










































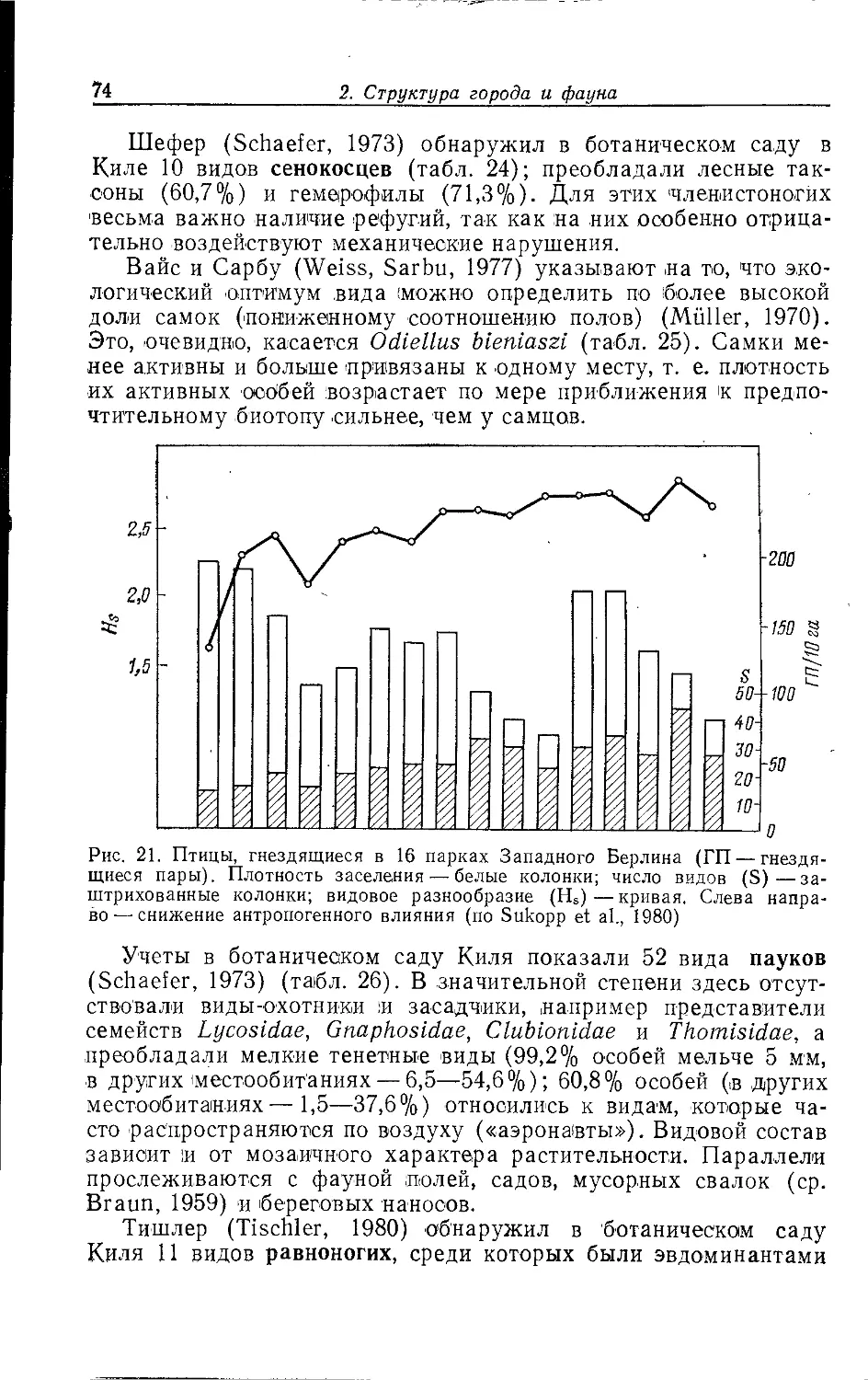



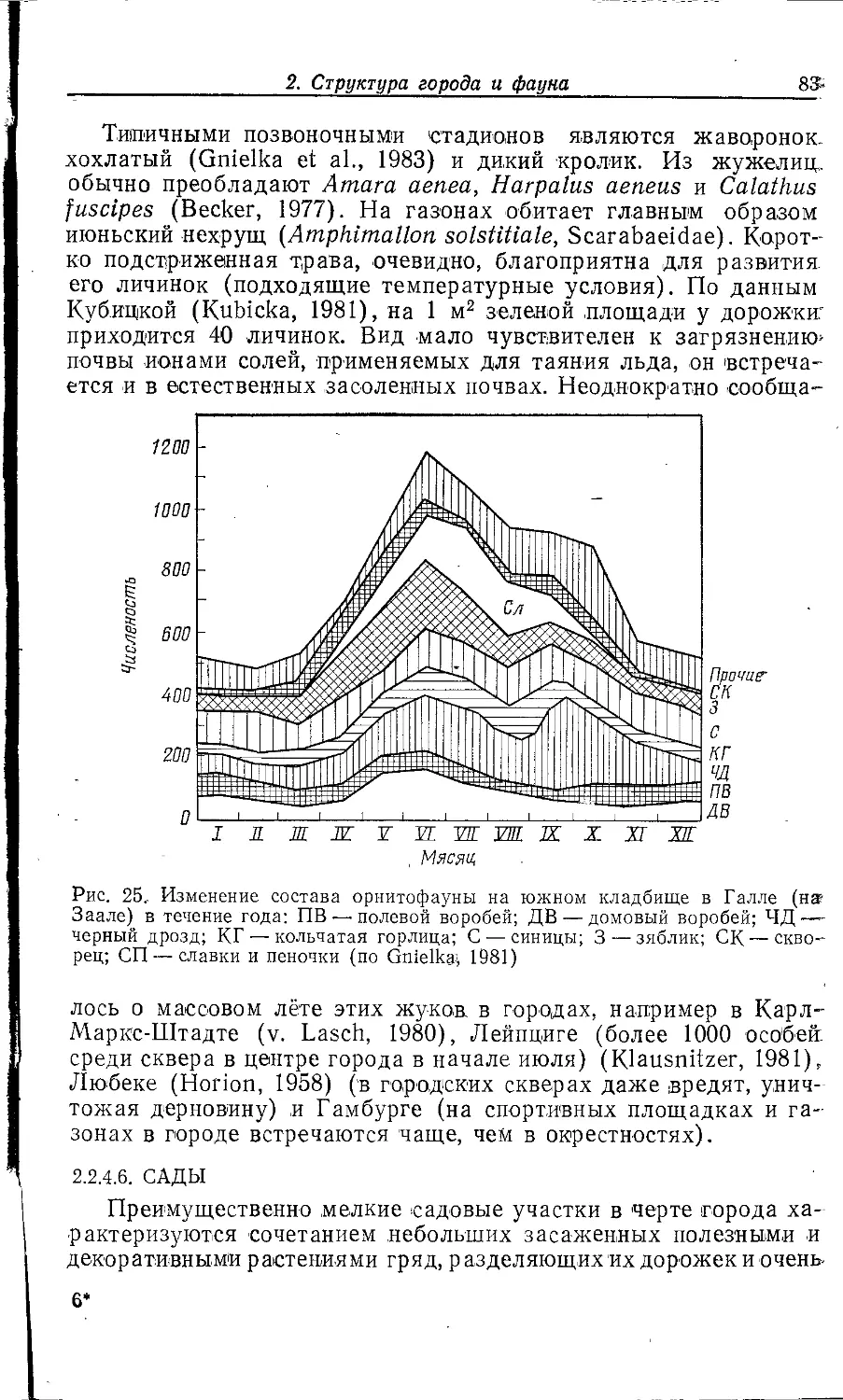
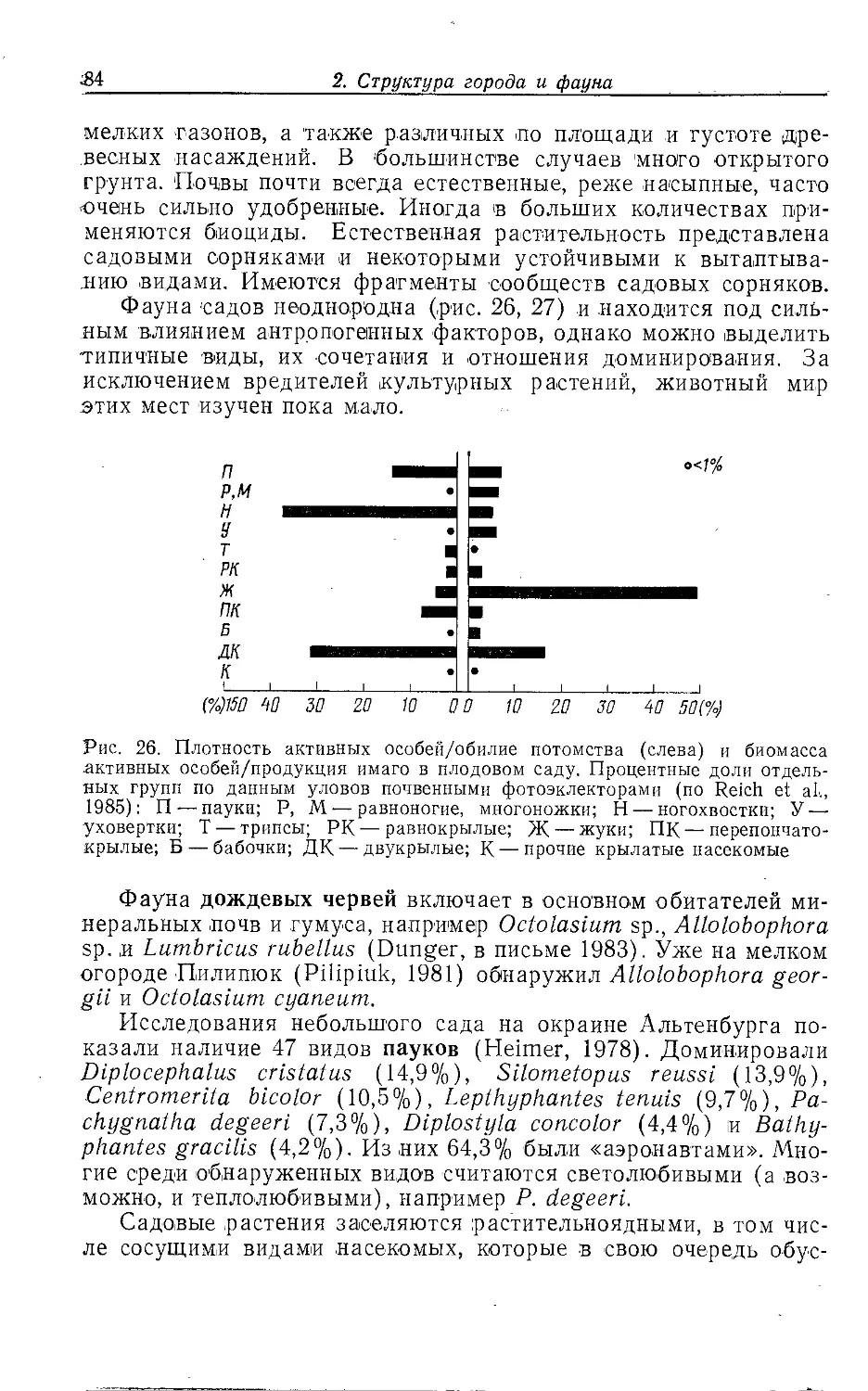
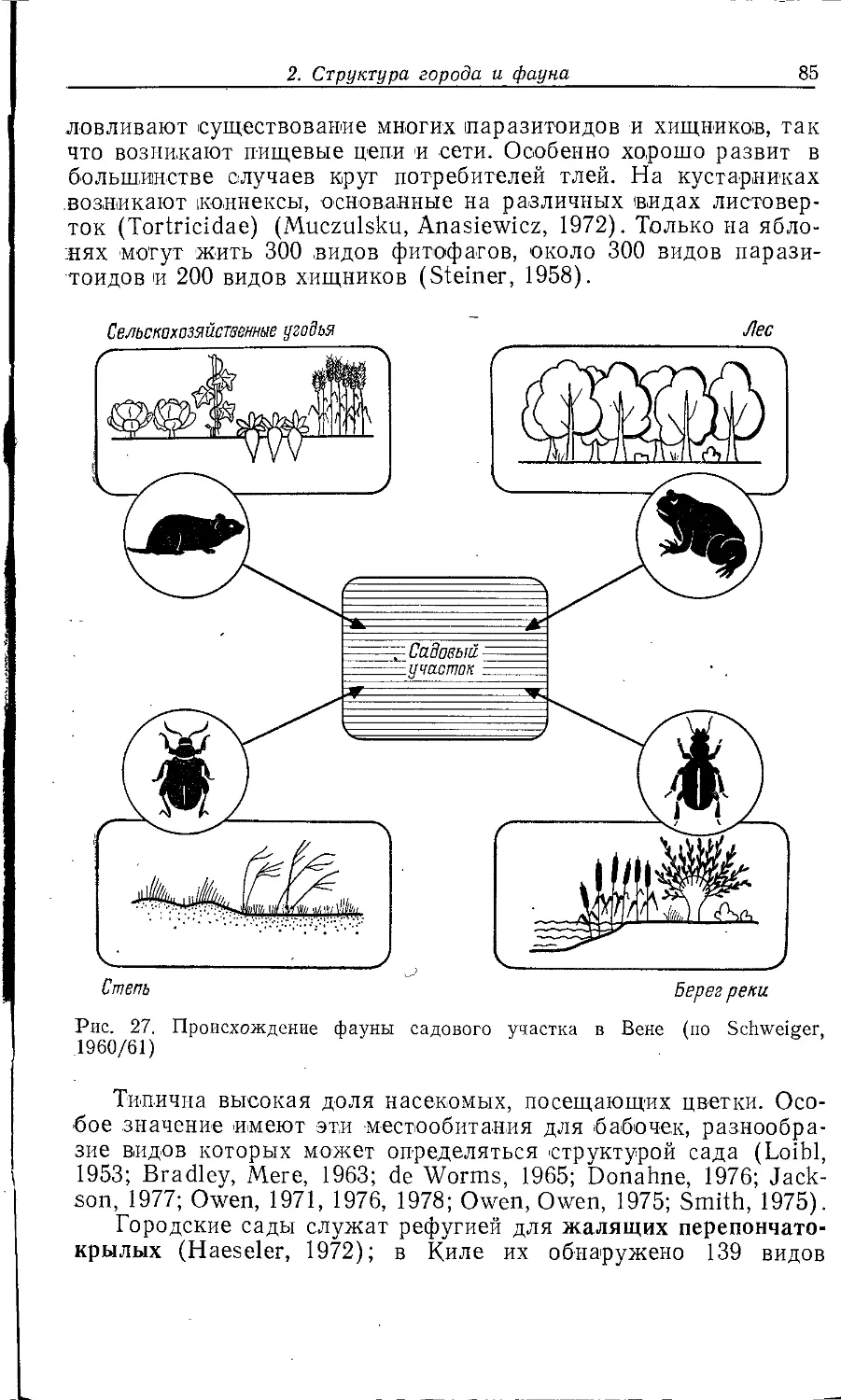
















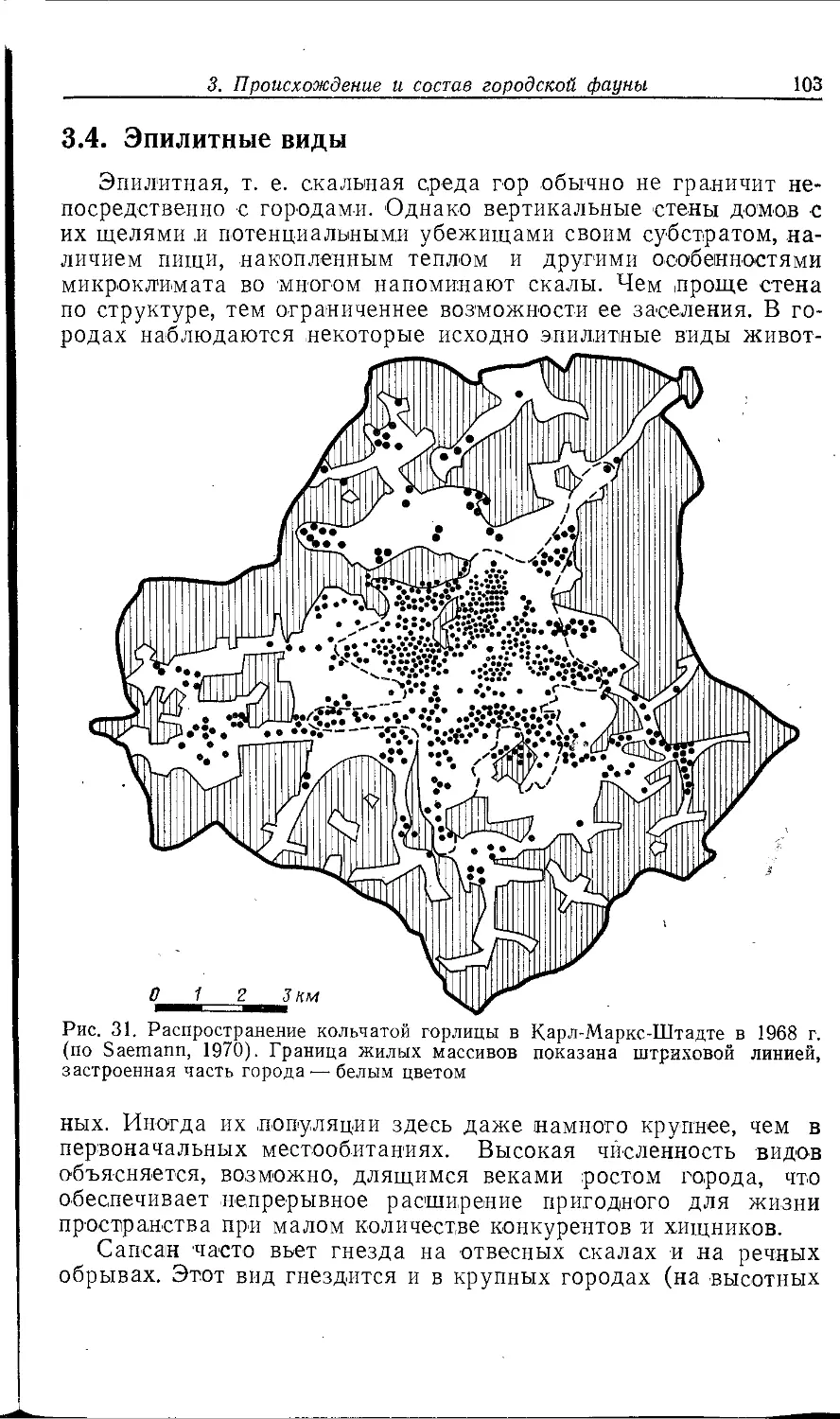




















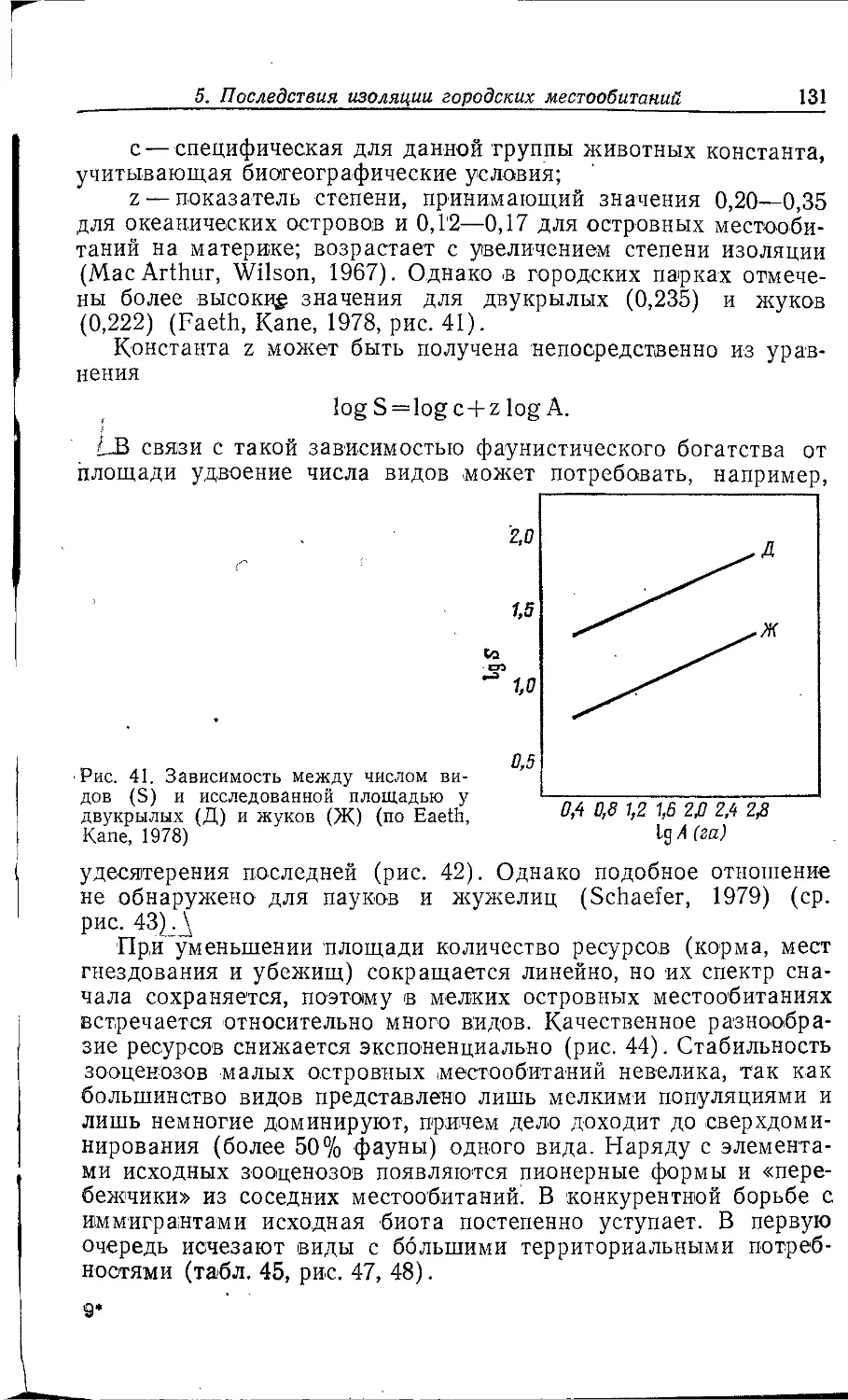
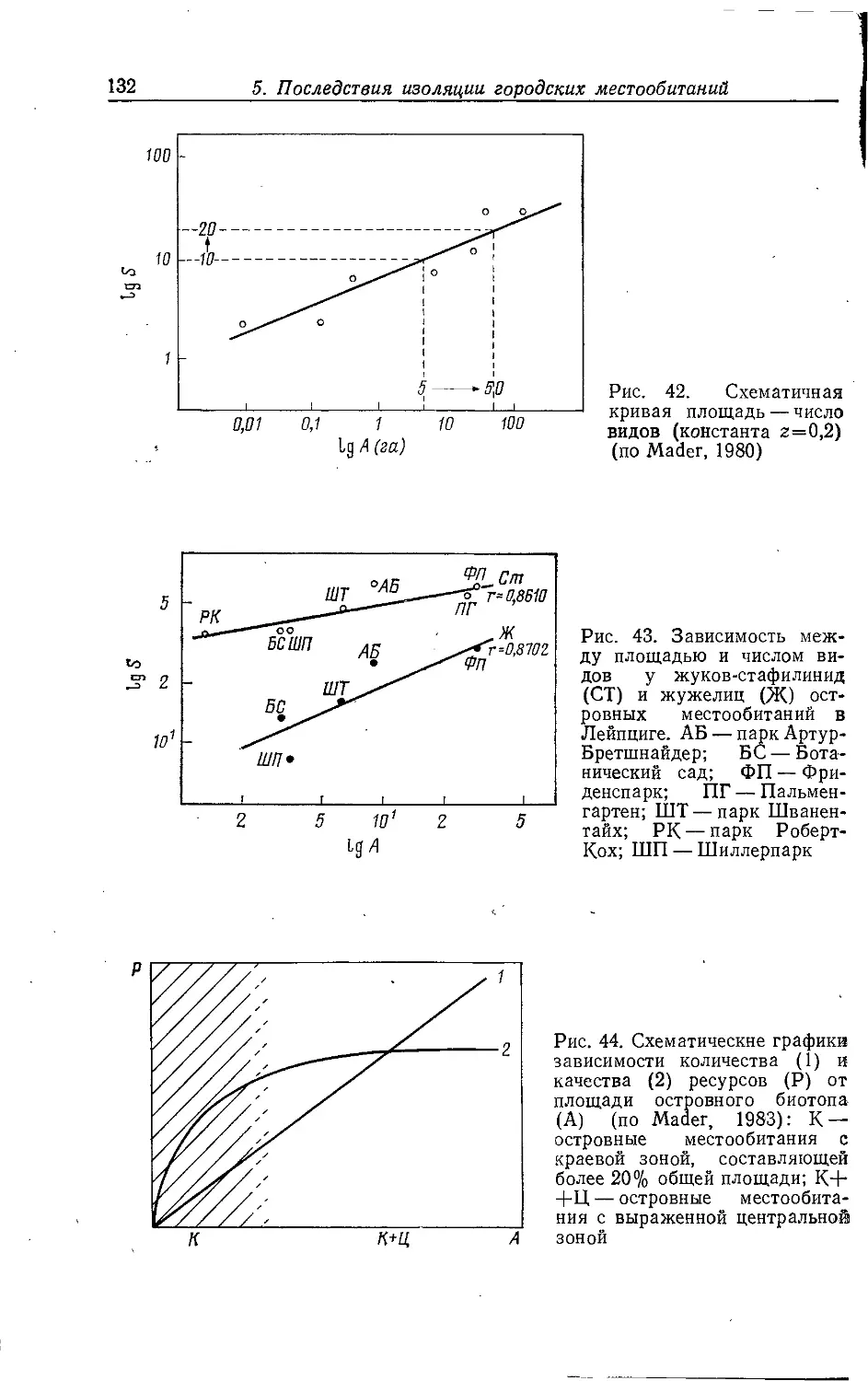
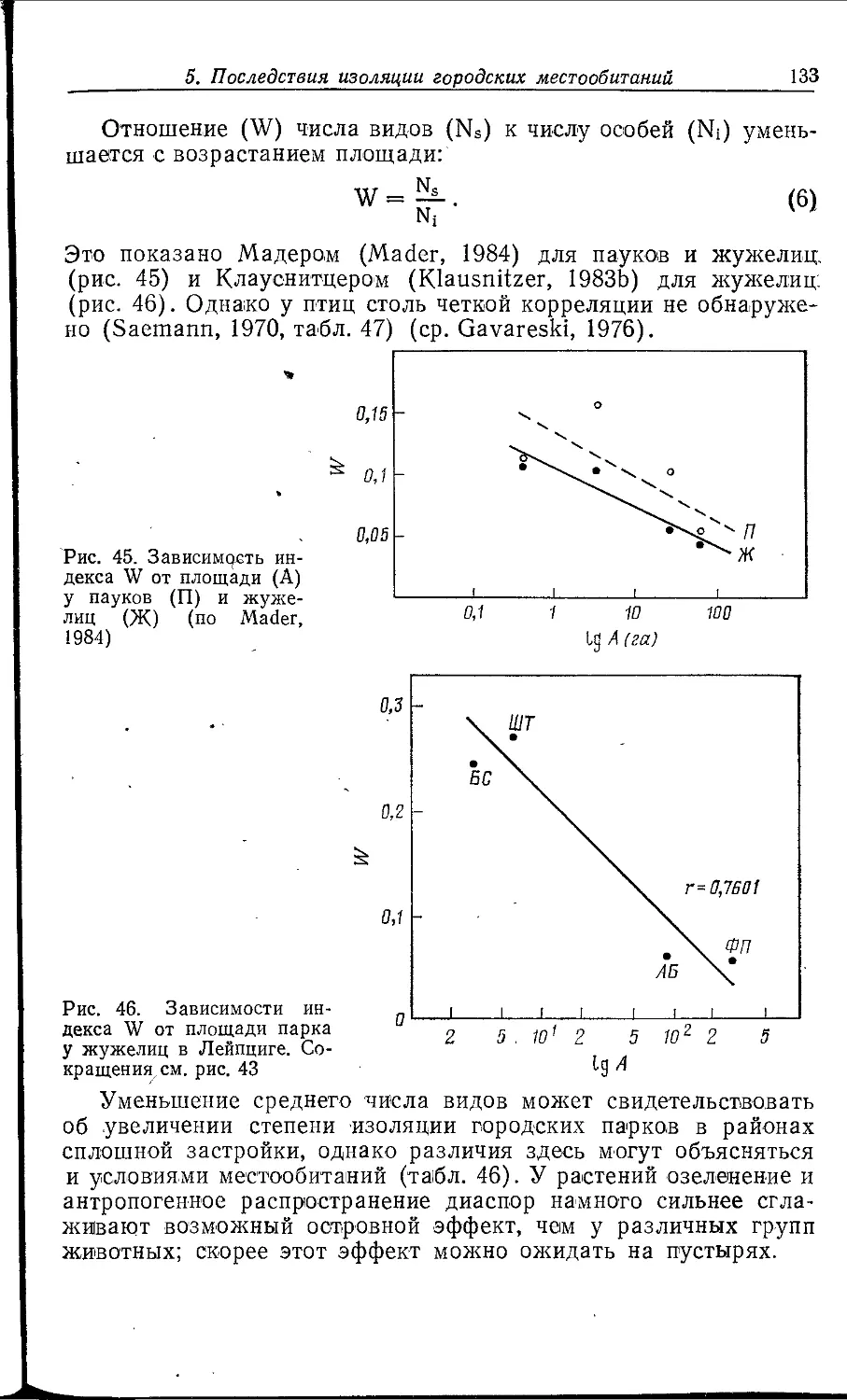
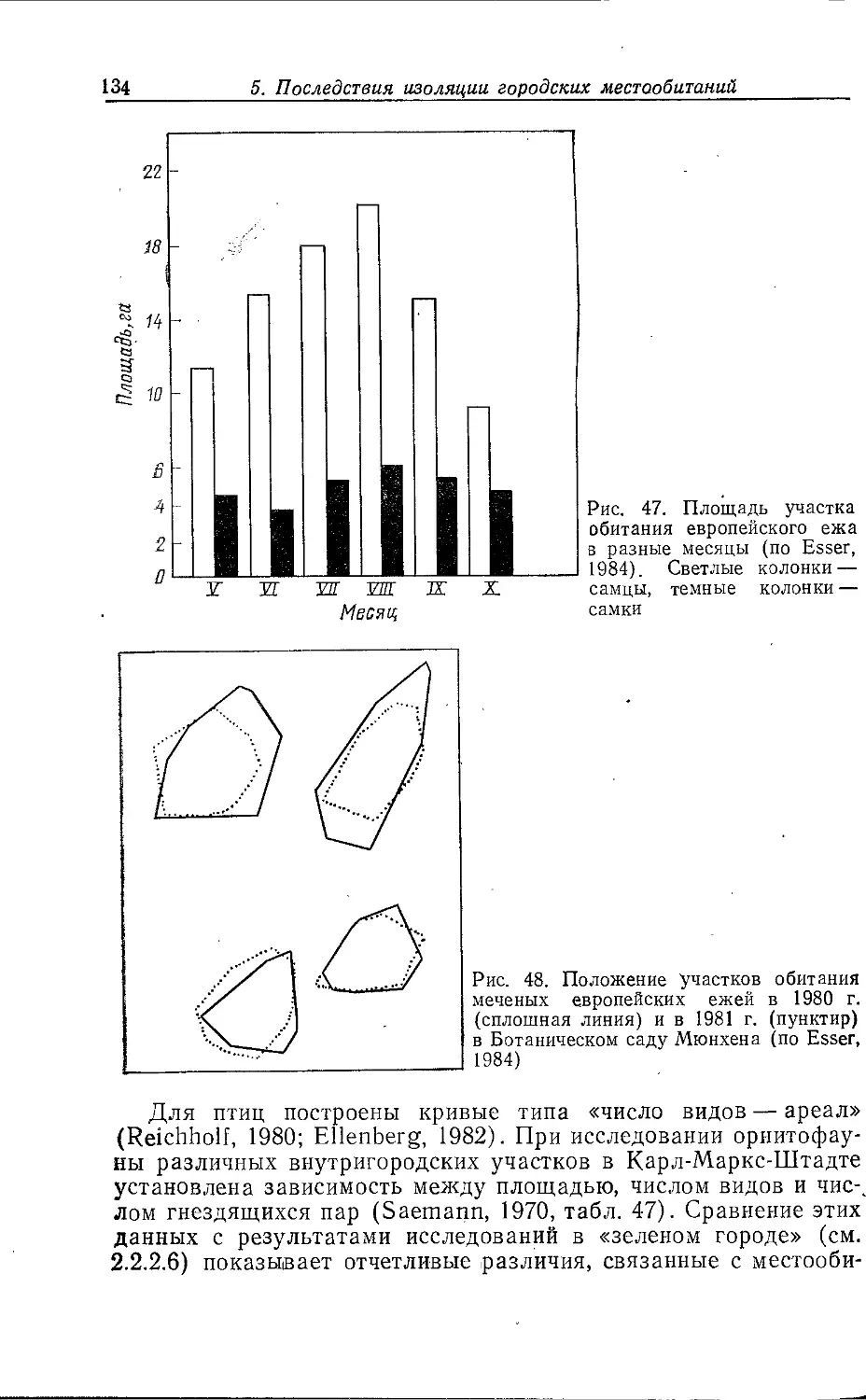












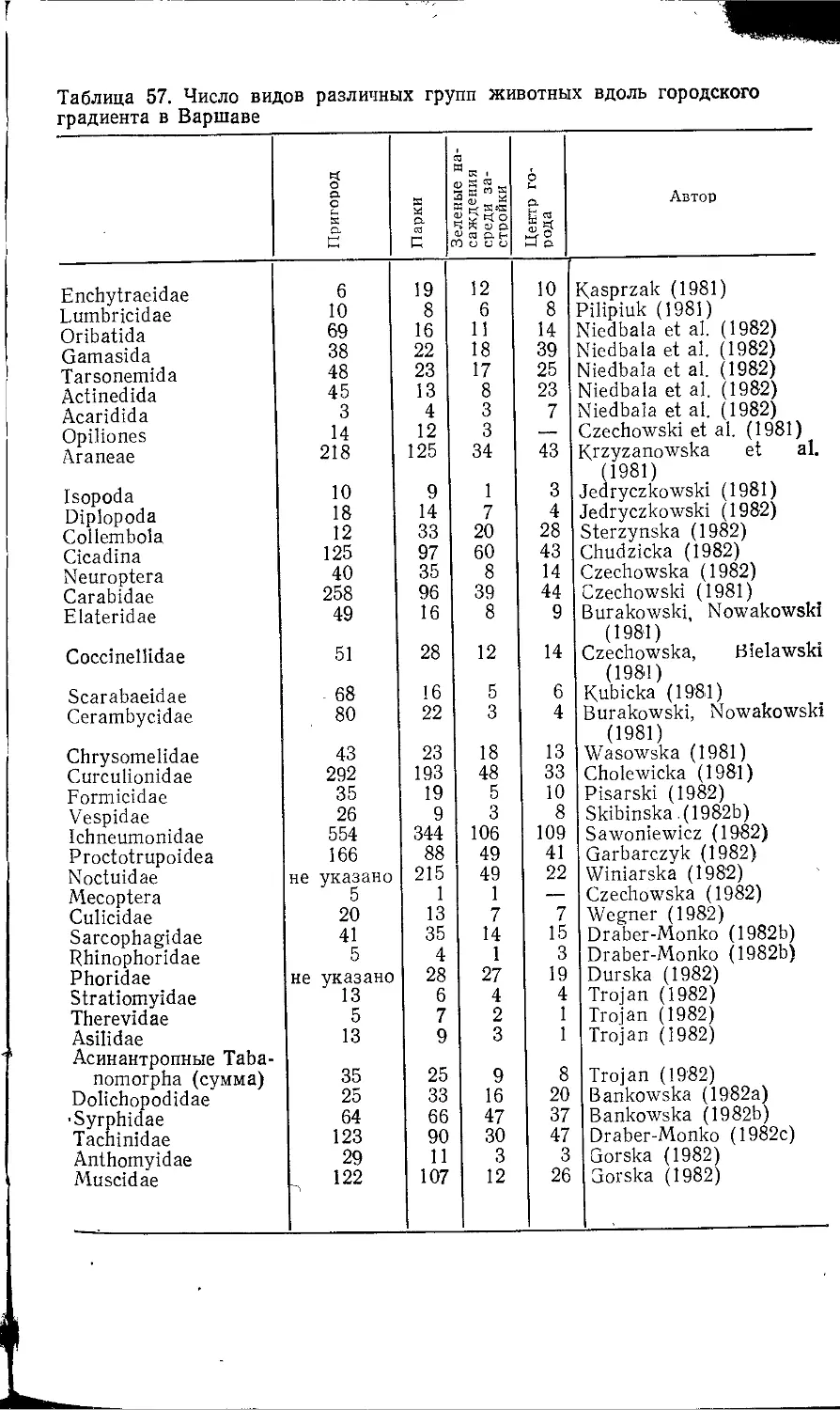
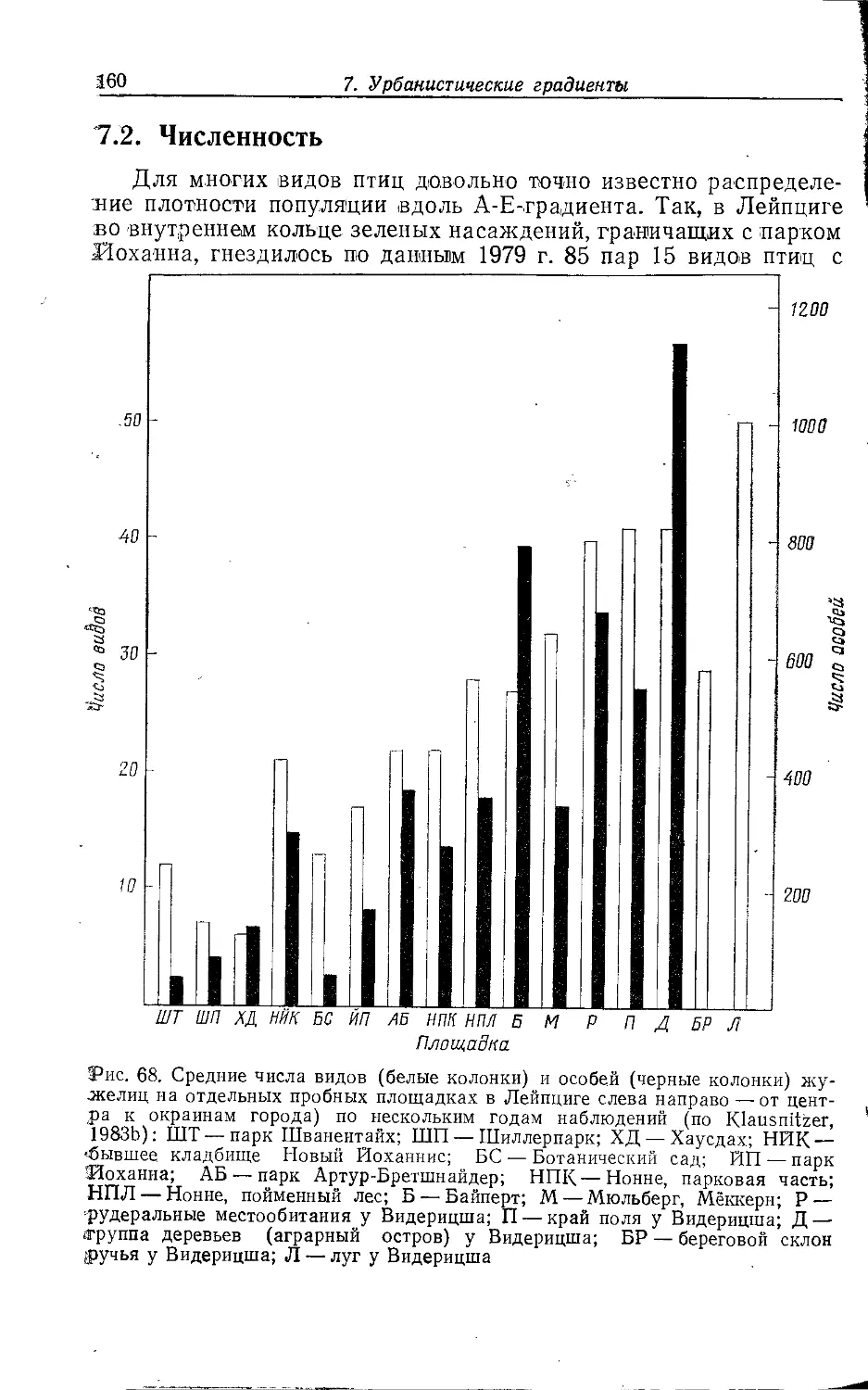
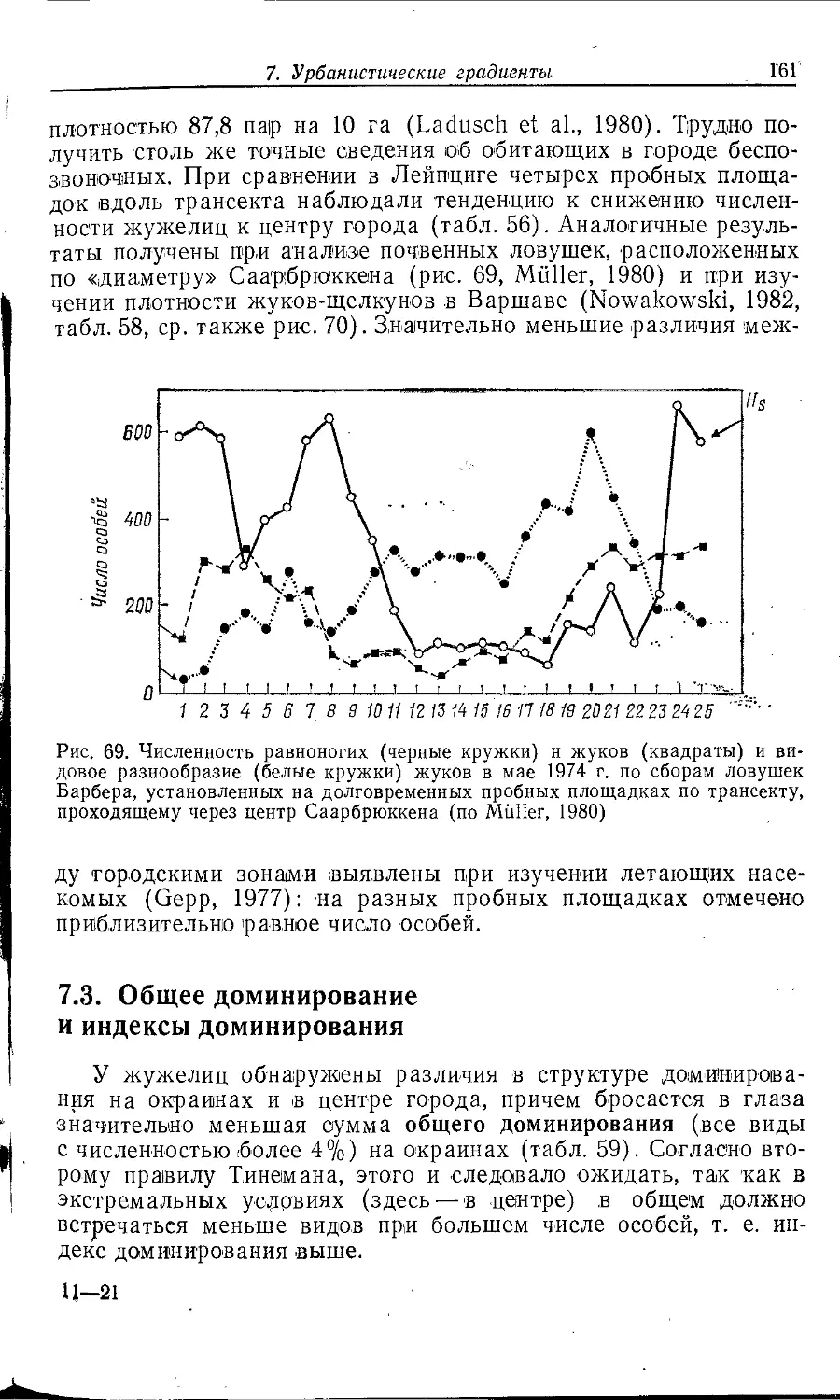
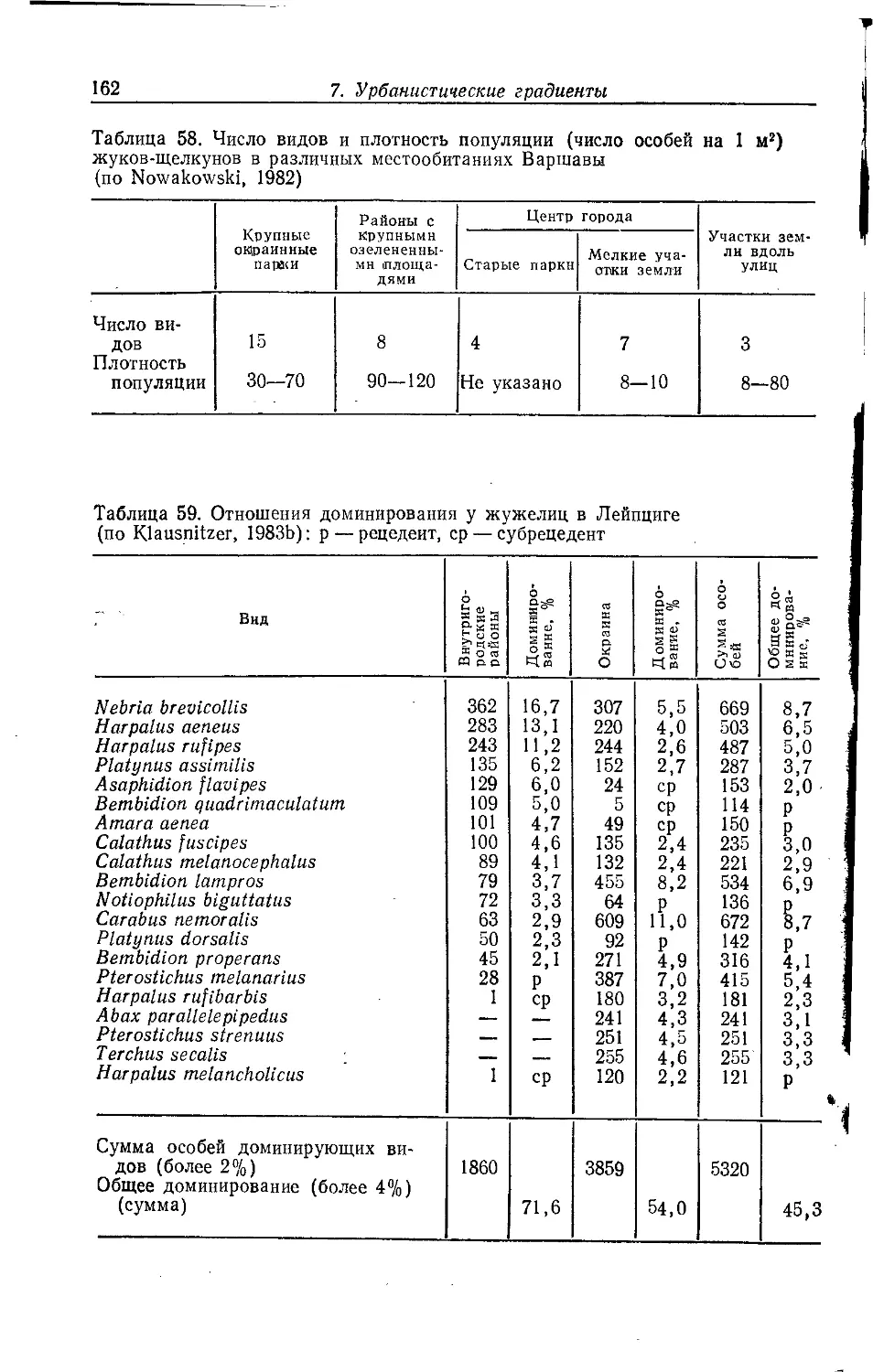
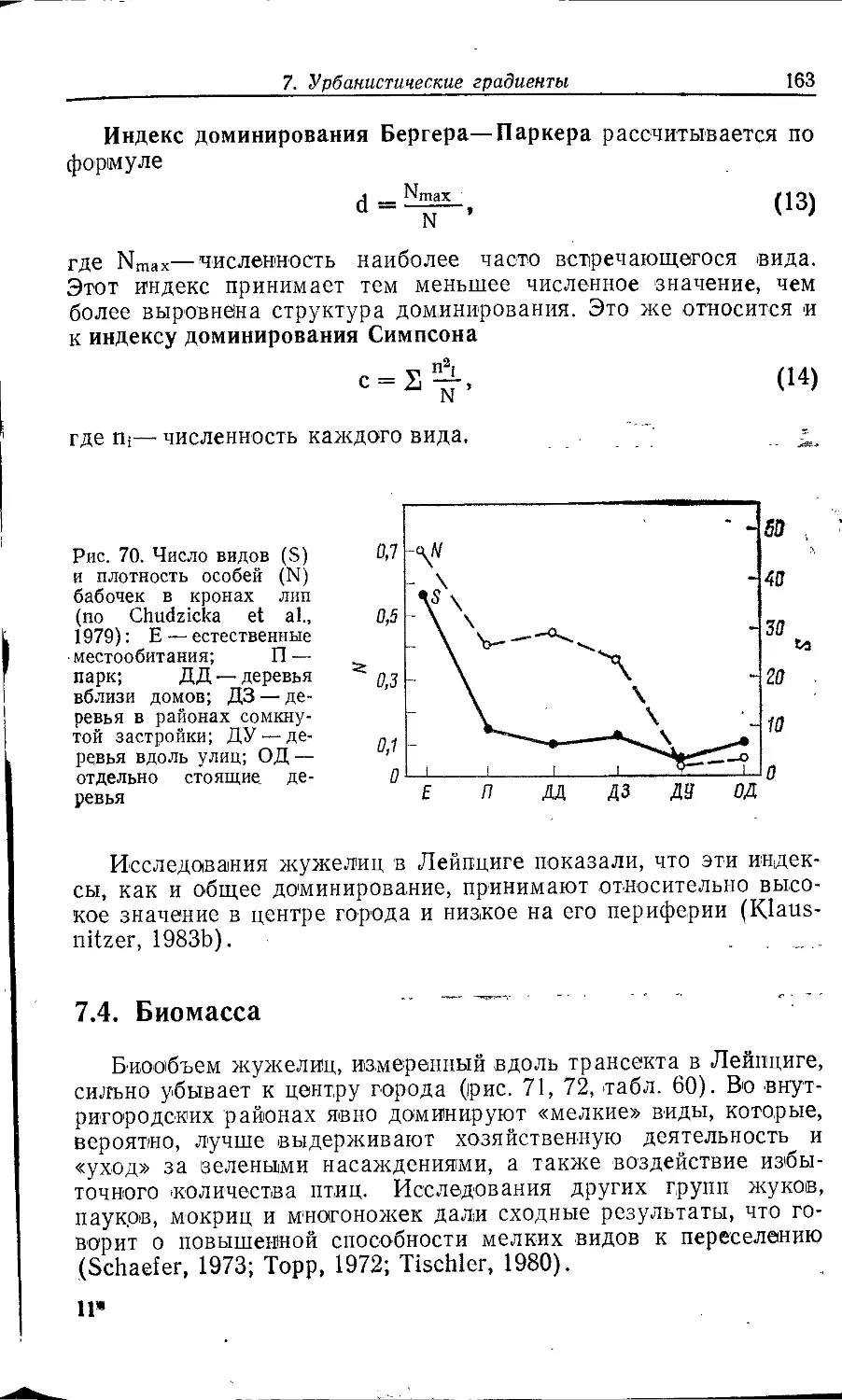
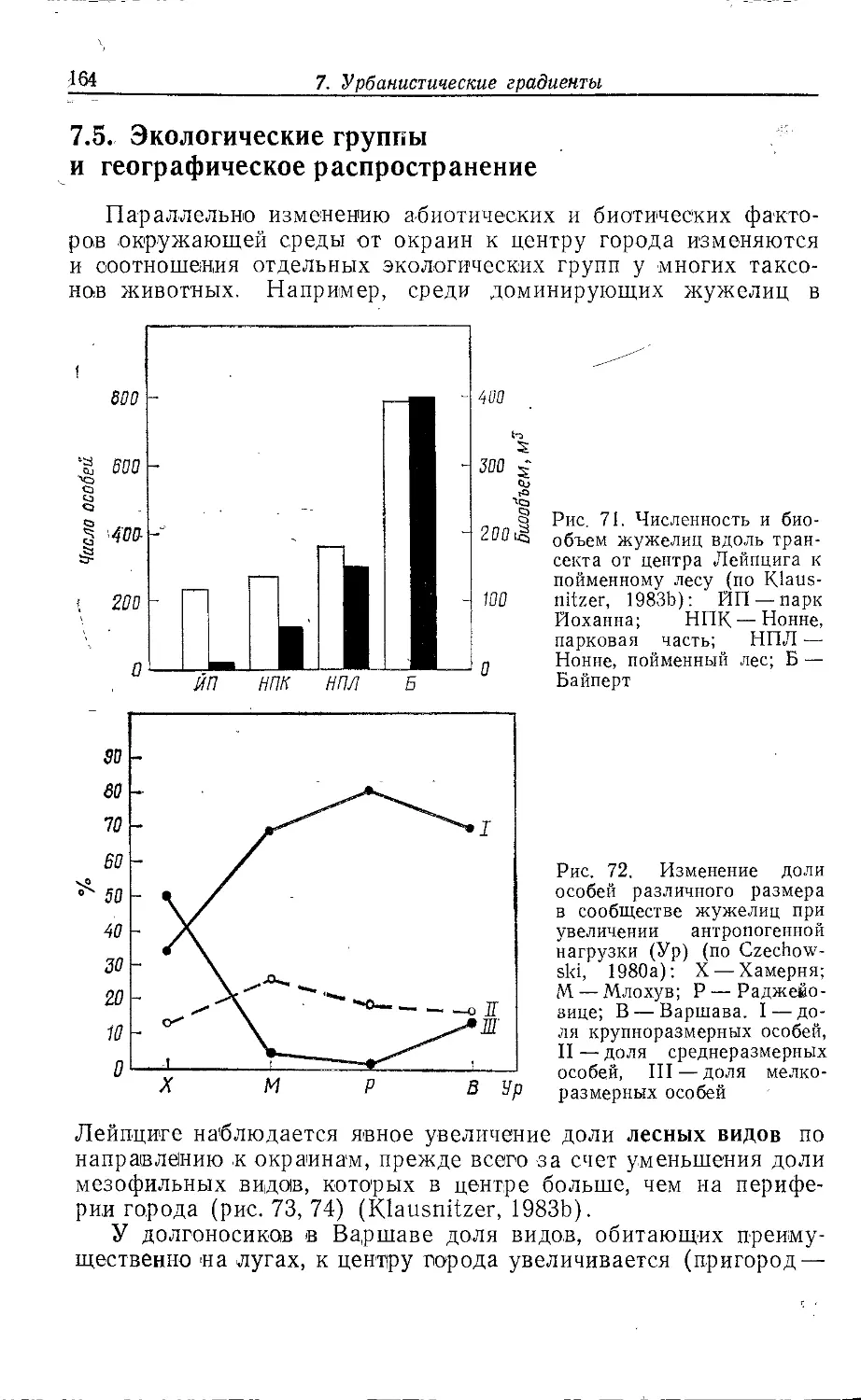
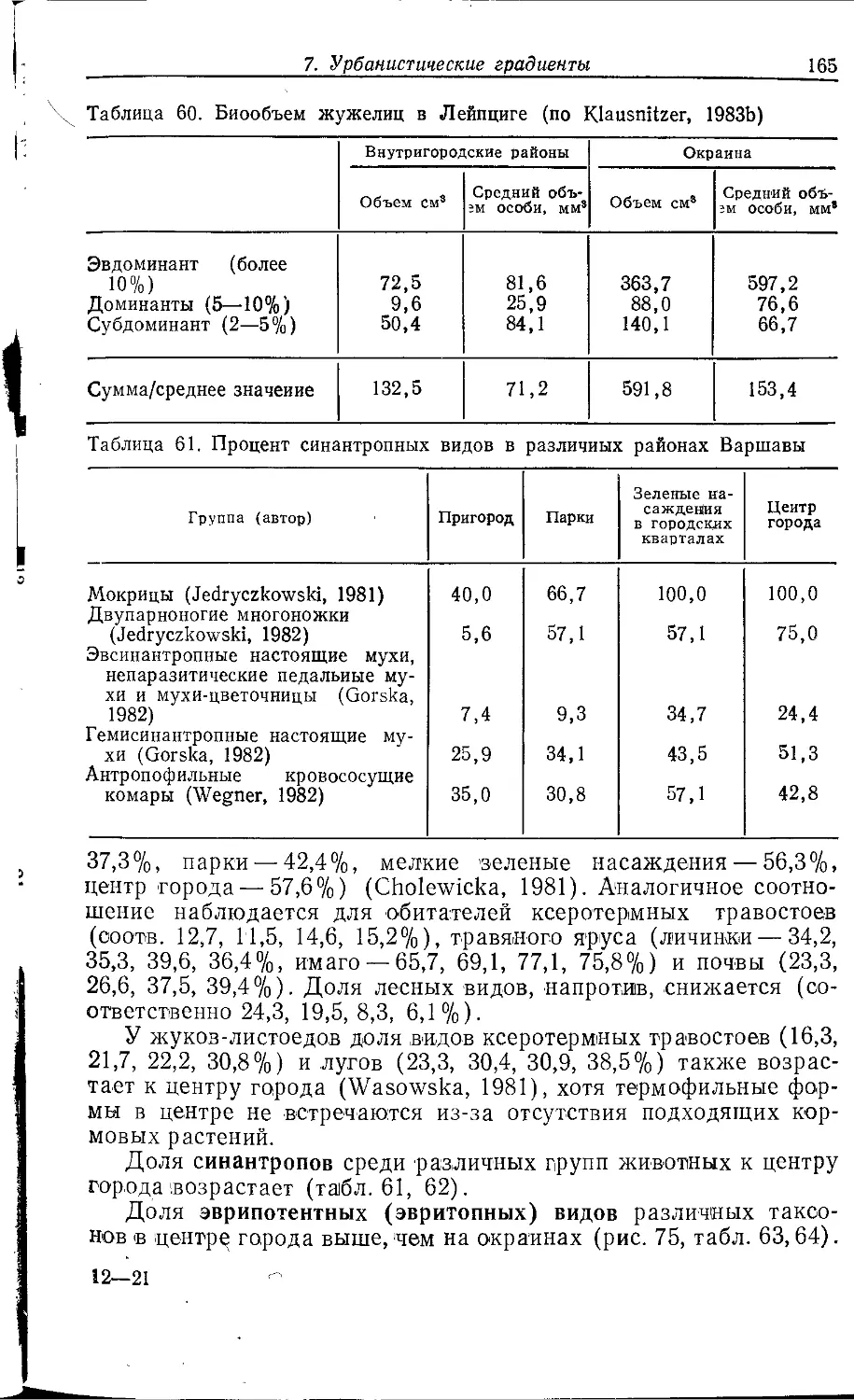
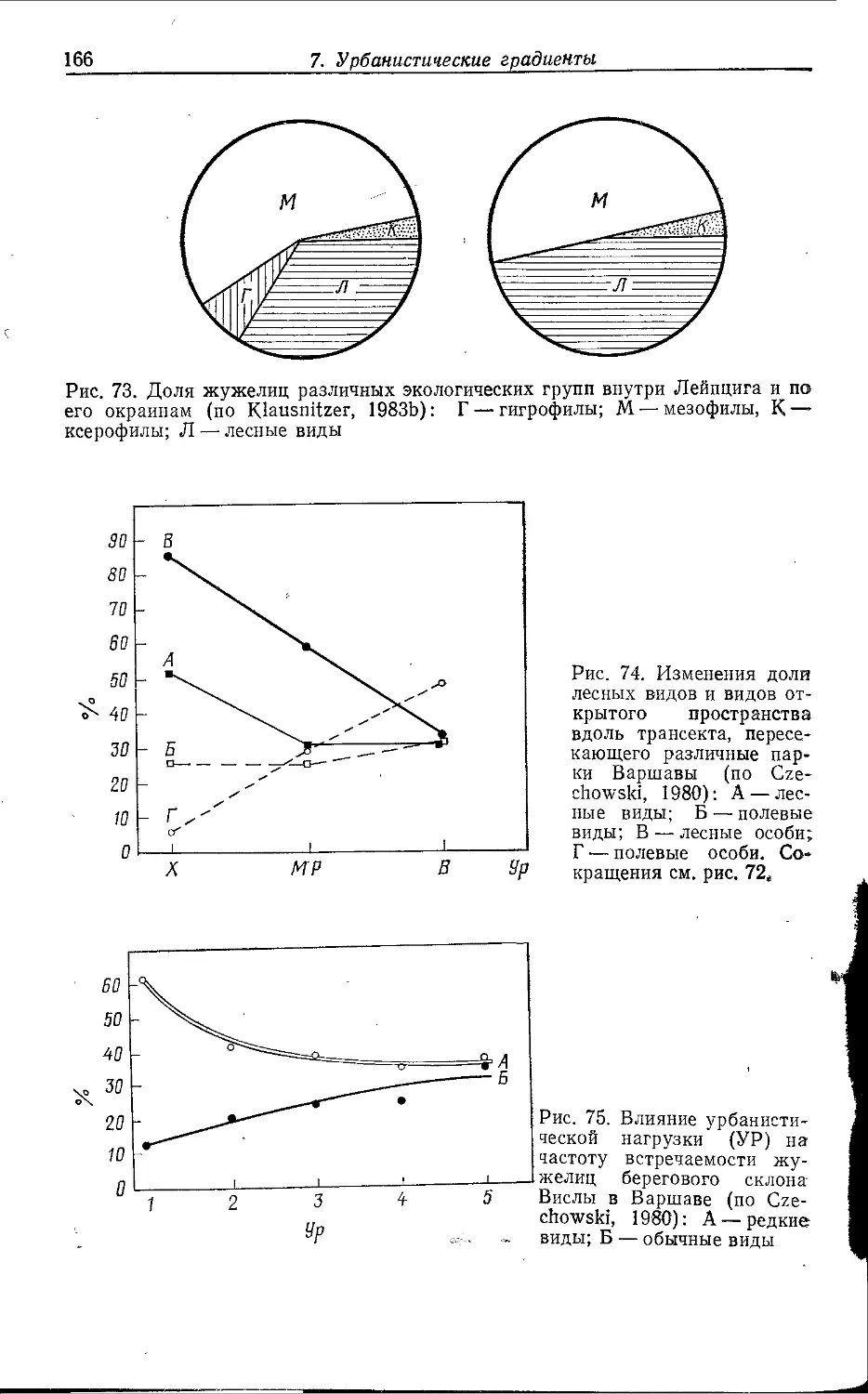




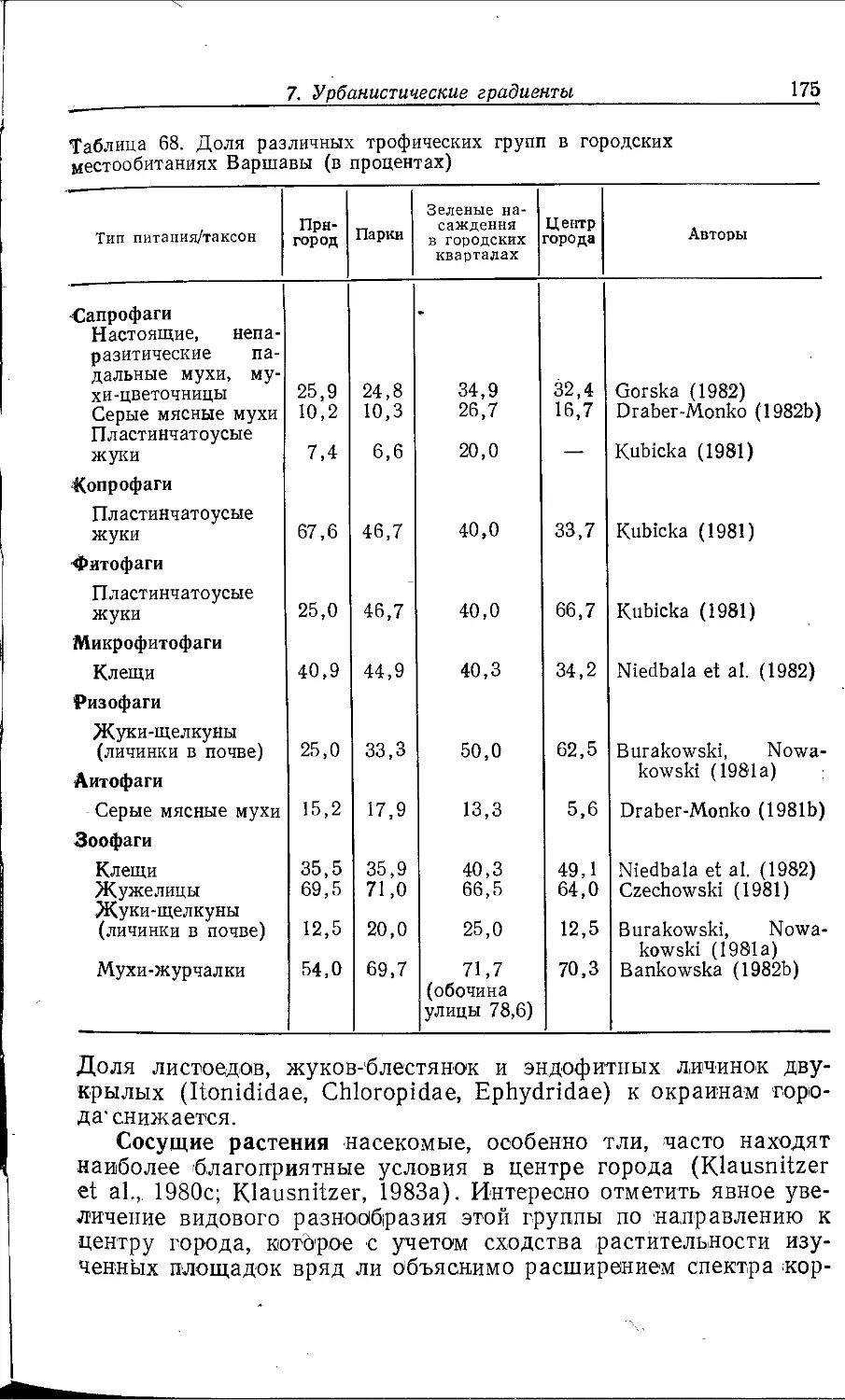
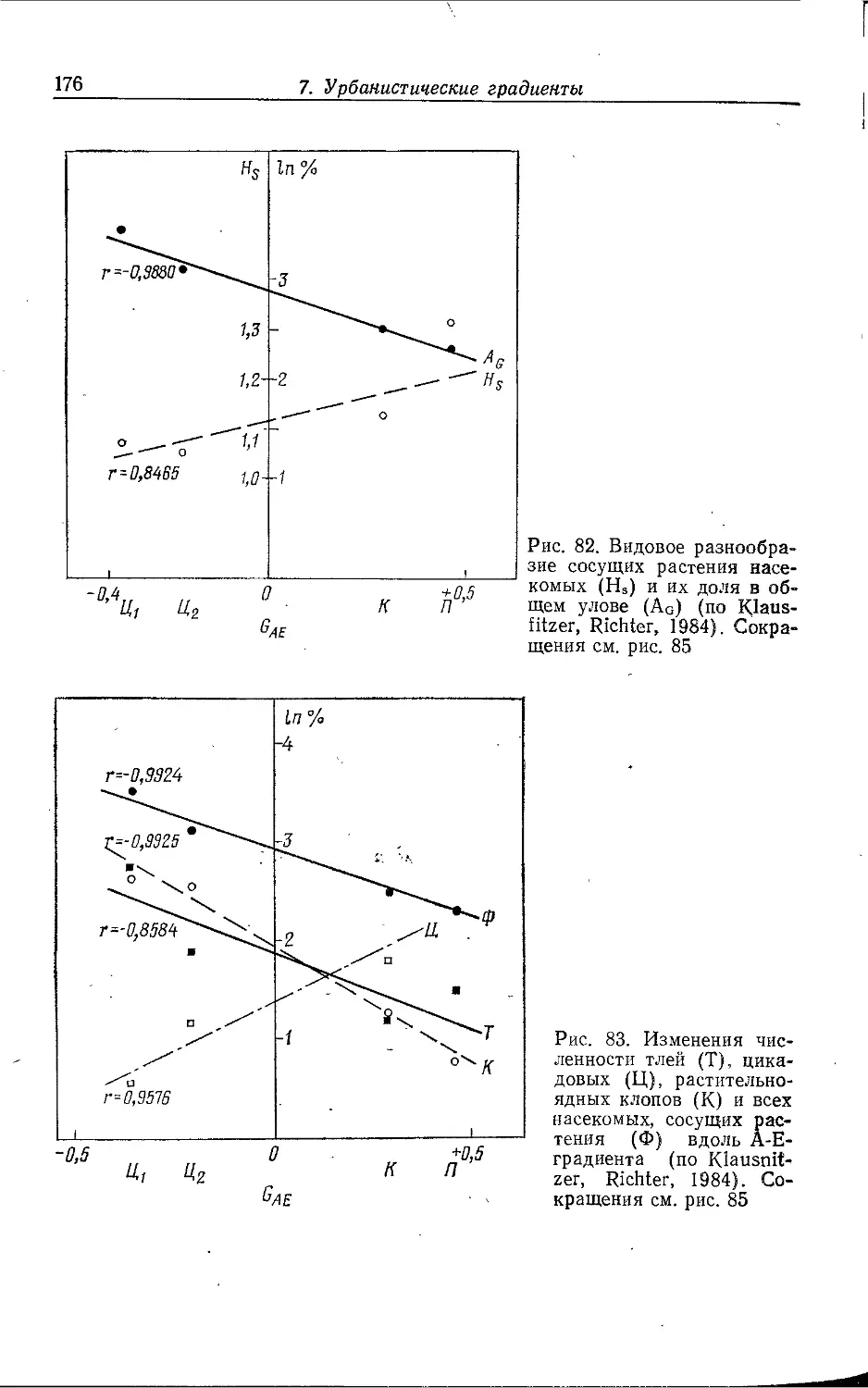

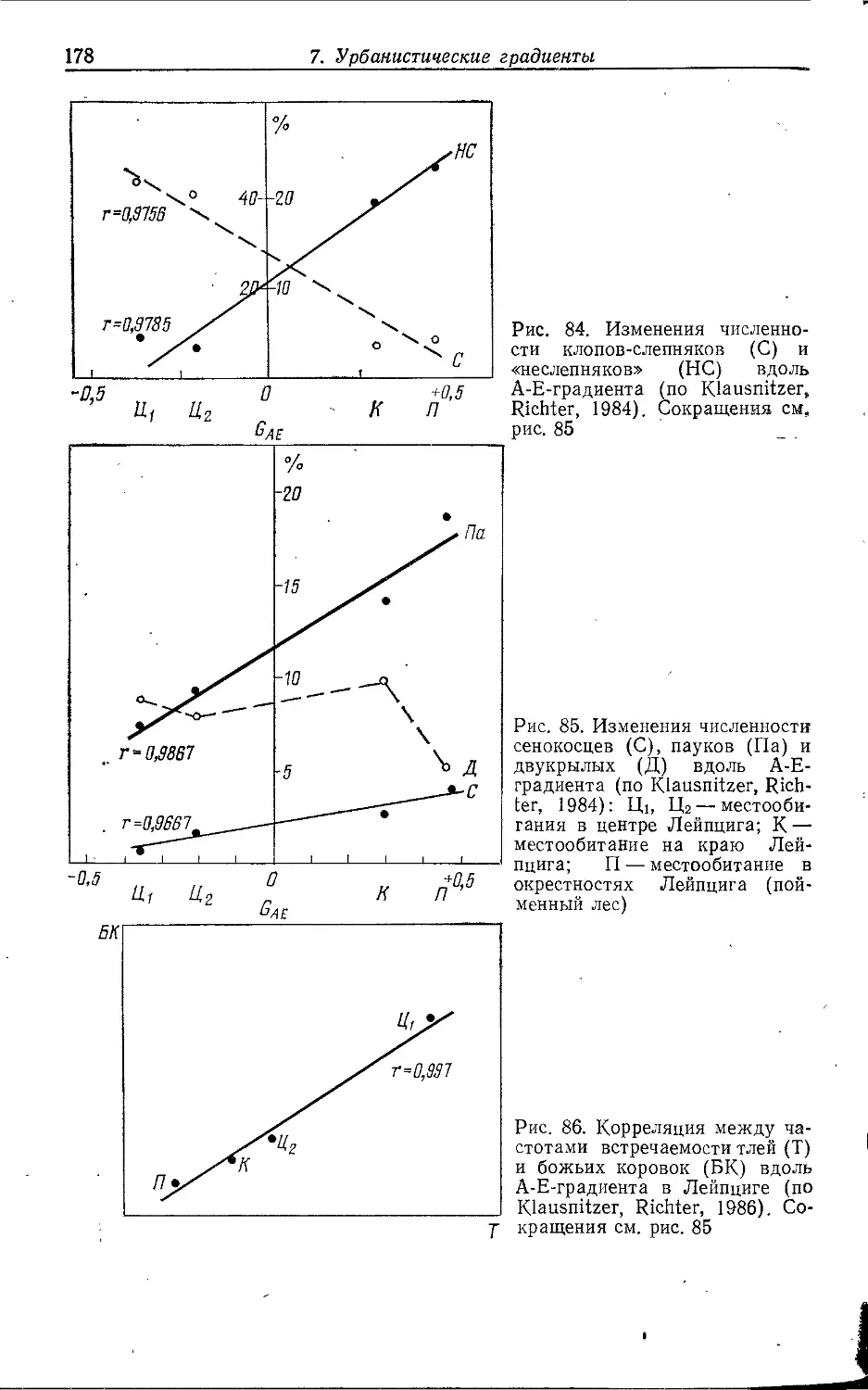

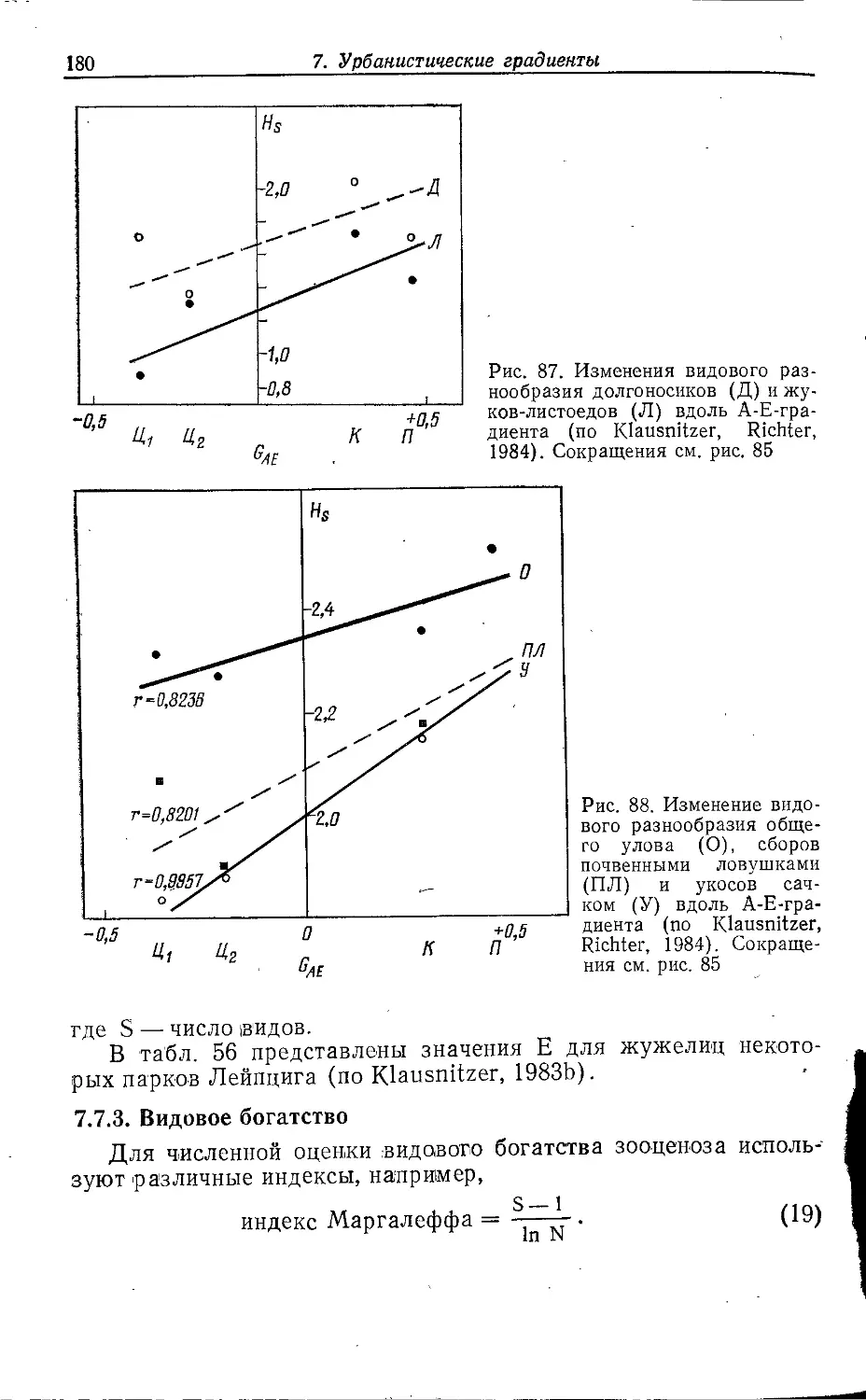






















































Author: Клауснитцер Б.
Tags: общая экология биоценология гидробиология биогеография экология и биогеография охрана живой природы экология зоология фауна города
ISBN: 5-03-001383-0
Year: 1990




Экология
ГОРОДСКОЙ
фауны
Ökologie der Großstadtfauna
Bernhard Klausnitzer, Leipzig
Mit 105 Abbildungen, 8 Tafeln und 78 Tabellen
VEB GUSTAV FISCHER VERLAG JENA. 1987
Б. КЛАУСНИТЦЕР
Экология
городской
фауны
Перевод с немецкого
И. В. Орловой, И. М. МароЕОй
*^ гор. КАЛИНИН \
М0
Москва «Мир» 1990
ББК 28.08
К47
УДК 574
Клауснитцер Б.
К47 Экология городской фауны: Пер. с нем. — М.: Мир,
1990.—246 с, ил.
ISBN 5-03-001383-0
в книге известного зоолога из ГДР, профессора ЛеЯпцигского университета
впервые обобщены сведения о всех группах животных, обитающих в крупных
городах. Основой обзора послужили данные изучения наземной фауны восьми
городов Европы: Варшавы, Вены, Гамбурга. Лейпцига и др. Наряду с
теоретическими обобщениями книга содержит ценный справочный материал, а также
рекомендации по охране многих представителей городской фауны.
Для зоологов, экологов, педагогов и лиц, занимающихся планировакнем
городских территорий.
1903040000—305
К J^^^,, " 107-90 ББК 28.08
041(01)—90
Научное издание
Берихард Клауснитцер
ЭКОЛОГИЯ городской фауны
Заведующий редакцией чл.-корр. АН СССР Т. М. Турпаев. Зам. зав. редакцией
М. Д. Гроздова. Научн. редактор Ю. Л. Амченков. Мл. редактор И. А. Деменцова.
Художник А. А, Лукьяненко. Художественные редакторы А. Я. Муснн, Л. М. Аленичева.
Технический редактор Т. А. Максимова. Корректор Л. Д. Панова
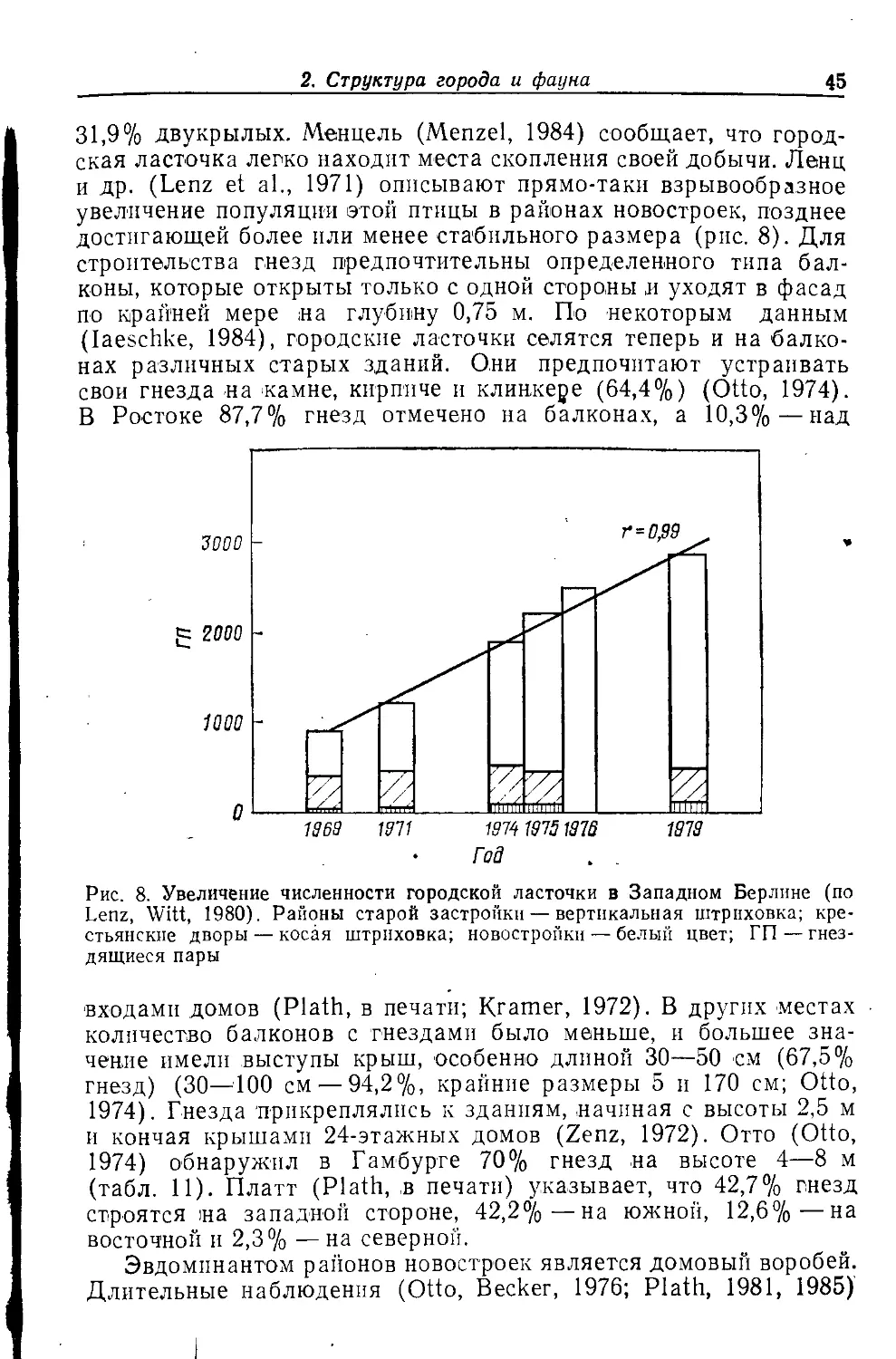
ИБ № 7167
Сдано в набор 03.01.90. Подписано к печати 28.04.90. Формат бОХЭО'/и. Бумага кн.-жури.
Печать высокая. Гарнитура Литературная. Объем 7,75 бум. л. Усл. печ. л. 15,5.
Усл. кр.-отт. 15,5. Уч.-изд. л. 15.84. Изд. № 4/6715. Тираж 6500 экз. Зак. 21,
Цена 2 р. 80 коп.
Издательство «Мир>
В/О «Совэкспорткнига» Государственного комитета СССР по печати.
129820, ГСП, Москва. 1-й Рижский пер., 2.
Московская типография № 11 Государственного комитета СССР по печати.
113105, Москва, Нагатинская ул. д. 1.
Редакция литературы по биологии
ISBN 5-03-001383-0 (русск.) © VEB Gustav Fisher Verlag Jena,
ISBN 3-334-00122-1 (нем.) 1987
© перевод на русский язык:
Орлова И. В., Марова И. М., 1990
ПРЕДИСЛОВИЕ
Книга Бернхарда Клауснитцера «Экология городской фауны»
не имеет аналогов среди русскоязычных публикаций. Это не
означает, что подобных работ в СССР не проводилось. Такие
наблюдения непрерывно десятилетиями ведутся в нашей стране
тысячами людей: паразитологами санэпидстанций, работниками
служб зеленых насаждений, парков, пригородных зон отдыха,
педагогами, студентами биологических и географических
факультетов, многочисленными натур(алистами-любителями и просто
любителями домашних животных, особенно собак и птиц.
Разумеется, существует и обширная информация о животных в
городах СССР, но, к сожалению, она не обобщена и никем не
сведена воедино. Правда, были публикации и довольно крупные,
например книга С. И. Огнева по млекопитающим Московской
губернии (1913), сборник «Животное население Москвы и
Подмосковья» (1967), другие сборники и статьи. Их наберется
несколько сотен лишь для Москвы, а ведь такая же богатая
информация собрана и опубликована практически по любому крупному
городу, причем по Ленинграду, Киеву, Харькову, Воронежу,
Ростову, Одессе, Днепропетровску, Минску существует, можно
сказать без преувеличения, целая литература. Об этом необходимо
напомнить читателю, так как автор использовал в своей работе
почти исключительно гермэноязычную литературу.
Полагаю, что нет нужды комментировать книгу Б.
Клауснитцера или давать ей какую-либо оценку — это сделает
самостоятельно каждый из читателей. Но нет сомнений, и об этом стоит
сказать, что перед нами итог огромного добросовестного и
вдумчивого труда опытного зоолога, работа, исключительно полезная
не только, а, мож^т быть, и не столько специалистам, но и
студентам биологам и географам, многочисленным любителям
природы.
Следует также сказать, что книга эта мол<ет представлять
немалый интерес и для экологов-урбанистов, поскольку экология
города у нас на глазах формируется в самостоятельную и
довольно специфическую область экологии. Об этом аспекте
использования сведений по животным в городской среде автор
почти ничего не говорит, а ведь дикие и синантропные животные
любого города помогают составить объективное впечатление и о
Предисловие
его санитарно-гигиеническом состоянии, и о качестве жизни его
населения, и о культуре отношения жителей к окружающей их
среде.'Анализ городской фауны позволяет получить объективную
информацию о то.м, насколько вредны для организмов
существующие в любом городе загрязнения воздуха, воды, почвы, зеленых
насаждений, насколько эффективны те или иные
природоохранные мероприятия, какова динамика экологической обстановки
города.
Обо всем этом можно сказать подробнее, так как именно
такие аспекты исследования городской фауны и составляют
основную его цель.[Города — сравнительно новая среда обитания
животных, весьма специфическая по всем своим параметрам.
Впервые они появились в эпоху перехода от первобытнообщинного к
рабовладельческому строю, т. е. в III—I тыс. до н. э., в
Месопотамии, Египте, Сирии, Малой Азии, Индии, Китае. По
археологическим данным, города с население.м 20—30 тыс. человек по тем
временам следует считать очень крупными.)Преимущества и
недостатки городской жизни известны каждому, но их восприятие,
как правило, предельно субъективно. Каждый город по многим
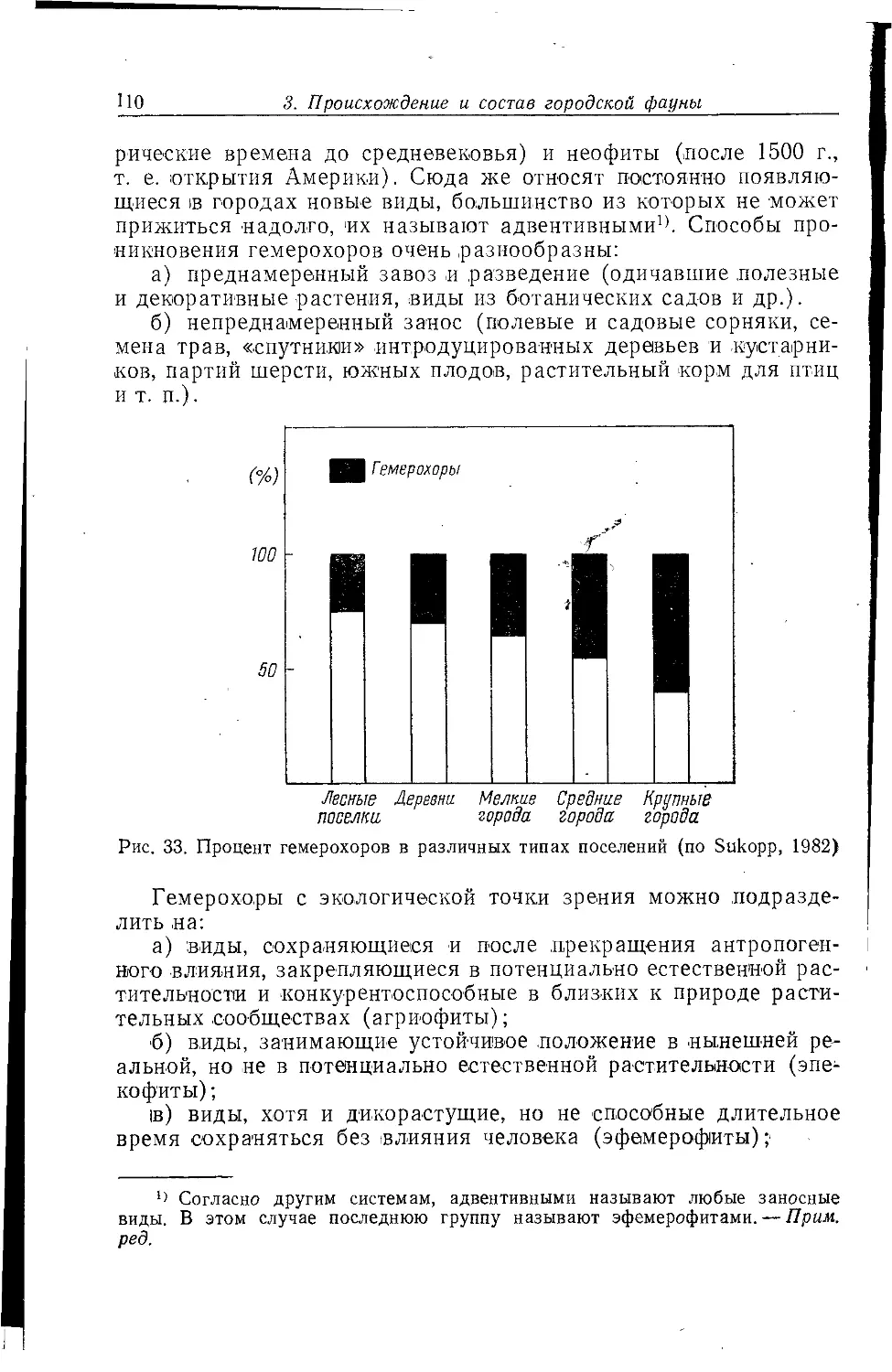
параметрам уникален, особенно в динамике, но ряд проблем
актуален для всех таких систем. Экологические проблемы городов
в капиталистическом мире подробно рассматриваются в книге
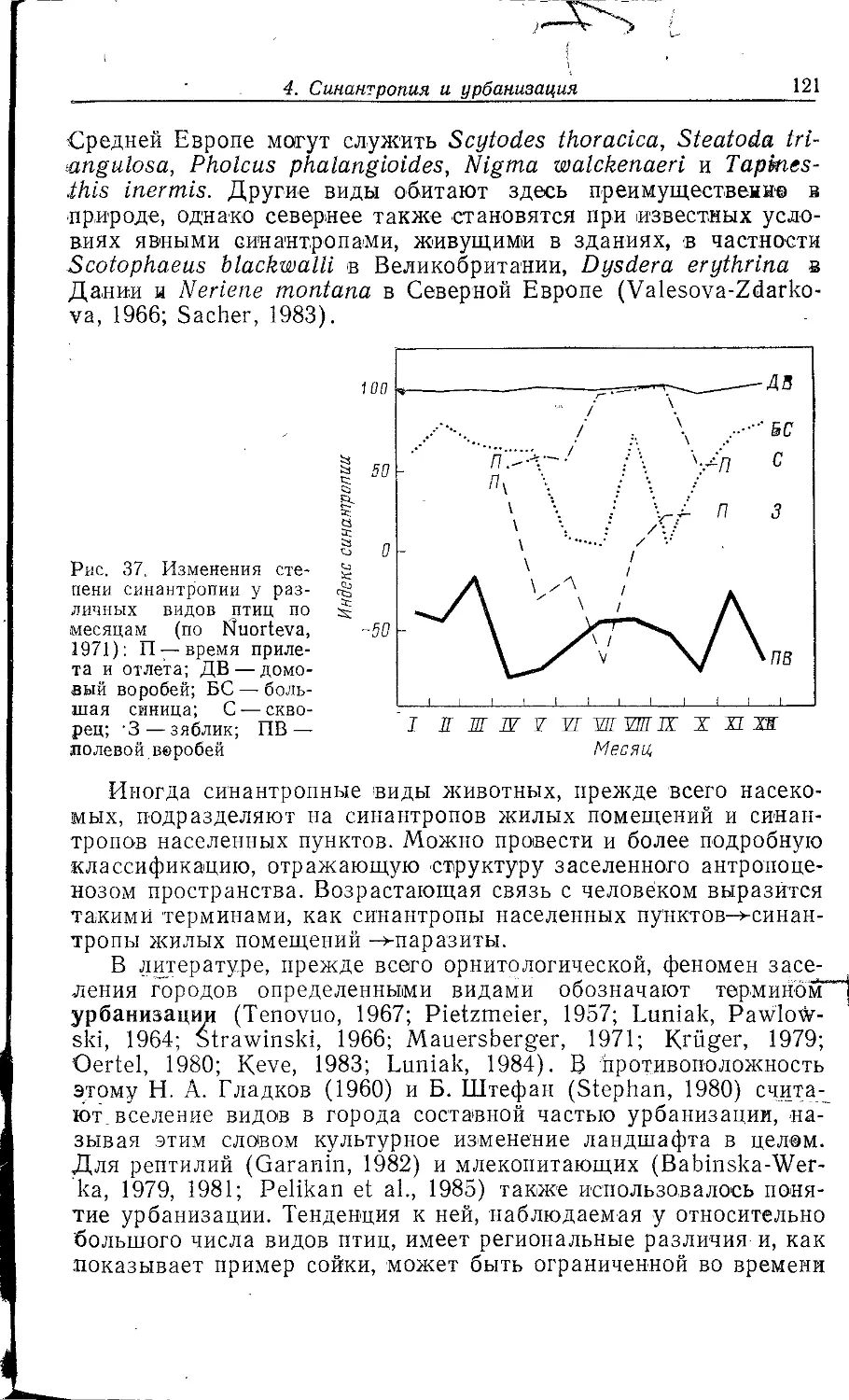
А. К. Быстровой (1980), а аналогичным проблемам
социалистического города посвящен специальный сборник статей «Город,
природа, человек» (1982), а также ряд книг по городским
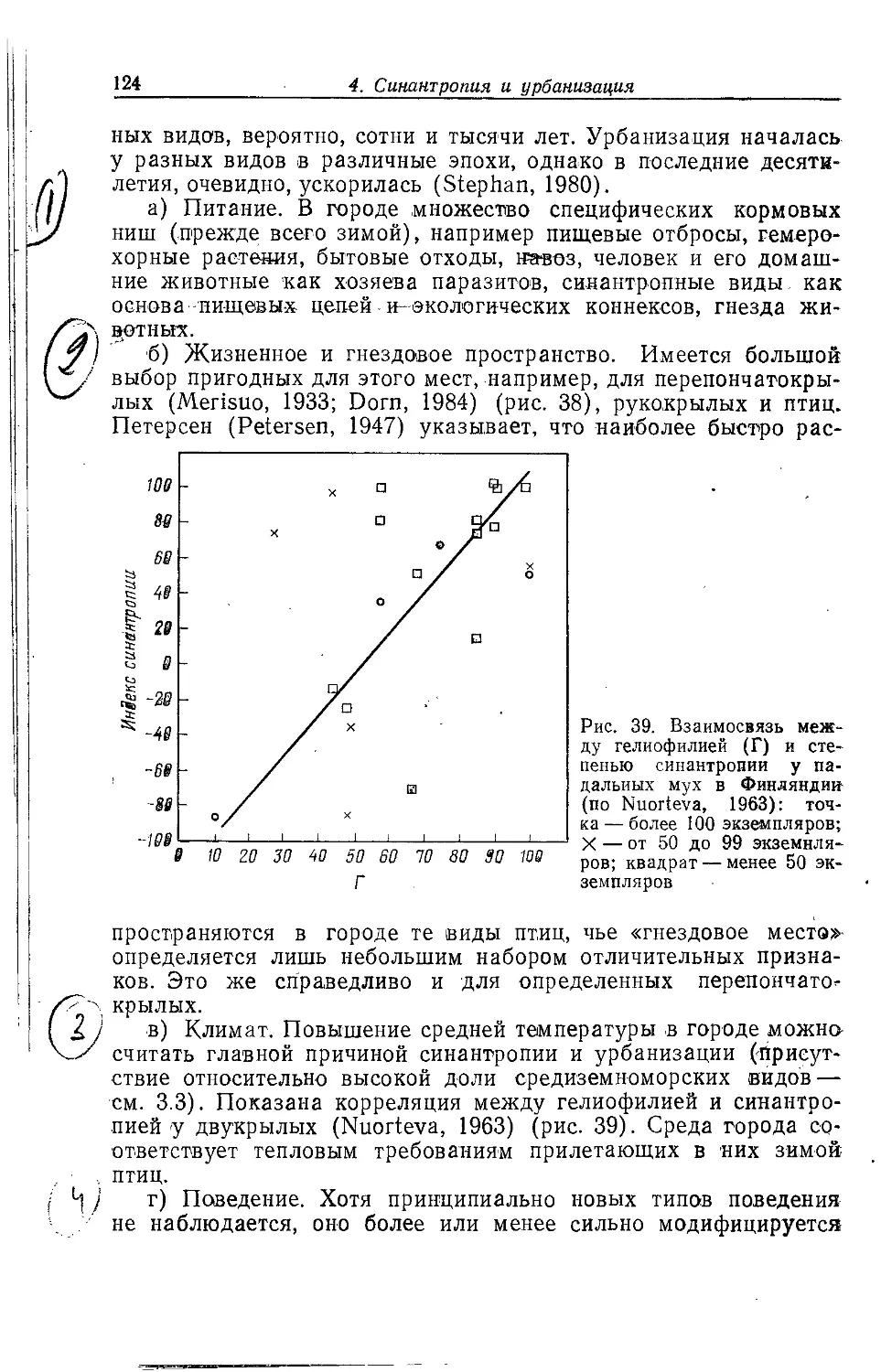
ландшафтам.
Для жизни животных в городе важны многие обстоятельства,
но на них всегда воздействует повышенная (на 1—2°С)
температура, загрязненность, «акустическое неблагополучие» среды,
запыленность атмосферы, совершенно отличный от природного
растительный мир. Особенностью городов является и обилие
домашних животных, а также хорошо приспособленных для жизни
и размножения в городе таких птиц, как вороны, галки, голуби,
чайки, которые определяют для многих других птиц и зверей
возможность или невозможность проникновения в городской
ландшафт. '
Известный зоолог Московского университета К. Н. Благоскло-
нов указывал на возникновение городских рас птиц,' у которых
по сравнению с исходными формами меняются и питаМе, и гнез-
достроительные инстинкты, и поведение в целом: перелетные
птицы становятся оседлыми, устраивают свои гнезда в самых
неожиданных местах, причем из необычных, неодинаковых в
разных городах материалов; хищники становятся санитарами
(мусорщиками), санитары — хищниками и т. д.-Сейчас орнитофауна
Москвы насчитывает 177 видов. В их числе такие редкие для
города птицы, как дубонос, чечевица, иволга, соловей, жаворо-
предисловие
нок, сова, сорока, майна, -кольчатая горлица... Эти животные —
тоже часть среды обитания, окружающей горожанина. А ведь
значительная часть населения (в Англии — 80%) проводите
городах основную часть своей жизни, и наличие островков зелени,
даже малых участков естественного ландшафта здесь трудно
переоценить.
Городская фауна — весьма существенный компонент
санитарной и эмоциональной среды человека, поэтому элементы
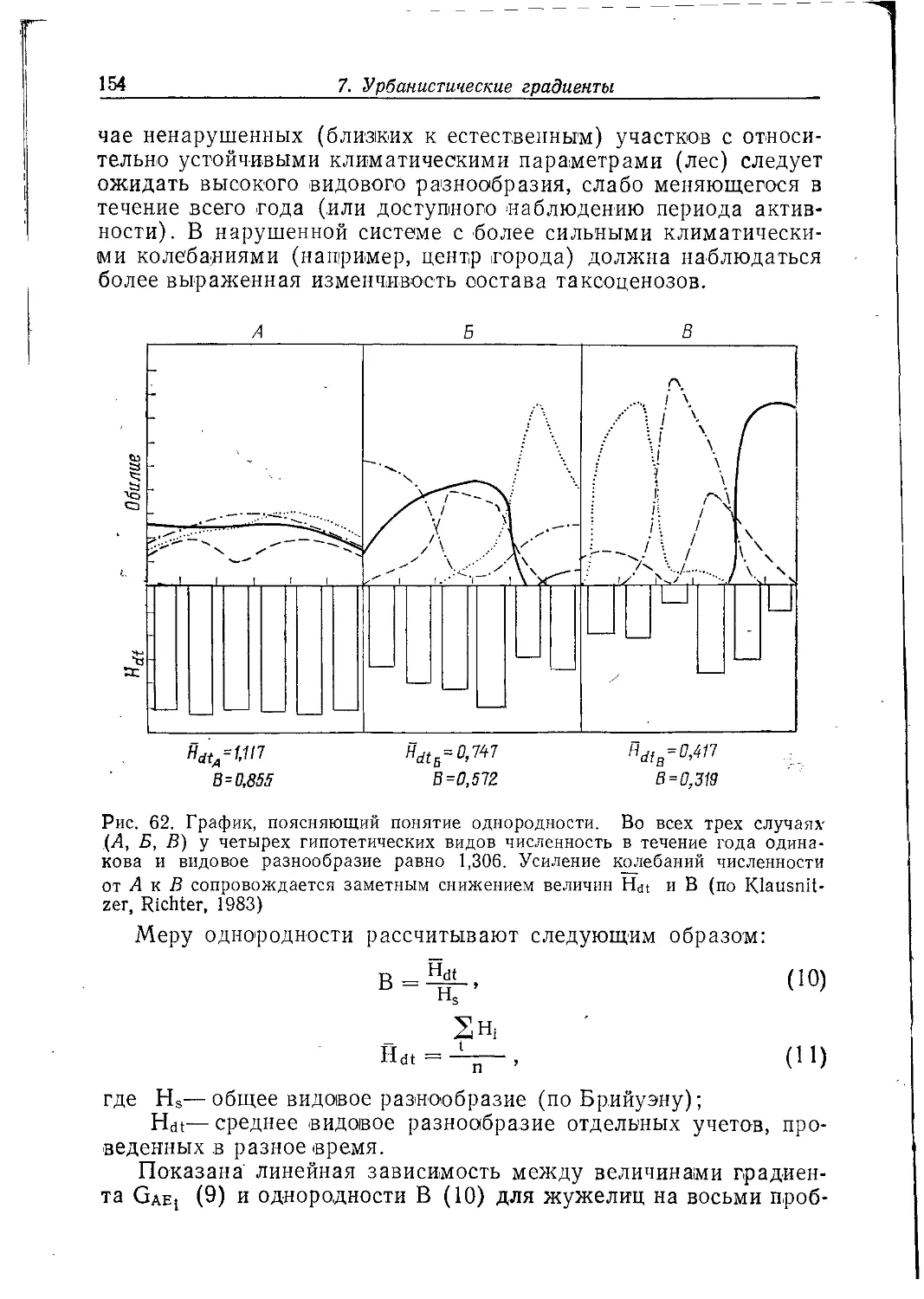
стихийности в ее формировании должны быть сведены до минимума.
Достижение гармонии в отношениях горожанина с его
«зоологическим окружением» — процесс сложный. Для этого нужна
повседневная конкретная работа экологической службы
каждого города, научно обоснованная система биотехнических
мероприятий.
Кроме таких экологических условий, как обеспеченность
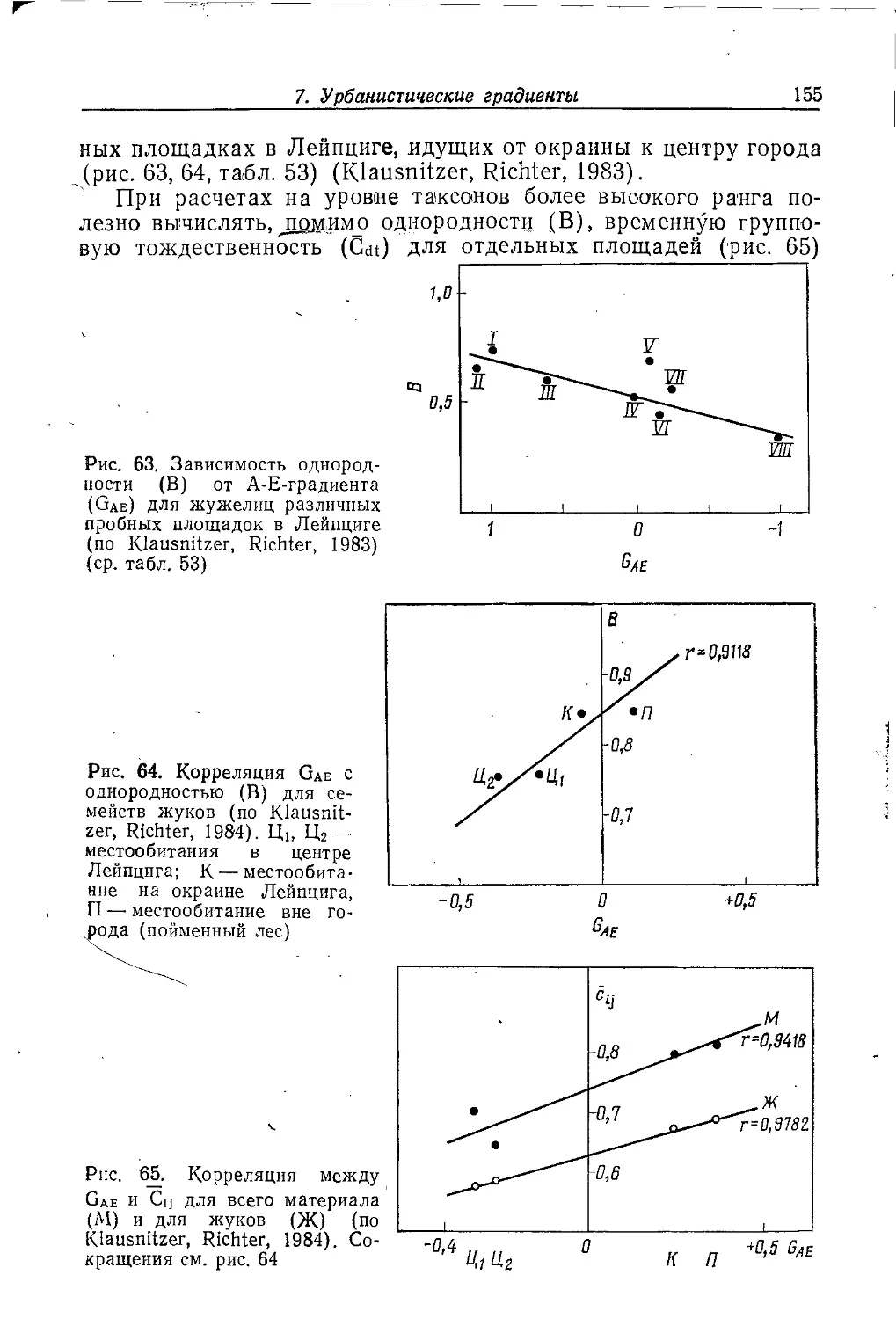
пищей или возможностями для размножения, в жизни
подавляющего большинства животных огромное значение имеет
использование занимаемого пространства, то, что Г. К. Нобль в 1939 г.
назвал «территориальностью», а В. X. Барт в 1943 г. оформил
в виде концепции «home ränge» или «хоминга». Под ними
подразумеваются особенности пространственного и временного
размещения животных в ландшафте. Очевидно, что городская среда
обитания является совершенно особой, эволюционно новой для
жизни любых видов животных, поскольку все они возникли
задолго до появления на планете городов. И далеко не все виды
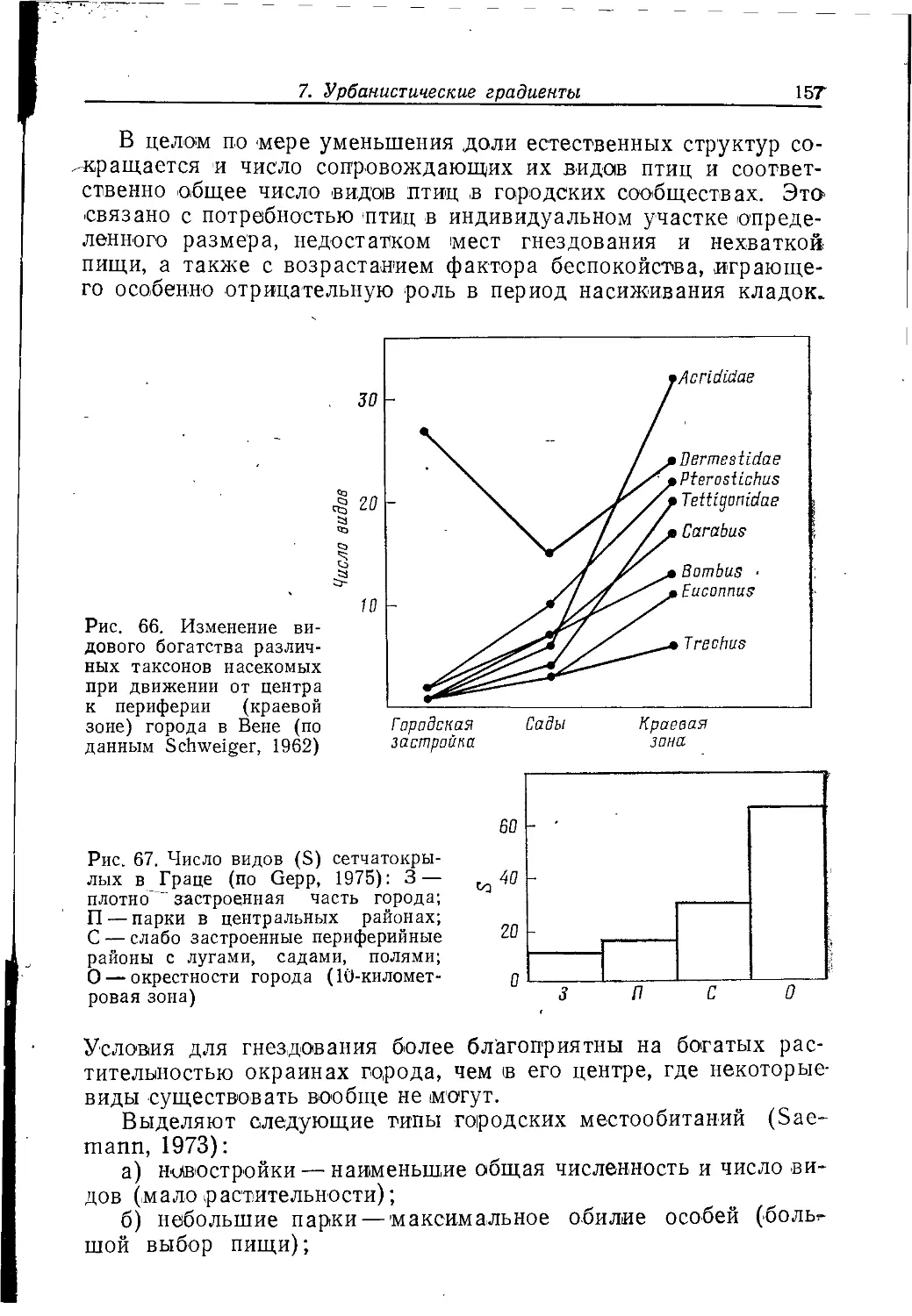
к ней могут приспособиться. Особенностью этой среды является
и ее строение из экотонов — переходных, промежуточных зон
между типичными ландшафтами. Экология города по сути
является экотональной экологией, концепции которой еще только
формируются.
Рассматривая экологические и градостроительные аспекты
среды, в которую должны «вписаться» дикие виды животных,
нельзя забывать и этносоциальную обстановку, которая может
стать определяющей для их жизни в городах, особенно для
таких заметных видов, как птицы, рептилии и млекопитающие
относительно крупных размеров.
Б. Клауснитцер совершенно не касается этого фактора,
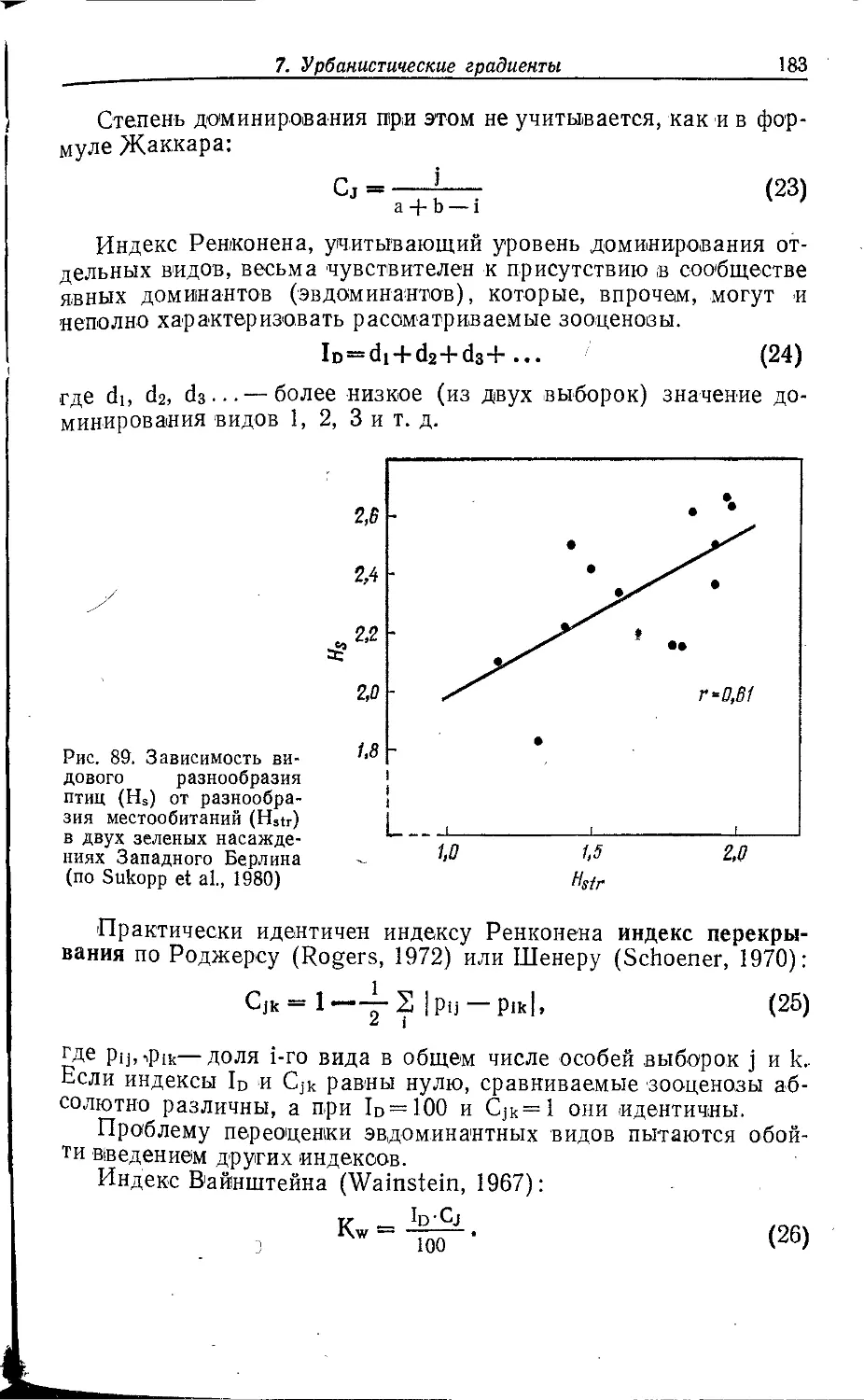
видимо, потому, что анализирует фауну городов Европы. Но в других
регионах мира отношение к городским животным совершенно
иное. В частности, в Индии в городах обитает поражающее
воображение количество птиц, зверей, ящериц, змей, которых в
большей мере охраняют религиозные обычаи, чем законы
государства. И совершенно иная картина в городах среди пустынь, где
большая часть населения сравнительно недавно занималась
отгонным животноводством и в обычаях и верованиях нет
традиций оберегать любых диких животных.
8 Предисловие
Способствует поддержанию разнообразия фауны города и
материальный уровень жизни людей. На моей памяти за
послевоенное время в Москве параллельно с ростом благосостояния шло
увеличение количества голубей, скворцов, а с 1973 г. на зимовку
стали оставаться и крупные стаи грачей. Лет 15 назад каждая
пролетная дикая утка в Москве была редкостью, а в 1986 г. их
насчитывалось здесь 14 тысяч, 'причем живущих оседло.
Проблемы городской экологии сегодня волнуют миллионы
людей. Настало время, говоря об окружающей среде, включать в
это понятие не только природу, но и городскую среду обитания
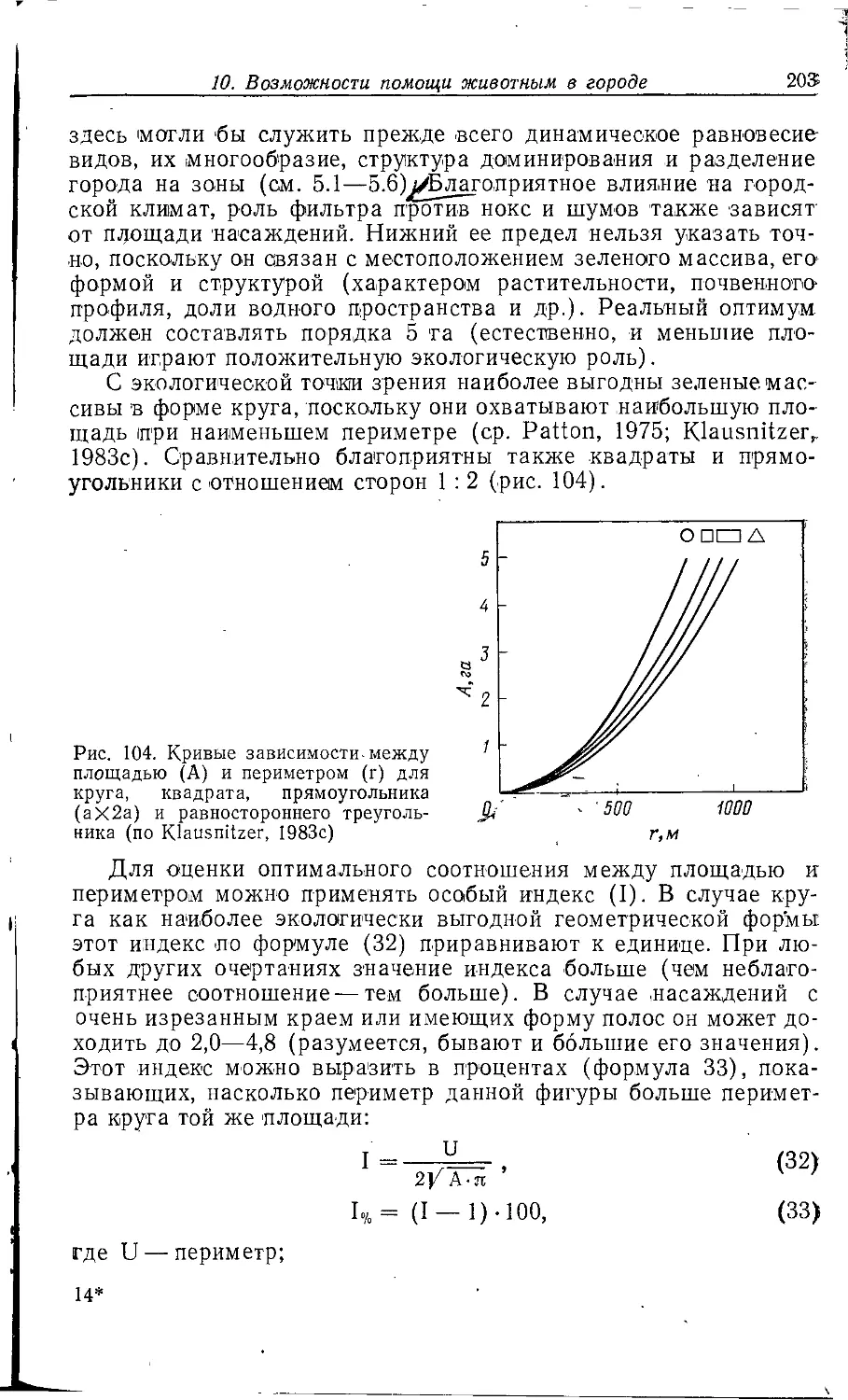
человека— наши жилища, места нашей работы и отдыха.
В лредставлениях многих до сих лор нередко отдается дань
традиционно урбанистическому видению города. Нагромождение
зданий, «cKaiMeHHbie джунгли», гигантская ярмарка или
гигантский вокзал-i:вот только некоторые из образов, которые обычно
возникают IB этой связи.
Именно поэтому лозунг «Города для людей» находит немало
приверженцев; он свидетельствует о росте внимания населения
к качеству жизни в современном городе. В этом направлении
советские архитекторы накопили немалый опыт. Упомянем
Международное совещание в Суздале в 1984 г. по программе
ЮНЕСКО «Человек и биосфера», один из 14 проектов которой
специально посвящен изучению городов и называется
«Экологические аспекты городских систем».
Широкую известность в СССР и за рубежом получила
программа «Экополис», которая посвящена экологическим
исследованиям г. Пущино — небольшого города Московской области.
Инициатором программы выступила лаборатория экологии и
охраны природы кафедры высших растений биологического
факультета МГУ, а руководство Научного центра биологических
исследований АН СССР в Пущине предложило свой город в
качестве полигона для совместной работы. В программе
«Экополис» целый ряд тем: от социально-психологических разработок
до изучения уровня городского шума и техногенных примесей в
дожде и снеге, от гибели животных на дорогах до аллергии
горожан при контакте с пыльцой цветущих растений.
По решению Пущпнского городского Совета народных
депутатов в 1982 г. в городе была учреждена система экологических
заказников для сохранения экологически важных природных
участков. Но в более крупных городах все эти проблемы решать
сложнее.
Независимо от принципов, которых придерживаются
приверженцы разных школ н направлений градостроительства, все они
выступают за значительное возрастание роли природных
элементов в структуре современного города. «Экогорода», в которых
гармонично сочетаются естественные ландшафтные структуры
Предисловие
С удобной для жизни и\работы человека застройкой и дорожной
сетью, стали мечтой многих градостроителей.
Развивается и наука об экологии города, а с 1975 г.
Международной ассоциацией экологов издается специальный журнал
«Urban Ecology» («Городская экология»).
Все эти вводные замечания, как я надеюсь, помогут читателю
лучше осознать, насколько ценна для практических прикладных
задач информация и методы анализа животного населения
городов, которые приведены в предлагаемой книге.
В истории человечества именно возникновение городов
привело к зарождению явления, которое было названо
«цивилизацией» и противопоставлялось таким понятиям как «варварство»
и «дикость». По мнению ряда исследователей, расцвет культуры
Древней Греции был обусловлен в первую очередь
распространением городского стиля жизни. Еще на заре нашей эры в эпоху
возникновения христианства безмятежная, счастливая, легкая
л^нзнь в представлении многих связывалась с жизнью именно в
городе, Б «Граде Божьем». Знаменитая книга Блаженного
Августина так и называлась «О граде Божьем».
Времена изменились. Город стал средоточием не только
культуры, новейшей техники, комфорта, индустрии услуг, по и
промышленных загрязнений, символом стрессов, сутолоки, суеты, «ас-
фальтовьгх пустынь». Вернуть нашим городам высокое «качество
жизни» для людей должны помочь не только новые формы
градостроительства, планирования городской среды, но и ее
гармоничное сочетание со средой естественной. А в этом деле не
последнее слово должны сказать экологи, в том числе и те,
которые изучают экологию животных крупных городов.
Д. А. Криволуцкий
Дополнительная литература
по экологии городской среды
Белоусов В. Н. Оздоровление городской среды — важнейшая
градостроительная задача. — М.: Знание, 1977.
Биоиндикация состояния окружающей среды' Москвы и Подмосковья. — М,;
Наука, 1982.
Бочкарева Т. В. Экологический джинн» урбанизации. — М.: Мысль, 1988.
Влияние промышленных предприятий на окружающую среду. — М.: Наука,
1987.
Гиренок Ф. И. Экология, цивилизация, ноосфера. — М.: Наука, 1987.
Глазычев В. Л. Социально-экологическая интерпретация городской среды. —
М.: Наука, 1984.
Город, природа, человек.—М.: Мысль, 1982.
Григорян А. Г. Ландшафт современного города, — М.: Стройиздат, 1986.
Животное население Москвы и Подмосковья. — М.: пзд-во МОИП, 1967.
Залесская Л. С. Курс ландшафтной архитектуры. — М.: Стройиздат, 1964.
Маергойз И. М. Географическое учение о городах. — М.: Наука, 1987.
Молодежь и экология Москвы. — М.: 1986.
Николаевская 3. А. Водоемы в ландшафте города. — М.: Стройиздат, 1975.
Новиков Э. Л. Город и природопользование. — Л.: Наука, 1986.
Окружающая среда крупного города. — Л.: Наука, 1988.
Огнев С. И. Fauna mosquensis. Опыт описания фауны Московской губернии.
1. Млекопитающие,—М.: изд-во Комиссии для исследования фауны
Московской губернии, 1913.
Природа и жилые районы городов. — М.: Стройиздат, 1986.
Растительность и животное население Москвы и Подмосковья. — М.: пзд-во
МГУ, 1978.
Формозов А. Н. Фауна. — В кн.: Природа Москвы и Подмосковья. — М. — Л.:
изд-во АН СССР, 1947.
Экология малого города. — Пущпно: изд-во Научного центра биологических
исследований АН СССР, 1987.
1. ПРЕДИСЛОВИЕ
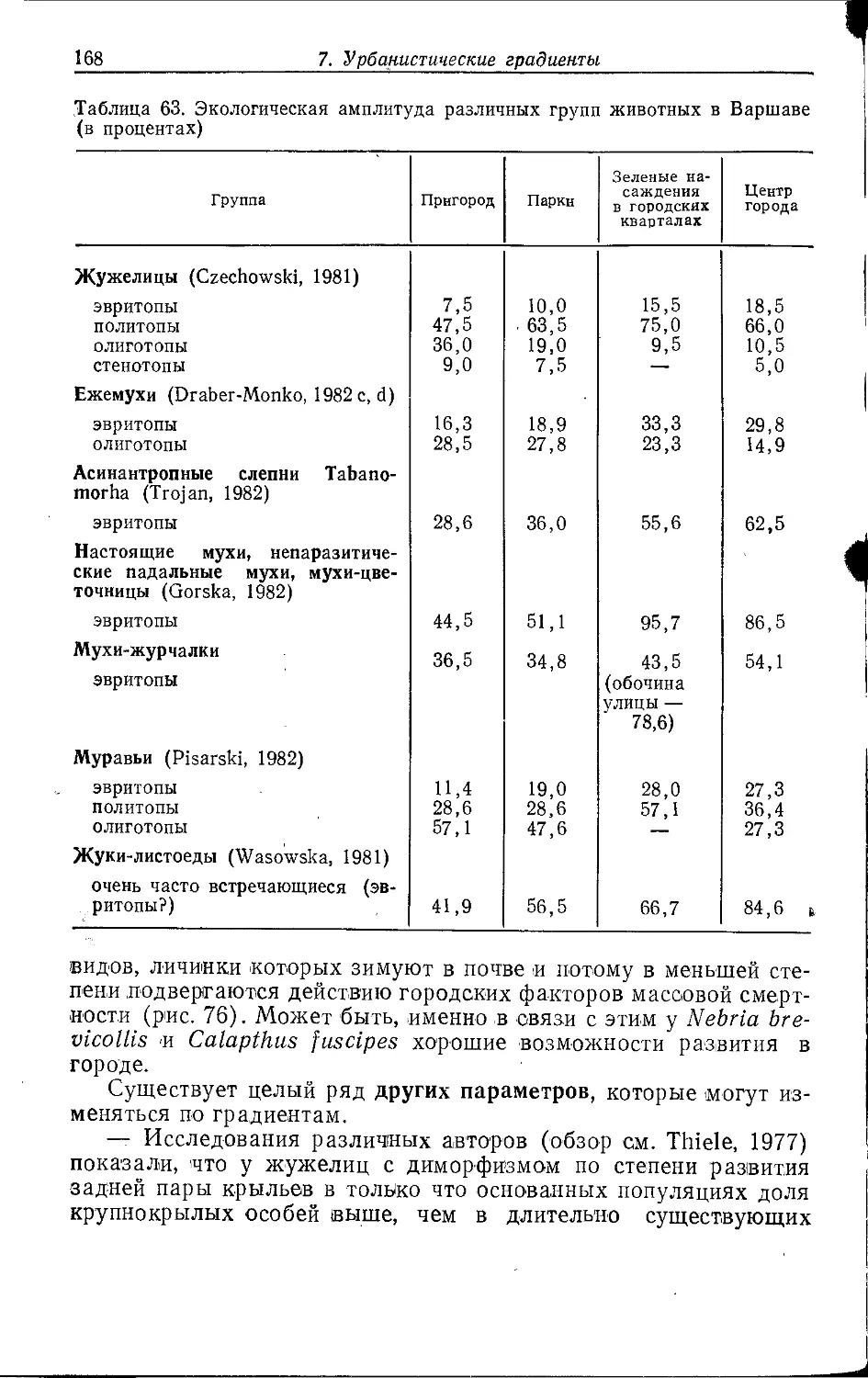
Одними из первых исследователей, более 30 лет назад
приступивших к углубленному изучению экологии животных в городах,
были Вильгельм Кюнельт, Вольфганг Тишлер и Герберт Вайднер.
С этого времени интерес экологов к «жизненному пространству
города» непрерывно рос. Следует, однако, подчеркнуть, что наше
зооэкологическое понимание городской среды по сравнению с
другими биотопами содержит еш,е много пробелов. Несмотря на
развитие в течение последних 30 лет исследований городских
экосистем, большинство экологов продолжала привлекать
естественная среда, изучением которой они занп1мались куда охотнее.
Информация по городским экосистемам оставалась, таким образом,
сравнительно фрагментарной, п о многих городах мы не знаем
почти ничего. К тому же зооэкологические исследования были
сосредоточены лишь на небольшом количестве объектов. И все-
таки сейчас в этой области накопилась уже довольно обширная
литература (2000 названий по Центральной Европе), так что
пришло время сделать некоторые обобщения, даже если они и
продемонстрируют больше пробелов в наших знаниях, чем
действительных закономерностей.
Данная работа является попыткой систематизации
имеющихся сведений, их сопоставления и, когда это возможно, обобщения.
За основу взяты данные прежде всего о Западном Берлине,
Граце (Австрия), Гамбурге (ФРГ), Киле (ФРГ), Лейпциге (ГДР),
Саарбрюккене (ФРГ), Варшаве (ПНР) и Вене (Австрия). О
городах, расположенных вне Центральной Европы, информации
мало. Кроме того, рассматриваются только наземные
местообитания, а пресноводные экосистемы опускаются.
Наличие новых, вряд ли существующих в природе
экологических ниш (особенно с точки зрения питания и структуры
занимаемого пространства) вместе с климатическими особенностями
привело к необычным сочетаниям факторов и формированию
особой фауны (специалисты, убиквисты, множество адвентивных
видов), а также специфических сообществ л<ивотных, связанных
новыми взаимоотношениями. Это относится как к городу в целом,
так, пожалуй даже в большей степени, к отдельньш его
местообитаниям. Многие экологические параметры закономерно изме-
^4-
12 /■ Предисловие
няются от периферии к центру города, при этом окраина в
некотором отношении нарушает линейность прослеживаемых
градиентов. Часто встречающееся в этой зоне необычайное богатство
видового состава заставляет задуматься о создании здесь
природоохранных территорий.
Многие виды и более высокие таксономические группы, а
также некоторые жизненные формы потенциально являются особо
пригодными для заселения города. Это подтверждается и эколо-
го-зоогеографичес'ким происхождением городакой фауны. Она
самобытна в количественном и качественном отношении, как и
фауна сухих лугов, верховых болот и других более пли менее
природных биотопов. Однако существенное своеобразие города
заключается в мозаичности часто совершенно противоположных
по характеру местообитаний. Внутри него близкие к природным
участки, дал^е небольшого размера, могут привести к появлению
видов, которых едва ли можно причислить к собственно
городской фауне.
Многие городские местообитания настолько резко
изолированы друг от друга транспортными путями и постройками, что их
мол<но рассматривать как островные и пытаться применять в
данном случае теоретические концепции островной
биогеографии. По мнению некоторых авторов, здесь наблюдаются даже
эволюционные процессы, так что вполне возмол^но (хотя, на наш
взгляд, и несколько преладевременно) говорить о городе как об
«экспериментальном поле эволюции». В этой связи особого
внимания заслул^ивают физиологические, морфологические и
экологические реакции л-снвотных на особенности городской среды, их
адаптации к ней и устойчивость к антропогенным нарушениям,
а таклче специфические факторы смертности в условиях города,
явления урбанизации и сннантропизацин (включения в новые
системы).
Для дальнейшего изучения городской фауны необходимо
прежде всего расшпрение наших фактических знаний. Поэтому
некоторые разделы (например, гл. 5, 7.7, 7.8, 8.1) следует
рассматривать как предварительные наброски, не претендующие на
полноту, не говоря уже об общей значимости сделанных выводов.
Возможно, они смогут послужить стимулом для аналогичной
обработки нового материала и позволят выявить некоторые другие
экологические закономерности городской среды.
Ряд авторов пытаются рассматривать весь город как единую
экосистему. Какими бы проблематичными ни были эти взгляды,
несомненно, что определенные экосистемы связаны с
различными местообитаниями города и в большей или меньшей степени
друг с другом. Трудность определения и описания системы в
целом заключается прелюде всего в существовании весьма
различных точек зрения да понятие «городская экология».
/. Предисловие 13
Именно в связи с тем, что для людей в городах общение с
природой имеет особую и все возрастающую важность,
экологические исследования городской фауны приобретают большое
практическое значение. Соответствующие данные необходимы для
планирования, наблюдения за окружающей средой, оценки
стабильности городских экосистем и снижения ущерба,
причиняемого животными. Получение как можно более точных знаний о
жизненном пространстве, в котором во всем мире с каждым
годом живет и работает все больше людей,— одна из
первоочередных задач экологии.
Бернхард Клауснитцер
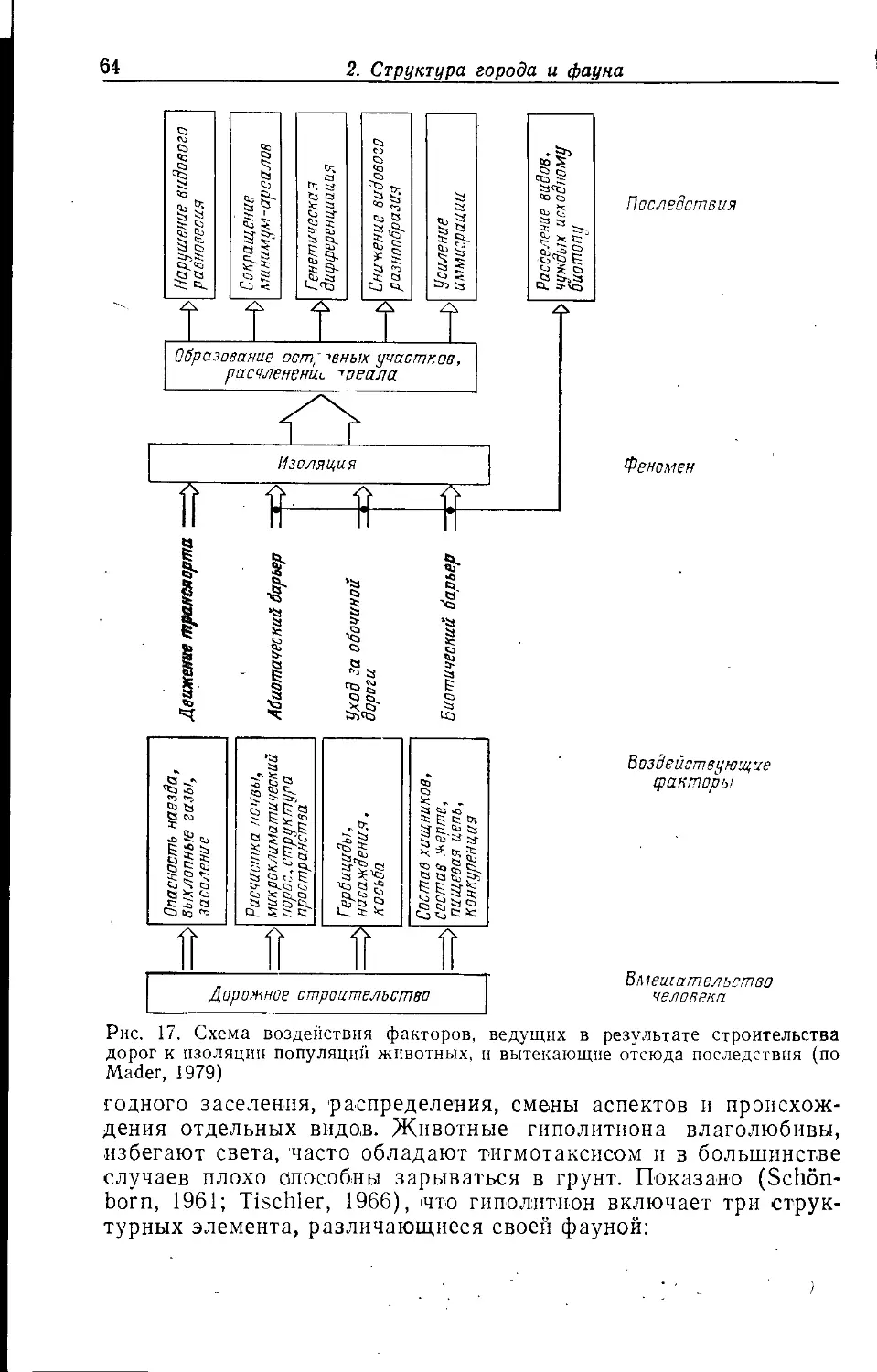
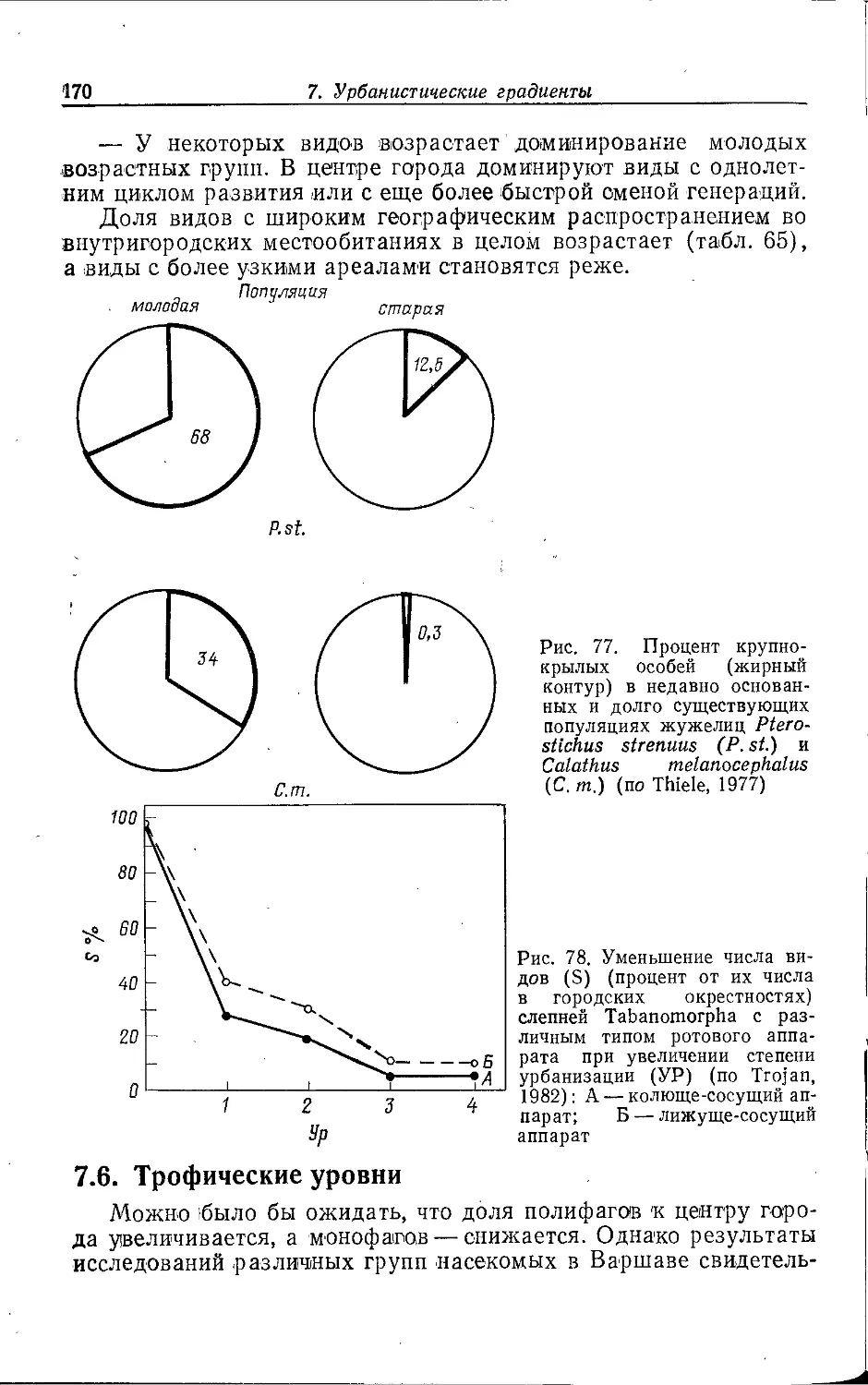
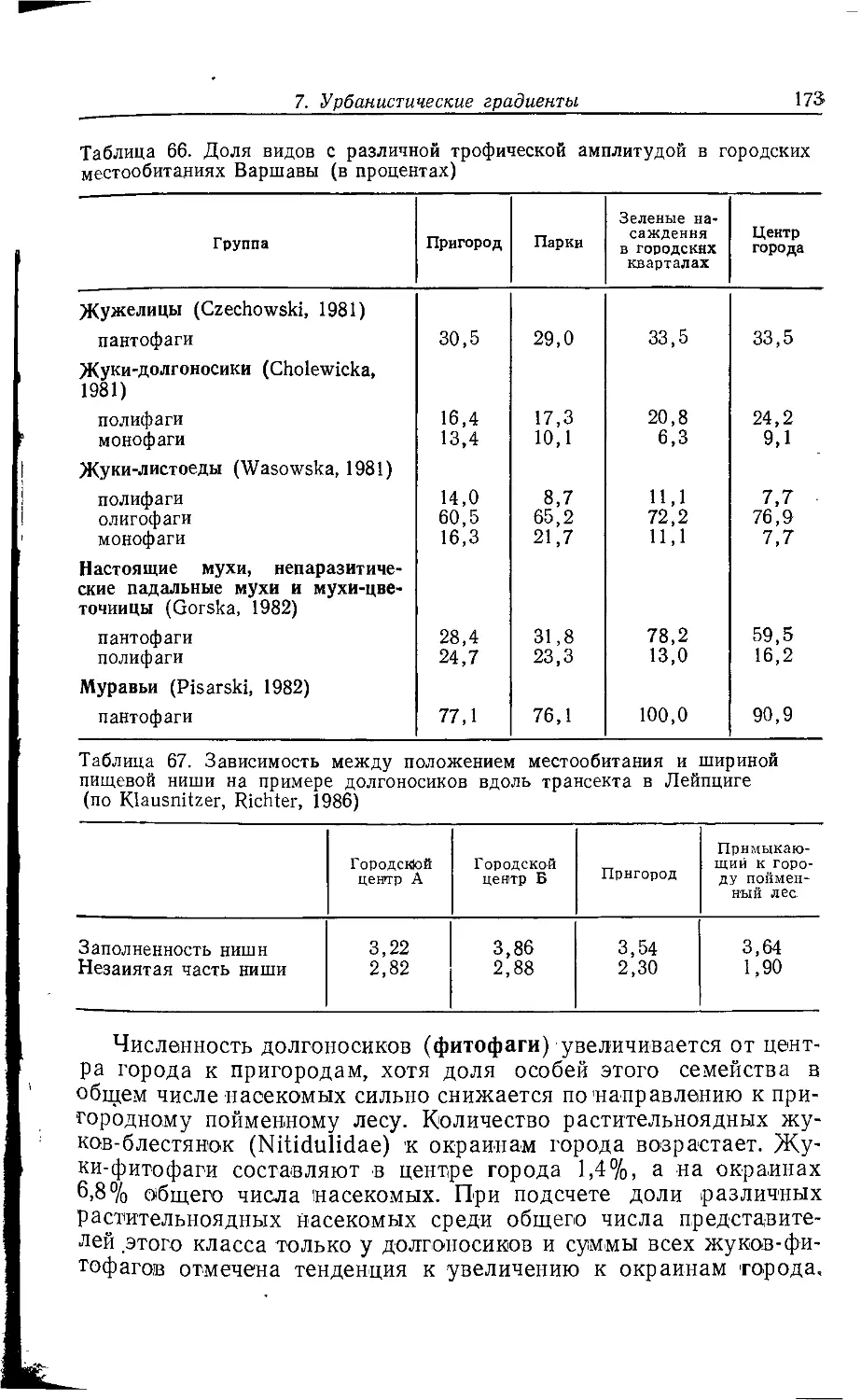
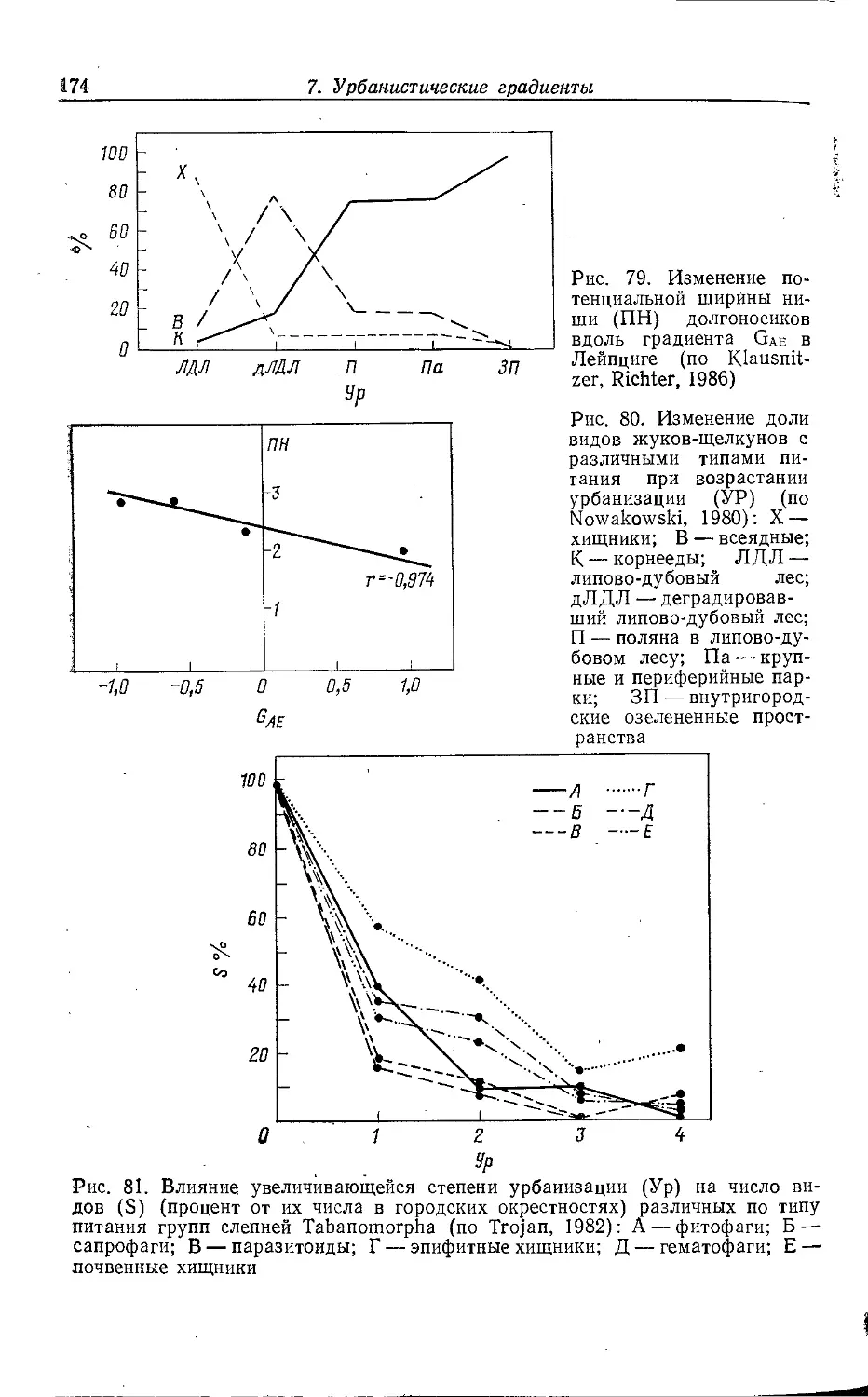
, 2. СТРУКТУРА ГОРОДА И ФАУНА
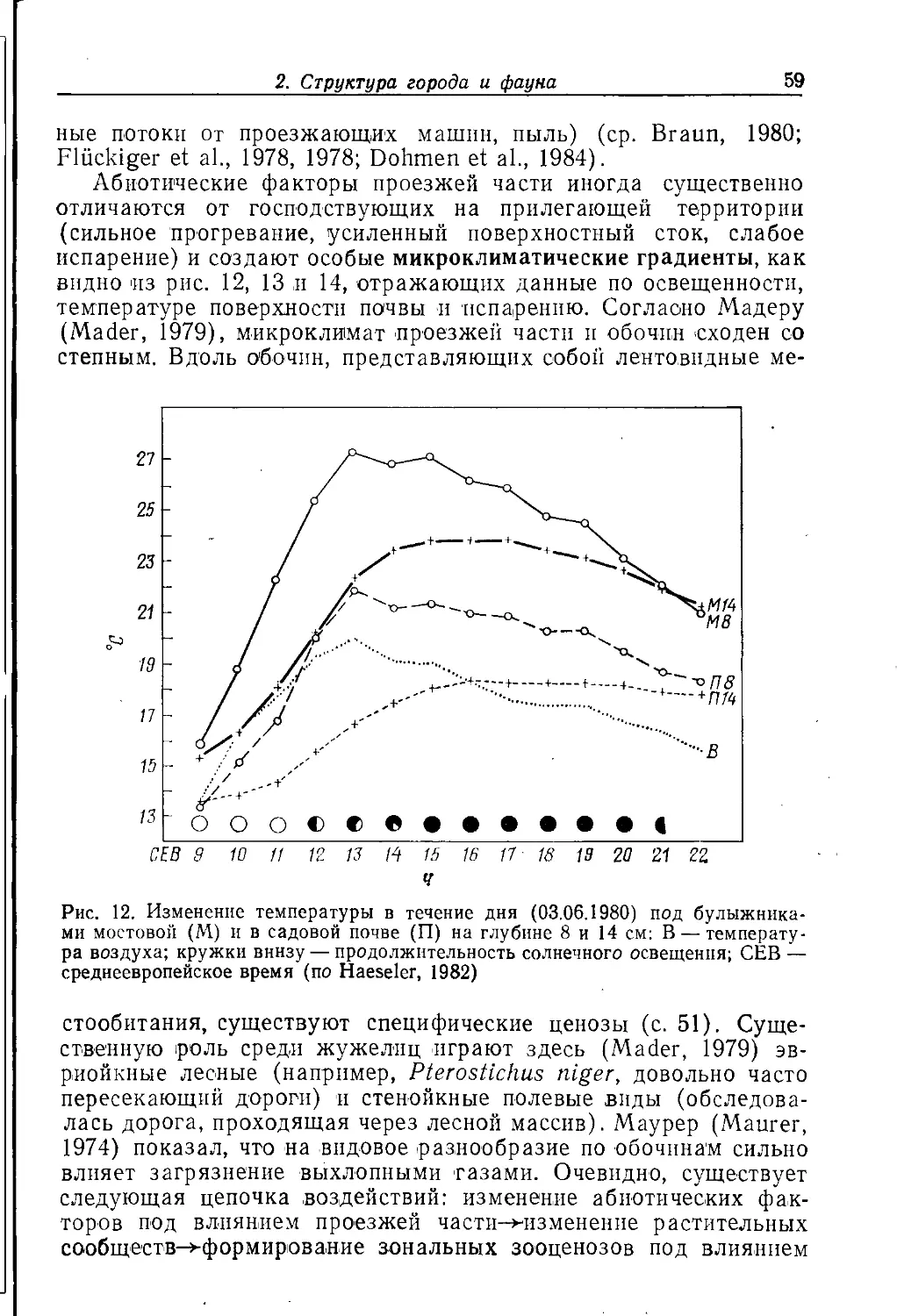
Крупный город часто представляет собой в историческом пла-
i не неустойчивую конечную стадию развития населенного пункта;
промежуточными этапами на этом пути были деревня, мелкий и
средний города. Его нельзя рассматривать как единую
экосистему; в большинстве случаев речь идет о мозаике разлпчны.х
биотопов (квазигомогенных элементарных систем). Экологическое
своеобразие отдельных городских местообитаний зависит прежде
всего от антропогенных форм их использования.
Соответствующие структурные единицы можно сравнивать между собой, по-
^скольку эти формы использования в различных городах схожи.
Схемы городского деления предлагались многими авторами.
В данной работе делается попытка обобщить взгляды,
изложенные в разных работах (Weidner, 1952; Sukopp, 1973, 1979, 1981,
1983а, b; Sukopp et al., 1980; Friese et al., 1976; Blume et al.,
1978; Kelcey, 1978; Brady et al., 1979; Bornkamm, 1980), но
прежде всего в статьях Клоца, Гутте и Клауснитцера (Klotz, Gutte,
Klausnitzer, 1984а, b).
Поч1вы, климат, антропогенные ноксы'', растительность,
другие пищевые ресурсы и наличие специфических структурных
элементов в отдельных городских местообитаниях имеют более или
менее сильные различия, что во многих случаях ведет к
образованию характерных зооценозов. Мозаичное распределение
местообитаний накладывается на более или менее выраженное
зонирование городской среды от центра к окраине. Поэтому в
зависимости от их расположения в одинаковых типах местообитаний
прослеживаются эколого-фаунистические различия, выявление
которых является главной предпосылкой для описания
городских градиентов (с. 151).
Городские местообитания мы подразделяем на две большие
группы: «строения» и «прочие наземные местообитания».
Пресноводные экосистемы нами не рассматриваются.
2.1. Строения
Строения (здесь — главным образом здания) как
местообитания, созданные человеком, несомненно, представляют собой
совершенно особые и отчасти новые для живого экологические
'> Ноксамп в спецпа.пьной литературе принято называть вредные для
живого агенты, как правило, загрязннтелп среды, биоциды и т. п. — Прим. ред.
2. Структура города и фауна 15
НИШИ. По структуре поверхности они сравнимы со скалами и тем
самым предоставляют подходящие условия (субстрат, трещины,
отверстия, пища, микроклимат) некоторым видам, исходно
обитавшим в скалистой местности. Следующей особенностью
является относительно высокая температура, которую внешние стены
могут приобретать в зависимости от экспозиции и солнечного
излучения и сохранять более или менее длительное время. Это
обеспечивает существование некоторых термофильных видов.
Дополнительные пиши возникают, если поверхность здания покрыта
растительностью.
В зависимости от использования можно выделить различные
типы строений: жнлые дома, служебные здания, складские
помещения, производственные строения и др. Для заселения
животными внутренних помещений важно прежде всего наличие там
специфических источников питания, а также температурный
режим и 1Влажность.
Эта группа местообитаний по сути идентична биотопу,
который Вайднер (Weidner, 1952) называет «культурной пустыней».
Он различает три биохора: здания с их обстанов'кой, хорпоцено-
зы хранящихся в зданиях запасов и .материалов и «стерильные
почвы» (см. 2.2.3). Понятие «биохор» нами не употребляется,
поскольку оно означает скорее значительно более мелкие
пространственные структуры.
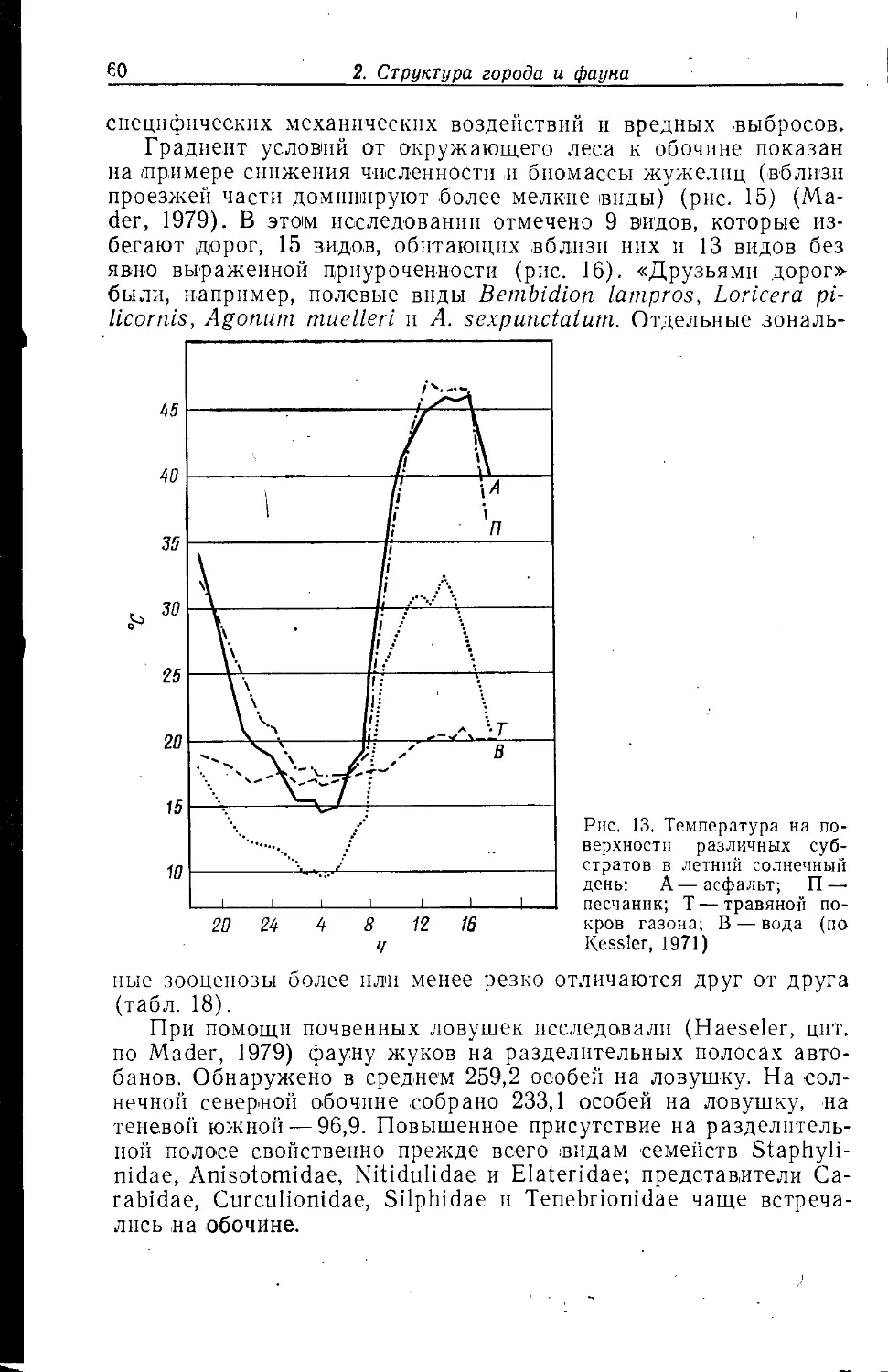
2.1.1 Непостоянно отапливаемые жилые дома
Характерным для этого типа зданий является часто
относительно сильная изменчивость температуры в помещении на
протяжении суток и года, а также соседнее расположение
отапливаемых и неотапливаемых в данный момент помещений. Это создает
прежде всего в средней части здания (на этажах) мозаику
различных жизненных условий. Внутри таких жилых домов
различают, как правило, три основных зоны: чердаки, этажи, подвалы.
2.1.1.1. ЧЕРДАКИ
Для чердаков характерны максимальные амплитуды
температурных колебаний в течение суток и года. Тепловой режим
очень существенно зависит от кровельного материала, формы
крыши и вентиляции. Под крышами из красной черепицы
температура летом может колебаться в течение суток от 12 до 42°С,
при толевом покрытии —от 16 до 42 °С. Чердаки в основном
сухие, лишь местами из-за повреждений крыши возникают более
или менее влажные условия, стимулирующие рост грибов.
Кроме этих грибов единственный постоянный источник питания для
животных — обработанная древесина различных пород, разного
16
2. Структура города ц фауна
возраста и состояния. По своей пространственной структуре
чердаки пригодны также для гнездования, ночевки и зимовки.
Дерево, а также повышенная температура создают
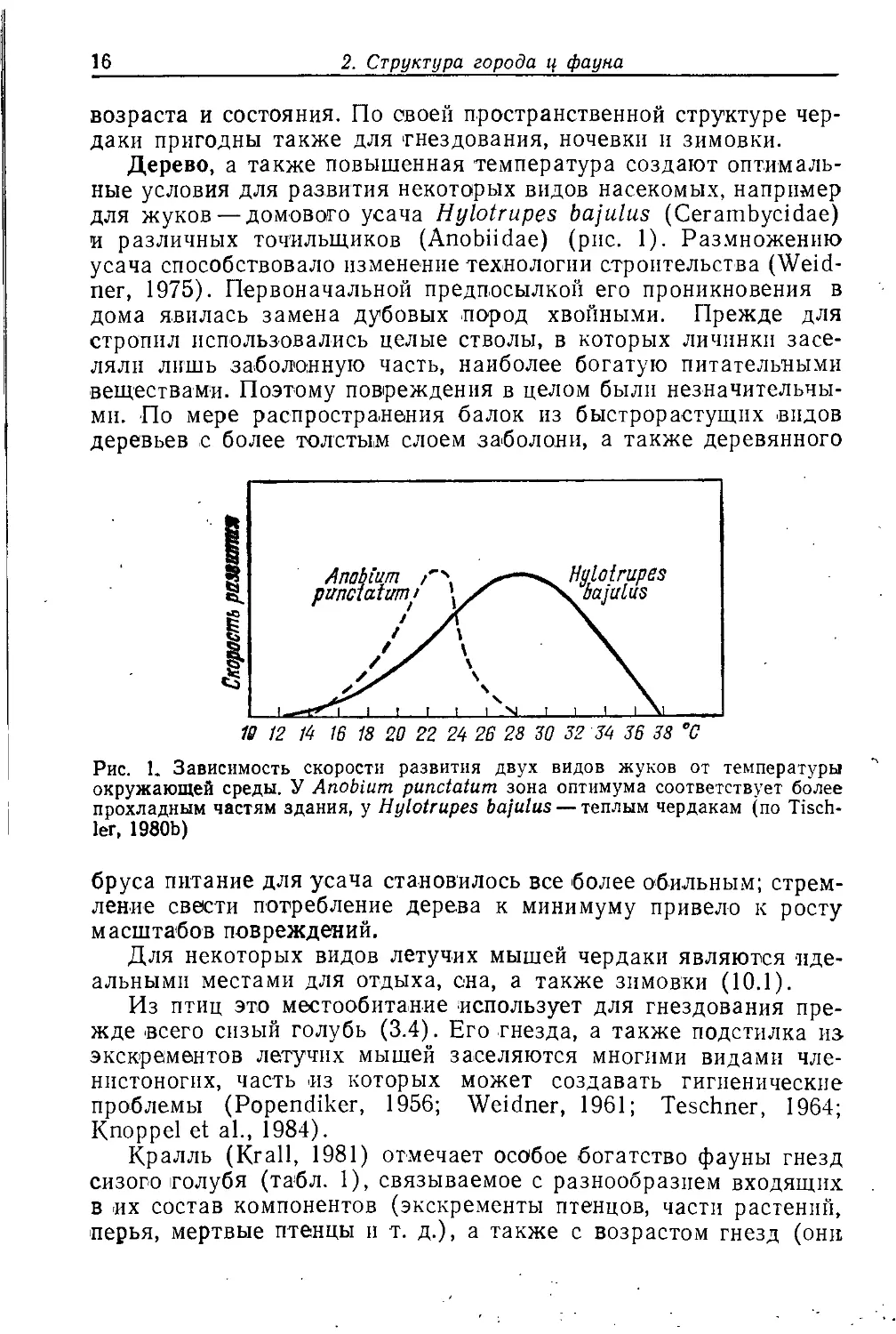
оптимальные условия для развития некоторых видов насекомых, например
для жуков — домового усача Hylotrupes bajulus (Cerambycidae)
и различных точильщиков (Anobiidae) (рис. 1). Раз.множению
усача способствовало изменение технологии строительства
(Weidner, 1975). Первоначальной предпосылкой его проникновения в
дома явилась замена дубовых пород хвойными. Прежде для
стропил использовалнсь целые стволы, в которых личинки
заселяли лишь заболонную часть, наиболее богатую питательными
веществами. Поэтому повреждения в целом были
незначительными. По мере распространения балок из быстрорастущих видов
деревьев с более толстым слоем заболони, а также деревянного
punctatumi
^ Hylotrupes
|^-Г>''| 1 ! I 1 1 4J
I 1 1 1 \i
IB 12 14 16 18 20 22 24 26 28 30 32 34 36 38 'С
Рис. 1. Зависимость скорости развития двух видов жуков от температуры
окружающей среды. У АпоЫит pundatum зона оптимума соответствует более
прохладным частям здания, у Hylotrupes bajulus — теплым чердакам (по
Tischler, 1980b)
бруса питание для усача становилось все более обильным;
стремление свести потребление дерева к минимуму привело к росту
масштабов повреждений.
Для некоторых видов летучих мышей чердаки являются
идеальными местами для отдыха, сна, а также зимовки (10.1).
Из птиц это местообитание использует для гнездования
прежде всего сизый голубь (3.4). Его гнезда, а также подстилка на
экскрементов летучих мышей заселяются многими видами
членистоногих, часть пз которых может создавать гигиенические
проблемы (Popendiker, 1956; Weidner, 1961; Teschner, 1964;
Knoppel et al., 1984).
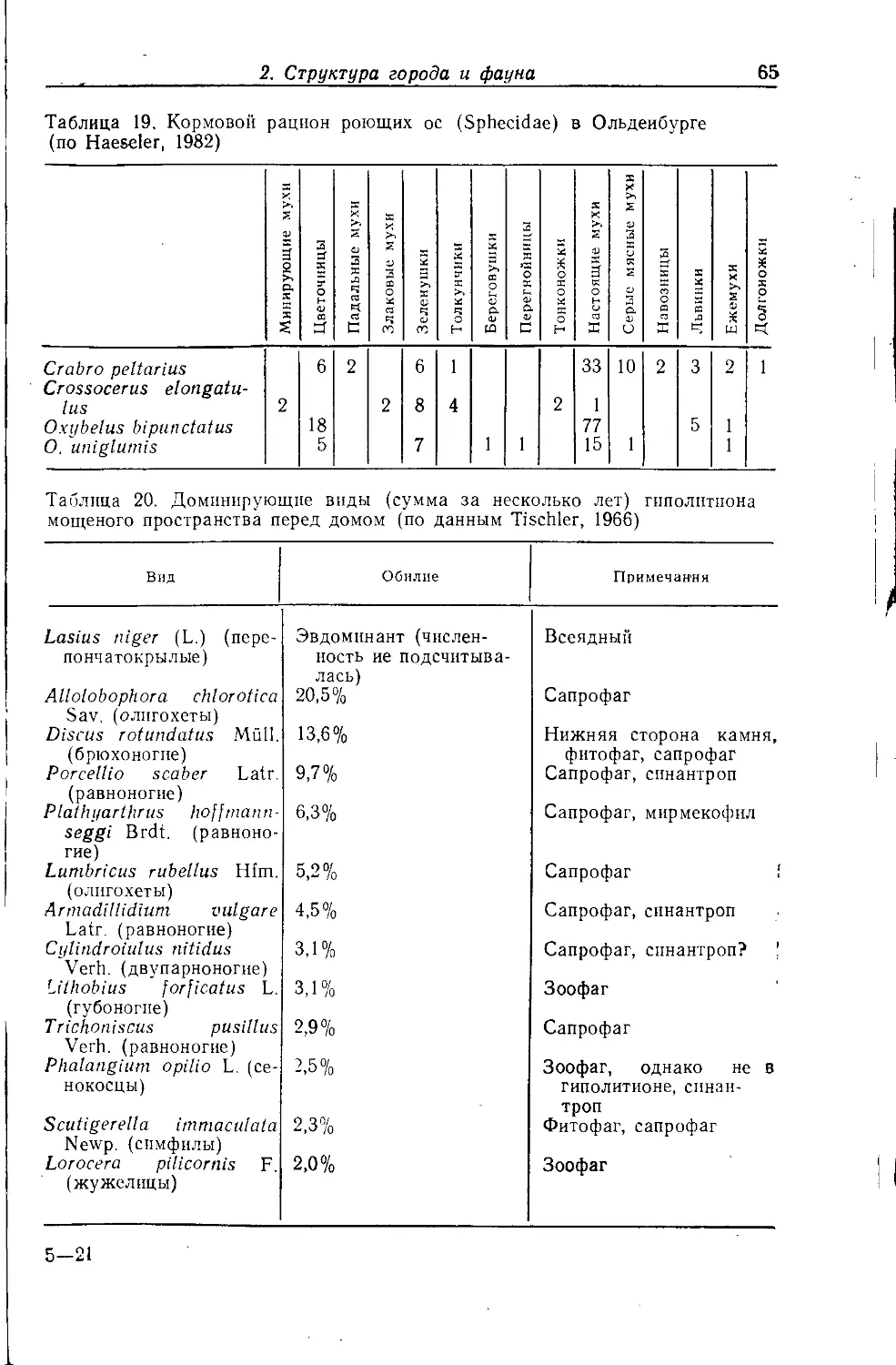
Кралль (Krall, 1981) отмечает особое богатство фауны гнезд
сизого голубя (табл. 1), связываемое с разнообразием входящих
в -их состав компонентов (экскременты птенцов, части растений,
перья, мертвые птенцы и т. д.), а также с возрастом гнезд (они
2. Структура города и фауна
17
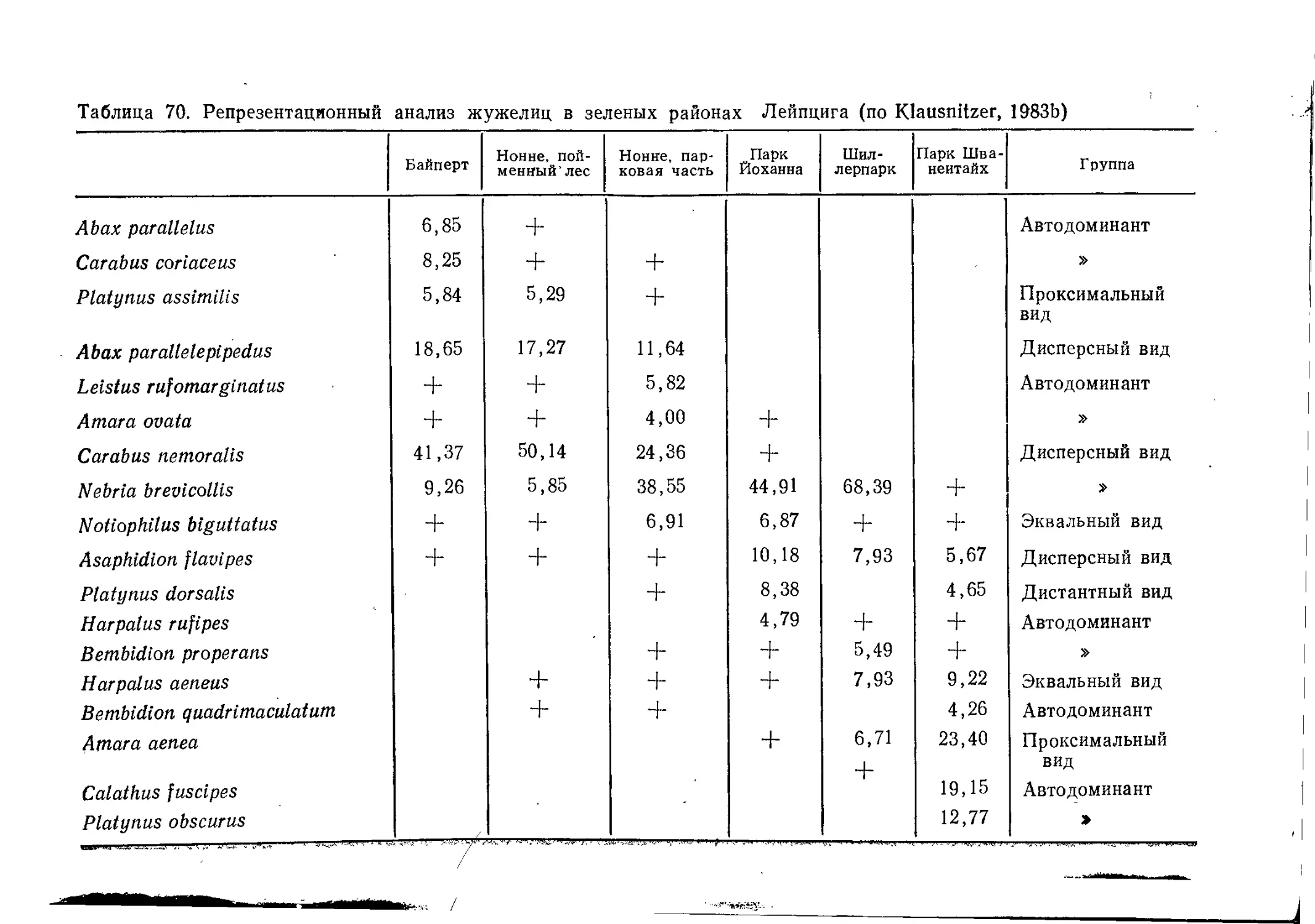
Таблица 1. Фауна гнезд сизого голубя (всего 20 гнезд) (по Krall, 1981)
Отряд
Жесткокрылые
Двукрылые
Перепончатокрылые
Чешуекрылые
Ногохвостки
Сеноеды
Пухоеды
Трипсы
Равнокрылые
Полужесткокрылые
, Блохи
Щетинохвостки
Кожистокрылые
^ Всего
Число
видов
Число
гнезд
Всего
особей
Имаго
Личинки
18
12
5
7
6
3
2
2
2
2
2
1
1
63
16
9
16
9
13
4
2
2
3
12
4
2
1515
2903
50
1938
2293
3924
15
И
13
3
241
632
31
353
239
50
27
5
1
11
2
67
20
13 569
775
1162
2664
1911
174
11
5925
Неполовозрелые
стадии
2288
3924
15
10.
632
6869
используются многократно). Обилие пищи, прежде всего
голубимого помета, обусловливает высокую численность видов.
Повышенной температурой (насиживание и присутствие птенцов)
Кралль объясняет высокое видовое разнообразие.
Главной причиной встречаемости доминирующих таксонов
являются трофические связи:
— кровь и детрит гнезда: блохи;
— плесень и детрит: ногохвостки, сеноеды, жуки-притворяшки;.
— остатки животных, кератин: кожееды, моли;
— экскременты: двукрылые;
— материал гнезда, пищевые остатки: бабочки семейства Оесо-
phoridae.
Регулярно зимуют на чердаках некоторые виды насекомых,
например крапивница {Aglais urticae) и дневной павлиний глаз
{Inachis io). Примечательно проникновение на зимовку в здания
златоглазки Chrysoperla сагпеа, прежде всего на чердаки и в
неотапливаемые комнаты (Gepp, 1967; Weidner, 1971).
Наступление диапаузы у этого вида сопровождается резким изменением
окраски (Мае Leod, 1967). Гепп (Gepp, 1975) указывает, что в
жилые помещения проникает и ряд других сетчатокрылых, в
частности Drepanopterix phalaenoides, Wesmaelius subnebulosus
(Micromus variegatus); он полагает, что причиной этого
являются неустойчивая погода и более постоянный климат внутри
зданий.
Некоторые виды божьих коровок зимуют в домах сообщест-
rfäim,(аГрёгац'115!.МИ)'; ^гГреЖДУ Всего это Oenopia conglobata с Ada-
ммвн
!_8 2. Структура города и фауна
На bipunctata. Они предпочитают щели в оконных рамах и
аналогичные пространства; преобладает всегда О. conglobata (40—
70-95%).
Характерным животным чердаков является (была?) черная
крыса, численность которой в последнее время во многих
городах снизилась. Кюнельт (Kühnelt, 1955) объясняет это тем, что
теперь на чердаках почти не хранятся продукты питания.
2.1.1.2. ЭТАЖИ
Фауна этажей (см. также 2.1.2) в первую очередь зависит от
наличия пищи, а также от структуры пространства, температуры
и влажности. Уже само расположение помещений обусловливает
различие температурных условий: южные комнаты теплее
северных, мансарды теплее комнат первого этажа, отдельно стоящие
до.ма холоднее домов, примыкающих друг к другу.
Экологически фауна этан\ей подразделяется на следующие
группы: вредители запасов, вредители материалов, паразиты
человека, паразиты домашних животных, обитатели домашней
пыли, обитатели плесени, фауна цветочных горшков п ко.мнат-
ных растений, синантропные двукрылые, пауки, случайно
проникающие (главным образом через окна) виды и «фауна
холодильников».
Вайднер (Weidner, 1952) приводит 308 видов специфических
домашних насекомых. Присутствие вредителей запасов в
квартирах зависит прежде всего от подходящего пищевого субстрата.
Их доля среди акклиматизировавшихся и заносных видов очень
высока (Weidner, 1939, 1958, 1962).
Решающим фактором для присутстви.- видов кроме наличия
пищи является также температура.
Например, постоянными обитателями домов стали некоторые
теплолюбивые виды щетинохвосток. Происходящая из
Средиземноморья и, В0.3М0ЖН0, также из Азии и ставшая ныне
космополитом сахарная чешуйница {Lepisma saccharina) широко
распространена в жилищах, складских помещениях, а также в гнездах
голубей и пчелиных ульях (Krall, 1981); встречается она и в
природе (обычна на юге Центральной Европы, в Юл<ной Европе)
(Samsinak, Stys, 1978). Бывают иногда случаи массового
размножения этого вида в домах (Samsinak, 1981). Основной пищей
чешунницы служат сахар, крахмал и другие углеводы,
извлекаемые насекомьгм также из проклеенной бумаги пли грязных
тряпок. Развитие вида идет при температуре 15—32 °С (оптимум —
28 °С) и относительной влажности воздуха более 75% (Crome,
1955; Laibach, 1952; Weidner, 1969, 1983b) (табл. 2).
С вредителями материалов ситуация похожа на описанную
выше. О паразитах человека и комнатных животных накоплена
2. Структура города и фауна
Таблица 2. Зависимость между температурой и продолжительностью
эмбрионального развития сахарной чешуйницы (по Laibach, 1952)
Температура, °С
35
30
25
20
15
14
13
Эмбриональное развитие,
дни
—
29
28,5
41,3
89,2
117
131,5
Таблица 3. Распределение трех видов пироглифидных клещей
Dermatophagoides pteronyssinus, D. farinae и Euroglyphus maynei (no Keil
1983)
Вид
Всего
обнаружено
в
процентах
Общее число
в пересчете
на I г пыли
в процентах
Процент от
общей
популяции
клещей
D.
pteronyssinus
4760
80,3
61 737
86,6
72,8
D. farinae
1067
18,0
8 552
12,0
10,0
E. maynei
100
1,7
999
1,4
1,2
Сумма
5927
100
71 288
100
84,0
обширная специальная литература (эта тема нами подробно не
рассматривается). Мало внимаяия уделялось тому факту, что
здесь возможно формирование простых трофических цепей,
например клоп редувий ряженый [Reduvlus personatus) питается
постельным клопом (Cimex lectularius) (фото 15, 16).
Весьма типичными и имеющими особое значение для зоны
этажей являются клещи домашней пыли (Abbot et al., 1981;
Andersen, 1984, 1985; Bronswijk, 1972, 1973, 1978; Hallas, Korsgaard,
1983; Karg, 1973, Keil, 1983; Keil, Rack, 1985; Koekkoek,
Bronswijk, 1972; Lustgraaf, 1978; Mumcouglu, 1975; Samsinäk et al.,
1972, 1978, 1979, 1982; Sesay, Dobson, 1972; Spieksma, Spieksma-
Boezeman, 1967; Vobräzkovä et al., 1979; Wharton, 1971, 1976).
Сегодня считается доказанным, что различные виды клещей,
прежде всего пироглифиды (семейство Pyroglyphidae), являются
составной частью 'ингаляционных аллергенов домашней нылп.
Среди них Euroglyphus maynei, Dermatophagoides pteronyssinus
н D. farinae (табл. 3). Кайл (Keil, 1983) при исследовании проб
пыли в квартирах Гамбурга выявил не менее 35 видов .клещей из
22 семейств. Пироглифиды состав.ляли 84% сборов, п особо
часто встречались D. pteronyssinus (73%) и D. farinae (10%).
Клещи скапливаются в основном в постелях, где встречаются
в сотни раз чаще, чем в остальной домашней пыли (Samsinäk
et al., 1982). Следующим местом их сосредоточения является
мягкая мебель. Пироглифиды питаются фрагментами кожи и во-
20
2. Структура города и фауна
лос, которых каждый взрослый человек теряет за неделю около
5 г. Клещк этого семейства тесно связаны с птичьими гнездами
и комнатными птицами. Распростраияются они, активно
передвигаясь или пассивно с использованием прел<де всего птиц и
воздушных потоков, а также самим человеком (например, их
систематически находят на состриженных волосах в парикмахерских).
Сильная зависимость клещей от относительно высокой влажности
I
I
I
Si
180
160
ш
120
WO
80
60
40-
20
•
-
•
•
f
•
-
•
»
«
•
•
• •
•
—
• •
•
-
•
*
t
•
*
■
•
V
•
•
• ••
• •
t
" *• *•
■
»*
# •
"h'
:■!>.
fill
0 10 20 30 40 SO 60
Пыль, мг/ЮОсм^
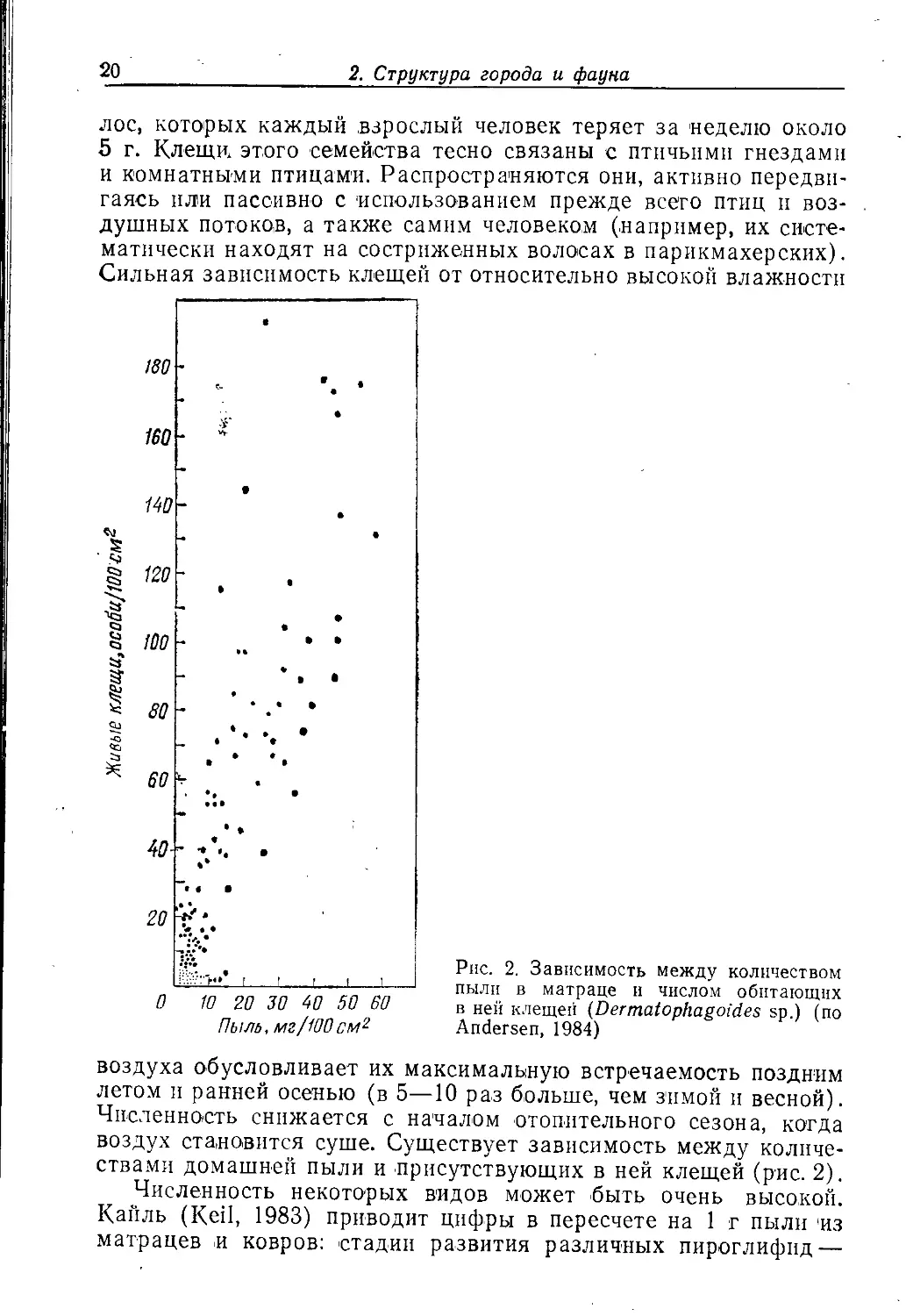
Рис. 2. Зависимость между количеством
пыли в матраце и числом обитающих
в ней клещей (Dermatophagoides sp.) (по
Andersen, 1984)
воздуха обусловливает их максимальную встречаемость поздним
летом и ранней осенью (в 5—10 раз больше, чем зимой и весной).
Численность снижается с началом отопительного сезона, когда
воздух становится суше. Существует зависимость между
количествами домашней пыли и присутствующих в ней клещей (рис. 2).
Численность некоторых видов может быть очень высокой.
Кайль (Keil, 1983) приводит цифры в пересчете на 1 г пыли из
матрацев и ковров: стадии развития различных пироглифид —
2. Структура города и фауна 21
35 823 (матрацы), 23 271 (ковры); взрослые особи D. pteronyssi-
nus соответственно — 5248 и 5313, D. farinae — 291 и 1 '70, Е. тау-
nei — 57 и 115.
Постоянными обитателями домов стали некоторые виды но-
гохвосток. Особенно часто встречается вид Seira domestica,
который можно было бы по праву считать «домашним»; он
является явным синантропом (родина — Средиземноморье). Часто в
домах наблюдается также Entomobrya marginata, что,
возможно, связано с наличием плесневых грибов. Известны случаи
массового размножения этого вида, в частности в зернохранилище
(Dunger, 1969).
На комнатных растениях поселяются различные насекомые,
в частности тли, белокрылки и кокциды; так, червец Pseudococ-
CUS adonidum, который сам в квартирах посещается муравьями,
обитает на различных кактусах. Поскольку в домах
предпочитают разводить вполне определенные виды растений, здесь
складывается и довольно устойчивое сообщество сосущих фитофагов.
Однако большинство видов комнатных растений практически
свободно от насекомых.
Даже небольшое количество почвы в цветочных горшках
достаточно для заселения некоторыми видами ногохвосток, а
также энхитреид (Moszynski, Urbanski, 1932). Среди ногохвосток
это, как правило, очень мелкие виды (0,8—2,0 мм), очевидно,
особенно хорошо приспособленные к такому специфическому
местообитанию. Одни из них встречаются регулярно (например,
Folsomia fimetaria, F. Candida, Neelus minimus, Sinella coeca,
Sminthurinus aureus, Sminthurides shoetti), другие от случая к
случаю.
Среди двукрылых ряд видов можно считать квартирными
синантропами (Tischler, 1950, Peters, 1959). Часть из них служит
пищей специфическим комнатным паукам (табл. 4).
Случайно проникающие через окна виды, конечно, не
относятся к типичной фауне этажей. Однако существует и более
регулярный приток фауны, меняющийся в зависимости от времени
года и суток. Комнатные пауки (фото 10), вероятно, частично
зависят от этого источника питания. Иногда проникновение
животных в дома особенно заметно. Так, сообщалось (Samsinak,
1981) о массовом появленш! в квартирах Allajulus londinensis.
Холодильники считаются непригодным для жизни местом,
однако черный садовый муравей (Lasius niger) (куколки и имаго —
дочерняя колония на верхнем крае двери) и рыжий таракан, или
прусак (Blattella germanica), встречаются в них все чаще
(Engelbrecht, 1981, Weidner, 1983с). При применении различных мер
борьбы с ними рыжий таракан укрывается прежде всего в
пустотах холодильников или на внутренней стороне их двери (Vater,
1979; Engelbrecht, 1981; Engelbrecht, Buske, 1983). Физиологиче-
и ^
в ч
л о
t- О
?J с
о л
Ja t- о
н ш
U о 01
о m 3
;5 д
as
03
0> о tH
" m -
m о .
с m
- л
ч m
ОЗ о
к к о,
я о а)
S Ч
X -
И я -
§ ч ч
с
л
а.
о
S
о
S
S
(U
СП
S
а.
и
о
CQ
га
3
S
о
я Ч - ш
э- о
СП
о S
' О- г-
pa ffl
о
g
S
S Ä
S
CD
CJ
О
С
SS
3
н
и
о
л
S-
CJ
О
S
к
о
Е
о
а.
К
X
О
£ 2
а о
а.
со
0)
к
с;
О.
X
£Х
X
а
>1
^1
S
С
О
о.
со
иг
(Г)
оа
и
га
о.
п
о
к
га
Н
и m к
Я _
я 3 S
я ^
от ^ S
W О) га
Р о U «
о
о
о я
CQ Я
ч
о
г- я
о; ^
53^
к ч
га
t4
3
к
к
си
си
га
си
к
га
СП
:а
ffl
та
о
и
Э
CQ
а
с
О к
о, <и
в
га"
S
та
со
н
о
Си СХ
3
СО
о
я
со О,
; о.
■ о
о
я
си
о
п
о
а
о
1£
g-3
о 5
Л
О. !-
5=5
О Я
я ч
(li
н
СО
о
!=:
С
С
Q,
о
О
о
f-
о.
а.
н
н
га
X
X
га
А
га
га
о
о
Ö
CJ
со
CQ
0^
^5
о
5 -J
Е
iO
со
- ^ ■—■
о
5 РЭ
•а
а:
! с 5
-rat;
;и 5
S j5 cl.'-
О =
,2 -J ■
с/) <УЗ
22
2. Структура города и фауна
23
скпе особенности этого теплолюбнвого вида субтропическо-тро-
пического происхождения легче понять, если учесть значительные
суточные колебания температуры в области его происхождения.
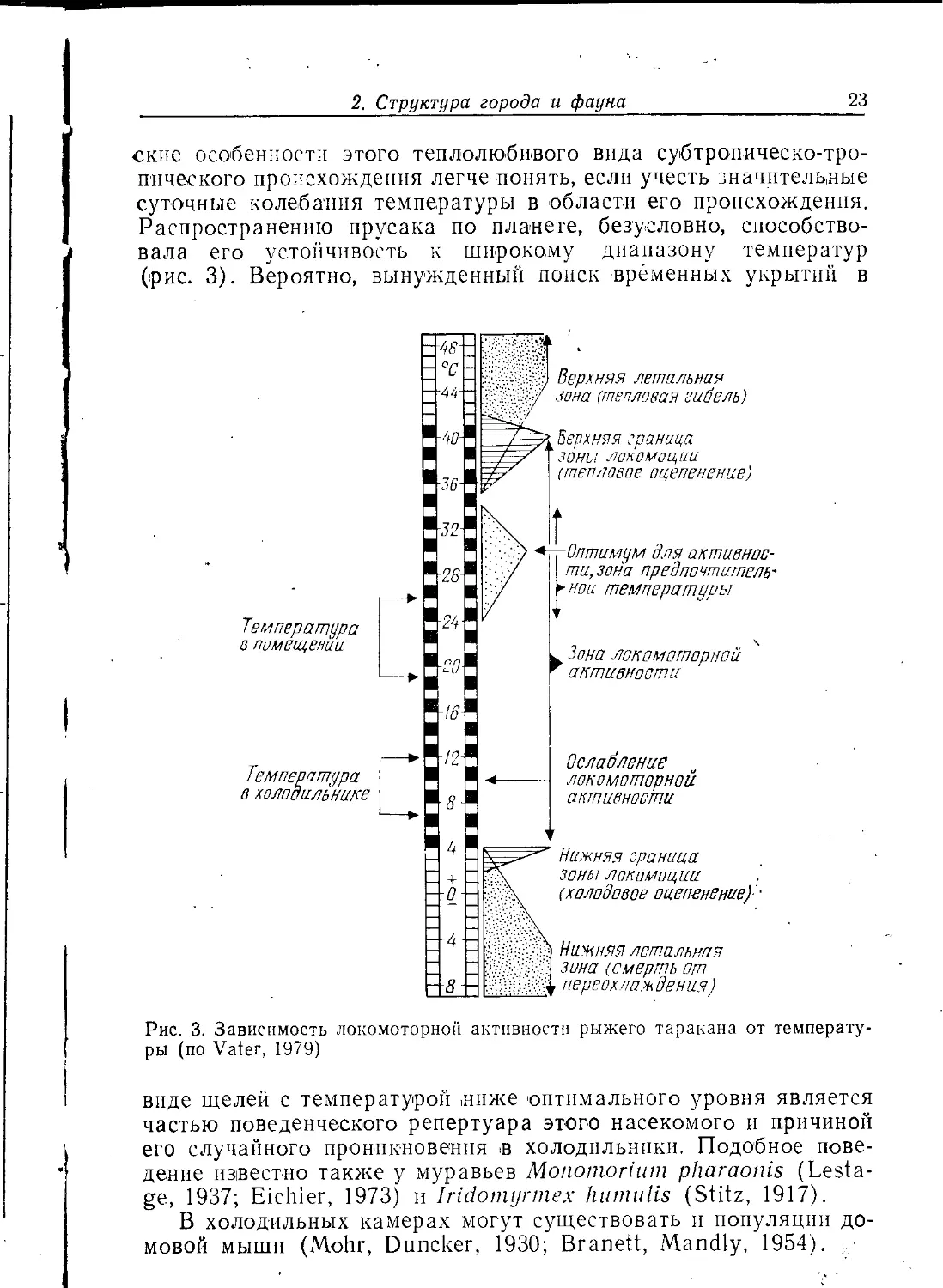
Распространению прусака по планете, безусловно,
способствовала его устойчивость к широкому диапазону температур
(рис. 3). Вероятно, вынужденный попек временных укрытий в
Температура
в помещении
Температура
в хояоаи/гьт.ке
I Верхняя метальная
jorn (тепловая гиаель)
Верхняя граница
зоны яакомащии
(тепловое ои,епенение)
Оптимум для
активности, за на преВпочтитель-
>-нои температуры
Зона локомоторной
активности
Ослабление
локомоторной
активности
Нижняя граница
зоны локомоции
(холодовое оцепенение) ■
Нижняя летальная
зона (смерть от
переохлаждения)
Рис. 3. Зависимость локомоторной активности рыжего таракана от
температуры (по Vater, 1979)
виде щелей с температурой ниже оптимального уровня является
частью поведенческого репертуара этого насекомого и причиной
его случайного проникновения в холодильники. Подобное
поведение известно также у муравьев Monomoriuin pharaonis
(Lestage, 1937; Eichler, 1973) и Iridomyrmex hiimulis (Stitz, 1917).
В холодильных камерах могут существовать и популяции
домовой мыши (Mohr, Duncker, 1930; Branett, Mandly, 1954).
24 2. Структура города и фауна
2.1.1.3. ПОДВАЛЫ
Присутствие специфических видов животных в подвалах
зависит прежде всего от абиотических факторов (темнота, высокая
относительная влажность воздуха, устойчивая низкая
температура), наличия пищи (хранящиеся продукты питания, дерево и.
плесневые грибы), структуры пространства и сравнительно
слабых .нарушений (Bender, 1941; Dahl, 1938; Hartmann, 1955;
Klausnitzer et al., 1984; Klippel, 1957; Kühnelt, 1955; Pax, Maschke,.
1935; Sacher, 1983; Stadler, 1962; Stellwag, 1924; Weidner, 1952).
Сюда идет постоянный приток фауны из соседних биотопов
(садов, дворов), в том числе вместе с заносимым людьми
материалом (картофелем, фруктами, деревом и т. п.). Собственно,
«подвальными» можно считать лишь некоторые из этих видов,
остальные отчасти являются их добычей пли используют подвалы для
зп.мовки.
Решающими факторами для поселения здесь пауков
являются температура и влажность воздуха (табл. 5). Большинство их
видов размножаются не в определенный сезон, а в течение всего
года (почти всегда можно обнаружить половозрелых осо'бей).
Однако у Amaurobius ferox размножение происходит главным
образом весной, у Pholcus phalangioldes максимальная
яйцекладка наблюдается в мае, а у Steatoda bipunctata, вероятно, два
основных копуляционных периода.
Из равноногих (мокриц) особенно часты в подвалах Oniscus
asellus и Porcellio scaber; они питаются в основном хранимыми
здесь овощами. Оба вида часто встречаются вместе, но первый,,
как правило, реже второго, поскольку имеет более узкий
диапазон устойчивости .к температуре и влажности. Однако при
наличии подходящих условий он мол^ет встречаться чаще Р. scaber.
Различные многоножки отмечаются более или менее
регулярно, например костянки Lithobius forficatus (заносится с
картофелем и овощами), L. melanops (таклче и в сухих помещениях),
кнвсяк пятнистый (Blaniulus guttulatus) и многосвяз Polydes-
mus inconstans (на запасах). С картофелем, овощами и досками
в подвалы попадают и другие кивсяки (luloidea), гломерисы
(Glomeridae) и многосвязы (Polydesmidae).
Одни виды ногохвосток попадаются в подвалах регулярно
(например, Hypogastrura assimilis), другие — временами.
Домовый сверчок {Acheta dotnesticus) живет преимущественно в
теплых, прежде всего в отапливаемых подвалах. Тля Rhopalosipho-
nius latysiphon питается в основном ростками картофеля и
способна к массовому размножению. Этот вид заселяет также
сельдерей, морковь и сладкий, или черный, корень {Scorzonera sp.).
В мае в колонии появляются крылатые формы, в остальное
время встречаются только бескрылые. Размножаются они исключи-
2. Структура города и фауна
25
Таблица 5. Пауки, обитающие в подвалах (по Sacher, 1983)
Вид
Аутэкология
Область происхождения,
прочие местообитания
Amaurobius ferox (Wal-
скепаег)
Pholcus phalangloides
(Fuesslin)
Physoeyclus simoni Ber-
land
Tegenaria dornestica
(Cierck)
T. ferruginea (Panzer)
Dysdera crocata
(C. L. Koch)
Meta menardi (Latreille)
M. merianae (Scopoli)
Lepthyphantes leprosus
(Ohlert) и L. nebulo-
sus (Sundevall)
Neriene montana (Cierck)
Steatoda bipunctaia (L.)
Nesticus celliilanus
(Cierck)
Liocranum rtipicola (Wal-
ckenaer)
Влажные до сырых
условия
Сухие до влажных
условия, теплые подвалы
с небольшими
температурными колебаниями
Тепло, сухо
Высокая влажность
воздуха, темнота,
прохлада (сходство с
пещерами)
Влажно
Влажно
Высокая влажность
Эврипотент, но не вла-
Г0-, а скорее
теплолюбивый
Высокая влажность
воздуха
Умеренная влажность
Под камнями
Космополит, другие
части зданий
Космополит; почва,
скотные дворы, сараи,
садовые домики, пещеры
Сараи, складские
помещения, амбары, кора,
скалы, деревья, пеще-
^ры
Почти космополит,
теплицы
Пещеры
Пещеры, норы
животных, каменная
облицовка берегов
Пещеры, скотные дворы,
прачечные
Теплицы, кора деревьев,
расселины скал
Все здание в целом,
кора деревьев,
расселины скал
Пещеры, влажные
природные
местообитания, на почве
Темные сухие комнаты,
под камнями, кора
деревьев
тельно партеногенетически. Этот вид был завезен в Европу из
Северной Америки.
Как примеры живущих в подвалах жуков можно привести
жужелиц Sphodrus lecoplithalmus и Pristonychus terricola, а
также чернотелок Blaps mortisaga (фото 13) (Klausnitzer, 1973),
В. mucronata и В. lethifera. Барндт (Barndt, 1981) считает
синантропами жужелиц Bembidion quinquestriatum (в земле, норах
мелких грызунов, подвалах, садах и на стенах домов) и Tachys
26
2. Структура города и фауна
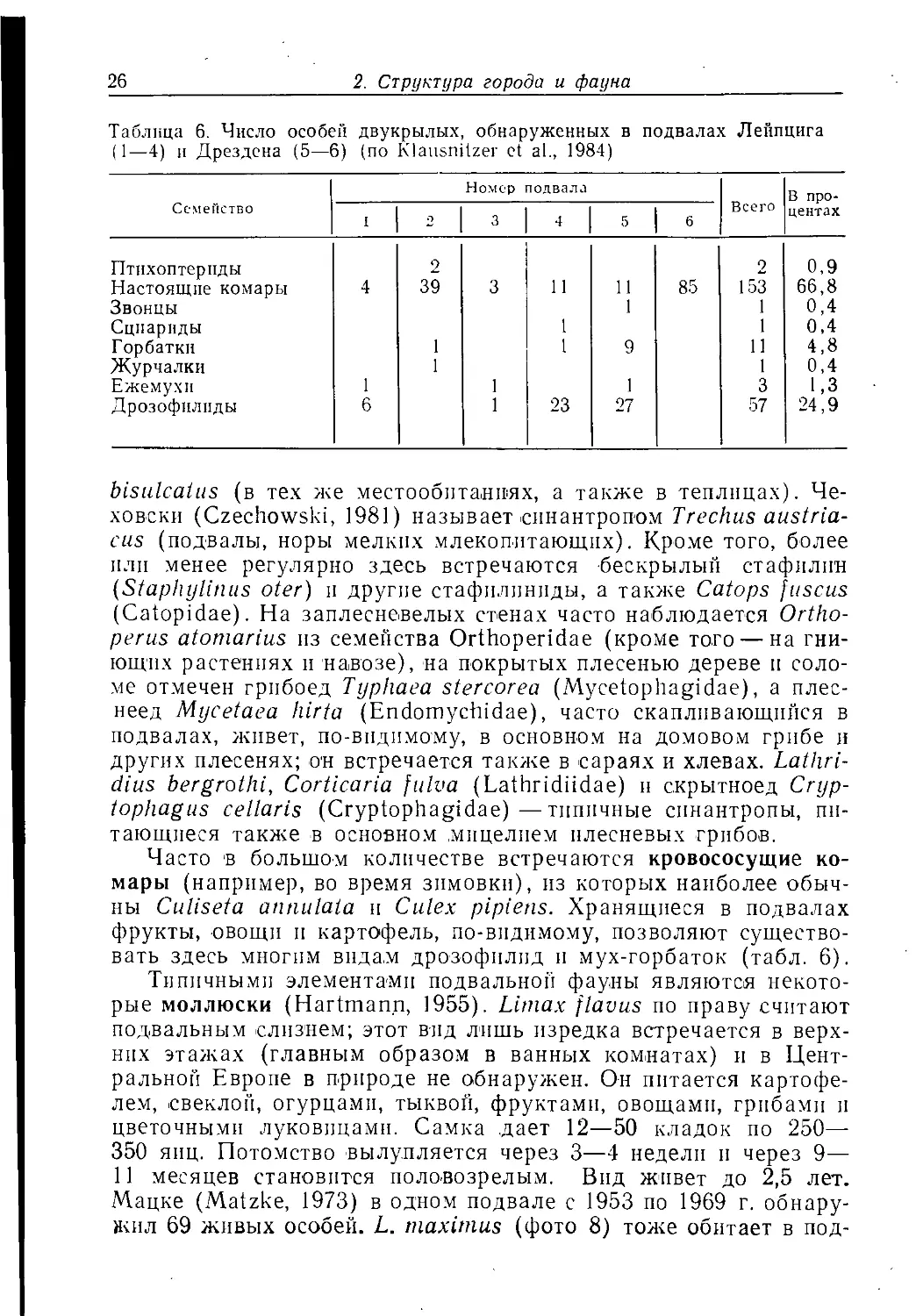
Таблица 6. Число особей двукрылых, обнаруженных в подвалах Лейпцига
(1—4) и Дрездена (5—6) (по Klausnilzer et al., 1984)
Семейство
Номер
тодвала
Всего
в
процентах
1
о
3
4
5
6
Птихоптериды
2
2
0,9
Настоящие комары
4
39
3
11
11
85
153
66,8
Звонцы
1
1
0,4
Сцпарпды
1
1
0,4
Горбатки
1
1
9
11
4,8
Журчалки
1
1
0,4
Ежемухп
1
1
1
3
1,3
Дрозофплпды
6
1
23
27
57
24,9
bisulcaius (в те.х же MecToooiiTaHiMX, а также в теплицах).
Чеховски (Czechowski, 1981) называет синантропом Trechus austria-
cus (подвалы, норы мелких млекопитающих). Кро.ме того, более
или менее регулярно здесь встречаются бескрылый стафилин
(Staphylinus oter) и другпе стафилнниды, а также Catops fiiscus
(Catopidae). На заплесневелых стенах часто наблюдается Ortho-
perus atomarius из семейства Orthoperidae (кроме того — на
гниющих растениях и на-возе), на покрытых плесенью дереве и
соломе от.мечен грибоед Typhaea stercorea (Mycetophagidae), а плес-
неед Mycetaea hirta (Endomychidae), часто скапливающийся в
подвалах, живет, по-види.мому, в основно.м на домовом грпбе и
других плесенях; он встречается также в сараях и хлевах. Lathri-
dius bergrothi, Corticaria fulva (Lathridiidae) и скрытноед Cryp-
tophagus cellaris (Cryptophagidae) — типичные синантропы,
питающиеся также в ochosho.m мицелием плесневых грибов.
Часто в большом количестве встречаются кровососущие
комары (например, во время зимовки), из которых наиболее
обычны Culiseta antiulaia и Culex pipiens. Хранящиеся в подвалах
фрукты, овощи и картофель, по-видимому, позволяют
существовать здесь многим видам дрозофилнд и мух-горбаток (табл. 6).
Типичными элементами подвальной фауны являются
некоторые моллюски (Hartman.n, 1955). Llmax flavus по праву считают
подвальным слизне.м; этот вид лишь изредка встречается в
верхних этажах (главным образом в ванных комнатах) и в
Центральной Европе в природе не обнаружен. Он питается
картофелем, свеклой, огурцами, тыквой, фруктами, овощами, грибами и
цветочными луковицами. Самка дает 12—50 кладок по 250—
350 яиц. Потомство вылупляется через 3—4 недели и через 9—
11 месяцев становится половозрелым. Вид живет до 2,5 лет.
Мацке (Matzke, 1973) в одном подвале с 1953 но 1969 г. обнару-
жпл 69 живых особей. L. maxirnus (фото 8) тоже обитает в под-
2. Структура города и фауна 27
валах, а кроме того, в теплицах п в природе. Этот слизень
питается фруктами, тыквой, капустой, .морковью, брюквой и
картофелем. Самка дает 13—370 кладок по 680—830 яиц. Мацке
(Matzke, 1979) обнаружил в 'подвале с 1955 по 1971 г. 81 живую
особь. Oxychilus cellarius регулярно встречается в подвалах, а
также в норах н в других природных местообитаниях.
Среди млекопитающих постоянные обитатели подвалов —
серая крыса и домовая мышь.
2.1.2. Постоянно отапливаемые здания
Большие города характеризуются прежде всего крупными
комплексами постоянно отапливаемых зданий, к которым
относятся, например, новые жилые районы, отели и больницы. Хотя
их температура иногда сильно различается, все же в помещении
она на протяжении всего года остается довольно высокой,
колебания ее незначительны, экстремальные значения отсутствуют,
а относительная влажность .воздуха в большинстве случаев явно
понижена. Часто в таких зданп1ях нет «настоящих» чердаков, а
подвалы обычно теплые и сухие. Поэтому мы рассмотрим здесь
только зону «этажей», фауна которой характеризуется видами,
как правило, отсутствующими в домах с непостоянным и печным
отоплением.
Некоторые животные встречакЬ^ся только в постоянно
отапливаемых зданиях, например-фараонов муравей (Monornoriuni plia-
raotüs) и термит Reticulifermes jlavipes (Becker, 1970; Weidner,
1978). О фараоновом муравье 'накоплены довольно подробные
данные (Eichler, 1962, 1962, 1962, 1962, 1973; Lestage, 1937, Britz,
1957). В связи с тем что те.мперату^Е(1и оптимум вида равен
32 °С, постоянно отапливаемые зданпя TjfiTä больниц,
хлебопекарен, ресторанов, отелей, закрытых бассейнов и оранжерей
пригодны для образования им гнезд, которые устраиваются в стенах
вблизи источников тепла. Крупные колонии могут насчитывать
до миллиона особей. Муравей тропического (восточного?)
происхождения Tapinoma melanocephalum был обнаружен в
теплицах, а в последнее время и в квартирах с центральным
отоплением (Scheurer, 1984).
Переоборудование непостоянно отапливаемых зданий в
постоянно отапливаемые влечет за собой глубокие изменения в
составе фауны. Например, развитие точильщика АпоЫит рипс-
tatum приостанавливается при включении центрального
отопления (Samsinäk, 1981). Постоянно отапливаемые здания
благоприятны для поселения ц непрерывной смены поколений
некоторых жуков-кожеедов, например Dermestes haemorrhoidales, Tro-
goderma angustum, Thylodrlas contractus, Attagenus fasciatus и
An ihren us scrophulariae.
28
2. Структура города а фауна
Таблица 7. Продолжительность развития платяной моли в зависимости
от температуры (по Titschack, 1925)
Температура, 'С
Продолжительность развития, дни
Самцы
Самки
15
187
196
20
123
135
25
72
89
30
62
73
Изменение численного соотношения в городах между черным
{Blatta orientalis) и рыл<им {Blattella germanica) тараканами
также объясняется сильным увеличением доли постоянно
отапливаемых зданий (Weidner, 1975): более теплый и сухой климат
в помещениях благоприятнее для второго вида, а кроме того,
его размножению способствуют общественные кухни.
Развитию платяной моли {Tineola bisselliella) также
благоприятствуют тепло 1и сухость; это насекомое стало домашним
после того, как было завезено в Европу предположительно из
Африки. В постоянно отапливаемых зданиях оно может давать
3—4 поколения в год, а в других условиях только 1—2 (табл. 7).
Наблюдающаяся с некоторого времени тенденция к снижению
численности этого вида объясняется прежде всего ростом
применения синтетических и специально обработанных текстильных
материалов, т. е. сокращением трофической базы.
2.1.3. Складские и некоторые производственные
помещения
Складские и производственные помещения более или менее
сильно различаются как по важнейшим абиотическим факторам,
так и по наличию пищи (свободных ниш), а соответственно и по
населяющей их фауне (рис. 4), которая отчасти сходна с фауной
жилых до.мов.
Некоторые типы зданий трудно поддаются классификации
(например, сараи и прачечные), однако могут служить
местообитанием для вполне определенных видов; это показал Захер
(Sacher, 1983) на примере пауков. В сараях встречаются Pholcus
opilionoides, Sosticus loricatus, Tegenaria domestica и Т. ferrugi-
nea. Первый упомянутый вид нуждается в относительно высокой
влажности воздуха. Dolichovespula saxonica из настоящих ос
также предпочитает строить гнезда в сараях. Прачечные часто
заселяются пауками Ph. opilionoides, Achaearanea tepidariorum,
Lepthyphantes leprosus и L. nebulosus.
2. Структура города и фауна
29
Мельницы, хранилища муки и зерна
Цахер (Zacher, 1938, 1939), .обследовав членистоногих на
мельницах и зернохранилищах в Гамбурге, обнаружил 407
видов (156 видов жуков, 83 — двукрылых,
51—перепончатокрылых, 36 — пауков, 26 — бабочек и др.) - Этот тип строений,
по-видимому, из всех описанных здесь, обладает самой специфичной
фауной, включающей высокую долю космополитов п постоянно
обогащающейся заносными видами. Некоторые виды насек01мых
из родов Sitophilus, Trogoderma, Ephestia и Sitotroga, а также
ряд их паразитов в природе больше не встречаются. Как мука,
так п обработанное зерно разных культур — местообитания во
многом самобытной фауны, которая наряду с паразитами может
Рнс. 4. Доля различных местообитаний
)ыжего и черного тараканов в округе
Лотсдам (по Engelbrecht, Buske, 1983):
1 — места общественного питания,
общественные, производственные и
административные помещения; 2 — торговые
предприятия; 3 — рестораны, отели, клубы;
4 — предприятия хлебопечения и других
отраслей пищевой промышленности; 5 —
жилища; 6 — детские учреждения; 7 —
учреждения здравоо.хранения;
8—прочие объекты
включать некоторых вторичных консументов, например жуков-
карапузиков (Histeridae) и стафилинид, питающихся
личинками двукрылых. Мельничные отходы заселяются
жуками-чернотелками и кожеедами. Кроме того, на мельницах и в амбарах
присутствуют микробпотоиы для лчпвотных, у которых зерновые
продукты не являются основной пищей.
Пекарни
Типичными животными пекарен уже в течение столетий
считаются рыжий и черный тараканы н домовый сверчок, а в
последнее время к ним прибавились фараонов муравей,
мельничная огневка, комплекс амбарных клещей {Acarus siro),
различные чернотелки н некоторые щетинохвостки. Для
существования этих видов наряду с пищей необходима прежде всего
высокая температура. Происходящая, по-видимому, пз
Средиземноморья щетинохвостка термобия домашняя (Thermobia do-
mestica) обитает почти исключительно в пекарнях, где питается
30
2. Структура города и фауна
мукой и мучными изделиями. Этот вид еще более теплолюбив,
чем са.харная чешуйнпца, и нуждается в температуре выше 24 °С
(Boettger, 1954; Sährhage, 1950, 1953, 1954; Kühnelt, 1970; Simon,
1957, 1958; Tischler, 1980b; Weidner, 1952, 1958).
Мясокомбинаты
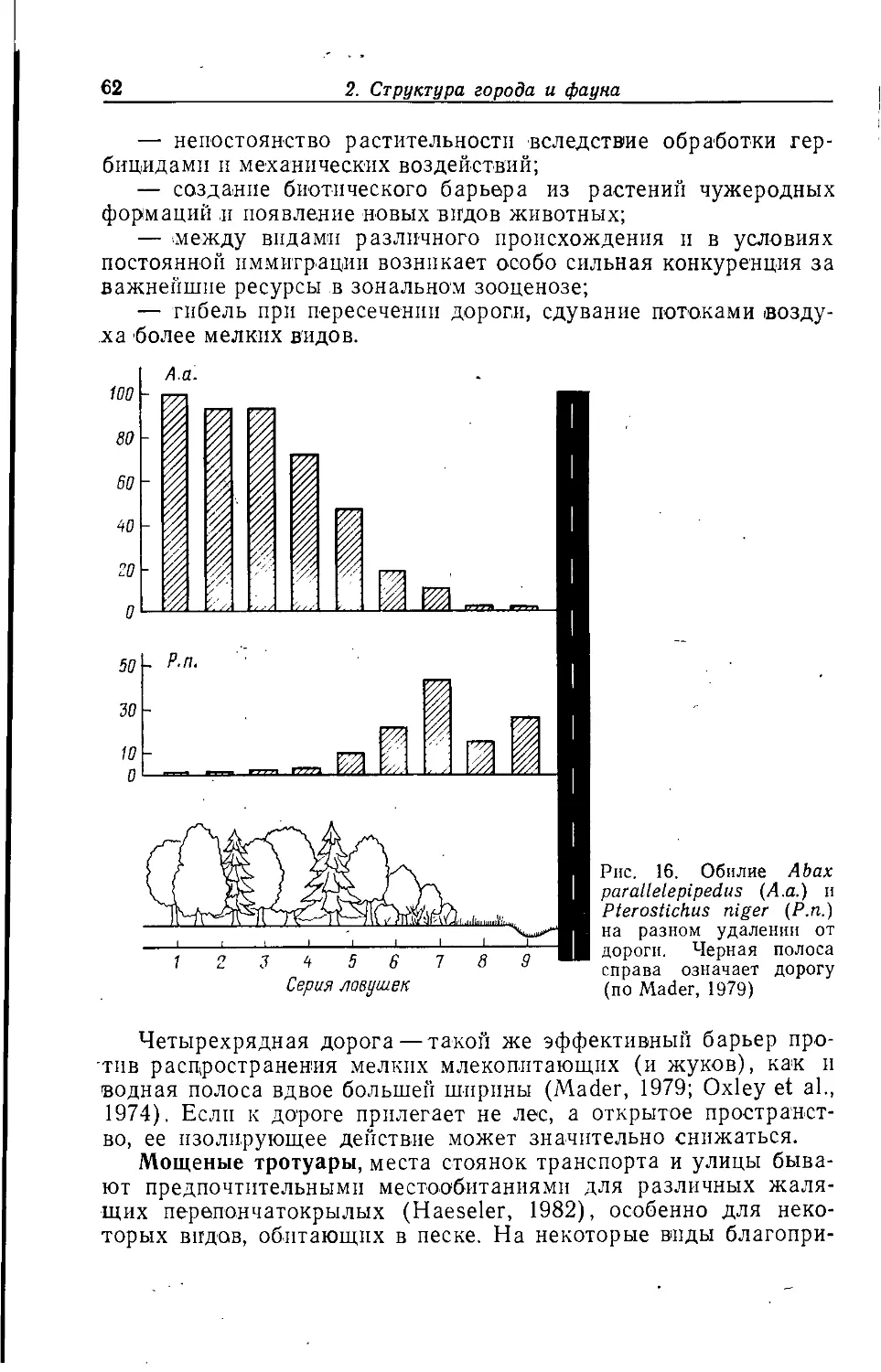
Ha мясоперерабатывающих предприятиях имеются прежде
всего два типа лищи: готовые продукты и отходы переработки.
Некоторые виды падальных и серых мясных мух (Calliphoridae
п Sarcophagidae) заселяют свежее мясо и поэтому постоянно
встречаются на скотобойнях (Green, 1951) и пищевых продуктах
220 у ^
200 Y
ту
I
I т
% 100
60
40
20
1
3 4 5 6
Рис. 5. Среднее количество яиц,
откладываемых самками комнатной
мухи при кормлении их разными
мясными продуктами (по Munzel, 1981):
1—солонина; 2 — грудинка; 3 —
вареная телячья колбаса; 4 — ветчина;
5 — говяжья колбаса; 6 — колбаски к
пиву; черный цвет — число
отложенных яиц; белый цвет — число яиц,
обнаруженных в яичниках
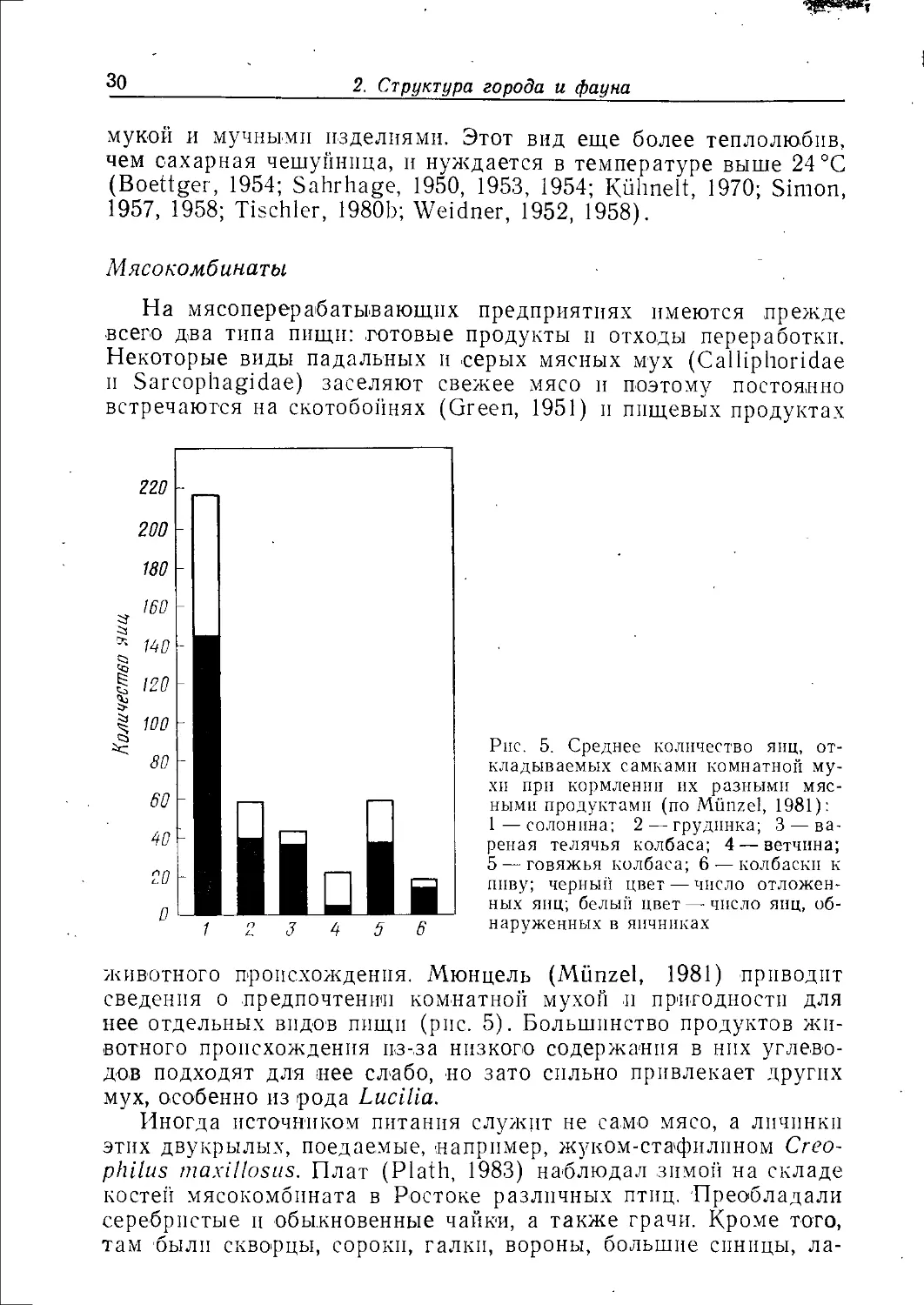
животного происхождения. Мюнцель (Munzel, 1981) приводит
сведения о предпочтении комнатной мухой и пригодности для
нее отдельных видов пищи (рис. 5). Большинство продуктов
животного происхождения из-за низкого содержания в нпх
углеводов подходят для нее слабо, но зато сильно привлекает других
мух, особенно из рода Lucllia.
Иногда источником питания служит не само мясо, а личинки
этих двукрылых, поедаемые, например, жуком-стафилпном Сгео-
philus maxillosus. Плат (Plath, 1983) наблюдал зимой на складе
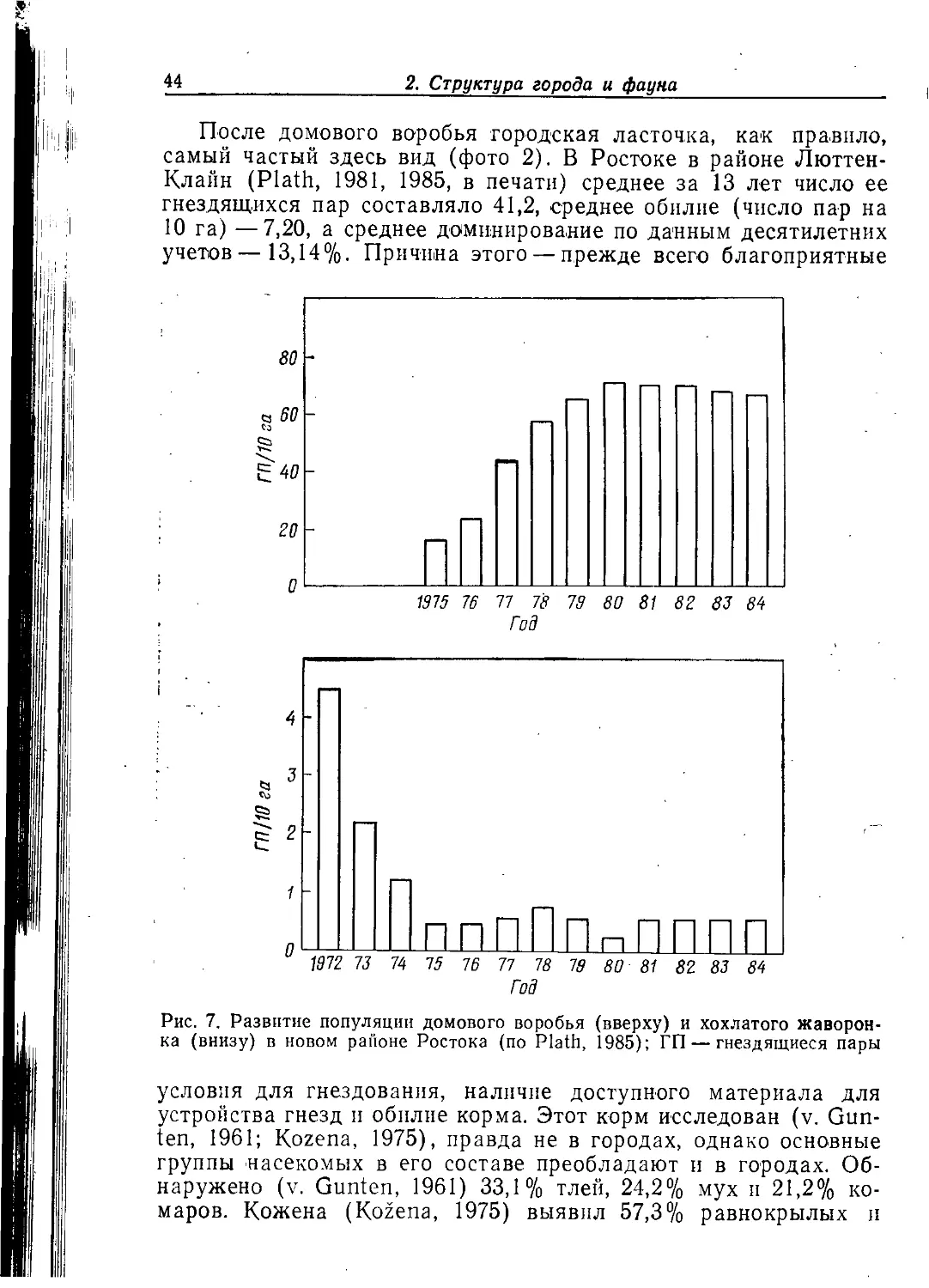
костей мясокомбината в Ростоке различных птиц. Преобладали
серебристые и обыкновенные чайки, а также грачи. Кроме того,
там были скворцы, сороки, галки, вороны, большие синицы, ла-
2. Структура города и фауна 31
Естественнонаучные коллекции и книгохранилища
В естественнонаучных коллекциях и библиотеках со старыми
книгами могут развиваться плесневые грибы. Они являются
пищевым субстратом прежде всего для некоторых сеноедов (см.
2.2.2.3) и ногохвостки Seira domestica. Эти насекомые, прежде
всего сеноеды, поедаются другими членистоногими, например
ложноскорпиопом Clielifer cancroides и пауком Oonops domesti-
cus. Последний вид длиной 1,5—2 мм, очевидно, живет главным
образом а этом местообитании.
Гербарии заселяются примерно таким же набором видов, что
и лекарственное сырье. Зоологические коллекции
(чучел,насекомых) — местообитания прежде всего кожеедов (Hämäiäinen,
Mannerkoski, 1984 и др.).
зоревки, черные дрозды и канюки обыкновенные, а также
единичные домовые и полевые воробьи, малиновки и лесные
завирушки.
Фрукты и овощи
С фруктами и овощами (в широком смысле) в основном в
начале XX в. завезены некоторые виды животных, среди которых
лишь малая часть смогла прижиться на более или менее
длительное время. Временно поселившимися видами являются,
например, уховертка ЕиЬогеШа peregrina, завезенная с
бразильским орехом из Бразилии, уховертка Marava arachidis, попавшая
в Европу с арахисом, копрой, каменным орехом, семенами
масличных (этот вид яйцеживородящий пли производит живых
личинок), жужелица Somotrichus unifasciatus, завезенная с
бразильским орехом из Бразилии (хищнпк вредителей запасов).
С бананами из Южной Америки завезли тараканов Panchlora
exoleta, Leucophaea maderae и Perlplaneta ausralastae (этот
вид — также и с другими фруктами), из которых прижился
последний.
При обследовании мух на овощных рынках Будапешта
обнаружены (Michalyi, 1966; Aradi, Michalyi, 1971) характерные
сочетания видов. Эвдоминантами этих местообптаний были
различные виды дрозофил.
Лекарственное сырье
Различное лекарственное сырье — под.ходящий пищевой
субстрат прежде всего для некоторых видов насекомых (Weidner,
1963, 1973).
-3^ 2. Структура города и фауна
Кожа, шкуры
На складах кож и шкур обитают главным образом некоторые
виды жуков-кожеедов.
Текстиль и мягкая мебель
Текстиль и мягкая мебель заселяются молями (Tineidae),
жуком Niptus hololeucus и кожеедами. Мебель иногда поражается
плесневыми грибам'и, на которых могут существовать некоторые
сеноеды, например Lepinotus patruelis.
Винные погреба
Винные погреба во многом отличаются от обычных подвалов
(зачастую в них еще более постоянная температура, более
высокая влажность воздуха и специфические пищевые субстраты,
в частности дерево, обросшее водорослями и грибами). Поэтому
здесь существуют особые виды (например, бабочка Dryadaula
pactolia) и виды с более широкой нишей типа бабочки Oenophila
v-flavum и других представителей семейств Tineidae, Oenophili-
dae и Geleciiidae, гусеницы которых питаются водорослями и
грибами на бочках, потолках и стенах (Bender, 1941; Stellwag,
1924). В основном в винных погребах встречается жук Orthope-
rus atomarius (Orthoperidae), поедающий плесень; здесь
отмечены различные виды дрозофил и грибной комар Масгосега sp.
2.2. Прочие наземные местообитания
Под этим названием мы объединяем все наземные
местообитания, расположенные снаружи помещений; к ним относятся так
называемая «внешняя оболочка» зданий и различные неозеле-
ненные и озелененные городские территории.
2.2.1. Внешняя оболочка зданий
2.2.1.1. ВНЕШНИЕ СТЕНЫ ДОМОВ
Озелененные стены домов имеют своеобразную фауну,
заметно отличающуюся от фауны голых стен. Особенность ее
заключается прежде всего в структуре пространства, создаваемой
вьющимися растениями. Например, среди этой растительности
(девичий виноград, виноград, кирказон крупнолистный, плющ)
обитает паук Nigma walclienaeri, растягивающий свои сети на
пластинках листьев. Подобным же образом ведет себя паук Dic-
tyna cirica, который встречается в основном спорадически, но
2. Структура города и фауна 33
иногда в громадном количестве (см. Billaudelle, 1957, Hertel,
1968).
Земан (Saemann, 1970) наблюдал гнездящегося снегиря на
зданиях с девичьим виноградом и плющом, а Фелленберг
(Fellenberg, 1966, 1968) — серую му.холовку на девичьем винограде.
Отмечались гнездования садовой горихвостки на плюще (Gniel-
ка et al., 1983), там же— зеленушка, а также колонии домового
воробья на плюще и винограде.
Неозелененные стены зданий заселяются эвсинантропными
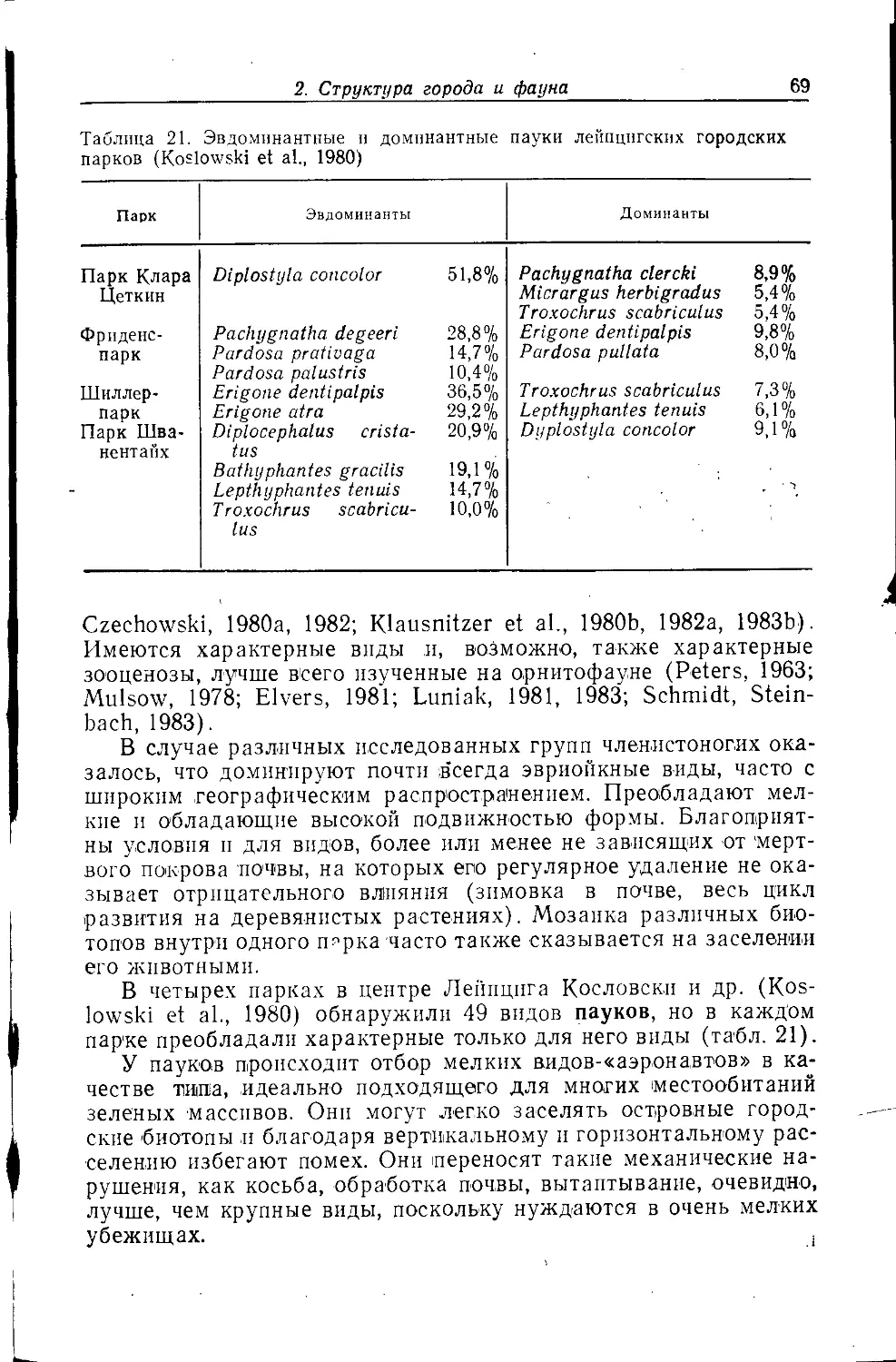
пауками Cyphereira ixobola и Zygiella x-notata (Sacher, 1983).
Гемисинантроиньши видами этого местообитания являются
Amaurobius similis, Наг pact еа rubicunda и Text rix denticulata
(этот вид также в зданиях, отверстиях и трещинах каменных
стен) ,Araneus diadematus, Zygiella atrica и Theridion melanurum.
Солнечные стены -и каменные ограды заселяются также геми-
синантропным пауком-скакуном Salticus scenicus, который
встречается преимущественно летом, иногда в больших количествах,
и в это же время достигает половой зрелости. Этот вид живет
также на скалах и каменистых откосах. На стенах встречается
и созревающий летом синантропный вид Sitticus pubescens,
обитающий, кроме того, в каменоломнях и на стволах деревьев.
Далее следует указать некоторые виды сенокосцев (см. 3.4). К
обитателям более или менее каменистых мест относятся сеноеды
Epipsocus lucifugus, Pseudopsocus meridionalis, Blaste quadrima-
culata, Neopsocus rhenanus, Psocus bipunctatus и Trichadenotec-
tum germanicum.
В заполненных раствором швах более старых домов
поселяется пчела-коллетес Colletes daviesanus. Шелоске (Scheloske,
1973) считает этот вид вредителем зданий и сообщает о его
колониях в стенах из песчаника; особое значение при этом имеют
их зернистость и твердость (см. Leclercq, 1964; Rode, 1962;
Scheloske, 1973; Mader, 1980, 1981, 1981, 1982). Первоначально этот
вид, как полагают Шелоске (1973) н Тишлер (Tischler, 1951),
жил в твердых отвесных стенках долин древних рек и
прибрежных обрывов, поэтому был иреадаптирован к поселению на
зданиях. Другие городские перепончатокрылые, возможно, также
происходят из таких природных биотопов (например, Hylaeus
pictipes, Lasioglossum morio, Diodontus trist is, Trypoxylon figulus,
T. clavicerum). Crossocerus elongatulus, если верить Леклерку
(Leclercq, 1964), обитает даже в бетонных стенах. Челюстями
эти виды выбирают выветрелый раствор; гнездовые ходы
приспособлены к структуре швов кирпичной стены. Colletes daviesanus
селится также в личиночных ходах домового дровосека {Hylo-
trupes bajulus) (Weidner, 1952).
Согласно Вайднеру (Weidner, 1952), в таком же
местообитании живут эвменовые осы Ancistrocerus parietinus и А. trlfascla-
3-21
34 2. Структура города и фауна
tus (Eumenidae). Оба вида вне города устрапвают гнезда в
полых стеблях растений и в ходах, прогрызенных в дереве, так
что ходы домового дровосека создают благоприятные условия
для их развития. Под.ходящие каменные деревянные и глиняные
стены заселяются пчелой-осмией Osmia rufa и различны.ми
видами пчел рода Hylaeus, которые наряду с пыльцой других цветов
собирают ее с очитков, особенно очитка едкого, часто
встречающегося в городах (цветочные ящики, трещины каменных стен,
крыши). Gsmia rufa, очевидно, весьма пластична в выборе мест
гнездования и кроме ходов домового дровосека заселяет и
другие трубчатые структуры. Виды рода Hylaeus гнездятся в
аналогичных местообитаниях, а также в глиняных стенах. В Киле
(ФРГ) этот вид строил свои гнезда даже на пятом этаже дома
(Haeseler, 1972). В цементных швах, трещинах стен и в старых
оконных рамах гнездятся Osmia caerulescens, Hylaeus communis
и Н. hyalinatus, причем последние виды на высоте до 16 м
(Haeseler, 1972). В городе особенно благоприятны условия, вероятно,
для пчелы-антофоры Anthophora acervorum и ее паразита Melec-
ta punctata, заселяющих старые гл1шяные стены и швы старых
домов. Chelostoma florisomne и Ch. distinctum размнол<аются в
старых заборах и в стенах сараев (прежде всего в отверстиях,
проделанных жуками). В трещинах каменных стен селятся
также роющие осы родов Liodontus и Crossocerus. Теплолюбивый
кожеед Trogoderma glabrum нуждается, по-видимому, в
дополнительном тепле солнечных стен домов, где он развивается в
гнездах одиночных пчел. Сходным образом л<нвет и кол-сеед Ме-
gatoma undata в гнездах Osmia rufa.
2.2.1.2. КРЫШИ
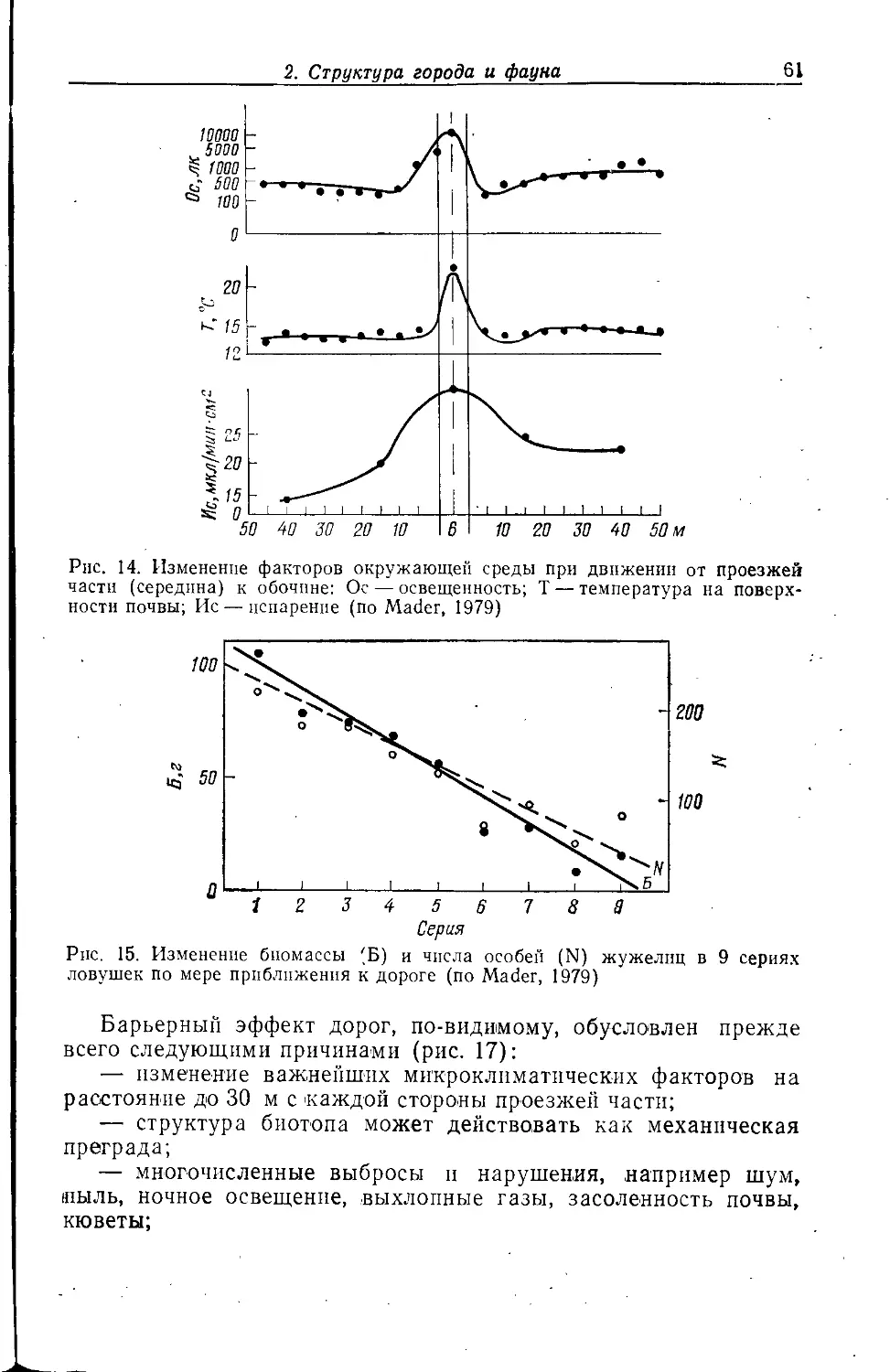
Указывалось (Bornkamm, 1961), что гравийно-битумные
кровли имеют свойственное только и.л{ растительное сообщество, так
что на них следует олчидать присутствия н особого ценоза
членистоногих.
Обследование гравийно-битумной плоской кровли в
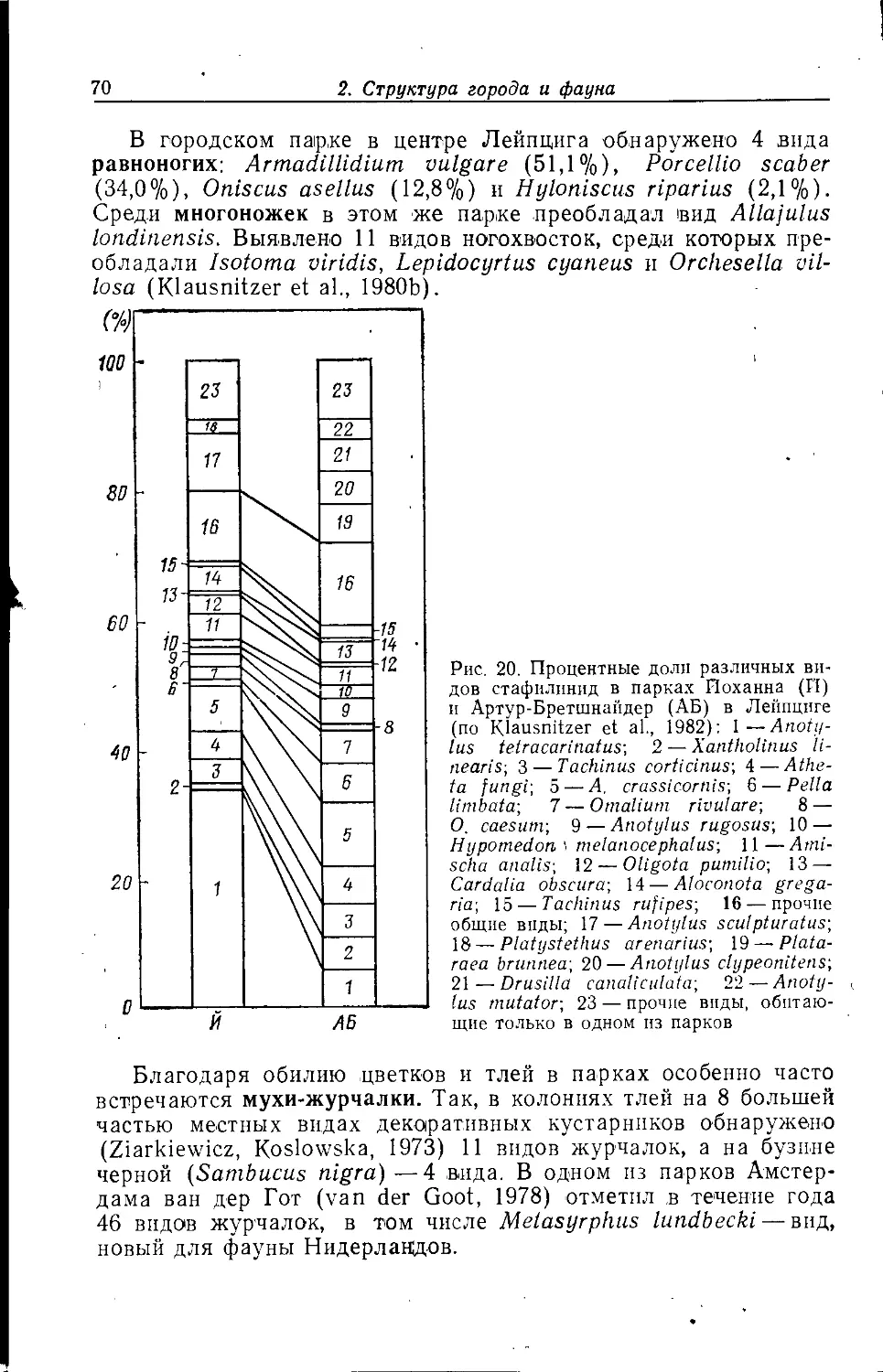
Лейпциге (Klausnitzer et al., 1980), заросшей в основном Sedum acre,
Ceratodon purpureus, Bryum caespiiicium и В. argenteum,
показало удивительно богатую фауну (рис. 6). Примечательно наличие
60 видов лчуков, в том числе 31 вида стафплинпд и 6 видов
жужелиц. Фаунистической особенностью были такие виды, как
Atheta aegra и Chilomorpha longitarsis. Среди л^уков
преобладали Simplocaria semistriata (27,2%, питается, в том числе и
личинки, мхом), Bembidion quadrimaculatum (21,8%, найдены так-
л<е личинки), Helophorus nubilis (5,8%, вероятно, поселению
благоприятен мох) и Xaniliolinus linearis (5,1%, обнаружены
также личинки). Наряду с жуками указаны фитофаги из цикадо-
2. Структура города и фауна
35
вых и клопов, причем иногда в больших количествах, гусеницы
бабочек-совок Triphaena pronuba ц Agrotis exclamationis, три
вида ногохвосток {Bourlettiella hortensis, Orcliesella cincta, En-
tomobrija multifasciata), более менее регулярно встречающихся
на отвалах и других находящихся под сильным антропогенным
влиянием в основном ксеротермных^ местообитаниях.
Среди пауков преобладали Erigone dentipalpis (40,7%),
Е. atra (23,8%) и Meioneta rurestris (22,6%). Большая часть
пауков на обследованной крыше относилась к ксенантропным
«аэронавтам».
Рис. 6. Доминирующие
обитатели битумно-гра-
вийной кровли в
Лейпциге (по Klausnitzer et al.,
1980)
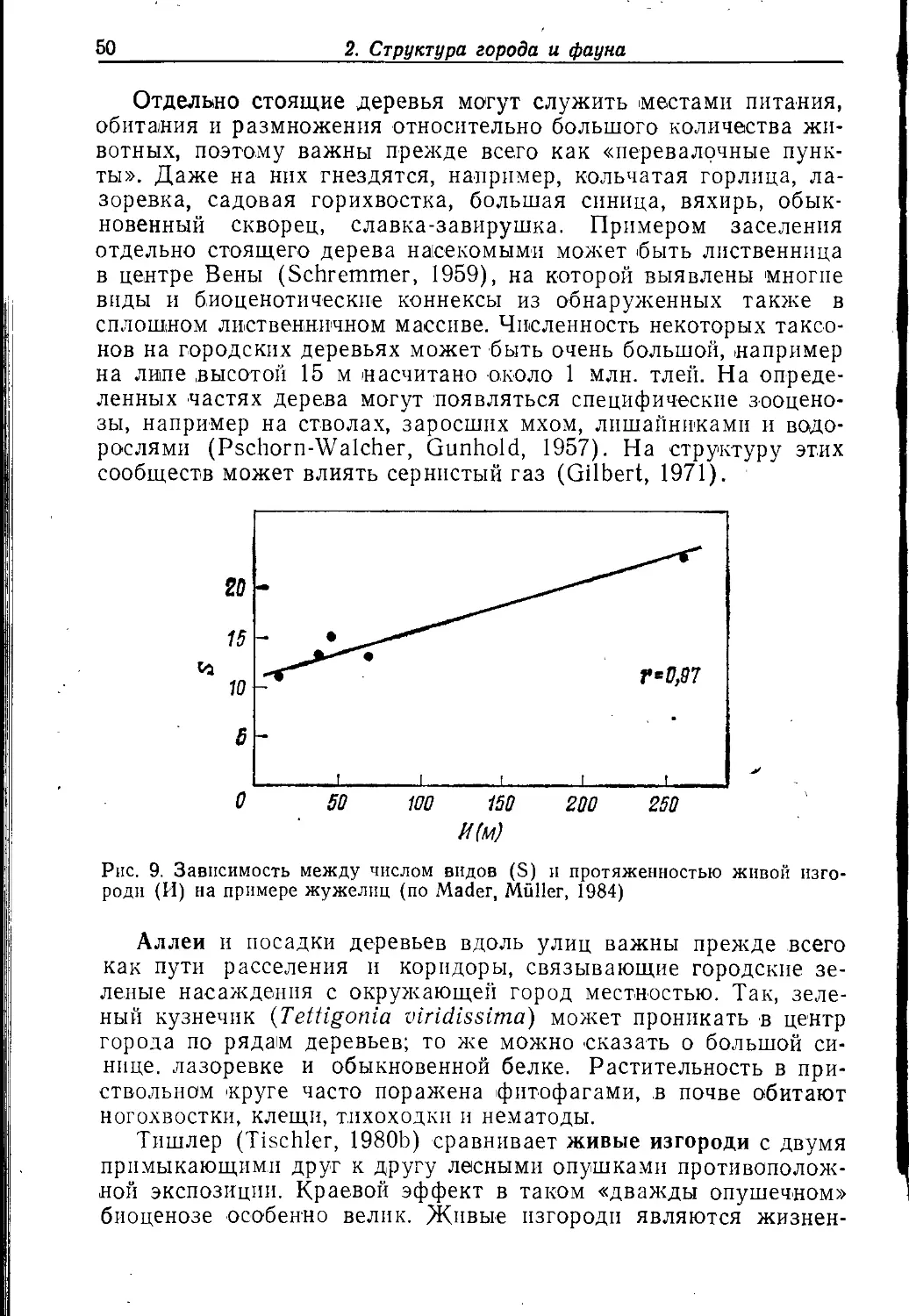
Под выступающими частями крыш находили гнезда ос Polis-
tes gallicus, P. nimpha и Dolichovespula saxonica. Ha плоских
крышах иногда гнездится хохлатый жаворонок.
В желобах крыш, вероятно, также присутствует собственный
ценоз, в котором изучены прежде всего тихоходки и коловратки.
2.2.1.3. БАЛКОНЫ
Балконы вследствие наличия на них различных видов
растений и определенных пространственных структур могут быть
местами концентрации некоторых видов животных. Здесь
гнездятся некоторые птицы: домовый воробей (Legen, 1972), зеленушка
(Grimm, Teis, 1972), деревенская ласточка и черный дрозд (Gni-
elka et al., 1983). Городская ласточка часто гнездится иод
балконом (см. 2.2.2.3). Наблюдалось, что балконные растения (на-
^ 2. Структура города и фауна
пример, очиток Sedum spectabile) служат кормом для
перепончатокрылых, селящихся в стенах домов (пчелы Hylaeus
communis, Н. hyalinatus) (Haeseler, 1972). Петере (Peters, 1971, 1973)
указывает, что цветочные ящики из пластмассы (полнстиролово-
го пенопласта) могут быть подходящим местом для размножения
роющих ос и других насекомых (ср. Becker, 1962; Kühne, 1969).
Было обнаружено прежде всего средне-южноевропенское
перепончатокрылое Ectemnius sexcinctus, широко
распространившееся и в ГДР (Oehlke, 1970); первоначально этот вид гнездился в
гнилом дереве (а таклче в стропилах, балках, оконных и дверных
рамах) (Peters, 1971), но полистироловый пенопласт, по-види-
мо.му, прекрасно заменяет ему дерево, поскольку цветочные
ящики заселяются не только единичными особями, но иногда и очень
большим их количеством. В систему ходов проникают на
зимовку уховертки, божьи коровки, хлебный трипе {Limothrips сегеа-
Иит), а также перепончатокрылое Pemphredon montanus.
Большеголовая оса {Crabro vagus) гнездится в косяках, перилах и
цветочных ящиках. Ее ходы длиной 4—5 см проходят
параллельно верхней поверхности доски. Обыкновенная уховертка (Forti-
cula auricularia) часто встречается на балконах под цветочными
горшками и в цветочных ящиках.
/
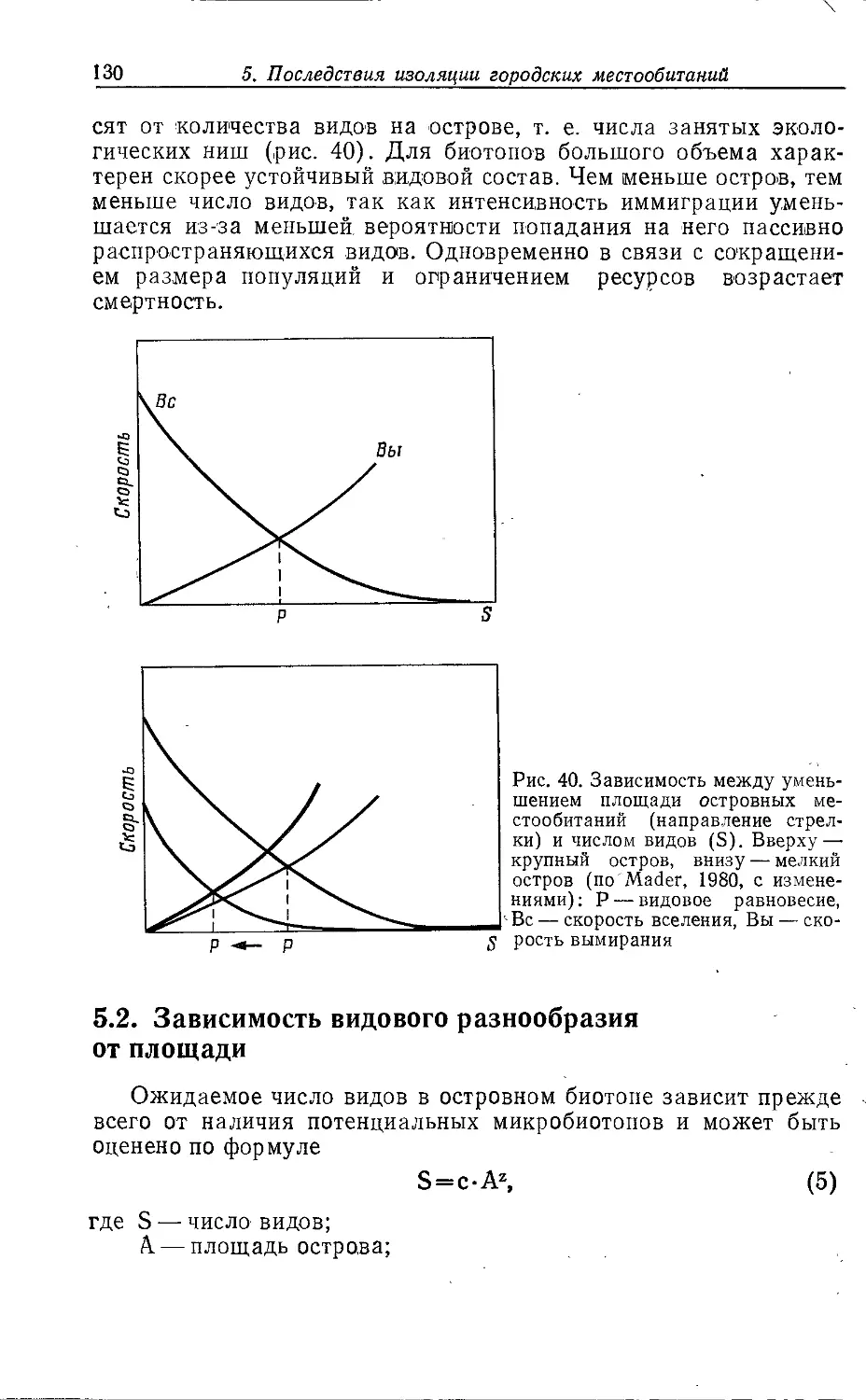
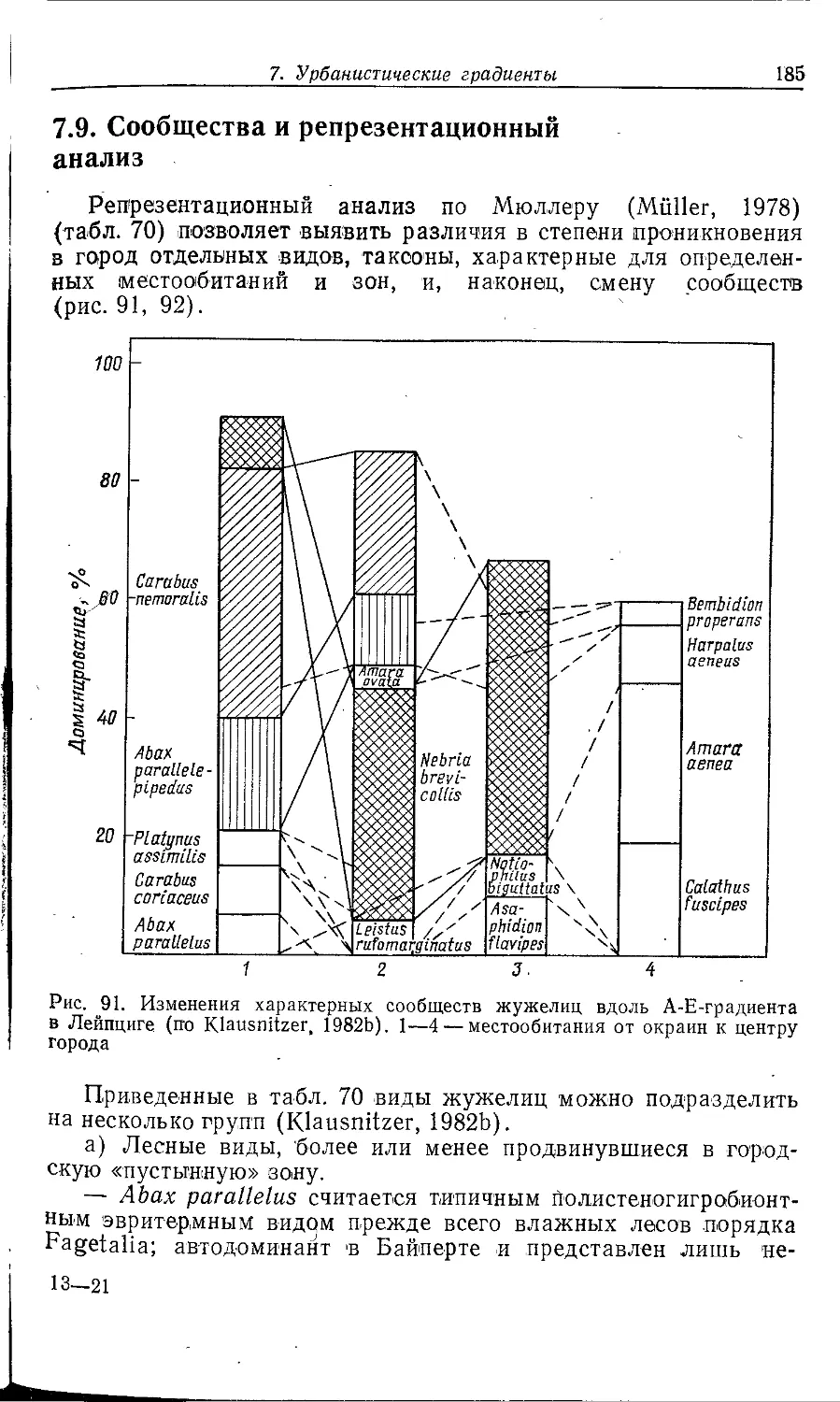
2.2.2. Застроенные территории
2.2.2.1. ГОРОДСКИЕ ЦЕНТРЫ /
Во многих городских центрах расположены исторические или
новые административные здания, например ратуши, башни,
церкви и высотные дома. Они часто окружены пешеходными зонами
и главными торговыми улицами. Степень застроенности
территории очень велика (свыше 80%). Оставшаяся свободной почва,
как правило, сильно эвтрофизирована и в большинстве случаев
представляет собой антропогенный насыпной покров из
искусственного или естественного субстрата.
Важные абиотические факторы окружающей среды
достигают своих экстремальных величин. Примечательна прежде всего
повышенная и относительно устойчивая температура (Eriksen,
1976), а также очень быстрый сток дождевой воды и связанная
с этим низкая относительная влажность воздуха. Воздух в
большинстве случаев загрязнен сернистым газом и пылью. Большую
роль играют выхлопные газы, приводящие к высокой
концентрации тяжелых металлов. Типична во многих случаях сильная
засоленность почвы.
Растительность развита в общем незначительно и
испытывает сильное антропогенное воздействие. Преобладают очень
ухоженные газоны, цветочные клумбы, вазоны и кустарниковые на-
/ ч
2. Структура города и фауна « 37
саждення. Деревья встречаются обычно отдельными
экземплярами, дикорастущие виды почти полностью отсутствуют. Иногда
на клумбах встречаются садовые сорняки. Отдельные
придорожные 'ПО экологии растения могут образовывать по краям газонов
сообщества из класса Plantaginetea majoris. На клумбах
преобладают травянистые декоративные растения. Адвентивные виды
в этом местообитании почти не встречаются.
На более высоких зданиях размножаются бывшие обитатели
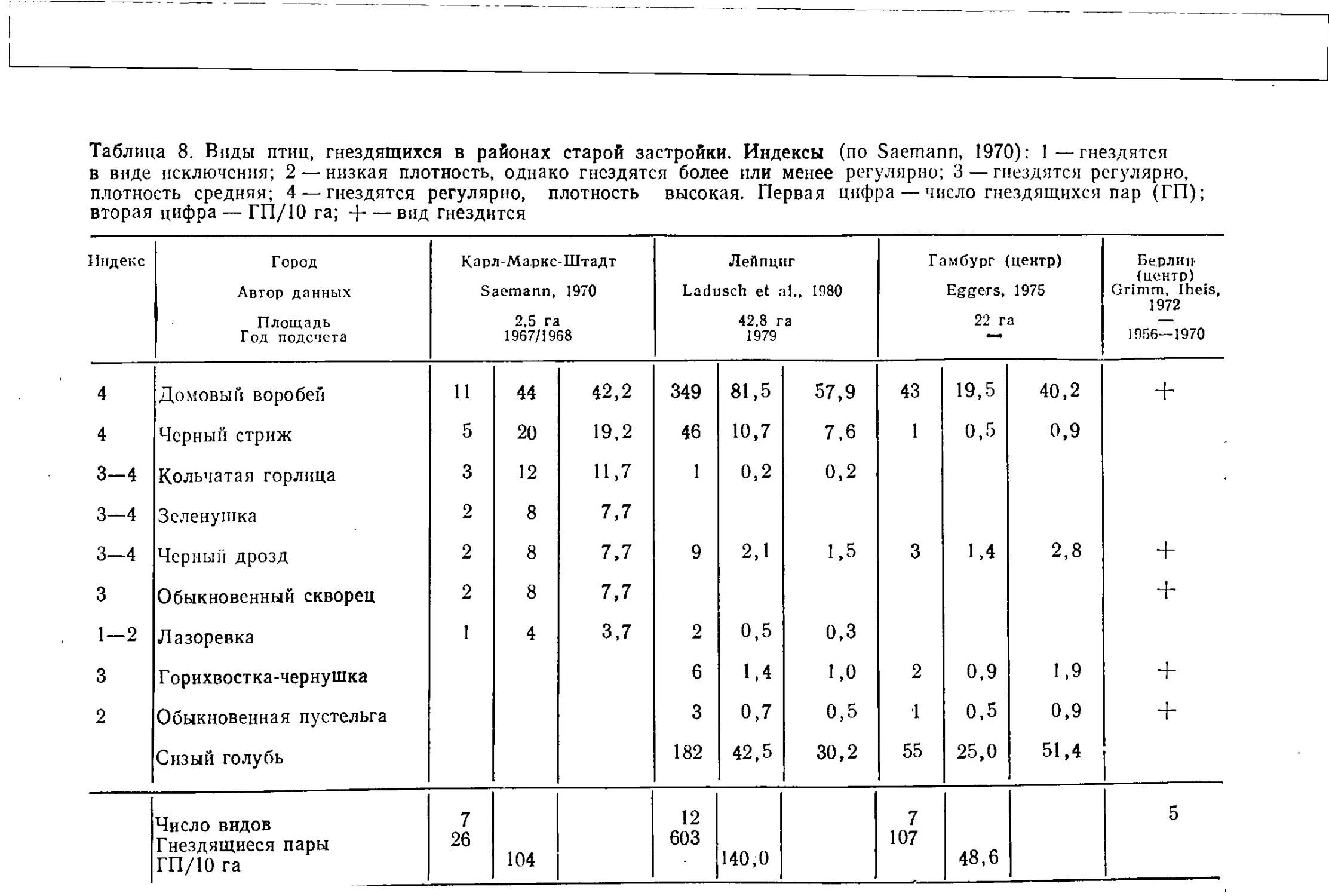
скал — галка (4), пустельга (4), черный стриж (3) и сизый
голубь, многочнсленен домовый воробей (3) (в скобках приведены
показатели постоянства по Saemann, 1970, см. табл. 8). Подсчет
гнездящихся птиц на деревенских церквях Мекленбурга показал,
что эти местообитания, где много отверстий и мало внешних
помех, можно рассматривать в экологическом плане как
«культурные скалы» (Plath, 1980). Из 15 обнаруженных видов
преобладали домовый воробей, сизый голубь и обыкновенный скворец.
В небольших группах деревьев гнездится кольчатая горлица (2).
На чердаках живут различные виды летучих мышей, прежде
всего позднш'! кожан. Целлик (Zöllick, 1980) наблюдал на церкви
Св. Николая в Висмаре — 800—900, а на церкви Св.
Варфоломея в Деммине — 3000 зимующих особей нетопыря-карлика.
В подвалах и канализационных системах встречается серая
крыса.
2.2.2.2. районы СТАРОЙ ЗАСТРОЙКИ
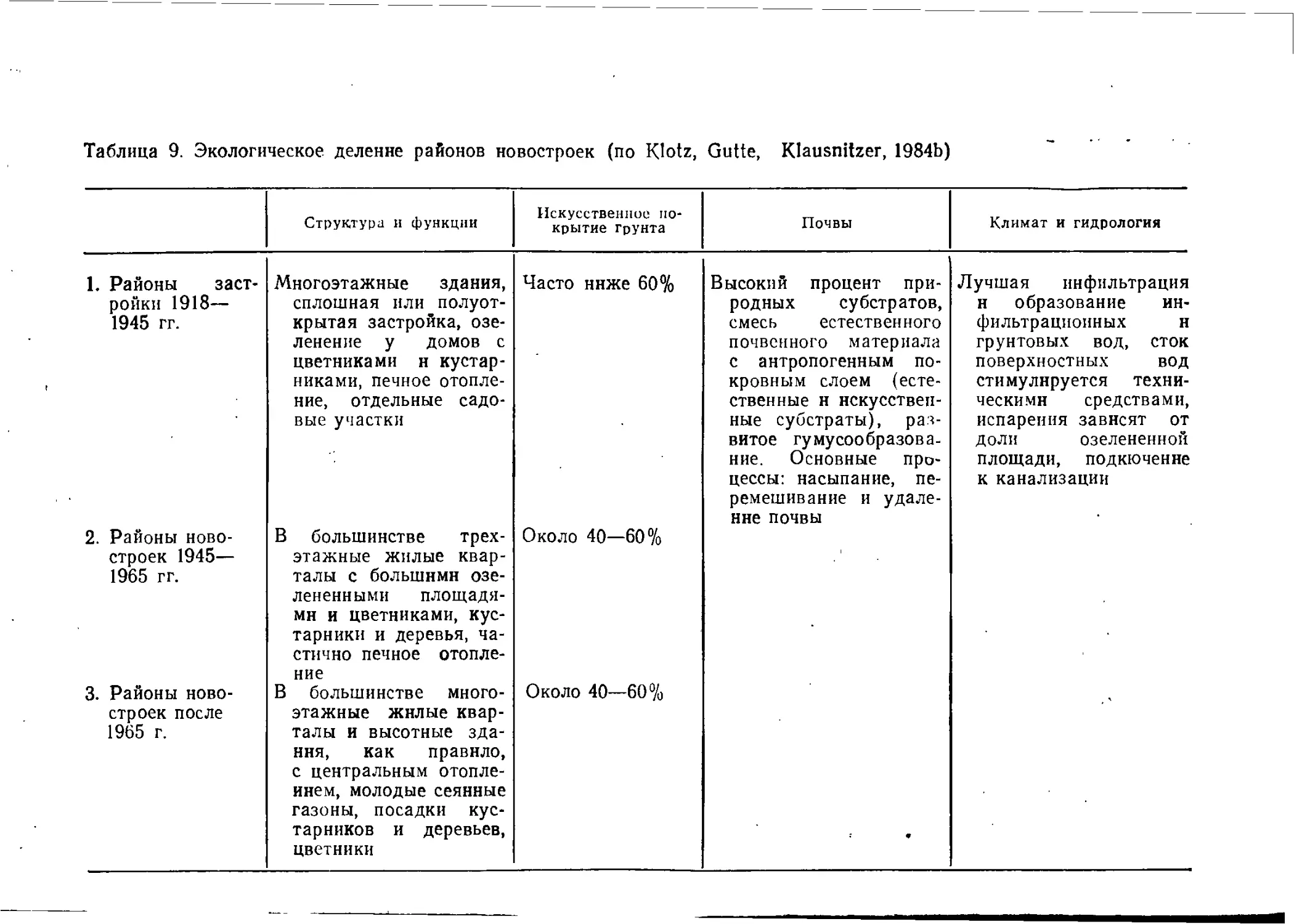
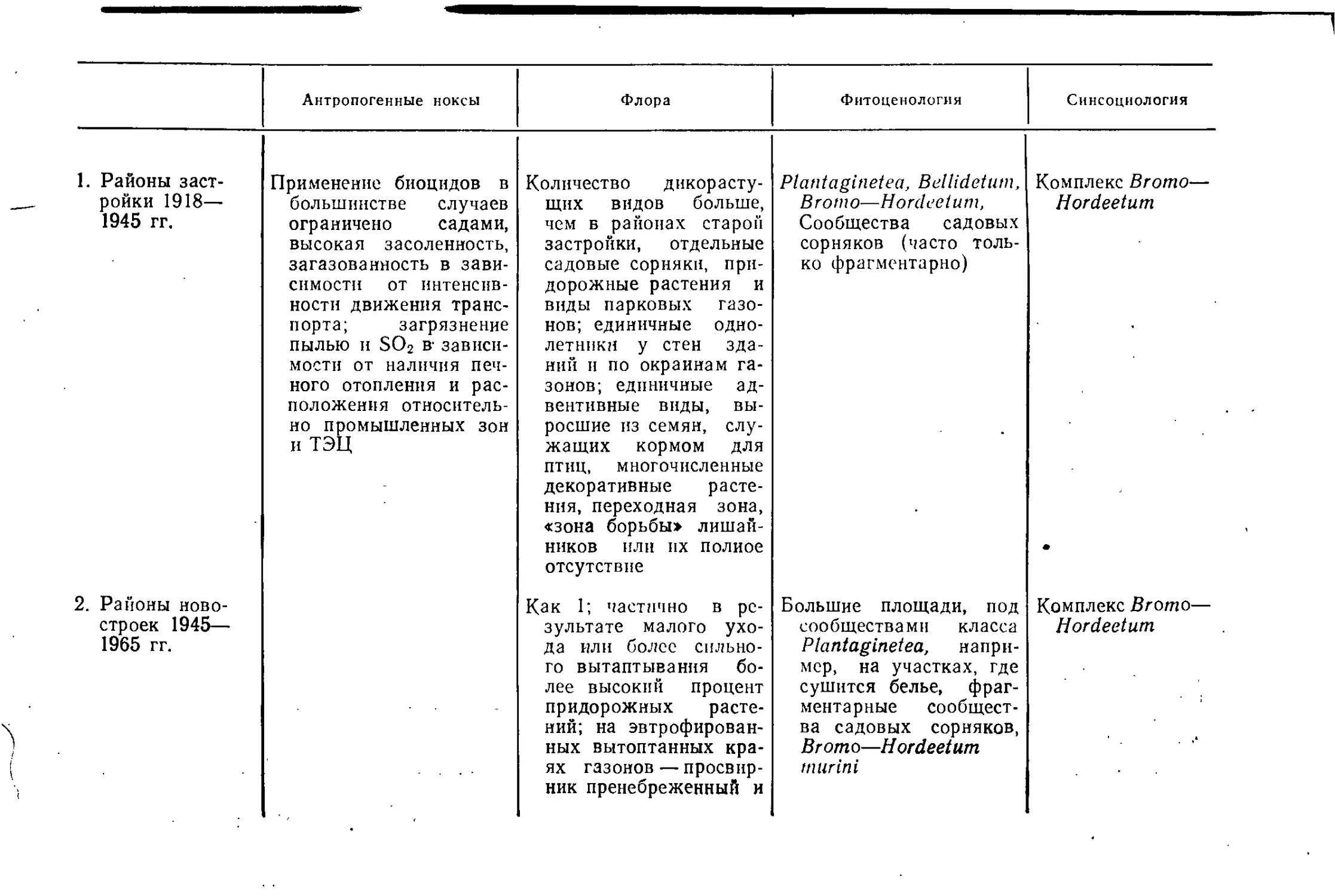
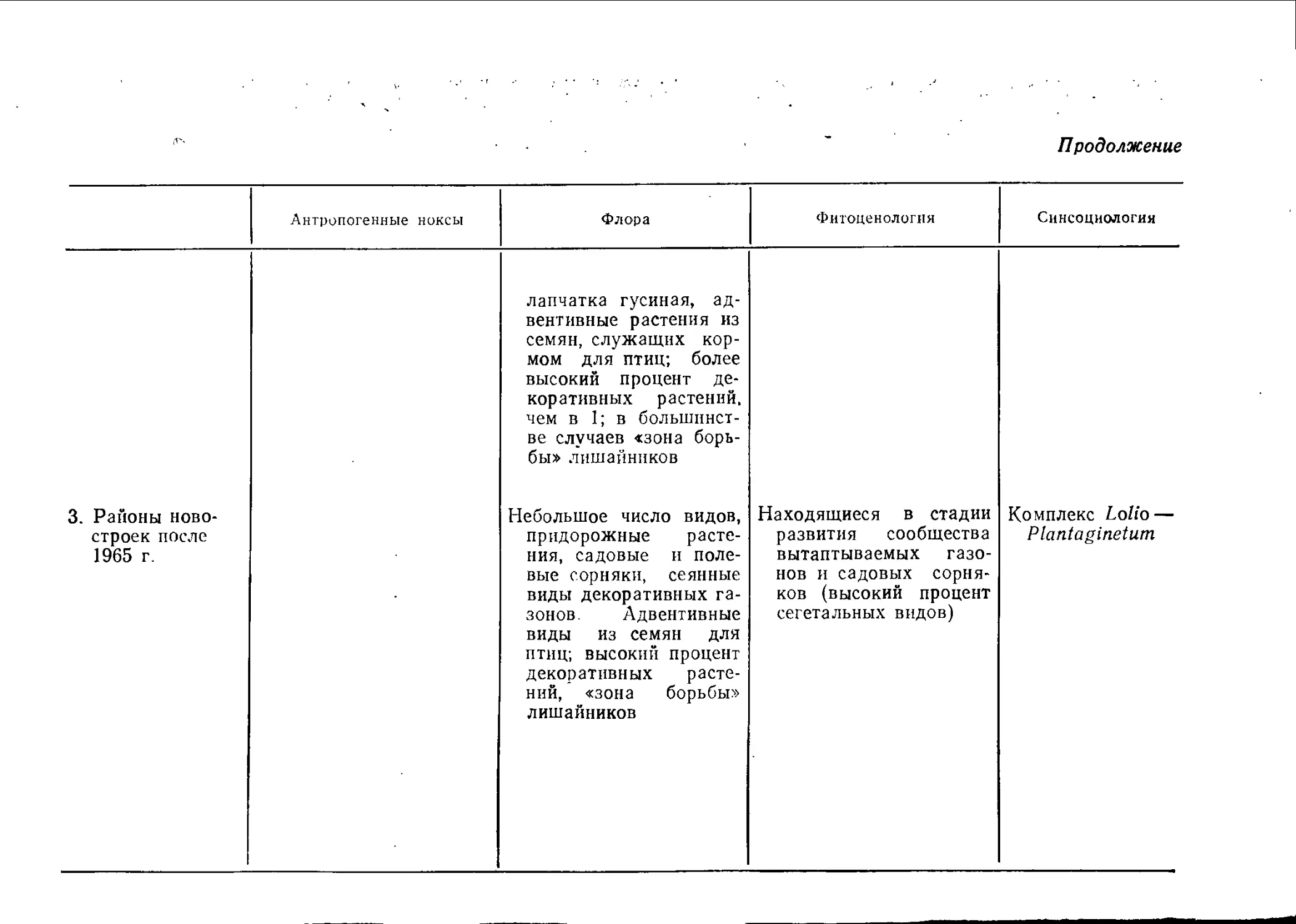
Районами старой застройки мы называем жилые кварталы,
возникшие до 1914 г. с двух-, трехэтажными (постройки до
1870 г.) или иятиэтажными зданиями (1870—1914 гг.), часто без
озелененных внутренних дворов.
Здесь характерны узкие улицы и сплошная застройка,
прерываемая развалинами или участками снесенных зданий, почти
нет свободных участков земли (занято до 80%). Почвы, климат
и влияние антропогенных нокс примерно такие же, как в центре
города (2.2.2.1).
Растительность отличается небольшой долей декоративных
растений. Число видов дикой флоры в общем незначительно,
только на развалинах и местах сноса оно увеличивается.
Плотная застройка обусловливает присутствие теневыносливых видов,
а высокая засоленность — наличие галофитов, напри.мер бескиль-
ницы расставленной. Встречаются адвентивные виды,
прорастающие из семян, используемых как корм для комнатных птиц. Во
внутренних дворах и на улицах растут некоторые породы
деревьев, например ясень обыкновенный, клен-явор и неофит айлант
высочайшш! Характерны некоторые виды птиц, гнездящиеся на
зданиях: домовый воробей (фото 7), сизый голубь, черный стриж,
ч а.
>> га
^ а
1 5 а
о
га 2 5
ill
о S-g-
я 4)
S к га
||£
■к
S га
§ а
!-
га "
" Ё л
о 3 о
o-S а:
S - g
" о §
^ !й с
2 я
а
к 5 к
СО Ж СП _1_
OJ СП CD '
U 5 U ^"
=f 1 1о
5 I
"lis.
га = Ö
Я щ о ts
= et X га
ч Е н а,
ю 5 о о
я Ч (-
Н а с (О
ш
et
га
О
Н
t-
э
.
с
а
с
я
«J
1
с;
о.
+
+ +
о
°1
о"
00
<N
1^
о
со —
о'
Ю TJ- t~. Ю
о" — о' 53
ст> «о —
(М со
см <N 1-
— 1Л CO <M <N (N —
о
о.
о
m
s
I
Ч
a.
о
3 <-
о
cx
о
CI
о
ч
5
о.
о
CS
3
X
X
CJ
ta
о
X
3
ю
о
о.
о
СП
га
с:
п
:<:
а
>>
а
о.
га
о
с
X
S
а.
о
и.
S
ш
СП
о
3
о
VC
>.
ч
о
с-
>а
Я
3
га
С
g 52
•Ф Tf
I I I
CO CO CO CO
—' CO (M
со
СП
3
3
о
и
га
Си
£ § °
" S I
« ° 5
b « "
»g5
Sog.
n ж g
о X 5
(_ a. ><
X 4> S
>, Ш s
CL о H
u с о
l°1
га a:
S f- 5
П R к
У о Ol
" n о
s s
= 9" -
^ ra -
о с Ч
Qj о о
в- s ч
S
3-
ü
га
с м
к й S
о- 2 о
" £
о 2 5
CQ
1 с S
га CJ
Х о;
rags
S к ш
(U о
р о о
о о 5
= а.£
я н- л
о S X
ш га (О
3- о
о а.
сих
S с^ га D о
а) га п о. с
" -S. s"
>.^хо^ со £
"
g ?
а о
S aj
О) S
О
с
g
п
о
в
о
•S
S
о
~ а
Ii
о
ю
га
о
to
I
о
^
о
ч
о
о
о
т
о
Оч н С)
Я О m
Я >.о
га ч
Ч о «-
W я fcd
•S
Я о
^ § ь
я - о
о. V
га Ч
га
я S 5
я я S
. га
Ol 1£
а
я
■о _
g S
о 2
я (О
и о. <и
<и га я
о, m о
я
о S
а я
^ е 3
S я 3
_ й ч
я ^ о
3 3 о
о *
о га
ь
СП
к Ö са i
ä ^§
5 Й я S
S Ю _ с
(U £ га H
Ч S H о
D. ra
ra i=C
ffi CT)
Ii
(D О
га s-
a. о
с „
CJ
Я
я
Э
gl
я
CQ
й 4)
а а
к а CU
я ОТ
§^
га
о я
Я
п я
. о а:
а « я
о
f2
со I
00
&< о.—
6 I
Я02
О о
Я о
Ii
со
2. Структура города и фауна _39
горихвостка-чернушка, а на озелененных участках гнездится
кольчатая горлица (табл. 8). Для зданий типичны серая крыса
и домовая мышь. Регулярно встречается каменная куница (Nicht,
1969).
2.2.2.3. районы НОВОСТРОЕК *
Районы новостроек особенно удобны дл^ наблюдений за
сукцессией городской фауны. Эти районы в зависимости от времени
их возникновения подразделяют на три группы (Klotz, Gutte,
Klausnitzer, 1984b) (табл. 9). В экологии животных еще так
много пробелов, что такое деление может быть обосновано
только частично. Для заселения н<пвотными новых районов важно их
окраинное положение по отношению к ранее застроенным
городским кварталам, более сильное влияние окрестностей и довольно
стандартный внешний облик, для которого характерны
прямолинейные ул1щы, проходящие через кварталы крупноблочных и
крупнопанельных зданий, и озелененные территории. В
различных городах здесь сходны сначала свободные экологические
ниши, что дает широкие возможности для сравнения.
Имеется целый ряд работ по заселению новостроек птицами
(Saemann, 1973; Hanoldt, 1973; Plath, 1976, 1981, 1985, в печати,
Gnielka et al., 1983). Совершенно необычные для них жизненные
пространства очень быстро заселяются определенными видами
(фото 4); при этом можно наблюдать некоторые общие
закономерности (частично — сукцессии):
— доминируют виды, гнездящиеся на зданиях; они
составляют в большинстве случаев свыше 90% общего числа
гнездящихся пар (табл. 10);
— по мере развития растительности возрастает количество
птиц, размножающихся в природе на кустарниках и деревьях;
— общее число видов, ,их число на 10 га, число гнездящихся
пар и плотность заселения увеличиваются с возрастом районов
(табл. 10);
— численность хохлатого жаворонка при этом уменьшается
(рис. 7), поскольку по мере застройки исчезают оптимальные для
него местообитания;
— городская ласточка очень быстро заселяет районы
новостроек, однако численность ее колеблется и впоследствии
уменьшается (Plath, 1985, в печати), поскольку:
а) по мере озеленения становится меньше материалов для
постройки гнезд,
б) развитие зелени на фасадах зданий мешает взлету птиц и
сокращает количество мест, пригодных для постройки гнезд,
в) жильцы из-за загрязнения балконов не допускают
присутствия гнезд.
s
s
3
Ii
5^
m
0 о
1 I Й
1 §
U Q.
m S
о is
uä га
« й-
я -е-
Ч га 4.
~ s а
i с
ч га
■ к S
о
с
га
sit
g о с
3 go
1 о
OS'S
IS
>>\о а)
Зой
§
>< о
о I
'"11
и Da с
га 1^
а, о
§»2
== Ol
о 3
о. а:
га ja
& S
В.
S
о о
m я
га CJ
t- о
cf га
ю я
о о
о к га
ш ш О-
£ S S Ö
с- я о га
g а :т m
и tu
X га m д
0.0-0 в-
о а
" ä о-я
„, я га S
ё я с с(
W 5
о о э ...
с< о. et я
га о я о
о Ч я В
га га Ч
4 t- га
" й
ТО UJ
5= к 3
О) К д
5 äs
Я Я n
я _ о
н « я
OJ я о
ч я m
3 >> ч 3
я S Чя
з" я" й 5
4 к о я
я S й 3-
ш а> Си о
U о
OJ СП * я
3 = й
" я g
Ра|я
и о. ä я
V «".в
CD
ч
о
ч
к
га
CD -г а
3 §ю
в 2 л
ill
§в§
<D я й
4 В W
Ч V
" m
ь
я ^
§&-
ООО
С1. X я
>> JJ
о с
в о
- га о
Р 2 3
то
~ н ■«
6 Ё
ю 5
я
о
2 в
в
га
W 9Я
3 я
S- в:
я о
га CD
t 3
3 ш
(С
S ^
О) я га
f-, га о.
•е-я
^ о я
й о. га
5«
о я
о в
о. в
я О)
CD
В CQ
I т
О * W
В.Я ><
о. я я
я я я
ad
XI я
в в
n ? I ё
5 S S =
= " CD CD
5"
9 Ö
£ о m
о Ö я
я = CD „
Я ^ 5
Sags
a О u 3
я
S
6
UJ
Ё
л,
m
я
О
ÖJ
га
о
га
с
со
я
о.
со
CD
(Г)
то
к
Я
ее
л
X
о,
(-.
CQ
f-
я
то
о
о
в
в
га
ю
о
CD
^
я
m
о
то
ж
-
а
Ч
н
о
о
о
то
а.
2
со
га
U.
о
й
Ö
ja
1
р
га
о
о
о
со
о
X
с
с:
я
о
о. CD
= g
§2
о о
вт
я" U
0 I
я 1Л
о
в а>
1 <u
,2 о ю
Ii
I
о ч
« о
о
О
к £ „
га О) S
а 5 Э И
I 1 " и
о.
о
ю
.о S
:s а
°1
ч с
с а
я га Д
ä S °
2
о g
и я ч
о 3
о ™
QJ CQ
S Е я
i^B
е[ « 5
О 2 Ч
га 3
га o-ffl
о о
о я
^ га
со со
о о
к 1«:
ЕС о с;
о f- ч
Ü °
к га д
а
X а
3 я
§ я 3
о 5 л
S я
га я я
S 5 S
raSgo
Ч а о S
о
•я а —
S S
м н а
о га
а 0-S
л о ф
а « л-
со S
" Я
CQ
ш ш
? а
ч
CD 3
.« UJ щ н
о
ч
CJ CJ CU
g 3 3
я и
<" ^ §
111
ю
я 5^
в; о
я «
о .
- а
-|§
К О
CQ и m
«я 5
sc ^
й) "
со о и Q
к 3 = m й
CQ Н ^ Д
га sS
зяё' ™
я м « я
я (- ш я я
а с =£ я ч
й ^-
3 ^= .
S 1^
lis
со
ю
00
00
о
1
ю
to
со
«3
CM
со
о
см
см
СП
CD
■*
со
см
-*
00
ю
<J5
CD
см"
о
со
со
ю
со
со
со
CN
о
1
00
со
•*
со
о
ю
со
ю
ю
CD
см"
СП
ю
CD
см"
о"
СП
со
см
со
со"
О)
со
со
со
о
ю
со
СП
S
00
со
а>
см
ю
оГ
о"
«
CM
00
■ч<
ю
со
см
■ф
со
Oi
см
со
со
со
гО
•*
о
1С
оГ
00
&.
00
о
00
•ф
со
со
щ.
со"
о
2
СП
СП
со
m
со
S
g
см
о
г-
а>
00
ел
о
00
оо
-
-
m
ю
со"
ю
CD
со
со
о
со
ш
см
со
со"
00
t~-
сп
00
о"
со
со
со
■ф
й
СП
CD
Ol
со
о
t--
■*
ю
ю
со
со
•ч-
оо'
^-
см
Ol
ю
оо
о"
ю
00
о
■ф
см
■ф
1
S
со
00
CD
00
см
со
о
СП
00
Ol
Ol"
о
со
см
СП
со
о
■ф
со"
ю
2
CD
со
00
см
см
ю
ю
00
со"
со
СП
ьО
о
иО
СП
1-.
Ol
ц.
со*
ю
со
а>
CD
см
со
<D
см
CD
о
см
2
со
со
00
Ol
со
ю"
см
е\.
см
ю
1
С5
Ol
CD
1
со
00
СП
N.
СП
QO
ш
см
1
со
о..
rt.
CD
Ol
1
оо
со
CM
О)
2
ю
см
1
с»-.
п.
ю
о
ст.
о
1
о-.
-
1
ю
CM
с
с
U
с
с
а"
сз
I-i
сч"
о
а
X
о
а.
cl;
GJ
ю
о
н
о,
о
о
cj
ю
а.
са
о
га
т
•S
R
cq
•S
я
о
и
CJ
3
о
о
S
о
cj
ч
о
ся
ч
3
ш
и
Си
X
3,
о
о
и
и
о
U
X
R
Я
Я
га
Я
о.
о
о
н
Я
а,
ti
н
о
я
о
ч
со
га
о.
VO
О
о
я
т
га
Си
я
CJ
о
си
S
X
д
э
CQ
а
с:.
га
к
CJ)
к
к Си
Ü
со
<и
S
CJ
я
и.
U
«=;
со
га
н
се;
Ü
S
2&
я 2
S «
g ё
I в
Н W ь>
о и g
с
44
2. Структура города и фауна
После домового воробья городская ласточка, как правило,
самый частый здесь вид (фото 2). В Ростоке в районе Люттен-
Клайн (Plath, 1981, 1985, в печати) среднее за 13 лет число ее
гнездящихся пар составляло 41,2, среднее обилие (число пар на
10 га) —7,20, а среднее доминирование по данным десятилетних
учетов—13,14%. Причина этого — прежде всего благоприятные
1975 76 77 78
Год
79 80 81 82 83 54
/972 73 74 75 76 77 78 7Э 80 81 52 83 84
Год
Рис. 7. Развитие популяции домового воробья (вверху) и хохлатого
жаворонка (внизу) в новом районе Ростока (по Plath, 1985); ГП — гнездящиеся пары
условия для гнездования, наличие доступного материала для
устройства гнезд и обилие корма. Этот корм исследован (v. Gun-
ten, 1961; Kozena, 1975), правда не в городах, однако основные
группы насекомых в его составе преобладают и в городах.
Обнаружено (V. Gunten, 1961) 33,1% тлей, 24,2% мух и 21,2%
комаров. Кожена (Kozena, 1975) выявил 57,3% равнокрылых и
2. Структура города и фауна
45
31,9% двукрылых. Менцель (Menzel, 1984) сообщает, что
городская ласточка легко находит места скопления своей добычи. Ленд
и др. (Lenz et al., 1971) описывают прямо-таки взрывообразное
увеличение популяции этой птицы в районах новостроек, позднее
достигающей более или менее стабильного размера (рис. 8). Для
строительства гнезд предпочтительны определенного типа
балконы, которые открыты только с одной стороны и уходят в фасад
по крайней мере на глубину 0,75 м. По некоторым данным
(laeschke, 1984), городские ласточки селятся теперь и на
балконах различных старых зданий. Они предпочитают устраивать
свои гнезда на 'камне, кирпиче и клинкере (64,4%) (Otto, 1974).
В Ростоке 87,7% гнезд отмечено на балкона.х, а 10,3%—над
3000 h
Р: 2000 h
1000 h
1369 1911
1974 1975 im
Год
1973
Рис. 8. Увеличение численности городской ласточки в Западном Берлине (по
Lenz, Witt, 1980). Районы старой застройки — вертикальная штриховка;
крестьянские дворы —косая штриховка; новостройки — белый цвет; ГП
—гнездящиеся пары
входами домов (Plath, в печати; Kramer, 1972). В других .местах
количество балконов с гнездами было меньше, и большее
значение имели выступы крыш, особенно длиной 30—50 см (67,5%
гнезд) (30—100 см — 94,2%, крайние размеры 5 и 170 см; Otto,
1974). Гнезда прикреплялись к зданиям, начиная с высоты 2,5 м
и кончая крышами 24-этажных домов (Zenz, 1972). Otto (Otto,
1974) обнаружил в Гамбурге 70% гнезд на высоте 4—8 м
(табл. 11). Платт (Plath, в печати) указывает, что 42,7% гнезд
строятся на западной стороне, 42,2%—на южной, 12,6%—на
восточной и 2,3% — на северной.
Эвдоминантом районов новостроек является домовый воробей.
Длительные наблюдения (Otto, Becker, 1976; Plath, 1981, 1985)
46
2. Структура города и фауна
Таблица 11. Распределение гнезд городской ласточки в районе
новостройки в Ростоке (по Plath, в печати)
Этаж
Число гнезд
в процентах
Вход В дом и нижний
39
5,6
этаж
1
38
5,4
2
125
17,8
3
177
25,2
4
156
22,3
5
156
22,3
6
1
0,1
7
1
0,1
8
2
0,3
9
4
0,6
10
2
0,3
показали, что после фазы быстрого роста плотностп его
популяции наступает некоторое ее снижение, и в конце концов
численность более-менее стабилизируется (рис. 7).
Типичными видами, появляющимися только по мере развития
деревьев и кустарников, являются славка-завирушка, черный
дрозд и кольчатая горлица. В новом районе Галле-Нойштадт
черный дрозд стал гнездиться только спустя 10 лет (Gnielka,
1983). Отмечалось за это же время увеличение численности
славки-завирушки.
Видовое разнообразие после основания района новостройки
сначала очень высокое, но со временем оно уменьшается и снова
возрастает параллельно развитию растительности (табл. 10).
Плат (Plath, 1985) отмечает после первоначально высокой
плотности новое сокращение популяции горихвостки-чернушки.
Причина этого, по-видимому, заключается в конкуренции за
места гнездования с домовым воробьем, чпсленность которого
одновременно значительно увеличивается. В Галле-Нойштадте эта
горихвостка все шире распространяется, становясь обычной здесь
птицей (Gnielka, 1983).
Сравнение данных шести учетов в районах новостроек Росто-
ка (Plath, 1981), Берлина (Otto, Becker, 1976), Карл-Маркс-
Штадта (Saemann, 1973), Хойерсверды (Krüger, 1973) и
Гамбурга (Mulsow, 1968; Hanoldt, 1973) повсюду показало наличие
потомства у домового воробья, хохлатого жаворонка, черного
дрозда и зеленушки. Более чем в 75% районов гнездились
славка-завирушка, горихвостка-чернушка, черный стриж и также,
вероятно, сизый голубь. В двух новых районах Карл-Маркс-
Штадта (Saemann, 1973) среднее обилие домового воробья
составляет 77,0%, черного стрижа —6,3%, черного дрозда —5,0%„
2. Структура города и фауна 47
Таблица 12. Сукцесспя членистоногих, питающихся плесневыми грибами,
в новостройках (по данным Rack, 1981). Виды перечислены в порядке
уменьшения влажности и, вероятно, в зависимости от изменения структуры
доминирования плесневых сообществ
1. Tyrophagus putrescenUa (клещи) Очень высокая
влажность, почти сырость
2. Виды Ameroseius (клещи)
3. LiposceUs sp. (сеноеды)
4. Willowsia buski (Lobbock) Lepidocyrtus cyaneus
(Tullberg) (ногохвостки)
5. Psyllipsocus ramburi (сеноеды)
зеленушки — 3,6%. Средняя плотность заселения (число
гнездящихся пар на 10 га) у тех же видов соответственно была 22,9,
1,9, 1,5 и 1,0.
По мере развития растительной сукцессии в районах
новостроек орннтоценоз становится все более похожим «а
характерный для так называемого «зеленого города» (Mulsow, 1976; Plath,
1981). Плат (Plath, 1981) отметил 18 видов птиц из 28
встречающихся в «зеленом городе», среди них все — первого 'класса
постоянства (от 75 до 100%) (Mulsow, 1976). В новом районе
Хойерсверды, существующем с 1950-х гг., обнаружено 20 видов
гнездящихся птиц (Krüger, 1973), в том числе много тех, что
гнездится в природе на кустарниках. Сравнение двух более
молодых и одного более старого районов новостроек в Карл-Маркс-
Штадте (Saemann, 1973) показало общую плотность 29,6 и
соответственно 84,8 гнездящихся пар на 10 га. Разница связана
прежде всего с наличием птиц, гнездящихся в природе на
кустарниках и деревьях, например черного дрозда, зеленушки,
кольчатой горлицы (обилие—12,1, 7,9, 12,9%; плотность заселения —
10,3, 6,7, 10,9 пар на 10 га).
Вскоре после постройки новых зданий в них активно или
пассивно проникают первые представители внутридомовой фауны.
Захер (Sacher, 1983) сообщает, что паук-сенокосец Pholcus
phalangloides появляется менее чем через год, а паук Tegenaria
atrica— через два года. Еще быстрее происходит заселение
клещами, сеноедами и различными жуками. По мере снижения
влажности сообщество этих грибоядных и частично плотоядных видов
изменяется (табл. 12). Как и в других вновь возникающих
местообитаниях, идет сукцессия. Иногда отмечаются ярко
выраженные вспышки размножения отдельных видов, численность
которых затем по мере высыхания зданий резко сокращается
(или они сменяются другими видами).
iB особенно больших количествах уже через два месяца после
заселения дома встречаются клещи рода Ameroseius (табл. 13).
Оптимальные условия в новостройках создаются, очевидно, для
клеща А. plumigerus; они обусловливают сильное размножение
48
2. Структура города и фауна
Таблица 13. Виды клещей в новостройка.х (по Rack, 1972)
Вид
Число
находок
Ameroseiidae
Ameroseius plumigerus (Oudemans)
A. pavidus (C. L. Koch)
A. ptumosus (Oudemans)
A. pseudoplumosus Rack
Aceosejidae
Proctolaelaps pygmaeus (Müller)
Phytoseiidae
Neoseiulus barkeri Hughes
Podocinidae
Lasioseius penicilliger (Berlese)
Laelapidae
Hypoaspis lubrica Voigts et Oudemans
Androlaelaps casalis (Berlese)
Haemogamasidae
Hae/nogamasus pontiger
Acaroidea
Tyrophagus putrescentiae (Schranck)
Glycyphagus domesticus (Degeer)
27
5
6
4
10
2
1
1
3
10
2
Пища
Грибы
Грибы
Грибы
Грибы
Грибы
Грибы ?
Грибы ?
Хищник ?
Грибы ?
Хищник ?
Грибы
Грибы
ЭТОГО вида, который в природе'массовыми скоплениями не
встречается (это же касается ВгуоЫа graniinum). Раннее отопление
при закрытых окнах приводит в связи с испарением воды из
каменных стен к высокой влажности воздуха, а это в свою
очередь— к образованию обширных пятен плесени на полу, мебели,
изоляции, обоях и т. д. Массовому появлению клещей
способствует партеногенетическое размножение. При этом времени
для образования крупных популяций видов, питающихся
плесневыми грибами, очевидно, слишком мало, поскольку весь цикл
развития до вспышки своей численности они должны успеть
пройти менее чем за 12 месяцев (ср. Rack, 1956, 1963а, 1963b,
1964, 1965, 1968, 1970, 1971, 1972, 1973, 1980, 1981, 1984).
Возникают их сообщества из 2—7 видов, проникающих из
окружающей среды и сначала относительно слабо заселяющих только что
построенные :здания; исключением является акарондный клещ
Tyrophagus putrescentiae.
Часто встречается также жук-плоскотелка Ahasverus adve-
пае — вредитель запасов, занесенный из тропиков с
заплесневелыми растительными продуктами. Он был обнаружен
(Engelbrecht, Buske, 1982) в 300 квартирах пяти городов округа
Потсдам, в большинстве случаев — уже в первые четыре недели после
вселения в квартиру. Причиной является наличие плесени. В этом
местообитании регулярно встречаются также виды семейства
Lathridiidae и сеноеды (например, Psyllipsocus ramburi, Dorypte-
2. Структура города и фауна 49
2.2.2.5. ОТДЕЛЬНЫЕ ЗЕЛЕНЫЕ НАСАЖДЕНИЯ
В понятие «отдельные зеленые насаждения» мы объединяем
одиночные деревья, аллеи и живые изгороди. Они интересны как
«перевалочные пункты»'' и экологические каналы, соединяющие
сплошные зеленые массивы, а также имеют эстетическое и
климатическое значение. '
Определенные свойства различных видов деревьев позволяют
им хорошо развиваться даже в центре городов. К таким породам
относятся, например, первоначальные обитатели ущелий, виды,
устойчивые к соли, применяемой для таяния льда, и другим
вредным факторам. С другой стороны, деревья в городских условиях
подвергаются множеству опасностей и стрессовых воздействий
(повреждения, инфекции, изменения почвы, плохое обеспечение
водой, загрязнение воздуха). ВысО'к процент «больных» деревьев;
они часто предрасположены к заселению ксилофагами,
короедами и сосущими насекомыми.
I
Так переведен термин Trittstein-Habitate (по-английски — stepping
stones), т. е. буквально местообитания, функционально соответствующие
камням для перехода через ручей (т, е. через непригодные для заселения
зоны) . — Прим. ред.
4—21 i
гух pallida п Lachestlla pedlcularia) (Peus, 1933; Rack, 1978).
Примером образования пищевой цепи могут служить клопы Em-
picoris culiciformis и Е. vagabunda, питающиеся сеноедами и
жуками.
2.2.2.4. ВАЗОНЫ С ЦВЕТАМИ
Во многих крупных городах по архитектурным
соображениям, особенно в центре, устанавливаются разного размера
бетонные емкости, которые засаживаются pa3iih4hbimh растениями.
Поскольку в течение года, как правило, проводят пересадку
(а иногда и ие один раз), эти вазоны становятся местами
постоянного проникновения в центр города различных видов
животных, которые заносятся вместе с посадочным материалом.
Некоторые из таких емкостей засаживаются также многолетними
кустарниками, так что становится возможным образование соб-*
ственного зооценоза. Определение животных пз почвенных
ловушек показывает, что широко распространены в основном виды,
считающиеся эврипотентными. В центре Лейпцига в таких
местах были обнаружены некоторые жуки, чаще всего жужелицы
Harpalus rufipes и Enicmus transversus (Klausnitzer, Lehnert,
1980).
50
2. Структура города и фауна
Отдельно стоящие деревья могут служить местами питания,
обитания и размножения относительно большого количества
животных, поэтому важны преладе всего как «перевалочные
пункты». Даже на них гнездятся, например, кольчатая горлица,
лазоревка, садовая горихвостка, большая синица, вяхирь,
обыкновенный скворец, славка-завирушка. Примером заселения
отдельно стоящего дерева насекомыми может (быть лиственница
в центре Вены (Schremmer, 1959), на которой выявлены многие
виды и биоценотические коннексы из обнаруженных также в
сплошном лиственничном массиве. Численность некоторых
таксонов на городских деревьях может быть очень большой, например
на липе высотой 15 м насчитано около 1 млн. тлей. На
определенных частях дерева могут появляться специфические
зооценозы, например на стволах, заросших мхом, лишайниками и
водорослями (Pschorn-Walcher, Gunhold, 1957). На структуру этих
сообществ может влиять сернистый газ (Gilbert, 1971).
100 т
И(м)
Рнс. 9. Зависимость между чис.пом видов (S) и протяженностью живой
нагороди (И) на примере жужелиц (по Mader, Müller, 1984)
Аллеи и посадки деревьев вдоль улиц важны прежде всего
как пути расселения и коридоры, связывающие городские
зеленые насаждения с окружающей город местностью. Так,
зеленый кузнечик {Tettigonia viridissima) может проникать в центр
города по рядам деревьев; то же можно сказать о большой
синице, лазоревке и обыкновенной белке. Растительность в
приствольном «руге часто поражена фитофагами, в почве О'битают
ногохвостки, клещи, тихоходки и нематоды.
Тишлер (Tischler, 1980b) сравнивает живые изгороди с двумя
примыкающими друг к другу лесными опушками
противоположной экспозиции. Краевой эффект в таком «дважды опушечном»
биоценозе особенно велик. Живые изгороди являются жизнен-
2. Структура города и фауна 51
2.2.3. Транспортные зоны
2.2.3.1. улицы, площади, мосты
Территория, отведенная транспорту: городские автобаны,
транзитные трассы, крупные автомобильные стоянки и т. д.—
характеризуется почти полным отсутствием открытой почвы.
Исходный грунт отсюда часто удален и заменен чужеродным
естественным и искусственным субстратами. Почва по обочинам
трансиортных путей уплотнена и часто более щелочная, чем
вдали от них. Растительность здесь и в других труднодоступных
непроезжих участках Tpaioc представлена средним количеством
видов (приблизительно 400, Sukopp et al., 1980; Way, 1977).
Соль, используемая для таяния льда, способствует
распространению бескильнпцы расставленной, а гербициды —
проникновению растений с глубокой корневой системой, например купыря
лесного, борщевика европейского (Heracleum spondijlium) и
осоки мохнатой; на городских окраинах здесь встречаются
обширные заросли лебеды продолговатолистной. На нескошенных
обочинах дорог разрастаются многолетние рудеральные растения.
Образуются краевые полосы бескильнпцы расставленной,
ассоциации Anthriscetum sylvestris (обедненная), Descurainio — Atri-
ным просрранством прежде всего жпвотных, требующих близких
к лесным условий. Мадер и Мюллер (Mader, Müller, 1984)
указывают на зависимость между протял<енностью живой изгороди
■и числом видов (на примере жужелиц) (рис. 9) (см. 5.2). Живые
изгороди иногда на очень маленьком пространстве располагают
разнообразнейшими пищевыми ресурсами как для фитофагов
(эврифагов и стенофагов), так и для хищников и паразитоидов;
велико также их структурное разнообразие (Forman, Baudry,
1984). Важнейшие функции живых изгородей, связанные с их
животным миром, приведены в табл. 14.
2.2.2.6. зеленый город
Клоц, Гутте и Клауснитцер (Klotz, Gutte, Klausnitzer, 1984b)
называют зеленым городом два типа местообитаний (табл. 15),
которые при изучении экологии животных не всегда отчетливо
разделялись пли вообще рассматривались вместе (Hobner, 1984).
Лучше всего нам пока известна их орнитофауна (табл. 16).
(Saemann, 1970, 1973; Mulsow, 1974, 1976; Luniak, 1980; Wal-
dert, 1982). Изучались также их жуки (Reich et al., 1985)
(рис. 26), жужелицы и журчалки (Jacob-Remacle, 1980), бабочки
(Loibl, 1953).
о
с
S
и
S
>>
•е-
X
3
S
о.
>,
(-
а.
ь
3
я:
щ X
с
3
3 я
ф сп
га
S-
i га-§ i х"
t=c „ о
я ca CJ
^ ^ ffl о ra
. о =f я
га S S «
b я я ra ;2
о CJ я и С
га
m a.
Яхо
я8
" Я-
= g
a.
ra S
O- m
CJ s
S
co eS
ra о
Q. CQ
Si ^
к X я
g.
S и !з
ill
о. я
я"
3 << -
^■^^
а. 1^ о S
я CD
И «а
к n
§1я
_ OJ м
~1э
öS.
' га u CR
я о Ä Ч
о о.
я с
3 сч
(L> о
^1я
<u л Ьй
а.ю в-
X
3
я
я
о;
о
с
о
о.
(- „
2 =^ •
га « со
в- х^
я g S
X к
3 - ч
я X га ш
§ 5 " ч
<u га
я f.
is
.. i>: a.
ra >. к
я ra о
о я „
>> о
со (U
о CJ
CQ га
¥ я
си —
§1
is
я
я
а
о
S
S
сч
2 .
я
•е-
я
£■
3
я
Q.
С
Я
S
о-
2
X
я
ш
га
я
СО
я
я
S
с
о.
о
о
я
я
>.
се
е
я 5 S
^ 5 "
я
я
я я я
& о. со
^ CD со
S о "
га » V
с с*
.я -g S
я« Ё "
Ч ~ га S
CJ h
я «
Q. га
я *iJ
о
X
3 =
IS
га о
ё ё я« ч
хО о
g
С
я
о
1,
ч
Ii
5 ^
4 о
я н
5 .0
S ч
X
3 _
S Ö
си « я
4 сп а:
о &■
я я о
к о о
о
3^ ■
т
о .я
4" X га
О.Я >,
S
Я я о
со о
° ^, X
Си
о
я я
я га о
га а. к
га ссз CU
&, н о
га t*j га
я g я_
к Я
4 о
r=t «11 о
5 1«
£ о о
S 'S
с?:
S о cf
X я Э
2 *
я п £
Ю о
я ч й
я я га § 3
я ь с
m
с
S
со
Я
В
я
о.
о
ч
S
S2
g £
я
з" S X
а а. я
g ё g
О <1>
я о " Р
52
2. Структура города и фауна ; 53
pliceium oblongifoliae, сообщество Artemisia vulgaris— Arrhena-
therum elatius.
Проезжая часть оказывает в основном три различных вида
воздействий на животный мир:
— животные погибают в результате движения транспорта
(см. 9.2) или высыхают на лишенных почвы транспортных
магистралях;
— она изолирует местообитания друг от друга и значительно
способствует приданию им островного характера (Mader, 1981b);
— транспортные выбросы и другие антропогенные факторы
(соль для посыпания дорог, гербициды) оказывают решающее
воздействие на фауну обочин, так что здесь могут возникать
совершенно новые ассоциации, так называемые антропогенные
зональные зооценозы (Tamm, 1976). Между ними и природными
переходными экосистемами, т. е. опушечными биотопами (эко-
тонами) с характерным краевым эффектом, имеются
существенные различия, например:
— предположительно другая структура сообществ;
— другая реакция на иммигрантов и вероятное отсутствие
прочных синэколотических связей, которые оказывают
стабилизирующее воздействие на граничащие друг с другом зональные
зооценозы;
— постоянное нарушающее влияние на сообщество ухода за
обоч1шами дорог (Sauer, 1977), которое нериодически
прерывает формирование зооценоза пз-за частого скашивания травы и
применения гербицидов.
С другой стороны, обочины представляют собой пути
проникновения эвритопных, а также ксеро-, гелио- и термофильных
форм (Way, 1977). Некоторые виды находят здесь
оптимальные жизненные условия вследствие обильной пищи в виде
мертвых позвоночных животных (Bergmann, 1974; Mader, 1979)
(рис. 10).
Дороги создают и другие трофические ниши, например, для
насекомоядных видов (трясогузки белой,
горихвостки-чернушки, домового воробья, обыкновенного скворца, ежа
обыкновенного), собирающих раздавленных насекомых с проезжей части, а
также для .видов, ловящих насекомых, используя привлекающий
эффект дороги (например, деревенской и городской ласточек,
черного стрижа), и для растительноядных, которые питаются
упавшими фруктами и семенами (например, домового воробья,
хохлатого жаворонка, черного дрозда, зеленушки).
Озеленение обочин дорог создает благоприятные условия для
некоторых фитофагов. Так, отмечено (Free et al., 1975) 67 видов
насекомых (прежде всего журчалки и настоящие пчелы),
посещающих цветки 39 видов растений на отрезке обочины длиной
200 м. Птицы (Haas, 1964; Bergmann, 1974) и млекоиитающне
о
3
О
S
3
S
sc
о
с
о
с
о
g
V
R
К
s
«о
о
о
. я « к f-
. ^ П я GJ
д sc t-
§ög = g
2.° "
Ii
я и
5 о « ^<
в S >Я R
•е- m 3 " g
о 9-
я я
SJ (1) й 5,«
g я я я fc
° в о н о
g- о я Ü ю
X
к
я
в
CJ
Ii
et m
О Я
с Ч
а 3
в в
я о
OJ ч
я го
н о
о •—'
а
я
Ig§
та I
В 7
о.
« га
га Ö
я к
га
о. о
2 e-oj
о
CJ
я
я
о"
га
о.
в
га
S я I
я (- ч
" ,s
ч ч
о о
Я °
я я
в
ч,
о в я
Я
■ О) я
о га
ag
га га
OJ
я в
а I
5 §
go
OJ OJ
S К
<u
to
^ i
в о
О <и
Ч Ч
С
га
В о
It
3 га
я о
о S
СП Ч
В га ,
I о '
о <и
g О S
g
I В dJ
ÜJ В
d *
я в OJ
, в а.4
Й «
1 га i_ я
I Оч о PJ
I о >, га
( м к о.
g ^ I
S OJ g
- о >о
о UJ
.«Ли
' о S
о в
s- я
Sil
га ^
_ Ч >,
S га ч
й о о
ia
m га
г- BJ д
>> в Д
Н Ч
га о
CJ о-ю
OJ о
я м ш
чё Si g §.i
^ а я я 'S
3 я я g о о
4 я ^ °Е ?
га гя
5 Sg-g g ö_
.-;f-3ras>>rara
ОЭовйЧЧга»
I
С
а
о
■^^ S
4 .=
я
5 а
о ^
о
о
я°-§
я М щ
о к Si
о 3
■■J
OJ и 3
3 3 £^
га ч Я
33
ь _
а. о -
е
я
а Ö
! я га
goj
! Л
; я с
: о о
; ч ч
Si га А
Й со
а"
« я «
§s
к о
OJ я о
° о 3
£5 °
S й X
н в «
а gg
S S ч
&i ч О
1 =
о с
а «
оа ^ v
о
а
о
о,
о
X
,3? s s
■5-5
§ g s
о
— О)
s S
а о,
та
'II
к " и,
я а
я о
0J с
-г: >.
is ч
я о
gel
а VO с
>> о £
я Ог=?>
а о я S
я О) m d
я S CD
4 с.
я «я
^ ™ я
5 1 я g л
Sis
с -е- [; \с а.
к
я
а.
8 1>
я
я о
Ь U
я
& я
о Ч
<u о
Ч я
•я
О)
я
т а
ч ч
я о
ovo
о
а
о
о <и
2 3
g g
^ 2
1"я
g а *
•я я
о « ю
я я я
0) о
\о о а. и
t, о =я
о о
s-я я
о й я
Ч СЯ «J
я я
J0 " |i
i""
Ч о
11
ч
о
я VO
£ » s я
а -я я
р <и =^ а)
1ч ^
£?и я
СЕЗ
2 " ö
й я
с ЯЮ о f- £
М 3
я о с
о §
Си я S
2 я 2
Km-*
га ск; «
3 я «
я ^ г? cu й
с я э С1
Is
н со
о го
§2
« 2 § .2 I
3 5-е-!=~
я s O.s а,
я о я ^ о
о ч я ■
о 1»
Sei
со ^
о
а 's."? о
C.1JJ
я
я
R 1
я 8S
и о
2 о-
с н
а. я о ^
S- н я я
я л 2 я ^ к о
s ч ч я и я x
ч 2 а^^я я э-
гййэзячяя
Ii
та CT)
X
SS"
о" g
к и 2
g S Ё
Е UJ
g о я
о «'Л
5
га
ь
S
к
о
с
CI,
о
£
о
ш
о
g
со
3
=f
к
i-
п
<и
3
в
о-
?i
га
Он
го
X
га
Я
I
го
н
'СО
ю с-
С-1 CT)
S с
m та
ч В
+ + + + +
+
+ +
— о—<tO — OOOOOOOTf'C'b-
со —г CD to" оо" cri см" о о о <><
Ot--00 — СЧ — tDCDC£>c£)CO_COO^
.*^C30-^lOC0t-^^ — ООО--—-
Г^ЮС001М1ЛЮ(МС>) — into
(М ^ _ _ CM
— CJOCDTflOCD-rJ^CMOCOCO
CM
Ю о ^ LO CM
fM ^ 00 CM tM
—■CDCN'MCDCMOOOOOOCOCOCMGO-^O ^"^^
Со"с5Г^1ЛЮС---^СМО!ГОСО — (MtM
CO-^OOCO-^OOCMt^CMCJ^CJ^crjr^-^O t-^co
ООООСОСГОЮЮЮСОСЗОСЮСЗО
CO —
OCO<Dt^t^----^t^^tOCO
in CO o" o" f-T t-^ о CO те CO
— CM —> — —
ao
O) CO
(M ^
CM Ю
— Ti-CX)COCOCMCM<MCO—"^—^
Ю
о
о.
о
n
.я
3 ,я
§ g
й о
о 0J
!Я
3
я
в
CL)
га
о
я
3
о
>в в
го
!<!
g
1 й ч
g о
со и ю
о.
о
а £
о. ю
1^
Си
S
R
га
о
го
!<:
го
га
а
н
о
о
о
га
!<:
X
о.
я
го
с
и
в
о
га
U
Я
к
со
к
га
го
го
га
га
\£
ю
я
^
я
о
в
о
о
к
^
га
я
о.
го
го
CJ
га
и
и
и
с (Г)
га
в
я й _
« S
= 1
ГО
к
го
к
и
ш
Sei
со со со
I 1 I
Tt"
со со (М со <м со
I I I I I
со ■*
со со <м см
I I 21 1
-«^ со — СО
с
3
о.
я я
р
в Н го
§§р
ч со:
о си
я яС
56
2. Структура города и фауна
57
(Heinrich, 1978) также могут использовать здешние пищевые
ресурсы.
Растительный мир обочпны испытывает влияние различных
антропогенных факторов, например соли для таяния (Rüge, 1972,
1974; Evers, 1971; Ernst, Feldmann, 1975), пыли, тяжелых
металлов, фотосмога (Steubing, Kirshbaum, 1976), свинца (Sommer
et al., 1971; Smith, 1971; Davies, Hohnes, 1972; Lerche, Brechle,
1974), кадмия и никеля (Lagerwerff, Specht, 1970). Эти
загрязнения могут использоваться некоторыми видами животных как
ресурсы, о чем свидетельствует присутствие здесь галофильных
видов (Maurer, 1974; Williamson, Evans, 1972; Gish, Christensen,
1973).
Дождевые черви {Lutnbricus terrestris) содержали в зоне
действия выхлопных газов свинца в 11 раз, кадмия в 5,9 раза, меди
в 5,5 раза и цинка в 2,7 раза больше, чем в контроле без таких
Рис. 10.
Распространение Phosphuga atrata в
раз.чичных зонах (W, I,
R, G, S) обочины дороги.
Черная полоса справа
означает дорогу (по
Mader, 1979); N —число
особей
30
20
10
W
R
Зона
G S
выбросов (рис. 11) (Czarnowska, lopkiewicz, 1980). У жужелицы
Pterostichus cupreus вблизи проезл<ей части содержание свинца
было в 7—8 раз выше, чем в ее окрестностях, однако у стафпли-
на Ocypüs olens подобной разницы не обнаружено (Geiler, 1976).
Кюнельт (Kühnelt, 1977) приводит содержание свинца 13,7 мг/1 г
сухой массы для равноногих Porcellio scaber и Armadillidiuin
vulgare и 7,9 мг/г для уховертки Forficula auricularia (на
городской окраине — 2,5 мг/г). Клаусен (Clausen, 1984) указывает на
то, что концентрация свинца у пауков Araneus umbratus четко
отражает содержание этого элемента в воздухе и коррелирует с
его содержанием в лишайнике Lecanora conizaeoides.
Отсутствие явной корреляции между содержанием свинца в почве и
плотностью проволочников (личинок жуков-щелкунов) отмечает
Новаковски (Nowakowski, 1982). Шмидт (Schmidt, 1981)
установил, что самцы ящерицы Lacerta muralis в Саарбрюккене
(ФРГ) содержали свинца и кадмия больше, чем самки, пр]1чем
с возрастом это расхождение между ними увеличивалось. У сам-
58
2. Структура города и фа у на
цов ящерицы живородящей [Lacerta vivipara) он обнаружил
отчетливые градиенты содержания тяжелых металлов в
зависимости от расстояния до проезжей части. В центральной части
Саарбрюккена у главного вокзала важнейшие группы животных
(многоножки, равноногие, дождевые черви и слизни) содержали
больше свинца и кадмия, чем питающаяся ими L. muralis
(Schmidt, 1981).
Браун и др. (Braun et al., 1981) наблюдали очень сильное
поражение яблоневой тлей {Aphis pomi) боярышника однопес-
тпчного на разделительной полосе и обочине автобана с очень
интенсивным движением (табл. 17).
I 12
I.
I
1
4h
i^—
of/ovsa
12
1
24
1
36
1
48
Цаалетоть от дороги, м
Рнс. 11. Содержание кадмия (млн-') в сухом веществе дождевых червей
и почвы на различно удаленных от дороги участках (США) (по Müller, 1981)
Причиной усиления присутствия яблоневой тли были наряду
с благоприятными температурными условиям,и биохимические
изменения растения-хозяина (изменение питания тли),
вызванные загрязнением на автобане (выхлопные газы, соль, воздущ-
Таблнца 17. Максимальная пораженность зеленой яблоневой тлей (Aphis
pomi) побега боярышника однопестичного (по Braun et al., 1981)
Год
Разделительная
полоса
Обочина
Контрольный
участок (удален от
проезжей часта
на 200 ы)
1979
1980
234
489
91
163
5
46
2. Структура города и фауна
59
ные потоки от проезжающих машин, пыль) (ср. Braun, 1980;
Flückiger et al., 1978, 1978; Dohmen et al., 1984).
Абиотические факторы проезжей части иногда существенно
отличаются от господствующих на прилегающей территории
(сильное прогревание, усиленный поверхностный сток, слабое
испарение) и создают особые микроклиматические градиенты, как
видно из рис. 12, 13 и 14, отражающих данные по освещенности,
температуре поверхности почвы и испарению. Согласно Мадеру
(Mader, 1979), микроклимат проезжей части и обочин сходен со
степным. Вдоль обочин, представляющих собой лентовидные ме-
27 -
25 -
23 -
21 -
13 -
п
15 -
13 -
CIB 9 10 11 IS 13 /4
15
ч
IB П - 18 13 го 21 22
Рис. 12. Изменение температуры в течение дня (03.06.1980) под
булыжниками мостовой (М) и в садовой почве (П) на глубине 8 и 14 см: В
—температура воздуха; кружки внизу — продолжительность солнечного освещения; СЕВ —
среднеевропейское время (по Haeseler, 1982)
стообитания, существуют специфические ценозы (с. 51).
Существенную роль среди жужелиц играют здесь (Mader, 1979) эв-
риойкные лесные (например, Pterostichus niger, довольно часто
пересекающий дороги) и стенойкные полевые виды
(обследовалась дорога, проходящая через лесной массив). Маурер (Maurer,
1974) показал, что на видовое разнообразие по обочинам сильно
влияет загрязнение выхлопными газами. Очевидно, существует
следующая цепочка воздействий: изменение абиотических
факторов под влиянием проезжей части->-нзменение растительных
сообществ-*-формирование зональных зооценозов под влиянием
60
2. Структура города и фауна
специфических механических воздействий и вредных выбросов.
Градиент условий от окружающего леса к обочине показан
на (примере снижения численности .и биомассы жужелиц (вблизи
проезжей части доминируют более мелкие виды) (рис. 15)
(Mader, 1979). В этом исследовании отмечено 9 видов, которые
избегают дорог, 15 видов, обитающих вблизи них и 13 видов без
явно выраженной приуроченности (рнс. 16). «Друзьями дорог»
были, например, полевые виды Bembidion lampros, Loricera pi-
licornis, Agonum muelleri и А. sexpunctatum. Отдельные зональ-
Рис. 13. Температура на по-
вер.хности различных
субстратов в летний солнечный
день: А—асфальт; П —
песчанпк; Т — травяной
покров газона; В — вода (по
V Kessler, 1971)
ные зооценозы более пли менее резко отличаются друг от друга
(табл. 18).
При помощи почвенных ловушек псследовали (Haeseler, цит.
по Mader, 1979) фауну жуков на разделительных полосах авто-
банов. Обнаружено в среднем 259,2 особей на ловушку. На
солнечной северной обочине собрано 233,1 особей на ловушку, на
теневой южной — 96,9. Повышенное присутствие на
разделительной полосе свойственно прежде всего видам семейств Staphyli-
nidae, Anisotomidae, Nitidulidae и Elateridae; представители Ca-
rabidae, Curculionidae, Silphidae и Tenebrionidae чаще
встречались на обочине.
2. Структура города и фауна
61
50 40 30 20 10
40 50 м
Piic. 14. Изменение факторов окружающей среды при движении от проезжей
части (середина) к обочине: Ос — освещенность; Т — температура на
поверхности почвы; Ис —испарение (по Mader, 1979)
A20Ü
5 6 7 8 3
Серия
Рис. 15. Изменение биомассы 'Б) и числа особей (N) жужелиц в 9 сериях
ловущек по мере приближения к дороге (по Mader, 1979)
Барьерный эффект дорог, по-видимому, обусловлен прежде
всего следующими причинами (рис. 17):
— пзменение важпейщих микроклиматических факторов на
расстояние до 30 м с 'каждой стороны проезжей части;
— структура биотопа может действовать как механическая
преграда;
— многочисленные выбросы и нарушения, например шум,
яыль, ночное освещение, выхлопные газы, засоленность почвы,
кюветы;
62
2. Структура города и фауна
— непостоянство растительности вследствие обработки
гербицидами и механических воздействий;
— создание биотического барьера из растений чужеродных
формаций и появление новых видов животных;
— .между видами различного происхождения и в условиях
постоянной иммиграции возникает особо сильная конкуренция за
важнейшие ресурсы в зональном зооценозе;
— гибель при пересечении дороги, сдувание потоками
воздуха более мелких видов.
4 5 Ä 7
Серия ловушек
Рнс. 16. Обилие АЬах
parallelepipedus (А.а.) и
Pterostichus niger (Р.п.)
на разном удалении от
дороги. Черная полоса
справа означает дорогу
(по Mader, 1979)
Четырехрядная дорога — такой же эффективный барьер
против расдространения мелких млекопитающих (и жуков), как и
водная полоса вдвое большей ширины (Mader, 1979; Oxley et al.,
1974). Если к дороге прилегает не лес, а открытое
пространство, ее изолирующее действие может значительно снижаться.
Мощеные тротуары, места стоянок транспорта и улицы
бывают предпочтительными местообитаниями для различных
жалящих перепончатокрылых (Haeseler, 1982), особенно для
некоторых видов, обитающих в песке. На некоторые виды благопри-
2. Структура города и фауна
63
Таблица 18. Сравнение ассоциаций различных зон (W, I, R, G, S), обочины
дороги (по Mader, 1979). (H-diff и Qs отрицательно коррелируют)
(г = -0,84, Р<0,01)
Зона
Q,
H-diff
Жужелнцы
Стафнлнннды
Жужелицы
СтафплиЕиды
W — I
88,1
77,2
0,01
0,12
I-r
78,3
61,8
0,05
0,16
r-g
62,2
12,0
0,23
0,64
g-s
32,8
18,2
0,09
0,65
ят«о влияет высокая температура, которая под уличным
покрытием значительно выше, чем в открытом грунте (рис. 12). По
сравнению с окружающей местностью здесь наблюдается
ускорение развития и более раннее появление насекомых, а иногда
частичное образование в течение года второго поколения. Кроме
того, по-видимому, благоприятное влияние испытывают здесь
виды со склонностью к о1бщественному образу жизни. В такп.х
местообитаниях отмечено 22 их вида (П — паразиты) из семейств
Formicidae (2), Mutillidae (1П), Pompilidae (1), Sphecidae (9),
Andrenidae (1), Anthophoridae (1П), Halictidae (3, ЗП) n Melit-
tidae (1) (Haeseler, 1982). Чаще всего встречались: Laslus niger,
Crabro peltarius, Crossocerus wesmaeli, Oxybelus bipunctatus,,
0. uniglumis, Andrena barbilabris, Nomada sheppardana (y
Lasioglossum sexstrigatum), Halictus rubicundus и Lasioglossum sex-
strigatum. У последнего вида было найдено 34 гнезда на 1 м-,
у С. peltarius — 31 (180 на 15 м^); минимальное расстояние
между гнездами составляло 1,4 см. Рудник (Rudnick, 1973)
наблюдал 35 гнезд песчаной осы Cerceris arenaria между булыжниками
мощеного тротуара на его отрезке длиной около 50 м, а Хезелер
(Haeseler, 1972) обнаружил в таком местообитании 13 гнезд этой
осы на площади 1 м^. Здесь же на 1 м^ было 86 гнезд Halictus
sexstrigatus (между плитами тротуара — 70) п 32 гнезда Andrena
barbilabris. Кормовой базой для перепончатокрылых этого
местообитания являются прежде всего плодовые деревья, а также
нектар других цветковых растений. Некоторые часто
встречающиеся виды роющих ос (Sphecidae) кормят своих личинок
определенными группами! двукрылых, которых в городе особенно
много (табл. 19). Видимые снаружи признаки гнезд более или менее
видоспецпфичны; расстояние между булыжниками и основным,
грунтом очень существенно влияет на видовой состав.
О фауне гиполитиона транспортных путей почти ничего не
известно. Только Тишлер (Tischler, 1966) провел анализ этого
местообитания на примере мощеного пространства перед домом,
обнаружив многообразную фауну (71 вид, не счптая клещей и
ногохвосток, табл. 20), и выявил некоторые закономерности еже-
6*
2. Структура города и фауна
II
5: ci.
Ii
Ii:
"5
14
Co
Образование ост;-'вньш участков,
расчленение -'реала
Изоляи,ия
Последствия
Феномен
то »5
л &
III
а is <о
с 5 а
СНГ) rrj
1
1
1
'gist
I'll
sill
^ 55
I
I
§ war
iiii
•
Дорожное строительства
Воздействующие
щапторы
вмешательство
человека
Рнс. 17. Схема воздействия факторов, ведущих в резу.аьтате строительства
дорог к изоляции популяций животных, и вытекающие отсюда последствия (по
Mader, 1979)
ГОДНОГО заселения, распределения, смены аспектов и
происхождения отдельных видов. Животные гиполитиона влаголюбивы,
избегают света, часто обладают тигмотаксисом и в большинстве
случаев плохо апособны зарываться в грунт. Показано
(Schönborn, 1961; Tischler, 1966), что гиполитион включает три
структурных элемента, различающиеся своей фауной:
2. Структура города и фауна
65
Таблица 19. Кормовой рацион роющих ос (Sphecidae) в Ольденбурге
(по Haeseler, 1982)
X
S
E
3
?^
а)
i
Q
>^
2
:i
ж
з:
■ aj
3
ja
3
g
D-
а
03
X
'S
о
1
X
и
3
X
S
X
к
О
o.
X
a;
о
ь
ё
Ч
га
«
о
iö
сз
ГО
X
m
о
н
о
CJ
а
Ol
о*
с
о
i<
X
Ö
н
о
н
X
о
&
О
о
ю
№
UJ
ш
Crabro peltarius
6
2
6
1
33
10
2
3
2
1
Crossocerus
elongatulus
2
2
8
4
2
1
Oxybelus bipunctatus
18
77
5
1
0. uniglumis
5
7
1
1
15
1
1
Таблица 20. Доминирующие виды (сумма за несколько лет) гиполитиона
мощеного пространства перед домом (по данным Tischler, 1966)
Вид
Обилие
Примечания
Lasius niger (L.)
(переЭвдоминант
(численВсеядный
пончатокрылые)
ность не подсчитыва-
лась)
Allolobophora chlorotica
20,5%
Сапрофаг
Sav. (олигохеты)
Discus rotundatus Müll.
13,6%
Нижняя сторона камня.
(брюхоногие)
фитофаг, сапрофаг
Porcellio scaber Latr.
9,7%
Сапрофаг, синантроп
(равноногие)
Plathyarthrus hoffmann-
6,3%
Сапрофаг, мирмекофил
seggi Brdt.
(равноногие)
Lumbricus rubellus Hfm.
5,2%
Сапрофаг !
(олиго.хеты)
Armadillidium vulgare
4,5%
Сапрофаг, синантроп
Latr. (равноногие)
Cijlindroiulus nitidus
3,1%
Сапрофаг, синантроп?
Verh. (двупарноногие)
Lithobius forficatus L.
3,1%
Зоофаг
(губоногие)
Trichoniscus pusillus
2,9%
Сапрофаг
Verh. (равноногие)
Phalangium opilio L.
(се2,5%
Зоофаг, однако не в
нокосцы)
гиполитионе,
синанScutigerella immaculata
2,3%
троп
Фитофаг, сапрофаг
Newp. (симфилы)
Lorocera pilicornis F.
2,0%
Зоофаг
(жужелгщы)
5-2
66 2. Структура города и фауна
2.2.3.2. ЖЕЛЕЗНОДОРОЖНЫЕ СООРУЖЕНИЯ
В зоне железнодорожных сооружений (рельсовый путь,
вокзалы, путевые сооружения), за исключением зданий, много
открытого грунта. Он характеризуется антропогенными
наслоениями камня, гравия, песка ai незначительной долей естественной
почвы. Наблюдается сильное поверхностное нагревание насыпей.
Часто применяются гербициды и соли для таяния, горючее и
смазочное масло образуют локальные загрязнения. Флора здесь
очень богата. Преобладают солеустойчивые однолетние и
теплолюбивые многолетние виды. Наряду с этим произрастает
много неофитов и адвентивных растений, например Salsola kali,
Amaranthus albus, Eragrostis poaeoides, Euphorbia virgata, виды
ослинника, Vulpius myuros, Brassica napus. Распространены
различные очитки. Виды с глубокой корневой системой могут
выживать после применения гербицидов. По бокам
железнодорожного полотна и на бездействующих путях появляются деревья
(тополь, роб'Иния, береза).
Данных по фауне этого 'местообитания сравнительно
немного. Гримм (Grimm, 1961, 1962) занимался птицами,
населяющими в Берлине вокзалы метро .и надземной железной дороги. В
последнем случае отмечено гнездование кольчатой горлицы (Ditt-
berner, Mönke, 1980). Каменка встречается на бедных
растительностью теплых путевых сооружениях (Gnielka et al., 1983).
Железнодорожные насыпи могут быть подходящими
местообитаниями для дикого кролика (Schweiger, 1960/61) и ящериц рода
Lacerta. Клауснитцер (Klausnitzer, 1957) наблюдал колонию пчел
Dasypoda plumipes возрастом не менее 50 лет на грузовой плат-
— нижняя сторона камня (литоф-нльные виды);
—: пустоты (место дневного отдыха или зимовки видов,
живущих на поверхности);
— грунт и детрит, который может отсутствовать или
.исчезать со временем (почвенные В1иды, часто сапрофаги).
Существуют также виды, не связанные ни с одним из этих
структурных элементов. Различные виды образуют между собой
определенные связи, прежде всего в форме пищевых цепей.
Городские мосты более или менее регулярно заселяются
некоторыми видами животных, напри-мер пауками Cyphepeira se-
ricata и Meta merianae, которые заселяют прежде всего мосты,
переброшенные через водные пространства (Sacher, 1983). В
Галле «а'блюдали (Gnielka et al., 1983) колонии городской ласточки
на стальной конструкции моста через реку Заале (1974 г.—
8 гнездящихся пар, 1975 г.— 14, 1977 г.— 50). Здесь же
гнездились трясогузка белая и домовый воробей.
2. Структура города и фауна
67
форме вокзала. Швайгер (Schweiger, 1960) обнаружил на
кустарниках железнодоролчной насыпи в Вене три вида саранчевых
(Acrididae), что свидетельствует о важности таких трасс как
путей имш1грацип. В старых гниющих железнодорожных шпалах
может жить жук-узкокрылка Nacerda ferruginea (Oedemeridae).
Сосна обыкновенная
очень
суаая
Ci/хая
Умеренно
сцхая
Умеренно
Света я
'Свеушя
Умеренно
влажная
Бла?нная
Умеренно
сырая
Сырая
Очень
сырая
Брина черная
/|
^ i:
Ива козья j:
(+Явор)
^ / г' ?.
i Тополь I i
V к J
Ильм
Соответствующие условия отсутствуют
Сильна Кислая Умеренно Слабо Ней- Щелочная
кислая кислая кислая
тральная
Рис, 18. Зависимость важьгйших древесных пород в центре Западного
Берлина от влажности и рН почвы (по Sukopp et al., 1980)
2.2.4. Озелененные площади
Озелененные площади в городе, несомненно, являются
центрами поселения животных. Обнаруженные здесь зооценозы
сравнительно близки к природным 41 часто богаты видами, поскольку
в этих местообитаниях во многих случаях имитируют, а ашогда
намеренно или ненамеренно сохраняют первоначальные
природные условия (островки леса) (pine. 18).
68
2. Структура города и фауна
2.2,4.1, ПАРКИ
Парки представляют собой комплекс сочетающихся в раз-
Л1ИЧНЫХ количественных отношениях газонов, кустарников,
деревьев и иасаичдений декоративных цветов. Обычно много
открытого грунта. Часто имеются естественные почвы, местами
насыпной естественный субстрат; в большинстве случаев
вносится умеренная доза удобрений. Парки оказывают смягчающее
воздействие на городской климат, выравнивают температуру,
осаждают ш связывают вещества, загрязняющие воздух. На
инфильтрацию воды их влияние почти не оказывается, но частично
повышается влажность. Биоциды и соли для таяния
применяются здесь лишь в небольших количествах. Видовое разнообразие
газонов зависит от частоты скашивания, характера почвы,
вытаптывания, возраста сообщества, удобрений п затенения. Доля ав-
Рнс. 19. Доля числа видов
разлпчны.х отрядов сухопутных
насекомых в парке Шванентайх в
Лейпциге по учетным данным в 1978 г.
Общее число впдов — 357
(подсчеты Klausnitzer et al., 1980b)
тохтонных видов определяется размером, возрастом газона и
уходом за ним. Часто встреч1аются интродуцированные деревья
и кустарники. Парки являются рефугиями для исчезающих
местных видов, а также археофитов и неофитов; кроме того, это
острова существования в городах лишайников. Доминирующее
газонное сообщество— Bellidetum; типичны также фитоценозы класса
Plantaginetea (в особенности ассоциация Lolio — Plantaginetum),
а также союза Arrhenatherion и опушечные сообщества; местами
встречаются близкие к природным заросли кустарников, в
ландшафтных парках возможны остатки естественных лесов.
В соответствии со структурой биотопа, составом
растительности 'И состоянием парка его фауна бывает относительно богата,
свидетельством чего является црежде всего наличие некоторых
групп членистоногих (рис. 19) (Loibl, 1953; Wlodarczyk, 1960;
2. Структура
города и
фауна
69
Таблица 21.
Эвдо.мпнантные п доминантные
пауки лейпцигскнх городских
парков (Koslowski et al, 1980)
Парк
Эвдрмннанты
Доминанты
Парк Клара
Diplostyla concolor
51,8%
Pachygnatha clercki
8,9%
Цеткин
Micrargus herbigradus
5,4%
Troxochrus scabriculus
5,4%
Фриденс-
Pachygnatha degeeri
28,8%
Erigone dentipalpis
9,8%
парк
Pardosa prafivaga
14,7%
Pardosa pullata
8,0%
Pardosa palustris
10,4%
7,3%
Шиллер-
Erigone dentipalpis
36,5%
Troxochrus scabriculus
парк
Erigone atra
29,2%
Lepthyphantes tenuis
6,1%
Парк Шва-
Diplocephalus crista-
20,9%
Dyplostyla concolor
9,1%
нентапх
tus
Bathyphantes gracilis
19,1%
Lepthyphantes tenuis
14,7%
Troxochrus scabricu-
10,0%
lus
Czechowski, 1980a, 1982; Klausnitzer et al., 1980b, 1982a, 1983b).
Имеются характерные виды ii, возможно, такл^е характерные
зооценозы, лучше в'сего изученные на орнитофауне (Peters, 1963;
Mulsow, 1978; Elvers, 1981; Luniak, 1981, 1983; Schmidt, Stein-
bach, 1983).
В случае различных исследованных групп членистоногих
оказалось, что доминируют почти всегда эвриойкные виды, часто с
широким географическим распростраиением. Преобладают
мелкие и обладающие высокой подвижностью формы. Благошрият-
ны условия и для видов, более или менее не зависящих от
мертвого покрова почвы, на которых его регулярное удаление не
оказывает отрицательного влияния (зимовка в почве, весь цикл
развития на деревянистых растениях). Мозаика различных
биотопов внутри одного п'-рка часто также сказывается на заселении
его лсивотными.
В четырех парках в центре Лейпцига Кословски и др.
(Koslowski et al., 1980) обнаружили 49 видов пауков, но в каждом
парке преобладали характерные только для него виды (табл. 21).
У пауков происходит отбор мелких в.идов-«аэронавтов» в
качестве типа, идеально подходящего для многих местообитаний
зеленых массивов. Они могут легко заселять островные
городские биотопы и благодаря вертикальному и горизонтальному
расселению избегают помех. Они переносят такие механические
нарушения, как косьба, обработка почвы, вытаптывание, очевидно,
лучше, чем крупные виды, поскольку нуждаются в очень мелких
убежищах. j
70
2. Структура города и фауна
В городском парке в центре Лейпцига обнаружено 4 вида
равноногих: Armadillidium vulgare (51,1%), Porcellio scaber
(34,0%), Oniscus asellus (12,8%) и Hyloniscus riparius (2,1%).
Среди многоножек в этом же парке преобладал вид Allajulus
londinensis. Выявлено 11 видов ногохвосток, среди которых
преобладали Isotoma viridis, Lepidocyrtus cyaneus и Orchesella vil-
losa (Klausnitzer et al., 1980b).
Рис. 20. Процентные до.чп различных
видов стафнлинпд в парках Иоханна (П)
и Артур-Бретшнайдер (АБ) в Лейпциге
(по Klausnitzer et al, 1982): I ~Anoty-
lus tetracarinatus; 2 — Xantholinus
linearis; 3 — Tachinus corticinus; 4 —
Atheta fungi; 5 — A. crassicornis; 6 — Pella
limhata; 7 — Omalium rivulare; 8 —
0. caesum; 9 — Anoiylus rugosus; 10 —
Hypomedon s melanocephalus; 11—Ami-
scha analis; 12 —Oligota pumilio; 13 —
Cardalia obscura; 14 — Aloconota grega-
ria; 15 — Tachinus rufipes; 16 — прочие
общие виды; 17 — Anotylus sculpturatus;
\Ъ — Platystethus arenarius; IQ — Plata-
raea brunnea; 20 — Anotylus clypeonitens;
21 — Drusilta canaliculata; 22
—Anotylus mutator; 23 — прочие виды,
обитающие только в одном из парков
Благодаря обилию цветков и тлей в парках особенно часто
встречаются мухи-журчалки. Так, в колониях тлей на 8 большей
частью местных видах декоративных кустарников обнаружено
(Ziarkiewicz, Koslowska, 1973) 11 видов журчалок, а на бузине
черной (Sambucus nigra) —4 вида. В одном из парков
Амстердама ван дер Гот (van der Goot, 1978) от.метил в течение года
46 видов журчалок, в том числе Meiasyrphus lundbecki — вид,
новый для фауны Нидерландов.
2. Структура города и фауна
71
Таблица 22. Эвдоминантные (эв) и доминантные (д) стафилиниды
в различных парках Лейпцига (по Klausnitzer et al., 1982). сд — субдоминант,
р — рецедент, ср — субрецедент, + — без количественных данных, И — парк
йоханна, АБ —парк Артур-Бретшнайдер, ПШ —парк Шванентайх
Вид
аб
ПШ
Anotylus sculpturatus (Qrav.)
А. clypeonitens (Fand.)
A. tetracarinatus (Block)
Xantholinus linearis (Ol.)
Tachinus corticinus (Qrav.)
Aloconota gregaria (Er.)
Amischa analis (Grav.)
Plataraea brunnea (F.)
Aiheta fungi (Grav.)
A. crassicornis (F.)
Pella limbata (Payk.)
эв
p
сд
сд
сд
сд
д
p
д
д
Д
Д
cp
сд
д
д
эв
д
+
+
+
+
+
+
в самом центре Лейпцига были обнаружены примечательные
в фаунистическом отношении виды стафилинид (Klausnitzer et al.,
1980а, 1980b, 1982). Здесь часто встречались виды, считающиеся
обычно «редкими», но наблюдались и обратные случаи. В парке
Шванентайх собрано 53 вида, в парках Иоханна и
Артур-Бретшнайдер—по 61. В каждо-м из двух последних парков
обнаружено 23 вида, (которых не было в других imecrax. Наряду с
качественными различиями наблюдались и значительные
количественные расхождения во встречаемости отдельных видов (рис. 20).
Преобладающее большинство обнаруженных стафилинид были
эвритопными или предпочптали открытую местность; типичные
лесные виды почти не встречались (табл. 22). Богач (Bohac,
1986) сообщил о преобладании в рудеральных биотопах Праги
видов с г-стратегией (независимая от плотности популяции
высокая смертность, сильные колебания плотности, быстрое
заселение новых территорий, короткий цикл развития), например, из
родов Oxyfelus и Aleochara. В двух парках Праги доминировали
(>5%) Omatium rivulare, Rugilus rufipes, Othius punctulatus,
Xantholinus linearis, Heterothops dissimills -и Atheta fungi (Bohac,
1985). ^
Орнитофауна парков имеет характерный видовой состав
(рис. 21, табл. 23). Луняк (Luniak, 1981) приводит 66 видов
гнездящихся птиц для парков Варшавы (в самом богатом ими парке
насчитывалось 29 видов, с плотностью 104 пары на 10 га). На
мелких парковых терр.итор'иях в центре города высокая плотность
достигается за счет в^идов-визитеров как во время гнездования
(до 570 особей на 10 га), так и зимой (до 709 особей и 192 кг
72
2. Структура города и фауна
Таблица 23. Виды птиц, гнездящихся в городских парках
Лейпцпг
Индекс
Вид
Внутреннее кольцо.
Фрнденспарк,
23,2 га
(по
Sae9,7 га (Ladusch
(Seidel, рукопись)
mann, 1970)
et
al., 1980)
%%
%%
4
Черный дрозд
22
22,7
25,9
30
12,5
31,6
4
Обыкновенный скворец
10
10,3
11,8
12
5,2
12,6
1 4
Зеленушка
3
3,1
3,5
8
3,4
8,4
4
Большая синица
6
6,2
7,1
8
3,4
8,4
4
Зяблик
1
1,0
1,2
4
1,7
4,2
5
Лазоревка
2
2,1
2,4
8
3,4
8,4
4
Вяхирь
10
10,3
11,8
3
1,3
3,2
1—4
Кольчатая горлица
7
7,2
8,2
1
0,4
1,1
1-2
Домовый воробей
3
3,1
3,5
2-4
Зеленая пересмешка
8
8,2
9,4
3-4
Серая мухоловка
2
0,9
2,1
2
Славка-завирушка
4
4,1
4,7
3-4
Певчий дрозд
2
0,9
2,1
3-4
Садовая горихвостка
2
2,1
2,4
1
0,4
1,1
3-4
Обыкновенный
поползень
2-3
Полевой воробей
Черноголовая славка
3
2
1,3
0,9
3,2
2,1
3-4
Садовая славка
1
1,0
1,2
2—4
Большой пестрый дятел
Зарянка
2
Канареечный вьюрок
4
1,7
4,2
1
Щегол
4
1,7
4,2
3
Камышевка
4
4,1
4,7
Пеночка-весничка
2—4
Пеночка-теньковка
4
Пеночка-трещотка
Галка
Обыкновенная пищуха
2
2,1
2,4
биомассы на 10 га). Разнообразие видов и плотность заселения
зависят прежде всего от структуры древостоя, возраста
насаждений, количества мест гнездования для птиц, гнездящихся в
дуплах, размеров и положения территории, наличия кустарникового
яруса, укрытий, зданий и водоемов, предоставляемого людьми
корма, растительной и животной пищи. Во время
вегетационного периода на открытых парковых площадях преобладают сизый
голубь, обыкновенный скворец и домовый воробей (визитеры,
добывающие корма). Зимой доминируют грач (фото 3), галка,
домовый воробей, сизый голубь и большая синица. В этот сезон
та'м, где человек регулярно подкармливает птиц, может скапли-
2. Структура города и фауна
73
(пояснения см. табл. 8)
Карл-Маркс-Штадт
Централер-Клайн парк,
6,5 га (Saemann, 1973)
%%
Шлостай.хпарк
10,7 га
%%
Гроспарк, 23.5 га
%%
18
27,7
20,5
39
28,0
20,5
15
6,4
7,5
9
13,8
10,2
11
10,3
7,5
46
19,6
23,1
23
35,4
26,1
28
26.2
19,2
2
0,9
1,0
2
3,1
2,3
4
3,7
2,7
15
6,4
7,5
2
2,1
2,3
17
15,9
11,6
14
6,0
7,0
2
3,1
2,3
8
7,5
5,5
13
5,5
6,5
1
1,5
1,1
2
1,9
1,4
4
1,7
2,0
20
30,8
22,7
18
16,8
12,3
6
9,2
6,8
5
4,7
3,4
2
0,9
1,0
1
1,5
1,1
3
2,8
2,1
1
0,4
0,5
1
1,5
1,1
2
1,9
1,4
2
0,9
1,0
1
1,5
1,1
2
1,9
1,4
1
0,9
0,7
5
2,1
2,5
3
1,3
1,5
1
1,5
1,1
3
2,8
2,1
4
1,7
2,0
4
3,7
2,7
5
2,1
2,5
6
2,6
3,0
1
0,9
0,1
4
1,7
2,0
10
4,3
5,0
8
3,4
4,0
7
3,0
3,5
7
3,0
3,5
4
1,7
2,0
ваться 83—148 кг п.х; биомассы на 10 га (сизый голубь, грач,
галка, домовый воробей), а в местах, бедных кормом — 20—25 кг
на 10 га (Luniak, 1981).
2.2.4.2, БОТАНИЧЕСКИЕ САДЫ
Ботанпческпе сады в зоожологпческом отношении мало
отличаются от парков. Однако особенностью их, по-впднмо'му,
являются большое число видов растений, среди которых много ин-
трудоцированных, на очень малой территории и высокое
разнообразие микробиотопов, из-за своих небольших размеров
непосредственно контактирующих друг с другом (множество границ
соприкосновения).
74
2. Структура города и фауна
Шефер (Schaefer, 1973) обнаружил в ботаническом саду в
Киле 10 видов сенокосцев (табл. 24); преобладали лесные
таксоны (60,7%) и гемерофилы (71,3%). Для этих 'членистоногих
весьма важно наличие :рефугий, так как на них особенно
отрицательно воздействуют механические нарушения.
Вайс и Сарбу (Weiss, Sarbu, 1977) указывают на то, что ако-
логический оптимум вида (можно определить по более высокой
доли самок (пониженному соотношению полов) (Müller, 1970).
Это, очевидно, касается Odiellus bieniaszi (табл. 25). Самки
менее активны и больше привязаны к одному месту, т. е. плотность
их активных особей возрастает по мере приближения к
предпочтительному биотопу 'Сильнее, чем у самцов.
Рис. 21. Птицы, гнездящиеся в 16 парках Западного Берлина (ГП —
гнездящиеся пары). Плотность заселения — белые колонки; число видов
(S)—заштрихованные колонки; видовое разнообразие (Hs) — кривая. Слева
направо—снижение антропогенного влияния (по Sukopp et al., 1980)
Учеты в ботаническом саду Киля показали 52 вида пауков
(Schaefer, 1973) (табл. 26). В значительной степени здесь отсут-
ствовал1и виды-охотники ,и засадчики, например представители
семейств Lycosldae, Gnaphosidae, Clubionidae и Thomisidae, а
преобладали мелкие тенетные виды (99,2% особей мельче 5 мм,
в других'местообитаниях — 6,5—54,6%); 60,8% особей (в других
местообитаниях—1,5—37,6%) относились к видам, которые
часто распространяются по воздуху («аэронавты»). Видовой состав
зависит и от мозаичного характера растительности. Параллели
прослеживаются с фауной полей, садов, мусорных свалок (ср.
Braun, 1959) и береговых наносов.
Тишлер (Tischler, 1980) обнаружил в ботаническом саду
Киля И видов равноногих, среди которых были эвдоминантами
2. Структура города и фауна
75
Таблица 24. Доминирующие сенокосцы ботанического сада в Киле
(по данным Schaefer, 1973)
Вид
Количество
в
процентах
Примечания
Lacinius ephippiatus (С. L. Koch)
164
35,2
Обитатель леса, гемерофил
Nemastoma lugubre (Müller)
104
22,3
Обитатель леса
OpiUo saxatilis (C. L. Koch)
60
12,9
Вид открытой местности,
гемерофил
OUgolophus tridens (C. L. Koch)
59
12,7
Эвриойкный вид, гемерофил
Phalangium opilio (L.)
49
10,5
Вид открытой местности,
гемерофил
OUgolophus hanseni (Kraepelin)
15
3,2
Обитатель леса и открытой
местности
Таблица 25. Сенокосцы ботанического сада города Яссы (по Weis,
Sarbu, 1977)
Вид
Число
особей
в
процентах
Самцы
Самки
Отношение
полов
Odlellus bleniaszi (Kulcz.)
Zacheus crista (Brülle)
Phalangium opilio L.
Opilio saxatilis C. L. Koch
Trogulus tricarinatus (L.)
550
271
35
23
19
61,2
30,2
3,9
2,6
2,1
150
56
20
10
270
49
15
9
5
0,56
1,14
1,33
1,11
898
Oniscus acellus (40,1%), Armadillidium vulgare (37,5%) и
Porcellio scaber (12,8%). Плотность заселения здесь гораздо выше,
чем ,в окрестностях города, причем плотность активных особей —
в 58 раз. В целом 8 видов (94,3% особей) —синантропы,
остальные считаются эвриойкными. Сравнителино часто встречается
паук Hyloniscus riparius, очевидно, благодаря очень мелким
размерам своего тела. '
В этом же саду Тишлер обнаружил 11 видов губоногих
многоножек. Эвдоминантами были гемисинантропный вид Lithobius
microps (67,6%) и многоножка-камнелаз {L. forficatus) (19,0%).
Плотность активных особей, как и у двупарноногих многоножек,
была почти в 4 раза выше, чем в схожих, но больших по
площади и однородных биотопах за городом. Ассоциация губоногих и
двупарноногих своеобразная и иная, чем в окрестностях города,
хотя биотопные фрагменты ботанического сада несколько
сходны с коренными местообитаниями этой местности. Особенно част
мелкий вид L. microps. Тишлер видит причину этого в легкости
заноса и слабой чувствительности к механическим нарушениям.
76
2. Структура города и фауна
Таблица 26. Доминирующие пауки ботанического сада в Киле (по данным
Schaefer, 1973)
Вид
Число
в
проособей
центах
Примечания
Diplocephalus crlstatus
(Black611
19,2
Вид открытой
местности, ге-
well)
мерофил, аэронавт
Stylophora concolor (Wider)
588
18,5
Лесной вид, гемерофил
Erigone dentipalpis (Wider)
447
14,0
Эвриойкный вид, гемерофил
(вряд ли встречается в
природных местообитаниях),
аэронавт
Diplocephalus latifrons
292
9,2
Лесной вид, гемерофил
(0. P. Cambrige)
Erigone atra (Blackwell)
290
9,1
Эвриойкный вид, гемерофил,
аэронавт
Oedothorax fuscus (Blackwell)
167
5,2
Обитатель берегов, аэронавт
Diplocephalus picinus
(Black156
4,9
Лесной вид
well)
Meioneta rurestris (C. L. Koch)
152
4,8
Вид открытой
местности, ге-
мерофил, аэронавт
2703
84,8
Процент видов
Процент
особей
Лесные
43,3
40,7
виды
Виды
от-
26,7
41,3
крытой
местности
Береговые
16,7
16,0
виды
Островное положение, очевидно, обусловило отсутствие видов,
чаще встречающихся в окрестностях города.
Фауна двупарноногих в этом же месте насчитывала 15 видов,
среди которых эвдоминантами были Allajulus occultus (22,9%)
(гемисинантроп), Polydesmus superus (22,8%), А. londinensis
(12%), Ophyiulus pilosus (11,4%) и Unciger foetidus (11,0%).
К 'более мелким видам А. occultus и Р. superus (Tischler, 1980)
принадлежало 45,7% особей.
Особенно ценным из немногих количественных учетов дожде-
-вых червей в черте города является проведенный Эссером (Esser,
;1984) (табл. 27). Он содержал прежде всего данные по количе-
.;ству дождевых червей, потенциально служащих пищей различным
:зоофага'М, в частности черному дрозду, 'Обыкновенному скворцу,
обыкновенному ежу и некоторым двукрылым.
3
я
со
сп
.—.с»
го
s о
о с
ю
■ си
w и
■го cu
« а
у
го
cu
ю
8 „
о я
tu о
-i
см^
то ^
и
ш о-
то cj
с»
-*
из
о"
со
см
о_
см"
см
см
о"
1
I
i
см
<м
о
<м
ю
о"
ю"
1
<м
о_
о"
1
cd
с£>
со__
1
1
см'
о"
ю
со"
ю"
о"
сз^
<м"
1
1
о"
ю
о"
cd
со
о
ю"
о
оо"
со
см"
1
1
00
t-"
со"
а?
ю
cj>"
ю
о"
2?
о"
I
о)
- о"
о_
ю"
ю
со"
о"
со"
со
со"
см
см
о
00
^"
1
о"
(м
о
со
со
ю
\®
оо
со
см
со
1
1
ю"
ю"
см
о"
иг"
ю"
1
1
со
cd
cd
см
со__
со
1л
см
1
1
1л
1л
со"
-*"
о
ю"
со"
1
1
о
ja
о
S
■-j
q
со
ä
о
_е
"о
о
со
сз
о
cj
ч
"о
g
-о
3
ä
е
сз
й
cü
I
s
s
ч
о
о
с
78 2. Структура города и фауна
Фауна жуков ботанического сада в Киле существенно
зависела от мозаики местообитаний исследованной местности, хотя
у некоторых очень часто встречающихся видов из-за их эвриойк-
ности (Практически невозможно было точно определять центры
распределения. Стафилиниды Lesteva longelytrata и Falagria
sulcata обнаружены, например, только на берегу пруда.
Дорн (Dorn, 1977), обследовав пчел ботанического сада в
Галле, обнаружил более 100 их одиночных диких видов.
Доминировали Hylaeus hyalinatus (11,8%), Colletes daviesanus (7,0%)^
я. communis (5,8%), Lasioglossum calceatum (5,8%), Osmya rufa
(5,6%), L. morio (4,9%) и Andrena minutula (4,2%). Банашак
(Banaszak, 1976) обнаружил в ботаническом саду Познани 67
видов пчел, пооещавщих цветки 55 видов растений.
2.2.4.3. ЗООПАРКИ
Разнообразные способы содержания животных в зоопарках^,
а также их в больщинстве случаев сходство с парками позволяют
. существовать там богатой *фауне. Так, Краль и Олекса (Kral,.
Olexa, 1975) характеризуют зоопарк в Праге как рефугию
редких видов, что в .первую очередь 'обусловлено его частично
степным характером. Примером могут служить ящерица Lacerta
viridis, прямокрылое Oecanthus pellucens (Oecanthidae),
многоножки Monotarsobius dubosqui (Chilopoda) и Symphyllelopsis
arvenorum (Symphyla), жуки Abraeus parvulus (Histeridae), Or-
thocerus clavicornis (Colydiidae) и Cylindromorphus bohemicus
Obenberger — terra typica! (Buprestidae) (отсюда описан этот
вид).
Некоторые виды в связи с особенностями содержания
животных наблюдаются особенно часто, например жуки Necrophorus
humator ч Оeceoptoma thoracica (Silphidae) (на остатках корма).
Частота встречаемости домового воробья может быть даже
избыточной (Gnielka et al., 1983).
Исследования фауны перепончатокрылых в зоопарке Берлина
(Dathe, 1960, 1971) показали присутствие 213 видов жалящих
(Aculeata) (рис. 22). Садовая терраса и окружающие стены
оказались местами концентрации многих видов. Абсолютным геме-
рофилом был вид роющих ос Cerceris rybensis. Обнаружены
также некоторые паразиты личинок этого вида — Omalus auratus,.
О. constrictus, Hedychrum gerstaeckeri и Chrysis cyanea (Chrysi-
doidea). otto Мюллер (Otto Müller) поймал на свет в зоопарке
Галле с 1971 по 1981 г. 592 вида бабочек (в письме 22.9 1982).
Зоологические сады считаются также местами 'концентрации я
проникновения адвентивных видов, что может отражаться на
составе паразитов содержащихся здесь животных.
Клещи Blattisocius tarsalis и В. keegani развиваются в
культурах насекомых и пищевых продуктах, пораженных насекомы-
2. Структура города и фауна
79
МИ (Rack, 1968). Paracarophenax dermestidarum поедает яйца в
культуре жуков, Percanestrinia blaptis — в культуре жуков-мед-
ляков рола Blaps. Hlstiostoma feroniarum встречается в
различных культурах насекомых, а Я. laboratorium в культуре дро- !
зофнл. -
2.2.4.4. КЛАДБИЩА
Кладбища во многих отнощениях существенно не отличаются
от парков. Особенностью их является эмоционально обусловлен- ■
ное отсутствие нарушений, что благоприятствует появлению не- :
которых видов и прежде всего позвоночных. Беспозвоночные в ]
этих местах до сих пор почти не изучались. Структура кладбищ \
определяется в основном количеством, размерами, расположени- J
40
1^0
Й 20
\
/
V
-4<^
/
/
S
•
ж
ш
ж
Мест
ж
ж к
Рис. 22. Изменение в течение года числа пчел (А) и роющих ос (S) в зоопарке 1
Берлина (по Dathe, 1971) |
ем и видом древесной растительности. Много открытого rpyHTa.j
Часто речь идет о естественных почвах, но с перемешанными го- \
ризонтами. Местами применяют в больших количествах биоциды \
(гербициды на дорожках, удрбрения). На кладбищах травяной'
покров очень разнообразен, встречаются отдельные теневыносли-:
вые сорняки, кустарники из нитрофильных опушечных видов и
декоративные растения. На сильно обработанных гербицидами
участках доминируют различные очитки. Среди деревьев преоб-'
ладают интродуцированные виды, в большинстве случаев много
хвойных. Существуют особые кладбищенские адвентивные виды.
Кладбища —это острова массового развития лишайников. Сточ-;
ки зрения растительности для них характерна ассоциация Belli-\
detum, а также опушечные сообщества, например, с фиалками?
Viola odorata и V. hirta, фитоценозы классов Plantaginetea (на-j
пример, Sagino-Bryetum) и Sedo-Scleranthetea (фрагменты). .!
В мелких водоемах может жить тритон обыкновенный; часто]
также встречаются обыкновенная жаба и ящерица прыткая.?
1по™п?е1ка m™aIm^T 197з" городских кладбищах (пояснения см. табл. 8)
Индекс
(по
Saemann, 1970)
Вид
4
4
1
3—4
3—4
4
2-3
3
4
2—3
4
4
2-3
3—4
3—4
3—4
3
3—4
2
4
Черный дрозд
Зеленушка
Домовый воробей
Кольчатая горлица
Певчий дрозд
Зяблик
Пеночка-теньковка
Лесной голубь
Обыкновенный
скворец
Полевой воробей
Лазоревка
Большая синица
Коноплянка
Садовая славка
Садовая горихвостка
Дрозд-рябинник
Лесная завирушка
Канареечный вьюрок
Зеленая пересмешка
Славка-завирушка
Карл-Маркс-Штадт
Колумбарий, 8,5 га
Rinnhofer (1965)
%%
Число видов
Гнездящиеся пары
(ГП)
ГП/Юга
19
54
8,2
9,4
8,2
4,7
4,7
3,5
1,2
2,4
5,9
2,4
2,4
2,4
1,2
1,2
1,2
1,2
1,2
63,5
Saemann (1973)
%%
13,0
14,8
13,0
7,4
7,4
5,6
1,9
3,7
9,3
3,7
3,7
3,7
1,9
1,9
1,9
1.9
1,9
2
2
2
2
23
70
11,8
10,6
9,4
5,9
4,7
4,7
3,5
3,5
2,4
2,4
2,4
2,4
1,2
2,4
1,2
2,4
2,4
2,4
2,4
Городское кладбище, 30,7 га
Rinnhofer (1965)
%%
14,3
12,9
11,4
7,1
5,7
5,7
4,3
4,3
2,9
2,9
2,9
2,9
1,4
2,9
1,4
2,9
2,9
2,9
2,9
82,4
35
22
5
20
12
12
2
10
2
15
10
3
8
21
2
6
4-
1
6
"32"
[218
Saemann (1973)
I %%
11,4
7,2
1,6
6,5
3,9
3,9
0,7
3,3
0,7
4,9
3,3
1,0
2,6
6,8
0,7
2,0
1,3
0,3
2,0
71,0
16,1
10,1
2,3
9,2
5,5
5,5
0,9
4,6
0,9
6,9
4,6
1,4
3,7
9,6
0,9
2,8
1,8
0,5
2,8
46
31
5
27
18
14
13
5
15
4
13
14
1
7
5
16
9
4
4
5
"Зб"
1297
15,0
10,1
1,6
8,8
5,9
4,6
4,2
1,6
4,9
1,3
4,2
4,6
0,3
2,3
1,6
5,2
2,9
1,3
1,3
1,6
96,7
15,5
10,4
1,7
9,1
6,1
4,7
4,4
1,7
5,1
1,3
4,4
4,7
0,3
2,4
1,7
5,4
3,0
1,3
1,3
1,7
Галле
Южное
кладбище, 24,0 га
(средние значения за
1965-1978 гг.)
Gnielka (1981)
I %%
57,5
45,6
0,3
34,6
4,9|
14,4
14,9|
11,0
29,1
26,1
12,
11,9]
0,6
4,2
5,0j
10,1
22,6
14,7
12,
2б"
|360,6
24,0
19,0
0,1
14,4
2,0
6,0
6,2
4,б|
12,
10,9]
5,0
5,0,
0,3|
1,8
2,1
4,2,
9,4|
6,1
5,0
15,9
12,6
0,1
9,6
1,4
4,0
4,1
3,1
8,1
7,2
3,4
3,3
0,2
1,2
1,4
2,8
6,3
4,1
3,4
150,3
2. Структура города и фауна
81.
Наряду с домашними кошками 'более или менее регулярно
наблюдаются каменная куница, горностай, ласка, обыкновенный еж,,
белобрюхая белозубка, дикий кролик, белка обыкновенная, мыш'и
полевая, лесная и желтогорлая, полевка обыкновенная (Elvers,.-
Elvers, 1984 и др.).
Количественные данные имеются в основном по птицам
(Schlegel, 1920; Rinnhofer, 1965; Gremple, 1966, 1973, 1982;
Saemann, 1973; Elvers, 1977; Gnielka, 1978, 1981; Dobberkau et al,
1979; Otto, 1983; Schmidt, Steinbach, 1983) (табл. 28, рис. 23, 24),
Колебания их численности в течение года исследовал Гнилька
(Gnielka, 1981) (рис. 25). Виды птиц, гнездящихся в природе на
деревьях и в дуплах, на кладбище так же разнообразны, как и в
ЭО
80 h
70 Ь
60 h
^ 50
g
зо\-
20 h
Wh
19В4
Рис. 23. Изменение числа гнездящихся пар доминирующих видов птиц на
южном кладбище в Галле (по данным Gnielka, 1981): Г —кольчатая горлица р
Д —черный дрозд; 3 —зеленущка; С — скворец; К — канареечный вьюрок;,
В — полевой воробей
6—21
«2
2. Структура города и фауна
Пригородных биотопах со схожей структурой (Otzen, Schaefer,
1980).
Немногие учеты членистоногих показывают ожидаемое
высокое разнообразие видов. Отмечалось (Strobl, Könecke, 1984)
46 видов дневных (бабочек, 61 вид шелкопрядов, 133 вида совок
и 108 видов пядениц. Такое видовое богатство объясняется
прежде всего обилием кормовых растений. Особым пищевым
ресурсом является старый хворост венков, в котором часто обитает
жук-плоскотелка Silvanoprus fagi (Cucujidae). На кладбищах
обычен жук Rhizophagus parallelocollis (Rhizophagidae). С
конца апреля до мая на памятниках и каменных стенах можно
наблюдать массовые его скопления, включающие тысячи особей.
Этот вид, вероятно, относится к трупной фауне и считается по-
W
Высота, м
Рис. 24. Высотное
распределение гнезд вяхиря
на южном кладбище в
Галле (на Заале) (по
Gnielka, 1976)
жирателем плесени (дерево гробов). Он явно ведет подземный
образ жизни и предпочитает темные местообитания типа
погребов, нор млекопитающих, глубоких «омпостных ям.
.2.2.4.5. СПОРТИВНЫЕ СООРУЖЕНИЯ
В этих местах наряду с территориями, лишенными
растительности, преобладают газоны и разных масштабов посадки
декоративных деревьев и кустарников. Открытого грунта много. Он
сильно перемешан, часто насыпной (иногда искусственные
покрытия). Естественные почвы сохраняются главным образом на
спортивных площадках городских окраин. Применяются биоциды
(гербициды и замедляющие рост вещества). Растительность бед-
'На видами, часто преобладают растения, устойчивые к
вытаптыванию. По склонам растут отдельные элементы суховатых лугов,
а по краям площадок — декоративные кустарники.
Распространены сообщества класса Plantaginetea, особенно ассоциация
LoLio-Plantaginetum, частично переходящая в Bellidetum.
2. Структура города и фауна
83.
Типичными позвоночными стадионов являются жаворонок-
хохлатый (Gnielka et al., 1983) и дикий кролик. Из жужелиц,
обычно преобладают Атага аепеа, Harpalus aeneus и Calathus
fuscipes (Becker, 1977). На газонах обитает главным образом
июньский нехрущ {Amphlmallon sotstitiale, Scarabaeidae). Корот-
ко подстриженная трава, очевидно, благоприятна для развития;
его личинок (подходящие температурные условия). По данным
Кубицкой (Kubicka, 1981), на 1 м^ зеленой площади у дорожки'
приходится 40 личинок. Вид мало чувствителен к загрязнению*
почвы ионами солей, применяемых для таяния льда, он
встречается и в естественных засоленных почвах. Неоднократно сообща-
1200 \-
1000 V
I
Прочие
■'К
ЕЖЖ¥ЖШШЖХ.ЛЖ
. Мясяи,
рис. 25, изменение состава орнитофауны на южном кладбище в галле (на»
Заале) в течение года: пв — полевой воробей; дв — домовый воробей; чд —
черный дрозд; кг — кольчатая горлица; с — синицы; 3 — зяблик; ck —
скворец; сп —славки и пеночки (по gnielka;, 1981)
ЛОСЬ О массовом лёте этих жуков, в городах, например в Карл-
Маркс-Штадте (v. Lasch, 1980), Лейпциге (более 1000 особек
среди сквера в центре города в начале июля) (Klausnitzer, 1981),
Любеке (Horton, 1958) (в городских скверах даже вредят,
уничтожая дерновину) и Гамбурге (на спортивных площадках и
газонах в городе встречаются чаще, чем в окрестностях).
2.2.4.6. сады
Преимущественно мелкие садовые участки в черте города
характеризуются сочетанием небольших засаженных полезными и
декоративными растениями гряд, разделяющих их дорожек и очень
2. Структура города и фауна
мелких газонов, а также различных по площади и густоте
древесных насаждений. В 'большинстве случаев много открытого
грунта. Почвы почти всегда естественные, реже насыпные, часто
.очень сильно удобренные. Иногда в больших количествах
применяются биоциды. Естественная растительность представлена
садовыми сорняками .и некоторыми устойчивыми к вытаптыва-
лию видахми. Имеются фрагменты сообществ садовых сорняков.
Фауна 'садов неоднородна (рис. 26, 27) и находится под
сильным влиянием антропогенных факторов, однако можно выделить
типичные виды, их сочетания и отношения доминирования. За
исключением вредителей культурных растений, животный мир
зтих мест изучен пока мало.
П
Р,М
Н
У
т
PK .
ж
ПК
Б
дк
к
о<1%
(%)т to 30 20 10 OD 10 20 30 4/7 50(7.)
Рис. 26. Плотность активных особей/обилие потомства (слева) и биомасса
активных особей/продукция имаго в плодовом саду. Процентные доли
отдельных групп по данным уловов почвенными фотоэклекторами (по Reich et al.,
1985): П —пауки; Р, М — равноногие, многоножки; Н — ногохвостки; У —
уховертки; Т —трипсы; PK — равнокрылые; Ж —жуки; ПК —
перепончатокрылые; Б —бабочки; ДК — двукрылые; К —прочие крылатые насекомые
Фауна дождевых червей включает в основном обитателей
минеральных .почв и гумуса, например Octolasium sp., Ällolobophora
sp. и Lumbricus rubellus (Dünger, в письме 1983). Уже на мелком
огороде Пилипюк (Pilipiuk, 1981) обнаружил Ällolobophora geor-
gü и Octolasium суапеит.
Исследования небольшого сада на окраине Альтенбурга
показали наличие 47 видов пауков (Heimer, 1978). Доминировали
Diplocephalus cristatus (14,9%), Silometopus reussi (13,9%),
Centromerita bicolor (10,5%), Lepthyphantes tenuis (9,7%), Pa-
chygnatha degeeri (7,3%), Diplostyla concolor (4,4%) и Bathy-
phantes gracilis (4,2%). Из них 64,3% были «аэронавтами».
Многие среди .О'бнаруженных видов считаются светолюбивыми (а
возможно, и теплолюбивыми), например Р. degeeri.
Садовые растения за.селяются растительноядными, в том
числе сосущим1И видам.и насекомых, которые в свою очередь обус-
2. Структура города и фауна
85
ловливают существовавие многих паразитоидов и хищников, так
что возникают пищевые цепи и сети. Особенно хорощо развит в
больщинстве случаев круг потребителей тлей. На кустарниках
.возникают коннексы, основ,анные на различных видах
листоверток (Tortricidae) (Muczulsku, Anasiewicz, 1972). Только на
яблонях могут жить 300 видов фитофагов, около 300 видов
паразитоидов и 200 видов хищников (Steiner, 1958).
Семьсшозяйсттные угодья
Степь
Берег реки
Рис. 27. Происхождение фауны садового участка в Вене (ио Schweiger,
1960/61)
Типична высокая доля насекомых, посещающих цветки.
Особое значение имеют эти местообитания для бабочек,
разнообразие видов которых может определяться структурой сада (Loibl,
1953; Bradley, Mere, 1963; de Worms, 1965; Donahne, 1976;
Jackson, 1977; Owen, 1971, 1976, 1978; Owen, Owen, 1975; Smith, 1975).
Городские сады служат рефугией для жалящих
перепончатокрылых (Haeseler, 1972); в Киле их обнаружено 139 видов
86
2. Структура города и фауна
Таблица 29. Доминирующие жалящие перепончатокрылые в городских садах
Киля (по Haeseler, 1972). ОС—обитатели отвесных стен, л—лесные виды„
О — виды открытой местности
Вид
Экологическая группа
Pemphredon shuckardi (А. Мог.)
Л
Pasialoecus tenuis А. Мог.
Л
Ectemnius sexcinctus (F.)
Л
Crossocerus elongatulus (vd. L.)
0
Hylaeus communis (Nyl.)
Л
H. hyalinatus (Sm.)
0
Andrena armata (Gmel.)
Л
A. barbilabris (Kir.)
о
A. helvola (L.)
л
A. nigroaenea (Kir.)
о
Lasioglossum morio (f.)
о
L. sexstrigatum Sch.
0
Osmia rufa (L.)
л
Chelostoma fuliginosum (Pz.)
л
Anthophora acervorum (L.)
ОС
Bambus hypnorum (L.)
л
(табл. 29), в том числе 62,4% с эндогейным, остальные —с гипо-
гейньш развитием. На газонах с редким травостоем и по их
краям предпочитают строить норки пчелы-андрены Andrena armata,
А. haemorrhoa, А. clarkella, А. nigroaenea, А. tibialis и А. jacobi.
По краю цветочных клумб гнездится А. varians, а в ее гнездах
паразитирует пчела-'бродяжка Nomada ruficornis.
Личинки жука-носорога {Oryctes nasicornis; Scarabaeidae) в
большинстве случаев развиваются в черте или вблизи
населенных пунктов в компосте, парниках, навозных кучах, навозных
ямах, среди древесных опилок, в гниющей соломе и аналогичных
местообитаниях, поэтому сады являются для них важной рефу-
гией. Иногда популяции этих жуков достигают большой
плотности, особенно в непосредственной близости от соответствующих
производственных площадей (огородов, мест изготовления
органических удобрений, бумаги и т. п.). Реннер (Renner, 1977)
обнаружил в компостных кучах 146 видов жесткокрылых, в том
числе 96 видов стафилинид. Иногда в таких местообитаниях
находят таксоны, считающиеся редкими, например Philontus adden-
dus, обитателя ходов дождевых червей (Веу, 1959).
Высокую биомассу (20—36 кг/100 м^) могут давать в садах
слизни рода Derocerus.
Особое внимание исследователи уделяли садовому орнитоце-
нозу (Zimmermann, 1967; Böhm, 1968; Gremple, 1968;
Winterbottom, 1971; Mulsow, 1976; Jacubiec, Bluj, 1977; Luniak, 1980; Wal-
dert, 1982; Dittberner et al., 1983; Gnielka et al., 1983, 1984). Его
2. Структура города и фауна
87
Таблица 30. Характерные виды садовых орнитоценозов (или «зеленого
города») (а —75—100% пробных площадей)
Mulsow
Waldert
Luniak
Gnielka
Saemann
Вид
(1976)
(1982)
(1980)
et al.
(1970)
(зеленый
город)
(1983)
Мелкие сады;
индекс
Домовый воробей
а
+
+
2
Черный дрозд
а
+
+
+
3
Обыкновенный скворец
а
+
3
Зеленущка
а
+
+
2
Большая синица
а
+
+
4
Лазоревка
а
+
4
Зяблик
а
+
3
Садовая горихвостка
а
3
Лесная завирушка
А
+
1
Славка-завирушка
а
4
Мухоловка-пеструшка
4
состав (табл. 30) аависит йрежде всего от развития
растительности и наличия мест, пригодных для гнездования. Согласно
Луняку (Luniak, 1980), самая богатая фауна наблюдается через
20 лет после закладки сада. Летом гнездится И—16 видов птиц
(83—118 пар на 10 га), зимой—11—14 видов (56—190 особей
на 10 га). На садовых участках под Берлином Диттбернер и др.
(Dittberner et al., 1983) обнаружили 129,1 пар на 10 га. В это
местообитание регулярно .проникает славка-завирушка,
единственный здесь представитель семейства славковых (Gnielka et al.,
1983).
В садах часто селятся водяная крыса и обыкновенный крот.
2.2.4.7. ТЕПЛИЦЫ \ 1:3 \
Под термином «теплицы» мы объединяем очень разные виды
сооружений, общим для которых является только изоляция от
внешней среды с помощью стекла или другого материала.
Поэтому здесь иногда наблюдаются значительные различия в
температурном режиме, водоснабжении, составе флоры и почвы,
характере использования и интенсивности обработки пестицидами.
В постоянно отапливаемых оранжереях ботанических садов с
долговременной относительно ненарушаемой растительностью и в
общем незначительным применением ядохимикатов могут
поселяться даже виды тропического про,исхождения. При сравнении
фаун сходно устроенных теплиц обнаруживается удивительная
близость их видового состава. Значительную долю видов можно
назвать «космополитами теплиц». Это предпочтительное место
поселения адвентивных видов. Ниже приводятся некоторые при-
2. Структура города и фауна
меры, свидетельствующие о маогообразии тепличной фауны, в
которой Айхлер (Eichler, 1952) насчитывает около 800 видов
(без простейших).
Пресноводная медуза Craspedacusta sowerbyi была занесена
из Бразилии в подогреваемые водоемы различных ботанических
садов, а затем расселилась и в природу >. В теплицах живут
завезенные, вероятно, из тропиков плоские черви Placocephalus
(Bipalium) kewensls и Rhynchodemus biUneatus. Шреммер
(Schremmer, 1954) обнаружил последний вид и в природе. Среди
кольчатых червей Eisenia japonica занесена из Японии, а Dend-
robaena veneta — из Средиземноморской области. j
Из теплиц ботанических садов Берлин-Далема и Берна,
орхидейных оранжерей близ Парижа и различных теплиц
Великобритании сообщалось о пауке Hasarius adansoni (Boettger, 1929;
Holzappel, 1932; Locket, Milljdge, 1951). Ha необычайно
массовое размножение паука Ostearius melanopigius в теплицах с
культурами томатов указывает Захер (Sacher, 1978).
Микроклиматические условия способствуют появлению популяций,
насчитывающих .многие тысячи особей (ср. также Braun, 1961).
Происходящий, возможно, из субтропиков или тропиков эвсинантроп-
ный .космополит Achaearanea tepidariorum является типичным
для теплиц пауком, считающимся пионером их заселения
(Boettger, 1932). Подходящие влажность и температуру он иногда
находит и в других местообитаниях, например в складских
помещениях, прачечных и на скотных дворах. Эвсинантроп Heteropoda
venatoria, возможно, относится к адвентивным видам; его
популяции существуют с 1923 г. в аквариумных помещениях
зоопарков Лейпцига, Берлина и Дрездена.
Два вида теплолюбивых равноногих — Porcellio laevls и Р. di-
latatus были завезены из Средиземноморской области и часто
встречаются в теплицах. Ендрычковски (Igdryczkowski, 1979)
обнаружил здесь 19 синантропных видов мокриц.
Среди ногохвосток наряду с .многочисленными
эврипотентными видами известны специфические обитатели теплиц, например
Slnella соеса, на что в особенности указывают Кассаньо и Руке
(Cassagnau, Rouqet, 1962) (из ботанического сада Тулузы).
Ногохвостки часто распространяются по .всему миру с клубнями
растений или .под корой древесных стволов (антропогенные
космополиты). Вайднер (Weidner, 1983b) приводит для Гамбурга 18
примеров такого рода.
В подогреваемых аквариумах удается разводить даже
тропические виды стрекоз; это показано на примере Ictinogomphus de-
В этой книге выражение «в природе» означает, как правило, не
естественный биотоп (в городской зоне Центральной Европы их практически не
осталось) , а местообитания под открытым небом. — Прим. ред.
2. Структура города и фауна 89
coratus melaenops (Lieftinck, 1978) из Малайзии. К условиям
теилиц успешно адаптировался космополитный оранжерейный
кузнечик {Tachycines asynamorus), отмечаемый с 1982 г. в
Гамбурге (Boettger, 1950, 1951; Eichler, 1952; Madel, 1937; Weiss,
1947). Кузнечик Phlugiola dahlemica в одной теплице
наблюдался более 20 лет (Eichler, 1939). На папоротниках отмечена тля
I diopter us nephrelepidis. В теплицах встречаются также тли Мас-
rosiphoniella oblonga, Macrosiphum euphorbiae, Aulacorthum cir-
cumflexum, A. sotani и Myzus ornatus.
К теплицам, a также квартирам приурочен ряд видов трипсов,
например Parthenothrips dracaenae, Hercinothrips femoralis, Не-
liothrips haemorrhoidalis, Chaetanaphothrips orchidii, Scirtothrips
longipennis и Leucothrips nigripennis (Morison, 1957; Pelican,
1954; Schliephake, 1984).
В теплицах ботанических садов обнаружен восточноазиатский
жук Ptitodactyla luteipes (Ptilodactylidae) (Eichler, 1938, 1952).
2.2.5. Пустыри
Этот тип биотопов включает в городах в основном ст'арые ру-
деральные участки, заросшие свалки и защитные полосы,
пользоваться которыми запрещено законом. Грунт, как правило,
открыт, а почвы по составу очень разные в зависимости от
первоначального использования места; они бывают как естественными,
так и антропогенными. Обычно имеется богатая рудеральная
флора. Большую роль играют виды, типичные для
первоначальной формы использования данной площади, например сегеталь-
ные, луговые растения сухих травостоев. Встречаются отдельные
деревья; в том числе ясень, горный и остролистный клены. На
пустырях растут лишь немногие адвентивные виды.
Преобладают сообщества классов Artemisietea и Agropyretea, а также
заросли рудеральных кустарников.
Среди членистоногих встречаются главным образом
ассоциации растительноядных, в том числе сосущих видов, часто с
необычайно большим их количеством на один вид растений: на
полыни обыкновенной и пижме отмечено примерно по 100 видов;
характерный вид для полыни обыкновенной —
муха-пестрокрылка Охупа parietina (Trypetidae) (Klausnitzer, 1968).
Характерными видами птиц этого местообитания являются
прежде всего болотная камышевка, затем хохлатый жаворонок,
серая славка и каменка (Rinnhofer, 1969; Rinnhofer, Saemann,
1968; Handke, 1982; Eggers, 1976) (табл. 31). Болотная
камышевка только в последние годы стала гемерофилом,
гнездящимся на участках, заросших сорняками. В городе этот вид заселяет
рудеральные местообитания, в первую очередь крупные заросли
крапивы двудомной, а также золотой розги, чертополоха и бодя-
90
2. Структура города и фауна
Таблица 31. Доминирующие виды птиц, гнездящихся на пустыре (52,5 га)
в Саарбрюккене (по Handke, 1982)
Вид
Гнездящиеся
пары
ГП/Ю га
Доминирование,
'%
Болотная камыщевка
16
3,0
15,4
Полевой жаворонок
12
2,3
11,5
Камыщевая овсянка
10
1,9
9,6
Желтая трясогузка/"
8
1,5
7,7
Серая славка
8
1,5
7,7
Черный дрозд
8
1,5
7,7
Луговой конек
5
1,0
4,8
Черноголовый чекан
5
1.0
4,8
28 видов
104
19,8
ка. По данным Земана (Saemann, 1970), камышевка в крапиве
достигает плотности 4 пары на 10 га. Требования к занимаемой
площади зачастую довольно незначительные, и наблюдались
даже 4 пары на 2 га пустыря.
Хохлатый жаворонок, проникший с юга, натурализовался в
северных районах Центральной Европы только в конце XIX в.
После второй мировой войны он был типичным видом руин и
развалин многих городов (Boettger, 1962). С исчезновением
таких местообитаний численность его заметно уменьшилась. Теперь
этот вид заселяет прежде всего пустыри, свалки, фабричные и
спортивные сооружения, конечные трамвайные «круги», крупные
стройки, мусорные кучи и даже зеленые разделительные полосы
проезжих дорог с интенсивным движением, а также плоские кры-
щ.и. Гнезда наблюдались и между железнодорожными колеями
около товарных станций. Повышенная встречаемость этих птиц
отмечена в некоторых районах новостроек (см. 2.2.2.3). Гнезда
за пределами городов почти не известны. В Галле .наблюдали
40—120 гнездящихся пар, в Галле-Нойштадте — 60—160
(Gnielka et al., 1983).
Зукопп и др. (Sukopp et al., 1980) обнаружили в ловушках
на городских пустырях лесную мышь (эвдоминант), полевку
обыкновенную, полевую и домовую мышь.
2.2.6. остатки негородских экосистем
В черте города могут в различной форме существовать остат-.
ки негородских экосистем (например, лесов, лесопосадок,
кустарников, рощ, полей, лугов, болот, прудов, озер, сухих травостоев).
Эти местообитания особенно важны как рефугии для заселения
2. Структура города и фауна ^ 91
городских озелененных территорий, «леревалочные пункты» и
связующие элементы. Здесь преобладают естественные открытые
почвы. Негородские биотопы [видоизменяются под влиянием
города; например, в пригородные леса ироникают нитрофильные
растения и неофиты (бузина черная, недотрога мелкодветковая,
черемуха поздняя, клен ясенелистный). Когда-то богатые видами
биоценозы более или менее обедняются.
Старые древесные насаждения, довольно часто
сохраняющиеся в городах, являются прежде всего местообитаниями животных
(особенно насекомых), которые уже почти не встречаются за
городом, т. е. близкие к природным городские биотопы
представляют собой их реликтовые местонахождения, иногда называемы-
)Ми даже «реликтами девственных лесов» (Korge, 1967). На
старых деревьях ботанического сада в Западном Берлине
обнаружены, например, такие редвие жуки, как Anitys rubens
(Anobiidae), Scydmaenus hellwigi (Scydmaenidae), Pycnomerus terebrans
(Colydiidae), Aderus oculatus (Aderidae). Жук Protaetia
aeruginosa (Scarabaeidae) встречается в трухе стволов старых дубов
(Horion, 1958). Естественно, вид не ограничен в своем
распространении только старыми городскими дубами, однако города и
прежде всего крупные парки являются для него важными
местообитаниями. Так, Швайгер (Schweiger, 1962) обнаружил это
жесткокрылое в городских парках Вены. В ГДР Гиварц (Schwartz,
1981) сообщает о его находках (провести границу ареала
нелегко), например, в различных частях Берлина, в Бранденбурге,
Галле, Кётеие, Дрездене и Майсене. Довольно часто этот вид
встречается в Лейпциге.
3. ПРОИСХОЖДЕНИЕ И СОСТАВ
ГОРОДСКОЙ ФАУНЫ
Городская фауна может включать как первоначально
существовавшие в данном месте виды, так и пp^ищeл.ьцeв (иммитран-
тов). Точные доказательства сохранения в городах полуляций,
присутствовавших еще до основания города, получить трудно, по
крайней мере для длительно существующих населенных пунктов.
Это означает, что их фауна, вероятно, большей частью состоит из
иммигрантов, связанных происхождением с аграрным
ландшафтом, эпилитными местообитаниями (скалы), троглобиотой (пеще^
ры в широком смысле слова, норы животных сублитион), лесами
и пустынями, т. е. безлесными сухими ландшафтами (здесь мы
не учитываем водных биотопов).
3.1. Реликтовые виды :
Хотя точные доказательства реликтового характера
популяций доступны лишь в очень редких случаях, м.ожно предполагать,
что часть видов сохранилась в городах еще с «догородских»
времен. Вероятно, это применимо, с одной стороны, к почвенным
формам, устойчивым к перемещениям грунта или обитающим в
местах, которые никогда не застраивались, а с другой стороны —
к обитателям деревьев (особенно старых), способным очень
долго существовать мелкими популяциями.
Примерами могут служить двухвостка Japyx braueri (Kühnelt,
1970) и пластинчатоусый жук Protaetia aeruginosa (см. 2.2.6).
Большего числа реликтов можно ожидать в остатках
негородских экосистем (например, бабочка Thaumetopoea pinivora, Skell,
1966).
3.2. Адвентивные виды
С иродовольствием, материалами, растениями и др. в города
постоянно заносились и заносятся различные животные. Если они
относятся к местной фауне, то этот процесс почти не заметен
(и затрудняет выявление реликтовых видов). Однако во многих
случаях речь идет о чужеродных элементах фауны, которые в
данном регионе обнаруживаются впервые именно в городах.
Прежде всего их находят в складах, амбарах, теплицах, на
вокзалах, в портах, на аэродромах. Только немногие из этих видов
3. Происхождение и состав городской фауны 9^
поселяются на некоторый срок и поэтому сравнимы с зфемеро-
фитами, многие же даже не замечаются зоологами. Однако
приток этот чрезвычайно интенсивен; достаточно представить себе,,
сколько особей завозится «епреднамеренно с прицепами к
легковым машинам, палаточным оборудованием и т. д. в первую
очередь из южных стран. Возможностью поселения обладают
главным образом виды, находящие свободные или слабо
используемые экологические ниши (пищевые продукты, материалы, геме-
рохоряые растения), соответствующие их требованиям.
Некоторые виды уже давно поселились на данной территории
и стали составной частью местной фауны. Строго говоря, их уже
нельзя назвать адвентивными. К сожалению, в отличие от
(растений время и происхождение их только в редких случаях точно
известны. Животных, -которых можно сравнивать с неофитамп
(например, американский таракан Periplaneta americana),
немного (если не учитывать завезенных из Америки в последние
десятилетия). Больше, вероятно, видов, аналогичных археофи-
там, т. е. попавших в данный район еще до средневековья
(например, рыжий и черный тараканы, домовой сверчок, сахарная-
чешуйница, постельный клоп) и уже причисляемых к
среднеевропейской фауне (см. 3.3). Только в отношении сравнительно
небольшого их числа можно говорить о сознательном завозе;
большинство же занесено непреднамеренно и активно
распространилось «само по себе» (Peters, 1956, 1961; Döhring, 1972, 1973;,
Metzger, Hanisch, 1979).
Довольно часто можно наблюдать адвентивных пауков,
долговременное лоселение которых остается под вопросом. Так,
сообщалось (Huhta, 1972) о популяции паука Loxosceles laeta из
Центральной и Южной Америки в Хельсинкском зоологическом-
институте. Поиски пассивно завезенных видов 'особенно
результативны в местах, постоянно связанных с международной торгов--
лей и транспортом (гавани, склады, товарные станции), а также
в крупных теплицах ботанических садов. \
Теплолюбивые щетинохвостки, например Lepismodes inquili-
nus и Ctenolepisma lineatum, завозятся прежде всего с
посадочным материалом или древесиной (Weidner, 1938b). Опять же
неясно, могут ли эти виды поселиться на длительное время.
Из нескольких завезенных в Гамбург видов длинноусых
прямокрылых не смог прижиться ни один, включая египетскую
кобылку {Anacridium aegyptium), хотя она каждый год
неоднократно «импортируется» (Weidner, 1981). Некоторые уховертки-
завозились с продукта1МИ питания, главным образом с орехами,
(см. 2.1.3).
Иногда заносятся сеноеды, в частности Psoquillamarginepunc-
tata из тропических зон Америки, Африки и Азии, Rhyopsocus-
speciophilus из неотропиков, Soa flaviterminata из тропиков (nai
^4
3. Происхождение и состав городской фауны
Таблица 32. Жуки-кожееды, занесенные в Центральную Европу
Вид
Область происхождения
.Dermestes haemorrholdalis Küster
D. carnivorus F.
D. peruvianas Cast.
Attagenus fasciatus (Thunb.)
Reesa vespulae Milliron
.Anthrenocerus australis (Hope)
Trogoderma angustum (Sol.)
~T. megatomoldes Reitter
T. inclusum Leconte
T. granarium Everts
.Anthrenus sp.
.Л. flavipes le Conte
Южная Франция (1937 г.-—первая
находка устойчивой колонии, Но-
rion, 1955)
Америка
Южная Америка
Космополит
Северная Америка (с 1963 г. в
Европе, Mehl, 1982)
Австралия (Англия, Нидерланды)
Южная Америка (Weidner, 1984;
история натурализации и
биология: Elbert, 1978; Frei, 1971;
Geisthardt, 1982; Kemper, Döhring; 1963;
Mehl, 1982; Mroczkowski, 1956,
1960; Philipp, 1967, 1968; Schmidt,
1960; Veerman, Schulten, 1977;
Wegelins, 1965; Wohlgemuth, 1975)
Центральная Америка
Южная Америка
Индия, почти космополит
Несколько видов (например, А. olgae
и А. polonlcus) в основном в
коллекциях насекомых, происхождение
в большинстве случаев неизвестно
Средиземноморье (Herfs, 1936)
•Орхидеях), Echmepteryx madagascariensis с Мадагаскара и Tri-
chopsocus acuminatus из Средиземноморья (последние два вида
'Обнаружены в теплицах). Мильднер (Miidner, 1983) сообщает о
появлении в Австрии адвентивного .клопа Corythucha ciliata из
•США под корой (во время зимовки) и на листьях платана
гибридного. Этот вид обнаружен также в Италии, Югославии, Венг-
.рии, ФРГ и Чехословакии.
Многие жуки-кожееды были в разное время (вероятно, и сто-
.летия тому назад) завезены в города Центральной Европы
(Hämäiäinen, Mannerkoski, 1984). Некоторые из них считаются
космополитами; область их происхождения часто неизвестна
(табл. 32). <ч
С фаунистической точки зрения особый интерес представляет
жук-водолюб Cryptopleurum subtilae (Hydrophilidae) из Японии,
«отмечаемый в Центральной Европе только с 1959 г. .и, очевидно,
-сейчас здесь распространяющийся. Из Средиземноморья
неоднократно завозился жук Zygia oblonga (Melyridae) (например, в
Гамбург), однако он не смог поселиться на длительное время.
,Жук Lophocaterus pusillus (Ostomidae) часто завозится с зер-
3. происхождение и состав городской фауны 95-
НОМ, рисом, земляными орехами, кунжутом, соевыми бобами,
мукой из Южной Азии и Южной Америки, однако также не
приживается. В Северной Америке родина жука-блестянки Glischrochi-
lus quadrisignatus, широко распространенного в Центральной.
Европе. Жук Adistemia watsoni (Lathridiidae) обнаружен в-
птичьем гнезде в Гамбурге (Lefkovitch, 1960). Жук Thorictodes
heydeni (Thorictidae) завезен с пшеницей, рисом, просом,
земляными орехами, птичьим кормом, семенами капокового дерева и
кунжутом в основном из Иидо-Малайской области. Вайднер
(Weidner, 1982) в своем обзоре жуков-усачей, завезенных в
Гамбург с деловой древесиной, называет, например, Plocaederus vi-
ridipennis (из Африки), Brasilianus lacordairei (из Южной
Америки), Cordylomera spinicornis (из Африки), Trachyderus striatus
(из Южной Америки). Вид Parandra brunnea из Неарктики
известен еще с 1916 г. на липах в центре Дрездена. Им было
заселено около 100 деревьев на 1 км^. Завезен он с дело,вой
древесиной или посадочным материалом (Nüssler, 1961). С материалом,,
в котором развиваются личинки, (плетеными изделиями из ивы,,
корзинами, обручами для бочек) из Северной Африки, Кавказа
или Малой Азии в Центральную Европу завезен усач Gracilia
minuta (Klausnitzer, Sander, 1981), расселившийся здесь
преимущественно в городах. Вайднер (Weidner, 1983а) проследнл-
историю распространения в Европе некоторых видов
долгоносиков из рода Sitophilus.
В Центральную Европу с юга распространяется и заселяет в-
основном города перепончатокрылое Sceiiphron sp. (Leclercq,.
Claparede, 1978; Vecht, 1984). To же происходит и с моллюском.;
Arion lusitanicum (Reischütz, 1984).
Блоха Ceratophyllus columbae считается адвентивным видом,,
проникшим в.города одновременно со своим хозяином — сизым
голубем. В голубиных гнездах здесь этот вид часто встречается;
в orpoiMHOM количестве (Krall, 1981).
Строго говоря, искусственно расселенные виды нельзя
причислять ,к адвентивным, однако между ними можно провести и-
некоторые лараллели, например часто незначительные
перспективы длительного выживания, необходимость подходящих
экологических ниш, а также большую роль случайностей. Так,
небольшие популяции юго-восточно- и западноевропейской ящерицы
Lacerta muralis, возникшие в результате выпуска животных,
сохраняются в городах иногда десятилетиями в местах, сходных с ее
природными местообитаниями (изобилующие трещинами
каменные стены, развалины, памятники, виноградники) (Mertens,
1917; Schiemenz, 1980). Иногда в городах выпускают черепах, и
те более или менее успешно приживаются. Это касается прежде-
всего европейской болотной черепахи, раньше встречавшейся в
Берлине и других городах Центральной Европы (она хорошо»
S3
3. Происхождение и состав городской фауны
Таблица 33. Кормовой рацион азиатского бурундука на главном кладбище
•Фрайбурга в разное время года. Сначала указан наиболее часто поедаемый
или запасаемый корм. Сверху вниз частота потребления снижается (по
■Geinitz, 1980)
Весна
Лето
Осень
"Семена липы
(прошлогодние)
Венчики отцветших
анютиных глазок
Цветки липы
«Семена вяза
Различные почки
Ростки пшеницы
Косточки вишни
Желуди (прошлогодние)
Мох
.Личинки тлей (на
листьях тополя)
Медвяная роса (?)
Косточки терна
Семена клена
Косточки вишни
Личинки тлей (на
листьях тополя)
Медвяная роса (?)
Цветки бегонии
Венчики отцветших
анютиных глазок
Мирабель
Лесная земляника
Плоды лавровишни
Черемуха обыкновенная
Семена жостера
Лютик ползучий
(семена)
Семена липы
Семена клена
Снежноягодник
Косточки вишни
Плоды шиповника
освоилась и в Граце), а также Pseudemys scripta elegans (из
Северной Америки, Nabrowsky в письме, 1983), средиземноморской
(из Юго-Заиадной и Юго-Восточной Европы, Северной Африки,
Передней Азии), балканской (из Южной Европы, Schweiger,
1960/61) я среднеазиатской (из Средней Азии) черепах.
На главном кладбище Фрайбурга с 1969 г. существует
популяция сибирского бурундука, насчитывающая 120—150 особей
(Geinitz, 1980). Для этого вида из Японии или Кореи условия во
■Фрайбурге, очевидно, настолько благоприятны, что возможно
длительное сохранение популяции. Летом во время жары
бурундук прячется в норы. Растительность этой территории
обеспечивает ему разнообразный и, вероятно, обильный корм (табл. 33),
к тому же животное прикармливают посетители. Основной пищей
служат семена липы и клена; семена липы он закапывает в
землю как запас на весну.
Изолированное положение кладбища в центре города
обусловливает островной эффект. У .небольшого количества особей,
поселившихся здесь первыми, по-видимому, не было эндопарази-
-тов или они исчезли из-за исходной .немногочисленности
популяции. Во всяком случае, Гайниц (Geinitz, 1980) паразитов не об^
яаружил, что является еще одним преимуществом этой
популяции.
3_ Происхождение и состав городской фауны 97
3.3. Средиземноморские виды
Некоторые -распространенные в Средиземноморье животные
привязаны к этой области лишь существующими там для них
оптимальными экологическими условиями. Когда непреодолимых
препятствий для расселения нет, они проявляют тенденцию «
расширению своего ареала и оседанию на новых территориях в
качестве адвентивных видов. Для существования их наряду с пищей
необходима в первую очередь мягкая с обильными осадками зима
и теплое, сухое, солнечное лето. Даже если сочетание этих
факторов встречается только на небольшом пространстве, возникает
возаможность активного или пассивного поселения
средиземноморских видов за пределами их сплошного ареала (Klausnitzer,
1982а). Эксклавами таких фаунистическпх элементов в
Центральной Европе часто служат южные склоны, котловины и
другие «островки тепла».
Меньше внимания обращалось в этой связи на города
Центральной Европы, особенно крупные (Eriksen, 1976), которые
вполне могут стать жизненным пространством для
средиземноморских видов, та,к как температура в них обычно выше, чем в
окрестностях. Так, Зукопп и др. (Sukopp et al., 1980)
определили, что среднегодовая температура ,в окрестностях Западного
Берлина равна 8,3—8,5 °С, а в егс центре — 9,9 °С, прячем
максимальная температура здесь выше на целых 7°С. Мюллер
(Müller, 1975) сообщает, что в центре Саарбрюккена (ФРГ)
тем.пература на 1,7 °С выше, чем в среднем по городу (9,8 °С).
Кюнельт (Kühnelt, 1970) и Тишлер (Tischler, 1980b) считают,
что городская температура в среднем на 1 °С выше сельской, а
Волькингер (Wolkinger, 1977) приводит как типичную для
многих городов разницу тем,пературы в 1,8 °С и пишет, что в
безветренные дни она может достигать и 10 °С.
Вторая особенность температурного режима внутри города —
значительно меньшие суточные и годовые колебания. Такая вы-
ровненность возникает в результате влияния нескольких
факторов. Так, в городе существуют в большинстве случаев
относительно устойчивые температурные слои, теплообмен между
которыми слабее, чем в окрестностях. Смог, хотя и снижает днем
солнечное излучение, ночью препятствует отдаче тепла. При
температурных инверсиях застой воздуха особенно заметен.
Наблюдается так называемый «эффект духовки», т. е. нагревание стен,
отражение ими тепла, измененная проводящая способ.ность
грунта, быстрый сток осадков и ослабленное в связи с этим
охлаждение. Кроме того, город сам производит тепло.
Сведения об отклонениях в городе других важных для среди- ,
земноморских видов абиотических факторов приведены в
табл. 34.
7-21
98
3. Происхождение и состав городской фауны
Таблица 34. Особенности абиотических факторов городской среды
(по Sucopp et al., 1980)
Параметры
Показатели
в городе по сравнению
с окрестностями
Осадки
Общая сумма (дождь)
На 5—10% больше
Дни с менее 5 мм
осадНа 10% больше
ков (дождь)
Снегопад
На 5% меньше
Относительная
влажЗимой
На 2% меньше
ность
Летом
На 8—10% меньше
Скорость ветра
Среднегодовая
На 10—20% меньше
Безветренное время
На 5—20% больше
Благоприятным для средиземноморских животных может
быть и то, 'ЧТО в городах по приведенным выше причинам
произрастает большое количество растений из Средиземноморья
(см. 3.7).
HoroxiBocTKa Seira domestica — средиземноморский вид
(Dunger, в письме 1983), встречающийся прежде всего внутри домов
(ом. 2.1.1.2 и 2.2.2.3). Кузнечик Месопета meridionale —
насекомое, распространенное, как сообщает Харц (Harz, 1957), в
Южной Франции, Италии, на юге Австрии и в Швейцарии (рис. 28).
С 1965 т., а, по данным Tröger (1981), местами даже с 1958 г.
он отмечен в ФРГ в теплых местообитаниях на юге Верхяерейн-
ской равнины (Фрайбургская котловина, Кайзерштул.ь) (v.
Heiversen, 1969) и в городах Гундельфинген, Фрайбург, Нойнлинден
(Кайзерштуль) и Фалькау (Шварцвальд). Ингриш (Ingrisch,
1980) называет еще Вену и Инсбрук, ссылаясь на Талера
(Thaler, 1977), а Трегер (Tröger, 1981) добавляет Оффенбург.
Примечательно, что npiH продвижении этого вида на север прежде
всего заселяются в основном города. Он отмечен, например, в
зеленых насаждениях (Thaler, 1977). Очевидно, многочисленная
популяция этого вида замечена на стене здания, заросшей Fal-
lopia aubertii (Heiversen, 1969) вместе с видом М. thalassinum.
Другие места городских находок включают виноградные
шпалеры, кусты роз, парки со старыми деревьями, аллеи в центре
города. Трегер (Tröger, 1981), отмечает, что М. meridionale за
пределами города почти всегда встречается одиночными особями.
По-видимо-му, при распространении этого вида решающую
роль играют автомобили; его регулярно замечали (v. Heiversen,
1969; Tröger, 1981) сидящим на транспортных средствах,
которые он не покидает даже на полном ходу.
Средиземноморский жук Amischa forcipata из стафилинид
был впервые обнаружен в ГДР в двух местах на территории
Лейпцига (Klausnitzer et al., 1980, 1982). Раньше северо-восточ-
3. происхождение и состав городской фауны
99
НОЙ границей его ареала считалась Австрия. Этот вид
распространился также в ФРГ (Рейнланд, Дюссельдорф) (Benick,
1967; Benick, Lohse, 1974).
Хорошо изучено расселение божьей коровки Epilachna argus
из единственного известного с 1954 г. маленького ареала в
районе Кнфхойзера, ГДР (рис. 29) (Dorn, 1963; Klausnitzer, 1965;
Dubberke, Creutzburg, 1970; Witsack, 1977; Heinicke, Klausnitzer,
1977). Во время проникновения в соседние районы (вероятно, по
железной дороге) основными местообитаниями на ее пути были
различные города, что объяснялось обилием в них переступня
(корм для жука) и, возможно, благоприятными местными кли-
Рис. 28. Распространение
кузнечика Меесопета
meridionale (по V. Helver-
sen, 1969; Tröger, 1981)
матическими условиями. Сначала вид был обнаружен в Вад-
Франкеихаузене, в 1967 г.—-в Гатерслебене, в 1968 г.— в Ашер-
слебене, затем Кведлинбурге (1969), Хальберштадте (1972) и
Иене (1978). Подобное наблюдалось ив Австрии (Kühnelt, 1981;
Christian, 1981).
Клингер (Klinger, 1985) обнаружил средиземноморский вид
усача Calamobius filum на газонах в центре Франкфурта-на-
Майне.
Бальдовски (Baldovski, в письме от 24.06.1982) наблюдал с
1979 г., очевидно, средиземноморский вид перепончатокрылых
Hylaeus punctatus на цветах нивяника, сныти, очитка едкого и др.
на территории города Герлица, а в 1985 г.— в Дрездене (в пись-
leo
3. Происхождение и состав городской фауны
ме от 9.09.1985). Оба города — единственные местонахождения
этого вида в ГДР и до сих пор самые северные из известных
пунктов его распространения. Муравей Нуроропега punctatissi-
та из Средиземноморья недавно обнаружен в Центральной
Европе; сначала он появился в городах ГДР — Берлине, Потсдаме,
Грайце, Галле и Лейпциге (Seifert, 1982). При постройке гнезд
вне зданий эти насекомые, вероятно, всегда нуждаются в
дополнительном тепле для зимовки.
Некоторые южноевропейские пауки селятся в центральноев-
ропейских городах, поскольку могут использовать запасы тепла
внутренних и внешних стен зданий (Sacher, 1983). Речь идет
прежде всего о видах Nigma walckenaeri (половозрелые особи
появляются только в августе), Dictyna civica и Zygiella x-notata
(тепло амбаров позволяет зимовать коконам).
— Гвсцаарствентя граница _
Рис. 29. Расселение божьей коровки Epilachna argus в ГДР
Klausnitzer, 1979)
(ио Heinicke.
Южный вид сенокосцев Nelima semproni (рис. 30) был
обнаружен только в городской черте Варшавы, в окрестностях горо^
да находок не было (Czechowski, Starega, 1977).
Из Средиземноморья происходят два вида равноногих
Porcellio laevis и Р. dilatatus, встречающиеся в Центральной Европе в
теплицах (см. 2.2.4.7). Эти и некоторые другие средиземномор-'
ские виды можно считать адвентивными. Средиземноморье —
один из важнейших иервичных ареалов таких элементов фауны.
Проникновение их в новые районы происходило, вероятно, уже
много столетий назад, поэтому сейчас множество исходно
средиземноморских В.ИДОВ можно отнести к городской фауне. В
качестве примеров можно назвать также виды Armadillidum vulgare
и Haplophthalmus danicus, из которых первый находит в горо-
3. Происхождение и состав городской фауны
101
дах, по-видимому, особо благоприятные условия. На севере
Центральной Европы оба считаются даже эвсинантропами (Tischler,
1980).
Кольчатый червь Allolobophora icterica из Юго-западной
Европы проник в Центральную Европу и встречается здесь, по всей
вероятности, только в городах (Höser, устное сообщение, 1984).
Происходящая, вероятно, из Средиземноморья и
встречающаяся там, по-видимому, в естественных условиях, сахарная че-
Рис. Зв. Распространение
сенокосца Nelima semproni
(по Czechowski, Starega,
1977)
шуйница [Leplsma saccharlna) широко распространена в
городах Центральной Европы (квартирный синантроп), однако в
природе здесь не встречается (по крайней мере в ГДР; Dunger,
в письме от 13.10.1981). Средиземноморская щетинохвостка
термобия домашняя [Thermobia domestica) может развиваться
только в пекарнях, поскольку нуждается в особенно высокой и
относительно постоянной температуре (см. 2.1.3). Черный
таракан происходит из Черноморской области и встречается на юго-
востоке Европы в природе. В центральноевропейских городах он
102 3. Происхождение и состав городской фауны
известен уже более 400 лет (Kühnelt, 1970). То же самое можно
сказать о рыжем таракане (его родина — Крым), который в
противоположность черному иногда наблюдается в природе даже в
ГДР. Домовый сверчок с естественным ареалом в южном
Средиземноморье и Западной Азии успешно расселился в централь-
ноевропейские города. Возможно, он был завезен сюда еще
римлянами (Weidner, 1972). В городах он все чаще снова
встречается вне домов, особенно летом — на улицах, в трещинах
каменных стен, возле труб центрального отопления, на мусорных
свалках (Haas, 1959, 1960; Weidner, 1972). Постельный клоп тоже
происходит из Средиземноморской области; полагают, что он
появился в Центральной Европе около 1000 лет назад (Weidner,
1952). В своем распространении постельный клоп, 'как и
большинство южных видов, не ограничивается только городами.
Естественное расширение ареалов также может быть
причиной появления средиземноморских видов (Neithammer, 1963).
С некоторой оговоркой здесь может быть приведен пример с
кольчатой горлицей. Первоначально область ее распространения
ограничивалась Передней Азией и областями к востоку от нее.
До XX в. этот вид встречался в Европе только местами на
Балканском полуострове. С 1930 г. наблюдается расширение его
ареала, который в 1932 г. захватил Венгрию, в 1936
г.—Чехословакию, затем Австрию (1938), Германию (1943), Северную
Италию (1944), Нидерланды (1947), Данию (1948), Швецию
(1949), Норвегию (1954) и Англию (1955). В процессе
расселения важную роль играли города, часто служившие приютом
крупным популяциям. Гримм (Grimm, 1953) сообщает, что этот вид
уже в 1953 г. наблюдали в Галле (ср. Gnielka, Wolter, 1970) и
йене. В 1950 г. кольчатая горлица впервые стала гнездиться в
Берлине, а в 1964 г. в Западном Берлине насчитывали по
меньшей мере 1432 пары этих птиц (Löschau, Lenz, 1967; Lenz, Witt,
1978). Самая высокая плотность заселения наблюдалась в
районах, застроенных виллами с садами и в зоне жилых кварталов
с редкими деревьями вдоль улиц (до 7 гнездовых участков на
10 га). В крупных зеленых массивах и остатках леса, а также в
районах города, где нет деревьев, этот вид почти не встречается.
В Карл-Маркс-Штадте первые гнезда были обнаружены в 1947 г.,
а в 1968 г. здесь гнездились уже 680 пар (Saemann, 1970)
(рис. 31). Здесь они также концентрировались в определенных
частях застроенной городской территории, откуда шло
дальнейшее заселение городских окраин и окрестностей (Saemann, 1969,
1975).
Сообщалось (Gnielka, 1981) о поразительном увеличении
численности кольчатой горлицы на кладбище в Галле, где этот вид
стал впервые гнездиться, по-видимому, в 1957 г. (рис. 23).
3. Происхождение и состав городской фауны
103
3.4. Эпилитные виды
Эпилитная, т. е. скальная среда гор обычно не граничит
непосредственно с городами. Однако вертикальные стены домов с
их щелями .и потенциальным.и убежищами своим субстратом,
наличием пищи, накопленным теплом и другими особенностями
микроклимата во многом напоминают скалы. Чем проще стена
по структуре, тем ограниченнее возможности ее заселения. В
городах наблюдаются некоторые исходно эпилитные виды живот-
Рис. 31. Распространение кольчатой горлицы в Карл-Маркс-Штадте в 1968 г.
(по Saemann, 1970). Граница жилых массивов показана штриховой линией,
застроенная часть города ■— белым цветом
ных. Иногда их популяции здесь даже намного крупнее, чем в
первоначальных местообитаниях. Высокая численность видов
объясняется, возможно, длящимся веками ростом города, что
обеспечивает непрерывное расширение пригодного для жизни
пространства при малом количестве конкурентов и хищников.
Сапсан часто вьет гнезда на отвесных скалах и на речных
обрывах. Этот вид гнездится и в крупных городах (на высотных
184 3. Происхождение и состав городской фауны
зданиях, башнях), где его основной пищей служат одичавш«е д©-
машние голуби. Гримм (Grimm, 1953) объясняет отсутствие саи-
саяа в Берлине .после второй мировой войны разрушением
многих высоких домов и малочисленностью голубей (ср. Scknurre,
1950). #*te».
Высокие здания подходят для гнездования обыкновенной
пустельги, которая считается по крайней мере отчасти обитателем
скал. Крампиц (Krampitz, 1949) сообщает, что участки
гнездования и охоты этого вида могут отстоять друг от друга на 2—3 км.
Согласно Рюмекопфу (Rümekopf, 1951), пустельга, гнездящаяся
в Лейпциге, питалась птенцами скворцов, воробьями, мелкими
млекопитающими и жуками. Земан (Saemann, 1969) упоминает
в качестве ее добычи кольчатую горлицу, а также домового
воробья, зеленушку и черного дрозда.
—Встречающиеся главным образом в городах очень многочис-
ленные колонии голубей ведут свое начало от сизого голубя и
по окраске часто более или менее схожи со своей предковой
формой (Обухова, Креславский, 1984, 1985). В природе сизый голубь
,гнездится почти исключительно в расселинах и углублениях
скал, а также на крутых морских берегах. Здания являются,
очевидно, оптимальным для него местом гнездования, поскольку
полностью отвечают унаследованному поведению. Сизые голубя
предпочитают высокие постройки типа церквей и башен, а также
развалины, жилые дома и опоры мостов. Для сооружения гнезд
особенно часто используются восточная и южная стороны. Во
многих городах существуют 'четко очерченные места скопления
голубей (для дикой формы также типично гнездование
колониями). Сегодня голуби обитают, по-видимому, во всех крупных
городах Европы и Аз'ии, некоторые из которых давно славятся
высокой численностью этих птиц, например Венеция и Лондон.
В Гамбурге обитает 150 000—^200 000 их особей (Krall, 1981).
«Завоевание» некоторых городов голубями произошло только в
последние десятилетия; в, Гамбурге их численность увеличилась
с 1959 г. в 10 раз (Krall, 1981). Земан (Saemann, 1970) считает
началом заселения 'одичавшими домашними голубями Карл-
Маркс-Штадта 1965 г.; в 1968 г. в центре города и в зоне жилых
'Кварталов их гнездилось уже 120 пар. Благоприятствовало
этому 'Прежде всего наличие корма.
■ Сипуху также считают изначально обитателем скал,
расселины которых она 'Использует для постройки гнезд. Здания, прежде
всего башни церквей 'И развалины,— предпочтительные места ее
'Гнездования в городах и в деревнях; для этого подходят прежде
всего ниши в каменных стенах, чердаки, колокольни и голубятни.
Гнезда этого вида иногда встречаются и в парках. 'В Меклен-
бурге в 1968/69 г. отмечено 144 гнезда (Krägenow, 1970), из
которых 60 располагались снаружи и внутри церквей. В городах
3. Происхождение и состав городской фауны 105
всновную пищу сипухи составляют мелкие грызуны, а также
летучие мыши (прежде всего у лётных отверстий) и воробьи.
В башне ратушп в Иене обнаружены остатки 128 жнвотных-
жертв (114 мелких млекопитающих и 14 птиц).
Черпыйстриж—характерная птица почти всех городов,
обитающая также в крупных деревнях. Предполагают, что
возрастание встречаемости этого вида в городах со времен средневековья
обусловлено изменением застройки. Основную пищу черного
стрижа составляют летающие насекомые (в первую очередь тли)
и примерно на 5% пауки-«аэронавты». Для выкармливания
птенцов ежедневно уничтожаются до 40 000 экземпляров жертв, чтэ
составляет по массе примерно 50 г (ср. Dathe, 1951)'.
Первоначальные места гнездования черного стрижа—скалы, хотя в Во-
сточво-й и Северной Европе он селится и в дуплах деревьев.
Стены башен, фабрик и высоких старых жилых зданий представляют
ему схожие условия (отверстия и трещины под выступами крыш).
Гнезда строятся, как правило, на высоте более 12 м. Решающее
значение имеет, вероятно, необходимая свободная высота
падения; ©птнмальное расстояние от земли до гнезда, по-видимому,
соста,вляет 10 м и более (вряд ли ниже 3 м). Важно также
наличие узких горизонтальных щелей для взлета птиц под скатоил
кровли. Новостройки, как правило, неудобны для гнездования..
В старых районах с по крайней мере трехэтажными зданиями
Земан (Saemann, 1970) отметил 20 гнездящихся пар на 10 га.
Для Галле (Gnielka et al., 1983) приводят 550—1800 пар.
Черный стриж селится в городе преимущественно колониями в 4—
10 пар. Как показывает кольцевание птиц, привязанность к
местам гнездования сохраняется десятилетиями (Gnielka et al.,
1983). Примечательны единовременный прилет и отлет птиц. Для
Карл-Маркс-Штадта, например, день прилета приходится в
среднем на 29 апреля. Отлет происходит чаще всего в конце июля —^
середине августа.
Белобрюхий стриж гнездится в природе на отвесных скалах
в высокогорье, ■но-вет.речается и в городах на высоких зданиях.
Предполагают, что деревенская ласточка стала синантропом
м.НОГО тысяч лет назад, сменив свои исходные местообитания
(вертикальные скалы и др.) на здания. Сегодня она живет почти
исключительно в таких условиях, однако чаще в деревнях, чем
в городах (даже на юртах). Деревенская ласточка характерна
для пригородов, а внутри города она гнездится большей частью
изолированно, вероятно, из-за удаленности от водоемов. Эти
гнезда часто размещаются у мест содержания животных,
например в зоопарках, у ветеринарных лечебниц, конюшен.
Деревенские ласточки встречаются даже в промышленных зданиях.
Требования их к типу сооружений высокие и в городах удовлётво-;
ряются редко. После периода гнездования лтиды. часто стаями
106 3. Происхождение и состав городской фауны
залетают в город, ловят на улицах и в парках насекомых и
греются на крышах. В большинстве городов их численность падает,
вероятно, в связи с сокращением поголовья скота. В Галле
(Gnielka et al., 1983) отмечено 600—2000 пар, а в Западном Берлине
250—500 (Bruch et al., 1978).
Городская ласточка в природных условиях гнездится на
скалах (Me^izel, 1984); .в качестве вторичных мест гнездовий она
использует определенные структуры зданий. В некоторых
городах наблюдалось взрывообразное увеличение ее численности.
Излюбленными Местами для поселения служат районы
новостроек (см. 2.2.2.3, рис. 8) (Lenz et al., 1971; Saemann, 1970;
Grimm, 1974; Plath, 1981; Gnielka et al., 1983; Menzel, 1984).
Галки в природе также гнездятся в расселинах скал и дуплах
деревьев. В городах они заселяют отверстия в высоких
строениях, развалинах, каменных стенах и т. д. По данным Земана
(Saemann, 1970) ,в Карл-Маркс-Гитадте 80% гнезд находились на
высоких зданиях (промышленных предприятиях, церквях) и
жилых домах (неоштукатуренные фронтоны, отверстия печных
труб) (см. Zimmermann, 1951). Большое количество галок
прилетает в города зимой вместе с грачами. Доля галок в таких
стаях составляет около 10—15%. В Галле зимует ежегодно
около 3000 их особей (Gnielka et al., 1983).
В пустотах и расселинах скал, а также на каменных осыпях
в горах гнездится горихвостка-чернушка. Такая экология стала
хорошей предпосылкой для заселения городских каменных
строений; она стала характерным видом различных городских
местообитаний и гнездится почти исключительно на зданиях (в
отверстиях стен); за пределами городской застройки обнаружено
только несколько гнезд (исключительно в деревнях).
Кроме названных птиц, можно привести несколько примеров
эпилитных видов города из других групп животных. Каменная
куница, (ПО-видимому, обитает в природе среди скал. Сейчас она
живет в основном вблизи или внутри населенных пунктов, вплоть
до центра крупных городов, где ее особенно часто встречают на
чердаках. Иногда к ним приурочены основные места ее
концентрации в урбанизированной местности (Nicht, 1969; Klawitter,
1979). В Западном Берлине на территории в 209 га обнаружены
следы 30 особей. Основные места охоты каменной куницы —
парки, кладбища, садовые участки, заброшенные деревенские
постройки (скотные дворы), дорожные строения и складские
помещения. Главная пища в городе — голуби, крысы и мыши.
Летучая мышь двухцветный кожан — исконный обитатель
скал (Bauer, 1954). В Европе вид обитает преимущественно (на
севере — почти исключительно) в городах и охотится над
крышами их центров (Ryberg, 1947; Natuschke, 1960). Нынешнее
широкое распространение этого пришельца с гор Юго-Западной
3. происхождение и состав городской фауны
107
Азии — результат сходства климатических условий (прежде все»
го континентальности) его естественных местообитаний и
городских центров, где, возможно, имеются и другие .благоприятные
для него факторы (Bauer, 1954). В Вене двухцветный кожан
отмечается главным образом на чердаках (Kühnelt, 1955).
Сенокосец Leiobunum limbatum (фото 9) широко
распространен 1в альпийской зо.не Швейцарии, Австрии, ФРГ и Италии, а за
ее пределами часто обнаруживается в городах, в частности ;в
Рис. Распространение
генокосца ОрШо canestrinil
в ГДР и Западном Берлине
(по Bliss, 1981) -—"^ Государственная граница ■
Гере, Галле, Берлине и Эберсвальде (Sacher, 1978). Присутствие
этого адвентивного вида на зданиях и внутри них, на каменных
стенах и т. д., вероятно, объясняется его приуроченностью в
природе к скальному биотопу. Аналогичная ситуация с сенокосцем
Opilio canestrinil также родом из Альп (и Апениияского
полуострова) (рис. 32), который обнаружен во многих местах ГДР
(иногда вместе с сенокасцем Phalangium opilio) ib перовую
очередь на стенах зданий и каменных оградах, а также на деревьях
в парке (Bliss, 1981; Martens, 1978; Thaler, 1979).
108
3. Происхождение и состав городской фауны
Таблица^ 35. Сравнение некоторых абиотических факторов среды пещер н
Температура
Свет
Влажность
Пещеры
Подвалы
Относительно постоянная, в
большинстве случаев низкая
От сплошной темноты
(внутри) до хорошей
освещенности у входа в пещеру
Относительно высокая
Относительно постоянная,
в большинстве случаев
низкая
От сплошной темноты (если
нет окон) до хорошей
освещенности (если имеются
окна)
Очень разная (в новых
зданиях меньше, чем в старых,
где в большинстве случаев
относительная влажность
более 80%). Зависит, например,
от отопления и вентиляции
Наружные стены домов — места посадки и отдыха многих
видов насекомых, прежде всего двукрылых, которые являются
идеальным источником питания для некоторых пауков,
обитавших изначально на скалах (см. 2.2.1.1), например для петрофи-
ла Salticus scenicus. Этот вид встречается иногда в огромных
количествах, используя также накопленное стенами тепло. Здесь
обитает Sitticus pubescens (тоже скальный охотник) и другие
пауки-скакуны (Salticidae).
Некоторые эпилитные брюхойагие преадаптированы для
жизни на каменных стенах и часто встречаются на развалинах
крепостей, а также в городах (Кегпеу et al., 1983). В качестве
примера можно назвать ClausiÜa bldentata, С. dubia, Balea perversa
И другие виды рода Balea, Helicigöna lapicida (Klausnitzer, 1965;
Streib, 1984).
3.5. Троглобионты
Экологические факторы подвалов, пещер (табл. 35) и
некоторых нор млекопитающих, несомненно, сходны, что отражается
и на составе их фауны. Часть подвальной бноты (см^ 2.1.1.3)
обнаружена в пещерах и норах. Аллохтонная органическая пища
в.:ПОдвалах в большинстве случаев значительно разнообразнее и
обильнее, чем в пещерах; к тому же часто имеются грибы. Если
в пещерах, большей частью преобладают детритофаги, то в
подвалах к ним добавляется большое число хищных видов.
Различные пауки, селящиеся в природных условиях в пеще-
||»-ах Счаще всего у входа в них),-а также мелких пустотах,
находят в сырых и прохладных.подвалах такие же или схожие уело-
3. происхождение и состав городской фауны
109
Таблица 36. Процент некоторых домашних животных в Саарбрюккене
яа 1976 г. (по Müller, 1981)
таксон
процент
1 таксон
процент
Рыбы (число
владель1 Млекопитающие
(53,50)
цев)
6,04
Собаки
22,20
Черепахи
5,56
Кошки
14,19
Птицы
(34,90)
Хомяки
7,86
Волнистые попугаи
16,87
Морские свинки
4,94
Канарейки
8,44
Мыши
4,31
Прочие попугаи
2,97
Прочие
6,62
1
вия. примерами могут служить Nesticus cellulanus, Meta menar-
di, M. merianae, Lepthyphantes nebulosus и L. leprosus (Sacher,
1983).
3.e. Домашние животные
Количество домашних животных, содержащихся в городах,
очень велико (табл. 36) и во многих местах увеличивается.
Некоторые из них (прежде всего собаки и кошки) становятся здесь
существенным экологическим фактором. Домашние животные
создают условия для существования большого числа паразитов,
например блох (на собаках) (Müller, 1986). Собачьи
экскременты, с одной стороны, имеют паразитолого-гигиеническое значение
(Kirchberg, 1951; Teschner, 1959; Berghausen, 1973; Kühlhorn,
1981), а с другой — являются трофическим фактором (в среднем
500 г в день иа особь). Многочисленные одичавшие кошки
выполняют в той или иной степени экологическую функцию
верховных хиитников. Вылетевшие из домов волнистые попугаи
составили 9% добычи сокола Falco subbuteo в Западном Берлине
(Fiuczynski, 1979). Количество кошек, как правило, к центру
города уменьшается,,^^^ собак увеличивается (Müller, 1981)-
В 1979 г. в столице ГДР насчитывалось около 40 000 собак и
60 000 кошек. В ФРГ отмечено примерно 3 млн. собак и 2,5 млн.
кошек, во Франции 7,4 млн. собак и столько же кошек.
3.7. Заселение животными гемерохорных
растений
Для городской флоры характерен высокий процент гемеро-
хоррв (ряс. 33). По времени пронииновения в синантропные
местообитания различают архёофиты (доисторические и раннеисто-
no
3. Происхождение и состав городской фауны
рические времена до средневековья) и неофиты (после 1500 г.,
т. е. открытия Америки). Сюда же относят постоянно
появляющиеся ib городах новые виды, большинство из которых не может
прижиться надолго, нх называют адвентивными^'. Способы про-
иикновения гемерохоров очень разнообразны:
а) преднамеренный завоз и разведение (одичавшие .полезные
и декоративные растения, виды из ботанических садов и др.).
б) непреднамеренный занос (полевые и садовые сорняки,
семена трав, «спутники» интродуцированных деревьев и
кустарников, партий шерсти, южных плодов, растительный корм для птиц
и т. п.).
■Лесные Деревни Мелкие Средние Крупные
посети города города города
Рис. 33. Процент гемерохоров в различных типах поселений (по Sukopp, 1982)
Гемерохоры с экологической точки зрения можно
подразделить на:
а) виды, сохраняющиеся и после .прекращения
антропогенного влияния, закрепляющиеся в потенциально естественной
растительности и конкурентоспособные в близких к природе
растительных сообществах (агриофиты);
б) виды, занимающие устойчивое .положение в нынешней
реальной, но не в потенциально естественной растительности (эпе-
кофиты);
1в) виды, хотя и дикорастущие, но не способные длительное
время сохраняться без влияния человека (эфемерофиты)
') Согласно другим системам, адвентивными называют любые заносные
виды. В этом случае последнюю группу называют эфемерофитами. — Прим.
ред.
3. происхождение и состав городской фауны ш
г) виды, встречающиеся только в разводимом состоянии
(культурные растения).
Многие из этих растений вообще не заселяются или
заселяются только немногими местными насекомыми-полифагами, так
что высокий процент гемерохоров ведет к обеднению энтомо-
фауны.
На примере оолинника двулетнего {Oenothera biennis) можно
рассмотреть некоторые особенности освоения гемерохоров
животными (Klausnitzer, Förster, 1984). Ряд ботаников полагают,
что все среднеевропейские виды Oenothera завезены из Северной
Америки или возникли в Центральной Европе в результате апо-'
микоиса. Ростански (Rostanski, 1968) считает автохтонами
Центральной Европы по крайней мере два вида — О. biennis и
О. rubricaulus.
Если завезенные из Северной Америки насекомые-фитофаги
в Центральной Европе живут на ослиннике, то частичная
европейская автохтония этих растений должна сочетаться с
заселением не всех их видов или предпочтением одних видов другим.
Если часть видов этого рода происходит из Северной
Америки, то на автохтонных видах Центральной Европы должна была
бы встречаться главным образом среднеевропейская энтомофау-'
на, в большей или меньшей степени переходящая на неофиты.
Самым четким у^казанием на происхождение было бы развитие
автохтонных монофагов только на автохтонных
среднеевропейских растениях. Правда, олигофаги, питающиеся сейчас в
Центральной Европе всеми видами этого рода, 500 лет назад могли
обитать только на исконно среднеовропейсних таксонах.
Если весь род происходит из Северной Америки, европейские
фитофаги должны реагировать на новые растения попытками
использовать новые экологические ниши. Это удалось бы в
первую очередь видам, живущим на близкородственных родах
растений, особенно на других представителях семейства кипрейных
{Ludwigia, Epilobium, Circaea). В этом случае фитофаги,
ограниченные только ослинником, должны были бы отсутствовать
или иметь североамериканокое происхождение.
Наши данные о заселении насекомыми ослинника двулетнего
дают следующую картину:
а) обнаружен единственный североамериканский вид тлей
Aphis oenotherae, олигофаг различных кипрейных (Müller, 1974,
1975; Klausnitzer, Forster, 1984);
б) все другие фитофаги, обитающие на ослиннике, в
Центральной Европе автохтонны. Их можно разделить на две группы
(Klausnitzer, Forster, 1984):
— олигофаги, основной пищей которых являются виды ЕрИо-
Ыит; и реже — Circaea, в большей или меньшей степени
переходящие на ослинник, например бабочки Proserpinus proserpina и
112
3. Происхождение и состав городской фауны
Mompha epilobiella, жуки НаШса oleracea и Adoxus obscurus.
Данных о монофагах, ограничивающихся только видами
ослинника, нет;
■— полифаги, которые наряду с другими кормовыми
растениями заселяют ослинник, например различные клопы, бабочки
Eupithecia castigata и Coleophora troglodytella, двукрылое Phyto-
myza atricornis, цикадовое Philaenus spumarius, тля Brachycau-
dus cardui.
Ha основании этих наверняка еще не полных данных о фауне
фитофагов можно сделать вывод, что ослинник двулетний —
завезенный вид. Неясность в вопросе о происхождении видов этого
рода, несомненно, .представляет собой особый случай; в других
приводимых ниже примерах происхождение кормовых растений
сомнений не вызывает.
пж
Цветки
СтеЁли
Листья
Норна
П'О
/7ж
п1
Г
Рис. 34. Число насекомых-фитофагов на различных частях пижмы (ПЖ)
и полыни обыкновенной (ПО) (по Klausnitzer, 1968): Б — бабочки; Ж —
жуки; Д — двукрылые; Т — тли
На 100 видах декоративных растений (Рарра, 1976) было об- 1
наружено 166 видов насекомых-фитофагов: 37 местных на мест- '
ных, 85 местных на местных и интродуцированных, 44 вида —
только на завезенных. Число 166 относительно невелико, если j
вспомнить, что на каждый сравнимый автохтонный вид в Цент- '
ральной Европе часто может приходиться 5—10 видов
фитофагов (Klausnitzer, 1977) (рис. 34). На деревьях-неофитах в
Англии обнаружена очень бедная энтомофауна (каштан — 5 видов;
конский каштан — 4; орех — 3; робиния — 1), а на дубе и
березе—более 200 видов насекомых (Southwood, 1961). Дорн (Dorn,
1982) указывает 42 вида диких пчел, опыляющих декоративные^!
растения, которые по большей части растут ил.и высаж1иваются ■
в городе.
С некоторыми растениями-гемерохорами были завезены тесно
связанные с ними виды насекомых, для других
насекомые-специалисты появились позже; иногда их считали адвентивным,и
видами. Вместе с сиренью обыкновенной из Восточного
Средиземноморья в Центральную Европу .проникла бабочка CaloptiUa sy-
ringella, сейчас широко распространенная в ГДР и всюДу здесь
3. Происхождение и состав городской фауны 1щ
часто встречающаяся (Borkowski, 1973; Рарра, 1976). В
городских парках она отмечена также на близкородственной сирени,
происходящей с Востока я яз Средиземноморья бирючине обьж-.
новенной (Patzak, в письме, 1981). Завезенный, вероятно, на
транспорте долгоносик Otiorhynchus stnreczynskii (фото 14) оби-,
тает в Центральной Европе только в населенных,пунктах (парт,
ки, сады, обочины дорог) на бирючине (его кормовое растение) ^
а также на сирени (Dieckmann, 1980). То же самое относится к
долгоносику О. dieckmanni, (Gospodar, Korge, 1982) и тле Myzus:
lygustri.
Листовертка Olethreutes siderana (Tortricidae) обитает на.
интродуцированных из Северной Америки двух видах спиреи;
(Spinaea douglasii и 5. saücifoUa). По данным ван Франкенхей-.
зена (van Frankenhuysen, 1981), она была впервые отмечена в_,
Голландии в 1975 г., а в 1980 г. встречалась там уже очень часто.
На различных видах спиреи живут также тли Acyrthosiphon ig-
notum {S.Xvanhouttei, S.Xarguta, S. solicifolia), Aphis spirae-:
phaga {S.Xvanhouttei, S.Xarguta) и Brachycaudus spiraeae-
{S. salicifolia), географическое происхождение которых неясно,
Еще в 1900 г. распространение средиземноморской бабочки
Phyllonoryder platani ограничивалось Южной Европой и Малой.
Азией. В последние 50 лет у нее наблюдалась все возрастающая,,
тенденция к расселению на север. Очевидно, вид активно
следовал за своим кормовым растением чинарой (Platanus orientalis).
Сначала эти бабочки, ло-видимому, заселили значительную часть,
Франции и Голландии (van Frankenhuyzen, 1983), а около 1920 г.
были замечены на юго-западе Гер(мании. Примерно в 1950 г. за^..
регистрированы их крупные скопления в районе Боденского -озе^
ра, близ Штутгарта я на равнине верхнего Рейна (Tiederaann,
1978). Наконец, стали известны, находки с юга Чехословакии,
в 1972 г.— в Польше (Borkowski, 1973), в 1973 г. в Западном
Берлине и в 1975 г. в Гамбурге (Tiedemann, .1978). В последние,
годы Ph. platani обнаружена в нескольких городах ГДР на
платане гибридном {Platanus hybrida). Повсюду численность этого,
вида относительно велика. Он наблюдался в Лейпциге (1978),
Галле (1980), Ашеролебене (1979), Магдебурге (1980) (Patzak,
в письме, 1981). В 1977 г. вид достиг Дании (Tiedemann, 1980).
На айланте высочайшем, неофите из Китая может селиться,
айлантовый шелкопряд {Philosamia cynthia], уже отмеченный в„
центре Вены (Kühnelt, 1955).
Вместе со своим кормовым растением гребенщиком Tamarix
tetranda в Центральную Европу из Средиземноморья проникла,
(понвидямому, пассивно) цикада Opsins stactogalus (она посели-,
лась также на мирикарии Myricaria germanica) из того же ее-
мейства. Гребенщик высаживают преимущественно в городах,
реже —в деревнях, где также находят эту цикаду, занесенную,;
8—21 ~
" x
'ft
to j
co
v, 00
?5 <» • -
i£ ^
<3) ol cd
— ^
«5 — -г:
^ - oj
к-a
о —' a>
tu
о ~ со
о. .
« Е-2
ся со
я j о.
•е- 3
0 .-0^
я со . .
s-co
о>
Й cj"~
я -с c
я
s
ш — <
1 с
5 ся ю
■га о
, р1<
га
га
со си t>
lis
э а
о) о
X га
и 2 »
s и
о сэ
= 2 „
3„
s3
11
£ со
о и
и к j3 ,
п <и ч
е * s t
га °м
о к
о. к
eg
га о
я
ч
се
е-
s
я
о
а.
а.
е
а
u
о к
— я 1я
~ я S
. се о.
сч r-f ccj
er
ё5а
№
со
3 а
й л cs
д ч я
э
яо,
я
к
S
а.
се
cf
•я
о)
я
со 05 — со
00 « со
05 cj5 05 ci
со <м —. о)
ю со со
<j> а>Oi (jiO^
ocmcoino)—■ —<
cdcocococdt--t~ 00
05 05 05 Oi 05 05 q> ob
ft
u
o
ю
co
я'
»я
. я
— я
со^
05
со
05 >
1я
я
в
со
а.
©
см"
ю
о)
.с^ 53
11
cu
.со
|о
о.
s
3
е
со
сз о)
s-2
■2 ьо
с
о
1эд
с
о,
>
x
со
s3
о
о
к
со
S
><
со
я
со
т
cp я
s &
и S
о=я
cq
к
со
я
5 я
s<
к
со
ч я
со со
со
сп
со
« я
со 5
ш s
о) ^
.ci cu
co
c3
"c3
3
co ,
q
1«
q
g «
:г о
2-3
3. Происхождение и состав городской фауны
115
вероятно, с посадочным материалом. В ГДР вид обнаружен в
Дрездене, Лейпциге, Галле (первое сообщение—15.08.1919),
Стендале и Ростоке. Он завезен также в Северную Америку
(Рарра, 1976). Иногда с лекарственным сырьем в Центральную
Европу попадал жук-грибоед Berginus tatnarisci из Средиземноморья,
обитающий там среди прочих растений и на гребенщиках.
Рис. 35. Распространение тли Impatientinum asiaticum в Европе (по Lampet.
1978)
Тли играют особенно больщую роль при заселении гемерохо^
ров (табл. 37). По данным Мюллера (Müller, 1986), в ГДР
обнаружен 21 вид тлей иноземного происхождения, отмечаемых %
городах на гамерохорах и сорняках. В некоторых случаях весьма,
вероятен их завоз самолетами, например, для Impatientinum asia^
ticum (рис. 35) и Uroleucon erigeronensis (Lampel, 1983).
8«
4. СИНАНТРОПИЯ И УРБАНИЗАЦИЯ
Синантропия и урбанизация (последнее слово употребляется
здесь только в своем частном значении как экологическое
понятие) — это биологические феномены, вызванные прежде всего
возникновением городов и тесно связанных с их строительством
и развитием. С появлением поселений уничтожаются природные
биоценозы и создаются новые со свободными и совершенно
своеобразными экологическими иишами, которые осваиваются
животными различного происхождения (обитателями скал, обрывов,
нор и пещер, древесины, семян и плодов, некрофагами, копро-
фагами и паразитами). Для некоторых синантропных видов,
например Sitophilus granarius (Weidner, 1983а), природных
популяций вообще неизвестно. Вероятно, синантропия возникла 6—
10 тыс. лет назад, однако в отдельных случаях она может иметь
и более древнее происхождение.
Часто различают виды, «избегающие» культурный ландшафт,
«индифферентные» к нему и являющиеся его «спутниками». Ге-
мерофобами называют виды, которые параллельно
антропогенным изменениям среды отступают в менее нарушенные области,
т. е. вытесняются туда (Schwerdtfeger, 1975; Schäfer, Tischler,
1983). Важнейшие местообитания, исчезающие из
урбанизированных областей, следующие (Mader, 1980):
— сырые или влажные участки (с застойным увлажнением);
— бедные иитательными веществами почвы и воды;
— участки с густой растительностью;
— древесина различной степени разрушения;
— скальные породы и продукты их выветривания;
— ложбины, низины, склоны и проч.
Гемеродиафорами называют виды, чье существование почти
или совершенно не зависит от антропогенного изменения ланд--
шафта (Schwerdtfeger, 1975).
Гемерофилы предпочитают местообитания, созданные
человеком, используя новые экологические ниши (климат, питание,
материалы, пространственную структуру). Еще до
возникновения постоянных поселений человека существовали его виды-
«спутники» (Povolny, 1963). С развитием населенных пунктов и
дальнейшим изменением ландшафта появились гемерофилы.
Среди видов, осевших в зоне поселений, были потенциальные
синантропы. Из видов, использующих другие антропогенно изме-
4. Синантропия и урбанизация И7
ненные ландшафты, возникли агробиоценозы (Povolny, 1963).
Отличие гемерофилов от синантропов состоит в том, что для
первых характерна только этолого-экологическая, но не биоденоти-
ческая зависимость от человека (Povolny, 1963). Обычно
понятие гемерофилии употребляется в узком смысле, поскольку в
противном случае почти все обитающие в Средней Европе виды
должны были бы считаться гемерофилами (ср. Warnecke, 1936).
Синантропию определяют по-разному (Peters, 1960; Povolny,
1962, 1963; Povolny, Sustek, 1982, 1985; Tischler, 1973). Здесь
вслед за Повольни (Povolny, 1963) мы используем как
решающий критерий принадлежность того или иного вида к антропо-
ценозу, считая антропоценоз системой взаимосвязей между чело-
.веком, домашними и синантропными животными. Ограничение
этого понятия «совместной жизнью неодомашненных животных с
человеком» неточно. Для характеристики синантропии
существенны два критерия (Schäfer, Tischler, 1983):
— спонтанное присутствие организмов в поселениях
человека без или против его воли;
— тесное сосуществование с человеком или зависимость от
•его деятельности.
«Спонтанность» .можно считать реализацией экологической
яиши. Термин «синантропия» должен означать связь с
человеком не только через пространственную структуру поселений, но
и через непосредственно зависимых от него домашних животны.х.
Различают следующие формы синантропии:
а) облигатная (звсинантропия): вид встречается как минимум
в одной из климатических зон только в антропогенных условиях,
а в пределе — только в зоне поселений человека (рис. 36);
б) факультативная (гемисинантропия, олигосинантропия):
виды имеют в зоне поселений человека оптимальные условия
существования, однако образуют популяции и вне антропоценозов,
т которых возможна иммиграция в природные биотопы;
в) непрерывная (перманентная): жизненный цикл вида
полностью протекает в антропоценозах;
г) временная (ксенантропия): виды находятся в антропоце-
:Нозах только в определенное время (например, в период зимов-
аси) или при определенных условиях, не образуя там
самовозобновляющейся популяции;
д) частичная: вид принадлежит к антропоценозу на опреде-
.ленной стадии своей жизнедеятельности (возможно, лишь часть
суток), а в остальное время входит в другие биоценозы. В
качестве примера можно привести городскую ласточку, гнездящуюся
на домах, но находящую корм (по крайней мере его часть) за
городом. Частичную синантропию легко спутать с временной, од-
лако следует учесть, что популяции частично синантропных видов
принадлежат к антропоценозу.
118
4. Синантропия и урбанизация
Ендрычковски (Jgdryczkowski, 1979) выделяет на примере
равноногих следующие понятия:
— синантропофилы — заносные виды, встречающиеся
преимущественно в зданиях и их ближайшем окружении;
— синантропобионты —заносные виды, встречающиеся
исключительно в теплицах и внутри зданий.
н
46
44
42
40
38
36
34
32
W
28
26
26
22
20
18
16
14
12
10
8
б
4
2
О
-
М.а.
-
LS. \
Се. \ \
\ \
-
Рис. 36. Эвсинантропные.
двукрылые (число особей на ловушку в-
день) по трансекту от центра
Варшавы (В) к ее окрестностям (О)
(по Gorska, 1979): N —число
особей; Се. — CaUiphora erythrocepha-
la; P.t. — Protophormia terraeno-
vae; L.s. — Lucilla sericata; M.a. —
Muscinai assimilis; F.c. — Fannia
canicularis"
Для характеристики синаятропии используют разнообразные
индексы. Индекс сннантропизацин (Ws) (J§dryczkowski, 1979)
означает долю синантропных видав в таксоценозе (табл. 38):
(I)
где Ls— число синантропных видов,
Lo— общее число видов.
Для численного выражения степени синантропии вида
используют (Nuorteva, 1963) индекс Si, позволяющий, в частности, точ-
4. Синантропия и урбанизация
Таблица 38. Индекс синантропи,зации (Ws) для таксоценозов мокриц
различных районов Варшавы (по Jgdryczkowski, 1980)
Меатошахождение
j Местонахождение
Урсынув
0,42
Университет
0,75
Аркадия
0,60
Цитадель
0,66
Улица Дольна
0,50
Улица Гданьска
0,66
Лазенки
0,75
Парк Каскада
0,60
Парк Культуры
0,60
Беляны
0,33
нее разделить облигатно и факультативно синантропные виды:
2а + b — 2с
(2)
где а — доля (%) особей видав урбанизированной области,
b — доля (%) особей того же вида в аграрной области,
с — доля (%) особей того же вида в биотопах, менее
затронутых антропогенным воздействием.
Этот индекс может иметь значения от -{-100 до —100, что
означает (Nuorteva, 1963):
; +100 — явное предпочтение плотно заселенных человеком
\/ мест;
/f75 —явное предпочтение заселенных человеком мест;
|/-|-50 — предпочтение заселенных человеком мест;
, О —независимость от поселений человека;
—25 — предпочтение незаселенных человеком областей;
—50 —избегание поселений человека.
Если понимать под'«независимостью от поселений человека»
равную долю рассм-атриваемого вида во всех трех
местообитаниях, этот индекс принимает значение + 16.7.
Удобство использования индекса синантропии для
характеристики поведения различных видов и его изменений в течение
года показано для птиц (табл. 39, рис. 37) (Nuorteva, 1971).
По мере приближения к границам ареала параллельно
увеличению суровости абиотических факторов часто наблюдается
возрастание синантропии, что характерно прежде всего для
видов южного происхождения. Из факультативной синантропия
становится о-блигатной, из временной — постоянной. Особенно
заметно ее увеличение к северу. Примером может служить муха
Lucilla sericata, у которой индекс синантропии достигает в
Венгрии 33, в Чехословакии 89, в Финляндии 98. Возрастание аинан-
тропии по направлению к северу обеспечивает существование
видов в тех областях, где иначе они вовсе не могли бы жить,
поэтому многие синантропные виды — космополиты. Решающим
I2i
4. Синантропия и урбанизация
Таблица. 39. Степень синантропии (Si) наиболее обычных видов птиц
в райвне Хельсинки (по Nuorteva, 1971)
Вид
Число
осов процентах
Индекс
сибей
нантропии
Сизый голубь
6758
35,5
+ 100
Домовый воробей
5171
27,2
+99
Черный стриж
343
1,8
+89
Белая трясогузка
269
1>4
+81
Обыкновенный скворец
399
2,1
+5в.
Большая синица
728
3,8
+53. .
Полевой жаворонок
177
0,9
+50'
Городская ласточка
91
0,5
+45.
Обыкновенная овсянка
386
2,0
+44
Луговой конек
97
0,5
+39
Деревенская ласточка
144
0,8
+29.
Дрозд-белобровик
148
0,8
+25..
Вяхиць
90
0,5
■_5.
Зяблик
1098
5,8
— 13
Сойка
99
0,5
—17
Чиж
230
1,2
—22
Снегирь
109
0,6
—28;
Певчий дрозд
341
1,8
—33,
Серая мухоловка
160
0,8
-34
Зарянка
199
1,0
—39
Лесной конек
396
2,1
—39
Черный дрозд
144
0,8
-44
Клесты
280
1,5
—44
Буроголовая гаичка
422
2,2
-56
Большой пестрый дятел
229
1,2
—58
Желтоголовый королек
389
2.0
—66
Рябчик
80
0,4
-81
Хохлатая синица
60
0,3
-95
фактором в этом случае считают более высокую температуру.
Показано также (особенно при изучении мусорных свалок), что
повышение питательности корма отчасти компенсирует
неблагоприятное воздействие других факторов.
Явление, принципиально сходное с возрастанием синантропии
к северу, наблюдается и в горах с увеличением высоты.
Известны также случаи возрастания синантропии как к северу, так и
к югу, например, для мух Lucitia caesar и Calllphora vicina
(Steinborn, 1981). Значения Si для последнего вида следующие:
север Финляндии — 95, Средняя Европа —43, Южная
Европа—87. ,
Некоторым паукам, распространенным на юге, свойственна
возрастание синантропии на северной границе их ареала.
Макроклимат уже не является лимитирующим фактором для видов,,
обитающих в (иди. на) ностройках. Характерными примерами в."
4. Синантропия и урбанизация
121
Средней Европе могут служить Scytodes thoracica, Steatoda tri-
langulosa, Pholcus phalangloides, Nigma walckenaeri и Tapkies-
ihis inermis. Другие виды обитают здесь преимуществеын© в
природе, однако севериее также становятся при известных
условиях явными синантропами, живущими в зданиях, в частности
Scotophaeus blackwalll в Великобритании, Di/safera erythrlna и
Дании и Neriene montana в Северной Европе (Valesova-Zdarko-
уа, 1966; Sacher, 1983).
100
Рис. 37. Изменения
степени синантропии у
различных видов птиц по
месяцам (по fJuorteva,
1971); П —время
прилета и отлета;
ДВ—домовый воробей; БС —
большая синица; С —
скворец; '3 — зяблик; ПВ —
полевой веробей
50
-50
-Дй
sc
с
\ \ 1 кг '
3
\ ■ / ■••
\
А \ /
/\ " ' Л
-
1
I I 1 1 1 1 1 1
1 1
j Ш Ш 7 Ш ШШЖ x хгж
Месяи,
Иногда синантропные виды животных, прежде всего
насекомых, подразделяют на синантропов жилых помещений и
синантропов населенных пунктов. Можно провести и более подробную
классификацию, отражающую 1структуру заселенного антропоце-
нозом пространства. Возрастающая связь с человеком выразится
такими терминами, как синантропы населенных пунктов->-синан-
тропы жилых помещений -^паразиты.
В литературе, прежде всего орнитологической, феномен
заселения городов определенными видами обозначают термином
урбанизации (Tenovuo, 1967; Pietzmeier, 1957; Luniak, Pawlo-#-
ski, 1964; Strawinski, 1966; Mauersberger, 1971; Krüger, 1979;
Oertel, 1980; Keve, 1983; Luniak, 1984). В йротивоположность
этому Н. А. Гладков (1960) и Б. Гитефан (Stephan, 1980) счита-
ютвселение видов в города составной частью урбанизации,
называя этим словом культурное изменение ландшафта в целэм.
Для рептилий (Garanin, 1982) и млекопитающих (Babinska-Wer-
ка, 1979, 1981; Pelikan et al., 1985) также использовалось
понятие урбанизации. Тенденция к ней, наблюдаемая у относительно
большого числа видов птиц, имеет региональные различия и, как
показывает пример сойки, может быть ограниченной во времени
/l22 4. Синантропия и урбанизация
Таблица 40. Распределение гнездящихся птиц Карл-Маркс-Штадта (ГДР)
по группам в зависимости от степени урбанизации (по Saemann, 1970),
1. Виды, освоившие город: гнездятся почти исключительно внутри поселений,
:! / жилых и промышленных комплексов. В других местообитаниях очень ред-
1,' ки или связаны с городскими биотопами
1.1. Гнездятся главным образом в центре города и в жилых кварталах:
галка, домовый воробей, сизый голубь, черный стриж, кольчатая
горлица, пустельга
1.2. Наибольшей плотности достигают в «зеленом городе», на окраинах и
Э пригородах: черный дрозд, полевой воробей, пеночка-пересмешка,
, канареечный вьюрок, серая мухоловка, зеленушка,
горихвостка-чернушка, деревенская ласточка
1.3. Гнездятся исключительно на городских пустырях: хохлатый
жаворонок
2. Виды, осваивающие город: гнездятся с большой плотностью и вне города.
Процесс урбанизации продвинулся в разной степени
2.1. Проникают до самых бедных растительных местообитаний:
обыкновенная лазоревка, большая синица, садовая горихвостка, вяхирь,
скворец, славка-завирушка
2.2. Встречаются в более озелененных местообитаниях: например, певчий
tyt^ дрозд, зяблик, сорока, поползень
' ' 2.3. Гнездятся в богатых растительностью городских местообитаниях: 20—
25 видов, например, большой пестрый дятел, зарянка, щегол
2.4. На городских пустырях и залежах: 8 видов, например, болотная
камышевка, серая славка, обыкновенная овсянка
2.5. Водоемы в черте города: кряква, лебедь-шипун, камышница, малый
зуек
3. Нейтральные виды: 27 видов, например, лесной конек, просянка
(табл. 40) (Heer, 1962; Küchler, 1951; Piechocki, 1956). По-влди-
мому, общепринятого определения урбанизации не существует»
однако могут быть выделены некоторые аспекты, достаточно
хорошо характеризующие .это явление. Решающим является
использование видом городской экологической ниши.
— Город служит (или становится) оптимальным биотопом
для данного вида, поэтому он встречается в нем либо
исключительно, либо с высоким обилием. Неоднородность городских
биотопов определяет основные зоны заселения. Некоторые типично
городские структурные элементы служат местами питания или
размножения. Доминируют закрытогнездящиеся виды (прежде
всего использующие пустоты построек) (75%), а кустарниковые
и древесные открытогнездящиеся виды составляют только 24%
(Saemann, 1968; Mulsow, 1971; Суг, Суг, 1979).
— Происходит расширение экологической амплитуды (Erz,
1964, 1966, 1967), например увеличение экологической
пластичности в новых субоптимальных биотопах (Oertel, 1980).
— Городские популяции относительно стабильны. Они
самовоспроизводятся и лишь в слабой степени обмениваются
особями с внегородской биотой (Schneider, 1982). Для существования
популяции регулярная иммиграция не обязательна.
4. Синантропия и урбанизация
123
— Развитие гонад и созревание гамет начинается раньше, за
счет чего удлиняется репродуктивный период (Seibert, Berthold,
1964 — данные по скворцу); продолжительность жизни
отдельных особей также увеличивается. С другой стороны, существуют
противоположные данные, т. е. наблюдались сокращение числа
яйцеклеток, повышенная гибель их самих и потомства,
нарушенное соотношенне полов (преобладание самцов).
— Происходит изменение поведения; примером может
служить зимнее пение черного дрозда, потенциально способного к
зимнему гнездованию (Oertel, 1979; Saernann, 1979; Grummt,
1962, 1979). У кольчатой горлицы наблюдалось до 2% зимних
гнездований (Saemann, 1975). Отмечались изменения в биологии
размножения закрытогнездящихся видов (Berresem et al., 1983,
1984).
Рис. 38. Местоположение гнезд жалящих перепончатокрылых в городских
домах и садах. По краям рисунка — расположение гнезд в (или на) структурных
частях первичных биотопов (по Haeseler, 1972): В3*-виды, гнездящиеся в
земле; НЗ — виды, гнездящиеся на земле; CT — гнезда в стенах домов; Д—'
гнезда на стенах домов; ДР — гнезда в древесине; Ч — гнезда на чердаках;
Ц — цементные постройки
— В пределах города происходит формирование новых
сообществ птиц (орнитоценозов)," не встречающихся вне города
(см; 7.9). g синантропии и. соответственно к урбанизации
(использованию городеких^эколотических ниш).„привели,..пять прин-
1(йпиально равнозначных комплексов причин (а—д, с. 124—126),
поэтому можно считать последнюю зарождающейся синантропи-
ей. За возникновением городских экологических ниш следует их
использование одновременно или попеременно с природными
(урбанизация), иногда ведущее к исключительно городскому
существованию (эвсинантропия). Важным моментом могло бы
стать отсутствующее сейчас ограничение понятия синантропии
городами. На примере некоторых видов птиц в настоящее время
прослеживаются процессы, длившиеся у большинства синантроп-
124
4. Синантропия и урбанизация
ных видав, вероятно, сотни и тысячи лет. Урбанизация началась
у разных видов (В различные эпохи, однако в последние
десятилетия, очевидно, ускорилась (Stephan, 1980).
а) Питание. В городе множество специфических кормовых
ниш (прежде всего зимой), например пищевые отбросы, гемеро-
хорные растения, бытовые отходы, ipasos, человек и его
домашние животные как хозяева паразитов, синантропные виды- как
основа пищевых цепей и-экологических коннексов, гнезда
животных.
б) Жизненное и гнездовое пространство. Имеется большой
выбор пригодных для этого мест, например, для
перепончатокрылых (Merisuo, 1933; Dorn, 1984) (рис. 38), рукокрылых и птиц.
Петерсен (Petersen, 1947) указывает, что наиболее быстро рас-
Рис. 39. Взаимосвязь
между гелиофилией (Г) и
степенью синантропии у па-
дальных мух в Финлянди»
(по Nuorteva, 1963):
точка — более 100 экземпляров;
X — от 50 до 99
экземпляров; квадрат — менее 50
экземпляров
прост.раняются в городе те виды птиц, чье «гнездовое места»
определяется лишь небольшим набором отличительных
признаков. Это же справедливо и для определенных перепончато^
крылых.
в) Климат. Повышение средней температуры в городе можно
считать главной причиной синантропии и урбанизации
(присутствие относительно высокой доли средиземноморских видов —
см. 3.3). Показана корреляция между гелиофилией и синантро-
пией у двукрылых (Nuorteva, 1963) (рис. 39). Среда города
соответствует тепловым требованиям прилетающих в них зимой
птиц.
г) Поведение. Хотя принципиально новых типов поведения
не наблюдается, оно более или менее сильно модифицируется
4. Синантропия и урбанизация
125
Таблица 41. Встречаемость синантропных видов мокриц в биотопах
с возрастающим антропогенным влиянием (Польша)
(по Jgdryczkowski, 1979, 1980, 1981)
Виды
о
Я
t-
(J
О
К
н
h
к
о
d
a
Z
t--
о
к
m
я
3
О
X CJ
all
S ffl о
S
<D
& ^
tu ja
V
Is
Ч s я
p.
oi
с S
Sie
Ligidium hypnorum (Cuv.)
+
+
+
Trichoniscus pusillus pusilus Bedt.
+
+
+
+
1
T. p. provisorius Rae.
+
+
4-
+
Hyloniscus riparius (C.L.K.)
+
Androniscus dentiger Verh.
+
+
+
Haplophthalmus danicus B.-L.
+
H. montivagus Verh.
+
+
Trichorkina tomentosa (B.-L.)
+
+
Platyarthrus hoffmannseggi Brdt.
+
+
Philoseia muscorum (Scop.)
+
+
+
+
Onisctts asellus L.
+
+
Cylisticus convexus (de Geer)
+
+
+
+
+
Protracheoniscus major (Dollf.)
+
+
+
+
+
Trachelipus rathkei ^rdt.)
+
+
T. waechtleri (Strouh.)
+
PorceUium conspersum (C.L.K.)
+
+
+
Metoponorthus pruinosus (Brdt.) -
+
Porcellio scaber Latr.
+
+
+
P. dilatatus Brdt.
+
+
+
P. laevis Latr.
+
P. spinicornis Say
+
i
+
+
+
Armadillidium nasatum B.-L.
+
f4. opacum (C.L.K.)
+
+
+
+
+
A. vulgare (Latr.)
+
. +
A. pulchellum (Zenck.)
+
Armadillio officinalis Dum.
Всего видов: 25
13
12
12
4
8
8
18
(cm. 8.3)^.-.например выбираются нетипичные места гнездования
(Gnielka et al., 1983, 1984; Grimm, 1953), научение ведет ,к
использованию новых кормовых ресурсов (открывание крышек
молочных бутылок, Thorpe, 1951) и определенных кормовых
пространств, привыканию к человеку и его технике (Stephan, 1980),
к уменьшению дистанции избегания. Непрерывный свет неоновых
ламп влияет на биоритмы, в частности, у ласточек (Semadam,
1966, 1967). Гримм (Grimm, 1953) указывает на повышение
порога оовещенности, необходимой для йгробуждения.
По-видимому, становятся возможными явления, подобные акселерации
{включая сходные с импринтингом), что объясняют лучшим пи-
126
4. Синантропия и урбанизация
Таблица 42. Эвсинантропные пауки (по Sacher, 1983)
Вид
Примечания
Nigma walckenaeri (Roewer)
■Dictyna civica (H. Lucas)
Cyphepeira ixobola (Thorell)
Zygiella x-notata (Clerck)
Amaurobius ferox (Walckenaer)
Oonops domesticus Dalmas
■Scytodes thoracica (Latreille)
Pholcus phalangioides (Fuesslin)
Physocyclus simoni Berland
Tegenaria domestica (Clerck)
Achaearanea tepidariorum (C. L. Koch)
Steatoda castanea (Clerck)
■S. grossa (C. L. Koch)
? S. triangulosa (Walckenaer)
■Sosticus loricatus (L. Koch)
Jieteropoda venatoria L.
Наружные части построек, в
зарослях растений; центр расселения —
Южная Европа.
Стены построек и каменные ограды
(пользуется накопленным ими
теплом); южноевропейский вид (Bil-
laudelle, 1957; Braun, 1952;
Hertel, 1969; Mader, 1952)
Ha постройках
Ha постройках
Подвалы
Между старыми бумагами и в
коллекциях
Квартиры (Gettmann, 1980)
Подвалы, квартиры
Подвалы
Постройки (все ярусы)
Оранжереи (заносный вид)
Аквариумные помещения
ские сады)
(зоологиче-
танием, удлинением действия света, изменением климата и
нарушением биоритмо^^ Изменения в поведении отмечаются не
только у птиц, но тий5<1чны и для перепончатокрылых
(наблюдались у 11 видов, Haeseler, ,1972), где они, возможно, приводят
к отбору (например, у Cro':ssQcerus elongatulus, Leclerq, 1964;
Haeseler, 1972). Жук-точильщй'к-,. ЛдоЬшт thomsoni и златка
Buprestis haemorrhoidalis изменяютспособ яйцекладки (в лесу —
на коре, в помещениях — на голой древесине).
д) Из(менения исходных биотопов. В оптимальных
загородных местообитаниях вследствие усиленного размножения может
возникнуть популяционный пресс, ведущий к заселению
субоптимальных городских биотопов С^дешапп, 1979). Причиной
последнего бывает также пространственное сокращение природных
местообитаний (Haeseler,-1-97-2).
Кроме того, в городе меньше влияние хищников и
относительно слабая конкуренция (Tomialojcr 1982). Некоторые виды
пользуются дополнительной защитой человека.
Преадаптированы к урбанизации прежде всего эвриойкные
виды с высокой плодовитостью (г-стратеги), хорошими
способностями к расселению и известной этологической пластичностью.
Синантропные виды известны почти во всех группах живот-
Таблица 43. Синантропные виды жуков-притворяшек (Ptinidae)
Вид
Биотоп (субстрат), происхождение
Gibblum psylloides (Czempinski)
Mezium affine Boieldieu
М. sulcatum F.
Trigonogenius globulus Solier
Sphaericus gibbioides Boieldieu
Niptus hololeucus Faldermann
Tipnus unlcolor (Pill.)
Paranlptus globulus (Illiger)
Ptinus raptor Sturm
P. teCtus Boieldieu
Ptinus pusillus Sturm
P. villiger Reitter
P. fur L.
P. clavipes Panzer
P. latro F.
Квартиры, склады (зерновые),
библиотеки, музеи, перекрытия старых
домов (Weidner, 1979)
Запасы (завезен из
Средиземноморья)
Запасы (завезен из
Средиземноморья)
Зерно, москательные тов'ары
(завезен из Южной Америки)
(Weidner, 1968, 1979)
Москательные товары (завезен из
Южной Европы) (Weidner, 1979;
Elbert, 1978)
Квартиры, склады (зерно, мука, рис,
овсяные хлопья, чай), перекрытия
старых домов
Сараи, помещения для домашних
животных (сено, солома), вне
построек (космополит, с 1861 г. в
Германии, по происхождению пон-
тийско-средиземноморский вид,
склонный к массовым
размножениям) (Weidner, 1975, 1979; Cymo-
гек, Koch, 1969; Döhring, 1970)
(Происходит из Северной Африки)
Дома (москательные товары, мука),
сараи, помещения для животных^
на каменных оградах
Дома, музеи (также в птичьих
гнездах), склады (зерно, мука,
отруби, рис, какао) (адвентивный вид
из Австралии, Тасмании и Новой
Зеландии, с 1916 г. в Германии,,
очевидно, имеет «склонность к
путешествиям») (Weidner, 1979)
Курятники, голубятни, сараи,
гаражи, склады (зерно)
Дома, склады, помещения для
животных
Квартиры с сырыми стенами, склады
(зерно), курятники, голубятни,
гаражи, птичьи гнезда (Weidner,
1979)
Дома, курятники, голубятни,
гаражи, птичьи гнезда, склады (зерно)
Дома, мельницы, пекарни, склады
(зерно)
127
128
4. Синантропия и урбанизация
Таблица 44. Различная степень синантропии подвидов домовой мыши
{ш Serafinski, 19S5)
Подвид
Область распространения
Степень синанжропин
Mtts musculus bactri-
Малая Азия
Вне домов
anus
Mus т. hortulanus
от Болгарии до
КазахСтепная зона, вне
достана
мов, сооружает
холмики для зимних запасов
Mus т. spicilegus
Юго-восточная Европа
В мягкие зимы вне
домов, в более суровые
Mus. т. musculus
холода в постройках
Север и восток
ЦентЗимой в постройках
Mus m. domesticus
ральной Европы
Эвсинантроп, если вне
домов, то толькв
вблизи них
ных (табл. 41—44), однако различные группы исследо^ваны в
этом отношении весьма неодинаково. Вместе с тем следует
отметить, что для ряда таксонов синантропия особенно характерна
(Klausnitzer, 1987).
К синантропам зданий относятся представители 15 (45,5%)
из 33 семейств пауков ГДР (Amaurobiidae, Dictynidae, Sicariidae,
Oonopidae, Pholcidae, Dysderidae, Agelenidae, Araneidae, Liny-
phiidae, Theridiidae, Nesticidae, Gnaphosidae, Liocranidae, Hete-
ropodidae, Salticidae). Среди примерно 800 видов пауков ГДР
44 являются эв- и гемисинантропами (5,5%). В отличие от
других групп животных причины поселения их в зданиях вряд ли
связаны с пищевыми потребностями. Синантропами становятся
прежде всего южные виды, обитатели пещер, трещин скал,
поверхности скал и древесных стволов (виды-охотники). У всех о^би-
тателей зданий растянутый период половой активности, причем
у некоторых оба пола способны размножаться в течение' всего
года (полностью ли исчезает периодичность размножения,
неясно; возможно, остается предпочтительный период копуляции).
К последним относятся, например, Nesticus cellulanus,
Achaearanea tepidariorum, Lepthyphantes leprosus, Meta menardi. Для
Meta merianae и Steatoda bipunctaia показаны различия в
фенологии размножения между городской и природной
популяциями (Sacher, 1983).
Из 25 синантропных видов равноногих Польши 13 —
космополиты, 6 — европейские, 3 — среднеевропейские, 1—палеаркти-
ческий, 1 — атлантический и 1 — неотролический (заносный вид
Trichorhina tomentosa). Удивительно много среди них обитателей
теплиц (18 видов) (Jgdryczkowski, 1979) (табл.41).
5. ПОСЛЕДСТВИЯ ИЗОЛЯЦИИ ГОРОДСКИХ
ОСТРОВНЫХ МЕСТООБИТАНИЙ
, Городские островные местообитания возникают в связи с
изолирующим эффектом улиц и сооружений, которые определяют
границы, размеры и форму ареалов. Примерами могут служить
изолированные парки, кладбища, небольшие сады, пустыри и
водоемы. Степень изоляции определяется прежде всего
удаленностью от сходного биотопа (Крупной площади и соотношением
площадей «острова» и «материка», с которого происходит
заселение. По Мак-Артуру и Уилсону (Мае Arthur, Wilson, 1967),
ni = c-wi2.e-4i/^, (3)
где ni—число иммигрантов на острове;
с —■ константа географических условий;
di— удаленность острова от источника биоты; '
wi—размер острова;
А — средний радиус расселения изучаемой группы животных.
Мадер (Mader, 1979) модифицировал это уравнение для
островных биотопов, ограниченных транспортными магистралями:
e-i/a = e-&"H, (4)
где V — компонента движения транспорта;
D — ширина разделяющей биотопы улицы;
К — специфическое для данной группы животных давление
конкуренции.
Многие авторы считают, что экосистемы городских островных
местообитаний в принципе сравнимы с экосистемами настоящих
островов (Мае Arthur, Wilson, 1967; Mader, 1979а, 1979b, 1980,
1983, 1984; Janzen, 1983 и др.). От последних они отличаются
значительно меньшей степенью изоляции, менее выраженными
границами и в большинстве случаев меньшим размером. Как
будет показано ниже, в обоих случаях наблюдаются,
по-видимому, сходные, иногда еще недостаточно изученные
закономерности. ^
5.1. Динамическое равновесие видов
Приток иммигрантов, т. е. направленное переселение и
случайный занос («принцип выстрела дробью»), компенсируется на
острове гибелью (эмиграцией) видов. Обе эти переменные зави-
Э—21
130
5. Последствия изоляции городских местообитаний
сят от количества видов на острове, т. е. числа занятых
экологических ниш (рис. 40). Для биотопов большого объема
характерен скорее устойчивый видовой состав. Чем меньше остров, тем
меньше число видов, так как интенсивность иммиграции
уменьшается из-за меньшей, вероятности попадания на него пассивно
распространяющихся видов. Одновременно в связи с
сокращением размера популяций и ограничением ресурсов возрастает
смертность.
1
I
\Вс
Вы
1 ^—
1 -~
Рис. 40. Зависимость между
уменьшением площади островных
местообитаний (направление
стрелки) и числом видов (S). Вверху —
крупный остров, внизу — мелкий
остров (по Mader, 1980, с
изменениями): Р — видовое равновесие,
'Вс — скорость вселения, Вы —
скорость вымирания
5.2. Зависимость видового разнообразия
от площади
Ожидаемое число видов в островном биотопе зависит прежде
всего от наличия потенциальных микробиотопов и может быть
оценено по формуле
S = c-A% (5)
где S — число видов;
к — площадь острова;
5. Последствия изоляции городских местообитаний
131
с—специфическая для данной труппы животных константа,
учитывающая биогеографические условия;
z — показатель степени, принимающий значения 0,20—0,35
для океанических островов и 0,Г2—0,17 для островных
местообитаний на материке; возрастает с увеличением степени изоляции
(Мае Arthur, Wilson, 1967). Однако в городских парках
отмечены более высоки^ значения для двукрылых (0,235) и жуков
(0,222) (Faeth, Капе, 1978, рис. 41).
Константа z может быть получена непосредственно из
уравнения
log S = logc+z log А.
Lb связи с такой зависимостью фаунистического богатства от
площади удвоение числа видов может потребовать, например,
Рис. 41. Зависимость между числом
видов (S) и исследованной площадью у
двукрылых (Д) и жуков (Ж) (по Eaeth,
Капе, 1978)
0,4 0,8 1,2 1,6 2Л 2,4 2,8
lg А (га)
удесятерения последней (рис. 42). Однако подобное отношение
не обнаружено для пауков и жужелиц (Schaefer, 1979) (ср.
рис. 43у\
При уменьшении площади количество ресурсов (корма, мест
гнездования и убежищ) сокращается линейно, но их спектр
сначала сохраняется, поэтому в мелких островных местообитаниях
встречается относительно много видов. Качественное
разнообразие ресурсов снижается экспоненциально (рис. 44). Стабильность
зооценозов малых островных местообитаний невелика, так как
большинатво видов представлено лишь мелкими популяциями и
лишь немногие доминируют, причем дело доходит до
сверхдоминирования (более 50% фауны) одного вида. Наряду с
элементами исходных зооценозов появляются пионерные формы и «пере-
беж1чики» из соседних местообитаний. В конкурентной борьбе с
иммигрантами исходная биота постепенно уступает. В первую
очередь исчезают виды с большими территориальными потреб-
ноотями (табл. 45, рис. 47, 48).
132
5. Последствия изоляции городских местообитаний
100 [■
0,1 1 10
Lg А (га)
Рис. 42. Схематичная
кривая площадь — число
видов (константа 2=0,2)
(по Mader, 1980)
Рис. 43. Зависимость
между площадью и числом
видов у жуков-стафилинид
(CT) и жужелиц (Ж)
островных местообитаний в
Лейпциге. АБ — парк Артур-
Бретщнайдер; ВС —
Ботанический сад; ФП — Фри-
денспарк; ПГ — Пальмен-
гартен; ШТ —парк
Шванентайх; PK — парк Роберт-
Кох; ШП — Шиллерпарк
Рис. 44. Схематические графики
зависимости количества ( ) и
качества (2) ресурсов (Р) от
площади островного биотопа
(А) (по Mader, 1983): К —
островные местообитания с
краевой зоной, составляющей
более 20% общей площади; К+
+Ц — островные
местообитания с выраженной центральной
5. Последствия изоляции городских местообитаний
133
Отношение (W) числа видов (Ns) к числу особей (Ni)
уменьшается с возрастанием площади:
Ni
(6)
Это показано Мадером (Mader, 1984) для пауков и жужелиц,
(рис. 45) и Клауснитцером (Klausnitzer, 1983b) для жужелиц;
(рис. 46). Однако у птиц столь четкой корреляции не
обнаружено (Saemann, 1970, табл. 47) (ср. Gavareski, 1976).
Рис. 45. Зависюи^сть
индекса W от площади (А)
у пауков (П) и
жужелиц (Ж) (по Mader,
1984)
о,г\-
Рис. 46. Зависимости
индекса W от площади парка
у жужелиц в Лейпциге.
Сокращения см. рис. 43
\ шт
х •
• \
БС \
\. г =0,7601
. \ ФП
АБ \
1 III
III 1
5 , Ю
10^ 2
2 5
lg Л
Уменьшение среднего числа видов может свидетельство,вать
об увеличении степени изоляции городских парков в районах
сплошной застройки, однако различия здесь могут объясняться
и условиями местообитаний (табл. 46). У растений озеленение и
антропогенное распространение диаспор намного сильнее
сглаживают возможный островной эффект, чем у различных групп
животных; скорее этот эффект можно ожидать на пустырях.
134
5. Последствия изоляции городских местообитаний
г Ж
Ш Ш Ж х.
Месяи,
Рис. 47. Площадь участка
обитания европейского ежа
в разные месяцы (по Esser,
1984). Светлые колонки —
самцы, темные колонки —
самки
Рис. 48. Положение участков обитания
меченых европейских ежей в 1980 г.
(сплошная линия) и в 1981 г. (пунктир)
в Ботаническом саду Мюнхена (по Esser,
1984)
Для птиц построены кривые типа «число видов — ареал»
(Reichholf, 1980; Ellenberg, 1982). При исследовании
орнитофауны различных внутригородских участков в Карл-Маркс-Штадте
установлена зависимость между площадью, числом видов и чис-,
лом гнездящихся пар (Saemarin, 1970, табл. 47). Сравнение этих
данных с результатами исследований в «зеленом городе» (см.
2.2.2.6) показывает отчетливые различия, связанные с местооби-
5. Последствия изоляции городских местообитаний
135
Таблица 45. Минимальные гнездовые территории некоторых видов птиц
в городе (по данным Saemann, 1970)
Минимальная -территория,
Виды
га
20
Зеленый дятел, пеночка-трещотка
5
Большой пестрый дятел
2
Зеленая пересмешка, славка-черноголовка,
бо1
лотная камышевка, европейский поползень
Зарянка, зяблик
0,5
Славка-завирушка, серая славка
Таблица 46. Зависимость между числом дикорастущих видов и размером
площади городских зеленых насаждений (по Sukopp, 1983b)
Местообитание
Площадь, га
Среднее
число видов
Среднее
число видов
на 1 га
Крупные парки
100
335
3,4
Городские парки в слабо
застроенных районах
15
155
10,3
Городские парки в районах
сплошной застройки
15
132
8,8
Мелкие зеленые насаждения в
слабо застроенных районах
1
100
100,0
Мелкие зеленые насаждения в
районах сплошной застройки
1
53
53,0
i
Таблица 47. Количество птиц различных пробных площадей
в Карл-Маркс-Штадте (по Saemann, 1970)
M.iCTHOCTb
Площадь,
Число
Число
виЧисло
W
ГП/га
га
видов
дов на 1 га
гнездящихся пар
(ГП)
■Центр города
0,9
5
5,6
14
0,18
15,6
Центр города
2,5
7
2,8
26
0,13
10,4
Центр города
4,0
11
2,8
32
0,17
8,0
«Зеленый город»
2,8
17
6,1
57
0,15
20,4
танием (меньшая изоляция, разница в ресурсах). Относительно
большее число видав и гнездящихся пар.отмечено на участках
наименьшей площади.
Для листовых минеров и галлообразователей найдена
зависимость между встречаемостью и величиной биотопа, а также
между числом видов и структурным разнообразием
растительности островного местообитания (Segebade, Schaefer, 1979).
136
5. Последствия изоляции городских местообитаний
5.3. Формирование краевых зон
с сильным антропогенным воздействием
Более крупные островные местообитания состоят из краевой
и центральной зон (рис. 49). Такое подразделение зависит от
размеров, формы и структуры острова. Чем он меньше, тем
больше относительная площадь краевой зоны. Когда ее воздействие
распространяется на весь остров и центральная зона исчезает,
зонирование теряет смысл. Лесное островное местообитание с
диаметром менее 80 м уже не может считаться особым зоотопом
для лесных животных (Mader, Mühlenberg, 1980). Краевая зона
испытывает особенно сильное антропогенное воздействие (авто-
Естествениые
[ракторы
Влияние краевой зоны
Излучение
АнтропогеннЫЕ
/ракторы
Механические
нарушения
Загрязнения
воздуха
О
Отражение
Ядовитые
вещества
Удоаршя
Рис. 49. Подразделение крупного островного местообитания на зоны (по
Mader, 1980)
мобильные выхлопные газы, хлориды, пыль, пестициды, механи-'
ческие нарушения, микроклиматические параметры). Глубина
проникновения внутрь «острова» этих агентов в большинстве слут
чаев довольно велика (в лесу—30—40 м. Mader, 1979).
5.4. Смещения в видовом спектре
краевой зоны
Изменение микроклимата и быстрое чередование
антропогенных нарушений ведут к эмиграции стенойкных видов.
Освобождающиеся экологические ниши занимаются эвриойкными
видами, часто достигающими высокой численности, гемерофилами
5. Последствия изоляции городских местообитаний
137
ИЛИ остаются свободными. Склонные к миграциям виды, спосо-б-
ные преодолевать крупные расстояния, имеют больше шансов на
поселение здесь, чем менее мобильные формы.
5.5. Смещения в видовом спектре
центральной зоны
Специализированные и особенно часто более крупные виды
вы.мнрают первыми. Освобо,дйвшиеся экологические ниши
занимают либо оппортунистические виды исходной биоты, либо им-
(%)\ (%)\ \Hs (%П
50 Y
12 3 4
501
? 2 J 4
0 ^
Гр ШП ШТ
(%)\ Ws \Hs W
5(7 h
12 3 4
50 h
50 h
12 3 4
П
ГД
Рис. 50. Связь распределения численности четырех наиболее часто
встречающихся видов жу_желиц и доли особей самого частого из них вида в общей
численности с показателем видового разнообразия (Hs). Гр — плоская гравий-
но-битумная кровля в центре Лейпцига; п, ГД, Л —поля, группы деревьев
и луга на окраине Лейпцига; остальцые сокращения см. рис. 43
138
5. Последствия изоляции городских местообитаний
(%)
50
Hs (%)
-
50
2
Л
50
-
\
f
0
1 ' 1
0
f г J 4
52
пг
Рис. 51. Графики, аналогичные приведенным на рис. 50, для жуков-стафили-
нид в Лейпциге. Условные обозначения см. рис. 43 и 50. Диаграммы Л и 5
составлены по исследованиям Мадера (Mader, 1980): Л — островное
местообитание; Б — исследованный район большой площади
мигранты. С сокращением центральной зоны ..всесильнее изменя- ■
ется плотность видов и структура доминирования. У пауков
островных местообитаний обнаружена конкуренция (Duffey, 1975).
5.6. Нарушение структуры доминирования
в островных местообитаниях
У^меньшение общего видового богатства и увеличение доли
часто встречающихся видов обычно ведет к снижению индекса
разнообразия. Вместе с низким индексом равномерности распре-
5. Последствия изоляции городских местообитаний
139
деления это указывает на нестабильность системы (рис. 50, 51).
Такие системы легче нарушаются внешними воздействиями и
испытывают наибольшую опасность исчезновения. Повысить
стабильность может увеличение способности организмов к
рассредоточению по вертикали и горизонтали (Schaefer, 1973), как
показано для ряда биохоров (морские выбросы, падаль,
экскременты и т. д.), фауна которых отличается именно такой
особенностью.
5.7. Дисгармония фауны
Полностью отсутствуют некоторые виды и более крупные
таксономические группы, встречающиеся в исходных
местообитаниях. Причина заключается в значительном удалении островов от
этих местообитаний, в величине угла вторжения (рис. 52),
различной способности животных к расселению и недостатке
определенных ресурсов (табл. 48).
Рис. 52. Угол
вторжения (а) и вероятность
заселения островного
местообитания (по Evers,
1977)
Наиболее приспособлены к заселению островных
местообитаний виды с большим количеством потомства, обладающего
широкими возможностями расселения (г-стратеги), поскольку
потери, связанные с вероятностью попадания на остров, очень
велики. Островная фауна обычно состоит, с одной стороны, из
крупных, активно внедряющихся видов, а с другой — из мелких,
пассивно заносимых.
Таблица/48. Группы животных, способные быстро заселять городские
островковые местообитания (А) и практически лишенные возможности
переселяться на такие «острова» (Б)
А
Б
,Пауки, тли,
птицы
• жуки-стафилиниды.
Брюхоногие моллюски, мокрицы,
многоножки, (сенокосцы), длинноусые
прямокрылые, амфибии, рептилии
140
5. Последствия изоляции городских местообитаний
Небольшие подвижность и радиус активности некоторых групп
животных препятствуют достижению ими новых островных
биотопов за счет собственной локомоции; особи погибают на пути к
ним (дождевые черви, брюхоногие моллюски, равноногие на
асфальте).
С увеличением возраста и вследствие этого дифференциации
островных местообитаний повышается богатство их фауны. На
начальной стадии здесь преобладают случайно занесенные виды,
и видовой состав сильно колеблется.
5.8. Различие фаун
Из-за различного запаса ресурсов, а также, возможно, и
вследствие случайного характера заселения между фаунами
островных местообитаний наблюдаются более или менее значитель-
Рис. 53. Эвдоминантные пауки
(названия видов см. табл. 21)
различных городских парков
Лейпцига (по данным
Koslowski et al., 1980). КЦ — парк
Клара-Цеткин; др. сокращения
см. рис. 43
ные различия (рис. 53, табл. 49). Мадер (Mader, 1980) считает'
маловероятными прогнозы об ожидаемом доминировании тех
или иных видов, придавая случайной компоненте большее
значение, чем принципу реализации экологической ниши.
Островная изоляция городских местообитаний может
существенно влиять на образование новых комбинаций видов, или
ассоциаций. Примеры здесь известны прежде всего из
фитоценологии, но сходные эффекты должны иметь место и у животных, на
что указывают исследования, проведенные на жужелицах, ста-
филинидах, пауках, сенокосцах, многоножках и равноногих в
Лейпциге (Klausnitzer, рукопись).
5. Последствия изоляции городских местообитаний
441
Таблица 49. Различия в распределении эвдоминантных (Э) и доминантных (Д)
видов жужелиц по внутригородским островным местообитаниям Лейпцига
(по Klausnitzer, 1983b)
Вид
Парк
ШваШиллер-
БотаничесФриденс-
нентайх
парк
кий сад
парк
Atnara aenea ^
Э
Д
Calathus fuscipes
Э
э
э
Platynus obscurus
Э!
Harpalus aeneus
Д
Д
д
Asaphidion flavipes
Д
д
Platynus dorsalis
Д!
Nebrla brevicollis
э
Д
э
Bembidion properans
Д!
Э!
B. lampros
Harpalus rufipes
Э!
Calathus melanocephalus
Э!
Notiophilus biguttatus
Э!
Anisodactylus binotatus
Э!
Amara familiar is
Э1
5.9. Генетическая дифференциация
Принцип основателя приводит к^повышенной частоте
определенных шмбинаций аллелей в изолированной популяции.
Вследствие этого проявляются определенные признаки и может
начаться процесс образования рас. Дифференциация касается
прежде iBcero видов, менее склонных к миграциям, для которых
граница местообитания служит почти непреодолимым препятствием.
В связи с действием принципа основателя и непрерывным
дрейфом генов естественные генетические процессы маскируются и
становятся малоэффективными. Мадер (Mader, 1980)
принимает за норму закрепления или потери отдельного признака (т. е.
исчезновения гетерозиготности по нему) 1/4 популяции за
поколение. Процессы видообразования должны происходить на
островах быстрее, так как прекращение межпопуляционного обмена
генами может ускорять генетическую дифференциацию. Благода ■
ря эффекту «разбавления» генофонда здесь при известных
обстоятельствах будут иметь успех аллели, которые не дали бы
преимуществ при полной панмиксии (подходят для городских
условий?).
Генетическая дифференциация в городе воз.можна у моллюска
Сераеа (различия в распределении типов полосатости, Müller,
1980), серой крысы (резистентность к кумарину. Bishop, Hartley,
1976), жужелиц (аллельные типы. Steiniger, 1978, рис. 54),
пяденицы Biston betularius (меланизм, Kettlewell, 1961, 1973), дро-
142
5. Последствия изоляции городских местообитаний
зофилы (расы, Franke, Ehler, 1978), полев-ой мыши (пропорции
черепа й частоты его неметрических варнантов, Sikorski, 1982а, Ь;
Klenke, 1986) (рис. 55).
5.10. Пути заселения островных
местообитаний
Воздушные течения переносят не только крылатых, но и
бескрылых насекомых -и других членистоногих. Способность к
пассивном^^, перемещению .по воздуху обеспечивается малым .весом,
опушением, шипами и собственными .крыльями.
Рис. 54. Частоты ал.челей в локусе Ahp-C пяти популяций Carabus neinoralis
в Саарбрюккене (по Müller, 1980): l~Ahp-0, 2 — Ähp-C^ Z — Ahp-C^, 4 —
Ahp-C\5 — Ahp-C^
оСЙ ofßK -fBC
54
56
58
ВО
62
Рис. 55. Результаты неэ.пементарного дискриминантного анализа для
возрастных классов V, и VI полевой мыши по промерам черепа (по Klenke, 1986):
СИ — кладбище Старый Йоханнис; ЮК — южное кладбище; БС —
ботанический сад. Круги отражают принадлежность к отдельным выборкам (F-кванти-
ли). Wi, Ws — векторы дискриминантного анализа
Миогие пауки достигают островных местообитаний как
«аэронавты» (по крайней мере на неполовозрелой стадии) (Duffey,
1956; Freemann, 1946; Knülle, 1954; Richter, 1970; Visse, van
Winger den, 1982), что объясняет быстрое заселение ими вновь воз-
5. Последствия изоляции городских местообитаний
143
никших местообитаний и высокое видовое разнообразие этой
группы в городах. Так, иа плоской кровле одного из домов в
Лейпциге (рис. 6) собрано 24 вида пауков, среди которых
эвдоминантами были аэронавты Erigone dentipalpis (40,7%), Е. atra
(23,8%) и Meioneta rurestris (22,6%) (Klausnitzer et al., 1980).
Виссеи ван Вингерден (Visse, van Wingerden, 1982) 29.08.1979—
29.11.1979 собрали на одной из крыш с помощью специального
приспособления 314 дрейфующих по воздуху пауков, из которых
83,9% относились к семейству Linyphiidae (67,1% —Erigone sp.,
прежде всего Е. atra и Е. dentipalpis, принем удивительно много
половозрелых особей). Над морскими островами существует
термически обусловленный восходящий ток воздуха, который может
"вести к «присасыванию» воздушного планктона (эффект ловчей
воронки) (Evers, 1977) (рис. 56). Аналогичное явление не иск-
Рис. 56. Воздушные потоки (тер-
мики) вокруг нагретой солнцем
постройки, которая в результате
может действовать как
воронкообразная ловушка (по Evers, 1977,
с изменениями)
лючено в случае более пли менее .изолированно расположенных
высоких зданий и способно влиять на заселение крыш. В этой
связи нелишне напомнить, что роза ветров внутри города сильно
зависит от застройки, и здесь могут возникать локальные ветры
(Sukopp et al., 1980).
В городские островные местообитания происходит активный
и пассивный занос животных людьми из самых разнообразных
климатических зон (см. 3.2). Преимущество его в том, что особи
непосредственно^попадают в подходящие для них местообитания
(складьС теплицы, вдания, (культуры кормовых растений), часто
находя в новых областях особенно благоприятные условия для
размножения (Weidner, 1970). Новые виды постоянно попадают
в городские островные местообитания также благодаря
озеленению и мероприятиям по благоустройству ландшафта.
6. ПИЩЕВЫЕ ЦЕПИ И СЕТИ
Естественные и городские экосистемы наряду с глубокими
экологическими различиями имеют и некоторые общие черты,
например наличие границ, самоподдержание за счет обмена
веществ и притока энергии, стабилизация за счет круговорота
энергии и тенденция перехода от экспансии к интенсивному росту
(табл. 50) (Kroske, 1981).
О городских пищевых цепях и сетях известно пока немного,
однако ясно, что паразиты и паразитоиды (возможно, в
противоположность хищникам) зависят в городе от присутствия своих
хозяев и иногда находятся с ними в таких же количественных
отношениях, что и в естественных биотопах. Однако доля
некоторых паразитоидов в городских местообитаниях, по-видимому,
ниже, чем в природе из-за большей их изоляции и меньшей
вероятности встречи с хозяином (хотя сравнительные данные для
такого заключения отсутствуют).
При изучении первичных и вторичных паразитоидов тлей в
Лейпциге из 10 000 зараженных особей 31 вида получено 4427 пе-,
репончатокрылых, из которых большинство — вторичные
паразитоиды (Klausnitzer, 1980b). Всего обнаружено И видов
вторичных и 31 вид первичных паразитоидов (рис. 57). Центром
одного из паразитоценотических коннексов является, например,
тля Microlophium evansi, живущая на крапиве двудомной
(рис. 58).
В большом количестве представлены во внутригородских
районах также некоторые хищники, прежде всего божьи коровки и
мухи-журчалки. В качестве примера паразитирования на
хищниках следует упомянуть двукрылое Phalacrotophora beroUnen-
sis. Этот вид паразитирует на куколках божьих коровок и
особенно часто — на Adalia bipunctata. Из более чем 1000 собранных
в Дрездене куколок А. bipunctata было поражено 1-3,4%: в 4,1%
отмечен вид Ph. berolinensis, в 9,3% — Tetrastichus neglectus
(перепончатокрылое) (Klausnitzer, 1969). Другие исследования
в центре города показали возможности развития там
паразитоидов яиц из рода Trichogramma (перепончатокрылые)
(Klausnitzer, 1980а).
Крылатые тли {Periphyllus и Drepanosiphum), обитающие
главным образом на клене, служат важными кормовыми
объектами некоторых типичных городских птиц, в частности черного
стрижа, деревенской и городской ласточек (Gnielka, 1981).
5
to
58
о
В
л
о
а
S
о
т
X
S
о
§
о
я
g
о,
I
et
О
§
I-,'
в
ш
о
о я к О)
« о га Ч
ш м о 5*
Ч о-
W ш 1=
г я ш
" Ё§
cd
g
Ё
О)
к
о
с
S
§
я
\D
О
О
«
о
а
ч
о
CD
о CTi
S
S си
о
са 5
о
I о ä
и
1^
... к о
S ш
н S к
га >. та
С1ч о СП
Si
о 3
t< сс
О
f- Ч
S „
cd л
t=( et
3 со к
d Ч ш
й о О)
g
™ 2 S
о « CU
4 s я
о,
о н
сь
ёЯ
я о
3 а,
о
|§
,й X
VO
3 i- 3
g S 8
о
f-
я
а.
с
о
я
в-
о
cd
ь
CJ
о
я
^1
Е- 3
Cd Ь
И cd
о ю
Q.. cd
я а.
Н CU
Cl, о.
О <D
Я Я
cd
ш
I
О)
я
g >,cd
' CU
-г си
о я g
S S 3
в я я
S « о
(Я о
я
я
cd cd
я§
я и
я Cd
Cd S
о §
я о s
" я ö
SSO
Чин
ra ,v
я
о
в
S
о
OJ
cd о
О. си
>> 2
CD Я
CU
Iii
ч
Я
S
cd
а.
я
с
>.
я
я
яг
я
я
Оч
с
н
S
о
о
и
S
>1
о
я
о
о
Он
га
ю
i|
II
S CD
О
S CU
s »*
5 я
s «
CQ
я
CU
s
>^
о
я
о
«
о
«
я
S
I
^o^S ч:
^ м о <u
1 cd
CU со
я я -я со
я CU й я
m - "
^ о
о-ю Cd m
о S о
Ml
S S о
s
05
I
Я
\о
о
_ё
S
со
Я cd
ES
я
я
я <а ^ ^
я н Б а.
со
се „
а. а-
га е(
5 о
Я я
1 §g
ш S я ч
Cd R 3 о
с-, я 03 я
Sou
= 4g
Й я Ш-&
со =я CU f;:
К fU i—I
к ^
О ^
ra
я
^ Ч „
g s §
о E OJ
я S и
Sis
g S §
a. 5 &
ra Ь l>i о
5 я Ш
1 II
Sil«
S Ч н л
к с
§
в-
2 Я
Йю
(.4 о
а.
1
я sS
S 5
S >>
:S3
1§
к
о
о- д
S 3 g
2 S g
Ül
о я о
н " «
га
я
Ö
о
с га
CU
о я
а. CJ
о
ю ч
Я О
Я
а"
VO
о
§
я
4 (U
5 Э
Is
t4 <n
ш
s
m
о
га
а.
к
CU
о
>^
а
о
о
UJ
О
я
CO
SI
со
га
к
s
я
я
Ч
О
О
И
о
CK
га
s
s
о
S
о
о
Я
о
<D
m
са
Я"
та
Он
о
я
О
о
о
К|
ч
QJ
га
CU
н
та
я
ч
S
я
о
я
га
о.
и
sE-
о О)
CU О-
m о
я S
га
и
CU
Я" «
« о
Е Н
>. о
о
ш
о
с
>1
о.
о
в
и
я
ч
о
о,
я
л
ч
я
\о
га
6
в
ся
о
в
о
CJ
о
я
и
03
=1
о
Я
я
'Я
в
о
в
я
10—21
146
6. Пищевые цепи и сети
В городах обитают также паразитоиды сетчатокрылых
насекомых; это показано для Conwetitzia psociformis [Aphanogonus
steinitzi) (Weidner, 1939, 1941) н Wesmaelius subnebulosus (Lip-
kow, 1969). Иногда они способны погасить локальные вспышки
массового размножения сетчатокрылых (Gepp, 1975).
Рис. 57. Результаты выведения паразитов тлей в центре Лейпцига (по
подсчетам Klausnitzer, 1980): P.c. — Ргаоп. volucre; A.hi. — Aphidius hieraciorum;
A.ho.—Aphidius hortensis; L.fa. — Lysiphlebus fabarum; L.fr. — Lysiphlebus
fritzmuelleri; T.a. — Trioxys acalephae, Oc—остальные виды; A.v. — Asaphes
vulgaris; P.m. — P achy neuron minutissimum; C.c. — Coruna clavata; A.a.—
Aphldencyrtus aphidivorus; D.c. — Dendrocerus carpenteri; С — Cynipidae
Вторичные
паразиты
Первичные
паразиты
Coruna
Asaphes_
Cynipidae
Dendrocerus
Pachyneuron
Тля
Microlophium evansi
Кормовое растение Urtica dioica
Рис. 58. Паразитоценотический коннекс тли Microlophium evansi в центре
Лейпцига (по Klausnitzer, 1983)
Около 70% зоофагов среди роющих ос охотятся на Hemiptera
и двукрылых; хищники прямокрылых из этой группы в черте
города отсутствуют. Добычей являются пауки (3 вида), сеноеды
(2), трипсы (3), настоящие клопы (1), цикадовые (6), тли (9),
жуки (2) и двукрылые (12) (табл. 51) (Haeseler, 1972). Рою-
6. Пищевые цепи и сети
147
Таблица 51. Кормовой рацион роющих ос в Ольденбурге (по Haeseler, 1982)
Вид
Семейство двукрылых
СумS
cf
и
&•
о
а
X
л
ч
5!
"й:
о
о:
о
к
tJ"
к
?^
S
3
2
=§
Щ
u
<D
о _^
6
К S
>^
=r
к
■ K
т
И
К
О
К
о
ма
н
t-
&
О) 3
а'
о ^
D. <!>
О
и
ä
С S
о
о
" н
а.
(u d
и к
гз
л
Crabro peltariuS'
6
2
6
1
33
10
2
3
2
1
66
Oxybelus blpun-
101
ctatus
18
—
—
—
—
—■
77
—
—
5
1
—
0. uniglumis
5
—
7
—•
1
1
15
1
—
—
1
—
31
Сумма
В процентах
29
14,5
2
1,0
13
6,6
1
0,5
1
0,5
1
0,5
125
63,1
11
5,6
2
1,0
8
4,0
4
2,0
1
0,5
198
щая оса Ectemnius sexcinctus находит в городе особенно
благоприятные условия в связи с питанием мухами-журчалками
(Peters, 1971).
Осы-блестянки (Chrysididae) представлены в Варшаве 10
видами (Skibinska, 1982а), из которых 7 обитают в парках и по 4
(прежде всего из рода Omalus) в центре города и в небольших
садах. Распространение этого семейства определяется
присутствием |ВИДов-хозяев. Обитающие в городских биотопах блестян-
ки паразитируют прежде всего на роющих осах.
В Варшаве обнаружен 381 вид наездников-ихневмонид из
перепончатокрылых (Ichneumonidae) (Sawoniewicz, 1982).
Доминируют паразитоиды листовых минеров и галлообразователей
(33%), затем следуют паразитоиды насекомых-экзофитофагов
(27%) и афидофагов (11%), прежде всего афидофагов-хищни-
ков (подсемейства Diplazontinae мух-журчалок, представленных
в городе тем же числом видов, что и вне города).
Йроктотрупоидные наездники (Proctotrupoidea,
перепончатокрылые) представлены в Варшаве 197 видами (Garbarczyk,
1982). В городских зеленых насаждениях отхмечены семейства Не-
loridae (3), Proctotrupidae (14), Diapriidae (39), Scelionidae (32)
и Platygastridae (19 видов). Доля паразитоидов жужелиц и
стафилинид довольно высока (18,7%), как и доля паразитоидов
минирующих насекомых и орехотворок (23,4%); в то же время доля
паразитоидов синантропных двукрылых низка.
Из паразитических падальных мух в Варшаве обитает 18
видов (Draber-Monko, 1982). Большая часть их (в среднем 76,5%;
в центре города даже 85,7%)—паразитоиды дождевых червей.
Другие виды поражают моллюсков и птиц.
Показано, что высокоспециализированные ежемухи (Tachini-
dae), например Medina separata, могут паразитировать на ши-
10*
148
6. Пищевые цепи и сети
Таблица 52. Паразитоиды (Hymenoptera) вредителей запасов
и синантропных мух
Хозяин
Паразитоид
Семейство
надсемейство
Ephestia kuehniella
Sitophilus, Stegobium
Lasioderma, Rhlzoderma,
Carpophilus
Oryzaephilus, Tribolium,
Sitophilus
Cryptolestes, Ahasverus
Tribolium confusum
Musca domestica
Sarcophaga
Bracon hebetor Say,
Plastonoxus westwoodi (Kiefer),
P. munroi Richards,
Holepyris hawaiiensis Ashmead,
Idechthis canescens
(Gravenhorst)
Lasiophagus distinguendus
(Förster)
Anisopteromalus calandrae
(Howard)
Cephalonomia tarsalis
(Ashmead)
Cephalonomia waterstoni Gahan
Rhabdepyris zeae Turner et Wa-
terston
Figiies striolatus Hartig,
Stenomalina muscarum (L.),
Figites scutellaris (Rossi),
F. consobrinus Giraud,
Brachymeria minuta (L.)
Braconidae
Bethylidae
Ichneumonidae
Pteromalidae
Pteromalidae
Bethylidae
Bethylidae
Bethylidae
Cynipoidea
Pteromalidae
Cynipoidea
Chalcididae
poKo распространенных в городе божьих коровках {Adalia
bipunctata, Oenopia conglobata) (Klausnitzer, 1969). Хотя
городские условия не особенно благоприятны для чешуекрылых, они
составляют 57,9% хозяев ежемух в Варшаве (Draber-Monko,
1981), причем можно проследить слабую тенденцию к
увеличению их доли по направлению к центру города. На втором месте
находятся паразитоиды жуков (в среднем 15,0%). Доля видов-
полифагов составляет в среднем 12,2%.
Основная часть серых мясных мух (Sarcophagidae) Варшавы
паразитирует на перепончатокрылых (30,2%), равноногих
(11,6%) и дождевых червях (11,6%) (Draber-Monko, 1982b).
Доля этих трех групп среди жертв возрастает по направлению к
центру города. Паразитоиды прямокрылых в городе
отсутствуют, а паразитоиды моллюсков встречаются лишь в
незначительном количестве.
Хозяевами перепончатокрылых паразитоидов являются
различные виды насекомых, постоянно обитающие в домах (прежде
всего вредители запасов и материалов). Их поражают,
например, представители Chalcidoidea, Cynipoidea, Evaniidae,
Ichneumonidae, Braconidae и Bethylidae (табл. 52). В оотеках
тараканов (фото 12) обнаружены Evania appendigaster (Cameron,
1957; Edmunds, 1953, 1954), Tetrastichus hagenowii (Cameron,
6. Пищевые цепи и сети
149
1955; Edmunds, 1955; Fleet, Frankie, 1975; Roth, Willis, 1954;
Vargas, Falles, 1974) и Comperia merceti (Gordh, 1973; Lawson, 1954).
Жук-кожеед Thytodrias contractus (Dermestidae) известен как
паразитоид черного таракана. Жук Rhipidius pectinicornis (Rhi-
piphoridae), паразитоид прусака, распространен, как и этот та-
Рис. 59. Рацион ящерицы Lacerta muralis по
данным содержимого желудков (по Sclimidt,
1981); П —пауки; Ж —жуки; Д —двукрылые;
О — олигохеты; М — муравьи; Пер — прочие
перепончатокрылые; Н — ногохвостки; Р —
равноногие; Мн — многоножки; С — сенокосцы
Рис. 60. 'Вторичый цикл развития (изменение хозяев в городских условиях)
бычьего цепня (по Engelbrecht, 1984): 1 —мясо, зараженное финками; 2 —
выход яиц и проглоттид; 3, 4, 5 — города; 6 — водоочистное устройство; 7 —
слив очищенных сточных вод; 8 — поедание скотом травы на полях орощения;
9 — загрязнение пастбищ при подтоплении; 10 — прямое заражение крупного
рогатого скота
ракан, космополитно, но очень редок. Личинки насекомых,
развивающиеся в древесине, могут быть поражены
перепончатокрылым Scleroderma domesticum (Bethylidae) (Kühne, Becker, 1974).
Виды подсемейства Diapriinae (Proctotrupoidea) паразитируют
на пупариях синантропных двукрылых (Teodorecsu, Ursu, 1979).
150 6. Пищевые цепи и сети
Существенные компоненты пищевых цепей и сетей, даже
внутри зданий,—пауки. Виды Zygiella x-notata, Cyphepelra
sericata и С. ixobola предпочитают изобилующие добычей участки
построек (иапример, пространства вокруг неоновых ламп,
привлекающие ночных бабочек). Паук Dysdera питается
обитающими в подвалах мокрицами, Pholcus phalangloides — тоже (до
25% пищи!) (Nyffeler, Benz, 1981). При обилии добычи (Tribo-
lulm destructor) наблюдались массовые скопления Pholcus
(Sacher, 1983). Паук Oonops domesticus питается сеноедами,
особенно LiposceUs dlvinatorius. Показано (Pötzsch, 1966), что
основной пищей паука Meta menardi служат мокрицы (27%),
жуки (29%), пауки (10%) и многоножки (11%).
Серьезные враги вяхиря— сорока, горностай и каменная
куница (Gnielka, 1978). Одичавшие городские кошки среди
прочего корма поедают прытких ящериц, которые, как и другие виды
Lacerta, сами находятся на вершине пищевой пирамиды (рис. 59).
Показана возможность использования пищевых цепей для
биоиндикации (например, для анализа на тяжелые металлы)
(Höffei, Müller, 1983; Müller, 1984).
Все сказанное выше о пищевых цепях и сетях относится
также к паразитирующим на человеке, домашних и диких
животных плоским червям (рис. 60), круглым червям, пиявкам, вшам
(Baum, 1968; Cerny, 1970; Mann, Ribbeck, 1973; Zlotorzycka,
1977a, b), клопам, мухам и блохам (Vater, 1984, 1985, 1986).
i
7. УРБАНИСТИЧЕСКИЕ ГРАДИЕНТЫ
Главная причина существования урбанистических (городских)
градиентов — зональность, определяемая городской застройкой
и характером использования пространства. Обычно выделяют
следующие зоны (от окрестностей с сельско- и лесохозяйствен-
ным землепользованием к центру города):
— окраина (свалки, зона отдыха с парками, садовыми
участками, спортивными площадками);
— свободная застройка:
• одно-двухсемейные коттеджи с небольшими садами;
• сплошная застройка кварталов с парками, скверами,
аллеями между ними и садами во дворах;
• сплошная застройка кварталов с озелененными
внутренними дворами;
— сплошная застройка (кварталы с мощеными или
бетонированными дворами, максимум—с отдельными деревьями и ру-
деральными участками).
Используют также несколько иное подразделение на зоны
(Sukopp, 1980):
а) преобладание открытых зеленых площадей (газоны и
пустыри, небольшие рощи и группы деревьев);
б) преобладание облесенных зеленых площадей (плотные —
разреженные заросли деревьев и кустарников);
в) асфальтированные, бетонированные, мощеные
пространства среди зеленых площадей (широкие улицы);
г) свободная застройка с высокой долей зеленых площадей
(равные площади зелени и застройки, трех-четырехэтажные
дома);
д) преобладание площадей с искусственным покрытием и
уплотненным грунтом и отдельных построек (товарные станции,
территории портов, заводские и фабричные районы);
е) плотная внутригородская застройка с ограниченным
озеленением (площадь 1—5 га с отдельными группами деревьев и
кустарников);
ж) плотная внутригородская застройка (четырех-шестиэтаж-
ные кварталы середины XIX в.).
Господствующие в пригородах поля по направлению к центру
города постепенно сменяются небольшими садовыми участками
и палисадниками (вплоть до ящиков и горшков с цветами) и
152
7. Урбанистические градиенты
клум[ба!ми разных типов (вплоть до растительности в бетонных
вазонах). Пропсходит омена полеводства садоводством. С
известными оговорками можно говорить о R-Н-градиенте (от
латинских слов rusticus — деревенский и hortus—^сад). Все
относящиеся' сюда местообитания имеют различные, но экологически
в обще4 одноплавовые характеристики. К ним относятся,
например, регулярная обработка и перемещение почвы, полная,
обычно ежегодная ротация, использование посадочного материала,
часто — закладка монокультур, довольно обильное удобрение,
использование пестицидов и полива.
Рис. 61. Снижение уровня загрязнения, создаваемого автомобилями в
зависимости от расстояния до края дороги (ио Sukopp et al., 1980)
По-видимому, существует также и R-М-градиент (от
латинских слов rupes — скала и murus—^стена), отражающий
увеличение «каменистости» ландщафта и реакцию на это фауны. Для
зонирования могут быть использованы сообщества птиц
(Saemann, 1970). На постройках небольшой высоты гнездятся белая
трясогузка, горихвостка-чернушка, деревенская ласточка.
Постройки средней высоты (трех-четырехэтажные жилые дома)
заселяют преимущественно черные стрил^и, а самые высокие
здания (башни, церкви, промышленные предприятия) — галки и
пустельги (см. 7.9). Возможно, существует также и С-С-градиент
(от caverna — пещера, и cella — камера), описывающий переход
от жизни в пещерах к заселению подвалов.
А-Е-градиент (arbor — дерево, eremus — пустыня)
характеризует переход от леса через многолетние луговые сообщества к
7. Урбанистические градиенты 153
V
Затем вычисляем значение градиента для отдельных
площадей:
Gae, = Ia,-Iep (9)
Кроме того, не лишена смысла оценка однородности
отдельных площадей во времени (рис. 62), т. е. относительного
постоянства ^азноо1бразших_биоть1 ib течение годового цикла. В слу-
каменистому ландшафту (Klausnitzer, 1982, 1983а).
Соответственно происходит увеличение жонтинентальности окружающей
среды.иЗ городах, находящихся в лесистых районах, этот
градиент существует почти всегда. Города пустынной зоны часто
имеют в центре наиболее крупные древесные насаждения,
поэтому в них прослеживается обратный Е-А-градиешс1
Важен также комплексный ноксовый градиент, который, хотя
и состоит из большого числа отдельных градиентов,
соответствующих каждой ноксе, должен, вероятно, рассматриваться как
единое целое (аддитивное, синергическое и антагонистическое
взаимодействия). В противоположность прочим он лишь в
редких случаях отражает непрерывную смену условий от периферии
к центру города (разрывы в зависимости от размещения
источников нокс) (рис. 61).
Наиболее детально исследован А-Е-градиент (Klausnitzer,
1983; Klausnitzer, Richter, 1983, 1984, 1986).
По уравнению
можно рассчитать «тождественность лесным видам» для всех
городских площадей; при этом Ру^выражает долю вида j на
данной площади, а Paj:—его долю на лесной площади (в принципе
аналогичные рассуждения применимы и для других градиентов).
Следует, однако, указать, что приведенное уравнение не
правомерно для слишком крутых градиентов.
С другой стороны, исходя из площади с наиболее низким
показателем «тождественности лесным видам», можно рассчитать
долю «пустынных» видов на всех площадях, причем, естественно,
для сравнения берется не вся городская «пустыня», а лишь
«наиболее пустынная» из исследованных площадей.
Аналогично формуле (7) получаем
154
7. Урбанистические градиенты
чае ненарушенных (близких к естественным) участков с
относительно устойчивыми климатическими параметрами (лес) следует
ожидать высокого видового разнообразия, слабо меняющегося в
течение всего года (или доступного наблюдению периода
активности). В нарушенной системе с более сильными
климатическими колеба^гиями (например, центр города) должна наблюдаться
более выраженная изменчивость состава таксоценозов.
Q=0,855
В =0,572
l^diB = 0,417
В = 0,319
Рис. 62. График, поясняющий понятие однородности. Во всех трех случаях-
(А, Б, В) у четырех гипотетических видов численность в течение года
одинакова и видовое разнообразие равно 1,306. Усиление колебаний численности
от Л к ß сопровождается заметным снижением величин Нл и В (по
Klausnitzer, Richter, 1983)
Меру однородности рассчитывают следующим образом:
Hdt
В =
Н,
2 Hi
Hdt =
(10)
(И)
где Hs—общее видовое разнообразие (по Брийуэну);
Hdt— среднее видовое разнообразие отдельных учетов,
проведенных в разное время.
Показана линейная зависимость между величинами
градиента Gaej (9) и одноро,дности В (10) для жужелиц на восьми проб-
7. Урбанистические градиент
155
ных площадках в Лейпциге, идущих от окраины к центру города
^(рис. 63, 64, табл. 53) (Klausnitzer, Richter, 1983).
При расчетах на уровне таксонов более высокого ранга
полезно вычислять, помимо однородности (В), временную группо-
_ _ 1 \ "' _'. о / /^г-\
Рис. 63. Зависимость
однородности (В) от А-Е-градиента
(Qae) для жужелиц различных
пробных площадок в Лейпциге
(по Klausnitzer, Richter, 1983)
(ср. табл. 53)
Рис. 64. Корреляция Gae с
однородностью (В) для
семейств жуков (по
Klausnitzer, Richter, 1984). Ц,, Us —
местообитания в центре
Лейпцига; К —
местообитание на окраине Лейпцига,
П —• местообитание вне
города (пойменный лес)
Рис. 65. Корреляция между
Gae и С|] для всего материала
(М) и для жуков (Ж) (по
Klausnitzer, Richter, 1984).
Сокращения см. рис. 64
I
•
F
оа
0,5
1 1
1 1 1
0 -1
й
-0,5
!
-0,7
г
-0,5 0 +0,5
1
JA
м 7
-0,8
1
-Ofy
156
7. Урбанистические градиенты
Таблица 53. Параметры А-Е-градиента на примере ассоциаций жужелиц
8 пробных площадок в Лейпциге с возрастающей урбанизацией
(I —VIII) (по Klausnitzer, Richter, 1983)
Площадка
'e
"dt
в
I
1,000
0,026
0,974
1,874
1,394
0,744
II
1,090
0,023
1,067
1,799
1,174
0,653
III
0,694
0,105
0,589
1,774
1,111
0,626
IV
0,243
0,260
-0,017
1,893
1,014
0,536
V
0,291
0,404
—0,113
2,038
1,430
0,702
VI
0,276
0,454
-0,178
1,098
0,493
0,449
VII
0,045
0,311
—0,266
2,423
1,376
0,566
VIII
0,014
1,000
—0,986
1,264
0,438
0,347
по формуле
Gdt =
к ■ к к
п 2 (Pkti — Pkti+i) + S (Pkti+i — Pkti+2) + 2 (Pktn+i -Pktn)
(12)
7.1. Число видов
Для многих групп животных показано уменьшение числа
видов от окрестностей к центру города (табл. 55, 56, 57) (Kühnelt,
1977; Klausnitzer, 1983а, Ь). Причинами можно считать прежде
всего сужение спектра кормовых растений, снижение
разнообразия местообитаний, а у некоторых групп — повышенную
смертность под воздействием антропогенных и структурных факторов.
Отмечается, напримор, обеднение фауны от окраин через
зеленые массивы до зоны сплошной застройки в Вене (рис. 66)
(Schweiger, 1962). Об уменьшении числа видов к центру города
свидетельствуют уловы летающих насекомых в палаточные
ловушки в предместьях, на окраинах и в центре города (Gepp,
1975, рис. 67), а также данные по минерам и галлообразователям
(Segebade, Schaefer, 1979).
Показано (Owen, 1978; Haeseler, 1972; Owen, Owen, 1975;
Klausnitzer, 1983 и др.), что число видов многих групп
членистоногих по окраинам города значительно выше, чем в его
окрестностях. Возможно, этот краевой эффект позволяет говорить об
экологической границе города. Аналогичное явление
наблюдается в случае цветковых растений и птиц (табл. 54, 55).
В Лейпциге обнаружено 175 видов жужелиц, из них 94 —
в зоне сплошной застройки и 168 — на окраине (Klausnitzer,
1983b, рис. 68).
7. Урбанистические градиенты
15Г
В целом по мере уменьшения доли естественных структур
сокращается и число сопровождающих их видов птиц и
соответственно общее число видов птиц в городских сообществах. Это
связано с потребностью 'птиц в индивидуальном участке
определенного размера, недостатком мест гнездования и нехваткой
пищи, а также с возрастанием фактора беспокойства,
играющего особенно отрицательную роль в период насиживания кладок.
Рис. 66. Изменение
видового богатства
различных таксонов насекомых
при движении от центра
к периферии (краевой
зоне) города в Вене (по
данным Schweiger, 1962)
20
/ тПегтвзШае
1 Pterostichus
yf / f Tettignnidae
1/ / » Carabus
/ ^ л Bambus •
// yfmiucannus
"pi/Trechus
Городская
застройка
Сады
60 b
Рис. 67. Число видов (S)
сетчатокрылых в Граце (по Gepp, 1975): 3 —
плотно" застроенная часть города;
П —парки в центральных районах;
С — слабо застроенные периферийные
районы с лугами, садами, полями;
О — окрестности города (Ш-километ-
ровая зона)
со
40
20
Краевая
зона
Условия для гнездования более благоприятны на богатых
растительностью окраинах города, чем в его центре, где некоторые-
виды существовать вообще не могут.
Выделяют следующие типы городских местообитаний
(Saemann, 1973):
а) ншвостройки — наименьшие общая численность и число
видов (мало растительности);
б) небольшие парки — максимальное обилие особей (боль1-
шой выбор пищи);
i58
7. Урбанистические градиенты
Таблица 54. Число видов растений в различных зонах города
(ио Sukopp et al., 1980)
Зона
Число видов
Зона
Число видов
Сплошная застройка в
Окраины города
440
центре
400
Окрестности города
250
Разреженная застройка
430
Таблица 55. Число гнездящихся видов птиц в различных городских зонах
Саарбрюккена (по Ellenberg, 1982)
Городская зона
На 25 га
На 1 кш'
На 80 га
Центр города
7,8+1,0
18,5+2,3
46
Внутренняя часть окраинной зоны
21,3±1,3
37,0+2,6
68
Наружная часть окраинной зоны
26,1±1,4
49,1±1,7
81
Таблица 56. Встречаемость жужелиц в различных местообитаниях Лейпцига
от окраин к центру города (по Klausnitzer, 1983b)
. Число
ЧисленБиообъем,
РазнообраРавномер-
Райою
видов
ность
смЗ
ем/числензие
яость
расность
пределения
Байперт (1980)
27
788
405,6
0,51
1,874
0,564
Нонне, пойменный
лес (1980)
28
359
150,9
0,42
1,799
0,564
Нонне, парк
(1980)
22
275
61,4
0,22
1,774
0,576
Парк Иоханна
(1980)
19
235
15,8
0,07
1,991
0,669
Шиллерпарк (1981)
8
137
9,2
0,07
0,569
0,289
Парк Шванентайх
(1981)
6
16
0,4
0,03
1,002
0,832
в) местообитания с хорошо структурированной
растительностью (зеленый город, крупные парки, предместья, кладбища) —
наибольшее число видов.
Земан (Saemann, 1970) указывает ряд закономерностей,
касающихся видового богатства сообществ птиц:
— возрастание плотности заселения и числа видов с
увеличением возраста застройки;
— чем больше возраст растительности и чем сильнее
выражена ее ярусность в слабо застроенных зонах, тем выше
плотность заселения и число видов;
— увеличение числа видов при возрастании площади
«однородного» биотопа или доли площади под растительностью.
Таблица 57. Число видов различных групп животных вдоль городского
g
a
о
к
к
а
о.
а
1^ я 2
3 к п 5
S и « о
4 Ä oj р
£
ig
Автор
с
С
Enchytraeidae
6
19
12
10
Kasprzak (1981)
Lumbricidae
10
8
6
8
14
Pilipiuk (1981)
Oribatida
69
16
11
Niedbala et al. (1982)
1 XkJ %.* \fL
Gamasida
38
22
18
39
Niedbala et al. (1982)
Tarsonemida
48
23
17
25
Niedbala et al. (1982)
Actinedida
45
13
8
23
Niedbala et al. (1982)
Acaridida
3
4
3
7
Niedbala et al. (1982)
Opiliones
Araneae
14
12
3
—
Czechowski et al. (1981)
218
125
34
43
Krzyzanowska et al.
(1981)
Isopoda
10
9
1
3
Jedryczkowski (1981)
Diplopoda
Collernbola
18
14
7
4
Jedryczkowski (1982)
12
33
20
28
Sterzynska (1982)
Cicadina
125
97
60
43
Chudzicka (1982)
Neuroptera
40
35
8
14
Czechowska (1982)
Carabidae
258
96
39
44
Czechowski (1981)
Elateridae
49
16
8
9
Burakowski, Nowakowski
(1981)
Coccinellidae
51
28
12
14
Czechowska, Bielawski
(1981)
Scarabaeidae
68
16
5
6
Kubicka (1981)
Cerambycidae
80
22
3
4
Burakowski, Nowakowski
(1981)
Chrysomelidae
43
23
18
13
Wasowska (1981)
Curculionidae
292
193
48
33
Cholewicka (1981)
Formicidae
35
19
5
10
Pisarski (1982)
Vespidae
26
9
3
8
Skibinska. (1982b)
Ichneumonidae
554
344
106
109
Sawoniewicz (1982)
Proctotrupoidea
166
88
49
41
Garbarczyk (1982)
Noctuidae
не указано
215
49
22
Winiarska (1982)
iVlecoptera
5
1
1
—
Czechowska (1982)
Culicidae
20
13
7
7
Wegner (1982)
Sarcophagidae
41
35
14
15
Draber-Monko (1982b)
Rhinophoridae
5
4
1
3
Draber-iVlonko (1982b)
Phoridae
не указано
28
27
19
Durska (1982)
Stratiomyidae
13
6
4
4
Trojan (1982)
Therevidae
5
7
2
1
Trojan (1982)
Asilidae
13
9
3
1
Trojan (1982)
Асинантропные Taba-
25
Trojan (1982)
nomorpha (сумма)
35
9
8
Dolichopodidae
25
33
16
20
Bankowska (1982a)
■Syrphidae
64
66
47
37
Bankowska (1982b)
Tachinidae
123
90
30
47
Draber-Monko (1982c)
Anthomyidae
29
11
3
3
Gorska (1982)
Muscidae
122
107
12
26
Gorska (1982)
S60
7. Урбанистические градиенты
7.2. Численность
Для многих видов птиц довольно точно известно
распределение плотности популяции вдоль А-Е-градиента. Так, в Лейпциге
во внутреннем кольце зеленых насаждений, граничащих с парком
Йоханна, гнездилось по данным 1979 г. 85 пар 15 видов птиц с
.50
40
J
30
20 V
10 ■
1
ШТ ШП лд ИЙК ВС йп
АБ ИПК НОЛ Б
Площадка
Н /200
п Ч ШО
А 800
М Р П Д БР л
600
400
200
S
рис. 68. Средние числа видов (белые колонки) и особей (черные колонки)
жужелиц на отдельных пробных площадках в Лейпциге слева направо — от
центра к окраинам города) по нескольким годам наблюдении (по Klausnitzer,
1983b): ШТ —парк Шванентайх; ШП — Шиллерпарк; ХД —Хаусдах; НИК —
«бывшее кладбище Новый Йоханнис; БС — Ботанический сад; ИП — парк
Иоханна; АБ — парк Артур-Бретшнайдер; НПК—Нонне, парковая часть;
НПЛ —Нонне, пойменный лес; Б —Байперт; М-Мюльберг, Мёккерн; Р —
^рудеральные местообитания у Видерицша; П — край поля у Видерицша; Д —
агруппа деревьев (аграрный остров) у Видерицша; БР — береговой склон
ручья у Видерицша; Л — луг у Видерицша
7. Урбанистические градиенты
161
ПЛОТНОСТЬЮ 87,8 naip на 10 га (Ladusch et al., 1980). Трудно
получить столь же точные сведения об обитающих в городе
беспозвоночных. При сравнении в Лейпциге четырех пробных
площадок вдоль трансекта наблюдали тенденцию к снижению
численности жужелиц к центру города (табл. 56). Аналогичные
результаты получены при анализе почвенных ловушек, расположенных
по «диаметру» Саарбрю!кквна (рис. 69, Müller, 1980) и при
изучении плотности жуков-щелкунов в Варшаве (Nowakowski, 1982,
табл. 58, ср. также рис. 70). Значительно меньшие различия меж-
1 2 3 4 5 В 7, 8 9 ЮН 12 1314 15 IS 17 18 19 2021 2223 24 25
Рис. 69. Численность равноногих (черные кружки) и жуков (квадраты) и
видовое разнообразие (белые кружки) жуков в мае 1974 г. по сборам ловушек
Барбера, установленных на долговременных пробных площадках по трансекту,
проходящему через центр Саарбрюккена (по Müller, 1980)
ду городскими зонами выявлены при изучении летающих насе-
КО.МЫХ (Gepp, 1977): на разных пробных площадках отмечено
приблизительно равное число особей.
7.3. Общее доминирование
и индексы доминирования
У жужелиц обнаружены различия в структуре
доминирования на окраинах и в центре города, причем бросается в глаза
значительно меньшая сумма общего доминирования (все виды
с численностью более 4%) на окраинах (табл. 59). Согласно
второму правилу Тинемана, этого и следовало ожидать, так как в
экстремальных усдовиях (здесь —в центре) в общем должно
встречаться меньше видов при большем числе особей, т. е.
индекс доминирования выше.
U-21
162
7. Урбанистические градиенты
Таблица 58. Число видов и плотность популяции (число особей на 1 м^)
жуков-щелкунов в различных местообитаниях Варшавы
(по Nowakowski, 1982)
Крупные
очраинные
параш
Районы с
Центр города
крупными
озелененными
(Площадями
Старые парки
Мелкие уча-
атки земли
Участки
земли вдоль
улиц
Число
видов
Плотность
популяции
15
30—70
8
90—120
4
Не указано
7
8-10
3
8-80
Таблица 59. Отношения доминирования у жужелиц в Лейпциге
(по Klausnitzer, 1983b): р — рецедент, ср — субрецедент
Ö
i
га
6
ё
Вид
s Я 3
too
ET '^'^
«SS
|s
R, й
ЕС
К
1
о
ц
R и
го
Ii"
Iii
Nebria brevicollis
362
16,7
307
5,5
669
8,7
Harpalus aeneus
283
13,1
220
4,0
503
6,5
Harpalus rufipes
243
11,2
244
2,6
487
5,0
Platynus assimilis
135
6,2
152
2,7
287
3,7
Asaphidion flavipes
129
6,0
24
ср
153
2,0
Bembidion quadrimaculatum
109
5,0
5
ср
114
p
Amara aenea
101
4,7
49
ср
150
p
Calathus fuscipes
100
4,6
135
2,4
235
3,0
Calathus melanocephalus
89
4,1
132
2,4
221
2,9
Bembidion lampros
79
3,7
455
8,2
534
6,9
Notiophilus biguttatus
72
3,3
64
р
136
p
Carabus nemoralis
63
2,9
609
11,0
672
8,7
Platynus dorsalis
50
2,3
92
р
142
p
Bembidion properans
45
2,1
271
4,9
316
4,1
Pterostichus melanarius
28
р
387
7,0
415
5,4
Harpalus rufibarbis
1
ср
180
3,2
181
2,3
Abax parallelepipedus
—
241
4,3
241
3,1
Pterostichus strenuus
251
4,5
251
3,3
Terchus secalis ;
255
4,6
255
3,3
Harpalus melanchollcus
1
ср
120
2,2
121
P
Сумма особей доминирующих
видов (более 2%)
1860
3859
5320
Общее доминирование (более 4%)
(сумма)
71,6
54,0
45,3
7. Урбанистические градиенты
163
Индекс доминирования Бергера—Паркера рассчитывается по
формуле
d = ^. (13)
где Nraax—числениость наиболее часто встречающегося .вида.
Этот индекс принимает тем меньшее численное значение, чем
более выровнена структура домпнирования. Это же относится и
к индексу доминирования Симпсона
с - У ^
где Hi— численность каждого вида.
(14)
Рис. 70. Число видов (S)
и плотность особей (N)
бабочек в кронах лип
(по Chudzicka et al.,
1979): Е — естественные
местообитания; П —
парк; ДД — деревья
вблизи домов; ДЗ —
деревья в районах
сомкнутой застройки; ДУ
—деревья вдоль улиц; ОД —
отдельно стоящие
деревья
Исследования жужелиц в Лейпциге показали, что эти
индексы, как и общее доминирование, принимают относительно
высокое значение в центре города и низкое на его периферии
(Klausnitzer, 1983b).
7.4. Биомасса
Биоо1бъем жужелиц, измеренный вдоль трансекта в Лейпциге,
сильно убывает к центру города (рис. 71, 72, табл. 60). Во внут-
ригородсних районах явно доминируют «мелкие» виды, которые,
вероятно, лучше выдерживают хозяйственную деятельность и
«уход» за зелеными насаждениями, а также воздействие
избыточного (количества птиц. Исследования других групп жуков,
пауков, мокриц и мпогоножек дали сходные результаты, что
говорит о повышенной способности мелких видов к переселению
(Schaefer, 1973; Topp, 1972; Tischler, 1980).
И»
464
7. Урбанистические градиенты
7.5. Экологические группы
и географическое распространение
Параллельно изменению абиотических и биотических
факторов окружающей среды от окраин к центру города изменяются
и соотношения отдельных экологических групп у многих
таксонов животных. Например, среди доминирующих жужелиц в
600
5 -т
{ 200
1
400
s
300 5
Си
^■5
А т
ИП НПК НПЛ
Рис. 71. Численность и
биообъем жужелиц вдоль
трансекта от центра Лейпцига к
пойменному лесу (ио
Klausnitzer, 1983b): ИП —парк
Иоханна; НПК —Нонне,
парковая часть; НПЛ —
Нонне, пойменный лес; Б —
Байперт
Рис. 72. Изменение доли
особей различного размера
в сообществе жужелиц при
увеличении антропогенной
нагрузки (Ур) (по
Czechowski, 1980а): X —Хамерня;
М — Млохув; Р — Раджейо-
8ице; В —Варшава. I
—доля крупноразмерных особей,
И—доля среднеразмерных
особей, П1—доля
мелкоразмерных особей
Лейпциге наблюдается явное увеличение доли лесных видов по
направланию к окраинам, прежде всего за счет уменьшения доли
мезофильных видав, которых в центре больше, чем на
периферии города (рис. 73, 74) (Klausnitzer, 1983b).
У долгоносиков в Варшаве доля видов, обитающих
преимущественно на лугах, к ценпру города увеличивается (пригород —
7. Урбанистические градиенты
165
Таблица 60. Биообъем жужелиц в Лейпциге (по Klausnitzer, 1983b)
Внутригородские районы
Окраина
Объем см'
Средний объ-
эм особи, мм'
Объем см®
Средний об-ь-
2м особи, мм'
Эвдоминант (более
10%)
Доминанты (5—10%)
Субдоминант (2—5%)
72,5
9,6
50,4
81,6
25,9
84,1
363,7
88,0
140,1
597,2
76,6
66,7
Сумма/среднее значение
132,5
71,2
591,8
153,4
Таблица 61. Процент синантропных видов в различных районах Варшавы
Группа (автор)
Пригород
Парки
Зеленые
насаждения
Б городских
кварталах
Центр
города
Мокрицы (Jedryczkowski, 1981)
40,0
66,7
100,0
100,0
Двупарноногие многоножки
(Jedryczkowski, 1982)
5,6
57,1
57,1
75,0
Эвсинантропные настоящие мухи.
непаразитические педальные
мухи и мухи-цветочницы (Gorska,
1982)
7,4
9,3
34,7
24,4
Гемисинантропные настоящие
му51,3
хи (Gorska, 1982)
25,9
34,1
43,5
Антропофильные кровососущие
30,8
57,1
42,8
комары (Wegner, 1982)
35,0
37,3%, парки —42,4%, мелкие зеленые насаждения — 56,3%,
центр города — 57,6%) (Ctiolewicka, 1981). Аналогичное
соотношение наблюдается для обитателей ксеротермных травостоев
(соотв. 12,7, 11,5, 14,6, 15,2%), травяного яруса (личинки — 34,2,
35,3, 39,6, 36,4%, имаго —65,7, 69,1, 77,1, 75,8%) и почвы (23,3,
26.6, 37,5, 39,4°/о). Доля лесных видов, напротив, снижается
(соответственно 24,3, 19,5, 8,3, 6,1%).
У жуков-листоедов доля видов ксеротермных травостоев (16,3,
21.7, 22,2, 30,8%) и лугов (23,3, 30,4, 30,9, 38,5%) также
возрастает к центру города (Wasowska, 1981), хотя термофильные
формы в центре не встречаются из-за отсутствия подходящих
кормовых растений.
Доля синантропов среди различных групп животных к центру
города возрастает (табл.61, 62).
Доля эврипотентных (эвритопных) видов различных
таксонов в центре города выше, чем на окраинах (рис. 75, табл. 63,64).
12—21
166
7. Урбанистические градиенты
Рис. 73. Доля жужелиц различных экологических групп внутри Лейпцига и по
его окраинам (по Klausnitzer, 1983b): Г —гигрофилы; М —мезофилы. К —
ксерофилы; Л — лесные виды
Рис. 74. Изменения доли
лесных видов и видов
открытого пространства
вдоль трансекта,
пересекающего различные
парки Варшавы (по
Czechowski, 1980): А
—лесные виды; Б — полевые
виды; В — лесные особи;
Г — полевые особи.
Сокращения см. рис. 72,
Рис. 75. Влияние
урбанистической нагрузки (УР) на
частоту встречаемости
жужелиц берегового склона
Вислы в Варшаве (по
Czechowski, 19Ш): А —редкие
виды; Б — обычные виды
7. Урбанистические градиенты
167
Таблица 62. Экологические группы пауков и сенокосцев Варшавы
(в процентах) (по Krzyzanowska et al., 1981; Chechowski et al., 1981)
Пригород
Парки
Зеленые
насаждения
в городских
кварталах
Центр
города
Пауки
эвритопные
5.1
7,3
14,7
13,9
политопные
20,2
23,2
35,3
23,3
олиготопные
56,0
56,8
44,1
44,2
стенотопные
18,8
12,8
5,9
18,6
очень сухолюбивые
3,7
3,2
2,9
2,3
сухолюбивые
36,7
32,8
32,4
32,6
умеренно сухолюбивые
6,4
7,2
—
2,3
умеренно влаголюбивые
11,0
11,2
11,8
18,6
влаголюбивые
32,6
40,0
50,0
32,6
очень влаголюбивые
4,6
1,6
—
—
сухо- и влаголюбивые
5,1
4,0
2,9
11,6
эпигейные
и гиполитные
57,3
55,2
64,7
60,5
обитатели травяного яруса
20,2
19,3
23,5
18,6
обитатели кустарникового яруса
13,8
18,4
8,8
9,3
обитатели древесных стволов и
деревянных оград
4,6
2,4
2,9
9,3
обитатели крон деревьев
4,2
4,8
—
2,3
асинантропы
91,3
84,7
74,9
67,4
гемисинантропы
5,5
10,5
17,7
16,3
эвсинантропы
. 3,2
4,8
2,9
16,3
Сенокосцы
гигрофилы
71,4
75,0
66,7
33,3
ксерофилы
28,6
25,0
33,3
66,7
эпигейные
35,7
50,0
66,7
50,0
древесные
57,1
50,0
33,3
50,0
асинантропные
42,9
50,0
33,3
—
синантропные
57,1
50,0
66,7
100,0
Олиготопные и стенотопные виды встречаются чем ближе к
центру города, тем реже. Это, например, четко продемонстрировано
для пауков (табл. 62). По требованиям к влажности биотопа
такой явной тенденции у пауков не прослеживается, но иногда
отмечается увеличение в центре доли эвригигробионтов. Доля
обитателей поверхности почвы н гиполитиона к центру города
возрастает, а видов кустарникового яруса убывает, доля гигрог
филов среди сенокосцев уменьшается, ксерофилов — возрастает
(табл. 62).
У ногохвосток эдафические (почвенные) виды внутрп города
оказываются в более благоприятных условиях, чем эпигейные
(наземные) (Sterzynska, 1982). Фауна жужелиц в центре города
характеризуется более высокой долей так называемых «осенних^»
12'
168
7. Урбанистические градиенты
Таблица 63. Экологическая амплитуда различных групп животных в Варшаве
(в процентах)
Группа
Пригород
Парки
Зеленые
насаждения
в городских
кварталах
Центр
города
Жужелицы (Czechowski, 1981)
эвритопы
политопы
олиготопы
стенотопы
7,5
47,5
36,0
9,0
10,0
63,5
19,0
7,5
15,5
75,0
9,5
18,5
66,0
10,5
5,0
Ежемухи (Draber-Monko, 1982 с, d)
эвритопы
олиготопы
16,3
28,5
18,9
27,8
33,3
23,3
29,8
14,9
Асинантропные слепни Tabano-
morha (Trojan, 1982)
эвритопы
28,6
36,0
55,6
62,5
Настоящие мухи,
непаразитические падальные мухи,
мухи-цветочницы (Gorska, 1982)
эвритопы
44,5
51,1
95,7
86,5
Мухи-журчалки
эвритопы
36,5
34,8
43,5
(обочина
улицы —
78,6)
54,1
Муравьи (Pisarski, 1982)
эвритопы ,
политопы
олиготопы
11,4
28,6
57,1
19,0
28,6
47,6
28,0
57,1
27,3
36,4
27,3
Жуки-листоеды (Wasowska, 1981)
очень часто встречающиеся
(эвритопы?)
41,9
56,5
66,7
84,6 к
ВИДОВ, личинки которых зимуют в почве и потому в меньшей
степени подвергаются действию городских факторов массовой
смертности (рис. 76). Может быть, именно в связи с этим у Nebria
brevicollis и Calapthus fuscipes хорошие возможности развития в
городе.
Существует целый ряд других параметров, которые могут
изменяться по градиентам.
— Исследования различных авторов (обзор см. Thiele, 1977)
показали, что у жужелиц с диморфизмом по степени развития
задней пары крыльев в толшо что основанных популяциях доля
крупнокрылых особей выше, чем в длительно существующих
7. Урбанистические градиенты
169
Таблица 64. Процент видов божьих коровок из различных экологических
групп в Варшаве (по Czechowska, Bielawski, 1981)
Экологическая группа
Пригород
Парки
Зеленые
насаждения
в городских
кварталах
Центр города.
Эвритопы
48,9
57,2
66,6
64,3
Олиготопы
14,3
10,7
16,7
14,S
Стенотопы
36,7
32,1
10,7
21,4
Убиквисты
12,2
17,9
33,3
28,6
«Лесные виды»
59,4
67,8
41,7
35,7
Обитатели открытых
14,3
25,0
35,7
мест
28,6
Ксерофилы
20,4
21,4
33,3
35,7
Мезофилы
51,0
64,3
66,7
64,3
Гигрофилы
28,6
14,3
(рис. 77). Подобного же соотношения можно ожидать и при
заселении ими городских местообитаний (ср. Stein, 1968, 1977)-.
В дольше существующих популяциях увеличивается процент бра-
хиптерии, т. е. «еспоообности к полету, при сохранении высокого
расселительпого потенциала.
— Доля видов с колюще-оооущим ротовым аппаратом у аси-
нантропных Tabanomorpha уменьшается к центру города
быстрее, чем доля видов с лижуще-сосущим аппаратом (рис. 78).
— По направлению к центру города возрастает
доминирование г-стратегав, в связи с чем в отдельных популяциях
уменьшается доля самок.
— Колебания размеров популяций у субдоминантов
усиливаются к центру города.
Рис. 76. Доля различных типов зимовки у жужелиц во внутригородских
районах (слева) и по окраинам (справа) Лейпцига (по Klausnitzer, 1983b): И —
имагинальная зимовка; (И) — имагинальная и частично личиночная зимовка;
Л — личиночная зимовка; (Л) — личиночная и частично имагинальная
зимовка
170
7. Урбанистические градиенты
— У некоторых видов возрастает' доминирование молодых
аозрастных групп. В центре города доминируют виды с
однолетним циклом развития .или с еще более 'быстрой сменой генераций.
Доля видов с широким географическим распространением во
внутригородских местообитаниях в целом возрастает (табл. 65),
а виды с более узкими ареалами становятся реже.
Популяция
момсая старая
P.St.
80
-.9 ВО
40
го
о
с. т.
V
V
\\
\ \
1 1 '
— об
-—
Ур
Рис. 77. Процент
крупнокрылых особей (жирный
контур) в недавно
основанных и долго существующих
популяциях жужелиц
Pterostichus strenuus (P. st.) и
Calathus melanocephalus
(С. т.) (по Thiele, 1977)
Рис. 78. Уменьшение числа
видов (S) (процент от их числа
в городских окрестностях)
слепней Tabanomorpha с
различным типом ротового
аппарата при увеличении степени
урбанизации (УР) (по Trojan,
1982): А — колюще-сосущий
аппарат; Б — лижуще-сосущий
аппарат
7.6. Трофические уровни
Можно было бы ожидать, что доля полифагов к центру
города увеличивается, а монофагов — снижается. Однако результаты
исследований различных групп насекомых в Варшаве свидетель-
Таблица 65. Зоогеографические элементы фауны Варшавы в процентах
Распространение
Пригород
Парки
Зеленые
насаждения в
городских
кварталах
Центр
города
Энхитреиды (Kasprzak, 1981)
космополиты
10,5
16,7
18,2
голарктические виды
50,0
36,8
41,7
45,5
европейские виды
50,0
47,4
41,7
27,3
«
Пауки (Krzyzanowska et al., 1981)
космополиты
0,5
—
2,9
6,9
голарктические виды
13,7
16,8
14,7
20,9
палеарктические виды
15,6
13,6
17,7
16,3
бореальные виды
0,9
—
—
—
европейско-сибирские виды
25,5
20,0
17,7
20,9
южноевропейско-сибирские
виды
4,6
5,6
8,8
6,9
европейские виды
36,2
41,6
38,2
27,9
субатлантические виды
0,5
0,8
—
—
средиземноморские виды
1,4
0,8
—
—
горные виды
0,5
0,8
—
—
Клещи (Niedbala et al., 1982)
европейские виды
35,5
37,2
40,3
44,8
Сенокосцы (Czechowski et al.,
1981)
голарктические виды
21,4
8,3
33,3
66,7
европейско-сибирские виды
7,1
8,3
33,3
33,3
европейские виды
71,5
83,4
33,3
—
Ногохвостки (Sterzynska, 1982)
космополиты
Не
ука25,0
33,3
40,8
зано
палеарктические виды
«
6,3
9,5
11,1
европейские виды
«
31,2
28,6
18,5
Цикадовые (Chudzicka, 1982)
голарктические виды
20,8
26,8
26,7
37,2
европейские виды
12,0
12,3
3,3
4,6
Сетчатокрылые и скорпионовые
мухи (Czechowska, 1982)
голарктические виды
22,5
28,6
37,5
33,3
европейские виды
37,5
28,6
12,5
16,7
Жужелицы (Czechowski, 1981)
голарктические виды
7,0
12,5
15,5
18,0
палеарктические виды
33,0
33,5
33,5
38,5
европейско-сибирские виды
29,5
36,5
43,5
34,0
вропейские виды
21,5
10,5
5,0
7,0
172
7. Урбанистические градиенты
Продолжение
Распространение
Пригород
Парки
Зеленые н'а-
сажден51я
в городских
кварталах
Центр
города
Жуки-листоеды (Wasowska, 1981)
палеарктические виды
европейско-сибирские виды
30,2
32,2
39,1
21,7
50,0
16,7
53,8
15,4
Жуки долгоносики (Cholewicka,
1981)
голарктические виды
8,2
10,1
16,7
21,2
Муравьи (Pisarski, 1982)
палеарктические виды
, европейские виды
28,6
20,0
33,3
19,0
57,1
28,6
45,4
27,3
Ежемухи (Draber-JV\onko, 1982с, d)
палеарктические виды
европейские виды
47,3
10,6
52,2
11,1
60,0
3,3
57,5
6,4
Настоящие мухи,
непаразитические падальные мухи,
мухи-цветочницы (Gorska, 1982)
космополиты
голарктические виды
палеарктические виды
европейские виды
6,4
30,3
25,9
36,4
5,4
34,1
25,5
31,8
21,8
52,1
17,4
8,7
16,2
54,0
13,5
13,5
ствуют о том, что такая тенденция наблюдается не всегда
(та'бл. 66). Правда, отнесение видов к определенным
трофическим группам затруднительно, поскольку сами подходы к
определению этого понятия могут значительно различаться
(Klausnitzer, 1983).
Выявлена взаимосвязь между шириной трофических ниш
фитофагов и различными стадиями сукцессии (Brown, Southwood,
1983), а также степенью урбанизации местообитания (у
долгоносиков в Лейпциге, рис. 79, табл. 67, Klausnitzer, Richter, 1986).
Кюнельт (Kühnelt, 1977) пишет, что в Ресльпарке в Вене
почти полностью отсутствуют группы видов, питающиеся
листьями деревьев, травянистыми растениями, навозом и падалью, а
также посетители цветов. Исследования в Лейпциге по'казали, что
различные таксоны неодинаково реагируют на степень
урбанизации в зависимости от своего положения в пищевой цепи
(Klausnitzer, Richter, 1984) (ср. также табл. 68, рис. 80, 81).
Численность мокриц (сапрофаги) снижается по направлению
к окраинам (86% особей отмечено в центре), а многоножек-сап-
рофагов, клещей и ногох1восток, напротив —к центру города.
7. Урбанистические градиенты
173
Таблица 66. Доля видов с различной трофической амплитудой в городских
местообита{1иях Варшавы (в процентах)
Группа
Пригород
Парки
Зеленые
насаждения
в городских
кварталах
Центр
города
Жужелицы (Czechowski, 1981)
пантофаги
30,5
29,0
33,5
33,5
Жуки-долгоносики (Cholewicka,
1981)
полифаги
монофаги
16,4
13,4
17,3
10,1
20,8
6,3
24,2
9,1
Жуки-листоеды (Wasowska, 1981)
полифаги
олигофаги
монофаги
14,0
60,5
16,3
8,7
65,2
21,7
11,1
72,2
11,1
7,7
76,9
7,7
Настоящие мухи,
непаразитические падальные мухи и
мухи-цветочницы (Gorska, 1982)
пантофаги
полифаги
28,4
24,7
31,8
23,3
78,2
13,0
59,5
16,2
Муравьи (Pisarski, 1982)
пантофаги
77,1
76,1
100,0
90,9
Таблица 67. Зависимость между положением местообитания и шириной
пищевой ниши на примере долгоносиков вдоль трансекта в Лейпциге
(по Klausnitzer, Richter, 1986)
ПримыкаюГородсйой
Городской
Пригород
щий к
гороцентр А
центр Б
ду
пойменный лес
Заполненность ниши
3,22
3,86
3,54
3,64
Незанятая часть ниши
2,82
2,88
2,30
1,90
Численность долгоносиков (фитофаги) увеличивается от
центра города к пригородам, хотя доля особей этого семейства в
общем числе насекомых сильно снижается по'направлению к
пригородному пойменному лесу. Количество растительноядных жу-
ков-блестямо« (Nitidulidae) к окраинам города возрастает.
Жуки-фитофаги составляют в центре города 1,4%, а на окраинах
6,8% общего числа насекомых. При подсчете доли различных
растительноядных Насекомых среди общего числа
представителей этого класса только у долгоносиков и суммы всех
жуков-фитофагов отмечена тенденция к увеличению к окраинам города.
174
7. Урбанистические градиенты
WD
80
ВО
40
20
О
- ^
—
\ / \ /
\/ \/
/\ /\
: / \ / V
« Т
ЛМ дЛМ
зп
Рис. 79. Изменение
потенциальной ширины
ниши (ПН) долгоносиков
вдоль градиента Gae в
Лейпциге (по
Klausnitzer, Richter, 1986)
Рис. 80. Изменение доли
видов жуков-щелкунов с
различными типами
питания при возрастании
урбанизации (УР) (по
Nowakowski, 1980): X —
хищники; В — всеядные;
К — корнееды; ЛДЛ —
липово-дубовый лес;
дЛДЛ —
деградировавший липово-дубовый лес;
П — поляна в
липово-дубовом лесу; Па—
крупные и периферийные
парки; ЗП —
внутригородские озелененные
пространства
Рис. 81. Влияние увеличивающейся степени урбанизации (Ур) на число
видов (S) (процент от их числа в городских окрестностях) различных по типу
питания групп слепней Tabanomorpha (по Trojan, 1982): А — фитофаги; Б —
сапрофаги; В — паразитоиды; Г — эпифитные хищники; Д — гематофаги; Е —
почвенные хищники
7. Урбанистические градиенты
175
Таблица 68. Доля различных трофических групп в городских
местообитаниях Варшавы (в процентах)
Зеленые
наЦентр
города
Тип питания/таксон
Пригород
Парки
саждения
в городских
кварталах
Авторы
Сапрофаги
Настоящие,
непаразитические
падальные мухи,
му32,4
хи-цветочницы
25,9
24,8
34,9
Gorska (1982)
Серые мясные мухи
10,2
10,3
26,7
16,7
Draber-Monko (1982b)
Пластинчатоусые
20,0
Kubicka (1981)
жуки
7,4
6,6
—
*;опрофаги
Пластинчатоусые
40,0
33,7
жуки
67,6
46,7
Kubicka (1981)
Фитофаги
Пластинчатоусые
25,0
жуки
46,7
40,0
66,7
Kubicka (1981)
Микрофитофаги
Клещи
40,9
44,9
40,3
34,2
Niedbala et al. (1982)
Ризофаги
Жуки-щелкуны
62,5
(личинки в почве)
25,0
33,3
50,0
Burakowski,
NowaАнтофаги
kowski (1981a)
Серые мясные мухи
15,2
17,9
13,3
5,6
Draber-Monko (1981b)
Зоофаги
Клещи
35,5
35,9
40,3
49,1
Niedbala et al. (1982)
Жужелицы
69,5
71,0
66,5
64,0
Czechowski (1981)
Жуки-щелкуны
12,5
(личинки в почве)
12,5
20,0
25,0
Burakowski,
Nowakowski (1981a)
Мухи-журчалки
54,0
69,7
71,7
(обочина
улицы 78,6)
70,3
Bankowska (1982b)
Доля листоедов, жуков-блестянок и эндофитных личинок
двукрылых (Itonididae, Chloropidae, Ephydridae) к окраинам
города-снижается.
Сосущие растения насекомые, особенно тли, часто находят
наиболее 'благоприятные условия в центре города (Klausnitzer
et al.„ 1980с; Klausnitzer, 1983а). Интересно отметить явное
увеличение видового разнообразия этой группы по направлению к
центру города, которое с учетом сходства растительности
изученных площадок вряд ли объяснимо расщирением спектра «ор-
176
7. Урбанистические градиенты
1п%
•
-3
ЬЗ
о
1,2-
-2
о
Г = 0,84В5
1,0-
-1
0
к
-0,5
О
К п
■0,5
Рис. 82. Видовое
разнообразие сосущих растения
насекомых (Hs) и их доля в
общем улове (Ag) (по Klaus-
fitzer, Richter, 1984).
Сокращения см. рис. 85
-4
r'-0,9924
1=-0,3325 ^'"'"■^
-J
-2
-i
r^0,S51B
1
1
Рис. 83. Изменения
численности тлей (Т),
цикадовых (Ц),
растительноядных клопов (К) и всех
насекомых, сосущих
растения (Ф) вдоль А-Е-
градиента (по
Klausnitzer, Richter, 1984).
Сокращения см. рис. 85
7. Урбанистические градиенты 177
иовых растений (рис. 82). В центре города отмечена высокая
численность таких насекомых, причем прослеживается отчетливая
тенденция к снижению их доли в общем улове насекомых к
окраинам города. Количество тлей, растительноядных клопов и
всех насекомых, питающихся соком растений снижается в этом
же направлении (рис. 83, 84). Такая же тенденция наблюдается
и у листоблощек (Psyllina), хотя их численность в
примыкающем к городу пойменном лесу выще, чем в пригороде.
Большинство цикадовых в отличие от других сосущих растения
насекомых обитает именно на окраинах.
Численность зоофагов, в частности пауков, сенокосцев, клопов
из семейств Nabidae и Anthocoridae, а также жуков-стафилинид
(не все они хищники), возрастает к окраинам города (рис. 85),
а у двукрылых, являющихся хищниками на стадии имаго
(прежде всего Empididae, а также Asilidae и Dolichopodidae), и
божьих коровок она, наоборот, в этом направлении уменьшается
(рис.86).
7.7. Синэкологические характеристики
7.7.1. видовое разнообразие
Видовое разнообразие (Shannon, 1948; Shannon, Weaver,
1962, или Brillouin, 1958), мера видовой неоднородности
сообщества, определяют по формулам
Hs (по Шеннону) =-2 (y)Ii(^)' ^^^^
где П(—число особей вида i (соответственно Ni, N2, ...);
N — общее число ооОбей;
S —число видов:
tt / с - ч Ii NiINz!... N,! .,с\
Hs (по Брииуэну)= n" ~^—n ^
При равном общем числе особей оба индекса возрастают при
увеличении числа видов и сближении их численностей, причем
индекс Шеннона принимает большие численные значения, чем
индекс Брийуэна. При неизменном соотношении численностей и
равном числе видов увеличение числа особей не влияет на
значение индекса Шеннона, тогда как индекс Брийуэна возрастает
(Pielou, 1966; Roth, 1976; Moss, 1978).
Многие авторы критически относятся к использованию
понятия видового разнообразия в качестве меры стабильности
сообщества (Leigh, 1965; Pianka, 1966; Hairston et al., 1968; Marga-
leff, 1969; Mac Arthur, 1975; Kushlan, 1976). Тиле и Вайс (Thiele,
178
7. Урбанистические градиенты
БК
Рис. 84. Изменения
численности клопов-слепняков (С) и
«неслепняков» (НС) вдоль
А-Е-градиента (по Klausnitzer,
Richter, 1984). Сокращения см,
рис. 85
%
-20
-15
^Х •
-10
--Л
\
.. Г=0,98В7
-5
\
*-с
1 ■ II 1
1
1
-—■ "т^
lilt
-о,д
4i Чг
0
Рис. 85. Изменения численности
сенокосцев (С), пауков (Па) и
двукрылых (д) вдоль А-Е-
градиента (по Klausnitzer,
Richter, 1984): Ui, Дг
—местообитания в центре Лейпцига; К —
местообитание на краю
Лейпцига; П — местообитание в
окрестностях Лейпцига
(пойменный лес)
'=0,931
/7^
Рис. 86. Корреляция между
частотами встречаемости тлей (Т)
и божьих коровок (БК) вдоль
А-Е-градиента в Лейпциге (по
Klausnitzer, Richter, 1986).
Сокращения см. рис. 85
I
7. Урбанистические градиенты 179
Weiss, 1976) рекомендуют в этой связи использовать индексы На
только при долговременных наблюдениях равных площадей в
условиях изменяющейся окружающей среды. Снижение Hs, по-
видимому, может указывать «а антропогенную нагрузку (Müller
et al., 1974; Nagel, 1977), так как в городе Hs обычно ниже, чем
в его окрестностях. Однако следует учитывать также спектры
видов и свойства биотопов (микроклимат, запас ресурсов).
Видовое разнообразие снижается в непосредственной близости от
проезжей части (Maurer, 1974; Mader, 1979). У жужелиц в
луговом сообществе (многолетние травы) на окраине Лейпцига,,
значение Hg равно 3,373, в полевой культуре — 2,397, на берегу
ручья —2,564, в рудеральном биотопе — 2,464, на мусорной
свалке—2,638 (Klausnitzer, 1983b).
Как видно из табл. 53, значения индекса видового
разнообразия пробных площадок вдоль А-Е-градиента в Лейпциге
значительно ниже и почти во всех случаях не достигают 2,0, причем
отмечена тенденция к уменьшению этого показателя к центру
города. На сухом лугу (аэродром Кёльн/Бонн) Hs = 0,584, а на
рудерализированном лугу — 0,763 (Becker, 1977). В городскою
парке видовое разнообразие значительно ниже (0,523—1,449),.
чём в пригородном лесу (Schäfer, Kock, 1979). Результаты
учетов жуков в Саарбрюккене (Klomann, 1977, 1978; Müller, 1980;.
Müller et al., 1974) также свидетельствуют о снижении Hs по
направлению к центру города (рис. 87, 88).
Аналогичные результаты получены и при орнитологических,
исследованиях (Elvers, 1978, 1980; Erdelen, 1977; Mulsow, 1980;:
Otto, 1983). Мульсов (Mulsow, 1980) и Otto (Otto, 1983)
рассчитали разнообразие групп птиц с различной экологией
гнездования. В небольших садах оно составило 0,719, на кладбищах —
0,956, в заповедниках —0,977—1,094 (Otto, 1983).
7.7.2. Равномерность распределения
Согласно Беццелю и Райххольфу (Bezzel, Reichholf, 1974),.
равномерность раопределения (выровненность) выражает
степень разнообразия. Этот показатель также может в ряде
случаев отражать ход сукцессии. Он рассчитывается по формуле
Е=-^, (17>
где Нтах—максимальное значение разнообразия при равном
числе особей всех видов.
Некоторые авторы определяют равномерность распределения!
следующим о1бразо.м:
180
7. Урбанистические градиенты
-2,0
° -Л
о
о
•
^— •
•
1
-1,0
-0,8
1
-0,д
4i Чг
+0,5
К П
"АС
Рис. 87. Изменения видового
разнообразия долгоносиков (Д) и
жуков-листоедов (Л) вдоль
А-Е-градиента (по Klausnitzer, Richter,
1984). Сокращения см. рис. 85
•
r-0,8238
-2,4 .
-2,2
•
ПЛ
у'
т=0,8201 у ^ у
4,0
г-0,095гЖ
1
-
-0,5 0
Ui U2 .
l'AE
+0,5
П
Рис. 88. Изменение
видового разнообразия
общего улова (О), сборов
почвенными ловушками
(ПЛ) и укосов
сачком (У) вдоль
А-Е-градиента (по Klausnitzer,
Richter, 1984).
Сокращения см. рис. 85
где S — ЧИСЛО 1в'идов.
В табл. 56 представлены значения Е для жужелиц
некоторых парков Лейпцига (по Klausnitzer, 1983b).
7.7.3. Видовое богатство
Для численной оценки видового богатства зооценоза
используют различные индексы, например,
индекс Маргалеффа = ^~ ^ .
In N
(19)
7. Урбанистические градиенты 181
Так называемое «внутриэкотопное» разнообразие в отличие от «меж-
экотопного» (fi) и «межобластного» {•{).—Прим. ред.
Чем больше видов, тем выше значение этого индекса; но
направлению к окраинам города оно возрастает. Исследования
жужелиц в Лейпциге показали, что в центре города видовое богатство
составляет 1,2—4,7, а на периферии — 5,6—7,3 (Klausnitzer,
1983b).
В качестве меры видового богатства используют также
понятие а-разнообразия'' (Williams, 1959):
S = aln —. (20)
а
Возрастание числа особей при неизменном числе видов ведет
к снижению значений индексов. Они могут и не отражать
изменений в структуре сообщества (распределения особей по видам
при постоянном числе последних).
7.7.4. Структурное разнообразие
Индекс структурного разнообразия (Hstr) можно вычислить
по,формуле Брийуэна (16) (Mader, 1979), подставив в нее, на-
np^iiмер, численные значения признаков местообитания из табл. 69.
Для нризнаков 1—4 и 7 количественная оценка производится не-
поаредственно путем подсчетов или измерений, а .для признаков
5'—6 и 8—10 используется шкала бонитировки. Этот индекс
применим для характеристики лространственной структуры
исследуемых площадок в парках, а также (в модифицированном виде)
на газонах и в рудеральных биотопах. В качестве примера на
рис. 89 показана взаимосвязь между видовым разнообразием
птиц и разнообразием их местообитаний (Sukopp et al., 1980).
H3tr = -—In fmu!Tn„!...m,3! (21)
M \ M! /'
где mi—численное значение признака i;
М —общее численное значение;
S — число признаков.
7.8. Индексы сходства
Для сравнения между собой зооценозов различных
городских местообитаний, а также для анализа их изменений от
центра к окраинам города используют различные индексы сходства
(Schaefer, 1973; Weiss, Sarbu, 1977; Klausnitzer, 1983b).
182
7. Урбанистические градиенты
Таблица 69. Элементы структурного разнообразия (по Mader, 1979)
Пробная
плоПризнак
Численная
щадка
(раоценка
диус
круга, м)
1. Общее число видов сосудистых растений
1/вид
1,5
2. Число деревянистых растений
1/экземпляр
1,5
3. Число деревьев с диаметром ствола более
10 см
1/экземпляр
5
4. Число многолетних травянистых растений
высотой более 20 см весной перед началом
1
вегетационного периода
5. Подлесок:
— обильный (более 5 ветвей на 1 м^)
10
— средний (2—5 ветвей на 1 м^)
5
5
— редкий (1 ветвь на 1 м^)
2
— отсутствует
—
6. Состав лесной подстилки:
10
— иглы хвойных деревьев и листва
1
— иглы хвойных деревьев или листва
5
— ни то, ни другое.
—
1
7. Глубина лесной подстилки в см
1/см
8. Вода (ручей, низина, сырое место)
на расстоянии:
10
— менее 2 м
—
— 2—5 м
5
— более 5 м
—
9. Древесные пни на расстоянии:
10
— менее 2 м
— 2-5 м
5
— более 5 м
—
10. Камни или скалы на расстоянии:
10
— менее 2 м
— 2-5 м
5
—
— более 5 м
—
Индекс сходства по Серенсену (Sorensen, 1948) 'рассчитыва-
■ ется по фор1муле
„ 2j
a-f b
(22)
где j — число видов, встречающихся одновременно в выборках
1 и 2; ■ I
а— число видов в выборке 1;
b —число видов в выборке 2.
Этот индекс выраж'ает в процентах сходство видового состава
биоценозов, например, на двух пробных площадках;
Qs=?=0 — ни одного общего вида на двух площадках;
Qs = 100 — все виды для двух площадок общие. ^ , ^
7. Урбанистические градиенты
183
Степень доминирования при этом не учитывается, как и в
формуле Жаккара:
j
Cj
(23)
a + b —i
Индекс Ренконена, учитывающий уровень доминирования
отдельных видов, весьма чувствителен к присутствию в сообществе
явных доминантой (звдоминантов), которые, впрочем, могут и
неполно характеризовать расаматриваемые зооценозы.
b=cli-t-d2+d3+... (24)
где dl, d2, ds... —более низкое (из двух выборок) значение
доминирования видов 1, 2, 3 и т. д.
Рис. 89. Зависимость
видового разнообразия
птиц (Hs) от
разнообразия местообитаний (Hstr)
в двух зеленых
насаждениях Западного Берлина
(по Sukopp et al, 1980)
Практически идентичен индексу Ренконена индекс
перекрывания по Роджерсу (Rogers, 1972) или Шенеру (Schöener, 1970):
Cjk= 1—^? IP'i~P'k|.
(25)
где pi],-Pik—доля i-го вида в общем числе особей выборок j и к.
Если индексы Ь и Cjk равны нулю, сравниваемые зооценозы
абсолютно различны, а при Ь = 100 и Cjk=l они идентичны.
Проблему переоценки эвдоминантных видов пытаются
обойти введением других индексов.
Индекс Вайнштейна (Wainstein, 1967):
100
Kw =
(26)
184
7. Урбанистические градиенты
Метрика Канберры (Lance, Williams, 1976), задаваемая
индексом
1 Л |П1] —П1к|
(27)
СМ= 1 —
•= ГГ1 (пц -f nik)
где Hi'— число особей i-ro вида;
S — число видов.
Выровненный индекс перекрывания (Klausnitzer, Richter,
1986):
CA = 1 —
i=l
qik
Sq.]
■^qik
i
(28)
Oh устраняет слабую нелинейность индекса СМ. Чем больше Hs,
тем меньше его отличие от Ь и Cjk (рис. 90).
0,5
Hi Н
0,808
К П Щ Hi К Л
0,727
0,574
0,681
0,523
0,397
üij
CA
Рис. 90. Сравнение результатов
вычисления двух различных
индексов сходства для одного и того же
материала •— общих сборов в
четырех местообитаниях вдоль А-Е-
градиента в Лейпциге (по
Klausnitzer, Richter, 1986) (ср. рис. 88)
Разница разнообразий (H-diff, MacArthur, 1965) отражает
различия в видовом составе и численности видов двух выборок.
Этот индекс позволяет количественно сравнить соседние
зооценозы Или островные местообитания с контрольными.
H-diff = 0 означает, что в обеих выборках виды и их
численность одинаковы.
Н, + Н2
H-diff = Ht
Pn + Pl2
2
Pu + Pl2
где Hl—разнообразие в выборке 1;
Ha— разнообразие в выборке 2;
Hi
(29)
(30)
(31)
7. Урбанистические градиенты
185
7.9. Сообщества и репрезентационный
анализ
Решрезентационный анализ по Мюллеру (Müller, 1978)
{табл. 70) позволяет выявить различия в степени проникновения
в город отдельных видов, таксоны, характерные для
определенных местообитаний и зон, и, наконец, смену сообществ
(рис. 91, 92).
100 \-
80
1
S
40
го
Carabus
■nemoralis
Abax
parallele-
pipedus
Platjjnus
assimilis
Carabus
coriaceus
Abax
paraUelus
Bembidion
properans
Harpalus
aeneus
Amara
aenea
Calathus
fuscipes
Рис. 91. Изменения характерных сообществ жужелиц вдоль А-Е-градиента
в Лейпциге (по Klausnitzer. 1982b). 1—4 — местообитания от окраин к центру
города
Приведенные, в табл. 70 виды жужелиц можно подразделить
на несколько групп (Klausnitzer, 1982b).
а) Лесные виды, более или менее продвинувшиеся в
городскую «пустынную» зону.
— Abax parallelus считается типичным полистеногигробионт-
ным эвритермным видрм прежде всего влажных лесов порядка
Fagetalia; автодоминант в Байперте и представлен лишь не-
13-21
СО
05
3
о
с
с
и
03
я
о
•S
о.
я
к
s
о»
S
ч
«
а
(Я
•S
S
в
S
о
в
Cf
со
н
и
Ö
«
S
ч
«
о. О)
3«
с се
да
5 s:
«
S
Ч
S
ш
!-|
я
о
я
Ш
«
я
на
я
Е-
=я
а
f-
я
я
я
о
я
Я
я
я
л
»S
са
>s
»S
аЯ
со
со
л
со
я
ч
а
я
я
3
К
=я
а
я
=я
я
ч
я
я
м
я
3
я
я
а
я
со
я
о
S
о
я
о
н
я
S
S
о '
Я
о.
о *
а.
.л
а.
я
о ^
л
о
я
о
л
=с
U
ш
1<
О)
ч
я
сС
ч
CJ
я
et
о
к
о
с
со
с
f-
о
со
о
о
о
t-f
Ü
о
CQ
о
о
н
со
о
ш
Е-
m
о.
R,
я
и
а
я
ш
п
о.
я
<:
с
Я
m
<
с(
<
т
<;
с
<
IM
to
о
ю
ю
(М
см
'S«
1—1
+
+
+ +
СП
со"
см
от
CM
05 СО
СО СП
00 + [>•"
CD
СП СО
CD- +
— 00 00 от
ОТ 00 со
+ + CD о" оо' Н Ь
+ + -
CD Ю
СО Ю
00 СО
СМ со
+ + + +
+ + ю
+
+ +
со 00 ю
+ от" +
S
а
а
S
<3
■5"
"а
а
о
Vi
3
s
3i
5
s
CO
1
—H
1
O)
-^и 1
1—1
1
(N
■ CO
^ 1
1
<N
00
(M
ai
^ 1
1
1
CO
Ю
Ю
о
^ 1
1
1.
CO
8
1
1
1
CO
я
s
C3 ■
m
о
о,
s
IS
s
о
Ч
ID
CJ
a
о
3
f-
я
Я
CS
и
«
«
s
S
S
о
Ч
о
о
et
я
о
S
<
и
О)
3
3
я
S
S-
S
я
к
к
ч
га
X
га
Е-
Он
«
ю
о
<и
S
S
m
а
m
|=С
В"
3
3
3
н
н
S
я
я
О)
0)
0)
S
S
к
ч
а
п.
о.
о.
о.
t-.
U
U
о
о
о
О
в
я
я
я
га
СЗ
га
га
о.
о.
о.
с
Е-
Н
3
3
Я
я
<и о
4 U
<D
II
е га
й о
5 »
Ч S
t|
я
й «
§1
я а> о
|„з5-ё-У&ё§
3 to ш .. й S m
h га о 'S я
3 Я f_ Я
« га S «я .я
^ &■« ь 3 3
»TS СГ W д
si
et
13*
188
7. Урбанистические градиенты
сколькими экземплярами в пойменном лесу Ноняе. Доля
биообъема 2,2%.
Carabus coriaceus — стенойкный вид влажных (мезофиль-
ных) лиственных лесов; автодоминант в Байперте, встречаемость
в Нонне ниже. Доля биообъема в Байперте 33,7%, в двух частях
Нонне 12,5% и 3,4% (доминирование 2,51 % и 0,36%).
— Platynus assimilis — холодо- и сухолюбивый вид прежде
всего пойменных лесов; в Байперте — высший траисгредиент, в
пойменном лесу Нонне — низший; встречается до парковой части
Нонне.
— АЬах parallelepipedus считается лолистеногигробионтным
мезоэвритермным видом лиственных лесов; частота
встречаемости убывает от Байперта к обеим частям Нонне. На фоне общего
МЛ
waloijius
marginaius 7В%
Prosternon
iesseilaium 10%
Athous
isubfuscus 10%
дЯЛЛ
П
Па
ЗП
Athous 77%
haernorrboidalis
Agrioies
sputator 75%
Aqriotes
sputator 80%
Agrioies
Agriotes
sputator 15%
Dalopius
warijinaiu's 4%
Agriotes
obscurus 10%
Agriotes
obscurus 7%
sputator SU-WOX
Рис. 92. Тенденции антропогенного изменения сообществ жуков-щелкунов при
возрастании урбанизации (по Nowakowski, 1982). Сокращения см. рис, 80.
снижения биообъема вдоль А-Е-градиента процентная доля в нем
этого вида возрастает, хотя его численность снижается.
, Местонахождение
Число особей
Биообъем, %
Байперт (Б)
147
11,3
Нонне, пойменный лес
(НПЛ)
124
12,8
Нонне, парк (НП)
64
16,2
— Carabus nemora/ts —по встречаемости почти
исключительно лесной вид, однако его экологическая амплитуда (эвригигро-
бионтность, эвритермия) значительно шире («гемерофильнее»),
чем у всех леречиоленных выше видов.
б) Относительно эврипотеятные виды, проникающие от
«древесных» окраин внутрь города и достигающие, возможно, за счет
конкуренции высокого уровня доминирования в средней части
А-Е-градиента, однако не доходящие до явно «пустынной»
городской зоны.
7. Урбанистические градиенты
189
Местонахождение
Число особей
Биообъем, %
Б
326
48,0
НПЛ
360
71,2
НП
134
6.5,2
Парк Йоханна (И)
7
26,4
— Nebria brevicollis считается мезоэвригигробионтным поли-
эвритермяым видом лиственных лесов. Хотя распространен
дисперсно, наблюдается увеличение встречаемости ближе к центру
города.
Местонахождение
Число особей
Биообъем, %
Б
73
1,3
НПЛ
42
1,0
НП
212
12,9
и
107
50,5
Шиллерпарк
116
94,3
в) Виды, сходные с представителями группы «б», но
достигающие городакой «пустынной» зоны.
— Asaphidion flavipes часто доминирует на полях, иногда
считается гигрофилом; дисперсно распространен от Байперта до
парка Шванентайх.
г) Виды, тяготеющие к «пустынной» части градиента; в
остальных зонах встречаются единично или отсутствуют.
—' Calathus fuscipes — эврипотентный мезофил открытых
пространств; доминант на полях и эвдоминант в травяных
сообществах аэропорта Кёльн/Бонн (Becker, 1977).
— Атага аепеа — вид, известный как явный гелиофил;
предпочитает очень сухие биотопы (часто поля под зерновыми).
— Harpalus aeneus — явно эврипотентный мезофил, часто
доминирующий на полях; в центральных парках является экваль-
ным траногредиентом, проникает в обе части Нонне.
Вдоль изученного градиента можно с некоторыми
оговорками выделить четыре сменяющих друг друга в пространстве
сообщества жужелиц (рис. 91).
Существование орнитоценозов сильно зависит от
определенной структуры местообитания, 'которая в свою очередь связана
с зонированием города. В городских условиях различают
следующие сообщества птиц (Saemann, 1970, ср. Mulsow, 1968):
190 7. Урбанистические градиенты
а) Орнитоценозы городоких построек:
—• Сообщество «галка—пуательга». Населяет самые высокие
постройки, например башни, церкви, высокие промышленные
сооружения, (МОСТЫ.
— Сообщество черного стрижа. Населяет преимущественно
строения средней высоты (трех-, в основном четырехэтажные
жилые дома).
— Сообщество «трясогузка — горихвостка-чернушка —
деревенская ласточка». Населяет преимущественно постройки еще
меньшей высоты (в основном двухэтажные жилые дома,
индивидуальные дома, хозяйственные строения и др.).
■б) орнитоценозы природных структур крупных городов:
— Сообщество «зеленушка — кольчатая горлица».
Предпочитает местообитания, в которых природные структуры (деревья,
кустарники) занимают до 50% общей площади, или же участки
природной структуры (площадью до 2 га) внутри застроенных
районов.
— Сообщество «канареечный вьюрок —садовая
горихвостка — зеленая пересмешка — славка-завирушка». Предпочитает
местообитания «зеленого города» с естественными структурами,
занимающими свыше 50% площади.
— Сообщество «серая славка — болотная камыщевка».
Заселяет естественные структуры с богатым травяным и менее
выраженным кустарниковым ярусами без деревьев, прежде всего
рудеральные местообита1нвя и другие участки с сильным
антропогенным нарушением.
— Сообщество хохлатого жаворонка. На сходных с предыду- i
щим сообществом площадях, однако с меньшим покрытием ниж- \
него яруса растительности. !
В орнитоценозах выделяют характерные и сопутствующие !
виды. \
8. РЕАКЦИИ ЖИВОТНЫХ НА СВОЕОБРАЗИЕ
ГОРОДСКИХ УСЛОВИЙ
8.1. Физиология
Изучение тлей (Aphis sambuci, А. fabae, Macrosiphoniella ar-
temisiae) показало, что в зависимости от влияния окружающей
среды изменяется удельная активность различных ферментов,
например малатдегидрогеназы (МДГ), глютаматоксалоацетат-
трансаминазы (ГОТ) и глутаматпируваттрансамияазы (ГПТ).
При уоилении антропогенной нагрузки активность ГОТ и ГПТ
явно возрастает (вероятно, под влиянием эвтрофизикации), а у
МДГ она снижается (Richter, Klausnitzer, 1980а, 1980b) (рис. 93,
94). Какие же ноксы влияют на МДГ? При изучении действия
различных веществ на активность трех названных ферментов in
vitro показано, что наиболее эффективным ингибитором
является SO2. Уже в концентрации порядка 1 моль/л SO2 ведет к
50Моному подавлению активности фермента (рис.95).
WÖ
50 h
A.S.
A.f.
Mm.
Г Р Л Э
Г И К Р Л 3
Г К Р Л
Рис. 93. Относительная удельная активность МДГ у тлей в местообитаниях
с различной антропогенной нагрузкой (возрастает от Г к Э): Г — Гримма;
М —Мелькау; К—Кульквиц; Р —Рета; Л — Лейпциг-Центр; Э —Эспенхайн
(по Richter, Klausnitzer, 1980а). A.s.—Aphis sambuci, A. f.— Aphis fabae,
M. a. — Macrosiphoniella artemisiae
192
8. Реакции животных на своеобразие городских условий
Ингйбирование МДГ и других NAD-зависимых ферментов
502(50з^~) доказано и для других организмов (Dohmen et al.,
~та84; Pfleiderer et al., 1956; Raabe, 1977; Schlee, 1977; Ziegler,
1973 и др.).
Рис. 94. Пункты измерения
активности МДГ у тли Aphis
sambuci во внутригородских
районах Лейпцига. Черный
кружок — 100% ингибирования (по
Richter, 1982)
I
Е
§
S
J
Ингйбирование 50 %
1 1 1 1 1
0,2
0,6
ммоль/л
Рис. 95. Ингйбирование МДГ у тли Aphis sambuci сульфидом (S^-),
сульфитом (SOa^-) и ртутью (Hg2+) (по Richter, Klausnitzer, 1980а)
Общему воздействию SO2, приводящему в числе прочего к
снижению фертильности (я соответственно рождаемости), несо-
М1ненно, подвержены многие биохимичеокие компоненты. Инакти-
вируются, например, такие белки с дисульфидными связями, как
каталазы и перокоидазы (Ziegler, 1973, 1975), может
непосредственно изменяться рН клетки.
8. Реакции животных на своеобразие городских условий
193
Примечательно повышение активности МДГ ,и фе-ртильности
при низких концентраЦ|Иях^уль4ш.т-1ИОН-9§ (Р'^^^- Возможно
совместное действие инг»бируюш,его и выравнивающего 1механиз-
мов для МДГ Aphis fabae (рис. 97). В последнем случае не
исключена индукция, поскольку химическое строение фермента
остается неизменным. Большое значение может иметь воздействие
Рис. 96. Корре.пяция
между фертильностью и
активностью МДГ после
воздействия различных
концентраций сульфит-
ионов на шесть
поколений тли Aphis fabae
(в контроле — 100%) (по
Richter,
1980b)
Klausnitzer,
Вторичная реакция
^•^§Ь1равни8ания (индукция)
^^^■-^^.^озраотаюш,ая
концентрация
" ~ ~ ^ ноксы
Прямое первичное ^
Фактическая удельная
активность
ингиаированое
фермента
Рис. 97. Гипотетический механизм влияния ноксы на активность фермента,
предложенный на основе экспериментальных данных -по сульфиту (SO2)
и МДГ (по Richter, Klausnitzer, 1980b)
SO2 через .растения. Как нитратная, так и в первом поколении
сульфитная их обработка почти всегда ведут к повышению фер-
тильности тлей. Это, вероятно, обусловлено среди прочего
повышением доступности питания при повреждении растений
(увеличение уровня свободных аминокислот). Установлено, что
содержание азота во флоэмном соке является лимитирующим
фактором питания тлей. Возможно, как повреждение (растений, так
194
8. Реакции животных на своеобразие городских условий
И улучшенное снабжение их азотом (эвтрофизация) ведут
прежде всего к общей активизации Обмена веществ у тлей. Прямое
ингйбирование его у насекомых, по-видимому, наступает
несколько позже, чем у растения, и этим мог бы объясняться сдвиг
максимума фертильности к более низким концентрациям сульфитов.
И напротив, нитратная подкормка, повышающая доступность
азота для растений, а вследствие этого и для тлей приводит к
более длительному устойчивому повыщению фертильности
последних.
-^•г-
Выравнивающая
активация
- [Aijph?
—Интенсивность
Зьтшя
Различные
механизмы
ингиаирования
Рис. 98. Гипотетический
механизм регуляции
дыхательной активности в условиях
ртутного загрязнения (по
Richter, Borneleit, 1984)
Бремя, МЕд.
Рис. 99. Зависимость индекса рождаемости от времени и относительной
концентрации ионов Cu2+ (по Walther et al., 1984а)
8. Реакции животных на своеобразие городских условий
195
Таблица 71. Морфологические реакции животных на своеобразие городских
местообитаний (по Klausnitzer, 1985)
Вид/таксономическая
группа
Реакция
Авторы
Aphis sambuci (тли)
Psyllipsocus ramburi
(сеноеды)
Охута parietina
(двукрылые)
Aphis fabae (тли)
Жужелицы
Чешуекрылые
Adalia bipunctaia (жуки)
Bombas hypnorum. Bom-
bus pratorum
(перепончатокрылые)
Cepaea hortensis
(моллюски)
Neottiura bimaculata
(пауки)
Orchesella villosa
(ногохвостки)
Drepanosiphum platanoi-
des (тли)
Птицы
Европейский еж
Сизый голубь
Размер тела, например,
длина сифона
Микроптерия
Размер пупария
Структура поверхности
тела
Полиморфизм аллелей,
скульптура покровов
Меланизм:
Меланизм
Меланизм
Типы полосатости
Окраска
Окраска
Окраска
Альбинизм
Альбинизм
Изменчивость, окраска
Richter et al., 1980
Weidner, 1969
Klausnitzer, Jacob, 1982
Klausnitzer, Jacob, 1982
Steiniger, 1978 (рис. 54)
Сводка в: Klausnitzer,
1985
Сводка в: Klausnitzer,
1985
Wagner, 1971
Müller et al., 1974;
Müller, 1980; Bartholome,
1974
Schäller, Peter (в печати)
Schäller, Peter (в печати)
Dixon, 1972
Grimm, 1953; Sage, 1962;
Saemann, 1970; Mayer,
1976; Grimm, 1985
Dittrich, 1980; Hutterer,
1980
Обухова, Креславский,
1984, 1985
Показано снижение интенсивности дыхания Aphis fabae под
влиянием ртути (рис. 98) (Richter, Borneleit, 1984).
Экспериментальное воздействие ионами металлов влияет на активность
кислой фоофатазы (Walther et al., 1984а) (рис. 99) и на
рождаемость А. fabae (Walther et al., 1984b) (рис. 100).
Показаны также физиологически обусловленные реакции
обитающих на коре микроартропод и моховых сообществ клещей на
загрязнение в0!здуха в городе (Gilbert, 1970, 1971; Lebrun, Т976а,
1976b; Vanek, 1967). Панцирный клещ Humerobates rostrolamel-
latus менее устойчив к действию SO2, чем лишайники, поэтому
является более высокочувствительным биоиндикатором (Lebrun
196
8. Реакции животных на своеобразие городских условий
1 г
Рис. 100. Зависимость
индекса рождаемости (1) и
активности кислой фосфата-
зы (2) тли Aphis fabae от
действия ноксы in vivo (по
Walther et al, 1984b)
Гнездо
Рис. 101. Положение гнезда кольчатой горлицы на
поперечных перекрытиях крыши у края
платформы трамвайной станции в Берлине (по Dittberner,
Möncke, 1980)
et al., 1977, 1978), однако известны и устойчивые к ноксам виды
(Andre, 1977).
8.2. Морфология
Ранее нами уже вкратце рассматривались (Klausnitzer in
Schubert, 1985) анатомо-морфологические реакции животных на
урбанизацию, поэтому вдесь мы лишь приводим табл. 71.
8.3. Этология
Указания на изменения в поведении животных в городе
многочисленны и разнообразны, особенно для птиц (табл. 72,
рис. 101).
8. Реакции животных на своеобразие городских условий
197
Таблица 72. Примеры особенностей поведения в городе
Вид
Поведение
Авторы
Большая синица
Вяхирь
Большая синица, обык-
. новенная лазоревка,
европейский поползень
Зяблик _ 3
Черный дрозд
Большая синица, белая
трясогузка
Обыкновенный скворец,,
черный дрозд .,if
Домовый воробей,
зеленушка
Домовый воробей,
деревенская ласточка 5
Пауки
Перепончатокрылые
АпоЫит thomsoni
(точильщики), Bypres-
tis haemorrhoidalis
(златки)
Серая крыса . .;
Способность к научению
(открывание молочных
бутылок)
Уменьшение «дистанции
бегства»
Кормление с руки
Возникновение
«диалектов»
Изменение стереотипа
гнездостроения,
поведения при гнездовании
Необычные места
гнездования
Необычные места
гнездования
Необычные места
гнездования
Раннее начало
гнездования (при
искусственном освещении)
Поведение при
размножении
Изменение
гнездостроения
Способ откладки яиц
Ритмика
Thorpe, 1951
Tomialojc, 1976
Gerlach, 1942';
Klausnitzer (неопубликовано)
Sick, 1989 '
Grimm, 1953
Gnielka et äl., 1983;
Hoesch, 1940
Peitzmeier, 1940
Saemann, 1970
Schob, 1983; Semadam,
1966, 1967; Saemann,
1970
Sacher, 1983
Leclerq, 1964; Haeseler,
1972
Weidner, 1975
Tischler, 1980b
9. УРБАНИСТИЧЕСКИЕ ФАКТОРЫ
СМЕРТНОСТИ
•По-видимому, не существует специфических для города
факторов смертности животных, однако несомненно, что некоторые
из них проявляются здесь особенно сильно. Ущерб, наносимый
зоопопуляциям путем изменения местообитаний, применения
пестицидов, интродукции гемерохорных растений («эффект ловущ-
ки», токсичность пыльцы и нектара), образования островных
местообитаний, антрапогенного влияния на конкуреицию и
хищничество здесь подро'бно не рассматриваются. Некоторые из этих
вопросов освещены в других разделах (см., например, гл. 10).
9.1. Техногенные и структурно
обусловленные факторы смертности
Соответствующие факторы (табл. 73) в основном являются
феноменами XX в., и животные не успели выработать к ним
никаких специальных адап! ащий.
9.2. Уличное движение
На проезжей части гибнут преимущественно следующие виды
и группы животных:
■— брюхоногие моллюски (недостаточная подвижность,
опасность высыхания);
— дождевые черви (недостаточная подвижность, опасность
высыхания);
— различные насекомые (привлечение теплом, освещенностью
и сухостью мостовой и т. д.);
— земноводные (недостаточная подвижность, пересечение
улиц при миграциях);
— черный дрозд, воробьи (малая высота полета, поиск
корма над проезжей частью);
— мыши (слабо развитое зрение в связи с ночным образом,
жизни, ослепление фарами);
—- землеройки (слабо развитое зрение в связи с ночным
образом жизни, ослепление фарами);
— летучие мыши: поздний горный .кожан и серая ночница
(зона охоты преимущественно на обочине улиц);
9. Урбанистические факторы смертности
199
Таблица 73. Техногенные и зависимые от структуры факторы смертности
(по Gepp, 1977, с дополнениями)
Фактор
Вид воздействия
Примеры
Скашивание,
выжигание травы
Строительные и
транспортные работы
Структура построек и
материала
Свойства материалов
Засасывающие и
нагнетающие воздух
устройства
Привлечение в
неподходящие условия
Прямое уничтожение
фауны травяного
яруса и поверхности
почвы, разрушение
местообитаний, сокращение
пищевых ресурсов
Строительные
котлованы и ямы как
ловушки, перемещения
грунта и выравнивание
почвы, заполнение
прудов
Одна, покрытия из
полимерных пленок,
чердаки как ловушки;
столкновение со
стеклянными фасадами,
проволокой, оградами;
высыхание при
пересечении участков с
искусственным покрытием
Прилипание к смоле и
свежей краске,
масляной пленке и
жидкостям, приманивающее
действие известковой
пыли
Воздушные фильтры
Источники света,
оптические обманы
(жестяные крыши
привлекают водных
насекомых) , выброшенные
бутылки и
пластмассовые коробки как
ловушки
Сокращение
численности ломкой веретеницы
После заполнения пруда
весной на поверхности
воды найдено 400 000
мертвых жуков
Златоглазки, дневные
бабочки, летучие
мыши, сипухи и др.
птицы (Schmidt, 1973),
улитки, дождевые
черви, мокрицы
Пчелы, двукрылые, жуки
Один воздушный фильтр
засасывает и убивает
за год 5 млн.
насекомых, мелкоячеистая
сетка уменьшает это
число до 20 ООО
Дефектная мощная
лампа за ночь может
уничтожить 100 ООО
насекомых (ср. Teschner,
1962; Stritt, 1971;
Joost, 1985)
— европейокий еж (реакция испуга, поиск пищи на мостовой);
— домашние кощки (узкое поле зрения, ослепление фарами).
За 38 месяцев на 19 204 км проезжей части найдено 704
погибших позвоночных, т. е. на каждые 27,3 км приходится в
течение года по 1 экземпляру (за первый квартал — одна особь иа
200
9. Урбанистические факторы смертности
86,2 км, за второй квартал —на 44,6 км, за третий — на 16,6 км,
за четвертый — на 26,1 км) (Н. Blümel, R. Blümel, 1980). Для
большинства позвоночных, за исключением европейского ежа,
максимальная смертность на дорогах приходится на летние
месяцы (рис. 102). Из 53 отмеченных видов наиболее часто гибли:
европейский еж (25,1%), полевой воробей (10,5%), домовый
воробей (9,9%), ваяц-русак (9,9%) и черный дрозд (4,8%).
Домовый воробей составляет 50% всех погибших позвоночных на
австрийских дорогах (Gepp, 1977). Изучены потери земноводных
(Arnold, 1982; Beebee, 1973; Heusser, 1973, 1974) и птиц (Haas,
1964; Lüpke, 1970). Согласно одному из проведенных
исследований, 49,4% всех погибших позвоночных составляли амфибии,
прежде всего обыкновенная жаба (Arnold, 1982). Булыжная
I ЕШШУИШШШХШШ
Месяц
Рис, 102. Число (п) ежемесячно погибающих позвоночных животных на
1000-километровом участке дороги (по Н. Blümel, R. Blümel, 1980): П —общее
число позвоночных; Пт — птицы; Е — ежи
мостовая и выбоины на дорогах уменьшают гибель амфибий, в то
время как влажный от дождя, теплый асфальт обусловливает
максимальную смертность.
В большом числе работ показано, что жертвами уличного
движения становятся прежде всего ежи (Berthoud, 1980, 1981;
Davies, 1957; Esser, Reichholf, 1980; Esser, 1984; Goeransson et
al., 1976; Hansen, 1969; Heinrich, 1978; Hodson, 1966; Knierer,
1967; Massey, 1972; Reichholf, 1983; Reichholf, Esser, 1981; Rettig,
1965; Schoenemann, 1977). Главный максимум их гибели
приходится на лето, меньший — на осень (рис. 103). Летний максимум
обусловлен прежде всего активностью самцов в период
спаривания, осенний — повышенной активностью, связанной с
накоплением на зиму жировых запасов, увеличением плотности
популяции и поиском мест зимовки. В населенных пунктах на дорогах
погибает больше ежей, чем в других местообитаниях, а в мелких
поселках — больше, чем е крупных городах, где такая смертность
9. Урбанистические факторы смертности
20!
Таблица 74. Вероятность гибели жужелиц при пересечении улицы
(по Mader, 1979)
Скорость пересечения
улицы
100% гибели при
АЬах ater (опыт)
Molops piceus
Pterostichus niger
6,6 см/с
3,9 см/с (по Thiele, 1977)
16,1 см/с (по Thiele,
1977)
792 автомобиля в час
468 автомобилей в час
1932 автомобиля в час
особенно велика на окраинах. В мелких поселках за год
подвергается наездам до 5 ежей на 1 км, вследствие чего их
популяции могут з'начительно сокращаться (5—20% особей погибает на*
дорогах) (Esser, 1984). На 75-километровом участке автострады
Мюнхен — Ингольштадт в мае — июне 1957 г. было задавлено
90 ежей (Grzimek, 1979). На 1000 км шоссейных дорог в Дании
в течение года находят 9345 погибших ежей. В ШлезвиГ'—Голь-,
штейне (ФРГ) как минимум 24 ООО ежей в год гибнет под
колесами транспорта (König, 1965).
30 [-
Рис. 103. Распределение по
месяцам ежей, раздавленных на
шоссе Мюнхен — Пассау в
течение года (по Reichholf, Esser,
1981)
20 h
Wh
Опыты, поставленные для изучения способности лесных видов*
жужелиц пересекать дороги, показали, что при определенных
положениях проезжей части, интенсивности движения, ширине
улицы и т. д. 'этих жуков ожидает 100%-пая вероятность наезда
(Mader, 1979), зависящая только от скорости передвижения
жужелиц и транапортной нагрузки, но не от ширины проезжей части
(табл. 74). Как показывают опыты с мечением и повторным
отловом жуков, многие виды, по-видимому, не пересекают улиц.
Доля успешных пересечений у лесных видов составила 2,3%, у
обитателей обочин — 22,4% (Mader, 1979).
В Австрии в течение года 14 триллионов насекомых
становятся жертвами уличного движения (Gepp, 1973, 1977). До 3000
насекомых на 1 км разбивается о ветровое стекло каждой
легковой машины. В зеленых насаждениях на 1 км пути за одно
воонресенье затаптывается до 1600 муравьев.
14—21
10. возможности помощи животным
в ГОРОДЕ
Помощь животным, присутствие которых желательно в
городе, маправлена прежде всего па охрану подходящих биотопов и
уход за ними (например, за сохранившимися лесными участками,
-старыми парками, пустырями). Выделение охраняемых
территорий в самом городе вполне возможно.
Зеленые массивы города выполняют множество более или
менее перекрывающихся функций:
— главная составная часть расчленяющей город системы
свободных пространств, расширение непроезжей части улиц;
— обеспечение отдыха людей (в зависимости от величины и
положения массива). Зеленые насаждения выполняют помимо
природной и собственные функции (обработка садовых участков,
использование растений);
— удовлетворение эстетических запросов путем
формирования цельных ландшафтов с участием в них растений как
элементов дизайна;
— сохранение и возобновление разнообразия флоры и
фауны, обеапечивающие прежде всего:
• стабильность (почва как дезактивирующая ноксы система,
противодействие размножению вредителей, сокращение затрат
по уходу за средой);
• охрану видов (многообразие дикой флоры и фауны, вплоть
до создания специальных заповедников);
• банки генов (например, сохранение редких видов прямо в
городах);
■ возможности исследования и народного образования;
• повышение эффективности отдыха;
• этические основы общежития;
— улучшение городского климата;
— дезактивация нокс и защита от загрязнения и шумов.
Среди этого многообразия функций следует выделить
создание благоприятных условий для животных, поддерживающее
разнообразие фауны и повышающее качество отдыха за счет
возможности зоологических наблюдений; кроме того, животные
повышают стабильность городских экосистем.
Величину городских островных местообитаний (зеленых
насаждений) нельзя проиввольно уменьшать, если требуется
одновременное выполнение всех перечисленных функций. Критериями
10. Возможности помойки животным в городе
20S
здесь могли бы служить прежде -всего динамическое равновесие
видов, их [Многообразие, структура доминирования и разделение
города на зоны (см. 5.1—5.6)^^лагоприятное влияние на
городской климат, роль фильтра против нокс и шумов также зависят
от пдощади насаждений. Нижний ее предел нельзя указать
точно, поскольку он связан с местоположением зеленого массива, его*
формой и структурой (характером растительности, почвенного-
профиля, доли водного пространства и др.). Реальный оптимум
должен составлять порядка 5 га (естественно, и меньшие
площади играют положительную экологическую роль).
С экологической точш зрения наиболее выгодны зеленые
массивы в форме круга, поскольку они охватывают наибольшую
площадь при наименьшем периметре (ср. Patton, 1975; Klausnitzer,.
1983с). Сравнительно благоприятны также квадраты и
прямоугольники с отношением сторон 1 : 2 (рис. 104).
Рис. 104. Кривые зависимости, между
площадью (А) и периметром (г) для
круга, квадрата, прямоугольника
(аХ2а) и равностороннего
треугольника (по Klausnitzer, 1983с)
Для оценки оптимального соотношения между площадью и
периметром можно применять особый индекс (I). В случае
круга как наиболее экологически выгодной геометрической формы
этот индекс по формуле (32) приравнивают к единице. При
любых других очертаниях значение индекса больше (чем
неблагоприятнее соотношение — тем больше). В случае насаждений с
очень изрезанным краем или имеющих форму полос он может
доходить до 2,0—4,8 (разумеется, бывают и большие его значения)..
Этот индекс можно выразить в процентах (формула 33),
показывающих, насколько периметр данной фигуры больше
периметра круга той же площади:
^ (32)
^ ' 2/А-я '
L,= (1-1)-100,
(33)
где U — периметр;
14*
504 10. Возможности помощи животным в городе
' А — площадь.
/ Для ослабления островного эффекта могут быть
предприняты следующие меры:
— предотвращение дальнейшего раздробления и измельчения
остро1вных местообитаний;
— сохранение вблизи города центральных зон естественных
биотопов (зеленый пояс);
С/ —/создание сети из отдельных «островов» и ее соединение с
центральными зона.ми крупной площади с помощью оплошных
«экологических коридоров» (например, аллей, живых изгородей)
и «перевалочных пунктов», которые обеспечивают постепенное
распространение животных из слишком удаленных от прочих в
противном случае исходных биотопов (мелких садов,
изолированных зеленых массивов и т. д.) (Mader, 1980, 1981а; Forman,
1983; Sukopp, Weiler, 1984)_/
Древесные насаждения должны удовлетворять следующим
параметрам:
— предпочтение местным видам с соответствующей
экологией;
— при закладке относительно протяженных насаждений —
обеспечение оптимальной ярусности; кроме того, важно
создание труднопроходимых участков;
—■ сохранение старых деревьев; из соображений безопасности
для людей следует удалять в первую очередь только склонные к
обламыванию ветви. Гнилые пни вследствие богатой
заселенности животными имеют важное экологическое значение.
Свободные от деревьев биотопы должны:
— предпочтительно создаваться в виде лугов с
дикорастущими цветами;
— скащиваться с разной интенсивностью;
— в особых случаях поддерживаться в состоянии, близком
к естественному или «предоставляться самим себе» (например,
свободные от растительности участки, площади с изреженной
растительностью, оползневые зоны).
Кроме того, на них:
— многообразие видав должно повышаться путем
целенаправленного разведения и устранения препятствий для пассивного
распростр анения;
— цветы для посадки следует выбирать не только с точки
зрения их декоративности, но и по экологическим показателям
с обеспечением максимальной непрерывности цветения видов,
дающих пыльцу и нектар, в течение вегетационного периода.
— необходимо создание благоприятных условий для
душистых растений;
— не следует рассматривать местные дикорастущие виды как
нежелательные.
10. Возможности помощи животным в городе 205
и
Озеленение зданий дает не только эстетический эффект, но
и заметно улучшает баланс излучения и энергии, что
предотвращает резкие порывы ветра. Нельзя недооценивать также роль
озеленения в связывании пыли. Для озеленения построек
пригодны прежде всего девичий виноград, плющ обыкновенный и
Fallopia aubertii. Следует обращать особое внимание на
озеленение зданий в районах новостроек, поскольку оно устраняет
характерный в этих зонах недостаток мест для гнездования птиц.
10.1. Млекопитающие
Некоторые виды зверей в городе должны охраняться
человеком. Это касается прежде всего летучих мышей, которые,
несмотря на все усилия пропаганды, не пользуются особой
любовью населения, хотя в их иаключительной полезности нет
никакого сомнения. Переходящее в синантропию предпочтение
некоторыми видами рукокрылых городских условий требует
преимущественной охраны этой группы животных именно в городах.
Охрана других видов, например ежей и белок, имеет у
горожан более благоприятную эмоциональную основу. То, что
следует охранять и каменных куниц как истребителей крыс и
голубей, воспринимается частью населения пока еще с трудом,
поскольку этим хищникам «инкриминируют» и перегрызенный
телевизионный кабель, и пропавшую курицу. Большинство
других живущих в городе млекопитающих либо лишь изредка
попадается на глаза (например, землеройки) и поэтому
охраняется фактически неосознаино (прежде всего за счет охраны их ме-
сгообитаний), либо является вредителями и преследуется (серые
крысы, домовые мыши, дикие кролики).
Гибель европейского ежа происходит главным образом на
дорогах (см. 9.2) и, кроме того, в результате сжигания куч
опавшей листвы, хвороста и сорной травы во многих садах поздней
осенью. Для охраны ежей можно предложить следующие меры:
— создание мест зимовки (сохраняющиеся в течение многих
лет кучи Х1вороста, водонепроницаемые деревянные ящики);
— размещение по обочинам дорог специальных ограждений
(в мелких населенных пунктах, по окраинам более крупных
поселений), за счет чего уровень смертности ежей можно снизить
на 80%;
— перенос куч опавших листьев в день намечаемого
сжигания в другое место сада;
— молодые ежи, не достигшие к осени (к концу октября)
массы тела 500—600 г (до 750 г), должны зимовать под
присмотром человека, так как на свободе они едва ли переживут
зиму. Этих животных следует держать при комнатной темпера-
206 10, Возможности помощи животным в городе
туре (около +20 °С) и хорошо кормить (вода! мясо, вареные
яйца, сырные корки, мучные и дождевые черви, слизни, мягкие
фрукты, витамины и минеральные соли) ср. Poduschka, 1980-,.
Lau, 1977). Следует избегать перекармливания (взрослый еж
весит около 1000 г). Размещать ежей нужно в картонных
коробках или деревянных ящиках, набитых листвой, сеном или мятой
газетной бумагой, что обеспечивает устройство убежища.
Убежище необходимо чистить. Весной (в конце апреля) ежей
следует выпустить на волю как можно дальше от улиц с
оживленным движением.
Если выпущенные ежи не приступают к размножению (этот
вопрос, впрочем, почти не исследован), они скорее препятствуют
развитию популяции, чем способствуют ему. Последнее
обстоятельство ведет к мнению, что обеспечивать зимовку ежей под
присмотром человека бессмысленно.
Важнейшее мероприятие — создание или сохранение крупных
местообитаний ежей, которые могут находиться в
непосредственной близости от человека. Сюда же относится повторное
заселение подходящих площадей, где ежи исчезли вследствие
островного эффекта, а также создание между «островами»
перевалочных пунктов».
По охране рукокрылых существует обширная литература
(Issl, Issl, 1955; Roer, 1968, 1971, 1979; Stratmann, 1971; Klawit-
ter, 1976; Blab, 1980; Heise, 1980; Höhne, 1981; Stutz, Haffner,
1984).
Важнейшие факторы, создающие угрозу для летучих мышей
в городе, следующие:
— исчезновение подходящих убежищ (особенно мест
зимовок и дневного отдыха):
• коренное изменение типа построек (исчезновение теплых
и темных легкодоступных чердачных помещений);
• исчезновение старых построек;
• перестройка и обновление зданий, заделывание летных
отверстий при санобработке и ремонте крыш;
• меры, принимаемые против одичавших домашних голубей
и действенные также в отношении рукокрылых;
• и без того редкие в городах стены из приро.дного камня с
широкимн щелями все больше вытесняются бесщелевыми
бетонными стенами (строительный бетон сам по себе летучими
мышами не избегается) (Deegen, 1983);
• вырубка дуплистых деревьев;
— отравление химической пропиткой и консервантами
кровли (Stebbings, 1971; Gottschalk, 1972; Schmidt, 1972);
— беспокойство;
— целенаправленное преследование и уничтожение;
— ограниченность пищевых ресурсов.
10. Возможности помощи животным в городе 207
10.2. Птицы
Плотность гнездования и видовое разнообразие птиц зависят
от: наличия мест гнездования, питания, меж- и внутривидовой
конкуренции. Целью мероприятий, проводимых прежде всего в
районах старой « новой застройки, является снижение «суммы
видовых потерь» до ее уровня в «зеленом городе». Важно
постоянно увеличивать количество мест гнездования (скворечники и
т. п.). В районах новостроек стоит проблема нех1ватки таких мест
для видов, гнездящихся на зданиях. Мендель (Menzel, 1984)
отмечает необходимость специального их обеспечения здесь для
деревенской ласточки (ср. табл. 75). Болотная камышевка и
серая славка могут гнездиться в зарослях крапивы. Следует
также охранять места ночевок птиц (деревья).
В то время как некоторые виды находят в городе избыток
корма (проблема голубей: Reinke, 1959; Berndt, 1970; Sixl, 1975;
Haag, 1985) и способны хорошо использовать специфические
городские пищевые ресурсы (сизый голубь, домовый воробей), для
других видов подходящая пища (по крайней мере периодически)
отсутствует. Кормушки компенсируют ее нехватку только в зим-
Особое внимание следует уделить таким возможным мерам
помощи летучим мышам в городах (с учетом видоспецифичных
потребностей), как:
— обеспечение убежищами:
• сохранение дуплистных деревьев в парках, аллеях, садах
и на кладбищах (проведение с ними оздоровительных
хирургических мероприятий);
• создание отверстий для проникновения на чердаки и в
другие пустоты зданий, например под обшивку фронтонов;
• навесы для летучих мышей на южных стенах вместо жалю-
зей и ставней;
' - устройство убежищ под коньками крыш;
• размещение ящиков для сна и размножения по
возможности небольшими группами (по 5—10 штук);
• специальные черепичные кровли, неудобные для голубей,
с отверстиями для влета летучих мышей;
• полые каменные блоки в подвалах;
— меньшее использование я1дов на крышах;
— забота об убежищах (с участием владельцев или
пользователей построек);
— уменьшение воздействия человека (санитарные обработки
и ремонт построек по возможности не во время спячки мышей,
продолжающейся с октября по март);
— разъяснительная работа.
208 10. Возможности помощи животным в городе
Горец птичий
Переступень двудомный
Пастушья сумка
Просвирник
Роза собачья
Малина, ежевика
Ослинник
Плющ
Пастернак посевной
Подмаренник цепкий
Бузина черная
Повой заборный
Дереза
Паслен черный
Льнянка обыкновенная
Подорожник
Лопух
Бодяк полевой
Латук дикий
Скерда зеленая
Костер бесплодный
Ежа сборная
нее время. Резерв для таких птиц составляют прежде всего
семена и плоды местных дикорастущих растений (табл. 76).
Так, например, ягоды черной бузины служат пищей кольчатой
горлице, лазоревке, дрозду-белобровику, певчему и черному
дроздам, садовой горихвостке, зарянке, славке-черноголовке, садовой
славке, славке-завирущке, мухоловке-пеструшке, зяблику.'Л-Ла-
честве примеров интродуцированных видов растений с
пригодными для питания птиц плодами можно назвать кизильник
(певчий и черный дрозды, зарянка, зеленушка) и яблоню Malus flo-
rlbunda (дрозд-белобровик, черный дрозд, славка-черноголовка,
зеленушка, снегирь, зяблик). Газоны имеют большое значение
для 'питания прежде всего таких видов, как черный и певчий
дрозды, галка, грач, скворец и зеленый дятел. Снижение доли
открытого грунта ведет к нехватке гнездового материала для
городской и деревенской ласточек.
10.3. Пресмыкающиеся
Рептилии, особенно ящерицы, могут быть сохранены в городе
(или поселены в нем) прежде всего при условии создания и со-
Таблица 75. Особенности зеленььх насаждений в районах новостроек,
облегчающие гнездование птиц (по Plath, 1981)
Растения, которые могут использоваться как места гнездования:
— хвойные деревья, клен, липа, рябина, рододендрон, можжевельник, тис
— кустарник высотой 2—3 м
— кустарники, расположенные в несколько рядов
— оставшиеся нетронутыми (старые) деревья и кустарники
Растения, которые практически не используются для гнездования:
— сирень, форзиция, тополя
■— кустарники высотой не более 1 м
— одиночные ряды кустарников
— кустарники в непосредственной близости от жилых домов
Таблица 76. Местные дикие растения в городе как пища для птиц (плоды
и семена)
10. Возможности помощи животным в городе
209
Таблица 77. Земноводные, отловленные 6—31 марта 1981 г. вдоль
250-метрового заграждения по краю улицы в Нидерлаузице (по Donat, 1982)
Вид
Число особей
В процентах
Обыкновенная жаба
Обыкновенный тритон
Травяная лягушка
Гребенчатый тритон
Прудовая лягушка
Остромордая лягушка
Обыкновенная чесночница
Обыкновенная квакша
2253
228
162
40
29
27
6
4
82,0
8,3
5,9
1,5
1,1
1,0
0,2
0,1
Всего 8 видов
2749
1
хранения благоприятных местообитаний (подходящие части
зеленых насаждений, старые каменные стены, альпийские горки).
10.4. Земноводные
Проведены многочисленные исследования (Heusser, 1968,
1970; Thielcke, 1975; Loos, 1980; Haenschke, 1982; Bregulla, 1983;
Kral et al., 1983; Loske, 1983; Steinberg, 1983; Griffiths, 1984; Kle-
wen et al., 1984; Thiesmeier, 1984), посвященные устройству и
оздоровлению в городах мелких водоемов, слун<ащих
местообитанием амфибий, прежде всего обыкновенного и гребенчатого
тритонов, чесночницы, обыкновенной и зеленой жаб, прудовой
лягушки. Важно при этом обеспечить поих берегам укрытия и
убежища. В ряде случаев удается переселение популяции
земноводных. Это не относится к видам с большим радиусом
активности (обыкновенная жаба, травяная лягушка), которые в массе
гибнут на улицах. Некоторый выход из положения составляет
способность головастиков к «запечатлению» особенностей
родного водоема. Обыкновенную жабу следует особенно внимательно
охранять от транспорта во время ее передвижения к местам
икрометания. Для этого сооружают заграждения, подземные
переходы и переносят жийбтных в подходящие биотопы (Bauer,
f977;üonat, 1982; Heusser, 1967, 1974; Jakobs, 1983) (табл. 77).
10.5. Моллюски
Содействие желательным в городе брюхоногим моллюскам,
прежде всего раковинным, заключается в защите^их
местообитаний (участков почвы). Наряду с этим должны специально
охраняться небольшие существующие в городах колонии.
210 10. Возможности помощи животным в городе
10.6. Длинноусые прямокрылые
Сохранению этих насекомых способствуют древесные и
кустарниковые насаждения с выраженной ярусностью. Важна связь
между городскими зелеными массивами и пригородными лесами,
так как многие виды нуждаются в пополнении поиуляций извне
(Martens, Gillandt, 1985).
10.7. Короткоусые прямокрылые
Эти насекомые встречаются прежде всего на редко (дважды
в год) скашиваемых газонах. «Кобылковые лужайки» можно
заложить в городе в теплых местах даже с нарушенной почвой.
Отдельные нескашиваемые участки должны быть оставлены в
качестве рефугий. Сохранение пустырей в черте города
обеспечивает существование большего числа видов (В. Klausnitzer,
и. Klausnitzer, 1982; Martens, Gillandt, 1985).
10.8. Перепончатокрылые
Создать благоприятные условия для этих насекомых в
городе можно путем устройства «гнездовых ящиков» (Friese, 1926;
Krombein, 1967; Peters, 1973; Preuss, 1980; Rühl, 1977).
Некоторые перепончатокрылые заселяют полости и ходы в древесине.
В садах и на балконах можно сделать, особенно в летнее время,
следующее:
— устроить «норки» диаметром 1—10 мм и длиной 5—10 см,
причем ходы различной величины должны располагаться
поблизости друг от друга;
— поместить на деревья, стены домов, заборы куски
расщепленных надвое буковых стволов длиной 50 см и диаметром
около 20 см с просверленными отверстиями (диаметром 2—10 см).
Размеры таких убежищ достаточны для обеспечения их
прочности и терморегуляции (исключение летнего перегрева);
—■ предоставить насеко.мым соломины или стебли с мягкой
сердцевиной (ежевика, малина, роза, бузина) длиной 10—15 см,
а также пластмассовые трубочки для коктейлей. Более прочные
стебли можно связать вместе, а соломины тугим пучком
вставить в консервную банку или же воткнуть одним концом в
гипсовый раствор.
Разработаны гнездовые ящики для шершней (\7. Hagen, 1982;
Müller, 1984); можно также изготовить искусственные гнезда для
шмелей (Preuss, 1980). Кроме того, необходимо сохранение
типичных мест гнездования перепончатокрылых, к которым
относятся глиняные стены, швы в 'каменных стенах, деревянные сараи,
тростниковые и соломенные крыши, газоны, необлицованные бе-
10. Возможности помощи животным в городе
211
Таблица 78. Виды, цветки которых предпочитают шмели (по Wagner, 1971)
Роза морщинистая
Бирючина обыкновенная
Клевер луговой
Горошек заборный
Рододендрон
Крыжовник
обыкновенный
Малина
ЯиЬив fruticosus
Клевер ползучий
Горошек мышиный
Горошек посевной
Bombus terrestris (L.), В. hypnorum
В. lucorum (L.)
В. lapidarius (L.), В. hortorum (L.), B.
distinguendus Morowitz, B. agrorum (F.), В. equestris (F.)
В. lapidarius (L.)
В. pratorum
В. agrorum (F.)
В. agrorum (F.), В. equestris (F.), В ruderarius
(Müller)
В. agrorum (F.)
реговые откосы, бревна, старые деревья (срубленные или упав-
шйе~стволы не уничтожаются, а лежат на месте 3—4 года). В
современных садах традиционные деревянные беседки все больше
заменяются бетонными, а деревянные ограды — проволочными,
что снижает выбор мест гнездования перепончатокрылых. Охра-
да колоний рациональнее, чем их создание (Blab, 1980).
Очень важно для перепончатокрылых наличие даюших нектар
и пыльцу цветков, прежде всего местных дикорастущих видов
(ясиостка, пикульник, клевер, саротамнус, бодяк). Нельзя
допускать возникновения «безнектарных» мест (табл. 78).
Следует также уделять внимание разведению растений,
связанных с медвяной росой (тли находят в городе весьма
благоприятные условия существования, что обусловливает появление ее
в изобилии). При закладке зеленых насаждений необходимо
учитывать виды, посещаемые пчелами (Dorn, 1984). На
существовании переиончатокрылых благоприятно сказывается также
сохранение пустырей (Preuss, 1980).
10.9. Чешуекрылые
Мероприятия по охране и разведению в городе дневных
бабочек действенны прежде всего при сохранении
соответствующих биотопов. Этому способствует также охрана растений, на
которых кормятся гусеницы, и постоянное наличие цветков,
дающих пищу имаго. На одной только крапиве двудомной живут
гусеницы изменчивой пестрокрыльницы, углокрыльницы С-'белое,
крапивницы, дневного павлиньего глаза, адмирала и других
видов. Различные подорожники служат кормовьши растениями для
гусениц более 40 видов. На одуванчике лекарственном также
отмечено свыше 40 видов. 1^1 еобходимо «целенаправленное
управление цветочными лужайками» на озелененных площадях] Они
212 10. Возможности помощи животным в городе
должны скашиваться только 1—2 раза в год, причем первый
раз — не раньше середины июня./Следует отказаться от
удобрений и особенно от применения гербицидов^^^Особую роль наряду с
газонами могут играть соответствующим образом оформленные
сады и сохранившиеся лесные участки. Возможна и
целенаправленная посадка кормовых растений, среди которых не только
буддлея (ср. Owen, Whiteway, 1980), но и, например, виды
лопуха, чертополоха, бодяка, синяка, цикория, кипрея, донника. Для
бабочек в городе полезны даже мелкие, изобилующие цветами
участки. Правда, и в этом случае существует нижний предел
площади (примерно 1 га). Такой участок может состоять и из не-
'скольких небольших площадок с дикорастущими видами,
пространственно связанных друг с другом./Особое значение имеют
края полей, канав, обочины дорог и т. п.Следует указать на не-
обходимость~сохранёния и создания зкивых изгородей, оообенно
с изрезанным краем и прогибами, ориентированными в южно.м
направлении. Кроме того, на их южной стороне должны быть
созданы широкие полосы дикорастущих видов (Owen, 1976; Gepp,
1981; Blad, Kudrna, 1982). Искусственное разведение бабочек с
успехом используется для сохранения отдельных их видов (Wei-
demann, 1982).
10.10. Почвенные животные
Условия для почвенной фауны можно создать уже
оставлением в зеленых насаждениях хотя бы части опавших листьев и
ветвей. Наряду с этим должны сохраняться рефугии с постоян-
ным запасом гумуса и hml,
10.11. Пауки
Рекомендуются следующие мероприятия по охране пауков в
городе (Schaefer, 1980): как можно большая
структурированность простраяспва, частичное сохранение листового опада,
отказ от__скашивания определенных^уч'асткав газонов, устройство
и сохранение небольших прудов с плоскими берегами и
естественных раститёлыШх сообществ.
п. ЗАКЛЮЧИТЕЛЬНЫЕ ЗАМЕЧАНИЯ
В настоящей книге намеренно не выделена специальная гла-^
ва, посвященная городским экосистемам. Порядок изложения
материала способствует пониманию структурных и функциональных,
свойств таких систем, что в известном смысле дает основу для
изучения последних (ср. рис. 105). Возможно, эта книга
послужит стимулом к дальнейшему развитию экологических
исследований в городах. Интересным, на мой взгляд, было бы
сравнение трудностей, возникающих при выявлении экологических
закономерностей в урбанизированной среде (где животные
существуют в экстремальных, часто совершенно новых для них
условиях) и в сбалансированных, часто тысячелетиями
складывавшихся природных сообществах.
Описок литературы из-за недостатка места включает только
работы, цитируемые и обсуждаемые в данной книге. В моей
книге об освоении животными городов (В. Klausnitzer,
«Verstädterung von Tieren» A. Ziemsen Verlag, Wittenberg Lutherstadt,.
1987) приведена библиография по экологии городской фауны,,
насчитывающая около 2000 названий.
В процессе работы автор получал советы и критические
замечания от целого ряда коллег, друзей и сотрудников и имел
счастливую возможность обмениваться с ними мнениями по
настоящей теме. Сердечную благодарность я бы хотел выразить
госпоже Сильвии Беме (Лейпциг) и господам Г. Бальдовски
(Герлиц), Г. Дате (Берлин), М. Дорну (Галле), В. Дунгеру
(Герлиц), И. Геппу (Грац), Г. Гримму (Берлин), П. Гутте и
Л. Кемпфе (Грайфсвальд), Е. Крайслю (Грац), В. Кюнельту
(Вена), М. Луняку (Варшава), Г. Мадеру (Бонн), Е. Мею (Ру-
дольштадт), Г. Мюллер-Моцфельду (Грайфсвальд), Г. Мюллеру
(Лейпциг), Г. И. Мюллеру (йена), П. Мюллеру (Саарбрюккен),
Г. Набровски (Берлин), П. Нагелю (Саарбрюккен), Ф.-Й. Обету
(Дрезден), Г. Оше (Фрайбург), Р. Пихоцки (Галле), Л. Плату
(Рост^?к), К. Рихтеру (Лейпциг), П. Захеру (Виттенберг-Лютер-
штадт), Г. Шименцу (Радебойль), Р. Шуберту (Галле), У. Зедла-
гу (Эберовальде), Б. Штефану (Берлин), М. Штуббе (Галле),
Г. Зукоппу (Западный Берлин), В. Тишлеру (Киль), В. Топну
(Кёльн), Г. Вайднеру (Гамбург) и Г. Вуоренринне (Эспо). Я
особенно благодарен также господам М. Ферстеру (Лейпциг) и
Г. Ринхоферу (Эберсвальде) за фотографии, любезно
предоставленные в мое распоряжение.
Госпоже Йоханне Шлютер (редактору издательства «Густав
Фишер», Йена) я глубоко благодарен за понимание и
любезность. Моя жена, Герта Клауснитцер помогала мне в период
написания этой книги, и здесь я хочу воспользоваться случаем,
чтобы сердечно поблагодарить ее за это.
Негородская
зкасистет
* Окраинный
жилой
комплекс
Т Протшлеи- Т Городской
ный район жилой
комплекс
Т ,, „ ,„„„я„ ^ Городской УПромышлен-^Сварка ^Окпаиииьш ^Негородская
Центр гороаа Милой ный район жилой экосистема
^ комплекс комплекс
Климат
- Излучение (общее)
- Время солнечного сияниям
-t°C (средне годовая)
- Vsempa (средне годовая)
- ОсаВпи (оцмма)
- Снегопад
-Оти.елажность воздуха
- Центры конденсации
-Туман (зима)
-Туман (Лето)
Грунтовые воды
Почвы
-Степень застроенности
-Антропогенные
изменения
(главные)
Растительность
-Покрытие
площади.
- Видовое разнообразие
-Даля неофитов
-Лишайники ' ■
-^на 15-20% меньихе
->-на 5-15% меньше
^ на 0,6 -! °С больше
-^на 20-30% меньше,
на 5-10% больше
—^ на 5 % меньше
-^на 2-8% меньше
^ в 10 раз больше
на 100% больше
^ на 30% больше
<20% —
Перемешивание Перемешивание с -
привнасом или выносам,
частичное погребение
естественной почвы
Число растительных
сообществ
(разнообразие зкотопав)
35%
■■■-^Максимум ■*
10-15% --
тш-^Зана борьбы —
-^Максимум -4^
Глубине
100%
Антропогенное
перекрывание искусствеиныти
и естественными
субстратами, перемешивание
1%
Минимим -
-20%
Отсутствие
Минимум
Перемешивание с -
привносом или выносам,
частичное погребение
естественной почвы
-<20%
Перемешивание
^■Максимум ■
W-1S%
ш Зона борьбы <
. Максимум-^
Рис. 105. Биом промышленно-городской экосистемы (по различным авторам, из Schubert, 1984)
Фото 1. Гнездо обыкновенной кряквы в башне костела на высоте 25 м (фото;
G. Rinnhofer)
Фото 2. Городская ласточка — характерная птица новостроек (фото G.
Rinnhofer)
'Фото 3. Кормежка грачей на городском газоне осенью (фото G. Rinnhofer)
Фото 4. Стая чечеток зимой в районе новостроек (фото G. Rinnhofer)
Фото 5. Стая озерных чаек в апреле на окраинной свалке (фото G. Rinnhofer)
*алЫ1е^'(фото Q R°nnhof™r)"""'^^^' ^^""^^ " лебедей-шипунов на канале в Эбврс-
15-21
Фото 7. Стайка домовых воробьев в центре города (фото G. Rinnhofer)
Фото 8. Слизень Limax maxitnus обитает в различных городских биотопах,
в том числе в подвалах (фото G. Rinnhofer)
Фото 9. Различные виды сенокосцев обитают в основном на стенах домов; на
фотографии Leiobunum limbatum (фото G. Rinnhofer)
Фото 10. Пара пауков {Pholcus opilionoides?); самка несет яйцевой пакет. этот
вид встречается преимущественно внутри зданий (фото М. Förster)
15*
.12
Фото 11. Ночные бабочки-монашенки, отдыхающие на фонарном столбе после
массового ночного лета на свет. Лето 1982 г. (фото G. Rinnhofer)
Фото 12. Перепончатокрылое Brachygaster minuta паразитирует на оотеках
тараканов, в том числе прусака (фото М. Förster)
1*'
13
i
14
Фото 13. Жук-чернотелка Blaps mortisaga и другие виды этого рода образуют
в подвалах стабильные популяции (фото М. Förster)
Фото 14. Повреждения долгоносиком Otiorhynchus smreczynskii листьев
бирючины (фото М. Förster)
Фото 15. Клоп-хищнец редувий ряженый обитает преимущественно в зданиях
(фото М. Förster)
Фото 16. Постельный клоп (фото М. Förster)
12. ЛИТЕРАТУРА
Обухова Н. Ю., Креславский А. Г.: Изменчивость и наследование окраски
у сизых голубей. Зоол. ж. 63 (1984): 233—244.
Обухова Н. Ю., Креславский А. Г.: Изменчивость окраски в городских
популяциях сизых голубей (Columba livia). Возможные механизмы
поддержания полиморфизма. Зоол. ж. 64 (1985): 1685—1694.
Andersen, А.: Abundance and spatial distribution of house-dust mites in their
natural environment (Acari: Sarcoptiformes & Trombidiformes). Ent. iVleddr.
(Copenhagen) 52 (1984): 25—32.
Aradi, M. P., Mihalyi, P.: Seasonal investigations of flies visiting foods markets
in Budapest. Acta Zool. Hung. 17 (1971): 1—10.
Babinska-Werka, J., Gliwicz, J., Goszczynski, J.: Synurbization processes in a
population of Apodenius agrarius U. Habitats of the striped field mouse.
Acta theriol. 24 (1979): 405—415.
Babinska-Werka, J., Gliwicz, J., Goszczynski, J.: Demographic processes in an
urban population of the striped field mouse. Acta theriol. 26 (1981): 275—
283.
Banaszak, J.: Apoidea (Hymenoptera) of Warsaw and Mazovia. Memorabilia
Zool. 36 (1982): 129—142.
Bankowska, R.: Dolichopodidae (Diptera) of Warsaw and Mazovia. Memorabilia
Zooi. 35 (1982a); 33—45.
Bankowska, R.: Hover Flies (Diptera, Syrphidae) of Warsaw and Mazovia.
Memorabilia Zool. 35 (1982b): 57—78.
Barndt, D.: Liste der Laufkäferarten von Berlin (West) mit Kennzeichnung und
Auswertung der verschollenen und gefährdeten Arten (Rote Liste). Ent. Bl.
Sonderheft 77 (1981): 3—35.
Barnett, S. A., Mandly, B. M.: Breeding of mies at -3°C. Nature 173 (1954):
355—356.
Baum, H.: Biologie und Ökologie der Amselfederläuse. Angew. Parasitol. 9
(1968): 129—175.
Becker, G.: Vergleichende Untersuchungen zur Ökologie einiger Reticulotermes-
Arten (Ins., Isopt.). Z. angew. Ent. 65 (1970): 183—216.
Becker, J.: Die Carabiden des Flughafens Köln/Bonn als Bioindikatoren für die
Belastung eines anthropogenen Ökosystems. Decheniana 20 (1977): 1—9.
Bender, E.: Untersuchungen zur Biologie und Morphologie der in Weinkellern
lebenden Kleinschmetterlinge. Z. angew. Ent. 27 (1941): 543—584.
Berndt, K. P.: Das Taubenproblem und Wege zu siener Lösung. Falke 17 (1970):
370-375.
Berressem, K. G., Beressem, H., K. H. Schmidt: Vergleich der Brutbiologie von
Höhlenbrütern in innerstädtischen und stadtfernen Biotopen. J. Ornithol. 124
(4) (1983): 431-446.
Berressem, K. G., Berressem, H., K. H. Schmidt: Existenzprobleme der Vögel in
innerstädtischen Lebensräumen. J. Ornithol. 125 (3) (1984).
Billaudelle, H.: Zur iBologie der Mauerspinne Dictyna civica (H. Luc.)
(Dictynidae, Araneida). Z. angew. Ent. 41 (1957): 475—512.
Blab, ].: Grundlagen für ein Fledermaus-Hilfsprogramm. Themen der Zeit, Heft 5,
Kilda-Verlag, Greven 1980.
Bliss, P.: Zur Verbreitung von Opilio ravennae Spoek in der DDR (Arachnida,
Opiliones). Faun. Abb. Mus. Tierk. Dresden 8 (1981): 87—90.
224 12. Литература
Blümel, Н., R. Blumel: Wirbeltiere als Opfer des Strassenverkehrs. Abb. Ber.
Naturkundemus. Görlitz 54 (1980): 19—24.
Boettger, C. R.: Die Gewächshausschrecl<e [Tactiyeines asynamorus Adel.). Abb.
Braunschweig, wiss. Ges. 2 (1950): 13—39.
Boettger, C. R.: Die Trümmerfauna Braunschweigs. Ein Beitrag zur Frage der
Auswirkung brachliegender Trümmergebiete auf die städtische Gemeinschaft
des Menschen Verb. 11. Int. Kongr. Ent. Wien 1960, 3 (1962): 195—200.
Borkowslti, A.: Arealausweitungen bei einigen minierenden Lepidopteren durch
anthropogene Pflanzenverbreitung. Bull. Ent. Pologne 43 (1973): 461—
467.
Braun, S., Flücliiger, W., Oertli, J. J.: Einfluss einer Autobahn auf den Befall
von Weissdorn {Crataegus monogyna) mit Aptiis pomi. Mitt. dtsch. Ges. allg.
angew. Ent. 3 (1981): 138—139
Bregulla, D.: Zur Situation von Amphibien in Ballungsräumen, am Beispiel der
Stadt Herne im Ruhrgebiet. Salamandra, Bonn, 19 (1983): 169—172.
Britz, L.: Ober die Stechmücken-Fauna (Dipt., Culicidae) des Stadtkreises
Leipzig. Z. angew. Zool. 42 (1955): 61—79.
Britz, L.: Die Pharao-Ameise und ihre Bekämpfung. Der Drogist 4 (1957):
21—22.
Burakowski, В., Nowakowski, E.: Click beetles (Col, Elateridae) of Warsaw and
Mazovia. Memorabilia Zool. 34 (1981a); 165—180.
Burakowski, В., Nowakowski, E.: Longicorns (Col, Cerambycidae) of Warsaw
and Mazovia. Memorabilia Zool. 34 (1981b): 199—218.
Cerny, v.: Die parasitischen Arthropoden der synanthropen Taubenvögel einer
Grossstadt. Angew. Parasitol. 11 (1970): 244—248.
Cholewicka, K.: Curculionids (Col, Curculionidae) of Wazsaw and Mazovia.
Memorabilia Zool 34 (1981): 235—258.
Chudzicka, £.: Auchenorryncha (Homoptera) of Warsaw and Mazovia. Memora-
biha Zool. 36 (1982): 143—164.
Chudzicka, E., Pisarski, В., Wegner, E.: Number compensation between
associations of insect fauna living in lime crowns in urban areas. Memorabilia
Zool. 32 (1972): 79—86.
Clausen, I. H. S.: Notes on the impact of air pollution (SO2 & Pb) on spider
(Araneae) populations in North Zealand, Denmark. Ent. Meddr.
(Copenhagen) 52 (1984); 33—39.
Cmoluch, Z.: Weevils (Col, Curculionidae) of town plants associations of Lub-
hn. Pol. Pismo Entomol. 42 (1972): 545—562.
Cyr, A., Cyr, J.: Welche Merkmale der Vegetation können einen Einfluss auf
Vogelgemeinscbaften haben? Vogelwelt 100 (1979): 165—181.
Czechowska, W.: Neuroptera and Mecoptera of Warsaw and Mazovia.
Memorabilia Zool. 36 (1982): 165—184.
Czechowska, W., Bielawski, R.: Coccinellids (Col, Coccinellidae) of Warsaw and
Mazovia. Memorabilia Zool. 34 (1981): 181—197.
Czechowski, W.: Influence of the manner of managing park areas and their
situation on the formation of the communities of carabid beetles (Col,
Carabidae). Fragm. Faun. 25 (1980a): 199—219.
Czechowski, W.: Carabids (Col, Carabidae) of the Vistula escarpment 1н
Warsaw. Fragm. Faun. 25 (1980b); 293—316.
Czechowski, W.: Carabids (Col, Carabidae) of Warsaw and Mazovia.
Memorabilia Zool. 34 (1981); 119—144.
Czeckomski, W.: Occurence of carabids (Col, Carabidae) in the urban greenery
of Warsaw according to the land utilization and cultivation Memorabilia
Zool. 39 il982); 3—108.
Czechowski, W., Kubicka, A., and Starega, W.: Harvestmen (.'Vrachnoidea,
Opiliones) of Warsaw and Mazovia. Memorabilia Zool 34 (1981); 111 — 118.
Czechowski, W., Starega, W.: Nelima semproni Szalay, 1951 (Opiliones. Gagrel-
lidae) in Warszawa. Fragm. Faun. 23 (1977); 13—24.
12. Литература 225
Dathe, Н. Н.: Zur Hymenopterenfauna im Tierparlt Berlin I. Milu 2 (1969):
430—443.
Dathe, H. H.: Zur Hymenopterenfauna im Tierpark Berlin II. Milu 3 (1971):
231—241.
Dittberner, H., Möntie, R.: Zum Brutvorkommen der Türkentaube auf Bahnhöfen
der Berliner Stadtbahn. Falke 27 (1980): 185—188.
Dittrich, L.: Die Vererbung des Albinismus beim Westeuropäischen Igel, Erina-
ceus europaeus Linne, 1758. Säugetierkdl. Mitt. 28 (1980): 281—286.
Dobberliau, Т., lander, G., Otto, W.: Untersuchungen zur Siedlungsdichte der
Brutvögel Berliner Friedhöfe 1972. Beitr. Vogelkd. 25 (1979): 129—166.
Döhring, E.; Zur Häufigkeit, hygienische Bedeutung und zum Fang sozialer
Faltenwespen in einer Qrossstadt. Z. angew. Entomol. 47 (I960): 67—79.
Döhring, E.: Vorkommen und Verbreitung von Schaben in der Bundesrepublik
Deutschland. Prakt. Schädlingsbek. 24 (1972): 29—35.
Döhring E.: Über Vorkommen von Schaben in der Bundesrepublik Deutschland.
Bundesgesundheitsbl. 16 (1973): 274—283.
Donat, R.: Amphibienschutz an Strassen. Feldherpetol. Mitt. 5 (1982): 10—11.
Dorn, M.: Ergebnisse faunistisch-ökologischer Untersuchungen an solitären
Apoidea (Hymenoptera) im Botanischen Garten der Martin-Luther-Universität
in Halle (Saale). I. Teil Hercynia N. F. Leipzig 14 (1977): 196—211.^
Dorn, M.: Das urbane Requisitenangebot und seine Nutzung durch solitäre
Apoidea. Tag.-ber. 2. Leipziger Symp. urbane Ökologie (1984): 53—55.
Draber-Monko, A.: Calliphoridae Parasitica (Diptera) of Warsaw and Mazovia.
Memorabilia Zool. 35 (1982a): 123—129.
Draber-Monko, A.: Sarcophagidae and Rhinophoridae (Diptera) of Warsaw and
Mazovia. Memorabilia Zool. 35 (1982b): 131-140.
Draber-Monko, A.: Tachinid Flies (Diptera, Tachinidae) of Warsaw and Mazovia.
Memorabilia Zool. 35 (1982c): 141—162.
Duffey, E.: Aerial dispersal in a known spider population. J. Anim. Ecol. 25
(1956): 85—111.
Durska, E.: Phoridae (Diptera) of Warsaw. Memorabilia Zool. 35 (1982): 47—
56.
Ebeling, W.: Urban entomology. Univ. Calif. Div. Agric. Sei., Berkeley,
California 1975.
Eggers, J.: Zur Siedlungsdichte der Hamburger Vogelwelt. Hamb. Avifaun. Beitr.
13 (1975): 13—73.
Eggers, ].: Zur Vogelwelt einer grossstädtischen Brachfläche (City Nord,
Hamburg). Hamb. Avifaun. Beitr. 14 (1976): 47—53.
Eichler, W.-D.: Die Tierwelt der Gewächshäuser. Leipzig 1952.
Eichler, W.-D.: Synanthrope Aspekte zur Ökologie der Pharaoameise. Dtsch. Ent.
Z. 20 (1973): 425—432.
Ellenberg, H.: Zur Erfassung und Bewertung einer ländlichen Lokal-Avifauna.
Ein Beispiel aus dem Saarland mit Bemerkungen zur Methodik. Faun.-flor.
Notizen Saarland 14 (1982): 111—126.
Elvers, H.: Die Brutvögel des Waldfriedhofes Heerstrasse 1974. Orn. Ber. f.
Berlin (West) 2 (1977): 139—150.
Elvers, H.: Die Brutvögel in den Grünanlagen von Berlin (West). Sitzungsber.
Ges. Naturforsch. Freunde zu Berlin (N. F.) 20/21 (1981): 107—124.
Elvers, H., Elvers, K.-L.: Verbreitung und Ökologie der Waldmaus (Apodemus
sylvaticus L.) in Berlin (West). Zool. Beitr. N. F. 28 (1984): 403—415.
Engelbrecht, H.: Taenia saginata, eine entdeckungsgeschichtliche und
ökologischepidemiologische Betrachtung. Angew. Parasitol. 25 (1984): 17—26.
Engelbrecht, H., Buske, M.: Zum Vorkommen des tropischen Schimmelplattkäfers
Ahasverus advena (Waltl, 1832) (CoL, Silvanidae) in modernen Wohnbau-
ten. Z. ges. Hyg. 28 (1982): 112—114.
Engelbrecht, H., Buske, M.: Ein Modell der komplexen Schaben-Grossraumbe-
^ kämpfung. II. Vorkommen und Verbreitung von Blattella germanica und
226 ]2. Литература
Blatta orientalis im Bezirk Potsdam (DDR)—eine Befallsanalyse. Angew
Parasitol. 24 (1983): 27—39.
Erdelen, M.: Zur Diversität von Vogelgemeinschaften. Charadrius 13 (1977):
1—7.
Eriksen, W.: Die städtische Wärmeinsel. Neuere Erkenntnisse zur Gliederung,
Genese und Bedeutung des innerstädtischen Temperaturfeldes. Geogr. Rdsch.
(Braunschweig) 9 (1976): 368—373.
Erz, W.: Populationsökologische Untersuuchungen an der Avifauna zweier
norddeutscher Grossstädte. Z. Wiss. Zool. 170 (1964): 1—111.
Erz, W.: Ecological principles in the urbanization of birds. Ostrich Suppl. 6
(1966): 357—363.
Erz, W.: Verstädterung unserer Vogelwelt. Umschau (Frankf.) 67, 85—88.
Esser, J.: Untersuchung zur Frage der Bestandsfährdung des Igels {Erinaceus
europaeus) in Bayern. Ber. ANL 8 (1984): 22—62.
Evers, A. M. ].: Faktoren zur Verbreitung kleiner Organismen. Jber. Naturwiss.
Ver. Wuppertal, 30. H. (1977): 116—122.
Faeth, H., Kane, Th. C: Urban biogeography. City parks as island for Diptera
and Coleoptera. Oecologia 32 (1978): 127—133.
forman, R. T. Т.: Corridors in a Landscape: Their ecological structure and
function. Ekologia (CSSR) 2 (1983): 375—387.
Forman, R. T. Т., Baudry, J.: Hedgerows and hedgerow networks in landscape
ecology. Environmental iHanagement 8 (1984): 495—510.
Frankenhuyzen, A. van: Olethreutes sideriana Treitschke, 1835 (Lep.,
Tortricidae). Ent. Berichten 41 (1981): 117—119.
Frankenhuyzen, A. van: Phyllonorycter platani (Staudinger, 1870) (Lep.: Gracil-
lariidae), een bladmineerder op Plataan in Nederland. Ent. Berichten 43
(1983): 19—25.
Frankie, G. W., Ehler, L. E.: Ecology of insect in urban environments. Ann. Rev.
Entomol. 23 (1978): 367—387.
Frankie, G. W., Koehler, C. S.: Perspectives in Urban Entomology. Academic
Press, New'York, San Francisco, London 1978.
Garanin W. !.: Die Urbanisation und die Herpetofauna. Vertebrata Hung. 21
(1982): 141 — 145.
Garbarczyk H : Proctotrupoidea (Hymenoptera) of Warsaw and JVlazovia. JVlemo-
rabilia Zool. 36 (1982): 41—60.
Gavareski, C. A.: Relation of park size and vegetation to urban bird population
in Seattle. Condor 18 (1976): 375—882.
Geinitz, Ch.: Beiträge zur Biologie des Streifenhörnchens (Eutamias Sibiriens
Laxmann, 1769) auf einem Friedhof in Freiburg (Süddeutschland). Z. f.
Säugetierkde. 45 (1980); 279-287.
Gepp, J.: Kraftfahrzeugverkehr und fliegende Insekten. Natur und Landschaft 59
(1973): 127—129.
Gepp, J.: Die Neuropteren von Graz: Ein Beitrag zur Kenntnis der
mitteleuropäischen Grossstadtfauna. Mitt. naturwiss. Ver. Steiermark 105 (1975):
265—278.
Gepp, /.: Technogene und strukturbedingte Dezimierungsfaktoren der
Stadttierwelt—ein Überblick. Stadtök. Tagungsber. 3. Fachtagung des Ludwig-Boltz-
mann-Inst. Graz (1977): 99—127.
Gepp ].: Programmrahmen für einen umfassenden Lepidopterenschutz. Beih.
Veröff. Naturschutz Landschaftspflege Bad.-Württ. 21 (1981); 191—216.
Gnielka R.: Brutstatistik einer Urbanen Population der Ringeltaube {Columba
palumbus). Orn. Jber. Mus. Hein 3 (1978); 31—42.
Gnielka, R.: Die Vögel des Südfriedhofs in Halle. Hercynia N. F. Leipzig 18
(1981): 134—184.
Gnielka, R., Liedel, K., Spreike, Т., Stiefel, A., Tauchnitz, H., Wischof, W.:
Avifauna von Halle und Umgebung. Natur und Nmwelt, Halle 2 (1984):
1—96.
12. Литература 227
Gnielka, R., Spreike, Т., Tauchnitz, H., Reuter, В.: Avifauna von Halle und
Umgebung. Natur und Umwelt, Halle 1 (1983): 1—96.
Gorska, D.: Communities of synanthropic flies (Diptera) in tbe Region of
Warsaw and Kalisz. Memorabilia Zool. 30 (1979): 3—26.
Gorska, D.: Anthomyidae, Muscidae and Non-Parasitic Calliphoridae (Calyptrata,
Diptera) of Warsaw and Mazovia. Memorabilia Zool. 35 (1982); 93—114.
Gospodar, и., Korge, H.: Die Berliner Population des Otiorhynchus (Arammich-
nus) dieckmanni Magnano. Entomol. Bl. 78 (1982); 7—14.
Grempe, G.: Die Entwicl<Iung des Brutvogelbestandes auf dem Alten Friedhof
(Lindenpark) in Rostock in den Jahren 1962 bis 1982. Natur und Umwelt,
Rostock (1982); 22—27.
Griffiths, R. Л.: Seasonal behavior and intrahabitat movements in an urban
population of smooth newts {Triturus vulgaris. Amphibia Salamandidae). J. Zool.
(Lond.) 2 (1984); 203, 241—252.
Grimm, H.: Die Grossstadt als Lebensraum der Vögel. Vogelschutz u.
Vogelforschung (1953): 41—57.
Grimm H.: Zur Vogelwelt der U- und S-Bahnhöfe Berlins. Wiss. Z. Univ. Leipzig
10, Ges. u. Sprachw. R. (1961); 237—241.
Grimm, H.: Die Vogelwelt der Grossstadt und der Industrielandschaft. Falke 16
(1969); 41—49.
Grimm, H.: Aussagen einer 30 jährigen Vogelartenstatistik zur Auswirkung
baulicher Veränderungen in Berlin-Stadtmitte 1954/82. Tag. ber. 2. Leipziger
Symp. urb. Ökologie (1984): 37—39.
Grimm. H., Theis, G.: Die Vogelarten in Berlin-Stadtmitte. Falke 19 (1972):
150—156.
Grimm, P.: Bemerkungen zum Auftreten von Albinismus und Leuzismus bei
einigen Vogelarten. Falke 32 (1985); 240—242.
Haag, D.: Ein Beitrag zur Oekologie der Stadttaube. Diss. Univ. Basel 1984.
Haas, W.: Verluste von Vögeln und Säugern auf Autostrassen. Orn. Mitt. 16
(1964); 245—250.
Haeseler, V.: Anthropogene Biotope (Kahlschlag, Kiesgrube, Stadtgärten) als
Refugien für Insekten, untersucht am Beispiel der Hymenoptera Aculeata.
Zool. Jb. Syst. Jena 99 (1972); 133—212.
Haeseler, V.: Ameisen, Wespen und Bienen als Bewohner gepflasterter
Bürgersteige, Parkplätze und Strassen (Hymenoptera, Aculeata). Drosera 1 (1982):
17—32.
Hämäläinen, M., Mannerkoski, /.; Occurence and distribution of dermestids (Col.,
Dermestidae) in Finland. Notulae Entomol. 64 (1984): 167—184.
Handke, K.: Ergebnisse einer Siedlungsdichte-Untersuchung der Brutvögel auf
einer 52,5 ha grossen Brachfläche in Saarbrücken, den St. Arnualer Wiesen.
■!> Faun.-flor. Notizen aus dem Saarland 14 (1982); 127—133.
Hanoldt, W.: Der Brutvogelbestand eines Neubauviertels in Hamburg-Meiendorf
im Jahre 1971. Hamb, avifaun. Beitr. 11 (1973): 17—20.
Harksen, E., Mönke, R., Schumann, H.: Faunistisch-Ökologische Untersuchungen
zur Stechmückenfauna Berlins (Diptera, Culicidae). Dtsch. Ent. z. N. f. 23
(1976): 367—406.
Hartmann, E.: Schnecken in Münsters Kellern und Gärten. Natur und Heimat
15 (1955): 33—47.
Heimer, S.: Zur Spinnenfauna eines Gartens am östlichen Stadtrand von
Altenburg. Abh. Ber. Naturkundl. Mus. «Mauritianum» Altenburg 10 (1978):
171—180.
Heinicke, W., Klausnitzer, В.: Ergebnisse bei der Erforschung der Insektenfauna
der Deutschen Demokratischen Republik. Ent. Ber. 21 (1977): 74—84.
Heiversen, 0. v.: Месопета meridionale (Costa, 1860) in der südlichen
Oberrhein-Ebene (Orth., Ensifera). Mitt. Dtsch. Ent. Ges. 28 (1969): 19—22.
Hobner, M.: Die Bedeutung von Kleingärten für die Fauna und das Stadtklima.
Berl. Naturschutzblätter 4 (1984): 96—100.
228 12. Литература
Höffei, /., Mummer, P.: Schwermetallrückstände in Honigbienen (Apis mellifi-
ca L.) in einem Ökosystem (Saarbrücken). Forum Städte-Hygiene 34 (1983):
191—193.
Holman, ].: Taxonomy and ecology of Impatientinum asiaticum Nevsky, an aphid
species recently introduced to Europe (Homoptera, Aphididae). Acta entom.
bohemoslov. 68 f 1971): 153—166.
Ingrisch, S.: Zur Orthopterenfauna der Stadt Glessen (Hessen) (Saltatoria, Der-
- maptera und Blattoptera). Ent. Ztschr. 90 (1980): 273—280.
lacob-Remacle, Л.: Dipteres Syrphides et Coleopteres Carabides piegds dans
trois jardins de Liege «Intra Muros». Bull. Soc. Roy. Sei. Liege 49 (1980):
378—385.
ledryczkowski, W.: Synantropijne röwnonogi ladowe (Isopoda, Oniscoidea)
Polski. Fragm. Faun. 25 (1979): 95—106.
ledryczkowski, W.: Röwnonogi ladowe (Isopoda, Oniscoidea) skarpy wislanej
w Warszawie. Fragm. Faun. 25 (1980): 317—324.
ledryczkowski, W.: Isopods (Isopoda) of Warsaw and Mazovia. Memorabilia
Zool. 34 (1981): 79—86.
ledryczowski, W.: Millipedes (Diplopoda) of Warsaw and Mazovia. Memorabilia
Zool. 36 (1982): 253—262.
loost, W.: Beleuchtete Fabrikfenster als Lichtfalle für Meconema thalassinum
(de Geer, 1773) sowie Bemerkungen zur Orthopteren-Fauna der Stadt Gotha.
Ent. Nachr. Ber. 29 (1985): 115—117.
Karg, W.: Haustaubmilben in der Deutschen Demokratischen Republik. Allergie
u. Immunologie 19 (1973): 81—85.
Kasprzak, K.: Enchytraeids (Oligochaeta, Enchytraeidae) of Warsaw and
Mazovia. Memorabiha Zool. 34 (1981): 59—67.
Keil, H.: Ökofaunistische Untersuchungen der Haustaubmilben in Hamburg. Ent.
Mitt. Zool. Mus. Hamburg 7 (1983): 343—386.
Keil, H., Rack, G.: Systematik, Morphologie und Ökologie von Milben (Acari)
in Häusern und Vorräten sowie Milben von medizinischer Bedeutung (Teil
VI). Haustaubmilben (Dermatophagoides) und Haustaubmilben-Allergie.
Prakt. Schädlingsbek. 37 (1985): 78—86.
Kessler, A.: Über den Tagesgang von Oberflächentemperaturen in der Bonner
Innenstadt an einem sommerlichen Strahlungstag. Erdkunde 25 (1971):
13—20.
Kettlewell, H. B. D.: The phenomen of industrial melanism in Lepidoptera. Ann.
Rev. Ent. 6 (1961): 245—262.
Kettlewell, H. B. D: The Evolution of Melanism. A Study of a Recurring
Necessity. Oxford: Clarendon 1973. 423 pp.
Keve, A.: Über die Urbanisation der Vögel. Beitr. Vogelkde 29 (1983): 191—
197.
Kirchberg, E.: Untersuchungen über die Fliegenfauna menschlicher Fäkalien. Z.
hyg. Zool. 39 (1951): 129—139.
Klausnitzer, В.: Zur Kenntnis der Entoniofauna von Tanacetum vulgare L. und
Artemisia vulgaris L. Wiss. Z. Techn. Univ. Dresden 17 (1968): 19—21.
Klausnitzer, В.: Evolution der Insekten als Einnischungsprozess bei
Angiospermen. Biol. Rdsch. 15 (1977): 366—377.
Klausnitzer, В.: Nachweis von Trichogramma embryophagum (Hartig) im
Stadtzentrum von Leipzig (Hym.). Ent. Nachr. 24 (1980a): 47—48.
Klausnitzer, В.: Blattlausparasiten aus dem Stadtzentrum von Leipzig. Wiss. Z.
KMU, Math.-Nat. Reihe 29 (1980b): 574—582.
Klausnitzer, В.: Grossstädte als Lebensraum für das mediterrane Faunenelement.
Ent. Nachr. u. Ber. 26 (1982a): 49—57.
Klausnitzer, В.: Zur Kenntnis urbancr Gradienten. Tag. ber. 1. Leipziger Symp.
Urb. Ökologie 1981 (1982b): 13—20.
Klausnitzer, В.: Zur Insektenfauna der Städte. Ent. Nachr. u. Ber. 27 (1983a):
49—59.
12. Литература 229
Klausnitzer, В.: Faunistisch-Ökologische Untersuchungen über die Laufkäfer (Col.,
Carabidae) des Stadtgebietes von Leipzig. Ent. Nachr. Ber. 27 (1983b)'
241—261.
Klausnitzer, В.: Bemerkungen zur Optimierung von Grösse und Form städtischer
Grüianlagen aus ökologischer Sicht. Landschaftsarch. 12 (1983c): 40—41.
Klausnitzer, В.: Wirkung anthropogener Stressoren auf
anatomisch-morphologische Strukturen bei Tieren. In: Schubert, R.: Bioindikation in terrestrischen
Ökosystemen. VEB Gustav Fischer Verlag, Jena 1985, S. 67—73.
Klausnitzer, В.: Verstädterung von Tieren. A. Ziemsen Verlag, Wittenberg 1987.
Klausnitzer, В., Bley, U., Joost, \X'.: Entomologische und arachnologische
Untersuchungen in städtischen Kellern von Leipzig und Dresden. Ent. Nachr. Ber.
28 (1984): 115—117.
Klausnitzer, В., Förster, G.: Zur Insektenbesiedlung von Oenottiera biennis
Linnaeus. Beitr. Ent. 34 (1984): 421—424.
Klausnitzer, В., Jacob, U.: Probleme der morphologischen Bioindikation bei
Tieren unter Urbanen Bedingungen. Biol. Rdsch. 20 (1982): 351—363.
Klausnitzer, В., Klausnitzer, U.: Bemerkungen zur Feldheuschrecken-Fauna des
Stadtzentrums von Leipzig (Caelifera, Acrididae). Ent. Nachr. u. Ber. 26
(1982): 133—134.
Klausnitzer, В., Köberleln, C., Köberlein, F., Vogel J., Uhllg, M.: Zur Staphy-
linidenfauna zweier Leipziger Stadtparks. Faun. Abh. Mus. Tierk. Dresden 9
(1982); 195—202.
Klausnitzer^ В., Richter, K-: Presence of an urban gradient demonstrated for
carabid associations. Oecologia 59 (1983): 79—82.
Klausnitzer, В., Richter, K-: Veränderungen tropischer Strukturen bei
unterschiedlich urban beeinflussten Arthropodengesellschaften. Tag. ber. 2.
Leipziger Symp. urb. Ökologie (1984); 19—24, 18.
Klausnitzer В., Richter, K.: Zur Struktur von unterschiedlich urban beeinflussten
Arthropodengesellschaften entlang des A-E-Gradienten. Tag. ber. XI. Int.
Symp. Enlomofaunistik Mitteleuropas Gotha 1986: im Druck.
Klausnitzer, В., Richter, K-, Lehneert, J.: Zur Insektenfauna der Parkanlage am
Schwanenteich im Zentrum von Leipzig. Hercynia N. F. 16, 213—224.
Klausnitzer, В., Richter, K., Pfüller, R.: Ökofaunistische Untersuchungen auf
einem Hausdach im Stadtzentrum von Leipzig. Wiss. Z. Karl-Marx-Univ.
Leipzig, Math.-Naturwiss. R. 29 (1980a); 629—638.
Klawitter, /.: Zur Verbreitung der Fledermäuse in Berlin (West) von 1945—
1976. Myotis 14 (1976): 3—14.
Klawilter, J.: Zum Vorkommen des Steinmarders in Berlin (West). Berl. Natur-
schutzbl. 23 (1979); 462—467.
Klenke, R.: Ökofaunistische Untersuchungen an den Kleinsäugerpopulationen
unterschiedlicher Habitatinsein in Leipzig. Wiss. Z. Karl-Marx-Univ. Lei-
* pzig, Math.-Nat. Reihe 35 (1986); 607—616.
Klewen, R., Knauf, A., Meinhold, A.: Vegetation und Amphibienfauna eines
Regenrückhaltebecl\ens in Duisburg — ein Beitrag zum Artenschutz in
Ballungräumen. Natur und Heimat Münster 44 (1984): 119—128.
Klinger, R.: Eine städtische Grünanlage in Frankfurt am Main; Nördlichste
Enl<Iave für den Getreidbockkäfer Calamobius filum Rossi (Col,
Cerambycidae). Mitt. int. ent. Ver. 10 (1985); 37—38.
Klippel, R.: Ökologische Untersuchungen an Arthropoden in Kellern Hamburger
Wohnhäuser. Ent. Mitt. Staatsinst. Zool. Mus. Hamburg 1, Nr. 9 (1957);
239—287.
Klomann, U.: Untersuchungen an Carabidenpopulationen auf immissionbelasteten
Standorten im Stadtverband Saarbrücken. Jahresber. Naturwiss. Ver.
Wuppertal, 31. H. (1978); 140—144.
Klöppel, R., Tscharntke, Т., Zucchi, H.: Vogelnester als Überwinterungsorte von
Insekten und Spinnen. Anz. Schädlingskd., Pflanzenschutz, Umweltschutz 57
(1984): 25—30.
230 12. Литература
Klotz, S., Gutte, P.. Klausnitzer, В.: Literaturübersicht: Charal\terisierung und
Gliederung urbaner Ökosysteme. Hercynia N. F. 21 (1984a): 218—234.
Klotz, S., Gutte, P., Klausnitzer, В.: Vorschlag einer Gliederung urbaner
Ökosysteme. Arch. f. Landschaftsforsch u. Naturschutz 24 (1984b): 153—156,
Tabelle.
König, D.: Der Igel, Erinaceus europaeus L., als Verkehrsopfer. Faun. Mitt. aus
Norddtschl. 2 (1985): 236—237.
Koslowski, R., Kuckelkorn, В., Pfäller, В., Pfüller R., Süssengut, C.:
Ökologischfaunistische Untersuchungen an Araneae in Grünanlagen Leipzigs. Wiss. Z.
Karl-Marx-Univ. Leipzig, Math.-Naturwiss. R. 29 (1980): 561—566.
Kral, В., Pellantova, 1., Kokes, /.: Amphibians and Reptiles of the Brno Urban
Agglomeration. Folia Zool. 32 (1983): 51—66.
Krall, S.: Ökofaunistische Untersuchungen der Insekten in Nestern der Stadttaube
(Columba livia domestica L.) unter besonderer Berücksichtigung schädlicher
und lästiger Arten. Ent. Mitt. Zool. Mus. Hamburg 7 (1981): 29—44.
Krüger, H.: Einige Bemerkungen zur Urbanisierung. Falke 26 (1979): 341—
343.
Krzyzanowska, E., Dziabaszewski, A., lackowska. В., W. Starega: Spiders
(Arachnoidea, Aranei) of Warsaw and Mazovia. Memorabilia Zool. 34
(1981): 87—110.
Kubicka, A.: Scarabaeids (Col, Scarabaeidae) of Warsaw and Mazovia.
Memorabiha Zool. 34 (1981): 145—164.
Kuhlhorn, F.: Über den Dipterenbefall von Hundekot und dessen hygienische
Bedeutung. Bundesgesundheitsbl. 24 (1981): 283—289.
Kühnelt, W.: Gesichtspunkte zur Beurteilung der Grossstadtfauna (mit
besonderer Berücksichtigung der Wiener Verhältnisse). Österr. Zool. Zschr. 6
(1955): 30-54.
Kühnelt, W.: Grundriss der Ökologie. VEB Gustav Fischer Verlag, Jena 1970.
Kühnelt, W.: Die Grünflächen der Städte und ihre Tierwelt. — In: Stadtökologie.
Tag. ber. 3. Fachtagung des Ludwig-Boltzmann-Institutes Graz, 69—77.
Ladusch, M., Lehnert, J., Strache, R.-R.: Die Brutvögel des Stadtkerns von
Leipzig im Jahre 1979. Wiss. Z. Karl-Marx-Univ. Leipzig, M. 29 (1980): 556
bis 560.
Laibach, E.: Lepisma saccharina L., das Silberfischchen. Z. hyg. Zool. 40 (1952):
431 bis 470.
Lampel, G.: Die Blattläuse (Aphidina) des Botanischen Gartens Freiburg/
Schweiz. Bull. Soc. Frib. Sc, Nat. 63 (1974): 59—137; 64 (1975): 125—184;
65 (1976): 197—255.
Lampel, G.: Impatientinum asiaticum News., 1929, eine asiatische Blattlausart,
neu im Botanischen Garten Freiburg/Schweiz. Bull. Soc. Frib. Sc. Nat. 67
(1978): 69—72.
Lampel, G.: Über einige Neo- und Xerophyten und ihre Blattläuse (Aphidina)
in der Schweiz. Mitt. Schweiz. Ent. Ges. 56 (1983): 212.
Lau, D.: Kurze Anleitung zur Pflege hilfsbedürftiger Igel. Tierpark-Merkblätter
Nr. 9 (1977). Berlin.
Leclerq, ]., Claparede, L.: Sceliphron caementarium (Drury) (Hymenoptera
Sphecidae) s'installe en Europe meridionale. Entomops (Nice) 47 (1978):
245—252.
Lenz, M.: Zur Problem der Erfassung von Brutvogelbeständen in Stadtbiotopen.
Vogelwelt 92 (1971): 41—52.
Lenz, M., Hindemith, I., Kroger, В.: Zum Brutvorkommen der Mehlschwalbe
(Delichon urbica) in West-BerUn 1969 und 1971. Vogelwelt 93 (1971): 161—
180.
Lenz, M., Witt, K.: Verbreitung der Türkentaube (Streptopelia decaocto) und
Ringeltaube (Columba palumbus) 1974 in zwei Berliner Bezirken. Vergleich
mit einer Zählung 1964. Orn. Ber. f. Berlin (West) 3 (1978); 171—188.
12. Литература 231
Lenz, М., Witt, К.: Das Brutvorkommen der Mehlschwalbe in Berlin (West)
1969—1979. Manuskript 1980.
Loibl, H.: Schmetterlinge in Hamburgs Gärten und Parks. Schmetterlinge als
Kulturfolger. Mitt. Faun. Arbgem. Schleswig-Holstein, Hamburg und
Lübeck N. F. 6 (1953): 38—45.
Lomolino, M. V.: Mammalian Island Biogeography; Effects of Area, Isolation
and Vagility. Oecologia 61 (1985): 376—382.
Loos, W.: Amphibien und Kleingewässer in der Stadt Kamen, Kreis Unna. Mitt.
Landesanstalt Ökol. NRW 5 (1980): 118—119.
Löschau, M., Lenz, M.: Zur Verbreitung der Türkentaube (Streptopelia decaocto)
in Gross-BerHn. J. Orn. 108 (1967); 51—64.
Loske, R.: Zur Situation der Kleingewässer und ihrer Amphibienfauna im Gebiet
der Stadt Lippstadt. Natur und Heimat 43 (1983); 97—128.
Luniak, M.: Birds of allotment gardens in Warsaw. Acta Orn. 17, Nr. 20 (1980):
297—317.
Luniak, M.: The birds of the park habitats in Warsaw. Acta Orn. 18 (1981);
335—374.
Luniak, M.: The avifauna of urban green areas in Poland and possibilities of
managing it. Acta Orn. 19 (1983): 3—61.
Luniak, M.: Preliminary results of the international inquiry on the composition
and recent changes of urban avifauna in Central and Eastern Europe. Tag.
ber. 2. Leipziger Symp. urb. Ökologie (1984); 33—36.
Luniak, M., Kalbarczyk, W., Pawlowski, W.: Vögel von Warszawa. Acta Orn. 8
(1964): 175—285.
Mac Arthur, R. H., Wilson, E. O.: The Theory of Island Biogeography.
Monographs in Population Biology. Princeton, N. J. (Ed.). Princeton Univ. Press
1967.
Mader, H.-J.: Die Isolationswirkung von Verkehrsstrassen auf Tierpopulationen
untersucht am Beispiel von Arthropoden und Kleinsäugern der
Waldbiozönose. Sehr. R. Landschaftspflege u. Naturschutz 19 (1979a), 131 S.
Mader, H.-J.: Biotopisolierung durch Strassenbau am Beispiel ausgewählyer
Arten —Folgerungen für die Trassenwahl. Ber. ANL 3 (1979b): 56—63.
Mader, H.-J.: Der Verinselung der Landschaft aus tierökologischer Sicht. Natur
und Landschaft 55 (1980); 91—96.
Mader, H.-J.: Untersuchungen zum Einfluss der FLächengrösse von
Inselbiotopen auf deren Funktion als Trittstein oder Refugium. Natur und Landschaft
56 (1981a): 235—242.
Mader, H.-J.: Der Konflikt Strasse — Tierwelt aus ökologischer Sicht. Sehr. R.
Landschaftspfl. Naturschutz H. 22 (1981b): 104 S.
Mader, H.-J.: Warum haben kleine Inselbiotope hohe Artenzahlen? Natur und
Landschaft 58 (1983); 367—370.
Mader H.-J.: Animal Habitat Isolation by Roads and Agricultural Fields.
Biological Conservation 29 (1984): 81—96.
Mader, H.-J., Mühlenberg, M.: Artenzusammensetzung und Ressourcenangebot
einer kleinflächigen Habitatinsel, untersucht am Beispiel der Carabidenfauna.
Pedobiologie 21 (1980): 46—59.
Mader, H.-J., Müller, K.: Der Zusammenhang zwischen Heckenlänge und
Artenvielfalt. Z. f. Kulturtechnik und Flurbereinigung 25 (1984); 282—293.
Mann, H.-G., Ribbeck, R.: Untersuchungen zum Vorkommen von Mallophagen
bei Haustauben, Columba livia domestica, speziell bei Sporttauben. Lounais-
Hämeen Luonto (Forssa) 46 (1973): 73—77.
Martens, J. M., Gillandt, L.: Schutzprogramm für Heuschrecken. Naturschutz und
Landschaftspflege Hamburg H. 10, 1985.
Matzke, M.: Landgastropoden innerhalb einer Grosstadt am Beispiel von
Halle an der Saale. Malak. Abh. Mus. Tierk. Dresden 4 (1973): 21—38.
232 }2. Литература
Matzke, М.: Landgastropoden innerhalb einer Kleinstadt am Beispiel von
Lichtenstein am Fusse des Westerzgebirges. Malak. Abb. Mus. Tierk. Dresden ß
(1979): 145—167.
Mauersberger, G.: Ökologische Probleme der. Urbanisierung. Falke 18 (1971):
72-82.
Maurer, R.: Die Vielfalt der Käfer- und Spinnenfauna des Wiesenbodens im
Einflussbereich von Verkehrsimmissionen. Oecologia (Berlin) 14 (1974):
327—351.
Mayer, G. ГЛ.: Ein Massenauftreten von Haussperlingsweisslingen (Passer
domesticus (L.)) in Linz. Naturkundl. Jahrb. Stadt Linz 22 (1976): 137—
152.
Meier, W.: Impatientinum asiaticum News., eine für Schweiz und für Westeuropa
neue Blattlausart. Mitt. Schweiz. Eni Ges. 50 (1977): 336.
Menzel, H.: Die Mehlschwalbe. Neue Brehm-Bücherei 548, Wittenberg 1984.
Merisuo, A. K.: Eine reichliche und interessante Sphecidenfauna (Hymenoptera)
mitten in der Stadt. Notul. ent. 13 (1933): 59—60.
Metzger, R., Hanisch, ].: Grösse und Verteilung einer Population von Blattella
germanica. Angew. Parasitol. 20 (1979): 193—202.
Mihalyi, E.: Flies visiting fruit and meat in an open-air market in Budapest.
Acta Zool. Hung. 7 (1966): 331—337.
Mildner, P.: Neues zur Kärntner Arthropodenfauna. Carinthia II 173/93 (1983):
137—141.
Moszynski, A., Urbanski, J.: Etude sur la faune des Serres de Poznan (Pologne)
Bull. Biol. Fr. Belg. 66 (1932): 45—76.
Müller, F. P.: Aphidologische Beobachtungen im Botanischen Garten der
Universität Rostock. Wiss. Z. Univ. Rostock, Math.-Nat. Reihe, 24 (1975): 811—
815.
Müller, F. P.. Buhr, H. ].: Hyadaspis tataricae (Ajzenberg), ein neuer Schädling
in Mitteleuropa an Lonlcera tatarica L. und einigen anderen Lonicera-Zier-
sträuchern. Anz. Schädlingskd. 38 (1965): 69—72.
Müller, G.: Der Sexual-Inde.x bei Carabiden als ökologisches Kriterium. Ent.
Ber. 14 (1970): 12—18.
Müller, ].: Das aktuelle Flohatrenspektrum (Siphonaptera) auf Stadthunden
Wiss. Z. Karl-Marx-Univ. Leipzig, Math.-Nat. R. 35 (1986): 653—658.
Müller, P.: Anpassung und Informationsgehalt von Tierpopulationen in Städten
Verh. Dtsch. Zool. Ges. Stuttgart (1980): 57—77.
Müller, P.: Arealsysteme und Biogeographie. Eugen Ulmer, Stuttgart 1981.
Müller, P.: Experimentelles Biomonitoring und Nahrungskettenanalysen in
Urbanen Ökosystemen. Tag. ber. 2. Leipziger Symp. urb. Ökologie (1984):
3-17.
Müller, P., Klomann, U., Nagel, P., Reis, H., Schäfer. A: Indikatorwert
unterschiedlicher biologischer Diversität im Verdichtungsraum von Saarbrücken.
Verb. Ges. Ökol. Erlangen, Junk, Den Hague (1974): 113—128.
Mulsow, R.: Untersuchungen zur Siedlungsdichte der Hamburger Vogelwelt. Abb.
Ver. Naturwiss. Ver. Hamburg N. F. 12 (1968): 123—188.
Mulsow, R.: Vogelbiotope und Siedlungsdichte in der Stadtlandschaft. Vogelwelt
. 92 (1971): 23—24.
Mulsow, R.: Der Sommervogelbestand 1971—1973 in einem Waldstück zwischen
Villenvierteln im Nordosten Hamburgs. Hamb, avifaun Beitr 12 (1974):
151—160.
Mulsow, R.: Die Avizönose der Gartenstadtzone. Vogelwelt 97 (1976): 55—68.
Mulsow, R.: Zur Struktur und Jahresdynamik der Parkvogelgemeinschaft in
Hamburg. Beitr. Avifauna Rheinl 11 (1978): 44—50.
'Munzel, M.: Vorkommen und Entwicklung von Fliegen (Diptera: Muscidae,
Calliphoridae) an Lebensmitteln tierischer Herkunft Z. angew Zool Berlin
68 (1981): 393—414.
12. Литература 23S
Nicht, М.: Ein Beitrag zum Vorkommen des Steinmarders, Maries foina (Erxle-
ben, 1770) in der Grossstadt (Magdeburg). Z. Jagdwiss. 15 (1969): 1—6.
Niedbala, W., Blaszak, C, Bloszyk, J., Kaliszewski, M., Kazmierski, A.: Soil mites
(Acari) of Warsaw and Mazovia. Memorabilia Zool. 36 (1982): 235—252.
Niethammer, G.: Die Einbürgerung von Säugetieren und Vögeln in Europa. Paul
Parey Hamburg, Berlin 1963.
Nowakowski, E.: Influence of urbanization on the structure of wireworn
communities (Col, Elateridae). Animals in urban environment, Proc. Symp.
Institute of Zoology, 1979, Warszawa (1982): 79—90.
Nuorteva, P.: Synanthropy of blowflies (Dipt., Calliphoridae) in Finland. Ann.
Ent. Fenn. 29 (1963): 1—49.
Nuorteva, P.: The synanthropy of birds as an expression of the ecological cycle
discorder caused by urbanization. Ann. Zool. Fenn. 8 (1971): 547—553.
Oertel, S.: Zu: «Einige Bemerkungen zur Urbanisierung». Falke 27 (1980): 230—
233.
Otto, W., Recker, W.: Zum Einfluss nistökologischer Faktoren auf die Abundanz
des Haussperlings in Berliner Neubauwohnvierteln. Falke 23 (1976): 330—
337.
Otzen, W., Schaefer, M.: Untersuchung:en zur Überwinterung von Arthropoden
inVogelnestern. Ein Beitrag zur Winterökologie. Zool. Jb. Syst. 107 (1980):
435—448.
Owen, D. F.: Conservation of butterflies in garden habitats. Environ Conserv. 3
(1976): 285—290.
Owen, D. F.: Insect diversity in an English suburban garden. In: Frankie, G. W.,
and Koehler, C. S. (Eds.): Perspectives in Urban Entomology. Academic-
Press, New York, San Francisco, London 1978, pp. 13—29.
Owen, D. F., Whiteway, W. R.: Buddleia davidii in Britain: history and
development of an associated fauna. Biol. Conserv. 17 (1980): 149—155.
Owen, J., Owen, D. F.: Suburban gaedens: England's most important nature
reserve? Environ Conserv. 2 (1975): 53—59.
Pappa, В.: Zierpflanzenschädlinge in und um Hamburg. Ent. Mitt. Zool. Mus.
Hamburg 5 (1976): 25—47.
Patton, D. R.: A diversity index for quantifying habitat «edge». Wildlife Soc
Bull. 3/4 (1975): 171 — 173.
Peitzmeier, I.: Zum Ursachenkomplex der Verstädterung der Vögel. Orn. Mitt. 9
(1957): 92—93.
Pelikan, J., Zejda, J., Homolka, M.: Mammals in the urban agglomeration of
Brno. Pfir. Prace ust. Ceskoslov. Akad. ved. v Brno 17, H. 9 (1983): 3—49.
Peters, D. St.: Ökologische Studien an Parkvögeln. Biol. Abb. 27/28 (1963):
1—44.
Peters, D. St.: Die Grabwespe Ectemnius sexcinctus als Bewohner von Kunststoff-
Blumenkästen. Natur und Museum 101 (1971): 26—31.
Peters, D. St.: «Nistkästen» für Insekten. Natur und Museum 103 (1973): 162—
165.
Peters, H.: Biologie einer Grossstadt. Verlag Honing, Heidelberg 1954.
Peters, H.: Das Auftreten der Braunbandschabe und anderer neuer Blattarien
in Deutschland. Städtehygiene 7 (1956): 40—42.
Peters, H.: Beitrag zur Biologie und Ökologie der synanthropen Dipteren einer
Grossstadt. Merkblätter, Beitr. Schädlingskd. 9 (Folge 1) (1959): 1—83.
Peters, H.: Über den Begriff Synanthropie. Z. angew. Zool. 47 (1960): 35—42.
Peters, H.: Die synanthropen Schaben Mitteleuropas (Gattungen Blatta, Blattella,
Peripllaneta und Supella). Merkbl. agnew. Parasitenk. Schädlingsbek 3
(1961) (Beilage Angew. Parasitol. 2).
Peus, F.: Flechtlinge als Wohnungs- und Materialschädlinge besonders in
Neubauten. Z. Gesendh. Techn. Städtehyg. 25 (1933): 483—490.
Pilipiuk, I.: Earthworms (Oligochaeta, Lumbricidae) of Warsaw and Mazovia
Memorabilia Zool. 34 (1981): 69—77.
16—21
234 12. Литература
Pisarski, В.: Ants (Hymenoptera, Formicoidea) of Warsaw and Mazovia.
Memorabilia Zool. 36 (1&82): 73—90.
Pisarski, В., Czechowski, W.: Influence de la pression urbane sur la myrmeco-
faune. Memorabilia Zool. 29 (1978): 109—128.
Plachter, H.: Tierbestände im Siedlungsbereich und ihre Erfassung im Rahmen
von Biotopkartierungen. Garten und Landschaft 90 (1980): 569—576.
Plath, L.: Neststandorte und Sidlungsdichte des Haussperlings (Passer
domesticus) in einem Rostocker Neubauwohngebiet. Orn. Rundbr. Meckl. NF 17
(1976): 13—15.
Plath, L.: Entwicklung und Wertung des Brutvogelbestandes eines Rostocker
Neubeuwohngebietes. Natur und Umwelt, Rostock (1981): 55—73.
Plath, L.: Die Besiedlung eines Neubauwohngebietes durch Vögel — Ergebnisse
ISjähriger Bestandserhebungen. Falke 32 (1985): 335—343.
Plath, L.: Zum. Brutvorkommen der Mehlschwalbe [Delichon urbica) in den
Neubaugebieten der Stadt Rostock. Beitr. Vogelkd. Im Druck.
Popendiker, K-: Die in Vogelnestern lebenden Insekten unter besonderer
Berücksichtigung der als Haus- und Vorratsschädlinge auftretenden Arten Mitt
Hamburg. Zool. Mus. Inst. 54 (1956): 49—127.
Povolny, D.: Versuch einer Klärung des Begriffs der Synanthropie von Tieren
Folia Zool 25 (1962): 105—112.
Povolny, D.: Einige Erwägungen über die Beziehungen zwischen den Begriffen
«synanthrop» und «Kulturfolger». Beitr. Ent. 13 (1963): 439—444.
Povolny, D., Sustek, Z.: An attempt at a methodical separation of the concept
«Synanthrope» and «Kulturfolger». Ekologia (CSSR) 1 (1982): 13—24.
Povolny, D.: Some considerations on animal synanthropy and its manifestations
on model groups Sarcophagidae (Diptera) and Carabidae (Coleoptera). Acta
Universitatis Agricult. 33 (1985): 175—199.
Preuss, G.: Voraussetzungen und Möglichkeiten für Hilfsmassnahmen zur
Erhaltung und Förderung von Stechimmen in der Bundesrepublik Deutschland.
Natur und Landschaft 55 (1980): 20—26.
Pschorn-Walcher, H., Gunhold, P.: Zur Kenntnis der Tiergemeinschaft in Moos-
und Flechtenrasen an Park- und Waldbäumen. Z. Morph. Ökol. Tiere 46
(1957): 342—354.
Rack, G.: Massenvorkommen von Neoseiulus barkeri Hughes, 1948 (Acarina,
Phytoseiidae) und sechs weiterer Milbenarten in einem Neubau. Anz. Schäd-
lingsk. 43 (1970): 184—186, Berlin.
Rack, G.: Milben in Neubauten. Prakt. Schädlingsbek. 23 (1971): 149—152,
Braunschweig.
Rack, G.: Weitere Beobachtungen über Milben in Neubauten mit Beschreibung
einer neuen Art, Ameroseius pseudoplumosus sp. n. (Acarina, Mesostigma-
ta). Ent. Mitt. Zool. Mus. Hamburg 4, Nr. 77 (1972): 249—257.
Rack, G.: Auftreten von Terpnacarus subterraneus Weis-Fogh, 1947 (Acarina,
Actinedida, Terpnacaridae) in einem neuen Wohnhaus in Süddeutschland.
Ent. Mitt. Zool. Mus. Hamb. 7 (^Sl): 3—9.
Rack, G.: Systematik, Morphologie und Biologie von Milben (Acari) in Häusern
und Vorräten sowie Milben von medizinischer Bedeutung. Teil III: Die
Modermilbe Tyrophagus putrescentiae (Schrank, 1781) und andere Milben
als Verursacher von Neubauplagen. Prakt. Schädlingsbek. 36 (1984): 13—16.
Reich, M., Roth, M., Majzlan, 0.: Die Coleopteren-Zönose im Ökosystem
«Obstgarten — Eklektorfauna. Jber. naturwiss. Ver. Wuppertal 38 (1985): 20—23.
Reichholf, J., Esser, ].: Daten zur Mortalität des Igels [Erinaceus europaeus),
verursacht durch den Strassenverkehr. Z. Säugetierkunde 46 (1981): 216—
222.
Reinke, E. M.: Die verwilderten Haustauben in Hamburg. Z. angew Zool. 46
(1959): 285—301.
Reischütz, P. L.: Zum massenhaften Auftreten von Arion lusltanlcus Mabille in
den Jahren \Ш und 1983. Mitt. Zool, Ges. Braunau 4 (1984): 253—254.
12. Литература 235
Rettig, К.: Tierverluste auf Autostrassen. Orn. Mitt. 17 (1965): 234—244.
Richter, C. J. J.: Aerial dispersal in relation to habitat in eight wolf spider species.
(Pardosa, Araneae, Lycosidae). Oecologia (Berlin) 5 ("1970): 2PP —214.
Richter, Beitrag zur Aphidenfauna des Grossstadtraumes von Leipzig
(Homoptera, Aphidina). Ent. Nachr. 23 (1979): 49—54.
Richter, K-: Beeinflussung der MDH-Aktivität von Blattläusen durch anthropo-^
gene Umweltfaktoren, insbesondere SO2 (SOa^-). Tagungsber. 1. Leipz.
Symp. urb. Okol. 1981 (1982): 21—24.
Richter, K., Borneleit, F.: Zur Atmungsaktivität von Aphis fabae Scopoli (Insecta,,
Homoptera) unter Quecksilberbelastung. Wiss. Z. Karl-Marx-Univ. Leipzig,.
Math.-Naturwiss. R. 33 (1984): 166—169.
Richter, K., Klausnitzer, В.: Ziim Einfluss ausgewählter anthropogener Noxen
auf NAD-abhängige Malatdehydrogenase und Transaminasen aus Aphis
sambuci L. und einigen anderen Blattläusen. Wiss. Z. Karl-Marx-Univ.
Leipzig, Math.-Naturwiss. R. 29 (1980a): 611—619.
Richter, K., Klausnitzer, В.: Experimentelle Untersuchungen zum Einfluss von,
S02(SÖ32") auf Fertilität und MDH-Aktivität von Aphis fabae Scop. (Horn.,
Aphidina). Wiss. Z. Karl-Marx-Univ. Leipzig, iVlath.-Naturwiss. R. 29-
(1980b): 620—626.
Rinnhofer, G.: Die Vogelwelt eines Grossstadtfriedhofes am Fusse des Erzgebir-^
ges. Zool. Abb. Staatl. Mus. Tierk. Dresden 28 (1965): 1—55.
Rinnhofer, G.: Köderfang auf Ruderalstellen. Ent. Nachr. 11 (1967): 18—24.
Rinnhofer, G.: Beobachtungen an der Vogelwelt eines Grossstadtruderalgeländes.
Hercynia N. F. 6 (1969): 1—35.
Roer, H.: Fledermauskasten — Vogelnistkasten — Kombinierter
Vogel-Fledermauskasten. Myotis 6 (1968): 10—15.
Roer, H.: Gittertore und Nistkästen als wichtige Hilfsmittel zur Erhaltung der
in ihrem Bestand bedrohten europäischen Fledermäuse. Decheniana Beh. 1&
(1971): 109—113.
Sacher, F.: Zum Vorkommen des Weberknechtes Leiobunum limbatum L. koch
in Gera. Veröff. Mus. Gera, Naturwiss. R. (1978): 77—78.
Sacher, P.: Spinnen (Araneae) an und in Gebäuden. — Versuch einer Analyse
der synanthropen Spinnenfauna in der DDR. Ent. Nachr. Ber. 27 (1983):
97—104, 141—152, 197—204.
Saemann, D.: Zur Typisierung städtischer Lebensräume in Hinblick auf avifauni-
stische Untersuchungen. Mitt. IG Avifauna DDR 1 (1968): 81—88.
Saeemünn, D.: Der Brutbestand der Türkentaube, Streptopelia decaocto, in Karl-
Marx-Stadt nach 20jähriger Siedlungszeit; Falke 16 (1969): 188—191.
Saemann, D.: Die Brutvogelfauna einer sächsischen Grossstadt. Veröff. Mus.
Naturkde. Karl-Marx-Stadt 5 (1970): 21—85.
Saemann, D.: Untersuchungen zur Siedlungsdichte der Vögel in verschiedenen
Grossstadthabitaten. Mitt. IG Avifauna DDR 6 (1973): 3—24.
Saemann, D.: Studien an einer Grossstadtpopulation der Türkentaube
Streptopelia decaocto im Süden der DDR. Hercynia N. F. 12 (1975): 361—368.
Saemann, D.: Ein Beitrag zur Brutphänologie der Amsel, Turdus merula, in der
Grossstadt. Actitis 17 (1979): 3—14.
Sage, B. L.: Albinism and melanism in birds. British Birds 55 (1962): 201—-
225.
Sahrhage, D.: Ökologische Untersuchungen am Ofenfischchen, Thermobia
domestica (Packard) und Silberfischchen, Lepisma saccharina L. Z. angew. Ent.
35 (1954): 495—499.
Samsinäk, K.: Ober einige in Häusern lästige Arthropodenarten. Anz.
Schädlingskde., Pflanzenschutz, Umweltschutz 54 (1981): 120—122.
Sawoniewicz, /.: Ichneumonidae (Hymenoptera) of Warsaw and Mazovia.
Memorabilia Zool. 36 (1982): 5—40.
16,
236 12. Литература
Schaefer, М.: Welche Faktoren beeinflussen die Existenzmöglichkeit von
Arthropoden eines Stadtparks — untersucht am Beispiel der Spinnen (Araneida)
und Weberknechte (Opihonida)? Faun. — ökol. Mitt. 4 (1973): 305—318.
Schaefer, M.: Gedanken zum Schutz der Spinnen. Natur und Landschaft 55
(1980): 36—38.
Schaefer, M., Kock, K.: Zur Ökologie der Arthropodenfauna einer Stadtlandschaft
und ihrer Umgebung. I. Laufkäfer (Carabidae) und Spinnen (Araneida).
Anz. Schädlingskd., Pflanzenschutz, Umweltschutz 52 (1979): 85—90.
Scheurer, S.: Erstnachweis des Hygienschädlings Tapinoma melanocephalum
(Hymenoptera, Formicidae) in der DDR. Angew. Parasitol. 25 (1984): 96—
99.
Schlegel, R.: Die Brutvögel der Leipziger Friedhöfe. Mitt. Sachs. Heimatsch. 9
(1920): 111—119.
Schlegel, R.: Die im Stadtgebiet Leipzig brütenden Vögel. Leipzig 1922.
Schliephake, G.: Thripse (Thysanoptera) als Gewächshausschädlinge. Ent. Nachr.
Ber. 28 (1984): 189—194.
Schmidt, J.: Blei- und Cadmium-Rückstände bei inner- und ausserstädtischen
Lacerta-Populationen. Verh. Ges. Ökologie (1980) 9 (1981): 297—300.
Schmidt, K. H., Steinbach, ].: Niedriger Bruterfolg der Kohlmeise (Parus major)
in städtischen Parks und Fridhöfen. J. Ornithol. 124 (1983): 81—83.
Schneider, N.: Les psocopteres du Grand-Duche de Luxembourg. I. Faunistique
et ecologie des especes domicoles. Bull. Ann. Soc. R. Belg. Ent. 113 (1977):
210—222.
Schneider, N.: Les psocopteres du Grand-Duche de Luxembourg. III. Faunistique
et ecologie des especes urbaines. Bull. Ann. Soc. R. belg. Ent. 118 (1982):
131—144.
Schneider, W.: Rückblick auf eine dreissigj ährige ununterbrochene Beobachtung
einer örtlichen Starpopulation im Leipziger Raum. Beitr. Vogelkd. 28 (1982):
207—221.
Schremmer, F.: Freilandfund der Landplanarie Rhynchodemus billneatus Metsch.
(zugleich ein Beitrag zur Fauna des Arkadenhofes der Wiener Universität).
Verh. Zool.-Bot. Ges. Wien 94 (1954).
Schremmer, F.: Beobachtungen und Untersuchungen über die Insektenfauna der
Lärche (Larix decidua) im östlichen Randgebiet ihrer natürlichen
Verbreitung mit besonderer Berücksichtigung einer Grossstadtlärche Z. angew.
Ent. 45 (1959): 1—48, 113—153.
Schubert, R.: Lehrbuch der Ökologie. VEB Gustav Fischer Verlag, Jena 1984.
Schwarz, A.: Zum gegenwärtigen Vorkommen von Protaetia aeruginosa (Drury)
in der DDR. Faun. Abh. Mus. Tierk. Dresden 9 (1981).
Schwarz, R.: Biologisch-ökologische Untersuchungen über die Blattläuse der
Unkraut- und Ruderalflora Berlins (Homoptera, Aphidina). Beitr. Ent. 9
(1959): 473—506.
Schweiger, H.: Versuch einer zoogeographischen Gliederung der rezenten Fauna
des Wiener Stadtgebietes. Österr. Zool. Z. 4 (1953): 556—586.
Schweiger, H.: Die Vertebratenfauna des Wiener Stadtgebietes und ihre
Probleme. Jubiläums-Jahrbuch, Österr. Arbeitskreis f. Wildtierforsch. (1960/61):
137—153.
Schweiger, H.: Die Insektenfauna des Wiener Stadtgebietes als Beispiel einer
kontinentalen Grossstadtfauna. 11 Int. Kongr. Ent. Wien 1960, 3 (1962):
184—193.
Segebade, R., Schaefer, M.: Zur Ökologie der Arthropodenfauna einer Stadtland-
scheft und ihrer Umgebung. II. Pfkanzengallen und Pflanzenminen. Anz.
, Schädlingskd., Pflanzenschutz, Umweltschutz 52 (1979); 117—121.
Seifert, В.: Hypoponera punctatissima (Roger) (Hym., Formicidae) eine
interessante Ameisenart in menschlichen Siedlungsgebieten). Ent. Nachr. Ber. 26
(1982): 173—175.
12. Литература 237
Serafinski, W.: The subspecies diiferentiation of the Central European house
mouse {Mus musculus L.) in the light of their ecology and morphology.
Ekol. Pol. A, 13 (1965): 305-348.
Sikorski, M. D.: Non-metrical divergence of isolated populations of Apodemus
agrarius in urban areas. Acta Theriol. 27 (1982a): 169—180.
Sikorski, M. D.: Craniometric variation of Apodemus agrarius (Pallas, 1771) in
urban green areas. Acta Theriol. 27 (1982b): 71—81.
Simberloff, D.: Experimental zoogeography of islands. Effects of island size.
Ecology 57 (1976): 629—648.
Simberloff, D., Wilson, E. O.: Experimental zoogeography of islands. The
colonization of empty islands. Ecology 50 (1969): 278—295.
Sixl, W.: Zum Problem der verwilderten Stadttaube. Mitt. Abt. Zool.
Landesmus. Joanneum (Graz) 1 (1975): 87—97.
Shell, J.: Thaumetopoea pinivora Tr. im Stadtgebiet von Dresden. Ent. Nachr.
10 (1966): 164—166.
Skibinska, е.: Chrisididae (Hymenoptera) of Warsaw and Mazovia. Memorabilia
Zool. 36 (1982a): 61—72.
Skibinska, E.: Wasps (Hymenoptera, Vespidae) of Warsaw and Mazovia.
Memorabilia Zool. 36 (1982b); 91—102.
Skibinska, E.: Sphecidae (Hymenoptera) of Warsaw and Mazovia. Memorabilia
Zool 36 (1982c); 103—127.
Southwood, Т.: The number of species of insects associated with various trees.
J. Anim. Ecol. 30 (1961); 1—8.
Stadler, H.: Haus-, Keller- und Stallspinnen. Nachr. naturw. Mus.
Aschaffenburg 69 (1962); 1—24.
Stein, W.: Der Einfluss des Flügeldimorphismus auf die Ausbreitung von Curcu-
lioniden-Arten. Z. angew. Entom. 61 (1968): 442—445.
Stein, W.: Die Beziehungen zwischen Biotop-Alter und Auftreten der Kurzflüge-
ligkeit bei Populationen dimorpher Rüsselkäfer-Arten (Col., Curculionidae).
Z. angew. Entomol 83 (1977): 37—39.
Steiniger, H.: Genetische Variabilität bei Carabidae-Populationen inner- und aus-
serstädtischer Standorte (Coleoptera). Diss. Univ. Saarbrücken 1978.
Stellwag, F.: Die Fauna tiefer Weinkeller. Archiv Naturgesch. 90 (1924): 130—
149.
Stephan, В.: Veränderungen der Fauna am Beispiel der Verstädterung von
Vögeln. Naumann, Festschrift (1980); 102—115.
Sterzynska, M.: Springtails (Collembola) of Warsaw and Mazovia. Memorabilia
Zool. 36 (1982): 217—234.
Strawinski, S.: Die Vogelverstädterung vom ökologischen Standpunkt. Orn. Mitt.
18 (1960); 72—74.
Stbeib, U.: Verbreitungsmuster rezenter Schnecken im Stadtgebiet von Mainz
(Mollusca: Gastropoda). Mainzer Naturw. Archiv 22 (1984): 149—209.
Stritt, W.: Wartehäuschen als Lichtfalle für Hautflüger. Dtsch. Ent. Z. 18 (1971):
99—112.
Strobl, P., Könecke, F.-W.: Die Schmetterlinge des Stendaler Friedhofes und
dessen Umgebung. Ent. Nachr. Ber. 28 (1984); 9—11.
Stutz, H., Haffner, M.: Richtlinien für die Erhaltung und Neuschaffung von
Flede'rmausquartieren in und an Gebäuden. Aktiver Fledermausschutz 3
(1984); FEBEX, Zürich.
Sukopp, H.: ökologische Charakteristika von Grossstädten. In; Grundriss der
Stadtplanung. Hannover 1983a.
Sukopp, H.: Ökologische Charalderistika der Grossstadt. Tag^ ber. 1. Leipziger
Symp. urb. Ökologie 1981 (1983b).
Sukopp, H., Blume, H.-P., Elvers, H., Horbert, M.: Beiträge zur Stadtökologie von
Berhn (West). Lanschaftsentw. u. Umwehforsch. Nr. 3 (1980), Techn. Univ.
Berlin.
238 12. Литература
Sukopp, Н., Weiler, S.: Vernetzte Biotopsysteme. In: Artenund Biotopschutz,
Aufbau eines vernetzten Biotopsystems. Hrsg.: JVlin. f. Soziales, Gesundh.
und Umwelt Rheinland-Pfalz. Fachtagung 1984.
Szelegiewicz, H.: Vier für die Fauna Polens neue Blattlausarten (Homoptera,
Aphidoidea). Bull. Acad. Polon. sei., Ser. sei. biol., Cl. II, 20 (1972): 229—
231.
Tamm, K..: Choriozönosen auf und an Autostrassen. Versuch einer
Charakterisierung. Naturwiss. Rdsch. 29 (1976): 197—202.
Tenovuo, R.: Zur Urbanisierung der Vögel in Finnland. Ann. Zool. Fenn. 4
(1967): 33—44.
Teschner, D.: Hausfliegen als Fäkalienbesucher im Stadtgebiet. Z. angew. Zool.
46 (1959): 358—363.
Teschner, D.: Fliegen einer Hamburger Wohnung und in Hamburg
neuaufgefundene Fliegenarten (Diptera). Ent. Mitt. Zool. Staatsinst. Zool. Mus.
Hamburg 37, 2 (1962): 221—282.
Teschner, D.: Die Bedeutung der Nester verwilderter Tauben in Grossstädten.
Anz. Schädlingsk. 37 (1964): 40—43.
Thiele, H. U.: Carabid Beetles in their Environments. Springer-Verlag, Berlin,
Heidelberg, New York 1977.
Thiesmeier, В.: Die Amphibien und ihre Lebensräume in Bochum — Beitrag zum
Amphibienschutz in der Grossstadt. Dortmunder Beitr. Landeskde., naturwiss.
Mitt. Dortmund 18 (1984): 17—46.
Thorpe, W.: The learning abihties of birds. Ibis 93 (1951): 1—52, 252—296.
Tischler, W.: Biozönotische Untersuchungen bei Hausfliegen. Z. angew, Ent. 32
(1950): 195—207.
Tischler, W.: Ein biozönotischer Beitrag zur Besiedlung von Steilwänden. Verh.
Dt. Zool. Ges. Marburg 1950, 214—229 (1951).
Tischler, W.: Biozönotische Untersuchungen an Ruderalstellen. Zool. Jb. Syst.
81 (1952): 122—174.
Tischler, W.: Untersuchungen über das Hypolithion einer Hausterrasse. Pedobiol.
6 (1966): 13—26.
Tischler, W.: Ecology of arthropod fauna in manmade habitats: The problem of
synanthropy. Zool. Anz. 191 (1973): 157—161.
Tischler, W.: Asseln (Isopoda) und Tausendfüsser (Myripoda) eines Stadtparks
im Vergleich mit der Umgebung der Stadt: z um Problem der
Urbanbiologie. Drosera 80 (1980a): 41—52.
Tischker, W.: Biologie der Kulturlandschaft. Eine Einführung. Gustav Fischer
Verlag Stuttgart, New York 1980b.
Titschack, E.: Untersuchungen über den Temperatureinfluss auf die Kleidermotte
(Tineola bisselliella Hum.). Z. wiss. Zool. 124 (1925): 213—251.
Tomialojc, L.: The Urban Population of the Woodpigeon Columba palumbus
Linnaeus, 1758, in Europe —its Origin, Increasee and Distribution. Acta
Zool. Cracov 21 (1976): 585—632.
Tomialojc, L.: Urbanization of birds and the preypredator relations. Animals in
urban environment, Symp. Institute of Zool. 1979, Warszawa (1982): 131—
139.
Topp, W.: Die Besiedlung eines Stadtparks durch Käfer. Pedobiol. 12 (1972):
336—346.
Tormola,T., V aninen, L: Leaf hopper (Homoptera, Auchunorrhyncha) communities
in 2 urban lawns in Central Finland. Ann. Entomol. Fenn. 49 (1983): III—
114.
Trojan, F.: Diptera Tabanomorpha of Warsaw and Mazovia. Memorabilia Zool.
35 (1982): 3—31.
Urbahn, е.: Zunahme von Melanismus-Beobachtungen bei Makrolepidopteren
Europas. Mitt. Münch. Ent. Ges. 61 (1971): 1—15.
Valesova-Zdarkova, E.: Synanthrope Spinnen in der Tschechoslowakei (Arach.,
Araneae). Senckenb. biol. 47 (1966): 73—75.
12. Литература 239
Vater, G.: Hausschaben (Blattella germanica) in Kühlschrank. Angew. Parasitol.
20 (1979): 147—154.
Vater, G.: Flöhe (Siphonaptera) beim IVlenschen. Befundanalyse 1961 bis 1983
im Bezirk Leipzig (DDR). Teil IL Räumliche und zeitliche Verteilung.
Angew. Parasitol. 26 (1985); 27—38.
Vater, G.: Gesundheitsschädlinge in Städten. "Wiss. Z. Karl-iVlarx-Univ. Leipzig,
iVlath.-Nat. R. 35 (1986): 627—639.
Vater, G., Vater, A.: Flöhe (Siphonaptera) beim iVlenschen. Befundanalyse 1961
bis 1983 im Bezirk Leipzig (DDR). Teil I: Arten, Befallsquellen und
Ausbreitung. Angew. Parasitol. 25 (1984): 148—156.
Vecht, J. van der: Die orientalische Mauerwespe Sceiiphron curvatum (Smith,
1870) in der Steiermark, Österreich (Hymenoptera, Sphecidae). Entomo-
fauna 5 (1984): 213—219.
Vepsäläinen, K., Pisarski, В.: The structure of urban ant communities along
a geographical gradient from North Finland to Poland. Animals in urbcn
environment, Symp. Institute of Zool., 1979, Warszawa (1982): 155—168.
Visse, G., van Wingerden, W. K. R. E.: Aerial dispersal of spiders in a city. In:
Bornkamm, R., Lee, J. A., and Seaward, M. R. D.: Urban Ecology. Symp.
Berhn 1980, Oxford, London, Edinburgh, Boston, Melbourne (1982): 344.
Wagner, R.: Die Veränderung der Hummelfauna Cuxhavens in diesem
Jahrhundert. Der Versuch einer Deutung. Ent. Mitt. Zool. Mus. Hamburg 4 (1971):
207—232.
Walther, H., Klausnitzer, В., Richter, K.: Beeinflussung der Natalität von Aphis
fabae Scopoli (Inst., Homoptera) durch anthropogene Noxen, insbesondere
Schwermetalle. Zool. Anz. 212 (1984a): 26—34.
Walther, H., Klausnitzer, В., Richter, K-: Zum Einfluss ausgewählter
anthropogener Noxen auf die saure Phosphatase (E.C.3.1.3.2.) eines clonierten
Stammes von Aphis fabae Scopoli (Ins., Homoptera). Wiss. Z. Karl-Marx-Univ.
Leipzig, Math.-Nat. R. 33 (1984b): 170—177.
Warnecke, G.: Kulturfolger unter den Lepidopteren. Mitt. Münch. Ent. Ges. 25,
(1936): 61—66.
Wasowska, M.: Chrysomelids (Coleoptera, Chrysomelidae) of Warsaw arid
Mazovia. Memorabilia Zool. 34 (1981); 219—233.
Way, J. М.: Roadside verges and conservation in Britain: A review. Biol. Con-
serv. 12 (1977): 65—74.
Wegner, E.: Mosquitoes (Diptera, Culicidae) of Warsaw and Mazovia.
Memorabilia Zool. 36 (1982): 201—216.
Weidner, H.: Die Grossstadt als Lebensraum der Insekten. Verh. 7. Int. Kongr.
Ent. Berlin 2 (1939); 1348—1361.
Weidner, H.: Die Insekten der «Kulturwüste». Mitt. Hamb. Zool. Mus. 51 (1952):
89—173.
Weidner, H.: Die Entstehung der Hausinsekten. Z. angew. Ent. 42 (1958): 429—
447.
Weidner, H.: Die Niststätten verwilderter Tauben als Reservoire für
Ungezieferplagen. Städtehyg. 12 (1961): 91—94.
Weidner, H.: Die Entwicklung der Arthropodenfauna im Stadtgebiet Hamburg.
Verh. 11. Int. Kong. Ent. Wien 1960, 3 (1962): 175—184.
Weidner, H.: Schädlinge an Arzneidrogen und Gewürzen im Hamburg. Beitr.
Ent. 13 (1963): 527—545.
Weidner, H.: Zur Bedeutung der Verschleppung für die Verbreitung der Insekten.
Z. angew. Ent. Hamburg 65 (1970): 334—338.
Weidner, H.: Insekten als Schädlinge an Drogen in Apotheken und Drogerien.
Anz. Schädlingsk. Pflanzensch. 46 (1973): 177—180.
Weidner, H.: Häufigkeitsschwankungen bei Hausinsekten in Abhängigkeit von
Hausbau und Wohnweise der Menschen Der prakt. Schädlingsbek. 27 (1975):
54—6$.
240 12. Литература
Weidner, Н.: Die Gelbfüssige Bodentermite Reiiculotermes flavipes (Kollar, 1837)
in Hamburg (Isoptera). Ent. Mitt. Zool. Mus. Hamburg 6 (1978): 49—
180.
Weidner, H.: Einschleppung von Heuschrecken (Saltatoria und Phasmida) nach
Hamburg. Anz. Schädlingskde., Pflanzenschutz, Umweltschutz 54 (1981):
65—67.
Weidner, H.: Nach Hamburg eingeschleppte Cerambycidae (Coleptera). Anz.
Schädlingskde., Pflanzenschutz, Umweltschutz 55 (1982): 113—118.
Weidner, H.: Herkunft einiger in Mitteleuropa vorkommender Vorratsschädlinge:
1. die Sitophilus-Arten (Col., Curculionidae). Mitt. int. Ent. Ver.
Frankfurt/M. 8 (1983a): 1 — 17.
Weidner, H.: Nach Hamburg eingeschleppte CoUembola und Zygentoma. Anz.
Schädlingskde., Pflanzenschutz, Umweltschutz 56 (1983b): 10,5—107.
Weidner, H.: Neue Untersuchungen über Vorkommen und Verbreitung der
Schaden in der DDR und einige Bemerkungen über die Häufigkeitszunahme der
Deutschen Schabe. Prakt. Schädlingsbek. 35 (1983c): 151—153.
Weiss, I., Sarbu, S. I.: Zur Kenntnis der Spinnen und Weberknechte des
Botanischen Gartens lasi. Studii si Comunicari, St. nat. 21 (1977): 225—243.
Wendland, V.: Die Wirbeltiere Westberlins. Sitz. ber. Ges. Naturf. Freunde zn
Berlin (N. F.) 11 (1971): 4—128.
Winiarska, G.: Noctuida (Noctuidae, Lepidoptera) of Warsaw and Mazovia.
Memorabiha Zool. 36 (1982): 185—200.
Wlodarczyk, ].: Psocoptera in the parks of Lodz. Fragm. Faun. Warszawa 8
(1960): 469—483.
Zacher, F.: Die Gliedertiere (Arthropoda) der Mühlen und Getreidespeicher in
Deutschland. Mitt. Ges. Vorratsschutz, Sonderh. (1938): 1—48.
Ziarkiewicz, Т., Kozlowska, A.: Materialen zur Kenntnis der
Artenzusammensetzung von den Schwebfliegen (Syrphidae, Diptera), die auf den
Ziersträuchern in Blattlauskolonien auftreten, Pol. Pismo Ent. 43 (1973): 621 —
626.
Zlotorzycka, /.: Observations on the differentiation of mallophagan fauna on
birds in and outside towns. Wiadomosci parazytol. (Wroclaw) 23 (1977a):
223—226.
Zlotorzycka, J.: Parasites of wild birds and mammals, in conditions of anthro-
popressure. Wiadomosci parazytol. (Wroclaw) 23 (1977b): 195—202.
Zöllick, H.: Notizen zur Invasion von Zwergfledermäusen aus dem Stadtgebiet
Rostock — Pipistrellus pipistrellus (Schreber, 1774), Natur und Umwelt,
Beitr. aus dem Bezirk Rostock, H. 1 (1980): 65—67.
Zwölfer, H.: Die Bewertung von Hecken aus tierökologischer Sicht. Laufener
Seminarbeitr. 5/82, Akad. f. Natursch. u. Landschaftspflege (1982): 130—
134.
ПРЕДМЕТНЫЙ УКАЗАТЕЛЬ
Адмирал 211
Аллеи 50
Амфибии 200, 209
Анализ репрезентационный 185
Антофоры 34
Бабочки 85, 111, 112
— дневные 82, 211
Балконы 35, 36, 45
Белки 81
Белозубки 81
Белокрылки 21
Биохоры 15
Блестяики 95, 173, 175
Блохи 17, 95, 109
Божьи коровки 17, 36, 99, 144, 148,
177
Брюхоногие 108
Бурундук 96
I
Вазоны 49
Виды
— адвентивные 92, 93, 94
—^оппортунистические 137
— средиземноморские 96, 97, 98, 99,
101, 102, 108
— термофильные 15
Водолюбы 94
Воробей домовый 31, 35, 37, 44, 45,
46, 53, 66, 72, 73, 104, 200, 207
Ворона 30
Вредители запасов 18
— материалов 18
Вши 150
Вяхирь 50, 72, 82, 150
Галка ЭО, 37, 73, 10, 190, 208
Галлообразователи 135, 156
Голубь сизый 16, 17, 37, 46, 72, 73,
104, 207
Горлица кольчатая 37, 39, 46, 47, 50,
66, 102, 190, 208
Горихвостка
— чернушка 39, 46, 53, 106, 190
— садовая 33, 50, 190, 208
Горностай 81, 150
Градиенты
— комплексный коксовый 153
— микроклиматический 59
— R—Н 152
— R—М 152
— А—Е 152, 153, 154, 160, 179, 188
— С—С 152
Грач 72, 73, 208
Двукрылые 17, 21, 45, 63, 76, 112,
144, 146, 177
Доминирование общее 161
Дровосек домашний 33
Дрозд
— белобровик 208
— певчий 208
— черный 31, 35, 46, 47, 53, 1»8, 200,
208
Дрозофилы 31, 32, 141, 142
Долгоносики 113, 164, 173
Дятел зеленый 208
Еж европейский 53, 81, 199, 208, 205
206
Ежемухи 147
Жаба
— зеленая 209
— обыкновенная 79, 200, 209
Жаворонок хохлатый 39, 46, 53, 83,
89, 90, 109
Животные домашние 109
Жужелицы 25, 34, 49, 51, 57, 59, 60,
83, 133, 140, 141, 154, 156, 161, 163,
164, 167, 168, 179, 185, 201
242
Предметный указатель
Жуки 25, 47, 49, 51, 60, 78, 79, 91,
112, 146, 163
Журчалки 51, 53, 70, 147
Завирушка лесная 31
Зарянка 208
Заяц-русак 200
Зеленушка 33, 35, 46, 47, 53, 190
Земноводные 198, 200
Златки 126
Златоглазки 17
Зоны
— краевые 136, 137
— центральные 136, 137
— транспортные 51
Зоопарки 78
Изгороди живые 50
Индекс
— Бергера — Паркера 163
— Брийуэна 177
— Вайнштейна 183
— перекрывания 183
— Ренконена 183
— Симпсона 163
— синантропизации 118, 119
— Шеннона 177
Камышевка болотная 89, 190, 207
Канюк обыкновенный 31
Кладбища 79, 179
Клещи 47, 78, 195
— домашней пыли 19
— чесоточные 29
Клопы 19, 49, 93, 102, 112, 177
Книгохранилица 31
Кобылка египетская 93
Кожан
— поздний 37, 198
— двухцветный 106, 107
Кожееды 27, 31, 32, 94, 149
Кошки 109, 199
Комары 26, 44
Кролик дикий 66, 81, 83, 205
Крот обыкновенный 87
Крыса
— черная 18
— серая 37, 39, 141, 205
— водяная 87
Крыши 34, 35
Кузнечик зеленый 50
Куница каменная 39, 81, 106, 150, 205
Лазоревка обыкновенная 30, 31, 50,
208
Ласка 81
Ласточка
— городская 35, 39, 44, 45, 56, 66,
106, 208
— деревенская 35, 53, 105, 106, 190.
207, 208
Листоблошки 177
Листоеды 163, 175
Листовертки 85, 113
«Лужайки кобылковые» 210
Малатдегидрогеназа 191, 192, 193
Мельницы 29
Многоножки 24, 70, 75, 140, 150, 163
Мокрицы 88, 150, 163
Моллюски 26, 147, 209
— брюхоногие 140, 198
Моль платяная 28
Мосты 51, 66
Муравьи 21, 29, 201
Мухи 44, 119, 120
Мыши летучие 16, 105, 199, 205, 206,
207
Мышь
— домовая 23, 39, 90, 205
— желтогорлая 81
— лесная 81
— полевая 81, 90, 142
Мясокомбинаты 30
Наездники 147
Насаждения
— древесные 204
— зеленые 49, 50
Ногохвостки 17, 21, 167
Ноксы 14, 153, 191, 196, 202. 203
Осы 33, 36, 63, 78, 146, 147
Озеленение зданий 205
Павлиний глаз 17, 211
Парки 68, 69, 70, 71
Пауки 21, 24, 33, 35, 57, 66, 69, 74,
84, 100, 105, 108, 121, 128, 133, 140,
142, 146, 150, 163, 167, 177, 212
Перепончатокрылые 210, 211
Пироглифиды 19, 20
Погреба винные 32
Подвалы 24, 25
Полевка обыкновенная 81, 90
Пустельга обыкновенная 104, 190
Пустыри 89, 90
Пчелы 78, 86, 112, 211
Пяденицы 82, 141
Разнообразие
— видовое 177
— структурное 181
Районы
I
предметный указатель
243
— старой застройки 37
— новостроек 39, 46
Редувий ряженый 19
Сады 73, 82, 83, 84
Сверчок домовый 24, 29, 93, 102
Сеноеды 17, 31, 33, 47, 48, 49, 93, 146,
150
Сенокосцы 33, 74, 100, 107, 140, 167,
177
Сетчатокрылые 17, 146
Синица большая 30, 50, 72
Сипуха 104, 105
Скворец обыкновенный 30, 37, 50, 53,
72, 208
Славки
— завирушка 46, 50, 87, 190, 208
— серая 89, 190, 207, 208
— Черноголовка 208
Снегирь 33, 208
Совки 35, 82
Сооружения железнодорожные 66, 67
Сорока 30, 150
Стафилиниды 29, 34, 71, 140, 177
Стриж черный 37, 46, 53, 105, 190
— белобрюхий 105
Таракан
— рыжий, прусак 21, 28, 29, 93, 102,
149
— черный 28, 29, 93, 101, 149
Теплицы 87, 88
Тли 21, 44, 58, 112, ИЗ, 144, 146, 175,
177, 190, 193, 194, 211
Точильщики 16, 126
Трипсы 36, 89, 146
Тритон обыкновенный 79, 209
Трясогузка белая 53, 66, 190
Усачи 16, 95
Холодильники 21, 23
Центры городские 36, 37
Цикадовые 34, 35, 112, 146, 147
Черви дождевые 57, 76, 84, 140, 147,
148, 198
Чердаки 15, 16, 17, 18
Черепахи 95, 96
Чернотелки 25, 29
Шелкопряды 82, 113
Шершни 210
Щелкуны 161
Щетинохвостки 18, 29, 93
Этажи 18, 19
Эффект
— барьерный 61
— краевой 156
Ящерицы 208
— живородящая 58
— прыткая 79
«Ящики гнездовые» 210 "
УКАЗАТЕЛЬ ЛАТИНСКИХ НАЗВАНИЙ
АЬах parallelus— 185
— parallelepipedus—188
Abraeus parvulus — 78
Acarus slro — 29
Achaearanea tepidariorum — 28,
128
Acheta aegra — 34
— domestica — 24
Acrididae— 67
Adalia bipunctata—-18, 144, 148
Aderus oculatus — 91
Adistemia watsoni — 95
Adoxus obscurus — 112
Agelenidae — 128
Aglais urticae— 17
Agriotis exclamationis — 35
Agonum muelleri — 60
— sexpunctatum — 60
Ahasverus advenae — 48
Allajulus londinensis — 21, 70, 76
— occultus — 76
Ällolobophora georgii — 84
— let erica — 101
Amara aenea —83, 189
Amaurobiidae— 128
Amaurobius ferox — 24
— similis — 33
Ameroseius — 47
— plumigerus — 47
Amischa forcipata — 48
Amphimallon solstitiale — 83
Anacridium aegyptium — 93
Ancistrocerus parietlnus — 33
— trlfasciatus — 33, 34
Andrena armata — 86
— barbilabris —63
— clarliella — 86
— haemorrhoa — 86
— jacobi — 86
— minutula — 78
— nigroaenea — 86
— tibialis —86
— varians — 86
Andrenidae — 63
Anisotomidae — 60
Anitys rubens— 91
Anobiidae — 16
Anobium punctatum — 27
— thomsoni — 126
Anthocoridae— 177
Anthophora acervorum — 34
Anthophoridae — 63
Anthrenus scrophulariae — 27
Aphis fabae— 191, 193, 195
— pomi —58
— oenotherae — 111
— sambuci — 191
Arion lusitanicum — 95
Armadillidium vulgare —57, 70, 75,
100
Araneidae— 128
Araneus diadematus — 33
— umbratls — 57
Asilidae — 177
Bembidion lampros — 60
— quinquestriatum — 25
— quadrimaculatum — 3^
Bethylidae — 148
Blston betularis— 141
Blaps lethifera — 25
— mortisaga — 25
— mucronata — 25
Blatta orientalis — 28
Blattella germanica — 21, 28
Braconidae — 148
Calathus fuscipes —83, 168,
Calllphora vicina — 120
CalUphoridae —30
Carabidae —60
Carabus coriaceus — 188
— nemoralis— 188
Cepaea — 141
Cerceris rybensis — 78
Chalcidoidea — 148
89
Указатель латинских названий
245
Chelifer cancroides — 31
Chloropidae—175
Chrysididae— 147
Cimex lectularius — 19
Clubionidae — 74
Colletes daviesianus — 33, 78
Crabro vagus — 36
— peltarius — 63
Craspedacusta sowerbyi — 88
Crossocerus —33, 34, 126
Culex pipiens — 26
Culiseta annulata — 26
Cyphepeira ixobola — 33, 150
— sericata — 66, 150
Dermatophagoides farinae—19, 21
— pteronyssinus— 19, 21
Dermestes haemorrhoidales — 27
Dictyna civica —32, 100
Dictynidae — 128
Dolichopodidae — 177
Dolichovespula saxonica — 28, 35
Dorypteryx pallida — 48, 49
Ectemnius sexcinctus — 36, 147
Elateridae — 60
Empididae — 177
Ephydridae — 175
Erigone atra —35, 143
— dentipalpis — 35, 143
Evaniidae — 148
Falagria sulcata — 78
Falco subbuteo — 109
Folsomia Candida — 21
— fimetaria — 21
Forficula auricularia — 36, 57
Formicidae — 63
Glomeridae — 24
Gijaphosidae —74, 128
Halictidae —63
Haltica oleracea — 112
Harpalus aeneus — 83, 189
— rufipes —
Heloridae— 147
Heteropodidae — 128
Histeridae —29
Humerobates rostrolamellatus — 195
Hylaeus — 34
— communis — 34, 36, 78
— hyalinatus — 34, 36, 78
— pictipes —33
— punctatus — 99
Hyloniscus riparius — 70, 75
Hylotrupus bajulus — 16, 33
Ichneumonidae—147,148
Ictinogomphus decoratus malaenops —
88, 89
Idiopterus nephrelepidus ■— 89
Inachis io — 17
Iridomyrniex hamulus — 23
Isotoma viridis — 70
Itonididae — 175
luloidea — 24
Japyx braueri — 92
Lacerta muralis — 57, 58, 95
— viridis — 78
— vivipara — 58
Lasioglossum calceatum — 78
— morio — 33, 78
— sexstrigatum — 63
Lasius niger —21, 63
Lathridiidae — 48
Lathridius bergrothi — 26
Lepisma saccharina— 18, 101
Lepthyphantes leprosus — 28, 109, 128
— nebulosus —28, 109
— tenuis — 84
Limax flavus — 26
— maximus — 26
Linyphiidae — 128, 143
Liocranidae — 128
Lithobius forficatus — 24, 75
— melanops — 24
— mlcrops — 75
Lucilla — 30
— caesar — 120
— sericata — 119
Lumbricus rubellus — 84
— terrestris — 57
Lycosidae — 74
Marava arachidis — 31
Meioneta rurestris— 35, 143
Melittidae — 63
Meconema meridionale — 98
' — thalassinum — 98
Meta menardi — 109, 128, 150
— merianae —66, 109, 128
Micromus variegatus—17
Monomorium pharaonis — 23, 27
Monotarsobius dubosqui — 78
JV1.utillidae — 63
Nabidae—177
Nacerda ferruginea — 67
Nebria brevicollis—168
Necrophorus humator — 78
Neelus minimus — 21
Nelima semproni—100
246
Указатель латинских названий
Neriene montana— 121
Nesticidae— 128
Nesticus cellulanus—m, 128
Nigma walckenaeri —32, 100. 121
Niptus hololeucus — 32
Nitidulidae —60
Nomada ruficornis — 86
— sheppardana — 63
Oecophoridae — 17
Oenophila v-flavum — 32
Oenophilidae —32
Oenopia conglobata—17, 18, 148
Oonopidae— 128
Oonops domesticus — 31, 150
Opillo canestrinii — 107
Orthoperus atomarius — 26, 32
Osmia caerulescens — 34
— rufa — 34, 78
Oxyna parietina — 89
Pachygnatha degeeri — 84
Parandra brunnea — 95
Pemphredon montanuus — 36
Periplaneta americana — 93
— australasiae — 3\
Phalanglum opillo — 107
Philaenus spumarius — 112
Philontus addendus — 86
Pholcidae— 128
Pholcus opilionoides — 28
— phalangioides — 24, 47, 121, 150
Platygastridae — 147
Polistes galllcus — 35
— nimpha —35
Polydesmus inconstans — 24
— superus — 76
Pompilidae — 63
Porcellio dilatatus — 88, 100
— laevis —88, 100
— scaber — 24, 57, 70, 75
Proctotrupidae— 147
Proctotrupoidea — 147, 149
Proserpinus proserpina — 111
Protaetia aeruginosa — 91, 92
Pseudococcus adonidum — 21
Psyllina—m
Pterostichus cupreus — 57
— niger — 59
Pyroglyphidae — 19
Reduvius per sonatas — 19
Reticulotermes flavipes — 27
Salticidae — 128
Salticus scenic us — 33, 108
Sarcophagidae —30, 148
Scelionidae— 147
Sceiiphron — 95
Seira domestica — 21, 31, 98
Sicariidae— 128
Silphidae — 60
Simplocaria semistriata — 34
Sinella coeca —21, 88
Sitophilus — 29, 95
— granarius — 116
Sitotroga — 29
Sitticus pubescens —33, 108
Sivanoprus fagi — 82
Sphecidae — 63
Sphodrus lecophthalmus — 25
Slaphylinidae — 60
Steatoda bipunctata — 24, 128
— triangulosa — 121
Tachinidae— 147
Tachycines asynamorus — 89
Tachys blsulcatus — 25, 26
Tapinoma melanocephalum — 27
Tegenaria atrica — 47
— domestica —18
— ferruginea —28
Tenebrionidae — 60
Tettigonia viridissima — 50
Textrlx denticulata — 33
Thaumetopoea pinivora — 92
Theridiidae— 128
Theridion melanurum — 33
Thermobia domestica — 29, 101
Thomisidae — 74
Thorictodes heydeni — 95
Thylodrias contractus — 27
Tineidae —32
Tineola bisselliella — 28
Tortricidae — 85
Trachyderus striatus ■— 95
Trechus austriacus — 26
Trilobium destructor — 150
Triphaena pronuba — 35
Trogoderma — 29
— angustum — 27
— glabrum — 34
Trypoxylon clavicerum — 33
— figulus — 33
Typhaea stercorea — 26
Tyrophagus putescentlae — 48
Unciger foetldus — 76
Wesmaelius subnebulosus— 17, 146
Xantholinus linearis — 34, 71
Zygia oblonga — 94
Zygiella atrica — 33
— x-notata —33, 100, 150
ОГЛАВЛЕНИЕ
предисловие ^
Дополнительная литература по экологии городской среды ... 10
1. предисловие (перевод и. в. орловой) И
2. структура города и фауна 14
2 1. Строения 14
2.1.1. Непостоянно отапливаемые жилые дома 15
2.1.2. Постоянно отапливаемые здания 27
2.1.3. Складские и некоторые производственные помещения . 28
2.2. Прочие наземные местообитания 32
2.2.1. Внещняя оболочка зданий 32
2.2.2. Застроенные территории 36
2.2.3. Транспортные зоны 51
22.4. Озелененные площади 67
2.2.5. Пустыри 89
' 2.2.6. Остатки негородских экосистем 90
3. Происхождение и состав городской фауны 92
3.1. Реликтовые виды 92
3.2. Адвентивные виды 92
3.3. Средиземноморские виды 97
3.4. Эпилитные виды 103
3.5. Троглобионты 108
3.6. Домашние животные 109
3.7 Заселение животными гемерохорных растений 109
4. синантропия и урбанизация {перевод И. М. Маровой) . 116
5. Последствия изоляции городских островных местообитаний 129
5.1. Динамическое равновесие видов 129
If 5.2. Зависимость видового разнообразия от площади .... 130
5.3. Формирование краевых зон с сильным антропогенным
воздействием 136
5.4. Смещения в видовом спектре краевой зоны 136
5.5. Смещения в видовом спектре центральной зоны 137
5.6. Нарушение структуры доминирования в островных
местообитаниях 138
5.7. Дисгармония фауны 139
5.8. Различие фаун 140
5.9. Генетическая дифференциация 141
5.10. Пути заселения островных местообитаний 142
6. пищевые цепи и сети 144
7. урбанистические градиенты 151
7.1. Число видов 156
7.2. Численность 160
^48
7.3. Общее доминирование и индексы доминирования . . . . 161
7.4. Биомасса 163
7.5. Экологические группы и географическое распространение . . 164
7.6. Трофические уровни 170
7.7. Синэкологические характеристики 177
7.7.1. Видовое разнообразие 177
7.7.2. Равномерность распределения 179
7.7.3. Видовое богатство 180
7.7.4. Структурное разнообразие 181
7.8. Индексы сходства 181
7.9. Сообщества и репрезентационный анализ 185
8. Реакции животных на своеобразие городских условий . . . . 191]
8.1. Физиология 191
82. Морфология 196
8.3. Этология 196
9. урбанистические факторы смертности 198
9.1. Техногенные и структурно обусловленные факторы смертности 198
9.2. Уличное движение 198
10. возможности помощи животным в городе 202
10.1 Млекопитающие 205
.10.2. Птицы 207
10.3. Пресмыкающиеся 208
10.4. Земноводные 209
10.5. Моллюски 209
10.6. Длинноусые прямокрылые 210
10.7. Короткоусые прямокрылые 210
10.8. Перепончатокрылые 210
10.9. Чещуекрылые . 211
10.10. Почвенные животные 212
10.11. Пауки 212
11. заключительные замечания 213
12. литература 223
предметный указатель 241
указатель латинских названий 244