/




















































































































































































































































































































































































































































































































































































































































Text
АКАДЕМИЯ НАУК СССР
УСТАНОВКА ТЕЛА
ЭКСПЕРИМЕНТАЛЬНО-ФИЗИОЛОГИЧЕСКИЕ ИССЛЕДОВАНИЯ ОТДЕЛЬНЫХ ОПРЕДЕЛЯЮЩИХ УСТАНОВКУ ТЕЛА РЕФЛЕКСОВ, ИХ ВЗАИМНЫХ ВЛИЯНИЙ И ИХ РАССТРОЙСТВ
Перевод с немецкого
И. Г. БАУЭР, Н. Н. БЕНУА и К. Г. ЛЕБЕНТРАУ Под редакцией
Э. Ш. АЙРАПЕТЬЯНЦА ж В. А. КИСЛЯКОВА
ИЗДАТЕЛЬСТВО АКАДЕМИИ НАУК СССР
Москва 1 9 6 2 Ленинград
МАГНУС
KORPERSTELLUNG
EXPERIMENTELL-PHYSIOLOGISCHE UNTERSUCHUNGEN ÜBER DIE EINZELNEN BEI DER KÖRPERSTELLUNG IN TÄTIGKEIT TRETENDEN REFLEXE,
ÜBER IHR ZUSAMMENWIRKEN UND IHRE STÖRUNGEN
von
R. MAGNUS
BERLIN
VERLAG VON JULIUS SPRINGER 1924
ОТ РЕДАКЦИИ
По всеобщему мировому признанию труд известного голландского физиолога Р. Магнуса «Установка тела» оценивается как классическое произведение, в котором изложена одна из существенных систем рефлекторной деятельности животного организма. Имя знаменитого ученого ставится рядом с тремя другими основоположниками современной разработки законов деятельности центральной нервной системы — И. П. Павлова, Н. Е. Введенского, Ч. С. Шеррингтона.
Рудольф Магнус принадлежит к той плеяде великих учителей физиологии XIX и начала XX в., жизнь которых неотделима от создания целых областей науки. Однажды увлекшая экспериментальная задача ничем не может быть вытеснена, пока предмет исканий не будет представлен как стройная грандиозная система жизнедеятельности цельного организма. Магнус по существу за 10—15 лет создал новое представление об одной из важнейших функций организма на основе богатейшего физиологического и клинического материала, добытого преимущественно своей личной работой и в творческом соавторстве с небольшим (по нашим временам) числом сотрудников. Многие из них впоследствии стали выдающимися деятелями науки (де Клейн, Радемакер, де Бурле, ван дер Гуве и др.).
Вместе с этим Магнус был исследователем с широкими научными интересами и, несмотря на казалось бы специальную врачебную подготовку, являл собой тип ученого с общебиологической эрудицией, естествоиспытателя по методам познания природы, по всестороннему охвату предмета изучения.
Открытия Магнуса и его сотрудников показали роль стволовой части головного мозга в равновесии тела. Магнус создал учение о рефлексах, обеспечивающих нормальное положение тела в пространстве, и вскрыл причины, вызывающие их нарушения. Результаты исследований автора книги «Установка тела» представляют фундаментальное значение не только для успешного развития самой физиологии, но и для многих сопредельных с физиологией областей медицины, психологии, биологии и, в частности, для зарождающегося раздела естествознания — космической биологии и физиологии.
Благодаря трудам лаборатории Магнуса наука приобрела точные знания о природе основных актов локомоторной деятельности (стояние, ходьба), о центральных аппаратах, ведающих координацией частей тела по отношению друг к другу, о роли лабиринтов внутреннего уха в управлении мышцами конечностей, шейными и глазными мышцами.
Магнус описал разнообразные структуры тонических рефлексов: Haltung или Stehreflexe — рефлексы положения, позы тела, Stellreflexe — установочные рефлексы. Исследования Магнуса положили основу дальнейшему изучению стато-кинетических рефлексов, раздражителями которых являются изменения скорости движения тела (положительные и отрицательные ускорения). Все основные опыты и закономерности, описанные Магнусом, вошли в учебники, и знания, вытекающие из этих
2
От редакции
исследований, продолжают служить живым источником в разработке новых теоретических и прикладных задач физиологии.
Книга Р. Магнуса «Körperstellung» вышла на немецком языке в 1924 г. Ныне это библиографическая редкость. Не во всех старых и крупных библиотеках она имеется, не говоря уже о том, что не все современные ученые имеют ее в личном пользовании. Что же касается медицинской и биологической научной молодежи, то можно быть уверенным, что вряд ли •большинство из них имело возможность держать ее в руках.
Таким образом, издание труда Магнуса на русском языке позволит ознакомиться с его фактами и идеями широкому кругу специалистов.
Рудольф Магнус прожил 54 года (1873—1927). За этот краткий ~для экспериментатора срок жизни он успел оставить такое научное наследство, которое позволило, увы- посмертно, и современникам, и потомкам отнести его имя к классикам науки. В его деятельности поражает фантастическая продуктивичсть, темпы творчества, неутомимость в постановке непрекращающихся опытов, потребность ежедневного письменного осмысливания материалов для подготовки к публикации. Достаточно сказать, что, помимо капитального труда «Körperstellung», он опубликовал около 400 статей. Исследовательская деятельность Магнуса ^сочеталась со служебно-педагогической работой, вначале ассистента, ‘«а затем заведующего кафедрой фармакологии. Магнус любил педагогическую деятельность, он был блестящим лектором. Магнус был жизнерадостным человеком, отцом большого семейства.
По воспоминаниям его коллег и учеников, умер Магнус «знаменитым, но бедным человеком» (в Швейцарии).
Рудольф Магнус, немец по происхождению, родился в Брауншвейге, закончил высшее образование в Гейдельберге (1898), работал вначале ва физиологической кафедре у проф. Вильгельма Кюне, затем после оставления при университете был зачислен на должность ассистента по кафедре фармакологии у проф. Готлиба, где и работал в течение 10 лет. Однако уже в эти годы фармакологические и физиологические интересы молодого ученого тесно переплетаются. Он выступает еще молодым ученым на IV (Кембриджском) Международном конгрессе физиологов с докладом о микромоментальной фотографии изменений зрачка рыб и лягушек при действии света. Два сезона работает на неаполитанской станции, изучая зрительный аппарат у осьминога. Много лет посвятил Магнус вопросам обмена воды в организме, функции почек, вместе с Шефером в 1900 г. открывает диуретическое действие экстракта гипофиза, детально исследует моторную функцию кишечника, применяет рентгеновское просвечивание кишечника. В этот же период он сотрудничает в первоклассных лабораториях: его учителем и другом был Икскюль, вместе с Ленглеем работает в Кембридже (1905), с Шеррингтоном — в Ливерпуле (1908). Магнус пробует свои силы в самых различных областях биологии. Его энергия-, его исследовательская страсть, к чему бы они ни прикасались, оставляют глубокий след в науке. Однако пока все это можно назвать талантливыми этюдами на разнообразные темы. Это еще не концентрация энергии для грандиозного полотна. Талант Магнуса по совокупности обстоятельств — научных, материальных, личных — как бы приторможен. Он не имел своей кафедры, был лишен возможности самостоятельного развертывания задуманных исследований. И это происходило из-за того, что на родине не сумели увидеть в Магнусе нечто новое в науке — оригинальное, самобытное, перспективное, и вся истинная слава Магнуса -родилась в Голландии. Он с радостью принял предложение занять кафедру фармакологии в Утрехте и с 1908 г. почти до последних дней своей
жизни работал в этом очень милом, тихом^ университетском городке. Здесь появились первые исследования по мёханизмам локомоторного^ акта (1909), навеянные еще следами сотрудничества с Шеррингтономг здесь же начались и завершились собственные, уже новаторские искания,, вылившиеся в учение о системе рефлексов и закономерностей в деятельности центральной нервной системы.
Научные труды Магнуса были высоко оценены советской физиологией.
В «Лекциях о работе больших полушарий головного мозга» И. П. Павлов писал: «. . .благодаря работам Магнуса и Клейна (Magnus und de Kleyn) мы знаем, что и стояние, и ходьба, и вообще уравновешивание тела в пространстве — рефлексы» (Поли. собр. соч., т. IV, М.—Л., 1951 ^ стр. 26).
А. А. Ухтомский сразу же по выходе в свет «Körperstellung» стал горячим пропагандистом этого труда, увлеченно излагая суть открытий Магнуса студентам на лекциях и ученикам и сотрудникам на семинарах. В своих «Очерках по физиологии нервной системы» он подверг специальному рассмотрению шейные и лабиринтные рефлексы.
С глубоким знанием исследований Магнуса подробно и систематично,, с иллюстрацией оригинальных рисунков излагает И. С. Беритов основные положения «Körperstellung» и работы сотрудников Магнуса в своем учебнике «Общая физиология мышечной и нервной системы» (т. II). Впервые в советской печати в год смерти Магнуса (1927) вышла обширная! статья А. Ф. Самойлова, лучшего знатока работ утрехтской лаборатории. В столь же основательной по содержанию, сколь и доступной по форме- изложения статье в строгой последовательности описываются опыты лаборатории Магнуса. Можно сказать, что здесь счастливым образом сочетались энциклопедические знания, высокий талант педагога с обстоятельствами, давшими А. Ф. Самойлову возможность длительное время знакомиться с лабораторией своего друга, лично видеть знаменитые эксперименты непосредственно «в руках» Магнуса и слышать их толкование иа уст самого автора. А. Ф. Самойлов пророчески предсказал перспективы развития трудов И. П. Павлова и Магнуса и как настоящий и дально*- видный теоретик современной физиологии усмотрел общую стратегическую* линию в обеих научных школах. «Павлов и Магнус шли в своих работах с разных концов центральной нервной системы, и в их продвижении вперед уже намечено место встречи. Невольно напрашивается сравнение с двумя партиями рабочих, прорывающих туннель с двух сторон горного массива. Они уже слышат удары молотков противоположной стороны.. Еще один удар — и они подадут друг другу руки». Так заканчивается: блестящее обозрение трудов Магнуса в статье А. Ф. Самойлова. Мы сочли в высшей степени полезным предоставить возможность читателям Магнуса, может быть вновь, а может быть впервые, ознакомиться со< статьей А. Ф. Самойлова, опубликовав ее в виде приложения к русскому изданию «Körperstellung».
Кто знаком с немецким подлинником, тот согласится с нами, насколько* нелегок был перевод специализированного языка, стиля изложения,, необычных терминов, натуральных описаний наблюдаемых явлений щ наконец, необыкновенных образных средств, когда исчерпывается физиологическая научная лексика для обозначения совершенно конкретной позы животного. Прежде всего затруднения начались с перевода названия книги «Körperstellung». Наиболее адекватным мы признали перевод «Установка тела». Однако избрав эти слова для перевода с немецкого «Körperstellung», мы, естественно, в отличие от А. Ф. Самойлова и Stellreflexe перевели не как «рефлекс выпрямления», а как «установочный:
4
От редакции
рефлекс». Таких затруднений в процессе работы было немало, нет надобности их перечислять, но в конце концов мы приходили к убеждению, что задача решена правильно. К сожалению, по техническим причинам (из-за отсутствия оригинала), только по рисункам книги, не удалось обеспечить репродукцию стереоскопических снимков. Необходимо также оговорить, что в немецкий подлинник вкралось некоторое число опечаток (и смысловых, и цифровых, и корректорских), в ряде случаев искажающих смысл. Мы считали необходимым внести в русское издание исправления в точном соответствии с сутью магнусовского изложения.
По-разному идет разработка проблем, вытекающих из трудов Магнуса: одни развиваются относительно широко, другие — все еще медленно. В Советском Союзе значительно продвинулось изучение тех вопросов, которые связаны с корковой регуляцией функций и применением метода условных рефлексов. Мы нашли уместным в «Приложении» в виде послесловия кратко изложить накопленный за последние три десятилетия фактический материал, относящийся к проблеме «Кора больших полушарий и функции вестибулярного аппарата», снабдив статью соответствующей библиографией. Выбор темы объясняется также и тем, что пишущие эти строки заняты специальной разработкой указанных вопросов.
Главы между переводчиками были распределены следующим образом: И. Г. Бауэр — предисловие и главы I, VIII, IX, XI, XII, И. Г. Бауэр и К. Г. Лебентрау — главы II—IV, Н. Н. Бенуа — главы V—VII, X.
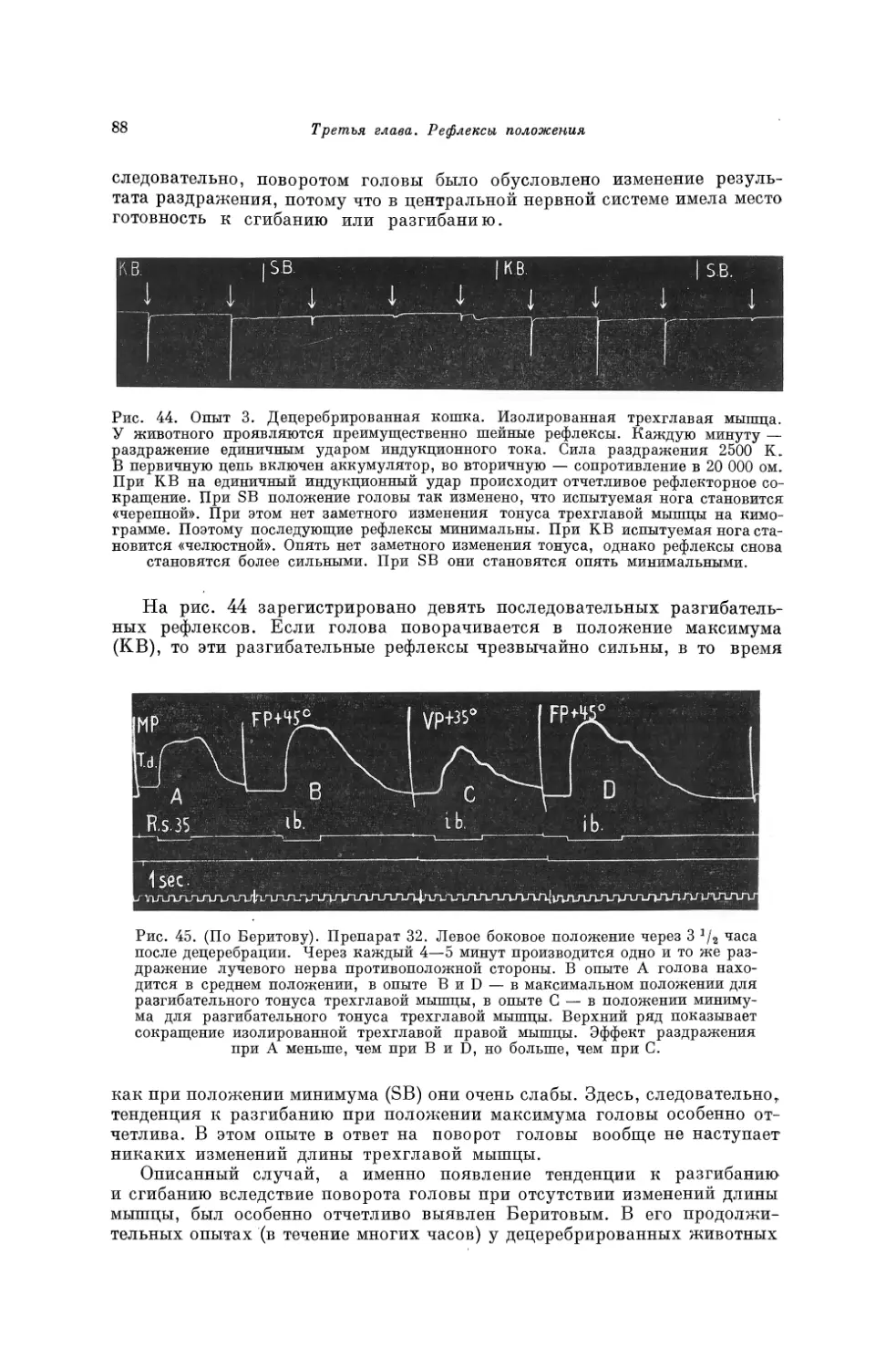
Рукопись перевода любезно прочитали профессора Я. А. Винников, А. В. Лебединский, М. А. Панкратов и высказали ряд ценных замечаний и рекомендаций. Мы приносим им искреннюю благодарность.
Работа над переводом книги и подготовка ее к изданию заняла несколько лет, теперь в связи с выходом монографии Р. Магнуса «Körperstellung» на русском языке мы, естественно, переживаем большое удовлетворение и уверены, что она займет подобающее ей место в личной библиотеке советского физиолога, клинициста, биолога, психолога. Один из нас всякий раз взволнованно вспоминает то огромное впечатление, которое много лет назад оказали на него демонстрации опытов Магнуса, осуществленные бывшими сотрудниками Магнуса, профессорами де Клейном в Утрехте и Радемакером в Лейдене. Точно, четко, безотказно принимало заданные позы тело кошки и кролика, как послушная сложная машина.
Можно не сомневаться, что вдумчивый советский читатель классического произведения Магнуса непременно будет увлечен и очарован постепенной, строго последовательной расшифровкой закономерностей, лежащих в основе самых разнообразных установок тела в пространстве. Он сможет на примере магнусовских рефлексов увидеть ту универсальную роль, которую осуществляют на разных уровнях сложные рефлекторные механизмы в гармонично функционирующем высшем животном организме. Мы надеемся, что вслед за монографией Р. Магнуса будут изданы на русском языке и другие классические произведения корифеев мировой физиологии, в том числе и Ч. С. Шеррингтона.
Э. Айрапетъянц, В. Кисляков.
с. Павлово (Колтуши), 1962 г.
ПРЕДИСЛОВИЕ
Изложенные в этой книге исследования были вызваны одним случайным наблюдением, сделанным мною в марте 1909 г. У децеребрированной кошки с перерезанным в области грудных позвонков спинным мозгом наблюдалось сильнейшее судорожное состояние разгибателей передних конечностей после того, как ее переложили из бокового положения на спину. При систематическом исследовании подобных реакций были найдены некоторые закономерности рефлексов положения. В связи с этим были проведены другие работы, которые постепенно выяснили очень сложную и точно согласованную систему рефлексов установки тела.
В течение ряда лет Фармакологическим институтом в Утрехте было выпущено настолько большое количество отдельных работ, посвященных этой проблеме, что разобраться в них нелегко. К тому же в то время, когда писались первые работы, многое было еще неизвестным и стало понятным лишь в дальнейшем. Поэтому один и тот же физиологический механизм анализировался в работах, далеко отстоящих друг от друга во времени.
Все эти исследования никоим образом нельзя считать законченными, в чем можно убедиться при чтении этой книги. Но они теперь настолько продвинулись, что видны основные линии проблемы. Поэтому я считаю своевременным в наглядном виде дать единое представление об основных установленных фактах так, чтобы и не специалисты физиологи могли разобраться в этом вопросе и притом так обстоятельно, чтобы читателю не нужно было прибегать к опубликованным ранее статьям.
Я не мог бы выполнить эту работу в таком объеме и охватить столь различные направления, если бы меня не поддержали старшие и младшие товарищи по специальности. Прежде всего нужно назвать следующие имена: Дюссер де Баренн (Dusser de Barenne), Бийльсма (Bijlsma), де Хаас -(de Haas), в. д. Хове (v. d. Hoeve), Гофманн (Hoffmann), Ионкхоф (Jonk- boff), Костер (Koster), Лильестранд (Liljestrand), Оорт (Oort), Раде- макер (Rademaker), Сосэн (Socin), Стенверс (Stenvers), Сторм ван Лееу- вен (Storm van Leeuwen), Ферстеег (Versteegh), Вейланд (Weiland), Вольф (Wolf). Форма совместной научной работы, как она развилась в Голландии и прежде всего в Утрехте, оказалась необходимой основой для такого рода исследований.
Среди своих помощников я прежде всего обязан выразить благодарность следующим.
Д-ру де Клейну, который почти сразу после начала опытов над рефлексами положения поступил ассистентом в Фармакологический институт и с тех пор непрерывно занимался этими исследованиями, так что постепенно между нами установилось тесное научное сотрудничество. В течение ряда лет мы совместно обсудили все основные вопросы. Я старался в дальнейшем изложении отмечать участие Клейна в отдельных исследованиях, но во многих местах, где его имя не упоминается, он также принимал участие в проводимых работах.
Проф. Винклер охотно делился своими глубокими знаниями о тончайшем строении центральной нервной системы и, несмотря на большую
6
Предисловие
перегруженность работой, всегда находил время, чтобы приготовить препараты оперированного мозга и зарисовать их. От него я получил много сведений по анатомии, его объективное описание результатов операций дало нам возможность сделать многие выводы.
Д-р де Бурле со своими сотрудниками установил при помощи трудоемких исследований расположение и структуру отдельных отделов лабиринта, прежде всего отолитовых пятен у различных млекопитающих, и разработал соответствующую методику, так что для теорий, относящихся к функции отолитов ит. п., была создана прочная анатомическая база.
Сделаем еще одно существенное замечание: очень трудно описать установку тела и трудно ее изобразить. Поэтому я поместил в книге много стереоскопических снимков. И тот, кто хочет более глубоко вникнуть в изложение, должен вооружиться стереоскопом. Консультация с многими специалистами показала, что этот метод весьма полезен.
Курсивные цифры в скобках, например {14), относятся к списку утрехтских работ. Остальная цитированная литература приведена в конце книги в алфавитном порядке. Работы одного и того же автора различаются цифрами в скобках, например ц(4). Составление полной сводки литературы не лреду сматрив а лось.
Утрехт, декабрь 1923.
Р. Магнус.
Первая глава
ОБЩИЙ ОБЗОР
В этой книге излагается учение об установке тела главным образом: у млекопитающих.
В неживой природе эта проблема сравнительно проста. Если знать положение центра тяжести по отношению к опорной плоскости, то этим уже решен вопрос, находится ли тело в равновесии и является ли это равновесие устойчивым или неустойчивым. Если тела плавают в воде или свободно падают в воздухе, то положение тела в пространстве можно определить при помощи соотношения между центром тяжести тела и центром тяжести массы вытесненного воздуха или воды, учитывая также сопротивление воздуха. Равновесие некоторых низших животных, как это, в частности, показал Бете (Bethe), также подчиняется этим простым физическим закономерностям.
Встречаются животные без статолитов, которые после выключения глаз всегда плывут в воде в правильном положении, так как они могут ориентироваться благодаря форме своего тела и положению центра тяжести. Захлороформированные шмели и стрекозы падают всегда на землю в нормальном положении.
У большинства же организмов и в особенности у высших животных эта проблема гораздо сложнее. Если попытаться поставить скелет человека на землю в вертикальном положении, то он просто упадет, и то же самое- случится, если проделать этот опыт с трупом человека, который еще не успел окоченеть. Кости, суставы, связки и все мускулы не в состоянии поддержать тело в нормальном положении. Для этого необходимо, чтобы мышцы были живыми и подчинялись центральной нервной системе. Но одной связи мускулов со спинным мозгом для этого недостаточно. Нам известно из исследований Гольца [Goltz (2)] и Шеррингтона [Sherrington (5)], что если перерезать собаке спинной мозг на уровне середины грудной части и после этого подождать, пока исчезнут все шоковые явления, то окажется, что животное способно выполнять задней половиной своего тела всевозможные сложные движения. Если животное вертикально подвесить головой вниз или держать его за голову и хвост так, чтобы позвоночный столб находился в горизонтальном положении, то задние конечности совершают ритмические и координированные движения бега, которые тормозятся при раздражении чувствительных нервных окончаний. Щипок лапы вызывает на соответствующей стороне рефлекс сгибания, благодаря которому лапа отстраняется от вредного раздражения, а на противоположной стороне конечность вытягивается. Если раздражать пальцем кожу живота, то животное начинает совершать вполне координированные чесательные движения. При соответствующем положении задних конечностей животного и поднятом хвосте со слизистой оболочки прямой кишки вызываются нормальные дефекационные движения. У таких животных можно наблюдать еще много других хорошо координированных рефлекторных движений. То же самое можно видеть у кошки, если перевязать у нее по способу
Первая глава. Общий обзор
:8
Шеррингтона четыре большие мозговые артерии и выключить этим головной мозг вместе с продолговатым мозгом. У такого спинального животного имеются всевозможные сложные рефлексы, которые вполне сходны с движениями, производимыми нормальными животными в ответ на подобные же раздражения. Мускулатура таких спинномозговых животных никоим образом не лишена тонуса. Еще Брондгеест (Brondgeest) показал, что обычно мышцы находятся в определенном среднем напряжении, зависящем от рефлекторных влияний. Благодаря работам Шеррингтона (9) мы знаем, что главный источник этого тонуса надо искать в проприоцептивных возбуждениях, которые исходят из самих же мышц. Выяснилось, однако, что, кроме возбуждений от сокращающихся мышц, на центры мускулатуры влияет еще ряд других чувствительных раздражений и этим вызывается более или менее сильное длительное возбуждение (тонус).
Рис. 1.
Но, несмотря на все это, млекопитающее после отделения спинного тиозга от головного не может стоять, оно просто падает (рис. 1, а).
Правда, Филипсон (Philippson) опубликовал кинематографические снимки собак, у которых он перерезал спинной мозг в грудной области и которые могли не только стоять, но бегать и галопировать. Однако вылепилось, что здесь налицо кажущееся явление, так как известно, что через некоторое время собаки приспосабливаются держать заднюю половину тела горизонтально при определенном положении головы и сильном сокращении плечевой мускулатуры, так что задние ноги иногда как бы стоят на полу и таким образом, свешиваясь с задней половины тела, выполняют простые рефлекторные движения бега; это выглядит так, как будто животные могут стоять на задних ногах. Но многочисленные наблюдения на собаках с перерезанным спинным мозгом, многие из которых жили по нескольку лет, показали, что задние ноги не приобретают настоящего статического тонуса и не в состоянии сами выдерживать вес задней половины тела.
Как показал Шеррингтон (1, 5, 9), животное способно стоять самостоятельно только тогда, когда спинной мозг остается в связи с продолговатым мозгом. Если млекопитающему перерезать мозговой ствол в области крыши мозжечка и этим отделить таламус и часть среднего мозга от спинного и продолговатого мозга, то наступает так называемая децеребрационная ригидность (рис. 1, Ъ). Для того чтобы вызвать это явление, достаточно наличия продолговатого мозга. После поперечного разреза, который проходит через продолговатый мозг перед самым входом 8-го нерва, наблюдается сильная децеребрационная ригидность. Точное расположение центров, необходимых для появления ригидности, нужно еще установить.
При децеребрационной ригидности определенная группа мышц тела находится в состоянии сильнейшего тонического сокращения. Это те мышцы, функция которых в течение жизни состоит в том, чтобы противо¬
Первая глава• Общий обзор
9
стоять силе тяжести, именно — разгибатели конечностей, мышцы, поднимающие затылок, разгибатели спины, мышцы, поднимающие хвост и мускулы, смыкающие нижнюю челюсть. Антагонисты этих мышц, в противоположность этому, находятся в расслабленном состоянии. Во многих случаях у них совсем нет тонуса, а если есть, то очень незначительный.
Шеррингтон показал, что главный, но не единственный источник этого тонуса лежит в проприоцептивных чувствительных нервах мышц и притом в проприоцепторах тех мышц, которые находятся в состоянии ригидности. И в этом случае видно, что, кроме этих проприоцепторов, на появление тонуса при децеребрационной ригидности влияют еще другие чувствительные нервы. Но, кроме того, существенным условием для возникновения ригидности является еще целостность вышеназванныхцентровв продолговатом мозгу. Шеррингтон (9) назвал состояние децеребрационной ригидности «рефлекторным стоянием». Но это «стояние» чисто пассивное: животное стоит, если его поставят, но тотчас же падает, если его слегка толкнуть, и не в состоянии подняться из лежачего положения.
Если же в мозговом стволе, кроме продолговатого, сохраняется еще неповрежденным средний мозг, то картина сразу меняется (рис. 1, Ь).
Во-первых, исчезает децеребрационная ригидность, отпадает одностороннее преобладание разгибателей и вместо этого наступает «нормальное» распределение тонуса между разгибательными и сгибательными мышцами, т. е. как раз то, что имеет место у нормального интактного животного. Децеребрационная ригидность является как бы карикатурой на стояние, причем разгибатели находятся в преувеличенном тонусе, в то время как у животного с сохраненным средним мозгом разгибатели находятся в точном соответствии с весом тела и, кроме того, не только разгибатели, но и сгибатели имеют свой нормальный тонус.
Таким образом, распределение напряжения во всей мускулатуре тела становится точно таким, как и у интактного животного; при этом отсутствует чрезмерное преобладание тонуса разгибательных мышц (мышц, поддерживающих позу стояния).
Но имеется еще второе изменение, обусловленное сохранением среднего мозга. В то время как децеребрированное животное способно только к пассивному стоянию и не может подняться из лежачего положения, мезэнцефалическое животное может самостоятельно принять нормальную позу. Значит, животное теперь выполняет то, что у децеребрированного животного должен был делать экспериментатор. Оно способно рефлекторно принять из всех неестественных положений основное положение и притом выполняет это вполне уверенно. Если сравнить животное с неповрежденным средним мозгом с нормальным, неоперированным животным, то вряд ли при первом взгляде можно установить между ними разницу. Мезэнцефалическое животное выполняет движения бега и прыжка с нормальной быстротой и уверенностью; ему не хватает только спонтанных движений, и каждый раз требуется внешнее раздражение, для того чтобы животное, которое ведет себя как автомат, пришло в движение. При сохраненном же большом мозге могут выполняться спонтанные движения. Другие различия между интактным и таламическим животным, которые связаны с наличием большого мозга, будут рассматриваться позднее.
На рис. 1 изображены три вышеописанных типа животных. Слева находится спинальное животное: кошка, у которой под эфирным наркозом были перевязаны сонные и позвоночные артерии и таким образом отключен головной мозг, включая продолговатый мозг. У животного искусственно поддерживается дыхание, и его согревают; при исследовании у него были обнаружены сгибательный рефлекс, перекрестный разгибательный рефлекс,
10
Первая глава. Общий обзор
коленный рефлекс, чесательные движения, хвостовые рефлексы и т. д~ Животное, как это видно из рисунка, не может стоять; хотя конечности: и обладают определенным тонусом, но они не способны нести вес тела. Справа находится децеребрированное животное (В): кошка, у которой: после перевязки сонных артерий была произведена трепанация черепа и. перерезка ствола мозга в области крыши мозжечка* Животное после операции отогрели, и вскоре у него начала развиваться децеребрационная ригидность. Из рисунка видно, что голова поддерживается зажимом, но' тело животного держится на передних и задних конечностях. В других: случаях даже нет необходимости закреплять голову, и такое децеребриро- ванное животное стоит с незакрепленной головой на четырех ногах, пока! оно не потеряет равновесия и не упадет. В середине находится животное с сохраненным средним мозгом (С): кролик, у которого накануне вечером был удален головной мозг до таламуса. Целесообразно для таких опытов оставлять таламус, так как при этом остается неповрежденным центр теплорегуляции и животному легче сохранить жизнь. Для выполнения функций стояния и установки тела основным условием является сохранность среднего мозга. Кролик сидит в нормальном положении, и если его' положить на один или другой бок, он моментально снова садится и остается затем сидеть в нормальном положении, изображенном на рисунке.
Поддержание положения тела и равновесия обусловливается и сохраняется афферентными возбуждениями, которые исходят от различных органов. Спрашивается, не можем ли мы изучать эти афферентные возбуждения: так, как мы привыкли это делать, например при зрительных и слуховых возбуждениях, т. е. посредством субъективного анализа исходящих от них ощущений. Однако если этот способ дал чрезвычайно важные и точные результаты в физиологической оптике и акустике, то при изучении равновесия тела он совершенно непригоден. Причина этого различия заключается в том, что нашим зрительным и слуховым ощущениям соответствуют определенные участки коры и поэтому в данном случае мы можем использовать зрительные и слуховые восприятия как определенные категории. В противоположность этому у нас нет участка коры для статических восприятий. Правда, мы осознаем ряд афферентных возбуждений из различных частей нашего тела и обладаем также очень точным чувством положения наших конечностей, но мы должны выводить наше суждение о положении тела и равновесии из ряда различных вторичных афферентных возбуждений, которые доставляются нам от лабиринтов, мышц и суставов, органов осязания и ощущения давления, от глаз. Эти возбуждения, как: определенные компоненты, часто остаются подпороговыми, не доходя до сознания. Поэтому мы часто входим в заблуждение и никоим образом не можем посредством субъективного анализа определить участие отдельных органов чувств в общей статической деятельности.
Успешное изучение различных положений тела и равновесия возможно только объективным путем, особенно на животных. Нужно найти ряд объективных, поддающихся исследованию рефлексов. Только таким образом можно достигнуть успеха и свести эту сложную деятельность в систему закономерных рефлексов и реакций, которые связаны с функцией определенных центральных групп.
При ближайшем изучении выяснилось, что речь идет об очень,сложной совместной деятельности разнообразных органов чувств, причем различные центральные группы объединяют отдельные возбуждения. Мускулатура- тела участвует в реакциях в различных комбинациях, причем можно выделить несколько разнородных групп рефлексов. Для того чтобы упростить
Статические рефлексы,. Положение тела
п
эти представления, нужно дать краткий обзор важнейших участвующих здесь рефлексов.
Прежде всего нам нужно установить разницу между поведением тела в покое и при движениях. Те рефлексы, которые обусловливают и сохраняют положение тела и равновесие при спокойном лежании, стоянии и сидении в различных положениях, мы можем обозначить как статические рефлексы. Те же рефлексы, при помощи которых животное совершает активные и пассивные движения и которые частично компенсируют последствия этих перемещений, мы назовем стато-кинетическими.
Статические рефлексы делятся в свою очередь на две большие группы. При абсолютном покое животное принимает определенную «позу» (Haltung), т. е. определенное положение отдельных участков тела по отношению друг к другу, обусловленное закономерным распределением напряжения по всей мускулатуре и тонической фиксацией в различных суставах. Те статические рефлексы, которые обусловливают положение или позу тела, мы можем обозначить как рефлексы стояния (Stehreflexe).
Во вторую группу статических рефлексов следует отнести те рефлексы, благодаря которым тело способно принимать нормальное положение из различных неестественных положений. Таким образом, здесь идет речь о способности животного самостоятельно вставать, и потому эти рефлексы называют установочными рефлексами (Stellreflexe).
I. СТАТИЧЕСКИЕ РЕФЛЕКСЫ
А. Положение тела (рефлексы стояния)
1. Влияние положения головы на позу
При исследовании рефлексов стояния оказалось целесообразным предварительно выключить установочные рефлексы, т. е. производить опыты на децеребрированном животном. На последнем удалось установить, что, придавая голове определенную позу, можно управлять положением тела и распределением напряжения во всей мускулатуре тела. Если голове животного придавать то или другое положение, то происходят два различных процесса. Во-первых, изменяется положение головы по отношению к телу, и, во-вторых, изменяется положение головы в пространстве; в обоих случаях вызываются различные группы рефлексов.
•а) Тонические шейные рефлексы, влияющие на положение конечностей
В зависимости от установки головы по отношению к телу (повороты, вращение, поднимание, опускание головы и т. д.) меняется и положение шеи. Это вызывает рефлекторные тонические влияния на мускулатуру конечностей, которые у децеребрированного животного проявляются в увеличении или уменьшении тонуса мышц разгибателей. Эти рефлексы подчиняются точно определенным законам. Такое измененное распределение напряжения в разгибателях конечностей сохраняется до тех пор, пока голова занимает определенное положение по отношению к туловищу: при изменении положения головы относительно туловища сейчас же возникает другое распределение напряжения. Выяснилось, что в большинстве случаев при изменениях положения головы по отношению к телу конечности правой и левой стороны или задние и передние конечности реагируют противоположно.
12
Первая глава. Общий обзор
б) Тонические лабиринтные рефлексы, влияющие на мускулатуру тела
Если голова животного не изменяет своего положения относительно тела, но ей придают различные положения в пространстве, то и этим вызываются тонические рефлексы на мускулатуру тела, которые длятся до тех пор, пока голова сохраняет одно и то же положение в пространстве. Изменение тонуса вызывается тем, что голова и лабиринты, как это установлено точным анализом, изменяют свой наклон по отношению к горизонтальной плоскости. На децеребрированном животном эти рефлексы выявляются точно так же, как и тонические шейные рефлексы, и главным образом на тех мышцах, которые находятся в состоянии тонуса при деце- ребрационной ригидности Шеррингтона. Эти мышцы имеют максимальный тонус при вполне определенном положении головы в пространстве и минимальный при повороте головы на 180°.
Тонические лабиринтные рефлексы мышц, конечностей. В этих тонических лабиринтных рефлексах участвуют прежде всего мышцы конечностей, так же как и при тонических шейных рефлексах; при изменение положения головы децеребрированного животного в пространстве можно видеть отчетливые изменения тонуса мышц разгибателей четырех конечностей. При этом разгибатели всех конечностей реагируют всегда одинаковым образом.
Тонические лабиринтные рефлексы мускулатуры шеи и туловища. Не только мышцы конечностей, но и вся мускулатура туловища находится в отношении распределения тонуса под влиянием лабиринтов. Из опытов выяснилось, что лабиринты оказывают сильнейшее влияние на шейную мускулатуру и что это влияние у определенного рода животных ограничивается шеей, в то время как у других видов животных лабиринты влияют и на остальную мускулатуру туловища. Из дальнейшего будет видно, что целесообразно отделить эти тонические лабиринтные рефлексы мускулатуры шеи и туловища от рефлексов мышц конечностей. Это станет ясно, когда мы будем обсуждать двустороннее влияние лабиринтов на мускулатуру обеих сторон тела. Однако в отношении этих двух мышечных групп речь идет о длительных влияниях очень сходного характера.
в) Взаимодействие тонических шейных и лабиринтных рефлексов
Если изменять положение головы при различных положениях тела в пространстве, то тонические шейные и лабиринтные рефлексы должны различным образом комбинироваться друг с другом. Это становится ясным при рассмотрении следующего примера: если при положении тела на боку поднимать и опускать голову (дорсальное и вентральное сгибание),, положение головы по отношению к горизонтальной плоскости не изменяется, и потому при этих различных положениях головы шейные рефлексы проявляются только в изменениях тонуса конечностей. Если же поднимать и опускать голову в нормальном положении животного или когда оно лежит на спине, то положение головы изменяется и по отношению к горизонтальной плоскости, и поэтому теперь, кроме тех же шейных рефлексов, вызываются тонические лабиринтные рефлексы мышц конечностей.
Эти группы рефлексов (тонические шейные и тонические лабиринтные) взаимодействуют таким образом, что или их влияние суммируется и тогда мышца находится в максимальном тонусе, или мышца расслабляется, если влияние шейных и лабиринтных рефлексов уменьшает ее тонус, или же напряжение мышцы не изменяется, если влияние шейных и лабиринтных рефлексов взаимно уничтожается.
Статические рефлексы. Положение тела
13’.
г) Непрямое влияние лабиринтов на мускулатуру конечностей посредством шейных рефлексов
Выше уже было упомянуто, что существуют тонические лабиринтные' рефлексы шейной мускулатуры. Если голова животного меняет свое положение в пространстве, то наблюдается изменение напряжения шейных, мышц. Эти тонические изменения мышц шеи могут в свою очередь снова, вызывать тонические рефлексы мышц конечностей, и этим создается двойное воздействие лабиринтов на мускулатуру конечностей: 1) прямое воздействие благодаря тоническим лабиринтным рефлексам на конечности;,
2) непрямое воздействие, при котором лабиринты сперва действуют на>. мышцы шеи и этим вызывают тонические рефлексы конечностей и таким образом влияют на напряжение мускулатуры конечностей. Оба эти воздействия на мышцы конечностей также суммируются алгебраически.
Таким путем у животных, которые лишены больших полушарий головного мозга, а также среднего мозга, создается очень запутанное отношение между положением головы и распределением напряжения во всей мускулатуре тела. И действительно, можно, изменяя положение головы, придавать телу животного ряд самых различных положений, которые обусловливаются вышеупомянутыми различными рефлексами позы. Все это становится еще более сложным, если еще участвуют средний мозг и кора больших полушарий, но основой таких различных, крайне выразительных положений и поз животных и человека, встречающихся в жизни, которые нас так восхищают в художественных и скульптурных произведениях, в конечном счете являются закономерности, создаваемые взаимодействием рефлексов позы.
2. Прямые влияния на положение тела
При изучении рефлексов позы выяснилось, что главные влияния на распределение напряжения в мускулатуре тела исходят от головы и что влияния от других частей тела хотя и отступают на задний план, но не полностью. Стенверс (Stenvers) в психиатрической клинике в Утрехте показал на человеке, что при вращении таза, когда пациент лежит на спине, вызываются закономерные повороты головы и глаз. Однако эти наблюдения до сих пор остались единичными. Во всяком случае можно сказать, что влияния других частей тела на распределение тонуса во всей мускулатуре значительно меньше, чем влияния головы.
3. Непрямое влияние на положение тела
Все факторы, которые влияют на положение головы, должны поэтому косвенно вызывать изменение положения тела. Если, например, поставить миску с молоком перед кошкой, стоящей на полу, то животное будет сидеть с опущенной вниз головой, и вследствие этого мышцы передних конечностей, поддерживающие позу стояния (разгибатели), будут иметь меньший тонус, в то время как тонус в задних конечностях из-за противоположного влияния шейных и лабиринтных рефлексов станет средним. Вследствие этого кошка всей передней частью тела приблизится к полу и примет положение, которое мы видим у пьющего животного.
Если, наоборот, держать высоко в воздухе кусок мяса, то животное повернет голову за пищей вверх, т. е. дорсально, и этим вызовет тоническое
;14
Первая глава. Общий обзор
разгибание своих передних ног, что приведет к поднятию передней половины тела, в то время как задняя половина тела мало изменит свое положение. Вследствие этого тело животного подастся вперед и рот приблизится к пище.
Если кошка стоит посреди комнаты и сбоку вдоль стены бежит мышь, то животное поворачивает голову по направлению услышанного шума. Вследствие этого увеличивается тонус тех конечностей, в сторону которых повернута голова животного. Поворот головы вызывает большее смещение центра тяжести по направлению поворота. Но, несмотря на это, животное не падает, потому что увеличивается тонус разгибателей в передней ноге, в сторону которой повернута голова. Теперь эта нога в состоянии вынести увеличенную нагрузку, вызванную смещением центра тяжести. Далее выяснилось, что если животное вслед за этим побежит, то первый шаг делает разгруженная нога. Следовательно, слуховым раздражением, идущим со стороны, были вызваны, во-первых, движения головы, во-вторых, изменение распределения напряжения, которое приспосабливается к перемещению центра тяжести, и, в-третьих, готовность мускулатуры конечностей к бегу или прыжку.
Эти примеры показывают, что раздражения, поступающие через телерецепторы (глаз, ухо и нос) в центральную нервную систему, вызывают движения головы, которые в свою очередь обусловливают соответствующее положение тела животного и создают условия для движений, вызванных афферентными возбуждениями.
Уже это краткое обозрение показывает, что речь идет о чрезвычайно гармоничном совместном влиянии различных тонических рефлексов, благодаря которым взаимное расположение отдельных частей тела и положение тела в целом приспосабливаются к различным условиям.
Б. Компенсаторные положения глаз
Компенсаторные положения глаз можно считать особым случаем рефлексов положения. Распределение напряжения в мышцах глаз также подчиняется совместному влиянию тонических лабиринтных и тонических шейных рефлексов. Эти компенсаторные положения глаз у человека и у животных, глаза которых расположены фронтально, играют незначительную роль, но все же можно точно доказать их наличие. У животных с фронтально расположенными глазами поле зрения перекрывается в наибольшей степени и вследствие этого положение обоих глаз по отношению друг к другу контролируется главным образом зрительно. Поэтому сами зрительные возбуждения «заботятся» (благодаря чувствительности мышц) о том, чтобы изображения на обеих сетчатках соответствовали друг другу и при изменении положения тела правильно совпадали. Иначе обстоит дело у животных с боковым зрением, как например у кроликов и морских свинок, у которых поля зрения не перекрываются или перекрываются только частично. У них не может осуществляться совместная работа этих полей зрительным путем, и поэтому у таких животных выработалась сложная система рефлексов положения, «заботящаяся» о том, чтобы при различных положениях головы в пространстве и относительно тела животного зрительные впечатления от обоих глаз оставались закономерно согласованными друг с другом.
Такие рефлексы, как уже было указано выше, можно обнаружить также и у человека и у животных с фронтально поставленными глазами, но по своему значению они отступают на задний план; поэтому целесообразнее изучать эти рефлексы у кроликов и морских свинок.
Статические рефлексы. Компенсаторные положения глаз
15
1. Влияние тонических лабиринтных рефлексов
на глаза
Каждому положению головы в пространстве соответствует определенное положение.глаз в орбитах. На эти различные положения глаз влияет взаимодействие вертикальных и круговых движений.
Вертикальные движения лучше всего изучать (на кроликах и морских свинках), если исходить из нормального положения головы с горизонтально расположенной ротовой щелью и производить вращение по затылочноносовой оси. Если, например, повернуть голову так, чтобы правый глаз двигался книзу, то правое глазное яблоко отклоняется вверх, а левый (находящийся наверху) глаз отклоняется вниз. Это отклонение становится наибольшим приблизительно тогда, когда голова достигает бокового положения. При обратном повороте головы, левым глазом вниз, глаза совершают обратное отклонение, так что теперь правое глазное яблоко отклоняется вниз, а левое — вверх. При других положениях головы в пространстве также имеются вертикальные отклонения глаз, но они не достигают максимума, как это наблюдается при боковом положении головы. Так как при правом боковом положении головы правый глаз отводится вверх, а левый вниз, то, следовательно, до некоторой степени глаз «старается» сохранить свое положение относительно горизонтальной линии.
Круговые движения лучше всего изучать, если исходить из нормального положения и вращать голову 1 вокруг височной оси, т. е. мордой вверх или вниз. Если морда поднимается вверх, то оба глаза также выполняют круговые движения в том же направлении, причем верхний полюс роговицы поворачивается вперед к носу. И наоборот — верхний полюс роговицы обоих глаз идет назад, если морду поворачивают вниз. И в этом случае глаза также стремятся при изменяющихся положениях головы сохранить неизменное положение по отношению к горизонту, что им и в, этом случае полностью не удается. Максимум кругового движения в обоих направлениях достигается в том случае, если голова стоит вертикально мордой вверх или вниз; при всех остальных . положениях головы в пространстве осуществляются незначительные круговые движения.
Комбинации вертикальных и круговых движений лучше всего удается изучить, если поместить голову в боковое положение и поворачивать ее вокруг горизонтально расположенной дорсо-вентральной оси. При исходном положении (боковое положение) вертикальное отклонение бывает максимальным. Если теперь повернуть голову мордой вверх, то достигает максимума круговое движение. Поворот до противоположного бокового положения вызывает максимальное вертикальное отклонение в противоположном направлении. Затем голова приводится в вертикальное положение мордой вниз, при этом круговое движение в противоположном направлении становится максимальным; в конце концов голова возвращается в исходное положение с наибольшим вертикальным отклонением. При различных поворотах головы положение глаз изменяется, при этом их движения слагаются из комбинации вертикальных и круговых движений, которые ограничены в ту и другую сторону разными максимальными отклонениями.
Если производить движения головы, находящейся в любом положении в пространстве, в горизонтальной плоскости, то по отношению к горизонту положение головы и лабиринтов не изменяется. Значит, горизонтальными движениями головы нельзя вызвать тонические лабиринтные рефлексы
1 Не меняя положения головы относительно туловища.
16
Первая глава. Общий обзор
глаз. С этим согласуется факт, что при тонических лабиринтных- рефлексах глаз горизонтальные отклонения не играют роли и что для этой группы рефлексов не удалось выявить участие горизонтальных прямых мышц глаза — наружной и внутренней. Компенсаторцые положения глаз, вызываемые лабиринтами, обусловлены в основном действием только обеих косых мышц глаз.
При различных положениях головы в пространстве тонические лабиринтные рефлексы вызывают соответствующие движения глаз, благодаря которым глаза «стараются» сохранить до некоторой степени свое прежнее положение в пространстве. Но количественные исследования показали, что эта цель не достигается одними лабиринтными рефлексами. Для этого должна присоединиться еще другая группа возбуждений.
2. Влияние тонических шейных рефлексов
н а г л а з а
Эти рефлексы были сперва установлены Барани [Bäräny (1)], а точными знаниями о них мы обязаны де Клейну [de Kleyn (53)]. При их описании мы исходим из нормального положения животного. Если, например, голова поворачивается влево так, что левое ухо приближается к левому плечу, то оба глаза, вследствие тонического шейного рефлекса, двигаются вправо; это значит, что правый глаз движется по направлению к виску, а левый — по направлению к носу. Здесь речь идет о влиянии на прямую наружную и внутреннюю мышцы. Если повернуть голову влево, то левый глаз перемещается книзу, а правый кверху, тогда нижний (левый) глаз отклоняется дорсально, а верхний (правый) — вентрально. Значит, при этих обстоятельствах из-за влияния тонических шейных рефлексов происходит отклонение глаз в таком же порядке, как оно вызывается и лабиринтами. Если поднимать или опускать голову, то последуют круговые движения глаз. Если морда поднимается вверх, то оба глаза верхним полюсом роговицы перекатываются вперед, если же морда опускается, то оба глаза поворачиваются назад. И здесь реакция глаз такая же, как и та, которая вызывается лабиринтами.
Эти положения глаз остаются до тех пор неизменными, пока голова находится в упомянутом положении по отношению к туловищу. При сравнении тонических шейных рефлексов глаз с реакциями глаз, вызванными лабиринтами, выяснилось, что при обеих группах рефлексов происходят вертикальные отклонения и круговые движения глаз и что в противоположность этому горизонтальные отклонения глаз вызываются исключительно с шеи.1
Тонические шейные рефлексы, изменяющие положение глаз, также способствуют такой установке глаз, при которой они в некоторой степени стремятся сохранить свое положение в пространстве при изменении положения головы. Но здесь обнаружилось, что тонические шейные рефлексы сами по себе не в состоянии при изменении положения головы произвести полную компенсацию положения глаз.
3. Совместное действие тонических лабиринтных ишейных рефлексов на глаза
Для исследования влияния тонических лабиринтных рефлексов на глаза следует исключить движения шеи. Лучше всего закрепить иссле¬
1 Дальше мы увидим, что при стато-кинетических лабиринтных рефлексах глаз большую роль играют горизонтальные отклонения, при статических же рефлексах горизонтальные отклонения не вызываются лабиринтами.
Статические рефлексы. Компенсаторные положения глаз
17
дуемое животное на специальном станке (рис. 70, стр. 124). Затем последний приводят в различное положение в пространстве, причем, конечно, положение головы относительно тела не изменяется. Для изолированного исследования шейных рефлексов нужно препятствовать появлению тонических лабиринтных рефлексов, влияющих на глаза. Этого можно достичь или удалением обоих лабиринтов, или тем, что во время всего исследования положение головы остается неизменным в пространстве и, таким образом, тело животного перемещается относительно неподвижно установленной головы. При свободных движениях и положениях животного яти два условия неосуществимы, и поэтому влияние лабиринтных и шейных рефлексов на глаза всегда суммируется. В то время как каждая из названных групп рефлексов не в состоянии в отдельности полностью компенсировать движения глаз, этого удается в некоторой степени достигнуть совместным действием обеих групп. Как показал де Клейн (53, 65) для круговых движений и вертикальных отклонений, глаза правильно ориентируются в пространстве при нормальном исходном положении животного и при движениях головы, которые животное обычно производит в жизни; поэтому при таких движениях головы положение полей зрения не изменяется и изображения на сетчатке остаются неподвижными. Важно то, что это происходит одновременно для обоих глаз и что поэтому у животных с латерально расположенными глазами правая и левая половины их зрительного поля должны и при длительных движениях соответствовать друг другу. У таких животных благодаря совместному действию шейных и лабиринтных рефлексов достигается тот же эффект, какой наблюдается у человека и животных с фронтально расположенными глазами вследствие взаимного оптического контроля зрительных полей. Как было уже упомянуто, и у последних удается обнаружить тонические шейные и лабиринтные рефлексы, изменяющие положение глаз, но их значение отодвинуто на задний план из-за оптического контроля положения глаз.
4. Совместное влияние компенсаторных положений глаз и стато-кинетических рефлексов на глаза
Но этим не исчерпывается деятельность этого тонкого установочного аппарата. В дальнейшем мы увидим, что при повороте головы в пространстве вызываются стато-кинетические рефлексы глаз, так называемые вращательные реакции глаз, которые также приводят к вертикальным отклонениям, круговым движениям и горизонтальным отклонениям глаз. Эти реакции осуществляются при нормальном положении животного таким образом, что определенное движение головы (поднятие, опускание, наклон и поворот) вызывает движение глаз в том направлении, в котором они удерживаются в дальнейшем статическими рефлексами (тоническими шейными и лабиринтными рефлексами), так что глаза управляются самим началом движения и поворачиваются в необходимую сторону.
И наоборот, компенсаторное положение глаз может оказать влияние на направление вызываемых лабиринтами глазных вращательных реакций. Именно из-за того, что при компенсаторном положении глаз осуществляются круговые движения и вертикальные отклонения, точки прикрепления глазных мышц к глазному яблоку будут смещены по отношению к орбите. И. если теперь с лабиринтов вызывается стато-кинетический рефлекс определенной пары глазных мышц, то может случиться, например, что вместо горизонтального отклонения произойдет косое или вертикальное отклонение глаза просто по той причине, что вследствие кругового
2 Р. Магнус
18
Первая глава. Общий обзор
движения точки прикрепления прямой наружной и прямой внутренней мышц лежат теперь в глазнице не в горизонтальном, а скорее в вертикальном направлении.
Так благодаря лабиринтным и шейным рефлексам выполняется чрезвычайно сложная взаимная работа глазных мышц и закономерная совместная деятельность обоих глаз, которая осуществляет правильное видение как при движении, так и в,покое и при различных физиологически возможных положениях головы относительно тела и в пространстве. Уже из этого ясно, что при патологических изменениях этого механизма должны наступить отчетливые нарушения в зрительной ориентировке, и очень возможно, что это играет существенную роль и в патологии человека.
В. Установочные рефлексы
В начале этой главы было показано, что если у млекопитающих, кроме центров в продолговатом мозгу, сохранен еще неповрежденный средний мозг, то животное способно самостоятельно вставать. Децеребрированное животное стоит, если его поставить, и падает, если ему дать толчок, но оно не в состоянии само подняться. Животное же с сохраненным средним мозгом, в противоположность этому, способно рефлекторно принять из каждого неестественного положения совершенно точно нормальное положение, которое потом может служить исходной точкой для всевозможных произвольных и-рефлекторных движений. Дальнейший анализ показал, что в этой установочной функции участвует ряд различных рефлексов и что различные органы восприятия служат исходным пунктом для установочных рефлексов.
1. Лабиринтные установочные рефлексы
головы
Для исследования этих рефлексов нужно держать животное свободно в воздухе — так, чтобы не было никакого соприкосновения тела с подставкой. Кроме того, нужно у кошек, собак и обезьян исключить зрительные возбуждения. Это можно произвести или при помощи колпака, надеваемого на голову, или путем удаления больших полушарий. Наоборот, у кроликов и морских свинок не нужно устранять зрительные импульсы, так как'выяснилось, что животные без лабиринтов, с надетым на голову колпаком или без него, или же животные с работоспособными полушариями или без них, но после экстирпации лабиринтов ведут себя при свободном нахождении в воздухе совершенно одинаково.
Если взять кролика или морскую свинку за таз и держать их в воздухе в нормальном положении, то передняя половина тела и голова стоят правильно, причем последняя находится в нормальном положении, теменем вверх и с ротовой щелью, опущенной немного ниже горизонтали. Если, исходя из этого положения, так поворачивать таз, чтобы крестец стоял вертикально (оральным концом вверх — висячее положение головой вверх), то голова остается в нормальном положении. Если повернуть таз вокруг битемпоральной оси так, чтобы голова была внизу (висячее положение головой вниз), то положение головы в пространстве не меняется или меняется, очень мало, ротовая щель сохраняет свой наклон к горизонту или очень незначительно отклоняется в вертикальном направлении. В то время как положение таза в пространстве изменилось на 180°, положение ротовой щели осталось или то же самое, или сдвинулось очень ненамного. В первом случае (висячее положение головой вверх) это происходит бла-
Статические рефлексы. Установочные рефлексы
19
годаря вентральному перегибу; при висячем положении головой вниз — напротив, из-за сильного дорсального перегиба головы. Если привести таз в боковое положение, то голова благодаря повороту передней половины тела приводится в нормальное положение и таз можно вращать в воздухе из одного бокового положения в другое на 180° без того, чтобы положение головы изменялось в пространстве. Кажется, что голову держит в пространстве какая-то «магическая» сила: этот опыт является чрезвычайно
наглядным. При помещении животного в положение на спину голова также приводится в нормальное положение, а именно — вентральным наклоном передней половины тела так, что голова приближается к животу, или же спиральным вращением передней половины тела на 180°.
Подобные рефлексы также можно обнаружить при исследовании кошек и собак с головным колпаком или обезьян с зашитыми веками; животные при этом находятся в воздухе в свободном состоянии.
После удаления обоих лабиринтов эти рефлексы отсутствуют. Теперь животных можно держать при вышеуказанных условиях в любом положении в воздухе без того, чтобы голова переходила в нормальное положение. Получаем спинное, боковое положение головы и т. д. Животные полностью дезориентированы в пространстве.
2. Влияние установочных рефлексов тела
на голову
Если кролика или морскую свинку после двустороннего выключения лабиринтов свободно держать в боковом положении в воздухе, то голова также перейдет в боковое положение. Но как только животное положат боком на пол, голова сейчас же поворачивается и переходит в нормальное положение. Этот рефлекс вызывается прикосновением тела к подстилке и основывается на асимметричном возбуждении чувствительных нервов тела вследствие давления тела на подстилку. Это можно доказать, если компенсировать асимметричное давление, положив на верхнюю боковую поверхность тела доску с грузом. Вслед за этим голова сейчас же переходит в боковое положение; если удалить доску, то голова снова поворачивается в нормальное положение.
Этими двумя группами рефлексов (лабиринтные установочные рефлексы и установочные рефлексы тела) обеспечивается то, что голова всегда приводится в нормальное положение. Как только последнее осуществилось, а тело еще не находится в нормальном положении, происходит перекручивание шеи. Этим вызываются следующие рефлексы.
3. Шейные установочные рефлексы
Эти рефлексы приводят к тому, что сперва передняя половина тела, а затем также и задняя половина тела животного следуют заданному положению головы и таким путем наконец все тело животного приходит в нормальное положение. Значит, здесь речь идет о цепном рефлексе, при котором сперва, благодаря своеобразному перекручиванию шеи, передняя часть тела, а затем, вследствие поворота поясничной части позвоночника, и задняя половина тела приводятся в правильное положение. Несмотря на эту цепную реакцию, ради простоты нужно употреблять общее название шейных установочных рефлексов.
Все до сих пор описанные установочные рефлексы действовали сперва на голову, которая сразу приводилась в нормальное положение. К этому затем вторично присоединялась правильная установка тела.
2*
20
Первая глава. Общий обзор
4. Влияние установочных рефлексов тела
на тело
Однако правильное положение тела не полностью зависит от головы. Если положить кролика в боковом положении на стол и голову держать также в боковом положении, то часто при этих условиях животное поднимается правильным образом. Особенно четко можно обнаружить этот рефлекс, если употреблять грубую подстилку (например, плетеную циновку). При этих условиях животное садится, противодействуя шейному установочному рефлексу, который стремится удержать тело в боковом положении. Действующие раздражения и в этом случае вызываются асимметричным соприкосновением тела животного с подстилкой. Это можно доказать наложением доски с грузом на верхнюю боковую часть тела.
Из этого следует, что как голова, так и тело приводятся в нормальное положение двойным механизмом рефлексов: голова — лабиринтными
установочными рефлексами и установочными рефлексами тела на тело, тело же — шейными установочными рефлексами и установочными рефлексами тела на тело.
5. Оптические установочные рефлексы
Центры четырех названных рефлексов находятся в общей функциональной группе в среднем мозгу и в области моста. У собак, кошек и обезьян, кроме того, присоединяется еще пятая группа рефлексов: оптические установочные рефлексы, для функционирования которых требуется наличие неповрежденной коры больших полушарий (вероятно, зрительная область коры). Собаки, кошки и обезьяны ведут себя после двусторонней экстирпации больших полушарий так же, как кролики и морские свинки. Если имеются большие полушария" то в их поведении принимают участие и вышеназванные установочные рефлексы. Если удалить у этих животных оба лабиринта и через короткое время после операции держать их свободно в воздухе, то голова, лишенная ориентации, принимает любое положение в пространстве. Но через несколько дней животные «выучиваются применять» свои глаза для ориентировки и можно отчетливо видеть, что если какой-нибудь предмет (корм, экспериментатор или предметы в комнате) фиксируется животным, то голова при этом приходит в нормальное положение. Еще через несколько дней животные обычно уже хорошо развивают эту способность, и если их теперь (т. е. с неповрежденными большими полушариями и открытыми глазами) исследовать в воздухе в свободном положении, то голова сразу же после фиксации какого-нибудь предмета приводится в нормальное положение. То, что здесь действительно речь идет об установочных рефлексах, вытекает из того, что после надевания колпака на голову эта способность сразу исчезает и животные не способны ориентироваться в воздухе, так же как кролики и морские свинки, лишенные лабиринтов. Оптические установочные рефлексы действуют также сперва на голову. К этому присоединяются потом вышеупомянутые шейные установочные рефлексы, которые вторично приводят тело в правильное положение.
Итак, видно, что особенно важные для животного функции установки тела обеспечиваются совместным влиянием разнородных рефлексов. Из всех неестественных положений в пространстве тело животного автоматически принимает нормальное исходное положение еще до совершения произвольного или рефлекторного движения.
Стато-кинетические рефлексы. Вращательные реащии
21
И. СТАТО-КИНЕТИЧЕСКИЕ РЕФЛЕКСЫ
Описанные статические рефлексы являются рефлексами положения. Рефлексы положения длятся до тех пор, пока голова находится в соответствующем положении в пространстве или относительно тела. Установочные рефлексы действуют и тогда, когда животное в совершенно спокойном состоянии находится в неправильном исходном положении и благодаря установочным рефлексам достигает нормального положения.
В противоположность этому стато-кинетические рефлексы вызываются движениями. Поскольку здесь лабиринты являются рецепторами для этих рефлексов, удалось выяснить, что раздражителем является не само движение как таковое, а изменение движения. Эти стато-кинетические рефлексы вызываются или угловыми ускорениями, или линейными ускорениями, притом как положительными, так и отрицательными.
А, Вращательные реакции
Лабиринтные рефлексы, вызываемые угловыми ускорениями, настолько общеизвестны и из-за их большого клинического значения так всесторонне изучены, что здесь можно только кратко остановиться на них.
1. Вращательные реакции головы
Если, например, посадить животное в нормальное положение на вертящийся диск так, чтобы позвоночник имел направление радиуса, а голова была направлена наружу, то при вращении животного по направлению его правой стороны тела голова поворачивается влево (вращательная реакция головы). После прекращения вращения наступает поворот головы вправо (поствращательная реакция головы). Как при вращательной, так и при поствращательной реакции может наступить нистагм головы, который своим быстрым компонентом всегда действует в направлении, противоположном вращательной реакции. Изменением положения животного и направления вращения можно также достичь вертикальных и ротаторных отклонений головы с соответствующим нистагмом.
2. Глазные вращательные реакции
Если, например, производить на кролике вышеописанный опыт на вращающемся диске, то при вращении вправо оба глаза отводятся влево, т. е. левый глаз движется по направлению виска, правый глаз — по направлению носа (вращательная реакция глаз), причем вращательный нистагм действует обратным образом. После прекращения вращения левый глаз движется к' носу, правый глаз — к виску (поствращательная реакция глаз). Поствращательный нистагм ^лаз действует своим быстрым компонентом в обратном порядке. Ротаторные отклонения можно получить, если вращать животное в боковом положении, вертикальные — при висячем положении головой вниз или вверх. Общим всегда является следующее правило: если вращают животное вокруг вертикально стоящей в пространстве оси, то отклонение глаз следует в горизонтальной плоскости и при этом в таком направлении, что глаза стремятся сохранить свое первоначальное положение в этой плоскости.
Описанные вращательные реакции головы и глаз являются в общем компенсаторными; это значит,, что в начале движения голова и глаза двигаются таким образом, что оптические изображения по возможности
22
Первая глава. Общий обзор
сохраняются. Уже выше обращалось внимание на то, что благодаря этим вращательным реакциям глаза при нормальном положении животного приводятся именно в то положение, в котором они позже удерживаются статическими рефлексами (компенсаторное положение глаз).
Кроме того, указывалось, что компенсаторные движения глаз влияют ца направление вращательной реакции глаз только в таком пределе, в каком может изменяться положение глазного яблока в глазнице. Этим также определяется направление вращательных реакций.
3. Вращательные реакции конечностей и туловища
Кроме описанных вращательных реакций головы и глаз, имеются такие же реакции и мускулатуры тела. Существование влияний такого рода вытекает из многочисленных клинических опытов. Например, при так называемой указательной пробе предварительное вращение влияет на направление произвольного движения, выполненного после вращения. Но условия этого опыта довольно сложны и пока не поддаются простому анализу. Мах (Mach), а также Барани (Вагапу), Райх (Reich) и Ротфельд (Roth- feld) описали у кролика после вращения реакции движения и падения, при которых, по всей вероятности, имеются и прямые лабиринтные влияния на мускулатуру тела. Непосредственному наблюдению поддаются вращательные реакции четырех конечностей и туловища у обезьян; на них удалось показать, что здесь речь идет о рефлексах, вызываемых прямо лабиринтами. То, что и у человека имеются в наличии такие рефлексы, вытекает из самонаблюдений при автомобильной езде: при каждом крутом повороте появляется очень ясное противодействующее движение в поясничной области, которое не зависит от движения головы и глаз и которое едва подавляется произвольно.
Б. Реакции на прогрессивные движения
Можно обнаружить следующие вызванные с лабиринтов стато-кинетические реакции на линейное ускорение.
1. На голову
Эти реакции лучше всего поддаются исследованию у собаки при вертикальных движениях вверх и вниз.
2. Влияние прогрессивных движений на конечности
Для наглядности здесь названы только две из этих реакций.
а) Лифтная реакция. Животное сажают на доску. Если теперь производить вертикальное движение вверх, то сначала передние ноги сгибаются, а в конце движения они, наоборот, разгибаются. При вертикальном движении вниз разгибание следует сначала, а сгибание конечностей в конце движения,
б) Готовность к прыжку. Если держать морскую свинку в висячем положении головой вниз и производить незначительное верти¬
Стато-кинетические рефлексы. Реакции на движения частей тела
23
кальное движение также вниз, то передние конечности двигаются по направлению рта и при этом животное принимает положение, которое способствует при падении на пол принятию веса всего тела. Для того чтобы вызвать эту реакцию, достаточно совсем незначительное вертикальное движение.
Все описанные реакции на прогрессивное движение отсутствуют после удаления обоих лабиринтов.
Влияния небольших прогрессивных движений на глаза описаны Флай- шем (Fleisch).
В. Реакции на движения отдельных частей тела
Описанные до сих пор стато-кинетические рефлексы вызваны положительными или отрицательными ускорениями головы, которые приводят к возбуждению лабиринтов. Но и при движениях других частей тела возникают реакции, которые, поскольку они вызывают или поддерживают положение тела, можно причислить к стато-кинетическим. Они, естественно, вызываются не с лабиринтов, а в первую очередь с самих двигающихся частей тела.
Достаточен один пример для того, чтобы уточнить, о чем здесь идет речь.
Если у собаки, стоящей на четырех ногах, ущипнуть палец задней ноги, то она притягивается к телу сгибательным рефлексом той же стороны. Животное стоит только на трех ногах, и задняя половина тела, которая до этого поддерживалась двумя задними ногами, теперь держится только на одной ноге. Это осуществляется тем, что одновременно увеличивается тонус в мышцах — разгибателях этой задней ноги. Здесь появляется вместе со сгибательным рефлексом (скажем, левой) ноги перекрестный раз- гибательный рефлекс правой задней ноги [Goltz und Freusberg, Gergens, Philippson, Sherrington (9)]. Одновременно из-за усиленной нагрузки правой задней ноги вызываются мышечные рефлексы (проприоцептивные рефлексы), которые ведут к усилению напряжения мышц — разгибателей этой ноги [Шеррингтон (8)]. Этим достигается то, что задняя половина тела не опускается и что правая задняя нога в состоянии сразу же нести увеличенную нагрузку и препятствовать падению животного.
Эта реакция вызывается и управляется движением (сгибанием) левой задней ноги и может поэтому считаться стато-кинетической реакцией. Но она сохраняется до тех пор, пока левая нога остается согнутой, и поэтому к стато-кинетической реакции присоединяется соответствующая статическая реакция.
Такие длительные влияния наблюдаются у спинальной собаки. Иногда, например, можно наблюдать, что у собаки с перерезанным спинным мозгом, которая вначале находилась в прекрасном состоянии и у которой имелись всевозможные сложные рефлексы задней половины тела, развивается язва на одной из задних ног (например, левой). Тогда больная нога притягивается к телу, т. е. находится в длительном сгибательном состоянии, в то время как здоровая (правая) задняя нога удерживается в состоянии разгибания (Шеррингтон). Итак, уже у спинальной собаки из-за такой язвы имеется реакция, которую можно видеть и у интактной собаки с язвой. Что здесь речь идет не только об анатомически вызванных сокращениях, вытекает из того, что вышеописанный вынужденные положения сейчас же прекращаются, если язву анестезировать кокаином или новокаином. Тогда сразу могут снова возникнуть нормальные движения ходьбы
24
Вторая глава. Переключение
и другие симметричные реакции на обеих конечностях (собственные наблюдения).
Посредством таких длительных влияний у неоперированного животного, находящегося в положении стоя с согнутой левой ногой, правая нога получает необходимое для сохранения положения тела увеличение тонуса.
Описанный пример достаточен, чтобы пояснить, что при движениях и при определенных положениях отдельных конечностей появляются реакции отчасти стато-кинетического, отчасти статического характера, имеющие свои центры в соответствующих сегментах спинного мозга, которые также способствуют сохранению положения тела.
Известно, что жизненно необходимые функции обычно обеспечиваются разнообразно. Так, для переваривания белка имеются различные ферменты в желудочном, в панкреатическом и кишечном соках, способные взаимно заменять друг друга; переваривание углеводов обеспечивается слюнюй и панкреатическим соком. И в центральной нервной системе известны многочисленные подобные примеры. Следует только напомнить о ритмических движениях ходьбы, вызванных посредством взаимодействия центрального [Graham-Brown (3)] и периферического [Sherrington (9)1 возбуждения. Функция положения тела принадлежит к наиболее застрахованным, четким функциям центральной нервной системы. Мы едва ли знаем другой пример взаимной деятельности такого большого количества разнородных рефлексов для осуществления единой конечной цели. Статокинетические и статические реакции, как это особенно четко удалось установить на примере глаз, взаимно дополняют друг друга: благодаря кинетическим реакциям выполняются движения, приводящие отдельные части тела в такое положение, в котором они потом удерживаются статическими рефлексами. Рефлексами стояния и установочными рефлексами осуществляется принятие основного положения, а также принятие и сохранение определенного положения. Важно то, что при этих реакциях голова, в которой расположены телерецепторы — глаза, уши и орган обоняния, — играет такую ведущую (хотя и не исключительную) роль. Таким образом, осуществляется то, что уже при дистантных раздражениях тело может принять соответствующее положение, которое часто является и защитным положением.
В этом кратком обзоре мы узнали о самых главных рефлексах, участвующих в создании положения тела; в дальнейшем будет проведен анализ возникновения каждой отдельной группы рефлексов.
Вторая глава ПЕРЕКЛЮЧЕНИЕ
Прежде чем перейти к подробному описанию различных рефлекторных групп и специальному анализу отдельных рефлексов, надо обсудить еще одну проблему общего значения, а именно вопрос о том, каким образом определенное положение тела при определенном распределении напряжения во всей мускулатуре влияет на различные виды реакций тела животного при разных раздражениях и при различных рефлексах. Уже повседневный опыт учит, что одно и то же раздражение при различных позах и положениях тела вовсе не всегда вызывает одну и ту же реакцию. Сама положение тела, следовательно, обусловливает смену реакций на одинаковые раздражения.
Вторая глава. П ер включение
25
При обсуждении этих отношений нецелесообразно исходить из постановки вопроса — почему рефлекс, в общем протекающий закономерно и одинаково при определенном положении тела, вдруг протекает иначе, а сперва надо задать вопрос, чем же обусловлено, что вообще локальное раздражение вызывает в определенном смысле локальную реакцию.
Если отравить животное стрихнином с целью повышения рефлекторной возбудимости, то можно привести в состояние возбуждения всю мускулатуру тела раздражением любого чувствительного нерва или нервного окончания. Значит, в этих условиях каждый афферентный чувствительный путь связывается функционально с каждым эфферентным моторным нервом и центральная нервная система представлена как диффузная сеть, в которой приходящие возбуждения могут передаваться на все моторные центры. Исходя из этого факта, возникает вопрос, чем же обусловлено, что при нормальных условиях возбуждение распространяется в центральной нервной системе высших животных не диффузно, ибо при обычных рефлексах приводятся в деятельное состояние только отдельные нервные образования и ограниченное количество мышц тела.
О законах, которые при этом действуют, мы знаем по более старым исследованиям Гольца и особенно по новым исследованиям Шеррингтона. Задачей этой книги не является подробное описание этих закономерностей. Шеррингтон сделал это в своей монографии «The integrative action of the nervous system». Там показывается, что процессы торможения играют большую роль в ограничении распространения возбуждения по центральной нервной системе. При возбуждении определенных центральных групп благодаря «одновременной индукции» уменьшается возбудимость соседних центральных групп, а после завершения рефлекса последовательная индукция оказывает противоположное влияние на возбудимость. Из «принципа общего пути» следует, что один и тот же моторный путь может быть одновременно использован только одним типом рефлекса. Вследствие этого на одной конечности одновременно может проявляться или чесательный, или длительный сгибательный, или же перекрестный разгибательный рефлекс. Как только «общий путь» занимается определенным рефлексом, он тотчас блокируется для других, частично антагонистических рефлексов. Большое значение имеет и рефрактерный период, во время которого центр, пришедший в состояние возбуждения, становится невозбудимым и может отвечать на новые раздражения только после уменьшения процесса возбуждения. Как раз для ритмических движений особенно важен рефрактерный период. Все эти и еще многие другие процессы регулируют закономерное распространение возбуждения в центральной нервной системе, препятствуют тому, чтобы при отдельном раздражении в нормальных условиях приходили в возбуждение все моторные центры, и делают возможным использование центральной нервной системы для самых разнообразных реакций.
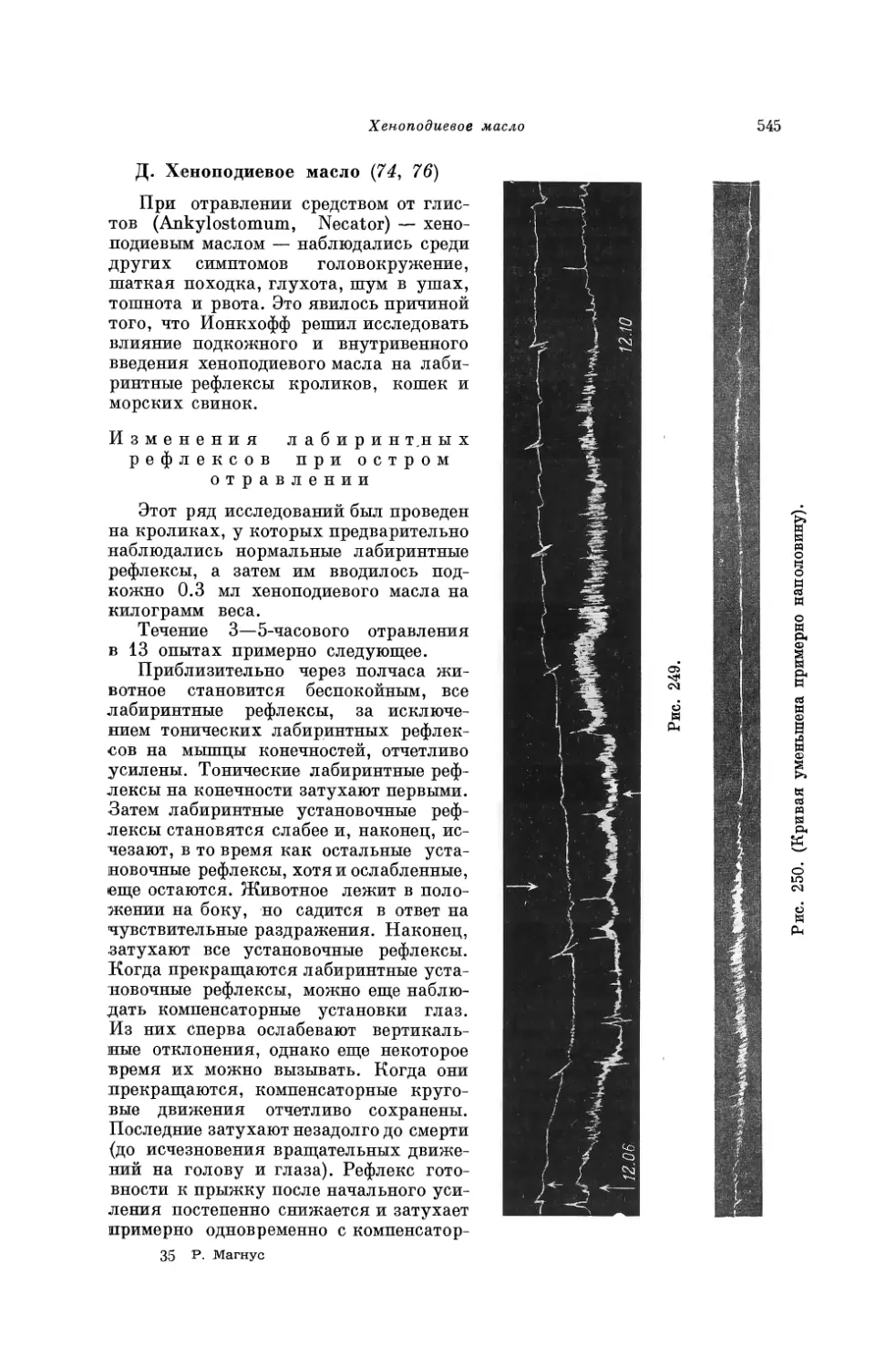
Из множества закономерностей подробно здесь описывается только один исключительный случай, имеющий значение для положения тела. Оказалось, что и состояние периферии тела влияет на распределение возбудимости в центральной нервной системе, другими словами, что данной положение и поза тела оказывают закономерное влияние на реакции центрального органа. Наши исследования в этой области можно связать с опытами, которые провели Икскюль (Uexküll) и Иордан (Jordan) на беспозвоночных. Оказалось, что у целого ряда беспозвоночных с более простым строением тела возбуждение распространяется в диффузной нервной сети по относительно простым законам и что возбудимость центров, находится под влиянием растяжения соответствующих мышц^ Центры растя¬
26
Вторая глава. Переключение
нутых мышц, по выражению Икскюля, «включены» для данного раздражения, и поэтому любое чувствительное раздражение приводит у этих животных в состояние сокращения только растянутые мышцы.
Для доказательства Икскюль (1) описывает следующий опыт на офиурах (ophiuroidea):
Офиура принадлежит к морским звездам. От центральной части тела, в которой находятся органы пищеварения, отходят пять длинных, круглых, очень подвижных лучей. Луч состоит в основном из многих костистых позвонков, которые, подобно столбику монет, сидят друг на друге. Они соприкасаются друг с другом своими центральными частями и связаны мышцами (рис. 2).
Основные движения — боковые, поэтому каждое отдельное сочленение позвонка можно рассматривать как двуплечий рычаг. Эти многочисленные рычаги взаимодействуют так, что вследствие сближения плеч рычагов на одной стороне плечи на другой
расходятся. Следовательно, при каждом боковом сгибании луча сближаются плечи рычагов вогнутой стороны, а на выпуклой стороне расходятся; это влечет за собой растяжение мышц на выпуклой стороне луча и сокращение мышц на вогнутой.
Нервная сеть не распространяется по всему лучу, а образует обособленный радиальный нерв, связывающий между собой все позвонки на нижней стороне луча.
Эти пять радиальных нервов входят в нервное кольцо, которое находится в центральной части тела и окружает оральное отверстие.
Основной опыт производится следующим образом. У офиуры удаляют все лучи, за исключением одного. Затем обнажают нервное кольцо и перерезают его на противоположной оставшемуся лучу стороне. Таким образом получают осевой нерв, который держится справа и слева, так сказать, двумя «вожжами». Раздражение нервного кольца индукционным током всегда вызывает при нормальном положении луча (т. е. когда луч висит перпендикулярно вниз) его сокращение в сторону раздражения. Таким путем мы узнаем, что все мышцы одной и той же стороны луча имеют между собой нервную связь.
Теперь центральная часть тела офиуры прикрепляется на штативе так, что луч сгибается под собственной тяжестью и свисает (рис. 3). Мышцы позвонков теперь растянуты на выпуклой стороне, а на нижней вогнутой стороне укорочены.
Раздражение на такой луч воздействует совершенно по-другому. Направление, из которого идет возбуждение, не является больше единственным компонентом, влияющим на сокращение мышц. При раздражении — безразлично какой половины нервного кольца (Rj или Яц) — теперь преимущественно отвечают растянутые мышцы, и луч сгибается вверх. Следовательно, растяжение мышц повлияло на исход возбуждения, направляя его к растянутым мышцам.
Таким образом, основной закон распространения возбуждения гласит: возбуждение в нервной сети всегда движется по направлению к растянутым мышцам. Для офиуры этого закона достаточно, чтобы вызвать у нее ритмические движения, возможность передвижения всей офиуры и уход от определенного раздражения.
Этот опыт, как сообщает Икскюль (2), удается не во всех 100%, так как результат раздражения обусловлен противодействием двух компонентов:
1) большей готовностью мускулатуры к сокращению на раздраженной стороне и 2) большей готовностью мускулатуры к сокращению на растянутой стороне.
В качестве следующего примера Икскюль приводит поведение иглы морского ежа. Морские ежи состоят из шаровидного известкового панциря, в котором заключены внутренности. Поверхность панциря покрыта многочисленными бугорками, служащими для прикрепления игл. Основание каждой иглы имеет суставную ямку, и иглы могут вращаться в разных направлениях. Сочленение иглы охвачено суставной сумкой, состоящей из мышечных волокон. В эпидермисе, близко над мышцами, лежит кольцо ганглионарных клеток, посылающих к нижележащим мышцам короткие центробежные нервы. Кольцо ганглионарных клеток связано с многочисленными нервными «сетями, распространяющимися по всей поверхности морского ежа. Более быстрая
Вторая глава. Переключение
27
связь обеспечивается особыми путями — радиальными нервами, находящимися внутри панциря и связывающими нервные сети разных сторон друг с другом.
Для эксперимента лучше всего пользоваться маленькой стеклянной трубкой, которая вдавливается между иглами, придерживающими ее.
Если теперь раздражать в любом месте •эпидермис, лучше всего легким похлопыванием, то стеклянная трубка начнет самым кратким путем передвигаться к месту раздражения. При этом замечательно, что в движение приходят только те иглы,которые непосредственно соприкасаются с трубкой, все остальные остаются неподвижными.
Из других опытов мы знаем, что нервные сети расположены так, что мышцы игл, лежащие в одном направлении, всегда связаны с одной и той же нервной сетью. Поэтому мы можем схематически себе представить условия, при которых происходит передвижение трубки.
На рис. 4 представлены три иглы морского ежа (И, В ж С). Нервная сеть связывает левые стороны игл друг с другом. Допустим, эта нервная сеть приводится в возбуждение (от радиальных нервов). Пусть возбуждение исходит из места раздражения (R), лежащего по ходу нервной сети влево. Стеклянная трубка лежит между В ж С. Возбуждение оставляет А жВ полностью в покое, действие его сказывается только на игле С. Растянутые давлением стеклянной трубки мышцы С сокращаются, и игда С придвигает теперь трубку к В. Игла В поддается, как это и следовало ожидать, слабому давлению, сгибается в сторону А ж создает таким образом возможность стеклянной трубке двигаться дальше. Стеклянная трубка, продвигаемая иглой С, попадает через В к игле А . Теперь поднимается В ж находится в таком положении, в каком находилась игла С в начале опыта. Игла В со своей стороны придавливает теперь трубку к игле А, и повторяется то же самое, пока стеклянная трубка не попадает наконец к месту раздражения. При этом в передвижении
принимают участие только те иглы, ** мышцы которых растянуты и возбуж¬
дены. Возбуждение снова распространяется по растянутым" мышцам, и опять только растянутые мышцы «включены» для возбуждения и придают возбуждению направление, по которому оно должно распространяться.
Если морской еж находится на земле и ему встретится где-нибудь раздражитель, то он будет как бы передвигать землю к месту раздражения, как перед тем стеклянную трубку, т. е. он сам будет, выполняя ритмические движения иглами, удаляться от раздражения.
Рис. 4. (По Икскюлю). Оба эти опыта показывают,
в чем сущность закона растяжения Икскюля и каким образом растяжение мышц и положение соответствующих частей тела определяют направление возбуждения в диффузных нервных системах.
Икскюль и Иордан считают, что у морских ежей и морских звезд отсутствуют афферентные чувствительные нервы мышц и что описанные реакции разыгрываются в простой системе: центр, нерв, мышцы. Что у таких животных растяжение мускулатуры действительно действует на центр, яснее
28
Вторая глава. • Переключение
р
#4
V
Рис. 5.
всего видно из так называемого опыта Иордана на половинном животном! (Halbtierversuch). Если разделить пополам в длину улитку (Helix) так,, чтобы животное состояло из правой и левой половин тела, связанных, между собой только педальным ганглием (рис. 5, Р), и растягивать небольшим весом правую половину тела (К) в длину, а левую половину- (L) соединить с регистрирующим аппаратом, то можно увидеть, что при растяжении R L расслабляется. Одновременно происходит изменение рефлекторной возбудимости L. Изменение длины и рефлекторной возбудимости L доказывает, что в соответствующем центре Р произошел сдвиг, который- мог быть вызван растяжением i?. Так как обе половины тела связаны между собой только центром Р, то растяжение мышц у этих животных действительно влияет на состояние центра.
Если бы удалось доказать такие же закономерности,, как у беспозвоночных, и в центральной нервной системе высших позвоночных, то это имело бы решающее значение для выяснения влияния положения тела на реакции животных. Действительно, и у млекопитающих удалось показать соответствующие отношения (1).. В доказательство этого приводим некоторые примеры.
Если собаку с перерезанным в нижней грудной области спинным мозгом и полностью поправившуюся от операционного шока положить на спину и ударить каким-нибудь предметом — например, тыльной стороной карманного ножа — по сухожилию колена, то, кроме коленного рефлекса раздражаемой стороны, будет двигаться и другая, перекрестная нога; такие- движения обычно называют перекрестными коленными рефлексами. Этот рефлекс обычно вызывает разгибание колена, а иногда и бедра. Выяснилось, что движения перекрестной ноги в этом рефлексе совершенно' закономерно зависят от ее исходного положения. Если нога согнута в бедре, колене и голеностопном суставе, то происходит разгибание. На рис. 6 представлен ряд кинематографических снимков, которые были полученыг на собаке через 7 недель после операции. На первом снимке собака лежит на спине на коленях служителя. Левая рука экспериментатора держит правую (на снимке впереди) заднюю ногу в согнутом во всех трех суставах положении. Правая рука поднята для удара по левому коленному сухожилию (на снимке сзади). Удар происходит на втором снимке; на третьем снимке видно уже начало реакции, разгибание бедра. Четвертая, пятая и шестая картины показывают дальнейшее осуществление полного разгибания. В этом опыте правая нога, разумеется, слегка фиксируется рукой в исходном положении, так что наступающее сильное рефлекторное движение легко может преодолеть сопротивление. Совершенно противоположный результат получается, если нога находилась вначале в разогнутом положении. Серия снимков на рис. 7 показывает, что теперь происходит сгибание ноги: после раздражения (удар по левому колену) правая нога сгибается в бедре, колене и голеностопном суставе.
Как видно, при одном и том же раздражении проявляются совершенно- разные реакции в зависимости от того положения, которое имела до этого- конечность («извращение рефлекса» — «Reflexumkehr»). Исследования Шеррингтона (9) наглядно показали движения мышц при сгибании и разгибании ног. При сгибании происходит сокращение системы «сгибателей», и в это время антагонисты — «разгибатели» — расслабляются (реципрок- ная иннервация). При разгибании, наоборот, происходит сокращение «разгибателей» и торможение «сгибателей». Если, следовательно, одно и
Прав ая нога согнута в бедре, колене и ступне.
Правая нога разогнута в бедре, колене и ступне.
Удар по левому коленному сухожилию.
Удар по левому коленному сухожилию.
Начинающееся разгибание правого бедра.
Разгибание правого бедра, колена и ноги.
f Сгибание правого бедра, колена и ступни.
Рис. 6. Собака «А» (оперирована 6 мая 1909 г. Перерезка спинного мозга на уровне 12-го грудного позвонка). Опыт 23 июня 1909 г.: разгибание согнутой правой ноги при ударе по левому коленному сухожилию.
Рис. 7. Собака «А». Опыт от 19 июня 1909 г. Сгибание разогнутой правой ноги при ударе по левому коленному сухожилию.
30
Вторая глава. Переключение
то же раздражение вызывает один раз сгибание, другой раз — разгибание,, то в нервных центрах спинного мозга должно происходить каждый раз
переключение, вызванное изменением положения конечности. На рис. 8 приведена схема, на которой изображены как целое центры разгибателей и сгибателей. Для успеха опыта не является существенным, что реагирую-
Рис. 9. Собака Спот (оперирована 3 октября 1904 г. Перерезка спинного мозга на уровне 10—11-го грудного сегмента). Опыт 26 марта 1908 г. Регистрация движений левой ноги. При разгибании рычажок поднимается вверх, при сгибании— вниз. Раздражение — удар по правому коленному сухожилию. Кривую следует читать справа налево. Время дано в секундах. Нога в согнутом положении (XX). а—g — семь следующих друг за другом разгибательных рефлексов, причем нога постепенно все больше переходит в положение разгибания.
щая нога придерживается искусственно в определенном положении. Если животное случайно держит свою ногу спокойно в разогнутом или согнутом положении, то получаются те же самые результаты.
На рис. 9 и 10 изображены записи, которые я снял на оперированных Шеррингтоном собаках в его лаборатории в Ливерпуле. Лапа животного
Вторая глава. Переключение
Ъ\
была соединена нитью с рычагом для регистрации. При разгибании ноги рычаг поднимался вверх, при сгибании опускался вниз. На рис. 9 изображены семь последовательных разгибательных рефлексов согнутой конечности, на рис. 10 — восемь последовательных сгибательных рефлексов разогнутой конечности. Раздражением в каждом случае был удар по правому коленному сухожилию.
Из рис. 9 видно, что при каждом отдельном рефлексе нога сразу же из положения покоя (сгибания) производит сильное разгибание. Из рис. 10, видно, что нога из положения покоя (разгибания) производит сразу же. сильное сгибание.
Рис. 10. Собака Спот. Опыт 26 марта 1908 г. Регистрация движений левой ноги. При разгибании рычажок поднимается вверх, при сгибании — вниз. Раздражение — удар по правому коленному сухожилию. Кривую следует читать справа налево. Время дано в секундах. Нога в положении разгибания (X). а—h — восемь следующих друг за другом сгибательных рефлексов.
Стало быть, описанное чередование рефлексов не обусловливается тем, что в случае максимального разгибания ноги дополнительно присоеди-- няется еще незначительное, для простого глаза незаметное разгибание, за которым вторично следует сгибание. То же самое можно получить и при измерении латентных периодов. Латентный период рефлекса при разогну- той ноге, наверно, не короче, чем в том случае, когда исходным положением ноги является максимальное сгибание или среднее положение при сгибании.
Так как мускулатура задней ноги собаки довольно сложна, то очень важно, что можно вызвать описанное влияние на направление рефлекса и в том случае, когда изменяется положение ноги только в бедре, при разогнутом коленей голеностопном суставе. Если разогнутая нога максимально, сгибается в бедре, то происходит разгибание бедра; если нога разгибается в бедре, то происходит сгибание. При аддукции ноги в тазобедренном суставе происходит абдукция, при абдукции ноги — аддукция. Следовав тельно, при разных положениях тазобедренного сустава «включаются» или «выключаются» центры четырех разных мышечных групп для одного и того же афферентного импульса. Эта последовательность четырех разных реакций в тазобедренном суставе очень удобна для демонстрации вышеописанных закономерностей, так как она несомненно является самым простым, и самым наглядным примером. -
32
Вторая глава. Переключение
Если при исходном положении ноги по-разному комбинировать сгибание и разгибание в трех больших суставах, то оказывается, что самое большое влияние на результирующее движение имеет положение бедра по отношению к туловищу (к тазобедренному суставу) и что положение коленного сустава имеет уже второстепенное значение, а положение голеностопного сустава вообще не оказывает большого влияния.
Значительное чередование рефлекторных движений вызывается в большинстве случаев только изменением положения тазобедренного сустава.
Для других перекрестных рефлексов можно доказать то же самое чередование. Так, например, это имеет место при перекрестном разгибательном
Рис. 11а. Рис. 116.
Рис. И. Диаграмма центрального переключения при вызове левостороннего сгибательного рефлекса. Рис. 11а — правая нога согнута. Рис.
116 — правая нога разогнута, а — афферентный путь с левой лапы;
В — центры сгибательных мышц задних ног с соответствующими эфферентными путями; S — центры разгибательных мышц с соответствующими эфферентными путями. + обозначает, что идущее по афферентному пути раздражение возбуждающим образом влияет на соответствующий центр, —
тормозящим образом.
рефлексе, при перекрестном чесательном рефлексе и при проприоцептивных рефлексах мышц. На рис. 11 изображена схема перекрестного (разги- бательного) рефлекса при различном положении ноги.
Если при перекрестном коленном рефлексе результат раздражения неоспоримо определен исходным положением ноги, то центральная связь между односторонним сгибательным и перекрестным разгибательным рефлексами более прочна, так что этот последний не во всех случаях поддается чередованию при измененном исходном положении ноги-исполнителя (ноги-эффектора); однако переключение ясно можно видеть в большинстве случаев на хорошо возбудимых животных. Это последнее наблюдение показывает, что причину измененной реакции ноги-эффектора при разных исходных положениях нельзя искать в анатомически преформированной связи, а что здесь речь идет о функциональных изменениях в центральном органе. То же самое вытекает из того факта, что описанное извращение рефлекса можно наблюдать только у совершенно здоровых животных. Маленькие изъязвления на конечностях уже достаточны, чтобы подавить этот феномен. Необходимо также, чтобы животное оправилось от шока, наступившего после пересечения спинного мозга. У взрослых животных на эту поправку часто требуется 4 недели, хотя уже через несколько дней после операции можно вызвать перекрестные рефлексы. Однако в течение первых недель в большинстве случаев не удается установить даже малейшего признака такого рода направления рефлекса, вызванного исходным положением конечности.
Вторая глава. Переключение
33
Шеррингтон (2) уже раньше наблюдал влияние начального положения на исход раздражения при неперекрестных рефлексах, однако доказать это здесь гораздо труднее, и в общем невозможно наблюдать такую регулярность, как при перекрестных рефлексах.
Описанный феномен основывается не на механических свойствах самих конечностей, а на измененном состоянии центральной нервной системы (спинного мозга). Изменение положения и установки конечностей вызывает переключение моторных центров отдельных мышц и мышечных групп, возбудимость которых изменяется так же, как и значение (смысл реакции: возбуждение или торможение). Регуляция эта чрезвычайно тонка. Мы узнаем при этом, что спинной мозг в каждый момент отражает положение и установку разных частей тела и всего тела. Каждому положению тела соответствует определенное распределение возбудимости и наиболее легко доступных путей в центральной нервной системе. Тело само настраивает правильным образом свой центральный орган.
Хотя на ноге собаки с ее сложно расположенной мускулатурой ц можно демонстрировать переключение, все-таки желательно показать все это на анатомически более простых объектах. Таким объектом является хвост кошки (2). Он представляет собой всесторонне подвижный и расчлененный стержень, отдельные части которого — позвонки хвоста — связаны между собой мышцами, выполняющими функции разгибателя (экстензора или леватора), сгибателя (флексора или депрессора) и абдуктора. Первые позвонки хвоста и благодаря этому корень хвоста связаны мышцами этих четырех групп с туловищем. Местом прикрепления их является таз, крестец, бедра и поясничная часть позвоночного столба. Корень хвоста у кошки снаружи не виден. Пятый позвонок хвоста находится в вертикальном положении над анусом, и только шестой позвонок лежит в свободной, снаружи поднимающейся подвижной части хвоста животного. Двигательные центры хвоста находятся в крестцовых и в непосредственно прилегающих к ним поясничных и копчиковых сегментах спинного мозга.
Чтобы получить хорошие рефлексы хвоста, кошку сперва децеребрируют при глубоком эфирном или хлороформном наркозе (перерезка мозгового ствола на уровне tentorium cerebelli). После развития децеребрационной ригидности, когда становится возможно наблюдать на конечностях односторонние и перекрестные рефлексы, производится перерезка спинного мозга на уровне 12-го грудного позвонка (уже без наркоза). Шеррингтон показал, что если производить эту операцию на децеребрированных животных, то шок не появляется. Сразу же после перерезки можно доказать наличие рефлексов задних ног и хвоста, которые обычно в первые часы после перерезки еще усиливаются. Этот препарат очень удобен для проведения опытов.
Если держать кошку в висячем положении головой вверх свободно в воздухе так, что хвост перпендикулярно висит вниз, — то при прикосновении к кончику хвоста вызываются рефлекторные движения хвоста в любую сторону. При этом не имеет значения, раздражается ли конец хвоста слева или справа, спереди или сзади. Направление полученного при таком положении рефлекса совершенно неопределенно.
Эта картина сразу изменяется, как только животное помещают в боковое положение и хвост свисает вниз через край стола, так что основание хвоста сильно изогнуто в одну сторону и сам хвост висит почти перпендикулярно вниз. Теперь прикосновение к кончику хвоста вызывает регулярные движения хвоста вверх. На рис. 12 изображены два ряда кинематограмм, которые были получены непосредственно одна за другой 3 Р. Магнус
34
Вторая глава. Переключение
на том же животном в правом (а) и левом боковом положении (б) и наглядно поясняют этот процесс.
В обоих случаях видно, как движение начинается с основания хвоста, т. е. на самом растянутом сегменте, и распространяется затем таким образом, что в каждый следующий момент двигается вверх та часть хвоста,
Раздражение кончика хвоста боковым щипком.
Раздражение кончика хвоста при дорсо- вентральном щипке.
Начинающаяся реакция: поднятие основания хвоста вследствие сокращения левосторонней отводящей мышцы. Сильнейший изгиб в проксимальной трети хвоста.
Начинающаяся реакция: поднятие ос¬
нования хвоста вследствие сокращения правосторонней приводящей мышцы. Сильнейший изгиб одной трети хвоста.
Основание хвоста поднимается дальше, проксимальная треть хвоста поднята (значит, двигается влево). Сильнейший изгиб в середине хвоста.
Основание хвоста поднимается дальше, середина хвоста также поднята. Изгиб кончика хвоста.
Хвост разогнут и in toto направлен влево, теперь также произошло сгибание середины хвоста.
Хвост разогнут и in toto направлен вправо.
Рис, 12а, Рис. 126.
Рис. 12. Децеребрированная кошка. Перерезка спинного мозга у 12-го грудного позвонка. Через 2 часа произведен опыт. Кинематографические снимки. (На том же животном была получена хронофотограмма, рис. 14). Рис. 12а — правое боковое положение, хвост висит через край стола вниз, так что левая сторона основания хвоста растянута. Рис. 126 — левое боковое положение, хвост висит через край стола вниз, так что растянута правая сторона основания хвоста.
которая в предыдущий момент была сильнее всего изогнута. Движение всегда направлено в сторону наибольшего изгиба. У сильно возбудимых животных хвост часто поднимается даже выше горизонтальной линии. При этом совершенно безразлично, прикасаются ли пальцами к волоскам кончика хвоста или же щиплют кончик хвоста в дорсо-вентральном направлении или в боковом, — всегда вызывается движение основания хвоста вверх., Эту же закономерность удается доказать, если перед тем пере¬
Вторая глава. Переключение
35
резать депрессоры хвоста, которые при боковом положении животного также должны отклонять хвост от среднего положения. Если при боковом положении животного пальцем поднимать хвост вверх, как это показано на рис. 13, то у основания хвоста будут растянуты правые абдукторы, а в середине хвоста — левые. Вследствие этого исход раздражения на прикосновение к кончику хвоста теперь определен не ясно — или основание хвоста может опускаться, или же дистальная половина хвоста поднимется. Если положить хвост по всей его длине на доску и поднимать его таким образом выше горизонтальной плоскости, оставляя при этом основание хвоста свободным, то на прикосновение к кончику хвоста последует сильное движение основания вверх (поворот вправо). Этот последний опыт показывает, что описанное переключение вызывается не боковым положением животного, т. е. асимметричным раздражением чувствительных нервов тела вследствие давления на подставку, а измененным положением хвоста.
То же самое можно доказать следующим опытом: животное сажают в нормальном положении на стол и опускают хвост через край стола вниз. Задние ноги расположены симметрично в согнутом положении. Теперь поворачивают переднюю часть тела в боковое положение, так что животное лежит одним плечом на столе. Так как спинной мозг перерезан на уровне 12-го грудного позвонка, то отсутствуют все нервные связи передней части тела, приведенной в боковое положение, с двигательными центрами хвоста. Однако поворотом передней части тела вызывается асимметричное растяжение абдукторов обеих сторон хвоста, и этого достаточно, чтобы определилось направление рефлекса хвоста. Хвост двигается всегда в ту сторону, в которую направлены передние ноги животного. На рис. 14 изображено правое и левое боковое положение передней части тела животного. У этой кошки на спине от поясничной части позвоночного столба до основания хвоста была чернилами намечена черная линия. При помощи этой линии можно было увидеть, что абдукторы основания хвоста на стороне, обращенной к передним ногам, находились в состоянии растяжения. В эту сторону закономерно и вызывалась реакция хвоста. Этот опыт показывает, что описанное переключение обусловливается только асимметричным растяжением мускулатуры хвоста.
Если держать животное вертикально головой вниз, то при вертикальном перегибе хвоста происходит рефлекторное его поднятие, если же хвост перевешивается в дорсальную сторону, то, напротив, наблюдаются движения хвоста в вентральном направлении. Следовательно, исходящее с кончика хвоста раздражение можно направлять, в зависимости от исходного положения хвоста, с четырех различных мышечных групп в центры. В тех случаях, когда в ответ на раздражение кончика хвоста происходит только движение хвоста, а задние ноги почти не участвуют в этом рефлексе, направление движения хвоста совершенно ясно обусловлено описанным переключением и имеет место в 100% всех опытов. Хвост кошки, таким образом, является более наглядным объектом для демонстрации этих явлений, чем луч офиуры, у которого имеет значение положение луча и сторона, на которую наносится раздражение. Это последнее влияние отпадает при раздражении хвоста кошки, и реакция зависит только от исходного положения органа.
Опыты с раздражением хвоста кошки приводят к таким же выводам, как и уже описанные опыты на ноге собаки. Если раздражается кончик
я*
36
Вторая глава. Переключение
хвоста, то возбуждение, идущее по чувствительным нервам в спинной мозг, может избирать разные пути, как это показали опыты с симметричным положением хвоста. Но как только хвост загибают в сторону, в центральной нервной системе происходит переключение, которое заставляет возбу-
Раздражение кончика хвоста легким прикосновением к шерсти на самом кончике.
Раздражение кончика хвоста боковым щипком.
Начинающаяся реакция: сгиба¬
ние основания хвоста влево.
Продолжающееся смещение хвоста у влево, причем движение идет от основания к кончику хвоста.
Реакция: хвост дви- ' гается влево.
i
Рис. 14а. Рис. 146.
Рис. 14. Та же кошка, что и на рис. 12. 2 часа после перерезки спинного мозга. Кинематографические снимки. Рис. 14а — хвост висит через край стола вниз, задняя часть тела животного лежит на обеих задних лапах, животом на столе, передняя часть тела повернута так, что правая лопатка — на столе, передние лапы направлены влево; вследствие этого происходит растяжение левых отводящих мышц хвоста. Рис. 146 — то же положение, что и на рис. 14а, только передняя часть тела повернута вправо и лежит левой лопаткой на столе, вследствие чего происходит растяжение правых отводящих
мышц хвоста.
ждение избирать из возможных путей только один, а именно путь к центрам таких мышц, которые сильнее всего растянуты. Можно совершенно произвольно направлять раздражения в разные центры мышц хвоста. Это влияние длится столько времени, сколько животное находится в соответствующем положении. Следовательно, переключение является тоническим изменением состояния в центральной нервной системе, обусловленным положением тела животного.
Вторая глава. Переключение
37
Спрашивается теперь, каким путем влияет изменение положения и установка определенной части тела на распределение возбудимости в центральной нервной системе, другими словами — как «узнает» центральная нервная система, что на периферии произошло изменение (3). Как уже выше было отмечено, Икскюль и Иордан считают, что у беспозвоночных переключение осуществляется без посредничества чувствительных нервов и разыгрывается в системе центр—нерв—мышца. У млекопитающих же можно было предположить, что переключение осуществляется посредством чувствительных нервов, т. е. является по происхождению рефлекторным. Чтобы это доказать для ног собаки, у интактных собак были с одной стороны интрадурально перерезаны люмбальные и сакральные задние корешки. Затем после выздоровления животного на деафферентированной ноге восстанавливалась хорошая двигательная активность (участие этой ноги в разнообразных рефлексах,и движениях). У мышц этой ноги развивался также определенный тонус. Точная проверка показала,, что вся задняя нога соответствующей стороны, а также соответствующая половина хвоста, промежности, спины и живота были лишены чувствительности.
У одной из этих собак, согласно Винклеру 1 (Winkler), самыми верхними перерезанными корешками были второй люмбальный и, вероятно, первый люмбальный корешки, а у другой собаки — первый люмбальный и, вероятно, 13-й торакальный корешки. Во всей деафферентированной области нельзя было найти островки с сохраненной чувствительностью, так что у этих животных вся задняя нога и прилегающие к ней части туловища действительно были денервированы. Так как, по опытам Шеррингтона (12), первый люмбальный корешок не принимает участия в двигательной иннервации ноги и, по тому же автору, двигательные и чувствительные нервы мышц выходят через передние и задние корешки того же сегмента, то получается, что операцией была денервирована и вся мускулатура ноги. После перерезки задних корешков была произведена перерезка спинного мозга на уровне 11-го или 13-го грудных позвонков. Затем выждали, пока пройдут шоковые явления, восстановится способность производить обеими задними ногами движения и станет возможно вызывать с чувствительных мест тела все сгибательные и разгибательные рефлексы на деафферентированной ноге. На обеих собаках в течение 4—5 месяцев было произведено несколько сотен отдельных опытов, таких же, какие были описаны выше на спинномозговых собаках. Результат был такой, что ни в одном случае нельзя было доказать какое-либо влияние исходного положения на направление рефлекторного движения этой денервированной ноги. Все феномены переключения окончательно исчезли.
То же самое можно было показать на хвосте кошки. Для этого у не- 'скольких интактных кошек с обеих сторон интрадурально перерезали задние корешки, начиная с 5-го люмбального сегмента вниз, причем по возможности сохранялись самые последние копчиковые нити корешков. Так как Шеррингтон (12) при известных условиях наблюдал у кошки еще при раздражении 7-го поясничного корешка (верхняя граница) движения основания хвоста в сторону раздражения, можно с точностью принять, что вследствие этой операции денервируется вся мускулатура основания хвоста. После заживления раны и полного восстановления движений хвоста точно определялось выпадение чувствительности. Выпадение чувствительности в трех опытах наблюдалось в промежности, вульве, анусе, в прилегающих частях живота, спины, бедра и в проксимальной половине хвоста. У двух животных были нечувствительны и обе лапы. В соответствии
1 Устное сообщение. См. также: Winkler u. van Rynberk (5).
38
Вторая глава. Переключение
с результатами опытов Мерцбахера (Merzbacher) на собаке и у наших животных отчетливо восстановился тонус хвоста, который был активен и сильно двигался во все стороны. В другом опыте весь хвост, включая и его конец, был нечувствительным. В третьем случае только основание хвоста оказалось нечувствительным. Затем животных децеребрировали и перерезали спинной мозг известным способом в области 12—13-го торакальных позвонков. После этого исследовали вышеописанные переключения на хвосте и получили такие результаты: хвост рефлекторно сильно двигался во все стороны, но направление движения было совершенно незакономерным, и было невозможно с точностью предсказать направление вызываемых рефлекторных движений, как это можно было сделать в нормальных опытах. В опыте, в котором было лишено чувствительности только основание хвоста, изменение его положения ни в коей мере не влияло на исход раздражения, в то время как изменение положения середины хвоста вызывало отчетливое переключение. Следовательно, перерезкой соответствующих задних корешков и на хвосте кошки полностью устраняется влияние исходного положения, хотя двигательная активность исследованных конечностей не была нарушена операцией перерезки задних корешков.
Для того чтобы дальше исследовать, какие чувствительные нервы передают в центральную нервную систему необходимые для переключения импульсы, следует все чувствительные нервы разделить на три функциональные группы: кожной чувствительности, суставной чувствительности и, наконец, все остальные чувствительные волокна. Для исключения суставной чувствительности спинномозговой собаке вводился стоваин или новокаин с примесью адреналина в три больших сустава одной задней ноги (тазобедренный, коленный и голеностопный суставы). После этой инъекции сохранились без изменения все переключения. В других опытах анестезировалась обильной инфильтрацией новокаин-супраренина кожа всей задней ноги (за исключением возвышения мышц большого пальца ноги), а также кожа живота, спины и промежности, и после этого можно было ясно показать реакции переключения. Комбинация анестезии кожи с инъекцией трех больших суставов также ничего не изменила. В других опытах инъецировались только тазобедренный сустав и смещающаяся при изменении положения тазобедренного сустава кожа. Оказалось, что реакции переключения не изменились и при перемене положения ноги в тазобедренном суставе. Из всех этих опытов вытекает, что ни суставная, ни кожная чувствительность не являются необходимыми для осуществления переключения. Итак, мы приходим к выводу, что для переключения необходимы проприоцептивные нервы, т. е. чувствительные нервы мышц, сухожилий и фасций. Самую большую роль при этом играют, вероятно, чувствительные нервы мышц. Следовательно, при переключениях дело идет о рефлексах, которые вызываются самими мышцами и передаются (по чувствительным нервам) центральным органам. Пока эти пути функционируют, возможно и переключение в вышеописанных случаях. Если эти пути разъединяются, прекращается описанный феномен. Этим и подтверждается вышеуказанный вывод, что место нахождения переключения надо искать в центральной нервной системе.
Влияние положения и установки конечности на направленность последующих рефлекторных актов является тоническим; оно продолжается, пока животное или соответствующие конечности находятся в заданном положении, и все равно, когда вызывают рефлекс — сразу же, или через 5 минут, или через полчаса. Возбуждение, вызванное положением конечности, в свою очередь совсем не должно вызывать появление
Вторая глава. Переключение
39
каких-либо рефлекторных движений. Влияния, о которых здесь идет речь, юпособствуют только тому, что последующий импульс идет по совершенно определенному пути, возбуждает определенные центры и, напротив, минует такие центры, в которые он тоже мог бы притекать. Дело идет не о движениях или изменениях тонуса мускулатуры, а исключительно о «переключениях» в центральном органе. Говоря образно: на сортировочной станции переводятся только стрелки, чтобы следующий поезд мог идти по правильному пути. Эти переведения стрелок носят характер постоянных реакций.
Шеррингтон (9) объяснил значение вышеописанных реакций переключения для осуществления ритмических движений ходьбы. Он пришел к выводу, что переключения поддерживают ритмические движения, но не являются исключительной причиной их осуществления. Это вытекает уже из того факта, что после перерезки задних корешков.у спинномозговой собаки еще возможны попеременные движения ходьбы. Однако ясно, что если при движениях ходьбы одна нога переходит в растянутое положение, то она этим уже «включается» для последующего сгибательного движения, и наоборот, если сгибательное движение полностью совершилось, то нога этим уже приходит в готовность для последующего разгибательного движения. Итак, переключения при попеременных ритмических рефлексах поддерживают чередование возбуждения между антагонистическими группами мышц.
Однако переключения должны функционировать во всех реакциях равновесия организма как поддерживающий механизм. Если, например, тело почти падает в одну сторону, то мышцы противоположной стороны, как правило, растягиваются. Если теперь включается, чтобы препятствовать падению, любая реакция равновесия, по ходу которой сокращаются растянутые мышцы, то рефлекс находит центры тех мышц, которые он должен возбуждать, уже во «включенном» состоянии. Однако и в этом случае речь идет только о поддерживающем механизме.
Описанные выше переключения используются практической ортопедией при пересадках сухожилий. Известно, что функция парализованной мышцы часто должна быть замещена функцией своего антагониста. Можно даже мышцу или ее сухожилие оперативным путем разделить в длину пополам, после чего одна половина сохраняет старую функцию, а другая приобретает функцию антагониста (деление функции), — например, это производилось на передней большеберцовой, разгибателе пальцев и на икроножной мышце. Этот процесс лучше всего можно наблюдать, если часть сухожилия трехглавой мышцы пересадить на двуглавую мышцу плеча. Кодивилла (Codivilla) наблюдал после вывиха сухожилия малоберцовой кости действие мышц как экстензоров ноги. Но как только они оперативным путем были обратно приведены в их нормальное положение, они иннервировались как флексоры. Иногда пересаженная мышца начинает сразу после снятия перевязки функционировать по-новому. Подобные результаты получил Марина (Marina) при пересаживании мышц глаз на обезьянах. Он пересаживал прямую верхнюю мышцу концом прикрепления на место перерезанной прямой внутренней или же косую верхнюю на место внутренней, или менял места прикрепления наружной и внутренней. После этого сохранились в норме и произвольные движения и направление вращательного нистагма, хотя замененные мышцы иннервировались разными нервами — п. п. oculomotorius, trochlearis et abducens. Вероятно, все это основывается на процессах переключения, причем мышца в новом положении действует, как и раньше, правильно влияя с помощью своих афферентных нервов соответственно новым условиям
40
Вторая глава. Переключение
на возбудимость своих двигательных центров. Этим она частично навязывает своему центру другую функциональную роль, частично создает условия, благодаря которым в течение последующего лечения головной мозг и высшие органы чувств могли бы тренировкой улучшить этот механизм.
До сих пор все описанные случаи переключения подчинялись правилу Икскюля, по которому центры растянутых мышц «включены» для последующего возбуждения и легче всего отвечают при следующем рефлексе. Однако высокоразвитая центральная нервная система млекопитающих отнюдь не связана этим простым правилом. Последующие наблюдения показывают, что имеется ряд переключений, не связанных с растяжением мышц. Хорошим примером этого является чесательный рефлекс у спинномозговой собаки (4). Чесательный рефлекс был описан Гольцем (Goltz) и его учениками (см. Freusberg) и прежде всего был подробно изучен Шерринг- тоном (7). Этот рефлекс у собак легко можно вызвать раздражением кожи туловища с перерезанным в нижнем грудном отделе спинным мозгом. У некоторых собак рефлексогенная зона заходит даже на обе задние конечности до голеностопного сустава. Этот рефлекс легко вызывается и раздражением кожи слабым индукционным током.
Если положить такую собаку на спину, то рефлекс появляется на обеих сторонах. При раздражении правой стороны живота делает чесателъные движения правая, при раздражении левой стороны живота — левая задняя конечность. Только иногда при определенных условиях в положении животного на спине наблюдается двустороннее чесание.
При разгибании и отведении в сторону ноги на раздражаемой стороне чесание производит нога противоположной стороны. Если, следовательно, при положении животного на спине разгибать и абдуцировать правую ногу и потом раздражать правую сторону живота, то чесательный рефлекс осуществляется левой ногой. Влияние разгибания и отведения правой задней конечности угасает, если перерезаются на уровне люмбального и сакрального отделов спинного мозга задние корешки правой стороны, и наоборот, оно сохраняется, если суставы и кожа анестезируются новокаином. Следовательно, переключение вызывается чувствительными нервами мышц. Чесательный рефлекс начинается тоническим сгибанием до этого разогнутой ноги, причем прежде всего должен сгибаться тазобедренный сустав. Разгибанием и отведением ноги растягиваются как раз те мышцы, которые при чесательном рефлексе должны отвечать первыми, и все-таки соответствующая нога «выключена» для чесательного рефлекса. В этом случае переключение не подчиняется правилу Икскюля, центры растянутых мышц не становятся доступными возбуждению, а наоборот, выключаются.
Описанная реакция играет, вероятно, при нормальном чесании животных определенную роль. Шеррингтон показал, что при чесательном рефлексе контралатеральная нога обычно растянута и отведена в сторону. Уже этим, следовательно, она исключается для рефлекса, так что чесание почти всегда должно происходить односторонне.
Если перевернуть лежащую на спине собаку, у которой чесательный рефлекс происходит односторонне, на бок, то отношение изменится. Как уже наблюдал в лаборатории Гольца Гергенс (Gergens), теперь при любых условиях производит акт чесания всегда верхняя нога независимо от того, с какой стороны тела прикладывается раздражение: находящаяся ниже нога для чесательного рефлекса исключена. Значит, в зависимости от того, на какой стороне наносят раздражение, вызывается односторонний или перекрестный чесательный рефлекс. Гергенс считал, что ниженаходящаясн задняя нога не может выполнять чесательного рефлекса просто из-за механического препятствия. Что это не так, показывает
Вторая глава. Переключение
41
следующее наблюдение: с находящейся внизу задней ноги легко можно вызвать всевозможные другие рефлексы, как например сгибательный, коленный и различные перекрестные рефлексы, иногда даже движения ходьбы, И только чесательный рефлекс не осуществляется на этой нижней задней ноге. Место нахождения этого переключения расположено в центральной нервной системе. Это вытекает уже из следующего соображения: если животное лежит, например, на правом боку, то чесание всегда производит левая задняя нога независимо от того, где раздражается кожа — на правой или левой стороне тела. Таким образом, акт чесания левой задней ногой может быть вызван со всех соответствующих чувствительных путей.
Если же, с другой стороны, раздражать определенное место кожи, то можно с этого места, в зависимости от того, лежит ли собака на левом или правом боку, вызывать чесание той или другой ногой. Уже из этого факта следует, что место переключения должно находиться там, где пересекаются все эти пути, т. е. в самом спинном мозгу. То же самое вытекает из разных других наблюдений. Так, например, исходное положение верхней или нижней задних лап не имеет никакого влияния на переключение чесателъного рефлекса. Только их нельзя разгибать и отводить в сторону, так как при этом они исключаются для чесателъного рефлекса. Также не имеет значения и изогнутость позвоночного столба при соответствующем боковом положении (изогнут ли он вверх или вниз, что легко можно вызвать, подложив подушку).
Переключение осуществляется в результате одностороннего соприкосновения тела животного с подставкой. Когда собака лежит на правом боку на столе или на коленях служителя, то при всех обстоятельствах чесание производит вышележащая левая задняя лапа. Если теперь взять животное рукой за хвост, то при этом ничего не изменяется. Когда приподнимают животное и сгибают позвоночный столб в другую сторону, то переключение остается неизменным. Но как только задняя половина тела животного лишается соприкосновения с подставкой, переключение прекращается и чесательный рефлекс опять происходит строго на той же стороне (как при положении на спине). То, что в результате давления подставки на бок туловища действительно осуществляется переключение, вытекает из следующего: если держать животное за туловище в области плечевого пояса и за хвост в воздухе, то давлением ладони на вышенаходящийся бок тела можно вызвать обратное переключение, при котором верхняя лапа оказывается «включенной» и чесательный рефлекс осуществляется только нижней лапой. Для этого достаточно также сделать складку на коже и сдавить ее. Очень хорошо можно продемонстрировать это обратное переключение, если поддерживать животное рукой в области плечевого пояса и за складку кожи в поясничной области в боковом положении в воздухе. Последнее наблюдение показывает, что в возникновении реакции во всяком случае участвуют нервы кожи, воспринимающие давление. Анестезия кожи новокаином, однако, .полностью не снимает такого переключения, так что, вероятно, здесь участвуют и более глубоко лежащие нервные окончания чувства давления.
Таким образом, описанное переключение не вызывается самим исполнительным органом и прежде всего не вызывается мышцами. Растяжение или расслабление мышц не играет при этом никакой роли. Перерезка задних корешков на одной стороне снимает это переключение, и именно таким образом, что давление на денервированную сторону никак не влияет на переключение чесательного рефлекса. В то же время переключение на денервированную лапу можно, наоборот, вызвать с абсолютной точностью. Следовательно, чувствительность лапы-эффектора не играет никакой роли.
42
Вторая глава. Переключение
Значение имеет только чувствительность той части тела, которая лежит на подставке.
При описанном переключении речь идет не о простом торможении. Можно допустить, что в случае, когда у животного, лежащего на правом боку, раздражается кожа той же стороны, чесательный рефлекс по какой- либо причине оказывается заторможенным и появление такого же рефлекса на противоположной стороне обусловлено чисто непроизвольным усилением раздражения. Против подобного допущения говорит, во-первых, тот факт, что при усилении раздражения кожи в положении животного на спине чесание на противоположной стороне никогда не вызывается, а имеет место только его усиление на стороне раздражения. Кроме того, точное определение порога раздражения индукционным током показало, что при различных боковых положениях животного с одного и того же места кожи можно вызвать при одинаковом пороге раздражения как односторонний, так и перекрестный чесательный рефлекс. Таким образом, при одном и том же пороге возможно распространение раздражения как к правой, так и к левой ноге из одного и того же места. В этих случаях дело идет именно о переключении, т. е. о направлении возбуждения по тому или иному пути, а не о выпадении последствия одного раздражения вследствие торможения и проявлении другого раздражения вследствие усиления и распространения этого последнего в центральной нервной системе.
Такие определения порога раздражения вообще имеют значение лишь тогда, когда приходится решать, идет ли вообще речь о переключении.
Описанное здесь переключение также является постоянной реакцией. При этом не имеет значения, вызывается ли чесательный рефлекс сразу же после поворота животного на бок или через полчаса.
Из описанных обоих влияний, переключающих чесательный рефлекс, разгибание и отведение лапы являются наиболее сильными. Если при положении животного на боку разогнуть и отвести в сторону верхнюю лапу, которая, собственно говоря, должна была осуществлять чесание, то чесание производит нижняя лапа. Таким образом, этот опыт показывает, что нижняя лапа отнюдь не имеет механических препятствий к осуществлению чесательного рефлекса и что при определенных условиях она действительно может произвести чесание.
Эти переключения чесательного рефлекса на спинномозговой собаке бесспорны и удаются в 100% всех опытов. Следовательно, в данном случае, где в результате определенного раздражения могут возникать разные реакции, удалось так же, как и при рефлексах хвоста кошки, полностью найти правило, от которого зависит это различие. Однако в описанном случае это правило не является законом растяжения Икскюля. Влияние переключения в этом случае вообще не исходит от той конечности, которая выполняет рефлекторные движения, а исходит из более отдаленного места тела, и растяжение мышц при этом не играет никакой роли. Описанные опыты показывают, что спинной мозг млекопитающего обладает гораздо более разнообразными возможностями переключения, чем простые нервные системы беспозвоночных. Не только афферентные нервы, возбуждающиеся растяжением мышц, но и другие чувствительные пути отдаленных мест тела возбуждаются положением, давлением и, вероятно, еще другими влияниями и воздействуют переключающим образом на центральную нервную систему, т. е. они определяют тот путь, по которому распространяются позже поступающие в центральную нервную систему возбуждения. Таким образом, центральная нервная система отражает в каждый данный момент состояние тела, его позу, положение его конечностей, соприкосновение с внешним миром; поэтому становится понятным, почему нервная си-
Влияние тонических рефлексов шеи на конечности
43
етема при самых различных условиях реагирует так разнообразно, но в то же время совершенно закономерно.
Как в случаях, где переключение осуществлялось благодаря разному исходному положению реагирующей конечности, так и в последнем описанном случае, в котором переключение зависело от положения тела животного и от его соприкосновения с полом, становится понятным, какое большое влияние должна иметь установка тела на протекание самых разных движений. Каждому определенному положению тела соответствует определенное распределение возбуждения в центральной нервной системе и установка стрелок, как на сортировочной станции, что дает возможность .поступающим возбуждениям идти по определенным путям и направляться к эффекторам. Итак, уже теперь становится ясным, каким путем установка тела как таковая должна действовать переключающим образом, и в дальнейшем изложении мы должны опять поставить себе вопрос, имеет ли переключение место при разных рёфлексах, вызванных определенной установкой тела. Поэтому, прежде чем перейти к описанию отдельных реакций, нужно было более подробно изложить учение о переключениях.
Третья глава
РЕФЛЕКСЫ ПОЛОЖЕНИЯ (HALTUNG — STEHREFLEXE)
После краткого обзора интересующей нас области и после внесения самых основных понятий мы теперь переходим к изучению отдельных рефлексов, обеспечивающих положение тела. При этом прежде всего будут рассмотрены рефлексы положения.
Во введении уже упоминалось, что наиболее существенные рефлексы положения или стояния вызываются различными установками головы. При этом имеют значение два конкурирующих влияния: шейные тонические и лабиринтные тонические рефлексы. Мы начнем с тонических рефлексов шеи. I.I. ВЛИЯНИЕ ТОНИЧЕСКИХ РЕФЛЕКСОВ ШЕИ НА КОНЕЧНОСТИ
Эти рефлексы, как мы еще увидим, можно исследовать и у животных с нормальной деятельностью головного мозга. Однако для установления точных закономерностей их проявления лучше исключить возможность произвольных движений, вызвав у животных децеребрационную ригидность. Поэтому удобнее всего исследовать тонические шейные рефлексы на децеребрированных животных. Для того чтобы при различных положениях головы не помешали лабиринтные рефлексы, следует исключить деятельность лабиринтов. Это лучше всего достигается их двусторонней экстирпацией. После этого исследования можно проводить или непосредственно после вмешательства, или по прошествии некоторого времени.
Для техники лабиринтэктомии при исследовании шейных рефлексов особенно важной является сохранность мест прикрепления мышц шеи на черепе. Для этой цели де Клейном (8) был разработан метод подхода к лабиринту у морских свинок, кроликов, кошек и собак со стороны буллы (bulla), полукруглого выступа у основания черепа, при вскрытии которого сразу обнажается среднее ухо. Основания черепа можно достигнуть тупым путем через кожный разрез медиально от угла нижней челюсти и обнажить буллу, не повреждая при этом мышц и не нарушая их иннервации. После операции подвижность головы и шеи остается без изменения.
Однако у обезьян, как и у человека, лабиринтэктомия производится со стороны сосцевидного отростка.
44
Третья глава. Рефлексы положения
Вместо хирургической операции можно впрыскивать в преддверье через овальное окно 209о-й раствор кокаина и этим выключать без явлений раздражения деятельность лабиринта. Инъекция производится или через отверстие буллы, или через барабанную перепонку — у кошек, а также у кроликов (менее удобно); при небольшой тренировке легко удается попадать кончиком иглы в овальное окно. При этом, конечно,, полностью нарушается функция лабиринта. Впрочем, у морской свинки можно выключить деятельность лабиринтов и временно, впрыскивая кокаин только в среднее ухо: яд диффундирует в лабиринт и выключает его полностью на несколько часов.. На следующее утро деятельность лабиринта на инъецированной стороне чаще всего снова нормальна.
При исследовании децеребрированных животных после двустороннего' выключения лабиринтов наблюдается полная независимость тонуса конечностей от положения головы в пространстве. При этих обстоятельствах можно исследовать влияние тонических рефлексов шеи на мускулатуру туловища и в нашем специальном случае — изолированно на конечности.
При различных изменениях положения головы вызываются теперь совершенно закономерные и отчетливые реакции.
Поворот головы. У морских свинок, кроликов, кошек и собак поворотом головы называется движение вокруг оси морда—затылочное отверстие; у человека и обезьян — напротив, вращение вокруг оси темя— затылочное отверстие. Общим у всех видов животных является то, что повороты осуществляются в атланто-эпистрофейном суставе и вокруг бородки эпистрофея. Некоторые трудности возникают при обозначении направления поворота. У подопытных животных, к которым экспериментатор подходит спереди, под поворотом вправо понимают обычно вращение головы, при котором правое ухо двигается в вентральном надрвле- н и и. Если, однако, вращение головы вправо осуществляется человеком, причем его нос приближается к правому плечу, то левое ухо двигается в вентральном направлении. Поэтому при переносе результатов опытов на животных в клинику могут возникать недоразумения, вследствие чего необходимо точно указывать, что понимается под поворотом вправо или влево. Во избежание путаницы в нашей лаборатории оказался пригодным метод обозначения, исключающий всякое недоразумение: мы обозначаем конечности той стороны, в которую поворачивается нижняя челюсть или нос животного или пациента, как «челюстные ноги» (Kieferbeine — КВ), а конечности противоположной стороны, куда поворачивается затылок, — как «черепные ноги» (Schädelbeine — SB).
Для всех видов животных и для человека установлено простое правило: при повороте головы тонус разгибательной мускулатуры повышается в «челюстных ногах» и уменьшается в «черепных ногах», причем передние и задние конечности реагируют при этом всегда одинаково. Если, таким образом, у децеребрированной кошки после двусторонней лабиринтэктомии поворачивать голову вправо, т. е. так, что правое ухо направляется вентрально, то левые конечности являются «челюстными ногами» и тонус разгибательной мускулатуры повышается в левой передней и левой задней ногах, в то время как тонус разгибательной мускулатуры в правой передней и правой задней ногах уменьшается. Если у человека, у которого обнаруживаются тонические шейные рефлексы, поворачивать голову влево — так, что нос приближается к левому плечу,, то левые конечности становятся «челюстными ногами», а -правые конечности — «затылочными ногами», и поэтому повышается разгибательный тонус в левой руке и в левой ноге, а в правой руке и в правой ноге уменьшается.
Влияние тонических рефлексов шеи на конечности
45
У разных подопытных животных можно видеть, что не имеет значения, находится ли голова перед началом вращения в нормальном положении или она была сильно откинута дорсально или вентрально. Можно фиксировать рукой поперечные отростки 4-го и 5-го шейных позвонков и произвести поворот только в верхнем шейном отделе позвоночного столба, тогда реакция вызывается вращением атланта относительно эпистрофея.
Наклон головы. Наклоном головы называется движение, которое осуществляется в шейном отделе позвоночного столба вокруг дорсо- вентральной оси. У морских свинок, кошек, кроликов и собак голова при этом движется вокруг дорсо-вентральной оси, у обезьян и человека — напротив, вокруг оси, которая проходит параллельно линии glabella— затылочный бугор (наклон головы). Обозначение направления наклона у всех животных одинаково: при наклоне влево левое ухо приближается к левому плечу, при наклоне вправо правое ухо приближается к правому плечу. У морских свинок, кроликов, собак и кошек правые конечности, например, при наклоне вправо можно обозначать как «челюстные конечности», а левые — как «затылочные конечности»; у человека и обезьяны, однако, это ведет к недоразумениям.
Если у децеребрированных животных наклонять голову вправо, то тонус мышц-разгибателей правой передней и задней ноги увеличи- нается, в то время как тонус мышц-разгибателей левой передней и задней ноги ослабляется. При наклоне влево наблюдаются обратные отношения. У кроликов, кошек, собак и морских свинок можно, например, при найлоне головы вправо правые конечности обозначать как «челюстные конечности», а левые — как «затылочные», и тогда получается для наклона такое же правило, как и для поворота: «челюстные ноги» разгибаются, а тонус «затылочных» понижается. Это имеет некоторое значение для запоминания. У обезьян, однако, такое обозначение применять нельзя. Тут •следует только сказать, что на стороне, куда наклоняется голова, наступает усиление разгибательного тонуса конечностей. Если у животного в положении стоя (кошки) голова наклоняется, например, вправо, то вследствие этого смещения перемещается вправо и центр тяжести всего животного.
Закономерно связанное с наклоном головы вправо усиление разгибательного тонуса в правой передней ноге соответствует перемещению центра тяжести, и правая передняя лапа оказывается в состоянии выдержать этот увеличенный вес. Вследствие этого усиленного разгибания цравой передней лапы одновременно перемещается влево и туловище (в сторону черепа), и таким образом более или менее уравновешивается перемещение центра тяжести.
Если у децеребрированного животного, которое в нормальном положении стоит на четырех лапах, вследствие наклонения головы слишком сильно ослабляется разгибательный тонус конечностей на стороне черепа, животное может упасть в эту сторону.
Для исхода реакции не имеет значения, при каком положении головы (максимальном сгибании головы дорсально или вентрально) производится наклон ее. Можно рукой фиксировать в середине позвоночный столб и производить наклон только в передней части шеи.
Насколько необходимо в каждом отдельном случае указывать, что понимается под «поворотом» и «наклонением» и под «вправо» и «влево», — вытекает из следующего: если пользоваться вышеуказанным способом обозначения, то поворот вправо и наклон вправо у собаки, кошки, кролика и морской свинки действуют на тонус конечностей диаметрально противоположно, потому что при повороте вправо «челюстными конеч¬
46
Третья глава. Рефлексы положения
ностями» становятся левые лапы, а при наклоне вправо «челюстными конечностями» становятся правые лапы.
Поднимание и опускание головы. При поднимании и опускании голова вращается вокруг битемпоральной оси. При поднимании происходит движение головы в дорсальном, при опускании — в вентральном направлении.
У всех исследованных видов животных разгибательный тонус обеих передних лап ослабляется при опускании головы, а при поднимании, напротив, усиливается. При этом имеет значение движение в середине* шейного отдела позвоночного столба. Если голова поднимается или опускается исключительно в атланто-затылочном суставе, то в большинстве* случаев всякие влияния на разгибательный тонус отсутствуют, только у собак в некоторых случаях можно было вызвать отчетливую реакцию.
У кошек, собак, морских свинок и обезьян задние лапы реагируют1 противоположно передним, т. е. при поднимании головы они сгибаются,, а при опускании головы разгибаются. Только кролик ведет себя по- другому: у него задние лапы действуют так же, как и передние.
Реакция задних лап у собак и у кроликов часто сильнее передних, у кошек, напротив, в большинстве случаев слабее.
У большинства обследованных животных надо точно отличать так называемый vertebra-prominens-рефлекс1 от влияния поднимания или опускания головы. Первый вызывается или давлением на остистые отростки нижнего шейного или верхнего грудного позвонков, или перемещением шеи в самых нижних суставах вентрально по направлению к туловищу. Это ведет к равномерному расслаблению всех четырех ног, при этом передние ноги расслабляются сильнее задних. В результате этого, если исходить из нормального положения животного, оно наконец оказывается лежащим головой, грудью и животом на полу. Рефлекс с 7-го шейного позвонка отличается от влияния опускания головы тем, что у кошки,, собаки, морской свинки и обезьяны в ответ на опускание головы в середине шейного отдела позвоночного столба задние ноги разгибаются, в то время как при рефлексе с 7-го шейного позвонка происходит одновременно расслабление всех четырех конечностей.
Вопрос о том, где начинаются описанные тонические шейные рефлексы — в мышцах или в суставах шеи, до сих пор не решен, потому что при сложном анатомическом расположении шейной мускулатуры и шейных суставов их невозможно отделить друг от друга на живом, способном к реакциям животном. Однако афферентные пути можно установить. После двусторонней перерезки задних шейных корешков (1—3-я пары) поворот, наклон, поднимание и опускание головы у кошки перестают влиять на тонус конечностей, в та время как у таких животных можно вызвать vertebra-prominens-рефлекс. Однако в одном случае у кролика после двусторонней перерезки трех верхних шейных задних корешков еще можно было наблюдать слабую реакцию в ответ на поворот головы, так что у таких животных при вызове тонических шейных рефлексов, вероятно, могут принимать участие и задние корешки 4-й пары шейных нервов.
Для силы реакции не имеет значения, были ли произведены движения шеи пассивно или животное двигало свою голову активно в этом же направлении. Это впервые было доказано Дюссер де Варенном [Dusser de Barenne (1)], который приготовил на кошке двусторонний vastocrureus-
1 В дальнейшем этот рефлекс иногда будет называться рефлексом с 7-го шейного позвонка. (Ред.)'
Влияние тонических лабиринтных рефлексов на конечности
47
препарат, исключив путем перерезки моторную иннервацию всех идущих к задним ногам мышц и оставив интактной на обеих сторонах только vastocrureus, т. е. ту часть четырехглавой мышцы, которая начинается на бедре и заканчивается на голени. При выходе из наркоза кошка активно двигала головой, при этом оказалось, что правый и левый vastocrureus в ответ на поворот, наклон, поднимание и опускание головы реагировали как обычно.
На человеке можно установить то же самое. Ниже мы приводим пример того, что в некоторых патологических случаях при мозговых заболеваниях человека можно вызвать типичные шейные рефлексы. Доктору Стен- версу (Stenvers) удалось в одном таком случае (утрехтская педиатрическая клиника) при помощи кинематографических съемок установить, что при активном повороте головы «челюстная рука» вытягивалась, а «черепная рука» сгибалась — точно так же, как это произошло при пассивном повороте головы.
Центры тонических шейных рефлексов лежат в верхнем шейном отделе спинного мозга (18). Если у децеребрированной кошки сделать после экстирпации мозжечка фронтальный разрез через продолговатый мозг непосредственно позади места выхода 8-го нерва, то вначале еще сохраняется хорошая децеребрационная ригидность. Так как, однако,8-йнерв отделен, то тоническое лабиринтное рефлекторное влияние на конечности не может осуществиться. На таких препаратах появление тонических шейных рефлексов при повороте, наклоне, поднимании и опускании головы можно еще показать с большой ясностью. То же самое наблюдается, когда поперечный разрез производится каудальнее через «писчее перо». В этом случае удается получить типичную реакцию на поворот и наклон головы и вызвать vertebra-prominens-рефлекс.
Когда производится поперечный разрез непосредственно позади места входа 1-й пары шейных нервов, то поворот и наклон оказываются еще явно действующими, но реакция становится немного слабее. То же самое имеет место, если поперечный разрез производится приблизительно в середине между 1-м и 2-м корешками шейных нервов. Однако после разреза непосредственно позади места входа 2-й пары шейных нервов реакция на поворот и наклон прекращается почти полностью. Только в одном из семи опытов еще наблюдалась незначительная реакция, которая прекратилась только после разреза позади С3. В большинстве случаев, однако, достаточно сделать поперечный разрез позади С2, чтобы исчезли шейные рефлексы на поворот и наклон головы. Следовательно, центры тонических шейных рефлексов лежат в двух верхних шейных сегментах. Афферентные пути проходят у кошки (за исключением путей для vertebra-prominens-рефлекса) через три верхних шейных задних корешка.
II. ВЛИЯНИЕ ТОНИЧЕСКИХ ЛАБИРИНТНЫХ РЕФЛЕКСОВ НА КОНЕЧНОСТИ (7, 9)
Для изолированного исследования тонических шейных рефлексов необходимо исключить лабиринтные рефлексы. И наоборот, для исследования тонических лабиринтных рефлексов следует исключить шейные рефлексы.
Это достигается:
1. Просто тем, что при изменениях положения исключаются все движения шеи; т. е., например, при перекладывании животного из положения на животе в положение на спину нужно тщательно следить за тем, чтобы ротовая щель не изменила своей оси по отношению к туловищу животного.
48
Третья глава. Рефлексы положения
Этот прием очень часто применяется в обычных лабораторных условиях, и он производится тем легче, чем мельче исследуемое животное. Однако при самых первых исследованиях свойств тонических лабиринтных рефлексов этот прием оказался недостаточным потому, что всегда неизбежны незначительные движения шеи.
2. На основании вышеизложенных результатов исследования у кошек можно перерезать три верхних шейных задних корешка (у кроликов, как уже было сказано выше, этого недостаточно). Этим выключаются вызываемые вращением и наклонением, подниманием и опусканием головы шейные рефлексы на конечности, но при этом сохраняется vertebra- prominens-рефлекс, что может привести к ошибкам в опыте, если на это не обратить особого внимания.
3. Лучше всего положить голову, шею и грудь децеребрированных животных в гипс так, чтобы никакие движения шеи не были возможны.
Для этой цели животное после развития децеребрационной ригидности и перерезки спинного мозга в нижнем грудном отделе кладут спиной книзу на соответствующей ширины свинцовую пластинку, которая идет приблизительно от лобных бугров черепа до 10-го грудного позвонка. На этой пластинке животное фиксируется при помощи гипсовой повязки, охватывающей грудь, шею, голову и оставляющей свободными трахейную канюлю и передние конечности до плеч. Теперь контролируют, стоит ли голова симметрично по отношению к туловищу, и накладывают вторую гипсовую повязку, при помощи которой с правой и левой сторон прикрепляют к грудной клетке, шее и голове две узкие свинцовые пластинки, стоящие почти перпендикулярно к первой дорсальной пластинке. Гибкие свинцовые пластинки можно предварительно примерить к формам тела. На все это накладывается еще третья повязка. При таком приеме животное может дышать беспрепятственно, так как экскурсии диафрагмы и брюшной стенки ничем не ограничены. После застывания гипса передняя половина тела животного настолько иммобилизована, что можно быть уверенным, что каждое изменение положения животного не вызывает ни малейшего движения в суставах шеи. При помощи простого угольника и ватерпаса можно измерить наклон плоскости ротовой щели по отношению к горизонтальной плоскости. Если требуется поставить опыт и на задних конечностях, то спинной мозг не перерезается и применяется такая дорсальная свинцовая пластинка, которая доходит до корня хвоста. При помощи третьей повязки эта пластинка прикрепляется к тазу и бедрам. При этом надо следить за тем, чтобы живот остался свободным, иначе создается затруднение дыханию. Само собой разумеется, что конечности должны свободно выступать из гипсового панциря. Поразительным и неожиданным было то, что эта повязка, покрывающая большую часть поверхности кожи животного, почти не препятствовала развитию децеребрационной ригидности. При этом наблюдается хороший разгибательный тонус передних конечностей.
Этот же прием применялся с успехом и на собаках, кроликах и обезьянах. На конечности, которые вследствие децеребрационной ригидности сильно растянуты, можно, смотря по обстоятельствам, прикреплять резиновые постромки, которые заходят за спину животного и стремятся согнуть конечности. Тогда особенно отчетливо выявляется изменение тонуса в мышцах-разгибателях конечности (рис. 16).
Можно кратко и просто резюмировать правила для тонических лабиринтных рефлексов, влияющих на мышцы конечностей. Все четыре конечности при этом реагируют однотипно. Есть одно и только одно положение головы в пространстве, при котором наступает максимум разги- бательного тонуса четырех конечностей, и одно и только одно положение головы в пространстве, при котором имеется его минимум. Положение максимум и положение минимум отличаются друг от друга углом в 180°. У до сих пор исследованных видов млекопитающих в большинстве случаев положение максимум наблюдалось при положении головы на затылке и немного приподнятой ротовой щели.
Об обозначениях различных положений тела животного и головы в пространстве надо сказать следующее.
Положение на спине, положение на животе или на ногах, правое и левое боковое положение — ясные обозначения. При висячем положении головой вниз позвоночный
Влияние тонических лабиринтных рефлексов на конечности
49
столб расположен вертикально, также и при висячем положении головой вверх. В первом случае голова образует нижний, в последнем случае верхний конец тела. Для описания положения головы в пространстве определяют угол, который образуется плоскостью ротовой щели при закрытой ротовой полости с горизонтальной плоскостью. Если при исходном положении головы затылком вниз и горизонтальном расположении ротовой щели (0°) поворачивать животное вокруг битемпоральной оси, то получают положения головы, показанные на рис. 15, которые определяются величиной угла в градусах. При поднимании морды она проходит ряд положений: +45°, +90°, +135°. Потом голова приходит в положение 180° с горизонтальной ротовой щелью и теменем вверх; после этого следует положение —135°, это то положение, в котором кошка держит голову в обычной жизни; 90° — это положение с ротовой щелью, расположенной вертикально, и при дальнейшем вращении голова возвращается через —45° в исходное положение. Исходя из каждого из этих положений, можно поворачивать голову в боковое положение на определенное количество градусов. Очевидно, что таким образом можно количественно определить различные положения головы в пространстве. Такое же изображение можно сделать ^ также для кроликов и для других видов животных. Ротовая щель оказалась удобным показателем для определения положения головы.
У большинства животных положение для максимального влияния тонических лабиринтных рефлексов на конечности находится в области между 0 и +45°, чаще всего при +45°. Соответственно этому «минимальное» положение находится между 180 и —135°, чаще всего при —135°.
При перемещении кошек из по- Рис. 15.
ложения головы теменем вниз (0°)
до тех пор, пока ротовая щель не достигает 45°, происходит максимальное разгибание конечностей, которые делаются совсем ригидными. Если это движение продолжается так, что морда направляется вертикально вверх (+90°), то тонус снова немного снижается и продолжает снижаться, когда достигается положение +135 и 180°. При —135° имеется минимум тонуса. Конечности в этом положении можно легко сгибать и они часто совершенно вялые. В это время спина находится наверху, живот внизу, конец хвоста животного выше головы.
Если это перемещение продолжать, то тонус постепенно снова повышается, что обычно и видно при положении —90°. При —45° снова имеется сильный разгибательный тонус, который при достижении исходного положения (положения на спине) продолжает повышаться и достигает вновь максимума при +45°.
Из двух промежуточных положений (+135° и —45°) тонус в последнем положении явно сильнее, чем в первом. В общем можно сказать, что в положениях от —80 через 0 до +90° разгибательный тонус сильнее, чем в положениях от +100 через 180 до —100°. При +90° тонус всегда значительно сильнее, чем при —90°. Рис. 16 демонстрирует разгибание передних конечностей загипсованной кошки при переходе положения головы от -110 до 55°.
Если в качестве исходного берется положение * животного на боку, при котором имеет место средний тонус, и при этом перемещают животное вокруг оси, параллельной позвоночному столбу, до тех пор, пока оно не достигает положения на спине, то тонус повышается; если же животное перемещать в положение на живот, то он снижается. В зависимости от 4 Р. Магнус
50
Третья глава. Рефлексы положения
исходного положения, которое придается голове по отношению к оси позвоночного столба, она может проходить или не проходить положения максимум и минимум для тонических лабиринтных рефлексов.
Специальные контрольные опыты показали, что эти реакции действительно зависят от положения головы в пространстве. Также можно показать, что эти реакции не вызываются изменением положения конечностей. Если при разных перемещениях положение головы по отношению к горизонтальной плоскости не изменяется, то и никакие тонические лабиринтные рефлексы не осуществляются. При положении животного на животе или на спине можно, следовательно, произвести поворот вокруг (перпендикулярно стоящей) дорсо-вен- тральной оси, а при положении животного на боку — поворот вокруг (также перпендикулярно стоящей) фронтальной оси, что не влияет при этом на тонус конечностей.
У децеребрированных кошек все чистые прогрессивные сдвижения также не имеют никакого влияния на тонус. Так как прогрессивными движениями можно вызвать лабиринтные рефлексы, изменяющие положение конечностей (см. введение), то это отсутствие влияния на
Рис. 16. Из кинематографического снимка. Кошка де- церебрирована при хлороформном наркозе. Через 1/2 часа перерезка спинного мозга на уровне 12-го грудного позвонка. После этого у животного помещается в гипс передняя часть тела до 9-го грудного позвонка. Ротовая щель примерно на 30° повернута вертикально по отношению к позвоночнику. На рис. 16, а животное держат свободно в воздухе. Голова, шея и грудь в гипсовой повязке, передние ноги свободны. Передние лапы притянуты к шее резиновыми повязками. Разгибатель- ный тонус передних ног проявляется, таким образом, в натяжении этих повязок и измеряется углом в локте. На рис. 16, а конец головы опущен, так что ротовая щель находится в положении 110°. Животное находится довольно продолжительное время в этом положении: тонус передних ног незначителен, угол в локте равен примерно 11°. После этого животное поворачивают в воздухе во- круг фронтальной оси, пока спина не будет внизу, конец головы поднят, и ротовая щель находится в положении +55°, значит, близко к положению максимума лабиринтных рефлексов. Через 3 сек. сделан рис. 16, в. Разгибательный тонус передних ног не изменился, угол в локте составляет 95°. Через 3.3 сек. начинается медленное сильное разгибание ног, которое в целом длится 5 сек., пока не достигается максимальный тонус. Это остается так долго, пока животное находится в этом положении- Рис. 16, с сделан через 0.5 сек. после завершения разгибания. Вследствие увеличения тонуса мышц-сгибателей резиновые повязки растянуты. Пальцы находятся гораздо выше, чем на рис. 16, в, а угол локтя составляет 150°. Очень характерным является длительный латентный период реакции, который начинается через 7.5 сек. после окончания поворота и заканчивается через 12.5 сек. после начала поворота.
тонус, очевидно, связано с тем, что рефлексы на прогрессивные движения очень чувствительны к операционному шоку. Для наших целей это имеет то преимущество, что нам эти реакции не мешают при исследовании тонических лабиринтных рефлексов на децеребрированных животных.
Если у загипсованных животных при помощи кокаина или хирургическим путем выключают лабиринты, то после этого все реакции на изменение положения исчезают. При этом чаще всего наблюдается средняя величина тонуса, которая слабее бывшего максимума, но значительно силь-
Влияние тонических лабиринтных, рефлексов на конечности
51
нее бывшего минимума. У таких животных исчезают также шейные и лабиринтные рефлексы.
Вышеописанные реакции обусловлены только положением головы в пространстве. Они не вызываются угловыми ускорениями при перемещениях, так как тогда было бы такое положение тела, при котором вращение в горизонтальной плоскости вызывало бы эти реакции. Но это вряд ли имеет место. Что речь идет о настоящих рефлексах положения вытекает и из того, что для окончательного состояния тонуса конечностей не имеет значения, достигается ли определенное положение головы в пространстве вращением в сагиттальной, фронтальной или в другой плоскости и происходит ли вращение, ведущее к окончательному положению, в том или обратном направлении. Вращательные реакции при этом не участвуют.1 Дальше можно показать, что соответствующее состояние тонуса конечностей (у загипсованных животных) продолжается до тех пор* пока не изменится положение головы в пространстве. При определении максимума и минимума тонуса выяснилось, что у разных подопытных животных наблюдаются значительные индивидуальные вариации. На возбудимых животных положение максимума и минимума можно установить с точностью до 5° (иногда до 10°). Но обычно наблюдаемые вариации выпадают из этого предела точности.
Количество опытов
Максимум
Минимум
15
Кошка (18 опытов) +45°
1
со
СП
о-
1
0 до +40°
+135 до —135°
1
+10°
—170°
1
0 до —10°
180 до +170°
4
Кролик (9 опытов) 0°
180°
1
+20°
—160°
1
0 до +45°
Ó
ю
со
1
о
te
о
00
ттН
3
+45°
—135°
2
Собака (4 опыта) +45°
—135°
1
0°
180°
1
+90°
— 90°
2
Обезьяна (2 опыта) +45°
—135°
Среди этих 33 опытов у кошки нашли один раз максимум между 0 и —10°, у собаки один раз при +90°. В 31 случае максимум лежал между1 0 и 45°, в том числе в 20 случаях точно при +45° (±5°). Все вариации (за одним исключением у собаки) находятся в положении около 0°. Мы возвратимся к этим отношениям, когда речь будет идти о зависимости тонических лабиринтных рефлексов от определенных частей вестибулярного аппарата (отолиты).
Афферентным нервом тонических лабиринтных рефлексов является,, конечно, 8-й нерв. Экстирпация лабиринтов или перевязка 8-го нерва
1 Магнус имеет в виду реакции на угловые ускорения. (Ред.)
4*
52
Третья глава. Рефлексы положения
полностью снимает эти рефлексы. Опыты с последовательными перерезками на центральной нервной системе децеребрированных животных показали, что полностью можно удалить средний мозг и мозжечок, не влияя на осуществление этих рефлексов; кроме того, они наблюдаются также и тогда, когда поперечный разрез через продолговатый мозг производится непосредственно перед вступлением в него 8-го нерва.
Центры тонических лабиринтных рефлексов, изменяющих положение конечностей, расположены, следовательно, каудально от вступления 8-го нерва в продолговатый мозг. Это вытекает из трех совпадающих опытов на кошках (18) и из трех — на кроликах (37).
Так как при тонических шейных рефлексах конечности чаще всего реагируют противоположно (исключениями являются лишь рефлексы с 7-го шейного позвонка и реакция на поднимание и опускание головы у кроликов), в то время как при тонических лабиринтных рефлексах все четыре конечности реагируют однотипно, то в случае сомнения можно использовать это для различения шейных рефлексов от лабиринтных.
Влияние тонических лабиринтных рефлексов на конечности после одностороннего выключения лабиринта (7, 9, 15)
А priori можно было ожидать, что после одностороннего выключения лабиринта описанные лабиринтные рефлексы должны были бы измениться в том смысле, что теперь реагировали бы только конечности одной стороны тела или по крайней мере реакция на одной стороне была бы слабее, чем на другой. В опытах, однако, выяснилось, что достаточно одного лабиринта, чтобы его влияние проявлялось в одинаковом смысле и с одинаковой силой на конечностях как правой, так и левой стодюны тела. Постоянные результаты получились в пяти опытах на кошках, через 17, 21, 28, 31 и 140 дней после односторонней экстирпации лабиринтов (кошки были загипсованы после децеребрации). Внимательное наблюдение в это время за животными позволило установить явления, характерные для одностороннего выключения лабиринта. После децеребрации на обеих сторонах тела наблюдалась одинаковая децеребрационная ригидность. После ее проявления и после перерезки спинного мозга в нижнем грудном отделе на животных накладывался гипс. Реакции на изменения положения в пространстве были такие же, как они описаны выше для животных с двумя интактными лабиринтами. Никогда не наблюдалось разницы в силе реакций правой и левой передней конечности, хотя, конечно, на это обращалось особое внимание.
В другом опыте децеребрировалась нормальная кошка и после перерезки спинного мозга загипсовывалась. У нее наблюдались все реакции, которые были описаны в предыдущем разделе: максимальный тонус оказался при +45°, минимальный — при —135°. После этого левый лабиринт выключался кокаином через окошечко в гипсовой повязке. Тонус левой передней конечности после этого был явно немного слабее, чем на правой стороне. Эта левая конечность, однако, реагировала на каждое изменение положения животного в пространстве точно таким же образом и с такой же силой, как и первая.
Следовательно, достаточно одного лабиринта, чтобы иметь одинаковое влияние на тонус конечностей обеих сторон тела.
Такое же наблюдение было сделано на пяти кроликах соответственно через 5, 7, 7, 18 и 24 дня после односторонней экстирпации лабиринта и на одной децеребрированной собаке, у которой один лабиринт был
Общий характер изменений тонуса при рефлексах положения
53
выключен кокаином. Такое же поведение можно было наблюдать и у обезьян.
Интересно то, что оставшийся лабиринт вызывает изменения тонуса одинаковой величины на конечностях обеих сторон тела независимо от того, имелся ли одинаковый исходный тонус на правых и левых конечностях (через продолжительное время после лабиринтэктомии) или же конечности на стороне лабиринтэктомии были более ослаблены (непосредственно после лабиринтэктомии). Уже из этого вытекает, что временное одностороннее ослабление конечностей после односторонней лабиринтэктомии нельзя объяснить выпадением влияния тонических лабиринтных рефлексов на мышцы конечностей.
III. ОБЩИЙ ХАРАКТЕР ИЗМЕНЕНИЙ ТОНУСА ПРИ РЕФЛЕКСАХ ПОЛОЖЕНИЯ
Если тонус конечностей усиливается, то это чаще всего вызывает — как при тонических шейных, так и при лабиринтных рефлексах — сильное разгибание конечностей, при котором преодолеваются значительные сопротивления. Если нагрузить конечность или изолированную разги- бательную мышцу постоянным грузом, то наблюдается явное укорочение разгибательных мышц. С другой стороны, требуется нагрузка с большим весом, чтобы снова привести мышцу к ее исходной длине. Следовательно, при постоянной нагрузке наступает укорочение мышцы, а при постоянной ее длине — усиление напряжения.
При уменьшении тонуса можно наблюдать противоположное явление. Уменьшается тонус разгибателей, причем при той же нагрузке мышца удлиняется и при той же длине она имеет меньшее напряжение. При известных условиях может произойти полное расслабление разгибательных мышц.
В плечевом и локтевом суставах передней ноги и в бедренном и коленном суставах задней ноги всегда имеется более сильная реакция. Реакция в голеностопном суставе и суставах пальцев гораздо слабее, но и тут иногда можно наблюдать разгибание и растопыривание пальцев, а у ко шек и выпускание когтей.
У животных с более слабой децеребрационной ригидностью реакцию можно определить таким образом: прежде всего разгибается в локте конечность (например, передняя) и затем определяется тот угол, при котором впервые почувствуется сопротивление при пассивном сгибании При этом получаются очень точные данные, так как при пассивном сгибании вдруг при определенном положении наступает как бы «защелкивание» разгибательной мышцы, после чего при дальнейшем сгибании приходится преодолевать некоторое сопротивление. Этот момент, например на локте, при максимальном тонусе наступает при угле 160°, при минимальном тонусе — при 90°. При значительном уменьшении тонуса иногда вообще больше не замечается никакого сопротивления.
При выраженной децеребрационной ригидности сокращены только разгибательные мышцы. Поэтому результат изменения положения головы на таких животных яснее всего проявляется на разгибательных мышцах. При графической записи изменений длины сгибательных и разгибательных мышц одной и той же конечности отклонения можно наблюдать только на кривой разгибателей, в то время как кривая сгибателей не изменяется. Следовательно,у децеребрированного животного для описанных тонических шейных и лабиринтных рефлексов часто «включены» только разгибательные мышцы. Иногда можно получить результаты, которые говорят об
54
Третья глава. Рефлексы положения
участии сгибательных мышц в вышеописанных рафлексах. Если у животных со значительными изменениями тонуса вызвать, например, минимум разгибательного тонуса соответствующим движением головы, при котором происходит полное ослабление экстензоров, то при этом иногда наблюдается активное сгибание. Это сгибание можно обнаружить по сопротивлению, которое оказывает соответствующая конечность пассивному разгибанию. В особенно демонстративном опыте животное держали в положении на животе в воздухе, а затем ранее вентрально поднятая голова опускалась. Это вызвало непосредственное уменьшение имевшегося до того разгибательного тонуса, но ноги при этом не повисли вниз по закону тяжести, а прижались активным сгибанием к туловищу. И эти сгибания выражены сильнее в локте, чем в запястном суставе. Итак, и для вышеописанных рефлексов имеет значение закон реципрокной иннервации [Шеррингтон (5)]. Торможение разгибательного тонуса сопровождается сокращением сгибателей, а усиление разгибательного
Рис. 17. (Опыт Беритова). Кошка, эфирный наркоз. Перерезка всех мышц, соединяющих лопатку и плечевую кость с телом, и всех нервов плечевого сплетения, за исключением лучевого и кожно-мышечного. Кроме того, перерезаются
m. anconaeus и мышечная ветвь лучевого нерва. Поверхностный лучевой нерв от- препарован для раздражения. Ампутация предплечья в локтевом суставе. Фиксация лопатки, головы и дистального конца плечевой кости. После этого децеребрация. Конец эфирного наркоза. Верхняя кривая — регистрация движения латеральной и медиальной головки, трехглавой мышцы, средняя — длинной головки трехглавой мышцы, нижняя — двуглавой мышцы. Кошка находится в боковом положении, регистрируемая нога наверху. В начале кривой голова находится в среднем положении. При КВ голова поворачивается теменем вниз, нижней челюстью вверх, регистрируемая нога становится «челюстной». Происходят сокращение трехглавой мышцы (верхняя и средняя кривые) и расслабление двуглавой мышцы соответственно разгибанию регистрируемой ноги. Это разгибание временно прерывается неполным сгибанием (торможение трехглавой мышцы и сокращение плечевой мышцы). Примерно через 13 сек. голову поворачивают теменем вверх (SB), после чего происходят расслабление трехглавой мышцы и сокращение плечевой, которая постепенно возвращается в исходное положение. При КВ голову снова поворачивают теменем вниз, следствием чего является сокращение трехглавой мышцы и торможение двуглавой. При SB голова поворачивается теменем вверх, на верхней кривой видно расслабление трехглавой мышцы, на нижней — возвращение плечевой мышцы в исходное положение. Время в секундах.
тонуса — затормаживанием сгибателей. Участие сгибателей можно доказать в соответствующих случаях как для лабиринтных, так и для шейных рефлексов.
На рис. 17 видна графическая запись сокращений мышц плеча с реципрокной иннервацией при тонических шейных и лабиринтных рефлек¬
Общий характер изменений тонуса при рефлексах положения
55
сах. Этот опыт был поставлен Беритовым в Утрехтском фармакологическом институте. Верхняя кривая — это реакция латеральной и медиальной головок трехглавой мышцы, средняя кривая — реакция длинной головки трехглавой мышцы, т. е. в обоих случаях разгибателей локтя; нижняя кривая — реакция плечевой мышцы, т. е. сгибателя конечности в локте. Мышцы были изолированы и соединены с рычажками. Опыт начинался при среднем положении головы. Затем голова поворачивалась так, что нога, у которой регистрировалось сокращение мышц, становилась «челюстной ногой» (КВ). После этого наступает сокращение разных частей трехглавой
Рис. 18. (По Беритову). Кошка. Децеребрация и удаление мозжечка. Опыт производился через 2*/2 часа после операции. Животное лежит на левом боку. Верхняя кривая соответствует правой плечевой мышце, нижняя — правой трехглавой. При КВ голова вращается теменем вниз, регистрируемая нога при этом становится «челюстной» и наступает сокращение трехглавой мышцы, в то время как плечевая мышца остается в покое. При SB голову поворачивают теме- нём вверх, трехглавая мышца после этого ослабляется. Этот же опыт повторяется еще раз. Наконец, при MS голову снова вращают, приводя ее в среднее положение. Во время опыта изменяется только положение трехглавой мышцы, в то время как плечевая мышца (сгибатель) не принимает участия в реакции.
мышцы, прерывающееся временно частичным ослаблением. Но одновременно с этим происходит расслабление плечевой мышцы.
Приблизительно через 13 секунд голову начинают поворачивать так, что регистрируемая нога становится «черепной» (SB). После этого происходит расслабление трехглавой и сокращение плечевой мышцы, которая возвращается к исходному положению. Через некоторое время голова поворачивается еще раз так, что регистрируемая нога становится «челюстной» (КВ); при этом явно видно сокращение трехглавой и расслабление двуглавой мышцы. Если голову повторно вращать так, что регистрируемая нога становится «черепной ногой» (SB), то расслабляется трехглавая мышца (что особенно хорошо видно на верхней кривой), а плечевая мышца возвращается в исходное положение.
При продолжении своих опытов в Петербурге Беритов (1) получил ряд хороших кривых. В его опытах, обычно в первые часы после децереб- рации, наблюдалась ярко выраженная децеребрационная ригидность, при которой мышцы-разгибатели были сокращены, а мышцы-сгибатели совершенно расслаблены. При вызове тонических шейных и лабиринтных рефлексов реагировали только разгибательные мышцы, в то время как мышцы-сгибатели оставались в состоянии расслабления (рис. 18).
56
Третья глава. Рефлексы положения
В условиях опытов Беритова (вероятно, очень высокая децеребрация) эта закономерность изменялась в течение последующих часов. Тонус разгибателей уменьшился, и одновременно с этим выступил более или менее выраженный тонус сгибателей. В этом состоянии в реакциях на вращение головы стали участвовать как сгибатели, так и разгибатели. Это явление изображено на рис. 19.
Но и в тех случаях, когда после децеребрации ригидность хорошо развита и вследствие этого тонические лабиринтные и шейные рефлексы влияют только на разгибатели, можно наблюдать реципрокные отношения при раздражении чувствительного нерва на регистрируемой лапе (появлс-
Рис. 19. (По Беритову). Правое боковое положение. 13 часов после децеребрации. У этого животного отпрепарированы на левой задней ноге vastocrureus (разгибатель колена) и semitendinosus (сгибатель колена). Верхняя кривая соответствует vastocrureus, нижняя — semitendinosus. При SB голову вращают теменем вверх, при КВ — теменем вниз, притом так, что по возможности достигается положение максимума и минимума для лабиринтных рефлексов. Видно, что при В каждый раз разгибатель расслабляется, а сгибатель сокращается, при КВ — разгибатель сокращается, а расслабляется сгибатель. В конце опыта голову приводят в среднее положение.
ние сгибательного рефлекса). Это наглядно поясняется опытом Беритова в Утрехтской лаборатории (рис. 20). Верхняя кривая записывается от изолированной трехглавой мышцы (разгибатель), нижняя — от изолированной плечевой мышцы (сгибатель). Голова животного находится в среднем полоя^ении. Затем центральный конец лучевого нерва регистрируемой лапы раздражается индукционным током. В ответ на это осуществляется сгибательный рефлекс на раздражаемой стороне (а), который вызывает сокращение плечевой мышцы и очень незначительное расслабление трехглавой. Затем голова яшвотного поворачивается резко дорсаль- но (Ъ). Это приводит к тоническому шейному рефлексу передних ног, благодаря которому обе ноги разгибаются. На кривой это можно наблюдать в виде сильного сокращения трехглавой мышцы и наступающего одновременно с этим торможения сокращения плечевой мышцы. Из обоих конкурирующих влияний, по-видимому, постепенно начинает преобладать влияние от раздражения индукционным током, и вследствие этого сокращение трехглавой и торможение плечевой мышц прекращаются. Наконец, голову возвращают в исходное положение (с), и раздражение нерва индукционным током прекращается (d).
В отдельных опытах Беритова напряжение разгибателей полностью прекращалось и в течение многочасовых опытов развивалось напряяшние
Общий характер изменений тонуса при рефлексах положения
57
сгибателей. После этого разгибательные мышцы больше не участвовали в тонических лабиринтных и шейных рефлексах и реакция наблюдалась только в сгибательных мышцах.
По опытам Ионкхофа [Jonkhoff {70)], проведенным в нашей лаборатории, это последнее состояние можно по произволу вызывать на каждом децеребрированном животном путем отравления пикротоксином. Пикро- токсин обладает свойством превращать после децеребрации напряжение разгибателей в напряжение сгибателей. Это видно на рис. 21, а и б.
Па рис. 22 и 23 (по Ионкхофу) графически изображены реакции двуглавой (верхняя кривая) и трехглавой мышц (нижняя кривая).
Время дано в секундах. На рис. 22 видна нормальная реакция. При повороте головы теменем вниз (КВ) сокращается трехглавая мышца верхней лапы при положении животного на боку, в то время как двуглавая мышца слегка затормаживается. При обратном повороте в среднее положение (MS) сокращение трехглавой мышцы прекращается.
После интравенозного введения 1 мг на 1 кг пикротоксина картина совершенно изменяется. В соответствии с вышеописанным максимальным тонусом разгибателей теперь при отравлении пикротоксином (рис. 23) расслабленная трехглавая мышца вообще больше не принимает участия в реакциях на поворот головы. Возбуждение же двуглавой мышцы, наоборот, выступает теперь на передний план.
В то время как раньше при повороте головы теменем вниз (КВ) происходило сильное разгибание, теперь при таком же положении головы реакция совсем не наступает. Наоборот, максимум подъема кривой наблюдается в том случае, если голова поворачивается теменем вверх (SB). При этом происходит максимальное сокращение двуглавой мышцы, продолжаю- рис 2о
щееся все время, пока голова удерживается в соответствующем положении. Особенно отчетливым это становится при повторном повороте головы. Если после этого голова возвращается в среднее положение (MS), то сокращение двуглавой мышцы немедленно прекращается. На верхней кривой видны 6 судорожных сокращений вследствие отравления пикротоксином.
Интересно рассмотреть влияние отравления. В то время как до отравления, как и у всех обычных децеребрированных животных, при положении головы с наклоном в +45° происходило сокращение трехглавой мышцы, а двуглавая мышца находилась в покое или в состоянии легкого расслабления, теперь при положении головы в —135° наступало максимальное сокращение двуглавой мышцы, а трехглавая находилась в покое. Другими словами, в то время как раньше тонические шейные и лабиринтные рефлексы выражались в изменениях разгибательного тонуса, теперь выявляются исключительно изменения сгибательного тонуса, и этот сгибательный тонус имеет свой максимум при таких положениях головы в пространстве и по отношению к телу, при которых разгибательный
58
Третья глава. Рефлексы положения
тонус имеет свой минимум, и наоборот; следовательно, при положениях головы в минимуме на кривых видны самые сильные реакции.
Все описанные наблюдения, однако, не могут скрыть (завуалировать) основное наблюдение, а именно, что у децеребрированного животного
Рис. 21а. Рис. 216.
Рис. 21а. 10 ч. 40 м. Децеребрированную кошку держат в положении на спине, ротовая щель примерно в горизонтальном положении, значит, почти в положении максимума для лабиринтных рефлексов на мышцы конечностей. Передние ноги показывают сильный разгибательный тонус, задние ноги не участвуют в реакции.
Рис. 216. 12 ч. 30 м. После введения 1.2 мг на килограмм веса пик- ротоксина разгибательный тонус децеребрированной кошки превратился в сгибательный тонус. Животное держат в положении на животе в воздухе, примерно в положении минимума для лабиринтных рефлексов. Передние ноги сильно согнуты, противодействуя силе тяжести. Виден сильный сгибательный тонус передних ног, которые активно согнуты в колене и в голеностопном суставе.
в реакциях участвуют преимущественно разгибательные мышцы. То, что их надо рассматривать и при физиологических условиях как самые
важные исполнительные органы при тонических шейных и лабиринтных рефлексах, вытекает уже из того, что они как раз являются «мышцами стояния», которые обусловливают и поддерживают главным образом установку и положение тела. Влияние положения головы на позу тела, следовательно, обусловливается преимущественно зависимостью тонуса разгиба- аельных мышц (мышц стояния) от рецепторов лабиринтов и шеи.
Тонические шейные и лабиринтные рефлексы влияют еще и в том случае на мышцы конечностей, если перерезаны и соответствующие задние корешки. У одной кошки под глубоким хлороформным наркозом вскрывался канал спинного мозга в области затылка и на левой стороне экстрадурально перерезались задние корешки с С5 до Th2. Последующее вскрытие Рис> 22. показало полноту перерезки. Затем животное
децеребрировалось, и через 35 минут перерезался спинной мозг на уровне 11-го грудного позвонка. Правая передняя нога находилась в состоянии сильной ригидности, в левой ригидность была намного слабее. Однако при поднятии и
Общий характер изменений тонуса при рефлексах положения
59
опускании головы в положении животного на спине обе ноги реагировали одинаково. Когда ротовая щель была повернута на 45° над горизонтальной плоскостью, тогда тонус в обеих ногах, особенно в локте, был максимальным (причем на левой ноге он всегда был гораздо слабее, чем на правой). При изменении положения головы в дорсальном или вентральном направлении тонус уменьшался. В основном здесь можно говорить о лабиринтных рефлексах. Следовательно, при децеребрационной ригидности еще можно изменением положения головы влиять на тонус конечности, афферентные пути которой перерезаны. То же самое удалось установить Шеррингтону (11) на изолированном vastocrureus-препарате у декапити- рованной кошки, у которой также были перерезаны соответствующие зад-
Рис. 23.
ние корешки. Поворот головы вызывал типичные реакции. Итак, вызванное оцределенным положением головы возбуждение надо рассматривать как самостоятельный источник для тонуса конечностей.
Можно устранить и симпатическую иннервацию поперечнополосатых мышц, не изменяя при этом влияния тонических шейных и лабиринтных рефлексов. Это вытекает из одного наблюдения, сделанного Дюссер де Варенном, который у кошек за 3—6 недель до опыта экстирпировал на одной стороне симпатическую цепочку живота от Lx до L7 или Sx и после этого приготовил двусторонний vastocrureus-препарат. На обеих сторонах можно было одинаково хорошо наблюдать влияние тонических шейных и лабиринтных рефлексов на vastocrureus-препарат. УУ децеребрированных животных латентные периоды тонических шейных рефлексов короткие и чаще всего лежат в пределах от х/3 до 1 секунды. Только в исключительно редких случаях наблюдаются латентные периоды до 6 секунд. Напротив, лабиринтные рефлексы имеют очень непостоянные латентные периоды, которые колеблются в пределах от /4 до 23 секунд. В опыте, изображенном на рис. 16, латентный период лабиринтных рефлексов составлял IV2 секунд.
Для того чтобы отчетливее себе представить форму изменений тонуса, приведем еще некоторые кривые.
Рис. 24 показывает движение правой передней ноги у незагипсованной кошки в положении на спине. Правое плечо зафиксировано в перпендикулярном положении при помощи зажима. Правое предплечье свободно, на правой лапе висит груз в 100 г, при помощи которого стремятся согнуть
60
Третья глава. Рефлексы положения
ногу в локтевом суставе. Лапа соединена с легким пишущим рычагому разгибание ноги вызывает движение рычага вниз. В начале кривой ротовая щель повернута на 30° над горизонтальной плоскостью, голова, таким образом, находится близко от положения максимума лабиринтных рефлексов.
При 1 голова сильно наклоняется вентрально (140°), так что морда оказывается между передними ногами. В результате происходит ослабление тонуса, которое вначале протекает очень быстро, потом медленнее,, пока наконец линия не приблизится к горизонтали. Видно, что здесь
Рис. 24. Опыт 62. Кошка децеребрирована под хлороформным наркозом. Через полчаса перерезка спинного мозга у 12-го грудного позвонка. Животное лежит в положении на спине на подогретом столе, задние ноги крепко привязаны, голова удерживается головодержателем, при его помощи голова может быть фиксирована в различных положениях. Плечо так фиксировано, что сгибание и разгибание локтя могут происходить беспрепятственно и плечо стоит вертикально. Правая лапа имеет груз в 100 г, который стремится согнуть локоть. Кроме того, правая лапа связана с рычажком. Увеличение через рычаг троекратное, нагрузка рычага 1.5 г. Разгибание локтя вызывает опускание рычага. Описание опыта дано в тексте.
речь идет о длительной реакции, которая продолжается до тех пор, пока голова находится в этом положении (1/4 минуты). После возвращения головы в максимальное положение (2) сразу же опять появляется разгибательный тонус и удерживается 23/4 минуты. При 3 голова перемещается из максимального положения в дорсальном направлении (ротовая щель повернута на —20° ниже горизонтали), и сразу же тонус опять уменьшается, но не так сильно, как прежде, в положении при сильнейшем вентральном сгибании головы. Следовательно, кривая показывает, как на измененное положение головы нога реагирует новым тоническим положением покоя.
Но есть еще другой тип реакции, который также можно наблюдать более или менее часто и который иллюстрируется рис. 25.
Кошка находится на правом боку. Регистрируются движения находящейся сверху левой ноги. Разгибание вызывает движение рычага вверх. В начале опыта голова расположена симметрично по отношению к туловищу (9). При 10 — голова наклоняется влево, так что морда смодрит вверх и левая нога становится челюстной ногой. Вследствие этого наступает большое усиление тонуса. Однако, в нр хгивоположность ранее описанному опыту, это усиление происходит иначе и нога не сразу принимает новое положение тонического равновесия. В начале реакции происходит очень сильное ■ разгибание, настолько сильное, что рычаг достигает наи-
Общий характер изменений тонуса при рефлексах положения
61
высшей точки подъема. После этого тонус опять медленно уменьшается и только через две минуты устанавливается уровень тонуса, который лежит гораздо выше исходного. Эта форма реакции характеризуется, следовательно, тем, что нога сначала выполняет чрезмерно сильное движение и только после этого принимает окончательную позу. В этом случае положение головы удерживалось в течение 5 минут. Потом голова наклонилась в обратном направлении, что привело к сильному уменьшению тонуса, которое в этом случае произошло почти так же, как ив опыте № 62 (рис. 24).
Обе реакции, следовательно, можно иногда наблюдать в одном и том же опыте. При 12 — голова возвращается снова в среднее положение и конечность опять принимает приблизительно свое исходное положение.
Как в этом, так и во многих других опытах можно показать, что если голова поддерживается от 5 до 10 минут в определенном положении, то и тонус конечности так же долго бывает высоким или низким.
Если изменяется положение головы, то этим устраняется и тоническая реакция. Часто (не всегда) можно увидеть, что нога после этого принимает ту же самую исходную позицию.
Иногда можно наблюдать, что конечность при неизменном положении головы сохраняет в течение часа тонус без изменений и что рычаг при неподвижном барабане кимографа не изменяет положения. Однако в случае изменения положения головы тонус сейчас же реагирует соответствующим образом. Следовательно, при описанных рефлексах речь идет о тонических длительных реакциях необычной силы и длительности.
Уже тут бросается в глаза, что описанные реакции очень мало утомляются. Позже будет показано, что эта неутомляемость может длиться целыми днями, неделями и месяцами. Обычные фазные рефлексы и спонтанные движения, записываемые эргографом Моссо, чрезвычайно быстро утомляются. По наблюдениям Шеррингтона (5, 7), многие спинномозговые рефлексы, например чесательный, также быстро утомляются. Однако здесь описанные реакции можно практически рассматривать как неутомимые.
Даже если исключаются антагонисты и, следовательно, работает только одна изолированная разгибательная мышца, например трехглавая, то изменения тонуса, вызванные шейными и лабиринтными рефлексами, носят характер длительных реакций, что и показано на рис. 26.
Рис. 25. Опыт 68. Кошка децеребрирована под хлороформным наркозом. Через полчаса перерезка спинного мозга у 12-го грудного позвонка. Животное лежит на правом боку. Левое плечо фиксировано зажимом. К левой лапе прикреплен груз в 100 г, который стремится согнуть локоть. При разгибании поднимается рычаг. Увеличение в 1.5 раза. Время в минутах. Описание опыта дано в тексте.
62
Третья глава. Рефлексы положения
На этом рисунке видны движения изолированной правой трехглавой мышцы децеребрированной кошки, находящейся в положении на левом боку. В начале рисунка (1) имеет место средний тонус трехглавой мышцы. При 2 голова поворачивается так, что темя находится внизу, а челюсть наверху. В ответ на это сразу происходит тоническое разгибание в локте, вызванное сокращением трехглавой мышцы, которое длится все время, пока голова удерживается в этом положении (приблизительно 70 секунд).
Рис. 26. Опыт 11. Кошка децеребрирована. Изоляция правой трехглавой мышцы. Нагрузка на правую лапу в 20 г стремится согнуть локоть. Увеличение в 3 раза. При разгибании локтя рычаг поднимается вверх. Время в секундах. Животное в левом боковом положении. Искусственное дыхание. Описание опыта дано в тексте.
При 3 голова поворачивается так, что темя находится наверху, а челюсть внизу. Трехглавая мышца сразу же расслабляется, и рычаг доходит до верхнего края бумаги. Все приведенные примеры показывают, что при тонических шейных и лабиринтных рефлексах мышца при постоянном весе принимает разную длину. Иначе говоря, речь идет о положениях покоя.
В опытах с сильно выраженными рефлексами может случиться, что тоническое состояние конечности прерывается приступами чередующихся шагательных движений (7, 19). Эти движения никогда не осуществляются при минимальном тонусе конечностей, проявляясь только при среднем и максимальном уровне тонуса. Их можно в любое время затормозить, даже тогда, когда они очень сильно выражены, понижая соответствующим изменением положения головы разгибательный тонус. Если максимальный тонус вызывается только или преимущественно лабиринтными импульсами и обе передние ноги растягиваются при этом в одинаковой степени,
Комбинации шейных и лабиринтных рефлексов
63
то нельзя точно определить ногу, которая начинает шагательные движения. Но когда имеется неодинаковое влияние на ноги, вызванное поворотом или наклоном (головы), то наблюдается следующее явление: первый шаг вперед делает всегда «черепная нога», т. е. нога с более низким тонусом, причем после этого сгибается локоть.
IV. КОМБИНАЦИИ ШЕЙНЫХ И ЛАБИРИНТНЫХ РЕФЛЕКСОВ
Как выше уже было сказано, для исследования лабиринтных рефлексов надо исключить шейные рефлексы и для исследования шейных рефлексов надо препятствовать осуществлению лабиринтных реакций. Если это не делать, то всегда надо учитывать взаимодействие шейных и лабиринтных рефлексов. Как это происходит в отдельных случаях, описывается дальше.
Анализ был полностью проведен на децеребрированной кошке (7), у которой были изучены тонические реакции конечностей при всех возможных положениях головы, при самых разнообразных положениях тела в пространстве. Все наблюдаемые во многих тысячах случаев реакции можно было объяснить взаимодействием шейных и лабиринтных рефлексов. При этом можно установить следующее правило: влияние возбуждений, исходящих из шеи и лабиринтов, на разные мышечные группы суммируется алгебраически. Следовательно, сильное возбуждение наступает тогда, когда тонус определенной мышцы повышается импульсами, исходящими как из шеи, так и из лабиринтов. Мышца эта расслабляется, когда тонус ее уменьшается под влиянием шейных и лабиринтных импульсов. Состояние ее не изменяется, когда тонус в одинаковой степени повышается лабиринтными импульсами и уменьшается шейными.
Тонические лабиринтные рефлекторные влияния на конечности осуществляются тогда, когда изменяется положение головы в горизонтальной плоскости. Поэтому при любом положении тела в пространстве можно найти такое движение головы, при котором не осуществляются лабиринтные рефлексы, а только шейные; это происходит тогда, когда не изменяется положение головы в горизонтальной плоскости, т. е. если при положении животного на боку производится поднимание и опускание головы или наклон головы при положении на спине, а также при нормальном положении животного с горизонтальной ротовой щелью или при повороте головы при висячем положении головой вниз или вверх.
С другой стороны, было показано, что поднимание и опускание головы в атланто-окципитальном суставе (при фиксации середины шеи и нижних шейных позвонков) не вызывают у кошки никаких заметных шейных рефлексов, так что, следовательно, при таком движении выступают только лабиринтные рефлексы.
Если производится поднимание и опускание головы в атланто-окципитальном суставе при положении животного на боку, то ни шейные, ни лабиринтные рефлексы не появляются. Это движение головы поэтому можно использовать для того, чтобы в сомнительных случаях установить, имеют ли влияние на исход опыта другие рефлексы у животного, например прикосновение к голове или захват кожи и т. д.
При всех остальных движениях головы и положениях тела речь идет всегда о комбинации шейных и лабиринтных рефлексов.
Исследование и объяснение отдельных случаев осложняется еще тем, что у разных животных шейные и лабиринтные рефлексы развиты не одинаково. Таким образом, у некоторых животных преобладают шейные рефлексы, у других лабиринтные, в то время как у третьих обе эти группы
64
Третья глава. Рефлексы положения
рефлексов выражены приблизительно в одинаковой степени. Все это касается не только децеребрированных животных, но и интактных с функционирующим большим мозгом (16).
В этой книге не описываются все возможные комбинации. Для этого следует читать отдельные работы (7, 9). Здесь же достаточно описать только самые важные различия. Изложение в основном ограничивается фактами, полученными на кошках.
1. Положение на спине
а) Поднимание, опускание. При изолированном влиянии лабиринтных рефлексов максимум для тонуса всех четырех конечностей лежит при положении головы +45° (при положении головы затылком вниз ротовая щель немного поднята над горизонтальной плоскостью) или при чередующихся положениях головы между +45 и 0°.
Шейными рефлексами вызывается при дорсальном наклонении головы усиление разгибательного тонуса передних ног и уменьшение тонуса задних, в то время как вентральный наклон головы вызывает ослабление разгибательного тонуса передних ног и усиление тонуса задних.
Комбинированное действие лабиринтных и шейных рефлексов на передние и задние конечности децеребрированных кошек видно из следующей таблицы (опыт 86).
Положение
головы
Передние конечности
Задние конечности
+ 135°
Тонус минимальный; ак- тЕЕвное сгибание.
Тонус немного понижен.
+ 90°
Тонус минимальный; активное сгибание.
Тонус сильный.
+ 45°
Тонус максимальный.
Тонус максимальный.
— 45°
Тонус еще сильный.
Тонус минимальный; активное сгибание.
i
со
О
О
Тонус понижен.
Тонус минимальный; активное сгибание.
При положении головы +45° тонус в передних и задних ногах максимальный, голова находится в максимальном положении для лабиринтных рефлексов. Если теперь ротовая щель опускается ниже горизонтали и голова при этом сгибается дорсально, то лабиринтные рефлексы уменьшаются, но шейные рефлексы для передних ног увеличиваются, в то время как для задних ног они уменьшаются. В результате при положении головы —45° еще наблюдается сильный тонус в передних ногах, в то время как в задних ногах наблюдается минимальный разгибательный тонус и значительный сгибательный. При положении головы —90°, при котором ротовая щель направляется вертикально вниз, понижается и тонус передних ног, потому что ослабляются лабиринтные рефлексы, однако это понижение происходит не так сильно, как в задних ногах, из-за того что передние конечности еще тонизируются со стороны шеи. Обратная картина наблюдается при вентральном наклонении головы. Если голова наклоняется вентрально настолько сильно, что морда оказывается в вертикальном положении (+90°), тогда тонус передних ног снижается вследствие уменьшения влияния от лабиринтов и шеи, разгибательный тонус на передних ногах становится минимальным и наступает их активное сгибание. Но так как задние конечности при этом еще тонизируются со стороны шеи, то их разгибательный тонус остается сильным. Он только тогда немного уменьшается, когда голова сгибается максимально в вентральном направлении (+135°), так как лабиринтные рефлексы теперь сильно ослабевают.
Этот пример иллюстрирует обычное поведение животных, у которых преобладают лабиринтные рефлексы; их передние и задние ноги реаги-
Комбинации шейных и лабиринтных рефлексов
65
руют более однотипно, в то время как у животных с преобладанием шейных рефлексов передние и задние ноги реагируют не одинаково. В вышеописанном случае, при положении головы +90° (морда направлена вертикально вверх), передние ноги активно сгибались, задние ноги активно разгибались, в то время как при —45° (ротовая щель опущена ниже горизонтальной плоскости) наблюдались об р атные отношения.
Поведение децеребрированной кошки с преобладанием лабиринтных рефлексов и с перерезанным спинным мозгом в грудной области зарегистрировано на кинематографическом снимке (рис. 27).
При максимальном положении для лабиринтных рефлексов (+45°) — на рис. 27, а ж с — передние ноги выпрямлены. Как при сильнейшем вентральном (рис. 27, è), так и при дорсальном наклоне (рис. 27, d) разгибательный тонус передних ног уменьшается. Следовательно, в этом случае действие шейных рефлексов гораздо слабее, чем лабиринтных, иначе на рис. 27, d должно было бы произойти разгибание передних ног.
Рис. 27. Из ряда кинематографических снимков. Кошка децеребрирована под хлороформным наркозом. Через 3.5 часа перерезка спинного мозга у 12-го грудного позвонка. Через полчаса киносъемка. Снятый на этом же рисунке метроном дает 60 колебаний в минуту.
Рис. 27, а — положение на спине, положение головы + 45°. Передние ноги разогнуты. Затем сильнейшее вентральное сгибание головы, пока она не достигнет положения +135°.
Через 0.4 сек. после этого начинается расслабление передних ног. Еще через 5 сек. сделан рис. 27, Ь: передние ноги согнуты. Через
0.5 сек. начинается дорсальное сгибание головы, посредством которого она снова приводится в положение максимума лабиринтных рефлексов (+45°). Через 0.3 сек. начинают разгибаться передние ноги. Через 7 сек. сделан рис. 27, с: максимальное разгибание передних ног. Еще через 1 сек. происходит дальнейшее сгибание головы, вследствие чего она приводится в положение —45°. Через 0.4 сек. после этого начинается расслабление передних ног. Через 2 сек. сделан рис. 27, <7, на котором видно сильнейшее сгибание передних ног.
б) Наклон. При наклоне головы в положении на спине положение ротовой щели не изменяется по отношению к горизонтали. Вследствие этого проявляются не лабиринтные рефлексы, а только шейные. Следовательно, результат наклона головы определенный: каждый раз обе «челюстные ноги» разгибаются сильнее, а разгибательный тонус «черепных ног» понижается.
в) Поворот. При положении на спине голова находится для лабиринтных рефлексов в положении максимума. Следовательно, при повороте головы должен уменьшаться тонус разгибателей всех четырех конечностей. Шейные рефлексы, наоборот, обусловливают усиление разгибатель-
5 Р. Магнус
66
Третья глава. Рефлексы положения
ного тонуса на обеих «челюстных ногах» и уменьшение его на «черепных ногах» (рис. 28). При комбинации шейных и лабиринтных рефлексов происходит суммация лабиринтных и шейных влияний на «черепные ноги», и вследствие этого разги-
Среднее по- батвЛЬНЫЙ ТОНуС В НИХ лошение ВСегда уменьшается.
Рис. 28. Из кинематографической серии снимков. Кошка децеребрирована под хлороформным наркозом. Спинной мозг не перерезан. Сфотографировано через 1.5 часа. Рис. Поворот 28, а: положение на спине,
симметричное положение головы. Ротовая щель примерно при+30°. Передние ноги равномерно разогнуты. Рис. 28, Ъ: поворот головы так, что темя направлено к зрителю (поворот вправо). Находящаяся спереди правая нога является «черепной», поэтому сгибается, Наклон левая нога разогнута сильнее.
Рис. 28, с: голова наклонена так, что морда отведена от зрителя (наклон влево). Находящаяся впереди правая нога является «черепной», поэтому сгибается. Левая нога — «челюстная» и поэтому разгибается. (Белые лапы ретушированы черными контурными линиями для того, чтобы они лучше выделялись на белом фоне.)
В «челюстных ногах», наоборот, наблюдаются не одинаковые эффекты: При сильных шейных и лабиринтных рефлексах разгибательный тонус остается неизменным, при преобладании лабиринтных рефлексов он уменьшается, при преобладании шейных — усиливается.
2. Нормальное положение
а) Поднимание — опускание. При нормальном положении животного, когда ротовая щель опущена немного ниже горизонтальной плоскости, голова находится в положении минимума для лабиринтных рефлексов. Следовательно, в случае поднимания головы тонус всех четырех конечностей должен усиливаться благодаря лабиринтным рефлексам. К этому присоединяется еще влияние тонических шейных рефлексов на передние ноги, вследствие чего весьма усиливается разгиба- тельный тонус передних ног. Это усиление тем больше, чем больше голова отклоняется дорсально. При опускании голова снова проходит положение минимума для лабиринтных рефлексов, при этом вентральное сгибание шеи обусловливает благодаря шейным рефлексам уменьшение тонуса передних ног, в результате чего опускание головы приводит к расслаблению разгибательных мышц передних ног (рис. 29).
Влияния шейных и лабиринтных рефлексов на задние ноги противодействуют друг другу. В случае преобладания лабиринтных рефлексов животное после подъема головы поднимается на задние ноги (хотя и ме-
Комбинации шейных и лабиринтных рефлексов
67
нее сильно, чем на передние), а при опускании головы животное немного приседает. Однако в случае преобладания шейных рефлексов животное при поднимании головы приседает, а при опускании головы поднимается. Хвост часто реагирует так же, как задние ноги. Если шейные и лабиринтные рефлексы уравновешены, то при поднимании и опускании головы изменяется только положение передней части тела.
У животных с отчетливо выраженными шейными и лабиринтными рефлексами в за- опускание
головы
висимости от того, производится ли движение головы в атланто-окципитальном суставе или в средних шейных суставах, можно наблюдать, что в первом случае появляются исключительно лаби-
Рис. 29. Из кинематографической головы серии снимков. Тот же опыт, что и на рис. 27. Рис. 29, а: кошка стоит на ногах, вес передней части тела удерживается передними ногами, голова только легко поддерживается рукой. Другая рука экспериментатора поддерживает заднюю часть тела за поясничную часть позвоночника. Положение головы —135°, разгибательный тонус передних ног незначительный. Передняя часть тела близка к полу. После этого голова сгибается дорсально, пока она не приходит в положение +105°. Продолжительность движения головы 1.3 сек. Через 1 сек. после этого передние ноги начинают сильно разгибаться и достигают в течение 2.5 сек. максимального разгибания. Через 1 сек. сделан рис. 29, Ъ: передняя часть тела сильно приподнята, передние ноги разогнуты, голова фиксируется в своем _ положении только указательным пальцем, так что весь вес ложится на передние ноги.
ринтные рефлексы, в последнем — комбинация лабиринтных и шейных рефлексов.
Когда производится движение в самых нижних шейных суставах, или когда перемещается весь шейный отдел позвоночного столба по отношению к грудной области в вентральном направлении, или же производится надавливание на область 7-го шейного позвонка, то получается максимальное уменьшение тонуса всех четырех ног, так что в результате животное ложится на живот с расслабленными конечностями (vertebra- prominens-рефлекс).
б) Наклон. При наклоне головы положение ротовой щели не изменяется по отношению к горизонтальной плоскости; вследствие этого на тонус конечностей могут влиять только шейные рефлексы. Тонус в «челюстных ногах» усиливается, в «черепных» уменьшается. Передние ноги реагируют сильнее, чем задние.
При наклоне головы (например, вправо) перемещение тяжелой головы в правую сторону приводит к перемещению центра тяжести. Вот почему разгибательный тонус передней правой ноги усиливается и вследствие этого способен противодействовать большей нагрузке. Разгибание правой передней ноги поддерживается, кроме того, перегибанием грудной области влево. Этим изгибом позвоночного столба уравновешивается перемещение центра тяжести. Если теперь такое животное после наклона головы начнет бегать, то точно известно, какая нога сделает первый шаг. Это всегда
5*
68
Третья глава. Рефлексы положения
будет «черепная нога», в то время как «челюстная нога» выполняет роль опоры (опорная нога).
в) Поворот. При нормальном положении животного голова находится в положении минимума для лабиринтных рефлексов. При повороте головы ее положение изменяется, при этом благодаря лабиринтным рефлексам все четыре конечности должны будут разгибаться сильнее. Шейные рефлексы действуют так, что разгибательный тонус «челюстных ног» усиливается, а «черепных» — уменьшается. Следовательно, «челюстные ноги» испытывают совместное влияние шейных и лабиринтных рефлексов и в каждом случае наступает сильное разгибание. На «черепных» же ногах влияние обеих рефлекторных групп противодействует друг другу и в итоге их эффекты взаимно нивелируют друг друга. В случае преобладания шейных рефлексов уменьшается разгибательный тонус. В случае преобладания лабиринтных рефлексов, наоборот, этот тонус усиливается. Если тонус «черепных ног» сильно уменьшается, то это может привести к падению животного.
Вышеописанное поведение наблюдается тогда, когда поворот и наклон головы начинаются при нормальной позе животного с обычным положением головы и с приблизительно горизонтально расположенной ротовой щелью. Если, напротив, голова сгибается максимально дорсально так, что морда стоит перпендикулярно вверх, то при повороте головы ротовая щель не изменяет своего положения относительно горизонтальной плоскости, и поэтому вызываются только шейные рефлексы, в то время как при наклоне головы вызывается комбинация шейных и лабиринтных рефлексов. При этом тонус на «черепных ногах» всегда уменьшается, на «челюстных» же цогах шейные и лабиринтные рефлексы противодействуют друг другу.
3. Положение на боку
а) Поднимани е—о пускание. Поднимание и опускание в положении на боку никаких лабиринтных рефлексов не вызывают. Это движение может служить проверкой шейных рефлексов при дорсальном и вентральном сгибании головы. Если производить движение исключительно в атланто-окципитальном суставе, то чаще всего шейные рефлексы не появляются, и это движение, следовательно, можно использовать для того, чтобы убедиться, что прикосновение к голове и перемещение на подставке никаких реакций у животного не вызывают. Если, наоборот, движения происходят в середине шеи, то дорсальное сгибание вызывает разгибание, а вентральное сгибание — расслабление передних ног. В задних ногах, наоборот, наблюдается уменьшение тонуса разгибательных мышц при дорсальном сгибании и усиление его при вентральном сгибании. Эти реакции проявляются отчетливо не во всех случаях.
Если надавить на остистые отростки самых нижних шейных позвонков и таким образом переместить их вентрально, то появляется рефлекс 7-го шейного позвонка и происходит расслабление всех четырех конечностей, причем сильнее на передних ногах, чем на задних.
б) Поворот. Так как при повороте головы в боковом положении передние и задние ноги реагируют одинаково, то здесь описываются только изменения, происходящие с передними ногами. Если поворачивать голову теменем вниз, то она оказывается в максимальном положении для лабиринтных рефлексов и обе передние ноги под влиянием лабиринтов разгибаются. Если, наоборот, голову поворачивать теменем вверх, то она оказывается в минимальном положении для лабиринтных рефлексов и обе передние ноги под влиянием лабиринтов расслабляются. Это поведение
Комбинации шейных и лабиринтных рефлексов
69
действительно наблюдается у животных с преобладанием лабиринтных рефлексов. В этих условиях решающим моментом является то, оказывается ли голова теменем вверх или вниз, и отсюда понятно, что один и тот же поворот головы (например, вращение вправо) при правом или левом боковом положении животного должен оказывать совершенно противоположное действие на конечности. Шейные рефлексы, наоборот, оказывают противоположное влияние на обе передние ноги. При вращении теменем вниз верхняя нога становится «челюстной» и разгибается, нижняя нога — «черепной» и расслабляется. При вращении теменем вверх верхняя нога становится «черепной» и расслабляется, нижняя нога — «челюстной» и разгибается. Следовательно, в этом случае реакция зависит от положения головы по отношению к туловищу и при преобладании шейных рефлексов один и тот же поворот головы (например, вращение вправо) должен при обоих боковых положениях дать один и тот же результат» Описанное поведение действительно наблюдается у животных с преобладанием шейных рефлексов.
Если же у исследуемого животного шейные и лабиринтные рефлексы выражены приблизительно одинаково, то их действие суммируется для верхней и нижней ноги таким образом, как это показано в нижеприведенной таблице.
Следовательно, если при положении животного на правом боку голова поворачивается теменем вниз, то на левой верхней ноге вследствие взаимодействия шейных и лабиринтных рефлексов происходит разгибание, в то время как тонус нижней ноги не изменяется. Если же, наоборот, поворачивать голову теменем вверх, то вследствие взаимодействия шейных и лабиринтных рефлексов на верхней ноге происходит расслабление, а тонус нижней ноги не изменяется. Таким образом, становится понятным, что при одинаково выраженных шейных и лабиринтных рефлексах поворот головы в положении на боку всегда вызывает реакцию только верхней ноги, в то время как на нижней ноге не обнаруживается никаких изменений или незначительное изменение тонуса. Значит, в зависимости от того, оказывается ли животное на правом или левом боку, можно обнаружить результат поворота головы на левой или правой передней ноге.
Это поведение ничего общего с «переключением» не имеет. Здесь дело не в том, что возбуждение при рефлексе направляется по тому или иному пути в спинном мозгу, а в .увеличении или уменьшении тонуса. При этом возбуждения встречаются в центрах соответствующих мышц конечностей и или суммируются, или вычитаются.
Если при положении животного на боку поворачивать голову теменем вниз, .то при таком положении ротовую щель можно поставить приблизительно горизонтально. В этом случае у большинства животных достигается максимум для лабиринтных рефлексов. Если произвести поворот головы так, что наступает последнее положение, то вероятность получения более сильных лабиринтных рефлексов больше, чем при повороте головы при простом горизонтальном положении ротовой щели.
Положение на правом боку
верхняя (левая) нога
нижняя
(правая)
нога
Поворот головы: темя вниз
Лабиринты Шея . . .
Результат
+
+
Разгибание
+
Без изменений
Поворот головы: темя вверх
Лабиринты
Шея
Результат
Расслабление
+
Без изменений
70
Третья глава. Рефлексы положения
Поворот головы в положении животного на боку является лучшим средством для того, чтобы на децеребрированных животных быстро убедиться, преобладают ли шейные или лабиринтные рефлексы. При различных реакциях передних ног преобладают шейные рефлексы, при одинаковых — лабиринтные; в то же время, если оба рефлекса выражены одинаково, реагирует только верхняя нога, а тонус нижней ноги остается неизменным.
в) Наклон. Реакции на наклон такие же, как и на поворот, но только они выражены немного слабее.
4. Висячее положение головой вниз
а) Поворот. Когда животное висит вертикально мордой вниз, то при повороте головы появляются не лабиринтные, а типичные шейные рефлексы.
б) Наклон. Когда голова висит вертикально мордой вниз, то она находится близко от минимального положения для лабиринтных рефлексов. Поэтому наклон головы должен привести благодаря лабиринтным рефлексам к усилению разгибательного тонуса всех четырех конечностей. Благодаря шейным рефлексам «челюстные ноги» разгибаются, в то время как «черепные ноги» расслабляются. При этом «челюстные ноги» всегда разгибаются, а «черепные ноги» реагируют по-разному. При преобладании лабиринтных рефлексов разгибательный тонус их усиливается, при преобладании шейных рефлексов он уменьшается.
Если при висячем положении головой вниз произвести наклон головы при максимальном дорсальном сгибании, то, конечно, будут вызваны только шейные рефлексы, так как теперь голова не изменяет своего положения относительно горизонтальной плоскости.
в) Поднимани е—о пускание. Здесь описывается только поведение передних ног. Когда голова висит мордой вниз, то разгибательный тонус конечностей под влиянием лабиринтов почти минимальный. При дорсальном сгибании головы она сперва попадает в положение максимума для лабиринтных рефлексов (—135°), однако при более сильном дорсальном сгибании тонус должен снова увеличиться, что обычно становится очевидным, как только морда поднимается над горизонтальной плоскостью. При вентральном сгибании головы, как только морда поднимается над горизонтальной плоскостью, также наступает разгибание под влиянием лабиринтов. Итак, при преобладании лабиринтных рефлексов в таком положении получается необычный результат: самые максимальные вентральные сгибания головы способствуют усилению разгибания передних ног. Если, кроме того, имеют место четкие шейные рефлексы, то последние усиливают описанную реакцию при дорсальном сгибании головы и ослабляют ее при вентральном сгибании.
Значит, усиление тонуса при максимальном вентральном сгибании в висячем положении головой вниз доказывает влияние сильных тонических лабиринтных рефлексов на конечности.
5. Висячее положение головой вверх
Децеребрированные животные обычно плохо переносят это положение, при нем легко наступает анемия продолговатого мозга с нарушениями дыхания. Хорошо, если исследование производится быстро и если при этом развитие анемии головы задерживается наложением тугой повязки на живот.
Если морда направлена вертикально вверх, то при повороте головы появляются только шейные рефлексы. Однако если голова удерживается
Комбинации шейных и лабиринтных рефлексов
71
в нормальном положении, то и наклоном головы вызываются только шейные рефлексы.
Если же производится наклон головы при вертикально вверх направленной морде, то всегда происходит расслабление «черепной ноги», так как шейные и лабиринтные рефлексы усиливают друг друга, в то время как поведение «челюстной ноги» зависит от того, преобладает ли расслабление, исходящее от лабиринтов, или усиление тонуса, исходящее от шеи.
За редким исключением, все данные, полученные на более чем ста подопытных животных, и тысячи отдельных наблюдений подчиняются вышеприведенным правилам. До сих пор исследованные другие виды животных (собаки, морские свинки, обезьяны) подчинялись таким же законам, которые были установлены выше для кошки. Только у кроликов наблюдается другое поведение. У них поднимание и опускание головы вызывают такое же влияние шейных рефлексов на задние ноги, как и на передние, в то время как у остальных животных задние ноги реагируют противоположно передним ногам.
Из описанного вытекает, что при разных положениях мускулатура тела устанавливается соответственно каждому положению головы и что при этом ведущую роль всегда играет голова, в то время как тело следует за ней. Нам почти не известен другой нервный механизм регуляции, при помощи которого вся мускулатура животного так объединялась бы воедино, как при помощи вышеописанных рефлексов. Так как при этом речь идет о длительных реакциях, то наблюдаются характерные положения тела, при которых голова и отдельные участки тела устанавливаются гармонически к различным положениям всего тела, именно так, как нам это известно из повседневных наблюдений и как это можно совершенно закономерно вызвать на децеребрированных животных.
Если имеется определенная степень децеребрационной ригидности и известно, какие преобладают у данных животных рефлексы (шейные или лабиринтные), — или они находятся в состоянии равновесия, — то при данном положении тела, в зависимости от положения головы, отчетливо определено распределение тонуса в мускулатуре конечностей (и, между прочим, также во всей мускулатуре тела).
В дальнейшем мы увидим, что если к центрам тонических шейных и лабиринтных рефлексов в верхней части шейного отдела спинного мозга и в продолговатом мозгу присоединяются еще центры среднего мозга, то отношения значительно усложняются. Однако и здесь удалось разобраться в сложных закономерностях. Само собой разумеется, что если к этому присоединится еще влияние произвольных движений со стороны коры головного мозга, то отношения становятся еще более трудно понимаемыми и едва ли они поддаются до сего времени полному обозрению.
Различие шейных и лабиринтных рефлексов
Здесь мы еще раз резюмируем, каким образом при исследовании разных животных можно в отдельном случае различать друг от друга шейные и лабиринтные рефлексы. Это особенно важно при клиническом обследовании человека.
Сначала надо убедиться в том, имеются ли вообще тонические рефлексы, рефлексы положения, или это рефлексы, вызванные движением. Следовательно, после каждого изменения положения головы надо подождать более или менее длительное время и затем посмотреть, каким является вызванное изменение установки — проходящим или постоянным. Только в последнем случае можно говорить о рефлексах положения.
72
Третья глава. Рефлексы положения
Наличие изолированных лабиринтных рефлексов лучше всего можно доказать на децеребрированных животных, находящихся в гипсе, у которых при разных положениях в пространстве не могут осуществляться никакие изменения положения головы по отношению к туловищу. Хорошим средством для различения шейных рефлексов от лабиринтных и для установления, преобладают ли у животного шейные или лабиринтные рефлексы, является поворот головы в положении животного на боку. Если при этом обе передние ноги реагируют одинаково, то преобладают лабиринтные рефлексы. Если они реагируют противоположно, то преобладают шейные рефлексы. Однако если реагирует только верхняя нога, в то время как положение нижней ноги не изменяется, то это значит, что шейные и лабиринтные рефлексы находятся в состоянии равновесия.
Сильно выраженные лабиринтные рефлексы имеют место и тогда, когда при висячем- положении головой вниз максимальное вентральное сгибание головы обусловливает разгибание передних лап.
На мелких животных особенно удобно перемещать голову и туловище без изменения их взаимного положения, из положения на животе в положение на спину. Если при этом наблюдается усиление разгибательного тонуса конечностей, то речь идет о лабиринтных рефлексах. Этот способ годен для ориентировочных опытов, но для точного доказательства надо предпочитать гипсовые повязки.
При каждом положении тела в пространстве имеется одно движение головы, при котором ротовая щель не изменяет отношения к горизонтальной плоскости и при котором, следовательно, лабиринтные рефлексы не появляются. Возможные изменения тонуса конечностей основываются тогда только на шейных рефлексах. Эти движения таковы: при положении на спине — и при нормальном положении — наклон головы; при висячем положении головой вниз — поворот головы; при положении на боку — поднимание и опускание головы. Если при нормальном положении голова направлена вследствие сильнейшего дорсального сгибания мордой вертикально вверх, то тогда поворот головы вызывает не лабиринтные рефлексы, а только шейные. Дальше, при положении на боку, можно вызвать изолированно У'еПеЬга-рготшепз-рефлекс (с 7-го шейного позвонка) давлением на остистые отростки нижних шейных позвонков.
Шейные рефлексы можно продемонстрировать изолированно и таким образом: голова фиксируется, ее положение в пространстве вообще не изменяется, и движения производятся только одним телом.
У. ТОНИЧЕСКИЕ ШЕЙНЫЕ И ЛАБИРИНТНЫЕ РЕФЛЕКСЫ У НОРМАЛЬНЫХ ЖИВОТНЫХ
До сих пор описанные наблюдения проводились на децеребрированных животных. Спрашивается, насколько можно эти же рефлексы показать и на нормальных животных с сохраненным мозговым стволом и большим мозгом.Что это действительно имеет место,учат нас следующие наблюдения (24).
На рис. 30 представлен нормальный кролик в обычном положении. Голова с ротовой щелью опущена немного ниже горизонтали, передние ноги согнуты.
Из рис. 31 видно, что если голову животного сгибать дорсально (поднимание головы), то передние ноги автоматически разгибаются, так что- туловище поднимается и «следует» за положением головы.
То же самое для кошки показывают оба следующих рисунка (16).
Из рис. 32 видно, что животное, глядя на мясо, лежащее на полу, сгибает голову .вентрально. В соответствии с этим передние ноги согнуты
Тонические шейные и лабиринтные рефлексы у нормальных животных
73
так, что передняя часть туловища находится близко к полу. Животное принимает такое же положение, когда оно пьет из стоящей на полу миски молоко и при этом наклоняется над миской.
На рис. 33 держат мясо высоко в воздухе, животное подняло голову, и вследствие этого наступило сильное разгибание передних ног с подни-
Рис. 30. Нормальный кролик свободно сидит с опущенной головой и согнутыми передними ногами.
Рис. 31. Тот же кролик с приподнятой головой. Передние ноги разогнуты, передняя часть тела активно поднята, спина поднимается спереди назад. Большой палец экспериментатора находится под углом нижней челюсти, чтобы поддержать голову в ее положении. Вес тела распределяется только на конечности животного.
манием передеи части туловища.
Положение задних ног мало изменилось, потому что здесь шейные и лабиринтные рефлексы противодействуют друг другу (в то время как они в передних ногах усиливают друг друга). Из этого рисунка мы можем заключить, что у этой подопытной кошки должно быть хорошо выражено влияние шейных рефлексов на задние ноги при поднимании и опускании головы.
Рис. 32. Нормальная кошка. Реак- Рис. 33. Нормальная кошка. Реакция передних ног на опускание ция передних ног на поднимание
головы (на полу лежит мясо). головы (мясо держат в воздухе).
Те же самые рефлексы можно видеть, если наблюдать бегающих по улице собак. Когда животные опускают голову, то передние ноги сгибаются в плече и в локте. Если животное поднимает морду, то оно бегает с выпрямленным плечом и локтями и высоко поднятой передней частью
74
Третья глава. Рефлексы положения
туловища. То же самое можно наблюдать у стоящих и бегающих лошадей и коров. При этом только нельзя забывать, что влияние тонических шейных и лабиринтных рефлексов на мышцы конечностей сильнее всего выступает в суставах плеча и локтя и меньше в голеностопном суставе и что у названных животных плечо и локоть лежат ближе к туловищу, в то время как свободно видимыми являются голеностопный сустав и суставы пальцев. В зоологическом саду также можно сделать многочисленные наблюдения над шейными и лабиринтными рефлексами у самых разнообразных видов животных.
Когда кошка собирается залезть в шкаф, она прижимает голову к полу и вызывает при этом посредством вентрального переме-
Рис. 34. Тот же кролик, что и на рис. 30 и 31, положен на спину, так что голова, туловище направлены точно дорсальной стороной вниз. После этого голова вращается вправо, это значит, что правое ухо движется вентрально, правый глаз смотрит вверх. Грудь остается в прежнем положении. Левая передняя нога — «челюстная» и разгибается, правая передняя нога — «черепная» и сгибается. Таз поворачивается в направлении, противоположном движению головы, так что правая задняя нога лежит внизу, левая задняя нога находится наверху. Тело животного вследствие этого повернуто винтообразно. Различие в тонусе задних ног на этом снимке распознать нельзя.
щения шеи по отношению к туловищу уеПеЬга-ргопипепзрефлекс, при котором расслабляются все четыре лапы и животное совершенно плоско лежит животом на полу.
Выше говорилось уже о том, что если кошка, стоящая в нормальном положении, наклоняет голову вправо и перемещает при этом свой центр тяжести вправо, то становится видным разгибание правой передней ноги, которое противодействует этому перемещению центра тяжести, в то время как в левой передней ноге разгибательный тонус уменьшается, левая нога часто поднимается; и если потом животное начинает передвигаться, то первый шаг всегда делается левой ногой. Подобное явление можно наблюдать у белых медведей, когда они ходят по клетке то в одну, то в другую сторону.
Ниже будет описано, что у кроликов и других видов животных после односторонней лабиринтэктомии наступает закономерный поворот (и наклон) головы. В дальнейшем мы покажем, что этим поворотом головы вызывается типичный шейный рефлекс, который ведет к усилению тонуса конечностей на стороне сохраненного лабиринта и к уменьшению тонуса на стороне, лишенной лабиринта. Разница в тонусе исчезает, если голову приводят в нормальное положение. Это доказывает, что она обусловливается тоническим шейным рефлексом на конечности, который у животных с сохраненным большим мозгом проявляется самым типичным образом и при известных условиях может наблюдаться месяцами и годами.1
На рис. 34 виден тот же самый кролик, что и на рис. 30 и 31, в положении на спине. Голова повернута направо, т. е. правый глаз смотрит вверх
1 Здесь исключается одновременно появляющаяся проходящая потеря тонуса конечностей на стороне с выключенным лабиринтом, которая является прямым следствием отсутствия лабиринта.
Тонические шейные и лабиринтные рефлексы у нормальных животных
75
ii в вентральную сторону животного. Вследствие этого благодаря тоническому шейному рефлексу левая передняя нога является «челюстной ногой» и разгибается, в то время как правая передняя нога является «черепной ногой» и сгибается. Разница в тонусе задних ног не видна. Поворот головы в обратную сторону показан на рис. 35. Теперь левый глаз животного смотрит вверх и в вентральную сторону животного. Вследствие этого правая передняя нога является «челюстной» и разгибается, в то время как левая передняя нога является «черепной» и сгибается. Следовательно, у интактных кроликов с сохраненным большим мозгом влияние тонического шейного рефлекса на конечности в ответ на поворот можно показать с большой ясностью и уверенностью.
У нормальных морских свинок с интактным большим мозгом удается иногда при перекладывании животного из положения на животе на спину,
Рис. 35. Тот же кролик был положен на спину, так что голо- sa, грудь и таз направлены точно дорсальной стороной вниз.
После этого голова вращается влево, это значит, что левое ухо движется вентрально, левый глаз смотрит вверх. (Голова
здесь закрыта рукой экспериментатора). Грудь остается лежать в прежнем положении. Правая передняя нога — «челюстная» и разгибается, левая передняя нога — «черепная» и сгибается. Таз вращается так, что левая ягодица лежит снизу и правая задняя нога больше выступает наверх. Вследствие этого тело животного винтообразно скручено. На этом снимке нельзя распознать различие тонуса задних ног. Все положение тела является зеркальным отражением положения на рис. 34.
когда положение головы по отношению к телу не изменяется, показать тоническое разгибание передних ног (лабиринтные рефлексы). Это, однако, можно вызвать у нормальных животных только в меньшинстве случаев, в то время как у децеребрированных морских свинок эта реакция нсегда происходит с особенной силой.
Описанные до сих пор наблюдения показывают, что у названных видов животных можно и при наличии большого мозга показать как тонические шейные, так и лабиринтные рефлексы. У обезьян (59) это становится уже труднее. В то время как у децеребрированной обезьяны выявляется весьма четкое влияние тонических шейных и лабиринтных рефлексов на конечности, которое подчиняется вышеописанным закономерностям и которое можно вызвать даже при наркозе, у нормального бодрого животного б большинстве случаев вызвать эти реакции не удается. Они маскируются деятельностью верхних частей ствола головного мозга, которая проявляется в живых спонтанных движениях, в реакциях оптического происхождения и т. д. Однако у одной обезьяны после двусторонней лабиринт- эктомии при закрытых глазах удалось вызвать дорсальным сгибанием головы типичное разгибание передних ног, которое у сидящего животного привело к так называемому Adorantenstellung поднятых лап. Это наблюдение показывает, что у обезьян с интактным большим мозгом имеются названные рефлексы, хотя узнать их при простом наблюдении чаще всего невозможно. В дальнейшем мы увидим, что у человека наблюдается такаяже картина и что у взрослых тонические шейные и лабиринтные рефлексы становятся очевидными только в патологических случаях, при которых деятельность вышележащих частей мозга более или менее повреждена.
76
Третья глава. Рефлексы положения
Влияние тонических шейных и лабиринтных рефлексов на дыхание уток
Чрезвычайно интересным в этом отношении наблюдением мы обязаны Хаксли (Huxley) и Ноэлю Патону [Noel Paton (1, 2)]. Они исследовали остановку дыхания, наступающую у уток при нырянии и сопровождающуюся сильным замедлением пульса. В соответствии с более старыми наблюдениями эти авторы тоже нашли, что рефлекторная остановка дыхания при нырянии вызывается соприкосновением поверхности головы с водою. Они, кроме того, установили, что это в значительной степени поддерживается тоническими шейными и лабиринтными рефлексами. Эти наблюдения проводились частично на децеребрированных, частично на «загипнотизированных» с завязанными глазами утках.
а) Тонические лабиринтные рефлексы. При нормальном положении головы с горизонтальной щелью клюва у животных апноэ не наблюдается, но зато его удается вызвать при положении головы теменем вниз и при близких к этому положениях. У животных с хорошо выраженными лабиринтными рефлексами одного этого положения уже достаточно для остановки дыхания. При положениях головы, средних между крайними положениями, лабиринты также вызывают рефлекторную остановку дыхания. Это влияние лабиринтов устраняется их экстирпацией или кокаинизацией.
б) Тонические шейные рефлексы. Если у уток шея вытягивается из нормального изогнутого положения вперед — так, что шея и голова образуют продолжение оси тела, — и если производится дальнейшее дорсальное сгибание, то тонические шейные рефлексы вызывают рефлекторную остановку дыхания. Эту реакцию можно вызывать от всех частей шеи: от атланто-затылочного сустава, от середины шеи или от грудной части шеи.
После выключения лабиринтов можно при всех положениях головы в пространстве вызвать вытягиванием (разгибанием) шеи рефлекторную остановку дыхания; этого, однако, нельзя осуществить при действующих лабиринтных рефлексах, если голова находится в нормальном положении. Но при положении головы теменем вниз апноэ можно вызвать с легкостью. Это же отчетливо достигается при промежуточных положениях головы.
Ноэль Патон дальше установил, что у большинства уток преобладают лабиринтные рефлексы, но что, кроме того, у многих наблюдаются отчетливые шейные рефлексы, которые у отдельных животных так же сильно выражены, как и лабиринтные рефлексы. Следовательно, оказалось, что у уток, так же как и у млекопитающих, встречаются индивидуальные различия в относительной силе шейных и лабиринтных рефлексов. При естественном нырянии животных соприкосновение головы с водою, положение головы в пространстве и положение шеи должны взаимодействовать для осуществления очень важной для жизни животных остановки дыхания при нырянии.
Эти интересные наблюдения показывают, что при нырянии птиц используется точно такой же рецепторный аппарат (лабиринты и шея) и что он работает таким же образом, как и у млекопитающих. Только рефлекторная реакция совершенно другая. Те же рецепторы, связанные у названных птиц с центрами дыхания и блуждающего нерва, у млекопитающих связаны с центрами мускулатуры тела. Здесь, следовательно, видно, как один и тот же рецепторный аппарат у разных групп животных может быть использован для совершенно разных рефлекторных реакций.
Влияние тонических лабиринтных рефлексов на шейные мышцы
77
VI. ВЛИЯНИЕ ТОНИЧЕСКИХ ЛАБИРИНТНЫХ РЕФЛЕКСОВ НА ШЕЙНЫЕ МЫШЦЫ (10)
Теперь мы вернемся к млекопитающим. До сих пор описывалось нлияние тонических шейных и лабиринтных рефлексов исключительно на мускулатуру конечностей. Но, кроме того, можно доказать влияние лабиринтов на шейную мускулатуру (а у некоторых видов животных и на остальную мускулатуру туловища). Исследование лучше всего проводить на децеребрированных животных, у которых проверяется способность к сопротивлению шеи пассивным движениям в разных направлениях.
После двусторонней лабиринтэктомии не наблюдается разницы в тонусе мускулатуры шеи в зависимости от различных положений головы в пространстве. Однако если имеются отчетливые лабиринтные рефлексы, то оказывается, что тонус мышц, поднимающих голову, т. е. тонус мышц, способствующих дорсальному наклону головы, максимален в затылочном положении головы с немного приподнятой над горизонталью ротовой щелью, причем этот тонус наблюдается во всех мышцах, поднимающих голову. Эти мышцы у децеребрированных животных при таком положении головы обычно не имеют никакого тонуса.
При противоположном положении головы, т. е. в нормальном положении с немного опущенной ниже горизонтали ротовой щелью, мышцы, поднимающие голову, имеют минимальный тонус, хотя у децеребрированных животных этот тонус все еще заметен; в это время тонус в мышцах, нагибающих голову, заметно усиливается; его легко можно обнаружить по сопротивлению дорсальному наклону головы.
Положение максимумов и минимумов для тонических лабиринтных рефлексов на мышцы шеи оказывается, следовательно, таким же, как и для лабиринтных рефлексов на мускулатуру конечностей. Оно характеризуется теми же индивидуальными вариациями (0 и +45°, 180 и —135°). При этом оказывается, что у каждого животного положение максимума и минимума для рефлексов на мускулатуру конечностей и на мускулатуру шеи одинаково.
При всех остальных положениях головы в пространстве величина тонуса затылочных мышц лежит между названными крайними точками. Следовательно, можно сказать, что тонус мышц, поднимающих голову, изменяется так же, как и тонус разгибательных мышц конечностей. Имеется только одна характерная разница между обеими группами рефлексов.
Как раньше уже было сказано, один лабиринт может рефлекторно изменять тонус мускулатуры конечностей обеих сторон тела. Если у де- церебрированного или у нормального животного выключить лабиринт, то часто находят (однако не всегда), что ноги той стороны тела, на которой отсутствует лабиринт, более вялы, чем ноги другой стороны. Эта разница в тонусе является проходящей и может вообще отсутствовать. Однако имеет ли место такая разница или нет, в обоих случаях сохраненный лабиринт способен при изменении положения головы в пространстве влиять в одинаковом смысле и с одинаковой силой на тонус всех конечностей. Один лабиринт, следовательно, имеет влияние на тонус конечностей обеих сторон тела.
Однако в опытах с тоническими лабиринтными рефлексами на шейную мускулатуру отчетливо выяснилось, что отношение лабиринтов к мускулатуре шеи является другим. Один лабиринт влияет исключительно или по крайней мере преобладающим образом на шейную мускулатуру только одной стороны.
78
Третья глава. Рефлексы положения
При лабиринтэктомии у децеребрированного животного или при выключении лабиринта кокаином вскоре становится заметным известное отклонение головы; голова поворачивается и наклоняется в сторону отсутствующего лабиринта. Сразу же после операции у кошек и собак преобладает наклон головы, в то время как у кроликов более сильно выражен поворот (причем в течение нескольких дней после операции у недецеребри- рованных кошек и собак также преобладает поворот). Этот поворот головы после односторонней лабиринтэктомии мы обозначили — по причинам, о которых будет говориться дальше, —как «основной поворот». Его можно показать и на децеребрированных животных, так как он зависит от центров, лежащих в продолговатом мозгу позади уровня входа в него 8-го нерва. Если исследовать у животного, лишенного на одной стороне лабиринта, изменение тонуса затылочных мышц, то сперва можно установить, что в положении максимума (0 до +45°) сгибание и перемещение головы вентрально все еще наталкиваются на более сильное сопротивление, чем в положении минимума. Эта разница отчетлива и в том случае, если сохраняется лабиринтное отклонение головы, а также если корригируют поворот и наклон головы и приводят голову в симметричное положение относительно туловища, при этом наблюдается отчетливое мышечное сопротивление. Если, например, экстирпирован правый лабиринт и голова вследствие этого повернута и наклонена вправо, то при повороте и наклоне головы влево можно заметить сильное сопротивление тонически укороченных мышц, поворачивающих и наклоняющих затылок вправо. И наоборот, нет никакого сопротивления при наклоне вправо и очень незначительное сопротивление при повороте вправо. Таким образом, на обеих сторонах шеи появляется разница в тонусе затылочных мышц.
Если теперь поместить такое животное в различные положения в пространстве, то можно заметить, что тонус прежде расслабленных мышц, (при правосторонней лабиринтэктомии это мышцы, поворачивающие и наклоняющие влево) при этом не изменяется. Как при положении минимума, так и при положении максимума головы их тонус остается минимальным (в нашем примере поворот и наклон вправо при всех положениях головы в пространстве не наталкиваются ни на какое сопротивление)..
По-другому изменяется тонус тех мышц, которые после лабиринтэктомии сокращаются (в нашем примере это мышцы, поворачивающие и наклоняющие голову вправо).
В многочисленных опытах на кошках, собаках и кроликах можно было показать, что тонус этих мышц в положении максимума головы (от О1 до +45°) больше, чем в положении минимума (от 180 до —134°). Следовательно, в положении животного на спине корреляция отклонения головы после лабиринтэктомии наталкивается на более сильное сопротивление, чем в положении на ногах. Те изменения тонуса, которые при этом, можно наблюдать, никогда не были такими значительными, как в описанном выше случае для мышц, поднимающих и опускающих голову при наличии обоих лабиринтов. Эти изменения в удачных опытах выступают настолько отчетливо, что в правильности этих данных нет сомнений. При положении минимума мышц головы односторонний тонус мышц, наклоняющих и поворачивающих голову, полностью никогда не исчезает; следовательно, отклонение головы остается и в положении животного на ногах. Только разница в тонусе затылочных мышц обеих сторон в положении максимума гораздо больше, чем в положении минимума, так как напряжение мышц, которые после односторонней лабиринтэктомии остаются вялыми, заметно не изменяется при разных положениях головы в пространстве. Такое поведение можно наблюдать не только у децеребриро-
Влияние тонических лабиринтных рефлексов на шейные мышцы
79
ванных животных, но и у животных с односторонне разрушенным лабиринтом и с сохраненным большим мозгом. У кроликов с односторонне разрушенным лабиринтом при сильно выраженном повороте головы описанные явления лучше всего можно проиллюстрировать таким образом: животное держат в положении на спине в воздухе, поддерживая одной рукой позвоночный столб, а другой охватывая шею и голову, и следят при этом за тем, чтобы сгибание (дорсо-вентральное) головы и шеи по отношению к туловищу не изменилось; затем поворот головы можно скорри- гировать, вращая голову до тех пор, пока не почувствуется хорошо выраженное эластичное сопротивление. Если после этого, ничего не изменяя в положении животного, поворачивать его постепенно через положение на боку в положение на ноги, то чувствуется, как эластичное сопротивление на шее уменьшается уже при положении на боку и достигает минимума в положении на ногах. Если вращать животное обратно в положение на спину, то чувствуется возврат эластичного сопротивления. Результаты исследований видны на следующей таблице.
Децеребрированный кролик. Левый лабиринт выключен 20%-м кокаином
Сопротивление головы по отношению к смещениям
Положение на спине +45°
Положение на боку
Положение на ногах —135°
Дорсальное перемещение. . . .
Ноль.
Отчетливо.
Вентральное перемещение. . . .
Сильно.
—
Слабо.
Наклон влево
Ноль.
Ноль.
Ноль.
Наклон вправо
Сильно.
Средне.
Слабо.
Поворот влево
Слабо.
—
Слабо.
Поворот вправо
Отчетливо.
Слабее.
До сих пор было невозможно определить отдельные мышцы шеи, вступающие в деятельность при поднимании, опускании, повороте и наклоне в разных направлениях. Если мы ранее для простоты изложения говорили о мышцах, поднимающих, опускающих, наклоняющих и поворачивающих голову, так, как будто речь при этом шла о разных мышцах, то сейчас мы должны указать на то, что в этих движениях часто участвуют одни и те же мышцы.
Из этого можно заключить об одностороннем влиянии каждого лабиринта на шейные мышцы. При односторонней лабиринтэктомии определенные группы шейных мышц утрачивают свой тонус, который у животных с сохраненным большим мозгом не восстанавливается даже через недели и месяцы. Таким образом, влияние одного лабиринта на шейные мышцы остается односторонним и в течение долгого времени не наступает никакой компенсации путем замещения функций одного лабиринта другим.
После односторонней лабиринтэктомии не только шея, но и туловище спирально изгибается. Если выпрямить голову по отношению к грудной части тела и таким образом устранить поворот шеи, то у морских свинок, собак и обезьян это спиральное вращение туловища исчезает. Следовательно, оно вторично зависит от поворота шеи. Однако у кроликов и кошек после выпрямления головы поворот туловища частично еще остается. Значит, здесь, кроме шейного рефлекса, осталось еще непосредственное влияние целого лабиринта на мускулатуру тела. У этих животных влия¬
80
Третья глава• Рефлексы положения
ние лабиринтов распространяется не только на шейную мускулатуру, но и на мускулатуру всего туловища.
Поворот головы в случае односторонней лабиринтэктомии происходит и после перерезки трех пар верхних шейных задних корешков. Таким образом, проприоцептивные рефлексы здесь не участвуют и речь идет о прямом влиянии лабиринтов на мускулатуру.
Центры тонических лабиринтных рефлексов лежат ниже уровня входа 8-го нерва в продолговатый мозг и функционируют еще и после перерезки прямо перед его входом.
Из этих наблюдений вытекает, что каждый лабиринт функционально связан с шейными мышцами (у некоторых животных и с мышцами туловища) с одной стороны тела. Под «шейными мышцами одной стороны тела» нельзя понимать мышцы только правой или только левой стороны тела, но следует иметь в виду мышцы, вызывающие наклон и поворот вправо или наклон и поворот влево, причем неизвестно, на какой (анатомической) стороне тела расположены эти мышцы, а также находятся ли мышцы, наклоняющие вправо, как на правой, так и на левой стороне шеи. Если оба лабиринта интактны, то тонус мышц, цоворачивающих и наклоняющих голову вправо и влево, на шее изменяется в одинаковой степени. Это, наверное, должно приводить к увеличению и уменьшению степени фиксации шеи, а может быть, и к дорсальному или вентральному сгибанию.
Разностороннюю связь лабиринтов с мышцами конечностей и шейными мышцами можно наглядно продемонстрировать следующим сравнением. В экипаж впрягается пара лошадей, которые представляют мышцы конечностей правой и левой стороны тела. На облучке сидят два кучера, соответственно обоим лабиринтам; каждый кучер имеет в руках вожжи для обеих лошадей. Как оба лабиринта при разных положениях головы изменяют свое положение по отношению к горизонтальной плоскости всегда одинаковым образом, так и оба кучера выполняют одни и те же движения вожжами, и упряжка поэтому остается в полном порядке. Если один кучер удаляется с облучка, то другой в состоянии управлять двойной упряжкой совершенно нормальным образом.
Для выяснения влияния тонических лабиринтных рефлексов на затылочные мышцы мы должны, напротив, предполагать, что левый кучер держит вожжи левой лошади, а правый кучер — правой. Как оба лабиринта при разных положениях головы изменяют свое положение по отношению к горизонтальной плоскости всегда одинаковым образом, так и каждый кучер будет управлять своей лошадью, как и его сосед. Если, однако, удаляется один кучер, то его лошадь идет теперь без вожжей. На нее с облучка уже невозможно больше повлиять, и она пассивно следует за движением своего партнера, в то время как другой кучер продолжает управлять своей лошадью нормальным образом.
Из вышеназванных фактов следуют новые отношения между лабиринтами и мускулатурой тела. Лабиринты имеют не только непосредственное (прямое) влияние на распределение тонуса в мышцах конечностей, но воздействуют и на шею, и со стороны шеи опять вторично вызываются тонические шейные рефлексы, изменяющие тонус мускулатуры конечностей и туловища. Каждое изменение тонуса затылочной мускулатуры должно вызвать соответствующее рефлекторное изменение в тонусе мускулатуры конечностей. Таким образом, выходит очень сложная связь головы с конечностями (и также с туловищем), благодаря чему устанавливается внутренний контакт между распределением тонуса всей мускулатуры тела и положением1 головы.
Влияние положения тела на движения
81*
VII. ВЛИЯНИЕ ПОЛОЖЕНИЯ ТЕЛА НА ДВИЖЕНИЯ
Мы познакомились с закономерностями, благодаря которым положение головы управляет распределением тонуса во всей мускулатуре тела при помощи центров, лежащих позади уровня входа 8-го нерва. Таким образом, установка тела должна, со своей стороны, влиять на произвольные и рефлекторные движения. Каждой данной установке тела соответствуют определенная рефлекторная готовность и определенное распределение возбудимости во всей центральной нервной системе для различных рефлексов, так что при изменяющихся условиях установки тела с большей или меньшей легкостью может быть вызван то один, то другой рефлекс и одно и то же раздражение может вызывать разные реакции животного.
Для изучения закономерностей, играющих при этом ведущую роль, полезно упростить условия опыта и исследовать изменения рефлекторной деятельности на отдельной мышце или на изолированной группе мышц при разных положениях головы. Такие эксперименты впервые систематически были проведены Сосэном и Шторм ван Лееувеном [Socin u. Storm van Leeuwen, {19)]. При этом были исследованы рефлексы на изолированной трехглавой мышце, разгибающей локоть у децеребрированной кошки.
При этом все мышцы, которые связывают лопатку с туловищем, и все нервы плечевого сплетения, за исключением радиального, были перерезаны. Только трехглавая мышца (разгибающая локоть) осталась в связи с центральной нервной системой. Далее перерезались все остальные мышцы или все соответствующие двигательные нервы. Чувствительная ветвь лучевого нерва служила для рефлекторного раздражения. Животное находилось в положении на боку; передняя нога, с которой осуществлялась регистрация, лежала сверху, так что при повороте головы, вследствие комбинации шейных и лабиринтных рефлексов, должны были появляться особенно сильные реакции. Исследовалось влияние поворота головы и vertebra-prominens-рефлекса (рефлекс с 7-го шейного позвонка). На кривой SB («черепная нога») обозначено такое положение головы, при котором темя находится наверху, а нижняя челюсть внизу, — следовательно, положение минимума для лабиринтных рефлексов. Буквами КВ («челюстная нога») обозначено такое положение головы, при котором темя находится внизу, а челюсть наверху, т. е. положение максимума для влияния лабиринтных рефлексов на разгибательные мышцы.
Согласно опытам Шеррингтона (10), при раздражении лучевого нерва слабым током появляется на той же стороне разгибательный рефлекс, при более сильном раздражении лучевого нерва появляется сгибательный рефлекс. В соответствующих опытах можно найти такую силу раздражений, при которой разгибательный или сгибательный рефлексы появляются в зависимости от внешних условий. Беритов (1, 2) расширил эти опыты, изолируя одновременно с мышцей-разгибателем антагонистическую мышцу-сгибатель и регистрируя изменения ее длины. Он использовал в качестве разгибателя на задней лапе vastocrureus и в качестве сгибателя m. semitendinosus, на передней ноге в качестве разгибателя трехглавую мышцу, а двуглавую в качестве сгибателя, рефлексы он вызывал раздражением чувствительного нерва как гомолатеральной ноги, так и контрлатеральной. Беритов вел свои опыты много часов подряд и при этом наблюдал, что с течением времени у децеребрированных животных появлялся активный сгибательный тонус. Таким образом, он мог видеть результаты поворотов головы как при преобладании разгибательного тонуса, так и при преобладании сгибательного тонуса и при промежуточных состояниях.
Хотя эти влияния поворотов головы на сгибательные и разгибательные рефлексы многообразны, все же и здесь удалось установить основные 6 Р. Магнус
82
Третья глава. Рефлексы положения
закономерности. Результат зависит от того, появились ли в регистрируемых мышцах при изменении положения головы изменения тонуса или нет.
а) Влияние установки головы на проявление рефлекса в условиях, когда имеют место видимые изменения длины мышц, обусловленные тоническими, шейными и лабиринтными рефлексами
Если при изменениях положения головы изменяется длина регистрируемых мышц, то при этом должны произойти ранее описанные переключения по правилу Икскюля, по которому центры растянутых мышц должны быть «включены» для рефлекторного раздражения. Другими словами, на максимально разогнутой конечности с большей степенью вероятности могут быть вызваны сгибательные рефлексы, на согнутой конечности — разгибательные рефлексы.
Соответственно этому в опытах Сосэна и Шторм ван Лееувена отмечалось, что при высоком разгибательном тонусе рефлекторное торможение трехглавой мышцы было,,выражено отчетливо, рефлекторное же возбуждение было выражено слабо; наоборот, при низком разгибательном тонусе рефлекторное торможение было выражено слабо, возбуждение же, напротив, было весьма отчетливым. Для наглядности мы приводим следующие кривые из - их работы.
На этой, как и на последующих кривых, зарегистрированы гомолатеральные рефлексы на изолированной трехглавой мышце левой передней ноги децеребрирован- ной кошки, находящейся на правом боку. Установка рычага всегда так избрана, что движение рычага вниз означает сокращение мышцы, а движение рычага вверх — ее расслабление. Соответственно этому при сильном тонусе трехглавой мышцы рычаг устанавливается на более низком уровне, чем при более слабом тонусе. Если изменения тонуса вызываются вращением головы, то обозначение КВ означает, что голова поворачивается так, что верхняя нога (рефлексы которой регистрируются) становится «челюстной» ногой, т. е. в ней усиливается разгибательный тонус, в то время как при SB верхняя нога становится «черепной» ногой, — следовательно, имеет уменьшенный разгибательный тонус. Если изменения тонуса вызываются через рефлекс с 7-го шейного позвонка, то это каждый раз отмечается под соответствующими кривыми. Все рефлексы вызываются раздражением гомолатеральной ветви лучевого нерва. Раздражение наносится фарадическим током или отдельными индукционными ударами. Интервалы между двумя раздражениями составляют всегда 1 минуту. Непосредственно перед раздражением кимограф переводится на быстрый ход. Момент начала рефлекса на кривых отмечен стрелкой и, кроме того, чаще всего виден на нижней сигнальной линии.
На рис. 36 показано шесть последовательных односторонних разги- бательных рефлексов, при разгибании рычаг опускается вниз. Вначале голова находится в положении максимума (КВ), трехглавая мышца укорочена, разгибательные рефлексы незначительны, хотя мышца еще способна к дальнейшему сокращению. При SB голова переводится в положение минимума, трехглавая мышца расслабляется и разгибательные рефлексы значительно усиливаются. При последнем рефлексе мышца вновь сократилась и разгибательный рефлекс уменьшился.
На рис. 37 зарегистрированы пять последовательных односторонних сгибательных рефлексов: первые три — при максимальном положении головы, последние два — при минимальном. Видно, что сгибательные рефлексы становятся тем сильнее, чем интенсивнее было до этого сокращение трехглавой мышцы. При положении КВ они по мере расслабления трехглавой мышцы уменьшаются, при положении SB они по мере сокращения трехглавой мышцы увеличиваются. Разница величин рефлекса в зависимости от исходного состояния мышцы видна очень хорошо. В дан¬
Влияние положения тела на движения
83
ном случае расслабление мышцы во время рефлекторных ответов не было максимальным и оставалась возможность ее дальнейшего расслабления.
На рис. 38 показаны односторонние сгибательные рефлексы. Вначале, при положении головы КВ, трехглавая мышца укорочена и сгибательные
Рис. 36. Опыт 17. (Уменьшено на 3/4). Децеребрированная кошка. Изолированная трехглавая мышца. Животное проявляет преимущественно шейные рефлексы. Раздражение каждую минуту единичным индукционным ударом. Сила раздражения 2500 К. В первичной цепи — аккумулятор, во вторичной цепи — сопротивление 20 000 ом. При КВ имеется довольно сильный разгибательный тонус трехглавой мышцы (нижний уровень кривой), наблюдаются три маленьких сокращения. При SB голова поворачивается так, что испытуемая нога становится «черепной». Тонус трехглавой мышцы падает, первое рефлекторное сокращение гораздо больше, чем вызванное при положении КВ. После раздражения тонус снова не падает, но кривая устанавливается на более низком уровне. Вследствие этого следующий рефлекс слабее, уровень остается опять низким, поэтому следующий за этим рефлекс еще меньше.
рефлексы поэтому очень отчетливы, при положении SB наступает расслабление трехглавой мышцы и на этой мышце теперь вообще невозможно
Рис. 37. Опыт 12. (Уменьшено на 1/2). Децеребрированная кошка. Изолированная: трехглавая мышца. Каждую минуту подается короткое фарадическое раздражение. Сила раздражения 500 К. В первичной цепи аккумулятор, во вторичной цепи не имеется особого сопротивления. Животное проявляет преимущественно лабиринтные рефлексы. При КВ сильное рефлекторное торможение. После рефлекса мышца вновь не восстанавливает своего полного тонуса, и рычаг устанавливается поэтому на более высоком уровне. Следующий рефлекс вследствие этого немного меньше, а следующий же за этим еще меньше. Тогда при SB вследствие поворота головы тонус трехглавой мышцы падает еще больше. Первый следующий за этим рефлекс очень мал. Потом тонус снова немного увеличивается после каждого рефлекса. В связи с этим рефлексы становятся снова более сильными.
вызвать рефлексы торможения, несмотря на то что мышца была способна к дальнейшему расслаблению.
На рис. 39 сила раздражения подобрана так, что могли проявляться или разгибательные или сгибательные рефлексы. В начале кривой трехглавая мышца (при положении SB) расслаблена, и поэтому проявляются разгибательные рефлексы, после этого трехглавая мышца (при положении КВ) сокращается, и теперь на то же раздражение выявляются сгибаем
84
Третья глава. Рефлексы положения
тельные рефлексы, голова снова переводится в положение SB, трехглавая мышца расслабляется и наблюдаются разгибательные рефлексы (изменение рефлекса зависит от длины мышцы).
Рис. 38. Опыт 14. (Уменьшено на 3/4). Децеребрированыая кошка. Изолированная трехглавая мышца. Каждую минуту производят фарадическое раздражение. Животное проявляет преимущественно лабиринтные рефлексы. При КВ зарегистрированы три отчетливых рефлекторных торможения. При SB нога отчетливо реагирует падением тонуса. На последующие три раздражения нет никакой реакции в исследуемой ноге. После изменения положения головы (на кривой этого уже не видно) снова наблюдалось сильное увеличение тонуса и вследствие этого отчетливое рефлекторное торможение. Это торможение имело ту же величину, что и предыдущее. При положении КВ наступают шагательные движения, которые удается затормозить поворотом головы.
На рис. 40 показано своеобразное изменение рефлекса вследствие влияния vertebra-prominens-рефлекса. Вначале трехглавая мышца сокращена, в ответ на раздражение лучевого нерва происходит односторонний сгиба-
Рис. 39. (Уменьшено на 1/2). Децеребрированная кошка. Изолированная трехглавая мышца. У животного проявляются преимущественно лабиринтные рефлексы. Каждую минуту подаются короткие фарадические раздражения 2500 К. В первичную цепь включено сопротивление в 20 000 ом. При SB на фарадическое раздражение наступают три отчетливых рефлекторных сокращения (рычаг опускается). При КВ сильно увеличивается тонус трехглавой мышцы. На то же раздражение тогда следует отчетливое рефлекторное торможение (рычаг поднимается). После каждого рефлекса тонус немного падает. Тогда следующий рефлекс немного меньше, чем предыдущий. При SB снова сильно спадает тонус трехглавой мышцы. Следующие три рефлекса —
опять рефлекторные сокращения.
тельный рефлекс. Давление на 7-й шейный позвонок вызывает расслабление трехглавой мышцы. Рефлекторное раздражение лучевого нерва вызывает теперь разгибательный рефлекс (при с). При d давление на 7-й шейный позвонок заканчивается, тонус трехглавой мышцы снова усиливается, и теперь происходит (при е) рефлекторное торможение тонуса трехглавой мышцы (сгибательный рефлекс).
Все эти кривые наглядно поясняют один и тот же закон: что положение соответствующей конечности имеет переключающее влияние на рефлекторный процесс в зависимости от длины мышцы. Когда разгибатель- ная мышца сокращена, то ее можно только незначительно возбудить, но сильно затормозить. Когда разгибательная мышца расслаблена, тогда
Влияние положения тела на движения
85
ее можно сильно возбудить, но незначительно затормозить. При сильном разгибательном тонусе конечности вызываются преимущественно сгибательные рефлексы, при слабом разгибательном тонусе — разгибательные рефлексы.
Рис. 40. Опыт 7. (Не уменьшено). Условия опыта такие же, как и на рис. 39. При а (при среднем положении головы) на фарадическое раздражение происходит отчетливое рефлекторное торможение. При Ъ вызывается отчетливый vertebra-prominens- рефлекс (поднятие рычага). При с в ответ на фарадическое раздражение той же силы, как при а, наблюдается отчетливое рефлекторное сокращение (опускание рычага). При d прекращается давление на 7-й шейный позвонок. Следующий рефлекс при е — снова торможение.
б) Влияние положения головы на проявление рефлекса, когда тонические шейные и лабиринтные рефлексы не вызывают заметных изменений тонуса
мышц тела
Если изменения положения головы не вызывают никаких видимых изменений тонуса мускулатуры конечностей, тогда не могут осуществляться и переключения. Если установка тела не изменяется, то должны прекратиться и все вторичные, обусловленные ею рефлекторные влияния с периферии. При этих условиях лучше всего можно изучать, какое влияние имеет положение головы как таковое на рефлекторную возбудимость центров, расположенных в спинном мозгу, и на остальные центры нервной системы. Те опыты, в которых изменения положения головы не вызывают никаких изменений тонуса конечностей, надо с экспериментально-технической точки зрения рассматривать как неудачные опыты. Но именно потому, что не происходят никакие переключения, они особенно поучительно разъясняют влияние, которое оказывают тонические шейные и лабиринтные рефлексы непосредственно на распространение возбудимости в центральной нервной системе.
При таких условиях выяснилось следующее. Если изменением положения головы не вызывается ожидаемая реакция и в мышцах конечностей не происходит никакого изменения тонуса, то последующими чувствительными возбуждениями часто можно еще вызвать первоначально предполагаемое изменение тонуса. Если, например, было выбрано положение го- ловчы, которое вообще должно было привести к разгибанию исследуемой ноги, то последующим афферентным раздражением с особенной легкостью часто можно вызвать разгибательный рефлекс. В центральной нервной системе изменением положения головы до некоторой степени была вызвана готовность и тенденция к разгибанию, которая обнаруживалась при последующем рефлексе. Если голову установить в такое положение,
86
Третья глава. Рефлексы положения
что ожидается расслабление исследуемой ноги, то центры настраиваются на готовность к сгибанию, и этим облегчается последующий вызов сгибательных рефлексов.
Следующие примеры подтверждают только что сказанное.
Рис. 41. Опыт 15. (Не уменьшено). Децеребрированная кошка. Изолированная трехглавая мышца. У животного проявляются преимущественно шейные рефлексы. Каждую минуту подаются короткие фарадические раздражения. Сила раздражения 100 К. В первичной цепи — аккумулятор, во вторичной цепи особого сопротивления не включено. При SB зарегистрированы два рефлекторных раздражения. При КВ испытуемая нога становится «челюстной». Сперва не наблюдается никаких видимых изменений тонуса. Первый рефлекс в этом положении меньше, чем в предыдущем положении SB. Верхняя сигнальная линия — время в секундах. Перед началом раздражения кимограф включен на быстрый ход.
На рис. 41 голова находится сначала в положении минимума и поворачивается при КВ в положение максимума. Длина трехглавой мышцы при этом не изменяется. Отчетливо видно, что гомолатеральный сгибательный рефлекс гораздо сильнее выражен в положении минимума, при ко-
Рис. 42. Опыт 13. (Уменьшено на 3/4). Децеребрированная кошка. Изолированная трехглавая мышца. Животное проявляет преимущественно шейные рефлексы. Каждую минуту подаются короткие фарадические раздражения. Сила раздражения 1500 К. В первичную цепь включен аккумулятор. При изменении положения головы наступают видимые изменения тонуса трехглавой мышцы. При КВ три раза были вызваны торможения примерно одинаковой интенсивности. Теперь при повороте головы при а на более высоком уровне рычага происходит меньшее торможение. При Ъ уровень рычага немного опустился и рефлекторное торможение стало больше. При с уровень все еще немного выше, чем при КВ, но, несмотря на это, вызванный здесь рефлекс почти в два раза больше, чем при
положении КВ.
тором, как выше сказано, в центральной нервной системе осуществляется тенденция к сгибанию, чем в положении максимума, при котором имеет место тенденция к разгибанию.
Влияние, которое в этом случае вызывается поворотом головы, прямо противоположно тому, каким оно должно было бы быть при переключении.
Влияние положения тела на движения
87
Если при КВ, когда голова повернута в положение максимума, трехгла- вал мышца действительно сократилась бы, то этим должен был бы усилиться и сгибательный рефлекс; но так как никакого сокращения трехглавой мышцы не наступает, то измененное влияние на центральную нервную систему становится теперь очевидным.
Это взаимное противодействие между переключением и тенденцией и сгибанию отчетливо видно на рис. 42. Здесь в начале опыта голова находится в положении максимума (КВ), при этом вызываются три гомолатеральных сгибательных рефлекса, которые незначительны по величине, потому что положением головы обусловлена тенденция к разгибанию.
Рис. 43. Опыт 5. (Не уменьшено). Децеребрированная кошка. Изолированная трехглавая мышца. Каждую минуту подаются короткие фарадические раздражения. В первичную цепь включен аккумулятор, во вторичную — сопротивление в 20 000 ом. Сила раздражения 1250 К. При SB раздражение вызывает рефлекторное торможение. При КВ испытуемая нога становится «челюстной». В трехглавой мышце немного увеличивается тонус. Уровень рычага спадает. Следующее раздражение не вызывает никакой реакции. Тонус трехглавой мышцы постепенно падает и становится таким же или немного меньше, как при положении SB. Следующий рефлекс — отчетливое рефлекторное сокращение. Уровень снова поднимается. Третий рефлекс при положении КВ — снова сокращение.
При SB голова поворачивается в положение минимума для разгибатель- ного тонуса, и при с видно, что теперь, вследствие тенденции к сгибанию, огибательный рефлекс на одно и то же раздражение стал гораздо сильнее. При с именно трехглавая мышца приобрела первоначальную длину. При а ж Ъ трехглавая мышца, наоборот, еще расслаблена. Вследствие этого переключения сгибательные рефлексы при а и Ъ значительно меньше, чем при с. Таким образом, при а влияние тенденции к сгибанию немного компенсировано наступающим переключением, обусловленным изменением мышцы.
Вторая часть кривой на рис. 43 соответствует положению головы в максимуме (КВ), и поэтому преобладает тенденция к разгибанию. Го- молатеральный разгибательный рефлекс сильно выражен при с; при b, где мышца более сокращена, он немного слабее, а при а, где имеет место еще большее сокращение трехглавой мышцы, разгибательный рефлекс равен нулю. В этом случае влияние переключения видно отчетливо. Вначале голова находится в положении минимума (SB) и поэтому преобладает тенденция к сгибанию. Длина мышцы соответствует при этом ее длине в участке кривой между а и Ъ, Следовательно, если бы голова находилась в положении максимума, можно было бы ожидать более слабого разгибательного рефлекса. Вместо этого в ответ на точно такое же раздражение наступает теперь сгибательный рефлекс. В этом случае,
88
Третья глава. Рефлексы положения
следовательно, поворотом головы было обусловлено изменение результата раздражения, потому что в центральной нервной системе имела место готовность к сгибанию или разгибанию.
Рис. 44. Опыт 3. Децеребрированная кошка. Изолированная трехглавая мышца. У животного проявляются преимущественно шейные рефлексы. Каждую минуту — раздражение единичным ударом индукционного тока. Сила раздражения 2500 К. В первичную цепь включен аккумулятор, во вторичную — сопротивление в 20 000 ом. При КВ на единичный индукционный удар происходит отчетливое рефлекторное сокращение. При SB положение головы так изменено, что испытуемая нога становится «черепной». При этом нет заметного изменения тонуса трехглавой мышцы на кимо- грамме. Поэтому последующие рефлексы минимальны. При КВ испытуемая нога становится «челюстной». Опять нет заметного изменения тонуса, однако рефлексы снова становятся более сильными. При SB они становятся опять минимальными.
На рис. 44 зарегистрировано девять последовательных разгибатель- ных рефлексов. Если голова поворачивается в положение максимума (КВ), то эти разгибательные рефлексы чрезвычайно сильны, в то время
Рис. 45. (По Беритову). Препарат 32. Левое боковое положение через 3 J/2 часа после децеребрации. Через каждый 4—5 минут производится одно и то же раздражение лучевого нерва противоположной стороны. В опыте А голова находится в среднем положении, в опыте В и D — в максимальном положении для разгибательного тонуса трехглавой мышцы, в опыте С — в положении минимума для разгибательного тонуса трехглавой мышцы. Верхний ряд показывает сокращение изолированной трехглавой правой мышцы. Эффект раздражения при А меньше, чем при В и D, но больше, чем при С.
как при положении минимума (SB) они очень слабы. Здесь, следовательног тенденция к разгибанию при положении максимума головы особенно отчетлива. В этом опыте в ответ на поворот головы вообще не наступает никаких изменений длины трехглавой мышцы.
Описанный случай, а именно появление тенденции к разгибанию и сгибанию вследствие поворота головы при отсутствии изменений длины мышцы, был особенно отчетливо выявлен Беритовым. В его продолжительных опытах (в течение многих часов) у децеребрированных животных
Влияние положения тела на движения
89
можно было видеть все степени разгибательного тонуса, промежуточного состояния и сгибательного тонуса, и чаще всего рефлексы у них были одновременно зарегистрированы на сгибательных и разгибательных мышцах. Мы приведем еще некоторые примеры из работы Беритова (2).
На рис. 45 зарегистрированы четыре последовательных перекрестных разгибательных рефлекса правой трехглавой мышцы кошки. При А голова находится в промежуточном положении. При В и Д — в положении максимума для лабиринтных рефлексов, поэтому преобладает тенденция к разгибанию и разгибательный рефлекс отчетливо повышен и растянут во времени. При G голова находится в положении минимума, и разгиба-
Рис. 46. (По Беритову). Децеребрированная кошка № 58. Правое боковое положение через 12х/4 часа после операции. Верхняя кривая — левый vastocrureus, вторая кривая — левый semitendinosus. Раздражение правого малоберцового нерва. Время в секундах. При А голова находится в максимальном положении для разгибательного тонуса, при В — в положении минимума для разгибательного тонуса. При В наступает долго длящееся тоническое сгибание.
тельный рефлекс поэтому весьма ослаблен. В положении максимума тенденция к разгибанию наибольшая, в положении минимума — наименьшая.
На рис. 46 (верхняя линия) зарегистрированы изменения длины vastocrureus’а, внизу — изменения длины полусухожильной мышцы. Вначале (А) голова находится в положении максимума для разгибательного тонуса. Преобладает тенденция к разгибанию. В ответ на раздражение
n. peroneus противоположной стороны происходит сильное сокращение разгибательной мышцы (vastocrureus), которое продолжается дольше, чем раздражение, в то время как m. semitendinosus остается в покое. После этого голову поворачивают в положение минимума для разгибательного тонуса (VP). Vastocrureus расслабляется, a m. semitendinosus кратковременно сокращается. В ответ на раздражение правого n. peroneus теперь происходит значительно более слабое сокращение vastocrureus’а, которое с прекращением раздражения сразу же исчезает. M. semitendinosus остается во время раздражения в покое, однако после прекращения раздражения эта мышца очень сильно тонически сокращается, что продолжается длительное время. В опыте А отчетливо проявляется тенденция к разгибанию, в опыте В — тенденция к сгибанию. При А отчетливо видно длительное последействие сокращения vastocrureus’а после прекращения раздражения, мышца вообще не возвращается к своей первоначальной длине, пока голова при VP не примет другого положения.
90
Третья глава. Рефлексы положения
Беритов опубликовал еще целый ряд кривых, на которых отчетливо видно, что при преобладании тенденции к разгибанию разгибательные мышцы обнаруживают длительное тоническое последействие, в то время как при преобладании тенденции к сгибанию это тоническое последействие на разгибателях отсутствует и обнаруживается при определенных условиях на сгибателях.
На рис. 47 зарегистрированы реакции плечевой (верхняя линия) и трехглавой мышц (внизу) при раздражении обоих лучевых нервов. При А голова находится в положении минимума для разгибательного
тонуса, вследствие этого преобладает готовность к сгибанию. Во время раздражения контралатерального лучевого нерва (разги- бательный рефлекс) происходит только незначительное движение. Как только к этому присоединяется раздражение гомолатераль- ного лучевого нерва, происходит сильное сокращение сгибателя. Обратное соотношение видно при В, где преобладает готовность
Рис. 47. (По Беритову). Препарат 28. Левое боковое положение, через 10.5 часов после децеребрации. Верхняя кривая — правая плечевая мышца, вторая кривая — правая трехглавая мышца. Первый сигнал (R. d.) — раздражение правого лучевого нерва, второй сигнал (R. s.) — раздражение левого лучевого нерва. Время — 0.2 секунды. При А голова находится в минимальном положении для разгибательного тонуса, при В — в максимальном положении. В обоих случаях раздражается сперва контрлатеральный лучевой нерв, потом при продолжающемся его раздражении присоединяют раздражение лучевого нерва той же стороны. Раздражение первого заканчивается раньше. Раздражение производилось каждый раз через 6 минут после ■ соответствующего
поворота головы.
мышцы к разгибанию. Теперь раздражение контралатерального лучевого нерва вызывает сильное сокращение разгибателя и только незначительно влияет на сгибательную мускулатуру.
И совершенно индифферентные раздражения могут воздействовать на сгибатель или разгибатель в зависимости от положения головы (рис. 48).
На рис. 48 изображен опыт, в котором животное производит ритмические движения конечностями синхронно с дыханием. Когда голова поворачивается в положение минимума для разгибательного тонуса, так что преобладает готовность к сгибанию (А), тогда выступают ритмические сокращения только двуглавой мышцы, в то время как в положении максимума головы для разгибательного тонуса (В) двуглавая мышца остается в покое, а трехглавая — производит ритмические движения. В этом опыте были выключены лабиринты, так что речь идет исключительно о влиянии тонических шейных рефлексов.
Все эти наблюдения показывают одно, а именно — что при изменении положения головы в нервных центрах мускулатуры тела устанавливается определенная тенденция к движению. Последующее афферентное раздражение находит тогда центры в состоянии некоторой «готовности», так что рефлекс или облегчается или затрудняется. Следовательно, каждое положение тела сопровождается определенным распределением «готовности» к рефлексу Bi центральной нервной системе.
Влияние положения тела на движения
91
Эти экспериментальные данные дают следующие представления о влиянии установки тела на движение.
Если тоническими шейными и лабиринтными рефлексами действительно вызывается новое положение тела, то этим обеспечивается переключение, которое обусловливается, во-первых, тем, что при новом положении тела определенные группы мышц растянутый вследствие этого соответствующие им центры находятся, по правилу Икскюля, в состоянии повышенной рефлекторной возбудимости. Во-вторых, тело при каждом новом положении в определенных местах будет соприкасаться с полом и этим будут вызываться переключающие влияния, точно так же, как это раньше было показано на спинномозговой собаке, находящейся в положении на боку, где возможно было осуществление переключения чесательного рефлекса раздражением кожных нервов. Следовательно, во всех этих случаях при определенном положении тела вызываются афферентные возбуждения, частично проприоцептивного, частично экстероцептивного происхождения, которые действуют в центральной нервной системе переключающим
Рис. 48. (По Беритову). Децеребрированный препарат 35. Лабиринты экстирпированы и выключены кокаином. Левое боковое положение через 3 часа после операции. В опыте А голова находится в минимальном положении для разги- бательного тонуса, у препарата тенденция к сгибанию. При В голова находится в максимальном
положении для разгибательного тонуса, у препарата тенденция к разгибанию. В первом случае трехглавая мышца находится в покое {нижняя кривая), двуглавая мышца сокращается ритмично, синхронно дыханию {верхняя кривая). При В двуглавая мышца находится в покое, а трехглавая ритмически сокращается одновременно с дыханием.
образом. Они заставляют новое приходящее возбуждение идти по определенным путям к определенным центрам и обусловливают степень готовности к осуществлению рефлекса в разных центрах. Во всех этих случаях речь идет о влиянии афферентных возбуждений, которые обусловливаются новым положением тела.
Кроме того, тонические шейные и лабиринтные рефлексы непосредственно влияют на распределение возбудимости в центральной нервной системе. Это становится очевидным прежде всего тогда, когда но каким- либо причинам при определенном положении головы не осуществляется соответствующая установка тела и поэтому не могут проявиться вышеназванные переключения. Тогда имеют место только эти центральные влияния. В результате этого в центральной нервной системе происходит соответственно положению головы и изменение распределения возбудимости. В одних конечностях или в их центрах появляется тенденция к сгибанию, в других — к разгибанию и т. д. Если теперь какое-либо раздражение поступает в центральную нервную систему, то оно находит одни центры «включенными», другие «выключенными» или более или менее возбудимыми и может поэтому вызвать такую же реакцию, какую вообще должны вызывать тонические шейные или лабиринтные рефлексы. Таким образом, одно и то же раздражение может вызвать в зависимости от положения тела или сильную, или слабую реакцию, или даже ее извра¬
92
Третья глава. Рефлексы положения
щение. Даже индифферентные раздражения, вообще не приводящие к изменениям положения, могут, таким образом, вызвать новые установки тела.
Беритов (2) попытался объяснить только что названные процессы переключениями. Но мне кажется это недостаточно доказанным; речь могла бы идти только об алгебраической суммации возбуждений в определенном отдельном центре, поступающих туда частично от лабиринтов и шеи, частично от других афферентных нервов.
Поэтому при разных положениях тела совместно проявляются как периферические переключающие влияния, так и изменения возбудимости центральных образований. Это изменяет, распределение рефлекторной возбудимости в центральной нервной системе и тем самым обусловливает реакции, соответствующие принятым установкам тела. И в этом случае выходит, что центральная нервная система не представляет собой неподвижного соединения определенных рефлекторных центров, а приспосабливается целесообразно к данным состояниям тела, так что каждой установке тела соответствует определенная готовность к осуществлению рефлекса и распределение рефлекторной возбудимости. Основные действующие здесь закономерности были, как показано выше, выяснены, но потребуется еще много работы, чтобы исследовать, какой вид будут принимать эти явления в каждом отдельном случае.
Следует только обсудить еще один пункт, о котором частично уже говорилось выше, а именно — закономерности, выступающие при беге и ходьбе.
Когда животное находится в нормальном положении и наклоняет голову, то передняя «челюстная нога» становится опорной ногой и передняя «черепная нога» всегда делает первый шаг. Локомоция начинается, следовательно, всегда с той конечности, в которой наблюдается наименьший разгибательный тонус. Если приготовить на децеребрированной кошке вышеописанным образом изолированный препарат трехглавой мышцы и переместить животное в положение на бок изолированной трехглавой мышцой вверх (нижележащая нога остается интактной), то на таких животных можно иногда наблюдать спонтанные шагательные движения. У животных с преобладанием шейных рефлексов не имеет значения, в каком положении находится голова. Все равно, находится ли темя наверху или внизу; в одной из передних ног будет максимальный, в другой передней ноге — минимальный тонус, и поэтому животное будет в состоянии произвести первый шаг. По-другому обстоит дело у животных с преобладанием лабиринтных рефлексов. Если у таких животных вращать голову теменем вверх, тогда обе ноги будут иметь минимальный разгибательный тонус и шагательные движения сразу же затормаживаются. Если вращать голову теменем вниз, тогда разгибательный тонус в обеих передних ногах становится максимальным щ в ответ на это сразу же выступают сильные шагательные движения (рис. 49).
Следовательно, в этих случаях шагательные движения начинаются только тогда, когда в обеих передних ногах тонус максимален. То же самое часто можно наблюдать, когда животных перемещают в положение на спину. Если голова находится в положении максимума для разгиба- тельного тонуса передних ног, то они часто выполняют сильные шагательные движения, которые можно сразу затормозить, переместив голову любым образом в такое положение, при котором уменьшается разгибательный тонус в передних ногах.
Описанное здесь влияние установки тела на движения имеет большое значение при обсуждении исследований всех рефлексов на животных,
Тонические шейные и лабиринтные рефлексы у человека
93
у которых верхняя часть шейного отдела спинного мозга и продолговатый мозг с лабиринтами или без лабиринтов остались в функциональной связи со спинным мозгом, — следовательно, прежде всего во всех опытах на децеребрированных животных. При обсуждении последствий рефлекторных влияний на таких животных часто не обращали внимания на одновременно наступающие изменения положения головы. При исследовании результатов раздражения мозгового ствола, коры больших полушарий и т. д. также надо всегда принимать во внимание возможные реакции головы.
При изучении литературы часто можно встретить исследования, в которых описываются реакции, являющиеся просто следствиями изменен-
Рис. 49. Опыт 7. Децеребрированная кошка. Изолированная трехглавая мышца. У животного наблюдаются преимущественно лабиринтные рефлексы. При положении КВ отчетливо проявляются альтернирующие движения ходьбы обеих передних ног. Эти движения верхней ноги зарегистрированы. Вследствие поворота головы [верхняя нога становится «черепной» (при SB)] затормаживаются движения ходьбы обеих ног.
ного положения головы, — реакции, вообще не обусловленные как таковые раздражением, которое дает экспериментатор. Или можно считать, что определенным раздражением вызываются движения конечностей и одновременно изменяется положение головы. Что эти явления могут находиться в причинной связи между собой, часто еще не известно исследователям. Желательно в будущем обратить больше внимания на эти явления; тогда станет ясно, что часто наблюдаемые изменения установки тела и движений тела являются следствиями одновременно вызванных шейных и лабиринтных рефлексов.
Симонелли (Simonelli) недавно показал при анализе принудительных движений после экстирпации задней доли мозжечка, что сложные процессы движения можно понимать как простые следствия определенных положений головы.
VIII. ТОНИЧЕСКИЕ ШЕЙНЫЕ И ЛАБИРИНТНЫЕ' РЕФЛЕКСЫ У ЧЕЛОВЕКА (7, 12 22) УУ здорового взрослого человека в обыкновенной жизни не представляется возможным обнаружить тонические шейные и лабиринтные рефлексы. Выше говорилось уже о том, что чаще всего эти рефлексы невозможно наблюдать и у нормальных обезьян и что тонические шейные рефлексы обнаруживаются у них только в исключительных случаях (лаби- ринтэктомия, зашивание глазных век). Что у взрослого человека эти явления должны быть гораздо сложнее, чем у до сих пор исследованных четвероногих животных, вытекает уже из того, что у человека с появлением прямой походки выступают и иные закономерности, так как балансирование на одной или двух ногах требует иных условий, чем стояние на
94
Третья глава. Рефлексы положения
четырех конечностях. Кроме того, у человека руки далеко не в такой степени влияют на положение тела, как передние конечности у животных.
У человеческого же плода, возможно, и будут наблюдаться тонические шейные рефлексы. Минковский [Minkowski (2)] наблюдал у плода человека 3—5 месяцев (от 8.5 до 23 см длины), который он получил при кесаревом сечении под местной анестезией, реакции при вращении головы по отношению к туловищу. Руки при этом реагировали реципрокно, хотя и довольно неправильно. Это были длительные реакции, которые сохранялись до тех пор, пока не изменялось положение головы. Так, например, при повороте головы вправо обнаруживалось отведение правой и приведение левой руки. Следовательно, речь идет, вероятно, о тонических шейных рефлексах. Тонические лабиринтные рефлексы на мышцы тела у зародыша человека Минковский не мог наблюдать. Однако при движениях головы в пространстве, — например, при перекладывании плода на спину или, наоборот, при поднимании в сидячее положение, — можно было наблюдать проходящие симметричные движения рук и ног, иногда ритмического характера, которые, вероятно, как будет показано ниже, представляют собой реакций’ с полукружных каналов. Полукружные каналы и отолитовый аппарат у плода человека такого возраста достаточно развиты.
В собственных наблюдениях над 26 новорожденными, которые были исследованы частично через несколько часов после рождения, частично через ЗУ2 месяца, тонические шейные рефлексы в ответ на вращение головы никогда не наблюдались. Исследование, кроме того, очень затруднялось многочисленными спонтанными движениями новорожденных.
Тонические шейные и лабиринтные рефлексы у человека отчетливо“ выступают только при патологических условиях. В настоящее время известны многочисленные наблюдения над тоническими шейными рефлексами. Однако тонические лабиринтные рефлексы были установлены только в редких хорошо изученных случаях. Недавно Уелш [Walche (1)] опубликовал более точные наблюдения над этими рефлексами. Эти тонические лабиринтные рефлексы следует строго отличать от реакций с полукружных каналов на конечности, и в каждом отдельном случае надо убедиться в том, что это действительно рефлексы положения, длительные реакции и. что соответствующие рефлексы не вызываются движениями.
Влияние тонических шейных рефлексов на конечности
Вначале мы приведем более подробно некоторые примеры из клинических наблюдений.
Первое наблюдение было сделано в Утрехтской неврологической клинике, проф. Хейльброннером [Heilbronner (7)].
Речь идет о шестилетней девочке, принятой 12 января 1910 г. Клинический диагноз: гидроцефалия, вероятно вторичная после опухоли мозжечка. До 4 лет ребенок был нормальным. Затем заболел, появились судороги с потерей сознания. С пятилетнего возраста — паралич, потеря речи, позднее исчез слух. Спустя еще полгода стало очевидным увеличение размера головы. Резко выражена гидроцефалия. Атрофические застойные соски. Ребенок ни на что не реагирует. Калорический нистагм нормален. При обычном положении в кровати конечности гипертоничны. Имеются рефлексы! Бабинского и Оппенгейма. Ребенка кормят через эзофаготомический зонд.
19 января. Ребенок лежит на спине, голова повернута вправо. На рис. 50 сзади видна громадная водянка, лицо повернуто направо, оба плеча поддерживаются так, что они стоят перпендикулярно, предплечья вяло свисают вперед, кисти рук лежат на. груди. Теперь голову пассивно поворачивают лицом влево вокруг оси: темя—шейная.
Тонические шейные и лабиринтные рефлексы у человека
95
Рис. 50.
часть позвоночного столба. В ответ на это происходит медленное, сильное тоническое разгибание левого локтя (рис. 51).
Предплечье стоит перпендикулярно, кисть руки активно и сильно сгибается дор- сально. Пассивное сгибание в локте встречает значительное сопротивление. Одновременно появляется разгибательный тонус в левом колене. При повороте головы в исходное положение (рис. 50) левое предплечье сейчас же снова вяло падает на грудь. Эта реакция проявляется с механической правильностью, и ее можно продемонстрировать в клинике.
20 января. Более точное исследование, причем за разгибательный тонус всегда принимается такое состояние, при котором в ответ на пассивное сгибание имеет место отчетливое сопротивление; сгибательный тонус характеризуется сопротивлением пассивному разгибанию. Ребенок лежит на спине, голова немного повернута вправо.
Глаза смотрят направо вниз. Правое колено и правое бедро атоничны; левое бедро также атонично. В левом колене и в левом локте незначительный, но отчетливый разгибательный тонус. Левое плечо и вся левая рука вялые. Поворот головы влево вызывает тоническое разгибание левого локтя с пронацией и разгибательным тонусом руки («челюстная рука»), более слабое разгибание правого локтя. Теперь голова удерживается в течение 5 минут в этом положении лицом влево; тонус в правом локте исчезает через Уч—1 минуту, но левая рука все время сильно растянута, только через 4 минуты
происходит короткое сгибательное движение, и вслед за этим рука сразу возвращается в разогнутое положение. Через 5 минут голова снова возвращается в среднее положение. Левое предплечье сразу вяло падает вниз. Левая кисть и плечо также становятся вялыми. Поворот головы вправо оказывается недейственным. Наклон головы (около сагиттальной оси) в направлении правого или левого плеча тоже оказывается недейственным, также и сгибание в вентральном направлении. После 5 минут лежания на левом боку в левой руке имеет место отчетливый разгибательный тонус; спустя еще 5 минут только голова возвращается в исходное положение, так что затылок ложится на стол (туловище остается на левом боку); после этого разгибательный тонус левой руки сразу же уменьшается. Теперь, когда голова находится в покое, туловище поворачивается на спину, при этом левая рука снова переходит в разогнутое состояние.
22 января. Продолжение. Ребенок лежит на спине, ноги атоничны.
Поведение ног. Вращение головы лицом влево обусловливает появление отчетливого разгибательного тонуса в левой рис четырехглавой мышце («челюстная нога»),
который в понимании Шеррингтона (8) «пластичен». Через некоторое время происходит спонтанное разгибание левой руки и ноги. После возвращения головы в среднее положение это разгибание и пластический тонус четырехглавой мышцы исчезают. Голова повернута вправо, неуверенная реакция, возможен незначительный сгибательный тонус в колене. Голова снова возвращена в среднее положение: кратковременное, но сильное разгибание левого колена, которое после этого опять расслабляется. Голова повернута влево: сперва незначительное, но отчетливое усиление тонуса четырехглавой мышцы, через некоторое время разгибание левого колена. При возвращении в среднее положение тонус опять исчезает.
96
Третья глава. Рефлексы положения
Поведение рук. Голова повернута влево: разгибание левого локтя, тонус «пластичен». При обратном вращении в среднее положение — расслабление левого локтя. Голова повернута вправо: в обоих локтях незначительный сгибательный тонус, исчезающий при обратном повороте головы.
27 января. Производится укол в мозолистое тело, при этом удаляется 15 мл жидкости (Prof. Lameris).
2 февраля. Если ребенка положить на спину, голова спонтанно поворачивается и наклоняется вправо, левая рука переходит в супинацию, после чего происходит сгибание в локте и сгибание пальцев. Поворот головы влево вызывает торможение сгибательного тонуса в левой руке, тоническое разгибание в левом локте, в то время как правый локоть тонически сгибается.
Начиная с 8 февраля, реакции конечностей в ответ на поворот головы становятся непостоянными. 14 июня родители берут ребенка домой, он умирает 25 ноября. Вскрытие не производится.
Заключение. Речь идет о ребенке с крайне выраженной гидроцефалией, у которого деятельность большого мозга почти полностью выключена. Поворот головы лицом влево вызывает сильное тоническое разгибание левой руки и ноги, сильнее всего разгибание выражено в левом локте. Иногда (не всегда) можно констатировать одновременное сгибание и правой руки. Реакция продолжается и при 5-минутном испытании до тех пор, пока не изменится положение головы; при возвращении головы в исходное положение сразу же исчезает разгибательный тонус на левой, стороне. Этот рефлекс является тоническим шейным рефлексом потому, что он проявляется независимо от положения головы в пространстве всякий раз, когда изменяется положение головы по отношению к туловищу или когда изменяется положение туловища по отношению к голове в названном направлении. Реакция подчиняется правилу шейных рефлексов, согласно которому усиление разгибательного тонуса всегда должно обнаруживаться на «челюстной ноге» или «челюстной руке». В этом случае очень отчетливо было видно, что торможение разгибательного тонуса, часто сопровождалось появлением активного сгибательного тануса.
В дополнение можно привести еще второй случай из этой же клиники (7).
Трехлетний мальчик. Принят 27 октября 1910 г. Клинический анализ: опухоль мозжечка. Гидроцефалия. Ребенок родился нормально, научился нормально ходить и начал говорить. Заболел в феврале, когда началась рвота, ухудшение ходьбы, недержание мочи и кала. После этого — спастический парез обеих ног и увеличение объема головы.
27 октября. Гидроцефалия крайней степени (окружность 55.1 см). Ходит очень плохо. Атаксия. При ходьбе нуждается в поддержке, так как в противном случае у него подгибаются колени или он падает в ту или другую сторону. Коленные рефлексы повышены. Атаксические толчкообразные произвольные движения рук. Интеллект понижен. Двусторонние атрофические застойные соски. Промывание ушей холодной водой влечет за собой характерный нистагм глаз.
3 ноября. Положение на спине. Разгибательный тонус обеих колен; при поднимании бедра голень держится в воздухе в ригидном состоянии, при пассивных движениях голень падает вниз, а затем несколько раз активно разгибается.
На правой ноге отмечается слабое сопротивление флексоров, более сильное в экстензорах. В четырехглавой мышце наблюдается пластический тонус, исчезающий при быстром форсированном сгибании и после этого постепенно возвращающийся.
В каждом положении нога «дергается» («schnappt»), после чего в этом положении голень спокойно удерживается в воздухе мышцами-разгибателями.
Если поддерживать правое бедро так, что видны движения голени, то поворот головы лицом влево обусловливает уменьшение тонуса четырехглавой мышцы справа («черепная нога»). Поворот головы лицом вправо обусловливает усиление правостороннего тонуса четырехглавой мышцы («челюстная нога»), обе реакции происходят с очень большим латентным периодом. Кроме того, если ноги остаются спокойно лежать на подстилке, поворот лица вправо обусловливает отведение правого бедра, а поворот влево — приведение левого бедра.
Тонические шейные и лабиринтные рефлексы у человека
97
4 ноября. Укол в мозолистое тело. При этом выделяется под давлением 75 мл жидкости.
Заключение. У ребенка с резко выраженной гидроцефалией имеет место спастический парез обеих ног. Особенно на правой ноге отмечается отчетливый экстенсорный и пластический тонус четырехглавой мышцы. При повороте головы тонус четырехглавой мышцы правой
Рис. 52.
ноги увеличивается, если она является «челюстной ногой», и затормаживается, если она является «черепной ногой». Кроме того, при повороте головы происходит приведение «челюстной ноги». В этом случае речь идет о шейных рефлексах.
Рис. 53.
В противоположность ранЦе описанному случаю здесь эти рефлексы выступают у ребенка, который еще выполняет произвольные движения и у которого сохранились еще определенные признаки интеллекта.
Случай 3. Родильный дом в Утрехте, проф. Каувер.
Новорожденная девочка. Вес 3.5 кг. Клинический диагноз: церебральные кровоизлияния вследствие искусственных родов.
Родилась И сентября 1911 г.
Placenta praevia, кровотечение, комбинированный поворот по Бракстону Хиксу, извлечение. Только через 3/4 часа ребенок начинает хорошо дышать. Глотательные и сосательные рефлексы отсутствуют; ребенка кормят при помощи зонда через нос. Ясно выраженный разгибательный тонус всех четырех конечностей с бросающимся в глаза дрожанием. Временами руки производят чередующиеся разгибательные и сгибательные движения, которые сопровождаются дрожанием; тризм.
20 сентября 1911 г. Положение на спине (рис. 52, ребенок сфотографирован сверху). Видно разгибание конечностей. Разгибание ног совершенно ненормально для новорожденного. Все четыре конечности оказывают явное сопротивление пассивному сгибанию (разгибательный тонус). Если поворачивать голову лицом влево (рис. 53 показы-
7 Р. Магнус
98
Третья глава. Рефлексы положения
вает ребенка сбоку, рис. 54 — более сверху), то левая рука разгибается сильнее и совершенно ригидна («челюстная» рука), правая рука («черепная» рука) переходит- в согнутое положение, особенно в локте; разгибательный тонус (сопротивление, которое оказывается пассивному сгибанию) исчезает, вместо этого появляется сильный сгибательный тонус и, наконец, правая кисть доходит до правого уха. Левая нога («челюстная») остается разогнутой, правая нога («черепная») слегка сгибается в бедре-
Рис. 54.
и колене (рис. 53), а также в голеностопном суставе (рис. 54). Реакция тоническая и продолжается до тех пор, пока голова остается в этом положении. При обратном повороте в среднее положение конечности снова занимают исходное положение.
При поворачивании головы вправо (рис. 55 снят сверху) изменения в положении ног незначительны. В левой руке уменьшается разгибательный тонус, вместо этого
Рис. 55.
появляется сгибательный тонус (сопротивление, оказываемое разгибанию), рука переходит в согнутое положение («черепная» рука), в то время как правая рука («челюстная») тонически разгибается. Реакция на поворот головы вправо слабее, чем реакция на поворот влево.
Результат поворота головы остается таким же и тогда, когда ребенок находится в положении на правом боку или в висячем положении головой вниз. Рис. 56 показывает ребенка в последнем положении. Видно, что лицо активно повернуто вправо и правая рука максимально разогнута. Эти реакции не зависят от положения головы в пространстве, они не являются лабиринтными рефлексами, а вызываются изменением
Тонические шейные и лабиринтные рефлексы у человека
99
положения головы по отношению к туловищу; следовательно, они являются шейными рефлексами и в этом случае подчиняются найденному в опытах на животных правилу: «черепные» конечности обнаруживают уменьшение, а «челюстные» — увеличение разгибательного тонуса.
Кроме этих тонических шейных рефлексов, ребенок обнаруживает еще другую реакцию. А именно: если ребенка из сидячего положения поместить в положение лежа или если выполнять это движение при положении тела на спине одной только головой, то происходит симметричное отведение обеих рук, причем отводятся плечи, локти разгибаются и иногда широко расставляются пальцы. Сначала эта реакция носит тонический характер, но через некоторое время прекращается. Этот рефлекс не зависит от изменения положения головы по отношению к туловищу и вызывается изменением положения головы в пространстве. Он не является ни рефлексом положения, ни тоническим лабиринтным рефлексом, но зависит от движения перекладывания ребенка и носит преходящий характер. Наличие этого последнего рефлекса можно было доказать и у нормальных грудных детей, только с тем различием, что реакция протекала тут еще быстрее, чем у данного ребенка. Этот рефлекс в последнее время был подробно изучен Моро и Фрейденбергом (Moro и.
Freudenberg). Речь здесь идет, вероятнее всего, о реакции, вызванной с полукружных каналов.
Тонические шейные рефлексы на конечности можно было показать 27 сентября, 11 октября и 8 ноября 1911 г. Затем состояние ребенка ухудшилось. Ребенок умер 20 декабря 1911 г. После смерти была произведена люмбальная пункция, при этом выделилось под давлением 20 мл жидкости. В люмбальный мешок введен формалин.
В скрытие. Оба желудочка сильно расширены.
С обеих сторон очаги размягчения в области чечевичного ядра, в мазке которого видны микроскопические зернистые клетки. В очагах наблюдаются пятна, имеющие вид старых кровоизлияний.
Заключение. У новорожденного ребенка с двусторонними кровоизлияниями и позже с очагами размягчения в области чечевичного ядра наблюдаются спастические явления на конечностях, нарушения глотания и дру- рис
гие церебральные симптомы. На поворот головы ребенок обнаруживает выраженные шейные
рефлексы, причем в «челюстной» руке и в «челюстной» ноге имеется сильный разгибательный тонус, в то время как в «черепной» руке и в «черепной» ноге разгибательный тонус затормаживается и вместо этого появляется сгибательный тонус. Реакция происходит независимо от положения головы в пространстве и вызывается соответствующим изменением положения головы по отношению к туловищу. На многочисленных нормальных новорожденных эти рефлексы не наблюдались.
Кроме того, ребенок обнаруживал реакции со стороны полукружных каналов (проходящие лабиринтные рефлексы на движение), если голова перемещалась из вертикального в горизонтальное положение. Рефлекс этот характеризуется расхождением обеих рук. Эту реакцию можно показать и на многочисленных нормальных грудных детях в первые месяцы их жизни, в то время как на некоторых недоношенных детях ее наблюдать не удалось.
Случай 4. R. de G., 9 лет, психиатрическая больница в Хееренло, д-ра Шеурер и Дюпон [Scheurer u. Dupont (12)]. 22 октября 1912 г. Анамнез: нормальные роды. У новорожденного отчетливая мышечная слабость. Световосприятие не удается наблюдать. На психические раздражения ребенок никогда не реагировал. Уже больше года находится в больнице. Слабый,
7*
100
Третья глава. Рефлексы положения
беспомощный ребенок, со спастически-паретическими конечностями и некоординированными глазными движениями, не реагирует на психические раздражения, на свет и на обычный шум, лежит спокойно и не издает никаких членораздельных звуков. Дефекация и мочеиспускание непроизвольны. Самостоятельно есть не может. Создается впечатление, что ребенок слеп. Зрачковая реакция на свет отсутствует. Соски зрительного нерва атрофичны. Спонтанный горизонтальный, иногда ротаторный нистагм. Парез
Рис. 57.
правой отводящей мышцы глаза. Вероятно, ребенок глухой (никакой реакции на камертон, окрик и т. д.).
Невозможно вызвать калорические лабиринтные рефлексы. При движениях головы не наблюдается отчетливой девиации глаз. Переход из лежачего в сидячее положение или из положения на боку в положение на спине не оказывает никакого влияния
Рис. 58.
на тонус конечностей, если при этом не изменять положения головы по отношению к туловищу. Хватательный рефлекс сохранен.
Следующие шейные рефлексы можно вызвать независимо от положения тела.
При вращении головы влево увеличивается разгибательный тонус в левом колене и локте («челюстные ноги»), справа («черепные ноги») он уменьшается, как это видно на рис. 57.
При повороте головы вправо имеют место обратные явления: разгибательный тонус в колене и в локте справа увеличивается, а слева уменьшается. На рис. 58 это показано для рук, на рис. 59 — для ног. При исследовании этих изменений тонуса конечностей отчетливо чувствовалось, что при растяжении тонус разгибателей усиливался и одновременно уменьшался тонус сгибателей, в то время как при обратной реакции, в случае расслабления конечностей, тонус разгибателей уменьшался и одновременно выступало активное сокращение мышц сгибателей. Об этих явлениях судили по сопротивлению, которое оказывается пассивному сгибанию (мерило для тонуса разгибательных мышц) или пассивному разгибанию (мерило для тонуса сгибательных
Тонические шейные и лабиринтные рефлексы у человека
101
мышц). На одновременно снятых кинематографических снимках, во время которых поддерживались плечи и бедра, видно, как при чередовании поворотов головы то влево, то вправо машинообразно поднимаются и опускаются предплечья и голени.
Наклон головы (вращение вокруг сагиттальной оси головы, при котором одно ухо приближается к плечу) вправо и влево также вызывает реакции. При наклоне вправо (правое ухо приближается к плечу) конечности левой стороны разгибаются, а конечности правой стороны расслабляются. При наклоне влево, т. е. когда левое ухо приближается к плечу, конечности правой стороны разгибаются, а левые конечности расслабляются.
Поднимание и опускание головы отчетливого влияния на тонус конечностей не имеет.
Заключение. У ребенка с выраженной идиотией, у которого не обнаруживалось следов каких-либо высших функций головного мозга,
Рис. 59.
который слеп и глух, изменения положения головы в пространстве не вызывают каких-либо изменений тонуса конечностей; напротив, эти реакции можно вызвать, изменяя положение головы по отношению к туловищу независимо от положения тела. Эти шейные рефлексы являются длительными реакциями.
Реакции на поворот головы происходят в этом случае точно таким же образом, как в двух до сих пор описанных случаях и в опытах на животных. Этот случай интересен и потому, что здесь в ответ на наклон головы появлялись тонические рефлексы на конечности. Наклон вправо вызывал разгибательный тонус на левой и расслабление на правой стороне. У животных, наоборот, в ответ на наклон вправо происходит усиление разгибательного тонуса на правой и уменьшение его на левой стороне.
Такая же реакция на наклон головы была описана еще в другом случае Броувером (1).
Случай 5. Речь идет о нормально рожденной девочке (5 октября 1913 г.), у которой после третьего месяца жизни постепенно исчезли нормальные движения. Через некоторое время руки и ноги стали ригидными. Эту девочку мы могли более подробно исследовать 26 ноября 1913 г. Зрачки на свет реагируют, ребенок не слеп, руки и ноги аддуци- рованы и почти полностью разогнуты, предплечья чрезмерно пронированы, ноги сильно дорсально согнуты, во всех конечностях ригидность высшей степени с сильным спазмом приводящих мышц. Ригидность затылка и опистотонус. Коленный рефлекс повышен. Симптомы Бабинского и Оппенгейма положительны.
У ребенка обнаруживаются типичные тонические шейные рефлексы на руки и ноги при повороте головы. При наклоне головы, т. е. когда голова двигается таким образом, что одно ухо приближается к плечу, наблюдаются совершенно аналогичные рефлекторные движения. Когда правое ухо приближается к плечу, конечности левой стороны растягиваются, на правой стороне расслабляются, при поворачивании влево происходит обратное. При поднимании и опускании головы не наблюдается сгибания или разгибания конечностей.
102
Третья глава. Рефлексы положения
No
Тонические шейные
Автор
Диагноз
рефлексы
Лабиринтные рефлексы и др.
7 Магнус, де Клейн. Pflügers Arch, f. d. ges.
Physiol., 145, 539, 1912.
8 Бейланд.
Münch, med. Wochenschr., 1912> стр. 253У.
Ребенок, 9 месяцев. Гнойный энцефалит. (Больница д-ра Бое- ла, Утрехт).
Типичные шейные рефлексы на руки и ноги (сгибатели и разгибатели ре- ципрокны).
Рефлекс с полукружных каналов при перекладывании ребенка (в положении на спине длительная реакция отведения вытянутых рук: тонические лабиринтные рефлексы).
Гумма мозжечка (и паралич?). (Мед. клиника Киль).
При коме (паралитический приступ?) типичные шейные рефлексы на руки, которые исчезают при возвращении сознания.
9
10
И
Ребенок, 16 месяцев. Амавротическая идиотия. (Невролог, клиника проф. Хейльброннера, Утрехт).
Типичные шейные рефлексы на руки и ноги при повороте головы (с ре- ципрокной иннервацией).
Магнус, де Клейн. Pflügers, Arch, f. d. ges. Physiol., 160,429, 1915.
Де Бруин. Ned. maandschr. у. Verlosen vrouwenziekten, 3, 593.
Боеме и Вей- ланд.
Zeitschr. f. d. ges. Nenrol. u.Psychiatrie, 44, 94, 1918.
Ребенок, 15 месяцев. Амавротическая идиотия.
Типичные шейные рефлексы на руки (менее отчетливо на ноги) при повороте головы при различных положениях. Наклон не действует. Дорсальное сгибание головы при положении на спине и боку вызывает длительную реакцию разгибательного тонуса обеих рук.
Реакция полукружных каналов при перекладывании ребенка. Типичные лабиринтные рефлексы на руки и ноги. Лабиринты пе- рзраздражены.
При положении на спине руки и ноги имеют больший тонус, чем в положении сидя (лабиринтный рефлекс?). Дорсальный наклон головы вызывает рефлекторную ригидность затылка и описто- тонус (лабиринтный рефлекс? ),— не ясно, наступает ли все это только в положении на спине.
78 лет, мужчина. Артериосклероз. Почти полный стеноз обеих сонных артерий, тромбоз сонной внутренней артерии. Размягчение ствола мозга и большого мозга (вскрытие).
Типичные шейные рефлексы на правую руку и обе ноги при повороте головы.
Тонические шейные и лабиринтные рефлексы у человека
103
№
Автор
Диагноз
Тонические шейные рефлексы
Лабиринтные рефлексы и др.
12
Те же.
56 лет, женщина. Слева — субдуральная гематома. Кровоизлияние в левую затылочную долю и во все три черепные ямки (вскрытие).
Типичные шейные рефлексы на обе руки (появление и исчезновение активного сгибательного тонуса) при повороте головы.
Вентральное сгибание головы вызывает сгибание обеих ног, иногда и правой руки (шейные или лабиринтные рефлексы?).
43
Те же.
39 лет, мужчина. Справа — гемо- плегия после сифилиса. Апоплексия.
Правые конечности спастические и неподвижные. Поворот головы вправо и влево вызывает типичные шейные рефлексы (сгиб, и разгиб, на правой руке и ноге), сгибание головы при положении на спине без результатов.
Поднимание туловища и вентральное сгибание головы вызывают сгибательный тонус, обратное помещение в положение лежа — ра згибате льный тонус в правых конечностях (лабиринтный рефлекс).
14
Долингер. Zeitschr. f. Kinderheilk., 22, 167, 1919.
Мальчик, 5 месяцев. Амавротическая идиотия. (Дом Августы - Виктории, Берлин, проф. Ланг- штейн).
Шейные рефлексы при" повороте головы на руки и ноги с активным сгибанием и разгибанием.
В положении на спине на чувствительное раздражение — типичная ригидная установка (лабиринтный рефлекс??).
15
Письменное сообщение проф. Л. Боумана и д-ра Т. М. Мес- дага.
Женщина. Автомобильная катастрофа — гладкий поперечный разрыв среднего мозга до моста (вскрытие).
Типичные шейные рефлексы на поворот головы, кома.
16
Ионкхофф. Ned. tijdschr. у. geneesk., 1920, I, стр. 307.
17 лет, девушка. Эпилепсия. Состояние кровоизлияния в центр, извилину и правый боковой желудочек.
Во время и после эпилептических припадков шейные рефлексы при повороте головы, которые отсутствуют при полном сознании.
16а
Уелш. Lancet, 1923, II, 644.
i
33 года, девушка. Медиальная опухоль между ножками мозга над гипофизом (вскрытие).
Двусторонняя гемиплегия. Децереб- рационная ригидность. Типичные шейные рефлексы на все 4 , конечности при повороте головы. Зависимость рефлекса Бабинского от положения головы.
104
Третья глава. Рефлексы положения
Подробное анатомическое исследование по Броуверу показало, что у этого ребенка был менингоэнцефалит, вследствие которого были почти полностью выведены из строя кора большого мозга и кора мозжечка. Кроме того, в вентральной части моста имелись очаги. Подробнее апатомическое описание можно посмотреть в опубликованной Броуве- ром работе.
Ниже я еще остановлюсь на наблюдениях Симонса (Simons), который в некоторых случаях наблюдал противоположное влияние поворачивания головы у человека. На обезьянах и остальных до сих пор исследованных млекопитающих также можно было наблюдать противоположное поведение. Поэтому потребуются еще дальнейшие наблюдения, чтобы
установить, почему у человека реакции на поворачивание головы (наклон головы к плечу) так непостоянны.
Случай 6. Пятимесячный ребенок. Университетская клиника, Утрехт, проф. Хавершмидт (Haverschmidt).
Рис. 60а.
Ребенок родился нормальным, с 2% месяцев залихорадил. Появились судороги и парез правой стороны тела.
На рис. 60а (кинематографические снимки докт. Стенверса) виден ребенок в положении на спине (снят сверху слева). Ребенок самостоятельно повернул голову влево, вследствие этого левая рука разогнута, правая рука согнута. Также разогнута левая нога и согнута правая нога. На рис. 606 видно, что ребенок активно поворачивает голову вправо, вследствие этого левая рука согнута, а правая разогнута. Обе ноги разогнуты.
Интересным в этом случае является то, что здесь имеют место типичные реакции рук на вращение головы при спонтанных движениях головы ребенка вправо и влево, следовательно — при активных движенияхг а не только при пассивных. Реакции рук такие же, как и при пассивных движениях головы. Реакции ног у этого ребенка осуществлялись не так регулярно, как реакции рук, и были нарушены тем, что ребенок производил беспорядочные движения ногами.
До сих пор мне были известны из литературы всего 14 случаев (из них 6 с описанием вскрытия), в которых можно было доказать тонцческие шейные рефлексы на конечности. Однако по устному сообщению д-ра Стенверса такие рефлексы наблюдались в гораздо большем количестве случаев.
В таблице на стр.102—103 объединены описанные в литературе случаи,, на которых я выше подробно не останавливался. Я не привожу наблюдений Симонса и Уелша (Simons u. Walshe), так как на них следует остановиться особо.
Из клиники Винклера нам стали известны, благодаря любезному сообщению д-ра Стенверса, следующие еще не опубликованные случаи:
Из этой таблицы (стр. 105) видно, что проявление тонических шейных рефлексов на конечности у человека не характерно для какой-либо определенной формы заболевания. Существенное значение при этом, вероятно, имеет выключение определенных мозговых путей или определенных высших отделов мозга (децеребрация). О каких путях здесь идет речь, на сегодняшний день еще полностью не ясно. Симонс на основе своих наблюдений полагает, что существенное значение имеет выключение пирамидных
Тонические шейные и лабиринтные рефлексы у человека
105
№
Пациент
Диагноз
Шейные
рефлексы
17
Г. С., ж., 29 лет.
Правосторонняя гемиплегия после удаления большой внутренней опухоли в левой париетальной области (мозга).
Очень отчетливые шейные рефлексы на парализованной стороне.
18
И. В. М., м., 31 год.
Левосторонняя гемиплегия. Посттравматический энцефалит.
Очень отчетливые шейные рефлексы на парализованной стороне (киносъемка).
19
Т. Г., ж., 28 лет.
Правосторонняя гемиплегия. Глиома мозга (оперирована).
Очень отчетливые шейные рефлексы на парализованной стороне.
20
И. В., ж., 48 лет.
Левосторонняя гемиплегия. Саркома (оперирован).
Отчетливые шейные рефлексы на парализованной стороне.
21
Э. Д. Е., м., 76 лет.
Правосторонняя гемиплегия. Паралич взора вверх. Очаг не известен. (Вскрытие не производилось).
Отчетливые шейные рефлексы обеих сторон. (Пациент сонлив, сознание отсутствовало полностью).
22
А. В. О., ж., 14 лет.
Опухоль медиальной линии, выше тенториума, растущая преимущественно влево.
Весьма отчетливые шейные рефлексы на обеих сторонах.
23
А. Д., ж., 14 лет.
Опухоль в области эпифиза.
Непостоянны. При ухудшении состояния на время становятся отчетливыми.
24
И. де С., ж., 12 лет.
Опухоль медиальной линии (желудочек?).
Во время инсульта с нарушением сознания отчетливы
25
И. Ц., м., 17 лет.
Атетоз после энцефалита.
Шейные рефлексы имеются (частые интеркуррентные движения являются помехой).
26
А. X., 3 года.
Энцефалит.
Шейные рефлексы появляются во время постэпилептической потери сознания.
27
Ц. Ф., 12 лет.
Энцефалит.
Видны на киносъемке во время припадка.
28
Л. Ц., 6 лет
Диффузное заболевание мозга. Опухоли.
Во время припадка типичные положения.
путей. Эти наблюдения позволяют сделать заключение о том, что центры тонических шейных рефлексов расположены приблизительно так же, как и у животных, т. е. в верхнем шейном отделе спинного мозга. Во всяком случае, из наблюдений Броувера вытекает, что большой мозг не нужен для этих рефлексов, а из случая № 16 Уелша видно, что может отсутствовать и промежуточный мозг; случай № 15 показывает, что рефлексы имеются и после перерезки мозгового ствола на уровне тенториума; наблюдение Броувера (№ 5) показывает, что может отсутствовать вся кора мозжечка, и, кроме того, это наблюдение показывает, что и очаги в области моста не препятствуют проявлению этих рефлексов.
Наблюдение № 8 Вейланда показывает, что в одном из случаев мозгового сифилиса во время комы после (паралитического?) приступа появлялись тонические шейные рефлексы. После выхода из комы и с
106
Третья глава. Рефлексы положения
постепенным возвращением сознания тонические шейные рефлексы исчезали. Подобные йаблюдения (появление рефлексов при нарушении сознания и во время припадков и их исчезновение по мере улучшения общего состояния организма, возвращения сознания и прекращения припадка) были сделаны Стенверсом и Ионкхофом.
До сих пор клиницисты для вызывания шейных рефлексов использовали поворот головы. При этом в исключительных случаях реагируют все четыре конечности, чаще только некоторые из них или же одна конечность. Если не все конечности участвуют в реакции, тогда обычно рефлексы проявляются на более или менее спастических конечностях. В известных условиях спазмы этих конечностей проявляются только при определенных положениях головы.
Как показывает наблюдение над случаем 2, рефлексы могут отчетливо выявляться и при наличии произвольных движений. Но по легко понятным причинам для решения вопроса о том, идет ли речь действительно о тонических шейных рефлексах, конечно, лучше, если никакие произвольные движения не мешают наблюдениям.
Характерным для шейных рефлексов является то, что реакции не зависят от положения головы в пространстве, т. е. что они появляются при положении на спине, при сидении и стоянии, при положении на боку и в известных условиях даже при положении на животе. Эти рефлексы зависят от положения головы по отношению к туловищу, и их можно, следовательно, вызвать при определенных условиях и при фиксированной голове одними движениями туловища.
Если при повороте головы в положении на спине реакции правых конечностей противоположны реакциям левых, то речь идет об истинных шейных рефлексах. Если реакция отчетлива только на одной конечности и при положении пациента на спине в ответ на повороты вправо и влево происходят противоположные реакции, то и здесь речь идет об истинных шейных рефлексах. Всегда следует обращать внимание на длительность реакций и на то, что они продолжаются до тех пор, пока голова остается в соответствующем положении. Сюда нельзя отнести преходящие рефлексы. От характера клинического случая зависит, проявляются ли рефлексы преимущественно на сгибателях, или на разгибателях, или на обеих мышечных группах. Обычно тонические шейные рефлексы отчетливее всего выявляются на таких мышцах, которые уже до этого обладали наиболее сильным тонусом. В конечностях с разгибательными спазмами они поэтому выражаются преимущественно как изменения разги- бательного тонуса, при сократившемся сгибателе — как изменения сгибательного тонуса, в то время как при промежуточном положении конечностей при одном положении головы могут выступать преимущественно разгибательные спазмы, а при противоположном положении головы — сгибательные спазмы.
Во многих случаях при определенных положениях головы в определенных мышечных группах можно доказать пластический тонус в понимании Шеррингтона (8). Если, например, конечность является «челюстной ногой», то у разгибательных мышц обнаруживается пластический тонус, причем соответствующие мышечные группы могут в любом положении «включиться» и зафиксировать соответствующий сустав в этом положении; однако если голова поворачивается в другую сторону, то тонус исчезает и фиксация сустава больше не осуществляется.
Как было уже упомянуто, в нашем распоряжении находятся преимущественно клинические наблюдения о результатах поворота головы. Гораздо меньше сообщений имеется о результатах наклона головы.
Тонические шейные и лабиринтные рефлексы у человека
107
У человека мы в двух случаях наблюдали противоположную реакцию по сравнению с животными. Симонсу же удалось получить в отдельных случаях такой же результат наклона головы, как и нам в наших экспериментальных исследованиях; недавно мы смогли это подтвердить в случае, подобном описанному Симонсом. Необходимо большее количество исследований на человеке, прежде чем высказывать что-нибудь определенное о тех условиях, от которых зависит характер тонических шейных рефлексов при наклоне головы у человека.
Гораздо меньше наши знания о последствиях поднимания и опускания головы. Де Бруин наблюдал в случае (№ 10) амавротической идиотии в ответ на дорсальное сгибание головы появление разгибательного тонуса обеих рук, который держался до тех пор, пока положение головы не изменилось. Эта реакция наступала как при положении пациента на спине, так и при положении на боку. Отсюда можно заключить, что здесь речь шла о тонических шейных рефлексах. Симонс при дорсальном наклоне головы также наблюдал шейные рефлексы. Некоторые авторы (Боеме, Вейланд, Фреуденберг) полагают, что и симптом Брудзинского (сгибание рук и ног при сгибании головы вперед в положении на спине) является тоническим шейным рефлексом. По данным Фреуденберга, этот рефлекс у грудных детей проявляется нормально в первые месяцы жизни. Позже его можно наблюдать только при менингите, энцефалите, рахите и т. д. Симонс не считает, что здесь речь идет о тоническом шейном рефлексе, однако до сих пор он не дает определенных объяснений.
Заранее можно сказать, что вентральное сгибание головы благодаря тоническому шейному рефлексу может вызвать реакцию как при симптоме Брудзинского, однако этим не утверждается, что во всех случаях проявления рефлексов Брудзинского причиной является действительно тонический шейный рефлекс.
Симптом же Кернига (сгибание ног при приподнимании туловища) не имеет отношения к тоническому шейному рефлексу, потому что он осуществляется как после перерезки спинного мозга (Боеме и Вейланд), так и при сгибании бедра в направлении к лежащему туловищу.
Описанные Бонди реакции конечностей на движение головы также являются тоническими шейными рефлексами, потому что они представляют собой преходящие и кратковременные движения.
Благодаря исследованиям Симонса возникла новая точка зрения на значение тонических шейных рефлексов у человека (2, 2), Он наблюдал раненых, страдающих гемиплегией, у которых обнаружил сопутствующие движения на парализованной стороне и разные реакции паретических конечностей в зависимости от положения головы. Подробные исследования на 250 больных с гемиплегией и на многочисленных других неврологических больных показали, что часто можно констатировать отчетливое влияние положения головы на сопутствующие движения. На таких пациентах следует сперва установить, имеют ли вообще место сопутствующие движения; это лучше всего следует таким образом: пациенту дают производить любые активные движения на здоровой или на одной из паретических конечностей — например, сильное надавливание на динамометр. При разных положениях головы — повороте, наклоне, поднимании и опускании — должны вызываться подобные сопутствующие движения. Проверка производится в положении лежа, сидя, стоя и при ходьбе.
108
Третья глава. Рефлексы положения
Оказалось, что в зависимости от положения головы сопутствующие^ движения паретических конечностей бывают разными: так, наблюдалось сгибание или разгибание, приведение или отведение, усиление или уменьшение сгибательного или разгибательного тонуса.
При таком способе проверки получают гораздо больше положительных результатов, чем в случае простого пассивного изменения положения головы пациента, — например, поворота головы. Симонс наблюдал влияние положения головы на сопутствующие движения только у больных гемиплегией и у больных с нарушением пирамидных путей в спин-
Рис. бСИа. (По Симонсу). Левосторонний тяжелый гемипарез вследствие прострела головы (из киносъемки). Исходное положение. Правая рука держит свободно эластическую металлическую гильзу. Левой, пораженной парезом ноге, пассивно придано указанное на рисунке положение, чтобы сделать более наглядным разгибательный тонус. Вследствие паралича и тяжелого пареза соответствующих мышц левая нога падает без поддержки в бедре из приведенного здесь исходного положения в сторону (наружу). Поэтому исследователь поддерживает слегка колено в сагиттальной плоскости до вступления в действие рефлекторного тонуса. Исходное положение в следующих исследованиях такое же, как и на данном рисунке.
ном мозгу. Среди них можно было установить положительный результат в % всех случаев. Если у таких пациентов удавалось вызвать пассивными изменениями положения головы непосредственные тонические шейные рефлексы, то они всегда оказывались более слабыми, чем при сопутствующих движениях. Реакции отсутствовали при всех экстрапирамидальных заболеваниях, — так, например, при paralysis agitans (дрожательный паралич) они, по Симонсу, связаны с нарушением пирамидных путей. Во время эпилептического припадка также можно было установить влияние положения головы на конечности.
При этих исследованиях Симонс отметил, что в ответ на такой поворот головы при сопутствующих движениях в «челюстной ноге» и в «челюстной руке» наблюдалось разгибание или усиление разгибательного тонуса, чаще всего сопровождаемое приведением, в то время как в «черепной ноге и руке» наблюдалось сгибание или усиление сгибательного тонуса и отведение (рис. 601 а—в). При вентральном сгибании головы в большинстве случаев происходило усиление разгибательного тонуса рук, при дорсальном сгибании — усиление сгибательного тонуса рук. В меньшинстве случаев обнаруживалась наблюдаемая и в опытах на животных реакция: разгибание передних конечностей при дорсальном сгибании головы и сгибание их при вентральном сгибании, без видимой причины для этих различий. При наклоне головы разгибание происходило на той стороне, на которой ухо приближалось к плечу, на противоположной стороне — сгибание или усиление сгибательного тонуса, т. е. точно такое же пове-
Тонические шейные и лабиринтные рефлексы у человека
109
щение, какое было обнаружено в экспериментах на животных (противо- лоложное этому поведение в двух клинических случаях на человеке
Рис. 601 б. Голова повернута вправо. Вызов гемиплегического сопутствующего движения вследствие сильнейшего сжатия кулака вокруг металлической гильзы. Положение покоя руки (рис. 60 ха) нарушается из-за боковой абдукции плеча, сгибания под острым углом локтя, поднятия руки, сжимания кулака. Сгибательный тонус руки очень сильный, одновременно видно напряжение трехглавой мышцы. В ноге сильное сгибание колена и бедра при легкой пронации бедра. Сгибательный тонус непреодолим (особенно в колене) по отношению пассивного разгибания, это заснято в следующих, здесь не представленных кинематограммах. Поднятие и аддукция ноги при супинации, поднятие пальцев. Отчетливо видно сопутствующее движение здоровой ноги.
Рис. 601 в. Голова повернута влево. Исходное положение такое же, как и на рис. 601 а. Следует обратить внимание на напряжение лица, разгибательный тонус, на приведение и легкий поворот вовнутрь плеча, разгибание руки, сжатие кулака. Лучезапястный сустав — близко к симфизу. Сильное разгибание ноги в колене и бедре с сильной внутренней ротацией, опускание ноги, довольно сильное поднятие пальцев. Одновременное напряжение двуглавой мышцы в плече не видно на рисунке.
было описано на стр. 101). В наблюдениях Симонса реакции рук чаще всего были сильнее, чем реакции ног.
Исходное положение конечности имеет мало значения для реакции при сопутствующих движениях, и почти не имеет значения, находилась ли конечность до этого в согнутом или в разогнутом положении; однако
HO
Третья глава. Рефлексы положения
относительно отведения и приведения можно было установить переключающее влияние исходного положения.
Описанные реакции являются шейными рефлексами и не зависят от положения головы в пространстве.
Эти рефлексы наблюдались не только на больных с гемиплегией в результате травм, но и у больных со спастическими явлениями после сифилиса, эмболии, энцефалита, повреждений шейного отдела спинного мозга, грубого двустороннего повреждения или выключения большого мозга (децеребрация).
Очень отчетливо было и влияние положения головы в таких случаях на ходьбу и позу животного. Влияние же на мускулатуру туловища уменьшалось.
Активные и пассивные движения головы действовали в том же смысле, только последние чаще всего слабее.
Недавно такие же наблюдения над влиянием положения головы на сопутствующие движения сделал Уелш (1) на больных спастической гемиплегией. По его данным, сопутствующие движения появляются только на спастических конечностях. Самыми действенными раздражителями для них являются сильные тонические произвольные сокращения — например, сжимание кулака, и даже в том случае, когда он не сильно сжимается. Латентный период составляет гЛ—2 секунды. Поворот головы в начале исследования пациента чаще всего не вызывает изменения тонуса и положения конечностей, однако через полчаса исследования эти изменения вызываются почти во всех случаях.
Эти наблюдения Симонса и Уелша имеют большое теоретическое и практическое значение — последнее потому, что этим чрезвычайно увеличивается возможность доказать наличие тонических шейных рефлексов, теоретически же по следующей причине: в наблюдениях Симонса, по моему мнению, речь идет о таких же отношениях, какие были выявлены в опытах Сосэна и Шторм ван Лееувена {19) и Беритова. В названных опытах на животных исследовался ряд рефлексов на изолированной трехглавой, двуглавой, полусухожильной мышцах и vastocru- reus-препарате под влиянием различных рефлекторных возбуждений. Было установлено, что если при изменении положения головы соответствующие изменения тонуса, вызываемые шейными и лабиринтными рефлексами, не проявлялись в регистрируемых конечностях и поэтому переключающее влияние оказалось не действующим, центры в спинном мозгу все же настраиваются определенным образом к разгибанию или к сгибанию. То же самое происходит в опытах Симонса. И здесь на соответствующих пациентах изменением положения головы вначале не вызывается изменение спастико-паретических конечностей. Однако, если теперь в этих конечностях вызвать любым способом (например, тем, что другая конечность выполняет энергичный произвольный акт) сопутствующее движение, это последнее оказывается таким, каким оно должно было бы быть после первоначального изменения положения головы. Другими словами, произвольная иннервация сделала то, что уже до этого должно было сделать положение головы. Этим определенным положением головы была в разных конечностях вызвана тенденция к разгибанию, к сгибанию, к приведению или отведению, которая в действительности проявилась только теперь при последующих сопутствующих движениях.
Сосэн и Шторм ван Лееувен и Беритов раздражали афферентные нервы конечностей. Беритову удалось показать, что иногда и возбуждения, исходящие из дыхательного центра, вызывают эти реакции. В кли¬
Тонические шейные и лабиринтные рефлексы у человека
111
нических наблюдениях Симонса речь идет о произвольной иннервации непарализованных конечностей, при которой сопутствующие движения появляются на паретических конечностях; на эти движения влияет закономерным образом определенное положение головы. Уелш полагает, что сопутствующие движения при этом вызываются проприоцептивными рефлексами от произвольно сокращенных мышц (например, при сжимании кулака). Но все это требует дальнейших доказательств.
Недавно Стенверс сделал в психоневрологической клинике относящиеся сюда наблюдения. Речь идет о ребенке с опухолью мозга. Поворот головы не вызывал спонтанного изменения положения рук. Но как только при повернутой вправо или влево голове на подошву наносилось сильное чувствительное раздражение, происходило сгибание (если соответствующая рука была «черепной») и разгибание (если она была «челюстной»). Следовательно, и в этом случае одно и то же чувствительное раздражение в зависимости от положения головы вызывало противоположные рефлексы. В реагирующей руке положением головы была обусловлена тенденция к разгибанию или сгибанию. В приведенном на стр. 103 случае опухоли в промежуточном мозгу Уелш (2) нашел отчетливое влияние поворота головы на рефлекс Бабинского, и действительными при этом оказались те же правила. Недавно Уелш (3) наблюдал у больного гемиплегией обратимость гомолатерального сгибательного рефлекса, вызванную поворотом головы.
Следует ожидать, что таким образом удастся сделать многочисленные наблюдения влияния тонических шейных рефлексов на рефлекторную готовность конечностей.
Тонические лабиринтные рефлексы у человека
В то время как мы имеем большое количество наблюдений тонических шейных рефлексов у человека, непосредственные тонические лабиринтные рефлексы на мускулатуру туловища человека отмечены лишь в небольшом количестве хорошо изученных случаев. Если наблюдать эти рефлексы, то, во-первых, надо исключить влияние шейных тонических рефлексов и, во-вторых, надо доказать, что наблюдаемые изменения тонуса мускулатуры являются продолжительными реакциями и что они вызываются определенным положением головы или же лабиринтов в пространстве и не зависят от движения.
Первое более подробное наблюдение было сделано на одном ребенке в Утрехтской психоневрологической клинике [проф. Хейльброннер (22)J.
Случай 1. Амавротическая идиотия (см. выше, № 9). М. К., 16-месячный мальчик, родился нормальным и в первые месяцы при грудном кормлении развивался нормально; вначале видел. В возрасте 3 месяцев поведение его стало отличаться от поведения нормальных детей. Он все время лежал спокойно, не играл. Затем стал отставать в умственном развитии, не научился ни стоять, ни ходить, ни говорить. В телесном отношении он развивался приблизительно так же, как и нормальные дети. Мать не может сказать определенно, видит ли он в данный момент. У ребенка 5 нормальных сестер и 1 нормальный брат, преждевременно рожденные близнецы умерли во время родов, один ребенок умер 8 лет тому назад, он был' парализован и слеп, точно так же как этот ребенок.
Отец и мать здоровы, по крови не родственны, оба евреи. Родители матери из Германии, отец из Голландии. Все братья отца и их дети здоровы.
Состояние. С 19 мая по 1 июля 1914 года. — Офтальмологическое обследование. Гладкое дно на обеих сторонах дает типичную картину амавротической идиотии, темно-красное круглое пятно в области макул,. окруженное широким бело-серым кольцом, а в остальном глазное дно нормально. Зрачки деформированы, реагируют на свет содружественно. Правый зрачок всегда
112
Третья глава. Рефлексы положения
немного шире левого. Движения глаз сохранены. Несомненно, ребенок видит, на показ блестящих предметов открывает рот и производит сосательные движения. Горизонтальный нистагм обоих глаз, когда ребенок смотрит вправо и влево. Нистагм имеет медленный и быстрый компоненты, последний чаще всего влево, реже вправо. Интенсивность нистагма меняется в разные дни, иногда при взгляде вперед он отсутствует. Перед припадками, описываемыми ниже, нистагм всегда становится сильнее. Корнеальный рефлекс на обеих сторонах сохранен, но снижен.
Данные терапевтического обследования. Для своего возраста ребенок мал. Небольшие рахитические изменения костей. Фронто-окципитальная окружность головы 45% см. Сердце в норме. Моча без белка и сахара. Реакция Вассермана отрицательна. Температура нормальная, в некоторые дни — повышение температуры до 3b° С; у ребенка недержание мочи, стул нормален; бронхи без особенностей. Пациент лежит спокойно, апатично, не кричит и не плачет, как это делают нормальные дети. Единственным признаком реагирования, который иногда можно у него наблюдать, является чуть заметная улыбка. Введенная в рот жидкая пища легко проглатывается. Несмотря на хороший аппетит, ребенок все же теряет в весе. При повышении температуры, все ухудшающемся пульсе и при очень частом дыхании он умирает 1 июля 1914 г. Секция не производилась.
Неврологическое обследование. На конечностях заметны легкие спазмы разной степени. Симптом Хвостека на обеих сторонах положителен, на левой стороне сильнее, чем на правой, особенно в области средних ветвей лицевого нерва. Брюшные рефлексы сохранены. Коленный и ахиллов рефлексы живые; клонуса нет. Подошвенный рефлекс с медиального края стопы плантарный, с латерального края иногда и дорсальный. Рефлекс Оппенгейма плантарен.
На левой стороне все эти рефлексы выражены сильнее, чем на правой. Лицо в покое симметрично.
Во время пребывания ребенка в клинике у него наблюдалось несколько эпилептиформных припадков, которые начинались рвотой. Эти припадки можно характеризовать как клонические; часты ритмические подергивания лица и конечностей, когда конечность только редко двигается как целое, так что возникает впечатление произвольных движений. В большинстве случаев речь идет о подергивании отдельных грудных мышц, причем каждой свойствен свой ритм. Приходит в действие разнообразная комбинация мышечных групп, затем они успокаиваются, в то время как другие охватываются судорогами, так что возникает весьма разнообразная картина судорожного состояния. Даже отдельные мышечные группы лица сокращаются в разном ритме. Один из этих припадков продолжался 3 часа. Ребенок во время припадков бледен, иногда цианотичен. Пульс сильно учащен, 140—160 в минуту. Дыхание не учащено. После припадка цвет лица ребенка становится обычным. Он выглядит хорошо и спокойно засыпает.
Отологическое обследование. Барабанная перепонка нормальна. Сильная чувствительность к громким звукам. При сильных шумах пациент вдруг вытягивает все четыре конечности. Кажущиеся спонтанными движения появляются почти исключительно на акустические раздражения. Эта повышенная чувствительность к звукам прогрессивно усиливается до момента смерти. При вызове калорического и вращательного нистагма лабиринт оказывался значительно гиперчувствительным; появлялся сильный ротаторный и горизонтальный нистагм.
Ринологическое обследование. Патологических изменений не обнаружено.
Речь идет о типичном случае прогрессирующей амавротической идиотии у 16-месячного мальчика, у которого наблюдалось почти полное выключение нормальных функций большого мозга. Иногда появлялись судороги, вероятно кортикального происхождения. Имели место чрезвычайно сильные рефлексы с лабиринта на глаза. На конечностях можно было отметить легкие спазмы.
Исследования пациента 18 и 26 июня дали следующие результаты, которые были зафиксированы двумя кинематографическими съемками: при повороте головы в положении на спине ребенок обнаруживал типичные тонические шейные рефлексы на руки и ноги обеих сторон; кроме того, наблюдался тот же самый рефлекс полукружных каналов при перемещении из сидячего в лежачее положение (и из положения сидя в положение стоя), как это было описано выше в случае 3.
У ребенка имелся в положении на спине разгибательный тонус четырех конечностей. При перемещении его из положения на спине в бо¬
Тонические шейные и лабиринтные рефлексы у человека
ИЗ
ковое положение или на живот, — причем надо было тщательно следить за тем, чтобы не изменилось положение головы по отношению к туло- тзищу, —разгибание конечностей исчезало. Если, наоборот, переместить ребенка из положения на животе или на боку в положение на спине, чо снова происходило сильное разгибание четырех конечностей.
Эти реакции зависят не от изменений положения головы по отношению к туловищу, а от изменений положения головы в пространстве, т. е. они являются лабиринтными рефлексами. Их можно было регулярно и точно вызывать у ребенка.
После получения этих результатов возник вопрос — можно ли установить одно и только одно положение головы в пространстве, при котором разгибательный тонус конечностей максимален, и только одно положение в пространстве, при котором он минимален, точно так же, как это наблюдалось в наших опытах на животных. Если это так, тогда было бы можно установить положение головы для этого максимума и минимума. В опытах на животных для проверки этих отношений мы полностью исключали все шейные рефлексы, вызываемые движениями головы но отношению к туловищу, — гипсовой повязкой были зафиксированы голова, шея и туловище животного, и тогда тело Рис- 61.
как единое целое помещалось
в разных положениях в пространстве, при этом разгибательный тонус конечностей изменялся закономерным образом. Этот прием, конечно, нельзя было применить к пациенту. Но мы достигли цели тем, что тщательно обили войлоком большую доску, положили на нее ребенка на спину и зафиксировали голову и грудь бинтом. Таким образом, ребенок лежал в удобном положении, при котором голова по отношению к туловищу была достаточно зафиксирована, не мешая дыханию пациента и т. д.
Если поднять доску головным концом вверх, пока позвоночный столб не встанет вертикально (рис. 61 и 62), то активного движения конечностей не происходит. Пассивное сгибание в локте (рис. 61), в бедре или в колене (рис. 62) не встречает отчетливого сопротивления. Если теперь опустить очень медленно и постепенно головной конец, поддерживая при этом в течение некоторого времени доску в определенном положении (для того чтобы избегнуть всех лабиринтных рефлексов, которые могли быть вызваны угловым ускорением), то обнаруживается сгибание и разгибание рук до того момента, когда позвоночный столб окажется в горизонтальном положении, в то время как тонически растянутые ноги при помощи легкого сгибания в бедре отрываются от подстилки. При горизонтальном положении позвоночного столба разгибание конечностей усиливается (рис. 63). Пассивное сгибание в локтевом и коленном суставах теперь встречает сильное сопротивление. Если задержать ребенка в этом положении длительный срок, то через некоторое время разгибательный тонус отчетливо уменьшается, не исчезая, однако, полностью. Более 8 Р. Магнус
114
Третья глава. Рефлексы положения
того, остается постоянное тоническое разгибание в левом локте и в левом колене. Если опустить головной конец доски ниже (рис. 64) — до того- момента, когда позвоночный столб образует с горизонтальной плоскостью угол в 45°, — то тоническое разгибание конечности становится максимальным. Однако держать ребенка продолжительное время в этом положении нельзя, так как он становится беспокойным.
Затем головной конец опять поднимается. Как только голова поднята приблизительно на 45а над горизонтальной плоскостью,, разгибание конечностей уменьшается, но сгибание в коленном суставе все еще встречает заметное сопротивление. При продолжении вращения доски разгибательный тонус уменьшается и полностью исчезает,, когда позвоночный столб переходит вертикальное положение Рис> 62, на 20—30° (рис. 65). В этом
положении осуществляется активное сгибание в локте.
Для того чтобы еще точнее определить положение максимума и минимума, ребенка поворачивают вокруг битемпоральной оси на полный круг. При этом разгибательный тонус четырех конечностей становится максимальным, если головной конец при положении пациента на спине опускается на 45° под горизонтальную плоскость (рис. 64). Он остается еще сильным, когда голова находится перпендикулярно вниз. При продолжении вращения он затем уменьшается и, вероятно, достигает минимума, когда тело находится головой вверх и головной конец опускается вперед от 0 до 45°
(рис. 65).
Если повернуть ребенка из положения на спине вокруг позвоночного столба в положение на животе, то при этом разгибательный тонус конечностей постепенно уменьшается.
Если, наоборот, повернуть его Рис- 63.
из положения на животе в положение на спину, то происходит опять тоническое разгибание.
При всех этих движениях постоянно следят за тем, чтобы не изменилось положение головы по отношению к туловищу.
Этими опытами, которые были несколько раз повторены с одними и теми же результатами, было установлено, что для лабиринтных рефлексов на конечности у нашего пациента существует одно и только одно положение головы в пространстве, при котором тонус разгибательных мышц
Тонические шейные и лабиринтные рефлексы у человека
115
всех четырех конечностей максимален, и одно и только одно, при котором он становится минимальным. Оба положения отличаются друг от друга на 180°. Следовательно, тут действуют те же законы, что и у исследованных животных. Спрашивается, совпадает ли положение максимума и минимума у нашего пациента с тем, которое найдено у животных. У них, по ранее установленным результатам, разги- бательный тонус конечностей становится максимальным, если при положении животного на спине ротовая щель поднимается приблизительно на 45° над горизонтальной плоскостью.
Если посмотреть на рис. 64, то видно (если использовать положение ротовой щели как линию ориентации), что у человека можно найти такое же положение для максимума. На рис. 65
также видно, что у человека, как Рис. 64.
и у животного в положении минимума, ротовая щель опущена приблизительно на 45° ниже горизонтальной плоскости.
Далее, из наблюдений на этом пациенте следует, что описанные рефлексы действительно являются рефлексами положения и не вызываются
угловыми ускорениями. Это вытекает, во-первых, из того факта, что для вызова этих реакций движения головы или тела отнюдь не должны быть быстрыми, а что их, напротив, лучше всего можно изучить, когда они выполняются очень медленно и с перерывами. Но совершенно определенно это вытекает из того факта, что тоническая реакция конечностей, относящаяся к каждому положению головы, происходит совершенно одинаково, независимо от того, поворотом в какую сторону было РИС. 65. достигнуто данное положение
головы. Приближаемся ли мы к положению максимума (рис. 64) с помощью поворотов вокруг битем- поральной оси 'из положения на спине или из положения на животе, или мы приближаемся к нему из положения на животе поворотом вокруг позвоночного столба, всегда будет происходить максимальное разгибание конечностей. Весь этот цикл разгибания и сгибания конечностей появляется и тогда, когда пациента поворачивают из положения на спине равномерным вращением вокруг битемпоральной оси на 360°.
Из этого вытекает, что действительно положение головы или лабиринтов в пространстве обусловливает описанные тонические реакции.
я*
116
Третья глава. Рефлексы положения
Если ребенок помещался в описанное положение максимума для разгибательного тонуса конечностей, то сначала появлялось сильное тоническое разгибание рук и ног. Через некоторое время интенсивность равгибательного тонуса уменьшалась, однако не исчезая полностью. Более того, в левом локте и в обоих коленях можно было отметить тоническое разгибание. Нам хотелось положить ребенка на длительное время в положении на спине в кровать, ножной конец которой был поднят, для того чтобы установить, имеется ли у конечностей при таком положении продолжающийся часами или днями разгибательный тонус. К сожалению, наши намерения нельзя было осуществить, потому что пациенту это положение явно было неудобно и потому что прогрессирующая болезнь не позволила осуществить такую проверку. Можно было установить только то, что когда ножной конец кровати был на минуту поднят, разгибательный тонус конечностей в это время был заметно повышенным.
Поэтому наблюдение на следующем больном является очень важным, щк-жак здесь удалось доказать, что и при длительном неизменном положении сохраняются изменения тонуса в соответствующей конечности.
Случай 2. Клиника проф. Хавершмидта 1 (см. стр. 104, № 6).
Речь идет о ребенке, родившемся 1 июня 1921 г., который был первые 2 месяца здоров, заболел в возрасте 2У2 месяцев с повышением температуры; через 3 недели у него появились судороги. После этого наступил проходящий правосторонний гемипарез. При исследовании поражал маленький размер черепа.
У ребенка обнаружились при повороте головы типичные шейные рефлексы на обе руки, которые одинаково проявлялись ‘ при активных и пассивных движениях головы.
При исследовании 6 ноября были обнаружены положительные вращательные реакции глаз и вращательный нистагм в обе стороны, также были обнаружены вращательные реакции головы. Тонические шейные рефлексы на глаза отсутствовали.
Ребенок, как и в случае 1, был зафиксирован на обитой войлоком доске, причем с помощью подушек было достигнуто то, что голова не могла изменить свое положение относительно туловища.
Когда ребенок находился на горизонтально лежащей доске в положении на спине, обе руки были согнуты, иногда спонтанно сгибались и ноги, Если ножной конец доски поднимался на 70°, то через 10—15 секунд происходило сильное тоническое разгибание правой руки, причем рука была дорсально флексирована. В левой руке и в обеих ногах никакого изменения не появилось.
После этого доска помещалась горизонтально, и через несколько секунд правая рука опять занимала исходное сгибательное положение.
Доска поставлена вертикально, голова вверху: при этом обе руки были согнуты, кисти рук находились у подбородка и оставались так 5 минут при неизменном положении доски (фотография). После этого нижний конец доски опять был поднят на 70°, и вначале изменения тонуса конечностей не наблюдалось, потому что ребенок заснул и конечности были расслаблены. Как только, однако, ребенок проснулся, рука опять сразу же разогнулась и оставалась в таком состоянии 6 минут без изменения при постоянном положении доски (фото).
При горизонтальном положении на спине сразу же вновь наступило сгибание рук с отчетливым активным сгибательным тонусом в обоих локтях.
После того как ножной конец подняли на 60°, на левой стороне сгибательный тонус сохраняется, справа, напротив, сгибательный тонус
1 Этот случай будет подробно опубликован доктором Карстексом и д-ром Стен- версом.
Тонические шейные и лабиринтные рефлексы у человека
117
исчезает, и появляется очень сильный разгибательный тонус в локте и в кисти, пальцы сильно сжимаются в кулак. В локте наблюдается сильное сопротивление при пассивном состоянии. В мышцах нет отчетливого изменения.
Этот случай дополняет предыдущий желаемым образом и показывает, что при этих лабиринтных рефлексах на конечности речь действительно идет о постоянных реакциях. Интерес представляет и то, что положения максимума и минимума в обоих случаях приблизительно одинаковы.
В качестве приложения к этому мы приведем некоторые примеры из литературы, где мы с более или менее большой вероятностью можем предполагать присутствие тонических лабиринтных рефлексов.
Случай 3 (см. выше, стр. 103, № 14). Ребенок с амавротической идиотией (Dol- linger).
В положении на спине в ответ на любое афферентное раздражение происходит типичное разгибание, причем ноги разогнуты и руки очень медленно и дрожа поднимаются почти в сагиттальной плоскости до перпендикулярного положения и даже дальше, после чего они как будто тетанически окостеневают и остаются в таком положении или обнаруживают незначительные колебания. Через %—2 минуты они медленно опускаются и ребенок возвращается в свое летаргическое состояние.
Как мне сообщил автор, разгибательные спазмы рук в основном проявлялись в положении на спине или при поднятии нижнего конца кровати на 45°; при положении на животе они не появились. Таким образом, становится вероятным, что и здесь играют роль тонические лабиринтные рефлексы на конечности, благодаря которым конечности при положении ребенка на спине оказываются в состоянии разгибательной готовности. Поэтому любое чувствительное раздражение в состоянии вызвать названные разгибательные спазмы. Акустически ребенок был гиперчувствителен.
Случай 4 (см. выше, стр. 102, № 7).
Ребенок нз городской больницы (д-р Bosscha).
Гнойный менингит. При перемещении из положения сидя в положение лежа появляется неоднократно описанный рефлекс полукружных каналов, при котором руки разводятся и переходят в разгибательное положение. Если оставить ребенка на спине, то описанное положение рук остается в виде правосторонней абдукции тонически сильно разогнутых конечностей, и поэтому ребенка находят в кровати на спине всегда в этом положении. При сидении сначала также происходит реакция полукружных каналов, а после этого руки вяло падают вниз.
И тут, вероятно, имеет место тоническое влияние лабиринтов, благодаря которому при положении на спине поддерживается постоянно разгибание и сгибание рук, в то время как при сидении они (руки) расслабляются.
Случай 5 (см. Böhme и. Weiland).
Мужчина 48 лет. Кровоизлияние в правый зрительный бугор и в правый боковой желудочек. Все желудочки заполнены кровью. Старый очаг в правом полушарии мозжечка (вскрытие).
У пациента обнаруживается сильнейшая децеребрационная ригидность во всех четырех конечностях. Нет отчетливых шейных рефлексов. Вентральное сгибание головы в положении на спине не вызывает никаких изменений. Однако, когда пациента сажают на кровати и затем наклоняют голову вентрально, происходит отчетливое уменьшение разгибательных спазмов в руках и ногах, так что теперь их можно легко пассивно сгибать. В положении на спине разгибательные спазмы появляются вновь.
В этом случае, следовательно, можно показать, что вентральное сгибание головы не вызывает никаких шейных рефлексов, потому что ее движение при положении пациента на спине остается без последствий. Тот же наклон головы ведет, однако, в прямо сидячем положении
118
Третья глава. Рефлексы положения
к расслаблению конечностей. При этом голова попадает в вышеописанное положение минимума для тонических лабиринтных рефлексов на разги- бательные мышцы конечностей, и в соответствии с этим разгибательный смазм в руках и ногах уменьшается.
Случай 6 (Боеме и Вейланд) (см. стр. 103, № 13).
Мужчина 39 лет. Апоплексия после сифилиса. Правосторонняя гемиплегия. Шейные рефлексы на правую руку и правую ногу имеются.
Наклон головы в положении на спине остается без последствий. При поднимании туловища и вентральном наклоне головы появляется сгибательный тонус в правой руке и правой ноге. Обратное помещение пациента на спину вызывает, наоборот, разгибательный тонус в обеих названных конечностях.
И в этом случае, следовательно, не представляется возможным с помощью вентрального сгибания головы в положении на спине вызвать шейный рефлекс, в то время как при изменении положения всего туловища названное движение вызывает сгибательный тонус. Как только голова попадает в положение минимума для тонических лабиринтных рефлексов, правая рука,и правая нога переходят в согнутое положение, в то время как при положении на спине приближение головы к положению максимума вызывает разгибательные спазмы.
Случай 7 (Броувер) (см. стр. 101, № 5).
Девочка, 1 год. Менингоэнцефалит с выключением большого мозга и коры мозжечка. Децеребрационная ригидность, шейные рефлексы на поворот и наклон головы. Ребенок обнаруживает в положении на спине ригидность затылка и опистотонус. Последние исчезают при переходе в сидячее положение, при этом голова наклоняется вперед.
В этом случае, следовательно, можно доказать типичные тонические лабиринтные рефлексы на шейные мышцы, причем положение минимума совпадает с положением, найденным при тонических лабиринтных рефлексах на мышцы конечностей.
Бруннер недавно описал некоторые случаи острого одностороннего заболевания лабиринта, при котором появилась «кривая шея», вызванная односторонним сокращением грудино-ключично-сосковой мышцы. Это сокращение появлялось или односторонне, или перекрестно, что автор объясняет явлением возбуждения или параличом воспаленного лабиринта. Квике [Quix (1)] уже раньше указал на случай врожденной кривошеи, в котором лабиринт одной стороны не был возбудимым.
Эти случаи делают вероятным предположение, что у человека влияние лабиринтов на шейную мускулатуру является таким же односторонним, как это было выше описано для животных.
В самое последнее время наши знания были расширены и в этой области исследованиями Уелша (1). Он нашел у больного гемиплегией в положении на животе активный сгибательный тонус в локте, который уменьшался в положении на спине. У другого такого же больного он наблюдал появление сильного разгибания в локте, как только пациент перемещался в положение на спину.
Далее Уелш обнаружил и влияние положения головы в пространстве на сопутствующие движения. Так, например, в спастичной руке при стоянии пациента произошло сгибание в локте; при положении на спине — разгибание, пронация и незначительное движение вперед; в положении на животе — сильное сгибание в локте и отведение в плече.
Очень хорошо можно было показать и комбинацию тонических шейных и лабиринтных рефлексов.
Тонические шейные и лабиринтные рефлексы у человека
119
Соответствующие движения спастической правой руки. С — сгибание, Р — разгибание
Положение головы
Поло¬
жение
стоя
Поло¬
жение
на
спине
Поло¬
жение
на
живо¬
те
Пра¬
вое
боко¬
вое
поло¬
жение
Левое
боковое
поло¬
жение
Повернута вправо
Р
Р
(Р)
Р
Р
Среднее положение
(С)
(Р)
С
С
С
Повернута влево
С
С
С
Р
С
Следует обратить внимание на то, что при среднем положении головы и положении на спине происходит разгибание, в положении на животе — «сгибание и что при голове, повернутой влево в правом боковом положении происходит разгибание, в левом боковом положении — сгибание. Это можно объяснить правилами для шейных и лабиринтных рефлексов.
Оказалось, что у этого же больного в разные дни иногда преобладали то шейные, то лабиринтные рефлексы.
Из предыдущего вытекает, что и у человека можно при подходящих условиях определенно Показать присутствие тонических лабиринтных рефлексов.
И у человека имеются положения головы в пространстве, при которых тонические лабиринтные рефлексы на разгибательные мышцы имеют свой максимум, и положение, отличающееся от предыдущего на 180°, при котором они имеют свой минимум. Представляет интерес то, что при этом плоскость ротовой щели лежит подобно плоскости у животных. Если пользоваться теми я^е обозначениями, что и у животных, то и у человека положение максимума находится при +45°, положение минимума—при —135°. Эти данные до сих пор приблизительны, точнее их определение можно предпринять только тогда, когда будет исследовано больше больных.
У человека также речь идет о рефлексах положения в чистом их виде. Тоническое влияние лабиринтов продолжается до тех пор, пока голова находится в соответствующем положении в пространстве. И в этом случае следует обратить внимание на то, что нри исследовании нельзя путать тонические лабиринтные рефлексы с рефлексами на движение (реакциями полукружных каналов).
Влияние лабиринтов у человека распространяется на четыре конечности, на шейные мышцы и специально на грудино-ключично-сосковую мышцу. Однако не во всех случаях можно показать такое влияние на все -эти мышцы, точно так же как при тонических шейных рефлексах случается, что реагирует только одна или две конечности, и это преимущественно те, которые раньше уже были спастическими.
Возможно, эти наблюдения приобретут практическое значение. Имеющие место спазмы в соответствующих случаях можно будет уменьшить или устранить соответствующим положением пациента; например, при разгибательных спазмах именно положение на спине оказывает неблагоприятное влияние, в то время как при другом положении эти «спазмы могут пройти. И в работе Симонса (2) указывается на подобные явления.
Имеют ли описанные тонические лабиринтные рефлексы у здорового человека и при интактной центральной нервной системе вообще влияние на положение тела и на распределение тонуса в мускулатуре, или эти
120
Третья глава« Рефлексы положения
влияния обнаруживаются только у больных — выяснение этого вопроса потребует еще дальнейших исследований.
У животного после симметричной перерезки мозгового ствола тонические шейные и лабиринтные рефлексы на конечности всегда отчетливы и определенны. В исследованиях на больных людях, у которых анатомические изменения в центральной нервной системе не являются такими простыми, были сделаны некоторые наблюдения, как это выше упомянуто, при которых исход рефлекса как раз оказался противоположным ожидаемому. Например, в нескольких случаях (случай 4, стр. 99, и случай 5, стр. 101) при наклоне было найдено, что конечности на стороне, на которой ухо приближалось к плечу, сгибались, конечности же на другой стороне разгибались, в то время как в исследованиях Симонса конечности на стороне, на которой ухо приближалось к плечу, разгибались и конечности другой стороны сгибались. Далее Симонс указывает на то, что в большинстве случаев при вентральном наклоне головы руки сгибаются, при дорсальном наклоне разгибаются, в то время как положение ног иногда изменяется в обратном направлении.
Недавно в клинике Винклера наблюдался случай опухоли в области: эпифиза, когда имели место тонические лабиринтные рефлексы на конечности. Стенверс и де Клейн установили, что при положении максимума, соответственно рис. 64, появился сгибательный тонус рук, в то время как руки переходили в разогнутое положение тогда, когда головной конец доски был поднят над горизонтальной плоскостью. Это наблюдение, однако, не являлось постоянным, спустя несколько дней тонические лабиринтные рефлексы больше не наблюдались.
Надо, следовательно, считаться с тем, что иногда у человека при патологических условиях происходит своеобразное изменение тонических шейных и лабиринтных рефлексов. Небезынтересно указать на то, чта подобные наблюдения были сделаны и на животных. Минковский (1} разрушал у обезьян сначала слева восходящую и верхнюю теменную- извилину коры большого мозга, через год он произвел эту же операцию на правой стороне и удалил при этом и лобную долю. Через 22 месяца у этого животного были экстирпированы правая восходящая фронтальная извилина и соседняя часть фронтальной доли. Начиная с 10-го дня после операции наблюдалась синергия между положением головы и положением конечностей, которая оставалась постоянной не менее 6 месяцев. Когда животное в ответ на любое раздражение поворачивало голову так, что морда была направлена к левому плечу, происходило сгибание (аддукция) левой парализованной руки и в более слабой степени— ноги. При повороте головы к правому плечу наступало легкое разгибание левой руки. Здесь, следовательно, реакция на поворот головы оказалась противоположной ожидаемой — и именно после асимметричного нарушения коры большого мозга у обезьян.
Ротфельд (6) указывает, что у кролика после частичной перерезки затылочных мышц наблюдались анормальные рефлексы при изменении положения головы. Он отметил, что у кролика после перерезки дорсальных шейных мышц голова падает вентрально на подстилку и что в связи с этим передние лапы тонически разгибаются вперед. Если перерезать, кроме дорсальных шейных мышц, на обеих сторонах боковые мышцы; (главным образом лестничные), то голова сильно наклоняется вентрально, производя при этом качательные боковые движения.
К спиральному повороту тела присоединяется, кроме того, еще своеобразное положение четырех ног, противоположное тому положению, которое имеет место при обычном шейном рефлексе. Конечности на «че¬
Четвертая глава. Компенсаторные положения глаз
121
репной стороне» разгибаются и на «челюстной» сгибаются. Указание Ротфельда на то, что и у нормального кролика в ответ на поворот максимально наклоненной головы получается подобное изменение рефлексов, я подтвердить не мог.
В обоих случаях речь идет о тонических шейных рефлексах. Исследование показывает, что и у животных при определенных экспериментальных условиях влияние тонических шейных рефлексов на конечности может быть противоположным. Более точного анализа до сих пор не сделано, и поэтому мы не можем сказать, какие условия являются для этого необходимыми. Понятно, что когда нарушения в центральной нервной системе человека являются несимметричными и расположены в других местах по сравнению с нашими обычными экспериментальными вмешательствами, изменение рефлексов может также иметь место.
Четвертая глава
КОМПЕНСАТОРНЫЕ ПОЛОЖЕНИЯ ГЛАЗ
Компенсаторные положения глаз наблюдаются у всех исследованных до настоящего времени млекопитающих, они также были изучены у человека.1 Однако только у животных с боковым расположением глаз они выражены отчетливо и поэтому у них количественно точнее изучены, лучше же всего — у таких животных, которые вообще не имеют или имеют незначительные произвольные движения глаз [Бартельс "(3)]. Поэтому они лучше всего поддаются исследованию у кроликов и морских свинок.
Альбрехт фон Граэфе (A. v. Graefe) исследовал более точно компенсаторные круговые движения глаз у кроликов, однако он еще не провел резкого разграничения между вращательными реакциями и рефлексами положения. Он наблюдал их и у слепых животных, а также нашел сохраненными после перерезки прямых мышц глаза, в то время как после перерезки косых мышц глаза они исчезали. Бреуер [Breuer (1)] первый провел отчетливое различие между компенсаторными положениями и вращательными реакциями глаз. Он же первый выявил зависимость обеих реакций от лабиринтов. Он нашел у слепых людей компенсаторные вертикальные отклонения и обратил внимание на то, что соответствующие компенсаторные положения глаз наступают неизменно, независимо от того, с какой стороны достигается определенное положение головы. В следующей работе (2) он относит их к деятельности отолитов (utriculus). Хёгиес (Högyes) позднее описал отклонения глаз после односторонней экстирпании лабиринтов и пришел к заключению, что нормальное положение глаз зависит от суммы тонических влияний от обоих лабиринтов на мышцы каждого глаза. Вертикальное отклонение при боковом положении головы обусловлено деятельностью находящегося внизу лабиринта, однако он не отделяет постоянные положения глаз от вращательных реакций. Изменения отклонений при различных характерных положениях головы были произведены Хёгиесом и Кубо (Kubo). Ротфельд (4) определил с помощью измерения в пределах одного квадранта (90°) круговые движения глаз при вентральном наклоне головы. Однако
1 И у остальных позвоночных, начиная от рыб, наблюдаются компенсаторные положения глаз.
122
Четвертая глава. Компенсаторные положения глаз
б ряде опытов, очевидно, примешивались шейные рефлексы на глаза. Первые измерения компенсаторных положений глаз были сделаны ьан дер Хове и де Клейном [von der Hoeve u. de Kleyn (27)].
При компенсаторных положениях глаз речь идет и о взаимодействии тонических лабиринтных и шейных рефлексов. При изолированном исследовании лабиринтных рефлексов следует исключить шейные рефлексы и, кроме того, прежде чем определить постоянное положение глаз, которое относится к определенному положению головы в пространстве, нужно следить за тем, чтобы отсутствовали все реакции на движения, т. е. все вращательные реакции глаз.
I. ТОНИЧЕСКИЕ ЛАБИРИНТНЫЕ РЕФЛЕКСЫ НА ГЛАЗА (27, 38)
Своими исследованиями ван дер Хове и де Клейн хотели установить, соответствует ли компенсаторному положению глаз при любом положении головы в пространстве также определенное положение глаза в орбите; далее, имеется ли одно или несколько положений максимума и минимума для этих постоянных рефлексов на глаза; и, в-третьих, каково влияние одного лабиринта на оба глаза.
Техника исследования следующая: глаз исследуемого кролика ко- каинизировался и на роговице выжигался крест, одна линия которого заканчивалась поперечной чертой, а Рис. 66. другая — точкой так, чтобы поло¬
жение креста можно было узнать после поворота (+). Если производится однократное наблюдение, тогда более целесообразно выжечь этот крест нагретым латунным штампом путем легкого прикосновения. Это почти не вызывает раздражения, и поэтому сразу же можно вести наблюдение за глазами; сморщивание роговицы оказывается настолько поверхностным, что через несколько дней крест исчезает, не оставляя никаких повреждений. Если производятся постоянные наблюдения на одном и том же животном, то требуется крест с постоянным положением на роговице, и тогда лучше всего вытравить его при помощи кристалла уксуснокислого овинца. После этого наступает временное раздражение глаза, так что наблюдение можно производить только через несколько дней. Но этим достигается то преимущество, что крест остается на роговице неизменным.
Для того чтобы не вызывать шейных рефлексов на глаза, нельзя в течение всего исследования изменять положение головы по отношению к туловищу. Поэтому кролика прикрепляют к доске го л ово держателем и, кроме того, фиксируют туловище неподвижно на доске с помощью завязок. К головодержателю (рис. 66) прикрепляется четырехугольная рамка, которая находится непосредственно перед исследуемым глазом. Эта рамка с двух сторон имеет тонкие проволочки, которые фотографируются как устойчивая координационная система. Сам глаз держится открытым при помопщ веко держателя. Если теперь сфотографировать глаз, то получается следующая картина (рис. 67).
Определением угла а и расстояния ab и ас можно совершенно точно определить положение глаза (см. рис. 68).
Фотографический снимок глаза при различных положениях был сделан при помощи маленького киноаппарата Эрнемана, который неподвижно фиксировался на той же доске, на которой находился кролик, так что в течение целой серии снимков расстояние от аппарата до глаза и до системы координат оставалось неизменным. Только таким образом можно получить точные измерения. При каждом снимке рядом с глазом прикреплялся листок с номером, что давало возможность знать потом для каждого снимка •соответствующее положение животного в пространстве. Полученные снимки затем увеличивались на белом экране при помощи проекционного аппарата и измерялись.
Так как животное и фотографический аппарат были неподвижно зафиксированы на доске, то оказалось возможным сравнить непосредственно друг с другом снимки,
Тонические лабиринтные рефлексы на глаза
123
полученные при различных положениях доски (т. е. головы) в пространстве. Чтобы поместить животное в любое необходимое положение в пространстве, поступали следующим образом: доска (см. рис. 69) pqrs закреплялась в рамке PQRS так, что доску можно ‘было вращать вокруг оси tu, а доску и рамку вместе — вокруг оси iт, во внешней рамке АВСВ\ как на рамке PQRS, так и на рамке AB CD были прикреплены угломеры, при помощи их можно было отсчитывать величину поворота вокруг обеих осей. Если животное зафиксировано на доске в положении на животе, то вращение вокруг оси vw вызывает поворот животного вокруг битемпоральной оси, в то время как вращение доски вокруг оси tu — поворот животного вокруг окципитально-каудальной оси.
Если же, наоборот, привести животно е при помощи поворота доски на 90° вокруг tu в боковое положение и зафиксировать его в этом положении в рамке PQRS, то вращение рамки вокруг оси то дает поворот животного вокруг дорсо- вентральной оси. При помощи комбинаций поворотов вокруг оси tu И VW можно привести животное в любое необходимое положение в пространстве.
При описании последующих опытов будут употребляться следующие обозначения.
Вращение I: животное в положении на животе, ротовая щель горизонтальна, вращение животного вокруг битемпоральной оси; направление вращения — головой вниз, хвостом вверх.
Вращение II: животное в положении на животе, ротовая щель горизонтальна, вращение животного вокруг окципитально-каудальной оси; направление вращения — исследуемым глазом вниз. У
Вращение III: животное в положении на боку, исследуемый глаз — вверху, ротовая щель вертикальна; направление вращения — мордой вниз.
При исследовании в течение каждого вращения делалось 25 снимков.
Рис. 67.
Рис. 68.
Первый снимок изображает исходное положение (например, при вращении I жи- ioiHoe в положении на животе), затем после поворота на 15° (при вращении I ротовая щель на 15° ниже горизонтальной линии) производился второй снимок, после 30° — третий снимок и т. д., пока на снимке № 25 — после вращения на 360° — не достигалось исходного положения. Таким образом, только с одного глаза можно получить после вращения по трем вышеназванным способам 75 снимков, но так как нужно было снимать и промежуточные положения, количество снимков глаза достигало 100.
После каждого изменения положения животного, конечно, надо было подождать, пока не затихнут вращательные реакции, но все же этот ряд снимков можно было произвести один за другим на одном и том же животном, не слишком его утомляя.
124
Четвертая глава. Компенсаторные положения глаз
Рис. 70 изображает снимок всей установки для опытов. При измерении снимков и изображении результатов на графиках оказалось, что при различных положениях головы в пространстве регулярные горизонтальные отклонения глаз не появляются, для вращательных и вертикальных отклонений глаз, напротив, получились очень отчетливые кривые* которые следует обсудить особо.
Круговые движения
Рис. 71 изображает характер круговых движений глаз у кролика. Расстояние между двумя горизонтальными линиями соответствует одному круговому движению на 10°. Подъем кривой означает вращение глаза верхним полюсом назад. При вращении I морда животного опускается вниз от нормального положения головы с горизонтальной ротовой щелью, при этом глаз двигается назад и максимум кругового движения глаза достигается тогда, когда голова висит вертикально вниз. При дальнейшем вращении этот максимум пока сохраняется (приблизительно до Рис* 70* 150°); но как только голова приближа¬
ется к положению на спине, происходит быстрое изменение положения глаз, так что между 165 и 210° глаза переходят (Rollstellung) из максимального заднего положения в максимальное переднее положение. В этом же положении глаз остается и при вращении животного через вертикальное положение мордой вверх, чтобы потом, при возвращении в нормальное положение, медленно вернуться к исходному. Характерной является сильная асимметрия кривой, которая показывает, что положение глаз при вращении через нормальное положение изменяется постепенно, однако при вращении через положение на спине оно изменяется быстро. Если же, наоборот, вращать голову животного из нормального положения вокруг сагиттальной оси (вращение II), то при этом движения глаз очень незначительны. При вращении же III снова наблюдается энергичное поворачивание глаз. При этом исходным положением является положение на боку. Из этого положения голова поворачивается на 90°, пока не оказывается мордой вниз; тогда поворот глаза назад становится максимальным. При дальнейшем вращении на 90° голова находится в противоположном положении на боку. При вращении на 270° голова оказывается мордой вверх, и глаз поворачивается теперь максимально вперед. При возвращении в исходное положение (на боку) глаз также возвращается снова в исходное положение.
При переходе тела от одного положения в другое при вращении I, II и III оба глаза всегда производят согласованные движения, т. е. всегда вместе переводятся или вперед, или назад.
Из рис. 71 следует, что действительно каждому положению головыв пространстве соответствует совершенно закономерное круговое движение глаза. Притом именно это положение головы в пространстве обусловливает соответствующее положение глаз. Все равно, с какой бы стороны ни дости-
Тонические лабиринтные рефлексы на глаза
125
галось определенное положение головы, всегда устанавливается одно и то же закономерное положение глаза. Уже из этого видно, что рефлексы на движения здесь не мешают. Отсюда следует, что если при разных вращениях достигается одинаковое положение головы, то всегда можно установить и одинаковое положение глаз. Это является очень важным для контролирования снимков, потому что на разных точках кривой должны появляться одинаковые положения глаз. Так, например, нормальное положение головы имеет место при вращении I (снимок 1 и 25) и при вращении II (снимок 1 и 25). Видно, что при этом имеются почти одинаковые круговые движения глаз. Положение головы на спине имеет место при вращении I (снимок 13) и при вращении II (снимок 13). Голова стоит мордой вертикально вниз при вращении I (№ 7) и при вращении III (№ 7). Голова стоит вертикально мордой вверх при вращении I (№ 19) и при вращении III (№ 19).
12 3 k Е 6 7 8 9 10 11 12 13 1k 15 16 17 18 19 20 21 22 23 2k 25 О IS 30 k5 60 75 9010512013515016S180 Ï952102252k0255270285300315330345360°
Положение на боку с заснятым на фото глазом вверх имеет место при вращении II (№ 19) и при вращении III (№ 1); положение на боку с заснятым на фото глазом вниз при вращении II (№ 7) и при вращении III (№ 13). Видно, что названные точки дают почти полное совпадение круговых движений, так что нет сомнения в достоверности кривых.
Когда один снимок был сделан вскоре после вращения (конечно, после затухания вращательной реакции), а второй снимок — при неизменном положении животного через 10 минут, то глаза оказались в одинаковом положении.
Всего было поставлено шесть удачных опытов. На рис. 72 представлены средние данные из пяти или шести опытов.1 Как видно, средняя кривая (60) почти полностью соответствует кривой, изображенной на рис. 71. Прежде всего, положения максимума и минимума являются одинаковыми и отчетливо имеет место асимметрия при вращении I.
Следовательно, из этих измерений вытекает, что действительно имеют место два положения головы в пространстве, при которых круговые движения глаз вперед или назад имеют свой максимум. Глаз максимально перемещается назад, когда голова находится мордой вниз, и это максимальное круговое движение остается в пределах довольно большой области, которая распространяется при вращении I от 90 до 135°, т. е. от такого положения, при котором морда повернута вертикально вниз, до такого по¬
1 Вращение I является средним из шести, вращение II и III — средним из пяти опытов.
126
Четвертая глава. Компенсаторные положения глаз
ложения, которое отличается от него на 45° в направлении запрокидывания головы. Даже до того, когда достигается положение мордой вниз и& положения головы на боку (вращение III), это максимальное круговой движение остается некоторое время неизменным (от 75 до 120°).
Максимальный поворот глаза верхним полюсом роговицы вперед можно увидеть, когда голова стоит вертикально мордой вверх, а именно— опять в пределах довольно большой области, которая распространяется при вращении I от 225 до 300° с абсолютным максимумом приблизительно около 255°. При вращении III максимум полностью не достигается (при 270°).
Абсолютная величина круговых движений, т. е. разница между двумя максимумами, составляла в шести опытах 87, 87, 88, 91, 99 и 100°. У морских свинок наблюдаются такие же величины круговых движений.
1 2 3 Ч S 6 7 8 9 10 11 1Z 13 П 15 16 17 18 19 20 21 22 23 2k 25
Измерение кругового движения на обоих глазах у одного и того же животного в один и тот же день дало 91 и 99°. В разные дни абсолютные данные были немножко отличными друг от друга; форма кривых, однако, была всегда одинакова, и максимумы также наблюдались всегда на одном и том же месте.
Непосредственно после маркировки роговицы уксуснокислым свинцом опыт проводить нельзя, потому что вызванное этим раздражение оказывает значительное тормозящее влияние на поворот глаза. В опыте непосредственно после маркировки максимальное перекатывание глаза показывало только 26°, в то же время как приблизительно через 14 дней этот же глаз обнаруживал максимум при 99°. Поэтому окончательные опыты всегда ставились только через несколько дней после отметки уксуснокислым свинцом, когда все явления раздражения глаза полностью исчезали.
Вертикальные отклонения
Измерялось расстояние от середины роговицы до рамки, т. е. ab (рис. 68). В последующем это расстояние будет обозначаться как высотная разность. Оно велико, когда глаз идет вверх, и мало, когда глаз идет вниз, при нормальном положении оно составляет среднюю величину.
При отметке надо очень следить за тем, чтобы крест отпечатался совершенно в середине роговицы, потому что если этого не будет, то уже при простом вращательном движении получится различие в расстоянии (как это видно из рис. 73, ad и се).
Поэтому намечают не простой крест, а показанную на стр. 122 ( + ) фигуру. Если расстояние измерять всегда только от а, Ъ и с или a', V и с' до¬
Тонические лабиринтные рефлексы на глаза
127
рамки, то сразу можно определить, находится ли точка Ъ в середине роговицы или нет.
В качестве примера характера вертикальных отклонений могут служить кривые, приведенные на рис. 74. Видно, что кривые вертикальных отклонений идут совершенно по-другому, чем кривые круговых движений. В то же время, если при вращении I круговое движение было максимальным, вертикальное отклонение при этом является очень незначительным. Максимальное вертикальное отклонение появляется, напротив, при вращении II, при котором почти не возникает круговых движений. Если исходить от нормального положения головы с горизонтальной ротовой щелью и вращать голову исследуемым глазом вниз, то последний отклоняется в орбите вверх и это отклонение достигает максимума при положении головы на боку (90°), который остается почти неизменным до 150°, т. ,е. до тех пор, пока голова из положения на боку не приблизится на 60° к положению на затылке. Теперь при вращении через положение на затылке происходит быстрое изменение, так что противоположное отклонение глаза в орбите вниз достигается приблизительно при 210° и остается до 285°. После этого глаз возвращается в исходное по-
/ 2 3 4 5 6 7 в 9 10 11 11 13 П 15 16 17 18 19 20 21 22 23 2Ь 25 0 15 30 45 60 7590 1051201351501651801952102252*Ю2552702853003/53303*536(Г
>
*
f—к
И
у
c—
S
►--4
A
Щ.
г*
1
/
f
\
k
N
\
[
i
г .
Ш.
\
'+1Г
/
t
\
ke
\
к
1
t
\
'4
k
У
у
\
А-
/
N
\
)
l
—*
Г—
/
/
N
Г-*
\
*
у
П
/
\
*
Д
{Л
t
/
ƒ
\
;
L_
k
\
\
/
s
u__
Ь'4
/
tL
»
—^'
о--«
4--H*
*-•4
i
Рис. 74.
ложение. Следовательно, при вращении II обнаруживается сильная асимметрия. При вращении III глаз максимально отклонен уже в исходном положений. Если животное вращать из исходного положения (положение на боку) через вертикальное положение мордой вниз в противоположное положение на боку, то при этом достигается другой абсолютный максимум. При вращении III один максимум находится от 345 до 45° и другой от 150 до 240°. И в этом случае выходит, что имеется одно положение головы в пространстве, при котором глаз максимально отклонен вверх, и другое положение головы в пространстве, при котором он отклонен максимально вниз. Оба эти положения являются боковыми положениями головы. Однако так же, как при круговых движениях, оказывается* что максимумы отклонения не связаны с отдельными, строго ограниченными положениями головы в пространстве, а находятся в определенных областях. Поэтому и понятно, что кривые не имеют ни одного пика максимума, а обнаруживают плато, над которым незначительно возвышается абсолютный максимум.
128
Четвертая глава. Компенсаторные положения глаз
Точность определения вытекает из совпадающих результатов на следующих контрольных пунктах. Нормальное положение — вращение I (№ 1 и 25), вращение II (№ 1 и 25). Морда вниз — вращение I (№ 7), вращение III (№ 7). Морда вверх — вращение I (№ 19), вращение III (№ 19). Положение на спине — вращение I (№ 13), вращение II (№ 13 (здесь кривые совпадают не точно при 180°, а приблизительно при 190°)* Положение на боку с исследуемым глазом наверху — вращение II (№ 19), вращение III (№ 1). Положение на боку с исследуемым глазом внизу — вращение II (№ 7), вращение III (№ 13).
Следовательно, положение максимумов для вертикальных отклонений является совершенно другим, чем положение максимумов для круговых движений. При вертикальных отклонениях оно наблюдается при боковом положении головы, при круговых движениях — при вертикальном положении головы мордой вверх или вниз. Таким образом, речь идет о двух независимых переменных величинах, действующих на глаз, которые должны при каждом положении головы в пространстве комбинироваться друг с другом разным образом. Каждому положению головы соответствует определенная комбинация кругового движения и вертикального отклонения глаза, в то время как боковые отклонения при компенсаторных положениях глаз, поскольку они вызываются лабиринтами, вероятно не играют закономерной роли.
Если сравнить вертикальные отклонения обоих глаз при разных положениях головы в пространстве, то можно найти, что они перемещаются в противоположных направлениях. Если при определенном положении правый глаз отклоняется вниз, то левый глаз смотрит вверх, и наоборот. Это значит, что отношения здесь другие, чем при круговых движениях, которые происходят у обоих глаз однозначно. И благодаря этому обнаруживаются своеобразные взаимоотношения положения глаз. При асимметричных положениях головы один глаз смотрит вниз, другой вверх, но оба смотрят одновременно вперед (или повернуты назад). Но все эти сложные положения полностью можно отнести к взаимодействию обоих вышеописанных отклонений.
Влияние тонических лабиринтных рефлексов на отдельные мышцы глаз
При помощи описанных измерений узнают, конечно, только зависимость положения глаза от лабиринтов. Для понимания функции лабиринтов, однако, необходимо знать влияние положения головы в пространстве, т. е. влияние лабиринтов на каждую отдельную мышцу глаза (38).
Так как при разных положениях головы в пространстве вертикальные и круговые движения часто комбинируются друг с другом и так как при этом места прикрепления мышц перемещаются трудно обозреваемым образом, то а priori не является само собой разумеющимся, что когда глазное яблоко перемещено максимально вверх или вниз или когда оно максимально повернуто, то тогда и соответствующие мышцы глаз (прямые и косые) достигают максимума своего сокращения.
Поэтому надо еще исследовать, при каком положении головы в пространстве отдельные мышцы глаз сокращаются максимально и минимально. Только после этого можно обсудить зависимость тонических лабиринтных рефлексов на глазные мышцы от определенных структур вестибулярного органа, и только тогда можно объяснить разные компенсаторные положения глаз совместным действием определенных лабиринтных рефлексов на отдельные глазные мышцы.
Тонические лабиринтные рефлексы на глаза
129
Так как исследованиями ван дер Хове и де Клейна были точно установлены положения глаз для любого положения головы в пространстве, то для этого больше ничего не требовалось, кроме конструирования подходящей модели глаза. В этой модели глазное яблоко надо поставить в разные положения и затем точно измерить длину шести глазных мышц для каждого отдельного положения глаз.
Анатомическое расположение глазных мышц у кролика было описано с приложением рисунков Веселым [Wessely (2)], однако цифровые данные не были приведены. Поэтому мы точно определили положение глазных мышц и их длину у разных кроликов. В основу конструкции глазной модели были положены следующие данные.
Глазное яблоко имеет диаметр 17.5 мм, край роговицы образует круг диаметром 14.5 мм; этот круг лежит приблизительно в 4 мм от вершины роговицы. Линия, которая связывает место прикрепления прямых глазных мьппц с вершиной роговицы, образует с глазной осью угол в 15°, который'сзади открыт. Центральная точка прикрепления верхней и нижней прямых глазных мышц расположена в 3 мм от края роговицы. Центральная точка прикрепления обеих боковых глазных мышц (прямой наружной и прямой внутренней) расположена в 7 мм от края роговицы. Центральная точка прикрепления верхней косой глазной мышцы расположена в 5 мм от края роговицы и приблизительно на 3.5 мм дальше окципитально, чем центральная точка прикрепления прямой верхней мышцы. Длина косой верхней мышцы составляет приблизительно 15.5 мм от блока до ее прикрепления. Эта мышца образует с глазной осью угол в 62°. Нижняя мышца прикрепляется отступя 4.5 мм от края роговицы и на 4 мм окципитально от медиальной линии; длина этой мышцы составляет 19.5 мм; она образует с глазной осью угол в 72°.
Так как одним моим коллегой были выражены сомнения в правильности нашей модели глаза и так как верхней косой мышце было придано другое положение, мы в настоящее время проверили эти измерения еще на трех глазах и могли подтвердить правильность модели. Итак, для угла между верхней косой мышцей и верхней прямой мы нашли на трех глазах следующие данные: 78—83, 74 и 77°. Центральная точка прикрепления верхней прямой мышцы была расположена в 2.7, 3, 2 мм от края роговицы. Два измерения были проделаны на мертвых животных, одно на наркотизо- ванном животном. Исходное положение глаза было зафиксировано выжиганием крестика на роговице.
Наша модель, следовательно, передает, по нашим опытам, действительные отношения на глазе кролика. Если, однако, при препаровке глазное яблоко сильно вытаскивается наружу, то угол между верхней косой и верхней прямой мышцами становится более острым. По исходным данным, Имхоф, механик института Ф. А. Ц., сконструировал приведенную на рис. 75 модель правого глаза кролика.
Глазное яблоко было сделано из деревянного шарика. Он может, вращаться в суставе, находящемся на конце горизонтальной палочки, которая лежит на линии, связывающей исходное место прямых глазных мышц с Центральной точкой глаз, и которая привинчена к вертикальной доске. Параллельно этой палочке на подставке модели нарисована пунктирная линия; сплошная линия на подставке изображает положение глазной о<?и при нормальном положении глаза.
Глазные мышцы в модели представлены крепкими нитками. Они закреплены на глазном яблоке соответственно их местам выхода, проходят через петли и держатся натянутыми с помощью грузов. Верхняя косая мышца представлена от trochlea до bulbus. Все глазные мышцы (нитки) разделены на участки, составляющие Ц4 см, эти участки окрашены разными красками, так что легко можно отсчитывать целые сантиметры. Длину нитей при разных положениях глаз можно определить с точностью до 1 мм (это в действительности соответствует 1/ъ мм).
Край роговицы представлен черной линией, которая разделена на участки по 10°. Кроме того, на модели нарисованы перпендикулярная и горизонтальная медианы роговицы, которые разделены на отдельные участки в 3 мм.
Перед роговицей находится четырехугольная рамка, которая крепко связана с подставкой. В рамке натянут тонкий крестик.
Если работать с этой моделью, то сначала горизонтальную метку роговицы следует установить горизонтально, затем глазную ось установить параллельно черной линии на основной пластине (глазная ось при нормальном положении). Глаз находится в нормальном положении. Затем рамка устанавливается так. что крестик совпадает с меридианами роговицы. Теперь для каждого положения головы в пространстве глазное яблоко приводится в соответствующее положение по вышеприведенным кривым (рис. 71 — для круговых движений, рис. 74 — для вертикальных отклонений).
9 Р. Магнус
130
Четвертая глава. Компенсаторные положения глаз
На рис. 71 каждая горизонтальная линия соответствует круговому движению в 10°, на рис. 74 — вертикальному перемещению на одно деление линии роговицы модели. Затем отсчитывается длина шести глазных мышц и вносится в таблицу.
При каждом из трех проделанных вращений в пространстве ван дер Хове и де Клейном были сделаны 25 отдельных определений через каждые 15° вращения. Всего, следовательно, было сделано 75 разных определений. Эти 75 положений действительно были установлены на модели, и каждый раз измерялась длина шести глазных мышц. Результаты были объединены в таблицах (Pflügers Arch, f. d. ges. Physiol., Bd. 178, стр. 183, 1920), которые здесь не приводятся. Для того чтобы из полученных данных
можно было сделать заключение о влиянии лабиринтов, следует ввести определенные поправки, необходимость которых вытекает из следующего.
Если глаз, исходя из нормального положения, выполняет круговые движения, вызванные сокращением косых глазных мышц, не испытывая при этом вертикальных отклонений, то этим круговым движением перемещается место прикрепления прямых глазных мышц на глазном яблоке и пассивно изменяется длина прямых глазных мышц. Если к определенному круговому движению присоединяется вертикальное перемещение глазного яблока, то сокращение прямых глазных мышц происходит не из той длины, которую они имеют при нормальном положении глазного яблока, а из измененной длины, которая получилась в результате кругового движения (сокращения косых глазных мышц). Поэтому если глаз выполнил круговое движение, то при этом полученную длину прямых глазных мышц надо поправить на такую величину, которая соответствует длине пассивного растяжения или сокращения, вызванного круговым движением. Для того чтобы установить эти данные на модели, глазное яблоко сперва было поставлено в нормальное поло- Рис. 75. жение, а затем производились такого рода круго¬
вые движения, при которых длина верхней косой мышцы изменялась ступенчато на 4—11 мм. Таким образом, возникшие без вертикального перемещения глазного яблока изменения длины верхней и нижней прямых мышц были измерены и различия в длине этих мышц при нормальном положении глазного яблока были объединены в одной таблице. Эта таблица находится на стр. 185 вышеназванной работы и показывает, насколько следует увеличить или уменьшить длину верхней и нижней прямых мышц вследствие изменения длины верхней косой мышцы, вызванного круговым движением.
Таким же образом надо поправить результаты измерения для верхней и нижней косых мышц при разных вертикальных отклонениях глаза. Если глазное яблоко выполняет без круговых движений вертикальные перемещения, вызванные сокращением верхней и нижней прямых мышц, то этим точные прикрепления косых глазных мышц пассивно смещаются и исходная длина изменяется. Исходя из нормального положения на модели, были произведены вертикальные перемещения глазного яблока без круговых движений и измерена соответствующая длина косых глазных мышц. Полученные данные находятся на стр. 186 вышеназванной работы.
На основе этих таблиц были исправлены результаты измерения для верхней и нижней прямых глазных мышц и для обеих косых глазных мышц, полученные результаты этой поправки представлены на графике (рис. 76—79).
Изображение изменения длины внутренней и наружной прямых мышц является ненужным, потому что невозможно было установить закономерные боковые перемещения глаз при изменениях положения головы в пространстве.
На кривых (рис. 76—79) дана исправленная длина верхней и нижней прямых и обеих косых мышц в мм при трех произведенных вращениях головы в пространстве (ван дер Хове и де Клейном). Для верхней косой мышцы приведено только расстояние от блока до места прикрепления на глазном яблоке. Из этих кривых вытекает следующее.
Кривые для верхней косой мышцы (рис. 76) представляют собой почти точное и зеркальное изображение кривых для нижней косой мышцы (рис. 77); это значит, что обе мышцы функционируют как чистые антагонисты: если одна удлиняется, то другая сокращается, и наоборот.
Тонические лабиринтные рефлексы на глава
131
Точно так же кривая для нижней прямой (рис. 78) представляет собой почти точное зеркальное изображение кривой для верхней прямой мышцы
/77/77
(рис. 79), так что эти обе мышцы функционируют при тонических лабиринтных рефлексах на глаза как чистые антагонисты.
Если сравнить кривые для косых глазных мышц (рис. 76, 77) с кривыми, полученными ван дер Хове и де Клейном для круговых движений глаза
Рис. 77. Нижняя косая (корригировано).
(рис. 71), то оказывается, что они совпадают в главных пунктах. Главным образом существенно не изменилось положение максимума и минимума; круговые движения глаза и сокращения косых глазных мышц приблизительно сильнее всего тогда, когда голова стоит вертикально мордой или вверх, или вниз.
Так же совпадают кривые для прямых глазных мышц (рис. 78 и 79) с кривыми, найденными де Клейном и ван дер Хове для вертикальных перемещений глаза (рис. 74).
9*
132
Четвертая глава. Компенсаторные положения глаз
Только для вращения III форма кривой на рис. 78 и 79 более острая, чем на кривых ван дер Хове и де Клейна.
Однако положение максимумов и минимумов существенно не изменилось. Максимум сокращения верхней и нижней прямых глазных мышц находится приблизительно при положении головы на боку. Если сравнить
О 15 30 45 60 75 901051201351501651801952102252*02552702853003153303*5360° Рис. 78. Нижняя прямая (корригировано).
кривые для косых мышц с кривыми для прямых, то вытекает следующее: при
вращении I ( ) реагируют очень сильно косые, в то время как прямые
мышцы почти никакого движения не выполняют, — т. е. при таком вращении обнаруживаются преимущественно круговые движения глаза без
тт 28
26
2*
22
20
О 15 30 45 60 75 90105120135150165180195210 2252*0 255 270285300315 3303*5360°
Рис. 79. Верхняя прямая (корригировано).
вертикальных перемещений. Наоборот, при вращении II ( )
косые мышцы почти совсем не реагируют, в то время как верхняя и нижняя прямые мышцы выполняют интенсивные движения, — т. е. при таком вращении обнаруживаются преимущественно вертикальные перемещения глаза
без кругового движения; Только при вращении III ( )
в тонических рефлексах на глаза участвуют как косые, так и прямые (верхняя и нижняя) мышцы. Следовательно, оказывается, что при вращении I лабиринты воздействуют почти исключительно на косые, при вращении II — почти исключительно на прямые (верхнюю и нижнюю) и при вращении III — на все четыре глазные мышцы.
Тьчный ход кривых мы здесь описывать не будем. Это будет иметь место только при обсуждении зависимости тонических лабиринтных рефлексов
Тонические лабиринтные рефлексы на глаза
133
на глаза от определенных лабиринтных структур и прежде всего оД отолитов.
Точность измерений получается при сравнении приведенных на стр. 125 и 128 контрольных точек на разных кривых. Во всех этих положениях у всех измеренных глазных мышц имеется почти одна и та же длина.
Если резюмировать результаты измерений длины глазных мышц и определений положения глаз в орбитах, то о тонических лабиринтных рефлексах на глазные мышцы можно сказать следующее: у кролика каждому положению головы в пространстве соответствует определенная степень сокращения глазных мышц и определенное положение глаз, которое продолжается до тех пор, пока не изменится положение головы в пространстве.
В этих тонических лабиринтных рефлексах на глаза у кролика наружная и внутренняя прямые мышцы участвуют в незначительной степени. Главным образом речь идет о влиянии прямых мышц, верхней и нижней, которые обусловливают вертикальное отклонение глаз, и обеих косых, обусловливающих круговое движение. Обе прямые мышцы ведут себя здесь как антагонисты: когда одна мышца сокращается, другая удлиняется. Косые мышцы также ведут себя как антагонисты. Напротив, изменения в длине прямых мышц могут комбинироваться в разных соотношениях с изменениями длины косых. Обе эти группы мышц функционируют, следовательно, независимо друг от друга, хотя обе они зависят от лабиринтов. Если голова находится в вертикальном положении мордой вверх, то обе косые верхние мышцы (правая и левая) находятся в состоянии сильнейшего сокращения, обе косые нижние — в состоянии сильнейшего расслабления. Оба глаза тогда оказываются перемещенными верхним краем роговицы вперед.
Если голова находится в вертикальном положении мордой вниз, то обе косые верхние мышцы оказываются в состоянии наибольшего расслабления, обе косые нижние — в состоянии сильнейшего сокращения. Оба глаза тогда оказываются перемещенными верхним краем роговицы назад.
При всех остальных положениях головы в пространстве степень сокращения косых мышц глаза находится между этими крайними положениями. Всегда при этом оба глаза реагируют синхронно.
Если голова находится в положении на правом боку, то левая нижняя и правая верхняя прямые мышцы находятся в состоянии сильнейшего укорочения, левая верхняя и правая нижняя прямые мышцы — в состоянии наибольшего удлинения. Правый глаз тогда максимально отведен вверх, а левый глаз — максимально вниз.
Если голова находится в положении на левом боку, то правая нижняя и левая верхняя прямые мышцы находятся в состоянии сильнейшего укорочения, правая верхняя и левая нижняя прямые — в состоянии наибольшего удлинения. Правый глаз тогда оказывается отведенным максимально вниз, левый глаз — максимально вверх.
При всех остальных положениях головы в пространстве степень сокращения прямых верхних и нижних мышц глаз лежит между отмеченными крайними положениями. Оба глаза всегда реагируют противоположно направленными вертикальными отклонениями. Верхняя прямая мышца одной стороны и нижняя прямая другой стороны реагируют при этом одинаково.
Если голова находится сначала в нормальном положении, а затем поворачивается вокруг битемпоральной оси на 360°, то реагируют главным образом косые мышцы и глаза переводятся в том же направлении.
134
Четвертая глава,. Компенсаторные положения глаз
Если голова сначала находится в нормальном положении и затем поворачивается вокруг затылочно-носовой оси на 360°, то при этом реагируют главным образом верхняя и нижняя прямые мышцы, глаза выполняют вертикальные отклонения (в противоположном направлении).
Если голова находится сперва в положении на боку и поворачивается затем вокруг вентро-дорсальной оси на 360°, то реагируют обе группы мышц и положения глаз являются результирующей одинаковых круговых движений и противоположно направленных вертикальных отклонений.
Если объяснить, каким образом осуществляются вышеописанные тонические рефлексы на прямые верхние и нижние и на обе косые мышцы, тогда надо знать 1) «нулевое положение» глаз, т. е. такое положение, которое принимают глаза без всякого лабиринтного влияния, и 2) характер компенсаторных положений глаз над влиянием только одного лабиринта, т. е. после односторонней экстирпации лабиринта.
Только тогда, когда будут располагать всеми этими данными, можно ответить на вопрос, как осуществляются в результате взаимодействия лабиринтов описанные положения глаз.
Положение глаз после двусторонней экстирпации лабиринта
После двусторонней экстирпации лабиринта полностью отсутствуют компенсаторные положения глаз при изменениях положения головы в пространстве. Пока положение головы по отношению к туловищу не изменяется, положение глазного яблока в орбите оказывается неизменным.
Рис. 80 а. Левый глаз до экстирпации лабиринтов, голова в нормальном положении (—175°). На этом и последующих снимках сфотографирована нитка с грузом в виде отвеса.
Рис. 80 б. Тот же глаз через два дня после двусторонней экстирпации лабиринтов. Положение глаз то же самое. Круговое движение не наблюдалось после экстирпации лабиринтов.
Это с особенной легкостью можно наблюдать у кролика, у которого почти не осуществляются произвольные содружественные движения глаз (Blickbewegungen).
После двусторонней лабиринтэктомии оба глаза стоят симметрично, один стоит не выше другого, и оба не имеют вертикальных отклонений вниз и вверх. Этот факт особенно отчетливо можно наблюдать тогда, когда сначала экстирпируется один лабиринт, вследствие чего происходит очень сильное вертикальное отклонение обоих глаз. Если позднее экстирпиро- вать, и другой лабиринт и подождать до тех пор, пока не затихнут острые явления после экстирпации лабиринта, то оба глаза опять стоят в перво¬
Тонические лабиринтные рефлексы на глаза
135
начальном среднем положении. Такое поведение вполне соответствует ожидаемому.
Как обстоит дело с поворотом глаз после двусторонней лабиринтэктомии, а priori с уверенностью сказать нельзя. Предполагалось, что нулевое положение глаз будет находиться при максимальном наклоне вперед или назад или что оно лежит в среднем положении между этими двумя крайними положениями. В действительности оказалось, что имеет место последний случай. Для того чтобы установить это (50), у трех кроликов выжигались на обоих глазах, под местной анестезией, крестики на роговице, один конец которых отмечался поперечной черточкой. Положение глаз фо¬
тографировалось перед и после двусторонней лабиринтэктомии при разных положениях головы в пространстве. Результаты представлены на рис. 80.
После этих измерений мы знаем положение покоя глаза, которое он принимает, если лишен влияния лабиринтов. Положение покоя глаза — это симметричное положение глаз при нормальном положении головы с ротовой щелью, немножко опущенной ниже горизонтальной плоскости.
Из этого нулевого положения возбуждения, исходящие из лабиринтов при других положениях головы в пространстве, вызывают компенсаторные положения глаз.
Положение глаз после односторонней экстирпации лабиринта (15, 27, 50)
После односторонней лабиринтэктомии при определенных положениях головы в пространстве обнаруживаются очень сильные отклонения глаз, так что иногда роговица полностью скрывается и остается видным только белок глаза. При таких обстоятельствах фотографические снимки по вышеописанному способу должны отразить роговицу и отмеченный крестик на ней в сильно уменьшенном виде, и измерения по¬
Рис. 80 в. До экстирпации лабиринтов. Голова мордой вверх. Сравнение с рис. 80а показывает, что глаз передвинулся верхним полюсом роговицы в сторону носа (сокращение верх-
Рис. 80 г. Тот же глаз через два дня после двусторонней экстирпации лабиринтов. Глаз остановился в нормальном положении (ср. с рис. 80а и 806). Сравнение с рис. 80в показывает, что при интактных лабиринтах верхняя косая сокращается.
ней косой).
136
Четвертая глава. Компенсаторные положения, глаз
этому сопровождаются более крупными ошибками, чем измерения у животных с интактными лабиринтами. К этому, далее, присоединяется то, что кролики после односторонней экстирпации лабиринта обнаруживают сильные вращения головы, которые насильственно задерживаются фиксацией животного на станке для фотографирования. При этом создается неудобное положение головы, которому животное иногда сопротивляется, так что снимкам мешают промежуточные спонтанные движения всего животного. Таким образом получается, что измерения после фотографирования сопровождаются ошибками, и поэтому они могут только приблизительно передать действительные соотношения. Но все-таки существенные закономерности можно узнать и из таких снимков. Для того чтобы получить абсолютные данные, надо повторить эти определения более точным методом.
Если экстирпировать у одного кролика лабиринт, — допустим, правый, — и подождать некоторое время, пока не затихнут первые острые
Рис. 80 д. До экстирпации лабиринтов. Голова повернута мордой вниз. Сравнение с рис. 80а показывает, что глаз верхним полюсом роговицы переместился по направлению к затылку (сокращение нижней косой).
следствия операции, прежде всего сильный нистагм, и пока немного не уменьшится девиация (отклонения глаз), то можно наблюдать следующие постоянные явления.
Когда , животное сидит свободно на полу, то его голова более или менее (часто на 90°) оказывается повернутой вправо. При этом левый глаз, который смотрит вверх, не отклоняется или же отклоняется вертик
Рис. 80 е. Тот же глаз через два дня после двусторонней экстирпации лабиринтов. Глаз остановился в нормальном положении (ср. с рис. 80а и 80Ь). Сравнение с рис. 80е показывает, что при интактных лабиринтах сокращается нижняя косая мышца.
То же положение глаз, что на рис. 80b, 80d, 80е после экстирпации лабиринтов, было установлено у того же животного после смерти. Правый глаз был таким же, как и левый. Опыты на двух других кроликах показали те же результаты.
яо—дорсально; нижний (правый)
глаз часто оказывается слегка повернутым в вентральном направлении. Это же положение глаз обнаруживается и тогда, когда поворот го¬
ловы по отношению к телу оказывается компенсированным, т. е. когда животное кладут на правый бок (голову и тело). При таком положении головы в пространстве вертикальное отклонение после правосторонней лабиринтэктомии является минимальным. При всех остальных положениях головы в пространстве обнаруживаются более сильные отклонения.
Если, наоборот, повернуть голову в противоположную (левую) сторону, то обнаруживается максимальное тоническое вертикальное отклоне-
Тонические лабиринтные рефлексы на глаза
137
ние обоих глаз, которое оказывается более сильным на той стороне, на которой отсутствует лабиринт. В этом примере правый глаз отклонен вентрально и слегка вперед, а левый глаз — дорсально. Это отклонение может быть таким сильным, что в щели между веками можно видеть чаще всего только белок глаза. Оба глаза, следовательно, обнаруживают противоположную девиацию: один отклонен вентрально, другой дорсально. В этом положении глаз почти ничего не изменится и тогда, когда в описанное боковое положение, в котором находится голова, приводится и тело (60). Из этого следует, если исходить из среднего положения глаз, что после односторонней лабиринтэктомии при разных положениях головы в пространстве обнаруживается только вертикальная девиация глаз в одном направлении. Если голова находится в таком боковом положении, что сохраненный лабиринт оказывается наверху, то это отклонение глаз будет нулевым или минимальным. Если, напротив, голова находится в таком боковом положении, при, котором сохраненный лабиринт лежит внизу, то отклонение глаз оказывается максимальным ц именно таким, что верхняя прямая мышца одноименной стороны и нижняя прямая мышца противоположной стороны максимально сокращаются. Во всех остальных положениях головы в пространстве наблюдается вертикальное отклонение, лежащее между этими двумя крайностями. Оба глаза при этом всегда обнаруживают противоположные отклонения от нулевого положения.
Положение глаз при интактных лабиринтах
Эти замечания достаточны для того, чтобы объяснить вертикальные отклонения обоих глаз при интактных лабиринтах (38, 50). По¬
смотрим, например, соотношение при вращении II вокруг сагиттальной оси (нормальное положение—правое боковое положение — положение на спине—левое боковое положение—нормальное положение). В правом боковом положении левый лабиринт находится наверху и поэтому не имеет влияния или имеет только минимальное влияние на верхнюю и нижнюю прямые мышцы обоих глаз. Правый лабиринт находится внизу и вследствие этого имеет максимальное влияние, благодаря которому правая прямая верхняя и левая прямая нижняя мышцы сокращаются максимально. Правый глаз, таким образом, отклонен максимально вверх, левый — максимально вниз. Если голову повернуть в нормальное положение или в положение на спину, то возбуждение правого лабиринта уменьшается, левого — увеличивается до тех пор, пока оба не уравновесятся, и благодаря их симметричному влиянию на прямые мышцы обеих сторон глаз приходит в среднее положение. Если повернуть голову в левое боковое положение, то правый лабиринт полностью или почти не оказывает влияния, левый максимально возбужден, вследствие чего левая верхняя и правая нижняя прямые мышцы оказываются максимально сокращенными. Левый глаз поэтому отклонен дорсально, правый глаз — вентрально.
При вращении I вокруг битемпоральной оси не происходит почти никаких вертикальных отклонений глаз, потому что оба лабиринта при этом все время находятся в симметричном положении. При вращении III (исходное положение боковое, вращение вокруг дорсо-вентральной оси), наоборот, должны осуществиться максимальные вертикальные отклонения, потому что в исходном положении голова находится мордой вниз (оба лабиринта стоят симметрично), затем голова снова приводится в боковое положение с максимальным отклонением глаз, после чего голова направляется мордой вверх, причем лабиринты опять удерживают друг
138
Четвертая глава. Компенсаторные положения глаз
друга в равновесии, и наконец голова возвращается в боковое положение с максимальным отклонением глаз. Из этого следует, что действительно возможно объяснить вертикальные отклонения обоих глаз при сохраненных лабиринтах влиянием суммы возбуждений со стороны обоих лабиринтов на верхнюю и нижнюю прямые мышцы обоих глаз. Объяснение более тонких подробностей на кривых можно дать только тогда, когда будет обсуждена зависимость вертикальных отклонений от определенных отолитов.
Характер круговых движений. В то время как после односторонней лабиринтэктомии имеет место односторонняя девиация
/ 2 3 Ь 5 6 7 8 9 10 11 12 13 Щ 18 16 17 18 19 20 2122 23 29 2S О 15 30 95 50 75 90105120135150165180195210225290255270285300315330395360е
Рис. 81..
глаз в вертикальном направлении, девиация по окружности не наступает. В присутствии только одного лабиринта оба глаза при разных положениях головы в пространстве всегда совершают одноименные круговые движения, только величина экскурсий составляет половину той, которая наблюдается при сохраненных обоих лабиринтах (27). Следовательно, один лабиринт оказывается в состоянии однородно влиять на оба глаза, причем у некоторых животных лабиринт сильнее влияет на одноименный глаз, у других — на противоположный, иногда же невозможно заметить разницу в величине круговых движений обоих глаз. Максимум для круговых движений вперед и назад лежит при таких же положениях головы в пространстве, которые можно установить и при целостности обоих лабиринтов.
Если определить на основе измерений, сделанных ван дер Хове и де Клейном (27) для круговых движений у кроликов, лишенных лабиринта на одной стороне, влияние, которое каждый отдельный лабиринт оказывает на один глаз при разных положениях головы в пространстве, то можно суперпонировать результаты влияния, исходящие из правого и левого лабиринтов, и тогда получить кривые, соответствующие тем, которые представлены на рис. 71.
У одного и того же кролика удалось фотографически определить круговое движение до и после односторонней лабиринтэктомии. На рис. 81 прерывистая линия изображает круговое движение левого глаза при вращении I, сплошная линия — круговое движение глаза, найденное в одном опыте после односторонней лабиринтэктомии сложением влияний правого и левого лабиринтов. Видно, что и эти две кривые удивительным образом совпадают. Следовательно, и здесь удается круговые движения глаз у интактных кроликов свести к суперпозиции тех влияний, которые осуществляются каждым отдельным лабиринтом.
Тонические лабиринтные рефлексы на глаза
139
Так как согласно наблюдениям, приведенным выше, нулевое положение глаза есть в то же время и его среднее положение и так как для осуществления кругового поворота в обоих направлениях, а также в направлении вперед и назад обоих глаз достаточно одного лабиринта, то каждый лабиринт должен быть связан с верхней и нижней косыми мышцами обоих глаз. Таким образом, один лабиринт вызывает максимальный поворот благодаря сокращению обеих нижних косых мышц, если голова повернута вертикально мордой вниз. Наоборот, один лабиринт вызывает максимальный поворот благодаря сокращению обеих верхних косых мышц, если голова стоит вертикально мордой вверх. Величина сокращений косых глазных мышц составляет при одном лабиринте приблизительно только половину той, которая имеет место при наличии обоих лабиринтов. При нормальном положении головы возбуждение верхней и нижней косых мышц удерживается в равновесии.
Против такого взгляда можно сделать только одно возражение. До сих пор мы всегда исследовали компенсаторные круговые движения глаз с интактными верхней, так и нижней косыми мышцами. G самого начала не исключено, что положение максимумов станет другим, если исследовать влияние лабиринтов только на одну косую мышцу после исключения ее антагониста, иначе говоря — что степень кругового движения определяется не только сокращением одной косой мышцы с одновременным пропорциональным расслаблением ее антагониста, но что при определенных положениях головы сокращаются, вероятно, обе косые мышцы одновременно. Что это в действительности не имеет места, доказывается со всей очевидностью опытами, в которых мы у одного и того же животного на правом глазу экстирпировали верхнюю косую и на левом глазу — нижнюю косую мышцы от основания (от блока) до места прикрепления их на глазном яблоке (50). Через день после операции на обеих анестезированных роговицах были выжжены кресты, после чего животное было прикреплено к доске с ориентировочной рамкой (рис. 70, стр. 124), затем было произведено вращение I, ичерез каждые 30° были измерены круговые движения на обоих глазах. После этого животное забили, и на секции было установлено отсутствие экстирпированных глазных мышц при интактности остальных.1
Результат измерений был следующий (см. таблицу на стр. 140). Если представить эти круговые движения в виде кривых, то получается, что как для косой верхней, так и для косой нижней мышц ход кривой тот же, каким он был получен для глаза с интактными обеими косыми мышцами (рис. 71 и 72, вращение I) и каким он был представлен (рис. 76 и 77, вращение I) при изучении длины обеих косых глазных мышц в случае, когда обе они были интактны.
Результат этих исследований никакого сомнения не оставляет.
Максимум сокращения для косой верхней мышцы находится при таком положении головы, когда она приблизительно повернута вверх, и для косой нижней — когда голова находится приблизительно мордой вниз.
В этом опыте интересно, что после выпадения функции верхней косой мышцы глаз все-таки в состоянии верхним полюсом роговицы поворачиваться вперед, а после выпадения функции нижней косой мышцы верхним
1 За основу этих опытов можно принять старое положение Граэфе о том, что у кролика после полной экстирпации обеих косых мышц при сохраненных прямых круговые движения оказываются уничтоженными и что, напротив, после полной экстирпации четырех прямых при интактных косых мышцах они оказываются сохраненными.
140
Четвертая глава. Компенсаторные положения глаз
жениях головы совершают
Левый глаз: косая верхняя мышца интактна, косая нияшяя перерезана
Правый глаз: косая нижняя мышца интактна, косая верхняя перерезана
положе¬
ние
головы
враща¬
тельное
движение
глаза
положе¬
ние
головы
враща¬
тельное
движение
глаза
0°
87°
0°
87°
30
65
30
82
60
57
60
55
90
44
90
29
120
40
120
27
150
40
150
31
180
53
180
73
210
89
210
93
240
92
240
97
270
93
270
97
300
93
300
97
330
87
330
92
360
' 86
360
87
полюсом роговицы поворачиваться назад и что оба глаза при всех поло-
такие круговые движения, как если бы функции обеих косых мышц остались неизменными.
Компенсаторные положения глаз у кролика сохраняются и после перерезки ствола на уровне переднего края среднего мозга, когда ядра глазных мышц остаются интактными. Центральные пути, которые должны иметься у кролика для объяснения описанного изменения тонических лабиринтных рефлексов на глаза, схематически представ лены на рис. 82. Сплошные линии представляют собой пути для прямой верхней и нижней мышц глаза, а прерывистые линии— для косых мышц. Пути от правого лабиринта нарисованы жирной линией, а от левого — тонкой линией. Каждая из четырех косых мышц подвергается влиянию обоих лабиринтов, каждая из обеих прямых мышц (верхняя и нижняя) подвергается влиянию только одного лабиринта. Один лабиринт влияет на все четыре косые и только на верхнюю прямую гомолатеральной и на нижнюю прямую мышцу противоположной стороны. Внутренняя и наружная мышцы при тонических лабиринтных рефлексах заметно не сокращаются.
Следовательно, речь здесь идет о двух взаимодействующих друг с другом двигательных системах, из которых одна вызывает вертикальное от- кл онение, другая—круговое движение
При разных положениях головы по отношению к горизонтальной плоскости глаза стараются устанавливаться благодаря исходящим от лабиринтов тоническим возбуждениям каждый раз таким образом, чтобы положение сетчатки по отношению к внешней среде оставалось неизменным. Однако эта цель из-за влияния тонических лабиринтных рефлексов на глазные мышцы никогда полностью не достигается.
Если, например, исходя из нормального положения, повернуть голову вокруг битемпоральной оси на 60° мордой вниз, то глаза, как показывает рис. 71, поворачиваются на 38° верхним полюсом роговицы назад. Горизонтальный меридиан сетчатки, таким образом, поворачивается на 22° по отношению к горизонту. Компенсация неполная.
0Ы. inf. O^Ti
Reet. sup. Q
Гр--0 0ЫЖ 1 i i
I j Rect.sup.
Рис.
Влияние тонических шейных рефлексов на глаза
141
То же самое можно найти при вертикальных отклонениях. Если, например, исходя из нормального положения, повернуть голову вокруг назально-окципитальной оси (вращение II) на 45° вбок, так что левый глаз оказывается наверху, то он поворачивается в орбите приблизительно на 20° вниз, т. е. вентрально. Здесь также компенсация неполная и линия взора поднимается приблизительно на 25° над горизонтом.
Если, исходя из нормального положения, повернуть голову в горизонтальной плоскости вправо или влево, то глаза вообще не осуществляют компенсаторных установок, исходящих из лабиринтов. Лабиринтный механизм, следовательно, у кроликов сам по себе неспособен поддерживать глаза в неизменном положении по отношению к пространству при разных новых положениях головы. Здесь присоединяются еще тонические шейные рефлексы на глаза.
II. ВЛИЯНИЕ ТОНИЧЕСКИХ ШЕЙНЫХ РЕФЛЕКСОВ НА ГЛАЗА (53)
То, что изменения положения головы по отношению к телу могут вызвать изменения положения глаз, наблюдал уже Стевенсон (1892) в лаборатории Эвальда у собаки. Он не смог, однако, объяснить свое наблюдение.
Эти рефлексы были обнаружены только в 1907 г. Барани (1) на кролике, у которого при зафиксированной голове при движениях туловища наблюдались изменения положения глаз. Так как, однако, реакции получались разные в зависимости от исходного положения головы, Барани не смог окончательно доказать предположение, что здесь речь идет о шейных рефлексах. Удовлетворительное объяснение было дано только благодаря опытам де Клейна (53), который показал, что в наблюдениях Барани речь шла о суперпозиции тонических шейных и лабиринтных рефлексов на глаза.
При изучении тонических шейных рефлексов лучше всего исходить из опытов на делабиринтированных животных, в которых вызванные лабиринтами компенсаторные положения глаза не могут помешать результатам. При таких обстоятельствах шейные рефлексы у кролика осуществляются совершенно постоянно и независимо от положения головы в пространстве. Опыты показали, что каждому положению головы относительно туловища соответствует определенное положение глазного яблока в орбите.
Если исходить из положения кролика, фиксированного на животе, с головой в нормальном положении, то движение тела по отношению к фиксированной голове может вызывать следующие тонические изменения положения глаза.
1. Если повернуть тело животного в горизонтальной плоскости вокруг дорсо-вентральной оси (наклон туловища), то можно увидеть, что тот глаз, к которому туловище приближается, отклоняется вперед (назально), другой глаз — назад (окципитально). Речь идет о реакции прямой наружной и внутренней мышц. На стороне поворота туловища сокращается прямая внутренняя мышца, на другой стороне — наружная (противоположная реакция глаз).
2. Если двигать туловище в сагиттальной плоскости и если его при этом вращать вокруг фронтальной оси (поднимание—опускание туловища), то при приближении спины животного к темени верхний полюс роговицы обоих глаз будет двигаться вперед. Если, наоборот, приблизить живот к нижней челюсти, то оба глаза верхним полюсом роговицы повернутся назад. Оба глаза, следовательно, выполняют одноименное движение, при котором действуют косые верхние и нижние мышцы.
142
Четвертая глава. Компенсаторные положения глаз
3. Если вращать туловище вокруг позвоночного столба, приняв его за ось (поворот туловища), то тот глаз, в сторону которого двигается спина, отводится вертикально вниз, а тот глаз, в сторону которого двигается живот, — вертикально вверх. В этом случае, следовательно, оба глаза двигаются в противоположном направлении. Речь идет о реакции прямой верхней и нижней мышц.
Вызванные таким образом отклонения глаз не очень сильно выражены и увеличиваются только при значительных (эксцессивных) поворотах туловища по отношению к голове.
Описанные наблюдения можно резюмировать в виде следующих общих правил:
1) глаза двигаются всегда в такой же плоскости, в которой двигается туловище;
2) направление движения глаз оказывается таким же, как и направление движения туловища.
Следовательно, оказывается, что тело как будто бы представляет собой рычаг, при помощи которого глаза двигаются в закрепленной голове в определенном направлении.
Эти же рефлексы можно, конечно, вызвать, если у делабиринтирован- ного кролика изменить положение головы по отношению к телу. Тогда движения глаза, как легко понять, являются компенсаторными, потому что туловище не двигается; глаза, следовательно, тоже стараются сохранить свое положение в пространстве, что им, однако, удается только частично. При наклоне и повороте соответствующие прямые мышцы обоих глаз всегда реагируют противоположно, при поднимании и опускании головы косые мышцы, наоборот, — всегда одинаково.
В то время как при тонических лабиринтных рефлексах на глаза на каждой стороне функционируют только четыре глазные мышцы (обе косые и прямые, верхняя и нижняя), то при тонических шейных рефлексах на глаза функционируют все шесть глазных мышц, т. е. и прямые наружная, и внутренняя. Обе последние мышцы подвергаются при компенсаторных положениях глаз влиянию со стороны шеи.
Тонические шейные рефлексы у кроликов с интактными лабиринтами. Барани наблюдал, что поворот туловища при нормальном положении головы вызывает девиацию глаз в направлении щели между веками, в то время как то же самое движение тела вызывает девиацию глаз вертикально к щели между веками, если голова висит вертикально мордой вниз.
Де Клейн (53) смог дать объяснение этому, на первый взгляд кажущемуся парадоксальным наблюдению. Рис. 83, а изображает положение левого глаза, когда животное находится на животе с горизонтальной ротовой щелью и симметричным положением головы по отношению к туловищу. Если повернуть туловище вокруг дорсо-вентральной оси в горизонтальной плоскости в сторону левого глаза, то левый глаз перемещается по направлению щели между веками к носу, потому что происходит сокращение прямой внутренней мышцы. Вследствие этого глаз достигает своего нового положения, изображенного на рис. 83, Ъ. Если же положение головы изменяется так, что морда находится перпендикулярно вниз, то положение глаза в орбите изменяется благодаря тоническим лабиринтным рефлексам. Глаз производит сильное круговое движение верхним по¬
<Ж> <£>
т int 777 ext \
с с"!*:
<ж> <?>
Рис. 83. Объяснения в тексте.
Влияние тонических шейных рефлексов на глаза
143
люсом в окципитальном направлении (рис. 83, с). Благодаря этому места прикрепления внутренней и наружной мышц должны переместиться в орбите. Если произвести точно такое же движение туловищем по отношению к голове, как до этого, то выступает точно такой же рефлекс, а именно — сокращение внутренней и расслабление наружной мышц. Результат движения глаз по отношению к орбите является, однако, совершенно другим. Глаз двигается теперь не в направлении щели между веками, а почти перпендикулярно к ней (рис. 83, d). Левый глаз достигает своего нового положения движением вперед вверх, правый вследствие этого же механизма — движением назад вниз.
На рис. 84,а—д даны фотографические снимки вышеописанных разных положений глаз у нормального кролика.
Так как тонические лабиринтные рефлексы иногда действуют так же, как и тонические шейные, а иногда в противоположном им направлении, то получается, как кажется, совершенно нерегулярный комплекс рефлексов, который на первый взгляд очень трудно разгадать.
Рис. 84 а. Животное в положении на животе, ротовая щель горизонтальна, туловище симметрично по отношению к голове. На роговице крест. Поперечная черта у заднего конца горизонтальной насечки.
Рис. 84 б. То же положение головы. Туловище повернуто, насколько возможно, вокруг дорсальной оси но направлению правого глаза. Левый глаз повернут назад но направлению глазной щели (движение по направлению короткой черты, нанесенной на роговице).
Рис. 84 в. То же положение головы. Туловище повернуто, насколько это возможно, вокруг своей дорсальной оси по направлению левого глаза. Левый глаз — вперед по направлению глазной щели (движение также по направлению короткой черты, нанесенной на роговице).
Однако вышеописанным способом проведенные наблюдения удалось полностью объяснить взаимодействием тонических шейных и лабиринтных рефлексов. Результаты совместного действия этих обеих рефлекторных групп оказались совершенно постоянными.
144
Четвертая глава. Компенсаторные положения глаз
Для того чтобы выявить афферентные нервы для тонических шейных рефлексов на мышцы глаз, у кролика экстирпировались сперва оба лабиринта и затем, спустя несколько дней, исследовались шейные рефлексы на глаза. Затем перерезались на обеих сторонах задние корешки обоих верхних шейных нервов. В трех опытах после этого шейные рефлексы на глаза полностью исчезали, в двух других случаях они оказались сильно ослабленными, но их можно было еще частично вызвать. Из этого можно заключить, что рефлекторная дуга для тонических шейных рефлексов на глаза преимущественно проходит по чувствительным корешкам 1-го и 2-го шейных сегментов, но что у некоторых кроликов 3-й шейный сегмент также содержит центрипетальные волокна для этих рефлексов. Выше было сказано, что пути для тонических шейных рефлексов на мышцы конечно-
Рис. 84 г. Голова мордой вертикально вниз. Туловище повернуто, насколько возможно, вокруг своей дорсо-вентральной оси по направлению правого глаза. Движение глаза опять происходит по направлению короткой черты креста на роговице, однако на этот раз не назад по направлению глазной щели, а вниз назад из-за кругового вращения, вызванного тоническими лабиринтными рефлексами.
Рис. 84 д. Голова мордой вертикально вниз. Туловище повернуто, насколько возможно, вокруг своей дорсо- вентральной оси по направлению левого глаза. Движение глаза опять происходит по направлению короткой черты креста на роговице, однако теперь не вперед по направлению глазной щели, а вверх вперед.
стей проходят через задние корешки от С1 до С3, у кроликов незначительная часть может проходить и
через С4. Начальные пути для тонических шейных рефлексов на мышцы глаз выходят, таким образом, приблизительно из одних и тех же частей шеи.
Реципрокная иннервация. Известно, что Шеррингтон (5) показал, что движения глаз происходят благодаря реципрокной иннервации антагонистических мышц глаз, т. е. что сокращение агониста сопровождается расслаблением его антагониста, что смог показать и Бартельс (1) при вестибулярных реакциях глаз — нистагме.
То же самое можно особенно легко показать при тонических шейных рефлексах на мышцы глаз, потому что при этом можно фиксировать голову, в то время как при регистрации тонических лабиринтных рефлексов голову все время нужно помещать в другое положение в пространстве.
Для этой цели у кроликов были перевязаны под эфирным наркозом сонные артерии и перерезаны блуждающие нервы, были отпрепарованы внутренняя и наружная пря¬
Совместное действие шейных и лабиринтных рефлексов на глаза
145
мые мышцы одного глаза, в месте прикрепления этих мышц к глазному яблоку были привязаны нитки и затем мышцы отсекались от глазного яблока. Затем экстирпирова- лись глазное яблоко и остальные мышцы глаз и нитки соединялись с пишущими рычагами, как это делали в своих опытах Тополянский и Бартелье. Опыты продолжались иногда под эфирным наркозом, иногда же производилась экстирпация обоих полушарий (таламическое животное). Пример опыта приведен на рис. 85. Наверху записаны сокращения прямой внутренней, внизу — прямой наружной мышц левого глаза.
1. Исходное положение: животное в положении на животе, ротовая щель горизонтальна, туловище симметрично по отношению к голове (нормальное положение).
2. Из этого положения туловище поворачивалось как можно дальше вокруг дорсо-вентральной оси в направлении левого глаза (поворачивание в направлении OS). При этом наступает отчетливое сокращение внутренней
Рис. 85. Верхняя кривая — внутренняя прямая мышца; нижняя кривая — наружная
прямая мышца.
и отчетливое расслабление наружной прямой мышц. Эти состояния сохраняются до тех пор, пока туловище остается в этом положении.
3. Туловище возвращается в нормальное положение, причем наступает расслабление внутренней мышцы и сокращение наружной.
4. Туловище поворачивается вокруг своей дорсо-вентральной оси как можно дальше в направлении правого глаза (поворачивание туловища в направлении OD), при этом наступает дальнейшее расслабление внутренней и сокращение наружной прямых мышц, тонический характер которых отчетливо виден.
5. Туловище находится опять в нормальном положении. При этом сокращается внутренняя мышца и расслабляется внешняя, так что мышцы находятся опять в таком же состоянии, в котором они находились в начале опыта.
Опыт был повторен пять раз с принципиально таким же результатом. Эксперименты показывают, что реципрокная иннервация происходит и при этих реакциях компенсаторных положений глаз.
III. СОВМЕСТНОЕ ДЕЙСТВИЕ ТОНИЧЕСКИХ ШЕЙНЫХ И ЛАБИРИНТНЫХ РЕФЛЕКСОВ НА ГЛАЗА
На рис. 86 схематически изображены пути для тонических шейных и лабиринтных рефлексов на глаза. Пути для тонических лабиринтных рефлексов нарисованы жирной линией, а для шейных рефлексов — тонкой. Для упрощения показаны только связи с одной стороны. Пути для вертикальных отклонений отмечены сплошной линией, для круговых движений — прерывистой, для горизонтальных отклонений — пунктиром. На схеме видно, что связи от лабиринтов и шейных рефлексов 10 Р. Магнус
146
Четвертая глава. Компенсаторные положения глаз
к мышцам для вертикальных отклонений расположены по одному и тому же принципу, точно так же, как и пути от лабиринта и от шеи к косым глазным мышцам. Мышцы для горизонтального отклонения (наружная и внутренняя прямые мышцы), напротив, иннервируются только от шеи.1
В предшествующих разделах было показано, что (при нормальном положении туловища) как тонические лабиринтные рефлексы, так и тонические шейные рефлексы на глазные мышцы «стараются» каждый по-своему
сохранить положение глаза в про- ^ странстве при изменениях поло¬
жения головы в пространстве или по отношению к туловищу, однако это не удается в отдельности ни лабиринтам, ни шее. Необходимо еще убедиться, что сохранение определенного положения глаза действительно достигается взаимодействием этих обеих рефлекторных групп.
До сих пор это исследовал де Клейн (53) для круговых движений, которые наступают, если, исходя из нормального положения животного, поднимают или опускают голову вокруг битемпораль- ной оси.
В этом случае обе рефлекторные группы влияют одновременно: тонические лабиринтные рефлексы — потому что изменяется положение головы в пространстве, тонические шейные рефлексы — потому что изменяется положение головы по отношению к туловищу.
Рис. 87 наглядно иллюстрирует подобный опыт.
На роговице кролика после предварительной кокаинизации выжигался крестик. Перед глазами фиксировалась изображенная на затем фотографировался глаз с рамкой в раз-
Рис. 86. Схема центральной связи при компенсаторных положениях глаз у кролика. Все толстые линии — лабиринтные рефлексы; все тонкие линии — шейные рефлексы. Сплошная линия — вертикальные отклонения, пунктирная — горизонтальные отклонения, штриховая — круговые движения.
рис. 66 маленькая рамка ных положениях.
На кривой каждая горизонтальная линия соответствует одному круговому движению на 10°.
На рис. 87а сплошной линией изображено круговое движение при поднимании и опускании головы, прерывистой линией — круговое движение,
1 Эта схема передает действительные отношения только в грубых чертах. Во- первых, нарисованы только связи от лабиринтов и от шеи к таким мышцам, которые при соответствующих рефлексах приходят в состояние сокращения (а не торможения). Во-вторых, надо отметить, что нервные связи при шейных рефлексах, вероятно, в действительности гораздо сложнее. Если мы, например, поворачиваем шею направо, то в состояние возбуждения приходят не только рецепторы одной стороны шеи, но, вероятно, обеих сторон. Несмотря на это, в схеме нарисованы только односторонние связи. Перекрещивания путей могут быть в действительности гораздо сложнее, чем нарисованное на этой схеме. Однако этого рисунка достаточно, чтобы сделать наглядными важнейшие физиологические данные.
Совместное действие шейных и лабиринтных рефлексов на глаза
147
наступающее только вследствие тонических лабиринтных рефлексов, которое определялось ранее описанным способом (стр. 123) на вращающейся доске. Заштрихованная часть на кривой изображает те круговые движения, появление которых надо приписывать одному только влиянию шейных рефлексов.
Позднее у этого же подопытного животного были перерезаны чувствительные корешки 1-го и 2-го шейных нервов. На рис. 876 видно, что появляющиеся при поднимании и опускании головы круговые движения являются приблизительно такими же, как те, которые наблюдаются при тонических лабиринтных рефлексах (рис. 87а). Маленький заштрихованный участок, который еще виден на этой кривой (шейные рефлексы), доказы-
0 10 20 30 40 50 60 70 0 10 20 30 40 50 60 70
Опускание головы Опускание головы
Рис. 87а. Рис. 876.
вает, что у этого подопытного животного незначительную роль при шейных рефлексах играют и чувствительные корешки от С3.
При более подробном рассмотрении рис. 87а можно видеть следующее.
Начало опыта — 0°. Животное в положении на животе. Ротовая щель горизонтальна.
Опускание головы.
Опускание головы на 10°: круговое движение на 10°
» » » 20°: » » » 20° и т. д. до
» » » 70°: » » » 70°.
Отсюда видно, что при опускании головы до 70° ниже горизонтальной плоскости положение глаза в пространстве остается совершенно неизменным. Глаз делает круговое движение на столько же градусов (верхним полюсом назад), на сколько градусов опускается голова ниже горизонтальной плоскости.
Поднимание головы.
Поднимание головы на 10°: круговое движение на 10°
» » 20°: » » » 20° и т. д. до
» » 60°: » » » 37°
10*
148
Четвертая глава. Компенсаторные положения глаз
Из этого следует, что при поднимании головы положение глаз в пространстве остается постоянным только до 10° выше горизонтальной плоскости.
В этом опыте с фотографированием производилось опускание головы только до 70°, поднимание головы только до 60°. В пяти других опытах опускание производилось до 90° и поднимание приблизительно до 80° (исходя из горизонтального положения ротовой щели), причем круговые движения определялись при помощи угломера визуально.
При этом было найдено постоянное явление, что глаз сохраняет свое положение в пространстве неизменным до опускания головы на 90° ниже горизонтальной плоскости и до поднимания на 10° выше горизонтальной плоскости.
Если принять во внимание, что кролики при нормальном сидении держат свою голову опущенной приблизительно на 35° ниже горизонтальной плоскости, то из этого следует, что животные в обычной жизни могут придавать своей голове разные положения по вертикальной плоскости в довольно широких пределах (вниз приблизительно на 55° и вверх приблизительно на 45°), отчего положение глаз в пространстве и вследствие этого поля зрения никак не изменяются.
Этот факт был установлен уже Барани (5). Однако он предполагал, что тут дело касалось исключительно лабиринтных рефлексов, в то время как в действительности тонические шейные рефлексы, как это показывает рис. 87а, оказывают существенное воздействие. Отсутствие изменения в поле зрения при разных положениях головы зависит от совместного влияния тонических лабиринтных и тонических шейных рефлексов, а не от одних тонических лабиринтных рефлексов.
Принципиально те же самые факты были обнаружены де Клейном при поворотах и наклонах головы (65). Для этой цели на кокаинизированную роговицу прикреплялась металлическая чашечка, приблизительно соответствующая выпуклости роговицы глаза. Эта чашечка имела три отверстия и была прикреплена к роговице тонкими шелковыми нитями. На наружной стороне чашки в перпендикулярном положении имелась тонкая легкая проволочка длиною в 24 мм, конец которой был изогнут под прямым углом. По главной части этой проволочки можно увидеть вертикальные отклонения, а по изогнутому концу — круговые движения. Находящийся в нормальном положении кролик (не привязанный за лапы) закрепляется головой в держателе Чермака так, чтобы ротовая щель была опущена приблизительно на 35° ниже горизонтальной плоскости. На штанге голово- держателя, являющейся приблизительным продолжением окципитальноназальной оси головы животного, находится прочно фиксированный угломер, передним — лот, так что каждый градус вращения головы можно отсчитывать по угломеру. При нормальном положении головы между наблюдателем и животным (лицо наблюдателя находится напротив носа животного) закрепляется на штативе проволочка, которая устанавливается строго параллельно метке на роговице. Если производить повороты головы вправо или влево, то нетрудно узнать, до какого градуса можно вращать голову, чтобы метка на роговице не потеряла своего параллельного положения по отношению к проволочке, прикрепленной к штативу (визирная линия). Таким образом, получалось, что в семи опытах надо было вращать голову от 17 до 25° вправо или влево (среднее 21°), для того чтобы удержать глаз, в неизменном положении в пространстве.
Таким же образом в пяти опытах было установлено неизменное положение глаз при наклоне головы (наклон головы вправо и влево в горизон¬
Совместное действие шейных и лабиринтных рефлексов на глаза
149
тальной плоскости). При этом оказалось, что положение глаза при наклоне вправо или влево от 12 до 24° оставалось еще неизменным (среднее 17°).
Итак, если кролик при исходном нормальном положении поднимает, опускает, поворачивает или наклоняет голову, то его глаза остаются правильно ориентированными в пространстве, пока движения головы не превышают определенных пределов. Эти пределы являются следующими: для поднимания головы 45°, для опускания — 55°, для наклона — 17° и для поворота — 21° в каждую сторону. В целом, таким образом, кролик может производить движения головой вверх и вниз в пределах 100°, повороты в пределах 50° и наклоны в размере 44°, без изменения поля зрения.
Определения для наклона и поворота головы дают минимальные величины, так как прикрепление метки на роговице вызывает в опытах различной степени торможение рефлекса, так что у свободно сидящего животного с неповрежденными глазами эти величины могут оказаться несколько большими. Это тонкое совместное влияние тонических лабиринтных и шейных рефлексов на мышцы глаз наблюдается прежде всего при нормальном положении животного, особенно если учесть, что при движении головы поля зрения обоих глаз остаются как бы направленными в одно место. В других положениях обе рефлекторные группы могут прямо-таки противодействовать друг другу. Например, если кролик находится в положении на спине и таким образом поворачивает свою голову, что правый глаз оказывается наверху, то шейный рефлекс стремится удержать положение глаза по отношению к горизонтальной плоскости, т. е. глаз отклоняется дорсально, в то время как тонический лабиринтный рефлекс, наоборот, двигает глаз вентрально. Вопросы, каким образом оба эти фактора при различных положениях животного в пространстве перемещают глазное яблоко то в одну, то в другую сторону и в каких положениях наступает еще компенсация отклонений головы благодаря тоническим рефлексам, требуют дальнейшей разработки.
Совместным влиянием шейных и лабиринтных рефлексов при разных положениях головы в пространстве достигается положение покоя (основное полоя^ение) глаза. Как было уже упомянуто, произвольные движения взора у кролика осуществляются в весьма незначительном объеме. У других видов животных эти движения глаз играют большую роль и вызываются соответствующими положениями головы. По прекращении их глаз всегда возвращается в основное положение, относящееся к данному положению головы. Тому факту, что тоническими шейными и лабиринтными рефлексами обусловливаются положения покоя глаз, соответствует тот общеизвестный опыт, что движениями, приводящими к компенсаторному положению глаз, нистагм (быстрая фаза) не вызывается. Противоположное этому положение можно наблюдать при реакциях глаз на вращения головы или реакциях на промывание холодной или теплой водой. При этом глаза перемещаются от своего основного положения (вращательная реакция или калорическая реакция) и возвращаются затем при быстрой фазе нистагма более или менее полностью в положение покоя, для того чтобы снова переместиться благодаря влиянию, исходящему из полукружных каналов, в положение, отличающееся от основного положения глаз (медленная фаза). При оптическом нистагме (железнодорожный нистагм) глаз также постоянно перемещается из своего положения покоя. Так как компенсаторные положения глаз являются основными положениями, то благодаря этому отсутствует как бы «принуждение» для быстрой фазы ни стагма.
150
Четвертая глава. Компенсаторные положения глаз
IV. СОВМЕСТНОЕ ВЛИЯНИЕ КОМПЕНСАТОРНЫХ ПОЛОЖЕНИИ ГЛАЗ И РЕАКЦИЙ ПОЛУКРУЖНЫХ КАНАЛОВ
Если животное совершает движения головою, то оно этим перемещает голову из одного положения покоя в другое. Обоим положениям головы соответствуют определенные компенсаторные положения глаз. Однако вместе с этим совершается (вращательное) движение головы, которое обусловливает (временную) вращательную реакцию глаз. В это время и сразу после движения должно проявиться совместное влияние компенсаторных положений глаз и вращательных реакций.
Когда кролик сидит в нормальном положении и поднимает голову, то этим вращением головы вокруг битемпоральной оси вызывается реакция полукружных каналов, благодаря которой глаз перемещается верхним краем роговицы вперед, т. е. в таком же направлении, в котором он затем фиксируется комбинацией тонических шейных и лабиринтных рефлексов. При опускании головы благодаря реакции полукружных каналов также вызывается перемещение глаз в обратном направлении, вследствие чего глаз также перемещается в положение, которое затем соответствует последующему компенсаторному положению глаз.
То же самое относится к вращению головы вокруг назально-окципитальной оси; при этом один глаз перемещается благодаря реакции полукружных каналов вверх, а другой — вниз, в том направлении, при котором он затем фиксируется благодаря компенсаторным положениям глаз.
Если животное совершает наклон головы, то полукружные каналы обусловливают движения глаз в направлении глазной щели и именно в том же направлении, в котором глаз затем фиксируется тоническими шейными рефлексами.
Следовательно, при таких движениях головы, которые совершаются из исходного нормального положения, гармонично совместно действуют рефлексы полукружных каналов и тонические шейные и лабиринтные рефлексы. Вращательными движениями головы вызывается реакция полукружных каналов, благодаря которой глаз перемещается в таком направлении, в котором он затем фиксируется тоническими шейными и лабиринтными рефлексами. Тут перед нами имеется установочный аппарат чрезвычайной тонкости и быстроты.
После только что сказанного о совместном действии шейных и лабиринтных рефлексов становится понятным, что эта гармония не имеет места при всех исходных положениях в пространстве. Правда, реакции полукружных каналов и тонические шейные рефлексы при всех положениях тела действуют совместно, однако реакции полукружных каналов протекают, — например, при движениях головы, которые животное выполняет в положении на спине, — в противоположном направлении по сравнению с тоническими лабиринтными рефлексами на мышцы глаз.
Как взаимодействуют друг с другом эти три рефлекторные группы при других исходных положениях тела, следует еще установить в каждом отдельном случае.
У. КОМПЕНСАТОРНЫЕ ПОЛОЖЕНИЯ ГЛАЗ У ДРУГИХ МЛЕКОПИТАЮЩИХ
И У ЧЕЛОВЕКА
Компенсаторные положения глаз неоднократно исследовались у рыб, амфибий, рептилий и птиц; у первых названных видов животных они были установлены особенно подробно; например, в последнее время они были изучены у рыб при помощи фотографических методов Бенжамином
Компенсаторные положения глаз у млекопитающих и человека
151
(Benjamins). Заслуживает упоминания, что Лион (Lyon) уже в 1899 г. смог отметить, что сгибание туловища по направлению зафиксированной головы приводит к отклонению глаз. Хотя, как известно, у рыб нет шеи, в этом случае несомненно надо увидеть аналогию с тоническими шейными рефлексами на глаза. Дальше, однако, мы примем во внимание только то, что мы знаем о компенсаторных положениях глаз у млекопитающих.
I. У морских свинок имеются такие же отношения, как и у кролика. Как направление отклонения глаз, так и размер его, и положение максимума и минимума у них приблизительно такие же, как и у кролика. Бартельс (3) указал на то, что у морской свинки имеются и тонические шейные рефлексы на глаза.
II. Гораздо менее точно мы знаем об этих отношениях у кошки, у которой глаза расположены в черепе довольно фронтально. В этом случае количественные исследования полностью отсутствуют, и в качественном отношении мы располагаем только очень скудными знаниями.
После односторонней экстирпации лабиринта (15) в первые два дня можно установить только незначительное горизонтальное отклонение и вращение глаза верхним полюсом роговицы в оперированную сторону. Глаз оперированной стороны отклонен вентрально, а другой — дорсально. Отклонение глаз чаще всего сильнее на оперированной стороне. Кроме того, глаза обнаруживают нистагм в противоположном направлении относительно отклонения, который, однако, быстро исчезает. Так как речь идет о быстро проходящих явлениях, то по легко понятным причинам нельзя с полной достоверностью утверждать, что эти отклонения глаз после односторонней лабиринтэктомии действительно обусловливаются компенсаторными положениями глаз.
По аналогии с наблюдениями на собаках, которые приводятся ниже, можно, вероятно, эти вертикальные отклонения отнести к таким компенсаторным положениям глаз. Кроме того, об этом свидетельствует еще следующее.
Если кошку после односторонней лабиринтэктомии поставить в нормальное положение и затем повернуть голову на 90° в сторону операции, то наблюдается уменьшение девиации глаз и очень сильный нистагм. Однако при вращении головы в другую сторону наступает усиление девиации и очень незначительный нистагм; все это наблюдается только в первые дни после операции. Здесь, следовательно, положение головы имеет ■отчетливое влияние на отклонение глаз и на осуществление или неосуществление нистагма. Однако при этом нельзя сказать с полной уверенностью, идет ли здесь речь о тонических лабиринтных или шейных рефлексах.1
Эти отношения нуждаются в новых исследованиях на основе наших теперешних знаний о различии между тоническими шейными и тоническими .лабиринтными рефлексами, с одной стороны, и тоническими лабиринтными рефлексами и реакциями полукружных каналов, с другой стороны. Из этого, однако, видно, как трудно из случайных наблюдений, — например, после односторонней лабиринтэктомии, — сделать заключение о влиянии тонических шейных и лабиринтных рефлексов на глаза. У интактного бодрого животного эти положения глаз вообще невозможно исследовать, потому что осуществляются слишком живые спонтанные движения глаз во все ■стороны. Наблюдения в таком случае следовало проводить или в наркозе, или после экстирпации большого мозга.
1 Соответствующие наблюдения на собаке говорят за тонические лабиринтные рефлексы.
152
Четвертая глава. Компенсаторные положения глав
III. У собаки компенсаторные положения глаз выражены во всяком случае более отчетливо, чем у кошки.
Если поместить нормальную собаку в положение на боку, то видно, что глаз, находящийся наверху, отклонен вентрально. Это является при положении головы на боку нормальным направлением взора животного. Названное отклонение наблюдается, однако, и тогда, когда глаза прикрывают рукой или если их рассматривают через трубку, которая не позволяет собаке видеть. Это отклонение также имеет место у таламической собаки после полной двусторонней экстирпации большого мозга. Такие животные больше не обладают способностью оптической установки глаз.
После односторонней экстирпации лабиринта (15) в течение 3—4 дней оба глаза отклонены в сторону операции (на оперированной стороне — сильнее). Кроме того, глаз оперированной стороны отклонен вентрально, а другой стороны — дорсально. Это вертикальное отклонение максимально при таком положении головы на боку, при котором интактный лабиринт находится внизу.
Положение глаз при боковом положении головы и девиация глаз после односторонней лабиринтэктомии хорошо совпадают по отношению к вертикальному отклонению. Каждый лабиринт обусловливает вертикальное отклонение контрлатерального глаза вниз и ипселатерального — вверх; лабиринт максимально возбужден, если он при боковом положении головы находится внизу. Наблюдаемое после односторонней экстирпации лабиринта горизонтальное отклонение глаз в сторону операции, однако, вряд ли можно отнести к тоническим лабиринтным рефлексам на глаза, так как ведь при последних не отмечается никаких горизонтальных отклонений. Здесь речь идет, вероятно, о последствиях перерезки нервов полукружных каналов.
Кроме того, у собаки были установлены тонические шейные рефлексы на глаза. Как уже было упомянуто в лаборатории Эвальда, Стевенсон наблюдал у собаки, которая находилась в нормальном положении на столе, в ответ на нагибание (наклон?) головы вправо — перемещение глаз влево. Однако в то время эти реакции еще не обозначались как шейные рефлексы.
Можно, конечно, с полной уверенностью сказать, что у собаки имеются компенсаторные положения глаз и что в этом участвуют как тонические лабиринтные, так и шейные рефлексы. Более точные количественные' определения отсутствуют; прежде всего надо обратить внимание еще на круговые движения.
IV. У обезьян (59) компенсаторные положения глаз полностью исследовались в качественном отношении. Однако если наблюдать интактную нормальную обезьяну с ее чрезвычайно живыми движениями глаз, то компенсаторных положений глаз нельзя установить. Только более точные исследования выявили, что и у обезьян имеются эти рефлексы.
а) Тонические лабиринтные рефлексы на глаза лучше всего можно исследовать во время просыпания после глубокого эфирного или хлороформного наркоза. В определенных стадиях наркоза их можно показать с большой достоверностью.1
1. Вертикальные отклонения. Если животйое медленно перевести в положение на спине, не изменяя при этом положения головы по отношению к туловищу, то становится очевидным, что оба глазных яблока равномерно отклоняются каудально сокращением нижних прямых мышц. Если, напротив, поместить животное в положение на животе так,
1 В недавно описанных Грехем Броуном (4) отклонениях глаз при наркозе речь, вероятно, шла це о компенсаторных положениях глаз, а о вращательных реакциях.
Компенсаторные положения глаз у млекопитающих и человека
153
что морда направлена вниз, то глазные яблоки отклоняются однозначно фронтально благодаря сокращению верхней прямой мышцы. В обоих случаях сокращаются одноименные мышцы правого и левого глаза одновременно и однозначно. Оба названные положения головы в пространстве* при которых ротовая щель в одном случае направлена вертикально вверх, а в другом вертикально вниз, являются приблизительными положениями максимума для вертикального отклонения глаз у обезьян.
2. Круговые движения. Круговые движения выражены сильнее всего при положении головы на боку. Их размер незначителен, так что их едва можно заметить, если до наблюдения не выжигался на роговице крест. У обезьян имеются почти такие же закономерности, как у человека.
Если переместить голову из нормального положения в боковое, то у обоих глаз компенсаторные круговые движения происходят однозначно и именно так, что верхний край роговицы вращается по направлению к потолку комнаты. Из анатомического расположения косых глазных мышц следует, что при этом на верхнем глазу сокращается косая нижняя и на нижнем глазу косая верхняя мышцы; следовательно, на правом и левом глазах одноименные косые мышцы реагируют противоположно, благодаря чему оба глаза выполняют однозначные круговые движения.
Из сказанного следует, что у об.езьяны с фронтально расположенными глазами имеются иные связи афферентных лабиринтных путей с ядрами мышц глаз, чем у кролика. Если у обоих животных голова направлена мордой вертикально вниз, то у кролика под однородным влиянием находятся косые мышцы обоих глаз, а у обезьян, напротив, под тем же влиянием находятся прямая верхняя и прямая нижняя мышцы обоих глаз.
При боковом положении головы у кролика лабиринты, напротив, имеют противоположные влияния на прямую верхнюю и нижнюю мышцы, у обезьян же они имеют противоположные влияния на косые мышцы.
Таким образом, у кролика и у обезьяны обе косые и обе прямые (верхняя и нижняя) мышцы глаз до известной степени поменялись ролями. Наоборот, при одних и тех же положениях головы в пространстве как у кролика, так и у обезьяны всякий раз наступают одни и те же (голова +90° или голова —90°) или противоположные (голова в боковом положении) реакции.
Эти различия можно увидеть при сравнении жирно нарисованных путей на рис. 86 и 88. У обезьяны также нельзя вызвать горизонтальное отклонение глаз как постоянный рефлекс тонического характера (компенсаторные положения глаз).
После односторонней лабиринтэктомии у обезьяны, также как у кролика, глаза оказываются отклоненными в течение нескольких дней в оперированную сторону. Как выше было уже отмечено для кошки и собаки, речь идет, вероятно, не об односторонних тонических лабиринтных рефлексах на глаза, а о временных последствиях перерезки нервов полукружных каналов.
Бартельс однажды наблюдал после односторонней экстирпации лабиринта у обезьян вертикальное отклонение глаз. И у одного из моих подопытных животных глаз оперированной стороны стоял в течение двух дней немного ниже, чем другой. Возможно, что это обусловлено тоническими лабиринтными рефлексами.
б) Тонические шейные рефлексы на глаза.
1. Вертикальные отклонения. Если поместить животное в прямое (вертикальное) положение головой вверх и фиксировать голову в нормальном положении, то движениями туловища вокруг
154
Четвертая глава. Компенсаторные положения глаз
битемпоральной оси в сагиттальной плоскости можно вызвать вертикальные отклонения глаз. Если приблизить спину животного к темени, то оба глаза перемещаются однозначно вентрально. Если приблизить живот животного к носу, то оба глаза перемещаются фронтально. Речь тут идет о реакции верхней и нижней прямых мышц; одноименные мышцы обоих глаз реаги-
L
ОЫ. inf. 0=
Reel sup. D^i
- int. □-ii- ^ и
Ы D-J'
R.
О 0Ы. inf.
■O Rect.sup. •• int. '• inf.
Ill
OU.sup O Rect.ext □
NO«
Obl.sup.IV
руют однозначно.
У сидящего животного можно при соответствующих положениях головы наблюдать комбинации шейных и лабиринтных рефлексов. Когда
животное сидит в вертикальном положении головой вверх, тогда при вентральном сгибании головы (опускание головы) оба глаза идут вверх, при дорсальном (поднимание головы) — вниз. Девиация глаз в этом случае является компенсаторной для движения головы.
2. Круговые движения. Поместим обезьяну в прямосидячее положение головой вверх и зафиксируем голову в нормальном положении. Теперь можно вызвать круговые движения глаз движениями тела вокруг дорсо- вентральной оси в фронтальной плоскости (наклон туловища). Если, например, наклоняют туловище так, что правое плечо приближается к правому уху,то оба глаза поворачиваются в одном направлении (верхним краем роговицы влево), т. е. направление их наклона то же, что и у туловища. У правого глаза сокращается косая верхняя, у левого — косая нижняя мышцы. Следовательно, одноименные косые глазные мышцы реагируют на обоих глазах противоположно. Размах этих круговых движений не очень велик, но все-таки его можно с достоверностью увидеть.
Совместное действие тонических шейных и лабиринтных рефлексов на повороты глаз имеет место в том случае, если придать животному вертикальное положение головой вверх и наклонять голову к правому или левому плечу. При этом появляются круговые движения, противоположные направлению наклона головы. И здесь речь идет о постоянных положениях. Размах незначителен.
3. Горизонтальные отклонения глаз в виде постоянного положения вызываются только влиянием с шеи. По моим опытам, это удается менее хорошо в наркозе. Это возможно было установить с большой отчетливостью у двух таламических обезьян после полной двусторонней экстирпации большого мозга. У одного из этих животных до этого были удалены оба лабиринта. В описанных случаях голова была зафиксирована в нормальном положении. В противоположность вышеописанному для
Тонические шейные рефлексы на глаза
Laby¬
rinth
Рис. 88. Схема центральной связи при компенсаторных положениях глаз у обезьяны. Толстые линии — лабиринтные рефлексы, тонкие линии — шейные рефлексы. Штриховая линия — вертикальные отклонения, сплошная — круговые вращения, пунктирная — горизонтальные отклонения.
Компенсаторные положения глаз у млекопитающих и человека
155
круговых движений тело находилось не в вертикальном положении, а в горизонтальном, так что исходное положение обезьяны приблизительно соответствовало нормальному положению кролика. Туловище вращалось в горизонтальной плоскости вокруг дорсо-вентральной оси. В этом случае, следовательно, исходное положение являлось другим, чем в случае круговых движений. Если приблизить правое плечо к правому уху, то оба глаза идут горизонтально влево. При этом сокращаются левая прямая наружная и правая прямая внутренняя мышцы. Таким образом, одноименные глазные мышцы, правые и левые, обеспечивают противоположные реакции. В обоих случаях речь идет об очень отчетливых отклонениях глаз, которые сохранялись до тех пор, пока туловище не изменяло своего положения по отношению к голове.1
На рис. 88 изображены связи одного лабиринта с ядрами глазных мышц и упрощенные односторонние пути тонических шейных рефлексов у обезьяны, и потому этот рисунок следует сравнить с рис. 86 для кролика. У обеих групп животных тонические горизонтальные отклонения глаз вызываются исключительно с шеи благодаря сопряженному сокращению прямой наружной и прямой внутренней мышц. У обоих видов животных пути от лабиринта к шее и к мышцам обоих глаз организованы по одному и тому же принципу, поэтому и в схемах они изображены в виде параллельных линий. У обезьяны распределение путей косой верхней и нижней мышц является таким же, как и у кролика для прямой верхней и нижней мышц, в то время как нервные связи от прямой верхней и наружной мышц у обезьяны соответствуют нервным путям косой верхней и нижней мышц у кролика.
В течение жизни эти рефлексы у здоровой обезьяны имеют небольшое значение. Правильная установка глаз контролируется прежде всего тем, что глаза расположены в одной фронтальной плоскости и что поля зрения большей частью совпадают друг с другом. Следовательно, положение глаз регулируется главным образом оптическим путем. Компенсаторные положения глаз нужны, вероятно, только для того, чтобы поддержать в очень незначительной степени постоянство полей зрения при движениях головы и туловища.
У. И учеловека несомненно имеются тонические шейные и лабиринтные рефлексы на глаза; однако они, так же как у обезьян, очень незначительны, и, кроме того, до сих пор полностью не исследованы.
В большинстве случаев исследователи не делали различия между шейными и лабиринтными рефлексами, потому что движения головы производились по отношению к неподвижному туловищу. Далее, прежние исследователи часто не делали никакого различия между вращательными реакциями и компенсаторными постоянными положениями.
1. Круговые движения. Вероятно, Джон Гунтер был первым, кто при наклоне головы в сторону плеча наблюдал круговые движения глаз. Вероятно, здесь речь главным образом шла об оптических реакциях и о лабиринтных вращательных реакциях. Четкие различия
1 Если зафиксировать голову в нормальном положении и позвоночный столб поставить вертикально, то поворот туловища обусловливает круговое движение глаз, если же позвоночный столб стоит горизонтально, то появляются горизонтальные отклонения. В обоих случаях движения шеи различны. Уже по этой причине вызванные этими движениями тонические рефлексы на глазные мышцы могли бы быть разными. Может быть, при этом определенную роль играют еще и «переключения». У кролика эти отношения проще. Здесь отклонение туловища всегда обусловливает горизонтальные смещения, а поворот туловища — вертикальные смещения, независимо от того, был ли позвоночный столб при нормальном положении зафиксированной головы направлен первоначально горизонтально или вертикально.
156
Четвертая глава. Компенсаторные положения глаз
между компенсаторными положениями глаз, которые оказываются постоянными, и вращательными реакциями, оказывающимися временными, делает Брейер (1). Он так же, как и А. Нагель, показал, что компенсаторные изменения положения глаз при изменении положения головы в пространстве наступают и тогда, когда не изменяется положение головы по отношению к туловищу. Здесь, вероятно, играют роль лабиринтные рефлексы. Действуют ли у человека шейные рефлексы в одинаковом направлении — следует еще установить.
Величина (постоянного) компенсаторного кругового движения глаз при наклоне головы к плечу видна из следующих таблиц.
(По Доту) Таблица 1
Наклон головы |
15°
25°
35°
45°
55°
65°
Вращение по:
Мулдеру
3°
4°
5°'
5.5°
5.5°
6°
Кюстеру
4
6
6.5
7
8
9
Скребицкому
2
2.6
4.2
5.5
6.8
7.7
(По В. Нагелю) Таблица 2
Поворот голо¬
вы
10°
20°
30°
40°
50°
60°
70°
80°
90°
100°
Угол вращения .
1.3
3.8
5.2
5.4
6.3
6.7
6.8
8.0
8.1
8.6
Компенсирова¬
лось поворотом головы . .
1
1
1
1
1
1
1
1
1
1
7.7
5.2
5.8
7.4
7.9
9
10.3
10
11.1
1
11.8
Следовательно, у человека даже при комбинации тонических шейных и лабиринтных рефлексов поворот глаз при наклонении головы к плечу компенсируется только в очень незначительной степени. По наблюдениям Анжиера (Angier), оба глаза перемещаются при этом одинаково сильно; они перемещаются верхним краем роговицы в одинаковом направлении, одноименные косые реагируют при этом противоположно. Компенсаторные круговые движения являются такими же, как и у обезьян.
2. Вертикальные отклонения. Еще Брейер наблюдал, что у слепых при поднимании и опускании головы осуществляются компенсаторные вертикальные отклонения глаз — именно таким образом, как и в случае круговых движений, но при более сильном наклоне головы. Прямая верхняя и внутренняя мышцы обоих глаз реагируют при этом одинаково, так же как и у обезьян. В. Нагель утверждает, что такое вертикальное отклонение наблюдалось и у маленьких детей и что он мог их установить у взрослых в затемненной комнате при помощи мгновенного освещения. Он делает следующие замечания: «Если чередовать при закрытых глазах прямое положение с положением наклона вперед и в это время ощупывать слегка приложенными пальцами веки, то отчетливо чувствуется, как при наклоне вперед роговица перемещается вверх, при выпрямлении— вниз».
Клинические наблюдения показывают, что при таких реакциях определенную роль играют тонические шейные рефлексы. Де Клейн и Стен- верс (67) наблюдали такой случай, когда при поднимании и опускании го-
Компенсаторные положения глаз у млекопитающих и человека
157
ловы наступали компенсаторные вертикальные отклонения глаз, которые отсутствовали при действии только тонических лабиринтных рефлексов. Пациент при этом прочно фиксировался на носилках в разных положениях в пространстве. Активные оптические глазные движения устранялись путем надевания сильных двояковыпуклых очков. В этом случае исключительное внимание обращалось и на то, что речь шла не о вращательных реакциях, а о настоящих постоянных положениях. Следовательно, при
Рис. 88ха.
этом с достоверностью были доказаны тонические шейные рефлексы на глаза при вертикальных компенсаторных положениях глаз. Требуются дальнейшие исследования, чтобы выяснить, насколько в таких вертикальных реакциях участвуют и тонические лабиринтные рефлексы.
3. Горизонтальные отклонения. Барани (6) отметил, что если у новорожденных (в первые два дня жизни) и у недоношенных детей (7—8 мес.) при положении на спине зафиксировать голову и затем по-
Рис. 881б.
вернуть туловище влево вокруг вертикальной оси тела на 90° (не меньше!), то постоянно наблюдается горизонтальное отклонение обоих глаз влево. В этом случае речь должна идти о чистых шейных рефлексах.
В одном неопубликованном случае (ребенок с большой мозговой опухолью) де Клейн и Стенверс также могли вызвать горизонтальное отклонение глаз вращением туловища при зафиксированной голове (рис. 88*а и 8Кб).
На рис. 88ха этот ребенок виден в правом боковом положении. Пра- выи глаз находится в среднем положении, левый — во внутреннем глазном углу. Если голову держали без изменений в правом боковом положении, а туловище ребенка поворачивали в положение на спину (рис. 88хб), то правый глаз находился во внутреннем глазном углу, левый глаз — в среднем положении. Следовательно, оба глаза перемещались в орбите при неподвижной голове в таком же направлении, в котором вращалось туловище. Речь идет о постоянных реакциях благодаря тоническим шейным рефлексам на оба глаза.
158
Пятая глава. Установочные рефлексы
Это все, что до сих пор известно о компенсаторных положениях глаз у человека, но уже эти скудные наблюдения показывают, что у человека имеются такие же рефлексы, как и у обезьяны, и что, вероятно, имеются как тонические лабиринтные, так и шейные рефлексы на глаза. У взрослого здорового, бодрого человека можно показать только круговые движения при наклоне головы на бок.
По Нагелю, вертикальные отклонения появляются в темноте — следовательно, тогда, когда воздействие оптической установки глаз отсутствует.
Отчетливые лабиринтные рефлексы выявляются при круговых движениях глаз, когда голова находится в боковом положении. Отчетливые шейные рефлексы видны при вертикальных отведениях глаз во время поднятия и опускания головы, при горизонтальных отведениях у новорожденных и у больных при отклонении туловища.
Весь этот вопрос нуждается в новом систематическом исследовании. Более достоверные наблюдения можно, наверное, делать иногда при хирургическом наркозе у детей, которых без всяких затруднений можно поместить в разные положения в пространстве. Исследование нужно при этом проводить так же полно, как это было описано выше для обезьяны.
Приведенные в этой главе факты показывают, что у всех до сих пор исследованных млекопитающих тонические шейные и лабиринтные рефлексы имеются“ в принципиально одинаковой форме. Однако разные виды животных в зависимости от анатомического расположения глаз и совершенства оптического установочного аппарата используют эти рефлексы по-разному.
Пятая глава
УСТАНОВОЧНЫЕ РЕФЛЕКСЫ (STELLREFLEXE)
Как было уже описано в третьей главе, положение тела у децеребриро- ванного животного управляется исключительно рефлексами положения (Haltungsreflexe), т. е. тоническими шейными и лабиринтными рефлексами на мускулатуру тела, благодаря чему распределение напряжения в отдельных частях тела приводится в полное соответствие с положением головы. Коль скоро, помимо центров в верхнем отделе спинного и продолговатого мозга, необходимых для осуществления этих рефлексов, имеется еще и функционирующий средний мозг, то на этой основе надстраивается новая система рефлексов — установочные рефлексы, позволяющие животному самостоятельно принимать и удерживать нормальное положение.
То, что животное без большого мозга и с более или менее интактным стволом мозга в состоянии стоять и бегать, установлено уже давно известными наблюдениями Мажанди (Magendie), Лонге (Longet), Шиффа (Schiff), Вульпиана (Vulpian), Христиани (Christiani) и Мунка [Munk (1)] на кролике и знаменитыми исследованиями Гольца (Goltz) на его бесполушар- ной собаке, которые в последнее время подтверждены Ротманом (Roth- mann) и Дюссер де Варенном [Dusser de Barenne (3)]. Проанализировать, каким образом подобное животное оказывается в состоянии принять и удерживать свое нормальное положение, оказалось лучше всего на кролике, так как у этого животного шоковые явления после полной экстирпации большого мозга выражены весьма незначительно, а кроме того, у него отсутствуют оптические установочные рефлексы. К тому же такие же установочные рефлексы можно наблюдать у кошки, собаки, обезьяны и у интактной морской свинки.
Пятая глава. Установочные рефлексы
159
Техника экстирпации б о льшо г о мозга у кроликов
При операциях применяется методика (24), описанная Морита (Morita). При этом череп трепанируется справа и слева, после чего крыша черепа удаляется с обеих сторон, причем в середине оставляется костная перемычка. Эта последняя защищает от давления и повреждения лежащий под ней продольный синус, а в дальнейшем — четверохолмие и зрительный бугор. В переднем и боковом направлениях отверстие делается возможно более широким; в заднем направлении можно идти лишь настолько, чтобы не поранить поперечный синус. Затем вскрывается твердая мозговая оболочка на одной стороне, полушарие большого мозга отделяется сбоку и сзади от черепа маленьким ватным тампоном или узким, тупым, специально изогнутым шпателем и, наконец, выталкивается через отверстие наружу; при этом свод (fornix) сдвигается вперед от таламуса. Этим самым вся боковая поверхность ствола обнажается. Далее отыскивается поверхность зрительных бугров, большой мозг и полосатое тело отделяются разрезом, проходящим дорсально в борозде между таламусом и полосатым телом, вентрально — тотчас перед зрительным трактом в направлении кпереди, медиально — назад и латерально. Весь большой мозг, включая обонятельную долю и полосатое тело, удаляется при этом в виде одной или двух порций. Подобная же операция производится затем и на другой стороне. Оба разреза соединяются в lamina terminalis (замыкающей пластинке). Зрительные нервы при желании можно без труда перерезать под контролем глаза. Наоборот, если желательно сохранить их, рекомендуется оставить на вентральной стороне каждого зрительного тракта кусочек большого мозга величиной примерно с половину вишневой косточки, принадлежащий к медиальному отделу gyrus piriformis и заключающий в себе окончания аммонова рога, subiculum cornu Ammonis и, может быть, также связи taenia semicircularis с миндалевидным ядром [ср.: атлас Винклера (Winkler) (2) и Поттера (Potter), табл. 12]. Названные отделы большого мозга относятся к обонятельному аппарату, который у оперированных животных вследствие удаления обонятельных луковиц не функционирует. Поэтому, как видно и из наблюдений над животными, наличие этих отделов не искажает функциональных результатов операции. При этом щадится зрительный тракт. Впрочем, в большинстве опытов большой мозг удаляется тотально.
По окончании операции неповрежденный ствол лежит на основании черепа. Удаляются возможные сгустки крови; образовавшаяся полость может (но не обязательно) быть уменьшена с помощью стерильной ваты; кожа зашивается. Кровотечение в большинстве случаев весьма незначительно. Из черепномозговых нервов повреждаются обонятельный и (при намеренной перерезке) зрительный. Другие черепномозговые нервы остаются неповрежденными. При желании сохранить животных живыми более долгое время и учитывая опасность кровотечения, рекомендуется первое исследование производить лишь на следующий день.
Животные, оперированные таким способом, выживали до 11 дней. Так как они самостоятельно пищу не брали, им ежедневно через желудочный зонд вводилось 50—100 мл молока. После гибели каждое животное вскрывалось. Признаков раздражения мозга не отмечено ни в одном случае. Для анатомического исследования препараты помещались в формалин.
В интересах наглядности результатов приведены стереоскопические снимки двух препаратов.
Опыт 29. Кролик, 800 г. 3 мая 1915 г. Эфирно-хлороформный наркоз. Экстирпация большого мозга выше таламуса по методике Морита. Животное живет 2 дня и подвергается в связи с этим более длительному исследованию.
Секция. Большой мозг полностью отсутствует. Левый зрительный нерв перерезан. Правый зрительный нерв интактен. Черепномозговые нервы — 3—12-я пары — с обеих сторон интактны. Четверохолмие и зрительные бугры сохранены. Базальные ганглии отсутствуют. Кпереди от таламуса, между колоннами свода, осталось около 3 мм ткани. В полости черепа лишь минимальное кровотечение. Признаков раздражения (препарата) нет. При извлечении боковая часть правого * полушария мозжечка оторвана.
На рис. 89 препарат виден сверху: мозжечок, перед ним узкие задние и мощные передние двухолмия. Зрительные бугры оканчиваются сбоку — в латеральных коленчатых телах — и оставляют в середине между собой третий желудочек, в который можно заглянуть сверху. Кпереди от последнего сохранился узкий кусочек области конечной пластинки. Сбоку от передней стенки левого переднего холма четверохолмия видно левое внутреннее коленчатое тело.
На рис. 90 тот же препарат виден сбоку. Можно узнать продолговатый мозг, мозжечок, переднее и заднее двухолмия и таламус. От заднего двухолмия проходит
160
Пятая глава. Установочные рефлексы
ниже переднего двухолмия к медиальному коленчатому телу ручка заднего двухолмия. Частично видна поверхность разреза таламуса.
На рис. 91 препарат представлен с вентральной стороны. Виден продолговатый мозг, трапециевидное тело с пирамидами, перед ними — мост, по сторонам — полушария мозжечка. Видны ножки мозга, выходящие из моста. Между ними — сосочковое тело и перед ними—воронка. Спереди к хиазме с двух сторон подходит зрительный тракт, откуда выступают две культи зрительных нервов. Перед хиазмой еще сохранился остаток из области конечной пластинки.
Опыт 49. Кролик. 17 мая 9 1915 г. Хлороформный наркоз при помощи искусственного дыхания. 8 Экстирпация большого мозга по 5 методу Морита, тотчас выше таламуса. Зрительные нервы приподняты и перерезаны. Трахея за-
1 Рис. 89а. 1 — червь мозжечка; 2 — полушария мозжечка; 3 — заднее двухолмие; 4 — переднее двухолмие; 5 — медиальное коленчатое телое; 6 — зрительные бугры; 7 — третий желудочек; 8 — латеральное коленчатое тело; 9 — конечная пластинка.
Рис. 89. Рис. 89а.
шита. Животное выжило двое суток и погибло от пневмонии.
Секция. Мозг без признаков раздражения. Зрительные бугры и четверохол мия интактны. Большой мозг удален полностью. Зрительные нервы перерезаны
Черепномозговые нервы—3—12-я пары — интактны. Разрез проходит с обеих сторон по дорсальной стороне, непосредственно по переднему краю таламуса; в середине— незначительное количество оставшейся ткани конечной пластинки. * 1Рис. 90а. 1 — продолговатый
мозг; 2 — мозжечок; 3 — заднее двухолмие; 4 — переднее двухолмие; 5 — ручка заднего двухолмия; 6 — медиальное коленчатое тело; 7 — зрительный бугор.
2 5 4
Рис. 90. Рис. 90а.
Вентрально разрез идет справа — сразу перед зрительным трактом, слева — на
1 мм кпереди.
На рис. 92 препарат показан сбоку. Виден продолговатый мозг, мозжечок, переднее двухолмие. Ниже к внутреннему коленчатому телу тянется правая ручка четверохолмия. Перед последним лежит наружное коленчатое тело в виде бокового деривата таламуса, который интактен. Видна часть поверхности разреза.
Это животное при жизни было сфотографировано (рис. 98 и 115, стр. 173 и 188).
Профессор Винклер произвел микроскопическое исследование серийных срезов препаратов животного, которое жило после операции 5 дней и непрерывно находилось под наблюдением. При этом установлено следующее.
Опыт 20а. Кролик. 24 марта 1915 г. Эфирный наркоз. Экстирпация большого мозга и базальных ганглиев по методу Морита. Зрительные бугры и четверохолмия интактны. Зрительные нервы сохранены.
Животное прожило 5 дней.и в конце концов погибло от пневмонии. У него отлично осуществлялись все рефлексы, которые будут описаны в дальнейшем, особенно все установочные. Зрачковая реакция и рефлекторные смыкания век при засвечивании имели место отчетливо лишь слева.
Секция. Препарат без признаков раздражения. Большой мозг удален полностью. Черепномозговые нервы — 2—12-я пары — с обеих сторон интактны. Четверохолмие и зрйтельные бугры сохранены. Базальные ганглии отсутствуют.
Пятая глава. Установочные рефлексы
161
Микроскопическое исследование. Полушария большого мозга, а также вентральные доли полностью отсутствуют. От большого мозга, однако, сохранилась гипоталамическая часть переднего мозга, лежащая под передней спайкой. От свода осталась еще частица его передней спайки. От хвостатого ядра с обеих сторон остались самые медиальные участки.
Таламус сохранен с обеих сторон. Справа он остался полностью неповрежденным. Слева разрез направляется через латеральную часть таламуса так, что в плоскости рис. 14 атласа Винклера—Поттера (2) латеральная часть наружного коленчатого тела оказывается удаленной, а в плоскости рис. 15 названного атласа оказываются задетыми основания ножки и субталамическая область, * 10Рис. 91а. 1 — продолговатый
мозг; 2 — трапециевидное тело; 3 — пирамида; 4 — мост; 5 — полушарие мозжечка; 6 — ножка мозга; 7 — сосочковое тело; 8 — воронка; 9 — зрительный тракт;
10 — хиазма; 11 — зрительные нервы; 12 — остатки от области конечной пластики.
Рис. 91.
Рис. 91а.
вплоть до вентральной медуллярной пластинки. Вследствие этого оказывается поврежденным зрительное сияние слева, выпадает зрачковая реакция, а реакции смыкания век при засвете правого глаза получают свое объяснение.
Соседние с линией разреза участки решетки (gitterschecht) (табл. 12, F. г. а. и F. г. Ъ.) и клетки в наиболее передних ядрах таламуса дегенерировали (табл. 12, А. Ь. и менее сильно А. а.) слева сильнее, чем справа.
Рис. 92.
Рис. 92а. 1 — продолговатый
мозг; 2 — мозжечок; 3 — переднее двухолмие; 4 — ручка заднего двухолмия; 5 — медиальное коленчатое тело; 6 — зрительный бугор; 7 — латеральное коленчатое тело.
Остальные составные части промежутрчного мозга остались неповрежденными.
Средний мозг и лежащие за ним части‘мозга совершенно интактны.
В опытах с удалением мозга до уровня) четверохолмия я пользовался тем же методом Морита, как и., в случае экстирпации большого мозга выше таламуса. Только в этом случае ствол разъединяется разрезом, идущим на дорсальной стороне на 0—2 мм кпереди от переднего края четверохолмия, латерально — через или позади внутреннего коленчатого тела, на вентральной поверхности — через ножку мозга по заднему краю сосочкового тела или поперек его. Разрез пересекает ствол в плоскости, лежащей между плоскостями рис. 16 и 17 атласа Винклера—Поттера. Направление разреза примерно такое же, как на упомянутых рисунках. При этом в известных условиях щадятся и расположенные наиболее каудально отделы промежуточного мозга (каудальный конец сосочкового тела, внутреннее коленчатое тело полностью или в каудальной части), красное ядро покрышки остается сохраненным до переднего края. По существу, дело идет И Р. Магнус
162
Пятая глава. Установочные рефлексы
о животном, у которого средний мозг сохранен полностью, а от промежуточного мозга остались только каудальные отделы различной протяженности.
Эффект операции представлен на снимках (рис. 93—95).
Опыт 31. Кролик, 1000 г. 5 мая 1915 г. Хлороформный наркоз* Сонные артерии перевязаны, блуждающие нервы перерезаны. Экстирпация большого мозга и таламуса по методу Морита. Разрез проходит тотчас впереди четверохолмия, косо- кпереди и вниз. Почти никакого кровотечения. Кролик жил 2 суток.
Рис. 93а. Препарат показан с дорсальной стороны. Видны: мозжечок (1), переднее двухолмие (2) и кпереди от него — на протяжении 1—2 мм остатки зрительных бугров (3).
Секция. На поверхности разреза свернувшаяся жровь. Полость пуста. Обонятельные и зрительные нервы отсутствуют. Глазодвигательные и остальные черепные- нервы интактны. Разрез проходит по дорсальной поверхности (рис. 93) симметрично на 1—2 мм кпереди от переднего четверохолмия; по вентральной поверхности (рис. 95) — непосредственно позади заднего края сосочкового тела, через ножку мозга. На левой стороне (рис. 94) он проходит в 2 мм кпереди от заднего- края внутреннего сосочкового тела, справа — точно по его заднему краю.
Если опыты ставить под точным контролем секционных данных, обе операции осуществляются без труда; все, что при этом делается, хорошо видно.
К номенклатуре. В прежних исследованиях дело касалось но большей части децеребриро-
Рис. 94а. Сделан с левой стороны. Ясно видны: продолговатый мозг (1), левое полушарие мозжечка (2), переднее двухолмие (3) и ниже него ручка заднего двухолмия (4), идущая к медиальному коленчатому телу, задняя половина которого (5) сохранилась.
Рис. 94.
3 2
Рис. 94а.
ванных животных; лишь в специальных целях у отдельных животных удалялся мозжечок или продолговатый мозг. В экспериментах, описанных здесь и в дальнейшем, оставлялись функционально связанными со спинным и продолговатым мозгом различные более высокие и более низкие отделы мозга. Для обозначения операции, произведенной в каждом (отдельном) случае, желательно иметь краткую и четкую номенклатуру, позволяющую избегать ошибок или обстоятельных описаний. Поэтому я предлагаю называть препарат по наиболее орально расположенному отделу центральной нервной системы, который остался в функциональной связи с более глубоко лежащими образованиями. Соответственно этому декапитированное животное обозначалось бы как спинальное (Halsmarktier), децеребрированное — как церебеллярно-бульбарное, resp. мозжечково-мостовое животное. Смотря по обстоятельствам можно говорить.
Общее поведение таламических и мезенцефалических животных
163
также о безмозжечковом, бульбарном животном и т. д. Эксперименты, описанные в следующем разделе, выполнены на диэнцефалическом, resp. таламическом и четверохолмном кролике. Вместо «четверохолмный кролик» можно говорить «ме- зендефалический кролик», не забывая, однако, что наиболее каудальные отделы промежуточного мозга остаются при этом неудаленными. * УРис. 95а. Препарат показан с вентральной стороны. Видны: продолговатый мозг (1), трапециевидное тело (2) с пирамидой (5), мост (4), полушария мозжечка (5). Впереди моста лежат ножки мозга (6). Сосочковое тело отсутствует. Ср. рис. 91.
Рис. 95. Рис. 95а.
I. ОБЩЕЕ ПОВЕДЕНИЕ ТАЛАМИЧЕСКИХ И МЕЗЕНЦЕФАЛИЧЕСКИХ
ЖИВОТНЫХ
1. Кролик. Уже через короткое время после операции животное предпринимает попытки сесть. По прошествии от получаса до нескольких часов и во всяком случае на следующий день оно подобно интактному животному сидит совершенно нормально и всякий раз, будучи выведено из этого положения, например в положение на боку или на спине, вновь принимает нормальное положение и позу. Голова и туловище стоят симметрично, голова держится свободно, при этом ротовая щель наклонена кпереди обычно под углом в 20—40°, живот находится выше пола, передние и задние ноги согнуты в нормальном положении, животные сидят как бы на корточках (Hockstellung). Спонтанных движений, приводящих к перемене места, почти никогда не бывает. Если они все же наступают, то при этом часто удается установить наличие внешнего раздражения, вызвавшего эти движения. По большей части животное часами находится в клетке на одном и том же месте. При поверхностном наблюдении отличить такое животное от нормального невозможно.
Так как центры регуляции температуры тела расположены в промежуточном мозгу (Isenschmid u. Kerl; Isenschmid и. Schnitzler), температура тела поддерживается на нормальном уровне (в прямой кишке она равна 37.8 и 38.2°).
У животных с интактными зрительными нервами весьма отчетливо осуществляется зрачковая реакция. Точно так же при интенсивном освещении, особенно на солнце, наступает энергичное рефлекторное смыкание век. Но эти оптические рефлексы оказываются единственными, которые возможно наблюдать у животных. Даже при самом энергичном освещении не вызываются никакие рефлексы мышц шеи, туловища и конечностей. Точно так же быстрое приближение руки к глазу или быстрые движения руки в поле зрения не вызывают никакой реакции животного. Если по какой-то причине животное меняет место, оно при этом не в состоянии избежать препятствий на своем пути, на что ранее уже указывал Мунк (Munk).
У таламического кролика с перерезанными зрительными нервами зрачковый рефлекс и рефлекс смыкания век, естественно, отсутствуют. Во всем остальном установить различие в поведении животных с перере-
11*
164
Пятая глава. Установочные рефлексы
занными и сохраненными зрительными нервами невозможно. У кролика способности к поддержанию нормального положения тела и к активному принятию сидячего положения из нормальных положений не зависят от оптических путей, и у животных с перерезанным зрительным нервом они реализуются с не меньшей быстротой, чем при сохранных зрительных нервах. Все другие, близкие к рассмотренным реакции оказываются при перерезанном зрительном нерве также неизмененными.
Никакие рефлексы на Мускулатуру шеи и тела не вызываются также и при пассивных движениях глаз. Для этой цели лучше всего, не повреждая передней камеры глаза, укрепить при помощи тонкой иглы на роговице (Homhartscheitel) с обеих сторон тонкие шелковые нити. С помощью этих нитей можно осуществлять одно- и двусторонние согласованные конвергирующие и дивергирующие движения глаз. Если для выключения возможных в этом случае тригеминальных рефлексов роговица была предварительно анестезирована кокаином, то реакции животного на отведения глаз не наступает. Также не удается вызвать рефлекторные движения при растягивании отдельных, глазных мышц. Таким образом, проприоцептивные рефлексы с мышц глаза на остальную мускулатуру тела, по крайней мере в описанных до сих пор условиях, не выявлялись.
При прикосновении к роговице или переднему углу глаза наступает живой рефлекс смыкания век.'
Сами по себе движения глаз вполне нормальны. В отдельных, хотя и редких, случаях наступают, по-видимому, спонтанные движения глаз. Эти взаимоотношения лучше всего рассматривать при исследовании известных лабиринтных рефлексов на глаза. При этом было установлено, что как подъем и опускание, движения внутрь и кнаружи, так и все круговые движения глаз производятся точно так же, как и у нормальных животных.
Реакции на хлопок (Schallreaktionen) (на хлопанье в ладоши на значительном расстоянии) наблюдались в разнообразных модификациях. Это были: прядение ушами, резкое вздрагивание головы (Zucken), поднимание головы с дополнительными движениями ног, один или несколько нормальных шагов животного, сидевшего до этого спокойно, осуществление правильного прыжка, иногда с дополнительной пробежкой, смыкание век и пр. Таким образом, в ответ на возбуждение слухового нерва таламического кролика активируются самые различные моторные аппараты и этим путем таламическое животное может быть приведено в движение с помощью дистантного рецептора (орган слуха), что при зрительном раздражении, как показано выше, получить не удавалось.
Чрезвычайно удобны для наблюдения рефлексы приема пищи. Правда, ни в одном случае спонтанного приема пищи отмечено не было, так как обонятельный нерв был разрушен, а с помощью глаз животные не в состоянии различать предметы. Однако как только касаются любым предметом (палец, кусок репы) нижней губы хорошо возбудимого животного, его голова благодаря движению шеи энергично проталкивается вперед, вследствие чего зубы плотно прижимаются к приложенному предмету. Если кусочек репы приложить к резцам, животное грызет и откусывает часть кусочка, после чего следует нормальное пережевывание и проглатывание. Если кусок репы попадает в рот, начинаются сильные жевательные движения, длящиеся до тех пор, пока все не оказывается пережеванным и проглоченным. Изо рта при этом ничего не теряется. Во время жевания язык иногда высовывается и облизывает морду. Если запихнуть кусок репы за щеку, он скоро оказывается разжеванным и проглоченным. Энергичные жевательные движения наступают всякий раз, когда между верх¬
Общее поведение таламических и мезенцефалических животных
165
ней и нижней челюстями в рот просовывается палец. Жевательные движения при этом настолько мощны, что приходится остерегаться, дабы не быть укушенным. В отдельных случаях спонтанные жевательные движения можно наблюдать и у спокойно сидящего животного. Однажды животные уселись на задние лапы и передними чистили мордочки.
Уже прежние исследователи обращали внимание на интенсивные «псевдоаффективные» рефлексы у кроликов, лишенных большого мозга [Шеррингтон (5)]. Наклонность к подобным рефлексам у различных животных сильно изменяется. В ответ на каждое достаточно сильное афферентное раздражение у животных развиваются сильные подергивания (Strampeln); они пытаются убегать, наступают рефлекторные остановки дыхания или продолжительное, затрудненное, стенозированное дыхание с гортанным свистом, или животное начинает сильно кричать, что часто продолжается весьма и весьма долгое время. В результате этих «псевдоаффективных» рефлексов отдельные животные оказываются настолько измученными, что эксперименты с ними надолго прерываются.
Раздражая обычно спокойно сидящее животное, можно побудить его к бегу и к прыжкам. Для этого пригодны все возможные раздражения, как например звуковые, или симметричное давление на обе задние лапы, или щипок хвоста. В ответ на это животное проделывает один или несколько вполне нормальных скачков, продолжая затем, если оно не натолкнулось на препятствие, сидеть спокойно. Возбудимые животные пробегают с десяток и более метров. В ответ на сильные раздражения наступают настоящие прыжки, причем животное всякий раз приземляется весьма уверенно, не переворачивается и оказывается в состоянии удерживать вполне симметричное положение в пространстве. Если позволить такому животному выскочить из клетки или соскочить со стола на пол, то и в этом случае при приземлении лапы располагаются на полу правильно и животное не падает. Способность соблюдать равновесие налицо, таким образом, не только при сидении, но и при беге и прыжках.
Если тело животного пассивно передвинуто вперед или назад, оно производит правильные движения ходьбы по полу как передними, так и задними конечностями.
Одним из наиболее бросающихся в глаза различий между таламическим и децеребрированным (бульбарно-мостовым) кроликом является отсутствие у первого децеребрационной ригидности. У децеребрированного животного те мышцы, которые при стоянии противодействуют силе тяжести, т. е. разгибатели конечностей и спины, а также мышцы, поднимающие шею и хвост, находятся в состоянии тонического сокращения [Шеррингтон (1)], в то время как их антагонисты (сгибатели конечностей и мышцы, изгибающие шею, туловище и хвост в вентральном направлении) или полностью атоничны, или тонус их весьма незначителен.
Эта децеребрационная ригидность у таламического кролика наиболее выражена во время пробуждения после наркоза, хотя в едва заметных чертах ее можно наблюдать и у нормального животного. Как только животное немного отдохнуло, состояние его мускулатуры таково, что его проще всего обозначить как «нормальное». Это -значит: мышцы, противодействующие силе тяжести, хорошо тонизированы и точно в той степени, чтобы обеспечивать стояние животного; одностороннего преобладания тонуса этих мышц нет, их антагонисты также тонически иннервированы. При пассивных движениях одинаково хорошо ощущается противодействие как при разгибающих, так и при сгибающих движениях, конечности и спина не жестки, как (это бывает) у децеребрированного животного, но ведут себя как у интактного животного. У таламического животного отсут¬
166
Пятая глава. Установочные рефлексы
ствует и характерная для децеребрированного животного поза с максимально вытянутыми конечностями, опистотонусоми затылком, притянутым к спине.
Отсюда следует, что, как уже указывал Тиле (Thiele), децеребрацион- ная ригидность у кролика не обусловлена отделением центров, расположенных в продолговатом мозгу, от большого мозга, но наступает тогда, когда эти центры лишаются связи с приборами, лежащими в стволе мозга кпереди от моста. Точное расположение этих центров будет рассмотрено в дальнейшем (глава десятая). Реципрокная иннервация мускулатуры у таламического животного оказывается хорошо «сбалансированной», тогда как у децеребрированного животного напряжение разгибателей преобладает за счет такового сгибающих мышц. Поэтому вместе с Шерринг- тоном (9) вполне возможно обозначить децеребрационную ригидность как «рефлекторное стояние», однако вследствие одностороннего преобладания одной группы мышц оно представляет собой только карикатуру на стояние. Напротив, таламическое животное стоит нормально.
Тонические шейные и лабиринтные рефлексы на мышцы конечностей и туловища (рефлексы позы), которые у децеребрированных животных обнаруживаются при перемене положения головы по отношению к туловищу и в пространстве, осуществляются, как показано выше, и у интактного кролика. Само собой разумеется, что рефлексы эти имеются и у таламического, кролика. Так как распределение тонуса у последнего идентично таковому у нормального животного, то шейные и лабиринтные рефлексы ведут себя у обоих животных также одинаково. Поднимая голову, животное становится на передние конечности; при опускании головы передние конечности подгибаются. Точно так же при поворотах головы в положении животного на спине «челюстные конечности» вытягиваются, а «черепные конечности» сгибаются. На таламическом животном гораздо легче, чем на децеребрированном, установить, что при снижении тонуса разгибателей тонус сгибающих мышц повышается, и наоборот. Реципрокная иннервация при этих рефлексах, таким образом, весьма отчетлива. Если придать голове положение, при котором тонус разгибающих мышц одной конечности понижается, очень часто можно наблюдать ее сильное активное сокращение. Особенно отчетливо это видно при поворачивании головы в боковом положении (животного). Если голова поворачивается челюстью вниз, передние ноги по большей части активно сгибаются и подтягиваются к передней части тела (лабиринтный рефлекс); этот рефлекс играет роль при принятии животным сидячего положения из лежачего положения на боку, что будет точнее описано далее.
Сюда же относится реакция, которую можно обозначить как рефлекс прыжка (Sprungreflex). Если поднять голову сидящего таламического животного и с силой прижать ее к спине или если поднять переднюю часть тела животного так, чтобы позвоночник оказался в положении вертикально вверх, сгибая при этом голову дорсально, то животное, энергично разгибая одновременно обе задние ноги, нередко производит правильный прыжок. Рефлекс вызывается тогда, когда благодаря определенной комбинации шейных и лабиринтных рефлексов задние конечности приобретают сильный разгибательный тонус. Именно это имеет место, когда голова сидящего животного энергично сгибается в дорсальном направлении; в этих условиях шейный рефлекс на поднятие головы суммируется с лабиринтным рефлексом, так как голова приводится в максимальное положение для проявления разгибательного тонуса. У животных с преобладающими шейными рефлексами и у животных после двусторонней экстирпации лабиринтов рефлекс прыжка в ответ на дорсальное сгибание головы осуществляется с равным успехом во всех положениях в пространстве. Наоборот, у животных с преоб¬
Общее поведение таламических и мезенцефалических животных
167
ладающими лабиринтными рефлексами рефлекс прыжка отчетливее всего выявляется, если голова дорсально сгибается у животного, которое сидит или передняя часть тела которого приподнята, так как при этом голова приближается к максимальному положению. В этих условиях в боковом положении рефлекс выражен слабее, а в положении на спине — отсутствует. Несомненно, рефлекс облегчается при сгибании и нагрузке задних конечностей, однако рефлекс может наступать и без нагрузки на задние конечности. Из последнего факта следует, что рефлекс прыжка не может быть безоговорочно идентифицирован с экстензорным толчком (Extenser- stof), который Шеррингтон наблюдал у спинальной собаки при надавливании на пальцы энергично согнутой задней конечности. У таламического кролика рефлекс прыжка может быть получен и без раздражения подошвы.
Часто рефлекс не исчерпывается энергичным выпрямлением задних конечностей, но одновременно с ним передние конечности совершают альтернирующие движения бега. Рефлекс прыжка значительно лучше изучать на кошках, которые нередко совершают высокие прыжки вверх.
Из приведенного следует, что путем простого изменения положения головы можно заставить двигаться спокойно сидящее таламическое животное. У децеребрированного животного можно при этом наблюдать описанные на стр. 63 движения бега, когда разгибательный тонус конечностей благодаря соответствующему положению головы становится максимальным. Конечно, до настоящего бега дело при этом никогда не доходит, так как равновесие тела не соблюдается и животное тотчас падает.
В случае же таламического животного, благодаря тому что центры раз- тибателей конечностей «снабжаются тонусом», становится возможным осуществление истинных прыжков, бега и скачков. Этого удается достичь посредством описанных поворотов головы.
В этом находит также свое объяснение старое наблюдение Христиани (Christiani), что таламические кролики способны «перепрыгнуть через препятствие». По данным Мунка, это может осуществиться только тогда, когда животные на полной скорости устремляются по направлению к препятствию. Тогда голова вследствие препятствия оказывается пассивно поднятой и за этим следует рефлекс прыжка.
Исключительно живыми оказываются у таламического кролика вращательные реакции головы и глаз. В согласии с более ранними данными Флу- ранса (Flourens), Хёгиеса (Högyes), Эвальда (Ewald), Барани [Bäräny (2)], Бауэра и Лендлера (Bauer u. Leidler) и в противоположность Бартельсу и Розенфельду (Bartels u. Rosenfeld) оказывается возможным получить живой и быстрый компонент головного и глазного нистагма; он наступает независимо от большого мозга.
Также нормально осуществляются у таламического кролика компенсаторные установки глаз.
Приведенные до сих пор описания показывают, какими разнообразными и сложными функциями обладает таламический кролик. Эти функции могут осуществляться, во-первых, потому, что в реакциях принимают участие, исключая разрушенные обонятельные, все (остальные) черепномозговые нервы и, во-вторых, потому, что центры ствола, расположенные впереди моста, обеспечивают нормальное распределение тонуса мускулатуры тела, а также возможность принять и поддерживать положение тела в покое и при движениях.
Поведение четверохолмного кролика можно передать несколькими словами. Если исключить зрачковый рефлекс и смыкание век
168
Пятая глава. Установочные рефлексы
в ответ на сильное освещение, а также теплорегуляцию, эти животные спо- собны осуществлять все функции в том виде, как это описано выше для таламического кролика. В частности, возможность поддерживать нормальное положение тела полностью сохранена. Децеребрационная ригидность отсутствует или заметна только непосредственно после пробуждения от наркоза и шока. Тонус мускулатуры тела очень скоро становится «нормальным».
Разгибатели конечностей не преобладают над сгибателями. Сидение и поза идентичны таковым интактного животного. Если засунуть палец в рот животного, то проявляется жевательный рефлекс. Часто имеют место выраженные псевдоаффективные рефлексы (крик при щипке лапы, шумное стенозированное дыхание). В ответ на акустические раздражения (хлопок в ладоши) реакция отчетлива. Имеется рефлекс смыкания век. В ответ на энергичное дорсальное сгибание головы наступает рефлекс прыжка. Тонические шейные и лабиринтные рефлексы на мышцы шеи и конечностей проявляются у четверохолмного кролика точно так же, как это описано для таламического животного. Имеются так же реакции поворота головы и глаз, глазной и головной ротаторный нистагм и постнистагм. То же относится и к компенсаторным положениям глаз.
Как и у мезенцефалического кролика, у таламического (животного) можно отметить легкое нарушение чувства положения передних конечностей. Если-одну из передних лап установить на полу тыльной ее стороной,, то корреляция происходит не так скоро, как у нормального животного. В большинстве случаев лапа в конце концов устанавливается правильно, подошвой вниз. Напротив, стояние на тыльной стороне задней лапы корригируется немедленно.
2. Таламическая собака и таламическая кошка. Известные наблюдения Гольца [Goltz (3)] на собаке без большого мозга, подтвержденные во всех существенных чертах Ротманом (Roth- mann) и в справедливости которых на собаке без большого мозга Дюссер де Баренна [Düsser de Barenne (3)] убедился я сам (фотографии ствола мозга этого животного см.: Pflüger’s Archives, 1920, Bd. 180, S. 78), показывают, что поведение собаки без большого мозга в основном совпадает с поведением таламического кролика. Это же справедливо, как показал Дюссер де Баренн, и для таламической кошки. Оба животных вскоре после операции в состоянии самостоятельно усаживаться из лежачего положения и спустя короткое время после пробуждения от наркоза бегать. Однако- у животных обоих видов шок длится дольше, чем у кролика, так что улучшение состояния может отмечаться в течение 10—14 дней после операции. Животные сидят в нормальном положении, совершают побежки в ответ на раздражение, но, в противоположность кролику, у них отмечается также и «спонтанный» бег, особенно когда они голодны. В этом случае появление движений бега следует отнести за счет внутренних раздражений. Температура тела нормальная, наблюдаются зрачковая реакция и смыкание век при освещении. Однако в ответ на самые различные оптические воздействия, кроме обоих упомянутых рефлексов, не появляется никаких других реакций. Кошки, спустя некоторое время после операции, оказываются в состоянии с большой ловкостью избегать препятствия. Здесь установлено, что это обеспечивается уже самым легким прикосновением усов со- встреченным препятствием, что ведет к движениям головы и согласованным с ними движениям тела. Как и у кролика, рефлексы на голову и тело не вызываются пассивными движениями глаз, resp. глазных мышц. Как было установлено при исследованиях реакций вращения, движения глаз, во всех направлениях нормальны.
Общее поведение таламических и мезенцефалических животных
169
В ответ на акустические раздражения следуют движения ушами, в более редких случаях — бег, так что у этих животных при отсутствии большого мозга может вступать в действие механизм движения с названного дистантного рецептора. Рефлексы принятия пищи выражены хорошо.
Например, если кошку окунуть мордой в миску, то она может вполне удовлетворительно пить, при этом совершая нормальные движения языком, облизывая нос. Если голову животного отвести от миски, то спустя некоторое время оно может снова самостоятельно опустить ее туда. Животные также лижут себе лапы, в состоянии чесаться, однако случаетсяг что вместо лапы они лижут стол, а при чесательных движениях задевают не те части тела, которые раздражаются.
Псевдоаффективные рефлексы выражены весьма отчетливо; крик, мимика боли, защитные движения, изменения дыхания отмечены очень часто.
Рефлекс прыжка, особенно интенсивный у кошек, уже упоминался выше.
Как и у кролика, у таламической собаки и таламической кошки отсутствует децеребрационная ригидность; тонус мускулатуры тела обнаруживает нормальное распределение.
Бег осуществляется почти нормально, и только в отдельных случаях у животных обнаруживается петушиная походка; через несколько недель после операции, преодолевая препятствия, они перестают падать. Из повреждений имеют место: отсутствие мунковского рефлекса прикосновения, отсутствие истинного чувства локализации на коже, что устанавливается применением болевых раздражений, вызыванием чесательного рефлекса и т. д. Эти животные медленнее корригируют положение лап на полу, чем нормальные.
Как уже известно, кролик, собака и кошка после полного удаления большого мозга обнаруживают весьма значительные возможности в осуществлении даже сложных двигательных комплексов. Однако животные ведут себя подобно настоящим автоматам, поэтому они особенно подходят для изучения тех рефлексов, которые приводят к принятию и поддержанию положения тела.
3. Таламическая обезьяна. В то время как можно составить себе достаточное суждение о функциях мозгового ствола после удаления большого мозга у трех названных видов животных, у обезьяны это, к сожалению, не так. Никто еще не наблюдал обезьяну, свободную от явлений (послеоперационного) шока. Карплус (Karplus) и Креиндль (Kreindl) произвели обстоятельнейшие исследования обезьян, которые после двусторонней экстирпации большого мозга выживали до 26 дней. Сам я прибавлю только два наблюдения в остром опыте, когда животные жили более 36 час. В обоих случаях было установлено наличие примерно тех же функций, что и в опытах венских исследователей.
Животное может (самостоятельно) сесть из бокового положения, может стоять, если его поддерживать за одну руку, но не в состоянии бегать и лазать. Температура его тела нормальная. В ответ на световое раздражение из оптических реакций наступает только зрачковая. Имеются нормальные координированные движения глаз. В ответ на звуковые раздражения следуют: рефлекс мускулатуры рта, движения глаз и век, вздрагивание всего тела и т. д. Имеются рефлексы жевания и глотания. Выявляются также и псевдоаффективные рефлексы. Распределение тонуса мускулатуры нормальное, децеребрационная ригидность отсутствует. Животные в состоянии чесаться, обнаруживают, если в руку им положить палец, сильный хватательный рефлекс и при поддержке за обе руки в состоянии путем мощного подтягивания на руках приводить и удерживать голову и тело
170
Пятая глава. Установочные рефлексы
в нормальном положении. Кроме того, как это будет позднее рассмотрено, они обладают весьма выраженным установочным рефлексом.
Так как избежать шока у этих животных до настоящего времени невозможно, остается неизвестным, может ли обезьяна, помимо этих отдельных функций, идентичных функциям, наблюдавшимся у собаки, кошки и кролика, образовывать (zusammensetzen) также сложные двигательные комплексы, какие описывались для этих животных, и что в действительности в состоянии выполнить таламическая обезьяна в процессе ходьбы, бега, прыганья и лазанья.
Полные экстирпации большого мозга у антропоидов, насколько мне известно, до сих пор не производились. Хотя и имеются единичные наблюдения над патологическими случаями полной утраты большого мозга у человека, мы не вправе делать из патологических случаев подобного рода далеко идущие выводы в отношении максимума функционирования мозгового ствола человека. Этот вопрос, стало быть, пока остается открытым.
II. АНАЛИЗ УСТАНОВОЧНЫХ РЕФЛЕКСОВ {24)
Как уже упоминалось, после того как явления шока после операции полностью прошли, таламический кролик обладает способностью принимать нормальное положение и восстанавливать его всякий раз, когда он из этого положения выведен. Правильное положение поддерживается также во время бега или прыжков, так что животное не переворачивается или, если это случилось, снова быстро поднимается.
В дальнейшем будет сделана попытка проанализировать, каким образом это происходит, какие рецепторные органы и какие центральные отделы вступают при этом в действие и каким образом различные механизмы действуют совместно.
Уже Лонге (Longei) показал, что кролик после экстирпации большого мозга еще в состоянии стоять и—в ответ на раздражение — нормально бегать и прыгать. Шиф (Schiff) подтвердил эти данные и, кроме того, нашел, что кролик без большого мозга в состоянии бегать и после экстирпации глаз. Вулышан (Vulpian) заметил, что если животных положить на бок или на спину, они тотчас, снова усаживаются в нормальное положение.
Наблюдения в период шока
Если после операции таламическое животное исследовать в течение длительного времени, то усаживание и удерживание положения тела происходят с такой быстротой и уверенностью, что нельзя сказать многого относительно механизма этих реакций в результате простого наблюдения. Наоборот, весьма плодотворным оказывается прослеживание постепенного совершенствования способности принимать правильное положение тела по мере пробуждения после наркоза и исчезновения явлений шока.
Тотчас после операции животное лежит в боковом положении и никакое раздражение не в состоянии заставить его сесть. Спустя некоторое время можно видеть, что в ответ на любое раздражение (например, щипок хвоста) голова поворачивается из бокового положения сначала лишь незначительно, в дальнейшем все лучше, пока не примет в пространстве «нормального положения» (темя вверх, нижняя часть вниз, ротовая щель наклонена кпереди примерно на 30°). Тело при этом продолжает находиться в боковом положении. По прекращении раздражения голова снова возвращается в боковое положение.
Несколько позже дело не ограничивается одной реакцией головы. В ответ на раздражение животного раньше всего, поворачиваясь, прини¬
Анализ установочных рефлексов
171
Рис. 96. Тот же таламический кролик, что и на рис. 99. Нормальная посадка.
мает «нормальное положение» голова; к этому, однако, присоединяются поднимание тела и усаживание. В отдельных случаях можно установить, что это последнее происходит последовательно, когда при непрекращаю- щемся раздражении раньше поднимаются грудная клетка и передние лапы, а только потом задняя половина тела и задние конечности (иногда несколько толчкообразно). Обычно животное остается сидящим и после прекращения раздражения.
В дальнейшем, чтобы вызвать эти реакции, необходимы раздражения все меньшей интенсивности, и в конце концов экспериментатору уже совсем не приходится пользоваться искусственными раздражителями. В этом случае также удается установить, что раньше всего наступает стадия, когда лежащее па полу в боковом положении животное приводит в нормальное положение только голову, поворачивая ее, в то время как тело остается еще в лежачем положении.
В заключение наступает окончательное состояние, когда животное всякий раз «спонтанно» поднимается из бокового положения и усаживается при нормальном положении головы и тела (рис. 96).
Здесь в отдельных случаях, как и в промежуточной стадии, можно установить, что и поднимание, и усаживание осуществляются только головой и передней частью тела, в то время как задняя половина следует за передней только после раздражения (щипок хвоста или лапы), или же реакция наступает настолько медленно, что можно видеть, как первой устанавливается голова, к чему присоединяется затем реакция туловища.
Вся описанная здесь последовательность событий развивается в течение нескольких часов и заканчивается самое позднее на следующий день.
Эти наблюдения показывают, что процесс усаживания (принятия сидячего положения) происходит таким образом, что прежде всего в нормальное положение приводится голова, а затем туловище.
То, что у таламического животного приведение головы в правильное положение в пространстве рефлекторно ведет к принятию сидячего положения, показать легко. Если по какой-либо причине, например во время шока, голова животного, лежащего на боку, не повернута в нормальное положение, часто достаточно, захватив голову рукой, пассивно повернуть ее в это положение, чтобы туловище тотчас оказалось в сидячем положении. (Если оно не наступает спонтанно, можно форсировать это щипком хвоста). Наоборот, правильно сидящее таламическое животное можно привести в боковое положение, если, захватив голову, повернуть ее вокруг сагиттальной оси на 90° (рис. 97).
Рис. 97. То же таламическое животное, что и на рис. 99. До этого животное сидело в нормальном положении (ер. рис. 96). Затем рукой голова была повернута вправо. Туловище последовало за ней и также приняло боковое положение.
172
Пятая глава. Установочные рефлексы
Если децеребрированное животное (мосто-мозжечковое, мозжечково-бульбарное, безмозжечковое бульбарное), которое находится в состоянии децере- брационной ригидности, повернуть в нормальное положение, усаживания тела не наступает. Наблюдаются только описанные выше изменения тонуса конечностей. Разгибательный тонус передних конечностей понижается, тонус нижних конечностей. ведет себя по-разному, смотря по тому, преобладают ли шейные или лабиринтные рефлексы. Поворота передней части тела отчетливо не отмечается. У таламического животного без децеребрационной ригидности в ответ на поворот головы в нормальное положение происходит активное сгибание обеих передних конечностей (лабиринтный рефлекс), которые притягиваются под переднюю часть тела. Это является следствием; иного, более «нормального» распределения тонуса сгибающих и разгибающих мышц передних конечностей. Помимо этого наступает поворот передней части тела, который рефлекторно присоединяется к повороту шеи. Подобное же имеет место в случае поворота поясничной части позвоночника и реакции задних конечностей.
Первые простые наблюдения, таким образом, уже показали, что при пробуждении из наркоза и выходе из шока животное прежде всего пытается привести в нормальное положение голову, к чему в дальнейшем присоединяется приведение тела в сидячее положение.
Обзор установочных рефлексов
Как уже упоминалось, способность таламического кролика принимать и поддерживать нормальное положение тела обеспечивается совокупным действием следующих рефлексов.
1) Лабиринтный установочный рефлекс наг о- л о в у. При любом положении тела голова приводится в нормальное положение благодаря рефлексу с лабиринтов.
2) Установочные рефлексы, на голову вследствие асимметричного раздражения поверхности тела. Если тело лежит на полу в асимметричном положении, например на боку, то благодаря асимметричному возбуждению чувствительных кожных нервов осуществляется рефлекторный поворот головы в нормальное положение.
3) Шейные рефлексы. Когда голова находится в нормальном положении, а тело еще нет, то вследствие анормального положения шеи (поворот, растяжение, сгибание) приводится в действие рефлекс, благодаря которому часть позвоночника, расположенная каудально, устанавливается по отношению к голове в правильное и симметричное положение. Этот рефлекс развертывается вдоль позвоночника спереди назад.
4) Установочные рефлексы на тело вследствие асимметричного раздражения поверхности тела. Названные до сих пор три группы рефлексов обнаруживаются легко и весьма отчетливо. Помимо* этого, при известных условиях отмечен еще один рефлекс, благодаря которому тело и в том случае, когда голова не находится в нормальном положении, может быть установлено правильно. Возбуждения, вызывающие этот рефлекс, возникают при асимметричном раздражении поверхности' тела.
5) Оптические установочные рефлексы у кролика* не играют никакой роли.
Теперь эти отдельные рефлексы следует охарактеризовать более, точно.
Лабиринтные установочные рефлексы на голову
Так как, согласно только что приведенному обзору, у таламического животного соприкосновение с полом вызывает ряд установочных рефлексов, рецепторами которых следует считать чувствительные нервы тела,
Анализ установочных рефлексов
173
то рефлексы, исходящие из лабиринтов, можно лучше всего и без помех изучать тогда, когда исключается какое бы то ни было соприкосновение животного с полом, т. е. когда животное свободно висит в воздухе. Несколько фотографий сделают явления, наблюдающиеся при этих обстоятельствах, более ясными, чем длинные описания.
Рис. 98. 17 июня 1915 г. Кролик. Удалены большие полушария, перерезаны оба зрительных нерва.
18 июня. Животное свободно удерживают рукой в воздухе за таз таким образом, что задняя половина тела находится в правом боковом положении. Вследствие поворота позвоночника (главным образом середины тела) голова и передняя часть туловища приводятся в нормальное положение (лабиринтный установочный рефлекс на голову с присоединением шейного установочного рефлекса).
Секция: четверохолмие и зрительные бугры интактны; разрез проходит спереди от зрительного бугра; большие полушария полностью отсутствуют. Зрительные нервы перерезаны. Черепные нервы 3—12-й пары с обеих сторон интактны.
На рис. 98 и 99 видно, что если задняя часть тела таламического кролика удерживается в воздухе в боковом положении, то голова поворачивается в нормальное положение.
Эта реакция у всех таламических животных весьма отчетлива. По большей части к этому присоединяется шейный рефлекс, благодаря которому
Рис. 99. Опыт 27 мая 1915 г. Кролик. Большие полушария удалены до зрительных бугров (см. также рис. 96
и 97).
29 мая. Животное свободно удерживают за таз в воздухе таким образом, что задняя часть тела находится в правом боковом положении. При помощи поворота позвоночника, главным образом среднего отдела тела, голова и передняя часть туловища приводятся в нормальное положение (лабиринтный установочный рефлекс на голову с присоединившимся шейным установочным рефлексом).
'Секция. Четверохолмие и зрительные бугры интактны. Разрез проходит кпереди от зрительных бугров. От больших полушарий слева, вентрально от зритель- лого тракта, остался небольшой участок грушевидной извилины. Нервы глазных мышц и зрительные нервы интактны.
грудь, следуя за головой, также поворачивается в правильное полоя^ение. В отдельных случаях за этим даже наступает дополнительный поворот таза, так что животное, преодолевая сопротивление держащей руки, «переворачивается», пока все тело не примет в воздухе правильного положения.
Стремление правильно установить голову в пространстве настолько сильно, что таз животного можно перекрутить в воздухе из одного бокового положения в другое без того, чтобы при этом изменилось положение головы. При этом животное оборачивается в воздухе как бы вокруг своей головы, прочно удерживаемой (в определенном положении) лабиринтным установочным рефлексом; исключительно наглядная демонстрация.
При этом голова удерживается по большей части таким образом, что плоскость ротовой щели наклонена ниже горизонтальной плоскости примерно под углом в 30°.
Сравнение рис. 98 и 99 показывает, что реакция наступает совершенно одинаково независимо от того, интактны или перерезаны зрительные
174
Пятая глава. Установочные рефлексы
нервы. Это соответствует сообщенному выше факту, что у таламического' кролика оптические реакции полностью отсутствуют (кроме зрачковой реакции и реакции смыкания век при освещении).
Если животное удерживать за таз в воздухе в затылочном положении,, то голова поворачивается в сторону. Этот поворот составляет в большинстве случаев 90°, но может достигать 135 и даже 180°, так что голова принимает полностью нормальное положение. Иногда голова поворачивается на 90° и затем, оборачиваясь кверху, еще больше приближается к нормальному положению.
К повороту головы присоединяется (рис. 100) соответствующий поворот передней половины тела/ Если при положении животного на спине, когда таз принял положение, изображенное на рис. 100' (левое ухо повернуто книзу), последний повернут в воздухе в противоположном направлении (левая ягодица вниз), голова удерживается в своем положении в пространстве до тех пор, пока вращение оси- тела не окажется слишком сильным. Когда же это наступает, голова внезапно, толчком, поворачивается таким образом, что правое ухо оказывается внизу. Эта реакция также очень показательна и может быть использована для доказательства наличия лабиринтного установочного рефлекса.
Часто голова устанавливается иа затылочного положения в прямое и другим путем, а именно: посредством сильного вентрального сгибания шеи голова из затылочного приводится в нормальное положение, к чему присоединяется вентральное сгибание передней половины тела, так что грудь и голова, двигаясь в вентральном направлении, соприкасаются, в результате чего голова оказывается во вполне нормальном положении (рис. 101).
Если уложить животное на стол на спину, то на чувствительные нервы тела асимметричные возбуждения действия не оказывают. Лабиринтный рефлекс на голову поэтому можно исследовать и при положении животного на спине. Вследствие этого рефлекса сначала голова, передняя часть тела и далее задняя половина принимают боковое положение, и уже из этого последнего животное поднимается в нормальное положение.
Если животное держать в воздухе позвоночником, расположенным вертикально, и головным концом кверху, наступает вентральный наклон головы до тех пор, пока последняя не окажется в нормальном положении. К этому присоединяется, если животное удерживать за таз, вентральное сгибание грудного и поясничного отделов позвоночника, так что правильно устанавливается также и передняя часть тела. Рис. 101 показывает, что эта реакция осуществляется настолько легко, что даже если крестец удерживается не вентрально, но почти в горизонтальном положении (в положении на спине), голова еще принимает в пространстве нормальное положением
Рис. 100. Тот же кролик, что и на рис. 99. 29 мая 1915 г. Животное удерживают рукой за таз в положении на спине. Голова поворачивается в левое боковое положение. К этому присоединяется поворот передней части тела (лабиринтный установочный рефлекс на голову с присоединившимся шейным установочным рефлексом).
Анализ установочных рефлексов
175.
Если держать таз животного в воздухе в перевернутом положении,, то проявление этого вентрального сгибания или описанного выше поворота головы и передней половины тела (рис. 100) зависит от «случайных»- обстоятельств.
Рис. 101. Тот же кролик, что и на рис. 99. 29 мая 1915 г. Животное удерживают рукой за таз свободно в воздухе так, что позвоночник находится почти в горизонтальном положении (30°). Голова приведена в нормальное положение, а позвоночник настолько сильно изогнут вентрально, что грудь оказывается стоящей правильно (лабиринтный установочный рефлекс на голову с присоединившимся шейным установочным рефлексом).
Если животное удерживается за таз, а голова его свешивается вниз; при отвесном положении позвоночника (рис. 102), то голова путем дорсального нагибания приводится в нормальное положение.
В течение короткого времени после операции животному это удается произвести не в полной мере, так что ротовая щель еще образует с горизонтальной линией угол в 45°. Постепенно рефлекс осуществляется во все
большем объеме, пока наконец в указанном положении голова не устанавливается в нормальном положении с ротовой щелью, опущенной под углом в 10— 20-30°.
Рис. 102. Тот же кролик, что и на рис. 99. 29 мая 1915 г. Животное удерживают за таз свободно в воздухе так, что позвоночник стоит вертикально, а голова обращена вниз. При помощи дорсального сгибания голова приводится в нормальное положение (лабиринтный установочный рефлекс на голову). Дорсальное сгибание головы посредством тонического шейного' рефлекса вызывает разгибание передних конечностей. На фотографии видно, как при прыжке вверх голова благодаря совместному действию названных рефлексов приводится в правильное положение и в то же время передние ноги приобретают способность принять на себя вес тела.
Если животное первоначально удерживалось головой вверх (рис. 101)^ и таз постепенно поворачивался вокруг поперечной оси на 180°, пока но достигалось положение головой вниз (рис. 102), можно было наблюдать, что голова не совсем участвует или только очень незначительно участвует1 в этом повороте задней половины тела. В отдельных случаях морда опускается при этом книзу примерно на 30°; в других — вообще не опускается.. Здесь можно поворачивать тело в воздухе вокруг головы, стоящей в постоянном положении в пространстве.
Все эти положения в пространстве, как они описаны до сих пор, можно. также исследовать, если захватить животное не за таз, аза задние конеч¬
176
Пятая глава. Установочные рефлексы
ности, дистально от тазобедренного сочленения. Тогда в установочном рефлексе обычно принимает участие и задняя половина тела. Последний способ, однако, применим не всегда по той причине, что многие животные при попытке держать их за задние конечности начинают биться и производят прыгательные движения.
Рис. 103 демонстрирует животное, удерживаемое в воздухе за задние конечности, причем голова и тело находятся в нормальном положении.
Описанные до сих пор лабиринтные рефлексы, благодаря которым голова всякий раз приводится в нормальное положение, после чего в направлении, определенном движением головы, следует также и тело, естественно обеспечивают то, что таламическое животное оказывается в состоянии, прыгая в воздухе (при беге, при выскакивании из клетки или соскакивании со стола на пол), неизменно принять на полу нормальное положение.
Животные без лабиринтов эту способность утрачивают непосредственно после операции и должны с трудом снова обучаться этому с помощью других механизмов. Рис. 102 показывает, как талами-
Рис. 103. Тот же кролик, что и на рис. 99.
29 мая 1915 г. Животное свободно удерживают рукой в воздухе за задние конечности. Голова и туловище находятся в нормальном положении.
ческий кролик в висячем положении головой вниз в результате лабиринтного рефлекса на голову и присоединяющегося к нему шейного рефлекса на передние конечности приводится в то же положение, которое животное должно принять при прыжке вниз. Голова защищается от удара о пол, и передние конечности благодаря тоническому разгибанию в состоянии принять на себя вес приземляющегося тела.
То, что описанные до сих пор лабиринтные установочные рефлексы на голову действительно исходят из лабиринтов, доказывается, как это будет видно из следующего отдела, тем, что после удаления лабиринтов они исчезают.
Наблюдения на безлабиринтном таламическом
кролике
Все экстирпации лабиринтов произведены де Клейном (de Kleyn). У двух животных удаление обоих лабиринтов было предпринято накануне; после этого животные спокойно сидели, отдыхая после операции. Через 24 часа я удалил большие полушария выше таламуса. У восьми остальных животных экстирпация большого мозга производилась непосредственно после удаления лабиринтов. В одном, более позднем случае выключение лабиринта осуществлялось не хирургическим путем, а с помощью впрыскивания кокаина со стороны буллы.
Общее поведение безлабиринтного таламического кролика не отличается сколько-нибудь существенно от такового «нормального» таламического животного, как оно описано выше. Животные сохраняют способность принимать нормальное полояюние, сидят сгорбившись на земле, даже когда спустя короткое время после операции тонус затылочных мышц у них вследствие удаления лабиринтов ниже, чем при интактных лабиринтах. Несмотря на это, через некоторое время к ним снова возвращается способность держать голову в прямом положении (рис. 104).
Децеребрационная ригидность отсутствует, распределение тонуса в мускулатуре тела «нормально». При засовывании пальца в рот животного
Анализ установочных рефлексов
177
наступают жевательные движения. Рефлекс смыкания век и другие рефлексы лицевого нерва в большинстве случаев отсутствуют, так как у кролика во время операции на лабиринте n. facialis обычно разрушается. Если же его удается сохранить, рефлекс смыкания век и другие рефлексы имеют место. Рефлекс прыжка на энергичное дорсальное сгибание головы проявляется отчетливо, но у безлабиринтных животных он не зависит от положения тела в пространстве.
В любом положении тела, если голова сильно прижимается к спине, задние конечности производят мощное движение прыжка, а передние осуществляют движения бега.
Рис. 104. 1 июня 1915 г. Кролик. Двусторонняя экстирпация лабиринтов.
Экстирпация большого мозга до уровня зрительных бугров.
Снимок сделан через 4 часа после операции. Сидит на столе, голову держит правильно. Секция. Четверохолмие и зрительные бугры интактны. Базальные узлы отсутствуют. Разрез проходит дорсально кпереди от зрительных бугров. В середине виден остаток области конечной пластинки шириной в 1—2 мм. На правой стороне остался совсем маленький участок грушевидной доли на зрительном тракте. В остальном большой мозг удален полностью. Зрительные нервы перерезаны. Нервы глазных мышц
интактны.
Все лабиринтные рефлексы, естественно, полностью отсутствуют. Изменение положения головы в пространстве не вызывает более никаких изменений тонуса мускулатуры шеи и конечностей. Все реакции поворота также отсутствуют.
Для наших целей важно то, что после удаления обоих лабиринтов описанные в предыдущем отделе лабиринтные установочные рефлексы полностью упраздняются. Это иллюстрируется следующими стереоскопическими рисунками.
На рис. 105 видно, что если удерживать тело лабиринтэктомированного таламиче-
Рис. 105. Тот же лабиринтэктомированный таламический кролик, что и на рис. 104. Животное за таз свободно удерживают в воздухе рукой таким образом, что тело находится в правом боковом положении. Передняя часть тела и голова (ср. уши) находятся также в правом боковом положении; передняячасть тела, шея и голова вследствие силы тяжести опустились вниз. Следовательно, голова и передняя часть тела не приняли нормального положения в пространстве.
Сравнение с рис. 98 и 99 показывает, что после экстирпации лабиринтов отсутствует лабиринтный установочный рефлекс на голову. Вследствие этого выпадает и шейный
рефлекс.
•ского животного в воздухе в боковом положении, то голова в нормальное положение не поворачивается. Поэтому положения изображенного на рис. 98 и 99 и характерного для находящегося в боковом положении таламического животного с интактными лабиринтами не наблюдается. Голова и тело прежде всего благодаря силе тяжести свисают вниз. Если повернуть заднюю половину тела из одного бокового положения 12 Р. Магнус
178
Пятая глава. Установочные рефлексы
в другое, голова пассивно следует за этим движением и не удерживается принудительно, как при интактных лабиринтах в нормальном положении.
Так как лабиринтный установочный рефлекс на голову отсутствует, присоединяющийся к нему установочный шейный рефлекс, благодаря ко-
Рис. 106. 25 июня 1915. Кролик. Хлороформный наркоз. Сонные артерии перевязаны. Блуждающие нервы перерезаны. Двусторонняя экстирпация лабиринтов, далее — экстирпация большого мозга кпереди от зрительных бугров. Зрительные нервы приподняты и перерезаны.
Снимок через 5 У2 часов после операции. Животное удерживают рукой за таз на столе в положении на спине. Голова и передняя часть тела находятся в том же положении, т. е. повернуты на бок.
Сравнение с рис. 100 показывает, что после экстирпации лабиринтов отсутствует лабиринтный установочный рефлекс на голову. Вследствие этого выпадает и шейный установочный рефлекс.
Секция. Почти никакого внутричерепного кровотечения. Зрительные нервы перерезаны; нервы глазных мышц интактны. Четверохолмие и зрительные бугры ин- тактны. Разрез проходит дорсально кпереди от зрительных бугров, позади конечной пластинки. Вентрально он идет сразу впереди хиазмы. С каждой стороны на зрительном бугре осталось по небольшому кусочку грушевидной извилины, которые не связаны с препаратом.
торому грудь устанавливается в направлении, заданном головой, также не наступает.
Рис. 106 показывает, что если таламическое безлабиринтное животное удерживать за таз в положении на спине — безразлично происходит эта на столе или в воздухе, — голова и передняя половина тела не поворачиваются в сторону.
Больше не предпринимаются попытки принять сидячее положение из полностью симметричного положения на спине. Если кролика перевернуть из положения на спине на один или другой бок, то голова пассивно следует за этим движением.
Если безлабиринтное таламическое животное подвесить головой вверх (рис. 107), то голова не приводится путем вентрального сгибания в нормальное положение, как это бывает при интактных лабиринтах, а просто свисает назад и пригибается в дорсальном направлении.
Рис. 107. Тот же лишенный лабиринтов таламический кролик, что и на рис. 104. Животное рукой удерживают за позвоночник свободно в воздухе головой вверх.
Голова опустилась назад, теменем книзу. Голова и передняя часть тела не приняли в пространстве нормального положения.
Сравнение с рис. 101 показывает, что лабиринтный установочный рефлекс на голову и присоединяющийся к нему шейный рефлекс отсутствуют.
Вследствие этого соответствующее вентральное сгибание передней части тела также отсутствует и животное остается в своем ненормальном положении, не будучи в состоянии корригировать последнее.
Анализ установочных рефлексов
179
В случае висячего положения головой вниз (рис. 108) голова не приводится, как у животных с интактными лабиринтами (ср. рис. 102), в нормальное положение посредством дорсального сгибания, а попросту свешивается, следуя силе тяжести, мордой отвесно вниз.
Если таламическое животное, лишенное лабиринтов, привести сначала в висячее положение головой вверх, а затем повернуть таз вокруг поперечной оси на 180°, пока животное не окажется висящим головой вниз, то голова чисто пассивно следует этим движениям и никаких попыток удержать голову в пространстве в определенном положении отметить невозможно.
Точно так же, если таламическое животное удерживать в воздухе во всевозмож-
Рис. 108. Тот же лишенный лабиринтов таламический кролик, что и на рис. 104. Животное удерживают рукой свободно в воздухе за таз так, что позвоночник стоит отвесно, а голова обращена вниз. Голова пассивно свешивается мордой вертикально вниз, стало быть, не приводится в нормальное положение; лабиринтный установочный рефлекс, как показывает сравнение с рис. 102, отсутствует.
Так как голова спокойно свисает вниз, не наступает и тонического разгибания передних конечностей, «поза прыжка» отсутствует.
ных положениях не за таз, а за задние конечности, то все лабиринтные и примыкающие к ним шейные установочные рефлексы на переднюю половину тела отсутствуют.
Описанные взаимоотношения во всех опытах с экстирпацией обоих лабиринтов были всегда одни и те же. Отсюда следует, что описанные до
сих пор установочные рефлексы на голову, которые проявляются во время наблюдения животного в воздухе, действительно исходят из лабиринтов.
Рис. 109. Кролик, 5У2 недель после двусторонней экстирпации лабиринтов. Большой мозг интактен. Животное удерживают в воздухе в положении на спине. Лабиринтные установочные рефлексы отсутствуют. Никаких оптических установочных рефлексов.
Все описанные до сих пор наблюдения над лабиринтными установочными рефлексами на голову у таламического кролика могут быть воспроизведены точно таким же образом и с теми же результатами и на кролике с интактным большим мозгом. На нормальном кролике можно показать, кроме того, что после двусторонней экстирпации лабиринтов установочные рефлексы на голову при исследовании в воздухе полностью выпадают. Это обнаруживается не только непосредственно после операции, но остается неизменном в течение длительного времени. Еще недавно мне удалось поставить опыты на кролике, у которого за 5.5 недель перед этим де Клей-
12 15
180
Пятая глава. Установочные рефлексы
ном была произведена двусторонняя экстирпация лабиринтов.1 После такой операции животные обычно погибают, так как при этом с обеих сторон повреждаются лицевые нервы, вследствие чего животные не в состоянии осуществлять акт еды. У только что упомянутого животного, однако, оба лицевых нерва остались неповрежденными, поэтому акт еды был возможен. У этого животного отсутствовали все установочные лабиринтные рефлексы на голову в точности так, как это было описано выше для таламического кролика без лабиринтов (ср. рис. 105—108). Рис. 109 показывает это животное в воздухе в боковом положении. Отсюда следует вывод, важный для наших дальнейших рассуждений, а именно, что у кролика с интактным большим мозгом оптические установочные рефлексы отсутствуют и что животные после полной ликвидации всех шоковых явлений и в течение недель не научаются использовать свои глаза для ориентирования головы в пространстве. Стало быть, для демонстрации лабиринтного установочного рефлекса на голову можно без риска ошибиться использовать интактного кролика, не надевая ему на голову специальной шапочки.
У морской свинки лабиринтные установочные рефлексы ведут себя так же, как у кролика.
Лабиринтные установочные рефлексы у кошки и у собаки
Таламическая кошка, прооперированная Дюссер де Варенном в два приема, уже через полчаса после последней операции проделала несколько шагов с головой, повернутой влево. Спустя четыре часа она уже сидела в клетке, вполне удовлетворительно усаживалась из лежачего положения на , левом боку, причем начинала движение голова; на следующее утро вполне удавалась правильная установка головы из лежачего положения на правом боку, но не удавалось еще принятие сидячего положения. Примерно через 22 часа после последней операции можно было наблюдать усаживание из лежачего положения на правом боку и побежку в несколько шагов без раздражения извне. Через два дня животное сидело, удерживая голову в прямом положении; из лежачего положения на левом или правом боку усаживание осуществлялось немедленно, причем движение начиналось с головы. Спустя 6—7 недель наблюдения явления шока проходили полностью и достигался максимальный объем функций (ср. подробный протокол Дюссер де Баренна).
У оперированной Дюссер де Варенном собаки без большого мозга {41) через 21 час после удаления второго полушария можно было в ответ на раздражение получить правильную установку головы из левого или правого боковых положений, а из лежачего положения на левом боку — усаживание передней половины тела. Собака могла стоять и бегать, однако опрокидывалась чаще вправо, но тотчас снова поднималась. Она натыкалась на все препятствия и уклонялась по дуге в левую сторону. Через четыре дня внезапно стало удаваться вставание из лежачего положения на обеих сторонах. Скоро прекратились и круговые движения.
Как у таламической кошки, так и у таламической собаки во время исследований при свободном пребывании в воздухе удалось установить (41), что у них в точности так же, как это было описано выше для кролика, голова приводилась в нормальное положение из любых положений в про¬
1 На одной стороне операция не была радикальной, так что после нее сохранились слабо выраженные реакции. Наоборот, все отолитовые реакции, а следовательно, и установочные лабиринтные рефлексы, о которых здесь говорится, длительно отсутствовали.
Анализ установочных рефлексов
181
странстве. При этом можно передвигать тело вокруг прочно установленной головы во всех направлениях. После экстирпации обоих лабиринтов лабиринтные установочные рефлексы пропадают. У таламической кошки и таламической собаки оптических установочных рефлексов нет.
Если же исследовать кошку и собаку с интактным большим мозгом, оптические установочные рефлексы, как будет показано далее, у них выявляются. Эти рефлексы следует выключать, надевая на голову (специальную) шапочку. Тогда обнаруживается, что собака и кошка ведут себя в воздухе совершенно так же, как таламический кролик. После двусторонней лабиринтэктомии они утрачивают способность правильно устанавливать голову в пространстве, оказываясь совершенно дезориентированными (см. рис. 127—131).
Лабиринтные установочные рефлексы у обезьян (59)
У обезьян оптические установочные рефлексы также сильно развиты. При желании изучать установочные лабиринтные рефлексы обезьяны с сохранным большим мозгом в условиях свободного пребывания в воздухе следует закрывать глаза животного, что лучше всего удается, если под легким хлор- этиловым наркозом наложить на веки несколько поверхностных скобяных швов. Тогда все лабиринтные установочные рефлексы выявляются с максимальной ясностью.
Точные описания здесь излишни, так так их можно найти в Pilüger’s Arch. 1923,
Bd. 193, S. 415. Поведение обезьян в точности соответствует таковому кроликов.
Рис. НО. Macacus Cynomolgus. Таламическая обезьяна, 4У2 часа после операции. Таз удерживают в воздухе в правом боковом положении (задние ноги
пассивно свешиваются вниз). Грудь на 45°, голована 60—70° повернуты в направлении к нормальному положению. Лабиринтный установочный рефлекс.
Лабиринтные установочные рефлексы исследовались у двух таламических обезьян. При этом выяснилось, что, как только исчезали первые явления послеоперационного шока, лабиринтные рефлексы уже в день операции отчетливо проявлялись.
Если животное удерживалось за таз в нормальном положении в воздухе, то голова приводилась в нормальное положение. Из бокового положения голова более или менее интенсивно поворачивалась в направлении нормального положения (рис. 110), так что у двух животных оно было достигнуто полностью, у третьего — не завершено на 45°. Если тело животного поворачивалось в воздухе из одного положения на боку в противоположное, можно было ясно видеть, что благодаря описанному рефлексу голова с большим или меньшим успехом удерживается в нормальном положении. При «стоячем» положении тела в воздухе голова удерживается в нормальном положении. При свисании головой вниз отчетливо выявлялось дорсальное пригибание головы; ротовая щель располагалась или вертикально книзу, или поворачивалась из этого положения еще на 10э по направлению к нормальному. При переходе из висячего положения го-
182
Пятая глава. Установочные рефлексы
ловой вверх в висячее положение головой вниз, когда таз поворачивается на 180°, голова производила поворот только на 45°. Если животные удерживались в положении на спине, голова благодаря вент}эальному пригибанию приводилась в нормальное положение, причем ротовая складка располагалась горизонтально или несколько выше горизонтали. Так как у таламической обезьяны оптические установочные рефлексы отсутствуют, этот эксперимент возможно осуществить и без закрывания глаз животному.
Из сказанного следует, что у таламической обезьяны лабиринтные установочные рефлексы на голову выявляются с исключительной отчетливостью.
Наоборот, если безлабиринтную таламическую обезьяну удерживать в воздухе в боковом положении, то голова находится в том же положении (рис. 111) так, что упомянутый выше поворот головы из бокового положе-
Рис. 111. Macacus Rhesus. Полная экстирпация через 34 дня после двусторонней экстирпации лабиринтов. Животное удерживают рукой в воздухе за таз и передние ноги в левом боковом положении. Голова в левом боковом положении и, следуя силе тяжести, только немного опустилась вниз. Отсутствуют лабиринтные установочные рефлексы на голову.
ния в направлении к нормальному положению маскируется последствиями действия силы тяжести.
Точно так же у обезьян с сохранным большим мозгом можно показать, что после двусторонней экстирпации лабиринтов лабиринтные установочные рефлексы на голову полностью отсутствуют. Это лучше всего наблюдать на животных, глаза которых закрыты. Это было проделано на безла- биринтной обезьяне спустя 14 дней и 1^ месяца после экстирпации второго лабиринта. Если такое животное удерживать в воздухе в различных положениях, то выясняется, что голова его не принимает больше постоянного положения в пространстве.
Так, например, при нормальном положении тела в воздухе голова может свисать вертикально мордой вниз или же находиться в правом или левом боковом положении, или даже пригибаться дорсально. Если животное удерживается в воздухе в боковом положении, то голова оказывается в боковом положении и, следуя только направлению силы тяжести, несколько свешивается нижним ухом книзу. В «стоячем» (aufrecht) положении тела в воздухе голова может или удерживаться в нормальном положении, или же находиться в правом или левом боковом положении, причем это ее положение не корригируется. Если тело животного свисает головой вниз, последняя в большинстве случаев пригибается вентрально, так что ротовая складка располагается горизонтально или даже несколько кверху, или же голое а свешивается мордой вниз. Если животное удерживается в воздухе в положении на спине, голова находится или в затылочном положении, или в правом или левом боковом. Однако чаще всего она оказывается в затылочном положении.
Отсюда следует, что после утраты лабиринтов и при закрытых глазах обезьяна, свободно удерживаемая в воздухе, не в состоянии более ориентировать свою голову в пространстве.
Описанные наблюдения показывают, что лабиринтные установочные рефлексы на голову у кролика, морской свинки, кошки, собаки и обезьяны выявляются совершенно одинаковым образом и что у животных названных видов они подчинены совершенно одинаковым законам.
Анализ установочных рефлексов
183
Переворачивание при свободном падении
Эти лабиринтные установочные рефлексы на голову играют при свободном падении (66) решающую роль. Если кошек, кроликов или обезьян свободно удерживать в воздухе в положении на спине, а затем позволить им падать вниз, они, как известно, переворачиваются в воздухе и, приземляясь, правильно становятся на ноги. Реакция наступает с исключительной быстротой и уверенностью. Она связана с интактностью лабиринтов. Как у интактной, так и у таламической кошки, а также у интактной обезьяны удалось показать, что после двусторонней экстирпации лабиринтов у этих животных переворачивание в воздухе становится невозможным. Они, как мешок, шлепаются (plumpsen) на пол — независимо от (исходного) положения на спине или на боку— и не в состоянии более так перевернуться в воздухе, чтобы принять правильное положение — конечно- отями вниз. Реакция отсутствует также и у децеребрированного животного с интактными лабиринтами. Уже отсюда становится вероятным, что для осуществления реакции необходим средний мозг. Это превращается в уверенность после наблюдения, показавшего, что таламическое животное еще обладает способностью переворачиваться [наблюдения над таламической кошкой (41) Дюссер де Баренна]. Поскольку для осуществления рефлекса необходимы как лабиринты, так и средний мозг, близким истинному является заключение, что дело идет о действии лабиринтного установочного рефлекса на голову. Что в действительности дело обстоит именно так, выясняется из изучения кинематографических снимков. Как известно, Марей (Магеу) изготовил серийные фотографии кошек и кроликов. Они показывают, что реакция начинается с поворота головы. Это можно с отчетливостью проследить на ряде кинематографических снимков, которые сделаны мною с падающей кошки. На рис. 112 приводятся два примера. В серии А на первом кадре животное удерживается в воздухе в положении на спине. На втором кадре оно отпущено; на третьем видно, как голова поворачивается в направлении от наблюдателя назад, в то время как грудь и передние ноги находятся еще в положении на спине. На четвертом кадре голова повернулась на 90° и начинает также поворачиваться грудь, в то время как положение таза остается еще без изменений. Поворачивание головы и передней половины тела на следующих кадрах продолжается. На пятом и шестом снимках видно, что к повороту передней части тела присоединяется поворот таза, который завершается к девятому кадру. Подобное же иллюстрирует и серия В. На втором кадре животное отпускается, и уже здесь видно, как начинается поворачивание головы по направлению к наблюдателю, на третьем, четвертом и пятом снимках поворот постепенно становится все более сильным. На снимках со второго по четвертый, на которых поворот головы ясно виден, грудной отдел туловища еще находится в положении (передними) конечностями вверх, и только на пятом снимке поворачивается вперед и грудная клетка. На седьмом снимке видно, что голова и грудь повернулись уже значительно, в то время как таз еще отчетливо отстает и лишь на последнем снимке достигает земли также в нормальном положении. Таким образом, в случае реакции при свободном падении дело идет о лабиринтном установочном рефлексе на голову, благодаря которому голова поворачивается по направлению к нормальному положению. К этому присоединяется шейный установочный рефлекс, в результате чего тело следует за головой и таким образом, что раньше следует грудной отдел и только затем таз. Происходит своего рода штопорообразное движение животного в пространстве, начинающееся с головы.
Рис. 112
Анализ установочных рефлексов
185
Поворот головы по отношению к телу должен, кроме того, вызвать тонические шейные рефлексы. Эти последние можно видеть в серии А на седьмом, восьмом и девятом кадрах, где изображено разгибание передних конечностей, наступившее благодаря повороту головы; то же можно видеть на рисунках с седьмого по девятый серии ВА
Дальнейшее прямолинейное смещение головы в пространстве при свободном падении следует объяснять лабиринтным рефлексом на прогрессивное движение. Как будет подробно описано далее (восьмая глава), ускорение лабиринта в дорсальном направлении ведет к сгибанию передних конечностей, тогда как ускорение лабиринта в вентральном направлении вызывает их разгибание. В соответствии с этим находится также и то, что в начале движения, когда голова находится строго в затылочном положении или близко к нему, наступает сгибание передних конечностей, как это ясно видно на втором и третьем кадрах серии А и третьем, четвертом и пятом кадрах серии j5. Как только голова завершила свой поворот в пространстве, лабиринты при продолжающемся свободном падении в воздухе смещаются в противоположном направлении, в результате чего происходит соответствующее разгибание передних конечностей, которые можно увидеть на шестом—девятом кадрах серии А и седьмом—десятом кадрах серии В.
Из описанного следует, что реакция при свободном падении базируется, по существу, на лабиринтном установочном рефлексе на голову, к которому присоединяется соответствующий шейный рефлекс на остальное тело. Помимо этого, отмечаются еще тонические шейные рефлексы на конечности и реакции на прогрессивные движения. Эти последние в конце движения приводят к тому, что конечности тонически разгибаются, оказываясь благодаря этому в состоянии целесообразно распределить и принять вес тела, когда последнее достигает земли.
Можно видеть, что эту замечательную способность животных, свободно падая, оказываться на земле в нормальном положении удается свести к взаимодействию известных рефлекторных реакций.
Установочные рефлексы тела на голову
Возвратимся снова к изложению наших наблюдений над лишенными лабиринта таламическими кроликами. Как описано выше, безлабиринтное таламическое животное уже не в состоянии принять в воздухе правильное положение. Однако стоит только животному прийти в соприкосновение с опорной поверхностью (с полом), как эти отношения меняются.
Если удерживать безлабиринтное таламическое животное за таз сначала в боковом положении в воздухе, то, как видно из рис. 105 (стр. 177), голова и передняя половина туловища остаются в таком же положении. Никаких попыток привести голову в нормальное положение не предпринимается. Однако как только при неизмененном положении таза положить животное на стол, голова тотчас же принимает нормальное положение. Это осуществляется или путем простого поворота, или же сначала морда, пригибаясь вентрально, оказывается между обеими передними лапами, откуда затем поворачивается в нормальное положение. К этому движению головы присоединяется далее обычный шейный рефлекс, вследствие которого, еще в то время пока таз удерживается в боковом положении, передняя 11 Тонический рефлекс на передней конечности, возникающий при падении головой вниз как следствие лабиринтного установочного рефлекса, обсуждался уже в пояснении к рис. 102.
186
Пятая глава. Установочные рефлексы
половина тела поворачивается в сидячее положение. Рис. ИЗ позволяет видеть окончательный результат.
Эта реакция наступает тотчас после того, как тело животного коснулось пола (или стола, или какой-либо другой поверхности). На рис. ИЗ видно, что эта реакция осуществляется, когда не голова, а только туловище ле- ж итна столе. Отсюда напрашивается заключение, что дело в данном случае идет о рефлексе, который вызывается благодаря одностороннему возбуждению чувствительных (давление) нервов тела.
Рис. 113. Тот же лишенный лабиринтов таламический кролик, что и на рис. 106. Животное сначала удерживали в воздухе в правом боковом положении. Голова и передняя часть тела также находились в боковом положении (ср. рис. 105), так как лабиринтный рефлекс отсутствует. Затем животное было уложено на стол в правом боковом положении. Голова выступала за край стола. При этом сначала повернулась влево, т. е. в нормальное положение (действие одностороннего давления на тело плоскости опоры), голова, за ней последовала передняя половина тела, в то время как задняя часть его фиксировалась рукой в правом боковом положении.
Справедливость этого соображения легко доказать. Если поворот головы в нормальное положение вызывается односторонним давлением плоскости опоры на поверхность тела, то этому можно воспрепятствовать, оказывая примерно такое же давление на боковую поверхность тела, обращенную кверху. Именно это имеет место в действительности. Если в то время, как лабиринтэктомированное таламическое животное лежит на столе в положении на боку, на поверхность тела, обращенную кверху, наложить дощечку, например крышку от, ящика для сигар, а сверху поместить груз в 1 кг, то голова в нормальное положение более не приводится, а остается в боковом положении, как если бы животное свободно удерживалось в воздухе. (Этот простой и весьма наглядный опыт в дальнейшем
Рис. 114. Тот же лишенный лабиринта таламический кролик, что и на рис. 106 и ИЗ. Животное лежит на левом боку на столе. Голова выступает за край стола. На правую сторону
тела, обращенную вверх, наложена дощечка с грузом в 1 кг, придерживаемая рукой («проба с дощечкой»). Голова в результате этого остается в левом боковом положении и в нормальное положение не приводится. (Действие одностороннего давления плоскости опоры компенсировано давлением дощечки).
будет обозначаться как «проба с доской»). На рис. 114 изображены результаты опыта.
Однако как только груженая дощечка убирается, голова, поворачиваясь, тотчас снова принимает нормальное положение, а за этим поворотом следует и туловище.
Из изложенного мы познакомились с новым фактором, благодаря которому лежащее на боку животное рефлекторно поднимается в сидячее положение. В противоположность лабиринтному установочному реф-
Анализ установочных рефлексов
187
лексу, который обеспечивает правильную ориентацию животного и в воздухе, для только что описанного установочного рефлекса (установочный рефлекс на голову благодаря асимметричному раздражению поверхности тела, кратко — «установочный рефлекс на голову») необходимо соприкосновение (тела) с плоскостью опоры.
При более близком рассмотрении выяснилось, что речь идет о весьма действенной реакции. Самые различные односторонние раздражения в состоянии вызвать у лабиринтэктомированного таламического животного поворот или отклонение головы в противоположную сторону. Так, при одностороннем прикосновении к роговице или к углу глаза голова отворачивается в противоположном направлении. Если удерживать животное за таз в нормальном положении в воздухе и ущипнуть при этом переднюю лапу, голова отклоняется или поворачивается в противоположную относительно раздражаемой конечности сторону. Тот же опыт можно соединить с пробой с доской. Если уложить лишенное лабиринтов животное на стол в положении на боку и на поверхность тела, обращенную кверху, поместить дощечку с грузом, то голова остается в боковом положении (рис. 114). Если теперь ущипнуть нижележащую переднюю лапку, то голова тотчас же поворачивается в нормальное положение и после прекращения раздражения возвращается обратно. Если, наоборот, ущипнуть вышележащую переднюю лапку, голова поворачивается теменем книзу. В этом случае давление плоскости опоры скомпенсировано давлением наложенной доски, вследствие чего одностороннее раздражение лапки в состоянии оказать свое направляющее влияние на голову. Если животное удерживать за таз в нормальном положении в воздухе, то можно широко захватить кожу одной стороны тела, щипать и сдавливать ее: голова в ответ на это будет поворачиваться и отклоняться в противоположную сторону.
Из опыта со щипком, помимо прочего, следует, что для ориентирующего влияния на голову одностороннее раздражение туловища животного не является единственной причиной и что то же самое может быть вызвано асимметричным раздражением конечностей. Это обстоятельство важно для понимания равновесия стоящего животного.
Если соприкосновение с плоскостью опоры не асимметрично, а симметрично, то в обычных условиях1 поворота или отклонения головы не происходит. Отсюда следует, что если лабиринтэктомированное таламическое животное уложить на стол в положении на спине (рис. 106), то голова остается в затылочном положении, теменем вниз, так же как если бы животное удерживалось в воздухе. Поворот головы из положения на спине осуществим только в том случае, если возможен лабиринтный установочный рефлекс.
В какой степени описанный «установочный рефлекс тела на голову» в состоянии привести в правильное положение тело лабиринтэктомированного таламического кролика, легче всего установить, наблюдая этих животных, когда они пробуждаются после операционного шока и постепенно делают попытки подняться.
Непосредственно после операции животное лежит на боку и в ответ на самые разнообразные раздражения не предпринимает попыток привести голову в нормальное положение. Спустя некоторое время в ответ на симметричное раздражение (щипок хвоста) наступает простое барахтанье
1 После односторонней экстирпации лабиринта дело обстоит иначе.
188
Пятая глава. Установочные рефлексы
(zappeln) в боковом положении. Напротив, если ущипнуть нижнюю (не верхнюю!) переднюю ногу, то голова поворачивается в нормальное положение. Вначале на щипок нижележащей передней ноги реагирует только голова, позднее к этому присоединяются поднимание и усаживание передней половины тела, а в конце концов — также и задней его половины. Животное таким образом или приходит к положению правильно сидящего в нормальной позе, или перекатывается через брюхо на противоположный бок. Когда оно при этом оказывается в боковом положении, то снова перекатывается в первоначальное положение. Может получиться так, что в течение некоторого времени животное переваливается через брюхо с одного бока на другой. Перекатывания через спину не бывает никогда; во всех случаях раньше всего приводится в нормальное положение голова, вслед за чем перекатывается через живот туловище. На последующей стадии пробуждения после шока эта реакция наступает не только в ответ на щипок нижерасполо- женной передней ноги; достаточно симметричного раздражения щипком хвоста для того, чтобы параллельно с односторонним давлением со стороны плоскости опоры осуществились поворот головы и нормаль-
Рис. 115. Тот же таламический кролик с перерезанным зрительным нервом и интактным лабиринтому что и на рис. 98. Животное лежит на столе на левом боку, голова выступает за край стола. На сторону тела, обращенную кверху, наложена дощечка с грузом, которая придерживается пальцем. Голова поворачивается вправо, т. е. в нормальное положение (несмотря на то, что действие одностороннего давления со стороны плоскости опоры компенсируется давлением дощечки, лабиринтный рефлекс осуществляется) .
ное положение и переваливание через брюхо на противоположную сторону. Для этого не требуется никаких искусственных раздражений. Как только животное оказывается в боковом положении, голова поворачивается в нормальное положение, и тело следует за ней. Первое время при этом еще происходит описанное выше переваливание через брюхо с боку на бок, которое обычно приводит к правильной симметричной посадке в нормальном положении. В конце концов животное оказывается в состоянии прямо усаживаться на столе из положения на боку и больше не переворачивается. Если на этой стадии еще раз оставить его в лежачем положении на боку, то легкого стука по столу оказывается в большинстве случаев достаточно, чтобы осуществилось усаживание. Усевшись, животное нередко покачивает головой и телом из стороны в сторону, как это имеет место и у кроликов с интактным большим мозгом в первые дни после двусторонней экстирпации лабиринтов. Как только доска с грузом помещается на обращенную вверх сторону тела животного, лежащего на столе в боковом положении, все упомянутые реакции, благодаря которым голова и тело перемещаются в нормальное положение, исчезают. Наоборот, после удаления доски реакция усаживания тотчас появляется снова.
Проба с доской, разумеется, должна иметь иные последствия у таламического кролика с интактными лабиринтами. Если такое животное уложить на столе в боковом положении и компенсировать давление со стороны стола посредством доски с грузом (рис. 115), голова уверенно поворачивается в нормальное положение, так как лабиринтный установочный рефлекс остался функционирующим.
Анализ установочных рефлексов
189
Из этого опыта, между прочим, видно, что накладывание доски не ведет к общему торможению установочного рефлекса и что поэтому нельзя объяснять выпадение установочного рефлекса у лабиринтэктомированного животного при пробе с доской общим торможением рефлекса.
Все наблюдения на лабиринтэктомированном таламическом животном могут быть воспроизведены также и на кролике с интактным большим мозгом после экстирпации обоих лабиринтов. В воздухе животные совершенно дезориентированы; однако, едва коснувшись пола, они оказываются в состоянии приводить голову в нормальное положение и, кроме того, нормально садиться и бегать.
У кошки установочные рефлексы тела на голову также наблюдаются с большой отчетливостью. В качестве примера могут служить наблюдения {41) Дюссер де Баренна на таламических кошках после экстирпации лабиринтов.
Кошка 1. Спустя полчаса животное пытается привести голову, поворачивая ее, из лежачего левого бокового положения в нормальное; из лежачего правого бокового положения оно приводит голову, поворачивая ее, в нормальное положение, за этим следует усаживание тела, которое тут же опрокидывается в левую сторону.
Через 4 часа. Голова повернута влево; животное опрокидывается влево, после чего оно приводит голову в нормальное положение, поворачивая ее вправо, и усаживается. Из правого бокового положения — тот же результат, что и через первые часы после последней операции. Животное многократно переворачивается через брюхо то вправо, то влево, причем голова всегда предшествует туловищу.
Через 24 часа. При пробе с доской на полу в правом и левом боковых положениях голова остается в соответствующем положении; однако как только доска убирается, голова тотчас принимает нормальное положение и вслед за этим усаживается туловище. В левом и правом боковых положениях в воздухе голова находится в боковом положении, но как только животное укладывается на пол в одно из двух боковых положений, голова тотчас принимает нормальное положение.
Кошка 2. Через 5 часов. Животное часто опрокидывается влево и, наконец, усаживается из левого бокового положения. Из правого бокового положения на столе голова поворачивается на 45° по направлению к нормальному положению. Если при боковом положении животного в воздухе голова находится в боковом положении, то она приводится в нормальное тотчас, как только животное укладывают на стол. При пробе с доской в обоих положениях голова находится в боковом положении, чтобы принять нормальное положение тотчас после удаления доски.
Через 24 часа. Животное усаживается на пол, при этом опрокидывается в левую сторону, но тотчас же поднимается из этого бокового положения, причем голова идет раньше туловища; как только принята нормальная поза, животное опрокидывается вправо. Из этого положения оно снова усаживается, причем голова опять идет первой, достигая нормального положения, и затем снова опрокидывается в левую сторону. Так происходит много раз подряд, т. е. животное непрерывно перекатывается через брюхо с боку на бок, каждый раз проходя через нормальное положение, причем первой всегда движется голова.
Через 48 часов. Во всех опытах точно такие же результаты, что вчера.
Напротив, у обезьян (59) лабиринтные установочные рефлексы развиты •слабо.
При пробуждении после наркоза у обезьяны в отдельных случаях можно установить стадию, когда лабиринтные установочные рефлексы в воздухе еще не осуществляются, но голова приводится в нормальное положение тотчас после того, как животное укладывается на стол в боковое положение. То же самое удавалось видеть у таламических обезьян спустя 13/4 часа после операции, лабиринтный установочный рефлекс к этому времени был еще отрицательным, в то время как при боковом положении (животного) голова поворачивалась в нормальное положение почти полностью. У ла- биринтэктомированной обезьяны с интактным большим мозгом и при закрытых глазах толова при положении тела на боку поворачивалась в направлении к нормальному положению.
Однако все эти наблюдения представляют собой лишь единичные находки. В общем и целом установочные рефлексы с тела на голову у обезь¬
190
Пятая глава. Установочные рефлексы
яны развиты гораздо слабее других установочных рефлексов и играют в сохранении прямого положения тела лишь подчиненную роль.
Сообщенные до сих пор наблюдения над животными с интактными лабиринтами при удерживании первых в воздухе и над лабиринтэктомиро- ванными животными в воздухе и на столе (как с интактным, так и с удаленным большим мозгом) показали прежде всего, что, для того чтобы повернуть голову в «нормальное положение», необходима совместная работа двух групп рефлексов. Это, во-первых, лабиринтный установочный рефлекс и, во-вторых, установочный рефлекс с тела, который осуществляется благодаря асимметричному раздражению поверхности тела. В условиях свободного пребывания в воздухе, после выключения оптических влияний, направляющие влияния на голову могут исходить только из лабиринтов. Как только животное приходит в соприкосновение с плоскостью опоры к возбуждениям, исходящим из лабиринтов, присоединяются еще раздражения с нервов туловища, воспринимающих давление, и с нервов конечностей. Как те, так и другие правильно устанавливают голову в пространстве. Далее, тело следует в направлении движения головы и также достигает нормальной посаДки. О других факторах, играющих при этом роль, будет сообщено более подробно далее.
Установочные рефлексы на тело
1.Шейные установочные рефлексы (24)
При описании поведения таламического кролика с интактными лабиринтами, постепенно пробуждающегося после наркоза и шока, уже указывалось (стр. 170), что раньше всего в нормальное положение приводится голова и что только вслед за этой «правильной установкой» головы следует присоединяющееся усаживание туловища. Зависимость положения тела от нормального положения головы доказывается тем, что телу таламического животного, лежащего на боку после операции, можно придать сидячее положение, пассивно повернув ему голову рукой в нормальное положение. С другой стороны, если таламическое животное сидит в правильном нормальном положении, тело может быть тотчас же приведено в боковое положение, если повернуть голову рукой в это положение (рис. 97, стр. 171).
После пробуждения (животного) от шока можно установить, что это влияние головы на тело происходит спереди назад. На определенной стадии, если голова активно или пассивно повернута из бокового положения в нормальное, в сидячее положение приводится только передняя часть тела, в то время как задняя его половина остается в лежачем положении. Постепенно, по мере улучшения состояния животного, рефлекс распространяется и на заднюю половину тела и все животное принимает нормальную сидячую позу.
Этот рефлекс наступает и тогда, когда голова и тело животного не соприкасаются с плоскостью опоры. На рис. 98 и 99 видно, что когда таз свободно удерживается в воздухе, то благодаря лабиринтному установочному рефлексу в нормальное положение поворачивается не только голова, но за этим поворотом следует и передняя половина тела с передними конечностями, принимая в воздухе также нормальное положение. При этом вне вращения остается только область поясничного отдела туловища, так как таз прочно удерживается рукой. Однако очень сильные животные в состоянии продолжить эту реакцию также и в воздухе и, преодолевая
Анализ установочных, рефлексов
191
сопротивление держащей руки, резким толчком «провернуть» заднюю половину тела в нормальное положение.
Тот же рефлекс можно видеть на рис. 100 (стр. 174), где животное повернуло голову из бокового положения в сторону и передняя часть тела, последовав за этим поворотом, также лежит на боку, в то время как таэ удерживается в положении на спине.
Описанная реакция, как легко убедиться, не наступает вследствие вращательного движения (угловое ускорение) головы, но обусловлена только ее положением. Она наступает с той же скоростью и тогда, когда голова уже в течение продолжительного времени находилась в повернутом по отношению к туловищу положении, и фиксированное при этом туловище более не удерживается. Таким образом, дело здесь не в реакциях с полукружных каналов.
Если учесть еще тот факт, что наступает одна и та же реакция, передней и задней половины тела вне зависимости от того, приведена ли голова из нормального положения в боковое или из бокового в нормальное, или из затылочного в боковое положение, то оказывается, что о тоническом лабиринтном рефлексе вообще нельзя говорить. Это доказывается еще и тем,
Рис. 116. Кролик находится в положении на спине, так что голова, грудь и таз дорсальной стороны обращены точно вниз. Голова поворачивается при этом вправо, т. е. правое ухо движется в вентральном направлении, правый глаз смотрит вверх. Грудь остается лежать в прежнем положении. Левая передняя конечность является «челюстной» и вытягивается, правая передняя конечность («черепная») сгибается. Таз поворачивается в противоположном голове направлении, так что левая ягодица оказывается внизу, левая задняя нога — наверху. Вследствие этого тело животного скручено- штопорообразно. Различия тонуса задних конечностей на этом снимке видеть нельзя.
что у лишенного лабиринтов таламического животного имеется такой же рефлекс. Его можно привести из нормального положения в боковое, поворачивая в это положение голову, а переводя ее из бокового в нормальное положение, можно заставить животное тотчас принять сидячее положение. В этом случае также можно установить, что на более ранней стадии после операции в ответ на правильную установку головы прежде всего наступает усаживание передней половины тела, тогда как позднее правильное «сгорбленное положение» принимает все тело с передними и задними конечностями.
Все эти наблюдения с неизбежностью приводят к заключению, что- именно поворот шеи обусловливает последующий поворот передней части тела, в свою очередь ведущий к повороту и задней его половины. Поэтому рефлекс был обозначен как «шейный установочный рефлекс». Благодаря этому рефлексу, после того как голова приняла нормальное положение- вследствие лабиринтного установочного рефлекса и асимметричного давления со стороны плоскости опоры, тело следует за головой и также принимает нормальное положение.
Этот шейный установочный рефлекс, вызванный поворотом головы,, легко может быть получен у интактных животных, если в положении животного на спине поворачивать его голову. Тогда, помимо соответствующих тонических шейных рефлексов на конечности, можно получить отчетливый поворот туловища благодаря шейному установочному рефлексу.
192
Пятая глаеа% Установочные рефлексы
На рис. 116 изображено животное, лежащее на спине, голова которого вправо (т. е. правое ухо поворачивается в направлении вентральной стороны тела). Различие тонуса передних конечностей выступает отчетливо: левая нога является «челюстной» и вытягивается сильнее, чем правая «черепная». Но помимо тонического рефлекса на конечности, виден также и поворот туловища. Поворот головы вызывает типичное изменение положения таза, которое осуществляется благодаря изменению осевой мускулатуры (Stammesmuskulatur), особенно поясничного ее отдела. Вследствие этого задняя половина тела поворачивается в направлении, противоположном движению шеи. В то время как правый глаз смотрит вверх, правая ягодица находится внизу и лежит на столе, а левая задняя нога — наверху. Эта реакция отчетливо видна и после экстирпации обоих лабиринтов. Поясничный отдел позвоночника продолжает поворот туловища, начавшийся с шеи, так что весь позвоночник спиралеобразно изгибается. Если бы таз был (при этом) фиксирован,
Рис. 117. Тот же таламический кролик, что и на рис. 99. Животное сначала сидело в нормальном положении (рис. 96). Затем при помощи поворота головы в левую сторону было приведено в левое боковое положение (рис. 97). Голова удерживалась в левом боковом положении и в то же время несколько приподнималась. После этого тело животного раздражалось (щипок хвоста). При этом туловище принимало нормальное сидячее положение, в то время как голова оставалась фиксированной в левом боковом положении.
грудной отдел вследствие поворота поясничного отдела следовал бы за поворотом шеи.
Этот шейный установочный рефлекс играет важную роль при установке тела в определенное положение и при движениях перекатывания кролика после односторонней экстирпации лабиринта, когда этот рефлекс появляется вследствие длительного поворота головы (после операции), а благодаря коррекции этого ненормального положения, как это будет подробно описано далее, может в любой момент стать менее интенсивным.
Этот же рефлекс участвует и тогда, когда животное принимает нормальное положение после положения на боку или на спине.
На остальные шейные установочные рефлексы здесь следует только кратко указать. Намечается правило, что у таламического кролика тело, принимая нормальное положение, следует направлению движения головы. Так, например, на рис. 101 (стр. 175), где таз удерживается спинной стороной книзу, видно, что посредством вентрального пригибания в нормальное положение приходит не только голова, но что этому движению следует также и передняя половина тела, в процессе чего производится сильное вентральное изгибание поясничного отдела позвоночника. Если сравнить с этим рис. 102 (стр. 175), то можно видеть, что при повороте таза вокруг поперечной оси до тех пор, пока животное не повиснет головой книзу, вследствие дорсального сгибания шеи не только голова удерживается в нормальном положении, но наступает также сильное вентральное изгибание тазового отдела позвоночника, приводящее к легкому дорсальному изгибу в грудном его отделе.
Анализ установочных рефлексов
193
Шейные установочные рефлексы занимают среди установочных рефлексов особое положение постольку, поскольку центры их лежат в мозговом стволе несколько более кзади, чем центры остальных установочных рефлексов. В то время как все центры последних доходят до переднего края среднего мозга, шейные установочные рефлексы часто сохраняются еще и после удаления значительной части среднего мозга. Радемакер (Rademaker) (см. десятую главу) наблюдал еще шейные установочные рефлексы у кролика и после разреза через задний край заднего двухолмия до середины моста. Однако при этом выясняется также, что центры шейных установочных рефлексов совершенно иные, чем центры тонических шейных рефлексов на конечности, которые, как показано выше, лежат в верхнем шейном отделе спинного мозга.
Точно такие же рефлексы обнаруживаются у таламической кошки и таламической собаки {41), поэтому точное описание их является излишним. У интактной обезьяны в состоянии бодрствования и на определенных стадиях наркоза, как и у таламической обезьяны, имеются шейные установочные рефлексы. Если поворачивать голову в положении животного на спине, то можно видеть, что в реакции таза хвост также принимает участие. У кошки и обезьяны удалось показать, что шейные установочные рефлексы имеются и после двухсторонней экстирпации лабиринтов.
Выше (стр. 183) было определенно показано, что эти шейные установочные рефлексы играют важную роль при переворачивании животного во время свободного падения в воздухе.
2. Установочные рефлексы тела на тело
До сих пор описанные рефлексы, благодаря которым кролик в состоянии принимать нормальное положение тела, имеют однородный механизм постольку, поскольку прежде всего в нормальное положение приводится голова (исходит ли это из лабиринтов или обусловлено асимметричным раздражением поверхности тела), а затем к этому присоединяется соответствующий шейный установочный рефлекс, вследствие которого и тело приводится в правильное сидячее положение.
Повседневный опыт при наблюдении нормальных животных учит, что этим вся совокупность действующих установочных рефлексов не исчерпывается. Ведь сидящее животное в состоянии свободно двигать головой во все стороны и придавать ей различные положения в пространстве, не опрокидываясь при этом. Следовательно, должен существовать еще механизм, с помощью которого тело приводится в сидячее положение или удерживается также и в том случае, когда голова не находится в нормальном положении.
То, что такой механизм существует, с особенной наглядностью видно на кроликах после односторонней экстирпации лабиринта, у которых голова оказывается длительное время повернутой в сторону отсутствующего лабиринта и тело которых тем не менее в состоянии принять правильное, хотя и несколько повернутое в сторону сидячее положение. Способность сидеть не утрачивается у них и после закрывания глаз.
У таламического кролика подобный, не зависящий от головы установочный рефлекс также может быть обнаружен.
Если спустя более длительное время после операции, когда явления шока преодолены, таламических кроликов с помощью пассивного поворота головы привести в боковое положение (рис. 97, стр. 171), то в отдельных случаях можно видеть, что тело удерживает это положение недолго. Особенно при фиксации головы рукой в боковом положении не непосредственно на полу, но несколько выше пола (рис. 117), часто происходит
13 Р. Магнус
194
Пятая глава. Установочные рефлексы
внезапное усаживание туловища, несмотря на то, что голова находится не в нормальном, а в боковом положении.
При этом раздражение обеспечивается благодаря асимметричному возбуждению чувствительных нервов тела вследствие давления со стороны плоскости опоры. Это следует, во-первых, из того, что описанная реакция наблюдается и у лабиринтэктомированного таламического кролика, и, во-вторых, из того, что наступлению ее можно с определенностью воспрепятствовать, если на сторону тела, обращенную вверх, наложить дощечку с грузом в 1 кг и этим компенсировать одностороннее давление плоскости опоры на сторону тела, обращенную вниз.
Смотря по степени отдыха животных после шока, рефлекс возникает в ответ на раздражения различной интенсивности. Когда животное приведено в боковое положение посредством поворота головы в сторону, часто оказывается необходимым усилить одностороннее раздражение стороны тела, обращенной книзу, тем, что дополнительно произвести щипок нижерасположенных передней или задней конечностей. При этом туловище усаживается, в то время как щипок одной из вышерасположенных конечностей ведет лишь к барахтанью на боку. В других случаях, как например у таламического кролика, изображенного на рис. 117, тело вне раздражения лежит на боку; достаточно, однако, простого симметричного раздражения (щипок хвоста), чтобы реализовалось одностороннее давление плоскости опоры. В благоприятных случаях не требуется никакого экстренного раздражения и тело непосредственно принимает правильное сидячее положение, несмотря на боковое положение головы.
Причина, по которой этот рефлекс непостоянен или приводится в действие специальным раздражением, отчасти лежит несомненно в том, что в описанных условиях опыта он должен проявляться, противодействуя шейному установочному рефлексу, который стремится удержать тело в боковом положении. Поэтому его удается демонстрировать только у хорошо возбудимых животных.
У нормальных кроликов с сохранным большим мозгом этот рефлекс почти всегда обнаруживается с большой отчетливостью.
Из изложенного следует, что у животного, лежащего на полу в боковом положении, одностороннее давление плоскости опоры на тело оказывает двойственное действие. Во-первых, благодаря ему голова рефлекторно поворачивается в нормальное положение и этот поворот головы в свою очередь вызывает при посредничестве шейных установочных рефлексов приведение туловища в сидячее положение. Во-вторых, благодаря прямому рефлексу тело само приводится в это же положение.
Такой же рефлекс с большой отчетливостью обнаруживается у таламической кошки и таламической собаки (41). У лабиринтэктомированной таламической кошки он вызывался также быстро.
У обезьян установочный рефлекс тела на тело тоже развит очень сильно. Легчайшего соприкосновения с полом или с решеткой потолка клетки во время лазанья оказывается достаточно для того, чтобы оказать выправляющее (richtend) влияние на тело, что особенно отчетливо выражено у лабиринтэктомированных животных.
Наблюдения за обезьянами следует рассмотреть несколько подробнее, так как у таламической обезьяны в состоянии шока развитие этого рефлекса протекает с особенной отчетливостью.
Если такое животное уложить на столе в боковом положении, то нижележащая задняя конечность выпрямляется, лежащая выше — сгибается; на той же стороне совершается поворот таза в направлении к нормальному положению, что ясно видно на рис. 118 и 119.
Анализ установочных рефлексов
195
У мартышковых (Cercopithecus), помимо прочего, в установочном рефлексе тела также принимает участие хвост, причем он перебрасывается в вентральном направлении и таким образом перемещает центр тяжести в направлении необходимого поворота. Наоборот, у макаки хвост не реагирует, это обстоятельство согласуется с тем, что макака и в интактном состоянии, принимая и удерживая нормальное положение тела, мало пользуется хвостом. Названная реакция у децеребрированно- го животного отсутствует, но имеется у таламической обезьяны.
Рис. 118. Cercopithecus pentaurista. Таламическая обезьяна. 4% часа после операции. Начинающееся усаживание на столе из левого
бокового положения. Левая задняя нога выпрямлена, правые передняя и задняя ноги согнуты. Хвост перекинут вправо. Поворот тела в нормальное положение. Голова еще отчасти в боковом положении.
Таким образом, и в этом случае мы сможем локализовать положение центров в среднем мозгу. Реакция не зависит от положения головы в пространстве и появляется как при боковом, так и при нормальном положениях головы. Рецепторы для рефлекса располагаются сбоку на туловище
и на бедре, и необходимо довольно обширное асимметричное возбуждение рецепторов. Простого надавливания на бедро или только на боковую поверхность туловища недостаточно.
При пробуждении после наркоза в боковом положении животно-
Рис. 119. Macacus Cynomolgus. Таламическая обезьяна. 4У2 часа после операции. Начинающееся усаживание на столе из левого бокового положения. Левая задняя нога разогнута, правые передняя и задняя ноги согнуты. Голова и туловище перевернуты в направлении к нормальному положению. Хвост в реакции усаживания не участвует.
го реакция усаживания у обезьяны вначале не проявляется. Далее, раньше прочего, наступает выпрямление одной и сгибание другой задней конечностей, без других изменений положения тела на столе. По мере дальнейшего исчезновения наркоза таз поворачивается в нормальное положение, в то время как плечи еще лежат. В этой стадии иногда производится поворот головы по направлению к нормальному положению (или с лабиринтов или благодаря установочному рефлексу с тела). Далее, к начальной реакции задних конечностей присоединяется перемена положения передних конечностей и в конце концов наступает переворачивание в нормальное положение всего тела, т. е. реакция усаживания, которая осуществляется еще настолько медленно и постепенно, что без труда можно наблюдать ее шаг за шагом в различном темпе.
Чем полнее пробудилось животное после наркоза, тем быстрее разворачивается вся последовательность явлений, так что при усаживании неудерживаемого животного преемственности отдельных актов установить невозможно. Если такое животное уложить на столе на бок и удерживать
13*
196
Пятая глава. Установочные рефлексы
голову в боковом положении, тело приводится в сидячее положение с большой быстротой, но и здесь еще можно чаще всего увидеть, что реакция начинается с заднего конца тела и что таким образом начинают движение задние конечности. В конце концов у нормального животного усаживание тела происходит молниеносно и тогда, когда голова удерживается в боковом положении, так что различить отдельные этапы этой реакции уже не представляется возможным. Помимо столь определенно выраженной реакции задних конечностей у таламической обезьяны, у обезьяны с сохраненным большим мозгом выявляется еще и обратная реакция, когда раньше сгибается и подтягивается под туловище нижерасположенная задняя конечность, за этим следует поворот таза. Задняя конечность, расположенная сверху, выпрямляется и отводится в сторону, оказываясь таким образом подошвой на столе. Теперь животное этой стоящей ногой перемещает таз, в то время как первоначально нижележавшая задняя нога вытягивается сильнее, вследствие чего таз принимает нормальное положение. У интактных животных можно видеть обе эти реакции сменяющими друг друга, хотя до сих пор не удалось установить причину, благодаря которой доминирует то одна, то другая из этих реакций.
После двусторонней экстирпации лабиринтов описанный установочный рефлекс тела на тело естественно остается без изменения и закрывание глаз также не вызывает никакой разницы.
На обезьянах с обоими удаленными лабиринтами и при закрытых глазах можно было показать еще следующий прекрасный установочный рефлекс. На 14-й день после экстирпации лабиринтов сшивались веки. Если животное удерживалось за хвост в воздухе головой вниз, то тело свешивалось вертикально вниз и голова в большинстве случаев находилась в затылочном положении. Если теперь животное опускать вниз, то на прогрессивные движения никакой реакции не наступает.
Как только темя животного касалось пола, голова и тело тотчас принимали нормальное положение и именно таким образом, что раньше выпрямлялась и касалась пола одна рука, к чему в дальнейшем присоединялась реакция остального тела. Внимательное наблюдение на другой день показало, что когда животное касается теменем пола, сначала одна, например правая, рука вытягивается по направлению к полу; как только она коснулась пола, осуществился рефлекс усаживания, во время чего голова повернулась также в правую сторону, вслед за чем животное перевело на эту сторону и тело, а другая, левая рука произвела шагатель- ное движение. Таким образом были произведены прежде всего выпрямление правой руки, прикосновение руки к полу, дорсальное нагибание головы, переход головы в нормальное положение, поворот и отклонение головы вправо, первое шагательное движение левой рукой, поворот животного вправо. Эта последовательность рефлексов воспроизводилась в одном и том же виде многократно и закономерно. Самым существенным при этом является несомненно асимметричное возбуждение, обусловленное соприкосновением одной руки с полом.
Роль установочных рефлексов тела выявляется весьма отчетливо, если наблюдать лабиринтэктомированную обезьяну в клетке. В то время как удерживаемое в воздухе животное полностью дезориентировано, по полу клетки оно передвигается с большой уверенностью и ловкостью. Однако ловкость животного становится еще большей, если оно забирается на решетчатую крышу клетки. Очевидно, возбуждений, исходящих из четырех рук, достаточно для того, чтобы точно ориентировать голову и тело животного. Правда, в этом случае (лазание по крыше) голова Находится в положении, отличающемся от нормального на 180°, и почти всегда
Анализ установочных рефлексов
197
удерживается в затылочном положении. Однако движения животного исключительно уверенны и быстры, и если после операции прошло некоторое время, животное никогда не оступается и не падает.
Заключение
Из предыдущего следует, что у всех таламических животных, исследованных до сих пор, взаимодействует целый ряд различных рефлексов, позволяющих животному принимать и удерживать нормальное положение тела. Оптические возбуждения при этом существенной роли не играют. Скорее всего существуют две группы раздражений, которые главным образом вызывают описанные реакции: во-первых, лабиринтные раздражения и, во-вторых, возбуждения, зависящие от асимметричного раздражения чувствительных нервов тела со стороны плоскости опоры, когда тело выведено из нормального положения.
Благодаря этим обоим различным «раздражениям положения» (Lagereize) голова рефлекторно устанавливается в нормальном положении. Чтобы в то же положение привести туловище и конечности, необходимо снова одновременное действие двух факторов. Во-первых, если голова находится, а туловище еще не приняло нормального положения, имеющееся при этом ненормальное положение шеи вызывает рефлекс, который приводит тело в нормальное положение. И, во-вторых, асимметричное раздражение чувствительных нервов тела со стороны плоскости опоры непосредственно вынуждает тело принять нормальное положение. Одностороннее давление плоскости опоры на тело обусловливает нормальное положение, стало быть, двояким образом: во-первых, влияя на правильность положения головы и, во-вторых, непосредственно приводя тело в сидячее положение.
В этом случае, как и во многих других, когда речь идет о жизненно важных функциях тела, появляется «забота», чтобы осуществление функции страховалось дублированием ее и чтобы при заболевании или выпадении одного механизма, имелся бы другой, который обеспечивал бы осуществление этой функции — в нашем случае — положения тела.
Если таламическое животное свободно удерживается в воздухе, то только одни лабиринты в состоянии поддерживать нормальное положение тела. Если же таламическое животное находится на земле, все вышеназванные рефлексы, действуя совместно, гарантируют сохранение нормального положения.
Четыре названных рефлекса положения осуществляются у таламического животного одновременно, давая возможность при всех обстоятельствах принимать и удерживать нормальное положение.
Теперь встает вопрос, действительно ли описанные до сих пор четыре группы рефлексов, обнаруженные в приведенных опытах у таламических животных, являются также и единственно существенными, совместное действие которых обеспечивает нормальное положение животного, или же мы вправе ожидать, что у бесполушарных животных могут быть обнаружены еще и другие установочные рефлексы. Следующее соображение говорит за то, что установочные рефлексы таламического животного проделанными исследованиями в существе своем действительно исчерпываются. Известно, что лабиринтэктомированные животные в воде оказываются полностью дезориентированными, тогда как в обычных условиях (т. е. при соприкосновении с плоскостью опоры) они поддерживают положение тела с большой уверенностью. Вода окружает все тело оболочкой, равномерно действующей на его поверхность, и тогда асимметричное
198
Пятая глава. Установочные рефлексы
возбуждение чувствительных нервов тела при различных положениях (его) в пространстве не может осуществиться. У лабиринтэктомирован- ного животного в этих условиях отнимается всякая возможность принять нормальное положение, и оно беспомощно идет ко дну. Отсюда с большой вероятностью следует, что после выключения раздражений из лабиринтов и асимметричных возбуждений, возникающих вследствие соприкосновения с плоскостью опоры, действительно не существует других, сколько- нибудь действенных механизмов, гарантирующих нормальное положение тела.
Уже неоднократно указывалось на то, что интактный кролик с сохранным большим мозгом не отличается в отношении своих установочных рефлексов от таламического кролика, так что приведенное выше описание остается верным и для нормального животного. Интактный кролик не имеет оптических установочных рефлексов и не использует впечатлений своих глаз для того, чтобы принимать и удерживать положение тела. У филогенетических более высоко стоящих животных дело обстоит иначе.
Расположёние центров установочных рефлексов
Большинство описанных до сих пор наблюдений было получено на таламических кроликах, так как теплорегуляция у этих животных не нарушается, благодаря чему их без труда удается сохранить живыми в течение нескольких дней. Таким образом, оказывается возможным по миновании всех явлений шока повторно изучать в течение длительного времени функции каждого отдельного животного. При этом, однако, возникает вопрос, для всех ли описанных выше рефлексов у таламического животного, прежде всего для рефлексов установочных, необходимо наличие сохранной центральной нервной системы вплоть до передней стенки таламуса или для этого уже достаточно более каудально расположенных центральных образований. Для решения этого вопроса были поставлены опыты на четверохолмных (мезенцефалическое животное) и децеребрирован- ных (мосто-мозжечковое и мозжечково-бульбарное животное) кроликах, о которых будет сообщено в дальнейшем.
Выяснилось, что за осуществление установочного рефлекса следует считать ответственным главным образом средний мозг.
1. Наблюдения за м е з е н ц е ф а л ич е с ким кроликом
Ранее мною было осуществлено семь опытов (24), в которых мозговой ствол перерезался по переднему краю среднего мозга. Методика операции и направление разреза были описанывыше (стр. 161—162). Препарат изображен на рис. 93—95. Наблюдения над большинством животных производились только в день операции. Многочисленные дальнейшие исследования в последнее время произведены Радемакером (Rademaker).
В целях упрощения изложения вначале будут приведены сокращенные протоколы двух наиболее удачных старых опытов. Результаты наблюдений над остальными животными в основных чертах совпадают с приводимыми.
Опыт 31. Кролик, 1000 г. 5 мая 1915 г. Хлороформный наркоз. Перевязаны сонные артерии. Блуждающие нервы перерезаны. Большой мозг и зрительные бугры удалены. Разрез проходит тотчас перед четверохолмием, косо вперед и вниз. Кровотечение минимально.
10 ч. 45 м. Окончание операции. Вскоре после этого самостоятельное дыхание.
И ч. 45 м. Сидит симметрично, поднимает голову с пола. Будучи приведен в боковое положениё, продолжает лежать и делает только слабые попытки повернуть го¬
Анализ установочных рефлексов
199
лову в нормальное положение. Если голова пассивно приводится в нормальное положение, туловище остается лежать на боку. Однако если при этом ущипнуть одну из четырех конечностей, тело принимает сидячее положение. Если животное удерживать в висячем положении в воздухе головой вверх или головой вниз, последняя поворачивается в нормальное положение. Если животное усадить на пол и ущипнуть за обе задние ноги, оно делает несколько нормальных прыжков и затем хорошо сидит. На вертящейся плоскости — очень тонкие реакции поворота головы и постреакции поворота головы, реакции поворота глаз, смотря по направлению вращения — в каудальном, назальном, дорсальном и вентральном направлениях. Компенсаторные повороты глаз при различных положениях головы в пространстве. Двусторонний рефлекс смыкания век. При поднимании головы наступает выпрямление, при опускании — сгибание передних конечностей. Температура 34° С. Трахеальная канюля удалена, трахея зашита. При шпике лапки сильный крик.
2 ч. 30 м. Животное хорошо сидит в клетке. Правильное дыхание. Будучи посажено на пол, в ответ на раздражение скачет до половины экспериментального зала и затем спокойно сидит, долго кричит, появляется слышимое стенозированное дыхание, далее ведет себя снова нормально. На вращающейся плоскости, помимо реакций, наблюдавшихся в И ч. 45 м., глазной нистагм и глазной поствращательный нистагм.
В висячем положении в воздухе головой вниз или вверх голова приводится полностью в нормальное положение. В боковом положении это не вполне удается.
На полу сидит безупречно, голова и брюхо выше пола. Из бокового положения усаживается теперь самостоятельно.
Если при боковом положении животного на столе на сторону тела, обращенную вверх, положить дощечку с грузом, голова, несмотря на это, принимает нормальное положение.
Если удерживать животное за задние ноги свободно в воздухе в «ножном положении» (Fußstellung) или в боковом положении, голова устанавливается в пространстве правильно. Никакой децеребрационной ригидности. Распределение тонуса в мускулатуре конечностей «нормально».
4 часа. Если животное свободно удерживать в воздухе за таз (ножное положение, оба висячих и боковое положения), то голова приходит в нормальное положение. Из затылочного голова поворачивается в сторону. Жевательный рефлекс, если палец засовывается в рот животному, слабо положителен.
4 ч. 30 м. Спонтанно усаживается из бокового положения. Если, как исключение, усаживание осуществляется несовершенно (т. е. только при участии головы и передних конечностей), то задние конечности принимают сидячее положение в ответ на любое раздражение.
В ответ на раздражение прыгает по комнате. Если при этом животное падает на бок, оно тотчас же спонтанно снова принимает сидячее положение.
Животное живет еще и на следующий день и обнаруживает в основном те же реакции. Утром 7 мая оно погибло.
Секционные данные (стр. 162). Зарисовки препарата — рис. 93—95. Разрез проходит дорсально — на 1—2 мм кпереди от переднего двухолмия, вентрально — позади заднего края сосочкового тела, на левой стороне — на 2 мм перед задним краем медиального коленчатого тела, на правой — точно по заднему краю медиального коленчатого тела. Плоскость разреза оказывается между плоскостями рис. 16 и 17 атласа Винклера—Поттера (2).
Опыт 36. Кролик, 1200 г. 12 мая 1915 г. Хлороформный наркоз. Сонные артерии перевязаны. Блуждающие нервы интактны. Большой мозг и таламус удалены. Разрез проходит на 1—2 мм перед четверохолмием, косо вперед и вниз. Кровотечение незначительно.
9 ч. 50 м. Окончание операции. Спонтанное дыхание. Децеребрационная ригидность едва проявляется. Удалена трахеальная канюля. Трахея зашита.
10 ч. 35 м. Состояние хорошее. Никакой децеребрационной ригидности. Распределение тонуса в мускулатуре тела «нормально»,.
При лежачем положении животного на столе спонтанно или при прикосновении происходит устанавливание головы в правильное положение, а в отдельных случаях и усаживание передней половины тела. Если животное удерживается в боковом положении в воздухе, тоже наступает легкий поворот головы по направлению к нормальному положению. В положении животного на спине голова также поворачивается в сторону.
Если в боковом положении животного голову пассивно привести в нормальное положение, туловище усаживается сначала передней, затем задней частью. В ответ на поднимание головы наступает разгибание, на опускание головы — сгибание конечностей. Поворачивание головы в лежачем положении вызывает отчетливые тонические шейные рефлексы на передние конечности; поворачивание головы в боковом положении тела — тонические лабиринтные рефлексы на передние конечности.
200
Пятая глава. Установочные рефлексы
При пассивном сгибании головы вокруг сагиттальной оси наступает соответствующие компенсаторные положения глаз. На вращающейся поверхности — очень легкая вращательная и послевращательная реакции головы, вращательная и после- вращательная реакции глаз, вращательный и послевращательный глазной нистагм. Выраженные псевдоаффективные рефлексы (крик). Двусторонний рефлекс смыкания век.
10 ч. 50 м. Если животное свободно удерживается в воздухе за таз или задние ноги, правильное устанавливание головы наступает теперь весьма отчетливо.
Если голову удерживать на столе в боковом положении, туловище остается в боковом положении; если отпустить голову, наступает правильное устанавливание головы и усаживание передней части тела.
И ч. 10 м. Лабиринтные установочные рефлексы отчетливы при всех положениях в воздухе.
Жевательный рефлекс при засовывании животному пальца в рот имеется.
11 ч. 40 м. Если животное усадить на пол и ущипнуть его за задние конечности, оно безупречно совершает побежку по комнате и затем сидит спокойно.
Спонтанно осуществляет усаживание из бокового положения передней и задней частью тела.
2 ч. 15 м. Очень хорошо спонтанно усаживается из бокового положения. Лабиринтные установочные рефлексы при всевозможных положениях тела в воздухе теперь вполне выявляются. Из затылочного положения голова поворачивается более чем просто в сторону.
Если на столе в положении животного на спине голову его удерживать в боковом положении, туловище также остается в боковом положении. Если теперь ущипнуть одну из задних конечностей, то туловище (несмотря на боковое положение головы) принимает сидячее положение; щипок задней конечности, лежащей ниже, действует при этом более надежно, чем конечности, лежащей выше.
Поднимание и опускание головы оказывают весьма действенное влияние на тонус передних конечностей. Поворот головы в положении животного на спине ведет к интенсивным шейным тоническим рефлексам на передние конечности. Поворот головы в боковом положении вызывает комбинацию тонических шейных и лабиринтных рефлексов, причем последние преобладают. Если при положении животного на боку повернуть ему голову челюстью вниз, наступает активное движение передних конечностей.
Если при боковом положении (животного) на столе поместить на сторону тела, обращенную кверху, дощечку с грузом, голова, несмотря на это, приходит в нормальное положение и в том случае, когда она выступает за пределами поверхности стола.
Если животное свободно удерживать в воздухе в боковом положении за таз, голова находится в нормальном положении, тогда как грудь повернута на 45°; передняя «челюстная» конечность при этом выпрямлена сильнее, чем передняя «черепная».
4 часа. Если уложить животное на пол в положении на спине, оно тотчас переворачивается в нормальное сидячее положение. Распределение тонуса мускулатуры конечностей «нормально». Температура 35° С.
Результат. Поведение точно такое же, как у таламического животного (исключая оптические рефлексы и теплорегуляцпю).
Опыт продолжается, в процессе чего при помощи удаления среднего мозга животное децеребрируется.
Секция. Первый разрез проходит по дорсальной стороне слева на 1 мм, справа — на 1.5 мм кпереди от переднего края четверохолмия. На вентральной стороне он проходит на 2 мм кпереди от заднего края сосочкового тела. Латерально он проходит справа на 1.5 мм, слева — тотчас перед передним краем внутреннего коленчатого тела.
Разрез в этом опыте проходит, следовательно, несколько оральнее, чем в опыте 31. Он укладывается примерно в плоскость рис. 16 атласа Винклера—Поттера.
Приведенные протоколы и остальные опыты, находясь между собой в полном соответствии, учат, что, исключая оптические рефлексы и тепло- регуляцию, четверохолмный кролик еще способен к тем же функциям, какие описаны для таламического кролика. В частности, способность удерживать нормальное положение тела еще полностью сохранена.
В деталях установлено следующее.
Сразу после операции животное лежит на боку, однако вскоре начинает поворачивать голову в нормальное положение, сначала в ответ на раздражение, позднее спонтанно, сначала только несовершенно, позднее —
Анализ установочных рефлексов
201
вполне успешно. К этому через некоторое время присоединяется усаживание тела, что происходит сначала в ответ на раздражение, позднее — спонтанно. В промежуточной стадии часто наступают устанавливание головы прямо и усаживание передней половины тела, тогда как задняя его половина еще продолжает лежать в боковом положении и принимает сидячее положение только в ответ на раздражение.
В конце концов животное сидит совершенно нормально, голова в нормальном положении; затылочные мышцы удерживают голову, тонус конечностей достаточен, чтобы удерживать брюхо' нормальным образом над землей.
Спонтанные движения спокойно сидящего животного настолько же редки, как у животного таламического. На любое раздражение (щипок хвоста или лапы) четверохолмное животное или делает шаг, или производит ряд правильных прыжков, пробегает в различных случаях разное расстояние. Во время прыганья и беганья равновесие тела поддерживается вполне совершенно. Также и при соскакивании со стола на пол одно из животных правильно приземлилось на ноги и тотчас же побежало дальше.
Лабиринтные установочные рефлексы у четверохолмного животного развиты совершенно так же, как у таламического. Если животное в различных положениях удерживать в воздухе, голова его всегда движется по направлению к нормальному положению. Повторного описания не требуется, достаточной является ссылка на рис. 98—102. Если четверохолмное животное удерживать в боковом положении в воздухе за таз или за задние ноги и переворачивать его из одного бокового положения в другое, голова прочно удерживается в пространстве. Так же обстоит дело, если переворачивать животное в воздухе из положения головой вверх в положение головой вниз. Если животное уложить на стол в боковом положении и на сторону тела, обращенную кверху, поместить дощечку с грузом, голова приводится в нормальное положение так, как это изображено на рис. 115 (стр. 188) для таламического животного.
Установочный рефлекс с тела на голову после асимметричного раздражения поверхности тела у четверохолмного животного выражен также хорошо. Если животное в течение недолгого времени лежит на столе в боковом положении, оказывается достаточным щипка нижерасположенной передней конечности, чтобы вызвать поворот головы в нормальное положение. Если животное фиксировать в нормальном положении в воздухе и ущипнуть при этом одну из передних конечностей, также происходит поворот головы в другую сторону.1
К правильной установке головы благодаря посредничеству шейных установочных рефлексов присоединяется установка нормального положения тела. Шейные установочные рефлексы лучше всего видны при исследовании животного в воздухе. Например, если удерживать в воздухе четверохолмного кролика в боковом положении, то голова стоит в нормальном положении и передняя половина тела поворачивается по направлению к нормальному положению примерно на 45° или более (ср. рис. 99, стр. 173). Если спустя некоторое время после операции лабиринтные установочные рефлексы на голову еще отсутствуют и животное спокойно лежит на столе на боку, часто достаточно пассивно привести голову в нормальное положение, чтобы тотчас вызвать усаживание передней части тела или всего животного.
1 Полную эффективность установочных рефлексов с тела на голову можно обнаружить у мезенцефалического животного, лишенного лабиринтов (Радемакер).
202
Пятая глава. Установочные рефлексы
Точно так же у четверохолмного животного можно обнаружить установочный рефлекс на тело вследствие асимметричного раздражения его поверхности. Если, поворачивая голову в сторону, привести животное, сидящее в нормальном положении, в боковое положение (рис. 97, стр. 171), то часто оказывается возможным, ущипнув нижележащую переднюю конечность, вызвать усаживание туловища, несмотря на боковое положение головы (рис. 117, стр. 192).
Отсюда следует, что у четверохолмного животного можно обнаружить все установочные рефлексы, которые найдены у таламического кролика и которые проявляются таким же образом, способствуя поддержанию нормального положения тела. В отдельных исследованиях обычно видно, что у четверохолмного животного установочные реакции осуществляются менее быстро и менее уверенно, чем у животного таламического. Я, однако, не думаю, что из этого можно заключить, что будто промежуточный мозг у кролика принимает участие в осуществлении установочных рефлексов. Ведь таламические животные находились под наблюдением всегда несколько дней, тогда как четверохолмные животные, с единственным исключением (опыт 31, стр. 198), могли быть исследованными только в день операции, а в день операции и таламические животные обнаруживают не лучшие показатели. Впрочем, опыт 36 (стр. 199) показывает, до какой точности могут развиваться установочные рефлексы также и у четверохолмного животного.
Опыты позволяют поэтому сделать вывод, что способность принимать и удерживать нормальное положение тела сохраняется у кролика после удаления мозга до переднего края среднего мозга.
Этим опровергается мнение Лонгэ и Христиани (Longet u. Christiani), что центры, обеспечивающие поддержание равновесия тела у кролика, лежат в промежуточном мозге. Об опытах, устанавливающих более точную локализацию центров, будет сообщено в десятой главе.
2f Наблюдения за децеребрированными кроликами
(мосто-мозжечковое животное и мосто-бульбарное
животное)
После отделения среднего мозга от продолговатого и моста способность принимать и поддерживать положение тела исчезает. После отделения среднего мозга животные обнаруживают, во-первых, децеребра- ционную ригидность, во-вторых, описанные в третьей главе тонические шейные и лабиринтные рефлексы на мускулатуру тела, реакции поворота головы, в отдельных случаях также вращательный нистагм головы.
Напротив, установочные рефлексы у них отсутствуют — способность принимать и удерживать нормальное положение тела утеряна. Конечно, децеребрированное животное можно поставить на его тонически выпрямленные ноги, после чего в течение некоторого времени оно остается стоящим, пока само или в результате легкого толчка не опрокинется и не останется лежать. Оно не в состоянии противодействовать и воспрепятствовать падению с помощью движений или изменений положения. Оно не пытается освободиться из бокового или из других ненормальных.положений, а остается в этих положениях, пока не будет из них выведено. Бег и прыжки невозможны, хотя координированные движения четырех конечностей осуществимы и последние вследствие децеребрационной ригидности в состоянии выдержать вес тела, однако животное после первого шага или прыжка опрокидывается и только безрезультатно барахтается в боковом положении.
Анализ установочных рефлексов
203
При более подробном исследовании выясняется, что такое поведение обусловлено тем, что у децеребрированного кролика отсутствуют установочные рефлексы.
Ниже это будет обосновано полнее.
а) Лабиринтные установочные рефлексы
Поведение децеребрированных кроликов, которые свободно удерживаются в воздухе в различных положениях, показано на следующих рисунках.
На рис. 120 видно, что если задняя часть тела децеребрированного кролика удерживается в воздухе в боковом положении, то голова в нормальное положение не поворачивается.
Вследствие этого не наступает характерной позы, какая обнаруживается у таламического кролика в таком же положении его в воздухе (рис. 98 и 99, стр. 173).
Голова и передняя часть тела, подчиняясь силе тяжести, просто свисают вниз.
Если заднюю часть тела повернуть из одного бокового положения в другое, голова пассивно следует этому движению и не удерживается принудительно, как у таламического животного, в нормальном положении.
Рис. 120. Опыт № 45. Децеребрированный кролик (мозжечково-бульбарное животное). Животное рукой свободно удерживают в воздухе за таз.
Голова и передние лапы находятся также в боковом положении. Голова и шея, следуя силе тяжести, свисают книзу.
Голова и передняя часть тела не приняли в воздухе нормального положения.
Сравнение с рис. 98 и 99 показывает, что после децеребрации лабиринтный установочный рефлекс на голову отсутствует. Положение то же, что на рис. 105, где изображено лишенное лабиринтов таламическое животное.
Секция. После рассечения мозжечка по длине видно, что четверохолмия полностью отсутствуют. Разрез проходит дорсально с обеих сторон кпереди от средних мозжечковых ножек. Вентрально он идет впереди трапециевидного тела. Справа мост отсутствует; слева задние отделы его сохранились на протяжении до 3 мм. 8-е нервы
интактны.
Из рис. 121 видно, что если исследовать децеребрированного кролика в положении на спине в воздухе или на столе, голова и передняя половина тела остаются в этом положении и не поворачиваются, как у таламического животного, в сторону (ср. рис. 100, стр. 174). Лабиринтный установочный рефлекс на голову также отсутствует. Если повернуть децеребри- рованное животное из положения на спине на один или другой бок, голова пассивно следует этому движению.
Рис. 122 показывает децеребрированного кролика в подвешенном положении головой вверх. Животное за таз удерживается в воздухе. Голова запала назад и пригнулась дорсально. Лабиринтный установочный рефлекс, который у таламического животного приводит голову в нормальное положение, отсутствует.
На рис. 123 показан висящий головой вниз кролик, децеребрированный кпереди от заднего двухолмия и кпереди от моста. Голова свисает мордой отвесно вниз и не приводится, как у таламического животного, в нормаль¬
204
Пятая глава. Установочные рефлексы
ное положение (рис. 102, стр. 175). Лабиринтный установочный рефлекс отсутствует.
Если перевернуть децеребрированного кролика из висячего положения головой вверх (рис. 122) в висячее положение головой вниз (рис. 123)г
голова пассивно следует этому движению и не- удерживается в нормальном положении.
Рисунки 120—123 свидетельствуют о том, что у децеребрированного кролика лабиринтные установочные рефлексы отсутствуют.
При сравнении этих рисунков с рис. 106— 108, где мы видим безлабиринтных таламических животных, заметно, что после обеих операций при исследовании в воздухе обнаруживается точно такое же поведение; рисунки, изображающие соответствующие друг другу поло-
Рис. 121. Тот же децеребрированный кролик (мозжечково-бульбарное животное), что и на рис. 120. Животное рукой удерживают за таз в воздухе в положении на спине. Голова и передняя части тела также находятся в этом положении, т. е. не повернулись в сторону. Сравнение с рис. 100 показывает, что после деце- ребрации лабиринтный установочный рефлекс на голову отсутствует. Положение — то же, что на рис. 106, где изображено- лишенное лабиринтов таламическое животное.
жения, почти полностью идентичны.1 Причина одна и та же: отсутствие лабиринтных установочных рефлексов на голову. Только у безлаб>иринт- ного таламического животного это обусловлено удалением воспринимающего органа; у децеребрированного животного — удалением центров, необходимых для осуществления рефлекса.
Центры лабиринтного установочного рефлекса лежат в среднем мозге.
Рис. 122. Тот же децеребрированный кролик (мозжечково-бульбарное животное), что и на рис. 120. Животное рукой свободно удерживают в воздухе за таз, головой кверху. Голова запрокинута назад, теменем вниз, т. е. не удерживается в нормальном положении.
Сравнение с рис. 101 показывает, что после децеребрации установочный лабиринтный рефлекс на голову
отсутствует. Положение то же, что и на рис. 107, сделанном с таламического животного, лишенного лабиринтов.
б) Установочные -рефлексы тела на голову
Если положить на стол лабиринтэктомированного таламического кролика, у которого (при исследовании) в воздухе не обнаруживается никаких
1 Только на рис. 120—123 у децеребрированного животного ригидность передних конечностей выражена отчетливей, чем у таламического животного (рис. 105-108).
Анализ установочных рефлексов
205
установочных рефлексов, то благодаря асимметричному раздражению поверхности тела он тотчас приводит голову в нормальное положение {рис. 113, стр. 186) и затем усаживается. Только когда одностороннее раздражение плоскости опоры компенсируется наложенной дощечкой, голова и тело остаются в боковом положении (рис. 114, стр. 186).
Наоборот, если на стол уложить в боковом положении децеребрированное животное, оно остается спокойно лежащим в этом положении и не делает попытки повернуть голову в нормальное положение (рис. 124).
Отсутствует установочный рефлекс тела на асимметричное раздражение поверхности тела.
Отсюда следует, что установочный рефлекс с тела на голову обусловлен сохранностью среднего мозга.
Рис. 123. Опыт 44. Кролик. Децеребрация до уровня заднего двухолмия. Животное рукой свободно удерживают за позвоночник в воздухе, головным концом вниз. Голова свешивается, следуя силе тяжести, вниз, т. е. в нормальное положение не приводится.
Сравнение с рис. 102 показывает, что после деце- ребрации лабиринтный установочный рефлекс на голову отсутствует. Положение то же, что на рис. 108, сделанном с таламического животного, лишенного лабиринтов. Секция. Разрез проходит на дорсальной стороне кпереди от заднего двухолмия (справа осталась еще незначительная полоска переднего двухолмия), на вентральной стороне — по переднему краю моста. 8-е нервы интактны.
После выключения среднего мозга кролик оказывался не в состоянии, находясь на столе в боковом положении, повернуть голову в нормальное положение. В ответ на асимметричное раздражение (щипок хвоста) рефлекс не появлялся. Щипок нижерасположенной передней конечности еще вызывает поворот или наклон головы в другую сторону, когда сохранены са-
Рис. 124. Тот же децеребрированный кролик (мозжечково-бульбарное животное), что и на рис. 120. Животное лежит на столе в правом боковом положении. Голова и передняя часть тела также находятся в правом боковом положении, т. е. не повернулись в направлении к нормальному положению. Установочный рефлекс отсутствует. Положение то же, что и на рис. 114, где у таламического животного вследствие экстирпации лабиринтов устранен лабиринтный установочный рефлекс, но имеется рефлекс в результате асимметричного раздражения поверхности тела. В случае, изображенном на рис. 124, дело идет, наоборот, об исчезновении установочного рефлекса, так как соответствующие центры в среднем мозгу удалены.
мые задние отделы среднего мозга или даже передний край области моста. После разреза позади переднего края моста исчезает и этот рефлекс.
Если объединить результаты данного и предыдущего разделов, то оказывается, что в среднем мозгу имеется аппарат, регулирующий напря-
206
Пятая глава. Установочные рефлексы
жение шейной мускулатуры и обеспечивающий тем самым удерживание головы в нормальном положении и обратное возвращение ее, если она из этого положения выведена. Начальным толчком для этого могут явиться два рода афферентных возбуждений: раздражения лабиринтов и чувствительных нервов тела.
в) Установочные рефлексы с тела на тело
Как видно из рис. 117, стр. 192, таламическое и мезенцефалическое животные способны привести свое тело в правильное сидячее положение и тогда, когда голова не находится в нормальном положении, а повернута, например, на 90° в сторону. Реакция начинается благодаря асимметричному раздражению поверхности тела давлением плоскости опоры.
У децеребрированного кролика мне ни в одном случае не удалось обнаружить этого рефлекса и тогда, когда животное раздражалось путем щипка хвоста или одной из лап.
Несмотря на то что этот рефлекс не наступает строго постоянно и у таламического животйого, учитывая достаточное количество наблюдений на децеребрированных животных, правомерно заключить, что осуществление установочных рефлексов с тела на тело вследствие асимметричного раздражения поверхности тела также связано с наличием среднего мозга.
г) Шейные установочные рефлексы
Если при совместном действии различных установочных рефлексов голова таламического или четверохолмного животного из какого бы то ни было ненормального положения приводится в нормальное положение, сюда присоединяется шейный рефлекс, благодаря чему тело следует за головой и таким образом достигает нормального положения. Этот шейный рефлекс удобно изучать в процессе усаживания животного из бокового положения. При этом прежде всего в нормальное положение (т. е. челюстью вниз) поворачивается голова, чем обусловливается активное сгибание передних ног (тонический лабиринтный рефлекс на мышцы конечностей, поддержанный для вышерасноложенной передней ноги тоническим шейным рефлексом на мускулатуру конечностей); одновременно происходит поворот позвоночника, вследствие чего передняя часть тела поворачивается в направлении к нормальному положению (рис. 98 и 99, стр. 173). То и другое приводит к усаживанию передней части тела, а к этому присоединяется усаживание задней половины, часто путем весьма быстрого рывка. Активное сгибание передних ног в ответ на поворот головы в нормальное положение основано на обычном тоническом лабиринтном и шейном рефлексах на мускулатуру конечностей, которые контролируются центрами, расположенными каудально от уровня вступления nervi octavi. У таламического и мезенцефалического кролика эти рефлексы приводят к активному сгибанию передних конечностей, так как распределение тонуса в них «нормально», а разгибающие мышцы не преобладают над сгибающими. У децеребрированного животного, наоборот, разгибатели конечностей находятся в состоянии децеребрационной ригидности, сгибатели же не тонизированы совсем или тонизированы слабо, и если голову пассивно повернуть из бокового положения в нормальное, то и теперь разгибательный тонус остается более высоким, но в отдельных случаях в нижележащих передних конечностях уменьшается, однако обе ноги чаще остаются вытянутыми, препятствуя тем самым усаживанию передней половины тела. (Только в исключительных случаях и у децеребиррованного животного в от¬
Анализ установочных рефлексов
207
вет на поворачивание головы из бокового положения наступает активное сгибание передних ног, которое никогда не бывает полным).
Таким образом, поведение передних конечностей во время усаживания из бокового положения таламического и четверохолмного животных принципиально совершенно то же, что и у децеребрированного животного; дело идет об обычных рефлексах положения, которые в последнем случае вследствие сильной децеребрационной ригидности должны противодействовать усаживанию животного, вместо того чтобы способствовать ему, как это имеет место у таламического животного. Остальные установочные рефлексы для объяснения этой реакции не нужны.
Отсюда возник вопрос, не является ли поворот позвоночника, наступающий вследствие предшествующего поворота головы по отношению к туловищу, простым рефлексом положения, который имеется также и у децеребрированного животного, или же дело идет об отдельном «установочном» рефлексе.
Во всех прежних опытах на децеребрированных кошках и кроликах никогда не удавалось вызвать поворачивания позвоночника в ответ на поворот головы. Выраженная реакция уже и при тогдашних наблюдениях определенно бы обратила на себя внимание. Более слабые движения, возможно, могли оказаться просмотренными. Выше упоминалось, что у нормальных недецеребрированных кроликов и кошек в ответ на поворот головы в затылочное положение наступает поворот позвоночника, ведущий к поворачиванию таза в противоположном направлении и идентичный шейным установочным рефлексам. Ранее уже говорилось, что шейные установочные рефлексы после удаления среднего мозга у кролика могут еще иметь место и что их центры расположены тотчас позади заднего двухолмия, т. е. во всяком случае значительно далее в оральном направлении, чем центры тонических шейных рефлексов на конечности. Отсюда следует, что шейные установочные рефлексы следует четко отличать от шейных тонических рефлексов и что, с другой стороны, центры их расположены несколько каудальнее, чем центры остальных установочных рефлексов, которые достигают и области моста.
Сообщенные данные делают понятным, почему децеребрированное животное не в состоянии принимать и поддерживать нормальное положение и вследствие чего оно не в состоянии бегать и прыгать. У него отсутствуют все описанные установочные рефлексы, центры которых лежат в среднем мозгу (центры шейных установочных рефлексов — в области моста).
Следует еще упомянуть только один пункт. Как указывалось выше, децеребрированное животное обладает весьма живыми и быстрыми реакциями поворачивания. Несмотря на это, оно не в состоянии поддерживать равновесие. Эти рефлексы не могут предохранить животное от падения и не имеют, таким образом, решающего значения для поддержания нормального положения тела.
Конечно, весьма вероятно, что реакции поворачивания способствуют установочной функции. Когда животное стоит на полу в нормальном положении и благодаря какому-либо воздействию опрокидывается вправо или влево, то вследствие падения вправо наступает поворотная реакция головы влево, стремящаяся удержать голову в нормальном положении. Это означает, что реакция поворачивания обеспечивает поворот головы в том направлении, в котором голова будет удержана благодаря последующему установочному рефлексу (или лабиринтному установочному рефлексу, или установочному рефлексу с тела на голову). Здесь, таким образом, мы имеем те же отношения, с которыми познакомились ранее при уста¬
208
Пятая глава• Установочные рефлексы
новках глаз. Реакции поворачивания обусловливают движение головы в определенном направлении, приводя ее в положение, которое соответствует таковому, наступившему благодаря установочному рефлексу. Мы снова видим, что определенные рефлексы положения могут вызываться соответствующими реакциями поворачивания.
Однако, как показывают наблюдения над децеребрированными животными, сами по себе реакции поворачивания осуществить функцию установки не в состоянии.
В противоположность реакциям поворачивания установочные рефлексы являются истинными рефлексами положения. Безразлично, приводится ли животное в ненормальное положение, или долгое время находится в этом положении, или в течение долгого времени ранее спокойно удерживалось в нем; как только животное перестает удерживаться, оно усаживается. Таким образом, установочные рефлексы наступают вследствие ненормального положения.
Оптические установочные рефлексы (43)
Как уже неоднократно упоминалось, оптические рефлексы отсутствуют у всех таламических животных и, кроме того, у кролика с интактным большим мозгом. Наоборот, у интактных кошки, собаки и обезьяны они имеются. У последних глаза принимают участие в осуществлении принятия нормального положения.
Чтобы исследовать эти взаимоотношения, следует свободно удерживать животное в воздухе, ибо только тогда избегается соприкосновение его с плоскостью опоры и установочные рефлексы с тела на голову и на тело не могут осуществиться. При этих обстоятельствах животное прежде всего предоставлено своим лабиринтным установочным рефлексам, а если экстирпировать лабиринты, получение всех установочных рефлексов у бесполушарных собаки, кошки и у кролика с интактным большим или без большого мозга становится невозможным. При исследовании лишенных лабиринтов, но обладающих большим мозгом собак и кошек, свободно удерживаемых в воздухе, выяснилось, что у этих животных еще имеются установочные рефлексы, благодаря которым они в состоянии придавать своему телу правильное положение в пространстве. Эти установочные рефлексы начинаются с глаз.
Для придания наглядности этому факту следует прежде всего подробно рассмотреть наблюдения на щенке.
В интактном состоянии до экстирпации лабиринтов животное в разные дни свободно держалось в воздухе и исследовалось на наличие лабиринтных установочных рефлексов. При этом глаза сразу же прикрывались специальной шапочкой.
Если животное свободно удерживать в воздухе за таз в нормальном положении, голова его также находится в нормальном положении. При правом и левом боковых положениях таза голова поворачивается почти (примерно до 30°) в нормальное положение. При положении таза «на спине» голова приводится в нормальное положение благодаря тому, что передняя часть тела животного, особенно шея и верхний отдел груди, сильно сгибаются в вентральном направлении; в других случаях голова ориентируется нормально вследствие того, что передняя часть тела животного описывает спиральный поворот на 180°. В подвешенном положении головой вверх голова стоит в нормальном положении. В подвешенном положении головой вниз последняя свешивается мордой отвесно вниз; шея отчетливо согнута дорсально.
Анализ установочных рефлексов
209
При исследовании без шапочки голова животного принимает примерно те же положения; только в том случае, когда таз удерживается в боковом, положении, голова полностью поворачивается в нормальное положение.
Установочные рефлексы этого животного в воздухе сфотографированы в опытах с надетой шапочкой и без нее.
22 сентября 1919 г. де Клейном производится двусторонняя экстирпация лабиринтов.
23 сентября 1919 г. в 9 ч. 00 м. животное держит голову прямо, нистагма не обнаруживается.
Исследование установочных рефлексов в воздухе без шапочки (т. е. с открытыми глазами) показывает, что в этом случае установочные рефлексы
Рис. 125. Щенок в день двусторонней экстирпации лабиринтов. Таз свободно удерживается в воздухе. Голова находится в боковом положении. Установочный рефлекс на голову отсутствует.
Рис. 126. Та же собака в день двусторонней экстирпации лабиринтов. Таз свободно удерживается в воздухе в опрокинутом положении. Голова находится в затылочном положении. Установочный рефлекс на голову отсутствует.
у животного полностью отсутствуют. В висячем положении головой вверх последняя стоит в правом боковом положении; в висячем положении головой вниз последняя находится в затылочном положении; при боковом положении таза голова также держится в боковом положении (рис. 125); в положении животного на спине и голова находится в затылочном положении (рис. 126).
Таким образом, исследование показывает, что в день лабиринтэктомии установочные рефлексы в воздухе у животного отсутствуют и что глаза в этом случае также не помогают.
Однако, уже через два дня, 25 сентября дело обстоит иначе. Отчетливо видно, что теперь животное в состоянии, если не полностью привести, то по крайней мере повернуть голову в нормальное положение. Во время исследования становится ясным, что животное осуществляет это путем фиксации глазами. Прежде всего голова движется в нормальное положение всякий раз, когда животное увидит какой-либо предмет. В висячем положении головой вниз последняя во всяком случае еще находится в затылочном положении; в висячем положении головой вверх, наоборот, уже делаются попытки пригнуть голову вентрально и установить ее правильно. В положении на спине предпринимаются попытки придать голове нормальное положение при помощи вентрального пригибания, а при положении на боку голова поворачивается в направлении нормального положения, но не в состоянии еще установиться вполне правильно.
14 Р. Магнус
210
Пятая глава. Установочные рефлексы
29 сентября в сидячем положении животное держит голову совершенна прямо и не обнаруживает нистагма. Бегая по клетке, оно описывает большие круги в правом или левом направлениях, что после двусторонней лабиринтэктомии является правилом. Исследование установочных рефлексов в воздухе без шапочки, т. е. при открытых глазах, показывает следующее. При правом боковом положении тела голова благодаря повороту или наклону влево устанавливается правильно; при левом боковом положении тела голова устанавливается правильно благодаря повороту вправо. Результат, однако, бывает несколько переменчивым. При висячем положении тела головой вниз последняя сначала находится в затылочном
Рис. 127. Та же собака в шапочке через 16 дней после двусторонней экстирпации лабиринтов. Животное свободно удерживают в воздухе за таз в боковом положении. Голова находится в боковом положении. Установочный рефлекс на голову отсутствует.
Рис. 128. Та же собака через 16 дней после двусторонней экстирпации лабиринтов в шапочке. Таз свободно удерживается в воздухе в опрокинутом положении. Голова находится в затылочном положении. Установочный рефлекс на голову отсутствует.
положении, а затем благодаря поворачиванию влево приводится в боковое, а иногда и в полностью нормальное положение. При висячем положении тела головой вверх животное глядит влево или вправо, причем голова зачастую стоит в среднем положении и при отклонении вправо или влево устанавливается в пространстве почти правильно.
При этом исследовании становится ясным, что животное приводит голову в правильное положение благодаря тому, что фиксирует всякий предмет, который попадает в его поле зрения, например руку экспериментатора, карандаш, пищу и т. д.
Способность животного правильно устанавливать голову в пространстве в то время, когда оно свободно удерживается в воздухе, исчезает тотчас и окончательно, как только глаза прикрываются специальной шапочкой. В этом случае при правом положении тела голова находится в правом боковом положении, при левом боковом — в левом боковом или затылочном, при положении на спине — в затылочном, при висячем положении тела головой вниз — в затылочном; при висячем положении головой вверх — в правом или левом боковых положениях.
8 октября. Животное, удерживаемое свободно в воздухе, уже научилось с помощью глаз приводить голову в нормальное положение при всех положения?: тела в пространстве.
Анализ установочных рефлексов
211
Сделаны две серии снимков: одна — со специальной шапочкой, другая — без нее. Животное с одетой шапочкой оказывается полностью дезориентированным в пространстве. Результат исследования подобен таковому от 29 сентября: в правом боковом положении тела голова находится в том же положении (рис. 127); при левом положении тела голова находится в левом боковом положении; при положении на спине голова стоит в затылочном положении (рис. 128); при свешивании тела головой вниз» последняя находится в затылочном положении. Если шапочку снять,, картина совершенно меняется. При боковом положении тела голова приводится полностью в нормальное положение (рис. 129). При свисании тела
Рис. 129. Та же собака через 16 дней после двусторонней экстирпации лабиринтов без шапочки. Задняя половина тела свободно удерживается в воздухе в правом боковом положении. Голова при помощи поворота приведена полностью в нормальное положение. Передняя часть тела последовала за поворотом головы. Оптический установочный рефлекс на голову с присоединившимся шейным установочным рефлексом.
Рис. 130. Та же собака через 16- дней после двусторонней экстирпации лабиринтов без шапочки- Задняя часть тела свободно удерживается в воздухе в опрокинутом положении. Голова приведена в нормальное положение, животное фиксирует глазами фотографа. Передняя часть тела последовала за головой и перевернулась, в вентральном направлении. Оптический установочный рефлекс на голову с присоединяющимся шейным рефлексом.
головой вниз наступает сильное дорсальное пригибание головы, морда*, поднимается и голова приходит в нормальное положение. Это в особенности отчетливо выявляется тогда, когда животное фиксирует глазами какой-либо предмет. При висячем положении тела головой вверх последняя стоит в строго нормальном положении; при положении на спине передняя часть тела сгибается вентрально, голова оказывается в нормальном положении и животное фиксирует окружающее с большой живостью (рис. 130).
Таким образом, этот опыт показал, что тотчас после экстирпации лабиринтов собака в воздухе оказывается полностью дезориентированной, но что в течение немногих дней она научается использовать глаза и с их помощью приводить голову в нормальное положение. Этот процесс начинается уже спустя два или три дня после операции. Через неделю он еще не вполне закончен; однако спустя 14 дней животное оказывалось в состоянии, находясь в воздухе, привести голову в полностью нормальное положение. Совершенно то же было констатировано у многих других, собак.
212
Пятая глава. Установочные рефлексы
Заслуживает упоминания, что у собаки, у которой Дюссер де Баренн 4 декабря 1918 г. экстирпировал большую часть мозжечка, так что на последующей секции были обнаружены только передний отдел и по сторонам от продолговатого мозга незначительные остатки червя, также можно было наблюдать развитие оптических установочных рефлексов. 3 марта 1919 г. де Клейном у той же собаки был удален правый, а 4 апреля 1919 г. — левый лабиринты. В опытах 23 апреля и 2 и 26 мая животное, удерживаемое в воздухе с завязанными глазами, не обнаруживало никаких следов установочных рефлексов. Наоборот, при открытых глазах поведение этого животного было идентично поведению собаки, описанной выше. В обоих боковых положениях, в висячем положении головой вверх и в затылочном положении голова устанавливалась совершенно прямо; в висячем положении головой вниз шейная часть позвоночника сильно изгибалась в дорсальном направлении, так что голова двигалась в направлении к нормальному положению. Развитие оптических установочных рефлексов у собаки имеет место и после выключения большей части мозжечка.
У кошки результаты наблюдений над оптическими установочными рефлексами те же, что у собаки. Для этих опытов следует отбирать небольших, смирных животных, так как в большинстве случаев кошки, если держать их свободно в воздухе в различных положениях, оказываются весьма беспокойными и вследствие этого мешают наблюдениям. Следующий экспериментальный пример пояснит дело.
Небольшая кошка при исследовании с шапочкой, удерживаемая свободно в воздухе, обнаруживает отчетливые лабиринтные установочные рефлексы на голову. В висячем положении головой вниз шея сильно изгибается дорсально и ротовая щель устанавливается вертикально вниз. При висячем положении тела головой вверх последняя стоит полностью в нормальном положении. При положении животного на спине голова вследствие вентрального сгибания передней части тела приводится в нормальное положение. При боковом положении тела голова, поворачиваясь, приводится в нормальное положение как из положения на правом, так и из положения на левом боку. 23 октября были сделаны стереоскопические снимки во всех этих положениях в воздухе с надетой шапочкой. В этот же день де Клейном произведена двусторонняя экстирпация лабиринтов.
Уже через два дня удается показать, что оптические установочные рефлексы образовались. При исследовании с шапочкой в положении тела на спине и в висячем положении головой вниз или вверх последняя находится в затылочном положении, при боковом положении тела — в боковом положении. Наоборот, при исследовании в воздухе без шапочки, т. е. с открытыми глазами, при боковом положении таза голова поворачивается в направлении к нормальному положению, которого она, однако, вполне не достигает.
При положении на спине имеет место сильное вентральное сгибание передней части тела или же голова и передняя половина тела поворачиваются спирально, так что голова становится в приблизительно нормальное положение. При висячем положении головой вниз голова посредством дорсального сгибания движется в направлении к нормальному положению; при положении головой вверх последняя стоит приблизительно нормально.
, 3 ноября исследование установочных рефлексов в воздухе с шапочкой показало
то же, что исследование от 25 октября. Без шапочки, с открытыми глазами, голова приводится в нормальное положение при боковом положении тела, если животное фиксирует глазами какой-либо предмет. В положении на спине и в висячем положении головой вверх или вниз последняя приводится полностью в нормальное положение.1 5 ноября — те же результаты: никаких рефлексов в воздухе с применением шапочки, отчетливые установочные рефлексы в воздухе без применения шапочки, с открытыми глазами, причем ясно видно, что оптические рефлексы осуществляются главным образом посредством фиксации предметов глазами. Если сначала держать
1 При висячем положении головой вниз это удается не всем нормальным и ла- биринтэктомированным животным. При этом шея сгибается дорсально, но ротовая щель не приходит в совершенно нормальное положение.
Анализ установочных рефлексов
213
животное с надетой шапочкой в воздухе в боковом положении, Волова находится в том же положении. Если теперь снять шапочку, голова путем поворота или наклона тотчас приводится в нормальное положение. Рис. 131 показывает кошку в шапочке в висячем положении тела головой вниз. Видно полузатылочное положение головы.
На рис. 132 то же животное находится в том же положении без шапочки; голова теперь сильно пригнута дорсально, и на рисунке видно, что животное фиксирует (глазами) находящуюся перед ним руку экспериментатора.
Эти наблюдения были повторены на серии кошек и собак и давали постоянно одни и те же результаты.
У обезьян (59) также имеются живые оптические установочные рефлексы. Уже простое наблюдение над животными показывает, что го-
Рис. 131. Кошка через 13 дней после двусторонней экстирпации лабиринтов в шлемике. Висячее положение головой вниз. Голова находится в полузатылоч- ном положении. Установочный рефлекс на голову отсутствует.
лова их постоянно совершает «рассматривающие» движения (Blickbewegungen), ориентируясь при этом определенным образом по видимым предметам, например по экспериментатору, и что к этому присоединяются изменения установки всего тела. Значение этих оптических установочных рефлексов выступает особенно отчетливо, если сравнивать поведение двусторонне лабиринтэктомированных обезьян в воздухе с открытыми и с закрытыми глазами. При закрытых глазах животное полностью дезориентировано и принимает любое положение в пространстве. Если, наоборот, глаза открываются, то в спокойном положении (поза покоя — Ruhelage), когда животное смотрит на какой-либо предмет, определенного положения головы также не отмечается. Как только какой-нибудь предмет возбуждает внимание животного, будь то экспериментатор или входящее в комнату другое лицо, муха или еда, тотчас следует реакция головы, которая, если животное свободно удерживается в воздухе, заключается в повороте головы в направлении к нормальному положению. Таким путем голова может быть повернута в нормальное положение в боковом, висячем головой вверх или вниз, на спине и других положениях тела, а также и тогда, когда выключены лабиринтные установочные рефлексы. Однако всегда можно наблюдать, что эта реакция длится лишь столько времени, пока активизировано оптическое внимание жи-
Рис. 132. Та же кошка в тот же день без шлемика. Висит в воздухе головой вниз. Дорсальное огибание шеи (и поворот влево), вследствие чего голова приближается к нормальному положению. Животное фиксирует глазами находящуюся перед ним руку экспериментатора. Оптический установочный рефлекс на голову.
1214
Пятая глава. Установочные рефлексы
квотного; как только последнее выключается голова снова возвращается в первоначальное исходное положение, которое в отсутствие лабиринтных установочных рефлексов оказывается положением или на боку, или жа спине, или каким-либо иным ненормальным положением.
Суммируя эти наблюдения, можно заключить следующее.
Бесполушарные кошки, собаки и обезьяны обладают теми же четырьмя группами установочных рефлексов, которые были подробно описаны для кроликов. В воздухе животные предоставлены исключительно .лабиринтным установочным рефлексам на голову и примыкающим к ним затем шейным рефлексам. Если у подобных животных с обеих сторон :экстирпируются лабиринты, они оказываются в воздухе полностью дезориентированными.
Наоборот, кошки, собаки и обезьяны, не лишенные большого мозга, ведут себя иначе. Для ориентации в пространстве они имеют возможность привлечь глаза. Это можно показать, исследуя их после двусторонней лабиринтэктомии в воздухе. Животные научаются использовать свои глаза, а спустя более или менее продолжительное время у них вырабатывается гвозможность уже и без лабиринтов приводить голову полностью в нормальное положение при различных положениях тела в пространстве. Прямое наблюдение учит, что при этом животные используют глаза и что оптические установочные рефлексы присоединяются к фиксированию глазами предметов внешнего мира. Если таких животных исследовать с шапочкой или без шапочки, то можно с уверенностью выключать оптические установочные рефлексы или снова получать их.
Из того факта, что описанные установочные рефлексы развиваются только у животных с сохраненным большим мозгом, следует, что оптические установочные рефлексы связаны с наличием коры больших полушарий головного мозга. Последнее является вместе с тем и необходимостью, так как у бесполушарных кошек, собак и обезьян не обнаруживаются никакие оптические реакции, кроме зрачкового рефлекса и рефлекса смыкания век при засвете глаз.
Интересно различие собаки, кошки и обезьяны, с одной стороны, т кролика — с другой. Интактный кролик с сохранным большим мозгом же имеет никаких оптических установочных рефлексов и в своем поведении (в том, что касается установочных реакций) ничем не отличается от таламического кролика. Это значит, что у кролика аппарат стояния и установки ограничен исключительно стволом мозга; наоборот, у собаки, кошки и обезьяны играют роль связи с корой больших полушарий, жо всей вероятности — со зрительной областью коры. Действительно ли жаличия одной зрительной области коры достаточно для осуществления оптических установочных рефлексов, разумеется, должно быть выявлено специальными исследованиями. Из того факта, что непосредственно после экстирпации лабиринтов кошки и собаки первое время несколько дезориентированы в воздухе, вытекает, что эти животные в период их нормального существования, пока они обладают интактными лабиринтами, основательно используют эти сенсорные органы для ориентировки в пространстве и что они тогда только привлекают для ориентирования глаза, когда лабиринты выходят из строя. Наблюдать этот процесс с отчетливостью можно в течение первых дней после операции. Имеющиеся сведения не позволяют еще решить, как в этом отношении дело обстоит у обезьян. У одной из наблюдавшихся мною обезьян уже в день экстирпации второго лабиринта обнаружились оптические установочные рефлексы. Они, однако, значительно улучшились в течение последующих дней. Поэтому нельзя решить вопрос, играют ли оптические установочные
Анализ установочных рефлексов
215
рефлексы у интактной обезьяны с сохранными лабиринтами значительную роль при наличии оживленных движений глаз. Это, пожалуй, вероятно и тем в большей мере, — как это наблюдается у обезьян и после односторонней экстирпации лабиринтов, — что оптические установочные рефлексы мешают типичной установке головы, которая наступает после односторонней экстирпации лабиринта при различных положениях тела в воздухе. Как только внимание животного возбуждено, голова его в большей или меньшей степени возвращается из данного в нормаль- шов положение. Это также говорит о том, что обезьяна использует опти- 1 неские установочные рефлексы и тогда, когда она не лишилась еще обоих .лабиринтов.
В то время как центры всех описанных до сих пор рефлексов положения и установки лежат в самых высоких областях шейного отдела спинного мозга, оптические установочные рефлексы оказываются исключением: они проходят через кору полушарий. В дальнейших экспериментах, которые будут посвящены вопросу, в какой мере аппарат стояния и установки может приводиться в действие из коры, эти рефлексы, вероятно, будут играть важную роль.
Влияние установочных рефлексов на другие реакции
У децеребрированного животного с данной интенсивностью шейных ж лабиринтных рефлексов положение тела при определенной установке головы и поза тела определены раз навсегда. Интактное животное, наоборот, может принимать и другие положения. Поэтому возможности реагирования благодаря наличию высших отделов мозга должны быть более сложными. Такого рода усложнение создается, например, вследствие включения установочных рефлексов. Более определенного анализа изменений отдельных рефлексов, движений и положений, обусловленных этим обстоятельством, здесь не дается. Достаточным будет показать па единственном примере, как проявляется это усложнение.
На рис. 122 (стр. 204) виден децеребрированный кролик в висячем положении головой вверх. Голова пригнута дорсально и достигает максимального положения для тонических лабиринтных рефлексов. Вследствие этого подниматели затылка иннервированы тонически, т. е. голова фиксирована в этом положении, разгибательный тонус передних и задних конечностей высок и еще более усиливается шейным тоническим рефлексом (вследствие дорсального пригибания головы). Животное вынуждено оставаться в этом положении, пока оно удерживается рукой ъ висячем положении. На рис. 101 (стр. 175) виден таламический кролик, который удерживается в воздухе в таком же положении. Здесь в действие нключился лабиринтный установочный рефлекс, преодолел тонический лабиринтный рефлекс на разгибатели затылка, вызвал вентральное нагибание головы, к чему присоединился шейный установочный рефлекс, который привел в нормальное положение всю переднюю половину тела. Конечный результат, таким образом, — совершенно иное по сравнению с децеребрированным животным положение, причем установочный рефлекс снимает тонический лабиринтный рефлекс на шею. После экстирпации лабиринтов (рис. 107, стр. 178) и таламический кролик принимает в воздухе положение, похожее на положение децеребрированного животного, только конечности не обнаруживают децеребрационной ригидности и поэтому согнуты.
Этот пример показывает, как вследствие наличия среднего мозга и установочных рефлексов способ реагирования децеребрированного животного изменяется и усложняется.
216
Пятая глава. Установочные рефлексы
На таламических и четверохолмных кроликах может быть хорошо изучена еще одна группа усложнений.
Во второй главе на спинальной кошке и спинальной собаке было показано, что, смотря по положению и позе, которые придаются всему животному или какому-нибудь его'члену (нога, хвост), одно и то же раздражение вызывает совершенно разные реакции. После различных длительных сенсорных раздражений (проприоцептивные возбуждения, раздражения давлением) возможно получать изменения центрального органа, благодаря которому определенное возбуждение будет вынуждено приходить один раз к одному, другой раз к другому центру. Так, например, изменяя положение спинальной собаки, можно добиться, что чесатель- ный рефлекс произвольно наступит либо гомо-, либо гетеролатерально. Центральные изменения такого рода были обозначены как «переключения».
Подобное удается наблюдать также и у таламического или четверохолмного кролика. Если такое животное, когда оно сидит на полу в нормальном положении, сильно ущипнуть за хвост, т. е. нанести строго симметричное раздражение, оно начинает прыгать и бегать. Таким образом возникает ритмичное движение конечностей. Если же в силу какой- либо причины животное лежит на боку и не усаживается, то же самое раздражение вызывает не движения бега, а поворачивание головы в нормальное положение. Это значит, что в положении на правом боку стимулируются левые ротаторы затылка, а в положении на левом — правые. Таким образом, одно и то же раздражение в зависимости от положения и позы животного приводит к возбуждению совершенно различных мышц.
Еще показательнее следующий опыт. Если таламический или четверохолмный кролик находится еще в настолько глубоком шоке, что оказывается не в состоянии самостоятельно садиться и приводить голову в нормальное положение, то, раздражая нижерасположенную переднюю конечность, возможно получить поворот головы в этом положении. Если исследовать животное в боковом положении в воздухе или на столе, с компенсацией давления со стороны плоскости опоры наложенной дощечкой с грузом, раздражение вышерасположенной передней конечности постоянно вызывает поворот головы теменем вниз. Если же животное лежит на столе без дощечки, в ответ на щипок вышерасположенной передней конечности часто (не всегда) наступает поворот головы в нормальное положение. В этом случае асимметричное раздражение поверхности тела давлением плоскости опоры привело к тому, что даже в ответ на такое интенсивное раздражение, как сильный щипок передней лапки, наступает реакция, полностью противоположная таковой в отсутствие асимметричного длительного раздражения.
Вследствие различного положения животного в центральной нервной системе создается состояние, когда любые раздражения, которые в других условиях не могут действовать как «установочные раздражения», приводят установочный аппарат в активное состояние. Животное таким образом своим предшествовавшим положением приводится в определенную «установочную готовность», когда любое индифферентное раздражение достаточно для реализации функции установки. В определенных случаях при этом речь может идти об алгебраическом суммировании двух взаимонезависимых возбуждений. Последний описанный опыт, в котором щипок вышележащей передней конечности вызывал поворот головы в нормальное положение, однако, показывает, что здесь должны играть роль также и переключения. В этом случае приложенное раздражение вызывает реакцию, точно противоположную той, которая имеет
Анализ установочных рефлексов
217
место, если вследствие позиции животного оно не приведено в определенную «установочную готовность». Возбуждение вынуждено влиять в центральной нервной системе на иные центры, чем в нормальных условиях.
Более точный анализ всех этих явлений, который еще будет проделан, сулит интересные разъяснения. Однако уже теперь можно сказать следующее: при наличии среднего мозга и установочных рефлексов простые рефлексы положения из центров верхнего шейного отдела спинного и продолговатого мозга изменяются, а в определенных случаях подавляются. Если животное благодаря функционированию своих установочных рефлексов приняло определенное полоя^ение и позу, существующее при этом распределение тонуса мускулатуры его тела должно оказывать переключающее действие и влиять на последующие реакции. То же должно происходить благодаря влиянию возбуждения поверхностной и глубокой чувствительности тела при определенном положении тела вследствие соприкосновения его с плоскостью опоры. Если тело животного по какой-либо причине еще не находится в нормальном положении, то, несмотря на это, имеет место определенная «установочная готовность», благодаря чему индифферентные раздражения или раздражения, которые собственно должны вызывать противоположную реакцию, безразлично являются ли они сами по себе слабыми или очень сильными, активируют теперь установочную функцию и побуждают животное к нормальному усаживанию и, может быть, присоединяющимся к этому бегу и прыжкам.
Шестая глава
ПОСЛЕДСТВИЯ ОДНОСТОРОННЕЙ ЭКСТИРПАЦИИ ЛАБИРИНТА
Описанные в предыдущей главе факты достаточны в качестве отправных для исследования состояний после односторонней экстирпации лабиринта.
В прошлом эта операция производилась почти исключительно для того, чтобы узнать что-либо новое в отношении деятельности лабиринта. Не говоря о том, что по существу дела желание исследовать функцию того, что разрушается, является сомнительным, последствия вмешательства подобного рода настолько сложны, что выводы из них зачастую делались неверные.
Если убрать лабиринт, то в каждой из вышеописанных групп рефлексов остаются с обеих сторон рефлексы, не зависящие от лабиринтов. Кроме того, могут сказываться односторонние влияния оставшегося лабиринта, а также последствия поворота шеи, которые могут выявиться в виде асимметричных рефлексов позы, шейных установочных рефлексов и шейных тонических рефлексов на глаза. К этому добавляется то, что действие нормальных двусторонних рефлексов нелабиринтного происхождения также должны сказаться на туловище, которое изогнуто вследствие односторонней утраты лабиринта. Если учесть, что названные отдельные компоненты картины симптомов у различных видов животных и в различное время после операции выражены неодинаково сильно и что непосредственно после операции к явлениям выпадения присоединяются еще и явления раздражения, то становится понятной необходимость подробного экспериментального анализа, прежде чем удастся распутать и понять этот сложный симптомокомплекс.
218
Шестая глава. Последствия односторонней экстирпации лабиринта
Выяснилось, например, что у всех исследованных видов животных часть последствий удаления лабиринта можно тотчас исключить, коррелируя ненормальное положение головы по отношению к туловищу. Другая часть этих последствий остается и при прямо поставленной голове. Благодаря этому стало возможно простым способом отличать шейные рефлексы от прямых последствий выпадения лабиринта.
Для того чтобы отделить лабиринтные установочные рефлексы от установочных рефлексов тела, необходимо исследовать животных попеременно в воздухе и на земле и т. д.
Наблюдения, которые описаны в дальнейшем, проведены на кроликах, морских свинках, кошках, собаках и обезьянах. У каждого из этих видов животных названные группы явлений комбинируются различным образом: шейные рефлексы, лабиринтные влияния, установочные рефлексы тела, оптические впечатления и т. д. — и имеют для положения тела неодинаковое значение. Поэтому необходимо было предпринять наблюдения над каждым видом в отдельности и, только сравнивая результаты, перейти к заключениям общего характера.
Анализ нарушений,'' наступавших после односторонней экстирпации лабиринта, должен быть приведен в этой главе в той мере, в какой это позволяет сделать современное состояние знаний, с учетом опасности, что изложение будет растянутым и на первый взгляд теряющимся в частностях. Однако однажды должно быть показано на отдельном примере, насколько сложны существующие отношения и насколько возможно в настоящее время расшифровать эти отношения. Для некоторых запросов клиники при этом могут быть установлены путеводные вехи.
Так как животные, лишенные лабиринта на одной стороне, находятся под наблюдением в течение месяцев, можно дать довольно полный обзор исходных состояний после удаления одного лабиринта у различных видов. Для лучшего понимания дела картина симптомов всякий раз должна быть описана (возможно) более точно, несмотря на то, что выше уже приводился ряд скрупулезных исследований, которые будут дополнены лишь в отдельных пунктах.
Наиболее подробно должно быть описано поведение кроликов, тогда в случае других животных потребуется выделять только самые существенные различия.
В течение двенадцати лет в Утрехтской лаборатории ставились все новые и новые исследования над животными с одним удаленным лабиринтом, в процессе чего, по мере успехов в изучении нормальных лабиринтных, шейных и других установочных рефлексов тела, понимание поведения оперированного животного, естественно, также прогрессировало. В то время как в соответствии со сказанным в первоначальных сообщениях должен был последовательно излагаться произведенный анализ, здесь представляется целесообразным более синтетический подход и построение картины симптомов из рефлексов, описанных в предшествовавших главах. Изложение станет благодаря этому более понятным. Тот, кто интересуется развитием детализации вопроса в эксперименте, может обратиться к оригинальным работам.
Методика
Если исследовать влияние экстирпации лабиринта на положение тела, следует прежде всего требовать, чтобы во время операции не были повреждены прикрепления шейных мышц на голове и чтобы вследствие операции не было никаких изменений в положении головы. С учетом этого основного обстоятельства де Клейном (8) в Утрехтском фармакологическом институте была разработана техника экстирпации
Шестая глава. Последствия односторонней экстирпации лабиринта
219
лабиринта у различных животных, которая в дальнейшем будет кратко описана. Проще всего анатомические соотношения выглядят у кошки.1
Животное наркотизируется эфиром, фиксируется в положении на спине, волосы на шее коротко выстригаются, операционное поле смазывается настойкой йода. Кожный разрез 2 см, идет параллельно позвоночнику через верхушку bullae osseae, которая легко прощупывается (в форме полушара, медиально и каудально от угла челюсти). Разрез начинается орально на высоте угла челюсти.
В глубине разреза тотчас виднеется v. maxillaris externa, в которую с медиальной стороны впадают одна или две ветви. Тупым путем с медиальной стороны вены и орально от боковых ветвей осуществляется доступ в глубину, в щель между подчелюстной железой и двубрюшной мышцей, которая широким брюшком проходит над буллой и осторожно отодвигается пальцем в медиальном направлении. Тем самым
обнажается место деления общей сонной артерии и становится видной наружная сонная артерия, которая, пульсируя, косо пересекает буллу. Каудально от последней тупым путем проходят через ткань в направлении на буллу и обнажают ее. Отслаивая надкостницу, бескровно можно получить широкое отверстие, в основании которого лежит большая часть буллы. Тупые крючки в течение операции удерживают операционное поле, ограниченное медиально двубрюшной мышцей, латерально — подчелюстной железой и орально — наружной сонной артерией. Чтобы с максимальной уверенностью избежать повреждения сонной артерии, желательно использовать лобную лампу. Если последнее все же будет допущено, кровоточащее место принимается тампоном, а через специальный маленький разрез кожи рядом с трахеей отыскивается и перевязывается общая сонная артерия, после чего кровотечение останавливается. При этом доступе вне поля зрения оказываются не только v. jugularis interna, но и блуждающий нерв, языкоглоточный нерв и шейный симпатикус.
Теперь происходит собственно экстирпация лабиринта, которая лучше всего иллюстрируется относящимися сюда стереоскопическими фотографиями.
С помощью маленького долота булла вскрывается в середине, а отверстие расширяется до величины, позволяющей хорошо осмотреть содержимое буллы.
К рис. 133.2 После вскрытия буллы круглое отверстие лежит свободно, более или менее прикрытое костной перегородкой, которая встречается у кошек (не у кроликов и не у собак) и делит среднее ухо на два отдела. Часть круглого отверстия, которая прикрЪгта перегородкой, бывает различной величины; чаще отверстие лежит почти свободно, в других случаях видна только небольшая его часть. Поэтому до- 1 21 Методика является модификацией приема Эвальда (Ewald); см.: Эвальд, стр.196. Операционные методики у собак.
2 Вследствие увеличения на фотографиях удаление от буллы выглядит больше, чем это необходимо длд операции. Фотоснимки сделаны с того же черепа, на котором лоследовательно производилась операция. Лупа Цейсса—Дрюннера. Объектив (55). Увеличение примерно в 3 раза.
Рис. 133.
Рис. 133а. 1 — круглое отверстие; 2 — перегородка.
220
Шестая глава. Последствия односторонней экстирпации лабиринта
лотом и кусачками 1 перегородка убирается настолько, что все круглое отверстие оказывается свободным и становится возможным осмотр среднего уха.
Перегородка берет свое начало от стенки bulla ossea и дорсально достигает среднего уха, оставляя здесь маленькую щель. Если во время операции разрушить эту
перегородку лишь настолько, чтобы хорошо осмотреть круглое отверстие, создается выгодное положение, так как именно эта перегородка закрывает барабанную перепонку и слуховые косточки во время операции, которая теперь становится легко
осуществимой без повреждения ба- д рабанной перепонки.
/ К рис. 134. Соседняя с круглым
отверстием часть мыса скалывается \ долотом, и таким путем открываются
улитка и преддверие. На рисунки , очень отчетливо видна улитка с ба-
Р __ ^10 зальными отверстиями Scala tym-
/ ч pani и Scala vestibuli и приоткры-
3--t- \_]j тое преддверие (ср. рис. 1346).
Sv К рис. 135. Теперь наступает
S-~\- главная часть операции. Пинцетом
g—_Ч JJ/. очищаются преддверие и ампулы по-
-13 лукружных каналов, которые хоро-
y. --Щ шо видны. Всю улитку лучше всего
'1/5
J Рис. 1346. 1 — наружный полукруж¬
ный канал; 2 — ямка стремечка;.
3 — задний полукружный канал;
4 _ наружный полукружный канал;
5 __ объединенные верхний и нижний полукружные каналы; 6 — преддверие* 7 — барабанная лестница; 8 —
наружная ампула; 9 — лимфатический проток; 10 — верхняя ампула; 11 — стенка между мышцей, напрягающей барабанную перепонку, ямкой и преддверием; 12 — задняя ампула; 13 — улитка; 14 — молоточек; 15 — лестница преддверия.
До X. Джейну (Jayne): Анатомия млекопитающих, ч. 1, 1898.
очистить острым инструментом, удалив долотом костную стенку между Scala tympani и преддверием на таком протяжении, чтобы весь лабиринт в конце концов превратился в полость. ii Эвальд, стр. 198.
Шестая глава. Последствия односторонней экстирпации лабиринта
221
Особое внимание следует обращать на то, чтобы перепончатый лабиринт был удален полностью и блестящая поверхность костной стенки виднелась бы на всем ее протяжении, что осуществимо только при использовании мощной лобной лампы. На дне полости отыскивается внутренний слуховой проход, заполненный 8-м нервом. Вначале проход выдалбливался, а 8-й нерв основательно разрушался небольшим крючком. Этот метод применять не рекомендуется. При долблении проход обычно весьма сильно кровоточит и в дальнейшем при вскрытии часто обнаруживались субдуральные кровоизлияния. Кроме того, во время выдалбливания нередко все окружающие образования также раскалываются, причем дополнительные повреждения почти неизбежны. В дальнейшем постоянно отыскивался и выделялся только проход, после чего в интактном проходе крючком осторожно обходился 8-й нерв. Этим путем можно произвести операцию почти без кровотечения.
Из рис. 135 ясны топографические взаимоотношения. Более толстый зонд находится в слуховом отверстии. В отверстие, над предыдущим (Aquaeductus cochleae),
2
'3
вставлены щупики. Ампула заднего канала на рисунке не видна, так как оказалось невозможным сфотографировать все детали сразу. В действительности эта ампула хорошо видна: она находится несколько латеральнее и каудальнее от общего отверстия верхнего и заднего каналов.
К рис. 136. Чрезвычайно важным является тот факт, что при экстирпации лабиринта у кошек можно избежать повреждения п. facialis. На рисунке ясно виден путь этого нерва. В проходе торчит самый толстый щуп. Отверстия, обзначенные на рис. 135 щупами, закрашены черными чернилами. Путь канала лицевого нерва отмечен черной ниткой.
По завершении операции раньше всего накладывается глубокий шов, фиксирующий m. digastricus к латеральной стенке раны. При этом следует остерегаться ранения слюнной железы. В кожный шов лучше всего захватить поверхностный слой шейной мускулатуры, обращая при этом внимание, чтобы швы не прижали вену. Смочить швы настойкой йода. Снятие швов на 5-й день.
В этих опытах, таким образом, вычищаются: улитка, преддверие и ампулы полукружных каналов. Во внутреннем слуховом проходе выделяется 8-й нерв и старательно обходится крючком. Напротив, сами полукружные каналы не экстирпируются. Барабанная перепонка и лицевой нерв не повреждаются, мышцы, двигающие шею, остаются нетронутыми.1
Техника операции на собаке точно такая же, как на кошке. Единственная разница состоит в том, что у собаки костная перегородка в булле, которая у кошки де¬
Рис. 135.
так же как и в два отверстия полукружных каналов (общее отверстие верхнего и заднего полукружных каналов и ампула верхнего канала и наружного канала),
Рис. 135а. 1 — внутренний слуховой проход с зондом в нем; 2 — зонд в водопроводе улитки; 3 — зонд в объединенных верхнем и заднем полукружных каналах; 4 — зонд в наружной и верхнее ампуле.
1 На вскрытии у животных, которые выживали после операции в течение долгого времени, всякий раз обнаруживалась атрофия 8-го нерва. Особенно отчетливо эта атрофия видна при сравнении обеих сторон у животных, оперированных на одной стороне.
222
Шестая глава# Последствия односторонней экстирпации лабиринта
лит среднее ухо на две части, отсутствует. Барабанная перепонка, в противоположность тому, что бывает в случае использования методики Эвальда, остается неповрежденной.
Примерно те же отношения имеют место и у кролика. Требуется, однако, несколько больший опыт, так как работать приходится на довольно большой глубине. Только лицевой нерв при этом сохранить не удается; при вычищении преддверия он, как правило, повреждается в тот момент, когда начинается полное выделение участков с устьями полукружных каналов и культи 8-го нерва, что необходимо для обеспечения контроля полного удаления лабиринта. Во всех случаях удалялись улитка, преддверие и ампулы полукружных каналов и выделялся ствол 8-го нерва. У кроликов также отсутствует костная перегородка в булле, которая имеется у кошек. Барабанная перепонка остается неповрежденной, заживление протекает асептически.
Рис. 136. Рис. 136а. 1 — задняя ампула; 2 — объединен¬
ные верхний и задний полукружные каналы; 3 — водопровод улитки; 4 — путь лицевого нерва; 5 — зонд во внутреннем слуховом проходе.
Для сохранения лицевого нерва следует воспользоваться методикой, разработанной Винклером [Winkler (1)], когда производится большой кожный разрез и m. biventer отсекается в точке прикрепления.
У морских свинок экстирпация лабиринта в булле осуществляется таким же способом, как у кроликов.
У обезьяны, в противоположность названным до сих пор животным, булла отсутствует. Поэтому операцию следует производить таким же способом, как и у человека, т. е. со стороны сосцевидного отростка. Более полное описание излишне, так как все соотношения весьма близки к таковым у человека.
Вместо хирургической операции можно, следуя опыту Кёнига (König), осуществить выключение лабиринта путем впрыскивания кокаина. Для этой цели у деце- ребрированной кошки вышеописанным способом инъецируется булла. Однако экстирпация лабиринта продолжается не далее этапа, изображенного на рис. 134, т. е. до сдалбливания отделов мыса, соседних с круглым отверстием, и открытия улитки и преддверия. Затем обычным шприцем Провачека в преддверие и улитку впрыскивается примерно 0.2 мл 20%-го раствора кокаина или после вскрытия буллы раствор через круглое отверстие вводится непосредственно в лабиринт. Для этих целей раствор кокаина подкрашивался метиленовой синькой. На вскрытии постоянно устанавливалось, что синяя жидкость попадала только в лабиринт, никогда не обнаруживаясь ни суб-, ни эпидурально. Почти непосредственно после впрыскивания проявляются признаки выключения лабиринта. Иногда до наступления паралича наблюдается быстро проходящее раздражение инъецированного лабиринта. Тотальное выключэ- ние лабиринта после впрыскивания длится несколько часов, чего вполне достаточно для решения большинства поставленных задач.
У недецеребрированных кошек также можно с помощью кокаина надолго выключить лабиринт, для чего, однако, необходим опыт. Под наркозом шприцем при хорошем освещении, используя ушную воронку, из слухового прохода в лабиринт впры¬
Последствия односторонней экстирпации лабиринта у кролика
223
скивается раствор кокаина. При этом барабанную перепонку прокалывают и, идя от заднего дорсального края костного слухового прохода, ищут иглой овальное отверстие, затем проходят в лабиринт и производят инъекцию. При пробуждении от наркоза у животного обнаруживаются типичные явления, как и после экстирпации лабиринта. Полного восстановления при этом по большей части уже не наступает, так как при этом способе лабиринт, естественно, остается поврежденным более или менее длительное время.
Гораздо проще временно выключить лабиринт у морских свинок, введя кокаин в среднее ухо по способу ван Россема (van Rossem). Иглой шприца через наружный слуховой проход достигают барабанной перепонки, для чего необходим известный опыт. Затем барабанную перепонку прокалывают и стараются ощутить прикосновение кончика инъекционной иглы к костной стенке среднего уха. Теперь сразу инъецируют 0.1 мл 5%-го раствора кокаина в среднее ухо. Спустя короткое время деятельность лабиринта на стороне инъекции выключается. Иногда параличу предшествуют преходящие явления раздражения. Выключение длится весь день и в большинстве случаев полностью исчезает на следующее утро. Способ, таким образом, очень удобен для временного выключения; у одного и того же животного можно последовательно выключать сначала правый, а затем левый лабиринты и далее снова получать нормальную функцию обоих лабиринтов. Для кролика этот способ неприменим.
В некоторых случаях на морских свинках применялось также выключение лабиринта хлороформом по способу Броун-Секара (Brown-Séquard). При этом наружный слуховой проход многократно заполняется хлороформом, после чего спустя некоторое время можно наблюдать двигательные последствия выключения лабиринта. Опыт в большинстве случаев проходит удачно, однако не всегда. Выключение лабиринта полное, но не во всех случаях достаточно длительное; у многих животных через некоторое время появляются лабиринтные рефлексы.
I. ПОСЛЕДСТВИЯ ОДНОСТОРОННЕЙ ЭКСТИРПАЦИИ ЛАБИРИНТА У КРОЛИКА
Общее поведение кроликов после одностороннего выключения лабиринтов (15)
Поведение кролика, лишенного лабиринта с одной стороны, наиболее полно изучено и описано Винклером (1). Поэтому здесь мы и ссылаемся на его .описания. В качестве типичного примера следует привести сокращенный протокол опыта с одним из наших подопытных животных.
Кролик XV. 30 ноября 1912 г. Эфирный наркоз. Левосторонняя экстирпация лабиринта. Улитка и преддверие вылущены, отверстия полукружных каналов и внутреннего слухового прохода выделены и открыты.
При пробуждении от наркоза левый глаз повернут вентрально, правый — дор- сально, имеется сильный нистагм, причем быстрый компонент на правом глазу направлен вентрально, на левом глазу — дорсально. Если подвесить животное головой вниз, голова отклоняется и поворачивается влево, грудь по отношению к тазу также повернута примерно на 30°. Несколько позже при том же положении тела нижняя апертура грудной клетки поворачивается по отношению к тазу влево на 45°, верхняя апертура — на 70°, голова — на 90°, кроме того, она отклонена влево на 45—70°. Обе правые конечности, особенно правая передняя, сильно разогнуты и отведены; тонус обеих левых значительно ниже. Животное типичным образом перекатывается налево через всю комнату.
2 декабря. Животное сидит в своей клетке прямо, голова повернута влево на 45° (левое ухо — вентрально), правая передняя нога разогнута и отведена, левая — расслаблена и согнута. В клетке животное не переворачивается, однако, будучи извлечено оттуда, после того как его усаживают на пол и наносят раздражения, оно перекатывается через (всю) комнату. Затем оно усаживается, делает несколько шагов, которые хорошо ему удаются, но неоднократно падает при этом на левую сторону, однако уже не переворачивается после падения. При свешивании головой вниз нижняя апертура грудной клетки поворачивается на 45°, верхняя апертура — на 90°, голова — на 120° по отношению к тазу; кроме того, она отклонена на 30° влево. Отклонение глаз не изменяется, нистагм еще имеет место.
4 декабря. Животное больше не опрокидывается. В клетке сидит с головой, повернутой на 45°. При этом положении головы отмечается еще назначительная девиация находящегося выше правого глаза. Нистагма нет. Правые конечности, в особенности передняя, разогнуты и отведены. Голова может отклоняться в обе стороны
224
Шестая глава. Последствия односторонней экстирпации лабиринта
(причем всегда остается повернутой влево). Животное в состоянии бежать вправо и влево, однако чаще бежит влево, описывая круги по направлению часовой стрелки.
7 декабря. При сидении голова повернута на 70° и не наклонена. Отклонение глаз не наблюдается, или оно минимально. Нистагма нет.
14 декабря. При сидении голова повернута на 90° и поэтому находится слева от животного. При таком положении головы отклонения глаз нет. Нистагма нет. Правая передняя лапа отчетливо вытянута и отведена, левая — согнута. В положении сидя грудная клетка слегка наклонена влево. Все тело на столе легче сдвинуть влево, чем вправо. Если животное положить на левый бок, то иногда оно еще раз перекатывается через спину и снова принимает сидячее положение; однако кролик может также сесть и прямо из левого бокового положения, не переворачиваясь через спину. При висячем положении головой вниз нижняя апертура грудной клетки повернута относительно таза на 45°, верхняя апертура — на 90°, голова — на 160°; кроме того, голова наклонена на 30°; правая передняя нога сильно вытянута.
14 января 1913 г. Во время сидения голова повернута на 90°. Девиация глаз при этом незначительна или отсутствует. Нистагма нет. Обе правые ноги разогнуты и отведены. Поворот тела в отдельных случаях увеличивается, достигая 135°. Таким образом, вследствие перевешивания головы животное валится в левую сторону и однократно переворачивается при этом через спину. Однако животное в состоянии непосредственно из левого бокового принять сидячее положение. При висячем положении головой вниз повернуты влево по отношению к тазу нижняя апертура грудной клетки на 45°, верхняя — на 90°, голова — на 170°; голова отклонена влево на 45°, правая передняя конечность разогнута. Животное в состоянии вполне удовлетворительно пробежать через всю комнату.
18 февраля. В основном состояние не изменилось. Только животное сидит теперь, держа передние лапы симметрично, так что разгибания и отведения правой конечности Теперь обычно не наблюдается. Наоборот, в висячем положении головой вниз сгибание передней лапы весьма отчетливо. Грудь во время сидения животного все же наклонена несколько влево.
12 марта. В висячем положении головой вниз нижняя апертура груди повернута по отношению к тазу на 30°, верхняя — на 60°, голова — на 110—135°; голова наклонена влево на 45°; правая нога сильно разогнута. Наоборот, при сидячем положении ноги удерживаются полностью симметрично; грудной отдел тела свисает несколько влево.
В дальнейшем состояние больше не меняется; в мае животное свободно бегает, самостоятельно отыскивает пищу. Поворот головы и груди по отношению к тазу остается без перемен; разгибание передней правой конечности в положении головой вниз весьма отчетливо, но при сидении хорошо компенсируется. Перекатываний через спину больше не бывает. При беге держится совершенно прямо.
При обзоре динамики явлений после односторонней экстирпации лабиринта у кролика можно вместе с Винклером выделить две группы симптомов: прямые и отдаленные последствия операции.
К прямым последствиям, которые отмечаются только в течение короткого времени после операции, следует отнести, помимо шока после очевидного тяжелого вмешательства, нистагм и движения перекатывания. Чем лучше и осторожнее произведена операция, тем более короткое время имеют место оба эти симптома. В большинстве случаев их можно было наблюдать лишь в продолжение одного-двух дней; после отдельных, протекавших особенно гладко операций спонтанные движения перекатывания констатировались лишь в течение немногих часов; они могут и вовсе отсутствовать. Наоборот, в отдельных случаях выраженные движения переворачивания имели место на протяжении трех, а нистагм — даже 4—8 дней. Как это отмечал и Винклер, оба феномена не прекращаются сразу, но, не наступая более спонтанно, могут быть вызваны путем возбуждения животного, например при придании ему анормального положения. Это в особенности относится к движениям перекатывания.
В качестве отдаленных последствий следует учитывать: спиральный поворот всего тела, прежде всего головы, по отношению к груди, а далее и груди по отношению к тазу. В наиболее чистом виде этот поворот можно наблюдать, если подвесить животное головой вниз. В первые дни или недели после операции этот поворот отчетливо увеличивается и дли¬
Последствия односторонней экстирпации лабиринта у кролика
225
тельное время остается в качестве крайне выраженного явления. К отдаленным же последствиям принадлежит и различие тонуса правых и левых конечностей. Ноги на стороне, где отсутствует лабиринт, долее остаются расслабленными; конечности противоположной стороны, наоборот, отчетливо разогнуты. Это различие тонуса еще можно уловить, исследуя животное в висячем положении головой вниз или положении на спине. В сидячем положении животное спустя несколько месяцев научается корригировать это ненормальное положение, так что вновь оно обнаруживается тЬлько при определенных условиях. Осуществление этой компенсации (ибо речь идет именно о таковой) будет обсуждаться позднее. К отдаленным последствиям операции принадлежит также отклонение глаз. Из протокола, приведенного выше, следует, что девиация у свободно сидящего с повернутой головой животного через некоторое время уменьшается и наблюдается только как непостоянное и слабо выраженное явление. Однако ее можно вызвать и показать снова всякий раз при измененном положении тела.
Таким образом, возникает вопрос о внутренней взаимообусловленности и взаимной противоречивости симптомов. Какие явления следует считать прямыми последствиями утраты лабиринта и какие являются вторичными, вызванными прежде всего поворотом шеи, который следует рассматривать как компенсаторную реакцию животного при ненормальной позиции тела? Какую роль играют нелабиринтные установочные рефлексы при этих компенсациях? Насколько существенно для компенсации участие глаз и т. д.? Следующие разделы должны ответить на эти вопросы.
Основной поворот (Grunddrehung)
Если сразу после экстирпации лабиринта кролик пробуждается от наркоза, голова его повернута и склонена в сторону экстирпированного лабиринта. Таким образом, после левосторонней экстирпации лабиринта голова повернута так, что левое ухо стоит вентрально, правое — дор- •сально. Одновременно мордочка приближается к левому плечу.
Если животное в состоянии садиться сразу после операции, то в большинстве случаев голова повернута примерно на 45°, однако наблюдаются повороты и на 20, и на 90°. Наклон головы имеет различную степень выраженности; иногда он может отсутствовать, в других случаях — составлять 110 и более градусов.
У сидящего животного с сохранным большим мозгом в осуществлении этого положения головы принимают участие установочные рефлексы тела на голову и лабиринтные установочные рефлексы. Поворот и наклон остаются и тогда, когда животное вслед за этим децеребрируется, и наступают также, когда односторонняя экстирпация лабиринта производится на ранее децеребрированном животном. Так как при этом удаляются средний мозг, а вместе с ним и центры установочных рефлексов, то выходит, что поворот и наклон осуществляются не в зависимости от центров установки, а благодаря действию тонических лабиринтных рефлексов на мышцы шеи, исходящих из оставшегося лабиринта. Как подробно рассмотрено в третьей главе, оставшийся лабиринт односторонне влияет на тонус затылочных мышц. После удаления лабиринта исчезает тонус мышц шеи, которые поворачивают и наклоняют шею в сторону. Тоническое влияние оставшегося лабиринта на соответствующие мышцы затылка имеет место при всех положениях головы в пространстве; оно, однако, сильнее всего, когда голова находится в затылочном положении, а мордочка поднята на 0—45° над горизонтальной плоскостью (положение 15 Р. Магнус
226
Шестая глава. Последствия односторонней экстирпации лабиринта
головы от 0 до +45°). Оно слабее всего, когда голова находится в нормальном положении, а мордочка по отношению к горизонтали опущена на О—45° (положение головы от —180 до —135°). Этот «основной поворот» (и наклон) головы имеет место в точно таком же виде и сохраняется дни, недели или месяцы, если сначала производится экстирпация лабиринтаг а затем животное подвергается децеребрации. Он также совершенно не зависит от глаз. Центры этого тонического рефлекса лежат в продолговатом мозгу и не простираются кпереди далее плоскости вступления в мозг 8-го нерва. Поворот головы после односторонней экстирпации лабиринта или после односторонней перерезки 8-го нерва, таким образом, может быть еще обнаружен и после удаления среднего мозга, моста, мозжечка и передней половины продолговатого мозга.
Для понимания последующего целесообразно сначала описать изменения положения головы при различных положениях тела у децеребри- рованного кролика, у которого отсутствует средний мозг, а стало быть, и установочные рефлексы и который обнаруживает только «основной поворот» головы. Следующий пример сделает сказанное более доступным.
Кролик. 8 апреля 1918 г. Правосторонняя экстирпация лабиринта, произведенная дн Клейном. Животное исследуется в день операции и 25 апреля, обнаруживает нормальные установочные рефлексы, как они наблюдаются у животных только с одним лабиринтом (см. далее). 25 апреля у кролика в сидячем положении на полу голова повернута на 90°, при нормальном положении животного в воздухе — на 180°, в висячем положении головой вниз — на 180° вправо.
26 апреля 1918 г. 10 ч. 20 м. Под неглубоким хлороформным наркозом перевязаны сонные артерии и перерезаны блуждающие нервы. Децеребрация, после которой довольно сильное кровотечение. Животное удерживается тазом вверх, чтобы препятствовать отеканию крови к продолговатому мозгу. Тотчас — отчетливая децереб- рационная ригидность всех четырех конечностей. Спонтанное дыхание. Кровотечение прекратилось.
Животное исследовано через короткое время после операции и в 10 ч. 55 м. При этом установлено следующее.
В нормальном положении в воздухе: голова повернута вправо на 90°.
В правом боковом положении в воздухе: голова повернута вправо на 90°.
В левом боковом положении в воздухе: голова повернута вправо на 90°.
В висячем положении головой вверх: голова в правом боковом положении.
В висячем положении головой вниз: голова повернута на 90° вправо. Ротовая щель и сагиттальная ось головы наклонены по отношению к горизонтали на 45°.
В положении на спине в воздухе: голова повернута на 45° вправо (вследствие силы тяжести полный поворот на 90° затруднен). Животное не предпринимает никаких попыток привести голову в боковое положение к животу.
Поворот головы в затылочное положение вызывает интенсивные шейные рефлексы на конечности, но не установочные шейные рефлексы на таз.
Поворот головы в боковое положение вызывает отчетливые лабиринтные рефлексы на передние ноги.
Боковое положение на столе: установочного рефлекса на голову нет. При устанавливании головы прямо тело остается лежать.
Перекладывание ног из положения «на спине» без изменения положения головы по отношению к туловищу ведет к отчетливому увеличению тонуса передних конечностей.
Положение на спине с головой, установленной прямо: правая передняя нога несколько более расслаблена, чем левая.
Имеются реакция поворота головы, последовательная реакция поворота головы, реакция горизонтального поворота глаз и горизонтальный нистагм, последовательная реакция поворота глаз и постнистагм, характеризующие типичное состояние после утраты правого лабиринта.
Животное забивается. На секции обнаружен гладкий симметричный разрез через средний мозг, дорсально — посередине через переднее двухолмие, вентрально — на 2 мм впереди переднего края моста; латерально — через ручки заднего двухолмия.
Заключение. У децеребрированного кролика все установочные рефлексы отсутствуют. Тонические шейные и лабиринтные рефлексы на мышцы конечностей
Последствия односторонней экстирпации лабиринта у кролика
227
имеются, равно как и реакции поворота головы и глаз. Вследствие утраты лабиринта с одной стороны голова повернута в сторону операции («основной поворот»). Этот поворот примерно одинаково выражен при всех положениях тела в пространстве. Только при положении в воздухе «на спине» он уменьшен вследствие влияния на голову силы тяжести.
Эти взаимоотношения типичны для децеребрированного кролика с одним утраченным лабиринтом. В отдельных случаях поворот может быть выражен еще сильнее. Обычно можно показать, что противодействие в случае поворачивания головы в правильное положение оказывается сильнее тогда, когда последняя находится в затылочном, а не в нормальном положении.
«Основной поворот» после односторонней экстирпации лабиринта характерен тем, что со временем он увеличивается. Если лабиринт удаляется у кролика с сохранным большим мозгом, то сразу после вмешательства голова при различных наклонах повернута по большей части только на 45° (20—90°). Спустя различное время, иногда уже через несколько дней, а в других случаях только через несколько недель, поворот достигает максимума в 90—135°, а иногда голова повернута даже на 180°. Эвальд подобное соотношение видел у голубей; здесь также поворот головы с течением времени увеличивается (см.: Эвальд, рис. 4—И).
«Основной поворот» не зависит от глаз. При закрывании глаз он не изменяется, осуществляясь также и после децеребрации или экстирпации глазных яблок. Мало зависит он и от других частей тела, кроме лабиринта. Таким образом, здесь мы имеем дело с первичным длительным результатом утраты лабиринта. Основной поворот наступает и после предварительной перерезки трех верхних пар шейных задних корешков.
Описанные отношения в дальнейшем остаются, по существу, неизменными. И через 6 месяцев после правосторонней экстирпации лабиринта поворачивание головы влево при положении животного на спине встречает значительно большее противодействие, чем при положении на животе, тогда как противодействие поворачиванию вправо одинаково невелико при всех положениях головы в пространстве. Таким образом, влияние одного лабиринта на мышцы шеи остается односторонним в течение продолжительного времени.
Основной поворот имеет место со всеми своими особенностями и у кролика с интактной центральной нервной системой.
Поворот туловища
Помимо поворота головы, у односторонне лабиринтэктомированного кролика, находящегося в сидячем положении, часто (не всегда) можно отметить поворот туловища, когда при прямосидящем тазе грудь животного перегибается в сторону операции или лопатка оперированной стороны тела лежит на полу (ср., например, рис. 150 и 151, стр. 248 и 249—250). У сидящего животного, однако, трудно установить, обусловлено ли это ненормальное положение поворотом туловища или потерей тонуса передних конечностей оперированной стороны. Поэтому поворот туловища исследуется иначе.
Для этой цели животное удерживается за поясничную часть позвоночника на уровне пахового сгиба и ему придается висячее положение. Тогда наступившие после экстирпации лабиринта изменения положения головы, шеи и туловища можно изучать в чистом виде, вне мешающих влияний тонуса конечностей на положение. Сила тяжести действует на брюшную часть, грудь, шею и голову в одном направлении, поэтому не оказывает препятствующего влияния, какое должно наступать, если захватить животное за грудной отдел и исследовать в висячем положении головой вверх.1
При этом положении животного установочные рефлексы не нарушаются.
1 Имеющие место при этом последнем положении животного аномалии положения головы н шеи подробно описаны Винклером (1), стр. 22 и след.
15*
228
Шестая глава. Последствия односторонней экстирпации лабиринта
Как показывают рис. 137—139, в висячрм положении головой вниз спиральный поворот всего тела виден с особенной отчетливостью.
На рис. 139, например, дорсальная сторона таза обращена вправо, дорсальная сторона головы — влево. На всех трех рисунках видно, что поворот начинается уже в поясничной части позвоночника и продолжается вплоть до головы. Грудь поэтому принимает примерно промежуточное
положение между тазом и головой.
Чтобы дать более точное описание этой аномалии положения, целесообразно измерить поворот, который производит дорсо-вентральная ось тела в грудном, шейном и головном отделах по отношению к дорсо-вентральной оси таза. Дорсо- вентральную часть тела животного, удерживаемого в воздухе, можно ориентировать по стоя-
Рис. 137. Кролик III. Левосторонняя экстирпация лабиринта 23 октября 1912 г. Сфотографирован 2 ноября, т. е. спустя 10 дней после операции. Животное удерживается в висячем положении головой вниз, рука держит поясничную часть позвоночника на высоте сгиба бедра. Таз дорсальной стороной обращен вправо, задние ноги — влево. Наоборот, грудной отдел вентральной стороной направлен кпереди и несколько влево, правая передняя нога вытянута, левая — согнута. Голова обращена вентральной (нижнечелюстной) стороной вправо и вперед так, что левая щека сильнее повернута к наблюдателю. Кроме того, голова отчетливо отклонена влево. Левое ухо свисает
вниз (паралич лицевого нерва).
нию корня хвоста, зада и задних ног. Затем на вентральной стороне нащупывается мечевидный отросток, на дорсальной — остистый отросток последнего грудного позвонка; соединяющая их линия является дорсо-вентральной осью нижней апертуры грудной клетки. Такую же ось можно установить и для верхней апертуры грудной клетки, нащупав рукоятку грудины и остистый отросток первого грудного позвонка. Дорсо-вентральная ось головы также легко определяется непосредственно. Если, кроме того, определить еще и степень наклона головы, т. е. угол, который образует сагиттальная ось головы (морда—большое затылочное отверстие) и вертикальная ось, то можно дать достаточно точное описание анормального взаиморасположения отделов тела каждого животного в этом положении, а также сравнить данные, полученные на различных животных и в разное время.
Во время наблюдений на различных животных в периоды до полугода после операции установлено следующее.
Что касается наклона головы, то он, как было упомянуто, отмечается у сидящего животного только в первые дни после операции. Напротив, у висящего кролика он постоянно и отчетливо заметен также и спустя полгода. Обычно он составляет около 30—45°, реже больше. Вероятно, что в сидячем положении наклон головы корригируется животным. х
Для поворота тела были получены следующие величины: тотчас
после операции по отношению к тазу нижняя апертура грудной клетки повернута по большей части на 30—45°, верхняя апертура — на 45—70°, голова — на 80—135°. Этот поворот в течение последующих дней и недель увеличивается и постепенно достигает максимума. У отдельных животных это происходит уже через несколько дней, у других — может длиться до 4 недель. Когда этот максимум достигнут, поворот нижней апертуры грудной клетки составяет 45—90°, верхней — от 60 до 135°, головы — 1JÖ—1800. Эти цифры колеблются в указанных выше пределах от живот¬
Последствия односторонней экстирпации лабиринта у кролика
229
ного к животному, ото дня ко дню. Дальнейшие наблюдения показывают, что этот интенсивный поворот всего тела является стойким последствием операции. Он никогда не исчезает полностью, а при исследовании животных в висячем положении, не исчезает даже и частично. Достигнутый максимум чаще остается в упомянутых пределах без изменений. Таким образом, если со временем сидящее животное постепенно уменьшает поворот головы, то речь здесь идет не об уменьшении влияния лабиринта, так как исследование в висячем положении показывает, что это влияние остается без изменения, а о других компен-
Рис. 138. Тот же кролик, что и на рис. 137, сфотографированный с другой стороны. Левосторонняя экстирпация лабиринта 23 октября 1912 года. Сфотографирован 2 ноября, т. е. 10 дней спустя после операции, непосредственно после снимка, приведенного на рис. 137. Животное удерживается в воздухе в висячем положении головой вниз. Таз обращен дорсальной стороной влево, задние ноги — вправо. Грудь дорсальной стороной направлена вперед, видна (вытянутая) правая передняя нога. Голова повернута еще сильнее, так, что правая щека направлена к наблюдателю. Кроме того, голова отклонена так, что левая щека животного приближена к левому плечу, правая — обращена вниз. Левое ухо (паралич лицевого нерва) отвисает в сторону. Девиация правого глаза в дорсальном направлении отчетлива: виден нижний белый
край склеры.
сирующих группах рефлексов, из которых наиболее существенную роль играют установочные рефлексы туловища на туловище и на голову.
Ранее было показано, что у нормального кролика рефлекторно благодаря повороту шеи можно вызвать спиральный поворот туловища, который наступает и после экстирпации обоих лабиринтов и должен быть отнесен за счет шейного установочного рефлекса. На-
Рис. 139. Кролик II. Левосторонняя экстирпация лабиринта 19 октября 1912 г. Сфотографирован 20 января 1913 г., т. е. через 3 месяца после операции. Висячее положение головой вниз. Таз дорсальной стороной обращен вправо и несколько книзу, задние ноги — налево и несколько вперед. Эпига- стриум обращен вперед, вентральная сторона грудного отдела — кпереди и вправо. Более сильное сгибание правой передней ноги на этом снимке не видно. Голова повернута еще сильнее, так что вентральная сторона (сторона нижней челюсти) направлена вправо, темя же — влево. Кроме того, голова наклонена так, что морда приближена к наблюдателю, а левая щека обращена кверху. Левый глаз отведен вентрально так, что в глазной щели виден почти исключительно верхний край склеры, третье веко опущено. Левое ухо (паралич лицевого нерва)
свисает вниз.
правление поворота туловища совпадает с поворотом, который наблюдается у кролика после односторонней экстирпации лабиринта. Отсюда уже заранее , представляется вероятным, что этот шейный рефлекс имеет
230
Шестая глава. Последствия односторонней экстирпации лабиринта
значение и при аномалии установки после утраты лабиринта, и возникает даже вопрос: имеет ли поворот туловища после утраты одного лабиринта у кролика отношение к лабиринту вообще, и не вызывывается ли он скорее поворотом шеи? Если поворот туловища осуществляется только с шеи, он должен производиться в обратном направлении после приведения шеи в правильное положение. Позднее будет показано, что у морской свинки и у собаки поворот туловища действительно исчезает, если голову привести по отношению к грудному отделу в правильное положение. Однако у кролика дело обстоит не так. Скорее в данном случае основной поворот туловища складывается с проявлениями названных шейных рефлексов.
Если кролика после односторонней экстирпации лабиринта привести в висячее положение головой вниз и определить поворот нижней и верхней апертур грудной клетки и головы по отношению к тазу, то видно, что, тотчас после того как поворот головы скорригирован по отношению к грудному отделу, поворот туловища уменьшается, но исчезает полностью только в небольшом числе случаев. По большей части грудной отдел остается по отношению к тазу повернутым на 20—45°.
Различные кролики ведут себя во время этих опытов по-разному. У некоторых животных после коррекции положения головы имеется незначительный поворот или же совсем нет никакого поворота туловища, у других, наоборот, обнаруживается поворот грудного отдела тела по отношению к тазу на 45° после того, как голова приведена относительно груди в правильное положение. У одних животных отчетливее выражено влияние шейных установочных рефлексов; у других — лабиринтный основной поворот. Это наблюдение вполне соответствует данным, полученным на децеребрированных животных. То же Ноэль Патон (1) получил и на утках.
Нередко оказывается, что в первые дни или недели после операции при прямо стоящей голове поворот туловища почти или полностью исчезает, в то время как спустя недели или месяцы установление головы прямо уже не позволяет полностью снять поворот туловища. Это согласуется с упомянутым выше наблюдением, что вообще основной поворот после операции становится все более интенсивным и постепенно достигает максимума.
Если голову кролика, лишенного одного лабиринта и подвешенного головой вниз, установить относительно грудного отдела прямо (и если после этого еще сохраняется поворот грудного отдела по отношению к тазу), то можно поворачивать голову еще дальше и привести ее в положение, противоположное исходному относительно грудного отдела. Этим возбуждается шейный рефлекс на мускулатуру туловища в противоположном направлении, который оказывается в состоянии компенсировать излишек поворота так, что теперь грудной отдел стоит относительно таза прямо. В большинстве случаев для этого необходимо повернуть голову на 30—45° в противоположном направлении. Таким путем можно использовать ту степень поворота шеи, какая необходима для установки грудного отдела прямо, чтобы измерить величину влияния лабиринта, которое при этом компенсируется. Если теперь поворачивать голову еще дальше, наступает поворот туловища в противоположном направлении. В соответствии с этим у каждого кролика, лишенного одного лабиринта, путем более или менее интенсивного поворота головы в направлении, противоположном спонтанному ее повороту, можно снять поворот туловища и даже вызвать поворот его в противоположном направлении. Таким образом, хотя шею нельзя считать исключительно ответственной
Последствия односторонней экстирпации лабиринта у кролика
231
за поворот туловища, но она в значительной степени определяет этот акт. Влияние сохраняется недели и месяцы. Дело идет, таким образом, исключительно о тонических рефлексах.
С другой стороны, в согласии с изложенным оказывается, что прямое влияние лабиринта у кролика не ограничивается одной мускулатурой шеи, но что (по крайней мере у этих животных) действие одностороннего выпадения лабиринта может быть доказано и для мышц туловища. И здесь дело идет о длительных последствиях, о тонических иннервациях.
Описанные выше наблюдения над подвешенным кроликом подтверждаются и расширяются с помощью подобных экспериментов в положении животного на спине.
Если кролика без одного лабиринта уложить на спину так, чтобы дорсо- вентральная ось грудного отдела стояла бы вертикально, и прочно удерживать этот отдел в таком положении рукой, то поворот как головы, так и таза по отношению к грудному отделу выражен отчетливо. После правосторонней экстирпации лабиринта правый глаз глядит вверх,темя обращено вправо. Наоборот, правая защечная область прилегает к столу, левая — направлена кверху. Таким образом, животное находится в положении, похожем на изображенное на рис. 116, стр. 191, где нормальный кролик представлен в положении на спине с головой, повернутой вправо. Тем самым активирован шейный рефлекс, который обусловил поворот таза. Уже при этом становится ясно,что и у односторонне лабиринтэкто- мированного кролика поворот шеи должен вызвать такой поворот таза. Если теперь голове односторонне лабиринтэктомированного животного придать по отношению к фиксированному рукой грудному отделу прямое положение, то можно видеть, что поворот таза относительно грудного отдела уменьшается, но в противоположность животному с интактными лабиринтами в большинстве случаев полностью не исчезает. Чаще всего остается поворот таза на 20—45°. Чтобы этот последний выровнять полностью, голова должна быть повернута на 20—45° в противоположном направлении (при правосторонней экстирпации лабиринта влево). При поворачивании головы еще дальше происходит поворот таза в противоположном направлении. Таким путем можно, например, добиться, чтобы после правосторонней экстирпации лабиринта в положении животного на спине при поворачивании головы влево левая защечная область оказалась бы внизу.
Таким образом, этц опыты также приводят к заключению, что поворот туловища у кролика после односторонней утраты лабиринта является результатом двух влияний, которые складываются друг с другом, а именно: во-первых, описанного шейного рефлекса и, во-вторых, прямого одностороннего действия оставшегося лабиринта на мускулатуру туловища.
Опыты в положении животного на спине потому так особенно наглядны, что в процессе их можно удобно фиксировать грудной отдел в этом положении и затем измерять повороты как головы, так и таза по отношению к прочно установленному грудному отделу.
При исследовании в положении на спине оказывается также, что у некоторых кроликов лабиринт, у других — шейные установочные рефлексы оказывают большее влияние на поворот туловища.
Приведенные до сих пор наблюдения приводят к следующим заключениям.
Одностороннее выпадение лабиринта у кролика первично ведет к появлению односторонних влияний на осевую мускулатуру, представляющих
232
Шестая глава. Последствия односторонней экстирпации лабиринта
собой длительное действие, интенсивность которого после операции увеличивается.
При этом осуществляются: .
1) очень сильно выраженный поворот шеи;
2) наклон шеи (менее интенсивно);
3) не очень интенсивный основной поворот туловища вплоть до таза,, вследствие чего все тело животного поворачивается спирально.
Поворот шеи вторично вызывает интенсивный поворот туловища в том* же направлении, что и поворот, зависящий от лабиринтов, благодаря чему последний значительно увеличивается (шейный установочный рефлекс).
Поворот туловища в большинстве случаев может быть частично уменьшен при помощи устанавливания головы прямо относительно туловища. Для полного снятия поворота голову следует продолжать поворачивать дальше, на другую (т. е. противоположную спонтанному повороту) сторону. Еще более сильный поворот головы в другую сторону вызывает* поворот туловища в противоположном направлении.
Как было показано выше, оставшийся лабиринт господствует, смотря по его положению в пространстве, с помощью рефлекторных механизмов, над состоянием тонуса мускулатуры шеи одной стороны. Шейная мускулатура другой стороны,1 даже спустя длительное время после операции,, не подпадает под контроль оставшегося лабиринта.
Лабиринтные установочные рефлексы на голову после односторонней экстирпации лабиринта
(52)
До сих пор в качестве первичного следствия односторонней экстирпации лабиринта мы рассматривали основной поворот головы, который осуществляется благодаря посредничеству центров заднего отдела продолговатого мозга. К этому присоединяются основной поворот туловища и, кроме того, шейный установочный рефлекс, который усиливает основной поворот и осуществляется центрами области моста.
Кроме того, у животных с неповрежденной центральной нервной системой средний мозг также интактен, а потому должны осуществляться также и установочные рефлексы. В нашем случае наиболее важную роль при этом, конечно, играют лабиринтные установочные рефлексы, исходящие из оставшегося лабиринта. Таким образом, для понимания всего' симптомокомплекса оказывается необходимым раньше всего выделить установочные рефлексы, исходящие из оставшегося лабиринта.
Результат этого анализа следующий: в то время как при обоих интактных лабиринтах установочные лабиринтные рефлексы всегда приводят голову в нормальное положение, те же рефлексы после односторонней экстирпации лабиринта поворачивают голову интактным лабиринтом кверху. Изучение этого лабиринтного установочного рефлекса осложняется тем, что последний оказывает влияние не только на голову, находящуюся в нормальном положении, но также и на голову, которая в результате описанного основного поворота уже оказывается повернутой. На основной поворот накладывается влияние лабиринтного установочного рефлекса после односторонней экстирпации лабиринта, что усиливает*
1 Под шейной мускулатурой одной и другой сторон здесь понимается та часть мышц шеи, которая после экстирпации лабиринта приходит в тоническое напряжение или resp. расслабляется.
Последствия односторонней экстирпации лабиринта у кролика
233
или ослабляет этот поворот, смотря по тому, какое исходное положение принимает в пространстве тело животного.
Если желательно сделать влияние лабиринтного установочного рефлекса на голову особенно демонстративным, то лучше всего ставить опыты именно в первые дни после экстирпации лабиринта, так как в это время основной поворот еще не достиг слишком значительного развития и может быть с легкостью преодолен установочными рефлексами, в то время как сильно выраженный основной поворот уменьшает действие установочных рефлексов.
В свое время мне удалось произвести исследования установочных рефлексов на двенадцати кроликах, у которых де Клейном была произведена правосторонняя экстирпация лабиринта. После операции у животных обнаружилось типичное поведение; кроме того, радикальность операции всякий раз контролировалась на секции. Лишь у одного из этих животных при повторном исследовании были обнаружены некоторые отклонения положений. У того же животного на секции было обнаружено нагноение правой буллы, чем и объяснялись отклонения. В дальнейших исследованиях это животное не принималось во внимание.
Поведение животных изучалось при интактном большом мозге. Два животных исследовались с завязанными глазами, чтобы исключить влияние на их поведение оптических раздражений. У пяти кроликов на 5-й, 11-й, 28-й и 30-й день после экстирпации лабиринтов удалялся большой мозг. После этого продолжалось исследование таламического животного. Радикальность операции контролировалась на секции.
Выяснилось, что динамика установочных рефлексов у животного с интактным большим мозгом и у таламического животного в существенных чертах одна и та же, так что изложение результатов обеих серий наблюдений в дальнейшем может быть объединено. Равным образом у кролика с интактным большим мозгом во время исследования установочных рефлексов при закрытых или при открытых глазах существенных различий не наблюдается; у животных этого вида в поддержании нормального положения тела глаза играют лишь незначительную роль, которая в дальнейшем еще будет обсуждаться. Если только средний мозг сохраняет функционирующие связи с остальным стволом, лабиринтные установочные рефлексы осуществляются типичным образом. Чтобы выключить остальные установочные рефлексы, животные должны исследоваться в воздухе.
Динамика лабиринтных установочных рефлексов должна быть представлена с помощью стереоскопических снимков, сделанных с кролика через 3 дня после правосторонней экстирпации лабиринта. Как упоминалось, общим результатом исследований был факт, что лабиринтный установочный рефлекс всегда стремится установить голову таким образом, чтобы интактный лабиринт находился сверху и голова лежала в боковом положении. Смотря по тому, в каком положении находится животное в пространстве, этот рефлекс различным образом комбинируется с «основным поворотом».
1. Нормальное положение в воздухе. Если животное свободно удерживается рукой в воздухе за тазовую область (рис. 140), то видно, что голова повернута примерно на 90° вправо и удерживается в правом боковом положении левым глазом кверху. В этой позиции основной поворот и лабиринтный установочный рефлекс действуют сообща.
В большом числе случаев в первое время после операции голова повернута точно на 90°. Позднее, когда основной поворот усиливается,
234
Шестая глава. Последствия односторонней экстирпации лабиринта
голова может повернуться еще сильнее, иногда на 135° и даже на 180°, так что оказывается в затылочном положении. В отдельных случаях наблюдалось, что сначала голова вследствие основного поворота была повернута на 135—160°, а затем вследствие лабиринтного установочного рефлекса медленно подвигалась обратно в боковое положение, в каковом и оставалась.
Если основной поворот выражен сильно, то по большей части отмечается еще и поворот грудной клетки по отношению к тазу. Если желательно выключить влияние последнего на положение головы, то следует держать животное в воздухе, ухватив его за кожу спины, тогда поворот
Рис. 140. Рис. 141.
головы составляет даже при сильном основном повороте обычно только 90°, в редких случаях: — 135°.
Таламическое животное ведет себя точно так же, как животное с интактным большим мозгом. Закрывание глаз ничего в положении тела не изменяет.
2. Положение на спине в воздухе. В положении на спине основной поворот и лабиринтный установочный рефлекс противодействуют один другому. Благодаря основному повороту голова (после правосторонней экстирпации лабиринта) приводится в левое боковое положение. Лабиринтный установочный рефлекс стремится привести голову в правое боковое положение. В результате это положение оказывается для животного весьма неприятным, и энергичными движениями оно пытается выйти из него. Как интактное, так и таламическое животное в этом положении производят головой и передней половиной тела кружащие движения, а именно — повороты в левую сторону,1 которые продолжаются до тех пор, пока животному не удается принять весьма примечательное положение, которое соответствует обоим рефлексам.
Результат представлен на рис. 141. Грудной отдел повернут в направлении основного поворота относительно таза таким образом, что передние лапы направлены влево, а голова вследствие сильнейшего отклонения в сторону шеи прижата к вертикальной стороне животного так, что прилежит к брюху в правом боковом положении. Левый глаз направлен кверху, ротовая щель стоит почти вертикально, уши свешиваются к правой стороне тела.
Как только животное выходит из этого покоящегося положения, снова начинаются кружащие движения передней половины тела влево, что
1 Т. е. повороты в направлении, обратном катательным движениям (см. далее).
Последствия односторонней экстирпации лабиринта у кролика
235
длится до тех пор, пока голова снова не прижимается к животу кролика в правом боковом положении. Это поведение установлено для всех без исключения исследованных животных как без, так и при наличии большого мозга вне зависимости от времени, которое прошло от экстирпации лабиринта до постановки эксперимента.
3. Правое и левое боковое положение в воздухе. Если животное, у которого утрачен правый лабиринт, удерживается в воздухе в правом боковом положении (т. е. правой стороной тела вниз), то голова в случае, если никакого поворота ее не наблюдается, находится в правильном положении, в которое ее приводит лабиринтный установочный рефлекс. Основной поворот, однако, вращает голову из этого положения и стремится повернуть ее в затылочное (или даже превышающее его по углу поворота) положение. Оба рефлекса таким
Рис. 142. Рис. 143.
образом противодействуют друг другу. Лабиринтный рефлекс уменьшает основной поворот, а при известных обстоятельствах может даже вовсе его снять, но никогда не может повернуть голову дальше, чем в правое боковое положение.
Иное имеет место при левом боковом положении. Основной поворот ориентирует голову по направлению к нормальному положению. Лабиринтный установочный рефлекс точно так же стремится освободить голову из левого бокового положения. Оба рефлекса, таким образом, суммируются, и наступает особенно интенсивный поворот головы.
Э^то можно видеть на рис. 142 и 143.
ЕГа рис. 142 животное удерживается в воздухе за четыре ноги в правом боковом положении. Голова несколько опущена вниз, однако в общем повернута вправо не больше, чем на 45°, так что правый глаз обращен кверху. В других случаях, которые также могут быть фиксированы на фотографиях, голова стоит точно в плоскости всего животного в правом боковом положении, так что какой бы то ни было поворот головы относительно тела отсутствует.
На рис. 143 животное удерживается в воздухе за четыре ноги в левом боковом положении. Голова так сильно повернута и отклонена вправо, что правый глаз (также как на рис. 142) обращен кверху. Морда несколько опущена вниз. Последнее не является правилом. У сильных неутомленных животных морда может оказаться приподнятой почти до горизонтальной линии.
При сравнении рис. 142 и 143 видно, что, несмотря на то, что в обоих случаях тело животного находится в положениях, отличающихся одно от другого на 180°, положение головы разнится только на 45—60°. Это
236
Шестая глава. Последствия односторонней экстирпации лабиринта
происходит потому, что в одном случае лабиринтный установочный; рефлекс увеличивает основной поворот, в другом — уменьшает его.
Сравнение положения головы при левом и правом боковых положениях тела в воздухе является (в совокупности с исследованием в спинальном положении) лучшим способом убедиться в наличии действующего лабиринтного установочного рефлекса после односторонней экстирпации лабиринта.
В отдельных случаях получаются различные картины, смотря по тому, сильно или слабо выражен основной поворот. Если основной поворот выражен лишь незначительно, что в большинстве случаев и имеет место в первые дни после операции, то в правом боковом положении голова оказывается повернутой на 45—0°, так как лабиринтный установочный рефлекс совсем или в значительной степени уменьшает основной поворот. В левом боковом положении при этом имеет место основной поворот на 90—135°, только в исключительных случаях — на 180°. Наоборот, если основной поворот интенсивен, что по большей части имеет место спустя 2—4 недели, то в правом боковом положении обнаруживается поворот головы на 90° (голова в положении на спине), в левом боковом положении,, наоборот, — поворот на 180° (голова в правом боковом положении). При этом имеют место всевозможные промежуточные состояния. Однако- отчетливая, разница в степени поворота головы в левом и правом боковых положениях тела обнаруживается неизменно.
Иногда можно наблюдать, если животное находится в воздухе в боковом положении, как сначала после основного поворота голова поворачивается примерно на 45°, а затем медленно (уже вследствие лабиринтного установочного рефлекса) идет обратно до достижения полного бокового положения.
Различия между животными с имеющимся или удаленным большим мозгом установить не удалось. После завязывания глаз реакция животного также не меняется.
Если в боковом положении животные удерживаются в воздухе не за четыре ноги, как на рис. 142 и 143, а за таз, то можно установить, как при сильном или слабом повороте головы грудной отдел в правом боковом положении только немного, в левом боковом — весьма значительно повернут относительно таза (шейный установочный рефлекс).
4. Висячее положение головой вверх. Если животное ухватить за кожу спины и удержать так, чтобы позвоночник был расположен отвесно, головным концом кверху, то голова свисает в правую сторону и остается в правом боковом положении, левым глазом кверху, с вертикально расположенными ротовой щелью и горизонтальносагиттальной осью (рис. 144).
Это положение головы можно было обнаружить почти у всех животных. Оно возникает вследствие взаимодействия основного поворота и лабиринтного установочного рефлекса. Последний обеспечивает установку головы в правом боковом положении. Только когда основной поворот очень интенсивен, голова наклоняется на бок более, чем на 90°; в исключительных случаях этот поворот может спустя 3—4 недели после операции составить почти 180°. Это положение, однако, неприятно животному; нередко оно производит круговые движения головой и предпринимаем попытки придать голове правое боковое положение.
5. Висячее положение головой вниз. Это положение единственное, в котором лабиринтный установочный рефлекс не можем быть использован вполне и действительно уступает основному повороту.
Последствия односторонней экстирпации лабиринта у кролика
237
Поэтому именно это положение наиболее удобно для исследования динамики основного поворота у недецеребрированного животного.
Если ухватить животное за тазовую область и дать ему повиснуть головой вниз, все тело животного, как это подробно описывалось ранее, перекручивается спирально в сторону отсутствующего лабиринта на ‘90—180°, смотря по степени выраженности основного поворота. Кроме того, голова отклонена в направлении отсутствующего лабиринта на угол до 45° и сагиттальная ось головы наклонена примерно на 45° вниз. Изображения этой позы см. на рис. 137—-139, стр. 228—229.
Таким образом, после правосторонней экстирпации лабиринта голова стоит наполовину в положении на спине, наполовину — в левом боковом, с правым глазом выше левого.
Рис. 144. Рис. 145.
Это положение для животного исключительно неприятно, и в большинстве случаев, оно тотчас же производит передней частью тела энергичные круговые движения влево, пока наконец не обессиливает.
Если животное пытается освободить свою голову из столь неблагоприятного для лабиринтного установочного рефлекса положения, для этого ему остаются два пути, которые в действительности и используются. Животное может увеличивать наклон головы, пока, наконец, левый глаз не окажется обращенным не вниз, а в сторону. Для этого требуется значительное мышечное напряжение, и поза может сохраняться только в течение недолгого времени. Действительно, в отдельных случаях мне удавалось наблюдать эту реакцию у кроликов, в особенности у таламических животных; она может быть отмечена и у таламических кошек. Чаще же лабиринтный установочный рефлекс проявляется лишь в том, что наклон головы вправо уменьшается. До истинного наклона влево дело доходит лишь в редких случаях, да и тогда только ненадолго. Чаще же, наоборот, в первые дни после экстирпации лабиринта, когда основной поворот еще незначителен, при висячем положении головой вниз наклон вправо не выражен и голова животного свешивается вниз, почти вертикально (рис. 145).
Если голова должна была бы приводиться в правое боковое положение, животному пришлось бы, находясь в висячем положении головой вниз, отклонить свою голову влево, а это, очевидно, не может осуществиться без преодоления основного поворота и в тем меньшей степени,
238
Шестая глава. Последствия односторонней экстирпации лабиринта
что основной поворот как раз при затылочном положении головы выражен всего сильнее, так как соответствующий тонический лабиринтный рефлекс на мышцы затылка в этом положении достигает своего максимума.
Именно это положение у кролика является единственным, при котором лабиринтный установочный рефлекс играет лишь незначительную роль.
Однако во всех остальных положениях при наличии только одного лабиринта голова благодаря лабиринтному установочному рефлексу всегда приближается к такому положению, когда интактный лабиринт оказывается наверху. Если мы по-прежнему воспользуемся примером с правосторонней экстирпацией лабиринта, то лабиринтный установочный рефлекс будет стремиться привести голову в правое боковое положение, в котором интактный левый лабиринт находится наверху. В этом положении и голова, и животное успокаиваются. Таким образом, исходящие из лабиринтов установочные раздражения в этом положении минимальны. Во всех других положениях головы в пространстве установочные импульсы более сильные и максимальны тогда, когда интактный лабиринт при боковом положении головы оказывается внизу. В этих условиях на голове сказываются влияния мощных рефлексов, что ведет к тому, что голова оказывается возвращенной в положение покоя (в нашем примере — правое боковое положение). Если в сцлу каких-либо причин это оказывается неосуществимым (висячее положение головой вниз, фиксация животного), наступают энергичные оборонительные реакции, по большей части движения по кругу передней половины тела.
Противоположным лабиринтным установочным рефлексам, которые осуществляются через средний мозг, является основной поворот, центры которого лежат более каудально и который при других положениях головы оказывается максимальным и придает определенное положение в пространстве не голове, как лабиринтные установочные рефлексы, а туловищу. Правда, интенсивность поворота зависит от положения головы в пространстве, однако основной поворот постоянно приводит к поворачиванию головы в направлении движения туловища. Таким образом, оба рефлекса должны суммироваться или вычитаться, смотря по положению животного в пространстве, совершенно различным образом, потому что результат оказывается довольно сложным. Так как оба рефлекса исходят из лабиринта, рассмотреть их раздельно оказалось возможным, только доказав, что центры их имеют различное положение в центральной нервной системе и поэтому могут быть разделены оперативным путем.
Благодаря тому, что окончательное положение головы животного в воздухе определяется совместным действием обоих рефлексов, из которых основной поворот у различных животных и в разное время не является одинаково интенсивным, максимальные и минимальные положения для лабиринтных установочных рефлексов не могут быть определены с точностью до небольших степеней. Лишь в общем можно сказать, что в минимальном положении голова оказывается в боковом положении интактным лабиринтом кверху, в максимальном положении, наоборот, — на противоположной стороне. Соответствуют ли истинные максимальные и минимальные положения головы точно боковому положению тела или отклоняются от него в различных направлениях на 20— 30°, с уверенностью сказать, наблюдая животное, лишенное одного лабиринта, нельзя.
Как бы там ни было, наблюдения, описанные в этом разделе, учат, что в случае, если тело животного находится в пространстве в нормаль¬
Последствия односторонней экстирпации лабиринта у кролика
239
ном положении, то односторонний лабиринтный, установочный рефлекс направляет голову в боковое положение, при котором интактный лабиринт оказывается наверху. Таким образом, окончательный результат выглядит так, как если бы имел место один только основной поворот средней степени. Если, наоборот, тело находится в иных, чем нормальное, положениях, основной поворот и односторонний лабиринтный установочный рефлекс часто должны действовать не согласованно, но взаимно противодействуя. Становится поэтому понятным, почему животные, лишенные лабиринта с одной стороны, при различных положениях тела в пространстве придают такие исключительно неправильные положения голове и, как следствие этого, всему телу.
Установочные рефлексы тела на голову
Если после экстирпации лабиринта с одной стороны кролик исследуется не в воздухе, а сидит или лежит на полу, то к влияниям, описанным до сих пор, присоединяются установочные рефлексы с тела, из которых здесь прежде всего должны быть рассмотрены установочные рефлексы с тела на голову. Как было показано в предыдущей главе, асимметричное возбуждение чувствительных нервов тела вызывает поворачивание головы в нормальное наложение. Тот же рефлекс можно показать, если животное без одного лабиринта сначала подержать в боковом положении в воздухе, определить степень поворота его головы и затем уложить на стол. Если в левое боковое положение на стол укладывается животное, у которого отсутствует правый лабиринт, то сильно выраженный уже в воздухе поворот головы усиливается еще более и голова (если этого не было еще в воздухе) может оказаться полностью переведенной в правое боковое положение. Наоборот, если животное укладывать на стол в правом боковом положении, часто основной поворот становится еще меньше, чем поворот, обусловленный лабиринтным установочным рефлексом, и голова может оказаться повернутой влево (чего один лабиринтный установочный рефлекс никогда не мог бы вызвать), приближаясь, таким образом, к нормальному положению (рис. 146). До этого животное удерживалось в воздухе в правом боковом положении. Вследствие лабиринтного установочного рефлекса основной поворот оказался полностью компенсированным так, что животное держало свою голову в воздухе точно в правом боковом положении, т. е. голова не была больше повернутой по отношению к телу. При этом, как только животное было уложено на стрл в правом боковом положении, голова оказалась повернутой влево (рис. 146) и примерно на 45° приблизилась к нормальному положению. Как говорилось, этот рефлекс наступает в правом боковом положении на столе не во всех без исключения случаях, хотя у ряда животных мог быть установлен с определенностью.
Если животное, лишенное правого лабиринта, захватить за правую сторону тела и свободно удерживать в воздухе, то вследствие этого асимметричного раздражения поворот головы вправо уменьшается и животное ведет себя спокойно. Наоборот, если захватить его за кожу левой стороны тела, то поворот головы вправо увеличится и, помимо этого, животное обычно производит передней половиной тела оживленные кружащие движения, которые можно тотчас же прекратить, захватив рукой кожу правой стороны тела.
Интересно также следующее наблюдение, которое может быть сделано на многих животных в различное время после операции. На рис. 140 изображено животное, лишенное правого лабиринта, удерживаемое за
240
Шестая глава. Последствия односторонней экстирпации лабиринта
тазовую область в воздухе в нормальном положении. Голова по отношению к груди повернута на 90° и находится в правом боковом положении. Затем животное усаживают на стол, при этом поворот головы постепенно уменьшается и в конце концов составляет только 45° (рис. 147).
Опыт можно поставить и таким образом, чтобы сначала удерживать животное в воздухе в нормальном положении за кожу спины для того, чтобы быть уверенным, что грудной отдел стоит в воздухе симметрично в нормальном положении, и после определения поворота головы усаживать животное на землю. И в этом случае часто можно видеть наступающее уменьшение поворота головы. Если основной поворот выражен незначительно, то может случиться, что, сидя на полу, животное вообще не держит более голову повернутой. При сильно выраженном основном
Рис. 146. Рис. 147.
повороте, когда животное в воздухе держит голову повернутой на 180°, при сидении также часто можно видеть, как наступает уменьшение поворота до 90°. Опыт удается на таламическом животном, а также не нарушается у животного с интактным большим мозгом, если завязать животному глаза.
Установочные рефлексы тела по прошествии некоторого времени усиливаются. На этих рефлексах основан тот факт, что через различные сроки (после 1—3 дней в одних случаях и только после 1—4 недель — в других) поворот головы во время сидения исчезает полностью, в то время как в висячем положении головой вниз он может обнаруживаться в течение длительного времени. В дальнейшем поворот головы часто также уменьшается. Выше упоминалось, что после экстирпации лабиринта основной поворот понемногу увеличивается. Даже через несколько месяцев, как это можно показать в исследовании в висячем положении головой вниз, он не становится меньше. При сидении животного на полу можно нередко наблюдать, ,как по прошествии примерно 3—4 месяцев поворот головы снова уменьшается, так что никогда не превышает 90°, составляя обычно лишь 45° или меньше. Объяснение этому лежит в постепенно наступающем усилении установочных рефлексов тела на голову.
Этот рефлекс приводится в действие тем, что животное соприкасается с полом своими четырьмя лапами. Установочные рефлексы тела на голову, описанные в предшествующей главе, наблюдались у животных, у которых оба лабиринта были интактными или у которых оба лабиринта отсутствовали. У них голова стоит симметрично. Установочные рефлексы у них осуществляются благодаря асимметричному раздражению нервов тела при симметричном стоянии головы. В описанных здесь опытах,
Последствия односторонней экстирпации лабиринта у крол ика
241
наоборот, рефлекс приводится в действие благодаря симметричному раздражению тела при асимметричном (повернутом) стоянии головы. В этом случае дело идет о «переключении» (Schaltung), наличие которого в осуществлении установочных рефлексов было уже показано в предыдущей главе. Вследствие поворота головы мышцы обеих половин шеи находятся в состоянии различного напряжения, и, следовательно, в состоянии различной рефлекторной готовности. Если теперь с конечностей обеих сторон, как только они коснутся пола, начнут поступать возбуждения, наступит сокращение тех мышц шеи, которые благодаря повороту находятся в состоянии большего растяжения, и поворот головы уменьшается.
Опыт представляется важным, ибо показывает, что возбуждения, ведущие к установочным реакциям, достигают центральной нервной системы не только тогда, когда нервы тела раздражаются асимметрично, но также и тогда, когда возникают симметричные раздражения. Последние не в состоянии, во всяком случае при симметричном стоянии головы, т. е. у нормальных животных, мобилизовать установочные реакции.
ь Тонус конечностей (15)
Всем исследователям, которые до сих пор занимались последствиями односторонней экстирпации лабиринтов у кролика, бросалось в глаза значительное различие тонуса и установки конечностей обеих половин тела. После утраты, например левого лабиринта, обе левые конечности сидящего животного расслаблены и согнуты, обе правые — тонически вытянуты, а правая передняя конечность принимает особенно характерное разогнуто-отведенное положение, которое можно видеть на рис. 150а—с и 151а.
Возникает вопрос, является ли это различие тонуса прямым следствием выпадения одного лабиринта или оно вторично обусловлено поворотом шеи. Ведь, как было показано ранее, поворот головы, наступающий после экстирпации лабиринта, должен обусловить наличие как раз такого тонуса ног, какой наблюдается в действительности. Далее будет показано, что у кролика комбинируются друг с другом оба влияния — как утраты лабиринта, так и поворота шеи, — и что участие их в окончательном результате различно в разные сроки после операции.
Если распределение тонуса конечностей исследовать без помех в чис- стом виде, то лучше не исходить из наблюдений над сидящим животным, так как у этого животного на состояние конечностей влияет не только тонус их мускулатуры, но также и установка туловища, положение лопатки и пр. Лучше для этих целей привести животное в положение на спине. Тогда тонус разгибающих мышц передних и задних конечностей можно видеть непосредственно и без труда измерить разгибательный тонус путем противодействия пассивному сгибанию, сгибательный — пассивному разгибанию всей конечности или отдельного сустава.
В дальнейшем для упрощения изложения будет описываться поведение животного после экстирпации левого лабиринта. После утраты правого лабиринта отношения являются зеркальным отображением изменений.
Если такого кролика уложить на спину, голова его повернута налево, т. е. левый глаз смотрит вверх, а нижняя челюсть — вправо (рис. 148, стр. 242). Положение туловища описано в предыдущем отделе. Правые передняя и задняя нога разогнуты, обе левые вытянуты в меньшей степени, а иногда даже активно согнуты. В одних случаях различие тонуса 16 Р. Магнус
242
Шестая глава# Последствия односторонней экстирпации лабиринта
справа и слева сильнее выражено на передних ногах, в других — на задних. То же различие сохраняется, если, не изменяя положения головы относительно туловища, перевернуть животное на правую или левую сторону, сняв тем самым влияние положения головы в пространстве. Описанное различие тонуса выражено отчетливо уже сразу после операции, спустя месяцы оно также еще очень заметно.
Если удерживать туловища этого животного в положении на спине, а голову по отношению к грудному отделу поставить прямо, чтобы темя было обращено книзу, а нижняя челюсть кверху, при достижении чего приходится преодолеть довольно сильное противодействие со стороны
затылочных мышц, то различие тонуса правых и левых конечностей станет во всяком случае меньшим. Сразу после операции левые конечности и при прямо поставленной голове остаются вытянутыми немного меньше, чем правые. Однако через один или несколько дней, устанавливая голову прямо, можно весьма значительно уменьшить различие тону-
Рис. 148. Кролик XV. (Сокращенный протокол см. настр. 223). Левосторонняя экстирпация лабиринта 30 ноября 1912 г. Фотоснимок 20 января 1913 г., т. е. спустя 7% недель после операции. Животное лежит в положении на спине; голова вследствие экстирпации лабиринта повернута влево так, что левый глаз обращен кверху, темя — влево, нижняя челюсть направлена вправо. Таким образом, правые ноги являются «челюстными»^ левые — «черепными». Ясно видно, что как передние, так и задние левые нога вытянуты значительно меньше, чем правые.
са, не уничтожая его до конца. Отсюда следует, что на этой стадии разгибание правых ног, сгибание или соответственно менее значительное разгибание левых обусловливаются шейным рефлексом только частично, так как после прекращения поворачивания шеи еще сохраняется некоторое, хотя и уменьшенное различие тонуса справа и слева. Дело идет здесь о непосредственном результате выпадения лабиринта. Это проявляется в том, что ноги на стороне утраченного лабиринта обладают меньшим разгибательным тонусом, чем на стороне интактного лабиринта. На это влияние накладывается эффект поворота шеи, который согласно приведенному выше правилу действует таким образом, что тонус «челюстной» ноги (справа) повышается, а тонус «черепной» ноги (слева) становится ниже.
Если на одном и том же животном этот опыт (устанавливание головы прямо в положении на спине) повторять вначале ежедневно, а позднее еженедельно, то можно наблюдать, что после прекращения поворачивания головы различие тонуса справа и слева понемногу становится все меньше и меньше. У некоторых животных оно становится едва заметным вначале на задних ногах, у других — на передних и через различное по длительности время, в среднем около 8 недель, полностью исчезает. В одном случае, до тех пор пока не было достигнуто это состояние, прошло 10 недель, в другом — только 5/4 недель. По прошествии этого времени различия тонуса между правыми и левыми конечностями установить уже не удается. Теперь более сильное разгибание правых конечностей, которое всегда сохраняется в качестве длительного последствия операцви.
Последствия односторонней экстирпации лабиринта у кролика
243
если уложить животное на спину и не корригировать поворота его головы, обеспечивается исключительно упоминавшимся шейным рефлексом.
Отсюда следует, что у кролика после утраты лабиринта с одной стороны поворот шеи имеет своим следствием тонический рефлекс на конечности, который, согласно выведенному ранее правилу, обусловливает уменьшение разгибательного тонуса конечностей на стороне операции и увеличение разгибательного тонуса конечностей на противоположной стороне тела. Это влияние сохраняется на долгий срок (отмечалось спустя год и более). Дело идет, таким образом, о настоящем постоянном рефлексе, на действие которого прежде всего накладываются влияния односторонней утраты лабиринта (точнее — влияния оставшегося лабиринта), действующие в том же направлении, но понемногу ослабевающие и по прошествии 2 месяцев полностью исчезающие. Таким образом, если прямое влияние выпадения лабиринта на мышцы шеи и туловища оказывается длительным, то это влияние на мышцы конеч-
Рис. 149. Тот же кролик, сфотографированный сразу же повторно (снимок сделан более слева). Животное лежит в положении на спине, но голова повернута в другую сторону, что удалось после преодоления отчетливо выраженного противодействия шейных мышц. Нижняя челюсть обращена влево,
темя — вправо; левые ноги стали «челюстными», правые — «черепными». Вследствие этого левые передняя и задняя ноги сильно разогнуты, обе правые, которые на рис. 148 были разогнуты, наоборот, сильно согнуты. [При голове, установленной прямо (темя внизу, нижняя челюсть наверху), никакого различия в разгибатель- ной установке и тонусе между правыми и левыми конечностями не выявлялось].
ностей ограничено во времени. Позднее для других видов животных будет показано, что у них прямое влияние выпадения лабиринта на мышцы конечностей еще менее продолжительно.
Эти заключения подтверждаются результатами, которые получаются, если в положении животного на спине сообщить голове его поворот, обратный таковому грудного отдела.
Рис. 149 демонстрирует результат подобного опыта спустя 7% недель после экстирпации лабиринта у того же кролика, который непосредственно перед тем был показан на рис. 148. Рука экспериментатора придала голове направление, противоположное туловищу: правый глаз обращен вверх, нижняя челюсть — влево, темя — вправо. Можно видеть, как вследствие измененного положения головы распределение тонуса правых и левых конечностей стало совершенно иным. Обе левые ноги разогнуты, правые согнуты. Животное, у которого утрачен левый лабиринт, теперь устанавливает свои конечности в точности так, как животное, у которого был удален правый лабиринт. Сравнение обоих снимков (148 и 149) показывает, что поворот головы в противоположную сторону (рис. 149) определил разгибание левых конечностей по меньшей мере в той же степени, в какой правые конечности разогнуты, если голова повернута влево (рис. 148). Таким образом, видно, что на данной стадии распределение тонуса ног на левой и правой сторонах тела обусловлено исключительно шейными рефлексами.
Иначе обстоит дело тотчас после операции, когда прямое влияние односторонней утраты лабиринта выражено еще сильно. Если сразу
16*
244
Шестая глава. Последствия односторонней экстирпации лабиринта
после операции в период пробуждения после наркоза уложить кролика на спину, то в случае левосторонней операции левые конечности оказываются расслабленными. При устанавливании головы прямо положение, как указывалось выше, особенно еще не изменяется. Если повернуть голову вправо, то в ряде случаев благодаря повороту шеи удается преодолеть прямое влияние лабиринта, так что левые конечности разгибаются сильнее правых. В других случаях после поворачивания головы вправо тонус конечностей обеих сторон тела становится примерно одинаковым, а иногда тонус конечностей на левой стороне оказывается ниже, чем на правой. Таким образом, тотчас после операции прямое влияние утраты лабиринта еще настолько сильно, что оказывается в состоянии уравновешивать противоположно направленный шейный рефлекс на поворот головы вправо и даже одолевать его действие. Однако это положение меняется очень скоро: часто уже на другой день, а самое позднее через 6 дней прямое влияние лабиринта ослабляется настолько, что в ответ на поворот головы вправо левые конечности оказываются разогнутыми сильнее, нем правые; таким образом, шейные рефлексы преобладают и в таком ниде удерживаются длительное время.
В отдельных опытах постепенное развитие этого явления можно было наблюдать особенно отчетливо. При максимальном повороте головы непосредственно после операции левые конечности оказывались более расслабленными, на второй день имели неодинаковый тонус с обеих сторон, на третий день тонус задних конечностей был еще одинаков, тогда как спереди можно было уже установить более сильное разгибание слева, а на пятый день чрезмерная компенсация была вполне отчетливой и левые конечности как спереди, так и сзади при поворачивании головы вправо разгибались сильнее правых. Однако в ряде опытов, как было упомянуто, эта чрезмерная компенсация могла быть отмечена уже непосредственно после операции.
Отсюда следует, что спустя уже немного дней после операции шейные рефлексы значительно преобладают над прямым влиянием утраты лабиринта на тонус конечностей. Это лабиринтное влияние особенно сильно уменьшается в первые дни и (как это следует из наблюдений с установкой головы в прямое положение) спустя в среднем 8 недель полностью исчезает. Однако решающую роль в контроле над напряжением конечностей приобретает очень скоро после операции шея; поворотами которой главным образом и определяется это напряжение. Из этого не наблюдалось ни одного исключения.
Как было показано в третьей главе, наклон головы (поворот вокруг оси темя—основание черепа) действует таким образом, что «челюстные» конечности, за которыми следует морда, разгибаются, а «черепные», наоборот, сгибаются. Влияние этого наклона является значительно меньшим, чем влияние поворота головы вокруг продольной оси тела. Это можно увидеть также на животных после односторонней экстирпации лабиринта. Если кролика спустя несколько месяцев после левосторонней экстирпации лабиринта уложить на спину, установить его голову по отношению к туловищу прямо и добиться этим путем, чтобы его конечности на правой и левой сторонах тела обладали одинаковым тонусом, то, отводя голову (вправо или влево), можно привести конечности «челюстной» стороны к разгибанию, а конечности «черепной» стороны — к сгибанию. Реакции при наклоне всегда слабее тех, которые получаются вследствие поворачивания головы. Если тот же опыт проделать спустя более короткое время после операции, когда при прямо стоящей голове левые конечности оказываются несколько более расслабленными, чем правые, то в отдельных случаях, отводя голову влево, это различие удается ликвидировать или даже сверхкомпенсировать. Это удается,
Последствия односторонней экстирпации лабиринта у кролика
245
однако, не всегда, так как реакции на отведение головы обычно выявляются только в довольно слабой степени. Выше было указано, что непосредственно после односторонней экстирпации лабиринта у кролика часто появляется сильно выраженное отведение головы в сторону операции, которое скоро становится все меньше, а у сидящего животного спустя немного дней исчезает совсем и в течение долгого времени наблюдается только при висячем положении головой вниз. Так как это отведение уже само по себе ослабевает по сравнению с поворотом, а кроме того, даже и сильное отведение головы способно влиять на тонус конечностей лишь незначительно, оказывается, что влиянием отведения головы на тонус конечностей после односторонней утраты лабиринта практически можно пренебречь.
В самом крайнем случае непосредственно после операции, когда поворот головы еще не так интенсивен, сильное отведение головы в сторону утраченного лабиринта может в' некоторой степени противодействовать расслаблению конечностей на этой стороне. Однако и здесь дело идет лишь о весьма незначительном влиянии, что видно, если скорригировать отведение головы.
В предыдущих отделах показано, что часть исходных состояний после односторонней экстирпации лабиринта можно устранить, если голову установить относительно груди в прямом положении. Из этого был сделан вывод, что эти состояния обусловлены шейными рефлексами. Если, например, через 8 недель после (левосторонней) операции в положении на спине, когда голова повернута влево, левые конечности расслаблены больше правых, тогда, как и при прямо поставленной голове, тонус конечностей одинаков с обеих сторон, и можно считать доказанным, что на этой стадии различие тонуса конечностей обеих сторон обусловлено исключительно шейным рефлексом.
Против этого можно было бы возразить следующее. После левосторонней экстирпации лабиринта голова длительно держится сильно повернутой в левую сторону. Если пожелать установить голову по отношению к грудному отделу прямо, то это оказывается возможным только после преодоления сильного противодействия, тогда как поворачивание головы влево наталкивается лишь на незначительное сопротивление. Поэтому можно считать, что поворот головы влево вызывает слабый, поворот вправо — интенсивный шейный рефлекс и что уже при устанавливании головы прямо и преодолении сильного мышечного противодействия активировался шейный рефлекс, по действию совпадавший с эффектом правого поворота у нормального животного. Если в дальнейшем, несмотря на это, тонус левых и правых ног оказался бы одинаковым, это могло бы иметь место по той причине, что лабиринтный рефлекс подействовал бы в направлении, обратном тому, которое непосредственно компенсировалось (гипотетическим) шейным рефлексом. Если бы это допущение оказалось правильным, то и по прошествии 8 недель прямое влияние лабиринта имело бы место, в то время как раньше мы пришли к заключению, что на этой стадии это влияние отсутствует.
Не говоря о том, что было бы удивительным, если бы оба рефлекса спустя 8 недель с таким постоянством уравновешивали друг друга и что тонус конечностей при прямо поставленной голове был бы строго одинаков, легко установить, что такое возражение неправомерно. А именно, если при голове, установленной прямо, имел бы место шейный рефлекс на конечности, который вызывался бы при преодолении противодействия одностороннего напряжения шейных мышц, интенсивность его должна была бы изменяться, если бы односторонне изменилось напряжение мышц шеи. Из опытов, описанных на стр. 78—79, выяснилось, что напряжение шейных мышц после утраты одного лабиринта зависит от положения головы в пространстве. Таким образом, на интенсивность гипотетического шейного рефлекса должно было бы сказываться влияние изменения положения головы в пространстве. Наоборот, согласно данным, приведенным на стр. 51, после односторонней экстирпации лабиринта тонус конечностей при различных положениях головы в пространстве остается неизменным. Отсюда проистекает возможность испытать экспериментально справедливость приведенного выше возражения. Если бы оба рефлекса точно уравновешивали друг друга при положении животного на спине, этого не могло бы получиться, например, при положении животного на ногах. Эту пробу мы предпринимали неоднократно. Кролик через 8 недель после операции приводился в положение на спине, а голова его по отношению к грудному отделу устанавливалась прямо. При этом тонус пра¬
246
Шестая глава. Последствия односторонней экстирпации лабиринту
вой и левой конечностей был примерно одинаковым. Если теперь все животное без изменения положения головы относительно грудного отдела переворачивалось из положения на спине и ставилось на ноги (спина кверху, позвоночник горизонтально), ничто от этого не менялось и тонус на обеих сторонах тела оставался совершенно одинаковым. Отсюда можно с уверенностью заключить, что возражение, обсуждавшееся выше, неправомерно и что на этой стадии различие тонуса конечностей обусловлено исключительно шейным рефлексом. Этим самым устанавливается- общая закономерность: те последствия утраты лабиринта, которые могут быть уменьшены посредством устанавливания шеи прямо, следует понимать как следствия поворота головы.
На рисунках 148 и 149 (положение на спине), 139 (висячее положение головой вниз), 140 (нормальная установка в воздухе) и 144 (висячее положение головой вверх) можно видеть одинаково убедительно, что, если животное не сидит на полу, а держит конечности свободно в воздухе, отведение и приведение конечностей по отношению к груди или тазу отсутствуют. Это не наступает ни в качестве прямого последствия выпадения лабиринта, ни благодаря шейным рефлексам. Если сидящее животное, что будет позднее показано, отводит конечности одной стороны и приводит конечности другой, то это основывается на вторичных влияниях, а именно: повороте грудного отдела относительно плоскости пола.
В то время как после односторонней экстирпации лабиринта все описанные изменения положения головы, туловища и конечностей оказывалось возможным объяснить и свести к известным шейным и лабиринтным рефлексам, это до сих пор невозможно сделать по отношению к непостоянной односторонней утрате тонуса конечностей, которая наступает в качестве прямого следствия выпадения лабиринта. Как было указано выше, одного лабиринта оказывается достаточно для того, чтобы одинаковым образом осуществлять тонические рефлексы положения на мышцы конечностей обеих половин тела. Это удалось установить с полнейшей определенностью при помощи опытов с децеребрированными животными. При этом не существенно, обладают ли конечности в данном случае одинаковым или разным тонусом на правой и левой сторонах тела. В обоих случаях изменение положения головы в пространстве вызывает изменения тонуса в одном направлении и одной интенсивности в разгибателях обеих половин тела. Значит, выпадение влияния этих тонических лабиринтных рефлексов на мышцы конечностей не может вызывать различия в тонусе конечностей правой и левой сторон. Каким образом это происходит после односторонней экстирпации лабиринта, до сих пор совершенно неясно. Примечательно уже то, что дело идет о преходящем влиянии. Все же у кролика и обезьяны различие тонуса после односторонней экстирпации лабиринта имеет место в течение ряда недель, у кошки и особенно у собаки — только несколько часов и то не всегда. У морских свинок после хирургического удаления лабиринта оно может быть отмечено в течение нескольких дней, отсутствуя, однако, в случае выключения одного лабиринта с помощью кокаина. Ни один из лабиринтных рефлексов, известных до сих пор, не может рассматриваться как ответственный за появление этого различия тонуса. Дальнейшими исследованиями предстоит установить, идет ли вообще речь о последствиях одностороннего выпадения деятельности отолитов или полукружных каналов или же причина изменений лежит в перерезке, т. е. в повреждении, ствола 8-го нерва, или же, может быть, во влияниях иных, чем вестибулярные.
По поводу динамики тонуса конечностей у кролика без одного лабиринта можно, суммируя, сказать следующее.
Последствия односторонней экстирпации лабиринта у кролика
247
Передняя и задняя ноги на стороне утраченного лабиринта длительное время обладают разгибательным тонусом по сравнению с конечностями другой стороны. Это различие тонуса зависит от двух разных факторов, действия которых суммируются.
1) От шейного рефлекса, который приводится в действие благодаря повороту головы по отношению к туловищу, а при установке головы прямо может быть уменьшенным. Так как поворот головы является длительным последствием операции, этот шейный рефлекс также постоянен.
2) От прямого последствия выпадения лабиринта, вследствие чего конечности на оперированной стороне приобретают более низкий разгибательный тонус. Вскоре после операции этот тонус весьма высок, затем быстро уменьшается и спустя в среднем 8 недель исчезает.
Отсюда следует, что при установлении головы по отношению к грудному отделу прямо сначала отмечается различие тонуса конечностей обеих сторон тела, тогда как спустя более длительное время это различие тонуса полностью снимается.
При поворачивании головы в обратном направлении конечностям па стороне утраченного лабиринта можно сообщить более высокий тонус, изменяя различие их тонуса в прямо противоположном направлении.
Стало быть, вскоре после операции влияние шейных рефлексов на тонус конечностей в значительной степени оказывается господствующим.
В то время как поворот шеи оказывает сильное влияние на тонус конечностей, действие отклонения головы значительно менее эффективно, и как последствие односторонней экстирпации лабиринта практически во внимание не принимается.
Положение тела при сидении (15)
Сообщенные до сих пор факты дают нам возможность проанализировать положение тела сидящего животного. При этом будут описаны отношения, какие имеют место после левосторонней экстирпации лабиринта.
Непосредственно после операции во время пробуждения от наркоза животное, если оно не производит движений перекатывания, лежит на стороне утраченного лабиринта, т. е. на левой стороне. Нередко уже спустя полчаса, самое позднее — на следующий день оно уже сидит в своей клетке, если только вследствие катания по клетке или исследования временно не обессилело.
Если до этого животное лежало на правой стороне, то способность к усаживанию понятна, так как сначала наступает левый поворот головы, благодаря чему голова движется по направлению к нормальному положению, после чего к этому вследствие шейных рефлексов может присоединиться усаживание. В случае левого бокового положения все установочные рефлексы на голову вместе с примыкающим шейным установочным рефлексом должны «отказать».1 Если, несмотря на это, усаживание тела все же происходит, то это основано на установочном рефлексе с туловища на туловище.
1 По меньшей мере, если установочный рефлекс с туловища на голову не проявляется настолько интенсивно, чтобы преодолеть лабиринтный установочный рефлекс и своими силами привести голову в нормальное положение. Это наблюдается -только в исключительно редких случаях.
Рис. 150а. Животное сидит свободно. Поворот головы влево, она лежит между лапами. Вследствие разгибания правой задней ноги таз свешивается влево. Поворот туловища обусловливает смещение левого плеча вниз, правого — вверх. Разгибание и сильное отведение правой передней ноги. Левая нога лежит под левым ухом.
Рис. 150в. Животному наложен головодержатель Чер- мака. Поворот головы и разгибание и отведение правой передней ноги видны отчетливо.
Рис. 1506. Положение, похожее на приведенное на рис. 150а. Голова лежит слева от животного. Позади разогнутой и отведенной правой передней ноги видна разогнутая правая задняя нога.
Рис. 150г. С помощью го ловодержателя Чермака поворот головы корригирован. Теперь голова стоит прямо по отношению к туловищу. Вследствие этого- ненормальное положение таза и конечностей исчезло, животное сидит подобно нормальному.
Рис. 150а—г. Кролик I. Левосторонняя экстирпация лабиринта 10’ октября 1912 г. Фотографические снимки 18 октября, т. е. через. 8 дней после операции.
Последствия односторонней экстирпации лабиринта у кролика
249
Как было описано выше, на таламическом или мезенцефалическом животном с обоими интактными или с обоими удаленными лабиринтами можно показать, что и тогда, когда голова не находится в нормальном положении, тело может быть приведено в нормальное сидячее положение, если (благодаря давлению плоскости опоры и т. п.) имеет место асимметричное возбуждение чувствительных нервов тела. На стр. 192 был, например, показан таламический кролик, голова которого фиксировалась в левом боковом положении и тело которого, несмотря на это, было приведено в сидячее положение.
Этот рефлекс — важнейший, благодаря.кото рому тело кролика после утраты одного лабиринта в состояни и ' принять сидячее по- * УРис. 151а. Животное, свободно сидящее с головодержателем Чермака.
Вследствие более сильного разгибания правой задней ноги таз несколько отвисает влево. Передняя часть тела животного лежит на левом плече, правое плечо обращено кверху.
Правая передняя нога разогнута и отведена, левая — согнута. Спиральный поворот всего тела отчетливо виден. Голова повернута вправо более чем на 90° и находится слева от животного; левый глаз обращен к потолку.
Рис. 151а—б. Кролик III. (То же животное, что и на рис. 137 и 138). Левосторонняя экстирпация лабиринта 23 октября 1912 г. Фотографический снимок 1 ноября, т. е. спустя 9 дней после операции.
ложение, несмотря на повернутое положение головы. После левосторонней утраты лабиринта установка животного, находящегося в левом боковом положении, обеспечивается исключительно этим установочным рефлексом на тело при условии, что основной поворот достаточно выражен, поэтому механизм, изображенный на рис. 146, в состоянии приблизить голову к нормальному положению.
У некоторых животных уже через 24 часа после осторожно проведенной экстирпации лабиринта можно отметить, что тело принимает сидячее положение, если голова фиксирована в правом или левом боковом положении, а спустя немного дней этой способностью обладают в полной мере все животные. После удаления большого мозга в большинстве случаев даже через несколько часов после операции рефлекс выражен максимально.
Положение, принимаемое во время сидения, описывалось уже много раз (например, Винклером (1), стр 29]; рисунки 150а—в и рис. 151, сделанные на 8-й или 9-й день после левосторонней операции, дают об этом хорошее представление. Голова повернута в сторону операции, а в первые дни после операции также и отклонена в том же направлений. После экстирпации левого лабиринта правый глаз обращен к потолку, голова лежит или между лапами, или слева. Правая передняя нога максимально разогнута, обладает интенсивным разгибательным тонусом, особенно в плечевом и локтевом суставах, держится в отведенном положении. Правая задняя нога обычно несколько отведена и разогнута. Левая нога
250
Шестая глава. Последствия односторонней экстирпации лабиринта
расслаблена, иногда даже активно согнута и чаще всего лежит под животным. В отдельных случаях передняя часть тела лежит на левой лопатке.
Через короткое время после операции, часто уже на следующий день, голова снова может быть активно поднята. Обе передние ноги оказываются вследствие этого (так же как это имеет место и у нормального животного) в состоянии повышенного разгибательного тонуса; однако различие тонуса правой и левой передних ног сохраняется: правая разогнута и отведена, левая—согнута еще сильнее.
Это ненормальное положение передней части тела вызывается действием следующих факторов.
1) Спиральным поворотом туловища (стр. 227 и след,). Этот поворот, как упомина-
Рис. 1516. Голову животного с помощью головодержателя установили прямо, ничего другого не изменяя. Вслед за этим животное тотчас же приняло нормальное положение; разгибание и отведение передней правой ноги исчезло; тело сидит прямо и правильно, оба плеча стоят симметрично. Снимок сделан более с правой стороны, чтобы лучше показать положение обеих передних ног.
лось, в первое время после операции увеличивается, чтобы затем остаться довольно постоянным. Эффект этого поворота может особенно сильно проявиться тогда, когда, как изображено на рис. 150, таз вследствие более сильного разгибания правой задней ноги отчетливо перевешивается в левую сторону. Тогда грудной отдел может оказаться повернутым на 45° и более. Поэтому проксимальная часть правой передней ноги перемещается вправо и вверх, и этим осуществляется ее отведенное положение. При этом левое плечо перемещается книзу.
Как указывалось, спиральный поворот туловища является частично прямым и отдаленным последствием выпадения лабиринта, частично — результатом поворота шеи. При устанавливании головы по отношению к грудному отделу прямо в соответствии с этим уменьшается поворот туловища, оставаясь, как правило, еще вполне отчетливым. Степень поворота туловища сидящего животного, которая обусловлена, между прочим, также и положением конечностей, колеблется. Задняя апертура груди в большинстве случаев повернута на 10—30°, передняя — на 20—45° в левую сторону.
2) Прямым последствием удаления лабиринта по отношению к тонусу конечностей, причем разгибательный тонус левых ног ниже, чем правых. Это воздействие сразу после операции оказывается сильнее всего, в течение нескольких дней значительно уменьшается для того, чтобы спустя примерно 2 месяца исчезнуть полностью.
У сидящего животного это влияние можно видеть, установив голову по отношению к груди прямо, и уравнять противодействие пассивному сгибанию в локтевом суставе на правой и левой сторонах. Вскоре после операции слева он значительно ниже, чем справа; через несколько дней различие выражено уже меньше; спустя еще более продолжительное время оно исчезает полностью.
3) Тоническими шейными рефлексами, которые мобилизуются вследствие поворота и отклонения шеи. Отклонение влево (левые конеч-
Последствия односторонней экстирпации лабиринта у кролика
251
пости становятся «челюстными» конечностями) сообщает «челюстным» ногам более высокий разгибательный тонус и тем самым противодействует описанному выше положении) тела и конечностей. Однако, как было указано, влияние отклонения головы на тонус конечностей является слишком незначительным для того, чтобы существенно изменить результат. Кроме того, по прошествии уже немногих дней после операции голова при сидении больше не отклоняется. Это исчезновение отклонения основывается, во всяком случае частично, на действии на голову установочного рефлекса с тела, так как при висячем положении головой вниз отклонение головы отмечается в течение длительного времени.
Напротив, поворот головы оказывает длительное и мощное влияние. Вследствие этого левые конечности приобретают пониженный, а правые — повышенный разгибательный тонус. Значение этого влияния можно исследовать, установив голову относительно туловища прямо, и наблюдать, какие аномалии положения можно уменьшить этим путем.
Оказывается, что влияние этих мобилизованных поворотом головы шейных тонических рефлексов весьма значительно отражается на аномалиях положения сидящего кролика, лишенного лабиринта с одной стороны. Поражающей оказывается картина, когда кролик, который только что сидел в позе, изображенной на рис. 150а—в и рис. 151а, при голове, установленной прямо, тотчас принимает почти нормальное положение (рис. 150г и 1516).
Животное при этом притягивает отведенные правые ноги, особенно заметно исчезает ненормальное стояние правой передней ноги, плечи стоят симметрично, короче говоря, животное сидит подобно нормальному, и необходимо более точное исследование, чтобы установить, что имеют место аномалии положения. Как только голова освобождается, правая нога сильно разгибается и отводится, а все прочие аномалии положения •становятся снова отчетливо заметными.
Этот простой, неизменно удающийся опыт является особенно подходящим для демонстрации преобладающего влияния, которое шейные рефлексы оказывают на возникновение аномалий положения после односторонней экстирпации лабиринта.
Когда голова такого кролика установлена прямо, на нем можно показать еще и следующие отклонения. Чаще всего грудной отдел отвисает в левую сторону (см. стр. 250). Кроме того, разгибательный тонус конечностей на левой стороне, особенно в первые дни после операции, несколько ниже, чем на правой. Правая передняя или задняя нога может остаться еще несколько разогнутой и отведенной, и при прямом исследовании разгибательного тонуса обнаруживается, что на правой стороне он немного выше. Как было сказано, это различие становится все меньше и спустя некоторое время совсем исчезает. Таким образом, у сидящего животного с прямо стоящей головой также может быть обнаружено непосредственное влияние утраты лабиринта на туловище и конечности. Речь идет о длительных влияниях на туловище и преходящих влияниях на конечности. Однако можно видеть, что эти влияния совершенно отступают на задний план по сравнению с преобладающим влиянием шейных рефлексов у сидящего животного.
Как указывалось выше, приведение конечностей на левой и отведение на правой стороне является следствием только поворота грудного, resp. тазового, отделов, а не прямым результатом выпадения лабиринта. Если таз или грудной отдел стоят прямо, отсутствуют также и отведение, и приведение конечностей.
Рис. 152а. Свободно сидящее животное. Никакого разгибания и отведения правой передней лапы, несмотря на сильный поворот головы влево.
Рис. 152в. Животному рукой прикрывают глаза. Тотчас же возвращаются характерные для односторонней утраты лабиринта разгибание и отведение правой передней лапы.
Рис. 1526. Голова животного пальцем немного приподнимается для того, чтобы показать положение передних конечностей. Можно видеть, что правая передняя лапа не разогнута и не отведена. Животное, в течение дня прыгающее в саду, в состоянии удерживать голову в этом положении и без поддержки.
Рис. 152г. Теперь, при закрытых глазах, голова рукой поворачивается вправо, что осуществляется при преодолении сильного противодействия шейных мышц. При этом левая нога выпрямляется и отводится, правая — сгибается, и животное принимает положение, как если бы у него был экстирпирован правый лабиринт.
Рис. 152а—г. Кролик XV. (То же животное, что и на рис. 148 и 149). Левосторонняя экстирпация лабиринта 30 ноября 1912 г. Снимки 29 апреля 1913 г., т. е. спустя примерно 5 месяцев после
операции.
Последствия односторонней экстирпации лабиринта у кролика
253
Когда животное сидит прямо или в первые дни после операции лежит на левом боку., можно, устанавливая голову животного прямо, тотчас привести его в сидячее положение.
Для того чтобы составить себе приблизительное представление о разгибательном тонусе и противодействующей способности конечностей свободно сидящего животного, можно усадить животное на негладкую поверхность, например на соломенный мат, и попытаться рукой сместить его тело вправо или влево. В случае свободно сидящего животного постоянно обнаруживается, что смещаемость влево отчетливее, чем вправо, так как правые ноги вследствие разгибания и отведения с большей силой противодействуют смещению туловища. Если проделать эту пробу, когда голова установлена прямо, обнаруживается, что в первые дни после операции животное легче сдвинуть влево, нежели вправо. Однако, как можно ожидать после изложенного, по прошествии некоторого времени это различие оказывается меньшим и далее исчезает совсем, так как к этому времени прямое влияние утраты лабиринта на конечности проходит.
Преобладающая роль, которую играют шейные рефлексы в определении установки кролика, лишенного одного лабиринта, раскрывается также и из того, что, производя обратный поворот головы, удается вызвать точно противоположное положение тела.
Рис. 152в и г иллюстрирует этот опыт. На рис. 152в животное, у которого за пять месяцев до этого был удален левый лабиринт, принимает {при закрытых глазах) характерное положение с головой, повернутой влево, и разгибанием и отведением правой передней ноги. На рис. 152г голова, несмотря на сильное противодействие шейных мышц, оказывается повернутой вправо и принимает, таким образом, положение, какое должно было иметь место у животного после правосторонней экстирпации лабиринта. Можно заметить, что вследствие того, что передние ноги животного приняли соответствующее положение, левая передняя нога разогнута и отведена, а правая—согнута.
Компенсация установки
В предыдущих отделах часто и по-разному указывалось на то, что с течением времени кролики выучиваются компенсировать при сидячем положении часть нарушений, наступающих после односторонней экстирпации лабиринта. Преимущественно речь здесь идет о более полном использовании установочных рефлексов с туловища и на голову. Так, при сидении уже спустя несколько дней животные не держат больше голову отклоненной, тогда как отклонение головы при висячем положении тела головой вниз может отмечаться долгое время (установочный рефлекс с тела на голову).1 Через 3—4 месяца и поворот головы во время сидения становится несколько меньше, в то время как в висячем положении он неизменно остается интенсивным (установочный рефлекс с туловища на голову). Улучшение сидения, несмотря на то, что голова остается повернутой, особенно бросающееся в глаза в первые дни после операции, основано на повышении участия установочных рефлексов с туловища на туловище.
Как уже говорилось при обсуждении оптических установочных рефлексов, влияние глаз на удерживание положения тела у кролика с удаленным с одной стороны лабиринтом удивительно невелико. Поэтому и положение головы у животных с открытыми или закрытыми глазами при различных положениях тела в пространстве в основном одно и то же. Таламические кролики, которые, кроме зрачкового рефлекса и рефлек¬
1 Дальнейшая причина уменьшения отклонения головы может быть рассмотрена только в девятой главе.
254
Шестая глава. Последствия односторонней экстирпации лабиринта
торного смыкания век при сильном освещении не обнаруживают вообщо никаких оптических рефлексов, не проявляют в своих установочных рефлексах никакого различия по сравнению с животными с интактным: большим мозгом и с открытыми глазами.
В некоторых немногочисленных случаях можно было наблюдать, что кролики, лишенные лабиринта с одной стороны, сидевшие в маленьких круглых клетках с закрытыми боковыми стенками, держали голову повернутой несколько более сильно, чем тогда, когда они свободно сидели на полу в середине комнаты и, стало быть, воспринимали оптические образы. Большого различия, однако, не было, и наблюдалось оно нечасто. Можно было бы думать, что дело идет здесь о небольшом исправляющем влиянии глаз на голову, но с уверенностью утверждать это я не осмеливаюсь. Выясняется, наоборот, что кролики без одного лабиринта медленнее научаются компенсировать ненормальное положение своих передних ног с помощью глаз после операции.
Если в первые месяцы после операции свободно сидящее животное поворачивает голову, а переднюю ногу на стороне интактного лабиринта разгибает и отводит, то с течением времени, хотя поворот шеи выражен в весьма сильной степени и часто достигает 90°, передние ноги не удерживаются более в столь ненормальном положении, как это было прежде. Иногда можно наблюдать, что при сидеции животное подтягивает отведенную (правую) ногу и некоторое время находится в этой позе. В конце концов это положение становится правилом, и тогда такое животное можно видеть сидящим или свободно прыгающим с повернутой шеей и симметрично расположенными передними ногами. У одного из наблюдавшихся нами животных это состояние наступило на 75-й, у другого — на 102-й день и долгое время оставалось без изменений. У третьего животного компенсация выработалась почти через два месяца.
Более точное исследование показало, что дело идет в данном случае о явлении компенсации при помощи глаз. Достаточно закрыть глаза животному, чтобы вызвать тотчас же прежнее положение конечностей. Это явление иллюстрирует рис. 152.
Снимок сделан спустя 5 месяцев после операции и изображает опыт, который к этому времени и далее ежедневно может быть повторен с одним и тем же постоянным результатом. На рис. 152а и б животное сидит с сильно, примерно на 90°, повернутой влево головой, держа передние ноги совершенно симметрично; во всяком случае разгибание и отведение правой передней ноги отсутствует. Рис. 1526 показывает, что можно также захватить голову животного без того, чтобы что-нибудь изменилось. Даже энергичное удерживание головы рукой никакого влияния не оказывает. Однако как только животному закрывают глаза, снова наступает- характерное положение. На рис. 152в ясно видны разгибание и отведение правой передней ноги при закрытых глазах. То, что поворот головы вправо вызывает зеркально противоположное положение передних ног, уже сообщалось выше.
Если с помощью закрывания глаз вызвано типичное положение передних ног (рис. 152в), то достаточно отвести руки от глаз животного, чтобы увидеть, как последнее тотчас же подтягивает свою правую переднюю ногу, вслед за чем оно принимает положение, изображенное на рис. 152 а или б.
То, что животное при прямо установленной голове и закрытых глазах держит свои конечности совершенно симметрично, с еще большей убедительностью было установлено в каждом отдельном случае.
Если такое животное, которое держит свои передние ноги совершенно' симметрично, привести в висячее положение головой вниз, т. е. в поло¬
Последствия односторонней экстирпации лабиринта у кролика
255
жение, для животного необычное, то, как говорилось выше, правая передняя нога снова энергично разгибается, несмотря на то, что глаза остаются открытыми.
В одном единственном случае подобная компенсация с помощью глаз могла быть отмечена уже в первые дни после операции; дело касалось животного, оперированного с особой осторожностью, у которого уже на следующий день не обнаруживалось нистагма, оставался только незначительный поворот головы при сидении и, наоборотг интенсивный в висячем положении. При сидении с открытыми глазами на не совершенно гладком полу животное держало передние ноги симметрично, в то время как на гладком полу или при закрытых глазах типичное положение передних ног было очевидным. Осуществленная в дальнейшем секция подтвердила радикальность операции. Дело шло в данном случае об исключительном явлении; у всех других животных описанная компенсация наступала только через несколько месяцев.
Так как у таламического животного, помимо зрачкового рефлекса и рефлекса смыкания век, всякие оптические реакции отсутствуют, можно считать, что компенсация эта осуществляется с помощью коры большого мозга. Дело идет при этом об одном из самых редких случаев, когда оказывается возможным показать влияние глаз на положение тела у кролика. В других случаях оптические установочные рефлексы, включая и положение тела кроликов с односторонне удаленным лабиринтом, не играют никакой или во всяком случае совершенно незначительную роль. Наиболее существенные компенсации осуществляются у кролика с помощью установочных рефлексов, т. е. без участия коры большого мозга, исключительно под контролем ствола.
В отдельных случаях на молодых кроликах было установлено, что уже спустя 1—2 дня после односторонней экстирпации лабиринта передняя нога противоположной стороны больше не удерживается в разогнутом и отведенном состояниях. Это не изменялось и при закрывании глаз. Так как компенсация наступала только при сидении, она, конечно, будет иметь место и после соприкосновения конечности с плоскостью опоры.
Заключение
Поза, какую принимает кролик с одним удаленным лабиринтом во* время сидения, может быть обусловлена изменениями тонуса мышц туловища и конечностей и установочными рефлексами.
Главное влияние на установку тела оказывает поворот шеи. Последний является результатом основного поворота и установочного лабиринтного рефлекса на голову и во время сидения уменьшается благодаря действию установочного рефлекса с тела на голову.
Если поворот шеи уменьшить, животное сидит почти нормально. Вследствие обратного поворота животному возможно придать положение, как если бы у него был удален другой лабиринт.
Поворот шеи воздействует на позу тела:
1) посредством тонических рефлексов на мышцы тела;
2) посредством шейных установочных рефлексов на грудной и тазовый отделы.
Менее значительное влияние на положение тела при сидении имеют:
1) длительный поворот туловища, вызванный непосредственно утратой лабиринта;
2) вызванное непосредственно утратой лабиринта временное различие тонуса конечностей обеих сторон;
3) тонический шейный рефлекс, который в первые дни после операции вызывается отклонением головы.
256
Шестая глава. Последствия односторонней экстирпации лабиринта
Для положения тела во время сидения весьма существенным является установочный рефлекс с туловища на туловище, который приводит тело к нормальному положению, несмотря на то, что так сильно повернута голова.
Спустя более длительное время после операции ненормальное положение передних ног может корригироваться с помощью зрения. Здесь принимает участие кора большого мозга.
В остальном установка тела у кролика после экстирпации лабиринта с одной стороны находится под контролем центров, лежащих в верхних отделах спинного мозга и ствола.
Движения перекатывания (Rollbewegungen)
Главный интерес всех исследователей, производивших одностороннюю экстирпацию лабиринта у кролика, вызывали энергичные движения перекатывания, которые животное производит по направлению к оперированной стороне. В точном экспериментальном анализе этого явления не было недостатка. Исследования Эвальда и Винклера (1) приводятся здесь в дословном изложении в качестве важнейших из этих опытов.
Эвальд (стр. 195) так кратко описывает свои данные, полученные на кролике:
«В делом эти животные не слишком пригодны для исследования нарушений лабиринтов. Здесь упоминаю о них только потому, что во всяком случае один симптом они обнаруживают так отчетливо, как этого нельзя увидеть ни у собаки, ни у кошки. Это — движения перекатывания спустя непродолжительное время после удаления одного лабиринта.
«Если такое животное положить на пол, оно катится через всю комнату, и причину этого можно установить с большой отчетливостью. Как у всех односторонне лабиринтэктомированных животных, конечности противоположной стороны у них разогнуты и отведены, но подвижны. Конечности на стороне операции согнуты и приведены, т. е. прилегают к телу. В то же время они неподвижны. Если поэтому посадить кролика на пол, тело его вследствие разгибания ног противоположной стороны наклоняется в направлении оперированной стороны и падает в эту сторону, так как не находит здесь опоры на притянутых к нему ногах. Лежа на спине, кролик использует ноги противоположной стороны, чтобы снова перевернуться, причем подтягивает их под тело, продолжая таким образом катящееся движение в том же направлении. Эти же конечности, как только достигнуто положение на животе, тотчас же опять разгибаются, и события повторяются снова и снова. Если наблюдать внимательно, то нетрудно заметить, что ноги на одноименной стороне во время движений перекатывания вообще не двигаются, но остаются спокойно притянутыми к телу. Следовательно, ноги противоположной стороны одни производят это движение».
Значительно подробнее изучал движения перекатывания Винклер. При этом он пришел к результатам, которые в некоторых наиболее важных пунктах были подтверждены позже:
«Как только животное (которое до этого было привязано и должно было поэтому держать свою голову прямо) было освобождено и встало на ноги или улеглось на оперированную сторону, появляются новые, бурные движения. Голова с исключительной силой поворачивается в сторону операции таким образом, что щека этой стороны оказывается на полу. Иногда поворот головы настолько велик, что дорсальная часть головы, поворачиваясь в сторону операции, касается пола. Одновременно с движением головы передняя конечность на стороне, противоположной оперированной, оказывается вытянутой и отведенной от туловища настолько, насколько это возможно. Животное царапает пол этой ногой, как бы стараясь поддержать себя этой ногой для того, чтобы предотвратить дальнейшее перекатывание. Обычно это ему не удается.
«Животное безрезультатно бьет по воздуху передней ногой на стороне, противоположной оперированной. Эта передняя нога, все еще максимально разогнутая и отведенная, поднимается все выше и выше, пока, наконец, не оказывается в верти¬
Последствия односторонней экстирпации лабиринта у кролика
257
кальном положении. Дорсальная часть головы в этот момент касается пола, так как поворот вверх и поднимание противоположного плеча, наступающие вслед за поворачиванием головы, есть причина движения передней ноги. Как только передняя нога пересекла вертикальный уровень, появляется новое движение. Животное не в состоянии поддерживать заднюю часть тела в обычном положении, тем более, что передняя часть тела до сих пор была повернута. Это было так до тех пор, пока передняя нога не достигла вертикального уровня (или пока дорсальная часть головы не коснулась пола), однако поворачивание головы все продолжается. Теперь в определенный момент животное подворачивает заднюю часть тела, повертывая ее тем самым в оперированную сторону.
«Поступая таким образом, животное перекатывается вокруг продольной оси своего тела в направлении оперированной стороны, и это движение нередко можно видеть повторяющимся много раз подряд.
«Каждое переворачивание совершается в два темпа или скорее может быть разложено на два полупереворота. При первом из них голова и шея поворачиваются в сторону операции, противоположное плечо повернуто кверху, соответственно противоположная передняя нога разогнута и отведена и также повернута кверху. Поворот головы и шеи происходит до момента, пока не достигнуто положение, которое не позволяет поддерживать заднюю часть тела в ее исходной позиции по отношению к перекрученной передней части тела. Эта первая часть переворачивания, по-видимому, не зависит от воли животного, но кажется скорее непроизвольной, как неизбежный автоматизм. Голова и шея (и последовательно поднимающаяся противоположная нога) должны принять определенное положение в направлении оперированной стороны. Вторая же часть поворота имеет другой источник. Для животного оказывается невозможным поддерживать исходную позицию задней половины тела, когда передняя часть повернута так сильно, и, по-видимому, это движение зависит от воли животного (animal’s willing). Произвольно задняя часть перебрасывается в направлении оперированной стороны. . .
«Так как на стороне операции конечности остаются неактивными в продолжение поворота вокруг оси, они расслаблены. . .
«...Поэтому мы начнем с поворачивания тела вокруг его продольной оси в направлении оперированной стороны.
«Я уже показал, что голова и шея вскоре после операции оказываются повернутыми таким образом, что для нормального животного становится невозможным сидеть на полу. И это в тем большей мере, что поза головы и шеи не остается постоянной, а через определенные интервалы времени еще более изменяется.
«Если кролик обладал бы длинной и подвижной, как у голубя, шеей, которую можно было бы опустить вниз и уложить там, в то время, как голова повернута кверху, то, может быть, поворачивание могло бы быть, как это имеет место у голубя, приостановлено, а поворот головы мог бы осуществиться на 270 или даже на 360°.
«Но это невозможно у кроликов. Поэтому животное катится. Перекатывание животного вокруг его продольной оси осуществляется всегда в два этапа. Первый автоматический этап перекатывания тот же, что и у голубей. Голова энергично отбрасывается в сторону операции, поворачиваясь до тех пор, пока дорсальная поверхность ее не коснется пола. Голова при этом поворачивается на 180°. Как уже было описано, в этот момент верхняя конечность противоположной стороны, увлекаемая движением головы и шеи, максимально разгибается, отводится от тела и, цепляясь за пол, пытается предупредить дальнейшее поворачивание головы. Но если однажды голова повернулась дальше и дорсальная часть ее касается пола, а поворот превышает 180°, то помощь верхней конечности становится бесполезной. Сама она поворачивается кверху, и в момент, когда она достигает вертикальной плоскости (поворот головы достигает в это время 270°), мощным броском осуществляется второй этап поворота, животное перебрасывает заднюю часть тела энергичным произвольным движением. «Тот факт, что повороту тела вокруг его продольной оси постоянно предшествует весьма интенсивный поворот головы и шеи, поддерживает предположение, что механизм поворота может являться следствием автоматического начального поворота головы и шеи...
«...Когда девиация передней части тела оказывается настолько скорригированной, что сидение снова оказывается возможным, переворачивания прекращаются, и это является веским аргументом в пользу предположения, что переворачивание полностью зависит от интенсивности поворота головы и шеи.
«За это предположение говорит также и еще один аргумент. Перекатывающееся таким образом с крайней интенсивностью животное может быть немедленно избавлено от этих переворачиваний, если удалить и второй лабиринт. Посредством этой второй операции повернутое положение головы и шеи ликвидируется словно волшебной силой, а также исчезает и девиация глаз как следствие того же вмешательства».
17 Р. Магнус
258
Шестая глава. Последствия односторонней экстирпации лабиринта
Как можно видеть, Эвальд объясняет появление движений перекатывания главным образом установкой и движениями тонически разогнутых конечностей противоположной стороны. Такое представление, несомненно, чересчур упрощенно. Напротив, Винклер разлагает движение перекатывания на две фазы: в первой животное поворачивает свои голову и шею примерно на 180° и включает в движение разогнутую переднюю ногу противоположной стороны. Это движение продолжается до тех пор, пока животное окажется не в состоянии дольше сохранять равновесие. Тогда произвольным движением второй фазы оно перебрасывает себя и переводит тем самым свои задние конечности на другую сторону. Это описание также все еще слишком упрощает дело, но оно уже содержит два важных элемента, которые принимаются во внимание при объяснении явления перекатывания, а именно: первопричинное значение поворота головы и шеи и признание того, что переворачивание через спину следует рассматривать как самостоятельное ответное движение.
Совершенно невозможно уследить за движениями перекатывающегося кролика простым глазом. По мере упражнения, правда, вполне удается распознать отдельные характерные фазы движения, однако другие составляющие движения протекают настолько быстро, что нельзя разобраться в том, что происходит. Поэтому необходимо прибегнуть к помощи кинематографических снимков. Такого рода серии снимков, произведенные спереди, сзади и с обеих сторон, позволяют изучить явление во всех деталях.
Всякая попытка объяснить движения перекатывания должна исходить из факта, что кролик с односторонне удаленным лабиринтом сразу после операции не переворачивается непрерывно, но что движения эти наступают приступообразно. В паузах между приступами животное или лежит (в первые часы после операции) на стороне утраченного лабиринта, или (на первый или второй день после операции) может спокойно сидеть прямо; если оно свалилось набок, способно даже снова подняться и продолжать сидеть в положении, описанном выше. Нередко спонтанно или в ответ на какое-либо раздражение, которое наносится в процессе исследования животного, наступает приступ движений перекатывания, который может прогнать животное через всю комнату и обычно прекращается, пока одна из стен комнаты не окажется непреодолимым препятствием для дальйейшего перекатывания. Стало быть, животное в состоянии сидеть совершенно нормально, и, несмотря на это, в промежутках оно перекатывается. Значит, помимо различия тонуса конечностей и ненормального поворота головы, шеи и туловища, должно добавляться что-то еще, чтобы вызвать движения перекатывания.
При внимательном рассматривании уже простым глазом можно увидеть, в чем здесь дело. Если посмотреть на катящееся по полу животное сверху, то можно заметить, что ноги при этом попеременно то сгибаются, то разгибаются и что производится это с весьма большой силой. Другими словами, животное бежит. При этом позвоночник также попеременно
Рис. 153—158. Кролик, левосторонняя экстирпация лабиринта 29 июня 1912 г. Кинематографические снимки движений перекатывания через небольшой промежуток после пробуждения от эфирного наркоза. Из каждой негативной пленки был изготовлен позитив, а затем негатив и соответствующий позитив проецировались рядом на большой лист чертежной бумаги. Затем негатив тщательно срисовывался, причем находящийся рядом позитив использовался в качестве постоянного контроля.
Размеры оригинальных снимков — 10 : 15 мм.
Рис. 153.
17*
260 Шестая глава. Последствия односторонней окстирпации лабиринта
разгибается и сгибается, а тело животного приобретает при взгляде сверху выгнутость то вправо, то влево.
Действительно, при изучении кинематограммы выяснилось, что здесь заключается единственное объяснение движений перекатывания.
Дело идет о весьма энергичных движениях бега животного, голова которого повернута спирально и которое вследствие этого движется не вперед, но в процессе бега «ввинчивается» в пространство.
Детали происходящего лучше всего иллюстрируются снимками.
На рис. 153 можно видеть кролика после левосторонней операции, который катится прямо на наблюдателя. Кадр al показывает, что передняя часть тела животного находится в сидячем положении, в то время как задняя половина лежит на правом боку. Полный поворот заснят на 20 кадрах (до кадра с4). Если сначала проследить за движениями задних ног, то видно, что они при этом дважды полностью разгибаются и сгибаются. Первое разгибание начинается на аЗ и достигает своего максимума на а5. Затем задние ноги снова сгибаются (а8 и Ы). Второе разгибание начинается на Ь2 и достигает максимума на Ъ4 и Ъ5. С Ь7 задние ноги снова сгибаются до сЗ. С с4 начинается разгибание для следующего перекатывания (максимум на с8). Передние ноги ведут себя совершенно так же. Первое сгибание видно на al, первое разгибание — на ai и а5, второе сгибание — на а8 и Ы, второе разгибание — на а5\ на cl и с2 передние ноги снова согнуты, далее начинается следующее перекатывание. Если при этом наблюдать за позвоночником, то он вначале (al) оказывается согнутым, животное в сгорбленном положении. Одновременно 6 разгибанием ног наступает максимальное разгибание (дорсальная флексия) позвоночника, которое достигает своего максимума на а4 и а5. На а8 и Ъ1 позвоночник снова согнут вентрально, на а4 и Ь5 — разогнут, на сЗ — согнут и т. д. Другими словами, чтобы один раз обернуться вокруг своей продольной оси, животное произвело два полных прыжка. Один прыжок привел его из правого бокового положения через живот в левое боковое положение; второй прыжок — из последнего положения через спину снова в правое боковое положение.
Когда подобные прыжки совершает нормальный кролик, он продвигается при этом вперед. Животное с утраченным лабиринтом, наоборот, продвигается вперед лишь очень мало и вместо этого переворачивается вокруг своей продольной оси. Каким образом это осуществляется, лучше всего видно на серийных снимках, на которых животное сфотографировано спереди (рис. 154). Тогда видно, что паузы между отдельными прыжками, т. е. время, в течение которого животное находится в одном из боковых положений, оно использует для того, чтобы снова восстановить спиральный поворот своего тела, который во время прыжка оказывался частично или полностью снятым. Только тогда, когда спиральный поворот снова наступил, совершается следующий прыжок.
На кадре а5 рис. 154 животное достигло правого бокового положения после того, как оно на al—а4 перевернулось через спину. Теперь оно лежит на своем правом плече и правой защечной области, задние ноги лежат слева от животного. В этом положении они остаются до кадров сЗ и с4. За это время, однако, голова повернулась настолько далеко, что кверху оказывается обращен уже не левый, а правый глаз; точно так же повернулся грудной отдел. При этом на голову действуют в совокупности основной поворот, лабиринтный установочный рефлекс и установочный рефлекс с тела, поэтому (на кадрах а8—Ъ4) голова переходит из бокового положения в прямое, причем передняя часть тела вследствие шейного установочного рефлекса переваливается через лежащие слева передние
Fixe. 154.
262 Шестая глава. Последствия односторонней экстирпации лабиринта
ноги, а голова также поворачивается влево, так что все тело принимает положение выгнутой стороной влево. Поворачивание передней части тела, однако, продолжается далее (основной поворот и лабиринтный установочный рефлекс), левый глаз оказывается внизу, и при повороте шеи осуществляется относящийся сюда тонический шейный рефлекс на правую переднюю ногу («челюстная» нога), которая тонически разгибается (<Ь7—с5), в то время как левая передняя нога («черепная» нога) тонизирована в меньшей степени (с2 и сЗ). На этой стадии животное лежит левым плечом вперед, правой защечной областью назад. На рис. 155 (кадры с5—с7) это характерное перекручивание (Verdrehung) видно весьма отчетливо.
Только тогда, когда эта стадия достигнута, осуществляется следующий прыжок, во время которого голова поднимается, позвоночник сгибается дорсально и все четыре ноги разгибаются. Этот прыжок перебрасывает животное на другую сторону.
Серийные снимки (рис. 154) позволяют отлично рассмотреть отдельное элементы этого движения. На кадрах ряда Ъ тело животного вогнуто влево, а голова и грудь осуществляют описанный выше поворот; позвоночник флексирован вентрально. В то время как в первый момент животное спокойно удерживает это положение, голова описанным выше образом поворачивается левым глазом вниз. Что происходит в результате этого с позвоночником, можно видеть, если сравнить кадр ЪЗ с кадром сЗ. В течение всего времени голова оставалась с левой стороны животного, но благодаря тому, что она повернулась, шейный отдел позвоночника должен был перейти из положения вентральной в положение дорсальной флексии. Значит, разгибание шейной части позвоночника причинно связано с этим поворотом. Те же отношения проявляются и в грудном отделе. Последний остается вогнутым влево. Но так как он поворачивается, то переходит из вентрального в дорсальное сгибание (Ъ6—с5). Таким образом, поворот передней половины тела сопряжен с разгибанием позвоночника до тех пор, пока сохраняется вогнутость животного влево. Поворот шеи, как показано выше, является прямым последствием выпадения лабиринта. То же самое верно и для поворота туловища. Однако если поворачивание шеи однажды началось, оно должно, как указано выше, усилить поворот туловища. Если животное прыгает и при этом разгибает позвоночник, этим предопределяется следующий момент, действующий в том же направлении. Таким образом, в это мгновение все содействует усилению общего эффекта. Кадры c4—d2 показывают только собственно прыжок. Разгибание позвоночника делается постепенно максимальным и распространяется на заднюю половину тела. Поскольку вогнутость тела влево сохраняется, животное при этом переворачивается на другую сторону. Однако сначала задние ноги обращены еще влево (с4). Они только тогда переходят на другую сторону, когда за собственно прыжком следует разгибание ног. Это разгибание задних ног видно на кадрах с4—с8. На с8 все животное представляет собой вогнутую дорсально дугу. Только верхняя брюшная область еще лежит на плоскости опоры, и животному нужно перевалиться на левую сторону из-за веса его головы.1 Таким образом, задние ноги попадают с одной стороны на другую, в процессе чего они сильно разгибаются назад, и уже движением, следующим за этим (dl и d2), тазовый отдел переваливается на другую сторону. Видно, что сначала правая передняя нога оказывается тонически разогнутой (с4), что вследствие этого левая передняя нога делает первый «шаг»
1 То же снято на рис. 158, Ъ6—Ъ8 сзади.
Последствия односторонней экстирпации лабиринта у кролика
263
{с5 и сб), а вслед за ней идет вперед и правая передняя нога (с7 и с5). Эта последовательность соответствует произведенным ранее исследованиям на децеребрированных животных, когда у них при повернутой голове наступают движения бега и «черепная» конечность, тонус которой ниже, постоянно начинает это движение. На кадре с8 прыжок выражен полно. На dl животное уже лежит на другой стороне. Теперь позвоночник снова согнут в вентральном направлении, передние и задние ноги оказываются на полу и на dé — согнуты. Следующая фаза движения перекатывания состоит в том, что вновь формируется поворот передней части тела относительно задней его части.
Рис. 155 отчетливо показывает эти события. Особенно ясно можно видеть переворачивание головы на другую сторону, распространяющееся спереди назад разгибание позвоночника, прыжок с разгибанием задних ног, тоническое разгибание правой передней ноги, «выступание» левой передней ногой и заваливание (Herumfallen) всего тела влево (кадры c3—d3). Рис. 156 (кадры Ъ5—с2) показывает приблизительно то же, но только с примыкающим сюда вентральным резким сгибанием разогнутого позвоночника, причем голова и вся передняя половина тела поднимаются в воздух (сЗ—dl), что носит характер прыжка. Рис. 157 показывает ту же «фазу спереди (а1—а8) и слева (с1—с4).
Если на рис. 154 рассмотреть кадры сЗ—d2, на рис. 155 — кадры dl—d3, на рис. 157 — кадры е2—е4, то тотчас станет очевидным, что левая передняя нога не остается при перекатывании совершенно спокойной и неподвижной, как думали предыдущие наблюдатели, а, наоборот, производит весьма энергичные движения. То же относится и к левой задней ноге. Без этого переворачивание на другую сторону оказалось бы неосуществимым. Причина, в силу которой при обычном наблюдении это движение ускользает от наблюдателя, лежит в исключительной кратковременности этой фазы. Например, на рис. 157 уже на кадре Ъ8 животное лежит в левом боковом положении, оставаясь в нем (задней частью тела) до d7, т. е. на 16 кадрах, в то время как собственно «прыжок» совершается от d8 до еЗ, т. е. на 4 кадрах. Если кадры показать кинематографически, чо можно увидеть, что движение это осуществляется исключительно быстрым рывком (Ruck), и разглядеть при этом какие-либо детали совершенно невозможно.
Мы проследили на рис. 154 движение перекатывания до момента, когда животное (на кадре d4) лежало после прыжка на левом боку и тело -его переходило из положения дорсальной в положение вентральной флексии со сгибанием ног. Благодаря этой вентральной флексии голова, если смотреть от наблюдателя, перебросилась справа налево (d2—d6). Одновременно с этим происходит и восстановление поворота головы, шеи и грудного отдела по отношению к заднему отделу тела. На кадре d2 обращен вниз левый глаз, на кадре d6 — темя. Подобное же движение представлено на рис. 153, кадры а5—d2. На а5 левый глаз, а начиная с а7 правый глаз направлены на наблюдателя, в то время как животное задней частью остается еще лежащим в боковом положении, передняя часть тела переворачивается на спину. На рис. 156 (кадры а4—а8) этот поворот тела виден более отчетливо,
Таким образом, повторяется последовательность, которая имела место выше, при положении животного на противоположном боку. На рис. 156 {кадры а4 и аб) животное оказывается в левом боковом положении с вентрально согнутым туловищем, а если смотреть от наблюдателя, образует дугу, вогнутую влево. Эту установку оно первое время удерживает. Когда же оно поворачивает переднюю часть своего тела, сгибание позво-
Рис. 155.
Рис. 156.
Рис. 157.
Рис. 158.
268
Шестая глава. Последствия односторонней экстирпации лабиринта
ночника в переднем отделе должно превратиться в разгибание (а7, а8 и Ы). Этим самым дается начало следующему движению прыжка,которое осуществляется через спину. Разгибание позвоночника распространяется дальше кзади, задние ноги разгибаются назад, передние — вперед (рис. 153, кадры Ъ3—Ъ5), и теперь, как только поворот передней части тела достиг определенной степени, происходит переваливание всего животного на другую сторону.
Рис. 159. Тот же кролик, что и на рис. 116, уложен на спину так, что голова, грудь и таз дорсальной стороной обращены строго вниз. Затем голова поворачивается в левую сторону, т. е. левое ухо перемещается в вентральном направлении, левый глаз обращен вверх (голова на рисунке закрыта рукой экспериментатора). Грудной отдел остается в прежнем положении. Правая передняя нога оказывается «челюстной» и разгибается, левая («черепная») — сгибается. Таз поворачивается в направлении, обратном, чем голова, таким образом, что левая ягодица лежит внизу, а правая задняя нога перемещается больше вверх. Вследствие этого тело животного поворачивается винтообразно. Различия тонуса задних ног на этом снимке отметить нельзя. Все положение является зеркальным отражением положения, изображенного на рис. 116.
Это движение имеет в основе шейный установочный рефлекс и может быть вызвано в положении на спине у любого кролика. Если, например, уложить нормального кролика симметрично в положение на спине и повернуть ему, как показано на рис. 159, голову влево, то, как описано выше, наступает спиральный поворот всего тела. В то время как голова левой щекой обращена вверх, тазовый отдел левой ягодицей обращен вниз. Если теперь все животное перевернуть через спину немного вправо, то
Рис. 160. Тот же кролик в положении на спине, с головой, повернутой влево (левый глаз кверху).
Положение животного отличается от такового, изображенного на рис. 159, только тем, что животное все в целом несколько перевернуто направо так,
что дорсо-вентральная ось грудного отдела теперь стоит не отвесно, как на рис. 159, а косо, поэтому правая лопатка лежит на столе, левая — обращена несколько вверх. Вследствие этого изменилось стояние таза. Этот последний перевалился на другую сторону: правая ягодица лежит теперь скорее несколько глубже, а поясничная часть позвоночника вогнута вправо. Поэтому тело животного больше не повернуто винтообразно, но образует дугу, открытую в «челюстную» сторону. Различия тонуса конечностей подобны таковым на рис. 159. Передняя правая нога («челюстная») максимально разогнута, левая («черепная») — сильно согнута. На этом рисунке хорошо видно также различие тонуса задних ног. Правая задняя нога сильно разогнута, левая —
согнута.
можно получить позу, при которой таз уже нельзя удержать в первоначальном положении, он переваливается на другую сторону так, что левая ягодица обращена книзу, а поясничный отдел позвоночника оказывается вогнутым вправо. Тело животного больше уже не изогнуто спирально, но образует дугу, изогнутую вправо (рис. 160). Описанное движение таза иногда осуществляется быстрым толчком. Если поворот животного продолжить, то последнее в конце концов оказывается лежащим на пра-
Последствия односторонней экстирпации лабиринта у кролика
269
ьом боку. Переворачивание таза протекает одинаково при согнутых или разогнутых задних ногах животного. После экстирпации обоих лабиринтов наступает точно такое же движение.
Если определить положение дорсо-вентральной оси грудного отдела, при котором совершается переход таза из одного положения в другое, то оказывается, что до тех пор пока при голове, повернутой в сторону, ж при перекатывании через спину левая лопатка находится внизу, таз повернут в направлении, обратном повороту головы. Переход у некоторых животных совершается тогда, когда дорсо-вентральная ось грудного отдела образует с вертикалью угол в 45°.
Переброс таза при этом переходе является исключительно мощной реакцией. В отдельных случаях таз может повернуться на 180°. При сравнении рис. 159 и 160 можно заметить, что стояние таза различается примерно на 90°.
Точно такое же наблюдение можно сделать, если по прошествии любого времени после операции и периода катательных движений кролика, лишенного одного лабиринта, уложить на спину и дать ему перевернуться через спину в сторону нормального лабиринта. Теперь не нужно приводить голову в повернутое относительно груди положение, так как голова самостоятельно поворачивалась вследствие операции. При этом движении спиральный поворот туловища сохраняется. Таким образом, после левосторонней экстирпации лабиринта голова повернута влево, левое ухо — вентрально, левая ягодица лежит на столе. Однако как только грудной отдел своей дорсо-вентральной осью пересек вертикаль, — а у многих животных только после того, как вертикаль перейдена на 30—45°, — таз перебрасывается, правая ягодица оказывается внизу, позвоночник становится вогнутым вправо. Движение это в точности повторяет движение нормального кролика с насильственно повернутой головой. В обоих случаях эта реакция представляется чисто механической, причем таз с момента достижения определенного положения передней частью тела не в состоянии дольше удерживать спиральную изогнутость относительно грудного отдела и, следуя силе тяжести и тяге односторонне напряженных мышц, переваливается (herumklappen), чтобы принять согнутое положение вогнутостью в направлении «челюстной» стороны.
Если голову односторонне лабиринтэктомированного животного сильно повернуть влево, то оказывается возможным, как говорилось выше, вызвать противоположный поворот таза по отношению к грудному отделу. В этом случае удается вызвать точное зеркальное воспроизведение только что описанной реакции тазовой области, когда животное переворачивается через спину справа налево. При этом сначала спирально изгибается туловище, так что лежащей на столе оказывается правая ягодица, и затем, следуя повороту грудного отдела, переваливается влево, после чего и поясничная часть позвоночника оказывается вогнутой влево.
Точно такие же движения производятся и при перекатывании,' когда животное переворачивается через спину. Наиболее демонстративно они выглядят на серийных снимках, на которых животное снято сзади (рис. 158). Впрочем, на рис. 153—157 они хорошо видны. На рис. 158 к моменту, фиксированному на кадре cl, животное вследствие «прыжка» оказалось в левом боковом положении. К с2 уже начинается поворот передней части тела. На сЗ правая передняя нога пересекла вертикаль, на сб видны обе передние ноги, а дорсо-вентральная ось грудного отдела перешла вертикаль. На сб левая ягодица еще лежит на столе, на с7 и с8 происходит переворачивание таза, на d2 правая ягодица внизу; животное приняло правое боковое положение. На с2 задние ноги еще
270 Шестая глава. Последствия односторонней экстирпации лабиринта
согнуты. На сЗ правая задняя нога («челюстная») разогнута в коленном и голеностопном суставах, вторая нога следует за ней, на с5 начинается разгибание бедра, на сб — разгибание поясничной части позвоночника* и животное переворачивается через спину в полностью разогнутом состоянии. Это движение совершается быстрым рывком. При переворачивании поясничного отдела позвоночника дело идет о чисто пассивном движении повернутого таза, который перетягивается силой тяжести и напряжением мышц другой стороны. Однако в процессе переворачивания это движение подкрепляется мощным разгибанием ног и позвоночника. Винклер также выделил эту фазу движения как нечто особое и рассматривал ее как произвольное движение животного, когда тело его вследствие перекатывания оказывается приведенным в неудобное положение.
В принципе я соглашаюсь с Винклером; нет необходимости после приведенных опытов на нормальных животных усматривать в этом движении произвольную реакцию. Таламические животные с полностью удаленным большим мозгом, когда они переворачиваются после экстирпации лабиринта на одной стороне, тоже обнаруживают эту фазу и так же отчетливо, как и кролик без одного лабиринта с интактными большими полушариями.
Мы хотели бы снова рассмотреть те положения, которые принимает животное, прежде чем перевернется через спину, т. е. в нашем примере — левое боковое положение (рис. 155, al). Кролик, не производящий ката- тельных движений и подвергавшийся экстирпации лабиринта уже много дней назад, в состоянии из этого положения принять с помощью установочных рефлексов с туловища сидячее положение. Непосредственно после операции этот рефлекс еще силен,и животное, если оно не катится, остается лежать на левом боку, что соответствует в покое лабиринтному установочному рефлексу на голову.
При переворачивании через спину, наоборот, постепенное усиление основного поворота головы описанным своеобразным путем комбинируется с шейным установочным рефлексом и с движением прыжка, которое ведет к дорсальному сгибанию головы и передней части тела и в конце концов перебрасывает животное в правое боковое положение (рис. 155, bl; рис. 158, Ъ2). Именно с этого положения начиналось на стр. 262 описание движений перекатывания. Теперь начинается следующий ряд движений перекатывания точно таким же образом, причем сначала животное восстанавливает поворот передней части тела, а затем снова «прыжком» переворачивается через спину; таким образом дело продолжается до тех пор, пока какое-нибудь внешнее препятствие или утомление не остановят животное. Поскольку в обоих боковых положениях животное сгибает позвоночник вентрально, то получается весьма примечательное зрелище, если на катящееся животное смотреть сверху, так как все тело его болтается из стороны в сторону, становясь при этом вогнутым то вправо, то влево. Переход между этими положениями осуществляется всякий раз через одну из двух «фаз прыжка», из которых обе производятся с большой скоростью, так что простым глазом можно различить только самые разгибания, но не то, каким путем они осуществляются.
Согласно принятому здесь, основанному на кинематографической регистрации представлению, движения перекатывания у кролика после односторонней экстирпации лабиринта являют собой движения бега и прыжка животных, тело которых вследствие операции повернуто спирально и которые поэтому «ввинчиваются» в пространство. Этих движений бега и прыжка для возникновения истинного перекатывания еще не¬
Последствия односторонней экстирпации лабиринта у кролика
271
достаточно. Прежде всего, перекатывание через брюхо может осуществляться только тогда, когда позвоночник и задние ноги оказываются вытянутыми. В противном случае задние ноги не могут быть переправлены на другую сторону. Конечно, может случиться, что и спокойно сидящее животное по какой-либо причине опрокинется на сторону утраченного лабиринта, перевернет после этого свое тело и благодаря этому перекатится через спину. Однако когда оно окажется на другой стороне, оно обычно снова усаживается; в других случаях это анормальное положение служит раздражением, вызывающим движение правильного прыжка.3
При таком понимании перекатываний старый спорный вопрос — является ли перекатывание следствием раздражения или следствием выпадения — утрачивает противоречивый характер. В действительности оно и то, й другое. Спиральный поворот всего тела есть явление выпадения, с одной стороны, прямо обусловленное утратой лабиринта, с другой — вторично обусловленное поворотом шеи, который в свою очередь является снова прямым следствием выпадения. Наоборот, движения бега и прыжка, как и у каждого нормального животного, наступают в ответ на раздражение. При этом движения перекатывания тем интенсивнее и длятся тем дольше, чем больше пострадал при экстирпации ствол 8-го нерва, чем более обильным было наступившее кровотечение и т. д. По мере приобретения опыта во время операции перекатывания после нее все уменьшаются; можно прооперировать животных, которые после операции не совершают вовсе или производят лишь немногочисленные катательные движения и уже через несколько часов сидят спокойно. Для этого во время удаления преддверия следует по возможности меньше травмировать ствол 8-го нерва. Во время последующих операций обычно оказывалось возможным предсказать, будет ли животное перекатываться интенсивно или только незначительно. Таким образом, главное раздражение, вызывающее движения бега и прыжка, представляет собой операционная рана. Однако это раздражение может быть усилено самыми различными путями с других точек тела. Каждое прикосновение или исследование животного в состоянии вызвать приступ катательных движений. После сказанного становится понятным, почему животное может то сидеть спокойно, то вскоре после этого кататься по всей комнате.
В пользу того, что в процессе перекатывания осуществляется пусть даже весьма незначительное продвижение всего тела вперед, говорит сравнение кадров а5 и с8 на рис. 153 и al и d7 на рис. 154.
Если, как было показано выше, движения перекатывания действительно можно представить себе как движения бега и прыжка у животных, тело которых спирально повернуто, то препятствовать или ограничивать движения перекатывания должно не только то, что 8-й нерв во время операции максимально щадится, но также уменьшение или упразднение спирального поворота тела. Именно это и имеет место в действительности. Любой, даже самый сильный приступ движений перекаты- 11 Мы многократно видели, что односторонне лабиринтэктомированный кролик через 1—2 месяца после операции, когда период движений перекатывания давно миновал, а поворот головы достиг своего максимума, снова начинал переворачиваться через спину. Если такое животное посадить на пол, поворот головы его понемногу увеличивается и спустя 1—2 минуты составляет 135°. При этом передняя «челюстная» конечность сильно разгибается и отводится, что вместе с перевешиванием головы животного вызывает падение на сторону утраченного лабиринта. Далее кролик один раз переворачивается через спину на другую сторону и потом спокойно усаживается. Теперь снова может увеличиться поворот головы, а вся история повторится много раз. Через спину всегда производится только одно переворачивание, и животное затем снова сидит (ср, протокол на стр. 223—224).
272
Шестая глава. Последствия односторонней экстирпации лабиринта
вания можно прервать, установив голову прямо относительно груди. В большинстве случаев животное при этом усаживается прямо и больше не переворачивается или продолжает движения бега и выбрасывания ног (Strampel), но последние не ведут к перекатываниям, а позволяют животному скакать прямо вперед. Как только голова отпускается, начинаются сильнейшие движения перекатывания.
Как было показано выше, благодаря установке головы прямо уменьшался и поворот грудного отдела относительно таза, а кроме того, понижалось различие тонуса конечностей обеих сторон. Все это действует в том же направлении, что и снятие поворота головы, с тем результатом, что теперь движения бега и прыжка не в состоянии больше вызвать ка- тательных движений.
В согласии с этим находится наблюдение, что кролик, у которого перерезаны верхние шейные корешки, после односторонней экстирпации лабиринта, как показывает следующий экспериментальный пример. {16), никаких движений перекатывания не обнаруживает.
Кролик. 25 сентября 1913 г. Перерезка трех верхних шейных задних корешков с обеих сторон. На следующий день животное сидит безупречно прямо, только с незначительно качающейся головой. В дальнейшем оно ежедневно по нескольку часов бегает по двору. Спустя 25 дней (20 октября) было обнаружено следующее. Животное сидит нормально, только иногда передние ноги еще держатся разогнутыми. При беге и прыжках незначительная неловкость; в нормальном положении пассивное сгибание и опускание головы не вызывают отчетливых тонических шейных рефлексов на передние ноги, которые, однако, животное вполне способно активно разгибать и сгибать. В положении на спине в ответ на поворот головы появляются слабые, но несомненные тонические шейные рефлексы (уменьшение разгибательного тонуса в передней «черепной» ноге, увеличение — в «челюстной» ноге). Задние ноги реагируют лишь очень слабо. Наоборот, влияние поворота головы на туловище (шейные установочные рефлексы) отсутствует.
Таким образом, у этого кролика перерезка трех пар верхних шейных задних корешков не оказалась достаточной для того, чтобы тонические шейные рефлексы на конечности полностью исчезли. Последние только значительно снизились; при повороте оказывалось еще возможным влиять на тонус передних ног, в то время как при поднимании и опускании головы отчетливых шейных рефлексов более не наблюдалось. Шейные установочные рефлексы на мускулатуру туловища полностью отсутствовали.
31 октября де Клейном был экстирпирован левый лабиринт. После обнажения и освобождения отверстий полукружных каналов пинцетом был обойден 8-й нерв во внутреннем слуховом проходе и намеренно несколько потревожен. По многочисленным нашим наблюдениям, нормальный кролик после такой операции, наверное, станет перекатываться. Такие движения у этого животного с ослабленными шейными рефлексами отсутствовали.
Через час после операции животное хорошо сидит на полу прямо, голова повернута на 45° влево, настолько же она отведена, горизонтальный нистагм головы, правая передняя нога разогнута и отведена. То же остается при установлении головы прямо. С момента пробуждения от наркоза животное находилось под длительным наблюдением и ни разу не переворачивалось. Конечно, приступообразно оно совершает интенсивные движения бега и прыжка, но эти движения никогда не ведут к перекатываниям, а наоборот, благодаря этим движениям животное или продвигается вперед, или падает на бок и тотчас же снова усаживается.
На следующий день в основном наблюдается то же самое. Голова теперь повернута на 90° влево. При сидении левая нога разогнута и отведена. При установлении головы прямо и при переворачивании головы (до 45°) вправо положение правой передней ноги не изменяется. Животное не перекатывается. Если оно падает на левую сторону, то способно без труда подняться снова. В висячем положении головой вниз грудной отдел повернут по отношению к тазу на 30°, верхняя апертура груди — на 45°, голова — на 60°. При устанавливании головы прямо поворот груди не становится меньше. В положении на спине тонус левой ноги значительно ниже, чем правой. При установке головы прямо это различие изменяется. Животное погибло через 6 дней после экстирпации лабиринта. Таким образом, исключение влияния шейных установочных рефлексов путем перерезки задних шейных корешков является достаточным для того, чтобы воспрепятствовать возникновению у этих животных движений перекатывания.
Последствия односторонней экстирпации лабиринта у кролика
273
Значительно труднее при помощи поворачивания головы заставить перекатываться в другую сторону интактного кролика, у которого эк- стирпирован один лабиринт. Это удается только в небольшом числе случаев. Причину этого установить легко. Так как в первые дни после операции, устанавливая голову прямо, можно уменьшить поворот туловища и различие тонуса конечностей только отчасти, то необходим значительный поворот головы, чтобы полностью компенсировать аномалию установки. Поэтому и в случае более сильного поворота головы в другую сторону противоположный поворот туловища и грудного отдела может наступить лишь в умеренной степени, так что это оказывается недостаточным, чтобы осуществилось перекатывание тела в другую сторону.
В заключение следует рассмотреть еще только один вопрос: возможно ли побудить к движениям перекатывания здорового, обладающего обоими лабиринтами кролика, если поворачивать его голову? Как указывалось выше, каждого нормального кррлика, если поворачивать его голову, можно заставить перевернуться из одного бокового положения через спину в другое боковое положение, в процессе чего животное производит точно такие же движения, что и перекатывающийся однолабиринтный кролик. После того как нормальное животное перевернулось через спину, оно обычно тотчас снова усаживается и далее остается в нормальном сгорбленном положении. Только тогда, когда животное по случайному стечению обстоятельств как раз в этот же момент совершает прыжок или рывок, оно перекатывается также и через брюхо и может произвести множество завершенных движений перекатывания одно за другим. Нам неоднократно удавалось отчетливо получить это на нормальных кроликах; это совпадение, однако не всякий раз вызывается произвольно, отчего этот опыт не пригоден для демонстраций.
В предыдущем отделе на основании ряда серийных снимков оказалось возможным дать анализ движений перекатывания кролика, лишенного одного лабиринта. Дело идет о движениях прыжка и бега у животных, тело которых вследствие операции приобрело спиральную изогнутость и которые поэтому прыгают не вперед, но ввинчиваются в пространство. Полный оборот вокруг продольной оси осуществляется посредством двух прыжков, из которых один переворачивает животное через брюхо, другой же—через спину. При этом всякий раз конечности и позвоночник разгибаются. Вследствие этих прыжков животное оказывается попеременно то в правом, то в левом боковом положениях, причем оно сгибает свои ноги и придает позвоночнику вентральный изгиб. Как только одно или другое боковое положение достигнуто, поворачиваться начинает передняя часть тела, благодаря чему утраченный во время прыжка поворот туловища восстанавливается.
Перекатывание наступает приступами, между которыми животное может сидеть спокойно.
Перекатывание через спину можно имитировать у нормального кролика, передвинув его с предварительно повернутой головой из одного бокового положения через спину в другое боковое положение. При этом возникает совершенно такое же перебрасывание таза и задних ног, как у перекатывающегося безлабиринтного кролика.
Устанавливанием головы прямо движения перекатывания можно тотчас прервать.
Специальный поворот тела, необходимый для осуществления перекатывания, по данным предыдущих разделов, следует представлять себе как явление выпадения, приступы движений бега — как явления раздражения.
18 Р. Магнус
274
Шестая глава. Последствия односторонней экстирпации лабиринта
Отклонение глаз и нистагм (15, 27, 38, 82Ъ)
Как только после односторонней экстирпации лабиринта начинает проходить действие глубокого наркоза, наступает установленное всеми авторами отклонение глаз. Глаз оперированной стороны отклоняется в вентральном направлении (книзу) и несколько вперед; глаз на другой стороне — дорсально (вверх) и иногда несколько кзади. Одновременно начинается сильный нистагм, во время чего глаза, достигнув максимума отклонения, быстрым толчком передвигаются обратно по направлению к среднему положению. Затем глаза снова медленным движением переходят в положение отведения, чтобы тотчас же новым нистагматическим движением оказаться отброшенными назад. Отклонение глаз совершается таким образом на обеих сторонах в направлении удаленного лабиринта; быстрые нистагматические движения — в направлении неоперированной стороны. Это описание действительно для кроликов, когда они после операции лежат на боку или, пробудившись после наркоза, приняли сидячее положение. Тогда они держат голову повернутой и наклоненной в сторону операции, т. е. морда обращена в сторону операции и ухо оперированной стороны благодаря повороту головы подвинуто в вентральном направлении. При этом у сидящего животного глаз оперированной стороны обращен к полу, глаз нормальной стороны — к потолку.
В течение первых дней девиация глаз несколько уменьшается до степени, которая в дальнейшим остается без изменений в качестве длительного последствия операции. Это состояние выше уже было подробно описано. Если спустя некоторое время после операции животное спокойно сидит в клетке и держит голову повернутой на 70—90° в направлении оперированной стороны, глаз неоперированной стороны стоит или совершенно нормально, или все еще несколько отклонен кверху (дорсально). Глаз на оперированной стороне, обращенный к полу, напротив, на этой стадии еще отчетливо, хотя и не очень сильно, отклоняется вентрально. Вообще, как наблюдали разные исследователи, отклонение глаза оперированной стороны обычно бывает сильнее, чем на интактной стороне. В этом боковом положении головы интактным лабиринтом кверху вертикальное отведение глаз минимально. Как только голове придают другое положение в пространстве (сагиттальная ось — горизонтально, интактный лабиринт — внизу), отведение глаз становится максимальным и может оказаться настолько интенсивным, что в разрезе век остается только белая склера. Глаз на оперированной стороне в этом случае отведен максимально в вентральном направлении и несколько вперед, другой глаз — максимально дорсально (иногда несколько впереди, иногда — несколько кзади). При всех других положениях головы в пространстве величины вертикального отклонения глаз лежат между этими двумя пределами. Девиация глаз всегда удерживается до тех пор, пока голова находится в соответствующем положении.
В разделе о компенсаторных установках глаз отклонение глаз после односторонней экстирпации лабиринта было уже объяснено действием на глаза тонического лабиринтного рефлекса, исходящего из оставшегося лабиринта.
Оставшийся лабиринт воздействует на одноименные косые мышцы обоих глаз и таким образом на отведение обоих глаз в одном направлении, а также на верхнюю прямую мышцу гомолатерального и на нижнюю прямую гетеролатерального глаза, обусловливая отклонение глаз по вер тикали в противоположных направлениях.
Последствия односторонней экстирпации лабиринта у кролика
275
Оставшийся лабиринт вызывает максимальное отклонение глаз из нормального положения по вертикали в том случае, если он при боковом положении головы оказывается внизу, тогда верхняя прямая мышца одноименной и нижняя прямая перекрестной стороны находятся в состоянии наибольшего укорочения. Наоборот, если при боковом положении головы интактный лабиринт окажется сверху, то его влияние на названные мышцы незначительно и глаз или вовсе не отклонен в вертикальном направлении, или отклонен незначительно.
Оставшийся лабиринт вызывает максимальный поворот обоих глаз вследствие сокращения обеих нижних косых мышц, тогда как голова стоит вертикально мордой вниз. И наоборот, лабиринт вызывает максимальный поворот при сокращении обеих верхних косых мыщц в том случае, если голова расположена вертикально мордой вверх. Интенсивность поворотов при наличии только одного лабиринта составляет примерно половину таковой при обоих интактных лабиринтах (в среднем около 45°).
Поскольку оба лабиринта воздействуют на вертикальные глазные мышцы в противоположном направлении, то после выпадения одного лабиринта дело должно окончиться описанным вертикальным поворотом. Наоборот, косые глазные мышцы находятся под однозначным влиянием обоих лабиринтов, вследствие чего после выпадения одного лабиринта не создается основания для появления ротаторной девиации.
В то время как оставшаяся после односторонней экстирпации лабиринта девиация глаз в своем главном направлении постоянна, направление нистагма, имеющего место в первые дни после операции, непостоянно и индивидуально различно. В большинстве случаев главное направление быстрой фазы нистагма на оперированной стороне — вперед и вверх; на интактной стороне — назад и вниз. Правда, может оказаться, что быстрая фаза нистагма на оперированной стороне направляется горизонтально вперед, на противоположной же стороне — горизонтально назад. Отчего зависят эти различия, до сих пор с полной уверенностью сказать нельзя. Один момент во всяком случае определяется установкой головы и вызванным этим круговым поворотом. Если, например, при установке головы а нистагм направлен (schlägt) вертикально вниз, то при положении головы в, если вследствие последнего произведен круговой поворот на 45°, тот же нистагм может иметь направление косо вниз и назад и т. д. Дальнейшие причины этих взаимоотношений могут быть обсуждены только позднее (см. стр. 289 и след.).
Как бы там ни было, но во всяком случае выяснилось, что величиной нистагма можно управлять, смотря по положению, которое будет придано в пространстве голове. Как только голова или все животное поворачивается в боковое положение так, что интактный лабиринт оказывается внизу и вертикальное отклонение глаз вследствие этого становится максимальным, нистагм полностью или почти полностью исчезает. Наоборот, нистагм максимально интенсивен, когда отклонение глаз достигло минимума, т. е. при тех положениях головы, при которых интактный лабиринт находится сверху. При всех других положениях головы в пространстве отклонение глаз и нистагм имеют промежуточную величину. Если, например, голову сидящего животного установить прямо, так, чтобы темя находилось наверху, а оба глаза были направлены в стороны, нистагм уменьшается, в отдельных случаях он может исчезнуть совсем. Если же продолжить поворот головы до тех пор, пока глаз оперированной стороны не окажется наверху, то в ряде случаев нистагм снимается полностью, иногда же он едва заметен.
18*
276
Шестая глава. Последствия односторонней экстирпации лабиринта
Как известно, нистагм исчезает спустя различное время: иногда через 24 часа, в большинстве случаев — спустя 48 часов. В одном случае он наблюдался на третий, в другом — даже на четвертый день. В этих случаях нистагм можно обычно ненадолго вызвать снова, если раздражать животное, придавать ему различные положения и т. д. В конце концов у подобного животного спонтанный нистагм вызвать больше не удается.
Зависимость между поворотом головы и отклонением глаз
До сих пор в качестве прямых последствий односторонней лабиринтэк- томии изучали вертикальное отклонение глаз и поворот головы. Теперь перед нами встает вопрос: в какой степени оба эти симптома зависят один от другого и влияют друг на друга?
Как указывалось выше, поворот головы ведет благодаря тоническим шейным рефлексам к отклонению глаз в вертикальном направлении. Если после правосторонней экстирпации лабиринта животное сидит в нормальном положении и повернуло голову вправо, т. е. по направлению к правому боковому положению, то левый глаз вследствие этого отклоняется книзу, а правый глаз — кверху. Если животному придавать различные положения в пространстве, то, как было показано выше, степень поворота головы увеличивается или становится меньшей. Спрашивается, будет ли при этом существенно усиливаться или уменьшаться первичное, обусловленное выпадением лабиринта, отклонение глаз. Следующие протоколы опытов дают на это ясный ответ. 11) Серый кролик. 19 ноября 1921 г. Правосторонняя экстирпация лабиринта 22 ноября. В висячем положении головой вниз, основной поворот более чем на 90°. Во время сидения голова лишь незначительно (0—20°) повернута вправо. Если голова стоит в пространстве прямо, левый глаз несколько отведен вверх и вперед, правый — немного вниз. На левом глазу горизонтальный нистагм, направление которого назад (в сторону ушей), на правом глазу — вперед.
а) Голова строго в правом боковом положении, тело в нормальном положении: левый глаз отклонен несколько кпереди, но никакого следа отклонения вверх или вниз.
б) Голова в правом боковом положении, тело в том же положении: глаз стоит, как и в описании а.
в) Голова в правом боковом положении, тело из правого бокового повернуто в положение на спине: левый глаз несколько отклонен кверху, нижний край роговицы, однако, еще не свободен.
г) Голова в правом боковом положении, тело из нормального повернуто в левое боковое положение: левый глаз отходит немного книзу, склера не свободна. Последняя освобождается только тогда, когда тело поворачивается в положение на спине на 270° по отношению к голове.
Результат. Если исходить из спонтанного положения животного после односторонней экстирпации лабиринта, влияние поворота шеи до 90° на установку глаз отсутствует или бывает весьма незначительным. Только повороты шеи на 180—270° оказывают более сильное влияние.
2) Черный кролик. 19 ноября 1921 г. Правосторонняя экстирпация лабиринта 22 ноября. Время от времени садится или лежит в правом боковом положении, иногда движения перекатывания вправо. Нистагма нет.
а) Голова в правом боковом положении (ротовая щель — вертикально, назогокни- питальная ось — горизонтально), тело в нормальном положении. В щели век, даже при полном раскрытии, склеры не видно вовсе. При раздвигании век глаз едва отклонен вверх.
б) Голова в правом боковом положении, тело в том же положении: установка глаз едва изменилась.
в) Голова в правом боковом положении, тело повернуто сильнее, так что оказывается в положении на спине: только теперь рефлекс на глаза становится отчетливым и глаз отходит кверху.
г) Голова в правом боковом положении, тело повернуто в обратном направлении до достижения левого бокового положения: установка глаз по сравнению с описанной
Последствия односторонней экстирпации лабиринта у кролика
277
в пункте а едва изменилась. Только тогда, когда тело оказывается в положении на спине (т. е. повернутым по отношению к голове на 270°), глаз отходит книзу, но и тогда не слишком сильно.
д) Фото: животное сидит спокойно, тело поддерживается в нормальном положении, опираясь на таз, спонтанное положение головы (голова в правом боковом положении), ротовая щель спонтанно открыта. Снимок вертикально сверху: глаз не отклонен.
Результат. Повороты шеи в обоих направлениях из спонтанного положения до 90° отчетливого влияния не оказывают, только повороты на 180—270° вызывают заметное отклонение глаз.
Отсюда следует, что поворот шеи сам по себе влияет на вертикальное отклонение глаз лишь весьма незначительно и что поэтому установки глаз после односторонней лабирантэктомии в основном следует связывать с деятельностью оставшегося лабиринта.
Но с другой стороны, спрашивается, находится ли самое поворачивание головы под влиянием, исходящим со стороны глаз. Предположительно говоря, это могло бы осуществляться двумя различными путями: во-первых, поворот головы мог бы корригироваться с помощью глаз, во-вторых, поворот головы мог вызываться или усиливаться вследствие отклонения глаз.
Что касается первого пункта (коррекция поворота головы со стороны глаз), то выше было уже показано,что у кролика оптические установочные рефлексы не играют никакой роли и что поворот головы после односторонней экстирпации лабиринта при закрывании глаз не меняется. Если помещать кроликов в тесные клетки, куда свет попадает только сверху, они в небольшом числе случаев держат голову повернутой несколько больше, чем тогда, когда они в состоянии свободно передвигаться по комнате. Это непостоянное влияние лишь минимально. Стало быть, существенной корректировки поворота головы с помощью зрительных восприятий у кроликов нет.
Другой вопрос, не является ли поворот головы вторичным результатом отклонения глаз, мог появиться лишь в то время, когда осуществление поворота головы еще не было объяснено односторонним тоническим лабиринтным рефлексом на шейные мышцы и шейными установочными рефлексами. Несмотря на это, не лишено интереса коротко коснуться этого вопроса.
Когда у кролика, лишенного одного лабиринта, который спокойно сидит, корригируется анормальное положение головы, то, как было показано выше, этим самым вызывается отчетливое отклонение глаз. Если голову отпустить, так чтобы она могла возвратиться в свое повернутое положение, девиация глаз уменьшается или полностью исчезает. Именно исходя из этого, Винклер (1) поставил в свое время вопрос: не для того ли животное производит поворот шеи и головы, чтобы компенсировать отклонение глаз? С этой точки зрения первичным последствием утраты лабиринта являлась бы только девиация глаз, тогда как поворот головы был бы вторичным явлением, вызванным анормальным положением глаз. Сам Влнклер пробовал уже в то время решить этот вопрос экспериментально, надеясь путем перерезки у кролика различных глазных мышц вызвать соответствующую девиацию глазного яблока, чтобы наблюдать вслед за этим соответствующий поворот головы. Однако к намеченной цели эксперименты не привели, так как этим путем желательная девиация глаз не вызывалась.
То, что поворот головы и другие анормальные позы и движения тела не зависят от девиации глаз и вызываться последней не могут, поясняется следующим.
Прежде всего смещение зрительных образов не может явиться причиной какого- либо компенсаторного поворота головы, так как кролику с одним лабиринтом можно прикрыть оба глаза без того, чтобы положение головы изменилось.
Далее, и у децеребрированных кроликов после односторонней экстирпации лабиринта наступают типичный поворот и отклонение головы. При этом связи оптических путей с мостом, мозжечком, продолговатым мозгом и спинным мозгом отсутствуют. Тем не менее после экстирпации лабиринта типичное отведение головы имеет место.
Катательные движения после выключения оптических связей осуществляются типичным образом, что показали также и опыты Вейланда (9) с пересечением ствола мозга у кроликов.
278
Шестая глава. Последствия односторонней экстирпации лабиринта
У кролика, у которого в глубоком наркозе после левосторонней экстирпации лабиринта были энуклеированы оба глазных яблока, после пробуждения от наркоза голова была повернута на 90° и отклонена влево. Правая передняя нога имела сильный раз- гибательный тонус, левая — отчетливый сгибательный. При висячем положении головой вниз обнаружилось также, что грудной отдел повернулся относительно тазового на 45—90°. Кроме того, животное производило очень оживленные движения перекатывания, заснятые кинематографически. Снимки показали, что движения перекатывания производятся точно так же, как у животных без одного лабиринта с нормальной способностью зрения и интактными глазными яблоками. Таким образом, в этом опыте все аномалии положения и движения, а также поворот головы были зарегистрированы точно такими же, какими они были описаны выше для зрячих животных.
Из этих наблюдений с определенностью следует еще раз, что поворот головы и другие аномалии положения и движения головы, шеи, туловища и конечностей не вызываются вторично девиацией глаз, что они, следовательно, не являются компенсаторными реакциями по отношению отклонения глаз и этим отклонением существенно не усиливаются и не ослабляются.
В соответствии со сказанным при обсуждении глазных симптомов выяснилось:
что отклонение глаз после односторонней экстирпации лабиринта является следствием влияния тонических лабиринтных рефлексов на верхнюю и нижнюю прямые мышцы глаза, тогда как тонические рефлексы на косые мышцы девиации не вызывают;
что величина вертикального отклонения стоит в закономерной зависимости от положения головы в пространстве;
что величина эта максимальна, когда при боковом положении головы интактный лабиринт находится внизу, и минимальна, если он оказывается наверху;
что поворот шеи влияет на отклонение глаз лишь очень незначительно, даже если он достигает очень большой величины;
что отклонение глаз в свою очередь существенно не изменяет поворота головы, движения перекатывания и другие аномалии положения;
что это отклонение представляет собой стойкое последствие экстирпации лабиринта;
что нистагм, наоборот, наступает лишь временно и зависит от положения головы в пространстве, причем при максимальной девиации он исчезает, при минимальной — достигает максимума.
Центральные компенсации
Если у животных одновременно удалить оба лабиринта, то не наступает ни поворота головы, ни отклонения глаз, ни нистагма. Бехтерев (1) установил на собаке, что если оба лабиринта удалять двухмоментно и между двумя этапами дать пройти достаточно долгому времени, то после удаления второго лабиринта поворот головы, девиация глаз и нистагм имеют место так, словно первый лабиринт вовсе не удалялся. По единодушному мнению всех исследователей, дело здесь идет о центральной компенсации, постепенно появляющейся после удаления первого лабиринта. О возникновении и сущности этой компенсации подробнее будет сказано далее, при обсуждении исходных состояний односторонней экстирпации лабиринта у кошек. Здесь необходимо лишь сообщить, что явление это имеет место и у кролика. В последнем случае достаточно промежутка между двумя операциями в 5 дней, чтобы после удаления второго лабиринта с определенностью ждать появления отчетливого нистагма в том направле¬
Последствия односторонней экстирпации лабиринта у кролика
279
нии, в каком он наступил бы после удаления соответствующего лабиринта у интактного до того животного. Этот нистагм не исчезает после полного удаления большого мозга. Играет ли эта центральная компенсация существенную роль в постепенном выравнивании нарушений после односторонней экстирпации лабиринта также и у кролика, сказать с уверенностью до сих пор не представляется возможным.
Возникновение сколиоза у растущих животных
Если через небольшой промежуток времени после рождения у кролика удалить лабиринт и позволить животному расти дальше, то в результате длительного изогнутого положения развивается, сколиоз.
У кролика в возрасте нескольких недель де Клейном был экстирпиро- ван правый лабиринт. После этого вмешательства животное жило 8 меся-
Рис. 161. Скелет взрослого кролика, которому в раннем возрасте был удален правый лабиринт (объяснение в тексте).
Рис. 162. Грудная клетка того же кролика с вентраль ной стороны.
цев, превратилось во взрослого кролика и тогда подверглось экстирпации левого лабиринта. После его гибели доктор Бранд (Утрехт) снял шкуру и удалил внутренности, после чего животное, подвешенное за нос, было помещено в большой стеклянный сосуд с формалином таким образом, что могло свободно плавать в жидкости. Затем тщательно отпрепарировался скелет. Доктор Бранд уже сделал краткое сообщение по поводу обнаруженного им. Наиболее примечательными изменениями скелета оказались следующие: если захватить скелет за таз, дав ему повиснуть вниз головой, можно обнаружить отчетливое перекручивание его вправо. Точно так же
230
Шестая глава. Последствия односторонней экстирпации лабиринта
весь позвоночник вогнут вправо. Этот поворот кпереди усиливается (рис. 161).
Очень сильно выражен сколиоз грудной клетки. Грудная клетка уплощена в косом направлении (рис. 161 и 162). Если на грудной отдел взглянуть сзади, то ребра на левой стороне латерально от позвоночника изогнулись, тогда как с правой стороны стали более плоскими. Прикрепления срединных реберных хрящей к грудине на левой стороне образуют острый угол, открытый кзади, в то время как на правой стороне хрящ устанавливается по отношению к позвоночнику более отвесно. Меж-
Рис. 163. Шейная часть позвоночника и череп того же кролика. Вид сверху.
Рис. 164. Передняя Рис. 165. Таз того же кро- часть тела того же кро- лика справа,
лика с черепом. Вид спереди.
реберные промежутки, особенно между средними ребрами, на левой стороне значительно шире, чем на правой.
Вся шейная часть позвоночника вогнута вправо, особенно сильный поворот вправо отмечен у эпистрофея относительно 3-го шейного позвонка (рис. 161—163). Как видно из рис. 163, весь череп деформирован и при рассматривании сверху сильно вогнут вправо. Если держать череп таким образом, чтобы линия, связывающая затылочные мыщелки, была горизонтальной, то носовая перегородка и верхние резцы располагаются по линии, идущей косо слева направо и сверху вниз (рис. 164). Левая нижняя челюсть длиннее правой, в целом также вогнута вправо. Горизонтальный диаметр правой орбиты меньше такового левой. Поперечные отростки атланта асимметричны; правый длинен и узок, левый короток и широк. Оба они не лежат в одной плоскостй.
Таз также асимметричен (рис. 161 и 165). Передняя оральная часть таза относительно задней каудальной повернута вправо, т. е. правой стороной вентрально. Симфиз проходит косо сверху спереди вправо вниз. Запирательное отверстие в сагиттальном направлении на правой стороне длиннее, чем на левой.
В заключение мы обнаруживаем следующие изменения: 1) спиралепо- добный поворот всего животного — от таза через весь позвоночник до
Последствия односторонней экстирпации лабиринта у кролика
281
черепа; 2) поворот позвоночника вправо, который распространяется и на голову и в котором принимает участие также таз; 3) сколиотическое уплощение грудной клетки.
Это наблюдение, конечно, должно быть повторено на многих животных, а кроме того, изменения скелета должны быть изучены детально. Но уже данные, полученные на одном животном, говорят за то, что растущий скелет реагирует совершенно типичными сколиотическими изменениями на аномалии положения, наступающие при появлении после односторонней экстирпации лабиринта анормальных напряжений мускулатуры. Этот сколиоз вследствие вынужденного «лабиринтного» положения кролика находит свою аналогию в кривошее у человека. Здесь также дело приходит к искривлению позвоночника и типичным асимметриям черепа, которые, как показывают рисунки, приведенные в Эйленбургской энциклопедии Иоахимсталя (Ioachimsthal), обнаруживают большое сходство с изменениями, найденными у кролика. Эти аналогии доляшы быть изучены еще глубже. Далее, следует установить, могут ли односторонние заболевания лабиринта играть при кривошее у человека этиологическую роль. Во всяком случае можно считать уже установленным, что односторонняя экстирпация лабиринта у молодого кролика представляет собой простое средство, с помощью которого мы можем вызвать у этих животных выраженный сколиоз позвоночника.
Паралич симпатического нерва глаза на стороне операции
После экстирпации лабиринта у кролика наблюдаются симптомы паралича симпатического нерва глаза оперированной стороны: сужение глазной щели, оттянутая мигательная перепонка и сужение зрачка. Сужение зрачка было описано Камисом [Cannes (3)], сужение глазной щели—Винклером (1), оттянутость мигательной перепонки зарисована Винклером (стр. 19, рис. 6).
Такие же последствия экстирпации лабиринта наступают и у кошек (Камис). У последних де Клейн (11) показал, что паралич симпатического нерва связан с самой экстирпацией лабиринта как таковой, но обусловлен перерезкой постганглионарного симпатического пути, который, согласно исследованиям де Клейна, Сосэна [Socin (20)] и де Бюрле [de Burlet (21)], проходит через среднее ухо и потому более или менее сильно повреждается во время операции. Поэтому не будет ошибки в .допущении наличия тех же отношений у кролика, чтобы не относить паралич симпатической иннервации глаза, к которой присоединяется также расширение сосудов уха на стороне операции, к прямым последствиям выпадения лабиринта.
Заключение
Сложный симптомокомплекс, который появляется у кролика после односторонней экстирпации лабиринта, складывается (насколько сейчас это известно) из следующих составляющих.
I. Длительные влияния, которые исходят из оставшегося лабиринта.
А. Симметричные влияния.
1) Тонические лабиринтные рефлексы на мышцы конечностей имеют место в одинаковой мере на обеих сторонах тела.
2) Компенсаторные круговые повороты глаз сохраняются на обоих глазах, выраженные в одинаковой степени
282
Шестая глава. Последствия односторонней экстирпации лабиринта
и в неизменном направлении, но величина их примерно вполовину меньше.
Б. Асимметричные влияния.
1) Тонические лабиринтные рефлексы на мышцы шеи приводят к «основному повороту» шеи в направлении утраченного лабиринта, интенсивность этого поворота зависит от положения головы в пространстве: он максимален, когда темя обращено вниз; минимален, когда оно обращено кверху; при самом крайнем положении головы выражен весьма отчетливо. Его лучше всего исследовать в висячем положении животного головой вниз. С течением времени после операции он увеличивается.
2) Те же рефлексы принимают участие в отклонении головы в сторону утраченного лабиринта, которое в первые дни после операции уменьшается, но полностью не исчезает. Это отклонение лучше всего исследовать в висячем положении животного головой вниз.
3) Лабиринтный установочный рефлекс на голову увеличивает или уменьшает основной поворот, имея тенденцию при всех положениях тела в пространстве приводить годову всегда в такое положение, при котором интактный лабиринт оказывается сверху.
4) Тонические лабиринтные рефлексы на мышцы туловища приводят к «основному повороту» туловища, со временем все усиливающемуся (в сторону утраченного лабиринта). Этот поворот оказывается также и продолжением поворота шеи.Благодаря совместному действию обоих рефлексов все тело поворачивается спирально.
5) Вертикальное отведение глаз в сторону утраченного лабиринта обусловливается тоническим лабиринтным рефлексом на прямую верхнюю мышцу одноименной и нижнюю прямую мышцу противоположной стороны. Оно максимально при боковом положении головы, когда интактный лабиринт находится внизу, и отсутствует, когда интактный лабиринт находится наверху, т. е. при том положении головы, которое животное принимает, когда сидит.
II. Преходящие последствия односторонней эк- стирпациии лабиринта.
1) Снижение разгибательного тонуса конечностей на стороне утраты лабиринта вскоре после операции выражено сильно, но уже через несколько дней заметно меньше и спустя примерно 8 недель исчезает совсем. Это до сих пор не удавалось объяснить действием известного одностороннего лабиринтного рефлекса. Нельзя утверждать, обусловлено ли оно деятельностью интактного лабиринта или последствиями оперативного вмешательства на другой стороне.
2) Исчезающий по прошествии немногих дней н и с т ’а г м глаз сильнее всего, когда при боковом положении головы интактный лабиринт находится наверху, поэтому девиация глаз минимальна, а при противоположном боковом положении, когда девиация достигает максимума, исчезает. Направление нистагма не у всех животных одинаково.
III. Длительные последствия поворота шеи.
А. Поворот шеи вызывает.
Последствия односторонней экстирпации лабиринта у кролика
283
1) Посредством шейного рефлекса значительное усиление поворота туловища.
2) Посредством тонических шейных рефлексов на мышцы конечностей разгибание ног на стороне интактного лабиринта и уменьшение тонуса на стороне операции. Это различие тонуса в первые недели после вмешательства увеличивается вследствие упомянутого под цифрами II. 1 переходящего расслабления ног на стороне операции. Когда это прямое последствие выпадения лабиринта исчезает, длительное различие тонуса ног обусловливается тоническим шейным рефлексом.
3) Посредством тонических шейных рефлексов на глазные мышцы уменьшение вертикального отклонения, которое при поворотах шеи до 180° количественно может не приниматься во внимание, поэтому при естественных установках и позах животного существенного действия не оказывает.
4) Поворот головы оказывает на установку тела только относительно незначительное влияние.
Б. Действие шейных рефлексов можно выключить в любой момент,
устанавливая голову по отношению к грудному отделу прямо.
Тогда из асимметричных влияний остаются только прямые последствия выпадения лабиринта или рефлексы с интактного лабиринта.
Устанавливание головы прямо оказывает следующие действия.
1) Поворот туловища уменьшается, но не снимается полностью (I. Б. 4).
2) Разгибательный тонус конечностей (если после операции прошло более длительное время) становится одинаковым на обеих сторонах тела. По прошествии же после операции недолгого времени различие тонуса между конечностями обеих сторон при устанавливании головы прямо только становится меньше (Й. 1).
3) Анормальное положение тела при сидении кор- регируется почти полностью; остаются только незначительный поворот туловища и в первое время после операции — незначительное различие тонуса конечностей обеих сторон.
4) Движения перекатывания тотчас прекращаются.
5) Если кролику после утраты лабиринта с одной стороны повернуть голову в другую сторону, туловище также поворачивается в другую сторону (хотя и менее сильно, чем это бывает при исходном повороте туловища). Вскоре после операции различие тонуса конечностей с обеих сторон увеличивается; позднее, наоборот, вызывается даже противоположное различие тонуса конечностей и во время сидения наступает аномалия положения тела противоположного характера.
IV. Компенсации. Анормальная установка корригируется.
А. Во время сидения на полу.
1) Благодаря установочному рефлексу с туловища на голову. Вследствие этого поворот головы уменьшается и в редких случаях может совсем отсутствовать. Отклонение же головы исчезает в большинстве случаев совершенно.
2) Благодаря установочному рефлексу с туловища на туловище. Этот рефлекс вызывает усаживание туловища, несмотря на то, что голова повернута более или менее
284 Шестая глава. Последствия односторонней экстирпации лабиринта
сильно (до 180°). Он делает возможным усаживание животного, когда оно лежит на оперированной стороне.
3) Иногда уже при соприкосновении конечностей с полом анормальное положение их корригируется.
4) Спустя более длительное время после операции для этой же' цели используются глаза.
5) Тем не менее зрительные коррекции у кролика существенной роли, не играют.
Б. Посредством центральных компенсаций. Через некоторое время после операции развивается бехтеревская центральная компенсация. Если после этого удаляется второй лабиринт, появляются поворот головы и отклонение глаз в другую сторону. Еще достаточно не изучено, какую роль играет это центральное явление у кролика.
V. Движения перекатывания. Возбуждения, которые спустя
долгое время после операции исходят из раны, вероятно из ствола 8-го нерва, вызывают подкрепленные другими сенсорными раздражениями приступы' весьма мощных движений бега и прыжка. Так как тело животных повернуто спирально, то поступательного движения при этом не возникает, но животное ввинчивается в пространство. Это и есть движения перекатывания. Для полного поворота необходимы два прыжка. После каждого прыжка спиральный поворот тела восстанавливается. Участие отдельных рефлексов в этом явлении оказалось возможным выяснить с помощью динамических фотографий. Однократное перекатывание через спину может осуществляться и без движения прыжка.
VI. Если лабиринт удаляется у молодого животного, то в дальнейшем
вследствие анормального положения тела развивается типичный, сколиоз.
VII. Сужение глазной щели, оттягивание мигательной перепонки и миоз
на стороне операции с разрушением лабиринта никак не связаны. Они обусловлены перерывом постганглионарных симпатических, путей, которые проходят через среднее ухо (де Клейн).
Из всех исследованных до сих пор видов млекопитающих у кролика прямые последствия выпадения лабиринта выражены наиболее сильно. У других видов животных отношения эти более просты, поэтому могут* быть описаны более кратко. II.II. ОПЫТЫ НА МОРСКИХ СВИНКАХ (15, 82Ь)
Подробное описание последствий односторонней экстирпации лабиринта у морских свинок дал Дрейфус (1) из лаборатории Эвальда. Сразу после операции он видел появление симптомов, которые уже на следующий день в большей своей части исчезали. В качестве же длительного последствия он наблюдал прежде всего поворот головы в сторону операции. Наоборот, ван Россем видел вследствие применения им пасты (Arsen—Lysol), которую он закладывал в лабиринт, бурные симптомы,, не исчезавшие до самой гибели животного (самое длительное — через 9 дней). Наши данные в существенных пунктах совпадают с данными Дрейфуса.
В качестве примера может служить сокращенный протокол опытов на животном, выжившем дольше всех остальных.
Морская свинка IV. 3 декабря 1912 г. Экстирпация левого лабиринта под эфирным: наркозом. После пробуждения от наркоза голова повернута влево на 45°, голова и все-
Опыты на морских свинках
285
тело сильно изогнуты влево, так что позвоночник обращен вогнутостью влево и морда касается левой задней ноги. Отчетливое отклонение глаз в сторону операции, сильный нистагм. Животное перекатывается влево.
4 декабря. Нистагм еще наблюдается. Голова во время сидения на 30° повернута влево, величина ее наклона непостоянна. Вогнутость тела намного меньше. Перекатывания нет; наоборот, манежные движения влево. В висячем положении головой вниз грудь повернута по отношению к тазу на 45°, голова — более чем на 90°, на 20—30° наклонена влево. Правая передняя лапа разогнута.
5 декабря. При сидении голова повернута на 30°, более не наклонена; в круглой метке животное в состоянии бегать вдоль стены в обе стороны. Нистагма нет.
7 декабря. Сидит так же хорошо, как нормальное животное, только голова на 20° повернута налево. При этом положении головы девиация глаз весьма незначительна. Нистагма нет. Перекатывания нет. Из положения на спине животное в состоянии принять нормальное сгорбленное положение, переворачиваясь как через правую, так и через левую сторону. В висячем положении головой вниз нижняя апертура грудной клетки повернута относительно таза на 30°, верхняя — на 45°, голова — на 90°, однако наклона головы и грудной клетки уже нет.
12 декабря. Голова при сидении повернута на 60°, правая передняя нога разогнута и отведена. Задние ноги установлены нормально. Весьма незначительное отклонение глаз. Животное в состоянии бегать вокруг клетки вправо и влево.
18 февраля. При сидении голова повернута влево на 45°, грудь — на 20°. Правый глаз отклонен несколько кверху, левый — несколько вниз. Передние лапы держатся симметрично, однако если прикрыть животному глаза, правая передняя нога вытягивается и отводится. Смещаемость всего животного на плоскости опоры более выражена при смещении влево, нежели вправо. Животное оживленно бегает во всех направлениях, хорошо ест, сильно выросло. В висячем положении головой вниз нижняя апертура груди повернута по отношению к тазу на 30°, верхняя — на 60°, голова — на 90° влево. Голова и грудь не наклонены. Правая передняя нога разогнута.
Начиная с этого момента до 1 июля 1913 г. животное исследуется через каждые 14 дней без того, чтобы что-либо изменилось в его поведении.
В качестве непосредственных последствий операции следует рассматривать нистагм, движения перекатывания, сильную изогнутость всего тела в сторону утраченного лабиринта, манежные движения, а у некоторых животных также нистагм головы. Для демонстрации длительных (стойких) последствий лучше всего придать животному висячее положение головой вниз. При этом отчетливо видны девиация глаз, поворот головы и всего туловища в сторону операции и разгибание правой передней ноги.
Анализ взаимозависимости этих симптомов показал, что по существу эти отношения точно такие же, как у кролика. Лишь в отдельных пунктах были обнаружены отклонения, которые в дальнейшем будут рассмотрены более подробно.
Поворот головы, шеи и туловища
Если морскую свинку после односторонней экстирпации лабиринта исследовать после операции через несколько дней, недель, месяцев в висячем положении головой вниз, то тело ее точно так же, как это имеет место у кролика, поворачивается по спирали. Нижняя апертура грудной клетки повернута по отношению к тазу на 20—45° (в среднем на 30°), верхняя апертура — на 30—70° (в среднем на 45°), голова по большей части — на 90°. При этом передняя нога неоперированной стороны разогнута. Голова обычно свисает мордой прямо вниз, наклон головы отмечается только в первые дни после операции, он может составлять угол до 30°. Если теперь установить голову по отношению к груди прямо, спиральная изогнутость туловища исчезает и тело животного свисает вниз совершенно симметрично. При исследовании в положении на спине можно также показать, что в ответ на устанавливание головы по отношению к груди прямо спиральный поворот туловища исчезает и тело животного лежит совершенно симметрично.
286
Шестая глава. Последствия односторонней экстирпации лабиринта
Только у двух животных в первые два дня после операции при устанавливании головы прямо еще отмечался незначительный остаточный поворот туловища. У остальных животных этого не наблюдалось уже в первые дни после операции.
Таким образом, по сравнению с кроликом наблюдается значительно иное положение. В то время как у последнего при устанавливании головы прямо еще сохраняется отчетливо выраженный поворот таза, у морской свинки этого не бывает. У нее односторонние тонические лабиринтные рефлексы на мускулатуру туловища отсутствуют. Только поворот шеи оказывается прямым следствием удаления лабиринта или проявлением активности оставшегося лабиринта. Поворот остальных отделов туловища, наоборот, не является прямым результатом утраты лабиринта, но обусловлен поворотом шеи (шейный рефлекс).
Тонус конечностей
Если кролика без одного лабиринта уложить на пол, не корригируя поворота головы, то на „стороне утраченного лабиринта разгибательный тонус ног оказывается ниже, чем тонус ног противоположной стороны. По большей части различие тонуса на передних ногах больше, чем на задних, однако достаточно выражено и на последних. Это различие еще улавливается 7 месяцев после операции.
Если корригировать поворот головы морской свинки, устанавливая ео относительно туловища прямо, результат получается различный в зависимости от того, исследуется ли животное вскоре после операции или через некоторое время. Вскоре после операции различие тонуса конечностей обеих сторон сохраняется и в том случае, когда он вследствие поворота головы уменьшается. При этом для того чтобы уравнять разгибательный тонус конечностей обеих сторон тела, необходимо более или менее сильно повернуть голову в другую сторону, т. е. чрезмерно корригировать поворот- головы. Однако уже через несколько дней положение меняется; самое раннее на второй и самое позднее на шестой день при устанавливании головы прямо тонус конечностей обеих сторон одинаков. Это продолжается в течение месяцев. Если при этом поворачивать голову в другую сторону, то различие тонуса конечностей извращается, а ноги оперированной стороны приобретают более высокий тонус. Таким образом, на этой стадии разный тонус конечностей у морской свинки с одним лабиринтом обусловлен исключительно поворотом головы.
Итак, в то время как у кролика прямое влияние односторонней утраты: лабиринта на тонус конечностей отчетливо проявляется в течение двух месяцев, у морской свинки дело идет о скоро проходящем эффекте, который может быть отмечен лишь в течение немногих дней после операции. По прошествии этого срока различие тонуса конечностей обеих сторон обусловлено исключительно шейным тоническим рефлексом на мышцы конечностей.
Положение тела при сидении
У морской свинки на положение тела при сидении существенно влияют установочные рефлексы. Влияние лабиринтного установочного рефлекса на голову лучше всего исследовать, свободно удерживая животное в обоих боковых положениях в воздухе. После правосторонней экстирпации лабиринта в правом боковом положении в воздухе основной поворот уменьшен благодаря лабиринтному установочному рефлексу и при некоторых: условиях голова может устанавливаться в пространстве даже в, правом
Опыты на морских свинках
287
боковом положении. При исследовании в воздухе в левом боковом положении, наоборот, лабиринтный установочный рефлекс усиливает основной поворот, а поворот головы может достигать вследствие этого 180°. Влияние установочного рефлекса туловища на голову наиболее демонстративно тогда, когда сначала свинку удерживают в воздухе в нормальном положении. Если, например, свинка при этом повернула голову на 90°, то когда животное усаживают на пол, становится видно, как этот поворот уменьшается более или менее сильно, в среднем примерно на 45°. Установочный рефлекс с туловища на туловище очень отчетлив, когда животное, например спустя недолгое время после правосторонней экстирпации лабиринта, оказывается уже в состоянии усаживаться из правого бокового положения, несмотря на сильный поворот головы вправо.
Для дальнейшего описания позы сидящего животного целесообразно отдельно обсудить различные стадии после операции.
Тотчас после операции и на следующий день животное уже в состоянии принять сидячее положение из обоих боковых положений (установочный рефлекс с туловища на туловище), голова на 45° или более повернута в сторону операции (совместное действие основного поворота, лабиринтного установочного рефлекса и установочного рефлекса с тела на голову), грудь свешивается в ту же сторону (шейный установочный рефлекс, различие тонуса конечностей). Кроме того, голова и все туловище изогнуты в сторону операции так, что морда касается задней ноги. Передняя нога здоровой стороны сильно разогнута и отведена (тонический шейный рефлекс на мышцы конечностей, усиленный преходящими последствиями выпадения лабиринта, перевешивание груди).
Уже на этой стадии, устанавливая голову прямо, можно устранить аномалию положения животного, которая возникает, таким образом, вследствие отклонения головы. Поворот туловища, как показано выше, является почти исключительно результатом поворота шеи. Различие тонуса передних ног на этой стадии определяется не только поворотом головы; здесь также играет роль и прямое влияние утраты лабиринта, которое имеет (как это доказывается устанавливанием головы прямо) только относительное значение.
Уже Дрейфус обратил внимание на то, что сразу после операции именно передняя (а не задняя) нога здоровой стороны разогнута и отведена. Задняя же нога в отдельных случаях лежит под телом. Может оказаться даже, как мы это наблюдали в одном из случаев, что разогнутой и отведенной оказывается левая задняя нога оперированной стороны. Это кажущееся исключение объясняется просто. Оно обусловлено весьма сильным изгибанием животного и исчезает, если последнее корригируется. Благодаря этому изгибанию конечности оперированной стороны становятся «челюстными» и приобретают более высокий разгибательный тонус; этот эффект, таким образом, прямо противоположен повороту шеи и прямому влиянию выпадения лабиринта. Это влияние на передних ногах выражено настолько сильно, что они не могут быть чрезмерно компенсированы. На задних конечностях, наоборот, изогнутость иногда преобладает. Однако если изогнутость скорригировать, то разгибается задняя нога здоровой и сгибается задняя нога оперированной стороны.
Поворачивая голову в другую сторону, в большинстве случаев уже на этой стадии можно достичь разгибания и отведения передней ноги на оперированной стороне и сгибания на здоровой, когда животное принимает такое положение, как если бы у него был экстирпирован другой лабиринт.
288 Шестая глава. Последствия односторонней экстирпации лабиринта
Если морскую свинку исследовать через несколько дней или недель после операции, она оказывается в состоянии сидеть прямо, без изогнутости длинной оси тела (изогнутость исчезает через 2—5 дней). Но поворот головы остается. В среднем он составляет 45°, достигая в иные дни 80°. Грудь несколько перевешивает в сторону операции. Частично это объясняется поворотом туловища, частично же — различием тонуса передних ног. Наклон передней апертуры грудной клетки составляет обычно 20— 30°, достигая иногда 45°; нижняя апертура также может несколько (до 15°) наклоняться в сторону операции. Установкой головы в прямое положение это перевешивание груди тотчас устраняется, при этом поворот туловища, как и различие тонуса конечностей, уменьшается. Это различие у свободно сидящего животного на этой стадии еще весьма отчетливо. Особенно ясно видны разгибание и отведение передней ноги на здоровой стороне. На задних ногах различие тонуса меньше. Оно исчезает при установке головы в прямое положение, обе передние ноги становятся совершенно симметрично. При поворачивании головы в другую сторону наблюдается иное расположение передних ног: нога оперированной стороны оказывается разогнутой, нога на другой стороне — более согнутой. Если животное усадить на шероховатую поверхность, например на соломенный мат, то вследствие разгибания передней ноги до нормальной степени его труднее сдвинуть в эту сторону, чем в сторону утраченного лабиринта. Если голову установить прямо, то и смещаемость в обоих направлениях становится одинаковой. Таким образом, на этой стадии все аномалии положения туловища и конечностей обусловлены исключительно поворотом шеи.
Спустя некоторое время морская свинка корригирует ненормальное положение своих передних конечностей во время сидения точно так же, как это описано у кролика. При исследовании животного через несколько месяцев после операции оно еще держит голову повернутой примерно на 45°, грудь тоже перевешивает на сторону операции вследствие поворота туловища, обусловленного поворотом шеи, но передние ноги теперь держатся симметрично. В этой компенсации у кролика принимают участие глаза: если закрыть глаза животного, разгибание и отведение передней ноги здоровой стороны появляются снова; если глаза открыть, правая нога подтягивается и животное снова сидит, как нормальное. Так как на этой стадии при сидении с открытыми глазами передние ноги держатся симметрично, то и смещаемость свободно сидящего животного вправо или влево не так различается, как на более ранних стадиях.
Оказывается, следовательно, что поворот головы, который обнаруживается у сидящей морской свинки без одного лабиринта, зависит от совокупного действия основного поворота, одностороннего лабиринтного установочного рефлекса и установочного рефлекса с тела на голову. Положение тела почти исключительно зависит от влияния, которое поворот головы со своей стороны оказывает на тонус мышц конечностей и туловища, это влияние уменьшает действие установочного рефлекса с туловища на туловище. Поэтому прямая установка головы оказывает влияние на нормальное положение животного. Меньшим и преходящим оказывается влияние наклона головы, которое наблюдается на первых этапах после операции. Наоборот, незначительная разница тонуса конечностей, которая наступает непосредственно после операции как следствие односторонней утраты лабиринта и исчезает спустя несколько дней, является слишком слабой, чтобы оказать заметное влияние на положение тела во время сидения.
Опыты на морских свинках
289
Движения перекатывания
Движения перекатывания у морской свинки во всех существенных пунктах идентичны таковым у кролика. Интенсивность и длительность перекатывания изменяются у морской свинки весьма значительно, некоторые животные катаются в течение двух дней, другие — только сразу после операции; одни спонтанно перекатываются через всю комнату, другие — переворачиваются один или два раза только в ответ на раздражение. Как и у кролика, перекатывание наступает приступами, и животное в промежутках может сидеть совершенно спокойно в обычном сгорбленном положении. И только в ответ на раздражение наступают энергичные катательные движения.
Кинематографические снимки морской свинки с четырех сторон показали, что и здесь дело идет о движениях бега спирально повернутого животного и что для целого поворота необходимы два прыжка; один — через брюхо, другой — через спину. При этом всякий раз в боковом положении позвоночник изгибается в вентральном направлении, лапки сгибаются, поворот передней части тела относительно таза снова восстанавливается. При каждом из двух прыжков позвоночник разгибается, передние лапки идут вперед, задние — назад, и, точно как это делает кролик, животное перекладывается на другую сторону. В противоположность кролику у морской свинки при перекатывании наблюдаются следующие отличия: тело морской свинки короче и более сплющено, конечности короче. В силу последнего обстоятельства ей труднее, чем кролику, переместить задние ноги во время перекатывания через брюхо на другую сторону. Вследствие этого разгибание задних конечностей кзади может не быть таким резким, как у кролика. Тем не менее оно имеет место. Той фазой движения, которая у морской свинки осуществляется с наибольшей силой и скоростью, является не прыжок через брюхо, а прыжок через спину. Этот последний осуществляется в виде быстрого «толчка». Очевидно, это зависит от формы тела, которая именно это движение делает наиболее трудно осуществимым. То, что главное раздражение, вызывающее приступы движений бега и прыжка и приводящее при спиральном повороте тела животного к перекатыванию, исходит из самой операционной раны, главным образом из ствола 8-го нерва, становится вероятным, между прочим, и потому, что в опытах ван Россемса (van Rossems), который удалял лабиринт не хирургически, а с помощью прижигающей пасты, животные перекатывались до тех пор, пока не погибали.
Глазные симптомы
Глазные симптомы у морской свинки точно такие же, как у кролика, поэтому не требуют детального описания. Вертикальное отклонение глаз имеет то же направление и осуществляется таким же образом, как и у кролика. Исследуя морскую свинку спустя недели или месяцы при положении головы, когда интактный лабиринт оказывается наверху, обнаруживается, что при этой установке головы глаз оперированной стороны, направленный вниз, в большинстве случаев больше не отклоняется, в то время как направленный вверх глаз нормальной стороны несколько отводится дорсально так, что остается еще видимой узкий край белой склеры по краю нижнего века. Наоборот, если голове придать противоположное положение (интактный лабиринт внизу), то глаз оперированной стороны, направленный вверх, максимально отклоняется вентрально и вперед и настолько сильно, что в отдельных случаях в глазной щели:
19 Р. Магнус
290
Шестая глава. Последствия односторонней экстирпации лабиринта
виден один только белок. Глаз неоперированной стороны, направленный вниз, также отклоняется, но не в такой сильной степени, как первый. Он обращен дорсально и несколько вперед.
Нистагм сильнее всего непосредственно после операции, но вскоре становится меньше, чаще всего уже на следующий день он отчетливо уменьшается, а на 2—9-й день совершенно исчезает. Он находится в той же зависимости от положения головы в пространстве, которая описана для кролика.
В дополнение к эом у следует сообщить следующее.
Если у морской свинки, введя в среднее ухо 0.1 мл 5%-го кокаина, временно выключить лабиринт {61), последствия паралича лабиринта отчетливо наблюдаются около 5—8 минут и проявляются в повороте головы и типичном вертикальном отведении глаз (глаз инъецированной стороны движется вниз, глаз противоположной стороны — вверх). Во всех опытах этого рода с неизменным постоянством оказывалось, что сначала наступает только отведение без какого бы то ни было нистагма и лишь после примерно 20 минут возникал бурный нистагм, который в большинстве случаев удерживался в течение всего дня до вечера. Обычно на следующее утро последствия введения кокаина в среднее ухо исчезали. Попытка объяснить это наблюдение может быть сделана только в девятой главе. Здесь предположительно могут быть даны только следующие заключения: если при полном кокаиновом параличе лабиринт действительно нераздражим, что безусловно следует принять после всех наблюдений над животными, то это значит, что нистагм может наступать и после выключения лабиринта до полной утраты возбудимости. Далее, значительный интервал времени между наступлением девиации и нистагма делает вероятным предположение, что выключением (до полной утраты возбудимости) лабиринта другой стороны обусловлен скорее нистагм, чем девиация. Направление нистагма в сторону оперативно удаленного лабиринта также говорит за то, что раздражение наступает весьма быстро и что нистагм следует представлять себе как явление выпадения. Опыты, которые будут описаны позднее, приводят к представлению, что нистагм обусловливает одностороннее выключение аппаратов полукружных каналов, в то время как односторонний паралич отолитовых окончаний его не вызывает. Поэтому нистагм как симптом sui generis следовало бы строго отличать от остаточной девиации. За всеми подробностями отсылаем читателя к соображениям, изложенным в девятой главе.
Заключение
Анализ состояний, наступающих после односторонней экстирпации лабиринта у морской свинки и у кролика, приводит в основном к одинаковым результатам. Поэтому отсылаем читателя к стр. 281. Существенными же различиями являются следующие.
1) У морской свинки основной поворот туловища как прямое следствие одностороннего тонического рефлекса на мускулатуру туловища отсутствует.
2) Различие тонуса конечностей, обусловленное непосредственно удалением лабиринта, исчезает уже через несколько дней.
3) Установка головы в прямое положение относительно туловища полностью снимает различие тонуса конечностей и поворот туловища (уже после немногих дней) так, что животное сидит совершенно нормально. Поворот головы в другую сторону вызывает зеркально отраженную установку тела.
Опыты на кошках
291
4) Наступающий непосредственно после операции временный наклон головы у морской свинки оказывает более сильное влияние на установку тела, чем у кролика.
III. ОПЫТЫ НА КОШКАХ
Экстирпации лабиринтов у кошек были рассмотрены с различных сторон [Крейдель (1); Камис (3)], однако подробного описания последствий операции у этого животного авторами не дано. Вследствие изящества своих движений кошки весьма пригодны для наблюдений по поводу выпадения лабиринта. Наоборот, анализ симптомов удается на них значительно хуже, так как они позволяют осуществить установку головы в прямое положение, висячее положение головой вниз, исследование тонуса конечностей в положении на спине и т. п. значительно менее охотно, нежели кролик или морская свинка. Даже вполне ручные и привыкшие к лаборатории животные при исследованиях этого рода царапаются .Тем не менее на 13 животных (15) были проведены продолжительные наблюдения, а менее длительные опыты — более чем на 30 кошках. У одной кошки (41), у которой Дюссер де Баренн экстирпировал неоэнцефалон, 7 месяцев спустя была произведена односторонняя лабиринтэктомия, и животное в течение трех дней находилось под наблюдением, после чего был удален второй лабиринт. У другой кошки Дюссер де Баренн осуществил полную экстирпацию большого мозга; через 2% месяца после этого был удален один, через 4 дня — другой лабиринт. На обоих животных главным образом и исследовались установочные рефлексы после односторонней экстирпации лабиринта. Кроме того, на четырех кошках были изучены последствия односторонней экстирпации лабиринта (16) спустя 41, 164, 191 и 233 дня после предварительной перерезки трех самых верхних пар задних шейных корешков. Таким путем был собран довольно обширный материал.
Раньше всего в качестве примера приведем сокращенный протокол опыта на кошке после односторонней экстирпации лабиринта.
Кошка Лабу. 3 июля, 1911 г. Под атропин-эфиртгым наркозом экстирпация левого лабиринта. Обнажены три устья полукружных каналов и ствол 8-го нерва во внутреннем слуховом проходе. Через % часа после операции: горизонтальный нистагм вправо, незначительное отклонение глаз влево, движения против часовой стрелки, голова повернута влево. При этом морда иногда устанавливается прямо вперед, в большинстве же случаев имеется отклонение головы влево. Во время сидения качается. Некоторое горизонтальное качание головы. Левая нога при сидении несколько отодвинута. Наклонность к падению влево.
4 июля. Сидит в клетке прямо, только левая нога несколько отставлена. Голова повернута на 45° влево и, кроме того, наклонена влево. Позвоночник также изогнут влево. Пьет из миски, при этом все тело клонится влево, угрожая упасть в ту же сторону, что иногда и случается. При побежке вперед устанавливает ось тела правильно в направлении к пели; несмотря на это, отклоняется влево примерно на 45°. Потом прислоняется левым боком к стене. Если не имеет определенной цели, производит движения против часовой стрелки, причем позвоночник отчетливо вогнут влево. Спрыгивает с колен осторожно и при этом правильно падает на лапы, после чего сильно шатается. Левый зрачок уже правого. Нистагм вправо. Разгибательный тонус обеих левых ног несколько ниже, чем правых. Пассивные движения головы влево осуществляются легко, вправо же возможны лишь с преодолением сильного противодействия.
5 июля. Бегает вдоль стены, прижимаясь к ней левым боком. Может свободно выбежать из комнаты, но отклоняется при этом влево. Продолжает иногда падать влево. Движений по часовой стрелке больше нет. Легкий горизонтальный и ротаторный нистагм вправо. Голова повернута влево, только иногда отклонена влево. Позвоночник в области туловища более не изогнут.
6 июля. Ест мясо. Голова повернута влево, иногда также отклонена влево. Туловище не изогнуто. Во время бега иногда падает влево. Движения по часовой стрелке и манежные движения отсутствуют. Левый зрачок несколько уже правого. Обе левые
19*
292
Шестая глава. Последствия односторонней экстирпации -лабиринта
ноги оказывают пассивному сгибанию намного меньшее противодействие, чем правые. Иногда во время сидения левая нога отставлена. Во время еды поворот и наклон головы становится немного сильнее.
7 июля. Нистагма больше нет. Зрачки равномерные. При беге продолжает еще несколько отклоняться влево. Иногда также падает влево. Разгибательный тоунс левых ног отчетливо ниже, чем правых. В состоянии спуститься по лестнице.
8 июля. Выскакивает из клетки. Голова отчетливо повернута влево, почти не наклонена. При сидении ноги больше не расставляет. Животное удается легче сместить на плоскости опоры влево, нежели вправо. Во время бега больше не падает.
6 сентября. Сидит вполне правильно. Голова в состоянии поворачиваться во всех направлениях, однако во время сидения и бега по большей части повернута влево. Левая нога при сидении несколько отставлена в сторону. Ест и пьет хорошо. Нистагм, неравномерность зрачков и глазных щелей отсутствуют. К цели бежит уверенно и прямо. Однако в отдельных случаях при беге отклоняется примерно на 30° влево. Во время питья поворот и наклон головы отчетливо увеличивается, при этом сидит, расставив ноги также несколько шире. С уверенностью спрыгивает вниз с высоты 1 м 30 см, прыгая, однако, всегда отчетливо влево. Во время питья из миски тело легче сместить на плоскости опоры влево, нежели вправо.
24 октября. Голова повернута влево, не отклонена. Во время еды, питья и при виде мяса, которое держат перед животным, поворот отчетливо увеличивается. Может поворачивать голову во всех направлениях. Спрыгивает со шкафа двухметровой высоты, при этом не наблюдается никаких аномалий, кроме поворота головы. Сбегает вниз по лестнице по совершенно прямой линии. Вспрыгивает с пола на стул. Лишь во время очень быстрого бега иногда все еще отклоняется влево. Тело сидящего животного легче сдвинуть влево, чем вправо.
25 октября. Для исследования децеребрирована (см. стр. 51). На секции обнаружено полноё удаление левого лабиринта; левая булла пуста; ствол 8-го нерва слева значительно тоньше, чем справа. Мозг и оболочки без признаков раздражения.
Как легко можно заметить, сначала после операции имеют место весьма отчетливо выраженные явления; большая часть их, однако, постепенно проходит и через несколько месяцев животное снова ведет себя почти как нормальное, так что необходима особая внимательность, чтобы установить у него наличие последствий выпадения лабиринта. Вышеприведенный протокол дает хорошее представление о том, как постепенно происходит этот процесс. Далее приведен еще один протокол, по которому можно хорошо проследить поведение кошки в первые дни.
Картина симптомов у различных кошек сходна. Обнаруживаются лишь незначительные индивидуальные различия. Возникновение и взаимозависимость этих последствий в дальнейшем будут подробно исследованы.
Глазные симптомы
а) Паралич симпатического нерва
Явления временного паралича симпатического нерва на глазу оперированной стороны, которые наблюдал Камис (3) у кошек после экстирпации лабиринта (оттянутая мигательная перепонка, узкая глазная щель и узкий зрачок), согласно исследованиям де Клейна, Сосэна и де Бюрле (11, 20, 21), обусловлены тем, что постганглионарные симпатические пути глаза у кошек проходят через среднее ухо, поэтому во время операции в большей или меньшей степени повреждаются. Поэтому с выпадением собственно функции лабиринта эти симптомы не имеют ничего общего и должны рассматриваться как следствия операционной травмы.
б) Отклонение глаз и нистагм
В то время как у кролика и морской свинки девиация глаз принадлежит к наиболее выраженным длительным симптомам выпадения лабиринта, у кошек этот симптом совершенно отодвигается на задний план. Во-пер¬
Опыты на кошках
293
вых, он весьма скромно выражен и, во-вторых, недолговечен. Нужна особенная внимательность, чтобы установить наличие отведения глаз. Непосредственно после операции можно видеть, что оба глазных яблока отведены в сторону (горизонтально) в направлении отсутствующего лабиринта и что, кроме того, имеется ротаторная девиация, когда верхний край зрачка на обеих сторонах повернут в сторону операции. Помимо этого, глаз оперированной стороны отклоняется в вентральном, а другой глаз, — в дорсальном направлении. Отклонение на стороне операции сильнее, но и здесь имеет место лишь настолько, что вследствие горизонтальной девиации остается видимым белый край склеры глаза неопери- рованной стороны. Уже на второй день после операции девиация минимальна, а еще через несколько дней исчезает.
Сразу после операции можно видеть отчетливый нистагм. На первый взгляд кажется, что он не имеет никакого определенного направления, так как движения в одну и другую стороны не очень сильно отличаются по своей скорости. Только при ближайшем рассмотрении и в этом случае возможно различить, что движение в одном направлении совершается скорее, чем в противоположном. Нистагм осуществляется так же, как у кролика и у морской свинки, в направлении, противоположном отведению глаз, он горизонтален, с быстрым компонентом, направленным в здоровую сторону; ротаторный нистагмоидный толчок зрачка направлен в здоровую сторону. Этот нистагм хорошо выражен обычно только в первые 24 часа после операции, на второй день он совершенно исчезает. Только в одном случае на шестой и седьмой день можно было видеть незначительные следы нистагма. При исчезновении нистагма обычно исчезает и девиация глаз. У кошки также отмечается влияние установки головы на девиацию глаз и нистагм. Если голову зрячего животного повернуть в сторону операции (ухо оперированной стороны вентрально), пока ротовая щель не встанет вертикально, а интактный лабиринт не окажется наверху, то девиация глаз уменьшается, а нистагм становится максимальным. В случае поворота в противоположную сторону (интактный лабиринт оказывается внизу) отклонение глаз достигает наиболее высокой степени, а нистагм очень сильно уменьшается или вовсе исчезает. При промежуточном положении головы, а также в положении на спине девиация глаз и нистагм имеют среднюю степень интенсивности.
Уже в первые дни после операции, т. е. тогда, когда еще имеет место отклонение глаз, животное в состоянии поворачивать глазное яблоко во все стороны, включая и направление, противоположное направлению девиации. Именно поэтому установить наличие девиации так трудно. Через несколько дней движения глаз, по крайней мере насколько это можно установить, не прибегая к особенно тонким методам, не отличимы от таковых у нормальной кошки.
Положение головы, шеи и туловища
Непосредственно после операции голова повернута и отклонена в сторону операции. Сначала преобладает отклонение. Если односторонняя экстирпация лабиринта производится у децеребрированной кошки, также наступает отклонение или отклонение в комбинации с поворотом в сторону операции.
К отклонению головы часто присоединяется вследствие шейного установочного рефлекса отчетливое отклонение туловища в сторону операции, исчезающее на следующий день. Отклонение головы через 2—3 дня становится меньше и постепенно исчезает.
294 Шестая глава. Последствия односторонней экстирпации лабиринта
В дальнейшем временами оно снова появляется, особенно при отвлечении внимания (во время еды и питья). В этом постепенном уменьшении отклонения, по-видимому, принимает участие активация установочных рефлексов тела на голову.
Поворот же головы не исчезает. Он является стойким результатом операции и обусловлен основным поворотом вследствие односторонних тонических лабиринтных рефлексов на шейную мускулатуру. У сидящего животного он составляет не более 45°, по большей же части 20—30°. С течением времени он постепенно уменьшается до 10—30°. Поворот также становится отчетливее во время еды или тогда, когда животное предпринимает что-либо, привлекающее его внимание. Так, например, в момент осуществления прыжка поворот виден отчетливее.
Точно так же, как у кролика и морской свинки, к повороту шеи присоединяется соответствующий поворот всего туловища. У бегущего или стоящего животного поэтому видно перевешивание тела в сторону операции, однако вследствие того, что это перевешивание обусловлено не одним только поворотом туловища, но и различием тонуса конечностей, поворот тела лучше всего констатировать в висячем положении животного головой вниз. Это осуществимо только в случае молодых или очень ручных животных. Так, например, непосредственно после операции грудь оказалась, повернутой по отношению к тазу на 45°, голова — на 90°. У другой кошки спустя три дня после операции нижняя апертура грудной клетки была повернута относительно таза на 45°, верхняя — на 90° и голова — на 135°.
Этот поворот туловища точно так же, как и у кролика, возникает при взаимодействии двух влияний: 1) прямого основного поворота туловища и 2) шейного установочного рефлекса. Основной поворот туловища у кошек выражен особенно отчетливо. Если, например, у кошки в висячем положении головой вниз установить голову по отношению к туловищу прямо, то почти всегда отчетливый поворот туловища еще остается. Он исчез в одном случае только тогда, когда голова была повернута на 45° в другую сторону, т. е. он оказался полностью компенсированным шейным установочным рефлексом другого направления. Прямой основной поворот туловища лучше всего удавалось изолированно исследовать у кошек, у которых предварительно пересекались три верхних задних шейных корешка. У одного из таких животных тотчас после операции нижняя апертура, грудной клетки была повернута в сторону операции на 20°, верхняя — на 30°, голова — на 90° повернута и на 20° отклонена. Через 11 дней нижняя апертура была повернута на 20°, верхняя — на 45°, голова — на 70—90°. У другой такой же кошки поворот нижней апертуры грудной клетки составлял 45°, верхней — 90°, головы — 135°. Таким образом, спиральный поворот всего тела у кошки весьма интенсивен и тогда, когда шейный установочный рефлекс полностью выключен. У кошки без больших полушарий Дюссер де Баренн обнаружил через день после односторонней экстирпации лабиринта поворот головы на 90° и грудного отдела — на 45°. После того как голова по отношению к туловищу была установлена прямо, все еще оставался поворот груди на 15°. Из последнего наблюдения следует, что, помимо основного поворота, функционирует еще и шейный установочный рефлекс. То же следует из возможности компенсировать спиральный поворот туловища поворачиванием головы в обратную сторону. Таким образом, и у кошки, как и у кролика, поворот туловища обусловлен взаимодействием двух влияний: основного поворота и шейного установочного рефлекса.
Опыты на кошках
295
К действию основного поворота, как у кролика, присоединяется влияние лабиринтного установочного рефлекса на голову с оставшегося интактного лабиринта. Это влияние было более подробно исследовано Дюс- оер де Варенном на двух бесполушарных кошках после правосторонней экстирпации лабиринтов, которые легко позволяли придавать им любое положение в пространстве. Результат исследования был примерно таким же, как и результат, полученный в опытах на кролике.
Односторонний лабиринтный рефлекс стремится привести голову в положение, в котором интактный лабиринт находится наверху и вследствие этого при различных положениях животного в пространстве будет или усиливать, или ослаблять основной поворот. Так, например, различная степень поворота головы обнаруживалась при разном положении животного в воздухе. При правом боковом положении голова также находилась в боковом положении, т. е. она не была повернута по отношению к туловищу, в то время как при левом боковом положении тела голова поворачивалась в нормальное положение. В первом случае установочный рефлекс и основной поворот противодействуют друг другу, в последнем — усиливают друг друга. То же наблюдается, если уложить животное на стол в боковое положение и компенсировать влияние на голову установочных рефлексов с тела, накладывая на сторону тела, обращенную вверх, дощечку с грузом. В положении на спине животные были беспокойны и успокаивались только тогда, когда они сложным движением приводили голову в правое боковое положение на брюхе точно так, как это показано на рис. 141 для кролика. В висячем положении головой вниз голова вследствие основного поворота повернута вправо и, кроме того, отклонена вправо. Поэтому голова оказывается в том боковом положении, в котором установочные рефлексы проявляются сильнее всего. Одно из животных освободилось из этого неудобного положения при помощи поворота влево, другое — при помощи такого интенсивного поворота шеи и груди, что голова пришла полностью или почти в нормальное положение. Этих примеров достаточно для того, чтобы показать действительность лабиринтных установочных рефлексов у кошки без одного лабиринта. Более обстоятельное описание остальных положений приводить излишне, так как они соответствуют таковым у кролика.
Как только тело животного оказывается не в воздухе, а на земле, в действие вступают установочные рефлексы с тела на голову. Эти рефлексы оказалось легче всего изучать на двух бесполушарных кошках Баренна после экстирпации правых лабиринтов. Как говорилось выше, животные, находясь в правом боковом положении в воздухе, держали голову также в правом боковом положении (основной поворот минус лабиринтный установочный рефлекс на голову). Однако, как только их укладывали в правое боковое положение на столе, они поворачивали голову в нормальное положение. Последнее неосуществимо ни посредством основного поворота, ни вследствие лабиринтного установочного рефлекса. Дело идет здесь единственно об установочном рефлексе с тела на голову, который должен проявить свое действие как по отношению к основному повороту, так и по отношению к лабиринтному установочному рефлексу. К этому повороту головы в нормальное положение присоединялось усаживание тела (совместное действие шейного установочного рефлекса и установочного рефлекса с туловища на туловище).
То, что шейные установочные рефлексы у односторонне лишенной лабиринта кошки взаимодействуют, было уже показано при описании поворота туловища. Их можно обнаружить у спокойно сидящего живот¬
296
Шестая глава. Последствия односторонней экстирпации лабиринта
ного, когда при повороте головы вправо или влево удается переложить туловище в правое или левое боковое положение.
То, что при сидении кошки на полу установочные рефлексы с туловища также влияют на туловище, оказалось легко показать на бесполу- шарных кошках потому, что если удерживать голову в правом или левом боковом положении и затем, слегка подталкивая, сдвигать туловище относительно плоскости опоры, то тело принимает нормальное сидячее положение, несмотря на то, что голова удерживается в боковом положении (значение установочного рефлекса с туловища на туловище по сравнению с шейным установочным рефлексом).
Резюмируя, можно заключить, что после односторонней экстирпации лабиринта из интактного лабиринта исходят следующие влияния.
1. Тонические лабиринтные рефлексы на мышцы шеи. Эти рефлексы влияют на_ основной поворот (и (основное отклонение) головы.
2. Весьма отчетливые тонические лабиринтные рефлексы на мышцы туловища. Эти рефлексы влияют на основной поворот туловища.
3. Лабиринтные установочные рефлексы на голову. Эти рефлексы усиливают шли ослабляют, смотря по положению в пространстве, оснон- ной поворот.
Вследствие поворота шеи включаются асимметричные шейные рефлексы, которые усиливают основной поворот туловища и заставляют тело закономерно следовать за головой.
Этим асимметричным влиянием при соприкосновении с полом противодействуют:
1) установочный рефлекс с тела на голову;
2) установочный рефлекс с тела на тело.
Тонус конечностей
Получить впечатление о разгибательном тонусе ног у кошки без одного лабиринта лучше всего при испытании смещаемости тела вправо или влево у животного, сидящего- на неровной поверхности (соломенный мат). Животное старается противодействовать смещению, разгибая ноги на стороне, куда направлено движение. Это разгибание ощущается как противодействие боковому смещению и таким образом становится хорошим критерием существующего, даже незначительного, различия тонуса. При этом можно установить голову или повернуть ее в противоположную сторону и без труда наблюдать влияние этих манипуляций на тонус конечностей. Другим употребительным приемом служит сравнительное испытание противодействия пассивному сгибанию в обоих локтевых суставах. Однако многим кошкам не нравится, когда их укладывают на спину и непосредственно сравнивают тонус их ног. Но в отдельных случаях можно использовать и этот способ.
Сразу после пробуждения от наркоза у некоторых животных тонус конечностей, когда голова устанавливается по отношению к груди прямо г с обеих сторон одинаков. Прямого влияния выпадения лабиринта на тонус конечностей у них установить не удается. Наоборот, в большинстве случаев сразу после операции ноги на стороне утраченного лабиринта более расслаблены, чем на другой стороне (при голове, установленной прямо). Это различие почти всегда исчезает уже на следующий день, лишь в одном случае оказалось возможным отметить через 24 часа отчетливое, а на второй-четвертый день — минимальное различие тонуса конечностей при коррекции отклонения головы. После этого прямого влияния выпадения лабиринта на тонус конечностей больше не обнаруживалось. Это влияние, следовательно, всегда незначительно по длительности, выражено в большинстве случаев только очень мало и может отсутствовать полностью.
Опыты на кошках
297
То же самое можно установить, если децеребрировать нормальных кошек и затем экстирпировать у них или выключить с помощью кокаина один лабиринт. В 14 из 17 случаях после операции разгибательный тонус конечностей на стороне утраченного лабиринта был ниже, чем на другой стороне; в остальных трех случаях он был одинаковым. Исследовать тонус конечностей у децеребрированных животных легче, чем у нормальных, так как первые обычно не производят спонтанных движений.
В то время как прямое влияние выпадения лабиринта на тонус конечностей невелико и преходяще, влияние шейных рефлексов в той же мере отчетливо выражено. Здесь тонические рефлексы на конечности, вызванные поворотом шеи, являются наиболее важными, ибо, во-первых, поворот шеи является длительным следствием операции и, во-вторых, поворот оказывает более сильное влияние на конечности, чем отклонение.
Так как после односторонней утраты лабиринта голова повернута в сторону операции, то ноги этой стороны являются «черепными» ногами и разгибательный тонус их становится ниже, а ноги противоположной стороны («челюстные» ноги) тонизированы сильнее. Вследствие этого в любой момент после операции, когда животное сидит свободно, более высокий тонус обнаруживается на ногах здоровой стороны. Однако как только голову по отношению к груди устанавливают прямо, это различие исчезает (по крайней мере если исследование ведется не в самые первые дни после операции, когда вышеупомянутое прямое влияние лабиринта еще имеет место). При поворачивании головы в другую сторону животное становится легче сместить в здоровую сторону, т. е. животное ведет себя так, как будто у него был удален другой лабиринт. У одной маленькой кошки это влияние поворота головы на тонус ног с тем же эффектом удавалось исследовать и в положении на спине. Оказалось, что влияние на передние ноги выражено сильнее, чем на задние. Описанные изменения тонуса конечностей можно было видеть до 115-го дня после операции. Более длительные наблюдения не предпринимались.
Отклонение головы выражено более сильно лишь в первые дни после операции, поэтому оно только вначале способно оказывать влияние на тонус конечностей. Так как влияние это менее значительно, чем влияние поворота головы, то им чаще можно пренебречь. Однако именно у кошки влияние отклонения головы на тонус конечностей в отдельных случаях сразу после операции может оказываться отчетливо выраженным. Поскольку голова отклонена в сторону утраченного лабиринта и ноги оперированной стороны становятся «челюстными» ногами, конечности на стороне отсутствующего лабиринта при интенсивном отклонении головы приобретают более высокий тонус и поэтому отклонение головы должно действовать на конечности в противоположном направлении по сравнению с поворотом головы и прямым влиянием выпадения лабиринта. В одном случае даже сразу после операции ноги оперированной стороны были разогнуты сильнее, чем на здоровой, причем в то же время отмечено было сильное отклонение головы.
Кошка Ла-цэ. 23 сентября 1911 г. 9% часов. Экстирпация левого лабиринта. Три отверстия полукружных каналов и ствол 8-го нерва открыты. Лицевой нерв интактен. Сразу после операции голова сильно отклонена влево, неотчетливо повернута. Обе левые ноги характеризуются более сильным разгибательный тонусом, чем правые.
10% часов. Животное усаживается. Голова повернута влево, левая передняя нога более расслаблена, чем правая. Если голову установить точно прямо, различие уменьшается, однако левая передняя лапа все еще отчетливо более расслаблена, чем правая. Поворот головы влево увеличивает различие, при повороте вправо оно уменьшается до полного уравнивания тонуса. Если из симметричного положения голова отклоняется влево, тонус правой ноги падает; при отклонении головы вправо, что возможно только после преодоления сильного противодействия, понижается тонус левой ноги. Комбинация поворота и отклонения оказывает отчетливо противоположное действие.
298 Шестая глава. Последствия односторонней экстирпации лабиринта
24 сентября. Голова повернута и отклонена влево. Обе левые ноги обладают меньшим разгибательным тонусом, чем правые. При установке головы прямо различие тонуса конечностей справа и слева исчезает почти полностью.
Как можно видеть, дело идет только о кратковременном влиянии. То же самое в отдельных случаях можно установить и у децеребрирован- ных животных, у которых после экстирпации лабиринта отклонение головы часто выражено сильнее, чем, поворот.
Так, в четырех опытах на децеребрированных кошках удалось показать, что после удаления одного лабиринта тонус конечностей на стороне операции, когда голова установлена прямо, ниже, чем тонус конечностей противоположной стороны. Вследствие сильного отклонения головы это влияние компенсировалось, так что разгибательный тонус с обеих сторон становился одинаковым. В двух других опытах различие тонуса конечностей, которое имело место при установленной прямо голове, было даже чрезмерно компенсировано с помощью сильного поворота головы, так что ноги оперированной стороны были разогнуты сильнее, чем ноги нормальной стороны.
Влияние тонических шейных рефлексов особенно хорошо заметно при сравнении с таковщм у тех четырех кошек, у которых предварительно была произведена перерезка трех пар верхних шейных задних корешков. Сразу после экстирпации лабиринта у трех из четырех этих кошек разгибательный тонус ног на стороне операции был меньше, чем на нормальной стороне. Это различие у одного животного исчезло уже на следующий день, у другого — через два дня и у третьего — через пять дней.г По прошествии этого времени тонус конечностей на обеих сторонах тела был одинаковым. Ни в первые, ни в последующие дни поворот или отклонение головы в одну или в другую сторону не оказывали на тонус конечностей никакого влияния. После того как сразу после операции тонус конечностей стал одинаковым, различия в смещаемости животного вправо или влево более не отмечалось, а при повороте головы в обе стороны его вызвать не удавалось. Таким образом, у кошек без тонических шейных рефлексов сразу после операции отмечается непостоянное и скоропроходящее различие тонуса конечностей обеих сторон; после исчезновения этого различия тонус долгое время остается одинаковым с обеих сторон.
Как ясно из предыдущего, имеются следующие влияния, которые обусловливают наличие разгибательного тонуса конечностей после односторонней экстирпации лабиринта. Поворот головы посредством тонического шейного рефлекса способствует тому, что конечности оперированной стороны приобретают меньший разгибательный тонус, чем конечности нормальной стороны. Так как поворот шеи является длительным последствием операции, то и этот шейный рефлекс оказывается длительным. Непосредственно после операции у большинства животных это различие углубляется вследствие прямого влияния выпадения лабиринта, которое продолжается один или несколько дней. Если после операции голову сильно отклонить, то это влияние действует в отношении описанного различия тонуса в противоположном направлении и даже способно временно чрезмерно компенсировать это различие. Здесь также дело идет о быстро проходящем влиянии.
Положение тела и движение
Уже спустя час или несколько часов после операции животное без одного лабиринта сидит прямо. Бесполушарная кошка уже через полчаса после удаления лабиринта в состоянии усаживаться прямо. После утраты
Опыты на кошках
299
левого лабиринта усаживание из левого бокового положения обеспечивается совместным действием основного поворота й всех установочных рефлексов. Из правого бокового положения, наоборот, основной поворот ж лабиринтный установочный рефлекс на голову действуют не в одном направлении и, таким образом, препятствуют усаживанию. Последнее осуществляется только благодаря установочному рефлексу с туловища на голову, но прежде всего вследствие установочного рефлекса с туловища на туловище. Вначале животные еще опрокидываются в сторону операции, но тотчас же усаживаются снова. Уже вскоре после пробуждения от наркоза возможен также и бег.
Поворот и отклонение головы в сторону операции, отклонение туловища в ту же сторону и утрата тонуса на ногах на этой же стороне уже были описаны. При сидении потеря тонуса соответствующих ног прояв- .ляется в том, что животное широко расставляет эти ноги. Стало быть, в то время как кролик отводит конечности противоположной стороны, разгибательный тонус которых выше, у кошки расслабленные ноги оперированной стороны отставлены несколько более в сторону. Это нередко отчетливо видно на передней и задней ноге, иногда же только на задней. В течение первой недели после операции это можно видеть почти всегда. Позднее животные корригируют эту установку, и тогда сидение с расставленными ногами можно наблюдать только случайно. В первые дни после операции животные во время сидения и 6èra покачиваются из стороны в сторону, спустя 1—5 дней они снова сидят спокойно.
Пока отклонение головы и туловища выражено сильно, животное во время бега совершает движения по часовой стрелке в направлении оперированной стороны или, когда в беге принимают большее участие зад- :ние ноги, — манежные туры. Так как отклонение очень скоро уменьшается, то эти зависящие от него аномалии также вскоре прекращаются. Лишь у трех животных движения по часовой стрелке имели еще место после 24 часов, в двух случаях — по прошествии двух дней и в одном случае — через пять дней. После этого они больше не наблюдались.
Наоборот, другое нарушение движений постоянно и значительно более длительно. Это — отклонение в сторону операции во время бега вперед, что происходит весьма своеобразно. Когда животное бежит к какой-нибудь цели, оно устанавливает ось своего тела совершенно правильно в направлении к этой цели. Если оно бежит вперед, тело отклоняется ж направлении стороны операции без того, чтобы направление оси тела при этом изменялось. Если, например, тело кошки, оперированной на правой стороне, стоит в направлении ab и животное хочет бежать в &, тело его во время бега перемещается в направлении ас таким образом, что оказывается в положении а!Ь' и так дальше.
ъг
Причину этого явления нетрудно установить. Вследствие того что ноги на оперированной стороне обладают более низким тонусом и во время бега не в состоянии поддерживать тело так, как ноги здоровой стороны, тело и валится в эту сторону. В большинстве случаев конечности еще выдерживают вес тела, и бег продолжается. Таким образом, согласно этому представлению, боковое отклонение во время бега обусловлено €олее низким тонусом ног на стороне операции. Так как последний по
300
Шестая глава. Последствия односторонней экстирпации лабиринта
прошествии первых дней после операции обусловлен исключительно поворотом шеи, то отсюда следует, что и отклонение в сторону во время бега также следует рассматривать как результат поворота шеи.
Что это действительно так, видно из наблюдений на тех четырех кошках, у которых для выключения шейных рефлексов перерезались три пары верхних задних шейных корешков, после чего экстирпировался правый лабиринт. Опрокидывание во время бега тотчас после операции наблюдалось лишь у одной из этих кошек (когда еще можно было констатировать наличие различного тонуса конечностей). Уже на следующий день животное больше не опрокидывалось на бок. Другая из этих кошек уже через 6 часов после операции была в состоянии пройти по комнате без отчетливого отклонения вправо. Точно так же у всех четырех кошек не наблюдалось отклонения в сторону. Уже в первый день животные с их сильно повернутой головой могли бежать точно прямо. Если они пошатывались, то в обе стороны в одинаковой степени. У одной кошки наблюдалось спотыкание вправо только в первый день после операции (когда правые ноги были расслаблены еще более, чем левые), и только .однажды это повторилось позже. Во время бега вследствие поворота туловища грудной отдел отчетливо перевешивал в сторону операции. И несмотря на это, отклонения во время бега не было. На этом основании для возникновения отклонения и опрокидывания повороту туловища выше было приписано лишь вторичное значение. Кошка уже в первый день после правосторонней операции могла бежать несколько влево; спустя неделю она осуществляла даже круговые туры влево. При сидении характерная поза с широко расставленными ногами на оперированной стороне отсутствовала. Только у одной из кошек эта поза наблюдалась в первый день после операции, когда еще имело место различие тонуса как прямое следствие выпадения лабиринта.
У кошек с интактными шейными задними корешками значительно изменяется угол отклонения в стороны. Иногда он составляет только 20—30°, нередко 45°, но может достигать и 90°. В последнем случае наблюдается чисто боковое движение. Обычно в последнем случае продвижение вперед осуществляется таким образом, что животное сначала более или менее успешно делает несколько шагов вперед, затем, спотыкаясь, передвигается под углом в 90° в направлении оперированной стороны, после этого снова идет вперед, снова спотыкается и т. д. Получается своеобразное зигзагообразное движение к цели. Из этого описания следует, что временами такое животное в состоянии сделать несколько нормальных шагов вперед. Только с течением времени это происходит все чаще, а угол, под которым животное отклоняется во время бега, становится все меньше. Таким образом, бег постепенно делается лучше, пока, наконец, отклонение не появляется только в отдельных случаях, при очень быстром беге. Однако оно наблюдалось даже спустя 115 дней после операции. Как указывалось выше, к этому времени влияние поворота на тонус конечностей также еще имеет место.
Пока это боковое спотыкание еще сильно выражено, в отдельных случаях и во время бега дело доходит до настоящего опрокидывания в сторону операции, после чего животное тотчас же поднимается. В течение первых четырех дней после операции кошки (с интактными задними корешками) весьма нередко опрокидываются во время бега на бок. Позже это происходит все реже и реже, обычно только во время быстрого бега, и в конце концов более не наблюдается. Это опрокидывание также следует связать в основном со слабостью разгибательных мышц конечностей оперированной стороны и считать зависящим от поворота шеи. Поворот
Опыты на кошках
301
груди играет при этом меньшую роль, имея только вторичное значение, как выясняется из наблюдений над животными с перерезанными задними шейными корешками.
Постепенно животные снова выучиваются прыгать. Вначале они при этом опрокидываются в сторону операции, позднее при спрыгивании на пол отклоняются в оперированную сторону, наконец, прыжок становится почти нормальным, только случайное отклонение напоминает, что мы имеем дело с животным, имеющим только один лабиринт. Обычно уже через 24 часа кошка пытается соскочить с колен, однако, как правило, падает при этом. Через месяц три кошки уже выучились соскакивать, не падая, со шкафа высотой в 2 м. К этому же времени, а иногда и раньше они оказываются в состоянии вскочить с пола на стул. Бегание по лестнице сначала связано с трудностями, но по большей части уже спустя 4—11 дней животные способны сбегать вниз по лестнице, при этом они иногда спотыкаются и держатся-стенки той же стороны, на которой у них утрачен лабиринт. Таким образом, они предохраняют себя от отклонения в сторону и от опрокидывания. Но в конце концов они снова выучиваются спускаться по лестнице совершенно так же, как нормальные кошки.
Эти нарушения значительно менее выражены после выпадения тонических шейных рефлексов. Одна из кошек с перерезанными задними шейными корешками уже в день операции соскакивала со стола на пол, не падая при этом, расставляя ноги с обеих сторон, при приземлении раскачиваясь в обе стороны, но не отклоняясь в какую-либо сторону. Спустя недолгое время после этого она совершала прыжок в Уг м длиной со стола в клетку, не падая и не отклоняясь; на третий день она прыгала уже на расстояние 1 м. Другая кошка уже в первый день, не падая, соскакивала с наклоненного стула, на второй день — с обыкновенного стула, на седьмой день — со стола высотой в 1 м на пол и в этот же день была в состоянии спускаться и подниматься по лестнице.
Кошки без одного лабиринта с сохранными шейными рефлексами в первые дни после операции, пока отклонение и особенно опрокидывание в сторону операции еще сильно выражены, будучи посажены в комнате на пол, очень скоро находят стену комнаты и бегают потом вдоль этой стены, повернувшись к ней оперированной стороной. Так же как при беге по лестнице, они пытаются этим путем предохранить себя от падения в сторону. По истечении первой недели эта предосторожность уже не нужна.
Истинные движения перекатывания, какие описаны для кролика и морской свинки, у кошек после осторожно произведенного удаления лабиринта не имеют места. Наоборот, кошка, у которой де Клейн обнажил через буллу полость черепа и интракраниально перерезал 8-й нерв, в процессе чего было довольно сильное кровотечение, в течение четырех дней производила типичные движения перекатывания в сторону операции. И в этом случае движения перекатывания наступали приступообразно. Таким образом, для кошки необходимо весьма сильное раздражение вследствие операции, чтобы вызвать спиральный поворот тела, который обусловил бы появление типичных движений перекатывания.
В какой степени направление, в котором поворачиваются животные и в котором они отклоняются, зависит от установки головы, может быть показано весьма просто. Так, каждую кошку, лишенную одного лабиринта, которая при беге еще сильно отклоняется в сторону операции или даже падает в эту сторону и которая в состоянии с легкостью двигаться по часовой стрелке и совершать манежные движения в сторону операции, можно заставить поворачиваться в здоровую сторону. Для
302
Шестая глава. Последствия односторонней экстирпации лабиринта
этой цели над головой голодного животного держат в воздухе кусок мяса. Если теперь водить мясо в воздухе по кругу в сторону операции, кошка оказывается не в состоянии следовать за этим движением: она или опрокидывается, или безуспешно прыгает вверх, пытаясь достать мясо, или попросту убегает. Наоборот, если мясо вести в направлении здоровой стороны (т. е. после левосторонней экстирпации лабиринта в направлении часовой стрелки, если смотреть сверху), животное следует за этим движением и описывает при этом движение по часодой стрелке или очень ограниченные манежные движения в направлении здоровой стороны. Объяснение этому, на первый взгляд кажущемуся парадоксальным, явлению кроется в установке головы.
Голова, следуя за мясом, поднимается в воздухе (при этом обе передние ноги разгибаются) и сразу же поворачивается определенным, обусловленным выпадением лабиринта образом. Так, после левосторонней операции морда стоит в этом случае почти вертикально кверху, левое ухо обращено кпереди, правое — каудально. Следовательно, тыльная часть головы обращена влево, т. е. в сторону операции. Животное, таким образом, не может повернуться в сторону тыльной части головы, но может делать это всегда только в другую сторону. Если оно попытается повернуться в сторону затылка, то опрокинется или споткнется, а после этого скоро оставляет свою попытку. Наоборот, в противоположную сторону оно может поворачиваться без труда, особенно потому, что вследствие поднятия головы обе передние ноги приобретают сильный разгибатель- ный тонус и опасности опрокинуться на сторону операции нет. После левосторонней операции животное не в состоянии держать голову с поднятой мордой, повернутой в противоположном направлении. Если держать мясо менее высоко, в связи с чем голова животного не поднимается в дорсальном направлении настолько высоко, то оно может наклонить голову в оперированную сторону и следовать за мясом и тогда, когда голова повернута в воздухе в противоположном направлении. Эти отношения наблюдаются еще в течение нескольких месяцев после операции; обнаруживается своеобразная картина, когда животное, не проявляющее более почти никаких двигательных нарушений, предпринимает самые отчаянные попытки следовать за куском мяса, которым водят в воздухе, не будучи в состоянии этого осуществить.1 Нормальная кошка, естественно, способна следовать за движением мяса по кругу в любом направлении. При этом она поднимает морду кверху и наклоняет затем голову в направлении вращения. Для кошки с одним лабиринтом это невозможно. В целом нарушение движения зависит только от вынужденного положения головы.
Последствия односторонней утраты лабиринта у кошек без шейных рефлексов
В последних разделах уже неоднократно упоминались наблюдения над четырьмя кошками, у которых при помощи перерезки трех пар верхних шейных задних корешков были выключены шейные рефлексы. У этих кошек влияния поворота и наклона головы на положение тела, на поворот туловища, на тонус конечностей, на нарушения бега и так далее от¬
1 Ср. с этим описанное на стр. 234 поведение лишенных одного лабиринта кроликов, которые, будучи посаженными на пол, катятся в сторону утраченного лабиринта; в положении на спине в воздухе, наоборот, описывают передней частью тела кружащие движения в противоположном направлении.
Опыты на кошках
303
сутствуют. Таким образом, несмотря на то, что на этих животных произведены были два серьезных вмешательства, из которых каждое само по себе вызывает нарушения движения, симптомы после односторонней экстирпации лабиринта у животных с перерезанными шейными задними корешками не более, а менее значительны, чем у нормальных животных. Это положение является веским доводом в пользу того взгляда, что шейные рефлексы действительно обусловливают большую часть общих симптомов после односторонней экстирпации лабиринта. Хотя, согласно имеющемуся фактическому материалу, сомнений по этому поводу более не возникает и наиболее важные наблюдения уже были в деталях приведены выше, в дальнейшем в интересах большей рельефности общей картины приводятся в сопоставлении друг с другом протоколы опытов (16) на нормальной кошке и кошке с перерезанными тремя парами задних верхних шейных корешков, у которых де Клейном в один и тот же день был экстирпирован правый лабиринт. Поведение обеих кошек сравнивалось в течение долгого времени.
Б е л и а л ь (операция на задних корешках 23 марта и 5 апреля 1913 г.).
16 сентября 1913 г. Типичная правосторонняя экстирпация лабиринта. Внутренний слуховой проход и отверстия полукружных каналов открыты, ствол 8-го нерва обойден пинцетом. Лицевой нерв интактен (щадящая операция).
3/4 часа после операции; животное сидит. Голова немного наклонена вправо и немного повернута вправо. Движения по часовой стрелке вправо. В висячем положении головой вниз: грудь повернута по отношению к тазу примерно на 30°, голова — на 70°. При установке головы прямо и при сильном повороте головы, заходящем на другую сторону, поворот груди не изменяется. В положении на спине, когда голова установлена прямо, обе правые ноги обладают меньшим разгиба- тельным тонусом, чем обе левые ноги. Различие особенно отчетливо на передних ногах. При поворачивании головы шейный рефлекс на передние ноги не вызывается. На неровной поверхности кошку легче сместить вправо, чем влево; при голове, установленной прямо, это сохраняется. Отчетливы отклонения глаз вправо. Верхний край правого зрачка повернут вправо. Небольшой нистагм головы.
6 часов после операции. Идет по комнате, не падая, и без отчетливого отклонения вправо. Только если кошка сильно потрясет головой, она оступается при этом вправо. При устанавливании ног, как при повернутой, так и при прямо стоящей голове, разгибательный тонус правой передней ноги ниже, чем левой. Сидя держит ноги симметрично.
Ч е р н а я (нормальная кошка).
16 сентября 1913 г. Типичная правосторонняя экстирпация лабиринта. Внутренний слуховой проход и отверстия полукружных каналов открыты. Ствол 8-го нерва обойден пинцетом. Лицевой нерв интактен (щадящая операция).
1/4 часа после операции: действие наркоза окончилось. Голова повернута и наклонена вправо, девиация глаз вправо, нистагм влево, верхний край правого зрачка несколько отходит вправо. В висячем положении головой вниз грудь повернута на 45°, голова — на 90° вправо; наклонены в разной степени. При установлении головы относительно груди прямо поворот грудного отдела частично становится меньше (на 30°); чтобы грудь установилась прямо, нужно повернуть голову на 45°. В положении на спине, когда голова установлена прямо, разгибательный тонус правой передней ноги намного ниже, чем левой; сзади это различие меньше. При спонтанном повороте головы вправо различие тонуса передних ног становится больше; при повороте головы влево правая нога приобретает более высокий разгибательный тонус, чем левая. Животное легче сместить на плоскости опоры вправо, чем влево. При установлении головы прямо различие еще отчетливо, но значительно меньше.
6 часов после операции. Во время бега многократно падает вправо. Во время сидения туловище сильно перевешивает вправо; ноги держит симметрично.
304
Шестая глава. Последствия односторонней экстирпации лабиринта
17 сентября. Сидит прямо. Голова несколько повернута и наклонена в разной степени. Голова может быть наклоненной и в левую сторону. Иногда движения по часовой стрелке и манежные движения вправо. При сидении грудной отдел отчетливо перевешивает вправо. Перебегает комнату по прямой, при этом иногда даже чуть уклоняется влево. Голова еще немного качается. С косо поставленного стула соскакивает не падая, в состоянии сделать несколько шагов влево. Поза в висячем положении головой вниз, как накануне. Различие тонуса ног, смещаемость и глазные симптомы, как накануне.
18 сентября. С осторожностью бегает по прямой через всю комнату; в состоянии также бегать по кругу влево, направо не оступается. При беге ноги еще немного согнуты в коленях (как и до операции), но не расставлены широко. При сидении — никакого различия тонуса передних ног. Соскакивает со стула, делает при этом круговой поворот вправо и убегает в прямом направлении. Отчетливой разницы в смещаемости вправо или влево нет; ее нет также и при спонтанном повороте головы. Нарушений наблюдается неоспоримо меньше, чем у кошки Черная.
21 сентября. Бегает намного лучше (ноги несколько согнуты и расставлены) по прямой через всю комнату. Временами отклоняется во время бега вправо, но так же часто и влево. С пола влезает на кроличью клетку и оттуда на подоконник.
23 сентября. Один раз, отклонившись вправо, хорошо бежит по прямой. В состоянии выполнить полный поворот по кругу влево. Временами левым боком обращается к стене. Голова повернута вправо примерно на 20°, в отдельных случаях и наклонена; может наклоняться и влево. При соскакивании со стула и со стола иногда несколько отклоняется вправо.
24 сентября. Отклонения глаз и нистагма нет. Бегает, как накануне.
27 сентября. Вскакивает с пола на стул, со стола на подоконник и соскакивает со стола на пол, не оступаясь и не падая. Хорошо сбегает и взбегает но лестнице, не падая при этом. Однако посаженная косо на лестницу падает и скатывается вниз.
17 сентября. Сидит прямо. Голова повернута на 30° и наклонена в разной степени. Ноги при сидении симметричны. Пятится, когда бежит, часто падает на правую сторону. Грудь перевешивает на правую сторону. Голова еще покачивается. С поставленного косо стула сползает вперед, при этом сильно спотыкается вправо. В висячем положении головой вниз поза та же, что накануне. Различие тонуса на ногах, смещаемость и глазные симптомы, как накануне.
18 сентября. При сидении с головой, установленной прямо, различия тонуса на ногах нет. При поворачивании головы «челюстная» нога приобретает более высокий, «черепная» нога — менее высокий разгиба- тельный тонус. Смещаемость вправо больше, чем влево; когда голова установлена прямо, различие в сме- щаемости исчезает. При исследовании нередко оступается вправо. Со стула соскакивает, не падая, но после этого оступается вправо, проделывает несколько движений в направлении часовой стрелки и снова оступается.
21 сентября. Бегает очень мало, при этом повторно падает вправо. Нередки движения по часовой стрелке вправо.
23 сентября. Бег такой же, как 21 сентября. Лапы при сидении симметричны. Если подталкивать вперед, в отдельных случаях бежит вперед в косом направлении. Отклонения глаз и нистагма нет. При сидении различие тонуса ног и смещаемость, как 21 сентября. В висячем положении головой вниз грудь повернута примерно на 20°. При устанавливании головы прямо поворот уменьшается. С косо поставленного стула соскакивает или сваливается вниз и при этом оступается вправо. Вперед идет очень осторожно и правым боком неоднократно опирается на стену.
24 сентября. Во время бега спотыкается в правую сторону. Бегает вдоль стены правым боком к ней. Через комнату бежит наполовину косо вправо. С косо поставленного стула хорошо соскакивает.
27 сентября. Теперь хорошо бегает по прямой. На показываемое мясо идет, оступаясь вправо.
Опыты на кошках
305
4 октября. Изменений нет
13 октября. С большой уверенностью галопирует вверх по лестнице и затем быстро перебегает комнату по прямой, при этом один-два раза оступается.
20 октября. Достает себе мясо с решетчатой крыши клетки, по боковой стенке влезает вверх; двумя передними лапами удерживается за перекладину решетки крыши и забирает мясо ртом.
27 октября. Голова повернута вправо, иногда до 45°. Свободно становится на задние лапы. Спрыгивает вниз с крыши почти двухметровой клетки, на мгновение падает на бок и затем быстро убегает. Помимо повернутой вправо головы, никаких видимых последствий односторонней экстирпации лабиринта больше нет.
18 декабря. Голова повернута вправо на 30—45°. Бежит по прямой. На мясо, которое держат перед ней, поднимает голову, не разгибая передних лап. Может поднять голову на 90° без реакции передних ног. В висячем положении головой вниз последняя повернута на 45°, грудь — только немного. По лестницам хорошо бегает как вверх, так и вниз.
Вторая половина дня: экстирпация второго лабиринта.
4 октября. Бегает хорошо, оступаясь, однако, вправо. Соскакивает со стула, но и при этом оступается также вправо. С большой осторожностью сбегает вниз по лестнице.
13 октября. Хорошо соскакивает со стола.
20 октября. Хорошо соскакивает со стула.
27 октября. Состояние без перемен.
18 декабря. Голова повернута вправо. С уверенностью спрыгивает вниз со стула или подоконника. Хорошо бежит по комнате. На поднимание и опускание головы передние ноги реагируют быстро.
Вторая половина дня: экстирпация второго лабиринта.
Из этих протоколов и из наблюдений над остальными четырьмя кошками следует, что односторонняя экстирпация ^лабиринта у кошек без шейных рефлексов вызывает' меньше симптомов, чем у нормальных кошек. Конечно, все прямые последствия выпадения лабиринта (временное отклонение глаз и нистагм, временный наклон и длительный поворот шеи, длительный поворот туловища и скоро проходящая расслабленность ног на стороне операции) проявляются точно так же, как у нормальных кошек. Наоборот, все тонические шейные рефлексы, обусловленные временным наклоном и длительным поворотом шеи, у них отсутствуют. Вследствие этого стойкая разница в тонусе конечностей обеих сторон, которая у нормальных кошек вызывается поворотом шеи, также отсутствует. Поэтому кошки без шейных рефлексов не обнаруживают тенденции к падению или отклонению в сторону операции; сидя, они не отставляют ноги оперированной стороны и прыгают увереннее, чем нормальные кошки. Смещаемость на опорной плоскости (по прошествии первых послеоперационных дней) в обе стороны одинакова. Поворачивание и наклон головы не оказывают ни малейшего влияния на тонус конечностей и на поворот туловища, вследствие чего у этих животных устанавливанием головы прямо невозможно уменьшить эффекты экстирпации лабиринта. Ввиду того что шейные рефлексы также отсутствуют, поднимание и усаживание туловища обусловлены исключительно установочными рефлексами с туловища на туловище * надежная деятельность которых в этих экспериментах проявляется с максимальной отчетливостью.
Явления компенсации
В наблюдениях над кошками без одного лабиринта самым замечательным является то, что такая значительная часть нарушений, имеющих место непосредственно после операции, по прошествии нескольких не- 20 Р. Магнус
306 Шестая глава. Последствия односторонней экстирпации лабиринта
дель сглаживается, а спустя несколько недель остаются лишь незначительные явления выпадения. Понемногу животные «выучивались» снова бегать по прямой, не опрокидываясь на бок, держать голову совсем не наклоненной и менее повернутой, удовлетворительно сидеть, выполнять большие прыжки, бегать по лестнице и т. д.
Исчерпывающего анализа этих явлений компенсации до сих пор не сделано. Частично это объясняется тем, что к тому времени, когда мы проводили наблюдения на кошках без одного лабиринта (1911—1913), установочные рефлексы вообще не были известны, частично же тем, что и теперь еще не выяснено, каким образом большой мозг (как целое) и его отдельные поля воздействуют на различные стволовые центры установки тела. Влияние этого рода до сих пор разъяснено только для оптических установочных рефлексов. В настоящее время можно считать лишь, что в явлениях компенсации принимают участие: 1) установочные рефлексы с туловища, 2) оптические установочные рефлексы, 3) центральные подкорковые компенсации.
Благодаря установочным рефлексам с туловища на голову животное, когда оно лежит на стороне экстирпации лабиринта, оказывается в состоянии переместить голову в направлении к нормальному положению и в довершение этого принять сидячее положение. Установочные рефлексы с туловища на туловище делают для животного возможным удерживать тело более или менее прямо, несмотря на то что голова его повернута; они облегчают усаживание туловища, когда голова предварительно переместилась в том же направлении, а прежде всего и тогда, когда голова не находится в нормальном положении. Большое значение установочных рефлексов с туловища на туловище особенно отчетливо выступило у животных с перерезанными задними шейными корешками, у которых шейные рефлексы отсутствуют. У этих животных усаживание и удерживание головы прямо обусловлены одними установочными рефлексами с тела. У обеих бесполушарных кошек Баренна особенно хорошо можно было наблюдать, как в первые три дня после операции установочные рефлексы с туловища постепенно приобретали все большее значение и прочность. Если, например, у одной кошки, лежащей на столе на правом боку, через 17 часов после операции голова медленно приводилась в нормальное положение, вслед за чем следовало усаживание, то через 66 часов животное быстро усаживалось из бокового положение, причем голова шла вперед. Уменьшение наклона головы в первые дни после операции наиболее вероятно обусловлено частично установочным рефлексом с туловища на голову.
В наших ранних наблюдениях над кошками без одного лабиринта мы обратили внимание на существенное влияние глаз на положение и установку тела. По-видимому, положение конечностей корригируется кошкой не с помощью глаз. В первые дни после операции влияния глаз, очевидно, даже вредны, так как выяснилось, что едва заметные маятникообразные качания головы после надевания на голову шлемика уменьшались и часто вовсе исчезали, так что с высокой степенью вероятности можно считать, что маятникообразное качание головы и нистагм имеют общее происхождение. Исходя из того, что уже через несколько дней кошки после двусторонней экстирпации лабиринта выучиваются контролировать положение головы с помощью глаз, можно допустить, что это, несомненно, может иметь место и после односторонней экстирпации лабиринта. Специальных наблюдений по этому поводу еще не предпринималось, но тем не менее можно с уверенностью сказать, что перед прыжком животное использует глаза для направления и ориентирования тела.
Опыты на кошках
307
Систематические наблюдения за кошками без одного лабиринта в шле- мике и без шлемика при разнообразных положениях тела в воздухе и на земле должны еще быть проведены.
По-видимому, важную роль в явлениях компенсации играют изменения центральной иннервации. Первым это установил на собаках Бехтерев (1). Изменения после одномоментной двусторонней экстирпации лабиринтов, как и следует ожидать, строго симметричны. Если же у собаки сначала экстирпировать лабиринт на одной стороне тела, подождать, пока явления выпадения частично ликвидируются, и удалить второй лабиринт, то животное обнаруживает зеркально обратные нарушения по сравнению с последствиями после первой операции: отклонение глаз, нистагм, поворот головы, перекатывания и так далее происходят так, как если бы первый лабиринт не был вовсе экстирпирован. Так как поворот головы и отклонение глаз (по своему направлению) уже не могут обусловливаться влиянием лабиринта, то в интервале между двумя операциями животное, лишенное лабиринта с одной стороны, должно было изменить иннервацию таким образом, что удаление второго лабиринта (т. е. восстановление симметрии в лабиринтной иннервации) вызвало асимметричное нарушение. Именно такое толкование этому явлению дает Эвальд.
«После одностороннего удаления лабиринта (у собаки) возникают асимметричные симптомы, которые в значительной степени компенсируются викарными явлениями. Последние, таким образом, тоже асимметричны, поэтому, когда удаляется второй лабиринт (вследствие чего асимметрия первоначальных нарушений после первой операции, казалось, должна была бы ликвидироваться), асимметричные викарные влияния вызывают неравномерность в функции обеих половин тела, которая никогда не наблюдается после одностороннего удаления лабиринта» (стр. 201).
Отсюда следует, что при изучении возникновения этих викарных процессов следует удалять второй лабиринт в различные сроки после экстирпации первого, наблюдая при этом, в какой степени и на какой стороне тела наступят асимметричные нарушения. В процессе прежних наших опытов представлялась возможность у девяти кошек экстирпировать второй лабиринт на 4-й, 11-й, 17-й, 21-й, 22-й, 25-й, 31-й, 114-й и 164-й день после разрушения первого лабиринта. Четыре кошки находились после этого под длительным наблюдением, пять других вскоре после второй операции были децеребрированы, так что на них можно было увидеть явления, наступающие непосредственно после операции. Результат оказался следующий: через четыре дня после удаления
первого лабиринта экстирпация второго оказывала то же влияние, как в случае одновременной экстирпации обоих лабиринтов. Если интервал между обеими операциями составлял 11 дней, то после второй операции наблюдался типичный нистагм, но не возникало ни поворота, ни наклона головы. При интервале в три недели и более голова всегда (7 наблюдений) оказывалась повернутой или наклоненной в сторону второй операции. Кроме этого, возникал, конечно, и нистагм (у недецеребрированного животного). Различия в тонусе конечностей, когда голова была установлена прямо, наоборот, после второй операции констатировать с уверенностью было нельзя, хотя в двух случаях (через 31 и 114 дней) такое различие намечалось. Отсюда можно было бы заключить, что сначала изменяется иннервация глаз, а затем наступает изменение иннервации мышц, отклоняющих голову, в то время как конечности и туловище вовлекаются в происходящее в гораздо меньшей мере или совсем не вовлекаются.
20*
308 Шестая глава. Последствия односторонней экстирпации лабиринта
У обеих бесполушарных кошек Баренна второй лабиринт был удален через три и через четыре дня после экстирпации первого. У обоих животных появились как отклонение глаз и нистагм, так и поворот и нистагм головы. Девиация головы и глаз была в сторону лабиринта, оперированного во второй этап, нистагм — в сторону лабиринта, экстирпированного на первом этапе. Дело выглядит, таким образом, как будто описанная центральная компенсация после удаления большого мозга наступает скорее, чем при интактном мозге. Однако число наблюдений слишком невелико, чтобы можно было с уверенностью делать выводы. Для этого необходимы дальнейшие наблюдения на нормальных животных, у которых оба лабиринта удаляются один после другого по прошествии нескольких дней.
Глазной нистагм, наступающий после второй операции, длится только
1— 2 дня. У одной кошки удалось наблюдать, что наступивший после второй операции поворот головы в сторону последней операции в течение
2— 3 месяцев постепенно прошел и что вслед за этим голова удерживалась совершенно сцмметрично.
Что касается возникновения этой компенсации, то опыты на двух бесполушарных кошках учат, что в образовании ее ни кора большого мозга, ни глаза роли не играют. У упомянутых выше кошек, у которых были перерезаны три пары верхних задних шейных корешков, позднее также был экстирпирован второй лабиринт. Нистагм и отклонение головы изменялись точно так же, как только что было описано. Таким образом, перерезка соответствующих шейных задних корешков не препятствует возникновению компенсации отклонения головы, которое обеспечивается главным образом мускулатурой первых шейных сегментов.
Если лишенных одного лабиринта кошек децеребрировать по прошествии большого времени, а затем удалить у них второй лабиринт, то наклон (и поворот) головы в сторону второй операции происходит с большей уверенностью. Таким образом, центральная нервная система ниже четверохолмия должна претерпеть стойкие изменения, посредством которых осуществляется отклонение головы после удаления второго лабиринта.
Таким образом, измененная центральная иннервация может осуществляться после удаления больших полушарий, выключения оптических раздражений и перерезки верхних задних шейных корешков. Кроме того, она может обнаружиться после удаления больших полушарий, выключения глаз, экстирпации обоих лабиринтов и перерезки верхних шейных корешков. Возникновение ее вне всякого сомнения зависит от односторонних возбуждений, которые исходят из интактного лабиринта, с каковыми нелабиринтные возбуждения другой стороны уравновешиваются затем более или менее совершенно.
Если благодаря этому центральному переключению наступает частичная компенсация установки головы, различие тонуса конечностей, естественно, должно стать меньше, а этим самым уменьшатся нарушения движения и положения. То, что различие тонуса конечностей после операции действительно сокращается только с уменьшением поворота головы, оказалось возможным показать, исследуя боковую смещаемость тела; различие это снова становилось больше, когда исходный поворот головы восстанавливался. Отсюда становится понятным, что такие нарушения, как нарушения бега, сидения, которые у кошки обусловлены различием тонуса конечностей, могут быть таким путем постепенно уменьшены, т. е. компенсированы.
Опыты на кошках
309
В заключение следует еще раз подчеркнуть: я не считаю, что исчерпал этим все компенсаторные возможности. Без сомнения, дело обстоит значительно сложнее, чем это представлено здесь.
Заключение
У кошки из симптомокомплекса односторонней экстирпации лабиринта можно выделить следующие факторы.
I. Длительные влияния, исходящие из оставшегося лабиринта.
А. Симметричные влияния: тонические лаби¬
ринтные рефлексы на мышцы конечностей имеют место равномерно на обеих сторонах.
Б. Асимметричные влияния.
1. Тонические лабиринтные рефлексы нашейные мышцы приводят к «основному повороту» шеи, который зависит от установки головы в пространстве и достигает своего максимума, когда темя обращено вниз. В течение короткого времени после операции голова также наклонена в сторону операции, что с течением времени уменьшается.
2. Лабиринтный установочный рефлекс на голову увеличивает или уменьшает «основной поворот», постоянно стремясь установить голову в боковое положение интактным лабиринтом кверху.
3. Тонические лабиринтные рефлексы на мышцы туловища приводят к «основному повороту» туловища в стороны утраченного лабиринта.
II. Временные последствия одностороннегоуда-
ления лабиринта.
1. Девиация глаз в сторону операции с нистагмом 'в противоположном направлении. Если интактный лабиринт
находится внизу, девиация велика, а нистагм слаб; если он находится сверху, девиация мала, нистагм же выражен сильно. И то и другое через несколько дней исчезает.
2. Непостоянное, через несколько дней проходящее расслабление конечностей на стороне операции, которое (как и в случае кролика) пока объяснить действием определенного лабиринтного рефлекса не удается.
III. Последствия поворота шеи. Таковые отсутствуют
у животных, у которых предварительно перерезались три пары
задних верхних шейных корешков.
А. Длительные последствия.
1. Посредством шейных рефлексов усиливается поворот шеи.
2. Тонические шейные рефлексы на мускулатуру конечностей вызывают долго длящееся различие тонуса конечностей обеих сторон, вследствие чего ноги оперированной стороны приобретают пониженный, а ноги противоположной стороны — повышенный разгибательный тонус. Исключая самые первые дни, когда могут проявить свое действие прямые последствия выпадения лабиринта (cp. II, 2) и влияния наклона головы (cp. III, В. 1), различие тонуса конечностей обусловливается исключительно этим рефлексом.
3. К этому различию тонуса может быть сведен целый ряд нарушений движения, как например отставление ног оперированной стороны при сидении, опрокидывание на бок и отклонение в сто¬
310
Шестая глава. Последствия односторонней экстирпации лабиринта
рону операции во время бега и прыжка и многие другие. Поэтому у животных с перерезанными задними корешками эти симптомы отсутствуют.
Б. Временные последствия.
1. Временный наклон шеи, если он сильно выражен, может непосредственно после операции противодействовать описанному различию тонуса и даже на время перекомпенсировать его.
2. Отклонение шеи посредством шейного установочного рефлекса влияет в том же направлении на поворот туловища. И то и другое в совокупности составляют причину быстро проходящих движений по часовой стрелке и манежных движений в направлении оперированной стороны.
В. Установление головы прямо снимает упомянутые последствия поворота шеи. Вследствие этого оказывается:
1. Поворот туловища уменьшенным.
2. Тонус конечностей (не считая самых первых дней) одинаковым с обеих сторон.
У животных с перерезанными задними шейными корешками в ответ на устанавливание головы прямо изменений не наступает.
IV. Компенсации.
A. При сидении на полу.
1. Компенсация благодаря установочному рефлексу с туловища на голову. Вследствие этого поворот головы уменьшается.
2. Компенсация благодаря установочному рефлексу с туловища на туловище. Этот рефлекс приводит к усаживанию туловища, несмотря на поворот головы. У животных с выключенными шейными рефлексами тело приводится в нормальное положение только благодаря этому рефлексу.
Б. Компенсация при помощи глаз. Насколько велико влияние оптических установочных рефлексов после односторонней экстирпации лабиринта, следует установить в более обстоятельном исследовании.
B. Компенсация благодаря центральным механизмам. Это относится главным образом к измененной иннервации глазной и шейной мускулатуры. Последняя вторично изменяет также и те последствия выпадения лабиринта, которые обусловлены различием тонуса конечностей. Компенсация наступает также после экстирпации больших полушарий, выключения оптических раздражений и перерезки трех пар верхних шейных задних корешков. Она обнаруживается после удаления второго лабиринта.
V. Сужение глазной щели, выступание мигательной перепонки и миоз на стороне операции обусловлены перерывом постганглинарных симпатических путей в среднем ухе и с разрушением лабиринта ничего общего не имеют.
IV. ОПЫТЫ НА СОБАКАХ
Описание последствий одностороннего выключения лабиринта у собаки приведено у Бехтерева (1) и Камиса (2). Бехтерев перерезал слуховой нерв, к которому он шел через чешую затылочной.кости. Камис также шел от planum occipitale затылочной кости (на basis apophysis jugularis) и разрушал отсюда полукружные каналы и в некоторых случаях также улитку. Оба автора наблюдали сразу после операции бурные явления, которые мы видели при экстирпации лабиринта через буллу.
Опыты на собаках
311
В общем, односторонняя экстирпация лабиринта была произведена де Клейном у десяти собак (15). Пять из этих животных были децеребри- рованы до или после экстирпации, три других находились под наблюдением в течение 42, 56 и 67 дней, у двух из них после этого был удален второй лабиринт. У экспериментальных собак симптомы развивались совершенно одинаково. По существу, динамика их развития тождественна таковой у кошки, только обратное развитие весьма выраженных в первое время явлений выпадения происходит еще быстрее, чем у кошки. У двух собак, у которых Дюссер де Баренн неполностью экстирпировал мозжечок, был удален сначала один, затем второй лабиринт. Здесь внимание было обращено главным образом на состояние установочных рефлексов.
Чтобы стала ясной общая последовательность событий, следует привести выдержки из типичного протокола.
Черно-серая собака. 13 декабря 1912 г. Под эфирным наркозом экстирпация правого лабиринта, вылущивание преддверия, обнажение и очищение устий полукружных каналов. Внутренний слуховой проход осторожно вскрыт долотом. 8-й нерв обойден пинцетом.
И час. (У2 часа после операции). Лежит в клетке, перекатывается вправо (еще наполовину под наркозом). Голова повернута вправо на 30°, левая передняя нога разогнута и отведена. Посаженная на пол, многократно опрокидывается в правую сторону. Очень сильная девиация глаз, левый глаз обращен вверх назально, правый глаз — вниз кнаружи. Очень сильный нистагм. В положении на спине, когда голова установлена прямо, тонус обеих правых ног меньше тонуса левых. После полного пробуждения •от наркоза перекатываний больше нет.
12 час. Перекатываний нет. Голова повернута на 30° вправо. Движения по и против часовой стрелки. Правые ноги разогнуты сильнее левых.
4 ч. 30 м. Отклонение глаз вправо. Нистагм влево, голова на 10° повернута вправо, может также отклоняться влево. Бежит по комнате неуверенно, широко расставляя ноги, иногда падает вправо. Намек на движение по часовой стрелке и на манежные движения вправо; однако в состоянии бегать в любых направлениях. Иногда пробегает несколько шагов в передне-боковом направлении. При стоянии несколько покачивается. Маятникообразных движений головы нет. В положении на спине, если голова •стоит прямо, различия тонуса правых и левых конечностей нет. В висячем положении головой вниз нижняя апертура грудной клетки не повернута совсем, зато голова повернута относительно таза на 90°.
14 декабря. Беспокойно бегает по комнате, все еще несколько расставляя ноги, при этом иногда спотыкается вправо и немного отклоняется в ту же сторону. Голова на 30° повернута вправо, может отклоняться в любую сторону. Если перед животным держать мясо, то оно не в состоянии подняться на задние ноги. Однако потом благополучно съедает мясо. Девиация глаз вправо, нистагм меньше, чем накануне. Соскакивая ■со стула, не падает, но несколько отклоняется вправо.
16 декабря. Незначительное отклонение глаз, нистагма нет. В состоянии осуществлять движения глаз во всех направлениях. Выскакивает из клетки, при этом ударяется брюхом об пол, но не падает. Бегает много лучше, но еще неуверенно. Иногда •оступается вправо, но не падает. Удовлетворительно становится На задние ноги.
17 декабря. Бегает уверенно, не спотыкаясь, все еще несколько расставляя ноги. Соскакивает со стола, при этом передние ноги подгибаются, но животное не падает, девиация глаз исчезла.
20 декабря. Голова отчетливо повернута вправо. Бег весьма уверенный; только при быстром беге иногда еще несколько оступается вправо. Соскакивает со стола, не падая, только передние ноги при этом несколько подгибаются. При попытке сбежать вниз по лестнице падает вправо и затем через голову вниз. Не в состоянии также и подняться вверх по лестнице.
14 января 1913 г. Весьма оживлена. Голова повернута на 30° вправо; может наклониться в любом направлении. Отклонения глаз нет. Стоит на задних лапах, не падая^; ■соскакивает со стола, бежать вниз по лестнице боится. В висячем положении головой вниз нижняя апертура грудной клетки повернута вправо на 20°, верхняя — на 30—45°, голова — на 90°.
21 января. Осторожно и медленно, но не падая, сбегает по лестнице вниз. Может бежать и вверх. Голова при стоянии повернута вправо на 45°; грудной отдел несколько перевешивает в правую сторону.
312
Шестая глава. Последствия односторонней экстирпации лабиринта
28 января. Хорошо соскакивает со стола, однако при этом покачивается вправо. Если держать перед животным мясо, кошка становится на задние ноги; при этом голова и грудной отдел винтообразно повернуты вправо (голова до 90°).
7 февраля. С уверенностью сбегает вниз по лестнице. Можно констатировать еще следующие явления выпадения: при стоянии голова на 30° повернута вправо; грудной отдел передней частью еще немного перевешивает вправо. В висячем положении головой вниз нижняя апертура груди повернута вправо на 20°, верхняя — на 30°, голова — на 80°. Левая передняя нога на следующий день разогнута отчетливо сильнее, чем правая.
12 февраля (56 дней после экстирпации правого лабиринта). Экстирпация левого лабиринта. После этого наклон головы влево, девиация глаз влево. Нистагм вправо. Никакого поворота грудного отдела.
У двух других собак протекание симптомов было в основном таким же, только движений перекатывания у них не наблюдалось.
Глазные симптомы
Глазные симптомы выражены у собаки отчетливее, чем у кошки. При этом оба глаза отклоняются в сторону отсутствующего лабиринта, а глаз на оперированной стороне — сильнее, чем на противоположной. Глаз оперированной стороны, кроме того, девиирует вниз, глаз противоположной — кверху. Уже на следующий день после операции отклонение глаз меньше, на 3—4-й день — исчезает полностью. Нистагм своим быстрым компонентом направлен в сторону, противоположную девиации глаз. Непосредственно после операции он выражен очень сильно, через 24 часа — значительно слабее, а через 2—3 дня исчезает совсем. Однажды во время исследования животного посчастливилось снова выявить отсутствовавший нистагм на четвертый день. Девиация глаз сильна, если голова находится в боковом положении интактным лабиринтом вниз, слаба, если он находится наверху. Нистагм изменяется в противоположном направлении.
На первый или второй день после операции животное по большей части уже в состоянии осуществлять активные движения глазными яблоками во всех направлениях. Спустя 3—4 дня во время обычного исследования вообще невозможно обнаружить никаких нарушений движений глаз.
В противоположность кролику или морской свинке и в соответствии с данными, полученными на кошке, девиация глаз у собаки не является, следовательно, стойким результатом односторонней лабиринтэктомии.
Положение головы, шеи и туловища
Непосредственно после операции голова повернута и наклонена в сторону утраченного лабиринта. В одних случаях преобладает поворот, в других — наклон. Отклонение головы не зависит от глаз.
Наклон головы вскоре после операции становится менее интенсивным, через несколько дней исчезает совсем и впоследствии наблюдается только иногда, особенно когда тело животного оказывается в ненормальных положениях. Вскоре после операции голова может также активно отклоняться в другую сторону.
Поворот головы в первые дни операции становится сильнее и далее остается без изменений. При стоянии он составляет от 20 до 45°. Через некоторое время собака может повернуть голову произвольно также и в другую сторону. Однако при спокойном стоянии и беге девиация головы обнаруживается снова.
К этому повороту головы присоединяется соответствующий поворот всего туловища. Последнее у стоящего животного проявляется в том,
Опыты на собаках
313
что грудь несколько перевешивает в сторону операции, что становится особенно отчетливым во время быстрого бега. Однако лучше исследовать это в висячем положении животного головой вниз. Поворот туловища в первые дни после операции еще мал, затем становится более выраженным и остается таким долгое время. Так, например, у собаки, протокол опытов на которой был приведен выше, в первые четыре дня после операции нижняя апертура грудной клетки вообще еще не была повернута по отношению к тазу, а поворот верхней апертуры колебался между 0 и 20°. По прошествии же первой недели, наоборот, нижняя апертура грудной клетки оказалась повернутой относительно таза на 20—30°, верхняя — на 30—45°, голова — на 70—90°; этот поворот удерживался в течение всего наблюдения (56 дней). У другой собаки в первый день при исследовании в висячем положении головой вниз никакого поворота груди по отношению к тазу вообще не было установлено; через 3 дня он был уже вполне выраженным и к 62-му дню составлял для нижней апертуры грудной клетки 20—30°, для верхней — 35—40°, для головы — 70—90°. В то время как у кошки поворот туловища обусловлен главным образом зависящим от интактного лабиринта основным поворотом, у собаки мы находим иное. В отношении к повороту своего туловища собака ведет себя аналогично морской свинке. Если установить голову собаки по отношению к груди прямо, то и грудь относительно таза устанавливается прямо. Следовательно, поворот туловища в основном обусловлен шейным рефлексом, а прямое влияние лабиринта весьма незначительно, если оно существует вообще. Доказать наличие его во всяком случае не удавалось.
Среди многочисленных проб лишь один раз у одной собаки как будто наблюдался поворот туловища после устанавливания головы прямо. На^ другой день у того же животного таз при прямо стоящей голове также стоял относительно груди прямо. Подобный же результат дало исследование поворота туловища в положении животного на спине. Он был отчетливым при некорригированном повороте головы и исчезал при устанавливании головы прямо.
Поворачивание головы вправо или влево в положении животного на спине, если оно не производило при этом энергичных защитных движений, приводило к тем же результатам, какие выше описаны для кролика. Поворот головы влево (левое ухо в вентральном направлении) приводил, например, к повороту таза левой ягодицей вниз или к появлению вогнутости позвоночника вправо. Поворот вправо приводил к зеркально противоположному результату. Наклон головы в положении животного на спине вызывал появление вогнутости позвоночника в ту сторону, в которую была направлена морда.
У собаки имеются очень сильные лабиринтные рефлексы на голову, которые, смотря по положению животного в пространстве, усиливают или ослабляют основной поворот. Закономерности при этом точно те же, что и для кролика или кошки, так что более точное описание излишне. При исследовании следует надевать животному шлемик, чтобы выключать оптические установочные рефлексы. Например, после правосторонней экстирпации лабиринта, как при левом, так и при правом боковом положениях в воздухе, голова находится в правом боковом положении.
Маятникообразные движения и нистагм головы наблюдались только у одной собаки, и то лишь непосредственно после пробуждения от наркоза.
В качестве длительного последствия операции у собаки выступает поворот в сторону операции. Последний благодаря тоническому шейному рефлексу имеет своим результатом поворот туловища в том же направлении. Основной поворот туловища, непосредственно зависящий от оставшегося
314 Шестая глава. Последствия односторонней экстирпации лабиринта
лабиринта, обнаружить не удалось. Как времённое явление после операции наблюдался наклон головы в сторону операции. Отклонение головы не зависит от глаз. Лабиринтные установочные рефлексы оказывают на установку головы то же влияние, что и у лишенных лабиринтов кошек и кроликов.
Тонус конечностей
У одной собаки уже тотчас после пробуждения от наркоза тонус конечностей с обеих сторон при установленной прямо голове одинаков (испытание в положении на спине). Таким образом, здесь даже с самого начала прямого влияния выпадения лабиринта на тонус конечностей не отмечалось. Во всех других случаях, как у нормальных, так и у децереб- рированных животных, при голове, установленной прямо, конечности на стороне операции обладали меньшим разгибательным тонусом, чем на другой стороне. Это различие продолжалось всегда недолго. В случае, более точно описанном выше, оно исчезло уже через 6 часов; только однажды оно существовало 24 часа. Прямое влияние выпадения лабиринта на тонус конечностей не постоянно и не длительно (как у кошки).
По прошествии некоторого времени различия тонуса на конечностях обеих сторон обусловливаются исключительно отклонейием головы. Уже Бехтерев дал весьма наглядные описания сильного разгибания и отведения контралатеральных и сгибания и расслабления гомолатеральных конечностей, которые ему удалось наблюдать в течение короткого времени после операции. Когда животные отдыхали настолько, что могли свободно бегать по комнате, различия тонуса в обычных условиях более не отмечалось; однако оно отчетливо выступало при положении животного на спине или в висячем положении.
Отклонение головы обычно сильно выражено только непосредственно после операции. Поэтому только в этой стадии оно может оказывать отчетливое влияние на тонус конечностей. Как выяснено выше (стр. 297), влияние это противоположно влиянию поворота головы и при известных обстоятельствах может преодолеть последнее. Это наблюдалось у собак в двух случаях, когда непосредственно после операции имел место значительный наклон головы и почти не наблюдалось поворота головы. Вследствие этого передняя нога оперированной стороны оказалась в состоянии сильного тонического разгибания, в то время как нога противоположной стороны была активно согнута. Устанавливание головы прямо тотчас приводило к исчезновению этого различия. Спустя несколько часов в одном случае голова была повернута сильнее и вследствие этого и нога противоположной стороны оказалась тонически разогнутой.
Таким образом, у собаки после односторонней экстирпации лабиринта различие тонуса конечностей обусловливается совокупностью следующих влияний: поворот шеи благодаря тоническому шейному рефлексу вызывает увеличение разгибательного тонуса на противоположной стороне и уменьшение его на стороне операции. Речь идет о длительном действии. В первые часы после операции к этому добавляются еще два влияния: во-первых, непостоянное действие выпадения лабиринта само по себе (действует в том же направлении); во-вторых, непостоянное действие наклона головы (последнее действует в направлении, противоположном только что упомянутому шейному рефлексу). В качестве окончательного результата оказывается, что лишь в небольшом числе случаев и на короткое время ноги на стороне операции оказываются разогнутыми сильнее, в то время как ноги на противоположной стороне обладают большим разгибательным тонусом.
Опыты на собаках
315
Компенсация
Б явлениях компенсации у собаки принимают участие прежде всего установочные. рефлексы тела. Из них весьма отчетливы установочные рефлексы с тела на голову. Они уменьшают при сидении и беге поворот головы. Их можно демонстрировать различным способом. Если, например, животное без правого лабиринта держать в шлемике в воздухе в правом боковом положении, то голова устанавливается в правом боковом положении или (при сильном основном повороте) в затылочном положении. Если же животное в правом боковом положении уложить на стол, голова «совсем или частично (благодаря повороту и наклону влево) приводится в нормальное положение. Если затем на сторону тела, обращенную кверху, поместить дощечку с грузом, голова опять возвращается в первоначальное положение.
Установочные рефлексы с туловища на туловище позволяют телу принять сидячее положение, когда голова повернута в боковое положение. .Я не располагаю достаточными сведениями относительно интенсивности их проявления у собаки после односторонней экстирпации лабиринта. Ъо всяком случае у нормальной собаки они настолько живые, что, вероятно, играют значительную роль в выравнивании лабиринтных нарушений.
Весьма живыми оказываются и оптические установочные рефлексы, участвующие в явлениях компенсации. Нередко можно наблюдать, если держать лишенную одного лабиринта собаку в шлемике в висячем положении головой вверх, что голова устанавливается в боковом положении. Как только шлемик снимается, голова тотчас же принимает полностью или частично нормальное положение. Соответствующие отношения можно установить и при других положениях тела: например, при боковом положении с интактным лабиринтом наверху или при нормальном положении в воздухе. При беге и прыганье животное также использует глаза в качестве корригирующего фактора.
Как говорилось, центральные компенсации открыты у собаки Бехтеревым. У одной из двух наблюдавшихся нами собак, у которой через 56 дней после удаления одного лабиринта удалялся и другой, сразу после •операции появились отклонения головы, глаз и нистагм в направлении, противоположном направлению их после первой операции. У другой собаки интервал между двумя операциями был 67 дней. После второй операции наблюдались девиация глаз, нистагм и небольшой поворот головы в противоположном направлении. Без сомнения, и эта центральная компенсация принимает участие в быстром выравнивании нарушений у собаки.
Положение тела и движения
Аномалии установки и движения после односторонней экстирпации лабиринта у собаки очень похожи на таковые, подробно описанные в предыдущем отделе для кошки. Только выравнивание нарушений происходит еще более быстро и полно.
Движения перекатывания наблюдались только один раз, когда животное неполностью пробудилось от наркоза, во всех остальных случаях их не было. Аномалии установки головы, туловища и конечностей тотчас после операции уже были описаны. Когда собака после операции поднимается на ноги и начинает бегать по комнате (что может иметь место уже через час), она стоит, широко расставив ноги и покачиваясь, иногда падает в сторону утраченного лабиринта и во время бега несколько отклоняется в сторону. Движения по часовой стрелке и манежные движения были
316
Шестая глава. Последствия односторонней экстирпации лабиринта
выражены заметно лишь у одного животного в первые часы после операции, пока наклон головы и изгибание туловища были еще отчетливыми.
Боковое отклонение во время бега, так бросающееся в глаза у кошек, у собак в первый день после операции обычно еще заметно, на второй же день уже не наблюдается. Затем все менее выраженным становится широкое расставление ног при стоянии и исчезает покачивание.
При беге в течение первых дней собака еще нередко спотыкается,, опускаясь в сторону операции. Но уже через 4—6 дней это наблюдается значительно реже и преимущественно при быстром беге. Позднее это- совершенно исчезает. Опрокидывания в сторону операции (и во время бега) уже в день операции обычно не бывает. Если прибавить к этому, что спустя34—1 неделю собаки оказываются в состоянии стоять и «танцевать» на задних лапах, то становится очевидным, что все те нарушения при беги и стоянии, которые, как было показано выше у кошки, обусловлены-главным образом слабостью ног оперированной стороны, у собаки быстро исчезают. Другими словами, различие тонуса конечностей у собаки в висячем положении или в положении на спине, т. е. при ненормальных положениях тела, в продолжение еще многих месяцев при стоянии и беге недостаточно интенсивно для того, чтобы привести к появлению сильно выраженных нарушений движения. Ноги на оперированной стороне также не настолько бедны тонусом, чтобы не быть в состоянии выдерживать вес тела.
С этим согласуются и наблюдения над тем, как прыгает животное. Одна собака уже в первый день после удаления правого лабиринта удовлетворительно соскакивала со стула и уклонялась при этом несколько в сторону, на третий день она выскакивала из клетки, на четвертый — спрыгивала со стола, причем но? и ее слегка подгибались. Через 14 дней она без всяких нарушений соскакивала со стола и только потом немного покачивалась в сторону операции. Другая собака через 10 дней осторожно, но удовлетворительно соскакивала со стола на пол. Наибольшие трудности встречали собаки при беге по лестнице. У одной собаки прошел 21 день, у другой — 10 дней, прежде чем она снова выучилась хорошо это делать. Как и в наблюдениях Эвальда, собаки, если они во время этих упражнений однажды упали и сильно ушиблись, в дальнейшем обычно не расположены повторять эти процедуры. Это особенно отчетливо выявляется во время прыганья н бега по лестнице.
Через несколько месяцев, кроме поворота головы и, может быть, незначительного перевешивания грудного отдела в сторону операции, простое наблюдение над животными уже не позволяет установить никаких аномалий. Животных теперь приходится приводить в висячее положение головой вниз, чтобы по сильному повороту всего туловища и в некоторых случаях по разгибанию контралатеральной конечности убедиться, что дело идет об утрате лабиринта. То, что во время стояния и бега отклонения значительно уменьшаются, вполне можно объяснить особо деятельными установочными рефлексами.
Заключение
Симптомы, появляющиеся после односторонней экстирпации лабиринта у собаки, в наиболее существенных пунктах соответствуют таковым у кошки. Главнейшие различия следующие.
1. Обусловленного деятельностью оставшегося лабиринта основного поворота туловища выявить не удается. Имеющий место поворот туло-
Опыты на обезьянах
317
вища обусловлен одним шейным установочным рефлексом благодаря повороту шеи.
2. Расслабление конечностей оперированной стороны, обусловленное односторонней утратой лабиринта, непостоянно, и, если оно имеет место, то в течение нескольких часов проходит.
3. Выравнивание нарушений происходит у собаки особенно быстро. В этом принимают весьма заметное участие, помимо центральных изменений иннервации и установочных рефлексов тела, также и оптические установочные рефлексы.
У. ОПЫТЫ НА ОБЕЗЬЯНАХ (59)
Влияние односторонней экстирпации лабиринта на поведение обезьян изучалось на двух макаках, у которых де Клейном был удален через сосцевидный отросток левый лабиринт. Одно животное наблюдалось после этого в продолжение 22, другое — 38 дней. Затем у обеих обезьян был удален второй лабиринт.
В качестве примера общего поведения животных приведен сокращенный протокол опытов на обезьяне, у которой выявились более отчетливые явления, тогда как у другого животного, оперированного более осторожно, эти симптомы были выражены в значительно меньшей степени.
28 февраля 1921 г. обезьяна (Macacus rhesus) после пробуждения от наркоза была уложена на пол и перекатывалась, производя сильные движения бега в левую сторону (сторона операции), причем она продвигалась вперед в направлении своей головы, будучи в состоянии перейти из левого бокового положения в сидячее (чередование движений перекатывания и нормальных установочных рефлексов на голову). У обезьян, так же как это было показано выше для кролика, движения перекатывания наступают только тогда, когда животное делает попытку бежать. При усаживании и при движениях расслабленность левых конечностей, а также разгибание и отведение правых весьма отчетливы. Если голову установить по отношению к телу прямо, грудь при сидении на земле стоит прямо, но левая передняя нога несколько более расслаблена, чем правая. При повороте головы в левую сторону различие тонуса ног очень отчетливое. У посаженного в клетку животного появляются крайне интенсивные движения перекатывания, а затем — установочные рефлексы с тела на голову с усаживанием и сильным качанием головы. При этом обезьяна долгое время стукается головой о стеклянную стену клетки, а поэтому помещается в маленькую клетку. Здесь она сидит, широко расставив ноги и качаясь; голова наклонена и повернута примерно на 45°.
На следующее утро обезьяна сидит в клетке спокойно, голова сильно повернута, наклон ее меньше. Иногда на короткое время голова может оказаться в нормальном положении — при фиксации предметов глазами. Обезьяна сидит, широко расставив передние ноги, в состоянии также самостоятельно сидеть и на задних ногах, после чего однократно перекатывается влево, далее садится и снова многократно перекатывается в левую сторону. При выскакивании из клетки с шумом падает на пол и затем долгое время перекатывается по комнате, поочередно то катясь, то усаживаясь. Может сделать несколько прыжков по комнате, но при этом падает влево. Во время бега тело сильно клонится влево. Вскакивает на доску высотой около 40 см, но падает вниз и влево; влезает после этого, не падая, на крышу клетки высотой около 2 м; может также сделать прыжок в длину около 1 м. На решетчатой крыше клетки еще неловка, но не падает, при сидении покачивается, оступается, но сейчас же словно удерживается за брусья крыши. Может удовлетворительно сидеть в большой клетке, снизу забирается на решетчатую крышу, может пить из миски, причем сначала погружает туда всю голову, но после пьет спокойно. Спустя немного времени поедает в клетке репу, при этом сидит нормально при наличии отчетливого покачивания верхней части тела.
На следующий день уже в состоянии с ловкостью лазать, лишь один раз во время прыжка падает влево. Бежит по прямой. При сидении тело перевешивает в левую сторону. При улучшении движений передвигается вперед очень быстро. Уже через 6 дней вполне проворна. Примерно через месяц большая часть движений совершается безупречно, а к концу периода наблюдения, по-видимому, совершенно нормально; правда, служитель сообщает, что как исключение в некоторых случаях прыжок совершает неточно и падает влево.
318
Шестая глава. Последствия односторонней экстирпации лабиринта
У другой обезьяны через 14 дней после операции в отдельных случаях наблюдалось падение влево. Однако животное с большой легкостью забиралось на крышу клетки, через 3 дня со стола перепрыгивало в клетку, не падая при этом.
В частности, установлено следующее.
Глазные симптомы
У одного животного вскоре после пробуждения от наркоза в течение- недолгого времени имел место нистагм глаз влево (симптом раздражения), который скоро исчез. У другого животного тоже сразу же наступило отчетливое отклонение глаз влево с очень сильным нистагмом вправо. Нистагм строго горизонтальный. Отклонение глаз и нистагм имели место еще на следующий день, исчезнув только через 2—3 дня. С этого момента глаза стоят нормально и могут двигаться в любом направлении.
У одного животного левый глаз на второй и третий день после операции как будто* западал, т. е. отклонялся вертикально книзу. Впрочем, судить об этом нелегко, так как вследствие левостороннего паралича лицевого нерва обе глазные щели неодинаковы. У другой обезьяны подобного вертикального отклонения установить не удалось. Поэтому сказать, что это явление наступает как правило, нельзя.
Таким образом, девиация глаз у обезьяны, как у собаки и кошки, является быстро проходящим симптомом. Это соответствует сделанному раньше утверждению, что у этих животных компенсаторные установки глаз по сравнению с оптической установкой играют меньшую роль.
Положение головы, шеи и туловища
Благодаря тоническим влияниям на мышцы шеи, исходящим из оставшегося лабиринта, наступают основной поворот и наклон головы в сторону операции. На это накладывается лабиринтный установочный рефлекс, который стремится привести голову в боковое положение с интактным лабиринтом наверху. Затем наступает следующее.
При нормальном положении в воздухе и при открытых глазах голова или наклонялась в сторону отсутствующего лабиринта, или поворачивалась таким образом,, что ухо с отсутствующим лабиринтом оказывалось направленным вентрально. В большинстве случаев голова вследствие оптических установочных рефлексов удерживалась более или менее прямо. В боковом положении животного благодаря различным: комбинациям основного поворота и лабиринтных установочных рефлексов наблюдаются несимметричные положения ^обезьяны. После левосторонней экстирпация лабиринта при правом боковом положении животного в воздухе имеет место суммация основного поворота и лабиринтного установочного рефлекса, благодаря чему голова поворачивается из бокового в нормальное положение. То же можно было показать и на обеих обезьянах после закрывания им глаз. При левом боковом положении в воздухе, наоборот, основной поворот и лабиринтный установочный рефлекс действуют в противоположных направлениях и вследствие этого голова при закрытых глазах удерживается в левом боковом положении, т. е. не поворачивается. Эти: взаимоотношения отчетливо сохраняются также и через месяц после операции. При открытых глазах нередко имеет место поворачивание головы в направлении к нормальному и из левого бокового положения. Последнее, однако, всегда только временно. При прямом положении тела головой вверх последняя при открытых глазах не остается, повернутой и более или менее сильно наклоненной к плечу. При висячем положении головой вниз в большинстве случаев можно установить, что основной поворот, как это наблюдалось у кроликов и кошек, с течением времени увеличивается, так чт^> по истечении 2--3 недель составляет 70—90°. Поскольку в результате левосторонней утраты лабиринта голова вследствие основного поворота в общем имеет тенденцию* отклониться к левому плечу, то в висячем положении головой вниз последняя пришла бы в результате этого в правое боковое положение. Однако это боковое положение*
Опыты нй обезьянах
319
как раз обратно тому, в которое стремится привести голову лабиринтный установочный рефлекс. В итоге — борьба, в которой у обезьян нередко побеждает лабиринтный установочный рефлекс, поэтому в висячем положении головой вниз допускается поворот в одном направлении, наклон же головы (наклон к плечу) — в противоположном, так что голова находится в воздухе в левом боковом положении. При положении на спине в воздухе прежде всего идет борьба между основным поворотом и лабиринтным установочным рефлексом. Из исследований на кроликах выяснилось, что существует положение, в котором оба рефлекса действуют в одном направлении. Это положение, когда голова животного вследствие вентрального пригибания оказывается на брюхе животного, поэтому сторона отсутствующего лабиринта обращена вниз. Это своеобразное вынужденное положение, которое на рис. 141 приведено для кролика, в отдельных случаях принимает и обезьяна. В других случаях основной поворот приводит к правому боковому положению головы в пространстве. Оба положения, очевидно, неудобны для животного и приводят к беспокойству, верчению головой и т. п. Если вследствие какого-либо влияния присоединяется еще и оптический рефлекс, голова временно благодаря вентральному прогибанию может оказаться приведенной в нормальное положение.
У одного животного в день операции выявился отчетливый нистагм головы вправо, который на следующий день исчез. Наклон головы влево, т. е. тенденция головы приблизиться к левому плечу, у одного животного был весьма выражен в течение долгого времени, так что почти через месяц угол наклона составлял еще 45°. У другого он с самого начала был очень невелик, так как в свободном сидячем положении корригировался установочными рефлексами с тела на голову и оптическими установочными рефлексами.
Поворота туловища как прямого следствия выпадения лабиринта у обезьян не наблюдается. Если животное подвесить в вертикальном положении головой вниз и полностью корригировать поворот его головы, то грудь устанавливается тоже совершенно симметрично. Если отпустить голову так, что поворот появляется снова, грудь следует за головой; речь идет здесь о рефлексе, начинающемся с шеи. Действительно, у каждой нормальной обезьяны поворачиванием шеи можно вызвать поворот всего тела (включая хвост). Отсюда следует, что компенсирующие влияния, уменьшающие поворот головы, приводят также и к уменьшению поворота туловища.
Тонус конечностей
У обоих животных весьма отчетливо можно было показать одностороннюю утрату тонуса конечностей на стороне операции как прямое последствие выпадения лабиринта. Для этой цели животное в положении на спине удерживается в воздухе, а голову его устанавливают по отношению к телу прямо. У одного животного левая передняя нога была заметно более расслаблена и оставалась таковой примерно в течение месяца, тогда как на задних ногах различие было менее заметно. У другого животного это различие было весьма отчетливым на передних ногах на протяжении 38 дней и в последний день наблюдения было сильно выражено также и в наркозе. У этого животного можно было показать соответствующее различие и на задних ногах; последнее через 10 дней стало непостоянным, однако проявлялось время от времени, а на 38-й день было под наркозом еще весьма отчетливым.
Если экспериментатор намеренно не установит голову прямо относительно туловища, а позволит ей принять естественное положение, наступающее после экстирпации лабиринта, то вследствие тонического шейного рефлекса различие тонуса конечностей становится еще сильнее, так что при повернутой голове оно намного сильнее, чем тогда, когда голову устанавливают по отношению к туловищу прямо.
320 Шестая глава. Последствия односторонней экстирпации лабиринта
Явления компенсации
Бросается в глаза, насколько быстро и совершенно выравниваются у обезьян нарушения, наступающие после односторонней утраты лабиринта. Здесь принимают участие следующие механизмы.
1. Установочный рефлекс с тела на голову. Этот рефлекс развит у обезьян относительно слабо. Тем не менее иногда можно отметить его действие. Например, если обезьяну после левосторонней экстирпации лабиринта удерживать в воздухе в левом боковом положении, то голова (вследствие противоположного действия основного поворота и лабиринтного установочного рефлекса) оказывается в левом боковом положении. Если затем животное уложить на стол, голова повернется по направлению к нормальному положению (в первый день после операции), а спустя немного дней даже полностью оказывается в нормальном положении. (Правда, в этих опытах оптические рефлексы не выключались. Поскольку оптические раздражения действуют в воздухе и на столе одинаково интенсивно, наблюдение все же не теряет значения).
2. Установочные рефлексы с туловища на туловище. Эти рефлексы у обезьян развиты очень сильно и играют в процессе компенсации очень большую роль. Их постепенное усиление можно видеть из следующего наблюдения.
Левосторонняя экстирпация лабиринта вызывает поворот головы, вследствие этого получается, что в первые дни после операции усаживание при правом боковом положении удается лучше, чем при левом, потому что именно поворот головы препятствует усаживанию при одном боковом положении и помогает при другом. Однако уже через два дня это различие становится меньше и приблизительно после 11 дней полностью исчезает.
Установочные рефлексы с тела у обезьян осуществляются не только при соприкосновении туловища с полом, но и прежде всего рук. Отсюда проистекает то, что очень скоро после операции животные снова обнаруживают большую уверенность при лазанье.
3. Оптические установочные рефлексы. Эти рефлексы у обезьян выражены очень сильно и в широкой степени используются при коррекции положения головы. Благодаря их действию обезьяна, удерживаемая в воздухе в нормальном положении, часто держит голову совершенно прямо и, находясь в воздухе в левом боковом положении, поворачивает голову по направлению к нормальному положению (после левосторонней экстирпации лабиринта). При других позах и при спокойном сидении также можно установить, что, как только внимание животного возбуждено и какой-нибудь предмет фиксируется"глазами, голова в большей или меньшей степени поворачивается по направлению к нормальной установке, но снова оказывается в исходном положении, как только фиксация зрением прекращается.
4. Изменения центральной иннервации. Через 22—38 дней экстирпировался оставшийся лабиринт. После пробуждения от наркоза имело место отчетливое отклонение глаз вправо с очень сильным нистагмом влево.
У одного животного нистагм исчез уже во второй прловине дня, у другого — на следующий день еще имелся слабый нистагм влево, в то время как установка и движения глаз снова стали вполне нормальными. Голова непосредственно после операции также оказалась наклоненной вправо; у одного животного это отмечалось еще и во второй половине дня, у другого — отсутствовало уже на другой день. В висячем положении головою
Общий итог
321
вниз наклон головы вправо был выражен в день операции. У одного животного имел также место нистагм головы в левую сторону.
Различие тонуса конечностей также исчезало. У одного животного при пробуждении от наркоза правая нога была расслаблена более отчетливо, чем левая. Различие удерживалось до второй половины дня. У другого животного в день операции никакой разницы еще не наблюдалось, но со второго до пятого дня она отчетливо проявилась на передних ногах, а на пятый день — на задних. Позднее, наоборот, конечности правой стороны не были более расслаблены по сравнению с левыми.
Отсюда следует, что и у обезьян наступает описанная Бехтеревым центральная компенсация как со стороны глаз, так и со стороны головы (и конечностей?).1
Эти компенсации частью осуществляются корой больших полушарий, частью — стволом мозга. Кортикальными являются оптические установочные рефлексы, в то время как установочные рефлексы тела локализованы в среднем мозгу. После экспериментов на кошках следует признать вероятным предположение, что у обезьян центральная бехтеревская компенсация осуществляется субкортикально.
Заключение
В целом оказывается, что последствия утраты лабиринта у обезьян проявляются примерно так же, как и у собаки, только односторонняя потеря тонуса конечностей как прямое последствие выпадения лабиринта продолжается у обезьяны значительно дольше. Компенсация весьма совершенна и наступает быстро. При этом принимают участие прежде всего установочные рефлексы тела на тело и оптические установочные рефлексы.
Обезьяна без одного лабиринта отличается от кошки, помимо прочего, еще и отсутствием основного поворота туловища, так что поворот туловища осуществляется одними шейными установочными рефлексами.
VI. ОБЩИЙ ИТОГ
Изучение последствий односторонней экстирпации лабиринта у пяти различных видов теплокровных показало, что картина симптомов у всех животных в принципе одна и та же. Однако результат экстирпации у каждого животного, несмотря на это, различен.
Сначала обсудим положения, общие для всех животных.
Во всех случаях из сложного симптомокомплекса можно выделить следующие.
а) Прямые последствия удаления одного лабиринта, т. е. односторонние рефлексы, исходящие из интактного лабиринта. Среди них самый постоянный и самый богатый следствиями симптом — поворот головы.
б) Вторичные последствия поворота головы.
в) Симметричные, нелабиринтные рефлексы, обнаруживающиеся в той степени, в какой они воздействуют на повернутое тело.
г) Явления раздражения вследствие операции.
Последствия выпадения лабиринта у различных животных неодинаковы. Шейные же рефлексы, посредством которых вызываются вторичные
1 Последнее с уверенностью можно решить только после того, как будет выяснена причина одностороннего расслабления мускулатуры конечностей после удаления лабиринта.
21 Р. Магнус
322
Шестая глава. Последствия односторонней экстирпации лабиринта
последствия поворота головы, обнаруживают большее сходство. При их изучении оказалось следующее.
Выключение шейных рефлексов уменьшает последствия одностороннего удаления лабиринта. Это установлено прежде всего на кошках, у которых перерезались три пары верхних шейных задних корешков. В любое время у всех животных можно временно снять шейные рефлексы, установив голову по отношению к туловищу прямо. При этом из асимметричных влияний остаются только те, природа которых чисто лабиринтная. Лишь благодаря этому приему правильный анализ последствий односторонней утраты лабиринта и сделался возможным.
Одним из важных свойств шейных рефлексов является их длительность. Они держатся, не уменьшаясь в величине, в течение недель и месяцев. Это подтверждают опыты на кролике, который спустя много месяцев корригировал аномалии установки ног, обусловленные шейными рефлексами, но после закрывания ему глаз тотчас же принимал исходное вынужденное положение, показывая тем самым, что импульсы со стороны шеи продолжают действовать, не ослабляясь. Наблюдения над морскими свинками, собаками и обезьянами, у которых лабиринтный основной поворот туловища отсутствует, показали, что при исследовании в висячем положении головой вниз у этих животных и по прошествии месяцев влияние поворота шеи на мускулатуру туловища остается неослабленным. Хорошо и давно известно, что проприоцептивные импульсы, которые, по Бронд- гисту (Brondgeest), играют значительную роль в обеспечении мышечного тонуса, существуют в течение всей жизни. Однако тот факт, что в результате афферентных возбуждений, которые появляются благодаря определенному положению части тела, могут возникать анормальные положения на такие длительные сроки, поразителен. Значение этих данных для физиологии и патологии установки тела налицо. Речь идет о тонических рефлексах практически бесконечной длительности, которые можно рассматривать как неистощающиеся.
То, что в отношении шейных рефлексов для всех пяти исследованных видов животных речь идет, в сущности, об одних и тех же влияниях, в той мере, в какой эти влияния сказываются на последствиях односторонней экстирпации лабиринта, видно из таблицы I.
Таблица I
Кролик
Морская
свинка
Котка
Собака
Обезьяна
Поворот голо¬
Весьма
Весьма
Действует.
Весьма
Весьма
вы действует на туловище (шейный установочный рефлекс).
Поворот голо¬
отчетливо.
отчетливо.
отчетливо.
отчетливо.
Весьма
Весьма
Весьма
Отчетливо.
Отчетливо.
вы действует на конечно¬
отчетливо.
отчетливо
(особенно
отчетливо.
сти.
передние
ноги).
■Наклон головы
Мало.
Мало.
Мало.
Мало.
Мало.
действует на
конечности.
Влияние этих рефлексов на мышечный тонус у всех исследованных до сих пор животных идентично. В ответ на поворот и наклон головы раз- гибательный тонус повышается на «челюстной» ноге и уменьшается —
Общий итог
323
на «черепной». Тонус сгибательных мышц изменяется в обратном направлении: он повышается на «черепных» ногах и понижается на «челюстных». На мышцах плеча, локтя, бедра и колена эти изменения выражены сильнее, чем на сочленениях ступни. Кроме того, грудной и поясничный отделы позвоночника при повороте и наклоне головы в «челюстную» сторону становятся вогнутыми или поворачиваются таким образом, что туловище продолжает поворот шеи. Установочные шейные рефлексы, которые у нормального животного должны придавать телу симметричное положение, после утраты лабиринта приводят посредством вынужденного положения шеи также к повороту или изгибанию туловища.
Остальные шейные рефлексы, например влияние на тонус конечностей поднимания и опускания головы, у односторонне лабиринтэктомированных животных также имеют место и могут быть вызваны у них в любое время. Однако они не являются характерными симптомами односторонней утраты лабиринта.
Значение шейных рефлексов, вызванных анормальным стоянием головы (и у всех исследованных животных совершенно однотипных), для установки тела и моторики после удаления одного лабиринта у различных видов животных весьма и весьма различно. Стоит только сравнить кролика и собаку спустя несколько недель после операции. Кролик сидит в вынужденном положении с отведенными контралатеральными передними ногами, собака же почти во всех случаях бегает подобно нормальному животному, и требуется внимательное исследование, чтобы показать, что упомянутые рефлексы осуществляются и в этом случае. Нейтрализация шейных рефлексов путем установления головы прямо изменяет эту картину у кролика, словно по волшебству, и позволяет животному сидеть почти совершенно подобно нормальному, тогда как собака, двигавшаяся почти нормально и до подобной манипуляции, после нее едва ли сильно улучшает свои движения. Собака, несмотря на имеющуюся разницу тонуса и поворот туловища, может бегать, прыгать, взбегать по лестнице и т. д.; кролик компенсирует только часть нарушений, притом очень медленно и несовершенно.
Так как после удаления лабиринта поворот головы делается стойким, а наклон головы, по крайней мере у морской свинки, кошки и собаки, наоборот, быстро уменьшается, становится понятным,что шейные рефлексы, включенные наклоном головы, у только что названных животных бывают отчетливыми лишь на время, тогда как шейные рефлексы, вызванные поворотом, являются длительными последствиями операции.
Те нарушения, которые отмечены после устанавливания головы по отношению к шее прямо, должны в большей части рассматриваться как непосредственные результаты удаления лабиринта. Среди них у каждого исследованного животного можно различить две группы. Одна часть их представляет собой длительные последствия операции, которые не подвергаются обратному развитию. Другая часть появляется лишь на время и спустя часы, дни или недели уже не обнаруживается (табл. II, стр. 324 и 325).
При обсуждении непосредственных результатов выпадения лабиринта следует уточнить, насколько выявляются у оперированных животных известные до сих пор тонические рефлексы, исходящие из интактного лабиринта, и, наоборот, каким образом наблюдающиеся у животных симптомы могут быть сведены к известным лабиринтным рефлексам.
1. Тонические лабиринтные рефлексы на мышцы конечностей. Так как рефлексы из одного лабиринта воздействуют с одинаковой интенсивностью на конечности обеих сторон, после односторонней экстирпации лабиринта асимметричного влияния осуществляться не может.
21*
324
Шестая глава. Последствия односторонней экстирпации лабиринта
Прямые последствия выпадения л а
Эффекторный
орган
Рефлекс
Кролик
Шея.
Тонические лабиринтные рефлексы на мышцы шеи (максимум, когда тело обращено вниз).
Лабиринтный установочный рефлекс (минимум, когда интактный лабиринт при боковом положении головы обращен кверху).
Основной поворот, оперированная сторона вентрально. Наклон в оперированную сторону.
Голова по возможности в боковом положении. Интактный лабиринт наверху.
Туловище.
Тонические лабиринтные рефлексы на мышцы туловища.
Основной поворот.
Ноги.
Тонические ^ лабиринтные рефлексы на мышцы конечностей (один лабиринт действует на обе стороны тела одинаково).
о
?
Расслабление ног на стороне операции (8 недель).
Глаза.
Тонические лабиринтные рефлексы на глаза.
Ротаторный.
о
Вертикально(максимум, когда интактный лабиринт при боковом положении головы обращен вниз).
Глаз интактной стороны вверх; оперированной — вниз.
Девиация (с нервов полукружных каналов??).
Горизонтально.
Непостоянно.
Нистагм.
Противоположно девиации (иногда с горизонтальным компонентом).
Примечание. Стойкие последствия напечатаны курсивом, временные —
2. Тонические лабиринтные рефлексы на мышцы шеи. Они односторонние. Результатом их действия является основной поворот шеи, который имеет место у всех исследованных животных (ухо оперированной стороны поворачивается вентрально). Этот поворот — длительное следствие. У кролика удалось показать, что с течением времени он становится сильнее, точно так, как это видел Эвальд у голубей.
У животных всех видов вскоре после операции можно видеть наклон головы в сторону операции. Последнее у кроликов и обезьян, по крайней мере в анормальных положениях тела, наблюдается более длительное время,
Общий итог
325
Таблица II
биржнта у разных видов животных
Морская свинка
Кошка
Собака
Обезьяна
Основной поворот, оперированная сторона вентрально. Наклон в сторону операции.
Основной поворот. Оперированная сторона вентрально. Наклон в сторону операции.
Основной поворот. Оперированная сторона вентрально. Наклон в сторону операции.
Основной поворот, оперированная сторона вентрально. Наклон в сторону операции.
Голова по возможности в боковом положении, интактный лабиринт наверху.
(Непостоянно, весьма короткое время).
Основной поворот.
о
о
о
О
О
О
Расслабление ног на стороне операции (несколько дней; после кокаина исчезает).
Расслабление ног на стороне операции (непостоянно, на следующий день исчезает).
Расслабление ног на стороне операции (непостоянно и только на несколько часов).
Расслабление ног на стороне операции (более месяца).
о
Верхний край роговицы с обеих сторон в сторону операции.i
?
?
Г лаз интактной стороны вверх, оперированной — вниз.
Глаз интактной стороны вверх, оперированной — вниз.
Глаз интактной стороны вверх, оперированной— вниз.
?
Непостоянно.
В сторону операции.
В сторону операции.
В сторону операции.
Противоположно девиации (иногда с горизонтальным компонентом).
Противоположно девиации.
Противоположно
девиации.
Противоположно
девиации.
обычным шрифтом.
быстро проходит у морской свинки, кошки и собаки; при этом осуществляют свое действие частью лабиринтные установочные рефлексы, частью (при сидении на полу) — установочные рефлексы с тела на голову, а у обезьян также оптические установочные рефлексы.
Могло бы оказаться возможным, что при повороте и наклоне головы речь идет совсем не о двух реакциях шейных мышц, отличающихся одна от другой по существу, 11 Зависимость от тонических лабиринтных рефлексов на глаза неопределенная.
326 Шестая глава. Последствия односторонней экстирпации лабиринта
а о том, что одно и то же различие тонуса шейных мышц в зависимости от общего тонуса шейных мышц делает голову в одном случае более повернутой, в другом — более наклоненной. Однако возможным представляется и иное положение, а именно: более интенсивное и длительное влияние на мышцы атланто-эпистрофейного сочленения, чем на мышцы остальных шейных суставов. Ответить на это было бы возможно только после постановки сложных и детальных исследований на весьма сложно расположенных шейных мышцах. До сих пор такие исследования не начаты. Еще одно возможное предположение заключается в том, что наклон шеи осуществляется из других отделов лабиринта (полукружные каналы), чем поворот. К этому мы вернемся еще раз в девятой главе.
3. Лабиринтные установочные рефлексы на голову. Эти рефлексы также имеют место у всех исследованных животных. Они действуют в направлении, стремящемся привести голову в то боковое положение, при котором интактный лабиринт оказывается наверху. В зависимости от положения животного в пространстве происходит или усиление, или уменьшение основного поворота. Подробности детально описаны для кролика; то же относится и к остальным животным. Дело идет о длительном последствии операции.
4. Тонические лабиринтные рефлексы на мышцы тела. Эти рефлексы односторонние. Результат их действия — основной поворот туловища, который остается и после устанавливания головы по отношению к грудной клетке прямо. Он может быть обнаружен у кролика или у кошки, у морской свинки слаб, непостоянен и существует только весьма короткое время. У собаки и обезьяны его вообще не удалось обнаружить. Таким образом, у собаки, обезьяны и морской свинки отчетливо выраженный поворот туловища обусловлен исключительно (resp. преимущественно) шейным установочным рефлексом, у кошки — преимущественно тоническим лабиринтным рефлексом, у кролика — равномерным действием обеих групп рефлексов.
5. Компенсаторные установки глаз. При изучении отклонения глаз у кролика и морской свинки, с одной стороны, у кошки, собаки и обезьяны — с другой, обнаруживаются большие различия.
а) У кроликов и морских свинок оба лабиринта, если они интактны, вызывают повороты обоих глаз в одном направлении. Удаление одного лабиринта, таким образом, не вызывает ротаторного отведения глаз вследствие тонического лабиринтного рефлекса.
Наоборот, интактный лабиринт вызывает длительное вертикальное отклонение обоих глаз. Гомолатеральный глаз отклоняется дорсально, контралатеральный (т. е. глаз оперированной стороны) — вентрально. Эта девиация глаз максимальна, если при боковом положении головы интактный лабиринт находится внизу. Она нулевая или минимальная при противоположном боковом положении Речь идет о стойком результате операции.
Тонические шейные рефлексы (по крайней мере при поворотах шеи, не вызванных искусственно) не оказывают существенного влияния на девиацию глаз.
Вскоре после операции временно наступает нистагм, направленный или вертикально, в сторону, противоположную описанной выше девиации, или более горизонтально, в направлении интактного лабиринта. Последнее заставляет допустить возможность, что причиной этого является оперативное выключение полукружных каналов. За это говорит также то наблюдение, что после выключения при помощи кокаина одного лабиринта у свинки нистагм наступает всегда позже девиации. К этому мы также еще вернемся в девятой главе.
б) У кошек, собак и обезьян длительных отклонений глаз не наступает. Поэтому нельзя с такой же ясностью решить, в какой степени после одно¬
Общий итог
327
сторонней экстирпации лабиринта у этих животных оказывается существенным влияние тонических лабиринтных рефлексов на установку глаз.
У всех трех видов животных прежде всего наблюдаются временная горизонтальная девиация глаз в сторону отсутствующего лабиринта и нистагм в противоположную сторону. Тонические лабиринтные рефлексы не являются причиной этого изменения. Тонические шейные рефлексы на глаза, которые (рефлексы) вызываются поворотом шеи (обезьяна) или соответственно сгибанием ее (кошка, собака), должны были бы действовать в обратном направлении. Таким образом, представляется, что дело идет о последствиях односторонней перерезки нервов полукружных каналов.
Помимо этого, у кошки и собаки глаз на стороне интактного лабиринта отклонен кверху, а глаз на оперированной стороне — вниз. Эта девиация достигает максимума, когда интактный лабиринт при боковом положении головы находится внизу (в этом положении нистагм минимален). Возможно, речь идет здесь о тонических лабиринтных рефлексах на глаза, исходящих из интактного лабиринта. В этом случае следовало бы принять, что через несколько дней девиация компенсируется или оптически, или благодаря изменениям центральной иннервации (Бехтерев).
Относительно возникновения ротаторной девиации (у кошки) данных еще недостаточно.
в) Бурный временный нистагм у всех исследованных видов животных и преходящая девиация глаз у кошки, собаки и обезьяны послужили основанием предположить, что речь идет в данном случае о временном симптоме раздражения, обусловленном операцией; тем более что нистагм оказывается тем сильнее, чем грубее произведена операция. В противовес этому следует подчеркнуть, что направление девиации и нистагма соответствуют выпадению функции удаленного лабиринта. Лишь как исключение в течение недолгого времени после операции наблюдаются девиация и нистагм в направлении, соответствующем раздражению нервов удаленного лабиринта. При впрыскивании кокаина в отдельных случаях также можно наблюдать такую стадию раздражения, предшествующую выпадению. После этого появляется изменение направлений девиации и нистагма. (В случае преходящего раздражения 8-го нерва наступают сразу же поворот и наклон головы в направлении, обратном описанному выше).
6. Совершенно неясна в настоящее время причина временной утраты тонуса конечностей на стороне экстирпированного лабиринта. Одностороннее выпадение тонических лабиринтных рефлексов на конечности считать, ответственным за это явление (см. выше) нельзя по той причине, что при. этом каждый лабиринт влияет в одном направлении и одинаково сильно на конечности обеих сторон. Расслабление продолжается у кролика около двух месяцев, у обезьян — свыше месяца, у морской свинки — несколько дней, у кошек и собак оно непостоянно и по большей части продолжается лишь в течение нескольких часов. После выключения лабиринта кокаином у морской свинки оно в большинстве случаев не наблюдается. Еще не нполне определено, идет ли вообще при этом речь о вестибулярных, влияниях и, если это так, исходят ли эти влияния из оперированной или интактной стороны.
7. Определенно то, что триада симптомов симпатической природы, появляющаяся на глазу оперированной стороны (миоз, узкая глазная щель, оттянутая мигательная перепонка), не зависит от лабиринта и скорее обусловлена разрушением симпатических путей в среднем ухе.
Если рассмотреть таблицу II, можно увидеть, что прямые стойкие последствия, зависящие от лабиринта и наступающие после односторонней экстирпации лабиринта, обнаруживаются больше всего , у кролика,
328 Шестая глава. Последствия, односторонней экстирпации лабиринта
и менее у собаки. Этому соответствует то обстоятельство, что собаки способны компенсировать одностороннее выпадение лабиринта значительно более совершенно.
Далее можно видеть, что по мере восхождения по лестнице млекопитающих значение лабиринтов для установки тела уменьшается. У кроликов лабиринты господствуют над шеей, туловищем и глазами (не считая тонических лабиринтных рефлексов на мышцы конечностей, которые в симптоматике после одностороннего удаления лабиринтов не играют никакой роли). У собаки и обезьяны влияние на туловище уже не распространяется, тонические рефлексы на глаза утрачивают господствующую роль, а в качестве прямого следствия остается только основной поворот шеи. Похоже на то, что имеет место прогрессирующий процесс обратного развития вестибулярного механизма по мере формирования мозга.
У всех исследуемых видов животных можно было показать явления компенсации, которые уменьшали нарушения после односторонней утраты лабиринта. Эффективность ее, однако, не одинакова. С развитием мозга она повышается.
При этом принимают участие прежде всего аппараты установочных рефлексов, расположенные в среднем мозгу.
При определенных положениях тела уже лабиринтные установочные рефлексы, исходящие из интактного лабиринта, в состоянии уменьшить основной поворот; это имеет место, например, при боковом положении оперированной стороной вниз. При висячем положении головой вниз наклон головы нередко снимается благодаря лабиринтному установочному рефлексу.
Значительно более эффективны установочные рефлексы тела. У сидящего животного нормальные возбуждения исходят из обеих сторон тела (особенно из конечностей). Эти возбуждения воздействуют на тело, которое вследствие односторонней утраты лабиринта повернуто и отдельные группы мышц которого поэтому растянуты. Центры их находятся в состоянии повышенной возбудимости. Так, например, при сидении на полу поворот головы относительно грудного отдела оказывается меньше, чем при нормальном положении животного в воздухе (установочный рефлекс с тела на голову).
Эти рефлексы вступают в действие, во-первых, когда животное опрокинулось (в боковое положение). В этом случае голова (установочный рефлекс с тела на голову) поворачивается в направлении к нормальному положению и даже тогда, когда этот поворот должен осуществиться в направлении, противоположном основному повороту, а во-вторых (установочный рефлекс с тела на тело), тело усаживается и тогда, когда голова не может быть приведена в нормальное положение.
Установочные рефлексы тела вступают в действие всякий раз, когда животное соприкасается с полом. У обезьян действие этих рефлексов можно наблюдать во время лазанья, когда руки их обхватывают прутья решетки или другие предметы.
Независимо от соприкосновения с землей вступают в действие оптические установочные рефлексы, в осуществлении которых принимает участие кора больших полушарий. В момент зрительной фиксации они управляют поворотом головы, у собак и обезьян играют настолько важную роль, что некоторые нарушения вообще становятся отчетливыми только после надевания шлемика. У кошки они, по-видимому, являются менее важными, а у кроликов и морских свинок отсутствуют.
Два последних вида животных могут использовать глаза для коррекции анормальных установок ног.
Общий итог
329
Помимо этого, у всех видов млекопитающих развиваются открытые Бехтеревым изменения центральной иннервации, которые проявляются в том, что спустя некоторое время удаление второго лабиринта вызывает поворот головы и девиацию глаз в другую сторону. Испытывают ли также влияние центры конечностей, еще не вполне ясно. Эта компенсация осуществляется субкортикально, вне зависимости от глаз и афферентных проприоцептивных нервов соответствующих мышц (по крайней мере шейных мышц).
Кроме названных прямых последствий всех этих компенсаций, проявляется еще весьма важное вторичное влияние. А именно в той степени, в какой поворот и наклон головы уменьшаются благодаря установочным рефлексам и центральной компенсации, также уменьшаются тонические и установочные шейные рефлексы. Поэтому поворот туловища, различие тонуса конечностей, движение по часовой стрелке и манежные движения, опрокидывание и отклонение в сторону во время бега, движения перекатывания и прочее уже по этой причине становятся меньше или исчезают вовсе.
Уже упоминавшийся факт, что компенсация у обезьяны и собаки настолько совершеннее по сравнению с таковой кролика и морской свинки, объясняется, во-первых, тем, что у высших млекопитающих прямые последствия выпадения лабиринта, как было сказано, менее сильно выражены, поэтому легче преодолеваются. Во-вторых, не только у высших млекопитающих, особенно у обезьян и собак, но и у кошек имеются оптические установочные рефлексы, которые у кролика и морской свинки отсутствуют. В-третьих, значение установочных рефлексов тела в соответствии с уменьшением роли лабиринтных влияний повышается. Все это делает понятным то, что обезьяна и собака обнаруживают по сравнению с кроликом так мало стойких нарушений.
Прежние авторы использовали одностороннюю экстирпацию лабиринта для исследования функции этого органа. Анализ, приводимый в этой главе, показывает, что без предварительного знания нормальных лабиринтных рефлексов можно прийти к серьезной путанице и что значительная группа симптомов является вторичной, частично наступая вследствие тонических шейных рефлексов, частично вследствие действия компенсаторных механизмов. Этот результат является существенным и для других областей физиологии центральной нервной системы и органов чувств.
Здесь следует еще кратко упомянуть об установленном Эвальдом понятии «лабиринтотонуса». Большая заслуга Эвальда в том, что он подчеркивал зависимость мышечного тонуса от лабиринтов и обосновывал это многочисленными экспериментами. В настоящее время мы знаем, что эваль- довский «лабиринтотонус» в той мере, в какой он проявляется после односторонней экстирпации лабиринта, основан на трех полностью отделимых одно от другого влияниях.
1. На тонических лабиринтных рефлексах с интактного лабиринта на мышцы шеи, туловища и глаз, которые у различных видов животных развиты в весьма различной степени (табл. II).
2. На вторично приводимых в действие посредством поворота шеи тонических шейных рефлексах на конечности и на шейных установочных рефлексах. Эти весьма интенсивные влияния не зависят непосредственно от лабиринтов. При увеличивающемся основном повороте они могут с течением времени даже приобретать все большее значение.
3. На временном двустороннем расслаблении конечностей, которое у одних видов животных непостоянно и длится всего несколько часов, у других — несколько недель. Взаимосвязь этого явления с лабиринтами еще неясна.
330
Седьмая глава. Последствия двусторонней экстирпации лабиринтов
Поэтому я думаю, что теперь наступило время оставить собирательное понятие «лабиринтотонус» и в каждом случае заменять это понятие отдельными его элементами, тем более, что не лабиринтные влияния играют при этом такую значительную роль.
Из острых и скоропроходящих последствий экстирпации лабиринта о нистагме было уже сказано. Движения по часовой стрелке и манежные движения главным образом обусловлены изгибанием шеи и туловища и исчезают при компенсации последних. Боковое отклонение при беге в основном обусловлено односторонней утратой тонуса конечностей, поэтому также является временным. Перекатывание представляет собой следствие спирального поворота всего тела и наблюдается только, когда животное производит движение бега и прыжка (два прыжка на один поворот). Перекатывания бывают тем энергичнее, чем грубее выполнена операция. При осторожном оперировании у кролика можно ограничить эти движения до минимума, у кошек, собак и обезьян — избежать полностью. Даже у кролика после очень щадящей операции эти движения могут полностью отсутствовать. Исчерпывающий анализ движений перекатывания стал возможным благодаря кинематографии.
Седьмая глава
ПОСЛЕДСТВИЯ ДВУСТОРОННЕЙ ЭКСТИРПАЦИИ ЛАБИРИНТОВ
В третьей—пятой главах было показано, что для рефлексов положения, компенсаторных установок глаз и установочных рефлексов лабиринты играют роль исходных инстанций возбуждения этих реакций, хотя при этом функционируют и другие рецепторы, которые также приводят в действие рефлексы всех трех названных групп. Следовательно, лабиринты никогда не являются единственными источниками, вызывающими эти рефлексы. Таким образом, получается, что после двусторонней экстирпации лабиринтов ни одна из названных функций не выпадает полностью и что из сохранившихся органов чувств нелабиринтной природы исходят возбуждения, выравнивающие выпадение лабиринта и с течением времени способные более или менее обезвредить возникшие при этом нарушения. Пока эти нелабиринтные рефлексы были неизвестны, симптоматическая картина двусторонней экстирпации лабиринта была не совсем понятной, а сделав из поведения безлабиринтных животных сколько-нибудь определенные заключения относительно нормальной деятельности вестибулярного аппарата было невозможно. При удалении обоих лабиринтов непосредственно устанавливалось только то, как функционируют в связи с центральной нервной системой остальные рецепторы, а не функция лабиринтов.
И если в предыдущих отделах для отдельных групп рефлексов всякий раз описывалось состояние животных после двусторонней экстирпации лабиринтов, все же общая картина должна быть здесь описана и уточнена еще раз.
Точные описания поведения после двусторонней лабиринтэктомии ранее были сделаны Брейером (2) и Эвальдом для голубей и Бехтеревым (1) и Эвальдом — для собак. В нашем распоряжении имеются длительные собственные наблюдения на кошках, собаках и обезьянах, менее продолжительные — на кроликах и морских свинках. Раньше всего следует обратиться к поведению кошек и обезьян.
Опыты на кошках
331
Опыты на кошках
Для этого прежде всего можно привести наблюдения над двенадцатью кошками {16), у восьми из которых оба лабиринта были удалены одномоментно, у четырех — в два приема. У животных, оперированных в один прием, нарушения значительно тяжелее, тем не менее в большинстве случаев уже вскоре после операции они сидят в клетке прямо, живот распластывается на полу, голова удерживается прямо, но сильно качается в горизонтальном и вертикальном направлениях. Часто животные с громким стуком ударяются мордой о пол. Однако в промежутках между этими движениями голова может удерживаться прямо. Если животных взять из клетки и посадить на пол, они иногда сидят прямо, в других случаях падают на бок и затем снова усаживаются. В ответ на раздражение или спонтанно наступают характерные приступы, во время которых животные внезапно начинают дико прыгать, часто переворачиваются, бьются головой о пол, перекатываются и т. д. В первые дни после операции кошки не в состоянии бегать, но могут ползать вперед и назад. Особенно примечательно то, что первое время почти все животные ползают только в обратном направлении. Через 1—3 дня они начинают ползти и вперед, несколько позже пытаются бегать. При этом •они вначале помогают себе тем, что одним боком опираются о стену. В дальнейшем они бегают, широко расставляя ноги, почти касаясь животом пола (на согнутых ногах) и прикасаясь к земле большей частью подошвы задних ног, чем нормальное животное (медвежья походка). .Кроме того, походка колеблющаяся и неловкая. При сидении и стоянии на этой стадии животные также раскачиваются и туловищем. Противостоять этому они пытаются при помощи энергичного отведения ног, особенно передних.
Только по прошествии примерно недели бег становится несколько лучше, по крайней мере животное уже в состоянии сделать несколько шагов по комнате, не падая. Как только бег улучшается, обнаруживается весьма характерный симптом, который был установлен у всех кошек после удаления обоих лабиринтов и наблюдался после операции более года. Во время бега животное беспрестанно оглядывается влево и вправо. Большей частью голова поворачивается при каждом шаге. Вначале это выражено так сильно, что животные вынуждены производить полные или полукруговые повороты, в процессе чего к повороту головы присоединяется и соответствующий изгиб всего туловища (шейный установочный рефлекс). Поэтому сначала кошки почти совсем не бегают по комнате прямо, а только зигзагообразно. Лишь в том случае, когда они двигаются к определенной цели, направление выдерживается несколько лучше. Создается впечатление, будто кошка стремится оптическими восприятиями заменить слуховые восприятия, отсутствующие после операции.
Наступающие непосредственно после операции покачивание, маятникообразные движения головы и удары головой о пол уже в течение первых дней становятся менее интенсивными. Удары головой о пол обычно прекнащаются раньше всего. Сильные движения головы следует преимущественно представлять себе как следствие ошибочных впечатлений со стороны глаз, так как даже самые сильные маятникообразные движения головы и ее качание возможно полностью или в значительной степени снять, если закрыть глаза шлемиком (в то время как рот и ноздри остаются свободными). Как только шлемик удаляют, маятникообразные движения головы и ее качание тотчас появляются с той же
332 Седьмая глава. Последствия двусторонней экстирпации лабиринтов
Интенсивностью. Этот опыт настолько демонстративен и убедителен, что был зафиксирован кинематографически. Прекращение качаний головы при прикрывании глаз как будто не основано на том, что неудобный шлемик вызывает какое-то общее торможение рефлексов, так как в случае, если шлемик сидит плохо и неполностью закрывает глаза, качание головы продолжается без изменений. В дальнейшем в одном случае удалось у смирной кошки во время сильного качания головы двумя пальцами прикрыть веки и этим самым тотчас прекратить качание. Таким образом, это, очевидно, зависит от оптических воздействий (такой же опыт удается и у собаки). Через несколько дней маятникообразные движения уменьшаются. На этой стадии с большой уверенностью' можно вызвать приступ отчетливого качания головы всякий раз, когда перед животным держат молоко или мясо. При этом они, как безумные,, устремляются в горизонтальном или вертикальном направлении, натыкаются на молоко и разбрызгивают его по комнате. Только после закрывания глаз животные оказываются в состоянии пить удовлетворительно.
Вследствие этих ненормальных движений головы многие животные весьма стеснены при приеме пищи, поэтому в первые дни их следует кормить через зонд или давать им мелко нарубленное мясо, которое они в состоянии хорошо проглатывать. Однако две кошки могли при отчетливом качании головы уже через 48 часов пить молоко и есть кусочки мяса.
В течение второй недели спонтанное маятникообразное качание головы уже незначительно. Однако спустя и более длительное время можно его выявить, держа перед животным молоко и мясо. Это выявляется также и при спонтанных попытках к еде. К этому времени уже проходят и другие явления раздражения, неистовое беганье и пр., и животные в состоянии, пусть неуклюже, пробежать несколько шагов, не падая. Если же на их пути установить небольшие препятствия, то в большинстве случаев они не могут их преодолеть. Если в первые недели после операции кошку усадить на высоте 15—25 см на широкой горизонтальной доске, то она не в состоянии слезть оттуда обычным способом. Она сползает назад до края доски и затем сваливается вниз. Реже она делает попытку спуститься, направляясь вперед, но это редко удается. Кошки, лишенные лабиринта, будучи приведены в ненормальное положение, вообще ползут почти постоянно назад. Если животное установить таким образом, чтобы задние ноги стадии на доску, оно остается иногда в этом ненормальном положении до 10 минут, не изменяя его. С нижней ступени лестницы кошки также не могут сойти в первые дни, и только через 13 дней после операции одной из наших кошек удалось сбежать по нескольким ступеням. Однако после этого она упала.
У кошек с двусторонней лабиринтэктомией, произведенной в два приема, после второй операции обнаруживаются в принципе те же явления. Только явления раздражения менее интенсивны, приступы неистового беганья совершенно отсутствуют, качания и маятникообразные движения головы менее интенсивны и длительны, при беге, прыганье и беганье по лестнице отмечается меньше нарушений.
Наступающее после второй операции изменение центральной иннервации, которое ведет к временному нистагму и повороту головы, было описано в предыдущей главе (стр. 307).
С течением времени описанные симптомы двусторонней утраты лабиринта подвергаются обратному развитию. Глазные и головные симптомы исчезают полностью. Маятниковые движения головы в ответ на держание перед животным мяса сохраняются довольно долго, но в дальней¬
Опыты на кошках
333
шем также совсем исчезают. Бег по гладкому полу понемногу становится совершенно нормальным, только из-за частого оглядывания вправо и влево в отдельных случаях к нему присоединяется круговой поворот, в конце концов только зигзагообразный бег отличает этих животных от нормальных. Отсутствие лабиринтного установочного рефлекса по прошествии определенного времени перестает оказывать сколько-нибудь заметное влияние на изящество и ловкость движений.
Говоря о судьбах отдельных рефлексов, особенно в первые дни после операции, можно установить следующее.
После децеребрации из рефлексов положения отсутствуют тонические лабиринтные рефлексы на конечности, на мышцы головы и туловища. Вследствие этого тонус мускулатуры тела не зависит от положения головы в пространстве. Наоборот, тонические шейные рефлексы на мускулатуру конечностей остаются ненарушенными, поэтому, изменяя положение головы в пространстве относительно туловища, можно вызвать типичные изменения положения тела в пространстве.
У кошек, лишенных лабиринта, с интактным большим мозгом и у таламических кошек также можно с большой отчетливостью обнаружить тонические шейные рефлексы на конечности, например различие тонуса правых и левых конечностей в ответ на поворот головы, разгибание передних ног в ответ на поднимание головы и их расслабление в ответ на ее опускание. Особенно легко вызываются две последние реакции у свободно сидящего животного с сохранным большим мозгом в ответ на приближение мяса, наступая в этом случае с большой точностью и быстротой. Создается впечатление, что после утраты лабиринтных рефлексов тонические шейные рефлексы через некоторое время компенсаторно усиливаются (обратное имеет место после снятия тонических шейных рефлексов путем перерезки задних корешков; см. далее, стр. 337).
Судьба установочных рефлексов после утраты лабиринта уже описана для таламических кроликов в пятой главе. Последние в воздухе совершенно дезориентированы. Если положить их на стол, то сразу после операции они остаются лежать на боку. Через некоторое время становится отчетливым установочный рефлекс с тела на голову. Голова поворачивается в нормальное положение, к чему скоро (благодаря шейному установочному рефлексу и установочному рефлексу с тела на тело) присоединяется усаживание передней и позднее задней части тела. Нередко животное переворачивается при этом на бок, тогда наступает та же реакция в обратном направлении. Таким образом, получается, что животное в течение некоторого времени перекатывается через живот из стороны в сторону. В конце концов всякий раз наступает правильное усаживание из бокового положения, вслед за чем голова и тело сначала еще продолжают раскачиваться, а затем прекращается и это. Накладывание на обращенный кверху бок животного дощечки с грузом подавляет реакцию усаживания из бокового положения.
Совершенно то же было установлено у двух таламических кошек, лишенных лабиринта (47), как уже было описано на стр. 189. В воздухе они были полностью дезориентированы. На полу сначала в нормальное положение приводилась голова, а спустя некоторое время и тело. Здесь также было ясно выражено временное явление перекатывания через живот с одной стороны на другую. Испытывая в отдельности, можно было показать установочные рефлексы с тела на голову и на тело и шейные установочные рефлексы. Одна таламическая кошка уже через 5 часов после удаления второго лабиринта приняла сидячее положение,
334
Седьмая глава. Последствия двусторонней экстирпации лабиринтов
а через 72 часа могла в течение продолжительного времени стоять на ногаху не падая.
При соприкосновении с полом животное снова может стоять и ходить; в первое время после операции можно наблюдать, как установочные рефлексы тела постепенно становятся все более действенными. Опыты над таламическими кошками показывают, что для этого «выучивания» большой мозг не нужен, но что процесс (частично) разыгрывается в среднем мозгу.
Сначала спокойное состояние тела (у животного с нормальным мозгом) нарушается вследствие зрительных раздражений. Отсюда сильные приступы горизонтальных и вертикальных качаний головы и ударов головой о пол, которые после надевания шлемика или простого закрывания глаз прекращаются. У таламического животного качание головы значительно меньше, потому что при этом отсутствуют оптические восприятия.
Однако через небольшой промежуток времени у животных после двусторонней экстирпации лабиринтов в ориентировании принимают значительное участие и глаза. На стр. 208 было уже описано появление оптических установочных рефлексов при участии в этом большого мозга (рис. 131 и 132 на стр. 213). В то время как животные с закрытыми глазами остаются в воздухе по-прежнему дезориентированными, после удаления шлемика голова при всех возможных положениях ее в пространстве приводится в нормальное положение и прежде всего тогда, когда глаза фиксируют какой-нибудь предмет. Такое же наблюдение можно' было сделать на собаках (стр. 208). У них было особенно отчетливо видно, что зрительная ориентировка постепенно тренируется: в первый день после операции она отсутствовала, на третий день была установлена впервые и примерно за 14 дней оказалась вполне сформированной.
Благодаря взаимодействию установочных рефлексов тела и оптических установочных рефлексов осуществляется далеко идущая компенсация выпадения лабиринта. При этом голова и тело посредством тонических рефлексов на конечности и шейных установочных рефлексов оказываются объединенными в функциональное единство.
Выпадение тонических лабиринтных рефлексов на мышцы глаз ж возможную их компенсацию благодаря тоническим шейным рефлексам на глаза у кошек исследовать не удается, так как компенсаторные установки глаз вообще не играют у них большой роли.
То, что у животных без лабиринтов отсутствуют рефлексы с полукружных каналов, само собой понятно. Это прежде всего относится к реакциям поворота головы и глаз вместе с относящимся сюда нистагмом. Значительно большее значение для животных имеет утрата реакций на прогрессивные движения, прежде всего рефлекса «готовности к прыжку». Если вниз прыгает нормальная кошка, ее передние ноги тонически разгибаются и подвигаются вперед так, что при достижении пола они эластично подхватывают вес тела. После экстирпации лабиринта этот рефлекс отсутствует: животное, не притормаживая, резко ударяется телом и головой о пол и в определенных условиях может нанести себе повреждение. Отсюда у животных страх перед падением.
Точно так же после лабиринтэктомии, как правило, способность животных, свободно падая в воздухе, переворачиваться и приземляться всегда утрачивается. Эта способность основана на лабиринтном установочном рефлексе на голову (с присоединяющимся шейным установочным рефлексом), при этом проявляется и влияние прогрессивных реакций на ногио
Опыты на кошках
335
После сказанного становится понятным поведение животных в воздухе. Понемногу они снова научаются прыгать. Три из кошек, лишенных лабиринта, свободно прыгали на расстояние 34—2 м от стола до клетки. Перед этим было осуществлено точное ориентирование с помощью глаз. Сначала они производили слишком короткий прыжок и падали на пол, далее прыжок оказывался чересчур большим, и животные ударялись головой о расположенную напротив стеклянную стенку клетки. Здесь речь идет о горизонтальном прыжке. При прыжке вниз, наоборот, нарушение выступает значительно более отчетливо. В данном случае сказывается отсутствие рефлекса готовности к прыжку, вследствие чего животные наносят себе повреждения. Поэтому как только их усаживают на какое-либо возвышение (стул, стол, лестница), можно наблюдать удивительное беспокойство: они принимаются мяукать, совершают круговые туры или двигаются ползком назад, нередко падая в том же направлении вниз. Но и спрыгивание со стула или со стола в конце концов после длительного упражнения осваивается животными, хотя они при этом спрыгивают не так беззвучно, как нормальные. Сбегание вниз по лестнице также становится возможным. Однако даже тогда, когда животные могут совершать эти движения безупречно, сплошь и рядом случается, что они шлепаются (hinunterplumsen) со стола или скатываются с лестницы.
Явное нарушение проявляется, когда животные действительно падают с возвышения вследствие отсутствия рефлекса переворачивания. Для оптической ориентации время оказывается слишком коротким, поэтому они просто шлепаются на пол, сильно ударяясь спиной или головой, однако тотчас же снова поднимаются и убегают.
Чрезвычайно важно установить, что при длительных сокращениях сила мышц без сомнения не страдает. Производятся большие силовые акты, далекие прыжки и т. д., тело при стоянии и беге держится совершенно прямо. Об общей потере тонуса мускулатуры, которого следовало ожидать, в особенности исходя из представлений Эвальда, ничего больше сказать нельзя. К этому мы возвратимся еще раз в связи с другими явлениями.
Путем продолжающихся упражнений удается, следовательно, привести животных в такое состояние, что в обычных условиях и при выполнении определенных выученных движений они почти не отличаются от нормальных. Однако как только им предлагается какое-либо необычное задание или они оказываются в необычной ситуации, они снова полностью дезориентируются до тех пор, пока дальнейшее упражнение научит их преодолевать также и новую задачу. Если теперь перед животным поставить новую задачу такой же трудности, нарушение тотчас проявляется снова.
Так, одна из кошек хорошо поднималась и спускалась по лестнице, грациозно соскакивала со стола на пол и перепрыгивала по воздуху со стола в клетку, «служила» на разогнутых задних ногах, но тотчас же начинала дико носиться, когда ее помещали на решетчатую крышу клетки Ганса Майера для основного обмена, и, пятясь, падала оттуда на пол.
Другая кошка выучилась даже свободно стоять на задних вытянутых ногах и схватывать передними лапами помещенный высоко кусок мяса.
Поучительность этих наблюдений состоит в том, что становится возможным видеть, как посредством уже в большей части известных механизмов выпадение лабиринтной деятельности постепенно компенсируется настолько, что животные начинают вести себя подобно нормальным.
336
Седьмая глава. Последствия двусторонней экстирпации лабиринтов
Последствия двусторонней экстирпации лабиринтов у кошек, у которых предварительно перерезались задние шейные корешки (16)
После двусторонней экстирпации лабиринтов наряду с другими рефлексами сохраняются и рефлексы, осуществляющиеся с шеи, т. е. тонические шейные рефлексы на конечности и шейные установочные рефлексы. Если последние выключить, то из перечисленных здесь групп остаются только установочные рефлексы тела и оптические установочные рефлексы. Поэтому наблюдения над животными, оперированными таким образом, не лишены интереса. Здесь следует коротко привести их результаты.
Перерезка трех пар верхних шейных корешков у -кошек снимает тонические шейные рефлексы на мышцы конечностей, на поднимание, опускание, поворот и наклон головы, но оставляет ненарушенным vertebra-prominens-рефлекс. Передние ноги иннервируются, только начиная с Сб (С4). Таким образом, операция не затрагивает их.
У пяти кошек сначала перерезались три верхние пары задних шейных корешков. После 41—203 дней удалялся один лабиринт, а через некоторое время — второй. По прошествии различных сроков (до года) после этого животные продолжали еще находиться под наблюдением. У одной кошки двусторонняя экстирпация лабиринтов была произведена одномоментно, и сразу после этого животное было децеребрировано. О наблюдениях над четырьмя из этих животных после одностороннего удаления лабиринта сообщалось уже в предшествующей главе.
Прежде всего следует описать, какие последствия влечет за собой сама по себе перерезка задних шейных корешков.
Уже в первый день животное сидит прямо, в состоянии двигать головой во все стороны без признаков атаксии. Никакого качания головой также нет.
Таким образом, в то время как мышцы конечностей после перерезки их задних корешков производят атаксические движения, в случае шейных мышц дело обстоит иначе. Это тем более вероятно, что для контроля движений шеи используются также лабиринты. С этим хорошо согласуется тот факт, что после экстирпации лабиринтов появляется качание головы, которое, по-видимому, находится под влиянием глаз.
Движения передних ног только в первые дни несколько неловки, далее они становятся совершенно нормальными. Напротив, нарушения движений задних ног отчетливы. Последние во время стояния и во время бега оказываются согнутыми, так что живбт находится около пола и походка становится ползающей. Сначала покачивается задняя часть тела (отсутствие шейных установочных рефлексов), и животные спотыкаются задними ногами во время прыжков и бега по лестнице. Пон многу это выравнивается (установочный рефлекс тела на тело) и, наконец, остается только походка на согнутых задних ногах.
Последнее безусловно основывается на выпадении тонических шейных рефлексов на задние ноги, которые в норме во время бега (противоположно действию лабиринтов) разгибают задние ноги. После исчезновения этих рефлексов при нормальном положении головы остается только тонический лабиринтный рефлекс, сгибающий задние ноги (и в некоторых случаях с задней, интактной шейной части vertebra-pro- minens-рефл екс).
Влияние тонических шейных рефлексов на передние ноги лучше всего исследовать, если, держа мясо перед животным, заставить его
Опыты на кошках
337
Реакция
ПОЛНОСТЬЮ
совершенно
отсутствовала
частично отсутствовала
нормальная
Белая
?
?
С 69-го но 186-й день
Лабах
До 187-го дня
С 187-го до 198-го дня
—
Ахмед
» 11-го »
» 11 го » 40-го »
—
Белиаль
» 49-го »
» 49-го » 143-го »
—
Чернобелая
» 12-го » 20-го »
С 20-го по 184-й день
поднять или опустить морду. При этом у нормальных кошек (рис. 32 и 33, стр. 73) вследствие взаимодействия шейных и лабиринтных рефлексов в первом случае передние ноги разгибаются, в последнем — сгибаются. Эта реакция в первые дни или недели после перерезки задних корешков совсем или частично исчезает (рис. 166), но постепенно более или менее восстанавливается (прежде всего благодаря компенсаторному усилению тонических лабиринтных рефлексов) и у одной части животных снова может стать вполне нормальной, в то время как у других длительно остается нарушенной. Эти различия, по-видимому, основаны на том, что
Рис. 166. Белиаль. Перерезка I и II пар задних шейных корешков 23 марта 1913г. и III пары 3 апреля 1913 г. Снимок 26 апреля 1913 г. Вызывается поднимание головы, причем передние ноги остаются
согнутыми (ср. рис. 33).
у одних кошек сильнее выраженными оказываются шейные, у других — лабиринтные рефлексы.
Динамика этих реакций у пяти кошек видна из таблицы (стр. 337).
Таким образом, во всех случаях отчетливо видно наступление компенсаторных явлений. То, что эти влияния по большей части исходят из лабиринтов, видно из того, что достаточно удалить уже один лабиринт, чтобы реакции снова исчезли.
Смотря по тому, идет ли дело о животном с преобладающими шейными или лабиринтными рефлексами, длительные явления выпадения, которые всегда незначительны, выступают более или менее отчетливо. Кошка с преобладающими лабиринтными рефлексами (Ахмед) в конце концов не обнаруживала во время бега, прыганья, поднимания по лестнице никаких нарушений и прыгала на высоту 2х/2 м с немного меньшим изяществом и уверенностью по сравнению с нормальными кошками. Другая кошка с преобладающими шейными рефлексами (Белиаль), наоборот, долго бегала на согнутых ногах, волоча живот по полу, но также была в состоянии бегать по лестнице, спрыгивать со стола и т. д.
Таким образом, в каждом случае длительные нарушения после перерезки задних корешков верхнего шейного отдела спинного мозга оказываются очень легкими и весьма полно компенсируются.
Теперь можно обсудить те нарушения, которые наблюдались у трех кошек (Лабах, Ахмед и Белиаль) после двустороннего удаления лаби- 22 Р. Магнус
338
Седьмая глава. Последствия двусторонней экстирпации лабиринтов
ринтов. Лабах находилась под наблюдением свыше 10 месяцев, Ахмед — 4 месяца и Белиаль — 5 недель. Нарушения в первые дни после удаления второго лабиринта не отличались существенно от таковых, обнаруженных у нормальных кошек после удаления лабиринтов в два приема.
По прошествии более долгого времени после операции кошки, лишенные лабиринта, без шейных рефлексов все же отличаются от контрольных животных. У первых после затихания начальных бурных явлений ясно обнаруживается, что всякое влияние установки головы на тонус конечностей отсутствует (у них еще имеется только vertebra-
prominens-рефлекс). Поэтому, когда в силу какой-либо причины они становятся беспокойными, у них отмечается весьма отчетливая неловкость. В то время как у нормальных кошек без лабиринтов частые движения головой постоянно сопровождаются соответствующими изменениями тонуса конечностей, у животных без шейных рефлексов этого не бывает никогда. Самые обширные движения головой, которые наступали всегда, когда животные приводились в необычное положение (например, на ступеньке лестницы), не вызывали изменения установки конечностей. В особенности Лабах сидела с максимально поднятой и сильно повернутой головой, в то время как передние ноги оставались согнутыми. Подобного положения никогда не наблюдается у нормальных кошек. Как явствует из приведенных описаний, влияние установки головы на тонус конечностей после удаления обоих лабиринтов у нормальных кошек обнаружить всегда легко; при этом шейные рефлексы функционируют особенно отчетливо. С другой стороны, после выключения шейных рефлексов выпадение их через некоторое время более или менее полно компенсируется лабиринтами. Однако, если удаленными оказываются верхние задние шейные корешки и оба лабиринта, всякое прямое влияние установки головы на тонус конечностей долгое время отсутствует. Лучше всего это наблюдать, как сказано выше, если заставить животное поднимать и опускать голову, держа перед ним мясо. Тогда передние ноги не принимают участия в реакции: при поднятой голове они остаются согнутыми (рис. 167) или вытянутыми при опущенной голове, несмотря на то что животное совершенно без труда в состоянии сгибать и разгибать свои конечности. Иногда раньше поднимается голова, а передние ноги остаются согнутыми. Через некоторое время после этого (без изменения положения головы) передние ноги разгибаются, так что животное в достоянии мордой достать мясо. При исследовании тонуса конечностей путем поворота головы в положении животного на спине соответствующие изменения тонуса конечностей отсутствуют.
Весьма интересно было наблюдать, что, несмотря на это длительное нарушение, животные успешно стояли, сидели и бегали. Отсюда становится очевидным исключительное значение нелабиринтных установочных рефлексов. У животных отсутствуют все лабиринтные рефлексы, кроме того, тонические шейные рефлексы на конечности и шейные уста-
Рис. 167. Лабах. Снимок 4 апреля 1913 г., через 10 недель после экстирпации второго лабиринта. При сильном поднимании головы передние ноги остаются согнутыми.
Опыты на кошках
339
нов очные рефлексы. Однако у них имеются установочные рефлексы тела на голову и тело и оптические установочные рефлексы. Этих рефлексов достаточно, чтобы было возможным стояние и беганье. К сожалению, в то время, когда производились эти опыты (1913), отдельные установочные рефлексы еще не были известны и поэтому не было возможности установить, какую долю участия принимают обе группы установочных рефлексов тела и оптических установочных рефлексов в общей деятельности. Однако то, что совокупность их обеспечивает значительную часть функции установки, определенно известно. Интересно далее, что, несмотря на отсутствие шейных установочных рефлексов, совместная деятельность тела и головы все еще может так хорошо осуществляться.
В сложных функциях (прыжки, спускание по лестнице) эти животные также не отстают существенно от животных, лишенных лабиринта. Они, как и делабиринтированные животные, должны заново выучиваться всем сложным движениям. Конечно, все кошки не ведут себя при этом совершенно одинаково. Так, Лабах до последнего времени не могла спускаться и подниматься по лестнице, только в отдельных случаях медленно спускалась на одну ступеньку. Однако эта кошка была в состоянии перепрыгнуть расстояние в 2 м со стола в открытую клетку (дальше, чем любая из делабиринтированных кошек, находившихся под нашим наблюдением). Ахмед прыгал хуже, но очень хорошо бегал по лестнице. Во всяком случае можно с определенностью сказать, что хотя животные без шейных и лабиринтных рефлексов и двигаются менее изящно и с меньшей ловкостью, они тем не менее способны выучиться примерно таким же сложным движениям, как и кошки, лишенные лабиринтов.
В этом отношении существует только одно исключение. В первое время после экстирпации второго лабиринта все исследованные кошки не могли свободно стоять на задних лапах. Была сделана попытка выучить их также и этому. Оказалось, что кошки с интактными шейными рефлексами в конце концов снова выучивались стоять на задних лапах, кошки же без шейных рефлексов научиться не могли. Первые (с шейными рефлексами) сначала поднимались только на согнутых задних ногах, если высоко над их головой держали кусок мяса. Позднее им при этом удавалось разгибать задние ноги полностью. Одна из наших кошек была в состоянии, несмотря на отсутствие обоих лабиринтов, свободно «служить» на задних ногах. В конце концов Лабах и Ахмед (без шейных рефлексов) также смогли подняться на задние ноги. Они могли и стоять на разогнутых задних ногах, если при этом опирались передними ногами о стену, но этим животным никогда не удавалось свободно подняться на вытянутых задних ногах. Это различие особенно отчетливо выступило в поведении животных, когда их сажали в клетку для измерения основного обмена (Ганса Майера), в середине решетчатой крыши которой укреплялся кусок мяса. Крыша была настолько высокой, что кошка могла достать мясо только в том случае, если она на середине клетки свободно поднималась на разогнутые задние ноги. Делабиринтированным кошкам после того, как они в течение нескольких дней предпринимали соответствующие попытки, это удавалось всегда сразу, так что они овладевали мясом в течение нескольких секунд. Лабах же и Ахмед после попыток, длившихся месяцами, не смогли достать мяса этим способом. В конце концов они научились другому. По боковой стене клетки они влезали передними ногами наверх и стояли на вытянутых задних ногах, опираясь передними о боковую стенку. Затем глазами отыскивали место, где было подвешено мясо, и в этом направлении просто падали. При этом передними ногами они ударяли по воздуху. Обычно первое время им не удавалось заце¬
22*
340
Седьмая глава. Последствия двусторонней экстирпации лабиринтов
пить и сбить мясо, но они продолжали попытки, пока (иногда после чет- вертьчасового напряжения) им не удавалось таким способом завладеть лакомством. Неспособность встать на задние ноги находится в соответствии с наблюдением, когда перерезка задних шейных корешков уже сама по себе обеспечивает стояние и беганье на согнутых задних ногах.
В соответствии с тем, что было сказано выше об общем мышечном тонусе после двусторонней экстирпации лабиринтов, животные не страдают от общей утраты тонуса мускулатуры тела, если сверх этого перерезаются самые верхние задние шейные корешки. Наоборот, они производят свои движения с большой силой, и уже тот факт, что одно из наших животных могло прыгнуть на расстояние, равное двум метрам, причем толчок был дан одними задними ногами, показывает, что возможны силовые акты большой интенсивности. Не удалось также обнаружить какой бы то ни было слабости ног, спины и затылка и при соответствующих нагрузках. То, что разгибательная мускулатура ног была способна к интенсивным силовым актам, можно было показать при почесывании спины животного. Если пощекотать кошку между лопатками, наступает сильное тоническое разгибание передних ног, если почесать ее в области таза, то разгибаются задние ноги. Можно было затем с силой надавить на спину без того, чтобы ощущаемое противодействие было меньшим, чем у нормальных кошек. То обстоятельство, что и децеребрационная ригидность проявляется у кошек без шейных рефлексов и без лабиринтов с весьма большой интенсивностью, будет показано далее.
Кошки, у которых были перерезаны задние шейные корешки и удалены оба лабиринта, сначала обнаруживают те же явления, какие наблюдаются после двусторонней экстирпации лабиринтов. Кроме того, они движутся с меньшим изяществом, чем безлабиринтные нормальные кошки, и не могут свободно подниматься на задние ноги. Прямая зависимость тонуса мышц конечностей от установки у них головы (не считая vertebra-prominens-рефлекса) полностью и длительно отсутствует. Несмотря на это, они в состоянии выполнять более сложные движения (прыжки на далекие расстояния, сбегание вниз по лестнице и т. д.). При этом они привлекают, помимо установочных рефлексов с тела на голову и с тела на тело, оптические установочные рефлексы, в то время как шейные установочные рефлексы отсутствуют. В отношении общей потери тонуса мускулатуры тела у них ничего не обнаружено.
Опыты на трех животных (Лабах, Ахмед, Чернобелая) завершились тем, что кошки были децеребрированы в целях контроля, действительно ли тонические рефлексы, которые посредством изменения установки головы оказывают влияние на тонус конечностей, полностью отсутствовали вследствие операций на лабиринта^ и на задних корешках. У всех животных развилась отчетливая децеребрационная ригидность. У них также имелся отчетливый vertebra-prominens-рефлекс: при давлении на позвоночный столб на границе шейного и грудного отделов наступало сильное уменьшение разгибательного тонуса передних ног. Этим доказывается, что на тонус конечностей у животных рефлекторно влияют те пункты, которые локализуются вне области операции на задних корешках.
Затем проводились исследования (в положении животного на ногах, на боку и в положении на спине) влияния поднимания, опускания, поворота и наклона головы на разгибательный тонус передних ног. Эти
Опыты на кошках
341
опыты повторялись в течение последующих часов много раз. В результате оказалось следующее.
У Лабаха при исследовании в положении стоя, на спине, правом и левом боковых положениях и в висячем положении головой вниз ни путем поднимания и опускания, ни путем поворота и наклона головы не удалось выявить ни малейшего влияния на разгибательный тонус передних ног, хотя у животного имелась выраженная ригидность и при пробе vertebra-prominens-рефлекса быстро наступала реакция передних ног. В течение четырех часов после операции животное основательно исследовалось трижды, а результат был во всех случаях один и тот же. Так как у децеребрированного животного рефлексы вызываются обычно с большой отчетливостью и постоянством, можно утверждать, что у Лабаха не отмечалось ни малейшего следа влияния установки головы на тонус конечностей.
В том же направлении шли испытания у Ахмеда и Чернобелой. В различных положениях тела при помощи поднимания, опускания и наклона
Рис. 168. Лабах. 8 июля 1912 г. Перерезка трех верхних задних шейных корешков 27 января 1913 г. Правосторонняя экстирпация лабиринта 10 февраля 1913 г. Левосторонняя экстирпация лабиринта 22 декабря 1913 г.
Децеребрирован. Перерезка спинного мозга на уровне 11-го грудного позвонка. Вследствие этого задняя часть тела животного не участвует в децеребрационной ригидности и удерживается рукой. Передняя часть тела животного полностью опирается на сильно разогнутые передние ноги. Голова удерживается прямо благодаря тонически сокращенным мышцам затылка.
головы никакой реакции передних ног вызвать не удавалось. Поворачивание головы в большинстве положений также оказывалось безрезультатным. Только в некоторых исследованиях в боковом положении была установлена минимальная пограничная реакция (шейный рефлекс). В течение опыта нельзя было с уверенностью решить, сохрани- рись ли еще какие-либо следы шейного рефлекса или их не было совсем. На секции у Ахмеда было обнаружено около правого заднего корешка С2? У Чернобелой у правого заднего корешка С3 по тоненькой легко рвущейся нити; возможно, что именно ее следует считать ответственной за упомянутую сомнительную реакцию. На секции Лабаха не было обнаружено никаких неперерезанных волокон задних корешков.
Децеребрационная ригидность у всех трех животных была особенно сильной. Рис. 168 показывает, что Лабах, спинной мозг которого перерезался после децеребрации на уровне 11-го грудного позвонка, весьма хорошо «стоит» на задних ногах и выдерживает вес своего тела.
Голова также свободно удерживается, т. е. тонус затылочных мышц удовлетворителен. Последнее тем более примечательно, что удалены не только лабиринты, но и собственные проприоцептивные афферентные нервы затылочных мышц. При испытании с помощью давления рукой можно было убедиться в наличии сильного тонуса затылочных мышц. Это служит еще одним указанием на то, что тонус мышц может поддерживаться не только с соответствующей части тела, но и с других участков и даже тогда, когда лабиринты отсутствуют.
342
Седьмая глава. Последствия двусторонней экстирпации лабиринтов
В описании этих опытов на основании результатов, полученных до сих пор, принималось, что компенсация выпадения лабиринтных и шейных рефлексов при стоянии и беге осуществляется благодаря установочным рефлексам тела и оптическим установочным рефлексам. Первые локализуются в среднем мозгу, последние — в коре большого мозга. Конечно, не исключено, что и другие части коры больших полушарий (моторная зона) также участвуют в этом. Однако доказательств тому мы до сих пор не имеем. На основании наблюдений над таламическими животными без лабиринта следует принять, что в этой компенсации главная роль принадлежит упомянутым установочным рефлексам.
В заключение в качестве приложения приводится сокращенный протокол наиболее длительных (в общем 134 года) наблюдений над одним из животных.
Лабах. 8 июля 1912 г. Наркоз по Мельцеру с эфиром. Перерезка задних корешков от С2 — с обеих сторон снаружи позвоночного канала, от С3 и С, — с обеих сторон ин- традурально. Длительность операции 50 мин. Непосредственно после операции вызывался пателлярный рефлекс.
5 мин. после операции. Животное дышит самостоятельно. Сгибательный рефлекс конечностей отчетлив.
25 мин. после операции. Самостоятельно поднялась на четыре ноги. Свободно двигает шеей влево и вправо. Голову держит опущенной вниз. Нос на полу.
50 мин. после операции. Сидит в клетке как нормальная кошка с поднятой головой. Свободно двигает головой во всех направлениях.
9 июля 1912 г. Самостоятельно пьет. Сидит с поднятой головой и сильно согнутыми ногами. Из этого положения поднимается на передние ноги, при этом покачивается на задних ногах (полусогнутых). Задняя часть тела при ходьбе раскачивается. Передние ноги координированы хорошо. Атаксии шеи нет. Анормальные установки одной из задних ног тотчас же корригируются. Без нарушений перебирается бегом через порог высотой 5 см. Самостоятельно пьет молоко, но не ест.
10 июля 1912 г. Сидит с поднятой головой и согнутыми задними ногами. Во время бега ноги также сильно согнуты. В остальном производит совершенно нормальное впечатление. Перешагивает через газовую трубу высотой 10 см. Сидит на доске стола высотой 25 см, покачиваясь задней половиной тела, но соскакивает вниз хорошо, координированно. Позже то же осуществляет с высоты 50 см.
После полудня животное, ставшее после операции очень пугливым, исчезло.
12 июля 1912 г. Животное отыскалось наверху лестницы, т. е. после подъема по всем ее ступеням. Сидит нормально. Быстро перебегает комнату, при этом задние ноги полусогнуты.
13 июля 1912 г. Спрыгивает вниз со стола высотой 1 м. Передние ноги при этом хорошо принимают вес тела, но задние при соприкосновении с полом еще подгибаются, так что голень проезжается по полу. Однако животное не падает. Самостоятельно почти не пьет, не ест; кормится через зонд.
15 июля 1912 г. При беге еще несколько раскачивается задней частью тела, ест самостоятельно.
17 июля 1912 г. Соскакивает с плеча стоящего человека.
2 августа 1912 г. При беге задние ноги согнуты больше, чем у нормальной кошки. Задняя часть тела во время бега несколько раскачивается. В остальном бегает и прыгает с легкостью. С пола вскакивает на доску, на высоту м. Свободно может подняться на задние ноги.
19 сентября 1912 г. При опускании и поднимании, при повороте и наклоне головы соответствующих движений передних ног не наступает. Животное в состоянии полностью разгибать передние ноги, но не видно, чтобы этому разгибанию предшествовало бы соответствующее движение головы. Очевидно, у животного появляется зуд в голове; оно непрерывно трется задней частью головы о пол, причем голова максимально поворачивается и наклоняется без того, чтобы наступали соответствующие движения передних ног.
Достает мясо, которое держат над ней, свободно поднимается на задние ноги, во время чего согнутые передние ноги также поднимаются. Перепрыгивает со стола в клетку (1 м), при этом отталкивается от стола задними ногами.
30 сентября 1912 г. При поднимании головы в направлении мяса, которое держат перед ней, передние ноги остаются согнутыми; если мясо поднимают еще выше, животное встает на задние ноги, держа передние согнутыми (поза кенгуру). Полностью поднимается на задние ноги.
Опыты на кошках
343
19 октября 1912 г. В течение нескольких недель животное совершенно разодрало себе кожу задней части головы с обеих сторон.1 На голове эта рана теперь в основном зажила. Но обширная раневая поверхность еще сохраняется на правой стороне шеи и меньшая — там же слева, дорсально. Животное все еще чешет себе голову и шею. Подлезая под шкаф, обнаруживает отчетливый vertebra-prominens-рефлекс. Реакция передних ног на поднимание и опускание головы отсутствует. При взгляде вверх животное нередко принимает позу кенгуру.
15 января 1913 г. При сравнении с нормальной кошкой оказывается, что при поднимании и опускании головы соответствующие разгибание и сгибание передних ног осуществляются в меньшей степени и менее быстро, но полностью, по-видимому, не отсутствуют. Решить это с определенностью нелегко. Совершенно свободно животное может становиться на задние ноги, которые при этом максимально разогнуты.
26 января 1913 г. При поднимании и опускании головы передние ноги разгибаются и сгибаются, но реакция 15 января представляется более слабой и менее быстрой, чем у нормальной кошки.
27 января 1913 г. Правосторонняя экстирпация лабиринта (де Клейн). Полукружные каналы и внутренний слуховой проход открыты.
Вскоре после операции. Отчетливый нистагм в левую сторону. В висячем положения (головой вниз) грудь повернута относительно таза на 20°, верхняя апертура — на 30°, голова повернута на 90° и наклонена на 20°. Левая передняя нога разогнута несколько сильнее правой.
28 января 1913 г. Нистагм в левую сторону. Незначительная девиация глаз вправо. При сидении отчетливого отведения левой ноги нет. Маятникообразных движений головы нет. Бегает через всю комнату, широко расставляя ноги и не падая. Может бежать прямо, а также налево и направо. Один раз оступилась вправо, не падая. Может наклонять голову в обе стороны. Прыгает со стола в клетку (У2 м), в клетке немного раскачивается, но не падает. Спрыгивает вниз со стола, приземляется, широко расставив ноги и покачиваясь, но не падая.
При попытке пить и при показывании мяса покачивания головы и тела нет. Во время сидения тело несколько перевешивает вправо. Смещаемость вправо больше, чем влево.
31 января 1913 г. В клетке сидит спокойно. Голова повернута вправо на 30—45°. Нистагм влево не постоянен. Перепрыгивает со стола в клетку на расстояние больше 1 м. Бегает, все еще широко расставляя ноги и покачиваясь, многократно оступаясь вправо.
Висячее положение (головой вниз): грудь повернута на 45°, голова — на 90° и наклонена по-разному. При устанавливании головы прямо поворот груди совершенно не изменяется. Положение на спине (голова установлена прямо): тонус передних ног несколько ниже, чем задних. Сзади тонус с обеих сторон одинаков. Поворачивание головы не оказывает никакого влияния на тонус передних ног.
1 февраля 1913 г. Нистагма больше нет. Бегает, немного расставляя ноги. Оступается временами вправо, но только в тех случаях, когда этому предшествует качание головы.
5 февраля 1913 г. Бегая, еще едва заметно расставляет ноги; покачивается только в том случае, если двигает головой.
7 февраля 1913 г. Голова на 30° повернута вправо. Девиации глаз и нистагма нет.
Висячее положение (головой вниз): грудь повернута на 45°, голова — на 70°. Никакого различия тонуса правой и левой передних ног. В положении на спине, когда голова установлена прямо, тонус передних ног с обеих сторон одинаков. Поворачивание головы никакого влияния на тонус конечностей и на поворот таза не оказывает. Лазает и прыгает уверенно. Бегая, более не расставляет ноги широко.
10 февраля 1913 г. Типичная левосторонняя экстирпация лабиринта. Полукружные каналы и внутренний слуховой проход открыты.
Через 5 часов после операции. Сильное качание головы, особенно в вертикальном направлении. Отклонение глаз влево (незначительно, но отчетливо). Нистагм вправо. Левосторонний парез лицевого нерва. Голова стоит прямо.
Висячее положение (головой вниз): никакого поворота головы и груди.
Положение на спине (голова прямо): никакого различия тонуса передних ног. Сидит, широко расставив передние и задние ноги. При маятникообразных движениях головы мордой ударяется о пол. Посаженная на пол, производит движения головы по направлению и против часовой стрелки. Пятясь, ползет через всю комнату, живот над полом, но все же на согнутых ногах.
После надевания шлемика (который прикрывает глаза, но оставляет свободными ноздри) покачивание головы тотчас прекращается и снова начинается после того, как шлемик снят.
1 Гиперестезии и парестезии вследствие перерезки задних корешков. См. 16, стр. 164.
344
Седьмая глава. Последствия двусторонней экстирпации лабиринтов
Немного позже. Бежит через всю комнату, широко расставляя полусогнутые ноги, голова сильно раскачивается, но животное не падает.
11 февраля 1913 г. Маятникообразное раскачивание головы меньше, но еще отчетливо. Если поднести молоко, раскачивание усиливается, но не в большой степени. Самостоятельно не пьет и не ест. Однако нарубленное мясо, положенное в рот, проглатывает.
12 февраля 1913 г. В клетке сидит спокойно, без маятниковых движений или качаний головы. Может наклонить голову в любую сторону. Нет больше отчетливой девиации глаз и нистагма. Сидит и бегает, расставляя и сгибая ноги, волоча живот по полу. При поднесении мяса — все еще некоторое раскачивание головы. Со стула ве спрыгивает. При поднимании и опускании головы соответственного разгибания и сгибания конечностей не наступает.
17 февраля 1913 г. Спрыгивает со стула через голову, однако правильно приземляется на все четыре ноги. Бегает еще покачиваясь и на согнутых ногах. Во время бега пола касается большая часть подошвы задней лапы, чем у нормальной кошки (медвежья походка). Без труда корригирует анормальные положения передних ног. Пытается прыгнуть со стола в клетку (30 см), не допрыгивает и падает, крепко хватаясь за прутья решетки клетки.
21 февраля 1913 г. Раны от расчесов на шее (после того как регулярно обстригались когти всех лап) зажили. При поднимании и опускании головы передние ноги не участвуют в реакции. Перепрыгивает со стола в клетку (У2 м).
26 февраля 1913 г. При е,о;е голова больше не качается. На задние ноги подняться не может.
5 марта 1913 г. Еще имеется левосторонний парез лицевого нерва. Бегает намного лучше, но еще несколько расставляя задние ноги. Медвежья походка сзади едва заметна.
Поднимание и опускание головы и поворачивание головы в положении животного на спине на тонус конечностей влияния не оказывает.
7 марта 1913 г. Бегает гораздо лучше, еще слегка заметна медвежья походка, ноги несколько согнуты. Со стула соскочить не решается, падает назад вниз.
9 апреля 1913 г. При беге через комнату оглядывается вправо и влево. Часто производит повороты по кругу. При этих движениях головы конечности в реакции не участвуют. В состоянии прыгнуть со стола в клетку на расстояние 1 м, но, очевидно, ошибается в оценке расстояния и часто прыгает или слишком далеко, или слишком близко. Встать свободно на задние ноги не может. Если в середине майеровской клетки помещается мясо, то после нескольких напрасных попыток оказывается в состоянии достать его следующим образом: сначала передними ногами забирается по стеклянной стенке клетки наверх, затем в течение мгновения, стоя на задних ногах, падает вниз, пытаясь во время падения сбить мясо одной из передних ног, что в большинстве случаев, после нескольких попыток удается. Сбежать вниз по лестнице не может. Посаженная на лестницу, часто принимав! особенно характерную позу. При этом, хотя она сидит с максимально поднятой головой, передние ноги остаются совсем согнутыми так, как это никогда не наблюдается у нормальной кошки.
21 апреля 1913 г. Ненормальное положение показано на рис. 167, стр. 338.
23 апреля 1913 г. Бегает гораздо лучше, быстро и уверенно, несколько расставляя ноги.
26 апреля 1913 г. Снимок: голова высоко поднята по направлению к мясу без соответствующего разгибания передних ног.
13 мая 1913 г. Прыгает со стола в клетку на расстояние 1.5 м. При этом прыгает слишком высоко. Левый лицевой нерв все еще парализован. Посаженная на лестницу, не решается или не может спуститься или подняться, но достает кусок мяса, который лежит на ступеньку ниже. Посаженная на стул, начинает вертеться п «танцевать», а потом через голову сваливается вниз.
7 июня 1913 г. Во время бега все еще оглядывается вправо и влево. Остальных нарушений почти не видно.
Начиная с этой стадии состояние изменяется относительно мало.
18 декабря 1913 г. (за четыре дня до забивания). Иногда сидит в описанном ненормальном положении (рис. 167), иногда стоит с поднятой головой и согнутыми передними ногами. Часто наблюдалось, что при поднимании головы передние ноги остаются без изменений. Может с успехом произвольно сгибать и разгибать передние ноги. Временами разгибание ног совпадает с подниманием головы, а сгибание ног—с опусканием, однако никогда реакция не наступает так быстро, как у нормальной кошки, и в два темпа (реакция большого мозга?). При потирании спины между лопатками животное тонически вытягивает передние ноги, а при потирании спины, ближе кзади — задние ноги. Об общей потере тонуса мускулатуры тела не может быть и речи. Соскочить со стула не может. Падает через голову вниз. Посаженная на пол, бежит нормально, но постоянно оглядывается вправо и влево. Описанным способом достает мясо с решет¬
Опыты на обезьянах
345
чатой крыши клетки. Посаженная на решетчатую крышу клетки, балансирует довольно хорошо, но не как нормальная кошка. Не попадает лапами между прутьев, в конце концов передними ногами соскальзывает по боковой стенке клетки вниз, причем прочно держится задними ногами, затем отпускает их и падает на спину.
22 декабря 1913 г. Эфирно-хлороформный наркоз; трахеотомия. Сонные артерии перевязаны, блуждающие нервы перерезаны. Обнажение спинного мозга на уровне 11-го грудного позвонка. Децеребрация с временным зажатием позвоночных артерий. Весь мозг до четверохолмия удален. Тотчас с началом ригидности самостоятельное дыхание.
И час. 45 мин. Операция закончена.
11 час. 55 мин. Сильная ригидность передних ног. Ушной рефлекс слаб. Имеется пателлярный рефлекс.
12 час. Выраженная ригидность передних ног. На задних — сгибательный рефлекс. Свободно стоит на передних ногах, если задняя часть тела поддерживается. Свободно поднимает голову. Это положение — рис. 168.
Исследование в:
положении на ногах — поднимание и опускание головы. ) без влияния на тонус Поворот головы. J передних ног.
Vertebra-promiiiens-рефлекс слабо положителен
правом боковом — поворот головы, поднимание и опу- ) никакой реакции пе- положении скание, наклон. } редних ног.
Vertebra-prominens-рефлекс слабый
левом боковом положении
— поворот головы, поднимание и опускание. Наклон.
никакой реакции.
Vertebra-proniinens-рефлекс отчетлив
положении на спине — поворот головы, поднимание и) никакой реакции
опускание. Наклон. ] г ^
Повторное исследование в 2 час. 10 мин. и в 3 час. 35 мин. дало точно такой же результат, с той только разницей, что теперь vertebra-prominens-рефлекс мог быть получен отчетливо во всех положениях.
Животное забито.
Секция. Центральная нервная система без признаков раздражения. Передние корешки Сх и С3 еще сохранены. Передние корешки С2 — под вопросом. Задние корешки Cj и С2 перерезаны. Задние корешки С3 из-за спаек и рубцов трудно различимы. Оба лабиринта полностью вылущены.
Последствия двусторонней экстирпации лабиринтов у обезьян (59) УУ двух обезьян через 22 и 38 дней после удаления левого лабиринта де Клейном была осуществлена экстирпация правого лабиринта. Одно из животных на следующее утро было найдено мертвым, другое же находилось под наблюдением свыше двух месяцев, после чего опыт был завершен экстирпацией большого мозга (таламическая обезьяна).
Результаты в основном те же, что и в опытах на кошках, только, у обезьян динамика установочных рефлексов изучалась основ тельно и при интактной коре больших полушарий, в то время как эти рефлексы у безлабиринтных кошек до сих пор анализировались только после удаления большого мозга. Часть наблюдений уже была упомянута в различных разделах этой книги при обсуждении отдельных рефлексов, однако краткий обзор общих результатов не будет излишним.
Так как удаление правого лабиринта происходило спустя долгое время , после удаления левого, то проявлялась бехтеревская компенсация: продолжавшийся в течение нескольких дней наклон головы и го-
346
Седьмая глава. Последствия двусторонней экстирпации лабиринтов
ризонтальное отклонение глаз вправо с нистагмом влево при временном расслаблении правых конечностей. Животное, которое можно было наблюдать только в день операции, обнаруживало качание головы, будучи посаженным на пол, падало, затем могло снова усаживаться, ползло, расставляя ноги, причем тело перевешивало вправо. Во время бега оно спотыкалось и падало вправо, затем снова усаживалось, опираясь правой стороной тела о стену, обнаруживая склонность ползти назад. У другого животного общее поведение можно было хорошо наблюдать в течение двух месяцев. В день операции оно сидело, расставив ноги, качаясь, дважды перевернулось. После полудня оно оказалось уже в состоянии схватить рукой хлеб и съесть его. Обезьяна пила при качании головы, падала при прыжках, обнаруживала движения по часовой стрелке и при беге спотыкалась в правую сторону. Тем не менее она была в состоянии спокойно и уверенно повиснуть на решетчатой крыше клетки. На следующий день она сидела, покачиваясь, и оживленно осматривалась вправо и влево, ползла в направлении назад, обнаруживала движения по часовой стрелке, сидела, расставив передние и задние ноги, лазала еще неуклюже, но не падая. Сидя на решетке клетки, еще сильно покачивалась. Уже в этот день можно было наблюдать значительное развитие оптических рефлексов. Животное использовало глаза, чтобы ориентироваться и держаться прямо. Установочные рефлексы тела на тело играли при этом также очень большую роль. Через 5 дней обезьяна уже была в состоянии проворно влезть на решетчатую крышу клетки и, повиснув на ней, раскачиваться в обе стороны, во время чего голова устанавливалась во всевозможных положениях. Прыгая, животное еще продолжало спотыкаться. Один раз оно упало с решетки крыши клетки (с высоты двух метров) на землю не лапами, а головой, причем черепом сильно ударилось о пол. На шестой день животное лазало все еще неуклюже и с шумом падало с крыши на землю, бежало, расставляя ноги, волоча живот по полу. Через 19 дней способность лазать значительно улучшилась, однако на полу животное все еще было неуклюжим, падало вправо, а один раз даже через голову назад на спину. При падении с крыши оно падало на одну и ту же сторону.
В конце концов становится все яснее, что лазанье оказывается для животного делом намного более легким, чем беганье по земле. Через месяц оно влезало с пола на крышу и вскакивало на высоко расположенную доску, не спотыкаясь. Наоборот, будучи потревоженным, оно становилось весьма неловким и при соскакивании со стола на пол, причем со стуком падало прямо на голову. Спустя 36 дней оно было в состоянии перепрыгнуть с крыши одной клетки на крышу другой на расстояние 50 см, не падая. На полу оно иногда еще бежало задом наперед; прыгая, один раз упало на живот. На решетке крыши клетки оно гораздо более уверенно, ибо удерживается там всеми четырьмя руками. Падает с крыши (если его согнать) на спину, позднее один раз — ца голову. Через 50 дней быстро бегало по комнате, широко расставляя ноги и все еще несколько раскачиваясь, прыгало на 40 см с крыши на крышу, при этом два раза падало, но всегда удерживалось. Сидя, оживленно оглядывалось по сторонам. Следовательно, непрерывно получало зрительные впечатления. По прошествии 58 дней при соскакивании со стола все еще падало на бок, бегало по полу, расставляя передние ноги; влезая с пола на крышу клетки, соскальзывало, но не падало и затем старательно удерживалось на крыше четырьмя руками.
Таким образом, в течение этого времени животное снова обрело значительные двигательные способности. Даже прыжки стали удаваться
Опыты на обезьянах
347
в случаях, когда обезьяна перед прыжком хорошенько ориентировалась зрительно. Это происходило таким образом: перед прыжком животное точно оценивало глазами расстояние и направление и затем производило движение, которое после длительного упражнения удавалось ему с большой определенностью. Наоборот, животное не в состоянии ориентироваться в пространстве тогда, когда внезапно в воздухе производится неожиданное движение, что происходит, например, при свободном падении. Для оптического ориентирования в этом случае не остается времени. Животное падает вниз в любых положениях, так как у него отсутствуют лабиринтные установочные рефлексы на голову и вследствие этого примыкающий шейный рефлекс, поэтому оно не может правильно перевернуться в воздухе. Кроме того, у него отсутствует рефлекторная готовность к прыжку, благодаря которой вес тела, случайно оказавшегося в правильном положении на полу, был бы принят и удержан. Поэтому животное, словно мертвая масса, ударяется головой или телом о пол. При лазанье оно проявляет исключительную ловкость даже тогда, когда случайно оступается или поскальзывается. Однако на земле нарушения движения все еще более заметны, чем при лазанье. Покачивания здесь полностью не исчезали.
При повторных наблюдениях бросалось в глаза, что обезьяна, лишенная лабиринтов, имела весьма значительную мышечную силу и с большой энергией вырывалась из рук экспериментатора. Она была в состоянии совершать очень длинные прыжки и обнаруживала сильное сопротивление в руках и ногах. Выпадали только зависящие от лабиринта изменения тонуса. Таким образом, мнение, будто после двусторонней утраты лабиринта у обезьяны, как и у кошки, наступает длительное общее расслабление мускулатуры, можно спокойно передать в область преданий.
На тех же животных через 13 и 48 дней после экстирпации второго лабиринта было испытано закрывание глаз. Вследствие этого нарушения выступили гораздо более отчетливо. Животное стало гораздо осторожнее и более охотно висело без движения на решетчатой крыше клетки в положении на спине, когда голова находилась в затылочном положении. По земле обезьяна бегала, широко расставляя ноги, нащупывая дорогу, с манежными движениями, которые обусловливались поворотом головы. Если во время бега оно наталкивалось мордой на какое- нибудь препятствие, то ползло, пятясь назад. Дальнейшие наблюдения во время свободного бега и лазанья не проводились, так как я опасался, что животное при этом нанесет себе вред.
Обезьяны с интактными лабиринтами, глаза которых прикрывались, обнаруживали только весьма незначительные нарушения. Они оживленно бегали по комнате и на все натыкались; как только они набредали на какой-либо отвесно расположенный предмет, тотчас же с большой ловкостью влезали наверх, не падая при этом.
Большое значение оптических рефлексов у обезьян было уже описано на стр. 213. Голова оптически ориентируется по отношению к видимым предметам, к этому присоединяются затем соответствующие изменения положения остального тела. Это особенно отчетливо можно наблюдать при исследовании безлабиринтных обезьян в свободных положениях в воздухе, когда установочные рефлексы тела не могут принимать участия в реакциях. Реакция длится лишь до тех пор, пока возбуждено оптическое внимание животного. Как только это прекращается, голова снова принимает индифферентное исходное положение.
348
Седьмая глава. Последствия двусторонней экстирпации лабиринтов
Глаза используются обезьяной не только для того, чтобы влиять на положение тела и корригировать его, но также и для контроля движений тела. Это отчетливо можно проследить, например, при прыжках безлабиринтных обезьян.
Сильное развитие оптических влияний у обезьян видно также, если после двусторонней лабиринтэктомии продемонстрировать отсутствие вращательных реакций, исходящих из полукружных каналов на голову и глаза. При этом часто наблюдается, что голова поворачивается в направлении, противоположном тому, которое соответствовало бы повороту, обусловленному лабиринтной реакцией. Наблюдаются также отклонения глаз и нистагм. Причина этого в том, что во время вращения обезьяна старается фиксировать определенные предметы в пространстве и затем время от времени быстрым нистагмоидным движением снова приводит глаза в исходное положение (железнодорожный нистагм). Это можно доказать, сделав для животного фиксацию невозможной, набросив на голову самому себе и обезьяне большой платок. Тогда после экстирпации лабиринтов при вращениях никогда не наступает поворота головы и поворота глаз с нистагмом. После экстирпации большого мозга и в легком наркозе эта оптическая поворотная реакция также отсутствует.
Установочные рефлексы тела на голову у обезьян имеют второстепенное значение, однако их наличие у безлабиринтных обезьян можно показать, если прикрыть у них глаза и, захватив животное за нижнюю руку, фиксированную за его спиной, держать его в боковом положении в воздухе. Тогда голова также стоит в боковом положении. Как только животное в этом положении укладывается на стол, голова его движется по направлению к нормальному положению (опыты спустя 3 недели и 1/4 месяца после экстирпации лабиринтов).
Большее значение имеют у обезьян установочные рефлексы тела на тело. Эти рефлексы подробно описаны уже на стр. 194. У безлабиринтных животных достаточно легкого прикосновения к полу или (при лазанье) к решетке, чтобы тело выпрямилось. Развитие этого рефлекса можно с успехом проследить в первые часы после пробуждения от наркоза и после экстирпации большого мозга. При боковом положении тела на полу возбуждения исходят со стороны туловища и наружной поверхности верхней части бедра. Усаживание начинается с задней части тела, при этом первым в нормальное положение приходит таз, затем усаживается передняя часть тела. Если животное, находящееся в висячем положении головой вниз, прикасается к полу сначала теменем, то одна рука вытягивается вперед. Как только эта рука касается пола, наступает дорсальное пригибание головы с поворотом и наклоном в сторону опорной руки, в то время как другая рука делает первый шаг.
При лазанье по решетке крыши клетки (тайже с закрытыми глаз&ми) голова благодаря возбуждениям, исходящим из четырех рук, направлена строго симметрично по отношению к решетке. При этом животное находится в положении на спине, однако движется уверенно и, если его не подгонять, никогда не падает. Этот рефлекс делает возможным висение и лазанье в положении на спине и для нормальных обезьян с интактными лабиринтами и открытыми -глазами. Если одна рука выпускает прут решетки, установочный рефлекс с другой руки приводит к тому, что тело поворачивается по направлению к плоскости лазанья.
У лишенных лабиринтов таламических обезьян с большой отчетливостью могут быть обнаружены также* й шейные установочные рефлексы.
Через 64 дня после экстирпации -втброго лабиринта у одной обезьяны была произведена экстирпация большого мозга. После этого животное
Заключение
349
было в состоянии, когда его держали за одну или обе руки, стоять на задних ногах в обычной позе. Хватательный рефлекс был отчетлив. Животное свободно могло висеть на передних руках, причем принимало нормальную позу Тонус конечностей (так как децеребрационная ригидность скоро после операции прошла) был нормален.
Эти наблюдения на обезьянах показывают, до какой степени может компенсироваться выпадение функции лабиринтов и как при этом используются прежде всего установочные рефлексы тела, шейные и оптические установочные рефлексы. При их помощи не только во время сидения, но также и при прыжках и лазанье осуществляются поразительно сложные акты. Только в одном случае регуляция оказывается полностью несостоятельной. Это наступает при свободном (не производимом животным намеренно) падении в воздухе.
Заключение
Из описания, приведенного в этой главе, ясно видно, что картина симптомов, какой она складывается после двусторонней экстирпации лабиринтов, без предварительного точного знания лабиринтных рефлексов не может быть использована для суждения о нормальной деятельности вестибулярного аппарата. Конечно, при исследовании каждого отдельного лабиринтного рефлекса необходимо доказать его лабиринтный генез тем, что выключение лабиринта снимает этот рефлекс. Однако функции безлабиринтного животного обусловлены деятельностью нелабиринтных рефлексов, активность которых у таких животных можно изучать изолированно. Так как после утраты лабиринтов в известной степени сохраняются рефлексы положения, компенсаторные установки глаз и установочные рефлексы, сферу действия соответствующих лабиринтных рефлексов очертить определенно нельзя. В отношении рефлексов положения это возможно только, если предварительно снять тонические шейные рефлексы (с помощью перерезки задних корешков или гипсования); в отношении компенсаторных установок глаз — после выключения тонических шейных рефлексов на глаза и на оптический установочный механизм; в отношении установочных рефлексов — после устранения установочных рефлексов тела, установочных шейных и оптических рефлексов. Только в этих условиях, сравнивая животных с сохранными лабиринтами с животными без лабиринтов, можно установить что-то определенное относительно функции последних.
Существуют лишь два условия, при которых поведение животных определяется исключительно лабиринтами. Первое — это свободное падение в воздухе. При этом вследствие быстроты явления оптические установочные рефлексы не могут проявить своего действия, установочные рефлексы тела отсутствуют, переворачивание в воздухе выпадает (отсутствие лабиринтного установочного рефлекса), конечности при достижении пола не подхватывают веса тела (отсутствие «готовности к прыжку»), вследствие этого и по прошествии долгого времени после операции животное при падении оказывается таким же неуклюжим, как в первый день, и не гарантировано от болезненных ушибов и в известных случаях от повреждений в момент соприкосновения с полом. Во-вторых, нечто подобное имеет место во время купания. Для этих наблюдений наиболее подходящими оказываются собаки. Вода удерживает вес тела и тем самым уменьшает интенсивность установочных рефлексов тела. Оптические установочные рефлексы проявляться под водой не могут, и, если отсутствуют лабиринты, беспомощное животное
350
Седьмая глава. Последствия двусторонней экстирпации лабиринтов
тонет. Возможно ли постепенно так выдрессировать безлабиринтную собаку, чтобы она только с помощью своих оптических установочных рефлексов плавала бы спокойно и уверенно, без того чтобы неистово биться в воде, предстоит еще решить.
При всех остальных условиях после устранения деятельности лабиринтов в распоряжении животного находятся все другие механизмы, замещающие эту деятельность. До какой высокой степени это удается, показывают приведенные наблюдения на кошках и обезьянах. При этом с течением времени функция понемногу становится все более совершенной. В опытах на таламических кошках с определенностью установлено, что значительная доля этого приспособления («выучивания») обеспечивается субкортикальными образованиями, причем наиболее деятельными становились установочные рефлексы тела. Тонические шейные рефлексы на конечности также постепенно усиливались. Другая часть компенсации осуществляется с помощью коры большого мозга. Это устанавливается, во-первых, по постепенному образованию оптических установочных рефлексов, что особенно хорошо прослеживается на собаках. Во-вторых, как уже описал Эвальд, животные после нескольких падений становятся в своих движениях особенно осторожными. Вероятно, при этом действуют еще и другие компенсации.
В опытах с перерезанными задними шейными корешками у кошек, как и в наблюдениях над обезьянами, выявилось большое значение установочных рефлексов тела и оптических установочных рефлексов. Если с помощью закрывания глаз последние выключались, оставались в основном одни установочные рефлексы тела.
Уже в предыдущей главе указывалось на то, что чем выше млекопитающее стоит на филогенетической лестнице, тем меньше по сравнению с остальными установочными рефлексами значение лабиринтных установочных рефлексов. В опытах на кошках и обезьянах с обоими удаленными лабиринтами можно ясно видеть, насколько полно выпадение лабиринтов может замещаться установочными рефлексами тела. Благодаря этому становится понятным, почему лишенные лабиринта глухонемые люди обнаруживают такие относительно незначительные нарушения установки и движений (если они не оказываются под водой).
Опыты на кошках и обезьянах, точно совпадая, показывают, что удаление обоих лабиринтов не вызывает длительного понижения тонуса мышц. В отношении собаки можно сделать такое же заключение. Как статические функции, так и сила движений туловища и конечностей не слабее, чем у нормальных животных. Этому не следует удивляться. Ведь уже от Брондгиста (Brondgeest) мы знали, что мышечный тонус в основном поддерживается афферентными нервными влияниями соответствующей стороны тела и главным образом [Шеррингтон (5)] благодаря проприоцептивным импульсам самих соответствующих мышц (34). Помимо этого, действуют возбуждения с кожи и соответствующих областей другой стороны тела (Шеррингтон, Тренделенбург, собственные опыты). Влияние тонических шейных рефлексов на тонус конечностей подробно описано в третьей главе. Таким образом, если и в норме тонические рефлексы с лабиринтов влияют на большую часть мускулатуры, то само собой становится понятным, что после их выпадения достаточно других источников, чтобы предохранить мышцы от расслабления. В соответствии с этим Кольнер и Гофман нашли, что у кролика токи действия мышц глаза не обнаруживают после двусторонней утраты лабиринта ослабления или замедления тетануса покоя. В этой связи интересно наблюдение, что тонус шейных мышц не снижается, если,
Восьмая глава. Лабиринтные рефлексы на прогрессивные движения
351
кроме обоих лабиринтов, выключаются афферентные нервы шеи в виде трех верхних пар задних корешков. У децеребрированных животных (кошки, кролики, собаки) экстирпация обоих лабиринтов также не снимает децеребрационной ригидности независимо от того, предпринимается ли удаление сразу или спустя более или менее долгий срок после де- цереб рации.
Даже сразу после одномоментного удаления обоих лабиринтов у кошек не удается установить значительного снижения мышечного тонуса. Ноги могут интенсивно разгибаться, и различные движения (кивание головой, эпизодическое перекатывание и др.) выполняются с большой силой.
В общем оказывается, что выпадение обоих лабиринтов может компенсироваться вмешательством известных и доступных весьма детализированному исследованию в эксперименте механизмов.
Восьмая глава
ЛАБИРИНТНЫЕ РЕФЛЕКСЫ НА ПРОГРЕССИВНЫЕ ДВИЖЕНИЯ (51)
В девятой главе будут описаны исходящие из лабиринтов рефлексы положения, вызываемые отолитами и имеющие значение для установки тела, лабиринтные же рефлексы движения являются преимущественно реакциями с полукружных каналов. Несмотря на то что рефлексы движения имеют для установки тела в положении покоя только второстепенное значение и поэтому мало освещены в этой книге, все же необходимо подробнее остановиться на одной группе лабиринтных двигательных рефлексов, прежде чем разбирать вопрос о зависимости лабиринтных рефлексов от отолитов. Это и есть реакции на (прямолинейные) прогрессивные движения.
Причина этого заключается в том, что до сего времени отолитовые аппараты рассматривали как единственный источник вызываемых при этом возбуждений. До сих пор анализировались только ощущения, наступающие при прогрессивных движениях. Мах (1—3), Брейер (3) и другие показали, что при прогрессивных движениях лабиринтами воспринимаются только изменения скорости. Так как полукружные каналы служат для восприятия угловых ускорений и в них из-за их строения (схематически) по физическим законам при прямолинейных сдвигах не должны возникать токи жидкости или разница в давлении, то считали невозможным, чтобы полукружные каналы возбуждались при прогрессивных движениях, и на этом основании заключили, что отолиты следует рассматривать как место возникновения лабиринтных ощущений при прогрессивных движениях. Брейер это высказал уже определенно в своей последней большой работе (1891), и все остальные исследователи были с ним согласны. В дальнейшем будет показано, что анатомо-физические предположения здесь неверны и что аппарат полукружных каналов может очень хорошо реагировать на прямолинейные движения. Поэтому необходима экспериментальная проверка этого вопроса.
352 Восьмая глава. Лабиринтные рефлексы на прогрессивные движения
В то время как о других вызываемых лабиринтами реакциях и восприятиях имеется очень большая литература с огромным фактическим материалом, наши познания о реакциях на прямолинейные движения, исходящих из лабиринтов, до сего времени были довольно ограниченными. Это показывают обобщения В. Нагеля (2), Крейдля (2) и Барани(2). Особенно незначительны данные об объективных двигательных реакциях, которые вызываются у подопытных животных прогрессивными движениями.
Н. Ах (из лаборатории Эвальда) сообщает, что у лягушек, сидящих на стеклянной пластине, после быстрых движений вверх и вниз наблюдается реакция глазного яблока и смыкание век. Реакция эта появляется и при движениях вперед и назад, вправо и влево, правда в этом случае после прекращения движения. Она отсутствует при двусторонней лабиринтэктомии. Если удалялись с двух сторон отолиты, «в то время как сохранялась остальная перепончатая часть лабиринта», причем sacculus открывался часовым пинцетом и удалялась отолитовая масса (иногда даже с помощью тонкой водяной струи), то через 1—2 недели вращательные реакции были почти или совсем нормальными, зато отсутствовал или был очень ослабленным (!) рефлекс век. Из этого Ах сделал заключение, что смыкание век после прогрессивных движений вызываетсд отолитами, вследствие чего он увидел в своих опытах экспериментальное подтверждение теории Брейера. К сожалению, здесь отсутствует анатомический контроль, поэтому, с одной стороны, не установлена полнота удаления отолитов, с другой — не исключена возможная регенерация последних (через 1—2 недели). Поэтому требуется срочное повторение этих опытов с последующим микроскопическим контролем.
В. Мулдер провел в лаборатории Цваардемакера следующие наблюдения: если посадить в ящик морскую свинку и ее голову высунуть через отверстие наружу, то можно видеть при вертикальном движении вниз подъем головы, при движениях вверх — опускание головы. У этих рефлексов измерялось латентное время, но у животных не были удалены лабиринты, чтобы показать, что речь здесь идет о лабиринтных рефлексах. (Далее будет показано, что это действительно лабиринтные рефлексы).
Грэхем Броун (1) в лаборатории Эвальда прикреплял лягушек на конце рычага длиной в 2.7 м и, поворачивая его, передвигал лягушек назад и вперед. Он наблюдал изменения легочного дыхания, которые отсутствовали после двусторонней экстирпации лабиринтов, однако после удаления одних отолитовых мешочков это изменение дыхания еще наблюдалось, но только в том случае, если после этого сохранялись вращательные реакции. К сожалению, здесь отсутствует, во-первых, анатомический контроль удаления отолитов (по-видимому, неполное) и, во-вторых, доказательства, что вращение на длинном рычаге действует как прямолинейное прогрессивное раздражение, а не как вращательное раздражение. Однако Грэхем Броун пришел к правильному выводу, что и полукружные каналы могут возбуждаться прогрессивными движениями. Этот же автор (2) описал позже у морских свинок реакцию задних ног, когда животных передвигали прямолинейно, в различных направлениях при висячем положении головой вверх. Однако отсутствует доказательство, что здесь дело касается лабиринтных рефлексов.
По моим сведениям, это единственные объективные наблюдения рефлексов на мышцы тела, которые (исходят из лабиринтов?) появляются
Рефлексы у морских свинок
353
у животных на прогрессивные движения. Если хотят изучать лабиринтные рефлексы на прогрессивные движения у животных, то прежде всего требуется найти реакции, которые легко вызываются, всегда появляются и можно доказать, что они исходят из лабиринтов.
А. Лабиринтные рефлексы на прогрессивные движения у морских свинок
а) Лифтная реакция. Животное сидит в нормальном положении на горизонтальной доске. Если двигать доску вертикально вверх, то в начале движения передние ноги переходят в сильное сгибательное положение и голова приближается к подставке. После прекращения лифтного движения вверх передние ноги сильно разгибаются, иногда со значительным мышечным дрожанием, передняя часть тела поднимается, голова иногда сгибается дорсально. Если реакция развилась полностью, начинают участвовать также задние ноги, и животное стоит наконец на четырех тонически вытянутых конечностях, чтобы через некоторое время снова возвратиться в состояние покоя.
Противоположная реакция происходит при лифтном движении вниз. В начале движения конечности, главным образом передние, разгибаются, передняя часть тела поднимается. После прекращения движения передние ноги переходят в состояние сгибания, голова и передняя часть тела лежат на полу. Через некоторое время животное снова принимает нормальное по л ожение.
Лифтная реакция у морских свинок различно выражена по силе: у некоторых животных наблюдаются сильное тоническое сгибание и разгибание всех четырех конечностей, у других же — реакция ограничивается передними ногами, задние ноги и голова не участвуют. Однако у нормальных животных лифтная реакция никогда полностью не отсутствует. Иногда для вызова реакции достаточно медленного движения доской, в других же случаях необходимо быстрое движение вверх и вниз. После двусторонней экстирпации лабиринтов эта реакция полностью отсутствует. Правда, при быстрых движениях доски животные как бы подбрасываются в высоту или голова пассивно ударяется о подставку. Если посадить нормальную морскую свинку и свинку с экстирпирован- ными лабиринтами на доску и производить после этого лифтные движения, то можно ясно видеть, что у одного животного наступает активная лифтная реакция, у другого же — ее нет. Уже простое наблюдение показывает, что реакции наступают не просто на движение, а на изменение движения. Они появляются в начале и после окончания лифтного движения вверх и вниз.
Изменение тонуса конечностей при лифтной реакции не зависит от появляющихся при этом движений головы. Это вытекает из того, что реакция конечностей всегда гораздо сильнее, чем шеи, и что она и тогда имеет место, когда отсутствуют всякие реакции головы. Во-вторых, всегда появляется отчетливая реакция конечностей, если голову держать рукой и этим препятствовать каждому изменению положения головы по отношению к туловищу. Появление лифтной реакции не зависит от оптических возбуждений, она неизменно наступает, если закрыть глаза животного.
Реакция зависит от движения головы в дорсо-вентральном направлении. Она появляется и при других положениях животного в пространстве, если голова смещается с определенной скоростью в дорсо- вентральном направлении.
23 Р. Магнус
354 Восьмая глава. Лабиринтные рефлексы на прогрессивные движения
Если, например, поместить животное в висячее положение головой вверх и мордочку направить вертикально вверх (положение головы +90°), то тогда реакция вызывается движением животного в горизонтальной плоскости в вентральную и дорсальную стороны. При вентральном движении разгибание происходит в начале движения, а сгибание — после его прекращения. При дорсальном смещении сгибание наблюдается в начале, а разгибание конечностей после прекращения движения.
Если поместить животное в положение на спину, то при движении в дорсальную сторону (значит, вниз) наступает сгибание в начале и разгибание после прекращения движения. Смещение животного в вентральном направлении (значит, вверх) вызывает разгибание в начале и сгибание после прекращения движения. Из этого также вытекает, что здесь речь идет о лабиринтных реакциях, а не о пассивных бросаниях, выполняемых экспериментатором.
В общем можно сказать, что реакция наиболее сильно выражена тогда, когда животное нормально сидит на горизонтальной доске, что она, напротив, наиболее слабая при висячем положении животного головой вверх и при положении на спине, может быть потому, что это неудобные положения для животного, при которых рефлекс более или менее тормозится. Возможно поэтому реакция и вызывается с трудом, если животное находится в положении на боку.
Еще более слабая реакция наблюдается, если смещать голову не в вентро-дорсальном, а окципитально-назальном направлении в пространстве. Если, например, животное держать в висячем положении головой вверх и привести его голову в нормальное положение (180°), то при вентродорсальном перемещении животного (в горизонтальной плоскости) наступает только одна непостоянная реакция и притом такая, что в начале дорсального смещения проявляется разгибание конечностей; однако рефлекс все же гораздо слабее, чем при смещении головы в дорсо-вент- ральном направлении.
б) Растопыривание пальцев. Морскую свинку держат правой рукой со стороны спины под плечом, так что она свободно висит головой вверх. Голова находится при этом в нормальном положении. Пальцы обеих задних лап путем нежного поглаживания приводятся в соприкосновение друг с другом. Если теперь сделать легкое движение вниз, то пальцы сразу же расходятся. Эта реакция наблюдается не у всех животных, однако у большинства ее можно вызвать. Обычно достаточно совсем незначительного движения, чтобы вызвать реакцию, которая была уже описана Грэхемом Броуном (2).
Растопыривание пальцев наблюдается при начале движения. Если двигать животное в этом же положении вертикально вверх, то также отмечается растопыривание пальцев. Оно происходит в зависимости от чувствительности животного или в начале движения или только после прекращения его.
После экстирпации обоих лабиринтов растопыривание пальцев отсутствует. Если таких безла- биринтных животных очень резко опускать вниз, то у чувствительных животных может наступить в конце движения сгибательный рефлекс задних ног. Следовательно, он зависит не от лабиринтов, а как-то вызван внезапным толчком, может быть даже органами брюшной полости.
Описанное растопыривание пальцев наступает и при фиксированной голове, т. е. когда исключены движения шеи.
в) Готовность к прыжку. Если держать животное за таз: в висячем положении головой вниз, то голова сгибается дорсально
Рефлексы у морских свинок
355
вследствие лабиринтных установочных рефлексов, так что ротовая щель находится при этом в полунаклонном положении. Если теперь двигать животное вертикально вниз, то передние ноги в плечевом суставе идут вперед и передние конечности в целом разгибаются. Иногда наблюдается и растопыривание пальцев. Данная реакция должна привести к тому, чтобы животное было в состоянии при прыжке вниз удержать вес тела передними ногами, этим объясняется название — готовность к прыжку. Этот рефлекс чрезвычайно чувствителен и отчетливо появляется нри незначительном движении вниз. Он отсутствует при экстирпации обоих лабиринтов. Поэтому если безлабиринтное животное прыгает вниз со стула или шкафа, то при этом сильно ударяется о пол, а не эластично «улавливает» прыжок передними ногами, как нормальное животное; это обусловливается не общей потерей тонуса мышц, как предполагали прежние исследователи, а отсутствием этого рефлекса на прогрессивные движения вниз.
Реакция бывает и тогда, когда голова фиксирована и этим исключаются движения шеи, она наступает и при закрытых глазах.
Сгибание передних ног происходит в том случае, если животное двигать в том же положении вертикально вверх; тогда передние ноги в плечевом суставе идут назад и более или менее сгибаются.
Отчетливо удается установить, что готовность к прыжку при вертикальных движениях вниз наступает уже в начале движения. Речь идет, по-видимому, о той же реакции, как при лифтном движении вниз, которая вследствие положения животного на весу вниз головой проявляется главным образом на передних ногах.
г) Прочие рефлексы на прогрессивные движения. , У чувствительных морских свинок еще удалось установить различные реакции на прогрессивные движения в разных направлениях. Однако они или не достаточно отчетливы, чтобы могли быть использованы для точного исследования, или непостоянны. Бывает и так, что в отдельных случаях нельзя с уверенностью решить, вызываются ли они лабиринтами или другими органами чувств.
По этой же причине для наших целей не применима и реакция, демонстрированная Грэхемом Броуном. Если поддерживать животное под плечо в висячем положении головой вверх и затем двигать горизонтально вправо и влево, то последует асимметричная реакция задних ног. В большинстве случаев впереди идущая нога разгибается, отстающая сгибается. Однако эта реакция ни в коем случае не постоянна; кроме того, она при фиксации таза сильно затормаживается, этим также создается препятствие движениям подбрасывания задней части тела, которые/ в свою очередь могли бы вызвать рефлексы. Но я не сомневаюсь в том,, что эта ^реакция вызывается лабиринтами.
Наблюдающиеся при лифтной реакции движения мышц плеча можно отчетливо почувствовать, наложив на них палец, в виде мышечного напряжения (51). У безлабиринтных животных это в большинстве случаев не наступает. Имеются очень возбудимые морские свинки, и без лабиринтов проявляющие при лифтных движениях напряжение мышц посредством рефлексов с других рецепторов. Поэтому эта очень чув- ствительная реакция не подходит для решающих опытов.
Движения глаз у морских свинок , при прогрессивных движениях не удается наблюдать при простом рассматривании без точного измерения инструментами.
23*
356
Восьмая глава. Лабиринтные рефлексы на прогрессивные движения
Обобщая, можно сказать, что наиболее сильные реакции на прогрессивные движения происходят тогда, когда голова двигается в дорсо- вентральном направлении. Это отчетливо проявляется как при лифтной реакции, так и при растопыривании пальцев и рефлексе готовности к прыжку, в то время как смещения головы в окципито-назальном направлении вызывают только более слабые рефлексы. Какой из описанных рефлексов проявится в каждом отдельном случае, зависит от внешних причин: лифтная реакция, если животное сидит на подстилке, готовность к прыжку при свободном движении в воздухе, главным образом при висячем положении головой вниз, растопыривание пальцев — при висячем положении головой вверх.
Б. Кролик
У кролика лифтная реакция проявляется очень отчетливо. Если Животное посадить в нормальном положении на доску, то при движении вверх вначале наблюдается сгибание четырех конечностей, в конце движения — очень сильное разгибание с напряжением мышц. При этом животное часто поднимается с подстилки и стоит некоторое время на сильно разогнутых конечностях, пока оно снова не «осядет». При лифт- ном движении вниз происходят тоническое разгибание в начале и сгибание в конце прогрессивного движения. Эту реакцию можно вызвать и на передних ногах, если животное свободно держать в воздухе, поддерживая его со стороны живота. Тогда в начале прогрессивного движения вниз появляется разгибание передних ног (готовность к прыжку), в начале прогрессивного движения вверх — сгибание передних ног.
Если кролика держать за таз свободно в воздухе головой вниз, то при вертикальном движении вниз наблюдается, кроме того, еще своеобразная реакция задних ног. Последние в начале движения ра'згибаются в тазобедренном суставе назад (каудально), так что появляется рефлекс прыжка. При вертикальном движении вверх этот рефлекс прыжка наблюдается только после прекращения движения.
Все эти рефлексы появляются тогда, когда голова фиксируется по отношению тела так, чтобы исключались движения шеи, и тогда, когда закрыты глаза. Они полностью отсутствуют у кролика после двусторонней экстирпации лабиринтов.
Для демонстрации лифтной реакции лучше всего подходят кролики. И у кролика не удалось установить рефлексы на глаза при прогрессивных движениях с помощью простого рассматривания без особых приспособлений. Флейш доказал с помощью зеркальной регистрации у кроликов, у которых были выключены оптические впечатления пришиванием пелота на роговицу глаза, что при прямолинейном боковом смещении в горизонтальной плоскости привязанных на животе животных впереди идущий глаз в начале движения отклоняется вниз, после прекращения движения — вверх. Глаз противоположной стороны двигался в обратном направлении. Экскурсии настолько малы, что их можно было установить только таким образом, и они подавляются оптической фиксацией.
В. Кошка
Для исследования подходят ручные кошки, у которых можно закрывать глаза колпаком. У этих животных отчетливо проявляется лифтная реакция, притом точно таким же образом, как это было описано у морских
Рефлексы у собак
357
свинок и кроликов. В реакции всегда участвуют все четыре конечности. Она наступает и тогда, когда фиксирована голова, чем исключаются все движения шеи.
Если животное (можно и с фиксированной головой) держать за таз в висячем положении головой вниз, то на вертикальное движение вниз последует очень сильный рефлекс готовности к прыжку. Пальцы растопыриваются, передние конечности двигаются в плечевом суставе вперед, и одновременно сгибаются локти, так что они готовы принять вес тела при прыжке вниз. Реакция появляется в начале движения.
Если животное держать в висячем положении головой вверх или в нормальном положении в воздухе, то наблюдается растопыривание пальцев на всех конечностях при вертикальном движении вниз.
При свободном падении в воздухе описанные рефлексы становятся очень отчетливыми. На рис. 112 (стр. 184) показано, что пока животное падает теменем вниз, ноги сгибаются, однако они разгибаются, если голова повернулась и нижняя челюсть направлена вниз. При этом передние ноги переходят в готовность к прыжку, так что они при становлении на пол ловят вес тела, в то время как при отсутствии этих рефлексов животное ударяется головой о пол.
Г. Собака
У собаки, кроме до сих пор описанных реакций на конечности, появляются также отчетливые движения головы. Однако следует подчеркнуть, что реакции конечностей собак также являются прямым следствием возбуждения лабиринтов. Они наблюдаются неизменно, если голова фиксирована по отношению туловища и этим выключаются движения шеи. Все реакции на прогрессивные движения наблюдаются у собаки и после закрытия глаз колпаком. Они полностью отсутствуют после двусторонней экстирпации лабиринтов.
Лифтная реакция. Если собаку посадить в нормальном положении на доску и выполнять лифтные движения вверх, то в начале движения — сгибание лап и вентральное движение головы, в конце — разгибание лап и дорсальное движение головы. Если лифтная реакция производится вниз, то в начале наступает разгибание лап и дорсальное сгибание головы, в конце же — сгибание лап и вентральное сгибание головы. Реакция всегда сильнее на передних ногах, чем на задних.
Если собаку свободно держать в воздухе в висячем положении головой вверх, то при вертикальном движении вверх происходит в начале движения вентральное сгибание головы, в конце — дорсальное. Конечности в начале движения сгибаются, задние ноги, кроме того, приводятся (adduziert). Если собаку передвигают вертикально вниз, то в начале движения наблюдается дорсальное сгибание головы, в конце — вентральное. Конечности в начале движения разгибаются, пальцы растопыриваются, задние ноги, кроме того, отводятся. Если при висячем положении головой вверх собаку перемещать в сторону, то происходит поворот головы по направлению движешь который становится особенно отчетливым после прекращения движения. Если собаку в висячем положении головой вверх передвигать по направлению вперед (вентрально), то голова в начале движения идет вперед, в конце — сгибается назад.
358
Восьмая глава. Лабиринтные рефлексы на прогрессивные движения
Д. Обезьяна (59)
Для исследования этих реакций у обезьяны лифтная реакция мало пригодна, так как животные, если их поставить на доску и на ней поднимать в воздухе вертикально вверх и вниз, обычно предпринимают попытки к побегу. Однако у ручных животных удается доказать, что при лифтном движении вверх в конце движения происходит слабое разгибание конечностей. При лифтном движении вниз разгибание наблюдается в начале движения и видно отчетливее на передних ногах, чем на задних. Гораздо лучше удается исследовать реакции на прогрессивные движения, если животное свободно находится в воздухе (готовность к прыжку). Если держать животное в нормальном положении (позвоночник горизонтально) за кожу спины в воздухе и производить прогрессивное движение вниз, то конечности разгибаются и часто при этом растопыриваются пальцы. Следовательно, здесь реагируют все четыре ноги. Если же держать животное в поднятом положении головой вверх, то при вертикальном движении вниз реагируют только задние ноги и притом они разгибаются в тазобедренном, коленном и голеностопном суставах, одновременно растопыриваются пальцы. Таким образом, задние ноги приходят в готовность «поймать» вес падающего тела при соприкосновении с полом. Противоположное этому происходит тогда, когда животное висит головой вниз и производятся вертикальные движения вниз. В этом случае реагируют только передние ноги. Они также разгибаются, причем двигаются в направлении пола. Пальцы растопыриваются. И в этом случае конечности приготовляются в конце прыжка «поймать» вес тела.
Эта реакция проявляется также отчетливо, если зашиты веки; следовательно, она не зависит от оптических впечатлений. Речь идет об очень чувствительном рефлексе, для вызова которого достаточно незначительного вертикального движения. После экстирпации больших полушарий эта реакция также сохраняется, у одной таламической обезьяны она наблюдалась через 7 часов после операции.
После односторонней экстирпации лабиринтов реакции на прогрессивные движения сохраняются без изменения и появляются, насколько это удалось установить на двух животных, с одинаковой интенсивностью на правых и левых конечностях. Они появлялись и тогда, когда веки были зашиты. Однако эта реакция отсутствует после двусторонней экстирпации лабиринтов. Это отчетливо удается показать у безлабиринтных обезьян с закрытыми глазами. После экстирпации больших полушарий у безлабиринтных обезьян реакция на прогрессивные движения отсутствует. У интактных безлабиринтных животных с открытыми глазами в большинстве случаев при прогрессивных движениях реакции также не наблюдается. Только два раза у одного животного можно было видеть, что непосредственно перед соприкосновением с полом происходило растопыривание рук, обусловленное оптическим восприятием, так как при закрытых глазах этого не наблюдалось.
Наблюдения на свободно двигающейся и прыгающей безлабиринтной обезьяне, находящейся на исследовании в течение двух месяцев, показали, что отсутствие реакций на прогрессивные движения после выключения лабиринтов очень мешает животному и что замена этих реакций с помощью оптических впечатлений не достигает цели.
Поэтому когда безлабиринтная^ обезьяна в нормальном положении приземляется при прыжке, то она все же это совершает всегда с сильно слышимым ударом, так как конечности не могут принять правильное
Лабиринтные рефлексы после выключения отолитов
359
положение для «улавливания» веса падающего тела. Из этого видно, какое большое значение имеют для свободно живущих обезьян описанные рефлексы на прогрессивные движения.
Из этого обозрения вытекает, что у пяти обследованных видов животных очень отчетливо выступают реакции на прогрессивные движения, которые, как доказано исследованиями, вызываются с лабиринтов и хорошо поддаются дальнейшему анализу.
После перерезки соответствующих данной конечности задних корешков (34, 51) рефлексы на прогрессивные движения появляются еще отчетливее и в соответствии с имеющейся при этом атаксией — с утрированными экскурсиями. Это удались наглядно показать на кошках (лифт- ная реакция, готовность к прыжку) после односторонней перерезки от С7 до Th2 и С6 до Th2. Ввиду того что двуглавая мышца иннервируется главным образом от С8, трехглавая — от Th1? мышцы, двигающие локоть, по-видимому оказываются десенсибилизированными, поэтому очень доказательным является появление обильных движений в локтевом суставе.
После экстирпации больших полушарий (51) у кроликов, кошек, собак и обезьян прогрессивные реакции остаются в таком же объеме.
После (не совсем полной) экстирпации мозжечка (51) у собаки (Дюс- сер де Баренн) прогрессивные реакции на голову и конечности выражены очень сильно. Они появляются и после закрытия глаз. Реакция передних ног при этих обстоятельствах не зависит от движений головы.
Поведение животных после односторонней экстирпации лабиринта должно быть еще изучено. Точно установлено, что с одного лабиринта могут быть вызваны лифтная реакция и готовность к прыжку на конечностях обеих сторон тела. Вопрос о том, становятся ли реакции слабее или временно асимметричными, требует еще дальнейшего исследования. Так как результат исследования имеет большой теоретический интерес, то необходимы более расширенно поставленные опыты для окончательного обобщения.
ЛАБИРИНТНЫЕ РЕФЛЕКСЫ НА ПРОГРЕССИВНЫЕ ДВИЖЕНИЯ ПОСЛЕ ВЫКЛЮЧЕНИЯ ОТОЛИТОВ
В следующей главе при обсуждении функции отолитов будет показано, что у морской свинки после сильного центрифугирования вращательные реакции головы и глаз остаются без изменения, в то время как непосредственно после вмешательства исчезают все тонические лабиринтные рефлексы: тонические лабиринтные рефлексы на конечности, лабиринтные установочные рефлексы и компенсаторные положения глаз.
В тех случаях, когда тонические лабиринтные рефлексы отсутствовали несколько дней, а вращательные реакции оставались без изменения, микроскопическое исследование лабиринтов показало, что отолиты в большинстве случаев были отброшены или в отдельных случаях их функция выключалась кровоизлияниями. Из этого вытекает, что вращательные реакции в соответствии со всеми принятыми мнениями являются рефлексами полукружных каналов. Тонические же лабиринтные рефлексы вызываются с отолитов.
Предпринятые на этих животных исследования лабиринтных рефлексов на прогрессивные движения показали, что эти рефлексы ведут
360
Восьмая глава. Лабиринтные рефлексы на прогрессивные движения
себя так же, как и вращательные реакции, а не как тонические лабиринтные рефлексы. Из этого следует сделать вывод, что лабиринтные рефлексы на прогрессивные движения являются реакциями полукружных каналов и могут проявляться и после выключения функции отолитов.
Сразу после центрифугирования животного рефлексы на прогрессивные движения сохранены так же, как и вращательные реакции. Наблюдения на морских свинках показали, что сразу после центрифугирования полностью отсутствовали тонические лабиринтные рефлексы, а вращательные реакции и реакции на прогрессивные движения сохранялись. Примером могут служить следующие два протокола опытов.
Морская свинка № 38. 6 мая 1920 г. 10 час. До центрифугирования все лабиринтные рефлексы нормальны: как вращательные, так и рефлексы на прогрессивные движения (лифтная реакция, растопыривание пальцев и готовность к прыжку), а также тонические рефлексы на конечности, лабиринтные рефлексы выпрямления и компенсаторные положения глаз.
10 ч. 30 м. Центрифугирование в течение 1.5 мин. со скоростью 990 м в минуту.
11 час Исчезли все тонические лабиринтные рефлексы: тонические лабиринтные рефлексы на конечности, лабиринтные установочные рефлексы и компенсаторные положения глаз. Зато сохранились вращательные реакции головы и глаз; кроме того, удается наблюдать при прогрессивных движениях (вертикальное опускание) очень отчетливое растопыривание пальцев и готовность к прыжку.
Исследование с таким же результатом было повторено в И ч. 30 м. Теперь лифтная реакция явно положительна и тогда, когда голова фиксируется и глаза закрыты. После этого животное децеребрируют и показывают, что полностью отсутствуют тонические лабиринтные рефлексы на конечности.
Результат. После центрифугирования исчезли все тонические лабиринтные рефлексы, вращательные же реакции и реакции на прогрессивные движения в противоположность этому полностью сохранены.
Морская свинка № 41. Исследование 7 мая показало, что имеются все лабиринтные рефлексы. 8 мая, 9 ч. 30 м. — центрифугирование. 12 час. Наблюдаются вращательные реакции головы и глаз, при лифтном движении вниз отчетливы лифтная реакция и растопыривание пальцев. Отсутствуют рефлекс готовности к прыжку и все тонические лабиринтные рефлексы, слуховая реакция положительна. После децеребра- ции отсутствовали тонические лабиринтные рефлексы на конечности.
Результат. После децеребрации остались сохраненными вращательные реакции и реакции на прогрессивные движения, отсутствовали тонические лабиринтные рефлексы.
Из этих опытов вытекает, что после центрифугирования животных рефлексы на прогрессивные движения ведут себя так же, как вращательные реакции, и не так, как тонические лабиринтные рефлексы.
Как будет показано в следующей главе, у ряда животных через несколько дней снова восстанавливались тонические лабиринтные рефлексы. Микроскопическое исследование лабиринтов у этих животных показало, что отолиты находятся на своем месте и, следовательно, ма- кулы были функционально выключены после центрифугирования только временно из-за сотрясения специфически тяжелых отолитов. У других, же животных тонические лабиринтные рефлексы исчезают навсегда. При исследовании под микроскопом в этих случаях было обнаружено, что- отолиты или отброшены, или по крайней мере повреждены, или что на этих местах были кровоизлияния и таким образом анатомически выключена деятельность отолитов.
В тех случаях, в которых анатомически было. доказано выключение отолитов, реакции на прогрессивные движения сохранялись. Примером может служить морская свинка № 8, у которой особенно отчетливо была видна эта реакция. Для изучения подробностей мы ссылаемся на следующую главу.
Лабиринтные рефлексы после выключения отолитов
361
У морскойсвинки№ 8 проявляются до центрифугирования 10 февраля все лабиринтные рефлексы. В тот же день после центрифугирования вращательные реакции головы и глаз были положительными, рефлексы на прогрессивные движения (а именно: лифтная реакция и растопыривание пальцев) отчетливо выражены (рефлекс готовности к прыжку тогда не исследовался), зато исчезли все тонические рефлексы. На следующий день реакция на вращение как головы, так и глаз положительна, рефлексы на прогрессивные движения стали слабыми, тонические лабиринтные рефлексы полностью отсутствуют. 12 февраля вращательные реакции еще положительны, реакции на прогрессивные движения (лифтная реакция и растопыривание пальцев) выражены очень сильно, тонические рефлексы отсутствуют. После децеребрации также отсутствуют тонические лабиринтные рефлексы на конечности.
Микроскопическое исследование показывает, что все отолиты отброшены и аппарат полукружных каналов с обеих сторон остался интактным (см. рис. 201—207, стр. 409— 413).
Результат. После центрифугирования отолиты удалены, полукружные каналы остались интактными. Лабиринтные рефлексы на прогрессивные движения у этих животных сохранились.
Опыт на морской свинке № 35 показывает то же самое. После центрифугирования вращательные реакции сохранены, однако диагностически имеются нарушения со стороны полукружных каналов. Реакции на прогрессивные движения положительны, тонические рефлексы отсутствуют. Через два дня после центрифугирования наблюдаются реакции на прогрессивные движения, отсутствуют только лифтная реакция и тонические рефлексы. После децеребрации не удается установить никаких тонических лабиринтных рефлексов на конечности.
Микроскопические исследования показывают, что справа отсутствуют отолиты, слева они отчасти отсутствуют, отчасти повреждены и выключены кровоизлияниями.
Результат. После разрушения отолитов еще удается наблюдать рефлексы на прогрессивные движения.
Из этих и других опытов со всей отчетливостью видно, что лабиринтные рефлексы на прогрессивные движения: лифтная реакция, рефлекс готовности к прыжку и растопыривание пальцев у морской свинки — являются рефлексами полукружных каналов и могут наблюдаться после удаления отолитов. Так как эти рефлексы отсутствуют после экстирпации лабиринтов, можно сделать точный вывод, что здесь речь идет о реакциях полукружных каналов.
Другой вопрос: не исходят ли и из отолитов возбуждения, которые поддерживают эти лабиринтные рефлексы на прогрессивные движения. Мы, кроме того, считаем, что при определенном движении могут совместно проявляться (для достижения одной конечной цели) рефлексы как полукружных каналов, так и отолитов. Если, например, перевести голову животного из нормального положения в боковое, то этим движением прежде всего вызывается реакция полукружных каналов, благодаря которой глаза приходят в то положение, в котором они могут удерживаться и тоническими компенсаторными положениями глаз, вызванными с отолитов. Что-нибудь подобное можно себе, конечно, представить и при прогрессивных движениях.
Если, например, голова находится в нормальном положении, то отолиты utriculus’a стоят горизонтально и давят на чувствительные пятнышки. Если производить лифтное движение вниз, то отолиты должны в начале движения произвести прежде всего легкое оттягивание макул, а этим вызвать, как будет показано в следующей главе, разгибание передних конечностей. Зато в конце движения отолиты будут давить сильнее на макулы, чем обусловливается уменьшение разгибательного тонуса передних конечностей. При лифтном движении вверх происходят противоположные явления.
Вполне возможно, что при прогрессивных движениях отолиты также могут участвовать тем или другим образом. Однако несомненно, что после полного выключения деятельности отолитов у морской свинки
362
Восьмая глава. Лабиринтные рефлексы на прогрессивные движения
вышеописанные реакции остаются без изменения. Следовательно, здесь речь идет в основной своей части о реакциях полукружных каналов.
Флейш (Fleisch) признал наблюдаемые им при горизонтальных прогрессивных движениях вертикальные отклонения глаз за отолитовые рефлексы, не приводя для этого объяснения. Ему следовало бы показать, что эти рефлексы отсутствуют после повреждения отолитов.
Рис. 169.
Может ли аппарат полукружных каналов возбудиться прогрессивными движениями?
Известно общепринятое мнение, которое было еще высказано Махом и Брейером, что физически невозможно возбудить аппарат полукружных каналов прогрессивными движениями. Если шарообразное полое пространство, наполненное жидкостью, передвигать по прямой, то действительно токи жидкости не могут появиться. В существующих до настоящего времени предположениях всегда принималось, что лабиринты или аппараты полукружных каналов соответствуют этой простой модели. Однако это не так: аппарат полукружных каналов представляет собой сложную систему заполненных жидкостью трубочек, в котором находится перепончатый лабиринт, наполненный изнутри жидкостью и лежащий также в жидкости (см. следующую главу). Перепончатый аппарат полукружных каналов подвешен на плотной стенке костного лабиринта сложной системой нитей и мембран. Перилимфатическое пространство его отделено пограничной мембраной от перилимфатического пространства cochlea и sacculus’a [де Бурле (86)]. Самым важным для наших целей кажется то, что ни перилимфатическое, ни эндолимфати- ческое пространства «не закрыты намертво» от окружающей среды. Перилимфатическое пространство имеет связь со средним ухом, во-первых, через пограничную мембрану и, во-вторых, через овальное и круглое окно, они эластичны и податливы. Эндо лимфатическое пространство связано через эндолимфатический проток с эндолимфатическим мешочком в полости черепа, он является более или менее эластическим резервуаром, который сообщается с внутренним пространством полукружных каналов и остальным лабиринтом. Итак, мы имеем пространство с плотными стенками, заполненное жидкостью, у которого перилимфа связана с внешней средой через эластические окошечки, а эндолимфа — через эндолимфатические мешочки.
Проф. Орнштейн (Ornstein) и д-р Бургер (Burger) в Утрехте предприняли исследования по этому вопросу и построили модель (рис. 169). Результат был следующий. Модель костного аппарата полукружных каналов сделана из стекла (а), в ней подвешена модель перепончатого аппарата полукружных каналов из резины (6), в которой находится ампула из стекла (с) с эластичной купулой (d). Перилимфатическое пространство этой модели соединено с внешней средой через окошечко, закрытое эластичной резиновой мембраной (е), в то время как внутреннее пространство перепончатого лабиринта соединено с прикрепленным с внешней стороны стеклянной трубки резиновым мешочком (ƒ). Такая модель реагирует чрезвычайно отчетливо на прогрессивные движения коле¬
Анатомические замечания
363
баниями купулы, которые появляются как в начале, так и в конце движения. Кроме того, видно, что при этих прогрессивных движениях весь перепончатый лабиринт смещается в костных полукружных каналах. Такого рода смещения, конечно, предотвращаются в настоящих полукружных каналах благодаря тому, что перепончатый лабиринт висит :на вышеупомянутых нитях и мембранах. Вследствие этого при определенных обстоятельствах колебания купулы должны быть еще сильнее.
Эту модель, конечно, необходимо разработать дальше. Должны быть точно взяты действительные формы вестибулярного аппарата и его действительные размеры, точно учтены коэффициент эластичности и остальные физические константы отдельных частей внутреннего уха. Также, конечно, должна быть выработана физическая теория наблюдающихся при этом движений и сил. Однако уже теперь можно с уверенностью сказать, если принять, что перил имфатическое пространство соединено через пограничную мембрану и эластические окошечки со средним ухом, а эндолимфатическое пространство через эндолимфа- тический мешочек с полостью черепа, что имеются физические предпосылки, которые делают возможным реакцию аппарата полукружных каналов на прогрессивные движения.
Кроме этих опытов с моделями, для полного понимания прогрессивных реакций требуются еще наблюдения на животных с одним лабиринтом и, кроме того, точные опыты с вращательными реакциями на конечности, для того чтобы последние можно было бы связать с прогрессивными реакциями на конечности.
Однако наблюдения уже сейчас показывают, что у подопытных животных удавалось вызвать прямолинейными ускорениями ряд объективных рефлексов, исходящих из лабиринтов и остающихся после разрушения отолитовых мембран. Таким образом, выходит, что полукружные каналы первоначально принимают участие в их возникновении.
Девятая глава
ФУНКЦИЯ отолитов
А. Предварительные анатомические замечания
На рис. 170 представлена изготовленная де Бурле и де Клейном (de Burlet и. de Kleyn) восковая модель перепончатого лабиринта кролика в 20-кратном увеличении (23), установленная в правильном положении по отношению к сфотографированному рядом черепу кролика, ротовая щель которого опущена на 40° ниже горизонтали [соответственно данным де Бурле и Костера (84)]. При стереоскопическом рассматривании можно видеть три полукружных канала в их взаимно-перпендикулярном положении. Все они входят в utriculus. Горизонтальный полукружный канал расположен почти горизонтально, его ампула впереди. У переднего вертикального полукружного канала ампула также находится впереди, зато у заднего вертикального полукружного канала она сзади (позади середины горизонтального полукружного канала). Оба вертикальных полукружных канала соединяются сзади в общую ножку (crus commune), которая сверху также впадает в utriculus и не имеет ампулы. Рядом с общей ножкой виден восходящий эндолимфа¬
364
Девятая глава. Функция отолитов
тический проток (ductus endolymphaticus). Ниже utriculus’а висит (в виде черной массы) sacculus.
На рис. 171 видно схематическое изображение перепончатого лабиринта и его иннервации по Бурле. Utriculus расположен посередине,.
на его основании (заштриховано крестиком) находится отолит. У переднего конца (слева) впадают ампулы горизонтального и переднего вертикального полукружных каналов, сзади (справа) — ампула заднего вертикального полукружного канала. В ампуле crista и cupula обозначены схематически. В середину ut- riculus’a впадает общая ножка обоих вертикальных полукружных каналов. Под utriculus’oM расположен sacculus. Связь между ними осуществляется посредством эндолимфати- ческого протока. Последний пробуравливает каменистую часть височной кости и заканчивается расширением — эндо лимфатическим мешком, который расположен у задней поверхности каменистой части пирамиды височной кости, под наружной твердой оболочкой (dura mater) мозга. Он представляет собой расположенную вне костного лабиринта эластичную подушку, которая свободна
Рис. 170.
соединена с эндолимфатическими пространствами sacculus’a и utriculus’a. На физиологическое, значение, этого расположения уже указывалось выше (стр. 362). Отолит sacculus’a расположен на его медиальной стедке. Он состоит йз главной части (заштриховано светлее) и дорсальных
Анатомические замечания
365
отрезков (заштриховано темнее), расположенных орально и дорсально. От sacculus’a узкий ductus reuniens ведет к улитке.
Иннервация чувствительных образований осуществляется следующим образом: проксимальный конец ствола 8-го нерва (заштриховано) проходит ganglion proximale scarpae (1), разветвляется на ramus superior, идущую к чувствительному эпителию в ампулах горизонтального и переднего вертикального полукружного каналов, далее к чувствительному пятну utriculus’а и, кроме того, дает еще боковую ветвь (4) к чувствительному эпителию дорсального отрезка sacculus’a. Дорсальный отрезок [де Бурле (83)], как и его иннервация боковой ветвью ii. utricularis [Voit, Oort (55)1, были установлены на всех до сего времени обследованных видах млекопитающих. Дорсальный ствол 8-го нерва состоит прежде всего из ramus interior (черная), которая после прохождения ganglion distale scarpae (2) иннервирует чувствительное пятно главной части sacculus’a и ампулу заднего вертикального полукружного канала (5) и посылает соединительную ветвь к ramus cochlearis (3), слуховому нерву, идущему прямо к улитке. N. vestibularis состоит, та-
ЗРис. 172. (По де Бурле). Схема правого лабиринта морской свинки. Упрощено. Эндо лимфатическое пространство обозначено черным цветом.
Перилимфатическое пространство вокруг полукружных каналов и большей части utriculus’а пронизано -многочисленными соединительнотканными тяжами и отделено от нижнего перилим- ^атического пространства пограничной мембраной. Нижнее перилимфатическое пространство не содержит соединительнотканных тяжей. Овальное и круглое окошки как бы вставлены в нижнее перилимфатическое пространство.
ким образом, из проксимального ствола и ramus interior дистального •ствола 8-го нерва.
Перепончатый лабиринт, за исключением эндолимфатического мешочка, расположен в полости костного лабиринта, который он, однако, заполняет не полностью. Промежуточные пространства между обоими лабиринтами заполнены перилимфой. Перилимфатическое пространство имеет •с внешним миром очень эластичную связь: 1) тончайшими перилимфати- пескими протоками, заканчивающимися в подпаутинных пространствах мозга; 2) через овальное окно преддверия, расположенное напротив sacculus’a и запирающееся (подвижной) пластинкой стремечка; 3) через круглое окно, запирающееся membrana tympani secundaria и отделяющее rscala tympani от среднего уха.
Исследования де Бурле (86) на морских свинках показали, что перилимфатическое пространство разделено на верхнюю и нижнюю половины. В нижней части расположены sacculus и улитка. Здесь перилимфатическое пространство в основном заполнено только жидкостью. Верхняя часть отделяется от нижней эластичной пограничной мембраной и частично нижней поверхностью utriculus’a. Над этой перегородкой перилимфати- неское пространство пронизано сетью соединительнотканных тяжей, при помощи которых полукружные каналы и utriculus прикрепляются к стенкам костного лабиринта и которые препятствуют грубым смещениям. По-видимому, такое устройство имеет большое значение для деятельности вестибулярного аппарата при движениях (и различных положениях?). Рис. 172 дает схематическое изображение этого расположения.
366
Девятая глава. Функция отолитов
Взаимное расположение utriculus’a и sacculus’a с их чувствительным: эпителием можно видеть на фронтальном срезе левого лабиринта морской; свинки (рис. 203). Срез сделан спереди, слева — медиальная стенка преддверия, справа напротив — пластинка слуховой косточки. Между ними (пустое) перилимфатическое пространство преддверия. У медиальной стенки расположен (на срезе треугольной формы) sacculus, нижняя медиальная стенка которого несет чувствительный эпителий чувствительного пятна. Препарат пересекается горизонтально пограничной мембраной, на которой находится utriculus в виде почти овальной полости. Чувствительное- пятно расположено почти горизонтально на нижней поверхности utriculus’a. Соединительнотканные перекладины верхнего перилимфати-
ческого пространства видны над utriculus’oM и справа от него.
Форма отолитов изменчива у различных видов млекопитающих.
На рис. 173 представлено правое чувствительное пятно utriculus’a морской свинки (88), если смотреть сверху, в 40-кратном: увеличении. В действительности величина поверхности составляет
0.5 мм2. Она плоская, только передняя и примыкающая части медиального края изогнуты.
На рис. 174 показано правой чувствительное пятно sacculus’a морской свинки, если смотреть с латеральной стороны (88), в 40-кратном увеличении. Действительная величина поверхности также составляет только 0.5 мм2. Чувствительное пятно растянуто спереди назад, на переднем конце завернуто лате- рально под углом примерно в 160° и имеет наверху дорсальный отрезок, который наполовину повернут вниз и образует с остальным saccu- Ius’om угол в 155°, в середине — в 140°.
Недавно Гоффману под руководством Бурле удалось отпрепарировать in situ под цейссовской лупой отолиты различных млекопитающих. На рис. 175 виден правый отолит sacculus’a in situ, на переднем конце* можно распознать завернутый кверху дорсальный отрезок.
Для исследования тонических лабиринтных рефлексов необходимо более точно знать форму и расположение чувствительного пятна отолита в черепе. Де Бурле совместно со своими сотрудниками в течение ряда лег изучал эти условия. Исследования еще не закончены, но дали уже ряд, важных результатов. Техника в течение работы постепенно развивалась. Здесь будет сообщено только главное. Подробности даны в оригиналах (83—90).
Самым главным при таких топографически микроскопических работах является то, чтобы при предварительных обработках объектов не произошло бы смещения. Располагающим к этому обстоятельством является положение чувствительного пятна sacculus’a против медиальной костной лабиринтной стенки, чувствительного пятна utriculus’a — на пограничной мембране.
Череп животного перепиливается во фронтальной плоскости до височных костей. Получается костное кольцо, закрытое снизу основанием черепа, сверху — черепной крышкой, благодаря чему можно избежать.
Спереди
Медиально
Рис. 173. (По де Бурле). Правое чувствительное пятно utriculus’a морской свинки, видно сверху. Увеличение в 40 раз.
Анатомические замечания
3,67
смещения обеих височных костей 1 по отношению друг к другу. Фиксация и дальнейшая обработка производятся по методу Виттмаака, причем сроки пребывания в различных жидкостях у маленьких объектов (морские свинки) могут быть сокращены. Самым главным является заливка
d
В
а
cf
А
- Е
е
Рис. 174. (По де Бурле). Правое чувствительное пятно sacculus’а морской свинки (увеличено в 40 раз), а -г- вид сбоку; Ъ — в плоскости: АС DE — нижняя часть чувствительного пятна; ACD — главная часть sacculus’а; С DE — часть, отогнутая от медиальной плоскости в фронтальном направлении; abde — дорсальный отрезок; с — фронтальный разрез de — fg для демонстрации угла между дорсальным отрезком и главной частью; d — фронтальный разрез аЪс\ е — действительное расстояние точек А, В и Е при увеличении в 40 раз для демонстрации угла АВЕ.
препарата до декальцинации в целлоидин для того, чтобы не изменилась правильная топография после потери костями их твердости.
После этого надо позаботиться о том, чтобы после приготовления срезов все препараты могли бы быть правильно ориентированы по отношению друг к другу. Для этого служат направляющие каналы, которые до приготовления срезов делаются в целлоидиновом блоке и которые в целях измерения должны стоять точно перпендикулярно к поверхности среза. Блок в желаемом положении (в большинстве случаев направление срезов было по возможности фронтальным) устанавливается в микротом, а затем приступают к изготовлению срезов, но вначале только через целлоидин. Как только получена гладкая поверхность среза, на
1 У более крупных животных с твердым черепом основание черепа достаточно для прочного соединения обеих височных костей.
368
Девятая глава. Функция отолитов
нее накладывается плоскопараллельный медный блок толщиной в 1 % см, содержащий много ведущих каналов к основной площади, в которую входит полая игла с острым нижним краем. Если теперь сделать прокол иглой через каналы и целлоидиновый блок, причем, конечно, обойти заложенный в него препарат, то можно получить направляющие каналы точно перпендикулярно к поверхности среза. Они потом появляются в каждом срезе в виде круглых отверстий, которые дают возможность правильно ориентировать различные срезы по отношению друг к другу.
Рис. 175а. 1 — отолито- вая мембрана sacculus’а прилегает к медиальной костной стенке; 2 — ductus cochlearis; 3 — лестница преддверия;
4 — барабанная лестница; 5 — lamina spiralis ossea; 6 — костная стенка горизонтального Рис. 175. (По Гоффману). Правый ла- канала; 7 — полостьПос- биринт кролика, открыт латерально. culus’а.
Теоретически достаточно два направляющих канала, но в большинстве случаев их применялось больше, для лучшего контроля.
Так как речь идет не о гистологических тонкостях, а о топографической точности, срезы делаются по возможности толстыми: у кролика — 50—100 ц, у морской свинки — 50—60 ц. Тогда срезы легко поддаются полному сглаживанию и ложатся без складок; отолитовые мембраны лежат неповрежденными на сохраненном чувствительном эпителии. Если проецировать препараты один за другим на плоскость срисовыванием, то круги, соответствующие направляющим каналам, должны точно совпадать.
Эти серии срезов можно использовать для конструкции восковых моделей, причем направляющие каналы должны построиться вертикально. Такая модель представлена на рис. 170. Модели эти очень нужны для ориентации, но их нельзя использовать для точного измерения. Также невозможно измерить непосредственно на срезах углы, которые отолитовые макулы образуют друг по отношению друга и по отношению к медиальной плоскости. Это подтверждает простой пример. Представьте себе прямоугольный угол в комнате и проложите через него горизонтальную плоскость (т. е. плоскость, перпендикулярную линии среза обеих стен). Обе стены комнаты образуют в этой плоскости угол в 90°. Однако во всех других плоскостях, которые пройдут через угол, этот угол будет все мень-
Анатомические замечания
369
шим и под конец достигнет 0, если плоскость пойдет параллельно линии среза обеих стен комнаты. Следовательно, угол между двумя плоскостями в микроскопических препаратах только тогда можно будет измерить непосредственно, если препарат случайно срезан точно вертикально по отношению к линии среза обеих плоскостей. К этому еще присоединяется, что линии среза поверхности sacculus’a и urticulus’a проходят непараллельно, так что даже в самых благоприятных случаях нельзя будет измерить углы в одной серии срезов.
Задачу можно решить только математически. Де Бурле для этой цели пользовался преимущественно начертательной геометрией. Одна плоскость, параллельная направлению среза, служит первой проекционной поверхностью; другая, проложенная через оси двух направляющих каналов, стоит перпендикулярно на первой и служит второй проекционной поверхностью; третья — проводится перпендикулярно через направляющий канал к двум первым плоскостям. На эти плоскости надо спроецировать следующее.
Ik Медиальную плоскость черепа с расположенной в ней «линией основания черепа» (у кролика — линия, соединяющая середину межмыщелковой выемки с концом кости у переднего края основной части затылочной кости и проходящая по верхней поверхности основания черепа; у морской свинки эта же линия — на нижней поверхности основания черепа от середины межмыщелковой выемки до середины вентрального шва postsphenoideus с затылочной костью). Названные точки легко найти в сериях срезов.
2. Плоскости обеих макул utriculus’oB. Они, за исключением загнутого вперед и медиально края, являются почти плоскими поверхностями. Следовательно, если выбрать три хорошо определенные точки на каждом чувствительном пятне и через них провести плоскость, то ее следует рассматривать как плоскость utriculus’a.1
3. Плоскости обеих макул sacculus’oB. Как показывает рис. 174, это не так просто определить. Можно провести плоскость через точки ACD и таким путем определить положение главной части sacculus’a. Но, кроме того, надо определить положение дорсального отрезка в плоскости abde и можно еще провести плоскость через CDE. Даже это тройное деление еще недостаточно для сложной формы чувствительного пятна, так как главная часть еще повернута (торзионно) вокруг своей продольной оси, так что в основу конструкции положен средний наклон по отношению медиальной плоскости в дорсо-вентраль- ном направлении. Подробности приведены в работе де Бурле и де Хаас (88).
4. В случае надобности еще одну плоскость через каждый из шести полукружных каналов. Три точки можно найти в серии срезов: средние точки обоих мест входа полукружного канала в utriculus и в ампулу и среднюю точку в середине поперечного разреза полукружного канала. Через эти три точки можно проложить плоскость.
С помощью проекции этих трех площадей на три проекционные плоскости можно сконструировать углы, которые образуют отолитовые ма- кулы и полукружные каналы между собой и с медиальной плоскостью, и, кроме того, углы, которые образуют линии среза обеих плоскостей utriculus’oB и sacculus’oB, а также плоскости одноименных полукружных каналов. Таким образом достигается полное определение положения отдельных
1 Если необходимо, то можно проложить еще особые плоскости через края, загнутые вперед и медиально.
24 Р. Магнус
370
Девятая глава. Функция отолитов
частей лабиринта по отношению друг к другу и к черепу. Таким же путем определяется взаимное расположение отдельных частей отолита.
Что при помощи такого метода достигаются достаточно точные результаты, видно из следующего: де Бурле и де Хаас (89) резрезали у обезьяны левую височную кость сагиттально, правую (с основанием черепа) — фронтально. Из таблицы III видно (Macacus С), что определение углов, произведенное на основании этих серий, срезанных в совершенно различных направлениях, в основном совпадает и не дает больших отклонений,
какие, между прочим, имеются между правой и левой сторонами.
Проф. Орнштейн обратил внимание на то, что можно эту начертательную конструкцию заменить расчетным методом, который де Бурле использовал в трех сериях опытов на кроликах для контроля геометрических данных. Речь идет о системе формул, которая подробно изложена в работе де Бурле и Костер (84). Соответствие полученных на тех же сериях результатов обоих методов вполне удовлетворительное (см. таблицу с результатами измерения на кроликах).
Все данные измерений, поскольку они к настоящему времени закончены, обобщены в табл. I, Ia, II и III (стр. 371 — 375).
Чтобы сделать наглядным это топографическое изучение, в анатомической лаборатории де Бурле и его сотрудников в течение многих лет изготовлялся целый ряд различных моделей. Часть из них уже послужила основой для физиологических исследований; новейшие модели будут использованы при продолжении этой работы.
Показанная на рис. 170 модель является простой восковой конструкцией перепончатого лабиринта кролика, которая не пригодна для изучения положения отолитов в пространстве, так как на ней нельзя распознать положение чувствительного пятна.
Оорт построил модель, изображенную на рис. 176 (87). На ней череп кролика, укрепленный на длинном гвозде, который отображает линию основания черепа, дан в нормальном положении. Справа находится восковая модель utriculus’a, sacculus’a и примыкающих конечных частей полукружных каналов вместе с ампулами и эндолимфатическим протоком (в 30-кратном увеличении). На наружной стороне sacculus’a и utriculus’a в красках дано положение соответствующего чувствительного пятна. Эта восковая модель сконструирована по одной из серий табл. I и приведена в правильное положение к черепу и линии основания черепа. При этом использовались также данные измерения двух других серий. На левой стороне чувствительные пятна utriculus’a и sacculus’a изображены изолированно. Обе эти части, изготовленные в 30-кратном увеличении, смон-
Рис. 176. Модель sacculus’a, utriculus’a, начальных частей полукружных каналов кролика, а также чувствительное пятно отолитов при 30-кратном увеличении, ориентировано правильно по отношению черепа и основания черепа (по Оорту) (87).
вычер- вычи- вычер- вычи- | вычер- вычи-
чено слено чено слено чено слено
оооооооооооо
^СОО'Н'^ООТОСОООСОЮ
OSOOC^OSOSOOOOOOOO^stfOO
о о о о о о
^ СО СО с\1 -TH CD
О as со со оо
I I
о о *-Q о *Яо о ^ о . о о о о о *-Q О ^ о
asOO^HCMx^'T-iD^OOI^'asv^LOOOLO^HOOOCOCO^HOOOOCO asooc'-asascooooooo^p^ooooao <о> о со !>• со оо со со
CO'tHCO'vFO^ V V оооюсо
COCOCO'^CO^tr^CSI'dH's^s^v^
ОООООООООООО 1 1 CO'HCOOCO^'Ht'-b'HOCO 1 1
oooc^asasasasooooLOxPoo
sf СОхЧ|>ю CNJ СО СО ^Г1 оо 1 О О О О О О I I
1 о- со со со со со 1 1 ОЗОЗЮСОСЗОО
о о
00000*^0000^000 СМ^нч^гНЮЮтН|>^'Н0 1>ОЗСХ1
ооос—asasasasooooLOvfoooooo
о
100000^00 1 oqosvfcoc-cov^-^H oasLOc^-cNjooxfco
^гН
sjH ^ 00 Ю О СО СО СО V
юсо^со^-мрсчею ю см
ОООООООООООО 1 1 СО t-со юю-Н со ^ со 00 00 оо 1 1 csC'-iH-asasasasasos'stHcooo
00 sj* V со чн оо
ТГП-НОЗЮСОСЗ 1 О О О О О О 1 I 1 О- СО С" хр СО !>■ 1 1 OOsfMMOO
^ о о о о UJ. О о ^ о о о о “i о о о ^ о о о сооосоююмсоос^ооасосоооюьсосо^со^юо osC'-['-asasosasasas^cooooot^^-ioo^f<i>-c\]oococo
я
я
о
&
и
CÖ
и
Cd
Рн Ä
о
° Cd
Р м
S я ,
о Рн '
О Д
05 ^ О
cd
ffl cd
Cd PQ
P-I ф P Я
О Ü
cd Cd Й
и cd Ф Ph
я а
cd
cd
И
й
а ,
cd '
05
Я
H
Ph
О
OS
cd
И
»sl
td И g
и |g
SS* I
а
я
я
tc <
ф
Рн
О
О
Я , О '
о
я
д
я
cd
Р
й
Я < cd
а
к
н
Рн
ф
В
i
««
я
Я И
: Р te
я ®
fcC Рн
я ®
со К
н
н
Ü О Я о
О
я
Д 5Я О
Я
03
я
cd 03 р
н д
® Н
о Рн и Cd дз й Ф® н И § я И Р ф
ll IIs
g со И Р'”1
й я
М DO
S Pi
EH
я °
° ьз
со 5
я в ftg
Я
Н
О
О
я
о
о
я
д
**Н
о
я
й
я
i я а
cd
QQ
Р я я P« w
g£g
1 3 о
’f-H
§5 § 3
В 3В 3
g .2 £ “
О Eh О cd ^ cd - ю Р и
со
ф
Р !—1
II
И
р
со
ф < Рн
^ я * о
р
р
я
* Я Я
■н ^
i г i i и i и 11Т i i i i i i i i i i i
<м СО р
Р щ
g 3
р
о
tt
Й
ф
S
р
я
ф
Рн
ф
§
со
Я
Р
Р
Он
i>s
Р
Я
Я
Я
Я
р
ф
И
р я
Р СС
я ® §:£ я ® со R
й 03
Н Еч
о о
р р Я й р ffl cd
03 Сб Сй CD 0Q
© Рнф РнФ Я Д Д Д Д
a о о о о
о Я . о • о р Д
о
я
о ■ о
р
д
р 2 Р Я
я Д
Й й р 2 £ а '
ge
СО Рн
Я я
Рнй
POP Р Рн Р Я
te • ф
Рн
ф
В
й
н
о
р
Я
Р
р
Я . й Р Р Я
я
Й р : р
я : «=с
р
со
й
н
о
я ;
о
о
р
Я
Р
Р
Я i * PT ф Рн
и
р
я
о <
Рн
Р
Я
Я
р
й
о
я
р
1 я я я 1=3
р t>
2 р Я PQ Рн ф Я Р о о
со
р
й
Р i> со
я
Я н о о
м 8 ä 3 о о Я Я
я
_о
‘Ê-H
; р 1 р
я
^ tt' " ф Рн
и
24*
1 Этот угол открыт назад. 2 Этот угол открыт вперед, з Этот угол открыт кверху. 4 Этот угол открыт назад; он измеряется над линией основания черепа. 5 Этот угол открыт вперед; он измеряется над линией основания черепа. 6 Этот угол открыт назад; он измеряется над линией основания черепа. 7 Этот угол открыт вперед и вниз.
Для каждой серии приведены рядом результаты конструкции по методу начертательной геометрии и по расчетному Орнштейна, Отклонения между двумя рядами чрезвычайно незначительны и могут быть использованы для суждения о точности метода.
Таблица la
о о
о
о
о
ю
LO
о
о
о
о
о
о
о
о
о
о
ю
LO
sf
О-
о*
со
05
ю
«Ч-Н
ю
00
!>-
ю
ю
с—
05
CS]
00
ю
со
со
см
со
'гН
CNJ
со
ю
о
ft
1W
Я
Я
©
Ч
о
Сб Сб
В В
cd
сб
W
я
сб
Ч
ft
^0
я
я
te
я
й
ft
ф
я
я
о
сб
я
я
со
я
’со
г-<
я
"со
ф
в
я
я
ft
в
ф
я
о
я
05
я
о
о
я
я
'Й
4-Э
я
о
о
я
со
я
н
о
о
сб
ч
Рч
!>s
я
н
о
я
я
н
Ü
я
*
я
о
о
ч
я
я
я
я
я
я
я
я
я
я
я
я
я
я
я
я
я
я
я
te
ч
я
я
ф
ф
ч
ч
ft
В
ft
ft
CJ
о
ft
о
cd
Я
Я
Н
II!!! I
Я
н
о
о
я
о
о
Ч
И
Я
к
И
te
я
Ен
о
сб
Я
Я
сб
Я
Я
сб
Ч
ft
о
Я
Я
я
я
я
о
2 :В
* я
OS W
о о ft ft О О
и te
I Illi
сб
я
я
я
я
ы
я
я
в
я
я
я
я
я
я
я
я
ч
Рн
я
н
о
сб
я
я
я
я
te
ф
ft
о
я
я
я
я
1=2
я
В я о "и я я w я Й -2 о ч
со
я
я
н
о
Ä Ä
я
я
я
о
05
я
со
я
со
я
ф
я
ф
ф
ф
о
ft
я
ft
ft
ft
о
о
я
я
я
о
Ü
о
я
я
я
Ä
я
я
я
я
я
я
я
я
я:
Ä Я ft
я
я
я
я
я
я
я
я
я
я
я
я
я
я
я
ч
ф
ч
я
я
я
ч
ft
в
ft
1=3
t=3
я
4
ф
fct
о
5
я
я
я-
я
ft
н
о
я
о
я
я
ft
я
я
о
я
о
о
ч
я
й
о
ч
о
я
ф
ч
ft
>>
W
я
ft
>е<
я
яг
ф
я
я
я
4
ф
н
я
ft
Сб
я
tc
ф
ft
я
я
н
со
ф
я
я
сб
я
ф
5
я
ft
в
Морская свинка
Вперед открыты —1, 4, 6, 7, 8, 12, 15, 17, 18; назад открыты —13; вверх открыты —2, 5, 16; вверх латерально открыты вниз открыт—14; вниз латерально открыты — 10, 11; латерально открыт — 9.
Последняя графа этой таблицы содержит использованные для конструкции модели средней величины.
Обезьяны (Macacus)
*"4
а
а*
а
а
£■4
Ü
СО
Р
о
справа
(фрон¬
тально)
о
СО
05
О
С'-
о
со
00
о
со
со
о
[>-
о
05
ю
о
о
4t
•ч-Н
о
05
см
чгН
О
05
Ю
о
со
■ч—1
О
о
4t
о
О
-чЧ
о
05
Ю
о
4t
со
ев
о
ев
ей о о W —- в
о
о
о
О
0
О
о
F=i
. ч
г-
о
со
4t
о
05
05
ю
со
LO
4t
LO
4t
w
g н ч а
о S
■ч-Н
cd
м
о
о
О
о
о
о
о
о
о
о
о
о
о
О
О
о
W
cd
i>
СМ
4t
со
СМ
00
00
со
со
05
со
!>•
С"—
о
00
о
Рн
st
со
05
см
00
ю
со
LO
СО
Ю
О
см
о
ю
4t
Р
g
т-Н
'гН
-Г-Н
чН
нН
тН
■^Н
о
св
о
cd
О
о
О
О
о
о
о
о
о
о
О
О
О
о
о
о
ы
w
О"
СМ
см
о
см
со
со
чгН
00
4t
4t
ю
00
со
м
4t
со
05
см
00
ю
-чн
со
4t
СО
ю
О
см
о
4t
со
3
4гЧ
'ГН
-ч-Н
л
Н
О
CÖ
й
й
ев
И
PQ
ев
й
Рн
ев
И
ев
Рн
И
о
Р В ^ Р Р о
р й
ев
И
t>> 3
t£ ° _,
Й ев -
о со
S Э
ев
й
Ф
Рн
Ф
ев
й
Р
1>>
ев
И
И
В
§
й
В
Н
О
ев
й
й
ев
И
pq
ев
й
Р-»
О
й
Й
й
й
И
еС
ф
Рн
и
ев
ев
~оа
*со
р
р
jp
Р
н
Н
Р
н
о
о
О
о
о
о
о
О
В
о
Й
ев
И
H-q>
к
о
со
о
о
о
о
о
й
Р
й
Р
й
и
й
й
Н
О
ев
Й
И
й
й
Ен
о
ев
й
И
Й ■ Й
в
й
Й
й
и
ы
ев
te
«:
ф
й
и
ф
ев
ф
Рн
Рн
И
Рн
CJ
ев
й
Рн
о
Я
ев
й
о
Р
Рн
О
й
И
W
й
1=5
ев
И
й
й
И
ев
Й
О
W
й
и
W
й
1=3
н (М СО ю
Рн
О
Й
И
И
Н
1=3
Рн
ен
О
>й
Я
к
в
й
ев
о
Рн
В в:
в
о
Й
ев И В Й ев
й _
5 &
в В
й
й
и
ьс
в
о
й
ев
И
р
ев
й
Рн
о
ев
й
й
й
И
ЬС
й
ев W
_ Я Рн CÖ
Ф й
В Рн
Рн
О
й
в
и
й
1=3
cä
*со
^Р
"3
о
ев
о
ев
оа
со
^Р
Й
*Р
О
со
ф
О
о
ев
Рн
со
Ен
О
Р
н
>й
Ü
ев
Я
Й
И
Р
Й
Й
ев
ев
И
о
И
Рн
ев
О
й
tt
Рн
Рн
н
о
>й
Я
И
р
й
ев
о
Рн
О
fct
й
ев
Р
Р
й
ев
В
Ы
Ф
Рн
О
й
й
И
Таблица lil (продолжейие)
cd
§
а
Cd I 'n'
m ju 2 g О g
sei
L о
gg
2 Eh Ч а
ö E
cd
о
ю
со
о
ю
ю
о
ю
О
со
с—
о
00
00
о
со
со
о
05
о
о
о
о
>4
4P
со
00
’N
Я
H
4
cd
о
ft
о
fct
,3 ^
Д
H
о
cd
Ч
4
4
W
fct
a>
ft
Ф
Й
cd
4
4
4
Й
tC
a>
ft
CD
И
>>
fct
№
Ф
s
cd
W
©
ft
cd
4
t-
0>
cd
И
P
И
ч
©
w
т
Д
о
cd ^
W
о
со
CD
ft
H
о
Я
Ч
л
ч
cd
о
ft
о
fct
нр
н
о
о
т
о
о
Ч
Р
Ч
Ч
И
fct
и
«
cd
Ч
Р
Р
Ен
О
cd
Ч
К
cd
И
И
cd
Ч
р
cd
Ч
cd
со
ф
ft
о
ч
и
и
Ä
Ä
Ä
Ä
и
ч
об
05
о
см
со
’гН
чг-<
см
см
см
см
Вперед открыты —1, 4, 6, 7, 19, 20; вверх открыты — 2, 22; вверх и латерально открыты —3, 5, 12, 14, 16, 23; латерально и назад открыты — 9, 21; латерально и вниз открыты — 10, 11; вверх и назад открыт — 13; вперед и вниз открыт—15; латерально открыт — 17; назад открыт — 18.
У макаки G левая височная кость срезана сагиттально, правая — фронтально. Конструкция углов по геометрическому методу не дала никаких больших различий между правой и левой сторонами, что также было установлено у макаки В.
376
Девятая глава. Функция отолитов
тированы симметрично и расположены друг от друга на расстоянии* в 30 раз меньшем действительного. Эта модель послужила основой для маленькой ручной пробной модели, использованной для первого изучения функции отолитов и с которой (рис. 179—186) дан целый ряд изображений. Но здесь не учтены форма и точное положение дорсального отрезка (лоскута) sacculus’a.
Рис. 177 показывает одну из новейших моделей де Бурле и де Хааса, изображающую соотношения у морской свинки. В середине дан череп в нормальном положении. Косо вверх поставленные поверхности (внизу из жестщ наверху из стекла), которые срезаются над черепом, соответст-
Рис. 177. Модель черепа морской свинки с плоскостями utriculus’а и sacculus’а и чувствительных пятен utriculus’а и sacculus’а правильной формы и ориентировки (по де Бурле и де Хаасу) (88).
Рис. 178. Модель черепа обезьяны с плоскостями главной части передней части и медиального угла utriculus’а, а также главной части передней части и дорсального отрезка sacculus’а, с соответствующими углами при правильной ориентировке к черепу, без учета действительной величины площади чувствительного пятна (по де Бурле и де Хаасу).
вуют плоскости главной части sacculus’a. На них справа и слева приделаны чувствительные пятна (макулы) в их правильной форме и положении к черепу. Светлоокрашенная часть — дорсальный отрезок, иннервируемый ramus utricularis. Почти горизонтально справа и слева изображены плоскости главных частей utriculus’a. На них находятся модели чувствительного пятна utriculus’a правильной формы и ориентиров ки. Плоскости поставлены по средним цифрам, которые даны в т абл. II (последнее поле). На модели можно легко измерить углы названных главных плоскостей.
В последнее время де Бурле изготовил для морских свинок, кроликов, обезьян и человека моде i и, на которых отсутствуют чувствительные- пятна отолитов, но зато отдельно даны плоскости для главной и передней частей sacculus’a и дорсального отрезка, а также для главной и передней частей urticulus’a и для медиального угла. Рис. 178 изображает модель для обезьяны (Macacus) (89). Череп дан в нормальном положении. Большие, почти горизонтальные плоскости соответствуют главным частям utriculus’a. Спереди видны плоскости переднего края и медиального угла» Косо вве рх сходятся плоскости главных частей sacculus’a, спереди к ним
Отношение лабиринтных рефлексов положения к положению отолитов 377
и снизу — плоскость передней части sacculus’a, над ней — дорсальный отрезок. Все эти плоскости дают только направление, но не величину изображенных частей чувствительного пятна. Следовательно, они предназначены для более точных научных исследований, в то время как модель на рис. 176 больше подходит для демонстрации. Все углы могут быть измерены прямо на модели, так как каждый угол срезан, по крайней мере с одной стороны вертикально к линии среза обеих соответствующих плоскостей. Модель, можно поместить в любом положении в пространстве. В оригинале работы де Бурле и де Хааса даны для морских свинок и обезьян образцы срезов для этой модели вместе со всеми необходимыми цифрами, так что их можно самим сделать из картона.1
Из вышесказанного вытекает, что теперь возможно довольно хорошо представить себе форму и положение чувствительного пятна отолита в черепе и, таким образом, для физиологических исследований создана достаточно хорошая анатомическая основа. Изучение это будет и дальше продолжаться в различных направлениях в анатомической лаборатории в Утрехте.
Б. Отношение лабиринтных рефлексов положения к положению
отолитов (50)
О функции вестибулярного аппарата появились в последние два десятилетия различные обобщающие представления, из которых здесь прежде всего следует упомянуть взгляды Крейдля (2), Баранин Виттмаака (2). Из их работ и из отдельных статей выяснилось, что имеется почти общее мнение о том, что реакции на угловое ускорение (вращательная реакция головы, вращательная реакция глаз, ощущение вращения) вызываются с полукружных каналов, а именно с расположенных в ампулах cristae. Зато о значении отолитов господствует менее ясное представление и нет полного единства мнений. Брейер (3) в свое время приписывал отолитам те лабиринтные функции, обеспечение которых в полукружных каналах ему казалось не подходящим, а именно вызов реакций на прогрессивные движения и ощущения положения головы в пространстве. Далее Кубо (Kubo) на основании опытов на кроликах и рыбах стал рассматривать компенсаторные положения глаз как реакции отолитов. Барани (2) объяснял промахивание при изменениях положения головы в пространстве действием отолитов.
Благодаря основным опытам Эвальда и его учеников стало известно влияние лабиринтов на тонус поперечнополосатой мускулатуры. Вначале Эвальд ответственным за это считал аппарат полукружных каналов. Однако позже школа Эвальда Ах) стала также говорить об отолитовом тонусе как части лабиринтного тонуса. Но еще в 1906 г. Крейдль считал все это неясным понятием.
Осложнение стало еще больше вследствие того, что отдельные авторы думали, что угловым ускорением возбуждается не только аппарат полукружных каналов, но также и отолиты [Бартельс (1)].
В описанных в этой книге исследованиях я установил, что при положении тела, установочных функциях и компенсаторных положениях глаз лабиринты имеют не единственное, но все же очень важное значение. Из довольно сложных явлений удалось выделить ряд точно ограниченных тонических лабиринтных рефлексов, которые следует рассматривать
1 Кроме того, эти модели можно получить у служителей анатомической лаборатории в Утрехте.
378
Девятая глава. Функция отолитов
как первичные последствия раздражения лабиринтов (в противоположность другим рефлексам; связанным с лабиринтами опосредованно и вызываемым вторично вследствие измененного положения головы и т. п.). Все эти лабиринтные рефлексы имеют ту особенность, что они вызываются не движениями головы, а зависят от определенного положения головы в пространстве. Они тонические, т. е. длятся так долго, пока голова сохраняет определенное положение в пространстве. Их легко обследовать изолированно в опытах на животных.
Этими рефлексами являются следующие.
1. Тонические лабиринтные рефлексы на мышцы тела (рефлексы положения):
а) на конечности;
б) на шею (и туловище).
2. Лабиринтные установочные рефлексы.
3. Тонические лабиринтные рефлексы на мышцы глаз:
а) вертикальные отклонения;
б) круговые вращения.
Чтобы установить, какие части лабиринта следует принять во внимание для вызова этих тонических рефлексов положения, прежде всего были
установлены для всех тонических лабиринтных рефлексов те положения головы в пространстве, при которых соответствующий рефлекс имеет свой максимум и минимум как при интактных лабиринтах, так и при наличии только одного лабиринта. В дальнейшем поэтому должен обсуждаться вопрос: занимают ли отолиты определенное положение в пространстве при этих эмпирически найденных положениях максимума и минимума? Прежде всего разговор будет идти преимущественно об отношениях у кролика.
Рис. 179. Остальные животные будут прини¬
маться во внимание только попутно.
Для этих опытов своевременно (1921) была изготовлена вышеупомянутая маленькая ручная модель (рис. 179) при нормальном положении головы. Череп кролика несет в битемпоральном направлении на уровне глазной щели штангу, на обоих концах которой в увеличенном виде приделаны отолиты в правильном положении к черепу. Видно, что отолиты utriculus’a попадают почти в ту же плоскость и что они при таком положении головы стоят горизонтально. Сторона, на которой отолит находится на чувствительном пятне (она при этом положении головы является верхней), обозначена белыми точками на черном фоне. Значит, при таком положении головы отолит давит вертикально на свою подставку. Отолит sacculus’a находится медиально и под отолитом utriculus’a. Отолит находится латерально по отношению к своему чувствительному пятну. Эта сторона на модели заштрихована белыми линиями на черном фоне. Отолит sacculus’a не параллелен средней плоскости черепа. Он скорее проходит спереди латерально и по направлению назад медиально, так что оба отолита sacculus'oB образуют между собой угол в 461/2—64°. Кроме того, отолит sacculus’a наклонен сверху медиально и по направлению книзу латерально на 23—32°.
Отношение лабиринтных рефлексов положения к положению отолитов 379'
На этой модели передний угол отолита sacculns’a загнут латерально, так что он приближается к фронтальной плоскости. Этот угол на модели закрашен белым цветом. Точное положение передней части отолита sacculus’a и прежде всего дорсального отрезка тогда еще не было установлено. Так как в ближайшее время будут произведены в этом направлении еще более точные физиологические опыты, окончательные результаты которых надо еще подождать для дальнейших выводов, я дал без изменения старые рисунки. Следовательно, в дальнейшем надо постоянно помнить, что отношения дорсальных отрезков даны на рис. 179—189 неточно
На модели можно в любое время удалять вместе sacculus и utriculus и, таким образом, видеть отношения после односторонней экстирпации лабиринтов.
Подобная же модель была изготовлена также для положения полукружных каналов в черепе кроликов.
1. Тонические лабиринтные рефлексы на мышцы тела
а) Тонические лабиринтные рефлексы на конечности. В третьей главе было показано, что эти рефлексы лучше всего удается исследовать на децеребрированных животных, у которых мышцы-разгибатели конечностей находятся в состоянии децеребрационной ригидности. Тонус этих мышц-разгибателей зависит от лабиринтов. Имеется одно и только одно положение головы в пространстве, при котором этот тонус имеет свой максимум, и только одно положение, при котором он имеет свой минимум. Оба положения отличаются друг от друга на 180°. При всех других положениях головы в пространстве тонус мышц-разгибателей принимает величины, которые лежат между двумя крайними положениями. Тонус в мышцах-разгибателях четырех конечностей изменяется всегда в одном смысле при изменении положения головы в пространстве. Если положение головы (или лабиринтов) к горизонтальной плоскости не изменяется, то также не происходит никакого изменения тонуса мышц конечностей. Изменение тонуса длится до тех пор, пока голова находится в соответствующем положении. Следовательно, речь идет о чистых «рефлексах положения». При прогрессивных движениях и угловых ускорениях головы эти рефлексы не вызываются. Они отсутствуют после двусторонней экстирпации лабиринтов.
Способ изображения для различных симметричных положений головы в пространстве виден на рис. 180. Положение головы на спине с горизонтальной ротовой щелью обозначено как 0°, нормальное положение головы с горизонтальной ротовой щелью — 180°. Голова с вертикально вверх поднятой мордой +90°, голова с вертикально опущенной мордой —90°. Промежуточные положения головы видны на рисунке.
Положение максимума для тонических лабиринтных рефлексов на мышцы конечностей у кролика, если голова находится в симметричном положении на спине, колеблется индивидуально между 0 и —(—45°. Положение минимума при нормальном положении головы“ колеблется индивидуально между 180 и —135°.
Рис. 181 показывает модель отолита в положении максимума для тонических лабиринтных рефлексов на мышцы конечностей (—}—30°). Отолиты sacculus’a не имеют особо характерного положения, Оба отолита utriculus’а стоят горизонтально и притом так, что отолит висит на чувствительном пятне.
Рис. 179 показывает модель отолита в минимальном положении, отличающемся на 180° от предыдущего. И здесь отолит utriculus’a стоит горизонтально, но в этом случае он давит на чувствительное пятно.
380
Девятая глава. Функция отолитов
Таким образом, вытекает, что для обсуждаемых здесь рефлексов максимум возбуждения исходит от отолита, если он висит на чувствительном пятне. Этот вывод находится в противоречии с общим мнением, по которому решающим является давление отолита. Однако для последнего взгляда
Рис. 180. Схема положения головы при вращении вокруг битемпоральной оси с указанием соответствующего угла при каждом положении головы.
не приводились необходимые доказательства. В большинстве случаев его выводили а priori из анатомии отолитового органа.
Если речь шла при исследовании функции отолитов только лишь о тонических лабиринтных рефлексах на мышцы конечностей, то действительно имеется возможность привести в соответствие экспериментально полученные факты с ранее имевшимися взглядами на функцию отолитов. Выяснилось, что в этих рефлексах участвуют не только мышцы-разгибатели конечностей, но и мышцы-сгибатели и что при этом действителен закон реципрокнойиннервации; если увеличивается тонус разгибателей, то Рис. 181. тонус сгибателей уменьшается, и
наоборот. Вследствие этого тонус мышц-сгибателей на конечностях больше всего тогда, когда отолиты ut- riculus’a давят, и меньше всего — когда они висят. Однако эти рефлексы на мышцы-сгибатели у децеребрированного животного играют только второстепенную роль, их можно вызвать только в исключительных случаях и при применении особых условий опыта,1 в то время как рефлексы на мышцы-разгибатели чрезвычайно сильны и заметны. Никто из произ-
1 Например^ при отравлении пикротоксипом.
Отношение лабиринтных рефлексов положения к положению отолитов 381
водивших сам эти опыты не будет сомневаться в том, что рефлексы на разгибатели (которые так же, как «мышцы стояния», прежде всего могут быть под влиянием «статического» органа чувств) являются первичными. Этот предполагаемый вывод о том, что для тонических лабиринтных рефлексов на мышцы конечностей максимум возбуждения исходит из отолитов utriculus’а, если последние висят, подтверждается и дальше в этой главе тем, что при лабиринтных рефлексах и компенсаторных положениях глаз (вертикальные отклонения) удается доказать, что у них максимум возбуждения исходит из висящих отолитов.
Из девяти исследованных Вейландом (9) кроликов максимум был в трех случаях при +45°, один раз при +20°, один раз между 0 и 45° и четыре раза при 0°. Минималь-
Рис. 182.
Рис. 183.
яое положение во всех случаях было примерно на 180° отличным от положения максимума. С этими экспериментально найденными вариациями совпадают измерения де Бурле и Костера. Они нашли угол между линией основания черепа и линией среза чувствительного пятна utriculus’а в их трех сериях в 39, 31 и 63°. В первых двух случаях ротовая щель стоит при горизонтально лежащем utriculus’e примерно от +30 до +40°, зато в третьем случае почти горизонтально.1
Подобные вариации были найдены у кошек и собак. Среди 18 обследованных кошек максимум был 15 раз +45°, один раз между 0 и 40°, один раз при +10° и один раз между 0 и +10°. Минимум и здесь отличался всегда на 180° от этих показаний.
Среди четырех исследованных собак максимум был два раза при +45°, один раз при 0° и один раз при +90°. И здесь минимум всегда отличался на 180°. Число наблюдений на собаках слишком мало, чтобы из этого сделать заключение, потому что возможность вариаций у собак больше, чем у кошек и кроликов.
После односторонней экстирпации лабиринтов тонические лабиринтные рефлексы на мышцы конечностей не изменяются. Положение максимума и минимума остается неизмененным. Достаточен один лабиринт, чтобы вызвать изменения тонуса на конечностях обеих сторон тела. Этот факт делается совершенно понятным благодаря модели отолитов. Так как оба utriculus’a расположены почти в одной плоскости, то после односторонней экстирпации лабиринтов не должно наступить никакого изменения этих рефлексов. Рис. 182 показывает положение максимума после правосторонней экстирпации лабиринтов, рис. 183 — положение минимума. Из этого факта следует подтверждение вывода, что utriculus’bi, а не sacculus’bi являются местом вызова тонических лабиринтных рефлексов на мышцы конечностей. Если бы последнее имело место, то, оттого что чувствительные пятна sacculus’а расположены не в одной плоскости, должно было бы наступить изменение (асимметрия) рефлексов после односторонней экстирпации лабиринтов, как это дальше будет описываться для лабиринтных рефлексов и компенсаторных положений
1 На то, что и в положении полукружных каналов встречаются индивидуальные различия, указывают Шёнеман, Ротфельд [Schönemari, Rotbfeld (4)] и др.
382
Девятая глава. Функция отолитов
глаз (вертикальные отклонения). Из этого далее следует, что каждый utriculus должен находиться в функциональной связи с мышцами конечностей обеих сторон тела.
Рассмотрение этой первой группы рефлексов привело к целому ряду принципиально важных заключений о функции отолитов. Тонические лабиринтные рефлексы на мышцы конечностей являются рефлексами utriculus’a. Каждому положению utriculus’a в пространстве соответствует определенная степень тонуса в мышцах-разгибателях ног и прежде всего в разгибателях проксимальных участков конечностей. Тонус сгибателей (на неотравленных децеребрированных животных) находится гораздо меньше под этим влиянием, но ведет себя противоположно то- нусу разгибателей. Максимум тонуса разгибателей достигается во всех четырех конечностях, если отолиты utriculus’a лежат горизонтально, и тогда, когда отолиты висят на чувствительном пятне. Минимум тонуса разгибателей имеется тогда, когда отолиты давят на свою подставку. Натягивание и надавливание отолитов на волоски чувствительных клеток в чувствительном пятне вызывают сообразно этому тоническое возбуждение. Каждый отолит utriculus’a находится в связи с мышцами конечностей на обеих сторонах тела. Этим фактом и тем, что оба отолита utriculus’a расположены почти в одной плоскости, объясняется то, что после односторонней экстирпации лабиринтов тонические лабиринтные рефлексы на конечности обеих частей тела остаются без изменения. Экспериментально найденным вариациям в положении максимума и минимума для описанных лабиринтных рефлексов соответствуют анатомически найденные вариации в положении отолитов utriculus’ов.
В следующих разделах будет рассмотрено, насколько эти заключения поддаются подтверждению и насколько можно расширить исследования положений отолитов при других лабиринтных рефлексах.
У остальных обследованных млекопитающих наблюдаются, в сущности, те же отношения. Рис. 177 (морская свинка) и рис. 178 (обезьяна) показывают, что положение отолита utriculus’a такое же, как и у кролика. При исследовании тонических лабиринтных рефлексов на мышцы конечностей было найдено то же отношение максимума и минимума, как и у кролика.
б) Тонические лабиринтные рефлексы на мышцы шеи. Тонические лабиринтные рефлексы на мышцы затылка (так же, как и на мышцы конечностей) имеют центры в продолговатом мозгу позади места входа 8-го нерва и не должны быть спутаны с «лабиринтными установочными рефлексами на голову», которые имеют свои центры в среднем мозгу. Тонус мышц-разгибателей затылка (дорсальные сгибатели головы) имеет свой максимум, если голова находится в положении от 0 до +45° (рис. 181), минимум — при положении от 180 до —135° (рис. 179). В положении минимума у децеребрированного животного иногда наступает, хотя и незначительный, тонус сгибателей затылка. Следовательно, положения максимума и минимума такие же, как и для тонических лабиринтных рефлексов на мышцы конечностей. Из этого следует, что здесь речь идет также о рефлексах utriculus’a. В положении максимума отолиты utriculus’a висят, в положении минимума они давят.
В то время как после односторонней экстирпации лабиринтов тонические лабиринтные рефлексы на мышцы конечностей на обеих сторонах тела остаются без изменений, при тонических лабиринтных рефлексах на мышцы шеи это не имеет места. После односторонней экстирпации лабиринтов следует известный основной поворот (и наклон) головы в сто¬
Отношение лабиринтных рефлексов положения к положению отолитов 383
рону отсутствующего лабиринта. Последние с течением времени увелш- чиваются. Здесь выявляется, что мышцы затылка одной стороны, притом те, которые поворачивают шею в сторону сохраненного лабиринта, не подлежат влиянию лабиринта, в то время как у мышц затылка другой стороны тонические лабиринтные рефлексы остаются без изменения. Положение максимума такое же — от 0 до +45° (рис. 182), положение минимума от 180 до —135° (рис. 183). Из этого следует, что отолит utriculus’а находится в функциональной связи только с мышцами шеи одной стороны тела, в то время как он, по сообщенным данным предыдущих разделов, действует на мышцы конечностей обеих сторон тела. Это принципиальная разница между тоническими лабиринтными рефлексами на мышцы конечностей и шеи.
Из того факта, что оба отолита utriculus’a расположены почти в одной плоскости, еще не следует, что удаление одного отолита utriculus’a всегда должно пройти без всяких симптомов. Это, правда, имеет место при тонических рефлексах на мышцы конечностей, так как здесь один отолит utriculus’a равномерно влияет на мышцы конечностей правой и левой сторон тела. Однако это не имеет места при тонических лабиринтных рефлексах на мышцы шеи, так как один отолит utriculus’a влияет только на мышцы шеи одной стороны тела.
Как уже было указано при сложной анатомии мышц шеи, до сих пор было невозможно установить, какие именно мышцы шеи сохраняют свой тонус под влиянием только одного лабиринта. Под «мышцами шеи одной стороны тела» поэтому пока понимают те мышцы шеи, которые поворачивают или наклоняют шею в одну сторону, не исключая возможности, что, например, мышцы, поворачивающие шею вправо, также имеются на левой стороне тела, и наоборот. УУ кролика и кошки, но не у собаки, обезьяны и, по-видимому, у морской свинки, влияние лабиринтов распространяется и на мускулатуру туловища. И здесь оказалось, что один лабиринт влияет преимущественно на мышцы туловища одной стороны тела.
Связь каждого отолита utriculus’a с мышцами шеи (и туловища) одной стороны тела объясняется наступающим после односторонней экстирпации лабиринта «основным поворотом» шеи (и туловища), который в свою очередь снова вследствие шейных рефлексов вызывает такие вторичные последствия, которые были подробно проанализированы в шестой главе.
Согласно обсуждаемым в этом разделе данным, тонические лабиринтные рефлексы на мышцы шеи являются рефлексами utriculus’a. Каждому положению utriculus’a в пространстве соответствует определенная степень тонуса мышц шеи. Максимум тонуса мышц, поднимающих и поворачивающих затылок, достигается тогда, когда отолиты utriculus’a лежат горизонтально и висят на чувствительном пятне. Каждый отолит utriculus’a имеет функциональную связь с мышцами шеи только одной стороны тела (причем понятие «мышцы одной стороны тела» пока понимается функционально, а не анатомически). Этим объясняется поворот шеи после выпадения одного лабиринта.
У остальных исследованных млекопитающих наблюдаются те же отношения.
2. Лабиринтные установочные рефлексы
Среди установочных рефлексов, которые приводят к тому, что животное из ненормальных положений тела каждый раз рефлекторно снова принимает нормальное положение, «лабиринтные установочные реф-
384
Девятая глава. Функция отолитов
лексы на голову» занимают важное место. Их можно изучать изолированно на кроликах, если животное держать свободно в воздухе. Вследствие возбуждения лабиринтов голова из любого положения ставится в нормальное. Тогда туловище можно поворачивать во все стороны вокруг головы, находящейся в постоянном положении в пространстве. Лабиринтные установочные рефлексы отсутствуют после экстирпации лабиринтов.
Исследование установочных рефлексов у кролика после односторонней экстирпации лабиринтов показало, что действие установочных ла-
Рис. 184. Рис. 185.
биринтных рефлексов присоединяется к описанному выше и признанному за реакцию utriculis’a основному повороту головы и что голова стремится принять то положение на боку в пространстве, при котором сохраненный лабиринт окажется вверху. В этом положении исходящий из интактного лабиринта установочный рефлекс имеет свой минимум. Если же интактный лабиринт находится внизу, то установочный рефлекс имеет максимум.
Те лабиринтные установочные рефлексы на голову, благодаря которым голова при сохраненных обоих лабиринтах из асимметричного положения в пространстве приводится обратно в симметричное, можно объяснить совместным действием возбуждения обоих лабиринтов. Голова тогда приходит в такое положение покоя, когда возбуждение из обоих лабиринтов будет одинаковой силы. Как только симметричное положение головы нарушается, из находящегося внизу лабиринта исходит более сильное возбуждение, которое и вызывает возврат головы в нормальное положение.
Если при обсуждении роли отолитов при этих рефлексах исходить из отношений после правосторонней экстирпации лабиринтов, то максимум будет при положении головы на левом боку. Рис. 184 показывает,
Отношение лабиринтных рефлексов положения к положению отолитов 385
что при этом положении головы левый sacculus висит. При этих обстоятельствах вызываются сильные установочные рефлексы на голову, которые приводят к тому, что голова приводится обратно в положение минимума. Это правое боковое положение. Из рис. 185 видно, что в этом положении левый sacculus давит. Это — положение покоя головы после правосторонней экстирпации лабиринта, при котором не вызывается ни один установочный рефлекс на мышцы, поворачивающие шею.
На вышеприведенных стереоскопических снимках (рис. 140—143) видно, что положение покоя головы кролика после правосторонней экстирпации лабиринта в действительности не соответствует точному положению на правом боку, а что при этом морда немного висит вниз.
При этом положении головы левый sacculus лежит своей продольной осью действительно горизонтально, а не как на рис. 185 немного косо. Но этому совпадению очень большое значение нельзя придавать, так как голова при положении на боку уже вследствие тяжести немного оттягивается мордой вниз.
Следовательно, при лабиринтных установочных рефлексах после односторонней экстирпации лабиринтов удается доказать, что максимум возбуждения исходит из отолита, если он висит, и минимум, когда он давит. Этим подтверждается правильность вышеприведенного заключения.
Если оба лабиринта интактны и голова приведена в правое боковое положение (рис. 186), то левый sacculus находится в положении минимума и не может вызвать никакого действия.
Для правого sacculus’a имеется максимум, так как он висит. Он вызывает лабиринтный установочный рефлекс, благодаря которому, если бы имелся только один правый sacculus, голова была бы повернута в левое боковое положение. Но это не может произойти, так как с рис
увеличением поворота головы левый sacculus
все более удаляется из своего положения минимума, и вследствие этого все более и более возбуждаются мышцы, поворачивающие голову вправо. Как только голова приходит в нормальное положение (рис. 179), оба sacculu’sa стоят симметрично, двустороннее возбуждение поддерживает равновесие, а голова в этом положении находится в покое.
Как только голова удаляется из симметричного положения, увеличивается возбуждение вниз повернутого sacculus’a, в то время как возбуждение sacculus’a, находящегося вверху, уменьшается и голова благодаря этому приводится снова в симметричное положение.
Из сказанного вытекает, что положения максимума правого и левого лабиринтов для описанных лабиринтных установочных рефлексов, отличаются друг от друга примерно на 180°. Из этого следует, что здесь речь должна идти о рефлексах sacculus’a и что это не могут быть рефлексы utriculus’a, так как отолиты utriculus’a расположены почти в той же горизонтальной плоскости, в то время как отолиты sacculus’a образуют между собой угол в 120—140—150°. ! 1
25 Р. Магнус
386
Девятая глава. Функция отолитов
Такие же отношения были найдены у других обследованных млекопитающих. Следует сравнить раздел о лабиринтных установочных рефлексах и рис. 177—178 для морских свинок и обезьян.
Кроме до сих пор описанных установочных рефлексов с sacculus’а, благодаря которым голова из асимметричного положения в пространстве приводится в симметричное, могут, пожалуй, иметь место еще установочные рефлексы с utriculus’a, хотя их действие и не может демонстрироваться с такой же уверенностью. Как уже было сказано на стр. 175, голова приводится не только из асимметричного положения в симметричное, но она всегда ставится так, чтобы находиться в «нормальном положении», это значит: темя — вверху, челюсть — внизу, ротовая щель немного опущена ниже горизонтали. На модели отолита (рис. 179) можно видеть, что при этом нормальном положении sacculus’oB их продольная ось горизонтальна и что вследствие наклонного положения отолиты с обеих сторон немного давят на чувствительное пятно. Таким образом, этим может быть обусловлено как бы положение покоя обоих sacculus’oB, в то время как при всех других симметричных положениях головы, которые вызываются ее поворотом вокруг битемпоральной оси из нормального положения, это оптимальное положение sacculus’а не достигается. Все же различия в положении sacculus’а, которые здесь наблюдаются, сравнительно невелики, поэтому кажется сомнительным, являются ли они ответственными за сохранение нормального положения отолитов sacculus’a. Однако в норме (рис. 179) отолиты utriculus’a расположены горизонтально и давят на чувствительное пятно, т. е. они находятся в положении минимума; поэтому возможно, что и из utriculus’a исходят установочные рефлексы, благодаря которым голова всегда ставится так, что отолиты utriculus’a стоят горизонтально. То же самое наблюдается у других обследованных видов животных, как это наглядно показывают рис. 177 и 178 для морских свинок и обезьян. Следовательно, если придерживаться этого взгляда, то на sacculus возложена задача перевести голову из асимметричного положения в симметричное, в то время как utriculus выполняет функцию поддержания тела в нормальном положении.1 По другой теории, обе реакции исходят из sacculus’a. Которая из них правильна, можно решить только сравнительно-анатомическими исследованиями или опытами на животных с выключением sacculus’a или же utriculus’a. Эти опыты приводятся далее.
Обобщая, можно сказать, что те лабиринтные установочные рефлексы, благодаря которым голова из асимметричного положения приводится в симметричное, являются рефлексами sacculus’a. Для этих рефлексов удается доказать, что максимум возбуждения исходит из чувствительного пятна, когда отолит висит, и минимум, когда он давит. После односторонней экстирпации лабиринтов животное стремится поставить свою голову так, чтобы давил интактный отолит sacculus’a, т. е. голова поворачивается стороной с интактным лабиринтом вверх. Если оба лабиринта сохранены, то голова приходит в «нормальное положение» (к покою) тогда, когда оба sacculus’a стоят симметрично, поэтому исходящие от них возбуждения сохраняют равновесие. Если голова выводится из симметричного положения, то находящийся снизу sacculus
1 Против участия utriculus’а говорит факт, что после односторонней экстирпации лабиринтов четко не проявляется тенденция головы вернуться к нормальному положению, а голова в положении на боку с немного опущенной мордой приходит в состояние покоя. Но тогда только sacculus, а не utriculus находится в положении минимума. Однако - из этого не вытекают никакие непреодолимые затруднения.
Отношение лабиринтных рефлексов положения к положению отолитов 387
возбуждается больше, а этим голова приводится опять в нормальное положение. Связь sacculus’a с мышцами шеи проходит для этих установочных рефлексов через центры в среднем мозгу. Вызываются ли лабиринтные установочные рефлексы, благодаря которым симметрично стоящая голова приводится в «нормальное положение», с sacculus’a или с utriculus’а нельзя решить с уверенностью только с помощью модели отолитов.
3. Тонические лабиринтные рефлексы на мышцы глаз
Исследования компенсаторных положений глаз, которые, как известно, у кролика чрезвычайно выражены, привели к следующим результатам.
У кролика каждому положению головы в пространстве соответствует определенное сокращение его глазных мышц, благодаря этому определенное положение глаз длится до тех пор, пока голова сохраняет свое положение в пространстве.
В этих тонических лабиринтных рефлексах на глаза прямая наружная и внутренняя мышцы у кролика участвуют неодинаково. В основном речь идет о действии прямой верхней и нижней мышц, обусловливающих вертикальные отклонения глаз, и обеих косых мышц, вызывающих круговые движения. Обе прямые мышцы ведут себя как антагонисты: если одна укорачивается, другая — удлиняется. Косые мышцы также ведут себя как антагонисты. Но зато изменения длины прямых и косых мышц комбинируются в переменной степени. Следовательно, обе эти группы мышц функционируют независимо друг от друга (хотя они, конечно, вместе зависят от лабиринтов).
Если голова в начале находится в нормальном положении и затем поворачивается на 360° вокруг битемпоральной оси (вращение I), то при этом реагируют главным образом косые мышцы и глаза передвигаются (в том же направлении).
Если голова находится вначале в нормальном положении и затем поворачивается на 360° вокруг затылочно-носовой оси (вращение II), то при этом реагируют главным образом прямые верхняя и нижняя мышцы и глаза отводятся вертикально (в обратном направлении).
Если голова вначале находится в положении на боку и затем поворачивается на 360° вокруг вентро-дорсальной оси (вращение III), то реагируют обе группы мышц, вследствие чего положение глаз является результатом поворота глаз в ту же сторону и их вертикального отклонения в противоположную сторону.
У нормальных животных изменение положения глаз удается свести к сумме влияний, которые исходят из правого и левого лабиринтов на прямые верхнюю и нижнюю и косые верхнюю и нижнюю мышцы обоих глаз.
а) Вертикальные отклонения. Если голова находится в левом боковом положении, то правая прямая нижняя и левая прямая верхняя мышцы находятся в состоянии наибольшего укорочения, правая прямая верхняя и левая прямая нижняя мышцы — в состоянии наибольшего удлинения. Тогда правый глаз повернут максимально верх и левый глаз максимально вниз.
При всех других положениях головы в пространстве прямые верхняя и нижняя мышцы имеют те степени укорочения, которые, находятся между этими крайними состояниями. Оба глаза всегда реагируют вертикальным отклонением противоположного направления. Прямая верх¬
25*
388
Девятая глава. Функция отолитов
няя мышца одной стороны и прямая нижняя другой при этом реагируют в том же направлении.
После односторонней экстирпации лабиринтов оставшийся лабиринт вызывает наибольшее вертикальное отклонение обоих глаз от нормального положения, если он при боковом положении головы находится внизу. Тогда прямая верхняя той же стороны и прямая нижняя мышцы противоположной стороны находятся в состоянии наибольшего укорочения (максимальное положение). Тонического влияния оставшегося лабиринта на прямую верхнюю мышцу противоположной стороны и прямую нижнюю той же стороны установить не удалось.
При интактных лабиринтах при нормальном положении головы возбуждения правого и левого лабиринтов поддерживают равновесие прямых верхней и нижней мышц обоих глаз, так что тогда глаза находятся в нормальном положении.
На основании модели о роли отолитов при вертикальных отклонениях можно сказать следующее.
Так как положение максимума обоих лабиринтов отличается друг от друга почти на 180° (максимум правого лабиринта примерно при положении на правом боку, левого — примерно при положении на левом боку), то отсюда следует, что речь не может идти об рефлексах utriculus’а, так как тогда положение максимума было бы идентичным. Следовательно, это должны быть рефлексы sacculus’a. В положении максимума отолит sacculus’a висит (рис. 184), в положении минимума — он лежит (рис. 185).
После односторонней экстирпации лабиринтов вертикальное отклонение при положении максимума интактного лабиринта — максимально, при положении минимума — нулевое или очень незначительно. Это имеет также место, если отсутствует поворот шеи (см. стр. 276). Во всяком случае точно установлено, что в положении минимума интактного лабиринта не наблюдаются вертикальные отклонения в другую сторону. Из этого следует вывод, что действительно в положении минимума из отолитов sacculus’a не исходят или исходят очень незначительные возбуждения, зато в положении максимума — очень сильные возбуждения. И в этом случае (как при лабиринтных установочных рефлексах) удается доказать, что максимум возбуждения исходит из чувствительного пятна, если отолит висит, и минимум — если он давит. Следовательно, сделанные при изучении тонических лабиринтных рефлексов выводы о деятельности utriculus’a достоверны и для sacculus’a.
Каждое чувствительное пятно sacculus’a находится по крайней мере в функциональной связи с прямой верхней мышцей той же стороны и прямой нижней противоположной стороны. Совместным действием обоих sacculus’ов на прямые верхнюю и нижнюю мышцы обоих глаз объясняются вертикальные отклонения при различных положениях головы в пространстве.
При боковом положении (рис. 186) находящийся сверху sacculus пребывает в положении минимума и не влияет совсем или вызывает очень незначительное действие. Находящийся внизу sacculus тянет глаз, находящийся сверху, максимально в вертикальном направлении, глаз, находящийся снизу, — максимально дорсально. При нормальном положении головы (рис, 179) оба sacculus’a стоят симметрично, от них исходят одинаковые средней силы возбуждения, которые идут от каждого отолита sacculus’a к верхней мышце той же стороны и к нижней мышце противоположной стороны, поэтому глаза находятся в нормальном по¬
Отношение лабиринтных рефлексов положения к положению отолитов 389
ложении. В положении головы на спине (рис. 181) наблюдается такое же положение.
На рис. 187 представлены изменения длины прямой верхней мышцы у кролика с сохраненными двумя лабиринтами при различных положениях головы в пространстве.
Вращение I (сплошная линия). Исходное положение — нормальное (рис. 179). Поворот вокруг битемпоральной оси. Направление вращения — мордой вниз. Так как при этом повороте оба sacculus’a всегда остаются в симметричном положении, то почти совсем нет вертикального отклонения.
Вращение II (штрих-пунктир). Исходное положение — нормальное (рис. 179). Поворот вокруг затылочно-носовой оси. Направление вра-
/77/77
4
ч
N.
Dr. I
ƒƒ
III
/
Г*“
Чч
✓
у
*
/
ч
i у
N
Ч
'n
/
i
/
/
/
\
\
чж
4N
\
\
N
N
ч
\
\
ч,
у
г'
f
/
/
/
>
Ч
—
—
\
?д\———— i i i i I 1 -
1 2 3 9 5 6 7 8 9 10 11 12 13 П 15 16 17 1849 20 21 22 23 2425 О 15 30 45 60 75 9010512013515016518019521022529025527028530031533039536lf
Рис. 187. (Верхняя прямая мышца).
щения — исследуемый правый глаз двигается книзу. После поворота на 90° левый sacculus прошел положение минимума, правый — приближается к положению максимума (рис. 186). Вследствие этого правая верхняя мышца укорочена, правая нижняя — удлинена. При дальнейшем вращении на 110—120° правый sacculus приходит в положение максимума и отклонение глаз еще немного увеличивается. При повороте на 180° (положение на спине) оба sacculus’a стоят опять симметрично (рис. 181), поэтому глаза опять приходят в нормальное положение. При вращении на 240—250° левый sacculus приходит в максимум и правая нижняя мышца теперь укорочена, а верхняя — удлинена. При 290— 300° правый sacculus приходит в положение минимума. При дальнейшем повороте отклонение глаз постепенно идет в обратном направлении. На кривой видна сильная асимметрия. При повороте через темя (180°) происходит очень быстрое изменение длины обоих прямых мышц, в то время как при повороте через нижнюю челюсть (0 или 360°) это изменение идет очень медленно. Причина этого заключается в наклонном положении чувствительного пятна sacculus’a по отношению к средней плоскости. В нормальном положении вследствие этого наклона обоих отолитов sacculus’a имеется еще небольшое давление на чувствительное пятно. Поворот надо продолжать в обе стороны на 20—30°, пока один sacculus не встанет вертикально и вследствие этого не перестанет давить. Только при дальнейшем вращении наступают отвисание отолита sacculus’a и вследствие этого усиление его влияния, проявляющееся в более сильно вертикальном отклонении глаз. Таким образом, становится понятным, что при повороте через нижнюю челюсть положение глаз изменяется постепенно. Иначе обстоит дело, если голова находится в положении на спине (180°, рис. 181). Тогда оба sacculus’a немного свеши¬
390
Девятая глава. Функция отолитов
ваются со своих наклонно поставленных чувствительных пятен. Каждый небольшой поворот в ту или другую сторону приближает один из sacculus’ов к положению максимума, другой — к вертикальному положению, при котором он перестает висеть. Итак, незначительные повороты вызывают различные действия обоих sacculus’ов, поэтому имеются сильные вертикальные отклонения. Эти отношения можно еще уяснить следующим образом: правый sacculus стоит при вращении II в максимальном положении при 110—120°, левый — при 240—250°. Следовательно, разница поворотов между обоими положениями максимума составляет при повороте через темя только 130°, в то время как при повороте через нижнюю челюсть — 230°. Поэтому получается, что кривые для обоих прямых мышц при вращении через темя имеют очень крутой ход (особенно между 150 и 210°), в то время как при повороте через нижнюю че-
Рис. 188. Рис. 189.
л гость (от 315 через 360 до 90°) их ход изменяется постепенно. Это точно совпадает с наклонным положением чувствительного пятна sacculus’a и является дальнейшим подтверждением зависимости вертикальных отклонений головы животного от sacculus’a.
Вращение III (штриховая линия). Исходное положение — левое боковое положение, правый глаз вверху. Поворот вокруг вентро-дорсальной оси. Направление вращения — мордой вниз. При исходном положении правый глаз сильно отклонен вниз, левый sacculus перешел уже положение максимума, правый sacculus еще не пришел в положение минимума. Это достигается только поворотом на 23—32°. После вращения на 90° (мордой вниз) оба sacculus’a стоят симметрично (рис. 188), глаза находятся в нормальном положении. После поворота на 180° (правое боковое положение) левый sacculus прошел положение минимума, правый еще не достиг положения максимума (рис. 186). Правый глаз сильно отклонен вверх. При повороте на 170° (мордой вверх) оба sacculus’a снова стоят симметрично (рис. 189), глаза в нормальном положении и переходят при дальнейшем повороте до 360° в исходное положение.
И в этом случае, вероятно, из-за наклонного положения отолитов sacculus’a имеется асимметрия кривой. Однако она не очень выражена. Причина незначительной асимметрии кривой, может быть, заключается в том, что при вращении III плоскость вращения приближенно совпадает с продольным направлением отолитов sacculus’a, поэтому висячее положение отолитов sacculus’a маскируется загнутой передней частью sacculus’a. При повороте на 90° (морда внизу, рис. 188) давление отолита частично распространяется на эту переднюю часть, при вращении на 270° (морда вверху, рис. 189) отолит частично висит на ней. Так как передняя часть (с дорсальным отрезком) вследствие своего анатомического расположения механически изменяет влияния давления и отвисания отолита, то сказанное выше становится тогда понятным.
Отношение лабиринтных рефлексов положения к положению отолитов 391
Обобщая, можно сказать, что вертикальные отклонения при компенсаторных положениях глаз являются рефлексами sacculus’a и что каждое чувствительное пятно sacculus’a находится в функциональной связи с прямой верхней той же стороны и прямой нижней мышцей противоположной. Максимум возбуждения исходит из чувствительного эпителия, если отолит висит. Если отолит давит на чувствительное пятно, то возбуждения отсутствуют или минимальны. При симметричных положениях головы из обоих sacculus’oB исходят одинаковой силы возбуждения к прямым верхней и нижней мышцам обеих сторон, поэтому глаза стоят б орбите без вертикальных отклонений. Если же голова находится в несимметричном положении, то находящийся снизу sacculus приобретает инициативу и направленный вниз глаз отклоняется дорсально, глаз, направленный вверх,'— вентрально. При боковом положении головы или при положениях головы, при которых находящийся внизу отолит sacculus’a лежит горизонтально и висит на чувствительном пятне, отклонение глаз наиболее сильное. Это особенно отчетливо видно при вращении II. Вертикальные отклонения при различных положениях головы в пространстве, имеющиеся у кролика с интактными лабиринтами, объясняются совместным влиянием обоих чувствительных пятен sacculus’a па названные мышцы глаз. Особенности изменения длины прямых верхней и нижней мышц при изменениях положения головы в пространстве объясняются анатомическим расположением sacculus’a.
Эти утверждения должны быть еще дополнены точными количественными исследованиями вертикальных отклонений глаз при различных поворотах после односторонней экстирпации лабиринтов. Так как при .этом, особенно в положении максимума интактного отолита sacculus’a, появляется сильное вертикальное отклонение, то на сделанных до настоящего времени фотографиях роговица снята сильно уменьшенной и нельзя было измерить точный количественный ход отклонения. Поэтому эти опыты надо с улучшенной техникой повторить на кроликах -с односторонней экстирпацией лабиринта.
У морской свинки зависимость вертикального отклонения от положения отолитов в принципе такая же, как и у кролика.
У обезьяны же (и у человека) эти связи должны быть другими. Из сообщений на стр. 153 вытекает, что при обоих боковых положениях головы появляются такие же круговые движения глаз, причем нижняя косая мышца вышележащего глаза и косая верхняя нижележащего глаза сокращаются. Поэтому можно предполагать, что (при одинаковом модусе возбуждения чувствительного пятна отолита) каждая основная пасть sacculus’a находится в функциональной связи с косой верхней мышцей глаза той же стороны и косой нижней мышцей противоположного глаза и что эти мышцы сокращаются, если отолит висит на чувствительном пятне. Данная на рис. 178 модель отолита показывает, нто положение основных частей sacculus’a у обезьяны в основном совпадает с таковым у кролика и морских свинок.
б) Круговые движения. В то время как для до сих пор обсуждаемых лабиринтных рефлексов были определены сравнительно простые соотношения с положениями отолитов и поэтому могли быть сделаны довольно уверенные выводы об их зависимости от определенных частей отолитового аппарата, то это еще невозможно сделать для компенсаторных круговых движений. Имеющийся на сегодняшний день материал недостаточен, чтобы состояние сокращения косых мышц глаз (у кролика) свести к изменению длины только чувствительного пятна пли одной его части.
392
Девятая глава. Функция отолитов
В четвертой главе были описаны следующие факты.
Если голова находится мордой вверх примерно в зоне от + 60 да +120° (рис. 189) с относительным максимумом при +75°, то обе косые верхние мышцы (справа и слева) находятся в состоянии сильнейшего укорочения, обе косые нижние — в состоянии сильнейшей растянутости. Оба глаза повернулись верхним краем роговой оболочки вперед..
Если голова находится мордой вниз примерно в зоне от —90 до —15° (рис. 188) с относительным максимумом при —75°, то обе косые верхние мышцы находятся в состоянии наибольшей растянутости, обо косые нижние — в состоянии наибольшего укорочения. Оба глаза повернулись верхним краем роговой оболочки назад.
При всех других положениях головы в пространстве косые мышцы глаз укорачиваются в таких степенях, которые находятся между этими. крайними положениями. Положение покоя косых мышц (из опытов на трех кроликах) имеется при нормальном положении головы (—175°). В этом положении глаза стоят так же, как после двусторонней экстирпации лабиринтов.
Оба глаза реагируют при компенсаторных круговых движениях, перемещением в ту же сторону.
Круговые движения (при вращении I вокруг битемпоральной оси) изменяются несущественно, если перерезать косую верхнюю и нижнюю мышцы.
После односторонней экстирпации лабиринта движения обоих глаз качественно остаются без изменения. Один лабиринт действует одинаково на косые мышцы обоих глаз и на их движения. Оставшийся лабиринт вызывает самое большое перемещение обоих глаз вследствие сокращения обеих косых нижних мышц, если голова находится примерно вертикально мордой вниз. И наоборот, один лабиринт вызывает наибольшее перемещение обоих глаз вследствие сокращения двух косых верхних мышц, если голова находится примерно мордой вверх. Величина движений глаз при наличии только одного лабиринта примерно наполовину меньше, чем при интактных лабиринтах.
Для решения вопроса, какие положения отолитов вызывают компенсаторные перемещения глаз, имеет значение то, что после односторонней экстирпации лабиринтов положение максимума и минимума изменяется несущественно (в противоположность вертикальным отклонениям) и что лабиринт действует одинаково на одноименные косые мышцы обоих глаз.. Из этого можно заключить, если принять найденные до сих пор закономерности и для круговых движений, что положения отолитов правого* и левого уха примерно совпадают в одной и той же плоскости.
Для этого следует прежде всего принять во внимание чувствительные пятна обоих utriculus’oB. Однако если считать, что отолиты, вызывающие положение максимума и минимума, должны лежать горизонтально, то это не могут быть отолиты utriculus’a, так как они при вертикальном положении головы мордой вверх или вниз лежат не горизонтально, а приблизительно вертикально (рис. 188 и 189). При положении головы, в котором при вращении I (вокруг битемпоральной оси) глаза больше всего перемещаются вперед или назад от —90 до —15° и от +60 до +120°, они никогда не стоят горизонтально. Отолиты utriculus’a сами по себе не должны вызывать круговые движения глаз, так как максимум кругового движения тогда совпадал бы с минимумом тонических лабиринтных рефлексов на мышцы тела. Ио это не имеет места. По определениям Вейланда (.9), этот минимум расположен индивидуально между 0 и +45°. Далее при вращении II (вокруг затылочно¬
Отношение лабиринтных рефлексов положения к положению отолитов 393
носовой оси), при котором отолиты utriculus’а испытывают очень сильные изменения положения, должны были бы наступать сильные круговые движения, что вообще не имеет места (рис. 190). И круговые движения при вращении III не совпадают с изменениями положения отолитов utri- culus’a. Кроме того, необходимо учесть следующее. При нормальном положении головы влияние лабиринтов на обе косые мышцы наименьшее. С этим совпали бы горизонтальное положение отолитов utriculus’a и их давление на macula. Однако при положении головы (на спине), отличающемся на 180°, глаза опять-таки находятся в среднем положении. Что в этом случае косые верхняя и нижняя мышцы не сокращены максимально, видно из того, что глаза не отклонены в сторону носа; это подтверждается
12 3 9 5 6 7 8 9 10 11 12 13 19 15 16 17 18 19 20 21 2223 29 25 0 15 30 к5 60 75 90 105120135150165180195210225 290255270285300315 330395360'
А
*
Г
*4^
о
>
/
\
V
т{
/
Ч
I j
и
L
J
/
I
/
\
\
я*
.А
С—
V
Л
3
.А
Г
ц
i
fcid
Д
«-•н
«•-н
*—J
■
г
г
\
0
f ,
к
. ч
1 >
)
Г
Рис. 190. Кривые кругового движения глаз у кролика.
далее из опыта на животных с перерезкой одной косой мышцы — верхней или нижней. Однако при этом положении головы отолиты utriculus’a висят. Из этих умозаключений видно, что круговые движения глаз не определяются изменением положения чувствительного пятна utriculus’a.
Маловероятно, что в данном случае имеют значение основные части sacculus’а, так как они расположены не в одной плоскости и не стоят горизонтально при вертикальном положении головы.
Рис. 188 наглядно показывает положение чувствительного пятна utriculus’a и основных частей sacculus’a при положении головы —75°, когда оба глаза максимально переместились назад.
Чтобы избежать этих трудностей, я в моих первых опубликованных работах (50) высказал гипотезу, что на круговое движение влияют дорсальные отрезки sacculus’a, имеющие отдельную иннервацию через и. utricularis, и у кролика они так загнуты, что попадают почти в одну плоскость. Правда, здесь ’ сразу обнаружилась особенность, что тогда должен быть другой модус возбуждения, чем в до сих пор изученных случаях; тогда должны исходить возбуждения не только из висячего, но и из давящего отолита; при висячем положении должны были бы наиболее сильно сокращаться верхние косые мышцы, при давящем положении — нижние косые. Это уже затрудняет принятие гипотезы. Однако тогда еще не было известно точное положение дорсальных отрезков. Это последнее теперь выяснено де Б урле. Результат противоречит нашим прежним предположениям. Правда, оба дорсальных отрезка расположены почти в одной плоскости. Но они стоят горизонтально при положении головы примерно в +345 и —15°, а не при +75 или —75°. Это значит,.
394
Девятая глава. Функция отолитов
что они при принятых до сих пор модусах возбуждения не могут одни вызывать круговые движения.
Следовательно, раз чувствительные пятна utriculus’a, основные части sacculus’а и его дорсальные отрезки расположены таким образом, что не могут вызвать компенсаторных перемещений глаз, то надо думать {что маловероятно) о влиянии других частей отолитовых макул, а именно передней части utriculus’a или sacculus’a. Но и этого мало. Иначе, надо считать круговые движения зависимыми не только от одного места, а от совместного действия нескольких частей отолитового аппарата. В наших ранних работах уже указывалось на то, что чувствительное пятно utriculus’a и дорсальный отрезок sacculus’a, иннервируемые ramus utricularis, влияют на круговые движения глаз, но тогда это казалось маловероятным. Однако после того как оказалось, что одни дорсальные отрезки этого не могут вызывать, пришлось снова подумать об этой возможности.
Имеющийся до настоящего времени фактический материал недостаточен, чтобы решить вопрос при помощи добавочных данных о положении отолитовых макул или их отдельных частей. Для этого требуются новые опыты, при которых, во-первых, доляшы быть подтверждены измененными методами полученные до настоящего времени закономерности; во-вторых, должна быть исследована зависимость длины каждой косой мышцы от обоих лабйринтов в отдельности при различных положениях головы; в-третьих, надо привести голову (с помощью модели отолитов) во все те положения, при которых измеренные до сих пор плоскости отолитов принимают характерное (горизонтальное) положение; в-четвертых, надо отдельные части отолитового аппарата исключить из функциональной деятельности, другие же части привести в деятельное состояние. Следует ожидать, что таким путем удастся выяснить возникновение круговых движений. Однако при современном уровне наших знаний было бы лучше подождать результата новых опытов, чем затеряться в рассуждениях о различных возможностях.
У морской свинки имеются те же затруднения для объяснения круговых движений глаз, что и у кролика; у обезьяны же (и у человека) трудно объяснить вертикальные отклонения, здесь следует еще выяснить механизм их вызова в лабиринте.
4. Выводы о функции отолитовых аппаратов
В этом разделе будут изложены исследования, показывающие, какое положение занимают отолиты в черепе кролика и в пространстве, когда различные тонические лабиринтные рефлексы имеют максимум и минимум. Поэтому только здесь и можно решить, к каким выводам и предположениям приведет эта теория.
Этим путем можно прийти к следующему представлению: тонические лабиринтные рефлексы на мышцы тела (конечности, шеи и туловища) исходят из utriculus’a. Асимметричные лабиринтные рефлексы — во всяком случае от sacculus’a, причем еще остается неточно выясненным, исходят ли лабиринтные установочные рефлексы с utriculus’a или sacculus’a. Из компенсаторных положений глаз вертикальные отклонения во всяком случае зависят от основной части отолитов sacculus’a. Для круговых движений еще следует установить места их вызова.
О функциональном действии отолитовых аппаратов можно показать следующее.
Отношение лабиринтных рефлексов положения к положению отолитов 395
Из чувствительного пятна исходят длительные возбуждения, которые ъ зависимости от положения отолитов в пространстве имеют различную интенсивность и длятся постольку, поскольку отолит сохраняет свое положение по отношению к горизонтальной плоскости. При лабиринтных установочных рефлексах и вертикальных отклонениях глаз удается доказать для основных частей sacculus’а, что максимум возбуждения имеется тогда, когда отолит лежит горизонтально и висит на чувствительном пятне.
При тонических лабиринтных рефлексах на мышцы конечностей и шеи удалось установить то же и для utriculus’а.
G другой стороны, доказано, что при лабиринтных установочных рефлексах и вертикальных отклонениях глаз минимум возбуждения исходит из чувствительного пятна sacculus’а, если отолит лежит горизонтально и давит на чувствительное пятно. При тонических лабиринтных рефлексах на мышцы шеи и конечности то же самое удалось выяснить для utriculus’oB. При этом нельзя с уверенностью решить, возбуждены ли вообще при положении минимума макулы; может быть, они возбуждены шеоколько меньше, чем в положении максимума и в среднем положении, или же орган в положении минимума приходит в состояние покоя и не дает никаких возбуждений. Наблюдения над вертикальными отклонениями глаз после односторонней экстирпации лабиринтов подтверждают последнее предположение (основные части sacculus’а). Зато на основании исследований тонических рефлексов на мышцы шеи после односторонней экстирпации лабиринтов можно считать, что из utriculus’oB исходят возбуждения и при положении минимума, так как поворачивание головы сохраняется и в этом положении.
Если рассматривать отдельно тонические лабиринтные рефлексы ша мышцы тела, лабиринтные установочные рефлексы и вертикальные отклонения глаз, то можно прийти к следующему взгляду о деятельности отолитов: чувствительные пятна являются органами чувств, реагирующими на растяжение, и при этом от них исходят нервные импульсы, которые сохраняются без изменения до тех пор, пока отолит не изменит своего положения по отношению к горизонтали. При других положениях в пространстве эти импульсы тем меньше, чем больше отолит удаляется из «висячего» положения и приближается к «давящему» положению. Если отолит давит, возбуждение достигает минимума или становится нулевым.
В то время как эти взгляды на деятельность отолитов довольно удовлетворительно объясняют остальные тонические лабиринтные рефлексы, при изучении компенсаторных круговых движений возникают трудности, окончательное преодоление которых мы представляем дальнейшим экспериментальным исследованиям.
Вышеприведенные заключения о распределении функции между utriculus’aMH и sacculus’aMH и о центральных связях этих органов чувств прежде всего относятся к кролику. У морской свинки эти отношения в основном такие же. У остальных обследованных млекопитающих для тонических лабиринтных рефлексов на мышцы тела и для лабиринтных установочных рефлексов были найдены, по существу, такие же отношения. Компенсаторные же положения глаз у обезьян происходят иначе, чем у кролика. У обезьяны круговые движения зависят от основных частей sacculus’a, объяснение же вертикальных отклонений наталкивается здесь на те же трудности, как и при компенсаторных круговых движениях у кролика. Зависимость компенсаторных положений глаз от лабиринтов у собаки и кошки необходимо изучить еще подробнее. Однако следует
396
Девятая глава. Функция отолитов
подчеркнуть, что у остальных классов позвоночных животных, у которых с каждой стороны имеются три отолита, должно быть другое распределение функций, если при этом не принимать во внимание, что из отолитов, лагены исходят рефлексы, отсутствующие у млекопитающих. Поэтому пока нельзя переносить заключения о функции отолитов других классов позвоночных животных на млекопитающих, и наоборот. Попытки объяснить функцию отдельных отолитов у рыб были предприняты Кубо, Бенья- мином (Benjamins) и в недавнем времени Максвеллом (1, 2). Последние еще не привели к полностью разработанной и непротиворечивой теории о деятельности отолитов. Поэтому мы не проводим пока подробного обсуждения. Все же результаты Беньямина в основном совпадают с развитыми здесь взглядами.
Наконец, следует еще упомянуть, что различные положения максимума и минимума проверены для отдельных лабиринтных рефлексов не только на различных моделях отолитов, но и на моделях полукружных каналов, однако какую-либо закономерную связь между положением максимума и минимума тонических лабиринтных рефлексов и положением полукружных каналов или их ампул и cristae найти не удалось. Участие аппаратов полукружных каналов в этих рефлексах все же доказать не* удалось.
Один из приведенных здесь взглядов на функции отолитов предложен: Квиксом [Quix (2)] без экспериментальных данных на основе теоретических рассуждений и расчетов. Несостоятельность этой теории, ведущей: к многочисленным противоречиям с фактами, изложена в подробной: критической работе (60).
В. Изолированное выключение отолитов у морской свинки (52)
При обсуждениях в предыдущем разделе было сделано предположениег что лабиринтные рефлексы положения исходят из чувствительных пятевг utriculus’a, и на основе этого решалось, может ли быть найдена связь между положением отолитов в пространстве и экспериментально установленными положениями максимума и минимума этих рефлексов.
Теперь поднимается вопрос: может ли быть экспериментально доказано это предположение? Исследования лабиринтных рефлексов дали возможность различать две группы: 1) рефлексы положения, 2) рефлексы на движение. Если они действительно исходят от различных частей лабиринтов, то после выключения аппарата полукружных каналов рефлексы движения должны были бы отсутствовать, а рефлексы положения сохраниться, и наоборот: после выключения отолитов должны были бы отсутствовать рефлексы положения и сохраниться рефлексы движения. Но у теплокровных это наталкивается на очень большие технические трудности.1 Рис. 171 показывает, что эндолимфатические пространства аппарата полукружных каналов находятся в связи с utriculus’oM, а этот в свою очередь с sacculus’oM, поэтому при оперативном разрушении гребешков полукружных каналов едва ли удается избежать вскрытия utriculus’a и sacculus’а, в то время как при экстирпации чувствительных
1 У рыб, по-видимому, эти отношения более благоприятны, как вытекает из сообщений Лее, Кубо, Беньямина и Максвелла. Однако до сих пор опубликованные опыты не привели к одному окончательному результату. Даже если анализ у рыб был бы выполнен полностью, нельзя было бы его результаты так просто перенести на млекопитающих с их совсем отличным анатомическим расположением вестибулярного аппарата. Кроме того, лабиринтные рефлексы положения у рыб, за исключением компенсаторных, положений глаз (Беньямин), известны только в общих чертах.
Изолированное выключение отолитов у морской свинки
397
пятен отолитов неизбежен выход эндолимфы из полукружных каналов. Значит, в обоих случаях нельзя достичь никакого чистого экспериментального результата.
Другой возможностью было бы изолированное отделение ведущих и гребешкам или макулам нервов в височной кости без открытия лабиринтов. И это является тяжелой, до сих пор не решенной задачей, хотя предварительные работы ведутся сейчас де Клейном, Хоффманом и Верстеегом.
В то время как изолированное выключение гребешков полукружных каналов у теплокровных еще не проведено, выяснилась возможность выключить отолиты и оставить интактными аппараты полукружных паналов без вскрытия лабиринтов.
Необходимую для этого методику недавно привел Виттмаак [1909 (2)]. При помощи повторного непродолжительного, очень быстрого центрифугирования (примерно 2000 оборотов в минуту в продолжение 1/2—3/4 ми- путы) ему удалось у морских свинок отбросить отолиты, в то время как гребешки полукружных каналов остались полностью сохраненными. Однако он не провел подробнейшего физиологического исследования подопытных животных, так как имевшаяся тогда в наличии техника исследования была для этого недостаточной. Виттмаак только указывает, нто при калорическом исследовании его животных вместо быстрого ритми- неского нистагма наступал нистагм с грубыми и нерегулярными подерги- наниями, в то время как соответствующие отклонения глаз были нормальными или усиленными. Кроме этих малохарактерных явлений выпадения, у подопытных животных не обнаружено никаких нарушений со стороны отолитов и восприятия положения. Объяснение, которое Виттмаак дает этим неожиданным данным, не требует обсуждений, так как далее будет сообщено, что при дальнейших подробных исследованиях у таких животных удалось установить совсем другие нарушения. Виттмаак отмечает, что одна часть животных заболела гнойным воспалением среднего уха, вследствие чего нельзя было сделать связующие выводы.
Очень счастливой идеей у де Клейна оказалось привлечение опытной техники Виттмаака для ответа на поднятые здесь вопросы. Поэтому мы снова применили методику Виттмаака для изолированного выключения отолитов у морской свинки.
Если приведенный в прошлой главе взгляд на функцию отолитов был правильным, то в тех опытах, в которых при гистологическом исследовании отолиты были найдены оторванными, клинически не должна проявиться пи одна из функций отолитов. Напротив, очень возможно, что клинически не будут установлены никакие функции отолитов, в то время как гистологическое исследование покажет, что мембрана отолитов не оторвана или неполностью оторвана. Гистологическая сохранность мембраны отолитов ни в коем случае не исключает, что макула функционально тяжело повреждена центрифугированием.
Исследование проводилось следующим образом. У нормальных морских свинок по определенной схеме тщательно исследовались все до сих пор известные лабиринтные рефлексы. Такая схема, хорошо себя оправдавшая при многочисленных других исследованиях, приведена здесь.
Схема
Морская свинка № Число
Отметки
Исследование до и после центрифугирования Продолжительность Положение головы при центрифугировании Скорость центрифугирования
Положение спины » »
398
Девятая глава. Функция отолитов
Исследование
А. Рефлексы на движение:
1. Вращательные реакции
а) на голову
вращение вправо нистагм
вращение влево »
постреакция после вращения вправо »
постреакция после вращения влево »
б) на глаза1
вращение вправо нистагм
вращение влево »
постреакция после вращения вправо »
постреакция после вращения влево »
2. Прогрессивные движения
а)Лифтная реакция вверх Лифтная реакция вниз
б) Растопыривание пальцев при движении вниз
в) Готовность к прыжку Б. Рефлексы положения:
1. Тонические рефлексы на конечност и:2 Перекладывание из полржения на животе в положение на спину Поворот головы в боковом положении
2. Лабиринтные установочные рефлексы (в воздухе): Нормальное положение
Правое боковое положение Левое боковое положение Висячее положение головой вверх Ьисячее положение головой вниз Положение на спине
3. Компенсаторные положения глаз (следует подождать прекращения вращательных реакций!):
а) Вертикальные
правое боковое положение левое боковое положение
б) Вращательное движение
голова мордой вверх голова мордой вниз В. Можно исследовать:
1. Отклонение глаз (с нистагмом. . .) вертикально Имеется ли при этом круговое движение?
2. Поворот головы (с проходящим наклоном)?
При этом «основной поворот» (при висячем положении головой вниз)? Односторонние лабиринтные установочные рефлексы (сравнение обеих боковых положений) ?
3. Односторонняя потеря тонуса конечностей (только несколько дней подряд)?
Слуховая реакция
После этого животных под эфирным наркозом центрифугировали. Наркоз применялся по многим причинам. Во-первых, чтобы поберечь животных; во-вторых, потому что выяснилось, что при этом: опыты протекали с меньшими расстройствами, чем без эфирного наркоза. Вначале центрифуга имеет небольшую скорость, и тогда живот- 1 21 В этом ряду опытов обследовалась всегда только горизонтальная вращательная реакция и притом только качественная. Конечно, обращалось внимание и на сильные' количественные отклонения.
2 С уверенностью удается исследовать только децеребрированных морских сви¬
нок.
Изолированное выключение отолитов у морской свинки
399
ные без наркоза начинают сопротивляться этому движению; при этом они могут легко нанести себе повреждения.
Для опытов служила большая фабричная центрифуга, на которой точно друг против друга были прикреплены два ящичка. В них помещались закутанные в вату животные под глубоким эфирным наркозом. Изменением расстояния ящичков от центра можно было достичь желаемой скорости центрифугирования.
В течение исследования выяснилось, что при радиусе в 16% см достигалась скорость примерно 900—1000 м в минуту. Через 1Ц—2У2 минуты достигался ожидаемый результат. В противоположность опытам Виттмаака в большинстве случаев центрифугировали только один раз.
После центрифугирования часто необходимо было некоторое время делать животным искусственное дыхание. Только в очень редких случаях животные были мертвы после центрифугирования или имели кровохарканье, так что они были потом не пригодны для дальнейших опытов.
Сразу после центрифугирования у животных снова исследовались все лабиринтные рефлексы по приведенной выше схеме. На следующий день исследования повторялись. Таким путем можно по лучить, непрерывную картину заболевания животных.
Исходя из уже давно известной клинической диагностики полукружных каналов и, кроме того, принимая функцию различных отолитов так, как она была обоснована в предыдущем разделе, удалось поставить клинический диагноз о состоянии полукружных каналов utriculus’oB и sacculus’oB правой и левой сторон.
Затем животных децеребрировали под эфирным наркозом, чтобы точнее исследовать тонические рефлексы на конечности. Последние у недецеребри- рованных морских свинок большей частью нельзя установить с уверенностью. Таким образом, выяснилось, что у децеребрированных морских свинок во всех случаях имеются чрезвычайно сильные тонические лабиринтные рефлексы на конечности, так что их выпадение после повреждения отолитов можно с уверенностью экспериментально установить. После этого животные убивались.
Лабиринт сразу же фиксировался по методу Виттмаака. Исследование заканчивалось точным патологоанатомическим контролем в виде полных серий срезов лабиринтов.
Гистологическое исследование проводилось в анатомическом институте в Утрехте частично самим де Бурле, частично под его руководством.
При суждении о результатах анатомического исследования необходимо учесть следующее: если на серийных срезах надо точно установить, что отолиты остались или неизмененными на своих местах, или были отброшены, нужно иметь уверенность, что мембраны отолитов при изготовлении срезов или вообще при изготовлении препаратов не были смещены. Для этого имеются достаточные доказательства, полученные при изучении нормальных серий. Они показывают, что если этим способом хорошо фиксировать и делать срезы (что очень важно для результатов) не тоньше 50 jLi, то отолиты остаются на месте несмещенными. В редких случаях приподняты наружные края. У нецентрифугированных морских свинок полный отрыв никогда не наблюдался. Однако решающим является то, что диагноз отбрасывания отолитов вообще основывается не исключительно на отсутствии мембраны отолитов на макулах, а на доказательстве их присутствия в других местах внутри перепончатого лабиринта. Кроме того, недавно были произведены опыты, при которых головы морских свинок устанавливались и вращались в разных положениях в центрифуге. На основании этих исследований положения отолитов можно предварительно сказать, какие отолиты были отброшены и в каком
400
Девятая глава. Функция отолитов
направлении они двигались [подробности можно найти в работе де Бурле и де Хааса (88)]. Результат был следующий.
Опыт 1. Голова в центрифуге в нормальном положении (—135°) с радиальным направлением продольной оси, кончик носа направлен внутрь. Ось центрифуги вертикальна. Результат: мембраны отолитов in situ, отброшены только дорсальные части sacculus’а. Объяснение: при этом положении головы только дорсальные отрезки лежат так, что сила центрифуги полностью действует на них (рис. 179).
Опыт 2. Голова с радиально направленной продольной осью. Кончик носа направлен внутрь. Положение головы +120°. При этом положении головы отолиты utriculus’а очень легко отбрасываются, они должны быть отброшены в направлении передней вертикальной ампулы, в которой они действительно и были найдены. Отолиты sacculus’а также были отброшены, что соответствовало их наклонному положению.
Опыт 3. Положение головы +120°, тангенциально, левое ухо — наружу, кончик мордочки направлен на 15° вниз. Центрифуга вращается по часовой стрелке. Результат: левая мембрана utriculus’а отброшена в ампулу горизонтального по- лукружного канала, соответственно направлению центрифуги. Правая мембрана utriculus’а сдвинута немного медиально, соответственно направлению центрифуги, там она была задержана стенкой utriculus’а. Оба отолита sacculus’a in situ. Это соответствует на правой стороне ожидаемым результатам, так как мембрана при центрифугировании должна быть придавленной. Слева также ожидалось отбрасывание, но оно пе имело места.
Опыт 4. Голова центрифугировалась два раза, сперва как в опыте 3, затем как в опыте 2. Результат: обе мембраны sacculus’a отброшены, как в опыте 2. Левая мембрана utriculus’а отброшена в том же направлении, как и в опыте 3, — в ампулу горизонтального полукружного канала, т. е. как при первом центрифугировании. Правая мембрана utriculus’а оторвалась не полностью, осталась прикрепленной у латерального края чувствительного пятна, верхний край открепившейся части доходит до входа передней вертикальной ампулы. Это соответствует явлениям при первом центрифугировании (опыт 3) и отбрасыванию к передней вертикальной ампуле при втором центрифугировании (опыт 2).
Из этого вытекает, что результат центрифугирования при заданном положении головы можно до определенной степени предсказать^ При этом не только можно предсказать, что не должно произойти, но и с некоторой уверенностью сказать, какие мембраны отолитов и куда будут отброшены. Такие опыты необходимы как основа будущих экспериментов, при которых должна быть отброшена только одна часть отолитов. Там, где речь идет только об удалении или неудалении отолитов, совпадение ожидаемого и действительного результатов может быть использовано как основа достоверности при гистологических исследованиях.
Результаты экспериментов следующие.
1. Сразу после центрифугирования в большинстве случаев можно найти очень типичную картину, а именно: исчезли все тонические рефлексы положения, в то время как рефлексы движения, вращательные и постреакции, а также реакции на прогрессивные движения еще сохранены, даже в большинстве случаев усилены. Эта картина совпала в девяти особенно тщательно исследованных случаях.
2. В следующие дни можно наблюдать очень различные состояния.
A. У некоторых животных постепенно полностью восстанавливалась функция лабиринтов, так что под конец нельзя было найти никаких отличий от нормальных животных.
Б. У других же животных лабиринтные рефлексы не восстанавливаются. Тогда можно всегда найти картину, как это было только что описано, типичную для исследований сразу же после центрифугирования: исчезновение всех рефлексов положения и сохранение рефлексов движения.
B. У некоторых животных отмечается только частичное возвращение функций. При этом наблюдаются очень сложные картины, и в этих случаях надо ставить клинический диагноз о функционировании или нефункциони-
Изолированное выключение отолитов у морской свинки
401
ровании отдельных отолитов. Такие картины можно, как было сказано выше, вызвать произвольно.
В дальнейшем будут обсуждаться случаи А и Б, случаи В находятся еще в обработке; о них будет сказано позже.
Нижеследующие примеры приводятся из первой проведенной для этой цели серии опытов.
А. Животные, сохранившие после центрифугирования нормальные лабиринтные рефлексы или у которых они восстановились.
Из этой группы приведен анатомически точно исследованный случай. За исключением одного частично поврежденного, на что дальше будет обращено внимание, utriculus’a, все полукружные каналы и отолиты были найдены анатомически нормальными.
Морская свинка 2. 3 февраля 1919 г. Непосредственно до центрифугирования. Все лабиринтные рефлексы нормальны. Основной поворот в 30° был установлен только при висячем положении головой вниз.
Клинический диагноз: полукружные каналы нормальны; отолиты
нормальны, перевес правого utriculus’a по сравнению с левым.
3 февраля, 4 часа. Центрифугирование со скоростью 900 м в минуту. Продолжительность центрифугирования 1У2 минуты. Голова горизонтально, живот к центру.
Сразу после центрифугирования животное сидит спокойно, голова прямо, нет качания головы, нет спонтанного нистагма.
Вращательные реакции головы и глаз и постреакции нормальны.
Реакции на прогрессивные движения все отчетливы. Отсутствуют лабиринтные установочные рефлексы. При висячем положении головой вниз голова повернута влево на 45—90°, позже — направо, иногда и прямо.
Компенсаторные положения глаз: отсутствуют вертикальные отклонения, круговые движения примерно 60°.
4 февраля, 10 час. Все лабиринтные рефлексы вполне нормальны. При положении сидя или при висячем положении головой вниз голова стоит совсем симметрично.
Клинический диагноз: полукружные каналы интактны,
sacculus’bi интактны. Поскольку до центрифугирования имелся основной поворот влево, то на основании этого можно,заключить о перевесе функции правого utriculus’a. Так как после центрифугирования исчез основной поворот (функции utriculus’a стали одинаковыми), было принято (как возможное) частичное повреждение правого utriculus’a.
Результаты анатомических исследований. Аппарат полукружных каналов интактен. Оба sacculus’а интактны. На рис. 191 дан правый sacculus при сильном увеличении, виден чувствительный эпителий, а на нем интактная мембрана отолита.
На рис. 192 видна также интактная мембрана отолита левого sacculus’а при более слабом увеличении.
Правый utriculus: действительно частично поврежден.
На некоторых срезах можно найти мембрану отолитов почти нормальной, на других же срезах — наоборот, более или менее поврежденной. Часть мембраны, обращенная к полукружным каналам, лежит нормально на чувствительном эпителии, другая часть отброшена с подкладки. Рис. 193 дает картину среза в области сильнейшего повреждения.
Левый utriculus вполне нормален. Рис. 194 показывает его при большом увеличении; чувствительный эпителий покрыт нормальной мембраной отолитов.
Следовательно, в этом случае вполне совпадают клинический диагноз и анатомические данные.
Б. Животные, у которых после центрифугирования исчезли и длительно отсутствовали 26 Р. Магнус
-f
-2
Рис. 191. Морская свинка 2. Правый sacculus. 1 — мембрана отолита; 2 — чувствительный эпителий.
Рис. 192. Морская свинка 2. Левый sacculus. 1 — чувствительный эпителий с интактной мембраной отолита.
Рис. 193. Морская свинка 2. Правый utriculus. 1 тельный эпителий с мембраной; 2 — мембрана;
вительный эпителий без мембраны.
— чувстви- 3 — чувст-
Рис. 194. Морская свинка 2. Левый utriculus. 1 — мембрана отолита; 2 — чувствительный эпителий.
404
Девятая глава. Функция отолитов
рефлексы положения, в то время как рефлексы движения (вращательные реакции, постреакции и реакции на прогрессивные движения)
были сохранены.
Далее приведены протоколы опытов на трех первых животных этой группы. Одно из этих животных (морская свинка 1) центрифугировалось, когда нам еще не были известны рефлексы на прогрессивные движения.
Морская свинка 1. 23 мая 1918 г. Исследование до центрифугирования. Все лабиринтные рефлексы нормальны (прогрессивные реакции не обследованы).
23 мая, ЗУ2 часа. Центрифугирование со скоростью 660 м в минуту, в течение 1 У2 минут; сразу после этого еще одно центрифугирование со скоростью 900 м, в течение
1У2 мин.
Сразу после центрифугирования: горизонтальное покачивание головы.
4У2 часа. Голова при положении сидя в спокойном состоянии держится прямо, нет спонтанного нистагма глаз.
Имеют место вращательные и постреакции на голову и глаза, только поствращательная реакция головы не очень ясная.
Тонические лабиринтные рефлексы на конечности отсутствуют (зато наблюдаются шейные рефлексы).
Лабиринтные установочные рефлексы пол- Рис. 195. Слева — нормаль- ностыо отсутствуют.
ная морская свинка. Справа — Компенсаторные положения глаз полностью
морская свинка 1. отсутствуют.
24 мая, 11% час. Животное сидит вполне симметрично, спонтанный нистагм отсутствует. После бега несколько раз горизонтальное покачивание головы.
Положение такое же, как 24 мая, 4У2 часа.
2 часа. Животное сфотографировано, чтобы показать отсутствие лабиринтных установочных рефлексов в боковом положении (рис. 195).
4 часа. Вращательные реакции и постреакции на голову и глаза слабые.
25 мая, 9% часа. Вращательные и постреакции на голову и глаза отчетливы.
Тонические лабиринтные рефлексы на конечности, лабиринтные установочные рефлексы и компенсаторные положения глаз отсутствуют.
101/4 часа. Эфирный наркоз. Трахеотомия, перевязка сонных артерий. Блуждающие нервы перерезаны, децеребрация, отчетливая децеребрационная ригидность.
При перекладывании с живота в положение на спину: нет никакого следа тонических лабиринтных рефлексов на конечности.
Ii1/i часа. Децеребрационная ригидность средней силы. Поворот головы при положении на боку: отсутствие тонических лабиринтных рефлексов (зато отчетливы тонические шейные рефлексы).
Поворот головы в положении на спине: отчетливые шейные рефлексы.
Клинический анализ: на основании рефлексов движения и отсутствия лабиринтных рефлексов устанавливается диагноз — полукружные каналы интактны, все отолиты выключены.
Результаты анатомических исследований: полукружные каналы и гребешки интактны.
Правый sacculus: мембраны отолитов патологически изменены и отчасти плохо окрашены, но не отброшены (рис. 196).
Левый sacculus: мембраны отолитов полностью отброшены,
мембрана находится в углу sacculus’a (рис. 197).
Правый utriculus: мембрана отолитов полностью отброшена, лежит в ампуле горизонтального полукружного канала (рис. 196).
Левый utriculus: мембрана отолита полностью отброшена,
лежит в углу utriculus’a (рис. 197).
В этом случае анатомическое исследование выявило изменения, которые следовало ожидать по клинической картине. Теперь можно было из клини¬
Изолированное выключение отолитов у морской свинки
405
ческого исследования заключить, что и правая мембрана sacculus’a должна была быть отброшена, в то время как в действительности эта мембрана, правда, не была нормальной, но была найдена на своем месте. Конечно, возможно, что животное слишком рано было убито и что функция правого sacculus’a в последующие дни более или менее восстановилась бы. Как уже было выше сказано, сразу после центрифугирования можно найти временное выключение рефлексов положения, в то время как позже они в ряде случаев снова восстанавливались.
Однако не исключалась возможность и того, что мембрана вообще повреждалась центрифугированием, хотя гистологически это и не удавалось доказать.
Морская свинка 35. 31 марта 1919 г. Исследование до центрифугирования. Все лабиринтные рефлексы нормальные.
4 часа. Центрифугирование со скоростью 960 м в минуту, в течение 2% минут. Голова горизонтальна. Живот к центру.
1 апреля, 11 час. Вращательные реакции и постреакции на голову и глаза отчетливы. Прогрессивные рефлексы отчетливы.
Отсутствуют лабиринтные рефлексы на конечности.
Лабиринтные установочные рефлексы, по-видимому, полностью отсутствуют, может быть только небольшой след.
При положении сидя левый глаз отведен вниз, а правый — вверх.
2 апреля, И час. Наблюдаются вращательные реакции и постреакции.
Имеются рефлексы на прогрессивные движения.
Тонические лабиринтные рефлексы на конечности отсутствуют, но зато отчетливы тонические шейные рефлексы.
Полностью отсутствуют лабиринтные установочные рефлексы; незначительный основной поворот влево.
Компенсаторные положения глаз полностью отсутствуют. Отклонение глаз у животного в положении сидя такое же, как и 1 апреля,
4 апреля, 11 час. Имеются вращательные реакции и постреакции на голову и глаза, однако отмечена отчетливая разница между вращением вправо и влево. Картина совпадает с той, которая наблюдается у животного после левосторонней экстирпации лабиринта.
Рефлексы на прогрессивные движения очень отчетливы, только нет лифтной реакции.
Отсутствуют тонические лабиринтные рефлексы на конечности.
Полностью отсутствуют лабиринтные установочные рефлексы. Основной поворот исчез.
Компенсаторные положения глаз отсутствуют; при положении сидя у животного левый глаз немного отведен вниз, правый — немного вверх.
После этого эфирный наркоз, трахеотомия, перевязка сонных артерий, блуждающие нервы перерезаны; децеребрация с последующей отчетливой децеребрационной ригидностью.
Перекладывание из положения на животе на спину: нет и следа тонических лабиринтных рефлексов на конечности. Поворот головы при положении на боку: нет и следа тонических лабиринтных рефлексов, зато отчетливые шейные рефлексы.
Клинический диагноз: повреждение полукружных каналов. Вследствие разницы в реакции при вращении вправо и влево, причем была найдена картина такая же, как после левосторонней экстирпации лабиринта, можно было предположить возможное повреждение системы полукружных каналов левой стороны.
Sacculi и utriculi: на основании отсутствия всех тонических лабиринтных рефлексов следует признать выключение всех отолитов. Проходящий основной поворот влево и отклонение глаз, как после левосторонней экстирпации лабиринта, дают возможность заключить, что левый sacculus и utriculus больше никакого влияния не оказывают, в то время как правые мембраны отолитов хотя и не функционируют, однако все же легко раздражимы.
Результат анатомического исследования: в правых полукружных каналах незначительное кровоизлияние, слева
Рис. 196. Морская свинка 1. Правый лабиринт. 1 — эпителий utriculus’a без мембраны; 2 — эпителий sacculus’a с мембраной (измененной); 3 — мембрана utriculus’а в ампуле правого горизонтального полукружного канала.
Рис. 197. Морская свинка 1. Левый лабиринт. 1 — мембрана utriculus’a; 2 — эпителий utriculus’a без мембраны; 3 — эпителий sacculus’a без мембраны; 4 — мембрана sacculus’a.
Изолированное выключение отолитов у морской свинки
407
кровоизлияние в перилимфатическое пространство и в полукружные каналы.
Правый sacculus: мембрана отолита отброшена (рис. 198), находится в крайнем углу sacculus’a (рис. 199).
Левый sacculus: большое кровоизлияние в sacculus, мембрана отолита отброшена, находится в углу sacculus’a (рис. 200).
Правый utriculus: мембрана отолита отброшена (рис. 198), находится в заднем вертикальном полукружном канале (фотография не подошла к репродукции).
Левый utiiculus: сильное кровоизлияние в utriculus и его окружение. Мембрана отолита полностью разорвана, но не совсем отброшена (рис. 200).
И здесь имеется хорошее совпадение клинической и анатомической картин. Не говоря уже о разрушении мембран отолитов, большие кровоизлияния сами по себе достаточны, чтобы объяснить, что весь левый лабиринт исключен и клинически нельзя выявить функцию полукружных каналов и отолитов левой стороны.
Справа мембраны отолитов были отброшены, но оставшиеся сохраненными полукружные каналы объясняют клинически найденные рефлексы движения.
Морская свинка 8. 10 февраля 1919 г. Исследования до центрифугирования. Все лабиринтные рефлексы нормальные.
4 часа. Центрифугирование со скоростью 900 м в минуту в течение 2 минут. Голова горизонтально, живот к центру.
Сразу после центрифугирования: вращательные реакции и ностреакции на голову и глаза отчетливы.
Рефлексы на прогрессивные движения отчетливы.
Тонические лабиринтные рефлексы на конечности полностью отсутствуют (имеются тонические шейные рефлексы).
Лабиринтные установочные рефлексы отсутствуют. Компенсаторные положения глаз отсутствуют. Животное сидит вполне симметрично, нет спонтанного нистагма.
И февраля, 10 час. Положение такое же, как 10 февраля, только слабо выражены реакции на прогрессивные движения.
12 февраля, 10 час. Положение то же, однако сегодня реакции на прогрессивные движения очень сильно выражены.
И час. Эфирный наркоз. Трахеотомия, перевязка сонных артерий, перерезка блуждающих нервов. Децеребрация, очень отчетливая децеребрационная ригидность. Перекладывание с живота в положение на спину: нет и следа тонических лабиринтных рефлексов на конечности.
Поворот головы в положении на боку: нет и следа тонических лабиринтных рефлексов, зато сильные шейные рефлексы.
Клинический диагноз: полукружные каналы интактны.
Выключены отолиты sacculus’a и utriculus’a.
Результаты анатомического исследования: полукружные каналы и гребешки интактны.
Правый sacculus: мембрана отолита отброшена, находится в углу sacculus’a (рис. 201).
Левый sacculus: мембрана отолита отброшена, лежит в углу sacculus’a. На рис. 202 (левый sacculus) виден чувствительный эпителий, мембрана отолита отсутствует полностью, можно найти только отдельные зернышки в виде остатков мембраны (ср. рис. 191, где показана нормальная мембрана). На рис. 203 изображен левый sacculus при слабом увеличении. Видна отброшенная мембрана в углу sacculus’a.
Правый utriculus: мембрана отолита отброшена, найдена
в заднем вертикальном полукружном канале. На рис. 204 изображен чувствительный эпителий, на нем лежат отдельные зернышки, но нет
Рис. 198. Морская свинка 35. Правый лабиринт. 1 — эпителий sacculus’а без мембраны; 2 — эпителий utriculus’a без мембраны.
Рпс. 199. Морская свинка 35. Правый лабиринт. 1 — мембрана sacculus’a; 2 — utriculus.
Рис. 200. Морская свинка 35. Левый лабиринт. 1\— кровоизлияние в sacculus’е; 2 — эпителий sacculus’а; 3 — мембрана sacculus’a; 4 — кровоизлияние; 5 — разорванная мембрана utriculus’а; 6 — эпителий utriculus’а.
Рис. 201. Морская свинка 8. Правый лабиринт. 1 — эпителий utriculus’a без мембраны; 2 — мембрана sacculus’a; 3 — эпителий sacculus’a без мембраны.
Рис. 202. Морская свинка 8. Левый sacculus. 1 — эпите¬
лий sacculus’а без мембраны.
Рис. 203. Морская свинка 8. Левый лабиринт. 1 — utriculus; 2 — мембрана sacculus’а; 3 — эпителий sacculus’а без мембраны.
Рис. 204. Морская свинка 8. Правый utriculus. 1 — от¬
дельное зернышко мембраны; 2 — эпителий utriculus’а.
Рис. 205. Морская свинка 8. Правый задний полукружный канал. 1 — задний вертикальный полукружный канал; 2 — мембрана utriculus’а в заднем вертикальном полукружном канале.
412
Девятая глава. Функция отолитов
и следа от нормальной мембраны (ср. с рис. 194). Отброшенная мембрана находится в заднем вертикальном полукружном канале (рис. 205).
Левый utriculus: мембрана отолита отброшена (рис. 206),
находится в заднем вертикальном полукружном канале.
Рис. 207 показывает отброшенную мембрану в заднем вертикальном полукружном канале.
В этом совсем несложном случае наблюдается полное соответствие между клиническим и анатомическим диагнозами.
В дальнейшем изложении этой главы еще будет приведен подобный несложный случай, отличающийся от вышеприведенного примера тем, что он обследовался 11 дней после центрифугирования, без того чтобы изменился результат.
Обобщая эти опыты, можно сделать следующие выводы: центрифугированием по методу Виттмаака у морских свинок удалось во многих случаях вызвать состояние, при котором были нормальны лабиринтные рефлексы на движения (вращательные и постреакции на голову и глаза и реакции на прогрессивные движения), в то время как лабиринтные рефлексы положения отсутствовали.
Если животное оставить на несколько дней в живых и каждый день исследовать лабиринтные рефлексы, то можно наблюдать, что у ряда животных это состояние остается неизменным, в то время как у других тонические рефлексы положения частично, а иногда и полностью восстанавливаются.
Гистологические исследования показали в том случае, когда функции лабиринтов снова стали полностью нормальными, что все отолиты были на своих местах, за исключением отолитов utriculus’а, который был поврежден; эти данные полностью совпали с клиническими явлениями. Аппарат полукружных каналов оказался также сохраненным.
У одной морской свинки через два дня появились лабиринтные рефлексы на движения (вращательные и прогрессивные реакции), но зато у этой свинки не наблюдалось никаких лабиринтных рефлексов положения. При гистологических исследованиях обнаружено, что все четыре мембраны отолитов были отброшены и находились в другом месте лабиринта, в то время как гребешки полукружных каналов оказались интактными. То же самое было найдено у другого животного, у которого наблюдения могли длиться 11 дней после центрифугирования.
В двух других случаях, при которых после центрифугирования отсутствовали лабиринтные рефлексы положения, мембраны отолитов оказались также частично отброшенными, отчасти разорванными и вследствие кровоизлияния не функционировали. Только один отодит sacculus’а был на своем месте, показывая, однако, гистологические отклонения. Аппарат полукружных каналов у одного животного был интактным, у второго — очень сильно поврежден с другой стороны вследствие кровотечения, что полностью совпадало с клиническими явлениями.
Из этих фактов можно сделать следующие выводы.
1. Вращательные реакции и постреакции на голову и глаза являются рефлексами полукружных каналов; они могут нормально проявляться и при отсутствии мембраны отолитов.
2. То же самое действительно и для реакции на прогрессивные движения.
3. Лабиринтные рефлексы положения (тонические лабиринтные рефлексы на мышцы тела, лабиринтные установочные рефлексы и компенсаторные положения глаз), напротив, являются рефлексами отолитов, которые невозможно больше вызвать после разрушения мембраны отолитов.
Рис. 206. Морская свинка 8. Левый лабиринт. 1 — эпителий sacculus’a без мембраны; 2 — эпителий utriculus’a без мембраны (лишь отдельные зернышки).
Рис. 207. Морская свинка 8. Полукружные каналы слева. 1 — мембрана utriculus’a в левом заднем вертикальном полукружном канале; 2 — левый горизонтальный полукружный канал; 3 — левый задний вертикальный по- лукружный канал.
414
Девятая глава. Функция отолитов
4. Сразу после центрифугирования во многих случаях временно аппарат отолитов выключается из-за сильного механического воздействия на специфически более тяжелые мембраны отолитов. Он выключается настолько, что больше не вызываются рефлексы с отолитов даже тогда, когда мембрана осталась на своем месте. Функция затем восстанавливается через различное время.
5. Из этого следует, что рефлексы на движение (вращательные реакции и ностреакции на голову и глаза и на прогрессивные движения) вызываются с аппарата полукружных каналов. Для рефлексов же положения (тонические лабиринтные рефлексы на мышцы тела, установочные рефлексы и компенсаторные положения глаз) ответственным является аппарат отолитов.
6. Этим, конечно, не исключается, что и аппарат отолитов может возбуждаться определенными формами движения (центрифугированием, быстрыми прогрессивными движениями); так, например, этим объясняется участие отолита utriculus’а при лифтных реакциях вверх и вниз.
Г. Состояние возбуждения отолитовых макул (61, 64)
Так как лабиринтные рефлексы больше не вызываются после отбрасывания мембран отолитов, их следует рассматривать как рефлексы отолитов. Изменением положения головы (а этим и отолитов) в пространстве у животных, подвергшихся центрифугированию, больше не удается изменить состояние возбуждения чувствительного эпителия макул. Однако из этого не следует, что эпителий чувствительного пятна при этих условиях не находится в длительном и постоянном состоянии возбуждения. A priori это вполне возможно, так и сетчатка может быть длительно возбуждена без освещения. Однако после удаления мембраны отолитов это состояние возбуящения потом, при изменении положения головы в пространстве, не сможет измениться.
Если поставить вопрос, каким путем отолиты осуществляют изменение возбуждения в макулах, надо прежде всего знать, в каком состоянии находится чувствительный эпителий, если на него больше не воздействуют отолиты. A priori имеются две возможности: или чувствительный эпителий тогда находится в состоянии покоя и его деятельность вызывается только давлением и оттягиванием мембраны отолитов, или чувствительный эпителий может сам по себе вызывать длительное возбуждение, сила которого в зависимости от положения отолитов по отношению направления силы тяжести может увеличиваться или уменьшаться.
Для решения этого вопроса следует исходить из следующих предпосылок.
Если экспериментировать с лабиринтом нормальной морской свинки, то наступает описанное уже в шестой главе сложное состояние. При подробном анализе его вытекали следующие симптомы как прямые последствия одностороннего удаления отолитового аппарата (чувствительный эпителий+отолит) или, говоря точнее, как последствия одностороннего сохранения отолитового органа.
а) Основной поворот головы в сторону удаленного лабиринта.
б) Вертикальные отклонения глаз, причем глаз оперированной стороны отведен вниз, а другой стороны — вверх.
Вследствие основного поворота (и наклона) головы наступают вторичные изменения положения всего тела, различия в тонусе конечностей, движении глаз и т. д.
Состояние возбуждения отолитовых маку л
415
(Какая часть лабиринта ответственна за временное различие тонуса конечностей, которое имеется при прямо стоящей голове, до сих пор не известно. Поэтому этот симптом в дальнейшем не учитывается).
На основании этих фактов были проделаны следующие опыты.
Морские свинки центрифугировались по Виттмааку, а затем для дальнейшего наблюдения отбирались те животные, у которых продолжительное время отсутствовали лабиринтные рефлексы положения, в то время как имелись все рефлексы движения; другими словами, животные, у которых можно было предполагать, что отброшены все отолиты на обеих сторонах.
Чтобы по возможности исключить вызванное самим процессом центрифугирования раздражение или торможение чувствительного эпителия, животные регулярно обследовались. Уже очень скоро прекратились возможные симптомы раздражения после центрифугирования. Чтобы быть совсем уверенным, сам опыт проводился только через 7—9 дней после центрифугирования.
По истечении этого времени на одной стороне через барабанную перепонку вводится примерно 0.1 мл 5% -го кокаинового раствора в среднее ухо, чтобы полностью парализовать лабиринт на этой стороне.
Если чувствительный эпителий макул вследствие центрифугирования на неинъецированной стороне не возбужден, то после выключения другого лабиринта кокаином не должны наступать симптомы, исходящие из отолитовых макул (за исключением в крайнем случае возможного нистагма и поворота головы как следствия одностороннего выключения полукружных каналов).
Если же макулы будут находиться в длительном возбуждении после удаления отолитов, то выключение одного лабиринта кокаином должно выявить последствия возбуждения макул. Тогда должны наступить такие же симптомы, как после односторонней экстирпации лабиринтов у нецентрифугированных животных. Следовательно, исходящий из utriculus’а основной поворот головы в инъецированную сторону и обусловленные sacculus’ом вертикальные отклонения глаз (на инъецированной стороне — вниз, на другой — вверх) различаются только тем, что у центрифугированных животных эти симптомы не изменяются как у нормальных животных после односторонней экстирпации лабиринта в зависимости от положения головы в пространстве вследствие различного влияния отолитов интактной стороны, а остаются без изменения независимо от того, в какое положение привести голову животного в пространстве.
И это последнее обстоятельство действительно имеет место.
Всего было проведено 5 опытов с полностью совпадающими результатами. Примером служат следующие протоколы трех опытов.
Морская свинка R. 28 июня 1921 г. Все лабиринтные рефлексы нормальные.
Центрифугирование: голова вверх, живот к центру, длительность 2 минуты, скорость 1000 м в минуту.
2 июля 1921 г. Тонические лабиринтные рефлексы полностью отсутствуют.
4 июля 1921 г. Рефлексы с полукружных каналов: вращательные рефлексы вправо — положительны, влево — слабо выражены.
Прогрессивные реакции сомнительны или отсутствуют. Топические рефлексы полностью отсутствуют.
5 июля 1921 г. Рефлексы с полукружных каналов (также прогрессивные реакции) наблюдаются все, и они симметричны.
Тонические лабиринтные рефлексы: все отсутствуют. Животное сидит в симметричном положении. Нет отклонения глаз. В положении на спине с прямо поставленной головой нет отчетливого различия в тонусе конечностей.
416
Девятая глава. Функция отолитов
11ч. 39 м. 0.1 мл 5%-го кокаина вводится в левое среднее ухо.
11 ч. 41 м. Висячее положение в воздухе головой вниз: голова на 90° повернута вправо, при положении сидя правый глаз отведен вниз, левый глаз — вверх (значит, раздражение левого лабиринта!).
11 ч. 43 м. Висячее положение головой вниз: голова опять стоит симметрично.
11 ч. 47 м. Висячее положение головой вниз: голова на 20—30° повернута влево и немного наклонена влево. При положении сидя небольшой поворот головы влево, нет отчетливых отклонений глаз.
И ч. 49 м. Висячее положение головой вниз: голова на 45° повернута влево, животное при положении сидя падает на левую сторону. Если посадка головы прямая, отсутствует различие в тонусе конечностей. При спонтанной постановке головы сдвиг влево гораздо больше, чем вправо, животное сильно свешивается влево (начинающийся паралич левого лабиринта).
11 ч. 51 м. Висячее положение головой вниз: голова повернута влево на 70°, при положении сидя нистагм головы вправо. Перемещение животного экспериментатором на полу вызывает перекатывание влево. Отсутствуют отчетливые отклонения глаз.
11 ч. 54 м. Висячее положение головой вниз: голова повернута влево на 90°. Левый глаз отведен немного вниз, правый — вверх.
12 час. На левом глазу слабые нистагмические подергивания спереди вверх, на правом глазу — назад вниз. Отсутствуют изменения симптомов при изменении положения головы в пространстве (правое и левое боковое положения).
12 ч. 03 м. Сильный спонтанный нистагм в том же направлении, как и в 12 час.
12 ч. 06 м. Сильное отклонение глаз и нистагм, которые не изменяются при различных положениях головы в пространстве.
6 июля 1921 г. Рефлексы с полукружных каналов имеются все и симметричны.
Тонические рефлексы все отсутствуют. Асимметрия вследствие введения кокаина полностью исчезла.
12 час. Децеребрация. Выраженная децеребрационная ригидность.
При перекладывании с живота на спину нет и следа тонических лабиринтных рефлексов. При повороте головы в положении на боку типичные шейные рефлексы, отсутствуют тонические лабиринтные рефлексы.
Морская свинка S. 28 июня 1921 г. Все лабиринтные рефлексы нормаль¬
ные.
Центрифугирование головой вверх, живот к центру. Продолжительность 2 минуты. Скорость 1000 м в минуту.
4 июля 1921 г. Рефлексы с полукружных каналов асимметричны. Вращательные реакции на голову и глаза при вращении вправо — слабые, влево — сильные.
Прогрессивные реакции слабые. Отсутствует растопыривание пальцев.
Рефлексы положения отсутствуют. Незначительный основной поворот влево.
7 июля 1921 г. Рефлексы с полукружных каналов имеются и симметричны; прогрессивные реакции слабые.
Рефлексы положения отсутствуют. Нет больше основного поворота. Животное сидит симметрично, отсутствуют отклонения глаз.
При положении на спине и прямо поставленной голове: отсутствует разница в тонусе конечностей.
Введение 0.1 мл 5%-го кокаина в левое среднее ухо.
12 ч. 30 м. Висячее положение головой вниз: поворот головой вправо (раздражение левого лабиринта!).
12 ч. 30 1/2 м. Висячее положение головой вниз: голова поставлена прямо, больше не повернута.
12 ч. 31 м. Висячее положение головой вниз: голова повернута влево (обусловлено параличом левого лабиринта). *
12 ч. 31% м. Висячее положение головой вниз: голова повернута на 60° влево.
12 ч. 33 м. При положении животного сидя голова повернута и наклонена влево; при сидении — движение по часовой стрелке влево, отсутствует нистагм.
12 ч. 34 м. Левый глаз отведен вниз, правый — вверх. Нистагм отсутствует.
12 ч. 34% м. Сильное отклонение глаз, нистагма нет. Отсутствует разница в отклонении глаз при изменении положения головы в пространстве. При висячем положении головой вниз голова имеет поворот влево на 90°.
12 ч. 36 м. Правое боковое положение: голова в нор- 1 мальном положении.
Левое боковое положение: голова в положении на спине.
Положение на спине: голова в правом боковом положе- > нии. I
Висячее положение головой вверх: голова в левом бо- | новом положении. j
Отсутствует изменение поворота головы при различных положениях головы в пространстве.
Состояние возбуждения отолитовых маку л
417
12 ч. 38 м. Нистагма нет.
12 ч. 40 м. При вращении вправо и влево: вращательная реакция глаз и нистагм, вращательная реакция головы — положительная. На полу: движение по часовой стрелке влево; при ударе по ноге животное перекатывается один раз влево.
12 ч. 52 м. Отчетливое отклонение глаз, очень сильный спонтанный нистагм на левом глазу по направлению вперед вверх, на правом — назад вниз.
4 часа. При положении животного сидя голова имеет максимальный поворот и наклон влево, сильные движения перекатывания, сильный спонтанный нистагм.
8 июля 1921 г. Лабиринтные рефлексы положения полностью отсутствуют. Асимметрия предыдущего дня совсем исчезла.
9 июля 1921 г. У животного одышка. Децеребрация. Нет хорошей децеребрацион- ной ригидности. Совсем отсутствуют тонические лабиринтные рефлексы на конечности.
Морская свинка F. 28 мая 1921 г. Все лабиринтные рефлексы положительные.
Центрифугируют головой вверх, животом к центру. Продолжительность центрифугирования 2 минуты. Скорость 1000 м в минуту.
31 мая 1921 г. Рефлексы с полукружных каналов: вращательные и постреакции — положительны.
Прогрессивные реакции: лифтная реакция положительна, остальные прогрессивные реакции — слабые.
Отолитовые рефлексы отсутствуют.
2 июня 1921 г. Реакции с полукружных каналов положительные, только еще слабо выражено растопыривание пальцев.
Отолитовые рефлексы отсутствуют.
6 июня 1921 г. Рефлексы с полукружных каналов положительны.
Отолитовые рефлексы отсутствуют.
5 ч. 52 м. 0.05 мл 10%-го кокаина вводится через левую барабанную перепонку в среднее ухо.
6 час. При положении животного сидй с прямо поставленной головой правый глаз отклонен вверх, левый — вниз (начинающийся паралич левого лабиринта).
6 ч. 02 м. При положении сидя поворот головы немного влево. Само животное перевесилось влево. Висячее положение головой вниз: основной поворот на 90° влево.
6 ч. 06 м. При повороте влево слабая реакция поворота головы, отчетливая постреакция. При повороте вправо сильная вращательная реакция головы и отсутствие постреакции.
6 ч. 07 м. Вращательная реакция глаз при повороте вправо: отчетливая реакция с нистагмом, отсутствие постреакции. При повороте влево: вращательная реакция и постреакция.
6 ч. 10 м. Прогрессивные реакции: лифтная реакция неотчетлива. Готовность к прыжку — положительная.
Отолитовые рефлексы отсутствуют.
Положение головы в воздухе при:
правом боковом положении: голова в нормальном по¬
ложении, из-за основного поворота иногда висит вниз;
левом боковом положении: голова находится вследствие основного поворота в положении на затылке;
висячем положении головой вверх: голова в левом боковом положении, животное делается беспокойным (действие кокаина);
висячем положении головой вниз: поворот головы влево на 90°;
положении на спине: голова вследствие поворота влево находится в правом боковом положении, иногда в положении на затылке с наклоном влево.
• 6 ч. 18 м. При положении сидя с прямо поставленной головой правый глаз отклонен вперед, вверх, левый глаз — назад, вниз. Нистагм при этом в обратном направлении.
6 ч. 33 м. В правом боковом н зложении левый глаз отклонен назад, вниз. Нистагм в обратном направлении. Отведение глаз и нистагм левого глаза одинаковы по силе при левом и правом боковом положении головы, то же самое и для правого глаза.
7 июня 1921 г. Животное сидит в симметричном положении, отсутствуют отклонения глаз.
Реакции с полукружных каналов положительные.
Отолитовые рефлексы отсутствуют. Асимметричные явления после введения кокаина полностью отсутствуют.
8 июня 1921 г. Все рефлексы выражены так же, как и в предыдущий день. При положении животного сидя голова слегка повернута вправо. Но животное сидит симметрично, отсутствуют отведения глаз.
27 Р. Магнус
Следовательно, постоянный поворот головы, который сохраняется при изменении положения головы в пространстве.
418
Девятая глава. Функция отолитов
Анатомическое исследование де Бурле: все мембраны отолитов отброшены.
П р а вый sacculus: чувствительный эпителий без мембраны, мембрана отолита лежит свободно в полости sacculus’a, между эндолимфа тическим протоком и задним концом чувствительного эпителия.
П р а вый utriculus: чувствительный эпителий без мембраны,
мембрана отолита расположена между задним концом чувствительного пятна и входом общей ножки, совсем медиально.
Левый sacculus: чувствительный эпителий без мембраны,
мембрана отолита прилегает к латеральной стенке sacculus’a и по отношению чувствительного пятна сдвинута вверх.
Левый utriculus: чувствительный эпителий без мембраны,
медиально и кверху.
Эти опыты показывают, что чувствительный эпителий макул в течение одной недели после центрифугирования находится в постоянном состоянии возбуждения. Если временно исключить один лабиринт кокаином, то вследствие возбуждения макул неинъецированного лабиринта асимметричные симптомы появляются таким же образом, как и у нормальных животных после односторонней экстирпации лабиринта, однако с тем различием, что эти симптомы у центрифугированных и односторонне инъецированных кокаином животных не изменяются при изменении положения головы в пространстве.
Если подождать после центрифугирования больше недели, то станет очевидным, что это состояние возбуждения эпителия макул уже больше не зависит от центрифугирования. Поэтому следует чувствительному эпителию макул приписать свойство самому производить возбуждение, которое, если отсутствуют мембраны отолитов, имеет постоянную силу. Функция мембран отолитов должна заключаться в том, что они изменяют силу этого состояния возбуждения в эпителии в зависимости от того, оттягивают ли они макулу или же давят на нее.
Для иннервируемой г. saccularis основной части sacculus’a выше было приведено доказательство, что возбуждение возрастает при оттягивании благодаря отолитам и уменьшается при давлении. Для макул utriculus’а стал очевидным тот же механизм. Для тех частей лабиринта, которые вызывают круговые движения глаз, еще следует установить их способ действия. Следовательно, если здесь не откроются новые факты, можно будет сделать заключение, что задача отолитов состоит в том, чтобы в зависимости от их положения в пространстве усиливать путем оттягивания или уменьшать давлением возбуждение эпителия макул.
Таким образом, дано первое представление о деятельности отолитового аппарата.
Если развитый здесь взгляд является правильным, то мы этим снова приближаемся к выдвинутому Эвальдом учению о лабиринтном тонусе.1 Тогда из макулы, не заторможенной давлением отолитов, шли бы постоянные возбуждения к мышцам тела и глаз,1 2 пока исходящие из правого и левого лабиринтов раздражения взаимно не сняли или уничтожили бы друг друга в центральной нервной системе. Поэтому надо еще раз напомнить о факте, что после двусторонней экстирпации лабиринтов у млекопи-
1 При этом, конечно, нельзя забывать, что в лабиринтном тонусе Эвальда имеется много нелабиринтных элементов.
2 В описанных опытах проявлялись такие длительные влияния на мышцы шеи и мышцы, отводящие глаз вертикально. Следует еще проверить, действительно ли это- и для остальных мышц тела.
Анализ деятельности лабиринтов после введения кокаина
419
тающих не наступает общего расслабления мышц и что как тонус, так и способность к мышечной работе в течение времени не уменьшаются. Значит, лабиринты являются одним из многих источников тонуса и, наверное, они не самые главные. Поэтому их влияние при выключении заменяется деятельностью других органов. До какой степени это возможно, показывает наблюдение, что после двусторонней экстирпации лабиринтов и перерезки трех верхних шейных задних корешков сохраняется хороший постоянный тонус мышц шеи.
Д. Дальнейший анализ деятельности лабиринтов после введения кокаина. Состояние возбуждения гребешков полукружных каналов
(82Ь)
К приведенным выше наблюдениям присоединяются дальнейшие опыты с выключением лабиринтов путем введения кокаина, которые привели к интересным результатам.
Как уже было сказано на стр. 289, при опытах с введением кокаина в среднее ухо нормальных и центрифугированных морских свинок удается снова наблюдать, что (после того как прошло возможное раздражение инъецированного лабиринта и стали отчетливыми явления торможения) прежде всего наступает вертикальное отклонение глаз без какого-либо нистагма и что последний начинается позже, иногда только через 20 минут и больше. Так как, по прежним рассуждениям, отклонение следует рассматривать как следствие выключения sacculus’a, а наступающий тут же. основной поворот следует отнести за счет паралича utriculus’a, то из этого вытекает предположение, что появляющийся позже нистагм обусловлен выключением аппарата полукружных каналов. Правильность этого взгляда удается проверить экспериментально. Нужно доказать, что в первой стадии, когда еще нет нистагма, аппараты полукружных каналов на инъецированной стороне еще возбудимы (поворотом и калорически), в то время как с появлением нистагма они становятся невозбудимыми. Это в действительности и имеет место.
Однако при этих опытах выяснилось дальше, что чувствительное пятно sacculus’а и utriculus’а парализуется не одновременно, так что может наблюдаться стадия, при которой utriculus еще функционирует, в то время как sacculus уже парализован.
Описанные далее результаты получены на 19 удачно проведенных опытах на морских свинках. У 9 морских свинок лабиринты были сохранены, у 5 — один лабиринт был экстирпирован за 5 часов до исследования, у 5 — за 5 дней.
Если нормальной морский свинке, у которой были проведены по вышеприведенной схеме исследования на интактность всех лабиринтных рефлексов, ввести 0.1 мл 5%-го кокаина через барабанную перепонку в среднее ухо (например, левое), то через несколько минут (в среднем через 4—б мин.) ослабляются лабиринтные рефлексы на голову при левом боковом положении в воздухе, а затем совсем исчезают. Тогда животное держит голову в левом боковом положении, в то время как из правого бокового положения голова приводится в нормальное положение. Одновременно или сразу после этого появляется вертикальное отклонение глаз (без нистагма); левый глаз отведен вентрально, правый — дорсально, при правом боковом положении головы отклонение достигает максимума, при нормальном положении оно меньше, при левом боковом положении оно минимально или совсем отсутствует. Таким образом, отклонение происходит так же, как и при левосторонней экстирпации лабиринта.
27*
420
Девятая глава. Функция отолитов
Исходя из ранее изложенных рассуждений, одностороннее прекращение лабиринтных установочных -рефлексов при положении на боку и вертикальное отклонение глаз можно объяснить параличом sacculus’a на инъецированной стороне.
На этой стадии совсем еще не выявляется основной поворот, как это прежде всего удается установить в висячем положении головой вниз. Животное нормально сидит и способно бежать прямо, без каких-либо нарушений. Значит, utriculus еще не поврежден. Интересным является то, что лабиринтные установочные рефлексы при симметричном положении тела в этом состоянии еще нормальны. При висячем положении головой вверх голова приводится вентральным сгибанием в нормальное положение, при висячем положении головой вниз — дорсальным сгибанием. При положении на спине животное сгибает переднюю часть туловища вентрально, так что голова опять приходит в нормальное положение. Эти данные говорят за то, что, как уже предполагалось выше, симметричные лабиринтные установочные рефлексы исходят из utriculus’а. В этой стадии все вращательные реакции на голову и глаза с соответствующим нистагмом сохранены без изменения и симметричны.
Через различное время (7—14 минут, иногда и раньше) начинает появляться при висячем положении головой вниз основной поворот в сторону инъецированного уха (левого); вначале он незначителен, но скоро становится максимальным и достигает 90° и выше. Следовательно, теперь наступает паралич utriculus’a. При положении животного сидя также видны поворот головы влево и перевес тела в левую сторону. Но зато совсем нет наклона головы и туловища влево и животное способно ходить прямо, причем оно при сильном основном повороте может упасть на левую сторону, но затем снова поднимается. У свободно сидящего животного не наблюдается спонтанный нистагм головы или глаз.
На этой стадии интактность аппарата полукружных каналов можно доказать следующими опытами: вращательная реакция головы имеется при повороте вправо и влево и притом вполне симметричная. Также можно вызвать при вращении в обе стороны с одинаковой силой вращательную реакцию глаз с соответствующим нистагмом; это можно наблюдать при горизонтальных, вертикальных и ротаторных вращательных реакциях. При введении холодной воды в слуховой проход (левой) стороны, инъецированный кокаином, появляется типичный калорический нистагм, при котором в быстрой фазе левый глаз подергивается вперед, а правый — назад. Что он является нистагмом полукружных каналов, можно узнать по описанному Барани поведению: если придать голове положение мордой вниз, то нистагм изменяет свое направление в противоположную сторону. Опыты с калорическим нистагмом только тогда доказательны, когда нистагм снова прекращается после окончания промывания водой (а не переходит в появляющийся в следующей стадии спонтанный нистагм), что наблюдалось много раз.
Описываемые далее опыты, в которых экстирпировался сперва правый лабиринт, а затем выключался кокаином левый, дали возможность с еще большей уверенностью доказать, что на этой стадии сохраняется возбудимость полукружных каналов при вращательных движениях; здесь с уверенностью можно сказать, что правый лабиринт уже не может возбуждаться.
Через разное время (иногда очень быстро, иногда через 7—8 минут) присоединяется паралич полукружных каналов. Голова сначала незначительно, а затем все больше поворачивается влево, к этому присоединяется
Анализ деятельности лабиринтов после введения кокаина
421
поворот туловища влево. Вращательная реакция головы становится асимметричной, по направлению поворота головы ее сила остается без изменения; в обратном направлении (влево, если животное сидит мордой к центру вращения) она вначале ослаблена, затем исчезает. Спонтанный нистагм глаз появляется вначале в виде отдельных подергиваний с изменяющимся направлением, но вскоре он становится равномерным, сильным и в левом глазу принимает направление вперед вверх, а в правом — назад вниз. То, что это — нистагм паралича доказывается тем, что он, в противоположность калорическому нистагму, не изменяет направления, если голова находится в положении мордой вниз. Поворот вправо или влево влияет на этот спонтанный нистагм неодинаково, значит асимметрично. Теперь появляется и типичный спонтанный нистагм головы. Вследствие сильного поворота начинаются движения по направлению часовой стрелки, животные уже не могут ходить прямо.
Следовательно, при этих опытах на трех стадиях развивается типичная картина выключения лабиринта, как уже было описано в шестой главе. Сначала наступает паралич sacculus’a, характеризующийся односторонним исчезновением лабиринтных установочных рефлексов при боковом положении животного и вертикальным отклонением глаз, затем происходит паралич utriculus’а — основной поворот (без наклона), а под конец — паралич полукружных каналов — поворот головы и спонтанный нистагм головы и глаз.
Только один единственный симптом одностороннего выключения лабиринта никогда не наблюдался при всех этих опытах на морских свинках: односторонняя потеря тонуса конечностей. И после полного паралича лабиринта, где в одном случае был также затронут лицевой нерв, никогда не происходило ослабления тонуса конечностей левой стороны, если голова ставилась прямо по отношению тела. Следовательно, этот симптом, появляющийся почти постоянно после хирургического удаления лабиринта, остается и на сегодняшний день необъяснимым.
В частности, в этих опытах выявились следующие особенности.
Иногда наблюдается временное возбуждение, предшествующее кокаиновому параличу и продолжающееся только несколько минут. В этом случае (до паралича) наблюдается отведение глаз (sacculus) или основной поворот (utriculus) в противоположную сторону. В одном случае наблюдался паралич только sacculus’a, в то время как utriculus и полукружные каналы не были выключены. Время между отдельными стадиями очень варьирует. Один опыт не мог быть использован, так как стадии так быстро сменялись, что невозможно было провести исследование различных рефлексов. В других случаях развитие полного паралича лабиринта наступало через полчаса,, так что было возможно провести очень точные наблюдения. Один раз основной поворот- начал ся очень скоро (через 4 минуты), поэтому было трудно отличить первую стадию- от второй. В других же случаях стадии были резко ограничены.
Как уже было сказано, спонтанный нистагм не изменяется в противоположную- сторону после паралича полукружных каналов при изменении положения головы. Однако следует указать на то, что если исходить из нормального положения головы и привести ее в положение мордой вниз, то наступает компенсаторное круговое движение глаз, верхним полюсом роговицы вниз (исходя из интактного лабиринта другой стороны), вследствие чего направление нистагма немного изменяется; однако это нельзя путать с изменением направления нистагма по фазе.
Во всех случаях рефлексы с инъецированного уха на следующий день снова нормальны.
У отдельных животных после введения кокаина в среднее ухо не наступало выключения лабиринта. При вскрытии в этих случаях обнаруживалось воспаление среднего уха с эксудатом, этим объяснялось отсутствие ожидаемого действия.
Уже этот первый ряд опытов приводит ко многим интересным выводам, из которых необходимо остановиться на следующих.
422
Девятая глава. Функция отолитов
1. Разделение последствий выключения лабиринта на три резко отличающиеся группы. К первой относятся асимметричные лабиринтные установочные рефлексы и вертикальные отклонения глаз; этим подтверждается выше сделанное заключение, что они вызываются с одного и того же чувствительного пятна (с sacculus’а). Ко второй группе относятся основной поворот и симметричные установочные рефлексы. Так как основной поворот по вышеизложенным причинам следует отнести за счет utri- culus’a, то, по-видимому, последний решающим образом влияет и на симметричные установочные рефлексы. Третья группа: нистагм головы и глаз, а также и наклон головы у морской свинки вызываются с полукружных каналов. Этим дополняется проведенный в шестой главе анализ последствий односторонней экстирпации лабиринта. В то время как у кролика и обезьяны наклон головы является длительным последствием операции, в чем, может быть, принимают участие отолитовые аппараты, этот наклон головы у морской свинки, кошки и собаки является только преходящим явлением. Его следует (по опытам на морских свинках) рассматривать как симптом полукружных каналов.
Что касается нистагма головы и глаз после выключения лабиринта, проведенного без раздражения, то опыты показывают, что это выключение чувствительного пятна отолитов не вызывает нистагма и поэтому этот симптом зависит исключительно от деятельности полукружных каналов.
2. Если спонтанный нистагм не зависит от чувствительного пятна отолита и появляется после проведенного без раздражения выключения полукружных каналов, то из этого следует, что причина его возникновения лежит в возбуждении, исходящем из аппарата полукружных каналов интактной стороны. Нистагм наблюдается непрерывно (в первые дни после одностороннего выключения лабиринта), даже тогда, когда голова находится в полном покое. Этим напрашивается вывод, что чувствительный эпителий гребешков продуцирует в полукружных каналах длительное возбуждение и что задача cupulae заключается в том, чтобы ослабить или усилить длительное возбуждение эпителия гребешков, если cupulae приходят в движение или придавливаются эндолимфой полукружных каналов. Таким образом, мы приходим к тому же представлению для аппарата полукружных каналов, как это было выше развито для чувствительного пятна отолитов: эпителий, производящий длительное возбуждение, сила которого может быть изменена действием механических аппаратов. Возможно, что этим обусловливается чрезвычайно большая чувствительность рецепторных образований лабиринта. Механическое устройство в полукружных каналах такое, что изменения состояния возбуждения в чувствительных клетках проявляются только временно, на чувствительных же пятнах отолитов возможны длительные возбуждения. Рефлексы с полукружных каналов преходящие, в то время как с этих органов чувств к центрам идут постоянные возбуждения. Следовательно, и в этом случае мы опять приближаемся к представлениям Эвальда о тонусе лабиринтов.
3. Далее, бросающимся в глаза является то, что вызывающиеся у морской свинки симптомы с полукружных каналов при односторонней экстирпации лабиринтов являются все преходящими, в то время как симптомы отолитов остаются длительное время. Нистагм головы и глаз, наклон головы и тела, а также движения по часовой стрелке исчезают через несколько дней.
4. Последовательность, в которой чувствительные окончания парализуются кокаином, введенным через среднее ухо, соответствует их анатомическому расположению. На рис. 193 видно слева внизу пространство среднего уха, отделенное сравнительно легкой стенкой от овального окна и
Анализ деятельности лабиринтов после введения кокаина
423
пластинкой стремечка — от преддверия, на медиальной стенке которого расположен sacculus. Utriculus и полукружные каналы разделены пограничной мембраной (рис. 172) и этим лучше защищены. Может быть, тот факт, что кокаин влияет на чувствительное пятно utriculus’a раньше, чем на ампулы полукружных каналов, обусловливается тем, что utriculus лежит на пограничной мембране.
Сделанные здесь выводы были подтверждены следующими опытами и расширены в некоторых пунктах.
У пяти морских свинок был экстирпирован правый лабиринт, и через пять дней появилась типичная картина симптомов.
Голова повернута вправо, при положении животного сидя наклонена или вправо, или стоит прямо, но может быть наклонена и влево. Поворот головы асимметричен, так же как и горизонтальный поворот глаз с вращательным нистагмом. Наблюдаются вертикальные и ротаторные вращательные реакции глаз. Спонтанный нистагм отсутствует (или в некоторых случаях слабо выражен). Прогрессивные реакции положительны. Основной поворот на 90° вправо. Лабиринтные установочные рефлексы да ют-типичную картину: при правом боковом положении животного голова находится в правом положении на боку, при левом боковом положении — переходит в нормальное положение. Правый глаз направлен вниз (может быть, немного назад), левый глаз — вверх (может быть, немного вперед). Отклонение глаз при левом боковом положении максимально, при нормальном положении — незначительно, в правом боковом положении — минимально. Правая передняя нога при установке головы прямо более расслаблена, чем левая.
В одном опыте наблюдалось только изменение основного поворота по Бехтереву, в то время как не было видно извращения нистагма и изменения наклона головы. В этом случае на первой стадии паралича полукружных каналов исчезали горизонтальная вращательная реакция и правый наклон головы, вращательная реакция и поворот головы влево, в то время как вертикальная вращательная реакция глаз оставалась сильно выраженной, она исчезала только на второй стадии. В другом случае обратимость основного поворота наблюдалась на такой стадии, при которой вообще еще не имелось никаких симптомов паралича полукружных каналов. Зато в другом опыте не было изменения основного поворота, однако наблюдались обратный наклон головы, а также спонтанный нистагм глаз по Бехтереву. Все это также говорит о самостоятельности симптомов с полукружных каналов и utriculus’а, хотя наклон головы и основной поворот в этих опытах изменялись совершенно независимо друг от друга. Во всех случаях на следующий день действие кокаина полностью исчезало и левый лабиринт снова функционировал нормально.
Если теперь ввести 0.1 мл 5%-го кокаина в левое среднее ухо, то можно наблюдать, как последствия правосторонней экстирпации лабиринта € закономерной последовательностью исчезают соответственно постепенному параличу чувствительных образований в левом лабиринте.
Прежде всего наступает паралич sacculus’a. Вследствие этого при положении животного на правом боку в воздухе голова находится не в боковом положении (лабиринтный установочный рефлекс), а в положении на спине (основной поворот). Одновременно или сразу после этого уменьшается различие в вертикальном отклонении глаз при правом и левом боковых положениях, а затем и совсем исчезает. Глаза теперь находятся в нормальном положении, иногда остается еще очень незначительное отклонение глаз в первоначальном направлении (действие левых полукружных каналов). На этой стадии основной поворот остается еще неизменным при 90°, спонтанный нистагм отсутствует, вращательные реакции и наклон головы не изменяются. Затем начинает уменьшаться основной поворот и, наконец, совсем прекращается. Вследствие этого, так как выключены sacculus и utriculus, голова имеет боковое положение при обоих боковых положениях животного в воздухе. Левые полукружные каналы еще ин- тактны. Поэтому голова наклонена вправо (при отсутствии основного
.424
Девятая глава. Функция отолитов
поворота), вращательная реакция головы типично асимметричная, можно вызвать горизонтальные, вертикальные и ротаторные вращательные реакции глаз с нистагмом, как и до введения кокаина. Наконец, парализуются и полукружные каналы. Однако это, что очень интересно, происходит в два этапа. Сперва исчезает наклон головы вправо (и может вследствие компенсации Бехтерева перейти в наклон влево), горизонтальная вращательная реакция головы больше не вызывается, и в большинстве случаев появляется спонтанный нистагм Бехтерева в противоположном направлении. Однако на этой стадии вертикальная вращательная реакция глаз еще отчетливо выражена. Так как в горизонтальной вращательной реакции, по общему мнению, участвует горизонтальный полукружный канал, то эти данные можно объяснить только так, что вначале парализуются чувствительные образования горизонтального полукружного канала, в то время как один или оба вертикальных полукружных канала поражаются позже. Познания физиологии на сегодняшний день недостаточны, чтобы решить вопрос о сказанном. Наконец, парализуется весь аппарат полукружных каналов и животное ведет себя, как морская свинка после двусторонней тотальной экстирпации лабиринтов, только с той разницей, что не исчезает расслабление правой передней ноги.
Постепенный паралич аппарата полукружных каналов сопровождается типичным изменением нистагма Бехтерева. На первой стадии нистагм происходит больше горизонтально, во второй — вертикально. Например, у большинства животных он был на первой стадии в правую сторону назад ,вниз, в левую — горизонтально вперед, на второй стадии — вправо вертикально вниз, влево наверх вперед.
В опытах, в которых введение кокаина в левый лабиринт происходило уже через 4—5 часов после хирургической экстирпации, можно было очень -хорошо наблюдать последовательность исчезновения острых симптомов.
Сперва животные сидели с сильным наклоном головы и туловища влево, имели нистагм голоды влево, сильный основной поворот вправо, типичное отклонение глаз (влево-вверх-вперед, вправо-вниз, с нистагмом влево-вниз-назад, вправо-вверх-вперед), которое было максимальным при левом боковом положении животного и минимальным — при правом. При левом боковом положении в воздухе голова была в нормальном положении, при правом боковом положении в воздухе голова также была в положении на правом боку. Сильная асимметрия вращательной реакции головы и нистагма. Круговые движения вправо.
После введения кокаина в левое среднее ухо в одном типичном опыте исчезали сперва односторонние лабиринтные установочные рефлексы, так что при правостороннем положении тела голова находилась в положении на спине, и уменьшалась разница в величине отклонения глаз при правом и левом боковых положениях, затем она совсем исчезла. На этой стадии паралича sacculus'а имелись еще основной поворот, спонтанный нистагм, наклон головы, односторонняя асимметричная вращательная реакция головы и очень незначительное возвращение глаз в нормальное положение. Затем наступал паралич utriculus'a, который выражался в уменьшении основного поворота. Потом следовал паралич полукружных каналов: исчезали вращательная реакция головы, наклон головы, направление нистагма извращалось, — и все это вначале при прямо стоящей голове (без поворота и наклона). Только примерно через 10 минут происходило изменение основного поворота, после чего животное катилось влево.
Интересным было изменение нистагма при наступлении паралича полукружных каналов:
Анализ деятельности лабиринтов после введения кокаина
425
До паралича
Незадолго до исчезновения вращательной реакции головы После исчезновения вращательной реакции головы Через 6 минут
Вправо-вперед-вверх
» вверх (верт.).
» назад (гориз.)
» назад-вниз
влево-назад-вниз
» вниз (верт.)
» вперед (гориз.) » вперед-вверх
При наступлении паралича горизонтального полукружного канала диагональный нистагм становится чисто вертикальным. После полного паралича горизонтального полукружного канала измененное направление нистагма чисто горизонтальное, а при тотальном параличе лабиринтов становится наклонным, но в противоположном направлении, чем до введения кокаина.
Обусловливаются ли изменения нистагма, основного поворота и т. д., наблюдаемых в этих опытах через 4—5 часов после выключения лабиринта, центрально вызванной компенсацией Бехтерева или продолжающимся после операции раздражением правого ствола 8-го нерва, нельзя окончательно решить. В этом смысле должны влиять оба фактора.
Изменение направления спонтанного нистагма наблюдалось в трех из пяти опытов при появлении паралича полукружных каналов. Два раза наблюдался только совсем слабый обратный нистагм, начавшийся в одном опыте чисто горизонтально. Интересным было, что в этой последней серии опытов стадия паралича utriculus’а и стадия выключения полукружных каналов больше сливались вместе, чем в других экспериментах, где они резко разграничивались. Имеются разные возможности объяснения этого факта, но экспериментально это еще окончательно не проверено.
Описанные опыты с инъекцией кокаина в среднее ухо морской свинки показывают, что как при интактности другого лабиринта, так и при его предварительном оперативном удалении симптомы паралича кокаином развиваются закономерно и последовательно и что группы симптомов, наблюдающиеся при этом, соответствуют ранее установленным представлениям для отолитовых рефлексов. Кроме того, удается выделить еще одну группу симптомов и свести их к функции полукружных каналов.
Если лабиринт другой стороны сохранен, то кокаином вызываются соответствующие симптомы; если он удален, то эти симптомы исчезают. Следовательно, обе серии опытов являются зеркальным отображением.
Паралич sacculus’а вызывает при интактном втором лабиринте одностороннее прекращение асимметричных лабиринтных установочных рефлексов и вертикальное отклонение глаз без нистагма. Симметричные лабиринтные установочные рефлексы остаются сохраненными и зависят, по-видимому, от utriculus’a. Если второй лабиринт предварительно удален, то последствием паралича sacculus’a является исчезновение всех асимметричных установочных рефлексов и вертикального отклонения глаз (остается только очень маленькая часть, зависящая от полукружных каналов).
Паралич utriculus’a вызывает при интактном втором лабиринте основной поворот по направлению инъецированной стороны; при отсутствии же другого лабиринта основной поворот исчезает, а при известных обстоятельствах он направлен в противоположную сторону, что по Бехтереву является следствием раздражения при инъекции.
При сохранном втором лабиринте не появляются ни нистагм головы и глаз, ни наклон головы, пока полукружные каналы интактны и раздражаются вращением или вливанием воды в ухо. При наступлении паралича полукружных каналов наблюдаются наклон головы
426
Десятая глава. Центры установки тела
и спонтанный нистагм головы и глаз, причем вращательные реакции одновременно становятся асимметричными. При отсутствии другого лабиринта исчезают наклон головы, а также имеющееся иногда незначительное вертикальное отклонение глаз; вращательные реакции головы и глаз отрицательные, появляется спонтанный нистагм глаз в обратном направлении. При появлении паралича полукружных каналов удается установить, что прежде всего затухают горизонтальные вращательные реакции головы и глаз, вертикальные же вращательные реакции остаются дольше. Направление движения глаз при обратном нистагме говорит о том, что сперва парализуется горизонтальный, а затем вертикальный полукружный канал.
Ранее описанные опыты с параличом, вызванным кокаином на центрифугированных морских свинках, привели к выводу, что эпителий чувствительного пятна вызывает длительное возбуждение, сила которого изменяется оттягиванием или давлением отолитов.
Описанные в конце опыты делают в высшей степени достоверным, что чувствительный эпителий гребешков производит длительное возбуждение, сила которого изменяете# раздражением cupulae. Рефлексы с полукружных каналов вызывались бы тогда только изменением силы возбуждения эпителия гребешков, как это имело место при движениях или ускорениях головы.
В то время как в предыдущих главах в основном сообщались более или менее законченные результаты, исследование деятельности отолитов далеко еще не доведено до конца; вполне вероятно, что некоторые приведенные выше взгляды при дальнейших познаниях будут модифицированы. Однако, если это и будет иметь место, я считаю полученные на сегодняшний день результаты не излишними, так как они являются основой для дальнейших экспериментальных исследований этой до сих пор загадочной и поэтому привлекательной области.
Десятая глава
ЦЕНТРЫ УСТАНОВКИ ТЕЛА
В предыдущих разделах на уровне наших знаний того времени были описаны рефлексы, которые участвуют в установке тела. Насколько это возможно, до сих пор были описаны мышцы, которые участвуют в этих рефлексах; рецепторы, с которых они запускаются; афферентные пути, которые проводят импульсы в центральную нервную систему; законы, по которым различные рефлексы складываются в группы, накладываются одни на другие и взаимодействуют друг с другом; роль, которую они играют в нормальных и патологических установках и движениях животных. Остается еще описать те отделы центральной нервной системы, наличие которых совершенно необходимо для осуществления этих рефлексов. Хотя в предыдущих главах отдельные детали из этой области и были уже приведены, тем не менее обобщенное представление на основе фактических данных, изложенных до сих пор, представляется необходимым.
Уже a priori можно предположить, что для группы функций, где большое число различных рефлексов действует автоматически, в строго определенном единстве, должен существовать развитый центральный аппарат. Если до сих пор совершенно не удавалось раскрыть этот механизм во всех его деталях, тем не менее оказалось возможным определить положение и взаимное пр<остранственное расположение главнейших групп центров и
Десятая глава. Центры установки тела
427
тем самым по меньшей мере положить начало тому, чтобы связать отдельные анатомически известные ядра с установкой тела. Главную часть работы еще предстоит выполнить. Но установленное до сих пор может послужить в качестве фундамента для дальнейших экспериментальных исследований.
Если не учитывать оптических установочных рефлексов, для образования которых необходима кора большого мозга, то оказывается, что все описанные в этой книге рефлексы установки тела в полной мере присутствуют уже у таламического животного. В отношении рефлексов, исходящих из лабиринта, и некоторых других установочных реакций в этой главе будет показано, что они сохраняются и после удаления мозжечка. Отсюда следует, что центры для этих рефлексов лежат в стволе мозга. В действительности оказалось, что центральный аппарат установки располагается, начиная от самой верхней части шейного мозга, до самой передней части среднего мозга. В этом отделе территориально раздельно располагаются центры для различных групп рефлексов.
Если начинать с таламического животного, исследование расчленяется на следующие части.
1. Следует установить, какие рефлексы сохраняются после полного удаления мозжечка и, таким образом, имеют свои центры в мозговом стволе.
2. Следует выяснить путем постепенного, шаг за шагом, спереди назад отделения частей мозгового ствола, до какого уровня можно удалить ствол, не повредив определенного рефлекса. При этом центры должны лежать позади соответствующего разреза. Это исследование должно быть произведено для каждого отдельного рефлекса.
3. Определив этим путем уровень, на котором лежит определенный центр, можно произвести здесь различные локальные разрушения и установить, сохраняется ли при этом рефлекс или исчезает, и по этим результатам судить о зависимости рефлекса от определенных, анатомически известных ядер и путей.
4. Цель исследования — установить для каждого рефлекса центр или центры и совокупность афферентных и эфферентных путей.
Задачи, описанные в пунктах 1 и 2, в главных чертах решены. Третья проблема разрабатывается. Достижение цели, поставленной в пункте 4, конечно, еще очень далеко.
По поводу уверенности, с которой из опытов с оперативными вмешательствами и экстирпациями на центральной нервной системе могут быть сделаны заключения относительно локализации и функций определенных центров, необходимо сказать следующее.
Если у экспериментального животного после удаления части мозга (например, мозжечка или большого мозга) или после перерезки ствола можно показать, что определенный рефлекс совершенно не изменился, то отсюда с абсолютной достоверностью следует, что для осуществления данного рефлекса удаленная часть мозга не является необходимой и что соответствующие центры лежат в оставшейся части мозга. Если только физиологическое наблюдение производится точно, в этом случае достаточно одного или немногих опытов, чтобы с уверенностью сделать выводы.
Наоборот, если после поперечной перерезки или экстирпации определенный рефлекс отсутствует, нельзя с абсолютной уверенностью полагать, что необходимые для этого рефлекса центры удалены. Всегда остается возможность, что центры находятся в оставшейся части мозга, но настолько сильно повреждены во время оперативного вмешательства, что более не функционируют. Если вследствие кровотечений и т. п. оно так и слу¬
428
Десятая глава. Центры установки тела
чилось, это устанавливается последующим анатомическим исследованием. Более существенной помехой, однако, являются шок или явления диашизаг после оперативного вмешательства часть оставшихся центров даже без доказуемых анатомических изменений оказывается функционально поврежденной. Если при этом определенный рефлекс отсутствует, исследователь не знает, причина ли тому удаление или шок оставшегося центра. Этой неуверенности можно избежать, если поставить большее количество отдельных опытов, но прежде всего в том случае, если отбирать для этих опытов те виды животных, у которых, согласно имеющемуся опыту, явления шока невелики. По этой причине обезьяны, к сожалению, исключаются из большей части подобных экспериментов, так как уже исходный препарат (таламическая обезьяна) до сих пор не может быть получен свободным от явлений,шока. Это не удавалось полностью ни Карплусу и Крей- длю (Karplus u. Kreidl), ни мне самому (см. выше, стр. 169). Поэтому с уверенностью делать заключения можно только из тех опытов на обезьянах, в которых после экспериментального вмешательства исследуемый рефлекс оставался ненарушенным. В других случаях следует спуститься к более низко стоящим млекопитающим (кошки и кролики). Результаты в острых опытах несколько лучше у кроликов, чем у кошек. У первых часто уже сразу после операции, во время пробуждения от наркоза, можно наблюдать большую часть рефлексов. До какой степени это возможно, показывает следующий пример: в ядре глазодвигательного нерва, орально, лежат центры для зрачка и аккомодации, каудально от них — центры наружных глазных мышц. Радемакер (Rademaker) в отдельных опытах производил поперечные сечения, которые отделяли самый «кончик» ядра глазодвигательного нерва. У этих животных реакция зрачка на освещение угасала, в то время как компенсаторные установки глаз и реакции поворота глаз сохранялись без изменения — и, в частности, не только горизонтальные и ротаторные движения глаз, которые могли также вызываться блоковым или отводящим нервами, но и вертикальные движения, которые зависят только от глазодвигательного нерва. Отсюда видно, что ядро, передняя часть которого отрезана, у кролика может задним своим отделом обусловливать проявление неизмененных рефлексов.
С другой стороны, кошку (как и собаку) после более обширных вмешательств на центральной нервной системе легче сохранить живой, так что, имея с нею дело, можно дождаться, пока утихнут явления шока.
Весьма важно, что после поперечной перерезки позади среднего мозга шоковые явления незначительны и что,как это уже наблюдал Шеррингтон (5), после оперативного вмешательства на центральной нервной системе децеребрированных животных наблюдается вообще весьма незначительно выраженный шок.
Из этих фактов следует, что необходимо быть очень осторожным со всеми заключениями, которые основываются на отсутствии определенного рефлекса после операции на мозгу. Точно так же, работая с препаратом, у которого, согласно опыту, шоковые явления незначительны, возможно всегда прийти только к вероятным заключениям.
Во всех опытах, которые должны быть описаны в этой главе, старательно избегались односторонние перерезки и экстирпации. Операции были постоянно двусторонними, и животные избирались преимущественно только такие, у которых после операции не обнаруживалось асимметричных проявлений. Это чрезвычайно важно. Всякая асимметричная поза животного вызывает благодаря шейным рефлексам и т. д. такое множество сложных вторичных рефлексов, что толкование последствий первичного вмешательства затрудняется или становится невозможным.
Установочные рефлексы после удаления мозжечка
429
Опыты в продолжение последнего десятилетия развертывались следующим образом.
Прежде всего, у децеребрированных кошек и кроликов удалялся мозжечок, причем было обнаружено, что тонические шейные и лабиринтные рефлексы на мышцы тела оставались без изменения (18). У тех же животных делались затем поперечные разрезы мозгового ствола и определялось, на каком уровне продолговатого мозга и шейного отдела спинного мозга лежат центры названных рефлексов положения.
К этому примыкает более обширный ряд опытов (37), в которых было показано, что все известные до тех пор лабиринтные рефлексы после полного удаления мозжечка сохраняются. Опыты с реакциями поворачивания и компенсаторными установками глаз выполнены де Клейном.
Для завершения этих данных служат наблюдения на двух собаках, у которых Дюссер де Баренн неполностью удалил мозжечок. У этих собак, помимо прочих рефлексов, изучались также прогрессивные реакции (51) и установочные рефлексы тела.
Относительно уровня, до которого можно рассекать мозговой ствол, не уничтожая различных лабиринтных рефлексов и установочных рефлексов тела, на протяжении многих лет накопился значительный фактический материал. О протекании различных форм лабиринтного нистагма глаз говорят точные эксперименты де Клейна (63). В дополнение к этому Радемакер поставил на кошках две большие серии опытов, в которых ствол мозга поперечно пересекался на различных уровнях и затем, чисто клинически, устанавливалось, какие рефлексы оставались и какие исчезали. К этому примыкало анатомическое исследование на серийных срезах, так что для каждого рефлекса был получен поперечный срез, на уровне которого рефлекс еще имел место и ниже которого он отсутствовал.
Между прочим, этим способом был определен уровень для лабиринтных установочных рефлексов. Именно на этом уровне Радемакером была с большой точностью изучена локализация центра этих рефлексов. Здесь также находился центр для возникновения нормального распределения тонуса мускулатуры (уничтожение децеребрационной ригидности).
Последующее изложение хода этих исследований будет дополнять и завершать сказанное.
I. ЛАБИРИНТНЫЕ УСТАНОВОЧНЫЕ РЕФЛЕКСЫ
И УСТАНОВОЧНЫЕ РЕФЛЕКСЫ ТЕЛА ПОСЛЕ УДАЛЕНИЯ МОЗЖЕЧКА
(18, 37)
А. Методы исследования и анатомический контроль
Всего представлен 21 опыт на кошках и 17 опытов на кроликах. Все опыты на кошках выполнены мною; все опыты на кроликах (за исключением трех) — де Клейном. Обе серии опытов взаимно дополняют друг друга.
1. Оперативные методы
а) Экстирпация мозжечка у децеребрированных к о- ш е к. Под глубоким эфирным наркозом производится трахеотомия, и затем наркоз продолжается при искусственном дыхании. Перевязываются обе сонные артерии, блуждающие нервы перерезаются. Затем на уровне 12-го грудного позвонка спинной мозг временно обнажается и рана закрывается зажимами. Этим в дальнейшем течении опыта обеспечивается быстрая перерезка спинного мозга в том случае, если таковая потребуется для усиления рефлекса положения на передние ноги.
Череп освобождается, обе височные мышцы отсепаровываются и отводятся кзади, области прикрепления мышц на planum occipitale обнажаются до запирающей мемб¬
430
Десятая глава. Центры установки тела
раны — так что вся крыша черепа, от большого затылочного отверстия до глаз, оказывается обнаженной. После трепанации вся крыша черепа над большим мозгом удаляется костными щипцами, во время чего ассистент большим и указательным пальцами прижимает позвоночные артерии к позвоночнику между атлантом и эпистрофеем — так что вся последующая внутричерепная операция (по крайней мере у не слишком крупных кошек) может происходить совершенно бескровно. После вскрытия твердой мозговой оболочки ствол на уровне намета мозжечка поперечно перерезается шпателем и весь мозг, от заднего двухолмия, in toto вынимается. При этом следует постараться не оторвать твердую мозговую оболочку от основания черепа, ибо в противном случае после операции легко возникает кровотечение из надорванных артериальных сосудов.
Затем наркотизация прекращается и при продолжающемся прижатии позвоночных артерий удаляются палатка мозжечка и крыша черепа над мозжечком до большого затылочного отверстия. Латерально при этом доходят до каменистой части пирамиды.
Теперь мозжечок лежит совершенно свободно. Передний полюс червя путем осторожного отрывания мягкой и паутинной оболочек с заднего двухолмия отделяется, и тупой зонд проталкивается под мозжечком кзади, пока он не покажется в области писчего пера, дорсально от продолговатого мозга.
Для удаления мозжечка мною использовались различные приемы. Можно идти по зонду узким ножом и удалить с помощью двух горизонтальных разрезов, направленных вправо и влево, червь и дорсальные отделы. Затем удаляются оставшиеся вентральные части боковых отделов. Это весьма щадящий путь. Однако, поступая таким образом, обычно оставляют большую часть мозжечковых ножек, чем при других методах. Поэтому в ряде опытов мозжечок приподнимался на просунутом зонде до тех пор, пока натянувшиеся мозжечковые ножки не становились видными. Тогда эти последние перерезались ножом, направленным со стороны четвертого желудочка. Таким образом, мозжечок удаляется при этом целиком и анатомически результат получается наилучший. Однако при приподнимании мозжечка ножки более или менее надрываются, и, вероятно, именно это является причиной того, что у животных, оперированных этим способом, децеребрацион- ная ригидность развивалась не столь интенсивно, а то- Рис. 208. нические рефлексы не были столь живыми, как в осталь¬
ных экспериментах. Поэтому в последующих опытах сначала по зонду производилось расщепление мозжечка вдоль, обе половины отводились вправо и влево и после этого с медиальной стороны пересекались ножом хорошо видимые ножки. Этот способ — щадящий и дает очень хорошие анатомические результаты.
При таком способе оба 8-х нерва, так же как и оба тройничных и лицевых нерва, остаются незатронутыми. Латеральные области каменистой части височной кости лежат свободно. Дно четвертого желудочка полностью доступно для обозрения.
На рис. 208 приведен снимок оставшегося ствола мозга с интактным лабиринтом после децеребрации. Рис. 214 (стр. 444) изображает препарат, полученный в опыте 23, вместе с мозжечком, экстирпированным двумя частями.
Длительность всей операции от первого разреза кожи составляет 15—20 минут, собственно экстирпации мозжечка — 5—8 минут.
Временная компрессия позвоночных артерий теперь прекращается. Последующее кровотечение уменьшается тем, что животное в течение 15—20 минут держат вверх головой. Так как вся полость черепа пуста, кровяные сгустки обычно не приводят к явлениям сдавления. Так, только в одном опыте наступило умеренное сдавление вследствие коагулятов крови.
Поразительно, что после серьезной операции в непосредственной близости к продолговатому мозгу явления шока весьма невелики. В большинстве случаев уже в конце операции животные дышат самостоятельно. Децеребрационная ригидность передних и — часто — задних конечностей начинает развиваться тотчас же и, спустя х/4 час а, обычно бывает сильно выражена. Гомо- и контралатеральные рефлексы на
Установочные рефлексы после удаления мозжечка
431
конечностях получаются постоянно, корнеальный и ушной рефлексы часто могут быть получены спустя несколько минут. Таким образом, и для операций вблизи продолговатого мозга действительно установленное Шеррингтоном правило, что в случае, если эти операции производятся после децеребрации, шок выражен незначительно или не наступает вовсе.
После этого в ряде опытов последовательными фронтальными разрезами спереди назад удалялся ствол. При этом всякий раз снова временно клем- мировались позвоночные артерии и осторожно удалялись кровяные коагуляты, если они бывали обнаружены. После этого анатомически можно легко ориентироваться, измерить миллиметровой линейкой удаленность разреза от четверохолмия, писчего пера и т. д., проконтролировать полноту перерезки и т. д. 8-й нерв также может быть перерезан под контролем глаза. При фронтальных срезах в области шейного отдела спинного мозга костными щипцами удаляются дорсальные части верхних шейных позвонков.
б) У таламического кролика и децеребрированпого кролика. Под эфирным наркозом кролику делалась трахеотомия, налаживалось искусственное дыхание, перевязывались обе сонные артерии, перерезались блуждающие нервы и после этого, производи лась-трепанация-крыши черепа,_.затем_ .производилась децеребрация, причем разрез направлялся через ствол между передним и задним двухолмиями, или же экстирпировался большой мозг выше таламз^са. Применявшаяся при этом техника была подробно описана на стр. 159. Затем наркотизация эфиром прекращалась, чешуя затылочной кости удалялась вплоть до membrana atlanto-occi- pitalis, для избежания кровотечения позвоночные артерии временно зажимались ассистентом двумя пальцами между атлантом и эпистрофеем, мозжечок расщеплялся по средней линии, идя сзади под контролем пальца (причем следует обращать внимание на сохранность четверохолмия), обе половины мозжечка несколько раздвигались в стороны, и узким, изогнутым по плоскости инструментом, напоминающим шпатель, мозжечковые ножки перерезались на уровне четвертого желудочка. После этого обе половины мозжечка (включая каменистые дольки) легко вынимаются, и можно собственными глазами контролировать радикальность операции. Зажатие позвоночных артерий, которое продолжалось всего несколько минут, теперь прекращается, и рана закрывается мышечными и кожными швами. Кровотечение в большинстве случаев очень незначительно. Чтобы операция вполне удалась, важным условием является использование лобной лампы.
в) У интактных кошек. Чтобы удалить мозжечок у кошки с интактными большими полушариями и сохранить животное после операции живым, делалось следующее. После подкожного вспрыскивания 0.25 мг атропина применяется мельцеров- ский наркоз с эфиром (внутритрахеальное вдувание). Обе сонные артерии перевязываются, и затем переходят на хлороформный наркоз. Этим достигается уменьшение кровотечения из костей. Далее planum occipitale обнажается от атланта до затылочного гребня, и последний удаляется костными щипцами. После рассечения membrana atlanto-occipitalis костными и специальными щипцами («клюв попугая») средняя часть planum occipitale, по ширине примерно соответствующая червю мозжечка, удаляется. При этом можно идти в переднем направлении, не опасаясь вызвать кровотечение из синуса, так как у кошки поперечный синус лежит дальше кпереди, чем у собаки. Если позаботились о том, чтобы голова животного была расположена на некоторой высоте, кровотечение из костей по большей части бывает незначительным. Если оно усиливается, ассистент двумя пальцами зажимает позвоночную артерию между атлантом и эпистрофеем. Последнее почти всегда оказывается необходимым в момент собственно удаления мозжечка, чтобы последнее возможно было осуществить без всякого кровотечения и под хорошим контролем зрения, что для анатомической точности операции имеет весьма большое значение. Таким образом, при направленном освещении лобной лампой над продолговатым мозгом осторожно приподнимают задний полюс мозжечка узким, слегка изогнутым по плоскости инструментом, напоминающим шпатель, подходят под мозжечок, продвигают шпатель латерально от средней линии кпереди, пока он не упрется в костную палатку мозжечка, и разрезают мозжечок сначала на одной стороне, примерно на границе между средней его частью и боковым отделом, в дорсальном направлении. Затем шпатель заводится снова, и такой же разрез производится на другой стороне. Таким образом, нетрудно полностью вынуть весь средний отдел мозжечка, примерно соответствующий червю, и увидеть перед собой поверхность дна четвертого желудочка и задние холмы четверохолмия, в невредимости которых при этом можно убедиться. По сторонам видны ножки мозжечка, которые под контролем
432
Десятая глава. Центры установки тела
глаза перерезаются примерно на уровне дна четвертого желудочка. После этого легко могут быть удалены боковые отделы мозжечка, в большинстве случаев даже целиком,— и можно убедиться в том, достаточно ли полно произведена экстирпация, не повреждено ли место вступления в мозг 8-го нерва и были ли избегнуты прочие побочные повреждения. Затем сдавление позвоночных артерий, которые при некотором навыке в удалении мозжечка приходится пережимать лишь в течение нескольких минут, прекращается. В большинстве случаев кровотечение после этого оказывается поразительно небольшим. Далее следует весьма тщательное зашивание шейных мышц, которые также хорошо закрепляются — спереди до наружной части переднего края костной раны, чтобы обеспечить по возможности нормальные движения головы, что и наступает в действительности. После закрытия кожной раны наркоз прекращается, но на некоторое время животное остается лежать под трахеальным вдуванием и хорошо обогревается до тех пор, пока дыхание не станет снова интенсивным.
Преимущество этого способа состоит в том, что операция проходит без кровотечения и под контролем зрения, без зависимости от остановки дыхания, наступающей иногда (но далеко не во всех случаях) вследствие близости дыхательного центра, и в том также, что вагусные влияния оказываются выключенными.
При некотором упражнении операцию можно произвести в течение 15 минут. Собственно удаление мозжечка продолжается всего несколько минут.
г) Т от же способ был однажды с успехом применен к кролику. В этом случае при проталкивании шпателя под мозжечком вперед следует быть осторожным, так как намет мозжечка у кролика не является костным образованием, как это имеет место у кошки. После перерезки мозжечковых ножек и удаления боковых частей в нише каменистой части височной кости обычно остаются каменистые дольки (flocculi старых авторов), однако после перерезки ножек мозжечка они не имеют больше связи с мозговым стволом. Удалить их можно, но не необходимо.
2. Анатомический контроль
а) Предварительные замечания. Относительно тонкого анатомического строения ствола мозга у кошки и у кролика в настоящее время могут быть получены точные сведения, так как для обоих видов животных имеются отличные атласы Винклера и Поттера (2, 3). Эти атласы положены в основу последующих описаний. По поводу всех более тонких анатомических деталей ссылки будут сделаны на тот же источник. На рис. 209 воспроизведено уменьшенное изображение табл. 24 атласа кошки (3) и на рис. 210 и 211 — табл. 29 и 30 атласа кролика. Для мозжечка Винклер и Поттер пользуются терминологией Болька (Bolk).
Только один пункт нужно упомянуть особо, а именно значение термина «flocculus». В качестве flocculus’a старые авторы, а в последнее время Барани (4, 5), обозначали у кролика ту часть мозжечка, которая «помещается внутри капсулы, образованной тремя костными полукружными каналами и соединена с остальным мозжечком только тоненькой ножкой. К flocculus'y принадлежит еще маленькая долька, которая расположена уже вне этой капсулы, прилегая к мосту» (Барани). Об этой дольке в работах Барани и других не говорится. Эта последняя долька, которая на рис. 210 обозначена плюсом и которая непосредственно прилежит к месту входа 8-го нерва или, точнее, к вентральному ядру 8-го нерва, обозначается в данной работе исключительно как flocculus, в то время как заключенная в упомянутой костной капсуле долька мозга (++ на рис. 210 и 211) обозначается как lobulus petrosus.
Точно так же, вместе с Винклером и Поттером, мы называем flocculus’oM у кошки исключительно маленькую дольку (+на рис. 209), которая тесно прилежит к вентральному ядру 8-го нерва и слуховому бугорку, в то время как доли, лежащие латерально, мы обозначаем как каменистую дольку и петлевую дольку.
Таким образом, если в дальнейшем речь пойдет о том, что при экстирпации мозжечка flocculus оставлен на большем или меньшем протяжении, это относится исключительно к этой небольшой, часто видимой вообще
Рис. 209.
Рис. 210.
28 Р. Магнус
434
Десятая глава• Центры установки тела
только микроскопически, тесно прилегающей к месту вступления 8-го нерва пластинке, которую нельзя удалить без уверенности в целости части 8-го нерва и которая случайно экстирпируется только в отдельных опытах. Наоборот, во всех опытах на кроликах каменистая долька, а на кошках — и каменистая и петлевая дольки полностью удаляются.
Как видно на рис. 210, к flocculus’y кролика протягиваются мякотные волокна сверху от ножек мозжечка. Если эти ножки удалены на уровне четвертого желудочка, то можно принимать с достоверностью, что анатомическая связь остающейся пластинки flocculus’a с продолговатым мозгом
Рис. 211.
прервана. У кошки дело именно так и обстоит (рис. 209). Для извилин flocculus’a, лежащих латерально, это видно непосредственно. Однако и медиальная пластинка flocculus’a, обычно оставляемая при операции, несет связи с продолговатым мозгом в относительно дорсальной части ножки flocculus’a, которая накладывается дорсально на слуховые полоски. Рассмотрение табл. 24—26 атласа кошки показывает, что если операционный разрез проходит так, что зубчатое ядро удаляется, то связи ножки flocculus’a с продолговатым мозгом оказываются прерванными с наибольшей вероятностью.
На шести препаратах, изученных специально именно по серийным срезам или по результатам фронтальных срезов (три кошки и три кролика), анатомические связи оставшихся пластинок flocculus’a с продолговатым мозгом были нарушены. У большинства остальных, изученных только макроскопически, препаратов кролика и кошки следует принять с вероятностью, граничащей с достоверностью, что небольшие оставшиеся участки flocculus’a не находились больше в функциональной связи с продолговатым мозгом.
Поэтому, суммарно говоря, можно считать, что при операциях на мозжечке обычно остается небольшая часть flocculus’a, тесно прилегающая
Установочные рефлексы после удаления мозжечка
435
к месту вступления в мозг 8-го нерва. Полное и достоверное удаление этой части без сопутствующего повреждения 8-го нерва — задача достаточно трудная и осуществима только в отдельных случаях большой серии экспериментов. То, что пластинки flocculus’a не находились более в функциональной связи с продолговатым мозгом, можно было показать на шести тщательно изученных препаратах, однако и в остальных опытах, когда препарат исследовался только макроскопически, почти с полной уверенностью можно сказать, что в случае, если ножки мозжечка были перерезаны правильно, связь остатков flocculus’a с продолговатым мозгом уничтожалась.
Впрочем, шесть специально изученных препаратов позволяют нам в отношении большего числа испытывавшихся лабиринтных рефлексов 1 доказать также и анатомически (т. е. включая flocculus мозжечка) положение, что после тотального удаления мозжечка они удерживаются без изменения.
б) Анатомические исследования. Из двух препаратов серии децеребрированных кошек проф. Винклером изготовлены и зарисованы полные серии срезов (№№ I и II, опыты 22 и 23). Результат будет приведен на стр. 435 и 443. Три препарата той же серии опытов были подвергнуты Винклером микроскопическому исследованию.
Один препарат (№ III, опыт 12) экспериментальной серии на интактных кошках, который соответствует особенно удачному опыту, разрезан Винклером на серии и зарисован. Результат будет приведен на стр. 449.
Препарат (№ IV) кролика, у которого был удален мозжечок, а продолговатый мозг был перерезан тотчас у места вступления в мозг 8-го нерва, был изготовлен Винклером в виде серийных срезов и тщательно изучен, а два среза зарисованы. Результат будет приведен ниже, на стр. 454. Препараты трех кроликов, у которых большой мозг был экстирпирован выше таламуса и, кроме того, удалялся мозжечок, были разрезаны в виде тонких пластинок и описаны Винклером. Результат будет сообщен попутно с описанием экспериментов.
Далее была исследована центральная нервная система собаки, у^ которой Дюссер де Баренн (3) неполностью удалил мозжечок. Исследование произведено Винклером на серийных срезах. Результат будет сообщен на стр. 455.
Таким образом, заключению Винклера было представлено всего одиннадцать препаратов. Пять из них были представлены в виде серии и зарисованы, три исследовались по фронтальным срезам. Конечно, мне не нужно подчеркивать, насколько большой помощью для этих исследований явилась трудоемкая обработка анатомического материала специалистом, хорошо знающим свое дело.
Сверх перечисленного, профессор Броувер из Амстердама исследовал препарат собаки с неполно удаленным мозжечком (Дюссер де Баренн) (см. стр. 456).
Теперь следует привести описание серий срезов.
Препарат I (первая серия опытов, № 22; см. ниже, стр. 459).
Опыт XXII. 24 февраля 1914 г. Кошка. Эфирный наркоз, сонные артерии перевязаны, блуждающие нервы перерезаны. Обнажение спинного мозга на уровне 12-го грудного позвонка. Временное пережатие позвоночных артерий; дедеребрадия. Тотальная экстирпадия мозжечка после предварительного продольного расщепления.
1 Реакция поворота головы, постреакдия, реакция поворота и постреакция глаз, тонические лабиринтные рефлексы на конечности, лабиринтные установочные рефлексы, компенсаторные установки глаз, калорические реакции, а также наступающие после односторонней перерезки 8-го нерва изменения установки головы и глаз.
28*
436
Десятая глава. Центры установки тела
Окончание операции 9 ч. 55 мин. Сразу после операции ясная ригидность четырех ног, самостоятельное дыхание, рефлекс век.
10 ч. 29 мин. При исследовании рефлексов положения обнаруживается наличие интенсивных тонических шейных и лабиринтных рефлексов на передние ноги.
10 ч. 45 мин. Фронтальный разрез перед местом вступления 8-го нерва.
10 ч. 55 мин. Перерезка спинного мозга на уровне 12-го грудного позвонка. Отличная ригидность передних ног.
Боковое положение, поворачивание головы: если темя смотрит вниз, обе ноги разгибаются, если темя обращено вверх, обе ноги сгибаются (лабиринтные рефлексы).
Положение на спине, поднимание и опускание головы: разгибательный тонус передних ног максимален, когда морда поднята выше горизонтали на 45°; при дорсальном
пригибании головы (морда книзу) и при сильном вентральном нагибании головы (морда между передними ногами) тонус становится меньше (лабиринтные рефлексы).
При укладывании животного из положения на ногах в положение на спине, когда установка головы относительно туловища не изменяется, наступает сильное повышение тонуса (лабиринтный рефлекс).
Положение на спине, наклон головы: повышение тонуса «челюстной» ноги, понижение на «черепной» ноге (шейный рефлекс).
Положение на ногах, поворачивание головы с поднятой мордой: повышение тонуса «челюстной» ноги, понижение тонуса «черепной» ноги (шейный рефлекс).
Положение на ногах, поворачивание головы при горизонтальной ротовой щели: но- Рис. 212. вышение тонуса «челюстной»
ноги, понижение тонуса «черепной» ноги (шейный рефлекс).
Секция. Оба 8-х нерва интактны. Удален мозжечок. Часть его, которая охватывает заднее двухолмие, передний отдел четвертого желудочка и культи мозжечковых ножек, так же, как и оставшаяся часть ствола мозга, которой было достаточно для осуществления шейных и лабиринтных рефлексов, сфотографирована (рис. 212). Видны задняя половина дна четвертого желудочка, место возникновения 8-го нерва, ствол правого 8-го нерва, оба слуховых бугорка, поскольку они не перерезаны, и поверхность фронтального разреза.
Заключение проф. Винклера относительно микроскопического исследования препарата гласит следующее.
Рис. 212а. 1 —мозжечок, рассеченный вдоль, удален; 2 — поверхность фронтального разреза кпереди от возникновения 8-го нерва; 3 — заднее двухолмие; 4 — поверхности разреза отделенных ножек мозжечка; 5 — 8-й нерв; 6 — слуховой бугорок; 7 — слуховые полоски; 8 — дно четвертого желудочка; 9 — писчее перо;
10 — 1-й шейный нерв;
11 — 2-й шейный нерв.
Препарат состоит из четырех кусков: а и в — левая и правая половина мозжечка, удаленного при жизни; с —длинная часть спинного мозга с продолговатым мозгом и с частью моста. Оральный разрез макроскопически проходит следующим образом: слева и справа он идет не одинаково высоко, еще менее — вентрально и дорсально.
1. Справа разрез проходит (линия аа) вентрально на 2^—3 мм оральнее трапециевидного тела, через мост; отводящий нерв висит на препарате; по линии вв ла- терально свешиваются хорошо различимые на этом объекте лицевой нерв, оба корешка 8-го нерва и тройничный нерв; на уровне сс дорсально различим слуховой бугорок, разрез проходит точно на 1 мм оральнее последнего, затем несколько загибается кнутри и пересекает базальную стенку четвертого желудочка почти отвесно в направлении к шву.
Установочные рефлексы после удаления мозжечка
437
2. Слева линия аа: вентрально разрез лежит на 2%— 3 мм оральнее трапециевидного тела (отводящий нерв оторван), несколько дистальнее загибается и идет по линии вв латерально через тройничный нерв. Наоборот, лицевой нерв и оба корешка 8-х нервов висят на препарате. Таким образом, разрез лежит здесь слева примерно на 1 см дистальнее, чем справа, и достигает уровня сс дорсально как раз около слухового бугорка, направляется вдоль последнего, затем снова несколько загибается орально и разделяет базальную стенку по направлению к шву на одинаковой высоте слева и справа Наоборот, латерально он отклоняется слева более дистально. Особенно левая боковая поверхность моста лежит более каудально (дистально), чем правая латеральная поверхность моста.
d — объект оральнее верхнего; содержит мост, заднюю ножку и часть среднего мозга. Он ограничен двумя поверхностями разреза: дистальный разрез идентичен оральной поверхности разреза объекта с. Проксимальный разрез расположен косо.
Рис. 213 а.
1. Справа он идет (аа) вентрально через ножку мозга, затем загибается (вв) латерально через Laquear Reillii настолько далеко кзади, что дорсально (сс) он проходит примерно в 1У2—2 мм оральнее от заднего двухолмия, через переднее двухолмие.
2. С л е в а он пересекает (аа) препарат косо, так что достигает (вв) латеральной поверхности орально от заднего двухолмия. Он пересекает Laquear значительно ближе к мозговой ножке, чем справа, и достигает (сс) вентральной стороны, т. е. ножки мозга, снова на той же высоте или даже несколько оральнее, чем справа. На основании он проходит примерно на той же высоте.
Оба объекта, с ж d, затем прикладывались один к другому, спинной мозг отрезался и весь кусок c-\-d соединялся.
Слева, орально до слухового бугорка, висят обрывки, которые могут являться мозжечком (исследование, однако, показывает, что это не так; ср. ниже, разрез V).
Серия тщательно, насколько это возможно, разрезается в направлении ко шву. Однако само собой разумеется, что и в этом случае не следует ожидать, что левая и правая половины окажутся рассеченными на равной высоте. Склеивание оральной и каудальной частей также не может обеспечить точного совпадения того, что до этого лежало рядом. Окраска по Нисслю.
При последующем описании срезов в соответствии сцелями данной работы речь пойдет главным образом о дистальной части, так как после разреза тотчас у места вступления 8-го нерва тонические шейные и лабиринтные рефлексы на конечности еще не были пониженными.
Срез I (рис. 213а) — один из первых разрезов (из пластинки первой серии).
На правой стороне, пересеченной выше (слева перерезано и заднее двухолмие; справа — только переднее) децеребрационным разрезом, проходящим через моз¬
438
Десятая глава. Центры установки тела
говую ножку, сделан надрез. Крупноклеточная часть красного ядра получена на препарате без всякого кровоизлияния. Интактно также начало глазодвигательного нерва.
Срез II (рис. 2136) из пластинки второй серии.
Здесь начинается дистальный разрез, который идет на такой высоте, что затрагивается pes pedunculi в мосту. Часть моста, что было видно и макроскопически, висит на оральной части объекта. Задние двухолмия связаны поверх сильвиевого водопровода своей комиссурой. В заднем двухолмии много кровоизлияний.
Срез III (рис. 21 Зв) из пластинки четвертой серии.
Lobus anticus мозжечка между обоими холмами заднего двухолмия изолирована полностью и без малейшего повреждения лежащего ниже серого вещества tegmentum. Операционный разрез идет поперечно через мост. На проксимальной части висит остаток моста, вентрально от ganglion interpedunculare. Проксимальная и дистальная части
Рис. 213 б.
соприкасаются одна с другой, несомненно, не совсем правильно, так как на верхней части видно, что левая часть перерезана дистальнее, чем правая, в то время как на нижнем куске этого нет.
Кусок, свешивающийся с проксимальной части идентифицировать нелегко. Это, наверное, остатки ручек моста, но я не мог бы утверждать с уверенностью, что то, что здесь обозначено как pes pedunculi, не является уже частью чувствительной порции тройничного нерва. Образование это слишком повреждено.
Срез IV (рис. 213г) из пластинки шестой серии.
На верхней части различие между правой и левой сторонами исчезло. Кажется, будто левая сторона теперь задета более проксимально, чем правая. Это происходит, однако, вследствие того, что левый холм заднего двухолмия крупнее, чем правый. Латеральная петля с дорсальным и медиальным ядрами с обеих сторон сохранена (см. также срез III). N. n. trochleares выходят из мозга; их ядра также располагаются в проксимальном куске (на незарисованном срезе между II и III).
Операционный разрез проходит через вентральное ядро латеральной петли. Нижний (дистальный) кусок разрезан так, что слева лежит на более дистальном уровне, чем справа.
Срез V (рис. 21 Зд) из пластинки седьмой серии.
Верхняя часть разрезана слева несколько проксимальнее, чем справа. Нижняя часть слева — несколько дистальнее, чем справа.
Операционный разрез переходит слева через продолговатый мозг в разрез, посредством которого при экстирпации мозжечка были отрезаны ручки моста. Ножка мозжечка слева представляла собой лоскуток, который при макроскопическом рассматривании (см. выше, стр. 437) импонировал в качестве остатка мозжечка. В точке х начинается разрез, который пересекает справа brachium conjunctivum.
Рис. 213 в.
Рис. 213 г.
440
Десятая глава. Центры установки тела
На нижней (дистальной) части слева сидит 8-й нерв, начинается вентральное ядро 8-го нерва. Разрез идет через спинальную ветвь тройничного нерва. Справа из мозга выходит тройничный нерв. Слева в нижнем куске лежит интактная верхняя добавочная олива.
Рис. 213 д.
Рис. 213 е.
Срез VI (рис. 21 Зе) из пластинки двенадцатой серии.
Операционный разрез с обеих сторон переходит в разрезы, посредством которых: во время операции на мозжечке были перерезаны ручки моста, веревчатые тела и brachia conjunctiva.
Установочные рефлексы после удаления мозжечка
441
Слева разрез, идя вдоль интактного вентрального ядра 8-го нерва, полностью отсек flocculus. Справа, наоборот, небольшая часть flocculus’a (см. срез VII, рис. 213ж) осталась.
Рис. 213 ж.
Рис. 213 з.
На нижнем (дистальном) куске продолговатого мозга с обеих сторон лежат совершенно интактные верхние оливы, околооливарные ядра, nucleus trapezoides medialis, ventralis и lateralis.
На верхнем куске справа коротко отрезаны brachia conjuncbiva (точно так, как ручки моста на срезе V), но висит еще обрывок веревчатого тела, в котором можно найти (как на срезе VII) пару клеток зубчатого ядра.
Моторное ядро тройничного нерва лежит слева, частью в верхнем, частью в нижнем куске. Операционный разрез проходит, таким образом, слева как раз через это ядро.
442
Десятая глава. Центры установки тела
Срез VII (рис. 21 Зж) из пластинки четырнадцатой серии.
Операционный разрез через продолговатый мозг слева переходит в разрез, который при экстирпации мозжечка разделил веревчатые тела и brachia conjunctiva.
На нижнем (дистальном) куске справа располагается маленький кусочек flocculus’а (см. также срезы VI и VIII). Корешки лицевого и отводящего нервов выходят здесь из мозга. Справа начинается вступление в мозг 8-го нерва, а также тройничного нерва.
На верхнем куске справа висит частица черной субстанции мозжечка, выходящая из веревчатого тела по направлению к зубчатому ядру.
Срез VIII (рис. 21 Зз) из пластинки семнадцатой серии.
На этом срезе слева заканчивается операционный разрез через продолговатый мозг. Зато можно видеть, что разрез, который во время экстирпации мозжечка разделил веревчатые тела и brachia conjunctiva, прошел через дорсальное ядро 8-го нерва и частично разрушил его. Разрез проходит также через верхнюю часть ядра Дейтерса.
Рис. 213 и.
Справа на нижнем (дистальном) куске виден маленький кусочек flocculus’а, сидящий на вентральном ядре 8-го нерва. Путь, связывающий этот последний с продолговатым мозгом, как и на VI и VII срезах, перерезан.
Справа на нижнем куске видны неповрежденные вентральные ядра 8-го нерва, слева — вентральное ядро 8-го нерва и слуховой бугорок.
От верхнего проксимального куска справа виден конец. Он содержит клетки зубчатого ядра и кусочек пробковидного ядра, через которое проходят отчетливо видимые fibrae perforantes conjunctivi. Это самый крупный кусок мозжечка, который вообще можно найти в серии. Однако он лежит в верхнем куске.
Срез IX (рис. 21 Зи) из пластинки двадцать второй серии.
Операционный разрез через продолговатый мозг с обеих сторон закончился. При произведенном во время экстирпации мозжечка разделении веревчатых тел слева повреждены дорсальное ядро 8-го нерва и ядро Дейтерса, в то время как справа оба образования полностью интактны. Общий нисходящий дейтерсовский пучок с обеих сторон лежит в дистальном куске неповрежденным. Слуховой бугорок можно с обеих сторон видеть интактным.
Анатомическое заключение.
А. Первый децеребрирующий разрез через четверохолмие лежит проксимально от крупноклеточной части красного ядра, так что руброспиналь- ный пучок был в состоянии функционировать до 10 ч. 45 мин. без нарушений. Ядро Бехтерева с обеих сторон интактно лежит в проксимальном куске.
Б. Второй разрез проходит более дистально, 1) лежит проксимально от трапециевидного тела и всех расположенных там ядер (ядра олив, ядра трапециевидного тела); эти ядра могут функционировать без нарушений; 2) при этом ядра Дейтерса справа совершенно интактны, слева верхняя
Установочные рефлексы после удаления мозжечка
443
часть ядра отрезана; однако как справа, так и слева начало и остаток нисходящего дейтерсова пучка интактны, они также функционировали без нарушений; 3) вполне интактны справа и слева вентральное ядро 8-го нерва и слуховой бугорок, точно так же не повреждены с обеих сторон нисходящий корешок 8-го нерва и вступающие в мозг вестибулярные корешки;. слева отрезан кусочек латеральной части дорсального ядра 8-го нерва; справа это ядро не повреждено.
В. Мозжечок слева удален полностью. Справа на дистальном куске, в месте вступления 8-го нерва, остался малеький кусочек flocculus’а. Этот последний, однако, не состоит в функциональной связи с продолговатым мозгом, так как дорсально отходящий пучок перерезан. На проксимальном куске располагается пучок, собирающий волокна веревчатых тел и brachia conjunctiva к зубчатому и пробковидному ядрам.
Физиологическое заключение. Так как после второго разреза, расположенного более дистально, тотчас перед местом вступления 8-го нерва, тонические шейные и лабиринтные рефлексы на обе передние ноги сохранились, не став менее интенсивными, следует считать, что эти рефлексы могут осуществляться после полного удаления мозжечка, включая и ядра мозжечка. Далее следует, что для этих рефлексов все лежащие орально от линии второго разреза центральные ядра (включая и ядра Бехтерева) не являются необходимыми.
Препарат II (серия 1, № 23; см. ниже, стр. 460).
Опыт XXIII. 27 февраля 1914 г. Кошка. Эфирный наркоз. Сонные артерии перевязаны, двусторонняя ваготомия, обнажение спинного мозга на уровне 12-го грудного позвонка. Временное клеммирование позвоночных артерий. Децеребрация. Тотальное удаление мозжечка после предварительного расщепления. Ножки перерезаны с медиальной стороны.
10 ч. 30 мин. Окончание операции. Начинается ригидность.
И ч. Боковое положение, поворачивание головы: в положении теменем вниз обе передние ноги разогнуты, верхняя значительно сильнее нижней. В случае, если темя обращено вверх, обе передние ноги расслабляются, причем верхняя сильнее (шейные и лабиринтные рефлексы; последние преобладают).
Боковое положение, поворачивание головы: то же (шейные и лабиринтные рефлексы; последние преобладают).
Боковое положение, поднимание и опускание головы: без эффекта (шейные рефлексы при симметричных движениях головы отсутствуют).
Положение на спине, поднимание и опускание головы: передние ноги максимально разгибательно тонизированы, когда морда благодаря вентральному пригибанию головы оказывается между передними лапами и когда она, вследствие дорсального нагибания головы, направлена вниз (сильный лабиринтный рефлекс).
Положение на спине, поворачивание головы: уменьшение тонуса обеих передних ног, в «черепной» ноге больше (шейные и лабиринтные рефлексы, последние преобладают).
Положение на спине, наклоны головы: уменьшение тонуса «черепной» ноги, увеличение тонуса «челюстной» ноги (шейный рефлекс).
Положение на ногах, поднимание и опускание головы: при поднятой голове повышение, при опущенной — уменьшение тонуса разгибателей передних ног (преобладающий лабиринтный рефлекс).
При перекладывании всего животного из положения на животе на спину, причем положение головы по отношению к туловищу не изменяется, наступает сильное разгибание передних ног (лабиринтные рефлексы).
После перерезки спинного мозга на уровне 12-го грудного позвонка вся серия проб повторяется с прежним результатом. Так как в этом опыте лабиринтные рефлексы оказались особенно живыми, последующих фронтальных разрезов ствола не делалось, по животное было забито, ствол извлечен и сфотографирован (рис. 214).
Наверху виден продольно расщепленный мозжечок, внизу — ствол мозга. Деце- ребрирующий разрез направляется через середину переднего двухолмия. Ромбовидная
444
Десятая глава. Центры установки тела
ямка лежит свободно на всем протяжении от заднего двухолмия до писчего пера (calamus scriptorius). Справа сверху видны перерезанные правые ножки мозжечка. Поверхность разреза лежит на 1% мм выше уровня дна четвертого желудочка. Кау- дально от правой мозжечковой ножки перебрасывается на вентральную сторону правый 8-й нерв. Левые мозжечковые ножки скрыты в толще веществ продолговатого- мозга. Ствол левого 8-го нерва также был интактен, но вследствие легкой торзии препарата не виден на фотографии.
Про ф. Винклер сообщает следующее.
Серия срезов ствола мозга кошки охватывает 16 пластинок с непрерывным рядом- срезов 30 ц толщиной, начиная от области ядра лицевого нерва до переднего двухолмия. Часть срезов окрашивалась по Вейгерту, часть же — кармин-гематоксилином.
Рис. 214 и рис. 214а. 1 — мозжечок, расщепленный вдоль; 2 — поверхность разреза; 3 — заднее двухолмие; 4 — поверхности разреза ножек мозжечка;
5 — 8-й нерв; 7 — слуховые полоски; 8 — дно четвертого желудочка; 9 — писчее перо; 10 — 1-й шейный нерв; 11 — 2-й шейный нерв; 12 — 3-й шейный нерв.
Описание срезов приводится последовательно от задних к передним. Препараты зарисованы такими, какими они были видны под микроскопом. Поэтому правая и левая сторона переставлены — то, что на рисунке расположено справа, на препарате' лежит слева, и наоборот.
Срез I (рис. 215а) из пластинки 16, № 12.
Дистальный разрез серии проходит через продолговатый мозг через дистальный- конец ядра лицевого нерва. С левой стороны разрез пришелся по слуховому бугорку.
Этот разрез нигде не затрагивался на операции. Первые признаки этого появляются, на срезе XVIII пластинки 15.
Срез II (рис. 2156) из пластинки 15, № 26.
Этот разрез проходит через продолговатый мозг, середину ядра лицевого нерва и дистальный конец верхних олив. На правой стороне (слева на рисунке) вступает кохлеарный нерв и видны слуховой бугорок и вентральное ядро 8-го нерва. Из них возникают двумя ножками слуховые полоски Монакова. С левой стороны все это отрезана в соответствии с жирной линией на рисунке.
Срез III (рис. 215в) из пластинки 14, № 37.
Этот разрез проходит через середину верхних оливарных ядер и выходящих корешков отводящего нерва; проксимальный конец лицевого нерва также затрагивается разрезом.
Установочные рефлексы после удаления мозжечка
445
На правой стороне (на рисунке — слева) в мозг вступает вестибулярный нерв, видны также слуховой бугорок, вентральное ядро 8-го нерва и слуховые полоски. При экстирпации мозжечка разрез, проведенный справа, прошел прямо вдоль слуховых лолосок.
Слева, наоборот, все эти ядра, как и периферические, вступающие в мозг вестибулярные волокна (в точке 7), отрезаны. Разрез проходит поперек через овальную площадку веревчатого тела.
Срез IV (рис. 215г) из пластинки 13,
№ 42.
Этот разрез проходит через место выхода лицевого и отводящего нервов и располагается только немного более проксимально, чем предыдущий.
На правой стороне операционный развез слегка касается овальной площадки веревчатого тела, откуда он переходит на мозжечок.
На левой стороне отрезаны все латеральные ядра 8-го нерва, но видно, что вестибулярный нерв пощажен разрезом лосле его вступления в мозг, так что возможно, что вестибулярные волокна пере- Рис. 215а.
резаны не были.
СрезУ (рис. 215д) из пластинки 12, № 67.
Этот разрез проходит через проксимальный конец верхнего оливарного ядра и выходящие корешки лицевого нерва.
Справа во время экстирпации мозжечка отрезано веревчатое тело.
Справа (на рисунке — слева) около вентрального ядра 8-го нерва находится маленький кусочек flocculus’а, однако его связи с продолговатым мозгом перерезаны.
Слева операционный разрез проходит через веревчатое тело, отделяет flocculus и вентральное ядро 8-го нерва, которые поэтому на препарате отсутствуют.
Рис. 2156.
На самом разрезе с обеих сторон находятся большие кровоизлияния. Справа они расположены (заштрихованное поле) в tractus spinalis nervi V и в латеральных ядрах верхних олив. Слева они имеются в tractis spinalis nervi V. Трапециевидное тело и его перекрест, монаковский перекрест и поле, расположенное дорсально от ядер олив, кровоизлияниями не повреждены.
Ядро Бехтерева интактно с обеих сторон (главным образом оно лежит между V и VI разрезами).
Срез VI (рис. 215е) из пластинки 8, № 109.
Этот разрез проходит через ствол там, где brachium conjunctivum свободно выходит на поверхность. Brachia conjunctiva и ручки моста во время экстирпации мозжечка отрезаны с обеих сторон. С правой стороны имеется кровоизлияние, которое распространяется на область вентрального ядра латеральной петли и на эту петлю. В остальном этот срез не поврежден.
Срез VII (рис. 215ж) из пластинки 5, № 150.
Этот срез с обеих сторон расположен вне мозговой раны. Он идет несколько косо, так что правый холм заднего двухолмия уже пересечен, а слева, наоборот, еще не затронут. С обеих сторон в латеральной петле обнаруживается дорсальное ядро. Кроме незначительных остатков кровотечения в шве, этот срез не поврежден.
Рис. 215в.
Рис. 215г.
Рис. 215д.
Установочные рефлексы после удаления мозжечка
447
Анатомическое заключение.
А. Мозжечок с левой стороны удален полностью; справа сохранился небольшой кусочек flocculus’a, который удерживается на вентральном ядре 8-го нерва (срез V), но никакой связи с продолговатым мозгом не имеет.
Рис. 215е.
Рис. 215ж.
Б. Ядра мозжечка удалены полностью.
В. Справа оба корешка 8-го нерва во время операции не повреждены.
Г. Слева полностью удалены кохлеарный нерв, вентральное ядро 8-го нерва и слуховой бугорок. Дистальная часть вестибулярного нерва, вступающего в мозг (т. е. периферический корешок), перерезана более проксимально, но уже вступивший корешок сохранен, и так как разрез здесь идет как раз вдоль корешка, то возможно, что отдельные проксимальные вестибулярные волокна остались неповрежденными.
448
Десятая глава. Центры установки тела
Д. Относительно кровоизлияний на срезе V и VI нужно сказать следующее.
Кровоизлияние на левой стороне находится в tractus spinalis n. V и не захватывает вторичного пути 8-го нерва.
Кровоизлияние на правой стороне, наоборот, проникает в трапециевидное тело и ядра оливы (срез V) и даже в латеральную петлю и ее вентральное ядро (срез VI). В соответствии с этим кровоизлияние распространяется из латеральных ядер 8-го нерва (nucleus ventralis 8-го нерва и слуховой бугорок) на вторичные волокна 8-го нерва, возникающие на левой стороне после того, как они пересекли шов, и разрушает их.
Отсюда следует также, что слева nucleus ventralis 8-го нерва и слуховой бугорок полностью удалены и что на более высоком уровне возникающие здесь вторичные пути после перекреста их на правой стороне уничтожены кровоизлиянием. Наоборот, общее вступление правого 8-го нерва и всех принадлежащих его системе ядер и возникающие из них вторичные пути, как до, так и после их перекреста, остаются и не затронутыми и не поврежденными кровоизлиянием.
Пути, возникающие из ядра Дейтерса, интактны с обеих сторон.
Как было сказано выше, взгляд на снимок препарата (рис. 214) показывает также интактность области вступления 8-го нерва на правой стороне, в то время как слева операционный разрез проходит в толще вещества продолговатого мозга.
Физиологическое заключение. В третьей главе уже было показано, что одного лабиринта оказывается достаточным, чтобы вызвать тонические лабиринтные рефлексы одинаковой силы на конечности обеих половин тела. Описанный здесь препарат подтверждает с определенностью этот факт. В самом деле, весь левый 8-й нерв был уничтожен вплоть до немногих вступающих в мозг вестибулярных волокон, тогда как весь правый 8-й нерв полностью остался интактным. Несмотря на это, тонические лабиринтные рефлексы на обе передние ноги остались не уменьшенными по величине и силе. Кроме того, этот опыт показывает, что у деце- ребрированной кошки после полного удаления мозжечка, включая его ядра, имеет место выраженная децеребрационная ригидность и при изменении положения головы обнаруживаются отчетливые шейные рефлексы и высокие лабиринтные рефлексы на мускулатуру передних ног.
Препарат III (пятая серия опытов, № 12, см. ниже, стр. 466).
3 мая 1918 г. Кошка; подкожно 1 мг атропина. Эфирный наркоз с трахеальным вдуванием по Мельцеру. Сонные артерии перевязаны. После этого продолжение наркоза хлороформом. Обнаружение planum occipitale, удаление crista occipitalis, вскрытие planum occipitale примерно на ширину червя мозжечка. Зажатие вертебральных артерий. Без кровотечения удалена средняя часть мозжечка до четверохолмия. Горизонтальная перерезка ножек мозжечка, удаление боковых его ножек. Течение операции гладкое, тщательное зашивание мышц и кожи.
С 11 до 4 часов животное оставалось лежать под искусственным дыханием. После этого хорошее спонтанное дыхание, хороший пульс, отчетливые пателлярные рефлексы, легкий разгибательный тонус передних ног.
4 мая 1918 г. До полудня. Если животное укладывается на полу в правое или левое боковое положение, оно устанавливает голову прямо. Обнаруживает с несомненностью лабиринтные установочные рефлексы на голову. Удерживаемое в воздухе в боковом положении, стремится установить голову прямо, что обычно удается. В положении на спине в воздухе поворачивает голову вправо или влево. В висячем положении головой вверх последняя устанавливается в пространстве правильно. Только в висячем положении головой вниз темя обращено книзу вследствие операции на затылочных мышцах. В пробе с доской на столе в обоих боковых положениях голова устанавливается в пространстве правильно. Если при этом ущипнуть вышележащую переднюю ногу, голова остается в правильном положении.
Реакция поворота головы, реакция поворота глаз, последовательная реакция поворота глаз с обеих^сторон отчетливо положительны.
Установочные рефлексы после удаления мозжечка
449
Поворот головы в боковом положении вызывает тонические лабиринтные рефлексы на передние ноги.
4 мая 1918 г. Лабиринтные установочные рефлексы в воздухе в обоих боковых положениях, в висячем положении головой вверх и особенно в положении на спине весьма отчетливы; голова постоянно устанавливается в воздухе правильно.
Точно так же — при пробе с доской на столе.
Отчетливая реакция поворота головы.
5 мая 1918 г. Лабиринтные установочные реакции в воздухе: в обоих боковых положениях и в висячем положении головой вверх голова устанавливается в пространстве правильно. В положении на спине в воздухе голова сначала поворачивается на 90° в сторону и затем падает в вентральном направлении до тех пор, пока не установится правильно в пространстве ниже изгиба грудной части позвоночника. В висячем положении головой вниз отчетливой реакции
нет.
При пробе с доской голова устанавливается правильно.
При перекладывании из положения на животе в положение на спине отчетливые тонические лабиринтные рефлексы на передние ноги.
Отчетливые тонические шейные рефлексы на передние и задние конечности.
Реакция поворота головы и реакция последовательного поворота головы, реакция поворота глаз и последовательная реакция поворота глаз отчетливо положительны.
Делает напрасные попытки усаживаться из правого бокового положения; из левого бокового положения усаживается хорошо и остается сидеть спокойно.
Неправильную установку лап (всех четырех конечностей) не корригирует.
Если в нормальном положении живот- р
ного на полу ущипнуть кожу одной из сторон ^ис* Zlba'
тела, наклона головы в другую сторону не наступает.
Примечательно полное отсутствие всех явлений раздражения, которые являются правилом после двусторонней экстирпации лабиринтов.
Затем глаза завязываются повязкой. Однако и после этого все лабиринтные рефлексы в воздухе осуществляются так же безупречно, как и при открытых глазах; проба с доской с обеих сторон отчетливо положительна.
6 мая 1918 г. Утром найдена мертвой.
Секция. Причина смерти — правосторонняя пневмония. На дне четвертого желудочка лежит кровяной сгусток. Задняя черепная яма в большей части пуста, т. е. компрессии мозга нет. На основании черепа и в спинномозговом канале крови не обнаружено. Менингита нет. Рана без признаков раздражения. 8-й нерв с обеих сторон интактен. Мозг, in toto, помещенный в формалин, передан проф. Винклеру.
Данные проф. Винклера.
Переданный мне препарат был разрезан на серии и насчитывал примерно 700 срезов. Окраски по Вейгерт-Палю, кармином и гематоксилином. Первый срез, на котором становятся видны результаты операции, — № 524.
Срез I (рис. 216а), № 524.
Этот разрез проходит через продолговатый мозг там, где центральный канал, расширяясь, переходит в четвертый желудочек через расположенные одно над другим ядра 12-го и 10-го черепно-мозговых нервов. На оставшейся интактной крыше четвертого желудочка, особенно справа, находится тонкая пленка, состоявшая из фибрина и белых кровяных телец, в которой обнаруживаются многочисленные лакуны.
Срез II (рис. 2166), № 406.
Этот разрез проходит через продолговатый мозг приблизительно на уровне выхода языкоглоточного нерва. На крыше четвертого желудочка лежит tela chorioidea, еще не поврежденная. В четвертом желудочке находится сгусток, который состоит из фибрина и белых кровяных телец. Хотя этот сгусток по форме и напоминает uvula, это, однако, только сгусток, как по своему строению, так и по тому, что он лежит внутри четвертого желудочка. Сверху на продолговатом мозгу лежат погруженные в сгусток несколько включений (в точках айв), внутри которых с помощью окраски по Вейгерт- Палю можно еще обнаружить остатки нервных волокон. Эти мозжечковые включения лежат свободно, разделенные сгустком. Этот разрез приблизительно соотвеютвует табл. 27 атласа кошки.
29 Р. Магнус
450
Десятая глава. Центры установки тела
Срез III (рис. 216в), № 331.
Этот разрез проходит через продолговатый мозг в месте вступления дистального корешка 8-го нерва, через улитковый нерв, и идет через recessus lateralis. Четвертый желудочек оказывается разделенным на всем протяжении (включая и recessus lateralis) по ходу tela chorioidea. Здесь также обнаруживается небольшой сгусток. В четвертом желудочке лежит крупный сгусток с остатками мозжечка, которые полностью погружены в нем. Однако в значительной части этот сгусток состоит из крови,
Рис. 2166.
фибрина и лейкоцитов. Разрез проходит несколько дистальнее плоскости табл. 24 атласа кошки.
Срез IV (рис. 216г), № 233.
Разрез проходит через продолговатый мозг в месте выхода 6-го и 7-го нервов и через вестибулярный нерв. Четвертый желудочек покрыт передним мозговым парусом. Выше лежит язычок, который вследствие сильных кровотечений стал совершено не-
Рис. 216в.
узнаваемым. Далее лежит сгусток, помещающийся на отсеченных верхних, нижних и средних мозжечковых ножках.
На правой стороне еще свешиваются две пластинки flocculus. При рассечении ножек мозжечка образовалась трещина, проходящая через проксимальный конец вентрального ядра 8-го нерва и заполненная фибрином и кровью. Трещина эта проникает между областью мозга и самым медиальным отделом веревчатого тела, пересекает здесь мозг в поперечном направлении и продолжается в вентральном ядре 8-го нерва. Эта трещина отделяет оставшуюся часть flocculus’а от продолговатого мозга (см. также срез V).
Установочные рефлексы после удаления мозжечка
45 f
На левой стороне осталась только пластинка flocculus’а, но она полностью лишена связей с продолговатым мозгом.
Этот разрез соответствует табл. 23 атласа кошки. Для большей ясности здесь приведен еще разрез мозга нормальной кошки (рис. 216д), имеющий не вполне, но примерно»
Рис. 216г.
Рис. 216д.
то же направление, и лежащий между №№ 406 и 233, но ближе к 233. На рисунке обозначено приблизительное направление операционного разреза.
Срез У (рис. 216е), № 211.
Разрез проходит через область выхода тройничного нерва. Здесь также видно начало aquaeductus sylvii, прикрытое мозговым парусом, на котором лежит кровяной сгусток, имеющий форму язычка. Ножки отрезаны. Справа остались две пластинки fl ос-
29*
452
Десятая глава. Центры установки тела
culus’a. Упоминавшаяся выше трещина проходит здесь между brachium conjunctivum и brachium pontis и разъединяет вентральное ядро 8-го нерва Слева остались только отдельные пластинки flocculus. С обеих сторон всякая связь этих образований со стволом мозга прервана. Все, что лежит на ножках, представляет собой кровяной сгусток с отдельными отрывками мозжечка. Ядро Бехтерева с обеих сторон интактно.
Рис. 216е.
Рис. 216ж.
Срез лежит между табл. 23 и 22 атласа кошки. Для пояснения приводится рисунок мозгового среза нормальной кошки, который лежит между №№ 211 и 233, но ближе к 233, где дана приблизительная линия разреза (рис. 216ж).
Срез VI (рис. 216з), № 117.
Разрез проходит по середине моста через перекрест блоковых нервов в мозговом парусе. Этот интактный перекрест виден под сгустком.
Рйс. 216з.
Рис. 216и.
Рис. 216к.
454
Десятая глава. Центры установки тела
Справа совершенно свободно лежит небольшой кусочек lobulus ansatus мозжечка. Cl обеих сторон всякая связь этих образований со стволом мозга прервана.
Срез VII (рис. 216s), № 93.
Разрез проходит через дистальный конец заднего двухолмия. Правый corpus quadrigeminum posterior задет операционным разрезом. Справа лежат совершенно свободно и без связи со стволом мозга две пластинки lobulus ansatus.
С обеих сторон совершенно не повреждена боковая петля с находящимися в ней ядрами.
Этот срез проходит также выше плоскости табл. 22 атласа кошки.
Срез VIII (рис. 216к), № 15, является последним срезом, на котором видны некоторые следы после операции, а именно, небольшое кровоизлияние в левый холм заднего двухолмия. На разрезе VII и VIII находится кровоизлияние в aquaeductus, которое здесь лежит совершенно свободно.
Срез приблизительно соответствует табл. 20 атласа. (В остальном все срезы лежат iB ином направлении, чем рисунки атласа, и именно таким образом, что направление |разреза в атласе более косое, так что вентральные отделы оказываются расположенными более дистально, чем на сделанных здесь зарисовках с оперированной кошки).
Анатомическое заключение. Учитывая все приведенное, можно сказать, что мозжечок в соответствии с линиями разреза, изображенными на рис. 216д и ж, удалялся полностью, включая его ядра. Отдельные кусочки, как, например, flocculus и lobulus ansatus, которые еще обнаруживались, исключают всякую связь с продолговатым мозгом. Кроме того, в продолговатом мозгу не обнаружено ни одного кровоизлияния, которое оказало бы влияние на вторичные пути и ядра 8-го нерва, известные в настоящее время.
Единственным исключением является трещина, проходящая справа нерез проксимадьную часть вентрального ядра 8-го нерва (срез IV и V). Дистальный конец этого ядра и место возникновения трапециевидного тела при этом, однако, не повреждаются.
Физиологическое заключение. После полного удаления мозжечка у кошки констатируются .лабиринтные установочные рефлексы на голову, тонические шейные и лабиринтные рефлексы на мускулатуру конечностей, реакция поворота головы и последовательная реакция поворота головы, так же как реакция поворота глаз и последовательная реакция глаз.
Препарат IV (экспериментальная третья серия, № 3; см. ниже, стр. 463).
Кролик. Эфирный наркоз. Сонные артерии церевязаны, блуждающие нервы перерезаны. Децеребрация, удаление мозжечка при зажатии позвоночных артерий. Затем фронтальный разрез продолговатого мозга тотчас впереди от места вступления 8-го нерва.
После этого у животного обнаружены отчетливые тонические лабиринтные рефлексы на конечности, реакции поворота и последовательного поворота головы, так же, как после перерезки правого 8-го нерва — отчетливый поворот головы вправо.
Препарат был разрезан проф. Винклером на серии. Окраска кармином и гематоксилином.
Результаты исследований и зарисовок резюмированы проф. Винклером следующим образом.
С левой стороны операционный разрез идет тесно проксимально от места выхода жорешка лицевого нерва. Ядро Дейтерса на этой стороне полностью сохранно. На правой стороне операционный разрез проходит более проксимально, примерно через выходящий корешок тройничного нерва. Корешок лицевого нерва затрагивается разрезом, проходящим глубже, так что и на этой стороне ядро Дейтерса также остается полностью интактным. Таким образом, операционный разрез проходит несколько косо, почти точно между мостом и трапециевидным телом, на левой стороне ниже, справа же через место выхода тройничного нерва.
На правой стороне осталось несколько небольших пластинок flocculus’а, однако последние не сохранили здесь никакой функциональной связи со стволом мозга. Кроме того, отсутствует полностью весь мозжечок с его ядрами.
Установочные рефлексы после удаления мозжечка
455
Ядра Дейтерса и Бехтерева с обеих сторон интактны.
Справа при оперативном разъединении 8-го нерва повреждено его вентральное ядро. Слева вентральное ядро интактно и содержит лишь небольшие кровоизлияния, не имеющие существенного значения. Вход вестибулярного нерва не поврежден.
Кровоизлияния обнаружены по обеим сторонам от шва в заднем продолговатом пучке, не затрагивая, однако, его дорсального отдела.
Справа — несколько кровоизлияний, достигающих ядер верхних олив; однако эти последние не затронуты.
Слева — крупное кровоизлияние в tractus spinalis тройничного нерва, однако также оставляющее свободной верхнюю оливу. Далее влево — небольшое кровоизлияние в дорсальных слоях трапециевидного тела, оставляющее, однако, не задетым его вентральные слои.
Анатомическое заключение. Разрез через продолговатый мозг идет между трапециевидным телом и мостом с обеих сторон, перед интактными дейтерсовыми и бехтеревскими ядрами и входом в мозг 8-го нерва, который слева интактен, а справа был перерезан при исследовании его вентрального ядра. Мозжечок вместе с его ядрами удален полностью. Обе верхние оливы и вентральная часть трапециевидного тела интактны, задний продольный пучок с обеих сторон частично разрушен.
Физиологическое заключение. Тонические лабиринтные рефлексы на конечности, реакция поворота головы и последовательная реакция поворота головы, так же, как и‘поворот головы после односторонней перерезки 8-го нерва, у кролика после полного удаления мозжечка вместе с его ядрами сохранены. Для их осуществления достаточно центров, расположенных дистально от операционного разреза.
Приведенные в этом разделе анатомические исследования показывают» что у кошек и кроликов с помощью применявшихся способов полное удаление мозжечка вместе с его ядрами удается без труда, не считая небольших пластин fiocculu s'a, лежащих тотчас у места вступления в мозг 8-го нерва, гарантированное полное удаление которых
дело нелегкое и удается лишь случайно. Анатомические связи этих пластин fl ос с ul u s'a с продолговатым мозгом распространяются, однако, настолько дорсально, что во всех четырех описанных здесь случаях они полностью разрушались и в большом числе остальных он ыто в с весьма большой долей вероятности также
оказывались перерезанны ми.
Физиологические данные будут обсуждаться позже в связи с другими экспериментами.
Дополнительно здесь следует привести еще данные, полученные на двух •собаках, у которых Дюссер де Баренн (3) произвел неполную экстирпацию мозжечка.
Препарат V. 4 декабря 1918 г. Маленькая собачка. Дюссер де Варенном удален мозжечок.
Сначала животное находилось под наблюдением в лаборатории. Далее 22 марта де Клейн произвел правостороннюю экстирпацию лабиринта. 14 апреля был удален левый лабиринт. 27 мая 1919 г. животное было забито.
У животного были обнаружены типичные симптомы удаления мозжечка, совпадающие с классическими описаниями Лючиани и других. Из лабиринтных рефлексов были констатированы: реакция поворота и последовательного поворота головы, вращательный нистагм и поствращательный нистагм головы, реакция поворота и последовательного поворота глаз, нистагм глаз и постнистагм глаз, прогрессивные
45(3
Десятая глава. Центры установки тела
реакции — лифтреакция на голову и конечности (также и при исследовании со шлемиком), готовность к прыжку, тонические лабиринтные рефлексы на конечности (при повороте головы в боковое положение и при перекладывании из положения на животе на спину), тонические шейные рефлексы на конечности, установочные рефлексы, очень сильно выраженные, лабиринтные установочные рефлексы (со шлемиком),. компенсаторные установки глаз (в боковом положении). Установочные рефлексы тела на голову стали отчетливыми только после одно- и двусторонней экстирпации лабиринтов. После двусторонней экстирпации они сначала были вялыми, но спустя 12—18 дней вполне развились. В пространстве животное с надетым шлемиком было полностью дезориентировано; при соприкосновении с подлежащей плоскостью опоры, наоборот,, голова устанавливалась прямо, и к этому вследствие действия шейных рефлексов присоединялось усаживание тела. Оптические установочные рефлексы проявлялись отчетливо; наоборот, установочные рефлексы с тела на тело отсутствовали. Если животное лежало на столе в боковом положении и голова удерживалась в боковом положении, то путем энергичного встряхивания тела на плоскости опоры в разных направлениях удавалось привести к усаживанию последнего.
Анатомическое исследование мозга этого животного проф. Броувером в Амстердаме дало следующие результаты (51).
Макроскопически. Большая часть мозжечка удалена. На правой стороне еще сохранилась ткань, которая по виду напоминает formatio vermicularis. Далее сохранилось несколько пластинок передней доли непосредственно над задним двухолмием. На поверхности моста й на других участках мозга не отмечается ничего особенного.
Микроскопически. После помещения в парафин были изготовлены серийные срезы через продолговатый мозг и мозжечок. Окраска тионином. Серия начинается от шейной части спинного мозга и кончается областью ядер n. trochlearis. Оказалось, что от мозжечка сохранились следующие части.
а) Большая часть правого flocculus’a и paraflocculus’а.
б) Отдельные части правой lobus ansiformis (пластинки полушарий neocerebellum).
в) В черве — отдельные пластинки nodulus’а.
г) В черве — отдельные части lobus anterior, lobus simplex и несколько частиц lobus medius.
д) Совсем маленький кусочек левого flocculus’a.
е) Часть правого nucleus globosus.
ж) На обеих сторонах остатки мозгового вещества мозжечка.
В этих пластинках повсюду можно хорошо различить нормальное строение; lamina molecularis, lamina granulosa и положение клеток Пуркинье сохранено.
С обеих сторон висят остатки мозжечка вместе с продолговатым мозгом, так что вне всякого сомнения возбуждения из продолговатого мозга могли проводиться в оставшиеся части мозжечка.
В продолговатом мозгу обнаруживается значительная, но неполная дегенерация клеток в комплексе нижних олив, группах варолиева моста и боковых ядер.
Область ядер 8-го нерва отчетливой дегенерации клеток не обнаруживает.
Во всей серии не обнаружено кровоизлияний и воспалительных очагов.
Выв о д. После удаления большей части мозжечка все лабиринтные рефлексы, так же как и тонические рефлексы на конечности, шейные установочные рефлексы и установочные рефлексы тела на голову, проявляются отчетливо; наоборот, установочные рефлексы тела на тело отсутствуют. Однако, так как у этого животного осталась неудаленной немалая часть мозжечка, опыт этот сам по себе не может служить вполне удовлетворительным доказательством в пользу независимости названных рефлексов от мозжечка.
Препарат VI. 25 февраля 1919 г. Собака коричневой масти. Экстирпация мозжечка Дюссер де Варенном. Сначала животное находилось под непрерывным наблюдением в лаборатории до 6 ноября 1919 г. В этот день де Клейном удален левый лабиринт. 17 ноября животное погибло во время попытки удалить правый лабиринт.
У этого животного можно было обнаружить следующие лабиринтные установочные рефлексы и установочные рефлексы тела: реакцию поворота и последовательную' реакцию поворота головы, реакцию поворота глаз и последовательную реакцию поворота глаз, вращательный нистагм глаз, лифтреакцию на голову и передние конечности (в шлемике), готовность к прыжку, тонические лабиринтные рефлексы на конечности
Установочные рефлексы после удаления мозжечка
457
при перекладывании на спину и при поворачивании головы в боковое положение, лабиринтные установочные рефлексы (в шлемике), шейные установочные рефлексы, оптические установочные рефлексы, компенсаторные установки глаз в боковом положении. Кроме того, после левосторонней экстирпации лабиринта исключительно высоким оказался установочный рефлекс тела на голову при положении животного на левом боку (в шлемике). Наоборот, установочные рефлексы тела на тело отсутствовали вплоть до момента гибели животного или в крайнем случае только едва намечались. Кроме того, у данного и предыдущего животного после одностороннего удаления лабиринта отмечались еще следующие симптомы: поворот и наклон головы, перекатывание, девиация глаз и нистагм, понижение тонуса конечностей (наиболее отчетливо на стороне операции).
После экстирпации второго лабиринта у этой собаки весьма отчетливо проявилась также бехтеревская компенсация.
Анатомическое исследование, осуществленное проф. Винклером, показало следующее {51).
Нервная система собаки, у которой Дюссер де Баренн удалил мозжечок, была исследована мною (Винклер) на серийных тиониновых препаратах.
Обнаруживается довольно значительное расширение центрального канала спинного мозга. Вследствие этого в 3-м и 4-м сегментах шейного отдела спинного мозга образовалась довольно большая полость. Кроме того, обнаружена довольно значительная внутренняя водянка головного мозга.
При макроскопическом исследовании отмечается, что справа и слева латеральный и базальный отделы мозжечка, по-видимому, удалены полностью. Наоборот, справа простым глазом видно, что довольно значительная часть латерального и базального отделов мозжечка сохранились.
Просматривая серийные срезы, можно констатировать, что мозжечок, за исключением правой базальной латеральной части, действительно удален полностью. Слева же нож прошел идеально между передним холмом и вентральным ядром 8-го нерва, с одной стороны, и floccullus’oM — с другой, не повредив ни одного из названных ядер.
Далее, в продолговатом мозгу и в мосту случайных кровоизлияний не видно.
Справа еще сохранился кусок мозжечка, который находится в связи с мостом я продолговатым мозгом.
Первый висящий здесь кусочек flocculus’а найден там, где появляется слуховой бугорок. Вместе с ножкой flocculus’a последний лежит сверху на слуховом бугорке и вентральном ядре 8-го нерва, латерально — на отсеченном здесь веревчатом теле. Висящие здесь же корешки 8-го нерва (кохлеарный и вестибулярный нервы) совершенно не повреждены, без признаков дегенерации или атрофии и расходятся обычным путем. Та же картина наблюдается, естественно, и слева.
Слуховой бугорок с обеих сторон совершенно не изменен.
Вентральное ядро на дистальном и вентральном концах нормально. Из него исходит мощное трапециевидное тело, в котором группы ядер верхней оливы совершенно нормальны. Латеральная петля и лежащие в ней ядра тоже нормальны (конечно, так же обстоит дело и на левой стороне).
Наоборот, в проксимальной и дорсальной частях вентрального ядра 8-го нерва в четко ограниченном участке клетки отсутствуют (мозжечковая часть вентрального ядра). Слева это выражено несколько сильнее.
Nucleus triangularis невелик. Из него, в связи с почти полной атрофией продольных волокон в S. А. К. (Монаков), выпало множество крупных клеток (слева еще больше, чем справа). Вторичные волокна (fibrae perforantes) из nucleus triangularis к ядрам мозжечка погибли в этом ядре вместе со своими исходными клетками.
Ядро Дейтерса с обеих сторон развито вполне хорошо. Крупные клетки в этом ядре никак не изменены.
Висящий на продолговатом мозгу кусок правого flocculus cerebelli вскоре увеличивается на уровне, на котором обнаруживаются лицевой нерв и ядро отводящего нерва, и достигает своего наибольшего объема по отношению к дистальному концу моторного ядра тройничного нерва.
Здесь, помимо flocculus’a и paraflocculus’а, сохранилась также базальная часть lobus ansatus. Кроме того, довольно мощный связующий пучок соединяет прилежащий сюда кусок мозжечка как с внутренним отделом веревчатого тела, так и с ручкой моста.
Проксимальнее соответствующий кусочек мозжечка быстро уменьшается в объеме.
Анатомическое заключение. Мозжечок на левой стороне удален полностью; на правой не осталось ничего, кроме flocculus’a ра- raflocculus’a и базальных пластинок lobulus ansatus, которые сохранили свои связи со стволом. Первичные корешки 8-го нерва не повреждены. Первичные ядра 8-го нерва изменены настолько, что лежащие в них на-
458
Десятая глава. Центры установки тела
Таблица I
Опыты на децеребрированных кошках с последующей экстирпацией мозжечка
Ха
Дополнительная
реакция
Тонические лабиринтные рефлексы на конечности
Одного лабиринта достаточно для конечностей обеих сторон
Тонические лабиринтные рефлексы на
После односторонней экс-, тирпации лабиринта
Тонические шейные рефлексы на конечности
Секция
шею
туловище
поворот и наклон головы
одностороннее понижение тонуса
5
+
+
Децеребрирована через заднюю треть переднего двухолмия. Справа кое- что осталось от мозговой ножки. Четвертый желудочек лежит совершенно свободно. Мозжечок удален вместе с боковыми отделами. Мозжечковые ножки остались в виде культей, так что часть зубчатых ядер, возможно, еще сохранилась. Оба 8-х нерва интактны.
Макроскопический контроль Винклера.
6
+
+
Децеребрация от заднего двухолмия. Четвертый желудочек лежит совершенно свободно. Мозжечок удален вместе с боковыми частями. Слева полностью удалены также и ножки мозжечка; справа от них осталась культя, так что на правой стороне, возможно, частично сохранилось зубчатое ядро. Оба 8-х нерва интактны.
Макроскопический контроль Винклера.
8
+
+
Мозжечок полностью удален, так что на обеих сторонах остались лишь культи мозжечковых ножек. Ромбовидная ямка лежит свободно на всем протяжении (Магнус).
Установочные рефлексы после удаления мозжечка
459
Таблица I (продолжение)
Ж
Дополнительная
реакция
Тонические лабиринтные рефлексы на конечности
Одного лабиринта достаточно для конечностей обеих сторон
Тонические лабиринтные рефлексы на
После односторонней экстирпации лабиринта
Тонические шейные рефлексы на конечности
Секция
шею
туловище
поворот и наклон головы
одностороннее понижение тонуса
10
+
+
Мозжечок полностью отсутствует. Зубчатое ядро отсутствует с обеих сторон. Ядра Дейтерса с обеих сторон интактны. Децеребрация позади заднего двухолмия.
Макроскопический контроль Винклера.
Левый 8-й нерв перерезан
+
34
+
+
Мозжечок полностью удален. Мозжечковые ножки перерезаны точно на уровне ромбовидной ямки. Фронтальный разрез идет справа на 2, слева на 1 мм кпереди от вступления 8-го нерва, через слуховой бугорок (М а г- нус).
Поперечный разрез впереди от вступления в мозг 8-го нерва.
+
+
17
+
+
+
Мозжечок полностью удален. Экстрамедуллярные части мозжечковых ножек удалены.
Фронтальный разрез с обеих сторон проходит позади культей мозжечковых ножек, слева на 2 мм кпереди от вступления в мозг 8-го нерва, справа на 1 мм кпереди от вступления 8-го нерва, через слуховой бугорок (Магнус).
Левый 8-й нерв перерезан.
+
+
+
+
+
+
+
Поперечный раз- роз впереди от места вступления 8-го нерва.
+
+
+
+
+
+
+
22
Поперечный разрез впереди от места вступления 8-го нерва.
+
+
Рис. 212, стр. 436. Полное обследование на серийных срезах Винклером (см. выше, стр. 436). Препарат I. Вступление 8-го нерва, слуховой бугорок, вентральное ядро 8-го нерва, нисходящий корешок 8-го нерва, дей- терсов нисходящий тракт,
460
Десятая глава. Центры установки тела
Таблица I (продолжение)
№
Дополнительная
реакция
Тонические лабиринтные рефлексы на конечности
Одного лабиринта достаточно для конечностей обеих сторон
Тонические лабиринтные рефлексы на
После односторонней экстирпации
Тонические шейные рефлексы на конечности
Секция
шею
туловище
поворот и наклон головы
одностороннее понижение тонуса
(22)
вместе со своим началом, ядра трапециевидного тела и олив с обеих сторон интактны. Ядро Дейтерса справа совершенно интактно, слева несколько срезано в верхней части. Дорсальное ядро 8-го нерва справа интактно, слева латеральная часть его несколько срезана. Ядро Бехтерева отрезано. Мозжечок полностью удален.
23
+
(+)
См.
дан¬
ные
сек¬
ции.
+
Рис. 214, стр. 444. Полное обследование на серийных срезах Винклером (см. выше, стр. 444). Препарат II. Децеребра- ция через переднее двухолмие. Правый 8-й нерв со всеми центральными путями и ядрами интак- тен. Область вступления левого 8-го нерва полностью (может быть, исключая немногие проксимальные вестибулярные волокна) удалена. Относящиеся сюда оральные вторичные пути частично разрушены кровоизлиянием. Пути, исходящие из ядер Дейтерса, с обеих сторон интактны. Мозжечок удален полностью.
чальные клетки для вторичных мозжечковых систем перестали существовать. Однако вторичные системы 8-го нерва, направляющиеся к среднему мозгу, как и ядра Дейтерса, остались неизменными.
Физиологическое заключение. После полного слева и неполного справа удаления мозжечка у собаки сохранились все известные лабиринтные рефлексы и, кроме того, шейные установочные рефлексы
Установочные рефлексы после удаления мозжечка
461
и установочные рефлексы тела на голову, в то время как установочные рефлексы тела на тело отсутствовали. Симптомы односторонней экстирпации лабиринта появлялись в неизмененном виде i
Б. Результаты опытов
После того как в предыдущем разделе подробно описаны операционные методы, приведены протоколы опытов и результаты анатомических исследований, могут быть сообщены в краткой форме таблиц результаты самих опытов.
В таблицах + означает, что рефлекс имел место, — что он не вызывался.
Первая серия опытов
Децеребрированные кошки (табл. I, стр. 458—460).
Таблица охватывает 9 опытов. Протоколы опытов 22 и 23 находятся на стр. 435 и 443. Два препарата (опыты; 22 и 23) исследованы в виде серий срезов. Последующие препараты были макроскопически проконтролированы проф. Винклером.
Заключение. У децеребрированных кошек после удаления мозжечка вместе с ядрами и после фронтального разреза через продолговатый мозг, сразу перед вступлением в мозг 8-х нервов, в неизменном виде обнаруживаются тонические шейные и лабиринтные рефлексы на конечности и тонические лабиринтные рефлексы на шейные мышцы. У такого рода животных после односторонней перерезки 8-го нерва (точно так же, как у интактных животных после односторонней экстирпации лабиринта) происходит поворот и наклон головы в сторону операции и понижение тонуса конечностей одной стороны тела (также и при установлении головы прямо). Одного лабиринта у животных, оперированных таким образом, достаточно и для осуществления тонических лабиринтных рефлексов на мускулатуру конечностей обеих половин тела. Ядро Бехтерева для осуществления тонических лабиринтных рефлексов на мышцы конечностей не является необходимым.
Вторая серия опытов
Децеребрированные кролики (опыты де Клейна).
У трех кроликов после децеребрации удалялся мозжечок. У одного из этих животных дополнительно перерезался 8-й нерв. В результате оказалось следующее (табл. II, стр. 462).
Заключение. У децеребрированного кролика после экстирпации мозжечка (в связи с чем остатки flocculus’a, вероятно, не находятся больше в функциональной связи с продолговатым мозгом) реакция поворота головы, последовательная реакция поворота головы и наступающие после односторонней перерезки 8-го нерва поворот и наклон головы не изменяются.
Третья серия опытов
Децеребрированные кролики; поперечный разрез тотчас возле вступления в мозг 8-го нерва (опыты д е Клейна).
У трех кроликов после децеребрации был удален мозжечок, после чего произведен фронтальный разрез через продолговатый мозг тотчас возле вступления в мозг 8-го нерва. В опытах 1 и 2, помимо этого, был перерезан
462
Десятая глава. Центры установки тела
Таблица П
Опыты на децеребрированных кроликах с последующей экстирпацией мозжечка
№
Дополни¬
тельная
операция
Поворот и наклон головы после односторонней перерезки 8-го нерва
Реакция поворота головы
Последовательная реакция поворота головы
Секция
1
+
+
Децеребрация между передним и задним двухолмиями. Мозжечок удален полностью., кроме совсем небольших кусочков flocculus’a на 8-х нервах. Последние не повреждены.
2
■+
+
Как в «Ns 1.
3
Перерезка правого 8-го нерва
+
Не испытано.
Не испытано.
Как в «Ns 1.
Слева небольшой кусочек flocculus’а на месте вступления в мозг 8-го нерва. Справа 8-й нерв отсечен тотчас у продолговатого мозга.
правый 8-й нерв, в то время как в опыте 3 предварительно исследовались тонические лабиринтные рефлексы и реакция поворота головы и только в конце опыта перерезался. 8-й нерв справа. Препарат опыта 3 был исследован на серийных срезах проф. Винклером (ср. выше, стр. 454, препарат IV).
Заключение. У децеребрированных кроликов после экстирпации мозжечка, включая и его ядра, и после фронтального рассечения продол- говатого мозга тотчас возле вступления в мозг 8-го нерва обнаруживаются тонические лабиринтные рефлексы на конечности, а также реакция поворота головы и такая же последовательная реакция. После односторонней перерезки 8-го нерва наступает типичный поворот и наклон головы в сторону операции, можно еще обнаружить тонические лабиринтные рефлексы на конечности с обеих сторон тела, реакция поворота головы при повороте вправо и влево такая же, как у интактных животных после удаления лабиринта с одной стороны. Тонические рефлексы на конечности, реакция поворота головы и последовательная реакция поворота головы после частичного нарушения заднего продольного пучка также еще сохранились (табл. III, стр. 463).
Четвертая серия опытов
Таламические кролики (опыты де Клейна), См, табл. IV, стр. 464.
У четырех кроликов были удалены большие полушария выше таламусов, для того чтобы исследовать лабиринтные рефлексы на глаза, после чего удалялся мозжечок. В конце опыта у трех кроликов перерезался 8-й нерв, у одного кролика, кроме того, производился фронтальный раз-
Установочные рефлексы после удаления мозжечка
463
Таблица III
Опыты на кроликах, у которых после экстирпации мозжечка пересекался продолговатый мозг тотчас у входа
в мозг 8-го нерва
№
Дополнительная
операция
Тонический лабиринтный рефлекс на конечности
Реакция поворота головы
Последо¬
вательная
реакция
поворота
головы
Поворот и наклон головы после односторонней перерезки 8-го нерва
Секция
1
Перерезка правого 8-го нерва в начале опыта.
-f- (с обеих сто- рон).
При повороте налево при повороте направо —.
Это типичное поведение после правосторонней перерезки 8-го нерва.
Сомни¬
тельно.
+
Фронтальный разрез через продолговатый мозг возле вступления в мозг 8-го нерва. Мозжечок экстирпирован полностью. Оставались лишь небольшие кусочки flocculus’а на левом 8-м нерве.
2
Перерезка правого 8-го нерва в начале опыта.
+ (с обеих сто- рон).
Не испытано.
Не испытано.
+
Как в № 1.
3
Перерезка правого 8-го нерва в конце опыта.
+
+ (с обеих сто- рон).
+ (с обеих сто- рон).
+ (слабо).
Исследование на серийных срезах (Винклер) (см. выше, стр. 454, препарат IV). Фронтальный разрез через продолговатый мозг между трапециевидным телом и мостом. Дейтерсово и Бех- теревское ядра с обеих сторон ин- тактны. Задний продольный пучок частично разрушен кровоизлиянием. Мозжечок вместе с его ядрами удален полностью.
рез через продолговатый мозг тотчас выше места вступления 8-го нерва. Три препарата этой экспериментальной серии были исследованы проф. Винклером, который изготовил из них тонкие срезы и описал их.
Секционные данные опытов таблицы IV
№ 1 (данные проф. Винклера). Слева возле вступления в мозг 8-го нерва сохранился небольшой кусочек flocculus’а, который, возможно, еще находится в физиологической связи с продолговатым мозгом. Справа flocculus удален полностью. Слева поверх brachia conjunctiva лежит совершенно изолированно маленький кусочек мозжечка*
464
Десятая глава. Центры установки тела
Таблица IV
Опыты на кроликах, у которых экстирпировался головной мозг выше таламуса и затем мозжечок
№
Дополни¬
тельная
операция
Компенсаторные установки глаз
Калорическое отклонение глаз и нистагм
Реакция
поворота
глаз
Реакция поворота головы
После односторонней перерезки 8-го нерва
Секция
поворот
вертикальное отклонение
реакция и нистагм
последовательная реакция и постнистагм
реакция
последовательная реакция
поворот и наклон головы
отклонение глаз
нистагм
1
Перерезка 8-го нерва справа в конце опыта. ,
Справа 70°, слева 65°.
С'обеих сторон
С обеих сторон -f
Не исследовано.
+
+
+
+
+
+
См. стр. 463—465.
2
В конце опыта перерезка левого 8-го нерва и фронтальный разрез тотчас выше места вступления 8-го нерва в мозг.
С обеих сторон 30°.
С обеих сторон -|-
С обеих сторон +
+
+
+
+
(Фронтальный разрез выше 8-го нерва).
3
Перерезка левого 8-го нерва в конце опыта.
Справа 45°, слева 30°, позже с обеих сторон 35°.
С обеих сторон -{-
С обеих сторон + , слева слабее.
+
+
+
+
+
4
—
С обеих сторон 20°.
С обеих сторон -f
С обеих сторон -{-, справа слабее.
+
+
+
+
не находясь ни в какой связи со стволом. С обеих сторон отсечены холмы заднего двухолмия; переднее двухолмие разрушено кровоизлиянием. Большие полушария с обеих сторон удалены полностью. С обеих сторон разрез проходит через таламус. Справа в таламусе находится крупное кровоизлияние.
№ 2. Мозжечок удален полностью, лишь слева остался кусочек flocculus’а, более крупный, чем на других препаратах.
№ 3 (данные проф. Винклера). У основания дистального края четвертого желудочка дно его разрушено, справа — вплоть до сетевидной формации. На разрушенном дне, совершенно изолированные, лежат пластинки мозжечка, не связанные со стволом мозга.
Установочные рефлексы после удаления мозжечка
465
Слева и справа в области вступления 8-го нерва находятся отдельные небольшие пластинки flocculus’a. Физиологическая связь их с продолговатым мозгом, однако, полностью прервана. Слева отсечены веревчатое тело и боковая петля. Задний холм слева поверхностно поврежден кровоизлиянием, справа — ножом. Левый передний холм уничтожен, aquaeductus заполнен кровью; правый передний холм содержит отдельные кровоизлияния. Слева и справа срединные ядра таламуса разрушены кровоизлиянием; кроме того, слева полностью разрушено латеральное ядро и частично — вентральное ядро.
Слева разрез проходит через полосатое тело. Полушарие большого мозга удалено, остался только оральный конец аммонового рога, поврежденный кровоизлияниями. Правое полушарие удалено только частично. Позади остался базолатеральный кусок его, почти целиком — аммонов рог, у которого отсутствует только дорсальная часть. Впереди также находится большой кусок; сохранились медиальная (дорсально и вентрально от комиссуры) и полностью базальная стенка; удалена лишь часть дорсальной поверхности.
№ 4 (данные проф. Винклера). Возле вступления правого 8-го нерва находится небольшой кусочек flocculus’а, физиологические связи которого с продолговатым мозгом, однако, полностью прерваны. Слева полностью удален также и flocculus. В остальном от мозжечка не осталось никаких следов. Кровоизлияния в правый задний холм и в оба передние холма. Оба большие полушария отсутствуют, только слева спереди базально-медиально сохранился небольшой кусочек, справа — некоторая часть аммонового рога. Разрез, экстирпирующий большой мозг, проходит на обеих сторонах через таламус и стриатум.
Заключение. У таламического кролика после полного удаления мозжечка сохраняются компенсаторные установки глаз (вертикальные отведения и повороты по кругу), реакция поворота и последовательная реакция поворота глаз, нистагм глазных яблок, постнистагм, калорическое отклонение глаз с нистагмом, реакция поворота и последовательная реакция поворота головы. После односторонней перерезки 8-го нерва, как и у нормального животного, наступает поворот и наклон головы и отведение глаз (с нистагмом). Реакции глаз имеют место и тогда, когда четверохолмия (крыша среднего мозга) оказываются в значительной мере разрушенными вследствие оперативного вмешательства или кровоизлияния.
Компенсаторные повороты, которые у нормального кролика (см. выше, стр. 126) составляют примерно 90—100°, как видно из опыта 1, даже после серьезной операции экстирпации большого мозга и удаления мозжечка могут осуществляться почти с нормальной интенсивностью. По большей части (опыты 2—4) величина экскурсий снижена (20—45°). Так как мы знаем, что величина компенсаторных круговых поворотов при воздействии всех возможных тормозящих влияний значительно уменьшается, последнее не представляется неожиданным.
В качестве завершения этой серии опытов можно привести еще одно наблюдение над кроликом, у которого при сохранных полушариях был удален мозжечок и у которого тонические лабиринтные рефлексы на конечности, реакция поворота головы и нистагм головы, последовательная реакция поворота головы, реакция поворота глаз и нистагм глаз, последовательная реакция поворота глаз, постнистагм глаз и компенсаторные установки глаз могли быть установлены с большой отчетливостью. На секции (макроскопически) мозжечок оказался полностью удаленным, остатки flocculus’a простым глазом не определялись, четверохолмие было не повреждено. В дальнейшем у таламической кошки после удаления мозжечка имели место реакции поворота головы, последовательная реакция поворота головы, а также реакция поворота и последовательная реакция поворота глаз.
Пятая серия опытов
Нормальные кошки. Как уже упоминалось выше, у кроликов непосредственно после экстирпации мозжечка лабиринтные установочные рефлексы, несмотря на шок, оказываются отчетливо выраженными. 30 Р. Магнус
Таблица
tr
я
ноя91гфэ(1 энньоа -ohbxoä эинхнийид-вк
шлзхэинхэоп
Я ей н И Я
ф ф Я И S К й g.' о
И
о й ftg
® Й
Я
Н §
® §
К EH ft * Ф
2 я л н н 2 О й
И g й И
Я ф
и|
8 S
й £
СО 2
ы й
t>"3
I—J ф
я а Я И о ÏÏ ф 3 я в я й я “ о g я 2
0 «
° g ft ft
ST «
1 Ф
я 5
I S
Ф Й4 W
й а й
я я я я я я о Й И j§
н
ф W
Я ^ о
П лл я
м202
ф ч Э
■Щ о S
§ g g
s
."I
* ф я 2 я
Я Ф н Я о о И о и а К со и й 2 ф о а й
я я
:Я §
и § я
ф 5 °
к g й
Я о я fct £ й:
>S Ж О
о й
tf Я
оаиФяа^ял
П О Я ^ 3 н Я h
9‘?5sSgw§,§ £ 2 в й и £<
Н о я Я о S Й
ft t-i ft ^ о g Я
ф Ф о я !>,§ а
9,0 И 8 5 о о
§3*
■г О СО 1 Я
Ф § И g н U н о ^
р яяая^с000 ® и 5 ф з о й Wg=ggWgH^Ä |§ W Н й ^
Й cq я о
Я
я И а
О ж >■
FH Я Ж h
о а ь-, - £>2® §
.&h!s
3 й 3 я о 3 о g Я м со я a й Ë?S® я ° и
О Ф 2 н _
ж Я a
м Я <ZL, ф о я
за и К ж я я ‘ ы “ ® л ï я
ж. ,_,
а я
5 w
а ф ft Я н о Я Я ф
о Я о м Я Я йнй й
ЯЯьс§М§ф§
яяяойян^за а а йж 5 й
о а о ft я
ft° о ре; Я й
°s
1 ° н а й а я
ft н I гр о
» о к
°с S
Ф I—i Я
я я
В - О
я а 5
° я püp
Я Ф й
о а я ° а й ft® я
я ф а
я tO g
а я н
Я Q. ft Ф^ Я Я о ж н я s я о g я
^ Э я
tc g
. н Я г о —О ф оо о с* а —
- Ф^ Я Q. Н .
оо . о-я . а и и о нн 'ЯЯм о к а м я ^ 'ЯКа * н &W i—i • а ,п ф
рны*£ьоайо$.я
н н а ü 2^ я в я
°а& "|^и|
-§^2§Sös5§
я я 3 ^ и
3 я
Я g É
§SS<
а в о
Я Щ Н н ж Я о
g ° 2
ёю и
ж ft
JL • ф ^5 § e-jS И
ф
2 й §
м о §
г§
и&
ÏÏ Я ct
Ss
2 о g
^ я
О е£ ф я a
я gw
н °
=в a сЗ
Ф Я Н Ü ® Я
I I
о a ft я Я о a я 5 Й
. ЯС
л 2 я ft я я
te Я
ж5 Я
лйнН ф S ф Й 1 И
о g Н
°|§
н ft ft
5gS w
я ïï
S Ф я
Sh I p .«
я 2 за я а о
Я н я о О ж
ftp g ф
ft
я •
я 3 а я
*£ за я о н
w Э
Я й ф
ft я ф м
а о =я
ф за и я Э н
Я ж
а
Я а
ф
о ft о К
я ■ 3 wj • i-,
^°£ Фа ftp Я ^
° w й о Й й й я
я ч-^а^йжаж
2юНоР §-1фО й ^ Я '— ^ а ^ Я о. Е-1 нР -Я Я н л
и л В4^ и яй 2 ?
о к и -£ а g-н ё
§ я ф я я ^Я g. g.
2^йя&^ойоа о Ьй я Jo ° ® и и
g й а я 4 я й fto ф
о £ ®р ° SS
^Я й wo«« Я я
w QJ
Я й ж Я Н Я я я о а-
н й н я
2 о о ^
S3 Я ° 5
а н
g § н
£ й g °
о а Я я Я н Я н HOÏÏft О О Ь ф
и 2 § Я я и Q
Й S К .
й 2 О о а й Я
о й
н а я я
® ф о ьз
Я g g
а о ж Я я
Ж ■>Й
Н Я Я
в я я
Ф й ьс я а я ф tf я о Кр О ^ГП
£и
fto Я
Ж й Я
S я Я S И н
g я Я О о Я Я ж н
s я
Я в й Я я , я о а
о ф 2 я р4 2 •я а ^
Яр« й og а Я Я
ООН
рз 2
R
S& ь & §0 §В о
Н §
и §
g g
ж О
а и
41 rL
я Я Н я ф
о К
ф к
Й
5
Я ф
О Ж
я
о
О ЭЯ
ftg я в а я
s к
Я Я О Я й о
Ф Я
й 2
Я ft
о й
+
о
VO
а
й
+
g - а^ 93 m § g Я а Й ^ я я Ж Я ft Я я Я Я я Ж
+ к “ 2
CÖ
R
U
+
+
+ +
+ +
+
8шм -BJLodoaon оаончкэхваойэкооп
+
+
ешы Bxodoaon
+ +
+
iaaoifoj Bxodoaon оаоналэхваойэкооп
iaaoifoj Bxodoaon
ихоон
■ЬЭНОЯ "ВН ЫЭЯЭ1ГфЭ(1 9НН
-хнийидвк эиноэьинох
(инК)
BXHD0 чхоонакэхип'й'
+ +
+
+
+
+ +
оо о см
со см
ю со
Установочные рефлексы после удаления мозжечка
467
В предыдущей, четвертой серии опытов де Клейна только у двух животных были отмечены слабые следы этих рефлексов. Точно так же у кролика, у которого я удалил большие полушария и мозжечок, установочные рефлексы по крайней мере намечались, в то время как реакция поворота и нистагм глаз, компенсаторные установки глаз, реакция поворота и последовательного поворота головы были выражены отчетливо. На секции этих животных было макроскопически отмечено полное удаление больших полушарий и мозжечка и интактность четверохолмия.
Поэтому следующая серия опытов была поставлена на кошках, у которых удалялся мозжечок, но большие полушария не удалялись, после чего животные сохранялись живыми в течение нескольких дней. Результаты приводятся в табл. V. Подробный протокол анатомически строго проконтролированного опыта 12 приведен выше, на стр. 448.
Заключение. У кошек с сохранными большими полушариями после полного отделения мозжечка вместе с его ядрами можно видеть неизменными все лабиринтные установочные рефлексы, также и в том случае, когда влияние глаз выключается путем наложения на глаза повязки. В подтверждение предшествующих экспериментальных серий выяснилось, кроме того, что тонические лабиринтные рефлексы на конечности, реакции поворота и последовательного поворота головы, а также реакции глаз, нистагм и постнистагм глаз оказываются у этих животных неизмененными.
Общий итог
Описанные выше опыты показывают, что после удаления мозжечка с его ядрами все лабиринтные рефлексы сохраняются.
В отношении почти всех рефлексов приводится анатомическое доказательство того, что мозжечок удалялся полностью, т. е. включая сюда прилежащие к области вступления в мозг 8-го нерва пластинки flocculus’a. Ниже те рефлексы и реакции, для которых это доказательство приведено, напечатаны разрядкой (в решающем опыте). Для немногих остальных реакций имеется только макроскопический контроль, так что остается маловероятная возможность, что здесь небольшие участки flocculus’a остались в связи с продолговатым мозгом.
Реакции на прогрессивные движения в то время, когда ставились эти опыты, еще не были известны. Поэтому они были исследованы только на двух собаках Дюссер де Баренна с удаленным мозжечком. Так как при этом оставались неудаленными отдельные участки мозжечка, из этих опытов можно, строго говоря, заключить только, что прогрессивные реакции после удаления большей части мозжечка вместе с его ядрами сохраняются.
В связи с остальными приведенными в этой главе данными следует, однако, с большой долей уверенности принять, что и прогрессивные реакции не осуществляются через мозжечок и после удаления последнего остаются без изменения. Однако точное доказательство этого положения еще должно быть приведено.
В частности, после удаления мозжечка удалось показать наличие следующих лабиринтных рефлексов и реакций.
А. Двигательные рефлексы.
1. Поворотные реакции.
а) На шею: реакция поворота головы и последовательная реакция поворота головы [третья, се¬
30*
468
Десятая глава. Центры установки тела
рия опытов, опыт 3; четвертая серия опытов, опыт 4 (в опыте 3 только реакция поворота головы); пятая серия опытов, опыт 12].
Вращательный нистагм головы.
б) На глаза: реакция поворота глаз и последовательная реакция поворота глаз (четвертая серия опытов, опыты 3 и 4; пятая серия опытов, опыт 12). Вращательный нистагм глаз и постнистагм (пятая серия опытов, опыты 3 и 4).
2. Прогрессивные реакции (до сих пор наблюдались только у собаки с полностью удаленным мозжечком).
Б. Рефлексы положения.
1. Тонические рефлексы на мускулатуру тела.
а) На конечности (первая серия опытов, опыты 22 и 23; третья серия опытов, опыт 3; пятая серия опытов, опыт 12). При этом одного лабиринта достаточно для конечностей обеих сторон (первая серия опытов, опыт 23).
б) На шею и туловище.
2. Лабиринтные установочные рефлексы (пятая серия опытов, опыт 12).
3. Компенсаторные установки глаз.
а) Вертикальные отведения (четвертая серия опытов, опыты 3 и 4).
б) Круговые повороты (четвертая серия опытов, опыты 3 и 4).
В. Последствия односторонней перерезки 8-го
нерва.
1. Поворот и наклон головы (третья серия опытов, опыт 3; четвертая серия опытов, опыт 3).
2. Утрата тонуса на гомолатеральных конечностях (макроскопический контроль проф. Винклера в первой серии опытов, опыт 10).
3. Измененная установка глазе нистагмом (четвертая серия опытов, опыт 3).
Г. Калорические реакции: отклонение и нистагм (четвертая серия опытов, опыты 3 и 4).
Из других рефлексов, учитывающихся при установке тела после удаления мозжечка, имели место следующие.
Тонические шейные рефлексы на конечности.
Шейные установочные рефлексы.
Установочные рефлексы с тела на голову (до сих пор наблюдавшиеся только при неполном удалении мозжечка).
Оптические установочные рефлексы (до сих пор наблюдавшиеся лишь при неполном удалении мозжечка).
Наоборот, у обеих собак с неполностью удаленным мозжечком, находившихся под наблюдением в продолжение месяца, установочные рефлексы с тела отсутствовали или по меньшей мере оказывались нарушенными настолько, что показать их с уверенностью было невозможно. Этот пункт несомненно требует дальнейших исследований.
О динамике децеребрационной ригидности после удаления мозжечка можно сказать следующее. Уже в 1898 г. Шеррингтон (1) установил, что в отдельных случаях децеребрационная ригидность остается и после удаления мозжечка. Это имеет место в тех случаях, когда мозжечок удаляется без значительного кровотечения. Однако в 1914 г. из лаборатории Шерринг- тона вышла работа Уида (Weed), который пришел к противоположным вы¬
У с танов очные рефлексы после удаления мозжечка
469
водам. У децеребрированных кошек Уид дополнительно удалял мозжечок, в процессе чего последний приподнимался над дном четвертого желудочка и мозжечковые ножки перерезались. После этого децеребрационная ригидность исчезла в среднем через 20 минут; только в одном опыте ригидность не исчезала. В другом опыте сначала удалялся мозжечок, а затем животное децеребрировалось. Также и в этом случае ригидность наступала только на 1—2-й минуте, после чего животное расслаблялось. У одной кошки, кроме того, мозжечок был удален за четыре недели до децеребра- ции, которая вначале сопровождалась ригидностью, исчезнувшей спустя 2гЛ часа. Уид сообщает далее, что перерезка ствола тотчас позади заднего двухолмия сопровождается по истечении не более 5 минут полнейшим исчезновением децеребрационной ригидности. Из этих опытов Уид делает заключение, что главный центр децеребрационной ригидности лежит в среднем мозгу и, весьма вероятно, в красном ядре, а мозжечок составляет весьма важное, если не абсолютно необходимое соединительное звено в процессе возникновения децеребрационной ригидности.
Эти наблюдения и заключения Уида вышеприведенными опытами с удалением мозжечка у децеребрированных кошек не подтверждаются. Из них скорее следует, что как после полного удаления мозжечка, так и после удаления ствола вплоть до заднего двухолмия включительно может еще наступать интенсивная децеребрационная ригидность и что, следовательно, ни мозжечок, ни красное ядро не могут быть отнесены к центральным образованиям, необходимым для возникновения децеребрационной ригидности. Различие в результатах опытов, может быть, объясняется тем, что, за исключением двух опытов Уида, экстирпация мозжечка производилась спустя продолжительное время после децеребрации, и тем, что применявшаяся им техника не была достаточно щадящей.
Так как в части приведенных выше опытов предпринимались последующие удаления ствола мозга, причем децеребрационная ригидность в конце концов исчезала, здесь следует сообщить еще об одном опыте Беритова (18а), поставленном в 1918 г. в моем институте. Оказалось, что даже через 8 часов после децеребрации и полного удаления мозжечка может еще наблюдаться достаточно интенсивная ригидность. В другом опыте через 5% часов после удаления мозжечка и удаления ствола вплоть до задних холмов включительно имела место значительная ригидность.
Из этих наблюдений также следует, что мозжечок не может иметь значения для возникновения «статотонуса», которое приписывает мозжечку Эдингер.
Таким образом, в согласии с первоначальными данными Шеррингтона, оказывается, что после щадящей экстирпации мозжечка децеребрационная ригидность не исчезает.
Благодаря сообщенным сведениям, на основании тщательных физиологических наблюдений и авторитетного анатомического контроля было доказано, что все исследованные лабиринтные рефлексы и реакции после полного удаления мозжечка вместе с его ядрами сохраняются и что соответствующие лабиринтным рефлексам проводящие путине проходят черезмозжечок.
Этим, конечно, не сказано, что никакие исходящие из лабиринтов возбуждения при интактной центральной нервной системе не могут попасть
470
Десятая глава. Центры установки тела
в мозжечок и участвовать так или иначе во все еще неизвестных функциях этой части мозга. Это может быть исследовано только тогда, когда нормальная функция мозжечка будет более доступна физиологическому эксперименту, перспективы чего в настоящее время, несмотря на усилия многих исследователей, еще весьма неопределенны.
С другой стороны, остается вполне возможным, что импульсы из мозжечка достигают расположенных в мозговом стволе центров лабиринтных рефлексов и там оказывают усиливающее или тормозящее влияние на динамику лабиринтных рефлексов. В пользу этого говорят наблюдения Бауэра и Лейдлера (Bauer u. Leidler), которые после повреждений червя мозжечка обнаружили значительное усиление вращательных реакций глаз.
Все возможности подобного рода, однако, не противоречат заключению, что центры лабиринтных рефлексов лежат вне мозжечка и что поэтому следует окончательно оставить все еще широко распространенный взгляд, согласно которому мозжечок должен считаться центральным аппаратом для лабиринтов.
Уже Флуранс трактовал симптомы, наступающие вследствие поражений полукружных каналов, как мозжечковые, и многие исследователи, вслед за ним, думали так же, усматривая большое сходство явлений, обусловленных раздражением и удалением мозжечка и удалением и раздражением лабиринтов. По Феррье, анатомическим основанием влияния лабиринтов на равновесие является их связь с мозжечком. Лючиани принимает, что лабиринт осуществляет свое тоническое влияние на мышцы через посредство мозжечка. Стефани заключает, что деятельность мозжечка в значительной ее части, если не полностью, обусловлена импуль- сацией, которая опосредуется лабиринтами, и что описанный Лючиани церебеллярный тонус вызывается неакустической частью лабиринта. Бехтерев (2) видит в мозжечке орган поддержания равновесия, который свои периферические возбуждения частично получает из полукружных каналов. По Шеррингтону (3, 5), мозжечок является головным ганглием всех проприоцепторов и получает важные возбуждения из лабиринтов. Левандовский считает установленным, что «характер ориентирования посредством лабиринтов полностью соответствует характеру и назначению ориентирования посредством мозжечка»; связи мозжечка с областью окончания вестибулярного нерва остаются, однако, еще неизвестными. Барани (2) принимал, что весь мозжечок в целом находится под влиянием определенных вестибулярных раздражений, но что, однако, у человека влияние вестибулярного нерва на мозжечок оказывается лишь незначительным. После Барани (3) Рейх и Ротфельд, исходя из опыта на децеребрированном животном с неполностью (Ротфельд) удаленным мозжечком, заключили, что мозжечок определенно имеет какое-то отношение к лабиринтным поворотным реакциям, но что по крайней мере движения вперед и назад локализованы уже в продолговатом мозгу. Ротфельд (4, 5) утверждает, что у кролика мозжечковая кора не оказывает существенного влияния на реакции поворота, однако указывает, что ядра мозжечка, в особенности ядро покрышки, принимают в этом большое участие.
Все эти предположения экспериментами, приведенными в этой главе, лишаются фактических оснований. Как было здесь показано, деятельность лабиринтов протекает независимо от участия мозжечка. Теперь надо исследовать деятельность мозжечка вне участия лабиринтных влияний. Только после решения этой задачи можно с определенной надеждой на успех подойти к вопросу, в каких взаимоотношениях находится деятельность мозжечка и центров лабиринтных рефлексов.
Установочные рефлексы после удаления мозжечка
471
В этой связи следует указать на то, что в литературе уже имеется ряд данных, указывающих на далеко идущую взаимонезависимость лабиринтных рефлексов и мозжечка. В 1873 г. Лёвенберг (Löwenberg) показал, что у голубей реакции полукружных каналов (движения головы при раздражении лабиринтов) сохраняются после удаления мозжечка. В 1880 г. Шпамер (Spamer) наблюдал у голубей после почти полного удаления мозжечка типичные реакции, наступавшие в ответ на локальное раздражение лабиринтов гальваническим током. Хёгиес (Högyes) в 1881 г. наблюдал у кроликов после неполного удаления мозжечка сохранившуюся реакцию поворота глаз, а также вращательный нистагм глаз и высказал на основании этого мнение, что эти лабиринтные рефлексы с мозжечком ничего общего не имеют. Прежде всего, однако, здесь следует упомянуть о работе Богумила Ланге (Bogumil Lange, 1891) из лаборатории Гольца и Эвальда, который удалял у голубей более 2/3 мозжечка и производил одно- или двустороннее разрушение или пломбирование полукружных каналов. По Ланге, симптомы удаления мозжечка и выключения лабиринтов абсолютно различны. После удаления мозжечка односторонняя экстирпация лабиринта вызывает те же типичные последствия, что и у нормальных животных; с другой стороны, удаление мозжечка после двусторонней экстирпации лабиринтов вызывает симптомы, типичные уже для этого случая. Таким образом, удаление одного органа не препятствует возникновению симптомов после утраты другого. Мозжечок и лабиринты могут взаимно друг друга компенсировать. В опытах, описанных выше, также обращало на себя внимание, что после удаления мозжечка (у животных с сохраненными большими полушариями, пятая серия опытов) обычных симптомов двустороннего удаления лабиринтов никогда не наблюдалось: у кошек и кроликов не появлялось отклонения глаз, нистагма и вообще никаких явлений раздражения, как это обычно бывает после операций на лабиринтах. Качания и маятникообразные движения головой, удары мордой о пол, приступы бурных движений, ползание пятясь назад и все остальные симптомы, которые описаны в седьмой главе в качестве начальных последствий двусторонней утраты лабиринтов у кошек, здесь отсутствовали.
Далее, Бейер и Левандовский после двусторонней экстирпации лабиринтов нашли, что, повреждая мозжечок, можно вызвать дополнительно типичные насильственные движения. Наконец, Вильсон и Пайк, раздражая у собаки полукружные каналы горячей водой и удаляя один лабиринт после экстирпации мозжечка, получали отклонение глаз и нистагм. Как и Ланге, они указывают на различие симптомов утраты мозжечка от таковых экстирпации лабиринтов.
Насколько мне известно, о том, подтверждалась ли полнота удаления мозжечка микроскопически, до сих пор ничего не сообщалось.
Чрезвычайно важным является сообщение Лючиани, что безмозжеч- ковые собаки хорошо плавают, и; сообщение Ланге, что безмозжечковые голуби хорошо летают. Причина этого лежит (по крайней мере для собак; относительно голубей собственного опыта у меня нет) в том, что собака во время плавания ориентируется исключительно с помощью лабиринтных установочных рефлексов, в то время как установочные рефлексы с туловища не имеют возможности проявиться. Таким образом, если безмозжечковые собаки способны плавать, это доказывает, что установочные лабиринтные рефлексы у них функционируют успешно.
Наконец, различные исследователи установили, что отдельные функции лабиринтов сохраняются после частичного удаления мозжечка. Так, Кубо нашел после частичного удаления мозжечка и специально flocculus’a калорический нистагм сохранным. Бауэр и Лейдлер видели, что после
472
Десятая глава. Центры установки тела
удаления полушарий мозжечка нистагм глаз не изменялся, а после удаления червя — даже усиливался. Барани, Рейх и Ротфельд удаляли большую часть мозжечка, — вероятно, однако, оставляя его ядра, — и наблюдали сохранность лабиринтных реакций поворота головы вперед и назад.
Однако, как было сказано выше, несмотря на различные сообщения подобного рода, представление о существовании непосредственного контакта функций лабиринтов и мозжечка остается еще весьма широко распространенным. В настоящее же время для лабиринтных и некоторых других рефлексов, участвующих в установке тела, показано, что в противоположность этому представлению они совершенно не зависят от мозжечка.
И. ПОЛОЖЕНИЕ ЦЕНТРОВ УСТАНОВКИ ТЕЛА И ЛАБИРИНТНЫХ РЕФЛЕКСОВ В МОЗГОВОМ СТВОЛЕ
Благодаря опытам Мажанди, Лонже, Шиффа, Вульпиана, Христианин X. Мунка, Феррье, Гуддена, Морита и других исследователей на кроликах и Гольца, X. Мунка и Ротманна на собаках, а также благодаря описанным в пятой главе наблюдениям на таламических животных, мы знаем, что способность к стоянию и бегу после удаления больших полушарий и базальных ганглиев сохраняется. Ближайший анализ показал, что у таламического животного имеются все описанные в этой книге реакции и рефлексы с единственным исключением оптических рефлексов, пути которых проходят через большие полушария. Описанные в предыдущем разделе эксперименты учат, что удаление мозжечка влияет только на установочные рефлексы с туловища на голову, тогда как все остальные рефлексы установки тела и лабиринтные рефлексы могут осуществляться без участия мозжечка. Отсюда вытекает необходимое заключение, что центры и пути для всех обсуждавшихся здесь реакций, — за исключением, может' быть, установочных рефлексов тела на тело (см. ниже), — лежат в мозговом стволе.
В дальнейшем следует обобщить то, что мы знаем в результате произведенных до настоящего времени многолетних экспериментальных исследований и из литературных источников, имеющихся в нашем распоряжении, о точном расположении центров отдельных рефлексов в мозговом стволе.
В 1861 г. Флуранс показал, что после экстирпации больших полушарий у голубей и кроликов явления, наступившие после перерезки отдельных полукружных каналов, не исчезают. В 1873 г. Левенберг наблюдал, что у голубей после экстирпации большого’ мозга сохраняются движения головы в ответ на раздражения полукружных каналов, но исчезают после удаления зрительных бугров. В 1881 г. Хёгиес сообщил, что реакция поворота и последовательного поворота глаз, вращательный нистагм и постнистагм глаз у кролика после удаления больших полушарий и зрительных бугров остаются без изменений. После поперечной перерезки продолговатого мозга, приблизительно на уровне нижней границы ядер 8-го нерва, он наблюдал сохранение нистагма и лабиринтных реакций поворота глаз. В 1884 г. Люксингер (Luchsinger) нашел, что у лягушки реакция поворота головы сохраняется после удаления больших полушарий и среднего мозга, а в 1887 г. Шрадер (Schrader) показал, что реакция поворота головы у лягушки сохраняется после разреза передней части продолговатого мозга и исчезает после рассечения его позади вступления тройничного нерва.
Результаты, описанные в дальнейшем, частично основываются на многочисленных собственных наблюдениях на таламических и мезенцефали- ческих животных (24), на децеребрированных животных с мозжечком и без мозжечка (18 и 37), на животных, у которых продолговатый мозг и самая верхняя часть шейного отдела спинного мозга поперечно перерезалась на различных уровнях (18); далее, на приведенных выше второй и третьей
Положение центров установки тела в мозговом стволе
473
сериях опытов (стр. 461) де Клейна и на его же исследовании калорических реакций глаз после поперечной перерезки ствола на различных уровнях (63). Далее, они основываются на двух обширных сериях экспериментов Радемакера, который производил у кроликов и кошек поперечные разрезы ствола на различных уровнях между задним таламусом и передним отделом продолговатого мозга. Существенно, что в значительном числе этих опытов имеется анатомическое исследование на сериях срезов, с помощью чего с определенностью устанавливается, что именно было
Striatum Thalamus \'~ \
Corp. quadrig Corp, quadrig
Ag.Sylüii PPst Substgnsea\ fN/ centralis \ . ‘
s Nucl.oculomot.
Corpus _ \
callosunt's. '■
Fornix —1
s Nucltrochlearls
' , Nuci abducens
U С.П CM
Rad.77. Corpus h/em N.ru- Pons Corpus trapezoides
optici mamillare оси- ber lomotorius
Рис. 217. Кролик. Схема поперечных разрезов через ствол, после которых отдельные рефлексы еще имели место. Линия I — нормальное распределение тонуса, лабиринтные установочные рефлексы, установочные рефлексы с туловища на туловище; линия II — децеребрационная ригидность (повышение нормального распределения тонуса, лабиринтных установочных рефлексов и установочных рефлексов туловища); линия III — прогрессивные реакции, шейные установочные рефлексы; линия IV — тонические лабиринтные рефлексы на мускулатуру тела, реакция поворота головы и та же последовательная реакция — головной, поствращательный нистагм(разрез несколько
более орально).
удалено и что оставлено и имели ли место сопутствующие повреждения нервов или кровоизлияния.
Если пренебречь некоторыми частностями, которые ниже будут описаны более обстоятельно, то в качестве наиболее существенного результата можно рассматривать положение, что центры установки тела и лабиринтных рефлексов расположены в стволе ввиде трех больших функциональных групп.
1. От области вступления в мозг вестибулярного нерва кзади до верхней части шейного отдела спинного мозга: центры лабиринтных и шейных рефлексов на всю мускулатуру тела, за исключением установочных рефлексов.
2. Между вступлением 8-го нерва и ядрами глазных мышц: центры всех лабиринтных рефлексов на глаза.
3. В среднем мозгу: центры установки (только центры шейных установочных рефлексов достигают продолговатого мозга и даже заходят в передний отдел его).
Теперь следует описать детали. Для пояснения могут служить рис. 217, 218 (кролик) и 219 (кошка), представляющие сагиттальные разрезы через мозг, на которых изображены отдельные области (вступление 8-го нерва,
Рис. 218. Кролик (реакция поворота глаз). Линия I — компенсаторное вертикальное отклонение, реакция вертикального поворота глаз и та же последовательная реакция — вертикальный нистагм глаз (разрез несколько более орально); линия II — ротаторный нистагм глаз и постнистагм; линия III — компенсаторный поворот глаз; линия IV — реакция горизонтального полукружного канала с ни- с т агмом (к а дорическим).
Согр.
Согр.
Рис. 219. Кошка. Линия I — готовность к прыжку, установочные рефлексы с туловища на туловище; линия II — нормальное распределение тонуса, лабиринтные установочные рефлексы, лифтная реакция; линия III — децеребрационная ригидность (повышение нормально распределенного тонуса и лабиринтных установочных рефлексов); линия IV — шейные установочные рефлексы; линия V — тонические рефлексы на мускулатуру конечностей, реакция поворота и последовательная реакция поворота головы, односторонняя утрата тонуса конечностей после перерезки 8-го нервана одной стороне; линия VI — тонические шейные рефлексы на конечности; линия VII — повышение тонических шейных рефлексов на конечности.
Положение центров установки тела в мозговом стволе
475
начало глазодвигательного нерва, ядра глазных мышц, красное ядро, мозжечковые ножки и т. д.). Топография отдельных поперечных разрезов, таким образом, уясняется наиболее легко.
А. Децеребрационная ригидность и нормальное распределение тонуса
Согласно первоначальным положениям Шеррингтона (1), децеребрационная ригидность наступает после поперечного разреза через средний мозг между передним и задним двухолмиями в плоскости намета мозжечка. Базетту и Пенфилду удалось сохранить децеребрированных кошек живыми в течение трех недель, доказав тем самым, что состояние животных не обусловлено раздражающим действием операционного разреза, так как раздраженные пути за это время дегенерировали, но объясняется деятельностью центров, лежащих в оставшемся мозгу. То, что децеребрационная ригидность не зависит от мозжечка, было показано выше (стр. 469).
Все исследователи согласны в том, что у таламического животного децеребрационной ригидности не бывает. Я могу подтвердить это собственными наблюдениями над кроликами, кошками, собаками и обезьянами. Относительно положения центров, удаление которых вызывает появление ригидности, мнения расходятся. Большинство авторов локализуют эти центры, судя по данным Тиле, в каудальном отделе таламуса. В противоположность этому выше, в пятой главе, было показано, что у мезенце- фалического кролика, у которого разрезом выше четверохолмия полностью удалялись оба зрительных бугра, еще обнаруживалось нормальное распределение тонуса.
Точные определения границ Радемакером показали следующее.
У кошки (рис. 219) после разрезов дорсально через наиболее каудально расположенные области таламуса, вентрально через передний край сосочкового тела (линия I) никогда не наступала ригидность, но всегда имело место нормальное распределение тонуса мускулатуры тела. После разреза дорсально по переднему краю переднего двухолмия, вентрально через переднюю часть задней мозговой ножки, перед вступлением глазодвигательного нерва (линия /ƒ), в половине случаев имело место нормальное распределение тонуса и в половине — легкая ригидность. Как говорилось выше (стр. 427), когда реакция (в данном случае — нормальное распределение тонуса) имеет место, ее следует связывать с деятельностью центров, лежащих позади от разреза. Центры нормального распределения тонуса, таким образом, лежат у кошки позади линии II. Наоборот, во всех случаях после поперечного разреза дорсально кпереди от задней трети переднего двухолмия, вентрально кпереди от переднего края моста (линия III) нормальное распределение тонуса исчезало и наступала сильная децере- брационная ригидность. Взгляд на рис. 219 показывает, что у кошки децеребрационная ригидность наступает сразу после удаления центров, лежащих в мозговом стволе на уровне крупноклеточного отдела красного ядра и ядра глазодвигательного нерва.
Еще точнее определялась эта граница у кролика (рис. 217). После разреза, проходящего дорсально через середину переднего двухолмия и вентрально через середину ножки мозга позади сосочкового тела и выхода глазодвигательного нерва (линия ƒ), распределение тонуса было во всех случаях нормальным. При прохождении же разреза дорсально через заднюю часть переднего двухолмия и вентрально впереди моста и позади выхода глазодвигательного нерва (линия II) всегда наступала децеребрационная ригидность. Следовательно, децеребрационная ригидность у кро¬
476
Десятая глава. Центры установки тела
лика обусловлена удалением центров мозгового ствола, лежащих на уровне? красного ядра и середины переднего четверохолмия.
Строго говоря, с абсолютной точностью установлено лишь положение' переднего разреза (линия //, рис. 219 — у кошки; линия ƒ, рис. 217 — у кролика). Но полностью невозможно предположить, что задний разрез (линия III — у кошки, линия II — у кролика) располагается несколько более каудально, если принять во внимание, что именно эти разрезы могут вызвать шок оставшейся части ствола на уровне, непосредственно примыкающем к разрезу, вследствие чего могут оказаться выключенными и неудаленные центры. Однако такое предположение маловероятно, во-первых, потому, что децеребрационная ригидность и нормальное распределение тонуса, особенно у кошек и у кроликов, являются состояниями, на которых влияние шока сказывается незначительно; во-вторых, потому, что описанный эффект удавалось получить во всех без исключения случаях; и в-третьих, потому, что, как будет описано ниже, Раде- макеру действительно удалось анатомически установить на указанном уровне существование центра, наличие которого обусловливает нормальное распределение тонуса, отсутствие же — децеребрационную ригидность.
Таким образом, мы знаем, что центры, наличие которых обусловливает0 появление децеребрационной ригидности, следует искать в продолговатом мозгу, не включая сюда мозжечок, позади уровней, о коих речь шла до сих пор. Точное их положение еще не известно. Шеррингтон в разное время высказывал по этому поводу различные мнения. В 1910 г. он локализовал (9) эти уровни между передним двухолмием и задним краем моста.
В описанном выше опыте Беритова (стр. 469) у кошки после удаления ствола мозга до уровня позади заднего двухолмия в течение 5% часов, наблюдалась хорошо выраженная децеребрационная ригидность. В отдельных опытах на кроликах, после поперечного разреза дорсально позади заднего двухолмия, вентрально справа на 1 мм, слева на 3 мм кпереди от трапециевидного тела, хорошо выраженная децеребрационная ригидность имела место в течение 2 часов (до окончания опыта); в другом опыте, после разреза, проходящего дорсально позади заднего четверохолмия и приблизительно в 2 мм впереди слуховых бугорков и вентрально позади моста, по переднему краю трапециевидного тела, сильная ригидность наблюдалась в продолжение 1% часов (до конца опыта). В этом опыте была удалена по крайней мере передняя четверть продолговатого мозга.
Отсюда следует, что центры децеребрационной ригидности расположены в продолговатом мозгу. Более точная локализация должна быть еще установлена, и прежде всего — уровень, до которого, удалив продолговатый мозг, тем не менее возможно получить длительно продолжающуюся ригидность.
Если у кошки после предварительного удаления мозжечка продолговатый мозг перерезается рядом последовательных, следующих спереди назад фронтальных разрезов, то децеребрационная ригидность понемногу становится все менее интенсивной. Опыты подобного рода я производил до сих пор (для определения положения центров рефлексов позы) только- таким образом, что разрезы следовали один за другим через 5—10 минут,, так как при этом каждый разрез действует как раздражение, вызывая на некоторое время повышение тонуса мышц конечностей, после чего можно быстро произвести испытание на наличие тонических и лабиринтных рефлексов, так как подобные фронтальные разрезы на децеребрированных животных не вызывают ни малейшего шока. Во время этих опытов после поперечного разреза тотчас кпереди от вступления 8-го нерва ригидность была еще хорошо выражена; даже после разреза через писчее перо она была
Положение центров установки тела в мозговом стволе
477
♦еще отчетлива; разрез на 5% мм кзади вызвал появление незначительного, ла 11 мм — весьма незначительного разгибательного тонуса конечностей. Так как у животного (декапитированного по Шеррингтону), у которого разрез проходит примерно через писчее перо, спустя короткое время ригидность не наступает, центры последней позволительно локализовать только в области продолговатого мозга, не определяя их расположения более точно. Все гипотезы относительно того, о каких ядрах при этом идет цело, будучи рассмотрены со всех сторон, представляются преждевременными.
Б. Центры рефлексов положения и лабиринтных рефлексов на мускулатуру тела
1. Рефлексы положения
Протокол опыта в этом направлении с изображениями препарата и обстоятельным исследованием на серийных срезах приведен выше, на стр. 435 {препарат I, опыт 22). После поперечного разреза продолговатого мозга (тотчас кпереди от вступления 8-х нервов), оставившего ядро Дейтерса справа полностью интактным и отделившего самую верхнюю его часть •слева, можно было, изменяя положение головы животного, получить еще живые шейные и лабиринтные рефлексы на передние конечности.
Следующий пример иллюстрирует сказанное.
Опыт 14. 6 февраля 1914 г. Кошка, 1300 г. Эфирный наркоз. Перевязка сонных артерий. Перерезка блуждающих нервов. Обнажение спинного мозга на уровне 12-го грудного позвонка. Децеребрация при временном пережатии позвоночных артерий, тотальное удаление мозжечка и боковых отделов одним куском. Длительность всей операции — 15 минут. Экстирпация мозжечка — 5 минут. Непосредственно после этого в 10 ч. 20 мин. началась децеребрационная ригидность.
10 ч. 30 мин. Сильная ригидность всех четырех конечностей. Испытание реакций на изменение положения головы показывает, что на передних и задних ногах преобладают шейные рефлексы, но что при этом лабиринтные рефлексы проявляются также отчетливо.
11. ч. 15 мин. Производится фронтальный разрез позади четверохолмия. После этого шейные и лабиринтные рефлексы отчетливы.
И ч. 25 мин. Производится фронтальный разрез оральнее вступления в мозг «8-х нервов, проходящий справа на 2 мм, слева на 1 мм впереди вступления 8-го нерва через слуховые бугорки. Оба 8-х нерва интактны. Исследование тонических рефлексов положения показало следующее.
Боковое положение, поворот головы: повышение тонуса в «челюстной ноге», понижение в «черепной ноге». Нижележащая нога реагирует несколько слабее (преобладают шейные рефлексы; менее выражены лабиринтные рефлексы).
Положение на спине, поднимание и опускание: сильнейший разгибательный тонус передних ног при поднимании морды на 45° выше горизонтали. Тонус отчетливо падает, когда голова пригибается дорсально, пока морда не установится обращенной вниз; тонус максимален, когда голова сильно пригнута вентрально, так что морда оказывается между передними ногами (лабиринтные рефлексы).
Положение на спине, поворачивание головы: понижение тонуса в обеих передних ногах, в «черепной ноге» более сильное (лабиринтные и шейные рефлексы).
Висячее положение головой вниз, поднимание и опускание головы: тонус передних ног самый низкий, когда морда свешивается вертикально вниз; он повышается, когда вследствие вентрального или дорсального нагибания ротовая щель устанавливается выше горизонтали. Более сильное повышение тонуса наступает при вентральном нагибании головы (лабиринтные рефлексы).
11 ч. 35 мин. Фронтальный разрез позади 8-го нерва, на 2% мм кпереди от писчего пера. После этого все лабиринтные рефлексы исчезают. Ни при поворачивании головы в боковом положении животного, ни при поднимании, опускании или поворачивании головы в положении на спине не удается обнаружить и следов лабиринтных рефлексов. В ответ на поворачивание головы в боковом положении животного появляются «чистые» шейные рефлексы (увеличение тонуса в «челюстной ноге» и уменьшение в «черепной»). В положении на спине при помощи поднимания и опускания головы
478
Десятая глава. Центры установки тела
удается получить также одни шейные рефлексы (повышение разгибательного тонуса передних ног при дорсальном нагибании головы, понижение — при вентральном). При поворачивании головы в положении животного на спине появляются также только шейные рефлексы (повышение тонуса в «челюстной ноге» и понижение в «черепной»). По этой причине при дальнейшем продолжении опыта исследование рефлексов головы производилось только в боковом положении животного.
И ч. 45 мин. Фронтальный разрез через писчее перо. После этого еще имеется отчетливая ригидность передних ног. В боковом положении в ответ на поворот и наклон головы выступают еще очень отчетливые шейные рефлексы передних ног (повышение тонуса на «челюстной ноге» и понижение на «черепной»). Vertebra-prominens-рефлекс (понижение тонуса передних ног при вентральном смещении самого нижнего шейного позвонка) весьма живой.
И ч. 50 мин. Фронтальный разрез на 5% мм позади писчего пера, тотчас позади выхода первого шейного корешка. После этого ригидность передних ног незначительна. В ответ на поворот головы в боковом положении животного, однако, наступают еще отчетливые, хотя и не слишком сильные шейные рефлексы (повышение тонуса в «челюстной ноге»; понижение в «черепной»). Равным образом, при наклоне головы появляются еще отчетливые шейные рефлексы.
11 ч. 55 мин. Фронтальный разрез на 5% мм кзади от начала второго шейного корешка. После этого экстензорная ригидность весьма незначительна. При повороте и наклоне головы в боковом положении животного — слабые, однако же совершенно отчетливые и несомненно шейные рефлексы.
12 ч. 02 мин. Фронтальный разрез еще более кзади, У2 мм оральнее начала второго шейного корешка. После этого ригидность передних ног весьма мала. При поворачивании и наклоне головы никаких рефлексов на передние ноги более не вызывается.
Результат. У децеребрированной кошки мозжечок удаляется полностью и мозговой ствол перерезается тотчас спереди вступающего в мозг 8-го нерва. После этого могут быть вызваны тонические шейные и лабиринтные рефлексы. После перерезки позади 8-х нервов лабиринтные рефлексы исчезают. Шейные же рефлексы на передние конечности сохраняются очень хорошо. Специально это было показано для реакций на поворот, наклон, поднимание и опускание головы и для vertebra-prominens- рефлекса. После поперечного сечения мозга через писчее перо шейные рефлексы еще весьма отчетливы; после перерезки шейного отдела спинного мозга позади выхода первых шейных нервов реакция передних ног на поворачивание и наклон головы ослаблена, но проявляется еще отчетливо и исчезает только после разреза, проходящего в 34 мм впереди места выхода С2.
а) Тонические шейные рефлексы на конечности
Афферентные пути, если отвлечься от vertebra-prominens-рефлекса, проходят у кошки в составе трех, у кролика — в составе четырех пар верхних шейных корешков. Эфферентные пути идут к центрам мускулатуры конечностей обеих сторон в шейном и поясничном утолщениях спинного мозга.
Относительно расположения центров этих рефлексов в семи опытах Hä кошках установлено следующее.
Во всех опытах шейный отдел спинного мозга можно было перерезать вплоть до места выхода Сг, не вызвав нарушения шейных рефлексов на поворачивание головы. При перерезке мозга между С2 и С2 в одном опыте реакция на передние конечности исчезла; в трех опытах она оставалась еще отчетливой, хотя и ослабленной. Перерезка на уровне С2 или ниже С2 приводила к повышению шейных рефлексов на поворот и наклон головы. Лишь в одном опыте оказалось нелегким установить с абсолютной уверенностью, имела ли еще место минимальная реакция после поперечного разреза точно позади С2, которая в этом случае исчезла только после проведения разреза позади С3. Судя по моим собственным данным, в боль¬
Положение центров установки тела в мозговом стволе
479
шей части случаев поперечного разреза позади С2 достаточно для того, чтобы исчезли шейные рефлексы на поворачивание и наклон головы.
Следовательно, центры тонических шейных рефлексов на мускулатуру конечностей у кошек расположены в двух верхних шейных сегментах спинного мозга.
б) Тонические лабиринтные рефлексы на конечности
Эти рефлексы начинаются с maculae utriculi. Афферентные пути проходят через ramus utricularis к продолговатому мозгу. Каждая macula utriculi находится в функциональной связи с центрами мускулатуры конечностей обеих сторон тела, расположенными в шейном и поясничном утолщениях спинного мозга.
Тонические лабиринтные рефлексы на конечности сохраняются после поперечной перерезки тотчас возле вступления в продолговатый мозг 8-го нерва. В различных опытах разрезы отстояли от места вступления 8-го нерва на 1—2 мм и частично проходили через слуховой бугорок. В опыте 22, тщательно обработанном анатомически, ядро Бехтерева было удалено с обеих сторон, ядро Дейтерса справа было интактно, слева часть его верхушки была отсечена. Начало и остаток нисходящего пучка Дейтерса были на обеих сторонах интактны, так же как .и вентральное ядро 8-го нерва и слуховой бугорок. Слева оказался отсеченным кусочек латерального отдела дорсального ядра 8-го нерва. Мозжечок и его ядра отсутствуют.
В опыте 23 на децеребрированной кошке без перерезки продолговатого мозга во время экстирпации мозжечка с левой стороны оказались полностью удаленными ядра кохлеарного нерва, вентральное ядро 8-го нерва и слуховой бугорок, только некоторые проксимальные вестибулярные волокна, возможно, остались неповрежденными. На правой стороне оказались неповрежденными оба корешка 8-го нерва и его ядра. Пути, возникающие в ядрах Дейтерса, с обеих сторон интактны. У этого животного были высокие тонические лабиринтные рефлексы на конечности обеих половин тела.
В опыте 17 (16 февраля 1914 г.) поперечный разрез прошел слева в 2 мм, справа — в 1 мм кпереди от места вступления в мозг 8-го нерва; кроме того, оказался перерезанным левый 8-й нерв. У животных имелись отчетливые тонические лабиринтные рефлексы на обе передние ноги.
Отсюда следует, что центры тонических лабиринтных рефлексов на мускулатуру конечностей лежат ниже поперечного разреза, проведенного через продолговатый мозг в области вступления в него 8-го нерва (рис. 219, линия F), и что область вступления 8-го нерва вместе с вентральным ядром, 8-ми нервами и слуховым бугорком одной стороны могут быть полностью удалены без нарушения перехода возбуждений с 8-го нерва одной стороны на обе половины тела. Ядро Бехтерева для названных рефлексов не является необходимым.
Результаты, полученные на кошках, подтверждаются данными опытов на кроликах (рис. 217, линия IV). Третья серия опытов (стр. 461) показывает, что после поперечного разреза тотчас выше вступления 8-го нерва, тонические лабиринтные рефлексы на конечности сохраняются. На тщательно изученном анатомически препарате IV ядра Дейтерса и Бехтерева были с обеих сторон интактны, задние продольные пучки — частично разрушены кровоизлияниями на обеих сторонах. У кролика рефлексы на обе половины тела также оказались сохраненными после того, как один из стволов 8-го нерва был перерезан тотчас выше вступления его в мозг.
480
Десятая глава. Центры установки тела
в) Тонические лабиринтные рефлексы на шейные мышцы
Эти рефлексы также исходят из maculae utriculi и проходят в составе rami utriculi в продолговатый мозг. Каждая macula связана функционально с мышцами шеи только одной половины тела (в физиологическом смысле, как описано на стр. 383). У кроликов и кошек афферентные связи, проходя через шейный отдел спинного мозга, достигают центров мускулатуры тела.
Расположение центров этих рефлексов характеризуется тем же уровнем, что и центров тонических лабиринтных рефлексов на мышцы конечностей. У кошки (опыт 17) рефлексы на мускулатуру туловища сохраняются после поперечной перерезки мозга на 1—2 мм выше вступления 8-го нерва; у кролика то же было обнаружено после дорсальной перерезки тотчас выше средних мозжечковых ножек и вентрально — позади моста.
Поворот (основной поворот) и наклон головы, обусловленные действием этих рефлексов и наступающие у кошки после односторонней перерезки 8-го нерва, имеют место и после такой же перерезки (опыт 17) выше 8-го нерва; у кролика то же имеет место после поперечной перерезки тотчас выше вступления в мозг этого нерва (см. выше, табл. III, стр. 463).
Отсюда следует, таким образом, что и эти центры лежат в плоскости, расположенной выше вступления в мозг 8-го нерва (рис. 217, линия IV; рис. 219, линия V).
г) Односторонняя утрата тонуса после односторонней перерезки
8-го нерва
Это до настоящего времени не объяснявшееся выпадением одного определенного лабиринта явление, которое у различных животных проявляется с разной интенсивностью и длительностью, обнаруживается у кошки еще после поперечной перерезки на 1—2 мм кпереди от места вступления в мозг 8-го нерва (опыт 17, рис. 219, линия F).
2. Вращательные реакции и поствращательные
реакции головы
Эти рефлексы начинаются в гребешках полукружных каналов, и путь их проходит через ампуллярные ветви к продолговатому мозгу. Эфферентные ветви отсюда идут к центрам шейной мускулатуры, а у человека и обезьян, кроме того, еще к мускулатуре туловища.
По данным третьей серии опытов (см. выше, стр. 461), центры этих рефлексов у кролика лежат каудальнее поперечного разреза, проходящего впереди вступления 8-го нерва в продолговатый мозг. Реакция поворота головы после этой операции также сохраняется. Задний продольный пучок может быть частично разрушен без того, чтобы исчезли эти рефлёксы.
Эти данные соответствуют более ранним результатам, согласно которым у кролика после фронтального разреза вблизи средних мозжечковых ножек, позади моста и впереди трапециевидного тела, реакции поворота сохраняются. При повороте в нормальном положении появлялся наклон головы в сторону, при повороте, когда морда удерживается поднятой вертикально, наступал, в зависимости от направления вращения, наклон головы вправо или влево или же дорсальное или вентральное нагибание головы.
Нистагм головы после операций на центральной нервной системе, при наркозе, отравлениях и т. д. устранить много легче, чем реакции ново¬
Положение центров установки тела в мозговом стволе
481
рота. Поэтому со всеми заключениями относительно локализации нистагма следует быть особенно осторожным, пока дело идет об экспериментах, в которых нистагм отсутствовал. У кролика можно было показать вращательный нистагм головы после разреза выше мозжечковых ножек, позади заднего двухолмия и впереди моста. Особенно отчетливый вращательный нистагм головы обнаружился у только что упомянутого животного, у которого разрез проходил впереди средних мозжечковых ножек, позади моста и трапециевидного тела. Мозжечок может отсутствовать. После этого становится вероятным, что для нистагма головы не следует предполагать существование специальных центров, но что и в этом случае оказывается достаточно аппаратов, лежащих позади плоскости вступления 8-х нервов.
Данными, изложенными выше, опровергаются утверждения Ротфельда (7), согласно которым центры реакции поворота головы у кролика лежат в промежуточном мозге, а центры нистагма — в лобном отделе (!). Точно так же опровергается и дальнейшее сообщение того же автора (8) о том, что Центры поствращательной реакции головы лежат в среднем мозгу, а центры ноствращательного нистагма головы — в таламусе. Ротфельд допускал даже раздельное существование центров реакции поворота и последовательного поворота головы. Для этого не существует никаких оснований. Из приведенного примера отчетливо видно, к каким неправильным выводам можно прийти, если основывать свои заключения на опытах, в которых определенный рефлекс*отсутст- вовал.
У кошки шок выражен сильнее. В этом случае вращательный нистагм головы можно показать только после разреза поверх заднего двухолмия и впереди моста.
Таким образом, здесь мы отмечаем многократно подтверждавшееся положение, что в случае, когда дело идет о рефлексах, чувствительных к шоку, уровень разреза у кошки локализуется значительно более орально, чем у кролика, в то время как в случае рефлексов, резистентных к шоку, наоборот, уровни у обоих видов животных лежат почти на одной и той же высоте. В последнем случае сказанное можно использовать в качестве довода в пользу правильности полученного уровня, так как в силу того, что у кошки разрез оказывается проходящим более орально, чем у кролика, возможен шок, и истинное положение центров следует принимать как более каудальное, чем это следует из расположения экспериментально осуществленного разреза.
3. Реакции на прогрессивные движения
Последнее замечание особенно существенно также для прогрессивных реакций. Последние у всех видов животных весьма чувствительны к послеоперационному шоку, к наркозу, отравлению и т. д.
Судя по данным опытов с центрифугированием, эти реакции начинаются в гребешках полукружных каналов и, возможно, кроме того, в отолитовых аппаратах.
В реакциях принимают участие также мышцы конечностей и шеи.
В согласии с данными относительно положения центров лабиринтных рефлексов на мышцы тела заранее представляется вероятным, что центры прогрессивных реакций также лежат позади поперечного разреза, проведенного выше вступления в мозг 8-го нерва.
В действительности, однако, обнаруживаются уровни, расположенные значительно более орально.
Радемакер видел у кролика положительную лифтреакцию и готовность к прыжку после фронтального разреза (рис. 217, линия ƒƒƒ), дорсально проходящего позади заднего двухолмия, через средние мозжечковые ножки
31 Р. Магнус
482
Десятая глава. Центры установки тела
(ручки моста) и в середине через мост, причем задний полюс ядра блокового нерва сохранился.
Наоборот, у кошки лифтреакция имела место только после разреза (рис. 219, линия 77), проходящего от передней стенки переднего двухолмия к передней четверти ножки мозга, а готовность к прыжку — даже только после разреза дорсально через самую заднюю часть таламуса, вентрально через переднюю стенку сосочкового тела (рис. 219, линия I).
Так как у кошки были обнаружены уровни, расположенные значительно более орально, следует быть чрезвычайно сдержанным с выводами относительно положения центров. По крайней мере вероятно, что и у кролика они лежат более каудально, чем это следует из экспериментов, и представляется все-таки возможным, что они лежат в задней половине продолговатого мозга вместе с центрами лабиринтных рефлексов на мышцы тела.
В. Центры рефлексов на глаза
Все данные, которые будут описаны ниже, получены на кроликах, у которых глазные реакции проявляются наиболее отчетливо, а соответствующие обстоятельства могут поэтому быть изучены наиболее основательно.
1. Компенсаторные установки глаз а) Тонические лабиринтные рефлексы на глаза
а) Вертикальные отклонения. Эти отклонения у кролика осуществляются с основного отдела macula sacculus. Афферентный путь проходит через ramus saccularis. Каждая macula sacculus находится в связи с обоими ядрами глазодвигательного нерва, а именно с центром прямой верхней мышцы своей стороны и нижней прямой противоположной стороны. Таким образом, уже заранее можно с уверенностью сказать, что для осуществления этих рефлексов необходима сохранность области,, расположенной между вступлением 8-го и ядрами глазодвигательного нервов.
Радемакер обнаружил, что компенсаторные вертикальные отклонения глаз у кролика сохраняются после фронтального разреза, который проходит дорсально через середину переднего двухолмия, вентрально через середину ножки мозга, тотчас впереди вступления глазодвигательного нерва (рис. 218, линия I). Разрез отделил верхушку ядра глазодвигательного нерва. Обширное повреждение четверохолмия осталось без последствий. Вертикальное отклонение глаз после односторонней перерезки 8-го нерва у таламических животных выступает весьма отчетливо. Следует ожидать, что они имеют место и у мезенцефалических животных.
ß) Круговые повороты. Места возникновения этого рефлекса в отолитовомаппарате еще неизвестны. Каждый лабиринту кролика находится в двусторонней связи с нижней косой мышцей в ядре глазодвигательного нерва и с верхней косой мышцей в ядре блокового нерва. Компенсаторные круговые повороты глаза можно еще показать после перерезки нижней косой мышцы, т. е. повороты эти обеспечиваются деятельностью одной только верхней косой мышцы, иннервируемой из ядер блокового Нерва.
В согласии с этими данными находится тот факт, что после разреза, который, начавшись дорсально между передним и задним двухолмиями (рис. 218, линия III), прошел на одной стороне через границу между ядрами глазодвигательного и блокового нервов, а на другой стороне отде-
Положение центров установки тела в мозговом стволе
483
лил большую часть ядра блокового нерва и вышел вентрально через середину моста, на глазу, иннервируемом сохранившимся ядром блокового нерва противоположной стороны, имеются отчетливые компенсаторные круговые повороты (Радемакер). Конечно, разрез должен быть проведен таким образом, чтобы перекрест блоковых нервов остался неповрежденным.
Наблюдения над вертикальными и ротаторными компенсаторными отклонениями глаз показывают, что для их осуществления не требуется ничего, кроме сохранности уровня, на котором лежит ядро данной глазной мышцы. Шок, наступающий после операционного разреза, в случае этих рефлексов минимален. Описанный последним опыт на ядре блокового нерва показывает далее, что для ротаторных компенсаторных установок одного глаза достаточно участия одного глаза и что реакция осуществляется еще и тогда, когда ядро антагонистов (в данном случае — нижней косой мышцы) отсечено. Следовательно, центральный аппарат построен весьма просто.
б) Тонические шейные рефлексы на глаза
Афферентные пути проходят у кролика в составе двух самых верхних и, кроме того, в незначительном количестве в составе третьей пары задних шейных корешков. Пути достигают всех ядер глазных мышц — от ядра отводящего нерва до ядра глазодвигательного нерва (нижняя косая мышца). Таким образом, в то время как из лабиринтов следует импульсация к четырем ядрам с каждой стороны, с рецепторов шеи при компенсаторных установках глаз приводятся в деятельное состояние все шесть ядер глазных мышц.
Тонические шейные рефлексы у таламических животных, по наблюдениям де Клейна, сохраняются. Дальнейшие опыты с более каудально произведенными разрезами до сих пор еще только предстоит осуществить.
2. Реакция с полукружных каналов на глаза
(Реакции ^поворота, постреакции поворота, вращательный нистагм и постнистагм, калорическая реакция и калорический нистагм)
Эти рефлексы возникают в гребешках полукружных каналов. Центры лежат между вступлением 8-го нерва (Хёгиес перерезал продолговатый мозг в каудальной части) и ядрами соответствующих глазных мышц. В противоположность отолитовым реакциям, с полукружных каналов могут приводиться в деятельное состояние все шесть пар глазных мышц.
То, что вращательная реакция и постреакция глаз, вращательный нистагм и постнистагм остаются неизменными после удаления большого мозга и таламусов, установил уже Хёгиес. Бауэр и Лейдлер так же, как Барани, РейхиРотфельд, подтвердили это наблюдение. Кубо (Kubo) показал, что калорический нистагм после удаления больших полушарий сохраняется. Действительно, у мезенцефалического кролика вертикальные, ротаторные и горизонтальные поворотные реакции и последовательные реакции, нистагмы и постнистагмы оказываются сохраненными. Более точные локализации устанавливались Радемакером.
Вертикальная реакция и постреакция поворота глазу кролика сохранилась 1 (рис. 218, линия I) после фрон¬
1 Так как в этих экспериментах животные поворачивались таким образом, что морда была направлена вертикально вверх или вниз, должны быть поставлены еще контрольные опыты, для того чтобы исключить предположение, что дело не идет о реакциях наружной мышцы вследствие компенсаторных .ротаций.
31*
48.4
Десятая .глава. Центры установки тела
тального разреза, проходящего дорсально через середину переднего двухолмия, вентрально через середину ножки мозга перед выходом глазодвигательного нерва и отсекающего верхушку ядра этого нерва.
Вертикальны йнистагм глаз еще обнаруживался после разреза, который проходил дорсально через передний край переднего двухолмия, вентрально через самую заднюю часть сосочкового тела, т. е. несколько кпереди от верхушки ядра глазодвигательного нерва (рис. 218, несколько оральнее линии /).
Реакция ротаторного поворота глаз имелась у того же животного (см. выше, стр. 483), у которого имела место также ротаторная компенсирующая установка на одном глазу. Разрез (рис. 218, линия III) прошел на одной стороне точно между ядрами глазодвигательного и бокового нервов и на другой стороне отделил большую часть ядра блокового нерва.
Ротаторный нистагм и постнистагм глаз имели место после разреза (рис. 218, линия //), который прошел дорсально через заднюю часть переднего двухолмия, вентрально точно через место выхода глазодвигательного нерва, перерезая ядро последнего.
Все названные до сих пор реакции сохраняются после основательных повреждений четверохолмия.
То, что после удаления ядер глазодвигательного и блокового нервов могут вследствие возбуждения одного ядра отводящего нерва осуществляться реакции с полукружных каналов с нистагмом, доказывают опыты де Клейна с калорическим раздражением.
В этих экспериментах экстирпировалось глазное яблоко на одной стороне, наружная прямая мышцы изолировалась и соединялась с записывающим рычажком. После этого вскрывалась крыша черепа, большой мозг удалялся, а ствол мозга перерезался точно позади заднего двухолмия, причем отводящий нерв, принадлежащий изолированной наружной мышце глаза, заботливо щадился. Далее попеременно в оба слуховых прохода вводилась холодная вода, для того чтобы вызвать появление нистагма в обоих направлениях.
Кролик F. Изоляция левой наружной мышцы, удаление большого мозга путем перерезки ствола позади четверохолмия. Орошение левого слухового прохода холодной водой вызвало очень отчетливый нистагм (медленные сокращения и быстрые расслабления). Введение воды в правый слуховой проход вызвало незначительное сокращение мышцы без нистагма (парадоксальная реакция: вместо первичного расслабления в этом случае наступило сокращение).
Анатомическое исследование про ф. Винклера. Поперечный разрез ствола проходит позади заднего четверохолмия; ядра обоих блоковых нервов и обоих глазодвигательных оказались полностью удаленными.
Кролик G. Изоляция левой наружной косой мышцы, удаление большого мозга; поперечный разрез через ствол в самом заднем отделе четверохолмия. Введение холодной воды в левый слуховой проход вызвало отчетливый нистагм (медленные сокращения и быстрые расслабления) (рис. 220а).
Введение воды в правый слуховой проход также привело к нистагму (медленные расслабления и быстрые сокращения) (рис. 2206).
Анатомическое исследование проф. Винклера. Разрез через ствол проходит через самый задний отдел четверохолмия; ядра блокового и отводящего нервов на обеих сторонах удалены.
Де Клейн показал, далее, что у мезенцефалического животного после калорического раздражения правого и левого уха на изолированной наружной прямой мышце глаза наблюдается как девиация, так и нистагм в том случае, когда нервы всех остальных глазных мышц, за исключением соответствующего отводящего нерва, а также тройничные нервы на обеих сторонах перерезаны. Точно так же парализация новокаином проприоцеп¬
Положение центров установки тела в мозговом стволе
485
тивных нервных окончаний в изолированной наружной мышце не тормозит калорического нистагма (быстрой и медленной фазы). Таким образом, дело идет о чисто центральном явлении, для которого в качестве эфферентного нерва достаточно нерва одной глазной мышцы, а в качестве ядра (ср. также опыт Радемакера на ядре блокового нерва) — ядра глазодвигательного нерва одной стороны.
В соответствии с этим оказывается, что и для реакций с полукружных каналов на глазные мышцы имеется относительно просто организованный центральный аппарат и что ядра отдельных глазных мышц можно возбуждать с лабиринта только до тех пор, пока они остаются в связи с областью вступления 8-го нерва. В отношении ядер глазодвигательного и блокового нервов можно показать, что в простых реакциях поворота аппараты, располагающиеся орально от этих ядер, не участвуют.
То же с большой долей вероятности следует принять и в отношении быстрой фазы нистагма.
Если вспомнить, что по общепринятым представлениям нистагм является более чувствительным к действию наркоза и операционного шока, чем реакция поворота, достаточно неожиданным представляется то, что ротаторный нистагм после поперечного разреза через ядро глазодвигательного нерва и горизонтальный нистагм даже через короткий промежуток времени после перерезки ядер глазодвигательного и блокового нервов могут еще сохраняться. Этим уже далеко не в первый раз отвергается не только старое, установленное Бартельсом (1) и Розенфельдом и снова повторенное Омом мнение, будто для осуществления быстрой фазы нистагма необходимо наличие больших полушарий, но и лишается правдоподобности представляемое Барани учение о «надъядерном центре взгляда», который должен реализовать быструю фазу нистагма. Если, как будет показано здесь, быстрая фаза нистагма приурочена к деятельности каждого из трех ядер глазных мышц (по меньшей мере к деятельности окружения каждого из них), тогда экспериментальные и клинические данные в отношении того, что при заболеваниях, операционном шоке, наркозе и т. п. реакция поворота и калорическая реакция глаз сохраняется, в то время как быстрый нистагм исчезает, легче объяснить функциональным повреждением соответствующих центров, чем существованием названных «надъядерных центров взгляда», которые, согласно приведенным здесь экспериментальным данным, должны быть в определенной степени внедренными в зону ядра каждой мышцы, для чего отсутствуют какие бы то ни было анатомические основания.
Рис. 220а. Рис. 2206.
486
Десятая глава. Центры установки тела
Описанные результаты существенно упрощают во всяком случае предстоящее точное анатомо-физиологическое изучение лабиринтных глазных рефлексов.
Г. Установочные рефлексы
1. Лабиринтные установочные рефлексы
Те лабиринтные рефлексы, которые возвращают голову из асимметричных положений в симметричное, с обеих сторон исходят из основного отдела sacculus’a и передаются в центральную нервную систему через ramus saccularis.
В случае лабиринтных установочных рефлексов, вероятно, участвуют возбуждения, исходящие из maculae utriculi, достигающие ствола через rami utriculares и ведущие к тому, что голова из всевозможных симметричных положений принимает именно нормальное положение с горизонтально стоящими отолитами utriculi.
Так как лабиринтные установочные рефлексы (см. пятую главу) имеются у мезенцефалического животного и отсутствуют у децеребрирован- ного, центры их должны лежать в среднем мозгу. Афферентные пути проходят через ствол, а не через мозжечок. Эфферентные пути идут из среднего мозга через ствол в шейный отдел спинного мозга, к центрам мышц шеи, которые поднимают, поворачивают и наклоняют голову.
Более точное расположение центров лабиринтных установочных рефлексов определяется данными многочисленных опытов Радемакера.
У кролика (рис. 217) установочные рефлексы сохраняются после разреза, который дорсально проходит через середину переднего двухолмия, вентрально — через середину ножки мозга, спереди от. выхода глазодвигательных нервов и через самую вершину мелкоклеточного отдела красного ядра (линия 7). В собственном опыте (55а) после разреза, который на дорсальной стороне справа прошел через середину, слева — через заднюю треть переднего двухолмия, а на вентральной стороне прошел по самому заднему краю сосочкового тела, еще имелся лабиринтный установочный рефлекс на голову в боковом положении животного (однако не на спине и не в обоих висячих положениях). Этот разрез также должен был пройти через верхушку красного ядра.
Наоборот, лабиринтные установочные рефлексы отсутствуют после разрезов (линия 77), которые проходили дорсально через заднюю часть переднего двухолмия, вентрально впереди переднего края моста и позади выхода глазодвигательного нерва. Эти разрезы идут точно по заднему краю красного ядра и через ядро глазодвигательного нерва.
Центры лабиринтных установочных рефлексов, таким образом, лежат у кролика в среднем мозгу на уровне красного ядра. Передний разрез (установочные рефлексы сохранены) абсолютно доказателен, задний разрез (установочные рефлексы отсутствуют) доказателен лишь с учетом ограничений, которые обсуждались на стр. 428. Ниже будет показано, что центры лабиринтных установочных рефлексов действительно лежат на этом уровне.
У кошки (рис. 219) лабиринтные рефлексы в 50% опытов имели место после разреза (линия 77), который дорсально проходил по переднему краю переднего двухолмия, вентрально оставил интактной % ножки мозга, располагался впереди ядра глазодвигательного нерва и крупноклеточного отдела красного ядра, отделив часть и мелкоклеточного его отдела.
Наоборот, лабиринтные рефлексы постоянно отсутствуют после разреза (линия III), проходящего дорсально через заднюю треть переднего дву¬
Положение центров установки тела в мозговом стволе
487
холмия, вентрально впереди переднего края моста и позади выхода глазодвигательного нерва; этот разрез проходит через задний край красного ядра.
Таким образом, у кошки центры лабиринтных установочных рефлексов лежат в среднем мозгу, на уровне ядра глазодвигательного нерва и крупноклеточного отдела красного ядра.
Если сравнить эти данные относительно положения центров лабиринтных установочных рефлексов у кошек и у кроликов с данными, приведенными выше для центров, с удалением которых связано наступление деце- ребрационной ригидности и появление нормального распределения тонуса мускулатуры тела, то оказывается, что уровни этих функций совпадают. Позже будет показано, что в действительности ответственным за обе функции следует считать одно ядро.
2. Установочные рефлексы тела на голову
Афферентные возбуждения для этих рефлексов возникают благодаря •соприкосновению тела с плоскостью опоры. Это было показано для боковой поверхности туловища, конечностей (прежде всего подошв и ладоней) и поверхности головы. Пути эти поднимаются в спинном мозгу, проходят (по крайней мере частично) не через мозжечок, но непосредственно через ствол в шейном отделе спинного мозга вперед к центрам, откуда через шейную часть ствола проходят к ядрам нервов шейных мышц.
Ко времени написания нижеследующей части работы опыты Радемакера, направленные к точному определению локализации центров этих рефлексов, еще не были закончены. Эксперименты проводятся в течение длительного времени, так как должны выполняться на лишенных лабиринтов животных, у которых лабиринтные рефлексы отсутствуют.
До сих пор твердо установлено, что установочные рефлексы тела на голову у таламических животных, лишенных лабиринта, имеются и являются весьма живыми. Они отсутствуют после всех поперечных разрезов, которые снимают и лабиринтные рефлексы (рис. 217, линия II — у кроликов; рис. 219, линия III — у кошек). В соответствии, с этим их центры следовало бы искать на уровне центров лабиринтных установочных рефлексов или оральнее в таламусе.
Что центры лежат приблизительно на том же уровне, что и центры лабиринтных рефлексов, становится вероятным благодаря следующемлг опыту. У мезенцефалического кролика (спустя короткое время после удаления таламуса, когда лабиринтные рефлексы еще не успели восстановиться), уложенного на столе в боковом положении, щипок нижележащей передней ноги приводит к устанавливанию головы в правильное положение (три опыта). В одном же случае для этого оказалось достаточным даже щипка вышележащей передней ноги.
Отсюда следовало бы заключить, что искомые центры лежат в среднем мозгу впереди линии II (кролик) и линии III (кошка). Однако для окончательного установления уровня следует дождаться результатов экспериментов на животных, лишенных лабиринтов, у которых прошли явления операционного шока и у которых после этого можно исследовать установочные рефлексы тела на голову.
3. Установочные рефлексы тела на тело
Местом возникновения этих рефлексов являются чувствительные аппараты туловища и конечностей. После наблюдений на двух собаках с почти полностью удаленным мозжечком (см. выше, стр. 455) становится вероят¬
488
Десятая глава. Центры установки тела
ным, что пути идут через мозжечок. Предположительно говоря, в данном случае дело идет об афферентных путях. Эфферентные пути идут через ствол мозга в спинной мозг к центрам конечностей и туловища.
По данным опытов Радемакера, установочные рефлексы тела на тело у кролика (рис. 217) после разреза (линия /), проходящего дорсально по границе передней и средней трети переднего двухолмия, вентрально по заднему полюсу сосочкового тела, сохраняются. Они исчезают после разрезов, соответствующих линии II.
У кошки (рис. 219) они сохраняются после разреза (линия I) дорсально по заднему краю таламуса, вентрально через хиазму.
Таким образом, центры установочных рефлексов тела на тело следует искать на уровне красного ядра.
4. Шейные установочные рефлексы
Эти рефлексы берут свое начало в проприоцепторах мышц или суставов шеи, а так как дело идет здесь о цепных рефлексах, то и в проприоцепторах туловища, особенно' поясничной области. Пути идут через спинной мозг, наверх, непосредственно в ствол. Они не проходят через мозжечок. Эфферентные пути проходят в спинном мозгу, особенно к центрам осевой мускулатуры, от шеи до таза.
Центры этих рефлексов лежат более каудально, чем остальные центры установки.
У кошек Радемакер наблюдал усаживание передней части тела из бокового положения в случае, когда голова пассивно поворачивалась в нормальное положение, и, кроме того, поворот таза в положение на спине, когда поворачивается голова, после разреза, который проходил дорсально позади заднего двухолмия, вентрально через середину моста (рис. 217, линия III). Я сам после разреза позади заднего двухолмия и перед трапециевидным телом наблюдал исчезновение шейных рефлексов.
У кошки поворот таза в положении животного на спине, когда голова поворачивается пассивно, можно еще показать после разреза, который проходит дорсально через границу между передним и задним двухолмиями, вентрально через переднюю часть моста (рис. 219, линия IV). Усаживания тела из бокового положения после разрезов в этой области у кошек наблюдать нельзя, так как разгибание передних ног вследствие сильной децереб- рационной ригидности чисто механически делает эту реакцию невозможной.
Центры шейных установочных рефлексов лежат, таким образом, в области моста. Функционально они принадлежат с остальными центрами переднему отделу среднего мозга. От центров рецепторов шеи, вызывающих рефлексы положения и тонические шейные рефлексы, они отделены также и анатомически.
Дополнительно здесь следует указать еще и на то, что мезенцефаличе- ское животное, у которого поперечный разрез проходит через ствол по переднему краю четверохолмия и позади сосочкового тела, полностью лишено как базальных ганглиев, так и striatum’a и pallidum’a. У такого животного не бывает контрактур, ригидности, треморов, хореи или атетоза. Распределение тонуса у него нормально, посадка нормальна. Оно нормально бежит и прыгает в ответ на нормальное раздражение. Какие бы то ни было симптомы, которые в свое время объяснялись многими клиницистами как следствие выпадения функции базальных ганглиев, полностью отсутствуют. Здесь не место обсуждать вопрос, на чем основано это кажущееся противоречие между наблюдениями над оперированными живот¬
Положение центров установки тела в мозговом стволе
489
ными с полностью удаленными названными отделами мозга и больным человеком с более или менее сохранным мозгом, но у которого отсутствуют базальные ганглии и, в известных условиях, еще и другие центры'ствола и других отделов центральной нервной системы.
Д. Заключение
Результатом проделанных до сих пор исследований является положение, что в стволе мозга, начиная от шейного отдела спинного мозга до среднего мозга, располагается сложно построенный центральный аппарат, контролирующий общую установку тела. Он объединяет мускулатуру всего тела в функциональные комплексы.
Различные сегменты центральной нервной системы управляют этого рода реакциями для каждого отдела тела. Однако уже спинной мозг как целое способен к сложным комплексным деятельностям. Спинальная собака способна совершать обеими задними ногами упорядоченные движения. В чесательном рефлексе принимает участие не только чешущая нога, но и изгибается позвоночник и другая нога принимает соответствующее покоящееся положение. При движениях дефекации ноги и хвост совершают координированные движения и т. д.
Шеррингтон своими исследованиями децеребрационной ригидности, которую он понимал как «рефлекторное стояние» (9), показал, что дистальная часть ствола мозга приводит в действие «мускулатуру стояния» как определенную функциональную группу.
Над этим в стволе мозга надстраивается, далее, аппарат, который управляет всем тем, что связано с установкой тела. Какая совершенная функция складывается в результате этого, известно из наблюдений над беспо- лушарными собаками, которые были классически описаны Гольцем.
Это является основанием, на котором кора большого мозга, словно на рояле, разыгрывает сложнейшие мелодии, согласно законам, которые частью известны и которые теперь могут быть исследованы с точки зрения новых вопросов.
Центральный аппарат в мозговом стволе организован в виде трех функциональных групп и, таким образом, расчленен иначе, чем то соответствовало бы рецепторным органам, возбуждение которых оказывает влияние на установку тела. Эти рецепторы суть, во-первых, макулы отолитов, которые определяют положение головы в пространстве, далее — аппараты полукружных каналов, отвечающие на ускорение в пространстве, проприоцепторы, доставляющие сведения от различных частей тела противоположной стороны, экстероцепторы, сигнализирующие о положении тела и отдельных его частей относительно непосредственного их окружения, и, наконец, дистантные рецепторы, опосредующие отношение тела, и прежде всего головы, к удаленным источникам раздражения. Все эти отношения — положение и движение в пространстве, расположение частей тела относительно друг друга, установка головы относительно ближайшего и удаленного окружения (окружающий мир), — подчиняясь известным законам, оказывают через посредство афферентных нервов длительное влияние на установку тела. Реакции возникают неизбежно и не являются произвольными.
В центре следует территориально выделить следующие группы механизмов установки тела.
1. Все рефлексы на мышцы тела: проприоцепторы, прежде всего шеи, и отолиты управляют положением. Аппараты полукружных каналов ведают реакциями на прямолинейное и вращательное ускорение.
490
Десятая глава. Центры установки тела
2. Все рефлексы на глаза: отолиты и проприоцепторы шеи сообщают глазам их закономерные покоящиеся положения относительно пространства и, в зависимости от положения головы, по отношению к телу. Аппараты полукружных каналов обусловливают движения глаз при поворотах тела в пространстве в противоположном направлении. Первый из названных механизмов по своему значению отступает назад, как только становится возможной оптическая ориентация глаз относительно окружения.
3. Все установочные рефлексы: нормальная установка головы обеспечивается благодаря возбуждениям, исходящим из отолитов, экстерорецеп- торов тела и проприоцепторов. Телерецепторы (глаза) действуют в том же направлении через посредство коры большого мозга.
Попутно с этими, известными по меньшей мере в основных чертах, механизмами функционируют и другие аппараты центральной нервной системы, деятельность которых еще не удается объяснить достаточно ясно. В первую очередь таким аппаратом является мозжечок, о котором мы знаем, что выпадение его вызывает столь характерные, но тем в меньшей степени поддающиеся объяснению нарушения, положительные же функции которого до сих пор почти неизвестны. Далее, таким аппаратом являются базальные ганглии, заболевания которых и отношение к патологическим симптомам именно в последнее время привлекают большое внимание клиницистов и анатомов, и безусловно еще и другие системы.
То, что оказалось возможным понять деятельность аппаратов установки тела в стволе мозга настолько, насколько это сделано к настоящему времени, в немалой степени объясняется направлением исследования, которое было принято при этом и которое отлично от обычных путей. Со времен Фритча и Гитцига исследователи деятельности мозга поступали таким образом, что начиная с коры большого мозга, путем частичных или тотальных экстирпаций снимали деятельность коры и таким образом постепенно доходили до ствола, причем многочисленные опыты с экстирпациями предпринимались с целью узнать что-либо о функции экстирпированных отделов мозга. Даже бесполушарная собака зачастую использовалась для того, чтобы узнать что-либо о физиологической деятельности коры. В исследованиях, о которых сообщает эта книга, планомерно применялся обратный путь исследования. Известные функции спинного мозга служили в качестве исходного пункта. Далее было установлено, какие функции приобретает спинной мозг, когда он находится в связи с продолговатым мозгом. После того как это было выяснено, оказалось возможным распространить изучение на средний мозг, причем в качестве нового функционального приобретения оказались распределение тонуса и функция установки. Наконец, было показано сохранение главнейших из найденных функций после удаления мозжечка и таким путем подтверждена локализация их в стволе мозга. Я думаю, что этот путь, хотя и более долгий и трудоемкий, является в той же степени и более надежным. Если одновременно с появлением нового отдела мозга выступает и новая функция, с большей долей вероятности можно локализовать эту функцию в соответствующей части мозга, в то время как, идя сверху вниз, никогда нельзя быть уверенным, является ли отсутствие данной функции следствием удаления определенного отдела мозга или результатом какого-либо другого операционного повреждения.
Полученные до сих пор данные относительно центров и их положения в определенных отделах мозга, относительно афферентных и эфферентных путей, рецепторных органов и эффекторных аппаратов могут быть представлены в виде простых схем, как это изображено в четвертой главе (стр. 146, рис. 86) для компенсаторных установок глаз. Аналогичные схемы для лабораторного пользования я изготовил для рефлексов на мускулатуру тела
Значение красного ядра для установочных рефлексов
491
и установочных рефлексов. От воспроизведения их здесь мы отказались; они имеют только относительную ценность в качестве основы для последующих экспериментальных исследований, и каждый в состоянии по мере надобности изготовить их самостоятельно. Для большинства рефлексов еще нельзя показать, какие анатомически известные структуры (ядра) соответствуют физиологическим, лежащим на определенном уровне, центрам, по каким анатомически известным путям пробегают афферентные и эфферентные возбуждения и из каких нейронов составляются эти пути. Для некоторых рефлексов еще не известно даже, проходят ли пути одно- или двусторонне, пересекаются ли они и где происходит этот перекрест.
Таким образом, предстоит еще проделать большую работу до той поры, пока строение центральных аппаратов установки тела станет известно во всех подробностях. При этом физиологические эксперименты и точное анатомическое исследование должны идти рука об руку. В качестве первого шага на этом нелегком, но весьма перспективном пути следует рассматривать серию экспериментальных исследований Радемакером центров нормального распределения тонуса и установочных рефлексов, — исследований, которые к моменту написания этих строк хотя и не были еще завершены, но уже установили целый ряд важных данных. Некоторые из них могут быть сообщены ниже.
III. ЗНАЧЕНИЕ КРАСНОГО ЯДРА ДЛЯ НОРМАЛЬНОГО РАСПРЕДЕЛЕНИЯ ТОНУСА И ДЛЯ УСТАНОВОЧНЫХ РЕФЛЕКСОВ (80, 81, 82а)
В предыдущем разделе было показано, что те центры, наличие которых обусловливает «нормальное распределение тонуса», отсутствие деце- ребрационной ригидности и которые необходимы для осуществления лабиринтных установочных рефлексов, лежат, по крайней мере в основной их части, в среднем мозгу, позади плоскости, соответствующей у кролика линии / рис. 217, а у кошки — линии II рис. 219.
Далее, с большой долей достоверности установлено, что центры этих функций лежат в среднем мозгу впереди плоскости, обозначенной на рис. 217 линией II (у кролика)
и на рис. 219 линией/// (у кошки). рис> 221.
Таким образом, пласт среднего
мозга, в котором следует искать соответствующие центры, имеет в толщину 1.5 мм у кролика и самое большее 3 мм у кошки. Он лежит на уровне, который включает в себя переднее двухолмие, красное ядро (у кошки — его крупноклеточный отдел) и место выхода глазодвигательного нерва. Далее следовало установить, в какой части поперечного разреза локализованы названные функции. Опыты в этом направлении были поставлены Радемакером на кроликах.
На рис. 221 изображены два таламических кролика, из которых левый сидит нормально. У него имелись все лабиринтные установочные рефлексы, тонус был распределен правильно и не было никаких следов де- церебрационной ригидности. Кролик, находящийся справа, лежит на боку. У него обнаружилась отчетливая децеребрационная ригидность, в особенности на правой передней ноге и обеих задних, не было никаких следов
492
Десятая глава. Центры установки тела
лабиринтных установочных рефлексов и установочных рефлексов тела на тело. Установочные рефлексы тела на голову проявлялись лишь едва заметно: в ответ на раздражение животное, лежащее на столе, слегка пово- чивало голову из обоих боковых положений по направлению к нормальному.
У этих таламических животных была произведена частичная перерезка среднего мозга. После введения небольшого узкого ножичка в боковую поверхность мозга у одного животного (на рисунке — слева) был произведен разрез дорсальной половины, у другого (на рисунке — справа) — вентральной половины мозга. Точный путь разреза (рис. 222) был установлен на серии срезов.
У животного, находящегося слева, с сохраненными установочными рефлексами и нормально распределенным тонусом, разрез прошел от области ядра блокового нерва в дорсальном направлении и через заднюю треть переднего двухолмия (линия /). У животного, находящегося справа, у которого имеется отчетливо выраженная децеребрационная ригидность и отсутствуют лабиринтные установочные рефлексы и установочные рефлексы с тела на тело, разрез прошел (линия II) от вершины ядра блокового нерва в вентральном направлении, через задний отдел крупноклеточного красного ядра и через место выхода глазодвигательного нерва.
Согласно данным этого двойного опыта, подтвержденным многочисленными другими экспериментами, центры должны располагаться в вентральной половине среднего мозга.
То, что дорсальная половина среднего мозга не имеет существенного значения для нормального распределения тонуса и установочных лабиринтных рефлексов, вытекает и из следующих опытов (рис. 223).
У таламического кролика («Nanel»), у которого дорсальная половина среднего мозга была разделена разрезом, вышедшим наружу позади заднего двухолмия, были обнаружены нормальное распределение тонуса, все установочные лабиринтные рефлексы и отсутствие децеребрационной ригидности; не было только установочных рефлексов тела на тело. Другой таламический кролик («Mata Biroe»), у которого горизонтально направленным разрезом была удалена вся крыша среднего мозга, держал себя в отношении распределения тонуса, установочных рефлексов и всех испытанных у него лабиринтных рефлексов совершенно как нормальное животное.
Corpus Thala- Согрдиа- Subst. grisea triatum mus dm ant centralis
11 V \
Corpus mamillare Nemocumolotorius /V. ruber Рис. 222.
Значение красного ядра для установочных рефлексов
493
Отсюда следует, что центры, снимающие децеребрационную ригидность, обусловливающие нормальное распределение тонуса и контролирующие лабиринтные установочные рефлексы, лежат в вентральной половине среднего мозга, в области, описанной выше (рис. 217, между линиями /и II).
Как показывает табл. XIX атласа кролика Винклера—Поттера, так же как и рис. 213а (стр. 437) и рис. 224 (стр. 494), на этом уровне, помимо глазодвигательного нерва, лежат красное ядро, черная субстанция, ganglion interpedunculare и разные другие ядра. Наибольшее значение имеет красное ядро.
Поэтому следующей задачей было — вылущить красное ядро с обеих сторон и исследовать затем распределение тонуса и установочные рефлексы.
Рис. 223.
Так как с обеих сторон красное ядро лежит в толще среднего мозга и обладает удлиненно овальной формой, с длинной осью в назо-окципитальном направлении, изолированное двустороннее разрушение его без нанесения сопровождающих повреждений затруднительно.
Поэтому подобное вмешательство в качестве типового в этом направлении не является подходящим. С другой стороны, представилось возможным перерезать эфферентный путь этого ядра 1 и исследовать влияние последствий этого вмешательства на рефлексы. Эфферентным путем является рубро-спинальный тракт, который сразу после выхода из ядра пересекает среднюю линию в вентральном перекрестке покрышки (Форель) (рис. 213а). Форелевский перекрест содержит по крайней мере большую часть рубро- спинального пути, а может быть даже все рубро-спинальные волокна. Перерезка их на высоте каудального конца красных ядер на обеих сторонах снимает, таким образом, связь этих последних со спинным мозгом — полностью или в значительнейшей ее части.
Обозначения расположения видны из рис. 224, на котором показаны с обеих сторон холмы переднего двухолмия, далее вниз — сияние (Fontänenstrahlung) Мейнерта, сильвиев водопровод, ядро глазодвигательного нерва с корешками, задний продольный пучок, гудденовский тракт, красное ядро, форелевский перекрест, медиальная петля, черная субстанция, ножка мозга и corpus interpedunculare. Измерения показывают, что красное ядро у кролика лежит на расстоянии 3 мм от вентральной поверхности и 7 мм от дорсальной поверхности (между холмами верхнего двухолмия).
1 Первым указанием на целесообразность такого подхода я обязан проф. Хельду в Лейпциге.
494
Десятая глава. Центры установки тела
Симптом (выпадение рефлекса) может быть отнесен за счет рассечения рубро-спинального тракта только в том случае, когда разрез по средней линии с вентральной или дорсальной сторон прямо к форелевскому пере-
Corpus inter- lemniscus pedunculare medialis
Рис. 224.
кресту оставляет, однако, его интактным, еще не вызывая симптома, в то время как последний появляется, когда разрез несколько углубляется,
Кро¬
лик
1
Глубина разреза в мм
Лаби¬
ринтные
устано¬
вочные
рефлексы
Установочные рефлексы тела на тело
Децере-
брацион-
ная
ригид¬
ность
F
2 Va
+
S
2 Va
+
—
н
2 Va
+
+
—
I
2 V2
+
+
—
р
2 Va
+
—
Q
3 Va
—
—
+
в
3 Va
—
—
+
S
3 Va
—
—
+
т
3 Va
—
—
+
вследствие чего перекрест пересекается.
Если принять во внимание, что при введении ножа в центральную нервную систему окружающая ткань всегда несколько смещается в направлении разреза, можно ожидать, что при введении ножа с вентральной стороны на глубину 134 мм перекрест остается неповрежденным и пересекается при продвижении ножа на 3 34 мм.
Результаты девяти опытов подобного рода, в которых у таламического кролика нож соответствую¬
щей формы вводился с вентральной стороны по возможности строго медиально на глубину 2/4 и 334 мм
в средний мозг на уровне каудального конца красного ядра, видны из следующей таблицы.
У кроликов- F, G и Р установочные рефлексы тела на тело не исследовались. Шейные установочные рефлексы животных были у всех.
Значение красного ядра для установочных рефлексов
495
Таблица охватывает все опыты, в которых операционное поле во время операции было хорошо доступно для обозрения и поэтому не наступало сильных асимметричных проявлений. «Глубина разреза» приведена согласно намеченной глубине, т. е. с выбором применявшегося ножа.
По отчетливости результатов эти опыты не оставляют желать ничего большего. Во всех случаях после погружения ножа на 2г12 мм наблюдалось нормальное распределение тонуса и отсутствие децеребрацион- ной ригидности. Лабиринтные рефлексы не уменьшались в двух случаях,
Рис. 225.
которые впоследствии были подвергнуты исследованию; установочные рефлексы тела на тело также имелись. Наоборот, после погружения кожа на 31/2 мм во всех случаях отсутствовало нормальное распределение тонуса, наступала отчетливая децеребрационная ригидность, лабиринтные установочные рефлексы и установочные рефлексы тела на тело исчезали.
До сих пор на полных сериях срезов изучены препараты шести животных: G, Н, I (234 мм) и Q, В, S (3/4 мм). Результат может быть демонстрирован тремя следующими примерами. Данные остальных случаев те же.
Кролик I. Таламическое животное. Намеченная глубина разреза— 234 мм. У животного отсутствуют всякие признаки децеребрационной ригидности; распределение тонуса совершенно нормально. Животное сидело и прыгало нормально, обнаруживало все лабиринтные рефлексы на голову и установочные рефлексы тела на тело (и, конечно, также шейные рефлексы) и было забито через 4 часа после операции.
Рис. 225 показывает, что надрез проходит в /4 мм от средней линии и своей нижней частью затрагивает вентральный край форелевского пере-
496
Десятая глава. Центры установки тела
креста, в который и проникает. Выход глазодвигательного нерва с обеих сторон интактен, так же как и (что видно из всей серии срезов) перекрест соединительных ножек, перекрест Мейнерта, ядра глазодвигательного нерва, оба красных ядра, черная субстанция на обеих сторонах и пирамидные пути; Конечно, поскольку дело идет о таламическом животном, то пирамидные пути во время экстирпации большого мозга перерезаны на более высоком уровне.
Кролик В. Таламическое животное. Намеченная глубина введения ножа — 3/4 мм. У животного обнаружилась длительная децеребра-
Fasciculus longitvd. post. ~
Nucleus оси- lomotorius
Corp guadrig.ant
Substantia grisea centralis Aq.Sylvii Radix mesencephalic, n.trig.
Corp. genu- culatummed decussatio Meynert Brachium conjunctivum Lemniscus medialis Pes pedunculi
Nucleus ruber
Decussatio Forel
Рис. 226.
ционная ригидность; оно лежало на боку. Все установочные рефлексы (конечно, за исключением шейных установочных рефлексов) были сняты. Животное было забито через 2/4 часа после операции.
На рис. 226 видно, что разрез расположен строго медиально. Он пересекает форелевский перекрест. Просмотр всей серии показывает, что перекрест соединительных ножек задет разрезом по самому краю и остается почти не поврежденным. Интактны, далее, с обеих сторон: красные ядра, ядро глазодвигательного нерва, черная субстанция, пирамидный путь (последний перерезан на более высоком уровне во время экстирпации большого мозга).
Точно так же у кролика S, у которого было намечено введение ножа ::а глубину 3/4 мм и у которого после этого исчезло нормальное распределение тонуса, появилась сильная децеребрационная ригидность, исчезли лабиринтные установочные рефлексы на туловище, был найден строго медиальный разрез через ganglion interpedunculare, который точно перерезал форелевский перекрест.
Данные, полученные на кроликах G и Н, соответствовали таковым у животного 1, а данные кролика Q — данным В и S.
Значение красного ядра для установочных рефлексов
497
Эти определенные результаты будут дополнены опытами с введениями ножа в средний мозг с дорсальной стороны. Так как расстояние до форе- левского перекреста здесь больше, с большей легкостью может иметь место отклонение разреза в сторону. Поэтому в данном случае можно привести лишь те опыты, в которых исследование серийных срезов уже проделано.
Кролик Z. Таламическое животное. Глубина введения — 6 мм. У животного отсутствует децеребрационная ригидность; распределение
— Corp.guadrig.ant
Radix mesencephalic. п. trigemini
Nuci. oculomotorius Fasc.longitud.post
Lemniscus lateralis Radix n.oculomot. Nucleus ruber Decussatio Forel Lemniscus medialis Corp. interpedunculare
Pes pedunculi cerebri
Рис. 227.
тонуса нормально. Сидит, бегает и прыгает совершенно нормально. Лабиринтные установочные рефлексы всегда положительны, как и установочные рефлексы тела на тело и шейные установочные рефлексы. Забито через час.
Разрез (рис. 227) проходит справа от средней линии, через сильвиев водопровод, в котором находится кровь; затем он направляется несколько вправо от средней линии, по латеральному краю ядра глазодвигательного нерва, в котором находится несколько кровоизлияний. Он заканчивается у дорсо-медиального угла правого красного ядра, которое совершенно интактно (ср. микрофотографию конечной части разреза на рис. 228). Форелевский перекрест интактен, так же как и перекрест соединительных ножек (виден на более каудальном разрезе). Перекрест Мейнерта перерезан.
Кролик X. Таламическое животное. Намеченная глубина разреза — 9 мм. У животного обнаружилась очень сильная децеребрационная ригидность. Лабиринтные установочные рефлексы и установочные рефлексы тела на тело полностью отсутствуют. Установочные рефлексы тела на голову, напротив, имеются в обоих боковых положениях (в правом боковом положении спонтанно, в левом — в ответ на раздражение).
32 Р. Магнус
498
Десятая глава. Центры установки тела
Шейные установочные рефлексы положительны. Компенсаторные установки глаз и реакции поворота глаз положительны. Забито через 3 часа.
Рис. 228.
Разрез' (Stichkanal, рис. 229) идет точно медиально между двумя ядрами глазодвигательного нерва и между обоими красными ядрами, через форелев-
ский перекрест и заканчивается в ganglion interpedunculare. Перекрест соединительных ножек перерезан, равно как и мейнертовский перекрест. Задний продольный пучок с обеих сторон интактен. Все структуры, лежащие вне медиальной плоскости, полностью сохранны.
Значение красного ядра для установочных рефлексов
499
Если суммировать данные всех этих опытов, то оказывается, что разрезы среднего мозга на уровне каудального полюса красного ядра, проходящие в медиальной плоскости или близко около нее по вентральной или дорсальной стороне на различной глубине, перерезающие перекресты, расположенные оральнее перекреста соединительных ножек, не нарушают нормального распределения тонуса, лабиринтных установочных рефлексов и установочных рефлексов тела на тело
Рис. 230.
до тех пор, пока остается неповрежденным фо- релевский перекрест (и вместе с ним — рубро- спинальный путь). Наоборот, при перерезке форелевского перекреста на этом уровне наступает децеребрационная ригидность и пропадают лабиринтные установочные рефлексы на голову, а также установочные рефлексы тела на тело.
Этот факт удалось весьма ярко продемонстрировать в следующем остром опыте.
У кролика удалялись оба больших полушария. Начиная от передней поверхности разреза выше таламуса в срединной плоскости, с помощью иглы была протянута в каудальном направлении, дорсальнее красного ядра, тонкая шелковая нить. Каудальнее красного ядра нить изгибалась в вентральном направлении и у переднего края моста выходила наружу. Оба конца нити соединялись в петлю. Положение нити изображено на рис. 230.
Животное обнаруживало поведение обычного таламического животного. У него было отмечено нормальное распределение тонуса, отсутствовала децеребрационная ригидность, сидело нормально, обладало хорошими лабиринтными рефлексами, установочными рефлексами тела на тело и шейными установочными рефлексами. На рис. 231 изображено животное в этом состоянии.
Теперь зажим кожной раны временно снимался и (через 65 минут после операции) нить натягивалась в переднем направлении настолько, чтобы
32*
500
Десятая глава. Центры установки тела
перекрест Фореля оказался перерезанным. Результат был исключительно показательным (рис. 232): животное тотчас же упало на бок, появилась сильнейшая децеребрационная ригидность, от лабиринтных установочных рефлексов и установочных рефлексов тела на тело не осталось и следов. Фотографический снимок (сверху) сделан спустя 234 часа. К этим исследованиям, после того как было твердо установлено значение форелевского перекреста, примыкают дальнейшие опыты по двустороннему разрушению красного ядра с помощью надрезов по обеим сторонам среднего мозга. G этой целью у таламического кролика ствол на одной стороне слегка приподнимался до тех пор, пока не становился видимым глазодвигательный нерв. Место введения ножа лежит на уровне выхода глазодвигательного нерва. Несколько дорсально от ножки мозга нож вводился на глубину около 4 мм, и после этого носик его перемещался несколько раз вперед
Рис. 231. Рис. 232.
и назад в назо-окципиальном направлении. Эта операция производилась на обеих сторонах.
Из большой серии опытов прежде всего обратили на себя внимание два животных, у которых операция считалась прошедшей особенно удачно. Поразительным образом одно из животных обнаружило совершенно нормальное поведение, в то время как второе было ригидным и лежало на боку. Поэтому препараты от этих кроликов были разрезаны на серии раньше других. Результат оказался следующий.
Кролик М. Состояние после операции: абсолютно никакой ригидности, нормальное распределение тонуса. Лабиринтные установочные рефлексы во всех положениях положительны. Установочные рефлексы тела на тело отсутствуют. Шейные установочные рефлексы (на переднюю половину тела и таз) положительны. Животное сидит передней половиной тела правильно. Время от времени пытается привести в сидячее положение и заднюю половину тела, однако скоро опрокидывается на спину. Забито спустя примерно 3 часа.
Микроскопическое исследование. На рис. 233 видно, что каналы от введения ножа располагаются на. уровне форелевского перекреста, каудально от красного ядра. Разрез на левой стороне рисунка проходит косо, в дорсальном направлении к нижнему краю ядра глазодвигательного нерва, и не переходит через среднюю линию. Разрез на правой стороне рисунка идет в горизонтальном направлении дорсаль- нее форелевского перекреста и пересекает среднюю линию. Разрез на обеих
Значение красного ядра для установочных рефлексов
501
сторонах повреждает церебеллярно-рубральный путь орально от перекреста соединительных ножек мозжечка.
Рисунок 234 изображает разрез, который расположен более назально, чем на рис. 233. На этом разрезе видно максимальное поражение красного ядра. На правой стороне разрез доходит вплоть до области медиальной петли и остается латеральнее красного ядра. Слева разрез проходит ла- терально от красного ядра. Однако вентро-латеральную (крупноклеточ-
Subst. grisea centralis
Lemniscus
lateralis
decussatio
Forel
Lemnicus
medialis
Corpus inter pedunculare
ную) область красного ядра поглотило [angefressen] кровоизлияние. В большей же части ядро осталось интактным. Черная субстанция слева повреждена. Пирамидные пути в области среднего мозга не затронуты.
Таким образом, в этом случае красные ядра на одной стороне полностью, на другой стороне в значительной части остались интактными. Неповрежденным остался также и рубро-спи- нальный путь.
Кролик О. Состояние после операции: децеребрационная ригидность на задних ногах интенсивна, на передних умеренна. Лабиринтные установочные рефлексы и установочные рефлексы тела на тело отсутствуют. Установочные рефлексы на голову в обоих боковых положениях (без раздражения) выражены слабо. Шейные установочные рефлексы положительны. Забит спустя примерно 3 часа. .
Микроскопическое исследование. На рис. 235 изображен разрез, на котором сохранилась максимальная часть красного
Corp. guadrig {ant Ag. Syluii
/
Tractus mes - encepbalicus n.V
Nucleus Ш
Fasciculus longitud. post
Lemniscus 'lateralis
Formatio reticularis
Lemniscus
medialis
Pes pedunculi
Substantia nigra
Aquaeductus Sylvii Corp.quadrig.ant
Lemniscus
lateralis
Lemniscus
medialis
Granglion Ln terpedun- culare
Subst. grisea centralis
Nucleus
oculomoto¬
rius
lemniscus
lateralis
Fasciculus
longitud.
post
Corp.geniculatum mediale Nucleus ruber Lemniscus meäialus
Substantia nigra Pes pedunculi
Pes pedunculi
Radices n. Linker Stich- oculomotorii kanal
Рис. 234»
Corpus qua dri* nost.
Nucleus
ruber
Lemniscus
medialis
Pes pedunculi
Pons
Corp. qua- drlg.ant.
■Radix mesencephalica 77. trig
Corp.quadrig.
post
Nucleus III Fasciculus longitudinalis post.
Pes pedunculi
-Pons
Corpus interpedunculare
Рис. 235.
Значение красного ядра для установочных рефлексов
503
ядра. Разрез на левой стороне рисунка проникает в красное ядро, разрушает его почти полностью и оставляет интактной только небольшую вентро-латеральную группу ядер (крупноклеточного отдела). На правой стороне красное ядро полностью разрушено. Имеется кровоизлияние на месте перекреста Фореля, которое на более каудально расположенном уровне совершенно разрушено кровоизлияниями.
Интактны с обеих сторон: черная субстанция, медиальная петля, перекрест соединительных ножек мозжечка (в последних только слева совсем небольшое кровоизлияние). Пирамидный путь в толще среднего мозга не задет. В перекресте Мейнерта — большое кровоизлияние.
Таким образом, в этом случае красное ядро на одной стороне разрушено полностью, на другой стороне от него осталась лишь небольшая часть. Перекрест Фореля также разрушен.
Оба опыта зеркально дополняют друг друга и показывают, что при сохранном красном ядре распределение тонуса нормально, лабиринтные установочные рефлексы и установочные рефлексы тела на тело имеются, в то время как после разрушения красного ядра и форелевского перекреста эти реакции отсутствуют.
В качестве дополнения следует привести здесь еще один эксперимент.
Кролик «Уротропин». Большой мозг оставлен интактным. После вскрытия черепа задний полюс большого мозга приподнимается и с одной стороны, каудальнее уровня красного ядра, делается латеральный разрез среднего мозга через среднюю линию и кнаружи. Животное в течение 9 дней оставляется живым и затем забивается. Так как большие полушария и пирамидные пути остаются неповрежденными, появляется лишь незначительная дедеребрационная ригидность. Отмечаются некоторые асимметричные явления: основной поворот вправо, постоянное лежание на правом боку.
Лабиринтные установочные рефлексы и установочные рефлексы тела на тело отрицательны. Установочные рефлексы с тела на голову в правом боковом положении животного отсутствуют, в левом маскируются, так как голова вследствие основного поворота движется в нормальное положение. Шейные установочные рефлексы положительны.
Гистологическое исследование: разрез расположен каудально от красного ядра.
На рис. 236 видны максимальные повреждения несколько каудальнее разреза. Рубро-спинальный путь с обеих сторон полностью перерезан каудальнее форелев- ского перекреста. Сетчатая субстанция дегенерирована. С обеих сторон интактны: лучистость и перекрест Мейнерта, перекрест пирамидных путей (только несколько поврежденный слева), черная субстанция, пирамидные пути, медиальная петля, задний продольный пучок.
В этом опыте также отмечается исчезновение лабиринтных установочных рефлексов и установочных рефлексов тела на тело в сочетании с разрушением рубро-спинальных путей. Уровень повреждения располагается более каудально, чем в предыдущих экспериментах.
Если мы теперь обобщим результаты всех этих опытов и целого ряда экспериментов, не описанных здесь, мы сможем обсудить заключения относительно положения центров лабиринтных установочных рефлексов и нормального распределения тонуса, так как во всех случаях вмешательства на среднем мозге у таламических животных было установлено, что животные с нормальным распределением тонуса и без децеребрационной ригидности обладают также и интактными лабиринтными рефлексами, в то время как децеребрационная ригидность постоянно сочетается с исчезновением лабиринтных установочных рефлексов. Определение уровня показало, что центры лабиринтных установочных рефлексов и нормального тонуса лежат на одном и том же поперечном сечении среднего мозга, на котором находится и крупноклеточное красное ядро, и что их следует
504
Десятая глава. Центры установки тела
искать в вентральной половине (вентральнее ядра глазодвигательного нерва) этого сечения. Вентральный и дорсальный разрезы срединной плоскости среднего мозга не снимают названных функций до тех пор, пока форелевский перекрест остается интактным, и даже тогда, когда разрезы проходят совсем близко около перекреста. Наоборот, если форелевский перекрест перерезается на уровне заднего конца красного ядра, там, где перекрещиваются рубро-спинальные пути, наступает децеребрационная ригидность и пропадают лабиринтные установочные рефлексы. Боковые
Рис. 236.
разрезы среднего мозга, проходящие дорсально от красных ядер и оставляющие эти ядра, подобно форелевскому перекресту, интактными, влияния на нормальное распределение тонуса и лабиринтные установочные рефлексы не оказывают.
Отсюда с определенностью следует, что путь этих рефлексов пересекает среднюю линию на высоте заднего конца красного ядра и что центры этих рефлексов лежат на уровне красного ядра в вентральной части поперечного разреза спинного мозга. Пути, которые на этом уровне перекрещиваются в форелевском перекресте,1 возникают в красном ядре. Отсюда следует, что красное ядро является центром лабиринтных установочных рефлексов и нормального распределения тонуса мускулатуры тела, благодаря деятельности этого ядра снимается децеребрационная ригидность. Этим самым для названных функций установлены ядро и эфферентные пути, и, с другой стороны, для образования,1 2 настолько ясно
1 Здесь, как и везде в этом отделе, я строго придерживаюсь способа описания рисунков атласа Винклера—Поттера.
2 Относительно анатомии красного ядра см. прежде всего большую монографию Монакова.
Значение красного ядра для установочных рефлексов
505
выраженного анатомически, установлена совершенно определенная физиологическая функция. Определения уровня на кошке делают вероятным предположение, что крупноклеточная часть красного ядра достаточна для обеспечения обсуждаемых реакций.
Помимо этого, в опытах на таламических животных с сохранными лабиринтными установочными рефлексами и нормальным распределением тонуса установлено, что разрушение следующих ядер и путей не нарушает нормального осуществления обеих названных функций и что эти ядра и пути не являются необходимыми для лабиринтных установочных рефлексов и нормального распределения тонуса:
пирамидного пути (у таламического животного перерезан); перекреста соединительных ножек мозжечка (см., например, у кролика М);
всех перекрестов (кроме фореяевского) на уровне крупноклеточного красного ядра (дорсальные и вентральные разрезы среднего мозга); всей покрышки вместе с четверохолмием (рис. 223); всех путей из покрышки: лучистость и перекрест Мейнерта, текто- понтинные и тёкто-бульбарные пути (рис. 222 и 223); медиальной петли (например, кролик М); вероятно, также — черной субстанции.
Радемакер у многих кроликов производил по обеим сторонам вентро- латеральные надрезы среднего мозга, которые разрушали черную субстанцию, но должны были оставить интактными красные ядра и рубро- спинальный путь. У животных имелись нормальное распределение тонуса и интактные лабиринтные установочные рефлексы. Анатомическое исследование еще впереди. Далее, сюда же может быть отнесено и то, что у бес- полушарной собаки Гольца, у которой не было децеребрационной ригидности, черная субстанция дегенерировала (Гордон Холмс) и что у одной из кошек (собственное наблюдение), у которой после экстирпации больших полушарий не было следов децеребрационной ригидности и нарушения распределения тонуса, а также отмечались исключительно симметричные лабиринтные установочные рефлексы, согласно заключению Винклера, на стороне операции имела место гибель всех клеток черной субстанции.
В связи с этим естественно указать, что, хотя все названные анатомические структуры и не являются необходимыми для названных функций, при известных условиях (например, в качестве афферентных путей) эти образования могут воздействовать на красное ядро, приобретая тем самым влияние на распределение тонуса.
Ведь нельзя забывать, что до настоящего времени для нормального распределения тонуса установлены только ядро и эфферентный путь. Что касается афферентных путей, мы знаем, что путь для лабиринтных рефлексов во всяком случае проходит от места вступления 8-го нерва через ствол и далее к красному ядру и не делает отклонений через субстанции, лежащие в стволе более орально. Точное расположение этого пути еще, однако, не известно.
Нуждается ли красное ядро, вообще говоря, в афферентных путях для того, чтобы тормозить децеребрационную ригидность и вызывать нормальное распределение тонуса, или оно может работать «автоматически» — сказать еще нельзя. Так как лишенное лабиринтов животное не ригидно, красное ядро во всяком случае еще способно осуществлять нормальное распределение тонуса и после выпадения афферентных лабиринтных возбуждений. Роль остальных афферентных раздражений еще следует установить.
506
Десятая глава. Центры установки тела
Кроме того, еще предстоит изучить, каким образом и в каком месте красное ядро через посредство рубро-спинального пути снимает децеребрацион- ную ригидность. Говоря а priori, существуют две возможности. Одна из них — от красного ядра проходит прямой путь к «центру ригидности» в продолговатом мозгу, — путь, через который этот центр тормозится. Против этого говорит личное сообщение проф. Винклера о том, что, по его представлению, рубро-спинальный путь, проходя через продолговатый мозг, не отдает или, во всяком случае, отдает весьма мало коллатералей. Колье и Бузар описывают, правда, коллатерали к мозжечку, которые, однако, не могут рассматриваться как имеющие отношение к обсуждаемой здесь функции. Во всяком случае, торможение «центра ригидности» красным ядром не могло бы быть совершенным, так как в противном случае таламическое животное должно было бы вести себя в отношении мускулатуры тела подобно спинальному животному, чего не бывает.
Поэтому представляется более вероятной другая возможность — что влияние через рубро-спинальные пути осуществляется непосредственно на центры различных уровней спинного мозга, для чего существует анатомическая предпосылка в виде самой длины рубро-спинального пути. Дело представлялось бы тогда таким образом, что, например, центры спинного мозга одной конечности возбуждаются рефлекторно, через корешки, входящие в мозг на данном уровне, и посылают импульсы к центрам сгибателей и разгибателей. Кроме того, на том же уровне оканчивается путь из центров продолговатого мозга, которые, действуя на мозг изолированно, вызывают децеребрационную ригидность. Таким образом, этот путь смещает распределение тонуса на соответствующем уровне спинного мозга в пользу разгибающих мышц. Если же, помимо этого, к тому же уровню спинного мозга по рубро-спинальному пути проходят импульсы от красного ядра, то, согласно этому представлению, распределение тонуса снова оказывается сдвинутым в сторону сгибателей. До некоторой степени центры спинного мозга находятся под двумя влияниями: разги- бательным влиянием, которое оказывает продолговатый мозг, и сгибательным влиянием, идущим от красного ядра. У таламического животного в результате этих обоих влияний наступает равномерное распределение тонуса иннервируемой конечности.
Однако, кроме того, у животного с интактным большим мозгом того же уровня достигают возбуждения, идущие по пирамидному пути, которые, как показывают опыты с раздражением большого мозга, вызывают преимущественно сгибательные движения. Благодаря этому становится понятным интересное противоречие между поведением интактного и таламического животных после перерыва рубро-спинального пути. После этого вмешательства лабиринтные установочные рефлексы и установочные рефлексы с тела на тело у обоих животных отсутствуют. Как уже говорилось, у таламического животного наступает сильная децеребрационная ригидность. Наоборот, у животного с сохранным большим мозгом (кролик, кошка) после перерезки форелевского перекреста или рубро-спинальных путей как сразу, так и в течение ряда дней после операции хотя и имеется ригидность, однако она выражена далеко не так сильно, как у таламического животного, — очевидно, по той причине, что связь коры и спинного мозга через посредство пирамидного пути сохраняется. Этот последний в качестве «сгибательных вожжей» воздействует на центры различных уровней спинного мозга, так что при выключении рубро-спинальных влияний все еще сохраняется иннервация, которая способна — хотя и не полностью — парализовать влияние центров продолговатого мозга.
Значение красного ядра для установочных рефлексов
507
Таким образом, одного рубро-спинального пути достаточно, чтобы устранить децеребрационную ригидность и вызвать нормальное распределение тонуса. Один пирамидный путь снимает ригидность лишь частично. Оба вместе обусловливают у животного с интактной центральной нервной системой нормальное распределение тонуса. Если оба отсутствуют (перерезка перекреста Форели у таламического животного), наступает максимальная ригидность. Если, при сохранном большом мозге, оба вместе перестают функционировать, то также наступает максимальная ригидность, как это видно из работы Экономо и Карплюсса, когда сопоставляются следующие три наблюдения. Кошки после двусторонней перерезки ножки мозга (пирамидный путь) и черной субстанции уже на следующий день способны нормально бегать. Макака после такой же операции может сидеть спустя 5 дней. Наоборот, кошка после того же вмешательства, сопровождавшегося перерезкой рубро-спинального перекреста, остается лежать на боку в продолжение 13 дней, причем имеется ригидность. Один перерыв пирамидного пути, как показывают наблюдения над таламическими животными, ригидности не вызывает. Для антропоидов то же самое следует из опытов Лейтона и Шеррингтона.
Все эти наблюдения, взятые вместе, с достаточной ясностью свидетельствуют о том, что перерезка рубро-спинального пути при интактном большом мозге не вызывает такой сильной ригидности, как у таламического животного. Является ли развитое здесь представление о значении взаимодействия различных нисходящих пучков в спинном мозгу для возникновения децеребрационной ригидности правильным — должно быть, конечно, показано экспериментально; в настоящее время его следует рассматривать как рабочую гипотезу.
Центры установочных рефлексов тела на тело, согласно прежним представлениям, располагаются на уровне красного ядра. Эти рефлексы как у таламического животного, так и при сохранном большом мозге исчезают во всех случаях после перерезки рубро-спинального пути в форелев- ском перекресте или каудальнее его.
В соответствии с этим красное ядро следует рассматривать как центр установочных рефлексов на туловище. Судя по определениям уровня у кошки (рис. 219, линия ƒ), следует думать о том, что отграничение в переднем (оральном) направлении не является таким определенным, как в случае лабиринтных установочных рефлексов, и что мелкоклеточная часть красного ядра может принимать участие в этих реакциях. Наблюдения на двух собаках с почти полностью удаленным мозжечком делают вероятным предположение, что афферентный путь этих рефлексов проходит через мозжечок. В согласии с этим находится и то обстоятельство, что у кролика М, у которого церебелло-рубральный путь был поврежден оральнее перекреста brachia conjunctiva, установочные рефлексы тела на тело (при сохранности лабиринтных установочных рефлексов) отсутствовали. Другие опыты также показательны в этом отношении. Однако следует подождать последующих экспериментов и прежде всего данных о многочисленных сериях срезов, раньше чем утверждать что-либо определенное относительно расположения афферентных путей установочных рефлексов тела на тело.
Поразительным оказался факт, что установочные рефлексы тела на голову имеют другие центры, чем лабиринтные установочные рефлексы, с одной стороны, и установочные рефлексы тела на тело, с другой. Перерезка форелевского перекреста и даже полная перерезка среднего мозга по средней линии на уровне заднего края красного ядра этих рефлексов не снимает. Даже у кролика О (см. стр. 502, рис. 235), у которого красные
508
Десятая глава. Центры установки тела
ядра и форелевский перекрест были почти полностью разрушены боковыми надрезами и кровоизлияниями, голова из обоих боковых положений на столе еще слабо поворачивалась по направлению к нормальному положению.
Из этого следует, что для осуществления установочных рефлексов тела на голову красные ядра не необходимы. Центры лежат или на том же уровне, что и красные ядра, или, может быть, несколько более каудально. Еще предстоит установить, какое из ядер является центральным пунктом для этих рефлексов.
После всех вмешательств, описанных в данном разделе, сохранялись только шейные рефлексы, чего и следовало ожидать после предыдущих определений уровней. Эти рефлексы к красному ядру отношения не имеют.
Опыты, описанные в этой главе, показывают, что сделан первый шаг на пути анатомо-физиологической расшифровки центрального аппарата установки тела. Вместе с данными относительно общего расположения центров и путей в отдельных отрезках ствола удалось получить сведения относительно функции или части функции по крайней мере одного анатомически известного ядра и установить анатомическое положение центров некоторых физиологически известных функций. Использованные при этом приемы оказались целесообразными; надо надеяться, что результаты аргументированы достаточно убедительно для того, чтобы послужить основой для дальнейших экспериментов, и что последующими опытами они будут расширены, но не опровергнуты.
Наконец, еще только одно методическое замечание. Все заключения этой главы основаны на симметричных вмешательствах на центральной нервной системе. Я считаю это исключительно важным. Коль скоро осуществляются односторонние экстирпации, перерезки, надрезы (или раздражения), создаются непредвиденные и часто почти необъяснимые осложнения. Следует принять во внимание, что уже одностороннее удаление лежащего за пределами центральной нервной системы рецепторного органа, каким является лабиринт, вызывает картину симптомов, в которой прямые последствия утраты лабиринта настолько перекрываются и усложняются вторичными шейными рефлексами и действием симметричных рефлексов, что без анализа, основанного на предшествующих сведениях обо всех рассматриваемых рефлексах, понимание происходящего становится невозможным. Насколько же это должно усугубляться после односторонних операций на центральной нервной системе, где причиняемые функциональные нарушения заранее не известны и, кроме того, существующие взаимоотношения значительно более сложны! Легко убедиться, как трудно после односторонних перерезок среднего мозга, которые вызывают повороты и изгибания туловища, исследовать различные установочные рефлексы, даже после установления головы прямо, и делать достоверные заключения по наличию или отсутствию этих рефлексов только с одной стороны. Например, после удаления всей крыши среднего мозга все лабиринтные установочные рефлексы, как и бег и сидение, остаются нормальными. Однако, если подобную экстирпацию произвести лишь с одной стороны, наступают поворот головы, изгибание позвоночника и значительные нарушения бега и сидения, и исследователю, даже знакомому со всеми рассматриваемыми рефлексами, не удастся установить, являются ли лабиринтные рефлексы данного животного односторонними или двусторонними. Если в основном осуществлять только симметричные вмешательства и животных, у которых после этого обнаруживаются асим-
Одиннадцатая глава. Действие ядов
500
метричные симптомы, при построении заключений не учитывать, то хотя и приходится ставить большее число опытов и продвигаться вперед медленнее, однако продвижение в этих условиях является более уверенным. Причина, по какой, несмотря на бесчисленные осуществленные в течение последнего десятилетия операции, физиологический результат даже в отдаленной степени не соответствует затраченным усилиям, лежит в значительной мере в том, что вмешательства на центральном (в остальном неповрежденном) органе были односторонними.1
Одиннадцатая глава
ДЕЙСТВИЕ ЯДОВ
В предыдущих главах я не останавливался на действии ядов на установку тела и лабиринтные рефлексы. Это имело свое обоснование. Если необходимо обследовать действие такого сложного, тесным образом взаимодействующего механизма различных рефлексов, то для его анализа не подходят яды и лекарства. Вызываемые ядами симптомы зависят от двух неизвестных: во-первых, от неизвестного механизма рефлексов и, во-вторых, от неизвестного действия ядов на последние. Поэтому никогда нельзя получить точные и определенные результаты. Применение ядов для анализа неизвестных физиологических явлений является ошибочным. К сожалению, в последнее время это часто повторялось.
Только после того, как в основных чертах был выяснен механизм установки тела, стало возможным исследовать влияние ядов на эти функции. Теперь мы стоим на твердой почве и можем в отдельных случаях решить, какая группа рефлексов, какие центры, какая часть периферического органа чувств и так далее подверглись действию яда.
Опыты при этом проводились двумя путями.
Во-первых, исследовалось общее действие ядов и прежде всего устанавливалась последовательность, в какой различные рефлексы установки тела затрагиваются постепенно возрастающим отравлением. Подобный ряд наблюдений был сделан Верстеегом (Versteegh) для наркотиков и Ионкхоффом (Jonkhoff) — для различных возбуждающих средств.
Во-вторых, внимательно отнеслись к анализу отдельных компонентов действия ядов. Над действием стрихнина провели совместные наблюдения Вольф (Wolf) и Ионкхофф, над действием хинакетона — Билзма (Bilsma) и Вер- стеег, а подробный анализ мест приложения, довольно сложно действующего на реакцию глаз, и нистагм никотина сделали де Клейн и Верстеег.
Таким образом, сейчас имеется довольно обширный фактический материал. В виде главного результата пока можно привести то, что выявилась чрезвычайная специфичность влияния исследуемых ядов на систему установки тела и лабиринтные рефлексы. Различные рефлексы и центры затрагиваются каждым отдельным ядом с определенной закономерной последовательностью, которая, в свою очередь, для каждой группы ядов является совершенно другой; вследствие этого наблюдаются чрезвычайно изменчивые клинические картины отравления. Едва ли имеется лучший пример для точной локализации действия каждого яда в различных отделах центральной нервной системы, чем группа рефлексов установки тела и лабиринтных рефлексов.
1 Это замечание относится только к физиологическим исследованиям. Для анатомического определения путей и ядер односторонние вмешательства, конечно, весьма полезны.
510
Одиннадцатая глава. Действие ядов
В то время как раньше при отравлении (и при заболеваниях) только в общих чертах можно было говорить о нарушениях координации, о принятии бокового положения животным, о судорогах и т. п., то теперь возможен их точный анализ. Как будет видно дальше, во многих случаях удается показать, какие определенные группы рефлексов угнетаются или же усиливаются; таким образом, мы можем теперь в основном понять изменчивые состояния установки тела при отравлениях.
Я предполагаю, что при этом выявится также та или другая параллель с естественно встречающимися состояниями болезни и что выбранный здесь путь исследований окажется плодотворным и при анализе нарушений установки тела и равновесия у человека, как только у него будут полностью изучены нормальные рефлексы установки тела, как это удалось сделать на животных.
В приводимых далее наблюдениях действия ядов принимались во внимание не только собственно рефлексы установки тела, но и реакции полукружных каналов. Хотя последнее и выходит за рамки этой книги, следует все же сообщить об этом основное, так как благодаря этому ярче выступает специфичность различных ядов.
А. Наркотики жирного ряда
(эфир, хлороформ, уретан, паральдегид, алкоголь)
Изменения различных рефлексов и последовательность их появления изучались на кроликах и обезьянах при выходе из эфирного или хлороформного наркоза. У кроликов уретан вводился внутривенно от 0.25 до 1.75 г на килограмм веса. Паральдегид вводился per os от 1 до 4 мл на килограмм веса. Алкоголь — от 3 до 8 мл на килограмм в 10%-м разведении.
В общих чертах можно сказать, что реакции на прогрессивные движения являются самыми чувствительными (так же как у обезьян вращательные реакции на конечности и таз); после этого исчезают установочные рефлексы, в то время как компенсаторные установки глаз и вращательные реакции на голову и глаза являются более устойчивыми. Нистагм головы и глаз всегда исчезает раньше, чем вращательная реакция головы и глаз. У обезьян можно вызвать при глубоком наркозе, если имеется ригидность от наркоза, отчетливые рефлексы положения (тонические шейные и лабиринтные рефлексы на конечности).
В частности, наблюдаются отличия между различными наркотиками.
В большинстве случаев у обезьяны (59) удается установить при глубоком эфирном или хлороформном наркозе, если развилась выраженная ригидность от наркоза, отчетливые тонические лабиринтные рефлексы на мышцы тела при изменении положения головы в пространстве и тонические шейные рефлексы на мышцы конечностей при изменении положения головы по отношению к туловищу. В этом случае также сильно проявляются лабиринтные вращательные реакции на голову и глаза (рефлексы полукружных каналов), в то время как нистагм еще отсутствует. Вращательный нистагм головы и глаз наступает только при дальнейшем уменьшении наркоза. При глубоком наркозе наблюдаются также компенсаторные установки глаз (тонические лабиринтные и шейные рефлексы на мышцы глаз), а иногда они появляются уже до вращательного нистагма. Но нельзя смешивать вращательный нистагм с так называемым нистагмом наркоза (горизонтальным и вертикальным), который может появляться в покое и не зависит от лабиринтов, так как он наблюдается и у безлабиринтных
Наркотики жирного ряда (эфир, хлороформ и др.)
511
обезьян. Он вызывается воздействием наркотика на центральную нервную систему и прекращается при выходе из наркоза.
Если нормальную обезьяну подвергнуть очень глубокому наркозу, при котором не появляются вращательные реакции на конечности и таз, но зато наблюдаются рефлексы положения и вращательные реакции на голову, и в этом состоянии поворачивать животное в воздухе, то наступит поворот головы. Эта реакция вызовет соответствующие тонические шейные рефлексы, благодаря которым передняя («челюстная») лапа разгибается, но из-за отсутствия вращательной реакции она не будет отводиться. Следовательно, наркоз обусловливает у обезьяны модификацию вращательной реакции.
Все это удается установить в такой стадии, при которой еще никак не обнаруживаются установочные рефлексы. Последние снова появляются только при дальнейшем выходе из наркоза и притом таким образом, что сперва появляются установочные рефлексы на голову. Иногда установочные рефлексы тела на голову проявляются до лабиринтных установочных рефлексов. Тогда голова животного в воздухе дезориентирована (например, при положении на боку), в то время как в нормальном положении она поворачивается, как только животное кладут на стол. У безлабиринтных животных установочный рефлекс тела на голову также проявляется раньше шейных установочных рефлексов и установочных рефлексов тела на тело. Лабиринтные установочные рефлексы (у обезьян с интактными лабиринтами) появляются только немного позднее и при выходе из наркоза развиваются быстро, так что животное из всех положений в воздухе приводит голову в нормальное состояние. Шейные установочные рефлексы вначале еще отсутствуют, и тело поэтому не следует за головой. Но вскоре появляется и эта реакция. Под конец постепенно становятся снова нормальными и установочные рефлексы тела на тело. Правда, при боковом положении на столе можно большей частью видеть разгибание одной и сгибание другой задней конечности, возможно и слабое вращение таза до появления рефлексов выпрямления на голову. Однако дело не доходит до того, чтобы животное оказалось в состоянии сесть.
Постепенное появление установочных рефлексов тела на тело и завершение акта сидения при пробуждении от наркоза описаны более подробно в пятой главе.
После полного развития установочных рефлексов реакции на прогрессивные движения и прежде всего «готовность к прыжку» еще не вызываются. Тогда может случиться, что животное, у которого почти не проявляются никакие нарушения, при прыжке со стола сильно ударяется головой об пол. Прогрессивные реакции и вращательные реакции на конечности и таз восстанавливаются только через продолжительное время после появления установочных рефлексов.
а) Эфир и хлороформ
Верстеег наблюдал у кролика при выходе из эфирного наркоза следующую последовательность восстановления рефлексов.
1. Пателлярный и сгибательный рефлексы.
2. Отклонение глаз (горизонтально, ротаторно и вертикально) при вращении.
3. Отклонение глаз после вращения.
4. Горизонтальный вращательный нистагм глаз.
5. Компенсаторные круговые движения глаз.
512
Одиннадцатая глава. Действие ядов
6. Вертикальный и ротаторный нистагм глаз.
7. Поствращательный нистагм глаз.
8. Компенсаторные вертикальные отклонения глаз.
9. Лабиринтные установочные рефлексы на голову.
10. Установочные рефлексы тела на голову.
11. Шейные установочные рефлексы на переднюю часть тела.
12. Установочные рефлексы тела на тело.
13. Лифтная реакция.
14. Готовность к прыжку.
15. Шейные установочные рефлексы на заднюю часть тела.
Тонические шейные и лабиринтные рефлексы часто наблюдались в такой стадии, при которой отсутствовали все установочные рефлексы. Наркозный нистагм был установлен во многих случаях.
В общем можно сказать, что рефлексы при хлороформном наркозе изменяются примерно таким же образом, как и при эфирном.
При этих наблюдениях заслуживает внимания то, что из компенсаторных лабиринтных установок глаз круговые движения восстанавливаются раньше, чем вертикальные отклонения, и что горизонтальный нистагм глаз появляется раньше, чем вертикальный и ротаторный. Последнее уже наблюдал Ротфельд (2). Далее надо сказать, что шейные установочные рефлексы на переднюю часть тела развиваются раньше, чем на заднюю. Ротфельд (1) описал тонические шейные рефлексы на конечности у кролика при глубоком хлороформном наркозе.
б) Уретан
Верстеег нашел для уретана, переходя от более легкого к более глубокому наркозу, следующую последовательность исчезновения рефлексов.
1. Прогрессивные реакции.
2. Шейные установочные рефлексы на заднюю часть тела.
3. Установочные рефлексы тела на тело.
4. Ротаторный и вертикальный вращательный нистагм глаз.
5. Шейные установочные рефлексы на переднюю часть тела.
6. Вращательная реакция головы.
7. Установочные рефлексы тела на голову.
8. Горизонтальный вращательный нистагм глаз.
9. Лабиринтные установочные рефлексы на голову.
10. Вращательная реакция глаз (горизонтальная, ротаторная и вертикальная).
11. Компенсаторные установки глаз (вертикально и ротаторно).
12. Корнеальный рефлекс.
13. Пателлярный и сгибательный рефлексы.
Иногда находили тонические шейные и лабиринтные рефлексы на мышцы тела в стадии, при которой отсутствовали установочные рефлексы. Спонтанный наркозный нистагм не принимался в расчет.
При этом бросается в глаза, что и здесь, как при эфирном наркозе, горизонтальный вращательный нистагм глаз более устойчив, чем ротаторный и вертикальный. Оба последних вида нистагма исчезают раньше, чем лабиринтные рефлексы (при действии эфира — наоборот). И отклонения головы при вращении прекращаются относительно раньше.
в) Паральдегид
Паральдегид при усиливающейся глубине наркоза вызывает следующую последовательность исчезновения рефлексов (Versteegh).
Наркотики жирного ряда (эфир, хлороформ и др.)
513
1. Шейные установочные рефлексы на заднюю часть тела.
2. Прогрессивные реакции.
3. Установочные рефлексы тела на тело.
4. Шейные установочные рефлексы на переднюю часть тела.
5. Вертикальный вращательный нистагм глаз (вращательная реакция сохранена).
6. Ротаторный вращательный нистагм глаз (вращательная реакция сохранена).
7. Лабиринтные установочные рефлексы на голову.
8. Установочные рефлексы тела на голову.
9. Вращательные реакции головы.
10. Компенсаторные установки глаз (вертикально).
11. Горизонтальный вращательный нистагм глаз.
12. Корнеальный рефлекс.
13. Вращательная реакция глаз (горизонтально, вертикально и ро- таторно).
14. Компенсаторные круговые движения глаз.
15. Пателлярный и сгибательный рефлексы.
До сих пор в исследуемых стадиях паральдегидного наркоза не удалось установить достоверных тонических шейных и лабиринтных рефлексов на мышцы тела.
Часто наблюдался спонтанный наркозный нистагм.
Заслуживает внимания то, что, как и при эфире, компенсаторные круговые движения более устойчивы, чем вертикальные отклонения глаз, в то время как, наоборот, из разных видов вращательного нистагма, как уже установил Ротфельд (2), сперва исчезает вертикальный, затем ротаторный и под конец горизонтальный. Причем вращательные реакции глаз при этом остаются сохраненными. Вращательная реакция головы зато прекращается раньше. Установочные рефлексы на голову исчезают после вертикального и ротаторного, но до горизонтального вращательного нистагма*
г) Алкоголь
Популярным объяснением нарушения равновесия, положения и движения действием алкоголя является повреждение функции мозжечка. Однако, по моему мнению, для этого объяснения нет достаточного основания.
Ротфельд (3) установил при остром отравлении алкоголем кролика следующие нарушения в аппарате глазных мышц.
1. Исчезание вертикального компенсаторного положения глаз при сохраненной вертикальной вращательной реакции глаз.
2. Исчезание вертикального и ротаторного вращательного нистагма до затухания горизонтального, в то время как вертикальные и ротаторные вращательные реакции глаз остаются сохраненными.
3. Горизонтальный наркозный нистагм в покое при боковом положении головы.
Верстеег обнаружил у кроликов в основном то же поведение исследованных им рефлексов, как и после остальных наркотиков жирного ряда. В частности, он смог установить следующее.
Маленькие дозы (3 мл алкоголя на килограмм в 10%-м разведении per os) не изменяют лабиринтных и установочных рефлексов тела и не влияют на внешнее поведение животного. Прекращаются только реакции на прогрессивные движения (лифтная реакция, готовность к прыжку).
33 Р. Магнус
514
Одиннадцатая глава. Действие ядов
Опыт VIII. Кролик, 1.2 кг.
9 ч. 55 м. 5 мл алкоголя на килограмм per os.
10 ч. 45 м. Животное сидит немного вяло в нормальном положении, при щипании хвоста животное делает несколько неуверенных, хотя и нормальных прыжков и при этом часто падает на бок.
При исследовании наблюдалось следующее.
Установочные рефлексы тела на тело выражены хорошо. Лабиринтные установочные рефлексы на голову также положительны. Шейные установочные рефлексы на тело полностью исчезли.
Интенсивность лифтной реакции сильно снижена.
Все остальные рефлексы нормальны.
Опыт VII. Кролик весом 1.45 кг.
10 ч. 45 м. 5 мл алкоголя на килограмм per os.
11 ч. 00 м. Животное сидит нормально и спокойно. Нормально прыгает.
При исследовании установочных рефлексов оказалось, что нет и следа от шейных установочных рефлексов, которые до этого отчетливо проявлялись. Остальные установочные рефлексы нормальны.
Кроме прогрессивных реакций, наблюдаются все остальные рефлексы.
Дозы в 5 мл на килограмм уже вызывают отчетливые явления, но животные полностью поправляются.
В обоих опытах, каК и в целом ряде других исследований, было обнаружено, что шейные установочные рефлексы парализуются избирательно, в то время как другие установочные рефлексы сохраняются еще без изменения. Следовательно, в то время как голова и тело в отдельности сами по себе ставятся правильно, отсутствует связующее промежуточное звено, которое заставляет тело всегда правильным образом следовать за головой. Каждая из обеих частей как бы идет своей дорогой. Как немного пьяному человеку, так и животному еще легко выполнять нормальный бег (опыт VII), причем установочные рефлексы на голову и тело сохраняют равновесие; однако в большинстве случаев впоследствии уже отчетливо наступают нарушения ходьбы от опьянения (опыт VIII).
В редких случаях уже на этой стадии можно наблюдать нарушение установочных рефлексов тела на тело.
Опыт VI. Кролик, 1.42 кг.
10 ч. 00 м. 5 мл алкоголя на килограмм per os.
10 ч. 30 м. Животное начинает качаться и почти все время лежит задней частью тела на боку. Иногда оно прыгает и бегает, однако все это выполняется некоординированно.
Лифтная реакция исчезла, пателлярный и сгибательные рефлексы выражены отчетливо.
Лабиринтные установочные рефлексы на голову положительны. Если голову держать в боковом положении и трясти тело на подставке, то животное поднимается с трудом; установочные рефлексы тела на тело слабы, но еще сохранены. Шейные установочные рефлексы задней части тела нельзя уже вызвать поворотом головы при положении животного на спине с фиксированной верхней частью тела.
Если же держать животное за таз в боковом положении в воздухе, то голова благодаря лабиринтным установочным рефлексам становится прямо и передняя часть тела следует за этим движением.
Из этих двух наблюдений можно заключить, что шейный установочный рефлекс влияет только на переднюю часть тела.
Следовательно, у этих животных лабиринтный установочный рефлекс на голову был нормальным, причем этот цепной рефлекс распространялся только на переднюю часть и уже не действовал на заднюю часть тела. Установочные рефлексы тела на тело и шейные установочные рефлексы были явно нарушены. Из-за этого создалось особо сильное нарушение равновесия задней части тела.
Очень часто удается наблюдать, что животное в покое лежит задней частью тела на боку, в то время как голова и передняя часть тела на-
Наркотики жирмого ряда (афир, хлороформ и др.)
515
ходятся в нормальном положении. Если раздражать животное (щипанием хвоста), то поднимается и задняя пасть тела в позу «сидения», но через некоторое время снова падает. При дальнейшем развитии наркоза все тело оказывается на боку, только голова более или менее поворачивается в нормальное положение. В этом состоянии исследование в воздухе показывает, что лабиринтные установочные рефлексы на голову еще сохранены. Наконец, голова валится на бок и исчезают все установочные рефлексы. Иногда при затухании лабиринтных установочных рефлексов еще удается наблюдать слабые установочные рефлексы тела на голову.
Следующий пример с немного большей дозой алкоголя показывает, что последний рефлекс иногда может сохраниться дольше, чем лабиринтные установочные рефлексы.
Опыт XV. Кролик, 2 кг.
14 ч. 48 м. 7 мл алкоголя на килограмм per os.
15 ч. 05 м. Животное находится в положении на боку,, голова также на боку,, причем находящийся наверху глаз отклонен вниз (компенсаторная установка глаз).
Имеются шейный установочный рефлекс, лабиринтный установочный рефлекс, установочный рефлекс тела на тело, готовность к прыжку.
Установочный рефлекс тела на голову слабо положительный;, если животное спокойно, то голова находится в положении на боку, но если энергично потрясти животное над полом, то последует поворот головы в нормальное положение. Если раздражать животное щипанием хвоста, когда оно спокойно лежит на боку, то голова также принимает правильное положение.
15 ч. 15 м. Положение животного, как в 15 ч. 05 м.
Установочный рефлекс тела на голову полностью,исчез. Компенсаторная установка глаз, пателлярный рефлекс, сгибательный рефлекс, вращательная реакция глаз и нистагм положительны.
15 ч. 45 м. Нет никаких существенных изменений.
16 ч. 20 м. То же самое. Резко положительный вращательцый нистагм.
16 ч. 25 м У животного опистотонус, оно делает шагательные движения при положении на боку. Остальное без изменения.
16 ч. 45 м. То же самое.
На следующий день животное опять нормально.
На другом кролике при исключении лабиринтных установочных рефлексов вследствие двусторонней лабиринтэктомии было, еще показано,, что установочный рефлекс тела на голову действительно, более устойчив к алкоголю, чем шейные установочные рефлексы и установочные рефлексы тела на тело. Животное лежало на боку; если его в таком положении потрясти, то оно поднимало только голову.
Когда исчезли все установочные рефлексы, можно было наблюдать еще компенсаторную установку глаз, вращательную реакцию глаз и нистагм, а также корнеальный рефлекс, сгибательный и пателлярный рефлексы.
Следовательно, при отравлении кролика алкоголем удается констатировать постепенное нарушение установки тела и движений и связать это с поведением отдельных установочных рефлексов.
На первой стадии все установочные рефлексы нормальны, из остальных исследуемых рефлексов отсутствуют только прогрессивные реакции. Подопытные животные не проявляют в покое и при движении на полу никаких существенных нарушений.
На второй стадии исчезают шейные установочные рефлексы, притом сперва на заднюю, а потом на переднюю часть тела. Затем голова и тело устанавливаются независимо друг от друга, но животное сидит еще нормально, однако при ходьбе движения большей частью становятся уже неуверенными и животные часто падают на бок (походка пьяного).
На третьей стадии исчезают установочные рефлексы тела на тело и животное уже не в состоянии держать тело в нормальном положении.
33*
516
Одиннадцатая глава. Действие ядов
Только голова еще стоит прямо благодаря совместному действию лабиринтных установочных рефлексов и установочных рефлексов тела на голову. Из обоих последних рефлексов иногда лабиринтный установочный рефлекс исчезает раньше установочного рефлекса тела на голову, который остается слабо положительным.
На четвертой стадии все установочные рефлексы исчезают. Животное лежит беспомощно на боку и не в состоянии при раздражении привести в нормальное положение хотя бы одну часть своего тела.
Уже изучение этой первой группы действия ядов дает ряд интересных фактов.
Различные рефлексы исчезают со строго равномерной последовательностью. У разных наркотиков мы находим в основном одинаковое действие, но в частностях — характерные отличия. Из лабиринтных рефлексов выпадают сперва не рефлексы полукружных каналов и потом рефлексы отолитов (или наоборот), как это имело бы место при действии на периферический орган чувств, а яды проникают в Центральную нервную систему и парализуют избирательно отдельные центры.
Последовательность в основном такая: 1) прогрессивные реакции (у обезьяны поворот таза и верхних конечностей), 2) установочные рефлексы, 3) в различной последовательности компенсаторные установки глаз, вращательные реакции на голову и глаза, тонические шейные и лабиринтные рефлексы на мышцы тела.
Вращательный нистагм всегда затухает раньше вращательной реакции, так что на определенной стадии остается только медленная фаза вращательной реакции головы и глаз.
Из вращательных нистагмов глаз горизонтальный является самым устойчивым, вертикальный — самым чувствительным.
Из компенсаторных установок глаз ротаторные устойчивее вертикальных (паральдегид, эфир).
Из установочных рефлексов наиболее устойчивы рефлексы на голову, чем на тело.
Особенно легко прекращаются при алкоголе шейные установочные рефлексы; связь этого исчезновения с нарушением движения у пьяных уже обсуждалась.
Б. Стрихнин
Совсем другая картина наблюдается при отравлении таким сильным возбуждающим средством, как стрихнин. Ионкхофф (69) исследовал общее поведение лабиринтных рефлексов у кроликов при внутривенной инъекции.
Дозы в 0.01 мг на килограмм вызывают легкое, в 0.02 мг — отчетливое повышение лабиринтных рефлексов. После 0.05 мг в общем отмечается повышенная возбудимость рефлексов и усиление сухожильных рефлексов. Дозы от 0.15 до 0.5 мг вызывают беспокойство и легкие судороги, 0.7—1 мг —тяжелые общие судороги в положении на боку и остановку дыхания.
До введения рекомендуется вставить животным трахеальную канюлю, чтобы в любое время можно было бы спасти животное искусственным дыханием при судорогах, сопровождающихся остановкой дыхания.
Стрихнин
517
Общее течение отравления стрихнином
Изменение лабиринтных рефлексов при увеличивающемся стрихнин- ном отравлении видно из следующих двух протоколов и таблиц, причем первые получены на таламическом животном, а вторые — на интактном кролике.1
Опыт I. 1 апреля 1919 г. (см. табл. I).
Кролик, 1.76 кг. Предыдущее исследование выявило наличие всех лабиринтных рефлексов.
9 ч. 15 м. Эфирный наркоз. Вставление канюли в трахею. Перевязка сонных артерий, ваготомия, экстирпация мозга до таламуса, незначительная потеря крови.
9 ч. 40 м. Конец операции. Прекращение эфирного наркоза. Спонтанное дыхание.
10 ч. 40 м. Исследование лабиринтных рефлексов. Имеются рефлексы положения. Правда, лабиринтные установочные рефлексы не очень подвижны, при висячем положении головой вниз голова находится под углом 90°. Наблюдаются лабиринтные рефлексы на движения, не удалось только установить вращательный нистагм головы и постнистагм.
Введение внутривенно 0.01 мг азотнокислого стрихнина.
10 ч. 50 м. При висячем положении головой вниз голова отводится немного дор- сально относительно нормального положения. Другие лабиринтные установочные рефлексы и компенсаторные установки глаз также подвижнее, чем до введения стрихнина. Рефлексы на движение не изменены, только вращательные реакции глаз, по-видимому, немного живее.
Еще раз вводится 0.01 мг стрихнина на килограмм.
10 ч. 55 м. Лабиринтные рефлексы теперь уже явно усилены. Лабиринтные установочные рефлексы очень отчетливы, компенсаторные установки глаз чрезвычайно сильно выражены.
Вращательная реакция головы и постреакция живые, наблюдаются также вращательный нистагм головы и постнистагм.
Все вращательные реакции глаз живые, наблюдается лифтная реакция, рефлекс же готовности к прыжку не отчетлив.
Еще раз вводится внутривенно 0.05 мг стрихнина на килограмм.
11 ч. 05 м. Поведение лабиринтных рефлексов то же самое, как в 10 ч. 55 м., отсутствует только готовность к прыжку.
Введение 0.1 мг стрихнина на килограмм.
11 ч. 10 м. Положение то же. Введение 0.2 мг стрихнина на килограмм.
И ч. 20 м. Вращательный нистагм головы менее живой, исчез поствращательный нистагм головы. Вращательный нистагм глаз и постнистагм теперь не очень живые.
Введение 0.3 мг стрихнина на килограмм.
11 ч. 30 м. Наблюдаются все лабиринтные рефлексы положения. Компенсаторные установки глаз выражены особенно отчетливо.
Вращательные реакции головы и глаз и постреакции теперь не очень живые, исчезли вращательный нистагм головы и глаз и постнистагм. Еще имеется лифтная реакция, рефлекс готовности к прыжку уже нельзя вызвать. Судорог еще нет.
11ч. 40 м. Наступил первый сильный общий приступ судорог с остановкой дыхания.
Во время этого приступа исчезли все лабиринтные рефлексы, за исключением компенсаторных установок глаз (вертикально и ротаторно), которые были даже очень живыми.
Вскоре после этого вращательные реакции головы и глаз оказались слабо выраженными, отсутствуют вращательный нистагм головы и глаз, а также лифтная реакция.
Тонические лабиринтные рефлексы на конечности не отчетливы.
12 ч. 20 м. Компенсаторные установки глаз еще очень живые, вращательные реакции головы и глаз и последующие реакции слабо выра кены, все остальные лабиринтные рефлексы исчезли.
12 ч. 40 м. Компенсаторные установки глаз еще отчетливы, все остальные лабиринтные рефлексы исчезли.
У животного непрерывные сильные судороги до самой смерти (в час дня).
Вскрытие. Полностью отсутствует головной мозг. Таламус и четверохолмие сохранены, в мосте и у основания никаких кровоизлияний.
1 В этих и последующих протоколах и таблицах, если нет другого примечания, под «компенсаторными установками глаз» понимаются только вертикальные отклонения.
518
Одиннадцатая глава. Действие ядов
Таблица I
Таламический кролик, 1.76 кг
Рефлексы положения
Рефлексы на движение
Примечания
д
д
п Ь
W
03§
Д
Д й
03 Д
и а щ W ft X E-* а £ft Д Е-1 ВД О
§
^ а о о
|1 Д а>
£
Д
а аз
»3
\о a д о Ч и
§
к
аз
4 >8< аз Д
ft Ei
аз о
5 Д К д о 03
2 и
(Г О § *
i
д
g,
03 ft
Я 2 д5
ft
О со Д м Д м О
аз а д В к й я и ч о о д к д д
вращательные
реакции
реакции на прогрессивные движения
головы
глаз
и и й д
в
Д аз Ч ft
о к д 1
oft
Ен В 2 К
к
в
дг
к
д
03
ft
&
Ен
О
§
к
д
н
к
д
аз
ft
я
ä
О
Д
д
1 IY 1919
+
Преобладание лабиринтных рефлексов.
+
4-
4-
4-
4-
+
4-
Слуховая реакция. +
9 ч. 45 м.
Конец операции. Закончился эфирный наркоз. Животное дышит
спонтанно.
10 ч. 40 м.
0.01
+
Преобладание лабиринтных рефлексов.
+
+
—
+
+
+
4-
Слуховая реакция?
10 ч. 50 м.
0.01
+
То же.
+
+
+
—
+
4-
+
+
То же -f-
10 ч. 55 м.
0.05
+
»
+
+
+
+
+
+
4-
+
+
4-
?
» +
И ч. 05 м.
0.1
4-
»
+
+
+
+
+
+
+
+
+
+
—
* 4-
И ч. 10 м.
0.2
+
»
+
4-
4-
4-
+
+
+
4-
4-
+
—
» +
И ч. 20 м.
0.3
+
»
+
+
+
4-
т
+
+
4-
+
—
» +
И ч. 30 im.
+
»
++
+
—
4-
—
+
—
» ?
11 ч. 40 м.
—
Т ?
+
+
+
—
т
—
—
—
Общие судорожные приступы. Имеются компенсаторные круговые движения.
12 ч. 20 м.
—
—
+
+
+
—
+
—
—
—
Слуховая реакция. —
12 ч. 40 м.
—
—
+
+
—
—
—
—
—
—
То же. —
13 ч. 00 м.
Смерть. Вскрытие: не обнаружено никаких макроскопических отклонений.
-f- обозначает нормальные лабиринтные рефлексы; + ослабленные лабиринтные рефлексы; ^ повышенные лабиринтные рефлексы; ^ очень сильно выраженные лабиринтные рефлексы; — отсутствие лабиринтных рефлексов.
Стрихнин,
519
Таблица II
Таламический кролик, J .62 кг
Дозы стрихнина (внутривенно) (мг)
Рефлексы положения
Рефлексы на движение
Примечания
i
о
ам
|3
^я
О м
Я <х>
я а Я 2
я Я ю я Я о Я я
Я
о
К
ф
п
в< о Я ft
о Q5 О
ц
о 2 2 Я
ц
о w н Я
1
£я
и
2 ft
Я 2 и «,
ft
О СО
й Ч Я м о
2 я я я я Л Я « ч о о я к я я
вращательные
движения
реакции на прогрессивные
движения
головы
глаз
и g
Ё ft Я а>
Ч а
§1
§ä
н Я о
и Я
я
я
ft
о
ft
Я
Рн
я
В
О
§
§
ft
§
о
ft
6
6и
о
«
5 IV 1919, 9 час.
+
Преобладают лабиринтные рефлексы.
+
+
+
+
+
+
+
Слуховая реакция. -f-
10 ч. 30 м.
+
То же.
+
+
+
+
+
+
+
После наркоза слуховая реакция. -f-
10 ч. 40 м.
0.05
И ч. 10 м.
+
То же.
+
+
+
+
+
+
+
+
+
+
+
+
Оживленные защитные рефлексы.
И ч. 45 м.
0.05
+
»
+
+
+
+
+
+
+
+
+
+
+
+
То же.
12 ч. 30 м.
0.05
+
»
+
+
+
+
+
+
+
+
+
+
+
+
»
14 ч. 15 м.
0.4
+
»
+
+
+
+
+
+
+
+
+
+
+
+
Легкие судороги.
14 ч. 30 м.
0.4
+
»
+
+
+
+
+
+
+
+
То же.
14 ч. 45 м.
0.4
+
»
+
+
+
+
+
+
+
+
»
14 ч. 55 м.
+
—
4- +'
+
+
+
+
—
—
Сильные судороги.
15 ч. 00 м.
—
—
Остановка дыхания.
15 ч. 02 м.
—
—
+
+
Искусственное
дыхание.
15 ч. 04 м.
—
—
'+
+
+
—
+
—
—
—
Животное снова дышит спонтанно.
15 ч. 10 м.
+
?
+
+
+
+
+
+
—
?
15 ч. 40 м.
—
—
+
+
—
—
—
—
—
?
Сильные общие судороги.
15 ч. 45 м.
|Смерть. Вскрытие: макроскопически нет
никаких
отклонении.
520.
Одиннадцатая глава. Действие ядов
Опыт И. 5 апреля 1919 г. (см. табл. II).
Кролик, 1.62 кг. Все лабиринтные рефлексы в норме.
9 ч. 30 м. Эфирный наркоз. Трахеальная канюля, затем выключение эфирного наркоза.
10 ч. 30 м. Все лабиринтные рефлексы снова в норме.
10 ч. 40 м. 0.05 мг на килограмм азотнокислого стрихнина вводится внутривенно.
11 ч. 10 м. Отчетливые лабиринтные рефлексы. Ощутимы тонические лабиринтные рефлексы на конечности. Компенсаторные установки глаз очень выражены. Вращательные реакции очень живые. Реакции на прогрессивные движения не очень отчетливы. Животное очень восприимчиво к раздражениям. Сухожильные рефлексы более живые, чем до введения стрихнина.
11 ч. 45 м. Введено 0.05 мг на килограмм азотнокислого стрихнина внутривенно. Положение то же.
12 ч. 30 м. 0.05 мг на килограмм азотнокислого стрихнина внутривенно. Положение то же.
14 ч. 00 м. Без изменений.
14 ч. 15 м. У животного наблюдаются легкие судороги и мышечное беспокойство. Состояние лабиринтных рефлексов, как в И ч. 10 м.
0.4 мг на килограмм стрихнина введено внутривенно.
14 ч. 30 м. Животное беспокойно, отмечаются легкие судороги. Наблюдаются все- лабиринтные рефлексы положения и движения. Компенсаторные установки глаз являются единственными лабиринтными рефлексами, которые отчетливо усилились-
Еще раз 0.4 мг стрихнина на килограмм внутривенно.
14 ч. 45 м. Положение то же. Еще раз 0.4 мг внутривенно.
14 ч. 55 м. Лабиринтные установочные рефлексы едва проявляются. Отсутствуют лабиринтныё рефлексы на конечности. Компенсаторные установки глаз чрезвычайно- живые. Вращательные реакции головы и глаз и постреакции еще наблюдаются. Вращательные постреакции глаз слабо выражены. Вращательный нистагм головы и глаз- иногда слабо проявляется, нистагм теперь не очень отчетливо распознается. Исчезли рефлексы на прогрессивные движения. У животного постоянное мышечное беспокойство.
Это состояние переходит в сильный приступ судорог с остановкой дыхания.
15 час. Судороги. Исчезли все лабиринтные рефлексы. Остановка дыхания. После нескольких минут искусственного дыхания животное вновь начинает спонтанно дышать и одновременно восстанавливаются и компенсаторные установки глаз. Через несколько минут снова наблюдаются вращательные реакции головы и глаз и постреакция, хотя и очень слабые. Отсутствует поствращательная реакция глаз.
15 ч. 10 м. Только в положении на боку в воздухе можно видеть незначительный поворот головы в направлении нормального положения. Отсутствуют остальные- лабиринтные установочные рефлексы и установочные рефлексы тела на голову. Тонические лабиринтные рефлексы на мышцы тела вызываются неотчетливо, зато компенсаторные установки глаз очень отчетливы. Снова наблюдаются вращательные реакции головы и глаз; реакции на прогрессивные движения отсутствуют или в крайнем случаи только намечаются.
Животное дышит спонтанно, ощутимо постоянное «напряжение» мышц как при спокойном положении животного, так и при движении.
15 ч. 40 м. Незадолго до общего приступа судорог исчезли все лабиринтные рефлексы, кроме компенсаторных установок глаз.
Во время этих судорог животное погибает.
В дальнейшем опыте, который здесь подробно не описывается, кролику ввели внутривенно 0.9 мг стрихнина на килограмм веса. Последовал общий приступ судорог с остановкой дыхания. Во время этих судорог отсутствовали все лабиринтные рефлексы. Сразу с началом спонтанного дыхания наблюдались очень сильные компенсаторные установки глаз (вертикально), в то время как остальные лабиринтные рефлексы еще- отсутствовали. Затем появлялись вращательные реакции головы, а вращательные реакции глаз были выражены очень слабо. Нистагм головы и глаз отсутствовал. Также- отсутствовали установочные рефлексы и реакции на прогрессивные движения. В последующих стадиях вращательные реакции глаз становились отчетливыми и можно было наблюдать лабиринтные установочные рефлексы. Через короткое время после этого восстановились и тонические лабиринтные рефлексы на мускулатуру тела, вращательные реакции головы и глаз стали сверх нормы живыми, появился нистагм головы и глаз. Одновременно наблюдались и реакции на прогрессивные движения. Через 1—2 часа были введены внутривенно еще 3 дозы по 0.2, 0.3 и 0.4 мг стрихнина. После этого снова последовал общий приступ судорог, после которого можно было доказать восстановление лабиринтных рефлексов приблизительно в той же последовательности г как это было описано выше.
Стрихнин
521
Обобщая эти опыты, можно сделать вывод, что стрихнин действует с определенной последовательностью на различные лабиринтные рефлексы. Уже такие маленькие дозы, как 1/100—1/50 мг на килограмм, отчетливо усиливают у кролика лабиринтные рефлексы. Только реакции на прогрессивные движения не показали никакого усиления при отравлении стрихнином. После 0.4—0.5 мг на килограмм веса исчезло усиление вращательных реакций (вращательные реакции головы и глаз). При несколько больших дозах могут уже отсутствовать вращательный нистагм головы и глаз и соответствующие постнистагмы, в то время как реакции поворота головы и глаз, так же как и установочные рефлексы, еще наблюдаются.
Установочные рефлексы сохраняются до появления общего судорожного приступа. Он обычно наступает у кролика при дозах от 0.7 мг или немного выше. Тогда животное падает на бок и установочные рефлексы исчезают. Отсутствует нистагм головы и глаз, нё удается выявить прогрессивные реакции. Однако после этих доз могут еще наблюдаться слабые тонические лабиринтные рефлексы на конечности и реакции поворота головы и глаз, а компенсаторные установки глаз еще очень живые.
При исследовании отдельных установочных рефлексов выявилось, что установочные рефлексы тела на голову большей частью немного устойчивее при стрихнинном отравлении, чем лабиринтные установочные рефлексы. Но разница не очень велика.
В одном опыте, кроме того, удалось показать, что тонические шейные рефлексы на мышцы глаз держатся примерно столько же, сколько и реакции поворота головы и глаз.
Больше всего при стрихнинном отравлении бросается в глаза то, что в общем усилении лабиринтных рефлексов, которое наступает после самых малых доз стрихнина, не участвуют или очень мало участвуют реакции на прогрессивные движения; что нистагм головы и глаз исчезает в одно и то же время, тогда как реакции поворота головы и глаз еще очень живые; что установочные рефлексы исчезают одновременно с появлением больших судорожных приступов; что, наконец, после больших доз исчезают все лабиринтные рефлексы, за исключением компенсаторных установок глаз, которые остаются до самого конца очень живыми и только, когда наступает остановка дыхания, исчезают вследствие удушья. На рис. 247 (стр. 535) изображены вертикальные компенсаторные установки глаз у кролика в стадии судорог при боковом положении головы.
Следовательно, мы имеем здесь снова характерный пример того, что отдельная группа лабиринтных рефлексов имеет избирательную способность к сопротивлению против определенного яда.
Влияние стрихнина на рефлексы положения
В связи с этими общими установками следует сообщить еще о некоторых наблюдениях, которые относятся к особым свойствам отдельных лабиринтных рефлексов при отравлении стрихнином, прежде всего к тоническим шейным и лабиринтным рефлексам на мышцы конечностей.
Мы обязаны Шеррингтону (4—6) знанием того, что стрихнин глубоко нарушает координацию рефлексов спинного мозга, а именно, дозы стрихнина, не вызывающие еще судорог (0.08—0.1 мг на килограмм веса у кошки), превращают целый ряд рефлексов торможения в рефлексы возбуждения. Чем больше повышаются дозы стрихнина, тем больше рефлексы торможения «извращаются». Например, на vastocrureus-препарате
522
Одиннадцатая глава. Действие ядов
нормального децеребрированного животного получают при раздражении какого-нибудь афферентного ипсилатерального нерва задней ноги торможение тонуса четырехглавой мышцы. При стрихнинном отравлении наблюдается «извращение» эффекта раздражения, в результате этого наступает сокращение мышцы. При раздражении бедренной ветви седалищного нерва извращение наступает только после немного больших доз стрихнина, чем при раздражении других афферентных нервов ноги. Чтобы получить это извращение с коры мозга (разгибание вместо сгибания ноги), требуются еще большие количества стрихнина, которые уже одновременно вызывают у животных судорожные приступы.1 Еще больше нужно повысить дозы, если хотят получить с определенных мест коры мозга вместо открытия рта закрытие его. И при раздражении одного и того же чувствительного нерва Шеррингтон находил отличия, если он применял различные виды раздражения. Так, иногда извращение уже наблюдалось при грубом механическом раздражении, если за медленным раздражением следовало еще торможение. Одновременно выяснился интересный факт, что при отравлении стрихнином снимается реципрокная иннервация и что при вызове одного из названных рефлексов одновременно наблюдается и сокращение антагонистических мышц, в то время как в норме всегда одна мышца расслабляется, если сокращается ее антагонист. Соответствующее извращение вазомоторных рефлексов под влиянием стрихнина было найдено Бейлиссом (Bayliss), дыхательного рефлекса — Зееманом (Seemann).
На одной лекции при демонстрации «стрихнинного извращения» на кошке я сделал попутно наблюдения, что после дозы стрихнина, которая привела при раздражении малоберцового нерва к отчетливому извращению эффекта на vastocrureus’e той же стороны, тонические рефлексы с лабиринтов и шеи на передние ноги остались полностью без изменения и все торможения особенно хорошо поддавались демонстрации. В соответствии с этими данными я вместе с Вольфом (13) исследовал, как ведут себя тонические шейные и лабиринтные рефлексы при отравлении стрихнином. Всего было проведено 11 опытов на децеребрированных кошках. В части опытов изолировались вышеописанным способом или vastocrureus, или трехглавая мышца. В результате этих опытов оказалось, что даже дозами стрихнина, вызывающими судороги, не удается «извратить» шейные и лабиринтные рефлексы.
Азотнокислый стрихнин вводится внутривенно. Доза в 0.08 мг на килограмм веса вызывает в большинстве случаев значительное увеличение ригидности на децеребрированных кошках, после введения 0.13 мг наблюдалась повышенная возбудимость рефлексов, после 0.16 мг у животного наступали, по-видимому, «спонтанные» вздрагивания, 0.3 мг вызывали во всех случаях сильные судорожные приступы. Часто были действенными и меньшие дозы. Так, мы наблюдали один раз типичный приступ судорог уже после введения 0.08 мг.
В опытах, где исследовались реакции изолированного vastocrureus’a, мы всегда раздражали и центральный отрезок малоберцового нерва, чтобы установить появление извращений вызванных при этом рефлексов. В опытах с изолированной трехглавой мышцей для этой цели служил центральный конец лучевого нерва. В соответствии с данными Шеррингтона и Соу- тона [Sowton (10)] мы нашли, что при применении минимальных фарадических раздражений попутно и без отравления стрихнином вызывается
1 Р. Магнус, по-видимому, подразумевает недецеребрированное животное, т. е. с сохраненной корой (Ред.).
Стрихнин
523
рефлекторное возбуждение (вместо торможения) малоберцового нерва тгли лучевого нерва той же стороны. Более сильные фарадические раздражения, наоборот, вызывают всегда рефлекторное торможение исследуемых изолированных мышц-разгибателей. Во всех случаях была достаточна доза в 0.08 мг стрихнина на килограмм веса, чтобы извратить этот эффект фарадического раздражения названных нервов и вызвать возбуждение. Но зато для извращения рефлексов после механического раздражения нервов потребовались большие дозы стрихнина (0.18—0.23 мг на килограмм). После доз от 0.09 до 0.15 мг мы могли видеть, что на растягивание нерва следовало возбуждение, на перевязывание последнего, наоборот, наступало торможение изолированной мышцы-разгибателя.
Во всех наших опытах нам никогда не удавалось получить какое-либо извращение шейных и лабиринтных рефлексов, несмотря на то что мы повышали дозы до 0.44 мг на килограмм веса, после чего наступали такие сильные общие судороги, что исследование рефлексов было едва возможным.
Несколько протоколов и рисунков приводятся здесь для подтверждения сказанного.
Кошка, 1800 г. Хлороформный наркоз, перевязка сонных артерий. Перерезка п. vagi, канюля в яремную вену для инъекции.
10 ч. 20 м. Децеребрация при глубоком наркозе. Прекращение хлороформного наркоза.
10 ч. 25 м. Начинающаяся децеребрационная ригидность.
10 ч. 45 м. Положение на боку. Если голову повернуть теменем вниз, то увеличивается тонус разгибателей обеих передних конечностей, особенно у ноги, находящейся сверху. Если повернуть голову теменем вверх, то тонус разгибателей обеих передних конечностей уменьшается, причем у верхней ноги уменьшается сильнее (комбинация шейных и лабиринтных рефлексов).
Поднимание и опускание головы при положении на боку не влечет изменений.
Давление на остистые отростки последнего шейного и первого грудного позвонка вызывает расслабление обеих передних конечностей: шейный рефлекс.
И ч. 16 м. 0.2 мг стрихнина (0.11 мг на килограмм). Сразу же увеличение тонуса разгибателей передних конечностей. Поворот головы в положении на боку действует, как и раньше; только увеличение тонуса и расслабление проявляются болеС' отчетливо. Давление на остистые отростки последнего шейного и первого грудного позвонка вызывает сильное расслабление передних конечностей. Ecjp все животное, не изменяя положения головы по отношению туловища, повернуть из положения на бойу в положение на спину, то увеличивается тонус разгибателей передних ног; если таким же способом повернуть животное на бок, то тонус разгибателей опять уменьшается (лабиринтные рефлексы). Поднимание и опускание головы в положении на спине оказывает сильное влияние: тонус разгибателей сильнее всего, если ротовая щель поднята жад горизонталью на 45°; он снижается, если ротовая щель опускается ниже горизонтали или если голова так сильно сгибается к туловищу, что морда оказывается между лапами (комбинация шейных и лабиринтных рефлексов). Поворот головы при положении на спине вызывает увеличение тонуса в «черепной» ноге и расслабление в «челюстной» (шейные рефлексы). При стоянии на ногах опускание головы вызывает расслабление, поднимание — разгибание передних ног (комбинация шейных и лабиринтных рефлексов.)
11 ч. 32 м. 0.2 мг стрихнина внутривенно (всего 0.22 мг на килограмм). После этого сильные спонтанные подергивания животного.
11 ч. 35 м. Шейные и лабиринтные рефлексы на передние конечности исследуются снова. Все они остались без изменения, но стали более отчетливыми. Особенно отчетливы уменьшение тонуса и расслабление. Теперь поднимание головы в положении на боку вызывает разгибание, опускание — расслабление передних конечностей (шейный рефлекс).
11 ч. 43 м. Внутривенно 0.2 мг стрихнина (всего 0.33 мг на килограмм).
И ч. 44 м. Общий стрихнинный судорожный приступ.
11 ч. 50 м. Животное лежит и непрерывно подергивается. Во всех положениях животного можно наблюдать неизмененные шейные и лабиринтные рефлексы. Торможение выступает так же отчетливо, как и возбуждение.
11 ч. 55 м. Внутривенно 0.2 мг стрихнина (всего 0.44 мг на килограмм веса).
11 ч. 56 м. У животного сильнейшие судороги.
524
Одиннадцатая глава. Действие ядов
12 ч. 10 м. Шейные и лабиринтные рефлексы стали слабее, но их характер не изменился. Поднимание и опускание головы при стоянии на ногах и поворот головы при положении на боку имеют пока еще то же действие, как в 11 ч. 16 м. При повороте головы в положении на боку ясно чувствуется, как при повороте головы теменем вверх, мышцы-разгибатели верхней ноги расслабляются.
Этот опыт показывает, что дозы стрихнина, вызывающие сильнейшие судороги, не приводят к извращению шейных и лабиринтных рефлексов
и что особенно все торможения остаются без изменения. Судя по этому и другим опытам,, стрихнин даже является превосходным средством, вызывающим в малых дозах (0.08— 0.1 мг на килограмм веса) при демонстрации отчетливые шейные и лабиринтные рефлексы у животных, у которых они по какой-либо причине развиты слабо.
Рис. 237 показывает, что иногда удается временно затормозить сильные стрихнинные судороги переводом головы в другое положение.
Кошка, вес 1360 г. Изоляция левого vastocru- reus’a. Сокращение мышцы вызывает опускание записывающего рычага. В начале опыта фарадическое раздражение левого малоберцового нерва вызывает расслабление vastocrureus’a. После внутривенного- введения 0.15 мг стрихнина на килограмм веса животного раздражение левого малоберцового нерва вызывает сокращение vastocmreus’a. Затем дозу стрихнина увеличивают до 0.34 мг на килограмм, после чего наступают сильные судороги. Верхняя часть тела животного лежит в положении на боку. Кривая 1 показывает судорожный приступ, при котором vastocrureus проявляет значительное усиление тонуса и быстрое движение. Полная противоположность чесательному рефлексу. При 2 голову поворачивают теменем вверх, и сразу же прекращаются судороги, а мышца расслабляется. После этого голову вновь поворачивают в исходное положение, и тотчас же появляются судороги, чтобы Рис. 237. (Наполовину умень- спонтанно (3) заглохнуть. При 4 начинается новый шеЯо). судорожный приступ, который снова прерывается (5),
так как голову снова поворачивают теменем вверх. Судороги прекращаются, мышца расслабляется. После этого голову опять поворачивают в исходное положение, и снова начинаются судороги, как будто их и не прерывали, чтобы при 6 спонтанно затухнуть.
Рис. 238 показывает, что после введения дозы стрихнина, вызывающей, сильные судороги и способной вызвать извращение эффекта раздражения лучевого нерва (фарадическое и механическое) на изолированной трехглаво^ мышце, шейные и лабиринтные рефлексы остаются без изменения,.
Кошка, 3950 г. Изолирование правой трехглавой мышцы. Препаровка для раздражения правого лучевого нерва у локтя. Затем децеребрация. При графической регистрации животное находится в положении на левом боку; правая лапа с грузом в 50 г, который стремится согнуть локоть. Разгибание локтя вызывает поднятие рычага вверх. Время в секундах.
Рис. 238а. При 3 фарадическое раздражение центрального конца лучевого' нерва. Следует расслабление трехглавой мышцы. После этого животному вводится внутривенно 0.13 мг стрихнина на килограмм веса. Видны спонтанные подергивания. Рис. ,2386. После фарадического раздражения (96) лучевого нерва наступает сокращение трехглавой мышцы. Рис. 238с. Механическое раздраже-
Стрихнин
525
ние трехглавой мышцы вследствие растягивания нерва (99) вызывает расслабление мышцы. Животное получило всего 0.23 мг стрихнина на килограмм веса. После этого наступили сильные судороги. Рис. 238d. Перевя-
СГ
Ъ Cd
Рис. 238а—е. (Наполовину уменьшено).
е
зывание нерва (102) и (рис. 238е) растягивание нерва (118) вызывают сокращение трехглавой мышцы. Значит, теперь для всех видов раздражения лучевого нерва имеется извращение. Но несмотря на это, действие поворота головы на тонус трехглавой мышцы остается совсем без изменения. (Рис.
238/). В начале кривой — трехглавая мышца находится в «спонтанном» сокращении. При 104 голова животного, находящегося в положении на боку, поворачивается теменем вверх. Сразу же расслабляется трехглавая мышца. При 105 голову поворачивают теменем вниз и происходит сокращение. При 106 голову снова поворачивают теменем вверх, после чего сразу же следует расслабление. Оно прерывается, если (107) голова приводится в среднее положение, и снова усиливается, когда (108) темя поворачивают опять вверх. При 109 голова опять находится в среднем положении; трехглавая мышца приобретает после этого средний тонус, который сразу же повышается (110), если голову поворачивают теменем вниз. При 111 видно сильное торможение тонуса, после того как темя снова очутилось наверху. До и после графической регистрации этих рефлексов наступали сильные судорожные приступы.
Рефлекс с 7-го шейного позвонка также не поддается извращению. Например, у одного животного после внутривенного введения 0.3 мг стрихнина на килограмм веса, что вызвало рис. 238/.
сильные судороги, давлением в области
границы шейных и грудных позвонков удалось вызвать расслабление va- stocrureus’a, точно такое же, как и до отравления стрихнином. Как только прекращалось давление, кончалось и расслабление.
526
Одиннадцатая глава. Действие ядов
При вызове всех этих рефлексов во время стрихнинных судорог нужно действовать осторожно и голову поворачивать медленно и спокойно. Грубыми и резкими движениями: сотрясается все животное, поэтому вызываются не поддающиеся контролю рефлексы. Еще лучше, если взять голову в руки и сперва немного подождать, пока пройдут вызванные этим рефлексы, а только потом начать поворот или наклон головы. Только таким путем можно получить чистые и точные результаты.
Приведенные выше примеры показывают, что даже дозы стрихнина* вызывающие сильные судороги и «извращающие» большое число других тормозных рефлексов, не в состоянии превратить в возбуждение исходящие с шеи и лабиринтов тонические тормозные рефлексы на мышцы конечностей. Реципрокная иннервация этих рефлексов также остается незатронутой во время отравления стрихнином. Мы в различных случаях могли наблюдать, что при определенных движениях головы — одновременно с торможением тонуса разгибателей — появлялось повышение тонуса сгибателей на исследуемой конечности. В соответствии с цитированным выше положением Шеррингтона получается удивительная картина нарушения координации в центральной цервной системе при отравлении стрихнином. При определенных средних дозах стрихнина нормально вызываемое торможение при раздражении определенных нервов переходит в возбуждение, в то время как тормозные рефлексы с других эфферентных нервов остаются неизменными. Чем больше увеличивать дозу стрихнина, тем больше торможений превращается в возбуждения. Однако имеются рефлекторные торможения, которые остаются и после самых больших доз стрихнина. При этом одна и та же мышца может реагировать на одно раздражение возбуждением (вместо нормального торможения), на другое раздражение — неизмененным торможением. При определенных рефлексах отравлением стрихнина снимается реципрокная иннервация мышц конечностей, при других рефлексах эти же мышцы конечностей имеют ненарушенную реципрокную иннервацию.
Объяснение этих фактов при современном состоянии наших знаний о способе и месте действия стрихнина в центральной нервной системе наталкивается еще на большие трудности.
Кашни (Cushny) пытался свести наблюдаемое Шеррингтоном снятие реципрокпой иннервации к тому, что при отравлении стрихнином слабые раздражения становятся максимальными или сверхмаксимальными. Однако это объяснение едва ли применимо для только что описанных наблюдений. Если уже До отравления стрихнином видны чрезвычайно сильные тонические реакции конечностей на изменение положения головы, то вызванные рефлексы едва ли можно принять за «слабые». По крайней мере невероятно, что описанное поведение можно объяснить простым количественным различием.
Влияние стрихнина на рефлексы глаз
Подобные же положения можно применить и для мышц глаз (69). Это вытекает из наблюдения за компенсаторными установками глаз. Как уже было сказано* эти последние сохраняются и после самых больших доз стрихнина до самой смерти, и все это время они чрезвычайно подвижны. Если кролика при наибольшем отравлении положить, например, на левый бок (рис. 247), то находящийся сверху правый глаз будет отведен максимально вниз, т. е. вентрально. Это вызвано, так же как и у неотравленного животного, сокращением прямой нижней мышцы с одновременным расслаблением прямой верхней мышцы глаза. Если бы у этих двух мышц прекратилась реципрокная иннервация и вследствие этого обе мышцы
Стрихнин
527
сократились бы, то глаз находился бы в среднем положении и не был бы максимально отклонен.
То же самое было установлено при вызове калорической реакции на мышцы глаз. Следующий опыт является примером этого.
5 мая 1919 г. Кролик, 3.1 кг. Лабиринты калорически хорошо раздражаются.
9 час. Канюля вставлена в трахею, внутренняя и внешняя прямые мышцы левого глаза отпрепарованы. Места прикрепления мышц на глазном яблоке свободно отпрепа- ровываются и перерезаются, концы мышц прикрепляются ниткой к двум регистрирующим рычажкам. Глаз экстирпируют. Во время всего опыта легкий эфирный наркоз (1/10). На верхней кривой регистрируется прямая внутренняя мышца, на нижней — прямая наружная мышца.
Рис. 239.
10 ч. 30 м. Промывание левого слухового прохода водой 14°. Наружная прямая мышца отчетливо сокращается с хорошим равномерным нистагмом. Внутренняя прямая мышца расслабляется очень мало и дает едва заметный нистагм.
10 ч. 30 м. (рис. 239). При знаке + вводится внутривенно 0.34 мг стрихнина на килограмм веса.
Примерно через 50 сек. наступает общий судорожный приступ. Обе мышцы глаза сразу одновременно удивительно одинаково сокращаются (а), однако возможно, что это сокращение симулируется сильным общим судорожным движением всего животного. Затем внутренняя мышца начинает расслабляться, а наружная — сокращаться. При стрелке вверх начинается промывание левого слухового прохода водой 14°. После этого следует медленное максимальное сокращение наружной прямой мышцы, причем в поднимающейся части кривой видно несколько вялых ударов нистагма. Одновременно- расслабляется внутренняя прямая мышца. Обе кривые являются полным зеркальным отражением. При второй стрелке вверх прекращается введение воды. Кривые постепенно приходят в исходное положение.
Следовательно, если исходить от первого молниеносного сокращения* то видно, что как при общих судорогах, так и при калорической реакции мышцы глаз полностью сохраняют реципрокную иннервацию и сокращение наружной мышцы сопровождается зеркальным отражением расслабления внутренней прямой мышцы глаз.
То же самое было получено на 20 мин. позже, при повторном введении воды в слуховой проход.
Далее на кривой видно, что на определенной стадии отравления стрихнином еще сохранено отклонение глаз, вызванное с полукружных каналов, в то время как нистагм (в этом случае но крайней мере большей частью) исчез.
В другом опыте после внутривенного введения г/5 мг стрихнина на килограмм веса удалось установить несколько раз то же самое.
В следующем опыте наблюдаются чрезвычайно долгое последействие при калорическом раздражении вестибулярного аппарата и, кроме того*
528
Одиннадцатая глава. Действие ядов
постепенное исчезновение нистагма, так что, отклонение глаз.
Опыт 4. 15 апреля 1919 г. (рис. 240—243).
наконец, остается только
Кролик, 1.6 кг. Лабиринты калорически хорошо раздражаются.
9 ч. 05 м. Эфирный наркоз, вставление канюли в трахею. Мышцы глаз не препарируются, а регистрируются только горизонтальные движения глазного яблока в целом. В середине кокаинизированной роговой оболочки левого глаза прикрепляется нитка и соединяется с пишущим рычажком. Поднятие рычага вверх соответствует горизонтальному движению глаза в темпоральном направлении. Наркоз окончен. Во время всего опыта сохраняется искусственное дыхание.
10 ч. 05 м. Введение в левый слуховой проход воды (f). Отклонение левого глаза происходит темпорально с нистагмом в обратном направлении (см. норму, рис. 240). Следует заметить, что непосредственно после окончания введения воды (f) отклонение идет обратно, а глаз быстро возвращается в исходное положение.
10 ч. 10 Mï Введение воды в правый слуховой проход, нормальное отклонение глаз в направлении к носу с нистагмом.
10 ч. 18 м. Внутривенное введение 0.5 мг стрихнина на килограмм веса. Сразу после введения стрихнина наступают судороги глазных мышц.
10 ч. 20 м. Введение воды в левый слуховой проход. Никаких отклонений. Через одну минуту наступают мелкиё быстрые нистагмоподобные движения глаза. После окончания промывания водой, напротив, вскоре наступает максимальное отклонение глаз в темпоральном направлении, но без какого-либо следа нистагма. Это сильное отклонение продолжается примерно одну минуту, и затем глаз медленно идет обратно при сопутствующих судорогах глазных мышц и дрожании.
10 ч. 41 м.*То же самое. И теперь отклонение происходит не во время калорического раздражения, а только после его. прекращения.
11 ч. 00 м. Введение воды в левый слуховой проход. Сперва никаких отклонений нет, нистагм отсутствует. Примерно после одноминутного введения воды возникает судорожный приступ нистагма; в это время отклонение медленно увеличивается. Это отклонение продолжает увеличиваться и после прекращения промывания водой. Отклонение длится примерно три минуты, после чего постепенно идет обратно, сопровождаясь нистагмом переменной интенсивности.
11 ч. 20 м. Внутривенное введение 0.4 мг стрихнина на килограмм.
И ч 25 м. Введение воды в левый слуховой проход (рис. 241). Кривая была уже и до того довольно беспокойной. При первой стрелке вверх начинается введение воды. Глаз сразу же отклоняется темпорально, и одновременно наступает сильный нистагм. Но этот нистагм является преходящим, и за ним следует очень сильное отклонение без нистагма. Потом снова начинается нистагм, причем отклонение несколько уменьшается. После прекращения введения, при второй стрелке вверх, наступает чрезвычайно сильное, не прерываемое более нистагмом отклонение глаз. Кривая носит неравномерный характер из-за постоянного «беспокойства» мышц. Отклонение продолжается примерно три минуты, а затем возвращается к норме, сопровождаясь начавшимися в это время сильными приступами нистагма.
11 ч. 34 м. Лабиринтные установочные рефлексы и установочные рефлексы тела на голову отсутствуют; равным образом отсутствуют тонические лабиринтные рефлексы на конечности и реакции на прогрессивные движения. Компенсаторные установки глаз еще очень подвижны; все вращательные реакции, за исключением постнистагма, сохранены.
И ч. 48 м. Внутривенное введение 0.3 мг азотнокислого стрихнина на килограмм.
11 ч. 50 м. Промывание правого слухового прохода. Наблюдается отклонение глаз, но нистагм отсутствует.
12 ч. 00 м. Промывание правого слухового прохода. Отсутствуют отклонение и нистагм.
12 ч. 05 м. Промывание левого слухового прохода. Отсутствуют отклонение и нистагм.
12 ч. 45 м. Промывание праврго слухового прохода. Наблюдаются отклонение глаза и нистагм. Нистагм снова быстро прекращается, в то время как отклонение сохраняется продолжительное время.
12 ч. 55 м. Промывание левого слухового прохода. Только после одноминутного промывания наступает отклонение глаза без нистагма. Это отклонение длится очень долго и время от времени прерывается судорогами мышц в противоположном направлении.
В дальнейшем устанавливается изменчивая картина. Иногда нет ни отклонения, ни нистагма, иногда наблюдается одно лишь отклонение, а затем снова отклонение с сильным нистагмом.
о
Vf
CM
о
И
P-i
ЧИ
CM
о
И
Р-,
34 Р. Магнус
Рис. 242. Рис. 243.
530
Одиннадцатая глава. Действие ядов
13 ч. 40 м. Из лабиринтных рефлексов наблюдаются только компенсаторные установки глаз и вращательные реакции головы и глаз.
14 ч. 10 м. Промывание левого слухового прохода (t) (рис. 242). Теперь наступает очень сильное отклонение без какого-либо следа нистагма; длится оно еще долгое время
и после прекращения введения воды (при второй стрелке).
14 ч. 30 м. Промывание левого слухового прохода (рис. 243). Появляется сильное отклонение глаза. На максимуме последнего наступает смерть животного.
В скрытие: никаких отклонений от нормы не обнаружено.
В ряде случаев после введения стрихнина удалось еще, кроме того, наблюдать отчетливое извращение рефлексов. Тогда при промывании слухового прохода происходило отклонение глаз в обратном направлении с соответствующим нистагмом. Примером этого служит опыт V от 29 апреля 1919 г., представленный на рис. 244.
Кролик, 1.3 кг. Лабиринты хорошо воспринимают калорическое раздражение.
14 ч. 30 м. Эфирный наркоз. Канюля вставлена в трахею. Кокаинизация роговой оболочки левого глаза, к середине которой прикрепляется нитка, привязанная к рычагу. Поднятие рычага соответствует горизонтальному отклонению глаза в направлении к виску. Во время всего опыта сохраняется искусственное дыхание. Легкий эфирный наркоз (1/10), который по контрольным опытам не влияет ни на калорическую реакцию глаз, ни на течение отравления.
Животному вводится внутривенно в 15 ч. 40 м. 0.5 мг и ^ в 16 ч. 00 м. 0.3 мг азотнокислого стрихнина на килограмм веса. ^ 17 ч. 16 м. (рис. 244). Кривая отражает еще до промыва¬
ем ния водой мышечные подергивания. Промывание левого уха на- ^ чинается с первой стрелки вверх. Сразу после начала промывания прекращается сокращение мышц. Между ажЪ кривая поднимается вверх соответственно отклонению глаза в височном направлении, как это было в норме при введении воды в левое ухо. Одновременно появляется неравномерный нистагм в противоположном направлении. При Ъ линия кривой вдруг без видимой причины идет вниз, это значит, что глаз отклоняется в направлении к носу, что было отчетливо видно в этом опыте и простым глазом. Теперь нистагм происходит в направлении виска. Следовательно, наступило извращение как отклонения, так и нистагма. При второй стрелке прекращается введение воды. После этого отклонение снова возвращается обратно. Такого рода извращение рефлекса наблюдалось много раз.
Например, в опыте от 6 мая 1919 г., в котором отдельно двумя рычагами регистрировались движения наружной и внутренней прямых мышц левого глаза. После внутривенного введения 0.5 мг азотнокислого стрихнина сначала наблюдался нормальный результат раздражения, : после промывания водой правого уха внутренняя прямая мышца отчетливо сократилась, наружная же проявила соответствующее реципрокное торможение. Одновременно был виден нистагм в обратном направлении. Однако через 80 мин. после введения стрихнина стал наблюдаться противоположный результат. После промывания водой правого уха торможение было видно на внутренней мышце, а наружная мышца была в стадии сокращения соответственно отклонению глаза в височном направлении, т. е. противоположно тому, что должно было бы быть после промывания правого уха. Одновременно были видны нистагмоподобные сокращения обеих мышц в направлении, обратном отклонению.
Следовательно, в этом случае раздельной записью движения обеих антагонистических мышц удалось показать, что здесь речь идет о дейст¬
Стрихнин
531
вительном извращении рефлексов, при котором осталась неизмененной реципрокная иннервация антагонистов.
Другой интересный пример извращения рефлексов после отравления стрихнином показывает следующий опыт, в котором наступивший после односторонней экстирпации лабиринта горизонтальный нистагм стал «обратным».
Опыт VI. 13 мая 1919 г. (рис. 245 и 246).
Кролик, 1.6 кг. Все лабиринтные рефлексы в норме.
9 ч. 30 м. Эфирный наркоз. Канюля вставлена в трахею. В яремную вену тоже вставляется канюля. Наружная и внутренняя прямые мышцы левого глаза свободно отпрепаровываются на дистальных местах прикрепления и соединяются нитками с ре-
Rect ext sin. * УSek
Reet. int. sin.
Рис. 245.
гистрирующими рычагами. Глаз экстирнируется. Верхняя линия записи идет от наружной, нижняя — от внутренней прямой мышцы. Подкожное введение 20 мг атропина.
Экстирпация левого лабиринта по де Клейну. Кролика привязывают на подогретом исследовательском столике, голова его находится в нормальном положении.
У животного соответственно левосторонней лабиринтэктомии появляется горизонтальное отклонение глаз влево с нистагмом вправо (рис. 245). На кривой видно, что наружная прямая мышца показывает медленное сокращение с быстрыми нистагмо- идными расслаблениями, в то время как внутренняя мышца, наоборот, медленно расслабляется с быстрым сокращением. Вводится внутривенно 0.6 мг азотнокислого стрихнина на килограмм веса тела. Это вызывает общие судорожные приступы. В И ч. 15 м. после одного из таких судорожных приступов появляется извращение нистагма (рис. 246). Теперь наружная мышца показывает медленное расслабление и быстрое сокращение, в то время как внутренняя медленно сокращается и быстро расслабляется.
Это соответствует горизонтальному нистагму влево.
Выводы. 1. Небольшие, вообще недейственные дозы стрихнина вызывают усиление всех лабиринтных рефлексов, за исключением реакций на прогрессивные движения.
2. При увеличении доз стрихнина в одно время исчезает вращательный нистагм головы и глаз, когда реакции поворота головы и глаз еще сохранены и иногда еще могут быть очень подвижными.
3. Установочные рефлексы исчезают при появлении общих судорог. Установочные рефлексы тела на голову при этом немного более устойчивы, чем лабиринтные установочные рефлексы. В этой стадии отсутствуют и прогрессивные реакции, зато сохранены еще реакции поворота, тонические шейные и лабиринтные рефлексы на мышцы конечностей и компенсаторные установки глаз.
4*
532
Одиннадцатая глава. Действие ядов
4. Из всех лабиринтных рефлексов самыми устойчивыми по отношению к отравлению стрихнином являются компенсаторные установки глаз. Они наблюдаются с большой отчетливостью до самого конца, пока не наступило удушье.
5. При нарастании степени отравления стрихнином до стадии судорог при тонических шейных и лабиринтных рефлексах на мышцы тела не происходит снятия реципрокной иннервации антагонистических мышц. Но затем эти же мышцы отчетливо показывают при других рефлексах снятие реципрокной иннервации.
6. Также и при компенсаторных установках глаз и калорических лабиринтных рефлексах на мышцы глаз остается сохраненной реципрок- ная иннервация в стадии судорог при отравлении стрихнином.
Rect. ext. sin.
Sek
Rect int. sin.
Рис. 246,
7. При калорических лабиринтных рефлексах на глаза удается наблюдать в стадии судорог длительное последействие в виде сохраняющегося нормального отклонения в течение нескольких минут.
8. Не удалось установить извращения ни тонических шейных и лабиринтных рефлексов на мышцы тела, ни компенсаторных установок глаз, но зато это было установлено в стадии судорог при калорических лабиринтных рефлексах на глаза и при нистагме после односторонней экстирпации лабиринтов.
9. Следовательно, и этот пример показывает, как чрезвычайно избирательно действуют яды на системы установки тела и лабиринтные рефлексы.
В. Пикротоксин
Действие пикротоксина на установку тела и лабиринтные рефлексы отчетливо отличается от действия стрихнина.
Пикротоксин является типичным ядом, вызывающим судороги, главным местом воздействия которого у кролика является ствол мозга, однако исследованиями Лухсингёра [Luchsinger (1)] и Готтлиба (Gottlieb) установлено, что он действует и на спинной мозг. Одновременно с судорогами наступает увеличение возбудимости рефлексов. Морита [Morita (2)] нашел, что у кроликов после экстирпации большого мозга появляются такие же пикротоксиновые судороги, как и у нормальных кроликов. Рое- бер (Roeber), Поллок и Холмес (Pollok) наблюдали сильное возбуждение центра блуждающего нерва, Поллок и Тредвей (Treadway) — возбуждение дыхательного центра еще до начала судорог. Все эти наблюдения относятся
Пикротоксин,
533
к кролику. Все подробности этого отравления описаны в работе П. Трен- дел енбурга.
Описанные здесь опыты были проведены на кроликах и кошках [Ионк- хофф (70)].
Общий ход течения отравления пикротоксином у кролика
Сперва описывается опыт, при котором ежедневно вводились внутривенно, начиная с самых малых доз, постепенно увеличивающиеся дозы пикротоксина.
Кролик, 1.3 кг. Все лабиринтные рефлексы в норме, только тонические лабиринтные рефлексы на мышцы конечностей неотчетливы.
5 июня 1919 г. 10 ч. 00 м. 0.05 мг пикротоксина на килограмм веса введено внутривенно.
Через час заметно усиление лабиринтных рефлексов, но они еще не достигли максимума. Повышение отчетливо заметно главным образом на компенсаторных установках глаз и на вращательных реакциях, но реакции на прогрессивные движения и лабиринтные установочные рефлексы на голову стали отчетливыми. Однако тонические лабиринтные рефлексы на мышцы тела еще плохо распознаваемы.
После обеда и вечером того же дня еще заметно усиление рефлексов.
6 июня, 10 час. Снова внутривенное введение 0.05 мг пикротоксина на килограмм веса. Через час повышение лабиринтных рефлексов гораздо отчетливее, чем накануне. Для сравнения при исследованиях на вращение наряду с животным, отравленным пикротоксином, вращают и нормальное животное. Установочные лабиринтные рефлексы на голову и компенсаторные установки глаз очень живые. Вращательные реакции головы и глаз и нистагм гораздо сильнее, чем до введения пикротоксина. Реакции на прогрессивные движения также отчетливее, чем у нормального животного. Такое значительное увеличение рефлексов остается в продолжение всего дня.
7 июня, 10 час. У животного все еще усиленные лабиринтные рефлексы. Еще раз вводится внутривенно 0.05 мг/кг пикротоксина. И после этого удается установить повышенную возбудимость лабиринтных рефлексов, особенно усилен теперь нистагм головы. Тонические лабиринтные рефлексы на конечности впервые стали отчетливыми.
8 июня. То же самое. 0.05 мг пикротоксина.
9 июня. После введения 0.1 мг пикротоксина наблюдается то же повышение рефлексов, особенно отчетлив вращательный нистагм головы и глаз.
11 июня. 0.2 мг на килограмм. То же самое состояние.
12 июня, 'i
13 июня. > 0.6 мг на килограмм. То же самое.
14 июня. J
16 июня, 10 час. Внутривенное введение 1 мг пикротоксина.
10 ч. 30 м. У животного учащенное дыхание, непрерывно двигает головой, раздвигает передние ноги, так что лежит животом на полу. Нельзя вызвать движения бега, потому невозможно наблюдать нарушения равновесия и акта ходьбы. Судорог еще нет.
Лабиринтные установочные рефлексы на голову имеются в правом и левом боковом положениях в воздухе, но уже слабее, чем у нормального животного. При висячем положении головой вверх голова еще приходит в нормальное положение. В положении на спине она висит вниз. При висячем положении головой вниз ротовая щель направлена вертикально. Следовательно, установочные рефлексы больше не наблюдаются.
Установочные рефлексы тела на голову, а также шейные установочные рефлексы еще отчетливы. Компенсаторные установки глаз еще наблюдаются, но они больше не увеличены. Тонические лабиринтные рефлексы на мышцы тела уже нельзя получить, но зато хорошо видны вращательные реакции головы и глаз, постреакции. Вращательный нистагм головы и глаз и постнистагм ослаблены. Лифтные реакции стали неясными, так же как и рефлекс готовности к прыжку.
17 июня. 10 час. Все лабиринтные рефлексы снова стали отчетливыми, даже повышенными, только тонические лабиринтные рефлексы на мышцы конечностей отсутствуют. Животное, по-видимому, полностью поправилось и при приближении скачет в сторону.
10 ч. 30 м. 1 мг пикротоксина на килограмм внутривенно.
10 ч. 45 м. Животное сидит в углу, передние ноги расставлены широко, голова и живот на полу, дыхание очень учащено. Время от времени покачивает головой.
534
Одиннадцатая глава. Действие ядов
10 ч. 50 м. Лабиринтные установочные рефлексы на голову полностью исчезают при положении на спине и висячем положении головой вниз; при висячем положении головой вверх, при положении на правом и левом боку иногда еще удается их наблюдать. В противоположность этому установочные рефлексы тела на голову и шейные установочные рефлексы очень отчетливы.
Компенсаторные установки глаз отсутствуют, наблюдаются вращательные реакции головы и постреакции, вращательный нистагм и постнистагм головы очень слабо выражены или отсутствуют.
Вращательные реакции глаз и постреакции слабо выражены, вращательный нистагм и постнистагм глаз отсутствуют или едва проявляются.
Лифтные реакции и готовность к прыжку вызвать уже не удается.
12 час. Компенсаторные установки глаз снова слабо положительны. Вращательные реакции стали еще слабее. Но в общем картина отравления та же самая, животное имеет больной вид, у него наблюдается диаррея, и оно сидит сжавшись в углу.
15 час. Так как животное больное, его убивают внутривенным вливанием 2 мг пик- ротоксина. Наступают судорожные приступы, вскоре полностью парализуются лабиринтные рефлексы, вращательные реакции головы остаются при этом дольше всего. Калорические лабиринтные рефлексы на этой стадии уже нельзя вызвать.
При вскрытии не найдено никаких отклонений от нормы.
Таким же образом было проведено еще пять опытов с аналогичными результатами.
Следующий опыт демонстрирует острое течение отравления после введения большой дозы пикротоксина. При этом на первый план больше всего выступает парализующее действие.
5 июня 1919 г. Кролик, 1.3 кг. Все лабиринтные рефлексы нормальны, только неотчетливы тонические лабиринтные рефлексы на мышцы тела.
10 час. 2 мг/кг пикротоксина внутривенно.
10 ч. 10 м. Животное сидит тихо и съежившись, при приближении не убегает, дыхание сильно учащено, так что его едва можно сосчитать. Покачивание головы, расставление передних ног, так что голова и живот лежат на полу.
Лабиринтные установочные рефлексы на голову отсутствуют при положении на спине и висячем положении головой вниз. При висячем положении головой вверх голова приводится в нормальное состояние. При положении на правом и левом боку еще имеются лабиринтные установочные рефлексы. Тонические лабиринтные рефлексы на мышцы тела неотчетливы, компенсаторные установки глаз очень живые.
Вращательные реакции головы живые, постреакции еще имеются. Вращательный нистагм головы и постнистагм очень слабо выражены или отсутствуют.
Можно вызвать вращательные реакции глаз и постреакции, вращательный нистагм глаз и постнистагм выражены слабо или отсутствуют.
Реакции на прогрессивные движения очень слабы или отсутствуют.
При поднимании животного с пола очень легко возникают тонические и клониче- ские судороги.
10 ч. 15 м. Лабиринтные установочные рефлексы на голову полностью отсутствуют. Шейные установочные рефлексы еще наблюдаются (способность сесть из положения на боку, если голову привести в нормальное положение). Установочные рефлексы тела на голову и тело имеют место, но значительно ослаблены. Тонические лабиринтные рефлексы на мышцы тела неотчетливы, компенсаторные установки глаз слабо выражены.
Вращательные реакции головы и глаз и постреакции отчетливы. Вращательный нистагм головы и глаз и постнистагм отсутствуют. Не удается вызвать реакции на прогрессивные движения.
У животного тонические и клонические судороги, но у него еще не было большого общего эпилептиформного судорожного приступа.
10 ч. 20 м. Лабиринтные установочные рефлексы полностью отсутствуют, в то время как наблюдаются другие установочные рефлексы. Компенсаторные установки глаз отсутствуют или выражены очень слабо, тонические лабиринтные рефлексы на мышцы тела неотчетливы.
Вращательные реакции головы и глаз и постреакции выражены еще отчетливо, зато отсутствуют вращательный нистагм головы и глаз и постнистагм. Реакции на прогрессивные движения не удается вызвать, только иногда наблюдается слабая лифтная реакция.
10 ч. 30 м. и 10 ч. 40 м. В основном та же картина.
10 ч. 50 м. Отсутствуют все лабиринтные рефлексы, за исключением вращательных реакций головы, которые слабо выражены.
Пикротоксин
535
У животного все время тонические и клоническпе судороги с вовлечением жевательных мышц и мышц языка; сильное слюноотделение. Поочередно появляются опи- стотонус и эмпростотонус. В 11 час. во время этих судорог произошла остановка дыхания. При вскрытии сердце еще билось. Сильная перистальтика кишечника. При вскрытии не были найдены отклонения от нормы.
Больше всего бросается в глаза при отравлении пикротоксином чрезвычайное усиление лабиринтных рефлексов при еще очень малых дозах пикротоксина (1/20 мг), которые обычно у животных не вызывают никаких других изменений. Это усиление лабиринтных рефлексов очень медленно возвращается к норме, еще через 24 часа после введения 1/20 мг на килограмм это усиление рефлекса отчетливо видно.
Два животных четыре дня подряд получали 1/20 мг пикротоксина, а затем еще дозу в 1 мг на килограмм. Они пол-
Рис. 247. Влияние стрихнина и пикротоксина на компенсаторные установки глаз (вертикальные отклонения). На рисунке — два одинаковых по весу кролика, белый и серый. В 12 часов белому кролику вводится 0.4 мг стрихнина на килограмм веса, серый получает внутривенно 2 мг пикротоксина на килограмм. Эти дозы примерно равны по
своему действию (вызов судорог). Обоим животным после этого и даже во время фотографирования делают искусственное дыхание, и они лежат рядом на столе в боковом положении (паралич установочных рефлексов). Через два часа после введения яда их фотографируют сверху, причем у обоих кроликов судороги были почти одинаковой силы. Белый кролик (стрихнин) показывает очень отчетливые компенсаторные вертикальные отклонения, склера хорошо видна в верхней половине глазной щели, глаз отведен вниз. У серого же кролика (пикротоксин) наблюдается, напротив, паралич компенсаторных установок глаз. Глаз занимает среднее положение, хотя
животное находится на боку.
ностью поправились, но через продолжительное время и очень медленно; в течение многих дней после введения пикротоксина было заметно отчетливое усиление лабиринтных рефлексов без каких-либо других симптомов.
Дозы в 1 мг на килограмм вызывали иногда при этих хронических отравлениях пикротоксином легкие судороги, иногда же их не было. Первым симптомом тяжелого отравления является учащение дыхания. Тогда животное сидит сжавшись в углу, и его уже нельзя заставить спасаться бегством. Передние ноги широко расставлены, чтобы получить большую площадь опоры, голова и живот лежат на полу. Иногда голова движется в ту и другую сторону.
Избирательное действие пикротоксина на лабиринтные рефлексы становится отчетливым и при исследовании установочных рефлексов. В то время когда лабиринтные установочные рефлексы на голову уже отсутствуют, установочные рефлексы тела на голову и на тело и шейные установочные рефлексы еще очень отчетливы.
После введения 1 мг пикротоксина это состояние, при котором парализованы некоторые лабиринтные рефлексы, длится при наличии легких судорог приблизительно от получаса до полутора часов, затем животные медленно поправляются и лабиринтные рефлексы после этого остаются усиленными много дней подряд.
536
Одиннадцатая глава. Действие ядов
ё ё - s а м ©*&
S§SSSS§1«
4 Is S |S3 i S 8
^-ч vo ff ä ft w н
§>§ § g ai©н ff § 0 i § к a« g-s
wgftff^ g g 5 g 5H § I ® m gxg ч 2 ft 1 S 0 &ю w .
SötsJOsHg.&9Hgg
S6 3 ö я s ° и s
И g Ьсб H S р о 2 a
« S «&“ « S * ° я
оЗн°пВ«онФ
§ § -§ н Ä e § I §
и 2 и H и f^Wgw
ft&gwag g g S öHgg9S^ ff
H I H m H t>i
S 5 S Р^И Й
ц Л H H H rt w Л r. л 4 H ü bS - о Й о
§<g ftg cdffi 2 g B >> ^ и ы 2 ff g 2 w m ^göjga*ftcd H Ï 4 Ü ? P ^ H h-r о
При постепенном увеличении отравления лабиринтные установочные рефлексы одновременно с компенсаторными установками глаз становятся слабее, но первые полностью парализуются немного раньше, чем компенсаторные установки глаз. Одновременно исчезают реакции на прогрессивные движения и вращательный нистагм головы и глаз. Затем прекращаются (вертикальные) компенсаторные установки глаз. Вращательные реакции на голову и глаза выражены дольше всего; только в самом конце прекращаются вращательные реакции глаз, в то время как вращательные реакции головы, хотя и сильно ослабленные, остаются до самой смерти.
О тонических лабиринтных рефлексах на мышцы тела, которые не удается так остро наблюдать у интактного животного, будет говориться далее при обсуждении опытов на децеребриро- ванных животных.
Влияние отравления стрихнином на компенсаторную установку глаз значительно отличается от влияния пик- ротоксина: в то время как она у отравленного стрихнином животного остается до смерти, при отравлении пик- ротоксином она парализуется значительно раньше (рис. 247).
Возбуждающее действие пикротоксина при исследовании калорического нистагма очень хорошо удается изобразить графически (рис. 248).
Влияние пикротоксина на децеребра- ционную ригидность и рефлексы положения
Уже в третьей главе сообщалось, что на децеребриро- ванном животном введением
H m \0 ^ g- ffeB« И ц g « «
В СО 'S f_| к VD Л о И
I s s а а - s _
>а«о&§°йин S н &&SIw й, S^BSgi&aS
§f§ И jäjxo § И
frBtiiwSo *
вдоВта3сйрн2^Н
« Й©со Я ® В °
® йЧЙМф
S m О S Н Н Ч И n ff
3 аз М Н Л г; jsj L_r
а.3 « ® В L
и н н- в Я »д
« m в е-я я нс в
м Ф W Г1 Cl,
«ч ft i
OüSI
° s
1 8§
I g.« я ° ^8 S'S'S в « s S s-в a,
S"äo»0o|So
4 Ш8£&ЗК
м Рч о
О § Ьч ° ®
■ню 6\® и „ - -Я
^ ф о 1 А Я РнД 1 О О ЕН 1-4
S &&д^8*ёйн
2 ДМ О ев в 2 И ® g
.£«фЗ§<*|
CQ У
В о Я 5 НЧ I S-H g М ftS я g
§ g« g,® § в % g
ft ф О о й ffl
И
S о“м|
- g 8 ääg § ft
cd°Wa>oJ2ffcö
й fto н ff” ff S
2 t=l о О и о
W
о
Я
ft
пикротоксина можно превратить разгибательную ригидность в сгибательную (рис. 21) и что тогда тонические лабиринтные рефлексы на мышцы конечностей проявляются не на разгибателях, а на сгибателях. Рис. 22 и 23 иллюстрируют это на
Пикротоксин
537
изолированных трехглавой и двуглавой мышцах децеребрированной кошки. На изменение положения головы разгибатель вообще не реагирует (трехглавая мышца), в то время как сгибатель (двуглавая мышца) показывает сильнейшие изменения тонуса, а именно: он расслабляется при «максимальном положения» головы (-j-45°), а при «минимальном положении» головы (—135°) сокращается наиболее сильно. При рефлексах положения изменяется положении конечностей так же, как до отравления, но поскольку теперь вместо разгибателей реагируют сгибатели, то самые большие сокращения и расслабления наступают при противоположном положении головы. Следовательно, первостепенным является вызванное введением пикротоксина изменение разгибательной ригидности в сгибательную. Изменение рефлексов положения является вынужденным следствием этого, деятельность лабиринтов (и шейных рецепторов) остается при этом неизмененной.
При спонтанных судорожных приступах после введения пикротоксина видно, что судороги появляются главным образом в мышцах-сгибателях, в то время как мышцы-разгибатели остаются в покое.
Следующий протокол показывает течение явлений на децеребрированной кошке, у которой не были отпрепарованы мышцы конечностей.
20 июня 1919 г. Кошка, 2 кг.
10 час. Эфирный наркоз, вставление канюли в трахею, искусственное дыхание, перевязка сонных артерий. Перерезка блуждающего нерва, децеребрация без удаления большого мозга из черепной коробки, незначительное кровотечение.
10 ч. 15 м. Конец операции.
10 ч. 40 м. Очень отчетливая децеребрационная ригидность. При положении на боку голова согнута максимально дорсально, передние лапы сильно вытянуты, в задних лапах отчетливо ощущается тонус четырехглавой мышцы.
При нормальном положении животного с горизонтальной ротовой щелью тонус передних конечностей незначителен, на задних конечностях отчетливее, чем на передних.
В положении животного на спине тонус шеи и передних ног максимален. Следовательно, имеются тонические лабиринтные рефлексы на передние конечности. Задние конечности не разогнуты (рис. 21а).
При повороте головы в положении животного на боку тонус обеих передних конечностей становится максимальным, если голова находится внизу, и — минимальным, если голова вверху. Значит, лабиринтные рефлексы превалируют над шейными рефлексами.
На задних конечностях шейные рефлексы выражены больше, чем лабиринтные.
При висячем положении головой вниз удается получить при повороте головы отчетливые шейные рефлексы на передние конечности.
11 ч. 5 м. То же самое.
11 ч. 10 м. Внутривенное введение 0.8 мг пикротоксина на килограмм веса тела.
11ч. 15 м. Нет отчетливого усиления лабиринтных рефлексов.
11 ч. 20 м. Внутривенное введение 0.4 мг пикротоксина на килограмм веса.
И ч. 21 м. Судорожный приступ с ригидностью затылка. Передние и задние конечности во время судорог сгибаются. При положении на спине тонус мышц-разгибателей больше, чем в нормальном положении, следовательно, имеются отчетливые лабиринтные рефлексы. В положении на боку отчетливый тонус мышц-сгибателей, который сильнее, чем тонус мышц-разгибателей. В положении на спине также превалирует тонус мышц-сгибателей, но при этом мышцы-разгибатели обладают еще довольно хорошим тонусом, который уменьшается при переходе в нормальное положение. Тонус мышц-сгибателей в положении на спине немного слабее, чем в нормальном положении.
В соответствии с этим после отравления пикротоксином появляется активный тонус мышц-сгибателей, который до того не наблюдался.
При повороте головы в положении животного на боку лабиринтные рефлексы на передние конечности были еще отчетливы. При этом тонус мышц-разгибателей был максимальным при повороте головы теменем вниз, а при повороте теменем вверх — минимальным. Противоположные этому изменения наблюдались у мышц-сгибателей, но различие было менее отчетливым, чем на мышцах-разгибателях.
Во время исследования у животного имелись судороги. Они проявлялись в разгибании конечностей, движениях бега, растопыривании пальцев, реакции затылка, при¬
538
Одиннадцатая глава. Действие ядов
чем у животного попеременно сменялись опистотонус и эмпростотонус. В положении на спине у животного главным образом проявлялись сильные движения бега. Тонические лабиринтные рефлексы на передние ноги очень отчетливы, как и тонические шейные рефлексы.
11 ч. 40 м. Сильные судороги и остановка дыхания.
11 ч. 50 м. У животного весьма отчетливые лабиринтные и шейные рефлексы на передние конечности. В положении на спине тонус имеется как у сгибателей, так и разгибателей. Первые имеют при этом положении минимум, последние — максимум тонуса. Передние конечности вытянуты как одно целое. В положении на животе тонус разгибателей очень незначительный, сгибателей — максимальный. Передние конечности судорожно подергиваются в состоянии сгибания. При переходе из положения на животе в положение на спине выявляется очень большое отличие в положении передних конечностей.
12 ч. 30 м. У животного сильные судороги, полностью исчез активный тонус разгибателей и сменился активным тонусом сгибателей. Децеребрационная ригидность имеет место, но перешла с мышц-разгибателей на мышцы-сгибатели, так что животное все время лежит в положении крайнего эмнростотонуса. В положении на боку ноги согнуты максимально, пальцы ног сильно втянуты, голова согнута в вентральном направлении. Опистотонус, который вначале еще сменялся эмпростотонусом, больше не появляется; только все время наблюдаемый тонус сгибателей то немного усиливается, то немного ослабляется. В положении животного на спине уже не удается установить тонус разгибателей, ноги остаются согнутыми. Однако тонус мышц-сгибателей все же меньше в положении на спине, чем в положении на животе. В положении на животе (рис. 216) передние конечности и голова сильно согнуты. Повышение тонуса на задних конечностях не очень сильное, а при положении на животе животное не способно поднять их вверх.
Влияние тонических лабиринтных рефлексов на мышцы конечностей проявляется вследствие этого совсем по-другому. В то время как у нормального децеребрированного животного под влиянием лабиринтов наступает активное усиление тонуса разгибателей, если голова находится в положении на спине (рис. 21а), то после отравления пикроток- сином, наоборот* наблюдается активное повышение тонуса мышц сгибателей, если голова находится теменем вверх (рис. 216).
13 ч. 30 м. Животное совершенно расслаблено, рефлексы отсутствуют.
В скрытие: разрез в мозгу проходит между передними и задними четверохолмиями. Нет никаких кровоизлияний у основания черепа. Дальнейшие отклонения от нормы отсутствуют.
Этот опыт много раз был повторен с одним и тем же результатом. У животных, у которых до введения пикротоксина можно было вызвать только слабые тонические лабиринтные рефлексы на мышцы конечностей, последние после введения пикротоксина стали гораздо более отчетливыми.
Заключение
Внутривенное вливание пикротоксина имеет специфическое действие на все лабиринтные рефлексы.
Уже маленькие дозы пикротоксина (от 1/20 мг на килограмм) вызывают отчетливое и длительно продолжающееся усиление лабиринтных рефлексов, хотя, между прочим, никаких других симптомов на животном установить не удалось.
Если животное отравлять каждый день увеличивающимися дозами пикротоксина (до 1 мг на килограмм веса), то наступают судороги и преходящие параличи. Но если затем прекратить вливание, эти симптомы вскоре проходят, только много дней подряд остается сильно повышенная возбудимость лабиринтных рефлексов, без каких-либо других явлений. Происходит полное восстановление до нормы.
Возбуждающее действие пикротоксина удается также весьма отчетливо наблюдать при калорических лабиринтных рефлексах. На отпрепарированных изолированных мышцах глаз после кратковременного расслабления заметно чрезвычайное усиление отклонения и нистагма при калорических раздражениях. И в этом случае у животного не наблюдается никаких других явлений отравления.
Камфара
539
После больших доз пикротоксина (1—2 мг на килограмм и больше) выступает на первый план его парализующее действие на лабиринтные рефлексы.
При этом дольше всего сохраняются, иногда даже до самой смерти, вращательные реакции головы и постреакции. Вращательные реакции глаз и постреакции очень долго сохраняются. Однако остальные лабиринтные рефлексы парализуются раньше, как например вращательный нистагм головы и глаз и постнистагм, реакции на прогрессивные движения, тонические лабиринтные рефлексы на конечности, компенсаторные установки глаз и лабиринтные установочные рефлексы. Последние относятся к тем лабиринтным рефлексам, которые первыми парализуются и которые исчезают раньше, чем другие установочные рефлексы. В то время как компенсаторные установки глаз после отравления стрихнином остаются без изменения до самой смерти, при отравлении пикротоксином они сравнительно рано парализуются.
Бросается в глаза, что при отравлении пикротоксином изменяется картина децеребрационной ригидности. Вместо максимального тонуса разгибателей у децеребрированного животного может наступить максимальный тонус мышц-сгибателей конечностей, затылка, туловища. В судорогах также участвуют преимущественно мышцы-сгибатели. Это удивительное положение сгибателей можно видеть и у животных с сохраненным большим мозгом.
В то время как у нормальных децеребрированных животных с максимальной ригидностью мышц-разгибателей последние преимущественно участвуют в тонических шейных и лабиринтных рефлексах, причем сгибатели остаются совсем расслабленными или проявляют только незначительное изменение реципрокного тонуса, то это все изменяется после введения пикротоксина. При наступившей преимущественной ригидности сгибателей в тонических шейных и лабиринтных рефлексах участвуют исключительно мышцы-сгибатели, а мышцы-разгибатели пребывают в покое. При этом мышцы-сгибатели приходят в максимальное сокращение, если голова приводится в такое положение, при котором у нормального децеребрированного животного мышцы-разгибатели имели бы минимальный тонус.
Описанное действие пикротоксина на распределение тонуса в мускулатуре тела и вызванная этим измененная реакция на различное положение головы объясняются вместе с парализующим действием пикротоксина на установочные рефлексы и повышением других рефлексов в стадии судорог своеобразной картиной отравления, которую можно наблюдать после вызывающих судороги доз пикротоксина.
Г. Камфара
Гофман (Hoffmann) и Видеман (Wiedemann) показали, что камфара у холоднокровных действует парализующим образом, у теплокровных, наоборот, вызывает эпилептиформные судороги благодаря возбуждению судорожного центра в продолговатом мозгу. Готтлиб (Gottlieb) видел на изолированном спинном мозге кошки повышение возбудимости рефлексов, при наличии связи с головным мозгом — судороги. Морита (2) констатировал выпадение судорог от камфары после экстирпации у кролика головного мозга.
Стросс (Stross) и Виховский (Wiechowski) выявили в согласии с прежними работами Харнака (Harnack) и Витковского (Witkowski) наркотические свойства камфары.
540
Одиннадцатая глава. Действие ядов
Лео (Leo) и Исаак (Isaak) показали, что можно быстро и отчетливо получить при внутривенном введении насыщенного камфарой раствора Рингера (1 : 600) возбуждающее действие, особенно судороги.
Камфару применяют против морской болезни. Ауербах (Auerbach) объясняет ее действие расширением кровеносных сосудов мозга и рекомендует смесь из камфары, бензойнокислой соли кофеина, натрия и диуретина, которая носит название эйталатин (Euthalattin). Другие средства против морской болезни также содержат камфару, так «Mothersills seasick remedy» содержит, кроме камфары, еще кофеин и бромистый калий.
Важным является то, что камфара легко окисляется в организме, соединяясь с глюкуроновой кислотой, легко выделяется из организма и таким путем быстро делается безвредной.
В опытах Ионкхоффа кролику вводились подкожно 10%-е камфарное масло или внутривенно насыщенный камфарой раствор Рингера. При ежедневном подкожном введении 500 мг камфары наступает отчетливое влияние на лабиринтные рефлексы, но без опасности для жизни. Острое отравление камфарой вызывается введением подкожно 1 г или внутривенным вливанием 10—40 мг в растворе Рингера. При опытах с дозами, вызывающими судороги, животным заранее вставляется канюля в трахею, чтобы иметь возможность быстро произвести искусственное дыхание.
Следующие протоколы опытов показывают наглядно течение отравления камфарой [Ионкхофф (72)].
Повторные подкожные отравления (таблица III)
Кролик, 1.95 кг. Наблюдаются все лабиринтные рефлексы.
23 сентября 1919 г. 14 час. 500 мг камфары в масле на килограмм подкожно.
16 час. Сильное общее повышение возбудимости рефлексов. Судорог нет. Лабиринтные установочные рефлексы ослаблены. Вращательные реакции головы и прогрессивные реакции усилены. Другие лабиринтные рефлексы отчетливы.
24 сентября. Вращательные реакции головы весьма живые.
10 час. 500 мг камфары на килограмм подкожно.
11 час. Повышение всех лабиринтных рефлексов, за исключением поствращательного нистагма головы и глаз.
15 час. Еще раз 500 мг камфары на килограмм подкожно. После этого та же картина.
26 сентября. Сильная общая повышенная возбудимость рефлексов. Лабиринтные рефлексы так же лабильны, как день тому назад.
10 час. 500 мг камфары на килограмм подкожно.
14 час. Все лабиринтные рефлексы усилены, за исключением поствращательного нистагма головы и глаз.
27 сентября. Животное заболело. Диаррея. Усилены только вращательные реакции головы и глаз и вращательный нистагм глаз. Все остальные лабиринтные рефлексы в норме.
9 час. 500 мг камфары на килограмм подкожно.
10 час. Лабиринтные установочные рефлексы очень слабые. Тонические лабиринтные рефлексы на конечности едва ощутимы. Вращательный нистагм головы и глаз не очень отчетлив. Остальные лабиринтные рефлексы в норме.
11 час. 500 мг камфары на килограмм подкожно. Вскоре после этого исчезают все лабиринтные рефлексы, причем последними затухают компенсаторные установки глаз и вращательные реакции головы и глаз.
12 час. Смерть. Вскрытие: между плевральными листками слизистая масса. Помутнение почек. Сердце увеличено.
Четыре следующих опыта показали в основном те же результаты.
Острое внутривенное отравление камфорой
30 сентября 1919 г. Кролик, 1.5 кг. Все лабиринтные рефлексы в норме (ср. табл. IV).
10 ч. И м. Внутривенно введено 20 мг камфары в растворе Рингера.
Камфара
541
Таблица III
Таламический кролик, 1.95 кг
Дозы камфары в растворе Рингера внутривенно (мг)
Рефлексы положения
Рефлексы на движение
Примечания
лабиринтные установочные рефлексы
тонические рефлексы на конечности
компенсаторные установки глаз (вертикально)
вращательные
реакции
реакции на прогрессивные
движения
головы
глаз
лифтная
реакция
готовность к прыжку
реакция
нистагм
реакция
нистагм
23 IX 1919
+
+
+
+
+
+
+
+
+
14 час.
500
16 »
+
+
+
++
+
+
+
+
+
+
+
24 IX 1919
+
+
+
+
++
+
+
+
+
+
Общая повышенная возбудимость рефлексов.
14 час.
500
16 »
+
+
+
+
+
+
++
++
++
++
+
+
+
+
25 IX 1919
+
+
+
+
+
+
++
++
++
++
+
+
+
+
10 час.
500
И »
+
+
+
+
+
+
+
+
++
++
++
+
+
+
+
15 »
500
17 »
+
+
+
+
+
+
++
++
++
+
+
+
+
+
+
26 IX 1919, 10 час.
500
+
+
+
+
+
+
++
++
++
++
+
+
+
+
27 IX 1919
9 час.
+
+
+
++
+
++
++
+
+
10 »
500
И »
+
+
+
+
т
+
+
+
И ч. 30 м.
500
12 час.
—
—
=F
+
—
т
—
—
—
Смерть вследствие остановки дыхания. За 1/4 часа до смерти были парализованы все лабиринтные рефлексы.
542
Одиннадцатая глава. Действие ядов
Таблица IV
Таламический кролик, 1.5 кг
Дозы камфары в растворе Рингера внутривен то (мг)
Рефлексы положения
[ Рефлексы на движение
Примечания
§
gï
^Я
а *
S»
11
'S *
сб о « Я
й
о
Я
О)
О» Я 0) о
Я и я к о
gig
SK
о W н Я
g
О 1
!*>Я
Ен
2 а
а
я£
а
О со Н Сб
W O 2 Я Я
В W я
О О ей Я Я И
вращательные реакции
реакции на прогрессивные движения
головы
глаз
я
ей
о
а
Я
сб
Я
Ен
0 Я
1 а
о W О Я
S3
Ен Я
2 я
и
я
а
я
Сб
о
а
§
и
г
о
Я
и
я
а
я
Сб
а
и
г
я
й
10 ч. 00 м.
+
+
+
+
+
+
+
+
+
10 ч. И м.
20
+ ?
+
—
—
—
—
—
10 ч. 20 м.
+
+
+
+
+
+
+
+
+
10 ч. 58 м.
+
—
+
+
+
+
•+
+
+
+
И ч. 10 м.
10
+
+
+
+
+
+
+
+
+
11 ч. 20 м.
10
+
=F
+
+
—
+
—
—
—
И ч. 25 м.
—
—
—
+
—
+
—
—
—
И ч. 30 м.
+
—
=F
+
—
+
—
+
+
11 ч. 40 м.
Т
+
+
+
+
+
+
—
14 час.
+
+
+
+
+
+
+
+
+
14 ч. 35 м.
10
—
+
+
—
+
+
—
16 час.
20
—
—
+
—
+
+
—
т
Судороги.
16 ч. 20 м.
—
—
т
+
—
—
—
—
16 ч. 24 м.
Смерть вследствие остановки дыхания. При вскрытии не обнаружено никаких отклонений.
Сразу после этого животное ложится на бок, легкие судороги мышц конечностей и глаз. Лабиринтные установочные рефлексы наряду с другими установочными рефлексами сильно ослаблены. Наблюдаются компенсаторные установки глаз. Тонические лабиринтные установочные рефлексы на конечности неотчетливы. Все вращательные реакции исчезли, из прогрессивных реакций только слабо выражена готовность к прыжку.
Камфара
543
10 ч. 20 м. Животное поправляется, сидит. Не переносит положения на боку. Опять наблюдаются все лабиринтные рефлексы, ослаблены только вращательный нистагм головы и глаз.
10 ч. 58 м. Установочные рефлексы, а также вращательный нистагм головы слабо выражены. Отсутствуют тонические лабиринтные рефлексы на конечности. Остальные лабиринтные рефлексы вызываются нормально. Вращательный нистагм глаз живой.
11 ч. 10 м. Внутривенно вводится 10 мг. Опять отчетливы лабиринтные установочные рефлексы. Тонические лабиринтные рефлексы на конечности, вращательный нистагм головы и прогрессивные реакции выражены слабо. Имеют место все остальные лабиринтные рефлексы.
11 ч. 20 м. Внутривенное введение 10 мг камфары; прекращаются прогрессивные реакции и вращательный нистагм. Вращательные реакции головы и глаз в норме. Все отолитовые рефлексы слабы.
11 ч. 25 м. Отсутствуют все лабиринтные рефлексы, за исключением очень слабых вращательных реакций головы и глаз. Установочные рефлексы тела и шейные установочные рефлексы слабо выражены, а лабиринтные установочные рефлексы отсутствуют. Компенсаторные установки глаз отсутствуют непродолжительное время, затем появляются снова. Спонтанные подергивания глазных мышц, которые принимаются иногда за нистагмоидные движения глаз.
Затем постепенная поправка животного. Вращательные реакции головы и глаз снова становятся отчетливыми (11 ч. 30 м.). Отолитовые рефлексы слабо положительны (И ч. 40 м.). Можно установить вращательный нистагм (11 ч. 40 м.). Прогрессивные реакции в 11 ч. 30 м. отчетливы, а в И ч. 40 м. — слабо выражены.
14 час. Все лабиринтные рефлексы опять восстановлены. Неясны только тониче- кие лабиринтные рефлексы на конечности и лифтная реакция.
14 ч. 35 м. 10 мг на килограмм. Сразу после этого снова явления паралича. Отчетливы только вращательные реакции головы и глаз. Слабо выражены установочные рефлексы, компенсаторные установки глаз, вращательный нистагм глаз, готовность к прыжку, остальные лабиринтные рефлексы отсутствуют.
16 час. 20 мг на килограмм. Слабо проявляются вращательные реакции головы и глаз и вращательный нистагм глаз, компенсаторные установки глаз и готовность к прыжку. Все остальные лабиринтные рефлексы отсутствуют.
16 ч. 20 м. Сильные судороги. Очень слабо проявляютсякомненсаторныеустановки глаз и вращательные реакции на голову и глаза. Полностью прекращены все остальные лабиринтные рефлексы.
16 ч. 24 м. Смерть вследствие остановки дыхания.
Опыт был многократно повторен с теми же результатами.
Графическое изображение калорического нистагма
Некоторые особенности можно видеть на следующих кривых калорического нистагма.
Кролик, 1.7 кг. Вставление канюли в трахею, искусственное дыхание. Движения яблока левого глаза передаются при помощи нитки, прикрепленной к кокаинизиро- ванной роговой оболочке, на регистрирующий рычажок. Горизонтальное движение глаза в направлении к виску вызывает поднятие кривой. Периодическое промывание водой (12°) левого уха.
Рис. 249. Нижняя кривая — 12 ч. 06 м. В 10 ч. 58 м. и И ч. 35 м. было введено подкожно 500 мг камфары в масле. При первой стрелке начинается промывание водой. Отклонение (кривая идет вниз) гораздо сильнее, чем в норме, при этом имеется нистагм. Он особенно отчетлив и долгое время усилен после второй стрелки, при которой заканчивалось промывание уха (увеличение отклонения и нистагма, долгое последействие).
Верхняя кривая — 14 ч. 30 м. В промежутке, т. е. в 14 ч. 10 м., еще раз было введено подкожно 500 мг на килограмм. При первой стрелке начинается введение воды. Сильное отклонение, маленький и медленный нистагм, который на самом глазу едва виден (отчетливое отклонение, ослабление нистагма).
Рис. 250. 15 час. Введение воды в ухо между первой и второй стрелками. Отклонение и нистагм снова стали живыми. После прекращения промывания водой нистагм постепенно затухает. Минутой позже (при +) наступают отклонение и нистагм в обратном направлении (усиление отклонения и нистагма с последующей реакцией в обратном направлении).
Рис. 251. 16 ч. 50 м. Между 15 ч. 30 м. и 16 ч. 20 м. введено еще 500 мг камфары на килограмм веса тела. Введение воды в ухо между двумя стрелками: сильное отклонение, отсутствие нистагма.
544
Одиннадцатая глава. Действие ядов
При исследовании отсутствуют все лабиринтные рефлексы, наблюдаются только вращательные реакции головы и глаз.
Следующие кривые получены в подобном же опыте с длительным промыванием водой (12°) слухового прохода левого уха. Движения яблока левого глаза в направлении к виску вызывают поднятие рычажка.
Рис. 252а. До введения камфары. Отклонения вследствие непрерывного промывания не наблюдаются. Распознать можно только величину и скорость нистагма.
Рис. 252g. После двухразового внутривенного введения 8 мг камфары на килограмм. Отчетливое усиление нистагма. (В остальном картина очень изменчива. Сразу после внутривенного введения каждый раз наступали явления паралича, которые затем снова переходили в возбуждение).
Рис. 252q. Еще раньше прекратили введение воды. При стрелке снова промывание холодной водой. После этого наступило сильное отклонение без всякого нистагма.
Заключение
Маленькие дозы камфары вызывают значительное повышение всех лабиринтных рефлексов, причем иногда больше оказывается влияние на одну, иногда на другую функциональную группу. Это влияние после внутривенного вливания 10 мг камфары быстро проходит, после 500 мг подкожно — длится более продолжительное время. Однако после ежедневных повторных подкожных введений возбуждение проходит через 48 часов.
Возбуждение установочных рефлексов камфарой проявлялось уже в старых опытах Готтлиба, который путем введения через рот камфары приводил кроликов в глубоком паральдегидовом наркозе снова в сидячее положение, причем сперва в нормальное положение приходила голова, затем уже тело.
При графической записи калорических рефлексов на глаза удается установить усиление отклонения глаз и нистагма, долгое последействие после прекращения калорического раздражения, а также проявление последействия в противоположном смысле после предварительной паузы.1
Большие дозы камфары действуют нарализующе. После введения подкожно 1 г явления паралича кратковременны, после чего при постепенном выведении яда присоединяется стадия возбуждения. 2% мг подкожно длительно парализуют лабиринтные рефлексы. Внутривенное введение 20 мг вызывает сначала паралич лабиринтных рефлексов, затем беспорядочно чередуются явления паралича и возбуждения, а под конец одно возбуждение. 40 мг действуют сильно нарализующе и вызывают смерть вследствие остановки дыхания*
Паралич распространяется также на все лабиринтные рефлексы. Иногда сильнее поражается одна группа, иногда — другая. Большей частью сперва исчезает вращательный нистагм головы и глаз, затем прогрессивные реакции, в то время как компенсаторные установки глаз (вертикально), и прежде всего вращательные реакции головы и глаз, имеют самую большую сопротивляемость к яду. Установочные рефлексы находятся примерно в середине по своей сопротивляемости. Лабиринтные установочные рефлексы чаще всего затухают раньше других установочных рефлексов. Нельзя найти ни одной группы рефлексов, которая, как например компенсаторные установки при отравлении стрихнином, оставалась бы очень лабильной до самой смерти.
При калорической реакции глаз удается графически записать исчезновение нистагма при сохраненном отклонении глаз.
1 Нечто подобное наблюдается у неотравленного животного только после очень длительного утомительного промывания (де Клейн и Верстеег). Наоборот, после камфары это наблюдается уже после недолго длящегося промывания.
Хеноподиевое масло
Д. Хеноподиевое масло (74, 76)
При отравлении средством от глистов (Ankylostomum, Necator) — хено- лодиевым маслом — наблюдались среди других симптомов головокружение, шаткая походка, глухота, шум в ушах, тошнота и рвота. Это явилось причиной того, что Ионкхофф решил исследовать влияние подкожного и внутривенного введения хеноподиевого масла на лабиринтные рефлексы кроликов, кошек и морских свинок.
Изменения лабирин т,н ы х рефлексов при остром отравлении
Этот ряд исследований был проведен на кроликах, у которых предварительно наблюдались нормальные лабиринтные рефлексы, а затем им вводилось подкожно 0.3 мл хеноподиевого масла на килограмм веса.
Течение 3—5-часового отравления в 13 опытах примерно следующее.
Приблизительно через полчаса животное становится беспокойным, все лабиринтные рефлексы, за исключением тонических лабиринтных рефлексов на мышцы конечностей, отчетливо усилены. Тонические лабиринтные рефлексы на конечности затухают первыми.
Затем лабиринтные установочные рефлексы становятся слабее и, наконец, исчезают, в то время как остальные установочные рефлексы, хотя и ослабленные, еще остаются. Животное лежит в положении на боку, но садится в ответ на чувствительные раздражения. Наконец, затухают все установочные рефлексы.
Когда прекращаются лабиринтные установочные рефлексы, можно еще наблюдать компенсаторные установки глаз.
Из них сперва ослабевают вертикальные отклонения, однако еще некоторое время их можно вызывать. Когда они прекращаются, компенсаторные круговые движения отчетливо сохранены. Последние затухают незадолго до смерти (до исчезновения вращательных движений на голову и глаза). Рефлекс готовности к прыжку после начального усиления постепенно снижается и затухает примерно одновременно с компенсатор-
35 Р. Магнус
545
В
Рн
Рис. 250. (Кривая уменьшена примерно наполовину).
546
Одиннадцатая глава. Действие ядов
ными вертикальными отклонениями. Более устойчивой является лифт- ная реакция. Она вначале очень подвижная, но затем ослабеваетг чтобы в стадии судорог снова усилиться и исчезает незадолго до* прекращения компенсаторных круговых движений. Вращательные реакции на голову и глаза вместе с соответствующим нистагмом становятся сильными и продолжительное время остаются такими.
Рис. 251.
Рис. 252.
При затухании компенсаторных вертикальных отклонений они еще живые. Нистагм головы и глаз бывает еще иногда после исчезновения установочных рефлексов особенно живым. Только в последней стадии отравления он слабеет. Наконец, при вращении остаются только максимальные судорогоподобные отклонения головы и глаз (с реакцией последействия). Незадолго до смерти исчезают сперва нистагм и лифтная реакция, затем компенсаторные круговые движения глаз, в то время как вращательные реакции головы и глаз остаются подвижными и часто проявляются до» самой смерти.
Общие явления возбуждения постепенно усиливаются. При затухании установочных рефлексов проявляются жевательные движения, при ослаблении компенсаторных движений глаз — подергивания мышц конечно-
Хеноподиевое масло
547
стей и глаз. Наконец, наступают общие судорожные приступы с опистото- нусом и преходящей остановкой дыхания.
Типичным для отравления хеноподиевым маслом является устойчивость к нему рефлексов полукружных каналов. Всегда имеется стадия, при которой реакции поворота головы и глаз (часто и нистагм), а также лифтная реакция отчетливо сохранены или даже усилены, в то время как тонические лабиринтные рефлексы на конечности, лабиринтные установочные рефлексы и компенсаторные вертикальные отклонения глаз исчезли, так что из отолитовых рефлексов остаются только одни компенсаторные круговые движения глаз.
Графическая запись калорических реакций
Большая устойчивость рефлексов с полукружных каналов к хенопо- диевому маслу проявляется и при исследованиях калорических лабиринтных рефлексов на глаза. Следующий пример это наглядно показывает.
Рис. 253.
19 октября 1919 г. Кролик. Графическая запись движений левой наружной прямой мышцы после экстирпации глазного яблока при длительном промывании водой (10°) левого слухового прохода. Сокращение мышцы вызывает подъем рычажка.
Рис. 253. 10 ч. 25 м. Первая часть кривой до стрелки показывает нормальный нистагм при продолжительном промывании. Мышца сокращена, быстрая фаза нистагма направлена вниз. Двумя стрелками показано внутривенное введение 0.07 мл хеноподиевого масла на килограмм (5%-я эмульсия). Сразу после инъекции отклонение уменьшается, нистагм сохранил только одну треть своей первоначальной величины. После этого отклонение быстро возвращается снова к прежнему уровню с преходящими нерегулярными уменьшениями.
Нистагм после введения хеноподие- р
вого масла остается уменьшенным и за- ис*
медленным.
Рис. 254. В начале этой кривой виден нистагм наружной прямой мышцы через 20 мин. после первого введения хеноподиевого масла. Если сравнить этот нистагм с нистагмом на рис. 253, то сразу видны уменьшение величины и, кроме того, появление постепенно проходящих уменьшений отклонений и нистагма, как и непосредственно после первого вливания. После второго введения хеноподиевого масла (0.07 мл на килограмм) (показано двумя стрелками) происходит еще большее спадение отклонения, чем после первого введения. Быстрая фаза нистагма после этого уменьшается и замедляется, в то время как отклонение не уменьшается.
35*
548
Одиннадцатая глава. Действие ядов
12 ч. и 12 ч. 30 м. Сделаны еще два вливания по 0.07 мл на килограмм.
Рис. 255. 13 час. Конечная стадия опыта. Нистагм в начале рисунка уже очень неравномерный. При первой стрелке прекращается длительное промывание уха водой. Отчетливо видно, как лабильно было отклонение, кривая идет медленно вниз, нистагм сразу же прекращается.
При второй стрелке снова начинается промывание водой (10°) левого слухового прохода. Сразу же появляется сильное калорическое отклонение с медленным нистаг-
Рис. 255.
мом. Двумя последними стрелками показано введение пятой порции хеноподиевого масла, сразу после этого животное погибает. Следовательно, калорические отклонения и нистагм остаются до самой смерти.
Влияние хеноподиевого масла на тонические лабиринтные рефлексы на мышцы конечностей у децеребрированных кошек
Примером четырех сходных между собой опытов является следующий протокол.
Опыт III. 14 октября 1919 г. Кролик, 1.5 кг.
9 ч. 30 м. Эфирный наркоз, трахеотомия, искусственное дыхание, перерезка блуждающих нервов, перевязка сонных артерий. Децеребрация в области tentorium cerebelli.
10 час. Конец операции. Наркоз окончен. Животное лежит на подогретом операционном столе.
И час. Выраженная децеребрационная ригидность. Тонические лабиринтные и шейные рефлексы очень отчетливы. Хорошо вызывается реакция поворота головы.
■ Корнеальный и пателлярный рефлексы живые.
11 ч. 20 м. Внутривенное введение 0.07 мл хеноподиевого масла в 5%-й эмульсии на килограмм.
И ч. 25 м. Тонические лабиринтные и шейные рефлексы, а также реакции поворота головы чрезвычайно подвижны.
И ч, 35 м. Еще раз 0.07 мл хеноподиевого масла на килограмм.
И ч. 40 м. Тонические лабиринтные и шейные рефлексы на конечности только слабо ощутимы, как при перемене положения с живота на спину, так и при повороте головы в положении на боку. Зато вращательная реакция головы еще очень живая.
Децеребрационная ригидность проявляется еще хорошо, пателлярный рефлекс и сгибательный рефлекс на щипание пальца — живые. Животное очень сильно реагирует на чувствительные раздражения. При щипании уха начинаются сильные разги- бательные движения ног.
12 час. Тонические шейные и лабиринтные рефлексы очень слабо выражены. Зато вращательные реакции головы отчетливы.
12 ч. 15 м. Еще раз внутривенно 0.07 мл хеноподиевого масла на килограмм.
12 ч. 20 м. Не ощущаются больше тонические лабиринтные рефлексы на конечности, но вращательные движения головы отчетливы.
12 ч. 35 м. Тонические лабиринтные рефлексы на конечности полностью исчезли как при перекладывании с живота на спину, так и при повороте головы в положении животного на боку. Также нельзя больше вызвать тонические шейные рефлексы на конечности. Однако вращательные реакции головы пока еще отчетливы. Общее состояние, рефлексы и тонус такие же, как в 11 ч. 40 м. Хорошая децеребрационная ригидность.
14 час. Тонические шейные и лабиринтные рефлексы исчезли. Реакции поворота головы проявляются только при быстром вращении животного.
' Биения сердца слабы и неравномерны. Животное убивается, i Bi с к р ы т и е: разрез в мозгу идет через переднее четверохолмие. Продолговатый мозг не поврежден. Других отклонений не обнаружено.
Хеноподиевое масло
549
Все опыты этого ряда исследований показали, что при отравлении хеноподиевым маслом тонические лабиринтные рефлексы на конечности исчезают гораздо раньше, чем реакции полукружных каналов (в этом случае вращательная реакция головы), что также совпадает с результатами опытов первого ряда исследований.
Действие хеноподиевого масла на последствия односторонней экстирпации лабиринта у кролика и морской свинки
Из симптомов, которые наступают после односторонней экстирпации лабиринтов, как показал проведенный в шестой главе анализ, можно рассматривать только одну часть как прямое следствие отсутствия лабиринта. Другие симптомы являются вторичными и отчасти зависят от шейных рефлексов, вызываемых поворотом головы. Прямым последствием выключения лабиринта у кроликов и морских свинок следует рассматривать вертикальные отклонения глаз, поворот (наклон) головы в сторону отсутствующего лабиринта, а у кролика, кроме того, отчасти поворот туловища. При этом они обусловливаются односторонним действием исходящих из сохраненного лабиринта отолитовых рефлексов. Вышеприведенные опыты показали, что хеноподиевое масло способно парализовать, за исключением компенсаторных круговых движений,, отолитовые рефлексы, в то время как рефлексы с полукружных каналов остаются сохраненными. Однако после односторонней экстирпации лабиринтов не наступают ротаторные принудительные движения глаз. Поэтому следовало ожидать, что станет возможным устранить хеноподиевым маслом последствия односторонней экстирпации лабиринтов, но в то время, когда еще сохранены рефлексы с полукружных каналов. Опыты на кроликах и морских свинках после односторонней экстирпации показали, что это в действительности имеет место.
Опыты производились через 8 и более дней после односторонней лаби- ринтэктомии у кроликов, когда проходили острые последствия экстирпации (движения перекатывания, нистагм головы и глаз) и когда были сильно развиты поворот головы и туловища, потеря тонуса конечностей на оперированной стороне, отклонение глаз, и животные в основном были вполне здоровы.
У морских свинок симптомы одностороннего выключения лабиринта вызывались тем, что в течение нескольких недель в слуховой проход вводили хлороформ. После этого наступали поворот головы, туловища, потерй тонуса конечностей на стороне выключения лабиринта и отклонение глаз. Отравление хеноподиевым маслом вызывалось подкожным введением 0.4 мл хеноподиевого масла на килограмм.
Результат опыта на кролике дан наглядно на рис. 256.
Кролик, 1.5 кг и контрольный кролик. Подопытное животное держат в правой, а контрольное — в левой руке. У обоих животных 17 октября 1919 г. был экстирпиро- ван правый лабиринт по де Клейну. После этого на обоих кроликах наблюдались совершенно одинаковые по силе симптомы, из которых движения перекатывания, нистагм головы и глаз быстро прошли, в то время как поворот и наклон головы, поворот туловища, потеря тонуса конечностей оперированной стороны и отклонение глаз были очень отчетливо выражены и оставались сохраненными.
22 октября подопытному животному ввели подкожно в 13 ч. 34 м. 0.4 мл хеноподиевого масла на килограмм. После этого снова наступили сильные явления раздражения: движения перекатывания, нистагм головы и глаз. В 15 ч. 15 м. было введено еще 0.04 мл внутривенно.
550
Одиннадцатая глава. Действие ядов
15 ч. 45 м. (рис. 256а). Поворот головы у инъецированного животного (в правой руке) при висячем положении головой вниз уменьшается (поворот головы попеременно изменяется между 70 и 45°).
В 16 ч. 15 м. был снят рис. 256Ь, поворот головы еще более уменьшился (примерно
30°).
В 16 ч. 45 м. был снят рис. 256с. Голова стоит теперь прямо. У контрольного животного поворот оказывается без изменений.
Отклонение глаз к этому времени тоже почти совсем пропало. Отолитовые рефлексы с неповрежденного лабиринта (за исключением тонических круговых движений) сняты или же минимальны.
Зато вращательные реакции, постреакции на голову и глаза, лифтная реакция еще очень живые, остальные рефлексы на прогрессивные движения отчетливы. Судорог еще нет в этой стадии.
Следовательно, из этих рисунков видно, как поворот головы после односторонней лабиринтэктомии и по- # следующего введения хеноподиевого
масла медленно снимается.
Еще отчетливее протекает этот опыт на морской свинке.
Опыт V. 23 октября 1919 г. (рис. 257).
Три морские свинки весом от 275 до 310 г. В течение трех недель до опыта животным закапывался хлороформ в слуховой проход, что вызвало симптомы одностороннего выключения лабиринта. У животных нет движений перекатывания и нистагма головы и глаз, но зато еще отчетливо сохранены поворот головы, туловища, потеря Ъ тонуса конечностями на стороне выключен¬
ного лабиринта и отклонение глаз
13 час. Подкожное введение 0.4 мл хеноподиевого масла на килограмм.
€
Рис. 256а—с
Рис. 257.
13 ч. 30 м. Сразу после введения и еще теперь продолжающиеся движения перекатывания, нистагм головы и глаз.
Поворот головы при висячем положении головой вниз отчетливо уменьшился после односторонней экстирпации лабиринтов. Вращательные реакции и рефлексы на прогрессивные движения очень живые.
14 час. Животные сидят, голова и туловище едва повернуты в сторону выключенного лабиринта. При приближении к ним животные убегают. Они больше не делают манежеподобных движений, как до введения хеноподиевого масла.
Хеноподиевое масло
551
При висячем положении головой вниз больше нельзя выявить поворот головы. Голова висит прямо вниз (рис. 257 показывает одно из этих животных — в левой руке— по сравнению с одним из контрольных животных — в правой руке, — которое не получило хеноподиевого масла и поэтому у него имеются симптомы односторонней экстирпации лабиринтов без изменения).
Поворот туловища так же, как различие в тонусе конечностей, отсутствует, отклонение обоих глаз минимально или тоже отсутствует.
Вращательные реакции головы и глаз и рефлексы на прогрессивные движения еще ‘Отчетливы.
14 ч. 30 м. Животные сидят сжавшись на полу. Волосы подняты; при положении животного сидя голова еще минимально повернута в сторону выключенного лабиринта. Лабиринтные рефлексы и последствия одностороннего выключения лабиринтов, такие же, как в 14 час.
15 час. При положении животного сидя голова держится совершенно прямо, животное лежит в боковом положении на столе. Как в правом, так и левом боковом положениях оно остается лежать некоторое время. При этом оно выполняет непрерывно шагательные движения всеми четырьмя конечностями. Эти шагательные движения изменяются при вращении на диске, причем они или прекращаются, или значительно усиливаются.
При висячем положении головой вниз не видно поворота головы. Исчезли все последствия одностороннего выключения лабиринтов, зато отчетливы реакции поворота и реакции на прогрессивные движения.
В таком состоянии животные остаются в течение 3 часов, только после этого наступали общие судорожные приступы с остановками дыхания. Реакции с полукружных каналов восстанавливались (вращательные реакции головы и глаз и лифтная реакция) тогда, когда уже не наблюдались никакие последствия одностороннего выключения лабиринтов. Тонические лабиринтные рефлексы на конечности, лабиринтные установочные рефлексы и вертикальные отклонения глаз не удавалось вызвать. Это состояние оставалось до смерти (19 часов).
Эти опыты показывают, что у кроликов и еще отчетливее у морских ■свинок удается снять подкожным введением 0.4 мл хеноподиевого масла на килограмм уже в ранней стадии отравления (через 2 часа после введения) последствия односторонней экстирпации лабиринта, в то время как рефлексы с полукружных каналов остаются еще 5—6 часов после введения хеноподиевого масла. Животные еще могут стоять на ногах и при приближении к ним спасаться бегством, хотя основной поворот при висячем положении головой вниз уже не вызывается.
В первые полчаса после подкожного введения можно было еще наблюдать отчетливые явления раздражения (движения перекатывания, нистагм и т. д.).
Заключение
Сразу после введения хеноподиевого масла можно в большинстве случаев видеть возбуждение лабиринтных рефлексов. Оно сильнее всего отражается на реакциях полукружных каналов и последствиях односторонней экстирпации лабиринтов.
Реакции на движения и калорические раздражения в течение отравления хеноподиевым маслом только в крайнем случае бывают кратковременно снятыми (во время судорожных приступов) и остаются до смерти. В последних стадиях отравления они очень живые. Даже нистагм может до смерти, или по крайней мере незадолго до смерти, сохраняться, но •становится тогда слабым и неравномерным.
Среди отолитовых рефлексов сперва парализуются тонические лабиринтные рефлексы на конечности, потом лабиринтные установочные рефлексы. Затем следуют компенсаторные вертикальные отклонения глаз, в то время как компенсаторные круговые движения остаются избирательно почти до самой смерти.
552
Одиннадцатая глава. Действие ядов
Лабиринтные установочные рефлексы на голову парализуются раньше,, чем шейные установочные рефлексы и установочные рефлексы тела на голову и тело.
У кролика удалось снять длительные последствия односторонней экстирпации лабиринта (отклонение глаз, поворот и наклон головы), в то время как реакции с полукружных каналов еще сохранялись. При этом очень хорошо удается проследить, как при возрастающем отравлении поворот и наклон головы постепенно уменьшаются, чтобы, наконец, совсем исчезнуть. На морской свинке удается еще скорее снять последствия одностороннего выключения лабиринта (после закапывания хлороформа в слуховой проход), чем на кроликах.
На поздних стадиях отравления вращательные реакции головы и глаз, и лифтная реакция имеют характер судорог, так что реакции становятся максимальными и длятся дольше, чем в норме.
Е. Хинин
Шарко (Charcot) рекомендовал хинин против болезни Меньера. О пользе его имеются разные суждения (cp. Voss). Корнинг (Corning) не нашел при исследовании на самом себе никакого действия против вращательного головокружения. Дрейфус [Dreyfuss (2)] исследовал влияние хинина на те вращательные реакции и те симптомы, которые наступали у морской свинки после одностороннего выключения хлороформом, и нашел отчетливое парализующее действие. Виттмаак (1) увидел при гистологических исследованиях предварительно тщательно обработанных лабиринтов отравленных хинином животных дегенеративные изменения ганглиозных клеток, которые сильнее проявились в спинальных ганглиях, чем в вестибулярных ганглиях. В гребешке и отолитовой макуле были также найдены дегенерации.
Ионкхофф исследовал влияние хинина на лабиринтные рефлексы у морских свинок, кроликов и кошек. Свои данные мы здесь передадим только вкратце. Подробности можно найти в диссертации Ионкхоффа (57).
У морских свинок дозы в 30—100 мг на килограмм, даже если их продолжительное время вводить ежедневно подкожно, вызывают удивительно- мало явлений. У животных нет нарушения акта ходьбы и равновесия. На отолитовые рефлексы тоже не оказывается никакого влияния. Лифтная реакция через некоторое время немного ослабевает. Вращательный нистагм глаз усиливается после малых доз, а после больших — ослабевает. Бросается в глаза значительное усиление вращательного нистагма головы. Быстрая фаза начинается уже тогда, когда последовало незначительное отклонение (медленная фаза), и часто заходит за среднюю линию, что неопытный наблюдатель ошибочно принимает за извращение вращательной реакции. Эти дозы очень непостоянно действуют на слуховую реакцию. У некоторых животных она снимается повторным введением 30 мг, у других же она остается и после введения 100 мг.
Большие дозы (200—400 мг на килограмм подкожно) вызывают уже тяжелые явления отравления. 200 мг переживаются, 400 мг — смертельны. При этом лабиринтные рефлексы постепенно парализуются. Раньше исчезают лабиринтные установочные рефлексы, затем установочные рефлексы тела. Наконец, остаются только вращательные реакции головы и глаз, их можно вызывать до самой смерти животного.
У децеребрированных кошек после внутримышечного введения 100 мг солянокислого хинина на килограмм сначала повышаются тонические лабиринтные рефлексы на конечности, затем исчезает децеребрационная
Хинакетоны
553-
ригидность, и теперь уже нельзя вызвать ни тонических шейных, ни лабиринтных рефлексов на мышцы конечностей.
У кроликов 20 мг не оказывают влияния на калорические реакции. 100 мг на килограмм подкожно или внутримышечно вызывают сперва отчетливое усиление калорического отклонения глаз и нистагм. После больших доз происходит паралич, причем нистагм затухает немного раньше, чем отклонение.
Наступающее после закапывания хлороформа в слуховой проход морской свинки явление одностороннего выключения лабиринтов (поворот и наклон головы, поворот туловища, потеря тонуса конечностей на стороне выключенного лабиринта, отклонение глаз с нистагмом, движения перекатывания) снимается или задерживается подкожным введением 200 мг хинина на килограмм, как это нашел и Дрейфус. 40 мг на килограмм не вызывают действия.
Следовательно, из этих опытов видно, что у данных видов животных, на которых проводились исследования, только очень большие дозы хинина, вызывающие явления отравления, оказывают сильное влияние на исследуемые лабиринтные рефлексы и что неядовитые дозы, по сути говоря, только усиливают вращательный нистагм головы. Итак, до сих пор проведенными исследованиями на животных не удалось установить экспериментальную основу терапевтического применения хинина при лабиринтных состояниях раздражения.
Ж# Хинакетоны
Хинакетоны образуются из хиноалколлоидов замещением группы НОН кислородом у углерода, соединяющего хинолиновую и лоипоновую группы (Rabe).
Моргенрот (Morgenroth) 1 увидел при исследовании действия гидро- хининона на мышей, что после доз в 375 мг на килограмм мыши становились на судорожно вытянутые конечности, затем падали на бок и до полного изнеможения катались по полу. Эти приступы движений перекатывания повторяются время от времени. Иногда животные делают длинные прыжки. Потом они лежат на спине или боку. Внутривенная инъекция 25 мг на килограмм после появления судорог и движений перекатывания становится смертельной. Хининон, по Моргенроту, обладает примерно той же ядовитостью и при внутривенном введении.
Развивая дальше эти наблюдения, Билзма и Верстеег (75) проделали дальнейшие исследования влияния кетонов на установку тела. Хининоьг и гидрохининон имеют одно и то же качественное действие. При подкожном отравлении мыши могут остаться в живых при 230—400 мг хининона на килограмм и 750 мг гидрохининона на килограмм. Движения перекатывания наступали после 350 мг хининона и 500 мг гидрохининона и более высоких доз.
Первыми всегда переставали действовать задние конечности. Это проявляется в походке с широко поставленными ногами и волочением задних конечностей. В то время как нормальная мышь способна удержаться на карандаше, находящемся в горизонтальном положении и постепенно вращающемся, отравленное животное в этой стадии падает с него. Одновременно и вскоре после отказа задних конечностей животные становятся очень беспокойными; затем наступают некоординированные движения головы, а еще через некоторое время — тонические судороги конечностей,
1 Устное сообщение.
-554
Одиннадцатая глава. Действие ядов
приходящих при этом в состояние разгибания. В это же время появляются нпистотонус и жевательные движения. Судороги становятся клоническими, иногда проявляются сильные периодические движения ходьбы. При этом животное поочередно катится то вправо, то влево. Вскоре после начала судорог животное уже не в состоянии принять нормальное положение. Оно лежит на боку. Иногда судороги начинаются таким большим прыжком, какой нормальные животные никогда не делают. Судорожные приступы начинаются или спонтанно, или на раздражение (щипание хвоста, удар по столу).
Для лучшего исследования движений перекатывания стали отравлять шорских свинок. Следующий опыт наглядно дает картину этого отравления.
Морская свинка IV. Вес 0.57 кг.
До введения были исследованы следующие рефлексы: лабиринтные установочные рефлексы, шейные установочные рефлексы, установочные рефлексы тела на тело, реакции поворота глаз и головы, компенсаторные установки глаз, слуховые реакции.
И ч. 35 м. Подкожная инъекция 240 мг 2% %-го солянокислого хинонина (420 мг на килограмм веса животного).
И ч. 45 м. Животное вцолне нормальное. Все вышеперечисленные рефлексы положительны.
И ч. 55 м. Тонические судороги разгибателей конечностей и опистотонус, пере- ;межающийся с движениями ходьбы.
Во время этих движений ходьбы животное несколько раз перекатывалось вправо. 1Если сразу после этого исследовать животное в висячем положении головой вниз, то • оказывается, что голова повернута вправо.
Нет больше спонтанного нистагма, вертикальных компенсаторных установок глаз. При вращении вправо наступает нистагм глаз, при вращении влево — только отклонение.
Установочные рефлексы тела на тело и шейные установочные рефлексы исчезли. .Лабиринтные установочные рефлексы, которые еще положительны, вскоре тоже исчезают.
12 ч. 10 м. Животное уже совсем находится в положении на боку. Конечности тонически разогнуты, у животного сильный опистотонус. Пальцы растопырены. Животное реагирует на кожное и слуховое раздражения. При висячем положении головой вниз последняя немного повернута вправо.
12 ч. 20 м. При висячем положении головой вниз голова повернута влево. Если теперь животное положить в нормальном положении на пол, то вдруг начинаются движения ходьбы, причем животное несколько раз катится влево. После этого оно снова обессиленное лежит в положении на боку.
12 ч. 30 м. Нет никаких существенных изменений.
14 ч. 15 м. Животное снова сидит в нормальном положении, но немного неуверенно. Вращательный нистагм глаз слабо выражен, все рефлексы положительны.
Пять следующих подопытных животных показали примерно те же явления. Лифтная реакция ослабевает уже до появления судорог. Установочные рефлексы вскоре после начала судорог становятся слабее и затем исчезают, так же как и компенсаторные установки глаз. Реакции поворота головы и глаз, наоборот, еще положительны, а относящийся к ним нистагм еще долго сохранен.
Удивительно то, что движения перекатывания в стадии судорог появляются всегда вместе с движениями ходьбы и что это движение всегда происходит в ту сторону, в которую случайно повернута голова. И после прекращения движений перекатывания голова находится в повернутом положении в сторону, в какую катилось животное. Следовательно, можно прийти к заключению, что, так же как при движениях перекатывания после односторонней экстирпации лабиринта, поворот головы является самым существенным. Но вопрос остается еще открытым: как объяснить этот поворот головы? Конечно, возможно, что при этом имеют значение лабиринты.
Никотин
555
Чтобы решить этот вопрос, начали наблюдать явления, наступающие после введения гидрохининона при двусторонней лабиринтэктомии у морских свинок. Здесь дан протокол подопытного животного.
Морская свинка VII. Вес 0.58 кг.
Все вышеназванные рефлексы положительны.
12 час. Двусторонняя экстирпация лабиринтов.
17 час. Лабиринтные рефлексы (лабиринтные установочные рефлексы, компенсаторные установки глаз, реакции поворота и нистагм глаз, реакции поворота и нистагм головы) отрицательны.
17 ч. 08 м. Подкожное введение 490 мг солянокислого гидрохинонина на килограмм веса тела.
17 ч. 16 м. Животное сидит спокойно и симметрично. Умеренное качание головой, как это почти всегда наблюдается после двусторонней экстирпации лабиринтов. Ноги согнуты, так что тело животного лежит на полу.
1.7 ч. 19 м. Животное делается беспокойным и ползает в разных направлениях по полу, причем голова и тело остаются в нормальном положении.
17 ч. 20 м. Начало судорог (опистотонус, разогнутое положение конечностей, особенно .передних, и жевательные движения). После этого животное делае сильные движения ходьбы, причем оно часто падает на бок, но сразу снова поднимается.
17 ч. 24 м. При положении животного сидя голова повернута немного влево. Животное несколько раз падает влево, но сразу поднимается, так что можно думать, что установочные рефлексы тела не повреждены.
17 ч. 25 м. Сильный опистотонус с последующим рефлексом прыжка, причем животное в воздухе переворачивается влево. После этого снова сидит симметрично.
17 ч. 26^м. Животное делает круговые движения перекатывания влево. Один раз ■оно покатилось вправо (при этих движениях перекатывания животное всегда один раз поворачивается вокруг своей продольной оси). Во время пауз покоя оно еще способно правильно сидеть, что указывает на интактность установочных рефлексов. Постепенно эти рефлексы ослабевают.
17 ч. 40 м. Теперь животное лежит на правом или левом боку, время от времени появляются приступы движений ходьбы; между этими приступами оно лежит с описто- тонусом и разогнутыми конечностями.
18 ч. 07 м. После сильного раздражения животное три раза подряд перекатывается влево, после чего оно обессиленное остается лежать.
19 ч. 10 м. Слабые движения ходьбы в положении на боку. На следующий день животное мертвое лежит в клетке.
Итак, из этого опыта видно, что наличия лабиринтов не требуется для появления движений перекатывания после введения гидрохининона и что вообще явления отравления гидрохининоном у животных без лабиринтов наступают так же, как у животных с интактными лабиринтами.
Таким образом, исследуемые хинакетоны вызывают у мышей и морских ■свинок отравление центральной нервной системы, сильные моторные явления раздражения, которые проявляются в судорогах, опистотонусе, положениях разгибания и движениях ходьбы. Время от времени появляются движения перекатывания, при этом только тогда, когда перед этим имелся поворот головы. Следовательно, так же как после односторонней экстирпации лабиринтов и при отравлении хинакетонами (например, при отравлениях стофаином), движения перекатывания обусловливаются приступами движений ходьбы при повернутой голове. Для этого не требуется наличия лабиринтов. В противоположность движениям перекатывания при односторонней экстирпации изменяется направление этих движений в зависимости от того, повернута ли голова вправо или влево.
3, Никотин
В описанных до сего времени исследованиях речь шла в основном об установлении последовательности, в которой различные участвующие при установке тела и лабиринтных рефлексах аппараты возбуждаются или парализуются употребленными здесь ядами. При этом «клиническая
556
Одиннадцатая глава. Действие ядов
картина» сводится более или менее полностью к выпадению или усилению определенных групп рефлексов при сохранности других рефлексов. Конечно, этим не исчерпывается задача. Надо также знать для каждого отдельного определенного яда и определенного рефлекса точное место воздействия яда. Такого рода анализ требует приближенного знания анатомического расположения центров отдельных рефлексов. Это задача довольно сложная, как это видно из десятой главы. Как сложны отношения и какие методы можно применить для решения вопросов, видно на примере действия никотина на вестибулярный нистагм. Де Клейн и Вер- стеег (73) подвергли это экспериментальному анализу.
Целью этого исследования было выработать такую методику, которая позволяла бы определить точную локализацию действия яда в рефлекторной дуге вестибулярного нистагма (лабиринт—область вестибулярных ядер—ядра мышц глаза—глазные мышцы). Опыты проводились на ваго- томированных таламических кроликах в условиях искусственного дыхания, у которых после экстирпации глазного яблока прямая наружная и прямая внутренняя мышцы были соединены с тончайшими регистрирующими рычажками.
а) Действие на глазную мышцу
На стороне отпрепарованных глазных мышц перерезается у основания черепа n. oculomotorius, так что внутренняя прямая мышца (в противоположность наружной) отделяется от центральной нервной системы.
М. reet. int.
М. reet. ext.
Рис. 258. Таламический кролик, 1.5 кг. Сонные артерии не перевязаны. Левый глазодвигательный нерв интра- краниально перерезан. Верхняя кривая — левая внутренняя прямая мышца; нижняя кривая — левая наружная прямая мышца. Введение внутривенно 0.3 мг никотина на килограмм веса. Сокращение обеих мышц.
Значит, реакция этой мышцы должна после введения яда основываться: на периферическом действии яда.
Рис. 258 показывает, что после внутривенного введения 0.3 мг на килограмм сокращаются обе мышцы, а также и отделенная от центра внутренняя прямая.
В 11 опытах выявилось, что нижняя граница для этого действия лежит при 0.25 мг на килограмм и что сокращение усиливается с увеличением дозы никотина. Следовательно, никотин обладает периферическим действием на мышцы глаз.
б) Действие на ядра глазных мышц
Веселый [Wessely (1)] показал, что снабжение кровью глазницы у кроликов осуществляется почти исключительно сонными артериями. Значит, перевязка сонных артерий препятствует попаданию яда в глаз¬
Никотин
557
ные мышцы при внутривенном его введении. Опыты проводились, как и в первой серии исследований, с перерезкой соответствующего n. оси- lomotorius'a. Кроме того, перевязывались сонные артерии. Если после введения яда сокращается наружная мышца, в то время как внутренняя пребывает в покое, то это доказывает центральное действие никотина. Если же сокращается и внутренняя прямая мышца, то выключение глазницы из круга кровообращения не удалось и опыт должен быть отброшен. Рис. 259 показывает, что после введения 1.5 мг на килограмм веса действительно сокращалась наружная прямая мышца, в то время как внутренняя мышца оставалась в покое. Итак, эта доза обладает прямым возбуждать . reet. int.
М. reet. ext.
Рис. 259. Таламический кролик, 1.9 кг. Сонные артерии перевязаны. Левый глазодвигательный нерв интракра- ниально перерезан. Верхняя кривая — левая внутренняя прямая мышца; нижняя кривая — левая наружная йря- мая мышца. Стрелка — введение внутривенно 1.5 мг никотина на килограмм веса.
ющим действием на центры нервов глазных мышц. Если глазодвигательный нерв не перерезать, то сокращается и внутренняя прямая мышца.
17 опытов позволили установить, что это действие проявляется только при дозе в 1 мг на килограмм веса животного. Следовательно, центры менее чувствительны, чем сами глазные мышцы. Лучшей дозой является 1.5 мг на килограмм. При 3 мг не наблюдалось никакого влияния, так как можно предполагать, что эта доза парализует ядра глазных мышц.
в) Действие на вестибулярную систему (лабиринты и область вестибулярных ядер)
До Сих пор описанные влияния наблюдались на глазных мышцах, находящихся в покое. Но чтобы исследовать с помощью реакций глазных мышц влияние на вестибулярную область яда, который сам не вызывает нистагма, нужно сперва его вызвать.
Думать об изолированном влиянии яда на вестибулярную систему можно только тогда, когда яд или вообще не обладает никаким действием на ядра глазных мышц, или действует на последние в больших дозах, чем на вестибулярную область. Периферическое действие на мышцы глаз можно исключить перевязкой сонных артерий.
Поэтому пришлось проводить исследование на таламических кроликах с перевязанными сонными артериями. На таких животных длительным промыванием слухового прохода холодной или теплой водой вызывался вестибулярный нистагм. Применяемые дозы никотина должны были
558
Одиннадцатая глава. Действие ядов
быть меньше 1 мг на килограмм, чтобы исключить действие яда на ядра глазных мышц.
Результат был следующий.
Дозы в 0.025 мг никотина на килограмм действия не оказывали. Дозы от 0.05 до 0.1 мг вызывали уменьшение быстрой фазы нистагма.
М. reet. int .
М. reet. ext.
Рис. 260. Таламический кролик, 4.2 кг. Сонные артерии перевязаны. Верхняя кривая — движение левой внутренней мышцы; нижняя кривая — наружной. Рычаги при сокращении мышц поднимаются вверх. Нистагм вследствие промывания правого слухового прохода теплой водой: внутренняя мышца производит медленные расслабления, с быстрыми сокращениями, наружная — медленные сокращения с быстрыми расслаблениями. При горизонтальном отрезке (см. рис.) внутривенно вводится 0.3 мг никотина на килограмм веса. Из-за общего вздрагивания животного обе кривые сдвигаются вверх. Вскоре после этого в обеих мышцах прекращается быстрая фаза, наружная мышца останавливается в состоянии сокращения, а внутренняя — в состоянии расслабления. Через короткий промежуток времени нистагм начинается снова.
Дозы от 0.2 до 0.75 мг останавливали обе глазные мышцы в отклоненном положении: в сокращенном состоянии мышцу, первично сокращенную
М. reet. int.
М. reet. ext.
Рис. 261. Таламический кролик, 1.46 кг. Сонные артерии не перевязаны. Верхняя линия — левая внутренняя; нижняя линия — левая наружная мышца. Нистагм появляется при промывании левого слухового прохода холодной водой, i—i — введение 0.3 мг никотина на килограмм веса вызывает ровное сокращение обеих главных мышц вследствие периферического действия никотина.
лабиринтным раздражением, и в расслабленном состоянии мышцу, первично расслабленную лабиринтным раздражением. Быстрая фаза исчезала (рис. 260).
Из этого можно сделать вывод о действии никотина на вестибулярную* область.
Никотин
559*
Если не перевязывать сонные артерии, то вследствие прямого периферического действия никотина (даже при малых дозах в 0.3 мг) сокращаются обе мышцы и не обнаруживается действия на вестибулярную область (рис. 261).
М. reet. int.
М. reet. ext.
Рис. 262. Таламический кролик, 1.53 кг. Сонные артерии перевязаны. Наверху — левая внутренняя мышца, внизу — левая наружная. Нистагм вследствие промывания левого слухового прохода холодной водой. Введение 1.5 мг никотина на килограмм веса вызывает сокращение обеих глазных мышц вследствие возбуждения ядер глазных мышц после прекращения нистагма.
Дозы от 1 мг на килограмм возбуждают, как это было показана в пункте б, ядра глазных мышц. Если вводить эти дозы при перевязанных сонных артериях во время вестибулярного нистагма,1 то обе мышцы за- стывают в сокращенном состоянии и не удается выявить действия никотина, на вестибулярную систему (рис. 262).
г) Действие только на область вестибулярных ядер
Для решения вопроса, действует ли яд в области вестибулярных ядер> или в лабиринте, можно еще исследовать его влияние на компенсаторный нистагм Бехтерева, который наступает, если через некоторое время после экстирпации первого также удаляют второй лабиринт. Тогда отсутствуют оба лабиринта и действие яда проявляется центрально в области ядер. Но если действие наступит при калорическом, а не при бехтеревском нистагме, то следует предположить прямое влияние яда на периферический орган чувств. Если будет найдено, что оба вида нистагма отвечают на. очень различные дозы яда, то очевидным является влияние как на лабиринты, так и на ядра.
И действительно, в четырех опытах на кроликах с двусторонней экстирпацией лабиринтов, у которых наблюдался компенсаторный нистагм Бехтерева, получилось, что внутривенное введение 0.1 мг никотина на килограмм, значит той же дозы, что и при калорическом нистагме, снимает быструю фазу нистагма и останавливает мышцы глаз в отклоненном состоянии (рис. 263).
Из этого можно сделать вывод, что применяемые дозы никотина действуют непосредственно в области вестибулярных ядер. Де Клейн и Вер-
1 В некоторых опытах для этого достаточно было 0.75 мг на килограмм веса животного.
560
Одиннадцатая глава. Действие ядов
стеег предполагают, что здесь речь идет о сильном возбуждающем действии. Более точная локализация этой области ядер пока еще невозможна. Изолированного периферического действия на лабиринт пока не удалось установить.
Анализ действия никотина на рефлекторную дугу при вестибулярном нистагме показал следующее.
Дозы от 0.25 мг на килограмм и выше приводят мышцы глаз в сокращение. Это действие перекрывает в общем остальные эффекты.
Если препятствовать проявлению периферического действия на мышцы, то после доз от 0.05—0.75 мг наблюдается влияние на область вестибу-
Рис. 263. Кролик 1.8 кг (правосторонняя экстирпация лабиринта 19 ноября 1921 г.), 24 ноября 1921 г. Таламическое животное. Сонные артерии перевязаны. Левосторонняя экстирпация лабиринтов. Верхняя кривая — прямая внутренняя; нижняя кривая — прямая наружная мышца. Наружная мышца сокращена и показывает быстрые расслабления. Внутренняя — расслаблена и дает быстрые сокращения. За 50 сек. до начала опыта введено 0.3 мг никотина на килограмм веса. Внутренняя прямая мышца расслабляется, наружная — через состояние сокращения приходит к покою.
лирных ядер, которое проявляется после малых доз в уменьшении, после больших — в исчезновении быстрых фаз нистагма и остановке в состоянии отклонения (сокращение одной мышцы при расслаблении ее антагониста), если до этого любым путем вызывался вестибулярный нистагм. Иначе действие не проявится.
Большие дозы никотина (0.75—2 мг), действие которых на периферические глазные мышцы исключено, возбуждают ядра глазных мышц, так что глазные мышцы сокращаются. Как только наступило это действие, уже больше не может проявиться влияние никотина на область вестибулярных ядер. Еще большие дозы парализуют ядра глазных мышц.
Приведенная дозировка действительна только для исследованного препарата никотина, так как существующие в продаже сорта никотина имеют различное действие.
И* Заключение
Сообщенные в этой главе наблюдения, несмотря на то что до настоящего времени исследовалось ограниченное количество ядов, дали запутанное обилие самых различных картин действия яда. Можно предполагать, что дальнейшая обработка этой области приведет к удивительным фактам. Однако уже теперь можно установить некоторые общие результаты.
Приходящие в действие при лабиринтных рефлексах и установке тела аппараты проявляют по отношению к разным ядрам чрезвычайно различную
Заключение
561
чувствительность. Причем не удается установить какую-нибудь общую последовательность восприимчивости по отношению ко всем ядам, наоборот, каждый отдельный яд действует своеобразным путем.
В то время как, например, прогрессивные реакции в общем очень чувствительны к наркотикам, при отравлении хеноподиевым маслом они сохраняются почти до самой смерти животного.
Компенсаторные вертикальные установки глаз затухают после введения пикротоксина в стадии судорог, после введения стрихнина они сохраняются дольше, чем любой лабиринтный рефлекс.
Вращательный нистагм головы и глаз может после введения камфары отсутствовать в стадии, когда еще имеются установочные рефлексы, в то время как они после введения наркотиков затухают гораздо позже установочных рефлексов, а после введения хеноподиевого масла сохраняются почти до самой смерти животного.
Самым примечательным является то, что внутри одной рефлекторной группы различные частные функции по-разному чувствительны к одному и тому же яду. Так, из различных форм вращательного нистагма глаз отдельными наркотиками сперва снимается вертикальный, затем ротаторный и только гораздо позднее горизонтальный, в то время как относящиеся сюда отклонения глаз остаются без изменения.
Из компенсаторных установок глаз вследствие действия хеноподиевого масла затухают сравнительно рано вертикальные, а ротаторные только лишь незадолго до смерти, в то время как при отравлении стрихнином, наоборот, — вертикальные остаются до самой смерти.
Из комплекса установочных рефлексов часто раньше всего выпадают лабиринтные установочные рефлексы (например, при действии пикротоксина), в то время как алкоголь избирательно затрагивает шейные установочные рефлексы и вследствие этого вызывает своеобразные нарушения движений.
Если принять еще во внимание, что отдельные яды, как пикротоксин, уже в дозах, когда они еще не Ьказывают никаких других действий, влияют на лабиринтные рефлексы, то можно понять, что трудно найти другую функциональную группу как эта, в которой так хорошо можно было бы демонстрировать избирательность и специфичность различных ядов.
До того как была хорошо изучена эта избирательность действия ядов, из отдельных случайных наблюдений иногда делались неправильные выводы. Так, Бартельс [Bartels (1)] и Розенфельд (Rosenfeld) предполагали из этого факта, что нистагм глаз при наркозе исчезает раньше, чем отклонение, что нистагм зависит от других частей мозга (кора большого мозга), чем отклонение. Этот взгляд опровергается успехами экстирпации головного мозга, после которой сохранялся нистагм. Это один из многочисленных примеров того, что яды не пригодны для анализа неизвестных физиологических механизмов.
Раньше приходилось довольствоваться общим установлением того, что определенный яд вызывал судороги, центральные параличи, нарушения равновесия и т. д. Приведенные в этой главе примеры показывают, что теперь мы двинулись вперед и можем указать частные механизмы, которые нарушены. Благодаря этому изучение таких нарушений приобретает новый интерес, и нет сомнения, что и клиника судорожных состояний человека сможет вынести из этого пользу. Во всяком случае изучение картины отравления на животных дает более глубокий взгляд на эти сложные процессы. Хинакетоны возбуждают судорожные движения ходьбы, которые только тогда приводят к поразительным движениям 36 Р. Магнус
562
Одиннадцатая глава. Действие ядов
перекатывания, когда голова при этом случайно повернута в ту или другую сторону. Характерная картина пикротоксиновых судорог определяется наступающей при этом ригидностью сгибателей, которая даже после де- церебрации заменяет ригидность разгибателей и закономерным образом изменяет течение рефлексов положения. При отравлении стрихнином,, наоборот, рефлексы положения не участвуют в прекращении координации в стадии судорог, так что тонические шейные и лабиринтные рефлексы на мышцы тела еще вызывают типичные реакции в то время, когда уже снята координация многочисленных других рефлексов. Внезапное паденио животных в определенных стадиях многочисленных отравлений основы« вается на исчезновении установочных рефлексов, которые парализуются с различной, но для каждого яда определенной последовательностью. После этого наступают очень различные состояния, в зависимости от которых животное или лежит расслабленно, или находится в состоянии окоченения, или же более или менее упорядоченного клонического возбуждения.
Так как центры обсуждаемых здесь рефлексов почти все без исключения находятся в стволе мозга, то понятно то большое значение, которое он имеет для осуществления наблюдаемых симптомов. Здесь речь идет о центрах, расположенных в среднем и продолговатом мозгу, которые необходимо привлечь для объяснения патологии различных картин отравления. Знатоку литературы по данному предмету ясно, что здесь прежде всего нельзя оставить в силе некоторые неправильно приписанные мозжечку зависимости. Например, большая роль, которую еще часто приписывают мозжечку при объяснении нарушения движений отравлением алкоголем, не соответствует и прямо противоположна нашим малым познаниям о нормальной деятельности этого органа. Характерным является еще следующее: Форссман нашел интересный факт, что введение малых доз бараньей крови, гемолизированной сывороткой кролика, морским свинкам в сонные артерии (по направлению аорты) вызывает своеобразные нарушения равновесия: поворот головы, манежеподобные движения,
перекатывание, вертикальные отклонения глаз, которые, по нашим знаниям на сегодняшний день, по-видимому, надо объяснять влиянием на центры в продолговатом мозгу (может быть, в среднем мозгу). Форссман объяснил это нарушениями в мозжечке. Фридбергер и Шродер (Friedberger и Schröder) исследовали гистологически мозг таких животных и действительно нашли очаги только в продолговатом мозгу, а не в большом мозгу и мозжечке. Но несмотря на это, они сделали следующий вывод: «Эта локализация изменений из-за теснейших и многочисленных связей продолговатого мозга с мозжечком клинически достаточно ясно объясняет симптомы во время жизни (!)». Однако в действительности их анатомическое исследование открыло правильную локализацию поражения.
Конечно, мы здесь находимся еще у истоков познания. Каждый прогресс нашего знания о расположении центральных аппаратов пойдет на пользу понимания токсикологических и патологических процессов. Однако теперь все же решено большое количество поставленных вопросов, и в целом ряде отдельных случаев положено начало для глубокого понимания тесного взаимодействия нарушений очень сложного симптомокомп- лекса.
Двенадцатая глава. Установочные рефлексы у новорожденных животных 563,
Двенадцатая глава
УСТАНОВОЧНЫЕ РЕФЛЕКСЫ ТЕЛА У НОВОРОЖДЕННЫХ
ЖИВОТНЫХ (79)
Если в лаборатории рождались животные, у них изучались важнейшие лабиринтные рефлексы и некоторые другие реакции, участвующие в установке тела. В общем, имеются записи протоколов опытов над более чем 15 кроликами (два помета), 18 кошками (шесть пометов), тремя собаками и двумя морскими свинками.
Подопытный материал достаточно велик для кроликов и кошек, чтобы е уверенностью характеризовать у них рефлексы, зато на собаках и особенно морских свинках требуется провести дополнительные исследования.
Обзор полученных до настоящего времени результатов приводится в таблице.
Бросается в глаза то, что хотя у всех новорожденных имелись отчетливые реакции поворота головы и, насколько это удалось исследовать, также реакции поворота глаз 1 (у морских свинок сразу после рождения, у кошек в конце беременности), у них отсутствовал вращательный нистагм головы и вращательный нистагм глаз. У кролика отсутствовали оба нистагма в течение 8 дней, через 18 дней они были положительными; у кошки они отсутствовали в течение 6 дней, на седьмой день можно было установить отчетливый вращательный нистагм глаз.
Это наблюдение совпадает с данными о том, что и при выходе из наркоза и из шока после операции вращательный нистагм восстанавливается позже, чем реакция поворота. Следовательно, и в индивидуальном развитии животного имеется стадия, при которой на раздражение полукружных каналов появляется только отклонение без всякого нистагма. По Бартельсу (2), преждевременно родившиеся дети имеют реакции поворота глаз, но у них отсутствует нистагм, зато у доношенных новорожденных наблюдается также вращательный нистагм.
Следующей группой реакций, развившихся сравнительно поздно, являются реакции на прогрессивные движения. Они наблюдались только у новорожденных морских свинок, у новорожденных кроликов и кошек они отсутствовали. Лифтная реакция у них появлялась раньше, чем рефлекс готовности к прыжку. Лифтная реакция развивалась у кроликов в течение 8, у кошек — 1—3 дней, рефлекс готовности к црыжку у кроликов развивался в течение 18 дней, у кошек — (6?) 20 дней. У собак через два дня после рождения рефлекс готовности к прыжку еще отсутствовал, однако лифтная реакция один раз как будто проявилась.1 2
Тонические лабиринтные рефлексы на конечности у недецеребрирован- ных животных не удается вызвать каждый раз, поэтому следует всегда проводить большое количество наблюдений. При этом оказалось, что эти рефлексы на кроликах, кошках и морских свинках вызываются сразу же после рождения. У некоторых котят они были чрезвычайно сильно выра-
1 Под реакциями поворота в отличие от нистагма понимается тоническое отклонение головы и глаз при вращении. — Ред.
2 М. Минковский (2) описывает у плода человека в различные сроки развития реакции поворота и прогрессивные реакции на конечности. Ван дер Ховен ф. Гендерен описывает случай водянки головного мозга у плода с сохраненным дистальным средним мозгом, включая клетки красного ядра, у которого в первые дни жизни наблюдались реакций поворота головы и глаз, слабый вращательный нистагм глаз и отчетливые прогрессивные реакции на руки.
36*
564 Двенадцатая глава. Установочные рефлексы у новорожденных животных
Установоч-
ные рефлексы
Я
Отолитовые рефлексы
тела на
я
©
я
Ч
>&
я
©
о
а
<D
Я
Я
о
и
компенсаторная установка глаз
О м
я В
Я и S о
ч
о
о
а
и
о
Я Я
©
Я
g
я К
&§ ю ? cd *
»a
Я
о
и
о
go
О
и
о
я
в
о
ь
о
И
я
зЯ
© м вП
CÖ
н
о й « О
!>»
О)
Я о н Ч
ge
Я си
Я
ч
о
Я
Я » Я з
Эй
Ä м
О 5ч
1
а о Й Cf
о
р*
Я
к
о
© я
Я 2
Я 5 4*0*
® я Я о
В»
Я
«I
о,
со
о
ч
о
Ч
зЯ
©
Я
vo 2
Ен
а
1
%
Ий
И
о
(и
8
0
CÖ Л Ч Я
©
И
a
о © и а
о cd Н Я
1
1 морская
4 часа.
+
+
+
01
~Г
+
+
свинка.
2
6 кроликов.
Новорож-
+
+
+
+
+
денные.
3
9 »
То же.
р
ф
ф
?
Ф
2
6 »
1 день.
3
9 »
То же.
+2
+
+
р
Ф
3
9 »
2 дня.
+
+
3
9 »
8 дней.
+
+
+
+
0
+
3
9 »
18 »
+
+
+
“Г
4
2 собаки.
2 дня.
ф
ф
+
+ 3
+
5
1 собака.
4-5
недель.
+
+
+
+
6а
4 кошачьих
Сразу.
+
0
+
0
+
ф
плода,
живые из матки, i
Через 4 часа.
ф
(+)8
ф
0
82—90 г. 1 1
1 день.
+
+
+
(+)
_i_
+
1
1
2 дня.
+
+
+
4~
+
66
3 кошачьих
’
Сразу.
+
0
+
0
0
0
0
“Г
плода
из матки, 108—122 г/
Через 6 час.
+
ф
ж
+
р
1 день.
+
+
?
2 дня.
ф
+
+
7
2 кошки.
Новорож¬
+
+
+
1
"Г
денные.
8
3 »
То же.
ф
*
8
3 »
1 день.
+
+
9
4 »
1 »
+
+
ф
+
+
10
2 »
1 »
ф
0
10
2 »
2 дня.
+
+
+ 3
10
2 »
3 »
+
ф
+ 5
10
2 »
4 »
+
+
i
10
2 »
6 дней.
ф
10
2 »
7 »
10
2 »
20 »
i В положения на боку—отрицательны. 2 У одного маленького экземпляра отри сомнительно. 6 У одного животного слабо положительны, у другого отрицательны, передняя часть — отсутствуют. Объяснения значений: ф очень сильно; +
Двенадцатая глава. Установочные рефлексы у новорожденных животных 565
Рефлексы полукружных каналов
Тонические шейные рефлексы на глаза
Переворачивание при падении
Состояние глаз
вращение
прогрессивные движения
на голову
на глаза
лифтная реакция
готовность к прыжку
растопыривание-
пальцев
реакция
нистагм
реакция
нистагм
+
0
+
0
+
+
+
Открыты.
0
0
0
0
Закрыты.
+
0
0
0
»
+
0
?
0
0
»
+
0
?
0
»
0
?
0
»
ч-
0
*
0
+
0
+
У 4 открыты.
+
+
+
+
+
+
Открыты.
+
0
?
0
Закрыты.
+
+
+
+
+
+
+
Открыты.
+
0
0
0
Закрыты.
+
0
»
+
0
0
0
»
+
0
(+)
0
0
0
0
»
Оперативно открыты.
+
0
+ 7
О7
0
0
0
То же
+
0
0
0
0
»
»
Закрыты.
0
»
+
0
»
*
0
+
(+)4
»
+
0
0
0
»
+
0
+
0
*(+)4
0
\ У одного животного один
*
0
+
0
+
0
0
i глаз оперативно от-
+
0
+
0
+
0
0
J крыт.
+
0
+
0
+
(+)6
+
+
+
+
цательны. з у одного экземпляра отрицательны. 4 Слабые. 5 У одного животного 7 Горизонтально, вертикально и ротаторно. 8 Задняя часть тела — рефлексы имеются, отчетливо; (+) слабо; ? сомнительно; Ü отрицательно.
566 Двенадцатая глава. Установочные рефлексы у новорожденных животных
жены. Обследованный на 2-й день после рождения щенок также имел эти рефлексы.
Тонические шейные рефлексы на конечности наблюдались у кошек и кроликов сразу после рождения, у собак — через 2 дня. Более ранние наблюдения на собаках не проводились, но весьма вероятно, что они также имеются сразу после рождения.
Установочные рефлексы у новорожденных обследованных здесь видов животных удивительно хорошо развиты.1 У одной морской свинки и у почти доношенного кошачьего плода (приплод 6 штук) отсутствовали лабиринтные установочные рефлексы на голову, в то время как установочные рефлексы тела на тело наблюдались отчетливо. Следовательно, животные могли при соприкосновении с полом привести голову в нормальное положение. И у двух кошачьих плодов (весом 91,и 97 г), которые родились преждевременно, во время определения титра дигиталиса у матери удалось вызвать установочные рефлексы тела на голову. Шейные установочные рефлексы имелись во всех описанных случаях. Лабиринтные установочные рефлексы на голову развивались у упомянутых кошачьих плодов (приплод 6 штук) уже через 6 часов и были на следующий день очень живыми. Они были обнаружены у всех спонтанно родившихся кроликов и кошек сразу после рождения и были также установлены через два дня после рождения у обследованных щенков. Установочные рефлексы тела на тело также в большинстве случаев обнаруживались сразу после рождения. У плодов кошек они отсутствовали сразу после изъятия, но уже через 6 часов были положительны. У приплода из 9 кроликов они сразу после рождения были сомнительными, зато на следующий день — отчетливыми, только у одного маленького экземпляра они отсутствовали. Через 24 часа они у него также были отчетливыми. Один кролик в возрасте 18 дней имел очень отчетливый рефлекс переворачивания при падении в воздухе.
У приплода из 9 кроликов наблюдалось развитие акта ходьбы. В день рождения животные, покачиваясь, очень неуклюже ползли на животе по земле, время от времени падали на бок, но опять быстро приподнимались. Через 24 часа они еще падали, касаясь животом земли, иногда падали на бок, но тотчас же принимали сидячее положение. Один раз одно из животных при падении перекатилось через спину. Через 2 дня животные все еще бегали, касаясь животом земли, падали на бок, в то время как голова оставалась в нормальном положении, тело сейчас же поднималось. В возрасте 8 дней животные свободно несли живот над полом. Походка была еще неуверенной, но они больше не падали. Через 18 дней после рождения животные могли бегать нормально.
Кошачьи плоды (приплод 6а на таблице) могли сразу после изъятия при щипании хвоста поднимать из бокового положения — правда, еще неуклюже — переднюю и заднюю часть тела. Через 6 часов они ползали неуклюже в нормальном положении на животе, но при этом почти не двигались вперед. Через 24 часа они могли немного поднимать голову, голова качалась, ползли на животе по полу вперед, но еще не в прямом направлении. Ноги при этом еще не выполняли правильных альтернированных движений ходьбы. Животные иногда падали на бок. Через 48 часов они ползли на животе, не падая на бок, могли переползать один через другого. Через 72 часа ползание было лучше, но еще не по прямому направлению, а поочередно влево и право, причем голова задавала на¬
1 Ландау недавно описал у новорожденного ребенка лабиринтные установочные рефлексы вместе с шейными установочными рефлексами.
Двенадцатая глава. Установочные рефлексы у новорожденных животных 567
правление. Они по нескольку раз падали задней частью тела на бок, но опять садились. Голова теперь поднималась активно. К сожалению, животным дольше не удалось сохранить жизнь.
Компенсаторные установки глаз имелись у морских свинок сразу после рождения. У кролика их можно было исследовать только через 8 дней. К этому времени удавалось наблюдать как тонические лабиринтные рефлексы, так и тонические шейные рефлексы на глаза.
В итоге можно сказать, что из обследованных видов животных у морской свинки, которая рождается с открытыми глазами, все эти рефлексы проявляются наиболее полно; у нее имеются уже реакции на прогрессивные движения, отсутствует только вращательный нистагм. Отсутствуют ли лабиринтные установочные рефлексы на голову у новорожденных морских свинок, следует еще точно установить. У остальных животных из обследованных рефлексов отсутствуют вращательный нистагм головы и глаз и реакции на прогрессивные движения. Среди последних лифтная реакция наблюдается раньше, чем рефлекс готовности к прыжку. Но в остальном кролики, кошки и собаки обладают довольно совершенными, необходимыми для установки тела лабиринтными рефлексами и остальными реакциями. Однако наблюдения над обучением кроликов и кошек ходьбе показывают, что животные в первые дни жизни должны сперва выучиться использовать эти рефлексы для сложной общей деятельности. У почти доношенных кошачьих плодов отсутствуют лабиринтные установочные рефлексы тела на тело, в то время как установочные рефлексы тела на голову и шейные установочные рефлексы развиты хорошо.
РАБОТЫ ИЗ ФАРМАКОЛОГИЧЕСКОГО ИНСТИТУТА ГОСУДАРСТВЕННОГО УНИВЕРСИТЕТА В УТРЕХТЕ
1. R. Magnus. Zur Regelung der Bewegungen durch das Zentralnervensystem.
I. Mitteilung. Pflügers Arch. f. d. ges. Physiol., Bd. 130, S. 219, 1909 (Aus dem physiologischen Institut der Universität Liverpool).
2. R. Magnus. Zur Regelung der Bewegungen durch das Zentralnervensystem.
II. Mitteilung. Pflügers Arch. f. d. ges. Physiol., Bd. 130, S. 253, 1909.
3. R. Magnus. Zur Regelung der Bewegungen durch das Zentralnervensystem.
III. Mitteilung. Pflügers Arch. f. d. ges. Physiol., Bd. 134, S. 545, 1910.
4. R. Magnus. Zur Regelung der Bewegungen durch das Zentralnervensystem.
IV. Mitteilung. Pflügers Arch. f. d. ges. Physiol., Bd. 134, S. 584, 1910.
5. R. Magnus. Experimentelles und Klinisches über tonische Reflexe. Hande¬
lingen van het XIII. Ned. Natuur- en Geneeskundig Congres. 20.—22. April 1911, S.317.
6. R. Magnus. Über die Beziehungen des Kopfes zu den Gliedern. (Festvortrag,.
gehalten am 25. November 1911 in der medizinischen Geséllschaft in Leiden). Münch, med. Wochenschr., 1912, S. 681.
7. R. Magnus und A. de К 1 e у n. Die Abhängigkeit des Tonus der Extremi¬
tätenmuskeln von der Kopfstellung. Pflügers Arch. f. d. ges. Physiol., Bd. 145,. S. 455, 1912.
8. A. de К 1 e у n. Zur Technik der Labyrinthexstirpation und Labyrinthausschal¬
tung bei Katzen. Pflügers Arch. f. d. ges. Physiol., Bd. 145, S. 549, 1912.
9. W. Weiland. Hals- und Labyrinthreflexe beim Kaninchen, ihr Einfluss auf
den Muskel tonus und die Stellung der Extremitäten. Pflügers Arch. f. d. ges. Physiol., Bd. 147, S. 1, 1912.
10. R. Magnus und A. de К 1 e у n. Die Abhängigkeit des Tonus der Nacken¬
muskeln von der Kopfstellung. Pflügers Arch. f. d. ges. Physiol., Bd. 147, S. 403,. 1912.
11. A. de К 1 e у n. Zur Kenntnis des Verlaufs der postganglionären Sympathicus-
bahnen für Pupillenerweiterung, Lidspaltöffnung und Retraktion der Nickhaut bei der Katze. Zentralbl. f. Physiol., Bd. 26, S. 4, 1912.
12. R. Magnus und A de К 1 e у n. Ein weiterer Fall von tonischen «Halsre¬
flexen» beim Menschen. Münch, med. Wochenschr., 1913, S. 2566.
13. R. Magnus und C. G. L. Wolf. Weitere Mitteilungen über den Einfluss
der Kopfstellung auf den Gliedertonus. Pflügers Arch. f. d. ges. Physiol., Bd. 149, S. 447, 1913.
14. R. Magnus und A. de К 1 e у n. Die Abhängigkeit der Körperstellung vom
Kopfstande beim normalen Kaninchen. Pflügers Arch. f. d. ges. Physiol., Bd. 154, S. 163, 1913.
15. R. Magnus und A. de К 1 e у n. Analyse der Folgezustände einseitiger La¬
byrinthexstirpation mit besonderer Berücksichtigung der Rolle der tonischen Halsreflexe. Pflügers Arch. f. d. ges. Physiol., Bd. 154, S. 178, 1913.
16. R. Magnus und W. Storm van Leeuwen. Die akuten und die dauern¬
den Folgen des Ausfalles der tonischen Hals- und Labyrinthreflexe. Pflügers Arch. f. d. ges. Physiol., Bd. 159, S. 157, 1914.
17. A. de К 1 e у n. Zur Analyse der Folgezustände einseitiger Labyrinthexstirpation
beim Frosch. Pflügers Arch. f. d. ges. Physiol., Bd. 159, S. 218, 1914.
18. R. Magnus. Welche Teile des Zentralnervensystems müssen für das Zustande¬
kommen der tonischen Hals- und Labyrinthreflexe auf die Körpermuskulatur vorhanden sein? Pflügers Arch. f. d. ges. Physiol., Bd. 159, S. 224, 1914.
18a. J. S. Beritoff und R. Magnus. Zusatz bei der Korrektur. Ebenda, S. 249.
19. Ch. S о c i n und W. Storm van Leeuwen. Über den Einfluss der Kopfstel¬
lung auf phasische Extremitätenreflexe. Pflügers Arch. f. d. ges. Physiol.,Bd.159, S. 251, 1914.
20. A. de Kleyn und Ch. S о c i n. Zur näheren Kenntnis des Verlaufs der post¬
ganglionären Sympathicusbahnen für Pupillenerweiterung, Lidspaltenöffnung
Литература
569
und Nickhautretraktion bei der Katze. Pflügers Arch. f. d. ges. Physiol., Bd. 160, S. 407, 1915.
21. H. M. de В u r 1 e t. Anatomische Bemerkungen zur vorhergehenden Arbeit von A. de Kleynund Gh. Socin. Pflügers Arch. f. d. ges. Physiol., Bd. 160, S. 416, 1915. 22- R. Magnus und A. de К 1 e у n. Weitere Beobachtungen über Hals- und Labyrinthreflexe auf die Gliedermuskeln des Menschen. Pflügers Arch. f. d. ges. Physiol., Bd 160, S. 429, 1915.
23. H. M. de В u r 1 e t und A. de К 1 e у n. Über den Stand der Otolithenmem-
branen beim Kaninchen. Pflügers Arch. f. d. ges. Physiol., Bd. 163, S. 321, 1916. (Aus dem anatomischen und dem pharmakologischen Institut der Reichsuniver=> sität Utrecht).
24. R. Magnus. Beiträge zum Problem der Körperstellung. I. Mitteilung. Stell¬
reflexe beim Zwischenhirn- und Mittelhirnkaninchen. Pflügers Arch. f. d. ges. Physiol., Bd. 163, S. 405, 1916.
25. A. de К 1 e у n en W. Storm van Leeuwen. Over vestibulaire oogreflexen.
I. Over de oorzaak van het ontstaan van den calorischen nystagmus. Verslagen koninklyke Akademie van Wetenschappen Amsterdam. Wis- en natuurkundige afdeeling, Bd. 26, S. 381, 1917. — Concerning vestibular Eye-reflexes. I. On the origin of caloric nystagmus. Koninklyke Akademie van Wetenschappen Amsterdam. Proceedings, Bd. 20, S. 622, 1917.
26 A. de К 1 e у n und W. Storm van Leeuwen. Über vestibuläre Augenreflexe. I. Über die Entstehungsursache des calorischen Nystagmus nach Versuchen an Katzen und Kaninchen, v. Graefes Arch. f. Ophth., Bd. 94, S. 316, 1917.
27. J. van der Hoeve und A. de К 1 e у n. Tonische Labyrinthreflexe auf die
Augen. Pflügers Arch. f. d. ges. Physiol., Bd..l69, S. 241, 1917.
28. A. de Kleyn und R. Tumbelaka. Über vestibuläre Augenreflexe. II.
Vestibuläre Augenreflexe bei totaler einseitiger Oculomotoriuslähmung, v. Graefes Arch. f. Ophth., Bd. 95, S. 314, 1918.
29. A. de Kleyn und R. Magnus. Sympathicuslähmung durch Abkühlung
des Mittelohres beim Ausspritzen des Gehörganges der Katze mit kaltem Wasser. v. Graefes Arch. f. Ophth., Bd. 96, S. 368, 1918.
30. R. Magnus. Tonische Hals- und Labyrinthreflexe auf die Körpermuskeln beim
decerebrierten Affen. Arch, néerland. de physiol, de l’homme et des anim., Bd. 2, S. 484, 1918.
31. A. de Kleyn. Actions réflexes du labyrinthe et du cou sur les muscles de Poeil.
Arch, néerland. de physiol, de l’homme et dés anim., Bd. 2, S. 644, 1918.
32. R. Magnus. Beiträge zum Problem der Körperstellung. II. Mitteilung. Stell¬
reflexe beim Kaninchen nach einseitiger Labyrinthexstirpation. Pflügers Arch, f. d. ges. Physiol., Bd. 174, S. 134, 1919.
33. G. Liljestrand und R. Magnus. Warum wird die lokale Muskelstarre
beim Wundstarrkrampf durch Novokain aufgehoben? Münch, med. Wochenschr., 1919, S. 551.
34. G. Liljestrand und R. Magnus. Über die Wirkung des Novocains
auf den normalen und den tetanusstarren Skelettmuskel und über die Entstehung der lokalen Muskelstarre beim Wundstarrkrampf. Pflügers Arch. f. d. ges. Physiol., Bd. 176, S. 168, 1919.
35. A. de Kleyn und R. Magnus. Kleinhirn, Hirnstamm und Labyrinthre¬
flexe. Münch, med. Wochenschr., 1919, S. 523.
36. R. Magnus und A. de Kleyn. Tonische Labyrinthreflexen op de oogspie¬
ren. Verslagen koninklyke Akademie van Wetenschappen Amsterdam. Wis- en natuurkundige afdeeling, Bd. 28, S. 129, 1919. — Tonic reflexes of the labyrinth on the eye-muscles. Koninklyke Akademie van Wetenschappen Amsterdam. Proceedings, Bd. 22, S. 242, 1919.
37. A. de Kleyn und R. Magnus. Über die Unabhängigkeit der Labyrinthre¬
flexe vom Kleinhirn und über die Lage der Zentren für Labyrinthreflexe im Hirnstamm. Pflügers Arch. f. d. ges. Physiol., Bd. 178, S. 124, 1920.
38. A. de Kleyn und R. Magnus. Tonische Labyrinthreflexe auf die Augen¬
muskeln. Pflügers Arch. f. d. ges. Physiol., Bd. 178, S. 179, 1920.
39. A. de Kleyn en G. R. J. Versteeg h. Over de al of niet labyrinthaire-
genese van den donkernystagmus by honden. Verslagen koninklyke Akademie van Wetenschappen Amsterdam. Wis- en natuurkundige afdeeling, Bd. 28, S. 253r
1919. — On the question whether or no darkness-nystagmus in dogs originates* in the labyrinth. Koninklyke Akademie van Wetenschappen Amsterdam. Proceedings, Bd. 22, S. 393, 1919.
40. A. de К 1 e у n und C. Versteeg h. Über die Unabhängigkeit des Dunkel¬
nystagmus der Hunde vom Labyrinth, v. Graefes Arch. f. Ophth., Bd. 101. S. 228, 1920.
570
Литература
41 J. G. Dusser de Barenne und R. Magnus. Beiträge zum Problem der Körperstellung. III. Mitteilung. Die Stellreflexe bei der grosshirnlosen Katze und dem grosshirnlosen Hunde. Pflügers Arch. f. d. ges. Physiol., Bd. 180, S. 75,
1920.
42. R. Magnus en A. de К 1 e у n. Optische «Stellreflexe» by den hond en by de
kat. Verslagen Koninklyke Akademie van Wetenschappen. Amsterdam. Wis- en natuurkundige afdeeling, Bd. 28, S. 670, 1920. — On optie «Stellreflexe» in the dog and in the cat. Koninklyke Akademie van Wetenschappen Amsterdam. Proceedings, Bd. 22, S. 948, 1920.
43. A. de К 1 e у n und R. Magnus. Beiträge zum Problem der Körperstellung.
IV. Mitteilung. Optische Stellreflexe bei Hund und Katze. Pflügers Arch. f. d. ges. Physiol., Bd. 180, S. 291, 1920.
44. A. de К 1 e у n. Tonische Labyrinth- en halsreflexen op de oogen. Verslagen Ko¬
ninklyke Akademie van Wetenschappen Amsterdam. Wis- en natuurkundige afdeeling, Bd. 28, S. 1223, 1920. — On the effect of tonic labyrinthine and cervical reflexes upon the eye-muscles. Koninklyke Akademie van Wetenschappen Amsterdam. Proceedings, Bd. 23, S. 509, 1920.
45. A. de К 1 e у n en W. Storm van Leeuwen. Vestibulaire oogreflexen. II.
De genese van den koudwaternystagmus by konynen. Verslagen Koninklyke Akademie van Wetenschappen Amsterdam. Wis- en natuurkundige afdeeling, Bd. 28, S. 721, 1920. — Concerning vestibular eye-reflexes. II. The genesis of coldwater nystagmus in rabbits. Koninklyke Akademie van Wetenschappen Amsterdam. Proceedings, Bd. 22, S. 713, 1920.
46. A. de К 1 e у n und G. R. J. Versteeg h. Über den Einfluss der Reizung
der Nasenschleimhaut auf den vestibulären Nystagmus beim Kaninchen. Arch, f. Laryngol. u. Rhinol., Bd. 33, S. 437, 1920.
47. R. Ma,gnus. Die Funktion der Otolithen. Ber. über d. ges. Physiol., Bd. 2,
S. 174. Aus dem Bericht über die Tagung der deutschen physiologischen Gesellschaft in Hamburg 26—28. Mai 1920.
48. A. de К 1 e у n. Folgen der isolierten Otolithenausschaltung. Ber. über d. ges.
Physiol., Bd. 2, S. 175. Aus dem Bericht über die Tagung der deutschen physiologischen Gesellschaft in Hamburg 26—28. Mai 1920.
49. R. Magnus en A. de К 1 e у n. De functie der otolithen. Verslagen konin¬
klyke Akademie van Wetenschappen Amsterdam. Wis- en natuurkundige afdeeling, Bd. 29, S. 375, 1920. — The function of the otolithes. Koninklyke Akademie van Wetenschappen Amsterdam. Proceedings, Bd. 23, S. 907, 1920.
50. A. de К 1 e у n und R. Magnus. Über die Funktion der Otolithen. I. Mit¬
teilung. Otolithenstand bei den tonischen Labyrinthreflexen. Pflügers Arch, f. d. ges. Physiol., Bd. 186, S. 6, 1921.
51. A. de Kleyn und R. Magnus. Labyrinthreflexe auf die Progressivbewe¬
gungen. Pflügers Arch. f. d. ges. Physiol., Bd. 186, S. 39, 1921.
52. A. de Kleyn und R. Magnus. Über die Funktion der Otolithen. II. Mit¬
teilung. Isolierte Otolithenausschaltung bei Meerschweinchen. Pflügers Arch, f. d. ges. Physiol., Bd. 186, S. 61, 1921.
53. A. de К 1 e у n. Tonische Labyrinth- und Halsreflexe auf die Augen. Pflügers Arch.
f. d. ges. Physiol., Bd. 186, S. 82, 1921.
54. G. Versteeg h. Über eine Methode zur Lokalisierung der Angriffspunkte ver¬
schiedener Arzneimittel auf das Vestibülarsystem. Verhandl. d. Gesellsch. dtsch. Hals-, Nasen- und Ohrenärzte, Bd. I, S. 350, 1921.
55. A. de К 1 e у n. Experimente über die schnelle Phase des vestibulären Nystagmus.
Verhandl. d. Gesellsch. dtsch. Hals-, Nasen- und Ohrenärzte, Bd. I, S. 353, 1921. ü6. A. de Kleyn. Experimenten over de snelle phase van den vestibulairen nystagmus by het konyn. Verslagen koninklyke Akademie van Wetenschappen Amsterdam. Wis- en natuurkundige afdeeling, Bd. 29, S. 1230, 1921. — Experiments on the quick component phase of vestibular nystagmus in the rabbit. Koninklyke Akademie van Wetenschappen Amsterdam. Proceedings, Bd. 23, S. 1357,
1921.
.57. D. J. J o n к h o f f. De invloed van eenige geneesmiddelen op de labyrinthrefle- xen van konynen, caviae en katten. Diss. Utrecht, 1921.
58. R. Magnus. Zur Pharmakologie der Körperstellung und der Labyrinthreflexe.
Verhandl. d. dtsch. pharmakolog. Gesellsch., Bd. 2, S.'8, 1921. — Siehe Schmiedebergs Archiv, Bd. 92, 1922.
59. R. Magnus. Körperstellung und Labyrinthreflexe beim Affen. Pflügers Arch.
f. d. ges. Physiol., Bd. 193, S. 396, .1922. e
60. A. de Kleyn und R. Magnus. Über die Funktion der Otolithen. III. Mit¬
teilung. Kritische Bemerkungen zur Otolithen theorie von Herrn F. H. Quix. Pflügers Arch. f. d. ges. Physiol., Bd. 194, S. 407, 1922.
Литература
571
61. R. Magnus und A. de К 1 e у n. Nadere bydrage tot de functie der otoli-
thenapparaten. Verslagen Koninklyke Akademie van Wetenschappen Amsterdam, Bd. 31, S. 184, 1922. — A further contribution concerning the function of the otolithic apparatus. Koninklyke Akademie van Wetenschappen Amsterdam. Proceedings, Bd. 25, S. 256, 1922.
'62. A. de K.l e у n und W. Storm van Leeuwen. Über vestibuläre Augenreflexe.
III. Über die Genese des Kaltwassernystagmus bei Kaninchen, v. Graefes Arch, f. Ophth., Bd. ..107, S. 109, 1922.
63. A. de К 1 e у n. Über vestibuläre Augenreflexe. IV. Experimentelle Untersuchun¬
gen über die schnelle Phase des vestibulären Nystagmus beim Kaninchen, v. Graefes Arch. f. Ophth., Bd. 107, S. 480, 1922.
64. R. Magnus. Otolithenfunktion und Körperstellung. Referatvortrag auf der
Leipziger Naturforscherversammlung 1922. Naturwissenschaften, 1922, S. 927.
65. A. de К 1 e у n. Recherches quantitatives sur les positions compensatoires de
l’oeil chez le lapin. Arch, neerland, de physiol, de l’homme et des anim., Bd. 7, S. 138, 1922.
66. R. Magnus. Wie sich die fallende Katze in der Luft umdreht. Arch, neerland.
de physiol, de l’homme et des anim., Bd. 7, S. 218, 1922.
67. A. de К 1 e у n en H. S t e n v e r s. Tonische labyrinthreflexen op de oogen
bij menschen. Nederlandsch. tijdschr. v. geneesk., 1922, I, S. 486.
68. R. Magnus. Beiträge zur Pharmakologie der Körperstellung und der Laby¬
rinthreflexe. I. Mitteilung. Vorbemerkungen. — Acta oto-laryngologica, Bd. 4, S. 21, 1922.
69. D. J. Jonkhoff. Beiträge zur Pharmakologie der Körperstellung und der
Labyrinthreflexe. II. Mitteilung. Strychnin. — Acta oto-laryngologica, Bd. 4, S. 174, 1922.
70. D. J. Jonkhoff. Beiträge zur Pharmakologie der Körperstellung und der
Labyrinthreflexe. III. Mitteilung. Pikrotoxin. Acta oto-laryngologica, Bd. 4, S. 265, 1922.
71. G. V e r s t e e g h. Beiträge zur Pharmakologie der Körperstellung und der La¬
byrinthreflexe. IV. Mitteilung. Der Einfluss des Alkohols auf die Stellreflexe. Acta oto-laryngologica, Bd. 4, S. 394, 1922.
72. D. J. Jonkhoff. Beiträge zur Pharmakologie der Körperstellung und der
' Labyrinthreflexe. V. Mitteilung. Gampher. Acta oto-laryngologica, Bd. 4,450,1922.
73. A. de Kleyn und G. Versteeg h. Beiträge zur Pharmakologie der Körper¬
stellung und der Labyrinthreflexe. VI. Mitteilung. Über eine Methode zur Lokalisierung der Angriffspunkte verschiedener Arzneimittel auf den vestibulären Nystagmus, mit besonderer Berücksichtigung der Wirkung von Nicotin. Pflügers Arch, f. d. ges. Physiol., Bd. 196, S. 331, 1922.
74. D. J. Jonkhoff. Beiträge zur Pharmakologie der Körperstellung und der
Labyrinthreflexe. VII. Mitteilung. Oleum Chenopodii. Pflügers Arch: f. d. ges. Physiol., Bd. 196, S. 571, 1922.
75. U. G. В у 1 s m a und G. Versteeg h. Beiträge zur Pharmakologie der Körper¬
stellung und der Labyrinthreflexe. VIII. Mitteilung. Vergiftung mit Chinaketonen mit besonderer Berücksichtigung der Rollbewegungen. Pflügers Arch, f. d. ges. Physiol., Bd. 197, S. 415, 1922.
76. R. Magnus. Beiträge zur Pharmakologie der Körperstellung und der Laby¬
rinthreflexe. IX. Mitteilung. Weitere Erfahrungen mit Oleum Chenopodii. Pflügers Arch. f. d. ges. Physiol., Bd. 198, S. 427, 1923.
77. A. de Kleyn. Statischer Sinn. Jahresbericht über die gesamte Physiologie. 1920.
Berlin, 1923, S. 300.
78. J. G. Dusser de Barenne und A. de К 1 e у n. Über vestibuläre Augen-
reflexe. V. Vestibularuntersuchungen nach Ausschaltung einer Grosshirnhemisphäre beim Kaninchen, v. Graefes Arch. f. Ophth., Bd. Ill, S. 374, 1923.
79. R. Magnus. Körperstellungsreflexe bei neugeborenen Tieren. Skandin. Arch. f.
Physiol., Bd. 63, S. 39, 1923.
.80. G. G. J. Rademaker. Der rote Kern, die normale Tonusverteilung und die Stellfunktion. Klin. Wochenschr., Bd. I, S. 404, 1923.
60a. R. Magnus und G. G. J. Rademaker. Die Bedeutung des roten Kernes für die Körperstellung. (Vorl. Mitt.) Schweiz. Arch. f. Neurol, u. Psych., Bd. 13, S. 408, 1923.
81. R. Magnus. Die Bedeutung des Hirnstammes für Muskeltonus und Körperstellung. Leyden-Vorlesung des Vereins für innere Medizin und Kinderheilkunde in Berlin am 19. März 1923. Dtsch. med. Wochenschr., 1923, S. 501.
62. R. Magnus und A. de Kleyn. Experimentelle Physiologie des Vestibular-
apparates bei Säugetieren mit Ausschluss des Menschen. Alexander-Marburg, Handbuch d. Neurologie des Ohres., Bd. I, 1, S. 465. Berlin u. Wien. 1923.
572
Литература
82а. G. G. J. Rademaker. De beteekenis der roode kernen en van het overige mesencephalon voor spiertonus, lichaamshouding en labyrinthaire reflexen, Diss. Utrecht, 1924.
82b. R. Magnus und A. de К 1 e у n. Bydrage tot de functie van het vestibulaire apparat. Verslagen Koninklyke Akademie van Wetenschappen Amsterdam. Wis- en natuurkundige afdeeling, Bd. 32, S. 961, 1923.
РАБОТЫ ИЗ АНАТОМИЧЕСКОГО ИНСТИТУТА ГОСУДАРСТВЕННОГО УНИВЕРСИТЕТА В УТРЕХТЕ
83. Н. М. de В и г 1 e t en J. J. Koster. Over de bepaling van den stand van
booggangs- en maculavlakken in den schedel. Verslagen Koninklyke Akademie van Wetenschappen Amsterdam. Wis- en natuurkundige afdeeling, Bd. 24, S. 1828, 1916.
84. H. M. de Burl et und J. J. Koster. Zur Bestimmung des Standes der
Bogengänge und der Maculae acusticae in Kaninchenschädel. Arch. f. Anat. (u. Physiol,) 1916, S. 59.
85. H. О о r t. Über die Verästelung des Nervus octavus bei Säugetieren. (Modell des
Utriculus und sacculus des Kaninchens). Anat. Anz., Bd. 51, S. 272, 1918.
86. H. M. de В u r 1 e t. Der Perilymphatische Raum des Meerschweinchenohres.
Anat. Апг.л Bd. 53, S/’ 302, 1920.
87. H. О о r t. Über ein Modell zur Demonstration der Stellung der Maculae acusticae
im Kaninchenschädel. Pflügers Arch. f. d. ges. Physiol., Bd. 186, S. 1, 1921.
88. H. M. de В u r 1 e t und J. H. de Haas. Die Stellung der Maculae acusticae
im Meerschweinchenschädel. Zeitschr. f. Anat. u. Entwicklungsgesetz, Bd. 68, S. 177, 1923.
89. H. M. de Burlet und J. H. de Haas. Die Stellung der Maculae acusticae
im Macacus-Schädel. Zeitschr. f. Anat. u. Entwicklungsgesetz, Bd. 71, S. 233, 1924.
90. H. M. de Burlet. Zur Innervation der Maculae sacculi bei Säugetieren.
Anatomischer Anzeiger., 1924.
ЛИТЕРАТУРА 1
Ach N. Über die Otholithenfunktion und den Labyrinthtonus. Pflügers Arch. f. d.
ges. Physiol., Bd. 86, S. 122, 1901.
Auerbach. Therapie d. Gegenw., 1913, S. 358 (Zit. nach Jonkhoff).
A u g i e r. Zeitschr. f. Psychol, und Physiol, d. Sinnesorg., Bd.37, 1905. (Zit. nach W. Nagel [60]).
Bäräny R. (1). Augenbewegungen durch Thoraxbewegungen ausgelöst. Zentralbl. f. Physiol., Bd. 20, S. 298, 1907.
— und K. Witmaak(2). Funktionelle Prüfung des Vestibularapparates. VerhandU
d. dtsch. otol. Ges., Bd. 20, 1911.
—, Z. Reich und J. Rothfeld(3). Experimentelle Untersuchungen über die vestibulären Reaktionsbewegungen an Tieren, insbesondere im Zustand der decerebrate rigidity (vorl. Mitt.). Neurol. Zentralbl., Bd. 31, S. 1139, 1912.
— (4). Untersuchungen über die Funktion des Flocculus beim Kaninchen. Jahrb. d. Psy¬
chiatrie u. Neurol., Bd. 36, S. 1, 1914.
— (5). Theoretisches zur Funktion der Bogengänge und speziell des Flocculus beim
Kaninchen. Nordisk tidskrift för Oto-Rhino-Laryngologi., Bd. 2, S. 458,. 1917.
— (6). Über einige Augen- und Halsmuskelreflexe bei Neugeborenen. Acta Oto-laryn-
gologica., Bd. I, S. 97, 1918.
Dusser de В ar enne J. G. (1). Nachweis, dass die Magnus-de Kleynschen Reflexe bei der erwachsenen Katze mit intakten Zentralnervensystem bei passiven und aktiven Kopf-resp. Halsbewegungen auf treten, und somit im normalen Leben der Tiere eine Rolle spielen. Fol. Neuro-Biol., Bd. 8, S. 413, 1914.
— (2). Recherches expér. sur les functions du système nerveux central, faites en parti¬
culier sur deux chats, dont le neopallium a été enlevé. Arch, néerland. de physiol, de l’homme et des anim., Bd. 4, S. 31, 1920.
1 Этот список литературы охватывает только работы, приведенные в данной книге. В случае, если приводится несколько работ одного автора, то здесь и в тексто они даны с нумерацией в круглых скобках.
Литература
573
— (3). Proefondervindelyke Physiologie van het zenuwstelsel. In Bouman-Brouwers
Leerboek der zenuwziekten., Bd. I, S. 402, 1923.
Bartels M. (1). Über Regulierung der Augenstellung durch den Ohrapparat. Mit. 1—4, V. Graefes Arch. f. Ophth., Bd. 76, S. 1, 1910; Bd. 77, S. 531, 1910; Bd. 78, S. 129, 1911; Bd. 80, S. 207, 1911. (Mit. 1 u. 4 mit Shin-Jzi-Ziba). Siehe besonders Mitt. 3: Kurven des Spannungszustandes einiger Augenmuskeln unter dem Einfluss der Ohrreflexe.
— (2). Über willkürliche und unwillkürliche Augenbewegungen. (Nystagmus der Blin¬
den, Proprioreflexe, Blickbewegungen der Tiere). Klin. Monatsbl. f. Augenheilk., Bd. 53, S. 358, 1914.
— (3). Aufgaben der vergleichenden Physiologie der Augenbewegungen, v. Graefes
Arch. f. Ophth., Bd. 101, S. 299, 1920.
Bauer J. und R. Leidler. Über den Einfluss der Ausschaltung verschiedener Hirnabschnitte auf die vestibulären Augenreflexe. Arb. a. d. Wiener neurol. Inst., Bd. 19, S. 155, 1911.
Bayliss W. M. On reciprocal innervation in vasomotor reflexes and the action of strychnine and of chloroform thereupon. Proc. of the roy. soc. of London., Bd. 80, S. 359, 1908.
Bazett H. G. und W. G. Penfield. A study of the Sherrington decerebrate animal in the chronic as well as the acute condition. Brain., Bd. 45, S. 185, 1922. Bechterew W. (1). Ergebnisse der Durchschnei dung des N. acusticus nebst Erörterung der Bedeutung der semicirculären Kanäle für das Körpergleichgewicht. Pflügers Arch. f. d. ges. Physiol., Bd. 30, S. 312, 1883.
— (2). Über die Verbindung der sog. peripheren Gleichgewichtsorgane mit dem Klein¬
hirn. Versuche mit Durchschneidung der Kleinhirnstiele. Pflügers Arch. f. d. ges. Physiol., Bd. 34, S. 362, 1884. Arch. f. Physiol., 1896, S. 105.
Benjamins G. E. Contribution ä la connaissance des reflexes toniques des muscles de l’oeil. Arch, neerland, de physiol, de l’homme et des anim., B. 2, S. 536, 1918. Siehe auch Nederlandsch tijdschr. v. geneesk. 1918, I, S. 1036.
Beritoff J.S. (l).On the reciprocal innervation in tonic reflexes from the labyrinths and the neck. Journ. of physiol., Bd. 49, S. 147, 1915.
— (2). On the mode of origination of labyrinthine and cervical tonic reflexes and
on their part in the reflexes reactions of the decerebrate preparation. Quart, journ. of exp...physiol., Bd. 9, S. 199, 1915.
В e t h e A. Über die Erhaltung des Gleichgewichts. Biol. Zentralbl., Jg. 14, N 3, 1894. Beyer H. und M. Lewandowsky. Experimentelle Untersuchungen am Vesti- bularapparat von Säugetieren. Pflügers Arch. f. d. ges. Physiol., 1906, S. 451. В i e h 1 K. Über die intrakranielle Durchtrennung des Nervus vestibuli und deren Folgen. Wien akad. Sitzungsber. math.-nat. KL, Bd. 109, III, S. 324, 1900. Böhme A. und W. Weiland. Einige Beobachtungen über die Magnusschen Hals- und Labyrinthreflexe beim Menschen. Zeitschr. f. d. ges. Neurol. u. Psy- chiatr.,.Bd. 44, S. 94, 1918.
Bondi S. Über refletorische Bewegungen bei Kopfwendung in cerebralen Affekt ionen. Wien. klin. Wochenschr., 1912, N 41.
Brand B. Lichaamshouding en wervelkolom. Nederlandsch tijdschr. v. geneesk., 1922, II, 2011.
Breuer J. (1). Über die Funktion der Bogengänge des Ohrlabyrinths. Wien. med. Jahrb., 1874, H. I.
— (2). Beitrag zur Lehre vom statischen Sinn (Gleichgewichtsorgan. Vestibularappa-
rat des Ohrlabyrinths). 2. Mitt. Wien. med. Jahrb., 1875, H. 1.
— (3). Über die Funktion der Otolithenapparate. Pflügers Arch. f. d. ges. Physiol.,
Bd. 48, S. 195, 1891.
Brondgeest P. Q. Onderzoekingen over den tonus der willekeurige spieren. Diss. Utrecht, 1860.
Brown T. Graham (1). Die Atembewegungen des Frosches und ihre Beeinflussung durch die nervösen Zentren und durch das Labyrinth. Pflügers Arch. f. d. ges. Physiol., Bd. 130, S. 193, 1909.
— (2). Studies in the reflexes of the guinea-pig. III. The effect of removal of the cortex
of one cerebral hemisphere. Quart, journ. of exp. physiol., Bd. 3, S. 139, 1910.
— (3). The intrinsic factors in the act of progression in the mammal. Proc. of the roy
soc. of London B, Bd. 84, S. 308, 1911.
— (4). Reflexe orientation of the optical axes and the influence upon it of the cerebral
cortex. Arch, néerland. de physiol, de l’homme et des anim., Bd. 7, S. 571,
1922.
Brown-Séquard G. E. Nouveaux faits relatifs ä Faction du chloroforme appli- quée ä la peripherie du Systeme nerveux (peau et conduit auditif externe). Cpt. rend des séances de la soc. de biol., Bd. 32, S. 383, 1881.
574
Литература
Brouwer В. (1). Über Meningo-Encephalitis und die Magnus-de Kleynschen Reflexe. Zeitschr. f. d. ges. Neurol, u. Psychiatrie,.Origin., Bd. 36, S. 161, 1917.
— (2). Examen anatomique du système nerveux central des deux chats décrits par
J. G. Dusser de Barenne. Arch, neerland, de physiol, de l’homme et des anim.„ Bd. 4, S. 124, 1919.
Brudzinski J. Un signe nouveau sur les membres inférieures dans les meningites chez les enfants (signe de la nuque). Arch, de méd. des enfants., Bd. 12, S. 745„ 1909.
Bruin J. de. Enkele neurologische gevallen uit de kinderpraktyk. II. Een gecompliceerd geval van idiotia amaurotica progressiva familiaris infantilis (Tay-Sachs). Nederlandsch maandschr. v. verloskunde, vrouwenziekten en kindergeneeskunde- Bd. 3, S. 593, 1914.
Brunner H. Zur Pathogenese der labyrinthär bedingten Stellungsanomalien des Kopfes und der Augen. (Ein Beitrag zur Pathologie des Statolithenapparates beim Menschen). Monatsschr. f. Ohrenheilk. u. Laryngo-Rhinol., Bd. 55, H. 4/5, 1921.
Camis M. (1). Contribution ä la Physiologie du labyrinthe. II. Une methode opé- ratoire pour la destruction des canaux demi-circulaires du chien. Arch. ital. de- biologie., Bd. 55, S. 180, 1911.
— (2). Contributi alia fisiologia del labirinto. III. Effetti della labirintectomia nel
cane particolarmente sulla innervazione vasomotoria. Folia neurobiologica, Bd. 6,. S. 138, 1912.
— (3). Contributi alia fisiologia del labirinto. Nota VI. Sulla miosi e sulla midriasi
paradossa nel gatto labirintectomizzato. Arch, di farmacol. sperim. e scienze aff., Bd. 12, 1911.
Charcot J. M. Guérison de la maladie de Meniere par le sulfate de quinine. Gaz. des hop. civ. et milit., 1875, S. 753.
Christiani A. Zur Physiologie des Gehirns. Berlin, 1885 (Siehe auch Pflügers Arch. f. d. ges. Physiol., 1884, S. 465).
Codivilla. Zit. nach 0. Vulpius. Die Behandlung der spinalen Kinderlähmung ~ Leipzig, 1910.
Collier J. und F. Buzzard. Descending mesencephalic tracts in cat, monkey and man. Brain, Bd. 2, S. 178, 1901.
Corning I. L. The nature and treatment of vertigo. Journ. Am eric. med. assoc.,. 1901, S. 722. — The suppression of rotatory vertigo, its bearing on the prevention and cure of seasickness. New York med. journ. a. med. record, Bd. 2, S. 297,. 1904. Zitiert nach Bäräny in Lewandowskis Handbuch der Neurologie., Bd. 3, II, S. 845, 872. Berlin, 1910.
C u s h n у A. R. Note on strychnine tetanus. Quart, journ. of exp. physiol., Bd. 12,. S. 153, 1919.
Dollinger A. Zur Klinik der infantilen Form der familiären amaurotischen Idiotie (Tay-Sachs). Zeitschr. f. Kinderheilk., Bd. 22, S. 167, 1919.
Dreyfuss R. (1). Experimenteller Beitrag zur Lehre von den nichtakustischen Funktionen des Ohrlabyrints. Pflügers Arch. f. d. ges. Physiol., Bd. 81, S. 604, 1900.
— (2). Über den Einfluss des Chinins auf das Tonuslabyrinth. Arch. f. Ohrenheilk.,.
Bd. 64, S. 49, 1905; Zeitschr. f. Ohrenheilk., Bd. 49, S. 343, 1905. v. Economo C. J. und J. P. Karplus. Zur Physiologie und Anatomie des Mittelhirns. Arch. f. Psychiatrie u. Nervenkrankh., Bd. 46, S. 275, 1910. Edinger L. Über das Kleinhirn und den Statotonus. Zentralbl. f. Physiol., Bd. 26,. S. 618, 1912.
Ewald J. R. Physiologische Untersuchungen über das Endorgan des Nervus octavus. Wiesbaden, 1892.
Ferrier D. The functions of the brain. 2 ed., London, 1886.
Fleisch A. Das Labyrinth als befcchleunigungsempfindendes Organ. Pflügers Arch., f. d. ges. Physiol., Bd. 195, S. 499, 1922.
Flourens P. Recherches expérimentales sur les propriétés et les fonctions du système nerveux. 2 ed., Paris, 1842.
Forssman J. Ein neues Krankheitsbild nach Seruminjektion. Biochem. Zeitschr., Bd. 110, S. 164, 1920.
Freusberg A. Reflexbewegungen beim Hunde. Pflügers Arch. f. d. ges. Physiol.,. Bd. 9, S. 358, 1874.
Friedberger E. und P. Schroeder. Histologische Veränderungen im Gehirn von Meerschweinchen und Kaninchen bei primärer Antiserumgiftigkeit und bei Einspritzung gef tiger Normalsera (carotal-zentraler Einspritzung des Serums). Zeitschr. f. d. ges. exp. Med., Bd. 26, S. 287, 1922.
Литература
575*
Gergens Е. Über gekreuzte Reflexe. Pflügers Arch. f. d. ges. Physiol., Bd. 14,. S. 340, 1877 [Goltz].
Goltz F. (1). Über die physiologische Bedeutung der Bogengänge des Ohrlabyrinthes. Pflügers Arch. f..d. ges. Physiol., Bd. 3, S. 172, 1870.
— und Freusberg (2). Über die Funktionen des Lendenmarkes des Hundes. Pflü¬
gers Arch. f. d. ges. Physiol., Bd. 8, S. 460, 1874.
— (3). Der Hund ohne Grosshirn. Pflügers Arch. f. d. ges. Physiol., Bd. 51, S. 570,
1892.
Gottlieb R. Studien über die Wirkung des Pikrotoxins. Schmiedebergs Arch., Bd. 30, S. 21, 1892.
V. G r a e f e A. Beiträge zur Physiologie und Pathologie der schiefen Augenmuskeln. V. Graefes Arch. f. Ophth., Bd. I, S. 1, 1854. v. Gudden B. Über die Frage der Lokalisation der Funktionen der Grosshirnrinde. Allg. Zeitschr. f. Psychiatrie u. psych, gerichtl. Med., Bd. 42, S. 478, 1886 und Ges. Abhandl., S. 42, Würzburg, 1889.
Harnack E. und L. Witkowski. Pharmakologische Untersuchungen über das Physostigmin. Schmiedebergs Arch., Bd. 5, S. 401, 1876.
Held H. Die anatomische Grundlage der Vestibularisfunktionen. Beitr. z. Anat.,.
Physiol., Pathol, u. Ther. d. Ohres usw., Bd. 19, S. 305, 1923.
H ö g у e s A. Über den Nervenmechanismus der assoziierten Augenbewegungen. Ann d. Akad. d. Wiss. Budapest, Bd. 10, 11, 14. 1881 ff. (übers, in Monatsschr. f. Ohren heilk. Bd. 46, S. 809, 1912).
Hoffmann. Beitrag zur Kenntnis der physiologischen Wirkungen der Carboisäure- und des Camphers. Diss. Dorpat, 1866. Schmiedebergs Arch., Bd. 67, S. 50, 1912. Holmes G. M. The nervous system of the dog without a forebrain. Journ. of physiol., Bd. 27, S. 1, 1901.
Hoven van Genderen W. J. v. d. Een geval van encephalocele posterior. Diss. Utrecht, 1920.
Hunter J. Observations on certain parts of the animal oeconomy. London, 1786,. S. 209.
Huxley F. M. On the reflex nature of apnoea in the duck in diving. II. Reflex postural apnoea. Quart. Journ. of exp. physiol., Bd. 6, S. 159, 1913.
J oachimsthal. Artikel «Torticollis» in Eulenburgs Realenzyklopädie d. ges._ Heilkunde. 4. Aufl. Berlin, 1913, Bd. 14, S. 538.
Jonkhoff D. J. Een geval van halsreflexen van Magnus en De Kleijn bij een mensch en haar belangrijkheid voor de prognose. Nederlandsch. tijdschr. v. ge- neesk. 1920, I, S. 307.
Jordan H. Untersuchungen zur Physiologie des Nervensystems bei Pulmonaten. II. Tonus und Erregbarkeit. Pflügers Arch. d. ges. Physiol., Bd. 110, S. 533,. 1905.
Isaak J. Untersuchungen zur Wirkung gesättigter wässeriger Campherlösung. Pflügers Arch. f. d. ges. Physiol., Bd. 153, S. 491, 1913.
Isenschmidt R. und L. К r e h 1. Über den Einfluss des Gehirns auf die Wärmeregulation. Schmiedebergs Arch., Bd. 70, S. 109, 1912.
Isenschmidt R. und W. Schnitzler. Beiträge zur Lokalisation des^ der Wärmeregulation vorstehenden Zentralapparates im Zwischenhirn. Schmiedebergs Arch., Bd. 76, S. 202, 1914.
Karplus J. P. und А. К r e i d 1. Über Totalexstirpation einer oder beider Gross— hirnhemisphären an Affen (Macacus Resus). Arch. f. [Anat. u.] Physiol., 1914r S. 155.
Köllner H. und P. Hoffmann. Der Einfluss des Vestibularapparates auf“ die Innervation der Augenmuskeln. Arch. f. Augenheilk., Bd. 90, S. 170, 1922. König Ch. J. Contribution ä l’étude expérimentale des canaux semicirculaires. These. Paris, 1897.
Kreidl A. (1). Gesellsch. d. Ärzte in Wien, 21. Dezember 1895 (vgl. Wien. klin. Wochenschr. 1896).
— (2). Die Funktion des Vestibularapparates. Ergehn, d. Physiol., Bd. 5, S. 572,1906.
К uh о J. Über die vom N. acusticus ausgelösten Augenbewegungen. Pflügers Arch.
f. d. ges. Physiol., Bd. 114, S. 143, 1906 und Bd. 115, S. 457, 1906. Landau A. Über einen tonischen Lagereflex beim älteren Säugling. Klin. Wochenschr., Bd. 2, S. 1253, 1923.
L a n g e, Bogumil. Inwieweit sind die Symptome, welche nach Zerstörung des Kleinhirns beobachtet werden, auf Verletzungen des Acusticus zurückzuführen. Pflügers Arch. f. d. ges. Physiol., Bd. 50, S. 615, 1891.
L e e F. S. A study of the sense of equilibrium in fishes I. Journ. of physiol., Bd. 15, S. 311, 1894.
576
Литература
Leidler R. Experimentelle Untersuchungen über das Endigungsgebiet des Nervus vestibularis. II. Arb. а. d. neurol. Inst. d. Wiener Univ., Bd. 21, S. 151, 1914. L e о H. cm.: J. Isaak.
Lewandowsky M. Die Funktionen des zentralen Nervensystems. S. 164, Jena, 1907.
L e у t о n A. S. F. und C. S. Sherrington. Observations on the excitable cortex of the chimpanze, orang-utan, gorilla. Quart, journ. of exp. physiol., Bd. 11, S. 135, 1917.
Löwenberg. Über die nach Durchschneidung der Bogengänge des Ohrlabyrinthes auf tretenden Bewegungsstörungen. Arch. f. Augen- u. Ohrenheilk., Bd. 3, S. 1,
Longet F. A. Anatomie und Physiologie des Nervensystems. Übersetzt von Hein. Bd. 1, S. 349—420, Leipzig, 1847.
Luchsinger В. (1). Zur Kenntnis der Funktionen des Rückenmarks. Pflügers Arch. f. d. ges. Physiol., Bd. 16, S. 310, 1878. Siehe auch ebenda, Bd. 22, S. 158, 1880.
— (2). Zur Lage der Gleichgewichtszentren. Pflügers Arch. f. d. ges. Physiol., Bd. 34,
S. 289, 1884.
Luciani L. Das Kleinhirn. Ergehn, d. Physiol., Bd. III, 2, S. 318, 1904.
Lyon P. Compensatory motions in fishes. Amer, journ. of physiol., Bd. 4, S. 77, 1901.
Magendie F. Physiologie. Übersetzt von Hofacker, Bd. 2, S. 246. Tübingen, 1826. Mach E. (1). Physikalische Versuche über den Gleichgewichtssinn des Menschen. Wien. akad. Sitzungsberichte III, Bd. 68, S. 124, 1873.
— (2). Grundlinien der Lehre von den Bewegungsempfindungen. Leipzig, 1875.
— (3). Beitrag zur Analyse der Empfindungen. Jena, 1886.
M ar ey E. J. Des mouvements que certains animaux exécutent pour retomber sur leurs pieds lorsqu’ils sont précipités d’un lieu élevé. Gpt. rend, hebdom. des séances de Pacad. des sciences, Bd. 119, S. 714, 1894. (Kinematogramme der fallenden Katze). Abbildungen des fallenden Kaninchens nach Marey bei Liesegang. Wissenschaftliche Photographie. Düsseldorf, 1920.
Mari n a A. Die Theorien über den Mechanismus der assoziierten Konvergenz- und Seitwärtsbewegungen, studiert auf der Grundlage experimenteller Forschungsergebnisse mittels Augenmuskel transplantation beim Affen. Dtsch. Zeitschr,, f. Ner- venheilk., Bd. 44, S. 138, 1912.
Maxwell S. S. (1). Labyrinth and equilibrium. I u. II. Journ. of general physiol., Bd. 2, S. 123, 349, 1919—20.
— (2). The equilibrium functions of the internal, ear. Science, Bd. 53, S. 423, 1921.
.M e r z b a c h e r L. Die Folgen der Durchschneidung der scnsibeln Wurzeln im unteren
Lendenmarke, im Sakralmark und in der Cauda equina des Hundes. Pflügers Arch, f. d. ges. Physiol., Bd. 92, S. 585, 1902.
Minkowski M. (1). Etude sur la Physiologie des circonvolutions rolandiques et pariétales. Schweiz. Arch. f. Neurol. u. Psychiat., Bd. 1, S. 389, 1917.
— (2). Sur les mouvements, les reflexes et les réactions musculaires du foetus humain
de 2 ä 5 mois et leurs relations avec le Systeme nerveux Foetal. Rev. neurol.,
Bd. 37, S. 1105, 1922.
■ Morita S. (1). Untersuchungen an grosshirnlosen Kaninchen. Schmiedebergs Arch., Bd. 78, S. 188, 1915.
— (2). Untersuchungen an grosshirnlosen Kaninchen. 2 Mitt. Die Wirkung verschiede¬
ner Krampfgifte. Schmiedebergs Arch., Bd. 78, S. 208, 1915.
Monakow G. v. Der rote Kern, die Haube und die Regio hypothalamica. Wiesbaden, 1910.
M o r o E. Das erste Trimenon. Münch, med. Wochenschr. 1918, S. 1147.— Freudenberg E. Der Morosche Umklammerungsreflex und das Brudzinskische Nackenzeichen als Reflexe des Säuglingsalters. Münch, med. Wochenschr., 1921, S. 1646.
Mulder M. E. Arch. f. vergl. Ophth., Bd. 21, S. 68, 1875.
Mulder W. Quantitative betrekking tusschen prikkel en effect by het statisch orgaan. Diss. Ütrecht, 1908.
.Munk H. (1). Über die zentralen Organe für Sehen und Hören bei Wirbeltieren.
V... Sitzungsber. d. preuss. Akad. d. Wiss., 1884, S. 549.
— (2). Über Grosshirnexstirpation beim Kaninchen. Pflügers Arch. f. Physiol., 1884,
S. 470.
Nagel A. Arch. f. vergl. Ophth., Bd. 17, S. 247, 1871.
Nagel W. A. (1). Über kompensatorische Raddrehungen der Augen. Zeitschr.
f. Psychol, u. Physiol, d. Sinnesorg, Bd. 12, S. 331, 1896.
— (2). Die Lage-, Beweguus-und Widerstandsempfindungen. Nagels Handb. d. Physiol., Bd. 3, S.. 771, 1905.
Литература
577
Ohm J. Klin. Monatsbl. f. Augenheilk., Bd. 59, S. 538. 1917.
Paton, D. N o el (1). The relative influence of the labyrinthine and cervical elements in the production of postural apnoea in the dnck. Quart, journ. of exp. physiol., Bd. 6, S. 197, 1913.
Paton, D. N о e 1 (2). Studies of the breathing mechanism of the duck in submergence. Proc. roy. philos. soc. of Glasgow, 1914.
Philippson M. L’autonomie et la centralisation dans le Systeme nerveux des animaux. Bruxelles, 1905.
P o 1 1 о к L. J. und W. H. Holmes. Arch, of int. med., Bd. 16, S. 213, 1915 (Zit. nach Jonkhoff.)
P о 11 о к L. J. und Treadway. Arch, of int med., Bd. 12, S. 445, 1913 (Zit. nach Jonkhoff.)
Q u i X F. H. (1). Metingen en beschouwingen over de otolithenfunctie. Nederlandsch tijdschr. V. geneesk., 1919,' I, S. 912.
— (2). Examen fonctionel de l’appareil otolithique. Verhandl. d. 10 internat. Otolo-
genkongresses, 1922. (Dort Verzeichnis aller Arbeiten des Verfassers).
Röber H. Über die physiologische Wirkung des Pikrotoxins. Arch. f. Anat. u. Physiol., 1869, S. 38.
Rosenfeld M. Über calorischen Nystagmus bei Gehirnkranken mit Bewusstseinsstörungen. Verhandl. c.Ges. dtsch. Naturforsch, u. Ärzte 82, Bd. 2, S. 278, 1911.— Das Verhalten des calorischen Nystagmus in der Chloroform-Äther-Narkose und im Morphiumscopolaminschlaf. Neurol. Zentralbl., Bd. 30, S. 238, 1911. Rossem A. v. Gewaarwordingen en reflexen, opgewekt vanuit de halfcirkelvormige kanalen. Onderzoekingen in het physiol, laborat, d. Utrechtsche hoogeschool V, Bd. 9, S. 151, 1908.
Rothfeld J. (1). Beitrag zur Kenntnis der Abhängigkeit des Tonus der Extremitätenmuskeln von der Kopfstellung. Versuche mit Narkose. Pflügers Arch. f. d. ges. Physiol., Bd. 148, S. 564, 1912.
— (2). Über die Wirkung einiger Körper aus der Gruppe des Chloroforms auf die vesti¬
bulären Augenreflexe. Pflügers Arch. f. d. ges. Physiol., Bd. 149, S. 435, 1913.
— (3). Über den Einfluss akuter und chronischer Alkoholvergiftung auf die vestibulären
Reaktionen. Arb. a. d. neurol. Inst. d. Wiener. Univ., Bd. 20, S. 89, 1913.
— (4). Die Physiologie des Bogengangsapparates. Verhandl. d. Ges. dtsch. Naturforsch.
u..Ärzte, Bd. 1, S. 30, 1913.
— (5). Über die Beeinflussung der vestibulären Reaktionsbewegungen durch experimen¬
telle Verletzungen der Medulla oblongata. Bull, de L’acad. des sciences, S. 74, Krakau, 1914.
— (6). Über den Einfluss der Kopfstellung auf die vestibulären Reaktionsbewegungen
der Tiere. Pflügers Arch. f. d. ges. Physiol., Bd. 159, S. 607, 1914.
— (7). Über den Einfluss des Stirnhirns auf die vestibulären Reaktionsbewegungen.
Autoreferat: Ronas Berichte, Bd. 5, S. 86, 1920.
— (8). Experimentelle Untersuchungen über den Einfluss der Grosshirnhemisphären,
des Mittel- und Zwischenhirns auf die vestibulären Reaktionsbewegungen. Pflügers Arch. f. d. ges. Physiol., Bd. 192, S. 272, 1921.
Rothmann M. Der Hund ohne Grosshirn. Neurol. Zentralbl., Bd. 28, S. 1045, 1909 und Bd. 31, S. 867, 1912.
S a s s а К. On the effects of constant galvanic currents upon the mammalian nerve- muscle and reflex preparations. Proc. of the roy. soc. of London, B., Bd. 92, S. 341, 1921.
Schiff J. M. Lehrbuch der Physiologie des Menschen, Bd. I, S. 331 ff, Lahr 1858—59. Schönemann A. Schläfenbein und Schädelbasis. Neue Denkschr. d. allg. Schweiz.
Ges. f. d. ges. Naturwis., Bd. 40, Abh. 3, 1906 (zit. nach Bäräny—Wittmaak). Schrader M. Zur Physiologie des Froschgehirns (Vorl. Mitt.) Pflügers Arch. f. d. ges. Physiol,, Bd. 41, S. 75, 1887.
Seemann J. Über die durch Strychnin hervorgerufene «Reflexumkehr» bei Atemreflexen. Zeitschr. f. Biol., Bd. 54, S. 153, 1910.
Sherrington C. S. (1). Decerebrate rigidity and reflex coordination of movements. Journ. of physiol., Bd. 22, S. 327, 1898.
— (2). On the innervation of antagonistic muscles. 6th note Proc. of the roy. soc. of
London, Bd. 66, S. 66, 1899.
— (3). In Schäfers Textbook of Physiol., Bd. 2, S, 905—909, 1900.
— (4). On reciprocal innervation of antagonistic muscles. 7. note. Proc. of the roy. soc.
of London, B., Bd. 76, S. 291, 1905.
— (5). The integrative action of the nervous system. London, 1906.
— (6). Strychnine and reflex inhibition of sceletal muscle. Journ. of physiol., Bd. 36,
S. 196, 1907.
37 P. Магнус
578
Литература
— (7). Observations on the scratch reflex in the spinal dog. Jonrn. of physiol., Bd. 34,
S. 1, 1908.
— (8). On plastic tonns and proprioceptic reflexes. Quart. Journ. of exp. physiol., Bd. 2,
S. 190, 1909.
— (9). Flexion-reflex of the limb, crossed extension-reflex and reflex stepping and stand¬
ing. Journ. of physiol., Bd. 40, S. 28, 1910.
— und S. G. M. Sowton (10). Reversal of reflex of an afferent nerve by altering the
character of the electric stimulus applied. Proc. of the roy. soc. of London, B., Bd. 83, S. 435, 1911.
(11). Further observations on the production of reflex stepping by combination of reflex excitation with reflex inhibition. Journ. of physiol., Bd. 47, S. 196r 1913.
— (12). Notes on the arragement of some motor fibres in the lumbosacral plexus. Journ.
of physiol., Bd. 13, S. 621, 1892.
Simonelli G.(l). Sulla funzione dei lobi medi del cerveletto. Nota I. li lobo posteriore etc. Arch, di fisiol., Bd. 19, S. 447, 1921.
— (2). Ricerche sui rapporti funzionali tra cerveletto e labirinto. Lo sperimentale 76,
fasc. 4, 1922.
Simons A. (1). Kopfhaltung und Muskeltonus. Sitzungsber. d. Berl. Ges. f. Psychiatrie u. Nervenkrankh. 3. Dezember 1919 und 12. Jan. 1920. Ref. ZentralbL f. Neurol., Bd. 39, S. 132 u. 256, 1920.
— (2). Kopfhaltung und Muskeltonus. Zeitschr. f. d. ges. Neurol, u. Psychiatrie., Bd. 80,
S. 499, 1923.
S p a m e r C. Experimenteller und kritischer Beitrag zur Physiologie der halbkreisförmigen Kanäle. Pflügers Arch. f. d. ges. Physiol., Bd. 21, S. 479, 1880.
S t e f a n i, zit. nach Luciani.
StenversH. W. Un «Stellreflex» du bassin chez l’homme. Arcb. néerland. de physiol. de l’homme et des anim., Bd. 2, S. 669, 1918 (Pekelharinglestschrift).
S t г о s s W. Beitrag zur Pharmakologie des Camphers. Schmiedebergs Arch., Bd. 95, S. 304, 1922. Siehe auch Verhandl. d. Dtsch. pharmakol. Ges., 1921, S. XXII.
Thiele F. H. On the afferent relationship of the optic thalamus and Deiters nucleus to the spinal cord with special reference to the cerebellar influx of Dr. Hughlings Jackson and the genesis of the decerebrate rigidity of Ord and Sherrington. Journ. of Physiol., Bd. 32, S. 358, 1905.
Topolanski A. Das Verhalten der Augenmuskeln bei zentraler Reizung. Das- Koordinationszentrum und die Bahnen für koordinierte Augenbewegungen, v. Grae- fes Arch. f. Ophth., Bd. 46, S. 452, 1898.
Trendelenburg P. Pikrotoxin. In Heffters Handbuch der experimentellen Pharmakologie, Bd. II, 1, S. 406, 1920.
Trendelenburg W. und A. Kühn. Zur Physiologie des Ohrlabyrinthes der Reptilien. Pflügers Arch. f. d. ges. Physiol., 1908, S. 160. v. Uexküll J. (1). Die ersten Ursachen des Rhythmus in der Tierreihe. Ergehn, d. Physiol., Bd. 3 (2), S. 1, 1904.
— (2). Ein Wort über die Schlangensterne. ZentralbL f. Physiol., Bd. 23, S. 1, 1909. Voit M. Zur,Frage der Verästelung des N. acusticus bei den Säugetieren Anat. Anz.,
Bd. 31, S. 635, 1907.
Voss O. Artikel «Menièresche Krankheit» in Eulenburgs Realenzyklopädie. 4. Aufl., Bd. 9, S. 385, 1910.
Vulpian A. Legons sur la Physiologie du système nerveux. S. 532—538. Paris, 1866.
Walshe F. M. R. (1). On certain tonic or postural reflexes in hemiplegia with special reference to the so-called «associated movements». Brain, Bd. 46, S. 1,
, 1.923.
— (2). A case of complete decerebrate rigidity in man. Lancet, 1923, II, S. 644.
— (3). On variations in the form of reflex movements, notably the Babinski plantar-
response under different degrees of spasticity and under the influence of Magnus and De Kleijn’s tonic neck reflex. Brain., Bd. 46, S. 281, 1923.
Weed L. H. Observations upon decerebrate rigidity. Journ. of. Physiol., Bd. 48, S. 205, 1914.
Weiland W. Münch, med. Wochenschr., 1912, S. 2539.
Wessely К. (1). Über den Einfluss der Carotisunterbindung auf die Blutversorgung des Auges; nach gemeinsam mit Herrn Wolf ausgeführten Untersuchungen. Ver- handl. d. Ges. dtsch. Naturforsch, u. Ärzte, 1908 (siehe auch Münch, med. Wochenschr. 1909, S. 688).
— (2). Über den Einfluss der Augenbewegungen auf den Augendruck. Arch. f. Augen-
heilk., Bd. 81, S. 111, 1916.
Литература
579
Wiedemann С. Beitrag zur Pharmakologie des Camphers. Schmiedebergs Arch., Bd. 6, S. 216, 1877.
Wilson T. G. und F. H. Pike. The effects of Stimulation and extirpation of the labyrinth of the ear and their relation to the motor system. Philosoph. Transactions roy. soc. B., Bd. 203, S. 127, 1913.
Winkler G. (1). The central course of the nervus octavus and its influence on motility. Yerhandl. d. koninkl. akad. v. wetensch. te Amsterdam, II, Bd. 14, N 1, 1907.
—1 und A. Potter (2). An anatomical guide to experimental researches on the rabbits brain. A series of 40 frontal sections. Amsterdam, 1911.
— und A. P о t t e r (3). An anatomical guide to experimental researches on the cats
brain. A series of 35 frontal sections. Amsterdam, 1914.
— (4). Anatomische aanteekeningen by de hersenen van een hond, by wien voor vyf
maanden het cerebellum is verwyderd. Nederlandsch. tijdschr. v. geneesk. 1920, 11, S. 958.
— und G. A. van Rijnberk (5). Experim. onderzoekingen over segmenteel-in-
nervatie van de huid van den hond. VI. Verslagen d. koninkl. akad. v. wetensch. te Amsterdam. Wis-en natuurk. afdeeling., Bd. 19, S. 307, 1910. Wittmaack K. (1). Beitrag zur Kenntnis des Chinins auf das Gehörorgan, II. Der Angriffspunkt des Chinins im Nervensystem des Gehörorgans. Pflügers Arch. f. d. ges. Physiol., Bd. 95, S. 234, 1903.
— (2). Über Veränderungen im inneren Ohr nach Rotationen. Verhandl. d. Dtsch. otol.
Ges., Bd. 18, S. 150, 1909.
Z о t h O. Augenbewegungen und Gesichtswahrnehmungen. Nagels Handb. d. Physiol., Bd. 3, S. 318, 1905.
37*
ПРИЛОЖЕНИЕ
А. Ф. Самойлов
РАБОТЫ Р. МАГНУСА И ЕГО ЛАБОРАТОРИЯ В УТРЕХТЕ1
Р. Магнус занимает кафедру фармакологии в Утрехте и является одним из крупнейших фармакологов нашего времени. Но внимание и почет, которыми теперь окружено его имя, находятся в связи не столько с его фармакологическими работами, сколько с его изумительными исследованиями из области физиологии центральной нервной системы.
Недавно, во время своего заграничного путешествия, я прожил некоторое время в Голландии и провел около месяца в лаборатории Магнуса. Под впечатлением этого посещения, под впечатлением личного знакомства с Магнусом и с его лабораторией мне хотелось бы дать изложение его учения о статических рефлексах.
Мне приходилось не раз бывать в Голландии. Всегда и неизменно, и раньше и теперь, у меня при посещении этой страны являлось одно и тоже чувство какого-то спокойствия, какого-то умиротворения. Вся жизнь течет здесь, кажется, так же спокойно, как спокойна вода в каналах, изрезывающих всю Голландию. Борьба с природой, от которой нужно было отвоевать эту ниже уровня моря находящуюся страну, закончилась, и теперь вся она, разделенная каналами на маленькие четырехугольники зеленых лугов, так называемые польдеры, освобождается от избытка воды при помощи тысячи ветряных мельниц, откачивающих воду в верхние каналы, отсюда в реку и в море. Голландия не боится засухи. Вследствие обильного орошения — богатые луга, отсюда прекрасный корм для скота, прекрасное молочное хозяйство. Отсюда голландский пейзаж с каналами, ветряными мельницами, коровами, хижинами, приютившимися у опушки высоких рощ, с богатым, сытым населением; отсюда и голландское спокойствие, голландский юмор,— словом все то, с чем нас знакомят хорошо всем известные жанровые картины голландских художников. Как красивы голландские города с их чистенькими домами, подъемными мостами, с их шпицами церквей и ратуш, куранты которых весело отбивают каждый час различные мелодии! Как красивы их пристани, загроможденные судами, полные жизни! И опять всюду благосостояние, богатство, сильное и крепкое. Когда видишь эту спокойную, богатую жизнь, то невольно спрашиваешь себя: откуда это благополучие? Кто должен заплатить своим трудом за это благополучие? На это нетрудно ответить: за это платят колонии.
В Голландии 6 миллионов жителей, и это число не уменьшается и не увеличивается, но народонаселение сильно возрастает. Кажущееся противоречие объясняется тем, что избыток населения переводится в колонии, которые и питают метрополию. Колонии — это источник богатств Голландии. Элемент менее талантливый, не умеющий в борьбе за жизнь
1 Доложено в Обществе врачей при Казанском университете (Успехи современной биологии, т. VI, вып. 1—2, 1927).
584
А. Ф. Самойлов
устроиться в Голландии, а также и элемент наиболее непокладистый, строптивый, переправляется в колонии, и число жителей голландской Индии растет не только из-за увеличения числа туземцев, но также и из-за притока голландцев из Европы. Но Голландия принимает к себе также людей из других стран, но лишь в виде отдельных единиц, притом людей, наиболее ценных, лучших, первых. Таким и является Р. Магнус.
Магнус не голландец, он приехал из Германии, где родился, вырос. «Когда, — передает он, — мне было уже 35 лет, на моих руках была семья, а перспектива получения кафедры была весьма сомнительна, то я чувствовал себя прескверно. Предложение Цваардемакера (Zvaardemaker), теперешнего профессора физиологии в Утрехте, конкурировать на кафедру фармакологии в Утрехтском университете давало луч надежды. Телеграмма с извещением о том, что я избран, встречена была мною с восторгом». Оказалось, однако, что фармакологической кафедры и лаборатории в Утрехте еще не было и нужно было организовать все дело с самого начала. Магнусу предлагали для лаборатории различные здания и среди них предлагали старую запущенную церковь, служившую в средние века, помимо другого, также больницей для чумных больных. Выбор Магнуса пал именно на эту церковь, которая по его планам была внутри перестроена и приспособлена для лаборатории. Лаборатория эта, сохранившая и до сих пор свои церковные, готические своды, высокие окна, производит несколько необычное впечатление. Магнус устроился в этой церкви очень хороша и счастливо и плодотворно проработал там теперь уже около 20 лет. К сожалению, в последнее время здание, по-видимому, отказывается служить и дает серьезные трещины, так что в различных местах установлены деревянные подпорки, число которых, увы, постепенно увеличивается.
Лаборатория Магнуса очень богата; здесь расходами не стесняются. Все, что нужно для работы, немедленно приобретается. Отличная механическая мастерская, искусные механики, благожелательно входящие в интересы лаборатории, хорошо вышколенные служители сильно облегчают работу. Каждую неделю привозят из Амстердама в лабораторию не менее 30 кошек; кроликов и собак имеется тоже сколько угодно, несмотря на то что животные стоят в Голландии дорого — значительно дороже, чем у нас.
При всем своем богатстве лаборатория Магнуса имеет один недостаток — она, несомненно, тесна. Изолироваться там нельзя. Предоставить занимающемуся комнату Магнус не может, он и сам не имеет своей комнаты, и всю административную и литературную часть своей работы ведет в небольшой библиотечной комнате, куда всякий из работающих в любое- время имеет доступ. Почти все работы экспериментального физиологического характера производятся, (в сущности, в одной большой комнате. Кто знает, может быть, это обстоятельство имело благоприятное влияние на ход работ этой лаборатории. Магнус окружил себя рядом прекрасных сотрудников, из которых многие занимают и сами теперь видное место в науке. Его идеями и направлением заинтересовались ученые пограничных с физиологией областей. В работах Магнуса принимали и принимают участие анатом де Бурлет (deBurlet), известный специалист-невропатолог Винклер (Winkler), отиатр и ближайший сотрудник Магнуса де Клейн (de Kleyn), офтальмолог ван дер Гуве (van der Hoove), затем его ближайшие ассистенты ла Гай, де Линд, Радемакер (La Науе, de Lind, Rademaker) и др. Все эти лица образуют одну дружную научную семью. Магнус неоднократно говорил мне, что только в такой атмосфере содружества и можно было1 плодотворно работать в этой тесной лаборатории.
Работы Р. Магнуса и его лаборатория в Утрехте
585
Как сказано было раньше, лабораторное здание пришло в ветхость. В скором времени оно будет совсем оставлено и лаборатория будет переведена в новое, только что выстроенное здание. Новая лаборатория Магнуса производит совсем иное впечатление. Это громадное, гигантское четырехэтажное здание, напоминающее по внешнему виду скорее фабрику, чем лабораторию. Она представляет собой, вероятно, самую большую из существующих университетских лабораторий. Расходы по постройке ее были не по силам даже богатой Голландии. Здание лаборатории, обошедшееся в 300 тысяч долларов, построено почти целиком на средства американского миллиардера Рокфеллера. Все новейшие усовершенствования техники, представляющие какую-нибудь лабораторную ценность, нашли себе применение в новой лаборатории Магнуса. Несмотря на самые лучшие возможности и перспективы работы в новом помещении, Магнус и его сотрудники с совершенно определенным сожалением и какой-то робостью покидают свою старую, развалившуюся церковь. Они озабочены мыслями о том, удастся ли им на новом месте, при новых возможностях новых заданиях, при новом широком размахе сохранить тот дружный, интимный и коллективный уклад лабораторной жизни, который давал им до сих пор столько духовных радостей и столько достижений. .
Изложение работ Магнуса представляет большие трудности. Правда, задача эта значительно облегчена самим Магнусом в том отношении, что работы его, касающиеся физиологии центральной нервной системы, — а именно об этих работах только я и хочу говорить, — объединены в его прекрасной большой монографии, вышедшей в свет два года тому назад.
Трудность этой темы заключается в том, что работа Магнуса не есть плод какого-нибудь одного удачного открытия, которое можно в кратких словах изложить. Работа Магнуса и его сотрудников есть результат долгого, двадцатилетнего упорного труда, где каждый шаг вперед давался после больших усилий. Все здание строилось, так сказать, из маленьких, трудно добываемых кирпичей. Чтобы уяснить ценность всего здания, нужно неминуемо остановиться и на описании отдельных кирпичей.
Позвольте начать несколько издалека.
35 лет назад известный германский физиолог Фридрих Гольц, профессор Страсбургского университета, произвел замечательную операцию на собаке. Он удалил у собаки в несколько приемов весь головной мозг, так что в черепе оставались только продолговатый мозг с мозжечком, четверохолмие, зрительные бугры и небольшие куски полосатого тела. Одна из оперированных собак прожила 18 месяцев, и затем была убита для анатомического исследования. Задача Гольца, конечно, заключалась в том, чтобы определить, какие функции собака потеряла, лишившись своих полушарий, и какие функции она сохранила. Гольц обладал совершенно исключительной наблюдательностью, экспериментальным талантом и большим литературным дарованием. Его физиологические труды читаются с истинным наслаждением. Но когда я несколько времени тому назад, приготовляясь к сегодняшнему вечеру, прочел опять его статью о собаке без головного мозга, то я увидел теперь особенно ясно, что Гольц не был в состоянии, несмотря на свои дарования, определить, что собака потеряла и что сохранила. К своей операции Гольц пришел не путем безусловно строгих логических следствий; эта операция носила в себе, несомненно, элемент тематической случайности. Гольц не нашел в физиологии своего времени готовыми те понятия, те термины, которыми можно было бы охарактеризовать потерянные и сохранившиеся функции собаки без полушарий, и потому задача, стоящая перед ним, была выше его сил.
.586
Д. Ф. Самойлов
Опыт Гольда в течение многих лет не повторялся, а если и повторялся, то в результате мы имели такие же случайные, эпизодические описания, какие давал Гольц. Но в последнее время операция удаления полушарий у животного повторяется очень часто, и на этот раз она является конечным звеном хорошо продуманной цепи изысканий.
После обширных многолетних трудов по условным рефлексам И. П. Павлова мы теперь можем сказать, что собака Гольца, лишившись полушарий, потеряла все приобретенные в течение жизни условные рефлексы и потеряла способность образовывать таковые. На вопрос, что сохранила собака, лишившись полушарий, мы находим ответ в трудах Р. Магнуса.
Понятие о рефлексе, которое ведет свое начало еще со времен Декарта, очевидно, было выбрано чрезвычайно удачно. Рефлекс, как ответ на раздражение в пределах рефлекторной дуги, есть поистине функциональный элемент, из которого складывается вся работа центральной нервной системы. Все современные исследования о функциях центральной нервной системы приводят нас неизменно так или иначе к одному и тому же заключению: животный организм есть рефлекторная ма¬
шина.
Представим себе, что мы перерезали у собаки спинной мозг. После того как нанесенная рана зарастет, когда все зарубцуется, перед нами животное, у которого нижний участок мозга разобщен от всей вышележащей центральной нервной системы, причем этот участок сохраняет способность рефлекторной передачи. Мы можем наблюдать на такой собаке самые разнообразные и даже весьма сложные рефлекторные явления. Если мы уколем кожу на пальце задней ноги собаки, то эта нога сейчас же даст сгибание и таким образом удалится от раздражающей причины, а другая задняя нога выпрямится. Мы имеем здесь флексорный рефлекс одной ноги и экстензорный — другой. Если мы поднимем собаку вверх так, чтобы ноги ее висели в воздухе, то они начнут производить движения, напоминающие ходьбу. Если мы будем легко почесывать определенные участки кожи, то начнется очень характерный и сложный рефлекс почесывания. Можно дальше наблюдать, что животное проделывает при дефекации движения задними конечностями совершенно так, как это делают здоровые собаки. Словом, на такой собаке мы с чрезвычайной легкостью можем получить целый- ряд очень сложных движений, способность к которым заложена в спинном мозгу. Одного только эта собака не в состоянии будет выполнить. Если мы поставим ее на пол так, чтобы все четыре конечности касались пола, то задние ее конечности не в состоянии будут нести груз тела и согнутей.
Большая заслуга в выяснении особенностей рефлекторных явлений принадлежит английскому ученому Шеррингтону, который впервые в обширных работах с чрезвычайной ясностью раскрыл существенную сторону рефлекторных движений конечностей. Он указал, что деятельность сгибающих и разгибающих мышц конечностей реципрокна, т. е., если сокращается флексор, то его антагонист, экстензор, расслабляется, и обратно: если экстензор сокращается, то флексор расслабляется. При этом существенно то, что при сокращении агониста антагонист не пассивно растягивается, а расслабляется активно вследствие угнетения тех моторных клеток спинного мозга, при помощи которых иннервируется антагонист. Поэтому если мы отделим сухожилия агониста и антагониста от мест их прикрепления, привяжем к ним нитки и соединим их с рычагами, то увидим следующее: при флексорном рефлексе флексор сократится, а экстензор растянется, один рычаг пойдет кверху, другой — книзу.
Работы Р. Магнуса, и его лаборатория в Утрехте
587
Это явление редипрокности оказалось явлением общим, и мы имеем основания принимать, что наши волевые движения также совершаются но этому типу.
По отношению к занимающей нас задаче заслуга Шеррингтона заключается в том, что он указал на одно замечательное явление, развивающееся на животном, когда перерезка совершена не в пределах спинного мозга, а производится на границе между продолговатым мозгом и четверохолмием. В результате такой перерезки получается животное, которое Шеррингтон назвал «децеребрированным» животным. Такую перерезку очень удобно производить на кошках, на кроликах, так как у них имеется костяная перегородка на границе между малым мозгом и гемисферами. Если через трепанационное отверстие у кошки провести шпатель, то можно без труда произвести перерезку, причем продолговатый мозг с мозжечком окажется отрезанным от передних частей головного мозга. Животное, конечно, не нуждается в искусственном дыхании, ибо его дыхательный центр не поврежден, и если сохранять его от потери теплоты, то можно производить опыты над таким децеребрированным животным в течение многих — 10, 20 и больше — часов. После того как наркоз прекращен и все следствия его прошли, мы наблюдаем на таком животном такое замечательное явление: все четыре его конечности оказываются вытянутыми. Но сокращены не только мышцы конечностей, сокращены, в сущности, все те экстензоры, которые помогают животному сопротивляться действию силы тяжести. У децеребрированной кошки оказывается приподнятым хвост, приподнята голова, нижняя челюсть притянута к верхней. Это тоническое сокращение экстензоров есть как бы выражение вековечной борьбы, которую ведет животное с принижающей его к земле силой земного притяжения. Описанное тоническое сокращение экстензоров Шеррингтон назвал «ригидностью децеребрированного животного». Эта ригидность так резко выражена, что если поставить животное на землю, то его вытянутые конечности в состоянии нести туловище, и такое животное способно стоять: оно стоит неподвижно, как истукан, как призрак. Оно не способно, однако, удерживать равновесие, и достаточно легкого толчка, чтобы децеребрирован- ная кошка упала. Сама по себе она не в состоянии подняться и остается в лежачем положении неопределенно долгое время. Ригидность экстензоров децеребрированной кошки есть явление рефлекторное, и Шерринг- тону мы обязаны выяснением того, откуда начинается этот рефлекс. Он начинается не с кожных чувствительных аппаратов: можно удалить
кожу у такой кошки без того, чтобы ригидность изменилась в каком- нибудь отношении. Рефлекс начинается от проприоцептивных раздражений чувствительных нервных окончаний, заложенных в самой мышце.
Мы все привыкли оценивать мышцу со стороны ее механических особенностей, со стороны ее сократительных свойств, и считаем мышцу нашим рабочим органом. Это правильная, но не полная оценка. Мышца есть не только рабочий орган, мышца есть вместе с тем и своеобразный орган чувств, рецептивный орган. Всякое вытяжение мышцы сопровождается механическим раздражением чувствительных окончаний, заложенных в мышце, и может служить начальным моментом для рефлекса. Чувствительные волокна, направляющиеся от мышц к центральной нервной системе, проходят, как и другие чувствительные волокна, через задние корешки, и поэтому мы можем перерезать чувствительные волокна мышцы, не тронувши ее двигательных волокон, и таким образом вскрыть механизм происхождения ригидности. Если мы перережем задние корешки, принимающие чувствительные волокна мышц задней конечности, то ригидность
588
А. Ф. Самойлов
децеребрированной кошки будет по-прежнему наблюдаться на всех экстензорах, за исключением лишь той ноги, чувствительные волокна мышц которой мы перережем.
Такое децеребрированное животное является исходным пунктом исследования Магнуса. Первое открытие, сделанное им случайно, заключалось в следующем: он заметил, что если у децеребрированной кошки, спокойно лежащей, повернуть голову, то степень сокращения мышц ее конечностей меняется и конечности примут иное положение. Это явление, оказавшееся при повторных пробах постоянным, явилось началом целого ряда дальнейших работ. Заслуги и талант Магнуса заключаются не в том, что он сделал это, в сущности, случайное.открытие, а в том, что он понял глубокий смысл и цену этого случайного открытия, которое, может быть, многие, имевшие в руках децеребрированное животное, тоже делали, но пропускали, не умея понять его значения. Талант Магнуса заключался дальше не только в том, что он сумел оценить глубокое значение своего открытия, но особенно в том, что он умел распутать все нити, которые сходятся в наблюденном им факте.
Когда поворачивают голову у децеребрированного животного, то конечности его меняют свое положение. Магнус спросил себя, какие движения конечностей наступают вслед за движением головы, имеется ли какая- нибудь закономерность, какая-нибудь определенная связь между положением головы и положением, которое принимают конечности; оказалось, что установить какую-нибудь закономерность на первых порах было совершенно невозможно.
Решение поставленного вопроса явилось лишь тогда, когда Магнус понял, что положение конечностей в результате изменения положения головы зависит от двух условий: 1) от того, как меняется положение головы вообще в пространстве и 2) как меняется положение головы по отношению к туловищу.
Для того чтобы изменить одно из этих условий, не изменяя другого, он в конце концов пришел к такой постановке опыта: на шею, голову и грудь децеребрированной кошки была наложена гипсовая повязка, которая раз навсегда фиксировала положение головы по отношению к туловищу. Если такое животное, лежащее на спине, двигать вокруг его дорсо-вентральной оси, то никаких изменений в положении конечностей не наблюдается. Если же движения производить вокруг других осей, например, вокруг битемпортальной оси, то мы замечаем, что максимум экстензорного сокращения всех четырех конечностей имеет место тогда, когда животное лежит на спине и его нижняя челюсть возвышается на 45° над горизонтом. Если же животное поворачивается на 180° спиной кверху и его челюсть находится на 45° ниже горизонта, то наблюдается минимум экстензорного сокращения конечностей.
Таким образом, положение головы в пространстве связано с определенным положением конечностей. Какова эта связь? Дальнейшие опыты показали, что изменение положения нижних конечностей вместе с изменением положения головы есть рефлекс, начало которого нужно искать в лабиринте внутреннего уха. Если децеребрированную кошку лишить лабиринтов, то описанное только что изменение положения конечностей больше не наблюдается. Де Клейн, специалист по ушным болезням, работающий в лаборатории Магнуса, выработал прекрасный и простой прием разрушения лабиринтов через так называемую bulla ossea. Всю операцию можно произвести в самое короткое время, причем животное не теряет ни капли крови и, кроме того, ни одна шейная мышца не повреждается при этом. Животное, лишенное обоих лабиринтов с загипсованной шеей,
Работы Р. Магнуса и его лаборатория в Утрехте
589
может быть поворачиваемо вокруг любой оси без того, чтобы конечности е_го обнаружили какое бы то ни было изменение первоначального положения. Магнус поставил себе вопрос: какая же часть лабиринта нужна для того, чтобы мог осуществиться описанный раньше тонический лабиринтовый рефлекс. На основании того, что мы знаем из физиологии внутреннего уха, можно было смело исключить улитку как аппарат, назначение которого связано с передачей звуковых колебаний. Вопрос мог быть только в том, имеют ли значение для тонических лабиринтовых рефлексов конечностей полукружные каналы или так называемые пятна с их отолитами в преддверии кожного лабиринта.
Для исследования этого вопроса Магнус воспользовался приемом, предложенным раньше Витмааком. Этот прием заключается в том, что животное подвергается центрифугированию. При этом вследствие центробежной силы отбрасываются в сторону отолиты, помещающиеся на волосках клеток пятен. Опыт показал,, что указанные раньше тонические лабиринтовые рефлексы конечностей пропадают у животных, когда у них нарушена целостность пятен. Дальше можно было спросить себя: какое из пятен имеет значение в настоящем случае — пятно утрикула или пятно сакула? С большой степенью вероятности Магнус заключил, что исходным моментом раздражения, ведущим к тоническому сокращению мышц конечностей, служит аппарат, заложенный в пятне утрикула. Это заключение основывается на анатомических данных. Утрехтский анатом Бурле, вошедший в интересы работ Магнуса, предпринял обширный ряд исследований для уяснения вопроса о том, как расположены в лабиринте кролика и кошки отолиты утрикула и сакула. Исследования Бурле, препараты и методику которого я имел возможность видеть, произведены с большой тщательностью. Бурле установил, что оба отолита утрикула лежат в одной плоскости и что в том положении, которое, по Магнусу, дает у кошки максимум сокращения экстензоров конечностей, а равно и в том положении, когда имеется минимум сокращений экстензоров, оба отолита утрикула лежат в горизонтальной плоскости. Мы уже видели раньше, что для того чтобы перевести кошку из положения максимума в положение минимума, нужно повернуть ее на 180°. Почему же, спрашивается, слуховые пятна утрикула с их отолитами при поворачивании на 180° могут превращать максимум в минимум? Как известно, кристаллы углекислой извести, составляющие отолит и погруженные в студенистую массу, покоятся на волосках клеток пятен. Оказалось, что в том положении головы, при котором имеется максимум сокращения экстензоров конечностей, отолит висит на волосках и, следовательно, растягивает их; когда же голову поворачивают на 180°, то отолит лежит на волосках, и в этом случае мы имеем минимум действия. Отсюда получилось представление о том, что раздражающим моментом, служащим началом для тонического лабиринтового рефлекса на конечности, является нагрузка отолитом волосков пятен утрикула. Здесь следует упомянуть, что в случае потери одного лабиринта тонические рефлексы при той же форме опыта с загипсованной шеей остаются неизмененными, так как оставшийся лабиринт обслуживает все четыре конечности. Мы должны себе, следовательно, представить, что возбуждения, начинаясь в лабиринтах, и именно в клетках пятен утрикула, бегут к центрам в продолговатый мозг, а отсюда от каждого из центров обеих сторон дальше через спинной мозг ко всем четырем конечностям.
На основании сказанного можно ожидать, что после удаления обоих лабиринтов тонические лабиринтовые рефлексы должны прекратиться, что на самом деле и имеет место. Децеребрированное животное без лабирин¬
590
Ал Ф. Самойлов
тов с загипсованной шеей может быть поворачиваемо вокруг какой угодно оси без того, чтобы положение конечностей как-нибудь изменялось.
Но как только мы снимем гипсовую повязку, то мы встретимся с новыми тоническими рефлексами на конечности в зависимости от того или иного положения головы по отношению к туловищу. Если у животного, лишенного обоих лабиринтов и не загипсованного, мы будем вращать голову и тем изменять положение головы относительно туловища, то заметим следующие изменения в положении конечностей: вращение головы вокруг битемпоральной оси с наклонением головы к вентральной стороне, т. е. вентральная флексия головы ведет к тому, что передние конечности сгибаются, а задние — растягиваются, и наоборот, дорсальная флексия1 ведет к тому, что передние конечности вытягиваются, а задние — сгибаются. Если будем поворачивать голову в стороны, то конечности той стороны, куда приближается челюсть, так называемые, по Магнусу, «челюстные конечности», вытягиваются, а конечности другой половины сгибаются. Это так называемые «тонические шейные рефлексы на конечности». Всякое изменение положения головы в отношении туловища вызывает механическое раздражение нервных окончаний, заложенных в шейных мышцах.
Эти проприоцептивные раздражения ведут к рефлексу на конечности. Поэтому если у децеребрированного животного перерезать первые три задние шейные корешка, то этим устраняется возможность шейных рефлексов на конечности.
Как сказано было раньше, для выяснения влияния лабиринтовых рефлексов мы фиксировали положение головы по отношению к туловищу при помощи гипсовой повязки; но вместо гипсовой повязки можно, конечно, как это на самом деле нередко и делается, применить перевязку первых трех шейных корешков, устраняющих возможность шейных рефлексов. Ясно, конечно, что, удалив оба лабиринта и перерезав задние три верхних шейных корешка, мы тем самым устраняем всякую возможность воздействия положения головы на положение конечностей.
Если у животного лабиринты не разрушены и задние корешки не перерезаны, то всякому положению головы соответствует совершенно определенное положение конечностей, и это положение есть результат суммирования воздействия как со стороны лабиринтов, так и шейных мышечных нервных окончаний. Следует еще раз отметить, что у нас речь идет не о рефлекторных движениях конечностей вследствие движения головы: в описанных опытах мы имеем дело с тоническими рефлексами, в результате которых определенному положению головы соответствует определенное положение конечностей, причем пока голова сохраняет свое положение неизмененным, положение конечностей остается тоже неизменным.
Нам нужно остановиться еще на одном обстоятельстве, весьма существенно усложняющем представленные выше рефлексы. Дело в том, что лабиринты управляют не только мышцами конечностей, но и шейными мышцами. После удаления одного из лабиринтов голова животного повернута в сторону. Так, например, если удален правый лабиринт, то голова повернута так, что челюсть приближается к левой стороне, а темя — к правой, и это изменение положения головы стойкое, оно не уменьшается с течением времени. Это изменение основано на том, что каждый из лабиринтов управляет шейными мышцами своей стороны, и поэтому если один лабиринт удален, то мышцы другой стороны берут перевес и голова оказывается повернутой. Но степень этого поворота
Работы Р Магнуса и его лаборатория в Утрехте
5:91
головы и тонического сокращения шейных мышц неодинакова при различных положениях головы в пространстве. Оказалось, что максимум тонуса при повороте имеет место как раз в том положении животного, которое дает и максимум вытяжения конечностей, а минимум тонуса шейных мышц одной стороны совпадает как раз с тем положением животного, которое дает минимум тонических лабиринтовых рефлексов на конечности. Отсюда на основании сказанного раньше следует, что воздействия каждого из лабиринтов на мышцы своей стороны осуществляются при помощи отягчения и растяжения отолитом волосков клеток утри- кула. Итак, каждый из лабиринтов может управлять рефлекторно конечностями обеих сторон, но воздействие его на шейные мышцы будет односторонним. Если, следовательно, лабиринт может влиять на мышцы шеи, то он посредственным путем может влиять также и на конечности; поэтому каждый из лабиринтов управляет конечностями непосредственно и посредственно.
Воздействие лабиринтов и шейных окончаний чувствительных нервов не ограничивается мышцами конечностей. И лабиринты, и чувствительные аппараты шейных мышц обнаруживают сильное действие также еще на один орган, приводимый в движение соответствующими мышцами; этот орган — глаз. Мышцы конечностей могут производить перемещения сочлененных друг с другом костей. Мышцы глаза, четыре прямые и две косые для каждого, способны изменять положение глаза. В лаборатории Магнуса были произведены исследования, которые показали, что со стороны лабиринтов и шейных мышечных окончаний идет рефлекторное управление мышцами глаза, так что каждому положению головы соответствует определенное положение глаза. Эти исследования производились при участии офтальмолога Ван дер Гуве.
Децеребрированный кролик прикреплялся к столику, который был подвешен по принципу Кардана так, что его можно было вращать в различных направлениях вокруг различных осей. Для того чтобы исключить изменение положения головы относительно туловища, применялась, как описано раньше, гипсовая повязка. К доске, на которой находилось животное, прикреплялась фотографическая камера, которая поэтому при всех вращениях доски не изменяла своего положения относительно животного. При помощи этой камеры фотографировался глаз в различных положениях животного. На роговой оболочке, анестизированной кокаином, предварительно выжигался крест, благодаря чему на фотографии глаза можно было видеть, как изменялось положение глаза. Вращение доски производилось вокруг избранной оси постепенно на 360°, причем при повороте на 15° доска останавливалась, производился снимок, вращали доску опять на 15°, и опять производился снимок; таким образомг для всего круга получалось 24 снимка, по которым можно было судить, как менялось положение глаза.
Это сложное, очень обстоятельное и очень точно проведенное исследование дало определенные и легко формулируемые результаты.
Если исходное положение животного есть положение спиной вверх с несколько наклоненной вниз головой, и мы станем вращать животное вокруг затылочно-носовой оси так, что правая сторона головы будет направляться книзу, то окажется, что правый глаз поднимется, а левый — опустится. Когда голова повернется таким образом на 90°, то поворот глаз приблизится к максимуму; при дальнейшем вращении животного, когда оно будет находиться спиной вниз и затылок будет смотреть книзу, глаза опять будут в нормальном положении. При дальнейшем повороте, когда левая сторона головы будет смотреть книзу, правый глаз опустится,
592
А. Ф. Самойлов
а левый — поднимется. Мы здесь имеем рефлексы лабиринта на мышцы прямую верхнюю и прямую нижнюю, причем сокращение мышц вызывает движение глаза в обратную сторону движению головы. Глаза как бы желают не участвовать в движении головы, а оставаться в первоначальном положении так, чтобы изображение обоих полей зрения, получающееся на сетчатой оболочке, не изменилось во время движения головы вокруг указанной оси, что, конечно, имеет значение для ориентировки животного при помощи зрительных ощущений. Если вращать животное вокруг битемпоральной оси, то получаются ротаторные движения глаза вокруг его оси, причем смысл этого движения такой же, т. е. глаз сохраняет свое положение, несмотря на движение головы. Таким образом, каждому положению головы в пространстве соответствует определенная степень сокращения глазных мышц. Управление этими сокращениями идет опять- таки со стороны лабиринта; доказательством этому служит то, что удаление лабиринтов устраняет описанные тонические рефлексы на глаз.
Однако и в данном случае управление мышцами глаза идет не только со стороны лабиринта. У децеребрированных животных с удаленными лабиринтами наклонение головы с приближением ее к туловищу при флек- сорном сокращении шейных мышц или при дорсальном флексорном сокращении, а также при повороте головы с приближением челюсти к одному или другому плечу получается движение глаз, и смысл этого движения опять заключается в том, что мышечные сокращения делают возможным сохранение первоначального положения глаза при изменении положения головы. Таким образом, эти так называемые компенсаторные движения глаза обеспечиваются рефлекторными механизмами, идущими со стороны лабиринтов и чувствительных окончаний, заложенных в шейных мышцах. При тех сравнительно небольших движениях головы, какие животное в нормальных условиях своей жизни производит, компенсаторные движения глаз совершенно компенсируют движения головы, благодаря чему изображения на сетчатой оболочке удерживаются на своих местах.
Рассмотренные нами тонические рефлексы со стороны лабиринтов на конечности, со стороны лабиринтов на шейные мышцы, со стороны проприоцептивных окончаний шейных мышц на конечности, наконец, тонические рефлексы со стороны лабиринтов и чувствительных аппаратов шеи на глазные мышцы были изучены на децеребрированных животных. Было бы, однако, ошибочно думать, что эти рефлексы имеют место исключительно на децеребрированных животных. Дело обстоит иначе. Указанные рефлексы, правда, могут быть наиболее легко демонстрированы на децеребрированных животных. У нормального животного рефлексы эти часто затушеваны, заслонены целым рядом других рефлексов, идущих со стороны верхних частей ствола головного мозга и гемисфер. Но при известных обстоятельствах можно и на нормальных животных наблюдать игру этих тонических рефлексов, и они проявляют себя с такой же ясностью, как и на децеребрированном животном.
Представим себе, что перед нами кошка, стоящая на своих четырех конечностях на полу. Пусть голова ее наклонена несколько книзу, образуя с горизонтом угол приблизительно в 45°. В таком случае ее конечности в отношении экстензорного сокращения находятся, как мы уже знаем, в минимуме и потому легко согнуты. Поднесем такой кошке сверху кусок мяса. Мясо раздражит телерецепторы, помещающиеся в голове кошки, т. е. в данном случае обонятельный и зрительный аппараты, и кошка подымет голову; но как только она подымет голову, сейчас же фатальным образом выпрямятся ее передние конечности и согнутся зад¬
Работы Р. Магнуса и его лаборатория в Утрехте
593
ние. Почему в данном случае ее передние конечности выпрямятся сильно? Это случится потому, что поворот головы кверху переведет в силу лабиринтовых рефлексов минимум тонического лабиринтового воздействия в максимум, и все четыре конечности должны будут выпрямиться. С другой же стороны, шейный рефлекс при повороте головы кверху поведет к тому, что передние конечности выпрямятся, а задние — согнутся. В результате мы получим сильнейшее выпрямление передних конечностей и сгиб задних. Наверное, многим покажется, что можно истолковать всю картину иначе. Они скажут: «Голова животного поднялась, оно увидело мясо; для того чтобы достигнуть мяса, кошка выпрямила свои конечности: тут нет фатального выпрямления, а есть благоразумное желани, со стороны животного при помощи выпрямления конечностей достигнуть мяса». Это, однако, совершенно не так. Во-первых, мы видели уже, что и у децеребрированного животного, которое вряд ли имеет «желания», поворот головы кверху тоже вызывает выпрямление передних конечностей. Дальше, если бы мы взяли кошку, у которой перерезаны задние корешки первых трех шейных нервов и удалены оба лабиринта, и приблизили сверху кусок мяса, то такая кошка подняла бы голову кверху, но ноги ее не выпрямились бы, потому что испорчен тот механизм, который фатально при поднятии головы кверху ведет к выпрямлению конечностей. Нужно при этом иметь в виду, что кошка с перерезанными верхними тремя шейными задними корешками и лишенная лабиринтов оправляется с течением времени настолько хорошо, что ее трудно бывает отличить сразу от совершенно нормального животного; она владеет своими конечностями с большим совершенством, но все же при поднятии головы у нее передние конечности не выпрямляются сейчас же. «Желание» в данном случае при испорченном механизме является весьма посредственным суррогатом; только через некоторое время, когда мясо продолжает раздражать кошку, ее конечности медленно выпрямляются.
Другой пример. Кошка видит мышь, бегущую по краю комнаты с левой стороны, и поворачивает туда голову. При этом повороте смещается центр тяжести тела кошки влево и фатальным образом выпрямляется ее левая передняя конечность и сгибается правая. Это есть типичный шейный тонический рефлекс. Челюстная конечность выпрямилась, а теменная — согнулась. Дальнейшее поведение кошки будет уже определяться не перечисленными тоническими рефлексами, а скорее условными рефлексами: если она сообразно данной ситуации направится к мыши, то она должна будет повернуться вокруг тонически вытянутой левой ноги и сделать первый шаг в сторону своей цели при помощи согнутой правой ноги.
Магнус указывает, что если внимательно наблюдать движения людей во время спортивных игр, если рассматривать картины, в которых правильно изображаются отдельные фазы каких-либо сложных движений тела, то мы в большинстве случаев сумеем расшифровать положение тела, зафиксированное в определенные моменты как результат тонических рефлексов положения, изученных на децеребрированных животных. Но, конечно, наслаивающиеся новые рефлексы верхних частей центральной нервной системы осложняют картину указанных рефлекторных положений. Кроме того, на человеке все указанные особенности тонических рефлексов не могут проявиться в той форме, в какой они изучены на четвероногих, ибо вертикальное положение тела человека и пользование только двумя конечностями при ходьбе требуют совершенно иных механизмов равновесия. Имеет значение и то обстоятельство, что передние конечности меньше участвуют в структуре положения, позе животного, а служат главным образом хватательными аппаратами.
38 Р. Магнус
594
А . Ф. Самойлов
Поэтому некоторые, например Минковский в Швейцарии, пытались на человеческих эмбрионах 3, 4, 5 месяцев, вырезанных под местной анестезией при кесаревом сечении из чрева матери, обнаружить тонические рефлексы Магнуса. Минковский утверждает, что в некоторых случаях ему удавалось доказать шейные рефлексы на передние конечности.
Если обратиться к патологическим случаям, то на людях можно воспроизвести многие из тех тонических рефлексов, какие мы имеем в случаях децеребрированных животных.
Прежде всего подходящими случаями для этой цели являются заболевания, ведущие в конце концов к водянке мозга. У гидроцефаликов, в особенности в тяжелых случаях, удается иногда наблюдать тонические рефлексы Магнуса на конечности. Впоследствии оказалось, что эти же рефлексы можно наблюдать и у гемиплегиков на их паретических конечностях, но здесь эти явления наблюдаются не в такой чистой форме * как у гидроцефаликов.
Симонс первый показал, что у гемиплегиков тонические рефлексы могут проявляться на фоне так называемых содружественных движений. Представим себе, что перёд нами правосторонний гемиплегик. Он лежит на спине, мы поворачиваем его голову в правую сторону и при этом не наблюдаем совершенно никаких изменений в положении конечностей. Но если теперь при повернутой вправо голове больного заставить произвести левой здоровой рукой усилие, то правые верхняя и нижняя конечности вытягиваются в сильнейшей экстензии. Если, наоборот, вначале повернуть голову влево, то усиленное движение левой здоровой рукой вызовет содружественное усилие правой больной стороны в виде флексии правых конечностей. Наблюдение Симонса имеет цену и теоретическую, и практическую. В теоретическом отношении здесь важно то, что если у гемиплегика поворот головы и не вызывает обычных тонических рефлексов непосредственно, то все же он производит какую-то перегруппировку в иннервационной картине спинного мозга. Эта перегруппировка ведет к тому, что наиболее легко совершается то движение парализованной конечности, которое соответствует обыкновенным тоническим лабиринтовым шейным рефлексам, и если каким-либо образом пустить в ход содружественное движение, то это именно будет то движение, которое соответствует схеме Магнуса для лабиринтовых и шейных рефлексов и уже потенциально приготовлено определенным поворотом головы. Подобные же отношения можно наблюдать и на децеребрированных животных. Русский физиолог Беритов впервые в лаборатории Магнуса заметил, что если поворот головы на животном не влияет почему-либо на положение конечностей, то стоит лишь вызвать какое-нибудь раздражение, например кожи у животного, чтобы сейчас же конечности приняли то положение, которое соответствует данному повороту головы. В практическом отношении указание Симонса имеет то значение, что оно открывает целую новую область для клинических исследований, дает новые черты в картине гемиплегии и новые диагностические возможности.
Та картина рефлексов, какую мы наблюдаем на децеребрированных животных, должна быть понимаема как сумма рефлексов, центры которых находятся в продолговатом мозгу. Правда, у децеребрированных животных сохранен также мозжечок, эта наиболее загадочная часть ствола центральной нервной системы. Можно было бы думать, что для получения описанных лабиринтовых и шейных рефлексов у децеребрированных животных требуется участие мозжечка, но это не так. Удаление мозжечка у децеребрированного животного не меняет системы описанных выше рефлексов, что особенно оттеняется Магнусом.
Работы Р. Магнуса и его лаборатория в Утрехте
595
Мы переходим к другому отделу тонических рефлексов, выяснение которых составляет особенную заслугу Магнуса; это так называемые Stellieflexe. Я не знаю русского перевода этого термина; я предложил бы перевести Stellreflexe — рефлексы выпрямления. Рефлексы выпрямления отличаются от описанных выше и названных Магнусом Haltungsreflexe; Haltungsreflexe я предлагаю перевести — рефлексы положения, позы тела.
Рефлексы выпрямления отсутствуют у децеребрированного животного. Как было указано, децеребрация производится при помощи разреза на границе между продолговатым мозгом и четверохолмием. Магнус хотел выяснить, что произойдет с рефлексами позы, положения, если разрез будет сделан несколько выше, именно на границе между четверохолмием и зрительными буграми. Результат получился поразительный. Когда после такой операции мы выждем прекращения действия наркоза, то прежде всего мы можем удостовериться, что ригидности в конечностях у такого животного нет. Конечности легко поддаются сгибанию и выпрямлению. Дальше мы с удивлением увидим, что через час-два после операции, а иногда и раньше, кошка обнаруживает усилие приподняться. Прежде всего голова ее, лежащая на боку, подымается. Это первое усилие может и не повести к тому, что вся кошка подымется; но мы увидим дальше, что последовательно вместе с повторными усилиями поднять выпрямленную голову будут подниматься и выпрямляться последующие части тела, пока в конце концов кошка встанет на ноги и начнет шагать. Ее ходьба на первых порах ненормальна в том отношении, что она ставит свои конечности не ладонной, а тыльной стороной; но это длится недолго, и тут же на наших глазах ее поступь становится совершенно правильной. Замечательно и то, что кошка при этом будет обходить предметы, и это, очевидно, производится при помощи длинных ее усов. Дотрагивание усами до какого-либо предмета изменяет направление движения. Так как у такой кошки верхний отдел ствола мозга заканчивается четверохолмием, то регулирование тепла нарушено, ибо тепловые центры находятся в зрительных буграх; Магнус поэтому изменил первоначальную операцию и делает разрез на границе между зрительными буграми и полосатыми телами. В результате мы имеем животное, которое Магнус называет Thalamustier (по-английски Thalamusanimal); англичане в честь Магнуса называют так оперированное животное—Magnusanimal. Совершенно ясно, конечно, что Magnusanimal — это есть то же животное, что и животное Гольца, о котором мы раньше говорили. Как определенно отражается уже в самом названии разность точек зрения Магнуса и Гольца! Гольц говорит о животном без головного мозга, а Магнус о том же животном говорит, как о таламус-животном: один напирает на то, что отсутствует у животного, а другой на то, что у животного еще осталось.
Конечно, кошка с зрительными буграми способна так же подымать голову, выпрямляться, становиться на конечности и ходить, как и кошка только с четверохолмием в качестве верхнего отдела мозга. Уже предшествовавшие труды Гольца, Чермака, Брейера сделали вероятным, что способность, животного держать голову в нормальном положении зависит от различных чувствительных аппаратов кожи, мышц, сухожилий и прежде всего от лабиринтов с их отолитами. Но Магнус первый выяснил в деталях всю систему рефлексов, при помощи которых животное становится на ноги с нормально установленной головой.
Прежде всего мы рассмотрим значение лабиринтов. Представим себе, что, произведя описанную только что операцию, мы берем животное
38*
596
А. Самойлов*
в руки, фиксируем рукой его таз и придаем тазу такое положение, что все животное оказывается в воздухе в лежачем положении на одной стороне. Если описанным образом держать животное, то голова его сейчас же выпрямляется, и оно сидит прямо. Если мы теперь, держа животное за таз, повернем его так, что сторона, которая была верхней, станет нижней, то прямо стоящая голова нисколько не изменит своего положения. Если бы мы заставили животное висеть в воздухе вертикально, то тоже голова его поднялась бы. Если бы мы держали его вертикально головой вверх, то опять-таки голова сохранила бы нормальное положение. Итак, к?к бы мы ни изменяли положение туловища животного, если оно находится в воздухе, положение головы было бы все равно нормальным.
После двустороннего удаления лабиринтов эта способность пропадает навсегда. Если мы держим такое животное в воздухе на боку, то и голова остается в боковом положении. Если мы держим животное в вертикальном положении головой вниз, то голова принимает то положение, какое соответствует действию силы тяжести. Точно так же теряется способность головы выпрямляться, если у нормального животного мы произведем перерезку в области передней трети среднего мозга, т. е. четверохолмий.
Как показывают опыты центрифугирования, способность выпрямления головы у таламус-животного связана именно с действием отолитов.
Много ясности вносит поведение животных Магнуса в том случае, когда удаляется только один лабиринт. Мы видели уже раньше, что одностороннее удаление лабиринта у децеребрированного животного ведет к проявлению тонического влияния здорового лабиринта на мышцы одной стороны шеи. Поворот головы при этом сохраняется вообще во всех положениях головы, но он имеет максимум, когда животное лежит на спине с приподнятой несколько головой. Центры этих тонических рефлексов лежат в продолговатом мозгу несколько позади от места выхода восьмых нервов. Этот поворот головы наблюдается также и у животных Магнуса, когда у них удаляют один лабиринт. Но в этом случае поворот головы, как тонический рефлекс со стороны лабиринта, комбинируется дальше со способностью животных Магнуса выпрямлять голову. Животное старается установить голову в таком положении, в котором сохраненный лабиринт находился бы наверху. Лабиринтовый рефлекс выпрямления в этом случае имеет минимум силы. Если же сохраненный лабиринт находится внизу, то лабиринтовый рефлекс имеет свой максимум и животное делает чрезвычайные усилия освободиться от этого ненормального положения и перевести голову в положение минимума. Мы не будем входить в интересные подробности тех положений головы, какие при этом могут иметь место. Для нас важнее оттенить, что устойчивое положение головы после удаления одного лабиринта, определяемое лабиринтовым рефлексом выпрямления, отличается от того, какое мы имеем при сохраненных двух лабиринтах. Отсюда можно заключить, что действующие в данном случае отолиты не лежат в одной плоскости. А так как отолиты утрикула именно лежат в одной плоскости, то следует заключить, что действующими отолитами являются в данном случае не отолиты утрикула, а отолиты сакула. Пусть у животного удален правый лабиринт, тогда максимум будет в том случае, когда животное лежит на левой стороне, ибо в этом случае здоровый левый лабиринт находится внизу. В этом случае, как оказывается, отолит сакула висит, растягивая волоски клеток пятна сакула, и животное делает сильнейшие усилия повернуть голову на правую сторону так, чтобы отолит не тянул волоски, а лежал бы на них. В тех случаях, когда оба лабиринта сохранены и го¬
Работы Р. Магнуса и его лаборатория в Утрехте
597
лова животного лежит на правой стороне, левый сакул находится в положении минимума и не вызывает никакого действия, а правый — находится в положении максимума, потому что он висит. Поэтому рефлекс выпрямления старается перевести голову на левую сторону, но этого осуществить никоим образом нельзя, потому что, как только голова начинает поворачиваться, чтобы повернуться на левую сторону, левый сакул удаляется из положения минимума и таким образом противодействует правому. Когда голова приходит в нормальное положение, оба сакула стоят симметрично; раздражения, идущие от обоих лабиринтов, приводят в конце концов к известному равновесию, и нормальное положение головы удерживается. Лишь только голова уклоняется от симметричного положения, как увеличивается раздражение со стороны нижележащего сакула и уменьшается раздражение со стороны вышележащего, благодаря чему голова приходит опять в симметричное положение.
Мы убедились, таким образом,' что ( лабиринты способны путем рефлекса у животного с сохраненными четверохолмиями выпрямлять и удерживать голову в нормальном положении. Как уже было сказано, после двустороннего удаления лабиринтов эта способность прекращается, и потому животное, если мы его держим в воздухе, не способно ориентировать правильно свою голову. Если все животное находится в воздухе в боковом положении, то и голова в боковом положении. Но удивительно то, что если мы такое животное без обоих лабиринтов, которое не способно ориентировать правильно свою голову в воздухе, положим на какую-нибудь твердую поверхность — на пол, на стол, то уже вследствие одного соприкосновения кожи туловища с предметом, на котором оно лежит, получается рефлекс, в результате которого голова приводится в нормальное положение. Что в самом деле в данном случае рефлекс вызывается несимметричным раздражением чувствительных нервов туловища, доказывается тем, что рефлекс этот можно легко устранить, если на верхнюю поверхность тела животного, ни с чем не соприкасающуюся, произвести какое-нибудь давление. Если, например, на животное положить доску, отягченную каким-либо грузом, то приподнятая голова падает и лежит на боку так же, как в том случае, когда все животное находится в воздухе. Этот второй рефлекс есть так называемый «рефлекс туловища на выпрямление головы».
При помощи двух описанных рефлексов, лабиринтного рефлекса выпрямления головы и рефлекса туловища на выпрямление головы, обеспечивается нормальное положение головы. Если голова имеет нормальное положение, а туловище при этом еще лежит на боку, то получается вращение шеи. Это вращение служит исходной точкой для третьего рефлекса, вследствие которого туловище вынуждено следовать за головой. Так как первичным моментом в данном случае служит ненормальное положение тела с поворотом шеи, то эти рефлексы, по Магнусу, называются «шейными рефлексами выпрямления». Ход проявления этих рефлексов обычно такой, что прежде всего передняя часть туловища следует за головой и вследствие этого получается скручивание поясничной части позвоночного столба, которое в свою очередь ведет к тому, что таз следует за передней частью туловища.
Описанные нами три рефлекса имеют в своем результате прежде всего выпрямление головы вследствие лабиринтового рефлекса, а за этим выпрямлением головы следует в силу шейных рефлексов последовательное выпрямление всего туловища.
Магнус показал еще и четвертую группу рефлексов, и эта четвертая группа состоит в следующем. Положим животное Магнуса на стол на бок
598
А. Ф< Самойлов
и фиксируем его голову в этом боковом положении; окажется, что в этом случае туловище приподнимается само, несмотря на то что голова находится в боковом положении. Если компенсировать опять асимметричное давление со стороны стола при помощи отягченной доски, положенной на верхнюю сторону животного, то рефлекс пропадает и животное остается целиком лежать на боку. Описанный «рефлекс туловища на выпрямление туловища» имеет место и после удаления обоих лабиринтов.
Все четыре указанных рефлекса в своей совокупности обеспечивают животному способность выпрямляться и сохранять нормальное положение. Воздействия, ведущие к рефлексам, имеют отношение к двум группам чувствительных органов: во-первых, к отолитам и, во-вторых, к чувствительным окончаниям нервов кожи тела. Вследствие раздражения указанных аппаратов прежде всего выпрямляется голова. Для того чтобы привести и туловище в нормальное положение, приводятся два различных механизма: во-первых, прямой рефлекс, вызываемый соприкосновением с поверхностью, на которой животное лежит, причем раздражаются чувствительные нервы кожи туловища; во-вторых, шейный рефлекс, назначение которого заключается в том, чтобы заставить туловище следовать за головой. Одностороннее соприкосновение с поверхностью действует двояким образом. Прежде всего оно выпрямляет голову, а затем заставляет и тело принять нормальное положение. Мы видим, что таким образом осуществление важных функций тела принимать нормальное положение, когда голова стоит прямо, и сохранять положение туловища, покоящегося на конечностях, обеспечивается во всех частях двояко, поэтому выпадение, например в случае болезни, одного механизма не ведет к потере способности принимать нормальное положение, так как эту функцию берет на себя другой механизм.
Описанные четыре группы рефлексов выпрямления мы встречаем на животных Магнуса, а также и на нормальных кроликах и морских свинках. У высших млекопитающих, как например у кошки, собаки, обезьяны, с неповрежденной центральной нервной системой мы наблюдаем еще другие рефлексы выпрямления, имеющие своим началом зрительный аппарат. Для того чтобы изучать эти рефлексы, мы исследуем животных, держа их на воздухе и лишив их предварительно лабиринтов. В этом случае, если глаза животных защищены повязкой, то реакции выпрямления оказываются потерянными. Если же глаза открыты, то эти животные способны придать своей голове правильное положение, если только они фиксируют глазами какой-нибудь заинтересовавший их предмет. Как только зрительное внимание прекращается, голова сейчас же принимает ненормальное положение. С точностью не установлено положение центров для этого рефлекса, но есть основание предполагать, что оно локализовано в зрительных долях гемисфер. В этой пятой группе рефлексов выпрямления намечается таким образом звено, связывающее рефлексы ствола с рефлексами гемисфер.
Итак, мы видели, как велика разница между децеребрированным животным, т. е. таким, у которого продолговатый мозг отделен от четверохолмия, и животным, у которого разрез сделан на границе между четверохолмием и зрительными буграми. С одной стороны, децеребрированное животное с его ригидностью, животное, лишенное способности сохранять нормальное положение тела, лишенное способности выпрямляться, и, с другой — животное Магнуса, не обнаруживающее никаких признаков ригидности, обладающее гармоническим распределением тонуса мышц, способное выпрямлять свое тело и держать голову в нормальном положении, способное производить нормальные движения ходьбы и др. Какая громадная разница! Эта разница окажется тем более поразительной, если
Работы Р. Магнуса и его лаборатория в Утрехте
599
мы дадим себе отчет в том, как ничтожен по своей величине кусочек центральной нервной системы, который составляет преимущество животного Магнуса перед децеребрированным животным. Все четверохолмие кошки равно по величине горошине. Этот комочек мозгового вещества служит источником всей той сложной иннервации, при помощи которой осуществляются все рефлексы выпрямления, явления ходьбы и даже прыжка.
Это сопоставление ничтожной величины мозгового вещества с совершенством сложного механизма, включенного в него, произвело на меня лично сильнейшее впечатление. Я пришел к мысли о том, что среди различных характерных черт животной машины, отличающих ее от машин, сделанных нашими руками, не отмечена еще до сих пор черта компактности живой машины. И в самом деле, в живой машине нет пустых мест, между тем как в наших машинах всюду и везде между отдельными частями их мы видим ничем не заполненные пространства. Это можно сказать и о паровой машине, и о пишущей машине, и о швейной машине и т. д. Задумываясь над этим обстоятельством, я пришел к заключению о том, что компактность живой машины имеет своим основанием, по всей вероятности, не только экономию места и материала, а более глубокие соотношения. Даже и в наших машинах в известных случаях мы стараемся достигнуть компактности. Так, например, в индукционной машине мы наматываем проволоку возможно плотнее, завиток к завитку, и это делается не из-за экономии места или материала, а в интересах лучшего использования электрической индукции.
Нужно думать, что компактность живой машины имеет тоже свои глубокие основания. В качестве одного такого основания я хотел бы указать на явления диффузии. Кровь питает наши ткани; чтобы питательный материал мог при помощи диффузии жидкости через капилляры достигать возможно скорее клеток, необходимо построить всю систему компактно. Чем компактность будет совершеннее, тем скорее, даже при сравнительно медленно протекающих процессах диффузии, может быть осуществлено воздействие какого-нибудь вещества, несущегося с потоком крови, на< клетки. Не нужно много распространяться о том, какое громадное значение может иметь эта компактность, когда дело касается воздействия гормонов на нервную систему. Пустые места, промежутки между отдельными элементами, были бы, конечно, в указанном смысле ошибкой конструкции механизма. Но возвратимся к Магнусу.
В его лаборатории молодой ученый Радемакер предпринял обширное исследование относительно более точного определения места центров для рефлексов выпрямления. На большом материале Радемакер производил поперечные разрезы у кошек и кроликов на стволе мозга на различных •его высотах и определял, какие рефлексы сохранялись и какие пропадали. Радемакер нашел, что перерезка ствола как раз впереди места выхождения третьего черепного нерва не устраняет нормальных лабиринтных рефлексов выпрямления туловища. Между тем как разрез, сделанный на несколько миллиметров позади от выхода третьих нервов, разрушает возможность упомянутых рефлексов. Полное удаление дорсальной половины •среднего мозга не уничтожает рефлексов выпрямления. Полное удаление мозжечка оставляет рефлексы выпрямления совершенно нетронутыми. ■Этими опытами было доказано, что центры рефлексов выпрямления лежат позади места выхода третьего нерва. Таким образом, получилось представление, что центры выпрямления находятся на уровне глазодвигательного нерва. Здесь как раз находится красное ядро, клетки которого дают начало так называемому рубро-спинальному тракту; на этом уровне этот пучок спускается книзу, давая так называемый перекрест Фореля.
600
А. Ф. Самойлов
Радемакер доказал, что расщепление четверохолмия в сагиттальной плоскости совершенно уничтожает рефлексы выпрямления. Поэтому Радемакер продолжал свои исследования в том направлении, что он делал со спинной и с брюшной стороны разрезы по срединной линии, стараясь оставить перекрест Фореля нетронутым. Такая операция не нарушала рефлексов выпрямления. Другие опыты показали, что разрушение на обеих сторонах substantia nigra и пирамидных пучков тоже не нарушает рефлексов выпрямления. Словом, все эти опыты доказали, что и у кошек, и у кроликов красное ядро представляет собой центры лабиринтовых рефлексов выпрямления и рефлексов выпрямления туловища. Центробежными путями этих рефлексов нужно считать рубро-спинальный тракт. Таким образом, миниатюрность механизма, обеспечивающего рефлексы выпрямления, еще больше усугубляется, ибо механизм сосредоточен не во всем четверохолмии, а сравнительно в небольшой его части, именно в красном ядре.
Удаление красного ядра у животных Магнуса ведет еще к одному знаменательному изменению: к появлению ригидности, которую
мы обычно наблюдаем у децеребрированного животного. Таким образом, мы приходим к заключению, что присутствие в животных Магнуса рубро- спинальных импульсов противодействует влиянию нижележащих областей продолговатого мозга, которые в тех случаях, когда их деятельность не контролируется верхними отделами, вызывают усиленный экстензор- ный тонус и уменьшение флексорного тонуса.
Здесь следует отметить одно очень важное обстоятельство. Перерезка рубро-спинального тракта у животных Магнуса, как мы видели выше, вызывает ригидность экстензоров. Замечательно, однако, то, что та же перерезка у цельного, нормального животного с сохраненными полушариями вызывает лишь очень слабую ригидность. Если же у такого нормального животного, помимо рубро-спинального пучка, перерезать еще и пирамидальные пучки, то тогда наступает ригидность в полной мере. Таким: образом, нужно заключить, что пирамидальный пучок имеет назначение, помимо других заданий, противодействовать тем влияниям, которые исходят из центров продолговатого мозга к спинному и вызывают ригидность. Можно'поэтому сказать, что возбудимость и тонус спинномозговых центров, управляются со стороны головного мозга, причем отсюда идут влиянияг отчасти увеличивающие экстензорный тонус, отчасти уменьшающие его, почему в результате и флексоры, и экстензоры получают свой надлежащий тонус.
В последнее время в лаборатории Магнуса определилось новое направление исследований. Это направление имело своим началом одно случайное наблюдение, которое было сделано на животном, лишенном мозжечка. Если такое животное лежит на боку, например на левой стороне, то правые его конечности не оказывают сопротивления пассивному сгибанию и растяжению. Но стоит только произвести сильную дорсальную флексию- стопы или кисти, как конечность сильно выпрямляется, и лишь с трудом можно заставить ногу согнуться в ее сочленениях. Конечность становится тогда вытянутой и превращается как бы в несгибаемую палку. Подобный же- результат достигается и иным путем. Допустим, что собака без мозжечка лежит на спине, а голова ее согнута по направлению к животу на 90°. Тогда задние конечности этой собаки согнуты во всех сочленениях. Стоит только дотронуться до какого-нибудь пальца ноги, как нога с силой выпрямляется. Если палец исследователя удаляется, то конечность животного не отрывается от пальца, а следует за ним сколько возможно так, как если бы палец играл роль магнита, притягивающего конечность. Поэтому Магнус назвал этот рефлекс «магнитной реакцией». Если
Работы Р. Магнуса и его лаборатория в Утрехте
601
удалить палец еще дальше, так что он оторвется от конечности, то вызванная экстензия сейчас же прекращается и конечность опять становится податливой для пассивных движений.
Описанные два рефлекса вытяжения конечностей при дорсо-флексии стопы и при легком дотрагивании до пальца особенно хорошо выражены у животных после удаления мозжечка. Однако эти же рефлексы можно в несколько менее резкой форме наблюдать у животных Магнуса, а также и на совершенно нормальных животных. Характерно для выпрямления и неподатливости конечностей в указанных рефлексах то, что сокращены не только экстензоры, как у децеребрированного животного, а также и флексоры. Здесь нет явления Шеррингтона, так называемой реципрокной иннервации, когда сокращение флексоров сопровождается угнетением экстензоров и обратно. В случае рефлексов, которые мы рассматриваем теперь, все мышцы вытянутых конечностей — и флексоры, и экстензоры — сокращены, и конечность превращается в неподатливую палку. Исходя из описанных явлений, Магнус приходит к заключению, что наши конечности играют вообще двойную роль: в известных случаях они представляют собой инструмент для передвижения, тогда мы наблюдаем их реципрок- ную иннервацию, они легко поддаются пассивному сгибанию и выпрямлению; в других же случаях задача конечностей не перемещаться, а служить лишь поддержкой для туловища, тогда вместо реципрокной иннервации мы наблюдаем сокращение флексоров и экстензоров, и конечность превращается в неподатливую, несгибающуюся колонну, несущую туловище. Такую реакцию Магнус называет «рефлексом поддержки». Эта реакция поддержки, несомненно, начинает собой новую главу в учении о рефлексах. Может показаться, что здесь нет ничего существенно нового: мы все знаем, что в известных случаях наши конечности превращаются в прочные не- сгибающиеся столбы, на которых покоится туловище. Заслуга Магнуса заключается в том, что он указал, каким способом эта реакция поддержки вызывается и чем она отличается от обычных рефлекторных реакций. То, что Магнус отметил новую черту в явлении, хорошо всем известном, и указал на принципиальную ценность этой черты, служит признаком большого таланта. Только большой талант умеет вскрыть в хорошо всем знакомом, банальном явлении нечто новое.
Укажем на один очень ценный пример статической реакции поддержки, которую Магнус назвал «шункель-рефлекс».
Это странное название взято им от названия деревенского танца, распространенного в Швейцарии под названием «шункель-вальс». Пара танцующих держит друг друга руками за плечи, перегибаясь всем туловищем на одну ногу и приподнимая другую ногу кверху. В определенный момент они перегибают туловище в другую сторону, как бы падая, и в это время нога, находящаяся в воздухе, выпрямляется, упирается в землю и поддерживает падающее на нее туловище, а прежде выпрямленная нога в это время сгибается, повисая в воздухе; так туловище танцующих как бы падает то в одну, то в другую сторону.
Описанное движение Магнус воспроизводит в опыте над животным таким образом.1
Представим себе, что собака стоит на столе и что ее правая передняя конечность, поддерживаемая нами, приподнята и согнута. Рука наблюдателя перемещает грудную клетку и все туловище животного в правую сторону. Тогда отчетливо чувствуется, как увеличивается экстензор-
1 Нужно прибавить, что опыт удается с цельным животным или с животным, у которого удален мозжечок или гемисферы.
602
А. Фш Самойлов
ный тонус локтя, и затем конечность сразу совершенно выпрямляется и становится способной поддерживать переднюю часть тела от падения на правую сторону. То же самое можно демонстрировать и на задней конечности и даже на нижней конечности человека. Представим себе, что кто-нибудь из нас стоит на одной ноге, допустим на правой, а затем перемещает свое туловище на левую сторону. Тогда неизбежно левая нога выпрямляется и удерживает туловище от падения. Конечно, многие подумают, что в этом нет ничего удивительного: мы по своей воле выпрямляем ногу, чтобы не упасть. Но это неправильно: здесь совершенно другое явление. Мы не падаем, и, помимо нашей воли, наоборот, удержать ногу от выпрямления, когда туловище падает в ее сторону, стоит нам чрезвычайных усилий. Выпрямление ноги, когда тяжесть туловища надвигается в ее сторону, не есть выражение нашего волевого влияния, а это есть результат рефлекса. Когда животное стоит на столе с приподнятой одной конечностью, например правой, и мы перемещаем туловище в правую сторону, то при этом растягиваются аддукторные мышцы той ноги, которая в данный момент поддерживает туловище, т. е. левой. Это растяжение аддукторов и есть начало рефлекса, реакции поддержки со стороны другой конечности. Стоит только перерезать задние корешки чувствительных нервов, снабжающих нервными окончаниями аддукторные мышцы, и этот рефлекс пропадает. Перемещение тяжести тела в сторону согнутой конечности не даст реакции поддержки. Без участия нервных окончаний в аддукторах нельзя танцевать «шункель-вальс».
Радемакер передавал мне, что ему удалось сделать одно очень интересное наблюдение над собакой, лишенной мозжечка, когда он исследовал реакцию поддержки на ее конечностях. Как уже упоминалось, реакция поддержки производилась таким образом: экспериментатор держал
в своих руках одну конечность Животного, стоящего на других трех конечностях, и затем наклонял животное в сторону этой конечности; вследствие такого наклонения получалась реакция поддержки, и конечность с силой вытягивалась. Радемакер подметил, что если часто повторять этот опыт, то животное вытягивает с силой конечность сейчас же, как только дотронешься до конечности. Очевидно, как справедливо думает Радемакер, мы имеем здесь дело с условным рефлексом. Можно себе представить, -сколько излишних затруднений могло бы встретиться на пути исследования реакции поддержки, если бы мы не имели уже учения об условных рефлексах и лишены были бы возможности учитывать участие условных рефлексов в той области, в которой можно ожидать лишь существования безусловных рефлексов.
Нужно думать, что описанная реакция поддержки, когда конечность превращается в колонну, поддерживающую туловище, имеет место и при акте ходьбы. При нашей ходьбе центр тяжести все время перемещается то в одну, то в другую сторону. Если центр тяжести переместился в правую сторону, то правая нога на почве реакции поддержки превращается в неподатливую подпорку, в которой все мышцы, экстензоры и флексоры сокращены. В то же время левая нога, не касаясь земли и не испытывая дорсальной флексии ступни, находится вне условий реакции поддержки и представляет собой инструмент для передвижения. Нога перемещается, подчиняясь закону реципрокной иннервации, и как только центр тяжести туловища переместится в левую сторону и аддукторные мышцы правой стороны вытянутся, то сейчас же левая нога начинает выпрямляться и, коснувшись земли и испытав при этом дорсальную флексию, превращается в свою очередь в подпирающую туловище колонну, а правая нога, оторвавшись от земли, превращается в инструмент передвижения и т. д.
Работы Р. Магнуса и его лаборатория в Утрехте
603
Если мы охватим мысленно еще раз весь представленный материал, то придем к заключению, что ствол мозга вместе со спинным мозгом является в плане работ Магнуса изумительным по своей сложности и точности работы инструментом, машиной, регулирующей сокращение наших мышц в интересах сохранения равновесия, сохранения положения тела, выпрямления его и производства всевозможных движений. В интересах животного его мышцы выполняют различные задания, и вся сложность механизма, диктующего задания, сосредоточена в указанных отделах центральной нервной системы. И вся эта сложность укладывается в простую формулу рефлекторного акта. Вся сложная машина представляется нам по праву как рефлекторная машина.
В цельном животном на эту машину безусловных рефлексов накладывается власть гемисфер, т. е. власть другой рефлекторной машины, машины условных рефлексов. Я упоминал вначале, что Гольц со своим опытом собаки без полушарий не решил задачи о функциях центральной нервной системы, а лишь поставил задачу. В продвижении этой задачи успех выпал на долю трудов Павлова и Магнуса и их многочисленных сотрудников. Стоит отметить, что Павлов и Магнус начали свои работы приблизительно четверть века тому назад, начали совершенно независимо друг от друга и пришли к определенным результатам почти одновременно., Год тому назад вышла большая монография Магнуса «Die Körperstellung», а на днях вышла в свет книга Павлова «Работа головного мозга», в которой наш маститый ученый дает систематическое изложение учения об условных рефлексах. Отрадное, возвышающее впечатление производят эти две книги. Павлов и Магнус шли в своих работах с разных концов центральной нервной системы, и в их продвижении вперед уже намечено место встречи. Невольно напрашивается сравнение с двумя партиями рабочих, прорывающих туннель с двух сторон горного массива. Они уже слышат удары молотков противоположной стороны. Еще один удар — и они подадут ДРУГ другу руки.
Э. III. Айрапетъянц и В. А. К и с л як о в
КОРА БОЛЬШИХ ПОЛУШАРИИ И ФУНКЦИЯ ВЕСТИБУЛЯРНОГО АППАРАТА
Деятельность неслуховой части лабиринта издавна привлекала внимание физиологов и клиницистов. За последние десятилетия в особенности интенсивно добывались факты и создавались представления о связях вестибулярного аппарата с различными отделами центральной нервной системы. В результате этих исследований накопился значительный материал, позволивший обнаружить закономерные взаимоотношения между корой головного мозга и разнообразными проявлениями вестибулярных реакций.
Изучение физиологии полукружных каналов началось с экспериментов французского исследователя Флюранса (Flourens, 1842). Он отметил значение полукружных каналов в координации движений, статике и поддержании равновесия тела в пространстве. Благодаря систематическим исследованиям голландского физиолога Магнуса и его сотрудников (Magnus, 1924) было установлено, что и отолитовый аппарат лабиринта выполняет аналогичную функцию. Таким образом, выяснилась роль вестибулярного аппарата в организме как органа равновесия.
Различные авторы посвятили много работ раскрытию механизма раздражения полукружных каналов (Goltz, 1870; Breuer, 1874; Grum- Brown, 1874; Mach, 1875; Ewald, 1892) и отолитового аппарата (Magnus, 1924; Quix, 1925). В результате этих исследований возникла теория, согласно которой раздражителем чувствительных окончаний нервов полукружных каналов является ток эндолимфы, возникающий при действие угловых ускорений обоих знаков. Нервные же окончания отолитового аппарата раздражаются при смещении отолитов, вызываемом прямолинейными ускорениями, центробежной силой и изменениями положения головы в пространстве.
Вопрос о взаимодействии отолитового аппарата и полукружных каналов лабиринта был поставлен и изучен у нас в стране В. И. Воячеком и К. Л. Хиловым с сотрудниками. В. И. Воячек в 1925 г. на I Всесоюзном съезде оториноларингологов высказал предположение о том, что отолитовый аппарат внутреннего уха, раздражаемый во время вращения центробежной силой, тормозит проявление реакций с полукружных каналов. В 1927 г. эта гипотеза была подтверждена К. Л. Хиловым (1952) в опытах на кошках и белых мышах. Затем Т. Д. Бахвалова (1950) представила новые экспериментальные материалы по этому вопросу. В. И. Воячек (1927) на основании опытов К. Л. Хилова ввел в клиническую практику пробу на отолитовую реакцию (ОР). Отмеченные работы показали, что полукружные каналы и отолитовый аппарат работают не изолированно, а в тесном функциональном взаимодействии.
Современные знания о рефлекторной деятельности центральной нервной системы основой своей имеют труды Шеррингтона, Магнуса, Сеченова и Павлова. Благодаря этим выдающимся исследователям была экспери-
Кора больших полушарий и функция вестибулярного аппарата
605
ментально изучена «система рефлексов в восходящем ряду» (А. А. Ухтомский).1 Магнусом и его сотрудниками были глубоко и всесторонне изучены центральные механизмы вестибулярных реакций (преимущественно отолитовых). Эти исследования раскрыли подкорковые пути лабиринтных рефлексов и связь их с другими рецепторными полями при формировании установочных реакций. Эксперименты Магнуса высоко оценил И. П. Павлов. Он подчеркивал глубокую связь между работами Шеррингтона и Магнуса и считал Магнуса продолжателем дела Шеррингтона. И. П. Павлов писал: «В восемнадцатом, девятнадцатом и двадцатом столетиях физиологи фактически и подробно использовали идею рефлекса, но только на низших отделах центральной нервной системы, однако все выше и выше поднимаясь по отделам ее пока, наконец, после классических работ Шеррингтона (Sherrington) над спинномозговыми рефлексами продолжатель его Магнус (Magnus) не доказал рефлекторного характера всех основных актов локомоторной деятельности. Таким образом, идея рефлекса с полным экспериментальным оправданием была приложена к центральной нервной системе почти вплоть до больших полушарий».1 2
Изгостно, что в работах Магнуса имеются высказывания о роли коры головного мозга в регуляции лабиринтных рефлексов. Однако затронутая проблема не оказалась предметом специальной разработки в упомянутых исследованиях. В книге Магнуса можно найти прекрасные описания установочных реакций у нормальных животных и человека, но он как бы в этом сложном поведении старается отыскать в отчетливом, неизменном виде те группы рефлексов, которые им были обнаружены на объектах, лишенных больших полушарий головного мозга. Магнус считал, что механизмы установочных реакций можно изучать только объективным путем, опираясь на идею рефлекса. Только этот путь, по Магнусу, позволяет выделить систему закономерных рефлексов, связанных с функцией определенных нервных центров. Но дальнейший ход его рассуждений приводит к выводу о том, что все, связанное с корой головного мозга, может быть изучено только субъективным методом. Субъективный анализ, давший много ценных сведений при изучении слуха и зрения, по мнению Магнуса, не мог быть применен для исследования статических реакций в связи с отсутствием «. . .участка коры для статических восприятий».3 Под названными участками Магнус, очевидно, понимал определенные корковые центры, подобные зрительным и слуховым, которые должны осуществлять статическую функцию организма, воспринимая импульсы от вестибулярного аппарата, проприоцепторов и т. д. Магнус и Павлов были современниками, и к тому времени, когда вышла в свет книга Магнуса (1924), наука в этом вопросе уже могла опираться на учение И. П. Павлова об условных рефлексах и анализаторах. Одним из важнейших результатов в учении об анализаторах явилось доказательство афферентной роли двигательной области коры головного мозга (Красногорский, 1911). Позднее Н. А. Поповым (1920) были поставлены эксперименты, положившие начало изучению методикой условных рефлексов функции вестибулярного аппарата. Таким образом, указанными работами устанавливалась анализаторная структура двух важнейших элементов, регулирующих равновесие тела — про-
1 А. А. Ухтомский, Собр. соч., т. V, стр. 228, Изд. ЛГУ, 1954.
2 И. П. Л а в л о в, Поли. собр. соч., т. IV, стр. 18—19, Изд. АН СССР, М.— Л., 1951
3 См. настоящее издание книги Р. Магнуса, стр. 10.
606
9. Ш. Айрапетъянц и В. А. Кисляков
приоцепции и вестибулярной рецепции, корковые проекции которых являются важнейшими компонентами «статической коры».
В лаборатории К. М. Быкова и его сотр. принцип условных рефлексов-, был широко использован для изучения анализаторных систем, обращенных к внутренней среде организма. Результаты этих исследований явились- важной основой учения о кортико-висцеральных влияниях (Быков, 1947).
В настоящее время, благодаря применению методики вызванных потенциалов и условнорефлекторной методики, в значительной степени расширились и пополнились наши сведения о кортикальных влияниях на функции организма и о различных афферентных проекциях в коре головного мозга (Айрапетъянц, 1955, 1960а; Черниговский, 1960; Гершуни,. 1962; Ades, 1959; Bartley, 1959; Chang, 1959; Gernandt, 1959; Nakahama, 1961)* He останавливаясь на всех вопросах из этой области, мы сосредоточим внимание на связях вестибулярного аппарата с корой головного мозга.
Изменения вестибулярных реакций
при экстирпациях коры головного мозга и различных воздействиях на нее
Среди отечественных ученых первым обратил внимание на зависимость вестибулярных реакций от деятельности больших полушарий у млекопитающих В. М. Бехтерев (Бехтерев, 1881, 1882, 1885, 1905). Разрабатывая проблему корковой компенсации нарушенных функций, Э. А. Асратян (1940, 1947) включил в сферу исследований и вестибулярный аппарат,, показав значение коры головного мозга в компенсации последствий разрушения лабиринтов.
При анализе кортикальных влияний на функцию вестибулярного аппарата различными авторами в качестве показателей использовались как соматические, так и вегетативные вестибулярные реакции. В исследованиях была установлена общая закономерность — зависимость вестибулярных рефлексов от коры головного мозга. При этом одни авторы отмечали преимущественно тормозные влияния, а другие — стимулирующие влияния коры на вестибулярные реакции. В работе А. Н. Крестовникова и А. И. Яроцкого (1938) исследовались влияния коры головного мозга на поствращательный нистагм у кроликов. Авторы наблюдали, что после удаления передних и средних отделов больших полушарий происходит увеличение продолжительности поствращательного нистагма, нарушение его ритма и появление спонтанных нистагматических движений глаз. Было отмечено, что удаление коры левого полушария вызывает растормаживание нистагма при остановке вращения вправо и, наоборот, после удаления коры правого полушария—при остановке вращения влево. В статье делается вывод о том, что кора головного мозга оказывает в норме тормозящее влияние на длительность нистагма и регуляторное — на его ритм. Фитцджеральд и Холпайс (Fitzgerald a. Hallpice, 1942) приводят факты Бауера и Лейд- лера, также отметивших растормаживающее влияние удаления одной гемисферы на поствращательный нистагм у кроликов. Аналогичные факты у кошек получены Бэлэчану-Столнич 'p (Balaceanu-Stolnici, 1953). В других исследованиях (Pice, 1917; de Barenne u. de Klein, 1923) было обнаружено растормаживающее влияние экстирпации одной гемисферы и передних отделов больших полушарий головного мозга у кроликов не только на поствращательный нистагм, но и на нистагм, вызываемый тепловыми и холодовыми раздражениями вестибулярного аппарата. Г. А. Образцова и 3. Д. Пигарева (1957) отметили, что экстирпация боль¬
Кора больших полушарий. и функция вестибулярного аппарата
607
ших полушарий у кроликов на 5—8-е сутки после рождения также вызывала усиление поствращательного нистагма. В далекие сроки после операции в ряде случаев у тех же кроликов наблюдалось угнетение нистагма. Исследования проводились в течение 8—12 месяцев после операции. Сравнительно-физиологическое изучение влияния экстирпации больших полушарий на вестибулярный нистагм проводилось Айви (Ivy, 1919). Как показали его опыты на лягушках, черепахах и птицах, удаление переднего мозга не нарушает вестибулярный нистагм. Однако другие авторы описали в своих работах растормаживание поствращательного нистагма и усиление качательных движений головы у голубей после экстирпации больших полушарий (Тюмянцев, 1926а, 19266; Борнгард, 1875). У млекопитающих животных (кроликов, кошек, собак) Айви наблюдал растормаживание вестибулярного поствращательного нистагма на остановку вращения в сторону, противоположную экстирпированному полушарию. В тех случаях,, когда вращение производилось в' направлении удаленного полушария, часто отмечалось ослабление постнистагма. Такая же закономерность выявилась и в опытах на собаках при частичном повреждении коры головного мозга одного полушария в двигательной и теменной областях. В работе И. Н. Пинчука (1948) в хронических опытах было показано, что экстирпация различных участков коры или полная декортикация животного приводит к угнетению вестибулярных реакций сроком до 45 дней. И. Н. Пинчук считает, что в нормальных физиологических условиях кора головного мозга повышает возбудимость вестибулярных центров. Примером зависимости результатов вестибулярных проб от функционального состояния коры головного мозга являются факты, описанные в клиническом сообщении И. Я. Калиновской и В. Е. Майорчик (1952).
Явление растормаживания вестибулярных реакций после выключения функции больших полушарий широко изучалось К. Л. Хиловым с сотрудниками в большой серии исследований, посвященной изучению роли коры в регуляции вестибуло-соматических и вестибуло-вегетативных реакций. Результаты этих исследований обобщены К. Л. Хиловым в специальных работах (Хилов, 1951, 1952) и в отдельных публикациях его сотрудников. Так, С. А. Злотников в опытах на кошках еще раз подтвердил тот факт, что экстирпация коры, замораживание ее хлорэтилом и угнетение наркотиками влияют на течение поствращательного нистагма, увеличивая его продолжительность. Другими сотрудниками К. Л. Хилова изучались, главным образом у людей, изменения моторной хронаксии под влиянием вестибулярных раздражений в норме и после приема хлоралгидрата, угнетающего кору головного мозга (А. С. Лунева), изменения вестибуло- вегетативных реакций в норме и при гипоксемии (А. Л. Фрейдович). В тех же условиях опыта, что и у А. С. Луневой, исследовались реакции сердечно-сосудистой системы (Т. Д. Бахвалова, Д. А. Гришина, А. П. Ярославский, Н. В. Павлова), изменения секреции желудочных желез (Т. А. Бахвалова, Н. А. Королева), слюнной секреции и ольфакторных реакций (Г. М. Мисевич, А. С. Минакова), изменения дыхания (А. П. Ярославский) и т. д. Во всех этих случаях имело место усиление вестибулярных реакций после угнетения коры головного мозга. Исходя из описанного фактического материала, К. Л. Хилов развивает концепцию о тормозящем влиянии коры на вестибулярные рефлексы в условиях нормы.
Крейндлер (Kieindler, 1955) в своей экспериментально-клинической работе уделил большое внимание влиянию эпилептических приступов на течение вестибулярных реакций. После электрошокового эпилептического припадка калорический нистагм удлиняется и становится более частым. У односторонне делабиринтированных кошек с компенсированными
€08
Э. Ш. Айрапетъянц и В* А. Кисляков
симптомами разрушения лабиринта эпилептический приступ вызывал декомпенсацию этих симптомов. Автор рассматривает электрошоковый эпилептический приступ как своеобразный способ функциональной декортикации. Отмечая тормозящее влияние коры на вестибулярные центры, Крейндлер высказывает мысль, что и подкорковые образования могут оказывать такие влияния на вестибулярный нистагм. Последующие опыты автора с сотрудниками выявили определенные корковые зоны, удаление которых способствует еще большей активации вестибулярного нистагма после эпилептического приступа. Некоторые из этих зон совпадают с открытыми А. В. Гервером (1899) глазодвигательными центрами в коре головного мозга у собак. Крейндлер обсуждает свои данные с точки зрения кортикального представительства вестибулярной функции у высших животных. В работе Кемпинского и Уорда (Kempinsky a. Ward, 1950) рассматривается участие ретикулярной субстанции ствола головного мозга во взаимодействии кортикальных и вестибулярных импульсов. Исходя из того факта, что в вестибулярном нерве все время наблюдается ритмическая импульсная активность, тонизирующая нервные центры, авторы пришли к мысли, что перерезка 8-го нерва кожет изменить состояние определенных нервных центров. Опыты показали, что в условиях пороговых и околопороговых раздражений моторной коры перерезка 8-го нерва приводила к угнетению или выпадению кортикально вызванных движений. Только усиление напряжения раздражающих кору электрических импульсов могло вновь восстановить двигательные реакции. Авторы относят эффект перерезки 8-го нерва за счет снижения влияний ретикулярной субстанции в результате прекращения потока вестибулярных импульсов. При рассмотрении этих опытов важно подчеркнуть то обстоятельство, что при определенных условиях вестибулярные импульсы поддерживают возбудимость корковых клеток, а прекращение вестибулярных импульсов приводит к снижению возбудимости корковых клеток.
К вопросу о локализации коркового конца вестибулярного анализатора
Одной из первых экспериментальных попыток установления корковой проекции вестибулярного аппарата является работа Шпигеля (Spiegel, 1932а, 19326), проведенная на кошках и собаках. Автор использовал в своих опытах способ повышения возбудимости коры больших полушарий стрихнином. После стрихнизации задних отделов эктосильвиевой и супрасиль- виевой извилин раздражение вестибулярного аппарата вращением стало вызывать у животных эпилептические приступы. На основании большого количества контрольных экспериментов Шпигель допустил возможность корковой проекции вестибулярного аппарата в дорсокаудальных участках височной области на границе с париетально-окципитальной областью коры головного мозга. Он также отметил, что при стрихнизации этой области эпилептические приступы могут быть вызваны сильными звуками. Аронсон (Aronson, 1933) в подобной же постановке опытов, предварительно разрушив лабиринт на одной стороне, наблюдал эпилептические конвульсии после вращения как при стрихнизации задних отделов височной области коры на стороне целого лабиринта, так и на стороне, разрушенного. Таким образом, Аронсон показал, что каждый из лабиринтов связан с височными долями коры обоих полушарий.
В последующих исследованиях Шпигеля (1934) ранее полученные факты были подтверждены новой методикой. Адекватное раздражение вестибулярного аппарата вызывало изменение электрической активности в пре¬
Кора больших полушарий и функция вестибулярного аппарата
609
делах задней эктоснльвиевой и супрасильвиевой извилин. Аналогичные данные были получены Жеребцовым (Gerebtzoff, 1940) и С. Н. Хечинаш- вили (1952а). Н. М. Асписов (1946) на основании электрофизиологических исследований делает заключение о том, что лабиринтные импульсы достигают коры головного мозга. Наибольшие изменения электрической активности он отметил в теменной области коры. В опытах Б. Н. Клосов- ского (1951), изучавшего закономерности циркуляции крови в мозгу, при раздражении вестибулярного аппарата у кроликов наблюдалось расширение артерий мягкой мозговой оболочки в лобной и теменной областях коры больших полушарий, а в опытах А. И. Науменко и В. С. Олисова (1959) на кошках — в височной и затылочной областях.
Решающие данные о локализации вестибулярной афферентной проекции в коре были получены методикой вызванных потенциалов. Первые опыты, поставленные Уолцл и Маунткасл (Walzl а. Mountcastle, 1949), показали, что вызванные потенциалы при электрическом раздражении вестибулярного нерва у кошек появляются в передней части супрасильвиевой борозды противоположного полушария. Часть этой зоны активировалась также при раздражении ипсилатерального нерва. К подобному выводу пришел и Кемпинский (Kempinsky, 1951). В отличие от названных авторов, Майкл и Эйде (Mickle a. Ades, 1952, 1954) отметили появление вызванных потенциалов только в контрлатеральном полушарии в передних участках супрасильвиевой и эктосильвиевой извилин. Андерсон и Гернандт (Andersson a. Gernandt, 1954), применив методику стрихнинных аппликаций, отдифференцировали в описанной вестибулярной зоне проекции нервов бокового и верхнего полукружных каналов, а также утрикулярного нерва. Новое подтверждение корковой проекции вестибулярной афферентной системы было получено Массопустом и Дейглем в опытах с раздражением медиального и спинального вестибулярных ядер (Massopust a. Daigle, 1960). Эти авторы зарегистрировали вызванные потенциалы почти в-той же зоне, что и Андерсон и Гернандт. Амплитуда потенциалов, а также зона активации на контрлатеральной стороне были больше, чем на ипсилатераль- ной. Однако в латентных периодах различие было незначительным. Оценивая значение кортикальной проекционной вестибулярной зоны, Руволд и Снайдер (Ruwald a. Snider, 1956) считают, что различение вестибулярных импульсов на кортикальном уровне создает предпосылки для более адекватных приспособлений, чем это возможно достигнуть путем подсознательного приспособления. Однако с таким утверждением не согласуются факты пространственной дезориентации, появления окуложиральных и окулогравитационных иллюзий и т. п. в тех случаях, когда вестибулярные импульсы доходят до сферы сознания человека. Заметим, что Руволд и Снайдер зарегистрировали вызванные потенциалы в описанной ранее вестибулярной корковой проекционной зоне при раздражении «вестибулярных отделов» мозжечка.
Итак, по данным, полученным методикой вызванных потенциалов, корковая проекционная вестибулярная зона находится в передних отделах височной коры, а по данным авторов, работавших другими методиками, — в задних отделах. Экстирпация передней височной коры у собак в наших опытах (Кисляков, 1962) и задней — у кошек в опытах Бома и Гернандта (Bohm a. Gernandt, 1951) в хронических условиях не приводила к нарушению статокинетических функций. Создавалось впечатление, что результаты острых и хронических экспериментов не совпадают. Это противоречие оказалось разрешенным в работе Л. С. Гамбаряна с сотр. (1962) в пользу фактов, полученных методикой вызванных потенциалов. Удаляя передние части эктосильвиевой и супрасильвиевой извилин у 39 Р. Магнус
610
Э. Ш. Айрапетъянц и В. А. Кисляков
собак с перекрестной ампутацией двух конечностей, авторы отметили у них нарушение локомоции и статокинетических функций. При разрушении коры задних участков названных извилин этих нарушений не отмечалось.
Материал по локализации вестибулярной функции в коре головного мозга человека имеется главным образом в клинических и клинико-физиологических исследованиях. В. В. Крамер и О. Г. Агеева-Майкова (1935) описали случай вестибулярного головокружения у больного с опухолью, локализованной в теменно-височной области коры правого полушария, в поле 21 Бродмана. Во время операции раздражение этого участка электрическим током вызывало головокружение. После удаления опухоли головокружение прекратилось. В более поздних работах О. Г. Агеева- Майкова (1946, 1950) отметила большое количество вестибулярных расстройств у больных с ранениями головного мозга при поражениях височной и теменной областей коры. Пенфильд и Эриксон (1949) считают, что «чувство равновесия» у человека связано с полем 22 коры мозга. Фитцджеральд и Холпайс (1942) наблюдали увеличение продолжительности калорического нистагма у 1СЬ больных с локализацией повреждения в височной области коры. В 10 другие случаях с повреждениями лобной, теменной, двигательной и затылочной областей коры усиления нистагма не было выявлено. Поражение височной доли может также сопровождаться спонтанным глазным нистагмом (Шилейко, 1940; Колдобский, 1950).
А. В. Жукович (1952), используя метод электроэнцефалографии, отметила изменения электрической активности при вестибулярных раздражениях в лобно-теменной и височной долях. Однако другие авторы (Жирмунская и Иоселевич, 1951) не считают возможным этим методом решить вопрос о корковой локализации вестибулярной рецепции у человека.
Изучение кортикальной регуляции вестибулярных реакций по методу условных рефлексов
Вопрос о вестибулярном аппарате как анализаторе впервые стал экспериментально разрабатываться по методу условных рефлексов Н. А. Поповым (1920). В его опытах вращение животного в специальной камере (условный раздражитель) подкреплялось вливанием в ротовую полость раствора соляной кислоты. У собак были выработаны условные рефлексы на направление и скорость вращения и соответствующие дифференцировки. В дальнейшем указанная работа была развита и дополнена экспериментами П. И. Одинца (1956) и В. В. Петелиной (1956). Для изучения вестибулярного анализатора у голубей Н. А. Попов применял электрооборонитель- ную методику. Вращение птицы подкреплялось раздражением током кожи лапы. После разрушения полукружных каналов выработанный условный вестибулярный рефлекс исчезал. У других голубей экстирпация больших полушарий также приводила к полному выпадению вестибулярного условного рефлекса (Попов, 1922, 1923, 1926; Попов и Черников, 1923).
В работе XI. И. Одинца (1953, 1956) вращение собак в камере производилось со скоростью 3—8° в сек. Во время вращения на кимографе непрерывно регистрировалось слюноотделение, дыхание, скорость вращения и т. п. Опыты были поставлены на 4 животных. У 2 из них условный рефлекс вырабатывался на начало вращения, а у 2 — на остановку. Для первого проявления условного рефлекса потребовалось 31—71 сочетание ускорения с кислотным подкреплением, а для упрочения рефлекса — 95—191 подкрепление. Дифференцировочным раздражителем являлось
Кора больших полушарий и функция вестибулярного аппарата
61.1
вращение животных без подкрепления в другую сторону. Полная диффе- ренцировка была получена на 11—20-м применении неподкрепляемого вращения. Наряду с дифференцировочным торможением П. И. Одинец исследовал также угашение условных вестибулярных рефлексов, ориентировочных вестибулярных реакций и условное торможение.1
Шпигель и Оппенгеймер (Spiegel а. Oppenheimer, 1939) изучали при помощи метода электрооборонительных условных рефлексов роль полукружных каналов в анализе угловых ускорений и роль отолитов в анализе положения тела в пространстве. В одной серии опытов изменение положения тела в пространстве по отношению к горизонтальной оси служило сигналом оборонительной реакции, в другой серии условным сигналом являлось угловое ускорение при вращении животного вокруг вертикальной оси. После двусторонней лабиринтэктомии условный рефлекс на изменение положения тела сохранился, проявляясь за счет входивших в комплекс кожных и мышечно-суставных раздражений; однако дифференциация направления движения временно нарушилась. Условный рефлекс на угловое ускорение в 2—3°/сек.2 и дифференциация направления вращения в этот период также отсутствовали. Через 2 месяца условный рефлекс стал проявляться на ускорения в 10°/сек.2. По мере увеличения срока с момента лабиринтэктомии стала восстанавливаться условная реакция на меньшие ускорения, но дифференциация направления вращения так и не восстановилась. Условные рефлексы на раздражение вестибулярного аппарата при пищевом подкреплении выработал С. Н. Хечинашвили (1952а). Раздражителем вестибулярного аппарата являлось качание камеры с находящейся в ней собакой. Условный пищевой двигательный рефлекс на сильные толчкообразные качания выработался на 7—15-м сочетаниях. Постепенно уменьшая амплитуду качаний камеры, удалось выработать условный рефлекс на колебания очень малой величины (до 2 мм). У без- лабиринтных собак условный рефлекс образовался только на колебания свыше 10 мм. В дальнейшем В. Г. Тевзадзе (1961) исследовал на собаках дифференцирование не только интенсивности, но и длительности лабиринтных раздражений. Сравнительное изучение условных вестибулярных рефлексов на вращательное и калорическое раздражение при пищевом подкреплении провел Г. В. Бакурадзе (1959). С. Н. Хечинашвили (1956,
1958) исследовал роль мозжечка в условных вестибулярных рефлексах. Как известно, вопрос о значении мозжечка в лабиринтных и шейных рефлексах был предметом специального изучения в лаборатории Магнуса. После полного удаления мозжечка эти рефлексы сохранялись. Сделав вывод о том, что основные пути шейных и лабиринтных рефлексов не проходят через мозжечок, Магнус с сотрудниками не закрыли возможность выявления между мозжечком и другими отделами мозга более тонких взаимоотношений. Современная нейрофизиология представила большое количество доказательств представительства афферентных проекций в мозжечке, а также влияний мозжечка на центры ствола головного мозга, в том числе и вестибулярные. Большой обзор по этому вопросу дан Доу и Мо- руцци (Dow a. Moruzzi, 1958). В опытах С. Н. Хечинашвили после разрушения «вестибулярной» флоккулонодулярной части мозжечка временно выпадали условные двигательно-пшцевые рефлексы, образованные на вестибулярные и вестибулярно-проприоцептивные раздражения. Со временем эти условные рефлексы восстанавливались. Автор рассматривает флок¬
1 Благодаря созданию специальных вращающихся кресел в клинике проф.. А. X. Миньковского и в лаборатории проф. А. В. Лебединского в настоящее время в Советском Союзе появилась возможность исследовать функцию вестибулярного анализатора в широком диапазоне угловых ускорений.
39*
612
Э. Ш. Айрапетъянц и В. А. Кисляков
кулонодулярную часть мозжечка как источник «облегчающих» влияний на центральные механизмы вестибулярных реакций.
Во всех перечисленных работах раздражение вестибулярного аппарата являлось условным сигналом пищевых и оборонительных реакций. Образование этих условных рефлексов служит одним из доказательств кортикальной регуляции вестибулярной функции, обеспечивающей в совокупности с другими системами правильную ориентировку организма в пространстве. Другим доказательством является возможность выработки условных рефлексов на основе безусловных вестибулярных реакций.
Лёвенбах и Гент (Löwenbach a. Gantt, 1940) выработали у собак условный звуковой рефлекс на основе двигательных реакций, возникающих при раздражении вестибулярного аппарата постоянным током. Раздражение вестибулярного аппарата гальваническим током производилось специальными электродами, вставленными в наружный слуховой проход правого и левого уха животного. Один электрод являлся анодом, другой — катодом. Анодическое отклонение головы собаки служило безусловной реакцией. После 8—10 сочетаний условного звукового сигнала (тон) с подкреплением током образовался условный рефлекс — поворот головы в сторону анода только при действии условного сигнала. Укрепление условного рефлекса произошло после 75—100 сочетаний. Другие авторы в качестве подкрепления применяли адекватное раздражение вестибулярного аппарата и вырабатывали условные рефлексы на базе вестибулярных вегетативных (Яроцкий, 1954; Рындина, 1955; Балковская, 1957), двигательных (Кисляков, 1953; Дмитриев, 1958; Кесарева, 1959; Balaceanu-Stolnici с соавт., 1954) и глазодвигательных реакций (Крестовников и Яроцкий, 1938; Миньковский, 1953; Будо, 1959; Фельдман и Фетисова, 1961).
Е. IL Кесарева (1959), пользуясь методикой миотонометрии и электромиографии, исследовала шейные и лабиринтные рефлексы, описанные Магнусом, у здоровых людей, а также изменения этих рефлексов под влиянием занятий спортом. А. С. Дмитриев (1958) провел подробное изучение условных и безусловных лабиринтных рефлексов при вращении и вертикальных перемещениях. Условные рефлексы при вертикальных перемещениях у кроликов описаны в статье П. Тулбаева (1959).
Одной из ваяшейших заслуг Магнуса является доказательство того положения, что функция равновесия на уровне ствола головного мозга обеспечивается взаимодействием целой группы специализированных рефлексов: проприоцептивных, кожных и лабиринтных.1 Исходя из этого, можно было предположить, что на кортикальном уровне функция равновесия обеспечивается комплексом анализаторов, главнейшими элементами которого являются вестибулярный и двигательный анализаторы (Кисляков, 1953; Айрапетъянц, 1955; Айрапетъянц и Кисляков, 1959а, 19596). Функция равновесия изучалась нами (Кисляков, 1953) на собаках по методике условных статокинетических рефлексов. Опыты показали, что после двустороннего разрушения лабиринтов ранее образованные условные статокинетические рефлексы сохраняются, а кроме того, они вырабатываются и у предварительно делабиринтированных животных. В условнорефлекторной деятельности собак, лишенных лабиринтов, наблюдались компенсаторные изменения, выражающиеся в усилении условных двигательных реакций или же в перестройке их структуры. Отмеченные факты приводили к выводу о том, что выпадение вестибулярных импульсов замещается остав¬
1 Следует отметить, что Г. Л. Комендантов описал тонические поясничные рефлесы на глаза и голову, а также шейные и поясничные рефлексы на третье веко у кроликов., которые пополняют перечень рефлексов, выявленных Р. Магнусом ссотр. (Физиол. журн. СССР, т. 31, № 1—2, стр. 62, 1945; т. 34, стр. 449, 1949).
Кора больших полушарий и функция вестибулярного аппарата
613
шимися анализаторными системами, которые продолжают обеспечивать функцию равновесия организма в новых условиях (Кисляков, 1957). В следующей серии опытов у делабиринтированных собак путем энуклеации выключалась зрительная рецепция. После выключения зрения ранее выработанные условные статокинетические рефлексы на кожные раздражители сохранялись, а также оказалось возможным образовать условный рефлекс и на новый интероцептивный раздражитель. Однако следует отметить, что выключение зрения отразилось на соотношении возбудительного и тормозного процессов в коре головного мозга, что проявлялось в растор- маживании дифференцировок и изменении условных положительных рефлексов.
На основании описанных опытов было сделано предположение о том, что после выключения вестибулярной, слуховой и зрительной рецепции условные статокинетические рефлексы осуществляются благодаря функционированию двигательного и других интактных анализаторов (Кисляков, 1956).
Опыты с экстирпацией сенсомоторной области коры головного мозга подтвердили достоверность этого предположения. Оказалось, что одностороннее удаление коры головного мозга в указанной зоне у собак без лабиринтов или без лабиринтов и без глаз вызвало выпадение из условной реакции тонких приспособительных компонентов, имевших важное значение для обеспечения устойчивой позы и равновесия животных при вращении (Кисляков, 1959). В опытах на собаках с двусторонне удаленной двигательной областью коры после разрушения лабиринтов наблюдались более сильные и длительные нарушения локомоции, функции равновесия и статокинетических условных рефлексов по сравнению с контрольными животными, у которых кора головного мозга удалялась в затылочных областях больших полушарий. Компенсация функции вестибулярного аппарата у собак без двигательной коры происходила значительно медленнее по сравнению с собаками без затылочных корковых областей. У последних этот процесс протекал так же, как и у животных с интактной корой (Кисляков, 1960).
В связи со сказанным следует отметить, что в опытах И. В. Филатова (1957) разрушение лабиринтов у собак без мозжечка сопровождалось очень тяжелыми и длительными расстройствами статокинетических функций.
На основании изложенного можно сделать заключение, что вестибулярные симптомы после разрушения лабиринтов проявляются сильнее и исчезают медленнее у животных с предварительным повреждением центральных аппаратов, непосредственно обеспечивающих двигательную функцию организма.
Одним из наиболее спорных вопросов является возможность условно- рефлекторного воспроизведения вестибулярного нистагма. Впервые механизм условного рефлекса был привлечен В. И. Воячеком (1915) для объяснения природы вестибулярного нистагма. Затем А. Н. Крестовников и А. И. Яроцкий (1938), наблюдая за влиянием вращательной тренировки на постнистагм, отметили, что обстановка опыта стала вызывать нистаг- моидные движения глаз и головы подопытных кроликов. М. С. Медве- довский и И. М. Невский (1940) вызвали нистагм глаз у человека внушением в гипнозе. Однако А. И. Яроцкий (1954) сообщил, что воспроизвести условнорефлекторным путем вестибулярный нистагм у человека ему не удалось. В работе же А. X. Миньковского (1953) была показана такая возможность. После опубликования статьи А. X. Миньковского возникла дискуссия, так как Б. С. Будо (1959) не смог подтвердить данных
614
Э• Ш. Айрапетъянц и В. А. Кисляков
А. X. Миньковского. Этот спор был разрешен опытами С. П. Фельдман иЕ. В. Фетисовой (1961), которые у части испытуемых воспроизвели вестибулярный нистагм, иллюзию противовращения и реакцию отклонения рук на условные звуковые и обонятельные раздражители. Эти интересные исследования требуют дальнейшего развития с целью более глубокого понимания полученных феноменов.
Большое значение имеет изучение развития вестибулярной функции в онтогенезе. Исследованию становления безусловных вестибулярных и установочных реакций были посвящены работы А. Я. Галебского (1929), Б. Н. Клосовского и И. Н. Касаткина (1935), Г. А. Павловой (1936), М. А. Минковского (1936), Г. А. Образцовой (1947) и др. Эти вопросы были выдвинуты Магнусом, но не получили в его лаборатории достаточно широкого развития. Благодаря систематическому изучению Г, А. Образцовой установочных рефлексов в эмбриональном и раннем постнатальном периодах онтогенеза вопросы, затронутые Магнусом в последней главе его книги, вылились в монографическое исследование (Образцова, 1961).
Условнорефлекторная регуляция вестибулярных реакций у молодых животных и детей в различной степени описана в работах В. М. Бехтерева и Н. И. Щелованова (1925), Ц. П. Немановой (1935, 1940), Ц. П. Немано- вой и А. М. Левиковой (1940), М. М. Кольцовой (1949), В. А. Трошихина (1952), Г. А. Образцовой (1959). Г. А. Образцова провела сравнительное изучение вестибулярных, слуховых и кожных условных рефлексов в раннем постнатальном онтогенезе у кроликов. Условные рефлексы на вестибулярное раздражение появлялись на 12—15-й день, а упрочивались на 17—19-й день жизни. По данным В. А. Трошихина (1952), вестибулярные условные рефлексы у щенков можно выработать на 10—12-й день жизни.
Значение вестибулярно-кинестетических импульсов для ориентировки животных и человека в пространстве в условиях свободного передвижения рассматривалось в работах И. С. Бериташвили с сотр. (Бериташвили,
1959) и Э. Ш. Айрапетьянца с сотр. (Айрапетьянц, 19606, 1961). На основании опытов с пищевыми двигательными условными рефлексами И. С. Бериташвили считает, что при выключении зрения только вестибулярный аппарат без участия проприоцепции позволяет животному правильно ориентироваться в пространстве, находить место подкрепления кормом и т. п.
Как показали исследования нашей лаборатории (Айрапетьянц с сотр.,
1960) , пространственная ориентировка животных осуществляется на основе взаимодействия анализаторных систем в результате комплексной работы анализаторов. Одним из важнейших факторов, обеспечивающих ориентировку организма в пространстве, является функциональное взаимодействие между зрительным, вестибулярным и двигательным анализаторами. Так, А. С. Батуев (1960), исследуя различные формы пищевых и оборонительных двигательных реакций, показал, что степень участия зрительного анализатора определяется формой двигательной условной реакции (прыжок, побежка по полу, переход по перекладине и т. п.), ее прочностью и биологическим значением, а также расстоянием, которое животному необходимо преодолеть с помощью двигательного акта. Значение вестибулярного анализатора в пространственной ориентировке зависит от формы двигательной реакции, а кожный и двигательный анализаторы, принимающие участие в любом двигательном акте, остаются основным интегрирующим компонентом.
В опытах К. Г. Лебентрау (1960) на обезьянах было показано, что пространственный анализ и синтез, нарушившиеся после поэтапного выключе-
Кора больших полушарий и функция вестибулярного аппарата
615
ни я зрения, обоняния, слуха и вестибулярной рецепции восстанавливаются на основе кожной и мышечной рецепции при введении в эксперимент дополнительных факторов, повышающих тонус коры головного мозга.
Закономерности комплексного регулирования установки тела в пространстве, обеспечения равновесия и нормального расположения частей тела относительно друг друга были открыты Магнусом в пределах стволовой части головного мозга. Исходя из современных данных, можно высказать предположение, что совокупность закономерностей формирования установки тела преобразуется на кортикальном уровне в механизмы дистантной динамической оценки расстояния, размеров и положения объектов и других пространственных факторов. Эти механизмы обеспечивают ■большую «. . .вариантность рефлекторного результата применительно к многообразию текущих условий среды» (А. А. Ухтомский).1 Проблема пространственной ориентировки требует дальнейшей комплексной разработки с широким использованием новейших способов исследования.
ЛИТЕРАТУРА
Агеева-Майкова О. Г. Ото-рино-неврологическая симптоматика при простых и комбинированных ранениях головного мозга. Вестн. оториноларингол., № 3, стр. 36, 1946.
Агеева-Майкова О. Г. Вестибулярные расстройства при различных локализациях травматических нарушений в мозгу. Тр. II съезда оториноларингол. УССР, стр. 275, 1950.
АйрапетьянцЭ. Ш. Новые данные по физиологии внешних и внутренних анализаторов. Журн. высш. нервн. деят., т. 5, в. 5, стр. 644, 1955.
Айрапетьянц Э. Ш. О принципе корковой проекции внутренних сигнализаций. Журн. высш. нервн. деят., т. 10, в. 3, стр. 360, 1960а.
Айрапетьянц Э. Ш. Опыт сравнительного изучения принципа замещаемости в межанализаторной интеграции. В кн. «Вопросы сравнительной физиологии анализаторов», в. 1, стр. 9, изд. ЛГУ, 19606.
АйрапетьянцЭ. Ш. К вопросу о функциональной структуре пространственного анализа. В кн. «Проблемы восприятия пространства и пространственных представлений», изд. АПН РСФСР, стр. 11, М., 1961.
Айрапетьянц Э. Ш., А. С. Батуев, В. А. КисляковиК. Лебент- р а у. О физиологических механизмах пространственного анализа. Физиол. журн. СССР, т. 46, № 8, 908, 1960.
АйрапетьянцЭ. Ш. и Кисляков В. А. К физиологии вестибулярного анализатора. В кн. «Некоторые вопросы современной физиологии», Сб., поев. 70-летию П. С. Купалова, стр. 71, 1959а.
Айрапетьянц Э. III. и В. А. Кисляков. К вопросу о функциональной структуре вестибулярного анализатора. Тр. У съезда отоларингологов СССР, стр. 43, 19596.
Асписов Н. М. Лабиринт и кора. Реферат доклада. Вестн. оториноларингол., № 2, стр. 61, 1946.
Асратян Э. А. Новые данные по пластиночности нервной системы. Бюлл. экспер. биол. и мед., т. 9, № 5, стр. 290, 1940.
Асратян Э. А. Кора больших полушарий и приспособительные явления в поврежденном организме. Сообщ. 5. Опыты с разрушением лабиринтов. Физиол. журн. СССР, т. 33, № 3, 289, 1947.
БакурадзеГ. В. К вопросу о вестибулярном условном рефлексе. Тр. V съезда отоларингол. СССР, стр. 211, Медгиз, 1959.
Б а л к о в с к а я Н. А. Вестибулярные сосудистые рефлексы в норме и при болезни Меньера. Вестн. оториноларингол., № 6, стр. 18, 1957.
Батуев А. С. О функциональной структуре двигательных актов в пространственной ориентировке животных. Автореф. дисс., Л., 1960.
Бахвалова Т. Д. О функциональной связи отолитового аппарата и полукружных каналов. Вестн. оториноларингол., № 5, стр. 35, 1950.
1 А. А. Ухтомский, Собр. соч., т. V, стр. 230, изд. ЛГУ, 1954.
616
Э. Ш. Айрапетъянц и В. А. Кисляков
БериташвилиИ. С. О нервных механизмах пространственной ориентации высших позвоночных животных. Тбилиси, 1959.
Бехтерев В. М. Экспериментальные исследования над круговыми движениями у животных. Еженед. клинич. газета, № 34, стр. 585, 1881.
Бехтерев В. М. Об отправлении полукружных каналов перепончатого лабиринта. Мед. библиотека, № 12, стр. 1, 1882.
Бехтерев В. М. О вынужденных движениях, обнаруживающихся при разрушении мозговой коры. Русск. медиц., № 1, стр. 6; № 3, стр. 54, 1885.
Бехтерев В. М. Основы учения о функциях мозга. СПб., 1905.
Бехтерев В. М.иН. И. Шелованов. К обоснованию генетической рефлексологии. В кн. «Новое в рефлексологии и физиологии нервной системы», стр. 116,
Борнгард А. Материалы для вопроса о значении полукружных каналов ушного- лабиринта. СПб., 1875.
Б у д о Б. С. К вопросу о выработке условных вестибулярных рефлексов, в частности] нистагма. Вести, оториноларингол., № 1, стр. 79, 1959.
Быков К. М. Кора головного мозга и внутренние органы. Медгиз, 1947.
Воячек В. И. Практические методы исследования лабиринтной функции. Вести ушн., нос., горл, болезн., июнь—июль, стр. 384; август—сентябрь, стр. 467,. 1915.
Воячек В. И. (1925). Цит. по К. Л. Хилову, 1952.
В оячек В. И. Современное состояние вопроса о физиологии и клинике вестибулярного аппарата. Шурн. ушн., нос. и горл, болезн., № 3—4, стр. 121, 1927.
ГалебскийА. Я. О вестибулярном нистагме у новорожденных. Журн. ушн.,. нос. и горл, болезн., № 9—10, стр. 623, 1929.
Гамбарян Л. С., Л. С. Гезалян, А. А. Гарибян, С. А. Айрапетян. О роли коркового отдела вестибулярного анализатора в механизмах статокинетической координации. Известия АН Армянской ССР, т. 15, № 4, стр. 59,. 1962.
Гервер А. В. О мозговых центрах движения глаз. СПб., 1899.
Гершуни Г. В. Оценка функционального значения электрических ответов слуховой системы. Физиолог, журн. СССР, т. 48, № 3, стр. 241, 1962.
Дмитриев А. С. Безусловные и условные лабиринтные тонические рефлексы у человека. Автореф. дисс., Минск, 1958.
ЖирмунскаяЕ.А. иФ. М. Иоселевич. Влияние вестибулярного раздражения на электрическую активность коры и базальных областей мозга. Вести., оториноларингол., № 2, стр. 40, 1951.
Жукович А. В. Вестибулярные влияния на электроэнцефалограмму человека.. Сб. тр. Ленингр. н.-и. инст. по болезн. уха, носа, горла и речи, т. 10, стр. 197, 1952.
Калиновская И. Я. и В. Е. Майорчик. К вопросу о корковой регуляции вестибулярных рефлексов человека. Вести, оториноларингол., № 3, стр. 11,. 1952.
Кесарева Е. П. Безусловные и условные тонические рефлексы в их связи с двигательной деятельностью человека. Автореф. дисс., Минск, 1959.
Кисляков В. А. Об условных двигательных рефлексах, образованных на основе- реакций, вызванных вращением животного. Тр. Инст. физиол. им. И. П. Павлова, т. 2, стр. 69, 1953.
Кисляков В. А. Об условных рефлексах на двигательные реакции, возникающие при вращении животного. Дисс., Л., 1953.
Кисляков В. А. Изменение условных статокинетических рефлексов после энуклеации у делабиринтированных собак. Журн. высш. нервн. деят., т. 6, в. 3, стр. 438, 1956.
Кисляков В. А. Изменение условных вестибулярно-двигательных рефлексов* у собак после лабиринтэктомии. Физиол. журн. СССР, т. 43, № 3, стр. 271, 1957.
Кисляков В. А. Изменение установочных реакций у собак при разрушении вестибулярного аппарата и экстирпации двигательной области коры головного- мозга. Тр. Инст. физиол. им. И. П. Павлова АН СССР, т. 8, стр. 39, 1959.
Кисляков В. А. К вопросу о взаимодействии вестибулярного и двигательного анализаторов в функции равновесия. В кн. «Вопросы сравнительной физиологии анализаторов», в. 1, стр. 137, изд. ЛГУ, 1960.
Кисляков В. А. О корковой конпенсации нарушений вестибулярной функции. Тр. Арханг. мед. инст., 1962.
Клосовский Б. Н. Циркуляция крови в мозгу. Медгиз. М., 1951.
Клосовский Б. Н. и И. Н. Касаткин. Развитие вестибулярного нистагма) у человека иживотных. Арх. биол. наук, т. 38, № 3, стр. 711, 1935.
Кора больших полушарий и функция вестибулярного аппарата
617
Колдобский А. М. К вопросу о локальных менинго-энцефалитах. Весты, оториноларингол., № 4, стр. 13, 1950.
Кольцова М. М. О возникновении и развитии второй сигнальной системы у ребенка. Тр. Физиол. инст. им. И. П. Павлова АН СССР, т. 4, стр. 49, 1949.
Крамер В. В. и О. Г. Агеева-Майкова. Существует ли вестибулярное головокружение кортикального происхождения? Советск, невропат., психиатр, и психолог., № 1, стр. 7, 1935.
Красногорский Н. И. О процессе задерживания и локализации кожного и двигательного анализаторов в коре больших полушарий у собак. СПб., 1911.
Крестовников А. Н. и А. И. Яроцкий. К вопросу о взаимоотношении между большими полушариями и вестибулярным аппаратом. Физиол. журн. СССР, т. 25, № 3, стр. 341, 1938.
Лебентрау К. Г. О заместительной функции анализаторов у обезьян. Автореф. дисс., Л., 1960.
МедведовскийМ. С. и И. М. Невский. К вопросу о влиянии коры больших полушарий головного мозга на вестибулярную функцию. В кн. «Вопросы: физиологии и патологии верхних дыхательных путей и уха», стр. 111, Л., 1940.
Минковский М. А. О шейных и лабиринтных рефлексах у человеческого зародыша. В кн.: Проблемы нервной физиологии и поведения, сб., посвящ. И. С. Бе- риташвили, Тифлис, 1936.
Миньковский А. X. Условнорефлекторный лабиринтный нистагм. Вестн. оториноларингол., № 1, стр. 28, 1953.
Науменко А. И. и В. С. О лисов. Влияние раздражения вестибулярного анализатора на мозговое кровообращение. Тр. V съезда оториноларингол. СССР, стр. 177, Медгиз, 1959.
Неманова Ц. П. Наиболее ранние положительные и отрицательные защитные иг пищевые условные рефлексы на вестибулярные раздражения у грудного ребенка. Вопр. педиатр., педол. и охр. матер, и дет., т. 7, № 4, стр. 278, 1935.
Неманова Ц. П. О порядке появления условных рефлексов с разных органов чувств у ребенка первых месяцев жизни. Сообщ. 1. Опыты с рефлексом на дуновение. Вонр. педиатр., недол. и охр. матер, и дет., т. 12, № 2—3, стр. 68.
Неманова Ц. П. и А. М. Левикова. О порядке появления условных рефлексов с разных органов чувств у ребенка первых месяцев. Сообщ. 2. Опыты с рефлексом на освещение. Вопр. педиатр., педол. и охр. матер, и дет., 12, № 5У стр. 202, 1940.
Образцова Г. А. Развитие вестибулярной функции в онтогенезе. Рефер. н.-и. работ за 1946 г. медико-биол. отд. АМН СССР,-в. 1, стр. 117, 1947.
Образцова Г. А. О возникновении вестибулярных условных рефлексов в онтогенезе у кролика. Научн. сообщ. Инст. физиол. им. И. П. Павлова АН СССР, в. 2, стр. 87, 1959.
Образцова Г. А. Формирование вестибулярной функции в онтогенезе. Изд^ АН СССР, М.—Л., 1961.
Образцова Г. А. и 3. Д. Пигарева. Влияние декортикации в раннем онтогенезе на вестибулярный нистагм и активность ферментных систем мозга. Физиол. журн. СССР, т. 43, № 6, стр. 503, 1957.
Одинец П. И. Методика условных рефлексов для исследования вестибулярного- аппарата. Физиол. журн. СССР, т. 39, № 3, стр. 367, 1953.
Одинец П. И. Условные рефлексы с вестибулярного анализатора. Автореф. дисс., Л., 1956.
Павлова Г. А. О развитии двигательных реакций с вестибулярного аппарата у теплокровных (кошек). Арх. биол. наук, т. 44, № 2, стр. 111, 1936.
Петелина В. В. К характеристике условных рефлексов вестибулярного анализатора. Ежегодник ИЭМ, стр. 72, Л., 1956.
Пинчук И. Н. К вопросу о вестибулярном центре в коре больших полушарий головного мозга у животных. Тез. докл. IV науч. сессии. Центр, н.-и. инст. по болез. уха, горла и носа, стр. 44, М., 1948.
Попов Н. А. К учению об анализаторе пространства. Новочеркасск, 1920.
Попов Н. А. Искусственные условные рефлексы у птиц и проблема анализатора пространства. Изв. Бакинск. гос. унив., № 2, стр. 105, 1922.
Попов Н. А. К вопросу о локализации условных рефлексов в центральной нервной системе у птиц. Невролог, записки, № 1, стр. 144, 1923.
Попов Н. А. Новые данные по физиологии пространственного чувства, полученные с применением объективных методов. Журн. ушн., нос. и горл, болезн., №№ 11 — 12, стр. 666, 1926.
Попов Н. А. и А. М. Черников.О дифференцировании внешних раздражений птицами (Дифференцировка звуков и направления перемещения тела в пространстве). Арх. теор. и практ. мед., т. 1, № 1—2, стр. 122, 1923.
Xi XI
'618
Э. Ш. Айрапетъянц и В. А. Кисляков
Рындина А. М. Изучение функции вестибулярного анализатора при помощи кожно-гальванического рефлекса. Автореф. дисс., Л., 1955.
Тевзадзе В. Г. О дифференцировании интенсивности и длительности лабиринтных раздражений. Журн. высш. нервн. деят., т. 11, в. 6, стр. 1099, 1961.
Трошихин В. А. Некоторые итоги изучения высшей нервной деятельности в онтогенезе. Журн. высш. нервн. деят., т. 2, в. 4, стр. 561, 1952.
Тулбаев П. О вестибулярно-двигательных условных рефлексах у нормальных и делабиринтированных кроликов. Тр. Инст. физио л. им. И. П. Павлова АН СССР, т. 8, стр. 207, 1959.
Тюмянцев Н. Ф. Влияние некоторых частей головного мозга на вращательный и поствращательный нистагм. Тр. II Всес. съезда физиол., стр. 130, Л., 1926а.
Тюмянцев Н. Ф. О влиянии некоторых частей головного мозга на вращательный и поствращательный нистагм. Русск. отоларингол., № 6, стр. 409, 19266.
Фельдман С. П. и Е. В. Фетисова. Значение условнорефлекторного компонента в генезе вестибулярных реакций, в частности нистагма. Вести, оторинола- рингол., № 6, стр. 55, 1961.
Филатов И. В. К вопросу о роли вестибулярного аппарата в стато-кинетической рецепторной системе. Сб. тр. Архангельск, гос. мед. инст., в. 15, стр. 22, 1957.
Хечинашвили С. Н. Электрофизиологический анализ некоторых вестибулярных рефлексов. Сб. труд., посвящ. В. В. Воронину, стр. 253, Тбилиси, 1952а.
Хечинашвили С. Н. Вестибулярная функция в свете учения И. П. Павлова. Грузмедизд., Тбилиси, 19526.
Хечинашвили С. Н. К вопросу о роли мозжечка в вестибулярных реакциях. В кн. «Проблемы современной физиологии нервной и мышечной системы», стр. 261, Тбилиси, 1956.
ечинашвилиС. Н. Вестибулярная функция. Тбилиси, 1958. илов К. Л. Кора головного мозга и вестибуло-соматические рефлексы. Вестн. оториноларингол., № 1, стр. 6, 1951.
X и л о в К. Л. Кора головного мозга в функции вестибулярного анализатора. Мед- гиз, 1952.
Черниговский В. Н. Интероцепторы. Медгиз, 1960.
Шилейко Э. И. Изменения слуха, лабиринтной и обонятельной функции при опухолях головного мозга. Вонр. нейрохир., т. 4, № 3, стр. 68, 1940.
Яроцкий А. И. Особенности образования вестибулярных условных рефлексов. Журн. высш. нервн. деят., № 4, в. 3, стр. 381, 1954.
Ades Н. W. Central auditory mechanisms. Handbook of Physiol., sect. I, Neurophysiol., V. 1, p. 585, 1959.
AnderssonA. а. В. E. Gernandt. Cortical projection of vestibular nerve in cat. Acta oto-laryngol., suppl. 116, v. 10, 1954.
Aronson L. The conduction of labyrinthine impulses to the cortex. J. nerv. a. ment. dis., v. 78, № 3, p. 250, 1933.
Balaceanu-Stolnici C. Relatüle aparatului vestibular cu scoarta cerebrala. Nota I, II Bull. Stiint. Acad. R. P. Romane, Sec. med., t. 5, № 2, 1953.
Balaceanu-Stolnici C., S. Falutz, N. Laurian. Relatüle aparatului vestibular cu scoarts cerebrala. Reflexe conditionale labirintice la pisici, Buil. Stiint. Acad. R. P. Romane, Sec. med., t. 6, № 3, 1954.
De Ba renne D. u. A. de Klein. Über vestibulare augenreflexe, v. Greafes Arch. f. Ophtalm., t. 3, № 3—4, s. 374, 1923.
Barley S. H. Central mechanisms of vision. Handbook of Physiol., sect. I, Neurophysiol., v. 1, p. 713, 1959.
Bohm E. a. B. E. Gernandt. Locomotion after Removal of Inferior Colliculi or Cortical Vestibular Projection Area. Acta Physiol. Scanol., v. 23, fasc. 4, p. 320, 1951.
Breuer J. Ueber die Function der Bogengänge des Ohrlabyrinthes, Med. Jahrb., H. I, S. 72, 1874.
Chang H. T. The evoked potencials. Handbook of Physiol., sect. I, Neurophysiol., u. 1, p. 299, 1959.
Dow S. a. G. Moruzzi. The Physiology and pathology of the cerebellum. Minneapolis, 1958.
Ewald R. Physiologische Untersuchungen ueber das Endorgan des Nervus Octavus. Wiesbaden, 1892.
Fitzgerald G. a. C. S. Hallpice. Studies in human Vestibular function.
I. Observation on the directional preponderance («Nystagmusbereitschaft») of caloric nystagmus resulting from cerebral lesions. Brain, v. 65, № 2, p. 115, 1942.
Flourens P. Recherches experimentales sur les propriétés et les fonctions du Systeme nerveux dans les animaux vertebres. Paris, 1842.
Gerebzoff 1V1. A. Reserches sur la projection corticale du labyrinthe. Arch. Int. Physiol, v. 50, № 1, p. 59, 1940.
Säg fl«
Кора больших полушарий и функция вестибулярного аппарата
619
Gern an dt В. Е. Vestibular mechanisms. Handbook of Physiol., sect. I, Neurophysiol., V. 1, p. 549, 1959.
Goltz F., Ueber die physiologische Bewegung der Bogengänge des Ohrlabyrinthes. Pflüg. Arch., Bd. 3, S. 172, 1870.
Grum-Brown A. On the sense of rotation and the anatomy and physiology of the sem circular canals of the internal ear. J. Anat. a. physiol., v. 8, p. 327, 1874.
Ivy A. C. Experimental studies on the brain Stem. II. Comparative study of the relation of the cerebral cortex to vestibular nystagmus. J. Comp. Neurol., v. 31, № 1, p. 1, 1919.
Kempinsky W. H. Cortical projection of vestibular a. facial nerves in cat. J. Neurophysiol., v. 14, № 3, p. 203, 1951.
Kempinsky W. H. a. A. A. Ward. Effect of section of vestibular nerve upon cortically induced movement in cat. J. Neurophysiol., v. 13, № 4, p. 295, 1950. reindler A. Epilepsia. Ed. acad. RPR, 1955 (русск. перевод, Медгиз, M., 1960). öwenbach H. u. W. Gantt. Conditioned vestibular reactions. Ber. über d.
gesamte Biol., Abt. B, t. 119, № 3—4, S. 271, 1940. ach E. Grundlinien der Lehre von den Bewegungsempfindungen. Leipzig, 1875. agnus R. Körperstellung. Berlin, 1924.
a s s о p u s t L. S. a. H. J. D a i gl e. Cortical projection of the medial and spinal vestibular nuclei in the cat. Expt. Neurol., v. 2, № 2, p. 179, 1960.
Mickle W. A. a. H. W. Ades. A composite sensory projection area in the cerebral cortex of the cat. Amer. J. Physiol., v. 170, № 3, p. 682, 1952.
M i c k 1 e W. A. a. H. W. Ades. Rostral projection pathway of the vestibular system. Amer. J. Physiol., v. 176, № 2, p. 243, 1954.
Nakahama H. Functional organization of somatic areas of the cerebral cortex. Internat. Rev. of Neurobiol., p. 187, 1961.
ПенфильдВ. и Г. Эриксон. Эпилепсия и мозговая локализация, Медгиз, М., 1949 (перев. с англ.).
Pice F. Н. The effect of decerebration upon the quick component of labyrinthine nystagmus. Proc. soc. exp. biol. a. med., v. 36, № 4, p. 563, 1917.
Q n i X F. H. The function of the vestibular organ a. the clinical examination of the otolithic apparatus. Laryng. a. otol., v. 40, № 7, p. 425; № 8, p. 444, 1925.
Ruwald M. N. a. R. S. Snider. Projection of vestibular areas of the cerebellum to the cerebrum. J. Comp. Neurol., v. 104, № 3, p. 387, 1956.
Spiegel E. A. The cortical centers of the labyrinth. J < nerv. a. ment. dis., v. 75, № 5, p. 504, 1932a.
Spiegel E. A. Rindenerregung (Auslösung epileptiformer Anfälle) durch Labyrinth- reisung. Versuch einer Localisation der corticalen Labyrinthzentren. Zeitschr. f. d. ges. Neurol, u. Phsychiatr., t. 138, № 2, S. 178, 19326.
Spiegel E. A. Labyrinth and Cortex. The electrencephalogram of the cortex in Stimulation of the labyrinth. Arch, neurol. a. psychiatr., v. 31, № 3, p. 469, 1934.
Spiegel E. A. a. M. Oppenheimer. Conditioned reactions to position and angular acceleration. Amer. J. Physiol., v. 125, № 2, p. 205, 1939.
Walzl E.M.a. V. Mountcastle. The projection of vestibular nerve to cerebral cortex of the cat. Amer. J. Physiol., v. 159, № 3, p. 595, 1949.
ОГЛАВЛЕНИЕ
От редакции ' 1'
Предисловие 5
Первая глава. Общий обзор 7
I. Статические рефлексы 11
A. Положение тела (рефлексы стояния) И
1. Влияние положения головы на позу 11
а) Тонические шейные рефлексы, влияющие на положение конечностей 11
б) Тонические лабиринтные рефлексы, влияющие на мускулатуру
тела 12
в) Взаимодействие’ тонических шейных и лабиринтных рефлексов 12
г) Непрямое влияние лабиринтов на мускулатуру конечностей
посредством шейных рефлексов 13
2. Прямые влияния на положение тела 13
3. Непрямое влияние на положение тела 13
Б. Компенсаторные положения глаз 14
1. Влияние тонических лабиринтных рефлексов на глаза .... 15
2. Влияние тонических шейных рефлексов на глаза 16
3. Совместное действие тонических лабиринтных и шейных рефлексов
на глаза 16
4. Совместное влияние компенсаторных положений глаз и стато-кинетических рефлексов на глаза 17
B. Установочные рефлексы 18
1. Лабиринтные установочные рефлексы головы 18
2. Влияние установочных рефлексов тела на голову 19
3. Шейные установочные рефлексы 19-
4. Влияние установочных рефлексов тела на тело 20
5. Оптические установочные рефлексы 20
II. Стато-кинетические рефлексы 21
A. Вращательные реакции 21
1. Вращательные реакции головы 21
2. Глазные вращательные реакции 21
3. Вращательные реакции конечностей и туловища 22
Б. Реакции на прогрессивные движения 22
1. На голову 22
2. Влияние прогрессивных движений на конечности 22
B. Реакции на движения отдельных частей тела 23
Вторая глава. Переключение 24
Третья глава. Рефлексы положения (Haltung—stehreflexe) 43
I. Влияние тонических рефлексов шеи на конечности 43
II. Влияние тонических лабиринтных рефлексов на конечности 47
Влияние тонических лабиринтных рефлексов на конечности после одностороннего выключения лабиринта 52
III. Общий характер изменений тонуса при рефлексах положения .... 53
IV. Комбинации шейных и лабиринтных рефлексов 63
1. Положение на спине 64
2. Нормальное положение 66
3. Положение на боку 68-
4. Висячее положение головой вниз 70
5. Висячее положение головой вверх 70
Различие шейных и лабиринтных рефлексов 71
V. Тонические шейные и лабиринтные рефлексы у нормальных животных 72 Влияние тонических шейных и лабиринтных рефлексов на дыхание уток 76
VI. Влияние тонических лабиринтных рефлексов на шейные мышцы . . 77
Оглавление 621
VII. Влияние положения тела на движения 81
а) Влияние установки головы на проявление рефлекса в условиях,
когда имеют место видимые изменения длины мышц, обусловленные тоническими шейными и лабиринтными рефлексами . . 82
б) Влияние положения головы на проявление рефлекса, когда тони¬
ческие шейные и лабиринтные рефлексы не вызывают заметных изменений тонуса мышц тела 85
VIII. Тонические шейные и лабиринтные рефлексы у человека 93
Влияние тонических шейных рефлексов на конечности 94
Тонические лабиринтные рефлесы у человека 111
Четвертая глава. Компенсаторные положения глаз 121
I. Тонические лабиринтные рефлексы на глаза 122
Круговые движения 124
Вертикальные отклонения 126
Влияние тонических лабиринтных рефлексов на отдельные мышцы
глаз 128
Положение глаз после двусторонней экстирпации лабиринта ... 134
Положение глаз после односторонней экстирпации лабиринта . . 135
Положение глаз при интактных лабиринтах 137
II. Влияние тонических шейных рефлексов на глаза 141
III. Совместное действие тонических шейных и лабиринтных рефлексов на
глаза 145
IV. Совместное влияние компенсаторных положений глаз и реакций полукружных каналов 150
V. Компенсаторные положения глаз у других млекопитающих и у человека 150
Пятая глава. Установочные рефлексы (Stellreflexe) 158
Техника экстирпации большого мозга у кроликов 159
I. Общее поведение таламических и мезенцефалических животных ... 163
II. Анализ установочных рефлексов 170
Наблюдения в период шока 170
Обзор установочных рефлексов 172
Лабиринтные установочные рефлексы на голову 172
Наблюдения на безлабиринтном таламическом кролике 176
Лабиринтные установочные рефлексы у кашки и у собаки .... 180
Лабиринтные установочные рефлексы у обезьян 181
Переворачивание при свободном падении 183
Установочные рефлексы тела на голову 185
Установочные рефлексы на тело 190
1. Шейные установочные рефлексы 190
2. Установочные рефлексы тела на тело 193
Заключение 197
Расположение центров установочных рефлексов 198
1. Наблюдения за мезенцефалическим кроликом 198
2. Наблюдения за децеребрированными кроликами (мосто-мозжеч¬
ковое животное и мосто-бульбарное животное) 202
а) Лабиринтные установочные рефлексы 203
б) Установочные рефлексы тела на голову 204
в) Установочные рефлексы с тела на тело 206
г) Шейные установочные рефлексы 206
Оптические установочные рефлексы 208
Влияние установочных рефлексов на другие реакции 215
Шестая глава. Последствия односторонней экстирпации лабиринта . . 217
Методика 218
L Последствия односторонней экстирпации лабиринта у кролика . . . 223
Общее поведение кроликов после одностороннего выключения лабиринтов 223
Основной поворот 225
Поворот туловища 227
Лабиринтные установочные рефлексы на голову после односторонней
экстирпации лабиринта 232
Установочные рефлексы тела на голову 239
Тонус конечностей 241
622
Оглавление
Положение тела при сидении 247
Компенсация установки 253
Заключение 255
Движения перекатывания 256
Отклонение глаз и нистагм 274
Зависимость между поворотом головы и отклонением глаз 276
Центральные компенсации 278
'Возникновение сколиоза у растущих животных 279
Паралич симпатического нерва глаза на стороне операции 281
Заключение 281
II. Опыты на морских свинках 284
Поворот головы, шеи и туловища 285
Тонус конечностей 286
Положение тела при сидении 286
Движения перекатывания 289
Глазные симптомы 289
Заключение 290
III. Опыты на кошках 291
Глазные симптомы 292
а) Паралич симпатического нерва 292
б) Отклонение глаз и нистагм 292
Положение головы, шец и туловища 293
Тонус конечностей 296
Положение тела и движение 298
Последствия односторонней утраты лабиринта у кошек без шейных
рефлексов 302
Явления компенсации 305
Заключение 309
IV. Опыты на собаках 310'
Глазные симптомы 312
Положение головы, шеи и туловища 312
Тонус конечностей 314
Компенсация 315
Положение тела и движение 315
Заключение 3J6
V. Опыты на обезьянах 317
Глазные симптомы 318
Положение головы, шеи и туловища 318
Тонус конечностей 319
Явления компенсации 320
Заключение 321
VI. Общий итог 321
Седьмая глава. Последствия двусторонней экстирпации лабиринтов 330
Опыты на кишках • 331
Последствия двусторонней экстирпации лабиринтов у кошек, у которых предварительно перерезались задние шейные корешки 336
Последствия двусторонней экстирпации лабиринтов у обезьян . . 345
Заключение 349
Восьмая глава. Лабиринтные рефлексы на прогрессивные движения 351
A. Лабиринтные рефлексыша прогрессивные движения у морских свинок 353
Б. Кролик 356
B. Кошка 356
Г. Собака 357
Д. Обезьяна 358
Лабиринтные рефлексы на прогрессивные движения после выключения отолитов 359
Может ли аппарат полукружных каналов возбудиться прогрессивными движениями? 362
Девятая глава. Функция отолитов 363
А. Предварительные анатомические замечания 363
Б. Отношение лабиринтных рефлексов положения к положению отолитов 377
1. Тонические лабиринтные рефлексы на мышцы тела 379
2. Лабиринтные установочные рефлексы 383
Оглавление 623
3. Тонические лабиринтные рефлексы на мышцы глаз 387
4. Выводы о функции отолитовых аппаратов 394
В. Изолированное выключение отолитов у морской свинки 396
Г. Состояние возбуждения отолитовых макул 414
Д. Дальнейший анализ деятельности лабиринтов после введения кокаина
Состояние возбуждения гребешков полукружных каналов 419
есятая глава. Центры установки тела 426
I. Лабиринтные установочные рефлексы и установочные рефлексы тела после
удаления мозжечка 429
А. Методы исследования и анатомический контроль 429
1. Оперативные методы 429
2. Анатомический контроль 432
Б. Результаты опытов 461
Первая серия опытов 461
Вторая серия опытов 461
Третья серия опытов 461
Четвертая серия опытов 462
Пятая серия опытов 465
Общий итог 467
II. Положение центров установки тела и лабиринтных рефлексов в мозговом
стволе 472
A. Децеребрационная ригидность и нормальное распределение тонуса 475
Б. Центры рефлексов положения и лабиринтных рефлексов на мускулатуру тела 477
1. Рефлексы положения 477
а) Тонические шейные рефлексы на конечности 478
б) Тонические лабиринтные рефлексы на конечности 479
в) Тонические лабиринтные рефлексы на шейные мышцы . . . 480
г) Односторонняя утрата тонуса после односторонней перерезки
8-го нерва 480
2. Вращательные реакции и поствращательные реакции головы . . 480
3. Реакции на прогрессивные движения 481
B. Центры рефлексов на глаза 482
1. Компенсаторные установки глаз 482
а) Тонические лабиринтные рефлексы на глаза 482
б) Тонические шейные рефлексы на глаза 483
2. Реакции с полукружных каналов на глаза 483
Г. Установочные рефлексы 486
1. Лабиринтные установочные рефлексы 486
2. Установочные рефлексы тела на голову 487
3. Установочные рефлексы тела на тело 487
4. Шейные установочные рефлексы 488
Д. Заключение 489
III. Значение красного ядра для нормального распределения тонуса и для
установочных рефлексов 491
[иннадцатая глава. Действие ядов 509
A. Наркотики жирного ряда (эфир, хлороформ, уретан, паральдегид,
алкоголь) 510
а) Эфир и хлороформ 511
б) Уретан 512
в) Паральдегид 512
г) Алкоголь 513
Б. Стрихнин 516
Общее течение отравления стрихнином 517
Влияние стрихнина на рефлексы положения 521
Влияние стрихнина на рефлексы глаз 526
B. Пикротоксин 532
Общий ход течения отравления пикротоксином у кролика . . . 533
Влияние пикротоксина на децеребрационную ригидность и рефлексы положения 536
Заключение 538
Г. Камфара 539
Повторные подкожные отравления 540
>624
Оглавление
Острое внутривенное отравление камфарой 540
Графическое изображение калорического нистагма 543
Заключение 544
Д. Хеноподиевое масло 545
Изменения лабиринтных рефлексов при остром отравлении . . 545
Графическая запись калорических реакций 547
Влияние хеноподиевого масла на тонические лабиринтные рефлексы на мышцы конечностей у децеребрированных кошек . . . 548
Действие хеноподиевого масла на последствия односторонней
экстирпации лабиринта у кролика и морской свинки .... 549
Заключение 551
Е. Хинин 552
Ж. Хинакетоны 553
3. Никотин 555
а) Действие на глазную мышцу 556
б) Действие на ядра глазных мышц 556
в) Действие на вестибулярную систему (лабиринты и область вестибулярных ядер) • . 557
г) Действие только на область вестибулярных ядер ...... 559
И. Заключение 560
Двенадцатая глава. Установочные рефлексы тела у новорожденных
животных 563
Работы из фармакологического Института государственного университета
в Утрехте 568
Работы из анатомического Института государственного университета в Утрехте 572
Литература 572
Приложение
А. Ф. Самойлов. Работы Р. Магнуса и его лаборатория в Утрехте .... 583
Э. Ш. А йрапетьянциВ. А. Кисляков. Кора больших полушарий
и функция вестибулярного аппарата 604
Рудольф Магнус
УСТАНОВКА ТЕЛА Утверждено к печати
Редколлегией серии «Классики науки» Академии наук СССР
Редактор Издательства 3. А. Васильева Художник М. Н. Свиньина Технический редактор М. Е. Зендель Корректоры Ж. Д. Панкратова, В. А. Пузиков и Г. И. Шер
Сдано в набор 15/Y 1962 г. Подписано к печати 8/VIII 1962 г. РИСО АН СССР 7а—1. Формат бумаги 70Х108Vie- Бум. л. 199/16. Печ. л. ■ 3978=53.5 уел. печ. л.+1 вкл. Уч.-изд. л. 55.56+ +1 вкл. (0.04). Изд. № 1839. Тип. зак. № 680. Тираж 2200.
Цена 4 р. 10 к.
Ленинградское отделение Издательства Академии наук СССР Ленинград, В-164, Менделеевская лин., д. 1
1-я тип. Издательства Академии наук СССР Ленинград, В-34, 9 линия, д. 12