/




















































































































































Text
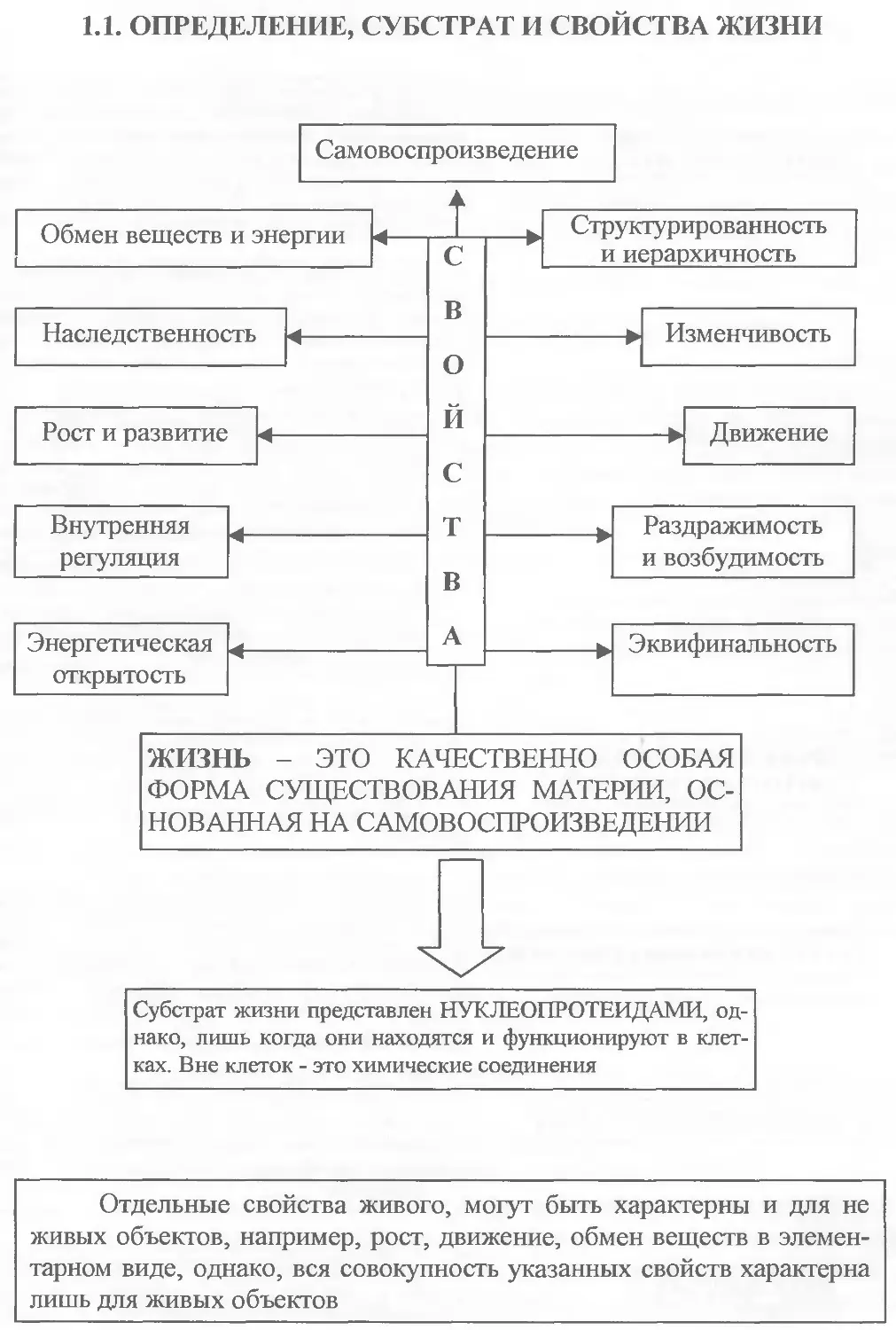
1.1. ОПРЕДЕЛЕНИЕ, СУБСТРАТ И СВОЙСТВА ЖИЗНИ
МОЛЕКУЛЯРНЫЙ
Молекулярный уровень представлен “биологическими молекулами”, основными из которых являются ДНК, три вида РНК и белки, определяющие потоки генетической информации в клетках. Основу жизни “биомолекулы” составляют лишь в клетке. Вне клетки это химические вещества.
КЛЕТОЧНЫЙ
Уровень представлен одноклеточными животными и растениями, а также клетками многоклеточных организмов. На клеточном уровне происходит сопряжение механизмов передачи генетической информации и превращения веществ и энергии. С этого уровня начинается собственно жизнь.
Сходные по размерам и функциям клетки составляют основу тканей. В разных тканях объединены клетки одного или нескольких видов, а также межклеточное вещество. У животных и человека выделяют четыре типа тканей: эпители-альную, соединительную, нервную и мышечную,
У многоклеточных организмов ткани формируют органы, объединенные в системы. Они обеспечивают важнейшие исполнительные и регуляторные функции, благодаря которым особи существуют как единое целое.
ОРГАНИЗМЕННЫЙ ИЛИ ОНТОГЕНЕТИЧЕСКИЙ
Одноклеточные и многоклеточные организмы представляют собой интегрированные единицы жизни. В ходе онтогенеза организмы реализуют наследственную информацию в конкретные биологические структуры и являются элементарным материалом для эволюционно-го процесса.
ПОПУЛЯЦИОННО-ВИДОВОЙ
Совокупность организмов одного вида, населяющих одну территорию и свободно скрещивающихся между собой, называют популяцией. Совокупность популяций составляет биологический вид. Элементарные эволюционные факторы вызывают эволюционно значимые изменения генофонда популяций и приводят к образо-ванию новых видов.
БИОГЕОЦЕНОТИЧЕСКИЙ
Реально вид всегда сосуществует в сообществе с представителями других видов и населяет территорию с конкретными абиотическими характеристиками, которые все вместе образуют устойчевые открытые системы - биогеоценозы.
БИОСФЕРНЫЙ
Комплекс биогеоценозов нашей планеты составляют биосферу - оболочку жизни. На этом уровне объединены все вещественно энергетические круговороты.
2.1. КЛЕТКА - ЭЛЕМЕНТАРНАЯ ЕДИНИЦА ЖИВОГО
КЛЕТКА - представляет собой обособленную наименьшую структуру, которой присуща вся совокупность свойств живого. В природе планеты клетке принадлежит роль элементарной структурной, функциональной и генетической единицы. Клетка составляет основу строения, жизнедеятельности и развития всех живых форм - одноклеточных, многоклеточных и даже неклеточных.
ИСТОРИЯ ОТКРЫТИЯ И ИЗУЧЕНИЯ КЛЕТОК
В 1665 году английский физик Р. Гук, рассматривая под микроскопом срез коры пробкового дерева и сердцевину бузины и камыша, обнаружил ячейки, напоминающие пчелиные соты, которые он назвал клетками (от латинского “cellula”). В дальнейшем во второй половине XVII века “клеточное” строение растений наблюдали итальянец М. Мальпиги и англичанин Н. Грю. Одноклеточные организмы -бактерии впервые обнаружил голландец А.Левенгук.
Чешский ученый Я. Пуркинье в 1825 году полужидкое студенистое образование назвал протоплазмой (от греческих слов protos — первый и plasma - образование).
Английский ботаник Б. Броун в 1831 году впервые открывает клеточное ядро.
В 1839 году немецкий зоолог Т.Шванн публикует книгу “Микроскопические исследования о соответствии в структуре и росте животных и растений”, в которой он впервые, опираясь на труды немецкого ботаника М. Шлейдена, сформулировал положения клеточной теории. В 1958 году была опубликована книга немецкого патолога Р. Вирхова “Целлюлярная патология”, дополнившая клеточную теорию положениями о соотношении клеток и многоклеточных структур.
СОВРЕМЕННЫЕ ПОЛОЖЕНИЯ КЛЕТОЧНОЙ ТЕОРИИ
1. Клетка является наименьшей единицей живого.
2. Клетки разных организмов сходны по своему строению.
3. Размножение клеток происходит путем деления исходной клетки.
4. Многоклеточные организмы представляют собой комплекс клеток и их производных, интегрированных в системы тканей и органов, связанные между собой межклеточными, гуморальными и нервными формами регуляции.
КЛЕТКИ
СРАВНИТЕЛЬНАЯ МОРФОФУНКЦИОНАЛЬНАЯ ХАРАКТЕРИСТИКА ПРОКАРИОТИЧЕСКИХ (БАКТЕРИАЛЬНЫХ) И ЭУКАРИОТИЧЕСКИХ (ЖИВОТНЫХ) КЛЕТОК
ПРИЗНАКИ ПРОКАРИОТЫ ЭУКАРИОТЫ
Функцию хранения и передачи наследственной информации выполняет... нуклеоид ядро
Ядерная оболочка нет есть
Число хромосом на клетку обычно 1 несколько
Форма хромосом кольцевая линейная
Оболочка клетки представлена ... Плазмалеммой, клеточной стенкой, капсулой Только плазмалеммой
Рибосомы есть есть
Размеры: рибосом; 70S 80S
большой субъединицы; 50S 60S
малой субъединицы. 30S 40S
Локализация рибосом только цитоплазма цитоплазма и эндоплазматический ретикулум
Митохондрии нет есть
Пластинчатый комплекс Гольджи нет есть
Лизосомы нет есть
Эндоплазматический ретикулум нет есть
Цитоплазматическая система микротрубочек возможно имеются у некоторых представителей есть
Диаметр жгутиков 0,01 - 0,02 мкм около 0,2 мкм
Деление простое митоз
Компартментализация нет есть
Нить
Длина 8 мкм
Кольца
Нить
Мезосома
Клеточная стенка
ТОЛЬКО ДЛЯ
Крюк Ось
КЛЕТОК
Жгутик
Липидные глобулы
Желатинозный слой
Фосфатная гранула
Белковая гранула
Малая суб-частица
для ОБОИХ ТИПОВ КЛЕТОК
ТОЛЬКО ДЛЯ - этого
ТИПА КЛЕТОК
Фотосинтезирующие мембраны (ламеллы)
Большая суб-частица
СИНЕ-ЗЕЛЕНАЯ ВОДОРОСЛЬ
БАКТЕРИЯ (3—4мкм) (грампо ложите льная)
Деухцепочечная ДНК (кольцевая)
Цитоплазма
Клеточная стенка
Белки
Периплазматнческое пространство
Плазматическая мембрана
Липидный бислой
Рибосомы
Полисомы
.Триплеты микротрубочек Окаймленная ямка Пиноцитозный пузырек
Комплекс Гольджи
► Десмосома
Лизосома —
Тонофиламенты
Жировав капля--
Центриоль
Плотный контакт -
| ЖИВОТНАЯ КЛЕТКА |
Окаймленный пузырек
Волокнистая область
Г падкий эндоплазматический ретикулум
Плазмодесма
Вакуоль
Комплекс Гольджи
ДНК
Хлоропласт
Г падкий эндоплазматический ретикулум
\ Шероховатый
\ эндоплазматический
\ ретикулум Рибосомы
(и полирибосомы)
Конденси' ровен ный хроматин
Плазматическая । '< мембрана
20-30нм
ТОЛЬКО ДЛЯ КЛЕТОК этого ТИПА
Ядерные поры
Ядрышко
для ОБОИХ " ТИПОВ
КЛЕТОК
Митохондрии
Клеточная стенка (слои целлюлозы)
ТОЛЬКО
для
>КЛЕТОК этого ТИПА
I РАСТИТЕЛЬНАЯ клетка ]
Фотосинтезирующие ламеллы
КЛЕТКА
ЦИТОПЛАЗМА
1) Ядерная оболочка (двумембранная),
2) Кариолимфа,
3) Хроматин (хромосомы),
4) ядрышко
мембранные
немембранные
Плазмалемма, митохондрии, эндоплазматический ретикулум, пластинчатый комплекс Гольджи, лизосомы, пероксисомы.
_____________L
Рибосомы, центриоли, микротрубочки, микрофиламенты, межуточные филаменты.
Трофические, секреторные, экскреторные, пигментные, неспецифические.
БЛАГОДАРЯ РАЗВЕТВЛЕННОЙ СЕТИ МЕМБРАННЫХ ОРГАНЕЛЛ ЭУКАРИОТИЧЕСКИЕ КЛЕТКИ РАЗБИТЫ НА ОТДЕЛЬНЫЕ УЧАСТКИ - КОМПАРТМЕНТЫ, ЧТО ПОЗВОЛЯЕТ РАЗГРАНИЧИТЬ МЕТАБОЛИЧЕСКИЕ И ИНФОРМАЦИОННЫЕ ПРОЦЕССЫ
ВО ВРЕМЕНИ И ПРОСТРАНСТВЕ
2.6 ПЛАЗМАТИЧЕСКАЯ МЕМБРАНА (ПЛАЗМАЛЕММА)
Плазмалемма - это барьер и, одновременно, связующее звено между цитоплазмой и окружающей клетку средой.
Каждому типу мембран присущи свои определенные липидные и белковые компоненты, однако основные структурные и функциональные особенности характерны как для плазмалеммы, так и для внутри-клеточных мембран.
ХИМИЧЕСКИЙ СОСТАВ МЕМБРАНЫ
БЕЛКИ 1) интегральные 2) полуинтегральные 3) поверхностные 4) примембранные ЛИПИДЫ 1) фосфолипиды 2) гликолипиды 3) холестерин УГЛЕВОДЫ (в составе гликолипидов и гликопротеинов) 1) сиаловые кислоты 2) глюкоза 3) галактоза
4) ацетилгалактозамин
СТРУКТУРА МЕМБРАНЫ БЫЛА ПРЕДЛОЖЕНА В 1971 ГОДУ НИКОЛСОНОМ И СИНГЕРОМ. В СООТВЕТСТВИИ С ЭТОЙ МОДЕЛЬЮ МЕМБРАНЫ СОСТОЯТ ИЗ НЕПРЕРЫВНОГО ДВОЙНОГО СЛОЯ ЛИПИДНЫХ МОЛЕКУЛ С ПОГРУЖЕННЫМИ В НЕГО БЕЛКАМИ
Наиболее “массовыми” липидами мембран являются фосфолипиды. У фосфолипидов две спиртовые группы глицерина образуют сложноэфирные связи с остатками высших жирных кислот, а третья - этерифицирова-на с каким либо фосфорорганическим соединением. Одна жирная кислота, как правило, насыщенная, а вторая - нет. На рисунке представлен фосфатидил холин. Его молекула состоит из диацилглицерина к которому по третьей спиртовой группе через ортофосфорную кислоту присоединена молекула холина.
Плазматические мембраны эукариот содержат большое количество холестерола. Холестерол придает жесткость и упругость мембранам.
Гликолипиды обнаруживаются только в наружном липидном слое мембраны и составляют около 5% всех его молекул. Их углеводные группы ориентированы к поверхности клетки. В животных клетках гликолипиды построены на основе церамида. Церамид образуется из аминоспирта сфингозина и жирной кислоты. Углеводы, например галактоза, присоединяются к спиртовой группе церамида. Гликолипиды делятся на полярные и нейтральные. На рисунке представлен глюкоцереброзид (справа) полученный присоединением к церамиду (слева) молекулы глюкозы.
Основные структурные особенности биологических мембран определяются липидами, но большинство специфических функций осуществляется белками. По характеру расположения в билипидном слое белки мембран можно разделить на четыре класса: 1) интегральные; 2) полу-интегральные; 3) поверхностные и 4) примембранные. По биологической роли белки мембран также можно подразделить на четыре основные группы: а) белки-ферменты; б) белки переносчики; в) рецепторные и г) структурные белки.
1) Интегральные белки пронизывают мембрану насквозь. Обычно они формируют ионные каналы и системы активного и облегченного транспорта. 2) Существование полуинтегральных белков признается не всеми авторами, их функция изучена в наименьшей степени. 3) Поверхностные белки в основном лежат на внутренней поверхности мембраны, не проникая в ее толщу. Они являются структурными и ферментными. Белки внешней поверхности обычно несут олиго- или полисахаридные фрагменты (элементы гликокаликса) и выполняют рецепторные функции. 4) Примембранные протеины, как правило, выполняют ферментативные функции.
На наружной поверхности мембран всех эукариотических клеток имеются углеводы. Они представлены олиго- и полисахаридными цепями, ковалентно присоединенными к мембранным белкам (гликопротеины) и к липидам (гликолипиды). Содержание этих углеводов по массе составляет от 2% до 10% от массы мембраны.
поверхностный
Углеводы, расположенные на поверхности мембраны образуют специфический слой, называемый гликокаликсом. В животных эукариотических клетках основными углеводами являются N-ацетилнейраминовая (сиаловая) кислота, N-ацетилгалактозамин, галактоза и глюкоза. Функция углеводов гликокаликса разнообразна и до конца невыяснена. Возможно, что некоторые углеводы принимают участие в процессах межклеточного узнавания. В частности они определяют антигенные свойства эритроцитов и формирование групп крови системы АВО.
Наиболее сложные гликолипиды — ганглиозиды, содержащие один или несколько остатков N-ацетилнейраминовой кислоты, несущей отрицательный заряд, могут выполнять рецепторную функцию. Установлено, что ганглиозид Gmi в эпителиальных клетках кишечника действует как рецептор для холерного токсина.
ГАНГЛИОЗИД GMi
1) N-ацетилнейраминовая кислота;
2) Галактоза;
3) Глюкоза;
4) N-ацетилгалактозамин.
Гидрофобные хвосты представлены церамидом.
2.10. ТРАНСПОРТ ВЕЩЕСТВ ЧЕРЕЗ МЕМБРАНУ
Транспортируемые
________________L____i______I__________________________________________
По другой классификации транспорт может быть индивидуальным (унипорт) или совместным (котранспорт). Последний представлен симпортом и антипортом.
Транслортируемм
УНИПОРТ
СИМПОРТ
АНТИПОРТ
котранспорт
2.11. СТРУКТУРА И ФУНКЦИЯ КЛЕТОЧНОГО ЯДРА
Ядро обеспечивает хранение, поддержание и реализацию наследственной информации. Основными структурными элементами ядра являются: 1) оболочка (двумембранная); 2) ядрышко; 3) кариоплазма; 4) хроматин (хромосомы). Ядерные мембраны отделяют содержимое ядра от цитоплазмы и их контакт происходит только через поры. Ядрышко осуществляет сборку рибосом. Кариоплазма (карио-лимфа) является внутренней средой ядра и по своей функции аналогична гиалоплазме. Хроматин представляет собой основной компонент ядра и определяет его главные функции.
Цитологически различают гетерохроматин и эухроматин. Гетерохроматическими называют участки, находящиеся в плотно упакованном, конденсированном состоянии. Деконденсированные зоны обозначают как эухроматин. В препаратах для световой микроскопии гетерохроматин окрашен более интенсивно, чем эухроматин. Гетерохроматин подразделяется на две фракции: конститутивный гетерохроматин и факультативный. Первый всегда плотно упакован. Второй может переходить в эухроматин. Свойства разных видов хроматина приведены ниже.
ХРОМАТИН
гетерохроматин
эухроматин
конститутивный
факультативный
1. во время интерфазы всегда остается конденсированным;
2. реплицируется в позднюю S-фазу;
3. скорость репликации выше, чем у эухроматина;
4. ассоциирован с небольшим количеством кислых белков;
5. не осуществляет транскрипции;
6. имеет большое число повторяющихся последовательностей нуклеотидов (сателлитная ДНК);
7. локализован в центромерных и теломерных участках хромосом, в области вторичных перетяжек.
1. во время интерфазы может оставаться конденсированным;
2. реплицируется в позднюю S-фазу;
3. скорость репликации выше, чем у эухроматина;
4. ассоциирован с небольшим количеством кислых белков;
5. не осуществляет транскрипции.
1. во время интерфазы деконденсирован;
2. реплицируется в раннюю S-фазу;
3. скорость репликации ниже, чем у гетерохроматина;
4. ассоциирован с большим количеством кислых белков, чем гетерохроматин;
5. активно транскрибируется;
6. имеет небольшое количество сателлитной ДНК.
Mb
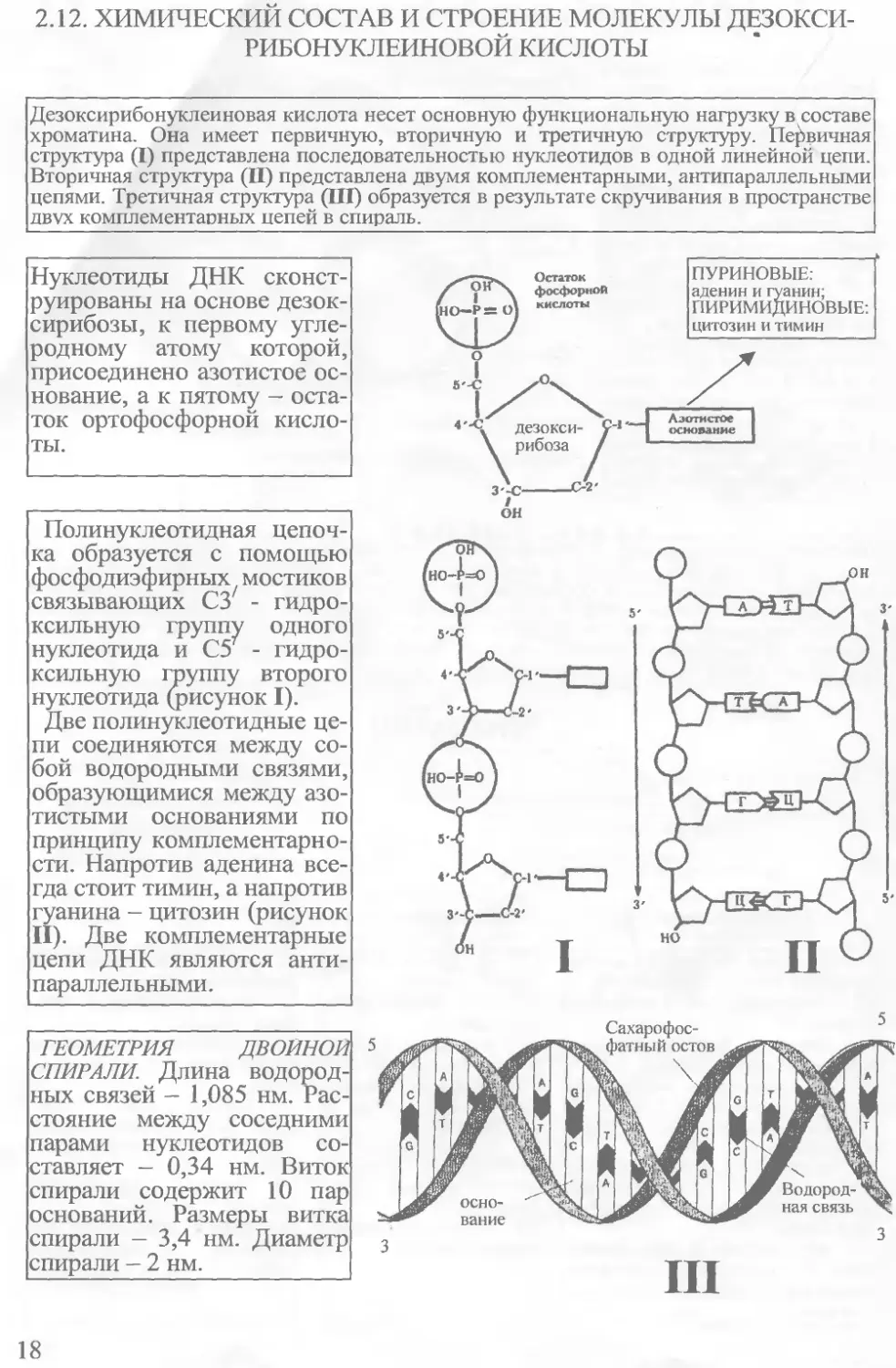
2.12. ХИМИЧЕСКИЙ СОСТАВ И СТРОЕНИЕ МОЛЕКУЛЫ ДЕЗОКСИРИБОНУКЛЕИНОВОЙ КИСЛОТЫ
Дезоксирибонуклеиновая кислота несет основную функциональную нагрузку в составе хроматина. Она имеет первичную, вторичную и третичную структуру. Первичная структура (I) представлена последовательностью нуклеотидов в одной линейной цепи. Вторичная структура (II) представлена двумя комплементарными, антипараллельными цепями. Третичная структура (III) образуется в результате скручивания в пространстве двух комплементаоных цепей в спираль.
Нуклеотиды ДНК сконструированы на основе дезоксирибозы, к первому углеродному атому которой, присоединено азотистое основание, а к пятому - остаток ортофосфорной кислоты.
Полинуклеотидная цепочка образуется с помощью фосфодиэфирных мостиков связывающих СЗ7 - гидроксильную группу одного нуклеотида и С5Г - гидроксильную группу второго нуклеотида (рисунок I).
Две полинуклеотидные цепи соединяются между собой водородными связями, образующимися между азотистыми основаниями по принципу комплементарно-сти. Напротив аденина всегда стоит тимин, а напротив гуанина - цитозин (рисунок II). Две комплементарные цепи ДНК являются антипараллельными .
ГЕОМЕТРИЯ ДВОЙНОЙ СПИРАЛИ. Длина водородных связей - 1,085 нм. Расстояние между соседними парами нуклеотидов составляет - 0,34 нм. Виток спирали содержит 10 пар оснований. Размеры витка спирали — 3,4 нм. Диаметр спирали - 2 нм.
III
2.13. СТРУКТУРНАЯ ОРГАНИЗАЦИЯ ХРОМАТИНА
Хроматин имеет сложный состав, основу, которого составляет нуклеопротеид - комплекс ДНК с гистонами. С помощью белков молекула ДНК подвергается многоступенчатому процессу компактизации (конденсации), что в зависимости от стадии клеточного цикла, позволяет наследственному материалу находится в состоянии эухрома-тина, гетерохроматина или конденсированной хромосомы.
На первом этапе молекула ДНК, диаметром 2 нм, образует комплекс с белковыми корами, состоящими из 8 молекул гистонов класса Нгд; Нгв; Нз и Н4. ДНК непосредственно “намотанная” на кору называется нуклеосомой. В контакте с каждым кором оказывается 150 - 200 пар нуклеотидов. Свободная от гистонов ДНК называется линкерной. В ее составе обычно находится от 15 до 100 пар нуклеотидов (в среднем 60). Структура, возникающая на первом этапе конденсации называется “нуклеосомная нить”. Ее диаметр составляет 11 нм и она в 7 раз короче первоначальной молекулы ДНК. (Рис. А и Б)
На втором этапе нуклеосомные нити сшиваются гистонами Hi с образованием хроматиновых фибрилл диаметром 30 нм. (Рис. В)
Далее хроматиновые фибриллы образуют серию петель размеры которых в диаметре достигают 300 нм. Такие структуры называют “интерфазными хромонемами”. В их образовании, по-видимому, принимают участие негистоновые белки. Одна хромонема содержит от 20 000 до 80 000 пар нуклеотидов. (Рис. Г).
Отдельные участки интерфазной хромонемы подвергаются дальнейшей компактизации с образованием структурных блоков (Рис. Д), входящих в состав метафазных хромосом (Рис. Е), видимых в световой микроскоп. Такие высококонденсированные участки в виде конститутивного гетерохроматина встречаются в области центромер хромосом, их теломерных участков и ин-тенсивно окрашиваются красителями.
2.14. МОРФОЛОГИЯ МЕТАФАЗНЫХ ХРОМОСОМ
Морфологию хромосом удобнее всего изучать в момент их наибольшей конденсации - в метафазу или раннюю анафазу митоза. В это время они представляют собой палочковидные структуры разной длины, с характерными морфологическими особенностями. Во всех хромосомах выделяют: 1) первичную перетяжку и 2) плечи. В зависимости от положения первичной перетяжки и, соответственно, размеров плеч, хромосомы делят на четыре группы: А) метацентрические (равноплечие); Б) субметацентрические (неравноплечие); В) акроцентрические (выраженнонерав-ноплечие и Г) телоцентрические (одноплечие). У человека в норме телоцентрический тип хромосом отсутствует. Длинное плечо хромосом обозначают буквой “q”, а короткое - “р”. Некоторые хромосомы, в области концевых участков (теломерных участков) имеют 3) вторичную перетяжку, которые отделяют короткие фрагменты - 4) спутники (сателлиты) хромосом.
В профазу и метафазу каждая хромосома состоит из двух сестринских хроматид. В анафазу и раннюю телофазу после разделения наследственного материала образуются однонитчатые дочерние хромосомы. На верхнем представлены хромосомы с еще не разошедшимися хроматидами. Каждая хроматида образована одной гигантской молекулой ДНК, упакованной с помощью гистоновых и негистоновых белков. В области вторичных перетяжек хромосом располагаются гены, ответственные за синтез рибосомных РНК. В интерфазу эти участки обнаруживаются в зоне ядрышек и поэтому их еще называют ядрышковыми организаторами. Теломерам хромосом (концевым участкам плечей) в последние годы придается особое значение. Высказана гипотеза, что толщина теломеров является определяющим фактором продолжительности жизни особей, в том числе и человека. Известно, что теломеры покрыты особым веществом, предупреждающим склеивание хромомсом, и как следствие, возникновение патологии не совместимой с жизнью. Показано, что при каждом делении клеток защитное вещество теломеров уменьшается и, со временем в клетках накапливаются опасные хромосомные мутации являющиеся причиной гибели особей.
2.15. ПОНЯТИЕ О КАРИОТИПЕ
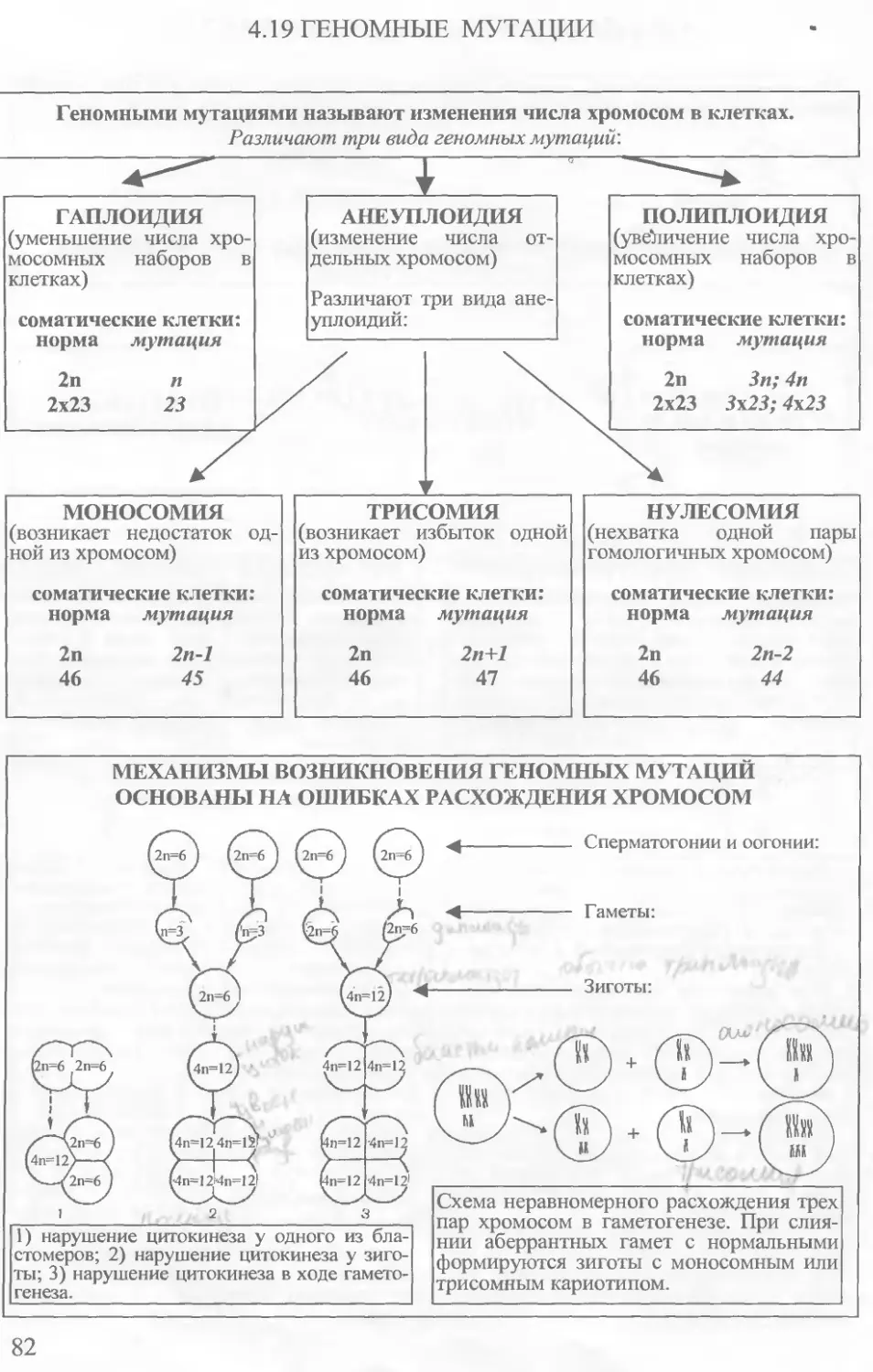
Кариотипом называют диплоидный набор хромосом клетки, характеризующийся их числом, величиной и формой. Термин введен в 1924 г. отечественным цитологом Г.А. Ливитским. Свойства кариотипа соответствуют четырем эмпирическим “правилам”.
ПРАВИЛА КАРИОТИПА
ПОСТОЯНСТВА
ПАРНОСТИ
ИНДИВИДУАЛЬНОСТИ
НЕПРЕРЫВНОСТИ
Число хромосом в клетках определенного вида всегда неизменно. Число хромосом — видовой признак. Эта особенность известна как правило постоянства числа хромосом.
В соматических клетках представителей любого биологического вида число хромосом четное, поскольку хромосомы составляют пары. Парные хромосомы называются гомологичными. Они совпадают по величине, форме, другим деталям строения, порядку расположения наследственного материала и его количеству, особенностям дифференциального окрашивания. Это правило справедливо для всех аутосом и гетеросом гомогаметно-го пола. Половые хромосомы гетерогаметного пола не совпадают по всем деталям строения и набору генов.
Негомологичные хромосомы всегда имеют морфологические и функциональные отличия. Следовательно, каждая пара хромосом характеризуется своими особенностями. В этом выражается правило индивидуальности хромосом.
При делении клеток число хромосом и их индивидуальность передается от одного поколения другому. В этом выражается правило непрерывности.
Гаплоидный набор хромосом в половых клетках обычно обозначают буквой п. Диплоидный набор соматических клеток организма записывают как 2п. При этом если возникает необходимость дифференцировать особи по полу, то к записи кариотипа добавляют символы половых хромосом. В соответствии с этим кариотип мужской и женской особи обозначают как 2n,XY и 2п,ХХ. Примеры хромосомных наборов некоторых организмов приведены в таблице.
Биологический вид Число хромосом Биологический вид Число хромосом
Малярийный плазмодий (Plasmodium malariae) 2 Гидра пресноводная (Hydra vulgaris) 32
Речной рак (Astacus fluviatillis) -116 о- Черный таракан (Blatta orientalis) 48
Комнатная муха (Musca domestica) 12 Лягушка зеленая (Rana esculenta) 26
Голубь сизый (Columbia livia) 80 Мышь домовая (Mus musculus) 40
Кролик дикий (Oryctolagus cuniculus) 44 Человек разумный (Homo sapiens) 46
2.16. КЛАССИФИКАЦИЯ ХРОМОСОМ ЧЕЛОВЕКА -
Первая международная (Денверская) классификация хромосом человека была создана в 1960 году. Согласно этой систематике учитываются линейные размеры аутосом и положение первичной перетяжки. Самая крупная хромосома имеет № 1, а самая мелкая - № 22. Дополнительными критериями являются наличие вторичных перетяжек и сателлитов. Согласно Денверской классификации хромосомы образуют 7 групп (А, В, С, D, Е, F,G). Гетеросомы выделяют отдельно.
Группа Номер хромосом Размер, мкм Характеристика
А 1-3 11-8,3 1 и 3 - крупные метацентрические, 2 - самая крупная субметацентрическая.
В 4, 5 7,7 Крупные субметацентрические
С 6-12 7,2-5,7 Средние субметацентрические
D 13-15 4,2 Средние акроцентрические
Е 16-18 3,6-3,2 Мелкие субметацентрические
F 19, 20 2,8 - 2,3 Мелкие метацентрические
G 21,22 2,3 Самые мелкие акроцентрические
На практике идентификация хромосом по указанным признакам имеет большие трудности. Фактически удается определить, к какой группе принадлежит та или иная хромосома, но ее конкретный номер обычно не поддается идентификации. Дальнейшее развитие классификация хромосом получила благодаря работам шведского генетика Касперссона, который к 1970 году разработал методику дифференциального окрашивания хромосом с помощью флуоресцентных красителей. В 1971 году его работы были положены в основу Парижской стандартизации хромосом человека.
Касперссон показал, что после обработки акрихин-ипритом, каждая хромосома не дает равномерного свечения. В хромосомах выделяют сильно флуоресцирующие участки, соответствующие гетерохроматиновым участкам и не флуоресцирующие зоны. Чередование светящихся и не светящихся фрагментов у каждой пары хромосом строго специфично. Сегодня разработано несколько методов выявления структурной неоднородности хромосом человека. Их сравнительный анализ показал, что одна и та же зона в хромосоме может быть светлой -неокрашенной или темной - окрашенной, но порядок расположения дисков идентичен при всех методах окраски. Следовательно, расположение дисков имеет закономерный характер, специфичный для каждой хромосомы.
1 2 3 4 8 6
7 8 9 10 11 12
X
22
2.17. КЛЕТОЧНЫЙ ЦИКЛ
Клеточным циклом называют последовательность событий от образования клетки до ее деления или гибели. Клеточный цикл любой клетки состоит из двух неравных по продолжительности периодов: митоза (или собственно деления) и интерфазы. Во время митоза (М-фаза) наследственный материал клетки делится строго пополам между двумя образующимися молодыми клетками. Интерфаза неоднородна по своим событиям и в ней выделяют фазы Gi, S, и G2.
Многие клетки сразу после образования подвергаются специализации и “выпадают” из клеточного цикла в фазу Go- Часть таких клеток (например, эритроциты человека) до самой гибели остаются в этой фазе, а некоторые (гепатоциты) могут возвращаться в клеточный цикл.
Фаза Gi называется постмитотической или пресинтетической. Ее основными событиями являются восстановление интерфазной ультраструктуры клетки, активизация синтеза белка и, как следствие, усиленный рост клетки. Готовясь к следующей фазе цикла, клетка образует нуклеотиды и ферменты репликации ДНК. Продолжительность Gi - периода у клеток млекопитающих в реальном времени обычно составляет от 1 до 34 часов.
S-фаза носит название синтетической. В это время в клетке происходит репликация ДНК. После завершения S-фазы, генетический материал в ядрах клеток удвоен и описывается формулой 2п4с. Наряду с ДНК в синтетическом периоде удваивается количество гистонов, интенсивно образуется РНК. Продолжительность S-периода интерфазы составляет от 6 до 10 часов.
Отрезок времени от окончания синтетического периода до начала митоза занимает фаза G2 или постсинтетический (предмитотический) период. В это время в цитоплазме клетки активизируется синтез РНК, тубулинов - белков микротрубочек, а также интенсифицируются процессы образования АТФ, необходимой для последующих этапов митоза. Длительность периода в реальном времени 2 до 5 часов.
Ключевыми периодами клеточного цикла являются S- и М- фазы. Их начала инициируются активатором S-фазы (А8Ф) и М-стимулирующим фактором (МСФ). К концу Gi периода в цитоплазме клетки нарабатывается А8Ф, который активирует начало репликации ДНК и исчезает к началу G2 -периода. МСФ появляется в цитоплазме к началу митоза и его выработка контролируется белком ииклином.
Время
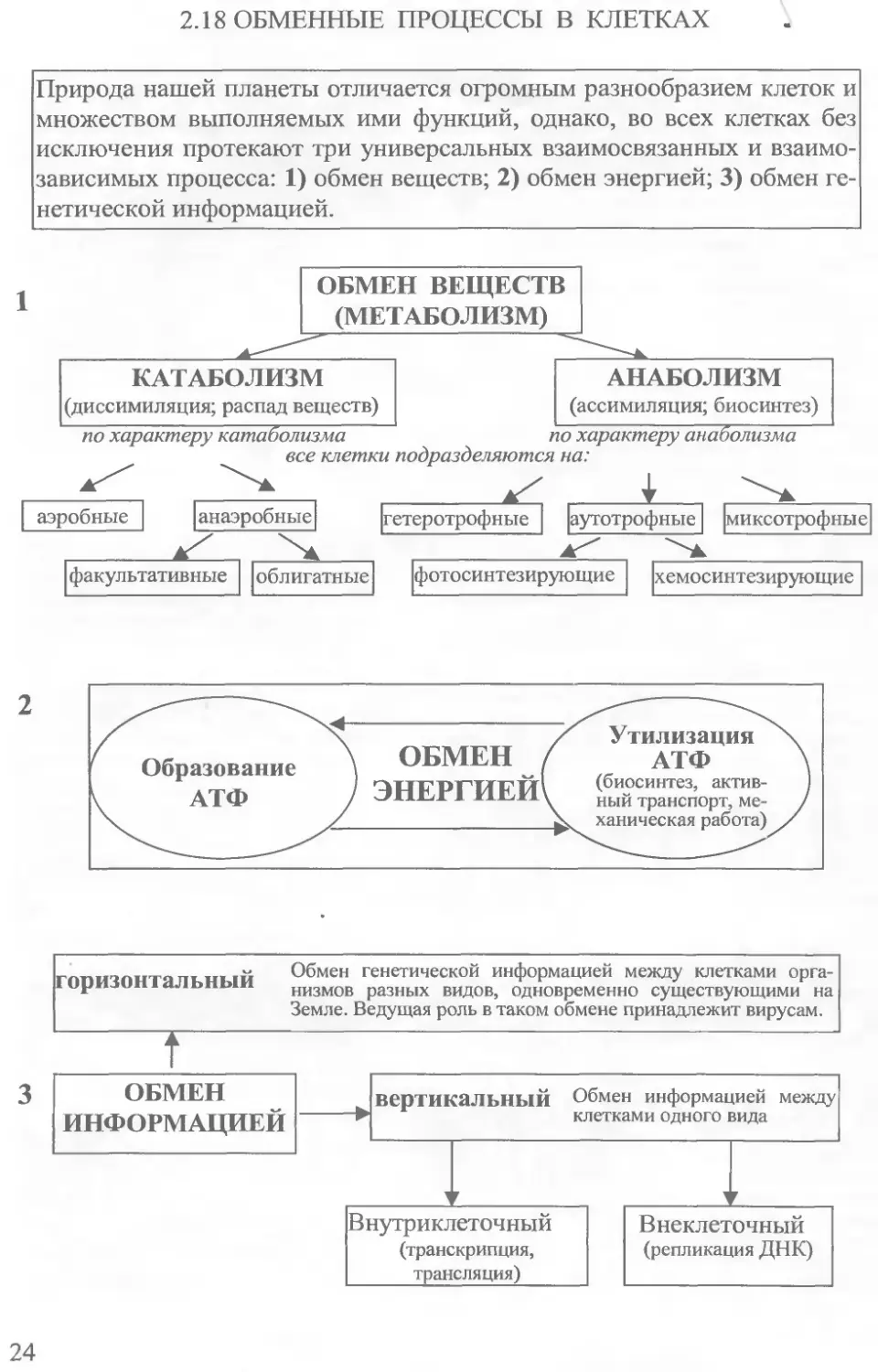
2.18 ОБМЕННЫЕ ПРОЦЕССЫ В КЛЕТКАХ
2.19 КАТАБОЛИЗМ БИОПОЛИМЕРОВ В КЛЕТКАХ
Распад основных пищевых веществ в клетках происходит в три стадии и сопряжен с образованием энергии в виде АТФ. Первая стадия не сопровождается непосредственным образованием АТФ и у многоклеточных организмов происходит в желудочно-кишечном тракте. Собственные отработанные компоненты клетки разрушаются лизосомами. Вторая стадия катаболизма протекает в цитоплазме клеток и обеспечивает небольшие количества метаболической энергии. В третью стадию завершается распад веществ до воды, углекислого газа, простых азотсодержащих соединений и происходит образование основного количества АТФ на кристах митохондрий.
Стадия 1.
Расщепление макромолекул на простые субъединицы
Стадия 2.
Расщепление простых субъединиц на ацетил-СоА, сопровождающееся образованием ограниченного количества ATOhNADH
Стадия 3.
При полном окислении ацетил-СоА до H2O и СО2 образуется большое ко-личство NADH, что обеспечивает синтез большого количества АТФ в дыхательной цепи митохондрий
2.20 АНАБОЛИЧЕСКИЕ ПРОЦЕССЫ В КЛЕТКАХ .
Синтез биологических полимеров, свойственных конкретным клеткам может происходить двумя принципиальными путями. 1) Наиболее выгодно белки, полисахариды и липиды синтезировать из соответствующих мономеров, образовавшихся при катаболизме пищевых компонентов. 2) При дефиците мономеров их можно образовать из промежуточных продуктов распада пищевых веществ. При биосинтезе расходуется большое количество молекул АТФ.
Первый принципиальный путь анаболизма
пищевые
характерные для конкретной клетки
Второй принципиальный путь анаболизма
пищевые
характерные для конкретной клетки
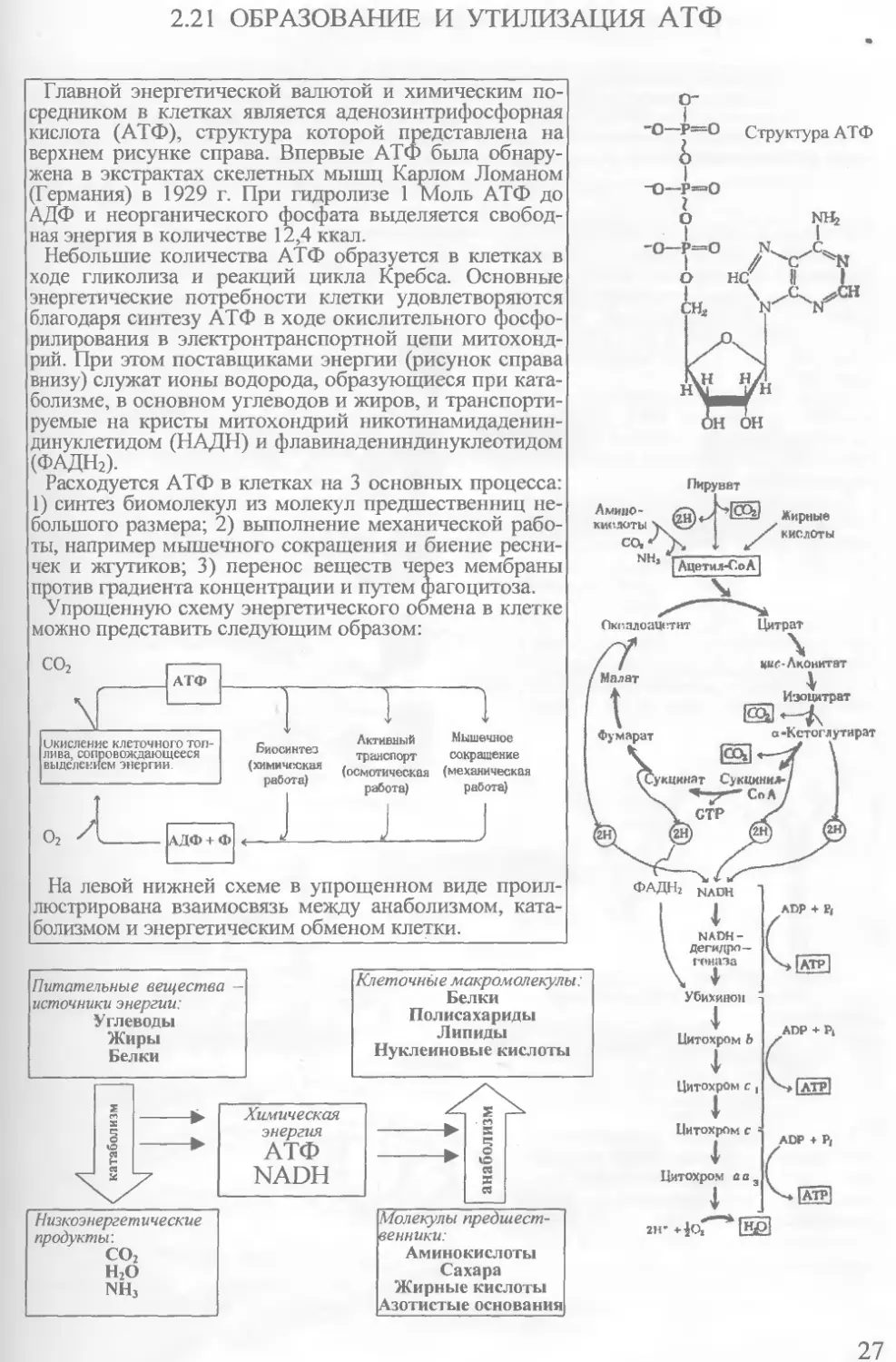
Главной энергетической валютой и химическим посредником в клетках является аденозинтрифосфорная кислота (АТФ), структура которой представлена на верхнем рисунке справа. Впервые АТФ была обнаружена в экстрактах скелетных мышц Карлом Ломаном (Германия) в 1929 г. При гидролизе 1 Моль АТФ до АДФ и неорганического фосфата выделяется свободная энергия в количестве 12,4 ккал.
Небольшие количества АТФ образуется в клетках в ходе гликолиза и реакций цикла Кребса. Основные энергетические потребности клетки удовлетворяются благодаря синтезу АТФ в ходе окислительного фосфорилирования в электронтранспортной цепи митохондрий. При этом поставщиками энергии (рисунок справа внизу) служат ионы водорода, образующиеся при катаболизме, в основном углеводов и жиров, и транспортируемые на кристы митохондрий никотинамидаденин-динуклетидом (НАДН) и флавинадениндинуклеотидом (ФАДН2).
Расходуется АТФ в клетках на 3 основных процесса: 1) синтез биомолекул из молекул предшественниц небольшого размера; 2) выполнение механической работы, например мышечного сокращения и биение ресничек и жгутиков; 3) перенос веществ через мембраны против градиента концентрации и путем фагоцитоза.
Упрощенную схему энергетического обмена в клетке можно представить следующим образом:
На левой нижней схеме в упрощенном виде проиллюстрирована взаимосвязь между анаболизмом, катаболизмом и энергетическим обменом клетки.
о-
-о—р—о ь I
-о—Р=О о
-О—р=о
Структура АТФ
МЧг
N С
НСГ II I
Я
СЦ.
н он
Пируват
(£Й)«> Жирные
< / кислоты
Аминокислоты > со,^ _
NH> ] Ацетил-СоА |
ОксалоаЦстит
Малат
Фумарат
укцииат
ФАДН2 MADH
Сукцинил-т— СоА
СТР
Цитрат
ituc-Лконитат
V
Иэоцитрат
а-Кстоглутират
ADP + ₽(
Убихинон
Цитохром Ь
Цитохром с ।
Цитохром с 1
Цитохром а а
2И’ 1Н1
АТР
ADP + Pt
ATPI
ADP + Р!
|АТР
2.22 РЕПЛИКАЦИЯ ДНК
Репликация (копирование) ДНК осуществляется в S-период интерфазы. Она происходит по полуконсервативной схеме и по принципу комплементарности. Это сложный процесс, в осуществлении которого принимает участие большое количество ферментов и белков. Начинается репликация с присоединения инициаторного белка к специфической последовательности нуклеотидов — точкам начала репликации.
К инициаторному белку присоединяется ДНК-геликаза, которая разрывает водородные связи между комплементарными цепями ДНК и образует репликационную вилку. Поддерживают структуру реплика-ционной вилки дестабилизирующие белки.
Далее праймаза синтезирует короткие фрагменты РНК, которые ДНК-полимераза - использует как затравку (праймер) для синтеза дочерних цепей ДНК.
Цепи ДНК антипараллельны. Поэтому учитывая, что ДНК-полимераза может вести синтез ДНК только от 5 к 3 концу, синтез лидирующей цепи осуществляется непрерывно (на ней образуется одна молекула РНК-затравки), а синтез отстающей цепи идет короткими фрагментами назад (фрагментами Оказаки). На отстающей цепи синтезируется множество РНК-затравок.
1) Отстающая цепь; 2) Лидирующая цепь; 3) Цепи ДНК, синтезированные последними.
В дальнейшем праймеры вырезаются ферментом ДНК-лигазой и на их место вшиваются дезоксирибонуклеотиды.
При образовании репликацион-ной вилки участок ДНК впереди вилки сильно закручивается и спутывается. Это напряжение ДНК снимается ферментом ДНК-топо-изомеразой, который работает по принципу “ножниц и клея”.
Точка начала репликации
Родительская спираль ДНК
Связывание инициаторного
3
5
Здесь спираль ДНК должна быстро вращаться
Матри ца для синтеза ведущей цепи
Матрица для синтеза отстающей цепи
З1
3
ДН К-пол имераза на ведущей цепи
Нов осин тезированная цепь ДНК
5
5
Транскрипция является первым этапом внутриклеточного потока наследственной информации и представляет собой перевод информации заключенной в ДНК на нуклеотидную последовательность матричной (информационной) РНК. Синтез мРНК осуществляется РНК-полимеразами. У эукариот существует 3 типа РНК-полимераз. В синтезе мРНК принимает участие РНК-полимераза II. Ферменты типа I и III синтезируют рибосомные и транспортные РНК. У прокариот все виды РНК образуются одним ферментом.
Наиболее хорошо изучена структура РНК-полимеразы Е. coli Она состоит из 5 белковых субъедениц, имеет формулу агрРосо и называется холофермент-РНК-полимераза. Фермент лишенный осубъединицы называется кор-фермент. Он способен катализировать большинство реакций, ведущих к образованию РНК, но не способен узнавать промоторные сайты ДНК.
Транскрипция это сложный процесс, в котором выделяют три основные стадии: 1) инициации, 2) элонгации (собственно транскрипция) и 3) терминации.
1) Молекула РНК-полимеразы способна присоединяться к специфическому участку ДНК, к так называемому промотору. В любом промоторе содержится специфическая последовательность нуклеотидов - старт сигнал. Этот сигнал (ТАТАТТ) обычно расположен на расстоянии примерно 10 оснований от того нуклеотида - с которого начинается транскрипция и, соответственно, его обозначают как 10-последовательность или Прибнов-Бокс.
Другая последовательность, длина которой равна девяти нуклеотидам, расположена на расстоянии 35 оснований до сайта инициации, ее называют 35 последовательностью. Она участвует в связывании РНК-полимеразы, которое предшествует перемещению фермента в Прибнов-бокс.
К премоторным участкам ДНК присоединяется холофермент. Затем после образования короткого фрагмента РНК (5-10 оснований) от холофермента отделяется о-субъединица и оставшийся кор-фермент осуществляет дальнейшие стадии транскрипции.
ДНК
Промоторт £ а
<—выше! ниже—>
Стартовая точка транскрипции |® х
б
транскрипции
Рисунок иллюстрирует “рабочий цикл” о-субъединицы. Кор-фермент связывается с субъ-диницей, образуя холофермент , который прочно связывается с промотором. После запуска ранскрипции о-субъединица отделяется, а кор-фермент катализирует элонгацию цепи.
Элонгация цепи мРНК происходит под влиянием кор-фермента. Он движется вдоль ДНК подобно застежке молнии, разрывает водородные связи между комплементарными основаниями, которые вновь образуются позади фермента, по мере прохождения участка в котором происходит спаривание оснований РНК с основаниями кодогенной цепи ДНК. Раскрытая кор-ферментом область простирается только на несколько пар оснований ДНК.
Терминация (прекращение роста) цепи мРНК происходит на специфических участках ДНК, называемых терминаторами. Эти участки обогащены GC- или АТ-парами нуклеотидов. Транскрипты после прохождения таких участков складываются с образованием шпилек разной длины.
С °и CAAUCAaOv'CC ииииииО
О- Зависимым р-Независимый
терминатор терминатор
Различают p-зависимую и р-не-зависимую терминацию транскрипции. При р-независимой терминации шпильки создают естественную помеху для РНК-полимеразы и транскрипция останавливается, после чего молекула РНК отрывается от матричной цепи ДНК вблизи U-богатого участка. При p-зависимом варианте р-белок, обладающий АТФ-азной активностью связывается с синтезируемой цепью РНК и перемещается по ней, за счет энергии гидролиза АТФ, в направлении 5-3' непосредственно за РНК-полимеразой. Если р-белок наталкивается на шпильку РНК, он останавливает полимеразу и вызывает отделение комплекса “мРНК-кор-фермент” от ДНК, после чего сам диссоциирует от этого комплекса.
РНК -полимераза транскрибирует ДНК
р-Белок присоединяется к 5'-концу цепи РНК
р-Белок перемещается по РНК за счет энергии гидролиза АТР
Полимераза останавливается в терминаторе под влиянием р-белка
Т ерминация транскрипции и отделение полимеразы и РНК
2.25 ТРАНСКРИПЦИЯ У ЭУКАРИОТ
Транскрипция у эукариот является более сложным процессом, чем - у прокариот. РНК полимераза II эукариот не может самостоятельно инициировать транскрипцию Для ее активирования необходимо большое число белков - факторов транскрипции. Факторы транскрипции к настоящему времени очищены и выделены. Всего их шесть: TFIIA; TFIIB; TFIID; TFIIE; TFIIF и TFIIH. Некоторые из них состоят из многих полипептидов. Например в состав TFIID входят белок ТВР и еще восемь молекул называемых TAF (ТВР - associated factors). В состав транскрипционного комплекса входят также белки SRB, которые связываются с большой субъединицей РНК-полимеразы II и помогают ей разрушить нуклеосомы и деконденсировать молекулу ДНК. Эту же функцию выполняют белки семейства SWI/SNF.
Процесс сборки комплекса показан на рисунке (а). Формирование белкового комплекса на премоторной последовательности начинается с того, что фактор TFIID связывается с ТАТА-последовательностью ДНК. После завершения сборки, до начала транскрипции РНК - полимераза II освобождается от всех факторов транскрипции. Ключевым в процессе инициации транскрипции является присоединение фактора TFIIH. Одна из его субъединиц, фосфорилирует молекулу РНК- полимеразы II, что и является толчком к началу процесса транскрипции.
Другой особенностью транскрипции у эукариот является наличие энхансерных - усиливающих (от англ, enhance) участков ДНК. Они часто расположены в тысячах пар нуклеотидов от промотора. Сейчас известно, что энхансерные участки связывают особые регуляторные белки, усиливающие процесс транскрипции. Примером энхансера может служить система GAL 4 -UAS, обнаруженная у дрожжей.
Точка начала транскрипции
РНК-транскрипт
2.26 ПОСТТРАНСКРИПЦИОННЫЕ ИЗМЕНЕНИЯ РНК
В ядерных генах эукариот, генах пластид и митохондрий, а также в некоторых генах вирусов участки ДНК, кодирующие структуру белков, не однородны. В них значащие участки (экзоны), действительно кодирующие последовательность аминокислот какого либо белка чередуются с интронами - инертными участками, не несущими информации. В связи с этим в ходе транскрипции вначале синтезируется молекула про-иРНК (первичный транскрипт). Такая молекула содержит информацию и об экзонах, и об интронах.
У низших эукариот, таких как дрожжи, 95% генов содержат только один экзон, значит, в подавляющей массе они не прерываются интоонами. У дрозофилы не имеет интронов только 17%, а у млекопитающих - лишь 6% генов. У последних число экзонов в некоторых генах может достигать 60. Общая длина всех интронов зачастую превышает суммарную длину экзонов. Например, из 7000 пар нуклеотидов гена овальбумина курицы на долю экзонов приходится всего 18/2 папы, то есть почти 75% длины ДНК составляют интроны.
Слева: количество экзонов в генах дрожжей, дрозофилы и млекопитающих.
Справа: размеры (в парах нуклеотидов) экзонных участков эукариот
В дальнейшем первичный транскрипт подвергается созреванию или процессингу. В ходе процессинга специальные ферменты вырезают из про-иРНК интронные участки. Далее оставшиеся экзонные участки сшиваются - этот процесс называют сплайсингом. В итоге образуется молекула зрелой иРНК, которая по своим размерам значительно меньше молекулы про-иРНК. Зрелая молекула иРНК является первичным продуктом действия генов. Через ядерные поры она выходит в цитоплазму и служит матрицей для формирования полипептидных цепей в рибосомах,
Экзон 1 Интрон 1 Экзон 2 Интрон 2 Экзон 3
мРНК
Белок
2.27 ОБЩИЕ ПРЕДСТАВЛЕНИЯ О ТРАНСЛЯЦИИ
Трансляция — это процесс декодирования мРНК, в результате которого информация с языка последовательности оснований мРНК переводится на язык аминокислотной последовательности белка. Синтез белка происходит на рибосомах. Рибосомы прокариот более мелкие и имеют константу седиментации 70S. Аналогичные рибосомы имеют митохондрии и пластиды эукариот. Константа седиментации цитоплазматических рибосом эукариот равна 80S. Каждая рибосома состоит из двух субъединиц. Их химический состав представлен на рисунке.
70S
Мол. масса 2500000
80S
1540 нуклеотидов
Мол. масса 900000
lespPHK
34 белка 21 белок
прокариотическая рибосома
нуклеотидов
*-49 белков
**33 белка
эукариотическая рибосома
В собранных рибосомах выделяют три функциональных центра. 1) центр связывания мРНК. 2) A-центр или аминоацильный центр к которому происходит присоединение ами-ноацил-тРНК (комплекса аминокислот с соответствующими тРНК). 3) P-центр или пеп-тидильный центр к которому присоединяется пептидил-тРНК (последняя тРНК с присоединенной к ней растущим полипептидным фрагментом).
Большая и малая частицы рибосомы несут различную нагрузку. Малая субъединица (4) отвечает за генетические (декодирующие) функции, а большая (5) - за биохимические (ферментативные).
Заказ №3775
33
2.28 ГЕНЕТИЧЕСКИЙ КОД
Структура белка зашифрована в последовательности нуклеотидов ДНК, а затем мРНК, называемой генетическим кодом. Содержание генетического кода представлено в таблице.
Первое положение (Б'-конец) Второе положение в кодоне T ретье положение (З'-конец)
ит C A G
U(T> UUU) UUCI UUA1 UUGJ Phe Leu UCU] ucc UCA UCGJ Ser UAUI UAC UAA) UAGI ►Tyr ►Терм UGUI^ UGCfCys UGA Терм. UGG Trp U(T> С A G
С CUU] cud CUA CUGJ Leu CCU] ccc CCA ccgJ Pro CAU1 CAC CAA CAG His Gin CGU] CGC CGAj CGGJ Arg U(T) C A G
А AUU1 AUC AUA ♦AUG lie Met ACU] ACC АСА acgJ Thr AAU1 AAC AAA] AAG Asn Lys AGU| AGCI AGA I AGGJ ► Ser Arg U(T) C A G
G GUU GUC GUA GUG Vai GCU1 GCC GCA gcgJ Ala GAU 1 GAC j GA Al GAG) Asp Glu GGU] GGC GGAi GGGJ Gly U(T) C A G
* Инициирующий кодон
СВОЙСТВА ГЕНЕТИЧЕСКОГО КОДА
Не перекры-ваемость - в последовательности оснований каждый нуклеотид входит лишь в какой-либо один кодон. Кодоны располагаются друг за другом без перерыва и запятых.
Триплетность - одна аминокислота задается последовательностью из трех нуклеотидов, называемой кодоном.
Универсальность - все живые организмы (вирусы, прокариоты и эукариоты) используют один и тот же генетический код.
Коллинеарность - последовательность триплетов (кодонов) полностью соответствует последовательности аминокислот в белке.
Вырожденность — большинство аминокислот кодируются более чем одним кодоном, поскольку на 20 аминокислот входящих в состав белков, приходится 43 = 64 кодона (три кодона обозначают конец синтеза, то есть являются термина-торами).
Считывание информации с мРНК на рибосомах происходит лишь в направлении от 5' к 3' концу. Рамка считывания задается первым основанием первого кодона. Обычно рамка считывания не меняется, но некоторые вирусы используют две или даже три рамки считывания и синтезируют разные белки. Это свидетельствует о нарушении такого свойства генетического кода как неперекрываемость. Свойство универсальности нарушается в генетическом коде митохондрий и дрожжей. В их мРНК триплет UGA является не терминирующим, а соответствующим триптофану. Кодоны AGA и AGC напротив -кодируют не аргинин, а являются терминаторами.
2.29 ИНИЦИАЦИЯ ТРАНСЛЯЦИИ
В ходе трансляции выделяют три последовательные стадии:
1) инициацию, 2) элонгацию и 3) терминацию синтеза полипептидной цепи.
Для начала инициации необходимо выполнение следующих условий: а) наличие соответствующих физико-химических условий в клетке (pH, температура, ионы Mg , К+ и др.); б) наличие ГТФ; в) наличие аминокислот ассоциированных с тРНК (аминоацил-тРНК); г) наличие рибосом и белковых факторов инициации трансляции; д) наличие мРНК.
Под инициацией трансляции понимают процесс формирования функционально активного комплекса рибосома-мРНК, содержащего метионил-тРНК (формилметионил-тРНК у прокариот) в P-участке рибосомы. Последовательность событий фазы инициации пред-ставлена ниже в виде схемы.
фмет-тРНК/мет-тРНК * 1
IF2/eIF2 . ГТФ
мРНК
fmet
fmet
мРНК
ГДФ + Фн
рибосома
finet
ГТФ, IF2
В начале несколько факторов инициации (от англ, initiation factors) стимулируют диссоциацию рибосомы на большую и малую субъединицы. У прокариот таких факторов сегодня известно 3, у эукариот (в обозначении добавляется префикс е) - 12. Важнейшим из них у любых клеток является IF3, который связывается с малой субъединицей. Далее судьба комплекса малой субъединицы и IF3 может идти в двух направлениях (на схеме левый и правый пути).
Либо вначале к комплексу присоединяется мРНК, а затем инициирующая аминоацил-тРНК (содержащая формилметионин у прокариот и метионин у эукариот), а также IF2 и ГТФ, либо наоборот - мРНК присоединяется последней. В любом случае после третьего этапа -образуется “инициаторный комплекс , состоящий из малой рибосомной субъединицы, мРНК, инициаторной аминоацил-тРНК (находящейся в P-участке), ГТФ и некоторых факторов инициации.
На четвертом этапе после удаления факторов инициации и гидролиза ГТФ присоединяется большая субъединица и формируется активная рибосома, содержащая в Р-участке инициаторную аминокислоту
Слева представлен рисунок иллюстрирующий события правого принципиального пути фазы инициации у прокариот.
Следует отметить, что в живой природе существует два способа поиска старт-сигнала на мРНК. У эукариот рибосомная частица, обычно сначала присоединяется к 5 концу мРНК, а затем движется вдоль цепи матрицы пока не встретит инициирующий кодон (терминальная инициация). Для реализации этого пути клетка имеет специальные мРНК-связывающие белки, помогающие связываться с матрицей и двигаться по ней, расщепляя АТФ. Внутренняя инициация используется прокариотами. В этом случае малая субъчастица рибосомы сразу ассоциирует с участком матрицы, содержащим старт-сигнал. В некоторых случаях эукариоты также используют внутренний путь инициации.
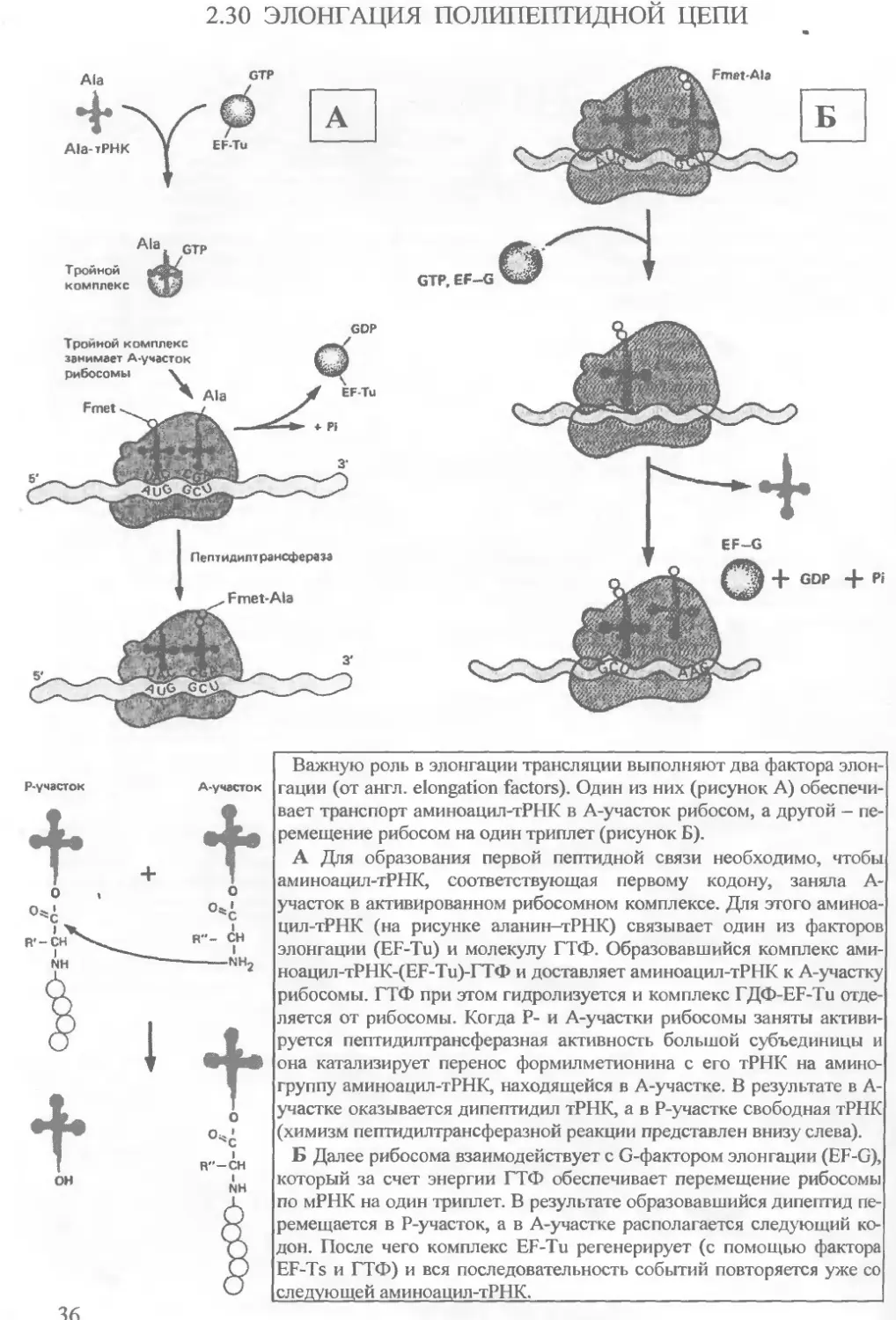
2.30 ЭЛОНГАЦИЯ ПОЛИПЕПТИДНОИ ЦЕПИ
Тройной комплекс
GOP
Пептидилт рансфера за
Р-участок
А-участок
Важную роль в элонгации трансляции выполняют два фактора элонгации (от англ, elongation factors). Один из них (рисунок А) обеспечивает транспорт аминоацил-тРНК в A-участок рибосом, а другой — перемещение рибосом на один триплет (рисунок Б).
А Для образования первой пептидной связи необходимо, чтобы аминоацил-тРНК, соответствующая первому кодону, заняла А-участок в активированном рибосомном комплексе. Для этого аминоацил-тРНК (на рисунке аланин—тРНК) связывает один из факторов элонгации (EF-Tu) и молекулу ГТФ. Образовавшийся комплекс ами-ноацил-тРНК-(ЕР-Ти)-ГТФ и доставляет аминоацил-тРНК к А-участку рибосомы. ГТФ при этом гидролизуется и комплекс ГДФ-EF-Tu отделяется от рибосомы. Когда Р- и A-участки рибосомы заняты активируется пептидилтрансферазная активность большой субъединицы и она катализирует перенос формилметионина с его тРНК на аминогруппу аминоацил-тРНК, находящейся в A-участке. В результате в А-участке оказывается дипептидил тРНК, а в P-участке свободная тРНК (химизм пептидилтрансферазной реакции представлен внизу слева).
Б Далее рибосома взаимодействует с G-фактором элонгации (EF-G), который за счет энергии ГТФ обеспечивает перемещение рибосомы по мРНК на один триплет. В результате образовавшийся дипептид перемещается в P-участок, а в A-участке располагается следующий кодон. После чего комплекс EF-Tu регенерирует (с помощью фактора EF-Ts и ГТФ) и вся последовательность событий повторяется уже со следующей аминоацил-тРНК.|
2.31 ТЕРМИНАЦИЯ БИОСИНТЕЗА БЕЛКА
Процесс последовательной трансляции кодонов в конце концов доходит до того момента, когда в A-участке оказывается один из трех терминирующих кодонов - UAG, UAA или UGA. Из-за отсутствия тРНК, отвечающих этим кодонам, по-липептидил-тРНК остается связанной с Р-участком.
Терминация биосинтеза полипептида и распад белоксинтези-рующего комплекса находится под управлением специфических факторов терминации или факторов освобождения. Сегодня известно 3 фактора освобождения (RF-1, RE-2 и RF-3). Их значение в процессе терминации различно. RF-1 и RF-2 связываются со стоп-кодоном, находящимся в A-участке, причем RF-1 узнает последовательность UAA или UAG, a RF-2 включается в том случае когда в А-участке оказывается UAA или UGA. Роль фактора RF-3 сегодня менее ясна. Считается, что он облегчает работу первых двух факторов.
После присоединения фактора освобождения изменяется активность пептидилтрансфераз-ного центра. Модифицированная пептидилтрансфераза присоединяет к пептидил-тРНК не свободную аминокислоту, а молекулу воды, что сопровождается гидролизом сложноэфирной связи между N-концевым карбоксилом полипептида и 3-концевым фрагментом последней тРНК. В результате синтезированная полипептидная последовательность отделяется от тРНК и происходит распад комплекса мРНК-рибосома-факто-ры терминации.
2.32 ОБЩИЕ ПРИНЦИПЫ РЕГУЛЯЦИИ ЭКСПРЕССИИ ГЕНОВ
Большая часть генома клеток многоклеточных организмов находится в неактивном (репрессированном) состоянии. Активно функционируют лишь 7 - 10% генов. Спектр функционирующих генов определяется типом клетки, стадией клеточного цикла и пе-риодом онтогенеза. Все функционирующие гены можно подразделить на две группы.
1) КОНСТИТУТИВНЫЕ
Гены определяющие синтез белков общего назначения (рибосомных, гистонов, тубулинов и т.д.). Транскрибирование этих генов определяется присоединением РНК-полимеразы к промоторам и, видимо, не подчиняется каким либо другим регулирующим воздействиям.
2) РЕГУЛИРУЕМЫЕ
Активность этих генов находится под контролем различных регулирующих факторов. Основными из них являются регуляторные белки.
РЕГУЛЯТОРНЫЕ БЕЛКИ
Являются продуктами действия генов-регуляторов. Связываясь с определенными участками ДНК способствуют или препятствуют присоединению РНК-полимеразы к промотору. Различают два типа регуляторов.
РЕПРЕССОРЫ
При контакте с ДНК занимают часть промотора или располагаются между промотором и структурной частью гена. Определяют негативный контроль экспрессии гена.
АКТИВАТОРЫ (АПОИНДУКТОРЫ)
При контакте с ДНК занимают область перед промотором (оператор) и облегчают связывание с ним РНК-полимеразы. Определяют позитивный контроль экспрессии гена.
Наряду с генетическими факторами в регуляции экспрессии генов на стадии транскрипции принимают участие негенетические факторы - эффекторы. К ним относят вещества небелковой природы, способные соединяться с белками регуляторами и изменять их сродство к промотору. Среди эффекторов различают коактиваторы (индукторы) и корепрессоры. Индукторы запускают транскрипцию, а корепрессоры препятствуют ее осуществлению.
Механизм действия индукторов состоит в инактивации (блокировании) белков —репрессоров, в результате чего они престают связываться с промотором или - во взаимодействии с апоиндукторами, что облегчает связывание РНК-полимеразы с промотором.
Корепрессоры или блокируют апоиндукторы, которые теряют способность связываться с оператором, или активируют репрессоры, находящиеся в неактивном состоянии.
2.33 РЕГУЛЯЦИЯ ЭКСПРЕССИИ ЛАКТОЗНОГО ОПЕРОНА
Область
РНК-полимераэа не может связаться с ДНК
РНК-лолимераза *-* в комплексе с ДНК
i мРНК v\Wv
Р ° Zhu.
Белок-репрессор/ в комплексе с ДНК
+Лактоза (L)
(Транскрипция не идет;
репрессор
Репрессор.
Покинувший
ДНК
ooL CSCSl
[Транскрипция]
Z Y А
На верхней части рисунка представлена структура лактозного оперона кишечной палочки. Он состоит из промотора (Р) - участка к которому присоединяется РНК-полимераза, оператора (О) - регуляторного участка способного присоединять белок-регулятор и трех структурных генов Z, Y и А кодирующих ферменты превращения молочного сахара (лактозы). Таким образом, три гена управляются одним оператором и транскрибируются с одной мРНК. Такая мРНК называется полицистронной или поли-генной. Следует отметить, что вне пределов оперона находится еще один ген (ген I), продуктом экспрессии которого является белок регулятор (репрессор), способный присоединяться к оператору и блокировать связывание РНК-полимеразы с промотором.
Добавление к среде где находятся кишечные палочки лактозы, вызывает примерно 100 кратное увеличение количества ферментов, метаболизирующих этот углевод в цитоплазме бактерий. Это ферменты, кодируемые генами Z, Y и А, соответственно 0-галак-тозидаза, (осуществляющая распад лактозы до галактозы и глюкозы), 0-галакто-зидпермеаза (регулирующая транспорт лактозы через мембрану) и тиогалактозидаце-тилтрансфераза (функция до конца не ясна). Такая активация метаболизма называется индукцией.
Механизм индукции представлен на нижней части рисунка. В среде бедной лактозой белок репрессор, экспрессируемый геном I, надежно блокирует транскрипцию структурных генов лактозного оперона. Появление во внешней среде лактозы в корне изменяет ситуацию. Белок репрессор также активно синтезируется геном I, однако лактоза присоединяется к репрессору, вследствие чего комплекс репрессор-лактоза диссоциирует от ДНК и освобождает промотор для присоединения РНК-полимеразы. Данный пример является примером негативной регуляции экспрессии генов.
Лактозный оперон может подвергаться и позитивной регуляции. Для его экспрессии, как и для экспрессии других индуцибельных опреонов, которые осуществляют контроль синтеза ферментов, участвующих в метаболизме сахаров, необходимо не только снять репрессию оперона, но и получить сигнал разрешающий экспрессию. Таким сигналом для 1ас-оперона служит комплекс циклического АМФ (цАМФ) с белком активатором катаболизма (САР от английского catabolite activator protein). Этот комплекс связывается со специфической последовательностью нуклеотидов, находящейся в самом начале промотора и вызывает усиление транскрипции в 50 раз. Возможно комплекс цАМФ-САР усиливает сродство РНК-полимеразы к промотору.
Промотор
!лс\
Область связывания САР-сАМР
Место присоединения PH К-лоли меразы
Оператор
bcZ
3.1 СПОСОБЫ РАЗМНОЖЕНИЯ ОРГАНИЗМОВ
Способность размножаться является одним из фундаментальных свойств живого. Биологический смысл размножения состоит в замещении погибших особей и в увеличении численности вида. Для живых существ характерно чрезвычайное разнообразие способов размножения и все их делят на две большие группы.
Различают бесполое и половое размножение.
Моноцитогенное
(одной клеткой)
Митоз
Простое деление
(прокариоты)
(эукариоты)
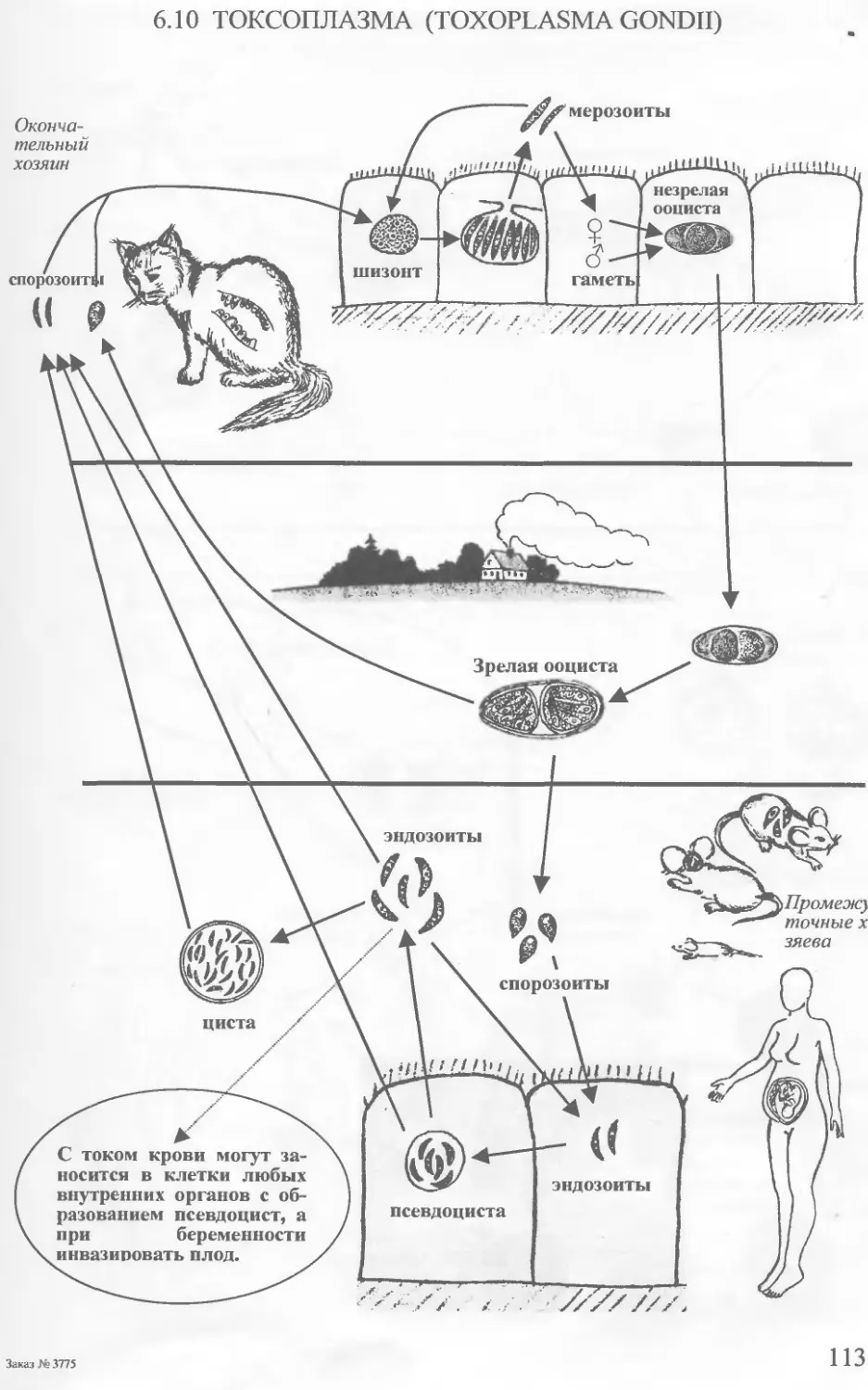
Шизогония (малярийные плазмодии) - - — “ ' , . - --— , -
Почкование(дрожжевые грибы)
Спорообразование
(папоротники)
Эндогония
(токсоплазма)
Бесполое размножение
Полицитогенное
(группой клеток)
Вегетативное размножение (растения)
Упорядоченное деление (морские звезды, кольчатые черви)
(плоские черви)
► Фрагментация
(пресноводная гидра)
Полиэмбриония (млекопитающие, человек)
Почкование
Половой процесс у одноклеточных
Конъюгация
(инфузории)
Копуляция
(споровики)
Половое размножение
С оплодотворением (большинство многоклеточных)
Половое размножение у многоклеточных
----^Гермафродитизм (сосальщики)
‘ > Партеногенез(пчелы, осы)
факультативный
облигатный
естественный
искусственный
3.2 МИТОТИЧЕСКОЕ ДЕЛЕНИЕ КЛЕТОК
Митоз представляет собой деление материнской клетки, при котором происходит равномерное распределение наследственного материала между образующимися дочерними клетками. Митоз непрерывный процесс, однако, в нем принято выделять несколько стадий: профазу, прометафазу, метафазу, анафазу и телофазу, заключитель-ным событием последней стадии является цитокинез — разделение цитоплазмы.
Ркпздвющмся ядрышко Цаитромцмс прикрепленными к
ней кинггохореми
Интактная я дернея оболочка
Плазмвтическвя мембран!
Конденсирующиеся хромосомы.
Цитоплазма
Образующееся биполярное веретено
Звезда, определяющая один из полюсов веретено
ПРОМЕТАФАЗА
Плазматическая мембрана
Полюсная микротрубочка Кинетохоры —
Кинетохорные микротрубочки
Астральная — микротрубочка
Полюс веретена
Случайно расположенные хромосомы беспорядочно движутся 'Фрагменты ядерной оболочки
'Полюс веретена
в которых две хроматиды соединены центромерой
Укорачивающаяся кинетохорная микротрубочка
Полюсы раздвигаются
Кинетохорные микротрубочки----
укорачиваются, по мере того как хроматида движется к полюсу
Удлиняющаяся-----
полюсная микротрубочка
цитрине:-!
Деконденсирующиеся хромосомы (хроматиды) без кинетохорных микротрубочек
Полюсная микротрубочка
Вокруг отдельных хромосом вновь образуется ядерная оболочка
Вновь образующееся ядрышко
Остаточное тельце — область перекрывания микротрубочек
Остатки полюсных микротрубочек веретена
Восстановление интерфазных микротрубочек, растущих от центросомы
Пара центриолей
Полная ядерная оболочка вокруг деконденсирующихся хромосом
Сократимое кольцо, образующее борозду деления
Основные события митоза: Профаза — конденсация хроматина и образование хромосом, состоящрЛс из двух хроматид, деструктурирование ядрышка, расхождение центриолей к полюсам, образование веретена деления; Прометафаза - дефрагментация ядерной оболочки, рост микротрубочек веретена и их прикрепление к кинетохорам хромосом; Метафаза - образование метафазной пластинки; Анафаза — движение сестринских хроматид к противоположным полюсам клетки; Телофаза — деконденсация хромосом, образование ядрышек и ядерных оболочек, разрушение веретена деления, завершение цитокинеза.
Биологический смысл митоза: 1) поддержание генетической стабильности; 2) механизм моноцитогенного бесполого размножения.
3.3 ЦИТОЛОГИЧЕСКИЕ ОСНОВЫ ГАМЕТОГЕНЕЗА
В основе полового размножения лежит слияние генетической информации мужской и женской гамет. Гаметы (половые клетки) вырабатываются в семенниках и яичниках в ходе гаметогенеза. Цитологической основой гаметогенеза является мейотическое деление или просто мейоз. Мейоз состоит из двух последовательных делений, которые обозначают как мейоз I и мейоз II. Перед первым делением клетки вступающей в мейоз в S-период интерфазы происходит репликация ДНК (хромосомно-хроматидный набор клетки после этого соответствует формуле 2п4с), перед вторым делением интерфаза выпадает. Оба деления мейоза состоят из профазы, метафазы, анафазы и телофазы, заканчивающейся цитокинезом. Профаза I включает такие стадии как лептотена, зиготена, па-хитена, диплотена и диакинез. События профазы I представлены ниже.
Рвкомбинаиионный уэалок
ЛЕПТОТЕНА. Хромосомы, изменив свою интерфазную конформацию, переходят в конденсированную форму. Каждая хромосома прикрепляется к ядерной мембране с помощью специализированной структуры - прикрепительного диска. Сестринские хроматиды очень тесно сближены и не различимы вплоть до поздней профазы. В цитоплазме начинается формирование веретена деления.
ЗИГОТЕНА. Моментом перехода лептотены в зи-готену считают начало синапсиса - тесной конъюгации двух гомологов. Обычно коньюгация начинается со сближения концов двух гомологичных хромосом на ядерной мембране, после чего соединение гомологов распространяется далее вдоль хромосом. При конъюгации каждый ген одной хромосомы приходит в соприкосновение с гомологичным ему геном другой хромосомы. Каждую пару хромосом, в профазе I, обычно называют бивалентом, но учитывая бихроматидный состав каждой хромосомы, гомологичные пары часто называют тетрадами.
ПАХИТЕНА. На стадии пахитены в продольной щели между гомологичными хромосомами появляются крупные рекомбинационные узелки, которым приписывают важную роль в обмене участками между хромосомами. Такие обмены приводят к перекрестам между двумя не сестринскими хроматидами. В пахитене эти перекресты еще не видны, но позднее они проявляются в виде хиазм. В обменах участвует по одной хроматиде каждой из двух спаренных хромосом.
ДИПЛОТЕНА. Стадия диплотены начинается с разделения конъюгировавших хромосом. Гомологичные хромосомы бивалента несколько отодвигаются друг от друга. Однако они еще связаны одной или несколькими хиазмами, т.е. местами, где произошел кроссинговер, происходит частичная деконденсация хромосом, осуществляется синтез мРНК.
ДИАКИНЕЗ. В эту стадию синтез РНК прекращается и хромосомы конденсируются, утолщаются и отделяются от ядерной мембраны. В стадию диаки-неза ясно видно, что каждый бивалент содержит четыре отдельные хроматиды, причем каждая пара сестринских хроматид соединена центромерой, тогда как несестринские хроматиды, претерпевшие кроссинговер, связаны хиазмами. Ядерная оболочка разрушается.
3.4 ЦИТОЛОГИЧЕСКИЕ ОСНОВЫ ГАМЕТОГЕНЕЗА (продолжение)
После окончания длительной профазы I два ядерных деления без разделяющего их периода синтеза ДНК доводят процесс мейоза до конца. Эти оставшиеся стадии обычно занимают не более 10 % всего времени, необходимого для мейоза. На схеме отслеживаются события мейоза после профазы I на примере одной пары гомологич-ных хромосом.
МЕТАФАЗА I. Нити веретена деления проникают в ядерное пространство и прикрепляются к центромерам хроматид. Биваленты выстраиваются в экваториальной плоскости, образуя метафаз-ную пластинку.
АНАФАЗА I. Центромеры каждого бивалента еще не делятся, но сестринские хроматиды уже не примыкают одна к другой. Нити веретена тянут гомологичные хромосомы к противоположным полюсам клетки.
ТЕЛОФАЗА I. Расхождение гомологов к противоположным полюсам означает завершение первого деления мейоза. Число хромосом в одном наборе стало вдвое меньше, но находящиеся на каждом полюсе хромосомы состоят из двух хроматид. Вследствие кроссинговера эти хроматиды генетически неидентичны. Нити веретена обычно исчезают. У животных на каждом полюсе вокруг хромосом формируется ядерная мембрана и образуется ядро. Параллельно происходит цитокинез.
ПРОФАЗА II. Происходят разрушение ядерной мембраны и исчезновение ядрышка, спирализация хромосом, перемещение центриолей к противоположным полюсам клеток и появление нитей веретена.
Метафаза II. Формируется метафаз-ная пластинка.
АНАФАЗА II. Нити веретена тянут за собой к противоположным полюсам клетки отделившиеся друг от друга се-стринские хроматиды.
ТЕЛОФАЗА II. Хромосомы деспира-лизуются. Нити веретена исчезают. Вокруг каждого ядра, которое содержит теперь гаплоидное число и хромосом и хроматид, вновь образуется ядерная мембрана. В результате последующего цитокинеза формируется четыре дочерние клетки.
Биологическое значение мейоза: 1) С его помощью формируются гаметы;
2) Он создает основу для генетической изменчивости.
3.5 СПЕРМАТОГЕНЕЗ
Формирование мужских и женских половых клеток называют гаметогенезом. Образование мужских гамет называют сперматогенезом, а образование женских гамет -оогенезом (овогенезом).
Образование спермиев происходит в семенниках (яичках). Весьма упрощенная схема строения яичка представлена на рисунке справа. В семенниках человека имеются многочисленные извитые семенные канальцы где и протекает сперматогенез.
Поперечный разрез семенного канальца
200
Базальная мембрана
митоз
Первое деление мейоза
Второе деление мейоза
Долька яичка
Сеть яичка
Белочная оболочка
Прямой семенной каналец
Извитой семенной каналец
1) головка придатка;
2) выносящие канальца;
3) основание придатка;
4) семявыносящий проток.
3
Ц— 4
Эпителий семенных канальцев состоит из нескольких слоев клеток. Выделяют наружний слой клеток, которые называются сперматого-ниями и примерно еще шесть слоев клеток, образующихся в результате многократных делений сперматогоний. Данные слои клеток соответствуют последовательным стадиям развития сперматозоидов.
сперматогонии
сперматоциты 1 -го попяпка
Сперматоциты 2-го порядка
МЕИОЗ I
МЕЙОЗII I
Л цитоплазматические мостики
сперматиды
дифференцирующиеся сперматиды
.остаточные тельца
Сперматогенез протекает в четыре стадии: размножения [1], роста [2], созревания [3] и формирования (спермиогенез) [4]. У человека он начинается лишь при достижении половой зрелости. Процесс образования спермия занимает 70 - 75 суток.
Фаза размножения начинается при достижении репродуктивного возраста. В наружних слоях эпителия извитых семенных канальцев начинаются многократные митотические деления сперматогоний.
Далее часть сперматогониев перемещается в следующую зону эпителия. Эти сперматогонии вступают в фазу роста. Они перестают делиться, увеличиваются в объеме и дифференцируются в сперматоциты первого порядка.
На стадии созревания сперматоциты первого порядка вступают в мейоз. Перед профазой мейоза в S-периоде у сперматоцитов первого порядка происходит удвоение ядерной ДНК. Следовательно, каждая клетка, вступающая в мейоз, является тетрап-лоидной (2п4С).
В результате первого деления мейоза образуются сперматоциты второго порядка, которые передвигаются еще ближе к просвету канальца. По размерам они меньше первичных сперматоцитов и содержат в два раза меньше ДНК (п2С).
Сперматоциты второго порядка претерпевают второе деление мейоза и превращаются в гаплоидные сперматиды (пС).
Сперматогонии вступают в стадию формирования. Спермиогенез происходит в боковых углублениях клеток Сертоли (поддерживающих клеток).
3.6 ООГЕНЕЗ
Образование женских гамет называют оогенезом (овогенезом). Оогенез происходит в яичниках и начинается уже в эмбриональном периоде, а продолжается до окончания репродуктивного возраста. Оогенез включает в себя три стадии: размножения, роста (малого и большого) и созревания.
ФАЗА СОБЫТИЯ ВИД ВИД СОБЫТИЯ
ОВОГЕНЕЗА ОВОГЕНЕЗА ФОЛЛИКУЛА КЛЕТОК МЕЙОЗА
У человека стадия размножения начинается на 2-3 месяце эмбрионального развития. Оогонии многократно делятся митозом, и их количество возрастает до 7-10 млн. К 7-8 месяцу эмбриогенеза стадия размножения заканчивается, и большая часть оогоний погибает.
В стадию малого роста (длится от 7-8 месяца эмбриогенеза до достижения репродуктивного возраста) сохранившиеся оогонии вступают в первое деление мейоза и преобразуются в ооциты I порядка. Мейоз приостанавливается на стадии диплоте-ны. Фаза малого роста у человека продолжается как минимум до полового созревания (12-13 лет). Некоторые ооциты I остаются в таком состоянии до последнего
репродуктивного цикла (45-50 лет).
В конце эмбриогенеза ооциты I приобретают оболочку из одного слоя фолликулярных клеток и формируют примордиальные фолликулы Некоторые ооциты I формируют несколько слоев фолликулярных клеток и соедини-тельно-тканную оболочку, преобразуясь в первичные
С наступлением половой зрелости под влиянием более высокой концентрации половых гормонов начинается период большого роста. У человека каждый месяц от 3 до 30 фолликулов увеличиваются в размерах. В одном (опережающем фолликуле) возрастает число фолликулярных клеток, между которыми появляются мелкие полости, заполненные жидкостью (вторичный фолликул). В ходе дальнейшего развития мелкие полости сливаются в одну большую, которая оттесняет ооцит I к стенке фолликула. Его окружает только тонкий слой фолликулярных клеток -лучистый венец (corona radiata). Такой фолликул называют зрелым (третичным или Граафовым пузырьком). В течение всего репродуктивного периода у человека образуется около 400 третичных фолликулов, а остальные погибают. В конце периода большого роста завершается первое деление мейоза, происходит разрыв Граафова пузырька и из него выходит ооцит второго порядка и первое направительное (или редукционное) тельце. Этот процесс называется овуляцией.
После внедрения сперматозоида в ооцит II начинается стадия созревания, в ходе которой завершается второе деление мейоза, отходит второе редукционное тельце и образуется зрелая яйцеклетка, в цитоплазме которой находится сперматозоид. Эта структура называется синкарион.
3.7. СТРОЕНИЕ СПЕРМАТОЗОИДА
Сперматозоид удивительным образом приспособлен к внесению своей генетической информации в яйцеклетку. Большая часть цитоплазмы спермия элиминирует при его созревании, и сохраняются только некоторые органеллы, видоизмененные для выполнения спермием основной функции. Морфологически в спермин выделяют головку, шейку и хвост.
головка
шейка
хвостик
Головка сперматозоида содержит гаплоидное ядро и акросому - видоизмененный комплекс Гольджи. Акросома содержит гиалуронидазу и другие протеазы, способные растворять оболочки яйцеклетки и помогающие сперматозоиду проникнуть в яйцеклетку при оплодотворении.
У многих видов, например у морских ежей, между ядром и акросомой имеется область, содержащая белок актин (на рисунке отсутствует). На ранних стадиях оплодотворения этот белок используется для образования пальцеобразного выроста. У таких видов молекулы на поверхности акросомного выроста участвуют в узнавании спермием и яйцом друг друга.
Шейка содержит пару центриолей, лежащих перпендикулярно друг другу. Одна из них (проксимальная) вносится спермием в цитоплазму яйцеклетки при оплодотворении и участвует в формировании сперматической звезды. Другая (дистальная) дает начало сократительной нити сперматозоида - аксонеме.
Передний отдел хвостика содержит большое число митохондрий, располагающихся спирально вокруг аксонемы. Митохондрии обеспечивают сократительную нить спермия метаболической энергией.
Средний и дистальный отделы хвостика содержат лишь аксонему. Ее строение типично для жгутиков эукариот и в поперечном разрезе представлено на нижних рисунках.
Аксонему образуют микротрубочки, отходящие от центриолей. Вокруг аксонемы располагаются девять плотных волокон, функция которых пока не ясна (рис А). Стержень аксонемы состоит из двух центральных одиночных микротрубочек, окруженных кольцом из девяти двойных микротрубочек (дублетов).
Одна микротрубочка каждого дублета имеет законченное строение и содержит 13 протофиламентов, тогда как вторая имеет С-образное строение и содержит только 11 протофиламентов (рис. Б). Протофиламенты химически представлены димерным белком тубулином.
С микротрубочками связан другой белок - динеин (рис. В). Этот белок может гидролизовать АТФ и преобразовывать химическую энергию в механическую, за счет которой осуществляется движение спермия.
Радиальная спица
микротрубочки
митохондрии
Головка спицы
Центральная микротрувочи
Центральная муфта
Динеиновые выступы
’\Дувлет микротрубочек
Рис. В.
Мостик между периферическими микротрубочками
3.8 ЯЙЦЕКЛЕТКА
Яйцеклетка в высшей степени дифференцирована для выполнения своей единственной функции -развития нового организма. В связи с этим она обладает уникальными особенностями строения:
1) Имеет большие размеры. V человека от 60 до 150 мкм, у лягушек и рыб от 1 до 2 мм, у птиц и рептилий достигает нескольких сантиметров.
2) Имеет запасы питательных веществ. Они сосредоточены в желтке. Желток представляет собой смесь веществ, основным компонентом которой является белок вителлогенин.
3) Содержит полный набор органоидов. В цитоплазме яйцеклеток содержатся все органоиды общего назначения, при этом имеется очень много рибосом и специальные морфогенетические факторы - молекулы направляющие дифференцировку клеток.
4) Периферический слой цитоплазмы содержит кортикальные гранулы. Это аналог акросомы сперматозоидов. Они принимают участие в формировании оболочки оплодотворения, которая надежно блокирует возможность полиспермии.
5) Окружена сложной системой оболочек, которые подразделяются на первичные, вторичные и третичные. Первичные - образованы преимущественно выделениями самой яйцеклетки. У млекопитающих и человека это блестящая оболочка (zona pel-lucida). Она обеспечивает видоспецифичность оплодотворения. Вторичные - образованы фолликулярными клетками. У млекопитающих и человека это лучистый венец (corona radiata). Третичные - формируются выделениями яйцеводов и у млекопитающих отсутствуют. У птиц представлены белочной, подскорлуповой и скорлу-повой оболочками.
ОСНОВНОЕ КОМПОНЕНТЫ ЯЙЦЕКЛЕТКИ
1. адро;
2. микроворсинки плазма-леммы;
3. лучистый венец;
4. отростки фолликулярных клеток лучистого венца;
5. кортикальные гранулы;
6. желточные включения;
7. блестящая оболочка;
8. рецепторы к сперматозоиду.
Классифицируют яйцеклетки учитывая количество желтка и его распределение в цитоплазме.
КЛАССИФИКАЦИЯ ЯЙЦЕКЛЕТОК
Полилецетальные
(многожелтковые)
Алецитальные (безжелтковые)
Олиголецитальные
(маложелтковые)
3.9. ОНТОГЕНЕЗ
Многоклеточные организмы не появляются полностью сформированными, а возникают в результате процесса прогрессивных изменений - развития. Развитие конкретной особи называют онтогенезом. История развития индивидуумов существенно зависит от их видовой принадлежности, однако у многоклеточных животных, размножающихся половым путем, онтогенез организма обязательно начинается с образования давших ему начало половых клеток и заканчивается смертью. Классификация видов онтогенеза при-ведена ниже.
Непрямой онтогенез наблюдается у земноводных, плоских червей, насекомых, у которых из яйца развиваются одна или несколько личиночных стадий, в дальнейшем дающих начало половозрелой особи.
Неличиночный онтогенез имеет место у пресмыкающихся, птиц и яйцекладущих млекопитающих. Они откладывают во внешнюю среду яйца, из которых выходит особь с течением времени достигающая половозрелого состояния.
Внутриутробый онтогенез характерен для плацентарных млекопитающих, в том числе и для человека.
ПЕРИОДИЗАЦИЯ ОНТОГЕНЕЗА
1 ПЕРИОД ПРОЭМБРИОНАЛЬНЫЙ.
Включает в себя сперматогенез и оогенез у родителей, завершающиеся образованием половых клеток дающих начало молодому организму.
2 ПЕРИОД ЭМБРИОНАЛЬНЫЙ (ПРЕНАТАЛЬНЫЙ).
Его событиями являются: 1) оплодотворение; 2) дробление; 3) гаструля-ция; 4) гисто- и органогенз; 5) рождение.
3 ПЕРИОД ПОСТЭМБРИОНАЛЬНЫЙ (ПОСТНАТАЛЬНЫИ).
У человека состоит из нескольких периодов: 1) ювенильного (до половой зрелости); 2) пубертатного (полового созревания); 3) зрелого;
4) старости; 5) смерти.
Оплодотворением называется слияние сперматозоида и яйцеклетки с образованием диплоидной зиготы. Выделяют четыре последовательные стадии оплодотворения:
1. Взаимное узнавание и контакт спермия и яйцеклетки;
2. Регуляция проникновения спермия в яйцо;
3. Слияние генетического материала спермия и яйца;
4. Активация метаболизма яйца для его вступления на путь развития.
Различают дистантные и контактные механизмы взаимодействия сперматозоида и яйцеклетки.
В основе дистантных взаимодействий лежат механизмы хемотаксиса. Миграция сперматозоидов человека к яйцеклетке направляется веществами небелковой и поли-пептидной природы. Эти вещества, выделяемые яйцеклеткой, называют гиногомонами. Сперматозоиды в свою очередь выделяют андрогомоны, которые способны модулировать активность своих собратьев.
Контактное взаимодействие видоспецифично и обеспечивается рецепторным аппаратом обеих гамет. Например, в сперматозоидах морского ежа содержится белок байн-дин, обеспечивающий прикрепление спермиев к яйцеклеткам только своего вида. Соответственно на оболочке яйца морского ежа имеется рецептор для байндина. Аналогичный механизм контактных взаимодействий гамет реализуется и у человека.
Следует отметить, что спермин человека не становятся активными, пока не пройдут капацитации т.е. не приобретут оплодотворяющей способности. Суть капацитации состоит в необходимости подвергнуть спермин воздействию выделений женских половых путей. Лишь после этого рецепторы спермиев становятся активными.
Углеводы, блокирующие рецепторы спермия
Спермий не прошедший капацитации
На рисунке:
NAr - N-ацетилглюкозаминовые остатки;
функции рецептора выполняет фермент гликозилтрансфераза, локализованный в мембране спермия.
У мыши и хомячка (по-видимому и у человека) после прикрепления спермия начинается акросомная реакция - выброс ферментов акросомы Ферменты изливаются на оболочки яйцеклетки и происходит это по механизмам экзоцитоза, что приводит к растворению оболочек и движению спермия в ооплазму за счет энергии биения жгутика.
Однако у некоторых видов (кролик) акросомная реакция происходит на некотором расстоянии от яйцеклетки и индуцируется растворимыми веществами, выделяемыми яйцом. У небольшого числа видов (морская свинка) акросомная реакция происходит в определенное время независимо от наличия яиц.
Заказ № 3775
К оболочке яйцеклетки одновременно прикрепляется множество сперматозоидов. Все они осуществляют ак-росомную реакцию и по законам конкуренции стремятся к проникновению в цитоплазму яйца (фото слева). Однако зигота любого организма должна быть диплоидной, для чего яйцеклетка может быть оплодотворена лишь одним спермием. В связи с этим природой предусмотрен механизм предотвращения полиспермии (полиспермией называют проникновение в ооплазму нескольких спермиев) как средство предупреждения полиплоидно-сти зиготы.
Таких механизмов сегодня известно два. Выделяют быстрый и медленный механизмы предотвращения полиспермии. Их сравнительная характеристика и свойства представлены ниже.
СРАВНИТЕЛЬНАЯ ХАРАКТЕРИСТИКА ОСНОВНЫХ МЕХАНИЗМОВ ПРЕДОТВРАЩЕНИЯ ПОЛИСПЕРМИИ
БЫСТРЫЙ МЕХАНИЗМ МЕДЛЕННЫЙ МЕХАНИЗМ
1) Реализуется за счет изменения электрического заряда на оолемме после проникновения первого сперматозоида. 1) Реализуется за счет экзоцитоза кортикальных гранул яйцеклетки и формирования оболочки оплодотворения.
2) Срабатывает быстро за десятые доли секунды. 2) Начинает работать через 30 — 40 секунд после проникновения сперматозоида.
3) Блокирует возможность полиспермии примерно на 60 секунд. 3) Навсегда блокирует возможность полиспермии.
4) Механизм известен для морских ежей, лягушек, пока не доказан для млекопитающих. 4) Механизм реализуется и у низших животных, и у млекопитающих.
Прикрепившийся спармий
На верхнем рисунке показан мембранный потенциал в яйце морского ежа до, и после оплодотворения. До добавления спермы внутренняя поверхность мембраны имеет более отрицательный потенциал, чем наружная. Через 0,1 сек после прикрепления спермия к оболочкам яйца потенциал сдвигается в направлении положительных величин.
На правом рисунке показан механизм кортикальной реакции у яйцеклетки морского ежа. Студенистая оболочка яйцеклетки морского ежа аналогична блестящей оболочке млекопитающих, рецепторы байндина - способны связываться с белком байндином, находящимся на мембране спермиев и, соответственно прикреплять сперматозоиды к оболочке яйцеклетки.
Секрет кортикальных гранул уплотняет ви-теллиновую оболочку яйца и разрушает рецепторы байндина, что делает невозможным проникновение следующих сперматозоидов в ооплазму.
Студенистая * оболочка
Рецептор байндина
Плазматическая ЬММбрвН* ЯЙЦД
Кортикальная
Гранула
Содержимое
ЭКЗОЦИТОЗ КОРТИКАЛЬНЫХ ГРАНУЛ (кортикагшмая реакция) >
гранул выходит в перивитеп— липовое пространство
Мембране кор ти калькой гранулы сливается с плазматической мембраной яйца
Измененный вителлиновый слой без рецепторов байндина (мембрана оплодотворения)
Мозаичная мембрана (продукт слияния мембран спермия, яйца и его кортикальных гранул)
Для того, чтобы оплодотворение привело к развитию яйца, должны произойти изменения в его цитоплазме. Их совокупность и составляют события заключительной стадии оплодотворения. Процессы активации метаболизма яйца подразделяют на ранние реакции и поздние реакции.
РАННИЕ РЕАКЦИИ
1) Повышение концентрации ионов кальция в цитоплпзме;
2) Повышение потребления кислорода;
3) Усиление синтеза НАДФ и, как следствие, липидов.
ПОЗДНИЕ РЕАКЦИИ
11 Повышение pH;
21 Активация синтеза ДНК;
3) Деблокирование имеющихся в цитоплазме яйцеклетки мРНК;
4) Активация синтеза белка.
Молекулярные механизмы приводящие к увеличению цитоплазматической концентрации ионов кальция и повышению pH приведены на рисунке.
Акросомв
Дмцилглииерол
Рецептор
Гипотетически акросомная молекула
Неактивна* фосфолипаза С
Активный переносчик ионов Na* м Н*
Фосфатидилинозитол-
-ВИСфОСфЭТ
Активная фосфолипаза С
Неактивный переносчик ионов Na*и Н*
ГТФ
АДФ
АТФ
11
Увеличение рн
Неактивная лротеинкиназа С
Активная протеинхиназа С
Неактивный ,
вело* G Гуанозин-5-дифосфат
Активный аелок 6
Ииозитолтрифосфат
1) Спермин взаимодействует с гипотетическим рецептором на мембране яйцеклетки. Рецептор активирует ГТФ-зави-симый G-белок. G-белок активирует фосфолипазу С, которая расщепляет один из фосфолипидов мембраны на два биологически активных вещества - инозитолтрифосфат (ИТФ) и диацилглице-рол (ДАТ).
2) ИТФ вызывает выход ионов кальция из эндоплазматического ретикулума, а ДАТ активирует протеинкиназу С, которая в свою очередь повышает активность КаТЕГ-обменника. Этот обменник удаляет из цитоплазмы ионы водорода в об-мен на ионы натрия.
Активация яйца
Высвобождение-
♦ Ci’+
Эндоплазматический ретикулум
3.14. ОБЩИЕ ПРЕДСТАВЛЕНИЯ О ДРОБЛЕНИИ
Следующим после оплодотворения этапом онтогенеза является дробление. Дробление это серия митотических делений яйцеклетки на клетки меньшего размера - бластомеры. Особенностями дробления является отсутствие в интерфазе стадии Gi - стадии роста и очень высокая скорость первых митозов. В ходе дробления формируется бластула, в клетках которой устанавливается нормальное ядерно-цитоплазматическое соотношение.
Характер дробления яйцеклетки зависит от нескольких факторов, но, главным образом, определяется количеством и положением желтка, то есть, типом яйцеклетки. Через зиготы, содержащие мало желтка, борозда дробления проходит полностью, а зиготы с большими запасами желтка делятся лишь частично. Классификация типов дробления.
Тип дробления Тип яйцеклетки Симметрия дробления Представители
Голобластическое (полное) Изолецитальные Радиальное Иглокожие, Ланцетник
Спиральное Моллюски, круглые и плоские черви
Билатеральное Асцидии
Асинхронное Млекопитающие, человек
Мезолецитальные Радиальное Амфибии
Меробластическое (частичное) Т ел о лецитал ьные Билатеральное Головоногие
Дискоидальное Рыбы, рептилии, птицы
Центролецитальные Поверхностное Членистоногие
Други классификации типов дробления, учитывающие размеры образующихся клеток (1) и синхронность процесса (2) представлены на нижней схеме.
РАВНОМЕРНОЕ
Все образующиеся бластомеры имеют равный размер.
НЕРАВНОМЕРНОЕ Образуются клетки большего и меньшего размера, которые называются макромеры и микромеры.
ДРОБЛЕНИЕ
СИНХРОННОЕ
Все бластомеры делятся одновременно, в связи с чем, число клеток в зародыше увеличивается в геометрической прогрессии.
АСИНХРОННОЕ
Разные бластомеры делятся в разное время, в связи с чем, в зародыше может находится нечетное число клеток.
3.15. РАДИАЛЬНОЕ ГОЛОБЛАСТИЧЕСКОЕ ДРОБЛЕНИЕ
Представлена последовательность митотических делений при голобластическом радиальном дроблении у иглокожих. На верхнем рисунке (А) приведен пример равномерного дробления у морского огурца (Synapta oigita). На нижнем рисунке (Б) пример неравномерного дробления у амфибий (Xenopus laevis).
Вегетативный полюс
Апикальная половина
Вегетативная половина
Апикальный полюс
Бластула Вегетативный полюс (в разрезе)
Яйцеклетки морского огурца изолецитальные, поэтому борозды деления разделяют зиготу на бластомеры равной величины. Яйцеклетки амфибий мезолецитальные и дробятся сложнее. Борозды дробления на нижнем рисунке обозначены римскими цифрами в порядке их появления. Желток, сконцентрированный на вегетативном полушарии, затрудняет дробление. Поэтому второе деление дробления у амфибий начинается раньше, чем заканчивается первое. Борозда третьего деления закладывается не экваториально, а со смещением к анимальному полюсу, который свободен от желтка. В результате последующих делений на анимальном полюсе образуется большое число микромеров, а на вегетативном -меньшее количество макромеров.
3.16 СПИРАЛЬНОЕ И БИЛАТЕРАЛЬНОЕ ГОЛОБЛАСТИЧЕСКОЕ ДРОБЛЕНИЕ
Спиральное голобластическое дробление характерно для кольчатых червей и не головоногих моллюсков. На рисунке приведен пример начальных этапов дробления у брюхоногого моллюска Trochus sp. Верхний ряд - вид со стороны анимального полюса, нижний ряд - вид с боку.
Первые два деления меридиональные и приводят к образованию четырех равных бластомеров. Экваториальная борозда смещается к анимальному полюсу поэтому после третьего деления образуется четыре макромера и четыре микромера, причем микромеры смещаются относительно своих сестринских макромеров либо по часовой, либо против часовой стрелки.
Билатеральное дробление встречается у асцидий. У этих животных первая - меридиональная плоскость деления закладывает единственную ось симметрии. Каждое последующее деление отклоняется либо от анимально-вегетативной оси, либо от плоскости экватора, поэтому формирующиеся микро- и макромеры лежат зеркально только относительно первой борозды деления. На рисунке представлен конечный результат такого дробления у Styela partita.
Вид со стороны вегетативного полюса
3.17 АСИНХРОННОЕ ГОЛОБЛАСТИЧЕСКОЕ ДРОБЛЕНИЕ У МЛЕКОПИТАЮЩИХ И ЧЕЛОВЕКА
Дробление у млекопитающих имеет ряд особенностей: 1) дробление одно из самых медленных; 2) дробление начинается через сутки после оплодотворения; 3) весь процесс дробления продолжается 3-4 суток; 4) одно деление длится 12-24 часа; 5) первые деления дробления протекают в маточных трубах; 6) дробление асинхронное и чередующееся; 7) на восьмиклеточной стадии происходит процесс компактизации.
Прозрачная оболочка
Натка
Морула
Яичник
Бластоциста
Ранняя стадий имплантации
2-клеточная ст
дро&лемил
Яйцевод
Первое деление
Плоскость деления Л
Плоскость деления I
Плоскость Плоскость деления! деления ПА
А. ИГЛОКОЖИЕ (морской еж)
Б. МЛЕКОПИТАЮЩИЕ (кролик)
Плоскость деления ПВ
Оплодотворение
Ридунок иллюстриру-ет, что оплодотворение у млекопитающих и человека происходит в маточных трубах, а дробление осуществляется при движении зиготы в матку. Имплантация происходит после образования бластулы типа бластоциста. Строение бластоцисты в поперечном разрезе представлено на нижнем рисунке.
Средний рисунок показывает одну из важнейших особенностей дробления млекопитающих. Видно, что у морского ежа оба бластомера, образовавшиеся после первого меридионального дробления, подвергаются следующему делению в меридиональной плоскости, проходящей перпендикулярно плоскости первого деления. У млекопитающих один бластомер делится меридионально, другой - экваториально. Такой тип дробления называется чередующимся и он сохраняется на протяжении всего процесса дробления.
Ранняя Компактный
8-клеточная стадия 8-клеточный
зародыш
А
Компак-тизация
Плотные контакты
Б
Внутренняя клетка
32-клеточная стадия (поперечный разрез)
Наружная клетка
Клетка трофовласт
Щелевые контакты
В
итация|’
Внутренняя клеточная Бластоциста масса
Полость Бластоцисты
На 8-ми клеточной стадии между бластомерами устанавливаются плотные контакты, на 16-ти клеточной -формируется морула состоящая из темных клеток (внутри) и светлых (снаружи). Далее в процессе кавитации зародыш образует бластоцель, заполненный жидкостью и формируется бластула.
При меробластическом дроблении большое количество желтка препятствует разделению всей цитоплазмы яйца за исключением небольшой ее части. При поверхностном дроблении от желтка свободен небольшой периферический слой цитоплазмы, а при дискоидальном - клеточные деления происходят на небольшом участке цитоплазмы на одном из полюсов (анимальном), расположенном над желточной массой. На рисунках приведены схемы дискоидального дробления у курицы (А) и поверхностного дробления у дрозофилы (Б).
На рисунке А видно, что процесс дробления затрагивает ограниченную часть цитоплазмы. Вначале образуется однослойная бластодерма, затем число клеточных слоев в дробящейся зоне цитоплазмы увеличивается и их становится три или четыре. Дис-коидальным этот тип дробления называется потому, что зона дробления по форме напоминает диск.
Рисунок Б иллюстрирует более сложные закономерности поверхностного дробления. При этом типе дробления обособление клеток происходит после многократного разделения ядер. Ядро зиготы претерпевает несколько митозов в центральной области яйца. Затем ядра, с окружающими их островками цитоплазмы (энергиды), мигрируют на периферию яйца, где митозы продолжаются с убывающей скоростью. Вокруг центрально расположенного желтка образуется синцитиальная бластодерма. Зародыш называется так потому, что все ядра (до 256 штук) окружены одной плазматической мембраной. Ядра мигрирующие к заднему полюсу в первую очередь окружаются самостоятельными мембранами. Их называют полярными клетками. В будущем они дадут начало половым клеткам взрослого организма. После образования полярных клеток и нескольких дальнейших делений ядер синцитиальной бластодермы, каждое ядро последней, вместе с островком цитоплазмы, окружается индивидуальной мембраной и становится клеточной бластодермой. Под бластодермой остаются запасы желтка.
мигрируют на периферию
Бластодерма, имеющая клеточное строение
Полярные клетки
Цифры над каждым зародышем соответствуют минутам после откладки яйца, а цифры внизу обозначают число имеющихся ядер.
3.19 ТИПЫ БЛАСТУЛ
Конечным результатом дробления является образование многоклеточной структуры -бластулы. Выделяют несколько типов бластул отличающихся некоторыми деталями строения. Слой бластомеров образующих бластулу называют бластодермой (1). В некоторых типах бластул имеется полость, заполненная жидкостью содержащей продукты жизнедеятельности бластомеров. Такая полость называется бластоцелем (2). Основные типы бластул и их структурные элементы представлены на рисунках.
При равномерном радиальном дроблении бластула принимает вид однослойного пузырька с большим бластоцелем. Такая бластула называется типичной или целобластулой.
В случае неравномерного радиального дробления телолецитальных яиц (например амфибий) бластодерма состоит из клеток разных размеров - мелких на анимальном полюсе и крупном на вегетативном. Бластоцель смещен к анимальному полюсу. Такая структура называется амфибластулой.
При дискоидальном дроблении возникает дискобластула, получившая свое название из-за небольшого участка цитоплазмы, подвергающегося дроблению и напоминающего по форме диск, расположенный на анимальном полюсе.
При поверхностном дроблении центральная часть зародыша заполнена желтком, а бластодерма состоит из одного слоя клеток. Такая бластула называется перибластулой.
Бластула с небольшим центрально расположенным бластоцелем называют стерробластулой.
Бластула млекопитающих и человека называется бластоцистой. Ее бластодерма (1) образована светлыми клетками и называется трофобластом. На внутренней поверхности одного из полюсов располагаются более темные клетки называемые внутренней клеточной массой или эмбриобластом (3).
3.20 ОБЩИЕ ПРЕДСТАВЛЕНИЯ О ГАСТРУЛЯЦИИ
Гаструляция представляет собой в высшей степени интегрированный процесс миграции клеток и тканей, приводящий к перераспределению клеток бластулы. Во время га-струляции клетки занимают новое положение и приобретают новых соседей В этот период устанавливается план строения многослойного тела животного. В ходе гаструля-ции формируется гаструла, в состав которой входят три зародышевых листка - наружняя эктодерма, внутренняя энтодерма и лежащая между ними мезодерма.
Инвагинация - впячивание участка бластодермы наподобии вдавливания внутрь стенки резинового мяча, когда на него нажимают
Ингрессия (иммиграция, выселение) - миграция клеток по отдельности из поверхностного слоя внутрь зародыша.
Эпиболия - (обрастание) движение эпителиальных пластов, которые распространяются как одно целое и окружают глубокие слои зародыша.
Инволюция - вворачивание внутрь зародыша увеличивающегося в размерах наружнего пласта клеток и его распространение по внутренней поверхности наружних клеток.
Деляминация - расщепление единого клеточного пласта на два более или менее параллельных.
целобластула
амфибластула
бластоциста
Исходный клеточный пласт
деляминация
Два образованных клеточных пласта
3.21 ПРИМЕРЫ ГАСТРУЛЯЦИИ
Анимальный полюс
Вегетативный полюс
энтодерма
На верхнем рисунке приведен пример га-струляции у морского ежа. Видно, что энтодерма закладывается путем инвагинации бластодермы. Мезодерма образуется путем миграци клеток, происходящих из микромеров бластулы.
На нижнем рисунке представлены этапы гаструляции у амфибий. На анимальном полюсе начинается интенсивное деление бластомеров (Б), приводящее к эпиболии бластодермы. Затем (В), сбоку от анимального полюса образуется бластопор (небольшое углубление) через который осуществляется инволюция нарастающей клеточной массы во внутрь зародыша, что приводит к смещению бластоцеля и образованию гастроцеля (В - Д). Мезодерма закладывается путем иммиграции (Е).
3.22 ГАСТРУЛЯЦИЯ У МЛЕКОПИТАЮЩИХ И ЧЕЛОВЕКА
Наиболее сложно гаструляция протекает у млекопитающих. Этот процесс подразделяют на две стадии. Первая протекает на 7 сутки после оплодотворения, вторая начинается на 14-15 сутки и заканчивается на 17 день. В промежутке между первой и второй стадиями гаструляции формируются провизорные органы и развивается трофобласт. События первой фазы отражены на рисунке.
Внутренняя клеточная масса (эмбриобласт) подвергается деляминации с образованием гипобласта и эпибласта (А, Б). Гипобласт дает начало —желточному мешку (В), а эпибласт совершает вторую деляминацию (Г) с образованием выстилки амниона и зародышевого эпибласта.
После завершения первой фазы трофобласт развивается с образованием цитотрофобласта (клеточного) и синцитиального трофобласта (Г). Син-цитиотрофобласт впоследствие (Д) даст начало хориону. Хорион вростая в стенку матки образует плаценту.
К началу второй фазы зародыш приобретает вид указанный на рисунке Д.
Желточный (париетальная
энтодерма)
События второй стадии гаструляции проиллюстрированы на рисунке (Е). В зародышевом эпибласте в области первичной полоски начинается интенсивная пролиферация клеток и их миграция на гипобласт. Нижний слой клеток даст начало энтодерме, средний слой - мезодерме, а сам зародышевый эпибласт по окончании гаструляции преобразуется в эктодерму.
3.23 ГИСТООРГАНОГЕНЕЗ
Дифференцировка зародышевых листков - эктодермы, мезодермы и энтодермы - с образованием тканей и органов называется гистоорганогенезом. Судьба зародышевых листков в упрощенном виде отражена на схеме.
1. эпидермис кожи и его производные (ногти, волосы, потовые и сальные железы, молочные железы, эмаль зубов),
2. Нервная система.
СОМИТЫ
1. дерматом -внутренний слой кожи (дерма);
2. миотом -мышечные ткани;
3. склеротом -все виды соединительной ткани.
НОЖКИ СОМИТА
1. нефрогоно-том - мочеполовая система.
БОКОВЫЕ
ПЛАСТИНЫ 1. спланхно-том — серозные оболчки (перикард, брыжейки, плевральные листки), сердце.
1. Эпителий желудочно-кишечного тракта,
2. Эпителий бронхо-легочной системы.
Образование органов - весьма сложный процесс. В их образовании принимают участие несколько зародышевых листков, поэтому правильнее говорить о преимущественной роли эктодермы, мезодермы или энтодермы в закладке того или иного органа. Механизмы органогенеза приведены на рисунке внизу на примере нейруляции - образования нервной системы.
Нейруляция начинается (А) с инвагинации нервной пластинки, выделяющейся на дорсальной эктодерме. Нервные валики, располагающиеся по краям нервной пластинки смыкаются с образованием нервной трубки. Клетки нервного валика преобразуются в клетки нервного гребня, дающие начало периферической нервной системе, а сама нервная трубка образует спинной и головной мозг. Этапы этого процесса представлены на рисунках (Б и В).
Эпидермис
Нервная трувка
4.24 ТЕРАТОЛОГИЯ
Различные факторы внешней среды способны нарушать нормальное течение эмбриогенеза и вызывать рождение детей с уродствами. Эти факторы называются тератогены. Наука, изучающая действие тератогенов, называется тератологией. Тератогены по своей природе можно подразделить на 3 группы.
ТЕРАТОГЕНЫ
ФИЗИЧЕСКИЕ
ХИМИЧЕСКИЕ
1) радиация;
2) удары, сотрясения;
3) опухоли, сдавливающие зародыш.
I) многие лекарства;
2) сырье и продукты производств;
3) сельхозхимия (пестициды, гербициды и т.д.);
4) бытовая химия.
БИОЛОГИЧЕСКИЕ
1) вирусы (краснухи, герпеса);
2) микроорганизмы (toxoplasma gondii).
В ходе эмбриогенеза имеются критические периоды развития, когда зародыш наиболее уязвим к действию тератогенов. Всего выделяют три критических периода. Первый — соответствует первой неделе беременности. В этот период тератогены могут нарушить процесс оплодотворения или дробления и их действие подчиняется правилу “все или ни чего”. Это значит, что при повреждении зиготы или большого числа бластомеров зародыш погибает, а при повреждении малого числа бластомеров он остается жизнеспособным за счет высокой регенераторной способности и ребенок рождается без аномалий. Второй критический период выделяют в интервале вторая - восьмая недели беременности. В этот период происходит гаструляция, гистогенез и начальные этапы органогенеза. Во второй критический период зародыш наиболее уязвим к действию тератогенов и большинство аномалий, проявляющихся в постнатальном онтогенезе “обязаны” своим появлением именно второму критическому периоду. Третий критический период захватывает 9-40 недели беременности, причем наиболее уязвимой фазой является 18-22 недели. В эти сроки наиболее активно формируется гуморальная система и закладывается биоэлектрическая активность головного мозга. Нарушение этих процессов тератогенами, может негативно сказаться не только на состоянии нервной и гуморальной систем, но и на строении и функции любого эффекторного органа, так как развитие последних происходит под нейрогумо-ральным контролем.
Пример тератогенного эффекта транквилизатора талидомида и зависимость его действия от сроков приема препарата проиллюстрированы на рисунках.
На левом рисунке отражена структура талидомида (А) и ребенок, (Б) родившийся у женщины, принимавшей талидомид во второй критический период. Рисунок (В)отражает зависимость эффекта от сроков приема.
Время, сут после последней менструации
Генетика - наука о закономерностях наследственности и изменчивости живых организме!
Наследственность - способность организмов передавать свои признаки и особенности развития последующим поколениям.
Изменчивость - способность организмов приобретать новые признаки и утрачивать прежние.
Под признаками понимают морфологические, биохимические, физиологические, патологические и другие свойства организмов, по которым одни особи отличаются от других. Примерами признаков являются цвет глаз, резус-группа, цвет кожи и.т.д. Признаки бывают альтернативные и не альтернативные.
Противоположные проявления одного и того же признака называются альтернативными, или взаимоисключающими, признаками. У одной особи обычно имеется только один альтернативный признак. Например, голубые глаза или карие, резус-положительная группа крови или резус-отрицательная, темный цвет кожи или светлый.
Разные или не альтернативные признаки - это признаки, которые могут одновременно присутствовать у одной и той же особи, то есть они не являются взаимоисключающими. Например, цвет глаз, группа крови, цвет кожи - это разные признаки. Совокупность разных признаков организма называют фенотипом.
Определяющим фактором развития любого признака организма является наследственный материал клеток, на реализацию которого оказывают существенное влияние факторы внешней среды. Различают три уровня организации наследственного миатериала.
Некоторые детали организация наследственного материала представлены на рисунке справа.
1. Пара гомологичных хромосом. Они имеют одинаковые морфологические характеристики (длину, положение центромеры) и содержат одни и те же гены. Одна из гомологичных хромосом является отцовской, а другая материнской.
2. Локус — место расположения конкретного гена в хромосоме. Оно постоянно и меняется только при некоторых мутациях (транслокации, инверсии).
3. Аллели — разновидности одного гена, определяющие развитие альтернативных признаков. Некоторые гены могут иметь множество аллелей (более двух), например гены, определяющие группы крови человека по системе АВО.
4. Гомозиготность говорит о присутствии в одинаковых локусах гомологичных хромосом одинаковых аллелей гена.
5. Гетерозиготность присутствие в гомологичных хромосомах разных аллелей гена.
Законы наследования признаков впервые установил австрийский монах Грегор Мендель. В 1865 году он сделал сообщение на заседании научного "Общества любителей естествознания" в городе Брно (ныне Чехия) на тему "Опыты над растительными гибридами". Его открытие не было понято современниками и позднее, в 1900 году трое ученых из Голландии, Германии и Австрии - Де Фриз, Коррейе и Чермак переоткрыли закономерности наследования выявленные Менде-лем и признали его приоритет.
Г. Мендель достиг успеха благодаря разработанному им гибридологическому методу анализа.
Основные положения этого метода:
В работе использовались только "чистые сорта" растений. В ряде потомств они не давали ни каких отклонений от стандартных признаков сорта - выражаясь современным языком Мендель исполь-зовал гомозиготные растения.
Учет проводился не по всем многообразным признакам у родителей, а лишь по некоторым альтернатив-ным признакам.
Применялся точный количественный учет в каждом ряду поколений и последующий математический анализ полученных результатов скрещивания.
Первый закон Г. Менделя называется законом “единообразия гибридов первого поколения”. Он гласит: при скрещивании гомозиготных особей с альтернативными признаками, все потомство единообразно по генотипу и фенотипу.
Первый закон Мендель вывел из простого наблюдения.
Для скрещивания он брал сорта гороха, имеющие альтернативные признаки. Например, одни растения имели только гладкие семена, а другие -только морщинистые. В результате мейоза родительские растения давали по одному сорту гамет. При скрещивании таких родительских форм все гибриды первого поколения имели только гладкие семена и по генотипу были гетерозиготными.
Скрещивание по одной паре альтернативных признаков называется моногибрид-ным, по нескольким признаком - полигибридным.
При записи родительские организмы обозначают буквой Р от латинского parents-родители. Знаком х обозначают скрещивание. Женская особь записывается симво-пом Венеры - $, а мужская - символом Марса - Гаплоидные гаметы обводятся кружком. В результате скрещивания формируются гибриды первого поколения Fi -от латинского fili — потомство.
Признак, проявившийся в первом поколении называют доминантным от латинского domine — господин. В нашем примере это гладкая форма семян гороха. Признак родителей не проявившийся в первом поколении называют рецессивным — от латинского recessivus- отступление - морщинистые семена гороха. Аллели доминантного признака обозначают заглавными буквами, а аллели рецессивного — строчными буквами.
Заказ № 3775
4.3 ВТОРОЙ И ТРЕТИЙ ЗАКОНЫ ГРЕГОРА МЕНДЕЛЯ
Второй закон Менделя называется закон расщепления. Он гласит, что при скрещивании двух гетерозиготных особей в потомстве наблюдается расщепление по генотипу в соотношении 1:2:1, а по фенотипу 3:1.
Второй закон вытекает из результатов скрещивания между собой гибридов первого поколения. Схема этого эксперимента Г. Менделя представлена на рисунке справа. Поскольку обе родительские формы являются гетерозиготными, и материнское и отцовское растения образуют по два типа гамет. В процессе мейоза в одну гамету уходит доминантный аллель Л, а в другую - рецессивный аллель а.
При оплодотворении отцовские гаметы имеют равную вероятность встречи с материнскими гаметами. Выявить все их возможные сочетания удобнее всего при построении решетки Пеннета. Для этого в верхней строке выписывают все типы женских гамет, а в левой колонке все типы мужских гамет. На пересечении получают все возможные сочетания интересующих нас генов в зиготе.
Анализ решетки показывает, что по генотипу в поколении F2 - I часть растений является гомозиготной по доминантному аллелю, 2 части - гетерозиготными и I часть - гомозиготной по рецессивному аллелю (расщепление 1:2:1). Фенотипически гомозиготные по доминантному аллелю растения и гетерозиготные растения имеют гладкие семена. Растения гомозиготные по рецессивному аллелю имеют морщинистые семена (расщепление 3:1).
Третий закон Менделя - закон независимого расщепления. Он гласит - гены определяющие формирование различных признаков, наследуются независимо друг от друга. Следовательно, этот закон справедлив для полигибридного скрещивания.
Для доказательства справедливости третьего закона проанализируем наследование двух признаков семян их формы (ГЛАДКИЕ и морщинистые) и цвета (ЖЕЛТЫЕ и зеленые). Доминантные признаки указаны заглавными буквами, зеленый горох обозначен черным.
При скрещивании гибридов первого поколения между собой во втором поколении появляется 4 категории потомков - расщепление по фенотипу 9:3:3:1. Рассмотрим механизм их появления. Родительские растения образуют по четыре типа гамет. Все возможные сочетания этих гамет, отражены в решетке Пеннета.
Р: в ААВВ
х $ aabb
Соотношение желтых семян к зеленым -12:4 или 3:1.
Соотношение гладких семян к морщинистым — 12:4 или 3:1.
Такое возможно лишь при независимом наследовании генов.
АВ АЬ аВ ab
АВ AABBQ) ААВЬ Q АаВВ АаВЬ Q
АЬ ААВЬ Q ААЬЬ ф АаВЬ АаЬЬ
аВ АаВВ Q АаВЬ 0^ ааВВ ^у^ ааВЬ ^у^
ab АаВЬ (2^ АаЬЬ ааВЬ ^у^ ааЬЬ
Большинство нормальных и патологических признаков человека наследуются в соответствии с законами Грегора Менделя. Такие признаки называются “менделирую-щими”. Однако часто наследование признаков отклоняется от законов Менделя. Причиной этому являются различные виды взаимодействия между генами, сцепление и неполная пенетрантность.
ВИДЫ ВЗАИМОДЕЙСТВИЯ МЕЖДУ ГЕНАМИ
АЛЛЕЛЬНЫМИ
НЕАЛЛЕЛЬНЫМИ
ы S д
Мд go It.
Eg s о 1=1
ы
S
ДИ ®о
os
о
ы
S s 3 а KS<
MSS Н t«S оэа
S >=1
S О
и ы е 6П «
о S к бп
к
ЫО
S о
§
*ы
9 g eg
§ ь. ы
S t? о
ДОМИНАНТНЫЙ
РЕЦЕССИВНЫЙ
К
СЦЕПЛЕНИЕ
АУТОСОМНОЕ
с половыми ХРОМОСОМИ
ПОЛНОЕ
ЧАСТИЧНОЕ
С Х-ХРОМОСОМОЙ
С У-ХРОМОСОМОЙ
Пенетрантность отражает долю особей, обладающих признаком от общего числа носителей гена, определяющих развитие этого признака. Пенетрантность выражается в процентах. Различают два вида пенетрантности.
ПЕНЕТРАНТНОСТЬ
ПОЛНАЯ
У всех особей в популяции ген обязательно пробивается в признак.
НЕПОЛНАЯ
У части особей ген блокируется и не пробивается в признак.
4.5 НЕПОЛНОЕ ДОМИНИРОВАНИЕ И ЛЕТАЛЬНЫЙ ЭФФЕКТ
Полное доминирование лежит в основе наследования признаков, подчиняющихся законам Г.Менделя. Все другие виды взаимодействия между аллельными и неаллельными генами могут сопровождаться модификацией формул менделевских расщеплений.
Неполным доминированием называют снижение фенотипической выраженности доминантного аллеля в присутсгвии рецессивного. Это объясняется тем, что и доминантный аллель и рецессивный активируют синтез белка или фермента определяющего развитие признака. Белок или фермент, опосредуемый рецессивным аллелем, несколько снижает активность продукта доминантного гена.
Примером неполного доминирования является наследование окраски цветов львиного зева. Если скрещивать между собой растение гомозиготное по доминантной красной окраске с рецессивной гомозиготой по белой окраске то в первом поколении все гибриды будут иметь промежуточную розовую окраску.
При скрещивании между собой гибридов первого поколения одна часть потомков будет иметь красные цветы (доминантные гомозиготы), две части будут с розовыми цветами (гетерозиготы) и одна гомозиготная по рецессивному аллелю часть будет иметь белые цветы.
Следовательно у гибридов второго поколения, вместо ожидаемого классического расщепления по фенотипу в соотношении 3 красных к 1 белому мы наблюдаем расщепление 1:2:1, совпадающее с расщеплением по генотипу.
Р: 3 АА х аа
Летальными называют гены, которые в гетерозиготном состоянии определяют развитие признака, а в гомозиготном состоянии оказываются смертельными для организма. Расщепление при моногибридном скрещивании в этом случае соответствует формуле 2:1.
Примером летального действия аллелей является наследование окраски шкурки у мышей.
У мышей ген желтой окраски (А), на рисунке он показан светлым цветом, доминантен по отношению к гену черной окраски шерсти (а). Оказалось, что желтые мыши всегда гетерозиготны, поскольку гомозиготы по доминантному аллелю погибают еще в эмбриональном периоде и никогда не рождаются.
Схема опыта по скрещиванию двух желтых мышей представлен на рисунке справа. Анализ решетки Пеннета, отражающей все разнообразие поколения Fi свидетельствует о том, что соотношение желтых и черных мышей составляет 2:1 как по генотипу, так и по фенотипу.
4.6. НАСЛЕДОВАНИЕ СЕРПОВИДНО-КЛЕТОЧНОЙ АНЕМИИ
Неполное доминирование и летальное действие генов у человека проявляется при наследовании серповидно-клеточной анемии. Для понимания этих закономерностей необходимо рассмотреть принципы структуры и генетического контроля синтеза гемоглобина человека.
Отличия между 0-цепями и S-цепями гемоглобина минимальны и касаются лишь 6
аминокислоты, гемоглобин a Val-His-Lcu-Thr-Pro Glu-Glu-Lys-гемотлобин s Val-His-Leu-Thr-Pro Val-Glu-Lys-
Замена глутаминовой кислоты на валин приводит изменению растворимости гемоглобина. В дезокси-форме гемоглобин S нерастворим и выпадает в осадок в результате чего форма эритроцита вместо нормальной округлой становится серповидной.
За синтез 0-цепи отвечает рецессивный аллель гена s, а за синтез S-цепи его доминантный аллель - S. Все гомозиготы по рецессивным аллелям (ss) это здоровые люди имеющие в крови округлые эритроциты с содержащимся в них гемоглобином А.
Оказалось, что если ребенок получает от родителей два доминантных аллеля S — то он погибает в пренатальный период или сразу после рождения. Следовательно, доминантные аллели в гомозиготном состоянии проявляют свойства летальных генов.
Гетерозиготы Ss имеют в крови и нормальные эритроциты и серповидные. Такие люди страдают серповидноклеточной анемией, так как половина их эритроцитов не способна транспортировать кислород. Наличие в крови больных гетерозигот и эритроцитов с гемоглобином А и серповидных эритроцитов с гемоглобином S свидетельствует о явлении неполного доминирования при наследовании серповидноклеточной анемии.
Р: 3 Аа х $ Аа болен больна
7 ~——-— А А а Аа Q
& АаО с А1 о с
4.7 МНОЖЕСТВЕННЫЙ алелизм
Многие гены имеют несколько аллелей, определяющих развитие одного признака. При этом каждая конкретная особь в популяции может быть носителем только двух из всех существующих аллелей. Подобный тип наследования носит название множественного аллелизма.
Одним из примеров служит серия аллелей гена, определяющего развитие окраски меха у кроликов. Известно 4 аллеля. Аллель дикого типа - агути (С+) доминантен по отношению к трем остальным: аллелю шиншиловой окраски (Сс1), аллелю гималайской окраски (Ch) и аллелю альбинизма (Са). Расположим их в порядке доминантности: С+ > Cch > Ch > Са. Возможные сочетания аллелей цвета меха в генотипе и фенотипы животных приведены в таблице._____
ГЕНОТИП ФЕНОТИП
с+с+, c+cch, c+ch, с+са Сс^Сс^ Qch^h chch,’chca саса Дикий тип (агути) Шиншилла Светло-серый Гималайский Альбинос
У человека по типу множественного аллелизма наследуются группы крови системы АВО. Существует четыре группы крови в этой системе. Установлено, что эритроциты человека содержат особые белки - агглютиногены. Всего их бывает два вида А и В. Люди, эритроциты которых не содержат агглютиногенов, имеют первую группу крови. Если в эритроцитах вырабатывается агглютиноген А - имеют вторую группу, для обладателей агглютиногенов В - характерна третья группа и если в эритроцитах человека содержатся оба агглютиногена формируется четвертая группа. За синтез агглютиногенов и следовательно за наследование этих групп крови ответственен ген, обозначаемый латинской буквой I. Существует три аллеля этого гена — два доминантных и один рецессивный. Все возможные сочетания аллелей гена I и формируемые при этом группы крови отражены в таблице.
-Группа крови генотип - 1(0) П(А) 1П(В) IV (AB)
гомозиготы ii IAIA IBIB -
гетерозиготы - IAi “TT“ IAIB
Анализ таблицы показывает, что аллели 1А и 1в являются доминантными в отношении рецессивного аллеля i. Однако аллели 1А и 1в проявляют свойство кодоминирова-ния по отношению друг к другу. Это свойство проявляется у лиц с четвертой группой крови. У них один доминантный аллель не подавляет действие другого и поэтому в эритроцитах происходит синтез обеих агглютиногенов. Возможные группы крови детей при различных сочетаниях генотипов родителей представлены в следующей таблице.
I(ii) II (1А1Аили IAi) in (IBIB или IBi) IV (IAIB)
I(ii) I I, II I, III п,ш
П (1А1А или IAi) I, п I, II I, П, in, IV П, III, IV
1П (1в1в или IBi) I, III I, П, III, IV I, III II, Ш, IV
IV ПА1В) II. Ill II. Ш. IV П. III. IV П. III. IV
4.8 . ДОМИНАНТНЫЙ ЭПИСТАЗ
Эпистазом называют подавление одного гена, другим неаллельныи ему геном. Гены подавители получили название эпистатических генов или генов супрессоров. Подавляемые гены называют гипостатическими генами.
ЭПИСТАЗ
ДОМИНАНТНЫЙ
При доминантном эпистазе один доминантный ген "А" подавляет действие другого доминантного гена "В". При этом у доминантных дигомозигот (ААВВ) не будет проявляться признак, контролируемый геном В.
РЕЦЕССИВНЫЙ
При рецессивном эпистазе действие доминантных аллелей “В” блокируется рецессивными аллелями гена “а . У обладателей генотипа ааВВ не будет проявляться признак В.
Явление доминантного эпистаза обнаруживается при изучении наследования серой и вороной окраски лошадей. Окраска у лошадей определяется несколькими неаллельными генами. Серый цвет определяется доминантным геном А, а вороной цвет -доминантным геном С. Гомозиготы по обеим рецессивным генам имеют рыжую окраску (на рисунке белую). Однако все особи, имеющие доминантные аллели С и хотя бы один доминантный аллель А - остаются серыми, поскольку ген серой окраски у лошадей является эпистатическим по отношению к гену вороной окраски.
Р: ? ААСС х в аасс
При скрещивании серых лошадей из поколения Fi между собой в F2 получим:
2 АС Ас аС ас
АС ААСС ААСс АаСС АаСс
Ас ААСс ААсс АаСс Аасс
аС АаСС АаСс ааСС ааСс
ас АаСс Аасс ааСс аасс
Таким образом, при доминантном эпистазе расщепление по фенотипу происходит в соотношении 12:3:1. Иногда рецессивная дигомозигота имеет такое же фенотипическое проявление как и доминантная дигомозигота. В этом случае формула приобретает вид 13:3.
4.9 РЕЦЕССИВНЫЙ ЭПИСТАЗ
Примером рецессивного эпистаза является подавление доминантных аллелей групп крови системы АВО неизвестным рецессивным аллелем в некоторых человеческих популяциях. Впервые это явление было обнаружено в окрестностях города Бомбея, поэтому оно было названо “Бомбейским феноменом”. Его механизм представлен ниже в виде схемы взаимодействия между эпистатическими и гипостатическими генами.
При Бомбейском феномене возможно неожиданное наследование групп крови системы АВО. Рассмотрим родословную семьи, у которой впервые было обнаружено данное явление сначала с обычных позиций (левый рисунок), а затем с учетом явления эпистаза (правый рисунок).
Феномен состоял в том, что у матери из второго поколения, имеющей первую группу крови (0), родилась дочь.с четвертой группой (АВ). Супруг женщины имел вторую группу крови (А). Если допустить, что доминантщле аллели 1А дочь получила от отца, то неясно происхождение доминантных аллелей I , поскольку ее мать в своем генотипе имеет только аллели i. Интересно, что вторая дочь имела первую группу.
Если учесть наличие у матери эпистатического гена, подавляющего в гомозиготном рецессивном состоянии (гг) доминантные аллели системы АВО, то механизм рождения в этой семье девочки с четвертой группой крови становится понятным. Мать девочек имеет в своем генотипе доминантные аллели I , но вследствие эпистаза они неактивны и фенотепически женщина имеет первую группу крови (0). Отец девочек, скорее всего гетерозиготен по эпистатическим генам, и первой своей дочери он точно передал доминантный аллель R, вследствие чего эпистаз у нее стал невозможным. Генотип второй дочери может быть самым разнообразным и, на данный момент, не ясен.
hi
й(0)
IAi(A)
—г—°
IBIB (В) или IBi(B)
-d
ii(0)
iiRr (0)
IAiRr (A)
°
IBIBRr (В) или IBiRr (B)
—О
IBirr (0)
hi
IAIB (AB)
ii(0)
IaIbRt(AB) iiRr (0) или ??rr (0)
и
и
4.10 КОМПЛЕМЕНТАРНОЕ ДЕЙСТВИЕ ГЕНОВ
Комплементарным называется взаимодействие, при котором признак проявляется лишь при наличии двух доминантных неаллельных генов, Причем каждый из этих ге-нов не имеет самостоятельного проявления.
Явление комплементарности хорошо изучено на примере наследования окраски зерен кукурузы. У кукурузы пурпурная окраска семян развивается только при встрече двух доминантных аллелей генов С и Р. Если в генотипе оказываются доминантные аллели только одного из указанных генов или все аллели оказываются рецессивными -то цвет зерен оказывается белым. То есть для развития пурпурной окраски семян у кукурузы необходимо сочетание двух доминантных неаллельных генов.
При скрещивании дигетерозиготных растений с пурпурными семенами у их потомков произойдет отклонение от известного менделевского расщепления по фенотипу 9:3:3:1. И вместо этого проявится соотношение 9:7 или 9:6:1.
Р: S СсРр х § СсРр
G: СР, Ср, cP, ср и СР, Ср, cP, ср
И:
Т: СР Ср cP ср
СР ССРР ССРр СсРР СсРр
Ср ССРр ССрр СсРр Ссрр
cP СсРР СсРр ссРР ссРр
ср СсРр Ссрр ссРр ссрр
Расщепление по фенотипу составляет: 9 (с пурпурной окраской семян) 7 (с неокрашенными семенами).
У человека комплементарным действием обладают гены пигментации волос. Ген М контролирует образование черного пигмента меланина. Причем этот ген представлен тремя аллелями Mb , Mbw и Мь . Первый аллель определяет синтез значительных количеств, второй промежуточных и третий малых количеств меланина. Другой ген R контролирует образование красного пигмента. Рецессивный аллель (Rj) определяет синтез малых количеств красного пигмента, а доминантный (R2) - больших количеств. Сочетания аллелей этих генов дают весь спектр окрасок волос человека.
R।Rj R 2
Светлый Шатен с Темно-
шатен рыжеватым рыжий
оттенком
Блондин Блондин с Яркорыжеватым рыжий оттенком
Шатен Каштановый Темно-рыжий
Темный Темный шатен 'Темный шатен (лоснящиеся шатен волосы) (волосы с глянцем)
Rl^2 R2R2
MbkMbk
Комплементарность действия проявляется в том, что при некоторых сочетаниях аллелей развиваются дополнительные признаки-такие как лоснящиеся или глянцевые волосы.
Все разнообразие оттенков нообразие оттенков волос представлено на схеме.
Брюнет Брюнет Брюнет (лоснящиеся (волосы волосы) с глянцем)
2
4.11 ПОЛИМЕРИЯ
О полимерном наследовании или полимерии говорят, когда один признак формируется под влиянием нескольких эквивалентных генов. Такие эквивалентные гены называют полимерными. Обозначают такие полимерные гены одной буквой, но приписывают индекс 1, 2 и т.д. - в зависимости от того, сколько пар полимерных аллелей участвует в развитии признака.
Полимерный тип наследования широко встречается в природе. Ему подчинена пере-дача по поколениям количественных признаков.
У овса красная окраска зерен определяется двумя полимерными генами. На развитие признака эти гены действуют в одном направлении. Доминантные аллели гена определяют синтез красного пигмента в зернах, а рецессивные не кодируют синтез пигмента. Следовательно, чем больше доминантных аллелей получит зигота, тем более окрашенными будут зерна у этого растения.
Р: с? К1К1К2К2 х ? kjk|k2k2
4 О
G: К,К2 kik2
Fi: К1к1Кгк2
2
Г: К,К2; K]k2; к,К2; к,к2
F2:
к,к2 К,к2 к,К2 kik2
К1К2 — ВД.КгЪ — К.Щг щ К1к]КгК2 ।—। К1к1Кгк2
№ — К.К.Кгкг [—। К1К1к2к2 □ К.к.Кгкг ।—। К1к1кгк2
kiK2 — К|к,К2К2 [—। №№ к.к^гКг ।—। к1к1кгК2
№ [—। К1к|К2кг ।— К|к]кгк2 ।— kikik2K2 k|kjk2k2
Примечания: |Н - ярко-красные семена;
^В- красные семена;
□ -розовые семена;
□— бледно-розовые семена;
□— неокрашенные семена.
Расщепление по фенотипу 15 - окрашенных частей: 1 неокрашенная часть.
Расщепление с учетом интенсивности окраски:
1 - ярко-красные : 4 - красные : 6 - розовые : 4 - бледно-розовые : 1 - неокрашенные.
Полимерия лежит в основе наследования количественных признаков и у человека, например, таких как масса тела, рост, величина артериального давления, пигментация кожи.
У европейцев кожа имеет светлый оттенок, поскольку в генотипе содержатся лишь рецессивные аллели полимерного гена Р (Р1Р1Р2Р2РзРзР4Р4)-
У негров имеются лишь доминантные аллели этого гена (Р1Р1Р2Р2Р3Р3Р4Р4) поэтому кожа имеет темный оттенок.
В браках европейцев и негров рождаются мулаты, являющиеся тетрагетерозиготами по гену Р (Р1Р1Р2Р2Р3Р3Р4Р4).
В браках мулатов возможно рождение детей с различными оттенками кожи от светлого до темного. Результат зависит от количества доминантных аллелей полученных при формировании зиготы.
4.12 ГЕНЕТИКА ПОЛА
Пол человека, как и любого другого организма, закладывается при формировании зиготы и механизм его определения является хромосомным. У большинства видов пол зависит от сочетания половых хромосом (гетеросом), у некоторых организмов половые хромосомы отсутствуют. Основные варианты хромосомного определения пола пред-ставлены в виде таблицы.
Хромосомные механизмы формирования пола
Представители гаметы генотип
<? 3 ?
Млекопитающие, дрозофила, человек Y или X X XY XX
Птицы, чешуекрылые (бабочки) X Y или X XX XY
Прямокрылые (кузнечики) X или 0 X ХО XX
Перепончатокрылые (пчелы, муравьи) п п п 2п
У человека, как и у большинства видов гетерогаметным является мужской пол, а го-могаметным - женский. Пол ребенка определяется, каким X- или Y- содержащим сперматозоидом будет оплодотворена яйцеклетка.
Р: $XY
Г: X, Y
Fi: XX 50%
$ XX
X
XY 50%
4.13 НАСЛЕДОВАНИЕ ПРИЗНАКОВ СЦЕПЛЕННЫХ С ПОЛОМ
Признаки, гены которых локализованы в половых хромосомах (гетеросомах) называются сцепленными с полом. Типы такого наследования представлены на схеме.
НАСЛЕДОВАНИЕ СЦЕПЛЕННОЕ С ПОЛОМ
С Х-ХРОМОСОМОЙ
С Y-ХРОМОСОМОЙ
Х-ДОМИНАНТНОЕ Х-РЕЦЕССИВНОЕ
Аллели генов могут быть локализованы в различных участках половых хромосом. В гетеросомах имеется участок структурно и функционально идентичный (2) для обеих хромосом. В то же время в Х-хромосо-ме имеется фрагмент (1) отсутствующий в Y- хромосоме и наоборот в мужской хромосоме есть участок (3), отсутствующий в женской хромосоме.
Закономерности наследования признаков сцепленных с полом существенным образом зависят от того, в каких участках гетеросом локализован ген, ответственный за развитие этого признака.
♦ Если ген расположен в участке 2, то характер наследования идентичен аутосомному;
♦ Если ген расположен в участке 1, то наследование осуществляется по X-доминантному или Х-рецессивному типу;
♦ Если ген находится в участке 3, наблюдается голандрический тип наследования при котором признаки передаются от отца только сыновьям.
Х-рецессивный тип наследования (на примере гемофилии)
Р: ?XHXh х ^XHY
здорова здоров
G: Хн, Xh Хн, Y
Fi: ХНХН, XHXh, XHY, XhY
В браке женщины, гетерозиготной по гену гемофилии и здорового, по этому признаку, мужчины в потомстве наблюдается расщепление Вероятность рожде-
ния у таких супругов здоровой гомозиготной девочки составляет 25%, здоровой девочки носительницы патологического гена - 25%, здорового мальчика - 25%, больного мальчика — также 25%.
Голандрический тип наследования (на примере ушного гипертрихоза)
Р: ?ХХ х dXYG
без признака с признаком
Г: X X, Yg
Fi: XX, XYg
В браках между женщинами (они никогда не имеют голандрических признаков) и мужчинами носителями такого признака все дочери рождаются без признака, а все сыновья несут признак отца.
Генотип мужчин, имеющих только одну копию гена в зоне Y-хромосомы, отсутствующей в Х-хромосоме называют геми-зиготным.
4.14 СЦЕПЛЕННОЕ НАСЛЕДОВАНИЕ ПРИЗНАКОВ
Различные (не альтернативные) признаки, гены которых находятся в одной хромосоме, называются сцепленными.
Выделяют два вида сцепления генов:
ПОЛНОЕ
Имеет место в том случае, когда два неаллельных гена расположены очень близко друг к другу и поэтому вероятность кроссинговера между ними ничтожно мала.
При полном сцеплении признаки передаются потомству всегда сцепленно.
ЧАСТИЧНОЕ
Наиболее частый вариант сцепления генов. Наблюдается когда между двумя генами имеется некоторое расстояние и, следовательно, высока вероятность кроссинговера.
При частичном сцеплении у части потомков наблюдаются комбинации родительских признаков.
У дрозофилы гены, отвечающие за цвет тела и форму крыльев расположены в одной паре гомологичных хромосом. Причем у самцов в гаметогенезе кроссинговер отсутствует и анализируемые гены проявляют свойства тесно сцепленных. У самок кроссинговер происходит и гены ведут себя как частично сцепленные.
Введем систему обозначения генов: А - нормальная длина крыльев;
а - короткие крылья;
В - серая окраска тела;
b - черная окраска тела.
При скрещивании дигетерозиготных самцов с гомозиготными по рецессивным аллелям самками в потомстве наблюдается расщепление в соотношении 1:1.
НАСЛЕДОВАНИЕ ПРИ ПОЛНОМ СЦЕПЛЕНИИ ГЕНОВ
При скрещивании дигетерозиготных самок с гомозиготными самцами вследствие кроссинговера появляется четыре сорта потомков. У части из них признаки сочетаются как у родителей, а у части - имеется комбинация родительских признаков.
Р: 2 АВ х r?ab
а-b “b
G: А В; а Ь; А Ь; а В а b
Fi: АВ А b а В а b
НАСЛЕДОВАНИЕ ПРИ НЕПОЛНОМ СЦЕПЛЕНИИ ГЕНОВ
Количественный учет потомков Fi при неполном сцеплении генов указывает, что соотношение фенотипов не соответствует формуле 1:1:1:1. Особей с родительскими (материнскими + отцовскими) признаками насчитывается 83%, а особей с комбинацией этих признаков - 17%. Следовательно, в ходе овогенеза образовалось 83% (41,5% + 41,5%) не кроссоверных гамет и 17% (8,5% + 8,5%) - кроссоверных гамет. Количество кроссоверных гамет определяется расстоянием между анализируемыми генами. В рассматриваемом примере расстояние между генами цвета тела и формы крыльев у самок дрозофилы составляет 17 морганид.
4.15 КАРТИРОВАНИЕ ХРОМОСОМ
Результаты анализирующего скрещивания при сцепленном наследовании дают возможность составить карту относительного расположения генов. При этом за единицу расстояния принимают 1% образования кроссоверных гамет. При расстоянии 50 мор-ганид и более признаки наследуются независимо, несмотря на локализацию генов в одной хромосоме.
При построении линейной карты хромосом учитывают, что гены лежат друг за другом в линейном порядке и не перекрываются. Если между генами А и В процент рекомбинаций составляет 5%, а между генами В и С - 7 %, то возможно два варианта их взаиморасположения:
57 1к 5
,-Л------А---Ч С 7—-----А
АВ С С А В
Для установления точного местоположения гена С необходимо знать процент перекреста между С и А. Если он равен 12%, то верна левая карта, а если он составляет 2% истинным является правый вариант карты.
Картирование хромосом человека, на основании гибридологического анализа невозможно (или является очень грубым), вследствие малого числа потомков в семьях и невозможности экспериментального (в частности анализирующего) скрещивания. Поэтому был разработан метод гибридизации соматических клеток.
Суть метода заключается в образовании гибридных соматических клеток мыши и человека. Особенностью этих гибридов является сохранение хромосомного набора мыши и потеря почти всех хромосом человека. Через несколько клеточных циклов в гибридах сохраняются лишь отдельные хромосомы человека (идеальным является вариант, когда сохраняется одна гомологичная пара).
ПОСЛЕДУЮЩИЙ АНАЛИЗ
Скнкврион
п=51- 57
50 мышиных +
1 -7 человеческих
Если последующий анализ гибридных клеток выявляет наличие, какого либо белка или фермента характерного для человека, то вывод очевиден — ген, ответственный за синтез этого белка локализуется в хромосоме, присутствующей в гибридных клетках. Номер этой хромосомы определяют методом дифференциального окрашивания.
Хромосомные делеции позволяют установить точное место локализации гена в хромосоме. Например, было показано, что ген кислой фосфатазы эритроцитов располагается во второй хромосоме. При делеции короткого плеча этой хромосомы в терминальном районе полосы 2р23 активность фермента в эритроцитах сохранялась. При делеции, затрагивающей проксимальный район полосы 2р23 —активность отсутствовала. Следовательно, ген кислой фосфатазы эритроцитов расположен в полосе 2р23. (центромеры указаны стрелками).
Изменчивостью называют свойство живых организмов приобретать и утрачивать признаки.
ИЗМЕНЧИВОСТЬ
ФЕНОТИПИЧЕСКАЯ
МОДИФИКАЦИОННАЯ
СЛУЧАЙНАЯ
ГЕНОМНЫЕ МУТАЦИИ
ХРОМОСОМНЫЕ МУТАЦИИ
ГЕННЫЕ МУТАЦИИ
1) моносомия
2) ТРИСОМИЯ
3) НУЛЕСОМИЯ
1) ДЕЛЕЦИИ
2) ДУПЛИКАЦИИ
3) ИНВЕРСИИ
1) РЕЦИПРОКНЫЕ 1) ДЕЛЕЦИИ
2) НЕРЕЦИПРОКНЫЕ 2) ДУПЛИКАЦИИ
3) ЦЕНТРИЧЕСКИЕ
Фенотипической называют изменчивость, которая возникает под действием факто-
ров среды (ФС) на продукты активности генов.
ГЕН------> ФЕРМЕНТ (БЕЛОК)
> ПРИЗНАК
ФЕНОТИПИЧЕСКАЯ ИЗМЕНЧИВОСТЬ
МОДИФИКАЦИОННАЯ
Возникает под влиянием одного известного фактора среды.
СЛУЧАЙНАЯ
Возникает под влиянием нескольких слабых по силе факторов среды или — под влиянием неизвестного фактора.
Диапазон изменчивости, в пределах которой под влиянием факторов среды один и тот же генотип дает различные фенотипы называется нормой реакции. В зависимости от широты нормы реакции признаки делятся на две группы:
ПРИЗНАКИ
ПЛАСТИЧНЫЕ
Имеют широкую норму реакции, то есть сильно зависят от факторов внешней среды.
НЕПЛАСТИЧНЫЕ
Имеют узкую норму реакции, следовательно, мало меняются под влиянием факторов внешней среды.
СВОЙСТВА МОДИФИКАЦИЙ
и сЗ а п
S
§ а <□ а (U S м а
Соматические мутации проявляются в признак непосредственно у той особи, у которой они возникли. Генеративные мутации в родительском поколении не проявляются, а реализуются в признак у потомков.
Если изменения наследственного материала происходят в соматических клетках зародыша, то после рождения часть клеток организма будет иметь нормальный наследственный материал, а часть клеток - измененный. Такие организмы называются мозаиками.
Изменения, возникшие в постнатальный период онтогенеза, могут быть причинами онкологических заболеваний, например рака кожи и ретинобластомы (опухоль сетчатки глаза).
Соматические мутации, количество которых накапливается с возрастом, по одной из гипотез, являются причиной старения и последующей смерти многоклеточных организмов.
Термин “мутация” впервые был введен Де Фризом для обозначения случайных изменений признаков у растений.
В зависимости от уровня повреждения наследственного материала различают генные, хромосомные и геномные мутации.
Частоту мутаций можно рассчитать по простой формуле : число случайных аномалий 2 х число обследованных
Факторы среды вызывающие мутации называют мутагенами.
Известны 3 механизма комбинатив-ной изменчивости:
1) Независимое и случайное расхождение хромосом при мейозе;
2) Случайное сочетание хромосом при оплодотворении;
3) Кроссинговер.
Комбинативной изменчивости принадлежит существенная роль в получении новых форм в дикой природе, новых сортов культурных растений и пород домашних животных.
Заказ № 3775
МЕХАНИЗМЫ ВОЗНИКНОВЕНИЯ ГЕНОМНЫХ МУТАЦИЙ ОСНОВАНЫ НА ОШИБКАХ РАСХОЖДЕНИЯ ХРОМОСОМ
1) нарушение цитокинеза у одного из бластомеров; 2) нарушение цитокинеза у зиготы; 3) нарушение цитокинеза в ходе гаметогенеза.
•4----------- Гаметы:
----------- Сперматогонии и оогонии:
Зиготы:
Схема неравномерного расхождения трех пар хромосом в гаметогенезе. При слиянии аберрантных гамет с нормальными формируются зиготы с моносомным или трисомным кариотипом.
4.20 ХРОМОСОМНЫЕ МУТАЦИИ
ИНВЕРСИИ
РЕЦИПРОКНЫЕ
НЕРЕЦИПРОКНЫЕ
ЦЕНТРИЧЕСКИЕ (робертсоновские)
ДЕЛЕЦИЯ (утрата фрагмента)
ДУПЛИКАЦИЯ
(дублирование фрагмента)
ИНВЕРСИЯ (изменение положения фрагмента)
норма мутация
норма мутация
РЕЦИПРОКНАЯ ТРАНСЛОКАЦИЯ (двусторонний обмен фрагментами хромосом)
НЕРЕЦИПРОКНАЯ ТРАНСЛОКАЦИЯ (односторонний обмен фрагментом хромосомы)
ЦЕНТРИЧЕСКАЯ ТРАНСЛОКАЦИЯ (соединение негомологичных хромосом в области центромер)
нормальные мутантные
нормальные
норма
механизм
мутация
ДЕЛЕЦИИ
ДУПЛИКАЦИИ
ЗАМЕНЫ ОСНОВАНИЙ
(Чаще приводят к изменению лишь одной аминокислоты в белке).
Норма Мутация
ДНК ГГГ-АТГ-ТТТТГ-АТГ-ТТТТ ГГГ-АЦГ-ЦЦГ-АТГ-ТТЦ
мРНК ЦЦЦ-УАЦ-ГГЦ-УАЦ-ААГ ЦЦЦ-УГЦ-ГГЦ-УАЦ-ААГ
Полипептид Про-тир-гли-тир-лиз Про-цис-гли-тир-лиз
ГЕННЫЕ ДЕЛЕЦИИ
(Могут приводить к радикальному изменению последовательности аминокислот).
Норма Мутация
ДНК ГГГ-АТГ-ЦЦГ-АТГ-ТТЦ... ГГГ-А..ГЦ-ЦГА-ТГТ-ТЦ,..
мРНК ЦЦЦ-УАЦ-ГГЦ-УАЦ-ААГ... ЦЦЦ-У..ЦГ-ГЦУ-АЦА-АГ,..
Полипептид Про-тир-гли-тир-лиз-... Про-сер-ала-тре-...
ГЕННЫЕ ДУПЛИКАЦИИ
(Также вызывают радикальное изменение последовательности аминокислот).
Норма Мутация
ДНК ГГГ-АТГ-ЦЦГ-АТГ-ТТЦ... ГГГ-АТТ-ГЦЦ-ГАТ-ГТТ-Ц...
мРНК ЦЦЦ-УАЦ-ГГЦ-УАЦ-ААГ... ЦЦЦ-УАА-ЦГГ-ЦУА-ЦАА-Г
Полипептид Про-тир-гли-тир-лиз-... Про-термин. Конец синтеза.
5.1 МЕТОДЫ ИССЛЕДОВАНИЯ ГЕНЕТИКИ ЧЕЛОВЕКА
Для изучения генетики человека неприменим гибридологический метод, разработан-ный Г. Менделем, в связи с чем используется ряд специальных методов.
Позволяет установить:
ГЕНЕАЛОГИЧЕСКИЙ Основан на составлении и анализе родословных ПОПУЛЯЦИОННО- СТАТИСТИЧЕСКИЙ Основан на законе Харди-Вайнберга БЛИЗНЕЦОВЫЙ Основан на выявлении внутри парного сходства в группах моно- и дизиготных близнецов ЦИТОГЕНЕТИЧЕСКИЙ Основан на анализе кариотипа клеток пациентов БИОХИМИЧЕСКИЙ Основан на качественном и количественном анализе активности продуктов генов ДЕРМАТОГЛИФИЧЕСКИЙ Основан на анализе кожного рисунка на ладонях и пальцах 1) является ли признак наследственным; 2) тип ниследования; 3) генотип членов семьи; 4) вероятность рождения больного ребенка; 5) пенетрантность признака. Позволяет установить генетическую структуру человеческих популяций: 1)частоту рецессивного и доминантного аллелей; 2) частоту доминантных гомозигот, гетерозигот и рецессивных гомозигот. Позволяет установить степень зависимости формирования конкретного признака от наследственных факторов и факторов среды. Позволяет выявлять геномные и хромосомные мутации и тем самым диагностировать хромосомную патологию, как в постнатальном, так и в пренатальном периоде онтогенеза. Позволяет: выявлять генные мутации и тем самым диагностировать генные заболевания. Позволяет подтверждать наличие хромосомной патологии.
При составлении родословных используют условные обозначения, предложенные Г. Юстом в 1931 году и применяют ряд правил.
Здоровая женщина
Здоровый мужчина
Пробанд
Пол неизвестен
Выкидыш
Медицинский аборт
Мертворождение
Брак
Внебрачная связь
Сибсы
Монозиготные близнецы
Кровнородственный брак
Повторный брак
Дизиготны близнецы
Бездетный брак
Бесплодие
Больные
гетерозиготные носители признака
1
ПРАВИЛА СОСТАВЛЕНИЯ РОДОСЛОВНОЙ:
1) Пробанд обозначается стрелкой;
2) Потомство одного поколения располагают в одном горизонтальном ряду в порядке рождения (слева направо) и обозначают арабскими цифрами;
3) Младшее поколение располагают под старшим и нумеруют римскими цифрами сверху вниз (по левому краю родословной);
4) Все линии вычерчивают строго параллельными и перпендикулярными;
5) Заканчивают родословную датой составления и личной подписью врача;
6) При необходимости к схеме прикладывают легенду (рукописное приложение), в! которой отмечают любую информацию, облегчающую последующий анализ1 родословной.
5.3 ГЕНЕАЛОГИЧЕСКИЙ МЕТОД (АНАЛИЗ РОДОСЛОВНЫХ)'
При анализе родословных в первую очередь определяют тип наследования признака, используя характерные признаки:
АУТОСОМНО-ДОМИНАНТНЫЙ С/ '
1)
2)
3)
4)
Признак встречается в каждом поколении, Болеют и мужчины и женщины в равной степени, Каждый больной ребенок имеет хотя бы одного больного родителя,
У здоровых родителей все дети здоровы.
В 9 10 11 12 13 14 1$ 16
Л
АУТОСОМНО-РЕЦЕССИВНЫЙ
1)
2)
3)
4)
Заболевание передается “по горизонтали” — болеют в основном сибсы, Болеют и мужчины и женщины в равной степени, Здоровые родители могут иметь больных детей В родословной встречаются кровнородственные браки.
(Нот
IV
2
5 в
9 10
* 1 2 3 4
1)
2)
3)
4)
5)
Х-ДОМИНАНТНЫЙ
Если болен отец, то все его дочери больны, а сыновья здоровы, Признак встречается в каждом поколении, Болеют и мужчины и женщины, но больных женщин примерно в 2 раза больше, Каждый больной ребенок имеет хотя бы одного больного родителя, У здоровых родителей все дети здоровы.
II
3
О 2
“• й □ й о X
о
д й
1)
2)
3)
4)
5)
Х-РЕЦЕССИВНЫЙ
Болеют в основном мужчины,
Болеют обычно мужчины - родственники пробанда по материнской линии, Сын никогда не наследует заболевание отца, От брака больных мужчин и здоровых женщин, все дети здоровы, но у дочерей могут быть больные сыновья,
Если пробанд женщина, ее отец и сыновья обязательно больны.
Y-СЦЕПЛЕННЫЙ
1)
2)
3)
Болеют только мужчины, Признак передается от отца к сыну, Если в браке больного отца и здоровой женщины рождаются только девочки, передача признака по поколениям прерывается.
5.4. ПОПУЛЯЦИОННО-СТАТИСТИЧЕСКИИ ’ И БЛИЗНЕЦОВЫЙ МЕТОДЫ
Популяционно-статистический метод позволяет выявить закономерности распределения генов и генотипов в популяциях. Эта закономерность была установлена в 1908 году английским математиком Дж. Харди и немецким врачом В. Вайнбергом независимо друг от друга. Поэтому закон стали называть их именами.
Закон справедлив для “идеальных популяций”, отвечающих следующим требованиям:
J Размеры популяции велики;
J Спаривание происходит случайным образом;
S Отсутствует мутационный процесс;
J Плодовитость особей с любыми генотипами одинакова;
Z Отсутствует миграция особей.
Закон Харди-Вайнберга включает два положения:
Сумма частот аллелей одного гена в идеальных популяциях - величина постоянная.
p+q=l где: р - частота доминантного аллеля; q — частота рецессивного аллеля.
Сумма частот доминантных гомозигот, гетерозигот и рецессивных гомозигот в идеальной популяции величина постоянная.
p2+2pq+q2=l где:
р - частота Доминантных гомозигот; 2pq - частота гетерозигот;
q - частота рецессивных гомозигот.
Сущность близнецового метода заключается в выявлении конкордантности (сходства) и дискордантности (различия) в группах монозиготных и дизиготных близнецов, что позволяет с помощью специальных формул оценить относительную роль наследственности и факторов среды в развитии конкретного признака. На практике чаще используют формулу Хольцингера:
П____
100%- Cdz ; Е=100% - Н где:
Н — коэффициент наследуемости;
Е — коэффициент влияния среды;
GnZ - конкордантность в группе монозиготных близнецов;
Cdz конкордантность в группе дизиготных близнецов.
Н>70%
Признак определяется наследственными факторами.
50%<Н<70%
Признак определяется и наследственными и средовыми факторами.
Н<50%
Признак определяется факторами среды.
5.5 ДЕРМАТОГЛИФИЧЕСКИЙ МЕТОД
Дерматоглифика (от греческого derma - кожа и gliphe - рисовать) - это изучение рельефа кожи на пальцах, ладонях и подошвенных поверхностях стоп. Метод используется в определении зиготности близнецов, в судебной медицине и криминалистике, как вспомогательный - при диагностике хромосомных заболеваний.
ДАКТИЛОСКОПИЯ
Изучение закономерностей папиллярных узоров.
ПАЛЬМОСКОПИЯ
Изучение ладонного рельефа.
ПЛАНТОСКОПИЯ
Изучение кожного узора поверхности стоп.
ВЫДЕЛЯЮТ ПЯТЬ ОСНОВНЫХ ТИПОВ ПАПИЛЛЯРНЫХ УЗОРОВ:
дуга петля переход от петли двойная петля завиток
к завитку
Папиллярные узоры индивидуальны и неповторимы (кроме однояйцевых близнецов). У монозиготных близнецов коэффициент конкордантности по этому признаку составляет 0,8 — 0,9. То есть не менее 8 пальцев рук имеют идентичные отпечатки. У дизи-готных близнецов конкордантность не превышает 0,5.
НАИБОЛЬШУЮ ДИАГНОСТИЧЕСКУЮ ЦЕННОСТЬ ПРЕДСТАВЛЯЕТ
ОПРЕДЕЛЕНИЕ ВЕЛИЧИНЫ УГЛА atd
У основания П, Ш, IV и V пальцев находятся пальцевые трирадиусы - точки где сходятся три разнонаправленных тока папиллярных линий. Их обозначают латинскими буквами a, b, с, d. Вблизи браслетной складки, отделяющей кисть от предплечья, по продольной линии, идущей от IV пястной кости, располагается главный (осевой) трирадиус -1. Если провести линии от трирадиусов а и d к t, то образуется ладонный угол adt, который в норме не превышает 57°.
Угол adt в норме и при хромосомных болезнях: 1 — синдром Патау; 2 — синдром Дауна; 3 - синдром Шерешевского-Тернера; 4 —здоровый человек ; 5 — синдром Клайнфелыпера.
5.6 ЦИТОГЕНЕТИЧЕСКИМ МЕТОД
С помощью цитогенетического метода устанавливают кариотип пациента или плода, на основании чего делают заключение о наличии хромосомной патологии. Метод включает несколько этапов:
1ЭТАП
Взятие материала
На практике для анализа обычно используют фибробласты, клетки костного мозга, амниотической жидкости, хориона. Наиболее удобны лейкоциты периферической крови.
2 ЭТАП
Культивирование клеточного материала
Клетки переносят в искусственную среду и добавляют стимуляторы митоза, например фитогемагглютинин.
ЗЭТАП
Накопление метафазных клеток
Остановка митозов всех клеток на стадии метафазы достигается добавлением в среду колхицина — вещества разрушающего нити веретена деления и препятствующего расхождению хромосом к противоположным полюсам.
4 ЭТАП
Гипотонизация клеток
Обработка клеток гипотоническим раствором сопровождается их набуханием, разрывом ядерной оболочки, потерей межхромосомных связей, что облегчает последующий анализ метафазной пластинки.
5 ЭТАП
Окрашивание препаратов
В зависимости от задач применяют простое (рутинное — по Романовскому — Гимза), дифференциальное или флуоресцентное окрашивание метафазных пластинок.
5.7 ЭКСПРЕСС - ЦИТОГЕНЕТИЧЕСКИЙ МЕТОД •
Экспресс цитогенетический метод применяется при диагностике хромосомных болезней, вызываемых числовыми нарушениями гетеросом. Кроме того, он находит широкое применение в судебной медицине и криминалистике когда по пятнам крови необходимо определить половую принадлежность.
Метод включает несколько этапов:
1ЭТАП
Взятие материала
На практике для анализа обычно используют клетки буккального эпителия или нейтрофилы периферической крови.
2 ЭТАП
Окрашивание препаратов
Используют простое (рутинное) окрашивание гематоксилином и другими основными красителями.
ЗЭТАП
Микроскопирование и анализ препаратов
Циагностика основана на обнаружении полового хроматина — конденсированной и инактивированной Х-хромосомы. В клетках буккального эпителия половой хроматин представлен в виде “телец Барра”, а в нейтрофилах — в виде “барабанных палочек ”.
СВЯЗЬ МЕЖДУ ЧИСЛОМ Х-ХРОМОСОМ, ЧИСЛОМ ТЕЛЕЦ БАРРА (I)
И ЧИСЛОМ “БАРАБАННЫХ ПАЛОЧЕК” (II)
Нормальный мужчина XY или
больная женщина ХО (синдром Шерешевского-Тернера)
Нормальная женщина XX
или больной мужчина XXY (синдром Клайнфельтера)
Больная женщина XXX (синдром трипло-Х) или больной мужчина XXXY (синдром Клайнфельтера)
Больная женщина ХХХХ (полисомия X) или больной мужчина XXXXY (синдром Клайнфельтера)
I II
5.8 ОБЩИЕ ПРЕДСТАВЛЕНИЯ О НАСЛЕДСТВЕННОЙ ПАТОЛОГИИ
В основе хромосомных болезней лежат геномные и хромосомные мутации.
Раличают две группы хромосомной патологии:
Полные формы
Мозаичные формы
В основе генных болезней лежат генные мутации.
Использую две классификации генных болезней:
Г енетическая:
Аутосомные болезни (доминантные, рецессивные), Сцепленные с полом (Х-доминантные, Х-рецессивные, Y-сцепленные).
Патогенетическая:
1) Болезни обмена веществ, 2) врожденные пороки развития, 3) комбинированные состояния.
Условием возникновения болезней с наследственной предрасположенностью является комплекс патологических генов и специфических условий среды.
Раличают две классификации.
Г енетическая:
1) Моногенные болезни (патология определяется одним геном), 2) Полигенные болезни (патология определяется группой генов).
Медико-практическая:
1) Врожденные пороки развития (Расщелина губы и неба, косолапость, гидроцефалия и др.),
2) Психические и нервные болезни (Шизофрения, маниакально-депрессивный психоз, рассеяный склероз, эпилепсия),
3) Соматические болезни среднего возраста (Ишемическая болезнь сердца, язвенная болезнь желудка и двенадцатиперстной кишки, гипертоническая болезнь, сахарный диабет, бронхиальная астма и др).
Генетические болезни соматических клеток развиваются вследствие хромосомных мутаций в соматических клетках, что в свою очередь вызывает активацию онкогенов и развитие злокачественных новообразований (ретинобластома, опухоль Вильмса). Если соматические мутации возникают в критическом периоде эмбриогенеза они вызывают врожденные пороки развития.
Болезни несовместимости матери и плода развиваются в результате иммунологической реакции матерей на антиген плода, который детерминирован аллелями отца (гемолитическая болезнь новорожденных при резус несовместимости).
5.9 СИНДРОМ ДАУНА
Известны 3 цитогенетических варианта не различающихся по своим клиническим проявлениям:
1) Простой трисомный вариант 47,XX (XY),21+. Составляет около 94% всех случаев.
2) Полная транслакоционная форма 46, XX(XY), t(21+15). Встречается в 4% случаев заболевания.
3) Мозаичная транслокационная форма 46,XX(XY)/46, XX(XY), t(21+15). Доля этого
варианта составляет 2% . частота
1:12
Следует отметить, что транслокации 21 хромосомы могут происходить с любыми акроцентрическими хромосомами группы D или G.
Среднестатистическая частота рождения детей составляет 1:700 - 1:800 существенно определяется возрастом матери.
1:45
1:1800 1;1300
1:100
1:1000 1:300
----------1---------1----------1---------г-
1 35 40 45 49
Возраст, матери, годы
ОСНОВНЫЕ ПРОЯВЛЕНИЯ СИНДРОМА ДАУНА
1. Низкий рост (100%),
2. Умственная отсталость (100%),
3. Плоское лицо (90%),
4. Уплощенный череп (81%),
5. Открытый рот (65%),
6. Раскосые (монголоидные) глаза (80%),
7. Широкая плоская переносица (40%),
8. Плоский затылок (78%),
9. Диспластичные уши (43%),
10. Врожденные пороки сердца (53%),
11. Пороки желудочно-кишечного тракта (23%),
12. Мышечная гипотония (8 0%),
13. Гиперподвижность суставов (80%),
14. Поперечная (“обезьянья”) складка на ладони (100%).
ДИАГНОСТИКА СИНДРОМА ДАУНА
1) Клинический метод,
2) Дерматоглифический метод, 3) Цитогенетический метод.
КЗДХХ «ГЙ/НКЙ
1 2 3 15 6
ЛХ ЛК ЛА ЛА XI ЛА
7 Я » 1П 11 12
АЛ а.-к.
13 И 15 16 17
ХК *А ла «.лл
18 10 20 22 21
5.10 СИНДРОМ ПАТАУ
Наиболее часто встречаются 2 цитогенетических варианта:
11 Простой трисомный вариант 47,XX(XY), 13+. Составляет 80-85% всех случаев.
2) Транслокационная форма 46, XX(XY), t(13+15). Встречается в 15-20% случаев.
Транслокации 13 хромосомы могут происходить с любыми акроцентрическими хромосомами группы D или G.
Среднестатистическая частота рождения детей составляет 1:5000 - 1:7000 и как и при синдроме Дауна существенно определяется возрастом матери.
ДИАГНОСТИКА СИНДРОМА ПАТАУ
1) Клинический метод,
2) Цитогенетический метод.
И 0 и
у 1____2____3 t *__4-5_,
А В
ь 6-Х-12 С ОдЛЙЛЛЛ XX XX ЛЛ у____13-15_t 10___17__18 у
О Е
ХХХХ ЛА ДА 4
v 19-20 t 21 22 Y
F G
5.11. СИНДРОМ ЭДВАРДСА
Наиболее часто встречаются 2 цитогенетических варианта:
1) Простой трисомный вариант 47,XX(XY), 18+. Составляет 90% всех случаев.
2) Мозаичная форма 46, XX(XY)/47,XX(XY), 18+. Встречается в 10% случаев.
Среднестатистическая частота рождения детей составляет 1:7000 и как и при синдроме Дауна существенно определяется возрастом матери.
ОСНОВНЫЕ ПРОЯВЛЕНИЯ
СИНДРОМА ЭДВАРДСА
1. Удлиненный череп (80%),
2. “Птичий” профиль лица (100%),
3. Короткие, горизонтально расположенные глазные щели (50%),
4. Низкопосаженные аномальной формы
ушные раковины (80%),
5. Задержка роста (100%),
6. Задержка психического развития (100%),
7. Крипторхизм (100%),
8. Наложение П пальца на Ш, а V на V (50%),
9. Дефект межжелудочковой перегородки или открытый боталлов проток (90%), Ю.Пороки развития почек (80%), 11 гипоплазия ногтей (50%).
ДИАГНОСТИКА СИНДРОМА ЭДВАРДСА
1) Клинический метод,
2) Цитогенетический метод. --->
On 8ш
1 ____2___3 ч__4-5__
/X В
v 6-X-12 s
С
ЛЬбйЛй хгг it вл»
ч 13-15 16 17 18
D Е
X X X X Л л лл
(__19-20_ ч 21 22 '
Ч~~ р 6
Причиной заболевания является делеция короткого плеча пятой хромосомы: 46,ХХ(Х¥), 5р- (85-90% всех случаев),
В 10 - 15 % случаев синдром связан с транслокацией теряемого пятой хромосомой фрагмента на одну из других хромосом.
Среднестатистическая частота рождения детей составляет 1:50000.
Соотношение полов при рождении М1:Ж1.
ОСНОВНЫЕ ПРОЯВЛЕНИЯ СИНДРОМА КОШАЧЬЕГО КРИКА
1. Специфический плач (98%),
2. Низкая масса тела при рождении (72%),
3. Отставание в росте (85%),
4. Микроцефалия (98%),
5. Умственная отсталость (100%),
6. Мышечная гипотония (60-80%),
7. Лунообразное лицо (70%),
8. Широкая переносица (84%),
9. Низкорасположенные и деформированные ушные раковины (85%),
10. Аномалии гортани (65%),
11. Антимонголоидный разрез глаз (85%),
12. Плоскостопие (75%),
13. Врожденные пороки сердца (30%).
Такие аномалии как “кошачий крик”, лунооб разное лицо, мышечная гипотония с возрастал могут исчезать
ДИАГНОСТИКА СИНДРОМА КОШАЧЬЕГО КРИКА
норма
1) Клинический метод,
2) Цитогенетический метод.
В метафазной пластинке обнаруживаются хромосомы №5 с укороченными р-плечами.
Причиной синдрома Шерешевского-Теренера является полная или частичная моносомия по Х-хромосоме. Различают три основных цитогенетических варианта синдрома, что определяет его большую клиническую вариабельность:
1) моносомия - 45X0 (встречается в 60% случаев заболевания);
2) частичная моносомия - 46ХХр- или 46XXq- (составляет 5% всех случаев);
3) мозаичная форма —46ХХ/45Х0 (10% случаев заболевания);
4) другие причины (изохромосома-Х, кольцевая Х-хромосома и др. - 25% случаев).
Частота рождения детей с синдромом Шерешевского-Тернера — 1:5000.
ОСНОВНЫЕ ПРОЯВЛЕНИЯ СИНДРОМА ШЕРЕШЕВСКОГО-ТЕРНЕРА
1. Фенотипически женщины,
2. Низкий рост - до 150 см (98%),
3. Короткая шея (71 %),
4. Крыловидные кожные складки на шее (56%),
5. Широкая грудная клетка (60%),
6. Половой инфантилизм (94%),
7. Аменорея (96%),
8. Бесплодие (99%),
9. Периферический лимфатический отек у новорожденных (40%),
10. Снижение слуха (52%),
11. Пороки сердца (27%),
12. Аномалии выделительной системы (38%),
13. Гиперпигментация кожи (60%),
14. Снижение умственного развития (16%).
ДИАГНОСТИКА СИНДРОМА ШЕРЕШЕВСКОГО-ТЕРНЕРА
1) Клинический метод,
2) Цитогенетический метод
К (1 п
1 2 3
Н Н Н I
4 5 6 X
>1 я
1 8
И
9
i 6
15
II It II
10 11 12
Тельца Барра в клетка слизистой ротовой полости (А) и “барабанные палочки” в ядрах лимфоцитов (Б) д g не выявляются. 19
«а
20
4 Л * Д 21 22
1аказ № 3775
97
5.14 СИНДРОМ КЛАЙНФЕЛЬТЕРА
Различают несколько цитогенетических вариантов синдрома::
1) “классическая форма” - 47,XXY (встречается в 80% случаев заболевания);
2) “редкая форма” - 48,XXXY;
3) мозаичная форма —46,XY/47,XXY (вместе со 2 вариантом составляют 20%).
4) разновидностью синдрома является вариант полисомии-Y - 47,XYY
Частота рождения детей с синдромом Шерешевского-Тернера — 1:1000 (мальчиков).
ОСНОВНЫЕ ПРОЯВЛЕНИЯ СИНДРОМА КЛАЙНФЕЛЬТЕРА
1. Фенотипически мужчины,
2. Признаки проявляются при достижении половозре-лости,
3. Высокий рост — более 170 см,
4. В детстве хрупкое телосложение,
5. У взрослых ожирение и оволосение по женскому типу,
6. Гинекомастия (50%),
7. Гипоплазия яичек и полового члена,
8. Снижение полового влечения, импотенция, бесплодие,
9. Слабоумие,
10. Склонность к алкоголизму, гомосексуализму и асоциальному поведению.
11. При варианте 47XYY интеллект соответствует низкой или средней норме, у части больных повышена агрессивность.
ДИАГНОСТИКА СИНДРОМА КЛАЙНФЕЛЬТЕРА
1) Клинический метод,
2) Цитогенетический метод
Ц К к
А
47,XXY
48,XXXY
>арра в клетка слизистой ротовой (А) и “барабанные палочки” в ядрах
Мйй
XX XX XX XX XX XX м
АЛ ЛА Ай
XX
JS
Тельца Барра в клетка слизистой ротовой полости ' лимфоцитов (Б) при двух формах синдрома Клайнфельтера.
XX
XX»
XXY
IX
5.15. ФЕНИЛКЕТОНУРИЯ
Причиной заболевания является мутация гена фенилаланингидроксилазы (ФАГ) -фермента, обеспечивающего превращение фенилаланина в тирозин.
Манифестация заболевания обычно происходит на 2-6 месяце после рождения. Наследование аутосомно-рецессивное.
ОСНОВНЫЕ ПРОЯВЛЕНИЯ ФЕНИЛКЕТОНУРИИ
повышенная нервно-мышечная возбудимость
судороги
слабоумие
радужной оболочки
>>волос
кожных покровов
ДИАГНОСТИКА
В основе диагностики лежит биохимический метод:
1) В моче больных детей обнаруживаются продукты метаболизма фенилаланина (фенилпируват, фениллактат, фенилацетат и др);
2) В крови больных детей обнаруживается повышенная концентрация фенилаланина.
ЛЕЧЕНИЕ
Главный способ лечения фенилкетонурии - диетотерапия, ограничивающая поступление в организм фенилаланина до минимальной возрастной потребности.
5.16. СИНДРОМ МАРФАНА
Причиной заболевания является мутация гена, ответственного за синтез фибриллина (ФБН) - белка соединительной ткани, придающего ей упругость и жесткость.
Наследование аутосомно-доминантное.
В норме
ФИБРИЛЛИН
Упругая соединительная ткань
ОСНОВНЫЕ ПРОЯВЛЕНИЯ СИНДРОМА МАРФАНА
1. Арахнодактилия (пучьи пальцы),
2. Высокий рост,
3. Непропорционально удлиненные конечности,
4. Гиперподвижность суставов,
5. Деформация передней стенки грудной клетки,
6. Плоскостопие,
7. Вывих хрусталика,
8. Пороки сердца,
9. Спонтанный пневмоторакс,
10. Аномалии развития нервной системы.
ДИАГНОСТИКА
Применяют клинический метод. Минимальными диагностическими признаками являются: высокий рост, арахнодактилия, гиперподвижность суставов, вывих хрусталика, аневризма аорты.
5.17 АРИТМИИ СЕРДЦА ПРИ СИНДРОМЕ УДЛИНЕНИЯ QT,
Ритмичность сердечных сокращений является одним из важнейших показателей гомеостаза. Аритмии сердца резко снижают качество жизни и часто являются причиной смерти. Механизмы возникновения аритмий весьма разнообразны, но при синдроме удлинения QT эта патология по своей природе мультифакториальна и возникает вследствие мутации белков калиевых каналов (БКК) кардиомиоцитов.
В норме
Ген БКК 11р15.5
НОРМАЛЬНАЯ СТРУКТУРА И СВОЙСТВА КАЛИЕВЫХ КАНАЛОВ КАРДИОМИОЦИТОВ
Нормальная продолжительность электрокардиографического комплекса QT.
Различают пренатальную (дородовую) и постнатальную (послеродовую) диагностику.
В основе постнатальной диагностики лежит клинический метод. Для уточнения диагноза используют цитогенетические, биохимические, дерматоглифические и молекулярно-генетические исследования.
Хорион- и плацентобиопсия
Взятие на анализ клеток хориона или плаценты.
Кордоцентез
Взятие на анализ крови (плазма и лимфоциты) из вен пуповины
Амниоцентез
Взятие на анализ клеток плода, находящихся в амниотической жидкости.
МЕХАНИЗМЫ РАЗВИТИЯ ПАТОЛОГИИ
ПРОЯВЛЕНИЯ (СИМПТОМЫ) ПАТОЛОГИИ
Возможна в отношении:
> Некоторых болезней с наследственной предрасположенностью;
> Некоторых генетических болезней соматических клеток.
ПРИНЦИП:
J Взятие клеток (например лимфоцитов при первичном иммунодефиците) и исправление в них генетического дефекта, являющегося причиной патологии;
J Размножение трансформированных клеток в культуре;
J Реимплантация трансформированных клеток.
Возможна в отношении:
> Генных болезней;
> Болезней с наследственной предрасположенностью;
> Генетических болезней соматических клеток;
> Болезней несовместимости матери и плода.
ПРИНЦИП:
Уменьшение в организме токсического субстрата реакции или напротив возмещение при его дефиците (диетотерапия, фармакотерапия, плазмо-ферез, гемосорбция);
•/ Тот же подход в отношении продуктов ферментативной реакции;
•/ Возмещение фермента или модификация ферментативной активности (витаминотерапия, фармакотерапия).
Возможна в отношении:
> Хромосомных болезней;
> Генных болезней;
> Болезней с наследственной предрасположенностью;
> Генетических болезней соматических клеток;
> Болезней несовместимости матери и плода.
ПРИНЦИП:
•/ Уменьшение выраженности основных симптомов заболевания (боль,судороги и другие проявления). С этой целью применяют фармакотерапию, климатотерапию, электротерапию, теплолечение и другие виды физиотерапии.
6.1 ХАРАКТЕРИСТИКА МЕЖВИДОВЫХ ВЗАИМООТНОШЕНИИ
6.2. ОСНОВНЫЕ ВИДЫ ПАРАЗИТИЗМА
3
И
Паразитом называют организм, использующий организм другого вида (хозяина) в качестве источника питания и среды обитания, чем приносит хозяину вред.
6.3. ХОЗЯЕВА И ПЕРЕНОСЧИКИ ПАРАЗИТОВ
МЕХАНИЧЕСКИЕ
> Переносят паразита в пространстве;
> В организме механического переносчика не происходит размножения и развития паразита;
> Один паразит может иметь механических переносчиков, относящихся к различным видам, родам и семействам;
> Не обеспечивают проникновение паразита в организм хозяина.
СПЕЦИФИЧЕСКИЕ
> Переносят паразита в пространстве;
> В организме специфического переносчика паразит проходит одну или несколько стадий развития;
> Круг специфических переносчиков паразита, как правило, ограничивается одним видом или родом.
> Обеспечивают проникновение паразита в организм хозяина трансмиссивными или контаминативными механизмами.
6.4 ВЗАИМООТНОШЕНИЯ В СИСТЕМЕ ПАРАЗИТ-ХОЗЯИН (НА УРОВНЕ ОСОБЕЙ)
хозяин
6.5. ЗАБОЛЕВАНИЯ, ВЫЗЫВАЕМЫЕ ПАРАЗИТАМИ
ИНФЕКЦИОННЫЕ
Вызываются:
♦ ВИРУСАМИ;
♦ БАКТЕРИЯМИ;
♦ ГРИБАМИ.
ИНВАЗИОННЫЕ
Вызываются:
♦ ПРОСТЕЙШИМИ;
♦ ГЕЛЬМИНТАМИ;
♦ ЧЛЕНИСТОНОГИМИ.
АНТРОПОНОЗЫ
Болезни свойственные только человеку.
АНТРОПОЗООНОЗЫ
Болезни свойственные и человеку и животным.
ЗООНОЗЫ
Болезни свойственные только животным.
АЛИМЕНТАРНЫЙ
Возбудитель проникает в организм хозяина с пищей.
АЭРОГЕННЫЙ
Возбудитель проникает в организм хозяина воздушнокапельным путем.
КОНТАКТНЫЙ
Возбудитель проникает в организм хозяина при контакте с больным, половым способом, при пользовании общим постельным бельем и средствами гигиены.
КОНТАМИНАТИВНЫЙ
Возбудитель проникает через микротрещины на коже при втирании гемолимфы членистоногих или их фекалий.
ТРАНСМИССИВНЫЙ
Возбудитель проникает в организм хозяина при укусе насекомым или клещом.
6.6. ПРИРОДНО-ОЧАГОВЫЕ ЗАБОЛЕВАНИЯ
Учение о природной очаговости заболеваний разработано отечественным паразитологом Е.Н. Павловским.
Циркулируют в природе не зависимо от человека.
Резервуаром служат дикие животные.
Болезнь встречается на определенной территории.
ПРИРОДНО-ОЧАГОВЫЕ ЗАБОЛЕВАНИЯ 1) трансмиссивные, 2) нетрансмиссивные).
ВОЗБУДИТЕЛЬ
I I КОМПЛЕКС
РЕЗЕРВУАР ПРИРОДНЫХ ______ I | УСЛОВИЙ
ПЕРЕНОСЧИК
(ДЛЯ ТРАНСМИССИВНЫХ БОЛЕЗНЕЙ)
МЕР
антилопы и носороги
Саванны Glossina morsitans
Африки
6.7. ПРОСТЕИШИЕ, ИМЕЮЩИЕ МЕДИЦИНСКОЕ ЗНАЧЕНИЕ
J Entamoeba histolytica, J Е. coli, J E. hartmanni, J E. gingivalis, J Negleria fowleri, J Acanthamoeba castellani.
Balantidium coli.
•J Toxoplasma gondii, J Plasmodium vi-vax, J Pl. malariae, J Pl. falciparum, J Pl. ovale, J Pneumocystis carinii,
J Sarcocyctis ho-mimis.
J Tripanosoma brucei gambiense, •J T.b.rhodesiense, •J T. crusi, J Leishmania tropica minor, J Leishmania tropica major, J L. mexicana, J L.donovani, L.brasiliensis,
J Lamblia in-testenalis, J Trichomonas-vaginalis,
J Tr. hominis, J Trtenax.
6.8 ДИЗЕНТИРИЙНАЯ АМЕБА (ENTAMOEBA HISTOLYTICA)
ЖИЗНЕННЫЙ ЦИКЛ ПРИ БЕССИМПТОМНОМ НОСИТЕЛЬСТВЕ
ЖИЗНЕННЫЙ ЦИКЛ У БОЛЬНОГО АМЕБИАЗОМ
6.9. БАЛАНТИДИИ (BALANTIDIUM COLI)
6.10 ТОКСОПЛАЗМА (TOXOPLASMA GONDII)
Окончательный хозяин
ооциста
шизонт
эндозоиты
циста
эндозоиты
СПОрОЗОИТ!}!
Заказ № 3775
С током крови могут заносится в клетки любых внутренних органов с образованием псевдоцист, а при беременности
инвазиоовать плод.
гаметь
алии
Зрелая ооциста
спорозоиты
псевдоциста
Промежу точные х зяева
113
11 МАЛЯРИЙНЫЕ ПЛАЗМОДИИ НА ПРИМЕРЕ PLASMODIUM VIVAX
Anopheles
Развитие ооцисты
спорозоиты
Взрослый трофозоит
оокинета
спорозоиты
Тканевые шизонты
шизонт
я
КРОВЯНАЯ ШИЗОГОНИЯ
ТКАНЕВАЯ ШИЗОГОНИЯ
Промежупг очный хозяин
Кольцевидный трофозоит
Тканевые * ( мерозоиты
Окончательный хозяин и специфический переносчик
зигота
макрогамета
Женский и мужской гаметоциты
АМЕТОГОНИЯ
амебовидный ^Л^ФОЗОИТ
СПОРОГОНИЯ
микрогаметы
Кровяные мерозоиты
Юный трофозоит
6.11 ГАМБИЙСКАЯ ТРИПАНОСОМА TRIPANOSOMA BRUCEI GAMBIENSE
6.12. РОДЕЗИЙСКАЯ ТРИПАНОСОМА TRIPANOSOMA BRUCEI RHODESIENSE
6.13 ЛАТИНОАМЕРИКАНСКАЯ ТРИПАНОСОМА TRIPANOSOMA CRUZI
Желудочно-кишечный тракт
род Triatoma род Panstrogylus
Трипаносомная форма
Критидиальная форма
Специфический переносчик
Лептомонадная Критидиальная форма форма
Лейшманиальная форма
Метациклическая форма
Лейшманиальная форма
Промежуточные ^резервуарные хозяева
0©% ° ($>с У ©О О*о О и
\ Трипаносомная1
\ форма ,
6.14 ВОЗБУДИТЕЛЬ КОЖНОГО ЛЕЙШМАНИОЗА НА ПРИМЕРЕ LEISHMANIA TROPICA
6.15 ВОЗБУДИТЕЛЬ ВИСЦЕРАЛЬНОГО ЛЕЙШМАНИОЗА НА ПРИМЕРЕ LEISHMANIA DONOVANI
6.16 ЛЯМБЛИЯ (LAMBLIAINTESTENALIS) И ТРИХОМОНАДА (TRICHOMONAS VAGINALIS)
БОЛЬНОЙ
Пользование общими предметами личной гигиены, постелью, бельем.
ЗДОРОВЫЙ
6.17.ПАРАЗИТИЧЕСКИЕ СОСАЛЬЩИКИ
ТИП: ПЛОСКИЕ ЧЕРВИ PLATHELMINTHES
КЛАСС: СОСАЛЬЩИКИ TREMATODA
КЛАСС: ЛЕНТОЧНЫЕ ЧЕРВИ CESTODA
ОБИТАЮЩИЕ В ЖЕЛУДОЧНО-КИШЕЧНОМ ТРАКТЕ
ОБИТАЮЩИЕ В КРОВЕНОСНЫХ СОСУДАХ
ЦИКЛ РАЗВИТИЯ СВЯЗАН С ВОДОЙ
ЦИКЛ РАЗВИТИЯ НЕ СВЯЗАН С ВОДОЙ
J Fasciola hepatica, у' F. gigantica, J Fasciolopsis buski.
J Schistosoma haematobium, J Sch. mansoni, J Sch. japonicum.
J Metagonimus yokogawai, J Nanophyetes salmincola, •J Opisthorchis fe-lineus, J O. Viverrini, J Clonorchis sinensis, J Paragonimus westermani.
•J Dicrocoelium lanceatum, J Eurytrema pancreaticum.
6.18 ПЕЧЕНОЧНЫЙ сосальщик (FASCIOLA HEPATICA)
марита
яйцо
мирацидии
спороциста
адолескарии
редия
церкарии
церкарии
Lyrnnea truncatula
Окончательный хозяин
Промежуточны хозяин
6.19 ФАСЦИОЛОПСИС (FASCIOLOPSIS BUSKI)
6.20 МОЧЕПОЛОВОЙ ШИСТОМАТОЗ (SCHISTOSOMA HAEMATOBIUM)
hhOlAt-WC
яйцо
мирацидий
спороциста I
спороциста II
церкарий
церкарий
Окончательный хозяин
Промежуточны хозяин
Роды Bullinus
Planorbis
6.20 КИШЕЧНЫЙ ШИСТОМАТОЗ
(SCHISTOSOMA MANSONI)
яйцо
мирацидии
спороциста I
спороциста II
церкарии
церкарии
Род Oncomelania
Окончательный хозяин
Промежуточны! хозяин
6.22 МЕТДГОНИМУС (METAGONIMUS YOKOGAWAI)
марита
яйцо
мирацидии
спороциста
метацеркарии
редия
церкарии
Род Melania
Окончательный хозяин
I промежуточный хозяин
Форель, сиг, лещ, сазан, каоась и до.
II промежуточный
. \ хозяин
6.23 НАНОФИЕТ (NANOPHYETES SALMINCOLA)
6.24 КОШАЧИЙ СОСАЛЬЩИК (OPISTHORCHIS FELINEUS)
6.25 КИТАЙСКАЯ ДВУУСТКА (CLONORCHIS SINENSIS) -
6.27 ЛАНЦЕТОВИДНЫЙ СОСАЛЬЩИК (DICROCOELIUM LANCEATUM)
6.28 ПАРАЗИТИЧЕСКИЕ ЦЕСТОДЫ
6.29 ЛЕНТЕЦ ШИРОКИЙ
6.30 БЫЧИЙ ЦЕПЕНЬ (TAENIARRHYNCHUS SAGINATUS)
6.31 СВИНОИ ЦЕПЕНЬ (TAENIA SOLIUM)
6.32 ЭХИНОКОКК (ECHINOCOCCUS GRANULOSUS)
6.33 АЛЬВЕОКОКК (ALVEOCOCCUS MULTILOKULARlS)
6.34 КАРЛИКОВЫЙ ЦЕПЕНЬ (HYMENOLEPIS NANA)
марита
яйцо
онкосфера
цистицеркоид
яйцо
онкосфера
цистицеркоид
Род: Tenebrio
Промежуточный хозяин
Окончательный (иногда промежуточный) хозяин
алой
6.35 КРУГЛЫЕ ЧЕРВИИ-ПАРАЗИТЫ ЧЕЛОВЕКА
ТИП: КРУГЛЫЕ ЧЕРВИ NEMATHELMINTHES
КЛАСС: СОБСТВЕННО КРУГЛЫЕ ЧЕРВИ NEMATODA
ГЕОГЕЛЬМИНТЫ
БИОГЕЛЬМИНТЫ
МИГРАНТЫ В ОРГАНИЗМЕ ЧЕЛОВЕКА
РАЗВИВАЮТСЯ С МИГРАЦИЕЙ
J Dracunculus medinensis, J Trichinella spiralis, Wuchereria bancrofti, J Brugia malai, Onchocerca volvulus, J Loa loa, Mansonella sp., Acanto-heilonema sp.
J Trichocephalus trichiurus, Enterobius ver-micularis.
J Ascaris lumbri-coides, Ankylostoma duodenale, J Necator ameri-canus, J Strongiloides stercoralis.
S Toxocara canis, J Toxocara leon-ina, J Anisacis sp.
6.36 РИШТА (DRACUNCULUS MEDINENSIS)
637 ТРИХИНЕЛЛА (TRICHINELLA SPIRALIS)
6.38 ФИЛЛЯРИИ НА ПРИМЕРЕ ВУХЕРЕРИИ
(WUCHERERIA BANCROFT!)
6.39 ВЛАСОГЛАВ (TRICHOCEPHALUS TRICHIURUS}
6.40 ОСТРИЦА ДЕТСКАЯ (ENTEROBIUS VERMICULARIS)
№ 3775
145
6.41 АСКАРИДА (ASCARIS LUMBRICOIDES)
6.42 АНКИЛОСТОМИДЫ (ANCILOSTOMA DUODENALE, NECATOR AMERICANUS)
6.43 УГРИЦА КИШЕЧНАЯ (STRONGILOIDES STERCORALIS)
1 1 11
филяриевидная личинка
Неблагоприятные условия внешней среды
рабдитовидная личинка
Оптимальные / условия внешней среды /
6.44 TOKCOKAPA (TOXOCARA CANIS)
Заказ № 3775
6.45 КЛЕЩИ, ИМЕЮЩИЕ МЕДИЦИНСКОЕ ЗНАЧЕНИЕ .
Надсемейство/ подотряд Семейство Представители Экологическая группа
1 Иксодовые (Ixodidae) Ixodes persulcatus, I. ricinus, Dermacentor pictus, D. marginatus, Hyalomma plumbeum, H. anatolicum. Временные эктопаразиты
Apra3OBbie(Argasidae) Omithodoros papillipes
2 Macronyssidae Omithonyssus bacoti
Dermanyssidae Dermanyssus gallinae, Allodermanyssus sanguineus.
3 Краснотелковые (T rombiculidae) Neotrombicula autumnalis, Leptotrombidiumakamushi.
Demodicidae Demodex folliculorum Постоянные эктопаразиты
4 Acaridiae Sarcoptes scabiei.
Tyrogliphus farinae, T. casei, Gliciphagus destractor, Dermatophagoides pteronissinus. Обитатели человеческого жилья
Ixodes ricin us $
Ixodes persulcatus
СХЕМА ЖИЗНЕННЫХ ЦИКЛОВ ОДНОХОЗЯИННЫХ (1), ДВУХХОЗЯИННЫХ (2) И ТРЕХХОЗЯИННЫХ (3) ИКСОДОВЫХ КЛЕЩЕЙ
О ---->L-
2
О —яйцо
L- личинка
N- нимфа
I - имаго
6Al ПАРАЗИТИЧЕСКИЕ ПРЕДСТАВИТЕЛИ РОДОВ DERMOCENTER И HYALOMMA
3 Dermacentor pictus $
Hyalomma plumbeum $
6.48 ПОСЕЛКОВЫЙ КЛЕЩ ORNITHODORUS PAPILLIPES
^ДДИЦИНСКОЕ ЗНАЧЕНИЙ
ЖИЗНЕННЫЙ ЦИКЛ НА ПРИМЕРЕ
ORNITHONYSSUS ВАСОТ
О - яйцо, L - личинка, Ni - протонимфа, N2C? - мужская дейтонимфа, N2$ - женская дейтонимфа, 3 - самец, 9 - самка.
паразитические личинки голодная и напитавшаяся
6.51 SARCOPTES SCABIEI И DEMODEX FOLLICULORUM -ПОСТОЯННЫЕ ЭКТОПАРАЗИТЫ ЧЕЛОВЕКА
имаго
личинка нимфа
ЖИЗНЕННЫЙ ЦИКЛ НА ПРИМЕРЕ sarcoptes scabiei
яйцо
Demodex folliculorum Sarcoptes scabiei
6.52 КЛЕЩИ-ОБИТАТЕЛИ ЧЕЛОВЕЧЕСКОГО ЖИЛЬЯ
ЖИЗНЕННЫЙ ЦИКЛ НА ПРИМЕРЕ
АМБАРНЫХ КЛЕЩЕЙ
6.53 НАСЕКОМЫЕ, ИМЕЮЩИЕ МЕДИЦИНСКОЕ ЗНАЧЕНИЕ
Отряд Семейство Представители Экологическая группа
1 Pulicidae Pulex irritans, Xenopsylla cheopis. Временные эктопаразиты.
4 Cimicidae Cimex lectularius.
Reduviidae Triatoma brasiliensis, Panstrogilus geniculatus.
3 Culicidae Culex pipiens, C. fatigans, Anopheles maculipennis, Aedes aegipti, A. polinesiensis, Mansonia sp.
Phlebotomidae Phlebotomus papatasii.
Simulidae Simulium damnosum, S. ochraceum, S. metallicum.
Ceratopogonidae Cullicoides pulicaris.
Tabanidae Chrisops dimidiata, C. silacea, Tabanus autumnalis.
Muscidae Stomoxis calcitrans, Glossina palpalis, G. morsitans.
Sarcophagidae Sarcophaga camaria, Wolfahrtia magnifica, Dermatobia hominis. Постоянные эктопаразиты.
Calliphoridae Calliphora eritrocephala, Lucilia caesar.
Hypodermatidae Hypoderma bovis
Gastrophilidae Gastrophylus intestinalis
Ostridae Ostrus ovis
Muscidae Musca domestica, Muscina stabulans. Синантропные виды не являющиеся паразитами.
5 Blattidae Blatta orientalis, Blattella germanica.
6 Tenebrionidae Tenebrio sp.
7 Formicidae Monomorium pharaonis
2 Pediculidae Pediculus humanus humanus, P. h. capitis, Phtirus pubis. Постоянные кровососущие паразиты.
Cimex lectularius
Triatoma sp.
постельный клоп поцелуйный клоп
Culex pipiens
Aedes aegypti
Anopheles maculipennis
личинка
маля-
и
ки
бру-
гиоза
malai)
Переносчики воз-
Переносчики воз-
Переносчики воз-
будителей вухе-
рериоза (Wuche-
rena bancrofu).
будителей
(Brugia
(Plasmodium
vivax, Р. malariae,
Р. falciparum,
Р. ovale).
Переносчики ви-
русов японского
Переносчики воз-
энцефалита
желтой лихорад-
будителен туля-
ремии (Francisella
tularensis).
ANOPHELES MACULIPENNIS
куколка
яйцо
имаго
6.57 МЕЛКИЕ КОМПОНЕНТЫ ГНУСА
Simulium sp.
Culicuides sp.
Phlebotomus sp.
'Эктопаразиты. Специфические переносчики возбудителей лейшманиозов (Leishmania sp.), вируса лихорадки пап-патачи, возбудителя бортенелеза (Вог-ДопеПа bacilliformis).
Эктопаразиты. Специфические переносчики возбудителя онхоцеркоза (Onchocerca volvulus).
Эктопаразиты. Специфические переносчики возбудителей некоторых филляриа-тозов - мансонеллеза и акантохейлонемато-за (Mansonella sp., Acantoheilonema sp.).
ЖИЗНЕННЫЕ ЦИКЛЫ
6.58 КРУПНЫЕ КОМПОНЕНТЫ ГНУСА - ПРЕДСТАВИТЕЛИ СЕМЕЙСТВ MUSCIDAE И TABANIDAE
Эктопаразиты. Специфические переносчики возбудителей африканской сонной болезни (Tripanoso-ma brucei gambiense и T. b. rhodesiense).
Эктопаразиты. Специфические переносчики возбудителей лоаоза (Loa 1оа). Механические переносчики возбудителей туляремии и сибирской язвы.
Эктопаразиты. Механические переносчики возбудителей туляремии, сибирской язвы и других инфекционных заболеваний.
Glossina sp.
Crisops sp.
Stomoxys calcitrans
ЖИЗНЕННЫЕ ЦИКЛЫ
6.59 НАСЕКОМЫЕ ТКАНЕВЫЕ И ПОЛОСТНЫЕ ЭНДОПАРАЗИТЫ
Wohlfartia magnifica
Вольфартова муха
пчеловидка обыкновенная
Fannia canicularis
Малая комнатная муха
Личинка Ostrus ovis овечьего полостного овода
Hypoderma bovis личинка и имаго бычьего подкожного овода
'L4IMUW
Gastrophylus intestinalis личинка и имаго лошадиного желудочного овода
ТКАНЕВЫЕ МИАЗЫ (ГЛАЗНЫЕ И НОСОГЛОТОЧНЫЕ)
Pediculus humanus humanus Phtirus pubis
Платяная вошь Лобковая вошь
Pediculus humanus capitis Головная вошь
ЖИЗНЕННЫЙ
Личинка
Яйцо .4
(гнида)
Имаго
►Нимфа
(2 поколения)
6.61 СИНАНТРОМНЫЕ НАСЕКОМЫЕ
НЕ ЯВЛЯЮЩИЕСЯ ПАРАЗИТАМИ
Sarcophaga carnaria Серая мясная муха
CaUiphora eritrocephala Lucilia caesar
Синяя мясная муха Зеленая падальная муха
Механические переносчики возбудителей инфекционных заболеваний и яиц гельминтов. Порча пищевых продуктов.
Monomorium pharaonis Муравей домовый
Tenebrio sp.
Мучной хрущ
Blattella germanica
Рыжий таракан
Blatta orientalis
Черный таракан
МОЛЕКУЛЯРНЫЙ
Молекулярный уровень представлен “биологическими молекулами”, основными из которых являются ДНК, три вида РНК и белки, определяющие потоки генетической информации в клетках. Основу жизни “биомолекулы” составляют лишь в клетке. Вне клетки это химические вещества.
КЛЕТОЧНЫЙ
Уровень представлен одноклеточными животными и растениями, а также клетками многоклеточных организмов. На клеточном уровне происходит сопряжение механизмов передачи генетической информации и превращения веществ и энергии. С этого уровня начинается собственно жизнь.
Сходные по размерам и функциям клетки составляют основу тканей. В разных тканях объединены клетки одного или нескольких видов, а также межклеточное вещество. У животных и человека выделяют четыре типа тканей: эпители-альную, соединительную, нервную и мышечную,
У многоклеточных организмов ткани формируют органы, объединенные в системы. Они обеспечивают важнейшие исполнительные и регуляторные функции, благодаря которым особи существуют как единое целое.
ОРГАНИЗМЕННЫЙ ИЛИ ОНТОГЕНЕТИЧЕСКИЙ
Одноклеточные и многоклеточные организмы представляют собой интегрированные единицы жизни. В ходе онтогенеза организмы реализуют наследственную информацию в конкретные биологические структуры и являются элементарным материалом для эволюционно-го процесса.
ПОПУЛЯЦИОННО-ВИДОВОЙ
Совокупность организмов одного вида, населяющих одну территорию и свободно скрещивающихся между собой, называют популяцией. Совокупность популяций составляет биологический вид. Элементарные эволюционные факторы вызывают эволюционно значимые изменения генофонда популяций и приводят к образо-ванию новых видов.
БИОГЕОЦЕНОТИЧЕСКИЙ
Реально вид всегда сосуществует в сообществе с представителями других видов и населяет территорию с конкретными абиотическими характеристиками, которые все вместе образуют устойчевые открытые системы - биогеоценозы.
БИОСФЕРНЫЙ
Комплекс биогеоценозов нашей планеты составляют биосферу - оболочку жизни. На этом уровне объединены все вещественно энергетические круговороты.