптации местных популяций ведут к увеличению генетического разнообразия вида.
Необходимы всесторонние и методически разнообразные конкретные исследования по выяснению механизмов и факторов, обусловливающих разные типы географической изменчивости. Только тогда станет понятной столь широкая вариабильность изменчивости и резко отличная форма ее проявления у разных видов птиц.
из
подвиды
Широко распространенные виды состоят из практически бесконечного числа локальных популяций, каждая из которых объединяет генетически различных особей. Совокупность локальных популяций, населяющих определенную часть ареала вида и таксономически, т. е. по ряду внешних диагностических признаков, Отличающихся от других популяций этого вида, называется подвидом (до сих пор иногда в качестве синонима используется термин «географическая раса»). Важно помнить, что подвид обычно состоит из многих локальных популяций, видимо, всегда хотя бы слегка отличающихся друг от друга генетически и фенотипически. Как правило, каждую локальную популяцию удается отнести к тому или иному подвиду, но для каждой отдельной особи это не всегда удается из-за индивидуальной изменчивости, иногда совпадающей с изменчивостью соседних подвидов. Поэтому при выделении подвидов большинство систематиков сейчас придерживается «правила 75%»: подвидом следует считать популяцию или группу популяций, не менее 75% особей которой отличимы от всех особей другого (ближайшего) подвида. Чем больше серии экземпляров, с которыми работает систематик, тем больше шансов выявить истинное положение.
Подвид занимает определенную территорию; два подвида не могут встречаться на одной и той же территории. Если эти популяции свободно скрещиваются, то данные «подвиды» выделены произвольно и требуют пересмотра. Если же между этими популяциями существует репродуктивная изоляция, то это самостоятельные виды. Между подвидами одного вида репродуктивной изоляции быть не может (немногие исключения в сложных таксономических случаях приведены ниже; см. кольцевые перекрывания,
Рис. 3. Зоны гибридизации между серыми и черными воронами в Палеарктике (по Майзе, 1928; упрощено);
/— ареал черных ворон группы согопе, 2— ареал серых ворон группы cornix, 3— зона гибридизации
14
стр. 19). Таким образом подвиды всегда аллопатричны, т. е. географически исключают друг друга, но обычно занимают смежные (соседние) территории. Подвиды более или менее постепенно переходят один в другой, образуя зону интерградации (перекрывания), где между двумя подвидами идет свободное скрещивание и многие особи имеют промежуточные признаки. Выявление границ подвидов — дело трудное, а сами границы в некоторой степени условны.
В некоторых случаях, при резких подвидовых отличиях, пограничная зона интерградации между подвидами выражена четко. Примером могут служить западные черный и серый подвиды обыкновенной вороны (Corvus согопе согопе и Corvus согопе comix), контактирующие в Западной Европе в узкой по сравнению с ареалами этих подвидов полосе шириной в 50—250 км (рис. 3). Спаривание (гибридизация) в зоне интерградации идет беспорядочно и здесь встречаются все возможные комбинации, родительских и промежуточных признаков. Такая же зона гибридизации (интерградации) между восточными подвидами вороны (серой Corvus согопе sharpii и черной Corvus согопе orientalis) проходит по казахстанским степям, северо-западному Алтаю, Прибалхашью и по долине Енисея. Полагают, что в период наибольшего оледенения ареал вороны был разделен на участки, где живущие в изоляции популяции образовали подвиды, резко разошедшиеся по морфологическим признакам, прежде всего по окраске и соответствующим генным комплексам. После отступания льдов, в послеледниковый период эти резко различающиеся внешне, но не выработавшие репродуктивной изоляции подвиды стали расселяться, образовав в районах стыков своих ареалов гибридные зоны. Относительную узость и устойчивость гибридных зон, видимо, можно объяснить тем, что отбор отбрасывает дисгармоничные комбинации генных комплексов в зоне гибридизации и отбор же препятствует распространению этих промежуточных комбинаций в смежные «чистые» подвидовые популяции (Майр, 1968). Многие авторы такие географические формы с четкими узкими зонами гибридизации рассматривают как самостоятельные виды; часть систематиков считает самостоятельными видами черную ц серую ворон, что кажется не вполне обоснованным и явно противоречит широкой концепции биологического вида.
КЛИНАЛЬНАЯ ИЗМЕНЧИВОСТЬ
Географическая изменчивость может проявляться не ступенчато, как в описанных выше случаях, а очень постепенно. При этом изучаемые таксономические признаки изменяются плавно и постепенно на значительном протяжении ареала вида; переходы между отдельными популяциями не улавливаются. Такая географическая изменчивость называется клинальной, а градиент изменчивости отдельных признаков — клиной (этот термин впервые ввел Д. Гексли, 1939). Клинальная изменчивость, видимо, может быть вызвана комплексом факторов. В ряде случаев постепенно изменяются факторы среды (например, климат). Потоки генов между близкими популяциями имеют тенденцию сглаживать межпопуляционные различия. Может быть, здесь сказывается эффект морфогенетического гомеостаза, скрывающего генные различия между популяциями, особенно при активном обмене генами между'ними. Майр (1968) характеризует клинальную изменчивость как результат взаимодействия двух противоречивых сил: отбора, стремящегося приспособить каждую популяцию к ее локальной среде, и потока генов, направленного на сохранение сходства всех популяций вида.
До сих пор клинальная изменчивость у птиц изучена очень мало, хотя она, видимо, достаточно широко распространена. Клинальная изменчивость отдельных признаков описана у ряда видов птиц семейства дронговых Dic-ruridae, у райской птицы Paradisaea apoda и немногих других видов. С запада на восток клинально изменяются размеры и пропорции клюва белоло
15
бого гуся Anser albifrons и гуменника Anser fabalis. Видимо, более или менее клинально изменяется окраска и пропорции у сорокопута жулана La-nius colurio (cristatus). Выявление и исследование случаев клинальной изменчивости — одна из насущных задач систематики.
МОНОТИПИЧЕСКИЕ И ПОЛИТИПИЧЕСКИЕ ВИДЫ
Разные виды могут, таким образом, по своей популяционной структуре резко отличаться. Виды, у которых все популяции по всему ареалу фенотипически однообразны, называются монотипическими. Политипическими называют виды, популяции которых проявляют географическую изменчивость и могут быть объединены в два или более подвидов. Среди воробьиных Палеарктики политипические виды составляют 66—70% (Хартерт, 1936; Вори, 1959). Среди птиц Северной Америки политипические виды составляют около 68%, среди птиц Новой Гвинеи — около 80% (Майр, 1968). В принимаемых сейчас примерно 8600 видах птиц выделяют около 28 000 подвидов, т. е. в среднем на вид приходится 3,3 подвида (в действительности больше, так как примерно около трети видов монотипические). Для разных семейств эти средние показатели очень различны; например, у ласточковых Hirundinidae на вид в среднем приходится 2,6 подвида, а у жаворонковых Alaudidae 5,1. Подвидовая изменчивость отдельных, даже близких видов варьирует в очень больших пределах. Обыкновенная сойка Garrulus glan-darts включает 26 подвидов (по мнению некоторых систематиков, 40), зяблик Frnigilla coelebs — 12 подвидов, домовый воробей Passer domesticus — 14 подвидов, полевой воробей Passer montanus — 6 подвидов, обыкновенная овсянка Etnberiza citrinella — 2 подвида, горная овсянка Emberiza cia — 13 подвидов, хохлатый жаворонок Galerida cristata — 37 подвидов, лесной жаворонок Lullula arborea — 2 подвида и ,т. п. Обычно у оседлых видов подвидовая (географическая) изменчивость выражена сильнее, чем у перелетных, у крупных птиц — слабее, чем у мелких. Однако эти обобщения имеют много исключений. Все это результат того, что изменчивость определяется сложным взаимодействием многих фактбров и естественно, что она очень отличается у разных родов и видов.
ГЕОГРАФИЧЕСКИЕ ИЗОЛЯТЫ
Ареал обычного политипического вида включает ареалы составляющих его более или менее различающихся популяций, которые группируются в подвиды или образуют клину. Между отдельными популяциями идет обмен генами. Но иногда популяции или группа популяций оказываются изолированными от остальной массы популяций своего вида внешними преградами, препятствующими или сильно затрудняющими обмен генов между ними. Такие популяции называют географическими и з о л я -т а м и. Географические изоляты возникают там, где экологические или географические условия определяют островное (пятнистое) распределение популяций данного вида. Особенно часто они встречаются на удаленных от Материка островах и на периферии ареала/ Могут образовываться географические изоляты и в результате изменения внешних условий (изменения растительности из-за повышения сухости климата, оледенения и т. д.) и при расселении отдельных популяций за пределы основного обитания вида.
Образование изолятов зависит не только от внешних условий, но и от характера изменчивости вида, способности его к расселению. Поэтому некоторые виды на обширных, зачастую достаточно резко изолированных или разобщенных территориях могут быть фенотипически однообразными, тогда как другие виды в сходных условиях образуют изоляты, вырабатывающие более или менее четкие морфологические, экологические, физиологи
16
ческие и другие отличия от остальных популяций своего вида. Скорость и степень обособления изолятов может варьировать в широких пределах; достаточно четко обособленные изоляты птиц на островах между Калимантаном и Сулавеси, видимо, имеют возраст менее 10 000 лет, подвиды птиц Евразии и Северной Америки, возникшие в периоды развития оледенения, значительно старше.
Судьба географических изолятов может быть различной (Майр, 1968). Они могут: 1) превратиться в самостоятельные виды, 2) существовать в изоляции длительное время, 3) восстановить контакт с основной частью популяций своего вида, образовав вторичную зону контакта, 4) полностью вымереть. Таксономическйй статус и степень фенотипических отличий изолятов обусловливаются длительностью изоляции, способностью изолированной популяции к изменчивости, характером факторов внешней среды в зоне изоляции и т. п. Одни изоляты фенотипически совершенно похожи на основную часть видовых популяций, другие отличаются столь резко, что многие исследователи рассматривают их, как самостоятельные виды, отличия третьих находятся на грани между видовыми и подвидовыми признаками и т. д. В ряде подобных случаев таксономическое положение изолятов оказывается спорным и вызывает резкие разногласия.
ВИДЫ или подвиды
Если географические изоляты выработали достаточно четкие морфологические, поведенческие или другие отличия и продолжают жить в изоляции, не контактируя с популяциями исходного вида, их обычно рассматривают как близкие, но самостоятельные виды с аллопатрическим распространением. В качестве примера можно привести тихоокеанскую ипатку Fra-tercula corniculata и атлантического тупика Fratercula arctica. При меньшей степени морфологического разобщения таксономическое положение подобных форм может быть спорным: часть систематиков рассматривает их как самостоятельные аллопатрические виды, а другие — как подвиды полити-пического вида (примеры приведены ниже).
Однако изоляты могут вновь восстанавливать контакт с основной частью популяций вида. Если ко времени восстановления контакта изолят резко не обособился от исходных популяций, то возникающая зона контактов (зона вторичной интерградации) выражена как обычная зона контактов между подвидами. Если же у изолята до вторичного контактирования выработались резкие различия (чаще морфологические), но не возникли механизмы репродуктивной изоляции, то зона вторичной интерградации обычно выражена достаточно резко. Примером такого рода, вероятно, могут быть уже упоминавшиеся зоны контакта серых и черных ворон (рис. 3).В подобных случаях некоторые систематики рассматривают вторично контактирующие формы как самостоятельные виды (хотя еще не окончательно обособившиеся), другие — как резко различающиеся подвиды, так как здесь отсутствует основной критерий вида — репродуктивная изоляция.
Таких спорных случаев в орнитологической таксономии довольно много; в нашей фауне их около 50. Если эти формы рассматриваются как самостоятельные виды, то подвидовое название второй формы становится ее видовым названием. Сопоставляемые формы могут входить в политипиче-ский вид с большим числом подвидов. Если указанным формам присваивается ранг самостоятельного вида, то тем самым исходный политипический вид разделяется на два (или больше) вида, а все остальные подвиды исходного политипического вида распределяются между вновь принятыми видами. Такое спорное положение занимают атлантическая Gavia arctica arctica L. и тихоокеанская G. arctica pacifica Lawrence чернозобые гагары, темноклювая Gavia immer immer Brunn и Светлоклю'вая G. immer adamsii Gray полярные гагары, азиатский Accipiter badius badius Gmelin и европейский
17
A. badius brevipes Severtzov тюбики, сапсан Falco peregrinus peregrinus Tunstall и пустынный сокол F. peregrinus pelegrinoides Temminck, европейский Falco vespertinus vespertinus L. и амурский F. vespertinus amuren-sis Radde кобчику, обыкновенный Alectoris graeca graeca Meisner и гималайский A. graeca chukar Gray кеклики, обыкновенный Coturnix coturnix co-turnix L. и японский C. coturnix japonica Temminck et Schlegel перепела, зуек галстучник Charadrius hiaticula hiaticula L., уссурийский зуек Ch. hiaticula placidus Gray и перепончатопалый галстучник Ch. hiaticula semi-palmatus Bonaparte, каспийский Charadrius asiaticus asiaticus Pallas и монгольский Ch. asiaticus veredus Gould длинноногие зуйки, американский Tringa (Heteroscelus} incana incana Gmelin и сибирский T. (IT). incana brevipes Vieillot пепельные улиты, эскимосский Nutnenius borealis borealis Forster и сибирский N. borealis minutus Gould кроншнепы малютки, атлантический Cepphus grylle grylle L. и тихоокеанский C. grylle columba Pallas чистики, южный Retupa zeylonensis zeylonensis Gmelin и северный /С. zeylonensis blakistoni Seebohm рыбные филины, обыкновенный Calandrella pispoletta (rufescens} pispoletta Pallas и манчжурский C. pispoletta cheleen-sis Swinhoe серые жаворонки, обыкновенный Corvus corax corax L. и пустынный C. corax ruficollis Lesson вороны, черная Corvus согопе согопе L. и серая С. согопе cornix L., обыкновенные вороны, обыкновенная Corvus monedula monedula L. и даурская С. monedula dauricus Pallas галки, европейская Acrocephalus a. arundinaceus L. и малоазиатская A. arundinaceus stentoreus Hemprich et Ehrenberg дроздовидные камышевки, европейская Sylvia cur-ruca curruca L., кашгарская S. curruca minula Hume и горная S. curruca al-thea Hume славки завирушки, японская Muscicapa narcissina narcissina Temminck и даурская M. narcissina zanthopygia Hay желтоспинные мухоловки, чернопегая каменка Oenanthe hispanica hispanica L. и каменка пле-шанка О. hispanica pleschanka Lepechin, западная Oenanthe lugens lugens Lichtenstein и восточная О. lugens finschi Heuglin черношейные каменки, бледный Turdus pallidus pallidus Gmelin, японский T. pallidus chrysolaus Temminck и оливковый T. pallidus obscurus Gmelin дрозды, рыжий Turdus naumanni naumanni Temminck и темный T. naumanni eunomus Temminck дрозды Науманна, американская Parus atricapillus atricapillus L., европейско-азиатская P. atricapillus montanus Baldenstein и тяньшанская P. atricapillus songarus Severtzov буроголовые гаички, белая Parus cyanus cya-nus Pallas и желтогрудая P. cyanus flavipectus Severtzov лазоревки, обыкновенная Parus major major L., бухарская P. major bokharensis Lichtenstein и японская P. major minor Temminck et Schlegel большие синицы, американский Sitta canadensis canadensis L., малоазиатский S. canadensis kruperi Pelzeln и китайский S. canadensis villosa Verreaux черноголовые поползни, домовый (городской) Passer domesticus domesticus L. и индийский P. domesticus indicus Jardine et Selby воробьи, европейский Mont if ringilia nivalis nivalis L. и азиатский ,M. nivalis alpicola Pallas снежные вьюрки, обыкновенный Carduelis carduelis carduelis L. и седоголовый C. carduelis caniceps Vigors щеглы, обыкновенная Acanthias (Carduelis} flammea flammea L. и толстоклювая (тундряная) A. flammea hornemanni Holboll чечетки, малая Erythrina (Caprodacus) rhodochlamys rhodochlamys Brandt и большая E. (C.) rhodochlamys grandis Bly th. розовые (арчевые) чечевицы, обыкновенный Pyrrhula pyrrhula purrhula L. и серый P. purrhula cinereacea Cabanis снегири.
НАДВИДЫ
Аллопатрические виды, морфологически и в других отношениях достаточно четко разошедшиеся (что позволяет предполагать существование между ними репродуктивной изоляции), но похожие по географическому размещению на политипические виды, Майр (1931, 1968) предлагает объединять
18
в надвиды (superspecies). Среди некоторых групп птиц они довольно обычны. Так, по Майру (1968), из распространенных в Юго-Восточной Азии 19 видов дронго (Dicruridae) 10 могут быть отнесены к 2 надвидам, включающим 4 и 6 аллопатрических островных видов; среди птиц Соломоновых островов в надвиды можно объединить 13,6% видов, а среди птиц Австралии — почти треть. Виды, включаемые в надвиды, Майр предлагает называть полувидами (semispecies). Этим подчеркивается, что между ними еще возможен, хотя и сильно затруднен, обмен генами, тогда как между истинными видами он отсутствует, а между подвидами осуществляется более или менее свободно. Употребление подобных группировок, видимо, полезно в эволюционных работах. Однако эти термины (надвид, полувид) не могут рассматриваться как номенклатурные категории; такого статуса они пока не получили.
Вторжение аллопатрических видов в ареал родительского или сестринского вида без гибридизации между ними свидетельствует о завершении процесса видообразования. Видимо, к подобным случаям относится перекрывание ареала белой трясогузки Motacilla alba с ареалом индийской трясогузки Mot. maderaspatensis в Кашмире и с япойской трясогузкой Mot. grandis на Хоккайдо и Хонсю; правда, некоторые систематики считают формы grandis и maderaspatensis подвидами Mot. alba. Более безупречны другие примеры. Вероятно, в ледниковый период (плейстоцен) языки ледников разделили родительскую популяцию пищухи Certhia sp. на западную и восточную группы. Развиваясь в изоляции, каждая из этих групп выработала резко и четко действующие механизмы репродуктивной изоляции, одновременно сохранив очень большое сходство фенотипов. Расселяясь после отступления ледника, эти изоляты встретились, и сейчас эти два самостоятельных вида — обыкновенная пищуха Certhia familiaris L. (восточная форма) и короткопалая пищуха С. brachydactyla Brehm (западная форма) — живут в сходных биотопах, не скрещиваясь, в обширной области перекрывания их ареалов: от Западной Европы до Альп, Закарпатья и Малой Азии. Видимо, совершенно аналогичен случай с южным Luscinia megarhynchos и обыкновенным L. luscinia соловьями, имеющими значительное перекрывание ареалов в Европе, и некоторыми другими видами.
КОЛЬЦЕВЫЕ ПЕРЕКРЫВАНИЯ АРЕАЛОВ
Примерно у десятка видов птиц с широкими ареалами и отчетливой политипичностью обнаружено перекрывание ареалов конечных звеньев (форм), что теперь часто называют кольцевым перекрытием ареалов: цепь интерградирующих подвидов образует кольцо или петлю, в которой концевые звенья (подвиды) перекрываются, населяя одну и ту же местность, но не скрещиваются в зоне перекрывания. Эти случаи можно трактовать как неполное или как незавершенное видообразование.
Обычно в качестве примера приводят большую синицу Parus major, распространенную по значительной части Евразии и образующую до 30 подвидов. Северные и" западные районы ареала вида занимают зеленоспинные и желтобрюхие подвиды группы major (рис. 4). Южную Азию населяют сероспинные и белобрюхие подвиды группы cinereus, а восток Азии — зеленоспинные и белобрюхие подвиды группы minor. Все эти три группы подвидов гибридизируют в зонах контактов, более или менее постепенно переходя одна в другую. Раньше полагали, что кольцевое перекрывание ареалов группы major и группы minor в бассейне Амура не сопровождается гибридизацией, поэтому последнюю группу подвидов иногда выделяли в самостоятельный вид Parus minor. Учитывая, что гибридизацию в бассейне Амура можно, вероятно, считать доказанной, видимо, правильнее относить все эти географические формы к одному большому политипическому виду Parus major (некоторые систематики дробят его на 3—4 вида).
19
Рис. 4.. Неполное видообразование у большой синицы Parus major (из Майр, 1968, по Делакуру и Вор и, 1950; с изменениями):
/— подвиды группы' major (зеленая спина, желтое брюшко, из Западной н Северной Евразии), 2— группы cinereus (серая спина, белое брюшко, из Южной Азии), 3— группы minor (зеленая спина, белое брюшко, из Восточной Азии); в зонах контактов образуются гибридные популяции: 4— в Иране, 6— в Южном Китае, 6— в бассейне Амура. Есть предположение, что ареалы групп major и minor в бассейне Амй>а перекрываются без гибридизации.
Рис. 5. Схема филогенетических отношений между географическими формами группы серебристая чайка — клуша (по Тимофееву — Ресовскому и Штреземанну, 1959). В Беломорско-Балтийском районе встречаются 4 формы: argentatus, antelius (heugli-ni), omissus, fuscus; из них наиболее обособлена клуша fuscus:
I— границы между группами форм, иногда выделяемых в самостоятельные виды (/— серебристая чайка, JZ— чайка хохотунья, III— клуша, IV— калифорнийская чайка), 2— предполагаемые пути возникновения и расселения отдельных форм
Бесспорнее другой пример кольцевого перекрывания ареалов: встречающиеся вместе на побережьях Балтийского и Белого морей и не образующие гибридов клуша Lotus fuscus и серебристая чайка L. argentatus могут рассматриваться как перекрывающиеся концевые звенья цепи подвидов одного вида, широко распространенного в умеренных и северных широтах Евразии и Северной Америки. По представлениям изучавших этот случай Е. Штреземанна и Н. В. Тимофеева-Ресовского (1947, 1959), можно предполагать, что в доледниковое время серебристые чайки занимали обширный ареал. В плейстоцене широко распространившиеся ледники разделили ареал этого вида на несколько изолятов. Весьма вероятно, что центром формирования современных форм была не подвергавшаяся оледенению Берингия (стык ареалов форм vegae и smithsonianus). Отсюда в межледниковые и послеледниковую эпохи чайки расселялись сложным путем (рис. 5): на восток по Северной Америке (thayeri — leucopterus; географически сейчас обособленная californicus; argenteus — argentatus; последняя форма, видимо, относительно недавно проникла из Северной Америки в Северную Европу); на запад по северным побережьям Евразии (birulae — taimyrensis — antelius); через Западную Сибирь на юг (cachinnans — ponticus — artneni-cus) и оттуда .на северо-запад (omissus) и через Средиземноморье в Атлантику (ponticus — michahellesi — atlanticus — britanicus — intermedins — fuscus). В результате в Беломоро-Балтийском районе сейчас встречаются четыре формы: fuscus, antelius (heuglini), omissus, argentatus (рис. 5), из которых первая (fuscus) морфологически и репродуктивно очень четко изолирована от остальных, а другие три морфологически отграничены довольно слабо, хотя гибрида между ними, видимо, довольно редки, и, кажется, намечается экологическая изоляция этих форм.
Поэтому при несомненной генетической близости всех упомянутых форм из-за перекрывания конечных звеньев и их репродуктивной изоляции в районе Балтика— Белое море сейчас обычно выделяют 2 вида: клушу L. fuscus (относя к нему формы fuscus, intermedium britanicus, иногда atlan-ticus) и серебристую чайку L. argentatus со всеми остальными формами. Штреземанн и Тимофеев-Ресовский предлагают объединить эти родственные формы (подвиды) в 4 самостоятельных вида: клушу L. fuscus, серебристую чайку L. argentatus, чайку хохотунью L. cachinnans и калифорнийскую чайку L. californicus (распределение подвидов между этими видами показано на рис. 5). Некоторые авторы доводят число выделяемых видов до 5 и даже 6.
Этот пример хорошо показывает один из возможных путей видообразования, когда образовавшиеся виды обособились еще не полностью. Подобный тщательный систематический анализ ряда других крупных политипи-ческих видов несомненно углубит представления о путях и формах эволюционного процесса.
ПРОИСХОЖДЕНИЕ НОВЫХ ВИДОВ
Объяснение происхождения видов — одна из основных проблем систематики. Состояние этой проблемы подробно изложено Э. Майром (1968).
С современной точки зрения, эволюционные изменения не обязательно приводят к увеличению числа видов. Например, изолированная популяция или группа популяций с течением времени может изменяться так, что вид «а» превратится постепенно в вид «d», пройдя эволюционные стадии видов «6» и «с» (рис. 6, Д). Такие эволюционные преобразования, видимо, свойственны видам, имеющим ограниченные ареалы, что облегчает широкое перекрестное скрещивание выживших в результате отбора особей. Поэтому непрерывное генетическое изменение популяций, слагающих данный вид, мо* жет не сопровождаться возникновением репродуктивной изоляции между
Рис. 6. Разные случаи видообразования по Майру (1968): А —изолированный остров, на котором вид «а» превращается со временем в вид «d>, последовательно проходя стадии видов <6» и «с»; Б — архипелаг из четырех островов, на которых вид «а» путем географического видообразования и колонизации образует 5 новых видов (и еще один — «6», вымирает), а на одном из островов сохраняется в неизмененном состоянии
22
ними и не приводит к дивергенции — разделению первоначального вида на два или несколько новых.
Как происходит увеличение числа видов? У высших животных наиболее вероятным сейчас представляется постепенное видообразование, являющееся результатом постепенной дивергенции (расхождения) популяций. Видимо, основной путь возникновения новых видов — это географическое видообразование. Популяция, попавшая в изолированные условия и не подвергающаяся поэтому притоку генов из соседних популяций, под воздействием определенных внешних факторов проходит основной этап видообразования: перестройку генного комплекса всей популяции. Эта перестройка постепенно приводит к выработке механизмов, репродуктивно изолирующих данную популяцию от других близких популяций. При этом иногда изолирующие механизмы могут вырабатываться раньше и скорее, чем приобретаются морфологические различия. Когда изоляция прекращается из-за устранения преграды (отступание ледника, облесение луговых участков, соединение острова с другим островом и т. п.) или впоследствии происходит расселение с колонизацией новых территорий, эти новые виды могут вновь проникнуть на территории, занятые предковым видом, но выработавшиеся в период обособленного существования механизмы репродуктивной изоляции приведут к тому, что они встретятся как совершенно самостоятельные виды с симпатрическим распространением (рис. 6, Б).
Вероятно, иногда можно говорить об усилении изолирующих механизмов при образовании области совместного обитания двух близких видов, возникшей при расселении обоих видов или одного из них. Сейчас общеизвестна широкая географическая изменчивость песни у многих видов птиц. В ряде случаев различия в песне у симпатрических (встречающихся в одном районе) популяций двух близких и сходных по внешнему облику видов значительно более резкие, чем у популяций этих же видов, географически не сталкивающихся. На всех территориях, где совместно встречаются пеночка весничка Phylloscopus trochilus (L.) и пеночка теньковка Phylloscopus colly-bitus (Vieill.), их песни резко отличаются. На Пиренейском полуострове пеночки веснички нет; песня живущего здесь испанского подвида пеночки теньковки Ph. collybitus ibericus Ticehurst похожа на песню веснички и явно отличается от песни других подвидов теньковки (Тайсхерст, 1938, из Майра, 1947).
Наконец, два (или даже больше) вида могут возникнуть из широко распространенного политипического вида, когда его конечные популяции достигли или почти достигли репродуктивной изоляции, а соединявшие их промежуточные популяции вымерли по каким-либо причинам: изменению среды обитания (вторжение ледника или морского залива, появление или исчезновение лесов на больших территориях из-за изменения климата и т. п.), может быть, из-за захватившей обширную территорию интенсивной эпизоотии и т. д.
В хорошо изолированных районах (горные хребты, острова и т. п.) иногда встречаются два или более близких вида широко распространенной группы (обычно рода), которая в любом другом районе представлена лишь одним видом. Такие случаи часто рассматривают как доказательство сим-патрического видообразования, т. е. дивергенцию одного вида на два в пределах одного и того же района. Ошибочность такой точки зрения была показана Э. Майром (1968) и другими исследователями; сейчас такие факты обычно объясняют иначе. Изолированный район был колонизирован группой особей, которая, развиваясь в изоляции, дивергировала от исходной родительской популяции и выработал^ изолирующие механизмы и экологическую обособленность к тому времени, когда в этот район вновь проникла следующая группа колонистов той же родительской формы. Такие двукратные и даже многократные волны нашествий выявляются при анализе состава фауны птиц ряда островов и даже континентов, например Австралии. Многократные переселения с острова на остров в пределах одного архипе
23
лага в некоторых случаях приводят к заметному увеличению числа видов и явному обогащению фауны. Сейчас не вызывает сомнения, что именно таким путем от 1—2 исходных предковых видов образовалось семейство (по представлениям части систематиков — подсемейство семейства Fringillidae) дарвиновых вьюрков Geospizidae на Галапагосских островах (14 видов, 21 подвид) и экологически и морфологически чрезвычайно разнообразное семейство гавайских цветочниц Drepanididae (45 подвидов, группируемых в 22 хороших вида).
Возможность симпатрического видообразования, т. е. возможность генетической дивергенции экологически специализированных популяций, населяющих одну и ту же территорию, без географической изоляции теоретически, видимо, допустима. Однако безупречных доказательств такого рода пока нет. Сейчас многими работами хорошо показано, что этому прежде всего препятствует расселение особей соседних, хотя экологически и разобщенных популяций, приводящее к генетической нивелировке подобных популяций и препятствующее видообразованию. Все факты, трактовавшиеся как случаи симпатрического видообразования, пока гораздо убедительнее и яснее объясняются с позиций географического видообразования (во всяком случае, применительно к птицам).
Образование нового вида путем слияния (гибридизации) двух ранее самостоятельных видов, видимо, в некоторых случаях возможно, но, вероятно, происходит крайне редко. Возможно, начало такого слияния видов представляет широкая гибридизация между двумя видами красноглазых тауи (Pipilo erythrophthalmus и Р. ocai) во многих районах Мексики, вызванная, как предполагает ряд исследователей, резким изменением экологических условий в результате расширения сельскохозяйственных угодий за последние 300—500 лет и сильного сокращения площади естественных местообитаний этих видов. Остальные описанные случаи «слияния» видов в действительности представляют собой полосы гибридизации подвидов, без должных обоснований рассматриваемых как самостоятельные виды (Майр, 1968).
В очень редких случаях изоляционные механизму между симпатри-ческими видами могут нарушаться в некоторых районах их совместного обитания.
Два близких вида воробьев — домовый Passer domesticus (L.) и испанский, или черногрудый Р. hispaniolensis (Temm.), — встречаются вместе на обширной территории в Южной Европе и Западной Азии (рис. 7), не скрещиваясь друг с другом. На большей части ареала домовые воробьи живут преимущественно в населенных пунктах, а черногрудые заселяют рощи и кустарниковые заросли в поймах рек и по берегам озер. В нескольких районах — Южной Италии, на островах Сицилия, Корсика, Крит и в Тунисе— оба вида живут в культурном ландшафте и между ними идет гибридизация. В гибридных популяциях фенотипически обычно представлены оба родительских вида и все возможные комбинации родительских признаков, что свидетельствует о неограниченном случайном спаривании. Однако в некоторых локальных районах (Южная Италия, оазисы на юге Туниса) закрепился один из промежуточных гибридных типов, а родительские типы исчезли (по Майзе, 1936, из Майра, 1968). Во влажных тропических лесах Африки обычны два вида райских мухоловок Terpsi phone ги-fiventer и Т. rufocinerea, а во вторичных и саванных лесах — третий вид Т. viridis. Встречаясь в одних и тех районах, каждый из видов придерживается своего биотопа, и гибридизация между ними не отмечена. Однако в северо-западной Анголе, в Западной Африке и Уганде, где в последнее время влажные леса были частично или полностью сведены, Т. viridis скрещивается с обеими видами, особенно с Т. rufiventer, местами образуя устойчивые гибридные популяции (Чэпен, 1948, из Майра, 1968).
Подобные случаи нарушения видовых изолирующих механизмов, видимо, определяются резкими изменениями экологических условий среды
24
Рис. 7. Распространение воробьев: домового Passer domesticus (L.) и черногрудого Р. hispaniolensis (Temm.) Майзе, 1936:
/ — ареал домового воробья, 2— черногрудого, 3— районы гибридизации этих видов
обитания, прошедшими в относительно короткое время преимущественно в результате хозяйственной деятельности человека. Резкие изменения среды обитания, видимо, нарушили видовые стереотипы поведения и тем самым способствовали ослаблению и нарушению механизмов видовой репродуктивной изоляции.
Случайная межвидовая гибридизация отмечена у довольно большого числа видов птиц. Она установлена у многих видов уток (внутриродовые и даже несколько случаев межродовой гибридизации), куриных (между глухарем и тетеревом, тетеревом и белой куропаткой, тетеревом и рябчиком, между американскими луговыми тетеревами Tytnpanuchus cupido и Т. phasianellus, между некоторыми видами фазанов), дятлов (в частности, для видов рода Dendrocopos), ряда видов колибри, многих видов воробьиных (между чижом и щеглом, домовым и полевым воробьем, между рядом видов райских птиц, манакинов и др.) и некоторых других видов.
Такие случайные гибриды чаще встречаются у видов, которые не образуют пар или образуют их только на короткий период спаривания: кратковременность контактов самца и самки увеличивает возможность «ошибки» в выборе партнера (утки, многие куриные, колибри и др.). Для многих родов райских птиц Paradiseidae характерны групповые тока самцов, на которых самки появляются на короткое время; строит гнездо и насиживает только самка. У большинства этих родов описаны межвидовые гибриды. У довольно многочисленных по числу особей видов рода Manucodia гибридов не обнаружено: у этого рода образуются пары, и самцы участвуют в насиживании и выкармливании птенцов. Случайная гибридизация чаще отмечается на периферии ареала одного из гибридизирующих видов, где популяция обычно сильно изрежена, и особи испытывают большие трудное^
25
ти в отыскании партнера своего вида. Видимо, этой же причиной и, одновременно, резким нарушением видового стереотипа поведения объясняются относительно частые случаи случайной межвидовой гибридизации в условиях неволи даже между видами, гибриды которых в природных условиях не встречены (некоторые виды фазанов, воробьиных и др.). Однако в целом случайные гибриды в природных условиях встречаются крайне редко. Например, за 50 лет из Новой Гвинеи вывезено более 100 тыс. шкурок райских птиц, среди которых выявлено лишь 30—50 гибридов (Майр, 1968).
Видимо, такие гибриды очень интенсивно отметаются отбором, и, вероятно, не играют никакой роли в эволюции видов.
ТАКСОНОМИЧЕСКИЕ КАТЕГОРИИ
И НЕКОТОРЫЕ ПРАВИЛА НОМЕНКЛАТУРЫ
Для построения системы необходимо установить таксономические группы разного ранга, распределив их в порядке иерархии, когда каждая вышестоящая группа включает одну или несколько более низких групп. Расположением групп в системе систематик старается отразить свои представления о их филогенетических (родственных) отношениях. Одновременно иерархия системы позволяет легче в ней ориентироваться, дает возможность довольно точно показать положение определенного организма.
Сейчас общеприняты следующие таксономические категории (перечислены в порядке от низших к высшим):
Подвид Вид
Подрод Род
Триба Подсемейство Семейство
— Subspecies
— Species — Subgenus — Genus
— Tribus (-ini)1
— Subfamilia (-inae)
— Familia (-idae)
Надсемейство— Superfamilia
Подотряд — Subordo
Отряд
Надотряд
Когорта Класс
(-oidea)
— Ordo (-formes)
— Superordo
— Cohors* * * * * 8
— Classis
Надкласс — Supercl assis
Подтип — Subphylum
Тип — Phylum
Царство — Regnum
Вид
Вид — основная систематическая категория. Начиная с К. Линнея, виду принято давать биноминальное научное название, т. е. обозначать его двумя словами (деревенская ласточка Hirundo rustica и т. п.), из которых первое — название рода, а второе обозначает именно данный вид и служит собственно видовым названием. Родовое название пишется с прописной буквы, а видовое — со строчной. В пределах одного рода не должно быть одинаковых видовых названий, а в разных родах использование одинаковых видовых названий допускается (большой пестрый дятел Dendrocopos
1 Здесь и ниже в скобках указано стандартное окончание названия данной кате-
гории (прибавляется к корню названия типового рода).
8 В большинстве систем класса птиц названия отрядов обычно оканчиваются на
formes: правила зоологической номенклатуры этого не требуют, но практически это
довольно удобно
8 В орнитологических системах обычно не применяется.
26
major, большая синица Paras major и т. д.) хотя, в общем, нежелательно, особенно в близких родах. Обычно принято, особенно в систематических работах, после видового названия ставить фамилию автора, впервые описавшего данный вид, и через запятую год публикации этого описания (Hirundo rustica Linnaeus, 1758; белая лазоревка Paras cyanas Pallas, 1770, и т. п.). В видовых названиях, впервые данных Линнеем, его фамилия обычно обозначается лишь одной заглавной буквой (L. вместо Linnaeus); фамилии других авторов чаще также обозначаются сокращенно одним или несколькими слогами, но так, чтобы данного автора нельзя было спутать с другим (например, But., вместо Buturlin, Lath., вместо Latham и т. п.).
Если данный вид первоначально включался в один род, а потом его перевели в другой, то его видовое название сохраняется, а фамилия автора, его описавшего, и год описания заключаются в круглые скобки; например, обыкновенная горлица Streptopelia tartar (L., 1758), так как Линней включал ее в род Colamba. Если при переводе вида в другой род оказывается, что там уже есть вид с таким же видовым названием, то более позднее (по дате описания) видовое название обязательно должно быть заменено.
Если один и тот же вид был описан двумя авторами под разными названиями, то действительным (валидным) названием согласно правилу приоритета признается то, которое было опубликовано раньше, а более позднее название считается синонимом и не употребляется. Этот же принцип применяется и к названиям всех других таксономических категорий.
Подвид и другие внутривидовые категории
Для обозначения индивидуальных отклонений (меланисты, альбиносы) Линней применял не имеющий точных критериев термин Varietas (разновидность). Постепенно накапливаются материалы по географической изменчивости, и уже в первой половине XIX в. становится очевидным противоречие между монотипическим пониманием вида у Линнея и его последователей и выявляющейся широкой географической изменчивостью. Его пытался решить Шлегель (1844), применяя для обозначения географических форм тройную номенклатуру. Однако это новшество не встретило поддержки и еще долго географические формы либо обозначались как разновидности (без особого названия), либо — чаще — описывались как самостоятельные виды (опубликованный Шарпом в 1909 г. список птиц мира включал более 19 тыс. видов). Но поиски путей Обозначения географической изменчивости продолжались. Клейншмидт (1900) предложил установить новую таксономическую категорию между родом и монотипическим линнеевским видом — «круг форм» Formenkreis, который бы объединял близкие, аллопатрически распространенные линнеевские виды. Ренш (1929) предложил географические формы объединять в политипический вид, называя его «кругом рас» Rassenkreis и противопоставляя его обычному монотипическому виду (Species).
Большинство систематиков предпочло пойти по другому пути: вместо введения новых терминов принять концепцию широкого биологического вида, который может изменяться во времени и в пространстве. Виды, изменяющиеся географически, стали подразделять на подвиды (иногда в качестве синонима «подвид» употребляется термин «географическая раса», что по ряду соображений мало желательно).
Подвид — единственная внутривидовая таксономическая категория, которая обозначается номенклатурно, т. е. получает собственное научное название. Подвиды обозначаются триноминальным названием (впервые ввел Шлегель, 1844), т. е. названием, состоящим из трех слов: первое — родовое, второе — видовое и третье — собственно подвидовое название, за которым ставится фамилия автора, описавшего или выделившего этот подвид и год публикации. Например, обыкновенная большая синица Paras major major
27
L., 1758; бухарская большая синица Parus major bokharensis Lichtenstein, 1823, и т. п.
Клинальная (постепенная) внутривидовая географическая изменчивость номенклатурно не обозначается. Нов практике систематических работ резко различающимся крайним популяциям клины иногда дают подвидовые названия. Установить даже приблизительные границы ареалов таких подвидов практически невозможно.
Подвиды состоят из локальных популяций, различающихся по средним значениям количественных признаков, стациальной приуроченности и некоторым тонким деталям биологии. Выявление и описание таких локальных популяций весьма полезно для систематических и экологических целей, для выяснения начальных этапов микроэволюции. Однако номенклатурно обозначать эти популяции, т. е. давать им таксономические названия, сейчас признано нецелесообразным, так как это крайне усложнило бы номенклатуру. Подобные локальные популяции обозначаются рядом терминов: дем, племя, экотип, экологическая раса, элементарная популяция и др. Практика использования этих названий, не имеющих номенклатурного статуса, еще не установилась и пока достаточно противоречива. Не получают специальных номенклатурных названий и гибриды; обычно их обозначают, используя названия обеих родительских форм. Например, гибрид между глухарем и тетеревом обозначается так: гибрид Tetrao urogallus X X Lyrurus tetrix.
Довольно часто в систематике используется нейтральный термин «группа». Этим термином иногда объединяют несколько близких родов в пределах большого семейства или несколько наиболее близких видов, в пределах большого рода. Более часто в группы объединяют подвиды широко распространенных, географически изменчивых политипических видов. Например, у большой синицы Parus major L. выделяют до 30 подвидов, которых часто объединяют в 3 группы (рис. 4). В этом случае группа именуется сочетанием двух слов: слова группа и названия наиболее характерного подвида, вокруг которого группируются остальные подвиды группы. Например, можно говорить применительно к большой синице: группа major, группа cinereus и т. п. Номенклатурного значения такие группы не имеют, но в ряде экологических и систематических исследований их применение оказывается полезным.
Высшие таксономические категории
К высшим таксономическим категориям относятся все категории выше вида: от рода до типа. Название любого высшего таксона должно состоять из одного слова, т. е. быть униноминальным, и нигде в системе не должно больше использоваться. Например, родовое название, использованное уже в системе насекомых или моллюсков, не должно использоваться в системе класса птиц и т. п.
Род
Сходные и близкие виды объединяются в род. Название рода входит в биноминальное название вида. Род — это систематическая категория, включающая один вид (монотипический род) или объединяющая несколько видов (политипический род), для которых предполагается общее происхождение, т. е. происхождение от одного общего предка. Один из видов, входящих в данный род, обозначается как типовой вид рода. Каждый род характеризуется определенным комплексом родовых признаков — морфологических, экологических, поведенческих и других, которые в той или иной степени свойственны всем видам данного рода. Обычно все виды, входящие в один
28
57 36
Рис. 8. Схема родственных отношений речных уток рода Anas (по Делакуру и Майру, 1945). Виды:
1— leucophrys, 2— waigiuensis, 3— angustirostris, 4— capensis, 5— punctata, 6— versicolor, 7— querquedula, 8—cyanopt era, 9— platalea, 10—aiscors, 11— rhynchotis, 12— clupeata, 13— smithl, 14— erythrorhynchos, 15— boha-rnensis, 16— georgica, 17— acuta, 18— falcata, 19— strepera, 20— america-na, 21— penelope, 22— sibilatrix, 23— flavirostris, 24— crecca, 25— fortno-sa, 26— aucklandica,- 27— castanea, 28— bernieri, 29— gibberifrons, 30— specularis, 31— sparsa, 32— undulata, 33— melleri, 34— fulvigula, 35— plathyrhynchos, 36— poecilorhyncha, 37—luzonica
род, занимают более или менее сходные экологические ниши. Как уже указывалось, вид отличается от любого другого вида репродуктивной изоляцией. Род отличается от любого другого рода комплексом характеризующих их признаков. Отличия этих комплексов признаков создают между родами разрывы. Характер и степень выражения разрывов между отдельными родами в разных группах птиц варьирует в значительной степени.
Объем родов может быть очень различным: от монотипических, включающих только один вид, как глупыш Fulmar us из трубконосых, люрик Plotus и топорик Lunda из чистиковых и многие др., до политипических, некоторые из которых объединяют десятки видов, например, род речных уток Anas включает 37 видов (рис, 8), род голубей Columba — 51 вид, род козодоев Caprimulgus — около 40 видов и т. п.
Иногда в пределах одного рода бывает целесообразно выделить часть видов, чтобы показать их более тесные и близкие родственные отношения по сравнению с остальными видами этого же рода. Таким группам присваивается ранг подрода. Подрод номенклатурно обозначается одним словом, которое пишется с прописной буквы и ставится в круглых скобках после родового названия; оно не входит в биноминальное название вида или триноминальное название подвида. Так, среди видов рода синица Parus иногда вьщеляют подроды гаичек Penthestes, черных синиц Peri par us, хохлатых синиц Lophophanes', в этом случае видовое название, например сероголо,-вой гаички, должно выглядеть так: Parus (Penthestes) cinctus Bodd., 1783.
Родственные аллопатрические популяции обычно рассматриваются как подвиды политипического вида. Однако в некоторых случаях они отли
29
чаются друг от друга достаточно резко и можно предполагать, что они почти достигли видового уровня, хотя между отдельными популяциями в местах их стыка возможен еще ограниченный обмен генов. Как уже указывалось выше, Майр (1931) предложил такую монофилитическую группу полностью или почти полностью аллопатрических форм называть надвидом (superspecies), а входящие в него популяции — почти сложившиеся виды — полувидами (semispecies). Эти категории пока не получили номенклатурного статуса (т. е. не получают специальных названий) и используются главным образом в исследованиях по зоогеографии и видообразованию.
Семейство
Семейство — систематическая категория, включающая один род или группу родов общего происхождения и отделенная от других семейств разрывом, т. е. более или менее явными различиями. Семейство получает название по типовому роду и имеет стандартное окончание —idae. Так, семейство синицевых по названию типового рода Parus получает наименование Paridae.
Большинство видов одного семейства характеризуется определенным морфологическим и экологическим сходством. Наиболее уклоняющиеся специализированные роды обычно (но не всегда) более или менее явными переходами связаны с типичными родами. Всем или преобладающему большинству видов семейства свойствен довольно сходный внешний облик, комплекс общих морфологических признаков и общие экологические особенности.
Для характеристики семейства и обособления его от других семейств стараются использовать большой комплекс разнообразных признаков: особенности скелета (пропорции конечностей, форма грудины, детали строения черепа, плечевого пояса и т. п.), мускулатуры, периферической кровеносной системы (характер ветвления сонных артерий и др.), расположение птери-лий и аптерий, размещение пуха, количество, форму и соотношение длины маховых перьев, ход линьки, форму, характер окраски и структуру скорлупы яиц, окраску птенцов, возрастные наряды и т. д. Для дополнительной характеристики используют и экологические признаки: сцособы и характер питания, тип гнездостроения, размеры кладки, характер позывных сигналов, тип токования, позы тока, отдыха, чесания, состав эктопаразитов и т. п. В последнее время, особенно в сложных и спорных случаях, предпринимаются попытки использовать различные физиологические и биохимические -критерии: состав гемоглобина, белковый состав сыворотки крови и мышц, фракции белка яиц, химический состав пигментов, иммунологические реакции и ряд других сложных показателей.
Разрывы между семействами обычно выражены более или менее четко, что позволяет при изучении местных фаун достаточно легко отличать друг от друга представителей разных семейств. Однако при изучении мировой фауны птиц приходится сталкиваться с тем, что имеются группы, представляющие как бы переходы между некоторыми семействами, что в значительной степени уничтожает разрывы между ними. Существование таких «переходных», или промежуточных, родов в тропиках вынуждает некоторых орнитологов объединять в одном громадном (около 1460 видов!) семействе Mus-cicapidae семейства мухоловковых Muscicapidae, славковых Sylviidae, дроздовых Turdidae, тимелиевых Timaliidae, толстоклювых синиц Paradoxor-nithidae и несколько других мелких семейств, а в объединенном семействе Fringillidae (около 550 видов!) — семейства ткачиковых Ploceidae, вьюрковых Fringillidae и овсянковых Emberizidae. При этом объединяемые семейства низводятся до ранга подсемейств. Подобная «объединительная» точка зрения пока не общепринята. Для ее обоснования необходимо дополнительное тщательное изучение с привлечением нового фактического мате
30
риала и с более углубленной и всесторонней его оценкой. Введение таких крупных объединений усложняет систему и существенно затрудняет ее практическое использование.
К. Линней группировал роды прямо в отряды, не применяя категории семейства. Это связано с тем, что среди всех животных он выделял лишь около 300 родов; известные ему 554 вида птиц он объединял в 65 родов. Резкое возрастание в XIX в. числа выделяемых родов вынудило систематиков ввести таксономическую категорию между родом и отрядом — семейство. Впервые этот термин, не отграничивая его четко от отряда, ввел Кювье (1798). Многие роды Линнея получили статус семейств. Сейчас в классе птиц принимают немногим более 200 семейств (около 170 включающих ныне живущие формы и около 40 представленных только ископаемыми формами).
При большом объеме семейства для удобства классификации и с целью лучшего выделения групп более близких родов сейчас обычно используют добавочные таксономические категории: подсемейства (название по типовому роду с окончанием -inae) и трибы (название по типовому роду с окончанием -ini). Хорошим примером их использования может служить система семейства утиных Anatidae (стр. 133).
Для отражения специфики филогенетических отношений в пределах отряда нередко используют промежуточные категории, объединяющие семейства, надсемейства (название по типовому семейству с окончанием -oidea, например надсемейство зимородковых Alcedinoidea) и Подотряды (окончание названия произвольное).
Если отряд подразделяется непосредственно на семейства, то это предполагает относительную четкость и простоту филогенетических отношений. Группировкой семейств в надсемейства и подотряды стараются показать большую разобщенность, большую степень дивергенции в пределах данного отряда.
Отряд
Отряды — главные филогенетические ветви класса. Каждый отряд характеризуется более или менее общим основным планом строения, в пределах которого вырабатываются иногда довольно разнообразные приспособительные модификации, характеризующие таксоны более низкого ранга (подотряды, надсемейства, семейства и т. д.). Отряд объединяет близкие, т. е. связанные родством, общим происхождением семейства. В системах некоторых классов, в том числе и в классе птиц, отряды группируются по родству в надотряды и в подклассы.
В орнитологической систематике почти общепринято придавать названию отряда определенное окончание formes (отряд воробьинообразных Passeriformes и т. п.). В целях классификации это очень удобно; сразу по названию можно представить ранг таксона. Однако в системах других классов этой практики применительно к отрядам придерживаются не всегда.
Для характеристики и обособления отрядов используют большой комплекс разнообразных признаков. Учитывают весь набор особенностей (признаков), используемых для характеристик семейств. Обращают внимание на основные черты пищеварительной системы (наличие зоба, степень развития мускульного желудка и слепых кишок, расположение петель кишечника), на строение верхней и, особенно, нижней гортани и т. п. Учитывают общий характер местообитаний, общий экологический облик, характер передвижения и другие особенности. Разрывы между частью отрядов птиц выражены достаточно резко, между другими — очень неясно. Следует отметить, что класс птиц в целом морфологически необычайно гомогенен (однообразен). Поэтому морфологическая обособленность между отрядами птиц выражена слабее, чем между отрядами других позвоночных животных.
31
РАСХОЖДЕНИЯ В ОБЪЕМЕ ТАКСОНОМИЧЕСКИХ КАТЕГОРИЙ
Объем и число видов птиц сейчас установлены более или менее четко. Подавляющее большинство систематиков-орнитологов считает, что в классе птиц около 8600 видов. Число спорных случаев (вид или подвид) относительно невелико и отражает объективно существующее в природе явление— случаи незавершенного видообразования.
Распределение видов по родам вызывает большие разногласия: по представлениям систематиков-«объединителей» все виды птиц должны быть распределены между 1500—2000 родов; однако некоторые систематики («крайние дробители») до сих пор признают до 7000—8000 родов! Большинство систематиков сейчас принимает около 2500 родов.
Разные взгляды и подходы к определению объема рода хорошо проиллюстрировали Майр, Линсли и Юзингер (1956): степень родства обычно выделяемых 37 видов речных уток рода Anas, основанную на характере их сходства (морфологического, экологического, этологического), графически можно изобразить в виде древа (рис. 8). Часть видов в пределах этого рода образует несколько групп, более близких между собой, чем с остальными видами (виды 32—37 группы крякв, виды 20—22 группы связей, виды 10— 13 группы широконосок и т. д.), другие виды более обцсоблены (1, 2, 18, 19, 30). Весь род достаточно четко обособлен от остальных родов три(5ы Anatini. Взаимоотношения видов, отражаемые этой схемой, практически не вызывают разногласий.
Систематики-«объединители» полагают, что степень родства между этими видами требует включения их в один род. Имеющиеся внутри рода группировки видов могут быть достаточно ясно охарактеризованы словесно или графически и не заслуживают номенклатурного выделения; в крайнем случае для выделения более четко выраженных внутриродовых групп можно использовать категорию «подрод». Систематики-«дробители» считают, что вышеприведенная схема дает основание разбить эту группу на 10— 12 родов, часть которых будет монотипична. Подобное деление, по их мнению, — единственный способ показа существования группиройок видов в этом объединении.
Подход «объединителей», видимо, и более объективен и более удобен практически. Такой позиции сейчас придерживается большинство систематиков (но, к сожалению, не все). Подобный подход значительно облегчает пользование системой (нужно запомнить одно родовое название, а не 10— 12), одновременно не затушевывая многообразных связей между видами и позволяя высказывать предположения о характере эволюции в пределах всей этой несомненно монофилетической, близкородственной группы видов. Подход «дробителей», наоборот, затушевывает несомненную близость всех этих видов и очень сильно усложняет ориентировку в системе (нужно запомнить значительно большее число родов).
Аналогичные расхождения, правда, не всегда столь резко выраженные, выявляются и при установлении объема других, более высоких таксономических категорий.
До сих пор не выработался единый взгляд на объем и количество отрядов птиц. Г. Гадов (1891, 1893) ныне живущих птиц группировал в ^отрядов, Е. Штреземанн относил их сначала к 48 отрядам (1927—1934), а позже— уже к 51 (1959); Уэтмор (1960) выделяет 33 отряда (из них 6 представлены только ископаемыми формами) и т. п. Это следствие в первую очередь недостаточности наших знаний: крайней скудности и отрывочности палеонтологических материалов, недостаточной морфологической и экологической изученности, особенно некоторых промежуточных и переходных групп и т. п. Несомненно, существенно сказывается и некоторый субъективизм в оценке уже известных.данных.
32
Разработка системы класса птиц продолжается, хотя и не очень энергично, и по мере накопления знаний существующие расхождения, несомненно, будут уменьшаться. Уже сейчас объем и таксономический статус многих групп более или менее установился и не вызывает особых дискуссий. Следует также отметить, что часть имеющихся расхождений носит в значительной степени формальный характер. Например, Уэтмор (1960), а вслед за ним К. А. Юдин (1965) и ряд других систематиков полагают, что кулики, чайки и чистиковые представляют, несомненно, родственную группу, что отражается объединением их в один отряд ржанкообразных Charadriifor-mes. Признавая несомненную близость и родство этих групп. Штреземанн (1927—1934) считал, что степень их разобщенности уже достигла отрядного уровня и потому придавал этим трем группам ранг самостоятельных отрядов. Позже Штреземанн (1959) объединил в один отряд чаек и куликов (отряд Laro — Limicolae), сохранив за чистиковыми ранг самостоятельного отряда, чем выразил свое мнение о большей близости чаек и куликов и о более резком обособлении от них чистиковых, но опять-таки не сомневался в родстве этих групп, поэтому в его системе они стоят рядом.
Таким образом, установление границ и объема высших систематических категорий до сих пор в некоторой степени носит субъективный характер. Однако любая систематическая категория имеет объективную основу, выраженную морфологическими и биологическими особенностями входящих в ее таксоны реальных видов. Майр (1971) вполне резонно указывает, что систематик не «создает» таксоны, а «открывает» группы, созданные эволюцией; он классифицирует организмы и популяции, независимо от него объективно существующие в природе. Тщательный всесторонний анализ накопленных сведений по морфологии, экологии и другим особенностям отдельных групп позволяет заметно уменьшить субъективизм в продолжающейся доработке и уточнении системы класса птиц. Хорошим примером такого анализа на уровне семейство — отряд может служить работа К. А. Юдина (1965) по филогении и классификации ржанкообразных.
В работе по уточнению и улучшению системы птиц следовало бы всегда помнить неоднократно высказывавшуюся рядом систематиков мысль о том, что классификация прежде всего должна быть наиболее полезной: соединять максимальное информационное содержание с наибольшей легкостью его отыскания. Излишнее дробительство, стремящееся отразить все возможные нюансы родства, неправомерно усложняет систему и крайне затрудняет отыскание нужной информации — выяснение положения в системе данного вида или любого другого таксона. Поэтому позиция умеренных «объединителей» представляется сейчас более полезной и плодотворной, чем деятельность крайних «дробителей». Это относится ко всем уровням системы — от подвида до класса.
НОМЕНКЛАТУРНЫЙ КОДЕКС
Практика работы систематиков уже в XVII—XIX йв. отчетливо показала, что во избежание путаницы в таксономических публикациях необходим какой-то общепринятый свод правил именования и объединения видов. Эту необходимость достаточно ясно чувствовал уже К. Линней, строго следовавший в своей работе определенным правилам: биноминальное обозначение вида, последовательное применение определенных таксономических категорий и т. п. В XIX в. стали появляться национальные кодексы (правила) зоологической номенклатуры: кодекс Стрикланда (1842) в Англии, кодекс Дэлла (1877) в США, кодексы Французского (1881) и Немецкого (1894) зоологических обществ и др. На I Международном зоологическом конгрессе в 1889 г. (Париж) был одобрен и принят кодекс Бланшара, заложивший основу современных международных номенклатурных правил. На III Международном зоологическом конгрессе (Лейден, 1895 г.) был соз
33
дан специальный Комитет по выработке правил номенклатуры. Этот Комитет выработал Международные правила зоологической номенклатуры, принятые V Международным зоологическим конгрессом в 1901 г. (Берлин). На следующем зоологическом Конгрессе Комитет был преобразован в Постоянную международную комиссию, задачей которой было рассматривать спорные случаи в разных разделах зоологической систематики, высказывать свое мнение по отдельным статьям правил, вырабатывать новые статьи и положения и т. п., способствуя этим стабильности номенклатуры. Результаты своей работы комиссия публиковала в нерегулярно издававшихся «Мнениях» и в «Декларациях», а позже в «Бюллетене зоологической номенклатуры».
В результате деятельности Постоянной международной комиссии по зоологической номенклатуре в послевоенные годы был подготовлен и в 1958 г. принят XV Международным зоологическим конгрессом в Лондоне «Международный кодекс зоологической номенклатуры», некоторые поправки к которому были дополнительно приняты XVI Международным зоологическим конгрессом (1963, Вашингтон).
«Международный кодекс зоологической номенклатуры» устанавливает основные принципы присвоения названий систематическим категориям (таксонам) от подвида до семейства включительно и ставит своей основной задачей способствовать максимальной универсальности, преемственности и стабильности зоологической номенклатуры. Замечания по номенклатуре, изложенные выше, основываются на положениях кодекса, но далеко не исчерпывают его содержания. Поэтому каждому, кто интересуется или занимается систематикой, необходимо внимательно ознакомиться с самим кодексом.
НЕКОТОРЫЕ ЗООГЕОГРАФИЧЕСКИЕ ПОНЯТИЯ,
БОЛЕЕ ЧАСТО ИСПОЛЬЗУЕМЫЕ В СИСТЕМАТИКЕ
Таксон любого ранга (подвид, вид, род, семейство, отряд), встречающийся только в одном определенном районе, называется эндемиком этого района. Например, краснозобая казарка Rufibrenta ruficollis (Pall.)— эндемик тундры и лесотундры Западной Сибири, кавказский тетерев Lyru-rus mlokosiewiczi (Tacz.) — эндемик Кавказа, семейство гавайских цветочниц Drepanididae — эндемик Гавайского архипелага, киви (виды, род, семейство, отряд) —эндемик Новой Зеландии, тупик Fratercula arctica (L.) — эндемик Северной Атлантики, а ипатка F. comiculata (Naum.) — Северной Пацифики.
Определение очагов (центров) возникновения и путей последующего расселения групп разного таксономического ранга — одна из задач зоогеографии. Но с этими вопросами приходится сталкиваться и систематику. Географический район, где предположительно возник тот или другой таксон, называют центром, иДи очагом, возникновения группы; эта группа для данного района автохтонная. Вероятным центром возникновения голубей считают Австралию, т. е. они, видимо, ее автохтоны; тетеревиные, вероятно, автохтоны Неарктики, а фазановые — Палеарктики.
В противоположность автохтонам, таксоны, проникшие в данный район с других территорий, называют иммигрантами.
Таксоны любого ранга от вида и выше, распространенные по всем материкам, называют космополитами. Как правило, чем ниже таксономический ранг группы, тем меньше (уже) область ее распространения— ареал. Но это общее правило имеет много исключений. Ареал рода уларов Tetraogallus ограничен горными системами Кавказа и Азии; род дроздов Turdus и род ласточек касаток Hirundo — практически космополиты. Очень широко — почти космополитически — могут быть распространены
34
Рис. 9. Разорванные ареалы:
/— голубой сороки Cycmoplca суета (Pall.), 2— горной чечетки Cannabina flavirostris (L.)
и некоторые виды. Например, скопа Pandion haliaetus (L.) встречается в подходящих местообитаниях почти по всем материкам (нет ее в Южной Америке). Очень широк ареал болотной совы и ряда других видов.
Узкие (малые по площади) ареалы обычно характерны для древних вымирающих форм (так называемые реликтовые ареалы, которые раньше были более широкими, например ареал африканского страуса) или, наоборот, для молодых, недавно возникших видов или более высоких таксонов (узкие ареалы уже упоминавшихся выше гавайских цветочниц или галапагосских вьюрков).
Ареалы, охватывающие все северные районы Европы, Азии и Северной Америки, называются кругоарктическими, или циркумполярными. Таковы ареалы пуночки Plectrophenax nivalis (L.) и лапландского подорожника Calcarius lapponicus (L.). Ареалы, охватывающие умеренные широты Европы, Азии и Северной Америки, называются кругобореальными, или циркумбореальными, Такие ареалы имеют ястребиная сова Surnia ulula (L.), щур Pinicola enucleator (L.) и ряд других видов. Ареалы, охватывающие большую часть Европы и Азии, называют палеарктическими.
В пределах своего ареала каждый вид обычно встречается не повсеместно, а лишь по подходящим биотопам, пятнисто, образуя так называемое «кружево» ареала. У некоторых видов заселяемые участки территории отстоят один от другого очень далеко. В этих случаях говорят о разорванном ареале. Такой разорванный ареал, например, имеет голубая сорока Cyanopica cyanus (Pall.), населяющая Пиренейский полуостров и Юго-Восточную Азию или живущая на трех изолированных участках на северо-западе Европы, на Кавказе и в прилегающих районах Малой Азии и Ирана и в Центральной Азии горная чечетка Cannabina flavirostris (L.) (рис. 9). Подобные разорванные ареалы свойственны и ряду других видов.
НЕКОТОРЫЕ МОРФОЛОГИЧЕСКИЕ ПРИЗНАКИ, ИСПОЛЬЗУЕМЫЕ В СИСТЕМАТИКЕ
В систематике, как сказано выше, широко используется очень большой набор морфологических признаков и особенностей строения. Поэтому систематик должен хорошо знать морфологические термины и понятия. Здесь упоминаются лишь некоторые, более часто употребляемые в орнитологической таксономии.
Типы расположения пальцев
Обычное для большинства птиц расположение пальцев — три вперед и один (первый) назад — называют анизодактильным типом лапы. У пингвинов, веслоногих, стрижей все четыре пальца более или менее отчетливо направлены вперед; такую лапу называют помпродак-тильной. У трогонов назад обращены I и II пальцы — это гетеро-дактильный тип лапы. У попугаев, кукушек, дятлов назад обращены I и IV пальцы; лапа такого типа называется зигодактильной.
Если основные фаланги направленных вперед пальцев — трех или двух — более или менее заметно срастаются роговыми покровами, то такое состояние называют синдактилией.
36
Типы скелетной основы ноздрей
Различают два типа ноздрей: голоринальный и схизо-ринальный (рис. 10). При го-лоринальном (округлом) типе ноздрей верхнечелюстной и межчелюстной отростки носовой кости разделены неглубоко, а широко закругленный угол их схождения, окаймляющий ноздрю сзади, лежит впереди от линии соприкосновения лобной и межчелюстной костей. Такой тип ноздрей характерен для пингвинов, гагар, трубконосых, куриных, веслоногих, пастушковых, дроф, кукушек, ракшеобразных, дятлообразных, воробьиных. При схизо-ринальном (щелевидном) типе ноздрей эти отростки носовой кости расщеплены глубоко и вершина образованного ими угла — задний край ноздри — очень остра или щелевидна
Рис. 10. Схема типов скелетной основы ноздрей. I — голоринальный (дрофа); II —схизоринальный (журавль):
/— носовая кость, 2— межчелюстная кость, 3— лобная кость; ноздри показаны черным цветом
и лежит на уровне лобно-межчелюст-
ного шва или даже заходит несколько назад. Схизориния характерна для ибисов, журавлей, большинства куликов, чаек, чистиковых, голубей,
рябков и некоторых других птиц.
Эти различия в скелетной основе ноздрей сказываются на характере
подвижности надклювья.
Кинетизм черепа и типы строения костного неба
Костное небо птиц подвижно. Оно образовано небными отростками межчелюстных и верхнечелюстных костей, сошником, небными и крыло-видными костями. Подвижные квадратные кости сочленяются с крыловидными костями, которые в свою очередь сочленяются с небными. Сочленение небных и крыловидных костей может скользить вдоль клювовидного отростка парасфеноида. Подвижность костного неба и истончение костей у основания верхней части надклювья в области «переносицы» (а иногда и образование здесь соединительнотканной перемычки— синдесмоза или даже настоящего сустава) обеспечивают кинетизм — способность надклювья смещаться вверх и вниз по отношению к мозговому отделу черепа. Для птиц характерна кинетичность двух типов (Юдин, 1965, 1970; Дзержинский, 1971): прокинетизм (гибкая зона лежит у основания надклювья) и ринхокинетизм (гибкая зона расположена на уровне носовых ям, верхняя челюсть обладает внутренней подвижностью). Кинетизм позволяет увеличить усилие на конце клюва, обеспечивает (вместе с дифференциацией челюстных мышц и связок) разнообразие производимых клювом движений, облегчает заглатывание добычи.
Выделяют 4 типа костного неба, различающихся по расположению образующих его костей (рис. 11):
1. Дромеогнатический тип. Небные и крыловидные кости неподвижно соединяются друг с другом; хорошо развиты базиптеригоид-
37
Рис. 11. Схема типов костного неба птиц. I — дромео-гнатический (нанду); II — схизогнатический (курица); III — десмогнатический (утка); IV — эгитогнати-ческий (ворона):
1— небные отростки верхнечелюстных костей, 2— сошник, 3— небные кости, 4~крыловидные кости, 5 — базиптеригоидное сочленение
ные сочленения; передний конец сошника обычно сливается с небными отростками верхнечелюстных костей, задний конец сошника расщеплен и его расходящиеся концы соединяются с небными и крыловидными костями, отделяя их от клювовидного отростка. Различные вариации такого типа костного неба характерны для тинаму и страусоподобных бескилевых птиц.
2. Схизогнатический тип. Небные отростки пра-вЬй и левой верхнечелюстных костей не соединяются друг с другом; небные и крыловидные кости подвижно сочленяются друг с другом и примыкают к клювовидному отростку; сошник спереди заострен и свободен —не соединяется с небными от
ростками верхнечелюстных костей. Различные вариации схизогнатического типа неба характерны для пингвинов, гагар, трубконосых, куриных, большинства журавлеобраз
ных, ржанкообразных, трогонов, большинства сов, козодоевых, колибри,
настоящих дятлов.
3. Десмогнатический тип. Небные отростки правой и левой верхнечелюстных костей сливаются друг с другом; сошник либо совсем редуцируется, либо узкий, палочковидный, задним концом иногда соединяющийся с небными костями и клювовидным отростком, а спереди — со слившимися небными отростками верхнечелюстных костей. Вариации такого типа неба свойственны веслоногим, голенастым, гусеобразным, дневным хищникам, некоторым козодоям, ракшеобразным, якамарам, пуховкам и туканам из дятлообразных.
4. Эгитогнатический тип. Тонкие удлиненные небные отростки верхнечелюстных костей подходят под сошник (если смотреть на череп снизу, то они нависают над сошником), но не соприкасаются ни с сошником, ни друг с другом; сошник широкий со свободным широким, иногда раздвоенным передним концом; задний его конец глубоко расщеплен и охватывает клювовидный отросток. Такой тип неба встречается у трехперсток, стрижей, бородатой и медоуказчиков из дятлообразных, почти у всех воробьиных.
Второй — четвертый типы неба иногда объединяют в группу неогнат
38
(новонебных), противопоставляя ее дромеогнатическому или палеогнати-ческому (древненебному) типу. Десмогнатический и эгитогнатический типы неба, видимо, возникли путем перестройки более древнего схизогнатического типа. Между всеми этими типами неба существуют более или менее выраженные переходы.
Базиптеригоидные сочленения. У страусоподобных птиц, тинаму, некоторых трубконосых, гусеобразных, сов и некоторых других птиц на боковых сторонах основной клиновидной кости расположены выросты — базиптеригоидные отростки; каждый из них образует скользящее сочленение с боковой поверхностью крыловидной кости соответствующей стороны. У пингвинов, чаек, чистиковых и некоторых других базиптеригоидные сочленения образуются в эмбриогенезе и полностью исчезают по мере роста птенцов.
У трубконосых, веслоногих, чаек и других птиц имеется стрепто-г н а т и з м: благодаря специальным устройствам ветви нижней челюсти (подклювья) могут выгибаться наружу, значительно увеличивая раскрытие ротового отверстия и обеспечивая заглатывание крупной добычи (Юдин, 1965, 1970).
Формула ножной мускулатуры
Детали строения мускулатуры разных частей тела широко используются в систематических целях. Гаррод (1873) ввел довольно часто теперь употребляемые буквенные обозначения некоторых мышц задней конечности: «А» — хвостовая часть m. caud-ilio-femo-ralis (m. adductor longus), «В»— подвздошная часть того же мускула, «X» — хвостовая часть ш. caud-ilio-flexorius (m. semitendino-sus), «У» — подвздошно-бедренная часть того же мускула (рис. 12). Если есть все порции этих мускулов, то ножная формула имеет вид «ЛВХК», а если какая-то из порций отсутствует, то из формулы исключается соответствующий индекс. Ножная мышечная формула гагар «ЛВХ», поганок — «ВХ», у сов, стрижей и некоторых других птиц — только «Л». Наличие или
Рис. 12. Схема расположения мышц, используемых для составления ножной формулы: А — хвостовая часть т. caud-ilio-femoralis, В — подвздошная часть того же мускула, X — хвостовая часть m. caud-ilio-flexorius, Y — подвздошно-бедренная часть того же мускула, а — m. ambiens
отсутствие отводящей мышцы ш.
ambiens обозначается в вышеописанной формуле знаком + или — . Так, он есть у гагар и отсутствует у поганок; их ножные формулы соответственно будут «ЛВХ4-» и «ВХ—».
Нижняя гортань
Обычно выделяют 3 основных типа нижней гортани птиц, каждый из которых в свою очередь может быть подразделен на группы. 1. Т р а х е о -бронхиальны й— голосовые перепонки и голосовые мышцы расположены на нижних кольцах трахеи и верхних кольцах бронхов; этот тип гортани свойствен большинству птиц. 2. Бронхиальный — голосовые
39
перепонки и мышцы расположены на верхних кольцах бронхов и не достигают нижних колец трахеи. Такая гортань описана у некоторых козодоев и кукушек. 3. Трахеальный — голосовые перепонки и мышцы расположены только на нижних утонченных кольцах трахеи; встречается у ряда южноамериканских кричащих воробьиных. У некоторых групп птиц нижняя гортань может быть промежуточного, переходного строения.
Типы оперения крыла
Большие верхние и нижние кроющие крыла прикрывают основания маховых перьев. Обычно число маховых и их больших кроющих строго совпадает (тинаму, кукушки, трогоны, дятлы, воробьинообразные). Такое крыло называют эутаксическим или квинтокубитальным. У гагар, трубконосых, голенастых, гусеобразных, дневных хищников, сов, козодоев между 4-м и 5-м второстепенными маховыми располагаются добавочные верхние и нижние кроющие. Такое крыло называют диастата к-сическим или аквинтокубитальным. В ряде отрядов (поганкообразные, курообразные, журавлеобразные, веслоногие, ржанкообразные, голубеобразные, попугаеобразные, ракшеобразные, стрижеобразные) встречаются оба типа крыла. Полагают, что эутаксия первична, а диа-статаксия вторична и развивается как приспособление, прежде всего облегчающее сильное складывание крыла (Штефан, 1970).
ИСТОРИЯ СТАНОВЛЕНИЯ СИСТЕМЫ КЛАССА ПТИЦ. СОВРЕМЕННЫЕ СИСТЕМЫ
Из многочисленных систем класса птиц, предлагавшихся разными авторами, здесь упоминаются лишь немногие, отражающие изменение подхода к изучаемому материалу и оказавшие большее влияние на развитие орнитологии.
Историю орнитологии, как и многих других разделов естественных наук, обычно и вполне обоснованна начинают с древнегреческого философа Аристотеля (384—322 гг. до нашей эры). Этот величайший мыслитель древности родился в г. Стагира в семье врача. В 17-летнем возрасте он отправился в г. Афины, где 20 лет был ближайшим учеником философа Платона. Позже в Течение 6 лет Аристотель был учителем и наставником будущего полководца Александра Македонского. В возрасте 48 лет Аристотель возвращается в Афины и открывает собственную школу Ликей. Во главе этого своеобразного университета Аристотель находился почти до самой смерти. Большинство известных сочинений Аристотеля, вероятно, представляют собой лекции, читавшиеся им в Ликеепо различным естественным и общественным циклам и охватившие все области знаний того времени.
Дошедшие до нас (видимо, не полностью, с более или менее значительными пропусками) 3 зоологических трактата — «История животных», «О частях животных»1 и «О возникновении животных» — представляют собой критическое обобщение всех известных Аристотелю сведений о различных животных: о их внешнем виде, анатомических особенностях, поведении, размножении и т. п. Практически это энциклопедия зоологических знаний того периода. При изложении и систематизации этих материалов Аристотель широко применяет сравнительный метод, пользуется принципом корреляции и аналогии, принципом общности всего живого и постепенного повышения организации. Следует отметить, что в области конкретных знаний, в том числе и в зоологии, Аристотель проявляет себя как материалист (хотя далеко не всегда последовательный).
Самостоятельной системы животного мира Аристотель не создал. Однако при чтении его зоологических сочинений отчетливо видно, что в,се известные ему животные (он упоминает около 540 видов) объединяются в несколько соподчиненных групп. Классификационная единица Аристотеля — род, который характеризуется многими признаками и объединяет
1 Аристотель. О частях животных. Биомедгиз, 1937.
Аристотель. О возникновении животных. Изд. АН СССР, М.—Л., 1940.
Aubert Н. und Wimmer Fr. Aristoteles Thierkunde. Bd.-l—2; Leipzig, 1868.
41
животных «по общности». В пределах каждого рода животные (виды) имеют «общую природу» и отличаются друг от друга размерами тела и внешним видом его отдельных частей, а также привычками и поведением. Судя по тексту, Аристотель различал несколько категорий рода: а) высший род (примерно соответствует классу), б) большой, или низший род (в современном понимании примерно соответствует отряду); в некоторых случаях род использовался им и примерно в современном понимании.
По выборке из зоологических трактатов Аристотеля классификация животного мира выглядит так (в скобках — обозначение группы в современном представлении):
А. Животные с кровью (Позвоночные):
Высшие роды: 1. Живородящие четвероногие, покрытые волосами
(Млекопитающие)
2. Яйцекладущие двуногие, с перьями, летают (Птицы)
3. Яйцекладущие четвероногие и безногие, дышат легкими (Пресмыкающиеся и Земноводные)
4. Яйцекладущие, живут в воде, дышат жабрами (Рыбы)
Б. Животные без крови (Беспозвоночные):
Высшие роды: 5. Мягкотелые, ноги на голове (Головоногие моллюски)
6. Мягкоскорлуповые, много ног (Ракообразные)
7. Черепокожие (Раковинные), без ног (Моллюски, Морские ежи)
8. Насекомые: тело с насечками (Насекомые, различные черви, в том числе и глисты)
9. Хаос (все остальные животные, не вошедшие в вышеуказанные группы)
Схема деления группы животных с кровью в общих чертах близка к современным представлениям об основных классах позвоночных животных (млекопитающие, птицы, рыбы); объединение пресмыкающихся и земноводных сохранялось еще до конца прошлого века). Интересно, что при описании строения скелета Аристотель подчеркивает, что только животные с кровью имеют хребет (т.,е. позвоночный столб).
В сочинениях Аристотеля упоминается около 160* птиц (видимо, он знал больше, так как не называются некоторые обычные в Греции виды), из которых по его описаниям более или менее точно сейчас можно узнать около 125 видов. Всех птиц он разделяет на 8 больших родов, примерно соответствующих рангу отрядов в последующих системах.
Классификация, птиц по Аристотелю (по выборке из его зоологических трактатов) выглядит следующим образом:
Высший род: Ornithes — Птицы
Большие роды:
1. Gampsonyches, или Sarcophaga, —Кривокоготные, или Мясоядные. Это группа хищных птиц, которых Аристотель подразделяет на две подгруппы:
а) Дневные птицы, к которым он относит орлов, ястребов, грифов, коршуна, соколов;
б) ночные птицы, к которым относятся филин, сычи, совы
2. Scolecophaga — Насекомоядные. Сюда он причисляет козодоя, трясогузку, конька, королька, пеночек, зяблика, воробья, зеленушку и др.
3. Acanthophaga — Растительноядные. К этой группе относятся мелкие воробьиные птицы, например коноплянка, (другие названия птиц этой группы сейчас трудно идентифицировать)
4. Schinopophaga — Фисташкоядные, но, видимо, можно понимать и как «долбящие». Сюда он относит дятлов (пестрого, зеленого) и некоторых воробьиных (пищуху, синицу)
5. Peristeroeide — Голубеобразные (клинтух, вяхирь, домашний голубь, горлица)
6. Parydra schizopoda — Водные раздельнопалые. К этой группе Аристотель причисляет разнообразных околоводных птиц: цапель, колпицу, куликов, скопу и, вероятно, орлана белохвоста и др.
7. Steganopoda — Перепончатопалые (водоплавающие), включающие уток, поганок, баклана, лебедя, гуся
8. Вагеа — Тяжелые, или Наземные. Эта группа включает куриных птиц: перепела, куропатку, кеклика, фазана
42
Так как Аристотель не ставил перед собой задачу создать всеобъемлющую классификацию птиц, то, естественно, часть упоминаемых видов не причисляется им ни к одной из вышеперечисленных групп (соловей, дрозды, ворон, щурка, попугай, дрофа). Часть названий птиц, относимых им к той или иной группе, до сих пор не удалось связать с определенными видами из-за отсутствия их описаний.
Эта классификация (которая для Аристотеля имела второстепенное, подчиненное значение) позволяет группировать известные ему факты «по общности», связывать некоторые особенности строения с привычками и поведением (у хищников быстрый полет и острое зрение, у водных птиц строение лапы приспособлено для плаванья и т. п.). Поэтому можно считать Аристотеля создателем первой системы всего животного мира и автором первой системы класса птиц.
Другие ученые древнего мира — Греции и Рима, писавшие о животных, для истории зоологии, в том числе и для орнитологии, имеют несравненно меньшее значение, так как в их сочинениях намного меньше фактического материала и он значительно хуже осмыслен и обобщен. Поэтому в истории науки Аристотеля вполне заслуженно называют «отцом биологии (зоологии)»; в его сочинениях истоки многих разделов современных естественных наук.
Первые века нашей эры характеризуются резким падением уровня знаний. И основная причина этого — не разгром античных государств ордами «варваров» (вандалы, готы и др.), а распространение в Южной Европе христианской религии. Нетерпимость «отцов церкви» к еретикам выражалась в разрушении древних храмов, в сжигании находившихся при них хранилищ рукописей, в физическом уничтожении людей, осмеливающихся хоть в чем-нибудь отклоняться от церковных догм. Так, в конце IV в. нашей эры была сожжена сокровищница античной мысли — Александрийская библиотека. Сочинения греков и римлян сохранились (часто в разрозненных отрывках) лишь в арабских странах: ислам несколько терпимее, чем христианская религия, относился к трудам инаковерующих. Часть античных сочинений была переведена на арабский язык. Развивались в арабских странах и науки: астрономия, математика, география и др.; при этом использовалось и античное наследство. Зоологией арабы интересовались мало, и сколько-нибудь существенного развития, по сравнению с Аристотелем, она не получила. Однако дошедшие до нас зоологические трактаты Аристотеля сохранились преимущественно в переложениях и переводах арабских ученых. В Европе же под защитой христианской церкви существовала и развивалась только одна «наука» — богословие.
Дальнейшее развитие и укрепление феодализма было связано с про-должавшимися около 200 лет крестовыми походами (1095—1270). Эти походы способствовали проникновению на христианский Запад культуры Востока. На латинском языке стали появляться переводы трудов арабских ученых и, преимущественно в их пересказах и переложениях, некоторые сочинения античных мыслителей. Так в Европе стали известны и зоологические трактаты Аристотеля. Появляются сочинения нескольких «святых отцов», которые описывают природу на основании фактов, почерпнутых из Аристотеля и других источников и обильно дополненных вымыслом (частично из сочинений древних, частично из сочинений более поздних времен) о различных «чудищах морских и земных», но в которых старательно выброшены все попытки объяснения природных явлений, к чему стре-: мились греки и что могло поколебать веру в бога. Такие «очищенные» сочинения в какой-то степени удовлетворяли интерес людей к окружающей их природе и в то же время не колебали церковных догм. Так в средне^ вековье сочинения Аристотеля начали свою вторую жизнь, но уже под защитой церкви. Естественно, что способствовать развитию зоологии они-не могли.
43
Единственным человеком, с именем которого в этот период действительно связано развитие орнитологии, был германский король (с 1220 г. — император) Фридрих II Гогенштауфен (1194—1250). Он не только покровительствовал наукам, но и сам изучал философию (греков и арабов), интересовался естественными науками, был инициатором одного из переводов сочинений Аристотеля с арабского языка на латинский. Особый интерес представляет законченный Фридрихом II примерно в 1247 г. трактат «Об искусстве охотиться с птицами» (De arte venandicum avibus). В отличие от других средневековых авторов и даже писателей эпохи Возрождения Фридрих II писал не по литературным источникам (среди которых основное место обычно занимали сочинения Аристотеля), а преимущественно по собственным наблюдениям. Он впервые описал пневматичность костей птиц и копчиковую железу, довольно детально охарактеризовал строение легких и гортани, дал описание перелетов и попытался объяснить их значение, впервые обратил внимание на географическую изменчивость птиц. В его сочинении есть сведения о питании, линьке, размножении, о зависимости скорости полета от размеров птиц и степени развития грудных мышц и т. п. При описании хищных птиц он не только приводит различные практические сведения, нужные охотящемуся с ловчими птицами, но дает описание внешнего вида и некоторых особенностей жизни и поведения, а также приводит их классификацию, которая очень близка к современной (деление на соколов и остальных дневных хищников, выделение среди последних ястребов, орлов, коршунов и т. д.). Пытался он объяснить и механику птичьего полета. Этот трактат среди других зоологических сочинений средневековья представляет совершенно исключительное явление. Более широко этот труд стал известен значительно позже, после того как он был напечатан в 1596 г. и, вторично, в 1788 г.
Охота с хищными птицами была широко распространена в средние века и сохраняла свое значение до XVIII в., а местами и до XIX в. О ней писали в XII—XVII вв. (в Китае первое сочинение о соколиной охоте появилось в VII в., в Персии — в 1145 г.). Однако действительно научных материалов эти трактаты о соколиной охоте содержали мало и, пожалуй, их значение было лишь в том, что они привлекали внимание к птицам; это в какой-то степени способствовало накоплению фактов и наблюдений.
Альберт Великий (род. в начале XIII в., умер в 1280 г.), доминиканский монах, пытался связать естественные науки с богословием и тем самым укрепить догмы церкви. Он выступал с лекциями и проповедями во многих городах Европы, привлекая большое число слушателей и широтой разбираемых вопросов и своим талантом оратора. В своих сочинениях он писал об астрономии и зоологии, о географии и философии. Его сочинение «О животных» (De animalibus; напечатано в 1478 г.) содержит некоторые сведения о птицах, взятые преимущественно из Аристотеля, с небольшими, обычно малоценными, собственными дополнениями и исправлениями. Популярность автора способствовала в какой-то степени привлечению внимания к естествознанию и помогала накоплению зоологических наблюдений. Однако в целом в период средневековья орнитология вперед продвинулась мало (и в основном лишь трудами Фридриха II).
XV—XVI вв. — эпоха Возрождения. Это период расцвета феодализма, период начала великих кругосветных путешествий (Колумб открыл Америку в 1492 г.), резко расширивших представление о земле и способствовавших развитию различных естественных наук (в первую очередь путем увеличения наблюдений, сбора коллекций в «заморских странах», привоза необычных животных и растений и т. п.). Развитию искусств (особенно литературы) и науки способствовало изобретение и довольно быстрое распространение книгопечатания (середина XV в.). Эта эпоха характеризуется и возрождением интереса к античному искусству и античной науке. Вновь переводятся на латинский язык сочинения греческих ученых, теперь уже в менее «исправленном» виде. Однако если Аристотеля
44
даже «не улучшенного», а более или менее подлинного церковь допускала, то за резкие отклонения от его взглядов грозил костер (Джордано Бруно был сожжен в 1600 г.). Но костры инквизиции не могли остановить начавшегося развития естественных наук. Однако в зоологии эти процессы пока проявляются слабо, идет преимущественно накопление новых, проверка ранее описанных фактов и постепенное освобождение от накопившихся зоологических мифов, выдумок и небылиц (например, зародыши гадюки пожирают внутренности своей матери; саламандра живет в огне и т. п.).
В середине XVI в. появляются крупные обобщающие орнитологические работы, заметно ускорившие развитие орнитологии.
Виллиам Турнер (1500—1568), англичанин, в 1554 г. в Кельне издал книгу «Avium praecipuarum, quarum apud Plinium et Aristotelem mentio est, brevis et succincta historia». («Тех птиц, которые у Плиния и Аристотеля упоминаются, краткая и сжатая история»), содержавшую первый по-настоящему критический обзор орнитологических материалов Аристотеля и других писателей древности. Хорошо зная птиц Англии и континентальной Европы, Турнер дал ясные и обстоятельные описания многих птиц, нечетко описанных его предшественниками, и в ряде случаев сопроводил их краткими заметками о их образе жизни (питание, способ добывания пищи и т. п.), обычности или редкости их в Англии и Германии и т. п.
Конрад Геснер (1516—1565), швейцарец, происходил из бедной семьи, но благодаря своей настойчивости смог окончить университет и в 22 года стал профессором греческого языка. Однако это его не увлекало и он стал врачом, одновременно занимаясь естественными науками. Знал много языков (более 10), что облегчало знакомство с литературой. Побывал во многих странах Европы, знакомясь с их природой. Написал ряд книг по ботанике, минералогии, литературе. Но длительную известность и славу ему принес многотомный труд (более 4000 страниц) «История животных» (Historia animalium), выходивший отдельными томами в 1551—1587 гг. (том I — млекопитающие, том II — яйцекладущие четвероногие — амфибии и рептилии, том III — птицы, том IV — рыбы и том V, опубликованный уже после смерти автора, насекомые). При выделении крупных групп Геснер применял классификацию Аристотеля. В посвященном птицам III томе (De Avium natura, 1555) на 800 страницах помещены изображения (гравюры на дереве) и описания 188 птиц, сделанных по одному плану; внешний вид и распространение, названия на разных языках, образ жизни и повадки, происхождение названия, использование этого вида в баснях, сказках, поговорках, гербах и т. п., подробный, практически исчерпывающий список сочинений, в которых упомянут или описан этот вид. При описании животных Геснер приводит не только критически оцениваемые литературные сведения, но и свои довольно многочисленные наблюдения.
Никакой классификации птиц (как и других описываемых групп) Геснер не дает: все животные располагаются в алфавитном порядке их названий.
Это сочинение Геснера представляет собой своеобразную зоологическую энциклопедию, собравшую сведения, накопленные почти за 2000 лет. Ясный язык, точность описаний животных, сопровождаемых рисунками (часть из них были вполне удовлетворительными), обилие фактического материала сделали все тома «Истории животных» очень популярными: они несколько раз переиздавались на разных европейских языках и вплоть до конца XVII в. служили настольной книгой любителей естествознания, привлекали внимание к изучению зоологии и способствовали дальнейшему накоплению фактического материала, в том числе и по орнитологии. Следует отметить исключительную работоспособность Геснера. Прожил он всего 49 лет. В 1565 г. в Цюрихе вспыхнула эпидемия чумы. Геснер бросил свои зоологические занятия и, работая как врач, пытался оказывать помощь в борьбе с эпидемией. Вскоре он сам заразился чумой и умер.
45
Отсутствие всякой классификации внутри крупных групп (перечисление животных по алфавиту) у Геснера, конечно, — шаг назад по сравнению даже с Аристотелем. Но этот недостаток с лихвой окупается (при недостаточности фактических материалов для построения системы) полнотой описаний, систематизацией всего известного материала и его критической (насколько это возможно при тогдашнем уровне знаний) оценкой. Все это обеспечивало дальнейшее развитие зоологии и было определенным этапом, подготавливающим возможность разработки системы животного мира, в том числе и системы птиц.
Альдрованди (1522—1605) из Болоньи написал три тома «Орнитологии» (1599—1603). Это тоже сводка — энциклопедия, в которой сильно чувствуется влияние Геснера. Однако Альдрованди был менее критичен, и в его работе значительно больше, чем у Геснера, мифов и небылиц. Положительным было описание довольно большого числа птиц из Азии, Африки, Америки по поступившим к нему коллекциям (например, туканов, птиц-носорогов, казуара и др.). Довольно тщательно описана мускулатура орла, язык зеленого дятла, трахея лебедя и т. п. При описании видов Альдрованди объединяет их в группы, уже применявшиеся Аристотелем, и выделяет новые: певчих птиц (зяблик, жаворонок, канарейка), птиц, купающихся в пыли и в воде (голуби, овсянки). Эта классификация весьма произвольна, и она не оказала заметного влияния на становление системы птиц.
Пьер Белон (1517—1564), француз, учился в университетах Германии и Италии, затем много путешествовал (Европа, Турция, Северная Африка), занимаясь изучением преимущественно птиц и, в меньшей степени, рыб и растений. Он напечатал описания своих путешествий, а также книги по птицам и рыбам. Полностью завершить обработку всех своих наблюдений он не смог. Для развития орнитологии особый интерес представляет его «L’ histoire de la nature des oyseaux, auec leurs descriptions, et naifs portraicts, retirez du naturel», Paris, 1555. В ней он описывает примерно 300 видов птиц. Как правило, описания оригинальны и точны, так как он пишет только о тех птицах, которых сам видел. При описании их биологии он сообщает свои наблюдения, а также довольно критически используемые литературные источники, включая Аристотеля (однако им использовано меньшее число источников, чем у Геснера).
Все описываемые виды на основе сходства образа жизни и некоторых особенностей внешнего облика Белон объединяет в 6 групп (в современном понимании в какой-то степени равноценных отрядам):
1. Хищные птицы (включая дневных и ночных хищников)
2. Водные птицы, имеющие плавательные перепонки (их он подразделяет на обитателей пресных и соленых водоемов)
3. Болотные (околоводные) птицы со свободными пальцами, т. е. без плавательных перепонок (цапли, кулики; сюда же отнесены зимородок и щурка)
4. Наземные птицы (страус, различные куриные, жаворонки и др.)
5. Крупные древесные и кустарниковые птицы
6. Мелкие древесные и кустарниковые птицы (сюда включены и ласточки)
Четкой характеристики отдельных групп он не дает. В целом классификация весьма схематична (основана в основном на различиях в характере местообитаний, отчасти на отличиях в питании) и очень похожа на классификацию Аристотеля (влияние великого грека на систематику птиц сохраняете^ практически до конца XVIII в.). Отличия сводятся лишь к изменению названий отдельных групп и к некоторой их переформировке, определяемой тем, что Белон в первую очередь основывался на сходстве местообитаний, а Аристотель — на сходстве в питании. Однако при отнесении определенной птицы к той или иной группе Белон больше, чем его предшественники, обращал внимание на многие внешние признаки — размеры и форму клюва и лап, отчасти окраску и характер оперения и т. п. Это полезное нововведение далее используется многими систематиками.
46
Особый интерес представляют анатомические разделы этого сочинения. Основываясь на собственных наблюдениях (вскрытия более 200 птиц), Белон дает сравнительноанатомические описания особенностей строения скелета и, менее подробно, некоторых других систем. При этом он пытается выявить гомологию (сходство) в строении скелета различных позвоночных животных (особенно птиц) и человека. В его книге приведен рисунок, на котором в одной позе рядом изображены скелеты птицы и человека; гомологичные части помечены одинаковыми обозначениями. Эти описания позволяют считать Белона вместе с Аристотелем и Фридрихом II основоположниками сравнительной анатомии. Но Белон не пытался использовать описанные им анатомические особенности для обоснования принятой им классификации птиц.
Все вышеперечисленные крупные работы середины XVI в. способствовали привлечению интереса к птицам и были известной базой, способствовавшей дальнейшему накоплению фактов; сходные процессы шли и в других разделах зоологии позвоночных. Появляются описания новых видов птиц не только европейских, но и привозимых из вновь открываемых стран. В начале XVI в. постепенно научились делать тушки и чучела. Это привело к созданию коллекций, что значительно облегчило и ускорило изучение птиц. Довольно много работ в конце XVI—XVII вв. посвящено описанию анатомии птиц.
В XVII в. развитию зоологии способствовало продолжающееся в широких масштабах освоение «заморских» территорий, откуда в Европу поступали все новые и новые коллекции. Усиливалось взаимное общение ученых: и путем улучшения и распространения книгопечатанья, и развития переписки (благодаря становлению почты), и заметно усилившегося личного общения. В этот период в разных городах и странах возникают различные научные кружки, некоторые из которых затем преобразуются в Академии — официальные объединения ученых, занимавшихся изучением и естественных наук (Академия рысей в Риме в 1590 г., Академия тайн природы в Неаполе в 1560 г., Английская Академия — Королевское общество — в 1645 г., Академия испытателей природы в Германии в 1652 г., впоследствии превратившаяся в Естественно-историческую Академию и т. п.). Образование академий, которые в конце XVII — начале XVIII вв. возникли во всех странах, сопровождалось организацией регулярных научных публикаций, опять-таки облегчавших общение и взаимное ознакомление с результатами исследований отдельных ученых. Все это наряду с расширением торгового обмена и развитием производственных отношений способствовало заметному ускорению развития всех естественных наук, в том числе и зоологии.
XVII в. характеризуется резким усилением анатомических исследований, в том числе и микроскопических, широким развитием дискуссий и экспериментальных проверок возможностей самозарождения (ранее считавшихся достоверными), выяснением роли яйца и сперматозоида в развитии и т. д. Одновременно продолжалось описание новых видов, накапливались сведения о лх распространении и биологии. Большое количество видов, которое теперь знали ученые, затрудняло ориентировку в материале. Явно назревала необходимость в системе, которая бы не только облегчала нахождение описания нужного вида, но и позволяла как-то обобщить известные сведения. В связи с этим во второй половине XVII в. ряд ученых пытается создать системы всего животного царства или его отдельных групп.
Из попыток создать систему всех животных следует отметить систему Рэя.
Джон Рэй (1628—1705) родился в Англии в семье кузнеца. Он сумел кончить Кэмбриджский университет и остался в нем преподавателем греческого языка, гуманитарных и математических наук. Здесь он подружился с сыном богатых родителей Фрэнсисом Виллоуби (1635—1672),
47
страстно увлеченным естественными науками. Видимо, под влиянием своего друга Рэй также увлекся естествознанием. Друзья решили составить новую классификацию растений и животных, в которой каждая группа сходных видов должна иметь достаточно четкие общие признаки. Рэй должен был заниматься ботаникой, Виллоуби — зоологией. Для сбора материала друзья отправились в длительное путешествие по Европе (1663— 1666); все расходы оплачивал Виллоуби. Вернувшись в Англию, они занялись обработкой материалов, причем оба были избраны членами Королевского общества (т. е. академиками). Однако вскоре Виллоуби умирает от лихорадки, перед смертью попросив друга завершить его работы. Рэй закончил и опубликовал «Орнитологию» и «Ихтиологию» Виллоуби, пополнив их своими материалами (поэтому их вполне можно считать соавторами).
Рэй в «Synopsis methodica animalium Quadrupedum et Serpenti generis», Londini, 1693 дает общую систему животных (в скобках — название групп в современном понимании):
Раздел I. Животные с кровью (красной, т. е. позвоночные)
1. Дыхание легочное
А. Сердце с двумя желудочками (теплокровные):
а) живородные (млекопитающие) водные (киты) наземные (остальные млекопитающие) б) яйцекладущие (птицы)
Б. Сердце с одним желудочком (амфибии + рептилии):
2. Дыхание жаберное (рыбы)
Раздел II. Животные без крови (беспозвоночные)
1. Большие (крупные)
А. Мягкотелые (головоногие)
Б. Раковинные (остальные моллюски)
В. Ракообразные
2. Маленькие (мелкие)
А. Насекомые (насекомые, черви, паукообразные и др.):
а) развитие без метаморфоза б) развитие с метаморфозом
Система Рэя практически полностью повторяет принципы классификации Аристотеля: то же деление на животных с кровью и без крови, практически те же группы и среди позвоночных, хотя для диагноза этих групп использован больший набор признаков: особенности дыхательной и кровеносной системы наряду с характером размножения, тогда как у Аристотеля преимущественно особенности размножения и покровов. Деление беспозвоночных еще менее удачно, чем у Аристотеля, хотя Рэй использует даже такой важный признак, как характер развития (в те времена беспозвоночные были изучены еще очень слабо, поэтому неудача их систематизации вполне закономерна). Однако важно подчеркнуть, что заслуга Рэя заключается в том, что впервые попытался дать именно систему всех животных, а не просто их «каталогизировать», что, по сути дела, делал Аристотель.
Очень велика заслуга Рэя в том, что он практически впервые охарактеризовал основную таксономическую категорию — вид. В его понимании вид — это совокупность особей, похожих друг на друга, как дети похожи на своих родителей. Это определение понятия «вид» в дальнейшем неоднократно менялось и расширялось и окончательно не сложилось еще и сейчас. Но оно было введено в обиход, и этим было положено начало систематики как самостоятельного раздела биологии. Четкого представления о более высоких таксономических категориях у Рэя нет. Иногда он упоминает собирательное понятие «род», не отличая его в практическом применении от неопределенного понятия «группа».
Из попыток построения системы птиц следует упомянуть две.
Вальтер Чарлетон (1619—1707), придворный врач английского короля
48
Карла II, в опубликованной в 1668 г. «Onomasticon Zooico» разделяет птиц по образу жизни на 2 группы, каждая из которых в свою очередь делится на соподчиненные группы:
А. Наземные птицы:
1. Мясоядные (орлы, ястреба, сорокопуты, соколы, кукушка, попугаи, вороны, совы, козодой, страусы)
2. Насекомоядные:
а) певчие (королек, славки, ласточки, синицы, трясогузки, соловей, удод и др.) б) не певчие (дятлы, поползень, вертишейка, пищуха, щурки)
3. Зерноядные:
а) купающиеся в пыли (куры, дрофа, авдотка и др.)
б) купающиеся в пыли и в воде (голуби, воробьи)
в) поющие (щегол, чиж, зяблик, мухоловка)
4. Ягодоядные (дрозды, скворец, дубонос)
Б. Водные птицы:
1. Перепончатопалые (т. е., имеющие выраженную плавательную перепонку между пальцами)
2. Раздельнопалые:
а) рыбоядные (аисты, цапли, фламинго, султанка, веретенник, кулик-сорока) б) насекомоядные (зуек, кроншнеп, коростель, травник, камышница, вальдшнеп, оляпка и др.)
в) травоядные (журавль)
Система Чарлетона представляет собой явный эклектизм: без всякого критического подхода перечисляются все группы, упоминавшиеся Аристотелем и Альдрованди. Основные критерии групп—поведение и характер питания — варьируют даже у близких видов, а иногда и у одного и того же вида (синицы и насекомоядные и зерноядные и т. п.), поэтому не пригодны в качестве единственных признаков. На внешние морфологические признаки он обращает мало внимания. Довольно много и фактических ошибок: насекомоядные мухоловки отнесены к зерноядным, а веретенник и кулик сорока — к рыбоядным и т. д.
Ответом на эту систему можно считать работу Виллоуби «Ornitho-logiae libri tres», Londini, 1676, окончательно оформленную, дополненную и опубликованную уже после его смерти Рэем. Здесь птицы разделяются также на 2 основные группы, каждая из которых в свою очередь разделяется на 2 и т. д. (дихотомический принцип деления):
А. Наземные птицы
1. Aves uncungues — птицы с сильно изогнутым клювом и когтями:
а) хищные птицы (совы и дневные хищники, сорокопуты) б) попугаи
2. Aves rectungues — птицы с относительно прямым клювом и когтями; дальнейшее деление этой группы идет в зависимости от величины и формы клюва; клюв округлый, клюв сжат с боков и т. д.
Б. Водные (и околоводные) птицы
3. Настоящие водные (дальнейшее деление по характеру питания, форме клюва, строению ног, по размерам тела)
4. Околоводные (журавли, кулики; дальнейшее деление также по характеру питания, форме клюва, строению ног, размерам тела)
Рэй в «Synopsis Methodica Avium», 1713, напечатанной тоже после смерти автора, приводит примерно такую же систему. В этих системах Виллоуби и Рэй еще более последовательно, чем Белон, используют в качестве систематических признаков особенности внешней морфологии (форма клюва и ног и т. п.). Очень важное нововведение, которое используется в систематике до сих пор, — короткие диагнозы, характеризующие каждую группу. Так, чайки: «Водные птицы, плавающие и летающие, пальцы соединены плавательными перепонками, четырехпалые, задний палец свободный, клюв сжат с боков, на конце острый и прямой». Хорошо видно стремление охарактеризовать группу кратко, но разносторонне, чтобы ее можно было отличить от других групп. Очень интересно, что выделяемые ими многие конечные (т. е. самые мелкие) группы по своему объему очень близки к современным таксономическим группам: вьюрки, вороновые,
49
куриные (включают дрофу), веслоногие, кулики, гусиные и утиные и т. п. В этом отчетливо проявилось знание материала, чутье и талант систематиков.
В книгах Виллоуби и Рэя собраны все имевшиеся тогда сведения о птицах: не только описание видов, их образа жизни и распространения, но также накопившиеся к тому времени материалы по анатомии и физиологии. Таким образом, это своеобразные орнитологические энциклопедии, обобщающие все знания о птицах конца XVII в. Эти сочинения способствовали дальнейшему прогрессу изучения птиц и подготовили почву для дальнейшего развития систематики.
XVIII в. характеризуется дальнейшим ростом техники, усложнением производственных отношений. Резко ускоряется развитие естественных наук, среди которых особенно Интенсивно развиваются астрономия, математика, оптика, механика. Заметно ослабевает влияние церкви на науку, хотя представление о неизменности природы после «божественного творения» еще остается основой мировоззрения даже передовых умов. Зология этого периода характеризуется более широким развертыванием сравнительноанатомических исследований, изучением особенностей эмбрионального развития (опять вспыхивают дискуссии о возможности самозарождения). Увеличиваются коллекции разных групп животных (отечественных и «заморских»). Накопление фактов требует их систематизации (классификации). Сейчас это чувствуется намного сильнее, чем раньше. Поток вновь описываемых новых видов столь интенсивен, что без классификации, без какой-то единой системы трудно разобраться в новых сведениях. Очень сказывается и отсутствие каких-то общепринятых правил наименования новых видов. Обычно, как это повелось еще с Древней Греции, вид старались назвать одним словом: синица, воробей и т. п. Если затем становился известным какой-то новый вид, но очень похожий на уже известный старый, его называли тем же словом, но с применением прилагательного: синица большая, малая и хохлатая, воробей домовый и полевой и т. п.
В некоторых случаях это название превращалось в длинную фразу. Так, обыкновенная пчела в одной из работ того периода получила такое название: Apis bubescens, thorace subgriseo, abdomine fusco, pedibus po-sticis glabris, utrinque margine ciliatis (пчела волосатая, головогрудь сероватая, брюшко бурое, задние ножки гладкие, остальные с ресничками; в то время научные сочинения писали только на латинском языке). Все это затрудняло работу и ученых и любителей, число которых, особенно интересовавшихся птицами и насекомыми, заметно возрастало.
Карл Линней (1707—1778) — шведский ученый, деятельность которого была важным этапом в развитии систематики как животных, так и растений. С очень посредственными успехами окончив гимназию, он начинает изучать в шведских университетах медицину и ботанику, особенно увлекаясь последней. Путешествие по Лапландии (1732) обогатило его ботанические знания. Весной 1735 г. он отправляется за границу и защищает там докторскую диссертацию «О лихорадке». Затем он посещает ряд европейских университетов (Лейден, Амстердам, Париж и др.), изучая там гербарии и, менее увлеченно, зоологические коллекции. В 1738 г. он возвращается в Швецию, работает практикующим врачом (не оставляя занятий ботаникой), в 1741 г. получает кафедру медицины университета в Упсала, но вскоре переходит на кафедру ботаники, которой заведует до конца жизни. Был одним из организаторов и первым президентом Шведской Академии наук.
К. Линней в 1735 г. опубликовал первое издание «Systema Naturae» («Система природы»), в которой дал классификацию и краткое описание минералов, растений и животных. Книга сразу выдвинула К. Линнея в ряды крупнейших ученых своего времени благодаря четкому, строгому применению относительно простых систематических принципов, фактически создавших основы научной систематики.
50
Систематические принципы Линнея в основном сводятся к тому, что устанавливаются четкие соподчиненные таксономические категории:
класс, отряд (порядок), род, вид.
Линней дает относительно четкое определение вида: его составляют особи, похожие друг на друга, как дети похожи на своих родителей. Соответственно взглядам своей эпохи Линней исходил из того, что все виды созданы творцом (богом) и неизменны. Однако к концу жизни, благодаря накопленному опыту и знаниям, Линней в некоторой степени отошел от этого, допуская, что новый вид может возникнуть при гибридизации двух старых видов, а виды одного рода ранее, видимо, составляли один вид.
Сходные виды объединяются в род. Сходные роды составляют отряд. Отряды объединяются в класс.
При обозначении видов Линней последовательно и твердо придерживался бинарной номенклатуры: наименование каждого вида обязательно состоит из двух слов: первого — наименования рода, второго — характеризующего именно данный вид. Например: Parus major — синица большая. Такое бинарное наименование очень удобно уже потому, что название вида сразу же дает возможность получить представление о близких видах. Неуклонное выполнение этого принципа показывало путь преодоления путаницы в обозначении вновь описываемых видов, упорядочивало работу систематика. Явно относящиеся к данному виду, но чем-то отличающиеся от массы других особей индивиды Линней называл разновидностями (вариациями). Он не придавал им особого таксономического значения и относил сюда индивидуальные отклонения (альбиносы, меланисты и т. п.) и немногие ему известные случаи географической изменчивости вида.
Используя эти принципы, Линней дал систему всего органического мира, изложенную в вышеупомянутом сочинении «Система природы». После выхода в свет первого издания Линней продолжал свою работу, улучшая и расширяя изложение, вводя новые виды. Всего при его жизни вышло 12 издании «Системы природы», завоевавших всеобщее признание. В зоологической практике за основу принимается 10-е издание «Системы природы», вышедшее в 1758 г. По этой системе вся вселенная составляет Империю, разделяющуюся на 3 Царства: Минералов, Растений и Животных.. Царство животных распадается на 6 классов: Млекопитающие (теплая кровь, живородящие, выкармливают детенышей молоком), Птицы (теплокровна, покрыты пером, откладывают яйца), Гады (кровь холодная, дышат легкими; относятся земноводные и пресмыкающиеся), Рыбы (кровь холодная, дышат жабрами), Насекомые (кровь белая, сердце без предсердий, щупальцы членистые) и Черви (щупальцы не членистые). В целом эта система животного мира похожа на систему Рея (и Аристотеля), но имеет более упорядоченный вид, благодаря применению единой крупной таксономической категории — класса. Очень неудачен объем класса Черви, куда относились все беспозвоночные, кроме членистоногих.
Изложение материалов по каждому классу построено по одной схеме. Дается краткая характеристика класса, перечень входящих в него отрядов с их краткой характеристикой, далее перечень родов каждого отряда с краткой характеристикой каждого рода и перечисление видов, причем за каждым видовым названием следует краткая характеристика (диагноз) вида, включающая несколько внешних признаков, распространение (очень общее) и часто ссылки на авторов, ранее писавших об этом виде.
Класс Птиц у Линнея включает 6 отрядов, 65 родов и 554 вида.
Класс Aves — Птицы
Отряд 1. Accipitres — хищные. Включает роды: Vultur (грифы)1, Falco (соколы, ястребы, коршуны, орлы, луни), Strix (совы, филины); Lantus (сорокопуты) Отряд 2. Picae —.сорочьи с родами: Corvus (вороновые), Coracias (сойки), Or lotus
1 В скобках ориентировочно показан объем принимаемых Линнеем родов. В современном понимании объем многих родов Линнея соответствует объему семейств.
51
(иволги), Sitta (поползни), Certhia (пищухи), Upupa (удоды), Paradisea (райские птицы), Picas (дятлы), Jynx (вертишейка), Ramphastos (туканы), Trochilus (колибри), Buceros (птицы носороги), Trogon (трогоны), Psittacus (попугаи), Cuculus (кукушки), Crotophaga (кукушки ани и др.), Merops (щурки), Alcedo (зимородки, тоди)
Отряд 3. Anseres — водоплавающие. Роды: Anas (утки, лебеди, гуси), Mergus (крохали), Alca (чистиковые), Pelecanus (бакланы, пеликаны), Phaeton (фаэтоны), Colymbus (гагары, поганки), Laras (чайки), Sterna (крачки), Rhyncops (водорезы), Diomedea (альбатросы), Procellaria (буревестники).
Отряд 4. Gralle — голенастые. Роды: Phoenicopterus (фламинго), Platalea (колпицы), Ardea (цапли, аисты, журавли), Psophia (журавли трубачи), Fulica (лысухи), Railas (пастушки, погоныши), Tringa (улиты и другие кулики), Recur-virostra (шилоклювки), Scolopax (вальдшнеп, бекасы, кроншнепы, плавунчики), Charadrius (зуйки, ржанки), Haematopus (кулики сороки), Otis (дрофы), Struthio (все страусовые)
Отряд 5. Gallinae — куриные. Роды: Phasianus (фазановые), Tetrao (тетеревиные и некоторые фазановые — куропатки, кеклики, перепел), Numida (цесарки), Pavo (павлины), Meleagris (индюки), Сгах (краксы, или гокко)
Отряд 6. Passeres — воробьиные. Роды: Loxia (клесты, дубоносы, зеленушки), Frin-gilla (вьюрки, чиж, воробьи и др.), Emberiza (овсянки), Hirundo (ласточки, стрижи), Tardus (дрозды), Muscicapa (мухоловки), Tangra (тангры), Parus (синицы), Motacilla (трясогузки, соловьи, славки), Alauda (жаворонки, коньки), Sturnus (скворцы), Columba (голуби), Caprimulgus (козодои)
В целом система класса птиц Линнея напоминает систему Виллоуби и Рэя, отчасти Белона. Положительно, кроме вышеуказанных общих принципов, стремление использовать для характеристики всех таксономических групп несколько признаков (но преимущественно или исключительно внешних: характер покровов, формы ног и клюва, расположение и форма ноздрей и т. п.), наличие диагнозов — кратких характеристик — для всех без исключения выделяемых групп, упоминание распространения в диагнозах вида и т. п.
Четкая форма, последовательность применения определенных принципов, систематизация накопленных к этому времени знаний о видовом разнообразии — вот что делало систему К. Линнея убедительной в глазах современников, привлекало внимание к зоологическим и ботаническим исследованиям, показывало, что нужно не только описывать новые виды, но одновременно и классифицировать их, строить систему. Все это способствовало не только дальнейшему расширению изучения птиц (как и других групп животных), но и развитию исследований по систематике.
В этом несомненная и большая заслуга Карла Линнея.
Бриссон (1723—1806), француз, написал работу, обобщающую сведения о видовом разнообразии птиц — 6-томную «Орнитологию». Она вышла в 1760 г. и имела большее значение, чем «Система природы» Линнея; здесь даются обычно четкие описания уже почти 1500 видов птиц, которые объединены в 115 родов. Опознаванию видов очень помогает прилагаемый атлас. Все роды распределяются по 26 отрядам, которые имеют только порядковые номера (т. е. цифровые, а не смысловые обозначения). Такой прием очень затрудняет и запоминание и пользование системой; дальнейшего развития он не нашел. При обозначении видов Бриссон последовательно не применял бинарной номенклатуры, что было шагом назад по сравнению с Линнеем, поэтому его видовые названия впоследствии при упорядочении систематических исследований не получили официального признания.
Очень большую роль в привлечении интереса к изучению птиц и в популяризации естествознания вообще, в конце XVIII—первой половине XIX вв. сыграла 10-томная «История птиц», написанная Бюффоном (1707— 1788). В ней было показано разнообразие мировой фауны птиц и обращалось внимание — в пределах знаний того времени — на их биологию.
Все орнитологические сведения, собранные академическими экспедициями Российской Академии наук в XVIII в., работавшими практически на всей территории нашей страны от Ледовитого океана до Каспия и от Белоруссии до Камчатки и Командорских островов, были обобщены академиком Петром Палласом в книге «Zoographia Rosso-Asiatica» (1811). В ней он
52
дал описание ряда новых видов (в книге дается перечень местных названий, описание внешних признаков, распространения, в очень общей форме, и отрывочных анатомических и биологических сведений для 425 видов птиц), расширил и уточнил родовую номенклатуру Линнея. Книга Палласа сохраняет свое значение и сейчас, позволяя, в частности, выявить ряд изменений в фауне птиц нашей страны за прошедшие 150—200 лет.
В конце XVIII — начале XIX вв. (послелиннеевский период) внимание многих, если не большинства, орнитологов было посвящено преимущественно описанию новых и переописанию старых, т. е. уже известных видов по правилам бинарной номенклатуры. Разобщенность ученых, чему способствовали длительные войны в Европе, привела к тому, что один и тот же вид часто получал разные названия (например, в Англии и Франции). Возникшая путаница с большим трудом была устранена — хотя ее отголоски весьма чувствуются и сейчас — лишь в конце XIX — начале XX вв., благодаря широким дискуссиям, приведшим в конечном счете к созданию «Международных правил зоологической номенклатуры». Разработка этих правил началась в середине XIX в., а дополнения и изменения в них вносятся и сейчас, после их утверждения периодически собирающимися Международными зоологическими конгрессами. Приложение общих правил зоологической номенклатуры к систематике птиц обсуждается на Международных орнитологических конгрессах.
С начала XIX в. начинают более быстро развиваться анатомия и физиология, и интерес к систематике вообще и к систематике птиц в частности уменьшается. Однако ряд ученых предпринимают попытки создания новых систем класса птиц.
Иллигер (liliger С. Prodromus systematis Mammalium et Avium, Berlin, 1811), подразделяя птиц на 7 отрядов, вводит новую таксономическую категорию — семейство (между отрядом и родом). Он выделял 41 семейство птиц, в которые группировал 147 родов.
Жорж Кювье (1769—1832) — крупнейший зоолог начала XIX в., сравнительный анатом, палеонтолог, создает свою систему животного мира, противопоставляя ее отвергавшейся им системе Линнея (Cuvier G. Le regne animal distribue d’apres son organisation, I—IV, Paris, 1817, 1829). На основании сходства и различия в строении скелета, Нервной и кровеносной систем, общего расположения органов Ж- Кювье делил всех животных на 4 группы (фактически типа, но этот термин высшей таксономической категории был предложен несколько позже учеником Кювье А. Блэквиллем).
1. Позвоночные Vertebrata — включали млекопитающих, птиц, гадов и рыб
2. Мягкотелые Mollusca — кроме моллюсков, объединяли оболочников и некоторых ракообразных
3. Членистые Articulata — объединяли настоящих членистоногих (ракообразных, паукообразных, насекомых), кольчатых червей, некоторых моллюсков 4. Лучистые животные Radiata — сюда относились все остальные беспозвоночные (иглокожие, плоские и круглые черви, кишечнополостные, простейшие), фактически это отряд Зоофитов в классе Червей Линнея
Все 4 группы в понимании Кювье резко обособлены и отражают 4 независимых «плана строения».
Класс Птиц Кювье делит на 6 отрядов, которые подразделяются на семейства. При этом он, как и предыдущие авторы, использует лишь внешние признаки (форму клюва и лап, некоторые особенности оперения и т. п.), совершенно не привлекая материалы сравнительной анатомии (одним из основателей которой совершенно справедливо его считают). Как и в системе Иллигера, у Кювье по сравнению с системой Линнея положительным было последовательное применение новой таксономической категории — семейства.
Меррем (В. Merrem. Tentamen systematis avium. Abhandlungen d.k. Akademie d. Wissenschaften, Berlin, 1813), впервые применяя для таксономии внутренние анатомические признаки, разделил птиц на 2 противостоя
53
щие группы (фактически на 2 подкласса) — бескилевых Ratitae и килевых Carinatae. Подавляющее большинство воробьиных птиц было выделено им в подгруппу певчих Oscines.
Вигоре (Vigors N. A. On the arrangement of the Genera of birds. Zoological Journal, 1826) предложил обозначать семейства птиц по названию типичного рода с окончанием -idae’: Parus — Paridae, Muscicapa — Musci-capidae и т. п.; впоследствии это предложение было закреплено правилами зоологической номенклатуры.
Нитцш (Ch. Nitzsch) впервые наиболее последовательно пытался расширить значение внутренних морфологических признаков для таксономии птиц. Он изучал строение носовых желез, некоторых отделов кровеносной и скелетной систем. В работе «Изучение общей сонной артерии птиц» (ОЬ-servationes de Avium arteria carotide communi. Halle, 1829) на основании этих исследований и, конечно, с учетом всех ранее используемых внешних признаков он предлагает свою систему птиц, ограничиваясь лишь перечнем групп и не давая их диагнозов.
I. Птицы килевые Aves carinatae
А. Килевые летающие птицы Aves carinatae aereae
1. Accipitrinae 2. Passerinae — дневные хищники, совы — воробьиные
3. Macrochires — длиннокрылые
4. Cuculinae — кукушки
5. Picinae — дятловые
6. Psittacinae — попугаи
7. Lipoglossae — удоды
8. Amphibolae — бананоеды
В. Килевые наземные птицы Aves carinatae terrestres
1. Columbinae — голубиные
2. Gallinaceae — куриные
С. Килевые водные птицы — Aves carinatae aquaticae
a) Пальцы не соединены плавательными перепонками Grallae:
1. Alectorides — дрофы
2. Gruinae — журавли
3. Fulicariae — лысухи и др.
4. Herodiae — цаплевые
5. Pelargi — аистовые
6. Odontoglossi — фламинго
7. Limicolae — кулики
6) Пальцы соединены плавательной перепонкой Palmatae:
8. Longipennes — чайковые
9. Nasutae — трубконосые
10. Unguirostres — гусеобразные
11. Steganopodes — веслоногие
12. Pygopodes — ныряющие
II. Птицы бескилевые Aves ratitae (все страусовые)
Позже Нитцш занялся тщательным изучением птерилографии птиц, т. е. выяснением особенностей размещения перьев по телу и различий в тонкой структуре пера. В сводной работе «Система птерилографии», которая вышла из печати через 3 года после смерти ее автора (Sysmem der Pterylo-graphie. Halle, 1840), он дает очень тщательное и детальное описание особенностей оперения по отдельным группам и видам (работа как сводка по птерилографии птиц не потеряла своего научного значения и сейчас). Здесь он дает несколько иной, чем в работе 1829 г., перечень групп, исходя из черт сходства и различия в птерилографии, хотя и не придает этим особенностям значения окончательных таксономических критериев, что видно хотя бы по отказу от разделения птиц на противостоящие группы килевых и бескилевых, хотя последние и по особенностям оперения резко отличаются от большинства других птиц.
Здесь птицы подразделяются на 8 групп (отрядов):
I. Хищные птицы Accipitrinae — дневные хищники, совы II. Певчие птицы (воробьиные) Passerinae
54
III. Дятловые птицы Picarinae 1. Macrochires —стрижи, колибри 2. Caprimulginae — козодои (по Нитцшу, ночные кукушки) 3. Todidae — ракши, момоты, тоди, якамары 4. Cuculinae — кукушки, медоуказчики, трогоны 5. Picinae — дятлы, туканы, пуховки 6. Psittacinae — попугаи 7. Lipoglossae — птицы носороги, удоды, зимородки 8. Amphibolae — бананоеды, птицы мыши, гоацин
IV. V. Голубеобразные Columbinae — голуби, рябки Куриные Gallinaceae 1. Tetraonidae —тетеревиные 2. Phasianidae — фазановые 3. Penelopidae — краксы 4. Cripturidae — сорные куры
VI. Бегающие птицы Currentes — все страусообразные
VII. Болотные птицы Grallae
1. Alectorides
2. Fulicariae
3. Herodii
4. Pelargi
5. Odontoglossae
6. Hemiglottidae
7. Limicolae
VIII. Плавающие птицы
1. Longipennes
2. Nasutae
3. Unguirostres
4. Steganopodes
5. Pigopodes
— паламедеи, дрофы, кариамы, журавли — лысухи и, видимо, другие пастушковые — цапли
— молотоглавы, аисты
— фламинго
— ибисы, колпицы
— кулики
Natatores
— чайковые
— трубконосые
— гусеобразные
— веслоногие
— поганки, гагары, чистики, пингвины
Несомненное плодотворное влияние работ Нитцша на последующее развитие систематики птиц заключается именно в попытке использовать для построения системы сочетание разнообразных как внешних, так и внутренних особенностей строения. Плодотворность такого подхода отчетливо выражается в том, что ряд выделенных Нитцшем групп птиц сохранился почти в неизменном виде в общепринятых современных системах птиц: объединение стрижей и колибри, голубей и рябков, выделение подгрупп (семейств) куриных, размещение по отдельным подразделениям гусеобразных, чайковых, трубконосых, попугаев, козодоев, воробьиных и т. д. В ряде позже создаваемых систем птиц можно более или менее явно увидеть плодотворное влияние Нитцща как по методике работы, так и по использованию конкретных результатов его сравнительноанатомических исследований.
В первой половине XIX в. публиковались и другие системы класса птиц: Виейльс (1816), Темминк (1820), Кабанис (1847), Бонапарт (1853) и др. Для этих авторов характерно стремление использовать при построении системы и для характеристик отдельных групп особенности распространения, отдельные черты внутреннего строения (но менее последовательное, чем у Нитцша). При этом постепенно уточнялся и улучшался объем родов и их группировок в семейства, что подготавливало почву для улучшения системы в будущем. Накопление фактического материала постепенно приводило ученых к представлениям о географической изменчивости ряда широко распространенных видов. Для обозначения таких географических вариаций Шлегель (Н. Schlegel. Kritische Ubersicht der europaischen Vogel, 1844) предложил обозначать их третьим латинским словом; например, одна из географических вариаций — в современном понимании подвид — большой синицы получает название Parus major major. Значительно позже это предложение вошло в зоологическую практику и применяется до сих пор.
Жан Батист Ламарк (1744—1829), француз, занимает совершенно особое место в развитии биологии. Он сформулировал в знаменитой «Философии зоологии» (1809) теорию эволюции органического мира, основывающейся на безграничной изменяемости видов: более высоко организованные формы произошли от менее организованных. При этом он пытался выяснить и причины эволюции. Здесь не место разбирать роль Ламарка в развитии эво
55
люционных идей, достоинства и недостатки его учения. Стоит лишь отметить, что эволюционные идеи Ламарка не нашли сторонников среди современников, хотя его конкретные работы, особенно по систематике и анатомии без-позвоночных животных, пользовались известностью. Ламарк впервые ввел употребление терминов «биология», «позвоночные» и «беспозвоночные».
Всех животных Ламарк делит на 2 группы: Беспозвоночных (10 классов вместо 2 у Линнея) и Позвоночных, включающих 4 класса: Рыб, Рептилий (земноводные и пресмыкающиеся), Птиц и Млекопитающих. При этом Ламарк предполагает, что Моллюски («при посредстве тех, которые остаются нам еще неизвестными») дали начало Рыбам, а от рыб произошли Рептилии (от рыб произошли лягушки, а от них — змеи, которые затем дали начало остальным отрядам рептилий). Птицы произошли от черепах, а Млекопитающие — от крокодилов (сначала возникли водные млекопитающие — амфибии — т. е. ластоногие, или тюлени; от травоядных тюленей произошли наземные копытные, от рыбоядных — наземные коготные; от тюленей же возникли и китообразные).
Класс Птиц Ламарк разделяет на 7 отрядов, пользуясь лишь внешними различиями и сходством в строении лап, клюва, крыльев, иногда размерами кладки и общим образом жизни:
Отряд 1. Лазуны (два пальца вперед, два — назад) — попугаи, трогоны, бананоеды, туканы, дятловые, кукушки
Отряд 2. Хищные — ночные (совы) и дневные
Отряд 3. Воробьиные — кроме собственно воробьиных в современном понимании, стрижи, колибри, козодои, зимородки, щурки, удоды и др.
Отряд 4. Голубиные — только голуби
Отряд 5. Куриные — кроме собственно куриных, дрофы и страусовые
Отряд 6. Голенастые (береговые птицы) — цапли, аисты, ибисы, кулики, пастушки Отряд 7. Лапчатоногие (плавающие птицы) — веслоногие, гусеобразные, чайковые, трубконосые, поганки, чистиковые, пингвины
Последовательность отрядов отражает последовательность их возникновения. Лазуны (сходные с хамелеонами из рептилий по расположению пальцев) — наиболее примитивный отряд. Первые 4, отряда — птенцовые, что тоже говорит о меньшем совершенстве их организации по сравнению с тремя последними отрядами выводковых птиц. Наиболее высоко организован отряд Лапчатоногих: не только летают, но плавают и ныряют. От Лапчатоногих произошли Однопроходные (ехидна и утконос) — ни птицы, ни млекопитающие; они рассматриваются Ламарком или как самостоятельный класс, или как дополнение к классу Птиц.
Особых преимуществ перед другими эта система птиц не имеет: при формировании групп (отрядов) использовано небольшое число внешних признаков, объем отрядов неравноценен, случаен и отражает лишь сходство, но не родственное, а преимуществнно конвергентное, т. е. вызванное приспособлением к сходным условиям жизни. Сам Ламарк не занимался изучением птиц и практически лишь перестроил то, что было более или менее принято в его время. Однако исторически весьма важно, что Ламарк впервые ясно сформулировал новые задачи системы: она должна отражать эволюцию всех групп и их происхождение, выражать естественные взаимоотношения организмов, а не служить лишь классификационным целям, создаваемым человеком для собственного удобства, чтобы удобнее ориентироваться в многообразии видов. К сожалению, общие недостатки эволюционных воззрений Ламарка, обусловленные в первую очередь недостатком фактического материала той эпохи, и активное противодействие многих именитых ученых (например, Кювье), привели к полному отрицанию учения Ламарка. Однако Ламарк, несмотря на все свои ошибки, был фактически первым, в столь ясной форме высказавшимся о эволюционном развитии органического мира и о необходимости разработки системы, отражающей эту эволюцию. «Теория Ламарка несовершенна, это первый набросок эволюционного учения, развитого позднее с большим успехом Ч. Дарвиным, но для своего
56
времени она была гигантским шагом вперед. И мы, люди XX века, рассматривающие мир с точки зрения диалектического его развития, будем ценить у Ламарка, этого несомненного борца за истину, не столько его ошибки, сколько его достижения» (Президент Академии наук СССР В. Л. Комаров1).
В ноябре 1859 г. вышла в свет книга Чарльза Дарвина «Происхождение видов путем естественного отбора или сохранение благоприятствующих пород в борьбе за жизнь». Оснащенная большим количеством фактов, тщательно и всесторонне рассматриваемых, охватывающих различные группы растений и животных, эта работа неоспоримо доказывала эволюцию органического мира, проходившую по собственным законам и не зависимую ни от «воли творца», ни от «мирового разума», показывала движущие силы (факторы) эволюции: изменчивость, наследственность и естественный отбор. Эволюционное учение Ч. Дарвина сначала встретило много противников, но многие крупные ученые мира приняли его и направили свою деятельность как на пропаганду и популяризацию эволюционного учения, так и на дальнейшее его развитие, проводя свои исследования с позиций эволюционизма. В России дарвинизм сразу же нашел многих сторонников как среди крупных ученых (А. П. Богданов, А. О. и В. О. Ковалевские, И. И. Мечников, М. А. Мензбир, Н. А. Северцов, И. М. Сеченов, К- А. Тимирязев и многие др.), так и среди публицистов (Д. И. Писарев).
Эволюционное учение сразу же поставило перед систематикой новые задачи. Систематические исследования должны были вскрыть, выяснить родственные отношения между отдельными группами, выяснить пути видообразования и на основе этого создать естественную систему, т. е. такую систему, которая реально отражала бы эволюцию и родственные отношения как отдельных групп (типы, классы, отряды), так и всего органического мира.
Для развития систематики в этом направлении очень важны были работы Э. Геккеля и Т. Гексли.
Эрнест Геккель (1834—1919), профессор в Иене, занимался изучением морских беспозвоночных. Приняв эволюционное учение Дарвина и ставя целью доказать его правильность, он опубликовал двухтомную «Общую морфологию организмов» (Е. Hackel. Generalle Morphologic der Organismen, Berlin, 1866J, где в популярной форме излагал идеи Дарвина, добавляя при этом отдельные идеи Ламарка и свои личные гипотезы и предположения, часто мало или совсем не обоснованные. Книга эта выдержала много изданий (каждое из них автором дополнялось й изменялось) и была переведена на многие языки мира. В этом сочинении Геккель впервые делает попытку изобразить эволюцию всего органического мира в виде «родословного древа», от основания ствола которого отходят 3 мощные ветви: растения, простейшие и многоклеточные животные. Каждая ветвь многократно ветвится, а характер отхождения ветвей отражает представления автора о родственных взаимоотношениях отдельных групп. Геккель впервые выделяет тип Хордовых, объединяя в нем оболочников, бесчерепных и позвоночных, разделяет позвоночных на 2 группы — анамний и амнйот. Система класса Птиц по Геккелю выглядит так:
1-й подкласс Saururae — ящерохвостые птицы (сюда он относил описанную к тому времени ископаемую птицу юрского периода Archaeopteryx)
2-й подкласс Ornithurae — веерохвостые птицы
I группа Autophagae (Nidifugae) — выводковые птицы Отряды: 1. Natatores — плавающие
2. Grallatores — голенастые
3. Rasores — куриные
4. Ineptae — дронты
5. Saurophalli -— трехпалые страусы (сюда включены и краксы из куриных)
1 Вступительная статья в кн. Ж. Б. Ламарк. Философия зоологии. Т. 1. М.-Л., 1935.
57
6. Apterygia — бескрылые (киви, моа)
7. Struthocameli— двухпалые страусы
II группа Paedotrophae Отряды: 1. Peristerae
2. Clamatores
3. Oscines
4. Scansores
5. Raptatores
(Insessores) — птенцовые птицы
— голуби
— кричащие воробьиные
— певчие воробьиные
— лазающие (попугаи, дятлы и др.)
— хищные птицы
Наиболее древняя группа веерохвостых птиц — выводковые птицы, а среди них — трехпалые страусы, давшие начало остальным отрядам выводковых птиц. От куриных птиц обособились голуби (группа птенцовых), давшие начало кричащим воробьиным и лазающим; от лазающих произошли певчие воробьиные, а от последних хищники.
Все эти филогенетические соображения Геккеля чисто умозрительны и не подкреплены анализом конкретных фактических материалов. Неудачно делецие всех веерохвостых птиц только по одному признаку — птенцовос-ти или выводковости, тем более, что вскоре выяснилось наличие постепенных переходов между этими' крайними группами. Искусственна и неудачна группировка по отрядам: объединение неродственных групп в отряды плавающих, голенастых, лазающих и т. п. Здесь явный шаг назад по сравнению, например, с системами Нитцша. Единственное, что сохранилось в последующих системах, — это деление птиц на 2 подкласса: ящёрохвостых и веерохвостых. Положительное влияние Геккеля на развитие систематики — в том числе и орнитологической — в подчеркнуто филогенетическом подходе к построению системы, в разработке полезного метода, позволяющего в наглядной графической форме изобразить предполагаемые филогенетические взаимоотношения отдельных групп.
Томас Гексли (1825—1895) занимался палеонтологией, сравнительной анатомией, эмбриологией, общими вопросами биологии. Был членом Королевского общества (Английская Академия наук) и его президентом. Широко эрудированный ученый и прекрасный оратор, он вошел в историю биологии как один из энергичнейших и страстных пропагандистов эволюционной теории Дарвина. На основании тщательного сравнительного изучения строения костного неба Т. Гексли попытался построить новую эволюционную систему класса птиц, изложенную им в работе «О классификации птиц и о таксономическом значении изменений в строении костного неба в этом классе» (Т. Н. Huxley. On the classification of Birds, and on the taxonomie value of the modification of certain of the cranial bones observable in that Class. Proc. Zool. Soc., London, 1867). Эта система выглядит так:
Класс Aves
Отряд I. Saururae — ящерохвостые, только ископаемые
Отряд II. Ratitae — бескилевые, в 5 групп выделены все 6 родов страусообразных птиц
Отряд III. Carinatae — килегрудые птицы
1-й подотряд Dromaeognathae — дромеогнаты (имеют дромеогнатический тип неба, одно семейство тинаму)
2-й подотряд Schizognathae — схизогнаты (схизогнатический или щелевидный тип неба), 15 семейств, объединяемых в 9 групп: кулики, журавли, пастушки, кариамы, дрофы, чайки, трубконосые, гагары, поганки, чистиковые, пингвины, трехперстки, рябки, голуби, куриные
3-й подотряд Desmognathae — десмогнаты (мосто-небные), 29 семейств, объединенных в 8 групп: гусеобразные, фламинго, цапли, аисты, ибисы, веслоногие, совы, дневные хищники, попугаи, • птицы мыши, бананоеды, дятловые, сизоворонковые, трогоны
4-й подотряд Aegithognathae — эгитогнаты (булаво-небные), две группы с несколькими семействами: колибри, стрижи, козодои, воробьиные.
Эта система сначала получила довольно широкое распространение, но быстро выявился ее основной недостаток: использование лишь одного таксономического признака (тип костного неба), который, как вскоре было установлено дальнейшими исследованиями, довольно сильно варьирует, и между отдельными типами существуют разнообразные переходы. Даже в пре
58
делах четких групп выявляются разные типы неба: например, у куликов — схизогнатический с явными переходами к десмогнатизму и эгитогнатизму, у дятловых — схизогнатический, десмогнатический и эгитогнатический, у краксов — десмогнатический, а у всех остальных куриных — схизогнатический. Это существенно нарушает стройность системы, делает ее искусственной. Очень большой объем основных таксономических групп (подотрядов), объединяющих множество разнообразных семейств, затрудняет пользование системой. Однако для диагнозов отрядов и подотрядов Гексли использовал и другие анатомические признаки, хотя основное значение придавал лишь костному небу.
Несмотря на эти недостатки, значение работы Гексли в дальнейшем улучшении системы класса птиц было очень большим. С полной очевидностью, очень ярко и выпукло было показано, что для построения естественной (филогенетической) системы необходимо тщательное изучение и широкое использование многих внутренних морфологических признаков. Этим было предопределено основное направление дальнейших систематических исследований. В ближайшие годы наряду с попытками строить эволюционные системы по старинке лишь по внешним признакам производились попытки построения систем с учетом накопившихся данных (оригинальных и литературных) по сравнительной анатомии птиц, включая многие признаки внутреннего строения (Карус, 1868; Гаррод, 1874; Склетер, 1880; Штейне-гер, 1885 и др.).
Макс Фюрбрингер (1846—1920) — профессор Амстердамского университета. Его многолетние исследования явились важным этапом в становлении современной системы птиц. Завершающим трудом была капитальнейшая монография «Исследования, морфологии и систематики птиц» (Unter-suchungen zur Morphologic und Systematik der Vogel. Theil 1—2, Amsterdam — Jena, 1888). Она включала более 1640 страниц большого формата текста, 66 страниц библиографии и 30 больших таблиц рисунков и схем. Ученик крупнейшего сравнительного анатома, основоположника филогенетической сравнительной анатомии Карла Гегенбауэра Фюрбрингер тщательно изучил многие анатомические особенности большого числа видов из всех основных групп птиц: плечевой пояс (размеры, положение, форма и другие особенности кар а ко и да, лопатки, вилочки, грудины и ее киля, суставной впадины и т. п.), скелет крыла, нервы, мускулы и связки плечевого пояса и передней конечности. При этом описываются вариации в строении деталей, проводится выяснение гомологии отдельных структур с рептилиями. Эти описания составляют содержание первой — специальной — части, занимающей 830 страниц текста и иллюстрированной 26 таблицами хорошо выполненных рисунков. Во второй — общей — части проводится обсуждение и сопоставление своих и литературных данных, высказываются соображения о таксономическом значении как изученных, так и других морфологических признаков (клюв, оперение, скелет, мускулатура, органы чувств, дыхания, пищеварения, кровообращения, размножения и т. д.). Далее дается морфологическая характеристика 73 семейств птиц (указываются черты сходства и различия каждого семейства с другими группами птиц) и затем излагается составленная на этой основе система класса.
Класс Aves — птицы
А. Подкласс Saururae — ящерохвостые
Отряд 1. Archornithes (включает только ископаемого Archaeopteryx)
Б. Подкласс Ornithurae — веерохвостые
Отряд 2. Struthiornithes— африканские страусы
Отряд 3. Rheornithes — американские страусы (нанду)
Отряд 4. Hippolectryornithes — австралийские страусы (казуары и эму) Промежуточный отряд 1. Aepyornithiformes —эпиорнисообразные (вымерли) Промежуточный отряд 2. Palamedeiformes — паламедееобразные Отряд 5. Pelargornithes — водные, околоводные и хищные птицы Подотряды: a) Anseriformes — гусеобразные
б) Podicipitiformes —поганкообразные (гагары, поган
59
ки, гесперорнисы — нелетающие ископаемые меловые зубастые птицы)
с) Ciconiiformes — айстообразные (фламинго, ибисы, аисты, молотоглавы, китоглавы, все дневные хищники, все веслоногие)
Промежуточный отряд 3. Procellariiformes — трубконосые Промежуточный отряд 4. Aptenodytiformes — пингвинообразные Промежуточный отряд 5. Ichthyornithiformes — ихтиорнисы (летающие ископаемые меловые зубастые птицы)
Отряд 6. Charadriornithes — куликообразные
Подотряды: a) Charadriiformes — куликообразные (кулики, чайки, . чистики, авдотки, дрофы)
б) Gruiformes — журавлеобразные (солнечные цап-
ли, все журавли, кариамы, кагу)
с) Ralliformes — пастушковые (собственно пастуш-
ки, лапчатоноги, мадагаскарские пайтушки, трехперстки} Отряд 7. Alectorornithes— курообразные птицы
Подотряды: a) Apterygiformes — киви (бескрылые)
б) Crypturiformes — тинаму
с) Galliformes — куриные (все группы в современ-
ном понимании, включая гоацина)
Промежуточный отряд 6. Columbiformes— голубеобразные (голуби, рябки} Промежуточный отряд 7. Psittaciformes— попугаеобразные
Отряд 8. Coracornithes — ракшеобразные
Подотряды: a) Coccygiformes — кукушкообразные (турако или ба-наноеды, кукушки, якамары)
б) Pico-Passeriformes — дятло-воробьинообразные (боро-дастики, туканы, медоуказчики, дятлы, все воробьиные, стрижи, колибри, птицы мыши, трогоны)
с) Halcyoniformes — зимородковые (зимородки, удоды, птицы носороги, щурки)
d) Coraciformes — ракши (собственно ракши, козодои, совы)
Фюрбрингер, стараясь более наглядно показать эволюционные отношения выделяемых таксономических групп, приводит в своей работе родословное древо (рис. 14), доводя ветвление до семейств, а в некоторых случаях —до подсемейств и даже крупных родов. Эта схема наряду с детальным обсуждением морфологических особенностей разных групп дает хорошую возможность представить генеалогические (филогенетические) отношения в пределах всего класса птиц. Отряды и промежуточные отряды — основные ветви этого родословного древа.
Громадный фактический материал, скрупулезность и тщательность его оценки, отчетливая филогенетичность (эволюционность) созданной системы — все это сделало труд М. Фюрбрингера классическим и определило последующее развитие систематики птиц. Многие представления Фюрбрингера о генеалогических отношениях отдельных групп птиц в почти неизмененном или малоизмененном виде сохранились и приняты и в наши дни. До сих пор полностью сохраняет свое значение и весь фактический материал этой сводки.
Из недостатков системы Фюрбрингера, затруднявших ее применение и измененных работами последующих систематиков, следует отметить ее чисто внешнюю сложность (основные и промежуточные отряды) и большой объем основных отрядов, включающих очень различные по внешнему виду и экологии крупные группы. Например, объединены в один отряд Pelar-gornithes гусеобразные, гагары, поганки, аистообразные, дневные хищники, веслоногие. Очень большой объем отрядов упрощает и затушевывает выдвинутую самим же Фюрбрингером идею о многосторонности связей отдельных групп. Палеонтологические материалы также пока не оправдывают таких крупных объединений (это было бы целесообразно лишь при наличии ископаемых форм, действительно связывающих, объединяющих эти современные группы).
Ганс Гадов (Hans Gadow, 1855—1928) — другой ученик Гегенбауэра, написал — почти одновременно с работой Фюрбрингера монографию, по
60
священную построению системы класса птиц. В первой части этого труда (Bronn’s «Klassen und Ordnungen des Thierreichs», Bd. VI, Abt. IV, Vogel. I. Anatomischer Theil. Leipzig, 1891) около 1000 стр. текста, иллюстрированного 59 таблицами анатомических рисунков, дается сравнительноанатомическое описание практически всех систем органов птиц: отделов скелета, мускулатуры, головного и спинного мозга, важнейших нервов, органов чувств, покровов (включая сравнительные данные по линьке в разных группах), пищеварительной, дыхательной (с учетом вариаций в строении нижней гортани), кровеносной, мочеполовой систем. В заключительных разделах приводятся данные по особенностям эмбрионального развития (особенно детально разбирается развитие скелета).
Во второй части (Bronn’s «Klassen und Ordnungen des Thierreichs», Bd. VI, Abt. IV, Vogel. II. Systeniatischer Theil. Leipzig, 1893, 303 стр. текста) Гадов приводит системы класса птиц предыдущих авторов, начиная с Линнея, затем дает перечень признаков, используемых для характеристики отдельных групп птиц (при этом он охватывает больший набор морфологических признаков, чем Фюрбрингер, и, кроме этого, привлекает эмбриологические материалы, данные по характеру питания и т. д.) и далее характеризует принимаемые им подразделения класса. При этом дается подробная, преимущественно морфологическая характеристика отряда, а затем входящих в него подотрядов и семейств; при описании семейств указывается географическое распространение отдельных родов. Система Гадова выглядит следующим образом:
Класс Aves — птицы
1-й подкласс Archaeornithes — древние птицы, ископаемые Archaeopteryx и Archaeor-nis (юра)
2-й подкласс Neornithes — настоящие птицы
I надотряд Ratitae — бескилевые, все ныне живущие и ископаемые страусо-подобные птицы
II надотряд Odontolcae — зубастые птицы, включает только гесперорнисов — ископаемых нелетающих зубастых птиц мелового периода
III надотряд Carinatae — килегрудые птицы
1-й отряд Ichthyornithes — ихтиорнисы, ископаемые летающие зубастые птицы мелового периода
oTpHA-Colymbiformes — гагарообразные
Подотряды: a) Colymbi — гагары
б) Podicipedes — поганки
отряд Sphenisciformes — пингвинообразные
отряд Procellariiformes — буревестникообразные, или трубконосые отряд Ciconiiformes — аистообразные
Подотряды: a) Steganopodes
б) Ardeae
в) Ciconiae
г) Phoenicopteri Anseriformes — гусеобразные Подотряды: a) Palamedeae
б) Anseres
Falconiformes — соколообразные
Подотряды: a) Cathartae — американские грифы
б) Accipitres — остальные дневные хищники Tinamiformes — тинамуобразные Galliformes Подотряды:
2-й
3-й
4-й
5-й
6-й
7-й
8-й
9-й
отряд
отряд
отряд отряд
— веслоногие
— цапли
— аисты, ибисы
— фламинго
— паламедеи — гусиные
— курообразные
a) Mesites (Mesoenades) — мадагаскарские пастушки б) Turnices — трехперстки
в) Galli — куриные
г) Opisthocomi — гоацины
— журавлеобразные, на подотряды не разделен, включает семейства пастушков, журавлей, кариам, дроф, кагу, солнечных цапель, лапчатоногов
11-й отряд Charadriiformes — куликообразные Подотряды: a) Limicolae
б) Lari в) Pterocles г) Columbae
10-й отряд Gruiformes
— кулики
— чайковые (чайки и чистики)
— рябки
, — голуби
12-й отряд Cuculiformes — кукушкообразные
61
Подотряды: a) Cuculi — кукушки (кукушки и банано-
еды или турако)
б) Psittaci — попугаи
13-й отряд Coraciiformes — ракшеобразные
Подотряды: a) Coraciae — ракши, (ракши, момоты, зимородки, то ди, щурки, удоды, птицы носороги)
б) Striges в) Caprimulgi г) Cypseli д) Colii е) Trogones ж) Pici — совы — козодои — стрижи (стрижи и колибри) — птицы мыши — трогоны — дятлообразные
14-й отряд Passeriformes — воробьинообразные
Подотряды: a) Passeres anisomyodi— рогоклювы и кричащие воробьиные
б) Passeres diacromyodi — птицы лиры и певчие воробьиные
Г. Гадов, стремясь показать филогенетические взаимоотношения между отрядами надотряда килегрудых птиц,' группирует их в 2 ветви (brigade) — 2 основных направления эволюции, каждая из которых в свою очередь подразделяется на 2 подгруппы (legion), объединяющие родственные отряды:
I ветвь. 1. Colymbomorphae — поганко-гагарообразные
Отряды: Ichthyornithes
Colymbiformes
Sphenisciformes
Procellariiformes
2. Pelargomorphae — водные, околоводные и хищные Отряды: Ciconiiformes
Anseriformes Falconiformes
II ветвь. 1. Alectoromorphae — курообразные
Отряды: Tinamiformes
Galliformes
Gruiformes
Charadriiformes
2. Coraciomorphae — ракшеобразные
Отряды: Cuculiformes
Coraciiformes
Passeriformes
Системы M. Фюрбрингера и Г. Гадова в общих чертах довольно близки, что, несомненно, определяется как общим, отчетливо эволюционным подходом к построению системы, так и сходством используемого фактического материала. Следует еще раз подчеркнуть, что обе эти работы до сих пор не потеряли своего большого научного значения как разносторонние морфологические сводки. Наиболее резкое принципиальное отличие — мнение о страусоподобных (бескилевых) птицах. Фюрбрингер помещает отдельные группы бескилевых птиц в разных подразделениях своей системы, подчеркивая этим их полифилетичность. Объединяя всех страусоподобных птиц в один надотряд бескилевых, Гадов показывает, что они представляют естественную, по сути дела монофилитическую группу. В ряде случаев у Гадова изменен, по сравнению с Фюрбрингером, порядок отдельных групп птиц, что отражает несколько иные представления о их филогенетических взаимоотношениях: например, сближение рябков и голубей с куликообразными, а не с курообразными, как у Фюрбрингера, включение дроф в журавлеобразных, а не в куликообразных и т. д. В какой-то степени эти различия определяются большим объемом использованного Гадовым морфологического материала, а с другой стороны — несколько иной его оценкой. Большая внешняя простота и стройность системы, выражающаяся в уменьшении объема особенно громоздких отрядов Фюрбрингера и в исключении градации промежуточных отрядов, очень сильно облегчают пользование системой и ее запоминание. Все это привело к тому, что система Гадова быстро завоевала расположение систематиков-орнитологов: в первой трети XX в. она
62
была наиболее распространена и общепринята. Некоторые систематики пользуются ею и сейчас.
В первой половине XX в. (а особенно в первые 20 лет) основное внимание систематиков-орнитологов было обращено на работу по низшим таксономическим категориям: видам (и подвидам), родам и отчасти семействам. Позже и вплоть до наших дней много исследователей-систематиков занимались изучением таксономических отношений в пределах различных отрядов птиц; упомянуть хотя бы часть из них здесь нет возможности. Результаты этих исследований, конечно, сказались и на общей системе класса: в перераспределении отдельных групп между отрядами, в изменении объема отрядов. Работ, посвященных пересмотру всей системы класса птиц, было относительно немного. Ниже упомянуты лишь те, которые получили большее распространение.
Практически все авторы, предлагавшие новые системы, обычно используют в качестве основы обширные морфологические материалы Гадова и Фюрбрингера, иногда несколько иначе их оценивая и лишь дополняют результатами более поздних исследований (только Ферхейен основывается преимущественно на оригинальных исследованиях). Палеонтологические данные по птицам до сих пор крайне отрывочны и малочисленны, поэтому их роль в построении системы и в выяснении родственных отношений между отдельными группами птиц крайне незначительна. Как и раньше, основной фактический материал для построения системы дает всестороннее изучение рецентных (современных) групп и видов.
А. Уэтмор, американский орнитолог, в 1930 г. выступил с новой системой класса птиц (Wetmore A. A systematic classification for the birds on the world. Proc. U. S. Nat. Mus., vol. 76, art. 24, Jan. 8, 1930, pp. 1—8). Позже он несколько раз ее уточнял и слегка видоизменял (1934, 1940, 1951). Последнее издание системы, вновь несколько пересмотренной в соответствии с новыми данными (в том,числе и палеонтологическими), опубликовано в 1960 г. (A classification for the birds on the world. Smithsonian Miscellaneous Collections, vol. 139, N 11, Washington, 1960, pp. 1—37).
Класс Aves — птицы
Подкласс Archaepmithes — древние или ящерохвостые птицы
1-й отряд Archaepterygiformes — ящерохвостые птицы, включают ископаемых (юра) Archaeopteryx и Archaeornis
Подкласс Neornithes — настоящие, или веерохвостые, птицы
I надотряд Odontognathae — зубастые птицы
2-й отряд Hesperornithiformes — гесперорнисообразные, ископаемые зубастые нелетающие птицы мелового периода
II надотряд Ichthyornithes ихтиорнисы
3-й отряд Ichthyornithiformfes — ихтиорнисообразные, ископаемые зубастые летающие птицы мелового периода
III надотряд Impennes — плавающие
4-й отряд Sphenisciformes — пингвинообразные
IV надотряд Neognathae — новонебные или типичные птицы
5-й отряд Struthioniformes — страусообразные
6-й отряд Rheiformes — нандуобразные
7-й отряд Casuariiformes — казуарообразные
8-й отряд Aepyornithiformes — эпиорнисообразные, страусо-или па-стушковоподобные птицы Мадагаскара и Северной Африки, полностью вымершие уже в историческое время
9-й отряд Dinornithiformes — моаобразные, страусоподобные птицы Новой Зеландии, полностью вымершие в историческое время
10-й отряд Apterygiformes — кивиобразные
11-й отряд Tinamiformes—тинамуобразные
12-й отряд Gaviiformes — гагарообразные
13-й отряд Podicipediformes — поганкообразные
14-й отряд Trocellariiformes — буревестникообразные, или трубконосые 15-й отряд Pelecaniformes — пеликанообразные, или веслоногие
(фаетоны, пеликаны, олуши, бакланы, фрегаты)
16-й отряд Ciconiiformes — аистообразные, или голенастые (цап-
ли, аисты, ибисы, фламинго)
63
17.-й отряд Ansenformes — гусеобразные (паламедеи и собствен-
но гусеобразные, утки, гуси, лебеди)
18-й отряд Falconiformes — соколообразные
19-й отряд Galliformes — курообразные
20-й отряд Gruiformes — журавлеобразные (мадагаскарские
пастушки, трехперстки, журавли, пастушки, лапчатоноги,
кагу, солнечные uai 21-й отряд Diatrymiformes
цы)
22-й отряд Charadriiformes чистики)
23-й отряд Columbiformes
24-й отряд Psittaciformes
25-й отряд Cuculiformes
турако, и кукушки)
26-й отряд Strigifbrmes
27-й отряд Caprimulgiformes
28-й отряд Apodiformes
29-й отряд Coliiformes
30-й отряд Trogoniformes
31-й отряд Coraciiformes
моты, щурки, ракши 32-й отряд Piciformes
медоуказчики, дятлы) 33-й отряд Passeriformes
:и, кариамы, дрофы)
— диатримообразные (ископаемые пти-
— куликообразные (кулики, чайки,
— голубеобразные (рябки, голуби)
— попугаеобразные
— кукушкообразные (бананоеды, или
— совообразные
— козодоеобразцые
— стрижеобразные (стрижи, колибри)
— птицы-мыши
— трогонообразные
— ракшеобразные (зимородки, тоди, мо-удоды, птицы носороги)
— дятлообразные (якамары, пуховки,
— воробьинообразные
По сути дела, это дальнейшее развитие и усовершенствование системы Гадова и одновременно системы Фюрбрингера. По сравнению с Гадовым у Уэтмора изменен порядок отрядов (особенно в начале системы), что отражает несколько иные взгляды на взаимоотношения этих групп. Увеличено число отрядов: разделены на самостоятельные отряды несколько крупных отрядов Гадова. Так, из аистообразных в самостоятельный отряд выделены пеликанообразные, из кукушкообразных — попугаи, из куликообразных — голубеобразные, все подотряды большого отряда ракшеобпазных по Гадову выделены в ранг самостоятельных отрядов. Произведено перемещение отдельных групп: например, мадагаскарские пастушки и трехперстки из куриных переведены в отряд журавлеобразных. Более детально разработана Уэтмором система подразделений внутри отрядов (выделение подотрядов, надсемейств и т. п.). Резкое отличие от Гадова и сходство с Фюрбрингером в том, что Уэтмор отряды страуеоподобных птиц не объединяет в обособленную самостоятельную группу (надотряд), чем подчеркивается их полифиле-тическое происхождение. Выделением в самостоятельный надотряд пингвинов Уэтмор показывает их в известной степени обособленное положение от других групп птиц, что подтверждается палеонтологическими и морфологическими исследованиями последнего времени.
Большая внешняя простота и логичность системы Уэтмора определили ее широкое распространение. XI Международный орнитологический Конгресс в Базеле (1954 г.) рекомендовал систему Уэтмора как стандарт для фаунистических и экологических публикаций, т. е. в тех случаях, когда в работе перечисляются виды разных таксономических групп, при этом перечислении следует придерживаться объема и той последовательности отрядов и более низких таксономических групп, которая принята Уэтмором. Соблюдение этой рекомендации значительно облегчает пользование подобными работами. Совершенно естественно, что подобная рекомендация вовсе не означает, что невозможны иные точки зрения на систему в целом или на отдельные ее части, что создана идеальная система, не нуждающаяся в дальнейшей разработке. Это означает лишь, что на современном уровне знаний система Уэтмора соответствует или близка к взглядам и представлениям большинства (или многих) систематиков.
В 1951 г. с классификацией ныне живущих птиц (вымершие формы не учитывались) выступили Майр и Амадон (Mayr Е., Amadon D. A classification of recent birds. American Museum Novitates, N 1496, New York, 1951, pp. 1—42). Они принимают практически то же число отрядов, что и Уэт
64
мор (выделены в самостоятельный отряд фламинго, а киви объединены с моа), но меняют их порядок: пингвины стоят между тинаму и трубконосыми, хищные птицы — между веслоногими и аистообразными, кукушки — между куриными и журавлеобразными и т. д. Резко упрощен внешний вид системы: отряды сразу разделяютсй на семейства. Только в отряде воробьиных оставлены 4 подотряда (того же объема, что у Уэтмора) и выделены надсемейства. Принципиальных отличий, которые позволяли бы предпочесть эту систему системе Уэтмора, видимо, нет.
Штреземанн, немецкий орнитолог, проявил несколько иной подход к построению системы класса птиц. Он полагал, что Ч. Дарвин и современные систематики-эволюционисты недостаточно оценивали роль различных форм изоляции в видообразовании и в становлении высших таксономических групп. Для преодоления этого упущения в случае неясности филогенетических отношений изучаемых групп лучше каждую из них рассматривать как самостоятельный отряд. При этом следует учитывать не только морфологические критерии, но и биологические характеристики групп (экология, поведение, распространение и т. п.). Впервые эти принципы были применены им в 1934 г. (Stresemann Е. Aves. Kiikenthal — Krumbach's Handbuch der Zoologie. Bd. VII, Hf. 2, Berlin, 1927—1934, pp. 728—853), когда он всех современных птиц сгруппировал в 49 отрядов. Еще большее число отрядов— 51 было им предложено в 1959 г. (The Status of Avian Systematics and its •unsolved problems. The Auk, vol. 76, N 3, 1959, pp. 269 — 280). Ископаемые группы упоминаются Штреземанном лишь в том случае, когда они объединяются с ныне живущими (в скобках приводится обозначение соответствующих групп по системе Уэтмора, 1960):
Отряды: 1. Struthiones
2. Rheae
3. Casuarii
4. Aepyornithes
5. Apteryges
6. Cryniiiri
7. Galli
8. Opisthocomi
9. Turnices
10. Columbae
11. Pterocletes
12. Ralli
13. Heliornithes
14. Mesoenades
15. Jacanae
16. Thinocori —
17. Rhynocheti
18. Eurypygae
19. Cariamae
20. Psophiae
21. Grues
22. Otides
23. Laro-Limicolae
24. Alcae
25. Gaviae
26. Podicipedes
27. Sphenisci
28. Tubinares
29. Anseres
30. Anhimae
31. Steganoppdes
— африканские страусы (отряд Struthioniformes)
— американские страусы (отряд Rheiformes)
— казуары и эму (отряд Casuariiformes)
— эпиорнисы (отряд Aepyornithiformes)
— бескрылы (отряды Apterygiformes -f- Dinornithi-formes)
— скрытохвосты (отряд Tinamiformes)
— куриные (отряд Galliformes)
— гоацины (подотряд отряда Galliformes)
— трехперстки (подотряд отряда Gruiformes)
— голуби (подотряд отряда Columbiformes)
— рябки (подотряд отряда Columbiformes)
— пастушки (надсемейство Ralloidea подотряда Grues отряда Gruiformes)
— лапчатоноги (подотряд отряда Gruiformes)
— мадагаскарские пастушки (подотряд Mesitorni-thides отряда Gruiformes)
— яканы (надсемейство Jacanoldea подотряда Cha-radii отряда Charadriiformes)
— зобатые бегунки (надсемейство Thinocoroidea подотряда Charadrii отряда Charadriiformes)
— кагу (подотряд Gruiformes)
— солнечные цапли (подотряд отряда Gruiformes)
— кариамы (подотряд отряда Gruiformes)
— трубачи (семейство Psophiidae надсемейства Gruo-idea подотряда Grues отряда Gruiformes)
журавли (семейства Gruidae 4- Aramidae надсемейства Gruoidea подотряда Grues отряда Gruiformes)
— дрофы (подотряд отряда Gruiformes)
чайки и кулики (подотряды Charadrii 4- Lari отряда Charadriiformes)
— чистиковые (подотряд отряда Charadriiformes)
— гагары (отряд Gaviiformes)
— поганки (отряд Podicipediformes)
— пингвины (отряд Sphenisciformes)
— трубконосые (отряд Procellariiformes)
— гусеобразные (подотряд отряда Anseriformes)
— паламедеи (подотряд отряда Anseriformes)
— веслоногие (отряд Steganopodiformes или Peleca-niformes)
65
32. Phoenicopteri
33. Gressores
34. Accipitres
35. Musophagae
36. Cuculi
37. Psittaci
38. Striges
39. Caprimulgi
40. Coraciae
41. Ha Icy ones
42. Meropes
43. Momoti
44. Todi
45. Upupae
46. Trogones
47. Colii
48. Apodes
49. Trochili
50. Pici
51. Passeres
— фламинго (подотряд отряда Ciconiiformes)
— голенастые (подотряды Ardeae + Ba laenici piles +
4- Ciconiae отряда Ciconiiformes)
— дневные хищники (отряд Falconiformes)
— бананоеды или турако (подотряд отряда Cuculi -formes)
— кукушки (подотряд отряда Cuculiformes)
— попугаи (отряд Psittaciformes)
— совы (отряд Strigiformes)
— козодои (отряд Caprimulgiformes)
— ракши (семейства Coraciidae + Leptosomatidae +
+ Brachypteraciidae подотряда Coracii отряда Coracii -formes)
— зимородки (надсемейство Alcedinoidea подотряда Alcedines отряда Coraciiformes)
— щурки (подотряд отряда Coraciiformes)
— момоты (надсемейство Momotoidea подотряда Alcedines отряда Coraciiformes)
— тоди (надсемейство Todoidea подотряда Alcedines отряда Coracciiformes)
— удодовые (семейства Upupidae Phoeniculidae подотряда Coracii + подотряд Bucerotes отряда Coraciiformes)
— трогоны (отряд Trogoniformes)
— птицы-мыши (отряд Coliiformes)
— стрижи (подотряд отряда Apodiformes)
— колибри (подотряд отряда Apodiformes)
— дятлообразные (отряд Piciformes)
— воробьиные (отряд Passeriformes)
Наиболее резкое отличие системы Штреземанна от системы Уэтмора — меньший объем многих отрядов; почти все подотряды Уэтмора и часть надсемейств возведены в ранг отрядов. Несколько изменен порядок отрядов. Так, отряд куриных поставлен сразу за отрядами страусоподобных птиц и тинаму; это отражает взгляды ряда исследователей, полагающих, что куриные одна из наиболее примитивных групп современных птиц. Около куриных поставлены голуби и рябки; однако, по мнению Гадова, подтвержденному исследованиями К. А. Юдина (1965), они близки к куликам и чайкам и должны стоять около них. Все группы водных птиц, включая пингвинов, поставлены рядом, что вряд ли оправдано в свете современных представлений об эволюции птиц.
Для практического применения система Штреземанна весьма удобна. Однако, видимо, следует отдать предпочтение более сложно построенной системе Уэтмора, которая более наглядно отражает выявленные или предполагаемые родственные отношения отдельных групп, особенно в пределах выделяемых отрядов.
Обзор систем класса птиц, появившихся в последние десятилетия, равно как и результаты систематических ревизий отдельных групп, позволяет утверждать, что схема системы класса птиц, созданная трудами выдающихся исследователей М. Фюрбрингера и Г. Гадова, в общих чертах справедлива и в наше время. Все исследования последних семидесяти лет смогли лишь несколько перестроить ее, придать большую стройность, сделать более удобной для пользования. Различия между описанными выше новыми системами в общем невелики. Более или менее определился и не вызывает разногласий объем многих крупных групп (отряды, подотряды, многие семейства). Относительно однозначно представление о филогенетических отношениях ряда крупных таксонов. Сохраняющиеся различия и противоречия будут постепенно устраняться при дальнейшей разработке системы.
Наибольшие разногласия сейчас вызывает система отряда воробьинообразных, особенно подотряда певчих воробьиных Passeres (Oscines), объединяющего почти половину видов (около 4000) ныне живущих птиц. Эти разногласия касаются как объема части семейств, так и порядка их перечисления, что отражает разные представления о родственных отношениях между семействами.
66
По Уэтмору (1960), подотряд певчих воробьиных Passeres включает следующие семейства: Alaudidae — Hirundinidae — Dicruridae — Orioli-dae — Corvidae — Cracticidae —Grallinidae — Ptilonorhynchidae — Paradi-saeidae — Paridae — Sittidae — Hyposittidae — Certhiidae — Paradoxorni-thidae — Chamaeidae — Timaliidae — Campephagidae — Pycnonotidae — — Chloropsei dae — Cinclidae — Troglodytidae— Mimidae — Turdidae — — Zeledoniidae — Sylviidae — Regulidae — Muscicapidae — Prunellidae — — Motacillidae — Bombycillidae — Ptilogonatidae — Dulidae — Artami-dae — Vangidae — Laniidae — Prionopidae — Cyclarhidae — Vireolanii-dae — Callaeidae — Sturnidae — Meliphagidae — Nectariniidae — Dicaei-dae — Zosteropidae — Vireonidae — Coerebidae — Drepanididae — Paruli-dae — Ploceidae — Icteridae — Tersinidae — Thraupidae — Catamblyrhyn-chidae — Fringillidae (Fringillinae + Emberizinae).
Майр и Амадон (1951) дают иную систему подотряда певчих воробьиных Oscines: Alaudidae — Hirundinidae — Pycnonotidae — Irenidae —Cam-pephagidae — Muscicapidae (Muscicapinae, Timaliinae, Sylviinae, Maluri-nae, Turdinae, Miminae, Troglodytinae, Cinclinae) — Prunellidae —: Mota-eillidae — Laniidae — Prionopidae — Vangidae — Artamidae — Bombycilli-dae — Certhiidae — Sittidae — Paridae (Parinae, Remizinae, Aegithalinae) — — Dicaeidae — Nectariniidae — Meliphagidae — Zosteropidae — Vireonidae — Drepaniidae — Thraupidae (Parulinae, Coerebinae, Catamblyrhynch-inae, Thraupinae, Pyrrhuloxiinae) — Tersinidae — Fringillidae — Icteridae — Ploceidae — Sturnidae — Oriolidae — Dicruridae — Corvidae — Cracticidae — Grallinidae — Callaeidae — Ptilonorhynchidae — Paradisaeidae.
По сравнению с Уэтмором здесь сильно изменен порядок семейств, а также их объем. Так, семейство мухоловковых объединяет более 1460 видов и включает, кроме мухоловок, тимелий, славок и пеночек, дроздовых, крапивниковых, оляпок и некоторые другие группы птиц, большинство которых рассматриваются Уэтмором как самостоятельные семейства. Все семейства, кроме жаворонковых и ласточковых, Майр и Амадон объединяют в 9 групп (примитивные насекомоядные, нектарницы Старого Света, скворцы и тка-чиковые и т. д.), которые не получают самостоятельных названий и определенного таксономического ранга. Некоторые подсемейства они подразделяют на трибы.
Делакур и Вори (Delacour J., Vaurie Ch. A classification of the Oscines (Aves). Los Angelos County Museum Contributions in Science. N 16, 1957, 1—6) разделяют все семейства певчих воробьиных на 3 группы, устанавливая при этом свой порядок и объем семейств: 1. Alaudidae — Hirundinidae — Motacillidae — Prionopidae — Vangidae — Artamidae — Cracticidae — Laniidae — Oriolidae — Dicruridae — Crallinidae — Callaeidae — — Sturnidae — Corvidae — Paradisaeidae (Paradisaeinae, Ptilonorhynch-inae); 2. Bombycillidae — Campephagidae — Pycnonotidae — Irenidae — — Cinclidae — Troglodytidae — Mimidae — Prunellidae — Muscicapidae (Pachycephalinae, Sylviinae, Polioptilinae, Malurinae, Muscicapinae, Turdinae, Timaliinae, Paradoxornithinae) — Aegithalidae — Paridae (Parinae, Sittinae, Tichodromaininae) —Certhiidae — Salpornithidae; 3. Remizidae — — Dicaeidae — Nectariniidae — Zosteropidae — Meliphagidae — Ploceidae — Fringillidae — Emberizidae (Emberizinae, Thraupinae, Parulinae) — — Vireonidae — Drepaniidae — Icteridae.
XI Международный орнитологический конгресс (Базель, 1954), пытаясь сгладить существующие разногласия, создал специальный Международный комитет, поручив ему рассмотреть различные точки зрения на систему певчих воробьиных. После длительных обсуждений этот Комитет рекомендовал при экологических и фаунистических исследованиях следовать системе, предложенной Майром и Гринуэем: (Mayr Е., Greenway J. С. Sequence of Passerine Families (Aves). Breviora, 1956(58), 1—11) :
Alaudidae — Hirundinidae — Motacillidae — Campephagidae — Pycnonotidae— Irenidae — Laniidae — Prionopidae — Vangidae — Bombycilli-
67
dae — Ptilognathidae — Cinclidae — Troglodytidae — Mimidae — Prunell-idae — Muscicapidae (Turdinae, Timaliinae, Paradoxornithinae, Polioptil-inae, Sylviinae incl. Regulus, Malurinae, Muscicapinae, Monarchinae, Pachy-cephalinae) — Paridae — Sittidae (Hyposittinae, Neosittinae) — Certhi-idae — Dicaeidae — Nectariniidae — Zosteropidae — Meliphagidae — Embe-rizidae (Emberizinae, Cardinalinae, Thraupinae, Tersininae, Coerebinae) — Parulidae — Drepaniidae — Vireonidae — Fringillidae (Fringillinae, Car-duelinae) — Estrildidae — Ploceidae — Sturnidae — Oriolidae — Corvidae.
Однако эта система всеобщего признания пока не получила. Поэтому с сожалением приходится констатировать, что до сих пор не удалось выработать сколько-нибудь единообразного взгляда на систему певчих воробьиных. Совершенно очевидна срочная необходимость дальнейших тщательных исследований, которые позволили бы снять или хотя бы уменьшить существующие разногласия. Определяются они рядом причин. Прежде всего, на современном уровне знаний совершенно неясны родственные отношения и связи многих семейств. В тех же случаях, когда они более или менее очевидны, их трудно отразить в линейной системе.
Неупорядоченность и нестабильность системы очень затрудняют практическую работу, в том числе и пользование литературой, особенно фаунистической. В связи с этим высказывалось предложение (Моро, 1960 и др.) располагать в определенном порядке, отражающем филогенетические отношения, только отряды, а перечень семейств и родов давать в алфавитном порядке их названий. Сходные предложения высказывались и в других разделах систематики (для термитов и других насекомых, некоторых групп морских беспозвоночных и т. д.). Эти предложения пока не встретили поддержки со стороны систематиков.
Таким образом, существующие системы, видимо, можно пока рассматривать как предварительные схемы естественной системы класса птиц. Под естественной системой следует понимать такую систему, которая с достаточной степенью точности и достоверности отражает эволюцию и родственные отношения отдельных групп, и, одновременно, характеризует степень их разобщенности. Для создания такой системы необходимы дальнейшие исследования, углубленный сбор и накопление разнообразного таксономического материала: морфологического, палеонтологического, экологического, этологического, зоогеографического и т. п. При обработке и оценке этих материалов следует учитывать и данные других биологических наук: генетики, сравнительной физиологии и биохимии. Важны именно комплексные исследования, сравнивающие разнообразные показатели.
Подобные широкие таксономические исследования, видимо, целесообразно вести по отдельным группам (отрядам) птиц, чтобы иметь возможность провести ревизию соподчиненных таксономических категорий в пределах отдельных отрядов, а уже затем — сравнение и сопоставление отрядов. Успешность такого подхода хорошо демонстрирует работа К. А. Юдина (1965) по филогении и классификации отряда ржанкообразных.
Степень морфологических различий между разными отрядами птиц в общем относительно невелика и не сравнима d теми различиями, которые, например, существуют между отдельными отрядами млекопитающих; они находятся примерно на уровне морфологических различий между семействами млекопитающих. Даже различия между надотрядами птиц в общем меньше, чем различия между некоторыми отрядами млекопитающих (например, между насекомоядными и хищными и т. п.). В связи с этим часто говорят о необходимости изменения в системе птиц ранга таксонов: группы, именуемые отрядами, следует называть семействами и т. д., но это было бы не совсем верно. Система каждого класса, включая и ранг ее таксонов, должна отражать специфичность его эволюции. А специфичность эволюции класса птиц как раз заключается в том, что они, сохраняя большое морфологическое сходство с пресмыкающимися, обособились от последних преимущественно путем мелких, иногда частных морфологических перестроек, которые,
68
однако, привели к резким физиологическим и биологическим отличиям (например, резкое возрастание интенсивности обмена веществ, теплокровность, сложный уровень высшей нервной деятельности и т. п.). Естественно поэтому, что отдельные отряды птиц различаются, между собой лишь по мелким морфологическим признакам (и иногда не очень четко), а в пределах каждого отряда амплитуда этих различий еще меньше. Однако если рассматривать биологические характеристики отдельных отрядов (т. е. учитывать не только морфологические различия, а различия в распространении и биотипической приуроченности, характере и способе питания, поведении, особенностям размножения и т. п.), то видно, что различия между отдельными отрядами достаточно четкие и в общем соответствуют различиям между отрядами других классов позвоночных животных, а иногда и превосходят их (например, более отчетливы, чем различия между многими отрядами костистых рыб).
В последние несколько десятилетий в разных разделах систематики неоднократно раздавались критические голоса: при построении системы очень велика роль субъективного фактора. Из практически неограниченного числа признаков, характеризующих данную группу, систематик выбирает лишь немногие, и этот выбор субъективен. Устранение субъективности необходимо для построения естественной системы. Наиболее часто предлагается следующий путь: резко увеличивать количество и разнообразие используемых таксономических признаков, а при их обработке использовать количественные (статистические) методы.
Ферхейен, бельгиец, в орнитологии был наиболее последовательным представителем этого направления. Он отмечал, что для построения естественной системы необходимо основываться на целостном таксономическом потенциале организма, используя для анализа возможно большее число характерных сврйств (признаков): сравнение 100 свойств позволит лучше построить естественную систему, чем каталогизация птиц по немногим морфологическим ига этологическим признакам, которые исследователь выбирает как имеющие наибольшее’таксономическое значение. В своих работах по ревизии отдельных отрядов птиц Ферхейен использовал от 100 до 200 преимущественно морфологических и отчасти экологических и поведенческих признаков: число позвонков в разных отделах, характер грудины и костей поясов, пропорции конечностей, число маховых и рулевых, характер оперения (развитие птерилий, наличие или отсутствие пуха), некоторые особенности кровеносной и пищеварительной систем, число яиц и их окраска, состояние птенцов при вылуплении, биотопическая приуроченность, иногда характер и способ питания и т. п. Все эти признаки в той или иной степени обычно используются и другими систематиками. Основное отличие — в оценке этих признаков.
По Ферхейену, если у сравниваемых видов более 90% общих признаков, то они относятся к одному роду, более 70% — к одному семейству, более 60% — к одному подотряду и более 50% —к одному отряду. При этом сам исследователь никак признаки не оценивает, исходя из того, что эволюция птиц шла преимущественно путем «параморфоза», когда сходные структуры появляются в дивергирующих группах общего происхождения. Конвергенция — выработка сходных структур при сходных условиях существования в разных по происхождению группах — при эволюции птиц, по мнению Ферхейена, возникала редко.
Ферхейен обработал большое число видов всех, кроме воробьиных, отрядов птиц, опубликовал материалы по отдельным отрядам в 36 работах (1953—1961 гг.) и выступил с итоговой статьей «Новая классификация (не воробьиных птиц мира» (Verheyen R. A new classification for the Non—Passerine Birds of the World. Institut royal des Sciences naturelies de Belgique. Bulletin. T. XXXVII, N 27, Bruxelles, 1961, pp. 1—36).
При построении системы ископаемые группы птиц он не учитывал.
69
Класс Aves
Надотряд I. Dinornithes — моа
1-й отряд Apterygiformes— киви, или бескрылы
Надотряд II. Hygrornithes — водные птицы
2-й отряд Sphenisciformes— пингвины
3-й отряд Procellariiformes— трубконосые
4-й отряд Alciformes — чистиковые (помимо собственно чистиковых сюда включены гагары и из трубконосых — ныряющие буревестники)
Надотряд III. Limnornithes — болотные (околоводные) птицы
5-й отряд Pelecaniforiftes — пеликанообразные или веслоногие (включены пеликаны, олуши, бакланы)
Первая ветвь
6-й отряд Lariformes — чайки (кроме чаек, включены фрегаты и фаэтоны из отряда веслоногих)
7-й отряд Charadriiformes — кулики
8-й отряд Jacaniformes — яканы (включены яканы из куликов, мадагаскарские пастушки, кагу и солнечные цапли из журавлеобразных)
9-й отряд Podicipediformes — поганки
10-й отряд Ralliformes — пастушковые (исключенные из журавлеобразных пастушки и лапчатоноги)
Вторая ветвь
11-й отряд Ardeifotmes — цаплевые (цапли, китоглавы)
12-й отряд Ciconiiformes — аистообразные (аисты, ибисы, молотоглавы) 13-й oTpHflPhoenicopteriformes—фламинго
14-й отряд Anseriformes—гусеобразные (лебеди, гуси, утки)
15-й отрядAnhimiformes—паламедеи (выведены из гусеобразных)
Надотряд IV. Chamaeornithes — наземные птицы
Первая ветвь
16-й отряд Sfruthioniformes — страусообразные (африканские страусы, казуары, эму, нанду)
17-й отряд Galliformes — куриные (включены отряды тинаму и собственно куриных, трехперстки — из журавлеобразных)
Надотряд V. Dendrornithes — древесные птицы
Первая ветвь
20-й отряд Columbiformes — голубеобразные (голуби, рябки и — из куликов — зобатые бегунки)
21-й отряд Psittaciformes — попугаи
22-й отряд Coraciiformes — ракшеобразные (ракши, зимородки, тоди, момоты, птицы-носороги, а также стрижи и колибри)
23-й отряд Caprimulgiformes — козодои
(в полном объеме)
24-й отряд Strigiformes — совы (в полном объеме)
Вторая ветвь
18-й отряд Gruiformes — журавлеобразные (журавли, трубачи, дрофы)
19-й отряд
Cariamiformes — кариамы
Вторая ветвь
Falconiformes — дневные (в полном объеме)
25-й отряд хищники
26-й отрядCiiculiformeS — кукушкообразные (турако, кукушки)
27-й отряд Coliiformes — птицы-мыши
28-й отряд Piciformes — дятлообразные ( в полном объеме)
29-й отряд Passeriformes — воробьиные
Таким образом, выделяемые Ферхейеном 29 отрядов группируются в 5 надотрядов. Некоторые из отрядов Уэтмора сохранены в полном объеме (киви, пингвины, поганки, попугаи, козодои, дневные хищники, кукушки, птицы-мыши, дятловые, воробьиные), другие разделены или, наоборот, объединены. Из особенно резких различий с Уэтмором обращает внимание объединение в один отряд всех страусоподобных птиц (кроме киви), включение в один отряд чистиковых, гагар и ныряющих буревестников (последние — из трубконосых), создание «сборных» отрядов чаек, якан, куриных, включение в отряд ракшеобразных стрижей и колибри. В трех надотрядах введены добавочные категории — ветви, которыми автор подчеркивает, что эволюция в этих группах шла в двух разных направлениях.
Отчетливо видно, что надотряды объединяют птиц сходных местообитаний и примерно сходного образа жизни. Это хорошо видно уже из назва
70
ний надотрядов: водные, околоводные, наземные, древесные птицы. Также произошло и объединение в новые «сложные» отряды. Чистиковые птицы, ныряющие буревестники и, в меньшей степени, гагары используют при нырянии крылья; эта общая особенность вызвала сходство многих черт строения скелета и пропорций конечностей, мускулатуры и т. д. Чайки, фрегаты и фаэтоны кормятся, преимущественно летая разведывательным меневрен-ным полетом над водой и пикируя на поднявшуюся к поверхности добычу. Сходный способ охоты, конечно, связан со сходством многих черт строения. Это относится и к отряду якан, объединяющих преимущественно тех птиц, которые живут на сильно заросших водоемах.
Таким образом, система Ферхейена прежде всего экологическая. Построение таких систем допустимо и полезно, если стоит задача показать отношение птиц к разным типам местообитаний, характеру добывания пищи, способам передвижения. Такие экологические системы создавались и задолго до Ферхейена (например, Шульпин Л. М. Орнитология, 1940, стр. 194—210). Однако Ферхейен ставил себе совсем иную задачу —создать естественную, т. е. отражающую эволюцию и родственные отношения систему. А она не получилась... Это произошло потому, что он недооценивал значение конвергенции.
Сейчас хорошо известно, что конвергенция (т. е. выработка сходных черт строения как приспособлений к однотипным условиям обитания, у неродственных групп на разной исходной морфологической основе) в эволюции встречается довольно часто. На хорошем палеонтологическом материале это показано для ряда групп пресмыкающихся и млекопитающих. Нет никаких оснований считать, что птицы представляют исключение, тем более, что о конвергентном развитии отдельных групп птиц можно с достаточной уверенностью говорить по уже имеющимся сравнительноморфологическим материалам. Именно конвергенция дает возможность видам из разных по происхождению групп занять сходные экологические ниши (совы и дневные хищники, ласточки и стрижи, колибри и цветочницы и т. п.). Применимый Ферхе^еном чисто статистический метод без оценки таксономического и биологического значения используемых признаков приводит к тому, что в случаях глубокого конвергентного сходства эти признаки резко доминируют над различиями, определяемыми разным происхождением отдельных групп. Поэтому и происходит необоснованное объединение чаек и некоторых веслоногих, чистиковых и ныряющих буревестников и т. п.
Система Ферхейена — шаг назад по сравнению с системами Фюрбрингера, Гадова, Уэтмора и Штреземанна. Однако обширный фактический материал, полученный Ферхейеном и изложенный в его многочисленных работах, представляет несомненный интерес и с успехом может быть использован для характеристик и ревизии средних и низших таскономических категорий (подотряды, семейства, роды). Общая реакция орнитологов на появление системы Ферхейена была весьма сдержанной. Кажется, до сих пор не было опубликовано ни одного положительного отзыва.
ПРОИСХОЖДЕНИЕ И ЭВОЛЮЦИЯ КЛАССА ПТИЦ
Палеонтологические материалы по птицам, как уже указывалось, очень скудны и отрывочны. Поэтому говорить о предках птиц среди рептилий и о происхождении и взаимоотношениях современных групп птиц приходится преимущественно предположительно, основываясь на морфологических особенностях современных групп и, в очень малой степени, на отрывочных палеонтологических данных.
По современным представлениям, предками птиц были архозавры Аг-chosauria — обширная и очень разнообразная группа рептилий, господствовавшая в мезозое и включавшая различных динозавров, крокодилов, летающих ящеров. Непосредственных предков птиц, видимо, нужно искать
71
Рис. 13. Гипотетический предок птиц — предптица (по Хейльманну, 1926)
среди наиболее древней и примитивной группы архозавров — текодонтов или псевдозухий Thecodontia (Pseudosuchia), давших начало и остальным более высокоорганизованным группам архозавров. Псевдозухии по внешнему виду были похожи на ящериц. Они вели преимущественно наземный образ жизни, питались, видимо, разнообразной мелкой животной пищей. Задние ноги были несколько длиннее передних, и при быстром беге животные опирались о почву только задними конечностями. По многим морфологическим особенностям псевдозухии весьма сходны с птицами (детали строения черепа, таза, задних конечностей). Промежуточных форм, которые позволяли бы выяснить стадии развития оперения и выработку способности к полету, пока не найдено. Полагают, что переход к древесному образу жизни и приспособление к прыжкам с ветки на ветку сопровождался разрастанием и усложнением структуры роговых- чешуй на боках туловища и хвоста, по заднему краю передних и задних конечностей, что обеспечивало возможность планирования и удлинения прыжков (гипотетическая стадия предптицы, по Хейльманну, 1926, рис. 13).
Дальнейшая Специализация и отбор привели к развитию оперения крыла, обеспечившего возможность сна
чала планирующего, а -затем активного полета. Оперение, покрывающее все тело птиц, вероятно, развивалось не столько как приспособление к. полету (обеспечение обтекаемости тела), сколько как приспособление к термоизоляции; высказываются предположения, что оно могло сформироваться еще до выработки способности к полету. У некоторых псевдозухий были удлиненные роговые чешуи с четким продольным гребнем, от которого отходили мелкие поперечные ребрышки. Такие чешуйки, видимо, можно рассматривать как структуру, из которой путем расчленения могло возникнуть птичье перо.
Обособление птиц от пресмыкающихся, видимо, произошло уже в конце триаса — начале юры. (170—190 млн. лет назад). Однако ископаемых остатков птиц этого времени пока не найдено. Довольно хорошие остатки двух самых древних известных нам птиц были найдены в 60-х годах XIX в. у Золенгофена (Бавария) на разработках сланцевых песчаников, представляющих отложения мелководного залива юрского моря (возраст около 150 млн. лет). Их назвали Archaeopteryx lithographica и Archaeornis simensi. В 1956 г. там же (в радиусе до 300 м, но на иной глубине) найден значительно худшей сохранности отпечаток еще одной особи. Часть исследователей рассматривает эти находки как остатки трех разных видов (два вида рода Archaeopteryx и один рода Archaeornis), другие же считают представителями одного вида Archaeopteryx lithographica, объясняя различия между особями как индивидуальные, возрастные и половые. Вопрос этот нуждается в дополнительном изучении, но и сейчас совершенно очевидна очень боль
72
шая близость всех этих особей. Все они имеют хорошо развитое оперение крыльев, хвоста и тела, некоторые пропорции черепа ближе к птичьим, чем к рептильным, птичьи черты несут пояс передних конечностей и таз, задние конечности. Типично рептильные черты: нет рогового клюва, имеются слабые зубы, кисть несет первостепенные маховые, но пряжка еще не образовалась и есть три хорошо развитых подвижных пальца с крупными когтями, образование сложного крестца только началось (позвонки еще не срастаются, подвижные кости соединяются с небольшим числом позвонков), бедро причленяется под тупым углом (направлено несколько вбок), проксимальные тарзальные кости еще не приросли к концу большой берцовой кости, хорошо развита малая берцовая кость, не завершено образование цевки и т. п. По внешнему облику, ряду внутренних морфологических признаков и, насколько это можно предполагать сейчас, образу жизни архептерикс (и археорнис) — древние, примитивные, но птицы.
Подкласс настоящих, или веерохвостых, птиц Neornithes, видимо, — прямые потомки каких-то ящерохвостых птиц Archaeornithes. Некоторые исследователи (например, Гадов) считают за прямых предков птиц археоптерикса.и археорниса, другие (их, пожалуй, большинство) рассматривают группу археоптерикса как примитивную, но специализированную боковую и слепую ветвь, полагая, что предками современных птиц должны быть другие, пока еще не обнаруженные ящерохвостые птицы (в случае их обнаружения они, вероятно, образуют другой отряд этого подкласса).
Происхождение и взаимоотношения современных групп (отрядов) птиц остаются, как уже говорилось, за отсутствием достаточных палеонтологических материалов, как правило, очень гипотетическими. Наибольшее внимание удеЛил этому вопросу М. Фюрбрингер, заметно меньшее — Г. Гадов. Не внесли много нового и бесспорного в эту проблему и современные авторы, в той или иной степени касавшиеся филогении птиц. Поэтому здесь изложено представление о взаимоотношениях отдельных групп птиц преимущественно по Фю^брингеру, но в ряде случаев, где это нужно, внесены некоторые изменения (в частных случаях эти изменения специально не оговариваются).
Ископаемые остатки веерохвостых птиц обнаружены только в меловом периоде (их возраст примерно 80—90 млн. лет), т. е. они на несколько десятков миллионов лет моложе археоптериксов. По нескольким плохой сохранности остаткам описаны гесперорнисы — очень крупные (длиной более 100 см) птицы, внешне, видимо, несколько похожие на гагар или поганок, совершенно не летавшие (грудина без киля, от передней конечности сохранилось только маленькое тоненькое плечо), но, вероятно, они хорошо плавали и ныряли: челюсти их несли зубы. В меловых же отложениях найдены остатки других зубастых птиц — ихтиорнисов. Они имели хорошо развитый киль грудины, типичное птичье крыло и, видимо, обладали активным полетом. По размерам они были примерно с голубя. Родственные отношения зубастых меловых птиц с другими птицами очень неясны. Возможно, часть этих остатков относится к меловым динозаврам, а не птицам, но часть, несомненно, зубастые птицы. Фюрбрингер сближал гесперорнисов с гагарами и поганками, а их тиорнисов — с чайками, но при этом подчеркивал их очень давнюю обособленность.
Гадов гесперорнисов выделял в надотряд, показывая этим их обособленное положение, а ихтиорнисы как самостоятельный отряд (тоже обособленный) начинали перечень отрядов килегрудных птиц. Подчеркивая неясность происхождения (да и объем) этих групп, Уэтмор (1960) выделяет их в самостоятельные надотряды.
Из меловых отложений описано еще несколько семейств птиц (по обломкам челюстей и трубчатых костей), но их положение и внешний облик очень неясны: возможно, хотя бы часть этих находок — остатки рептилий. Встреченные в отложениях третичного периода кайнозойской эры (возраст менее 55 млн. лет) остатки птиц с большей или меньшей вероятностью
73
Halcyoniformes
Palllformes
Trogones
Churadriiformes (Laro-L imicolae)
Gruiformes
Hippolectryomithes
Struthlornttties
Archornithes
Colii
Mucrochires Pico-Passeres
Caprimulgi Coraciiformes Striges Cornell
Coccygiformes Psitt uniform es
Columbiformes Coracarnithes Charudriorniihes Culliformes
Crypturiformes
Apterygiformes
Aleciorornlihes
Ichtyor.nithiformes Aptenodytiformes Procellariiformes Steyunopodes Accipiteri
Pelurgo Herodii r—Phoenicopteri Ciconiiformes Podicipitiformes Anseriformes
Pelargornithes Palamedeiformes Rheornithes
Рис. 14. Филогенетические отношения в классе птиц (по Фюрбрингеру, 1888; упрощено)
могут быть отнесены к современным отрядам. Во всяком случае они принадлежат к типичным птицам, близким к тем или иным группам современных птиц. Судя по этим материалам, можно предполагать, что интенсивная адаптивная радиация птиц и становление большинства современных отрядов проходило в самом конце мелового периода мезозойской эры — в третичный период кайнозойской эры, т. е. в период примерно 70—40 млн. лет назад.
Гадов всех ныне живущих и ископаемых крупных нелетающих страусоподобных птиц выделял в надотряд бескилевых, подчеркивая этим их сходство и филогенетическое родство (т. е. монофилию), а также их определенную обособленность от остальных групп птиц. Среди килевых птиц наиболее близки к страусоподобным тинаму (скрытохвосты), куриные, журавлеобразные и близкие к ним группы. Фюрбрингер и большинство современных авторов (Уэтмор, Штреземанн и др.) считают страусоподобных птиц сборной полифилетической группой, разбиваемой на ряд самостоятельных отрядов, начинающих систему, что подразумевает их определенную примитивность и изолированность. Их общие сходные черты — результат конвергенции: крупные размеры, потеря способности к полету, приспособление к быстрому бегу и т. д.
По Гадову, эволюция остальных птиц шла в двух направлениях (две ветви, стр. 62), каждое из которых в свою очередь разделяется на 2 группы родственных отрядов. Поганко-гагарообразные (ихтиорнисы, поганки, гагары, пингвины и трубконосые) родственны группе Pelargomorphae (аистообразные, гусеобразные, хищники), а близкая к страусоподобным группа курообразных (тинаму, куриные, журавлеобразные, куликообразные) дала начало ракшеобразным (кукушки, ракши, воробьиные).
Эволюционные представления Фюрбрингера в общих чертах близки к этой схеме Гадова (о иных взглядах на страусоподобных говорилось выше), но он излагает свои представления с большей степенью детализации. Крупный ствол птиц — отряд Pelargornithes — делится на ряд
74
ветвей: гусеобразных, поганкообразных (включая поганок, гагар, геспе-рорнисов); от основания этих ветвей отходит еще одна мощная ветвь, которая, разделяясь, дает начало веслоногим, дневным хищникам и группам подотряда аистообразных (рис. 14).
Отряд паламедей Palamedeiformes занимает промежуточное положение между нанду и отрядом Pelargornithes. Промежуточные отряды пингвинов Aptenodytiformes и трубконосых Procellariiformes довольно обособлены; возможно, они имеют давние слабые связи друг с другом и, еще более отдаленные, с отрядами Pelargornithes и куликообразных Charadri-ornithes. Ихтиорнисы Ichthyornithiformes — тоже промежуточный отряд, но с несколько более выраженными давними связями с куликообразными. Отряд куликообразных Charadriornithes очень давними связями видимо может быть приближен к отряду ракшеобразных. Основные ветвления ствола куликообразных — подотряд Laro-Limicolae — кулики, чайки и чистики; к ним довольно близки зобатые бегунки и дрофы (переходной группой между куликами и дрофами служат авдотки). От основания ствола куликообразных обособились еще две крупные ветви: журавлеобразные Gruiformes и пастушковые Ralliformes. Журавлеобразные разделяются на несколько ветвей: собственно журавлиных (журавли, арамиды, трубачи, кариамы) и занимающих несколько обособленное положение солнечных цапель и кагу. Пастушковые подразделяются на родственные ветви: пастушков, лапчатоногих и стоящих несколько обособленно трехперсток и мадагаскарских пастушков.
Один из древних стволов родословного древа птиц — отряд курообраз-ных Alectorornithis. Наиболее примитивная ветвь — подотряд бескрылых, или киви Apterygiformes, включающий киви и моа; обе группы разошлись почти сразу после своего обособления. Общим происхождением с предыдущей ветвью, видимо, связаны тинаму (или скрытохвосты) Cryptu-riformes, в свою очередь через предков близкие к подотряду куриных Galliformes {практически в современном объеме). Отряд курообразных очень давними родственными ,связями, вероятно, более или менее близок к пастушковым Ralliformes, а через них и ко всему отряду куликообразных.
Промежуточные отряды голубеобразных Columbiformes (включающий рябков и голубей) и попугаев Psittaciformes занимают в системе обособленное положение. Вероятно, через очень давние отношения они как-то связаны с обширным отрядом ракшеобразных. По представлениям Гадова, голуби и рябки в ранге подотрядов входят в отряд куликообразных, а попугаи — тоже как подотряд — в отряд кукушкообразных. По сути дела сходной точки зрения придерживается и Уэтмор, ставящий отряд голубеобразных сразу же за отрядом куликообразных, а отряд попугаев — рядом с отрядом кукушек.
Обширный и разнообразный отряд Coracornithes своими истоками связан с предками отряда куликообразных. Основная и наиболее мощная ветвь — подотряд дятлов и воробьиных Pico — Passeriformes, которая затем разделяется на отдельные группы собственно воробьиных и дятловых. От основания этой ветви обособились Macrochires (стрижи, колибри) и птицы мыши Colii. Еще раньше от этой ветви обособляются трогоны Тго-gones. От самого основания ветви дятлово-воробьиных птиц обособляются и три других подотряда ракшеобразных. Подотряд кукушкообразных Coccygiformes отделяет ветвь якамар Galbulae (якамары и пуховки, а по Уэтмору —это подотряд отряда дятлов), а затем разделяется на кукушек Cuculidae и бананоедов Musophagidae. Подотряд ракшеобразных Coraciiformes разделяется на собственно ракш Coraciae и на вторую ветвь, вскоре делящуюся на сов Striges и козодоев Caprimulgi. Подотряд зимородковых На Icy oni formes (он, видимо, ближе к Pico — Passeriformes, чем к другим подотрядам), отделяя в самом начале боковую веточку то ди Todi (кроме тоди Фюрбрингер сюда включает и момотов), разделяется на 3 род-
75
Poenicopteri
Gressores
Galli X 1 CucM
Grues
LimicoLae - Lari
Anseres
Accipitres Steganopodes Tubinares
Sphenisct
Macrochires Ptci
Passeres
Dendrorncthes
Geornithes
Psittaci
Striges Caprimulgi
Trogones
'Coracii Colli
Рис. 15. Схема филогенетических отношений в классе птиц (по Берндту и Майзе, 1960)
ственные группы: зимородковых Halcyones (Alcedinidae), удодов Bucero-tes (удоды, птицы-носороги) и щурок Metopes.
Сопоставление филогенетических представлений Фюрбрингера со взглядами Гадова и Уэтмора (очередность и порядок перечисления отрядов) показывает значительное их совпадение; резкие отличия в положении и взаимоотношении отдельных групп указаны выше.
В последние годы попытку графического изображения филогенетических взаимоотношений между отдельными группами птиц предприняли немецкие орнитологи Берндт и Майзе в капитальной трехтомной сводке «Естественная история птиц» (Bernd R.,MeiseW. Naturge«5chichte derVogel, Kosmos, Stuttgart, 1960 (1962), v. 2, стр 668—673). Подкласс древних ящерохвостых птиц Palaeornithes дал начало подклассу новых веерохвостых птиц Neornithes, который включает 24 отряда, подразделяемых авторами обычно сразу на семейства; таксономические категории ранга подотряда и надсемейства используются, как правило, только в отряде воробьиных. Взаимоотношения между отрядами птиц, по Берндту и Майзе, выглядят следующим образом:
От древних птиц эволюция шла по двум направлениям (ветвям): ветви (надотряд) наземных и водных птиц Geornithes и ветви (надотряд) древесных птиц Dendrornithes. От самого начала ветви наземных и водных птиц (рис. 15) общим стволом отделились, позже разделившись, тинаму Crypturi и бескилевые птицы Ratitae, включая всех ныне живущих страусоподобных, в том числе и киви, и ископаемых моа и эпиорнисов. Далее обособляются куриные птицы Galli (в полном объеме отряда Уэтмора) и на этом же уровне, но самостоятельной ветвью j— гагарообразные Pygopodes (включающие ископаемых меловых зубастых птиц гесперорнисов, гагар и поганок). Все остальные отряды этой ветви (надотряда) имеют общее происхождение — представляют собой как бы мутовку стволов, отходящих от одного участка. Относительно тесно связаны друг с другом и несколько противопоставляются другим группам отряды, журавлеобразных (пастушки, мадагаскарские пастушки, солнечные цапли,лапчатоноги, кагу, настоящие журавли, арамы, трубачи, дрофы, кариамы, трехперстки; из ископаемых — диатримы и форораки Phororhacidae) и болотно- и чайкообразных птиц Limicolae — Lari (яканы, различные группы собственно куликов, чайки чистики и — из ископаемых — зубастые меловые птицы — ихтиорнисы). Далее от общего основания мутовки (т. е. имея общих предков) последова
76
тельно обособляются отряды гусеобразных Anseres (паламедеи и собственно гусеобразные), фламинго Phoenicopteri, голенастые Gressores, дневные хищники Accipitres и веслоногие Stegaribpodes. От форм, близких к предкам веслоногих, отделяется ветвь, которая затем разделяется на отряды трубконосых Tubinares и пингвинов Sphenisci.
Развитие второй ветви (надотряда) древесных птиц Dendrornithes начинается от каких-то предковых форм, видимо, более древних, чем предки первой ветви (первого надотряда). Сначала общим стволом обособляются 3 отряда: кукушкообразных Cuculi, голубей Columbae (голубки, рябки и дронты) и попугаев Psittaci. Следующая обособившаяся ветвь разделяется на 2 ответвления, каждое из которых образует по 2 отряда. Одно из них дает начало совам Striges и козодоям Caprimulgi, другое — трого-нам Trogones и ракшеобразным Coracii. Дальше от общего ствола также общей ветвью обособляются отряды птиц-мышей СоШ и стрижеобразных Macrochires (стрижи и колибри), самостоятельной ветвью отходит отряд дятлов Pici (в объеме Уэтмора), а основной ствол ветвится на многочисленные подразделения отряда воробьиных Passeres. Помимо этого, авторы дают гипотетические схемы, в которых пытаются представить филогенетические взаимоотношения в пределах нескольких отдельных отрядов (дневных хищников, веслоногих, гагарообразных и воробьиных).
Филогенетические схемы Берндта и Майзе ясны, наглядны и легко запоминаются. Однако они практически совсем не аргументированы, не обоснованы материалами и, видимо, в значительно меньшей степени отражают действительные родственные (филогенетические) отношения между отдельными группами птиц, чем соответствующие схемы Фюрбрингера. Теоретически маловероятно, что уже на начальных стадиях эволюции класса четко определилась последующая биотопическая специализация птиц, что выражается в изложенной схеме деления подкласса «новых птиц» только на 2 обособленных надотряда и в почти линейном, последовательном обособлении отрядов. Оценивая по современным формам присущий птицам характер эволюции, выражающийся в широкой адаптивной радиации в пределах отдельных групп (о чем кратко упоминалось выше), кажется более логичным предполагать, что подобная же широкая адаптивная радиация была характерна и для начальных фаз эволюции класса. Если это так, то можно говорить, что филогенетические схемы Фюрбрингера хотя бы внешне действительно отражают преобладание такого типа эволюции.
Сейчас еще рано говорить о вполне сложившейся системе класса. Однако системы Фюрбрингера, Гадова и Уэтмора, видимо, могут рассматриваться как хорошая основа для построения естественной системы. Их материалы дают и самое общее представление о филогенетических взаимоотношениях большинства групп. Для улучшения и уточнения системы класса птиц необходимы дальнейшие разнообразные исследования на всех таксономических уровнях (от подвидовых группировок до отрядов и надотрядов): морфологические, экологические, этологические, генетические. Возможно, в будущем ценные материалы даст применение биохимических и физиологических показателей; использование современных методик пока существенного вклада в таксономию птиц не внесло. Хочется надеяться, что увеличится и количество палеонтологических материалов, которые позволят более обоснованно обсуждать эволюцию птиц в целом и отдельных групп (отрядов, семейств).
СИСТЕМАТИЧЕСКИЙ ОБЗОР КЛАССА ПТИЦ AVES*
Царство животных Animalia
Подцарство многоклеточных Metazoa Тип хордовых Chordata Подтип позвоночных Vertebrata Группа амниота Amniota Класс птиц Aves
Птицы — яйцекладущие амниота. Череп с одним затылочным мыщелком, большими глазницами, с образующими клюв вытянутыми челюстями, обычно лишенными зубов, с подвижным небом и довольно большим объемом мозговой полости. Шейные позвонки гетероцельные. Грудина большая, плоская, у большинства видов с хорошо развитым килем. Передние конечности превратились в крылья и не участвуют в движении по суше. Образуется сложный крестец: последний грудной, поясничные, крестцовые и часть хвостовых позвонков сливаются в монолитное образование вместе с костями таза. Единственная опора при движении по суше — задние конечности. Малая берцовая кость более или менее редуцирована. Проксимальный ряд костей предплюсны прирастает к нижнему (дистальному) концу большой берцовой кости. Дистальный ряд костей предплюсны (тарзус) и все косточки плюсны (метатарзус) соединяются (у большинства — сливаются) в цевку (тарзометатарзус). Чаще три пальца направлены вперед, один — назад (он может редуцироваться). Скелет (особенно трубчатые кости) обычно сильно пневматичен.
Желудок делится на 2 отдела: железистый и мускульный. Имеется клоака. Мочевого пузыря нет. Функционируют только левый яичник и левый яйцевод. Сердце четырехкамерное. От левого желудочка отходит один ствол — правая дуга аорты. Легкие сильно губчатые, компактные; с ними связана сложная система воздушных мешков. Голосовой аппарат — нижняя гортань. Кожных желез практически нет: есть только одна крупная кожная копчиковая железа, лежащая над основанием хвоста; у части видов она отсутствует. Тело покрыто пером. Более длинные и прочные перья — маховые и рулевые — образуют крылья и хвост. Характерна гомойотерм-ность. Температура тела довольно высокая (37-42° С). Поведение сложное. Обычно хорошо выражена забота о потомстве: устройство гнезд, иногда очень сложных и искусно построенных, насиживание яиц, вскармливание и обогревание птенцов, их защита и т. п.
Класс птиц разделяется на 2 подкласса, объединяющих 33 отряда (из них 6 только с ископаемыми формами) и 213 семейств (из них 42 — ископа-
1 В этом обзоре автор придерживается системы Уэтмора (A. Wetmore. A classification for the birds of the world. Smithsonian Miscellaneous Collections, vol, 139, № 11, Washington, 1960, 1—37). Все отклонения оговорены fi тексте.
78
емые). Всего сейчас известно около 8600—8800 видов (включая вымерших в историческое время, но без ископаемых форм), группируемых в 2000— 2400 родов. Распространены практически по всему земному шару. На территории СССР отмечен 734—751 вид (расхождение за счет спорных случаев: вид, подвид?), принадлежащих к 284—301 роду из 75 семейств 18 отрядов. Несомненно, гнездится 650—665 видов, из которых 26 — эндемики: их гнездовой ареал не выходит за пределы нашей страны.
ПОДКЛАСС ARCHAEORN1THES —
ДРЕВНИЕ, ИЛИ ЯЩЕРОХВОСТЫЕ, ПТИЦЫ
Известны по отпечатку пера и трем разной сохранности отпечаткам скелета и оперения, найденным в илистых сланцах —отложениях залива моря юрского периода (примерный возраст 150—130 млн. лет) в районе Золспгофен, Бавария. Имели настоящее оперение, включая маховые и рулевые перья. В скелете много рептильных признаков (рис. 16). Хвост состоял примерно из 20 позвонков; на боковых поверхностях каждого из них крепились парные рулевые перья (отсюда и название подкласса — ящерохвостые, т. е. имеющие длинный рептильный хвост). Позвонки амфицельные. Тонкие ребра, видимо, не имели крючковидных отростков. Были брюшные ребра (не соединяются с грудиной и позвонками, лежат в брюшной стенке). Пояс передних конечностей по форме и составу похож на птичий;
Рис. 16. Отпечаток ящерохвостой птицы Archaeopteryx litographica Meyer
79
ключицы уже срослись в вилочку. Грудина разбита и о ее виде судить трудно. Кости передней конечности относительно слабее, чем у птиц. Проксимальный конец плеча напоминает плечо птицы; отчетливо развит боковой гребень, что говорит о довольно сильных мышцах, двигавших крыло. Метакарпальные кости тесно прилегают друг к другу, но еще не срослись. Три свободных, хорошо развитых пальца заканчиваются острыми когтями. Второстепенные маховые прикреплялись к локтевой кости, первостепенные (их не менее 10) — к метакарпальным костям и основным фалангам II и III пальцев. Сложный крестец развит слабо: тазовый пояс соединяется только с 4—6 позвонками (у современных птиц с 10—22). Лобковые кости направлены назад и, видимо, образовывали симфиз. Задняя конечность внешне напоминает птичью, но имеет ряд примитивных признаков. Бедро направлено несколько вбок, что больше похоже на положение бедра у рептилий. Малая берцовая кость такой же длины, как большая. Дистальные тарзальные косточки тесно прилегают к метатарзальным костям, но еще не слились в монолитную цевку. Три направленных вперед пальца развиты сильнее, чем единственный палец, направленный назад.
Череп сохранился только на одном отпечатке. В целом он рептильного типа, но с отчетливо выраженным утончением костей. Объем мозговой коробки невелик. Межчелюстная кость меньше верхнечелюстной. Образуется некоторое подобие клюва, но мощного рогового покрова (рамфотеки), видимо, еще нет. Верхняя и нижняя челюсти несут мелкие острые зубы, сидящие в альвеолах. Из признаков, напоминающих череп современных птиц, можно отметить: увеличенные по сравнению с рептилиями размеры глазниц, разделенных тонкой межглазничной перегородкой (тропибазальный тип черепа), смещение назад наружных носовых отверстий, редукция верхней дуги, форма общей височной ямы, слияние костей без образования швов.
По размерам эти животные были близки к современной сороке. Судя по многим деталям строения, летали они плохо и, видимо, были способны лишь планировать или перепархивать с дерева на дерево. По земле они, вероятно, передвигались на задних конечностях, а при перемещении в кронах и по стволам деревьев использовали и подвижные пальцы передних конечностей.
ОТРЯД 1. ARCHAEOPTERYGIFORMES
Подкласс ящерохвостых птиц включает один отряд с единственным семейством Archaeopterygidae и двумя родами: Archaeopteryx (два отпечатка) и Archaeornis (один отпечаток).
Сейчас большинство исследователей полагает, что все отпечатки относятся к одному виду Archaeopteryx lithographica М. Отличия между изученными экземплярами объясняют половой и возрастной изменчивостью.
ПОДКЛАСС NEORN1THES —НАСТОЯЩИЕ, ИЛИ ВЕЕРОХВОСТЫЕ, ПТИЦЫ
Объединяет всех современных и ископаемых птиц, для которых характерно резкое укорочение хвостового отдела позвоночника и слияние последних хвостовых позвонков в специальную косточку — пигостиль, к которой веером прикрепляются рулевые перья (поэтому птиц этого подкласса называют веерохвостыми). В передней конечности образовалась пряжка и отсутствуют свободноподвижные пальцы. Заметно редуцирована малая берцовая кость, образовалась монолитная цевка (тарзометатарзус), имеется типичный сложный крестец и т. д.
Подкласс разделяется на 4 надотряда, объединяющие 32 отряда.
80
Надотряд I. Odontognathae — зубастые птицы
Включает только один отряд ископаемых птиц.
ОТРЯД 2. HESPERORNITHIFORMES -ГЕСПЕРОРНИСООБРАЗНЫЕ
В отряде 2 рода: Hesperornis и Hargeria с 4 видами. Остатки этих птиц найдены в меловых отложениях Канзаса, США (примерный возраст 80 — 90 млн. лет). Эти крупные птицы (один вид длиной более метра), видимо, хорошо плавали и ныряли, а по внешнему облику ' несколько напоминали гагар или поганок (рис. 17). Мощные задние конечности имели 4 пальца, видимо, соединенных плавательной перепонкой или отороченных роговыми щитками, как у поганок. Чашечка сливается с мощным и длинным акромиальным отростком. Узкий таз типично птичьего типа. Способностью к полету они не обладали: грудина плоская, без киля; от сильно редуцированной передней конечности сохранилась маленькая тонкая плечевая кость. Череп по общей конфигурации похож на череп гагар, но резко отличается рядом особенностей: мал объем мозговой полости, верхняя и нижняя челюсти имеют зубы, расположенные в общем желобке. Предчелюстные кости покрыты роговым покровом и не несут зубов. Часть позвонков гетероцельного типа; хвостовой отдел сильно редуцирован и лишен пигостиля. Видимо, жиди по морским, побережьям, питаясь рыбой, которую добывали, плавая и ныряя. По суше, вероятно, передвигались с большим трудом.
Возможно, к этому же отряду следует отнести еще 3 рода (Enaliornis, Neogaeornis, Baptornis) с 4 видами, описанными по плохо сохранившимся фрагментарным остаткам из меловых отложений в США, Чили и Англии.
Рис. 17. Гесперорнис Hesperornis regalis Marsch. (реконструкция)
81
Надотряд II. Ichthyornithes — ихтиорнисы
Включает один отряд, объединяющий тоже только ископаемых птиц.
ОТРЯД 3. ICHTHYORNITHIFORMES— ИХТИОРНИСООБРАЗНЫЕ
Рис. 18. Ихтиорнис Ichthyornis victor Marsch. (реконструкция)
Два рода (Ichthyornis, Apatornis) с 8 видами, описанными по остаткам из верхнемеловых отложений в США. Это были, несомненно, летающие, но очень своеобразные птицы (рис. 18), размером примерно с голубя. Скелет крыла типичен для летающих птиц (образовалась пряжка, редуцировались пальцы и т. д.), грудина с хорошо развитым килем. Задние конечности по пропорциям и строению похожи на конечности чаек. Позвонки амфицельные. Хорошо развит пигостиль. Голова непропорционально большая. Объем мозговой полости невелик. Длинный клюв. Нижняя челюсть и верхнечелюстная кость (нр не предчелюстная)
несут конические, сидящие в альвеолах зубы. Видимо, жили по морским побережьям, питались рыбой, которую схватывали в поверхностных слоях воды при пикировании.
Надотряд III. Impennes — плавающие
Включает только один отряд — пингвинов.
ОТРЯД 4. SPHENISCIFORMES — ПИНГВИНООБРАЗНЫЕ
Птицы средней и крупной величины (вес от 1,5 до 40 кг), нелетающие но прекрасно плавающие и ныряющие (ныряют на глубину более 20 м развивая скорость до 36 км/ч). Крыло преобразовано в ласт — орган поступательного движения при плаваньи и нырянии. Сильно отнесенные назад короткие задние конечности выполняют при плаваньи и нырянии только функцию рулей. Три направленных вперед пальца соединены плавательной перепонкой; 1 палец очень мал и направлен не назад, а внутрь. На суше тело держат вертикально (рис. 19), передвигаясь мелкими шагами или короткими прыжками. При движении иногда ложатся грудью на лед и быстро скользят вперед, отталкиваясь ногами и ластами.
Из многих своеобразных морфологических особенностей обращает внимание большая подвижность туловищного отдела позвоночника (малоподвижного или совсем неподвижного у остальных птиц); крестец не срастается
82
с тазом, а соединяется с ним мощными связками. Грудина с высоким килем. Пояс передних конечностей мощный; лопатка в задней половине очень широкая. Все кости крыла сильно уплощены (занимают почти треть ширины крыла-ласта), особенно скелет кисти. Кости крыла малоподвижны во всех суставах, кроме плечевого. Все крыло покрыто плотными мелкими чешуеобразными перышками с длинными, погруженными в кожу очинами, что резко повышает упругость заднего края ласта. Мышцы дистальной части крыла развиты слабо (крыло почти не сгибается), а мышцы, двигающие крыло в плечевом суставе, очень мощны; при этом подключичная мышца (поднимающая крыло) развита почти столь же хорошо, как и большая грудная. Все эти особенности связаны с движением крыла в воде. В задней конечности своеобразна короткая широкая цевка, на которой не только отчетливо видны швы, но имеются отверстия — результат неполного слияния трех метатарзальных костей. Пнев-матичность костей развита слабо или вовсе не выражена.
Рамфотека клюва у части видов не сплошная, а состоит из нескольких щитков. Язык и небо покрыты направленными назад длинными ороговевшими сосочками, помогающими удерживать скользкую подвижную добычу. Пищевод сильно растяжим. Железистый желудок очень объемистый, мускульный желудок с толстыми стенками, но относительно невелик; кишечник длинный, слепые кишки очень малы. Фабрициева сумка сохраняется и у взрослых. Есть оперенная копчиковая железа. Контурное перо равномерно покрывает все тело (аптерий нет). Ствол пера уплощенный и широкий. Бородки второго порядка не несут крючочков, поэтому сцепленные опахала отсутствуют. Каждое перо снизу имеет мягкий добавочный пуховой стержень. Густо сидящие в коже, черепицеобразно налегающие друг на друга упругие перья образуют плотный покров, успешно противостоящий и проникновению воды, и раздуванию ветром. Теплозащитные свойства оперения дополняются сильным отложением жира в соединительнотканном слое кожи. Новое перо формируется в толще кожи, затем, проникая вершиной к основанию очина старого пера, выталкивает его. Линька проходит в сжатые сроки (10—20 дней) и охватывает одновременно большие участки тела. В период линьки птицы стоят на суше.
Окраска однообразна: у всех видов верх тела черный или серый, низ белый. У части видов на голове растут яркие украшающие перья. Полового морфизма окраски нет. Моногамы. Гнездятся более или менее крупными колониями: от десятков до сотен и даже до десятков тысяч пар. У двух самых крупных видов в кладке одно яйцо, у остальных 2. Насиживание от 30 (мелкие виды) до 64 дней (крупные виды). Яйца откладывают в короткие норы, в трещины и расщелины скал, под валуны или открыто, в небольшие углубления почвы. Некоторые виды устраивают из камешков примитивные гнезда. Насиживают и выкармливают оба партнера.
Особенно своеобразно размножение у видов, гнездящихся по берегам Антарктиды. Наиболее южногнездящиеся императорские пингвины Apte-nodytes forsteri Gr. появляются на местах гнездовых колоний еще зимой.
83
Ближайшие разводья в это время находятся в 100—150 км от колоний. Птицы имеют большие жировые отложения (до 5—10 кг). После образования пар и брачных игр начинается откладка яиц (обычно в мае). Самка откладывает одно крупное яйцо (весом до 450 г) и почти сразу же покидает колонию, уходя к морю на кормежку. Насиживает в течение 62—64 дней самец, который держит яйцо на лапах, прикрывая его сверху и с боков складкой кожи на нижней части брюха. Насиживающая птица перемещается вместе с яйцом короткими шажками, держа туловище вертикально. За период насиживания самец теряет до 30—45% веса. Самки возвращаются на колонию примерно через 2 месяца, очень жирными и с запасом корма в пищеводе (до 1 кг). К этому времени вылупляются птенцы, которых начинают обогревать самки; они же кормят их отрыжкой. Птенцы вылупляются зрячими, покрытыми густым, но коротким пухом. Самцы после вылупления птенцов уходят на кормежку к морю и возвращаются только через 1—1,5 месяца, также с кормом. Подросшие к этому времени птенцы сбиваются в плотные скопления —«детские сады», — включающие до нескольких сотен особей. Это позволяет им, обогревая друг друга, противостоять еще нередким в это время морозам и снежным бурям. Оба партнера поочередно ходят за кормом. Благодаря весеннему торошению и таянью льда расстояние до открытой воды резко сокращается, и взрослые птицы носят корм птенцам с интервалами в несколько дней.
У всех видов подросшие птенцы сменяют свой первый( эмбриональный) пуховой наряд на второй, и лишь спустя некоторое время у них начинает формироваться первый перьевой наряд. Лишь полностью оперившись, в возрасте 3 (мелкие виды) — 6 месяцев (крупные виды) птенцы покидают гнездовые колонии и начинают вести кочевую жизнь в море; к этому времени они достигают или почти достигают размеров и веса взрослых птиц.
Кормятся только в море, ловя мелких рыб, головоногих моллюсков и ракообразных. Распространены только в Южном полушарии (рис. 20). Большинство видов приурочено к островам Субантарктики, несколько видов населяет береговую кромку ледового щита Антарктики. С холодным течением Spheniscus detnersus (L.) проникает на север щгнездится на островах и кое-где на юго-западном побережье Африки (на север до 17° ю. ш.), два вида гнездятся,по западному побережью Южной Америки (на север до берегов Перу), a Spheniscus mendiculus S. гнездится на расположенных у самого экватора Галапагосских островах (к ним подходит холодное Гум-больдтово течение от Антарктиды). Самый мелкий вид EudyptulcrminorE. по южному побережью Австралии проникает на север до 38° ю. ш.
Половой зрелости мелкие виды достигают в возрасте около 2 лет, крупные — на 4—5-м году жизни.
В отряде единственное семейство Spheniscidae, включающее 16 видов в 6 ныне живущих родах: Spheniscus, Aptenodytes, Eudyptes, Eudyptula, Megadyptes, Pygoscelis. Ископаемые остатки типичных пингвинов обнаружены в Новой Зеландии, Австралии, южной Аргентине в эоценовых (возраст 60—40 млн. лет) и более молодых отложениях. Эти остатки группируют в 21 род с 32 видами; один из ископаемых видов достигал размеров человека. Из ныне живущих видов в ископаемом состоянии обнаружено 4.
Надотряд IV. Neognathae — новонебные, или типичные, птицы
Сюда относят всех остальных ныне живущих и ископаемых птиц, в целом (за исключением, пожалуй, нескольких отрядов страусоподобных птиц) более прогрессивного строения, более разнообразной экологической специализации. Большое разнообразие включаемых в этот надотряд групп очень затрудняет выбор признаков, которые в равной степени были бы свой
84
150° 120° 90° 60°к западу от Гринвича 0*к востоку от Гринвича 60° 90° 120° 150° 180° 150°
Рис. 20. Ареалы:
1— отряд пингвинообразных Sphenisciformes, 2— отряд гагарообразных Gavii formes, 3— отряд поганкообразных Podicipediformes
ственны всем входящим сюда 29 отрядам. Во всяком случае челюсти никогда не несут настоящих зубов. (иногда могут развиваться роговые зубчики), крылья не преобразуются в ласты; цевка удлиненная, без отверстий (без следов неполного слияния метатарзальных костей), линька пера по всему телу идет постепенно.
Все встречающиеся на территории СССР птицы относятся только к этому надотряду.
ОТРЯД 5. STRUTHIONIFORMES — СТРАУСООБРАЗНЫЕ
Рис. 21. Страус Struthio camelus L-
В отряде одно ныне живущее семейство Struthionidae с единственным видом Struthio camelus L. (рис. 21), распространенным в степях и полупустынях Африки (рис. 22); в Аравии сейчас, видимо, уже полностью истреблен. Самая крупная из ныне живущих птиц: вес 75—100 кг. Прекрасно бегает, развивая скорость свыше 50 км/ч, не летает.
Череп отличается от черепов других бескилевых птиц: сошник смещен вперед и соединяется только с небными отростками верхнечелюстных костей; небные кости палочковидны и причленяются к средней части крыловидных костей; хорошо развиты базиптерйгоидные сочленения. Скелет крыла типично птичьего типа, но сильно укорочен, особенно кисть; кисть и предплечье вместе короче плеча. Карпальные кости частично редуцированы. Сильно упрощен пояс передних конечностей: вилочка редуцировалась, коракоид и лопатка срастаются, грудина без следов киля. Лобковые кости образуют симфиз. Задние конечности очень мощны. Сохранились только 2 пальца: более короткий наружный (IV) и более длинный внутренний (III) палец, который заканчивается широким когтем, напоминающим копыто. На бегу ширина шага достигает 3—4 м. Сохранился маленький пигостиль (из 4 слившихся позвонков).
В связи с преимущественно растительным характером питания пищевод сильно растяжимый, а железистый желудок объемистый. Мускульный желудок невелик по объему, но имеет очень мощные мускульные стенки и толстую бугорчатую кутикулу, что обеспечивает перетирание грубой растительной пищи (вегетативные части растений, плоды в плотной, иногда колючей, скорлупе и т. п.). На границе железистого и мускульного желудка находится расширение — фактически вырост железистого желудка, но без желез, увеличивающее объем желудка и потому позволяющее одновременно заглатывать большое количество пищи. Кишечник очень длинный; он превышает длину тела примерно в 20 раз и достигает 14—15 м. Сильно развиты слепые кишки: длиной до 70 см. Относительно длинна и прямая кишка, в которой, видимо, как и в слепых кишках, идет переваривание целлюлозы за счет деятельности бак-
терий. В клоаке обособляет-
86
ся ее верхний отдел — caprodeum, — фактически выполняющий функцию мочевого пузыря. Есть сложно устроенный непарный совокупительный орган (penis). Копчиковая железа отсутствует.
Оперение равномерно покрывает все тело (аптерий нет). На груди есть участок голой утолщенной кожи («грудная мозоль»), на который птица опирается, лежа на земле. Перья рассучены (из-за отсутствия крючочков бородки не сцепляются в опахало), их опахала симметричны, добавочный стержень отсутствует; 16 первостепенных, 20—23 второстепенных маховых и до 50—60 рулевых также рассучены и выполняют только роль украшающих перьев. Резко выражен половой диморфизм: самец ярко-черный с белыми маховыми и хвостовыми перьями, самка одноцветная, коричневато-бурая.
Держатся обычно небольшими группами. У самца отчетливо выражен ток: он принимает разнообразные позы, машет крыльями, издает громкие звуки. Видимо, редко моногамы, чаще полигамы: с самцом держатся 2—5 самок, каждая из которых откладывает 10 яиц в общее гнездо. Яйца относительно мелкие (1,5—1,7% от веса самки): длина до 15—16 см и ширина около 12—13 см, вес 1,1—1,6 кг. Скорлупа блестящая, белого цвета. Самки по очереди насиживают общую кладку днем, самец — ночью. Насиживание продолжается 6—7 недель. Птенцы выводкового типа: вылупляются опушенными, зрячими и, обсохнув, покидают гнездо. Кормятся сами. С птенцами чаще держится самец. К 5 месяцам птенцы имеют перьевой наряд, похожий на наряд самки, а к 6—8 месяцам почти достигают размеров взрослых. Половозрелыми становятся в возрасте 3—4 лет.
Усиленное преследование со стороны человека (от страусов получают мясо и перья для украшений) привело к резкому сокращению численности. На юге Африки есть фермы, разводящие страусов.
Семейство Eleutherornithidae с одним видом, описанным из эоценовых отложений Швейцарии (примерный возраст 60—40 млн. лет), близко к настоящим страусам, но более примитивно.
Ископаемые остатки страусов (рис. 22) известны из Северной Африки, Греции, южной Украины, Ирана, МНР и Внутренней Монголии (где, возможно, жили еще во II—I в. до нашей эры), Северной Индии, Китая. С этой же территории описаны и другие ископаемые виды этого семейства (7—21 по разным источникам), относимые к миоцену — плейстоцену.
ОТРЯД 6. RHEIFORMES — НАНДУ ОБРАЗНЫЕ
В отряде единственное семейство Rheidae с 2 видами: Rhea americana L. — обыкновенный нанду (вес 20—25 кг) и Pterocnemia pennata Or. (Rhea darwini)— нанду Дарвина (примерно в 1,5 раза мельче), населяющие саваны и полупустыни Южной Америки (рис. 22). Быстро бегающие, не летающие птицы (рис. 23).
Череп дромеогнатический: сошник смещен назад и соединяется как с небными, так и с крыловидными костями; задние части небных костей расширены. Пояс передних конечностей упрощен, как ,и у страусов. Кисть и предплечье вместе несколько длиннее плеча. Сохраняются все карпальные косточки; I палец заканчивается коготком. Лобковые кости правой и левой сторон не соединяются друг с другом, а седалищные образуют симфиз. Мощные задние конечности имеют 3 направленных вперед пальца.
Питаются преимущественно мягкой растительной пищей. Мускульный желудок слабее, чем у страусов, и имеет менее мощную кутикулу. Кишечник превышает длину тела примерно в 8 раз; слепые кишки очень длинны и по объему почти равны всему остальному кишечнику. Хорошо развита и у взрослых фабрициева сумка, полость которой выполняет функцию мочевого пузыря. Есть непарный выворачивающийся penis. Копчиковая же-
87
Рис. 22. Ареалы страусоподобных птиц:
1— страуссобразные Struthioniformes, 2— места находок ископаемых страусов, 3—нандуобразные Rheiformes, 4— кивиобразные Apterygi formes и моаобразные Dinornithi-formes, 5— эму Dromiceidae, 6— казуаровые Casuaridae, 7— места находок остатков эпиориисообразных Aepyornithiformes
леза отсутствует. В нижней гортани есть трахейно-бронхиальные мышцы (их нет у африканского страуса).
Оперение равномерно покрывает все тело. Рассученные перья не несут добавочного стержня; 12 первостепенных и 16 второстепенных маховых выполняют роль только украшающих перьев и в значительной степени скрыты под более длинными кроющими крыла. По окраске самцы неотличимы от самок. Рулевые перья редуцированы; полностью исчез и пиго-стиль.
Полигамы. В период тока около самца группируются от 2 до 5—6 самок. Самец роет гнездовую лунку, в которую самки откладывают яйца. В полных кладках от 13 до 20—30 яиц (бывает и больше). Яйца у обыкновенного нанду белые или желтоватые (у нанду Дарвина — зеленоватые) размером 13—15 X 9—10 см, весом около 600—700 г (каждое яйцо 2,7— 3,9% веса самки). Насиживает только самец. Инкубация около 6 недель. Самец же водит выводок. В возрасте 5—6 месяцев птенцы достигают размеров взрослых птиц. Половозрелыми становятся в возрасте 2—3 лет. Усиленное преследбвание из-за мяса и перьев и сбор яиц привели к резкому сокращению численности; сохранились лишь в наиболее глухих районах.
Ископаемые остатки нанду известны из верхнего плиоцена северной Аргентины. В разных пунктах Южной Америки найдены фрагментарные остатки, которые относят к 4 видам двух родов (нижний эоцен, около 60 млн. лет).
ОТРЯД 7. CASUARIIFORMES— КАЗУАРООБРАЗНЫЕ
Нелетающие страусоподобные птицы Австралийской зоогеографической области, объединяемые в 2 ныне живущих и в одно ископаемое семейства.
Скелет неба напоминает таковой у нанду, но сошник еще более мощен и срастается с крупными небными отростками верхнечелюстных костей. Скелет крыла сильно редуцирован: общая длина кисти и предплечья равна длине плеча; I палец полностью редуцировался; все развивающиеся карпальные и метакарпальные кости сливаются вместе. Длина цевки (тарзо-метатарзус) резко превышает длину бедра. Сохраняются 3 направленных вперед пальца ноги; они несут сильные когти. Таз объемистый и широкий, концы седалищных и подвздошных костей срастаются.
Питаются преимущественно плодами, реже нежными зелеными частями растений, крупными насекомыми, мелкими позвоночными. Мускульный желудок имеет менее мощные стенки, чем у других страусоподобных, и невелик по объему. Кишечник недлинный, слепые кишки развиты слабо. Фабрициева сумка у взрослых редуцируется. Копчиковой железы нет. Есть непарный penis. В нижней гортани трахеально-бронхиальные мышцы отсутствуют.
Оперение равномерно покрывает все тело. Рассученные перья имеют добавочный стержень, по длине не уступающий основному. Полового мор?
89
Рис. 24. Казуар Casuarius unappendiculatus Blyth.
физма окраски нет. Рулевые перья полностью редуцированы; нет и пигостиля. Скорлупа яиц грубозернистая темно-зеленого, иногда с бурым оттенком цвета.
СЕМЕЙСТВО 1.
CASUARIIDAE — КАЗУАРОВЫЕ
Один род Casuarius с 3 (5?) видами (рис. 24), распространенными на Новой Гвинее и прилегающих островах, в северо-восточной Австралии (рис. 22). Населяют тропические леса с густым подлеском, реже кустарниковые заросли. Питаются преимущественно мясистыми плодами. Вес 80—90 кг. Окраска черная. На голове и
шее есть участки голой ярко окрашенной кожи (синей, красной, оранжевой). На го-
лове тонкостенный роговой вырост («шлем»). Клюв сжат с боков. Маховые сохранились в виде заостренных длиной до 50 см толстых роговых стержней (6—8), прикрепленных к костям кисти и предплечья. Коготь внутреннего пальца ноги очень длинный и острый — он используется для защиты. Кишечник всего в 3—4 раза длиннее тела. Держатся одиночками или небольшими группами. Самка откладывает в гнездо из мха и листьев от 3 до 5 зеленых яиц длиной до
15 и шириной до 10 см, ярко- или темно-зеленого цвета, весом около 600 г (примерно, 1,5% веса самки). Насиживает самец, инкубация 6—7 недель, самец же водит и выводок (возможно, иногда в насиживании участвует и самка).
Один из ныне живущих видов известен и в ископаемом состоянии из верхнеплейстоценовых отложений в пределах современного ареала.
СЕМЕЙСТВО 2.
DROMAIIDAE — ЭМУ
Один род Dromaius с единственным рецентным видом D. novae-hollandiae Lath. (рис. 25), населяющим степи и кустарниковые пустыни Австралии. В прошлом веке истреблен в Тасмании (см. рис. 22). Питаются ягодами, семенами, мелкими животными. Вес 40—55 кг. Окраска коричневато-бурая. «Шлема» и ярко окрашенных участков голой кожи на го
Рис. 25. Эму Dromaius novae-hollandiae Lath.
90
лове и шее нет. Клюв приплюснут сверху вниз (как у страуса и нанду).
Сохраняются остатки ключиц. В скелете кисти только одна метакарпальная кость и лишь один палец, несущий на конце коготок. Коготь внутреннего пальца ноги не отличается от когтей остальных пальцев. Кишечник в 7—8 раз длиннее тела: в питании в отличие от казуаров довольно большое значение имеют вегетативные части растений. На трахее расположен соединительнотканный вырост, наполняемый воздухом и служащий резонатором.
Вне сезона размножения держатся группами, иногда до нескольких десятков особей. Самка откладывает от 8 до 10 яиц (редко больше) размером примерно 13 X 9 см, темно-зеленого или зеленовато-бурого цвета. Насиживание продолжается около 8 недель. Насиживает самец, он же водит выводок. Окраска птенцов сероватая с темно-бурыми продольными полосами. Половозрелыми становятся после двух лет жизни. Местами приносят существенный ущерб, кормясь на полях созревающих зерновых и при этом сильно вытаптывая посевы. Поэтому в культурном ландшафте усиленно преследуются и уничтожаются.
В прошлом веке полностью истреблен живший на о-ве Кенгуру (южное побережье Австралии) более мелкий — размером с крупную дрофу — D. minor Sp.
По остаткам из отложений верхнего плейстоцена в пределах современного ареала описаны еще 2 вида.
СЕМЕЙСТВО 3. DROMORNITHIDAE
По находкам костей из плейстоценовых слоев Австралии описаны 2 рода (с одним видом в каждом) крупных примитивных страусоподобных птиц, почти вдвое крупнее по размерам, чем эму.
ОТРЯД 8. AEPYORNITHIFORMES — ЭПИОРНИСООБРАЗНЫЕ
Крупные страусо- или пастушковоподобные птицы из эоценовых-оли-гоценовых отложений Северной Африки и плейстоценовых-голоценовых отложений Мадагаскара (см. рис. 22). Наиболее крупный Aepyornis maximum Geoffr. достигал высоты около 3 м и весил, видимо, примерно 400 кг. Откладывал очень крупные яйца (34 X 22 см) объемом 9—12 л. Характерны тяжелые столбообразные трехпалые задние конечности с широкой уплощенной цевкой, широкий таз, широкая грудина без следов киля, широко расставленные коракоиды, очень сильно редуцированный скелет крыла и т. п. Сильнее, чем у других страусоподобных птиц, выражена пневма-тичность бедра и, видимо, тел позвонков. Вероятно, подобно казуарам, жили в сырых тропических лесах. Судя по остаткам костей и яиц, носящих следы обработки человеком, служили объектом охоты коренного населения Мадагаскара. Вымерли в историческое время: по мнению некоторых исследователей, в XVIII, а может быть даже и в XIX в. Вымиранию, вероятно, способствовало не только прямое преследование человеком, но и очень интенсивное уничтожение тропических лесов. Видимо, мадагаскарские эпиорнисы были прототипом характерных для арабских сказок гигантских птиц «рок» или «роух» (арабы посещали Мадагаскар уже в середине нашей эры).
Сейчас описано 4 рода с 9 видами. Взаимоотношения эпиорнисов с другими отрядами птиц неясны.
91
ОТРЯД 9. DINORNITHIFORMES — МОАОБРАЗНЫЕ
Страусоподобные вымершие птицы (рис. 26), населявшие оба острова Новой Зеландии (см. рис. 22). Описано более 20 видов, группируемых в 7—8 родов. Наиболее древние находки датируют миоценом (10—20 млн. лет). Высота мелких видов около 1 м, самого крупного Dinornis maximum Н.—около 3,1 м (вероятный вес — 400 кг). Характерны широкий уплощенный череп с коротким клювом, у части видов прямым, у других
Рис. 26. Примерные размеры некоторых моа (по Архей, 1941)
с небольшим крючком на конце. Грудина выпуклая, без следов киля. Сливающиеся вместе лопатка и коракоид очень малы и не образуют суставной впадины для плеча. Скелет крыла, видимо, полностью редуцирован. Таз широкий, очень мощный. Цевка относительно короткая (обычно короче бедра). Хорошо развиты 3 направленных вперед пальца; задний палец мал и у части видов исчез. Кольца трахеи окостеневали. Оперение очень похоже на оперение эму (обнаружены отдельные перья и обрывки кожи с перьями) и по расцветке и по структуре пера: хорошо развит добавочный стержень, перо рассученное. Яйца относительно мелкие: самые крупные из обнаруженных яиц были длиной до 20—25 см и шириной 15 — 17 см-, примерный вес около 2,5—3 кг (при сходных предельных размерах птиц яйца эпиорнисов были примерно в 4 раза крупнее).
Многочисленные материалы подтверждают, что не менее 10—13 видов жили в нашу эру. Об этом свидетельствуют остатки птиц (кости, перья, яйца) на месте стоянок охотников-маорийцев и в их захоронениях, а также частые упоминания об этих птицах .в легендах и преданиях. Видимо, вымирание этих своеобразных крупных нелетающих птиц было в основном определено прямым преследованием человеком. Большинство видов истреблено к середине XIV в. (люди появились в Новой Зеландии в IX—X вв.). На Южном острове в глухих лесах последние моа, вероятно, встречались даже в XVIII в., а может быть и позже (р. Megalapteryx).
Моа, видимо, близки к отряду бескрылых, или кивиобразных.
92
ОТРЯД 10. APTERYGIFORMES — КИВИОБРАЗНЫЕ, ИЛИ БЕСКРЫЛЫЕ
Мелкие нелетающие птицы Новой Зеландии (рис. 27). Череп по типу строения напоминает череп казуаров, но отличается рядом особенностей: сошник срастается с небными и крыловидными костями, не соединяясь с небными отростками верхних челюстей; хорошо развиты базиптеригоидные отростки. Клюв длинный и тонкий (напоминает клюв некоторых куликов), а у основания окружен длинными щетинками. Ноздри открываются на конце клюва (резкое отличие от всех остальных птиц). Грудина без киля. Скелет крыла сильно редуцирован: кисть и предплечье вместе вдвое короче плеча. Предплечье неподвижно соединено с пле-чом. Вилочки нет, лопатка и ко-ракоид срастаются друг с дру-гом. Таз узкий и длинный. Лобковые кости не срастаются. Пигостиль из 2—3 позвонков, но рулевые отсутствуют. Зад-ние конечности мощные; хоро-шо развиты направленные впе-ред 3 пальца, заканчивающие- х
ся крепкими ногтями. Задний и! WL
палец рудиментарен, но отчет-ливо заметен. Скелет не пневма-тичен.
Глаза ЯВНО подверглись ре- Рис. 27. Киви Apteryx haasti Potts, дукции (их диаметр всего около 8 мм\ это единственный случай столь сильного уменьшения глаз в классе птиц). Основной орган рецепции, видимо, обоняние: очень сильно развиты и обонятельные полости в основании клюва, и обонятельные доли переднего мозга. Язык короткий, несмотря на длинный клюв. Мускульный желудок относительно слабый. Кишечник довольно длинный с длинными узкими слепыми кишками. В стенке клоаки находится непарный совокупительный орган. Оперение буроватого цвета(окраска самцов и самок одинакова). Перья рассученные, без добавочных стержней, равномерно покрывающие все тело. Можно различить 13 слабо дифференцированных маховых перьев. В отличие от остальных страусообразных птиц есть копчиковая железа.
В отряде единственное семейство Apterygidae с одним родом Apteryx, объединяющим 3 вида: A. australis, A. haasti, A. oweni. Распространены в сырых лесах Новой Зеландии. Ведут сумеречный и ночной образ жизни. Питаются главным образом червями, наземными моллюсками, насекомыми, добывая их в верхнем слое почвы. Самки крупнее самцов (2,5—3 кг против 1,5—2 кг). В кладке 1 — редко 2 очень крупных яйца (размером до 13 X Х8 см, весом около 450 г (более 15% веса самки). Насиживает только самец — до 40—50 дней (по другим данным, до 70—80 дней). Птенцы выводковые, по окраске похожи на взрослых.
Молодых водит самец. Половозрелыми становятся только на 3—5-й год жизни.
Неумеренное преследование и резкое уменьшение площади лесов привели к сильному уменьшению численности киви. Сейчас охраняются.
Достоверные палеонтологические остатки из плейстоцена Новой Зеландии описаны как особый род с одним видом.
В ископаемом состоянии обнаружены и ныне живущие виды.
ОТРЯД 11. TIN AMIFORMES — ТИНАМУ ОБРАЗНЫЕ, ИЛИ СКРЫТОХВОСТЫЕ
моря), в лесах, кустарниковых
Рис. 28. Тинаму Rhynchotus rufescens (Temm.)
тонкий, изогнутый книзу. Шейных грудных спинную
Внешне похожи на куропаток (рис. 28). Птицы средней величины (вес 200 г— 1,5 кг), живущие на равнинах и в горах (до 4000 м над уровнем ' . . зарослях и в степях Средней и Южной
Америки (рис. 31) — от Мексики до Патагонии.
Череп дромеогнатического типа, но небные кости соприкасаются по средней линии друг с другом. Хорошо развиты базиптеригоидные отростки. Клюв удлиненный, слегка позвонков 16—18, 4—5 позвонков срастаются в кость. Пигостиль маленький (образован только тремя позвонками). Пояс передних конечностей типично птичьего типа. Узкая длинная грудина несет хорошо развитый киль и очень длинные узкие боковые отростки. Коракоид и лопатка соединяются малоподвижным суставом, как и у большинства ныне живущих птиц. Хорошо развита вилочка. Таз широкий. Задний палец ноги слабо развит или редуцируется. Две сонные артерии. Сильно развит объемистый зоб, мощный мускулистый желудок и большие объемистые слепые кишки. Имеется penis. Есть небольшая, слабо оперенная копчиковая железа. Оперение своеобразное: вершины бородок сливаются друг с другом, образуя «жесткое» опахало. Есть узкие аптерии. Пух- расположен только по птерилиям. Первостепенных маховых 10. Крылья короткие, широкие, эутаксические. Короткие рулевые (8 —12) полностью прикрыты кроющими хвоста. Окраска буроватая или коричневатая. Самки несколько крупнее самцов, иногда более ярко окрашены.
Наземные птицы, ведут преимущественно сумеречный или ночной образ жизни, хорошо бегают, взлетают неохотно. Оседлы. Часть видов, видимо, моногамы с равным соотношением полов. Откладывают от 1 до 6—10 яиц. Насиживает преимущественно самец. У ряда видов полиандрия: число самцов в популяции превышает число самок (например, у Crypturellus — в 4 раза). Гнездовый период таких видов обычно очень растянут. Самец занимает и охраняет гнездовой участок, на котором одна или несколько самок откладывают по нескольку яиц в одно примитивное гнездо на земле и затем переходят на участок другого самца, заполняя яйцами его гнездо. Насиживает эту «коллективную» кладку, состоящую из 6—12, а иногда и больше крупных, обычно ярко окрашенных яиц только самец. Очень редко покидая гнездо для кормежки, самец маскирует яйца, прикрывая их листьями и растительным мусором. Насиживание продолжается 16—20 дней. Птенцы выводкового типа. Выводок водит только самец, но птенцы вскоре — иногда уже через несколько дней — начинают вести самостоятельную жизнь. За сезон размножения самец насиживает одну кладку, но у некоторых видов, возможно, и две.
Пища разнообразная, растительная (семена, ягоды, вегетативные части растений) и животная (насекомые и другие почвенные беспозвоночные; крупные виды при случае ловят мелких лягушек, ящериц и т. п.).
94
В отряде единственное семейство Tinamidae с 9 родами: Tinamus, Са-lopezus, Crypturellus, Nothocercus, Nothoprocta, Nothura, Rhynchotus, Taoni-scus, Tinamotus, включающими 42 ныне живущих вида. Центр возникновения, видимо, — Южная Америка. В пределах современного ареала описано до 14 вымерших видов (наиболеедревние — из плиоцена). По очень давним родственным связям, вероятно, ближе всего к нанду. Многие черты сходства с куриными и пастушковыми, скорее всего, результат конвергенции.
ОТРЯД 12. GAVIIFORMES — ГАГАРООБРАЗНЫЕ
Водные птицы средней и крупной величины (вес от 1 до 5 кг), распространенные только в Северном полушарии (см. рис. 20). Перелетны.
Характерно удлиненное туловище, довольно длинная шея, тонкая, удлиненная голова с прямым тонким, острым клювом (рис. 29). Череп схизогнатического типа, базиптеригоидные сочленения отсутствуют. Ноздри голоринальные. Хорошо развиты надорбитальные носовые железы. Шейных позвонков 14—15. Спинная кость не образуется. Грудина удлиненная. Таз узкий и длинный, вертлужная впадина сдвинута назад. Ноги далеко отнесены назад, поэтому по суше практически ходить не могут. Чашечка сливается с мощным акромиальным отростком. Цевка сильно сплющена с боков. Направленные вперед 3 пальца соединены плавательной перепонкой, задний палец маленький. Очень сильна мускулатура задних конечностей. Ножная формула АВХ +. Имеются две сонные артерии. Пищевод сильно растяжим. Мускульный желудок объемист, но имеет относительно тонкие стенки и тонкую кутикулу. Слепые кишки очень малы. Копчиковая железа оперена.
Оперение плотное, густое. Короткие перья и пух имеют добавочный стержень. Аптерии узкие. Пух равномерно распределен по всему телу. Первостепенных маховых 11 (первое недоразвито). Крылья узкие и довольно короткие, эутаксические; 16—20 коротких рулевых. Окраска самцов и самок сходна; есть возрастная и сезонная смена окраски. При линьке все маховые выпадают одновременно.
Летают быстрым, но не маневренным полетом; взлетая, разбегаются по воде. Прекрасно плавают и ныряют; в толще воды двигаются с помощью ног и, видимо, иногда с помощью крыльев. Моногамы. Гнездятся по берегам озер, на болотах с участками чистой воды от умеренной зоны до побережья Ледовитого океана. Гнездо—лунка умятых растений со скудной подстилкой, устраивается у самой кромки воды. В гнездо вползают с воды. В кладке обычно 2 оливково-бурых, с неясным темным крапом яйца. На-
Рис. 29. Чернозобая гагара Gavia arctlca (L.) на гнезде
95
сиживают оба партнера. Инкубация около месяца. Вылупившийся птенец покрыт одноцветным темным пухом. Обсохнув, он может хорошо плавать и нырять. Родители кормят птенца до приобретения им способности к полету. Поэтому могут гнездиться на совершенно бескормных водоемах. Пуховых нарядов два. Птенцы полностью оперяются и становятся способными к полету примерно через 45 дней после вылупления.
Питаются преимущественно рыбой, которую ловят на глубинах до 25 м, возможно, и глубже; поедают и разнообразных водных беспозвоночных. Гнездящиеся вблизи морских побережий птицы летают за кормом на море. Зимуют обычно на морях умеренной зоны.
В отряде единственное семейство Gaviidae с одним родом Gavia и 3 видами, гнездящимися по водоемам тундровой и лесной зон Европы, Азии и Северной Америки; в СССР встречаются и гнездятся все 3 вида. По ископаемым остаткам из верхнего палеоцена (возраст не менее 60 млн. лет) и из более поздних слоев в пределах современного ареала описано 12 видов (в том числе и современных), относимых к 4 родам.
Древняя, относительно примитивная группа птиц, возможно, имеющая очень давние родственные связи с меловыми зубастыми птицами — геспе-рорнисами. Высказывалось предположение об очень давнем родстве гагар с куликообразными.
ОТРЯД 13. PODICIPEDIFORMES — ПОГАНКООБРДЗНЫЕ
Водные птицы мелких и средних размеров (вес от 150 а до 1,5 кг), широко распространенные почти по всему земному шару (см. рис. 20). Перелеты в северных районах, в тропиках оседлые или кочующие.
По внешнему облику напоминают гагар: вальковатое туловище, длинная шея, удлиненная голова с прямым зацстренным клювом (рис. 30). Череп схизогнатический, базиптеригоидные отростки отсутствуют, надорбитальные железы развиты слабо. Шейных позвонков 17—21; 3—4 грудных позвонка срастаются в спинную кость. Грудина широкая, относительно короткая. Таз очень узкий и длинный, ноги далеко отнесены назад. По суше практически ходить не могут. Коленная чашечка не сливается с длинным акромиальным отростком большой берцовой кости. Конец малой берцовой кости не доходит до дистальной трети большой берцовой кости. Цевка плос-
Рис. 30. Большая поганка Podiceps cristatus L., под крылом птенец
96
кая, очень сильно сжата с боков. Каждый из трех направленных вперед длинных пальцев окаймлен самостоятельной широкой кожистой лопастью и заканчивается широким плоским когтем, похожим на ноготь. Задний палец мал, но тоже окаймлен кожистой лопастью. Пневматичность скелета слабая; Очень сильна ножная мускулатура. Ножная формула ВХ —. Есть только левая сонная артерия. Пищевод сильно растяжим, но зоба нет. Желудок объемист, но с относительно тонкими стенками; отчетливо развит пилорический отдел. Слепые кишки очень малы. Копчиковая железа оперенная.
Оперение густое и плотное; перья короткие, имеют слабо развитый добавочный стержень. Аптерии узкие. Уздечка не оперена. Пух покрывает и птерилии и аптерии. Первостепенных маховых 12, первое рудиментарно. Крылья узкие, относительно короткие, диастатаксические (у Centropelma — эутаксические). Короткие мягкие рулевые полностью прикрыты кроющими хвоста. Самцы и самки окрашены сходно. Зимний наряд тусклее брачного; оперение молодых птиц по окраске отличимо от оперения взрослых. При линьке маховые выпадают почти одновременно.
Полет быстрый, прямолинейный, с частыми взмахами крыльев; взлет после разбега по воде. Прекрасно плавают, великолепно ныряют, оставаясь под водой, как и гагары, до 1 мин и даже, редко, до 3 мин. В толще воды передвигаются только с помощью ног. Моногамы. Гнездятся преимущественно по пресным водоемам с обильной водной и надводной растительностью. Гнездо плавучее: представляет кучу растительной ветоши, обычно расположенную на окраине куртины надводных растений. Довольно сложен ток: токуют оба партнера. Спаривание на гнезде. В кладке 4—6 светлых однотонных яиц. Насиживают оба родителя. Инкубация 20—25 дней. При уходе с гнезда кладка обычно прикрывается гнездовым материалом. Вылупляющиеся птенцы покрыты густым пухом; окраска сверху полосатая (только птенцы Aechtnophorus сверху одноцветные). Птенцы прекрасно плавают, а через несколько дней и ныряют. Греясь, забираются под крылья плавающей взрослой птицы, очень редко отдыхают в гнезде. Держатся с самцом и самкой. Птенцовый пуховой наряд один. Взрослые кормят птенцов; подросшие птенцы, видимо, часть пищи добывают сами. Полностью оперяются и становятся способными к полету через 4—5 недель после вылупления. Вскоре после взлета птенцы начинают вести самостоятельную жизнь.
Кормятся рыбой, различными водными беспозвоночными (особенно насекомыми и их личинками), земноводными и их личинками. Добычу ловят на поверхности и в толще воды, плавая и ныряя на глубину до 20—25 м. У взрослых птиц в пилорической части желудка обычно всегда находится комок перьев, функция которого до сих пор неясна. Полагают, что они облегчают формирование погадки (после отрыгивания погадки птица собирает с воды и вновь заглатывает попавшие в погадку перья) или выполняют роль своеобразного фильтра, не пропускающего в кишечник непереваренные кости и комочки пищи.
В отряде единственное семейство Podicipedidae с 4 родами и 18—20 видами (Podiceps с 13—15 видами, Aechmophorus с 2 видами, Centropelma — 1 вид, Podilymbus — 2 вида). Встречаются по водоемам почти всего земного шара, кроме Субарктики и Субантарктики (см. рис. 20), в Палеарктике только виды рода Podiceps. В СССР встречаются и гнездятся 5 видов. В пределах современного ареала обнаружены ископаемые остатки нескольких современных видов и 5 видов вымерших (часть из них входит в вымерший род). Наиболее древние находки — из нижнего миоцена. Центр возникновения отряда, видимо, — Северная Америка.
Раньше гагар и поганок объединяли в один отряд. Сейчас общепринято, что сходство этих групп определяется не родственными связями, а конвергенцией — приспособлениями к водному образу жизни, сходным характером питания.
97
ОТРЯД 14. PROCELLARIIFORMES (TUBINARES) -БУРЕВЕСТНИКООБРАЗНЫЕ, ИЛИ ТРУБКОНОСЫЕ
Морские птицы, связанные с сушей только в период размножения. Размеры от мелких до'крупных: вес от 20 г до 8—10 кг. Гнездятся по морским побережьям (рис. 31); большая часть видов — только в Южном полушарии. Вне периода размножения широко рассеиваются по морям и океанам; часть видов Южного полушария зимует в морях Северного полушария. По внешнему виду несколько напоминают чаек. Прекрасно летают, хорошо плавают; ныряют немногие виды.
Клюв с крючком на конце, покрыт сложной рамфотекой, состоящей из нескольких роговых пластинок. У некоторых планктоноядных форм (Ра-chyptila и др.) по краям надклювья развиваются роговые пластинки, функционально сходные с пластинками краев клюва уток; основание клюва расширено. Ноздри открываются в роговые трубочки; у альбатросов они лежат по бокам клюва, у других видов обе трубочки лежат рядом по хребту клюва или сливаются в одну трубочку, внутри разделенную вертикальной продольной перегородкой. Череп схизогнатического типа, у альбатросов проявляется тенденция к десмогнатизму. Базиптеригоидные сочленения есть или находятся в разной стадии исчезновения. Над орбитами отчетливые углубления для хорошо развитых надорбитальных носовых желез. Шейных позвонков 15. Грудина относительно широкая и короткая (только у ныряющих буревестников — узкая и длинная), с 1—2 парами вырезок по заднему краю (у качурок вырезок нет). Грудные позвонки в спинную кость обычно не сливаются. Вилочка мощная, часто срастающаяся с килем грудины. Скелет крыла удлинен; у крупных форм особенно заметно удлинение плеча. Таз удлиненный, относительно узкий. Ноги умеренной длины или короткие. В области коленного сустава гребень большой берцовой кости очень высок. Коленная чашечка есть или отсутствует. Три направленных вперед пальца соединены плавательной перепонкой. Задний палец мал или отсутствует. Обычно хорошо развита пневматичность скелета. Мускулатура крыла и груди очень мощная. Сонных артерий две. Пищевод сильно растяжим. Железистый желудок очень объемист; мускульный желудок относительно мал и имеет довольно слабые мускульные стенки. Кишечник относительно короткий, слепые кишки малы или отсутствуют. Копчиковая железа хорошо развита и оперена.
Оперение густое. Контурные перья довольно длинные и имеют побочный ствол (нет у альбатросов). Пух с побочным стволом, равномерно покрывает все тело. Крыло обычно узкое, длинное, острое, диастатаксическое. Первостепенных маховых 11, первое зачаточное. Второстепенных маховых от 10 (мелкие формы) до 37 (альбатросы). Рулевых 10—16. Обрез хвоста прямой, округлый, конический или вильчатый. Самцы и самки сходной окраски. У части видов возрастной морфизм окраски. Линька маховых идет постепенно, без потери способности к полету. Полет легкий, стремительный. Прекрасно выражена способность к планирующему полету при слабых неустойчивых воздушных потоках (завихрения над волнами). Своеобразен сесамовидный, сухожильный и мускульный аппараты, что позволяет (особенно у крупных видов) изменять положение кисти при фиксированном положении проксимальных отделов крыла.
Моногамы. Гнездятся обычно большими колониями по морским побережьям, некоторые виды — в удалении от моря, даже на несколько десятков километров. Гнездо со скудной выстилкой из растительной ветоши, местоположение резко отличается у разных видов: расположено открыто на ровной поверхности или на скалистых уступах, в трещинах скал или под камнями, в вырытых самими птицами норах. В кладке одно относительно крупное белое яйцо. Эмбриональное и постэмбриональное развитие медленное. Насиживание от 6 недель (мелкие виды) до 2 месяцев. Птенец вылупля-
98
150° 120° 90° 60°н западу от Гринвича (Ук востоку от Гринвича 60° 90° 120° 150° 180е 150°
Рис. 31. Ареалы:
/— отряд тинамуобразных Tinamiformes, 2— отряд трубконосых Procellariiformes, 3— семейство авдоток Burhinidae (подотряд дроф из журавлеобразных)
ется опушенным, но слепым; прозревает быстро. Насиживают и выкармливают оба партнера. У многих видов смена насиживающих птиц и кормление птенцов идет только ночью или в сумерках. Птенцы получают корм 1—2 раза в сутки у мелких и 2—3 раза в неделю у крупных видов. Несмотря на это, к концу послегнездового периода птенцы накапливают очень большие запасы жира (до 30% и более от веса тела). Гнездо покидают полностью оперенными, способными к полету через 2 (мелкие виды)— 6 (альбатросы) месяцев после вылупления. Обычно молодые кочуют отдельно от взрослых.
Пища — разнообразные морские беспозвоночные, рыба, падаль и отбросы с судов — собирается с поверхности воды и ловится в верхнем слое при пикировании. При защите взрослые и птенцы выбрызгивают на врага сильно пахнущую ворванью маслянистую жидкость, которая, видимо, частично состоит из переваренной пищи, частично — из маслянистого секрета, выделяемого стенками железистого желудка и, возможно, пищевода. В первые дни после вылупления взрослые птицы кормят птенцов этой же отрыжкой. Местами имеют промысловое значение: собирают яйца, добывают выросших птенцов, реже взрослых птиц. Из-за неумеренного промысла чис-
ленность многих видов резко сократилась, а некоторые находятся на грани исчезновения.
Отряд включают 81 вид, объединяемых в 23 рода и 4 семейства. В СССР встречаются 14 видов из трех семейств; гнездятся только 5 видов. Часть ныне живущих видов найдена и в ископаемом состоянии. По ископаемым остаткам, начиная с эоцена и позже, описано 19 видов и 4 рода.
Древняя группа, по очень давним родственным связям, видимо, ближе всего стоящая к веслоногим и, может быть, к пингвинам. Центр возникновения отряда — Южное йолушарие, возможно, Субантарктика.
СЕМЕЙСТВО 1. DIOMEDEIDAE — АЛЬБАТРОСОВЫЕ
Крупные птицы весом до 8—10 кг и размахом крыльев до 3—4 м (рис. 32). Носовые трубочки по бокам клюва. Второстепенных маховых не менее 30. Летают преимущественно парящим полетом. Не ныряют. По земле передвигаются довольно свободно, опираясь только на пальцы. Гнездятся колониями на открытых участках. Молодые птицы одевают взрослый наряд на 4—5-й год жизни, половозрелыми становятся после 5 лет.
В семействе единственный род Diomedea (включает подроды Thalas-sarche и Phoebetria, ранее принимавшиеся за самостоятельные роды) с 12 видами, гнездящимися на островах в субтропиках, тропиках и Субантарктике. В период кочевок у тихоокеанских берегов СССР встречаются 3 вида.
Рис. 32. Альбатрос Diomedea exulans L.
100
Наиболее древние находки — из среднего эоцена Нигерии. Описано 2 ископаемых рода.
СЕМЕЙСТВО 2. PROCELLARHDAE — БУРЕВЕСТНИКОВЫЕ
Птицы средней величины (рис. 33), весом в 300—800 г; лишь один вид Macronectes giganteus (Gm.) весит около 5 кг. Носовые трубочки находятся на гребне клюва; у многих видов они сливаются в одну общую трубочку, разделенную вертикальной перегородкой. Второстепенных маховых не более 25. Хорошо летают как активным, так и парящим полетом; некоторые виды могут с разлета нырять на глубину в несколько десятков сантиметров.
Рис. 33. Глупыш Fulmarus glacialis (L.), светлая фаза.
Задний палец очень мал. По земле передвигаются с трудом, опираясь на пальцы и цевку. Большинство видов гнездится в трещинах скал, под камнями, в норах, немногие — на открытых скалистых уступах. Молодые по окраске не отличаются от взрослых. Половозрелыми становятся на 2— 5-й год жизни.
Семейство Procellariidae включает 12 родов (Procellaria, Adamastor, Bulveria, Daption, Fulmarus, Halobaena, Macronectes, Pachyptila, Pago-droma, Pterodroma, Puffinus, Thalassoicd) c 47 видами, гнездящимися по морским побережьям преимущественно Южного полушария; в Северном полушарии гнездятся лишь немногие виды. В водах СССР отмечено 7 видов, из которых гнездятся только 2: глупыш Fulmarus glacialis (L.) и пестролицый буревестник Puffinus leucomelas (Temm.), который гнездится в небольшом числе и, возможно, нерегулярно на некоторых островах Японского моря.
В ископаемом состоянии в пределах современного ареала встречаются со среднего олигоцена (описано несколько ископаемых видов и 2 рода); в более поздних отложениях встречено и несколько ныне живущих видов.
СЕМЕЙСТВО 3. HYDROBATIDAE — КАЧУРКОВЫЕ
Мелкие птицы весом в 20—200 г, редко больше (рис. 34). Ноздри в одной трубочке, лежащей на гребне клюва. Второстепенных маховых 10— 15. Полет быстрый, маневренный, напоминающий полет стрижей или ласточек (во многих районах местное название — морские ласточки). Большую часть времени проводят в воздухе. Не ныряют. Пищу — мелких беспозвоночных — обычно склевывают на лету с поверхности воды и с гребней волн. Гнездятся в норах, в щелях под камнями как на обрывах морских побере-
101
Рис. 34. Сизая качурка Oceanodroma furcata Gm.
жий, так и в нескольких километрах от моря. Птенцы в гнезде остаются 2—2,5 месяца. Молодые по окраске похожи на взрослых. Половозрелыми, видимо, становятся к концу 2-го года жизни.
В семействе 9 родов (Hydrobates, Cymochorea, Fregetta, Garrodia, Halo-cyptena, Nesofregetta, Oceanites, Oceanodroma, Pelagodromd) c 18 ныне живущими видами и одним видом, вымершим в историческое время. Гнездятся преимущественно в Южном полушарии; только немногие виды в Северном. У побережий СССР встречено 4 вида двух родов; в ряде мест тихоокеанского побережья и прилегающих островов гнездятся 3 вида.
Наиболее древние ископаемые остатки датируются верхним миоценом (Калифорния).
СЕМЕЙСТВО 4. PELECANOIDIDAE — НЫРЯЮЩИЕ БУРЕВЕСТНИКИ
Мелкие птицы, вес 150—250 г (рис. 35). Ноздри в очень коротких тру-
бочках, расположенных у основания клюва и открывающихся кверху. Полет стремительный, прямолинейный. Прекрасно нцряют с разлета и с поверхности воды. В толще воды свободно перемещаются, работая крыльями («подводный по-
Рис. 35. Ныряющий буревестник Pelecanoi-des glorgicus Murphy
лет»). Поэтому по многим особенностям мускулатуры, скелета и, отчасти, внешнего вида напоминают чистиковых птиц (конвергенция).
Распространены только в Южном полушарии на север до юго-восточной Австралии, по восточному и западному побережьям Южной Америки — на север до Перу. Гнездятся в норах и под камнями. Биология изучена плохо. Питаются преимущественно ракообразными, меньше мелкой рыбой. Добычу ловят в толще воды при нырянии.
В семействе один род Ре-lecanoides с 4 видами. Вид, гнездящийся по западному берегу Южной Америки, най-
ден там же в плейстоценовых отложениях.
102
ОТРЯД 15. PELECANIFORMES (STEG ANOPOD IFORMES) -ПЕЛИКАНООБРАЗНЫЕ, ИЛИ ВЕСЛОНОГИЕ
Очень разнообразные по внешнему облику и экологии водные птицы средней и крупной величины весом от 400 г до 10—14 кг. Населяют морские побережья и крупные пресные водоемы почти всего земного шара, кроме Антарктики. Распространены очень спорадично. Перелетные или кочующие, в тропиках некоторые виды оседлы.
Клювы разнообразной формы: остроконечные, конические, удлиненные с большим крючком на конце, очень длинные, сильно уплощенные с небольшим крючком на конце. Рамфотека сложная, из нескольких роговых пластин, но у старых птиц они обычно сливаются в монолитный роговой покров. Ноздри не сквозные (за исключением фаэтонов). Череп переходного типа между десмогнатизмом и схизогнатизмом. Сошник есть только у фаэтонов и фрегатов; у остальных групп полностью редуцируется. Базиптеригоидных отростков нет. Надорбитальные железы малы, и на черепе для них образуются плохо заметные углубления. Шейных позвонков 12—20. Грудина мощная, широкая, с большим килем. Таз широкий или у хорошо ныряющих форм заметно суженый. Ноги короткие. Цевка не оперена (исключение — фрегаты). Все 4 пальца соединены общей плавательной перепонкой. Пневматичность скелета варьирует в разных группах. Мускулатура грудины и ног мощная. Язык обычно редуцирован. Пищевод и желудок сильно растяжимы. В железистом желудке обособлен пилорический отдел. Стенки мускульного желудка довольно слабые. Слепые кишки сильно редуцированы. Копчиковая железа обычно хорошо развита, оперена и имеет несколько выводных протоков.
Оперение густое, плотное. Побочного ствола контурных перьев нет или он слабо развит. Аптерии узкие. Пух равномерно покрывает все тело. Самцы и самки сходной окраски. У многих видов есть брачный наряд. Молодые птицы по окраске обычно отличаются от взрослых. Первостепенных маховых 11. Крыло диастатаксическое (у Nannopterum эутаксическое). Хвост различной формы и длины, образован 12—24 рулевыми. Смена маховых во всех группах, кроме змеешеек, идет постепенно, без потери способности к полету. Характер полета варьирует в разных группах. По земле ходят мало, медленно и неуклюже. Подавляющее большинство видов хорошо плавает. Способность к нырянию различна: например, бакланы и змеешейки ныряют превосходно, олуши — с разлета, большинство пеликанов совсем не могут нырять и т. п.
Моногамы. Гнездятся обычно большими колониями по берегам морей и внутренних водоемов; редко колонии бывают в нескольких километрах от воды. Гнездо — сцементированная пометом груда мелких веток, стеблей тростника, растительной ветоши, иногда водорослей — строится на земле, на уступах береговых обрывов, в заломах тростника, реже (бакланы, фрегаты) в развилках ветвей деревьев. Яйца относительно мелкие, с довольно рыхлым наружным известковым слоем, одноцветные, у фаэтонов пестрые. Размеры кладок варьируют у разных видов от 1 др 4—6 яиц, редко больше. Насиживание обычно начинается после откладки первого яйца и продолжается 4—7 недель. Насиживают оба партнера. Птенцы вылупляются слепыми и голыми. Глаза открываются на 3—5-й день, к 6—10-му дню жизни покрываются густым коротким пухом. В гнезде или около гнезда птенцы остаются до 6—15 недель, пока полностью не оперятся и не приобретут способность к полету. Кормят оба родителя, отрыгивая в открытые клювы птенцов полупереваренную пищу; подросшим птенцам приносят добычу в клюве. В жаркую погоду поят птенцов, принося им воду в клюве и пищеводе. Покинувшие гнездовья молодые птицы кочуют вместе со взрослыми. Полный брачный наряд одевают, достигнув половой зрелости, на 3—4-й год жизни. Пища животная: преимущественно рыба, в меньшей степени разнообразные
103
водные беспозвоночные. Способы охоты различны в разных группах.
В крупных, смешанных по видовому составу колониях веслоногих в сухих районах тропиков накапливается в больших количествах гуано (высохший помет и трупы погибших птенцов, перемешанные с подстилкой гнезд), которое используют как очень эффективное удобрение. В промышленных масштабах (десятки тысяч тонн ежегодно) добывается гуано на островах, принадлежащих Перу. Основные гуанонакопители здесь бурый пеликан Pelecanus occidentalis L., баклан Бунгенвиля Phalacrocorax bounga-invillei Less., олуша Sula variegata Tschudi (общая их численность на островах и побережье Перу около 15 млн. особей). В меньших масштабах добывают гуано и в некоторых других местах. В ряде районов местное значение имеет массовый сбор яиц и, в меньшей степени, добыча выросших птенцов и взрослых птиц (используются мясо и перо, иногда шкурки). Местами некоторые виды могут причинять небольшой ущерб, кормясь на рыборазводных прудах.
Отряд разделяется на 5 подотрядов (из них 2 включают только ископаемые формы), объединяющих 12 семейств, 6 семейств представлены только ископаемыми формами (17 родов и более 60 видов), а 6 остальных семейств включают 7—8 ныне живущих родов, объединяющих 50—52 вида (23 из них обнаружены и в ископаемых остатках). В СССР встречаются 11 видов из 4 родов, а гнездится 8 видов, представляющих 2 рода.
Веслоногие, или пеликанообразные, — одна из древних ветвей класса птиц, очень давними родственными связями, вероятно, близкая к голенастым и, возможно, к трубконосым.
ПОДОТРЯД 1. ODONTOPTERYGES —ЛОЖНОЗУБЫЕ ПТИЦЫ
По плохой сохранности остаткам черепов описаны два ископаемых семейства: Odontopterygidae с 1 родом и 1 видом из нижнего эоцена Южной Англии и Pseudodontornithidae с 2 родами и 2 видами из верхнего миоцена Калифорнии. Края рамфотеки зазубрены, несколько напоминают зубы. По морфологическим особенностям и внешнему облику, вероятно, походили на олуш или очень примитивных пеликанов. Может быть, были близки к предковым формам всего отряда.
ПОДОТРЯД 2. PHAETHONTES — ФАЭТОНОВЫЕ
По внешнему облику напоминают крупных крачек (рис. 36). Распространены по морским побережьям тропической зоны (рис. 37).
Клюв конический, слегка зазубренный по краям. Ноздри сквозные, с довольно крупными щелевидными наружными отверстиями. Сошник есть. Шейных позвонков 14—15. Грудные позвонки гетероцельные. Сонные артерии парные. Ноги короткие и слабые; по земле ходят плохо и мало. Довольно хорошо плавают. Крылья узкие и длинные. Полет стремительный, высокоманевренный; могут длительное время находиться в воздухе. Хвост из 12—14 рулевых с очень длинной центральной парой. В окраске сочетаются белый и черный цвета, молодые пестрые.
Гнездятся одиночными парами или группами из 3—6 пар. В расщелину скалы, под камень или у основания густого куста без подстилки откладывается единственное, пестрой окраски яйцо. Насиживание продолжается около 6 недель. Партнеры сменяются на гнезде через 2—5 суток. Птенцовый пух светло-серый. Кормят оба родителя. Птенец покидает гнездо активным полетом через 11—15 недель после вылупления. Периодичность гнездования 9—10 месяцев; пары, видимо, сохраняются длительное время.
Держатся в прибрежных участках моря, отлетая от берегов обычно не далее 150—200 км. Питаются рыбой, головоногими моллюсками, крупными
104
Рис. 36. Фаэтон Phaethon rubricauda Bodd.
ракообразными. Разыскивают пищу, летая над морем на высоте 10—20 м и стремительно пикируя на замеченную добычу. При пикировании могут погружаться в воду на глубину более 1 м. Удар о воду амортизируется хорошо развитыми подкожными воздушными полостями на нижней стороне тела. Иногда отбирают корм у других рыбоядных птиц: стремительно пикируя на летящую птицу, фаэтон заставляет ее отрыгнуть или бросить добычу, которую подхватывает налету.
СЕМЕЙСТВО 3. PHAETHONTIDAE — ФАЭТОНОВЫЕ
В подотряде единственное семейство, включающее только один род Phaethon с 3 видами. В наших водах не отмечены.
Вероятное место обособления семейства — тропики. По ряду признаков (сквозные ноздри, крупные наружные носовые щели, наличие сошника, относительно длинный язык, пестрая окраска яиц и др.) заметно отличаются от других групп веслоногих. Возможно, наиболее примитивны из всех ныне живущих семейств отряда.
Наиболее древняя находка (вид ископаемого рода) — из нижнего эоцена (возраст 50 —60 млн. лет) Англии.
ПОДОТРЯД 3. PELECANI — ПЕЛИКАНОВЫЕ
Птицы средней и крупной величины. Центральная группа отряда. Наружные щелевидные отверстия ноздрей очень малы или полностью зарастают. Внутренняя носовая перегородка хорошо развита (т. е. ноздри не сквозные). Язык очень мал и, видимо, не функционирует. Горловой мешок, участки около глаз и основания клюва обычно голые (не оперенные). Окраска яиц одноцветная. В подотряд включают 7 семейств (в том числе 3 только с ископаемыми формами), объединяемых в 2 надсемейства.
Всего в этот подотряд входят 42—44 ныне живущих вида, объединяемых в 5—родов. В фауне СССР подотряд представлен 10 видами 3 родов; гнездится 8 видов, отосящихся к 2 родам.
Надсемейство 1. Pelecanoidea— пеликановые
Включает птиц с обликом пеликанов. 2 семейства.
105
Рис. 37. Ареалы семейств пеликанообразных (веслоногих) птиц:
.* — фаэтоновые Paethontidae и фрегатовые Fregatidae (ареалы созпадают), 2—пеликановые Pelecanidae, 3— олушевые Sulidae
СЕМЕЙСТВО 4. PELECANIDAE— ПЕЛИКАНОВЫЕ
Крупные птицы (вес 7—14 кг) с очень длинным оригинальным клювом. Надклювье сильно уплощено и заканчивается острым крючком. Ветви под-клювья соединены легко растяжимой голой кожей, образующей горловой мешок. При широко открытом клюве ветви подклювья сильно изгибаются и расходятся, образуя как бы обруч, на котором растягивается горловой мешок; образуется своеобразный сачок (рис. 38), которым птица подхватывает крупную рыбу. Наружные отверстия ноздрей щелевидные, отчетливо заметные. Шейных позвонков 17. Гетероцельные грудные позвонки сливаются в спинную кость. Конец вилочки срастается с вершиной киля грудины. Скелет сильно пневматизирован; хорошо развиты и подкожные воздухоносные полости. Слепые кишки короткие (около 5 см длиной). Копчиковая железа крупная, с 6—8 выводными протоками. Голосовая мускулатура нижней гортани не развита. Сохраняется только левая сонная артерия.
Оперение относительно рыхлое, белое или бурое. Крылья широкие и длинные; у крупных видов размах крыльев до 3,5 м. Округлый хвост из 20—24 довольно мягких рулевых. Полет плавный, медленный; часто прибегают к парению. Хорошо плавают. Не ныряют; только бурый пеликан способен нырять с разлета.
Гнездятся колониями вблизи воды, на заломах тростника на мелководье, иногда на низких кустах и деревьях. Гнездо—большая куча небрежно набросанных водных и наземных растений. В кладке 2—4 белых, желтоватых или голубоватых яиц. Инкубация 5—6 недель; самка насиживает более интенсивно, чем самец. Птенцы покрываются белым или бурым пухом на 8—12-й день. К этому времени при опасности оставляют гнездо и стараются уплыть в заросли. К 20-му дню жизни начинает разворачиваться оперение на брюхе и крыльях. Взлетают в возрасте 2—2,5 месяцев. Перьевой наряд взрослых одевают на 3—4-й год жизни; в эти же сроки впервые приступают к размножению.
Питаются рыбой. Ловят ее, плавая, иногда с опущенной в воду головой. Вычерпывают рыбу раскрытым клювом, как сачком. Только один морской вид — бурый пеликан Pelecanus occiden'talis L., гнездящийся по островам у берегов Центральной и Южной Америки, охотится иначе. Птицы летают активным или планирующим полетом на высоте 3—20 м над морем и, заметив добычу, пикируют на нее, погружаясь в воду на глубину 1—2 м.
Рис. 38. Розовый пеликан Pelecanus onocrotalus L.
107
В семействе один род Pelecanus с 6 видами, встречающимися по пресным и соленым внутренним водоемам и морским побережьям тропиков и южных районов умеренной зоны. Распространение очень спорадичное (см. рис. 37). В южных районах СССР встречаются и гнездятся 2 вида.
В ископаемом состоянии настоящие пеликаны известны, начиная с нижнего эоцена (Франция, возраст около 50—60 млн. лет); более поздние находки — в разных частях современного ареала. Эти ископаемые остатки относят к 5 ныне живущим и к 10 ископаемым видам, распределяя их между существующим и ископаемым родами (Рrotopelecanus).
По фрагментарным остаткам из отложений нижнего миоцена (возраст 20—25 млн. лет) на западном и восточном побережье США описаны 2 рода с 2 видами, видимо, пеликаноподобных птиц, которые по некоторым деталям строения, возможно, приближаются к олушам и, может быть, к бакланам. Их обычно выделяют в самостоятельное семейство (5-е) — Cyphornithidae.
Надсемейство 2. Suloidea — олушевые
Отличаются от надсемейства пеликановых прежде всего иной формой клюва, более узкими щелевидными наружными носовыми отверстиями, иногда полностью зарастающими, опистоцельной формой грудных позвонков и некоторыми другими особенностями. Надсемейство объединяет 5 семейств, из них 2 включают только ископаемые формы.
СЕМЕЙСТВО 6. PELAGORNITHIDAE
По плечевой кости длиной в 58 см из среднего миоцена (Юго-Восточная Франция) описан единственный вид рода Pelagornis. Это очень крупная птица, по особенностям строения предположительно занимавшая промежуточное положение между пеликанами и олушами, но, видимо, более близкая к последним.
СЕМЕЙСТВО 7. SULIDAE — ОЛУШЕВЫЕ
Морские птицы (рис. 39) средней и крупной величины (вес 1,5—5 кг). Клюв удлиненный, конический. Края надклювья и подклювья слегка зазубрены. Могут очень широко открывать рот, заглатывая крупную добычу. Наружные отверстия ноздрей полностью зарастают. Шейных позвонков 16—18. Грудина и таз удлинены. Основание киля занимает только передние 2/3 грудины. Вершина вилочки соединена с вершиной киля связкой. Скелет сильно пневматичен; хорошо развиты подкожные воздухоносные полости. Слепые кишки очень малы. Копчиковая железа с 5 выводными протоками. Сохраняется только правая сбнная артерия. Голосовая мускулатура нижней гортани не развита. Оперение плотное, густое. У большинства видов окраска белая с черными маховыми, молодые бурые. Первостепенных маховых 10. Крыло длинное, относительно узкое. Длинный клиновидный хвост из 12—18 рулевых.
Гнездятся колониями в сотни и тысячи пар по склонам морских побережий, преимущественно на островах. Гнездо — небрежно набросанная куча водорослей и прибрежных растений. В кладке 1, реже 2 голубоватых или зеленоватых яйца. Насиживают оба партнера в течение 6—7 недель. Наседных пятен не образуется и насиживающая птица греет яйцо, прикрывая его плавательными перепонками лап. Вылупляются птенцы голые с черной кожей. Через несколько дней они покрываются белым или светлобуроватым пухом. Контурное перо начинает раскрываться только через 6—7 недель после вылупления. Полностью оперяются и становятся способными к полету в возрасте 9—12 недель.
Полет быстрый (до 60—90 км/ч), маневренный; легко переходят к планированию. Могут длительное время находиться в воздухе. Хорошо плава-
108
Рис. 39. Олуша Sula bassana L.
ют. Питаются преимущественно рыбой, меньше - крупными морскими беспозвоночными. Пищу разыскивают, паря или летая в 10—30 м над поверхностью моря и, заметив добычу, пикируют на нее, погружаясь в воду иногда на глубину до 5 м. Приспособления к такому типу ныряния с пикирования: плотное оперение, сильное развитие воздухоносных подкожных полостей, удлинение грудины и т. п.
В семействе 1 род Sula с 7—9 видами (иногда выделяют еще род Morus с 2 видами). Спорадично распространены по островам и морским побережьям тропиков и субтропиков (рис. 37). Только один вид Sula bossana L. проникает далеко на север и гнездится на Ньюфаундленде и в Исландии. В прибрежных водах СССР отмечено 2 случайно залетных вида.
Центр возникновения семейства, видимо, тропики. Наиболее древняя ископаемая находка — из олигоценовых отложений Франции (ископаемый вид Sula ronzoni). Остатки из более поздних отложений в разных районах современного ареала относят к 6 ныне живущим и 15 ископаемым видам.
СЕМЕЙСТВО 8. ELOPTERYGIDAE
По плохой сохранности разрозненным остаткам из верхнемедовых или нижнепалеоценовых отложений Румынии и остаткам из эоцена Франции и Англии описаны 3 монотипичных рода, включаемых в это семейство. Вероятно, могут рассматриваться как формы, занимающие промежуточное положение между олушами и бакланами.
СЕМЕЙСТВО 9. PHALACROCORACIDAE— БАКЛАНОВЫЕ
Длинношейные средних и крупных размеров (вес 800 г — 4 кг) водные птицы (рис. 40). Клюв относительно тонкий и длинный, с большим острым крючком на конце. Края надклювья и подклювья гладкие, режущие. Клюв может раскрываться очень широко, что облегчает заглатывание крупной добычи. У молодых птиц наружные отверстия ноздрей очень узкие, щелевидные; у старых птиц они полностью зарастают. К верхнезатылочной кости черепа причленяется длинная палочковидная косточка, на которой укрепляются добавочные порции мышц, закрывающих клюв. Эта особенность, наряду с режущими краями клюва и его острым крючком, позволяет удерживать пойманную большую и сильную рыбу. Шейных позвонков 18. Грудина широкая, таз суженный. Коленная чашечка имеет отверстие, через
109
Рис. 40. Стеллеров баклан Phalacrocorax perspicillatus Pall, с Командорских островов, вымер в середине XIX в.
которое проходит мышечное сухожилие. Малая берцовая Кость доходит до интертарзального сустава, что обеспечивает возможность вращательных движений лапы (важно при нырянии). Пнев-матичность скелета слабая. Подкожных воздухоносных полостей нет. Слепые кишки рудиментарны. Копчиковая железа небольшая, с двумя выводными протоками. Функционируют обе сонные артерии. Есть слабо развитая мускулатура нижней гортани.
Оперение плотное; контурные перья относительно короткие. Окраска обычно черная с металлическим отблеском, иногда более или менее развиты участки белого цвета. Окраска молодых более тусклая. Первостепенных маховых 11. Крылья относительно короткие и широкие, с тупыми вершинами. Хвост длинный, слабоступенчатый из 12 —14 жестких рулевых.
Гнездятся колониями от групп в несколько пар до десятков и сотен гнезд (иногда и больше).
Гнездо — груда веток, травы, водорослей, скрепленных пометом, — помещается на уступах скал, на заломах тростника, в кронах деревьев и прямо на земле. Обычно колонии размещаются на бере-
гах водоемов, но иногда бывают и в удалении от воды на 3—5 км. В кладке 3—4 белых с зеленоватым или голубоватым отливом яйца. Насиживание, в котором участвуют оба партнера, продолжается 4—7 недель. У вылупляющихся птенцов темно-бурая кожа. На 10—14-йдень жизни покрываются коротким, но густым темно-бурым или серым птенцовым пухом. Через 4—5
недель после вылупления бывают покрыты почти полностью раскрывающимся контурным пером, а в возрасте около 7 недель способны к полету. Обычно взрослые кормят молодых птиц еще 1—2 недели после вылета. Затем молодые кочуют вместе со взрослыми, самостоятельно добывая пищу. Полный взрослый наряд получают на 3 -4-й год жизни.
Полет довольно быстрый, но не маневренный. Прекрасно плавают и с поверхности воды ныряют. В толще воды двигаются сложными движениями ног; слегка полураскрытые крылья и жесткий хвост выполняют роль рулей глубины. Ныряют на глубину до 10 м и более. Обычная продолжительность ныряния 1—2 мин. Основу питания составляют рыбы длиной до 15—25 см, ловят и разнообразных крупных водных беспозвоночных (ракообразных, морских звезд и т. п.). Пищу добывают ныряя. После продолжительного ныряния часто садятся на берег, на выступающие из воды камни и сушат крылья. При кочевках держатся в прибрежных районах морей.
В семействе 2 рода: Phalacrocorax с 25 видами, спорадично распростра
ненными по морским побережьям и крупным пресноводным водоемам практически всего земного шара, кроме Арктики и Антарктики (рис. 41), и Nan-nopterum с 1 видом — галапагосским бакланом N. harrisi Roth., практиче-
ски утратившим способность к полету.
ПО
. 150° 120° 90° 60°к западу отГрииета 0°к востоку отГринвича60° 90° 12СР 150° 180° 150°
Рис. 41. Ареалы семейств пеликанообразных птиц:
1— баклановые Phalacrocoracidae, 2— змеешейковые Anhingidae
В СССР сейчас встречается и гнездится 6 видов бакланов. На Командорских островах жил крупный, видимо, очень плохо летавший Стелле-ров баклан Phalacrocorax perspicillaris Pall. (рис. 40), полностью вымерший в первой половине XIX в. из-за неумеренного промысла и, может быть, вспышки какой-то эпизоотии.
Ископаемые остаки бакланов обнаружены в разных районах современного ареала, начиная с верхнего палеоцена (Северная Америка, возраст примерно 60—70 млн. лет). По ним описаны 23 вымерших вида 3 родов. Из ныне живущих видов в ископаемом состоянии найдено 8. Вероятный центр возникновения — тропики, возможно, район Индийского океана.
СЕМЕЙСТВО 10. ANHINGIDAE — ЗМЕЕШЕЙКОВЫЕ
Близки к бакланам; напоминают их по внешнему облику (рис. 42) и образу жизни.
Клюв длинный, прямой, с конически заостренной вершиной. Края надклювья и подклювья слегка зазубрены. Наружные отверстия ноздрей в виде узких щелей. Нет сесамовидной косточки, примыкающей у бакланов к затылочному отделу черепа. Шея более длинная, чем у бакланов, и более подвижная. Шейных позвонков 20, тела позвонков удлиненные. Характер суставных поверхностей позвонков и сложная система мышц и связок обеспе-печивают возможность резкого выпрямления шеи. При этом голова мгновенно выбрасывается вперед, и добыча, как копьем, пронизывается клювом или схватывается. Сухожилие мускула не пронизывает коленную чашечку, как у бакланов, а проходит по глубокому желобку по ее наружной поверхности. В желудке четко обособлен пилорический отдел. Пищеварительные железы лежат в углублениях стенок объемистого железистого желудка. Копчиковая железа с двумя выводными протоками. Функционирует только левая сонная артерия. Мускулатура нижней гортани развита слабо.
Оперение плотное. Преобладает темная окраска. Первостепенных маховых И. Хвост еще длиннее, чем у бакланов, из 10—12 жестких рулевых. При линьке все маховые выпадают почти одновременно, и птица на некоторое время теряет способность к полету.
Гнездятся небольшими группами или одиночными парами по берегам озер и заводям крупных рек. Гнезда в зарослях тростников или в кустах у воды. В кладке 3—6 белых с зеленоватым отливом яиц. Птенцы вылупляются голыми, затем покрываются белым пухом. Гнездо покидают полностью оперенными в возрасте 6—8 недель. Взрослые кормят молодых еще 1—2 недели после вылета. Вне сезона размножения кочуют мелкими стайками по водоемам в районе гнездовий.
Полет бакланьего типа. Прекрасно плавают и ныряют. Охотящаяся птица либо подстерегает добычу, сидя у кромки воды, на нависающей над водой ветви и т. п., либо разыскивает ее, плавая по водоему так, что
Рис. 42. Змеешейка Anhinga все тело погружено в воду, и периодически anhinga (L.) ныряя. Основной корм — разнообразные ры
112
бы. Ловят различных водных беспозвоночных, земноводных и их личинок и т. п.
Один род Anhinga с 3 видами (иногда их рассматривают как географические формы одного вида), приуроченными к крупным внутренним водоемам тропической и субтропической зон (рис. 41).
По немногочисленным ископаемым остаткам из среднего эоцена и более поздних отложений как в пределах современного ареала, так и в Центральной Европе, описано 6 видов (из них 1 и ныне живущий), относимых к современному и одному ископаемому роду. Вероятный центр возникновения семейства — тропики.
Иногда бакланов и змеешеек объединяют в 1 семейство Phalacrocorac-idae с 2 подсемействами: Phalacrocoracinae и Anhinginae.
ПОДОТРЯД 4. FREGATAE — ФРЕГАТОВЫЕ
В подотряде единственное семейство.
СЕМЕЙСТВО И. FREGATIDAE— ФРЕГАТОВЫЕ
Довольно крупные (вес около 2 кг), длиннокрылые (размах крыльев до 2 м), длиннохвостые морские птицы (рис. 43). Клюв тонкий и длинный, с большим острым крючком на конце. Узкие щелевидные наружные носовые отверстия. Сошник есть. Шея относительно короткая, из 14—15 позвонков. Гетероцельные грудные позвонки не срастаются в спинную кость. Грудина широкая и короткая. Киль высокий, но короткий: его основание занимает лишь около половины длины грудины. Вилочка срастается с коракоидами, а ее вершина прирастает к вершине киля. Скелет крыла удлинен, особенно за счет кисти и меньше предплечья. Очень сильно развиты сесамовидные косточки, заклинивающие локтевой сустав, что позволяет без особой затраты мышечной энергии сохранять определенное положение крыла при парении. Таз широкий и короткий. Цевка короткая, расширенная, с неполностью слившимися метатарзалиями; оперена почти до пальцев. Пальцы длинные. Плавательная перепонка соединяет все 4 пальца, но развита слабо и не доходит до концевых фаланг. Пневматичность скелета очень высокая; подкожных воздухоносных полостей нет. Сонные артерии парные.
Оперение относительно рыхлое. Окраска обычно черная, с металлическим отблеском, у части видов с белыми пятнами. Подбородок, горло и область зоба голые, у самцов ярко-красные. В период тока область зоба раздувается в виде большого красного шара (рис. 43). Первостепенных маховых
30.
Рис 43. Токующий фрегат Fregata minor (Gm.)
113
11. Крыло узкое, длинное, с острой вершиной. Хвост очень длинный, резко вильчатый, из 12 рулевых.
Гнездятся колониями. Гнездо — груда сухих веточек и растительной ветоши — на уступах скал или на деревьях и крупных кустах. В кладке 1, редко 2 яйца. Насиживают и выкармливают оба партнера. Инкубация 6—7 недель. Примерно к 12—14-му дню жизни голый при вылуплении птенец покрывается светло-серым пухом. Полностью оперяются и становятся способными к полету только в возрасте 4—5 месяцев. По некоторым данным, взрослые, кочующие вместе с молодыми в районе колоний, продолжают кормить молодых еще 2—6 месяцев.
По земле ходят плохо и неохотно. На воду тоже садятся редко и плавают мало. Не ныряют. Полет быстрый, маневренный, преимущественно парящий. Способны длительное время находиться в воздухе. Для отдыха возвращаются на берег и усаживаются на скалах или ветвях деревьев. Питаются преимущественно летучими рыбами, которых подхватывают налету в воздухе. С налета схватывают выплывших на поверхность моллюсков и ракообразных. Нападают на других морских птиц, заставляя их отрыгивать проглоченную добычу; подхватывают ее налету.
В семействе 1 род Fregata с 5 видами, встречающимися преимущественно на островах тропической и субтропической зон (рис. 37). В прибрежные воды СССР возможны только редкие залеты; одна птица добыта у Хабаровска.
Фрегаты — довольно резко уклоняющаяся и наиболее приспособленная к воздушному образу жизни группа веслоногих. Палеонтологические данные отсутствуют. Центр возникновения, видимо, тропики.
ПОДОТРЯД 5. CLADORNITHES
По остаткам метатарзальных костей из нижнего олигоцена Огненной Земли описано 2 вида, относимых к 2 родам (Cladornis, Crusceduld), составляющим единственное семейство (12-е) подотряда Cladornithidae. Недостаточность материала вынуждает считать эту группу, равно как и ее положение в системе, очень сомнительной.
ОТРЯД 16. CICONIIFORMES (GRESSORES) — ЛИСТООБРАЗНЫЕ, ИЛИ ГОЛЕНАСТЫЕ
Разнообразные по величине (вес от 100 а до 6 кг\ высота стоящей птицы от 20 см до 1,5 м), обычно длинноклювые, длинношейные и длинноногие око-ловодные птицы с характерным обликом аиста, цапли или ибиса. Распространены почти по всему земному шару, кроме Арктики и Антарктики, приурочены к берегам водоемов и болотам; только одиночные виды проникают в сухие степи. В тропиках оседлы или кочуют, в умеренных широтах пере-летны.
Ноздри щелевидные. Череп десмогнатического типа; базиптеригоидные сочленения редуцируются. Шейных позвонков 16—20. Обычно образуется спинная кость. Грудина и таз широкие. Цевка и нижняя часть голени не оперены. Задний палец у большинства видов довольно длинный. Хорошо развита пневматичность скелета. Пищевод и желудок легко растяжимы. Есть желчный пузырь. Слепые кишки обычно рудиментарны. У части групп сохраняется рудимент пениса. Копчиковая железа голая или оперенная. Обычно две сонные артерии. Оперение относительно рыхлое. Контурные перья с пуховой частью у основания и с небольшим добавочным стержнем (нет у некоторых аистов). Уздечка или кожа вокруг глаз или и то и другое голые, не оперенные. Крыло диастатаксическое. Первостепенных маховых
114
10—12, рулевых 8—12. Смена маховых идет постепенно. Половой диморфизм окраски свойствен немногим видам. У части видов более или менее отчетливо выражено сезонное изменение окраски; в брачном наряде иногда развиваются украшающие перья. Окраска молодых обычно отличается от окраски взрослых.
Моногамы. Колониальные, реже одиночно гнездящиеся. Гнезда грубой, неряшливой постройки, располагаются в зарослях тростника и камыша, на кустах и деревьях, реже в траве на земле. Обычно самец носит материал, а самка строит гнездо. Кладка из 2—8 яиц. Насиживание начинается после откладки первого или второго яйца и продолжается от 2,5 (мелкие виды) до 4,5—5 недель. Птенцов выкармливают оба партнера. У многих видов взрослые носят птенцам воду. До подъема на крыло (в возрасте 4—10 недель, у аистов марабу 16—17 недель) птенцы либо остаются в гнезде (аисты), либо лазают в зарослях в районе гнезда. Наряд взрослых птиц одевают в возрасте 2—3 лет и тогда же становятся половозрелыми (мелкие виды, возможно, в возрасте около года).
Питаются разнообразными животными кормами. Способы добывания пищи отличаются в разных семействах. Летают медленно, активным и парящим полетом. В полете ноги вытянуты назад и далеко выдаются за хвост.
Некоторые виды служат объектом спортивной охоты. В 20-х годах XX в. из-за неумеренного преследования во многих районах резко сократилась численность ряда видов цапель, добываемых ради украшающих перьев («эгреток»), которые использовались для украшения дамских туалетов. Сейчас в большинстве стран мира продажа перьев запрещена. Местами могут причинять некоторый ущерб, поедая рыбу на рыборазводных прудах. Однако большинство видов приносит некоторую пользу, истребляя большое количество насекомых как вредителей растительности, так и хищников, поедающий икру и молодь рыб.
Отряд разделяется на 4 подотряда, включающих 11 семейств (из них 4 представлены только ископаемыми формами). Ныне живущие аистообразные подразделяются на 49 родов, объединяющих 118 видов. В СССР встречаются 23 вида 14 родов; несомненно гнездится 18 видов, относимых к 12 родам.
По ископаемым остаткам, иногда хорошей сохранности, начиная с эоцена в разных районах современного ареала описано 64 вида 21 рода. Из ныне живущих видов в ископаемом состоянии обнаружено 37. Очень давние родственные отношения выявляются между голенастыми и веслоногими. Более глубокие, но тоже очень давние и трудно выявляемые родственные с;вязи, видимо, сближают голенастых, гусеобразных и дневных хищников.
ПОДОТРЯД 1. ARDEAE — ЦАПЛЕВЫЕ
Клюв длинный, обычно прямой, к концу конически заостренный. Сошник более крупный, чем у других групп отряда. Тонкая длинная шея содержит 19—20 удлиненных шейных позвонков. По деталям строения шеи много общего с змеешейками: боковые движения шеи резко ограничены, но она легко и быстро сгибается и выпрямляется, что облегчает схватывание подвижной добычи и позволяет бить ее клювом, как копьем. Задний палец ноги почти такой же длины, как и передние. Когти длинные, острые. На внутренней стороне когтя среднего пальца зазубринки в виде гребенки (рис. 44). Пищевод и желудок сильно растяжимы. Четко обособлен пилорический отдел. Обычно есть только один сильно редуцированный слепой отросток. Нет никаких следов пениса. Копчиковая железа развита слабо; у части видов оперенная, а у других — голая. Обычно сохраняются обе сонные артерии (кроме Ixobrychus).
Оперение довольно рыхлое. Побочный стержень контурного пера хорошо развит. Редкий пух расположен только по аптериям. На отдельных
115
участках тела (на груди, надхвостье, иногда на брюхе) есть «пудретки» — участки очень ломкого, сильно ветвящегося и непрерывно растущего пуха. Вершины пушинок в области пуд-реток все время обламываются, образуя талькоподобную роговую пудру. Соскабливая гребешком когтя эту пудру, птица присыпает ею оперение, что несколько уменьшает смачиваемость пера. Первостепенных маховых 11, рулевых обычно 12. У многих видов в брачном наряде обоих полов есть украшающие перья (на голове,шее, спине и т. п.). У немногих видов — четкий половой диморфизм окраски. Молодые птицы в более тусклом пере, у части видов окраска молодых резко отлична от окраски взрослых.
Летают медленным, довольно тяжелым подетом. В полете вытягивают ноги назад, а голову, резко согнув шею, подтягивают к туловищу.
В подотряде 2 семейства.
СЕМЕЙСТВО 1. ARDEIDAE — ЦАПЛЕВЫЕ
Рис. 44. Серая цапля Ardea cinerea L. Сбо- По внешнему облику — ти-
ку-коготь среднего пальца с гребенкой пичные цапли (рис. 44). Разме-
ры от мелких (вес около 100 г) до крупных (до 3—4 кг).
Клюв прямой, конический, заметно сжатый с боков, заостренный. Края надклювья и подклювья режущие, иногда слегка зазубренные. Ноздри сквозные. Язык длинный, тонкий. Нижняя часть голени не опер.ена на значительном протяжении.
Гнездятся преимущественно колониально (от групп в несколько пар до сотен) или, реже, одиночно на деревьях, в кустах, в залитых водой тростниках и камышах, реже среди густых зарослей на земле. В кладке от 2 до 5 (у мелких видов — до 8) голубоватых или зеленоватых яиц. Насиживают оба партнера, но самка обычно интенсивнее. Инкубация от 16—17 (мелкие виды) до 28—30 дней. Птенцы вылупляются беспомощными, голыми или слабо опущенными, с уже прорезавшимися, но еще не полностью открытыми глазами. В возрасте около 2 недель птенцы уже ходят по гнезду, а при опасности уходят из него и затаиваются поблизости. В это же время у них начинает разворачиваться контурное перо. Полностью оперяются и приобретают способность к полету в возрасте 4—5 недель мелкие и 7—8 недель — крупные виды. Молодые стайных видов после вылета из гнезда кочуют вместе со взрослыми, у видов с одиночным образом жизни — держатся самостоятельно. Половозрелыми становятся в возрасте около 2—3 лет.
Животноядны. Крупные виды питаются преимущественно мелкой рыбой, земноводными, крупными насекомыми, при случае ловят мелких млекопитающих (мышевидных и даже сусликов). Обычный способ охоты — под-
116
150° 120° 90° 60 к западу от Гринвича 0°к востоку от Гринвича 60° 90° 120°. 150° 180° 150°
’уг чи ьи ои и OU CU аи I2U юи 1оО - ЬО
Рис. 45. Ареалы семейств аистообразных птиц:
1— цаплевые Ardeidae, 2—китоглавовые Balaenicipi tidae, 3— молотог лавовые Scopidae
карауливание добычи: птица неподвижно стоит на мелководье, приблизившаяся добыча схватывается клювом при молниеносном движении головы. Иногда бродят по мелководью или по траве, выпугивая добычу. У мелких видов способы охоты такие же, но питаются они преимущественно разнообразными беспозвоночными: насекомыми и их личинками, червями, моллюсками и т. п.
Часть видов активна ночью и в сумерках, другие ведут преимущественно дневной образ жизни.
В семействе Ardeidae 14 родов — Ardea, Agamia, Ardeola, Botaurus, Bubulcus, Bator ides, Egretta, Gorsachius, Hygranassa (Melanophoyx), Ixobry-chus, Nyctanassa, Nycticorax, Tigrosotna, Zebrilus,—объединяющих 62 вида. Цапли почти космополиты, не встречающиеся лишь в приполярных районах (рис. 45). За последние несколько десятилетий египетская цапля Bubulcus ibis (L.) широко расселилась по тропикам и субтропикам Старого Света, проникла в Америку и сейчас успешно там расселяется (юг США, Колумбия, Венёсуэла и т. д.). В СССР отмечены 16 видов, относимых к 9 родам; несомненно гнездятся 13 видов из 8 родов.
По ископаемым остаткам в пределах современного ареала описано 14 ископаемых видов, относимых к 7 родам. Наиболее древняя находка — цапля из нижнеэоценовых отложений Англии. В более поздних отложениях обнаружены остатки, относимые к 20 и ныне живущим видам. Вероятный центр возникновения — тропики.
СЕМЕЙСТВО 2. COCHLEARIIDAE — ЧЕЛНОКЛЮВОВЫЕ
Птицы средней величины (вес 600—800 а), ведущие преимущественно ночной образ жизни и отличающиеся от настоящих цапель некоторыми чертами строения (рис. 46). Клюв широкий, приплюснутый сверху вниз, очень объемистый (внешне несколько напоминает клюв китоглава; это конвергентное сходство). Ноздри не сквозные. Язык редуцирован. Хвост из 12 жестких рулевых. Голень почти вся оперена. Гнездятся на деревьях. В кладке 2—4 белых с коричневатым отливом яиц. Питаются рыбой, крабами, мелкими грызунами. Объемистый клюв при схватывании добычи, видимо, играет роль своеобразного сачка.
В семействе 1 род Cochlearius с 2 видами, распространенными по берегам лесных водоемов Центральной и Южной Америки.
Подотряд цаплевых, видимо, наиболее прогрессивная и наиболее уклонившаяся от предковых форм группа отряда. В ряде мелких черт строения проявляется некоторое сходство с веслоногими. Конечно, черт сходства с аистами много больше и они с несомненностью говорят о естественности всего отряда в целом.
Рис. 46. Челноклюв Cochlearius cochlearius (L.)
ПОДОТРЯД 2. BALAENICIPITES — КИТОГЛАВОВЫЕ
В подотряде единственное семейство.
U8
СЕМЕЙСТВО 3. BALAENICIPITIDAE — КИТОГЛАВОВЫЕ
Один вид — Balaeniceps гех Gid. (рис. 47). Населяет поросшие папирусом обширные болота верховьев Конго и Белого Нила (рис. 45). Крупная птица (вес 5—7 кг, зоологическая длина около 120 см) с большим широким массивным клювом.
По многим морфологическим особенностям занимает промежуточное положение между подотрядами цаплевых и аистовых; сохраняются и некоторые черты сходства с веслоногими. Объемистый клюв длинен и очень широк, приплюснут; на конце небольшой острый крючок. Края надклювья и подклювья острые. Ноздри не сквозные. Затылочных фонтанелей нет. Шейных позвонков 16 — 17. Конец вилочки срастается с килем грудины. Ноги длинные; направленные вперед 3 пальца очень длинные, задний заметно короче. Коготь среднего пальца без гребенки. Язык маленький. Железистый желудок очень объемистый, с хорошо обособленным пилорическим отделом. Мускульный желудок мал, со слабыми стенками. Слепая кишка од-
Рис. 47. Китоглав Balaeniceps rex Gid.
на, очень маленькая. Рудимента пениса нет. Копчиковая железа сильно редуцирована. Побочный стержень контурного пера очень короткий. Пух растет по всему телу. На спине есть 2 большие пудретки. Первостепенных маховых 11, рулевых 12. Окраска темная.
По походке и позам напоминает аистов, но охотится, как цапли, подкарауливая добычу. Питается преимущественно рыбой и различными земноводными. Объемистый клюв при схватывании добычи, видимо, играет роль своеобразного сачка. Летит, как цапля, согнув шею и подтянув голову с тяжелым клювом к туловищу. Гнездо — большая куча стеблей в зарослях тростника или папируса. В кладке обычно 2 довольно крупных голубовато-белых яйца. Вылупившиеся птенцы покрыты коротким серым пухом. Экология изучена плохо.
ПОДОТРЯД 3. CICONIAE — АИСТОВЫЕ
Объединяет разнообразных по величине птиц с общим обликом аистов или ибисов; некоторые представители внешне похожи на цапель. Шея несколько короче, чем у цапель, и не столь подвижна. В полете шея вытянута вперед; только марабу, с их очень большим, массивным клювом, в полете, как и цапли, сгибают шею и подтягивают голову к туловищу. Летают активным и парящим полетом.
119
В подотряд включают 3 надсемейства — молотоглавов, аистов и ибисов, в каждое из которых входят 1—2 семейства. Введением ранга надсемейств подчеркивают обособленность этих насомненно родственных групп. Всего в подотряде 48 видов, относимых к 30 родам. В СССР встречается 6 видов 4 родов, несомненно гнездится только 4 вида.
Надсемейство 1. Scopoidea — молотоглавовые
Сюда входит одно семейство.
СЕМЕЙСТВО 4. SCOP1DAE— МОЛОТОГЛАВОВЫЕ
Единственный вид—Scops umbretta Gm. (рис. 48). Населяют берега лесных рек и озер Мадагаскара и Африки южнее Сахары (см. рис. 45). По внешнему облику это небольшая цапля (длина 60 см). Как и китоглав, занимает промежуточное положение между цаплями и аистами, но ближе к
последним.
Клюв прямой, довольно длинный, очень сильно сжат с боков; на конце клюва маленький крючок. Ноздри не сквозные. В затылочном отделе черепа есть щелевидные фонтанели. Шейных позвонков 16. Между основаниями
передних пальцев ноги есть
Рис. 48. Молотоглав Scops umbretta Gm.
маленькие перепонки. В отличие от аистов коготь среднего пальца имеет гребенку. Когти всех пальцев длинные, заостренные. Язык маленький, треугольной формы. Слепые кишки сильно редуцированы. Копчиковая железа небольшая, оперенная. Функционируют обе сонные артерии. Есть подкожный шейный воздушный мешок. Мускулатура нижней гортани развита. Первостепенных маховых 10, рулевых 12. Общая окраска коричневато-бурая. На затылке хохол из одноцветных перьев. Пуд-
реток нет.
Гнездятся обычно одиночными парами, устраивая на деревьях или в кустах большое — в диаметре до 1—1,5 м — крытое гнездо, внутри которого размещены 2—3 соединяющиеся друг с другом камеры; узкий вход расположен сбоку. Гнездо строится из веток; стенки гнездовой (самой внутренней) камеры обычно промазываются грязью или илом. Строят гнездо оба партнера в течение 35—40 дней. Кладка из 3—6 чисто белых яиц. Насижи
вает преимущественно самка, самец в это время продолжает надстраивать и укреплять гнездо. Инкубация около 4 недель. Вылупляющиеся птенцы покрыты сероватым пухом. Оставляют они гнездо уже летными, в возрасте около 50 дней и больше к гнезду не возвращаются. Питаются разнообразной животной пищей, подкарауливая ее или медленно ходя по берегу.
Надсемейство 2. Ciconioidea — аистовые
В надсемействе единственное семейство.
120
СЕМЕЙСТВО 5. С ICO N1IDAE—АИСТОВЫЕ
Крупные птицы (вес до 5—6 кг) с характерным внешним обликом аиста (рис. 49).
Клюв длинный, прямой, к вершине заостренный. Сквозные ноздри. Затылочных фонтанелей нет. Шейных позвонков 17. Задний край грудины с одной парой вырезок. Конец вилочки подходит к вершине киля грудины и иногда с ним срастается. Первый йалец кисти несет небольшой коготок. Задний палец ноги заметно короче передних и прикреплен несколько выше. Между основаниями передних пальцев 'есть маленькие перепонки. Когти притупленные; коготь среднего пальца не несет гребенки. Язык маленький, треугольной формы. В желудке обособляется пилорический отдел. Сильно редуцированные слепые кишки парные. Есть рудимент пениса. Копчиковая железа оперена. Сонные артерии парные. На шее под кожей расположен разделенный на камеры воздушный мешок. У большинства видов нет голосовых мышц нижней гортани, а у части видов редуцирована и звуковая мембрана. Такие виды немые и издают звуки, лишь щелкая клювом (Ciconia и др.). У некоторых видов гортань имеет голосовую мускулатуру, и они способны издавать довольно сложные звуки; длинная трахея образует петли, играющие роль резонаторов.
Контурные перья обычно без побочных стволов. Пух распространен по птерилиям и аптериям. Пудреток нет. Украшающие перья отсутствуют. Первостепенных маховых 11—12, рулевых обычно 12.
Гнездятся чаще одиночными парами, реже гнезда отдельных пар расположены поблизости. Гнездо — куча небрежно набросанных веток и растительной ветоши. Его устраивают на крупных ветвях деревьев, на уступах скал, реже на земле в зарослях околоводной растительности. В кладке 3—5, редко до 8 белых яиц. Насиживают оба партнера, у части видов более ак
тивно насиживает самец. Инкубация около 4—6 недель. Вылупляющиеся птенцы зрячие, покрыты коротким и довольно редким сероватым пухом. У многих видов вскоре этот первичный пуховой наряд дополняется вырастающим более длинным и густым пухом. В гнезде остаются до приобретения способности к полету — до 60—70 дней (у марабу—до НО дней). После вылета птенцов держатся обычно семьями, которые иногда объединяются в более крупные стаи. Половозрелыми становятся в возрасте 3—4 лет.
Кормятся разнообразной животной пищей: амфибиями, рептилиями, моллюсками, червями, крупными насекомыми и их личинками, иногда ловят мелких грызунов. Пищу отыскивают, медленно бродя по лесным полянам, лугам, мелководным болотам, берегам водоемов. В лужах и на мелководье иногда ловят рыбу. Некоторые виды, например африканский и индийский марабу — Leptoptilos crumeniferus Less., L. dubius Gm., питаются и падалью — трупами
Рис. 49. Африканский марабу Leptoptilos crumeniferus (Less.)
121
Рис. 50. Ареалы семейств аистообразных птиц:
1— аистовые Ciconiidae, 2— ибисовые Threskiornithidae, 3— фламинговые Phoenicopteridae
крупных животных. В связи с этим у них особенно большой и мощный клюв, а голова и значительная часть шеи не оперены.
Семейство Ciconiidae включает 17 ныне живущих видов, объединяемых в 10 родов: Ciconia, Anastomus, Dissoura, Ephippiorhynchus, Ibis, Jabiru, Leptoptilos, Sphenorhynchys, Tantalus, Xenorhynchus. Широко распространены в умеренных и жарких широтах, но заселяют лишь небольшую часть Австралии и почти не проникли в Северную Америку (рис. 50). Роды Старого Света не проникли в Америку, американские роды не встречаются в Старом Свете. Из-за изменения ландшафтов и прямого преследования ареалы и численность многих видов сейчас сильно сократились.
В СССР встречаются и гнездятся 2 вида рода Ciconia (белый и черный аисты).
По ископаемым остаткам, начиная с олигоценовых отложений (Франция, Патагония), описано 20 ископаемых видов (относят к 9 ископаемым родам) и 7 видов, живущих и сейчас. Центр возникновения семейства, видимо, тропики или субтропики. Вероятно, центральная, наименее специализированная группа отряда.
Надсемейство 3. Threskiornithoidea — ибисовые
В надсемействе 2 семейства.
СЕМЕЙСТВО 6. THRESKIORNITHIDAE—ИБИСОВЫЕ
Единственное ныне живущее семейство включает птиц средней величины (вес 0,5—2 кг) с характерным обликом ибиса, длинношейные, с длинным, тонким, обычно изогнутым книзу клювом (рис. 51) или колпицы с плоским, лопатообразно расширяющимся на конце клювом (рис. 52). От щелевидных наружных отверстий ноздрей к концу клюва идут четкие борозды. На нижней челюсти хорошо развит задний угловой отросток рг. angularis posterior (как и у фламинго; у других групп голенастых его нет). Есть затылочные фонтанели. Шейных позвонков 17—18. По заднему краю грудины 2 пары вырезок. Конец вилочки обычно не доходит до вершины киля и соединен с ним эластичной связкой. Первый палец кисти не имеет коготка. Задний палец но
ги заметно короче передних, но прикрепляется на одном уровне с ними. Когти заостренные; коготь среднего пальца гладкий, без гребенки. Язык сильно редуцирован. Слепые кишки не длинные, но обычно функционирующие. Рудимента пениса нет. Копчиковая железа оперена. Сонные артерии парные. У большинства видов голосовая мускулатура нижней гортани развита слабо или отсутствует. У части видов отсутствует и голосовая мембрана, они немые (например, колпица). У немногих видов с хорошим голосом длинная трахея образует усиливающие звук пет-
Рис. 51. Священный ибис Threskiornis aethioplcus-(Lath.)
123
ли, лежащие в углублении грудины. Подкожный шейный воздушный мешок не развит. Контурные перья со слаборазвитым побочным стержнем. У многих видов голова и часть шеи голые, не оперенные. Пух растет по птерилиям и аптериям. Пудреток нет. Первостепенных маховых 11, рулевых обычно 12. Окраска разнообразная.
Гнездятся обычно колониально, часто вместе с цаплями. Гнездо — в виде небрежно набросанных веток и стеблей, помещается на ветвях деревьев и кустов, на заломах тростника, реже на уступах скал или на земле. В кладке 2—3, редко 4 белых или голубоватых яйца; у немногих видов окраска яйца слабо пятнистая. Насиживание
Рис. 52. Колпица Platalea leucorodia L.
около 4—5 недель. Вылупившиеся птенцы слепые (обычно прозревают в первые сутки после вылупления), покрытые беловатым, коричневатым или бурым пухом. Подросшие птенцы иногда уходят из гнезда и держатся поблизости в ветвях или в тростнике. Полностью оперяются и приобретают способность к полету на 6—7-й неделе жизни. Половозрелыми становятся через 2—3 года. Вне периода размножения обычно держатся стайками или мелкими группами.
Кормятся разнообразной животной пищей. Ибисы обычно разыскивают Пищу — червей, моллюсков, ракообразных, насекомых и их личинок — по берегам водоемов и на мелководье, зондируя почву своим длинным, изогнутым книзу клювом. Колпицы копаются своим лопаткообразным клювом в илу или ловят добычу (преимущественно ракообразных) в верхних слоях
воды.
В семействе Threskiornithidae 19 родов и 30 видов, распространенных преимущественно в тропиках и субтропиках (см. рис. 50). Это семейство обычно подразделяют на 2 подсемейства: а) ибисов Threskior-n i t h i и a e, включающее 24 вида 17 родов (Threskiornis, Bostrychia, Cercibis, Eudocimus, Geronticus, Hagedashia, Harpiprion, Lampribis< Lopho-tibis, Mesembrinibis, Nipponia, Phitnosus, Platibis, Plegadis, Pseudibis, Thaumatibis, Theristricus), и б) колпиц Plataleinaec2 родами и 6 видами (Platalea с 5 видами и Ajdja). Только один род Plegadis встречается и в Старом, и в Новом Свете (каравайка Р. falcinellus L. — практически космополит). Остальные роды распространены либо только в Старом, либо только в Новом Свете.
В СССР отмечено 4 вида 3 родов, несомненно гнездятся 2 вида 2 родов (каравайка и колпица).
Центр возникновения семейства — тропическая или субтропическая зоны.
Близки к аистам, но есть и отдельные черты сходства с фламинго. В пределах современного ареала ископаемые остатки ибисов известны с верхнего эоцена (Англия). Их относят к 7 ископаемым (3 рода) и 8 ныне живущим видам.
Может быть, к семейству ибисов следует отнести род Plegadornis, описанный по остаткам из верхнего мела Алабамы (США) и выделяемый в близкое к. ибисам семейство (7-е) Plegadornithidae.
124
ПОДОТРЯД 4. PHOENICOPTERI —ФЛАМИНГОВЫЕ
Ныне живущие виды фламинго объединены в семейство (8-е) Phoeni-copteridae.
СЕМЕЙСТВО 8. PHOENICOPTERIDAE — ФЛАМИНГОВЫЕ
Довольно крупные (вес 2,5—4,5 кг), длинношейные и длинноногие птицы (рис. 53).
Клюв массивный; примерно посредине он перегнут почти под прямым углом, и вершина клюва направлена вниз. По краям плоского надклювья и объемистого вогнутого подклювья, а так же по краям большого мясистого языка расположены роговые пластинки, образующие фильтровальный аппарат, функционально сходный с фильтровальным аппаратом клюва уток. Большая часть надклювья образована верхнечелюстной костью (у гусеобразных — предчелюстной). На заднем краю нижней челюсти хорошо развит рг. angularis posterior. Ноздри сквозные. Хорошо развиты надорбитальные носовые железы. Есть затылочные фонтанели. Шейных позвонков 19, из них последний входит в состав спинной кости. Строение грудины и таза, мускулатура крыла и ноги сходны с таковыми аистов. Ноги длинные; особенно длинна цевка. Направленные вперед относительно короткие пальцы соединены хорошо развитой плавательной перепонкой. Задний палец мал и прикрепляется выше, чем передние (у некоторых видов полностью редуцируется). Когти тупые. Пневматичность скелета хорошо развита. Есть разделенный на несколько камер подкожный шейный воздушный мешок. Мускульный желудок довольно объемист и имеет мощную мускулатуру. Имеются парные функционирующие слепые кишки. Есть рудимент пениса. Копчиковая железа довольно крупная, оперенная. Нижняя гортань трахеобронхиального типа. Голос напоминает гоготанье гусей.
Структура контурных перьев сходна с таковыми аистов. Окраска взрослых преимущественно белая, с более или менее развитым розовым оттенком, или красная. Она обусловлена пигментом ному пигменту ракообразных. Окраска молодых птиц бурая. Первостепенных маховых 12, рулевых 12—16. При линьке первостепенные маховые выпадают одновременно, и птицы на 2—3 недели теряют способность к полету. Полет активный, с довольно частыми взмахами коротких крыльев. Летят, вытянув шею вперед. Ведут дневной образ жизни, держатся стаями. В тропиках оседлы, в умеренных широтах перелетны.
Гнездятся колониями, обычно на мелководьях солоноватых водоемов и лагун. Отдельные колонии, например в некоторых районах Африки, даже сейчас насчитывают десятки и сотни тысяч гнездящихся пар. Гнезда располагаются в непосредственной близости одно от другого и, представляют собой столбообразные кучи из ила и ракушечника высотой до 50—60 см. В период насиживания водоемы мелеют, и гнезда могут оказаться на суше. Внезапньк^подъемы воды часто приводят к массовой гибели кладок. Из года в год гнездятся
астаксантином, близким к крас-
Рис. 53. Розовый фламинго Phoeni-сор terus ruber L.
125
обычно на одних и тех же участках, но гнезда ежегодно строят заново. В кладке 1—2, редко 3 относительно крупных яйца. В насиживании, которое продолжается немногим более месяца, участвуют оба партнера. Птенцы вылупляются зрячими, покрытыми густым коротким светлым пухом, с прямым клювом. Через несколько дней оставляют гнездо и бродят поблизости. В возрасте около 2 недель клюв начинает постепенно изгибаться. К началу второго месяца жизни птенцы одевают второй пуховой наряд, обычно серого цвета и лишь позже начинает развиваться оперение. Способными к полету становятся на 65—75-й день жизни; к этому времени полностью формируется фильтрационный аппарат клюва. Окончательный наряд взрослых одевают на 3-м году жизни, вероятно, одновременно с наступлением половозрелости (но по некоторым данным впервые приступают к размножению лишь на 5—6-й год жизни). Продолжительность жизни в природе не установлена; в неволе погибают довольно быстро)
Кормятся на мелководьях, фильтруя воду и ил. Питаются преимущественно разнообразными мелкими ракообразными, червями, моллюсками, планктонными водорослями. Птенцов выкармливают отрыжкой из пищевода, содержащей, помимо полупереваренной пищи, еще слизистый секрет стенок пищевода, выпот лимфы и крови. Подросшие птенцы ищут корм сами, но родители продолжают их кормить почти до взлета.
Семейство Phoenicopteridae включает 3 рода (Phoenicopterus, Phoeni-conaias, Phoenicoparrus) c 4—6 видами, очень спорадично гнездящимися в тропической и субтропической зонах и лишь кое-где проникающих в умеренные широты (см. рис. 50). В СССР встречается и в очень ограниченных районах гнездится только 1 широко распространенный вид Phoenicopterus ruber L.; его ареал практически совпадает с ареалом всего семейства.
Начиная с верхнего эоцена (Англия) и в более поздних отложениях в разных районах ареала семейства обнаружены остатки, относимые к 11 видам (в том числе 2 ныне живущих).
Кроме этого, к подотряду фламинго относят еще 3 давно вымерших семейства (9—11-е): Agnopteridae, Palaelodidae, Scaniornithidae. Включаемые сюда более 10 видов описаны по ископаемым остаткам разной сохранности из верхнего мела нижнего палеоцена США, нижнего эоцена — верхнего миоцена Европы (Англия, Франция, Германия, СССР). Это примитивные формы, видимо, сохранявшие большее, чем современные фламинго, сходство с аистами (более прямой или совсем прямой клюв, более короткие ноги с более длинными пальцами и т. п.).
Взгляды на положение группы фламинго в системе класса птиц весьма противоречивы. Отчетливое сходство с гусеобразными (строение клюва и языка, редукция заднего пальца и наличие плавательных перепонок, одновременная линька маховых и др.) позволяло включать их в отряд гусеобразных в ранге подотряда. С другой стороны, значительно большее сходство с аистами и колпицами (форма клюва птенцов при вылуплении, затылочные фонтанели, форма и строение грудины и таза, особенности мускулатуры и т. д.) давало основание включать фламинго в отряд аистообразных (голенастых) в ранге подотряда. Часть систематиков решали эту проблему, выделяя фламинго в самостоятельный отряд, стоящий между отрядами аистообразных и гусеобразных. Сейчас, пожалуй, более распространено мнение, что многие морфологические особенности фламинго говорят о несомненном их родстве, хотя и относительно давнем, с аистообразными. Черты сходства с гусеобразными рассматривают как конвергентно выработавшиеся адаптивные признаки. Кстати, о большей близости фламинго к аистообразным и несколько большем их удалении от гусеобразных говорят и результаты серологических и иммунологических исследований.
По представлениям многих систематикой (Фюрбрингер, Гадов и др.), аистообразные, гусеобразные, веслоногие и дневные хищники имеют очень давние, но несомненные родственные связи. Можно предполагать, что все эти группы птиц возникли от близко родственных предковых форм.
126
ОТРЯД 17. AN SERIFORMES— ГУСЕОБРАЗНЫЕ
Объединяет 2 довольно резко различающихся по особенностям строения и экологии подотряда: паламедей и собственно гусиных, поэтому общая характеристика отряда несколько нечеткая.
Размеры от мелких до крупных (вес от 200 а до 8 —10 кг). Большая часть клюва образована предчелюстными костями; верхнечелюстная кость мала. Череп десмогнатического типа. Хорошо развиты базиптергоидные сочленения и рг. angularis posterior нижней челюсти. Шейных позвонков 16— 25. Истинных ребер 6—7 пар. У рецентных форм по заднему краю грудины одна пара вырезок. Базальные концы коракоидов не соприкасаются. Мускульный желудок с мощными стенками и прочной кутикулой. Довольно хорошо развиты функционирующие парные слепые кишки. Желчный пузырь объемистый. Копчиковая железа есть. Сонные артерии парные. Есть 2 пары трахейно-грудинных мышц.
Контурные перья без побочного ствола или он сильно редуцирован. Шейная аптерия отсутствует. Пух обильно и равномерно покрывает все тело. Крыло диастатаксическое. Первостепенных маховых 11, рулевых 12—24. Преимущественно растительноядны, но часть более или менее животнояд-ны. Моногамия выражена у разных видов в очень различной степени. Птенцы в момент вылупления зрячие, хорошо опушенные. Кормятся птенцы сами; сопровождающая выводок взрослая птица их греет, охраняет и в какой-то степени помогает в поисках пищи.
Всего в отряде 43 рода и 151—154 вида.
ПОДОТРЯД 1. ANHIMAE — ПАЛАМЕДЕЕВЫЕ
Грузные крупные птицы (вес 2—4 кг), по внешнему облику несколько похожие на куриных (рис. 54). Клюв короткий, слегка изогнутый (похож на клюв куриных), с гладкими краями (без окаймляющих роговых пластинок или зубцов) и плотной рамфотекой. Затылочные фонтанели отсутствуют. Шейных позвонков 19—20. Ребра не имеют крючковидных отростков. На первой и второй метакарпальных костях расположены 2 мощных костных
Рис. 54. Рогатая паламедея Anhima cornuta (L.)
127
Рис. 55. Ареалы семейств гусеобразных птиц: 1— паламедеевые Anhimidae, 2— утиные Anatidae
шипа, покрытых плотными, с острой вершиной роговыми чехлами; их длина достигает 10 см. Птица бьет врага крылом, вооруженным таким сильным оружием. Ноги довольно длинные и толстые; нижняя часть голени не оперена. Направленные вперед 3 пальца ноги относительно длинные и не соединены плавательной перепонкой: она заметна лишь в основании пальцев. Довольно длинный задний палец находится на уровне передних. Скелет сильно пневматичен; под кожей есть воздушные полости. Подкожного жира нет. Железистый желудок объемистый; мускульный желудок мал, но с мощными стенками и прочной кутикулой. Пениса нет. Копчиковая железа маленькая, оперенная.
Контурное перо равномерно покрывает все тело: не выражены птери-лии и аптерии. Оперение не очень плотное. Крылья с относительно широкой вершиной. Линька маховых проходит постепенно, без потери способности к полету. Хвост длинный, из 12—14 рулевых. Самцы и самки окрашены сходно. Полет довольно быстрый, маневренный; во время тока совершают сложные воздушные маневры: подъем на большую высоту, парение с криком, пикирование и т. п. Хорошо ходят и бегают. Не плавают и не ныряют.
Моногамы. Гнездо — большая куча сухих стеблей — устраивается в густых зарослях надводных растений на мелководье крупных болот или на берегах водоемов. В кладке 2—6 желтовато-белых яиц. В насиживании, продолжающемся около 6 недель, участвуют оба партнера. Вылупившиеся птенцы покрыты сероватым пухом и по поведению похожи на гусят. Вне периода размножения держатся небольшими стайками. Оседлы. Питаются преимущественно зелеными частями растений; случайно поедают и разнообразных животных: насекомых, моллюсков, червей, земноводных и т. п.
В подотряде единственное семейство (1-е) Anhimidae с 2 родами (Anhi-та и Chauna) и 3 видами, распространенными в Южной Америке от Гвианы и Венесуэлу до Аргентины (рис. 55). Держатся по окраинам обширных болот, по берегам водоемов, по заболоченным опушкам леса.
В ископаемом состоянии не обнаружены. По Гадову (1891), из 40 рассматриваемых им морфологических признаков у паламедей и собственно гусиных сходны 30. Родство этих двух подотрядов очень давнее, но несомненное. Может быть, обособились от предковых форм вскоре после разделения на гусеобразных и аистообразных.
ПОДОТРЯД 2. ANSERES — ГУСИНЫЕ, ИЛИ ПЛАСТИНЧАТОКЛЮВЫЕ
Околоводные и водные птицы разнообразных размеров (вес от 200 г до 8—10 кг) с характерным обликом лебедей, гусей и уток.
Клюв обычно более или менее уплощенный, с утолщением рогового покрова на конце надклювья (ноготок) и относительно мягким роговым покровом на значительной части клюва, богатым осязательными тельцами. Края надклювья и подклювья несут поперечные роговые пластинки; вместе с пластинками, расположенными по бокам мясистого языка, они образуют фильтровальный аппарат. У некоторых видов (крохали Mergus) роговые пластинки краев клюва превращаются в острые, направленные назад роговые зубчики. У большинства видов отчетливо заметны затылочные фонта-нели. У многих видов (более или менее связанных с морями) хорошо развиты надорбитальные носовые железы. Шейных позвонков 16—25. Ребра несут хорошо развитые крючковидные отростки. На рудименте первого пальца крыла может быть коготок, у нескольких видов выдающийся из оперения. У большинства видов ноги относительно короткие. Направленные вперед 3 пальца обычно соединены хорошо развитой плавательной перепонкой. Задний палец мал, расположен выше уровня передних пальцев и имеет более или менее развитую собственную кожистую оторочку. Пневматичность скелета заметно варьирует у разных видов. Подковных воздушных полос
129
тей нет. При высокой упитанности под кожей откладываются слои жира.
Пищевод легко растяжим. Железистый желудок довольно мал. Мускульный желудок объемистый, с мощными стенками и очень плотной кутикулой. Размеры слепых кишок сильно варьируют: они велики у преимущественно растительноядных форм, малы у животноядных. У самцов в стенке клоаки есть длинный, выворачивающийся наружу пенис. Копчиковая железа крупная, оперенная. У многих видов уток в нижней гортани образуется окостеневающее вздутие — барабан, резонирующий звук. У других видов (лебеди, полулапчатый гусь) очень длинная трахея образует петли, лежащие перед грудиной, в полости киля или на поверхности грудных мышц; эти петли трахеи также выполняют функцию резонатора.
Контурные перья относительно короткие, с сильно изогнутыми стержнями и хорошо развитой пуховой частью. Оперение плотное, прилегающее. Есть широкие птерилии и узкие аптерии. Пух густо и равномерно покрывает все тело. Крылья заостренные. Линька маховых за немногими исключениями проходит одновременно, с потерей способности к полету на 2—5 недель. У многих видов резкий половой диморфизм: самцы крупнее самок и значительно ярче окрашены. В этом случае после спаривания и яйцекладки самцы линяют и одевают на короткое время послебрачный наряд, похожий по окраске на оперение самок. Вскоре опять начинается медленная линька, и уже осенью самцы приобретают брачный наряд. Молодые птицы у большинства видов по окраске похожи на самок; у немногих видов (лебеди и др.) молодые по окраске легко отличимы от взрослых.
Полет довольно быстрый (70—90 ot/ч), не маневренный; парящим полетом обычно не летают. По суше ходят и бегают переваливающейся походкой. Все виды хорошо плавают, многие легко ныряют.
Как правило, моногамы. У части видов (гуси, лебеди) пары сохраняются длительное время; насиживает только самка, а самец охраняет район гнезда. Выводок водят оба партнера. У многих уток пары образуются только на период спаривания и яйцекладки: насиживает и водит выводок только самка, а самцы после завершения яйцекладки собираются в стаи и откочевывают в районы линьки. После завершения кладки самка выщипывает с поверхности брюшка пух, выстилая им лоток гнезда и прикрывая яйца сверху при уходе на кормежку. Гнезда обычно размещаются вблизи водоемов на земле, часто под защитой кустика травы, кочки и т. п. Некоторые виды гнездятся в разнообразных укрытиях. Яйца обычно одноцветные: желтоватые, голубоватые, зеленоватые и т. п. У гусей и лебедей в кладке 4—6, у уток 5—12 яиц. Обсохнувшие птенцы’уже на следующий день после вылупления хорошо бегают и плавают, у многих видов легко ныряют. Покинув гнездо, выводок в него больше не возвращается. Корм птенцы добывают сами. Держащиеся с птенцами взрослые птицы (оба партнера или только самка) лишь водят выводок, защищают его и временами греют птенцов. На крыло поднимаются в возрасте 1—3 месяцев. Половозрелыми становятся,в возрасте около года, крупные виды в возрасте 2—4 лет. Вне сезона размножения обычно держатся стайками. В тропиках и субтропиках оседлые и кочующие, в умеренных и северных широтах перелетные.
Все виды — объекты спортивной, местами промысловой охоты. В большинстве районов мира в последнее время численность гусеобразных резко снижается из-за неумеренной добычи, изменения ландшафтов в местах гнездования и зимовок в результате хозяйственной деятельности человека, загрязнения воды и т. п.
Очень гомогенный подотряд, включающий только 1 ныне живущее семейство (2-е) Anatidae, чем подчеркивается однородность группы, с 41 родом и 148—151 ныне живущими видами, распространенными практически по всему земному шару (рис. 55). На территории СССР отмечено 55 видов, из которых несомненно гнездится 48 (при несколько иной оценке видовой самостоятельности встречается 58 и гнездится 50 видов).
130
Семейство утиных1 подразделяется на 3 подсемейства; роды крупных подсемейств группируются в трибы.
ПОДСЕМЕЙСТВО 1. ANSERANATINAE —ПОЛУЛАПЧАТЫЕ ГУСИ
В подсемействе единственный вид — полулапчатый гусь Anseranas semipalmata (Lath.), встречающийся в южной части Новой Гвинеи и северных районах Австралии.
Крупная птица (самец весит более 3,5 кг, самка около 2,5 кг) с длинной шеей, широкими крыльями, длинными ногами (рис. 56). Плавательные перепонки соединяют только основные фаланги направленных вперед пальцев. Цевка сетчатая. В году одна или две линьки. Смена маховых постепенная, без потери способности к полёту. Полового диморфизма окраски нет. По многим мелким деталям строения близки к гусям, но, видимо, более примитивны. Проявляются некоторые черты сходства с аистообразными и, более отчетливо, с паламедеями. Видимо, боковая ветвь, обособившаяся от предковых форм подотряда.
Держатся по заросшим берегам пресных и соленых водоемов, по обширным болотам. Летают мало. Обычно не плавают, не ныряют. Для отдыха часто садятся на деревья. Иногда гнездятся группами до нескольких десятков гнезд. Чаще при одном самце держатся две самки, откладывающие яйца в общее гнездо. Гнездо — большая куча грязи и растительного мусора — строится в зарослях у берега водоема. Каждая самка откладывает от 4 до 8 белых, относительно крупных яиц (вес каждого около 120 г). Насиживают и заботятся о птенцах и самки, и самец. Насиживание около 4 недель. На следующий день после вылупления птенцы покидают гнездо и кочуют вместе со взрослыми по прибрежным зарослям. Птенцы кормятся сами и, кроме того, их кормят взрослые птицы. Оперяются и взлетают в возрасте около 10 недель.
Питаются преимущественно растительной пищей: семенами, корневищами, луковицами, зелеными частями растений, прибрежными водорослями и т. п. Животную пищу поедают случайно.
Рис. 56. Полулапчатый гусь Anseranas semipalmata
____________ (Lath.)
1 Система семейства Anatidae дается по работе Delacour and Mayr. The Family Anatidae. Wilson Bulletin. Band. 57, 1945.
131
ПОДСЕМЕЙСТВО 2. ANSERINAE - ГУСИНЫЕ
Цевка сетчатая хотя бы спереди. У большинства видов только одна линька в году (послегнездовая), у немногих две: полная послегнездовая (летняя) и частичная осенне-зимняя. При летней линьке маховые сменяются одновременно с потерей способности к полету на 3—5 недель. Полового диморфизма окраски обычно нет. Зеркальца на крыле нет. Выводок водят оба партнера. Многие виды преимущественно растительноядны.
В подсемейство входят 5 родов с 29 видами, объединяемые в 2 трибы. Распространены повсюду, кроме Антарктики. В СССР встречаются 16 видов лишь одной трибы, из которых 14 несомненно гнездятся.
1 .Триба. Dendrocygnini — древесные утки
Сюда включается только один род Dendrocygna с 8 видами, распространенными в тропиках Африки, Юго-Восточной Азии и Америки (к югу от
Рис. 57. Древесная утка Dendrocygna viduata (L.) (1/$ нат. вел.)
Мексики), есть в Австралии.
Птицы средних размеров (вес 0,5—1,5 кг), короткохвостые, относительно длинноногие, с несколько вертикальной постановкой туловища (рис. 57). Голос — своеобразный высокий свист. Держатся по мелководьям и берегам водоемов преимущественно в лесной зоне. Пища главным образом растительная; различные беспозвоночные в питании играют второстепенное значение. Гнездятся в лесу вблизи водоемощ Гнездо помещается укрыто: в дупле, в углублениях между корнями, в густых мутовках ветвей и т. п. Обычно пуховой выстилки не бывает. В кладке 8—12 яиц. Летают мало, прекрасно плавают и ныряют.
Рис. 58. Серый гусь Anser anser (L.) Г/в нат. вел.)
2 .Триба. Anserini — гуси и лебеди
Включает 21 вид 4 родов: Anser, Branta, Cygnus, Coscoro-ba. Роды Chen, Cygnopsis, Eu-labeia и Philacte сейчас обычно включают в род Anser (иногда сохраняя за ними ранг подрода); род Rufibrenta включается в род Branta. В СССР встречаются 16 видов 3 родов; достоверно гнездится 14 видов. Крас-нозобая казарка Branta ruficollis (Pall.) — наш эндемик, гнездится на Ямале и Таймыре.
Птицы средней и крупной величины (вес от 1 до 8—10 кг), распространенные практически
132
по всему земному шару, кроме Антарктики (и районов безводных пустынь). Имеют характерный облик гуся или лебедя (рис. 58).
Обычно живут на крупных водоемах. Пары сохраняются длительное время, иногда до гибели одного из партнеров. Гнезда либо на суше (тогда это лунка в земле, выстланная растительной ветошью и пухом), либо в зарослях на мелководье (большая куча стеблей с лункой на вершине, выстланной пухом). В кладке 3—5, редко 8 и больше яиц. Насиживает толька самка (у некоторых лебедей самец и самка); самец держится в районе гнезда и охраняет его. Выводок водят оба партнера. Семья сохраняется и после подъема молодых на крыло (у некоторых видов до конца зимы). У лебедей молодые по окраске отличаются от взрослых. Половозрелость наступает на 2—4-й год жизни.
Питаются преимущественно растительной пищей: семенами и зелеными частями наземных растений, побегами и прикорневыми частями водных растений ит. п.; животная пища обычно имеет второстепенное значение. Кормятся на суше и на водоемах. Как правило, не ныряют. Пищу на дне водоема добывают, погружая в воду шею, реже и переднюю часть туловища.
Все породы домашних гусей созданы человеком в результате одомашнивания, отбора и последующей гибридизации двух диких видов: серого гуся Anser anser (L.) и гуся сухоноса A. cygnoides (L.), и сейчас живущих на территории нашей страны.
ПОДСЕМЕЙСТВО 3. ANATINAE - УТИНЫЕ
Цевка спереди, хотя бы в нижней части, не сетчатая, а покрытая поперечно-вытянутыми роговыми пластинками. У всех видов в году 2 линьки: полная летняя (послегнездовая) и частичная осенне-зимняя. Сменяющиеся при полной линьке маховые выпадают одновременно, с потерей способности к полету на 2—т4 недели. Половой диморфизм у большинства видов хорошо выражен, самцы несколько крупнее самок и окрашены значительно ярче. Многие виды имеют на крыле зеркальце. У всех видов, кроме земляных уток, выводок водит только самка. Питание обычно смешанное; часть видов преимущественно животноядны.
В подсемейство утиных включают 118—121 вид 35 родов, распределяемых по 7 трибам. Распространены по подходящим местообитаниям практически всего земного шара, кроме ледового щита Антарктики. В СССР встречается 39 видов 13 родов; несомненно гнездится 34 вида. В нескольких случаях подвиды принимаются рядом систематиков за самостоятельные виды; тогда число встречающихся в СССР видов возрастает до 42, а гнездящихся до 36.
1 .Триба. Tadornini — земляные утки
К земляным уткам относят 20 видов, распределенных по 8 родам: Tadorna (Casarсо), Aloppchen, Cereopsis, Chloephaga, Cyanochen, Lophonetta, Neochen, Tachyeres. Распространены преимущественно в тропической и субтропической зонах всего земного шара. В СССР отмечено 3 вида 1 рода. Гнездятся в южных районах СССР 2 вида: пеганка Tadorna tadorna (L.) и красная утка, или огарь, Т. ferruginea Pall. Хохлатая пеганка Т. cristata (Kuroda) известна лишь по 4 коллекционным экземплярам (3 с Корейского полуострова и 1 —, из окрестностей г. Владивостока; рис. 59). Видимо, полностью исчезла в начале XX в., хотя иногда появляются сообщения о встречах этого вида, не подтвержденные коллекционным материалом; например, в 1964 г. самца и двух самок якобы видели в заливе Петра Великого.
Птицы весом от 1 до 3 кг. По внешнему облику представляют нечто среднее между гусями и утками (отсюда и иногда применяемое название по-лугуси). Клюв довольно короткий, иногда широкий, с хорошо развитым ноготком на конце. Относительно длинноноги. Задний палец мал и не не-
133
Рис. 59. Хохлатая пеганка Tadorna cristata (Kuroda)
сет кожистой оторочки. Половой диморфизм окраски выражен различно: у одних видов отсутствует, у других резкий.
Приурочены преимущественно к морским побережьям и берегам крупных, в том числе и солоноватых водоемов. Значительная часть видов гнездится в норах (либо вырытых в обрывах самими птицами, либо в норах млекопитающих, например сурков и др.), в промоинах берегов, расщелинах и т. п. (поэтому всю трибу называют земляными утками), часто на расстоянии нескольких километров от берегов ближайшего водоема. Часть видов гнездится на земле, под защитой куста, кочки и т. п. По земле ходят легко и быстро. Птенцы значительно лучше птенцов других групп уток ходят и бегают по суше, поэтому без особого труда переходят из гнезда на водоем и с одного водоема на другой. В глубине норы из растительной ветоши устраивается лоток гнезда; позже он обильно выстилается пухом. В кладке 8—12 яиц. Обычно насиживает только самка. Выводок водят оба партнера.
Питаются разнообразной животной и растительной пищей, которую собирают на мелководье, погружая в воду шею и переднюю часть туловища. Обычно не ныряют. Виды р. Tachyeres плохо или совсем не летают; они и виды р. Lophonetta хорошо ныряют. Виды родов Cereopsis и Chloephaga в Австралии и Южной Америке экологически замещают настоящих гусей и похожи на них по внешнему облику; они ведут более наземный образ жизни, преимущественно растительноядны. Во времена фараонов был одомашнен египетский гусь Alopochen aegyptiacus (L.); сейчас эта порода исчезла, хотя предковая форма в небольшом количестве продолжает жить в долине р. Нила.
2 .Триба. Anatini — речные утки
Сюда включают 6 родов — Anas, Hytnenolaimus, Malacorhynchus, Мег-ganetta, Rhodonessa, Stictonetta — c 42^видами; из них 37 относят к р. Anas, остальные роды монотипичные. В СССР встречаются 12 видов рода Anas’, достоверно гнездится 11 видов. Чирок клоктун— A. formosa эндемик, гнездящийся только в северо-восточной Сибири (от Енисея до Камчатки).
Типичные утки (рис. 60) мелкой и средней величины (вес 200 г — 1,5 кг, редко больше). Клюв довольно длинный и широкий, сильно уплощенный, с очень маленьким ноготком, занимающим лишь часть переднего конца надклювья. Роговые пластинки по краям клюва тонкие, довольно высокие, густо расположенные. Задний палец мал, его кожистая оторочка слабо развита. Резко выражен половой диморфизм окраски.
134
Рис. 60. Чирок клоктун Anas formosa Georgi
Встречаются на разнообразных, преимущественно пресных внутренних водоемах, хотя многие виды гнездятся и у солоноватых водоемов и по морским побережьям. Распространены практически по всему земному шару, кроме Антарктики. Пара сохраняется лишь на период спаривания и яйцекладки. Насиживает и водит выводок только самка. Гнездятся вблизи водоемов, обычно на земле, лишь изредка в дуплах, низ'йо над землей в развилках ветвей, в брошенных гнездах других птиц. Лоток выстилают растительной ветошью, а после завершения кладки — пухом. В кладке 6— 12 яиц. Насиживание 3—4 недели. Птенцы поднимаются на крыло в возрасте 1—2 месяцев.
Питание разнообразное. Процеживая воду и ил на мелководье, добывают мелких разнообразных беспозвоночных и водоросли; поедают проростки, клубеньки и мягкие корневища водных растений и т. п.; рыбу ловят лишь случайно. При добывании пищи не ныряют, погружая в воду лишь шею и переднюю часть туловища. Кормятся и на суше; собирают желуди, поедают пешую саранчу, упавшие колосья на полях и т. п.
Обыкновенная кряква — Anas platyrhynchos L. — родоначальник всех пород домашних уток Старого Света.
3 .Триба. Somaterini — гаги
Два рода (Somateria, Polysticta) с 4 видами, распространенными по морским побережьям преимущественно высоких широт северного полушария; гнездятся и по озерам вблизи морских побережий. В северных районах СССР встречаются и гнездятся все 4 вида.
По облику (рис. 61) —типичные утки. Вес от 0,5 до 3 кг. Клюв сильный, слабо уплощенный, с мощным и широким ноготком на конце; роговые пластинки по краям клюва грубые, не высокие. Задний палец окаймлен широкой кожистой лопастью.
Рис. 61. Самец обыкновенной гаги Somateria mollissima (L.)
1 Трибу Somaterini иногда включают в состав трибы морских уток Mergini.
135
Гнездо — лунка в земле, выстланная растительной ветошью и пухом, обычно под защитой кочки, камня и т. п. Иногда бывает групповое гнездование, при котором расстояние между гнездами может быть менее 1 м. В кладке 4—8 яиц. Насиживает только самка около 44/2 недель. Через несколько часов после вылупления самка уводит выводок на ближайший водоем. Обычно немного подросшие выводки с пресных озер перемещаются на морские побережья.
Пищу добывают преимущественно во время ныряния. Видимо, способны нырять на глубину более 10 м, находясь под водой 1—2 мин. Пища главным образом животная: больше всего поедают разнообразных моллюсков, в том числе и прикрепляющихся к скалам. Их отрывают при помощи мощного ноготка клюва; раздавливанию раковины помогают грубые пластинки краев клюва. Едят ракообразных, червей и т. п., иногда в небольшом количестве ловят мелкую рыбу, поедают водные растения (морскую траву, водоросли, на пресных водоемах — мягкие корневища, клубеньки, проростки).
В ряде районов (особенно в Исландии) до сих пор существует промысел гагачьего пуха (собирается пух из гнезд после вылупления птенцов).
4.Триба. Aythyini — нырковые утки
2 рода — Aythya, Netta — с 15 видами, распространенными преимущественно в умеренных и холодных широтах по всем материкам. В СССР встречаётся и гнездится 6 видов обоих родов.
Грузные, коротконогие утки (рис. 62); вес 0,5—1,5 кг. Клюв длинный, широкий, сильно уплощенный. Роговые пластинки по краям клюва высокие, часто расположенные. Короткий задний палец окаймлен широкой кожистой лопастью.
Гнездятся по берегам крупных озер с пресной и соленой водой и хорошо развитой водной и надводной растительностью. Гнезда скрыты среди растений и размещены вблизи воды; часто устраивают гнезда на сплавинах, заломах тростника и т. п. В особо благоприятных условиях отмечено групповое гнездование, когда гнезда расположены в нескольких метрах одно от другого. В кладке 6—10, иногда больше, яиц. Насиживает и водит выводок только самка. Инкубация около 4 недель. Птенцы начинают хорошо летать в возрасте 1,5—2 месяцев.
Пищу добывают преимущественно при нырянии, реже кормятся, опустив в воду шею и переднюю часть туловища. Питание разнообразное. Часть видов преимущественно растительноядна: поедают водоросли, проростки,
Рис. 62. Самец хохлатой чернети Aythya fuligula (L.)
136
клубеньки, корневища, семена и нежные листья различных водных растений. Другие виды преимущественно животноядны; питаются главным образом водными личинками насекомых, моллюсками, ракообразными, червями; мелкую рыбу поедают более или менее случайно.
5 .Триба. Cairini — шипуны
Объединяет 7 родов — Cairina, Aix, Amazonetta, Chenonetta, Nettapus, Ple-ctropterus, Sarkidiornis, — c 12 видами, распростра
Рис. 63. Самец мандаринки Aix galericulata L.
ненными преимущественно
в теплых широтах Афри-
ки, Юго-Восточной Азии, Австралии, Северной и Южной Америки. В СССР гнездится (в Приморье) только 1 вид — мандаринка Aix galericulata L. (рис. 63).
В русской литературе часть видов иногда называют древесными утками (Aix, Amazonetta, Chenonetta, Nettapus), так как они обычно гнездятся в дуплах, для отдыха садятся на крупные ветви деревьев. Остальные виды именуют мускусными утками (Cairina), гребенчатыми утками (Sarkidiornis) и шпорцевыми гусями (Plectropterus, рис. 64). У последних довольно
Рис. 64. Шпорцевый гусь Plectropterus gambensis (L.), наверху сгиб крыла с когтем
137
большой коготь первого пальца кисти заметно выдается из оперения. Южноамериканская мускусная утка одомашнена; выведено несколько культурных пород.
Птицы с типичным обликом коротконогих уток (первые 5 родов) или более длинноногие, несколько похожие на гусей (Plectropterus, Sarkidiornis). Размеры от мелких до крупных: вес от 0,5 до 5 кг и даже более. Задний палец не несет широкой кожистой оторочки.
Чаще приурочены к берегам лесных ручьев, рек и озер; встречаются и на больших озерах с обильной надводной растительностью. Обычно пары распадаются к началу насиживания. Некоторые виды, например южноамериканская мускусная утка Cairina moschata (L.), — настоящие полига-мы. Мелкие виды гнездятся в дуплах, брошенных гнездах других птиц, более крупные — на земле среди густой растительности.
Питание обычно смешанное: на берегах водоемов и на мелководье поедают разнообразную животную и растительную пищу. При кормежке обычно не ныряют, опуская в воду лишь шею и переднюю половину туловища.
6 .Триба. Mergini — морские утки и крохали
Включает 16—19 видов 6 родов: Mergus, Bucephala, Clangula, Camptor-hynchus (C. labradornis Gm. — лабрадорская утка, вымерла в конце XIX в.), Histrionicus, Melanitta. Населяют берега крупных пресных водоемов и побережья морей преимущественно умеренных и высоких широт северного полушария. Лишь 1 вид крохалей обитает в Южной Америке, а другой жил на Ауклендских островах к югу от Новой Зеландии (сейчас, видимо, вымер).
На территории СССР встречается 12—15 видов 5 родов, несомненно гнездится 9—11 видов (расхождения в количестве видов определяются тем, что часть систематиков выделяет в самостоятельные виды формы, которые другие считают лишь подвидами, например Melanitta fusca deglandi, М. nigra atnericand).
По внешнему облику типичные утки. Вес от 0,5 до 2 кг. Клювы разнообразной формы: сильно уплощенные и расширенные (турпаны), относительно короткие, слабо уплощенные (гоголи, морянки, каменушки), удлиненные цилиндрические с острыми мелкими роговыми зубчиками по краям надклювья и подклювья, с большим ноготком на конце клюва (крохали, рис. 65). Задний палец несет широкую кожистую оторочку.
Пары сохраняются только на период кладки. Часть видов гнездится укрыто: в дуплах, промоинах и углублениях между корнями деревьев, в расщелинах скал, под камнями; остальные — в кочках, куртинах около-водных растений и т. п. Насиживает и водит, выводок только самка. Моло-
Рис. 65. Самец большого крохаля Mergus merganser L.
138
дые становятся половозрелыми в возрасте 1—2 лет. Прекрасно ныряют; добычу обычно ловят в толще воды и на дне водоемов. Пища преимущественно животная. Крохали питаются главным образом мелкой рыбой. Другие виды в большей степени питаются различными водными беспозвоночными (насекомыми, особенно их водными личинками, ракообразными, моллюсками, червями, при случае ловят и мелкую рыбу).
7 . Триба. Oxyurini— савки
Сюда входят 9 видов 4 родов — Oxyura, Biziura, Heteronetta, Thalas-somis, приуроченных преимущественно к солоноватым водоемам и морским побережьям тропической зоны. Немногие виды проникают на водоемы засушливых районов умеренных широт Европы, Азии, Австралии, Северной Америки. В южных засушливых районах СССР встречается и гнездится 1 вид — савка Oxyura leucocephala (Scop.).
Большеголовые, относительно короткошейные утки (рис. 66) средней величины (вес около 1 кг, у немногих видов чуть больше). Клюв умеренной длины, расширенный, с хорошо развитыми роговыми пластинками по краям. Задний палец с широкой кожистой оторочкой. Рулевые в отличие от других уток очень узкие, удлиненные, жесткие; кроющие перья прикрывают лишь основную треть хвоста. Хвост резко ступенчатый. При плаванья хвост либо лежит на поверхности воды, либо поднят почти вертикально. По земле ходят мало и неуклюже. Прекрасно плавают и ныряют.
Гнезда строят на сплавинах и заломах в густых зарослях надводных растений. Скорлупа яиц грубозернистая, обычно белая. Насиживает только самка. У южноамериканской черноголовой утки Heteronetta atricapilla Мегг. выражен гнездовой паразитизм: самка подкладывает яйца в гнезда других видов уток, чаек, пастушковых птиц. Вылупившийся птенец, обсохнув в гнезде, сразу начинает вести самостоятельную жизнь.
Пищу собирают на поверхности и в толще воды. Поедают водоросли, проростки, клубеньки, свежие листья и семена многих водных и надводных растений, ловят ракообразных, моллюсков, червей и других водных беспозвоночных, при случае мелкую рыбешку.
Довольно многочисленные, но в большинстве плохой сохранности ископаемые остатки семейства утиных в пределах современного ареала известны с конца мелового периода (сомнительные находки) и из более поздних отложений. По ним описано около 160 ископаемых видов, распределяемых между ныне живущими подсемействами и трибами.
Рис. 66. Самец савки Oxyura leucocepahla (Scop )
139
Помимо семейства Anatidae в подотряде гусеобразных Уэтмор выделяет еще 1 семейство (3-е) Paranyrocidae с 1 ископаемым видом, описанным по остаткам из нижнего миоцена Бразилии. Видимо, это какая-то примитивная нырковая утка. Вероятно, было бы правильнее придать этой пока моно-типичной ископаемой группе ранг трибы, близкой к трибам морских и нырковых уток, и включить ее в подсемейство Anatinae.
Тесные родственные связи всех групп в пределах подотряда гусиных совершенно несомненны. Также очевидны явные, хотя и более отдаленные, родственные связи между подотрядами гусиных и паламедей, что особенно отчетливо подчеркивается многими чертами сходства паламедей с полулап-чатым гусем и, в несколько меньшей степени, с другими подсемействами гусиных. Многие детали морфологического строения позволяют предполагать очень давнюю близость этого древнего отряда с аистообразными, веслоногими и дневными хищными птицами.
ОТРЯД 18. FALCONIFORMES —СОКОЛООБРАЗНЫЕ, ИЛИ ДНЕВНЫЕ ХИЩНЫЕ ПТИЦЫ
Отряд объединяет птиц разнообразной величины с характерным обликом хищника. Вес от 60 г до 12 кг. Населяют самые разнообразные ландшафты и распространены практически по всему земному шару, кроме Антарктики и некоторых океанических островов. Оседлые или кочующие, в умеренных и высоких широтах — кочующие или перелетные.
Клюв довольно короткий, но высокий у основания, с острыми режущими краями и резко загнутым книзу острым крючком на конце надклювья. Основание надклювья покрыто восковицей — участком голой, иногда ярко-окрашенной кожи, на которой расположены наружные отверстия ноздрей. Череп голоринальный, обычно десмогнатического типа; в онтогенезе проходит схизогнатическую стадию, которая сохраняется у части видов и во взрослом состоянии. Шея из 14—17 позвонков. Грудина длинная и широкая, с высоким гребнем. Коракоиды широкие. Мощная, сильно изогнутая вилочка. Ребра несут крепкие крючковидные отростки. Очень мощный скелет конечностей. Самый длинный элемент крыла — предплечье; иногда почти такую же длину имеет кисть. Таз широкий, прочный. Ноги у большинства видов умеренной длины. Всегда неоперенные длинные пальцы заканчиваются крепкими, обычно очень сильно изогнутыми и острыми когтями. Пневматичность скелета хорошо выражена. Очень мощна мускулатура груди и задних конечностей.
Сердце крупное. Функционируют две сонные артерии. Головной мозг относительно велик. Глаза очень крупные (около 1% от веса тела). Слух хороший; у части видов есть зачаток наружного уха в виде кожистой складки у края наружного слухового прохода. Язык крупный, мясистый. Пищевод легко растяжим. Зоб хорошо развит. Железистый желудок объемистый. Мускульный желудок легко растяжим, стенки его слабые. Слепые кишки рудиментарны или их нет. Копчиковая железа небольшая.
Контурные перья с хорошо развитой пуховой частью, обычно довольно плотно прилегающие. Пух растет равномерно по всему телу. У нескольких родов есть пудретки. Крыло диастатаксическое. Первостепенных маховых обычно 10; у части видов есть скрытый кроющими рудимент первого махового. Рулевых 12—14. Смена маховых идет постепенно, без заметного ухудшения способности к полету. В году обычно одна полная линька (после-гнездовая). Как правило, самки заметно крупнее самцов; у части видов окраска самцов более яркая, чем у самок. У многих видов есть возрастное изменение окраски. Большинство видов обладает стремительным, маневрен
140
ным полетом; многие способны к длительному парению. По земле обычно ходят относительно редко.
Моногамы. У многих видов пары сохраняются длительное время, иногда до гибели одного из партнеров. Немногие гнездятся группами; у большинства видов пары гнездятся изолированно и имеют хорошо выраженные гнездовые участки. Часто одно гнездо используется несколько лет. Гнездо довольно примитивное: обычно это просто куча беспорядочно набросанных веток. Гнезда размещаются на деревьях, уступах скал, в заломах тростников, на земле. Иногда занимают гнезда других птиц. У многих видов хорошо ^выражен ток: брачный полет обоих партнеров со сложными эволюциями в воздухе, с громкими криками. Крупные виды откладывают 1—2, мелкие до 4—7 яиц. У видов с большими ареалами обычно выражена географическая изменчивость размеров кладки: на севере средние размеры кладки несколько больше, чем на юге. При недостатке кормов у многих видов уменьшаются размеры кладки; в бескормном году птицы иногда совсем не приступают к размножению. Яйца откладываются с интервалом в 2—3 суток. Насиживание обычно начинается после откладки первого яйца. Поэтому и вылупление птенцов идет не одновременно. Насиживают оба партнера, но самец насиживает меньше. У многих видов самец носит корм насиживающей самке. Продолжительность насиживания 4—8 недель. Вылупляющиеся птенцы зрячие, покрытые пухом (у американских грифов — голые), но терморегуляция устанавливается медленно, и птенцы нуждаются в обогреве. Из-за разновозрастности птенцов при недостатке кормов младшие часто гибнут от голода или их съедают более крупные птенцы (гнездовой каннибализм). Выкармливают оба партнера. Птенцы покидают гнездо полностью оперенными и способными к полету в возрасте 1,5—2 (мелкие виды)— 3 и даже 4 месяцев (крупные виды). Некоторое время после вылета родители продолжает кормить птенцов. Окончательный наряд взрослых молодые у мелких видов одевают в возрасте около года, у крупных видов в возрасте 3—4 лет. В это же время становятся половозрелыми.
Способы охоты и состав пищи у разных видов широко варьируют. У части видов выработалась довольно узкая пищевая специализация. Подавляющее большинство видов питается только животной пищей. Лишь несколько видов более или менее растительноядны; поедают различные плоды. Обычно активны днем; немногие виды активны и в сумерках. Многие полезны истреблением вредителей (грызуны, насекомые и т. п.) и как санитары (поедают падаль). Даже хищники, охотящиеся на промысловые виды, оказываются полезными, устраняя из популяции больных и ослабевших особей. Сейчас во многих странах большинство видов хищных птиц охраняются специальным законодательством.
Отряд дневных хищных птиц включает 81 род с 274 ныне живущими видами и подразделяется на 2 подотряда, объединяющие 7 семейств (из них 2 включают только ископаемые формы). В СССР встречаются представители только 1 подотряда соколиных: 50 видов из 20 родов. Доказано гнездование 47 видов.
По ископаемым остаткам, начиная с нижнего эоцена, из разных пунктов современного ареала отряда описан 91 ископаемый вид; их относят к 38 родам. В ископаемых остатках обнаружено также 58 из ныне живущих видов. Судя по сравнительноморфологическому материалу, дневные хищники имеют очень давние родственные связи (и, следовательно, более или менее общее происхождение) с аистообразными, среди которых наибольшие черты сходства улавливаются с аистами; установлены также очень давние родственные связи с веслоногими и гусеобразными. Эти черты сходства с другими отрядами более отчетливо проявляются в строении представителей подотряда американских грифов. Более молодая группа дневных хищных птиц подотряд соколиных — сохраняет многие черты сходства с американскими грифами, но в значительной степени утрачивает сходство с другими более или менее близкими отрядами.
141
150° 120° S0° 60° 30° 0° 30° 60° 90° 120° 150° 180° 150°
Рис. 67. Ареалы семейств дневных хищных птиц:
1— американские грифы Cathartidae, 2— птица секретарь Sagittariidае, 3—ястребинные Accipitridae
ПОДОТРЯД 1. CATHARTAE — АМЕРИКАНСКИЕ ГРИФЫ
Крупные хищники (вес от 4 до 9—12 кг) южных районов Северной и почти всей Южной Америки (рис. 67). Голова и часть шеи голые, не оперенные; у части видов на этих участках кожа ярко окрашенная, иногда имеются причудливые кожные выросты (рис. 68). В черепе характерны функционирующие базиптеригоид-ные сочленения. Ноздри сквозные. Большой объем носовых полостей и крупные обонятельные доли головного мозга косвенно свидетельствуют о хорошем развитии обоняния. По заднему краю грудины две пары вырезок. Длинные передние пальцы ног заканчиваются тупыми, слабо изогнутыми когтями; задний палец короткий и прикрепляется к цевке несколько выше уровня передних. Нижняя гортань с замкнутыми бронхиальными кольцами и без специальной голосовой мускулатуры. Птицы немые, звуков не издают. В желудке более или менее четко обособлен пилорический отдел. Сле-
Рис. 68. Королевский гриф Sarcorhamphus papa (L.)
пые кишки полностью редуцированы. Копчико-
вая железа не оперена. Контурные перья без следов побочных стержней. Включает 2 ископаемых и 1 ныне живущее семейство.
СЕМЕЙСТВО 1. NEOCATHARTIDAE
Из отложений верхнего эоцена Северной Америки описан 1 вид, видимо, весьма примитивного грифа, по особенностям скелета весьма близкого к аистам. Вероятно, летал плохо, но хорошо бегал. Возможно, довольно близок к предковым формам подотряда. Иногда это семейство выделяют в надсемейство Neocathartoidea.
СЕМЕЙСТВО 2. CATHARTIDAE — АМЕРИКАНСКИЕ ГРИФЫ
Включает 6 видов, относимых к 5 родам: Cathartes, Coragyps, Gytnno-gyps, Sarcoramphus, Vultur.
Моногамы. Гнезда обычно на уступах скал, обрывах, реже на деревьях. В кладке 2—3 яйца, чаще белых, иногда со слабым крапом. Насиживание около 8 недель. Птенцы вылупляются зрячие, но совершенно голые; лишь потом одевают птенцовый пух. Взрослые приносят корм птенцам в зобу.
Питаются различной падалью, которую разыскивают в продолжительном парящем полете. Живую добычу практически не ловят. На трупах крупных животных собираются группами. Летают хорошо, особенно парящим полетом. Довольно легко ходят по земле. Самые крупные из ныне живущих видов — калифорнийский кондор Gymnogyps californianus (Shaw.) и андский кондор Vultur gryphus L. весом 12 кг и до 3 м в размахе крыльев. Сейчас численность большинства видов катастрофически сократилась. Так, калифорнийского кондора в 1965 г. насчитывали 30—35 экз.
По ископаемым остаткам из отложений среднего эоцена в разных пунктах современного ареала и из Западной Европы описано 7 родов с 13 видами. В пределах современного ареала обнаружены и остатки 4 ныне живущих видов. Место возникновения, видимо, Северная Америка (хотя находки во Франции более древние).
143
СЕМЕЙСТВО 3. TERATORNITHIDAE
По остаткам из верхнего плейстоцена Северной Америки (Флорида, Калифорния) описано 2 рода с 3 крупными видами, вероятно, близкими к современным американским грифам. Один из них, видимо, имел до 4 м в размахе и, возможно, весил до 20 кг.
Семейства Cathartidae и Teratornithidae иногда объединяют в надсемейство Cathartoidea.
ПОДОТРЯД 2. FALCONES —СОКОЛИНЫЕ
Очень различные по величине (вес 50 г — 12 кг) хищные птицы, распространенные практически повсеместно, кроме Антарктики.
Базиптеригоидные сочленения, как правило, функционируют только у маленьких птенцов и редуцируются у взрослых. Ноздри несквозные. По заднему краю грудины только одна пара вырезок, обычно полностью зарастающих у взрослых. Задний палец хорошо развит и прикрепляется на одном уровне с передними пальцами. Почти у всех видов когти сильно изогнутые, очень острые. Нижняя гортань с незамкнутыми бронхиальными кольцами и хорошо развитой парой голосовых мышц. Пилорическая часть желудка не обособлена. Слепые кишки очень малы, но полностью не редуцируются. Копчиковая железа оперена. Контурные перья имеют, хотя и слаборазвитый, побочный стержень (у скопы только на нижней стороне тела). Птенцы вылупляются зрячими, покрытыми густым пухом, который через 1,5—2 недели заменяется вторым пуховым нарядом, и лишь после этого начинает развиваться оперение. Пищу птенцам обычно приносят в клюве или в когтях.
Подотряд соколиных разделяют на два надсемейства (птиц секретарей и соколиных) с 4 семействами, объединяющими 76 родов и 268 ныне живущих видов. В СССР встречается 50 видов из 20 родов; достоверно гнездится 47 видов.
Надсемейство 1. Sagittarioidea — птицы секретари
Включает одно семейство.
СЕМЕЙСТВО 4. SAGITTARIIDAE— ПТИЦЫ СЕКРЕТАРИ
Единственный ныне живущий вид — птица секретарь Sagittarius ser-pentarius Mill. (рис. 69). Обитает в степных и лесостепных районах Африки (рис. 67). По внешнему виду несколько напоминает кариам, отличаясь от остальных видов подотряда очень длинными ногами (особенно удлинена цевка) и длинным хвостом. Базиптеригоидные сочленения функционируют и у взрослых. Шейных позвонков 15. Грудина к заднему краю резко суживается. Вершина вилочки срастается с вершиной киля грудины. Относительно короткие пальцы несут острые, сильно изогнутые когти.
Прекрасно летают. Легко и быстро ходят и бегают. Охотятся обычно «пешком», хватая выпугнутых из травы рептилий (в том числе и ядовитых змей), насекомых, мелких млекопитающих. Гнездо на деревьях или крупных кустах. В кладке 2, редко 3 яйца. Насиживает самка, около 6—7 недель. Кормят птенцов оба родителя. Птенцы вылетают из гнезд через 10— И недель после вылупления. Взятые из гнезд птенцы хорошо привыкают к людям. Их часто держат в африканских деревнях: они поедают заползающих в деревни змей и защищают домашних птиц от нападения пернатых хищников.
Предполагаемый центр возникновения семейства — Африка. По фрагментарным остаткам из верхнего эоцена или нижнего миоцена Франции опи-
144
Рис. 69. Птица секретарь Sagittarius serpentarius Mill.
саны 2 вида (р. Amphiserpentarius), видимо, близких современным секретарям, но отличавшихся более короткими и толстыми цевками и некоторыми другими особенностями. Может быть, однако, что эти ископаемые виды близки к кариамам. По ряду особенностей птицы секретари занимают промежуточное положение между американкими грифами и соколиными; несомненна их большая близость к последним.
Надсемейство 2. Fal соло idea — соколиные
По внешнему облику типичные дневные хищные птицы типа орла, грифа, ястреба, сокола и т. п.
Базиптеригоидные сочленения функционируют у птенцов и редуцируются у взрослых. Предлобная кость образует большой надглазничный отросток. Вершина вилочки соединяется с вершиной киля грудины эластичной связкой. К заднему краю грудина не суживается. Ноги относительно короткие. Пальцы ног довольно длинные.
Надсемейство включает 3 семейства.
СЕМЕЙСТВО 5. ACCIPITRIDAE — ЯСТРЕБИНЫЕ
Размеры от мелких до крупных: вес от 50 г до 12 кг. Клюв без отчетливо выраженного добавочного предвершинного зубца (рис. 70, /). На небе йет продольного гребня. Шейных позвонков 14 (у грифов 17). Грудные позвонки обычно не срастаются в спинную кость. Мембрана в нижней гортани развита слабо. Линька первостепенных маховых идет от 11-го к 1-му или (орлы, грифы) полицентрично. У некоторых грифов есть пудретки.
Распространены практически по всему земному шару, кроме Антарктики, некоторых океанических островов и совершенно безводных участков пустынь (см. рис. 67).
В семейство включают 208 ныне живущих видов, объединяемых в 63 рода. В СССР отмечено 37 видов 18 родов; несомненно гнездится 34 вида. Большой объем семейства вынуждает подразделять его на подсемейства;
145
Рис. 70. Головы ястреба тетеревятника Accipiter gentilis L. (?) и сапсана Falco pere-grinus Tunst. (II; хорошо виден предвершинный зубец)
из нескольких схем такого деления приведем одну, более распространенную.
Подсемейства (с перечнем входящих в них родов):
1. дымчатые коршуны Elaninae: Elanus, Chelictinia, Gampsonyx, Machaerhamphus;
2) осоеды Perniпае: Pern is, Aviceda, Chondroh ierax, Elanoides, Henicopernis, Leptodon;
3) коршуны и орланы Milvinae: Milvus, Haliaeetus, Haliastur, Hamirostra, Harpagus, Ichthyophaga, Ictinia, Lophoictinia, Rostrha-mus;
4) ястребы Accipitrinae: Acciplter, Eryt-hrotriorchis, Kaupifalco, Megatriorchis, Melierax, Urotriorchis;
5) канюки, или сарычи, Buteoninae: Buteo, Busarelleus Butastur, Buteogallus, Heterospizias, Leucopternis, Parabuteo;
6) орлы Aquilinae: Aquila (рис. 71), Harpia, Harpyhaliaetus, Harpyopsis, Hiera-eetus, Ictinaetus, Morphnus, Oroaetus, Pithe-cophaga, Polmaetus, Spizaetus, Spizastur, Stephanoaetus;
7) грифы Старого Света СураёНпае: Gy-
paetus, Aegypius, Gypoh ierax, Gyps, Necrosyrtes, Neophron, Pseudogyps, Sarcogyps, Trigonoceps, Torgos\
8) змееяды СлгсаёПпае: Circdetus, Dryotriorchis, Eutriorchis, Spilornis, Terathop ius\
9) луни Circinae: Circus, Geranospiza, Poly boro ides (Gymnogenus'}.
По ископаемым остаткам, начиная с нижнего эоцена, описано 57 видов, относимых к 24 ископаемым родам. Из ныне живущих видов в ископаемом состоянии обнаружено 40.
Ястребиные — центральное семейство подотряда соколиных, эволюи-ровавшее путем широкой адаптивной радиации. Заселяют самые разнообразные ландшафты: леса, горы, тундры, степи, пустыни. Гнезда — в зависимости от характера ландшафта и от видовой специфики — на деревьях и кустах, уступах скал и обрывов, в заломах тростников, на земле.
Довольно резко различаются по пищевой специализации и способам охоты. Практически все виды животноядны: лишь африканский пальмовый
Рис. 71. Беркут Aquila chrysaetos (L.)
146
Рис. 72. Скопа Pandion haliaetus (L.)
гриф Gypohierax angolensis L. питается преимущественно плодами нескольких видов пальм. Орлы, ястребы, луни нападают главным образом на относительно крупную подвижную добычу (птицы, млекопитающие). Канюки ловят мышевидных грызунов, насекомых, земноводных, рептилий. Осоеды питаются различными насекомыми, в том числе и личинками ос, которых добывают, раскапывая их гнезда. Коршуны — полифаги, поедающие насекомых, грызунов, слетков птиц, разнообразную падаль (в том числе и снулую рыбу) и т. д. Грифы питаются преимущественно или исключительно падалью. Для некоторых видов характерна очень узкая пищевая специализация: американский коршун слизнеед Rostrhamus sociable is (V.) питается почти исключительно моллюсками, гарпия Harpia harpy la (L.)—обезьянами и ленивцами и т. п.
Способы охоты взаимосвязаны с пищевой специализацией. Грифы разыскивают падаль, паря на большой высоте. Многие орлы, коршуны, канюки, паря в воздухе, высматривают добычу и стремительно пикируют на нее. Луни выискивают добычу, летая медленным бесшумным полетом низко над землей. Некоторые орлы подкарауливают добычу, неподвижно сидя на земле в колонии грызунов. Ястребы выслеживают добычу, сидя в укрытии (в кроне дерева на опушке и т. п.) или стремительно пролетая вдоль опушек и лесных полян, бросаются на выпугнутых животных. Многие виды ловят насекомых, медленно бродя по земле. Для видов с узким набором кормов (стенофагов) характерны относительно постоянные, мало варьирующие способы охоты. Виды с широким набором кормов (полифаги) используют более разнообразные способы охоты. Например, коршуны высматривают добычу, паря в воздухе и пикируя на нее, ходят по земле, схватывая выпугнутых насекомых, иногда сидят у нор, ожидая выхода грызунов и т. п. Крупная добыча схватывается лапами и убивается проколом когтей. Обычно ведут одиночный образ жизни. Падальники (некрофаги) и полифаги (коршуны) иногда держатся и гнездятся группами.
СЕМЕЙСТВО 6. PANDIONIDAE — СКОП ИНЫЕ
Монотипичное семейство, включающее единственный вид — скопу Pandion haliaetus (L.) (рис. 72), спорадично распространенную по крупным внутренним водоемам и морским побережьям многих районов земного шара (рис. 73). Северные популяции перелетные, южные — оседлые.
Близка к семейству ястребиных (иногда включают в него в ранге подсемейства), но занимает несколько обособленное положение по своеобразной экологической специализации и ряду мелких деталей строения. Шейных позвонков 15.
147
Рис. 73. Ареалы семейств дневных хищных птиц:
1— соколиные Falconidae, 2— скопиные Pandionidae
Наружный палец ноги очень подвижен и может быть направлен либо вперед, либо назад. Когти сильно изогнутые, тонкие и острые. Роговой покров нижней поверхности пальцев имеет вид острых шипи-ков, облегчающих удерживание рыбы. Только на нижней стороне тела контурные перья имеют небольшой побочный ствол. По некоторым особенностям строения (характер птерилозиса, строение сгибателей пальцев ноги и др.) похожа на американских грифов.
Встречается по берегам богатых рыбой водоемов с относительно прозрачной водой. Питается почти исключительно крупной рыбой, которую высматривает, паря над водой и иногда задерживаясь на одном месте в трепещущем полете; стремительно пикирует на замеченную в верхних слоях воды добычу. За счет энергии пикирования может погрузиться в воду. Добычу схватывает когтями. Гнезда вблизи водоемов на вершинах одиночных деревьев, на уступах береговых обрывов и т. п.
СЕМЕЙСТВО 7. FALCONIDAE—СОКОЛИНЫЕ
Птицы мелкой и средней величины; вес от 50 г до 2—3 кг. На надклювье четко развит предвершинный зубец (см. рис. 70, II). На твердом небе отчетливо выражен продольный гребень. Шейных позвонков 15. Грудные позвонки срастаются в спинную кость. В нижней гортани хорошо развита мембрана. Линька первостепенных маховых обычно идет от 6—7-го к 1-му и потом к 10-му.
По характеру распространения — почти космополиты (рис. 73).
Семейство объединяет 58 ныне живущих видов, распределяемых по 11 родам. В СССР встречается и гнездится 12 видов, сейчас обычно относимых к 1 роду Falco. Часто внутри семейства соколов выделяют 3 подсемейства: лесные соколы, каракары, настоящие соколы.
Лесные соколы Herpetotherinae, с 2 родами Herpetotheres и М icrastur. По внешнему облику и экологии похожи на типичных ястребов. Приспособления к лесному образу жизни: характерны короткие закругленные крылья, длинный хвост. Полет быстрый, маневренный, легко летают в чаще ветвей. Парящим полетом и на дальние дистанции обычно не летают. Питаются преимущественно позвоночными, которых ловят из засады или,
Рис. 74. Сокол сапсан Falco peregrinus Tunst.
149
застав врасплох, при вылете из-за прикрытия. Населяют тропические и субтропические леса Америки и Юго-Восточной Азии.
2. Каракары, или соколы универсалы, Daptriinae объединяют 4 рода: Daptrius, Milvago, Phalcobaenus, Polyborus (в последний род включают в качестве подродов Ibycter и Сагасага). Населяют открытые пространства и опушки лесов всей Южной Америки и юг Северной Америки. Питаются самой разнообразной животной пищей (позвоночные, насекомые и др.), поедают отбросы и падаль; несколько видов в качестве второстепенных, а иногда и основных кормов используют плоды. Способы охоты разнообразны: парение и пикирование на добычу, преследование ее в воздухе, подкарауливание, преследование пешком по земле и т. п. Некоторые виды при обилии корма могут держаться небольшими стайками. Активный полет с относительно медленными взмахами крыльев, часто парят. Хорошо бегают по земле.
3. Настоящие соколы Falconinae (рис. 74) объединяют 5 родов: Falco (более 30 видов; сейчас в этот род в ранге подродов включают ранее выделявшиеся роды — Aesalon, Cerchneis, Erythropus, Ну patriarch is, Harpe, leracidea, и др.), Microhierax, Nesierax, Poliohierax, Spiziapterax. Последние 4 рода — соколы карлики, распространены в тропиках Старого и Нового Света, род Falco — почти космополит.
Населяют разнообразные, преимущественно более или менее открытые ландшафты; охотно держатся по опушкам лесов. Питаются разнообразной животной пищей; на падаль обычно внимания не обращают. Мелкие соколки — преимущественно энтомофаги, более крупные ловят разнообразных животных: наземных позвоночных, крупных насекомых. Иногда добыча может превышать по весу добывшего ее хищника. Основной способ охоты: стремительный полет — бросок, иногда со значительного расстояния, на бегущую или летящую добычу. Полет стремительный, высоко маневренный, с частыми ударами крыльев, временами кратковременным скольжением — планированием. Почти все виды хорошо ходят по земле. Мелкую добычу хватают клювом. Крупная добыча с разлета убивается или оглушается ударом когтей задних пальцев; оглушенная жертва умерщвляется перекусыванием шеи или затылочного отдела черепа. Некоторые, преимущественно насекомоядные виды могут гнездиться небольшими группами.
Относительно молодое семейство, обособившееся от ястребиных. По ископаемым остаткам, в большинстве плохой сохранности, описано 15 вымерших видов (наиболее древний — из верхнего олигоцена Патагонии), видимо, довольно близких к современным формам. Из ныне живущих видов в ископаемом состоянии обнаружено 12.
ОТРЯД 19. GALLIFORMES —КУРООБРАЗНЫЕ
Очень разнообразные по размерам (вес от 45 г до 8—10 кг), наземные или наземно-древесные птицы характерного куриного облика. Населяют разнообразные ландшафты почти всего земного шара, кроме Антарктики и некоторых океанических островов. Оседлые или кочующие: часть видов в умеренных и северных широтах перелетные. Почти все виды — объекты спортивной, местами и промысловой охоты. В связи с резким изменением ландшафтов и неумеренной охотой численность многих видов сильно уменьшилась, ареалы — сократились.
Клюв короткий, выпуклый, с более или менее крючковато загнутой тупой вершиной надклювья. Череп схизогнатического типа, со слабым палочковидным сошником и коротким клювовидным отростком парасфеноида. Ноздри сквозные, голоринальные. Шейных позвонков 16—19. Есть спинная кость. По заднему краю грудины обычно две пары глубоких вырезок. Задний палец ноги есть, хотя может быть очень маленьким. Ноги сильные. Четко обособлен от пищевода объемистый зоб. Слепые кишки более или
150
менее развиты. Есть желчный пузырь. Нижняя гортань трахео-бронхиаль-ного или трахеального типа. У большинства групп две сонные артерии.
Оперение плотное. Контурное перо с более или менее развитым побочным стержнем. Первостепенных маховых 10. Крылья короткие, широкие. Рулевых от 10 до 24. У многих видов четко выражен половой диморфизм: самцы крупнее самок, окраска ярче.
Отряд включает 85 родов с 253 ныне живущими видами, объединяемые в 8 семейств. Выделяют 2 подотряда, отчетливо различных по морфологическим и экологическим особенностям.
Куриные, видимо, древняя, довольно четко обособленная группа птиц, сохраняющая в какой-то степени известную примитивность. Родственные связи с другими отрядами птиц неясны. Возможно, по очень давним родственным отношениям они в какой-то степени близки к отряду журавлеобразных. Ископаемые остатки из мезозоя отсутствуют, но есть из эоцена.
ПОДОТРЯД 1. GALLI — КУРИНЫЕ
Размеры от мелких до крупных (вес от 45 г до 8—10 кг). Есть функционирующие базиптеригоидные сочленения. Шейных позвонков 16. Спинная кость образуется слиянием последнего шейного и трех грудных позвонков. Ребра с хорошо развитыми крючковидными отростками. Киль грудины хорошо развит. Цевка спереди покрыта двумя рядами поперечно вытянутых роговых щитков. Зоб объемистый, с тонкими стенками. Мускульный желудок с очень мощными стенками и жесткой, прочной кутикулой. Слепые кишки функционирующие. В кольцах трахеи часто развиваются окостенения.
Побочный стержень контурного пера обычно хорошо развит. Пух растет только по аптериям. Рулевых от 12 до 24. Крыло диастатаксическое (исключение — сорные куры). У многих видов отчетливо развит половой диморфизм. Взаимоотношения полов при размножении разнообразны: от типичной моногамии до чаще встречающейся полигамии. Соотношение полов примерно одинаково; лишь у немногих видов в естественных популяциях выявлено преобладание самок (павлины). Насиживание начинается после откладки всех яиц. Птенцы типично выводкового типа: вылупляются зрячими, хорошо опушенными, обсохнув, покидают гнездо и бродят с самкой, (реже — с обоими родителями), кормятся сами.
Подотряд куриных разделяется на 2 надсемейства и 7 семейств (одно представлено только ископаемой формой) с 84 родами и 252 ныне живущими видами. Куриные, встречающиеся в СССР, относятся только к этому подотряду; их 20—21 вид из 12 родов.
Надсемейство 1. Cracoidea — краксовые
Объединяет, видимо, наиболее примитивных куриных птиц. Шпор нет. Задний палец расположен на одном уровне с передними. Копчиковая железа обычно голая. Побочный ствол контурного пера развит слабо. Птенцы вылупляются с уже пробившими кожу и иногда даже начинающими раскрываться пейьками маховых.
В это надсемейство включают 3 семейства.
СЕМЕЙСТВО 1. MEGAPODIIDAE — БОЛЬШЕНОГИЕ, ИЛИ СОРНЫЕ, КУРЫ
Птицы средней величины (вес 0,5—2 кг), ведут наземный образ жизни, распространены на островах Малайского архипелага (включая Филиппины) и в Австралии (рис. 75).
Коренастые, плотного сложения, довольно высоконогие, часто длиннохвостые птицы куриного облика (рис. 76). Скелет слабо пневматичен.
151
Рис. 75. Ареалы семейств курообразных птиц:
1— большеногие Megapodiidae, 2— краксовые Gracidae, <3— тетеревиные Tetraonidae, 4—цесарковые Numididae
Рис. 76. Сорная курица Alectura lathami (L.)
Ноги сильные. Трахея не образует петель. Имеется только левая сонная артерия. На голове есть участки голой кожи. Окраска чаше тусклая. У 4 родов крыло эутаксическое, у 3 диастатаксическое. Рулевых 12—18. Семейство включает 11 (иногда выделяют 16) видов, относимых к 7 родам: Megapod las, Aepypodius, Alectura (Catheturus), Eulipoa, Leipoa, Megacephalon, Talegalla.
Очень своеобразно размножение. Более детально оно изучено у глазчатой курицы Leipoa ocellata Gid. (вес около 1,5 кг), живущей в полупустынных районах Юго-Западной Австралии. В легко дренируемом песчаном грунте самец роет яму, 3—5 м в диаметре. Затем он заполняет яму растительным мусором (листья, трава) и засыпает сверху грунтом. Высота кучи может достигать 1 м (рис. 77). Постепенно за счет гниения органического вещества температура внутри кучи повышается. Когда она достигает 29— 30° (через 1—3 месяца после постройки гнезда), начинается кладка. Самец роет узкий ход, доходящий до органического вещества в центре кучи, на дно которого самка откладывает крупное яйцо (до 10—15% от веса самки); ход засыпается. Следующие яйца самка откладывает с интервалом в 5—9 дней; всего может снести до 20—25 яиц. Кладка растягивается на несколько месяцев. Гнездо функционирует как инкубатор 6—9 месяцев. Самец неотлучно держится у гнезда, проверяя его температуру (видимо, языком или голыми участками кожи на голове) и регулируя ее: при жаркой солнечной погоде увеличивает слой грунта над гнездом; при повышении температуры
Рис. 77. Схема гнезда глазчатой сорной курицы Leipoa ocellata Gid.
153
внутри гнезда раскапывает утром и вечером верхний слой, охлаждая кладку; при снижении температуры раскапывает кладку в середине дня, обеспечивая обогрев яиц лучами солнца и, наоборот, усиленно засыпает ее в плохую погоду. Поэтому внутри гнезда постоянно поддерживается довольно высокая температура (порядка 34—40°). Инкубация каждого яйца продолжается от 50 до 90 дней.
Виды родов Megapod ius, Talegalla и некоторых других устраивают подобные же гнезда или, не роя ямы, сгребают большие кучи растительного мусора (диаметром до 3—5 м, высотой до 1—1,5 м), внутрь которых откладывают яйца. Другие (Megacephalon, Eulipoa) закапывают яйца в песок на прибрежных отмелях, где нужное для развития зародыша тепло получается за счет нагревания почвы солнцем. Часть видов, живущих в районах действующих вулканов, закапывает яйца на их склонах в участках с относительно постоянной температурой; температуру в гнезде они, видимо, не регулируют.
Вылупившийся из яйца птенец покрыт коротким пухом, имеет маленькие крылышки и способен немного перепархивать. Птенцы выбираются из гнезда сами (у части видов, возможно, с помощью взрослых птиц, раскапывающих гнездо над пищащим птенцом) и сразу же начинают вести самостоятельную жизнь, скрываясь в кустарниковых зарослях. Лишь достигнув размеров взрослых птиц, в возрасте около 3 месяцев, молодые объединяются в небольшие стайки. Взрослые держатся парами или небольшими группами. Летают мало и неохотно. От опасности стараются убежать, скрываясь в зарослях. Кормятся на земле разнообразной растительной и животной пищей.
Предположительно семейство сорных кур возникло в Австралии. Ископаемые остатки — 2 вида одного рода — найдены в Австралии в относительно молодых отложениях (плейстоцен) и близки к современным.
СЕМЕЙСТВО 2. GALLI NULOIDIDAE
Сюда относят 1 вид, описанный по фрагментарным остаткам из среднего эоцена Северной Америки. По ряду признаков, видимо, занимает промежуточное положение между тетеревиными и краксами, но, вероятно, ближе к последним.
СЕМЕЙСТВО 3. CRACIDAE — КРАКСОВЫЕ
Лесные птицы средних и крупных размеров (вес от 0,5 до 4 кг), распространенные в Центральной и Южной Америке (см. рис. 75).
Скелет сильно пневматичен. У самцов трахея удлинена и образует петли. Есть 3 пары наружных мышц нижней гортани. Сонных артерий две. Есть пенис. В оперении, особенно у самцов, обычны участки черного цвета с ярким металлическим отливом. У части видов на голове хохол из высоких вертикально торчащих закрученных перьев (рис. 78).
Ведут наземно-древесный образ жизни. Моногамы. Гнездо — кучка прутьев в развилке ветвей в кроне высокого дерева; строят оба партнера. В кладке 2, редко 3 крупных яйца. Насиживает только самка; инкубация около 4 недель. Птенцы вылупляются зрячими, хорошо опушенными, с пробившими кожу пеньками маховых. Несколько дней держатся в кроне дерева-в районе гнезда. Оба родителя носят им корм (червей, моллюсков, насекомых, ягоды). В возрасте 5—7 дней птенцы уже могут перепархивать и начинают перемещаться вместе с родителями, самостоятельно питаясь. Полово-зрелость наступает в возрасте около 2 лет. Корм собирают в кронах деревьев, ходя по толстым наклонным ветвям, и на земле; в лесной подстилке роются , как куры. Питаются плодами, семенами, различными беспозвоночными.
В семействе 39 видов, объединяемых в 11 родов: С г ах, Aburria, Chamae-petes, Mitu, Nothocrax, Oreophasis, Or tai is, Pauxi, Penelope, Penelopina,
154
Рис. 78. Хохлатый кракс Crax alector L.
Pipile. Центр возникновения семейства, видимо, Южная Америка. По остаткам из нижнего одигоцена и более поздних отложений в пределах современного ареала и вне его (на юге США) описано 6 видов, относимых к 4 родам. В плейстоценовых отложениях обнаружены остатки 3 ныне живущих видов.
Надсемейство 2. Phasianoidea — фазановые
Объединяет основную массу видов куриных птиц и, видимо, представляет относительно более молодую, прогрессивную ветвь отряда.
Грудина имеет длинные боковые отростки и несет глубокие вырезки по заднему краю. Задний палец короткий и расположен несколько выше, чем передние. У части видов на цевке развивается шпора. Функционируют обе сонные артерии. Пениса нет. Копчиковая железа оперена. Побочный ствол контурного пера хорошо развит. Самцы крупнее самок и у многих видов значительно более яркой окраски. Птенцы вылупляются зрячими и хорошо опушенными; пеньки маховых пробивают кожу через несколько дней после вылупления. Птенцы кормятся сами. Водящие выводок взрослые птицы обогревают и защищают птенцов, помогают им в поисках пищи (раскапывают муравейники и т. п.). Оперившиеся птенцы по окраске похожи на самок.
Надсемейство фазановых объединяет 4 семейства.
СЕМЕЙСТВО 4. TETRAONIDAE — ТЕТЕРЕВИНЫЕ
Птицы средней и крупной величины (вес от 300 г до 4—5 кг), распространенные в умеренных и северных широтах Европы, Азии, Северной Америки (см. рис. 75). Ведут наземный или наземно-древесный образ жизни. Оседлы; немногие виды кочующие или перелетные.
От близких семейств отличаются немногими признаками. Ноздри прикрыты перьями. Цевка оперена полностью или хотя бы в верхней половине. У многих видов в зимнем наряде пальцы оперены (лапа превращается в «лыжу», позволяющую ходить по снегу) или по краям пальцев развиваются
155
Рис. 79. Рябчик Tetrastes bonasia (L ) Сбоку его лапа; хорошо видны роговые выросты по бокам пальцев
роГовые выросты — бахромки (рис. 79), которые увеличивают площадь опоры и, возможно, уменьшают скольжение на промерзших ветвях. Обитатели лесов; немногие виды живут в тундре, лесостепи и степи. Лесные виды кормятся на земле и на деревьях (особенно зимой).
У всех видов хорошо проявляется токовое поведение. Гнездо — ямка на земле, скудно выстланная растительной ветошью. В кладке 6—12 яиц. Часть видов моногамы, но насиживает обычно только самка (белая куропатка, рябчик, дикуша и др.); водить выводок могут оба партнера. Другие виды (глухари, тетерева, луговые тетерева) — типичные полигамы, совсем не образующие пар; спаривание проходит на токовище или поблизости от него. Строит гнездо, насиживает кладку и водит выводок только самка. Инкубация 3—4 недели. Через несколько дней после вылупления пеньки маховых пробивают кожу и начинают разворачиваться; в возрасте 7—10 дней птенцы уже способны при опасности взлетать невысоко на ветку и там затаиваться. В возрасте около месяца, достигнув лишь примерно половины размеров самки, молодые уже довольно хорошо летают. Через 2—3 месяца После вылупления достигают размеров взрослых птиц. Половозрелыми становятся в возрасте 1—2 лет.
Вне сезона размножения держатся небольшими группами или стаями, иногда состоящими из нескольких десятков птиц. При опасности убегают или затаиваются. Взлетают неохотно и, пролетев небольшое расстояние, садятся. Преимущественно растительноядны с резкой сезонной сменой питания: летом едят зеленые части растений, ягоды, семена, насекомых, зимой — почки и кору молодых побегов деревьев и кустарников, хвою. Зимой ночуют в снегу.
Семейство тетеревиных объединяет 18 видов, распределяемых между 41 родами: Tetrao, Bonasa, Canachltes, Centrocercus, Dendragapus, Falci-pennis, Lagopus, Lyrurus, Pedioecetes, Tetrastes, Tytnpanuchus. В СССР встречается и гнездится 8 видов 5 родов.
Центр возникновения семейства — предположительно Северная Америка. Остатки 9 ископаемых и 9 ныне живущих видов обнаружены в разных районах современного ареала в нижнем эоцене и более поздних слоях.
156
СЕМЕЙСТВО 5. PHASIANIDAE— ФАЗАНОВЫЕ
Центральное семейство отряда с большим число видов, различных по внешнему облику (рис. 80) и размерам (вес 45 а — 5 кг). Почти космополиты, не проникающие лишь в высокие широты Северного полушария (в тундру и северную часть тайги), Антарктику и на удаленные от материков океанические острова (рис. 81).
Ноздри не прикрыты перьями. Цевка не оперена. По краям пальцев никогда не развивается роговая бахрома. Населяют самые разнообразные ландшафты: леса разных типов, степи и пустыни, горы вплоть до альпийских лугов. Ведут наземный образ жизни; только немногие виды при опасности садятся на деревья или ночуют в кронах.
Для многих видов характерно сложное токовое поведение. Взаимоотношения полов в период размножения весьма разнообразны. Многие виды — моногамы, но пары сохраняются только на период яйцекладки или, значительно реже, на весь период размножения. Часть видов — полигамы. Гнездо — небольшая ямка в земле со скудной выстилкой. В кладке 5—10, у некоторых видов до 15—20 яиц. Насиживает, как правило, только самка. Инкубация 2,5 (мелкие виды) — 5 недель. В вождении выводка у немногих видов участвует и самец. В возрасте 2 (мелкие виды ) — 4 (крупные виды) недели птенцы, лишь немного превысив четверть веса взрослых, способны перепархивать или немного летать. Формирование оперения заканчивается в возрасте 1—2 месяцев. Мелкие виды становятся половозрелыми в возрасте менее года, крупные — в 2—3 года.
Вне сезона размножения держатся группами. При опасности стараются затаиваться или убежать. Взлетают неохотно, при крайней опасности. По земле ходят легко, быстро бегают. Кормятся только на земле. Преимущественно растительноядны (семена, ягоды, зеленые части растений). Но, при возможности, поедают и животную пищу (насекомые и их личинки, черви, моллюски и др.). Птенцы особенно в первые дни жизни кормятся преимущественно различными наземными беспозвоночными.
Семейство фазановых объединяет 175 видов, распределяемых по 48 родам. Наиболее разнообразен видовой состав фазановых в Юго-Восточной Азии. В СССР гнездятся 12—13 видов (13, если европейского Coturnix coturnix coturnix L. и дальневосточного С. с. japonicus Temm. перепелов считать за самостоятельные виды), относимых к 7 родам. У части видов (например, некоторых фазанов) очень резко выражена географическая изменчивость окраски.
Рис. 80. Самка и самец обыкновенного фазана Phasianus colchicus L.
157
150° 120° 90° 60° 30° 0° 30° 60° 90° 120° 150° 180° 150°
Рис. 81. Ареалы семейств куриных птиц:
/— фазановые Phasianidae, 2— ивдейковые Meleagrididae, 3 — гоациновые Opi sthocomidae
Родственные отношения внутри семейства достаточно не выяснены. Поэтому в чисто практических целях роды часто объединяют в несколько групп, не придавая им определенного таксономического ранга:
американские перепела — Callipepla, Colinus, Cyrtonyx, Dactylortyx, Dendrortyx, Lophortyx, Odontophorus, Oreortyx, Philortyx, Rhynchortyx\
перепела Старого Света — Anurophasis, Coturnix, Excalf actor ia, Mar-gar operd ix, Sу no iceus;
куропатки, кеклики, улары: Alectoris, Ammoperdix, Arborophila, Bambusicola, Caloperdix, Francolinus, Galloperdix, Haematortyx, Lerwa, Melanoperdix, Ophrysia, Perdicula, Perdix, Ptilopachus, Rhizothera, Rollulus, Tetraogallus',
аргусы, павлины, фазаны, настоящие куры, трагопаны — Afropavo, Argusianus, Catreus, Chrysolophus, Crossoptilon, Gallus, Ithaginus, Lopho-phorus, Lophura, Pavo, Phasianus, Polyplectron, Pucrasia, Rheinartia, Syr-mat icus, Trogopan.
Центр возникновения семейства — видимо, южные районы Палеарктики. Ископаемые остатки известны из верхнего эоцена (Франция) и более поздних отложений. Их относят к 46 ископаемым и 19 ныне живущим видам.
Все породы домашних кур созданы в результате одомашнивания красной кустарниковой, или банкивской, курицы Gallus gallus (L.), живущей и сейчас в диком состоянии во многих районах Юго-Восточной Азии. Полу-одомашнены и разводятся в декоративных целях павлин Pavo cristatus L. и многие виды фазанов. Недавно одомашнен и разводится как лабораторная птица и пищевой объект японский перепел Coturnix japonicus Temm. (чаще рассматривается как подвид обычного перепела С. coturnix japonicus).
СЕМЕЙСТВО 6. NUMIDIDAE — ЦЕСАРКОВЫЕ
По внешнму облику и размерам похожи на кур (вес около 1 кг). Характерна крапчатость пера: четкие белые пятна на темном фоне (рис. 82). На голове и шее есть голые участки кожи, часто яркой окраски. Полового ди
морфизма нет. Распространены только Живут по опушкам и лесным полянам, в зарослях кустарников на равнине и на склонах гор. Ведут наземный образ жизни. Прекрасно бегают. Летают плохо и мало; при опасности взлетают и садятся на деревья или на обломки скал. Моногамы. Гнездо на земле в укрытии. В кладке 8—12, по некоторым данным до 20 яиц с очень толстой и прочной скорлупой. Насиживает только самка, но птенцов водят, вероятно, оба партнера. Питаются разнообразной растительной пищей и мелкими беспозвоночными. Держатся мелкими стайками.
Семейство включает 7 видов, распределяемых между 5 родами: Numida, Acryllium, Agelastes, Gut-tera, Phasidus. Сейчас распространены только в Африке, но раньше ареал был больше: в верхнем плейстоцене Чехословакии обнаружены остатки, которые относят
в Африке (рис. 75).
Рис. 82. Цесарка Numida tnelagris (L.)
159
к ныне живущему виду — обыкновенной цесарке Numida meleagris L. Этот вид сейчас одомашнен и разводится во многих странах; выведены культурные породы с весом птиц до 3—4 кг и хорошей яйценоскостью.
СЕМЕЙСТВО 7. MELEAGRIDIDAE — ИНДЕЙКОВЫЕ
Крупные (вес 5—10 кг) птицы типичного облика индейки (рис. 83), распространенные в южных районах Северной и в Центральной Америке (рис. 81). Сейчас ареал сильно сократился из-за изменения ландшафтов и неумеренной охоты. Обитают преимущественно в лесостепных районах.
Самцы значительно крупнее самок и более яркой окраски. На голове есть голые участки кожи и голые кожистые выросты. Хорошо развиты шпоры. Полигамы. Около токующего самца группируется гарем из нескольких самок. В кладке 8—15 яиц. Иногда две самки несутся в одном гнезде. Гнездо на земле, под защитой кустарников. Насиживает только самка около 4 недель. Выводок водит тоже только самка. Кормятся на земле разнообразной растительной пищей; животные корма (насекомые, черви, амфибии, рептилии и т. п.) поедаются случайно. Хорошо ходят и бегают. Летают медленно и неохотно. Вне сезона размножения держатся группами, самцы обычно отдельно от самок.
Два монотипичных рода: Meleagris и Agr iochar is. Формировались, видимо, в южных районах Северной Америки. В пределах современного ареала ископаемые остатки известны с олигоцена; их относят к 7 ископаемым видам. В плейстоцене найден и современный вид. В результате одомашнивания и последующего отбора были выведены разнообразные культурные породы домашних индеек.
160
ПОДОТРЯД 2. OPISTHOCOMI — ГОАЦИНОВЫЕ
Включает только одно семейство
СЕМЕЙСТВО 8. OPISTHOCOMIDAE — ГОАЦИНОВЫЕ
Единственный ныне живущий вид — Opisthocomus hoazin (Mil 11.) (рис. 84). Населяет влажные леса бассейна Амазонки (рис. 81).
Вес около 800 г. Сильно выражена подвижность надклювья (изгиб у переднего конца лобных костей). Базиптеригоидных сочленений нет. Края рогового надклювья слегка зубчатые. Шейных позвонков 16—19. Спинная кость образуется слиянием двух шейных и одного грудного позвонков. Ребра не несут крючковидных отростков. Киль развит только в задней полови-
не грудины и имеет расширение на вершине; над ним на коже находится мозолистое утолщение, на которое птица опирается при отдыхе. Цевка сетчатая. Задний палец довольно длинный и расположен на одном уровне с передними. Зоб очень объемистый, заходит на переднюю часть грудины (где не развит киль). Зоб имеет мускулистые стенки и в нем происходит частичное перетирание пищи. Мускульный желудок относительно тонкостенный, невелик по объему. Слепые кишки короткие. Трахеальные кольца окостеневают слабо. Оперение рыхлое. Пух есть на птерилиях и аптериях. Первостепенных маховых и рулевых по 10. Крыло эутаксическое. На голове хохол. Окраска самцов и самок сходна.
Держатся по опушкам влажных тропических лесов, зарослям тростника и камыша по берегам водоемов. Взаимоотношения полов при размно
жении не выяснены: вероятно, моногамы, но, видимо, встречаются и случаи полигамии. Гнездо — кучка веток в развилке сучков или залом тростника — помещается обыч-
но над водой. Гнездятся иногда мелкими группами и даже скоплениями в несколько десятков пар. В кладке 2—4 яйца. Птенцы вылупляются почти голыми; на хорошо развитых и подвижных концевых фалангах первого и второго пальцев крыла развиты когти. При опасности птенцы убегают из гнезда, используя при движении в ветвях клюв, передние и задние конечности. Упавшие в воду птенцы хорошо плавают и даже ныряют, быстро скрываясь в зарослях. Последовательно сменяются два пуховых наряда и лишь после этого начинает развиваться оперение. Птенцов взрослые длительное время выкармливают отрыжкой из зоба, затем они пос
161
тепенно переходят на самостоятельное питание. Взрослые кормятся в кронах деревьев и кустарников преимущественно листьями, в меньшей степени — плодами и семенами. Вне сезона размножения держатся стайками по 10—40 птиц.
Своеобразие строения и экологии гоацинов обусловливает выделение этого вида в отдельный подотряд. Видимо, они обособились от очень примитивных куриных птиц. Некоторые черты сходства с кукушкообразными обычно объясняют конвергенцией: приспособлением к древесному образу жизни.
Остатки, относимые к ископаемому роду, обнаружены в миоценовых отложениях Южной Америки. Остатки другого ископаемого рода описаны из верхнего эоцена — нижнего олигоцена Франции; плохая сохранность делает отнесение их к подотряду гоацинов весьма предположительным.
ОТРЯД 20. GRUIFORMES —ЖУРАВЛЕОБРАЗНЫЕ
Включаемые сюда виды очень разнообразны по величине (вес от 30 г до 16 кг), морфологическим и экологическим особенностям. Система отряда очень сложна: 8 подотрядов и 22 семейства (9 семейств представлены только ископаемыми формами). Это отражает сильную разобщенность отдельных групп и их сложную эволюцию, поэтому довольно трудно дать общую характеристику отряда. Всего в отряде 84 рода с 193 видами, ныне живущими или исчезнувшими в историческое время. В СССР встречается 23 вида из 14 родов — представители 5 семейств и 3 подотрядов. Достоверно гнездится 22 вида.
Преимущественно это наземные птицы, часть из которых заселила болота или перешла к более или менее водному образу жизни. Череп схизогнатический, в некоторых группах с переходом к десмогнатизму или эгито-гнатизму. Базиптеригоидные сочленения обычно отсутствуют. Шейных позвонков 15—20. По заднему краю грудины одна пара вырезок (у некоторых отсутствует). Базальные концы коракоидов при сочленении с грудиной не соприкасаются друг с другом. Гортань трахео-бронхиального типа. Зоба нет. Пищевод сильно растяжим и иногда имеет не ограниченное четко расширение (дрофы). Мускульный желудок с мощными стенками и прочной кутикулой. Слепые кишки разной длины, но всегда функционирующие. Есть желчный пузырь. У многих групп копчиковая железа оперена.Контурное перо обычно имеет побочный стержень. Первостепенных маховых 10—11, рулевых 8—20. Большинство видов моногамы. Гнездо обычно на земле; немногие виды гнездятся на деревьях. Птенцы выводкового типа.
Для каждой из входящих в этот отряд групп характерен именно комплекс всех или большинства перечисленных признаков. Отряд журавлеобразных — древняя группа. Отдаленные родственные связи, вероятно, сближают журавлеобразных с предками ржанкообразных (куликообразных). Еще более давнее родство возможно с предками куриных птиц.
Родственные взаимоотношения в пределах отряда неясны. Видимо, каждый из подотрядов — потомки общей предковой группы, более или менее сильно обособившиеся в результате специализации и занявшие определенные жизненные ниши. Некоторые систематики и сейчас рассматривают все подотряды журавлеобразных птиц как близкие, но самостоятельные отряды.
ПОДОТРЯД 1. MESITORNITHIDES —МАДАГАСКАРСКИЕ ПАСТУШКИ
Включает только одно семейство.
162
СЕМЕЙСТВО 1. MESITORNITHIDAE (MESOENATIDAE)
Объединяет 2 рода Mesitornis (Mesoenas) и Manias и 3 вида, встречающиеся только на Мадагаскаре. По внешнему облику напоминают каких-то длиннохвостых короткокрылых воробьиных птиц (рис. 85) величиной с крупного дрозда.
Рис. 85. Мадагаскарский пастушок Manias benschi Oust.
Клюв удлиненный, суживающийся к концу и более или менее изогнутый книзу. Череп схизогнатический, без базиптеригоидных сочленений и затылочных фонтанелей; сходен с черепом настоящих журавлей. Шейных позвонков 17. По заднему краю грудины пара вырезок. Коракоиды длинные и тонкие. Вилочка полностью редуцирована. Ноги относительно короткие, с длинными пальцами; хорошо развит задний палец. Гортань с парой трахеобронхиальных мускулов. Слепые кишки длинные. Копчиковой железы нет. Контурные перья без побочных стержней. Есть 5 пудреток. Первостепенных маховых 10, рулевых 16. Крыло эутаксическое.
Обитают преимущественно в саваннах, придерживаясь рощ и кустарниковых зарослей. Кормятся на земле. Хорошо бегают. Летают мало; взлетают неохотно. Поедают различных наземных беспозвоночных. Видимо, моногамы, но у Monias с одним самцом иногда держатся 2—3 самки. Примитивное гнездо из растительной ветоши на земле или в развилке ветвей низких кустов. Кладка из 2—3 пятнистых яиц. Птенцы вылупляются покрытые черным или буро-коричневым пухом; вскоре после вылупления оставляют гнездо. Возможны 2 кладки в год. Ископаемые остатки неизвестны. Видимо, древняя, хорошо обособившаяся боковая ветвь журавлеобразных. Прослеживается сходство по многим мелким морфологическим признакам с трехперстками и журавлями; менее явное — с солнечными цаплями и кагу.
ПОДОТРЯД 2. TURNICES —ТРЕХПЕРСТКИ
Мелкие птицы (вес 30—100 г), по внешнему облику похожие на перепелов (рис. 86, 87). Череп схизогнатического типа с переходом к эгитогнатиз-му. Есть функционирующиебазиптеригоидныесочленения. Ноздри не сквозные, схизоринальные. Образуется длинная спинная кость (сливается по 9 позвонков), но последний шейный позвонок в нее не входит. По заднему краю грудины пара вырезок. Сильно растяжим начальный участок пищевода, играющий у самок во время тока роль резонатора. Мускульный желудок мощный. Слепые кишки длинные. Трахеальные кольца не окостеневают. У самок в нижней части трахеи есть заметное расширение, также выполняющее роль резонатора. Токовые крики издают только самки. Хорошо раз
163
виты побочные стержни контурных перьев. Первостепенных маховых 10, рулевых 12. Копчиковая железа оперена.
Обитатели степных и луговых участков преимущественно в теплых широтах Африки, Азии, Австралии, в ограниченных участках Южной Испании, Сардинии. Самки несколько крупнее самцов и более яркой окраски. Видимо, у всех или по крайней мере у большинства видов выражена полиандрия: самки малочисленнее самцов. Самка токует и спаривается с самцом, затем откладывает 4 яйца (у Ortyxelus^—только 2), которые насиживает самец. Самка опять начинает токовать, спаривается другим самцом, откладывает вторую кладку, которую начинает насиживать второй самец, и т. д. Одна самка, вероятно, может отложить до 3—5 кладок. Кончившие кладку самки собираются в стайки, кочуют. Самцы насиживают 12—13 дней, а затем водят птенцов; первую неделю кормят их из клюва. В возрасте 7—10 дней птенцы уже могут перепархивать и начинают кормиться сами. Через 25—28 дней после вылупления птенцы хорошо летают и полностью самостоятельны, а через 6—7 недель ни по виду, ни по размерам неотличимы от взрослых. В тропиках у некоторых видов молодые становятся половозрелыми в возрасте 4—5 месяцев.
Дневные птицы. Кормятся только на земле, преимущественно растительной пищей (семена, вегетативные части растений); животные корма поедают случайно. Вне сезона размножения держатся маленькими стайками. Оседлые или кочующие, немногие перелетные.
Подотряд трехперсток включает 2 семейства с 3 родами и 15 видами. В СССР встречается и гнездится 1 вид.
СЕМЕЙСТВО 2. TURNICIDAE — ТРЕХПЕРСТКОВЫЕ
Задний палец полностью редуцирован. Ножная формула AXY. Есть только левая сонная артерия. Крыло эутаксическое. Яйца овальные. Сюда включают 14 видов 2 родов: Ortyxelus с 1 видом, живущим в Южном Судане, на севере Кении и Уганды, и Turnix с 13 видами, распространенными в Африке, Азии, Австралии (рис. 88). Один вид — Turnix
tanki Bly th. (рис. 86)—встречается и гнездится в южной части Приморского края.
СЕМЕЙСТВО 3. PEDIONOMIDAE — АВСТРАЛИЙСКИЕ СТРАННИКИ
Задний палец есть, хотя и мал. Ножная формула ABXY. Функционируют обе сонные артерии,
Рис. 86. Пятнистая трехперстка Turnix tanki Blyth.
Рис. 87. Астралийский странник Pedionomus torquatus (Gid.)
164
Рис. 88. Ареалы семейств журавлеобразных птиц:
1 — мадагаскарские пастушки Mesitornithidae, 2— трехперстковые Turnicidae (ареал в Африке и Европе не показан), 3— австралийские странники Pedionomidae, 4— настоящие журавли Gruidae, 5 — пастушковые журавли Arainidae, 6 — трубачи Psophiidae
но левая развита слабее правой. Крыло диастатаксическое. Яйца грушевидной формы. В это семейство включают единственный вид Pedionomus torquatus Gid. (рис. 87), встречающийся только в юго-западной Австралии (рис. 88). По биологии сходен с обыкновенными трехперстками, но встречается в более сухих ландшафтах.
Отчетливые различия между двумя семействами трехперсток, проявляющиеся во многих деталях строения, свидетельствуют о древности всего подотряда. Очевидные черты сходства сближают трехперсток с мадагаскарскими пастушками и подотрядом журавлей (с пастушками и собственно журавлями). Сходство с куриными, видимо, конвергентное.
ПОДОТРЯД 3. GRUES —ЖУРАВЛИНЫЕ
Объединяет довольно разнообразных птиц с внешним обликом журавлей (рис. 89) или пастушков (рис. 92). Размеры от мелких до крупных: вес от 30 г до 15 кг. Клюв удлиненный, конусообразный или короткий, сжатый с боков. Ноздри сквозные. Череп схизогнатический. Функционирующих ба-зиптеригоидных сочленений нет. Нижняя часть голени не оперена. Задний палец укорочен, но всегда есть; обычно он прикрепляется несколько выше, чем передние пальцы. Сохраняются обе сонные артерии. Побочный стержень контурного пера развит относительно слабо. Пух растет по птерилиям и аптериям. Копчиковая железа оперена. Рулевых 8—14, первостепенных маховых 10—11. Линька маховых у многих видов проходит почти одновременно, и птицы на короткое время (1—2 недели) теряют способность к полету. Полового диморфизма окраски нет; самцы немного крупнее самок. Возрастные отличия в окраске выражены слабо.
Подотряд журавлиных — центральная группа отряда — разделяется на 2 надсемейства. Надсемейство пастушковых имеет более примитивные особенности и, видимо, в большей степени сохраняет черты строения предков. Надсемейство журавлей более специализировано. В подотряде 6 семейств (2 только с ископаемыми формами), с 58 родами, 137 видами, ныне живущими или вымершими в историческое время. В СССР встречаются 18 видов 9 родов; достоверно гнездится 17 видов.
Надсемейство 1. Gruoidea — журавлиные
Птицы средней и крупной величины (вес 1—15 кг) с более или менее явным обликом журавля (рис. 89—91). Длинноклювые (за исключением трубачей), длинношейные птицы на высоких ногах. Часть грудных позвонков срастается в спинную кость. Задний край грудины без вырезок. Лобковые и седалищные кости срастаются друг с другом. Трахея обычно удлиненная, образует петли. Третьестепенные маховые длинные и при сложенном крыле достигают его вершины или выходят за нее.
Надсемейство включает 4 семейства (одно ископаемое) с 7 родами и 18 ныне живущими видами. В СССР встречается 7 видов только семейства настоящих журавлей.
СЕМЕЙСТВО 4. GERANOIDIDAE
По фрагментам задних конечностей из отложений нижнего эоцена Северной Америки описан один вид журавлеподобной птицы, выделяемый в это семейство.
СЕМЕЙСТВО 5. GRUIDAE — НАСТОЯЩИЕ ЖУРАВЛИ
Птицы средних и крупных размеров, с прямым длинным копьевидным клювом, длинной шеей, высоконогие (рис. 89). Спорадично, но широко распространены по земному шару; не проникают в Южную Америку, Антарк
166
тику и на многие океанические острова (рис. 88). В северных и умеренных широтах перелетные, в тропиках оседлые или кочующие.
На голове у многих видов есть голые, часто яркой окраски участки кожи или украшающие перья. Ноздри схизоринальные. Шейных позвонков 19—20. Грудина удлиненная. Вершина вилоч--ки у многих видов срастается с вершиной киля. 7 пар ребер. Ножная формула ABXY или BXY, редко ВХ. У большинства видов петли трахеи лежат в полости грудины и ее киля. Издают громкие курлыкающие крики. Крыло диастата ксичес кое. Первостепенных маховых 11, рулевых 12.
Населяют обширные болота
и луга, немногие виды — степи. Пары сохраняются много лет. Размножению предшествует сложный ток: групповые и парные «танцы». Гнездо — небрежно набросанная большая куча растительной ветоши. Кладка из 1т—2, редко 3 пятнистых яиц. Насиживает, ви-
Рис. 89. Венценосный журавль Balearica pavonina (L.)
димо, преимущественно самка. Самец держится в районе гнезда и предупреждает о появлении опасности. Инкубация около 4—5 недель. Птенцы покрыты коричневатым или желтовато-бурым пухом. Почти сразу после вылупления птенцы покидают гнездо и кочуют с родителями. В возрасте 4—7 недель полностью оперяются, на 1—2 недели позже приобретают способность к полету. Кочующие семьи вскоре объединяются в стаи. Молодые становятся половозрелыми на 2—3-е лето жизни.
Прекрасно ходят и бегают. Никогда не садятся на деревья. Полет спокойный, медленный. Кормятся только на земле. Пища преимущественно растительная: всходы, молодые побеги, корневища и луковицы, семена и ягоды. Попутно, иногда в значительном количестве, поедают и разнообразных мелких животных (насекомых, червей, моллюсков, амфибий, ящериц и т. п.).
Семейство настоящих журавлей включает 14 видов, относимых к 5 родам: Grus (10 видов), Anthropoides, Balearica, Bugranus, Tetrapteryx. В СССР гнездится 7 видов 2 родов. Стерх, или белый журавль, Grus leuco-geranus Pall, гнездится только на территории нашей страны в низовьях р. Оби и в Яно-Индигирской тундре (северо-восточная Якутия), а у 4 видов (красавки, черного, даурского и маньчжурского журавлей) в СССР расположена основная часть ареала. Численность некоторых видов очень мала:
так, всего из нескольких десятков птиц состоит вся популяция американского G. americana (L.) и маньчжурского G. japonensis (Mull.) журавлей; общая численность стерха, видимо, не превышает 300—400 пар. Во многих странах, в том числе и в СССР, охота на журавлей запрещена.
Семейство, видимо, возникло в Палеарктике, хотя самые ранние находки (нижний эоцен) сделаны в США (Вайоминг). По ископаемым остаткам разной сохранности описано 11 родов и 24 вида; в палеонтологических материалах обнаружены остатки 7 ныне живущих видов.
167
СЕМЕЙСТВО 6. ARAMIDAE— АРАМОВЫЕ, ИЛИ ПАСТУШКОВЫЕ, ЖУРАВЛИ
Семейство монотипическое: сюда входит только 1 вид — бразильская арама, или пастушковый журавль, Aratnus guarauna (L.), распространенный от юга США до Аргентины (рис. 88). По внешнему облику напоминает крупного пастушка (вес около 1,5 кг; рис. 90).
Клюв длинный, слегка изогнутый книзу. Ноздри схизоринальные. Шейных позвонков 17. Вилочка на конце не имеет расширения и не срастается с килем. Ребер 7 пар. Ножная формула BXY. Пальцы ног, особенно передние, удлиненные. Петля трахеи лежит в нижней части шеи и не входит в грудину. Голос громкий, каркающий. Крыло диастатаксическое. Первостепенных маховых 10, рулевых 12.
Живут по обширным лесным болотам. Объемистое гнездо — куча веток и сухих стеблей — обычно устраивается в развилке крупных ветвей на дереве, реже на сухом месте на земле. В кладке 4—8 пестрых яиц. Насиживают оба родителя. Птенцы вылупляются покрытые черным пухом, зрячие. Более недели остаются в гнезде и выкармливаются родителями; потом оставляют гнездо и кочуют со взрослыми, питаясь уже самостоятельно. Активны днем и в сумерках. Хорошо бегают, могут плавать. Взлетают неохотно. Полет медленный, тяжелый. Питаются преимущественно животной пищей, особенно разнообразными моллюсками. Вытаскивая моллюска, иногда придерживают раковину лапой.
Ископаемые остатки (6 видов 5 родов) описаны из нижнего олигоцена и более поздних слоев центральных районов США и юга Южной Америки, т. е. вне современного ареала. Вероятно, это семейство можно считать более примитивным, чем семейство настоящих журавлей. Часть признаков, сближающих арам с пастушками, возможно, выработалась конвергентно.
168
СЕМЕЙСТВО 7. Р SOPH П DAE — ТРУБАЧИ
Рис. 91. Трубач Рsophia crepitans L.
Включает только 1 род Рsophia с 3 видами (рис. 91), населяющими влажные тропические леса Южной Америки (рис. 88). Птицы средней величины (вес около 1 кг), внешне похожие на пастушков или на куриных. Клюв короткий, выпуклый по гребню надклювья. Ноздри голоринальные. Шейных позвонков 17—18. Ребер 8 пар. Формула ноги BXY. Передние пальцы ног удлиненные. Трахея образует петли которые лежат под кожей, не заходя в полость грудины. Издают громкие трубные звуки. Первостепенных маховых и рулевых по 10. Крыло эутаксиче-ское. На груди и шее есть металлический зеленовато-фиолетовый отблеск.
Перед началом гнездования характерен групповой ток: «танцы», сопровождаемые громкими криками.
Видимо, моногамы. Гнездо устраивают в развилке крупных ветвей или в больших дуплах. В кладке 6—10 яиц белого цвета, без пестрин. Насиживает, кажется, только самка. Птенцы покрыты коричневым пухом, на спине светлые полосы. Почти сразу после вылупления птенцы покидают гнездо; видимо, собирают корм сами и, кроме того, первое время подкармливаются родителями. Держатся обычно группами и стаями до нескольких десятков птиц. Кормятся на земле, поедая семена, ягоды, насекомых и других беспозвоночных. Хорошо бегают. При опасности затаиваются или убегают, иногда взлетают на деревья. Полет медленный, тяжелый.
Ископаемые остатки не обнаружены. По особенностям строения несомненно близки к журавлям. Однако проявляются и некоторые черты сходства с кариамами.
Надсемейство 2. Railoidea — пастушковые
Околоводные птицы мелкой и средней величины (вес от 30 а до 1,5—2 кг), относительно короткошейные (рис. 92). Большинство видов коротконоги, хотя есть и довольно длинноногие формы.
Клювы варьируют от удлиненных, довольно тонких, иногда слабо изогнутых, до коротких и высоких, сжатых с боков. У ряда видов на лбу голая, яркая кожная бляшка. Ноздри голоринальные. Шейных позвонков 14—15. Грудные позвонки не сливаются в спинную кость. Вилочка не доходит до вершины киля. По заднему краю грудины пара глубоких вырезок. Ребер 6—7 пар. Лобковая и седалищная кости не срастаются друг с другом. Ноги сильные. Ножная формула ABXY. Передние пальцы удлиненные. Трахея короткая и не образует петель. Крыло короткое, диастатаксическое, с 11 первостепенными маховыми; у утративших способность к полету некоторых островных видов крыло эутаксическое с 9—8 маховыми. Третьестепенные маховые при сложенном крыле не достигают его вершины. Хвост короткий, из 8—14 мягких рулевых. Половой диморфизм окраски свойствен немногим видам.
Надсемейство объединяет 2 семейства.
169
Рис. 92. Султанка Porphyria poliocephalus (Lath.)
СЕМЕЙСТВО 8. ORTHOCNEMIDAE
Сюда входят описанные по ископаемым остаткам из олигоцена Европы 2 рода и 7 видов пастушкообразных птиц, видимо, близких к настоящим пастушкам.
СЕМЕЙСТВО 9. RALLIDAE— ПАСТУШКОВЫЕ
Характеристика надсемейства целиком приложима и к семейству пастушковых. Распространены в тропических и умеренных широтах всего земного шара (рис. 93). Северные формы перелетны, южные оседлые или кочующие.
Несмотря на относительное обилие видов и большие вариации в размерах, морфологически и экологически семейство довольно однородно. Населяют заросли по берегам водоемов, болота, влажные луга, реже сырые леса. Некоторые виды хорошо плавают и ныряют; каждый палец у них имеет фестончатую кожистую оторочку. Большинство быстро бегает, ловко пробираясь среди густых зарослей. Взлетают неохотно; полет прямолинейный, довольно тяжелый.
Гнездо на земле, в густых зарослях; некоторые виды строят плавающие гнезда в надводных зарослях. В кладке чаще 6—12 яиц, светлых с крапом. У части видов в южных районах, видимо, две кладки в сезон размножения. Насиживают оба партнера. Инкубация 2—3 недели. Птенцы покрыты густым, обычно черным пухом. Почти сразу после вылупления способны бегать и прятаться, у плавающих видов нырять и плавать. Выводок водят оба родителя; иногда возвращаются для отдыха в гнездо. Птенцы, видимо, кормятся сами и, кроме того, их подкармливают взрослые. Полностью оперяются и становятся способными к полету, достигнув размеров взрослых. Половозрелыми становятся в возрасте 1—2 лет.
170
Питаются на земле и на сплавинах разнообразной животной (различные беспозвоночные) и растительной пищей (проростки, семена и т. п.). Плавающие и ныряющие виды поедают водоросли, проростки и зеленые части водных растений, иногда — видимо, в небольших количествах — ловят мелкую рыбу и водных беспозвоночных. У многих в сумерках и ночью активность увеличивается. Вне сезона размножения большинство живет одиночками или мелкими группами; немногие виды держатся стаями.
Семейство пастушковых включает 119 видов, обычно относимых к 51 роду (у разных авторов от 46 до 60 родов): Rallus (23 вида), Amaurornis, Апи-rolitnnas, Aphanapteryx, Aptornis, Aramides, Aramidopsis, Atlantisia, Canirallus, Coturnicops, Crex, Cyanolimnas, Diaphorapteryx, Dryolimnas, Fu-lica, Gallicrex, Gallinula, Gallirallus, Habropteryx, Habroptila, Himantor-nis, Laterallus, Limnocorax, Megacrex, Micropygia, Neocrex, Nesoclopeus, Nesotrochis, Notornis, Pardirallus, Poliolimnas, Porphyria, Porphyriops, Porphyriornis, Porphyrula, Porzana, Rallicula, Rallina, Rougetius, Sarthru-ra, Tricholimnas и др.
В СССР отмечено 11 видов 7 родов; несомненно гнездится 10 видов.
Более 10 видов, преимущественно плохо летавших или не летавших и живших главным образом на островах, по разным причинам полностью вымерли в историческое время (прямое преследование человеком, изменение ландшафтов, завоз хищников и крыс и т. п.). Часть видов сейчас находится под угрозой быстрого исчезновения.
По остаткам из верхнего палеоцена и более поздних отложений в пределах современного ареала описано 46 видов 24 родов. В отложениях из плиоцена и более поздних слоев найдены остатки 23 ныне живущих видов.
Из всех групп отряда журавлеобразных пастушковые ближе всего стоят к журавлям; проявляются некоторые черты сходства и с лапчатоногими.
ПОДОТРЯД 4. HELIORNITHES — ЛАПЧАТОНОГИЕ
Сюда включают птиц, по внешнему виду напоминающих и пастушков, поганок (рис. 94). Они составляют единственное семейство.
СЕМЕЙСТВО 10. HELIORNITHIDAE — ЛАПЧАТОНОГИЕ
Включает только 3 монотипических рода: Heliornis (Южная Америка), Heliopais (Южная Индия, Суматра) и Podica (Африка). Обитают в тропиках (рис. 93), на реках и больших озерах с густой береговой растительностью.
Клюв удлиненный, сжатый с боков. Голова небольшая. Шейных позвонков 14—15. Спинная кость не образуется. На расширенном заднем крае грудины одна пара вырезок. Расширенный конец вилочки прирастает к килю грудины. Ребер 6 пар. Лобковая и седалищная кости частично сливаются друг с другом. Каждый из направленных вперед трех пальцев имеет широкую кожистую оторочку. Оперение плотное (как у уток). Побочный ствол контурного пера развит или отсутствует (Podicd). Копчиковая железа оперена. Крыло эутаксическое. Первостепенных маховых 10, длинных и жестких рулевых 18. Самцы крупнее самок и более яркой окраски.
Гнездо — куча веток и сухой растительности — на полузатопленных кустах и деревьях. В кладке 2—5, редко до 7 яиц. Насиживают оба партнера. Птенцы вылупляются зрячими, покрыты серо-коричневым пухом, у Heliornis—голые. Довольно долго выкармливаются взрослыми. Есть указания, что птенцы вскоре покидают гнездо и перемещаются вместе с родителями, которые плавают, держа их под крыльями. По земле передвигаются довольно легко. Хорошо плавают; ныряют неохотно. При опасности птица бежит по воде или плывет, погрузив все туловище в воду, и старается скрыться в зарослях. Взлетают в крайнем случае; полет тяжелый. Корм собирают преимущественно на поверхности воды: водных насекомых, мол-
171
Рис. 93. Ареалы семейств журавлеобразных птиц:
1— пастушковые Rallidae, 2— лапчатоногие Не!iornithidae, 3— кагу Rhinochetidae, 4— кариамовые Cariamidae, 5— солнечные цапли Eurypygidae, 6— дрофиные Otididae
Рис. 94. Африканский лапчатоног Podica senegalensis Vieill.
люсков, ракообразных, земноводных, изредка ловят рыбу, возможно, используют и растительные корма.
Видимо, древняя, очень специализированная группа. Ископаемые формы не обнаружены. Много черт сходства с пастушковыми, меньше — с мадагаскарскими пастушками и солнечными цаплями. Сходство с поганками чисто конвергентное.
ПОДОТРЯД 5. RHINOCHETI — КАГУ
Сюда входит лишь одно семейство.
СЕМЕЙСТВО И. RHINOCHETIDAE — КАГУ
Единственный вид — Rhinochetus jubatus Verr. (рис. 95), встречается только на о. Новая Каледония (рис. 93). По внешнему виду и размерам кагу
несколько похож на квакву.
Клюв крепкий, заостренный. Ноздри не сквозные, схизоринальные; наружные отверстия ноздрей прикрыты кожистыми крышечками. Шейных позвонков 16. Спинная кость из 4 грудных позвонков. Задний край грудины без вырезок. Ребер 5 пар. Слепые кишки короткие, но функционирующие. Копчиковая железа голая. У контурных перьев сильно развит побочный стержень. Много мелких пудреток, разбросанных по телу. Крыло эутаксическое. Первостепенных маховых 10, рулевых 12. Самцы сходны с самками по размерам и окраске.
Держатся в сырых лесах с густым подлеском. Сохранились только в глухих районах
Рис. 95. Кагу Rhinochetus jubatus Verr.
173
гористой южной части острова. Примитивное гнездо устраивается на земле в густых зарослях. В кладке одно пестрое яйцо. Самец насиживает днем, самка — ночью. Инкубация 5—6 недель. Птенец покрыт бурым с желтоватыми пятнами пуховым нарядом. Гнездо покидает через 4—5 дней после вылупления и бродит с родителями, которые его кормят около 4 недель. Возможно, в году 2—3 цикла размножения. Активны преимущественно ночью. Вне сезона размножения держатся маленькими группами. Хорошо ходят и бегают. Летают плохо и очень неохотно. Питаются различными беспозвоночными, которых добывают, глубоко погружая клюв в почву, или собирают на поверхности.
Вместе с солнечной цаплей, с которой у кагу много общих черт, наиболее примитивны и, видимо, в большей степени, чем другие группы, сохраняют черты генерализованных предков всего отряда. Древность кагу проявляется также в сохранении некоторых признаков сходства с молото-главами из аистообразных. В ископаемых остатках не обнаружены.
ПОДОТРЯД 6. EURYPYGAE — СОЛНЕЧНЫЕ ЦАПЛИ
Сюда включают одно семейство.
СЕМЕЙСТВО 12. EURYPYGIDAE—СОЛНЕЧНЫЕ ЦАПЛИ
Единственный вид — Eurypyga hellos Pall. (рис. 96) населяет опушки заболоченных лесов и берега водоемов Центральной и Южной Америки (см. рис. 93). Похожа на маленькую, пестроокрашенную длиннохвостую цаплю. Солнечной цаплей названа потому, что любит греться на солнце.
Клюв тонкий, заостренный, прямой. Ноздри сквозные, схизориналь-ные. Шейных позвонков 18. Спинная кость из трех грудных позвонков. Грудина с парой вырезок по заднему краю. Голень и цевка удлиненные. Задний палец маленький. Слепые кишки рудиментарны. Побочный ствол контурного пера развит слабо. Есть многочисленные мелкие пудретки. Копчиковая железа голая. Крыло диастатаксическое. Первостепенных маховых 10, рулевых 12. Самцы и самки сходны по окраске и размерам.
Легко бегают, пробираясь сквозь заросли. Летают плохо и неохотно. Гнездо на земле или, чаще, в развилке ветвей низкого дерева или куста, из сучьев, листьев и сухой травы, скрепленных глиной или илом. Кладка из
Рис. 96. Солнечная цапля Eurypyga helios Pall.
174
2 пестрых яиц. Насиживают оба партнера около 4 недель. Вылупившиеся птенцы покрыты пестрым пухом; через несколько дней покидают гнездо и бродят с родителями. Видимо, до подъема на крыло выкармливаются взрослыми. Питаются солнечные цапли различными беспозвоночными, которых склевывают на почве и листве, мелкими рыбками и земноводными.
Древняя примитивная группа отряда, наиболее близкая к кагу. Много общих черт и с пастушковыми. Внешнее сходство с цаплевыми из аистообразных, видимо, в значительной степени конвергентное. Ископаемые остатки не обнаружены.
ПОДОТРЯД 7. CARIAMAE — КАРИ АМЫ
Включает довольно крупных, хорошо бегающих длинноногих птиц (рис. 97), по внешнему облику несколько похожих на птиц секретарей или дроф.
Клюв сильный, с загнутым книзу острым концом надклювья. Ноздри сквозные, голоринальные. Череп промежуточного типа между схизогнатическим и десмогнатическим. Шейных позвонков 14—15. На заднем крае грудины пара вырезок. Голень и цевка удлинены. Передние пальцы, особенно средний, длинные с острыми когтями; задний палец мал.
Подотряд объединяет 2 надсемейства.
Надсемейство 1. Phororhacoidea
Сюда включают около 40 видов 20 родов ископаемых птиц, описанных по разной сохранности остаткам из олигоценовых и миоценовых отложений юга Южной Америки. Эти виды относят к 5 семействам (13—17-е): Phororhacidae, Psilopteridae, Brontornithidae, Opisthodactylidae, Cunam-paiidae. По внешнему облику, видимо, походили на современных кариам, но отличались крупными размерами. Например, у Phororhacos longissimus длина черепа с клювом достигала 75 см. Все крупные формы, видимо, утратили способность к полету: скелет крыла явно подвержен редукции (укорочение всех отделов и т. д.). Видимо, это слепые ветви эволюции, обособившиеся от общих предковых форм подотряда.
Надсемейство 2. Cariamoidea — кариамовые
Скелетные особенности перечислены в характеристике подотряда. Хорошо развиты слепые кишки. Копчиковая железа голая. Контурные перья с развитым побочным стержнем. У основания надклювья пучок длинных щетинковидных перьев. Крыло эутаксическое. Первостепенных маховых 10, длинных рулевых 12.
В надсемейство входят 3 семейства; 2 из них представлены только ископаемыми формами.
СЕМЕЙСТВО 18. САRIAMIDAE—КАРИАМОВЫЕ
Характерные для семейства особенности приведены в характеристике подотряда и надсемейства. Сюда входят всего 2 вида птиц средних размеров (вес около 2 кг), относимых к двум монотипичным родам: Cariama (рис. 97) и Chunga. Населяют сухие лесостепи и степи с редкими кустами в Южной Америке (см. рис. 93).
Держатся парами или небольшими группами. Прекрасно бегают, легко скрываясь в высокой траве. На крыло поднимаются неохотно. Гнездо — кучка мелких веток — устраивается в кроне густого куста или невысокого дерева. В кладке обычно 2 белых с коричневато-красными пятнами яйца. Насиживают, видимо, оба партнера. Инкубация около 4 недель. Птенцы вылупляются зрячими, покрытыми длинным коричневато-желтым пухом.
175
Рис. 97. Кариама Cariama cristata L.
Через несколько дней покидают гнездо и кочуют вместе с родителями. Питаются главным образом рептилиями, убивая их сильным ударом лапы (помогает очень большой, сильно изогнутый и острый коготь внутреннего пальца), насекомыми, отчасти ягодами и. семенами.
Центр возникновения семейства — Южная Америка. В пределах современного ареала из нижнего олигоцена описан 1 из ныне живущих и 1 ископаемый вид. Кроме того, из отложений нижнего плиоцена Южной Америки описано 2 рода с несколькими видами кариамо-подобных птиц, которых относят к семейству (19-е) Hermosiorni-thidae, а из нижнеолигоценовых отложений Северной Америки (Колорадо, Небраска) — 1 род с 4 видами, выделяемый в семейство (20-е) Bathornithidae.
Все семейства подотряда кариам, вероятно, близки друг
к другу и, видимо, представляют результат адаптивной радиации исходной предковой группы. Морфологические особенности подотряда говорят об очень давних,
но, кажется, несомненных родственных связях с другими группами журавлеобразных, особенно с трубачами и дрофами. Сходство с птицей секретарем (из отряда дневных хищников) чисто конвергентное.
ПОДОТРЯД 8. OTIDES—ДРОФИНЫЕ
Объединяет хорошо бегающих птиц средних и крупных размеров (вес 0,5—15 кг). Населяют открытые ландшафты.
Ноздри сквозные, голоринальные. Череп схизогнатический. Шейных позвонков 16—17. По заднему краю грудины пара вырезок. Ножная формула BXY. Ноги трехпалые: передние пальцы укорочены, заднего нет. Гортань трахео-бронхиальная; голосовая мускулатура развита слабо или отсутствует. Хорошо развиты слепые кишки. У самцов есть рудиментарный пенис. Контурное перо с побочным стержнем. Пух (растет только на аптериях) и основания контурных перьев слущиваются, образуя обильную «пудру». Крыло диастатаксическое. Первостепенных маховых 10—И, рулевых 12—20. Линька маховых постепенная, без потери способности к полету.
Включает 2 семейства.
СЕМЕЙСТВО 21. OTIDIDAE — ДРОФИНЫЕ
Птицы средней и крупной величины (вес 1—16 кг). Населяют степи, полупустыни и некоторые пустыни Старого Света (см. рис. 93).
Коренастые птицы (рис, 98), по внешнему облику напоминающие куриных. Клюв короткий, слегка вздутый. Шейных позвонков 16—17. В основании пальцев ноги нет следов плавательной перепонки. Первостепенных
176
Рис. 98. Самец австралийской дрофы Ardeotis australis (Gr') во время тока; на заднем плане в спокойной позе
маховых 10. Рулевых 14—20. Копчиковая железа отсутствует. В большей или меньшей степени развит половой диморфизм: самцы обычно крупнее самок, несколько более яркой окраски, часто имеют украшающие перья. У самцов некоторых дроф ко времени наступления половой зрелости образуется подкожный горловой мешок, который сообщается с ротовой полостью. Во время тока он наполняется воздухом и сильно раздувается, резко изменяя общий облик птицы (рис. 98).
Живут в открытых ландшафтах, избегая участков с кустарниками. Хорошо ходят и бегают. При опасности убегают, иногда затаиваются. Полет тяжелый, но довольно быстрый. Держатся одиночками или стаями. Кочующие, в более северных районах перелетные. Активность дневная. Питаются разнообразной растительной и животной пищей: молодыми побегами, семенами, луковицами, насекомыми, мелкими рептилиями и т. п.
Характерно довольно сложное токовое поведение; у немногих видов — токовой полет. Часть видов моногамы, часть полигамы. Гнездо — небольшая лунка, слабо выстланная растительной ветошью — располагается на открытом месте среди травянистых растений. В кладке у крупных видов 1—3, у мелких 3—5 яиц. У моногамов насиживает самка, самец держится вблизи гнезда. Выводок водят оба партнера. У полигамов самцы обычно токуют поодиночке и собирают около себя иногда несколько самок. Насиживает и водит выводок только самка. Инкубация около 1 месяца. Птенцы вылупляются зрячими, хорошо опушенными (окраска пестрая, покровительственная); кормятся сами. Становятся способными к полету и почти достигают размеров взрослых в возрасте около 2 месяцев, у крупных видов — позже. Половозрелыми становятся в возрасте 2—3 лет.
В семействе 22 вида, обычно относимых к 11 родам: Otis, Afrotis, Ardeotis, Chlamydotis, Eupodotis, Houbaropsis, Lissotis, Lophotis, Neotis, Sypheo-tides, Tetrax.
177
Рис. 99. Авдотка Burhinus oedicnemus (L.)
В СССР гнездятся 3 вида трех родов. Ареал и численность всех видов в связи с неумеренным преследованием и хозяйственным освоением степей и полупустынь в последнее столетие резко сократились.
Ископаемые остатки известны из среднего эоцена (Германия) и более молодых отложений. Их относят к 3 ископаемым и 3 ныне живущим видам. Центр возникновения семейства, вероятно, степи Африки.
СЕМЕЙСТВО 22. BURHINIDAE*—АВДОТКОВЫЕ
Птицы средней величины (вес около 0,5 кг). Распространены в умеренных и жарких широтах Старого Света и в ограниченных рацонах Центральной и Южной Америки (см. рис. 31). По внешнему облику (рис. 99) напоминают куликов, по поведению во многом сходны с дрофами.
Клюв удлиненный (составляет более половины длины головы), крепкий. Ноздри расположены, примерно, посредине клюва. Большие глаза. Шейных позвонков 16. В основании пальцев слабо развита плавательная перепонка. Первостепенных маховых 11, рулевых 12—14. Окраска пестрая. Заметного полового диморфизма нет.
Активны преимущественно в сумерках. Живут в открытых ландшафтах: по участкам степи с изреженным травяным покровом, в полупустынях и пустынях, по песчаным отмелям крупных рек. Держатся одиночками и парами, осенью объединяются в небольшие стайки. Кочующие, в более северных районах перелетные. Прекрасно бегают, хорошо летают. Видимо, в основном животноядны (различные насекомые, моллюски, мелкие ящерицы и т. п.).
Моногамы. Гнездо — небольшая лунка в земле, иногда слабо выстланная растительной ветошью. В кладке 2, реже 3 яйца пятнистой окраски. Насиживают или оба партнера, или только самка, но самец держится в районе гнезда. Инкубация около 4 недель. Птенцы вылупляются зрячими, опушенными. Водят птенцов оба родителя. При опасности взрослая птица стара-
1 Уэтмор (I960) придает авдоткам ранг надсемейства и включает их в подотряд куликов отряда ржанкообразных. По Фюрбрингеру (1888), ряду других систематиков и по последним тщательным исследованиям К. А. Юдина (1965), авдотки имеют много общих черт с дрофами и должны объединяться с ними. Сходство с куликами у авдоток поверхностное и, вероятно, чисто конвергентное. Учитывая эти данные, видимо, необходимо внести эту поправку в систему Уэтмора и включить авдоток в подотряд дроф.
178
ется незаметно отбежать от гнезда или птенцов, а затем начинает энергично отводить. Молодые становятся способными к полету в возрасте около 1,5— 2 месяцев. У части видов, вероятно, 2 кладки в сезон размножения.
В семействе авдоток 9 видов, относимых к 3 родам: Burhinus, Esacus, Orthorhamphus (последние 2 рода монотипичны). В СССР встречается только 1 вид — авдотка Burhinus oedicnemus (L.).
Ископаемые остатки, относимые к одному современному и 2 ископаемым видам, обнаружены в нижнем миоцене (Франция) и в более поздних отложениях. Центр возникновения семейства, видимо, Старый Свет.
ОТРЯД 21. DIATRYMIFORMES — ДИАТРИМООБРАЗНЫЕ
Включает только ископаемые формы. По одному довольно хорошей сохранности скелету из отложений нижнего эоцена США (Вайоминг) и
отдельным костям из~ эоцена — палеоцена из разных пунктов Северной Америки и Европы (Англия, Франция) описано 7 родов и 13 видов крупных птиц, относимых к 2 семействам: Diatrymidae и Gastornithidae. Описанная по почти полному скелету Diatryma steini (рис. 100) — грузная птица, достигавшая высоты около 2 л; на относительно короткой шее была большая голова (длина черепа около 45 см) с мощным, сжатым с боков клювом и массивной нижней челюстью. Череп десмогнатического типа, голоринальный. Пояс передних конечностей и крыло заметно редуцированы;
Рис. 100. Диатрима Diatryma steini Matt, (реконструкция)
явно не могла летать.
Предполагают, что диатримообразные близки к отряду журавлеобразных и даже, может быть, представляют одну из его сильно обособившихся и затем вымерших ветвей.
ОТРЯД 22. CHARADRIIFORMES —РЖАНКООБРАЗНЫЕ
Птицы мелких и средних размеров (вес от 20 г до 3 кг) с характерным обликом куликов, чаек, чистиковых. Ведут преимущественно околоводный или водный образ жизни.
Число общих признаков для всех групп отряда относительно невелико, но в комплексе они довольно отчетливо обособляют ржанкообразных от других, даже родственных отрядов. Клювы очень разнообразной формы. Череп схизогнатический с развитым сошником; у некоторых групп более или менее выражен переход к эгитогнатизму. Функционирующие базиптеригоидные сочленения есть у птенцов; у части видов сохраняются и у взрослых. Обычно хорошо развиты надорбитальные железы. Шейных позвонков 12—16. Грудные позвонки чаще сливаются в спинную кость; у чистиковых она не образуется. На заднем крае грудины одна или две пары вырезок. Вилочка хорошо развита. Истинных ребер 5—8 пар. Цевка и нижний конец голени не оперены. Задний палец у всех групп, кроме якан, сильно редуцирован, у части видов полностью исчезает. Пневматичность скелета вы-
179
Рис. 101. Филогенетическое древо отряда ржанкообразных (по К. А. Юдину, 1965)
ражена относительно слабо. Гортань трахео-бронхиальная. Сонные артерии парные. Пищевод легко растяжим, зоба обычно нет. Мускульный желудок с мощными стенками. Слепые кишки развиты в разной степени. Есть желчный пузырь. Побочный стержень контурного пера обычно развит. Копчиковая железа оперена. Крылья заостренные, диастатаксические (исключение— белые ржанки). Первостепенных маховых 11; первое мало и часто плохо видно. Рулевых 12—18, редко больше. Пух растет по птерилиям и аптериям. В году две линьки: полная послегнездовая и частичная весенняя (предбрачная). Половой морфизм окраски свойствен немногим видам; у большинства хорошо выражено возрастное и сезонное ее изменение.
Большинство видов моногамы. Яйца обычно с окрашенной скорлупой и более или менее отчетливой пятнистостью. Птенцы выводкового типа, но у многих видов выкармливаются родителями до подъема на крыло. Распространены практически по всему земному шару.
Отряд ржанкообразных включает 3 подотряда с 17 семействами, объединяющими 287—289 видов, относимых к 87—113 родам.
В фауне СССР отмечено 132—133 вида 51—56 родов (несомненно гнездится 120—121 вид) из 10 семейств всех 3 подотрядов.
Журавлеобразные, видимо, могут рассматриваться как давние предки ржанкообразных. Примитивные генерализованные ржанкообразные дали начало голубеобразным. Некоторое сходство ржанкообразных с трубконосыми, вероятно, результат конвергентной эволюции. Внутри отряда ржанкообразных более примитивен подотряд куликов; от него обособились, эво-люируя по пути приспособления к водному образу жизни, подотряды чаек и чистиковых (рис. 101).
ПОДОТРЯД 1. CHARADRII — КУЛИ КОВЫЕ
Птицы мелких и средних размеров (вес от 20 г до 1 кг, редко немногим больше) с более или менее выраженным обликом куликов. Характерен око-ловодный образ жизни; лишь немногие виды освоили сухие местообитания.
180
150° 120° 90° 60°к западу от Гринвича 0 к востоку от Гринвта60° 90° 120° 150° 180° 150°
Рис. 102. Ареалы семейств ржанкообразных птиц:
1— яклновые Jacanldae, 2— цветные бекасы Rostratulidae, 3—кулики . ор жи Haematopodidae, 4—плавунчиковые Phalaropodidae
Клюв разнообразной формы. Ноздри сквозные или несквозные. Череп схизогнатический, иногда с тенденцией к эгитогнатизму. У части видов взрослые имеют функционирующие базиптеригоидные сочленения. Шейных позвонков 12—16. По заднему краю грудины чаще 2 пары вырезок. Истинных ребер 5—7 пар. Ножная формула ЛВЛТ4-; у некоторых родов ЛХУ4-. Ноги у большинства более или менее удлиненные. Слепые кишки развиты у одних и рудиментарны у других видов. Рулевых чаще 12—14, у некоторых видов (бекасы и др.) 14—18 и больше.
Бродят по мелководью или по суше, добывая пищу; некоторые виды склевывают добычу с поверхности воды, плавая. Прекрасно летают. Обычно моногамы (есть исключения). Кладки из 2—5 яиц, обычно пестрой окраски и резко, грушевидной формы, откладываемых прямо на землю в небольшую лунку или в небрежно высланный растительной ветошью лоток. У большинства видов птенцы оставляют гнездо сразу после вылупления. Выкармливают птенцов немногие виды.
Подотряд Куликовых разделяется на 6 надсемейств1, включающих 13 семейств (из них 2 только с ископаемыми формами) с 181 ныне живущим или вымершим в историческое время видом, распределяемых между 58— 83 родами. В СССР встречается 79 видов, относимых к 31—34 родам и 7 семействам двух надсемейств. Достоверно гнездится 72 вида.
Надсемейство 1. Jacanoidea — якановые
Включает только одно семейство.
СЕМЕЙСТВО I. JACANIDAE— ЯКАНОВЫЕ
Объединяет 7 видов, относимых к 6 родам: Jacana, Actophilornis, Hyd-rophasianus, Irediparra, Metopidius, Microparra. Обитают на сильно заросших пресных водоемах всей тропической зоны (рис. 102), включая ряд океанических островов. В СССР не встречаются.
Мелкие, стройные птицы (рис. 103), похожие на пастушков. Клюв уд-
Рис. 103. Якана Jacana jacana L.
линенный, слегка сжат с боков. Базиптеригоидные сочленения есть и у взрослых. Затылочных фонтанелей нет. Надорбитальные железы развиты слабо. Шейных позвонков 16. По заднему краю грудины одна пара вырезок. Ребер 5 пар. У обоих полов на кистевом сгибе крыла есть костная шпора, покрытая роговым чехлом. Ноги длинные. Передние и задние пальцы длинные, тонкие, с длинными, слабо изогнутыми когтями. Гортань с одной парой тра-хео-бронхиальных мышц. Слепые кишки очень малы. Побочный ствол контурного пера развит слабо. Окраска обычно
1 Уэтмор (1960) включает сюда еще 7-е надсемейство авдоток. В нашем обзоре они рассматриваются в подотряде дроф отряда журавлеобразных.
182
из сочетания разных оттенков черного и белого цвета; часто есть металлический отблеск. Самки немного крупнее самцов.
Живут на водоемах, густо поросших растительностью. Бегая по плавающим растениям, ловят насекомых, моллюсков, рачков; иногда едят семена и мягкие побеги. Хорошо плавают и ныряют. При опасности скры ваются в зарослях. Гнездо устраивают на плавающих растениях, реже в заломах среди зарослей надводных растений. В кладке обычно 4, реже 3—5 яиц. Часть видов моногамы, у которых насиживают и водят выводок оба пола или только самец. Насиживание около трех недель. У индийской Metopidius indicus (Lath.) два самца заняли и охраняли соседние участки. С обоими самцами спаривалась одна самка. Гнезда строили самцы; каждому помогала самка, отложившая в одно гнездо 3, а в другое — 4 яйца. Насиживали только самцы. У широко распространенной в Юго-Восточной Азии Hydrophasianus chirurgus (Scop.) около одной самки держатся 3—4 самца, занимающих смежные участки. Самка откладывает яйца в гнездо каждого самца. Предполагается, что самец иногда успевает вырастить 2 выводка. Самка за сезон откладывает 6—8 кладок. Птенцы вылупляются зрячими, хорошо опушенными, хорошо бегают и плавают. Кормятся сами, но у некоторых видов, возможно, их еще подкармливает самец. В возрасте 2—3 недель начинают вести самостоятельную жизнь. Оперение взрослых одевают к концу второго года жизни; в это же время становятся половозрелыми.
Яканы — рано обособившаяся ветвь древних ржанкообразных, занявших своеобразную жизненную нишу. Их древность подтверждается сохранением некоторых черт сходства с журавлеобразными. Ископаемые формы не найдены.
Надсемейство 2. Charadrioidea — собственно кулики
Птицы мелких и средних размеров с характерным обликом куликов.
Клюв разнообразной формы и длины. Ноздри схизоринальные, обычно сквозные. Часто сохраняются функционирующие базиптеригоидные сочленения. Есть затылочные фонтанели. Шейных позвонков 12—15. Истинных ребер 6—7 пар. У большинства видов по заднему краю грудины две пары вырезок, у части видов — одна пара. Шпора на кисти крыла есть только у немногих видов. Крылья чаще острые, довольно длинные. Передние пальцы ноги средней длины; задний палец очень мал или совсем отсутствует. В желудке выделяется пилорический отдел. Контурные перья с хорошо развитым побочным стволом. Половой и возрастной морфизм окраски резко выражен у немногих видов; чаще встречается сезонное изменение окраски.
Большинство видов моногамы; у немногих видов пары не образуются даже на короткое время (цветные бекасы, турухтан, бекасы, вальдшнеп). Почти у всех видов отчетливо выражены токовые явления. Некоторые виды гнездятся колониально. У большинства кладка — 4 резко грушевидных яйца. В умеренных и северных широтах перелетны, в тропиках оседлые или кочующие. Вне сезона размножения держатся группами или стаями, иногда насчитывающими сотни и тысячи особей; только немногие виды ведут одиночный образ жизни. Распространены повсеместно, кроме Антарктики.
Настоящие кулики — одна из основных ветвей отряда ржанкообразных, очень давно (в самом начале эволюции от генерализованных предков), видимо, обособившиеся от якан. Основное направление адаптивной радиации — приспособление к жизни в различных околоводных местообитаниях.
Надсемейство куликов подразделяется на 8 семейств (из них 2 представлены только ископаемыми формами), включающих 41—66 родов с 151 видом; несколько видов недавно вымерло. В СССР встречается 75 видов, включаемых в 29—32 рода 6 семейств. Достоверно гнездятся 69 видов.
183
СЕМЕЙСТВО 2. RHEGMINORNITHIDAE
По остаткам из нижнего миоцена Флориды (США) описан 1 вид, видимо, занимающий в известной степени промежуточное положение между яка-нами и настоящими куликами.
СЕМЕЙСТВО 3. ROSTRATULIDAE — ЦВЕТНЫЕ БЕКАСЫ
Сюда включают всего 2 вида: афро-азиатский Rostratula benghalensis (L.), распространен в тропиках Старого Света, в Австралии и Тасмании, а Nycticrypes semicollaris Vieill. — в Южной Америке (рис. 102). В Приморском крае добывался первый вид; гнездование не доказано.
По размерам и внешнему облику похожи на обыкновенного бекаса (рис. 104), но окраска более яркая. Клюв удлиненный, прямой. Крылья короткие и широкие. Первостепенных маховых 10 (первое сильно редуцировано), рулевых 14—16; хвост короткий. Пальцы ноги длинные и тонкие, следов плавательной перепонки нет; задний палец хорошо развит, хотя много короче передних. Четко обособлен зоб. Слепые кишки относительно длинные, функционирующие.
Ведут скрытный образ жизни, встречаются по болотам и сырым берегам водоемов. Активны преимущественно в сумерках и ночью. Кормятся среди растительности, реже на открытых отмелях. Пищу, видимо, добывают главным образом при зондировании клювом сырой почвы. Едят разнообразных почвенных и наземных беспозвоночных, иногда мелкие клубни и семена. Самки крупнее самцов и у афро-азиатского вида более ярко окрашены. Гнездо устраивается в зарослях на берегу или на заломах надводных зарослей. У южноамериканского вида в кладке 2 яйца; насиживают самец и самка. У австралийского подвида афро-азиатского цветного бекаса токующая самка совместно с самцом строит гнездо и откладывает обычно 4, редко 5—6 яиц, которые в течение 17—19 дней насиживает самец; он же водит выводок. У азиатского подвида после завершения первой кладки, которую насиживает самец, самка объединяется с другим самцом и откладывает вто-
Рис. 104. Цветной бекас Rostratula benghalensis (L.)
184
рую кладку, все заботы о которой падают на второго самца. Водящие выводок самцы не только греют птенцов, но и кормят их; постепенно птенцы начинают кормиться сами.
Цветные бекасы — древняя специализированная боковая ветвь настоящих куликов. Вероятно, возникли в тропиках Старого Света. По остаткам из среднего миоцена Германии описан вид, возможно, близкий к современным цветным бекасам.
СЕМЕЙСТВО 4. HAEMATOPODIDAE— КУЛИКИ СОРОКИ
Сюда входит 1 род Haematopus с 4 видами, широко, но очень спорадично распространенными по морским побережьям и кое-где по берегам крупных внутренних водоемов (рис. 102). В СССР встречается и гнездится только 1 вид (рис. 105)
Размеры средние (вес около 0,5 кг). Клюв длинный, почти прямой, сильно сжатый с боков. Хорошо развиты ямки для надорбитальных желез. Кисть короче предплечья. Ноги относительно короткие: цевка чуть длиннее бедра. Пальцы относительно короткие и толстые. Между основанием наружного и среднего пальцев есть плавательная перепонка; заднего пальца нет. Побочный стержень может достигать половины длины контурного пера. Окраска черная или из сочетания черного и белого. Полового морфизма нет; у молодых окраска более тусклая.
Хорошо бегают; могут плавать и даже нырять. Моногамы. Гнездятся отдельными парами или диффузными группами. Гнездо—мелкая лунка, выстланная растительной ветошью или камешками. В кладке 3, редко 2 или 4 яйца. Насиживают оба партнера около 4 недель. Пуховики держатся в районе гнезда. Взрослые их кормят около 3 недель, передавая пищу из клюва в клюв или бросая ее перед птенцом. Половая зрелость наступает в возрасте около 3 лет. Питаются разнообразной животной пищей, которую ловят на мелководье, и на суше, под камнями, в верхнем слое почвы и т. п. Поедают двустворчатых моллюсков: раковину открывают, вводя между створками уплощенный сбоку клюв. Редко ловят мелкую рыбу.
Специализированная ветвь куликов, видимо, близкая к ржанкам. Центр возникновения, возможно, Новый Свет. Наиболее ранние фрагментарные ископаемые остатки — из нижнего миоцена Северной Америки. Описано 2 ископаемых вида, а в более поздних слоях обнаружены остатки обыкновенного кулика сороки.
Рис. 105. Кулик сорока Haematopus ostralegus L.
185
СЕМЕЙСТВО 5. CHARADRIIDAE — РЖАНКОВЫЕ
Кулики мелкой и средней величины (вес 20—600 г) с относительно большой головой, короткими шеей и ногами, с характерным обликом зуйка или чибиса (рис. 106).
Клюв средней длины или короткий. Ротовое отверстие широкое. Ямки для надорбитальных желез развиты слабо, хотя сами железы крупные. Цевка широкая. Передние пальцы относительно короткие и толстые; между их основаниями может быть слабо развитая плавательная перепонка. Задний палец мал или его нет вовсе. Половой и возрастной диморфизм окраски выражен лишь у части видов.
Населяют преимущественно открытые, иногда сухие ландшафты: отмели по берегам морей и внутренних водоемов, тундры, луга и болота, степи, долины горных рекит. п. Некоторые виды более активны ночью. Гнездятся отдельными парами; некоторые виды— диффузными колониями. Гнездо — небольшая лунка, часто без всякой подстилки. В кладке обычно 4 яйца, у немногих видов 3 и даже 2. Насиживают оба партнера, у немногих видов — только самка, у хрустана Charadrius morinellus (L.) — самец. Инкубация 3—4 недели. Птенцы, видимо, с первых дней жизни кормятся сами; взрослые их только греют и охраняют, «отводя» приближающегося врага. У части видов (тулес, ржанка и др.) взрослые отлетают еще до подъема молодых на крыло. У других (чибисы) молодые птицы кочуют и отлетают вместе со взрослыми. Половозрелыми становятся в конце первого или второго года жизни. Пища — разнообразные беспозвоночные, которых собирают на поверхности почвы, в полосе заплеска или на мелководье. Характерно выпугивание прячущихся в грунте червей и насекомых частыми ударами лапки.
До сих пор нет единства взглядов на объем родов. Так, по мнению одних систематиков, все чибисы и близкие к ним виды должны объединяться в один род Vanellus, другие же распределяют их между 16—20 родами. Чис-
Рис. 106. Чибис Vanellus vanellus (L.)
186
Рис- 107. Ареалы семейств ржанкообразных птиц:
I— ржанкозыг Characlriidae, 2—бекасовые Scolopacidae
ло родов ржанок варьирует по разным представлениям от 8 до 12. Таким образом, в семействе от 9 до 32 родов: Charadrius, Afribyx, Anarhynchus, Апо-malophrus, Belonop terus, Chettusia, Elsyornis, Erythrogonys, Eudromias, Hemiparva, Hoplopterus, Hoploxypterus, Lobibyx, Lobipluvia, Lobivanel-lus, M icrosar cops, Oreopholus, Phegomis, Pluvial is, Pluvianellus, Pluvlo-rhynchus, Ptiloscelyx, Rogibyx, Sarciophorus, Squatarola, Stephanibyx, Thinorrtis, Tylibyx, Vanellus, Vanellochettusia, Xiphidiopterus, Zonibyx.
Всего в семейство вводит 60 видов (это число принимается большинством орнитологов), распространенных повсеместно, кроме Антарктики (рис. 107). Тяготеют к теплым широтам, в Арктику проникают немногие виды. В СССР встречается 18 видов, относимых к 7—10 родам; достоверно гнездится 16.
Вероятный центр возникновения — тропики Нового Света. Ископаемые остатки описаны из нижнего олигоцена Европы и из миоцена Северной Америки; их относят к 5 ископаемым и к 12 ныне живущим видам. Ржанковые — одна из основных ветвей подотряда куликов.
СЕМЕЙСТВО 6. SCOLOPACIDAE— БЕКАСОВЫЕ
Кулики различных размеров (вес от 20 г др 1 кг) и разнообразного облика (бекасы, песочники, кроншнепы, улиты и др.; рис. 108).
Клюв у большинства видов длинный, тонкий, иногда несколько расширен к вершине, у некоторых видов довольно сильно изогнут книзу, реже слегка загнут кверху. На конце клюва обычно много осязательных телец. У немногих видов клюв относительно короткий, сильный, конический. Ноги могут быть длинными или относительно короткими. Задний палец мал или отсутствует. У основания передних пальцев у некоторых видов развита плавательная перепонка. У части видов отчетливый возрастной и половой морфизм окраски; у других его нет.
Населяют преимущественно открытые пространства достаточного и избыточного увлажнения (не встречаются в сухих степях); немногие виды живут в сырых лесах, придерживаясь полян, долин ручьев и т. п. Большинство видов с дневной активностью, немногие с сумерочной и ночной. Большинство моногамы, немногие полигамы (турухтан, вальдшнеп). Обычно гнездятся одиночными парами, реже диффузными колониями. Гнездо — лунка в земле, со скудной выстилкой. В кладке, как правило, 4 яйца, в повторной кладке 3. Насиживают и водят птенцов оба пола (веретенники, кроншнепы, песочники) или только самка (турухтан, вальдшнеп и др.). Насиживание около 3 недель или чуть больше. Вылупившиеся пуховички кормятся сами. Начинают подлетывать в возрасте 2—3 недель. Половозре-
Рис. 108. Вальдшнеп Scolopax rusticola L.
188
лыми становятся в конце первого или второго (видимо, чаще) года жизни. Питаются разнообразной животной пищей: червями, моллюсками, ракообразными, насекомыми и их личинками; крупные виды иногда ловят мелкую рыбешку и земноводных. Добычу ловят на мелководье или на суше, зондируют клювом мягкую влажную почву. Некоторые виды (кроншнепы, веретенники и др.) охотно поедают ягоды.
Семейство объединяет 24—26 родов с 75 видами; по широте распространения (рис. 107) лишь немного уступают ржанковым. В отличие от последних большинство видов бекасовых тяготеет к умеренным и холодным широтам Северного полушария.
Обычно выделяют несколько подсемейств.
1. Песочников ые Calidritinae. Calidris (Erolia), Cro-cethia (иногда включают в первый род), Eurynorhynchus, Limicola, Micro-palma, Philomachus, Tryngites.
2. Улиты Tringinae. Tringa, Actitis (иногда включают в род Tringa'), Aechmorhynchus, Catopterophorus, Heteroscelus, Prosobonia, Xenus (Terekia).
3. Бекасов ые Scolopacinae. Scolopax, Chubbia, Caeno-corypha, Gallinago, Lymnocryptes, Philohela.
4. Веретенниковые Limosinae. Limosa, Bartramia, Mac-ror hampus (Limnodromus), Numenius.
5. Камнешарковые Arenariinae. Arenaria, Aphriza.
В СССР встречается 50 видов 16 родов (включен и эскимосский кроншнеп Numenius borealis (Forst.), видимо, полностью исчезнувший в последние годы); достоверно установлено гнездование 47 видов.
Как и ржанковые, бекасовые — одна из основных ветвей подотряда куликов. Ископаемые остатки известны из верхнего эоцена Европы (улиты) и Северной Америки (бекасы) и более поздних отложений. Описано 24 ископаемых вида!; в ископаемых остатках найден 31 из ныне живущих видов. Центр возникновения семейства не ясен. Предполагают, что улиты формировались в лесной зоне Евразии, песочники — в тундрах Старого и Нового Света, бекасовые — в Азии.
СЕМЕЙСТВО 7. RECURVIROSTRIDAE — ШИЛОКЛЮВКОВЫЕ
Сюда включают куликов характерного облика: шилоклювок (рис. 109), ходулочников, серпоклювов; вес 100—500 г.
Клюв длинный, тонкий, прямой (ходулочники), длинный, загнутый кверху, слегка уплощенный (шилоклювки) или длинный, крепкий, загнутый книзу (серпоклюв). Ноги средней длины (серпоклюв) или длинные (ши-
локлювки, особенно ходулочники); особенно удлинены цевка и голень. Перепонки между пальцами хорошо развиты (шилоклювки) или сильно редуцированы. Задний палец мал (шилоклювки, серпоклюв) или его нет вовсе. Оперение густое, плотноё. Хорошо развит пух, растущий по птерилиям и аптериям. Половой и сезонный морфизм окраски практически отсутствует; есть небольшие возрастные изменения.
Ходулочники и шилоклювки населяют открытые берега преимущественно соло
Рис. 109. Шилоклювка Recurvirostra avosetta L.
189
новатых водоемов теплых широт (рис. НО); серпоклювы держатся по каменистым долинам речек высокогорий Центральной Азии.
Моногамы. Ходулочники и шилоклювки гнездятся обычно группами, при низкой численности — отдельными парами; серпоклювы — отдельными парами. Гнездо на открытом месте близко от воды, выложено растительной ветошью или (серпоклюв) камешками. В кладке 3—4 яйца. Насиживают оба пола 3—4 недели. Защищая гнездо и птенцов, обычно энергично «отводят». Птенцов водят оба родителя. После подъема птенцов на крыло держатся семьями, потом объединяются в стайки. Шилоклювки хорошо плавают.
Активность дневная. Питаются разнообразной животной пищей — ракообразными, моллюсками, червями, насекомыми, очень редко — мелкими рыбешками, которых ловят в воде, бродя по мелководьям; реже охотятся на суше. Серпоклювы, отыскивая пищу, часто переворачивают камешки.
В семействе 7 видов, объединяемых в 4 рода: Recurvirostra, Cladorhyfichus, Н itnantopus, 1 b idorhyncha. В южных районах СССР встречаются и гнездятся 3 вида 3 родов.
Одна из боковых ветвей ржанковых, специализировавшихся на добывании пищи в воде. Остатки одного вида найдены в нижнем эоцене США; в более поздних отложениях в разных районах обнаружены еще 2 ныне живущих вида.
СЕМЕЙСТВО 8. PRESBYORNITHIDAE
По остаткам из эоценовых отложений в США (Юта) описан крупный кулик, возможно, приспособленный к водному образу жизни.
СЕМЕЙСТВО 9. PHALAROPODIDAE — ПЛАВУНЧИКОВЫЕ
В семейство входит один род Phalaropus с 3 видами мелких куличков (вес менее 100 г), распространенных только в Северном полушарии (см. рис. 102). Некоторые авторы каждый вид выделяют в самостоятельный род. В СССР встречается и гнездится 2 вида (рис. 111).
Клюв удлиненный, тонкий или уплощенный. Костные углубления для надорбитальных желез узкие и неглубокие, хотя железы очень велики и на две трети свободно нависают над орбитами. Грудина удлиненная. Ноги средней длины. Каждый из трех направленных вперед пальцев имеет кожистую оторочку. Задний палец мал; у одного вида его тоже окаймляет кожистая лопасть. Оперение плотное, густое. Побочный ствол контурного пера короткий, но с длинными бородками. Хорошо выражен сезонный и возрастной морфизм окраски; самки немного крупнее самцов и несколько ярче.
Гнездятся в северных районах по заболоченным берегам озер. На зимовках широко рассеиваются по океанам, проникая и в Южное полушарие; реже зимуют на внутренних водоемах. Полет быстрый. Очень хорошо бегают по суше, прекрасно плавают. Кормятся, бегая по берегам или плавая кругами на воде. Добычу — разнообразных мелких животных —склевывают с поверхности воды или земли, с растений. .
Токуют самки. Гнездо строят оба партнера. В кладке обычно 4 яйца. Закончив кладку, самки собираются в стайки, кочующие в районе гнездования и вскоре отлетающие к югу. Насиживают только самцы 19—21 день. Вылупившиеся и обсохшие птенцы бродят с самцом по берегам, уже с первых дней хорошо плавают; в редких случаях с птенцами встречали и самок. Молодые взлетают в возрасте 3—4 недель. Выводки быстро объединяются в кочующие стайки.
Плавунчики близки к бекасовым (улитам и песочникам) и, видимо, обособились от них, адаптируясь к питанию преимущественно планктонными организмами.
190
Рис. ПО. Ареалы семейств ржанкообразных птиц:
1— шилоклювковые Recurvirostridae, 2- рачьи ржанки Dromadidae, 3— тиркушковые Glareolidae, 4- зобатые бегунки Thinocoridae, 5— белые ржанки Chionididae
Рис. 111. Круглоносый плавунчик Phalaropus lobatus (L.) Сбоку его лапа:
на пальцах кожистые оторочки
Надсемейство 3. Dromadoidea — рачьи ржанки
В надсемейство входит лишь одно семейство.
СЕМЕЙСТВО 10. DROMADIDAE — РАЧЬИ РЖАНКИ
Единственный вид — Dromas ardeola Payk. (рис. 112), распространен по песчаным побережьям Красного моря и отдельным участкам берегов Индийского океана (рис. ПО).
Клюв мощный, слегка сжатый с боков, с острой вершиной. Ноздри сквозные. Надорбитальные железы велики. Функционирующих базипте-ригоидных сочленений нет. Ноги длинные. Основания передних пальцев соединены плавательной перепонкой; коготь среднего пальца гребенчатый. Задний палец маленький, но несет коготок. Окраска черная с белым. Полового морфизма нет. Весят около 300 г. Питаются главным образом крабами и моллюсками, которых ловят, медленно бродя по мелководью. Моногамы. Гнездятся обычно колониями. В песчаных дюнах побережья роют нору, иногда длиной до 2 и более метров, в конце которой откладыва-
ется одно очень крупное (до 25% веса самки) чисто белое яйцо. Птенцы вылупляются зрячими и покрытыми одноцветным серым пухом. В гнезде остаются длительное время; выкармливаются родителями.
В ископаемых остатках не обнаружены. Видимо, близки к тиркушкам и вместе о ними, вероятно, обособились от примитивных ржанок, которые дали начало и чайкам.
В своей системе после рачьих ржанок Уэтмор (1960) ставит надсемейство авдоток Burhinoidea (см. стр. 178).
Рис. 112. Рачья ржанка Dromas ardeola Payk.
192
Надсемейство 4. Glareoloidea — тиркушковые
Включают единственное семейство.
СЕМЕЙСТВО 11. GLAREOLIDAE— ТИРКУШКОВЫЕ
Объединяет тиркушек и бегунков. Размеры небольшие (вес 50—200 г). Клюв относительно короткий, с выпуклым надклювьем, кончающимся более или менее выраженным острым крючком. Ноздри не сквозные. Базип-теригоидных сочленений обычно нет. Крупные надорбитальные железы. У части видов — собственно тиркушек (рис. 113) — ноги короткие, задний палец есть, хотя и мал, крылья длинные и острые, хвост длинный, вильчатый; в полете напоминают ласточек. Другие виды — бегунки (рис. 114) — по внешнему виду напоминают ржанок: более длинные ноги с гребенчатым когтем среднего пальца, задний палец обычно отсутствует, крылья более короткие, короткий хвост прямо обрезан. В отличие от других семейств подотряда семейство тиркушковых характеризуются экологической неоднородностью: нет единого направления специализации.
Тиркушки (Glareola, Galachrysia, Stiltia) распространены преимущественно в степной зоне, местами проникают в полупустыни. Гнездятся на открытых ровных сухих или влажных участках вблизи водоемов, а иногда и на значительном удалении от них. Кормятся преимущественно крупными насекомыми, которых собирают, бегая по земле, или ловят в воздухе; способны налету схватывать добычу с поверхности воды. Часть видов гнездится колониально, другие одиночными парами. Гнездо — еле заметная лунка в земле. В кладке обычно 2—3, редко 4 яйца. Насиживают и водят птенцов, видимо, оба партнера. Птенцы покрыты густым крапчатым пухом. Первые 1—2 недели взрослые кормят птенцов, потом они постепенно привыкают сами ловить добычу.
Бегунки (Cursor i us, Peltohyas, Pluvianus, Rhynoptilus) населяют полупустынные и пустынные районы со скудной растительностью; немногие виды живут на больших песчаных речных отмелях. Кормятся различными беспозвоночными, собирая их на поверхности почвы; крупные виды ловят и мелких ящериц. Гнездо — ямка в земле без всякой выстилки. Кладки у разных видов варьируют от 1—2 до 4 яиц. Часть видов присыпает отложенные яйца песком. Насиживают оба пола. Некоторые виды насиживают только ночью, утром и вечером кладка обогревается солнцем, а днем птицы
Рис. 113. Луговая тиркушка Glareola pratincole (L.)
193
Рис. 114. Бегунок Cursorius cursor (Lath.)
стоят над кладкой, защищая ее от перегрева. Часть видов первое время кормит птенцов, у других они сразу после вылупления кормятся сами.
Всего 7 родов (перечёнь дан выше) с 16 видами, распространенными в жарких и, отчасти, умеренных широтах Старого Света (рис. 110). В СССР встречаются 4 вида 2 родов; достоверно гнездятся 3 вида.
Ископаемых остатков нет. Возможный центр возникновения — Северная Африка. Обособились от примитивных ржанок, приспосабливаясь к жизни в маловодных или безводных местообитаниях.
Надсемейство 5. Thinocoroidea — зобатые бегунки
Включает единственное семейство.
СЕМЕЙСТВО 12. TH INOCORIDAE — ЗОБАТЫЕ БЕГУНКИ
Объединяет всего 2 рода (Thitiocoriis, Attagis) с 4 видами, населяющими
холодные сухие горные и равнинные степи и полупустыни Южной Америки
(рис. НО). Птицы мелких и средних размеров (вес 100—400 г), по внешнему виду (рис. 115) напоминающие трехперсток.
Клюв короткий, слегка вздутый. Череп близок к эгитогнати-ческому типу, с коротким и широким сошником. Базиптериго-идных отростков нет. Ноздри не сквозные. Ребер 5 пар. Крылья длинные и острые; первостепенных маховых 10. Ноги относительно короткие. Плавательной перепонки нет. Задний палец мал. Зоб большой, круглый, четко обособленный, мускульный желудок мощный, функционирующие сле-
Рис. 115. Зобатый бегунок Trinocorus пые КИШКИ И прямая кишка ДЛИН-orbignyianus (Less) ные. Густой черный пух растет
194
только по аптериям. Половые различия в окраске выражены слабо или отсутствуют. Хорошо бегают. Полет быстрый, стремительный.
Питаются преимущественно разнообразными семенами, реже вегетативными частями растений. Редко и, видимо, случайно поедают наземных беспозвоночных. Регулярно летают на водопой, иногда за несколько десятков километров. Моногамы. Гнездо — ямка в земле, иногда слабо выстланная травинками. В кладке обычно 4 яйца, резко грушевидных, пятнистых. Экология изучена плохо. Вне сезона размножения кочуют небольшими стайками.
В ископаемом состоянии не обнаружены. Специализировались на питании растительной пищей; сильно обособившаяся ветвь куликов, видимо, блрзкая к настоящим и белым ржанкам.
Надсемейство 6. Chionidoidea — белые ржанки
Включает одно семейство.
СЕМЕЙСТВО 13. CHIONIDIDAE— БЕЛЫЕ РЖАНКИ
Единственный род Chionis (Chionarchus) с 2 видами. Снежно-белые птицы (рис. 116), по внешнему виду несколько похожие на короткокрылых чаек или голубей. Вес около 600 г. Живут по побережьям островов Антарктики и Субантарктики (рис. ПО).
Клюв относительно короткий, слегка вздутый. Над ноздрями роговые крышечки. Ноздри сквозные. Сильно развиты ямки для надорбитальных желез. Базиптеригоидных отростков нет. Туловище округлое. Ноги довольно короткие. Задний палец мал. Между передними пальцами нет и следов плавательной перепонки. Зоба нет. Слепые кишки довольно длинные. Оперение густое и плотное. Полового морфизма окраски нет.
Всеядны: ловят на литорали различных беспозвоночных и едят водоросли, таскают яйца и птенцов других птиц, поедают падаль, придерживая ее лапой. Гнездятся одиночными парами. Объемистое гнездо из сухих водорослей, перьев пингвинов, обломков раковин и другого мусора строится в расщелинах скал или под валунами. В кладке 2—3 голубоватых с пятнами яиц. Насиживают, видимо, оба партнера, около 4 недель. Птенцы вылупляются зрячими, в густом коричневом с черными пятнами пуховом наряде. В гнезде остаются длительное время, выкармливаются родителями, потом бродят с ними по литорали. Вне сезона размножения держатся маленьки-
Рис. 116. Белая ржанка Chionis alba Gm.
195
ми стайками. Оседлы; более южные популяции кочующие или перелетные. Плавают редко и неохотно.
Ископаемых остатков нет. Боковая, сильно специализированная ветвь куликов, вероятно, ближе всего стоящая к настоящим ржанкам и зобатым бегункам.
ПОДОТРЯД 2. LARI — ЧАЙКОВЫЕ
Водные птицы мелких и средних размеров (вес от 40 г до 3 кг) с характерным обликом чайки или крачки.
Клюв удлиненный, несколько сжатый с боков, с заостренной вершиной или крючком на конце. Ноздри сквозные, схизоринальные. Череп схизогнатический, с хорошо развитым сошником. Базиптеригоидных сочленений у взрослых нет. Ямки для надорбитальных желез большие. Шейных позвонков 15. Задний край хорошо развитой грудины с двумя парами вырезок (у поморников — одна пара). Истинных ребер 6—7 пар. Вилочка без резкого расширения на конце. Предплечье — наиболее длинный элемент скелета крыла. Таз широкий. Ноги умеренной длины. Передние пальцы соединены плавательной перепонкой. Задний палец мал и расположен выше уровня передних; у некоторых видов исчезает. Гортань трахео-бронхи-альная, с хорошо развитыми голосовыми мышцами. Пищевод сильно растяжим, но зоба обычно нет. Мускульный желудок с толстыми стенками. Слепые кишки рудиментарны (лучше развиты у поморников). Побочный ствол контурного пера заметен. Крылья длинные, острые. Первостепенных маховых 11, но первое обычно недоразвито. Рулевых 12. Полового морфизма окраски нет; возрастная, а иногда и сезонная смена окраски хорошо выражена.
Моногамы, обычно, с длительно сохраняющимися парами. В начальных фазах размножения характерно ритуальное кормление самки самцом. Большинство видов более или менее колониальны. Гнезда варьируют: от простой лунки в земле до большой груды растительной ветоши. Гнездо строится на земле или уступах скал, у некоторых тропических крачек в развилке ветвей. Кладка из 2—3 (реже из 1 или 4—5) яиц, пестро окрашенных и слабо грушевидных. Насиживают оба партнера. Птенцы вылупляются зрячими, хорошо опушенными; выкармливаются родителями вплоть до подъема на крыло и даже некоторое время после взлета. У мелких видов половозрелость наступает к концу первого года жизни, у более крупных — в возрасте 2, 3 и даже 4 лет. Пищу собирают в полете над водой и пикируя на замеченную добычу, бродя по суше или по мелководью и некоторые виды при плавании. Многие виды хорошо плавают. Ныряют на небольшую глубину только с пикирования. У всех видов свободный, маневренный полет. Распространены практически по всему земному шару. В высоких и умеренных широтах перелетные, в тропиках оседлые или кочующие.
Подотряд чайковых включает 3 семейства, объединяющих 85—86 видов 16—17 родов. В СССР встречается 35 видов, относимых к 10—11 родам 2 семейств; установлено гнездование 32 видов.
СЕМЕЙСТВО 14. STERCORARHDAE — ПОМОРИИ КОВШЕ
Птицы средних размеров (вес от 400 г до 2 кг) с темными участками в оперении (рис. 117). Рамфотека надклювья состоит из 4 пластин, обычно не сливающихся друг с другом. Клюв с резко выраженным крючком на конце. По заднему краю грудины одна пара вырезок. Формула мускулатуры ноги AXY-j-. Слепые кишки довольно развиты. Средние рулевые более или менее удлинены. Полет быстрый, легкий, маневренный. Хорошо плавают.
Распространены по морским побережьям и прилегающим участкам тундры высоких широт (рис. 118). Практически всеядны: ловят на суше и на воде
196
Рис. 117. Длиннохвостый поморник Strercorarius longicaudus Vieill.
разнообразных беспозвоночных, таскают яйца и птенцов, добывают мышевидных грызунов, едят падаль, ягоды. Довольно отчетливо выражен своеобразный паразитизм: стремительно пикируя, отбирают корм у многих птиц (чайки, крачки, чистиковые и др.), несущих его птенцам. Гнездятся отдельными парами. В кладке 2 яйца, которые лежат в небольшой, без всякой выстилки лунке. Насиживание около 4 недель. Птенцы в буром или коричневатом одноцветном пуху. Становятся лётными в возрасте около месяца. К размножению приступают в возрасте 2—5 лет. Вне сезона размножения держатся одиночками или маленькими группами по морским побережьям и в открытом море.
Семейство включает 2 рода: Stercorarius с 3 видами, распространенными в Северном полущарии, и Catharacta (иногда включают в первый род) с 1 видом, распространенным биполярно: в Арктике и в Антарктике (иногда северный и южный подвиды рассматривают как самостоятельные виды). В СССР встречаются 4 вида, достоверно гнездятся 3.
Видимо, наиболее древняя ветвь подотряда, хотя ископаемые остатки пока обнаружены лишь в плейстоценовых отложениях.
СЕМЕЙСТВО 15. LARIDAE— ЧАЙКОВЫЕ
Птицы мелких, средних и крупных размеров (вес от 40 г до 3 кг) с характерным обликом чайки или крачки (рис. 119).
Рамфотека надклювья сплошная. Клюв почти прямой, с острой вершиной (крачки) или с более или менее выраженным крючком на конце (чайки). По заднему краю грудины 2 пары вырезок. В окраске преобладают белые или светло-серые тона; лишь немногие виды имеют темную окраску.
Большинство хорошо плавает; лишь некоторые крачки плавают неохотно и не садятся на воду. Полет легкий, маневренный. Пикируя, могут хватать добычу не только с поверхности воды, но и на небольшой глубине. Большинство видов, помимо этого, могут собирать пищу, бродя по земле или по мелководью. Крупные виды практически всеядны: поедают рыбу, различных водных и наземных беспозвоночных, мелких грызунов, падаль, ягоды, таскают яйца и птенцов. Мелкие виды питаются преимущественно мелкой рыбешкой, насекомыми и другими беспозвоночными.
197
Рис. 118. Ареалы семейств ржанкосбразных птиц:
1— поморникозые Stercorariidae, 2— чайковые Laridae, 3— водорезовые Rynchopidae
Рис. 119. Пестроносая крачка Thalasseu& sandvicensis (Lath.)
Гнездятся обычно колониально. Гнезда на земле, уступах скал, сплавинах, реже на плавающих растениях; у единичных видов на деревьях. Размеры гнезда варьируют у разных видов и у одного вида в различных условиях: на сырых местах обычно гнезда больше. В кладке 2—3 пестрых яйца; у немногих видов только 1. Инкубация от 2 до 4 недель. Окраска птенцов пестрая. Через 1—2 дня после вылупления у большинства видов птенцы уже хорошо бегают и плавают, держатся в районе гнезда. Подъем на крыло в возрасте 2—3 недель у мелких и в возрасте 4—6 недель у крупных видов. Молодые птицы мелких видов приступают к размножению в возрасте 1—2 лет, крупные 3—5 лет. Вне периода размножения кочуют по берегам водоемов стайками разной величины, некоторые виды держатся в открытом море. Распространены практически по всему земному шару (рис. 118).
Семейство чайковых обычно разделяют на 2 подсемейства (иногда подсемейства возводят в ранг самостоятельных семейств).
1.Чайки Larina е. Клюв с более или менее выраженным крючком на конце, ноги средней длины, ножная формула AXY+, хорошо ходят, бегают и плавают; объединяют роды Lotus, Pagophila, Rissa, Rhodostethia, Хета (иногда часть этих родов включают в род Laru$.
2. Крачки Sterninae. Клюв с заостренной вершиной без крючка, крылья длиннее и острее, чем у чаек; некоторые виды плавают неохотно, формула мышц ноги ABXY-\-\ включают роды5/еттш, A nous, Chlidonias, Gelo-chelidon, Gygis, Hydroprogne, Larosterna, Phaetusa, Thalasseus (некоторые систематики принимают меньшее число родов); в высокие широты проникают немногие виды, большинство свойственно умеренным и тропическим широтам.
Всего в семействе чайковых 78 видов, относимых к 14 родам (иногда принимают меньшее число родов). В СССР встречается 31 вид 9 родов (21 вид чаек и 10 — крачек), достоверно гнездится 29 видов.
Это семейство —центральная ветвь подотряда, развивавшаяся путем адаптивной радиации в околоводных местообитаниях. Видимо, возникли в Северном полушарии, обособившись от примитивных генерализованных ржанок. Ископаемые остатки известны с нижнего эоцена (Англия). Описано 14 ископаемых видов; обнаружены остатки 17 ныне живущих видов.
СЕМЕЙСТВО 16. RYNCHOPIDAE— ВОДОРЕЗОВЫЕ
В семейство входит только один род Rynchops с 3 видами, распространенными в тропиках (рис. 118).
По внешнему виду водорезы (рис. 120) похожи на средних размеров крачек, но резко отличаются строением клюва. Он довольно высок и сильно сжат с боков; подклювье выступает вперед за конец надклювья. Формула
199
Рис. 120. Африканский водорез Rynchops flavirostris V.
мышц ногиЛВХУ —. Ноги короткие. Ходят мало. Крылья длинные. Хвост выемчатый. В окраске черный цвет сочетается с сизым и белым. Держатся в устьях рек и на больших внутренних водоемах. Пищу — мелкую рыбу и крупных беспозвоночных —добывают, летая низко над водой и выбрасывая встреченную добычу в воздух длинным подклювьем, где ее и хватают. Могут пикировать на добычу. Гнездятся колониями на отмелях. В кладке 4 яйца. Вне сезона размножения держатся стайками.
Ископаемых остатков нет. Близки к чайкам.
ПОДОТРЯД 3. ALCAE — ЧИСТИКОВЫЕ
Сюда входит только одно семейство.
СЕМЕЙСТВО 17. ALCIDAE — ЧИСТИКОВЫЕ
Типичные морские птицы, связанные с сушей только в период размножения. Размеры от мелких до средних (вес от 80 г до 1,2 кг).
Клюв более или менее уд-
Рис. 121. Бескрылая гагарка Pinguinus impennis (L.)
линенныи, конический или высокий, сильно Сжатый с боков, или короткий со слегка вздутым надклювьем. Ноздри сквозные, схизоринальные. Череп схизогнатический. Иногда сохраняются затылочные фонта-нели. Базиптеригоидные сочленения у взрослых не функционируют. Хорошо развиты ямки для надорбитальных желез. Шейных позвонков 15. Спинная кость не образуется. Грудина узкая и длинная, с хорошо развитым килем и одной парой вырезок по заднему краю. Вилочка с хорошо развитым расширением на конце. Истинных ребер 7—8 пар. Предплечье — наиболее короткий элемент скелета крыла. Ноги короткие, отнесенные назад. Кайры, гагарка, иногда и другие виды опираются при ходьбе на пальцы и цевку (рис. 121). Задний палец редуцирован. Передние пальцы соеди-
200
йены плавательной перепонкой. Ножная формула АВХ —, АХ —. Гортань трахео-бронхиальная со слабой голосовой мускулатурой. Большие воздушные мешки. Язык мускулистый, с многочисленными шипи-ками, помогающими удерживать добычу. У мелких видов в период размножения функционирует горловой мешок, в котором приносят птенцам корм. Мускульный желудок небольшого объема. Слепые кишки рудиментарны. Побочный стержень контурного пера развит слабо. Оперение плотное; число перьев на единицу поверхности примерно вдвое больше, чем у чаек. Крылья относительно короткие, узкие, острые. Первостепенных маховых 11, первое рудиментарно. Рулевых 12—18. Полового морфизма окраски нет. Сезонная и возрастная смена окраски выражена у многих видов. Часть видов по суше передвигается легко и свободно, другие с трудом. Полет быстрый, но не маневренный. Все виды прекрасно плавают и ныряют. В толще воды двигаются, работая крыльями (подводный полет).
Моногамы. Гнездятся более или менее крупными колониями, немногие виды одиночными парами. В кладке 1—2 относительно крупных яйца (более 10% веса самки). Обычно яйцо откладывается без всякой подстилки в укрытии: трещине скалы, под камнями, в норе, вырытой птицами в слое торфа; кайры откладывают единственное яйцо на открытых скалистых уступах; длинноклювый пыжик Brachyramphus marmoratus (Gm.) иногда гнездится на деревьях. Насиживают оба партнера. Инкубация 3—5 недель. Птенцы вылупляются зрячие, густо опушенные; окраска обычно более или менее одноцветная, не пестрая. У большинства видов птенцы остаются в гнезде 4—6 недель до подъема на крыло и выкармливаются родителями. У кайр и гагарки птенцы примерно через 3 недели после вылупления одевают промежуточный перьевой наряд — мезоптиль и, достигнув лишь */3 веса взрослых, покидают гнездовой уступ и уплывают с родителями в море; все их дальнейшее развитие идет на воде. Половозрелыми становятся, видимо, только в конце 2-го года жизни.
Вне сезона размножения держатся на воде у побережий и в открытом море, обычно стайками, реже поодиночке. Пища — разнообразные водные животные: мелкая рыба, ракообразные, моллюски, черви — добывается только в воде при плавании, чаще при нырянии. Способны нырять на глубину в несколько десятков метров.
Семейство чистиковых объединяет 21—22 ныне живущих вида, распределяемых по 13 родам: Alca, Aethia, Brachyramphus, Cepphus, Cerorhinca, Cyclorrhynchus, Endomychura, Fratercula, Lunda, Plotus (Plautus), Pty-choramphus, Synthlieoramphus, Uria. Распространены по морским побережьям северных и умеренных широт Северного полушария (рис. 123). В СССР встречается 18—19 видов 11 родов (часть систематиков выделяет 2 самостоятельных вида: атлантический чистик и тихоокеанский чистик). Достоверно установлено гнездование 16—17 видов. В первой половине XIX в. была истреблена жившая по побережьям Северной Атлантики бескрылая гагарка Pinguinus impennis (L.) (рис. 121), достигавшая веса 5 кг. Она полностью утратила способность к полету, что облегчило ее истребление. Последние особи добыты в 1844 г. на маленьком островке около Исландии.
Чистиковые обособились от чаек в результате приспособления к водному образу жизни. Сходство черт строения и экологии с гагарами и ныряющими буревестниками — результат конвергенции, главным образом использования крыльев при нырянии. Вероятный центр возникновения подотряда — побережья древнего Берингова моря. Ископаемые остатки плохой сохранности известны из нижнего эоцена и более поздних слоев. Их относят к 11 ископаемым и 9 ныне живущим видам. По остаткам из плиоценовых отложений Калифорнии описано 2 вида одного рода, который иногда выделяют в самостоятельное семейство Маисаllidae. Это крупные, видимо, нелетающие, птицы, вероятно, походили на кайр или на бескрылую гагарку, но имели более примитивные черты: меньшую уплощенность скелета крыла и т. п.
201
ж
* *
К. А. Юдин тщательно проанализировал ранее опубликованные материалы и, широко используя результаты оригинальных исследований (1965), предлагает несколько иной и, видимо, более обоснованный вариант системы отряда Charadriiformes:
1. Подотряд якан —Jacanae
Семейство Jacanidae
2. Подотряд куликов Limicolae
Семейство Rostratulidae
Семейство Charadriidae (по Е. В. Козловой, 1961, 1962), это семейство включает 11 подсемейств: Charadriinae, Vannellinae, Himantopinae, Ibidorhynchinae, Hae-matopinae, Tringinae, Phalaropinae, Arehariinae, Calidriinae, Scolopacinae, Li-mosinae
3. Подотряд чайко- и куликообразных Laro — Limicolae
Надсемейство тиркушковых Glareoloidea
Семейство Glareolidae
Семейство Pluvianidae
Надсемейство белых ржанок Chionidoidea
Семейство Chionididae
Семейство Thinocoridae
Надсемейство рачьих ржанок — Dromadidoidea
Семейство Dromadidae
Надсемейство чаек — Lari (Larioidea)
Семейство Stercorariidae
Семейство Laridae (включает 3 подсемейства: Larinae, Sterninae, Rynchopinea)1 Надсемейство чистиковых — Alcae (Alcidoidea)
Семейство Alcidae
По мнению К. А. Юдина, выделенные им подотряды соответствуют трем основным направлениям эволюции ржанкообразных. Яканы—наиболее ранняя ветвь отряда, еще сохраняющая ряд архаичных признаков, характерных для журавлей и дроф. Подотряд куликов — единая линия развития околоводных птиц с широкой адаптивной радиацией, что отражается делением сем. Charadriidae на 11 подсемейств. Цветные бекасы — более примитивная ветвь, резко обособленная от других куликов. Обособление от примитивных куликов подотряда Laro-Limicolae шло, прежде всего, по пути развития приспособлений^ обеспечивающих возможность питания более крупными пищевыми объектами. Первая ветвь, сохранившая многие предковые черты и выработавшая способность питаться крупными насекомыми, — надсемейство тиркушковых: сем. Pluvianidae сохранило связь с берегами водоемов-, а сем. Glareolidae перешло к охоте в воздухе (тиркушки) или к добыванию пищи на земле, но вдали от водоемов (бегунки). Своеобразные ограниченные жизненные ниши заняли надсемейства белых и рачьих ржанок.
Основная ветвь подотряда — надсемейство чаек — развивалась на берегах водоемов, вырабатывая черты водной специализации. Плаванье и полет наряду с перемещениями пешком стали основными способами добывания очень разнообразных пищевых объектов на побережьях и в верхнем слое воды. От предковых форм чаек обособились чистиковые птицы, выработавшие способность к нырянию, что позволило им добывать корм в толще воды. Филогенетические взаимоотношения внутри отряда ржанкообразных и его отношения с близкими отрядами птиц характеризует составленная К. А. Юдиным схема (см. рис. 101).
ОТРЯД 23. COLUMBIFORMES —ГОЛУБЕОБРАЗНЫЕ
Преимущественно растительноядные птицы мелких и средних, редко крупных размеров (вес от 30 г до 1 кг, редко до 3 кг) с характерным обликом голубей (рис. 125) или рябков (рис. 122).
202
Рис. 122. Саджа Syrrhaptes paradoxus (Pall.)
Ноздри не сквозные. Череп схизоринальный, схизогнатический; сошник обычно редуцируется (есть у дронтов). Есть базиптеригоидные сочленения (нет у дронтов). Надорбитальные железы малы или отсутствуют. Шейных позвонков 14—16. По заднему краю грудины две пары вырезок. У большинства форм ножная формула ABXYdz. Сонных артерий две. Хорошо развит зоб. Мускульный желудок обычно с мощными стенками. Копчиковая железа маленькая, не оперенная; у части видов нет вовсе. Контурные перья с хорошо развитой пуховой частью, на некоторых участках тела продуцирующей «пудру». Оперение плотное. Крылья удлиненные, заостренные. Первостепенных маховых 10—11, рулевых 12—20. Обычно самцы несколько крупнее самок и у части видов их окраска ярче. Возрастное и сезонное изменение окраски не отчетливо. Моногамы. Птенцы выводкового или птенцового типа. Распространены в тропических и умеренных широтах всего земного шара.
Отряд голубеобразных разделяется на 2 подотряда: рябков и голубей. Всего в отряде 310 видов, относимых к 49 родам. В СССР встречается 16 видов 5 родов; достоверно гнездятся 14 видов.
Предки голубеобразных — генерализованные ржанкообразные.
ПОДОТРЯД 1. РТЕ ROCLETES — РЯБКИ
Включает единственное семейство.
СЕМЕЙСТВО 1. PTEROCLIDAE — РЯБКОВЫЕ
Птицы небольшого размера (вес 300—500 г; рис. 122). Населяют сухие степи и пустыни Африки и Южной Азии (рис. 123).
Клюв маленький, со слегка вздутым надклювьем. Восковицы нет. Ноздри полуприкрыты кожистой складкой. Ноги короткие. Задний палец мал и расположен выше уровня передних пальцев; у части видов исчезает. Передние пальцы относительно короткие, у части видов срастающиеся основаниями. Зоб большой, без боковых выростов. Слепые кишки длинные, функционирующие. Желчный пузырь есть. Гортань трахео-бронхиальная, с симметричной голосовой мускулатурой. В контурном пере есть, хотя и слабо развитый, добавочный ствол. Редкий пух растет на птерилиях и аптериях. Первостепенных маховых 11, первое недоразвито. Крыло диастатаксическое. Рулевых 14—16, средние удлинненые. В окраске сверху преобладают желтоватые и охристые тона. Самцы окрашены ярче самок.
Полет быстрый, стремительный. Хорошо ходят, быстро бегают. Когда пьют, периодически поднимают голову вверх, в этот момент заглатывая воду. Держатся стайками и в гнездовое время. Обычно гнездятся рассеянными
203
Рис. 123. Ареалы:
/— чистиковые Alcidae (отряд ржанкообразных); голубеобразные: 2— рябковые Pteroclicjae, 3— голубиные Colurnbidae, 4— дронтовые Raphidae
группами. Гнездо — небольшая ямка в почве, иногда слабо выстланная травинками. В кладке обычно 3, реже 2 или 4 пестрых яйца. Насиживают оба пола около 4 недель. Птенцы вылупляются зрячими, покрытыми пестрым пухом. До подъема на крыло бродят в районе гнезда. Родители не только кормят птенцов, но и приносят им в зобе воду. По некоторым наблюдениям, взрослые рябки во время водопоя заходят в воду, так что оперение груди и брюшка сильно намокает. Когда птицы прилетают на гнездовье, часть воды сохраняется и птенцы, перебирая клювом оперение родителей, забирают эту влагу. Иногда гнездятся в десятках километров от водоемов; регулярно летают на водопой. Кормятся на земле, поедают семена, зеленые части растений, попутно ловят насекомых и других наземных беспозвоночных. Вне периода размножения кочуют большими стаями. Некоторые виды совершают непериодические вылеты за пределы своего ареала: например, обыкновенная саджа залетает в Западную Европу.
В семействе 16 видов в 2 родах: Pterocles (рябки, 14 видов) и Syrrha-ptes (саджи, 2 вида). В южных районах азиатской части СССР гнездятся 4 вида: 2 вида рябков и 2 вида садж.
Предполагаемый центр возникновения — Северная Африка. Ископаемые остатки верхнего эоцена и более поздних слоев (Франция, районы современного ареала) относят к ныне живущим родам.
ПОДОТРЯД 2. COLUMBAE — ГОЛУБИНЫЕ
Клюв более или менее удлиненный, несколько утолщенный в передней части. Ноздри не прикрыты кожистыми складками. Ноги длиннее, чем у рябков. Задний палец хорошо развит и расположен на одном уровне с передними. Зоб большой, развивается как два боковых выроста пищевода. Слепые кишки обычно рудиментарны или их нет вовсе. Желчный пузырь у многих видов не развит. У части видов нет копчиковой железы. Нижняя гортань с несимметрично расположенными мышцами.
Подотряд разделяется на 2 семейства.
СЕМЕЙСТВО 2. RAPHIDAE — ДРОНТОВЫЕ
Крупные (вес до 10 кг) нелетающие птицы (рис. 124), жившие на Мас каренских островах (Маврикий, Реюньон, Родригес) и полностью истреб
ленные в XVI—XVII вв. Всего 3 вида, относимых к 2 родам: Raphus (Didus) и Pezop-haps (на каждом острове встречался только 1 вид).
Клюв мощный, длинный, с большим вздутием и крючком на конце. Восковицы нет. Череп массивный. Базиптеригоидных сочленений нет. Грудина с очень низким килем; на ее заднем крае только одна пара вырезок. Скелет всех отделов крыла сильно укорочен. Кара-коиды и вилочка слабые. Таз и скелет задних конечностей мощные. Сохранилось 5—6 первостепенных маховых. Рулевые, видимо, отсутствовали. Оперение рыхлое. Лицевая часть головы не оперена.
Рис. 124. Дронт Raphus cucullatus L.
205
Вели наземный образ жизни. Судя по нахождению остатков скелетов, предпочитали болотистые участки. Питались разнообразной растительной пищей. Видимо, держались парами, охраняя свой участок. У дронта с о-ва Родригес на кисти есть округлые костные вздутия, которыми, как полагают, птица наносила удары другой особи, вторгшейся на ее участок. Рост птенцов продолжался несколько месяцев.
Обычно это семейство рассматривают как примитивную ветвь голубей, утратившую способность к полету в связи с островным распространением и увеличением размеров. Ископаемые остатки плейстоценового времени известны только с этих же островов. Иногда высказывается предположение, что дронты — сильно уклонившаяся ветвь не голубей, а пастушков и должны включаться в надсемейство Ralloidea.
СЕМЕЙСТВО 3. COLUMBIDAE — ГОЛУБИНЫЕ
Птицы обычно средней величины, но есть очень мелкие (горлицы весом около 30 г) и крупные (венценосный голубь до 3 кг; рис. 125).
Клюв тонкий, несколько удлиненный, с небольшим вздутием и крючком на конце. У основания надклювья есть восковица: участок голой кожи, у основания которой открываются ноздри. Грудина с высоким килем и обычно двумя парами вырезок по заднему краю. 3—5 грудных позвонков сливаются в спинную кость. Контурные перья без добавочного ствола, с сильно уплощенным основным стержнем и хорошо развитой пуховой частью. Обычно пуха нет, по всему телу есть длинные нитевидные пушинки. Первостепенных маховых 10—11. Крыло диастатаксическое или эутаксическое. Рулевых 12—14. Окраска разнообразная, часто с металлическим блеском. Полового морфизма окраски нет, возрастной выражен слабо.
Полет быстрый, довольно маневренный. Хорошо ходят, бегают плохо. Когда пьют, всасывают воду, не поднимая головы. Большинство видов гнездится одиночными парами, немногие разрозненными группами и даже настоящими колониями. Гнездо — рыхлая кучка прутиков в развилке
Рис. 125. Венценосный голубь Goura critsata (Pall.)
206
ветвей. Некоторые виды гнездятся в дуплах, расщелинах скал, под камнями, в промоинах оврагов, единичные виды — на земле. В кладке 2, реже 1 яйцо белого цвета (у немногих — кремового). Насиживают оба пола 2,5 (мелкие виды) 4 и даже 5 недель. Птенцы вылупляются слепыми, покрытыми редким нитевидным пухом. Кормят оба родителя так называемым «птичьим молочком». Эпителий стенок зоба взрослых птиц усиленно делится, происходит жировое перерождение и он отторгается; этой жидкой пищей птенцов выкармливают первое время, затем добавляются разбухшие в зобу семена. Птенцы растут быстро и в возрасте 2,5—4 недель приобретают способность к полету и покидают гнездо. Половозрелыми становятся в возрасте около года (венценосные голуби в возрасте около 2 лет). В теплых районах у многих видов в году 2—3 кладки.
Питаются разнообразной растительной пищей: семенами, ягодами, зелеными проростками растений и т. п. Охотно поедают раковины моллюсков, восполняя недостаток солей в растительной пище. Животные корма поедают случайно и в небольших количествах. Регулярно летают на водопой. Часть видов держится группами и в сезон размножения, в остальное время года — стайками. В умеренных широтах передетны, в тропиках — оседлые и кочующие. Многие виды — объект спортивной охоты.
Голуби — почти космополиты, населяющие умеренные и жаркие широты всего земного шара (рис. 123), включая и многие океанические острова.
В семейство включают 45 родов, из них 12 монотипичны: Columba (единственный род, встречающийся почти по всему ареалу семейства; включает 51 вид), Alectroenas, Aplopelia, Caloenas, Chalcopelia, Chalcophaps, Chloroe-nas, Cloravis, Columbina, Cryptophaps, Didunculus, Drepanoptila, Ducala (36 видов), Ectopistes, Gallicolumba (18 видов), Geopelia, Geotry gon, Goura, Gymnophaps, Hemiphaga, Henicophas, Leptotila (10 видов), Lopholaimus, Leucosarcia, Macropygia, Metriopetia, Microgoura, Ocyphaps, Oena, Otidi-phaps, Petrophassa, P hap it reran, Phaps, Ptilinopus (45 видов), Reinwardtoena, Scardafella, Sphenurus, Streptopelia (17 видов), Starnoenas, Treron (25 видов), Trugon, Turacoena, Tartar, Uropelia, Zenaida.
Большое число родов в пределах семейства заставляет группировать их в подсемейства, нечеткие межродовые отличия сильно затрудняют эту задачу. Поэтому число принимаемых подсемейств варьирует от 4 до 9. Система семейства нуждается в доработке. Всего в семейство голубей включают 285 ныне живущих видов; 6 видов истреблены в историческое время. Среди них и одна из ранее самых многочисленных птиц земли — странствующий голубь Ectopistes migratorius (L.), гнездившийся в Северной Америке колониями в миллионы пар; полностью истреблен в конце XIX в.
В фауне СССР отмечено 12 видов 3 родов (встречающегося в южной части Курильских островов зеленого голубя Sphenurus sieboldi (Temm.) сейчас обычно включают в род Treron), достоверно доказано гнездование 10 видов. В последние десятилетия по Европе, включая и западные районы СССР, энергично расселяется кольчатая горлица Streptopelia deccuocto (Friv.).
Ископаемые остатки (7 ископаемых и 27 ныне живущих видов) известны из миоцена Франции и более поздних отложений в разных районах современного ареала. Возможный центр возникновения — Австралия (по другим взглядам, Южная Америка).
ОТРЯД 24. PSITTACIFORMES — ПОПУГАЕОБРАЗНЫЕ
Включает 1 семейство Psittacidae.
СЕМЕЙСТВО 1. PSITTACIDAE — ПОПУГАЕВЫЕ
Преимущественно древесные птицы с характерным обликом попугая (рис. 126), мелких и средних размеров (вес от 10 а до 1 кг и несколько больше).
207
Рис. 126. Черный какаду Probosiger atterimus (Gm.)
Клюв массивный. Широкое и высокое у основания надклювье заканчивается длинным, заостренным крючком, нависающим над более коротким подклювьем. Подвижный клюв соединяется с лобными и носовыми костями суставом. Мощная жевательная мускулатура и скелетные особенности обеспечивают большую подвижность клюва и позволяют разгрызать и размельчать пищу. У основания надклювья развита восковица, на которой расположены наружные отверстия ноздрей. Череп голоринальный, десмогнатический, с редуцированным сошником и без базиптеригоидных сочленений. Надорбитальные железы малы. Голова относительно крупная. Шейных позвонков 14 (как исключение, бывает 13 или 15). По заднему краю грудины пара вырезок, у части видов зарастающих. Киль высокий. Вилочка без расширения на конце, у плохо летающих видов в разной степени редукции. Спинная кость не образуется. Истинных ребер 5—6 пар. Ноги сильные. Ножная формула AXY±. Цевка относительно короткая. Лапа зигодактильного типа; четвертый палец очень подвижен и может сильно отводиться в сторону. Когти острые.
Мускулистый язык хорошо развит, сильно варьирует по форме. Хорошо развит объемистый зоб. Между железистым и относительно небольшим мускулистым желудком расположен промежуточный отдел. Кишечник длинный, слепых кишок обычно нет. У большинства видов нет желчного пузыря. Копчиковая железа оперенная; у части видов развита хорошо, у части — сла
бо, у некоторых ее нет вовсе. Сонные артерии парные, иногда развиты в различной степени. Грудинно-трахеальные мышцы развиты слабо или отсутствуют; голосовая мускулатура развита хорошо. Головной мозг, особенно полушария, относительно крупный. Характерны высокие ассоциативные способности, хорошая память, способность к звукоподражанию. Некоторые виды обучаются произносить слова и короткие фразы. Побочный ствол контурного пера развит. Пух есть на птерилиях и аптериях. На боках и пояснице растет пух, продуцирующий «пудру». Первостепенных маховых 10 у (совиного попугая 9). Крыло диастатаксическое. Рулевых 12—14. У многих видов окраска яркая; часто развиваются хохлы, длинные хвосты. Самцы немного крупнее самок. Полового морфизма окраски обычно нет, а возрастной выражен слабо.
Большинство видов живет в кронах деревьев, немногие в кустарниковых зарослях; одиночные виды проникают в степи и полупустыни. В кронах иногда передвигаются очень своеобразно: вытягивая шею, хватают клювом ветку, повисают, затем захватывают ее лапами. Пища преимущественно или исключительно растительная: разнообразные плоды, семена, ягоды; некоторые мелкие виды поедают пыльцу и нектар, попутно ловя сидящих в цветках насекомых.
Моногамы. Обычно гнездятся пара от пары поблизости, иногда группами. Подавляющее большинство видов дуплогнездники. Некоторые виды гнездятся в расщелинах скал, углублениях между корнями и в норах.
208
150° 120° 90° 60 к западу от Гринвича 0°к востоку отГрипвичабО0 90° 120° 150° 180° 150°
Рис. 127. Ареалы семейств:
1— попугаевые Psittacidae (отряд попугаеобразных). 2— тураковые Musophagidae, 3—кукушковые Cuculidae (отряд кукушкообразных)
Немногие виды устраивают примитивное открытое гнездо в развилке ветвей. В кладке от 2—3 (крупные виды) до 5—8 округлых яиц с белой скорлупой. Насиживает самка; у части видов некоторое участие в насиживании принимает самец. Инкубация от 2 до 4 недель. Птенцы вылупляются слепыми и голыми. Последовательно развиваются два пуховых наряда и лишь после этого одевается первый перьевой наряд. Выкармливают оба родителя. Птенцы остаются в гнезде до приобретения способности к полету: до 5 недель у мелких видов и до 2—3 месяцев у крупных. У части мелких видов бывает 2—3 кладки в год, у крупных только одна. Оседлые или кочующие. Вне сезона размножения держатся стайками.
Отряд в целом довольно однообразен и морфологически и экологически. Это подчеркивают тем, что выделяют только 1 семейство (1-е) Psittacidae, объединяющее 75 родов с 324 видами, 19 видов полностью вымерли или истреблены в историческое время. Распространены в тропиках и, отчасти, в умеренных широтах (рис. 127). В СССР не встречаются.
В практических целях выделяют несколько (6—9) подсемейств. Наиболее часто принимают следующие подсемейства.
1. Nestorinae — несторы. Темноокрашенные попугаи Новой Зеландии и прилегающих островов. Единственный род Nestor с 3 видами, один из которых полностью истреблен. Иногда нападают на овец, вырывая у них со спины куски кожи с мясом и жиром. Истребляются.
2. Psittrichasinae — щетинкоголовые попугаи. Единственный монотипичный род Psittrichas, встречающийся только на Новой Гвинее. Размерами с грача, темной окраски, с широким, но низким клювом.
3. Kakatoeinae — какаду. 5 родов — Kakatoe, Callocepha-lon, Calyptorhynchus, Plyctolophus, Probosciger — c 17 видами, размерами от галки до ворона. Окраска яркая, но без зеленых тонов; часто хорошо развит хохол. Распространены в Австралии, Новой Гвинее, Восточной Индонезии, на Филлиппинах.
4. Micropsittinae — карликовые, или дятловые, попугаи. Единственный род — Micropsitta с 6 видами, населяющими Новую Гвинею и прилегающие острова. Мелкие птицы (длина около 10 см) преимущественно зеленой окраски. Хвост из жестких рулевых (похож на хвост дятлов). Могут лазить по вертикальным стволам деревьев.
5. Trichoglossinae — щеткоязычные попугаи. 14 родов — Trichoglossus, Chalcopsitta, Charmosyna, Domicella, Eos, Glossop-sitta, Lathamus, Neopsittacus, Opopsitta, Oreopsittacus, Phigys, Pseudeos, Psittaculirostris, Vini — c 62 видами, распространенными в Австралии, Новой Гвинее, Восточной Индонезии, на Филиппинах. Мелкой и средней величины, часто очень яркой окраски. У большинства видов язык оканчивается щеточкой сосочков. Питаются ягодами, плодами, семенами, пыльниками и пестиками цветков, сосут цветочный сок.
6. Strigopinae — совиные попугаи. Сюда включают один монотипичный род Strigops, живущий сейчас в сухих изреженных лесах только на Южном острове Новой Зеландии. Всюду редок. Ведет сумеречный и ночной образ жизни. Наземная птица, потерявшая способность к полету, хорошо ходит и бегает. Клюв короткий, с тупой вершиной. Крылья короткие, широкие. Оперение рыхлое, похожее на оперение сов. Окраска тусклая, зеленовато-бурая. Питается вегетативными частями растений, корешками, мхом, меньше ягодами и семенами. Днем прячется в нишах между корнями, под упавшими деревьями, в расщелинах скал, иногда в неглубоких норах. В этих же убежищах откладывает 2—4 яйца.
7. Psittacinae — настоящие попугаи. Самое большое подсемейство, включающее 234 вида, относимых к 52 родам: Psittacus, Agapomis, Alisterus, Amazona, Anodorhynchus, Aprosrpictus, Ara, Aratin-ga, Bolbopsittacus, Bolborhynchos, Brotogeris, Caracopsis, Conuropsis, Cyano-liseus, Cyanoramphus, Deroptyus, Enicognathus, Eunymphicus, Forpus, Geoff-
210
royus, Geopsittacus, Graydidasculus, Gypopsitta, Layardiella, Loriculus, Lorias, Melopsittacus, Myiopsitta, Nannopsittaca, Neophema, Neopsephotus, Northilia, Nymphicus, Pezoporus, Pionites, Pionopsitta, Pionus, Platycercus, Poicephalus, Polytelis, Prioniturus, Prosopeia, Psephotus, Psittacella, Psitta, cula, Psittinus, Purpureicephalus, Pyrrhura, Rhynchopsitta, Tanygnathus, Touit, Triclaria. Распространены практически по всему ареалу отряда. Птицы различных размеров, очень разнообразной, часто яркой контрастной окраски. Язык гладкий. Питаются разнообразными ягодами, плодами, семенами.
Вероятный центр возникновения отряда — Австралийская зоогеографическая область. Именно здесь обитает наибольшее число родов (около 60) и видов. Родов, распространенных одновременно в Старом и Новом Свете, нет. Ископаемые остатки немногочисленны (описано несколько видов); наиболее древняя находка — из нижнего миоцена Франции. Вероятно, можно говорить об очень давних родственных связях с ракшеобразными и кукушкообразными, но попугаи, видимо, отделились еще до обособления последних двух отрядов. Родственные отношения отдельных групп внутри отряда (семейства) не выяснены. Вполне возможно, что все подсемейства возникли путем адаптивной радиации от общих предковых форм.
ОТРЯД 25. CUCULIFORMES —КУКУШКООБРАЗНЫЕ
Преимущественно древесные птицы мелких и средних размеров (вес от 20 г до 1 кг).
Череп голоринальный, десмогнатический, без базиптеригоидных отростков. Ноздри не сквозные. Надорбитальных желез нет. Шейных позвонков 14—15. По заднему краю грудины две пары вырезок, во взрослом состоянии у части видов зарастающих или превращающихся в фонтанели. Истинных ребер 3—5 пар. Формула мускулатуры ноги ABXY + или, редко, AXY+. Лапа зигодактильная или переходного типа. Сонных артерий две. Гортань трахео-бронхиального или — у немногих родов — бронхиального типа. Желчный пузырь есть. Кожа тонкая. Пух есть только на аптериях. Первостепенных маховых 10. Крыло эутаксическое. Хвост обычно удлиненный, из 10 рулевых (у 2 родов кукушек — только из 8).
По характеру питания, как и по многим другим признакам, отчетливо разделяются на 2 группы: тураковые растительноядны, кукушки обычно животноядны. Моногамы или полигамы. Для всех видов характерен птенцовый тип развития. В тропиках оседлые или кочующие, в умеренных широтах перелетные.
Отряд кукушкообразных объединяет 43 рода с 148 видами. Разделяется на 2 подотряда: тураковых и настоящих кукушек. В СССР встречается 6, а гнездится 5 видов из подотряда кукушек.
ПОДОТРЯД 1. MUSOPHAGI — ТУРАКОВЫЕ, ИЛИ БАНАНОЕДЫ
В подотряд входит только одно семейство.
СЕМЕЙСТВО 1. MUSOPHAGIDAE — ТУРАКОВЫЕ
Свойственны тропическим лесам Африки (на Мадагаскаре нет; рис. 127). По поведению, отчасти и внешнему виду немного похожи на соек (рис. 128).
Птицы средних размеров (вес от 250 г до 1 кг). Клюв относительно короткий, слегка вздутый, напоминающий клюв куриных птиц. Края клюва зазубрены. Наружные отверстия ноздрей щелевидны и лежат ближе к концу клюва. Сошник редуцирован. Шейных позвонков 15. По заднему краю
211
Рис. 128. Турако Tauraco corythaix Wagl.
грудины две пары вырезок. Конец вилочки без расширения. Истинных ребер 5 пар. Четвертый палец очень подвижен, направлен вперед и может сильно отводиться вбок (переход к зигодактилии). Кишечник относительно короткий и широкий. Слепых кишок нет. Копчиковая железа оперена. Хорошо развит побочный стержень контурного пера. Окраска взрослых яркая (зеленая, синяя и т. п.). На маховых перьях есть участки красного цвета, который образуется специфическими пигментами, содержащими медь,— турацином и тураковердином, не встречающимся у птиц других групп. Полового диморфизма окраски нет. У многих видов на голове хохол.
Держатся в кронах деревьев, питаясь разнообразной растительной пищей: ягодами, плодами, цветками и почками деревьев. Бананы обычно не едят, поэтому название «бананоеды» неудачно.
Моногамы. Гнездо — рыхлая постройка из мелких веточек — помещается в развилке ветвей. В кладке 2 белых яйца. Инкубация около 18—20 дней. Птенцы вылупляются голыми, но уже через несколько дней покрываются густым коротким черным пухом. При опасности йокидают гнездо и прячутся в ветвях. У птенцов хорошо развит коготь первого пальца крыла, которым они цепляются за ветки (подобно птенцам гоацина). Начинают перепархивать с ветки на ветку еще полуоперенными, с остатками пуха, в возрасте 6-*-7 недель. Вне сезона размножения держатся мелкими стайками.
Семейство Musophagidae включает 5 родов — Musophaga, Corythaeola, Corythaixoides, Crinifer, Tauraco — c 18 видами.
В ископаемом состоянии неизвестны. Вероятный центр возникновения — Африка.
ПОДОТРЯД 2. CUCULI — КУКУШКОВЫЕ
Включает одно семейство.
СЕМЕЙСТВО 2. CUCULIDAE— КУКУШКОВЫЕ
Распространены практически по всем материкам, кроме Субарктики и Антарктики (рис. 127).
Кукушки (рис. 129) — птицы мелкой и средней величины (вес от 20 г почти до 1 кг). Клюв удлиненный, слегка изогнутый по хребту, с заостренным концом. Края клюва обычно гладкие. Ноздри округлые и лежат в основной половине надклювья. Сошник сохраняется, хотя у многих видов очень мал. Шейных позвонков 14. Вырезки по заднему краю грудины во взрослом состоянии зарастают или превращаются в фонтанели. Конец ви-
212
Рис. 129. Кукушка ани Сго-tophaga major Gm.
лочки с заметным расширением. Истинных ребер 3—4 пары. Лапа зигодактильная. Кишечник довольно длинный; слепые кишки есть, хотя и короткие. Копчиковая железа не оперена. Добавочный стержень контурного пера рудиментарен или его нет вовсе. Хохол на голове имеют единичные виды. Окраска обычно более или менее однотонная, у некоторых видов довольно яркая. Полового диморфизма окраски нет или он слабо выражен.
Большинство видов держится в кронах деревьев и кустарников, лишь изредка и ненадолго спускаясь на землю; ходят медленно. Немногие виды проникли в безлесные степи и полупустыни, где ведут наземный образ жизни; легко и быстро бегают. Все виды хорошо летают. Растительнояден только один род Eudynatnis (зазубрены края клюва). Все остальные кукушки животноядны. Подавляющее большинство видов питается различными насекомыми (взрослыми, гусеницами, личинками), которых собирают на ветвях, реже ловят в воздухе. Крупные виды, кроме насекомых, ловят ящериц, мелких млекопитающих, таскают яйца и птенцов. Некоторые виды в небольшом количестве поедают и растительную пищу (ягоды, мелкие плоды).
Примерно половина видов кукушек типичные моногамы, а у половины видов в той или иной степени выражен гнездовой паразитизм. Кукушки моногамы строят примитивное рыхлое гнездо на ветвях (у древесных видов) или на земле (у наземноживущих). В кладке 2—6 яиц, которые насиживают поочереди оба партнера. Насиживание продолжается около 18—20 дней. Птенцов кормят оба родителя. Постэмбриональное развитие относительно медленное. Птенцы оперяются и приобретают способность к полету в возрасте около месяца и более.
У видов с хорошо выраженным гнездовым паразитизмом относительные размеры яиц заметно уменьшаются, укорачивается период эмбрионального и постэмбрионального роста. Так, у обыкновенной кукушки продолжительность инкубации всего 12 дней, а в возрасте около 20 дней птенец уже вылетает из гнезда. Степень гнездового паразитизма у разных видов выражена различно. Некоторые виды (например, американские кукушки рода Соссу-zus) иногда строят гнезда сами, иногда занимают чужие, насиживают и выкармливают птенцов сами. Кукушки родов Crotophagav. Guira иногда гнездятся отдельными парами, занимая чужие гнезда, чаще же 2—3 пары строят общее гнездо, в которое все самки откладывают яйца; самки насиживают поочередно и совместно выкармливают птенцов. В то же время в находящихся поблизости гнездах других видов птиц часто находят подброшенные яйца этих кукушек, о которых они не заботятся; подкладывают они иногда яйца и в гнезда своего вида. Наконец, у видов с типичным гнездовым паразитизмом самка всегда подкладывает свое яйцо в гнезда других видов птиц,
никогда не пытается его насиживать и никогда не выкармливает птенца.
У видов с мало изменчивой окраской яиц круг птиц-хозяев, в гнезда которых подкладываются яйца, относительно невелик. Например, хохлатая кукушка Clamator glandaris (L.) подкладывает яйца лишь вороновым
213
птицам. Яйца обыкновенной кукушки Cucculus canorus L. находили в гнездах 150 видов птиц (в подавляющем большинстве, различных воробьиных). Но чаще всего яйца подкладываются в гнезда 20—30 видов мелких воробьиных птиц. Изменчивость окраски яиц обыкновенной кукушки очень велика, но каждая самка откладывает яйца только одного типа окраски. В одном и том же районе встречаются самки, откладывающие яйца разной окраски. В европейской части СССР наиболее обычны кукушки, несущие светлые с мелким крапом яйца (подкладывают их в гнезда белых трясогузок, чью окраску яиц они напоминают); светлые яйца с крупным крапом (их обычно находят в гнездах камышевок и славок) и голубые яйца (подкладывают в гнезда горихвосток и луговых чеканов, чьи яйца тоже голубые). Каждому району свойственно определенное соотношение различных биологических рас кукушек.
Семейство кукушек подразделяют на несколько подсемейств.
1. Настоящие кукушки Cuculinae, 47 видов, объединяемых в 16 родов: Cuculus (12 видов), Cacomantis, Caliechthrus, Cercococ-сух, Chalcites, Chrysococcyx, Clamator, Eudynamis, Microdynamis, Misoca-lius, Pachycoccyx, Penthoceryx, Rhamphomantis, Scythrops, Surniculus, Urodynamis.
2. Пестроклювые кукушки Phaenicophaeinae, 28 видов, распределяемых между 12 родами: Phaenicophaeus, Ceuthmoch-ares, Coccyzus, Dasylophus, Lepidogrammus, Playa, Rhamphococyx, Rhi-northa, Rhopodytes, Saurothera, Taccocua, Zanclostomus.
3. Кукушки личинкоед ы Crotophaginae, 4 вида 2 родов: Crotophaga и Guira.
4. Наземные (бегающие) кукушки Neomorphin а е, 13 видов 6 родов: Neomorphus, Carpococcyx, Dromococcyx, Geococcyx, Morococcyx, Taper а.
5. Мадагаскарские кукушки Couinae. Единственный род Соиа с 10 видами.
6 Шпорцевые кукушки Centropodinae. Включает лишь род Centropus, объединяющий 28 видов.
Всего в семействе кукушек 38 родов, объединяющих 130 видов. В фауне СССР встречается 6 видов 2 родов (только подсемейство Cuculinae); достоверно гнездится 5 видов рода Cuculus.
Ископаемые остатки кукушек немногочисленны (4 ископаемых и несколько видов ныне живущих). Наиболее древняя находка — из верхнего эоцена или нижнего олигоцена Франции. Центр возникновения, видимо, тропики Старого Света. И сейчас большинство видов свойственно тропикам; так, на о. Суматра встречается 14 видов. Ни один род не распространен одновременно в Старом и Новом Свете.
Очень давнее родство, видимо, сближает кукушкообразных с ракшеобразными; еще более отдаленные связи возможны с попугаями. Разобщение кукушек и тураковых очень давнее и достаточно резкое; это подчеркнуто делением отряда на 2 подотряда. Некоторое сходство тураковых с куриными (гоацин) большинство систематиков рассматривает как результат конвергенции (растительноядность, древесный образ жизни).
ОТРЯД 26. STRIGIFORMES — СОВООБРАЗНЫЕ
Преимущественно ночные хищные птицы с рыхлым пушистым оперением. Населяют самые различные ландшафты практически всего земного шара, кроме Антарктики. Размеры от мелких до крупных (вес от 50 г до 4—4,5 кг).
Череп голоринальный, схизогнатический; сошник обычно есть. Имеются функционирующие базиптеригоидные сочленения. Ноздри не сквозные. Надорбитальных желез нет. Клюв сильный с острым, круто изогнутым
214
крючком на конце и с режущими краями. В основании надклювья хорошо развита восковица, обычно прикрытая направленными вперед пучками жестких перьев, напоминающих вибриссы. Шейных позвонков 14. Шея умеренной длины, но очень подвижна: голова может поворачиваться на 180—270°. Грудные позвонки не сливаются в спинную кость. Грудина с большим килем и 1—2 парами вырезок по заднему краю. Самый длинный элемент скелета крыла — предплечье, плечо длиннее кисти. Очень мощная мускулатура ног, особенно сгибателей. Формула мускулатуры ноги упрощена: А — . Все четыре пальца ноги хорошо развиты и заканчиваются длинными, сильно изогнутыми острыми когтями; четвертый палец очень подвижен и может отводиться не только вбок, но и назад. На нижней поверхности пальцев расположены жесткие сосочки, помогающие удерживать добычу.
Глаза крупные, телескопической формы, направленные более или менее вперед. В связи с этим общее поле зрения относительно невелико (160— 180°), но резко возрастает поле бинокулярного зрения (до 60—70°). Очень хорошо развит слух. У части видов правая и левая ушные области довольно резко отличаются по размерам, что приводит к асимметрии черепа. По краю очень большого наружного слухового прохода есть складка кожи — зачаток наружного уха. У большинства видов нижняя гортань бронхиального типа с парой голосовых мышц и более или менее развитой наружной голосовой мембраной. Сонные артерии парные. Язык мясистый. Зоба нет, но пищевод сильно растяжим. Желудок объемистый; стенки мускульного желудка относительно тонкие. Хорошо развиты функционирующие слепые кишки, заканчивающиеся колбообразными вздутиями. Есть желчный пузырь.
Оперение Мягкое, рыхлое, пушистое. Контурные перья с хорошо развитой пуховой частью, без добавочного стержня. Оперение передней части головы образует лицевой диск, более или менее четко отграниченный от оперения боковых сторон головы несколькими рядами плотных мелких перышек. У большинства видов цевка оперена, у многих видов оперены и пальцы. Первостепенных маховых 10. Крыло диастатаксическое. Рулевых обычно 12. Маховые и рулевые относительно мягкие, «бархатистые», что делает полет совершенно бесшумным. Окраска обычно неяркая, маскирующая, из сочетания серых, бурых, охристых тонов. Половой и возрастной морфизм окраски обычно не выражен. Самцы, как правило, несколько мельче самок.
Активны преимущественно ночью или в сумерках. Населяют разнообразные ландшафты (степи, пустыни, горы, тундру), но большинство видов приурочено к лесным местообитаниям. Питаются животной пищей: крупными насекомыми, различными наземными позвоночными; несколько видов ловят на мелководье рыбу и крупных ракообразных. Падаль не едят. При ловле добычи ориентируются в основном на слух, отчасти на зрение. Возможность улавливания инфракрасного (теплового) излучения жертвы не доказана. Добычу подкарауливают или разыскивают, облетая охотничий участок. Реже охотятся «пешком». Добычу заглатывают целиком или — крупную — большими кусками. Кости перевариваются в желудке сов значительно хуже, чем у дневных хищных птиц. Непереваренные остатки пищи выбрасываются в виде погадок.
Моногамы. У многих видов пары, видимо, сохраняются длительное время. Гнездятся отдельными парами, обычно укрыто: в дуплах, брошенных гнездах других птиц, расщелинах скал, нишах между корнями, в норах крупных грызунов. Реже яйца откладывают на землю у основания кочки и т. п. Большинство видов гнезд не строит; у нас только болотная сова устраивает примитивное гнездо на земле, скрытое травянистой растительностью. Размеры кладки сильно варьируют в зависимости от количества корма; в малокормные годы возможно негнездование, в годы обилия пищи («мышиные годы») бывает иногда и 2 кладки. У крупных видов кладки меньше, чем у мелких: бывает от 1—2 до 10—13 яиц, чаще 3—5. Яйца белого цвета. Насиживает обычно только самка; самец носит ей корм. Насижива
215
ние начинается после откладки первого яйца (поэтому в гнездах птенцы разновозрастные) и продолжается около 3—5 недель. Развитие по птенцовому типу: птенцы вылупляются опушенными белым пухом, но слепые. В гнезде остаются до приобретения способности к полету: от 3 (мелкие виды) до 6 (крупные виды) недель. Корм птенцам носят оба партнера.
Оседлые или кочующие, немногие виды перелетные. Вне сезона размножения держатся поодиночке, редко небольшими, легко распадающимися группами. Подавляющее большинство видов сов полезны истреблением вредных животных: грызунов, крупных жуков и т. п.
Отряд сов включает 3 семейства (в одном только ископаемые формы), объединяющих 25—30 родов с 144 ныне живущими видами. В СССР встречается и гнездится 18 видов, относимых к 12 родам 2 семейств.
СЕМЕЙСТВО 1. PROTO ST RIGIDAE
По разной сохранности остаткам из нижнего или среднего эоцена США (Вайоминг) описано 4 вида, видимо, примитивных сов. Они и составляют это семейство.
СЕМЕЙСТВО 2. ТУТО NIDAE — СИПУХОВЫЕ
Отличаются от настоящих сов более стройным телом, не круглым, а суживающимся книзу «лицом» (рис. 130). Клюв относительно слабый. Ушные капсулы симметричны. По заднему краю грудины только одна пара вырезок, у взрослых иногда полностью зарастающих. Вилочка не пневматизирована. Цевка относительно длинная. Второй и третий пальцы ноги одинаковой длины; коготь третьего пальца зазубренный. Копчиковая железа оперена. Яйца овальной формы. В возрасте около 2 недель птенец сменяет эмбриональный пух на второй пуховой наряд, а затем у него развивается уже первый годовой перьевой наряд. Питаются землеройками, мелкими мышевидными грызунами, крупными насекомыми; изредка ловят мелких птиц. Оседлы.
Рис. 130. Сипуха Ту to alba (Scop.)
216
150° 120° 90° 60°к западу от ГринвичаО°к востоку ст Гринвича 60° .90° 120° 150° 180° 150° •
150°120°90°60°30°0°30°60° 90°120°150° 180° 150°
Рис. 131. Ареалы семейств:
1— сипуховые Tytonidae, 2— настоящие совы Strigidae (отряд сов), 3— трогоновые Trogonidae (отряд трогонообразных)
Семейство сипуховых объединяет И видов, относимых к 2 родам: Ту to (9 видов, распространены в жарком, отчасти умеренном поясе обоих полушарий; рис. 131) и Rhodilus (один вид в Юго-Восточной Азии; второй в Центральной Африке). В юго-западных районах европейской части СССР гнездится сипуха Tyto alba (Scop.).
По остаткам из миоцена и более поздних отложений в пределах современного ареала описано 6 ископаемых и один ныне живущий вид. Сипухо-вые, несомненно, близки к настоящим совам, но в то же время достаточно четко от них обособлены.
СЕМЕЙСТВО 3. STRIGIDAE— НАСТОЯЩИЕ СОВЫ
Рис. 132. Филин Bubo bubo (L.)
Большая голова с округлым лицевым диском (рис. 132). Клюв крепкий. У многих видов асимметрия ушных капсул приводит к более или менее выраженной асимметрии черепа. По заднему краю грудины две пары вырезок. Вилочка пневматизирована. Цевка относительно короткая. Третий палец ноги длиннее второго; его коготь гладкий. Копчиковая железа голйя. Яйца округлые, почти шаровидные. Примерно через 1,5—2 недели после вылупления птенцы сменяют эмбриональный пуховой наряд на промежуточный перьевой — мезоптиль, а вскоре после вылета из гнезда начинают линьку в первый годовой перьевой наряд (большие кроющие, маховые и рулевые не сменяются).
Семейство настоящих сов объединяет 133 вида (2 вымерли в историческое время), относимых к 23—28 родам: Strix, Aegolius, Asia, Athene, Bubo, Ciccaba, Glaucidium, Gymnoglaux, Jubula, Ketupa, Lophostrix, Micrat-hene, Mimizuka, Nesasio, Ninox, No-vipulsatrix, Nyctea, Otus, Pseudoscops, Pseudoptynx, Pulsatrix, Pyrrhoglaux, Rhinoptynx, Sceloglaux, Scotopelia, Speotyto, Surnia, Uroglaux. Практически космополиты (рис. 131), не заселившие лишь некоторые океанические острова и Антарктиду. Некоторые роды рапространены очень широко, хотя их ареалы несколько меньше ареала семейства. Так, Bubo, Glaucidium, Otus встречаются повсеместно, кроме Австралийской области. В СССР встречается и гнездится 17 видов настоящих сов, относимых к 11 родам.
Ископаемые остатки, датируемые верхним эоценом — нижним олиго-ценом и более поздними периодами, известны из многих районов современного ареала (наиболее ранние из Франции, в том числе и относимые к современным родам Bubo и Asia). Их относят к 21 ископаемому и к 26 ныне живущим видам (достоверность части определений сомнительна).
Центр возникновения сов, види-мо, Палеарктика. Сейчас большее видовое разнообразие характерно для тропиков. Обычно полагают, что совы—давно обособившийся отряд, вероятно, очень давними родственны
218
ми связями сближающийся с козодоеобразными и, в меньшей степени, с ракшеобразными. Сходство с дневными хищными птицами — результат конвергенции.
ОТРЯД 27. CAPRIMULGIFORMES — КОЗОДОЕОБРАЗНЫЕ
Птицы мелких и средних размеров (вес от 50 до 500—700 г), ведущие сумеречный или ночной образ жизни.
Клюв маленький, но разрез рта очень большой: челюстной сустав расположен позади глаза. В углах рта обычно развиты длинные упругие щетинки. Череп схизогнатический или десмогнатический; сошник есть. У многих видов есть функционирующие базиптеригоидные сочленения. Ноздри обычно не сквозные, голоринальные. Шейных позвонков 13—15. Спинная кость не образуется. По заднему краю грудины 1—2 пары вырезок. Истинных ребер 4 пары. Цевка короткая. Гортань бронхиальная или трахео-брон-хиальная. Язык плоский, обычно очень короткий. Зоба нет. Кишечник относительно короткий; слепые кишки длинные, со вздутиями на конце (как у сов). Желчный пузырь отсутствует лишь у единичных родов. Копчиковая железа голая, маленькая или ее нет вовсе. Кожа очень тонкая. Контурные перья мягкие, с развитой пуховой частью и обычно с добавочным стержнем. Пух растет только на аптериях. Первостепенных маховых 10. Крыло диа-статаксическое. Рулевых 10.. Маховые и рулевые «бархатистые», но не так отчетливо, как у сов. Окраска покровительственная буроватая или коричневатая, делающая неподвижно сидящих днем птиц мало заметными. Половой и возрастной морфизм окраски выражен слабо.
Хвост и крылья длинные. В полете несколько напоминают крупных ласточек или мелких ястребов. Полет бесшумный, маневренный. По земле большинство видов передвигается медленно, неуверенными прыжками. Пищу обычно хватают налету. Подавляющее, большинство видов животно-ядны. Моногамы. Для многих видов характерен своеобразный токовый полет с хлопаньем крыльев, криками или «урчаньем». В кладке 1—2, редко 3—4 яйца. Насиживают оба пола. Птенцы вылупляются зрячими. Родители выкармливают птенцов до подъема на крыло. Становятся половозрелыми в возрасте около года.
Населяют разнообразные местообитания: леса (тяготеют к опушкам и полянам), степи, горы, пустыни. Распространены преимущественно в тропиках; в умеренные широты проникают немногие виды. В тропиках оседлые или кочующие, в умеренных широтах перелетные.
Отряд козодоеобразных разделяется на 2 подотряда и 5 семейств, объединяющих 23 рода с 93 видами. В СССР встречаются и гнездятся 3 вида рода Caprimulgus.
ПОДОТРЯД 1. STEATO R NITH ES — ГУАХАРО, ИЛИ ЖИРЯКИ
Включает одно семейство.
СЕМЕЙСТВО 1. STEATORNITHIDAE — ГУАХАРО, ИЛИ ЖИРЯКИ
Единственный вид —Steatornis caripensis Humb. (рис. 133), распространен в горных тропических лесах севера Южной Америки и на некоторых прибрежных островах, включая о. Тринидад (рис. 134). Размеры довольно
219
Рис. 133. Гуахаро Steatornis caripensis Hamb.
крупные (вес около 400 г, длина около 45 см). Встречаются очень спорадично: только в районах, где есть глубокие пещеры. Оседлы.
Клюв сильный, с резко выраженным длинным крючком на конце надклювья и зубчатым выростом в конце подклювья. Межносовая перегородка окостеневает. Череп десмогнатический, с функционирующими базиптери-гоидными сочленениями. Шейных позвонков 15. Вырезки по заднему краю грудины неглубокие. Ноги более слабые, чем у остальных козодоев, но пальцы с крепкими острыми когтями (для удерживания на скалистых уступах). Две сонные артерии. Гортань бронхиальная. Копчиковая железа голая, маленькая. Оперение более плотное и менее «бархатистое», чем у остальных козодоев. Окраска коричневатая. Добавочного стержня контурного пера нет. Пудреток нет.
Видимо, исключительно растительноядны. Питаются разнообразными плодами (косточки выплевывают), ягодами и семенами, которые налету срывают с тонких ветвей. Кормятся ночью, день проводят в пещерах. Сидеть на ветвях и ходить почти не могут; сидят на уступах стен и на дне пещеры, опираясь о субстрат пальцами и цевкой.
Гнездятся большими скоплениями (раньше местами — десятками тысяч пар) в глубоких темных пещерах. Видимо, ориентируются с помощью звуковой локации. Пары, вероятно, сохраняются до гибели одного из партнеров. Гнездо, состоящее в основном из погадок (косточки плодов, оболочки семян и т. п.), помещается на уступе стен пещеры. В кладке 2—4 белых с небольшой пятнистостью яиц, откладываемых с интервалом в несколько дней. Насиживание начинается после откладки 1—2 яиц и продолжается 33—35 дней. Птенцы вылупляются голыми, но вскоре покрываются густым темным пухом. В первые 20—25 дней жизни усиленно обогреваются родителями. Постэмбриональное развитие медленное: гнездо покидают активным полетом в возрасте 3,5—4 месяцев. Птенцы накапливают очень большое количество подкожного и внутреннего жира, составляющего до 4/3 их веса. Местное население издавна ведет промысел птенцов, используя их жир в пищу. Многие колонии сейчас полностью истреблены.
В ископаемом состоянии не обнаружены. Вероятно, автохтоны тропиков Южной Америки. По многим мелким морфологическим особенностям (детали строения скелета, мускулатуры и т. п.) занимают как бы промежуточное положение между остальными козодоями и совами. Гуахаро, видимо, потомок одной из древнейших ветвей отряда, четко обособившийся от других групп козодоев.
220
Рис. 134. Ареалы семейств козодоеобразных птиц:
1—гуахаро Steatornithidae, 2—совиные козодои Podargidae, 3— исполинские козодои Nyctibiidae, 4—совиные лягушкороты Aegothelidae, Б—настоящие козодои Caprimulgidae
ПОДОТРЯД 2. CAPRIMULGI — КОЗОДОЕВЫЕ
Птицы мелких и средних размеров (вес от 50 до 500—700 г). Череп десмогнатический или схизогнатический. Шейных позвонков 13—14. Вырезки по заднему краю грудины обычно глубокие. Сонных артерий одна или две. Гортань бронхиальная или трахео-бронхиальная. Копчиковая железа маленькая, голая; может отсутствовать. Добавочный стержень контурного пера мал, но всегда заметен.
Животноядны; растительную пищу не едят. Не колониальны. Никогда не гнездятся в пещерах. Видимо, нет звуковой локации. Птенцы вылупляются покрытые густым пухом. Становятся способными к полету в возрасте менее 1,5 месяцев.
Подотряд включает 4 семейства с 22 родами и 92 видами. В СССР гнездятся только 3 вида 1 рода.
СЕМЕЙСТВО 2. PODARGIDAE — СОВИНЫЕ КОЗОДОИ
Всего 12 видов, объединяемых в 2 рода: Podargus (3 вида) и Batrachosto-mus (9 видов). Населяют тропические леса Юго-Восточной Азии, ряда островов Океании и Австралии, но не проникают в Новую Зеландию (рис. 134). Птицы средних размеров (рис. 135).
Клюв довольно массивный с крючком на конце надклювья. Ноздри у основания клюва полуприкрыты кожистыми складками. Носовая перегородка окостеневает. Череп десмогнатический с более или менее выраженным переходом к эгитогнатизму; функционирующих базиптеригоидных сочленений нет. Шейных позвонков 13—14. Задний край грудины с 2 парами вырезок. Лапы довольно сильные; могут охватывать ветви. Коготь среднего пальца гладкий. Четвертый палец состоит из 5 фаланг, очень подви-
Рис. 135. Совиный козодой Batrachostomus moniliger Bl. на гнезде
жен и может отводиться назад, каку сов. Сонная артерия одна. Нижняя гортань бронхиального типа. Копчиковая железа очень мала. В области надхвостья есть пудретки.
Питаются главным образом крупными насекомыми, которых ловят в воздухе или налету подхватывают с земли и ветвей; склевывая добычу, медленно ходят по земле и по толстым ветвям. Маленькое гнездо устраивают в развилке сучков или на толстой горизонтальной ветке. Выстилка гнезда—из порошковидных обломков пуха (е пудреток). В кладке 2—3 белых яйца. Насиживает, видимо, только самка, обычно сидящая на ветке и областью наседного пятна прикрывающая гнездо с яйцами. Инкубация около 1 месяца. Птенцы при вылуплении покрыты длинным белым пухом. Вскоре покидают гнездо и сидят на ветках рядом с ним. Способность к полету приобретают в возрасте около 1 месяца.
222
Вероятный центр возникновения — Австралия или Юго-Восточная Азия.
СЕМЕЙСТВО 3. NYCT1B11DAE— ИСПОЛИНСКИЕ КОЗОДОИ
Сюда включают только 1 род Nyctibius с 5 видами (длина 20—55 см), распространенными в тропических лесах Центральной и Южной Америки (рис. 134).
Клюв короткий, слабый, с острым крючком на конце. Ноздри открытые, расположены несколько вперед от основания клюва. Носовая перегородка не окостеневает. Череп схизогнатический, с функционирующими базиптеригоидными сочленениями. Шейных позвонков 14. По заднему краю грудины 2 пары мелких вырезок. Четвертый палец ноги с 5 фалангами. Коготь среднего пальца гладкий. Могут сидеть на ветке, охватывая ее пальцами. Сонная артерия одна. Гортань трахео-бронхиаль-ного типа. В области надхвостья большие пудрет-ки.
Экология изучена плохо. Пища — разнообразные крупные насекомые, добываемые в воздухе. Единственное белое яйцо откладывают в ямку с древесной трухой на конце сломанной ветви или высокого пня.
Насиживающая птица держит голову и тело вертикально; издали кажется продолжением ствола (рис. 136).
Остатки одного из ныне живущих видов найдены в плейстоценовых отложениях Бразилии. Видимо, автохтоны Южной Америки. По многим особенностям строения занимают промежуточное положение между настоящими и совиными козодоями.
Рис. 136. Насиживающий серый исполинский козодой Nycti-bius griseus Gm.
Рис. 137. Совиный лягушкорот Aegotheles cristatus (Wh.)
223
СЕМЕЙСТВО 4. AEGOTHELIDAE—СОВИНЫЕ ЛЯГУШКОРОТЫ
В семейство входит единственный род Aegotheles с 5 видами мелких козодоев (вес 50—100 г), распространенных в лесах Австралии, Новой Гвинеи и прилегающих островах (см. рис. 134). Близки к совиным козодоям.
Клюв короткий и слабый. Открытые ноздри расположены близко от конца клюва. Носовая перегородка не окостеневает. В углах рта длинные осязательные щетинки. Череп десмогнатический с функционирующими ба-зиптеригоидными сочленениями. Шейных позвонков 14. По заднему краю грудины 2 пары вырезок. Четвертый палец лапы из 5 фаланг. Сонных артерий две. Гортань бронхиальная. Слепых кишок нет. Пудреток нет.
Экология изучена плохо. Могут сидеть на ветвях; своеобразны по позам и общему облику (рис. 137). Животноядны. Днюют и гнездятся в естественных дуплах, на дно которых без всякой подстилки откладывают обычно 4 белых яйца. Центр возникновения семейства, вероятно, Новая Гвинея.
СЕМЕЙСТВО 5. CAPRIMULGIDAE — НАСТОЯЩИЕ КОЗОДОИ
Птицы мелких и средних размеров с характерным обликом козодоев (рис. 138), распространенные почти по всему земному шару, кроме Арктики, Новой Зеландии и Антарктики (см. рис. 134).
Короткий слабый клюв. Носовые отверстия в виде коротких мягких трубочек. Носовая перегородка не окостеневает. Череп схизогнатический (у Chordeiles — десмогнатический) с функционирующими базиптеригоид-ными сочленениями. Шейных позвонков 14. По заднему краю грудины одна пара вырезок. Ноги слабые, пальцы не могут охватывать тонкую ветку. Поэтому садятся только на толстые сучья, всегда располагая тело вдоль ветви, а не поперек, как остальные птицы. Коготь среднего пальца сбоку отчетливо зазубрен. IV палец образован 4 фалангами; не может отводиться вбок и назад. Нижняя гортань трахео-бронхиального или бронхиального типа. У единичных родов нет желчного пузыря. Копчиковая железа голая, небольшая. Побочный ствол контурного пера развит слабо. Пудреток нет.
Населяют разреженные леса или опушки и поляны густых лесов, степи, пустыни, горы. Охотятся только с наступлением сумерек; день проводят, затаившись на земле или на толстой горизонтальной ветви. Пища: разнообразные насекомые, которых ловят в воздухе. Вне сезона размножения держатся поодиночке. Южные виды оседлые или кочующие, обитатели умеренных широт перелетны. У американского белогорлого козодоя Phalaeno-ptilus nuttallii (Aud.) в северных частях ареала (юг США) у некоторых особей отмечена «зимняя спячка», продолжавшаяся до 85 дней; температура спящей птицы, забившейся в промоину в стенке обрыва, снижалась до 18— 19°. При резких падениях t° воздуха кратковременное оцепенение описано у обыкновенного козодоя Caprimulgus europeus L. и некоторых др .
Рис. 138. Буланый козодой Caprimtilgus aegyptius Licht.
224
В кладке 2 белых с размытым крапом яйца, откладываемых обычно прямо на землю. Развитие быстрре: инкубация около 3 недель, примерно через такое же время птенцы становятся способными к полету. Птенцы вылупляются зрячими, покрытыми густым пухом; их окраска защитная, пятнистая. Держатся, затаившись в районе «гнезда». Оба родителя некоторое время продолжают кормить уже летающих птенцов. У части видов в южных широтах, видимо, довольно часто бывает 2 кладки.
Семейство Caprimulgidae объединяет 70 видов, группируемых в 18 родов: Caprimulgus (почти космополитичен; включает около 40 видов), Chor-deiles, Eleothreptus, Eurostopodus, Hydropsalis, Lurocalis, Macrodipteryx, Macropsalis, Nyctidromus, Nyctiphrynus, Nyctiprogne, Otophones, Phala-enoptilus, Podager, Scotornis, Semeiophorus, Siphonorhis (S. americanus L. — ямайский козодой, недавно вымер), Uropsalis. Свойственны преимущественно тропикам; в умеренные широты проникают немногие виды. Наиболее разнообразна по числу видов фауна козодоев Нового Света. В СССР встречаются и гнездятся только 3 вида рода Caprimulgus.
Центральное, видимо, наиболее прогрессивное, семейство отряда. Вероятный центр возникновения — Новый Свет.
Ископаемые остатки немногочисленны (7 ныне живущих видов) и обнаружены лишь в поздних отложениях (наиболее ранние — из плейстоцена Румынии).
Весь отряд козодоеобразных довольно компактен и четко ограничен. Видимо, несомненные, но очень давние родственные связи прослеживаются с отрядом сов. В какой-то степени проявляются также очень давние связи с ракшеобразными и стрижеобразными. Вероятно, все эти отряды возникли от общей предковой группы примитивных птиц.
ОТРЯД 28. APODIFORMES —СТРИЖЕОБРАЗНЫЕ
Птицы очень маленьких или маленьких размерор (вес от 1,5 до 100 г), с характерным обликом стрижа или колибри (рис. 139, 141, 142).
Клювы разнообразной формы и размеров. Ноздри не сквозные, голоринальные. Череп схизогнатический или эгитогнатический; базиптеригоид-ные сочленения отсутствуют, сошник есть. Шейных позвонков 13—15. Истинных ребер 4 пары. Задний край грудины закругленный, без вырезок, но могут быть 1—2 фонтанели. Вилочка с небольшим расширением на конце (hypocleidium). Самый длинный отдел скелета крыла — кисть; плечо очень короткое и толстое, с сильно развитым боковым отростком. Ноги маленькие, относительно слабые. Формула мышц ноги упрощена: А—. По земле не ходят; ноги используются лишь как опора при сидении или для цепляния за края гнезда. Слепые кишки сильно редуцированы или полностью исчезают. Копчиковая железа голая. Нижняя гортань обычно трахео-бронхи-ального типа, с 1—2 парами голосовых мышц. Оперение довольно плотное. Контурное перо с более или менее заметным добавочным стержнем. Первостепенных маховых 10, коротких второстепенных 6—И, рулевых обычно 10. Крылья в разных родах диастатаксические или эутаксические.
Полет быстрый, стремительный, маневренный. Моногамы; у колибри пары образуются лишь на период спаривания. Строят гнезда, иногда довольно сложные. Кладка из 1—6 белых яиц. Насиживают кладку и выкармливают птенцов либо оба партнера, либо только самка. Птенцы вылупляются слепыми, голыми или очень слабо опушенными. В гнезде остаются, пока полностью не оперятся и не приобретут способность к полету. В тропиках оседлые или кочующие, в умеренных широтах перелетные (поздно прилетают, рано отлетают).
Отряд разделяется на 2 резко специализированных подотряда: стрижей и колибри. В отряде принимают 4 семейства (одно только с ископаемы
225
ми формами), объединяющие 378 видов, относимых к 109—129 родам. В СССР встречаются и гнездятся 5 видов лишь семейства настоящих стрижей.
ПОДОТРЯД 1. APODI —СТРИЖИ
Мелкие птицы (вес 20—100 г) с уплощенной головой, короткой шеей, очень длинными и узкими «саблевидными» крыльями (рис. 141).
Клюв короткий, сплющенный сверху вниз, в плане треугольный. Разрез рта большой: челюстной сустав находится позади глаза. Череп эгитогна-тический. Шейных позвонков 13 (у Chaetura 14). Строение лапы варьирует. Когти острые, изогнутые. Обычно сохраняется только левая сонная артерия (у Cypselold.es и некоторых Chaetura есть обе). Сильно развиты слюнные железы. Язык короткий, треугольный. Зоба нет. Есть желчный пузырь. Оперение довольно плотное. Добавочный стержень контурного пера развит. У взрослых на аптериях есть короткий пух. Второстепенных маховых 8—11. Окраска обычно из сочетания разных оттенков черного и белого цветов. Полового диморфизма окраски обычно нет, возрастной выражен слабо.
Активность дневная. Полет быстрый (до 120—170 км/ч}, стремительный, неутомимый. По земле практически не ходят. Все время проводят в воздухе, лишь изредка присаживаясь на уступ скалы, край гнездового дупла ит. п. По некоторым данным, негнездящиеся птицы некоторых видов (например, черный стриж) могут ночевать в воздухе, летая широкими кругами на высоте 1—3 км со скоростью в 40—60 км/ч.
Живут в горах, на равнинах, в лесах, поселениях человека. Более многочисленны в тропиках. В умеренные широты проникают немногие виды. Питаются разнообразными насекомыми, которых ловят в воздухе. Пьют налету, черпая воду подклювьем. Гнездятся группами или небольшими колониями. Строительный материал для гнезда подхватывают налету. В разных родах отличаются размещение и конструкция гнезда, размеры
кладки, которая варьирует от 1—2 до 4—6 белых яиц. Насиживают кладку и выкармливают птенцов оба родителя. Инкубация 2,5 — 3,5 недель. Постэмбриональное развитие медленное — до 1,5 месяца. Птенцы покидают гнездо активным полетом.
Подотряд стрижей включает 3 семейства (одно только с ископаемыми формами), объединяющие 58 ныне живущих видов, относимых к 9 родам.
Рис. 139. Хохлатый стриж Hemiprocne mystacea (Less.) у гнезда с яйцом
СЕМЕЙСТВО 1 AEG1ALORNITH1DAE
Включает 2 вида 1 рода, описанных по остаткам из
верхнего эоцена — нижнего олигоцена Франции.
226
СЕМЕЙСТВО 2. HEMIPROCNIDAE — ДРЕВЕСНЫЕ, ИЛИ ХОХЛАТЫЕ, СТРИЖИ
Объединяет 3 вида рода Hemiprocne (рис. 139), распространенных от Южной Индии до Новой Гвинеи (рис. 140). На голове развиваются яркие, чаще чисто белые украшающие перья. Когтя на первом пальце крыла нет. Лапа анизодактильная.
Экология изучена плохо. Маленькое гнездо в виде тоненькой чашечки из склеенных слюной кусочков коры и листьев прикрепляется к боковой поверхности ветки в вершине дерева. Лоточек выстилается тонкими травинками. Единственное яйцо прикрепляется к ветке и к стенкам гнезда ниточками засохшей слюны. Насиживающая птица сидит на ветке рядом с гнездом, прикасаясь к яйцу голой кожей наседного пятна. Вылупившийся птенец покрыт пухом. Немного подросший птенец выбирается из гнезда и сидит на ветке, тесно к ней прижимаясь.
В ископаемом состоянии не обнаружены. Уклоняющаяся, возможно, более примитивная ветвь стрижей, в какой-то мере сохраняющая черты предков (неспециализированная лапа, гнездование на ветвях и т. п.).
СЕМЕЙСТВО 3. APODIDAE— НАСТОЯЩИЕ СТРИЖИ
Птицы с типичным обликом стрижей (рис. 141), очень широко распространенные по земному шару (рис. 140).
На голове нет украшающих перьев. На первом пальце крыла развит роговой коготок. У большинства видов лапа помпродактильная (все 4 пальца направлены вперед); у видов рода Chaetura — анизодактильная (три пальца вперед, один назад), а у Cypsiurus — зигодактильная (назад направлены первый й четвертый пальцы).
Гнезда разных видов очень различны по конструкции и расположению. Стрижи родов Apus, Aeronautes , Chaetura в укрытии — дуплах, расщелинах скал, щелях в постройках и т. п. — строят рыхлое гнездо из тонких веточек, растительных волокон и перьев. Стрижи Tachornis и Panyptila из скрепленного слюной растительного пуха, травинок и листочков строят гнездо в виде трубы или мешка, открытого снизу и прикрепленного вершиной к уступу скалы или толстой ветке; яйца откладывают в своеобразный карманчик, построенный на внутренней стенке гнезда (рис. 141). Стрижи Cypsiurus маленькое чашеобразное гнездо приклеивают слюной к вертикальной поверхности пальмовых листьев, а к гнезду слюной приклеивают 1 или 2 маленьких яичка. Стрижи Cypseloides строят склеенные слюной лепные гнезда из грязи и растительных волокон, прикрепляя их на обрывах скал у воды. Гнездящиеся в пещерах Юго-Восточной Азии мелкие стрижи саланганы Collocalia устраивают гнезда из загустевающей на воздухе чистой слюны или из слюны с небольшим добавлением растительных волокон; эти гнезда, имеющие вид маленьких пергаментообразных чашечек, прикрепляются к верхним участкам стен пещер. Гнезда саланганов представляют объект промысла и используются в китайской кухне. В отличие от других стрижей гнездящиеся в пещерах саланганы способны к звуковой локации; они издают щелкающие звуки в пределах 1,5—5,5 кгц с интервалом около 0,25 сек. В кладке у Apus, Cypseloides, Collocalia, Cypsiurus 1—2 яйца, у Panyptila и Tachornis 3, редко 2, у Chaetura и A'eronautes 3—6. Насиживают кладку и выкармливают птенцов оба партнера.
При внезапных похолоданиях, когда исчезают летающие насекомые, гнездящиеся взрослые особи некоторых видов (например, черный стриж A. apus L.) отлетают к югу, а птенцы впадают в оцепенение и могут находиться в таком состоянии «мнимой смерти» до 5—8 дней. При возвращении родителей после наступившего потепления с началом обогрева птенцы «оживают», продолжая нормально развиваться; естественно, что общая продолжительность пребывания их в гнезде увеличивается.
227
Рис. 140. Ареалы семейств стрижеобразных птиц и птиц мышей:
1— хохлатые стрижи Hemiprocnidae, 2— настоящие стрижи Apodidae, 3— колибри Trochilidae, 4—птицы мыши Coliidae
<М
Рис. 141. Кайенский стриж Panyptilacayennensis (Gm.) около гнезда; справа — гнездо в разрезе
Семейство настоящих стрижей объединяет 55 видов, относимых к 8 родам: Apus (10 видов), Aeronautes (3 вида), Chaetura (17 видов; сюда включают и род Hirundapus, к которому относится гнездящийся в Приморье колючехвостый стриж), Collocalia (10 видов), Cypseloides (9 видов), Cypsiu-rus (1 вид), Panyptila (2 вида), Tachornis (3 вида). В СССР гнездится 5 видов 2 родов: Apus (4 вида) и Chaetura (Hirundapus).
Основное, наиболее специализированное семейство подотряда. Центр возникновения, видимо, Старый Свет. В ископаемом состоянии известны из нижнего эоцена — верхнего олигоцена (Франция) и из более молодых отложений в пределах современного ареала.
ПОДОТРЯД 2. TROCHILI — КОЛИБРИ
Включает единственное семейство.
СЕМЕЙСТВО 4. TROCH1LIDAE— КОЛИБРИ
Мелкие и очень мелкие птицы очень разнообразного облика (рис. 142) >весом от 1,6 г до 20 г; большинство видов меньше крапивника и весит 3—8 г. Распространены в Северной и Южной Америке и на прилегающих островах '(рис. 140). Подавляющее большинство видов встречается лишь в зоне тропи-.ков, но при этом многие виды летом живут в суровых условиях высокогорий
229
Рис. 142. Сосущий сок шпагоклювый колибри Ensifera ensifera (Boiss.)
Анд. Один вид распространен по всей Патагонии, 4 вида — на территории США и южных районов Канады, а один вид, населяющий юго-восток Аляски, заходит на север почти до Полярного круга. В жарких районах оседлы, в высокогорьях и в умеренных широтах перелетны.
Клюв длинный или очень длинный (у некоторых видов превышает длину тела), иногда более или менее заметно изогнутый книзу. Разрез рта небольшой; челюстной сустав располагается на уровне глаза или впереди него. Череп схизогнатический. Шейных позвонков 14—15. Когтя на первом пальце крыла нет. По сравнению со стрижами кисть еще более удлинена, а предплечье и плечо укорочены. Грудина и киль относительно большие. Цевка заметно укорочена. Лапа анизодактильного типа. Когти почти такой же длины, как пальцы. В целом скелет довольно мощный и прочный. Над-коракоидная мышца почти такой же величины, как большая грудная мышца (у стрижей в 5—6 раз меньше). Сохраняется только левая сонная артерия. Слюнные железы развиты много слабее, чем у стрижей. Язык у большинства видов мускулистый, очень подвижный; у многих видов его конец сворачивается в трубку при высасывании нектара из цветка. Взрослые колибри лишь у немногих видов имеют зоб, но он, кажется, есть у птенцов всех видов. Желчного пузыря нет. Оперение обычно довольно плотное, у части видов относительно рыхлое. Добавочный стержень контурного пера короткий. Пуха нет. Крылья относительно шире, чем у стрижей. Второстепенных маховых всего 6—7. Окраска самцов очень яркая, с металлическим блеском; часто развиваются украшающие перья: хохолки, воротнички, длинные рулевые, пушистые «манжеты» на лапках и т. п. Самки окрашены значительно тусклее. Окраска молодых похожа на окраску самок.
Активны преимущественно днем, но есть несколько видов с сумеречной активностью. Полет довольно быстрый (до 40—60, у некоторых видов до 90 км!ч), маневренный, с резким изменением направления. Способны длительное время держаться в воздухе на одном месте: колибри трепещет в воздухе около цветка, высасывая из него сок и выбирая насекомых. При этом крылья совершают до 50—80 взмахов в секунду! По земле обычно не ходят. Живущие в горах виды ловко лазают по скалам. Хорошо сидят на тонких ветвях и стеблях.
Населяют разнообразные местообитания. Немногие виды живут в густых и темных тропических лесах. Большинство видов приурочено к более
230
открытым, хорошо освещенным местообитаниям: опушкам или разреженным участкам леса, лесным полянам, зарослям кустарников, лугам разных типов, в том числе и альпийским; встречаются в степях и полупустынях. В горы проникают до 5—5,5 км над уровнем моря. Некоторые виды имеют очень ограниченные ареалы: отдельным горам Анд свойственны эндемичные виды. Распространение сильно специализированных видов — нек-тарофагов часто определяется распространением тех растений, на цветках которых они питаются.
Более крупные виды питаются насекомыми, которых они налету, в трепещущем полете выбирают из венчиков цветков, склевывают с листьев и ветвей. Мелкие виды подобно бабочкам высасывают нектар цветков, склевывают тычинки и мелких насекомых. Видимо, все виды могут ловить насекомых в воздухе. Потребность в корме высокая: до удвоенного веса своего тела в сутки (у мелких видов). Очень интенсивный обмен веществ, видимо, в первую очередь связан с высокими теплопотерями: односительно большая поверхность тела при малой его массе. В активном состоянии температура тела у разных видов равна 40—43° С. У сидящей птицы пульс до 500 ударов в минуту и до 180 дыханий в минуту; в полете эти показатели возрастают: пульс до 1200, дыхание — до 600 в минуту. Колибри, живущие в районах с резким колебанием суточных температур (высокогорья, умеренные широты), ночуют в гнездах. У севшей вечером в гнездо птицы температура тела снижается; когда она достигает 35—38° С, птичка впадает в оцепенение. Температура тела снижается до 15—20° С (по некоторым данным, даже до 10—12°) и лишь на 1—2° С превышает температуру в гнезде; пульс уменьшается до 100—50 ударов в минуту, становятся более редкими дыхательные движения. Такое оцепенение с резко сниженным уровнем обмена веществ дает возможность колибри провести ночь, экономно расходуя энергетические запасы; если бы не было оцепенения, то птичка, израсходовав все запасы на поддержание высокой температуры тела и не имея возможности их пополнить, быстро бы погибла от истощения. Утром птица просыпается, температура, пульс, дыхание в течение 20—30 мин вновь поднимаются до нормы, и она вновь становится активной, начиная свой день с поисков пищи.
У видов влажных жарких тропиков размножение идет в течение всего года, а у каждой самки бывает 2—3 выводка. У видов, населяющих районы с заметным сезонным изменением условий жизни, обычно один цикл размножения. Самцы токуют поодиночке, видимо, придерживаясь определенных участков: совершают токовые полеты, издают громкие отрывистые трели. У части видов отмечен групповой ток, когда несколько токующих самцов держатся вместе на одном небольшом участке. Самка одна строит гнездо, размещая его в развилке тонких веточек куста или дерева, а в горных районах — в узких расщелинах скал. Чашеобразное гнездо с глубоким лотком и толстыми стенками сплетается из тонких растительных волокон, растительного пуха, мха, шерсти; строительный материал часто оплетается паутиной, которую птичка собирает, обрывая тенета пауков. Постройка гнезда продолжается 15—20 дней. Построившая гнездо самка объединяется с самцом. Птички держатся вместе, совершают совместные токовые полеты, спариваются. После завершения кладки пара распадается; у некоторых видов она существует всего несколько часов.
В кладке обычно два, у мелких видов только одно белое яйцо. У колибри весом 1,6—1,8 г яйцо весит около 0,2 г. Насиживает только самка. Инкубационный период зависит от погодных условий и продолжается у разных видов от 14 до 20 и даже 25 дней. Птенцы вылупляются голыми, слепыми, с коротким клювиком, несколько напоминающим клюв стрижей. Кормит только самка, чередуя полеты за кормом с обогревом птенцов. Пищу приносит в ротовой полости и в пищеводе и отрыгивает ее в открытый клюв птенца. Постэмбриональное развитие медленное; оно несколько ускоряется при высоких температурах воздуха. Глаза открываются на 6—12-й день после
231
вылупления. В этом же возрасте начинает формироваться перьевой покров. Птенцы покидают гнездо активным полетом в возрасте 25—35 дней. Некоторое время после вылета (от 4 до 20 дней у разных видов) самка продолжает кормить птенцов.
Вне сезона размножения держатся поодиночке или мелкими рассеянными группами. Во время миграций некоторые виды способны пролетать без отдыха значительные расстояния. Так, рубиновогорлый колибри Archilochus colubris (L.) весом 2,5—3,5 а, весной и осенью пересекая Мексиканский залив, пролетает без отдыха со скоростью около 40—45 км!ч не менее 900 км над водой, расходуя накопленные запасы жира (полагают, что при обычном полете энергетические траты меньше, чем при трепещущем). В году одна полная линька, завершающаяся у оседлых видов в течение 2 месяцев; у перелетных видов линька растягивается на большее время.
В связи с резким изменением ландшафта во многих районах, а также неумеренным преследованием со стороны человека (добывают для изготовления различных украшений и сувениров) ареалы многих видов существенно сократились, а численность заметно уменьшилась.
Колибри — относительно однообразная группа. Поэтому деление семейства на более мелкие группы очень затруднено, хотя при очень большом объеме семейства это было бы весьма полезно в чисто практических целях. Сейчас выделяют 320 видов колибри, объединяемых в 100—120 родов: Тго-chilus, Acestrura, Adelomyia, Aglaeactis, Aglaiocercus, Agyrtria, Amazilia, Androdon, Anthracothorax, Archilochus, Avocettula, Boissonneaua, Calliplox, Calothorax, Calypte, Campylopteros, Chaetocercus, Chalcostigma, Chalybura, Chlorestes, Chlorostilbon, Chrysolampis, Chrysuronia, Cincludes, Coeligena, Cyanolesbia, Cyanthus, Discosura, Docimastes, Doricha, Dorifera, Ensifera, Eriocnemis, Eugenes, Eulampis, Eupetomena, Eutoxeres, Florisuga, Glaucis, Heliactin, Heliangelus, Heliomaster, Heliothrix, Hylocharis, Lafresnaya, Lampornis, Lesbia, Leucippus, Loddigesia, Lophornis, Melanotrochilus, Mellisuga, Metallura, Microstilbon, Myrmia, Ocreatus, Oreotrochilus, Oxy-pogon, Panterpe, Patagona, Phaethornis, Polyttnus, Popelairia, Pterophanes, Ramphomicron, Sappho, Selasphorus, Sephanoides, Stellula, Stephanoxis, Thalurania, Threnetes, Tilmatura, Topaza, Waldronia и др.
Колибри, видимо, возникли в тропиках Южной Америки. В верхнем плейстоцене Бразилии обнаружены остатки одного из ныне живущих видов.
Предполагается, что родство двух подотрядов — стрижей и колибри — несомненное, но давнее; колибри обособились от каких-то примитивных, но уже вполне сложившихся стрижей. Только Ферхейен (1956) полагает, что эти 2 группы птиц между собой совершенно не родственны: по его мнению, колибри близки к удодам, а стрижи— к ракшам.
Относительно родственных взаимоотношений всего отряда стрижеобразных единой точки зрения нет. Большинство систематиков полагают, что очень давними родственными отношениями стрижеобразные связаны с козодоеобразными. Однако высказывается предположение, что никаких родственных связей с козодоевыми у них нет, а они близки к примитивным воробьиным, и должны быть включены в отряд Passeriformes в качестве самостоятельных подотрядов стрижей и колибри. Решить это могут только дальнейшие исследования.
ОТРЯД 29. COLIIFORMES — ПТИЦЫ МЫШИ
Включает 1 семейство (1-е) Coliidae с одним родом Colius, объединяющим 6 видов, распространенных в Центральной и Южной Африке (рис. 140).
Мелкие (вес 40—50 а) длиннохвостые птицы (рис. 143). Клюв короткий, конический, слегка изогнутый к концу, с острой вершиной. Череп десмогнатический, с редуцированным сошником; базиптеригоидных сочленений нет. Ноздри не сквозные, голоринальные. Шейных позвонков 13. Грудина
232
Рис. 143. Птица мышь Colitis macrourus (L.)
Мускульный желудок слабый. Кишечник
с двумя парами глубоких вырезок по заднему краю. Вилочка с расширением на конце. Истинных ребер 4 пары. Первый и четвертый пальцы лапы очень подвижны и могут поворачиваться вперед и назад. Мускульная формула ноги AXY—. Гортань трахеобронхиальная, с двумя парами мускулов. Сонная артерия только левая. Язык плоский. Зоба нет. короткий и широкий. Слепые кишки отсутствуют. Есть желчный пузырь. Копчиковая железа оперенная. Кожа толстая. Оперение рыхлое. Хорошо развиты побочные стержни контурных перьев. Пуха нет. Первостепенных маховых 10. Крылья короткие, широкие, эутаксические. Длинный, резко ступенчатый хвост из 10 рулевых. Окраска коричневатая. На головке хохол из удлиненных, часто ярких перьев. Возрастной и половой морфизм окраски не отчетлив.
Населяют кустарники африканских саванн, проникают в полупустыни, встречаются по опушкам лесов и в культурном ландшафте. Птицами мышами названы потому, что шныряют в гуще кустов, редко показываясь на глаза. Взлетают неохотно. Питаются семенами, ягодами, почками и цветками; ловят и подвернувшихся насекомых, таскают яйца и птенцов мелких птиц. Моногамы. Пары сохраняются длительное время. Ток — ритмичные прыжки, сопровождаемые трескучими выкриками. Определенных сроков гнездования, видимо, нет: гнездятся при благоприятных погодных и кормовых условиях. Гнездо — чаша из стебельков и листьев, укрепляется в развилке ветвей. В кладке 2—5 яиц чисто белых или с крричневато-красноватым крапом. В году может быть несколько кладок. Насиживают оба партнера 12—14 дней. Птенцы вылупляются слепыми. На 3—4-й день жизни открываются глаза, а пеньки перьев пробивают кожу. Кормят оба родителя, всовывая корм в яркую — черную с оранжевым языком — ротовую полость птенца. Птенцы оставляют гнездо в возрасте 14—16 дней, достигнув примерно 2/3 веса взрослых. Родители кормят их еще 2—4 недели после вылета. Выводки не распадаются до наступления половой зрелости (до 10—12 месяцев).
Видимо, автохтоны Африки. Родственные отношения с другими отрядами неясны. Возможно, обособились от форм, позднее давших начало ко-зодоевым и стрижеобразным; может быть, есть очень давние связи с примитивными ракшеобразными.
ОТРЯД 30. TROGONIFORMES — ТРОГОНООБРАЗНЫЕ
По внешнему облику несколько напоминающие соек птицы тропических лесов Африки, Юго-Восточной Азии, Центральной и Южной Америки (см. рис. 131).
Вес 60—200 а. Клюв у большинства видов широкий у основания, крепкий, умеренной длины, со слабоизогнутой вершиной, иногда заканчивающейся небольшим крючком. Края клюва часто зазубренные. В углах рта — крупные щетинки. Ноздри голоринальные, несквозные. Череп схизогнатический с крупным сошником. У большинства видов есть функционирующие базиптеригоидные сочленения. Шейных позвонков 15. По заднему краю грудины 2 пары глубоких вырезок. Коракоиды на грудине соприкасаются. Вилочка с лопатообразным расширением на конце. Мускульная формула
233
Рис. 144. Квезал Pha-romachrus mocinno Lla-ve.
AX—. Цевка короткая, частично оперенная. Лапа гетеродактильная (единственный случай в-классе птиц): первый и второй пальцы направлены назад, третий и четвертый — вперед. Когти острые, крупные. Сонная артерия только левая. Гортань трахео-бронхиальная, со слабой мускулатурой. Язык короткий, слабо уплощенный. Зоба нет. Мускульный желудок тонкостенный, но большого объема. Слепые кишки короткие. Копчиковая железа не оперена. Кожа очень тонкая; перья держатся слабо. Оперение довольно рыхлое. Контурные перья с крупными побочными стволами. Пуха нет. Первостепенных маховых 10. Крыло эутаксическое. Хвост обычно длинный, из 12 рулевых. Иногда верхние кроющие хвоста длиннее рулевых (рис. 144). Окраска очень яркая, с металлическим отливом: преимущественно зеленая, часто с болыпимц участками синего, красного, желтого цветов. Самки несколько тусклее самцов, у молодых птиц окраска менее яркая.
Населяют тропические леса разных типов, многие виды освоили культурные ландшафты. Американские виды, видимо, питаются преимущественно мелкими плодами, которые срывают налету или сидя на ветке; реже кормятся на земле. Виды Старого Света питаются преимущественно насекомыми: обычно птица сидит на ветке и, взлетев, ловит в воздухе добычу; реже склевывают насекомых на ветвях и на земле; плоды, семена поедают редко. Моногамы. В насиживании и в выкармливании птенцов участвуют оба партнера. Яйца откладывают без подстилки в естественные дупла. В кладке 2—3, редко 4 одноцветных глянцевитых яйца: белых, коричневатых, светлоголубых или зеленоватых. Инкубация 17—20 дней. Птенцы вылупляются голыми и слепыми. Вылетают из гнезда пол-
ностью оперенными. Даже у растительноядных видов птенцы, хотя бы в первую половину гнездовой жизни, выкармливаются насекомыми.
В отряде единственное семейство Trogonidae. включающее 8 родов с 35 видами. В Америке встречается 21 вид 5 родов: Trogon (12 видов), Eupti-lotis, Pharomachrus (6 видов), Priotelus Temnotrogon. В Африке живут лишь 3 вида 2 родов: Apaloderma и Heterotrogon, в Азии — 11 видов рода Harpactes.
Вероятный центр возникновения — Новый Свет, хотя наиболее ранние из ископаемых остатков обнаружены в отложениях нижнего олигоцена — верхнего эоцена Франции. Описано 4 ископаемых вида. Видимо, можно говорить об отдаленном родстве трогонов, козодоевых, сов, птиц мышей и ракшеобразных. Предположительно трогоны обособились от ство-
ла, позднее давшего начало совам и козодоевым.
ОТРЯД 31. CORACIIFOPMES —РАКШЕОБРАЗНЫЕ
Отряд объединяет птиц, очень различных по величине (вес от 10 г до 4 кг), внешнему облику, экологии и другим особенностям.
234
Клювы разнообразной формы. Ноздри не сквозные, голоринальные. Череп десмогнатический, сошник мал или отсутствует, нет функционирующих базиптеригоидных сочленений. Надорбитальные железы не развиты. Шейных позвонков 13—15. По заднему краю грудины 1—2 пары вырезок, иногда превращающихся в фонтанели. Ребер 4—8 пар. Лапы анизодактильные. У многих видов более или менее отчетливая синдактилия: основные фаланги направленных вперед всех либо только второго— третьего или, редко, третьего — четвертого пальцев срастаются роговыми покровами. Цевка не оперена. Гортань трахео-бронхиальная; голосовых мышц 1—2 пары или их нет вовсе. (Зонные артерии парные, у части родов — только левая. Зоба нет. Оперение довольно плотное. Побочный стержень контурного пера более или менее развит или его нет. Первостепенных маховых 10—И, рулевых 10—12. Окраска обычно яркая, часто с металлическим отблеском, редко контрастная черно-белая. Половой и возрастной морфизм окраски выражен слабо или отсутствует. Разные группы резко отличаются по характеру питания и способам охоты. Обычно моногамы. Гнездятся укрыто одиночными парами, реже рассеянными группами и даже колониями. Почти всегда насиживают кладку и выкармливают оба партнера. Птенцы вылупляются слепыми, голыми или в редком пуху. Долго остаются в гнезде.
Отряд объединяет сильно разошедшиеся специализированные группы. Это подчеркивается сложной системой отряда, разделяемого на 4 подотряда. Всего в отряд включают 192—193 вида, объединяемых в 48—54 рода и 10 семейств. В СССР встречается 10 и достоверно гнездится 6 видов из 4 семейств. Подавляющее большинство видов свойственно тропикам; в умеренные широты проникают немногие виды. В тропиках оседлые или кочующие, в умеренных широтах — перелетные.
ПОДОТРЯД 1. ALCEDINES — ЗИМОРОДКОВЫЕ
Птицы мелких и средних размеров (вес от 10 до 500 а). Клюв длинный, прямой или слегка изогнутый, с острым концом. Сошник отсутствует или рудиментарен. Шейных позвонков 14—15. Вилочка без расширения на конце. Ноги слабые, относительно короткие. Сонных артерий две. Нижняя гортань с парой голосовых мышц. Желчного пузыря нет. Гнездятся в вырытых самими птицами норах, редко в дуплах.
Подотряд объединяет 99 видов, относимых к 19—23 родам, группируемых в 3 семейства. Подчеркивая разобщенность семейств, каждое из них включают в отдельное надсемейство. В СССР встречается 5 видов только 1 семейства зймородковых.
Надсемейство 1. Alcedinoidea— зимородковые
Сюда включают только одно семейство (1-е) Alcedinidae.
СЕМЕЙСТВО 1. ALCEDINIDAE—ЗИМОРОДКОВЫЕ
Птицы от 10 до 500 а с обликом зимородка (рис. 145): коренастые, большеголовые, с длинным клювом и коротким хвостом (исключение р. Tanysiptera), широко распространенные почти по всему земному шару (рис. 146). Наибольшее разнообразие видов — в тропиках Старого Света.
Клюв удлиненный, иногда с маленьким крючком на конце. Сошника нет. Шейных позвонков 14—15. Мускульная формула ноги АХ—. Язык короткий, плоский. Слепые кишки короткие, но функционирующие. Копчиковая железа оперенная или голая. Побочного стержня контурного пера нет. Пух растет по птерилиям и аптериям. Первостепенных маховых 11, у родов Alcedo и Tanysiptera 10. Крыло эутаксическое или, реже, диастатак-
235
Рис. 145. Обыкновенный зимородок Alcedo atthls (L.)
сическое. Хвост из 12 рулевых, короткий, прямо обрезанный; лишь у Tanysiptera средняя пара рулевых длиннее остальных в 2—3 раза. Окраска из сочетания черного и белого или яркая, из сочетания зеленого, синего, коричневого цветов с металлическим блеском. Самцы и самки сходной окраски, молодые птицы в более тусклом пере.
Полет стремительный, у многих маневренный. Обычно приурочены к берегам рек и ручьев; лишь немногие виды живут в лесах и кустарниковых зарослях далеко от воды. Многие мелкие виды исключительно насекомоядны. Высматривают добычу, сидя на сучке; взлетая, ловят подлетевшее насекомое, или, пикируя, хватают его на земле. Крупные-виды ловят насекомых, разнообразных рептилий, мелких грызунов, таскают яйца и птенцов других птиц. Примерно половина видов добывают пищу в поверхностных слоях воды: мелких рыбешек, водных насекомых и их личинок, ракообразных. Добычу высматривают, сидя на присаде или летая низко над водой, затем пикируют, хватая ее клювом. Виды рода Clytoceyx имеют короткий вздутый клюв и, вероятно, пользуются им, выкапывая земляных червей.
Гнездятся отдельными парами в норах, которые роют в обрывах с мягкой почвой. Некоторые виды делают норы в термитниках, немногие гнездятся в дуплах. Крупные виды откладывают 2—3, мелкие — 5—7 блестящих белых яиц. Инкубация 3—4 недели. У части видов 2 и даже 3 кладки в году. Яйца откладываются без подстилки, но постепенно на дне гнезда образуется слой мелких косточек и хитина из разрушившихся погадок. Птенцы оставляют гнездо полностью оперенными и способными к полету в возрасте около 4 недель у мелких и 5—6 недель — у крупных видов. Вне сезона размножения держатся одиночками или парами. Южные виды оседлые или, редко, кочующие; в умеренных широтах перелетные.
В семействе зимородковых 86 видов, группируемых в 12—16 родов: Alcedo (9 видов), Chloroceryla, Сегу la, Сеух, Cittura, Clytoceyx, Corythornis, Dacelo, Halcyon (около 40 видов), Ispidina, Lacedo, Megaceryla, Melidora, Mioceyx, Pelargopsis, Tanysiptera.
В СССР широко распространен обыкновенный зимородок Alcedo atthis (L.); 4 вида 2 родов отмечены как залетные.
Центр возникновения семейства, видимо, тропики Старого Света.
Надсемейство 2. Todoidea — тодиевые
Включает одно семейство.
СЕМЕЙСТВО 2. TODIDAE—ТОДИЕВЫЕ
Единственный род Todus с 5 видами. Мелкие птички (вес 10—15 г) ярко-зеленой окраски с красным пятном на горле, внешне немного- похожие на крапивников (рис. 147). Встречаются только на Больших Антильских островах (Куба, Гаити, Ямайка, и др.; рис. 146).
Клюв прямой и длинный, слегка уплощенный у основания. В углах рта длинные щетинки. Сошника нет. Мускульная формула ноги AXY—. Язык длинный, заостренный. Кишечник короткий. Короткие слепые кишки на концах имеют колбообразные вздутия. Желчного пузыря нет. Коп-
236
150° 120° 90Q 60°к'западу or Гринвича (ГквостокуогГринвича60° 90° 120° 150° 180° 150°
Рис. 146. Ареалы семейств ракшеобразных птиц:
/— зимородковые Alcedinidae, 2— тодиевые Todidae, 3— момотозые Momotidae, 4—щурковые Meropidae
Рис. 147. Тоди Todas todus (L.)
чиковая железа оперена. Пуха нет. Побочные стержни контурного пера рудиментарны. Первостепенных маховых 10. Крыло короткое, эутаксическое. Хвост короткий, прямо обрезанный, из 12 рулевых.
Живут оседло в лесах и кустарниковых зарослях, обычно вблизи ручьев и речек. Легко прыгают в ветвях. Кормятся насекомыми, которых чаще подстерегают, сидя на ветке, и ловят в коротком взлете; склевывают на ветвях и листьях. Иногда едят ягоды и семена. Держатся отдельными парами. Гнездо — нора глубиной до 60 см, вырытая птичками в обрыве, в вывороте дерна, в стенках ям и т. п. В кладке 2—3 белых яйца. Ископаемые остатки тоди не обнаружены.
Надсемейство 3. Мот о to idea — момотовые
Сюда входит единственное семейство (3-е) Momotidae.
СЕМЕЙСТВО 3. MOMOTIDAE — МОМОТОВЫЕ
Момоты (рис. 148) несколько напоминают щурок, по поведению — соек или сизоворонок. Размеры мелкие и средние (вес 50—400 а). Населяют тропические и субтропические леса Центральной и Южной Америки (рис. 146); нет их в Вест-Индии.
Клюв длинный, слегка изогнутый, с острой вершиной и зазубренными режущими краями. Сохраняется сильно редуцированный сошник. Мускульная формула ноги AXY—. Вырезки заднего края грудины часто превращаются в фонтанели. Язык длинный, с бахромой на конце. Мускульный желудок довольно мощный. Ки
шечник относительно длинный. Слепых кишок нет. Имеется желчный пузырь. Копчиковая железа голая или слабо оперенная. Побочные стержни контурного пера сильно редуцированы. Пуха нет. Первостепенных маховых 10. Крыло эутаксическое. Резко ступенчатый хвост из 12 рулевых. Средняя пара очень длинная; сама птица выщипывает на ней предвершинную часть опахал (рис. 148). Окраска яркая синяя, красная, зеленая с металлическим отблеском.
Хорошо летают. Держатся в зарослях кустарников или в нижнем ярусе леса. Кормятся различными насекомыми (преимущественно жуками), склевывая их с листьев и веток, или ловят, взлетая с присады. Иногда ловят мелких ящериц и змей, слетков мелких воробьиных птиц. Едят плоды. Гнездятся в норах, которые роют в обрывах ручьев и лесных оврагов
Рис. 148. Момот Momotus momota (L.)
238
(длина нор достигает 2—3 м). В кладке 3, редко 4 блестящих белых яйца. Насиживают 20—24 дня. Птенцы остаются в гнезде около месяца.
В семействе момотов всего 8 видов, относимых к 6 родам: Momotus, Aspatha, Baryphthengus, Electron, Eumomota, Hylomanes. Основываясь на находках плохой сохранности из среднего эоцена юга США, полагают, что центром возникновения семейства мог быть юг Северной Америки.
Весь подотряд зимородковых, видимо, обособился от примитивных ракш; сходство с щурками более отдаленное. Обособление семейств (надсемейств), вероятно, определялось в первую очередь пищевой специализацией и произошло очень давно (возможно, в нижнем эоцене).
ПОДОТРЯД 2. MEROPES—ЩУР КОВЫ Е
В этот подотряд включают одно семейство (4-е) Meropidae.
СЕМЕЙСТВО 4. MEROPIDAE — ЩУРКОВЫЕ
Мелкие птицы (вес 20—100 а) с обликом щурки (рис. 149), распространенные в тропиках и южных районах умеренных широт Старого Света, включая Мадагаскар и Австралию (рис. 146); в Новую Зеландию не проникают.
Клюв длинный, тонкий, слегка изогнутый книзу, с гладкими краями. Сошник сильно редуцирован. Шейных позвонков 14. Истинных ребер 4 пары. Ноги короткие, слабые. Мускульная формула ноги AXY—. Язык, заостренный, плоский; длина его варьирует. Слепые кишки довольно длинные. Желчный пузырь есть. Копчиковая железа голая. Обычно сохраняет-
ся только левая сонная артерия (у Nyctiornis — обе). Пуха нет. Контурные перья с коротким, но хорошо заметным добавочным стержнем. Первостепенных маховых 10. Крылья удлиненные, эутаксические. У многих видов средние рулевые заметно выдаются за обрез хвоста. Окраска яркая, иногда довольно пестрая.
По земле почти не ходят. Для отдыха садятся на ветки. Полет легкий, маневренный, могут планировать. Приурочены к открытым местообитаниям: степям, лугам, поймам рек. Держатся стайками. Кормятся преимущественно крупными насекомыми (пчелы, стрекозы, бабочки, жуки и др.), которых ловят в воздухе или склевывают налету с ветвей и листьев. Кормящаяся стайка обычно кружит в воздухе, меняя высоту и скорость полета. Местами причиняют ущерб, поедая пчел. Гнездятся группами в несколько пар или колониями, иногда в сотни пар. Роют норы длиной до 1 м и больше в обрывах или по склонам холмов. Рытье но
Рис. 149. Золотистая щурка Merops apia-ster L.
239
ры продолжается 2—3 недели. В конце норы на рыхлую подстилку из сухих травинок и хитина (разрушенные погадки) откладывается 2—5, реже до 7 белых яиц. Инкубация около 3 недель. Птенцы в возрасте около 1 месяца покидают нору активным полетом. В тропиках — кочующие, в умеренных широтах перелетные.
В семействе 25 видов, относимых к 7 родам: Мег ops, Aerops, Bombylo-tiax, Dicrocercus, Melittophagus, Moropogon, Nyctiornis. В южных районах СССР встречается и гнездится 2 вида рода Merops.
Вероятный центр возникновения — Африка. Остатки, обнаруженные в плейстоценовых отложениях, относят к современному виду. Имеют некоторые черты сходства с зимородками и удодами. Вероятно, обособились вместе с ними от примитивных ракшеобразных. Обособление подотряда шло по пути пищевой специализации — добыванию насекомых в воздухе в длительном активном полете.
ПОДОТРЯД 3. CORACII — РАКШЕВЫЕ
Птицы обычно яркой окраски, весом в 50—300 а, довольно разнообразного внешнего облика. Клюв грубый или удлиненный, тонкий. Сошник рудиментарен или его нет. Шейных позвонков 13—14. Ноги относительно длинные. Мускульная формула ноги AXY—. Крыло эутаксическое или диастатаксическое. Оперение плотное. Пуха нет. Гнездование укрытое.
Подотряд объединяет 24 вида 10 родов, распределяемых по 5 семействам. В СССР встречается и гнездится 3 вида 3 родов из 2 семейств.
СЕМЕЙСТВО 5. CORACIIDAE— НАСТОЯЩИЕ, ИЛИ ДРЕВЕСНЫЕ, РАКШИ
Рис. 150. Сизоворонка, или ракша Coracias garrulus L.
Крепкие коренастые птицы (рис. 150), вес 100—200 г. Распространены в Африке и Юго-Восточной Азии, немногие виды проникают в умеренные широты Евразии (рис. 151). Клюв крепкий, слабо изогнутый (похож на клюв вороновых), с острым концом; у широкоротое — широкий, с крючком на конце. Сошник рудиментарен. Очень длинны заднеглазничные отростки, слезная кость большая. Шейных позвонков 13—14. Истинных ребер 4—5. Язык длинный, уплощенный, с заостренной вершиной. Слепые кишки довольно длинные. Сонные артерии парные. Копчиковая железа голая. Побочный ствол контурного пера есть. Первостепенных маховых 10, рулевых 12. Окраска яркая, с преобладанием синих, голубых и рыжих цветов, у молодых более тусклая, чем у взрослых.
Распространены по речным долинам юга лесной зоны, в лесостепи, культурном ландшафте, безлесных степях с оврагами, предгорьях; немногие виды (широкороты) населяют высокоствольные леса. Полет
240
Рис. 151. Ареалы семейств:
/_ настоящие ракши Coraciidae, 2— хдодовые Upupidae, 3— птицы носороги Bucerotidae (отряд ракшеобразных). 4— якамаровые Galbulidae, 5— пуховковые Bucconidae-(отряд дятлообразных)
быстрый, свободный. Поземлепередвигаются довольно медленно. Питаются главным образом крупными насекомыми, которых ловят в воздухе, взлетая с присады, и собирают на земле и ветвях. Поедают моллюсков, червей и других беспозвоночных. Более крупные виды иногда ловят мышевидных грызунов, ящериц, таскают яйца и птенцов мелких воробьиных. Изредка поедают мелкие плоды, семена. Гнездятся одиночными парами. Характерен ток: полет с громкими криками. Кладка из 3—6 яиц, всегда в укрытии: в естественных дуплах, расщелинах скал, промоинах или вырытых птицами норах в обрывах. Инкубация около 3 недель. Через 25—30 дней после вылупления птенцы покидают гнездо активным полетом. Вне сезона размножения держатся мелкими группами или одиночками. Оседлые или кочующие, в умеренных широтах перелетные.
Семейство объединяет 11 видов, относимых к 2 родам: 8 видов сизоворонок рода Coracias и 3 вида широкоротое Eurystomus. В СССР гнездятся 2 вида: по одному виду каждого рода.
Ископаемые остатки известны из верхнего эоцена или нижнего олигоцена Франции, в более поздних слоях — из других районов современного ареала. Центр возникновения — Старый Свет, возможно Южная Европа. Сейчас наибольшее разнообразие видов в Африке. Видимо, одно из наиболее древних семейств подотряда.
СЕМЕЙСТВО 6. BRACHYPTERACIIDAE—ЗЕМЛЯНЫЕ РАКШИ
Похожи на сизоворонок, но отличаются большой приспособленностью к наземному образу жизни (рис. 152). Встречаются только на Мадагаскаре.
Клюв удлиненный. Ноги длинные; особенно удлинена цевка. Пальцы длинные, сильные. Когти второго и третьего пальцев повернуты внутрь, первого и четвертого — наружу. Синдактилия почти не выражена. Крылья относительно короткие, тупые. У части видов длинный «сорочий» хвост. Держатся по кустарниковым зарослям, опушкам леса, лесным полянам. Летают мало и неохотно. Кормятся на земле, разгребая ногами подстилку и выбирая из нее насекомых, червей, моллюсков; ловят мелких земноводных и пресмыкающихся. Экология изучена плохо. Гнездятся в коротких норах, в полостях между корнями и т. п.
В семействе всего 5 видов, относимых к 3 родам: Brachypteracias, Ate-lornis, Uratelornis. Иногда земляных ракш в ранге подсемейства включают в семейство древесных ракш.
Ископаемые остатки неизвестны.
Рис. 152. Земляная ракша Atelornis crossleyi Sh.
242
СЕМЕЙСТВО 7. LEPTOSOMAT1DAE — КУРОЛОВЫЕ
Выделяется единственный вид курол, или кирумбо, Leptosomus discolor Herm. (рис. 153), встречающийся только на Мадагаскаре и на Коморских островах.
Крупная ракша (вес 300 г). [Целевидные ноздри расположены почти посредине клюва и прикрыты роговыми крышечками. Своеобразна лапа: основные фаланги второго и третьего пальцев срастаются; четвертый палец свободен и подвижен: он направлен вперед, но может отворачиваться вбок и назад. Гортань бронхиального типа. Парные сон-
Рис. 153. Курол Leptosomus discolor Herm.
ные артерии расположены очень
поверхностно. Копчиковой желе-
зы нет. На пояснице парные пудретки. Рулевых 10. Отчетливый половой диморфизм: у самца резкий зеленовато-медный отблеск, самка тусклая.
Держатся в кронах деревьев, обычно по опушкам. Питаются различными насекомыми, особенно гусеницами, которых собирают на ветвях и листьях. Во время тока взлетают на большую высоту и с грубым свистом пикируют вниз. Кладка из 4—5 белых яиц в дупле.
Видимо, наиболее примитивное семейство отряда, сохраняющее некоторые черты сходства с кукушкообразными. В то же время, несомненно, близки к древесным и земляным ракшам.
СЕМЕЙСТВО 8. UPUPIDAE— УДОДОВЫЕ
В семейство включается единственный вид — удод Upupa epops L. (рис. 154), очень широко распространенный в Африке, Европе и Азии (рис. 151); встречается в
Рис. 154. Удод Upupa epops L.
южных, отчасти и в центральных районах СССР.
Вес около 70 г. Клюв длинный, тонкий, изогнутый книзу. Ноздри открытые. Сошника нет. Функционируют слабо развитые базиптеригоидные сочленения. Заглазничные отростки короткие. Шейных позвонков 14. По заднему краю грудины только пара вырезок, превращающихся в фонтанели. Ноги короткие, но сильные. Язык очень мал. Слепых кишок нет. Голосовые мышцы гортани не развиты. Пуха нет. Нет побочного стержня контурного пера. На голове хохол из пестрых перь
243
ев. Рулевых и первостепенных маховых по 10. Большая копчиковая железа оперена. У самок в период размножения и у подросших птенцов в ней образуется вонючая черновато-коричневая маслянистая жидкость, выбрызгиваемая при испуге; видимо, этот запах отпугивает мелких хищников. Гнездо всегда очень неприятно пахнет.
Полет довольно медленный, волнообразный. Хорошо ходят и бегают по земле. Легко прыгают в кронах деревьев. Встречаются в изреженных лесах, лесостепях и степях, полупустынях, предгорьях, в культурном ландшафте. Кормятся различными беспозвоночными, собирая их на земле, реже на ветвях и листьях, добывают пищу и в верхнем слое почвы, зондируя ее тонким клювом. Гнездятся отдельными парами, устраивая очень примитивное гнездо в естественных дуплах, трещинах скал и под камнями, в промоинах обрывов, брошенных норах грызунов, в щелях построек и т. п. В кладке 5—6 яиц (иногда до 12) матово-белых, с голубоватым или охристым налетом. Насиживает преимущественно самка; самец ее кормит. Инкубация 16—18 дней. Слепые птенцы покрыты редким рыжевато-серым волосовидным пухом. Выкармливают оба партнера, вкладывая насекомых в раскрытый рот птенца. Молодые вылетают в возрасте 22—25 дней. В южных районах, видимо, 2 кладки в году.
Ископаемые остатки современного вида встречаются в разных частях ареала, начиная с середины плейстоцена. Возник, видимо, в Северной Африке. Отдаленные, но несомненные признаки родства сближают удодов и ракш.
СЕМЕЙСТВО 9. PHOENICULIDAE — ДРЕВЕСНЫЕ УДОДЫ
Скрытно держащиеся мелкие птицы (вес 50—100 г) опушек тропических лесов и кустарниковых зарослей Африки.
Клюв длинный, тонкий, еще более резко, чем у обыкновенного удода, изогнутый книзу (рис. 155). Ноздри прикрыты роговыми крышечками. Функционирующих базиптеригоидных сочленений нет, хотя у части видов есть заметные базиптеригоидные отростки. Когти острые, сильно изогнутые. Копчиковая железа, как и у обыкновенного удода, выделяет маслянистую жидкость с резким неприятным запахом. Пуха нет. У контурных перьев есть маленькие побочные стволы. Хохла нет. Хвост из жестких рулевых, резко ступенчатый, его линька идет, как у дятлов: последней сменяется центральная пара. Окраска яркая, с металлическим отливом, клювы и лапы ярко-красные. Молодые окрашены более тускло.
Экологически похожи на дятлов и пищух. Питаются различными членистоногими, их личинками и яйцами. Разыскивают добычу в трещинах коры, лазая по ветвям и стволам (опираются на хвост). Гнездятся в естественных дуплах. В кладке 3 яйца: зеленоватых, голубоватых или коричневатых. Насиживает самка; самец ее кормит. Птенцов кормят оба партнера.
244
К древесным удодам относят 6 видов, распределяемых по 3 родам: Phoeniculus, Rhinopomastus, Scoptelus.
Вероятный центр возникновения — Африка. По остаткам из нижнего миоцена Франции описан ископаемый род с 1 видом. Таким образом, раньше ареал был, видимо, более широким. Несомненно, близки к настоящим удодам. Оба семейства имеют общих предков и их обособление — результат приспособления к наземному и древесному образу жизни.
ПОДОТРЯД 4. В UCE ROTES — ПТИЦЫ НОСОРОГИ
Включает только одно семейство (10-е) Bucerotidae.
СЕМЕЙСТВО 10. BUCEROTIDAE — ПТИЦЫ НОСОРОГИ
Это птицы весом от 150 г до 4 кг , населяющие тропические леса Африки и Юго-Восточной Азии (рис. 151).
Характерен длинный, толстый, изогнутый книзу клюв, у большинства видов с большим роговым выростом у основания надклювья (рис. 156). Несмотря на большие размеры клюва, он легкий, так как костная основа губчатая, а роговой вырост полый. Края клюва слегка зазубрены. Базип-теригоидных сочленений нет, хотя базиптеригоидные отростки имеются. Сошник отсутствует. Шейных позвонков 14—15; атлант и эпистрофей сливаются в единое костное образование. По заднему краю грудины пара неглубоких вырезок. Вилочка с расширением на конце. Кости конечностей очень пневматичны. Есть многочисленные подкожные воздушные полости. Мускульная формула ноги AXY—. Язык короткий. Пищевод сильно растяжим. Слепые кишки отсутствуют. Желчный пузырь есть. Сохраняется только левая сонная артерия (у Bucorvus — обе). Гортань трахео-бронхи-альная с одной-двумя парами голосовых мышц. Копчиковая железа оперена. Контурные перья без побочного ствола. Пуха нет. Первостепенных маховых 10. Крыло эутаксическое. Хвост длинный из 10 рулевых. Окраска черная или темная, иногда с белыми пятнами. У части видов на голове и шее есть участки голой яркой кожи. Самки чуть меньше самцов, с более коротким клювом и меньшим «рогом».
Ведут древесный образ жизни, кормятся в кронах, по земле передвигаются плохо. Только рогатые вороны (Bucorvus abissinicus Bodd.), живущие в разреженных лесах и зарослях кустарников, кормятся на земле, хорошо ходят и бегают. Практически всеядны: питаются разнообразными плодами и семенами, поедают насекомых, ловят мелких пресмыкающихся, слетков птиц и т. п. Благодаря длинному клюву и относительно длинной шее сидящая на толстой ветке тяжелая птица достает висящие на тонких веточках плоды. Пары у многих видов, видимо, сохраняются до гибели од-
Рис. 156. Большой носорог Buceros bicornis L.
245
ного из партнеров. Гнездятся в естественных дуплах, часто на большой высоте. Самец и самка уменьшают вход в дупло, замазывая его глиной, пометом, древесной трухой, сцементированными слюной. Затем самец продолжает носить строительный материал, а самка залезает в дупло и еще больше замуровывает вход, оставляя лишь щель, в которую может высунуть переднюю часть клюва. Рогатые вороны гнездятся в дуплах и расщелинах скал, входа не замазывают. У крупных видов в кладке 1—3, у мелких 3—5 белых яиц. Инкубация 1—1,5 месяца. Самец кормит насиживающую самку, а затем и появившихся птенцов. Самка в это время интенсивно линяет. У части видов через 2—3 недели после вылупления птенцов перелинявшая самка самостоятельно или с помощью самца расширяет щель и выходит из дупла; вход вновь замуровывается и остается щель, через которую самец и самка продолжают кормить птенцов. У других видов самка остается в гнезде до вылета птенцов, и всю семью кормит только самец. Постэмбриональное развитие медленное: у мелких видов птенцы вылетают из гнезда через 1,5—2 месяца после вылупления, у крупных через 3—4 месяца. В году может быть 2 кладки. Вне периода размножения держатся одиночками или мелкими группами. Оседлые или кочующие.
Семейство объединяет 44—45 видов, относимых к 12—14 родам: Ви-ceros, Aceros, Anorrhinus, Anthracoceros, Berenicornis, Bucorvus, Bycanistes, Ceratogymna, Penelopides, Ptilolaemus, Rhinoplax, Rhyticeros, Tockus, Tropicranus.
Центр возникновения, видимо, Африка. Ископаемые остатки обнаружены в слоях среднего эоцена Франции и Германии (точность определения сомнительна). Близки к удодам. Вероятно, обособились до их разделения на настоящих и древесных удодов. Более слабо выражено родство с другими группами ракшеобразных. Сходство с туканами (дятлообразные) конвергентное; оба семейства могут быть примером экологического викариата.
Отряд ракшеобразных в целом — достаточно четко обособленная группа, близкая к кукушкообразным и козодоеобразным. Обособившиеся от исходных форм примитивные ракшеобразные в ходе эволюции дали начало довольно близким друг другу семействам подотряда ракшевых и остальным подотрядам. При этом зимородковые и щурки ближе к настоящим ракшам, а птицы носороги — к семействам удодов.
ОТРЯД 32. PIC1FORMES — ДЯТЛООБРАЗНЫЕ
Мелкие и средние птицы (вес от 6 до 300 г), ведущие преимущественно древесный образ жизни.
Клюв прямой, долотообразный или к концу слегка изогнутый, разной длины. Ноздри голоринальные, не сквозные. Череп схизогнатический, десмогнатический или эгитогнатический. Сошник есть. Базиптеригоидные отростки отсутствуют. Шейных позвонков 14. Спинная кость не образуется. По заднему краю грудины обычно 2 пары вырезок, иногда превращающихся в фонтанели. Лапы зигодактильные; первый палец у отдельных видов исчезает. Мускульная формула ноги AXY — или АХ—. Язык короткий, умеренной длины или очень длинный и тонкий. Зоба нет. Кишечник относительно короткий. Слепые кишки функционируют или редуцируются. У части семейств отсутствует желчный пузырь. Копчиковая железа голая или оперенная. Гортань трахео-бронхиальная; голосовых мускулов одна пара или они не развиты. Оперение довольно рыхлое. Контурные перья имеют хорошо развитую пуховую часть и у части видов — побочный стер-жен ь. Пуха нет. Птерилии узкие, аптерии широкие. Первостепенных махов ых 9—11, второстепенных 10—12. Крыло эутаксическое. Рулевых 8— 12. Окраска разнообразная. Самцы и самки имеют сходную окраску или отличаются ее деталями. Молодые иногда окрашены более тускло.
246
Подавляющее большинство видов — обитатели лесов разного типа, проникают в сады и парки; немногие виды живут в степях. Питаются разнообразными насекомыми, их личинками и яйцами, поедают семена и ягоды. Корм обычно собирают в кронах и на стволах деревьев, реже кормятся на земле. Моногамы. Гнездятся в дуплах (естественных или выдолбленных самими птицами), некоторые виды роют гнездовые норы. В кладке от 2—5 до 10 яиц. Насиживают оба партнера, самка обычно более интенсивно. Птенцы вылупляются слепыми и у большинства видов голыми. У птенцов хорошо развиты пяточные мозоли. Выкармливают оба родителя. Птенцы покидают гнездо оперенными и способными к полету. Вне периода размножения держатся одиночками или мелкими группами. Оседлые или кочующие, немногие виды перелетные.
Отряд объединяет 379 видов, группируемых в 73—87 родов и относимых к 6 семействам 2 подотрядов. В СССР встречается и гнездится только 13 видов 1 семейства.
ПОДОТРЯД 1. GALBULAE — ПРИМИТИВНЫЕ ДЯТЛООБРАЗНЫЕ
Дятлообразные птицы, у которых признаки древесной специализации не выражены столь четко и многообразно, как у второго подотряда — настоящих дятлов. Клювы разнообразной формы. Череп десмогнатический или эгитогнатический. Язык не выдвигающийся.
Подотряд разделяется на 3 надсемейства, объединяющие 5 семейств с 37—42 родами и 170 видами. В СССР представители подотряда не встречаются.
Надсемейство 1. Galbuloidea— якамаровые
Череп десмогнатический, с небольшим парным сошником. Ключицы всегда сливаются в вилочку. Язык гладкий, почти доходит до конца ротовой полости. Слепые кишки функционирующие, с колбообразными вздутиями на концах. Желчного пузыря нет. Сонные артерии парные. Кожа тонкая. Первостепенных маховых 10, рулевых 12. Оседлы. Вне сезона размножения обычно держатся поодиночке.
Надсемейство якамар (местное бразильское название этих птиц) объединяет наиболее примитивных дятлообразных, еще сохраняющих черты сходства с ракшеобразными. Включает 45 видов 15 родов, распределяемых по 2 семействам. Населяют тропики Америки.
СЕМЕЙСТВО 1. GALBULIDAE — ЯКАМАРОВЫЕ
Мелкие птицы (вес 15—100 г) очень яркой, чаще зеленой окраски, обычно с ярким металлическим отблеском, внешне (рис. 157) немного похожие на щурок: населяют леса и кустарниковые заросли Центральной и Южной Америки (рис. 151).
Клюв длинный, постепенно утончающийся к концу, с более или менее заметными продольными гранями. Расширения на конце вилочки нет. Копчиковая железа голая. Побочный стержень кон
Рис. 157. Зеленая якамара Galbula gal-bula (L.)
247
турного пера есть. Хвост длинный, более или менее ступенчатый; резко укорочены крайние рулевые.
Питаются преимущественно крупными насекомыми, которых обычно ловят в воздухе: птица сидит на ветке и, быстро взлетев, ловит пролетающую добычу. Гнездятся в норах длиной в 30—50 см, которые роют в берегах ручьев и оврагов, насыпях дорог, термитниках. Кладка из 2—4 блестящих белых яиц. Насиживают 19—25 дней. Вылупляющиеся птенцы в отличие от остальных дятлообразных покрыты беловатым пухом. Молодые покидают нору на 20—26-й день жизни полностью оперенными и по окраске практически не отличаются от взрослых.
Семейство объединяет 15 видов, распределяемых в 5 родов: Galbula, Brachygalba, Galbalcyrhynchus, Jacamaralcyon, Jacamerops. В ископаемом состоянии не обнаружены. Центр возникновения, видимо, Южная Америка.
СЕМЕЙСТВО 2. BUCCONIDAE — ПУХОВКОВЫЕ, ИЛИ ЛЕНИВКОВЫЕ
Мелкие птицы (вес 20—100 г), по внешнему облику (рис. 158) несколько напоминающие сизоворонок. Обитают в лесах и кустарниковых зарослях тропической Америки; ареал практичес-
ки совпадает с ареалом якамар (рис. 151).
Клюв короче черепа, довольно толстый, слегка изогнутый книзу, часто с небольшим крючком на конце (напоминает клюв сизоворонок). У основания клюва длинные щетинки. На конце вилочки слабое расширение. Копчиковая железа обычно голая (оперена у нескольких видов). Оперение очень пушистое, рыхлое (отсюда и название семейства). Побочный стержень контурного пера не развит. Хвост умеренной длины, с прямым обрезом. Окраска темная или пестрая, обычно неяркая, без металлического блеска.
Питаются насекомыми, которых собирают, лениво прыгая в кронах или на зем-
Рис. 158. Пуховка Виссо tamatia ле’ Л0ВЯТ И В воздухе: сидящая птица Gm. взлетает, хватает подлетевшую добычу и
опять садится на приезду. Гнездятся в коротких норах, вырытых птицами в обрывах, в полостях термитников; иногда откладывают яйца в низко расположенные естественные дупла. В кладке 2—3 блестящих белых яйца, откладываемых на скудную подстилку из сухих травинок. Насиживают 2—3 недели. Птенцы вылупляются голыми. Вылетают из гнезда в возрасте около 3 недель.
30 видов пуховок распределяют по 10 родам: Виссо, Chelidoptera, На-paloptila, Hypnelus, Malacoptila, Micromonacha, Monasa, Nonnula, Nothar-chus, Nу status.
Центр возникновения, видимо, Южная Америка. Из ныне живущих видов 2 обнаружены в отложениях верхнего плейстоцена Бразилии.
Оба семейства надсемейства якамаровых — две близкие ветви, обособившиеся от предковых форм дятлообразных.
Надсемейство 2. Capitonoidea — примитивные дятловые
Череп эгитогнатический, иногда со следами десмогнатизма. Сошник непарный, обычно более или менее расщепленный. У части видов ключицы остаются свободными, у остальных сливаются в вилочку. Нижние кольца трахеи окостеневают и расширяются, образуя костный барабан. Язык до-
248
вольно короткий, не высовывающийся. Желчный пузырь есть. Копчиковая железа оперена. Хорошо заметен побочный стержень контурного пера. Первостепенных маховых 9—10, относительно короткий хвост из 10—12 рулевых.
В надсемействе примитивных дятлов признаки древесной специализации проявляются более отчетливо, чем в предыдущем надсемействе. Сюда входят 2 семейства — Capitonidae и Indicatoridae, объединяющие 88 видов 17—20 родов. Распространены в тропиках.
СЕМЕЙСТВО 3. CAPITONIDAE — БОРОДАТКОВЫЕ
Мелкие птицы (вес менее 100 г), обычно пестрой и яркой окраски
(рис. 159), по внешнему облику немного напоминающие сизоворонок. Насе-
ляют леса и кустарниковые заросли почти всей тропической зоны (рис. 160).
Клюв средней длины, слегка вздутый, с небольшим крючком на конце, у некоторых видов мощный, высокий и широкий. Иногда намечается продольная ребристость рамфотеки. Часто края клюва слегка зазубрены. У основания клюва сильно развиты щетинки, более или менее прикрывающие ноздри (отсюда и название семейства). Череп эгито-гнатический со следами десмогнатизма. Ключицы не сливаются в вилочку. Слепых кишок нет. Крылья тупые, первостепенных маховых 10. Хвост короткий, из 10 рулевых; крайняя пара заметно укорочена. Половой диморфизм есть у немногих видов.
Летают как дятлы. Хорошо лазают и прыгают в кронах, довольно легко двигаются по земле. Питаются разнообразными плодами, более или менее случайно поедают насекомых. Немногие виды преимущественно насекомоядны; корм собирают на ветвях и на земле, под отрываемой корой, роются в трухлявой древесине. Выдалбливают в гнилых стволах и пнях гнездовые дупла, некоторые
Рис. 159. Черноспинная боро-датка Trachyphonus vaillantii Ranz.
виды гнездятся в естественных дуплах и в норах, вырытых в обрывах или термитниках. Гнездятся отдельными парами, немногие мелкими группами. В кладке 2—5 блестящих белых яйца. Насиживают около 2 недель.
Птенцы остаются в гнезде около месяца. Вне сезона размножения держатся одиночками или мелкими группами. Оседлые.
Семейство бородатковых объединяет 76 видов, группируемых в 13— 16 родов: Capita, Buccanodon, Eubucco, Euphorbia, Gymnobucco, Lybius, Megalaima, Pogon lulus, Pogonornis, Psilopogon, Semnornis, Smilorhis, Stac-tolaema, Trachylaemus, Trachyphonus, Tricholaemus. Наибольшее число видов и родов встречается в Африке. Видов и родов, свойственных одновременно Старому и Новому свету, нет. Центр возникновения, видимо, Африка. Остатки одного из ныне живущих видов найдены в верхнем плейстоцене Бра-
зилии.
СЕМЕЙСТВО 4. INDICATORIDAE — МЕДОУКАЗЧИКОВЫЕ
Мелкие (длина 10—20 см) сероватые или зеленоватые птицы (рис. 161), внешним обликом несколько напоминающие синиц. Распространены в тропических лесах Африки и в Юго-Восточной Азии (рис. 160).
249
Рис. 160. Ареалы семейств дятлообразных птиц:
1— бородатковые Capitonidae, 2— медоуказчиковые Indicatoridae, 3— тукановые Ramphastidae, 4— дятловые PIcidae
Клюв крепкий, относительно небольшой, напоминающий клюв синиц. Щелевидные ноздри лежат почти у самого гребня клюва. Щетинок у основания клюва нет. Череп эгитогнатический. Ключицы срастаются в вилочку. Есть очень маленькие слепые кишки. Кожа относительно толстая (считают, что это защита от жала ос и пчел). Первостепенных маховых 9, у некоторых 10 (но первое тогда сильно укорочено). Крылья заостренные. Хвост прямо обрезанный или слабо ступенчатый из 10—12 рулевых. Половой диморфизм выражен слабо.
Полет быстрый, маневренный. Обычно держатся
Рис. 161. Большой медоуказчик Indicator indi-cator Sp.
одиночками в кронах, ловко
прыгая по ветвям. Кормятся разнообразными насекомыми, собирая их на ветвях или ловят в воздухе, взлетая с приезды; более или менее случайно поедают мелкие плоды. Многие, а может быть и все виды разыскивают гнезда ос и пчел, поедая личинок, мед и соты. Найдя недоступное дупло с сотами пчел, крутятся с криками поблизости, привлекая внимание хищников (например, медоедов) или человека (отсюда и название медоуказчики). Когда хищник или человек вскроют дупло, птички подбирают выпавших личинок, едят соты. Это единственная группа птиц, способная переваривать воск, благодаря своеобразной бактериальной флоре кишечника. В период размножения, подобно дятлам, барабанят клювом по сухим ветвям, с криком совершают токовой полет, издают громкие крики, сидя на ветви, и т. п. Видимо, все виды — типичные гнездовые паразиты. Гнезд не строят, яйца подкладывают в гнезда дятлов и бородаток, иногда яйца находят и в гнездах открыто гнездящихся мелких воробьиных (у белоглазок, славок, камышевок и др.). Насиживание и выкармливание осуществляется «приемными родителями» — хозяевами дупла. У вылупившегося птенца на конце клюва резко выражен крючок, помогающий ему выбрасывать из гнезда птенцов хозяев. К концу гнездового периода из-за неравномерного роста разных участков клюва крючок почти полностью исчезает, и клюв птенца становится похожим на клюв взрослой птицы.
12 видов медоуказчиков относят к 4 родам: Indicator, Melichneutes, Melignomon, Prodotiscus. 10 видов встречается в Африке, 2 — в Юго-Восточной Азии. Центр возникновения, видимо, Африка. Ископаемых остатков нет. Несомненно, близки к бородаткам, но в то же время достаточно четко от них отличаются.
Надсемейство 3. Ramphastoidea — тукановые, или перцееды
Сюда включают одно семейство (5-е) Ramphastidae.
СЕМЕЙСТВО 5. RAMPHASTIDAE—ТУКАНОВЫЕ
Птицы мелких и средних размеров (100—300 г) с большим длинным клювом (рис. 162); при длине 60 см длина клюва может достигать 20—23 см. Окраска клюва и оперения яркая, контрастная. Населяют тропические леса Центральной и Южной Америки (рис. 160).
251
Внешне необычайно мощный клюв относительно легок: его костная основа сильно губчатая. Края клюва зазубренные. Череп десмогнатический. Сошник не расщепленный. Ключицы не срастаются в вилочку. Язык не выдвижной, доходит до конца ротовой полости; его конец и края бахромчатые. Мускульный желудок объемистый, толстостенный. Кишечник короткий и широкий. Слепых кишок нет. Желчный пузырь большой. Копчиковая железа оперена. Нижние кольца трахеи образуют костный барабан. Функционирует
~ „ , , . , только левая сонная артерия. Побоч-
Рис. 162. Тукан Ramphastos artel Vig. HbIg ствол КОнтурного пера редуцирован. Уздечка и кожа вокруг глаза голые. Первостепенных маховых 11. Крылья тупые. Хвост прямой или слабо ступенчатый, из 10 рулевых, коротких или средней длины.
Полет дятлового типа. Держатся в кронах деревьев, легко лазая по ветвям. Подвижны и крикливы. Питаются преимущественно различными
плодами, при случае склевывают насекомых и моллюсков, иногда таскают из гнезд других птиц яйца и птенцов, ловят ящериц. Длинный клюв и удлиненная шея дают возможность туканам, сидя на прочной ветви, склевывать с тонких веточек ягоды; может быть, такой клюв облегчает защиту гнездового дупла. Спящая птица кладет клюв на плечо и сильно поднимает хвост, почти прикрывая им спину. Гнездятся в естественных дуплах, мелкие виды занимают брошенные дупла дятлов. В кладке 2—4 белых, относительно мелких яйца. Инкубация 2—3 недели. В насиживании, кроме родителей, иногда участвуют и другие особи, видимо, не вполне половозрелые. У вы-
лупившихся птенцов клювы относительно короткие, подклювье заметно длиннее и шире надклювья; это облегчает схватывание корма, бросаемого взрослыми в дупло. Постэмбриональное развитие медленное: лишь в возрасте около 3 недель у птенцов Прорезаются глаза, а покидают гнездо они лишь через 7—8 недель после вылупления. Кроме родителей, в выкармливании птенцов участвуют и другие взрослые особи (неполовозрелые или потерявшие кладки?): описан случай, когда в гнезде с 3 птенцами ночевало 5 взрослых птиц. Вылетевшие из гнезда молодые отличаются от взрослых более тусклой окраской клюва. Половозрелыми становятся, видимо, на 2— 3-м году жизни. Вне сезона размножения держатся маленькими стайками.
37 видов туканов относят к 5—7 родам: Ramphastos, Andigena, Aula-corhynchus, Baillonius, Beauharnaisius, Pteroglossus, Selenidera.
Центр возникновения, видимо, Южная Америка. Несомненно близки к бородаткам. Остатки 2 ныне живущих видов обнаружены в отложениях верхнего плейстоцена Бразилии.
ПОДОТРЯД 2. PICI — НАСТОЯЩИЕ ДЯТЛООБРАЗНЫЕ
Сюда входит одно семейство Picidae (6-е),
СЕМЕЙСТВО 6. PICIDAE — НАСТОЯЩИЕ ДЯТЛОВЫЕ
Мелкие и средние птицы (вес 6—300 г) с характерным обликом дятла (рис. 163). Широко распространены в умеренной и тропической зонах (рис. 160); нет в Австралии, Новой Зеландии, Новой Гвинее, на Мадагаскаре.
252
Клюв прямой, долотообразный, с продольными гранями. Череп схизогнатический. Сошник парный, сильно редуцированный. Ключицы срастаются в вилочку. Ноги короткие с сильными пальцами и острыми, изогнутыми когтями. Первый палец у отдельных видов редуцируется. Язык длинный, тонкий; ножки подъязычной кости очень длинные, и язык может сильно выдвигаться (почти на длину клюва). Конец языка заострен и несет острые, направленные назад щетинки. Сильно развиты нижнечелюстные слюнные железы. Липкая слюна тонкой пленкой покрывает язык. Птица выдвигает язык в узкий ход живущих в коре или древесине насекомых и добыча приклеивается к языку или накалывается на шипы. Желчный пузырь большой. Слепых кишок нет. Копчиковая железа оперенная, у немногих голая. Нижняя гортань без костного барабана с парой голосовых мышц. Есть только левая сойная артерия.
Кожа довольно толстая. Контурное перо со слабо развитым или полностью редуцированным побочным стержнем. У ряда видов пуховые части контурных перьев продуцируют роговую «пудру». Первостепенных маховых 10, рулевых 12 ( у Verreauxia мала и скрыта кроющими хвост довольно длинный,
Рис. 163. Черный дятел Dryocopus marti us (L.)
только 8), но наружная пара обычно очень хвоста. У подавляющего большинства видов резко конический; рулевые жесткие с прочным стержнем и острой вершиной. При линьке центральная, наиболее длинная
пара сменяется лишь тогда, когда остальные рулевые уже почти полностью вырастут. Окраска разнообразная, иногда яркая и контрастная. У части видов выражен половой и возрастной морфизм окраски.
Полет волнистый; обычно пролетают небольшое расстояние и садятся. Большинство видов легко передвигается по вертикальным стволам деревьев, цепляясь за неровности коры острыми когтями и опираясь на хвост. По земле передвигаются прыжками. Приурочены к лесам разных типов, немногие виды населяют безлесные степные пространства Африки и Америки. В питании преобладают разнообразные насекомые, их личинки и яйца. Собирают их на стволах и ветвях, иногда на земле, выдалбливают из коры и древесины. При недостатке насекомых поедают разнообразные семена; наши дятлы зимой выдалбливают семена из шишек хвойных деревьев. Вне сезона размножения держатся поодиночке. Оседлые или кочующие, немногие виды перелетные.
В период тока стучат клювом по сухим сучкам, издавая своеобразную «дробь». Обычно в гнилом или начинающем загнивать дереве ежегодно долбят новое гнездовое дупло; немногие виды занимают естественные или чужие брошенные дупла. В безлесных ландшафтах долбят норы в обрывах, термитниках. В кладке 2—8 блестящих белых яиц, откладываемых в дупло
253
без всякой подстилки. Насиживание продолжается 11—18 дней. Птенцы вылупляются голыми, у немногих видов слабо опушенными; хорошо развиты пяточные мозоли, а подклювье длиннее и шире надклювья. Оба родителя носят птенцам разнообразных насекомых. Птенцы покидают гнездо полностью оперенные и способные к полету в возрасте 19 (мелкие виды) — 30— 35 дней (крупные виды). Выводок 2—3 недели кочует вместе с родителями, продолжающими кормить птенцов, а затем распадается.
В семействе дятловых 36—45 родов и 209 видов. В СССР гнездится 13 видов, относимых к 5—6 родам. Иногда выделяют 2 подсемейства.
1. Вертишейки Jynginae. Хвост прямо обрезанный из мягких, не заостренных рулевых, язык гладкий, без щетинок на конце, гнездятся в естественных или брошенных дуплах других дятлов, сами дупел не выдалбливают. Включает только род Jynx с 2 видами (один широко распространен по Палеарктике, другой в Африке).
2. Настоящие дятлы Picinae. Хвост резко конический, из жестких заостренных рулевых, на конце языка есть щетинки. Гнездятся обычно в дуплах, которые долбят сами. Подсемейство объединяет 207 видов, включаемых в 35—44 рода: Picas, Asyndesmus, Blythipicus, Brachy-lophus, Campephilus, Campethera, Camponotus, Celeus, Ceophloeus, Chryso-colaptes, Chrysoptilus, Colaptes, Dendrocopos, Dendropicos, Desertipicus, Dinopium, Dryocopus, Gecinulus, Geocolaptes, Hemicircus, Ipophilus, Leuco-nerpes, Liopicus, Meiglyptes, Melanerpes, Mesopicos, Micropternus, Mulle-ripicus, Nesoceleus, Nesoctites, Phloeoceastes, Picoides, Piculus, Picumnus, Polipicus, Sasia, Sapheopipo, Scapaneus, Sphyrapicus, Thripias, Trichopicus, Veniliornis, Verreauxia, Xiphidiopicus.
Центр возникновения подотряда, видимо, Южная Америка, где и сейчас наиболее велико видовое разнообразие дятлов. Ископаемые остатки из нижнего миоцена (Франция) и более поздних отложений в разных пунктах современного ареала относят к 4 ископаемым и 20 ныне живущим видам. Вероятно, обособились от бородатковых.
В целом отряд дятлообразных обособился от примитивных ракшеобразных птиц, возможно, близких к предкам момотов и зимородков. В пределах отряда дятлообразных центральное семейство, вероятно, бородатковые Capitonidae, от предковых форм которых обособились якамары, туканы и настоящие дятлы. Медоуказчики отделились от бородаток, видимо, позже становления перечисленных групп.
ОТРЯД 33. PASSERIFORMES — ВОРОБЬИНООБРАЗНЫЕ
Птицы мелкой и средней величины (вес от 4 г до 1,5 кг) довольно разно образного внешнего облика. Распространены практически по всему земному шару, кроме Антарктики и некоторых мелких океанических островов, далеко удаленных от материков. Особенно разнообразен видовой состав воробьиных в тропических лесах.
Форма клюва р разных группах варьирует очень сильно. Череп обычно эгитогнатического типа (у немногих групп с переходом к схизогнатизму). Базиптеригоидных отростков нет. Сошник обычно более или менее отчетливо расщепленный. Надорбитальных желез нет. Шейных позвонков 14—15. Истинных ребер 4—6 пар. Ключицы сливаются в вилочку. По заднему краю грудины одна пара вырезок, иногда превращающихся в фонтанели (у единичных родов две пары вырезок). Все четыре пальца ноги относительно длинные, с острыми когтями; три пальца направлены вперед, один (первый) — назад. Язык хорошо развит. Зоба нет. Мускульный желудок небольшой, но с довольно мощными мускульными стенками. Слепые кишки обычно рудиментарны. Желчный пузырь у большинства видов есть. Копчиковая железа не оперена. Гортань трахео-бронхиальная, реже трахеальная.
254
В разных группах число голосовых мышц варьирует от 1 до 6—7 пар. Хорошо развита только левая сонная артерия.
Оперение довольно плотно прилегающее или рыхлое. Пуховая часть контурного пера обычно хорошо развита, побочный стёржень мал или полностью редуцируется. Пух редкий, расположен только по аптериям. Первостепенных маховых 9—11; у многих групп первое маховое мало или совсем редуцировано. Второстепенных маховых обычно 9. Крыло эутаксическое. Рулевых чаще 10—12, иногда 6—16. Окраска разнообразная: от скромной и незаметной до очень яркой, иногда с металлическим отблеском. У многих видов более или менее четко выражен половой и возрастной морфизм окраски, у части видов — сезонный. Обычно самцы несколько крупнее самок.
Большее число видов — обитатели разнообразных древесных и кустарниковых зарослей; часть из них проникает в открытые ландшафты. Меньшее число видов населяют только открытые ландшафты и ведут наземный образ жизни. Проникают высоко в горы, заселяют многие острова. Питание очень разнообразно. Поедают насекомых и других беспозвоночных, едят семена, плоды, некоторые виды питаются почти исключительно пыльниками и нектаром, есть и практически всеядные формы. Часть видов оседлые или кочующие даже в высоких широтах, другие перелетные.
Обычно моногамы: оба партнера в той или иной степени участвуют в насиживании кладки и выкармливании птенцов. Однако встречаются и случаи типичной полигамии. Гнезда на земле или в ветвях, дуплах и расщелинах скал, иногда в норах. Некоторые виды устраивают лепные гнезда. Размеры кладок очень варьируют: от 1 до 15—16 яиц, у большинства видов кладки обычно из 4—6—8 яиц. Развитие идет по чисто птенцовому типу: птенцы вылупляются голыми или очень слабо опушенными, слепыми. В гнезде остаются, пока не достигнут размеров взрослых и полностью оперятся. Родители кормят птенцов, вкладывая корм в рот, который голодный птенец очень широко открывает. У многих видов ротовая полость птенцов очень яркая, что служит стимулом для проявления инстинкта кормления у взрослых птиц. Обычно после вылета из гнезда птенцы некоторое время — от нескольких дней до нескольких недель у разных видов — держатся с родителями. Как правило, достигают половозрелости и приступают к размножению на следующее лето жизни, в возрасте немного меньше года.
Отряд воробьиных объединяет больше половины всех ныне живущих видов птиц. Он включает около 5100 видов (5035—5309), группируемых в 1080—1343 рода и относимых к 52—72 семействам (2 из них включают только ископаемые виды) и 4 подотрядам. В СССР встречается 317—329 видов воробьиных птиц только 1 подотряда певчих воробьиных.
Система отряда до сих пор разработана недостаточно. Этому мешает не только очень большой объем отряда, но и хорошо выраженная анатомическая его однородность. Выяснение родственных отношений сильно затруднено широко распространенным в отряде конвергентным развитием. Сказывается и недостаточная изученность многих групп. Ископаемые остатки воробьиных малочисленны и не помогают в построении системы. Самые древние остатки известны из верхнего эоцена. В верхнем олигоцене и миоцене уже находят остатки современных родов. Все это приводит к тому, что системы отряда, предлагаемые разными авторами, существенно различаются по числу и объему семейств, их расположению (см. стр. 66—68). Из-за большой морфологической однородности всех групп отряда для характеристики семейств приходится использовать преимущественно внешние признаки: общий облик, форму клюва, расположение щитков на цевке, некоторые особенности оперения, окраску яиц и т. д. Учитываются и экологические данные: биотопическая приуроченность, характер питания и гнездования, способы охоты. В меньшей степени используются черты внутреннего строения: детали строения нижней гортани, некоторые особенности мускулатуры конечностей и т. п. Несколько шире удается использовать морфологические особенности для характеристики высших подразделений
255
отряда — подотрядов. Подавляющее большинство систематиков выделяет в отряде воробьиных 4 подотряда: Eurylaimi, Tyranni, Menurae, Passeres (Oscines).
ПОДОТРЯД 1. EURYLAIMI— РОГОКЛЮВЫ
Подотряд рогоклювов включает единственное семейство.
СЕМЕЙСТВО 1. EURYLAIMIDAE— РОГОКЛЮВЫЕ
Коренастые лесные птицы (рис. 164) весом от 20 до 150 г, длиной1 13— 27 см, обычно яркой окраски (с преобладанием зеленого, иногда голубого). Внешне несколько похожи на ракш. Распространены в лесных районах Аф-
рики (кроме Мадагаскара) и Юго-Восточной Азии (рис. 165).
Морфологически довольно отчетливо отличаются от остальных воробьиных птиц. Клюв довольно мощный, уплощенный у основания, широкий и короткий (отсюда и название подотряда). Имеется лобно-носовое сочленение. Череп эгито-гнатический. Шейных позвонков 15. Истинных ребер 4 пары. Основные фаланги направленных вперед пальцев (особенно третьего и четвертого) срастаются друг с другом. Задняя сторона цевки покрыта многоугольными щитками (а не большими роговыми пластинками, как у остальных воробьиных). Трахео-бронхиальная ниж-
няя гортань имеет лишь одну пару слабо РИСа^0№па"“м.Гка“ЮВ Развитых голосовых мышц или они совсем не развиваются. Кажется, имеются парные сонные артерии (требует подтверждения). Первостепенных маховых 10—11. Рулевых обычно 12.
Живут в лесах, держатся в кронах, на земле кормятся редко. Питаются преимущественно насекомыми, отчасти плодами. Крупные виды иногда ловят ящериц и лягушек. Гнездо свивают из растительных волокон в виде сумки с длинным боковым входом — рукавом; оно висит на тонких ветвях. В кладке 2—6 яиц, у большинства видов пятнистых.
В семействе всего 14 видов, относимых к 8 родам. 4 вида 2 родов (Smi-thornis, Pseudocalyptomend) встречаются в Африке, 10 видов 6 родов (Еигу-laimus, Calyptomena, Corydon, Cymbirhynchus, Psarisomus, Serilophus) — в Юго-Восточной Азии.
Многие детали строения (число шейных позвонков, синдактилия, характер мускулатуры ног, особенности птерилозиса) сближают рогоклювов с ракшеобразными. Поэтому подотряд обычно рассматривается как наиболее примитивная группа отряда, занимающая в известной степени промежуточное положение между ракшеобразными и остальными группами воробьиных птиц.
1 Здесь и далее указывается зоологическая длина: от конца клюва до конца хвоста.
256
150° 120° 90° 60°к западу от Гринвича 0 к востоку отГринвича 60° 90° 120° 150° 180° 150°
Рис. 165. Ареалы семейств воробьиных птиц:
/—- рогоклювые Eurylaimidae, 2—птицы печники Furnariidae, 3— питтовые Pittidae, 4— филепиттовые Philepittidae, 5 • тиранновые Tyrannidae* 6 птицы лиры Menuri-dae, 7—кустарниковые птицы At г ichornit hidae
ПОДОТРЯД 2. TYRANNI (CLAMATO RES) — ТИРАН НОВЫЕ, ИЛИ КРИЧАЩИЕ ВОРОБЬИНЫЕ
Воробьиные птицы разнообразного внешнего облика и различных размеров (вес 4—800 г). Отличаются от остальных воробьиных преимущественно деталями строения нижней гортани: она трахеального или трахео-брон-хиального типа и имеет слабо развитую мускулатуру, обычно представленную лишь одной-двумя парами голосовых мышц. Издают разнообразные, иногда очень звучные крики, но настоящей сложной песни нет. Клювы разнообразной формы. Лобно-носового сочленения нет. Череп эгитогнатиче-ский. Шейных позвонков 14. Истинных ребер 4—5 пар. Задняя сторона цевки покрыта поперечными пластинками. Синдактилия выражена у немногих родов. Окраска разнообразная, часто яркая. У части видов хорошо выражен половой и возрастной морфизм окраски. По характеру питания семейства различны: есть преимущественно насекомоядные, есть и преимущественно растительноядные.
Распространены в тропиках Старого и Нового Света, относительно немного видов проникает в умеренные широты Америки. В Южной Америке по численности и видовому составу доминируют над певчими воробьиными примерно в соотношении 3 : 2. Оседлые или кочующие; немногие виды, проникающие в умеренные широты, перелетные.
По деталям строения нижней гортани, некоторым особенностям ветвления крупных кровеносных стволов (особенно сонных артерий), характеру мускулатуры конечностей, строению роговых щитков цевки, типу вырезок по заднему краю грудины, деталям костного неба, некоторым чертам экологии (типы гнезд, характер питания и т. п.) в подотряде выделяют ^семейств, обычно группируя их в 2 надсемейства. Различия между семействами очень нечеткие. Всего к подотряду относят 310—316 родов с 1077 —1084 видами. В СССР представители подотряда не встречаются.
Надсемейство 1. Furnarioidea — древолазы, муравьеловки, птицы печники и др.
Рис. 166. Красноватый древолаз Dendrocincla homochroa (V.)
Объединяет 5 семейств, распространенных только в Центральной и Южной Америке. Включает 527 видов, распределяемых по 136—137 родам.
СЕМЕЙСТВО 2. DENDROCOLAPTIDAE — ДРЕВОЛАЗОВ ЫЕ
Лесные птицы (длина 14—35 см), обычно сероватооливковой окраски с белыми или темными пестринами, иногда с красноватым или коричневатым отливом без металлического блеска. По внешнему облику (рис. 166) и повадками немного похожи на дятлов или пищух. Клюв прямой, мощный; у некоторых видов длинный, тонкий, иногда изогнутый, как у пищух. Гортан^ трахеального тийа с
258
двумя парами голосовых мышц. Рулевые перья с мощными уплощенными стержнями. Хвост похож на хвост дятлов и используется, как опора. Половой диморфизм обычно не выражен.
Держатся в кронах. На стволах и ветвях собирают насекомых, их яйца и личинок. Часть видов кормится и в кронах, и на земле, немногие виды — преимущественно на земле. Некоторые виды, добывая личинок и взрослых насекомых, могут немного долбить кору и гнилую древесину. Гнездо располагается в укрытии: в дупле (естественном или дятловом), узкой развилке ветвей, за отставшей корой и т. п. Яйца одноцветные белые или голубоватые. В кладке 2, редко 3 яйца. Инкубация около 2,5 недель. Птенцы вылетают из гнезда на 19—20-й день после вылупления. Вне сезона размножения держатся поодиночке, парами или стайками.
Распространены в Центральной и Южной Америке (в Венесуэле, например, встречается 23 вида). Известно 48 видов, относимых к 12—13 родам: Dendrocolaptes, Campylorhamphus, Deconychura, Dendrexetastes, Den-drocincla, Drymornis, Glyphorhynchus, Lepidocolaptes, Nasica, Sittasomus, X iphocolaptes, X iphorhynchus.
Центр возникновения, видимо, тропики Южной Америки.
СЕМЕЙСТВО 3. FURNARIIDAE — ПТИЦЫ ПЕЧНИКИ, ИЛИ ПТИЦЫ ГОНЧАРЫ
Большое семейство мелких и очень мелких птиц (длина 10—25 см), очень разнообразных по внешнему облику (рис. 167), повадкам и биотопи-ческой приуроченности. Распространены повсеместно в Центральной и Южной Америке, включая Огненную Землю (рис. 165). Многие похожи на мелких дроздов, каменок, жаворонков; другие напоминают славок, пищух, поползней, трясогусок. Клювы очень разнообразны: короткие и длинные, прямые; более или менее изогнутые, тонкие; конические и т. п. Нижняя гортань трахеального типа с двумя парами мышц. Хвосты разнообразной длины и формы: короткие и длинные, прямо обрезанные, округлые, заостренные, у нескольких видов лирообразные. Окраска чаще тусклая, буровато-коричневая; у некоторых видов на голове, горле, хвосте есть яркие цветные пятна.
Рис. 167. Печник Furnarius rufus (Gm.) около гнезда
259
Обитатели лесов, кустарниковых или тростниковых зарослей. Однако часть видов живет в открытых лугах и степях, на безлесных высокогорьях, некоторые виды приурочены к берегам горных речек и скалистым морским побережьям. Большинство видов — исключительно или преимущественно насекомоядны, часть видов более или менее регулярно использует растительные корма (мелкие плоды, семена). По характеру пищевой специализации и местам добывания пищи в семействе очень широкая радиация: многие виды — обитатели крон, кормящиеся на ветвях и вертикальных стволах; другие, также лесные виды, кормятся только на земле, выбирая пищу в лесной подстилке, и скрываются в кронах лишь при опасности. Виды, населяющие открытые местообитания, кормятся на земле.
По положению и характеру постройки гнезда крайне разнообразны. Виды рода Furnarius лепят из глины большие круглые гнезда (диаметром до 20—30 см) с боковым входом (рис. 167); иногда вход в гнездо идет через длинный глиняный туннель. Такой тип гнезда и послужил основанием для наименования семейства. Тяжелое гнездо помещается на земле, уступе скалы или у основания крупных горизонтальных ветвей дерева. Другие виды устраивают шарообразные гнезда с боковым входом из мелких веточек, сухой травы, листьев, помещая это крупное сооружение (в диаметре до 50 см и больше) в развилке ветвей, на заломах тростника. Некоторые виды гнездятся в расщелинах скал. Часть видов роет гнездовые норы. Наконец, у некоторых видов обычные открытые гнезда размещаются в развилке ветвей в кронах. В кладке 3—5, у некоторых видов до 9 обычно одноцветных (чаще белых) яиц. Насиживание 13—20 дней. Птенцы из гнезд вылетают через 13—29 дней после вылупления. Оседлые или кочующие; виды крайнего юга Южной Америки совершают короткие перелеты.
Входящие в семейство 215 видов группируются в 58 родов: Furnarius, Anabacerthia, Ancistrops, Anumbius, Aphrastura, Asthenes, Automolus, Berlepschia, Chilia, Cinclodes, Clibanornis, Cranioleuca, Coryphistera, Drioc-tistes, Geobates, Geositta, Heliobletus, Leptasthenura, Margarornis, Megaxe-nops, Metopothrix, Phacellodomus, Philydor, Phleocryptes, Premnoplex, Premnornis, Pseudocolap tes, Pseudoseisura, Pygarrhichas, Schizoeaca, Schoenio-phylax, Sclerurus, Siptornis, Spartonoica, Sylviorthorhynchus, Synallaxis, Thripodectes, Thripophaga, Upucerthia, Xenoctistes, Xenops и др. Некоторые систематики в пределах семейства выделяют 4 подсемейства. Центр возникновения, вероятно, тропики Южной Америки. В отложениях верхнего плейстоцена Бразилии обнаружены 2 ныне живущих вида.
СЕМЕЙСТВО 4. FORMICARHDAE—МУРАВЬЕЛОВКОВЫЕ
Мелкие (длина 10—34 см), преимущественно лесные птицы, довольно разнообразные по внешнему облику (рис. 168); похожи на каменок, короткохвостых дроздов, сорокопутов, крапивников. Населяют лесные и горные районы Центральной и Южной Америки.
Клюв чаще крепкий, удлиненный, иногда с очень маленьким крючком на конце. Ноги относительно длинные, сильные, с короткими когтями. Нижняя гортань трахеального типа с одной парой голосовых мышц. Крылья короткие, с округлой вершиной. Очень пушистое оперение надхвостья. Хвост короткий или средней длины. Окраска коричневатая, часто с пятнами черного, белого, оранжевого, оливкового цвета. Половой диморфизм окраски четко выражен лишь у немногих видов.
Населяют разного типа леса и кустарниковые заросли. Летают мало Некоторые виды кормятся преимущественно в кронах, большее число видов — на земле. Поедают разнообразных насекомых, особенно кочующих муравьев. Более крупные виды наряду с насекомыми используют и другие животные корма: таскают яйца и птенцов, ловят мелких ящериц и т. п. Гнездо — неглубокая чашечка из растительных волокон—располагается на земле или на низких ветвях. Яйца белые или кремовые, у большинства
260
видов с коричневатым крапом и пятнами. Насиживают 14— 17 дней. Рост птенцов быстрый: уже на 9—13-й день они вылетают из гнезд, а в возрасте около месяца начинают вести самостоятельную жизнь. Все виды оседлые, кочующие по небольшой территории в поисках корма. Обычно держатся парами и мелкими группами.
Семейство включает 224 вида, которые группируются в 53 рода. Более обычные роды: Formicarius, Batara,
Cractisus, Cercomacra, Chae- Рис. 168- Серая муравьеловка Formiciv^ra grisea maeza, Clytoctantes, Cymbila- (Bodd.)
Dysithamnus, Formi-civora, Grallaria, Gr altar i-
cula, Gymbilaimus, Gymnocichla, Gymnopithys, Hypocnemis, Hylophylax, Melanopareia, Microrhopias, Myrmeciza, Myrmecocichlia, Myrtnoborus, Myr-motherula, Neoctantes, Phaenostictus, Phlegopsis, Pithys, Pittasoma, Pyri-glena, Rhopornis, Sakesphorus, Sclateria, Tar ba, Terenura, Thamnocharis, Thamnophilus, Xenornis и др. Центр возникновения — тропики Южной Америки. Относительно поздние (плейстоцен) остатки одного ископаемого вида из Бразилии относят к этому семейству.
СЕМЕЙСТВО 5. CONOPOPHAGIDAE— ГУСЕНИЦЕЕДОВЫЕ
Мелкие (длина 10—14 см) лесные птицы (рис. 169), по повадкам и
внешнему виду несколько напоминающие лесного конька или зарянку. Распространены в тропиках Южной
Рис. 169. Золотистый гусеницеед Conapophaga aurita (V.)
Америки.
Клюв прямой, тонкий, но крепкий с небольшим крючком на конце. В отличие от других семейств у гусеницеедов череп схизогнатического типа. Нижняя гортань тра-хео-бронхиального типа с одной парой слабо развитых голосовых мышц. Окраска коричневатая с участками белого, красного, черного цветов. Самки тусклее самцов. Летают мало. По земле быстро передвигаются сильными прыжками.
Населяют тропические леса с густым подлеском и густые кустарниковые заросли. Кормятся чаще на зем-
ле различными насекомыми, их личинками и гусеницами, пауками и другими наземными беспозвоночными. Гнездо в виде глубокой чаши, иногда с крышкой и боковым входом; сплетается из растительных волокон на земле или в развилке у основания куста. В кладке всего 2 яйца, обычно
пятнистых.
Семейство объединяет 11 видов 2 родов: Conopophaga (9 видов) и Со-rythopis.
261
Рис. 170. Красногорлый топаколо Scelorchilus rubecula (Kitt.)
СЕМЕЙСТВО 6. RHINOCRYPTIDAE — ТОПАКОЛОВЫЕ
Мелкие птицы (длина 11—25 см, вес 14—160 г), по внешнему облику (рис. 170) и размерам похожи на крапивников, славок, дроздов. Распространены почти по всей Южной Америке (кроме районов заболоченных лесов) от Коста-Рики до Огненной Земли.
Крепкий прямой клюв к концу заострен. Ноздри прикрыты роговыми крышечками. По заднему краю грудины 2 пары вырезок. Грудная мускулатура относительно слабая. Ноги удлиненные, крепкие, с сильной мускулатурой. Нижняя гортань трахеального типа с одной парой голосовых мышц (у части видов они не развиты). Крылья короткие, округ-
лые. Окраска коричневатая, иногда с яркими цветными пятнами или полосами на голове, горле, груди и т. п. Половой диморфизм окраски выражен лишь у части видов.
Населяют леса с подлеском и густые кустарниковые заросли; некоторые виды проникают в травянистые степи и безлесные высокогорья. Летают мало и неохотно. Легко и быстро бегают по земле. Кормятся преимущественно на земле, иногда даже разгребая подстилку сильными ногами. Едят насекомых, пауков, червей, моллюсков; поедают ягоды и семена, сочные корешки. Гнездятся в коротких, вырытых самими птицами норах или (немногие виды) — в дуплах. Некоторые виды вьют крупные шарообразные гнезда, размещая их в густых мутовках ветвей в основании куста. В кладке 2—4, чаще белых яйца. Насиживают оба партнера.
В семейство включают 29 видов, относимых к 11 родам: Rhinocrypta, Acropternis, Eugralla, Liosceles, Melanopariea, Merulaxis, Psilorhampus, Pteroptochos, Scelorchilus, Scytalopus, Teledromas.
Центр возникновения, видимо, Южная Америка, хотя описанные из среднего эоцена Вайоминга (США) ископаемые остатки предположительно относят к этому семейству.
Надсемейство 2. Tyrannoidea — тиранновые
Сюда включают 8 семейств, из которых 5 встречаются только в Новом Свете, а 3 — только в Старом Свете. Всего в надсемействе 550—557 видов, относимых к 174—179 родам.
СЕМЕЙСТВО 7. COTINGIDAE— КОТИНГОВЫЕ
Объединяет очень разнообразных по величине (длина 7—50 см, размеры от чижа до вороны) и внешнему виду преимущественно лесных птиц Центральной и Южной Америки. Часть видов напоминает дроздов или крупных славок, другие не имеют аналогов в нашей фауне (рис. 171, 172).
Клюв сильный, слегка изогнутый к концу, с маленьким крючком на конце. Основание клюва широкое, зев большой. Ноздри открытые. Нижняя гортань трахео-бронхиального типа с двумя голосовыми мышцами, развитыми различно у разных видов. Голоса громкие, у некоторых видов с металлическим тоном. Окраска яркая: белая, зеленая, красная, голубая, черная или из сочетания этих цветов. В большей или меньшей степени вы-
262
Рис. 171. / — желтогрудая котинга Euchlornis rlefferi (Boiss.); II — голова трехусого кузнеца Procnias tricarunculatus (Verr.)
Рис. 172. Токующий самец зонтичной птицы Cephalopterus ог-natus Geoff.
ражен половой диморфизм. У многих видов развиты украшения в виде удлиненных рулевых, видоизмененных и удлиненных второстепенных маховых, сильно развитых хохлов на голове и т. п. У некоторых видов есть голые (рис. 171) или покрытые перьями кожные выросты (рис. 172). Во время тока они наполняются кровью и размеры их заметно увеличиваются.
Экология изучена плохо. Большинство видов — обитатели разных типов леса, постоянно держащиеся в кронах; некоторые виды приурочены к опушкам леса. Немногие виды населяют безлесные предгорья. Кормятся в кронах, реже — на земле. Питаются разнообразными мелкими плодами, в меньшей степени — семенами и различными беспозвоночными.
Большинство видов моногамы, но некоторые — полигамы. У многих видов сложное предбрачное поведение: токующие птицы издают громкие крики, топорщат оперение, поднимают кожные выросты и т. п. У скалистых петушков Rupicola самцы токуют группами на полянках среди скал; их ток очень похож на ток тетеревов. Расположение гнезд очень разнообразно. Часть видов гнездится в дуплах или вьет гнезда в расщелинах скал, другие устраивают в развилке ветвей объемистые закрытые гнезда с боковым входом или вьют обычные чашеобразные открытые гнезда в густых ветвях. У крупных видов в кладке 1—2, у мелких до 3—4 яиц. Яйца оливковые, зеленоватые или коричневатые, обычно с темным крапом. Насиживание, видимо, у большинства видов около 2,5—3 недель, птенцы вылетают примерно к 4-недельному возрасту. У части видов оба партнера строят гнездо, насиживают и выкармливают птенцов, у других самец не насиживает, но
263
помогает в постройке гнезда и выкармливании птенцов. У видов с резко выраженным половым диморфизмом вся забота о потомстве, вероятно, падает на самку. Оседлые или кочующие. Населяют Центральную Америку и тропические районы Южной Америки; один вид проник в южные районы США, один — на Ямайку.
В семействе 94 вида, объединяемые в 30—33 рода: Cotinga, Ampelion, Attila, Calyptura, Carpodectes, Cephalopterus, Euchlornis, Gytnnoderus, Haematoderus, Heliochera, lodopleura, Lipaugus, Pachyrhamphus, Perissocep-halus, Phibaltira, Phoenicircus, Platypsaris, Procnias, Pyroderus, Queru-la, Rupicola, Tityra, Tijuca, Xipholena, Zaratornis и др. Центр возникновения, видимо, тропики Южной Америки.
СЕМЕЙСТВО 8. PIPRIDAE — МАНАКИНОВЫЕ
Мелкие древесные птицы лесов и густых зарослей кустарников тропических районов Центральной и Южной Америки. По внешнему облику (рис. 173), размерам и поведению несколько напоминают синиц. Длина 8—16 см, вес 7—40 г.
Клюв короткий, но довольно сильный с маленьким крючочком на конце. Нижняя гортань трахео-бронхиальная; голосовая мембрана без хряща. Цевки длинные, лапки слабые; у большинства выражена синдактилия.
Хвост и крылья короткие. Обычен рез-
кий половой диморфизм. Окраска самцов яркая из сочетания черного, красного, желтого, синего, оранжевого, белого цветов. Иногда имеют хохлы, удлиненные маховые или рулевые. Самки однотонные: зеленоватые, буроватые, коричневатые.
Экология известна плохо. Кормятся в кронах деревьев и кустарников преимущественно разнообразными насекомыми, отчасти, видимо, семенами и мелкими ягодами. Вне размножения кочуют мелкими стайками. Видимо, у всех или большинства видов более или менее отчетливо выражен групповой ток: самцы сидят группами в 3—5 птиц на сухой ветке,
Рис. 173. Золотистый манакин издают громкие крики, подпрыгивают или Monacus vitellinus (Gid.) взлетают и вновь садятся на то же место и т. д. Спариваются с самками, подлетающими к месту тока. Кратковременность общения самца и самки, видимо, служит причиной относительно частых случаев гибридизации (межвидовой, реже межродовой). У видов,
экология которых изучена, строит гнездо, насиживает кладку и выкармливает птенцов только самка. Открытое сверху гнездо устраивается в развилке ветвей. В кладке обычно 2 яйца, чаще палевого цвета с отчетливым крапом. Насиживание плотное, продолжается до 3 недель, а птенцы покидают гнездо примерно через 2 недели после вылупления.
В семействе 61 вид, объединяемый в 21 род: Monacus, Chiroxiphia, Corapipo, Heterocercus, Machaerhopterus, Masius, Neopelma, Pipra, Piprites, Schiffornis, Teleonema, Tyranneutes и др.
Центр возникновения, видимо, тропики Южной Америки. Проявляются несомненные черты родства с котинговыми и тиранновыми.
СЕМЕЙСТВО 9. TYRANNIDAE— ТИРАННОВЫЕ
Большое семейство, объединяющее мелких птиц (вес от 5 до 80 г, длина от 7 до 27 см), очень разнообразных по внешнему облику (рис. 174) и экологии. Многие виды напоминают мухоловок, славок, пеночек, другие больше
264
похожи на каменок, мелких ракш и сорокопутов. Населяют только Новый Свет, встречаясь от северных районов Канады до Огненной Земли; есть ина многих прилегающих островах (рис. 165).
Клюв сильный, прямой, с небольшим крючком на конце. Цевка удлиненная, пальцы умеренной длины. Основные фаланги 3-го и 4-го пальцев срастаются. Нижняя гортань трахео-бронхиальная.с 1—2 парами голосовых мышц; голосовую мембрану поддерживают разрастания хряща. Крылья короткие и широкие или удлиненные, заостренные. Хвост умеренной длины, но у части видов длинный. Окраска разнообразна: однотонная
зеленоватая ИЛИ коричневатая, яркая рис. 174. Тиранн Tyrannus tyran-из сочетания желтого, красного, белого, nus (L.)
черного, синего цветов и т. п. У большинства видов половой диморфизм окраски отсутствует или выражен не ясно; у части видов — очень резкий.
Преимущественно насекомоядны, но при случае поедают ягоды и семена; у немногих видов преобладает растительная пища. Крупные виды иногда таскают яйца и птенцов других птиц. Большинство видов разыскивает корм в кронах деревьев и кустарников. Часто охотятся «по-мухоловочьи»: птица сидит на ветке и, взлетая, ловит близко подлетевшее насекомое. Относительно небольшое числб видов собирает корм на земле. Подавляющее большинство видов приурочено к лесам разных типов и кустарниковым зарослям. Немногие виды населяют открытые ландшафты: луга, степи, предгорья, полупустыни. Хорошо развито территориальное поведение. Гнезда разнообразны по типу и местоположению: открытые гнезда в развилке ветвей или у немногих видов — на земле; закрытые шарообразные гнезда с боковым входом на крупных ветвях и т. п. Немногие виды скрепляют растительную основу гнезда грязью. Некоторые виды гнездятся в дуплах. Насиживают оба партнера или только самка; в последнем случае самец охраняет гнездовой участок и кормит самку. В кладке у тропических видов 2—3 яйца, у видов умеренных широт 3—5, редко до 8. Яйца белые, кремовые или оливковые, с темным крапом. Инкубация 14—18 дней, редко больше. Птенцов выкармливают самец и самка. Птенцы вылетают из гнезд на 13—16-й день, у некоторых — на 20—23-й день жизни. Вне сезона размножения держатся стайками или одиночками.
В тропиках тиранны оседлые или кочующие, в умеренных широтах — перелетные.
Семейство включает 358—365 видов, группируемых в 114—116 родов, почти половина которых монотипична. Наиболее обычные роды: Tyrannus, Agriornis, Alectrurus, Arundinicola, Camptostoma, Colonia, Contopus, Elaenia, Empidonax, Empidonomus, Euscarthmus, Fluvicola, Gubernetes, Hirundinea, Knipolegus, Legatus, Lessonia, Lichenops, Machetornis, Megarhynchus, Musci-gralla, Muscisaxicola, Muscivora, Myiarchus, Myiobius, Myiodynastes, Myiozetetes, Neoxolmis, Nuttallornis, Ochthoeca, Onychorhynchus, Onco-stoma, Perissotricus, Phaeomyias, Phylloscartes, Pipromorpha, Pitangus, Pla-tyrinchus, Pyrocephalus, Rhynchocyclus, Sayornis, Serpophaga, Tachirnis, Tachuris, Todirostrum, Tolmomyias, Xolmis и др.
Центр возникновения, видимо, Южная Америка, где сейчас видовой состав тиранн наиболее разнообразен. Ископаемые остатки из верхнего плейстоцена, найденные в разных пунктах современного ареала, относят к 7 ныне живущим видам.
265
Рис. 175. Остроклюв Oxyruncus cristatus (Sw.)
Рис. 176. Красногрудый траворез Phytotoma rutila N.
СЕМЕЙСТВО 10. ОХУRUNCIDAE—ОСТРОКЛЮВЫЕ
Сюда включают единственный вид Oxyruncus cristatus Sw. (рис. 175), обитающий в лесах от Коста-Рики до Парагвая и Бразилии. Близок к тираннам (иногда включают в семейство тиранновых в ранге подсемейства). Отличается крепким, довольно длинным, коническим клювом и некоторыми другими особенностями. Общая длина около 17 см. Хорошо летают. Держатся в кронах поодиночке. Экология не выяснена. Питаются преимущественно мелкими плодами.
СЕМЕЙСТВО И. РНУТОТОМIDAE — ТРАВ0РЕ30ВЫЕ
Включает всего один род Phytotoma с 3 видами (рис. 176), распространенными в лесо-степных и степных предгорных и равнинных районах Южной Америки от Перу до Патагонии.
Размерами со скворца, коренастые. Клюв вьюркового типа: крепкий, высокий и широкий у основания, заостренный к концу. Края надклювья и под-клювья зазубренные,что облегчает срезание почек, листьев, молодых побегов. Цевка короткая; пальцы крепкие, довольно длинные. Окраска яркая, самки тусклее самцов. Питаются разнообразной растительной пищей: мелкими плодами, семенами, почками, молодыми листочками. Гнездо в виде небрежно построенной неглубокой чаши из тонких прутиков помещается в верхней части куста или невысокого дерева на развилке горизонтальной ветки. В кладке 2—4 зеленовато-голубых с коричневым крапом яйца. Вне сезона размножения кочуют стайками, иногда до нескольких десятков птиц.
СЕМЕЙСТВО 12. PITTIDAE— ПИТТОВЫЕ
Мелкие (длина от 15 до 28 см) короткохвостые наземные птицы (рис. 177) тропических лесов и кустарниковых зарослей Африки (нет на Мадагаскаре), Юго-Восточной Азии и Северной Австралии (см. рис. 164).
Сложение плотное. Клюв похож на клюв дроздов. Ноги длинные и сильные с довольно длинной цевкой и относительно короткими пальцами. Грудина с одной парой вырезок. Шейных позвонков 14—15. Нижняя гортань трахео-бронхиальная со слабо развитой парой голосовых мышц. Крылья короткие, широкие. Хвост очень короткий. Окраска яркая из сочетания зелёного, синего, красного, белого и др. цветов. У части видов половой диморфизм отсутствует, у других хорошо выражен. У всех видов отчетливый возрастной морфизм окраски. Летают плохо и неохотно. На ветки садятся редко. Держатся на земле в густых зарослях, передвигаясь резкими прыжками. Питаются разнообразными мелкими почвенными животными: на-
266
Рис. 177. Синегрудая питта Pitta baudi (Gid.)
Рис. 178. Новозеландский крапивник Acanthisitta chloris (Sp.)
секомыми и их личинками, червями, моллюсками и т. п. Видимо, моногамы. Гнездо в виде большого шара, сплетенного из тонких веточек и травы, с боковым входом помещается на земле (обычно у основания куста), пнях или низких горизонтальных ветвях. У африканских видов в кладке 2—3, у азиатских 4—5 яиц, блестящих, белых, с резким крапом. Птенцы становятся способными к полету уже в 2-недельном возрасте. Вне сезона размножения держатся поодиночке, реже группами в 2—4 птицы.
Семейство объединяет 25 видов, из которых 2 встречаются в тропической Африке, 3 в Северной Австралии, остальные в Юго-Восточной Азии. Все виды обычно относят к одному роду Pitta', иногда выделяют моноти-пичный род Anthocichla из Индии.
Центр возникновения, видимо, Юго-Восточная Азия.
СЕМЕЙСТВО 13. ACANTHISITTIDAE (XENICIDAE) — НОВОЗЕЛАНДСКИЕ КРАПИВНИКОВЫЕ
Миниатюрные (длина 7—10 см) короткохвостые птички (рис. 178), населяющие леса, кустарниковые заросли и скалистые обрывы Новой Зеландии и прилегающих островов. По повадкам, внешнему виду и экологии похожи на обыкновенных крапивников.
Клюв тонкий, прямой, довольно длинный. Нижняя гортань трахеобронхиальная с одной парой голосовых мышц. Крылья короткие, широкие. Хвост короткий. Ноги довольно длинные, с тонкими пальцами. Окраска зеленоватая или коричневатая; половой диморфизм не выражен или неотчетлив. Летают мало и неохотно. Насекомоядны. Пищу собирают в кронах деревьев и кустарников или — другие виды — на земле и скалистых обрывах. В щелях между камнями, в развилках корней или около упавших деревьев из травы и листьев строят большое шаровидное гнездо с боковым входом. Яйца белые. В кладке 1—2, у одного вида до 5 яиц.
Семейство включает всего 4 вида, относимых к 3 родам: Acanthisitta, Traversia (единственный среди воробьиных нелетающий вид; видимо, полностью истреблен в конце прошлого века домашними кошками), Xenicus (2 вида).
СЕМЕЙСТВО 14. PHILEPITTIDAE — МАДАГАСКАРСКИЕ ПИТТОВЫЕ
В семействе всего 2 рода (в каждом по 2 вида) Philepitta и Neodrepanis, резко отличающихся друг от друга по внешнему виду и населяющих влажные густые леса Мадагаскара. Ведут древесный образ жизни. Размеры мел-
267
Рис. 179. Бархатистая филепитта Philepitta costanea (Miill.)
репляют на низко расположенных
кие (длина 8—15сл«). Нижняя гортань трахео-бронхиальная, с сильно разросшимся верхним бронхиальным кольцом. Крылья и хвост короткие.
Мадагаскарские питты Philepitta крепкие, коренастые птицы с довольно мощным, сильно утончающимся к концу слабо изогнутым клювом (рис. 179). В окраске самцов преобладает черный, у самок зеленоватый цвет. Над глазом расположены яркие голые участки кожи. Грубо построенное висячее гнездо с боковым входом ук-ветвях. В кладке 1—3 яйца.
Насиживание продолжается около 4 недель, птенцы остаются в гнезде до 6 недель (эти данные требуют проверки). Питаются преимущественно мелкими плодами, собирая их в кронах и на земле.
Ложные нектарницы Neodrepanis имеют длинный, тонкий, изогнутый книзу клюв. Окраска яркая из сочетания желтого и голубого. Питаются преимущественно насекомыми: берут с листьев, достают их из трещин коры,
из венчиков цветков; возможно, склевывают пыльники и сосут цветочный сок. Раньше их относили к нектарницам Nectariniidae, с которыми они очень сходны по повадкам и внешнему облику, но отличаются по многим мелким морфологическим признакам.
Можно предполагать, что предковые формы кричащих воробьиных Tyrani близки к каким-то примитивным рогоклювам. В пределах подотряда, вероятно очень скоро обособились 2 ветви — надсемейства в излагаемой
системе,— адаптивная радиация которых привела к возникновению перечисленных семейств. В надсемействе Furnar io idea основные направления эволюции представлены семействами Furnariidae и Formicariidae, а в надсемействе Tyrannoidea — семейства Cotingidae и Tyrranidae.
ПОДОТРЯД 3. MENURAE — ПРИМИТИВНЫЕ ВОРОБЬИНЫЕ
Объединяет всего 4 вида, относимых к двум резко различающимся семействам, которые встречаются только в некоторых районах Австралии.
Череп не четко эгитогнатический: сошник спереди закруглен, без расщепа; небные отростки верхнечелюстных костей не окостеневают. Шейных позвонков 14. По заднему краю грудины одна пара вырезок. На цевке боковые роговые щитки расположены косо. Когти лапы слабо изогнуты. Гортань трахео-бронхиальная, с 2—3 парами хорошо развитых голосовых мышц. Язык стреловидный. Слепые кишки рудиментарны. Первостепенных маховых 11 (первое недоразвито), рулевых 12—16. Летают плохо и мало, хорошо ходят и бегают.
По морфологическим деталям проявляют некоторое сходство с примитивными дятлообразными и ракшеобразными. Видимо, представляют боковую ветвь, обособившуюся от примитивных предковых форм отряда.
СЕМЕЙСТВО 15. MENURIDAE— ПТИЦЫ ЛИРЫ
Внешне напоминают фазанов. Живут в густых лесах Юго-Восточной Австралии (см. рис. 165).
Вес 500—800 г. Клюв довольно длинный, слегка изогнутый к концу. На заднем крае грудины одна пара маленьких вырезок. На конце вилочки
268
не образуется расширения. Ноги сильные, с длинной цевкой и крепкими пальцами. В нижней гортани хорошо развиты 3 пары голосовых мышц. Контурное перо имеет сильно редуцированный побочный стержень. Пуха нет. Рулевых 16. У самки хвост удлинен, но рулевые имеют обычное строение. У самца они очень сильно удлинены и резко видоизменены: крайние рулевые имеют широкие опахала, а их концы изогнуты в сторону, остальные рулевые несут длинные, редко сидящие бородки, не сцепляющиеся в опахало (рис. 180). Во время тока самец поднимает и расправляет хвост, и он становится несколько похож на лиру. Общий тон окраски буровато-коричневый.
Живут во влажных лесах с густым подлеском. Быстро бегают, легко перепрыгивая через упавшие деревья; при прыжках иногда помогают взмахами крыльев. При опасности стараются убежать, а не взлететь. Ночуют в кронах деревьев. Кормятся на земле, разгребая ногами подстилку. Поедают различных наземных и почвенных беспозвоночных, в меньшей
степени используют растительные Рис 180 Птица Menura novaehollan. корма. Полигамы. Самцы токуют ^iae Lath.
поодиночке на маленьких лесных полянах: танцуют, совершая разнообразные движения хвостом, издают громкие крики. Хорошо имитируют голоса птиц и другие звуки (блеянье овцы, скрежет пилы и т. д.). Появившаяся на участке самка спаривается с самцом и где-то поблизости самостоятельно строит гнездо в виде большого шара с боковым входом. Оно помещается в развилке ветвей низко над землей, на пне, реже на земле в основании куста или в углублении между корнями. В кладке единственное яйцо сероватого цвета с маленькими крапинками. Насиживает только самка. Самец продолжает токовать и спариваться с другими самками. Инкубация продолжается 6—7 недель (по некоторым авторам, до 9 недель). Птенец вылупляется слепым и голым, но вскоре покрывается густым черным пухом. Глаза открываются в недельном возрасте. Самка кормит птенца различными беспозвоночными. Полностью оперенный птенец покидает гнездо в возрасте примерно 6 недель; остается с самкой еще 2—3 месяца. Вне сезона размножения держатся поодиночке. Очень пугливы и скрытны. Полный брачный наряд самец одевает, видимо, только на 3—4-й год жизни.
В семейство входит 1 род Menura с 2 видами, отличающимися по размерам и деталям окраски. Очень малочисленны, распределение спорадич-
ное.
Сходство с куриными чисто внешнее, конвергентное. Принадлежность птиц лир к воробьиным совершенно несомненна. Вполне очевидна и их близость со следующим семейством кустарниковых птиц.
269
СЕМЕЙСТВО 16.
ATRICHO R NITHI DAE — КУСТАРНИКОВЫЕ ПТИЦЫ
Мелкие, относительно длиннохвостые птицы (длина 17—22 см), по внешнему облику (рис. 181) напоминающие камышевок. Летают плохо и неохотно; хорошо бегают по земле, ловко лазают в ветвях кустов и деревьев. В семействе 1 род Atrichornis с 2 видами. А. rufescens (Ram.) распространен в кустарниковых зарослях, лесах и парках прибрежной части Юго-Восточной Австралии, A. clamosus (Gid.) встречался в кустарниках юго-запада Австралии (см. рис. 165) на площади не более 1000 км2; сейчас, видимо, уже полностью вымер.
Рис. 181. Кустарниковая птица Atric- Клюв ДОВОЛЬНО мощный, сла-hornis clamosus (Gid.) боконический. По заднему краю
грудины пара довольно глубоких вырезок. Конец сильно редуцированной вилочки с небольшим расширением. В нижней гортани хорошо развиты 2 пары голосовых мышц. Побочный стержень контурного пера полностью редуцируется. Крылья короткие и широкие; длина первостепенных маховых почти такая же, как второстепенных. Хвост ступенчатый, из 12 рулевых. Окраска коричневатая. Кормятся в зарослях кустов преимущественно на земле, разгребая сильными ногами подстилку, реже на ветвях. Поедают различных беспозвоночных и семена. Гнездо с боковым входом расположено на земле, обычно у основания куста или в куртине травы, реже на ветвях. В кладке 2 беловатых, с коричневым крапом яйца. Насиживает только самка.
ПОДОТРЯД 4. PASSERES (OSCINES) — ПЕВЧИЕ ВОРОБЬИНЫЕ
Подотряд включает подавляющее большинство видов отряда и почти половину всех ныне живущих видов птиц: 3944—4211 видов, относимых к 761—1017 родам и объединяемых в 33—54 семейства (еще 2 семейства включают только ископаемые формы). На территории СССР встречаются воробьиные только этого подотряда. Всего отмечено 317—329 видов, относимых к 112—123 родам 27 семейств. Достоверно гнездится 290—301 вид из 25 семейств.
Размеры птиц в этом подотряде от мелких до средних (вес от 4 г до 1,5 кг). Внешний облик, повадки, окраска очень разнообразны. Клювы различной формы и длины. Череп эгитогнатического типа с довольно большим сошником. Шейных позвонков 14. По заднему краю грудины одна, очень редко 2 пары вырезок; обычно они зарастают полностью или остается небольшое окошко. Ребер 5, реже 4 пары. Задний палец ноги, как правило, хорошо развит; синдактилия встречается редко. Нижняя гортань обычно трахеобронхиального типа; нижние кольца трахеи у многих видов срастаются в костный барабан. У большинства видов хорошо развиты 5—7 пар голосовых мышц, симметрично прикрепляющихся к концам бронхиальных полуколец. Издаваемые звуки очень разнообразны. Часть видов способна к
270
звуковому подражанию (пересмешиичество). Контурное перо с небольшим побочным стержнем или он может полностью редуцироваться. Пуховой покров редкий и развит только на аптериях; у части видов его нет вовсе. Первостепенных маховых 9—10—11; у части видов первое маховое недоразвито и плохо заметно, иногда совсем редуцировано. Рулевых 10—12 (у немногих видов больше или меньше). У части видов отчетливо выражен половой, возрастной и сезонный морфизм окраски, у других его нет.
Система подотряда нуждается в дальнейших исследованиях. Характеристики семейств основываются на внешних морфологических признаках и отчасти немногих экологических особенностях. Отличия между семействами иногда незначительны, что определяется не только их недостаточной изученностью, но и очень большой гомогенностью подотряда. Пока совершенно неясны родственные отношения между отдельными семейства' ми. Требует уточнения объем ряда семейств.
СЕМЕЙСТВО 17. ALAUDIDAE—ЖАВОРОНКОВЫЕ
Мелкие наземные птицы (длина 11—23 см, вес 15—70 г) с типичным обликом (рис. 182). Населяют преимущественно открытые ландшафты Старого и Нового света (рис. 183).
Клюв удлиненно конический, иногда тонкий. Цевка сзади округлая и покрыта многочисленными щитками (у всех остальных певчих воробьиных задний край цевки заострен). Пальцы умеренной длины, задний палец несет длинный, почти прямой коготь. Крылья заостренные, с 10 первостепенными маховыми. Хвост из 12 рулевых. Окраска обычно более или менее покровительственная, буровато-пестрая. У немногих видов выражен половой диморфизм окраски; почти у всех видов есть, но выражен слабо возрастной морфизм окраски. В году одна послегнездовая линька, но из-за обнашивания пера осенний наряд несколько отличается от весенне-летнего.
Населяют разнообразные открытые местообитания: тундры, луга, степи, пустыни, безлесные предгорья; немногие виды проникли в леса, но держатся обычно по опушкам и полянам. Хорошие певцы. Токующие самцы часто поют в воздухе, иногда при этом трепеща на одном месте. Полет быстрый. Гнездо вьют на земле, под защитой кочки, кустика травы и т. п. В кладке 3—5, редко до 8 светлых, с густым крапом яиц. Насиживает
обычно только самка 12 • кормят оба партнера. Молодые покидают гнездо полуоперенны-ми, еще не умея летать, в возрасте 9—11 дней; родители их кормят некоторое время и после подъема на крыло. У многих видов даже в умеренных широтах 2 кладки в сезон; ко второй приступают после подъема на крыло птенцов первого выводка. Кормятся на земле, поедая различных беспозвоночных (главным образом насекомых) и семена. Вне сезона размножения держатся стайками, иногда из нескольких десятков особей, реже мелкими группами или одиночками, кочуя в пределах гнездового ареала. Северные виды (и часть видов умеренных широт) перелетные.
16 дней. Вылупившихся птенцов
Рис. 182. Полевой жаворонок Alauda arve-nsis L.
271
150 120°90° ' - 60°30° 0°30° 60°30° 120° 150° 180° 150°
Рис. 183. Ареалы семейств певчих воробьиных:
1— жаворонковые Alaudidae, 2—районы акклиматизации жаворонков, 3— ласточковые Hii undinidae, 4— дронговые Ducruridae
Семейство Alaudidae включает 70—78 видов, распределяемых по 15 родам: Alauda (2 вида), Alaemon, Ammomanes, Calandrella (12 видов), Certhi-lauda, Chersophilus, Eremophila, Eremopteryx, Galerida, Heteromirafra, Lu-llula, Melanocorypha (6 видов), Mirafra (25 видов), Pseudalaemon, Rampho-corys. Особенно велико разнообразие видов жаворонков в Африке; часть видов свойственна Африке и Азии, несколько видов проникают в Европу, лишь 2 вида населяют Австралию и только 1 вид — рогатый жаворонок Eremophila alpestris L., населяющий тундры и горные .районы Старого Света, широко распространен в Северной Америке и образует небольшую изолированную популяцию в горах Колумбии. Некоторые виды акклиматизированы в ряде районов (рис. 183).
В СССР встречаются и гнездятся 13 видов 7 родов. Два вида — черный жаворонок Melanocorypha yeltoniensis (Forst.) и белокрылый М. leucoptera (Pall.) — эндемики СССР, их гнездовые ареалы ограничены степями При-каспия, Казахстана и Средней Азии.
Центр возникновения семейства, вероятно, Африка.
СЕМЕЙСТВО 18. PALAEOSPIZIDAE
Включает единственный вид Palaeospiza bella АП., описанный по хорошей сохранности остаткам, из среднего или верхнего олигоцена Колорадо (США). Возможно» занимает в какой-то степени промежуточное положение между котингами и певчими воробьиными.
СЕМЕЙСТВО 19. HIRUNDINIDAE—ЛАСТОЧКОВЫЕ
Мелкие (длина 9—23 см, вес 10—60 г), прекрасно летающие острокрылые птицы с типичным обликом (рис. 184). По характеру распространения почти космополиты (рис. 183), не проникшие лишь в Антарктику, Новую Зеландию, на мелкие океанические острова и в районы Арктики и Субарк-
тики.
Округлая, слегка уплощенная голова с коротким, довольно отчетливо уплощенным клювом; разрез рта очень большой (челюстной сустав на уровне глаза). Бронхиальные кольца более или менее замкнутые, а в области нижней гортани — полукольца. Слюнные железы очень крупные (увеличиваются перед гнездованием). Крылья длинные, заостренные, с 9 первостепенными маховыми. Хвост с более или менее резкой вырезкой. Ноги довольно слабые, с короткими цевками и тонкими пальцами. По земле ходят медленно и неохотно. Окраска темная сверху и светлая (белая) снизу. Половой и возрастной морфизм окраски выражен нерезко и не у всех видов. Обычно в году одна линька: полная послегнездовая.
Населяют преимущественно открытые ландшафты: долины рек на равнинах и в горах,
Рис. 184. Деревенская ласточка Hirundo rustica L.
273
обережья, опушки лесов; многие виды — синантропы. У самцов щебечущая песня. Гнезда большинства видов лепные, из комочков грязи, скрепляемых слюной и прикрепленных на скалистых обрывах, деревьях, постройках. Некоторые виды в обрывах роют норы, другие гнездятся в дуплах и расщелинах скал. У многих видов гнездование колониальное или групповое. Лоток гнезда всегда выстлан мягкой растительной ветошью и перьями.В кладке 3—7 яиц, обычно белых, иногда с крапом. Насиживание 13—20 дней. У многих видов самка насиживает интенсивнее самца. Птенцы вылупляются голыми или слабо опушенными. В гнезде остаются 3—5 недель. У некоторых видов — 2—3 кладки в сезон. Вне сезона размножения держатся стайками. В тропиках кочующие, в умеренных широтах перелетные. Практически исключительно насекомоядны. Добычу ловят в воздухе, реже склевывают налету насекомых, сидящих на ветвях, камнях и т п
Семейство Hirundinidae объединяет 75—79 видов, относимых к 19 родам: Hirundo (около 13 видов), Atticora, Cescropis, Cheramoeca, Delichort, Hylochelidon, Iridoprocne, Notiochelidon, Phedina, Petrochelidon (10 видов), Progne, P sal idopr оспе, Pseudochelidon, PseudohirundO, Ptyonoprogne, Riparia, Stelgidopteryx, Tachycineta.
Центр возникновения, видимо, Африка, где сейчас встречается большинство видов. Например, в Анголе гнездится 15 видов ласточек. В Южной Америке видовой состав ласточек более разнообразен, чем в Северной. Некоторые виды имеют очень большие ареалы: так, деревенская ласточка Hirundo rustica L. и береговая ласточка R. riparia (L.) широко распространены в Европе, Азии, Северной Америке, местами в Северной Африке. В СССР отмечено 9 видов 6 родов, несомненно гнездится 7 видов 4 родов.
СЕМЕЙСТВО 20. DUCRURIDAE— ДРОНГОВЫЕ
Рис. 185. Черный дронго Dicrurus macrocercus Vieill.
Обычно черные, с металлическим отблеском птицы (рис. 185) тропических лесов Африки, Мадагаскара, Южной Азии, Океании, Северной и Восточной Австралии (рис. 183). По повадкам несколько напоминают мухоловок.
Длина 18—36 см, вес 20—50 г. Клюв крепкий, слегка изогнутый, с небольшим крючком на конце. У основания клюва — направленные вперед щетинки. Первостепенных маховых 10. Крыло тупое. Ноги с сильными пальцами и короткой цевкой. Хвост довольно длинный, вильчатый или лирообразный, из 10 рулевых (у Chaetorhynchus из 12); у части видов крайние рулевые очень сильно удлинены. Самцы окрашены сходно с самками, молодые несколько светлее взрослых.
Держатся в кронах деревьев одиночками или парами. У самцов довольно сложная песня; у части видов хорошая способность к звукоподражанию. Чашеобразное гнездо укрепляется в развилке тонких густых ветвей; снаружи стенки иногда оплетаются паутиной. В кладке 3—4, реже 2—5 светлых или белых, обычно крапчатых яиц. Вне сезона размножения перелетные или
274
кочующие. Питаются преимущественно насекомыми, реже мелкими плодами, которые собирают в кронах деревьев и кустарников. Насекомых чаще ловят в воздухе: птица сидит на сухой ветке и, взлетая, ловит добычу.
Семейство объединяет 20—21 видов, сейчас обычно относимых к 2 родам: Chaetorhynchus (монотипичный род из Новой Гвинеи) и Dicrurus (включает ранее выделявшиеся роды Bhringia, Buchanga, Chaptia, Chibia, Disse-murus, Edolius, Ehringa).
В СССР в Приморском крае встречаются (и может быть изредка гнездятся) 2 вида.
Центр возникновения, видимо, Южная Азия. Очень разноречивы представления о родственных отношениях: их сближают с иволгами, мухоловками, сорокопутами, вороновыми и даже с райскими птицами.
СЕМЕЙСТВО 21. ORIOLIDAE — ИВОЛГОВЫЕ
Яркоокрашенные древесные птицы (рис. 186) Старого Света, распространенные по лесам разных типов в умеренных и тропических широтах Европы, Африки (нет на Мадагаскаре), Азии, Австралии (рис. 187). Размерами со скворца или крупного дрозда (вес 100—200 г).
Клюв довольно длинный, стройный, слегка изогнутый книзу. Наружные отверстия ноздрей открытые. Щетинок в углах рта нет. Крыло заостренное, удлиненное, с 10 первостепенными маховыми. Хвост средней длины, чаще прямо обрезанный, из 12 рулевых. У многих видов половой диморфизм окраски: самцы часто ярко-желтые с черными пятнами, самки и молодые — более тусклой желтовато-зеленоватой окраски; виды рода Irena с преобладанием голубого цвета.
Держатся преимущественно в кронах. Характерны звучная, флейтовая песня и резкие, грубые позывные крики. Гнездо в виде глубокой чаши подвешивается к горизонтальной развилке ветвей. В кладке 2—5 яиц, беловато-розовых с темным коричневым крапом. Насиживает преимущественно самка; самец кормит ее и изредка сменяет на гнезде. Инкубация около 2—3 недель. Птенцы покидают гнездо в возрасте около 2—3 недель и кочуют вместе со взрослыми. В умеренных широтах перелетные, в более южных оседлые или кочующие. Питаются разнообразными насекомыми, особенно гусеницами, собирая их в кронах, редко на земле. В качестве добавочных кормов охотно используют различные мелкие плоды и т. п.
В семейство включают 34—42 вида, объединяемых в 3 рода (Ortolas, Irena, Sphecotheres). В СССР гнездятся 2 вида типичного рода.
Центр возникновения, видимо, Юго-Восточная Азия. В Африке южнее Сахары встречается 6 видов, в Юго-Восточной Азии 15—20, в Австралии 8. Виды рода Irena иногда выделяют в самостоятельное семейство Irenidae.
СЕМЕЙСТВО 22. CORVIDAE — ВОРОНОВЫЕ
Очень различные по величине (вес 50 а — 1,5 кг), разнообразные по облику
Рис. 186. Черноголовая иволга Oriolus chi-nensis (L.)
275
Рис. 187. Ареалы семейств певчих воробьиных:
1— иволговые Oriolidae (ареал в Центральной и Южной Африке не дается), 2— вороновые Corvidae, 3— райские птицы Paradisaeidae
(рис. 188) широко распространенные птицы. Населяют самые разнообразные ландшафты (тундры, степи, пустыни, горы, поселения человека), но большинство видов приурочено к лесным местообитаниям. Семейство распространено почти по всему земному шару (рис. 187), кроме Антарктики, некоторых арктических районов, юга Южной Америки, центральной Австралии и части океанических островов; в Новой Зеландии акклиматизированы (здесь обнаружены субфоссильные остатки вороновых).
Обычно большеголовые, большеклювые, плотного сложения птицы. Клюв крепкий, довольно длинный, прямой или слабо изогнутый, часто с выпуклым коньком надклювья. Ноздри прикрыты густыми щетинкообразными перьями (исключение — взрослые грачи). Ноги сильные. Первостепенных маховых 10. Крыло у многих видов короткое и широкое, у части удлиненное, с заостренной вершиной. Хвост варьирует по длине, прямой или ступенчатый, из 12 рулевых. Оперение плотное, у части видов рыхлое. Окраска черная, черная с белым, серая, коричневатая, иногда с участками голубого цвета. Часто хорошо развит металлический отблеск. Самцы несколько крупнее самок, но по окраске не отличаются. Молодые в более тусклом пере. В году одна линька: полная послебрачная.
Гнездятся одиночными парами, часть видов группами или настоящими колониями. У некоторых видов пары сохраняются длительное время, иногда до гибели одного из партнеров. Гнезда на деревьях, в кустах, зарослях тростников, на скалах, в трещинах и промоинах обрывов, в дуплах, в постройках. Гнездо массивное, из веток, с мягкой выстилкой; у части видов гнезда с крышей и боковым входом. В кладке 2—4, у немногих видов до 7— 9 зеленоватых или голубоватых с коричневым крапом яиц. Инкубация 16— 22 дня. У одних видов попеременно насиживают оба партнера, у других насиживает преимущественно самка, а самец ее кормит, лишь изредка заменяя на гнезде. Выкармливают птенцов оба партнера. Птенцы вылетают через 3—5 недель после вылупления. Вне сезона размножения держатся группами или стаями, немногие виды поодиночке. Оседлые, кочующие, реже настоящие перелетные птицы. Подавляющее большинство видов всеядны: питаются самой разнообразной животной и растительной пищей, падалью и т. п. Многие крупные виды ведут себя как настоящие хищники, нападая на относительно крупных животных. Кормятся в кронах деревьев и на земле.
Вороновые — одна из групп птиц с наиболее высоким и сложным уровнем высшей нервной деятельности. Поэтому относительно легко приспосаб-
Рис. 188. Кедровка Nucifraga caryocatactes (Ь.)
277
ливаются к разнообразным условиям жизни; в частности, многие виды успешно освоили культурные ландшафты. Голос — карканье, громкие свистовые выкрики и т. п. Часть видов способна к звукоподражанию вплоть до имитации голоса человека.
Семейство вороновых включает 100—105 видов птиц, группируемых в 20—32 рода: Corvus (38 видов; Coloeus, Corvultur — синонимы), Aphelocoma, Boangerges, Calocitta, Cissa (К Uta), Cissopica, Cissolopha, Crypsirhina, Cya-nocitta, Cyanocorax, Cyanolyca, Cyanopica, Dendrocitta, Garrulus, Gymno-corvus, Gymnorhinus, Lalocitta, Nucifraga, Perisoreus (Cractes), Pica, Platy-tophus, Platysmurus, Podoces, Pseudopodoces, Psilorhinus, Ptilostomus, Pyrrhocorax (Graculus), Rhinocorax, Temnurus, Urocissa, Zavattariornis.
В СССР встречается и гнездится 14—16 видов из 8 родов. Саксаульная сойка Podoces panderi Fisch., гнездящаяся в наших среднеазиатских пустынях, вне СССР не гнездится; это эндемик Средней Азии.
СЕМЕЙСТВО 23. CRACTICIDAE— ФЛЕЙТОВЫЕ ПТИЦЫ
Коренастые крепкие птицы (рис. 189; длина 26—52 см, вес 100—350 г), населяющие лесные и степные районы Австралии, Тасмании, Новой Гвинеи и прилегающих островов; 2 вида акклиматизированы в Новой Зеландии. По внешнему облику похожи на вороновых.
Клюв сильный, удлиненно-конический, с маленьким крючком на конце надклювья. Открытые щелевидные наружные ноздри. Ноги сильные, с довольно длинной цевкой и крепкими пальцами. Крылья удлиненные, с острой вершиной. Окраска взрослых самцов и самок сходна и состоит из сочетания черного, серого и белого цветов; окраска молодых коричневатого тона. Хорошо поют; песня звучная, разнообразная (флейтовые птицы — местное название); у некоторых видов хорошо поют и самки. Гнезда в кронах деревьев и кустарников. В кладке 2—5 пятнистых яиц. Насиживает преимущественно самка; самец носит ей корм. Вылетевшие молодые иногда до следующего цикла гнездования держатся с родителями. Оседлые или мало кочующие. Кормятся на земле и в кронах деревьев и кустов, поедая разнообразных насекомых, плоды и семена; ловят мелких рептилий и млекопитающих, слетков птиц, грабят гнезда. Хорошо бегают и летают.
Семейство объединяет 10—11 видов, относимых к 3—4 родам: Cracticus, Gymnorhina, Strepera, Taraba.
Место возникновения,.вероятно, Австралия. Видимо, близки к дронго или к вороновым птицам.
Рис. 189. Белоспинная флейтовая птица Gymnorhina hypoleuca (Gid.)
278
Рис. 190. Сорочий жаворонок Grallina cyanoleuca (Lath.) около гнезда
СЕМЕЙСТВО 24. GRALLINIDAE — АВСТРАЛИЙСКИЕ СОРОЧЬИ ЖАВОРОНКИ
Птицы (рис. 190), внешне напоминающие некрупных вороновых или дроздовых (длина 18—45 см). Заселяют Австралию, один вид живет в горных районах Новой Гвинеи. Всего 4 вида в 3 родах: Grallina (2 вида), Сог-corax, Struthidea.
Клюв короткий, конический, слегка изогнутый или удлиненный, довольно тонкий, слабо или заметно изогнутый книзу. Ноздри не прикрыты перьями. Ноги довольно длинные; особенно заметно удлинена цевка. Отчетливый или слабо развитый половой и возрастной морфизм окраски, в которой преобладают черный, серый и белый цвета. Хорошие певцы; в период тока характерна звуковая перекличка партнеров. Все виды строят гнезда, слепленные из грязи или ила с добавлением сухих стебельков, и располагают их обычно на толстой горизонтальной ветке дерева или куста. Яйца варьирующей окраски, всегда с отчетливым крапом. Кладка из 2—5 яиц. В сезон размножения часто 2 кладки. Молодые одевают наряд взрослых в возрасте около 3 месяцев. У видов р. Grallina сложный групповой ток (издают разнообразные звуки при сложных телодвижениях, иногда взлетают с песней и т. д.) из года в год на одном и том же месте. Гнездятся отдельными парами, охраняя гнездовую территорию. Struthidea cinerea Gid. гнездятся группами: по нескольку гнезд на одном дереве. У Согсогах melanorhamphus (V.) в постройке гнезда обычно участвует несколько самок, продолжающих держаться около гнезда в период насиживания и выкармливания птенцов. В кладке бывает до 8 яиц. Предполагают, что либо в общее гнездо кладут по 1—2 яйца, все участвующие в постройке самки, либо только 1—2 «доминирующие» самки.
Встречаются в открытых местообитаниях с отдельными кустами и деревьями. Кормятся на земле и в ветвях, поедая разнообразных насекомых, семена, мелкие плоды. Вне сезона размножения держатся стаями, иногда в несколько сотен особей.
СЕМЕЙСТВО 25. PTILONORHYNCHIDAE — БЕСЕДКОВЫЕ ПТИЦЫ, ИЛИ ПТИЦЫ ШАЛАШНИКИ
По внешнему облику (рис. 191) и размерам напоминают дроздов (длина 25—40 см). Распространены в сырых прибрежных и горных лесах Новой Гвинеи и Австралии.
279
Клюв сильный, короткий или несколько удлиненный, слабо изогнутый, иногда с маленьким крючком на конце надклювья. Основание клюва обычно окаймлено мелкими бархатистыми перышками, прикрывающими наружные отверстия ноздрей или не доходящими до них. Первостепенных и второстепенных маховых по 10. Рулевых 12. Крылья и хвост относительно короткие. Ноги сильные. Окраска разнообразная: от яркой с металлическим отливом (сине-черной, желтой и т. п.) до покровительственной коричневатобурой; в последнем случае часто у самцов есть яркие хохлы. Половой диморфизм выражен более или менее отчетливо. Вылетевшие из гнезд птенцы окрашены как самки. Издают громкие крики, часть видов способна к пере-смешничеству.
Очень своеобразен ток. Задолго до начала размножения самец расчищает токовую площадку диаметром в 1—3 м и украшает ее цветными камешками, ракушками, косточками, яркими цветками и ягодами; увядшие цветки и ягоды периодически заменяются свежими (часть видов родов А Пи-roedus, Atnblyornis, Archboldia, Scenopoeets). Самцы других видов, расчистив площадку, строят на ней шалашик из веточек и стеблей травы высотой до 30—50 см и длиной стенок иногда почти до 1 л«.Стенки и пол шалаша, а также площадка перед ним украшаются камешками, раковинами, косточками, перьями, цветками и т. п. Самец постоянно держится около токовой площадки или шалаша, охраняя эту территорию. Перед началом размножения стайки самок распадаются, и самки появляются на участках самцов. Образовавшаяся пара держится преимущественно около токовой площадки или шалаша; здесь идут токовые танцы и спаривание. Затем самка где-то поблизости на дереве строит открытое гнездо, в которое откладывает обычно 2 (реже 1 или 3) яйца. У разных видов окраска яиц варьирует от однотонной до резко пятнистой. Насиживает кладку и выкармливает птенцов, видимо, только самка. У кошачьих птиц (они издают мяукающие звуки; роды А Пи-roedus, Scenopoeetes) самец помогает самке в выкармливании птенцов. У остальных видов самцы продолжают держаться у своих токовых площадок и
Рис. 191. Воротничковая беседковая птица Chlamydera maculata (Gid,); самец и самка у шалаша
280
шалашей. Возможно, у части видов есть полигения, и самец по очереди токует с несколькими самками.
Обычно оседлые или ограниченно кочующие птицы. Вне сезона размножения держатся небольшими стайками, кормясь на земле и в кронах. Пища разнообразная: мелкие плоды, семена, насекомые и их личинки, наземные моллюски, черви и т. п.
Семейство Ptilonorhynchidae включает 17 видов, относимых к 8 родам: Ptilonorhynchus, Ailuroedus, Atnblyornis, Archboldia, Chlamydera, Prio-nodura, Scenopoeetes, Sericulus. На Новой Гвинее встречаются виды 6 родов, в Австралии 5. Некоторые систематики включают беседковых птиц в ранге подсемейства в семейство райских птиц.
СЕМЕЙСТВО 26. PARADISAEIDAE — РАЙСКИЕ ПТИЦЫ
Разнообразны по внешнему облику (рис. 192), размеры — от большой синицы до галки. Очень подвижные птицы густых лесов Новой Гвинеи и прилегающих островов; несколько видов проникает в лесные районы Северной и Восточной Австралии (см. рис. 187).
Клювы разнообразной формы: от коротких, толстых, похожих на клювы вьюрков, до удлиненных, тонких, более или менее сильно изогнутых кни-
Рис. 192. Токующий самец большой райской птицы Paradisaea apoda L.
281
зу. Основание клюва покрыто короткими бархатистыми перышками, у части видов прикрывающими наружные отверстия ноздрей. Крылья относительно короткие и широкие. Первостепенных и второстепенных маховых по 10. Рулевых 12. Ноги довольно короткие. Задний палец обычно длиннее направленных вперед. У немногих видов самцы похожи на самок и имеют скромную окраску. Для подавляющего большинства видов характерен резко выраженный половой диморфизм. Окраска самок более или менее однотонная, с преобладанием бурого, коричневого, зеленоватого тонов. Окраска самцов очень яркая, с сильным развитием металлического блеска. Характерны украшающие перья, отличающиеся особенно яркой окраской, структурой, длиной. Они образуют хохлы, разной формы воротники, длинные пучки на крыльях, на боках тела, над хвостом и т. п. Часть рулевых иногда тоже превращается в украшающие перья (у одного вида средние рулевые превышают длину туловища более чем в 10 раз). Голос — хриплое карканье; в период тока обычно издают громкие резкие крики (у некоторых видов длинная трахея образует под кожей в области вилочки петлю, резонирующую звуки).
Немногие виды (Manucodia, Macgregoria, Phonygarntnus) co слабо выраженным половым диморфизмом — типичные моногамы: самец помогает самке в насиживании и выкармливании птенцов. Подавляющее большинство видов райских птиц характеризуется резким половым диморфизмом и типичной полигамией. Для самцов характерен групповой ток: до 10—20 самцов собираются в кроне одного дерева, принимают разнообразные позы, меняют положение украшающих перьев (рис. 192), издают громкие крики, взлетают, вновь садятся и т. п. Самки подлетают к месту тока и происходит случайное спаривание. Пары — даже на короткое время — не образуются. Кратковременность общения самца и самки приводит к относительно частой гибридизации (межвидовой и даже межродовой). Самка строит чашеобразное гнездо в густой кроне дерева или куста. Некоторые виды гнездятся в дуплах. В кладке 1—2 палевых или коричневатых яйца, с темными полосами или пятнами. Насиживает кладку и выкармливает птенцов только самка. Вне периода размножения держатся обычно поодиночке; у части видов самцы ведут одиночный образ жизни, а самки и молодые держатся мелкими стайками. Оседлы или кочуют в ограниченном районе.
Держатся в густых лесах и зарослях кустарников; часть видов свойственна только горным лесам. Кормятся в кронах; некоторые виды, подобно дятлам, собирают корм на стволах деревьев. Немногие виды собирают корм и на земле. Практически всеядны. Поедают различные плоды и семена; собирают на листве и ветвях насекомых и их личинок, пауков, моллюсков; при случае ловят мелких древесных лягушек и ящериц. Некоторые виды с тонкими изогнутыми клювами пьют нектар.
Семейство Paradisaeidae объединяет 39—43 вида, относимых к 20 родам: Paradisaea, Astrapia, Cicinnurus, Cnemophilus, Craspedophora, Diphyllodes, Drepanornis, Epimachus, Lophomia, Loria, Lycocorax, Macgregoria, Manu-codia, Paradigalla, Parotia, Phony gammas, Pteridophora, Ptiloris, Seleuci-dis, Semioptera.
Подавляющее большинство видов встречается только на Новой Гвинее и прилегающих островах; виды 2 родов проникают в Австралию и виды 2 родов — на Молуккские острова. У части видов очень маленькие ареалы (ограниченные площадью какого-нибудь небольшого острова). Издавна преследовались местными жителями: яркие перья использовали для украшений (символ власти вождя, ритуальные маски и прически и т. п.). В конце XVIII — начале XX вв. большое число шкурок отправлялось в Европу, где их употребляли для украшения женских нарядов. Все это привело к резкому падению численности и, видимо, к исчезновению ряда видов. Сейчас находятся под охраной, но ее действенность весьма относительна.
Райские птицы, несомненно, близки к беседковым птицам. Некоторые систематики объединяют эти 2 группы, присваивая им ранг подсемейств, в
282
общее семейство райских птиц Paradisaeidae. По многим чертам внутреннего строения райские птицы близки к вороновым Corvidae, что, видимо, свидетельствует о родстве, хотя и давнем.
СЕМЕЙСТВО 27. PARIDAE — СИНИЦЕВЫЕ
Очень подвижные мелкие птички (длина 10—20 см, вес 7—25 г) с характерным обликом синиц (рис. 193). Населяют леса разных типов, сады, пар
ки; часть видов по кустарниковым, очень немногие по камышовым зарослям проникают в открытые ландшафты. Распространены в Европе, Азии, Африке южнее Сахары (нет на Мадагаскаре) и в Северной Америке (рис. 194).
Клюв сильный, конический, со слабо изогнутым коньком. У основания надклювья растут густые перышки или щетинки, прикрывающие округлые ноздри. Крылья короткие, округлые, с 10 первостепенными маховыми, из которых первое сильно укорочено. Хвост средней длины, из 12 рулевых. Ноги сильные, пальцы относительно короткие; у части видов покровы основных фаланг более или менее срастаются. Когти крепкие, сильно изогнутые, острые; очень мощный коготь заднего пальца. Оперение мягкое, пушистое. Окраска самцов и самок сходна; молодые более тусклые. В году одна
линька: полная послегнездовая, у ремезов есть вторая частичная весенняя.
Все виды, кроме ремезов, гнездятся в дуплах (естественных или выдолбленных дятлами; некоторые виды в гнилых трухлявых пнях сами выдалбливают дупла), реже за отставшей корой, в густых мутовках ветвей, трещинах в обрывах, щелях построек и т. п. Гнездо устраивают из растительной ветоши, лоток выстилают тонкими стебельками, шерстью, перьями, паутиной и т. п. Ремезы Anthoscopus, Auriparus, Remiz из растительных волокон и пуха тополей ц ив плетут прочные мешкообразные гнезда с боковым входом, подвешивая их на тонких веточках, часто над водой. Кладки у большинства видов большие — до 6—10, даже до 16 яиц, только у некоторых тропических видов лишь из 3—5 яиц. Яйца белые или с отчетливым красноватым крапом. Насиживание начинается после завершения кладки и продолжается около 2 недель. Видимо, у всех видов насиживает только самка, самец ее кормит. Птенцов выкармливают оба партнера: в первые дни только нежной животной пищей (пауки, поденки и т. п.), потом самыми различными насекомыми, в том числе гусеницами, бабочками, жуками. Птенцы вылетают из гнезда в возрасте 2,5—3 недель. У части видов в сезон 2 кладки. Вне сезона размножения держатся стайками, часто вместе с другими видами синиц, с поползнями и др. Оседлые или кочующие, немногие виды настоя
щие перелетные.
Кормятся, внимательно Пища преимущественно животная, главным образом различные насекомые, их кладки и личинки, пауки и др. Поедают семена и ягоды. При обилии мелких семян некоторые виды делают запасы: прячут семена в трещины коры, под лишайники. Обычно эти запасы используют другие особи, попавшие в этот район уже в период недостатка кормов. Очень полезны истреблением вредных насекомых. Многие виды легко привлекают в культурный ландшафт развеской искусственных гнездовий.
обыскивая ветви деревьев и кустарников.
Рис. 193. Синица московка Parus ater L.
283
150° 120° 90° 60°к западу от Гринвича 0°к востоку от Гринвича60° 9СР 120° 150°________180°_________150°
Рис. 194. Ареалы семейств певчих воробьиных:
1— синицевые Paridae, 2— поползневые Sittidae, 3— пищуховые Certhiidae
Объем семейства по представлениям разных систематиков резко варьирует. Более часто сейчас в семейство Paridae вводят60—65 видов, относимых к 9 родам: Parus (объединяет до 27—29 видов; сюда в качестве подродов включают роды Cyanistes, Lophophanes, Penthestes, Periparus, Poecile), Anthoscopus, Auriparus, Cephalopyrus, Melanochlora, Psaltiparus, Psaltria, Remiz, Sylviparus.
Центр возникновения, видимо, Палеарктика. По ископаемым остаткам из верхнего эоцена (Франция) описан один ископаемый род. Остатки из более поздних отложений относят к 8 ныне живущим видам.
В СССР встречается и гнездится 13 видов 2 родов (Parus и Retniz).
СЕМЕЙСТВО 28. SITTIDAE — ПОПОЛЗНЕВЫЕ
Рис. 195. Поползень Sitta europaea L.
Мелкие птицы (длина 10—20 см-, вес 15—40 г), по внешнему облику похожие на поползней (рис. 195). Распространены в лесах разных типов, некоторые виды населяют безлесные скалистые участки в горах. Живут в Европе, Азии, на ряде островов Океании и в Северной Америке (рис. 194).
Голова довольно крупная. Клюв длинный, мощный у основания, с острой вершиной, прямой или чуть изогнутый. Ноздри прикрыты жесткими перышками. В углах рта есть короткие, направленные вперед щетинки. Крылья умеренной длины, с 10 первостепенными маховыми. Хвост короткий, из 12 рулевых. Лапы сильные, с крепкой цевкой, довольно длинными пальцами, несущими мощные, сильно изогнутые острые когти. Оперение довольно рыхлое. В году одна линька (полная послебрачная). Половой диморфизм окраски выражен слабо; молодые немного тусклее взрослых. В окраске верха преобладают голубовато-серые или голубовато-зеленые тона.
Гнездятся в укрытиях: дуплах, трещинах скал. Обычно уменьшают вход в укрытие, замазывая его грязью; некоторые горные виды делают закрытые с боковым входом гнезда из грязи. Внутри гнезда подстилка из шерсти, перьев (часто этот материал берут из разрушенных погадок хищных птиц), гнилой трухи, кусочков коры И'Т. п. Поползни рода Neositta в развилке ветвей строят открытые сверху толстостенные гнезда. Кладки из 4—8, редко больше белых яиц, обычно с красновато-коричневым крапом. Насиживает только самка. Инкубация 2— 2,5 недели. Птенцов кормят оба родителя обычно разнообразными насекомыми. Молодые вылетают примерно через 3 недели после вылупления. Вне сезона размножения часто присоединяются к стайкам синиц. Взрослые особи у многих видов оседлы и в течение круглого года держатся на своем гнездовом участке. Питаются пауками, насекомыми, их личинками и яйцами, разыскивая их в трещинах коры (легко лазают по вер-
285
тикальным стволам), пазухах листьев, трещинах скал. Поедают разнообразные семена и плоды. Скорлупу орехов раскалывают сильными ударами клюва, предварительно забив орех в какую-нибудь щель. При обилии кормов семена, орехи прячут в трещинах коры, под лишайниками и т. п.
Объем семейства по представлениям разных систематиков очень варьирует. Сейчас обычно сюда включают 23—35 видов, относимых к 3 родам: Sitta (16—18 видов), Neositta (6 видов), Daphoenositta (монотипичный род). В СССР встречается и гнездится 3—4 вида одного рода Sitta (малого и большого скалистых поползней часть систематиков считает за подвиды одного вида).
Поползни очень близки к семейству пищух Certhiidae, с которым нередко их объединяют (обе группы получают ранг подсемейств). Иногда в семейство поползневых включают все роды семейства пищух (т. е. стенолазов, австралийских пищух и др.), кроме номинального рода Certhia, образующего в этом случае монотипичное семейство.
СЕМЕЙСТВО 29. HYPOSITTIDAE — КРАСНОКЛЮВЫЕ ПОПОЛЗНИ
В это семейство выделяют единственный вид Hypositta coralliirostris Newt., населяющий леса Мадагаскара. По внешнему облику (рис. 196) похож на поползня.
Рис. 196. Красноклювый поползень Hypositta coralliirostris Newt.
Характерен рубиново-красный короткий клюв с небольшим крючком на конце и зазубринами по краю надклювья. Окраска из сочетания зеленого, голубого и черного цвета; самка бледнее. Кормятся в кронах. Не могут, как поползни, спускаться по стволу вниз, но ловко, как пищухи, поднимаются по стволам вверх.
Иногда включают в семейство Sittidae. Другие систематики вводят в семейство Vangidae.
СЕМЕЙСТВО 30. CERTHIIDAE — ПИЩУХОВЫЕ
Мелкие (длина 10—16 см, вес 10—30 а), стройные, подвижные птички (рис. 197). Населяют леса разных типов, реже скалистые безлесные участки гор Центральной Африки, Европы, Азии, Австралии, Северной Америки (рис. 194).
Клюв удлиненный, довольно тонкий, слабо или заметно изогнутый книзу. Ноздри щелевидные, сверху прикрыты роговыми крышечками. Щетинок в углах рта нет. Лапы крепкие с длинными пальцами и сильно изогнутыми острыми когтями. Коготь заднего пальца иногда длиннее пальца и
286
заметно длиннее остальных когтей. Крылья короткие, закругленные; первостепенных маховых 10. Рулевых 12. Хвост средней длины из обычных рулевых или удлиненный, конический из жестких (почти как у дятлов), заостренных рулевых. Оперение рыхлое, у части видов шелковистое. Половой диморфизм окраски не выражен; молодые похожи на взрослых.
Гнездятся в укрытиях: в дуплах, за отставшей корой, трещинах скал и т. п. Гнездо из растительной ветоши, лоток обычно мягкий, выложенный тоненькими стебельками, перьями, шерстью. В кладке 2—8 белых с крапом яиц. Насиживает, видимо, у всех видов только самка. Инкубация немногим более 2 недель. Птенцов кормят оба родителя. Молодые вылетают в возрасте около 2,5 недель Вне сезона размножения кочуют, обычно поодиночке; часто присоединяются к кочующим стайкам синиц. Кормятся насекомыми и другими беспозвоночными, собирая их на стволах и ветвях, на обрывистых скалах. Легко перемещаются (обычно снизу вверх) по вертикальным стволам. Лишь при бес-
Рис. 197. Обыкновенная пищуха Certhia familiar is L.
кормице поедают мелкие семена.
Объем семейства пищух у разных систематиков очень варьирует. Чаще сюда включают 15 видов 5 родов: Certhia (5 видов), CUmacteris (6 видов), Rhabdorrtis, Salpornis, Tichodroma. В СССР встречаются и гнездятся 4 вида: 3 вида пищух рода Certhia и единственный вид рода — краснокрылый стенолаз Tichodroma muraria (L.).
Иногда часть родов или все роды, кроме Certhia, из этого семейства изымают и включают в семейство поползневых. Тогда в семействе остается только род пищух Certhia с 5 видами. Некоторые систематики и пищух в ранге подсемейства включают в семейство поползневых Sittidae.
СЕМЕЙСТВО 31. PARADOXORNITHIDAE — ТОЛСТОКЛЮВЫЕ СИНИЦЫ
Мелкие длиннохвостые птички (вес 8—20 г, длина 10—20 см, причем примерно половина ее приходится на хвост; рис. 198), населяющие лесные опушки, кустарниковые, бамбуковые, тростниковые и камышовые заросли Европы и Азии (рис. 200). По внешнему облику , а часто и по повадкам похожи на синиц.
Клюв короткий, вздутый, без крючка на конце. Режущий край клюва волнистый. Ноздри прикрыты мелкими перышками или роговыми крышечками. Первостепенных маховых 10; первое очень короткое. Крыло короткое, закругленное. Хвост длинный, более или менее ступенчатый. Ноги сильные, довольно длинные. Когти не крупные, но сильно изогнутые. Оперение пушистое, рыхлое. Половой диморфизм окраски выражен лишь у части видов; молодые с более короткими хвостами и тусклее (а иногда темнее), чем взрослые. В году одна линька: полная послегнездовая.
В отличие от синиц, не гнездятся в дуплах. Гнездо помещается в развилке ветвей или в заломе стеблей тростника, реже среди густой травы на земле. Оно имеет вид очень глубокой, толстостенной чаши или рыхлого шара с боковым входом, сплетенного из мелких веточек, мха, сухих стеблей травы и т. п. Лоток выстлан мхом или растительным пухом (метелки тростника, рогоза и т. п.). В кладке 2—7, редко до 12—14 белых или голубоватых яиц
287
Рис. 198. Усатая синица Panurus biarmicus (L.)
ны длиннохвостая Aegithalos caudatus
с темным крапом (красного крапа, как у настоящих синиц, не бывает). Насиживают только самка или оба партнера. Инкубация немногим более 2 недель. Выкармливают птенцов оба партнера. Вылетевшие в возрасте 15—18 дней птенцы кочуют выводками, иногда объединяющимися в более крупные стайки. У некоторых видов 2 кладки в сезон. Преимущественно насекомоядны. Пищу (личинок насекомых и их кладки) добывают, раздавливая (разгрызая) крепкими клювиками прочные стебли, вытаскивают ее из трещин, из пазух листьев. Поедают и разнообразные мелкие семена.
Объем семейства и его положение в системе по разным источникам очень противоречиво. Чаще в семейство Paradoxorni-thidae включают 13—19 видов, объединяемых в 4—5 — родов: Paradoxomis (Cholomis), Aegit-halos, Conostoma, Panurus, Sut-hora (последний иногда включают в род Paradoxomis). В СССР гнездится 4 вида 4 родов. Довольно широко, распростране-(L.) и усатая Panurus biarmicus
(L.) синицы. В Южной части Приморского края встречается бурая сутора Suthora webbiana Gray., а на самом юге Приморского края на оз. Ханка в 1970 г. обнаружена небольшая гнездовая популяция толстоклювой синицы Paradoxomis heudei David.
Иногда семейство толстоклювых синиц в полном объеме включают в другие семейства (Paridae, Timaliidae, Muscicapidae) или исключают из него отдельные роды. Так, часть систематиков до сих пор включает длиннохвостых синиц Aegithalos в ранге подсемейства в семейство синицевых Paridae.
СЕМЕЙСТВО 32. CHAMAEIDAE — КРАПИВНИКОВЫЕ СИНИЦЫ Монотипичное семейство, включающее один вид Chamaea fasciata Gamb. (рис. 199), встречающийся по густым кустарниковым зарослям западного побережья США. Маленькая (длина около 15 см) длиннохвостая синич-
Рис. 199. Крапивниковая синица Cha-таеа fasciata Gamb.
288
Рис. 200. Ареалы семейств певчих воробьиных:
1— толггоклюзые синицы Paradoxornithidae, 2— тимелиевые Tlmaliidae, 3— личинкоедовые Campephagidae
ка, часто высоко поднимающая хвост (отсюда и название). Клюв конический, слабо вздутый у основания, удлиненный. Хвост слабо ступенчатый. Окраска коричневатая.
Глубокое чашеобразное гнездо строят в развилке густых ветвей низко от земли. Вкладке 3—5 яиц. Насиживают по очереди оба партнера. Инкубация около 16 дней. В возрасте 15—17 дней птенцы вылетают из гнезда и вместе с родителями остаются на гнездовом участке около 2 месяцев, потом начинают самостоятельно кочевать. Взрослые оседлы и держатся круглый год на своем участке. В сезон одна кладка. Питаются насекомыми, собирая их на ветвях, листьях и на земле, поедают семена и ягоды.
Иногда этот род включают в семейство толстоклювых синиц или, чаще, в семейство тимелиевых Timaliidae.
СЕМЕЙСТВО 33. TIMALIIDAE—ТИМЕЛИЕВЫЕ, ИЛИ КУСТАРНИЦЫ
Птицы мелких и средних размеров (длина 9—30 см, несколько видов чуть крупнее), по внешнему облику напоминающие славок, дроздов, соек (рис. 201). Распространены в тропиках Старого Света: Африке, Южной Азии, Океании, Австралии (рис. 200); нет в Новой Зеландии. Встречаются преимущественно в зарослях кустарников и в лесах разных типов в горах и на равнинах.
Форма и величина клюва сильно варьирует: у большинства удлиненный, крепкий, заостренный к концу клюв, у нескольких видов—очень длинный, тонкий, сильно изогнутый книзу. Обычно на конце клюва небольшой крючок, а перед ним слабозаметная выемка. Ноздри открытые или полуприкрытые щетинками и перышками. В углах рта жесткие щетинки. Крылья короткие, закругленные. Первостепенных маховых 10; первое не короче половины длины второго. Ноги сильные, довольно длинные. Пальцы крепкие, у части видов короткие. Задний палец примерно равен внутреннему. Когти крепкие, не удлиненные. Хвост из 12 рулевых. У подавляющего большинства видов он длиннее крыла, отчетливо ступенчатый с закругленной вершиной. У немногих видов хвост короткий. Оперение густое, довольно рыхлое. Окраска обычно коричневатая, часто с пестринами; у немногих видов яркая. Редко встречаются хохлы, ярко окрашенные участки голой кожи на голове и т. п. Самцы и самки имеют сходную окраску.
Ведут древесный и наземный образ жизни. Очень подвижные и бойкие, крикливые птицы. Летают неохотно; полет посредственный. Голос —чаще
Рис. 201. Кустарница Garrulax leucolophus (Hard.)
290
несложный свист. Немногие виды хорошие певцы. Гнездо рыхлое, чашеобразное, располагается низко над землей в густых ветвях куста или дерева, реже на земле среди густой травы. В кладке 2—6 яиц, одноцветных белых, голубоватых или пестрых. Насиживают и выкармливают птенцов, видимо, оба партнера. Вне сезона размножения держатся небольшими стайками. Оседлые или кочующие на небольшие расстояния. Кормятся в кронах и, чаще, на земле. Практически всеядны; поедают семена, мелкие плоды, различных беспозвоночных. Немногие виды исключительно насекомоядны.
В семейство Timaliidae включают 260—280 видов, группируемых в 52—57 родов; Timalia, Actinodura, Alcippe, Andropholus, Babax, Chrysotn-ma, Cinclosoma, Crateropus, Crocias, Cutia, Drymodes, Dumetia, Eupetes, Garrulax (27—32 вида), Gampsorhynchos, Heterophasia, Ifrita, Jabouilleia, Kenopia, Leiothrlx, Leonardina, Liocichla, Lioptilus, Macrotius, Malacocinc-la, M. alacopteron, Melampiita, Mesia, Microura, Minla, Moupinia, Myzor-nis, Napothera, Neomixis, Orthonyx, Pellorneum, Phyllanthus, Picathartes, Pnoepyga, Pomatorhinus, Pomatostomus, Proparus, Psophodes, Pteruthius, Ptilocichla, Ptyrticus, Rhopocichla, Rimator, Siva, Spelaeornis, Sphenocichla, Sphenostoma, Stachyris (24 вида), Turdoides (Turdinus), Trichastoma, Xiphi-rynchus, Yuhina.
Центр возникновения, видимо, Южная Азия. На территории СССР в южных районах Средней Азии гнездится один вид — полосатая кустарница Garrulax lineatus (Vig.).
Объем семейства и его положение в системе требуют уточнения. В семейство тимелиевых иногда включают (как подсемейства) толстоклювых синиц Paradoxornithidae и крапивниковых синиц Chamaeidae. С другой стороны, тимелиевых, как подсемейство, иногда вводят в широко понимаемое семейство мухоловковых Muscicapidae.
СЕМЕЙСТВО 34. CAMPEPHAGIDAE — ЛИЧИНКОЕДОВЫЕ
Древесные птицы размером от воробья до галки (длина 9—36 см), по внешнему облику (рис. 202) и повадкам несколько напоминающие мухоловок или сорокопутов. Распространены преимущественно в тропических широтах Старого Света, включая Мадагаскар и Австралию (рис. 200); нет в Новой Зеландии.
Клюв сильный, слабо изогнутый, приплюснутый у основания, с четко выраженным маленьким крючком на конце и маленькой предвершинной выемкой. Ноздри полностью или частично прикрыты щетинковидными перышками. В углах рта по нескольку щетинок. Крылья довольно длинные, со слабо заостренными вершинами. Первостепенных маховых 10; первое короткое. Ноги короткие: плюсна почти такой же длины, как средний палец. Когти короткие, сильно изогнутые. Оперение густое, довольно мягкое. Перья поясницы своеобразны: основная половина стержня жесткая, уплощенная, а верхняя мягкая и тонкая. Функциональное значение такого строения пера не выяснено. Хвост длиннее или чуть короче крыла, обычно ступенчатый. Окраска разнообразная, чаще из сочетания черных, серых, коричневых тонов, но есть и ярко окрашенные виды (черные
Рис. 202. Серый личинкоед Pericrocotus divaricatus Raffl.
291
с красным и т. п.). У большинства видов более или менее отчетливо выражен половой диморфизм окраски (самцы ярче); молодые похожи на самок.
Экология изучена плохо. Держатся в лесах, перелесках, куртинах кустарников; по участкам с деревьями и кустами проникают в горы. Гнездо в виде маленькой тонкостенной чаши укрепляется на тонких ветвях, обычно высоко над землей. Снаружи бывает замаскировано мхом, лишайниками, паутиной. Кладка из 2—5 яиц, пестрой окраски. Возможно, насиживает только самка. Птенцов выкармливают оба родителя. Вне сезона размножения держатся мелкими стайками. Оседлые или кочующие; виды умеренных широт перелетные. Хорошо летают. Ловко двигаются в густых ветвях. Питаются преимущественно насекомыми, которых ловят главным образом двумя способами: птица сидит на ветке и, заметив близко подлетевшее крупное насекомое, взлетает, ловит его в воздухе и вновь садится; прыгая по веткам, собирают корм, склевывая его с листьев, вытаскивая из расщелин коры ит. п.; значительно реже кормятся на земле. В качестве второстепенных кормов используют семена, мелкие плоды и т. п.
Семейство личинкоедовых объединяет 70—72 вида, группируемых в 9—11 родов: Campephaga (сюда включают и Lobotes), Campochaera, Chla-mydochaera, Coracina (обычно включают и Edolisoma, всего почти 50 видов), Hemipus, Lalagea, Pericrocotus, Pteropodocys, Tephrodortiis.
Центр возникновения, видимо, Индо-Австралийская область, где сейчас встречается наибольшее число видов.
На Дальнем Востоке Советского Союза (Приморье, Приамурье) встречается и гнездится один вид: серый личинкоед Pericrocotus roseus Vieil. (Р. divaricatus Raffl.).
СЕМЕЙСТВО 35. PyCNONOTIDAE— БУЛЬБУЛЕВЫЕ, ИЛИ КОРОТКОПАЛЫЕ, ДРОЗДЫ
Древесные и кустарниковые птицы (рис. 203) размером от воробья до дрозда (длина 14—28 см). По внешнему облику похожи на дроздов или славок. Распространены преимущественно в тропиках Африки, Азии, Се-
Рис. 203. Краснощекий бульбуль Pycnonotus jосоsus (G.)
верной Океании; не проникают на Новую Гвинею и в Австралию (рис. 204).
Клюв короткий или средней длины, довольно крепкий, слегка изогнутый по коньку, с маленьким крючком на конце. Овальные ноздри с нависающими крышечками. В углах рта хорошо развиты щетинки. Крылья относительно длинные, с округлой вершиной. Первое первостепенное маховое достигает примерно середины второго. Цевка и пальцы короткие. Кожа очень тонкая. Оперение мягкое, из длинных перьев. По всему телу есть тонкие длинные волосовидные перья, особенно многочисленные на затылке и задней стороне шеи. У части видов на голове хохолок. Окраска не яркая, из сочетания зеленых, жёлтых, коричневых тонов, иногда с белыми, черными, красными пятнами. Полового диморфизма окраски нет; у части видов самцы несколько крупнее самок. Молодые по окраске похожи на взрослых.
Населяют тропические леса и их опушки, рощи и куртины кустов в открытых
292
Рис. 204. Ареалы семейств певчих воробьиных:
1— короткопалые дрозды Pycnonotidae, 2— оляпковые Cinclidae, 3— крапивниковые Troglodytidae
ландшафтах и в горах, сады и парки. Гнезда открытые, чашеобразные помещаются среди густых ветвей невысоко над землей (1—3 м), лишь у некоторых видов в кронах высоких деревьев. В кладке 2—3, у немногих видов до 4—5 яиц. Их окраска белая или розоватая, с коричневым крапом. В период размножения часть видов очень скрытны, другие крикливы и держатся на виду. Некоторые виды очень хорошо поют. Вне сезона размножения держатся стайками. Оседлы, но виды, проникающие в горы или в умеренные широты, кочующие или перелетные. Летают хорошо. Очень подвижны. Питаются мелкими плодами, семенами; в качестве второстепенных кормов используют различных беспозвоночных, склевывая их на ветвях, листьях, реже на земле. Немногие виды преимущественно насекомоядны.
В семейство бульбулевых вводят 119 видов, объединяемых в 15—21 род: Pycnonotus (включает около 50 видов, в том числе и виды р. Molpaster и р. Microscelis), Andropadus (11 видов), Baeopogon, Bernieria, Bleda, Calyp-tocichla, Chlorocichla, Criniger (10 видов), Euptilosus, Hypsipetes, I ole, Ixonotus, Malta, Ncolestes, Nicator, Otocompsa, Phyllastrephus, Rubigula, Spizixos, Thescelocichla, Tricholestes. Большинство видов и родов населяют только Африку, 2—3 рода — Азию и 3 рода — встречаются в Африке и Азии. Вероятный центр возникновения — Африка.
В СССР встречаются 2 вида: Pycnonotus leucogenys (Gr.) отмечен на залетах в южных районах Средней Азии, a Pycnonotus (Microscelis) amauro-tis (Temm.), видимо, в небольшом количестве гнездится на Южном Сахалине и на острове Кунашир (юг Курильской гряды).
СЕМЕЙСТВО 36. PALAEOSCINIDAE
По остаткам из среднего миоцена Калифорнии описан род Palaeoscinis, возможно, близкий к короткопалым дроздам, или, по другим взглядам, к оляпкам и крапивникам. Пожалуй, выделение этого семейства малообоснованно.
СЕМЕЙСТВО 37. CHLOROPSEIDAE—ЛИСТОВКОВЫЕ
Мелкие древесные птицы (длина 10—22 см), встречающиеся в Юго-Восточной Азии (от Индии и Южного Китая до Филиппин). По внешнему виду (рис. 205) похожи на славок. По морфологическим особенностям и экологии близки к бульбулевым. Отличаются более стройным клювом с ме-
Рис. 205. Желтолобая листовка Chloropsis aurifrons (Т.)
нее развитым крючком на конце, большей длинноногостью, более острыми крыльями, отсутствием скоплений длинных волосовидных перьев на отдельных участках тела и другими деталями. Окраска ярко-зеленая с примесью других цветов; самцы ярче самок, реже половой диморфизм слабо выражен.
Подвижные птицы, держащиеся в кронах деревьев и кустов. Населяют леса разных типов, заросли кустарников, рощи, сады. Открытые, чашеобразные гнезда строят в развилках тонких ветвей. В кладке 2—4 пятнистых яйца. Вне сезона размножения держатся поодиночке или мелкими груп
294
пами; оседлы. Питаются в кронах, преимущественно мелкими плодами; меньший вес в питании занимают семена и различные насекомые, склевываемые с ветвей и листьев.
Семейство включает всего 12 видов 2 родов: Chloropsis (7 видов) и Aegi-thina (5 видов). Центр возникновения, видимо, Юго-Восточная Азия. Иногда эти два рода объединяют вместе с родом Irena, исключая его из семейства Oriolidae, в семейство Irenidae.
СЕМЕЙСТВО 38. CINCLIDAE — ОЛЯПКОВЫЕ
Семейство включает всего один род Cinclus с 4—5 видами, распространенными по берегам быстро текущих прозрачных ручьев и речек с каменистым дном в горных и предгорных районах Европы, Азии, запада Северной и Южной Америки; локально встречаются в немногих районах северо-западной Африки (рис. 204). По берегам водоемов с медленным течением и мутной водой не держатся. В СССР встречаются и гнездятся 2 вида.
Размерами с дрозда (вес 50—80 г), тело плотное, коренастое (рис. 206). Клюв прямой, удлиненный, довольно тонкий, без крючка на конце. Узкие щелевидные ноздри прикрыты сверху кожистыми крышечками. Щетинок в углах рта нет. Наружные ушные отверстия с кожистым клапаном. Скелет слабо пневматизирован. Крылья короткие, закругленные; первое первостепенное маховое не длиннее кроющих кисти. Ноги сильные, с несколько удлиненной цевкой и крепкими короткими пальцами. Никаких следов плавательных перепонок нет. Когти короткие, прочные. Хвост короткий, из 12 рулевых. Оперение густое, плотно прилегающее. В отличие от других воробьиных на аптериях хорошо развит пух. Окраска из сочетания белых и темно-бурых или темно-коричневых цветов. Самцы и самки окрашены сходно. В году одна линька (полная, послегнездовая). Молодые в первую осень жизни меняют пестроватый гнездовой наряд на окончательный.
Обычно имеют довольно четкие гнездовые участки. Большое шарообразное гнездо с боковым входом помещается около воды: у основания ствола, на уступе или в трещине скалы, в промоине берегового обрыва. Гнездо строится из мха, сухих листьев и травинок. В кладке 4—7 белых яиц. Насиживает преимущественно самка; птенцов в гнезде и некоторое время после вылета кормят оба родителя. Вне сезона размножения держатся поодиночке
Рис. 206. Оляпка Cinclus cinclus (L.)
295
вблизи гнездовых участков или откочевывают к незамерзающим речкам; северные популяции перелетные. Пища — преимущественно насекомые и их личинки, отчасти моллюски и ракообразные, которых собирают по берегам водоемов или на дне во время ныряния. Иногда в небольшом количестве ловят мелких рыбешек. Ныряющая птица держит крылья так, что течение прибивает ее ко дну; ныряют на глубину до 1—1,5 м и пробегают по дну до 10—20 м, оставаясь под водой до 15—20 сек. Когда птица складывает крылья, ее, как пробку, выбрасывает на поверхность, и она взлетает. Нырять в стоячую воду или в воду с медленным течением оляпки не могут. Полет быстрый, стремительный. Обычно летят низко над водой или берегом, следуя изгибам русла. Хорошо, легко бегают. Изредка плавают; легко взлетают с воды.
Единственная ныряющая группа воробьиных птиц. Центр возникновения предположительно Северная Америка, хотя палеонтологические остатки обыкновенной оляпки найдены в Австрии (верхний плейстоцен).
Оляпки близки к крапивникам; иногда эти семейства объединяют в общее семейство крапивниковых Troglodytidae, придавая им ранг подсемейств. Возможно, что более давние родственные связи есть между оляпками и дроздовыми.
СЕМЕЙСТВО 39. TROGLODYTIDAE — КРАПИВНИКОВЫЕ
Довольно коренастые (рис. 207) мелкие птички (длина 9—22 см), населяющие преимущественно леса с подлеском или кустарниковые заросли. Некоторые виды (и часть подвидов широко распространенных видов) живут на безлесных скалистых морских побережьях и в горах, в пустынях и полупустынях с чахлой кустарниковой растительностью или зарослями кактусов.
Клюв удлиненный, тонкий, заостренный, без крючка на конце, иногда несколько изогнутый книзу. Ноздри открытые, иногда с чуть нависающими над ними кожистыми крышечками. Щетинки в углах рта слабо развиты или их нет. Цевка длиннее пальцев. Задний и средний пальцы почти равны по длине. Когти большие, сильно изогнутые, острые. Крылья короткие, с закругленной вершиной. Первостепенных маховых 10; первое обычно не короче половины второго. Хвост из 12 рулевых, короче или — иногда — длиннее крыла, прямо обрезанный или ступенчатый. Птицы часто поднимают хвост кверху, иногда почти под прямым углом к телу. Оперение мягкое, густое, прилегающее. Окраска с пре-
Рис. 207. Крапивник Troglodytes troglodytes (L.)
обладанием коричневых тонов, на брюшной стороне более светлая. У части видов развита поперечная исчер-ченность перьев. Половой диморфизм выражен у немногих видов; обычно самец, самка и молодые птицы сходны по окраске. В году одна полная послегнездовая линька.
Очень подвижные птицы; несмотря на это многие виды держатся довольно скрытно. Летают хорошо, но обычно на короткие расстояния. Быстро и легко двигаются по земле и в кустах. У многих видов звонкая, мелодичная песня; поют и вне сезона размножения. У части видов хорошо поет и самка: партнеры на гнездовом участке исполняют своеобразный дуэт. Самец занимает большой гнездовой участок (площадью в несколько гектар)
296
и строит несколько «холостяцких» гнезд, в которых ночует. Большинство видов моногамы, но у части видов (а у некоторых лишь у отдельных популяций) более или менее выражена факультативная или постоянная полигамия: на участке самца поселяются 2, изредка даже 3 самки. Они достраивают «холостяцкие» гнезда и откладывают в них яйца. Гнезда по сравнению с размерами птиц очень массивны. Это толстостенная шарообразная постройка с боковым входом, свитая из стеблей травы, листьев папоротника, мха, тоненьких веточек. Гнездо помещается в густой кроне куста или невысокого дерева, в выворотах корней, в промоинах, в трещинах скал, иногда в больших дуплах или за отставшей корой. В тропиках в кладке 2—5 яиц, у более северных видов — до 8—10. Яйца белые или голубоватые, у части видов с темным крапом. Насиживает самка около 2 недель. В выкармливании птенцов участвует и самец, даже если у него на участке гнездится несколько самок. Птенцы вылетают из гнезда через 16—18 дней после вылупления и вскоре начинают вести самостоятельную жизнь. У некоторых видов в году 2 кладки.
Вне периода размножения держатся одиночками или парами. В тропиках оседлы; самец в течение всего года охраняет свой гнездовой участок. В умеренных широтах кочующие или перелетные. Питаются разнообразными беспозвоночными (насекомыми и их личинками, пауками, моллюсками, червями и т. п.), собирая их на земле и ветвях; на морских побережьях часто кормятся у заплеска волн, хватая выброшенных прибоем мелких ракообразных. В качестве второстепенных кормов используют ягоды, реже семена.
В семейство Troglodytidae включают 59—63 вида, объединяемых в 12—16 родов: Troglodytes, Campylopterus, Campylorhynchus, Cathereps, Cinnicerthia, Cistothorus, Ferminia, Hetiicorhina, Microcercuius, Salpinc-tes, Tematodytes, Thryomanes, Thryothorus, Uropsila и др. Практически все виды крапивниковых свойственны только Новому Свету — Северной и Южной Америке, причем большинство видов приурочено к тропикам. Так, на территории США встречается 10 видов, а в маленькой Коста-Рике — 22 вида крапивников. Лишь один вид — обыкновенный крапивник Troglodytes troglodytes (L.), живущий во многих районах Северной Америки, проник в Старый Свет, широко расселившись по Евразии (он встречается и на большей части территории СССР) и по некоторым районам Северной Африки (рис. 204). Для широко распространенных видов характерна клинальная изменчивость, но у части видов образуются более или менее дифференцированные географические формы, например, у обыкновеного крапивника выделяют до 38 подвидов.
Центр возникновения, видимо, Северная Америка. Ископаемые остатки обнаружены в отложениях среднего плейстоцена Флориды (США); их относят к одному ископаемому и к одному ныне живущему виду. Крапивни-ковые близки к оляпкам, с которыми их нередко объединяют в одно семейство.
СЕМЕЙСТВО 40. MIMIDAE— ПЕРЕСМЕШНИКОВЫЕ
Птицы размером со скворца или дрозда (длина 20—30 см), со стройным телом и длинным хвостом (рис. 208) внешне, пожалуй, похожие на крупных камышевок или дроздов. Распространены только в Новом Свете: от южных районов Канады до юга Южной Америки.
Клюв стройный, умеренной длины, обычно слабо изогнутый, но у части видов сильно удлинен и изогнут (похож на клюв пищух). Крылья короткие, широкие. Лапы сильные, с длинными, довольно толстыми цевками и крепкими длинными пальцами; когти крепкие, слабо изогнутые. Оперение рыхлое, мягкое. Окраска коричневатых или сероватых тонов, иногда с крапом. Полового морфизма окраски нет; у части видов самцы немного крупнее самок. Молодые похожи на взрослых.
297
Рис. 208. Коричневый пересмешник Тoxostoma rufum (Т.)
Большинство видов предпочитает открытые пространства (степи, полупустыни, культурный ландшафт) с одиночными деревьями и куртинами кустов; встречаются по опушечным участкам, реже в глубине леса. Многие виды — хорошие певцы с очень развитым подражанием и имитацией разнообразных звуков. Поют и вне сезона размножения. У части видов хорошо поют и самки. Открытое чашеобразное гнездо помещается в кроне дерева или куста невысоко над землей (0,5—3 м), редко на земле. У некоторых видов в году 2, иногда даже 3 кладки. Яйца голубоватые, зеленоватые или беловатые, с крапом или без него. В кладке 2—7 яиц. Насиживает преимущественно самка (у части видов только самка). Инкубация около 2 недель. Птенцов выкармливают оба партнера. Молодые оставляют гнездо в возрасте около 2 недель. Вне сезона размножения держатся парами на гнездовых участили кочуют поодиночке. Кормятся в
ках (оседлые виды теплых широт) кронах и на земле. Практически всеядны: пищей служат ягоды, семена, различные мелкие беспозвоночные.
Семейство объединяет 30—31 видов, распределяемых по 13 родам: Mimas (10 видов), Allenia, Cinclocerthia, Donacobius, Dumetella, Margarops, Melanopttla, Melanotis, Mimodes, Nesomimus, Oreoscoptes, Ramphacinclus, Toxostoma (10 видов). 6 из ныне живущих видов найдены в отложениях верхнего плейстоцена. Центр возникновения, видимо, Северная Америка, хотя сейчас большее число видов свойственно Центральной и Южной Америке (в США встречается 10 видов).
Семейство Mimidae близко по многим деталям строения к дроздовым Turdidae и к крапивниковым Troglodytidae.
СЕМЕЙСТВО 41. TURDIDAE— ДРОЗДОВЫЕ
Наземные и древесные птицы размером от синиц до дрозда (длина 10— 40 см, вес 10—200 г), с обликом дрозда (рис. 209) каменки, чекана, соловья, горихвостки и т. п. Распространение почти космополитическое (рис. 210): отсутствуют только, в Антарктике, в наиболее суровых районах Арктики, части Австралии и на немногих океанических островах; отсутствовали в Новой Зеландии, но сейчас там акклиматизировано несколько видов.
Тело стройное, несколько удлиненное. Голова небольшая. Клюв сильный, но тонкий; слегка изогнутый, он несколько сжат с боков, конек надклювья выпуклый, ноздри открытые. Щетинки в углах рта развиты слабо или их нет. Крылья умеренной длины, со слабо заостренными вершинами. Первостепенных маховых 10, из них первое укорочено, мало выдается из-под кроющих кисти или совсем закрыто ими. Хвост различной длины, но не бывает очень коротким. Обычно он прямо обрезан, реже выемчатый или ступенчатый. Рулевых 12, у немногих видов 14. Цевка удлиненная, но не толстая. Щитки, покрывающие цевку спереди, обычно полностью или частично сливаются. Пальцы крепкие; задний палец короче среднего. Оперение довольно плотно прилегающее или рыхлое. Окраска разнообразная: от скромной до яркой. У многих видов резкий половой диморфизм окраски;
298
Рис. 209. Дрозд рябинник Turdus pilaris L.
самцы немного крупнее самок. Гнездовой наряд всегда с пестринами, птенцы его сменяют во время осенней линьки. У видов с резким половым диморфизмом самцы участвуют в размножении в возрасте около года в оперении, похожем по окраске на самку; полный самцовый наряд одевают уже после первого размножения. У взрослых одна линька в году — полная послегнез-довая; хорошо выраженная у части видов брачная окраска образуется без линьки в результате снашивания вершин пера в период зимовки.
Живые, подвижные птицы. Большинство с дневной активностью, некоторые с сумеречной. Заселяют разнообразные ландшафты. Большинство видов населяет леса самых различных типов и кустарниковые заросли; другие виды встречаются по безлесным районам: в горах, тундре, степях, пустынях, на морских побережьях. Самцы многих видов хорошо поют. Гнезда чашеобразные, иногда с обмазкой стенок грязью или скрепленной слюной древесной трухой. Помещаются в развилке ветвей на деревьях и кустарниках или на земле, под прикрытием травы или кочки. Некоторые виды устраивают гнезда укрыто: в расщелинах скал и под камнями, в норах грызунов. В постройке гнезда и в насиживании обычно большее участие принимает самка; у части видов, возможно, только самка. Яйца беловатой, голубоватой или зеленоватой окраски, одноцветные или пятнистые. В кладке 3—6 яиц, редко больше. У части видов 2 кладки в сезон. Инкубация около 2 недель. Птенцов выкармливают оба партнера, принося им различных беспозвоночных. Молодые вылетают из гнезда в возрасте около 2 недель и некоторое время держатся вместе с родителями. Подавляющее большинство видов перелетные или кочующие. Вне сезона размножения часть видов держится стайками,’другие поодиночке.
Питаются разнообразной животной пищей (насекомые и их личинки, наземные моллюски, черви и т. п.), которую собирают на земле и в ветвях. Поедают мелкие плоды, семена; часть видов вне сезона размножения преимущественно растительноядны.
Объем семейства в трактовке разных систематиков очень различен и нуждается в уточнении. Обычно сюда включают 297—303 вида (из них 2 недавно вымерли), относя их к 40—70 родам (часть родов, в том числе и некоторые из перечисленных ниже, часто сводят в синонимы): Turdus (56— 60 видов), Alethe, Brachypteryx, Calliope, Cathar us, Cercomela, Chaimarror-nis, Cichlherminia, Cochoa, Copsychus, Cossypha, Cyanosilvia, Dessonornis, Diplootocus, Enicurus, Erithacus, Geocichla (около 18 видов), Grandala, Hylocichla, Irania (Ixoreus), Kittacincla, Lamprolia, Larvivora, Luscinia, Microcichla, Monticola, Myadestes, Myophonus (Myiophoneus), Myrmecoci-
299
Рис. 210. Ареалы семейств певчих воробьиных:
1— дроздовые Turdidae, 2— славковые Sylviidae, 3— корольковые Regulidae
chia, Neocossyphus, Nesocichla, Oenanthe, Oreocincla, Phaeornis, Phoenicurus, Pseudoaedon, Psophocichla, Rhyacornis, Saxicola, Saxicoloides, Sialia, Sti-zorhina, Tarsi ger, Xenocopsychus, Zoothera и др. Некоторые роды, например Тurdus, распространены почти по всему ареалу семейства. У широко распространенных видов хорошо выражена географическая изменчивость.
В СССР встречается 50—54 вида, относимых к 12—19 родам. Достоверно гнездится 48—51 вид. Гнездовой ареал дрозда Науманна Turdus паи-manni Теппп. — тайга и лесотундра от Восточного Саяна до Камчатки и Шантарских островов — не выходит за пределы нашей страны.
Вероятный центр возникновения, видимо, Старый Свет. Ископаемые остатки 23 ныне живущих видов обнаружены в пределах современного ареала в верхнем плейстоцене.
Морфологически дроздовые нечетко обособлены от близких семейств— славковых, мухоловковые и некоторых др. Поэтому часть систематиков объединяет эти группы в ранге подсемейств в большое семейство мухоловковых (подробнее ниже).
СЕМЕЙСТВО 42. ZELEDONIIDAE — ВОДЯНЫЕ ДРОЗДЫ
Монотипичное семейство, включающее единственный вид Zeledonia coronata Ridgway, встречающийся в горных районах Коста-Рики и Панамы (Центральная Америка). От настоящих дроздов отличаются тем, что первое первостепенное маховое совсем крошечное, хвост из 10 рулевых, клюв короткий, заметно приплюснутый у основания, и другими мелкими деталями. Экология не изучена. Ряд систематиков включают его в семейство дроздовых в ранге подсемейства или трибы.
СЕМЕЙСТВО 43. SYLVIIDAE—СЛАВКОВЫЕ
Мелкие или очень мелкие подвижные птицы, населяющие леса разных типов, кустарниковые, тростниковые и травянистые заросли, безлесные участки гор, открытые ландшафты с изреженной растительностью (степи, полупустыни, пустыни). Длина от 7 до 28 см\ вес от 6 до 30—40 г. Распространены практически по всему земному шару (рис. 210), но подавляю
щее большинство видов свойственно Старому Свету.
Внешний облик довольно типичен: это славки, пеночки, камышевки (рис. 211) и т. п. Тело стройное, голова небольшая. Клюв умеренно длинный, слегка приплюснутый у основания, дальше сжатый с боков, тонкий, заостренный, иногда слабо изогнутый книзу. По коньку надклювья проходит более или менее отчетливый гребень. Ноздри открытые; над ними слегка нависают перепончатые крышечки. Щетинки в углах рта есть или отсутствуют. Ноги средней длины, пальцы длинные, тонкие, с острыми когтями. Цевка спереди покрыта несколькими щитками (у старых птиц иногда сливающимися). Крылья довольно короткие, чаще более или менее закругленные. Первостепенных маховых 10, первое ко
Рис. 211. Дроздовидная камышевка Acrocephalus arundinaceus (L.)
роче половины второго, иногда очень мало. Хвосты у разных родов резко
301
варьируют по длине и форме. Обрез хвоста прямой, закругленный или более или менее ступенчатый. Рулевых 12 (у немногих родов 10).
Оперение мягкое, приглаженное. Хохлов нет, хотя при возбуждении некоторые виды приподнимают перья на темени. У многих видов половой диморфизм проявляется только в размерах (самцы немного крупнее самок), у части видов самцы окрашены ярче. Окраска разнообразная, у большинства видов довольно скромная, с преобладанием зеленоватых, сероватых, коричневатых тонов. Некоторые тропические виды ярко окрашены. Молодые птицы в первом гнездовом наряде не бывают пятнистыми; у большинства видов они похожи по окраске на взрослых (при наличии полового диморфизма — на самок). В году 2 линьки: полная послегнездовая и частичная, редко тоже полная предбрачная (ранневесенняя). Молодые птицы весной следующего года жизни одевают наряд, неотличимый от наряда взрослых. Виды многих родов трудно различимы по внешним признакам, но обычно довольно хорошо отличаются по голосу (песня, позывы). Большинство видов — хорошие певцы (у немногих поют и самки). У некоторых видов хорошо выражена способность к звуковому подражанию (звуковая имитация); они включают в свою песню строфы из песен многих видов птиц.
Большинство ведет скрытный образ жизни, выдавая свое присутствие только голосом. Обычно активны днем, но часть видов в сумерках и ночью. Гнезда открытые чашеобразные или закрытые с боковым входом. У части видов гнездо сплетается очень искусно, у других небрежно, с просвечивающим дном и тонкими стенками. Гнезда помещаются на разной высоте, чаще низко над землей, в развилках ветвей деревьев и кустарников, на крепких стеблях травы или тростника; некоторые виды строят гнездо на земле, под защитой кочки или кустика травы. Окраска яиц варьирует очень сильно: она может быть однотонной белой, голубоватой, зеленоватой или с коричневой или красноватой пятнистостью. В кладке 4—6 яиц, редко больше или меньше. Насиживают оба партнера, реже только самка. Инкубация 11—14 дней. Птенцов выкармливают оба партнера. Молодые покидают гнездо в возрасте около 2 недель, короткое время после вылета кочуя вместе с родителями. В году обычно одна кладка, у части видов две. Вне сезона размножения держатся поодиночке, реже рассеянными небольшими группами. Перелетные, в тропиках кочующие, реже оседлые.
Кормятся в кронах, на стеблях травянистых растений, реже, взлетая, хватают близко подлетевших насекомых или собирают корм на земле. Питаются преимущественно разнообразными беспозвоночными (насекомые и их личинки, пауки и т. п.), поедают мелкие ягоды.
Объем семейства нуждается в уточнении: он сильно различается в понимании разных систематиков. Чаще в него включают 400—425 видов, объединяемых в 83—93 рода. Большой объем семейства вынуждает выделять подсемейства, которые морфологически различаются очень слабо. Обычно выделяют 3 подсемейства:
1. Sylviinae — славки Старого С в е т а; .330 — 332 вида, распределяемых по 55—65 родам: Sylvia (19 видов), Abroscopus, Acrocephalus (18 видов), Apalis (более 20 видов), Bawdier ia, Bradypterus (включает часть видов р. Tribura, всего более 20 видов), Calamocichla, Cettia (Horeites), Cisticola (около 75 видов), Dromaeocercus, Eremomela, Erythropygla (Agrobates, Cercotrichas), Herbivocula, Hippolais, Hylla, Locus tella, Lusciniola, Macrosphenus, Megalurus, Orthotomus, Phragmaticola, Phyllergates, Phylloscopus (сюда включают Oreopneuste и Reguloides\ более 30 видов), Prinia, Scepotnycter, Scotocerca, Seicercus, Sphenoeacus, Spil-optila, Sylvietta, Tesia (Oligura), Tribura, Trichocichla, Urosphena и др. Населяют Африку (где встречается не менее 150 видов), Европу, Азию, Океанию; немногие виды проникают в Австралию и один вид — Phylloscopus borealis (Blass.) — на Аляску. В СССР встречается только это подсемейство: 51 вид, относимый к 11—14 родам; достоверно гнездится 49 видов.
302
2. Malurinae — австралийские славки. Распространены в Австралии, Новой Гвинее, Новой Зеландии и на островах Океании. 80—82 вида мелких птиц славкового или крапивникового облика, распределяемых по 25 родам: Malurus, Acanthiza, Amytornis, Aphelocephala, Ca-lamanthus, Cerygone, Chenorhamphus, Chthonicola, Cincloramphus, Clyto-tnyias, Crateroscelis, Dasyornis, Epthianura, Eretniornis, Gerygone, Hylacola, Origtna, Pycnoptilus, Pyrrhalaetnus, Sericornis, Smicrornis, Stipiturus, To-dopsis, Triodia, Vitia.
3. Polioptilinae — комароловы; 11—10 видов славкоподобных птиц, распространенных в северной половине Южной Америки и южной части Северной Америки. Всего 3 рода: Polioptila, Microbates и Ramphocaenus (последние 2 монотипичны и встречаются только в тропиках).
Иногда в семейство славок включают корольков Regulidae. Слабая морфологическая и экологическая обособленность этой группы вынуждает некоторых систематиков объединять славковых вместе с дроздовыми и мухоловковыми в общее семейство мухоловковых Muscicapidae.
Центр возникновения славковых — видимо, Старый Свет. Ископаемые остатки в пределах современного ареала известны со среднего миоцена; их относят к 10 ныне живущим видам.
СЕМЕЙСТВО 44. REGULIDAE — КОРОЛЬКОВЫЕ
Мелкие, подвижные птички (длина 8—12 см, вес 5—8 г), по внешнему облику напоминающие синиц или пеночек (рис. 212). Распространены в Евразии, Северной Африке и Северной Америке (рис. 210).
Клюв относительно короткий, тонкий, чуть изогнутый к вершине, с едва намечающимся крючком на конце. Ноздря или прикрыта одиночным перышком— щетинкой или над ней нависает тоненькая кожистая крышечка. В углах рта есть щетинки. Ноги умеренной длины. Цевка спереди покрыта сплошной пластинкой (корольки) или отдельными щитками. Пальцы тонкие, с короткими, сильно изогнутыми острыми коготками. Крылья короткие, туповершинные, Первостепенных маховых 10. Хвост из 12 рулевых, по длине примерно равен крылу, обычно слабо выемчатый. Оперение мягкое, пушистое. Окраска довольно яркая; темя по цвету отличается от спины. Половой диморфизм окраски выражен лишь у части видов. Ни у птенцов, ни у взрослых на перьях нет четких пестрин. В году одна линька: полная послегнездовая.
Населяют леса, преимущественно хвойные на равнинах и леса и кустарниковые заросли в горах. Довольно скрытны. Строят гнездо в густых вет-
вях либо в форме глубокой, прочно свитой чашечки, либо шарообразное, с боковым входом. Лоток выстилается шерстью и подобранными перышками. В кладке 4—8 пестрых яиц. Инкубация около 2 недель. Птенцы вылетают из гнезда на 16—18-й день. Выводки широко кочуют, иногда присоединяясь к стайкам синиц. . Кормятся в кронах деревьев и кустов, склевывая пауков, насекомых, их яйца, личинок и куколок с ветвей, из трещин коры, с листьев. Зимой поедают и мелкие семена. На земле кормятся редко.
Рис. 212. Красноголовый королек Regulus ignicapillus (Temm.)
303
В семейство включают всего 5—6 видов, относимых к 3 родам. 3—4 вида корольков рода Regains распространены по лесам Евразии, Северной Африки и Северной Америки. Сюда же относят 2 монотипичных рода: Lepto-poecile живут в куртинах кустов по горам Средней и Центральной Азии, Lophobas ileus — в Тибете. Близость двух последних родов к королькам сомнительна; их положение в системе требует уточнения (иногда включают к славкам, тимелиям или синицам). Семейство корольков иногда включают в ранге подсемейства в семейство славок.
Вероятный центр обособления корольков — умеренные широты Евразии. В СССР встречаются и гнездятся 3 вида: 2 вида корольков и расписная синичка Leptopoecile sophiae Sev.
СЕМЕЙСТВО 45. MUSCICAPIDAE ~ МУХОЛОВКОВЫЕ
Мелкие птицы (длина 10—20 см, вес 8—30 г). Встречаются в лесах разных типов, реже в кустарниковых зарослях; некоторые виды — в культурных ландшафтах (сады, парки, населенные пункты). Распространены только в Старом Свете почти повсеместно (рис. 213;, кроме приполярных областей и обширных пустынь.
Сложение довольно плотное. Клюв короткий или средней длины, у основания широкий и обычно заметно уплощенный. Хребет надклювья четко выражен. На конце клюва есть маленький крючочек. У основания клюва — перышки с удлиненными стержнями, иногда частично прикрывающие широко расставленные ноздри. Как правило, хорошо развиты щетинки в углах рта. Ноги слабые. Тонкая цевка спереди покрыта несколькими поперечными щитками. Пальцы тонкие, с короткими коготками. Крылья
Рис. 213. Ареалы семейств певчих воробьиных: /— мухоловковые Muscicapidae, 2— завирушковые Prunellidae
304
удлиненные, слабо заостренные. Первостепенных маховых 10; первое часто сильно укорочено. Хвост из 12 рулевых, умеренной длины. Однако у некоторых видов рулевые, особенно средние, резко удлинены, и хвост превышает длину крыла в 3—5 раз (рис. 214). Иногда на голове есть хохолок из удлиненных перьев. Оперение приглаженное, мягкое. Окраска очень разнообразна: от однотонной буроватой до очень яркой. У многих видов резко выражен половой диморфизм окраски, у части видов его нет. Окраска птенцов в первом гнездовом наряде пятнистая или с чешуйчатым рисунком. Птенцы осенью сменяют мелкое перо и становятся похожими на самок. У ряда видов самцы одевают полный брачный наряд лишь на втором и даже третьем году жизни, а в размножении участвуют в возрасте около года. У
Рис. 214. Длиннохвостая мухоловка Terpsiphone paradisi (L.); самка и самец
части видов в году одна линька: полная послебрачная, у других еще есть частичная или полная предбрачная линька.
Часть видов придерживается опушек и разреженных участков леса, другие ведут скрытную жизнь, населяя густые насаждения. Некоторые виды поднимаются в горы до верхней границы леса. Моногамы; у немногих видов описана факультативная бигамия: на участке самца гнездятся две самки. Песня самцов довольно однообразная, иногда звучная. Часть видов строит открытые, иногда очень искусно свитые гнезда, укрепляя их в развилке ветвей куста или дерева. Другие виды гнездятся укрыто: за отставшей корой, в прогнивших пнях, дуплах, охотно занимают искусственные гнездовья. В кладке 2—8 , редко больше белых или голубоватых яиц, часто с хорошо развитым крапом. В году обычно одна кладка. У большинства видов насиживают кладку оба партнера, у немногих видов только самка. Инкубация 2,5—3 недели, столько же времени птенцы остаются в гнезде. Птенцов кормят оба партнера. Вылетевшие из гнезда птенцы непродолжительное время кочуют вместе с родителями, а затем выводки распадаются. Вне сезона размножения большинство видов держится поодиночке, реже небольшими группами. В умеренных широтах перелетные; довольно поздно прилетают и рано улетают.
Питаются преимущественно насекомыми, в меньшей степени другими мелкими беспозвоночными; иногда поедают ягоды. Обычный способ охоты:
305
птица сидит на ветке и, взлетая, ловит насекомое. При недостатке летающих насекомых кормятся, склевывая добычу на ветвях, листьях и земле.
Объем семейства и отдельных родов, перечень видов требует уточнения. Чаще сюда относят 378—398 видов, объединяемых в 57—82 рода: Muscica-ра (более 30 видов; обычно сюда включают и роды Alseonax, Cyanoptila, Ficedula, Hemichelidon, Poliomyias, Siphia, Xanthopygia или Zanthopygia), Anthipes, Arses, Artomyias, Batis, Bradornis, Chasiempis, Chelidorhynx, Collar icincla, Coracornis, Cyornis, Dendrob iastes, Diaphorophyia, Eopsaltria, Erannornis, Erythrocercus, Eulacestoma, Eumyias, Falcunculus, Fraseria, Hylocitrea, Hypothymis, Machaerirhynchus, Melaenornis, Microeca, Miro, Monarcha (19 видов), Muscicapella, Myiagra (11 видов), Myiolestes, Myopor-nis, Namibornis, Newtonia, Niltava, Ochromela, Oreoica, Pachycare, Pachy-cephala, Parisoma, Pedilorhynchus, Peltops, Petroica, Pitohui, Platysteira, Poecilodryas, Pomarea, Rhagalogus, Rhiomyias, Rhipidura (24 вида), Terpsi-phone (Tchitrea), Zeocephus и др. В пределах семейства мухоловковых выделяют 3—4 подсемейства (трибы, если все мухоловковые рассматриваются как подсемейства).
В СССР встречаются 14 видов 2 родов; гнездятся 13 видов.
Между мухоловковыми, славковыми, дроздовыми и некоторыми другими группами певчих воробьиных, как уже указывалось, много общего, а морфологические и экологические различия нечеткие. Учитывая это и, видимо, родство групп, их часто объединяют в общее семейство мухоловковых. Наибольший объем семейства мухоловковых предлагают Майр и Амадон (1951), включающие сюда следующие семейства, выделяемые Уэтмором (1960): мухоловковых Muscicapidae, тимелиевых Timaliidae, крапивниковых синиц Chaemaeidae, славковых Sylviidae, корольковых Regul-idae, дроздовых Turdidae, водяных дроздов Zeledoniidae, пересмеши и ко вых Mimidae, крапивниковых Troglodytidae, оляпковых Cinclidae. Подавляющее большинство перечисленных групп получает ранг подсемейств, а немногие — триб. Всего в это громадное семейство входит свыше 1460 видов, т. е. более трети всех певчих воробьиных. Такой объем семейства неудобен хотя бы из чисто практических соображений. Поэтому точка зрения Уэтмора кажется более приемлемой. Может быть, стоило бы, принимая схему Уэтмора, объединить все вышеперечисленные семейства в надсемейство мухоловковых, что позволило бы, не усложняя системы, подчеркнуть близость этих групп.
СЕМЕЙСТВО 46. PRUNELLIDAE — ЗАВИРУШКОВЫЕ
Очень скрытные, малозаметные птицы (рис. 215) размером примерно с воробья, живущие в лесах и кустарниковых зарослях на равнинах и в горах Европы и Азии и в небольшом участке в Северной Африке (рис. 213).
Рис. 215. Лесная завирушка Prunella modularis (L.)
306
Клюв небольшой (короче головы), прямой и тонкий; основание клюва широкое, но затем он быстро сужается; перед ноздрями ровная площадка. Над ноздрями слегка нависают кожистые крышечки. В углах рта короткие щетинки. Тело коренастое. Короткая цевка спереди покрыта нерезко разграниченными поперечными щитками. Пальцы довольно короткие, с не очень острыми когтями. Крылья закругленные. Первостепенных маховых 10; первое почти не выдается за кроющие кисти. Хвост из 12 рулевых, короче крыла, прямо обрезанный, слабо вырезанный или слегка закругленный. Оперение густое, довольно плотное. Окраска буроватых или охристых тонов; самки несколько тусклее самцов. Молодые пестрые. Одна линька в году (послебрачная).
Чашеобразное гнездо с плотными стенками помещается в густых ветвях, трещинах скал, под камнями. В кладке обычно 4—6 голубовато-зеленых яиц. У части видов 2 кладки в году. Инкубация около 2 недель, столько же времени птенцы остаются в гнезде. Вне сезона размножения держатся мелкими стайками. Перелетные или, на юге, кочующие. Кормятся в кронах деревьев и кустарников, чаще на земле. Пища — разнообразные мелкие беспозвоночные, семена, ягоды. Экологически близки к вьюрковым птицам.
Семейство включает 12 видов, относимых к 1—2 родам: Prunella и Lais-copus (этот род часто объединяют с первым). В СССР встречается и гнездится 8 видов; большинство из них приурочено к горным районам Средней Азии и Сибири.
СЕМЕЙСТВО 47. MOTACILLIDAE — ТРЯСОГУЗКОВЫЕ
Мелкие птицы (длина 12—23 см, вес 20—30 г) с характерным обликом трясогузки или конька (рис. 216). По характеру распространения почти космополиты: нет в Антарктике, в части Океании, спорадично встречаются в Северной Америке (рис. 218).
Стройные птицы с удлиненным телом и небольшой головой. Клюв шиловидный, прямой, почти равный длине головы. Ноздри открытые овальные; над ними слегка нависают небольшие кожистые крышечки. Щетинки
в углах клюва маленькие или их нет вовсе. Ноги длинные. Цевка и пальцы
тонкие. Задний край цевки заострен. Коготь заднего пальца длинный (не
короче пальца), слабо изогнутый. Крылья заостренные, первостепенных маховых 9 (первое совершенно редуцировано). Задние второстепенные маховые удлинены, но не достигают вершины крыла примерно на длину цевки. Хвост из 12 рулевых, равен крылу или длиннее его, закругленный или слабо выемчатый. Оперение мягкое, прилегающее. Окраска разнообразная, у многих видов скромная. На крайних рулевых белые поля. Половой диморфизм окраски чаще отсутствует; слабо выражен сезонный морфизм окраски. Молодые тусклее взрослых. В году 2 линьки: полная осенняя и частичная ранневесенняя.
Приурочены преимущественно к открытым местообитаниям в горах и на равнине: лугам, степям, тундрам, опушкам и разреженным участкам леса, берегам водоемов. Песня довольно простая; немногие виды поют в воздухе. Гнездо открытое, чаще на земле под защитой кустика травы или кочки; немногие виды гнездятся укрыто: в промоинах берегов, в дуплах, под камнями и т. п. Кладка из 4—6 светлых, обычно
Лесной конек
пестроокрашенных яиц. После вылета молодые рИСе 216.
кочуют вместе с родителями, потом выводки рас-
Anthus trivialis (L.)
307
падаются. Вне сезона размножения обычно держатся стайками. По земле ходят и бегают, а не прыгают. Питаются преимущественно различными мелкими беспозвоночными, собирая их на земле, листьях и траве (подпрыгивая), изредка даже в воздухе (подлетывая). Как второстепенные корма используются семена и ягоды.
Семейство включает 50—53 вида, объединяемых в 6—7 родов: Motacilla (сейчас включает и р. Budytes\ 10 видов), Anthus (около 30 видов), Dendro-nanthus, Macronyx (8 видов), Oreocorys, Tmetothylacus. Некоторые виды имеют большие ареалы: например, белая трясогузка Motacilla alba L. распространена в Европе, Азии, Африке и проникает на Аляску, горный конек Anthus spinoletta (L.) населяет многие горные районы Европы, Азии, Северной Америки.
Вероятный центр возникновения, видимо, Африка. Наиболее ранние палеонтологические находки известны из среднего миоцена Франции.
В СССР встречается 15—17 видов 3 родов; достоверно гнездятся 14— 16 видов. Сибирский конек Anthus gustavi Swinh. наш эндемик — населяет лесотундру от Печоры до Чукотки.
СЕМЕЙСТВО 48. BOMBYCILLIDAE — СВИРИСТЕЛЕВЫЕ
Коренастые птицы (рис. 217) размером примерно со скворца (вес 60— 70 г, длина 16—22 см). Распространены по таежным лесам Евразии и Северной Америки и в некоторых полупустынных районах Евразии (рис. 218).
Шея короткая, голова довольно крупная. Клюв короче головы, широкий у основания, выпуклый по гребню, слабо изогнутый, с маленьким крючком на конце. Ноздри частично прикрыты мелкими щетинковидными перышками или открытые. Ноги короткие, толстые и сильные. Цевка спереди покрыта отдельными поперечными щитками. Когти толстые, изогнутые. Крыло слабо заостренное; первостепенных маховых 10 (первое мало). Хвост относительно короткий (свиристели) или длинный (свиристелевые сорокопуты). Оперение густое, мягкое. У свиристелей на лбу длинные перья, которые птица может поднимать, как высокий хохол. Окраска красивых мягких тонов. Половой диморфизм окраски выражен сла,бо. Окраска молодых более тусклая; мелкое перо гнездового наряда они сменяют в первую осень жизни. У взрослых одна полная послегнездовая линька.
Гнездо открытое, чашеобразное, с толстыми стенками, укрепляется в развилке веток. Кладка из 3—7 пятнистых яиц. Насиживает кладку около 2 недель самка; самец ее кормит. Выкармливают птенцов оба партнера.
Выводки кочуют вместе со взрослыми, обычно объединяясь в большие стаи; направление кочевок и скорость перемещения определяются наличием пищи. В гнездовой период преимущественно насекомоядны. Корм собирают в кронах и на земле. Взлетая, ловят близко подлетевших насекомых. В остальное время года поедают насекомых, разнообразные ягоды, семена. Гнездящийся в нашей тайге обыкновенный свиристель Botnb-ycilla garrulus (L.) в годы обильного урожая рябины, калины, можжевельника зимует в области гнездования и в средней полосе; при недостатке корма откочевывает
Рис. 217. Обыкновенный свиристель Вот-bycilla garrulus (L.)
дальше на юг. Семейство
свиристелевых
308
150° 120° 90° 60°к западу от Гринвича 0°к востоку от Гринвича30° 60° 120° 150° 180° 150°
150° !20° 90° 6СР 30° 0° 30° 60° 90° 120° 150° 180° 150°
Рис. 218. Ареалы семейств певчих воробьиных:
1— трясогузковые Motacillidae, 2— свиристелевые Bombycillidae
включает всего 4 вида 2 родов. Их обычно разделяют на 2 подсемейства.
1. Bombycillinae — свиристелевые. Один род Вот-bycilla с тремя видами, населяющими таежные леса Евразии и Северной Америки. В СССР встречаются и гнездятся 2 вида.
2. Hypocoliinae — свиристелевые сорокопуты. Один монотипический род Hypocolius. Гнездятся в кустарниковых полупустынях Аравии, во время кочевок залетают в Северную Африку, Индию; залетные птицы добыты в Южной Туркмении.
Иногда эти подсемейства рассматриваются как самостоятельные семейства. С другой стороны, в семейство свиристелевых часто включают еще 2 подсемейства: Dulinae и Ptilogonatinae, которым Уэтмор придает ранг самостоятельных семейств.
СЕМЕЙСТВО 49. PTILOGONAT1DAE — ШЕЛКОВИСТЫЕ МУХОЛОВКИ
Рис. 219. Шелковистая мухоловка Phainopepla nitens (Sw.)
По общему облику и размерам более или менее похожи на свиристелей, но отличаются большей длиной хвоста (рис. 219). Распространены от юго-западных районов США до Панамы и Коста-Рики. Отличаются от свиристе-левых мелкими деталями.
Оперение шелковистое с блестящим отблеском. Половой диморфизм окраски выражен четко. Населяют кустарниковые заросли, опушки леса. Плоское гнездо помещается в развилке ветвей; видимо, его строит преимущественно самец. В кладке 3—4 пестрых яйца. Вне сезона размножения держатся мелкими группами. Кочуют на небольшие расстояния. Питаются ягодами и насекомыми. Корм собирают на ветвях, реже на земле. Насекомых ловят и в воздухе: взлетев, схватывают близ-
ко подлетевшее насекомое.
Семейство включает всего 4 вида, относимых к 3 родам: Ptilogonys с 2 видами и монотипичные роды Phainopepla и Phainopiila. Эту группу в ранге подсемейства часто включают в семейство свиристелевых.
СЕМЕЙСТВО 50. DUL1DAE— ПОЖИРАТЕЛИ ПАЛЬМОВЫХ СЕМЯН
Единственный вид Dulus dominicus (L.), встречающийся только на о. Гаити и маленьком о. Гонав. Размеры примерно со скворца. Отличаются от свиристелевых рядом мелких деталей. Клюв более широкий и крепкий. Ноздри открытые. Крылья короткие, тупые. Хвост длинный. Окраска зеленовато-коричневатая с бурыми пестринами. Гнездование своеобразное: несколько пар (до 10 и больше) на вершине высокой пальмы строят большое шарообразное гнездо диаметром до 90—100 см из веточек и травы. В толще его расположены гнезда отдельных пар; каждое имеет сбоку свой вход. В кладке 2—4 белых яйца с резким крапом и исчерченностью. Насиживают и кормят оба партнера. Держатся стайками. Пища, видимо, исключительно растительная: мелкие плоды, отчасти семена.
Вероятно, близки к свиристелевым. Иногда включаются в это семейство в ранге подсемейства.
310
СЕМЕЙСТВО 51. ARTAMIDAE — ЛАСТОЧКОВЫЕ СОРОКОПУТЫ
Семейство содержит только один род Artemus с 10 видами (иногда выделяют до 15 видов), распространенными в открытых ландшафтах с зарослями кустов и отдельных деревьев в Австралии (нет в Новой ^Зеландии), на островах Океании и в Юго-Восточной Азии.
Крепкого сложения птицы (рис. 220) размером с воробья или скворца (длина 15—20 см). Клюв
крепкий, средней длины, слегка Рис. 22°- Ласточковый сорокопут Ariamus изогнутый, с широким основанием superciliosus (Gid.)
и заостренным концом. Ноги короткие. Пальцы короткие, крепкие с короткими, слабо изогнутыми ногтями. Крылья из 10 первостепенных маховых (первое очень маленькое), длинные, заостреннные. Хвост средней длины. Оперение прилегающее. Окраска из сочетания черных, серых и белых цветов. Половой морфизм окраски выражен слабо, возрастной — более отчетлив. Характерны мелкие пудретки по бокам тела и в основании хвоста (единственный случай у воробьиных птиц).
Экология изучена слабо. Рыхлые гнезда из сухих травинок, мха, перьев, шерсти располагаются в развилке ветвей, на вершинах пней; некоторые виды строят гнезда в дуплах, расщелинах скал, промоинах береговых обрывов. Часто гнездятся группами. В кладке 2—4 беловатых или кремовых яйца, покрытых темным крапом. Обычно держатся стайками. Оседлы или кочуют на небольшие расстояния; немногие виды (Южная Австралия) перед етны. Полет быстрый, маневренный, способны парить. Питаются преимущественно насекомыми, которых или догоняют в воздухе, подобно ласточкам, или, как мухоловки, сидят на ветке и, взлетев, схватывают добычу. Реже кормятся в кронах, очень редко на земле.
Вероятно, близки к семейству сорокопутовых; высказывались соображения и о близости их со свиристелевыми.
Вероятный центр возникновения — Австралия.
СЕМЕЙСТВО 52. VANGIDAE — ВАНТОВЫЕ
Мелкие (длина 12—30 см) птицы размером от воробья до удода, преимущественно древесные. Распространены только на Мадагаскаре и прилегающих островах. Пр повадкам отчасти напоминают сорокопутов, поползней, крупных мухоловок.
Тело коренастое. Голова средней величины. Форма и размеры клюва у разных видов варьируют в очень широких пределах: от длинного мощного клюва с острым крючком на конце до короткого прямого или длинного, тонкого, сильно изогнутого и заостренного (рис. 221). Ноздри полуприкрыты нависающими кожистыми крышечками. Щетинки в углах рта хорошо развиты или их нет. Ноги средней длины с довольно длинными крепкими пальцами. Боковые поверхности цевки покрыты несколькими щитками. Когти слабо изогнутые. Крылья умеренной длины, тупые или слабо заостренные. Хвост средней длины. Оперение прилегающее. Окраска разнообразная: из сочетания черного и белого; черного, белого, голубого, коричневого и т. п. Половой диморфизм окраски выражен у немногих видов; молодые тусклее взрослых.
311
Рис. 221. Ванговые: / —толстоклювая ванга Aerocharis prevostii Less.; // — серпоклювая ванга Falculea palliata Geof.; HI — крючкоклювая ванга Vanga curvirostris (L.)
Экология изучена плохо. Держатся в лесах разных типов, реже в кустарниковых зарослях. Гнезда в развилке ветвей, часто высоко над землей. Кладки из 3—4 беловатых или зеленоватых яиц, испещренных коричневыми пятнами. Оседлы или кочуют в районе гнездования. Обычно держатся стайками от 3—4 до нескольких десятков особей. Кормятся преимущественно в кронах, немногие виды охотятся чаще на земле. Пища — разнообразные беспозвоночные (насекомое, пауки, моллюски), которых склевывают с ветвей и листьев; тонкоклювые виды, видимо, вытаскивают добычу из щелей коры и т. п. Виды, имеющие мощные клювы, поедают также плоды; при случае, видимо, грабят гнезда мелких птиц, хватают маленьких ящериц, мышей и т. п. Некоторые виды ловят и летающих насекомых.
Семейство объединяет И—14 видов, распределяемых между 9—И родами: Vanga, Aerocharis (Euryceros), Calicalicus, Cyanolanius, Falculea, Leptopterus, Oriolia, Schetba, Tylas, Xenopirostris.
Ванговые могут служить примером резкой адаптивной радиации небольшой группы, живущей в изолированных условиях. Особенно рельефно радиация проявляется в многообразии клювов, видимо, обеспечивающих видовую специфичность пищевых связей и, тем самым, уменьшающих конкуренцию между близкими видами, населяющими сходные или одни и те же биотопы на ограниченной территории.
Система семейства нуждается в уточнении: видимо, завышено число родов; возможно, некоторые роды заслуживают исключения из этой группы и т. п. Некоторые систематики полагают, что более или менее явные родственные отношения сближают ванговых с семейством сорокопутовых.
СЕМЕЙСТВО 53. LANIIDAE — СОРОКОПУТОВЫЕ
Птицы размером от воробья до дрозда (длина 15—30 см, у нескольких особенно длиннохвостых видов до 40—45 см-, вес 25—120 а). Распространены в Африке (нет на Мадагаскаре), Европе, Азии, на части островов Океании (нет в Австралии) и в Северной Америке (рис. 222).
Птицы плотного сложения (рис. 223). Сильный, сжатый с боков клюв немного короче головы. Надклювье заканчивается изогнутым острым зуб-
312
Рис. 222. Ареалы семейств певчих воробьиных:
1— сорокопутовые Laniidae, 2— скворцовые Sturnidae, 3— районы акклиматизации обыкновенного скворца Sturnus vulgaris L.
Рис. 223. Серый сорокопут Lanius excubitor L.
цом; обычно хорошо развит и предвершинный зубец (как у соколов). Ноздри открытые или полуприкрыты жесткими перышками. Щетинки в углах рта хорошо развиты. Ноги умеренной длины. Цевка крепкая, спереди покрыта раздельными щитками, а по бокам слившимися роговыми пластинками. Пальцы относительно короткие, с короткими, сильно изогнутыми когтями. Крылья короткие, с закругленной вершиной. Первостепенных маховых 10, первое часто лишь немного короче половины второго. Хвост из 12 рулевых, удлиненный или длинный, округлый, иногда ступенчатый. Оперение густое, довольно рыхлое. Окраска разнообразная, у многих видов из сочетания белого, серого, коричневого и черного цветов; у тропических видов более яркая. Самцы немного крупнее самок и у части видов ярче окрашены. Молодые похожи на самок, но отличаются более или менее развитой поперечной, исчерченностью; сменяют гнездовой наряд во время частичной осенней линьки. У видов с резким половым диморфизмом молодые самцы следующей весной, в возрасте 8—9 месяцев, проходят полную линьку, одевая брачный наряд. Взрослые особи многих видов линяют раз в году, осенью; у других видов есть и частичная предбрачная линька.
Густых лесов избегают: держатся по опушкам и изреженным участкам. Широко распространены по разнообразным открытым ландшафтам в горах и на равнинах: обязательно должны быть хотя бы отдельные кусты. Проникают в культурные ландшафты. Голос грубый, резкий. Многие виды способны к имитации песни других птиц. Возможно, с этим связано русское название сорокопут (плетет путы сорока птицам, т. е. приманивает их песней). Гнездо строят оба партнера, но степень их участия в работе различна у разных видов. Открытые чашеобразные гнезда укрепляются в развилке ветвей на разной высоте от земли. В кладке 4—6 пестрых яиц. Насиживают оба партнера или только самка — тогда самец носит ей корм. Инкубация около 2 недель или немного больше. Птенцов кормят оба партнера. Птенцы покидают гнездо в возрасте 2—2,5 недель, иногда с еще не полностью выросшими маховыми и рулевыми. Выводок держится с родителями, продолжающими кормить птенцов, 3—4 недели. Вне сезона размножения встречаются одиночками, реже парами или — у немногих видов — группами в 4—10 особей. В умеренных широтах перелетные, в южных оседлые или кочующие.
Сильные агрессивные птицы. Поедают преимущественно крупных насекомых (жуков, бабочек, саранчовых, стрекоз и т. п.), при случае ловят мелких ящериц, молодых мышат, т аскают яйца и птенцов из гнезд мелких птиц. Добычу обычно подкарауливают, сидя на кусте, столбе и т. п., и, взлетев, ловят в воздухе или на земле. Крупную добычу накалывают на
314
острый сухой сучок и потом разрывают клювом. Накалывают на сучки около гнезда и избыток добычи (жуков и т. п.).
В семейство Laniidae включают 66—69 видов, распределяемых по 9— 12 родам. Особенно велико разнообразие видов в Африке. Часто семейство сорокопутовых разделяют на 2 подсемейства.
1. Типичные сорокопутовые Laniinae;27 видов, относимых к 3 родам: 25 видов объединяет род Lanins, распространенный практически по всему ареалу семейства; в Северной Америке всего 2 вида, в том числе серый сорокопут L. excubitor L., широко распространенный по Евразии и Северной Африке. Сюда же включают 2 монотипичных африканских рода Corvinella и IJrolestes. В СССР 11 видов рода Lanius', достоверно установлено гнездование 10 видов.
2. Кустарниковые сорокопуты Malaconotinae; 39—42 вида объединяют в 6—9 родов: Malaconotus, Chlorophoneus, Dryos-copus (Chaunonotus), Laniarius, Laniturdus, Nilaus, Rhodophoneus, Tchagra (Antichronus), Telephorus. Распространены только в Африке. По внешнему облику — мелкие сорокопуты (размерами с воробья), часто очень ярко окрашенные.
Высказывалось предположение, что сходство этих двух подсемейств конвергентное, поэтому кустарниковых сорокопутов следует выделять в самостоятельное семейство.
Центр возникновения, вероятно, Палеарктика или Африка. Ископаемые остатки обнаруживают с нижнего миоцена (Франция); их относят к роду Lanius.
СЕМЕЙСТВО 54. PRIONOPIDAE — ЛЕСНЫЕ СОРОКОПУТЫ
По облику (рис. 224) похожи на мелких сорокопутов. Населяют леса и кустарниковые заросли Африки южнее Сахары. От настоящих сорокойу-тов отличаются мелкими деталями. У многих видов на голове хохол из перьев. Окраска яркая, сходная у самцов и самок.
Экология изучена плохо. Все сезоны года держатся стайками в 5—20 особей. Гнездятся в кронах. Часто гнезда нескольких пар строятся рядом, соприкасаясь стенками. Иногда 2—3 пары строят общее чашеобразное гнездо, в которое каждая самка откладывает от 2 до 6 яиц. Насиживают общую кладку совместно. Птенцов выкармливают все партнеры; иногда к ним присоединяются и другие взрослые особи, видимо, потерявшие кладки или по каким-то причинам не приступившие к размножению. Животноядны. Кормятся преимущественно в кронах. Взлетая, ловят насекомых в воздухе.
Семейство объединяет 9 видов 3 родов: Prionops, Europephalus, Sigmodus. Иногда сюда включают и монотипичный род Pityriasis (живет в болотистых лесах о. Калимантан); его родство с лесными сорокопутами у ряда систематиков вызывает сомнение.
Часто лесных сорокопутов в качестве подсемейства включают в семейство Laniidae. Полагают, что семейства Laniidae, Prionopidae и Vangidae в какой-то степени родственны. Однако эта точка зрения нуждается в добавочных обоснованиях.
Рис. 224. Желтоголовый лесной сорокопут? Prionops alberti Schout.
315
СЕМЕЙСТВО 55. CVCLARHIDAE — ПЕРЦЕВЫЕ СОРОКОПУТЫ
Единственный род Cyclarhis с 2 видами, встречающимися в лесах и культурном ландшафте Центральной Америки от Мексики до Эквадора. Яркой окраски небольшие птицы (длинаоколо 15 см), крепкого сложения, большеголовые. Клюв очень мощный с резко выраженным острым крючком на конце. Крылья короткие, полет довольно слабый. Оперение рыхлое. Голос мелодичный; напоминает свист нашей иволги. Чашеобразное гнездо подвешивается к тонким веткам в кроне. В кладке 2—3 пестрых яйца. Охотятся в кронах, реже на земле, поедая разнообразных насекомых и других беспозвоночных.
Иногда этот род включают в семейство Vireonidae в ранге подсемейства.
СЕМЕЙСТВО 56. VIREOLANIIDAE—СОРОКОПУТОВЫЕ ВИРЕОНЫ
Крепкого сложения мелкие птицы (длина около 15 см) преимущественно горных лесов тропической Америки. Клюв сильный с большим острым крючком на конце. Окраска яркая с преобладанием зеленого цвета. Экология не изучена. Держатся в кронах. Поедают различных беспозвоночных, особенно крупных насекомых, разыскивая их на ветвях и листьях. Держатся чаще парами.
Семейство включает 3—4 вида, относимых к 2 родам: Vireolanius и Smaragdolanius. Роды семейства Vireolaniidae наряду с семейством Cyclarhi-dae иногда включают в семейство виреоновых Vireonidae, хотя они заметно отличаются от последних рядом признаков, прежде всего-мощным клювом с крючком на конце.
СЕМЕЙСТВО 57. GALLAEIDAE— НОВОЗЕЛАНДСКИЕ СКВОРЦЫ
Птицы плотного сложения размером со скворца или крупного дрозда (длина 25—45 см). Населяют леса Новой Зеландии.
Рис. 225. Новозеландские скворцы: I — разноклювая гуйа Heteralocha acutirostris (Gid.) самец и самка; II — новозеландский скворец Callaeas cinerea Gm.
316
Клювы сильно варьируют по форме. У самца разноклювой гуйи крепкий, удлиненный и слегка изогнутый клюв; у самки много длиннее, тоньше и сильнее изогнут; у новозеландского скворца клюв короткий, широкий у основания и высокий (рис. 225). Ноздри открытые. В углах рта ярко окрашенные выросты кожи: большие у самцов, меньшие у самок и очень малы у молодых. Ноги крепкие с удлиненной цевкой и пальцами. Крылья закругленные; первое первостепенное маховое длиннее половины второго. Хвост длинный, закругленный. Окраска черная или коричневая, с металлическим блеском, сходная у самцов и самок. Молодые отличаются более тусклой окраской.
Птицы подвижные, крикливые. Гнездо в виде почти плоской платформы из веточек и травы, строят на горизонтальной развилке в кроне дерева; реже гнездятся в полудуплах или в промоинах обрывов. В кладке 2—4 крапчатых яйца. Инкубация продолжается около 3 недель; птенцы вылетают из гнезда в возрасте около 4 недель. Оседлы. Вне сезона размножения держатся парами или семьями. Кормятся в кронах и на земле. Питаются разнообразной животной и растительной пищей: насекомыми и другими беспозвоночными, которых собирают на земле, ветвях и листьях, вытаскивают из трухлявой древесины и из-под отставшей коры, едят мелкие плоды, молодые листья и проростки и т. п.
Семейство включает всего 3 вида, относимых к 3 монотипичным родам: Callaeas cinerea (Gm.), Creadion (Philesturnus) carunculatus (Gm.) и, видимо, полностью вымершую (истребленную) в начале нашего века разноклювую гуйю Heteralocha acutirostris (Gid.) Иногда сюда же включают еще 1 вид тоже монотипичного рода Turnagra capensis Sp. Обычно это семейство сближают с семейством скворцовых (Уэтмор и др.); однако высказывались предположения о близости их к беседковым птицам Ptilonorhynchidae или ласточковым сорокопутам Artamidae.
Остатки всех ныне живущих видов найдены в плейстоценовых отложениях Новой Зеландии.
СЕМЕЙСТВО 58. STURNIDAE — СКВОРЦОВЫЕ
Очень подвижные наземно-древесные птицы (рис. 226) размером от воробья до галки (длина 17—45 см, вес большинства видов 50—150 а). Свойственны преимущественно открытым ландшафтам: лесостепи, безлесным предгорным и горным районам; некоторые виды проникают в полупустыни; заселяют и опушки леса. Освоили самые разнообразные культурные ландшафты. Распространены только в Старом Свете; сейчас несколько видов, в том числе и обыкновенный скворец Sturnus vulgaris L., акклиматизированы в Северной Америке, Новой Зеландии (рис. 222) и на ряде океанических островов.
Телосложение плотное. Клюв примерно равен длине головы, крепкий, заостренный, слабо изогнутый книзу. Ноздри открытые; над ними нависают голые или оперенные кожистые крышечки. Щетинок в углах рта нет. Ноги сильные, умеренной длины. Цевка спереди покрыта грубыми поперечными щитками, а с боков — слившимися пластинками. Сильные пальцы заканчиваются крепкими, острыми когтями. Крылья умеренной длины, заостренные или округлые. Первостепенных маховых 10, первое редуцированно и прикрыто кроющими кисти. Хвост из 12 рулевых, различной длины. Оперение густое, плотное. Часть видов имеет на голове хохол из удлиненных перьев. У некоторых видов уздечка или пространство около глаза не оперены и ярко окрашены. Окраска оперения чаще с преобладанием черного, коричневого, белого, с сильным металлическим блеском. Некоторые виды окрашены в очень яркие цвета: красный, синий и др. Самцы чуть крупнее самок и лишь у немногих видов более яркой окраски. Гнездовой наряд тусклый, без металлического блеска; окончательный наряд молодые одевают в первую осень жизни. У взрослых одна линька в году: полная послегнездо-
317
Рис. 226. Маскаренский скворец Fregilupus varius (Bodd.); вымер в конце XIX — начале XX вв.
вая. Обычное для некоторых видов, в том числе и для обыкновенного скворца сезонное изменение окраски происходит без линьки, за счет снашивания вершин контурного пера в период зимовки.
Самцы хорошо поют; у некоторых видов хорошо развита способность к имитации. У части видов поют и самки. Для многих видов характерно групповое или колониальное гнездование. Обычно рыхлое гнездо устраивается в укрытии: в дупле, трещине скалы, промоине обрыва, щелях построек и т. п. Охотно заселяют искусственные гнездовья. Несколько видов в обрывах роют неглубокие норы. Немногие виды гнездятся открыто, устраивая шарообразное, небрежно построенное гнездо с боковым входом на земле,
в основании куста или в развилке ветвей на разной высоте. Яйца однотонные, беловатые, голубоватые или зеленоватые; у части видов с коричневыми и красноватыми пятнами. В кладке чаще 4—6 яиц, у некоторых тропических видов 2—4. Инкубация около 2—3 недель. Насиживают оба партнера или только самка; птенцов выкармливают оба родителя. Птенцы оставляют гнездо через 2—4 недели после вылупления. Выводки кочуют с родителями; отдельные семьи объединяются в стаи, иногда включающие тысячи особей. Молодые становятся половозрелыми в возрасте менее года, но у части видов, особенно самцы, лишь в возрасте около 2 лет. В умеренных широтах пере-
летные, в тропиках кочующие; реже оседлые.
Полет быстрый, легкий. Хорошо ходят и бегают по земле, ловко прыгают с ветки на ветку. Кормятся на земле, стеблях тростника, в кронах кустов и деревьев. Пища разнообразная: насекомые и их личинки, моллюски, черви и другие беспозвоночные, мелкие плоды, семена, проростки растений и т. п. Крупные виды иногда ловят мелких ящериц, таскают яйца и маленьких птенцов из гнезд других птиц. Полезны уничтожением массовых видов вредных насекомых. Во время кочевок могут местами причинять существенный ущерб, расклевывая виноград, вишни и др.
Семейство Sturnidae объединяет 103—110 видов (в том числе 4 островных вида, вымерших в последнее столетие), распределяемых по 25—36 родам (из них около 20 монотипичны): Sturnus (14—16 видов), Acridotheres, Ampeliceps, Aplonis, Basilornis, Buphagus (Textor), Charitornis, Cinnyricin-clus, Cosmopsarus, Creatophora, Enodes, Fregilupus (вымер), Gracula, Gracupica, Hartlaubius, Rittlitzia (вымер), Lamprocolius (12 видов), Lamprotor-nis, Leucopsar, Melanopyrrhus, Mino, Necropsar (вымер), Onychognathus (10 видов), Pastor, Poeoptera, Rhinopsar, Sarcops, Saroglossa, Scissirostrum, Spodiopsar, Spreo, Streptocitta, Sturnia, Temenuchus.Буйволовых скворцов Buphagus (2 вида) иногда выделяют в отдельное подсемейство. Особенно разнообразен видовой состав скворцов в Африке и, несколько меньше, в Юго-Восточной Азии. В Австралии, Океании и умеренных широтах Евразии встречаются лишь немногие виды. В СССР отмечено 8 видов 5 родов; достоверно гнездится 6 видов.
318
Центр возникновения семейства, вероятно, Африка. Ископаемые остатки, относимые к 3 ископаемым и 6 ныне живущим видам, обнаружены в отложениях эоцена и более поздних слоях в пределах современного ареала. Иногда говорят о давних родственных связях скворцовых с иволговыми или ткачи новыми.
СЕМЕЙСТВО 59. MELIPHAGIDAE — МЕДОСОСОВЫЕ
Мелкие древесные и кустарниковые птицы (рис. 227) Австралийско-Папуасской области, проникающие на север до ряда островов Индонезии и до Гавайских островов; только один род встречается в Африке. По внешнему облику напоминают то колибри, то славок, пеночек или синиц, немногие относительно крупные виды похожи на мелких вороновых. У большинства видов длина 10—15 см, у немногих до 30 см и даже до 45 см (в последнем случае длина хвоста около 30 см).
Стройные птички с небольшой головкой. У большинства видов клювы длинные, тонкие, более или менее заметно изогнутые книзу, со слабо зазубренными режущими краями. У части видов клюв короткий, утолщенный, слегка вздутый. Щетинки в углах рта слабо развиты или их нет. Язык длинный, уплощенный, у многих видов при сосании нектара сворачивающийся в трубочку: на конце и по бокам языка развита бахромка сосочков. Кишечник короткий и широкий. На внутренней поверхности желудка есть глубокая борозда, по которой нектар, не задерживаясь, сразу стекает в кишечник; твердая пища перетирается в же'лудке и лишь потом
переходит в кишечник.
Ноги умеренной длины с довольно сильными цевками и пальцами; когти короткие, острые, сильно изогнутые. Крылья притупленные с 10 первостепенными маховыми. Хвост умеренной длины, у немногих видов удлиненный и даже очень длинный; рулевых 12. Оперение прилегающее, часто яркое (черного, красного, желтого, синего цвета) с сильным металлическим блеском. У некоторых видов развиваются украшающие перья на голове, шее, груди, иногда на плечах. У части видов четкий половой диморфизм: самцы
окрашены значительно ярче самок; у других видов окраска обеих полов сходна. Молодые более тусклые.
Очень подвижные, крикливые и драчливые птицы, живущие в лесах разных типов (в том числе и мангровых), зарослях кустарников; немногие виды населяют травянистые заросли по приморским дюнам и большим болотам. Большинство имеет звучную музыкальную песню. Некоторые виды способны к имитации чужих песен. У части видов самки поют почти так же хорошо, как и самцы. Открытое сверху чашеобразное гнездо с рыхлыми
стенками и мягкой выстилкой строится в развилке ветвей. Немногие виды вьют гнездо в зарослях травы, прикрепляя его к стеблям; одиночные виды строят гнездо в дуплах. Гнезд на земле, видимо, не бывает. Яйца белые, зеленоватые или розоватые, с коричневым или красноватым крапом. В тропиках в кладке 1—2 яйца, в субтропиках и умеренных широтах 3—4, редко 5. У многих видов в сезон размножения 2 кладки. У части видов насиживают только самка, иногда она одна выкармливает птенцов; у других видов оба
партнера насиживают и кормят птен- рис 227. Кардиналовый медосос Му-цов. Вне сезона размножения мелки- zomela cardinalis Lath.
319
ми стайками кочуют по гнездовой области; немногие виды перелетные.
Легко и быстро летают, но обычно лишь на короткие расстояния: от цветка к цветку. Ловко лазают в чаще ветвей или по стеблям крупных травянистых растений. Некоторые виды питаются преимущественно нектаром цветков и насекомыми, которых выклевывают из венчиков. Ареалы таких видов определяются распространением нескольких растений (эвкалипты и др.). Эта группа видов — типичные экологические викариаты колибри в Старом Свете; имеют большое значение как опылители. Другие виды наряду с питанием нектаром склевывают насекомых с ветвей и листьев, разыскивают их в трещинах коры, ловят, взлетая с присады, близко подлетевших насекомых, едят мелкие плоды и т. п. Самые крупные виды экологически напоминают иволг или мелких вороновых: кормятся в кронах, меньше на земле, поедая ягоды, различных беспозвоночных, при случае грабят гнезда мелких птиц, ловят мелких ящериц и лягушек и т. п.
В семействе медососовых сейчас насчитывают 162 вида (еще несколько видов вымерли в последние 1—2 столетия), группируемых в 35—50 родов (из них не менее 14 монотипичны). Наиболее обычны роды: Meliphaga (36 видов), Acanthorhynchus, Anthochaera, Anthornis, Apalopteron, Chaeto-ptila, Certhionyx, Conopophila, Entomyzon, Gliciphila, Lichmera, Mano-rina, Melipotes, Melithreptus, Moho, Myza, Myzantha, Myzomela (24 вида), Notiomystis, Philemon (16 видов), Prosthemadera, Ptilotus, Ramsayornis, Stomiopera и др. Многие виды свойственны Австралии (из них 49 — эндемики этого континента); несколько меньшее число видов характерно для Новой Гвинеи, Новой Зеландии и прилегающих островов. В семейство медососовых включают в ранге подсемейства 2 вида рода Promerops, распространенных в Южной Африке (скромно окрашенные, длинноклювые птицы размером со скворца с очень длинным хвостом). Некоторые систематики считают, что сходство этого рода с типичными медососами определяется конвергенцией и его положение в системе требует уточнения. Один вид — новозеландский туй Prosthemadera novaeseelandiae (Gm.) — в пределах современного ареала найден в плейстоценовых отложениях. Вероятный центр обособления медососовых — Австралия.
СЕМЕЙСТВО 60. NECTARINIIDAE — НЕКТАРНИЦЕВЫЕ
Мелкие (длина 9—25 см, вес 6—20 г) подвижные птицы тропиков и субтропиков Старого Света. Распространены в Африке южнее Сахары, на Мадагаскаре, в Юго-Восточной Азии, включая Новую Гвинею и прилегающие острова; немногие виды проникают в северо-восточные районы Австралии.
По повадкам несколько напоминают мелких синиц или пеночек, стройного сложения (рис. 228). Клюв длиннее головы, тонкий, изогнутый книзу и сжатый с боков; края надклювья имеют мелкие насечки. Язык длинный, тонкий, с продольным желобком. Ноги средней длины, с тонкими цевками И тонкими острыми коготками. Относительно короткие, туповершинные крылья имеют 10 первостепенных маховых. Хвост средней длины; у самцов части видов средние рулевые очень длинные. У большинства видов резкий половой диморфизм окраски: самцы яркие, с сильным металлическим отблеском; самки имеют более тусклое оперение. У немногих видов самцы окрашены как самки. Молодые похожи на самок.
Многие виды — хорошие певцы. Моногамы. Висячее гнездо с боковым входом прикрепляется к тонким ветвям дерева или куста. Для маскировки в наружные стенки вплетают мох. листья, кусочки коры, паутину. Лоток выстилают растительным пухом. Немногие виды строят открытые гнезда, помещая их в густых развилках ветвей. В кладке 1—2, редко 3 пятнистых яйца, отличающихся по окраске у разных видов. Инкубация около 2 недель. Птенцы вылетают из гнезда на 17—19-й день жизни. У части видов в году 2 и даже 3 кладки. Вне сезона размножения держатся одиночками, парами
320
или мелкими стайками. В ряде районов (горы, субтропики) перелетны, в других оседлые или кочующие.
Населяют леса разных типов, открытые ландшафты с куртинами кустарников; поднимаются в горы до верхней границы кустарниковой растительности; встречаются в культурных ландшафтах. Перелетая от цветка к цветку, садятся на веточки, высасывают нектар и выклевывают из венчиков цветков насекомых. Иногда короткое время могут сосать нектар, держась в воздухе около цветка в порхающем полете. Распространение семейства ограничено районами с обилием продолжительное время
Рис. 228. Манильская нектарница Cinnyris sperata (Sp.)
цветущих растений; некоторые виды тесно связаны лишь с немногими растениями. Выполняют роль опылителей.
Взлетая с присады, ловят в воздухе
близко подлетевших насекомых. Иногда, видимо, поедают мелкие ягоды, поч-
ки, бутоны. Вместе с медососовыми являются экологическим викариатом колибри.
Семейство нектарницевых Nectariniidae включает 105—108 видов, относимых к 8—11 родам: Nectarinia (более 10 видов), Aethopyga (14 видов), Anthreptes (16 видов), Arachnothera, Chalcomitra, Cinnyris (35 видов), Cyanomitra, Dreptes, Hydydipna, Leptocoma, Sanchezia. Наиболее разнообразен видовой состав нектарниц в Африке (где, видимо, и возникло это семейство); на островах Малайского архипелага живет 25 видов, и только 13 видов проникают на северо-восток Австралии.
СЕМЕЙСТВО 61. DICAEIDAE — ЦВЕТОЕДОВЫЕ
Мелкие (длина 7—10 см\ лишь один вид размером с мелкого дрозда) подвижные лесные птицы, распространенные от Индии и Южного Китая до Австралии и Тасмании (на Новой Зеландии нет).
По внешнему облику напоминают синиц, сложение коренастое (рис. 229). Клюв короче головы, прямой, более или менее конический, с острой вершиной, но с довольно широким и высоким основанием. Края
Рис. 229. Красногрудый цветоед Dicaeum ignipectus (Hodg.)
надклювья иногда слабо зазубрены. Язык может сворачиваться в трубочку, на конце бахромчатый или раздвоенный. Крылья округлые, из 9—10 первостепенных маховых (первое короткое или полностью скрыто под кроющими). Хвост короткий или умеренной длины. Оперение густое, мягкое. У многих видов резко выражен половой диморфизм окраски: самцы яркие (сочетание красного, желтого, синего и т. п.), самки буроватые. Молодые похожи на самок. У части видов относительно скромно окрашенных, молодые и самки лишь немного тусклее самцов.
321
Хорошо поют немногие виды. Экология изучена плохо. Плетут закрытые висячие гнезда с боковым входом, напоминающие гнезда ремезов, прикрепляя их к тонким ветвям. Несколько видов гнездится в естественных дуплах. В кладке 2—4 белых или сероватых яйца, иногда со слабой пятнистостью. У части видов самец, видимо, не участвует в постройке гнезда и в насиживании. Кормят птенцов оба родителя. Вне сезона размножения держатся семьями или парами, реже поодиночке. Обычно оседлы; немногие виды кочующие. Населяют густые дождевые леса; некоторые виды живут в кустарниковых и бамбуковых зарослях, проникают в культурный ландшафт. Держатся преимущественно в кронах. Питаются мелкими плодами, бутонами, цветками; склевывают насекомых и пауков. Многие виды выполняют функцию опылителей.
В семейство Dicaeidae включают 54—56 видов, распределяемых по 7 родам: Dicaeum (33—35 видов), Melanocharis (4—5 видов), Oreocharis, Paramythia, Pardalotus (7 видов), Prionochilus (6 видов), Rhamphocharis. В Гималаях встречается лишь 2 вида, на Филиппинах — 13 (из них 11 — эндемики), на Новой Гвинее — 11 (все эндемики).
Вероятный центр возникновения — Малайская подобласть или северные районы Австралийской области (Новая Гвинея).
СЕМЕЙСТВО 62. ZOSTEROPIDAE — БЕЛОГЛАЗКОВЫЕ
Мелкие (длина 10—14 см, вес 8—15 г) птицы (рис. 230), свойственные тропикам Старого Света; немногие виды проникают и в умеренные широты. Распространены в Африке южнее Сахары, Южной Азии, на островах Океании, в Австралии и Новой Зеландии (рис. 231).
По облику и повадкам похожи на пеночек. Клюв короче головы, тонкий, шилообразный, слегка выпуклый по коньку надклювья. Над щелевидными ноздрями нависают кожистые крышечки. В углах рта есть слабо развитые щетинки. Язык на конце раздвоен. Ноги сильные. Коготь заднего пальца очень длинный и сильно изогнут. Крылья умеренной длины, округлые или слабо заостренные. Первостепенных маховых 9. Хвост из 12 рулевых, прямо обрезанный или слегка выемчатый. Оперение мягкое, прилегающее. Окраска с преобладанием зеленых или желтых тонов; брюшко светлое. Почти у всех видов вокруг глаза белое кольцо из коротких белых шелковистых перышек. Сезонного, полового и возрастного морфизма окраски практически нет. В году одна линька: полная послегнездовая.
У самцов многих видов звучная, довольно сложная песенка. Встречаются в лесах разных типов, в кустарниковых и бамбуковых зарослях, чаще вблизи водоемов. Проникают в горы до верхней границы кустарников, заселяют культурные ландшафты. Открытое, глубокое чашевидное гнездо,
строится в развилке веток дерева или куста, иногда довольно высоко над землей. Лоток выстилается пухом, шерстью и т. п.; в наружные стенки
Рис. 230. Буробокая белоглазка Zosterops erythropleura Swinh.
вплетают для маскировки мох, кусочки коры. Кладки из 2—5, чаще 2—3 белых, голубоватых или зеленоватых яиц, обычно одноцветных. Строят гнездо, насиживают кладку и выкармливают птенцов оба партнера. Инкубация продолжается около 11—13 дней, птенцы вылетают из гнезд на 10—14-й день жизни. Вне сезона размножения держатся стайками; часто присоединяются к стайкам других мелких птиц (синиц и др.). В умеренных, отчасти в субтро-
322
Рис. 231. Ареалы семейств певчих воробьиных:
1— белоглазковые Zosteropidae, 2— американские славковые Parulidae, 3— ткачиковые Ploceidae, 4— районы акклиматизации домового воробья Passer domesticus (L •)
пических широтах перелетны, в тропиках оседлые или кочующие.
Кормятся в кронах деревьев и кустов, бойко шныряя в густых ветвях. Питаются преимущественно различными мелкими беспозвоночными (насекомые и их личинки, пауки и т. п.), склевывая их с листьев и ветвей; поедают мелкие ягоды и, вероятно, различные семена.
В семейство Zosteropidae включают 79—85 видов, распределяемых по 7—12 родам: Zosterops (qkqjiq 60 видов), Chlorocharis, Lophozosterops, Ма-danga, Rukia, Speirops, Woodfordia и др. Большинство видов очень сходны, а все семейство в целом весьма однородно. Вероятный центр возникновения— Индо-Малайская область. В СССР на Дальнем Востоке (Приморье) встречается и гнездится один вид — буробокая белоглазка Zosterops егу-thropleura Swinh.
Семейства Meliphagidae, Nectariniidae, Dicaeidae и Zosteropidae близки друг к другу по многим морфо-экологическим особенностям и, возможно, близко родственны. Иногда их объединяют в общее надсемейство — нек-тарницевые Старого Света Nectarinioidea.
СЕМЕЙСТВО 63. VIREONIDAE— ВИРЕОНОВЫЕ
Мелкие (длина 10—18 см) древесные птицы Северной и Южной Америки. По внешнему облику (рис. 232) напоминают крупных славок. Клюв чуть короче головы, сильный, прямой; надклювье слегка изогнуто книзу и образует на конце небольшой крючок. Ноздри щелевидные. Щетинки в углах рта не развиты. Ноги сильные. Цевка спереди покрыта не сливающимися поперечными щитками. Основные фаланги направленных вперед пальцев иногда более или менее сливаются друг с другом. Крылья короткие, тупые. Первостепенных маховых 10; первое короткое. Хвост средней длины. Оперение мягкое. Окраска самцов и самок более или менее сходна; молодые более тусклые. В году одна линька.
Населяют леса разных типов и кустарниковые заросли на равнинах и в горах. Многие виды хорошие певцы; песня свистового типа. Чашеобразное, с глубоким лотком, открытое сверху гнездо подвешивается к горизонтальной развилке ветви дерева или куста. Яйца с более или менее развитым крапом. В умеренных широтах в кладке 3—5, в тропиках 2—3 яйца. У разных видов степень участия самца в постройке гнезда и насиживании варьирует; у некоторых видов самцы иногда поют, обогревая яйца. Инкубация 11—13 дней. Кормят птенцов оба партнера. Птенцы вылетают из гнезда в возрасте 11—15 дней. Взрослые птицы продолжают кормить птенцов еще 3—4 недели. Вне сезона размножения держатся поодиночке или мелкими группами. В умеренных широтах перелетные, в тропиках — оседлые или кочующие. Кормятся преимущественно в кронах, склевывая с ветвей и лис-стьев разнообразных беспозвоночных (насекомые и их личинки, пауки, слизни ит. п.), едят мелкие мягкие плоды.
Семейство включает 42 вида, распределяемые по 4—7 родам: Vireo (около 20 видов; в США встречаются 12 видов), Hylo-
Vе-
Рис. 232. Белобровый виреон Vireo altiloquus (V.)
324
philus (15 видов), Laletes, Lawrencia, Neochloe, Vireosylva.
Остатки двух ныне живущих видов в пределах современного ареала найдены в верхне-плейстоценовых отложениях. Вероятный центр возникновения — Северная Америка. Иногда виреонов включают как подсемейство в семейство сорокопутовых Laniidae или в семейство славок Нового Света Parulidae. Чаще считают самостоятельным семейством; некоторые черты сходства с сорокопутами и славками, видимо, конвергентны.
СЕМЕЙСТВО 64. COEREBIDAE—АМЕРИКАНСКИЕ ЦВЕТОЧНИЦЫ
Мелкие птицы (длина 8—15 см\ рис. 233) тропической Америки; распространены от Мексики до Аргентины. По повадкам напоминают мелких
синиц.
Клюв удлиненный, тонкий, с острой вершиной, слегка изогнутый книзу. Ноздри открытые. Язык с желобком, на вершине бахромчатый. Крылья короткие с тупой вершиной. Первостепенных маховых 10; первое очень короткое. Хвост из 12 мягких рулевых, короткий или средней длины. Оперение густое, мягкое. Окраска яркая из сочетания синего, красного, желтого, черного. У части видов самцы заметно ярче самок, у других видов диморфизма окраски нет.
Держатся по опушкам и разреженным участкам леса, по куртинам кустов в саванне, заселяют сады и парки. Гнезда открытые, чашеобразные, в развилке ветвей или подвешенные к тонкой ветке, закрытые грушевидные с боковым входом. Маскируя гнездо, вплетают в
его наружные стенки свисающие около гнезда листья, паутину, мох. В кладке 2—3 пятнистых яйца. Строит гнездо и насиживает, видимо, только самка: самец ее иногда кормит. У некоторых видов самец по-
близости строит более примитивное гнездо, в котором ночует. Инкубация 12—13 дней. Выкармливают птенцов оба партнера, принося им разнооб-
разных насекомых. Птенцы вылетают из гнезда в возрасте около 2 недель. Вне сезона размножения держатся парами или мелкими стайками. Оседлые или кочующие по ограниченной территории.
Подвижные птицы, неутомимо шныряющие в гуще ветвей или перелетающие от цветка к цветку. Садясь на ветку рядом с цветком, просовывают в венчик тонкий клюв, высасывают нектар и склевывают пыльцу и сидящих
в цветке насекомых. Расклевывают мягкие плоды (многие виды — спелые бананы и т. п.).
К американским цветочницам относят 36—38 видов, распределяемых
по 7—12 родам: Coereba, Certhida, Chlorophanes, Со-nirostrum, Cyanerpes, Dacnis, Diglossa (10 видов) и др. Объем семейства требует уточнения. Взаимоотношения американских цветочниц с другими группами воробьиных неясны. Их включают в качестве подсемейства в семейство танагр Thraupidae, иногда сближают с овсянками Emberizinae. Некоторые роды, например Certhida, включают иногда в семейство американских славок Parulidae.
Рис. 233. Красноногая цветочница Cynaerpes cyaneus (L.)
325
Рис. 234. Форма клюва у гавайских цветочниц: / — Psittirostra bailleui (Oust.); // — Hemignathus wilsoni (Rotsch.); Ill — H. procerus (Licht.); IV — Loxops coccinea (Gm.); V — Vesiiaria coccinea (Forst.)
СЕМЕЙСТВО 65. DREPANIDIDAE — ГАВАЙСКИЕ ЦВЕТОЧНИЦЫ
Мелкие птички (длина 10—20 см, вес 10—30 г) Гавайских островов. По поведению и по внешнему виду напоминают синиц, медососов, пищух, вьюрков, 'Попугайчиков. Разнообразие внешнего облика и деталей строения—следствие резкой адаптивной радиации семейства, проходившей на ограниченной территории.
Клювы разнообразны (рис. 234); их можно свести к двум типам. Виды, имеющие длинные, тонкие клювы, сильно изогнутые книзу, пьют нектар, ловят в цвет
ках мелких насекомых, высасывают сок из сочных плодов и т. п. Виды, для которых характерен клюв короткий, прямой, заостренный или короткий, высокий и широкий, часто с крючком на конце, питаются преимущественно семенами. У длинноклювых, пьющих нектар, язык длинный, тонкий, сворачивающийся в трубочку, — служит для сосания. У короткоклювых семеноядных форм язык короткий, иногда с плотным режущим краем — напоминает язык клестов (служит для обрезания летучек семян и т. п.). Крылья относительно короткие, с закругленными вершинами; первостепенных маховых 10, первое сильно укорочено. Хвост умеренной длины. Оперение прилегающее. Окраска разнообразная, однотонная или яркая: черная, зеленая, иногда красная и т. п. У части видов отчетливый половой диморфизм окраски, у других самцы похожи на самок. В году одна линька.
Чашеобразные гнезда укрепляют в развилке ветвей дерева или куста, иногда в густых зарослях травы. В кладке 2—3 белых, обычно с сильным крапом яйца. Населяют разного типа леса, отдельные рощи и заросли кустарников, проникают высоко в горы. Близкие виды отличаются по местообитаниям, пищевой специализации и другим чертам экологии. Ареалы очень ограничены: некоторые виды населяют лишь определенные ландшафты одного небольшого острова.
Семейство включает 21—22 вида, распределяемых по 7—11 родам: Drepanis (2 вида), Ciridops (1 вид), Hemignathus (3 вида), Himatione (1 вид), Loxops (1 вид), Рaimeria (1 вид), Paroreomyza (1 вид), Pseudonestor (1 вид), Psittirostra (6 видов), Vesiiaria (1—2 вида), Viridonia (3 вида). В настоящее, время не менее 7—8 видов полностью исчезли: вымерли из-за изменения местообитаний в результате хозяйственной деятельности или истреблены акклиматизированными видами (крысы, кошки и др.) и человеком (шкурки шли на украшения).
Предполагают, что семейство гавайских цветочниц возникло от вселившегося на острова и эволюционировавшего по нескольким направлениям какого-то одного американского вида овсянок, вьюрков, танагр или американских цветочниц. Адаптивная радиация потомков этого вселенца шла по пути пищевой специализации. В итоге образовались близкие виды, резко различающиеся друг от друга по внешнему облику (клювы, окраска) и повадкам, по предпочитаемым местообитаниям. Это один из ярких примеров интенсивного видообразования в условиях островной изоляции.
326
СЕМЕЙСТВО 66. PARULIDAE — АМЕРИКАНСКИЕ СЛАВКОВЫЕ, ИЛИ ЛЕСНЫЕ ПЕВУНЫ
Мелкие птицы (размеры от пеночки до воробья; длина 10—20 см, вес 10—30 г) американского континента, распространенные от тундр Северной Америки (немногие виды) до Аргентины (рис. 231). По внешнему облику (рис. 235) и повадкам напоминают славок и пеночек, мухоловок, некоторые виды — мелких дроздов.
Стройные птички. Клюв короче головы, тонкий и острый, прямой или слегка изогнутый, у некоторых видов у основания довольно широкий и приплюснутый. Над ноздрями нависают перепончатые крышечки. В углах рта есть жесткие щетинки; их длина и количество варьируют у разных видов. Ноги умеренной длины, сильные. Когти маленькие, сильно изогнутые; коготь заднего пальца длиннее остальных. Крылья умеренной длины с заостренными вершинами. Первостепенных маховых 9—10. Хвост из 12 рулевых умеренной длины (короче крыла). Оперение мягкое. Окраска разнообразная, у многих видов яркая, чаще из сочетания черного, желтого, зеленого, белого цветов; у немногих видов — красного, голубого и др. У видов умеренных и северных широт выражен половой и сезонный морфизм окраски: самцы ярче самок, а после полной послегнездовой линьки сходны с самками. Ранней весной после частичной линьки самцы вновь одевают брачный наряд. В тропиках у большинства видов сезонного и полового морфизма окраски нет, и в году, видимо, лишь одна линька (послегнездовая).
Населяют леса разных типов (в том числе и дождевые), отдельные рощицы и заросли кустарников на полях, лугах, болотах, некоторые виды проникают в полупустыни и горные районы. Большинство видов ведут древесный образ жизни, многие виды открытых ландшафтов держатся на земле, лишь изредка присаживаясь на ветви. Песня простая, только немногие виды хорошие певцы. Гнездо чашеобразное или шарообразное с боковым вхо--дом, устраивается на земле или в развилке ветвей, обычно невысоко над землей, но некоторые виды гнездятся в кронах высоких деревьев. Немногие устраивают гнездо в дуплах или полудуплах. Гнездо строит преимущественно самка. Яйца беловатые, с более или менее выраженным крапом. В тропиках в кладке 2—4, в более северных широтах 3—6 яиц. Засиживает, видимо, только самка — в тропиках 13—17 дней, в северных широтах — 11—13. Птенцов кормят оба партнера. Молодые вылетают из гнезд в возрасте 12—15 дней в тропиках и 9—12 дней в умеренных и северных широтах. Вне сезона размножения держатся мелкими группами и поодиночке. В умеренных и северных широтах перелетные, в тропиках — оседлые.
Кормятся различными мелкими беспозвоночными, склевывая их с ветвей и листьев, едят ягоды. Немногие виды ловят летающих насекомых, охотясь за ними «по-мухоловочьи».
Рис. 235. Желтая американская славка Dendroica petechia (L.)
327
В семейство Parulidae (раньше оно носило другие названия: Compso-thlypidae или Mniotiltidae) включают 108—113 видов, относимых к 18—26 родам: Parula (2 вида), Basileuterus (22—23 вида), Cardellina, Catharopeza, Chamaethlypis, Dendroica (26—27 видов), Ergaticus, Geothlypis (9 видов), Granatellus, Helmitheros, Icteria, Leucopeza, Limnothlypis, Mniotilta, Myio-borus (10 видов), Oporornis, Peucedramus, Protonotaria, Seiurus, Setophaga, Vermivora (11 видов), Wilsonia и др. В США живет 59 видов (из них 42 — эндемики Северной Америки), несколько эндемичных родов характерно для островов Вест-Индии. В СССР на Чукотке встречаются 2 вида 2 родов; характер пребывания неясен: залетные, или, может быть, в очень небольшом числе гнездящиеся.
Остатки 4 ныне живущих видов обнаружены в отложениях среднего плейстоцена. Вероятный центр обособления Северная Америка. Иногда американских славок в ранге подсемейства включают в семейство танагр Thraupidae.
СЕМЕЙСТВО 67. PLOCEIDAE—ТКАЧИКОВЫЕ
Большое семейство, объединяющее довольно разнообразных по внешнему облику мелких птиц размером от пеночки до дрозда (вес 7—100 а, длина 7—30 см, у некоторых видов с очень длинным хвостом — до 50— 60 см). Свойственны всему Старому Свету (рис. 231); акклиматизированный домовый воробей Passer domesticus (L.) широко расселился по Северной и Южной Америке, Юго-Восточной Австралии, Новой Зеландии и на ряде океанических островов. В некоторых районах акклиматизировано еще несколько видов ткачиковых.
Морфологически очень сходны с вьюрковыми, отличаются от них лишь мелкими деталями строения и экологии. Телосложение плотное, голова округлая, довольно большая, но часть видов со стройным телом и маленькой головкой. Форма клюва очень варьирует. Чаще клюв конический, иногда несколько удлиненный, дугообразно изогнутый по коньку, у основания вздутый или несколько приплюснутый; вершина острая, без крючка. У некоторых видов Мощный клюв похож на клюв вьюрков и даже дубоносов. Ноздри округлые, сдвинуты к гребню клюва, обычно открытые, изредка прикрытые мелкими перышками. Характерно (отличие ткачиковых от вьюрковых) наличие трех продольных валиков на небе. Ноги средней длины или короткие, сильные с крепкими пальцами, покрытыми грубыми щитками. Крылья средней длины с округлой или, редко, слегка заостренной вершиной. Первостепенных маховых 10; первое очень варьирует по длине: от короткого, скрытого кроющими кисти, до длинного, превышающего половину длины второго. Хвост из 12 рулевых, средней длины, прямо обрезанный или округлый; у немногих видов (вдовушки, некоторые ткачики) очень удлиненные и широкие все рулевые или только две средних пары образуют пышный, «фазаньего» типа хвост.
Оперение плотное. Окраска разнообразна. Живущие в Палеарктике виды имеют буроватую окраску с коричневыми, белыми и черцыми пятнами и пестринами. В тропиках большинство видов с яркой, контрастной окраской. У многих видов более или менее четко выражен половой, возрастной и сезонный морфизм окраски; самцы ярче самок. Такие виды обычно имеют 2 линьки в году: полную послегнездовую и частичную весеннюю (предбрачную). У других видов отчетливый половой и возрастной морфизм окраски, но сезонной смены нарядов не бывает. Наконец, у некоторых видов самцы чуть крупнее самок, но окрашены одинаково; молодые более тусклые. У таких видов в году только одна полная послегнездовая линька.
Подавляющее большинство ткачиковых свойственны тропикам и субтропикам; относительно немногие виды проникают в умеренные широты. Заселяют разнообразные ландшафты на равнинах и в горах: опушки и разреженные участки лесов разных типов (в глухие леса обычно не проникают),
328
степи, саванны, пустыни с изреженной кустарниковой растительностью, берега водоемов, заросли по болотам и т. п. Довольно много типичных синантропов, живущих в культурных ландшафтах и поселениях человека. По земле и в ветвях передвигаются быстрыми прыжками. Некоторые виды довольно хорошие летуны, другие летают плохо и неохотно.
Песня — чириканье; немногие виды хорошие певцы. Для многих видов характерно групповое гнездование: гнезда отдельных пар расположены близко одно от другого. Часть видов — настоящие колониальные птицы. Особенно ярким примером может быть живущий в саваннах Африки красноклювый ткачик Quelea quelea (L.), некоторые колонии которого занимают 20—30 га и насчитывают по несколько миллионов гнездящихся пар; на одном дереве бывает до 400—600 гнезд, а на 1 ж2 зарослей тростника до 30— 40 гнезд. Отношения полов в период размножения весьма разнообразны. Многие виды — типичные моногамы, у которых оба партнера в той или иной степени участвуют в постройке гнезда, насиживании кладки и выкармливании птенцов. У некоторых видов строят гнезда и насиживают только самки, выкармливают птенцов оба партнера. Наконец, есть и типичные полигамы (буйволовые птицы, некоторые ткачи), у которых при половозрелом самце держится несколько самок, каждая из которых несет все заботы по постройке гнезда, насиживанию и выкармливанию; кажется, иногда самец участвует в выкармливании птенцов. Несколько видов (ткач из рода Anomalospiza, вдовушки Viduinae) — типичные гнездовые паразиты, подкладывающие яйца в гнезда других птиц, обычно ткачиковых. Их птенцы не выбрасывают яйца и птенцов приемных родителей, как это делают птенцы кукушек.
Гнезда разнообразны. У типичных воробьев и ряда других видов рыхлое, толстое, небрежно построенное шарообразное гнездо помещается в густой развилке ветвей или укрытии (дупло, щель в скале или постройке, промоина обрыва и т. п.). Многие ткачи и вьюрковые ткачики из сухой травы искусно вьют прочные шарообразные или грушевидные гнезда с боковым входом — трубкой (ее длина иногда достигает 50 см); изнутри оно высти-
Рис. 236. Африканский общественный воробей Philetairus socius (Lath.); справа общее гнездовое сооружение, входы в индивидуальные гнезда снизу
329
лается мягкой растительной ветошью, шерстью, перьями. Гнездо подвешивают на тонких ветвях или на крепких стеблях высокой травы. Некоторые виды строят общественные гнезда. Так, у африканского общественного воробья (рис. 236) несколько десятков пар птиц (иногда до 200—400) на ветвях одинокого дерева или куста в саванне совместно строят толстую крышу из сухой травы, под которой тесно друг к другу располагаются индивидуальные гнезда отдельных пар, входы в которые находятся внизу. Такие общие «гнезда» достигают 5—7 м в диаметре и до 3 м высоты; они служат защитой и от ряда хищников и, что, видимо, важнее, от сильных порывов ветра и ливней. У многих видов гнезда используются несколько лет; перед каждым гнездованием их ремонтируют. Некоторые виды гнезд не строят, а занимают брошенные гнезда других птиц.
Размеры кладок сильно варьируют: в одном районе у части видов в кладке 2—3 яйца, а у других 5—7, редко до 9. Яйца либо одноцветные белые или зеленоватые, либо с более или менее развитым крапом. Очень варьирует у разных видов и продолжительность гнездового периода. Инкубация продолжается от 10 до 17 дней, птенцы вылетают из гнезд в возрасте 12—15 дней у одних видов и лишь на 20—28-й день жизни у других. После вылета взрослые некоторое время продолжают кормить птенцов. Потом выводки молодых объединяются в «молодежные» стайки, а взрослые приступают к следующему циклу размножения: у ряда видов как в тропиках, так и в умеренных широтах бывает 2, реже даже 3 кладки в году. Другие виды моноцикличны. У некоторых тропических видов молодые приступают к размножению в возрасте 4—5 месяцев, у большинства видов в 9—10 месяцев, а вдовушки, некоторые ткачи и вьюрковые ткачики, видимо, только а возрасте около 2 лет.
Вне сезона размножения держатся стайками или большими стаями иногда из десятков тысяч особей (красноклювый ткачик). Большинство видов оседлы и вне периода размножения кочуют в райойе гнездовий, у других протяженность кочевок или перелетов может достигать сотен километров. Домовый (городской) воробей Passer domesticus (L.) на большей части своего громадного ареала, охватывающего почти веку Евразию, оседлый или (в ряде северных районов) кочующий, но живущий в Средней Азии подвид Р. d. bactrianus Zar. (его вместе с некоторыми другими подвидами иногда выделяют в самостоятельный вид Passer indicus Jard.) — настоящая перелетная птица.
Большинство видов преимущественно зерноядны, хотя поедают и ягоды, разнообразных насекомых и других беспозвоночных, весной склевывают проростки, почки и бутоны. Многие виды выкармливают птенцов преимущественно или исключительно насекомыми. Немногие тропические виды в основном насекомоядны. При большой численности в некоторых районах могут быть более или менее серьезными вредителями. Так, в южных районах СССР местами причиняют существенный ущерб, поедая созревающие семена зерновых (просо, ячмень и др.) и расклевывая вишни, виноград, абрикосы и т. п. В саваннах Западной Африки по приблизительным подсчетам полуторамиллиардная популяция красноклювого ткачика местами, уничтожает до 50% урожая, поедая около 1 млн. т зерна в год. С этим видом приходится вести напряженную истребительную борьбу.
Семейство ткачиковых Ploceidae включает 247—272 вида, группируемых в 37—68 родов. Подавляющее большинство видов свойственно тропикам, почти 4/5 всех видов — обитатели Африки. В СССР встречается 11— 12 видов 5 родов; несомненно гнездится 10—11 видов (11, если среднеазиатскую форму домового воробья выделяют в самостоятельный вид).
Большое число видов и эколого-морфологическое разнообразие отдельных групп вынуждают выделять внутри семейства ткачиковых от 4 до 11 подсемейств. Чаще сейчас принимают следующие подсемейства.
1. Буйволовые птицы Bubalornithinae; 2—3 африканских вида в 2 родах: Bubalornis, Dinemellia.
330
2. Типичные воробьи Passerinae; 38—42 вида в 9—10 родах: Passer (15 видов), Carpospiza, Histurgops, Montif ringilia, Petronia, Philetairus, Plocepasser, Pseudonigrita, Pyrgilauda, Sporopipes. Распространены в Африке и Евразии, в СССР встречаются представители только этого подсемейства.
3. Типичные ткачикиР1осе!пае; 92—103 вида в 10—16 родах: Ploceus (57 видов), Amblyospiza, Anaplectes, Anomalospiza, Brachy-cape, Coliuspasser (7 видов), Drepanoplectes, Eupledes (11 видов), Foudia, Hyphanturgus Malimbus (10 видов), Melanoploceus, Neospiza (1 вид, жил на острове в Гвинейском заливе, вымер в конце XIX в.), Niobella, Quelea. Живут в Африке южнее Сахары и на Мадагаскаре, лишь 5—6 видов Ploceus населяют Индо-Малайскую область.
4. Вдовушки Viduinae, 8—9 видов в 1—5 родах: Vidua, Hypochera, Linura, Sieganura, Tetraenura. Встречаются только в Африке южнее. Сахары.
5. Вьюрковые ткачики Estrildinae; 107—115 видов в 15—35 родах: Eslrilda (10 видов) Aegintha, Aidemosyne, Amadina, Атап-dava, Baihilda, Clyiospiza, Cripiospiza, Erythrura, Euodice, Granaiina, Heieromunia, Hypargos, Lagonosiicta (9 видов), Lonchura (24 вида), Neo-chmia, Nesocharis, Nigrita, Oreostruthus, Oriygospiza, Parmoptila, Pholi-dornis, Pirenestes, Poephila, Pytilia, Radda, Rhamphostruthus, Spermesies, Spprmophaga, Sporaeginthus, Steganopleura, Taeniopygia, Uraeginihus, Zo-naeginthus. Распространены в Африке, на Мадагаскаре, в Юго-Восточной Азии, Океании, Австралии.
Иногда последние 2 подсемейства выделяют в самостоятельные семейства: вдовушек и вьюрковых ткачиков.
Центр возникновения, несомненно, Старый Свет, вероятнее всего — Африка. Немногочисленные палеонтологические остатки известны с нижнего миоцена; их относят к 2 ископаемым видам современных родов и к нескольким ныне живущим видам. Высказываются предположения о давней близости ткачиковых к скворцовым, иволговым и дронго; для подтверждения необходимы специальные исследования.
СЕМЕЙСТВО 68. ICTERIDAE — ТРУПИАЛОВЫЕ (АМЕРИКАНСКИЕ ИВОЛГИ, КАССИКИ)
Размеры от воробья до галки (вес 20—400 г, длина 15—55 см). По внешнему облику и повадкам очень разнообразны: напоминают иволг, скворцов, жаворонков, мелких вороновых. Крепкие, подвижные птицы (рис. 237), распространенные в Новом Свете от Аляски и Южной Канады до Огненной Земли и островов Вест-Индии.
Клювы разной длины: у немногих видов короткие, чаще удлиненные: чуть короче или немного длиннее головы. Обычно клюв конический с острой вершиной и четко выраженным гребнем на надклювье и подклювье, прямой или несколько изогнутый книзу. Над открытыми ноздрями нависают кожистые крышечки. Щетинок в углах рта нет. Ноги сильные. Цевка спереди покрыта грубыми поперечными пластинками. Покров основной фаланги среднего пальца срастается с покровом основной фаланги наружного (но не внутреннего) пальца. Крылья удлиненные, с заостренными вершинами. Первостепенных маховых 9; первое почти достигает вершины крыла. Хвост из 12 рулевых, средней длины, у некоторых видов длинный. Оперение прилегающее. У части видов окраска черная или черно-бурая с сильным металлическим блеском. У других яркая окраска из сочетания черного, желтого, красного, оранжевого цветов. У молодых заметна поперечная исчерченность, во взрослом состоянии сохраняющаяся лишь у немногих видов. У большинства видов более или менее выражен половой и возрастной морфизм: самцы крупнее самок (иногда почти в 2 раза) и ярче окрашены,
331
Рис. 237. Черногорлый трупиал Icterus gularis Wagl.
молодые похожи на самок, но тусклее. У немногих видов есть и сезонное изменение окраски.
Большинство видов связано с древесными и кустарниковыми зарослями и населяют леса разных типов, прерии, луга, болота, культурные ландшафты. Немногие виды ведут наземный образ жизни в открытых прериях, лугах, полях.
Многие — хорошие певцы; у некоторых видов поют и самки. Некоторые виды колониальны: десятки и сотни гнезд располагаются в непосредственной близости; в редких случаях в колониях бывает до нескольких тысяч гнезд. Взаимоотношения полов в период размножения весьма разнообразны. Большинство видов — типичные моногамы. Самец выбирает место гнезда, помогает самке его строить, кормит насиживающую самку; основные заботы по постройке гнезда и насиживанию падают на самку. Птенцов выкармливают оба партнера. Некоторые виды — полигамы: около самца группируется от 2 до 10 самок; каждая из них после спаривания строит гнездо на участке самца, самостоятельно насиживает кладку и выкармливает птенцов. Среди колониальных видов встречаются и моногамы, и полигамы. У некоторых видов в зависимости от соотношения полов в данной колонии может быть или моногамия, или полигамия.
Немногие виды открытое чашеобразное гнездо устраивают на земле под кустиком травы, у кочки и т. п.; большее число строит гнезда в кронах деревьев и кустов, в зарослях тростника и камыша, на высоких стеблях в густой траве. У таких видов гнезда двух типов: глубокая, открытая сверху чаша или прочно сплетенное удлиненно-грушевидное сооружение (длиной иногда до 80—100 см) с боковым входом, подвешенное к тонким ветвям, часто над водой. У черноголового кассика Psarocolius decumanus 10—15 самок и 1—2 самца в течение 20—30 дней строят общее гнездо, в которое затем каждая самка откладывает 1—2 яйца. Насиживают общую кладку и выкармливают птенцов совместно.
Яйца белые или зеленоватые, пятнистые. В тропиках в кладке 2—3, в умеренных широтах 4—6, редко 8 яиц. Насиживание продолжается И— 14 дней. Возраст, в котором птенцы вылетают из гнезда, варьирует у разных видов: 2, 3, даже 4—5 недель. У части видов в сезон 2, редко 3 кладки. У видов многих родов выражена первая степень гнездового паразитизма: они занимают брошенные гнезда других птиц, иногда выгоняют хозяев и занимают их гнездо. У Molothrus badius (V.) пара строит гнездо, насиживает
332
кладку, выкармливает птенцов. Остальные 3 вида этого рода и виды моно-типичных родов Psomocolax и Tangavius — типичные гнездовые паразиты подкладывающие свои яйца в гнезда других птиц (преимущественно близких видов). На этом вся их забота о потомстве кончается.
Полет быстрый. Легко передвигаются в кронах, лазают в густых зарослях тростника, многие охотно и легко бегают по земле. Кормятся на земле и в кронах; немногие — только на земле. Большинство видов всеядны с хорошо выраженной сезонной изменчивостью питания: поедают различных беспозвоночных и мелких позвоночных (лягушек, ящериц, мышевидных грызунов, таскают яйца и птенцов и т. п.), разнообразные семена, мелкие плоды. Местами причиняют ущерб посевам зерновых и фруктовым садам. Немногие виды почти исключительно насекомоядны. Держатся преимущественно стаями. В тропиках оседлые или кочующие, в северных районах перелетные.
Семейство трупиалов Icteridae объединяет 87—93 вида, группируемые в 22—35 родов; Icterus (около 30 видов), Agelaius (7 видов), A mblycercus, Amblyrhampus, Cacicus, Cassidix, Dives, Dolichonyx, Euphagus, Gymnast inops, Leistes, Molothrus (4 вида), Ocyalus, Ostinops, Pezites, Psarocolius, Pseudoleistes, Psomocolax, Ptiloxena, Quiscalus, Scaphidura, Sturnella,Tangavius, Xanthocephalus, Zarhynchus и др.
Вероятный центр возникновения — Южная Америка. Ископаемые остатки из среднего плейстоцена и более поздних отложений относят к 6 ископаемым и к 13 ныне живущим видам. Большинство современных видов свойственно тропикам, относительно немногие проникают в умеренные широты (в США встречается 20 видов; Полярного круга достигает лишь один). В СССР* как залетная птица встречен 1 вид (на Чукотке).
СЕМЕЙСТВО 69. TERSINIDAE — ЛАСТОЧКОВЫЕ ТАНАГРЫ
Монотипическое семейство, включающее единственный вид — ласточ-ковую танагру Tersina viridis (Ill.), распространенную в северной половине Южной Америки от Панамы до северной Аргентины.
Довольно коренастые птицы (рис. 238) длиной около 15 см и весом 30 г. Широкий у основания клюв сильно уплощен; на конце надклювья маленький крючок. Ноздри в коротких трубочках. В углах рта развиты щетинки. Ноги укороченные с тонкими цевками и пальцами. Крылья удлиненные, заостренные. Хвост выемчатый. Самец бирюзово-голубой с черным горлом и лбом, белым брюхом и черными полосами на боках, самка зеленая сверху и желтая снизу, с черной маской.
Широко кочуют. В период раз-
множения приурочены к осветленным лесам, в остальное время к открытым болотистым равнинам, опушкам лесов, садам. Песня примитивная. Пара активно охраняет гнездовой участок. Рыхлое гнездо строится в дупле дерева, под камнями, в промоинах обрывов, иногда птицы роют короткие норы или занимают пустые норы грызунов. В кладке 2—3 белых яйца. Насиживает только самка, около 15 дней. Птенцы остаются в гнезде 24 дня; их кормит самка и немного самец. У части пар 2 кладки в году. Молодые похожи по окраске на самок. Окончательный яркий наряд самцы одевают лишь на 3—4-й год жизни. Пища смешанная. Подоб-
Рис. 238. Ласточковая танагра Tersina viridis (Ill.)
333
но ласточкам ловят разнообразных насекомых в воздухе, склевывают их на ветвях и листьях. Поедают мелкие плоды, почки, бутоны; птица долго мнет их в клюве, как бы «пережевывая».
Остатки вида найдены в отложениях верхнего плейстоцена Бразилии. Иногда ласточковых танагр в качестве подсемейства или трибы присоединяют к овсянковым, реже к танаграм. Систематическое положение вида требует уточнения.
СЕМЕЙСТВО 70. THRAUPIDAE— ТАНАГРОВЫЕ
Подвижные, коренастые (рис. 239), мелкие (длина 8—30 см) древесные птицы Нового Света, распространенные от Южной Канады до Аргентины; обычны на островах Вест-Индии. По внешнему виду и повадкам напоминают овсянок и вьюрков (от которых нечетко обособлены и по морфологическим признакам).
Клюв конический, удлиненный (у части видов почти равен длине головы), у основания слегка вздутый, иногда слегка изогнутый книзу. На режущей стороне надклювья более или менее выражена пологая предвершинная вырезка; надклювье может заканчиваться слабо изогнутым крючком. Ноздри открытые. Цевки крепкие, средней длины. Пальцы и когти относительно короткие, но крепкие. Первостепенных маховых 9. Крылья заостренные или закругленные. Хвост умеренной длины, у немногих видов длинный ступенчатый. Оперение плотное, прилегающее. Окраска яркая: желтая, зеленая, красная, синяя или из сочетания этих цветов. У многих видов хорошо развит металлический отблеск. Обычно самцы ярч§ самок, реже окраска обоих полов сходна. У части видов после размноженйя самцы одевают наряд, похожий на оперение самок, а потом, после предбрачной линьки, — вновь брачный наряд. Молодые похожи на самок.
Основные местообитания — леса разных типов, кустарниковые заросли, сады и парки, заросли тростников по болотам и берегам водоемов, луга с куртинами кустов и т. п. Большинство видов — хорошие певцы. Открытые чашеобразные гнезда, нередко небрежно построенные, устраивают в развилке ветвей высоко над землей; реже строят в кронах низких деревьев и кустов, заломах тростника. Немногие виды строят закрытые шарообразные гнезда с боковым входом,' некоторые виды гнездятся в дуплах. Гнездо строят оба партнера или у части видов только самка. Некоторые виды занимают гнезда других птиц, выбрасывая из них яйца и птенцов. Яйца пятнистые, с белым, зеленоватым, голубоватым ил« кремовым фоном. У многих видов в кладке 2, редко 3 яйца, у других 4—5. Обычно насиживает только самка; самец поет на гнездовом участке и, у ряда видов, приносит самке корм. Инкубация 12—14 дней, редко больше. Вылупившихся птенцов греет только самка; кормят оба партнера. В выкармливании птенцов, кроме родителей, иногда участвуют и другие взрослые особи, видимо, по каким-то причинам не имеющие своих гнезд. Вылет птенцов из гнезда
Рис. 239. Алая танагра Piranga olivacea (Gm.)
334
у разных видов происходит на 10—13, 14—20, 17—24-й день жизни. Вне сезона размножения держатся одиночками или парами, немногие виды — стайками. Перелетные и кочующие, часть видов в тропиках оседлые. Многие виды ведут довольно скрытную жизнь.
Полет быстрый. Ловко двигаются в чаще ветвей, зарослях тростника и высокой травы. На земле обычно не кормятся. Питаются преимущественно сочными мягкими плодами, почками и бутонами. Наряду с этим, видимо, все виды в той или иной степени поедают насекомых, которых ловят в воздухе и склевывают с ветвей и листьев. Некоторые виды разоряют гнезда ос, поедая личинок. Местами могут причинять ущерб плантациям: расклевывают спелые бананы, апельсины и другие фрукты.
Семейство танагровых Thraupidae (=Tanagridae) объединяет 192—205 видов, группируемых в 48—61 род: Thraupis (8 видов), Anisognathus, Chlo-rochrysa, Chlorophonia, Chlorornis, Chlorospingtis, Cissopis, Eucometis, Eup-honia, Habia, Hemispingus (9 видов), Hemithraupis, Lanio, Nemosia, Orc-hesticus, Piranga (9 видов), Ramphocelus (7 видов), Rodinocichla, Sericos-sypha, Spindalis, Stephanophorus, Tachyphonus (9 видов), Tanagrella, Tangara (=Calospiza; около 45 видов), Thypopsis к Подавляющее большинство видов свойственно тропической зоне. На территории США встречается 5 видов.
Иногда танагровых как подсемейство включают в семейство овсянковых Emberizidae (широко понимаемое и объединяющее также Catamblyrhyn-chidae, Tersinidae, Coerebidae). Другие систематики придают семейству танагровых значительно больший объем и включают в него, кроме танагр, в качестве подсемейств Parulidae, Coerebidae, Catamblyrhynchidae, Pyrrhu-loxiinae (кардиналы; Уэтмор их объединяет с овсянками и вьюрками) и иногда Icteridae. Систематическое положение всех этих групп и характер взаимосвязей между ними требует уточнения.
СЕМЕЙСТВО 71. САТАМВЬУ RHY NCHIDAE — ПЛЮШЕВОГОЛОВЫЕ ВЬЮРКИ
Монотипичное семейство, включающее единственный вид Caiamblyr-hynchus diodetna (Lafr.), населяющий предгорные районы Анд от Колумбии и Венесуэлы до Боливии. По внешнему виду (рис. 240) похож на вьюрков. Длина 14—15 сж.
Клюв короткий, высокий, конический, слегка сжатый с боков. Крылья короткие, закругленные. Хвост средней длины с небольшой вырезкой. Сверху темный голубовато-серый, снизу каштаново-коричневый; лоб и передняя часть темени желтые, на затылке черное пятно. Перья лба и темени слегка приподняты, поэтому оперение головы имеет бархатистый вид (отсюда и название на всех европейских языках).
Экология неизвестна. Скрытные видимо, повсюду редкие птицы, дер-
жащиеся поодиночке или парами в кустарниковых зарослях, рощах, изре-женных лесах.
Систематическое положение неясно. Часто в ранге подсемейства включают в семейство танагровых или овсянковых.
СЕМЕЙСТВО 72. FRINGILLIDAE — ВЬЮРКОВЫЕ
Очень большое, нечетко еще установленное семейство, объединяющее мелких птиц размером от пеночки до дрозда (длина 8—25 см, вес 7—100 г),
Рис. 240. Плюшевоголовый вьюрок Catamblyrhynchus diadema (Lafr.)
335
150° 120° 90° 60° 30° cP 30° 60° 90° 120° 150° 180° 150°
Рис. 241. Ареалы подсемейств семейства вьюрковых Fringillidae:
/— овсянковые Emberizinae, 2— дарвиновы, или галапагосские, вьюрковые Geospizinae, 3— настоящие вьюрковые Fringillinae
довольно резко различающихся по внешнему виду. Занимают самые разнообразные местообитания на равнинах и в горах: тундры, леса, луга, степи, пустыни, скалистые горные склоны, болота и берега водоемов; многие виды живут в культурных ландшафтах. Распространены очень широко в Африке, Палеарктике, Америке (рис. 241); нет на Мадагаскаре, в Австралии, Новой Зеландии (там и на некоторых океанических островах акклиматизировано несколько видов); в Индо-Малайскую область проникают немногие виды, которые распространены очень локально.
Сложение плотное. Голова круглая, довольно большая, шея короткая. Клюв крепкий, за немногими исключениями более или менее конический, с острым концом, у основания широкий и высокий, иногда несколько вздутый. Ноздри частично прикрыты мелкими волосовидными перышками. Жестких щетинок в углах рта нет. Ноги умеренной длины, крепкие. Когти небольшие, сильно изогнутые. Крылья средней длины, округлые или слегка заостренные. Первостепенных маховых 9: первое скрыто под кроющими кисти и в расчет не принимается. Хвост из 12 рулевых, средней длины, реже короткий или удлиненный, прямо обрезанный, слабо округлый или выемчатый, редко ступенчатый. Оперение густое, мягкое или плотное, прилегающее. У большинства видов выражен половой диморфизм; самцы крупнее и ярче самок. Птенцы похожи на самок, но окрашены более тускло; в первую осень жизни они сменяют контурное перо годового наряда. В году одна полная послегнездовая линька.
У части видов яркий брачный наряд самцов образуется к весне за счет обнашивания кромок контурного пера осенне-зимнего наряда; у других видов есть частичная предбрачная линька.
Многие виды — хорошие певцы. Гнездятся одиночными парами, немногие виды группами. У некоторых, главным образом тропических видов изредка бывает бигамия: на участке самца гнездятся две самки. Гнезда открытые, чашеобразные, свитые из сухих травинок. Лоток выстилается мягкими стебельками, мхом, шерстью, перьями. Положение гнезд очень разнообразно: на земле, в расщелинах скал и под камнями, на разной высоте от земли на крепких стеблях травы, в развилках ветвей деревьев и кустарников. У одних видов гнездо строит только самка, у других ей помогает самец. Яйца с более или менее выраженной исчерченностью и пятнистостью. В тропиках в кладке 2—4 яйца, в умеренных и северных широтах 5—7, редко больше. Насиживает обычно только самка; но у части видов некоторое участие в насиживании принимает и самец. У многих видов (у всех?) самец кормит насиживающую самку. Инкубация 10—14 дней, у немногих до 17—18 дней. Птенцов выкармливают оба партнера. Молодые вылетают из гнезд на 10—20-й день жизни. Выводки держатся с родителями, затем объединяются в стайки. У части видов в году 2 кладки, у немногих видов в тропиках — 3 и даже 4. Молодые становятся половозрелыми и приступают к размножению обычно на следующее лето жизни.
Вне периода размножения держатся стайками, иногда в сотни особей. В умеренных и северных широтах и в высокогорьях — перелетные или широко кочующие, в тропиках оседлые или кочующие на небольшие расстояния. Часть видов ведет скрытный образ жизни.
Преимущественно зерноядны или семеноядны. Однако, видимо, все виды в большей или меньшей степени используют и многие другие корма: мелкие плоды, проростки, почки, различных мелких беспозвоночных и т. п. Птенцов многие виды выкармливают только насекомыми и пауками, другие, и птенцов кормят преимущественно мягкими (поспевающими) семенами. Кормятся на земле, травянистых растениях, в кронах деревьев и кустов.
Система семейства вьюрковых разработана слабо: четко не установлен объем семейства и его структура, много противоречивых представлений о положении отдельных родов и т. д. По излагаемой нами системе семейство включает 4 подсемейства, объединяющие 414—468 видов, группируемых
337
в 89—123 рода. Объем подсемейств варьирует в представлениях разных систематиков.
В СССР встречается 69—71 вид 28 родов только 2 подсемейств; несомненно гнездится 61—63 вида 22 родов.
ПОДСЕМЕЙСТВО 1. EMBERIZINAE — ОВСЯНКОВЫЕ
Отличаются от других вьюрковых несколько более тонким коническим клювом, обычно не вздутым у основания (рис. 242). Нижний режущий край надклювья образует две линии, смыкающиеся под тупым углом на уровне ноздрей. В этом месте при закрытом клюве края надклювья и подклювья не соприкасаются и образуется узкая щель. В окраске взрослых птиц обычно нет металлического блеска. В птенцовом наряде, как правило, выражена продольная исчерченность. Распространены почти во всему ареалу семейства (рис. 241).
Подсемейство объединяет 171—197 видов, распределяемых между 46—56 родами: Emberiza (34—37 видов, только Старый Свет), Aimophila (14 видов), Ammodramus, Ammospiza, Amphispiza, Arremon, (Arremonops), Atlaptes (27 видов), Calamospiza, Calcarius, Catamenia, Chlorura, Chondestes, Coryphospingus, Diuca, Fr ingillaria (9 видов), J unco, Melanodera, Melano-spiza, Melophus, Melospiza, Nesospiza, Oryzoborus, Passerculus, Passerella, Passer herbulus, Phrygilus, (9 видов), Pipilo, Plectrophenax, Pooecetes, Poo-spiza (13 видов), Rhynchophanes, Rowettia, Sicalis (10 видов), Spizella, Spo-rophila (28 видов), Tiaris, Volatinia, Zonotrichia и др.Среди родов довольно много монотипичных, но четко обособленных. Часть систематиков некоторые из родов (Melanospiza, Sicalis, Tiaris) включает в подсемейство кардиналовых или вьюрковых. Наиболее разнообразен видовой состав овсянок в Центральной и Южной Америке. В СССР встречается 34 вида 9 родов, достоверно гнездится 26 видов 3 родов. Два вида — желтобровая овсянка Emberiza chrysophrus Pall и рыжая овсянка Е. rutila Pall — эндемики, гнездящиеся только в пределах СССР. Гнездящиеся у нас пуночка Plectrophenax nivalis (L.) и лапландский подорожник Calcarius lapponicus (L.) населяют и северные районы Северной Америки.
Овсянковых часто рассматривают как самостоятельное семейство. В этом случае иногда в него включают в ранге подсемейств танагровых, плюшевоголовых вьюрков, американских цветочниц и кардиналовых. Высказывалось предположение, что овсянковые ближе к ткачиковым, чем к вьюрковым.
Рис. 242. Камышовая овсянка Emberiza schoeniclus L.
338
ПОДСЕМЕЙСТВО 2. GEOSPIZINAE — ДАРВИНОВЫ, ИЛИ ГАЛАПАГОССКИЕ, ВЬЮРКОВЫЕ
Сюда относят 14 видов, объединенных в 4 рода (подроды приведены в скобках): Geospiza (Castornis; 6 видов), Camarhynchus, (Cactospiza, Pla-tyspiza\ 6 видов), Certhidae (1 вид), Pinaroloxias (1 вид). Три первых рода встречаются только на Галапагосских островах (рис. 241), последний — лишь на Кокосовом острове. Обособились в результате адаптивной радиации в условиях географической изоляции от какого-то, видимо, южноамериканского вида.
Изучение этой группы позволяет представить один из возможных путей видообразования (Лэк, 1949; Майр, 1968). По внешнему облику, строению клюва, повадкам напоминают вьюрков, славок, пищух и даже мелких дятлов. Обособление видов шло преимущественно по линии пищевой специализации (питание семенами, мякотью кактусов и их цветками, различными насекомыми). Видовые различия сводятся прежде всего к форме клюва: массивному, вьюркового типа; толстому, слегка изогнутому; тонкому изогнутому. Один из видов — дятловый вьюрок Camarhynchus (Castospiza) pallidas Scl. — широко известен тем, что, держа в клюве длинную колючку или тонкую веточку, ковыряет ею в щелях или в ходах насекомых (рис. 243), выгоняя их наружу, где хватает клювом. Земляной вьюрок Geospiza diffi-cilis Sharpe часто садится на гнездящихся здесь олуш и ловит кровососущих мух. Видимо, таким путем у этого вида возник своеобразный паразитизм (не выяснено, свойствен он лишь отдельным особям или всей популяции; Бовман, 1965): вьюрок садится на спину олуши, клювом прокалывает ее кожу и слизывает выступившие капельки крови. Олуши на такое нападение не реагируют.
Видовая обрсобленность по местообитаниям и характеру гнездования выражена менее отчетливо. Самцы в период тока строят примитивные гнезда, иногда токуют перед чужими. Объемистое гнездо с крышей и боковым входом окончательно строит самка, размещая его в развилке ветвей. В кладке 4 яйца. Окраска самок буровато-коричневатая, самцы сходны с самками или отличаются более темной окраской (развивается не у всех самцов данного вида). По мнению Лэка (1949), основным видовым опознавательным признаком служит форма клюва.
Иногда дарвиновых вьюрков выделяют в самостоятельное семейство. Кто был предком дарвиновых вьюрков и к кому они ближе — к овсянковым,
Рис. 243. Дятловый вьюрок Camarhynchus (Cactospiza) pallidas Scl., выгоняющий колючкой насекомое из щели
339
Рис. 244. Кардинал Pyrrhulo-xia cardinalis (L.)
вьюрковым или кардиналовым, пока неясно. Высказывалось предположение о их близости к ткачиковым; это маловероятно по зоогеографическим соображениям (ткачиковых в Америке нет) и не обосновано морфологическими данными.
ПОДСЕМЕЙСТВО 3. PYRRHULOXIINAE — КАРДИНАЛОВ ЫЕ
Похожи нй вьюрков (рис. 244). По морфологическим особенностям занимают промежуточное положение между овсянковыми и вьюрковыми. В частности, нижний край надклювья имеет такой же излом, как у овсянок. Клюв более мощный, чем у овсянок, и часто вздут у основания. Встречаются только в Новом Свете от южных районов Канады и до юга Южной Америки; большее разнообразие видов в тропиках.
В подсемействе ПО—132 вида, объединяемых в 12—31 род: Рyrrhaloxia (Richmondena), Cardinalis, Caryothraustes, Cyanocomposa, Gui-raca, Hedymeles, Hesperiphona, Paroaria, Passer ina, Periporyhus, Pheucticus, Pitylus, Rho-dothraupis, Saltator, Spiza и др.
Объем подсемейства определен плохо. Иногда оставляют здесь только около 50 видов, переводя часть родов (Hesperiphona и др.) к вьюрковым и
к овсянковым. Неясны и взаимоотношения кардиналовых с другими группами. Некоторые систематики считают, что они более близки к овсянковым, другие говорят о их большей близости к настоящим вьюрковым. Наконец, некоторые авторы включают подсемейство кардиналовых в семейство тана
гровых.
ПОДСЕМЕЙСТВО 4. FRINGILLINAE — НАСТОЯЩИЕ ВЬЮРКОВЫЕ
Нижний обрез надклювья прямой или слабо дугообразный, без тупого излома. При закрытом клюве края надклювья и подклювья смыкаются плотно, не оставляя щели. Клюв крепкий, у части видов необычайно мощный. Например, у обыкновенного дубоноса (рис. 245), весящего около 50— 60 г, в основании клюва может создаваться давление до 25—60 кг (легко
разгрызает вишневые косточки). В окраске взрослых часто встречаются участки с металлическим блеском. В птенцовом наряде продольная исчерченность выражена слабо или ее нет вовсе.
К подсемейству настоящих вьюрковых относят 119—125 видов, группируемых в 27—32 рода: Frin-gilla (3 вида), Acanthis (включает р. Cannabina), Alar io, Bucanetes, Cardue-lis, Carpodacus (более 20 видов; включает pp. Eryth-
/Q.C.
Рис. 245. Обыкновенный дубонос Coccothraustes coccothraustes (L.)
340
rina и Rubicilla) Chaunoproctus (монотипичный род с о. Бонин, недавно вымер), Chloris (6 видов), Coccothraustes, Eophona, Haematospiza, Kozlowia, Leucosticte (Fringillauda), Linurgus, Loxia, Loxigilla, Loximitris, Melopyrrha, Mycerobas, Pinicola, Pyrrhospiza, Pyrrhula (6 видов), Rhodope-chys, Rhodospiza, Rhynchostruthus, Serinas (15 видов), Spinas (19 видов), Uragus, Warsanglia и др.
Объем подсемейства окончательно не установлен. Иногда некоторые роды (например, Loxigilla и др.) перемещают к овсянковым; другие систематики отдельные роды из овсянковых и кардиналовых переводят сюда. В подсемействе много монотипичных родов. В Палеарктике встречается 69—71 вид, при этом из 22 живущих здесь родов 13 — эндемики; в Африке живет 36 видов 9 родов, из них 6 — эндемики. В Новом Свете (обе Америки) встречаются лишь 25 видов 8 родов, но из них 6 видов 4 родов очень широко распространены и по Палеарктике; эндемичны лишь 2 рода. Сейчас некоторые виды акклиматизированы на Новой Зеландии, Кубе, Тайване и ряде мелких океанических островов. В СССР встречается и гнездится 35—37 видов 19 родов.
Часто настоящих вьюрковых рассматривают как самостоятельное семейство; при этом в него иногда включают — в ранге подсемейств — кардиналовых и дарвиновых вьюрков.
ЛИТЕРАТУРА
Бутурлин С. А., Дементьев Г. П. Полный определитель птиц СССР. М.—Л., 1934—1941. КОИЗ, т. 1—5.
Гладков Н. А., Дементьев Г. П., П т у ш е н к о Е. С., С у д и-ловская А. М., 1964. Определитель птиц СССР. М., «Высшая школа».
Жизнь животных. М., 1970. Ред. Гладков Н. А., Михеев А. В. «Просвещение», т. 5. Птицы.
Дарлингтон Ф. Зоогеография. М., «Прогресс», 1966.
Дементьев Г. П. Птицы. Руководство по зоологии. М.—Л., 1940. Изд-во АН СССР, т. 6.
Дементьев Г. П. Класс Aves. Птицы. Основы палеонтологии. Земноводные, пресмыкающиеся, птицы. Изд-во АН СССР, 1964.
Дементьев Г. П. Систематика птиц. Сб. «Современные проблемы орнитологии». Фрунзе, «Илим», 1965.
Дзержинский Ф. Я. Челюстной аппарат птиц. Итоги науки. Зоология позвоночных. 1969. М., ВИНИТИ, 1971.
Иванов А. И., Коз л.о в а Е. В., П о р т е н к о Л. А., Тугаринов А. Я. Птицы СССР. М.—Л., Изд-во АН СССР, ч. 1—2, 1951—1953.
Иванов А. И., Штегман Б. К. Краткий определитель птиц СССР. М.—Л., «Наука», 1964.
Козлова Е. В. Гагарообразные, трубконосые. Фауна СССР Птицы, т.1, вып. 3, 1947; Подотряд чистиковые. Фауна СССР. Птицы, т. 2, вып. 3; 1957, Ржанкообразные. Подотряд кулики. Фауна СССР. Птицы, т. 2, вып. 1, ч. 2 и 3. Изд-во АН СССР. М.—Л., 1961, 1962.
Курочкин Е. Н. Основные вопросы изучения ископаемых птиц. Итоги науки. Зоология позвоночных. 1970 М., ВИНИТИ, 1971.
Л э к Д. Дарвиновы вьюрки. М., Изд-во иностр, лит-ры. 1949.
Майр Э. Зоологический вид и эволюция. М., «Мир», 1968.
Майр Э. Принципы зоологической систематики. М., «Мир», 1971.
Майр Э., Л и нс л и Э., Юзингер Р. Методы и принципы зоологической систематики. М., Изд-во иностр, лит-ры, 1956.
Международный кодекс зоологической номенклатуры, принятый XV Международным зоологическим конгрессом. М.—Л., «Наука», 1966.
Портенко Л. А. Птицы СССР. ч. 3—4. М.—Л., Изд-во АН СССР, 1954, 1960.
Птицы Советского Союза. Ред. Дементьев Г. П. и Гладков Н. А. М., «Советская наука», 1951—1954, т. 1—6.
Степанян Л. С. Систематический список видов птиц фауны СССР. Сб. Фауна, экология и география животных. Уч. зап. № 362. М., Изд-во Моск. гос. пед. ин-та им. Ленина, 1969; Итоги науки. Зоология позвоночных, 1969. М., ВИНИТИ, 1971;
Тугаринов А. Я. Пластинчатоклювые. Фауна СССР. Птицы. М., 1941, т. 1, вып. 4; Веслоногие, аистообразные, фламинго. Фауна СССР. Птицы. М.—Л., Изд-во АН СССР, 1947, т. 1, вып. 3.
Флинт В. Е., Беме Р. Л., Костин Ю. В., Кузнецов А. А. Птицы СССР (справочник-определитель). М., «Мысль», 1968.
Штегман Б. К. Дневные хищники. Фауна СССР. Птицы. М.—Л., т. 1, вып. 5, 1937; Основы орнитогеографического деления Палеарктики. Фауна СССР. Птицы. М.—Л., Изд-во АН СССР, 1938, т. 1, вып. 2.
Ш у л ь п и н Л. М. Орнитология. Л., Изд-во ЛГУ, 1940.
Юдин К- А. Филогения и классификация ржанкообразных. Фауна СССР. Птицы. М., «Наука», 1965, т. 2. вып. 1, ч. 1; Биологическое значение и эволюция кине-тичности черепа птиц. Орнитологический сборник. Тр. ЗИН АН СССР. Л., «Наука», 1970, т. XLVII.
Berndt R., Meise W., Naturgeschichte der Vogel. Kosmos, Stuttgart. 1959—1965, Bd. 1—111.
H art erf E. Die Vogel der palaarktischen Fauna. Friedlander und Sohn. Berlin. 1910—1922, Bd. 1—3.
Hartert E., Steinbacher F. Die Vogel der palaarktischen Fauna. Erganzungsband. Friedlander und Sohn. Berlin, 1932—1938.
H e i 1 m a n n G. The origin of birds. London, 1926.
Peters J. L. Chek — List of birds of the world. Cambridge, Harvard University Press. 1931—1951, Vol. 1—7.
Stresemann E. Aves. Kukenthal’Handbuch der Zoologie. Krumbach, Berlin und Leipzig, 1927—1934. Bd. V 11, Hf. 2.; Die Entwiclung der Ornithologie von Aris-toteles bis zur Gegenwart. Berlin, 1951.
Traite de zoologie. Publie sous la direction de P. Grasse. Oiseaux. Masson, Paris, 1950, T. XV.
V a u r i e Ch., The birds of the Palearctic fauna. Passeriformes. Non Passeres. H. und G. Witherby limited. London. 1959, 1965.
342
ПРИЛОЖ ЕНИЕ
Родовой и видовой состав основных систематических групп птиц и состав фауны птиц СССР
Отряды, подотряды, семейства Число родов Число видов
всего в мире в СССР всего в мире встречается гнездится
в СССР в СССР
абс. % абс. %
1 2 3 4 5 6 7 8
1. Archaeopterygiformes + •
2. Hesperornithiformes 4"
3. Ichthyornithiformes '+•
4. Sphenisciformes 6 — 16 —
5. Struthioniformes 1 1
Struthionidae 1 — 1
Eleutherornithidae + •
6. Rhe iformes 1—2 . 2 ——
7. Casuariiformes ......... 2 ' 4—6
Casuariidae , . 1 — 3-5 — —
Dromaiidae 1 — 1 —
Dromornithidae +•
8. Aepyornithiformes
9. Dinornithiformes +•
10. Apterygiformes 1 — 3 — —“
11. Tinamiformes 9 — 42
12. Gaviiformes 1 1 3 3 100,0 3 100,0
13. Podicipediformes 4 1 18—20 5 20,0 5 20,0
14, Procellariiformes 23 5 81 14 17,2 5 6,1
Diomedeidae 1 1 12 3 25,0
Procellariidae 12 2 47 7 15,0 2 4,3
Hydrobatidae 9 2 18 4 22,2 3 16,6
Pelecanoididae . . .’ 1 4
15. Pelecaniformes 7-8 4 50-52 11 21,1 8 15,4
Odontopteryges
Phaethontes 1 ! 3 —
Pelecani 5—6 3 42—44 10 22,7 8 18,1
Pelecanidae 1 1 6 2 33,3 2 33,3
Cyphornithidae Pelagornithidae + •
+
Sulidae 1—2 1 7—9 2 22,2
Elopterygidae . + •
Phalacrocoracidae 2 1 26 6 23,1 6 23,1
Anhingidae 1 — 3 —
Fregatae 1 1 5 1 20,0 —
Cladornithes t-
16. Ciconiiformes 49 14 117— 119 23 19,5 18 15,2
Ardeae 15 9 64 16 25,0 13 20,0
Ardeidae 14 9 62 16 25,8 13 21,0
Cochleariidae 1 .— 2
Balaenicipites 1 — 1 — —
Ciconiae 30 4 48 6 12,5 4 8,8
Scopidae 1 — 1 — —
Ciconiidae 10 1 17 2 11,8 2 11,8
Threskiornithidae 19 3 30 4 13,3 2 6,6
Plegadornithidae •
Phoenicopteri 3 1 4—6 1 16,6 1 16,6
Phoenicopteridae 3 1 4—6 1 16,6 1 16,6
Agnopteridae +•
Palaelodidae •J"
Scaniornithidae
17. Anseriformes 43 16 151— 154 55—58 37,7 48—50 32,4
Anhimae 2 .— 3 —
An seres 41 16 148— 151 55—58 38,4 48—50 33,1
Anatidae 41 16 148— 151 55-58 38,4 48—50 33,1
343
1 2 3 4 5 6 7 8
Anseranatinae 1 1
Anserinae 5 3 29 16 55,2 14 48,2
Dendrocygni ni 1 —. 8
Anserini 4 3 21 16 76,1 14 66,6
Anatinae 35 13 118— 121 39—42 35,5 34—36 29,7
Tadornini • 8 1 20 3 15,0 2 10,0
Anatini 6 1 42 12 28,8 11 26,2
Somaterini 2 2 4 4 100,0 4 100,0
Aethyini 2 2 15 6 40,0 6 40,0
Cairini 7 1 12 1 8,3 1 8,3
Mergini 6 5 16—19 12—15 78,9 9—11 57,8
Oxyurini 4 1 9 1 11,1 1 11,1
Paranyrocidae + •
18. Falconiformes < 81 20 274 50 18,3 47 17,1
Cathartae ; 5 6
Neocathartidae +•
Cathartidae ’ 5 6
Teratornithidae t +•
Falcones ; 76 20 268 50 18,7 47 17,9
Sagittariidae 1 — 1 —
Accipitridae , 63 18 208 37 17,8 34 16,3
Pandionidae 1 1 1 1 100,0 1 100,0
Falconidae - 11 1 58 12 20,7 12 20,7
19. Galliformes 85 12 253 20—21 8,3 20—21 8,3
Galli 84 12 252 20—21 8,3 20—21 8,3
Megapodiidae GaTlinuloididae 7 J.. — 11 — —
Cracidae 11 — 39 -
Tetraonidae 11 5 18 8 44,4 8 44,4
Phasianidae 48 7 175 12—13 7,4 12—13 7,4
Numididae 5 — 7
Meleagrididae 2 — 2 — —
Opisthocomi 1 — 1 —
20. Gruiformes ’ 84 14 193 23 11,9 22 11,4
Mesitornithides 2 —. 3 —
Turnices > 3 1 15 1 6,6 1 6,6
Turnicidae • 2 1 14 1 7,1 1 7,1
Pedionomidae $ 1 — 1
Grues 58 9 137 18 13,1 17 12,5
Geranoididae +•
Gruidae : 5 2 14 7 50,0 7 50,0
Aramidae 1 — 1
Psophiidae 1 — 3 —
Orthocnemidae Rallidae Ji 7 119 11 9,2 10 8,4
Heliornithes 3 —, 3
Rhinocheti 1 1 —
Eurypygae ' 1 —. 1 —
Cariamae 2 2 —
Phororhacidae +•
Psilopteridae + •
Brontornithidae +•
Opisthodactylidae : + •
Cunampaiidae t-
Cariamidae : 2 — 2
Hermosiornithidae Bathornithidae + f
Otides 14 4 31 4 12,9 4 12,9
Otididae 11 3 22 3 13,6 3 13,6
Burhinidae 3 1 9 1 11,1 1 11,1
21. Diatrymiformes +•
22. Charadriiformes 87— 51—56 287— 132— 46,0 120— 41,9
113 289 133 121
Charadrii = 58—83 31—34 181 79 43,7 72 40,0
Jacanidae 6 —, 7
Rhegminornithidae i-
Rostratulidae ' 2 1 , 2 1 5C,0 ?
344
1 2 3 4 5 6 7 8
Haematopodidae • . 1 1 4 1 25,0 1 25,0
Charadriidae ........ 9—32 7—10 60 18 30,0 16 26,6
Scolopacidae 24—26 16 75 50 66,6 47 62,6
Recurvirostridae 4 3 7 3 42,8 3 42,8
Presbyornithidae ....... +•
Phalaropodidae 1 1 3 2 66,6 2 66,6
Dromadidae 1 1 -
Glareolidae 7 2 16 4 25,0 3 18,8
Thinocoridae 2 —. 4
Chionididae 1 2
Lari 16—17 10—11 85—86 35 40,7 32 37,2
Stercorariidae 1—2 1—2 4—5 4 80,0 3 60,0
Laridae 14 9 78 31 39,7 29 37,2
Rynchopidae 1 3
Alcae 13 11 21—22 18—19 86,4 16—17 77,3
23. Columbiformes 49 5 310 16 5,1 14 4,5
Pterocletes 2 2 16 4 25,0 4 25,0
Columbae '47 3 294 12 4,1 10 3,4
Raphidae 2+. 3f.
Columbidae 45 3 291 12 4,1 10 3,4
24. Psittaciformes 75 324
25. Cuculiformes 43 2 148 6 4,0 5 3,4
Musophagi 5 18
Cuculi 38 2 130 6 4,6 5 3,8
26. Strigiformes 25—30 12 144 18 12,5 18 12,5
Protostrigidae t-
Tytonidae 2 1 11 1 9,0 1 9,0
Strigidae 23—28 11 133 17 12,8 17 12,8
27. Caprimulgiformes 23 1 93 3 3,2 3 3j2
Steatornithes 1 1
Caprimulgi 22 1 92 3 3,3 3 3,3
Podargidae . . , 2, — 12 —
Nyctibiidae 1 —. 5
Aegothelidae 1 5
Caprimulgidae 18 1 70 3 4,3 3 4,3
28. Apodiformes 109— 2 378 5 1,3 5 1,3
129
Apodi 9 2 58 5 8,6 5 8,6
Aegialornithidae +•
Hemiprocnidae 1 — 3
Apodidae 8 2 55 5 9,1 5 9,1
Trochili 100— 320
120
29. Coliiformes 1 6
30. Trogoniformes 8 35
31. Coraciiformes 48—54 7 192- 10 5,2 6 3,1
193
Alcedines 19—23 3 99 5 5,0 1 1,0
Alcedinidae 12—16 3 86 5 5,8 1 1,1
Todidae 1 5
Momotidae 6 8
Meropes 7 1 25 2 8,0 2 8,0
Coracii 10 3 24 3 12,5 3 12,5
Coraciidae 2 2 11 2 18,1 2 18,1
Brachypteraciidae 3 — 5
Leptosomatidae 1 1
Upupidae 1 1 1 1 100,0 1 100,0
Pfioeniculidae 3 6
Bucerotes 12—14 44—45
32. Piciformes 73—87 5—6 379 13 3,4 13 3,4
Galbulae 37—42 170
Galbulidae 5 15
Bucconidae 10 30
Capitonidae 13—16 76
Indicatoridae 4 12
Ramphastidae 5—7 — 37 5,2
Pici 36—45 5—6 209 13 6,2 13
345
1 2 3 4 5 6 7 8
33. Passeriformes 1081— 112- 5035— 317— 6,2 290— 5,7
1343 123 5309 329 301
Eurylaimi 8 — 14 — —
Tyranni 310— 1077— .—
316 1084
Dendrocola ptidae ...... 12—13 — 48 —
Furnariidae 58 215 —
Formicariidae 53 — 224 —
Conopophagidae 2 — 11 —
Rhinocryptidae 11 29 —
Cotingidae 30—33 — 94 — И»
Pipridae 21 — 61 —
Tyrannidae . . . 114— 358—
116 365
Oxyruncidae 1 — 1 — —
Phytotomidae 1 —• 3 —
Pittidae 2 — 25 — —
Acanthisittidae 3 — 4 —. —
Philepittidae 2 — 4 — —
Menurae 2 — 4 —
Menuridae 1 — 2
Atrichornithidae 1 — 2 — -
Passeres (Osciens) 761— 112- 3944— 317— 7,8 290— 7,1
1017 123 4211 329 301
Alaudidae 15 7 70—78 13 16,6 13 16,6
Palaeospizidae +•
Hirundinidae 19 6 75—79 9 11,4 7 8,9
Dicruridae 2—6 1 20—21 2 9,5 ?
Oriolidae 3 1 34—42 2 4,8 2 4,8
Corvidae 20—32 8 100- 105 14—16 15,2 14—16 15,2
Cracticidae 3—4 — 10—11 — .
Grallinidae 3 — 4 —
Ptilonorhynchidae 8 — 17 — —
Paradisaeidae 20 — 39—43
Paridae 9 2 60—65 13 20,0 13 20,0
Sittidae - 3 1 23—25 3—4 116,0 3—4 16,0
Hyposittidae 1 — 1 —. —
Certhiidae 5 2 15 4 26,6 4 26,6
Paradoxornithidae 4—5 4 13—19 4 21,0 4 21,0
Chamaeidae 1 — 1 —
Timaliidae 52—57 1 260— 280 1 0,3 1 0,3
Campephagidae 9-11 1 70—72 1 1,4 1 1,4
Pycnonotidae 15-21 1 119 2 1,7 1 0,8
Palaeoscinidae +•
Chloropseidae 2 — 12 —
Cinclidae 1 1 4—5 2 40,0 2 40,C
Troglodytidae 12—16 1 59—63 1 1,6 1 1,6
Mimidae 13 — 30—31 —
Turdidae 40—70 12—19 297— 303 50—54 17,7 48—51 16,7
Zeiedoniidae 1 1 —
Sylviidae 83—93 11—14 400— 425 51 12,0 49 11,5
Regulidae ,. . . 3 2 5—6 3 50,0 3 50,0
Muscicapidae 57—82 2 378— 398 12 14 3,5 13 3,3
Prunellidae . 1—2 1—2 8 66,6 8 66,6
Motacillidae 6—7 3 50—53 15—17 32,1 14—16 30,2
Bombycillidae 2 2 4 3 75,0 2 50,0
Ptilogonatidae 3 — 4 — —
Dulidae 1 — 1 —
Artamidae 1 — 10—15 —
Vangidae 9—11 11—14 —— —-
Laniidae 9—12 1 66—69 11 15,9 10 14,5
346
1 2 3 4 5 6 7 8
Prionopidae’ 3—4 9—10
Cyclarhidae 1 _— 2
Vireolaniidae 2 3—4
Callaeidae 3—4 3—4
Sturnidae 25—36 5 103— 8 7,3 6 5,5
110
Meliphagidae 35—50 162
Nect’ariniidae 8—11 — 105— 108 —, —
Dicaeidae 7 54—56
Zosterop idae 7—12 1 79-85 1 1,2 1 1,2
Vireonidae 4—7 42
Coerebidae 7—12 36—38
Drepanididae 7—11 21—22 —
Parulidae 18—26 2 108- 2 1,8 ?
113
Ploceidae 37—68 5 247— 272 11—12 4,4 10—11 4,0
Icteridae 22—35 1 87—93 1 1,1
Tersinidae 1 1 —
Thraupidae 48—61 — 192— —-
205
Catamblyrhynchidae 1 — 1
Fringillidae 89- 28 414— 69—71 15,2 61—63 13,5
123 468
Emberizinae 46—56 9 171— 197 34 17,3 26 13,2
Geospizinae 4 — 14
Pyrrhuloxiinae Fringillinae 12-31 — 110— 132 — — —
27—32 19 119— 125 35—37 29,6 35—37 29,6
Всего 2019— 284- 8539— 734— 8,5 650-665 7,5
|2354 301 8827 751
Примечание. Если в отряд или подотряд входит только одно семейство, то его название не приводится. Роды и виды, представленные только ископаемыми формами, не включены. Приводимые данные отражают существующие разные точки зрения на число видов и родов в отдельных группах. При вычислении процента числа видов, встречающихся и гнездящихся иа территории СССР, от общего числа видов данной группы орались максимальные цифры. Для; семейства Anatidae дается перечень подсемейств и триб, для Fringillidae—подсемейства.
УКАЗАТЕЛЬ ЛАТИНСКИХ НАЗВАНИИ РОДОВ1
Abroscopus 302 Aburria 154 Acanthis 340 Acanthisitta 267 Acanthiza 303 Acanthorhynchus 320 Accipiter 146 Aceros 246 Acestrura 232 Acridotheres 318 Acrocephalus 302 Acropternis 262 Aery Ilium 159 Actinodura 291 Act it is 189 Actophilornis 182 Adamastor 101 Adelomyia 232 Aechmophorus 97 Aechmorhynchus 189 Aegithina 295 Aegintha 331 Aegithalos 288 Aegolius 218 Aegotheles 224 Aegypius 146 Aepyornis 91 Aepypodius 153 Aerocharis 312 Aeronautes 227, 229 Aerops 240 Aethia 201 Aethopyga 321 Aesalon 150 Afribyx 188 Afropavo 159 Afrotis 177 Agapornis 210 Agelaius 333 Aglaeactis 232, Aglaiocercus 232 Agelaius 333 Agrobates 302 Agamia 118 Agelastes 159 Agr iochar is 160 Agriornis 265 Agyrtria 232 Aidemosyne 331 Ailuroedus 280, 281 Aimophila 338 Aix 137
Aiaja 124 Alaemon 273
Alario 340 Alauda 273 Alca 201 A Ice do 235, 236 Alcippe 291 A lector is 159 AleCtroenas 207 Alectrurus 265 Alectura 153 A let he 299 Alisterus 210 Allenia 298 Alopochen 133, 134 Alseonax 306 Amadina 331 Amandava 331 Amaurornis 171 Amazilia 232 Amazona 210 Amazonetta 137 Amblycercus 333 Amblyornis 280, 281 AmWyospiza 331 AmVlyrhampus 333 Ammodramus 338 Ammomanes 273 Ammoperdix 159 Ammospiza 338 Ampeliceps 318 Ampelion 264 Amphiserpentarius 145 Amphispiza 338 Amytornis 303 Anabacerthia 260 Anaplectes 331 Anarhynchus 188 Anas 134 Anastomus 123 Ancistrops 260 Andigena 252 Androdon 232 Andropadus 294 Andropholus 291 Anhima 129
Anhinga 113 Anisognathus 335 Anodorhynchus 210 Anomalophrus 188 Anomalospiza 329, 331 Anorrhinus 246 Anous 199
Anser 132
Anser anas 131 Anthipes 306
1 Включены роды, упомянутые в систематическом обзоре класса птиц
348
Anthochaera 320 Anthocichla 267 Anthornis 320 Anthoscopus 283, 285 Anthracoceros 246 Anthracothorax 232 Anthreptes 321 Anthropoides 167 Anthus 308 Antichronus 315 Anumbius 260 Anurolimnas 171 Anurophasis 159 Apalis 302 Apaloderma 234 Apalopteron 320 Apatornis 82 Aphanapteryx 171 Aphelocoma 278 Apheloephala 303 Aphrastura 260 Aphriza 189 Aplonis 318 Aplopelia 207 Aprosmictus 210 Aptenodytes 84 Apteryx 93 Aptornis 171 Apus 227, 229 Aquila 146 Ara 210 Arachnothera 321 Aramidopsis 171 Aramides 171 Aramus 168 Aratinga 210 Arborophila 159 Archaeopteryx 80 Archaeornis 80 Archboldia 280, 281 Archilochus 232 Ardea 118 Ardeola 118 Ardeotis 177 Arenaria 198 Argusianus 159 Arre mon 338 Arremonops 338 Artamus 311 Artomyias 306 Arses 306 Arundinicola 265 Asio 218 Aspatha 239 Asthenes 260 Astra pia 282 Asyndesmus 254 Atelornis 242 Athene 218 Atlantisia 171 Atlaptes 338 Atrichornis 270 Attagis 194 Atticora 274 Attila 264 Aulacorhynchus 252 Auriparus 283, 285 Automolus 260 Aviceda 146 Avocettula 232 Aythya 136
Babax 291 Baeopogon 294 Baillonius 252 Balaeniceps 119 Balearica 167 Bambusicola 159 Baptornis 81 Bartramia 189 Baryphthengus 239 Basileuterus 328 Batara 261 Bathilda 331 Batis 306 Batrachostomus 222 Basilornis 318 Beauharnaisius 252 Belonopterus 188 Berenicomis 246 Berlepschia 260 Bernieria 294 Bhringia 275 Biziura 139 Bleda 294 Blythipicus 254 Boangerges 278 Boissonneaua 232 Bolbopsittacus 210 Bolborhynchos 210 Bombylonax 240 Bombycilla 310 Bonasa 156 Bostrychia 124 Botaurus 118 Bowdleria 302 Bradypterus 302 Brachycope 331 Brachygalba 248 Brachyldphus 254 Brachypteracias 242 Brachypteryx 299 Brachyramphus 201 Bradornis 306 Branta 132 Brotogeris 210 Bubalornis 330 Bubo 218 Bubulcus 118 Bucanetes 340 Buccanodon 249 Bucco 248 Bucephala 138 Buceros 245, 246 Buchanga 275 Bucorvus 245, 246 Budytes 308 Bulveria 101 Buphagus 318 Burganus 167 Burhinus 179 Busare ileus 146 Butastur 146 Buteo 146 Buteogallus 146 Butorides 118 Bycanistes 246
Cacicus 333
Cacomantis 214
Cactospiza 339
Caenocorypha 189
Cairina 137, 138
349
Calamanthus 303
Calamocichla 302
Calamospiza 338
Calandrella 273
Calcarius 338
Calicalicus 312
Calidris 189
Caliechthrus 214
Calliope 299
Callaeas 317
Callipepla 159
Calliphlox 232
Callocephalon 210
Calocitta 278
Caloenas 207
Caloperdix 159
Calopezus 95
Calospiza 335
Calothorax 232
Calypte 232
Calyptocichla 294
Calyptomena 256
Calyptorhynchus 210
Calyptura 264
Camarhynchus 339
Campephaga 292
Campephilus 254
Campethera 254
Campochaera . 292
Camponotus 254
Camptorhynchus 138
Camptostoma 265
Campylopteros 302
Campylopterus 297
Campylorhamphus 259
Campylorhynchus 297
Canachites 156
Canirallus 171
Cannabina 340
Capito 249
Caprimulgus 219, 224, 225
Carpodacus 340
Caracara 150
Caracopsis 210
Cardellina 328
Cardinalis 340
Carduelis 340
Cariama 175
Carpococcyx 214
Carpodectes 264
Carpospiza 331
Caryotnraustes 340
Casarca 133
Cassidix 333
Castornis 339
Casuarius 90
Catamblyrhynchus 335
Catamenia 338
Catharacta 197
Catharopeza 328
Cathartes 143
Catharus 299
Cathereps 297
Catheturus 153
Catopterophorus 189
Catreus 159
Celeus 254
Centrocercus 156
Centropeljna 97
Centropus 214
Ceophloeus 254
Cephalopterus 264
Cephalopyrus 285
Cepphus 201
Ceratogymna 246
Cercibis 124
Cerchneis 150
Cercococcyx 214
Cercomacra 261
Cercomela 299
Cercotrichas 302
Cereopsis 133, 134
Cerorhinca 201
Certhia 286, 287
Certhida 325
Certhidea 339
Certhilauda 273
Certhionyx 320
Cerygone 303
Ceryla 236
Cescropis 274
Cettia 302
Ceuthmochares 214
Ceyx 236
Chaemaeza 261
Chaetocercus 232
Chaetoptila 320
Chaetorhynchus 274, 275
Chaetura 226, 227, 229
Chaimarrornis 299
Cha kites 214
Chalcomitra 321
Chalcopelia 207
Chalcophaps 207
Chalcostigma 232
Chalcopsitta 210
Chalybura 232
Chamaea 288
Chamaepetes 154
Chamaethlypis 328
Charadrius 186, 188
Charitornis 318
Charmosyna 210
Chaptia 275
Chasiempis 306
Chauna 129
Chaunonotus 315
Chaunoproctus 341
Chelictinia 146
Chelidoptera 248
Chelidorhynx 306
Chen 132
Chenonetta 137
Chenorhamphus 303
Cheramoeca 274
Chersophilus 273
Chettusia 188
Chibia 275
Ch ilia 260
Chionarchus 195
Chionis 195
Chiroxiphia 264
Chlamydera 281
Chlamydochaera 292
Chlamydotis 177
Chlidonias 199
Chloephaga 133, 134
Chloravis 207
Chlorestes 232
Chloris 340
Chloroceryla 236
Chlorocharis 324
Chlorochrysa 335
Chlorocichla 294
350
Chloroenas 207 Chloephaga 133, 134 Chlorophanes 325 Chlorophoneus 315 Chlorophonia 335 Chloropsis 295 Chlorornis 335 Chlorospingus 335 Chlorostilbon 232 Chlorura 338 Cholornis 288 Chondestes 338 Chondrohierax 146 Chordeiles 224, 225 Chrysococcyx 214 Chrysocolaptes 254 Chrysolampis 232 Chrysolophus 159 Chrysomma 291 ChrysPptilus 254 Chrysuronia 232 Chthonicola 303 Chubbia 189 Chunga 175 Ciccaba 218 Cichlherminia 299 Cicinnurus 282 Ciconia 121, 123 Cinclocerthia 298 Cinclodes 260 Cincloramphus 303 Cinclosoma 291 Cincludes 232 Cinclus 295 Cinnicerthia 297 Cinnyricinclus 318 Cinnyris 321 Circaetus 146 Circus 146 Ciridops 326 Cisticola '302 Cistothorus 297 Cissa 278 Cissolopha 278 Cissopica 278 Cissopis 335 Cittura 236 Cladornis 114 Cladorhynchus 190 Clamator 213, 214 Clangula 138 Clibanornis 260 Climacteris 287 Cloravis 207 Clytoceyx 236 Clytoctantes 261 Clytomyias 303 Clytospiza 331 Cnemophilus 282 Coccothraustes 341 Coccyzus 213, 214 Cochlearius 118 Cochoa 299 Coeligena 232 Coereba 325 Colaptes 254 Colinus 159 Colius 232 Coliuspasser 331 Collocalia 227, 229 Colluricincla 306 Coloeus 278
Colonia 265
Columba 207
Columbina 207
Conirostrum 325
Conopophaga 261
Conopophila 320
Conostoma 288
Contopus 265
Conuropsis 210
Copsychus 299 Coracias 242 Coracina 292 Coracornis 306
Coragyps 143
Corapipo 264
Corcorax 279 Corvinella 315 Corvultur 273 Corvus 278 Corydon 256 Coryphistera 260 Coryphospingus 338 Corythaeola 212 Corythaixoides 212
Corythopis 261 Corythornis 236 Coscoroba 132
Cosmopsarus 318
Cossypha 299
Cotinga 264
Cotnurnicops 171
Coturnix 157, 159
Coua 214
Cractes 278
Cracticus 278
Cractisus 261 Cranioleuca 260 Crateropus 291 Crateroscelis 303 Craspedophora 282 Crax 154
Creadion 317
Creatophora 318
Crex 171
Crinifer 212
Criniger 294
Criptospiza 331
Crocethia 189
Crocias 291
Crossopt ilon 159
Crotophaga 213, 214
Cruschedula 114
Cryptophaps 207
Crypturellus 95
Crypsirhina 278 Cuculus 214 Cursor ius 193 Cutia 291 Cyanerpes 325 Cyanistes 285 Cyanochen 133 Cyanocitta 278 Cyanocomposa 340 Cyanocorax 278 Cyanolanius 312 Cyanolesbia 232 Cyanolimnas 171 Cyanoliseus 210 Cyanolyca 278 Cyanomitra 321 Cyanopica 278 Cyanoptila 306
351
Cyanoramphus 210
Cyanosilvia 299
Cyanthus 232
Cyclarhis 316
Cyclorrhynchus 201
Cygnopsis 132
Cygnus 132
Cymbilaimus 261
Cymbirhynchus 256
Cymochorea 102
Cyornis 306
Cypseloides 226, 227, 229
Cypsiurus 227, 229
Cyrtonyx 159
Dacelo 236 Dacnis 325 Dactylortyx 159 Daphoenositta 286 Daption 101 Daptrius 150 Dasylophus 214 Dasyornis 303 Deconychura 259 Delichon 274 Dendrexetastes 259 Dendrobiastes 306 Dendrocincla 259 Dendrocitta 278 Dendrocolaptes 259 Dendrocopos 254 Dendrocygna 132 Dendragapus 156 Dendroica 328 Dendronanthus 308 Dendropicos 254 Deroptyus 210 Dendrortyx 159 Desertipicus 254 Dessonornis 299 Diaphorapteryx 171 Diaphorophyia 306 Diatryma 179 Dicaeum 322 Dicrurus 275 Dicrocercus 240 Didunculus 207 Didus 205 Diglossa 325 Dinemellia 330 Dinopium 254 Dinornis 92 Diomedea 100 Diphyllodes 282 Diplootocus 299 Discosura 232 Dissemurus 275 Dissoura 123 Dives 333 Diuca 338 Docimastes 232 Dolichonyx 333 Domicella 210 Donacobius 298 Doricha 232 Dorifera 232 Drepanis 326 Drepanoplectes 331 Drepanoptila 207 Drepanornis 282 Dreptes 321
Drioctistes 260 Dromaeocercus 302 Dromaius 90 Dromas 192 Dromococcyx 214 Drymodes 291 Drymornis 259 Dryocopus 254 Dryolimnas 171 Dryotriorchis 146 Dryoscopus 315 Dulus 310 Ducula 207 Dumetella 298 Dumetia 291 Dysithamnus 261
Ectopistes 207
Edolisoma 292
Edolius 275
Egretta 118
Enringa 275
Elaenia 265
Elanoides 146
Elanus 146
Electron 239
Eleothreptus 225
Elsyornis 188
Emberiza 338
Empidonax 265
Empidonomus 265
Enaliornis 81
Endomychura 201
Enicognathus 210
Enicurus 299
Enodes 318
Ensifera 232
Entomyzon 320
Eophona 341
Eopsaltria 306
Eos 210
Ephippiorhynchus 123
Epimachus 282
Epthianura 303
Erannornis 306
Eremiornis 303
Eremomela 302
Eremophila 273
Eremopteryx 273
Ergaticus 328
Eriocnemis 232
Erithacus 299
Erolia 189
Erythrina 340
Erythrocercus 306
Erythrogonys 188
Erythropus 150
Erythropygia 302
Erythrotriorchis 146
Erythrura 331
Esacus 179
Estrilda 331
Eubucco 249
Euchlornis 264
Eucometis. 335
Eudocimus 124
Eudromias 188
Eudynamis 213, 214
Eudyptes 84
Eudyptula 84 Eugenes 232
352
Eugralla 262 Eulabeia 132 Eulacestoma 306 Eulampis 232 Eulipoa 153, 154 Eumomota 239 Eumyias 306 Eunymphicus 210 Euodice 331 Euphagus 333 Euphonia 335 Euphorbia 249 Eupetes 291 Eupetomena 232 Euplectes 331 Eupodotis 177 Euptilosus 294 Euptilotis 234 Eurocephalus 315 Eurostopodus 225 Euryceros 312 Eurylaimus 256 Eurynorhynchus 189 Eurypyga 174 Eurystomus 242 Euscarthmus 265 Eutoxeres' 232 Eutriorchis 146 Excalfactoria 159 Gecinulus 254 Gelochelidon 199 Geobates 260 Geocichla 299 Geococcyx 214 Geocolaptes 254 Geoffroyus 210 Geopelia 207 Geopsittacus 211 Geositta 260 Geospiza 339 Geothlypis 328 Geotrygon 207 Geranospiza 146 Geronticus 124 Gerygone 303 Glareola 193 Glaucidium 218 Glaucis 232 Gliciphila 320 Glossopsitta 210 Glyphorhynchus 259 Gorsachius 118 Goura 207 Gracula 318 Graculus 278 Gracupica 318 Grailaria 261 Grallaricula 261 Grallina 279
Falco 149, 150 Falcipennis 156 Falculea 312 Falcunculus 306 Ferminia 297 Ficedula 306 Florisuga 232 Fluvicola 265 Formicarius 261 Formicivora 261 Forpus 210 Foudia 331 Francolinus 159 Fraseria 306 Fratercula 201 Fregata 114 Frege tta 102 Fregilupus 318 Fringilla 340 Fringillaria 338 Fringillauda 341 Fulica 171 Fulmarus 101 Furnarius 260 Granatellus 328 Granatina 331 Grandala 299 Graydidasculus 211 Grus 167 Gubernetes 265 Gugis 199 Guira 213, 214 Guiraca 340 Guttera 159 Gymbilaimus 261 Gymhobucco 249 Gymnocichla 261 Gymnocorvus 278 Gymnoderus 264 Gymnogenus 146 Gymnoglaux 218 Gymnogyps 143 Gymnophaps 207 Gymnopithys 261 Gymnorhina 278 Gymnorhinus 278 Gymnostinops 333 Gypaetus 146
Galachrysia 193 Galbalcyrhynchus 248 Galbula 248 Galerida 273 Gallicolumba 207 Gallicrex 171 Gallinago 189 Gallinula 171 Gallirallus 171 Galloperdix 159 Gallus 159 Gampsonyx 146 Gampsorhynchos 291 Garrodia 102 Garrulax 291 Garr ulus 278 Gavia 96 Gypohierax 146 Gypopsitta 211 Gyps 146 Habia 335 Habropteryx 171 Habroptila 171 Haematoderus 264 Haematopus 185 Haematortyx 159 Haematospiza 341 Hagedashia 124 Halcyon 236 Haliaeetus 146 Haliastur 146 Halobaena 101 Halocyptena 102 Hamirosfra 146
353
Hargeria 81 Harpactes 234 Harpagus 146 Harpe 150 Harpia 146 Harpiprion 124 Harpyhaliaetus 146 Harpyopsis 146 Hartlaubius 318 Hedy me les 340 Heliactin 232 Heliangelus 232 HeliobTetus 260 Heliochera 264 Heliomaster 232 Heliopais 171 Heliornis 171 Heliothrix 232 Helmitheros 328 Hemichelidon 306 Hemicircus 254 Hemignathus 326 Hemiparva 188 Hemiphaga 207 Hemiprocne 227 Hemipus 292 Hemispingus 335 Hemitnraupis 335 Henicopernis 146 Henicophas 207 Henicorhina 297 Herbivocula 302 Herpetotheres 149 Hesperiphona 340 Hesperornis 81 Heterolocha 317 Heterocercus 264 Heteromirafra 273 Heteromunia 331 Heteronetta 139 Heterophasia 291 Heteroscelus 189 Heterospizias 146 Heterotrogon 234 Hieraeetus 146 Himantopus 190 Himantornis 171 Himatione 325 Hippolais 302 Hirundapus 229 Hirundinea 265 Hirundo 274 Histrionicus 138 Histurgops 331 Hoplopterus 188 Hoploxypterus 188 Horeites 302 Houbaropsis 177 Hydrobates 102 Hydrophasianus 182 Hydroprogne 199 HydropsaTis 225 Hydydipna 321 Hygranassa 118 HyTacola 303 Hylia 302 Hylocharis 232 Hylochelidon 274 Hylocichla 299 Hylocitrea 306 Hylomanes 239 Hylophilus 324
Hylophylax 261 Hymenolaimus 134 Hypargos 331 Hyphanturgus 331 Hypnelus 248 Hypochera 331 Hypocnemis 261 Hypocolius 310 Hypositta 286 Hypothymis 306 Hypotriorchis 150 Hypskpetes 294
Ibidorhyncha 190 Ibis 123
Ibycter 150 Ichthyophaga 146 Ichthyornis 82 Icteria 328 Icterus 333 Ictinaetus 146 Ictinia 146 leracidea 150 Ifrita 291 Indicator 251 lodopleura 264 lole 294
Ipophilus 254 Irania 299 Irediparra 182 Irena 275, 295 Iridoprocne 274 Ispidina 236 Ithaginus 159 Ixobrychus 115, 118 Ixonotus 294 Ixoreus 299
Jabiru 123 Jabouilleia 291 Jacamaralcyon 248 Jacamerops 248 Jacana 182 Jubula 218 Junco 338 Jynx 254
Kakatoe 210 Kaupifalco 146 Kenopia 291 Ketupa 218 Kitta 278 Kittacincla 299 Kittlitzia 318 Knipolegus 265 Kozlowia 341
Lacedo 236 Lafresnaya 232 Lagonosticta 331 Lagopus 156 Laiscopus 307 Lalagea 292 Lalocitta 278 Laletes 325 Lamprocolius 318 Lampornis 232 Lampribis 124 Lamprolia 299 Lamprotornis 318 Laniarius 315
354
Lanio 335 Laniturdus 315 Lanius 315 Larosterna 199 Larus 199 Larvivora 299, Laterallus 171 Lathamus 210 Lawrencia 325 Layardiella 211
Legatus 265 Leiothrix 291
Leipoa 153
Leistes 333
Leonardina 291
Lepido.colaptes 259
Lepidogrammus 214
Leptasthenura 260
Leptocoma 321
Leptodon 146
Leptopoecile 304
Leptopterus 312 Leptosomus 243 Leptotila 207 Leptoptilos 121 i 123 Lerwa 159 Lesbia 232 Lessonia 265 Leucippus 232 Leuconerpes 254 Leucopeza 328 Leucopsar 318 Leucopternis 146 Leucosarcia 207 Leucost icte 341 Lichenops 265 Lichmera 320 Limicola 189 Limnocorax 171 Limnodromus 189 Limnothlypis 328 Limosa 189 Linura 331 Linugus 341 Liocichla 291 Liopicus 254 Lioptilus 291 Liosceles 262 Lipaugus 264 Lissotis 177 Lobibux 188 Lobipluvia 188 Lobivanellus 188 Lobotes 292 Locustella 302 Loddigesia 232 Lonchura 331 Lophobasileus 304 Lophoictinia 146 Lopholaimus 207
Lophonetta 133, 134
Lophophanes 285
Lophornia 282 Lophophorus 159 Lophornis 232 Lophortyx 159 Lophostrix 218
Lophotibis 124
Lop hot is 177
Lophozosterops 324
Lophura 159
Loria 282
Loriculus 211 Lorius 211 Loxia 341 Loxigilla 341 Loximitris 341 Loxops 326 Lullula 273 Lunda 201 Lurocalis 225 Luscinia 299 Lusciniola 302 Lybius 249 Lycocorax 282 Lymnocryptes 189 Lyrurus 156
Macgregoria 282 Machaerhamphus 146 Machaerhopterus 264 Machaerirhynchus 306 Machetornis 265 Macropsalis 225 Macrorhampus 189 Macronectes 101 Macronus 291 Macronyx 308 Macrodipteryx 225 Macropygia 207 Macrosphenus 302 Madanga 324 Malacocincla 291 Malaconotus 315 Malacopteron 291 Malacoptila 248 Malacorhynchus 134 Malia 294 Malimbus 331 Malurus 303 Manorina 320 Manucodia 282 Margaroperdix 159 Margarops 298 Margarornis 260 Masius 264 Megacephalon 153, 154 Megaceryla 236 Megacrex 171 Megadyptes 84 Megalaima 249 Megalapteryx 92 Megalurus 302 Megapodius 153, 154 Megarhynchus 265 Megatriorchis 146 Megaxenops 260 Meiglyptes 254 Melaenornis 306 Melampitta 291 Melanerpes 254 Melanitta 138 Melanocharis 322 Melanochlora 285 Melanocorypha 273 Melanodera 338 Melanopareia 261 Melanopariea 262 Melanoperdix 159 Melanophoyx 118 Melanoploceus 331 Melanoptila 298 Melanopyrrhus 318 Melanospiza 338
355
Melanotis 298 Melanotrochilus 232 Meleagris 160 Melichneutes 251 Melidora 236 Melierax 146 Melignomon 251 Meliphaga 320 Melipotes 320 Melithreptus 320 Melittophagus 240 Mellisuga 232 Melophus 338 Melopsittacus 211 Melopyrrha 341 Melospiza 338 Menura 269 Merganetta 134 Mergus 129, 138 Merops 240 Merulaxis 262 Mesembrinibis 124 Mesia 291 Mesitornis 163 Mesoenas 163 Mesopicos 254 Metallura 232 Metopidius 182 Metopothrix 260 Metriopelia 207 Micrastur 149 Micrathene 218 Microbates 303 Microcerculus 297 Microcichla 299 Microdynamis 214 Microeca 306 Microgoura 207 Micron ierax 150 Microrhopias 261 Micromonacha 248 Micropalma 189 Microparra 182 Micropsitta 210 Micropternus 254 Micropygia 171 Microsarcops 188 Microscelis 294 Microstilbon 232 Microura 291 Milvago 150 Milvus 146 Mimizuka 218 Mimodes 298 Mimus 298 Minla 291 Mino 318 Mioceyx 236 Mirafra 273 Miro 306 Misocalius 214 Mitu 154 Mniotilta 328 Moho 320 Molothrus 333 Mol paster 294 Momotus 239 Monacus 264 Monarch a 306 Monasa 248 Monias 163 Monticola 299
Montifringilla 331 Morococcyx 214 Moropogon 240 Morphnus 146 Morns 109
Moupinia 291
Motacilla 308
Mulleripicus 254
Muscicapa 306 Muscicapella 306 MuseigraIla 265 Muscisaxicola 265
Muscivora 265
Musophaga 212
MyadestCs 299
Mycerobas 341
Myiagra 306
Myiarchus 265
Myiborus 328
Myiobius 265 Myiodynastes 265
Myiolestes 306 Myiophoneus 299 Myiozetetes 265 Myiopsitta 211
Myophonus 299
Myopornis 306
Myrmeciza 261
Myrmecocichla 299
Myrmecocichlia 261
Myrmia 232
Myrmoborus 261
Myrmotherula 261
Myza 320
Myzantha 320 Myzomela 320
Myzornis 291
Namibornis 306
Nannopsittaca 211
Nannopterum 103, 110
Napotnera 291
Nasica 259
Necropsar 318
Necrosyrtes 146
Nectarinia 321
Nemosia 335
Neochen 133
Neochloe 325
Neocossyphus 301
Neocrex 171
Neodrepanis 267, 268
Neogaeornis 81 Neochmia 331 Neoctantes 261 Neolestes 294
Neomixis 291
Neomorphus 214
Neopelma 264
Neophron 146
Neophema 211 Neopsephotus 211 Neopsittacus 210
Neositta 285, 286
Neospiza 331
Neotis 177
Neoxolmis 265
Nesasio 218
Nesierax 150 Nesoceleus 254 Nesocharis 331 Nesocichla 301
356
Nesoclopeus 171 Nesoctites 254 Nesofregetta 102 Nesomimus 298 Nesospiza 338 Nesotrochis 171 Nestor 210 Netta 136 Nettapus 137 Newtonia 306 Nicator 294 Nigrita 331 NiTaus 315 Niltava 306 Ninox 218 Niobella 331 Nipponia 124 Nonnula 248 Northilla 211 Notharchus 248 Nothocrax 154 Notiochelidon 274 Notiomystis 320 Nothocercus 95 Nothoprocta 95 Nothura 95 Notornis 171 Novipulsatrix 218 Nuciiraga 278 Numenius 189 Numida 159, 160 Nuttallornis 265 Nyctanassa 118 Nyctea 218 Nyctibius 223 Nycticorax 118 Nycticrypes 184 Nyctidromus 225 Nyctiornis 239, 240 Nyctiphrynus 225 Nyctiprogne 225 Nymphicus 211 Nystalus 248
Oceanites 102
Oceanodroma 102
Ochromela 306
Ochthoeca 265
Ocreatus 232
Ocyalus 333
Ocyphaps 207
Odontophorus 159
Oena 207
Oenanthe 301
Oligura 302
Oncostoma 265
Onychognathus 318
Onychorhynchus 265
Ophrysia 159
Opisthocomus 161
Opopsitta 210
Oporornis 328 Orchesticus 335 Oreocharis 322 Oreocincla 301
Oreocorys 308
Oreoica 306
Oreophasis 154
Oreopholus 188 Oreopneuste 302
Oreopsittacus 210
Oreortyx 159
Oreoscoptes 298
Oreostruthus 331
Qreotrochilus 232
Oriolia 312
Oriolus 275
Origma 303
Oroaetus 146
Ortalis 154
Orthonyx 291
Orthornamphus 179
Orthotomus 302
Ortygospiza 331
Ortyxelus 164
Oryzoborus 338
Ost inops 333
Otidiphaps 207
Otis 177
Otocompsa 294
Otophanes 225
Otus 218
Oxypogon 232
Oxyruncus 266
Oxyura 139
Pachycare 306
Pachycephala 306
Pachycoccyx 214
Pachyptila 98, 101
Pachyrhamphus 264
Pagodroma 101
Pagophila 199
Palaeoscinis 294
Palaeospiza 273
Palmeria 326
Pandion 147
Panterpe 232
Panurus 288
Panyptila 227, 229
Parabuteo 146
Paradigalla 282
Paradisaea 282
Paradoxomis 288
Paramythia 322
Pardalotus 322
Pardirallus 171
Parisoma 306
Parmoptila 331
Paroaria 340
Paroreomyza 326
Parotia 282
Parula 328
Parus 285
Passer 328, 330, 331
Passerculus 338
Passerella 338
Passerherbulus 338
Passerina 340
Pastor 318
Patagona 232
Pauxi 154
Pavo 159
Pedilorhynchus 306
Pedioecetes 156
Pedionomus 166
Pelagodroma 102
Pelagornis 108
Pelargopsis 236
Pelecanoides 102
Pelecanus 104, 107, 108
Pellorneum 291
357
Peltohyas 193 Peltops 306 Penelope 154 Penelopides 246 Penelopina 154 Penthestes 285 Penthoceryx 214 Perdicula 159 Perdix 159 Pericrocotus 292 Periparus 285 Periporyhus 340 Perisoreus 278 Perissocephalus 264 Perissotricus 265 Pernis 146 Petroche lidon 274 Petroica 306 Petronia 331 Petrophassa 207 Peucedramus 328 Pezites 333 Pezophaps 205 Pezoporus 211 Phacellodomus 260 Phaenicophaeus 214 Phaenostictus 261 Phaeomyias 265 Phaeornis 301 Phaethon 105 Phaethornis 232 Phaetusa 199 Phainopepla 310 Phainoptila 310 Phalacrocorax 104, 110 Phalaenoptilus 224, 225 Phalaropus 190 Phalcobaenus 150 Phapitreron 207 Phaps 207 Pharomachrus 234 Phasianus 159 Phasidus 159 Phedina 274 Phegornis 188 Pheucticus 340 Phibalura 264 Phigys 210 Philacte 132 Philemon 320 Philepitta 267, 268 Philesturnus 317 Philetairus 329, 331 Philohela 189 Philomachus 189 Philortyx 159 Philydor 260 Phimosus 124 Phlegopsis 261 Phleocryptes 260 Phoebetria 100 Phloeoceastes 254 Phoenicircus 264 Phoeniconaias 126 Phoenicoparrus 126 Phoenicopterus 126 Phoeniculus 245 Phoenicurus 301 Pholidornis 331 Phonygammus 282 Phororhacos 175 Phragmaticola 302
Phrygilus 338
Phyllanthus 291
Phyllastrephus 294
Phyllergates 302
Phylloscartes 265
Phylloscopus 302
Phytotoma 266
Piaya 214
Pica 278
Picathartes 291
Picoides 254
Piculus 254
Picumnus 254
Picus 254
Pinaroloxias 339
Pinicola 341
Pionites 211
Pionopsitta 211
Pionus 211
Pipile 155
Pipilo 338
Pipra 264
Piprites 264
Pipromorpha 265
Piranga 335
Pirenestes 331
Pitangus 265
Pithecophaga 146
Pithys 261
Pitohui 306
Pitta 267
Pittasoma 261
Pitylus 340
Pityriasis 315
Platalea 124
Platibis 124
Platycercus 211
Platylophus 278
Platypsaris 264
Platyrinchus 265
Platysmurus 278
Platyspiza 339
Platysteira 306
Plautus 201
Plectrophenax 338
Plectropterus 137, 138
Plegadis 124
Plegadornis 124
Plocepasser 331
Ploceus 331
Plotus 201
Pluvialis 188
Pluvianellus 188
Pluvianus 193
Pluviorhynchus 188
Plyctolophus 210
Pnoepyga 291
Podager 225
Podargus 222
Podica 171
Podiceps 97
Podilymbus 97
Podoces 278
Poecile 285
Poecilodryas 306
Poeoptera 318
Poepnila 331
Pogoniulus 249
Pogonornis 249
Poicephalus 211
Polionierax 150
Poliolimnas 171
358
Poliomyias 306 Polioptila 303 Polipicus 254 Polmaetus 146 Polyboroides 146 Polyborus 150 Polyplectron 159 Polysticta 135 Polytelis 211 Polytmus 232 Pomarea 306 Pomatorhinus 291 Pomatostomus 291 Pooecetes 338 Poospiza 338 Popelairia 232 Porphyrio 171 Porphyriops 171 Porphyriornis 171 Porphyrula 171 Porzana 171 Premnoplex 260 Premnornis 260 Prinia 302 Prioniturus 211 Prionochilus 322 Prionodura 281 Prionops 315 Priotelus 234 Probosciger 210 Procellaria 101 Procnias 264 Prodotiscus 251 Progne 274 Promerops 320 Proparus 291 Prosobonia 189 Prosopeia 211 Prosthemadera 320 Protonotaria 328 Protopelecanus 108 Prunella 307 Psalidoprocne 274 Psaltiparus 285 Psaltria 285 Psarisomus 256 Psarocolius 333 Psephotus 211 Pseudalaemon 273 Pseudeos 210 Pseudibis 124 Pseuddaedon 301 Pseudocalyptomena 256 Psepdocolaptes 260 Pseudochelidon 274 Pseudohirundo 274 Pseudogyps 146 Pseudoleistes 333 Pseudonestor 326 Pseudonigrita 331 Pseudopodoces 278 Pseudoptynx 218 Pseudoscops 218 Pseudoseisura 260 Psilopogon 249 Psilorhampus 262 Psilorhinus 278 Psittacella 211 Psittacula 211 Psittaculirostris 210 Psittacus 210 Psittinus 211
Psittirostra 326 Psittrichas 210 Psomocolax 333 Psophia 169 Psophocichla 301 Psophodes 291 Pteridophora 282 Pterocles 205 Pterocnemia 87 Pterodroma 101 Pteroglossus 252 Pterophanes 232 Pteropodocys 292 Pteroptochos 262 Pteruthius 291 Ptilinopus 207 Ptilocichla 291 Ptilogonys 310 Ptilolaemus 246 Ptilonorhynchus 281 Ptilopachus 159 Ptiloris 282 Ptiloscelyx 188 Ptilostomus 278 Ptilotus 320 Ptiloxena 333 Ptychoramphus 201 Ptyonoprogne 274 Ptyrticus 291 Pucrasia 159 Puffinus 101 Pulsatrix 218 Purpureicephalus 211 Pycnonotus 294 Pycnoptilus 303 Pygarrhichas 260 Pygoscelis 84 .Pyrgilauda 331 Pyriglena 261 Pyrocephalus 265 Pyroderus 264 Pyrrhalaemus 303 Pyrrhocorax 278 Pyrrhoglaux 218 Pyrrhospiza 341 Pyrrhula 341 Pyrrhuloxia 340 Pyrrhura 211 Pytilia 331
Quelea 229, 331 Querula 264 Quiscalus 333
Radda 331 Rallicula 171 Rallina 171 Rallus 171 Ramphacinclus 298 Ramphastos 252 Ramphocaenus 303 Ramphocelus 335 Ramphomicron 232 Ramsayornis 320 Raphus 205 Recurvirostra 190 Reguloides 302 Regulus 304 Reinwardtoena 207 . Remiz 283, 285 Rhabdornis 287
359
Rhagalogus 306 Rhamphocharis 322 Rhamphococcyx 214 Rampnocorys 273 Rhamphomantis 214 Rhamphostruthus 331 Rhea 87 Rheinartia 159 Rhinochetus 173 Rhinocorax 278 Rhinocrypta 262 Rhinoplax 246 Rhinopomastus 245 Rhinopsar 318 Rhinoptynx 218 Rhinortna 214 Rhiomyias 306 Rhipidura 306 Rhizothera 159 Rhodilus 218 Rhodonessa 134 Rhodopechys 341 Rhodophoneus 315 Rhodospiza 341 Rhodostethia 199 Rhodothraupis 340 Rhopocichla 291 Rhopodytes 214 Rhopornis 261 Rhyacornis 301 Rhynchocyclus 265 Rhynchophanes 338 Rhynchopsitta 211 Rhynchortyx 159 Rhynchostruthus 341 Rhynchotus 95 Rhynoptilus 193 Rhyticeros 246 Richamondena 340 Rimator 291 Riparia 274 Rissa 199 Rodinocichla 335 Rogibyx 188 Rollulus 159 Rostratula 184 Rostrhamus 146 Rougetius 171 Rowettia 338 Rubicilla 341 Rubigula 294 Rufibrenta 132 Rukia 324
Rupicola 264 Rynchops 199
Sagittarius 144 Sakesphorus 261 Salpinctes 297 Salpornis 287 Saltator 340 Sanchezia 321 Sapheopipo 254 Sappho 232 Sarciophorus 188 Sarcogyps 146 Sarcops 318 Sarcoramphus 143 Sarkidiornis 137, 138 Saroglossa 318 Sarthrura 171 Sasia 254
Saurothera 214
Saxicola 301
Saxicoloides 301
Sayornis 265
Scapaneus 254
Scaphidura 333
Scardafella 207
Sceloglaux 218
Scelorchilus 262
Scenopoeetes 280, 281
Scepomycter 302
Schetba 312
Schiffornis 264
Schizoeaca 260
Schoeniophylax 260
Scissirostrum 318
Sclateria 261
Sclerurus 260
Scolopax 189
Scops 120
Scoptelus 245
Scotocerca 302
Scotopelia 218
Scotornis 225
Scythrops 214
Scytalopus 262
Schetba 312
Seicercus 302
Seiurus 328
Selasphorus 232
Selenidera 252
Seleucidis 282
Semeiophorus 225
Semioptera 282
Semnornis 249
Sephanoides 232
Sericornis 303
Sericossypha 335
Sericulus 281
Serilophus 256
Serious 341
Serpophaga 265
Setopnaga 328
Sialia 301
Sicalis 338
Sigmodus 315
Siphia 306
Siphonorhis 225
Siptornis 260
Sitta 286
Sittasomus 259
Siva 291
Smaragdolanius 316
Smicronis 303
Smilorhis 249
Smithornis 256
Somateria 135
Spartonoica 260
Speirops 324
Spelaeornis 291
Speotyto 218
Spermestes 331
Spermophaga 331
Sphecotheres 275
Spheniscus 84
Sphenocichla 291
Sphenoeacus 302
Sphenorhynchus 123
Sphenostoma 291
Sphenurus 207
Sphyrapicus 254
Spiloptila 302
360
Spilornis 146 Spindalis 335 Spinus 341 Spiza 340 Spizaetus 146 Spizastur 146 Spizella 338 Spiziapterax 150 Spizixos 294 Spodiopsar 318 Sporaeginthus 331 Sporophila 338 Sporopipes 331 Spreo 318 Squatarola 188 Stachyris 291 Stactolaema 249 Starnoenas 207 Steatornis 219 Steganopleura 331 Steganura 331 Stelgidopteryx 274 Stellula 232 Stephanibux 188 Stephanoaetus 146 Stephanophorus 335 Stephanoxis 232 Stercorarius 197 Sterna 199 Stictonetta 134 Stiltia 193 Stipiturus 303 Stizorhina 301 Stomiopera 320 Strepera 278 Streptocitta 318 Streptopelia 207 Strigops 210 Strix 218 Struthidea 279 Struthio 86 Sturnella 333 Sturnia 318 Sturnus 318 Sula 104, 109 Surnia 218 Surniculus 214 Suthora 288 Sylvia 302 Sylvietta 302 Sylviorthorhynchus 260 Sylviparus 285 Synallaxis 260 Synoiclus 159 Synthliboramphus 201 Sypheotides 177 Syrmaticus 159- Syrrhaptes 205 Tantalus 123 Tanygnathus 211 Tanysiptera 235, 236 Taoniscus 95 Tapera 214 Taraba 278 Tarba 261 Tarsiger 301 Tauraco 212 Tchagra 315 Tchitrea 306 Teledromas 262 Teleonema 264 Telephorus 315 Tematodytes 297 Temenuchus 318 Temnotrogon 234 Temnurus 278 Tephrodornis 292 Terathopius 146 Terekia 189 Terenura 261 Terpsiphone 306 Tesia 302 Tersina 333 Tetraenura 331 Tetrao 156 Tetraogallus 159 Tetra pteryx 167 Tetrastes 156 Tetrax 177 Textor 318 Thalassarche 100 Thalasseus 199 Thalassoica 101 Thalassornis 139 Thalurania ,232 Thamnocharis 261 Thamnophilus 261 Thaumatibis 124 Theristricus 124 Thescelocichla 294 Thinocorus 194 Thinornis 188 Thraupis 335 Threnetes 232 Threskiornis 124 Thripias 254 Thripodectes 260 Thripophaga 260 Thryomanes 297 Thryothorus 297 Thypopsis 335 Tiaris 338 Tichodroma 287 Tigrosoma 118 Tijuca 264 Tilmatura 232 Timalia 291
Taccocua 214 Tachirnis 265 Tachornis 227, 229 Tachuris 265 Taehycineta 274 Tachyeres 133, 134 Tachyphonus 335 Tadorna 133 Taeniopygia 331 Talegalla 153, 154 Tanagrella 335 Tangara 335 Tangavius 333 Tinamotus 95 Tinamus 95 Tityra 264 Tmetothylacus 308 Tockus 246 Todirostrum 265 Todopsis 303 Todus 236 Tolmomyias 265 Topaza 232 Torgos 146 Touit 211 Toxostoma 298
361
Trachylaemus 249 Trachyphonus 249 Traversia 267 Treron 207 Tribura 302 Trichastoma 291 Trichocichla 302 Trichoglossus 210 Tricholaemus 249 Tricholestes 294 Tricholimnas 171 Trichopicus 254 Triclaria 211 Trigo noceps 146 Tringa 189 Triodia 303 Trochilus 232 Troglodytes 297 Trogon 234 Trogopan 159 Tropicranus 246 Trugon 207 Tryngites 189 Turacoena 207 Turdinus 291 Turdoides 291 Turdus 299, 301 Turnagra 317 Turnix 164 Turtur 207 Tylas 312 Tylibyx 188 Tympanuchus 156 Tyranneutes 264 Tyrannus 265 Tyto 218 Vanga 312 Veniliornis 254 Vermivora 328 Verreauxia 253, 254 Vestiaria 326 Vidua 331 Vini 210 Vireo 324 Vireolanius 316 Vireosylva 325 Viridonia 326 Vitia 303 Volatinia 338 Vultur 143 Waldronia 232 Warsanglia 341 Wilsonia 328 Woodfordia 324 Yuhina 291 Xanthocephalus 333 Xanthopygia 306 Xenicus 267 Xenocopsychus 301 Xenoctistes 260 Xenopirostris 312 Xenops 260 Xenorhynchus 123 Xenornis 261 Xenus 189 Хеша 199 Xiphidiopicus 254 Xiphidiopterus 188 Xiphirynchus 291
Upucerthia 260 Upupa 243 Uraeginthus 331 Uragus 341 Uratelornis 242 Uria 201 Urocissa 278 Urodynamis 214 Uroglaux 218 Urolestes 315 Uropelia 207 Uropsalis 225 Uropsila 297 Urotriorchis 146 Urosphena 302 Xipholena 264 Xiphocolaptes 259 Xiphorhynchus 259 Xolmis 265 Zaratornis 264 Zarhynchus 333 Zanclostomus 214 Zanthopygia 306 Zavattariornis 278 Zebrilus 118 Zeledonia 301 Zeocephus 306 Zenaida 207 Zonaeginthus 331 Zonibyx 188 Zonotrichia 338
Vanellochettusia 188 Vanellus 186, 188 Zoothera 301 Zosterops 324
ОГЛАВЛЕНИЕ
Стр.
Предисловие ............................................................... 3
Основные задачи и принципы систематики. Систематические категории. Правила зоологической номенклатуры ................................................ 5
Цели и задачи систематики ................................................. 5
Понятие о виде ............................................................ 7
Виды-двойники ........................................................... 8
Индивидуальная внутривидовая изменчивость ................................. 9
Географическая изменчивость . . . .-.................................... И
Подвиды................................................................... 14
Клинальная изменчивость................................................... 15
Монотипические и политипические виды.................................... 16
Географические изоляты ................................................... 16
Виды или подвиды •........................................................ 17
Надвиды................................................................... 18
Кольцевые перекрывания ареалов............................................ 19
Происхождение новых видов................................................. 22
Таксономические категории и некоторые правила номенклатуры .... 26
Вид .......................................................... . 26
Подвид и другие внутривидовые категории............................. 27
Высшие таксономические категории ................................... 28
Род ........................................................... , 28
Семейство .......................................................... 30
Отряд ,............................................................. 31
Расхождения в рбъеме таксономических категорий............................ 32
Номенклатурный кодекс .................................................... 33
Некоторые зоогеографические понятия, болеечасто Используемые в систематике 34
Некоторые морфологические признаки, используемые в систематике .... 36
Типы расположения пальцев . . . :................................... 36
Типы скелетной основы ноздрей . . .................................. 37
Кинетизм черепа и типы строения костного неба....................... 37
Формула ножной мускулатуры.......................................... 39
Нцжняя гортань ................................................ . 39
Типы оперения крыла ............................................... 40
История становления системы класса птиц. Современные системы ... 41
Происхождение и эволюция класса птиц................................... 71
Систематический обзор класса птиц Aves................................... 78
Подкласс Archaeornithes.— древние, или ящерохвостые, птицы................ 79
Отряд 1. Archaeopterygiformes....................................... 80
Подкласс Neornithes — настоящие, или веерохвостые, птицы.................. 80
Надотряд I. Odontognathae — зубастые птицы ............................... 81
Отряд 2. Hesperornithiformes — гесперорнисообразные................. 81
Надотряд II. Ichthyornithes — ихтиорнисы................................. 82
Отряд 3. Ichthyornithiformes — ихтиорнисообразные .'................ 82
Надотряд III. Impennes — плавающие ....................................... 82
Отряд 4. Sphenisciformes — пингвинообразные......................... 82
Надотряд IV. Neognathae — новонебные, или типичные, птицы................. 84
Отряд 5. Struthioniformes — страусообразные......................... 86
Отряд 6. Rheiformes— нандуобразные '................................ 87
Отряд 7. Casuariiformes — казуарообразные........................... 89
Семейство 1. Casuariidae — казуаровые............................ 90
Семейство 2. Dromaiidae — эму................................... 90
Семейство 3. Dromornithidae ...................................... 91
Отряд 8. Aepyornithiformes — эпиорнисообразные...................... 91
Отряд 9. Dinornithiformes — моаобразные < , . . . •. • • • • 92
Отряд 10. Apterygiformes — кивиобразные, или бескрылые ._ . . . . 93
Отряд 11. Tinamiformes — тинамуобразные, иди скрытохвостые ... 94
Отряд 12.* Gaviiformes— гагарообразные . 95
Отряд 13. Podicipediformes — поганкообразные........................ 96
363
Отряд 14. Procellariiformes (Tubinares) — буревестникообразные, или трубконосые .........................................................
Семейство 1. Diomedeidae — альбатросовые...........................
Семейство 2. Procell ariidae — буревестниковые.....................
Семейство 3. Hydrobatidae — качурковые.............................
Семейство 4. Pelecanoididae — ныряющие буревестники................
Отряд 15. Pelecaniformes (Steganopo diformes) — пеликанообразные,
или веслоногие ....................................................
Подотряд 1. Odontopteryges— ложнозубые птицы...........................
Подотряд 2. Phaethontes — фаэтоновые................................... .
Семейство 3. Phaethontidae — фаэтоновые............................
Подотряд 3. Pelecani — пеликановые.........................................
Надсемейство 1. Pelecanoidea — пеликановые/................................
Семейство 4. Pelecanidae — пеликановые.............................
Надсемейство 2. Suloidea — олушевые........................................
Семейство 6. Pelagornithidae...................................
Семейство 7. Sulidae — олушевые....................................
Семейство 8. Elopterygidae.........................................
Семейство 9. Phalacrocoracidae — баклановые........................
Семейство 10. Anhingidae — змеешейковые............................
Подотряд 4. Fregatae — фрегатовые ...................................
Семейство И. Fregafidae — фрегатовые.........................
Подотряд 5. Cladornithes ..................................................
Отряд 16. Ciconiiformes (Gressores) — аистообразные, или голенастые Подотряд 1. Ardeae — цаплевые..............................................
Семейство 1. Ardeidae — цаплевые...................................
Семейство 2. Cochleariidae — челноклювовые.........................
Подотряд 2. Balaenicipites — китоглавовые..................................
Семейство 3. Balaenicipitidae — китоглавовые.......................
Подотряд 3. Ciconiae — аистовые........................................ .
Надсемейство 1. Scopoidea — молотоглавовые.................................
Семейство 4. Scopidae — молотоглавовые.............................
Надсемейство 2. Ciconioidea — аистовые ....................................
Семейство 5. Ciconiidae — аистовые ................................
Надсемейство 3. Threskiornithdidea — ибисовые..............................
Семейство 6. Threskiornithidae — ибисовые..........................
Подотряд 4. Phoenicopteri — фламинговые ...................................
Семейство 8. Pnoenicopteridae — фламинговые.................. ...
Отряд 17. Anseriformes — гусеобразные ...........
Подотряд 1. Anhimae — паламедеевые.........................................
Подотряд 2. Anseres — гусиные, или пластинчатоклювые.......................
Подсемейство 1. Anseranatinae — полулапчатые гуси............................
Подсемейство 2. Anserinae — гусиные........................................
1. Триба. Dendrocygnini — древесные утки...................................
2. Триба. Anserini — гуси и лебеди.........................................
Подсемейство 3. Anatinae — утиные..........................................
1. Триба. Tadornini — земляные утки . . ..................................
2. Триба. Anatini — речные утки...........................................
3. Триба. Somaterini — гаги ..............................................
4. Триба. Aythyini — нырковые утки........................................
5. Триба. Cairini—шипуны..................................................
6. Триба. Mergini — морские утки и крохали................................
7. Триба. Oxyurini — савки................................................
Отряд 18. Falconiformes — соколообразные, или дневные хищные птицы .................................................................
Подотряд 1. Cathartae — американские грифы.................................
Семейство 1. Neocathartidae.......................................
Семейство 2. Cathartidae — американские грифы.....................
Семейство 3. Teratornithidae......................................
Подотряд 2. Falcones — соколиные...........................................
Надсемейство 1. Sagittarioidea — птицы секретари...........................
Семейство 4. Sagittariidae — птицы секретари . . . -..............
Надсемейство 2. Falconojdae — соколиные ...................................
Семейство 5. Accipitridae — ястребиные............................
Семейство 6. Pandionidae — скопиные...............................
Семейство 7. Falconidae — соколиные...............................
Отряд 19. Galliform^s — курообразные................................
Подотряд Д. Galli— куриные ................................................
Надсемейство 1. Cracoidea — краксовые..............................• . . .
Семейство 1. Megapodiidae — большеногие, или сорные, куры . . .
Семейство 2. Gallinuloididae......................................
Семейство 3. Cracidae — краксовые ................................
98 100 101 101
102
103 104 104
105 105 105
106 108 108
108
109
114
114
115
116
1\8 118
119
119
120
120
120
121
123
123
125
125
127
127
129
131
132
132
132
133
133
134
135
136
137
138
139
140
143
143
143
144
144
144
144
145
145
147
149
150
151
151
151
154
154
364
Надсемейство 2. Phasianoidea — фазановые..................................
Семейство 4. Tetraonidae — тетеревиные ..........
Семейство 5. Phasianidae — фазановые .............................
Семейство 6. Numididae — цесарковые ..............................
Семейство 7. Meleagrididae — индейковые...........................
Подотряд 2. Opisthocomi — гоациновые....................,.................
Семейство 8. Opisthocomidae — гоациновые..........................
Отряд 20. Gruiformes — журавлеобразные..............................
Подотряд 1. Mesitornithides — мадагаскарские пастушки.....................
Семейство 1. Mesitornithidae (Mesoenatidae).......................
Подотряд 2.'Turnices — трехперстки........................................
Семейство 2. Turnicidae — трех пер стковые .......................
Семейство 3. Pedionomidae — австралийские странники...............
Подотряд 3. Grues — журавлиные............................................
Надсемейство 1. Gruoidea — журавлиные.....................................
Семейство 4. Geranoididae.........................................
Семейство 5. Gruidae — настоящие журавли .........................
Семейство 6. Aramidae — Арамовые, или пастушковые, журавли . .
Семейство 7. Psophiidae — трубачи.................................
Надсемейство 2. Ralloidea — пастушковые ..................................
Семейство 8. Orthocnemidae........................................
Семейство 9. Rallidae — пастушковые...............................
Подотряд 4. Heliornithes — лапчатоногие...................................
Семейство 10. Heliornithidae — лапчатоногие ......................
Подотряд 5. Rhinocheti — кагу.............................................
Семейство 11. Rhinochetidae — кагу ...............................
Подотряд 6. Eurypygae — солнечные цапли...................................
Семейство 12. Eurypygidae — солнечные цапли.......................
Подотряд 7. Cariamae — кариамы............................................
Надсемейство 1. Phororhacoidea............................................
Надсемейство 2. Cariamoidea — кариамовые..................................
Семейство 18. Cariamidae — кариамовые.............................
Подотряд 8. Otides — дрофиные..........................................
Семейство 21. Otididae — дрофиные.................................
Семейство 22. Burhinidae — авдотковые ............................
Отряд 21. Diatrymiformes — диатримообразные.........................
Отряд 22. Charadriiformes — ржанкообразные..........................
Подотряд 1. Charadrii — куликовые.........................................
Надсемейство 1. Jacanodea — якановые......................................
Семейство 1. Jacanidae — якановые.................................
Надсемейство 2. Charadrioidea — собственно кулики.........................
Семейство 2. Rhegminornithidae....................................
Семейство 3. Rostratulidae — цветные бекасы.......................
Семейство 4. Haematopodidae — кулики сороки.......................
Семейство 5. Charadriidae —. ржанковые ...........................
Семейство 6. Scolopacidae — бекасовые ............................
Семейство 7. Recurvirostridae — шилоклювковые....................
Семейство 8. Presbyornithidae . ..................................
Семейство 9. Phalaropodidae — плавунчиковые.......................
Надсемейство 3. Dromadoidea — рачьи ржанки................................
Семейство 10. Dromadidae — рачьи ржанки..........................
Надсемейство 4. Glareoloidea — тиркушковые ...............................
Семейство 11. Glareolidae — тиркушковые...........................
Надсемейство 5. Thinocoroidea — зобатые бегунки ..........................
Семейство 12. Thinocoridae — зобатые бегунки.....................
Надсемейство 6. Chionidoidea — белые ржанки...............................
Семейство 13. Chionididae — белые ржанки.........................
Подотряд 2. Lari — чайковые...............................................
Семейство 14. Stercorariidae — поморниковые......................
Семейство 15. Laridae — чайковые . .............................
Семейство 16. Rynchopidae — водорезовые .........................
Подотряд 3. Alcae — чистиковые ...........................................
Семейство 17. Alcidae — чистиковые ..............................
Отряд 23. Columbiformes — голубеобразные ..........................
Подотряд 1. Pterocletes— рябки............................................
Семейство 1. Pteroclidae — рябковые .............................
Подотряд 2. Columbae — голубиные .........................................
Семейство 2. Raphidae — дронтовые................................
Семейство 3. Columbidae — голубиные..............................
Отряд 24. Psittaciformes — попугаеобразные ........................
Семейство 1. Psittacidae — попугаевые.............................
Отряд 25. Cuculiformes — кукушкообразные ..........................
155
155
157
159
160
161
161
162
162
163
163
164
164
166
166
166
166
168
169
169
170
170
171
171
173
173
174
174
175
175
175
175
176
176
178
179
179
180
182
182
183
184
184
185
186
188
189
190
190
192
192
193
193
194
194
195
195
196
196
197
199
200
200
202
203
203
205
205
206
207
207
211
365
Подотряд 1. Musophagi— тураковые, или бананоеды............................211
Семейство 1. Musophagidae— тураковые............................. 211
Подотряд 2. Cuculi — кукушковые ...........................................212
Семейство 2. Cuculidae — кукушковые................................212
Отряд 26. Strigiformes — совообразные ...............................214
Семейство 1. Protostrigidae .......................................216
Семейство 2. Tytonidae — сипуховые ................................216
Семейство 3. Strigidae — настоящие совы............................218
Отряд 27. Caprimulgiformes — козодоеобразные ........................219
Подотряд 1. Steatornithes — гуахаро, или жиряки............................219
Семейство 1. Steatornithidae — гуахаро, или жиряки.................219
Подотряд 2. Caprimulgi — козодбевые........................................222
Семейство 2. Podargidae — совиные козодои..........................222
Семейство 3. Nyctibiidae — исполинские козодои.....................223
Семейство 4. Aegothelidae — совиные лягушкороты....................224
Семейство 5. Caprimulgidae — настоящие козодои ....................224
Отряд 28. Apodiformes — стрижеобразные ..............................225
Подотряд 1. Apodi — стрижи ................................................226
Семейство 1. Aegialornithidae......................................226
Семейство 2. Hemiprocnidae, — древесные, или хохлатые, стрижи . . 227
Семейство 3. Apodidae — настоящие стрижи...........................227
Подотряд 2. Trochili — колибри ........................................... 229
Семейство 4. Trochilidae — колибри.................................229
Отряд 29. Coliiformes — птицы мыши...................................232
Отряд 30. Trogoniformes — трогонообразные ...........................233
Отряд 31. Coraciiformes — ракшеобразные .............................234
Подотряд 1. Alcedines — зимородковые.......................................235
Надсемейство 1. Alcedinoidea — зимородковые................................235
Семейство 1. Alcedinidae — зимородковые............................235
Надсемейство 2. Todoidea — тодиевые ..................................... 236
Семейство 2. Todidae — тодиевые....................................236
Надсемейство 3. Momotoidea — момотовые ....................................238
Семейство 3. Momotidae — момотовые.................................238
Подотряд 2. Meropes — щурковые ............................................239
Семейство 4. Meropidae — щурковые .................................239
Подотряд 3. Coracii — ракшевые.............................................240
Семейство 5. Coraciidae — настоящие, или древесные, ракши .... 240
Семейство 6. Brachypteraciidae — земляные ракши....................242
Семейство 7. Leptosomatidae — куроловые . . . .....................243
Семейство 8. Upupidae — удодовые....................'..............243
Семейство 9. Pnoeniculidae — древесные удоды ......................244
Подотряд 4. Bucerotes — птицы носороги.....................................245
Семейство 10. Bucerotidae — птицы носороги.........................245
Отряд 32- Piciformes — дятлообразные ............................... 246
Подотряд 1. Galbulae — примитивные дятлообразные...........................247
Надсемейство 1. Galbuloidea — якамаровые...................................247
Семейство 1. Galbulidae — якамаровые ..............................247
Семейство 2. Bucconidae — пуховковые, или ленивКовые...............248
Надсемейство 3. Capitonoidea — примитивные дятловые........................248
Семейство 3. Capitonidae — бородатковые ...........................249
Семейство 4. Indicatoridae — медоуказчиковые ......................249
Надсемейство 3. Ramphastoidea — тукановые, или перцееды....................251
Семейство 5. Ramphastidae — тукановые..............................251
Подотряд 2. Pici — настоящие дятлообразные..............................J 252
Семейство 6. Picidae — настоящие дятловые..........................252
Отряд 33. Passeriformes — воробьинообразные................. . . . 254
Подотряд 1. Eurylaimi — рогоклювы..........................................256
Семейство 1. Eurylaimidae — рогоклювые.............................256
Подотряд 2. Tyranni (Clamatores)—тиранновые, или кричащие воробьиные . 258
Надсемейство 1. Furnarioidea — древолазы, муравьеловки, птицы печники и др. 258
Семейство 2. Dendrocolaptidae — древолазовые ....................258
Семейство 3. Furnariidae . — птицы печники, или . птицы гончары 259
Семейство 4. Formicariidae — муравьеловковые.....................260
Семейство 5. Conopophagidae — гусеницеедов^е.....................261
Семейство 6. Rhinocryptidae — топаколовые........................262
Надсемейство 2. Tyrannoidea — тиранновые...................................262
Семейство 7. Cotingidae — котинговые...............................262
Семейство 8. Pipridae — манакиновые................................264
Семейство 9. Tyrannidae — тиранновые ..............................264
Семейство 10. Oxyruncidae — остроклювые ...........................266
Семейство 11. Phytotomidae — траворезовые..........................266
Семейство 12. Pittidae — питтовые..................................266
366
Семейство 13. Acanthisittidae (Xenicidae) — новозеландские крапивни-ковые .............................................................267
Семейство 14. Philepittidae — мадагаскарские питтовые..............267
Подотряд 3. Мепигае — примитивные воробьиные...............................268
Семейство 15. Menuridae — птицы лиры...............................268
Семейство 16. Atrichornithidae — кустарниковые птицы.............270
Подотряд 4. Passeres (Oscines) — певчие воробьиные.........................270
Семейство 17. Aleaudidae — жаворонкбвые...........................271
Семейство 18. Palaeospizidae......................................273
Семейство 19. Hirundinidae — ласточковые ..........................273
Семейство 20. Ducruridae — дронговые .............................274
Семейство 21. Oriolidae — иволговые...............................275
Семейство 22. Corvidae — вороновые ............................275
Семейство 23. Cracticidae — флейтовые птицы.......................278
Семейство 24. Grallinidae — австралийские сорочьи жаворонки . . 279
Семейство 25. Ptilonorhynchidae — беседковые птицы, или птицы шалашники ....................................................279
Семейство 26. Paradisaeidae — райские птицы........................281
Семейство 27. Paridae — синицевые..................................283
Семейство 28. Sittidae — поползневые ..............................285
Семейство 29. Hyposittidae — красноклювые поползни ..... 286
Семейство 30. Certhiidae — пищуховые...............................286
Семейство 31. Paradoxornithidae — толстоклювые синицы..............287
Семейство 32. Chamaeidae — крапивниковые синицы....................288
Семейство 33. Timaliidae — тимелиевые, или кустарницы..............290
Семейство 34. Campephagidae — личинкоедовые .......................291
Семейство 35. Pycnonotidae — бульбулевые, или короткопалые дрозды 292
Семейство 36. Palaeoscinidae ......................................294
Семейство 37. Chloropseidae — листовковые..........................294
Семейство 38. Cinclidae — оляпковые................................295
Семейство 39. Troglodytidae — крапивниковые........................296
Семейство 40. Mimidae — пересмешниковые............................297
Семейство 41. Turdidae —дроздовые..................................298
Семейство 42. Zeledoniidae — водяные дрозды........................301
Семейство 43. Sylviidae — славковые................................301
Семейство 44., Regulidae — корольковые.............................303
Семейство 45. Muscicapidae — мухоловковые . . .....................304
Семейство 46. Prunellidae — завирушковые...........................306
Семейство 47. Motacillidae — трясогузковые.........................307
Семейство 4$. Bombycillidae — свиристелевые . . ...................308
Семейство 49. Ptilogonatidae — шелковистые мухоловки...............310
Семейство 50. Dulidae — пожиратели пальмовых семян.................310
Семейство 51. Artamidae — ласточковые сорокопуты...................311
Семейство 52. Vangidae — ванговые..................................311
Семейство 53. Laniidae — сорокопутовые ............................312
Семейство 54. Prionopidae — лесные сорокопуты......................315
Семейство 55. Cyclarhidae — перцевые сорокопуты....................316
Семейство 56. Vireolaniidae — сорокопутовые виреоны................316
Семейство 57. Callaeidae — новозеландские скворцы..................316
Семейство 58. Sturnidae — скворцовые...............................317
Семейство 59. Meliphagidae — медососовые...........................319
Семейство 60. Nectariniidae — нектарницевые........................320
Семейство 61. Dicaeidae — цветоедовые..............................321
Семейство 62. Zosteropidae — белоглазкбвые.........................322
Семейство 63. Vireonidae — виреоновые .............................324
Семейство 64. Coerebidae — американские цветочницы.................325
Семейство 65. Drepanididae — гавайские цветочницы..................326
Семейство 66. Parulidae — американские славковые, или лесные певуны 327
Семейство 67. Ploceidae — ткачиковые ..............................328
Семейство 68. Icteridae—трупиаловые (американские иволги, кассики) 331
Семейство 69. Tersinidae — ласточковые танагры.....................333
Семейство 70. Thraupidae — танагровые..............................334
Семейство 71. Catamblyrhynchidae — плюшевоголовые вьюрки . . . 335
Семейство 72. Fringillidae — вьюрковые ............................335
Подсемейство 1. Emberizinae — овсянковые...................................338
Подсемейство 2. Geospizinae — дарвиновы, или галапагосские, вьюрковые . 339
Подсемейство 3. Pyrrhuloxiinae — кардиналовые..............................340
Подсемейство 4. Fringillinae — настоящие вьюрковые.........................340
Литература ................................................................342
Приложение 343
Указатель латинских названий родов ........................................348
367
НИКОЛАЙ НИКОЛАЕВИЧ КАРТАШЕВ
СИСТЕМАТИКА ПТИЦ
Редактор Л. П. Петровская Художники В. М. С мири и и Ю. М. Смири и Художественный редактор Т. А. Коненкова Технический редактор Г. Г. Киселева Корректор С. К. Марченко
Сдано в набор 5/Х—73 г. Подп. к печати 26/11—74 г. Формат 70xl08*/i« Бум. тнп. № 2. Объем 23 печ. л. Усл. п. л. 32,2.Уч.-изд. л. 31,63.Изд. № Е-228.Тираж 20 000 экз.
Цена 1 р. 46 к. Зак. 774
План выпуска литературы издательства «Высшая школа» (вузы и техникумы) на 1974 г. Позиция № 89.
Москва, К-51, Неглинная ул., д. 29/14, Издательство «Высшая школа»
Ярославский полиграфкомбинат «Союзполиграфпрома» при Государственном комитете Совета Министров СССР по делам издательств, полиграфии и книжной торговли.
Ярославль, ул. Свободы, 97.