/

































































































































































































































































































Text
АКАДЕМИЯ НА у К СССР
фауна
ссср
ДВУКРЫЛЫЕ
ТОМ
ХУШ
8ЫП.
ИЗДАТЕЛЬСТВО АКАДЕМИИ НАУК СССР
МОСКВА * ЛЕНИНГРАД
ЗООЛОГИЧЕСКИЙ ИНСТИТУТ АКАДЕМИИ НАУК СССР
НОВАЯ СЕРИЯ, № 45
ФАУНА СССР
Главный редактор акад. Е. Н. ПАВЛОВСКИЙ
Редактор А. А. ШТАКЕЛЬБЕРГ
НАСЕКОМЫЕ ДВУКРЫЛЫЕ
Том XVIII, вып. 4
А. С. ЗИМИН
СЕМ. MUSCIDAE. НАСТОЯЩИЕ МУХИ
(ТРИБЫ MUSCINI.STOMOXYDINI)
ИЗДАТЕЛЬСТВО АКАДЕМИИ НАУК СССР
МОСКВА • 195 1 • ЛЕНИНГРАД
Светлой памяти
Алексея Алексеевича
ЗАХВАТКИНА
ПРЕДИСЛОВИЕ
Предпринимая обработку данной группы двукрылых, мы рассма-
тривали ее как первый этап ревизии семейства настоящих мух в целом.
Выбор группы был продиктован не только ее положением в системе
семейства, но и тем огромным значением, которое имеют виды обеих триб
в эпидемиологии и ветеринарии. Последнему вопросу посвящена обшир-
ная литература как научного, так и учебного характера. В ней с доста-
точной убедительностью доказана причастность мух к распространению
возбудителей многих трансмиссивных заболеваний. Если интерес меди-
цинских и ветеринарных кругов еще не так давно ограничивался только
синантропными видами в узком смысле, то сейчас, с расширением наших
знаний в области этиологии некоторых заболеваний, он возрос и рас-
пространился и на формы двукрылых, обитающие вне населенных пунк-
тов. В соответствии с этим повысились требования и к работам по диагно-
стике видов, их экологии и географии.
В семействе настоящих мух разбираемая группа по своему эпидемио-
логическому значению является наиболее существенной и, несмотря на
это, очень слабо изученной.
Основой для систематической обработки послужили автору богатей-
шие коллекции Зоологического института Академии Наук СССР в Ленин-
граде, насчитывающие по данной группе двукрылых более И ООО экзем-
пляров, многочисленные сборы и коллекции других учреждений и от-
дельных энтомологов, а также собственные материалы автора, в частности
обширные личные сборы автора по преимагинальным фазам развития.
Описания морфологии взрослых насекомых и ранних фаз онтогенеза
даются в несколько расширенном виде в связи с трудностью определения
мух этой группы и сильной индивидуальной их изменчивостью. Принята
в расчет и вероятность ошибки при определении новых систематически
близких форм.
Расширение объема глав, касающихся значения видов группы и их
экологии, представлялось нам совершенно необходимым при характе-
ристике столь опасной в медико-санитарном и ветеринарном отношениях
группы, как синантропные двукрылые.
Недостоверность литературных сведений по морфологии личиноч-
ной фазы или краткость существующих описаний сузили возможность
использования этих данных в определительной таблице, тем более, что
последние составлены на основе иных отличительных признаков, чем
это принималось до настоящего времени. Более 80% описанных личиноч-
ных форм получены автором в лабораторных условиях- Часть видов ли-
j *
4
ПРЕДИСЛОВИЕ
чинок за полной невозможностью (по вышеуказанным мотивам) их поме-
щения в определитель описывается в видовых диагнозах.
За исключением небольшого числа форм, не дошедших в работу, мы
даем здесь обзор, в сущности говоря, всей палеарктической фауны по рас-
сматриваемым группам двукрылых. Приводимый для СССР список ви-
дов может считаться полным лишь на данном этапе исследования, т. к.
сборы из малоизвестных в диптерологическом отношении обширных про-
странств Советского Союза и, в частности, из Сибири и Средней Азии
могут обещать еще очень много новых форм.
За предоставление возможности обработки обширных коллекцион-
ных материалов Зоологического института Академии Наук СССР и за
многочисленные указания в работе автор глубоко признателен директору
Института академику Евгению Никаноровичу Павловскому.
С особо теплым чувством и благодарностью автор вспоминает здесь
Александра Александровича Штакельберга, советами, указаниями и по-
мощью которого он пользовался на протяжении почти 25 лет.
Л. Зимин.
СИСТЕМАТИЧЕСКИЙ УКАЗАТЕЛЬ ВИДОВ
Сем. Muscidae
1. Триба Muscini. Настоящие мухи
1. Род Orthellia Rob.-Desv.
1. Подрод Orthellia Rob.-Desv. s. str.
Стр.
1. Orth, (in sp.) caesarion Mg. ................................. 75
2. Orth, (in sp.) cornicina F.................................... 81
3. Orth, (in sp.) pacifica, sp. nova ............................ 85
2. Подрод Pyrelliomima, subgen. nov.
4. Orth. (Pyr.) latipalpis, sp. nova................................................. 87
2. Род Musca L.
1. M. domestica L................................................................ 96
2. M. nebulo F.................................................................. 118
3. M. sorbens Wd................................................................ 114
4. M. albina Wd................................................................. 120
5. M. vitripennis Mg............................................................ 122
б. M. osiris Wd................................................................. 125
7. M. tempestiva Fall........................................................... 128
8. M. lucidula Lw............................................................... 133
9. M. crassirostris Stein....................................................... 135
10. M. autumnalis Deg. ......................................................... 139
11. M. arnica, sp. nova ........................................................ 145
12. M. mesopotamiensis Patt..................................................... 146
13. M. larvipara Portsch........................................................ 147
14. M. convexifrons Thoms....................................................... 154
15. M. lusoria Wd. ............................................................. 156
16. M. xanthomelas Wd........................................................... 157
17. M. ventrosa Wd............................................................... 159
18. M. hervei Vill............................................................... 160
3. Род Pyreilia Rob.-Desv.
1. Подрод Pyreilia s. str.
1. P. (in sp.) cadaverina L................................................... 165
2. P. (in sp.) secunda, sp. nov............................................... 170
3. P. (in sp.) minuta, sp. nov................................................ 171
4. P. (in. sp.) aenea Zett..................................................... 171
2. Подрод Rypellia Mall.
5t P; (R.) semilutea Mall. ..................................................... 174
6
СИСТЕМАТИЧЕСКИЙ УКАЗАТЕЛЬ ВИДОВ
3. Подрод Dasyphorornima, subgen. nov.
6. Р. (D.) pavlovskii, sp. nov.................................................. 175
4. Род Dasyphora Rob.-Desv.
1. D. serena Mg................................................................. 181
2. D. cyanella Mg.............................................................. 183
3. D. eriophthalma Macq........................................................ 186
4. D. saltuum Rd............................................................... 187
5. D. asiatica Zim............................................................ 188
6. D. quadrisetosa, sp. nov. .................................................. 191
7. D. latifrons, sp. nov. ..................................................... 192
8. D. setitibia, sp. nov....................................................... 193
9. D. similis, sp. nov.......................................................... 194
10. D. paraversicolor, sp. nov................................................. 194
11. D. versicolor Mg........................................................... 196
12. D. gussakovskii Zim........................................................ 197
13. D. trichosterna, sp. nov................................................... 198
14. D. pratorum’Mg............................................................. 199
14a.D. pratorum meridionalis, subsp. nov......................................... 203
5. Род Morellia Rob.-Desv.
1. Подрод Morellia s. str.
1. M. (in sp.) hortorum Fall.................................................. 206
2. M. (in sp.) podagrica Lw................................................... 208
3. M. (in sp.) nilotica Lw................................................... 211
4. M. (in sp.) aenescens Rob.-Desv............................................ 211
2. Подрод Dasysterna, subgen. nov.
5. M. (D.) simplex Lw.......................................................... 213
6. M. (D.) simplicissima, sp. nov.............................................. 221
7. M. (D.) hortensia Wd........................................................ 221
6. Род Mesembrina Mg.
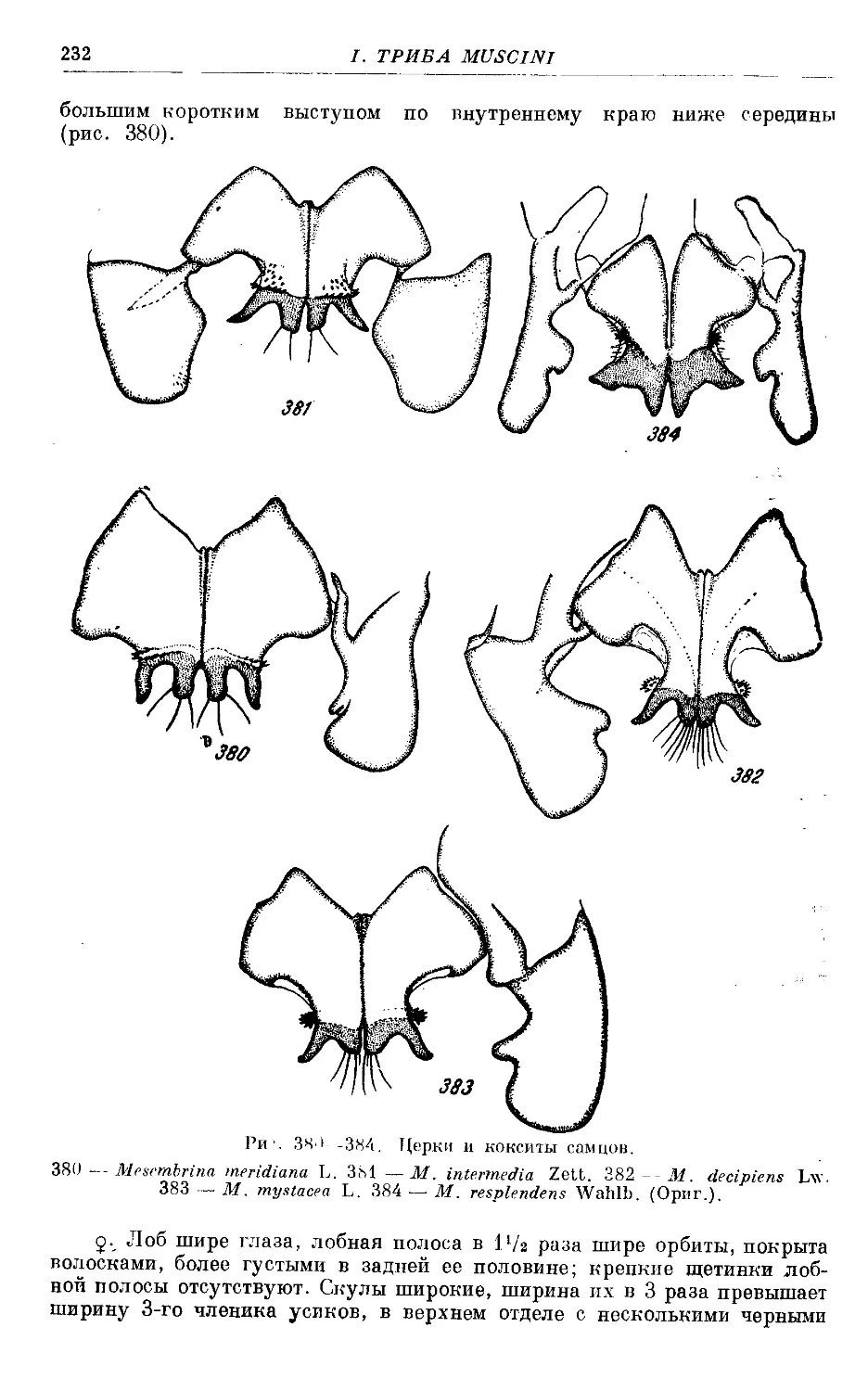
1. M. mystacea L............................................................... 224
2. M. decipiens Lw............................................................. 228
3. M. resplendens Wahlb........................................................ 228
4. M. meridiana L.............................................................. 230
5. M. intermedia Zett.......................................................... 236
6. M. gracilior, sp. nov....................................................... 238
7. M. montana, sp. nov......................................................... 239
2. Триба Stomoxydini. Жигалки.
7. Род Graphomyia Rob.-Desv.
1. G. maculata Scop......................................................... • 242
8. Род Stomoxys Geoffr.
1. St. calcitrans L............................................................ 250
9. Род Haematohia Rob.-Desv.
1. H. stimulans Mg.i........................................................... 259
2. H. atripalpis Bzz............................................................ 265
10. Род Lyperosia Rond.
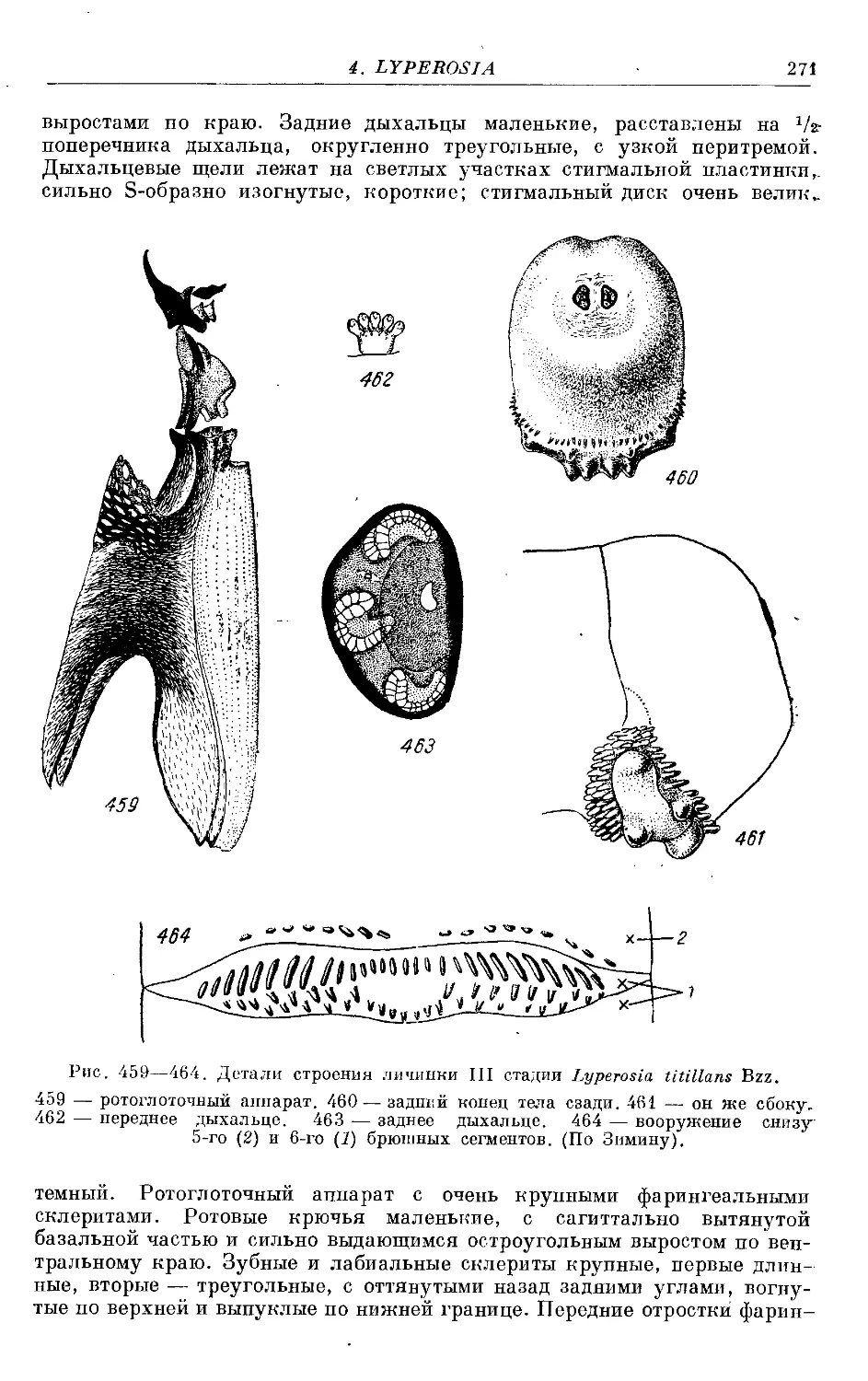
1. L. titillans Bzz............................................................. 269
2. L. minuta Bzz................................................................ 272
3. L. irritans L................................................................ 273
11. Род Acridomyia Stack.
1. A. sacharovi Stack....................................................... • 278
ВВЕДЕНИЕ
МОРФОЛОГИЯ!, ВЗРОСЛОГО НАСЕКОМОГО
Голова (рис. 1) соединена с грудью тонкой, короткой шеей, мяг-
кой у живых насекомых, обеспечивающей голове большую подвижность.
Ширина головы чуть меньше, равна или превышает ширину переднего
отдела груди. Обращенная вперед поверхность головы выпуклая, задняя,
именуемая затылком, то вогнутая, то плоская, то выпуклая в той или иной
степени. Боковые отделы головы заняты фасеточными глазами, у мерт-
вых насекомых красновато-коричневой окраски; они состоят из много-
численных фасеток, размер которых уменьшается сверху вниз, что осо-
бенно отчетливо выражено у самцов многих видов.
Фасеточные глаза видов трибы Muscini имеют округлый,
яйцевидный или эллипсоидный контур; у жигалок глаза удлиненно или
коротко почковидной формы (задняя граница глаз в той или иной степени
вогнутая). Как правило, у самцов фасетки глаз часто крупнее, нежели
у самок того же вида, а сами глаза занимают значительно ббльшую по-
верхность головы, чем у самок. У большинства видов поверхность глаз
лишена волосков; только у Dasyphora, Mesembrina и некоторых видов
Musca и Morellia она покрыта светлыми тонкими волосками; особенно
сильного развития они достигают у высокогорных и многих тропических
форм.
Пространство на верхней стороне головы между фасеточными глазами
у мух носит название лба. Этот термин по существу не верен, т. к. этот
отдел в действительности гомологичен темени насекомых других отрядов.
Мы ради удобства, однако, сохраняем его, т. к. замена этого термина
потребовала бы введения новой терминологии для многих склеритов.
Лоб состоит из нескольких хитиновых склеритов, более или менее
ясно отделенных швами друг от друга. Средняя часть лба образована
так называемой лобной полосой, обычно темноокрашенной
(черной, коричневой, красной). У самцов она значительн > уже, чем
у самок, что объясняется сильным развитием у первых фасеточных глаз,
нередко почти соприкасающихся друг с другом. Передний конец лобной
полосы упирается в поперечное узкое вдавление полулунной формы —
лунку. Она образуется после втягивания внутрь головы лоб-
ного пузыря, функционирующего в момент выхода мухи (Schizo-
phora) из ложного кокона и субстрата, в котором последний находился
(почва и т. п.). Задний конец лобной полосы соприкасается с небольшим
пологим возвышением — глазковым треугольником,
склеритом, несущим три простых глазка. По бокам от глазкового треуголь-
ника располагается с каждой стороны по одному мелкому склериту, не
имеющему резких границ, — теменному треугольнику.
8
ВВЕДЕНИЕ
Между внутренними краями глаз и краями лобной полосы располагаются
скуловые пластинки лба, или орбиты. Они представ-
лены узкими полосами и являются непосредственным продолжением скул.
С к у л И имеют вид узких полос, один край которых примыкает к перед-
ней границе фасеточных глаз, другой, внутренний, — к дуговидному
\ ____________ стш,
Рис. 1. Схема строения головы.
Части г о л о в ы: ус — усики; ар — ариста; фк— фронтокли-
пеус, лицо; щ — щеки; л — лунка; м— медиана; гл — фасеточные
глаза; ор — орбиты; ск— скулы; лги— лицевой дуговидный шов;
т — глазковый (теменной) треугольник с тремя простыми глаз-
ками; «к — вибриссальный киль; лп — лобная полоса. Щетин-
ки: лщ — лобные, или фронтоорбитальные; \глщ— глазковые;
ощ — орбитальные; стщ— стомальные; втщ— внутренние темен-
ные; нтщ — наружные теменные; вибр — вибриссы; рос — заглаз-
ничные. (Ориг.).
лицевому шву. Как орбиты, так и скулы у самцов уже, чем у самок.
Дуговидный лицевой шов, начинаясь вверху над усиками,
спускается вниз от боковых углов лунки по передней поверхности головы,
ограничивая собственно лицо, или фронтоклипеус. В верх-
нем отделе лица прикреплена пара усиков, или антенн. Нижняя граница
лица служит одновременно передним и нижним краем головы. На нижней
стороне головы выступает хоботок. Боковые края лица валикообраэно
приподняты и носят название вибриссаль ны х килей. Ниж-
ние концы килей несколько расширены, образуя утолщенные в и б р и с-
МОРФОЛОГИЯ ВЗРОСЛОГО НАСЕКОМОГО
сальные углы, несущие крепкие, обращенные вершинами друг
к другу щетинки. У некоторых видов лицо обладает продольным, сре-
динным возвышением в виде валика или киля, который, начинаясь между
основанием усиков, спускается вниз и, достигнув средины или нижней
трети лицевого щитка, сходит на нет. Наиболее сильного развития сре-
динный киль достигает у Graphomyia, где его верхний конец упирается
в лунку, широко раздвигая основание усиков. У большинства представи-
телей группы срединный киль отсутствует или развит слабо. Область
головы между нижним краем глаза и боковым нижним краем головы носит
название щек.
Передней границей щек служит легкое вдавление треугольной или
полулунной формы, так называемая медиана (.и), начинающаяся
вверху у нижнего конца лицевого дуговидного шва, и ограниченная
спереди вибриссальным килем, сверху — передним краем скулы, а сзади—
щекой. Задний отдел щеки переходит непосредственно в затылок, кото-
рый занимает всю заднюю поверхность головы.
Обращенная вниз сторона головы занята крупным отверстием,
к краям которого прикреплен хоботок с максиллярными щупальцами.
Придатки головы. В верхней части фронтоклипеуса при-
членены усики (антенны), являющиеся органами обоняния.
Усики — парные образования, симметрично расположенные по бокам
от средней вертикальной линии фронтоклипеуса. Каждый усик состоит
из 3 члеников, также подвижно соединенных друг с другом.
1-й (основной) членик усиков всегда небольших размеров. Он имеет
форму узкого колечка. 2-й — конусовидный, длиннее 1-го; по дорзаль-
ной стороне он разделен вдоль узкой, глубокой бороздкой; поверхность
его покрыта более или менее развитыми мелкими волосками и несет
сверху 1—3 щетинки. 3-й членик самый длинный, несколько сплюснутый
с боков; имеет форму вытянутого прямоугольника со сглаженными уг-
лами или почти квадрата. В спокойном состоянии 3-й членик прижат
к фронтоклипеусу. От основания 3-го членика отходит его придаток,
а р и с т а, которая состоит из двух или трех члеников и снабжена по
верхней и нижней (у большинства видов группы) или только верхней
(некоторые жигалки) стороне длинными волосками, придающими аристе
вид перышка. Среди Muscini и Stomoxydini нашей фауны известна лишь
одна форма (Acridomyia sacharovi), у которой длинные волоски на
аристе отсутствуют. Основание аристы, как правило, утолщено на боль-
шем или меньшем протяжении. Поверхность 3-го членика усиков покрыта
мелкими волосками, часть которых связана с нервными клетками и слу-
жит сензорными органами. Вместе с тем 3-й членик обладает обонятель-
ными ямками, расположенными обычно в основной части 3-го членика
книзу от места прикрепления аристы (рис. 3). Конфигурация усиков,
соотношение длины члеников, а также характер аристы, длина ее волос-
ков и тип распределения последних имеют значение в диагностике группы.
Хоботок (рис. 2) подвижно соединен при помощи мембраны
с нижними краями головной капсулы. В нем различают две основных
части, в совокупности составляющих коленообразно согнутый придаток.
Верхняя часть, или собственно хоботок (rostrum), конусовид-
ная, соединенная с головной капсулой своим расширенным основным
отделом, а нижним, узким дистальным, — с вершинной частью хоботка —
сосальцем (haustellum). В спокойном состоянии сосальце направ-
лено вершинным концом вперед, занимая, следовательно, горизонтальное-
положение. Основная часть хоботка несет пару максиллярных
10
ВВЕДЕНИЕ
щупалец; дистальная часть сосальца — две широких, мягких
л о н а с т и (labellae). Основной отдел хоботка состоит из внутренней
хитиновой рамы, служащей в качестве опорного элемента — fulcrum,
и наружной перепонки, образующей наружные стенки хоботка. Рама
в свою очередь слагается из двух вертикально стоящих хитиновых пла-
стинок, соединенных друг с другом системой хитиновых поперечных пере-
мычек. Задняя — более широкая, образует заднюю стенку fulcrum и
идет по всей длине последнего. Одновременно она служит и задней стенкой
глотки. Параллельно этой стенке располагается такая же длинная хити-
новая передняя пластинка, пред-
ставляющая в то же время переднюю
стенку пищевода.
Рис. 2. Схема строения хоботка
Musca.
а — аподема; р — ротовая полость;
са—'слюнная ампула; дек — дис-
кальный склерит с престомальными
зубцами; елпр — проток слюнных
желез; ф— фурка; фк— фулькрум
(по — передние; во — задние его от-
ростки); пел — подглоточник; сл—
сосательные лопасти; впл— верхняя
пластинка нижней губы; вг -|- нгл —
верхняя губа + надглоточник; п —
подбородок; пгц— пищевод (по —
передняя, зо— задняя стенки); гц —
максиллярные щупальцы; вп — ди-
стальные выступы подбородка; лт —
ложнотрахеи; т — торма.
новая лопасть, t о г m а, иногда
Рис. 3. Вертикальный разрез
через сензорную ямку на уси-
ках.
о 56 — обонятельный бугорок;
н — нервные клетки. (По Ли-
берману).
Верхний край боковых пластинок
fulcrum вогнутый, вследствие чего
образуются задне-верхние и перед-
не-верхние его отростки. Передние
отростки соединяются поперечной
перемычкой. В верхнем отделе на-
ружной перепонки по передней сто-
роне хоботка заключена тонкая хити-
разделенная вдоль на две обособленные
боковые части. Под наружными покровами боковых отделов хоботка за-
ключено с каждой стороны последнего по изогнутому, тонкому, длинному
хитиновому тяжу — аподеме, имеющей короткое расширение на
верхнем и короткий заостренный отросток на нижнем конце. Он достигает
более четкого выражения у некоторых Acalyptratae. К нижнему концу
аподемы подходит основание подглоточника и верхней губы, соединяясь
с ее концом при помощи перепонки.
В месте соединения хоботка и сосальца помещается мелкое полое
внутри хитиновое образование, веретеновидной или грушевидной
МОРФОЛОГИЯ ВЗРОСЛОГО НАСЕКОМОГО
11
формы — г и о ид (hyoid), представляющее собой ротовую полость. По
заднему краю гиоид соединен короткой мембраной с нижними краями
задней и передней стенки глотки, передним верхним краем — с основа-
нием нижней поверхности надглоточника, передним нижним — с основа-
нием верхней стороны подглоточника. В нижнем отделе сосальца раз-
личают: 1) верхнюю губу и 2) надглоточник, сращенные на вершине
и представляющие собой как бы одно образование (labrum-(-epipharynx),
3) подглоточник, 4) нижнюю губу, состоящую из верхней хитиновой
узкой пластинки — желобка — и широкой нижней, являющейся одно-
временно нижней наружной поверхностью сосальца и носящей название
подбородка. На дистальном конце нижняя губа несет две сосательных
лопасти.
Верхняя губа -(-надглоточник, как сказано, верх-
ним отделом проксимальной части соединены с нижним концом ападемы,
нижней стороной основания — с верхней стенкой гиоида (ротовой по-
лости). Он представлен вытянутым узким склеритом, заостренным на
конце; верхние и боковые его стенки ± выпуклые (в сечении представлены
полукругом или дугой), нижняя стенка — вогнутая, образующая жело-
бок, по которому проходит всасываемая мухой пища.
К нижним краям labrum-(-epipharynx примыкает подглоточ-
ник, а также сагиттально вытянутый и заостренный на вершине тон-
кий склерит. Он закрывает пищепроводный канал хоботка снизу, являясь
нижней внутренней стенкой последнего. Верхняя сторона основания
подглоточника сращена с нижним краем дистального конца ротовой по-
лости. Задний нижний конец (основание) подглоточника несколько вы-
ступает и принимает проток слюнных желез, проходящий позади фуль-
крума и далее через основание подглоточника по всей его длине в виде
тонкого канальца, открывающегося маленьким отверстием на его вершине.
Верхняя пластинка нижней губы — желобок — представлена длин-
ным хитиновым склеритом, идущим от основания сосальца до места при-
крепления сосательных лопастей по верхней стороне последнего. Верх-
няя желобовидно вогнутая сторона пластинки является вместилищем для
верхней губы с надглоточником и подглоточника. Боковые загнутые вверх
и вбок края желобка переходят в наружную перепонку хоботка, соединя-
ющую верхнюю пластинку нижней губы с краями ее нижней хитиновой
пластинки — подбородком. Основание желобка соединено с нижней
стороной основания подглоточника и дистальным концом хоботка. Вер-
шина, обычно более или менее выемчатая посредине, сочленяется при
посредстве дискального склерита с сосательными лопастями.
Подбородок шире верхней пластинки нижней губы, он обра-
зует нижнюю сторону сосальца и своими боковыми краями, загибаясь
на боковые отделы последнего, формирует боковые его стенки. У жига-
лок подбородок фактически образует почти всю наружную поверхность
сосальца. В этом случае мембрана, соединяющая края верхней (же-
лобка) и нижней (подбородка) пластинок нижней губы, сведена до мини-
мума. Задний край подбородка, не глубоко вогнутый или выпуклый,
соединен с задней мембранозной стенкой хоботка. Передний край б. м. глу-
боко выемчатый посредине и с выступами по бокам от нее. Выступы перед-
него края подбородка сочленены с подковообразным по форме, поперечно
расположенным хитиновым склеритом, носящим название вилки. Свобод-
ные концы вилки направлены вбок, вперед и вверх, охватывая основание
сосательных лопастей, с наружной перепончатой стенкой которых сращен
передний край вилки.
12
ВВЕДЕНИЕ
Рис. 4. Типы престомальных зубцов у кро-
волижущих и кровососущих видов Musca.
Кроволижущие: а —М. senior-
whitei Ptt.; б—М. domestica L.; в —
М. conducens. Кровососущие: е —
М. inferior St.; д — М. mesopotamiensis
Ptt.; е — М. crassirostris St. (По Пэттону).
Сосательные лопасти представляют собою производные
нижней губы. В них различают внутреннюю и наружную поверхности,
расположенные по бокам от пищепроводного канала и образующие своими
основаниями так называемый praestomum. Укрепляющим обра-
зованием для последнего служит дискальный склерит, боковые концы
которого, (или его боковые части в случае, если склерит раздвоен) распо-
лагаются в основании сосательных лопастей по бокам praestomum. Сними
соединены хитиновые мелкие склериты — предротовые зубцы
(рис. 4), при помощи которых осуществляется прокол тканей животного
или соскабливание мухой твердых питательных частиц. У кровососущих
форм (жигалок) престомальные зубцы достигают крупных размеров и
большой крепости. Нередко у одного и того же вида самцы снабжены
слабыми, а самки крепкими зуб-
цами. В роде Acridomyia самцы
вовсе лишены этого приспособле-
ния в связи с выключенной у них
функцией питания (Рукавишников,
1930). Не имеют крепких престо-
мальных зубцов и многие расти-
тельноядные мухи Graphomyia,
Mesembrina. У Musca domestica пре-
стомальные зубцы идут в несколь-
ко рядов, из которых некоторые,
развитые всегда слабее прочих,
прикреплены непосредственно к
внутренней перепонке сосательных
лопастей. На рис. 4 указаны раз-
личные типы престомальных зуб-
цов у кровососущих и кроволижу-
щих видов рода Musca. Нередко
каждый зубец в свою очередь бы-
вает зазубрен по свободному краю
или несет дополнительные зуб-
чики на наружной стороне.
Сосательные лопасти с внут-
ренней поверхности имеют тон-
кие продольные углубления — ка-
нальцы, именуемые псевдо-
трахеями. В поперечном сечении они представляются округлыми или
грушевидными. Внутренние стенки псевдотрахей по всей своейдлине укре-
плены тонкими склеротизованными полукольцами, концы которых совпа-
дают с наружными краями канальцев. У видов рода Musca каждое полу-
кольцо имеет один дуговидно раздвоенный, другой прямой конец. На
каждом крае канальца простые и раздвоенные концы хитиновых колец
перемежаются (рис. 5). При сокращении просвета псевдотрахей, противо-
положные концы хитиновых полуколец соприкасаются друг с другом.
Сензорные образования (исключая максиллярные щупальцы) рас-
полагаются на различных частях сосальца. Так, сосательные лопасти
имеют (Musca) на внутренней стороне сосцевидные выросты, представляю-
щие собою мелкие папиллы, пронизанные посредине тонким канальцем,
проходящим от вершины выроста внутрь и оканчивающимся груше-
видным комплексом сензорных клеток. Помимо того, внутренняя поверх-
ность лопастей может обладать мелкими плоскими волосками на 'псевдо-
МОРФОЛОГИЯ ВЗРОСЛОГО НАСЕКОМОГО
13
Рис. 5. Схема-
расположени я
хитиновых ко-
лец внутрилож-
нотрахеи на
сосательнойло-
пасти хоботка.
(По Хьюитту).
е н н ы х щ е-
трахеях. Они представляются в виде 2—3 мелких полых внутри щетино-
чек, выходящих из общего цилиндрического основания, связанного с нерв-
ными клетками. Им, как и сосцевидным выростам и волоскам на дорзаль-
ной стороне сосательных лопастей, приписывают функции вкусовых орга-
нов. Наружная сторона сосательных лопастей снабжена щетинистыми во-
лосками с проходящим внутри к нервным клеткам тонким канальцем.
Нервные клетки собраны у комнатной мухи группами по 6 клеток, состав-
ляющих в совокупности грушевидной формы образование, вершиной
своей соединенное с вкусовым волоском. Те же функции, как предпола-
гают, несут мелкие папиллы на нижней поверхности верхней губы и
волоски на внутренней поверхности передней стенки глотки. Сензорные
образования сосательных лопастей видны на рис. 6.
Щет инки головы (рис. 1) не достигают у Muscinae сильного
развития, за исключением разве некоторых групп, как вибриссы или вну-
тренние теменные щетинки. Как правило, на ногах у самцов
щетинки длиннее и толще, чем у самок. Редукция щетинок
особенно далеко зашла у рода Acridomyia (Stomoxydini).
Глазковый треугольник снабжен парой выделяю-
щихся по длине глазковых щетинок, или
щетинистых волосков, направленных вершинами вперед
и наружу; у рода Acridomyia эти щетинки едва разли-
чимы. На границе орбит и затылка расположены две
или одна пара щетинок — внутренние и на-
ружные теменные, причем первые из них обычно
развиты сильнее вторых. Наружные теменные щетин-
ки развиты лучше у самок, у самцов они часто имеют
характер волосков. У самцов Graphomyia остаются не-
доразвитыми и внутренние теменные, не превышающие
здесь длины заглазничных ресничек. Заглазничные
реснички идут правильным рядом параллельно зад-
нему краю глаз. Вблизи средней продольной линии го-
ловы, тотчас за глазковым треугольником, помещаются
две тонких, сближенных своими основаниями затем
тинки; в редких случаях (Acridomyia) они не развиты.
Лобная полоса у большинства видов группы лишена щетинок; лишь
у видов родов Mesembrina бывает развита одна пара в заднем отделе лоб-
ной полосы. Поверхность лобной полосы может быть голой или в большей
или меньшей степени опушенной (Mesembrina, Graphomyia). В последнем
случае волоски наиболее густо располагаются в боковых отделах полосы.
На орбитах или скуловых пластинках лба имеются лобные
или фронтоорбитальные щетинки. Расположенные на
внутренних краях орбит, они обращены своими вершинами навстречу
друг другу, как бы нависая над лобной полосой, а при узком лбе — пере-
крещиваясь своими концами. У самок некоторых видов может присут-
ствовать второй, добавочный ряд лобных щетинок в заднем отделе орбит.
Длина и толщина лобных щетинок убывает спереди назад. Ряды лобных
щетинок впереди заканчиваются на уровне прикрепления усиков, не пере-
ходя на скулы. Между рядом лобных щетинок и внутренним краем глаза
могут присутствовать орбитальные щетинки в количестве 1—
3 пар с каждой стороны. Для видов рода Morellia характерно наличие
сравнительно крепких орбитальных щетинок у обоих полов. У рода
Dasyphora и Pyreilia орбитальные щетинки развиты только у самок; то же
«амое имеет место и у представителей трибы Stomoxydini. Крайнюю сте-
14
ВВЕДЕНИЕ
пень редукции щетинок этой группы, когда они отсутствуют у обоих
полов, можно наблюдать у видов Orthellia и некоторых видов рода Musca.
Орбитальные щетинки, как правило, направлены вершинами вперед; из
известных нам в натуре родов лишь у Mesembrina орбитальные щетинки
обращены вершинами назад. Как исключение (Acridomyia), орбиты могут
быть вовсе без волосков. В большинстве же случаев они в той или иной
мере опушены.
Скулы на всем протяжении от лобных щетинок до нижней своей гра-
ницы обычно не имеют волосяного покрова. Только у Mesembrina meridi-
Рис. 7. Щетинки на верхней стороне
груди (схема).
акр — акростихальные; б— боковые; в—
вершинные; осн — основные; дц — дорзо-
центральные; диск — дискальные; пл —
плечевые; иа — интраалярные; зкр— за-
крыловые; зпл — заплечевые; пкр — пред-
крыловые; про — предосновные; нпл —
нотоплевральные; прш — предшовные;
пш — поперечный шов среднеспинки;
икр — надкрыловые; гц — щиток. (Ориг.),
Рис. 6. Поперечный разрез через
сосательную лопасть хоботка ком-
натной мухи.
<к — сензорные клетки; гк — гипо-
дермальные клетки; ев — сензорный
волосок; пт—псевдотрахеи на
нижней стороне лопасти; т — тра-
хеи. (По Хьюитту).
апа в вершинной трети скул присутствует группа щетинистых волосков.
Щеки и нижняя половина затылка покрыты то длинными, тонкими, то
короткими, густыми волосками. Верхний отдел затылка в укороченных
жестких волосках или с короткими щетиночками.
Вибриссальные углы имеют по одной крепкой саблевидно согнутой
щетинке — вибриссе. Вверх по вибриссальным килям от указанных
угловых вибрисс поднимаются в большинстве случаев несколькими не-
правильными рядами мелкие щетиночки, обращенные вершинами вниз.
В трибе Muscini ряды мелких щетиночек поднимаются до половины вы-
соты вибриссальных килей, тогда как у Stomoxydini они .развиты лишь
в нижней четверти последних. От угловых вибрисс назад следует ряд
ротовых щетинок, не во всех группах хорошо развитых.
МОРФОЛОГИЯ ВЗРОСЛОГО НАСЕКОМОГО
15
В грудном отделе (рис. 7, 8, 9) наиболее мощного развития достигает
средний (второй) сегмент — среднегрудь; ему принадлежит вся
верхняя и большая часть плевральных отделов груди. Передний грудной
сегмент образует поверхность груди, обращенную к голове. На боковую
сторону груди заходят лишь его узкие, вытянутые в дорзовентральном
направлении склериты —п роплевры, позади которых располагается
передняя пара дыхалец и плечевые бугорки. Дорзальная поверхность-
груди, как сказано, образована среднегрудным сегментом. При рассма-
тривании сверху (рис. 7, 8) дорзальная поверхность груди представляется
Рис. 8. Схема строения груди (вид сбоку).
Части груди: окр — основание крыла; mlt т2, т3 — тазики 1-й, 2-й, 3-й
пары ног; эт3 — эпимерит заднегруди; эс3 — эпистернит заднегруди; дпт —
дорзальная часть плевротергита; пл — плечевой бугорок; ж— жужжальце;
гпл—гипоплевра; мп л— мезоплевра; мф — мезофрагма; срсп — среднеспинка;
нпл — нотоплевра; ш— шов; зб— заднекрыловый бугорок; прпл — проплевра;
пск — постскутеллум; ппл — птероплевра; и д2 — переднее и заднее груд-
ные дыхальцы; щ — щиток; сквр — супрасквамальное ребро; ч— грудная че-
шуйка (вершинный ее отдел удален); ст. пл— стерноплевра; cj — 1-и стернит
брюшка; nij+2 — тергиты первый и второй; впт— вентральная часть плевро-
тергита. Волоски: гплв — гипоплевральные; пвгп — передне-верхние гипо-
плевральные; прпв — проплевральные. Щетинки, места прикрепления которых
отмечены светлыми кружками, именуются по несущему их склериту груди.
(Ориг.).
в виде пятиугольного вытянутого, слабо выпуклого склерита, рассечен-
ного бороздками на несколько отделов: плечевые и закрыл о-
вые бугорки, среднеспинку и щиток.
Среднеспинка в свою очередь разделяется поперечным швом
на передний и задний отделы (praescutum и scutum). Поперечный шов
спинки своими боковыми отделами упирается в сагиттально вытянутые,
неправильно треугольные участки, лежащие на границе среднеспинки-
16
ВВЕДЕНИЕ
и плевр, — нотоплевры, ниже которых располагается крупный
прямоугольный склерит — мезоплевра. Последняя является дор-
зальным эпистернитом среднегрудного сегмента. Нижний край мезоплевры
«ращен со стерноплеврами — треугольными склеритами, раз-
деляющими основания передней и средней пары ног. Стерноплевры пред-
оставляют собою вентральный эпистернит среднегруди. Стерноплевры
своими нижними отделами соприкасаются друг с другом и с задним кон-
цом переднегруди; последняя представляет собою стернит переднегруд-
ного кольца, расположенный между основаниями передних ног и имеющий
вид узкой или в той или иной мере расширенной на концах пластинки
(рис. 275). Передний отдел пластинки выпуклый, задний — вогнутый.
У переднего конца, кроме того, помещается небольшой склерит,
отвечающий престерниту.
Верхним отделом своего заднего края мезоплевры с помощью мел-
ких склеритов (parapterum 1, 2) соединены с основанием крыла, а его
нижней частью соприкасаются с дорзальным эпимеритом среднегруди или
птероплеврой. Снизу птероплевры граничат с задним верхним
отделом стерноплевр и передне-верхним углом гипоплевр; по-
следние представляют собою вентральный эпимерит среднегрудного сег-
мента. Задне-верхний угол гипоплевр подходит к заднему грудному
дыхальцу, а их верхний край упирается в вентральный отдел п л е в р о-
т е р г и т а. Нижний край гипоплевр соприкасается со склеритами задне-
груди, а задний их край — с эпистернитом и эпимеритом заднегруд-
ного кольца. Границы этого отдела плевральной области не всегда отчет-
ливы. Заднегрудные стерниты представлены узкой полоской, проходя-
щей вдоль верхней и передней границы тазика III пары ног, с которыми
они соединяются перепонкой.
Задний отдел среднеспинки — щиток — имеет треугольную или
полукруглую форму. Его верхняя сторона слабо выпуклая, реже — плос-
кая. Снизу к щитку подходит своим верхним краем медиотергит,
состоящий из трех частей: postscutellum, мезофрагмы
и боковой — плевротергита. Плевротергит не всегда ясным
швом или вдавлением делится на дорзальный и вентральный отделы;
первый лежит непосредственно под грудной чешуйкой, второй, именуе-
мый иногда метаплеврой, выпуклый и покрыт нежными, корот-
кими волосками.
Позади заднего дыхальца причленены к груди жужжальца,
мелкие булавовидные образования, в которых различают основание,
стебелек, или ножку, и булаву. В жужжальцах располагаются сен-
зорные органы.
В некоторых участках грудного отдела хитиновые склериты сра-
щены с соседними частями наружного склерита неподвижно. Это отчет-
ливо выражено в области дорзальных и плевральных склеритов передне-
грудного и заднегрудного кольца. Части среднегрудного сегмента
соединены преимущественно подвижно плотными эластичными перепон-
ками, обеспечивающими некоторое перемещение склеритов относительно
друг друга при работе грудных мышц. Ряд склеритов грудного отдела
образует выросты внутрь грудной полости. Внутренние выросты служат
местом прикрепления многих мышц груди, управляющих движением
крыльев и ног.
В разных отделах хитинового скелета груди можно обнаружить
небольшие мембранозные участки (под грудной чешуйкой вблизи
щитка, в области дыхалец).
МОРФОЛОГИЯ ВЗРОСЛОГО НАСЕКОМОГО
17
Грудной отдел покрыт волосками и щетинками, собранными
группами, рядами или располагающимися поодиночке. На среднеспинке
щетинки распределяются симметрично по обе стороны от средней про-
дольной ее линии. Ближайшими к продольной средней линии щетинками
являются акростихальные. Они представлены двумя сближен-
ными рядами, начинающимися от передней границы спинки и заканчи-
вающимися перед щитком. Нередко передние акростихальные щетинки
бывают неразвиты (Musca lucidula, Mesembrina и др.), почему их ряды
оказываются неполными. В некоторых случаях исчезают щетинки в сред-
нем отделе спинки (Orthellia, Mesembrina).
По отношению к поперечному шву спинки акростихальные щетинки
располагаются в числе 1—2—3 пар (иногда они отсутствуют вовсе) впереди
шва и 1—2—3—4 пар позади него; в редких случаях и зашовные акрости-
хальные щетинки могут отсутствовать.
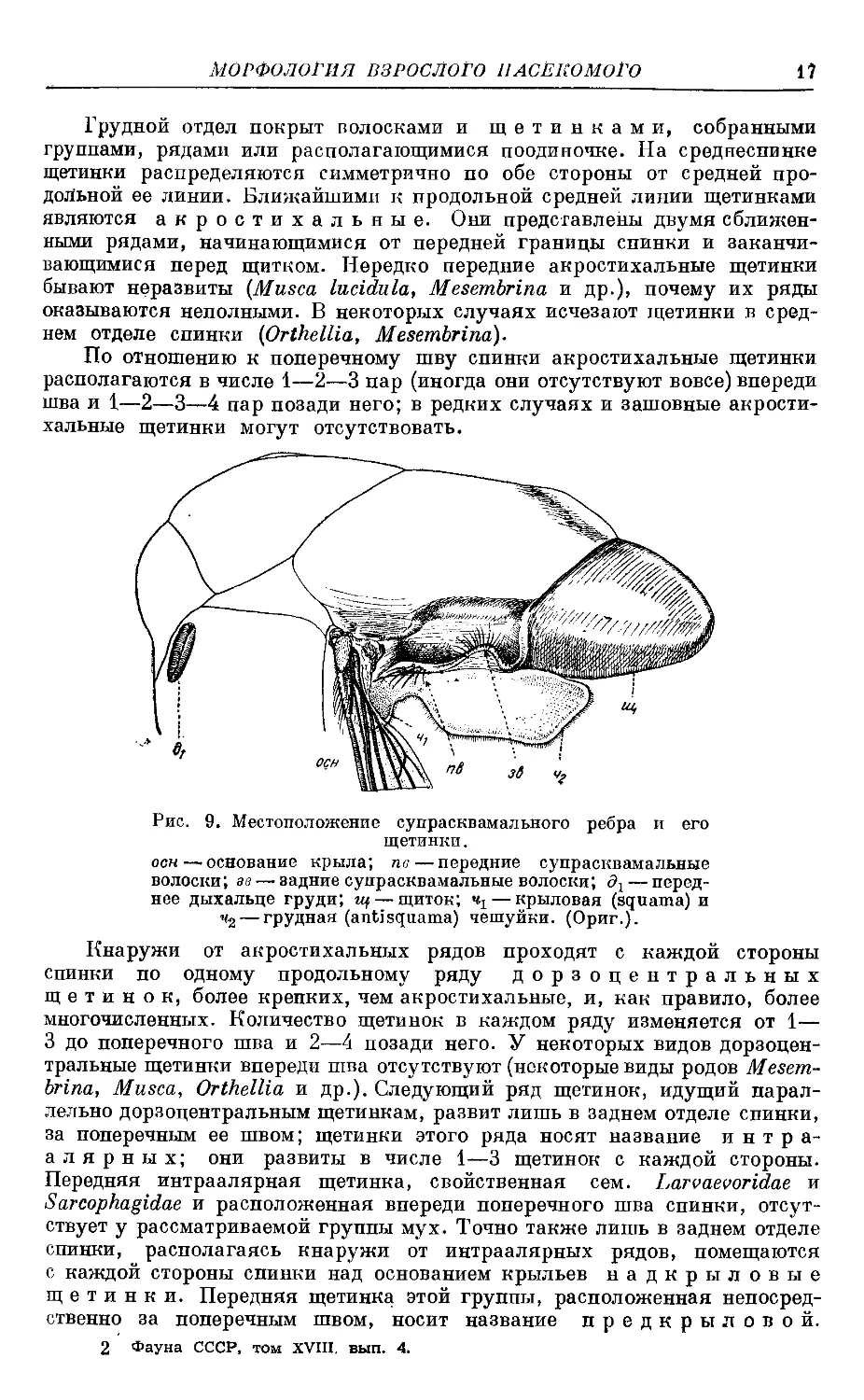
Рис. 9. Местоположение супрасквамального ребра и его
щетинки.
осн—основание крыла; пв — передние супрасквамальные
волоски; ав — задние супрасквамальные волоски; дг — перед-
нее дыхальце груди; щ — щиток; — крыловая (squama) и
Чз — грудная (antisquama) чешуйки. (Ориг.).
Кнаружи от акростихальных рядов проходят с каждой стороны
спинки по одному продольному ряду дорзоцентральных
щетинок, более крепких, чем акростихальные, и, как правило, более
многочисленных. Количество щетинок в каждом ряду изменяется от 1—
3 до поперечного шва и 2—4 позади него. У некоторых видов дорзоцен-
тральные щетинки впереди шва отсутствуют (некоторые виды родов Mesem-
brina, Musca, Orthellia и др.). Следующий ряд щетинок, идущий парал-
лельно дорзоцентральным щетинкам, развит лишь в заднем отделе спинки,
за поперечным ее швом; щетинки этого ряда носят название интра-
а л я р н ы х; они развиты в числе 1—3 щетинок с каждой стороны.
Передняя интраалярная щетинка, свойственная сем. Larvaevoridae и
Sarcophagidae и расположенная впереди поперечного шва спинки, отсут-
ствует у рассматриваемой группы мух. Точно также лишь в заднем отделе
спинки, располагаясь кнаружи от интраалярных рядов, помещаются
с каждой стороны спинки над основанием крыльев надкрыловые
щетинки. Передняя щетинка этой группы, расположенная непосред-
ственно за поперечным швом, носит название предкрыловой.
2 Фауна СССР, том XVIII. вып. 4.
18
ВВЕДЕНИЕ
За основанием крыльев на закрыловых бугорках помещены з а к р ы-
ловые щетинки, в количестве 1—2—3 с каждой стороны.
Су^прасквамальное ребро (рис. 9) у видов рода Orthellia,
как правило, а у некоторых Musca, как исключение, снабжено тонкими
щетиночками; различают переднюю и заднюю группы супрасквамальных
щетинок. Обе названных группы щетинок имеют большое значение в си-
стематике таких родов, как Musca, Orthellia. У большинства видов триб
Muscini и Stomoxydini супрасквамальные щетинки отсутствуют.
Передний отдел спинки, помимо акростихальных и дорзоцентральных,
несет на плечевых бугорках плечевые, заплечевые —
в числе 1—2 с каждой стороны (реже они отсутствуют) — и предшов-
ную щетинки. Последняя прикреплена в боковой части переднего
отдела среднеспинки перед поперечным ее швом.
Щетинки щитка располагаются по свободному его краю
и на поверхности симметрично по обе стороны от его продольной средней
линии. Вершина щитка снабжена 2, обычно сильно развитыми, вер-
шинными щетинками. По бокам от них с каждой стороны при-
крепляется по одной или по две боковых, или латеральных
щетинки, в большинстве случаев более коротких и тонких, чем вершин-
ные. Впереди боковых щетинок имеются основная, а несколько
впереди и ковнутри от них предосновная щетинка, последняя
часто слабо развита, а иногда и отсутствует вовсе.
Наименование щетинок боковых отделов груди (рис. 8) являются про-
изводным названий тех склеритов груди, на которых эти щетинки рас-
положены. Так, нотоплевры несут по 2—3 нотоплевральных
щетинки. Проплевра несет 1 или несколько щетинок в нижнем отделе
(над тазиками передних ног). Мезоплевры имеют 1 щетинку в передне-
верхнем углу и вертикальный ряд щетинок вдоль заднего шва склерита.
Стерноплевры несут щетинки как в нижних, так и в верхних своих отде-
лах; особенно существенным является количество и характер распо-
ложения стерноплевральных щетинок относительно
друг друга; наиболее обычным следует признать такое размещение щети-
нок, когда одна из трех стерноплевральных щетинок расположена в перед-
не-верхнем, а две (одна под другой) — в задне-верхнем углу стерноплевр;
такое расположение щетинок условно обозначается формулой 1 -|-2 (или
1 : 2). Реже (Dasyphora) количество щетинок увеличивается до четырех,
с расположением 1 —[-3, или, наоборот, уменьшается до 0-|-1 (Pyreilia),
1-|-1 (Mesembrina и др.), 0-|-2; Musca albina вообще лишена стерноплев-
ральных щетинок. Более постоянна хетотаксия птероплевры, дающая
признаки родового порядка, правда, пока мало разработанные.
Волосяной покров груди сравнительно редкий у большинства видов
группы. Наибольшего развития он достигает в роде Мesembrina, наимень-
шего — у Acridomyia sacharovi. Распределение волосков по поверхности
грудного отдела неравномерное. Участки грудной поверхности, наиболее
выступающие и не соприкасающиеся с соседними частями (ногами, голо-
вой, брюшком) тела, покрыты более густыми и обычно более длинными
волосками. В связи с этим переднее и заднее грудные кольца опушены
беднее среднегрудного кольца. Характер распределения волосков на скле-
ритах первого и третьего грудных сегментов является ценным системати-
ческим признаком. В особенности это относится к проплевре, переднегруди,
гипоплевре и метаплевре. Наличие или отсутствие волосков на этих
склеритах зачастую определяет не только видовую, но и родовую принад-
лежность. Волоски, имеющиеся на гипоплевре, могут быть разбиты па
МОРФОЛОГИЯ ВЗРОСЛОГО ИАСВКОМОГО
19
две самостоятельных группы соответственно с местом своего расположе-
ния; первая группа расположена в передневерхнем углу склерита
(Graphotnyia и др.), эта группа волосков отсутствует у многих родов,
в частности, у Mesembrina, Dasyphora и других. Вторая группа гипоплев-
ральных волосков находится впереди задних дыхалец (Graphomyia, РугеГ
lia, Musca)', у видов родов Orthellia, Mesembrina они нами не обнаружены.
Особое значение имеют мелкие волоски в нижнем заднем углу груд-
ного отдела — на эпимерах и эпистернах заднегрудного сегмента. Волоски
этой группы присутствуют у всех родов трибы Muscini и никогда не на-
блюдаются у представителей трибы Stomoxydini.
Опушенная проплевра — явление редкое, свойственное некоторым
видам рода Musca, а из жигалок — Haematobia, Haematobosca (в пони-
мании Мэллока) и Stomoxys. Весьма большим постоянством обладает
опушение переднегруди (prosternum). Волоски на переднегруди обнару-
жены у видов Dasyphora и Morellia, причем далеко не у всех видов, входя-
щих в названные роды. Наоборот, в роде Orthellia волоски на переднегруди
присутствуют у всех видов, являясь характерным родовым признаком.
Обращаясь к вопросу устойчивости хетотаксических и пилотакси-
ческих признаков, следует указать на то обстоятельство, что далеко не
все системы щетинок и волосков обладают одинаковой степенью посто-
янства у различных родов данной группы. В качестве примера можно при-
вести количество стерноилевральных щетинок у видов Musca. От вида
к виду признак варьирует от 0 до 4(1 -|-3), тогда как у видов рода Dasy-
phora он почти не претерпевает изменений, показывая стабильное число
14-3. В роде Dasyphora число щетинок щитка изменяется в пределах от
8 до 14, в то время как у видов Pyreilia оно дает значительно меньший
размах колебаний. Сходную картину можно наблюдать на примере дор-
зоцентральных щетинок в роде Orthellia, где наряду с сокращенным их
числом (одна пара) встречаются виды с 6 парами щетинок названной
группы. Эта же группа щетинок у рода Dasyphora является константной
(34-4)- Акростихальные щетинки, столь сильно колеблющиеся в числе
у родов Orthellia, Musca, Mesembrina и др., оказываются более стойкими
в пределах рода Morellia.
Что касается грудных дыхалец, то они много крупнее брюшных;
снаружи они имеют овальный, коротко или удлиненно треугольный кон-
тур; в дыхальцах различают узкую пластинку, образующую край
дыхальца — перитрему, от внутреннего края которой отходят
многочисленные столбики или выросты, покрытые простыми или развет-
вленными волосками (рис. 25—37); эти выросты заслоняют в той или иной
степени просвет дыхальцевого отверстия, почему им приписывается роль
фильтра. Относительные размеры дыхалец, конфигурация их, а также
характер выростов перитремы и волосков, имеют существенное значе-
ние в систематике группы.
Крылья (рис. 10) имеют форму вытянутого треугольника с ши-
роко закругленными углами. У основания крыла различают небольшие
лопасти — крылышко — прозрачную тонкую пластинку трапецие-
видных очертаний (кр), отделенную от крыла надрезом и являющуюся
югальной областью крыла, и две непрозрачные или полупрозрачные,
обычно белой, желтой или дымчатой окраски, округлые лопастинки
(рис. 66—68) — крыловую и грудную чешуйки; первая
меньше второй; она является непосредственным продолжением крылышка
и соединяет при его посредстве задний край крыловой пластинки с груд-
ной чешуйкой, срощенной по внутренней своей границе с грудным от-
2*
20
ВВЕДЕНИЕ
делом. При спокойном положении крыльев крыловая чешуйка распола-
гается над грудной. Характер прикрепления, конфигурации и относитель-
ные размеры подкрыловых чешуек, указывающие на особенности эволю-
ции органа полета, имеют большое значение и широко используются в си-
стематике Muscoidea.
Крылья стекловидно прозрачные или сероватые, нередко окрашены
{Mesembrina) в желтые или коричневатые тона.
По переднему краю крыла от основания до его вершины проходит
костальная жилка (с); вместе отделяясь от корня крыла и сле-
Рис. 10. Схема жилкования крыла.
Ж и л к и: ап — анальная; Ъс — базикоста; с — костальная; crv —
изгиб медиальной жилки; си, и си2 — первая и вторая кубитальные;
h — плечевая поперечная; т — медиальная; тси— медиокуби-
тальная поперечная; /•], r2+3, г4+5— первая, вторая и третья
ветви радиуса; гт — радиомедиальная поперечная; sc — субко-
стальная; tp(mcu) — задняя поперечная. Я чей к и: Bas^— пе-
редняя основная; Сг и С2— костальные 1-я и 2-я; Си2 (= Ап)—
2-я кубитальная или анальная; М1 {—Вая2)— первая медиальная
или задняя основная; М2 {=!>)— вторая медиальная или диско-
идальная; Blt R3, R& — радиальные 1-я, 3-я и 5-я; Sc — субко-
стальная; кр—крылышко, — крыловая чешуйка. (Ориг.).
дуя почти параллельно ей, идет тонкая субкостальная жилка
(sc), дистальным концом упирающаяся в костальную недалеко от средины
крыла. От субкостальной жилки далее к заднему краю крыла распола-
гаются 3 радиальных ветви, выходящие из основания крыла одним
общим стволом. Передняя более толстая — 1-я радиальная жилка (rj
короче остальных; ее дистальный конец дуговидно изогнут по направле-
нию к переднему краю крыла, вливаясь вблизи средины последнего
в костальную жилку недалеко от субкостальной жилки; вторая ветвь ра-
диуса — 2-я радиальная жилка (г2+3) длинная, вершинным концом сое-
диняется с костальной жилкой около вершины крыла; 3-я радиальная
жилка (г4+5) длиннее предыдущих, ее свободный конец достигает вер-
шины крыла, где сливается с костальной жилкой или с этой последней
-(-медиальной {Musca lucidula Lw.). Медиальная {т) продольная
жилка укрепляет срединный отдел крыла. Проксимальный ее конец со-
единяется близ основания крыла с нижележащей — кубитальной ветвью.
М0РФ0Л01 ИЯ ВЗРОСЛОГО НАСЕКОМОГО
21
Дистальный конец медиальной жилки перед крыловой вершиной обра-
зует 5-образный (Mesembrina), дуговидный (Pyreilia, Morellia, Dasyphora)
или угловидный изгиб (Musca, Orthellia). У представителей трибы
Stomoxydini дуговидный изгиб выражен слабо (Lyperosia, Stomoxys)', реже
(Acridomyia) медиальная жилка прямая на всем протяжении. От места
своего соединения с медиальной ветвью (от основания крыла) под острым
углом к этой последней отходит, направляясь к заднему краю крыла
первая кубитальная жилка (си1); к вершине жилка посте-
пенно утончается, переходя вблизи края крыла в складку. Последняя
жилка — анальная (ап), ее дистальный конец далеко не достигает
края крыла, более или менее резко обрываясь и не продолжаясь, как это
наблюдается у представителей семейства Anthomyidae, в виде тонкой
складки до заднего края крыла. Анальная жилка выходит из корня крыла
самостоятельной ветвью, иногда анастомозируя с основанием кубиталь-
ной жилки слабым расплывчатым утолщением мембраны крыла.
Дальнейшее укрепление крыловой мембраны происходит при помощи
поперечных жилок, количество которых у Muscinae равно пяти. Первая
из них — плечевая (h) поперечная жилка, соединяющая
костальную жилку с основанием субкостальной-^радиальной. Медиаль-
ная жилка связана с 3-й радиальной жилкой (г4+5) срединной
или радио медиальной поперечной жилкой (гт),
располагающейся в среднем отделе крыла. Связь кубитальной ветви
с медиальной осуществляется при посредстве двух поперечных жилок:
медиокубитальной (тси) в основной половине и задней поперечной жил-
кой (tp) в вершинной половине крыла. Последняя поперечная жилка —
вторая кубитальная (сгг2) — соединяет основания кубитальной и аналь-
ной ветвей.
Задний край крыла от вершины до его основания свободен от жилок,
он снабжен короткой бахромой из тонких густых волосков.
Большинство жилок голые, т. е. не несут на своей поверхности ни
волосков, ни щетинок; таковы субкостальная, вторая радиальная, меди-
альная, кубитальная, анальная и все поперечные жилки. Костальная
жилка всегда покрыта несколькими рядами коротких толстых щетинок.
Жилка г1 в редких случаях (Dasyphora) несет (рис. 300) по всей длине или
только в основном отделе короткие хорошо заметные при слабом увеличе-
нии щетинки. Жилка г4+Б, наоборот, почти всегда снабжена щетинками
(Dasyphora) или тонкими волосками при основании (Musca). Присутствие
волосков и щетинок может быть обнаружено на одной из сторон, или
одновременно с верхней и нижней поверхностей крыла. Чешуйки и кры-
лышко по свободным краям несут тонкие волоски.
В систематике Muscini и Stomoxydini большое значение имеют
характер изогнутости вершинного отдела медиальной жилки, соотноше-
ние длины участков костальной жилки, заключенных между точками
впадения в нее радиальных и медиальной жилок, пропорция отрезков
медиальной жилки, отграниченных rm, tp и вершиной крыла. Имеют
также значение в систематике степень выраженности и характер щети-
нок и волосков, покрывающих жилки крыла.
Участки крыла, ограниченные жилками, носят название ячеек. На-
именование ячей является производным названия ограничивающей их
(как правило, спереди, где это возможно) жилки. Поэтому первые две
ячейки, лежащие по переднему краю крыла, именуются как 1-я и 2-я
костальные (Ci, Cz); узкая ячейка позади субкостальной получает
название — субкостальной (Sc). Далее следуют: р а д и а л ь-
22
ВВЕДЕНИЕ
н а я первая (Ri), радиальная третья (Rs)* 1 радиаль-
ная пятая (Я5)2 (=Л-й заднекрайней прежних авторов). Центральная
часть крыла занята длинной второй медиальной (Ms) или
дискоидальной ячейкой, от которой к основанию крыла рас-
полагается первая медиальная ячейка (Mi). Непосредственно
позади нее помещается почти равная ей по размеру вторая куби-
тальная ячейка (анальная прежних авторов) (Сиг=Ап). К осно-
ванию крыла от ячейки Rs лежит узкая первая базальная
ячейка (Bas). Область крыла позади анальной жилки носит назва-
ние анальной лопасти.
Жужжальца (рис. 8) являются придатком заднегрудного
кольца, гомологичным задней паре крыльев. Жужжальца подвижно
причленены к плевральным и спинным склеритам задней груди. Состоят
они из утолщенного основания, тонкой, обычно цилиндрической ножки
и булавы. Основание жужжалец содержит в себе сензорные образования,
в том числе хордотональный орган.
Ноги имеют хорошо развитые тазики, из них передние дости-
гают наибольших размеров; передние тазики выпуклые по задней и во-
гнутые или плоские по передней стороне, имеют вид вытянутых сравни-
тельно массивных образований, подвижно сочлененных с переднегруд-
ным сегментом и неподвижно — с вертлугом. Аналогичный характер
наблюдается у средних и задних тазиков. Тазики задних ног достигают
наименьшего развития и представлены сравнительно коротким непра-
вильным в очертаниях кольцом. Тазики средних ног по своим размерам
занимают среднее положение между передними и задними. Бедра, в боль-
шей или меньшей степени сжатые с боков, как правило, постепенно и
слабо сужены к концам. Передние бедра толщей обычно несколько короче
бедер средней и задней пары ног. Вертлуги маленькие. Голени передних
ног прямые, более тонкие при основании чем у вершины. Сфедние голени
могут иметь ту же форму, что и передние (Musca, Graphomyia, Orthellia,
Stomoxys и др.), или обладать своеобразным строением, в котором можно
усматривать признаки не только видового (Mesembrina), но и родового
порядка. Мощного развития, сопровождающегося при этом сильной
искривленностью голеней, достигает вооружение ног в виде шипов, ще-
тинок, крепких волосков у самцов Mesembrina mystacea, М. decipiens.
Примером сильной искривленности могут служить голени самцов Morel-
lia aenescens. Некоторое сплющивание голеней (Morellia podagrica, М. hor-
torum) и разнообразие в характере хетотаксии обычно встречается только
среди представителей трибы Muscini. Виды трибы Stomoxydini в такой
мере не обладают этими чертами.
Лапка состоит из пяти члеников, подвижно сочлененных друг
с другом и с вершиной голени. 5-й членик снабжен на конце парой
крючковидно загнутых на вершине коготков. Под основанием каждого
коготка располагается по одной присоске, между которыми прикреплен?
дуговидно согнутая кверху, реже почти прямая, очень короткая, светло-
окрашенная щетинка — эмподий — рудимент средней (третьей) присоски.
У большинства видов Muscini щетинковидный эмподий весьма слабо раз-
вит, причем длина его часто не достигает половины длины присосок
1 Вторая радиальная ячейка ввиду слияния второй (га) и третьей (гз) радиаль-
ных жилок в одну радиальную ветвь, обозначаемую поэтому символом с2+3, исчезла.
1 Слияние 4-й и 5-й радиальных жилок в одну ветвь вызвало выпадение
находившейся между ними ячейки Ri.
МОРФОЛОГИЯ ВЗРОСЛОГО НАСЕКОМОГО
23
{Musca, Morellia, Dasyphora^, реже {Orthellia, Lyperosia) эмподий равен
длине присосок. Основание эмподия у некоторых видов покрыто тонкими,
светлыми волосками {Mesembrina и др.). 1-й членик лапки крупнее осталь-
ных; длина его составляет почти половину длины лапки. Длина члени-
ков убывает к дистальному концу лапки.
В строении ног наблюдается половой диморфизм, особенно резко
выраженный в родах Morellia, Mesembrina и у некоторых видов родов
Dasyphora и Lyperosia. Диморфизм сказывается в строении голеней
(изогнутость, сплющенность, сильное вооружение у самцов) и ла-
пок (выросты, опушение); иногда различия сводятся к увеличению
длины члеников лапок, присосок и коготков на лапках у самцов,
что свойственно всем без исключения видам группы {Muscini, Stomoxydini).
Хетотаксия ног. Все части ног покрыты короткими обычно
жесткими волосками, среди которых выделяются своей длиной или тол-
щиной щетинки и щетинистые волоски, располагающиеся поодиночке или
рядами. Так как щетинки ног располагаются продольными рядами, удобно
именовать ряды щетинок сообразно тем продольным поверхностям ноги,
на которой эти ряды расположены. Чтобы уяснить принцип такого обо-
значения, нужно представить себе бедро и голень в виде вытянутых вось-
мигранных призм. При разогнутом колене одна из продольных плоскостей
бедра и голени будет направлена вниз (вентрально). При согнутом колене
эти поверхности бедра и голени будут соприкасаться. Сторона, противо-
положная вентральной, при вытянутой ноге будет обращена кверху,
дорзально, почему эта плоскость носит название дорзальной; соответ-
ственно этому определяются плоскости, обращенные вперед и назад.
Плоскости, расположенные в промежутках между указанными выше
плоскостями, именуются сообразно названию двух соседних сторон ноги.
Таким образом, на ногах различаются следующие поверхности и одно-
именные щетинки: передние, задние, дорзальные, вентральные, передне-
дорзальные, заднедорзальные, передневентральные и задневентральные.
В систематике Muscini особенно широко используются щетинки го-
леней. В меньшей степени разработана и поэтому реже применяется хето-
таксия бедер. Передние голени вооружены слабее средних и задних. Им
свойственны в большинстве случаев лишь концевые и предвершинные
щетинки; в редких случаях {Morellia aenescens, Dasyphora setitibia) ще-
тинки могут присутствовать и в среднем их отделе. Вентральная поверх-
ность средних голеней у некоторых родов несет крепкую щетинку вблизи
средины {Dasyphora, Pyreilia)’, у большинства родов она не развита.
Переднедорзальные щетинки, представленные одной крепкой щетинкой
или рядом щетинок или щетинистых волосков, имеют значение в система-
тике Dasyphora. Задние и задневентральные или заднедорзальные ще-
тинки, присутствующие у большинства представителей группы, измен-
чивы в числе, почему в систематическом отношении оказываются мало
ценными. В этом смысле исключение представляют самцы Morellia, у ко-
торых эти группы щетинок достигают большого своеобразия и посто-
янства. Хетотаксия задних голеней, за исключением рода Morellia не
является устойчивой.
Задние голени снабжены щетинками и волосками на всех поверх-
ностях, кроме вентральной (исключение представляет триба Stomoxy-
dini, у которой хетотаксия ног слабо выражена).
Щетинки задних бедер в характере расположения довольно одно-
образны. Некоторое разнообразие в распределении щетинок наблюдается,
пожалуй, только у родов ^Morellia и Dasyphora.
24
ВВЕДЕНИЕ
Передние бедра обладают, как правило, тремя полными рядами
щетинок, а именно: задневентральным, заднедорзальным и дорзальным.
На средних бедрах ряды, обычно, не полные. Здесь могут присутствовать
вентральный, передний, переднедорзальный и передневентральный ряды,
которые, начинаясь от основания бедер, доходят до их средины. У вер-
шины бедер различаются заднедорзальный и дорзальный ряды щетинок.
Для задних бедер характерно присутствие передневентральных,
переднедорзальных и задневентральных лцетинок или волосков. Послед-
ние известны у представителей рода Dasyphora.
Брюшко (рис. 11) соединено с грудью плотной перепонкой, обес-
печивающей его подвижность. Брюшко состоит из 5 сегментов, соединен-
Рис. И. Схема строения брюшка самца ком-
натной мухи.
1-10— тергиты; I—VII—стерниты; IX—
9 — 9-й тергостернит;— мембрана, ({—цер-
ки; <fi — фаллозома (место прикрепления);
п —парамеры (место прикрепления); д — ды-
хальцы. (По Пэттону с изменениями).
ных межсегментными перепон-
ками. В свою очередь каждый
сегмент брюшка состоит из верх-
ней (тергит) и нижней (стернит)
пластинки. Тергиты развиты
сильнее стернитов и имеют вид
полуколец, тогда как стерниты
сокращены до степени узких, ±
прямоугольных или округлых
пластинок, расположенных в го-
ризонтальной плоскости одна за
другой снизу брюшка по его
средней продольной линии. Пер-
вый видимый тергит является
продуктом слияния 1-го и 2-го
тергитов, о чем можно судить по
наличию на каждом из его
концов двух дыхалец, вместо
одного; последнее свойственно
последующим сегментам. Ды-
хальцы первых пяти сегмен-
тов, как у трибы Muscini, так
и Stomoxydini, заложены в тер-
гитах. Тергиты на вентральной
стороне брюшка соединяются
своими концами со стернитами
широкой перепонкой, сильно
развитой как у Muscini, так
и Stomoxydini.
Брюшко Muscinae имеет
5—7 пар дыхалец.
Начиная с 6-го сегмента, происходит сильное видоизменение кон-
фигурации брюшных колец, смещение и редукция их. Эти дистальные
пять сегментов брюшка, из которых сформирован наружный гениталь-
ный аппарат насекомого, различно устроены у самцов и самок, почему
рассмотрение их морфологии проводится отдельно для каждого пола.
У самцов последние 5 сегментов, имеющие в совокупности вид
колбочки или груши, носят название гипопигия. Гипопигий
в спокойном состоянии несколько втянут в задний просвет 5-го тер-
гита и поддогнут книзу. Он соединен мембраной с задними краями
5-го тергита и V стернита. Дорзальная и боковые стороны гипопи-
ГРЯ (рис. 12, 13) образованы тремя узкими полукольцами, в строе-
МОРФОЛОГИЯ ВЗРОСЛОГО НАСЕКОМОГО
25
нии которых наблюдается асимметрия: левые края первых двух полу-
колец шире и спускаются по боковым сторонам гипопигия нижо, чем
правые их концы. Первый склерит является 6-м, второй (средний) —
7-м, а последний, ограничивающий сверху анальное отверстие и
перепонку, в которой оно расположено, — 10-м тергитом (П >ттон,
1932—1933). Тергиты 6-й и 7-й снабжены дыхальцами, расположен-
ными то в перепонке вблизи тергитов, то в самих тер:итах.
Лишь у Graphomyia дыхальцы 7-го тергита отсутствуют (гак у
многих Anthomyidae'), и, следовательно, гипопигий у видов этого
Рис. 12, 13. Схема строения конца брюшка самца.
12 — вид сзади, ао — анальное отверстие; к9— кокситы 9-го тергостернита; н —
парамеры; 4с—7с — стерниты с 4-го по 7-й; д5—5?—-дыхальца 5—7 сегментов;
5 т—Ю т — тергиты с 5-го по 10-й; 9тс — 9-й тергостернит; ф — фаллозома; ц —
церки. 13 — схема строения брюшка самца (вид сбоку); Обозначения как на рис. 12.
(Ориг.).
рода обладает только одной парой дыхалец, принадлежащих 6-му сег-
менту и заложенных в межсегментной перепонке вблизи переднего края
у концов 7-го сегмента. Положение дыхалец конечных сегментов г- раз-
личных родах не одинаково. У самцов Musca, Stomoxys, Haematobia 7-е
дыхальце заложено на крае 7-го тергита, причем у нижних его концов;
6-е дыхальце лежит ниже его и несколько позади в межсегментной пере-
понке, у Orthellia оба дыхальца сильно сближены, часто соприкасаясь друг
с другом; Pyreilia и Dasyphora имеют несколько расставленные, но все же
близко расположенные друг к другу дыхальцы. Расстояние 7-го дыхальца
от бокового края тергита различно с разных сторон тела: левое дыхальце
всегда несколько дальше отстоит от конца тергита нежели правое; это
особенно резко выражено у Mesembrina, где 7-е дыхальце лежит к тому же
далее от переднего края тергита; то же наблюдается у Polietes, Myicspila,
некоторых Phaonia и др. 10-й тергит не имеет дыхалец; своим передним
краем он соединен при помощи перепонки с задним краем 7-го тергита.
2fi
ВВЕДЕНИЕ
8-й тергит смещен на вентральную сторону гипопигия, тде, слившись
с одноименным стернитом, образовал так называемый 9-й тергосте р-
н и т (Awati, Patton). От заднего края 10-го тергита отходит широкая
перепонка, заключающая в себе анальное отверстие и своим свободным
краем срастающаяся с верхним и боковыми краями церок. Церки
соответствуют верхним форцепсам прежних авторов. Строение церок очень
разнообразно; они имеют парное строение, сказывающееся в наличии про-
дольного шва, рассекающего церки на две боковые, симметричные части.
Церки закрывают полость гипопигия сзади и в общем имеют вид хитино-
вой пластинки, то поперечно, то продольно вытянутой, и нередко снабжен-
ной симметрично расположенными по бокам от продольной средней ли-
нии выступами, отростками, шипами, щетинками и т. п. Дорзальный край
церок загнут внутрь; его можно наблюдать на препаратах в форме более
или менее широкой закраины. В некоторых случаях край бывает зазубрен
(Dasyphora, Pyreilia) или несет шипы.
Стерниты (рис. И, I—VII) располагаются на вентральной стороне
брюшка. I стернит передним краем соединен перепонкой с нижней поверх-
ностью заднего отдела груди и тазиками задних ног. Каждый из последу-
ющих стернитов соединен перепонкой с предыдущим стернитом. I стер-
нит имеет обычно полулунную форму; II, III и IV стерниты отличаются
от него, равно как и от V, по форме; они имеют характер удлиненных,
овальных или сравнительно широких прямоугольных пластинок. V стер-
нит всегда шире предыдущего, расширен назад и снабжен у большинства
видов Muscini и Stomoxydini по заднему краю двумя боковыми выростами;
задний край стернита посредине выемчатый. Боковые края V стернита,
как и предшествующих стернитов, сращены с концами соответствующих
тергитов. От заднего края V стернита отходит мембрана, заметно скле-
ротизованная в области выемки стернита; у Stomoxydini эта область до-
стигает наиболее сильного развития, приобретая иногда характер по-
перечно вытянутой и сочлененной с внутренними краями выемки V стер-
нита пластинки (рис. 11, 4, 5, 6). Значительной склеротизации достигает
мембрана и у некоторых Muscini (в частности, Dasyphora, большинство
которых, однако, имеет мембрану склеротизованную у заднего края
V стернита). Мембрана рассматривается нами как VI стернит. У всех из-
вестных нам видов группы мембрана эта от краев V стернита загнута
складкой внутрь гипопигия, и располагается, следовательно, под краями
срединной выемки V стернита. С ней соединен вершиной узкий непарный
саблевидный склерит, расширенным своим проксимальным концом по-
движно сочлененный с левым концом 7-го тергита. Саблевидный склерит
является VII стернитом (рис. И, VII).
Его проксимальная часть по заднему краю дает угловидно или полу-
кругло расширенную лопасть; весьма вероятно, что эта лопасть
является VIII стернитом, что, однако, требует подтверждения. Позади
мембрана VI стернита соединяется с угповидным или полулунным тон-
ким склеритом — 9-м тергостернитом; вершина его угла направлена впе-
ред (в сторону V стернита); от нее, расходясь вбок, отходят с каждой
стороны назад по одному узкому отростку; дистальные их концы под-
вижно сочленены с концами отростков кокситов; наружный край задних
отростков 9-го тергостернита подвижно сочленен с отростками 10-го тер-
гита. Между боковыми лопастями 9-го тергостернита расположена фал-
лозома и лежащие по бокам от нее парамеры (каждая состоящая из перед-
него и заднего отделов). Кокситы 9-г<5 тергостернита вторым своим от-
ростком соединяются с боковыми краями церок и задним краем 10-го
МОРФОЛОГИЯ ВЗРОСЛОГО НАСЕКОМОГО
27
Рис. 14. Строение фаллозомы
(схема).
а — аподема; пп — передний
отдел парамер; сч— склероти-
зованная часть penis; сп —
семенной проток; до — дор
зальный отросток фаллозомы;
мч—мембранозная часть pe-
nis; ап—задний отдел пара-
мер; п — penis; са — семенная
ампула; т— тека. (Ориг.).
penis, особенно в вершин-
тергита; соединение кокситов друг с другом осуществляется при помощи
прозрачной перепонки, натянутой между концами их отростков, которые
сращены с 9-м тергостернитом (рис. 11, 12, 13). Кокситы 9-го тергостер-
нита (рис. 12, 13), в дальнейшем изложении именуемые просто кокси-
тами, подобно церкам, имеют существенное значение в диагностике мно-
гих групп. У некоторых групп двукрылых, в частности, у рода Musca они
изменяются от вида к виду незначительно. Каждый коксит представляет
собою хитиновый склерит, то в виде широ-
кой лопасти, то в виде саблевидно или клю-
вовидно заостренного придатка, лежащего
с каждой стороны на боковых отделах гипо-
пигия.
Фаллозома (рис. 14) — частично
склеротизованное, частью мембранозное обра-
зование, расположенное с вентральной сто-
роны гипопигия между боковыми отделами
9-го тергостернита; по бокам от основания
фаллозомы- прикреплены парамеры, каждая
состоящая из двух частей — передней и зад-
ней. Парамеры также помещаются между от-
ростками 9-го тергостернита, примыкая на-
ружным краем основания к внутреннему
краю отростков 9-го тергостернита, а вну-
тренней его стороной — к основанию фалло-
зомы. Фаллозома слагается из следующих ча-
стей: 1 — собственно фаллозомы (penis — сово-
купительный орган), 2 — теки, 3 — отростка
фаллозомы (дорзальный отросток теки), 4 —
аподемы. Собственно фаллозома почти всегда
имеет склеротизованную основную (прокси-
мальную) и перепончатую дистальную части.
Основной отдел может иметь вид более или
менее длинной трубки (Musca, Stomoxys,
Haematobia) почти цилиндрической или ко-
нусовидной, резко отграниченной от мем-
бранозного отдела. У других форм (Orthel-
lia, Pyreilia, Dasyphora, Morellia, Mesem-
brina и др.) склеротизация основного отдела
фаллозомы выражена слабее.
Так, у Orthellia мембранозный отдел за-
нимает сравнительно небольшую часть вер-
шины и средины penis, тогда как у Musca
почти половина penis перепончатая. Стенки
ном отделе, могут быть вооружены то мелкими тонкими (Pyreilia, Das-
yphora, Morellia, Mesembrina), то крепкими, сильными шипами (Orthel-
lia). У Stomoxydini вершина penis несет поперечно расположенную
полосу, снабженную на концах шиповидными отростками. Исключением
является Acridomyia, где положение шипов и поперечной полосы иное,
а также Graphomyia (рис. 350, 472).
По дорзальной стороне penis соединяется узким или широким хитиновым
тяжем с проксимальной частью, при посредстве которой совокупительный
орган соединяется с парамерами и вентральной мембраной гипопигия. Эта
проксимальная часть — тека — имеет вид то узкой и короткой (Sto-
28
ВВЕДЕНИЕ
moxydini, Musca, Orthellia, Pyreilia и др.), то очень широкой, мас-
сивной {Graphomyia) трубки. Верхний ее край соединен тонкой,
обычно покрытой мелкими волосками, прозрачной мембраной с нижним
краем penis; нижний конец соединен с длинным тонким хитиновым тя-
жем, лежащим в полости гипопигия — аподемой, нижние края соединены
с мембраной гипопигия, соприкасаясь с основаниями парамер. От ниж-
него края теки отходит назад дорзальный отросток фаллозомы, сильно
развитый у Muscini и обычно едва заметный у Stomoxydini. От прокси-
мальной части аподемы отходят вперед узкие хитиновые тяжи, соединяю-
щие аподему с основаниями парамер.
Парамеры (forcipes inferiores-|-palpi genitalium auct.) состоят из двух
рядом стоящих и сочлененных основаниями переднего и заднего отделов.
Передние отделы парамер, как правило, развиты сильнее задних. Они
булавовидно, лопастевидно или клювовидно вытянутой формы, снабжены
при основании или в вершинной части щетинками или шипиками; иногда
их дистальные концы перепончаты и снабжены сензорными образованиями.
Задние отделы парамер обычно коротки, широки или представляются
в форме мелких конусовидных (некоторые виды рода Mused) выростов,
мембранозных на вершине. Как и передние отделы, они снабжены сен-
зорными образованиями. У Graphomyia maculata передние отделы пара-
мер практически отсутствуют, будучи слитыми с краями 9-го тергостернита.
С основанием фаллозомы связан маленький хитиновый склерит (се-
менная ампула), от которого направляется семеизвергательный канал
внутрь тела фаллозомы, открывающийся отверстием на дистальном конце
совокупительного органа.
Мужская половая система самцов (рис. 38) состоит
из пары семенников, имеющих различную форму в зависимости от видо-
вой и родовой принадлежности. У Musca они имеют в общем грушевидную
форму, с отходящими от суженного конца семепроводами. Вместе со
всеми прочими внутренними гениталиями семенники расположены в об-
ласти 5-го брюшного сегмента, помещаясь вентрально от прямой
кишки. Каждый семенник состоит из плотной хитиновой капсулы, содер-
жащей сперматозоиды, и покрывающей ее фолликулярной оболочки.
Семепроводы сравнительно коротки и соединяются в один общий семеиз-
вергательный канал; он образует несколько изгибов, расположенных
с левой стороны брюшка, утончается к дистальному концу и переходит в
правую часть брюшной полости, направляясь далее назад в область, огра-
ниченную 6-м и частично 7-м сегментами (тергитами). Здесь семеизвер-
гагельный канал дает грушевидной формы расширение, внутри которого
содержится особый склерит — apodema ejaculatorii; от нее канал в виде
тонкой трубки входит в тело фаллозомы, открываясь выходным отвер-
стием на вершине совокупительного органа.
Сегменты брюшка самок (рис. 15—24), начиная с 6-го, сильно умень-
шены в диаметре и в совокупности представляют втягивающийся в про-
свет 5-го брюшного сегмента яйцеклад.
Половое отверстие находится на вершине последнего, 10-го, сегмента
под анальным отверстием.
Яйцеклад образуется несколькими сегментами. По одним авторам
число сегментов, составляющих яйцеклад, равно пяти (с 6-го по 10-й),
по другим — четырем; мы придерживаемся первой точки зрения, прини-
мая за десятый сегмент мембранозное окончание яйцеклада.
Склериты сегментов яйцеклада в пределах рассматриваемой группы
редко имеют характер целых хитиновых колец; среди Muscinae всегда
МОРФОЛОГИЯ ВЗРОСЛОГО НАСЕКОМОГО
29
приходится сталкиваться с более или менее выраженной редукцией тер-
гитов и стернитов. Эти последние представлены чаще всего то узкими тя-
жами, то более широкими пластинками, лежащими в дорзальной и вен-
ДОЛ/: ’
24
Рис. 15—24. Строение яйцеклада.
15 —Dasyphora serena Mg. 16 — Orthellia cornicina L. 17 —
Stomoxys calcitrans L. 18 — Morellia hortorum Fall. 19 —
Dasyphora saltuum Rd. 20 — Graphomyia maculata Scop.
21—Mesembrina meridiana L. 22—24 — хитиновые шипики
на межсегментной перепонке яйцеклада: на 6-м сегменте
Orthellia cornicina F. (22); на 5-м сегменте Morellia horto-
rum Fall. (23); на 5-м, 6-м и 7-м сегментах Stomoxys calci-
trans L. (24). (Ориг.).
тральной мембране яйцеклада. Степени сплошных широких полуколец
стерниты и тергиты достигают лишь в яйцекладе Graphomyia (рис. 20),
в котором они представлены поперечно вытянутыми склеритами. Срав-
нительно мощные хитиновые тяжи наблюдаются в яйцекладе Mesembrir t
и Acridomyia, у которых, однако, они образуют не поперечные, а широ-
кие продольные пластинки.
Рис. 25—37. Заднегрудные дыхальцы.
25 — (левое) Polietes. 26 — (правое) Orthellia. 27 — (левое) Mesembrina. 28 — Gra-
phomyia. 29 — Scopeuma. 30 — Calliphora. 31 — Stomoxys. 32 — Hylemyia. 33 —
Musca. 34 — Morellia. 35 — Dasyphora. 36 —Pyreilia. 37 — Conops. (Opur )
МОРФОЛОГИЯ ВЗРОСЛОГО НАСЕКОМОГО
31
Отрезки яйцеклада, соответствующие 6-, 7-, 8- и 9-му сегментам, отде-
лены друг от друга межсегментной перепонкой, у большинства видов
покрытой волосками {Pyreilia cadaverina), шипиками {Orthellia, частично
Pyreilia) или нежными чешуйками {Haematobia)', эти особенности межсег-
ментной перепонки можно наблюдать у одних видов на всех ее частях,
у других — лишь на некоторых, вполне определенных ее участках.
Степень развития межсегментной перепонки неодинакова у разных
видов. То она очень сильно развита, представляя собою тонкую полупро-
зрачную трубку {Orthellia, Pyreilia), то редуцирована настолько, что
каждый сегмент яйцеклада принимает форму очень короткой трубки или
узкого кольца. Подобный тип строения наблюдается у многих живоро-
дящих видов. Варьируют в длине и склеротизованные части каждого сег-
мента яйцеклада и весь яйцеклад в целом. Особенно короток яйцеклад
у живородящих Musca larvipara, М. convexifrons, Dasyphora asiatica и
D. saltuum-, как упоминалось выше, наибольшего развития склеротизо-
ванные части сегментов яйцеклада достигают у Graphomyia. Dasyphora
saltuum (рис. 19) обладает, наоборот, наименее развитым скелетом яйце-
клада. В первом случае 6-й тергит представлен узким, загнутым на вен-
тральную сторону полукольцом. VI стернит имеет вид поперечно вытя-
нутой пластинки. Значительно слабее развиты твердые хитиновые части
6-го сегмента у трибы Stomoxydini {Stomoxys, Haematobia, Acridomyia),
у представителей которой тергит расщеплен вдоль на две широкие, лате-
рально смещенные пластинки; стернит имеет вид широкого, но непар-
ного склерита, лежащего по середине вентральной поверхности 6-го сег-
мента. Следующая ступень редукции хитиновых частей наблюдается
у Mesembrina meridiana, у которой тергит 6-го сегмента расщеплен вдоль
надвое; части его имеют вид широких прямоугольных полос; VI стернит
непарный, несколько уже их, но все же не имеет еще характера узкого
хитинового тяжа. У систематически более близких форм, как Pyreilia,
Orthellia, Musca, 6-й тергит представлен двумя узкими, длинными или
короткими, но тонкими, параллельно и близко друг к другу лежащими
тяжами, иногда (некоторые Musca и Orthellia) слитыми своими основными
отделами и в этом случае образующими вилку. Уклоняющимися от этого
обычного для группы типа формами являются живородящие Dasyphora,
где сокращение длины яйцеклада вызвало сокращение и хитинового ске-
лета. Здесь 6-й тергит иногда распознается по узким выступам тонкого
хитина, направленным от краев 5-го тергита по бокам яйцеклада; VI
стернит имеет форму маленького треугольника, заложенного в мембране
на вентральной поверхности яйцеклада.
Сходное строение с 6-м имеет следующий 7-й сегмент яйцеклада.
В нем наблюдаются иногда укорачивание стернита или редукция его
(некоторые Pyreilia). В редких случаях имеет место полная редукция
(некоторые Pyreilia) хитиновых тяжей (и тергитов и стернитов). У неко-
торых Dasyphora короткие латерально расположенные и имеющие вид
треугольников части тергита слиты с аналогичными частями следующего
(8-го) сегмента. 8-й сегмент отличается от предыдущего у большинства
форм {Orthellia, Musca, Pyreilia, Morellia) наличием парного склерита,
имеющего, подобно тергиту, два продольных хитиновых тяжа или пла-
стинки. У Stomoxydini {Stomoxys, Haematobia) стернит выражен хитиновой
(одной или двумя) пластинкой на дистальном крае сегмента. Dasyphora
имеет непарный стернит в форме поперечного маленького склерита.
9-й сегмент яйцеклада укреплен двумя хитиновыми пластинками
сверху (анальной) и снизу (субгенитальной); форма их то полукруглая или
32
ВВЕДЕНИЕ
овалыия, то квадратная или треугольная. По бокам сегмента ниже аналь-
ной пластинки причленено по одной анальной церке..
-я | (Дистальные края сегментов яйцеклада обладают мелкими пластин-
ками,
различного строения соединенными
с продольными хитиновыми
тяжами. Пластинки по-
Рис. 38—40.
крыты щетинками, до-
стигающими у некоторых
видов значительного раз-
вития ^Orthellia, некото-
рые Pyreilia, Morellia
и др.).
Сегменты яйцеклада
Muscini лишены дыха-
лец.
По отношению к дли-
не тела насекомого дли-
на яйцеклада неодина-
кова. У некоторых видов
Musca длина предельно
растянутого яйцеклада
равна длине тела. Наи-
меньшая длина яйцекла-
да свойственна живоро-
дящим формам; Dasyp-
hora saltuum, у ко-
торой длина яйцеклада
не превышает длины пя-
того тергита брюшка,
представляет собою в
этом отношении крайний
тип.
Половая си-
стема самок (рис.
39, 40) в схеме состоит
из следующих частей,
которые подвергаются
Рис. 38. Внутренний половой аппарат самца комнатной
мухи, сп — семенной проток; са — семенная ампула;
рж— ректальные железы; с — семенники; свпр— се-
мявыносящий проток. (По Хьюитту). Рис. 39. Вну-
тренний половой аппарат самки комнатной мухи, в —
влагалище; ж — придаточные железы половой сферы;
дж — полушаровидные придаточные железы половой
сферы; с — семеприемники; я — яйцевод. (По Хьюит-
ту). Рис. 40. Яйцеклад комнатной мухи, ц — церки;
п — межсегментная перепонка; с — стерниты; т — тер-
гиты; VI — IX — сегменты брюшка с 6-го по 9-й.
(По Хо).
весьма сильным измене-
ниям в своих деталях не
только в пределах дале-
ких групп, но и от ви-
да к виду, чте находится
в связи с характером
размножения вида и спо-
собом откладки яиц или
личинок.
Яичники распола-
гаются в заднем отделе
брюшной полости в числе
одной пары. Каждый яичник состоит из большего или меньшего коли-
чества яйцевых трубок. Каждая яйцевая трубка утончается к дисталь-
ному концу, который прикреплен тонкой нитью к внутренней стороне
спинной поверхности брюшка. Проксимальные концы трубок расширены
и открываются отверстием в яйцевод. Правый и левый яйцеводы со-
МОРФОЛОГИЯ ФАЗ РАЗВИТИЯ
33.
единяются в один общий проток — более широкую трубку, дистальный
конец которой образует влагалище, открывающееся отверстием на по-
следнем сегменте яйцеклада (под анальным отверстием). У жцйородящих
мух впереди влагалища общий проток яйцеводов расширен. Расширен-
ная часть протока носит название матки. Стенки последней в некоторых
случаях (у живородящих видов) образуют расширение, полость которого
предназначена для жизни и питания личинки.
В область матки или влагалища впадают придаточные железы, кото-
рых может быть 2 или 1 пара. Первая из них имеет характер тонких тру-
бок,. сильно суженных в проксимальном отделе. Вторая пара желез,’ об-
наруживаемая у некоторых Musca, имеет шаровидную или лопастевидную
форму. Вблизи придаточных желез от стенки влагалища отходит обычно
3 семеприемника. Они состоят из дистальной склеротизованной капсулы,
имеющей на вершине фолликулярную оболочку.
МОРФОЛОГИЯ ФАЗ РАЗВИТИЯ
За редкими исключениями (случаи живорождения) мухи триб Mus-
cini и Stomoxydini являются яйценесущими.
Яйцо (рис. 41—44) имеет эллипсоидные очертания, к одному полюсу
более заостренное, почти прямое или несколько изогнутое относительно
своей длинной оси; дорзальная сторона в этих случаях слабо .вогнута,
вентральная — дуговидно выпуклая. Дорзальная сторона несет два
почти параллельных друг другу рубчика, слабо выступающие над поверх-
ностью яйца. У некоторых форм {Graphomyia) вдоль боковых сторон
яйца бывают развиты узкие, тонкие лопасти, образующие равномерные
закругленные выросты по обе стороны полюсов яйца. Редко (например
у Musca autumnalis) на верхнем полюсе яйца имеется заостренный вырост
в виде крючка (рис. 44). Хорион яйца как в отношении толщины и скульп-
турных особенностей поверхности, так и в окраске сильно варьи-
рует. От очень тонкого и почти бесцветного, что наблюдается у некото-
рых видов Musca (группа М. domestica), хорион изменяется до упругого,
сравнительно толстого и темноокрашенного — рыжего, коричневатого.
Скульптурные особенности на поверхности хориона выражены обычно
слабо и становятся заметными лишь при сильном увеличении. В этом
случае поверхность яйца представляется покрытой мелкими ячейками.,
несколько напоминающими рисунок пчелиных сот, или же она обладает
тонкой исчерченностью. Сильно варьирует от вида к виду величина яиц,
причем между размерами яиц и тем количеством, которое откладывает
муха, наблюдается обратная зависимость: чем меньше продукция яиц
у мухи, тем крупнее ее яйца.
Стадии личинки различаются особенностями ротоглоточного аппа-
рата, дыхалец, размерами тела и его моторными приспособлениями. Тело
личинки (рис. 58) состоит из 1 головного, 3 грудных и 10 брюшных сег-
ментов. Оно постепенно сужено к головному концу и более или менее
расширено назад; только у Graphomyia личинка сужена к обоим
концам.
Головной сегмент, или псевдоцефал, двураздельный: передний конец
несколько выемчатый, благодаря чему боковые отделы головы имеют
форму округлых или цилиндрических выступов, каждый из которых
несет по одной антенне и максиллярному щупальцу (рис. 64).
3 Фауна СССР, том XVIII, вып. 4.
34
ВВЕДЕНИЕ
Количество сензорных образований на головном сегменте различно.
Максимальное число их указывается для личинок Musca domestica (Том-
сен, 1938).
дп до
Рис. 41—46.
Рис. 41—44. Яйца. 41 — Musca domestica L. 42 —
Mesembrina meridiana L. 43 — Lyperosia irritans L.
44 — Musca autumnalis Deg'. Рис. 45, 46. Передний
конец тела личинки (схема). 45— I стадии: ус —
усики; по — передние отростки фарингеального скле-
рита; дп — дорзальная перемычка; дс— срединный
дорзальный склерит; до — дорзальные отростки фарин-
геального склерита; гс— гипостомальный склерит;
лс — лабиальные склериты; мщ—>максиллярные щу-
пальцы; фс— фарингеальный склерит; во — вентраль-
ные отростки фарингеального склерита. 46 — Перед-
ний конец тела личинки II стадии: рк— ротовые
крючки; уг — углевидный склерит; фм— фарингеаль-
ная мембрана; дг—переднее дыхальце. Прочие обоз-
начения как на рис. 45 и 47. (Ориг.).
Ротовой аппарат личинок, хорошо просвечивающий через тонкие
покровы тела, состоит из 3 основных склеритов: 1) ротовых крючков,
МОРФОЛОГИЯ ФАЗ РАЗВИТИЯ 35
у личинок I стадии они заменены аналогичным по функции срединным
дорзальным склеритом; 2) гипостомального и 3) фарингеального (или
базального) склерита. Помимо перечисленных, может присутствовать
различное число дополнительных мелких склеритов, которые изменяются
в зависимости от вида и стадии личинки.
Каждый из основных склеритов состоит из двух (кроме срединного
дорзального у личинок I стадии) симметрично расположенных по обе
стороны от средней вертикальной плоскости тела личинки хитиновых
пластинок (рис. 45—47), более или менее параллельных друг другу и со-
единенных поперечными перемычками.
Ротовое отверстие, к просвету которого направлены концы ротовых
крючков, лежит на нижней стороне головного сегмента ближе к его осно-
ванию. Боковые наружные его стенки обычно покрыты бороздками,
радиально расходящимися от ротового отверстия по нижней стороне
головного сегмента и распространяющимися далее на боковые отделы
последнего (рис. 64).
Грудные дыхальцы появляются у личинок этой группы мух только
со II стадии. Они без труда обнаруживаются на границе 1-го и 2-го груд-
ных сегментов. Они состоят из широкого или узкого трубковидного осно-
вания, или атриума, который по своей функции является фильтрующей
камерой, и пальцевидных или сосцевидных выростов, выступающих на
свободном крае атриума в числе 4—7, реже 7—12 (Musca) или 16—18
(Mesembrina).
За редкими исключениями, грудные и брюшные сегменты тела личинки
по переднему или переднему и заднему краям вооружены кутикулярными
шиповидными выростами. В подавляющем большинстве случаев шипы
располагаются поперечными рядами, которые образуют присущий
каждому виду рисунок на поверхности сегментов. Характерно, что, кроме
триб Muscini, Stomoxydini, Phaoniini и Mydaeini, ни одно из подсемейств
и триб Muscidae, а также систематически близких семейств серии
не имеет в личиночной фазе (III стадии) такого расположения и такого
характера шиповидных выростов кутикулы. Именно этот признак в связи
с другими свидетельствует, с одной стороны, о генетической близости
Anthomyidae к Calliphoridae -\-Sarcophagidae -\-Larvaevoridae (частью)-фИса-
lypteratae (в широком смысле), с другой стороны, о более отдаленном
родстве Anthomyidae с Muscini и Stomoxydini, а также Fanniini. Черты
строения личинок Fanniini в совокупности с морфологическими особен-
ностями имагинальной фазы дают все основания считать эту т руппу дву-
крылых достаточно резко очерченной. Если представители триб Muscini,
Mydaeini и Phaoniini в фазе личинки обладают рядами крепких и крупных
кутикулярных шиповатых выростов на вентральной стороне брюшных
сегментов, то в пределах Acalypteratae, Calliphoridae, Sarcophagidae, Lar-
vaevoridae и им близких семейств личинки вооружены с вентральной
стороны лишь мелкими многорядными шипиками. Триба Stomoxydini,
близкая к Muscini по ряду морфологических признаков личиночной фазы,
характеризуется таким же порядком расположения крупных кутикуляр-
ных выростов, имеющих, однако, здесь не форму острых шипов, а корот-
ких ребер.
В разнообразных группах личинок шиповидные кутикулярные обра-
зования распределены по телу с известной закономерностью. Средним
типом в этом смысле может служить такой, где головной и грудные сег-
менты тела вооружены сильнее брюшных, кроме одного-двух последних,
точно так же имеющих сильное вооружение. Примерами такого типа воо-
3*
36
ВВЕДЕНИЕ
ружения могут служить личинки видов Musca. Сокращение кутикулярных
выростов осуществляется здесь за счет верхней и боковых поверхностей
срединных брюшных сегментов. Вооружение нижней стороны тела, несу-
щей у данной группы личинок преимущественно двигательные функции,
претерпевает меньшие изменения. Установлено, например, что в III ста-
дии личинки родов Orthellia, Musca, Pyreilia, Dasyphora и других почти
утрачивают шиповидные образования на дорзальной поверхности средин-
ных брюшных сегментов, в то время как первая стадия их личинок,
на долю которой падает процесс внедрения в субстрат, обладает шипо-
ватой скульптурой брюшных сегментов с боковых и верхней сторон.
В расположении и характере кутикулярных выростов личинок
можно наметить несколько типов. Первый, охватывающий трибы Muscini,
Stomoxydini, Mydaeini, Phaoniini, характеризуется присутствием на вен-
тральной стороне тела одного-двух поперечных рядов крупных зубцов,
окаймленных сзади (часто и спереди) несколькими рядами более мелких
зубчиков (обычно двух размеров). У Stomoxydini крупные шипы превра-
тились в ребра, не нарушающие, однако, характера расположения, свой-
ственного Muscini.
Второй тип характеризуется многорядными очень мелкими, почти
одинакового размера шипиками, образующими своеобразный узор
на валикообразных вентральных поперечных вздутиях тела личинки.
Примером такого типа могут быть личинки большинства Anthomyidae,
многие Сalliphoridae и Larvaevoridae.
Третий тип характеризуется наличием крупных редких плоских
зубцов, окруженных многими рядами очень мелких шипиков, как это
наблюдается у личинок Cypselidae. Сходный тип, но с удвоенным рядом
крупных, редких зубцов, окруженных рядами мелких, можно наблюдать
у личинок Sepsidae и некоторых других. Своеобразным отклонением
от этого типа является расположение кутикулярных выростов, встре-
чающееся у Milichiidac, их личинки на вентральных вздутиях снабжены
кривыми острыми зубцами, образующими волнообразную линию, впереди
которой расположен ряд более мелких острых крючьев; оба ряда окру-
жены мелкими зубчиками. У некоторых семейств шипы заменены много-
численными щетиночками (Cordyluridae). Особенное строение вентраль-
ных кутикулярных выростов имеют Fanniini, у которых параллельно
с пучками и рядами щетинок имеются выросты различного строения,
бородавки и т. п. Своеобразный тип вентральных кутикулярных выро-
стов наблюдается у Sarcophagidae, личинки которых вооружены много-
численными, беспорядочно разбросанными по поверхности тела, мелкими,
одинаковыми шипиками.
Порядок расположения шипообразных выростов, форма, размеры
и число их служат надежными признаками в систематике и могут быть
широко использованы с диагностическими целями. Задний конец личи-
нок обладает характерной для каждого вида формой и постоянными
скульптурными признаками. В разбираемой группе тело личинок утол-
щено в направлении спереди назад. Сзади оно округлое или плоское,
как бы обрубленное.
Задние дыхальцы расположены у личинок в средней части 8-го брюш-
ного сегмента. Представлены они парой округлых или почковидных,
обычно темно окрашенных образований. Каждое дыхадьце (рис. 53)
состоит из дыхальцевой пластинки (<Эи), окруженной по краю сильно
склеротизованной полоской — перитремой (и), не всегда отчетливо раз-
витой. У внутреннего края каждого дыхальца, что отчетливо выступает
an 9, Но
56
vna..ia—ы.
47 — Передний конец тела личинки III стадии: ас — зубной склерит; ап — вентраль-
ная перемычка фарингеального склерита; спр— общий проток слюнных желез.
Прочие обозначения как на рис. 45 и 46. 48 -- задний конец тела личинки: ап —
анальная пластинка; б — бугорки; д— задние дыхальцы. 49—51 — типы задних
дыхалец у личинки I стадии. 52 — заднее дыхальце личинки II стадии. 53 — заднее
дыхальце личинки III стадии: дщ — дыхальцевые щели; пак. — пери спи ракулярные
железы, п — перитрема; дп — дыхальцевая пластинка; сд— стигмальпый диск.
54 — схема строения дыхальцевой щели: п — перегородки; р — рама; а — зубцы.
55 — схема строения заднего дыхальца: дщ— дыхальцевая щель;фк—фильтрую-
щая камера; п — перитрема; di, дз— диски; т — хитиновый тяж; т.р — трахея.
56 — общий вид ложнококона: пд — передние дыхальцы личинки. 57 — дыхальце
куколки: р — рожки; вд — внутренние дыхальцы:фк — промежуточная фильтрую-
щая камера; вф — внутренняя фильтрующая камера (рис. 55 по Мейере, рис. 56, 57
по Томсону, рис. 54 по Зимину прочие—оригинальные).
38
ВВЕДЕНИЕ
у личинок III стадии, помещается округлый стигмальный диск (с<Э) («цен-
тральное тело» других авторов), неподвижно срощенный с дыхальцевой
пластинкой. От центра диска отходит для соединения с фильтрующей
камерой хитиновый тяж (рис. 55, т), фиксирующий положение дыхальца
и камеры в кутикуле личинки. Соответственно стадиям личинки дыхаль-
цевые пластинки имеют 1 (I стадия), 2 (II стадия) или 3 (III стадия) ды-
хальцевых щели. Дыхальцевые щели имеют вид змеевидно или S-образно
изогнутых просветов. Исключением являются Acridomyia, где дыхаль-
цевые щели представлены короткими полосками, расположенными как
у жигалок на светлых участках перитремы. Каждая из дыхальцевых
щелей у взрослых личинок состоит из наружной сильно склеротизован-
ной рамы (рис. 54, р), от которой внутрь просвета щели отходят тонкие
перегородки (рис. 54, п). У некоторых видов личинок от рамы могут
отходить также зубцы (з), нависающие над просветом щели. У взрослых
личинок можно обнаружить помимо округлых щелей мелкие просветы,
являющиеся выходными отверстиями периспиракулярных желез (рис. 53,
пж); вокруг этих светлых отметин обычно располагаются венцом упругие
волоски. Периспиракулярные железы с системой волосков являются,
как предполагают, приспособлением, препятствующим смачиванию ды-
хальцевых щелей водой и проникновению последней в дыхательные пути
личинок. Одно дыхальце у личинок последней стадии может располагать
двумя-тремя и большим количеством периспиракулярных желез.
Подобно тому, как это наблюдается у передних дыхалец, задние
дыхальцы обладают фильтрующей камерой (рис. 55, дбк), представленной
широкой и короткой трубкой, одним концом соединенной с дыхальцевой
пластинкой, другим — с трахейным стволом (рис. 55, тр).
По своим относительным размерам, взаиморасположению, характеру
дыхальцевых щелей и перитремы задние стигмы очень разнообразны,
что позволяет использовать их не только для установления стадии
личинки, но обычно и ее вида.
Головной сегмент (псевдоцефал) (рис. 64) личинки развит незначи-
тельно. Длина и диаметр сегментов возрастает от передних к задним
сегментам, достигая максимума в области 7-го и 8-го брюшных сегмен-
тов. Сегменты 9-й и 10-й развиты незначительно, смещены на вентральную
поверхность тела, вклиниваясь между нижним краем 8-го и задним ниж-
ним краем 7-го брюшного сегментов.
Каждый из сегментов тела, кроме головного и трех последних брюш-
ных, обладает на нижней стороне поперечным валикообразным вздутием,
расположенным у переднего края сегмента и снабженным многочислен-
ными и крепкими кутикулярными выростами. Задний конец личинки,
в частности, 8-й, 9-й и 10-й сегменты (все одновременно или лишь неко-
торые из них) снабжены крупными мягкими выростами кутикулы. Среди
Muscini и Stomoxydini такие крупные мясистые выросты всегда присут-
ствуют на 10-м, 9-м и, только как исключение, на 8-м брюшном сегменте.
У представителей семейств Sarcophagidae, Calliphoridae и большинства
Acalypteratae имеют выросты все 3 последних сегмента, причем здесь,
наоборот, их отсутствие на 8-м сегменте представляется редким явлением.
Сегмент 8-й (брюшной) в схеме может обладать 14 выростами: 2 дор-
зальными, 2 субдорзальными, 2 верхними и 2 нижними латеральными,
2 вентральными, 2 субвентральными и 2 паравентральными. Они рас-
пределяются кольцом вокруг задних стигм. Личинки Muscini и Stomo-
xydini, как правило, несут на 8-м брюшном сегменте лишь пологие воз-
вышения (рис. 199) и лишь в редких случаях мелкие выросты.
58
6g Cg
Рис. 58—64.
58 — общий вид мускопдной личинки III стадии: jE>i—5ц>— брюш-
ные сегменты с 1-го по 10-й; В — валикообразные вздутия; Г1—Гз —
грудные сегменты; ГЛ—главный сегмент. П — плевральные отделы тела;
С], С2 — переднее и заднее дыхальцы. 59 — строение усика личинки
Graphomyia. 60 — максрллярное щупальце личинки Graphomyia. 61 — ося-
зательный орган на псевдоподиях личинки Graphomyia. 62 — бугорки
ротовой полости. 63 — нижнегубное щупальце е сензорной щетинкой
у личинки того же вида. 64 — головной сегмент личинки комнатной
мухи: ус — усики; щ — максиллярные щупальцы; бг— б4— сензорные
бугорки; р — ротовое отверстие (Рис. 58 ориг., рис. 64 — по Томсену,
прочие — по Кейлину).
40
ВВЕДЕНИЕ
Сегмент 9-й имеет не более 5 выростов, у большинства видов разби-
раемой группы представленных коническими узкими возвышениями
вблизи анального отверстия. Среди этой категории выростов мы разли-
чаем 2 субанальных, 2 экстраанальных и 1 постанальный (рис. 270).
Максимальное число выростов 10-го сегмента равно 6: 2 аналь-
ных, 2 преанальных и 2 параанальных (рис. 85, 461).
10-й брюшной сегмент личинок Muscini и Stomoxydini очень мал.
Он имеет форму сравнительно небольшой, полулунной, округлой или
веретеновидной площадки — анальной пластинки, по середине которой
между двумя маленькими анальными бугорками открывается анальное
отверстие. Анальная пластинка имеет вид отчетливо отграниченной
области, форма и размеры которой константны в пределах вида. В неко-
торых случаях анальная цластинка {Morellia, Musca larvipara, М. auturn-
nalis) имеет вид поперечно вытянутой полоски, далеко заходящей на боко-
вые поверхности тела личинки. У Musca domestica и у целого ряда видов
Mydaeini и Phaoniini она представлена короткой площадкой на вентраль-
ной стороне тела.
Фаза личинки у Muscoidea имеет 3 стадии. Различие между личи-
ночными стадиями сказывается в строении дыхалец, устройстве рото-
глоточного аппарата, вооружении тела (в форме шипов, щетинок и их
размерах).
Личинка I стадии (рис. 45, 49, 50, 51). Парные ротовые крючки отсут-
ствуют, их заменяет не гомологичный им хитиновый непарный склерит —
срединный дорзальный склерит, имеющий листовидную или клиновидную
форму. Он помещается в головном сегменте личинки и направлен своим
заостренным дистальным концом к ротовому отверстию; этот склерит
является гомологом верхней губы. Задний конец срединного дорзального
склерита раздвоен. Каждый из двух концов направлен назад и обычно
несколько вбок, где с ними подвижно соединяются боковые части пар-
ного гипостомального склерита. Последний не всегда ясно выражен; он
имеет вид двух коротких хитиновых тяжей, соединенных друг с другом
более или менее широкой перемычкой. Задние концы гипостомального
склерита сращены со следующим — базальным или фарингеальным —
склеритом, состоящим из двух вертикально расположенных хитиновых
лопастей. В этой стадии он очень длинный, тонкий, с 4 длинными, узкими,
обычно заостренными задними отростками (дорзальными и вентральными).
Дорзальные отростки по дорзальным краям соединены друг с другом
узкой, иногда имеющей решетчатый вид, дорзальной перемычкой, в ред-
ких случаях последняя в среднем отделе плохо выражена. Передние
отростки базального склерита очень длинные, имеют характер сагит-
тально вытянутых тонких, почти прямых, хитиновых тяжей. С нижней
стороны к гипостомальному склериту подходит общий проток слюнных
желез.
Вентральные края базального склерита у некоторых видов бывают
ясно соединены со слабо склеротизованной лопастью, лежащей более
или менее параллельно нижней поверхности тела личинки и закрываю-
щей с вентральной стороны просвет между правой и левой пластинками
базального склерита. Эта лопасть, хорошо выраженная у большинства
личинок старших стадий, покрыта продольными так называемыми
фарингеальными реб ры ш к а м и, носит название базаль-
ной лопасти или фарингеальной мембраны. Фарингеальный склерит зало-
жен в области 1-го и 2-го грудных сегментов личинки. К вентральным
его отросткам подходит пищевод.
МОРФОЛОГИЯ ФАЗ РАЗВИТИЯ
41
Личинки I стадии метапнейстичны. Задние дыхальцы мелкие,
каждое снабжено одним маленьким, округлым, почковидным или сердце-
видным отверстием, нередко перегороженным узенькой перемычкой
на две части (рис. 49).
В I стадии личинки чаще всего не имеют того вооружения из шипи-
ков, которым обладают личинки всех последующих стадий. Нужно счи-
тать за правило, что у личинок, ведущих хищный образ жизни, воору-
жение из шипиков развито сильнее, нежели у копрофагов; особенно резко
это сказывается на первых (головном и грудных) сегментах тела. Антенны
и максиллярные щупальцы присутствуют в виде очень мелких образо-
ваний на вершине головного сегмента; последний обычно имеет более
или менее ясную двулопастную вершину (рис. 45).
Со II стадии (рис. 46) личинки становятся амфипнейстичными. Перед-
ние дыхальцы расположены на границе 1-го и 2-го грудного сегментов;
они представляют собою короткие выросты, заканчивающиеся рядом
пальцевидных или сосцевидных выступов.
Задние дыхальцы (фиг. 52), по сравнению с таковыми личинок I ста-
дии, достигают значительно больших размеров. Количество дыхальце-
вых отверстий равно 2; они слабо изогнуты и сравнительно широкие.
В строении ротоглоточного аппарата происходят глубокие измене-
ния. Это выражается в конфигурации хитиновых склеритов и в их коли-
честве (рис. 46). Срединный дорзальный склерит заменяется парными
ротовыми крючками, иногда сращенными в вершинных частях, распола-
гающимися под некоторым углом к продольной оси тела личинки. Обычно
это заостренные, реже тупые, саблевидно или серповидно изогнутые или
прямые, сравнительно короткие пластинки, подвижно сочлененные сво-.
ими основаниями друг с другом и с передними концами гипостомального
склерита и угловидным склеритом; последний, как правило, отсутствует
у хищных форм, но может быть обнаружен у копрофагов - (Musca, Sto-
moxys, Orthellia и др.). Непосредственно под ротовыми крючками поме-
щаются парные губные склериты, имеющие различное строение у разных
видов. Помимо губных склеритов нередко присутствуют добавочные скле-
риты, лежащие по бокам ротового отверстия впереди губных склеритов.
Что касается угловидного склерита, то он состоит из двух узких пласти-
нок, соединенных друг с другом своими дорзальными концами (Dasy-
phora') или имеющих вид самостоятельных мелких склеритов (Haematobid).
Угловидный склерит подвижно сочленен с основанием ротовых крючьев
и гипостома л ьным склеритом; последний у этой стадии личинок выражен
в виде двух нешироких, сагиттально вытянутых лопастинок, длина кото-
рых более чем в два раза превышает ширину. Задними концами гипосто-
мальный склерит сочленен с передними отростками фарингеального
(базального) склерита, более широкого и массивного в этой стадии. Перед-
ние отростки базального склерита широкие, не очень длинные; срединный
отдел склерита относительно шире и длиннее, в том же направлении
изменяются и задние отростки. Дорзальная перемычка, связывающая
дорзальные края склерита, по сравнению с телом последнего, кажется
маленькой; решетчатое строение перемычки выражено у личинок II ста-
дии резче. Передние отростки базального склерита иногда бывают соеди-
нены снизу поперечной вентральной перемычкой, хорошо заметной
у личинок Mesembrina и соответствующей, очевидно, мелкому попереч-
ному склериту у личинок Orthellia, не связанному здесь столь тесно
с концами фарингеального склерита. С переходом личинок во II стадию
увеличивается количество шипиков на теле.
и.
ВВЕДЕНИЕ
Личинки III стадии резко отличаются от личинок II стадии строе-
нием ротового аппарата (рис. 47), устройством задних дыхалец и рядом
других особенностей (вооружение тела, конфигурация анальной пла-
стинки и др.).
Дыхальцевые пластинки имеют по 3 (рис. 53), а не по 2, как во II ста-
дии, щели. В большинстве случаев каждая из дыхальцевых щелей пред-
ставляется в виде тонкой, светлой более или менее извитой линии {Musca,
Dasyphora, Mesembrina, Orthellia и др.). Значительно реже мы встречаем
на дыхальцевой пластинке вместо тонких линий широкие или узкие
слабо S-образно изогнутые просветы {Haematobia, рис. 452).
У личинок, близких к линке на III стадию, удается наблюдать одно-
временно 2 щели, свойственные II стадии, и под ними слабо намечаю-
щиеся 3 щели будущей III стадии.
Характер дыхальцевых щелей, степень изогнутости, число изгибов,
направление концов щелей по отношению к центру пластинки имеют
значение в систематике личинок. Не меньшую систематическую ценность
представляет собою строение и форма самих дыхальцевых пластинок,
расстояние между ними, степень развития перитремы.
Личинки III стадии обладают весьма мощным ротовым аппаратом
(рис. 47), резко отличным от такового у личинок предыдущей стадии.
Ротовые крючки, гипостомальный и фарингеальный склериты у III ста-
дии становятся много крупнее. Ротовые крючки делаются шире, осно-
вание их более массивно. Расширяются пластинки гипостомального
и фарингеального склеритов. У некоторых форм, ведущих не хищный
образ жизни, резче выступает фарингеальная мембрана; обычно она
несколько выступает позади вентральных отростков. Дорзальная пере-
мычка базального склерита, наоборот, становится более короткой и не
такой высокой.
Боковые лопасти фарингеального склерита образуют боковые стенки
глотки. Нижнюю стенку образует фарингеальная мембрана с ее продоль-
ными ребрышками, верхнюю — эластичная перегородка, боковые края
которой срощены с боковыми лопастями фарингеального склерита. Верх-
ние края последних являются местом прикрепления косых мышц глотки,
которые нижними своими концами срощены с перегородкой (верхней
стенкой глотки). При сокращении мышц перегородка подтягивается
кверху, расширяя полость фарингса. Нижняя стенка глотки в области
фарингеального склерита изнутри покрыта продольными ребрышками,
несущими на своем свободном верхнем крае по две тонкие перепончатые
полоски. При подтянутой кверху верхней стенке глотки полоски под-
няты кверху, при опущенной вниз верхней стенке глотки, полоски закры-
вают сверху просвет между ребрами, образуя, таким образом, род глухих
продольных канальцев.
Вблизи ротовых крючьев обнаруживается ряд парных хитиновых
образований, достигающих наибольшей сложности у хищных личинок,
как Graphomyia и др. У копрофагов устройство рото-глоточного аппарата
проще.
Для Muscini и Stomoxydini характерно присутствие зубных склери-
тов, помещающихся под основанием ротовых крючьев. Они имеют форму
треугольников. Нижние их углы только у хищных или полухищных форм
соединяются поперечной перемычкой, отсутствующей у копрофагов
(Кейлин, 1917; Томсен, 1938). Перемычка, получившая название вен-
тральной перемычки, хорошо видна (при рассматривании объекта снизу)
в виде узкой ленты, нередко зазубренной по краю. Помимо зубных скле-
СИСТЕМАТИЧЕСКОЕ ПОЛОЖЕНИЕ ГРУППЫ
43
Рис. 65. Куколка ком-
натной мухи, извлечен-
ная из ложнококона.
(По Хьюитту).
д—дыхальце; вг—верх-
няя губа; к — крылья;
не — нижняя губа; ус —
усики.
ритов в этом отделе ротового аппарата могут присутствовать добавочные
склериты, помещающиеся вентрально от ротовых крючков. Эти склериты
у Graphomyia достигают сравнительно большой длины и сложности строе-
ния. Только у одного известного нам вида группы, Graphomyia таси-
lata, личинки III стадии имеют сверх перечисленных еще один склерит,
срощенный с добавочными склеритами: так называемый задний тяж,
весьма характерный для представителей Phaoniini, в частности, для
личинок Phaonia, Muscina и им близких.
Куколка (рис. 65). По достижению предельной зрелости тело
личинки III стадии сокращается в длине и несколько утолщается, при-
обретая форму вытянутого бочоночка. Оболочка личинки становится
твердой. Затвердевшая оболочка личинки — ложнококон или пупа-
рий — имеет желтый или коричневый цвет. Ин-
тенсивность окраски ложнококона зависит не
только от вида личинки, но и от возраста ложно-
кокона. Недавно сформировавшиеся ложноко-
коны светлее старых. Являясь затвердевшей
шкуркой личинки III стадии, ложнококон сохра-
няет черты строения последней. На нем без труда
различимы сегментация, шипики, скульптура
поверхности, свойственные личинке, анальная
пластинка и задние дыхальцы. Новообразованием
являются лишь мелкие, часто конической формы,
выросты на границе 1-го и 2-го брюшных сегментов
ложнококона, напоминающие короткие рожки.
Они представляют собою наружные части грудных
дыхалец куколки, заключенной внутри ложно-
кокона (рис. 56, 57).
Вершины рожек (фиг. 57) пронизаны много-
численными мелкими дыхательными отверстиями,
число которых не одинаково у различных видов.
От стенок рожек отходит внутрь ложнококона тон-
костенная трубка, прямая или изогнутая, пред-
ставляющая собою промежуточную фильтрующую
камеру. Камера в основном отделе или у середины имеет на одной
из стенок расширение в форме площадки, несущего дыхательные папиллы,
число которых может варьировать у разных видов от нескольких штук
до нескольких сотен (Мейере, 1916; Томсон, 1937). От места расшире-
ния далее внутрь пупария отходит большей или меньшей длины цилин-
дрическая камера — внутренняя фильтрующая камера, переходящая
непосредственно в упругую трахею. Присутствие рожек на поверхно-
сти пупария обнаружено не у всех Schizophora и не у всех представи-
телей трибы Muscini.
СИСТЕМАТИЧЕСКОЕ ПОЛОЖЕНИЕ ГРУППЫ
Трибы Muscini и Stomoxydini представляют часть богатого видами
семейства Muscidae (Anthomyidae auct.).
Границы семейства впервые были определены Гнршнером (Girscbner,
1893, 1896), выделившим его из группы близких семейств (Cordyluridae,
Larvaevoridae). Основанием к разделению послужили хетотаксические
признаки, строение брюшка, жилкование крыла и строение его базаль-
ного отдела. В этом крупном комплексе двукрылых Гиршнер различал
ВВЕДЕНИЕ
2 подсемейства (Coenosiinae, Muscinae), причем последнее представляло
собою смесь из филогенетических различных категорий не меньшего
систематического значения, чем первое подсемейство автора.
Тогда же Брауэром и Бергенштаммом (Brauer u. Bergenstamm, 1893)
была предложена иная система Muscoidea. Отделив от Anthomyidae трибы
Muscini и Stomoxydini, авторы включили их вместе с семействами Calli-
phorida'1, Sarcophagidae, Larvaeooridae и другими в разряд Muscaria Schi-
zometopa. Трибы Muscini и Stomoxydini вошли совместно с сем. Callipho-
ridae в «Секцию Muscina», причем Stomoxydini (вместе с Glossina) соста-
вили «подсекцию Stomoxys», а триба Muscini (-\-Calliphoridae) — «подсек-
цию Musca». В этой системе секция Muscina занимала промежуточное
положение между секциями «Rhinia» и «Oestridae». Вследствие чисто фор-
мальной оценки некоторых признаков в подсекцию Musca попали роды
Muscina (Cyrtoneura) и Myiospila. Если с фактом сближения Callipho-
ridae с трибой Muscini можно условно согласиться, то нельзя того же
сделать в отношении систематического положения Stomoxydini. Эта
триба, четко охарактеризованная морфолого-анатомическими признаками
(строение головы, хоботка, глаз, генитального аппарата, хетотаксия тела,
строение личинок и т. д.), должна после отделения от Muscini -\-Calli-
phoiidae занимать самостоятельное положение, а не как часть одной
секции, общей с последними.
Принятое авторами положение Stomoxydini, с нашей точки зрения,
допустимо лишь при условии целостности сем. Muscidae, без присоедине-
ния последнего к Calliphoridae, или при условии выделения каллифор
в отдельную подсекцию. Подобная операция потребовала бы пересмотра
Mydaeini и Phaoniini, систематически близких Muscini.
Система Бецци и Штейна (Bezzi u. Stein, 1907) явилась некоторым
улучшением классификации Гиршнера в том отношении, что все сем.
Muscidae (включая и Anthomyidae) было поделено на 3 подсемейства;
триба Stomoxydini осталась в подсемействе Muscinae.
Разработанная Шнаблем и Дзедзицким (Schnabl u. Dziedzicki, 1911)
система Muscidae (Anthomyidae), основанная главным образом на строе-
нии копулятивного аппарата самцов, внесла много ошибочных положений
в систему Гиршнера. Соединив в одну группу трибы Stomoxydini и Mus-
cini, авторы отделили от последней наиболее близкие ей в систематиче-
ском отношении роды (Pyreilia, Orthellia, Dasyphora) в самостоятельную
группу (Ordo II: Muscinae polietes-formes). Совершенно формально в одну
категорию с Musca, Stomoxys и другими введена Atherigona, имеющая
больше общих черт с трибой Coenosiini.
В 1916 г. Штейн разделял семейство на 7 групп, соответствующих
трибам — Muscini, Stomoxydini, Phaoniini, Mydaeini,Fucelliini, Anthomyini
и Coenosiini. Однако и здесь на основе жилкования крыльев роды Mus-
cina и Myiospila вошли в трибу Muscini, что представляется ошибочным.
В своем труде, посвященном семейству Muscidae, Штейн (1917) трибу
Stomoxydini помещает в системе семейства первой. Сеги (Seguy, 1923)
повторяет в основных чертах систему Штейна (1916), увеличив объем
семейства присоединением оводов (Gasterophilinae). Включение оводов
в семейство Muscidae не представляется нам обоснованным.
Карль (Karl, 1928) делит европейских Muscidae на 6 подсемейств.
С его точки зрения Muscini и Stomoxydini образуют отдельное подсемей-
ство (Muscinae), куда они входят на положении триб.
Рингдаль (Ringdahl, 1929) разбивает семейство по степени развития
анальной жилки на два подсемейства — Muscinae (толстая, короткая,
СИСТЕМАТИЧЕСКОЕ ПОЛОЖЕНИЕ ГРУППЫ
45
не достигающая края крыла ап жилка) и Anthomyinae (ап жилка в виде
складки доходит до заднего края крыла), очевидно предусматривая в даль-
нейшем дробление подсемейств на трибы. Ни Cordyluridae, ни тем более
Calliphoridae и Gasterophilidae им в расчет не принимаются.
Своеобразного взгляда на систему Muscidae придерживается Таун-
сенд (Townsend, 1935). По этому автору, семейство Muscidae входит само-
стоятельным семейством в надсемейство Muscoidea вместо с сем. Glossi-
nidae. Семейство Muscidae автором разделено на большое количество
триб, среди которых Stomoxydini, Graphomyini, Mesembrinini и Mus-
cini в совокупности соответствуют нашим двум трибам. Трибы поделены
на множество родов, что создает значительные неудобства. Целая серия
вновь установленных автором родов являются монотипическими. Морфо-
логически хорошо очерченные роды Mesembrina, Morellia, Musca и дру-
гие разбиты на несколько мелких родов. Так, вместо одного рода Mesem-
brina автор устанавливает 3 рода; Morellia разбивается на 2 рода, а род
Musca на 13 родов. То же самое имеет место и в отношении Dasyphora.
По нашим представлениям ошибка системы Таунсенда заключается
в сближении Glossinidae и Muscidae', к этому вопросу мы еще вернемся
в дальнейшем. Недостатком системы, кроме того, является излишнее
дробление родов, для чего Таунсенд использовал второстепенные адап-
тивные (а нередко вторичные половые) признаки, упуская из вида
общность в строении многих систем органов имагинальной и личиночной
фаз.
Из вышеприведенного обзора основных работ по систематике группы
с очевидностью вытекает недостаточная определенность в целом ряде
вопросов. Остается неясным прежде всего положение семейства Muscidae
внутри серии Muscoidea, и, следовательно, филогенетические связи его
с другими семействами той же серии двукрылых; неопределенными пред-
ставляются и границы как всего семейства в целом, так и составляющих
его звеньев (подсемейств, триб).
До полной систематической обработки сем. Muscidae и близких к нему
групп, как Cordyluridae, Anthomyidae, Calliphoridae, Sarcophagidae, Lar-
vaeooridae (в широком смысле) и Gasterophilidae — Oestridae, точное опре-
деление систематических взаимосвязей между ними вообще неосуще-
ствимо.
При имеющемся материале мы, поставив перед собою более узкие
задачи, имеем возможность говорить лишь о тех ближайших системати-
ческих категориях, которые непосредственно связаны с трибами Muscini
и Stomoxydini. Условно мы принимаем сем. Muscidae в меньшем объеме,
чем понимали его Штейн и Карль, а именно мы выключаем из состава
семейства формы с длинной, доходящей до края крыла анальной жилкой.
Таким образом, в сем. Muscidae по нашим представлениям не входят
Anthomyinae и Eginiinae, которые мы объединяем в семейство Anthomyidae.
Сем. Muscidae нами подразделяется на трибы: Muscini, Stomoxydini,
Phaoniini, Mydaeini, Сoenosiini и условно—Fanniini, положение кото-
рых, вероятно, потребует пересмотра.
Первые две трибы, которым посвящена настоящая работа, пред-
ставляют собою морфологически четко отграниченные от ближайших
триб группы. Триба Muscini характеризуется: 1) развитыми грудными
чешуйками (рис. 66, 67), имеющими характер широких пластинок, 2) угло-
видно или дугообразно изогнутой по направлению к переднему краю
крыла жилкой т, 3) оперенной аристой и 4) наличием волосков на плев-
ральных склеритах заднегруди вблизи задних тазиков. Характерно для
46
ВВЕДЕНИЕ
трибы также отсутствие крепких хитиновых склеритов в дистальной,
мембранозной части penis, короткие церки самцов; 6-й и 7-й тергиты
самцов имеют вид узких полуколец, снабженных дыхальцами; брюшные
сегменты самок с 6-го по 9-й лишены дыхалец. У обоих полов первое
и второе дыхальце заключены в хитин тергита (1 -|—2).
Взрослые личинки имеют S-образно или многократно змеевидно
изогнутые дыхальцевые щели задних стигм; валикообразные поперечные
вздутия снизу брюшных сегментов несут один поперечный ряд крупных
зубцов, ограниченный сзади и в меньшей степени спереди рядами мелких
шипиков. Дополнительные склериты ротоглоточного аппарата неразвиты
(как правило). У личинок I стадии присутствует непарный срединный
дорзальный склерит.
Триба Stomoxydini характеризуется следующими признаками: 1) глаза
удлиненные с более или менее вогнутым задним краем, 2) затылок обычно
выпуклый в нижнем отделе, 3) хоботок тонкий, длинный, с маленькими
сосательными лопастями, 4) медиальная жилка на вершине дуговидно
изогнута в направлении костального края крыла, 5) грудная чешуйка
узкая (рис. 68), языковидная, как исключение — широкая или развита
в форме узкой поперечной полоски (Acridomyia)-, ариста перистая
(у Acridomyia — голая). Penis с двузубчатым поперечным образованием,
с склеротизованными продольными отростками на вершине; перки
широкие.
Взрослые личинки обладают S-образно изогнутыми щелями задних
дыхалец; вентральные вздутия брюшных сегментов с рядом косых круп-
ных ребер и мелкими зубцами позади и впереди них. Ротоглоточный
аппарат без дополнительных склеритов (исключение — Graphomyia).
В первой стадии личинки обладают срединным дорзальным склеритом.
В нашем представлении Phaoniini и Mydaeini филогенетически наибо-
лее близки друг другу. Современных Muscini, имеющих много общего
с Phaoniini, мы рассматриваем как производное ствола последних. Что
касается Stomoxydini, то современные их виды имеют больше сходства
с современными Mydaeini, почему нам кажется наиболее справедлиным
искать родоначальную форму современных жигалок среди предковых
видов Mydaeini.
Трибу Muscini сближает с Phaoniini строение генитального аппарата
самцов, присутствие дорзальной (точнее — задне-дорзальной) щетинки
на задних голенях, конфигурация грудных чешуек и характер рисунка
на спинке и брюшке (парные полосы или пятна). Mydaeini сближаются
с жигалками на основании следующих признаков: отсутствие дорзальной
щетинки на задних голенях, узкие грудные чешуйки, рисунок на спинке
и брюшке у Mydaeini часто характеризуется наличием непарного ряда
пятен или непарной полосы, что присуще Stomoxydini (кроме Acridomyia).
Как явствовало из нашего обзора, Muscidae и Anthomyidae объединя-
лись многими авторами в одно семейство (Muscidae). Мы считаем необ-
ходимым рассматривать их в качестве самостоятельных семейств на осно-
вании следующих признаков.
Muscidae
1. Анальная жилка не достигает зад-
него края крыла.
2. Стигмы на сегментах яйцеклада
отсутствуют.
3. Грудные чешуйки в подавляющем
большинстве случаев шире и много
длиннее крыловых чешуек.
Anthomyidae
la. Анальная жилка в виде складки
доходит до заднего края крыла.
2а. Одна или две пары дыхалец на сег-
ментах яйцеклада.
За. Грудные чешуйки не бывают широ-
кими и очень длинными.
СИСТЕМАТИЧЕСКОЕ ПОЛОЖЕНИЕ ГРУППЫ
47
4. Брюшко коническое или яйцевидное,
причлененное сравнительно высоко,
отчего мезофрагма оказывается су-
женной (поперечной).
5. Взрослые личинки без венца мягких
конических выростов на заднем конце
тела, или с остатками их; задние
стигмы часто с извилистыми дыхал ь-
цевыми щелями.
6. Брюшные сегменты зрелых личинок
снизу, по крайней мере, с одним
рядом крупных зубцов или валиков.
7. Личинки, сапрофаги, копрофаги, хищ-
ники, очень редко растительноядные.
4а. Брюшко, как правило, цилиндри-
ческое, низко причлененное к груди,
отчего мезофрагма выступает длин-
ным склеритом (продольно вогнутым).
5а. Взрослые личинки с длинными ко-
ническими выростами на заднем
конце тела; щели задних стигм
всегда прямые.
6а. Брюшные сегменты зрелых личинок
с многорядными мелкими шипиками.
7а. Личинки по преимуществу расти-
тельноядные, реже паразиты, хищ-
ники или копрофаги.
Если основываться на морфологии имагинальной фазы, то из се-
мейств тахиноидного комплекса наиболее близким к Muscidae является
сем. Calliphoridae', сближает их строение головы, жилкование крыльев,
строение фаллозомы, окраска тела. Что касается морфологии личинок,
то в ней мы не находим подтверждения этой близости. Строение личинок
Calliphoridae и Sarcophagidae указывают на родственные связи этих
семейств с семейством Anthomyidae (характер шипов на теле, присутствие
паростомального склерита в ротоглоточном аппарате, прямые щели зад-
них дыхалец, присутствие венца конических выростов на заднем конце
тела, парное во многих случаях строение ротовых крючьев у личинок
I стадии). Большинство перечисленных признаков отсутствует у личинок
Muscidae и многих Larvaevoridae. Мусцидный тип личинок с теми или
иными изменениями мы встречаем у семейств Muscidae, Larvaevoridae
и Oestridae. В то же время большое сходство в личиночной фазе обнару-
живается у семейств Cordyluridae—Anthomyidae—Calliphoridae—Sarcopha-
gidae. Близки к ним по морфологии и личинки некоторых семейств
Acalypti atae.
Таким'образом, анализ личиночных структур с полной очевидностью
подтверждает необходимость пересмотра общей системы Schizophora,
построенной без учета всех фаз онтогенеза и их биологии, а лишь на мор-
фологических особенностях окрыленных мух, что свидетельствует о ее
неестественности.
Триба Muscini содержит 6 палеарктических и 8 родов, распростра-
ненных в тропических странах Эфиопской, Ориентальной и Неотропи-
ческой областей (Dasymorellia Mall., Xenomorellia Mall., Fraseria Mall.,
Alluaudinella Giglio-Tos, Aethiopomyia Mall., Ochromusca Mall., Neo-
musca Mall.1
Центральным родом трибы представляется род Pyreilia R.-D. (с типом
aenea Zett.).
Генетически близок к Dasyphora род Morellia R.-D., еще сохраняю-
щий опушенность глаз, сходную хетотаксию грудного отдела и характер
строения фаллозомы; однако церки уже значительно изменяются, при-
обретая некоторое сходство с таковыми рода Mesembrina Mg., с которым
Morellia имеет большое сходство и в окраске тела.
У Mesembrina мы встречаем уже много общих черт и с трибой Phao-
niini и особенно с видами рода Polietes. Однако у Mesembrina чешуйки
1 Pyrellina Mall, (тип inventrix Walk.), обладающий узкими подкрыловыми
чешуйками, очевидно не должен входить в трибу. Положение родов Synthesiomyia
Вг. В., Reinwardtia Вг. В., Aethiopomyia Mall., Ochromusca Mall., Neomusca Mall.,
не вполне ясно без просмотра типов.
43
ВВЕДЕНИЕ
сохраняют тип, свойственный трибе Muscini, тогда как у Policies они
уже, как это свойственно типичным представителям трибы Phaoniini.
Генетическая близость Mesembrina к Policies подкрепляется особенностями
биологии, характером жилкования крыльев (широкая 7?б) и строением
личинок (передние дыхальцы со склеротизованным склеритом в атриуме).
Подрод Rypellia Mall, отражает в себе признаки родов Pyreilia
и Orthellia, но в большей степени рода Pyreilia R.-D. Так, при наличии
свойственных РугеШа признаков: дуговидный изгиб медиальной жилки,
отсутствие волосков на переднегруди и супрасквамальном ребре, узкий
лоб самца, узкие щеки, слабо выступающий вперед нижний край лица, —
виды Rypellia имеют длинное заднее дыхальце и широкую переднегрудь,
что присуще роду Orthellia. В роде Orthellia фаллозома несет крепкие
многочисленные шипы, которые лишь в незначительной степени развиты
у Pyreilia. По всем морфологическим особенностям, исключая угловид-
ный изгиб медиальной жилки, род РугеШа представляется генетически
более близким роду Musca L., нежели род Orthellia (строение генита-
лий 55, головы: лоб, скулы, щеки, глаза, медиана, хетотаксия ее и дру-
гие).
Род Graphomyia R.-D., мы относим к трибе Stomoxydini. С трибой
Stomoxydini род имеет много общего в строении глаз (длинные выем-
чатые по заднему краю), щек (узкие), головы (высоко приподнятый ниж-
ний край лица), отсутствие, как правило, передних стерноплевральных
щетинок; хорошо развитые хитиновые части 6—10 тергитов брюшка
самок, удлиненные церки самцов и характер темного рисунка, типич-
ный для многих жигалок.
По нашим представлениям, триба Stomoxydini в пределах Палеарк-
тики разделяется на три подтрибы: Graphomyiina (с одним родом), Sto-
moxydina (с 4 родами) и Acric.om.yina (с одним родом). В пределах Пале-
арктической области Stomoxydini представлены небольшим числом видов.
Некоторые их виды распространены всесветно. Наибольшего многообра-
зия группа достигает в Эфиопской и Ориентальной областях, где против
8 палеарктических видов мы имеем свыше 70 экзотических.
За исключением одного рода (Acridomyia), приспособившегося
к паразитическому образу жизни на саранче и в связи с этим значительно
отличающегося от остальных родов трибы, все известные нам палеаркти-
ческие представители родов трибы, относящиеся к родам Stomoxys Geoffr.,
Haematobia R.-D., Lyperosia Rd., представляют собою морфологически
и биологически резко очерченную группу.
Бецци (1911) в своем обзоре разбивает жигалок на 2 подсемейства:
Glossinae и Stomoxydinae, относя к последнему 6 родов. Достоинством
системы Бецци является отделение Glossina в качестве самостоятельной
группы. Пэттон (Patton 1933), включая род в подсемейство Muscinae,
относит его к трибе Muscini, наряду с родом Stomoxys. По Генделю (Hen-
del, 1936), Glossina составляет особое подсемейство Muscidae (Glossinae),
равно как и Stomoxydinae, при чем в последнее отнесено 2 трибы (Sto-
moxydini и Haematobiinae). Таунсенд выделил Glossina в особое семейство.
Принимая в качестве основы нижеперечисленные признаки, мы
считаем мух це-це в систематическом отношении далеко отстоящими
от сем. Muscidae и, наоборот, чрезвычайно близкими кровососкам Hippo-
boscidae.
Glossina сближается с Hippoboscidae на основе следующих морфоло-
гических признаков. 1) Сильно развитый вентральный отдел медиотер-
гита; 2) редуцированная срединная часть переднегруди, замененная мем-
Географическое распространение
49
браной; 3) сильно развитый основной стернит брюшка и редуцированные
срединные стерниты; 4) мощно развитая мембрана брюшка, снабженная
многочисленными волосками; 5) развитие стернитов 2-го и 3-го грудных
сегментов; 6) строение наружной поверхности фасеточных глаз; 7) харак-
тер устройства наружных гениталий самцов; 8) устройство внутренних
половых органов самцов; 9) сильно раздутое у основания сосальце;
10) разветвленность волосков аристы; 11) ясно расширенный у основа-
ния эмподий. Помимо этого виды рода Glossina, как и кровососки, отно-
сятся к категории куклородных двукрылых, что имеет свое отражение
и в морфологии внутренних половых органов самок и личиночной фазы.
Таким образом, с нашей точки зрения глоссины должны составлять
особое семейство — Glossinidae, близкое семейству Hippoboscidae. О фило-
генетической близости глоссин и жигалок можно говорить лишь
постольку, поскольку те и другие относятся к семействам мускоидных
двукрылых.
ГЕОГРАФИЧЕСКОЕ РАСПРОСТРАНЕНИЕ
Данные о географическом распространении группы сведены в виде
таблицы. В таблицу, где показано обилие видов группы по зоогеогра-
фическим областям, не включен ряд экзотических родов по той при-
чине, что принадлежность их к разбираемым здесь трибам вызывает
сомнение. К числу таких родов относятся: Aethiopomyia G. Т., Och.ro-
musca Mall., Reinwardtia Вт. В., Synthesiomyia Вт. В. Роды Muscina
R.-D., Myiospila Rd., Pyrellina Mall, при нашем толковании трибы
Muscini должны быть из нее исключены и присоединены к трибам Phao-
niini и Mydaeini.
В таблицу не включены сомнительные виды, относящиеся к родам
Musca, Orthellia, Pyreilia, Graphomyia, что заставляет считать приведен-
ные цифры до известной степени условными.
По неполным данным триба Muscini содержит, таким образом, около
350, а триба Stomoxydini около 75 видов.
Наибольшего видового многообразия та и другая трибы достигают
в Эфиопской и Индомалайской областях, где сосредоточено около 2/з всех
описанных видов группы. В Неарктике группа представлена наимень-
шим числом форм. Что касается эндемичных родов, то они в Неарктике
вообще отсутствуют, тогда как в Эфиопской, Австралийской, Индо-Ма-
лайской и Неотропической областях известно по 1 эндемичному роду.
После выяснения систематического положения упомянутых выше родов,
систематическое положение которых недостаточно выяснено, эти соотно-
шения могут несколько измениться. В Палеарктике эндемиком мы
должны считать род Dasyphora, представленный здесь 14 видами; правда,
один вид, характеризующийся большим сходством с видом рода Pyreilia,
особенно богато представленного в Неарктике, является общим с Север-
ной Америкой. Таково же распространение и рода Mesembrina.
Из 17 родов рассматриваемой группы 8 наиболее богатых по
видовому составу распространены по всему земному шару. Из прочих
родов Acridomyia свойственна одновременно Палеарктике и Северной
Америке; Mesembrina представлена 7 видами в Палеарктике и одним
общим с Палеарктикой видом — в Неарктической области; два вида
Mesembrina известны из Индо-Малайской области.
Таким образом, в Палеарктике насчитывается 6 родов трибы Muscini
и 5 — трибы Stomoxydini. Обильнее всех представлен род Musca, изве-
4 Фауна СССР, том XVIII, вып. 4,
50
ВВЕДЕНИЕ
Наименование родов и триб Палеарк- тическая Неаркти- ческая Эфиопская Индо-Ма- лайская и Австра- лийская Неотропи- ческая
Триба Muscini
1. Род Orthellia R.-D 4 2 35 34 11
2. » Musca L 19 21 25 32 2
3. » Pyreilia R.-D 6 4 24 15 16
4. » Dasyphora R.-D 14 1 — — —
5. » Morellia R.-D 9 2 15 8 16
6. » Xenomorellia Mall — — — — . 1
7. » Mesembrina Mg 7 1 — 2 —
8. » Fraseria Mall — — 1 —
9. » Alluaudinella G.-T — — 6 — —
10. » Neomusca Mall — — — — 2
Триба Stomoxydini
11. » Graphomyia R.-D 1 6 11 16 5
12. » Gordonia Mall — — — 1 (Австр.) —
13. » Stomoxys Geoffr 1 3 23 11 2
14. » Haemato^ia R.-D 3 2 6 9 3
15. » Lyperosia Rd 5 1 9 3 1
1 (Индо-
16. » Haematostoma Mall — — — Мал.) —
17. » Acridomyia Stack 1 2 — —
Итого: Muscini . . . . 59 31 105 92 48
» Stomoxydini . . 11 14 49 41 11
всего: . . . 70 45 154 133 59
ГЕОГРАФИЧЕСКОЕ РАСПРОСТРАНЕНИЕ
51
стный от крайних северных пределов области до южных ее границ, при-
чем в Арктическую подобласть проникает лишь один М. domestica. Энде-
мичными для области являются два вида. Из них один распространен
в Сибири на восток от р. Енисея и связан, видимо, с таежным ландшаф-
том {М. arnica, sp. nov.). Типичный средиземноморский вид — М. vitri-
pennis Mg., известный из южной и центральной Европы, а в пределах
СССР обнаруженный по побережью Черного моря и в югозападной
Украине, является также эндемичным для области видом; он не распро-
страняется в пределы восточно-средиземноморской провинции и связан,
очевидно, с условиями теплого и влажного климата. Близкий ему вид —
М. osiris Wd. обитает в условиях континентального климата и, невиди-
мому, представляет собою экологически весьма пластичную форму, изве-
стную как в Индии и Африке (в последней — повсеместно), так и повсюду
в степной зоне Палеарктики; он обычен в пустынях и нагорьях Азии
и сравнительно далеко проникает на север восточно-европейской провин-
ции (Кировская обл.); в Средней Азии вид заходит в горы на высоту
до 1000—2000 м.
Особый интерес представляют виды с явной адаптацией к суровому
климату песчаных пустынь — Musca albina Wd. и М. lucidula Lw.,
личинки которых найдены в норах грызунов и черепах. Эти виды не захо-
дят на север далее пустынь Средней Азии и южных границ западно-среди-
земноморской подобласти. Ксерофильный характер этих видов объясняет
нахождение их в Африке и отсутствие в условиях влажного и жаркого
климата Индо-Малайской области. Элементы фауны последней проса-
чиваются в Палеарктику в восточных ее частях (Средне- и Южнокитай-
ские и Амурская провинции). В СССР такие виды обнаружены в При-
морском крае {Musca convexifrons Thoms). Связанный с влажными тропи-
ками Musca sorbens Wd. — обычный синантропный вид в Индии и на
Малайском архипелаге — в Эфиопской области и в Среднеазиатских
пустынях придерживается оазисов; М. sorbens в кругу других смежных
с нею видов является единственной формой, проникающей в Палеарктику;
прочие распространены в Индо-Малайской области, Африке и Австра-
лии. Не имеет своих представителей в Палеарктике и группа опушенно-
глазых форм, как М. inferior St., М. lasiopa St. и другие, свойственные
лишь тропикам. Группы видов, родственных М. domestica L. и М. larvi-
para Portsch., в Палеарктике представлены бедно; первая одним видом,
вторая — тремя видами, в то время как в экваториальных странах
Африки и Азии эти группы насчитывают в совокупности более 20 форм.
Как известно, М. domestica распространена всесветно.
Orthellia R.-D. является родом явно тропического происхождения;
он представлен в Палеарктике всего 4, а в Северной Америке — 2 видами;
прочие виды рода свойственны тропическим странам Старого и Нового
Света. Распространение рода всесветное; таким же всесветным распро-
странением обладают и отдельные виды рода, в частности О. caesarian
Mg., встречающийся как за северным полярным кругом в Европейской
провинции Арктической области, так и у нкватора во всех частях света;
О. cornicina F. имеет, очевидно, циркумбореальный ареал, встречаясь
в Евразии и в Северной Америке. Группа тропических видов, характе-
ризующаяся слабым развитием щетинок на среднеспинке, в Палеарктике
представлена лишь на крайнем ее юговостоке, в СССР в Амурскую про-
винцию заходит один вид этой группы {О. pacifica, sp. п.). Подрод Pyrel-
liomima, subgen. nov. связывает фауну Индо-Малайской области и юга
Эфиопии; в СССР этот подрод пока не зарегистрирован.
4*
52
ВВЕДЕНИЕ
Pyreilia R.-D. обнаруживает сходный с предыдущим родом харак-
тер распространения. В Палеарктике известно всего 6, в Неарктической
области — 4 вида; остальные виды, составляющие около 80%, связаны
с тропиками, причем наиболее пышного расцвета род достигает в Эфиопии.
Из всех известных форм Р. cadaoerina обладает весьма широким ареалом,
захватывающим Евразию и Неарктику; близкие ей виды связаны с Афри-
кой. Р. aenea Ztt., в отличие -от большинства форм рода, преобладает
в таежной зоне, не встречаясь в условиях открытого степного ландшафта,
проникновение ее на юг происходит более интенсивно в условиях повы-
шенного рельефа. Эндемиками для Палеарктики являются Pyreilia (Dasy-
phoromima) pavlovskyi, sp. nov. (Приморье), Pyreilia (in sp.) minuta,
sp. nov., и P. (in sp.) secunda, sp. nov. Последние два вида известны
пока из Восточно-Средиземноморской и Нагорно-Туркестанской про-
винций.
Кроме Dasyphora serena Mg., обитающей в Северной Америке, род
Dasyphora распространен только в Палеарктике. Особенно богаты видами
рода Монголо-Тибетская и Нагорно-Туркестанская провинции, откуда
есть основания ожидать еще не мало новых форм. В Палеарктике широ-
кого распространения достигает сравнительно небольшое число форм.
Некоторые виды обитают лишь в условиях теплого и влажного климата
Средиземноморья (D. saltuum Rd.). Такие виды, как D. cyanella Mg., D. pra-
torum Mg., D. versicolor Mg., имеют более широкие ареалы, встречаясь
в Европейских провинциях Боревразийской подобласти и одновременно
в Средиземноморье. Для большинства видов характерен узкий геогра-
фический ареал, лежащий в области сухих центрально-азиатских наго-
рий и пустынь, где, очевидно, и возможно было закрепление за видами
рода такого признака, как живорождение. D. serena Mg. является един-
ственным видом рода, обладающим циркумбореальным ареалом п связан-
ным с лесным ландшафтом.
Род Morellia R.-D. отличается особым многообразием в Южной
Америке; близкий к нему род Xenomorellia Mall. — эндемик последней.
Несколько беднее фауна Morellia Эфиопской области, что, правда, может
быть объяснено недостаточной исследованностью. Особенно богатой ока-
зывается фауна горных областей южной и восточной Африки и Южной
Америки, где некоторые виды найдены только в горах на больших высо-
тах. В Палеарктике род Morellia представлен 9 видами, из которых два,
М. nilotica St. и М. hortensia Wd., являются общими с Эфиопской
областью. Обычные для области виды широко распространены от Скан-
динавии до южной Европы, Малой Азии и на восток до Монголии.
Из числа видов с узким географическим ареалом следует отметить М. sim-
plicissima, sp. nov., известного лишь из Сибири, и М. asetosa Ваг.,недавно
описанного из Югославии. Ближайший к Morellia род Xenomorellia Mall,
за пределами Неарктической области не известен.
Род Mesembrina достигает наибольшего видового многообразия
на северо-востоке Палеарктики (Восточная Сибирь); один вид рода яв-
ляется общим с Северной Америкой. Род связан с зоной хвойных и лист-
венных лесов, проникая на юге в горные районы. Наиболее эколо-
гически пластичным видом является М. meridiana L., известный из лес-
ных областей, степей Европы и гор Центральной Азии. Вместе с двумя
южно-китайскими видами М. meridiana L. составляет хорошо морфоло-
гически очерченный комплекс форм.
Близкий к Mesembrina род Fraseria Mall., которого составитель
не имел в натуре, известен из Индо-Малайской области.
ЭКОЛОГИЯ ГРУППЫ
53
Роды Alluaudinella G.-T. и Neomusca Mall, отсутствуют в Палеарк-
тике, первцй из них отмечен для Эфиопской, второй — для Неотропи-
ческой областей.
Род Graphomyia R.-D. наибольшего расцвета достигает в Индо-
Малайской и Эфиопской областях. В Палеарктику заходит один вид рода,
образующий здесь многочисленные уклоняющиеся формы; этот вид {G.
maculata Scop.) связывает Палеарктику с Эфиопской и Индо-Малайской
областями, равно как и со странами Нового Света. Род Gordonia Mall,
пока найден лишь в Австралийской области.
Нужно заметить, что в основном распространение видов трибы Sto-
moxydini сходно с распространением трибы Muscini. Область обитания
большинства жигалок ограничена тропическими странами, на долю
которых приходится 2/з мировой фауны Stomoxydini. Жигалки известны
из всех частей света, показывая особенно большое видовое разнообразие
в Эфиопской и Индо-Малайской областях. Лишь один из известных
из нашей фауны родов трибы обитает только в северном полушарии {Acri-
domyia) и один род эндемичен для Индо-Малайской области {Haemato-
storna Mall.).
Самым крупным родом трибы является Stomoxys Geoffr., наиболее
богато представленный в тропической Африке. В Палеарктику проникает
всего 1 вид этого рода {Stomoxys calcitrans L.), распространенный почти
по всему свету, исключая, правда, Сибирь. Бедно представлен род
в обеих Америках, из которых одна располагает одним, вторая— двумя
эндемичными видами, не считая обыкновенной жигалки. Индо-Малай-
ская область опять-таки, помимо осенней жигалки, обладает 10 видами,
часть их заходит в Эфиопскую область.
Роды Haematobia R.-D. и Lyperosia Rd. менее богаты видами.
В Палеарктике насчитывается 8 видов этих родов, среди них L. irritans L.
встречается также в Америке и Африке, а Н. stimulans L. одновременно
известна из Палеарктики и Неарктической области. Подобно тому, как
это наблюдается в отношении осенней жигалки, Н. stimulans и L. irri-
tans не встречаются в Сибири, за исключением Приморского края, куда
они заходят из Манчжурии; возможно, что приморские особи названных
видов представляют собою и самостоятельные формы, что можно уста-
новить лишь после сравнительного анализа более обширного материала.
Род Haematostoma Mall, в Палеарктику на заходит.
Последний род трибы — Acridomyia — до сего времени обнаружен
лишь на юге СССР и в Северной Америке. Эти данные не дают пока
ясного представления о широте ареала рода.
Биологически главная масса видов обеих триб связана с различными
млекопитающими, реже — беспозвоночными. Связь осуществляется либо
одновременно и в стадии взрослого насекомого и личинки, как, напри-
мер, в трибе Stomoxydini, либо только в стадии личинки, когда развитие
последних может проходить за счет помета животных {Dasyphora R.-D.).
Значительно реже встречаются среди видов группы хищные (в фазе
личинки) формы {Mesembrina Mg., Graphomyia R.-D.), или эндопаразиты,
как, например, Acridomyia Stack. Некоторые формы теснейшим образом
связаны с человеком.
ЭКОЛОГИЯ ГРУППЫ
Из форм, включенных в настоящую работу, многие виды являются
синантропами. Связь с самим человеком у одних видов почти повседнев-
ная и непосредственная, как, например, у комнатной мухи, у других она
54
ВВЕДЕНИЕ
осуществляется косвенным путем — через жилище, предметы обихода,
домашних животных и т. п.
В фазе личинки преобладающее число видов — копрофаги (Musca,
Orthellia, Dasyphora, Lyperosia и др.), причем для одних форм (Haema-
tobia, Lyperosia и др.) размножение возможно лишь при наличии помета
животных, в то время как другие, более пластичные виды, без видимого
для себя ущерба могут развиваться и в других веществах. Примером
этого Может служить комнатная муха, личинки которой удовлетворяются
любым гниющим веществом как растительного, так и животного проис-
хождения, живыми растительными и животными тканями, хотя и пред-
почитают помет животных всему остальному-
Некоторые виды рода Musca (как М. autumnalis Deg., larvipara
Portsch. и им близкие) явно предпочитают навоз жвачных животных,
никогда не встречаясь у лошадей; личинки М. tempestioa Fall, преобла-
дают в коровьем помете, но могут быть обнаружены в навозе и друтих
животных (лошадей, свиней).
Нужно отметить, наконец, и факультативных хищников, как
личинки Mesembrina или Graphomyia, или такие высоко специализиро-
ванные паразитические формы, как Acridomyia, живущие в фазе ли-
чинки в теле саранчи. Таким образом, главная масса форм разбирае-
мых триб в фазе личинки представляет собою копрофагов, роже —
сапрофагов и только незначительная их часть является хищниками или
паразитами.
В трибе Stomoxydini все палеарктические виды яйценесущи. Среди
Muscini встречаются, правда сравнительно редко, формы живородящие,
откладывающие личинок. Полное созревание яиц, а иногда и личинок,
протекает в теле самок, обладающих особым маткообразным расшире-
нием влагалища, отсутствующим у яйценесущих видов группы.
Степень зрелости эмбрионов в откладываемых яйцах у различных
видов неодинакова, вследствие чего период эмбрионального развития
имеет то большую, то меньшую длительность. В одних случаях от момента
откладки яйца мухой до вылупления из него личинки проходит 1—2
суток (некоторые Musca, Stomoxys), у других этот срок не превышает
нескольких часов. Переходную форму к живорождению представляют
собою виды Mesembrina (М. mystacea L.), у которых вылупление личи-
нок из яйца наблюдается почти тотчас же после того, как яйцо было
отложено на субстрат.
Продолжительность периода эмбрионального развития у одною
и того же вида варьирует в широких пределах в зависимости от условий
среды. Повышение температуры ускоряет развитие эмбрионов, правда,
до известного предела, за которым следует замедление развития или
остановка последнего. Увеличение влажности среды, окружающей яйцо,
выше оптимальной тормозит развитие зародыша, так же как и пониже-
ние ее, когда развитие эмбрионов может прекратиться совершенно. Харак-
терны для каждого вида не только оптимальные гигротермические усло-
вия, но и минимальные сроки, необходимые для того, чтобы завершилось
развитие в яйце.
Рождение живых личинок — явление сравнительно редкое среди
видов трибы Muscini, а у Stomoxydini оно не наблюдается вовсе. Явле-
ние живорождения свойственно некоторым видам рода Musca (М. larvi-
para, М. convexifrons, М. lusoria и ряду видов, несвойственных палеаркти-
ческой фауне), а также Dasyphora (D. pratorum, D. saltuum, D. asiatica).
Только что отложенная личинка может быть в I во II или даже в III
ЭКОЛОГИЯ ГРУППЫ
55
(D. pratorum) стадии. У видов Musca самка откладывает обычно личинку
I стадии, которая спустя короткое время линяет на II стадию. Среди
палеарктических Muscini только у некоторых форм Dasyphora личинка
почти полностью завершает цикл своего развития в теле мухи. В этом
отношении Dasyphora является как бы переходной формой к кукло-
родным двукрылым. Интересно указать, что живородящая в естествен-
ной обстановке Musca larvipara при воспитании ее в неволе нередко начи-
нает откладывать яйца. То же наблюдается и у Dasyphora asiatica (Зимин,
1944; Дербенева-Ухова. 1940).
Наблюдениями в природных условиях установлено, что живородя-
щие мухи трибы Muscini откладывают за раз обычно по одной личинке,
а не по несколько, как это свойственно многим живородящим сапрофа-
гам. Такой способ кладки является результатом последовательного созре-
вания половой продукции самок, когда полного развития каждое следую-
щее яйцо достигает лишь по откладке самкой личинки, причем созрева-
ние в одном яичнике зависит от степени зрелости яйца в другом. В срав-
нении с яйценесущими видами живородящие мухи значительно менее пло-
довиты й откладываемые личинки много крупнее тех, которые вылуп-
ляются из яиц у неживородящих видов (Порчинский, 1910; Дербенева-
Ухова, 1940).
Яйненесущие виды за раз откладывают по несколько яиц, при-
чем число последних в каждой кладке может достигать нескольких
десятков.
Некоторые синантропные мухи обладают весьма высокой плодови-
тостью. У комнатной мухи созревает одновременно около ста (от 46
до 150) яиц, вслед за откладкой которых идет созревание следующей
порции. Наряду с такой высокой плодовитостью, как у комнатной мухи,
способной за свою жизнь дать более 2000 яиц, мы встречаем крайне низ-
кие пределы, когда самка в течение жизни способна отложить не более
десятка яиц (Mesembrina) (Порчинский, 1910; Дербенева-Ухова, 1940).
Между сроками созревания каждой порции яиц у мух проходит
обычно несколько дней, причем, насколько нам известно, М. domestica
и в этом отношении оказывается на первом месте: каждая следующая пар-
тия яиц созревает при благоприятных условиях уже через 36 часов.
Характер пищи имеет первостепенное значение для созревания яиц.
Так, Дербенева-Ухова и Рубо, на примере с комнатной мухой, устано-
вили, что при углеводистой пище в отсутствии белков мухи остаются
бесплодными. Дербенева-Ухова (1935) экспериментальным путем пока-
зала, что при неполноценной пище самки развитие у нее яиц не доходит
до конца, и в этом случае фолликулы сходны с таковыми у мух, под-
вергавшихся голоданию. В этом отношении, примерно, одинаковая кар-
тина получается при кормлении мух только сахаром или только белком.
Лишь комбинированная по своему составу пища позволяет яичникам
мух развиваться вполне нормально.
Вмйсте с тем быстрота созревания яиц в сильной степени связана
с гигротермическим режимом среды, в которой находится самка. Высо-
кие температуры (при прочих равных условиях) способствуют ускоре-
нию созревания яиц, в то время как низкие — тормозят развитие.
Личинки всех известных нам видов данной группы имеют 3 стадии.
Продолжительность каждой предыдущей стадии короче последующей.
Сроки личиночного развития подвержены сильным колебаниям не только
от вида к виду, но и в пределах одного вида в зависимости от окружаю-
щих условий среды (температура, влажность, свет, пища).
56
ВВЕДЕНИЕ
Насколько сильно отрицательные условия сказываются на продол-
жительности личиночной фазы, можно видеть из того, что против мини-
мальной ее длительности, которая обеспечивается оптимумом вышена-
званных условий, сроки развития могут быть искусственно удлинены
в 6 и более раз (Stomoxys-, по Ныостиду) (при помощи пониженных тем-
ператур, увеличенной влажности и других факторов).
Еще в недавнее время особенное значение в борьбе с мухами при-
давалось влиянию высоких температур. В этом отношении могут пред-
ставить интерес исследования Томсена (1938) над различными фазами
развития комнатной мухи и осенней жигалки. Яйца комнатной мухи
особенно резистентны к высоким температурам при высокой влажности
(90—100%), тогда как понижение последней до 70% вызывает при тех
же температурах (16—40° С) увеличение процента смертности яиц. При
температуре ниже 9 и выше 43° С смертность эмбрионов при той же
влажности (от 70 до 100%) и длительности экспозиции вызывает 100%-ю
гибель яиц.
Под действием высоких температур (при 100% относительной влаж-
ности воздуха) гибель яиц наступает уже при 44° С через 15 минут, тогда
как за тот же период при 41—42° часть яиц (17—40%) выживает. При
одноминутной экспозиции достаточно 47° С, чтобы вызвать 100%-ю гибель
эмбрионов.
Среди личиночных стадий наиболее стойкими к высоким температу-
рам оказываются взрослые личинки (Тетеровская, 1944; Дербенева-Ухова,
1940). При этом смертность личинок каждой последующей стадии при
равной экспозиции наступает при все более высоких температурах.
Находясь в субстрате, личинки концентрируются в зоне оптималь-
ных температур и влажности и, обычно, избегают света. Нарушение
экологических условий в ту и другую сторону от оптимума вызывает
миграцию личинок, которая совершается иногда на сравнительно дале-
кое расстояние. Передвижение личинок может быть обусловлено и по-
исками подходящих мест для окукления, когда взрослые личинки распол-
заются от питавшего их субстрата иногда на 100 и более метров.
Окукление большинства видов происходит в почве на глубине не пре-
вышающей обычно 10—15 см. По отношению к некоторым видам известны
случаи окукления личинок в периферических частях питавшего их суб-
страта. Впрочем, многие синантропные виды показывают в этом отно-
шении достаточное непостоянство, что, очевидно находится в зависимости
от гигротермических условий окружающей среды. На примерах с ком-
натной мухой, жигалкой, И. tempestiva и другими, известно, что для
окукления личинки предпочитают прогреваемую и малоувлажненную
среду. Процесс формирования'пупария длится ;1—3 !часа.
Продолжительность фазы куколки, как и прочих преимагинальных
фаз, весьма изменчива. Кукол очный период обычно продолжается немного
дольше личиночной фазы, а у некоторых видов значительно превосходит
его по продолжительности; примером тому могут служить зимующие
куколки, продолжительность их жизни исчисляется несколькими ме-
сяцами.
Диапауза, свойственная, видимо, всем фазам развития мух, у тро-
пических форм падает на засушливый и наиболее жаркий период года.
Большинство видов группы зимует в фазе куколки, некоторые —
в имагинальной фазе. Наряду с этим известно, что у одного и того же
вида зимовка протекает то в одной, то в другой фазе развития. Здесь
следует указать также, что при благоприятных условиях размножение
ЭКОЛОГИЯ ГРУППЫ
мух может проходить безостановочно в течение круглого года, как, напри-
мер, у комнатной мухи.
Вылупление мух из пупария происходит под действием лобного
пузыря, функционирующего при посредстве кровяной жидкости. Лоб-
ный пузырь помогает насекомому выбраться и из субстрата, где нахо-
дится пупарий. Опытами установлено, что вылупившаяся из ложноко-
кона муха в состоянии преодолеть толщу песка до 60 см и более (Дербе-
нева-Ухова, 1943). В естественной обстановке для такой большой работы
необходимости не возникает, ибо окукление мух если и происходит
в почве, то на небольшой глубине.
С вылетом из пупария муха приступает к питанию. Мухи отыски-
вают источник питания при помощи органов обоняния; зрение играет
в отыскании пищи (равно как и субстрата для откладки яиц) подчинен-
ную роль (Кузина, 1938).
Характер веществ, идущих в пищу мухам, очень разнообразен.
Среди кровососущих жигалок, где основной пищей самок служит кровь
животных, самцы часто прибегают к растительной пище. В той же группе
мух самки паразитической Acridomyia питаются кровью саранчи, тогда
как у самцов функция питания, видимо, совершенно выключена (Рука-
вишников). Если не все, то большинство навозных мух питаются поме-
том животных, являясь в то же время и кроволижущими видами; среди
них самцы чаще самок могут быть отмечены за питанием на раститель-
ной пище. Главная масса синантропов обладает наиболее широким диапа-
зоном в выборе питательной среды, о чем легко судить по комнатной мухе.
Продолжительность жизни мух исчисляется днями, а если принять
во внимание зимовку или диапаузу, то и месяцами. При известном пище-
вом рационе жизнь мухи может быть продлена. Таким путем продолжи-
тельность жизни комнатной мухи, нормально исчисляющуюся двумя неде-
лями, удавалось в искусственных условиях продлить до 112 дней (без
диапаузы) (Кулагин, 1921).
Что касается активности мух, то она в сильной степени зависит от
условий среды (температура, влажность, освещение, ветер и т. п.). В этом
отношении каждый вид обладает своим преферендумом.
Нижний предел активности у известных с этой стороны синантроп-
ных видов расположен между 7—12° G, верхний, вызывающий уже силь-
ное беспокойство у насекомого, определяется 34—43° С. Нужно заме-
тить, что у одного и того же вида имеет место сильная индиви-
дуальная изменчивость в этом отношении; как общее правило, у видов
с южным географическим ареалом термический преферендум распола-
гается выше чем у северных форм. Что касается влияния влажности воз-
духа на активность имаго, то в этом отношении мы не располагаем соответ-
ствующими материалами.
У 'Свет весьма сильно действует на активность насекомого. В темноте
мухи остаются неподвижны, если термические условия нормальны. Повы-
шение температуры до критического уровня выводит мух из пассивного
состояния независимо от условий освещения. На крайние цвета спектра
реакция мух слабая; допускается, что некоторые из них мухи вообще
не улавливают и веДут себя как в темноте. Таково, например, действие
синей части спектра. Из наблюдений и опытов лабораторного характера
явствует, что различные виды обладают неодинаковой реакцией на пря-
мое солнечное освещение.
У дикоживущих форм и особенно у форм южного происхождения
(М. sorbens, М. osiris) отрицательный гелиотропизм обнаруживается лишь
58
ВВЕДЕНИЕ
при очень высоких температурах и то в очень неясной форме. У комнат-
ной мухи или осенней жигалки, тесно связанных с помещениями, наблю-
даются обратные соотношения: отрицательный гелиотропизм резко выра-
жен и проявляется при сравнительно низких температурах.
Соответсвенно отношению видов к гигротермическим условиям и со-
стоянию мест выплода размножение мух в продолжении теплого периода
года проходит то медленным, то ускоренным темпом, чем определяется
численность особей данного вида в природе. Одни виды характеризуются
подъемом кривой численности особей в сравнительно прохладный сезон,
у других вершина кривой размножения совпадает, наоборот, с наиболее
жарким периодом лета. И в этом отношении мы обнаруживали связь между
условиями, обеспечивающими максимальную численность особей дан-
ного вида в природе с характером его географического распространения.
Виды явно южного происхождения достигают в наших условиях наи-
большего расцвета в самый жаркий и обычно сухой период года. Север-
ные формы успешнее развиваются в весенний и осенний периоды; депрес-
сию в размножении вызывает у них знойный сезон.
ЭПИДЕМИОЛОГИЧЕСКОЕ И ХОЗЯЙСТВЕННОЕ ЗНАЧЕНИЕ ГРУППЫ
Виды трибы Muscini являются, как известно, распространителями
возбудителей острых кишечных заболеваний, как холера, дизентерия,
брюшной тиф. Возбудители этих болезней неоднократно были находимы,
как в кишечнике мух, так и на наружных частях тела. Соприкасаясь
с зараженными нечистотами, больным организмом или его выделениями,
мухи загрязняют наружные покровы своего тела, а насасываясь заражен-
ной пищей, вводят в кишечник огромное число патогенных микробов
(в кишечнике одной мухи удавалось находить до 660 миллионов бакте-
рий). Тысячи и десятки тысяч микробов муха носит и на внешней по-
верхности тела. Если представить себе, насколько велико количество
точек, с которыми муха при ее высокой подвижности войдет в сопри-
косновение после того, как она оказалась носительницей столь обиль-
ной бактериальной флоры, то станет понятным и размер дисперсии микро-
бов, осуществляемой одним лишь насекомым. Помножив его на общую
численность синантропных мух, убедимся в значительной роли мух в рас-
пространении инфекции. Отсюда становится также ясным, насколько
обширным может быть отрицательное значение синантропных двукрылых
при возникновении в населенном пункте крупного очага инфекции.
Причастность мух к распространению острых кишечных заболеваний,
помимо экспериментальных исследований, доказывается сопоставлением
характера кривых роста численности мух и роста кишечных заболева-
ний. Обычно последняя кривая запаздывает в сроках против первой,
оставаясь ей параллельной почти на всем протяжении. Предполагается
поэтому, что кривая заболеваемости связана с кривой численности мух.
Отсутствие синхронности в развитии эпидемий и размножении мух объяс-
няется периодом инкубации болезнетворных начал.
Представители трибы Stomoxydini как кровососы, невидимому, не
участвуют в разнесении кишечных инфекпий. Патогенное значение их
сводится к распространению возбудителей других тяжелых заболеваний
среди животных и населения, как сибирская язва, трипанозомозы. Им при-
писывают роль механических переносчиков туляремии, фрамбезии. Экспе-
риментально доказана возможность переноса жигалками вируса желтой
лихорадки среди обезьян. Сказанным далеко не исчерцьщается эпидемио-
ЭПИДЕМИОЛОГИЧЕСКОЕ И ХОЗЯЙСТВЕННОЕ ЗНАЧЕНИЕ ГРУППЫ 59
логическое значение жигалок; дополнительные данные приведены в опи-
сании биологии осенней жигалки.
Среди врачей офтальмологов уже сравнительно давно возникали
подозрения относительно того, что некоторые синантропные виды мух,
в том числе и комнатная муха, являются переносчиками вируса некото-
рых глазных болезней. Имелись в виду остро заразные болезни, как тра-
хома и эпидемический конъюнктивит.
Хьюитт и Пэттон считали несомненными переносчиками глазных
заболеваний комнатную и базарную муху. До недавних пор эти подо-
зрения одними принимались на веру без соответствующих эксперимен-
тальных доказательств, другими исследователями отрицались.
На юге эпидемический конъюнктивит имеет широкое распростране-
ние, проявляясь ежегодно в конце летнего сезона, т. е. в период массового
появления в природе синантропных мух. Для доказательства связи между
обоими явлениями автором были проведены специальные исследования
базарной мухи (Musca sorbens Wd.) и поставлены эксперименты с искус-
ственным заражением глаз человека при посредстве мух, которые дали
положительные результаты (Зимин, Парадоксов, Механикова, 1947).
Таким путем удалось экспериментально показать, что базарная муха
действительно переносит возбудителей острого эпидемического конъюнк-
тивита (палочки Кох-Викса) и способствует сильному развитию эпидемий
этого опасного заболевания.
Подозревали мух и в разнесении ими проказы (Павловский, 1948).
Опытным путем прямых доказательств по отношению к человеку полу-
чено не было. Что касается косвенных наблюдений, то они не дают пока
права снять существующие на этот счет подозрения.
Из общего количества болезней, которые могут разноситься мухами
(а известно около 50 таких заболеваний), изучалось менее половины.
В отношении прочих заболеваний экспериментальные данные отсутст-
вуют. Следует отметить, что в литературе описывались случаи обнаруже-
ния в теле мух возбудителей туберкулеза, дифтерии, оспы и т. д.
Область вредной деятельности синантропных двукрылых расши-
ряется еще способностью их транспортировать на себе яйца глист (ленте-
цов, нематод). По отношению к некоторым видам червей мухи являются
промежуточными хозяевами. Таковы, например, Habronema microstoma
Schneid., обитающая в стадии имаго в желудке лошади, а личиночную
стадию проводящая в теле осенней жигалки. Hymenolepis carioca (Magal-
haeus) обычная у домашней птицы, имеет в качестве промежуточного
хозяина ту же осеннюю жигалку. Передача большинства инвазий совер-
шается механическим путем.
Тропические виды жигалок служат транспортерами яиц мухи Der-
matobia hominis, помогая, таким образом, распространению потомства
последней среди животных. Помимо распространения заразных начал,
синантропные мухи нередко сами являются причиной заболевания
людей и животных, известного под названием миаза, когда личинки мух
попадают в тело человека или животного и питаются их тканями.
Десятки видов мух в фазе взрослого насекомого тесно связаны с на-
шими домашними животными. Независимо от возможного патогенного
значения мухи этой категории одним своим присутствием, не говоря уже
о кровососущих формах, причиняют животным сильное беспокойство,
что отрицательно сказывается на размерах молочной продукции и на
весе животных. Снижение молочной продукции может достигать по нека-
торым данным 50% нормального удоя.
60
ВВЕДЕНИЕ
Все вместе взятое дает право оценивать эту группу двукрылых как
одну из наиболее вредных для человека. Что касается полезной деятель-
ности некоторых видов, входящих в трибы Muscini и Stomoxydini, то
на фоне суммарной вредоносности она ничтожна. Можно указать на неко-
торую пользу от паразитической Acridomyia, несколько снижающей
численность саранчи в ее гнездилищах. Деятельность хищных личинок
таких мух, как Mesembrina, Graphomyia, мало ощутительна в связи
с ограниченным количеством мух в природе и сравнительной редкостью
их в пределах населенных пунктов. Попытки использования Mesembrina
путем колонизации на новой территории не оправдали возложенных на
них надежд.
В заключение укажем на опыты по использованию личинок комнат-
ной мухи в хирургии для лечения некоторых болезней. Как выяснено,
присутствие личинок на ранах животных и человека в некоторых слу-
чаях способствует более быстрому заживанию тканей, что ставится в зави-
симость от химического состава веществ, выделяемых мушиными личин-
ками (аллантоин).
МЕТОДЫ БОРЬБЫ С МУХАМИ
Борьбу с настоящими мухами и жигалками необходимо рассматри-
вать как неотъемлемую часть противоэпидемических и санитарно-гигие-
нических мероприятий.
Борьба с синантропными двукрылыми должна быть ориентирована
в двух направлениях: 1) уничтожение мух на разных стадиях их разви-
тия; 2) проведение профилактических мероприятий, имеющих целью
пресечение контакта окрыленных мух, с одной стороны, с очагами инфек-
ции, с другой — с самим человеком, продуктами питания и жилищем.
Истребительные мероприятия являются основой борьбы с перенос-
чиками, профилактические — дополнением, обязательным постольку, по-
скольку в подавляющем большинстве случаев истребительными меро-
приятиями не удается добиться полного уничтожения мух.
Мероприятия истребительные. Если не принимать
в расчет пастбищных видов, методика истребления которых оставляет
еще желать многого, то следует признать существующие в настоящее
время средства уничтожения синантропных жигалок и настояших мух
вполне надежными для достижения высокой смертности в популяциях
как личинок, так и окрыленных насекомых.
Казалось бы, что владея высоко эффективными средствами истребле-
ния, мы в праве ожидать во всех случаях их применения сокращения чис-
ленности синантропных мух до минимума, при котором роль мух как
переносчиков практически будет сводиться к нулю. В действительности
оказывается, что даже при условии использования в борьбе одних
и тех же высоко эффективных средств, результаты получаются подчас
совершенно неодинаковые. В большинстве примеров неудачные резуль-
таты борьбы являются прямым следствием недооценки значения биологи-
ческих особенностей синантропных видов мух — сроков развития, условий
и мест размножения, периодов активности насекомых, их миграций ит. д.
Итоговые результаты истребительной компании в сильной степени зави-
сят и от того, на каких фазах развития мух — ранних или имагиналь-
ной — сосредоточено внимание в борьбе. Наиболее целесообразной
является такая’Порганизация истребительных противомушиных меро-
приятий, при которой в действие параллельно введены и средства унич-
МЕТОДЫ БОРЬБЫ С МУХАМЙ В1
тожения окрыленных мух и средства истребления ранних стадий разви-
тия — яиц, личинок и куколок в местах выплода. Чтобы добиться высо-
ких результатов лишь методами уничтожения крылатой формы, нужно
подвергнуть обработке все без исключения постройки населенного
пункта сверху до низу, что по техническим возможностям и материаль-
ным затратам представляется явно неосуществимой задачей. С другой
стороны, подвергая дезинфекции только места выплода мух, мы никогда
не можем быть гарантированы в том, что обработаны все очаги, начиная
от зарегистрированных обледованием крупных скоплений навоза, отбро-
сов и т. д., и кончая мелкими скоплениями мусора и нечистот, возникаю-
щими порою неожиданно для обследователя, и продуцирующими нередко
огромное количество окрыленных мух.
Таким образом, только параллельная борьба со всеми фазами раз-
вития мух дает основание рассчитывать на успех (Павловский, 1948).
Этот принцип является первым условием организации противомушиных
мероприятий. Истребление ранних стадий развития и окрыленных мух
ведется различными отравляющими веществами и разными методами,
почему их мы рассматриваем обособленно друг от друга.
Методы борьбы с ранними стадиями раз-
вития мух. Основная масса синантропных двукрылых развивается
в крупных скоплениях навоза, в пищевых отбросах, в помойных ямах,
мусорных ящиках (на свалках) и в фекалиях человека. Все перечисленные
вещества содержат колоссальные количества яиц, личинок и куколок
мух. Истребление их может быть достигнуто различными путями.
Для ликвидации мух в навозе применяются специальные навозо-
хранилища, в которых взрослые личинки во время своих передвижений
для окукления попадают в специальный приемник, где они подлежат
уничтожению. При отсутствии навозохранилищ в скоплениях навоза,
путем утрамбовки его, создают условия для усиления ферментативного про-
цесса: развивающаяся при этом высокая температура убивает все пре-
имагинальные стадии мух. В местностях с сухим жарким климатом навоз
целесообразно просушивать на открытых площадках, хорошо прогревае-
мых солнцем. Навоз распределяется тонким слоем на земле в самое жар-
кое время дня и периодически перелопачивается до полной его просушки.
Для уничтожения мушиного выплода в выгребных ямах, уборных и по-
мойках с жидким содержанием, а также в пищевых отбросах мусорных
ящиков (в сельских местностях) рекомендуется периодическая обработка
их поверхности креолиновой эмульсией, нефтью или слабым раствором
карболовой кислоты. Высоким лярвицидным действием обладают также
каменноугольные масла среднелегких фракций (Павловский, 1948; Зимин
и Быховская, 1944). Заливка нечистот хлорно-известковым молоком
может быть рекомендована лишь в том случае, если применяется известь
с высоким содержанием хлора.
При правильной организапии работ и тщательной обработке мест
выплода можно в кратчайший срок добиться снижения численности мух
более чем на 90% даже в том случае, если мы не ведем борьбы с окрылен-
ными мухами. Обязательными условиями для этого являются следую-
щие: ^предварительное (до начала работ) и периодическое (в течение всего
теплого времени года) выявление и точная регистрация крупных и мелких
очагов выплода мух (уборные, мусорные ящики, помойки, свалки, скопле-
ния навоза на скотных дворах и т. д.); 2) дезинсекция всех без исключе-
ния мест выплода через промежутки времени, равные срокам развития
основных синантропных видов данной местности от фазы яйца до
62
ВВЕДЕНИЕ
стадии предкуколки; 3) тщательная обработка субстратов инсектицидами
или другими способами.
Выполнение данных строго обязательных условий возможно лишь
при ведении работ по графику, который позволяет учитывать рабочую
нагрузку персонала, без труда осуществлять контроль проводимых меро-
приятий и планировать расход ядоматериалов. В графике все места
выплода должны идти под постоянными номерами, а сроки дезинсекции
каждого очага должны быть расчитаны соответственно местным условиям
развития мух на весь теплый период года. Каждый дезинфектор отвечает
за своевременность и тщательность обработки определенной группы оча-
гов выплода (номеров). Таким образом, наиболее длительные проме-
жутки между обработками совпадут с ранне-летним и осенним перио-
дами, когда развитие мух идет замедленно, а самые короткие интервалы
совпадут с жарким периодом лета.
Стопроцентной гибели личинок, яиц и куколок мух в субстрате
можно добиться лишь в том случае, когда личинконосные слои послед-
него хорошо пропитаны инсектицидом.
Борьба с окрыленными мухами. Если борьба с ран-
ними стадиями сосредоточена сравнительно на небольшом количестве точек
(мест выплода), что является несомненным преимуществом этого метода,
то истребление окрыленных мух при широком масштабе работ охваты-
вает огромную площадь, включающую все жилые дома, помещения торго-
вой сети, фабрики, фермы, детские и лечебные учреждения и т. п. Для
уничтожения мух используются крайне разнообразные средства, начи-
ная от липкой бумаги, раствора формалина, мухоморов, мухоловок и кон-
чая сплошной обработкой внутренних, а то и наружных частей помещений
порошковидными (ДДТ, пиретрум) или эмульгированными (ДДТ и др.)
ядами. В борьбе с окрыленными мухами химические средства истребле-
ния составляют основу, механические способы в большинстве случаев
являются дополнительными. Ведущая роль последним отводится лишь
там, где по тем или иным соображениям недопустимо применение инсек-
тицидов.
Механические способы борьбы сводятся главным обра^
зом к вылавливанию мух ловушками различных систем и на липкую
бумагу. Чаще других применяемые мухоловки (стеклянные или сетча-
тые) основаны на принципе использования приманочных веществ
(кваса, молока и бродящих веществ), с помощью которых мухи привле-
каются в мухоловку. Устройство ловушек не позволяет попавшим в нее
насекомым выбраться обратно. Ловушки, устанавливаемые в стенках
стойл животных или навозоприемниках, действуют в силу положитель-
ного фототропизма мух, стремящихся из темных помещений к свету.
При надлежащих условиях ловушками можно уничтожить сотни, а при
высокой численности популяции насекомых и тысячи мух в сутки.
Липкие составы, изготовляемые обычно из канифоли в смеси с ка-
сторовым маслом и наносимые тонким слоем на листы плотной бумаги,
широко используются в быту. В липкий состав никаких привлекающих
мух веществ обычно не добавляется, и, следовательно, насекомые попа-
дают на липучку более или менее случайно. Чтобы повысить частоту
попадания насекомых, липучки следует раскладывать в местах, особенно
охотно посещаемых мухами. Так как в течение дня мухи концентри-
руются в различных частях жилища, целесообразно в соответствии с этим
перекладывать с места на место и липучки. Это замечание в известной
степени относится и к использованию мухоловок.
МЕТОДЫ БОРЬБЫ С МУХАМИ 83
Химические средства борьбы многообразны по
составу, своему действию на мух и местам применения.
Яды кишечного действия имеют широкое применение внутри глав-
ным образом жилых помещений. К ним относятся фтористый и кремне-
фтористый натрий, мышьяковистый натрий, раствор формалина и д у-
гие. Растворы фтористых и мышьяковистых соединений идут на изготов-
ление ядовитой бумаги («мухомор», «флорида») и широко используются
населением для борьбы с мухами, как и раствор формалина. Все пере-
численные растворы требуют добавления различных привлекающих мух
веществ (молока, сахара и др.).
Яды контактные, получившие широкую известность, в борьбе с му-
хами являются особенно перспективными и среди них особенно препарат
ддт.
В связи с введением ДДТ, пиретрум, сыгравший в свое время боль-
шую положительную роль в борьбе с вредными насекомыми, отошел на
задний план. В настоящее время пиретрум в виде порошка или вытяжки
(флицид) применяется значительно реже. Действующим началом
является пиретрум, вызывающий у мух явление временного паралича,
почему после применения пиретрума упавшие насекомые должны в крат-
кий срок сметаться и уничтожаться. Пиретрум не ядовит для людей
и позвоночных животных, почему применение его возможно без1 особых
ограничений.
Препарат ДДТ, стоящий сейчас в центре внимания исследователей
токсикологов и практиков и по существу заменивший все ранее извест-
ные средства борьбы с мухами, является действительно эффективным
и, что очень существенно, весьма стойким инсектицидом.
ДДТ, применяемый против мух преимущественно в виде порошко-
видных смесей (дусты «ДДТ», «Дуелит»), может быть использован и в вод-
ной эмульсии или суспензии.
ДДТ (дихлордифенилтрихлорэтан) — яд контактного действия,
смертельный для окрыленных мух. Опыты с личинками пока не дают
уверенности в целесообразности использования ДДТ для борьбы с ран-
ними стадиями мух.
Техника применения препарата сводится к опыливанию (с большим
или меньшим процентным содержанием действующего начала) или опры-
скиванию эмульсиями внутренних, а в случае необходимости и наружных
частей помещений. Мухи, входящие в соприкосновение с обработанной
ДДТ поверхностью, быстро погибают. Опытные данные показывают,
что ДДТ может быть с успехом применен и для борьбы с мухами, отро-
ждающимися в местах выплода и посещающими последние. Это дости-
гается соответствующей обработкой ближайших к данному очагу размно-
жения предметов (стены уборных, мусорных ящиков, помоек и др.) и по-
верхности почвы. Цель данного мероприятия заключается в том, чтобы,
с одной стороны, парализовать очаг размножения мух, минуя, следова-
тельно, обработку его ларвицидами, с другой стороны, — нарушить
контакт подлетающих насекомых с нечистотами.
Эта правильная в своей основе идея, к сожалению, не во всех слу-
чаях и не повсеместно может найти себе применение. Как установлено,
ДДТ токсичен для птиц, рыб и некоторых рептилий, а в чистом виде и в кол-
лоидальных растворах — для некоторых млекопитающихн в частности,
грызунов. Совершенно очевидно поэтому, что в сельских местностях
к повсеместному применению ДДТ следует относиться с большой осмот-
рительностью.
64'
ИЁЕДЕНЙЕ
Профилактические мероприятия ограничивают
возможность мух распространять заразные начала. В числе предупреди-
тельных средств используются отпугивающие вещества и приспособле-
ния, преграждающие или ограничивающие доступ мух к источникам
инфекции жилища человека и к продуктам питания, или, наконец, делаю-
щие помещения непривлекательными для мух.
Отпугивающие вещества, главным образом карболовая кислота,
применяется в дворовых уборных, помойках, мусорных ящиках.
Для преграждения мухам доступа в жилища и другие помещения
рекомендуется засетчивание окон, форточек, затемнение помещений
в часы наивысшей активности мух. Важным условием является хранение
съестных припасов в недоступных для мух местах.
ЛИТЕРАТУРА
Вайнштейн Б. А. и Р. А. Р о д о в а. 1940. Места выплода навозных мух
в условиях горного Таджикистана. Мед. паразитолог., IX, 4 : 364—368.
В а'н с к а я Р. А. 1941. Применение углекислого аммония для борьбы с Musca dome-
stica L. Мед. паразитолог., X, 5—6 : 562—567.
Дербенева-Ухова В. П. 1935. Влияние питания imago на развитие яични-
ков у Musca domestica L. Мед. паразитолог., IV, 5 : 394—403.
Дербенева-Ухова В.П. 1935. О количестве генераций у Musca domestica L.
Мед. паразитолог., IV, 5 : 404—407.
Дербенева-Ухова В.П. 1937. Экология личинок Musca domestica L. в при-
роде. Мед. паразитолог., IV, 3 : 409—417.
Дербенева-Ухова В.П. 1940. К экологии навозных мух в Кабарде. Мед.
паразитолог., IX, 4 : 323—339.
Дербенева-Ухова В. П. 1940. Влияние температуры на личинок Musca
domestica L. Мед. паразитолог., IX, 5 : 521—524.
Дербенева-Ухова В. П. 1940. Адаптация личинок Musca domestica L.
к высоким температурам. Мед. паразитолог., IX, 5 : 525—527.
Дербенева-Ухова В.П. 1941. К экологии Musca domestica L. и Muscina
stabulans Fall. Мед. паразитолог., X, 5 : 534—543.
Дербенева-Ухова В. П. 1943. Влияние влажности и трамбовки почвы
на выход imago Musca domestica L. на поверхность. Мед. паразитолог., XII,
3 : 72—76.
Дербенева-Ухова В.П. 1946. Некоторые данные по биологии мух в связи
с разработкой методов борьбы. Гигиена и санитария, 5 : 44—46.
Дербенева-Ухова В.П. 1947. Опыт применения препаратов ДДТ против
мух. Мед. паразитолог., XVI, 1 : 16—28.
Дербенева-Ухова В.П. 1947. Характер распределения мух по помеще-
ниям населенного пункта. Мед. паразитолог., XVI, 6 : 72—79.
Дербенева-Ухова В. П. 1948. Применение препарата ДДТ против мух
в Орехово-Зуеве в 1947 г. Мед. паразитолог., XVII, 1 : 34—45.
Дербенева-Ухова В. П. 1948. О сезонном ходе численности мух Musca
domestica L. Мед. паразитолог., XVII, 6 : 491—496.
Дербенева-Ухова В. П. 1949. Высшие двукрылые или круглошовные.
Учебн.мед. энтомолог, под ред. Беклемишева. Разд. Г : 357—400; фиг. 223—280.
Дербенева-Ухова В. П. иО. С. Кузина. 1938. Некоторые наблюде-
ния во время опыта борьбы с мухами на новостройке. Мед. паразитолог., VII,
3 : 399—405.
Егоров П. И. 1940. Опыт применения почвенной фумигации как средства унич-
тожения пупариев мух. Мед. паразитолог., IX, 5 : 528—530.
Зайцев Ф. А. 1947. К фауне домашних мух г. Тбилиси. Тр. Зоолог. Инет. АН
Груз. ССР, VII : 165—169.
Змеев Г. Я. 1944. Опыт оценки роли мушиного фактора в распространении дизен-
терии в южном Таджикистане. Пробл. кишечных инфекций: 95—117. Сталин-
аба д.
Змеев Г. Я. 4944. Опыт бактериологического обследования различных видов мух
с целью оценки их эпидемиологического значения. Там Же: 118—122.
Зимин Л. С. 1944. Базарная муха (Musca sorbens Wd.) в Таджикистане и возмож-
ная ее роль в распространении глазных заболеваний. Изв. Тадж. фил. АН
СССР, 5 : 144—151.
ЛИТЕРАТУРА
65
Зимин Л. С. 1944. Синантропные мухи Гиссарского района. Проблемы кишеч-
ных инфекций. Изд. НКЗдр. Таджикск. ССР, Сталинабад : 133—136.
Зимин Л. С. 1944. Сезонные и суточные колебания численности мух в жилых
помещениях в связи с температурой и влажностью. Там же : 167—176.
Зимин Л. С. 1944. Синантропные мухи южного Таджикистана. Там же : 177—192.
Зимин Л. С. 1945. Два новых вида рода Dasyphora R.-D. из Средней Азии. Энто-
молог. обозр., XXVII, 3—4 : 116—120.
Зимин Л. С. 1947. Эпидемический конъюнктивит в Таджикистане и роль базарной
мухи (Musca sorbens Wd.) в его распространении. Сб. раб. клин. глаз. бол.
Сталинабадск. мед. инет., Сталинабад : 11—21.
Зимин Л. С. 1948. Определитель личинок синантропных мух Таджикистана. Опре-
делители по фауне СССР, 28 : 113, 61 фиг.
Зимин Л. С. иИ. Е. Быховская. 1944. Средне-легкое каменноугольное
масло как лярвицид в борьбе с личинками синантропных мух. Изв. Таджикск.
фил. АН СССР, V : 152—154.
Зимин Л. С., Л. Ф. Парадоксов и В. Г. Механикова. 1947. Эпи-
демический конъюнктивит в Таджикистане и базарная муха (Musca sorbens
Wd.) как его переносчик. Таджикск. ГИЗ, Сталинабад : 54.
Зимин Л. С. и Т. О. Т е т е р о в с к а я. 1943. Сезонный ход численности комнат-
ной мухи в связи с развитием дизентерийных заболеваний в Таджикистане.
Мед. паразитолог., XII, 5 : 44—53.
Каландадзе Л. П. иС. В. Чилингарова. 1940. Результаты наблюде-
ний над комнатной мухой Musca vicina Macq. Мед. паразитолог., IX, 4 : 350—
354.
Каландадзе Л.П. иС.В. Чилингарова. 1940. Материалы к изучению
мух Грузии. Мед. паразитолог., IX, 5 : 518—520.
Квасникова П. А. 1931. Мухи жилых и хозяйственных построек человека
г. Томска. Изв. Томск. Гос. унив. : 88.
Клесов М. Д. 1949. Изучение нематоды Thelazia Rhodesi Desmarest, 1827. Докл.
АН СССР, LXVI, 2 ; 309—311.
Крастин Н. И. 1949. Расшифровка цикла развития нематоды Thelazia Rhodesi
(Desmarest), паразитирующей в глазах крупного рогатого скота. Докл. АН
СССР, XIV, 6 : 885—887.
Кузина О. С. 1936. Плодовитость и преимагинальная смертность мух (Musca
domestica L.). Мед. паразитолог., V, 3 : 329—339.
Кузина О.С. 1938. Выбор субстрата при откладке яиц комнатной мухой (Musca
domestica L.). Мед. паразитолог., VII, 2 : 244—257.
Кузина О. С. 1940. Роль органов чувств у Musca domestica L. при посещении
навоза и откладке яиц. Мед. паразитолог., IX, 4 : 340—349. »
Кулагин Н. М. 1916. Комнатная муха Musca domestica L. К вопросу о борьбе
с ней. Любит, прир., XI, № 3—4 : 93—100.
Кулагин Н.М. 1921. Комнатная муха (М. domestica L.) как разноситель зараз-
ных заболеваний, и способы борьбы с ней. Изд. Ком. Здравоохр. Москва :
30 стр.
Метелкин А. С. 1935. Значение мух в распространении кокцидиозов среди
животных и людей. Мед. паразитолог., IV, 1—2 : 75—82.
Набоков В. А. 1949. Медицинская дезинсекция: меры борьбы с мухами. Учеб-
ник мед. энтомолог., II, под ред. Беклемишева, ч. 1 : 124—129.
Набоков В. А. иН. А. Тибурская. 1935. О пргменении пиретрума
в борьбе с насекомыми. Мед. паразитолог., IV, 6 : 486—491.
Олсуфьев Н. Г. 1929. Этюды по паразитам азиатской саранчи (Locusta migra-
toria L.) из отряда двукрылых и их сверхпаразитам. Изв. прикл. энтомолог.,
IV, 1 : 61—119.
Павловский Е. Н. 1921. Мухи. Изд. НКЗдр. РСФСР.
Павловский Е. Н. 1921. Насекомые и заразные болезни человека. Москва :
48—65.
Павловский Е. Н. 1933. О применении личинок мух вместо хирургического
лечения. Природа, № 2 : 68.
Павловский Е.Н. 1948. Руководство по паразитологии человека. Изд. Акад.
Наук СССР, II : 860—914.
Петрищева П. А. 1932. К биологии комнатной мухи в условиях г. Самары.
Паразитолог, сб., III : 161—182,
Петрова Е. Ф. 1942. Синантропные мухи г. Алма-ата. Мед. паразитолог., XI,
2 : 86—93.
Петрова Е.Ф. 1948. Некоторые данные о комарах и мухах центральной части
пустыни Бетпак-дала. Мед. паразитолог. XVII, 6 : 527—531.
5 Фауна СССР, том XVIII, вып. 4.
66
ВВЕДЕНИЕ
П о д ъ я п о л ь с к а я В. П.п М. Н. Гнедина. 1934. О роли мух в эпидемио-
логии глистных заболевай й. Мед. паразитолог., III, 2 : 179—185.
П о р ч и н с к и й И. А. 1875. Materieux pour servir a une faune dipt^rologique de
la Russie. Tp. Русск. энтомолог, общ., XI, 1 : 27—36.
Порч и некий И. А. 1885. О различных формах размножения и о сокращенном
способе развития у некоторых обыкновеннейших видов мух. Тр. Русск. энто-
молог. общ., XIX : 210—244.
Нерчинский И. А. 1891—1892. Биология мясных и навозных видов мух. Тр.
Русск. энтомолог, общ. : 63—131.
П о р ч и н с к и й И. А. 1910. Осенняя жигалка, ее биология в связи с друг! ми
мухами и борьба с нею. Тр. Бюро энтомолог., VIII, 8 : 90, 13 фиг.
П о р ч и н с к и й И. А. 1911. Обыкновенная зубоножка, ее биология и истребле-
ние личинками ее личинок комнатной мухи. Тр. Бюро энтомолог., IX, 5 : 30,
23 фиг.
Пор ч и н с к и й И. А. 1913. Домовая муха (Muscina stabulans Fall.), ее значение
для человека и ее хозяйства и отношение ее к комнатной мухе. Тр. Бюро энто-
молог., X, 1 ; 39, 32 рис.
Романов А. П. 1940. Экология синантропных мух южного Таджикистана. Мед.
паразитолог., IX, 4 : 355—363.
Рукавишников Б. И. 1930. Материалы по изучению му х, паразитирующих
в личиночной и взрослой фазах саранчи (Locusta migratoria L.). Тр. защ. раст.,
сер. энтомолог., I, 1 : 205—225, фиг. 5—30.
Семенов В.Д. 1945. Наблюдения над сезонными изменениями численности ком-
натной мухи (Musca domestica L.) в связи с движением некоторых желудочно-
кишечных инфекций в Горьком. Мед. паразитолог., XIV, 6 : 50—54.
Смирнов Е. С. 1934. Исследования по экологии Mormoniella vitripennis, пара-
зита синантропных мух. Мед. паразитолог., III, 4 : 330—336.
Смирнов Е.С. 1937. Краткие итоги работ лаборатории по борьбе с мухами. Мед.
паразитолог., VI, 6 : 872—879.
Смирнов Е. С. 1940. Мушиная проблема в Таджикистане. Мед. паразитолог.,
IX, 5 : 515—517.
Смирнов Е. С. и О. С. Кузина. 1933. Экспериментально-экологические
исследования над паразитами мух. Зоолог, журн., XII, 4 : 96—109.
Тетеровская Т.О. 1944. Влияние температуры и влажности в местах выплода
на распространение в них личинок комнатной мухи. Тр. Инет, малярии и мед.
паразитолог. НКЗдр. Таджикск. ССР, I, Сталинабад : 49—55.
Чеботаревич Н. Н. 1937. О биологии комнатной мухи Musca vicina Macq.
и борьба с нею в Средней Азии. Тр. Сов. по изуч. произвол, сил, сер, турк-
' менская, 9 : 35—115.
Шта кельберг А. А. 1929. О новом паразите азиатской саранчи (Locusta migra-
toria L.) из сем. Muscidae. Изв. прикл. энтомолог., IV, 1 : 121—129.
Ш такел ьберг А. А. 1933. Определитель мух европейской части СССР. Ленин-
град—Москва.
Шура-Бура Б. Л. 1948. Метод количественного учета выживаемости патоген-
ных микробов в кишечнике комнатной мухи. Тр. Воен.-Мед. Акад. им. С. М.
Кирова, XXIV : 79—86.
Aubertin D. 1933. Notes on certain Species of the Genus Orthellia, with a Descrip-
tion of one new Species. Ann. Mag. Nat. Hist., ser. 10, XI : 139—144.
Austen E. E. 1909. New genera and species of Blood-sucking Muscidae from the
Ethiopian and Oriental Regions in the British Museum (Nat. Hist.). Ann. Mag.
Nat. Hist., ser. VIII, III : 285—299.
A w a t i P. R. 1915 (1916). Studies in flies. I. Chaetotaxy and pilotaxy of the Muscidae
and range of their variability in the same species. Indian Journ. medic. Res.,
Ill : 135—148, pl. 12—14.
Awati P. R. 1916. Studies in flies. II. Contributions to the study of specific
differences in the Genus Musca. Indian Joum. Med. Res., Ill : 510—529. pl.
35—50.
Awati P. R. 1917. Studies in flies. III. Classification of the genus Musca and descrip-
tion of the Indian species. Indian Journ. Med. Res., V : 160—191.
Banks N. 1912. The structure of certain Dipterous larvae with particular reference
to those in human foods. Li. St. Dep. Agric. Bur. Entom., Technic, series, No 22 :
44 pp.
В ez z i M. 1907. Die Gattungen der blutsaugenden Musciden. Zeitschr. Hymen, u-
Dipter., VII : 413—416.
В e z z i M. 1911. Etudes systematiques sur les Muscides hematophages du genre Lype-
rosia. Arch. Parasitol., XV : 110—143.
ЛИТЕРАТУРА
67
Brauer F. 1883. Die Zweifliigler d. Kaiserl. Mus. zu Wien, III. System. Studien
auf grundlage der Dipteren Larven nebst einer Zusammenstellung von Beispielen
aus der Literatur fiber derselben und Beschreibungen neuer Formen. Denkschr.
Kais. Akad. Wiss., Math.-Naturwiss. Classe, Wien, XL, VII : 100 pp., 5 pl.
Brauer F. & F. Bergenstamm. 1893. Die Zweifliigler d. Kaiser Mus. zu
Wien. III. Muscaria Schizometopa. Denkschr. Kais. Akad. Wiss. Wien., I : 86—90;
II : 89—91.
BrunettiE.1910. Revision of the Oriental blood-sucking Muscidae (Stomoxynae, Phi-
laematomyia Aust. a. Pristirhynchomyia, gen. nov.). Rec. Indian Mus., IV : 59—93.
De Mei j ere J. 1916. Beitrage zur Kenntniss der Dipterenlarven und Puppen. Zool.
Jahrb., Syst., XL : 177—322; Tafl. : 4—14.
D i n u 1 es c u G. 1930. Armature gftiitale male et female du Stomoxys calcitrans.
Ann. Parasitol., VIII : 61.
Enderlein G. 1924. Eine neue Blutsaugende Muscidae aus Afrika. Konowia, III :
51—52.
Enderlein G. 1933 (34). Dipterologica. I. Muscoidea, Muscidae, Muscinae. Sit-
zungsberichte Gesell. Naturforsch. Freunde, Berlin, III : 416—429.
Girschner E. 1893. Beitrage zur Systematik der Museiden. Berlin. Entomol.
Zeitschr., XXXVIII : 297—312, figs.
Girschner E. 1896. Eines neuen Musciden-System auf Grund der Thoracalbe-
borstung und der Segmentierungdes Hinterleibes. Illustr. Wochenschr. Ent., II :
11—15, 30—32, 61—64, 105—112.
Graham-Smith G.S. 1913. Flies in relation to disease. Nonbloodsucking flies.
Cambridge.
Graham-Smith G.S. 1916. Observations the habits and parasites of common
flies. Parasitology, VIII : 440—544.
II andlirsch A. 1906—1908. Die Fossilen Insecten und die Phylogenie der rezenten
Formen.
II e n d e 1 F. 1936. Dipteren — Fliegen. Handbuch der Zoologie. Insecta 2, IV, Heft 2,
Lief. 9—10, Figs. : 1853—2171.
Hewitt G. 1914. The Housefly, Musca domestica L. Cambridge.
Howard L. O. 1900. A contribution to the study of the insect fauna of human excre-
ment. Proc. Wash. Acad. Sci., II.
II о Ch ’ i. 1936. On the blood-sucking Muscidae of Peiping. Bull. Fan Memorial
Institute Biol., VI, No 5 : 185—206, 15 figs.
Huckett H.C. 1921. On the morphology of the ovipositor of certain Anthomyian
genera. Ann. Entom. Soc. Amer., XIV : 290—320, 8 pl.
Karl O. 1928. Die Tierwelt Deutschlands und der angrenzenden Meeresteile. 13. Zwei-
fliigler Oder Diptera, III, Muscidae : 1—232, 14 figs.
К e i 1 i n D. 1914. Les formes adaptives des larves des Anthomyides, Les Anthomyides
a larves carnivores. Bull. Soc. Entom. France : 496—501.
К e i 1 i n D. 1915. Recherches sur les larves des Dipteres Cyclorrhaphes. Bull. Sci.
France—Belgique, XL : 15—198, 16 pl.
К e i 1 i n D. 1916. Sur la vivarite des Dipteres et les larves des Dipteres vivipares.
Arch. Zool. Exp. Gen., LV : 393—415.
К e i 1 i n D. 1917. Recherches sur les Anthomyides a larves carnivores. Parasitology,
IX : 325—450, pl. V—XV.
Lamborn- W. A. 1936. The experimental transmission to man of Treponema per-
tenue by the fly Musca sorbens. Journ. trop. Med. Hyg., 39.
Lamborn W. A. 1937. The haematophagous Fly Musca sorbens Wd. in relation
to the transmission of Leprosy. Journ. trop. Med. Hyg., XL, No 4 : 37—42.
LorinczF. & G. Makar a. 1935. Investigations into Fly Density in Hungary
in the Years 1934-a. Quart. Bull. Hlth, Org. L. N., V, No 2 : 219—222,
figs.
Ma ckkerr as 1933. The Taxonomy of Lyperosia exigua De Meijere (Diptera, Mus-
cidae). Ann. Mag. Nat. Hist., 10, XI : 58—64.
Melvin R. 1934. Incubation period of eggs of certain muscoid flies at different con-
stant temperatures. Ann. Ent. Soc. Amer.. XXVII : 406—410.
Newstead R. 1906. On the life-history of Stomoxys calcitrans L. Journ. Econ.
Biol., I : 157—166; pl. XII.
NieschulzO. &duToit. R. 1933. Uber die Temperaturabhangigkeit der Akti-
vitat u. die Vorzugstemperatur von Stomoxys calcitrens, Musca vicina u. M. cras-
sirostris in Siidafrica. Centralbl. Bakter., II. Abt., 89, No 8—12 : 244—249.
Pandelle L. 1898. Etudes sur les Muscides de France. Rev. entom. XVII : 1—303.
Patton W. S. 1926. The Ethiopian species of the genus Musca Linnaeus. Bee. Indian
Mus., XXVIII : 29—52, pl. 1—4.
5*
68
ВВЕДЕНИЕ
Patton W. S. 1932. Studies on the higher Diptera of medical and veterinary impor-
tance : a revision of the species of the genus Musca based on a comparative study
of the male terjpinalia. I - The natural grouping of the species a. their relationship
to each other. Ann. trop. Med. Parasitol., XXVI : 347—405.
Patton W. S. 1933. Study on the higher Diptera of medical and veterinary impor-
tance: a revision of the genera of the tribe Muscini, subfamily Muscinae, based
on a comparative study of the male terminalia. I. The genus Musca L. Ann. trop.
Med. Parasitol., XXVII : 135—156.
Patton W. S. 1933. Study on the higher Dipt, of med. and veter. importance. II.
A practical guide to the Palearctic species. (Musca). Ann. trop. Med. Parasitol.,
XXVII : 327—345, 397—430.
Patton W. S. 1933. Study on the higher Dipt, of med. and veter. importance. II.
The genus Stomoxys Geoffroy (sens. lat.). Ann. trop. Med. Parasitol., XXVII :
501—537.
Patton W. S. 1936. Study on the higher Dipt, of med. and veter. importance. III.
A practical guide to the Ethiopian species. (Musca). Ann. trop. Med. Parasitol.,
XXX : 469—490.
Patton W. S. 1937. Study on the higher Dipt, of med. and veter. importance. IV.
A practical guide to the Oriental species (Musca). Ann. trop. Med. Parasitol.
XXXI : 127—140.
Patton W. S.&E.G. Gibbins. 1934. To же. III. The metallic Muscinae. Ann.
trop. Med. Parasitol., XXVIII : 571—578.
P e u s F. 1937. Zwei bisher unbekannte Arten der Stechfliegengattungen Haphospatha
End. aus Mitteleuropa (Dipt. Stomoxydidae). Zeitschr. angew. Entomol., XXIV :
150—154, 6 figs.
Ringdahl O. 1929. Bestimmungstabeller till svenska muscids slakten Muscinae.
Entomolog. Tidskrift. : 8—43 (sep.).
Schnabl J. 1906. Einige Worte uber die Terminologie der Beborstung der Dipte-
ren-Beine. Wiener. Entomolog. Zeitung, XXV : 269—272.
Schnabl J. &H. Dziedzicki. 1911. Die Anthomyiden. Nova Acta. Abh.
Kaiserl. Leop.-Carol. Deutsch. Akad. Naturforsch., Halle, XLV, No 2 : 1—301,
959 figs.
Seguy E. 1923. Faune de France. Dipteres Anthomyides. Encycloped. Entomol.,
ser. В., II : 1—390; 812 figs.
S ё g u у E. 1935. Etudes sur les Stomoxydines. Encycloped. Entomol., ser. В., VIII :
39—46.
Stein P. 1916. Anthomyiden Europas. Arch. Naturgesch., LXXXI (1915) : 1—224.
Stein P. 1917. Die Anthomyidengattungen der Welt, analytische Bearbeitung, nebst
einem kritisch systematischen Verzeichniss aller aussereuropaischen Arten. Arch.
Naturgesch., LXXXII, A : 85—178.
Thomsen M. 1938. Stuefluen (Musca domestica L.) og stikfluen (Stomoxys calci-
trans L.). 176-de Beretning Fors0gslaboratori et Kepenhavn: 352 pp, 13 pl, 107
text figs.
Thomsen M. & O. Hammer. 1936. The. breeding Media of some common flies.
Bull. Ent. Res., XXVII : 559—587, 4 figs.
Thomson R.G. 1937. Observations on the biology a. larves of the Anthomyidae.
Parasitology, XXIX : 278—358, 157 figs.
Townsend H. T. 1935. Manual of myiology, V : 1—64.
W i 1 h e 1 m i J. 1910. Zur Biologieder Kleinen Stechfliege Lyperosia irritans L. S : tz.-
ber. Gesell. Naturf. Ireunde Berlin : 179—194.
W i 1 h e 1 m i J. 1917. Die gemelne Stechfliege (Wadenstecher) Untersuchungen uber
die Biologie der Stomoxys calcitrans L. Zeitschr. angew. Entom., V : 2.
СПЕЦИАЛЬНАЯ ЧАСТЬ
ОПРЕДЕЛИТЕЛЬНАЯ ТАБЛИЦА ЛИЧИНОК III СТАДИИ 1
1 (2). Тело суженное к обоим концам, с продольными бороздками; зад-
ний конец тела с двумя выростами, несущими дыхальцы; на вен-
тральной стороне каждого сегмента имеется по два мясистых бу-
горка, покрытых изогнутыми шипами..............................
................................Graphomyia maculata Scop.
2 (1). Тело личинки сужено к переднему и утолщено к заднему концу,
последний без двух длинных дыхальценосных выростов посредине.
3 (4). Анальная пластинка очень длинная и узкая: отношение длины
к ширине равно 30 : 1; задняя сторона последнего сегмента в мел-
ких ямках, напоминает поверхность наперстка, задний конец тела
почти плоский с резким кантиком по краю; кутикула плотная,
передние края сегментов с многорядными плоскими, зазубрен-
ными на вершине хитиновыми шипи ками..........................
...................................... Morellia simplex Lw.
4 (3). Анальная пластинка шире, отношение длины ее к ширине состав-
ляет от 4 : 1 до 12 : 1; задняя сторона последнего сегмента без глу-
боких резких ямок и резкого кантика по краю, ± выпуклая.
5 (6). Передние дыхальцы с 16—18 длинными пальцевидными выро-
стами по краю, атриум передних стигм с треугольным черным хити-
новым затемнением посредине; длина тела 20 мм..................
..................................Mesembrina meridiana L.
б (5). Передние дыхальцы с 6—10, редко с 12—14 короткими выростами,
округлыми по краю: атриум передних стигм внутри без черного
хитинового склерита, длина тела до 16 мм.
7 (20). Задний конец тела внизу в области анальной пластинки не менее
чем с 5—7 мягкими выростами.
8 (15). Дыхальцевые щели короткие, S-образно изогнутые, располо-
жены на светлых участках дыхальцевых пластинок, последние
часто имеют ± треугольные очертания.
9 (12). Расстояние между задними стигмами в Р/г—2 раза превышает
поперечник одной стигмы, стигмы ± треугольных очертаний. Круп-
нее: 8—12 мм.
10 (И). Постанальный бугорок ясный, покрыт мелкими шипиками . . .
........................................Stomoxys calcitrans L.
И (10). Постанальный бугорок отсутствует......................
..................................Haematobia stimulans Mg.
12 (9). Расстояние между задними стигмами равно поперечнику дыхальца
или менее его; задние стигмы почковидной или треугольной формы.
Мельче: 5—7.0 мм.
1 В таблицу введены лишь виды, известные автору в натуре.
70
ОПРЕДЕЛИТЕЛЬНАЯ ТАБЛИЦА ЛИЧИНОК III СТАДИИ
13 (14). Задние дыхальцы расставлены на расстояние, равное попереч-
нику стигмы или чуть менее его, треугольных очертаний, мелкие;
параанальные бугорки отчетливые, образуют ясный угол на боко-
вых концах анальной пластинки...............Lyperosia titillans Bezzi
14 (13). Задние дыхальцы приближены друг к другу на г/4—х/з попереч-
ника стигмы, крупные, почковидные; параанальные бугорки не вы-
ражены.......................................Lyperosia irritans L.
15 (8). Дыхальцевые щели задних стигм многократно змеевидно изо-
гнуты, дыхальцы овальной или почковидной формы.
16 (17). Рядом с боковыми концами анальной пластинки располагаются
экстраанальные бугорки; анальная пластинка короткая, конец ее
далеко не достигает средины боковой поверхности тела личинки . .
.............................................Pyreilia cadaverina L.
17 (16). Экстраанальные бугорки отсутствуют, анальная пластинка длин-
ная, концы ее доходят до середины боковой поверхности тела
личинки.
18 (19). Анальная пластинка узкая с заостренными концами...........
.......................................Orthellia cornicina F.
19 (18). Анальная пластинка широкая . . . Orthellia caesarion Mg.
20 (7). Задний конец тела снизу в области анальной пластинки не более
чем с 4 мягкими бугорками (2 маленьких по бокам от анального
отверстия, а 2 — у заднего края анальной пластинки).
21 (22). Вокруг задних дыхалец располагается 8 мелких конических,
острых бугорочков; поверхность последнего сегмента в мелких
шипиках........................................Musca sorbens Wd.
22 (21). Мелкие конические острые бугорочки вокруг задних стигм и мел-
кие шипики на поверхности последнего сегмента отсутствуют.
23 (24). Субанальные бугорки значительно крупнее анальных. Позади
анального отверстия присутствует не более 10 очень мелких свет-
лых шипиков, под задними стигмами располагается две крупных
округлых выпуклости ................... . . Dasyphora asiatica Zim.
24 (23). Субанальные бугорки мельче, равны или едва крупнее анальных.
25 (26). Поверхность последнего сегмента позади анальной пластинки
между субанальными бугорками без мелких шипиков....................
......................................Musca tempestiva Fall.
26 (25). Позади анальной пластинки имеются мелкие светлые шипики.
27 (30). Анальная пластинка длинная, концы ее заходят на боковые
отделы сегмента до средней линии или далее нее; конпы анальной
пластинки с прямоугольным расширением; дыхальцевые щели тон-
кие, пятикратно извитые, концы всех щелей обычно направлены
внутрь дыхальцевой пластинки.
28 (29). Шипики вдоль задней границы анальной пластинки доходят
до концов последней; три продольных бороздки на дорзальной сто-
роне последнего сегмента с шипиками по краям.......................
...................................... Musca autumnalis Deg.
29 (28). Шипики вдоль задней границы анальной пластинки не дости-
гают концов последней; продольные бороздки на дорзальной сто-
роне последнего сегмента без шипиков по краю.......................
......................................Musca larvipara Portsch.
30 (27). Анальная пластинка короткая, концы ее не доходят до боковой
средней линии сегмента, округлые, или прямые без расширения;
дыхальцевые щели сравнительно широкие, не более чем с 4 изги-
бами.
ОПРЕДЕЛИТЕЛЬНАЯ ТАБЛИЦА ЛИЧИНОК Ill СТАДИИ
71
31 (34). Анальная пластинка вытянутая, узкая с округлыми концами;
шипики впереди и позади анальной пластинки располагаются в рай-
оне ее среднего отдела, не достигая ее концов.
32 (33). Шипиков позади анальной пластинки не более 10, расположен-
ных обычно в один неправильный ряд. . . . Musca lucidula Bezzi
33 (32). Шипики позади анальной пластинки в числе нескольких десят-
ков, расположены в несколько рядов..............Musca osiris Wd.
34 (31). Анальная пластинка очень короткая с прямыми концами, со
всех сторон окружена многочисленными мелкими светлыми шипи-
ками, среди которых отчетливо выделяется группа темных шипиков
между субанальными бугорками..................Musca domestica L.
ОПРЕДЕЛИТЕЛЬНАЯ ТАБЛИЦА ТРИБ ПО ВЗРОСЛОЙ ФАЗЕ
1 (2). Задний край глаз выпуклый; хоботок толстый, мясистый с широ-
кими сосательными лопастями (рис. 2); грудная чешуйка крыла
широкая, заходящая внутренним задним углом далее основания
щитка (рис. 66, 67); задние голени с дорзальной щетинкой . . . .
......................................................Muscini
Рис. 66—68. Форма грудной (ui) и крыловой (чг)
чешуек.
66 — Orthellia cornicina', 67 — Mesembrina meridiana.
68 — iS tom ox ys calcitrans'. 4i —чешуйка; — прогиво-
чешуйка. (Орвг.).
2 (1). Задний край глаз вогнутый (исключение — Acridomyia Stack.);
хоботок тонкий, обычно с маленькими сосательными лопастями;
грудная чешуйка (рис. 68) узкая, языковидная, без заднего внут-
реннего угла (исключение: Acridomyia и Graphomyia)', задние голени
без дорзальной щетинки .........................................Stomoxydini
72
J. ТРИБА MUSCINI
I. Триба Muscini
Грудные чешуйки сильно развиты, с лопастевидно расширенным
внутренним концом, соприкасающимся с основанием бокового отдела
щитка и нередко достигающим средины бокового края последнего (рис. 66,
67); усики с длинными (превышающими поперечник 3-го членика усиков)
волосками с обеих сторон; жилка т в вершинной части угловидно или
дуговидно изогнутая по направлению к вершине крыла; r4+s с волосками
при основании; гипоплевральные щетинки отсутствуют; эпимериты
и эпистерниты заднегруди (метаплевры над задними тазиками) обычно
с группой волосков (рис. 8). Щетинки груди сравнительно хорошо раз-
виты. Хоботок короткий, с толстым сосальцем и широкими сосатель-
ными лопастями.
Мейкие или средней величины мухи с голыми или опушенными гла-
зами с более узким, чем у самок, лбом самца; брюшко у обоих полов из 5
крупных (1—5) сегментов и 5 мелких, сильно измененных и образую-
щих гипопигий у <5 <5 и яйцеклад у gg. 6-й и 7-й тергиты самцов асиммет-
ричные: в левой половине развиты сильнее нежели в правой; 6-е и 7-е
дыхальцы брюшка у самцов имеются, у самок — отсутствуют. Фалло-
зома всегда с развитым дорзальным отростком. Церки самцов короткие,
не бывают глубоко разделенными снизу (как у Calliphoridae). Первые
две пары брюшных дыхалец заключены в хитиновую пластинку сегментов.
ОПРЕДЕЛИТЕЛЬНАЯ ТАБЛИЦА РОДОВ
1 (2). Жилка п сверху с несколькими короткими щетиночками в основ-
ной части; средние голени с крепкой вентральной щетинкой близ
средины; глаза в светлых волосках, или голые; переднегрудь широ-
кая, супрасквамальное ребро без щетинистых волосков; жилка т
в дистальном отрезке с мягким дуговидным изгибом. Тело отчетли-
вой металлически-зеленой, бронзовой или синей окраски . с более
или менее сильно развитым светлым налетом........................
................................. . . 4. Dasyphora R.-D.
2 (1). Жилка т-i сверху у основания без щетиночек.
3 (4). Щиток по свободному краю с 10—16 щетинками; гипоплевры,
метаплевры над задними тазиками, переднегрудь и супрасквамаль-
ное ребро без волосков; жилка т дуговидно изогнута в дистальном
отрезке. Крупные виды: 10—16 мм с яркожелтым основанием
крыльев; некоторые виды в густых волосках........................
.........................................6. Mesembrina Mg.
4 (3). Щиток с 8 щетинками по заднему краю; по крайней мере, мета-
плевры над задними тазиками с группой волосков; переднегрудь
и супрасквамальное ребро с волосками, или без них. Длина тела
от 2.5 до 9 мм.
5 (8). Вентральная щетинка на средних голенях близ их средины креп-
кая, длинная; щиток по краю с 6 щетинками (предосновные щетинки
отсутствуют), если с 8, — то тело металлически зеленой или синей
окраски.
6 (7). Супрасквамальное ребро в длинных, черных, щетинистых воло-
сках; переднегрудь широкая, с черными волосками по краям; задне-
грудные дыхальцы длинные, передние своим краем достигающие
передней границы вентральной части плевротергита.................
...........................................1. Orthellia R.-D.
I. ORTHELLIA
73
7 (6). Супрасквамальное ребро без длинных щетинистых волосков;
жилка т с мягким дуговидным изгибом в дистальном отрезке . . .
.....................................................3. РугеШа R.-D.
8 (5). Вентральная щетинка средних голеней отсутствует; щиток по
' краю с 8 щетинками (предосновные щетинки развиты и отчетливо
выделяются среди волосков щитка). Формы с бурой окраской
тела, в более или менее развитом светлом налете.
9 (10). Жилка т угловидным изгибом в дистальном отрезке (рис. 10),
если с дуговидным, то I стернит брюшка без волосков. Передне-
грудь широкая, всегда с волосками по краям. Голени самцов без
длинных щет шистых волосков или мелких шипиков (помимо обыч-
ных щетинок); наружные теменные щетинки самок крепкие. Тело
преимущественно бурой, реже зеленой окраски, боковые отделы
брюшка самцов часто желтые..............................2. Musca L.
Ю (9). Жилка т с мягким дуговидным изгибом в дистальном отрезке;
I стернит брюшка в щетинистых волосках; переднегрудь широкая,
без волосков, реже с волосками по краю. Голени самцов с более
или менее развитым дополнительным вооружением в виде мелких
шипиков, щетинок или длинных щетинистых волосков; наружные
теменные щетинки у обоих полов имеют вид тонких волосков. Тело
черное с легким синеватым оттенком и развитым светлым нале-
том .......................................... 5. Morellia R.-D.
1. Род ORTHELLIA R.-D.
Robineau-Desvoidy, 1863, Hist. Dipt., II :837; Malloch, 1923, Ann. Mag. Nat.
Hist., (9), XII : 505; 1925(9), XVI : 365; 1928, (10), 1:473; Aubertin, 1933,(10),
XI : 139; Karl, 1935, Arb. morph, taxon. Ent. : 34. — Pseudopyrellia Girschner, 1893,
Berl. ent. Zeitschr., XXXVIII : 306; Bezzi u. Stein, 1907, Catal. pal. Dipt., IV : 608;
Stein, 1916 : 18. — Cryptolucilia Brauer u. Bergenstamm, 1893, Denkschr. Ak. Wien,
LX : 206; Stein, 1918 : 105; S<5guy, 1923 : 359; Karl, 1928 : 18; —Scutellorthellia
Townsend, 1932, Journ. New York ent. Soc., XL : 439—479. — Stenomitra Enderlein,
1934, Sitzungsber. Gesell. Nat. Fr. Berl. : 416. — Anacrostichia Enderlein, 1934 : 417. —
Neomyia Walker (y Enderlein), 1934 : 418. — Agalmia Enderlein, 1934 : 421. -r- Com-
mosia Enderlein 1934, 1. c. : 421.
Тип рода Orthellia cornicina F.
Средней величины мухи с металлически зеленой, синей или фиоле-
товой окраской тела, обычно лишенные матового налета; лоб самца узкий
{Pyrelliomima) или в два раза шире 3-го членика усиков, у самки равен
или менее ширины глаза; лобная положа уже орбит {Orthellia s. str.) или
шире их {Pyrelliomima). В подроде Orthellia s. str. оба пола лишены направ-
ленных вперед крепких орбитальных щетинок; у подрода Pyrelliomima,
subg. п., орбитальные щетинки присутствуют у самок. Щеки широкие или
узкие {Pyrelliomima)', скулы узкие. Глаза у в микроскопических ред-
ких волосках, иногда исчезающих у этого пола (чаще у самок). Супра-
сквамальное ребро всегда, как и переднегрудь, в щетинистых темных воло-
сках; заднегрудное дыхальце крупное, поперечно вытянутое.
Акростихальные щетинки в числе 1 пары перед щитком (реже отсут-
ствуют), перед швом отсутствуют или {О. caesarion) имеются (1 пара);
дорзоцентральные щетинки весьма изменчивы в числе: от 0-J-1 до 24-
4-3; интраалярная щетинка 1 позади шва; надкрыловых — 2, из которых
задняя слабая; закрыловых — 3; щиток с 3 парами щетинок; пребазаль-
ные щетинки отсутствуют. Изгиб т обычно в виде угла, иногда несколько'
74
I. ТРИБА MUSCINI
сглаженного. Жилка г, без щетинок, r4+s со щетиночками сверху и воло-
сками снизу в основной половине первого отрезка жилки. Средние голени
с крепкой вентральной щетинкой; переднедорзальные щетинки обычно
отсутствуют; задние бедра по задневейтральной стороне с 1 щетинкой
у средины; положение видов, снабженных рядом щетинок на этом месте,
как, например, О. superba Karl, 1935, требует пересмотра. Коготки корот-
кие. Фаллозома с очень короткой текой и крупной, расширенной, ме-
стами вооруженной крепкими широкими зубцами мембранозной частью;
парамеры с широкими зубцами; парамеры с широким передним и узким,
заостренным к вершине задним отделом; дорзальный отросток хорошо
развит; церки широкие в основной половине, у середины суженные и на
вершине вновь слабо расширенные с двумя выростами, разделенными по
средней линии более или менее глубокой полукруглой выемкой.
Род Orthellia (в широком смысле) представляет собою один из наибо-
лее богатых видами родов трибы, в то же время в систематическом отно-
шении наименее удовлетворительно изученных. Краткие диагнозы преж-
них авторов, основанные по преимуществу на цветовых признаках, поро-
дили до чрезвычайности путанную синонимику видов, разобраться в кото-
рых возможно лишь после просмотра типов. В этом направлении много
ясности внесено работами Мэллока (Mallocb, 1923 и последующие) и Обер-
тин (Aubertin, 1933). Первый из названных авторов установил хорошие
отличительные признаки рода (хетотаксия супрасквамальных ребер
п переднегруди). В противовес Мэллоку, Эндерлейн (Enderlein, 1933,
1934) раздробил род на несколько родов, приняв в качестве одного из
основных признаков для этого число дорзоцентральных и акростихаль-
ных щетинок; в действительньсти та и другая группа щетинок (в особен-
ности первая) весьма изменчивы в пределах как раз рода Orthellia, при
существовании к тому же постепенных переходов между крайними фор-
мами. Именно поэтому система Эндерлейна не может быть названа
удачной.
Особенно пышного расцвета род Orthellia в широком смысле дости-
гает в тропических странах. По предварительным подсчетам, наибольшее
число форм обитает в Ориентальной и Эфиопской областях (около 34
видов); из Неотропической области известно 11 видов. В остальных обла-
стях фауна Orthellia бедна: из Палеарктики известно 4 вида, из Неаркти-
ческой области — 2 вида, общие с Палеарктикой.
Биология видов, кроме двух известных из Палеарктики, не изу-
чена. Что касается последних, то личинки их являются копробионтами.
Род Orthellia R.-D. мы разбиваем на 2 подрода.
ОПРЕДЕЛИТЕЛЬНАЯ ТАБЛИЦА ПОДРОДОВ,
1 (2). Самцы: ширина лба составляет от т/з до Уг ширины 3-го членика
усиков; щеки в 4—5 раз уже вертикального диаметра глаза; фа-
сетки в передне-верхнем отделе глаз очень крупные: диаметр фасе-
ток равен % поперечника скулы.
Самки: лобная полоса почти в 5 раз шире орбиты; ширина
щеки в 4—5 раз менее вертикального диаметра глаза; орбиталь-
ные щетинки развиты................2. Pyrelliomima, =ubg. nov.
2 (1). Самцы: ширина лба в 1)4—2 раза превышает поперечник 3-го
чл. усиков; поперечник щеки в 3 раза короче вертикального диа-
метра глаза; фасетки в передне-верхнем отделе глаз относительно
мелкие: диаметр фасеток в 5 раз короче диаметра скулы.
1. ORTHELLIA
75
Самки: лобная полоса уже орбиты или равна ей; щеки в 2% —3
раза уже вертикального поперечника глаза; орбитальные щетинки
отсутствуют................................Orthellia R.-D. (s. str.)
1. Подрод Orthellia R.-D.
Robineau-Desvoidy, 1863, Hist. Dipt., II : 837.
Тип подрода: Orthellia cornicina Fabricius (1781).
Диагноз подрода сходен с. диагнозом рода, за исключением призна-
ков. указанных для Pyrelliomima, subgen. nov.
Подрод включает главную массу видов рода. С достоверностью из-
вестны лишь 2 и под вопросом еще 2 вида, относящиеся к подроду Pyrel-
liomima mihi, почему сведения о географическом распространении под-
рода Orthellia и численности входящих в него видов почти полностью
совпадают с данными, приведенными для рода.
ОПРЕДЕЛИТЕЛЬНАЯ ТАБЛИЦА ВИДОВ
1 (2). Акростихальные щетинки впереди шва в числе 1 пары; дорзоцен-
тральных щетинок позади шва — 3, впереди него — 2..................
....................................*1. О. (in sp.) caesarion Mg.
2 (1). Акростихальные щетинки впереди шва отсутствуют.
3 (4). Дорзоцентральные щетинки перед поперечным швом отсутствуют;
если они развиты, то едва отличимы среди покрывающих спинку
волосков; позади шва две пары дорзопентральных щетинок . . .
......................................*3. О. (in sp.) pacifica, sp. n.
4 (3). Дорзоцентральные щетинки впереди поперечного шва сдинки
толстые, длинные в числе 2 пар, позади шва — 4 пар.................
................................... *2. О. (in sp.) cornicina F.
*1. Orthellia (in sp.) caesarion Mg. (рис. 69—71, 73, 75, 76—86).
Meigen, 1838, Syst. Beschr., V : 97 (Musca)', Girschner, 1893, Berl. Ent. Zeitsch.,
XXXVIII : 306 (Cryptolucilia asiatica Br.); Pandelle, 1898, Rev. ent., XVII : 38
(Lucilia cornicina Pand.); Schnabl u. Dziedzicki, 1911 : 229, figs. 645, 646 (Pseudopy-
rellia cornicina Schn.); Stein, 1916 : 18 (Pseudopyrellia)',—fennica Frey, 1909, Acta
Soc. Fauna Flor. Fenn,, 31, 9 : 9.
Биология. Thomson, 1938 : 319; Дербенева-Ухова, 1940, Мед. парази-
толог., IX : 337 (cornicina Fabr.0); Вайнштейн и Родова, 1940, Мед. паразитолог.,
IX . 366; Зимин, 1948 : 81, рис. 44.
Весьма близок к О. cornicina F., от которой отличается следующими
признаками.
Перед швом присутствует одна пара крепких, длинных акрости-
хальных щетинок; дорзопентральных щетинок позади шва — 3, а не 4
пары, как у О. cornicina. Лоб самца шире: ширина его равна двойной
ширине 3-го членика усиков; ширина щеки у самца составляет 2/б,
у самки — 2/з вертикального диаметра глаза. Тело короче: 4.5—8 мм.
Окраска обычно светлее, брюшко всегда с более отчетливо выраженным
бронзовым или золотистым оттенком, особенно сильно развитым на 5-м
тергите. Тека узкая, в 1% раза короче и почти в 4 раза уже дистальной
части фаллозомы; последняя бокаловидно расширена к вершине, сильно
склеротизована и покрыта многочисленными шипами, более мелкими
78
I. ТРИБА MUSCINI
нежными оороздками; хорион
Рис. 69. Самец Orthellia caesarion Mg.
(Ориг.).
в срединной части. Передняя часть парамер длинная, дуговидно изо-
гнутая, сильно расширена на вершине; задний отдел парамер в Р/г раза
короче переднего, постепенно сужен к вершине; дорзальный отросток
тонкий, длинный. Церки (рис. 75).
Яйцо неправильно веретеновидной формы, к одному полюсу сужено
несколько сильнее, нежели к другому, цвета слоновой кости или желто-
белое; по дорзальной стороне с 2 параллельными узко расставленными
ый, тонко шагренированный, без
ячеистой скульптуры. Длина
около 2 мм (от 1.9 до 2.2 мм).
Личинка I стадии крупная,
с суженным к переднему концу
телом; анальный сегмент в про-
филь полукруглый с бугорча-
тыми выступами в нижнем отделе,
из которых 2 располагаются по
бокам рядом с анальным бугорком
и 4 позади последнего. Брюшные
сегменты снизу по передним краям
с полосками из темных, крепких,
изогнутых шипов, заходящих на
боковую сторону сегментов до
конца нижней ее трети; на 2 брюш-
ном сегменте шипики образуют
одну узкую полоску, на сегментах
с 3-го по 8-й — по 3 поперечных по-
лоски, из которых передняя Уже
задних; 9-й сегмент имеет одну уз-
кую полоску из мелких шипиков
впереди анальных бугорков; 1-й
брюшной сегмент снабжен редко
расставленными шипиками снизу
по переднему краю; передние края
груднйх сегментов узко шагрени-
рованы. Личинка метапнейстична;
задние дыха льны с 1 округлым
и сердцевидным отверстием; пе-
ритрема сильно склеротизована.
Ротоглоточный аппарат (рис. 77, 78) обычного для рода Musca строения.
Фарингеальный склерит с дуговидными, узкими боковыми частями;
вентральные и дорзальные отростки заостренные, дорзальная перемычка
высокая, сравнительно широкая; передние отростки в виде двух тонких
длинных тяжей, соединенных с коротким гипостомальным склеритом;
последний слабо дуговидно изогнут и слегка утолщен в заднем отделе,
с вентральной стороны соединен широкой перемычкой; срединный дор-
зальный склерит широкий, коротко раздвоен сзади и здесь соединен
с частями гипостомального склерита; верхний его край слабо выгнут
у средины, в дистальной части дуговидно загнут вперед, образуя на вер-
шине крепкий, загнутый вперед и вниз зубеп; от основания последнего
передне-нижний край зазубрен (с 4—5 короткими угловидными высту-
пами, величина которых убывает книзу), оканчивается резким пойти
прямым углом внизу и переходит в вентральный край склерита. Боковые
стенки ротового отверстия укреплены плотными, сравнительно с родом
Рис. 70—76.
70 — передняя половина крыла Orthellia caesarion Mg. 71 — заднее ды-
хальпе Orthellia caesarion Mg. 72 — церки самца О. (Pyrelliomima)
latipalpis, sp. n. 73 — голова самца О. caesarion Mg. сбоку. 74 —
церки самца О. pacijica, sp. n. 75 — переднегрудь О. caesarion Mg.
76 — церки самца О. caesarion Mg. (Ориг.).
78
I. ТРИБА MUSCINI
Musca, узкими пластинками; впереди ротового отверстия проходит попе-
речный ряд тонких, узких слабо склеротизованных полосок. Длина тела
2.5—3.6 мм.
Личинка II стадии (рис. 79, 80) амфипнеистичиа; передние дыхальцы
с 7 лучами (реже с 6); задние дыхальцы с более или менее сильно склеро-
тизованной перитремой, имеют каждое по 2 коротких слабо изогнутых
щели; верхняя короче нижней. Тело лишено крепких шипиков; сег-
менты снизу по переднему краю с валикообразными поперечными утол-
Рпс. 77—80.
77 — срединный дорзальный склерит ли-
чинки Orthellia caesarion Mg. 78 — ротогло-
точный аппарат личинки 1 стадии О. caesa-
rion Mg. 79 — то же II стадии . 80 — задние
дыхальцы личинки'II стадии того же вида.
(Ориг.).
шениями; анальный сегмент сходен
с таковым личинки I стадии, но
отличается более широкой аналь-
ной пластинкой. Ротоглоточный
аппарат с прямыми и соединен-
ными на вершине ротовыми крюч-
ками, в основной половине не-
сколько расширенными, разделен-
ными и дуговидно изогнутыми;
угловидные склериты с более или
менее длинным, направленным
вперед от места изгиба острым
клиновидным отростком; верхний
конец склерита направлен назад
и несколько вверх. Гипостомаль-
ный склерит неправильных пря-
моугольных очертаний с длинным,
обращенным назад отростком, сое-
диняющимся с верхней частью пе-
редних отростков фарингеального
склерита. Фарингеальный склерит
длинный и относительно широкий,
вентральные его отростки коротко
косо срезаны на вершине, дорзаль-
ные— крыловидные, постепенно за-
остренные к дистальному концу;
дорзальная перемычка узкая у
места срощения ее с фарингеаль-
ным склеритом и полукругло рас-
ширена с каждой стороны в верх-
ней части, образуя здесь два на-
правленных вперед и два обра-
щенных назад выступа, верхний ее край слабо выдается над верхним
краем фарингеального склерита, перемычка с крупными отверстиями,
придающими ее стенкам решетчатый вид. Передние отростки удлинен-
ные, с оттянутым клиновидно вперед нижним дистальным углом: снизу
соединены вентральной перемычкой. Антенны и максиллярные шупальцы
ясные. Длина тела 3.8—4.7 мм.
Личинка III стадии (рис. 81—86) крупная, оливково-серая; кутикула
гладкая, блестящая, тонкая. Головной сегмент двураздельный спереди;
ротовые борозды покрывают нижнюю и боковые стороны сегмента; пре-
стомал ьные зубцы в виде светлых прижатых к телу угловидных пласти-
нок, идущих от краев ротового отверстия вбок; усики с длинным цилинд-
рическим 1-м й маленьким конусовидным 2-м члеником. 1-й грудной
сегмент вдоль всего переднего края в мелких многорядных шипиках,
1. ORTHELLIA
7!>
занимающих Vs—V* длины сегмента; по средней продольной линии на
спинной стороне сегмента короткая бороздка, иногда замененная круп-
ным пунктиром; прочая поверхность 1-го грудного сегмента гладкая, как
и последующие грудные и 1-й брюшной сегменты. Брюшные сегменты
со 2-го по 7-й снизу по переднему краю с валикообразными поперечными
возвышениями, усаженными шипами разного размера; наиболее мелкие
шипы расположены по заднему краю возвышений. Брюшные сегменты
со 2-го по 6-й снизу по бокам от средней продольной линии у заднего
края несут по группе бугорков (по 4—5 штук в каждой). 8-й брюшной
Рис. 81—86. Детали строения личинки III стадии Orthellia caesarion Mg.
81 — ротоглоточный аппарат. 82 — передние дыхальце. 83 — заднее дыхальце.
8 4 — задний конец тела личинки сбоку. 85 — он же сзади. 86 — вооружение
на нижней стороне 5-го (2) и 6-го (1) брюшных сегментов. (По Зимину).
сегмент чуть короче 7-го, сзади округленный; дорзальный край его вы-
дается над дорзальным краем 7-го сегмента, по дорзальной поверхности
с тремя продольными бороздками; задняя сторона вокруг дыхалец слабо-
вдавлена, в нижнем отделе под дыхальцами с 2 слабо выступающими
пологими бугорками; анальная пластинка широкая, длинная, своими
концами достигающая средней боковой продольной линии сегмента или
даже заходящая за нее; концы пластинки несколько сужены и слабо-
загнуты вперед. Вдоль переднего и заднего краев анальной пластинки
располагаются светлые шипы, достигающие ее боковых концов; шипики
вдоль заднего края заходят вверх несколько далее переднекрайних.
Нижняя сторона заднего конца тела несколько вздута с 7 бугорками
в перианальной области (2 преанальных, 2 анальных, 2 субанальных
SO
I. ТРИБА MUSCINI
и 1 постанальный); наибольшего развития достигают субанальные
бугорки, наименьшего — конусовидный, постанальный. Передние ды-
хальцы поперечные, с 7 округлыми выростами. Задние дыхальцы
крупные, расставленные на Уг ширины стигмы; перитрема широкая,
черная; стигмальный диск крупный, черный; стигмальная пластинка
желтовато-рыжая, дыхальцевые щели тонкие, многократно змеевидно
изогнутые; внутренние концы верхней и нижней щели обращены к стиг-
мальному диску, как и нижний конец средней щели, прочие концы направ-
лены наружу. Ротовой аппарат типа Musca, но с дополнительными скле-
ритами под ротовыми крючьями. Ротовые крючья длинные, левый короче
правого, слабо дуговидно изогнутые в вершинном отделе; основание
с сильно оттянутым назад вентральным углом; зубные склериты непра-
вильно треугольные; добавочные склериты в форме 2 узких волнисто
или дуговидно изогнутых тяжей. Гипостомальные склериты в 2 раза
короче ротовых крючьев, неправильных очертаний, широкие. Фарин-
геальные склериты с широкими задними и передними отростками; перед-
ние отростки выдвинуты вперед значительно дальше конца дорзальной
перемычки; вентральная перемычка узкая; дорзальная перемычка решет-
чатая с узкими длинными задними отростками, сращенными с дорзаль-
ными краями склерита; вентральные отростки с крупными угловидными
выступами по верхнему краю; дорзальные отростки крыловидные, дуго-
видно согнутые, к концу суженные, фарингеальная мембрана широкая.
Длина тела 11—13.3 мм; толщина у средины 1.7—2.4 мм.
Ложнококон красно-коричневый, ± блестящий, поперечно мелко
морщинистый снизу, с матовыми грубо шагренированными полосами
по передним краям сегментов; сегментация резко выражена. Передние
стигмы (рожки) крупные, черные. Анальная пластинка широкая в сре-
дине, к концам постепенно сужена. Крупные бугорки, характерные для
личинок, отсутствуют; задний конец ложнококона правильно, узко за-
круглен, передний, по сравнению с ним, слабо расширен. Длина 4.3—
5.2, диаметр 1.7—2.2 мм.
Распространение. В Палеарктике северная граница вида
проходит через скандинавские страны и Кольский полуостров, заходя
здесь за Северный Полярный круг. С продвижением на восток граница
ареала смещается к югу, проходя через Соловецкие острова, Онегу,
Усть-Цыльму (на Печоре), Березов (на Оби), Тобольск, Томск, Красно-
ярск, Иркутск и Якутск. Южная граница идет по северной Африке через
Малую Азию и Северный Китай до Японии (Нагасаки). Вид известен
за пределами Палеарктики из Северной (США, Мексика) и Южной Аме-
рики (Бразилия).
В СССР южная граница вида, по всей вероятности, совпадает с юж-
ной его границей, отступая от нее на север лишь в зоне степного и полу-
пустынного ландшафта. О- caesarion связан по преимуществу с лесной
(хвойные и лиственные леса) зоной, в южных широтах придерживаясь
горных областей.
Биология вида изучена недостаточно. Известно, что в своем размно-
жении вид связан с пометом животных (коров и лошадей), где про-
текает развитие личинок, являющихся копрофагами. По данным Дер-
беневой-Уховой (1940) для Кабарды и нашим наблюдениям в Средней
Азии, вид размножается в коровьем помете или в навозе этих жи-
вотных.
Окрыленные мухи растительноядны и копрофаги. Откладка яиц
производится на навоз. По подсчетам Дербеневой-Уховой (1940), каждая
1. ORTHELLIA
81
кладка содержит в среднем около 37 яиц. Максимальное количество мух
в природе падает на май и первую половину июня (Таджикистан).
*2. Orthellia (in sp.) cornicina F. (рис. 87—96, 102).
Fabricius, 1781, Spec. Ins., II : 438 (Musca); Pandelle, 1898, Rev. entom., XVII :
38 (Pyreilia)', Stein, 1916 : 18 (Pseudopyrellia)', Stein, 1917 : 105 (Cryptolucilia)', Mal-
loch, 1923, Ann. Mag. Nat. Hist, (9), XII : 518 (Orthellia),
Биология. Порчинский, 1910, Tp. Бюро энтом., VIII : 22, рис. 15—20;
Keilin, 1917, Parasitology, IX : 389, 422; Brues, 1902, Psyche, IX : 354; Bequaert,
1922, Psyche, XXIX : 89; Thomson, 1937 : 276 figs. 1, 2, 4, 5, 11, 12, 52, 53,
145 (Cryptolucilia); Дербенева-Ухова, 1940, Мед. паразитолог., IX : 337; Hand-
schin, 1934, Rev. Suisse Zool., 41 : 8.
Ширина лба самца в наиболее узком его месте вблизи средины в Р/г
(реже l3/.») раза шире 3-го членика усиков; орбиты в том же отделе в 1 %—
2 раза шире узкой бурой лобной полосы; скулы в верхнем отделе по ши-
рине равны 3-му членику усиков, книзу постепенно расширены; высота
щеки в 3—3/4 раза короче вертикального диаметра глаза; передний
верхний край щеки заходит углом выше нижнего уровня глаза; верх-
ний край рта выставлен вперед далее нежели лоб; вибриссальный угол
несколько приподнят над передним краем рта; глаза в редких светлых
волосках, заметных в нижнем отделе глаза, чаще у 33 чем у jj; усики
коричневато-бурые, 3-й членик в 2/4—3 раза длиннее 2-го; щупальцы
рыжевато-бурые, слабо расширены к вершине; подбородок длиною с усик,
блестящий, с зеленоватым металлическим отливом. Затылок и щеки
в тонких бурых волосках, орбиты в коротких редких волосках, распо-
юженных вблизи лобных щетинок и немного спускающихся на скулы,
заходя за линию передних лобных щетинок. Внутренние теменные ще-
тинки и вибриссы крепкие, длинные; лобные, кроме нескольких более
крепких передних пар, тонкие, короткие, как и глазковые щетинки.
Голова черно-бурая; орбиты, затылок и щеки металлически зеленые
или сине-зеленые, блестящие, лишены налета, исключая нижне наружный
край затылка, покрытый светлым налетом. Скулы в шелковисто белом
густом, лицо в тусклом, редком, сером налете. Грудь блестящая, метал-
лически зеленая или синяя; гипоплевры местами коричневые; коричне-
ватый налет груди слабо выражен (практически налет отсутствует).
Грудь в черных волосках; спинка с 2 узкими, слабо расходящимися назад
штрихами, иначе окрашенными, чем прочая поверхность спинки, и слабо
заметными. Переднегрудь в многочисленных длинных темных волосках;
суцрасквамальное ребро в бурых щетинистых волосках; проплевры голые
в центре; гипоплевра с одиночными волосками впереди заднего дыхальца
и более многочисленными темными волосками в нижнем отделе. Перед-
нее дыхальце бурое. Щетинки груди: акростихальных — 1 пара перед
щитком; дорзоцентральных — 2-J-4; интраалярных — 1; предкрыловых—
1; надкрыловых — 2 (из них задняя много короче и тоньше передней);
закрыловых — 3; плечевых — 3; заплечевых — 1; предшовных — 1; ното-
плевральных — 2; стерноплевральных — 1+2; на щитке 3 пары крепких
щетинок (вершинных, боковых, основных); предосновные щетинки отсут-
ствуют. Крылья сероватые с несколько более темным основанием. Жилки
рыжевато-желтые, при основании — коричневатые; г4+3 со щетинками
на верхней и волосками на нижней стороне, занимающими 2/з—3/« пер-
вого отрезка жилки. Чешуйки белые или желтовато-белые; жужжальца
буроватые, с желтоватыми отметинами при основании. Ноги коричнево-
5урые с металлическим оттенком на бедрах, особенно передней и задней
g Фауна СССР, том XVIII, вып. 4.
82
I. ТРИБА MUSCINI
пар ног. Передние голени по передне-дорзальному краю с рядом удли-
ненных щетинистых волосков, образующих род короткого гребня; неко-
торые волоски достигают крепости щетинок. Средние голени без передне-
дорзальных щетинок, с длинной крепкой вентральной (сдвинутой на
задне-вентральную сторону, почему она может быть названа задне-вен-
тральной) щетинкой. Задние бедра без длинных щетинистых волосков на
нижней стороне, исключая 1—,2 согнутых, тонких при основании бедер;
задне-вентральная поверхность бедер посредине с 1 крепкой, передне-
вентральная — с несколькими крепкими щетинками, сконцентрирован-
ными в вершинной половине бедра. Брюшко цвета груди, с легким серо-
ватым налетом на вентральной поверхности, покрыто короткими на верх-
ней и более тонкими и длинными на нижней стороне волосками; марги-
нальные щетинки короткие, тонкие, слабо выделяются среди прочих во-
лосков брюшка. Генитальный аппарат самца сохраняет тип строения
такового предыдущего вида, отличаясь сравнительно небольшими дета-
лями в строении церок, парамер и относительными размерами фалло-
зомы. V стернит по заднему краю выемчатый с боковыми выступами.
Ширина лба самки равна или чуть более ширины глаза; орбиты
выпуклые, в задних 2/з металлически зеленые, синие или с фиолето-
вым оттенком; в передней трети, как и скулы, покрыты светлым шелко-
висто блестящим налетом; в этом месте несколько вдавленные; лобная
полоса в 1 %—12/з раза уже орбиты, слабо расширена в переднем отделе,
рыжевато-коричнево-бурая, без волосков. Скулы в 13/4—2 раза шире
3-го членика усиков. Орбитальные щетинки отсутствуют. Наружные
и внутренние теменные щетинки и вибриссы крепкие. Окраска и хетотак-
сия как у самца. Длина тела 6.5—8.7.
Яйцо (рис. 94) очень сходно по строению с яйцом М. domestica, но
шире его; слабо вогнутое на дорзальной стороне, снабженное здесь 2 па-
раллельно-крайними нежными ребрышками. Длина 2 мм.
Личинка I стадии (рис. 87, 92) метапнейстична; с вентральной сто-
роны брюшных сегментов (от 2 до 7) располагаются по передним их краям
двойные полоски из мелких шипиков, доходящие до половины боковых
поверхностей каждого сегмента. Такие же полоски, но состоящие из
более мелких шипиков, имеются на переднем крае 3-го грудного и 1-го
брюшного сегментов. Анальная пластинка ясная, заходит на бочки сег-
мента далее средины. Базальный склерит ротоглоточного аппарата тонкий
с заостренными задними отростками; дорзальная перемычка хорошо
выражена в средней своей части; передние отростки, соединяющиеся
с гипостомальным склеритом, тонкие, длинные; срединный дорзальный
склерит узкий, в виде вытянутого листа; передний конец оттянут
несколько вниз и заострен; под ним располагается парный Т-образный
склерит (по Кейлину — гомолог нижней губы). Головной сегмент с парой
антенн и максиллярных щупалец. Задние стигмы сердцевидные со сре-
динной перегородкой (по Порчинскому). Длина личинки до 4 мм.
Личинка II стадии (рис. 88) амфипнейстична; передние дыхальцы
с 7 лучами; задние дыхальцы имеют по 2 изогнутых щели. Базальный скле-
рит ротоглоточного аппарата крупный, сагиттально вытянутый; вен-
тральные отростки шире и много длиннее дорзальных, от средины своего
дорзального края до вершины скошены; задние концы отростков заост-
рены. Дорзальная перемычка низкая, широкая сверху, крупно решет-
чатая; передние отростки базального склерита сравнительно длинные,
прямые, сочленяются с гипостомальным склеритом и поперечно располо-
женным вентральным склеритом. Гипостомальный склерит парный,
1. ORTHELLIA
83
в виде 2 длинных слабо изогнутых хитиновых тяжей. Передние их концы
соединены с угловидным склеритом, расщепленным снизу. Ротовые боко-
вые крючки короткие, тонкие, слабо дуговидно изогнутые; под ним рас-
положены два губных коротких, широких склерита, укрепляющих стенки
ротового отверстия.
Личинка III стадии (рис. 89, 90, 91, 93) имеет передние дыхальцы
с 7 лучами; задние дыхальцы имеют по 3 щели, изгиб которых почти
Рис. 87—96. Детали строения преимагинальных фаз Orthellia cornicina F.
87 — ротоглоточный аппарат личинки 1 стадии сбоку. 88 — то же II стадии. 89 —
то же III стадии. 90 — то же III стадии сверху. 91 — задние дыхальцы личинки
III стадии. 92 — задний конец тела личинки I стадии сбоку. 93 — анальная пла-
стинка личинки III стадии. 94 — яйцо. 95 — ложнококон. 96 — дыхальце куколки.
(По Томсону).,
в точности отвечает таковому личинок III стадии Musca, но более изо-
гнутые и расстояние между пластинками много короче, чем у личинок
домашней мухи. Промежуток между пластинками составляет всего 1/е—
Vs ширины дыхальца. По периферии дыхалец располагаются отверстия
периспиракулярных желез. Передние края брюшных сегментов_от 2 до
7 с нижней стороны несут двойные полоски из мелких шипиков. Аналь-
6*
84
I. ТРИБА MUSCINI
ная пластинка узкая, длинная с возвышающимися тотчас позади нее
3 анальными сосочками, направленными вниз и вбок. Тело личинки голу-
боватого, зеленоватого или бронзового оттенка, перед окуклением^цвет
личинки почти белый. Базальный склерит ротоглоточного аппарата
сзади глубоко выемчатый; дорзальная его перемычка грубо сетчатая,
с двулопастным краем спереди и двувильчатым отростком посредине зад-
него края; нижний край слабо склеротизован и ; снабжен продольными
фарингеальными ребрышками; передние отростки базального склерита
срощены с гипостомальным и вентрально_от задних его концов распо-
ложенным поперечновытянутым склеритом. Гипостомальный склерит
массивный Н-образный; ротовые крючья тесно сближены; они имеют
широкую базальную часть, снабженную снизу клиновидным выступом.
Зубные склериты соединены снизу, образуя вентральную перемычку.
Кпереди от зубных помещается пара мелких добавочных склеритов.
Головной сегмент с парой антенн и максиллярными щупальцами. Длина
тела 7—8 мм.
Ложнококон (рис. 95, 96) светлокоричневый, длиною 6—7 мм. Перед-
ние дыхальцы и задние ясно выступают; анальная пластинка резко очер-
чена.
Распространение. В пределах СССР распространен от
крайних западных областей на восток до Приморского края. В Средней
Азии известен из различных мест Таджикистана. Распространение в Си-
бири и Казахстане за малочисленностью материала не вполне ясно;
известны следующие места нахождения: Кокчетав, Боровое, Северный
Казахстан, Алтай, Томск, Красноярск, Березов на Оби, Иркутск, Яков-
левна Приморского края; в Европейской части СССР южная граница
проходит по линии Киев—Полтава—Харьков—Саратов—Кубань—Кавказ
(Железноводск, Кисловодск).Известен по всей Зап. Европе от Италии
до Скандинавии. Известен из Северной Монголии: Урга, ущ. Сюцзуктэ,
пров. Кентэй. Данные по Северной Америке требуют проверки. Вид из-
бегает открытого ландшафта, придерживаясь облесенных и горных обла-
стей.
Биология вида изучалась Порчинским, Томсоном и в отдель-
ных деталях Беквертом, Кейлиным, Вайнштейном, Родовой, Дербене-
вой-Уховой. За исключением последней, все авторы свидетельствуют
о том, что развитие мухи проходит исключительно в коровьем помете.
Дербенева-Ухова указывает (1940 : 337), что личинки в условиях Ка-
барды развиваются лишь в конском помете, что, очевидно, является для
вида „необычным. Яйца закладываются группами по 30—50 штук в по-
лость, сделанную в помете яйцекладом самки. Выход личинок из яиц
наступает при благоприятных условиях через сутки. В противном слу-
чае выход задерживается до 2—3 дней. В первой стадии, по свидетельству
Томсона, личинка пребывает около 18 часов; длительность II стадии ли-
чинки при средней суточной температуре 18° С составляет 48 часов,
с понижением температуры до 15° С развитие несколько задерживается.
В ложно-коконе муха находится от 10 до 20 дней. При условии теплой
погоды на развитие мухи требуется до 22 дней.
Подобно большинству калоядных личинок, личинки О. cornicina
подвергаются нападению хищников. Хьюитт (1914) указывает на унич-
тожение личинок зеленой коровницы личинками мухи Scopeuma ster-
corarium L. По нашим наблюдениям в Таджикистане личинки этого вида
уничтожаются личинками хищных мух: Anaclysta multipunctata Stein
и Myiospila meditabunda F.
1. ORTHELLIA
85
*3. Orthellia (in sp.) pacifica Zimin, sp. nov* (рис * 74, 97 ' 99, i04).
$. Лоб в 2 раза шире 3-го членика усиков, лобная полоса черная,
в наиболее узком месте имеет вид тонкой линии, слабо расширенной на
обоих концах; скулы в средней их части равны ширине 5-го членика уси-
ков книзу постепенно расширенные; высота щеки в 4 раза короче верти-
кального диаметра глаза. Глаза в едва заметных светлых волосках в ниж-
нем своем отделе. Усики бурые, 3-й членик в 2V1 раза длиннее 2-го; щу-
пальцы черно-бурые, слаборасширенные к дистальному концу; затылок
и щеки металлически зеленые или синезеленые; такой же окраски зад-
няя треть орбит и наружный, примыкающий к глазам их край; скулы
и внутренний край орбит около лобной полосы — черные, лакировано-
блестящие; скулы от уровня прикрепления усиков до нижней их границы
в шелковисто-блестящем белом налете. Внутренние теменные щетинки
и вибриссы длинные, крепкие; лобные щетинки (кроме передних не-
сколько утолщенных пар) короткие, как и затылочные щетинки, и имеют
вид щетинистых волосков; орбиты в редких, коротких, бурых волосках,
заходящих немного ниже передних лобных щетинок. Глаза в верхне-
переднем отделе со слабо увеличенными фасетками. Грудь темнозеленая
пли синевато-зеленая в черных, торчащих, коротких щетинистых воло-
сках. Переднее дыхальце бурое; переднегрудь расширенная кпереди,
в тонких бурых сравнительно длинных волосках; передние плевры
в центре без волосков; гипоплевра с группой волосков впереди заднего
дыхальца и в заднем нижнем углу; супрасквамальное ребро с длинными
бурыми щетинистыми волосками. Щетинки груди: акростихальные тон-
кие — 1 пара перед щитком; дорзоцентральных — 1 пара крепких и 1
слабо развитая (позади шва); интраалярных — 1 хорошо развитая; пред-
крыловых — 1, надкрыловых — 1, плечевых — 3; заплечевых — 1;
предшовных — 1; акростихальные щетинки перед швом отсутствуют,
как и дорзоцентральные; у некоторых экземпляров последние могут при-
сутствовать в виде 1 пары очень коротких тонких волосовидных щети-
ночек; стерноплевральных — 1-)-2; нотоплевральных — 2. На щитке:
вершинных — 2, боковых — 2, основных — 2, все щетинки щитка хорошо
развиты. Крылья бледно серые, при основании рыжеватые; жилки
рыжие, при основании буроватые; чешуйки бледнодымчатые; верхняя
чешуйка с белым пятном у наружного края. Жилка г4+5 снизу с воло-
сками. заходящими далеко от г—т сверху с мелкими щетинистыми воло-
сками лишь в основной половине первого отрезка жилки; жужжальца
желтые с буроватой булавой. Ноги толстые, чернобурые, передние бедра
по задней поверхности и задние по передней поверхности с зеленым ме-
таллическим отблеском. Переднедорзальные щетинки на средних голе-
нях отсутствуют; вентральная щетинка крепкая. Задние бедра по передне-
вентральной и задневентральной поверхности со щетинистыми воло-
сками, длина которых не превышает поперечника бедра. Коготки и при-
соски короче 5-го членика лапки. Брюшко цвета груди; выпуклое, лишено
светлого налета, в черных щетинистых волосках. Щетинки брюшка очень
короткие, длиной не превышающие 1/в длины тергита. Низ брюшка в более
густых удлиненных черных волосках. Церки расширены в основной
половине, далее сужены к вершине, на дистальном конце образуют две
широких лопасти, разделенные узким вырезом. Фаллозома с узкой
трубковидной текой и массивной вершинной частью, сильно склероти-
зованной и покрытой от средины до вершины многочисленными узкими
зубцами. Передняя часть парамер длинная, дуговидно изогнутая,
86
I. ТРИВА MUSCINI
расширенная на конце; задний отдел ее короче переднего, сужен к вер-
шине. Дорзальный отросток фаллозомы длинный, узкий, изогнутый;
аподема длинная, утолщенная на вершине.
Лоб самки равен или чуть шире глаза; орбиты широкие, металли-
чески зеленые или сине-зеленые; атласно-черная лобная полоса слабо
сужена в задней половине; ширина ее в Р/г—13/« раза менее ширины
орбиты; лоб с выпуклыми орбитами. Скулы блестяще черно-бурые в белом
налете; ширина их посредине в раза более ширины 3-го членика
усиков. Орбитальные щетинки отсутствуют; орбиты в черных, редких
щетинистых волосках, заходящих на скулы чуть ниже уровня лобных
щетинок. Теменные наружные и теменные внутренние щетинки и ви-
бриссы толстые, длинные. Затылок сильно выпуклый, как и щеки, метал-
лически зеленый или сине-зеленый. Чешуйки серовато-белые. В осталь-
ном окраска и хетотаксия как у самца. Длина тела 6—8 мм.
Распространение. В СССР известен с Дальнего Востока:
дер. Майхэ, Шкотовского р-на; Владивосток, Бухта Золотой Рог; Яков-
левка, Спасского р-на; Камень-Рыболов и сел. Троицкое на оз. Ханка;
ст. Пограничная; Сидеми, Нагорная в южной части Приморского края.
О. pacifica, sp. п., относится к обширной группе видов, распростра-
ненной в экваториальных странах Старого и Нового Света, которая
достигает наиболее пышного расцвета на островах Малайского архипе-
лага. Для группы является характерным слабое развитие хетотаксических
признаков, в частности щетинок среднеспинки. В пределах СССР пред-
ставитель этой группы видов рода отмечается впервые.
В систематическом отношении О. pacifica, sp. п., близка к ряду ви-
дов, от которых отличается следующими признаками. От О. bimaculata
St. — сплошным светлым налетом на лице и скулах, который у назван-
ного вида присутствует лишь в форме овальных пятен на скулах; от О. du-
Ыа Mall. — отсутствием очень крупных фасеток в верхнем отделе глаз
и темными ногами. Светлые крылья О. pacifica, sp. п., лишенные темных
отметин, отличают новый вид от О. chalybaea Wd. От недавно описан-
ных О- (Neomyia) diadema End., О. (N.) argentigena End., 0. (N.) lauta
End, — светлой окраской чешуек, темными жужжальцами, распределе-
нием светлого налета на скулах и окраской орбит. По системе Эндер-
лейна (1928), вид должен быть отнесен к его роду Neomyia, который,
-с нашей точки зрения, являетбя синонимом рода Orthellia R.-D., о чем
см. описание рода.
2. Подрод Pyrelliomima Zimin, subgen. nov.
(Рис. 72, 100, 101, 105)
Тип подрода: Orthellia (Pyrelliomima) latipalpis, sp. nov.
Глаза голые; жилка r4+5 co щетинками в передней половине основного
отрезка сверху и с волосками в том же месте жилки снизу; переднегрудь
широкая, как и супрасквамальное ребро в щетинистых волосках; задние
грудные дыхальцы длинные, поперечно вытянутые, как у О. cornicina
и О. caesarion', гипоплевры с волосками снизу и впереди дыхалец. Изгиб
т тупоугольный, на самом сгибе узко округленный; глаза $$ разделены
очень узким лбом с крупными фасетками вверху, у самки лобная полоса
очень широкая, орбиты с 3 изогнутыми вбок орбитальными щетинками,
в 5 раз уже лобной полосы, последняя с одиночными волосками в заднем
отделе; щеки почти в 5 раз короче вертикального диаметра глаза, перед-
ний их край заходит выше нижнего уровня глаз; щупальцы расширен-
1. ORTHELLIA
87
ные; скулы очень узкие. Спинка в редком налете; щетинки груди: акро-
стихальных — 1 пара перед щитком; дорзопентральных — 2-f-4 (у самца—
3); интраалярных — 1; предкрыловых — 1; надкрыловых — 2 (задняя
слабо развита), закрыловых — 3; плечевых — 3; заплечевых —1; ното-
плевральных — 2; стерноплевральных — l-f-З (у развита 4-я, распо-
ложенная ниже трех остальных, стоящих в ряд); на щитке 3 пары креп-
ких щетинок (вершинных, боковых, основных). Брюшко блестящее, без
налета.
По строению переднегруди, заднего дыхальца, опушению передне-
груди и супрасквамального ребра, а также по хетотаксии груди и щитка
Pyrelliomima наиболее близка в Orthellia R.-D. и рассматривается нами
как его подрод. С Pyreilia R.-D. подрод имеет общие черты в строении
головы: узкие скулы; орбиты, щеки и лоб сампа, очень узкие орбиты
самки.
С достоверностью известны 2 вида этого подрода. Под вопросом сюда
могут быть отнесены О. trispina Mall, и О. indica R.-D.
Распространение. Ориентальная область и юг Эфиоп-
ской области. Б пределы Палеарктики (Китай, Япония) заходит всего
1 вид. R СССР представители Pyrelliomima пока не известны.
4. Orthellia (Pyrelliomima) latipalpis Zimin, sp. nov. (рис. 72, 100,
101, 105).
Голова самца шире груди, большая; глаза сверху разделены узким
лбом, ширина которого в 3 раза уже 3-го членика усиков; в передне-верх-
нем отделе глаза фасетки сильно укрупнены; диаметр крупных фасеток
составляет почти половину ширины верхнего отдела скулы; усики длин-
ные, рыжевато-коричневые, с более светлым основанием 3-го и вершиной
2-го члеников; 3-й членик в ЗРг—4 раза длиннее 2-го; скулы очень узкие,
ширина их в среднем отделе составляет 8А ширины 3-го членика усиков;
щеки узкие, бурые, внизу красновато-коричневые, в редком сероватом
налете; щупальцы рыжевато-коричневые, сильно расширены на вершине;
наибольшая их ширина в 1*/г—РА раза превосходит ширину среднего
отдела скул. Бнутренние теменные щетинки и вибриссы крепкие; прочие
щетинки слабые, короткие. Грудь сине-зеленая, передняя половина спинки
буровато-бронзовая, в сероватом редком налете; задний отдел плевр корич-
неватый. Дорзопентральных щетинок позади шва — 3, причем передняя
щетинка далеко отстоиТ от поперечного шва. Церки расширены в основ-
ном отделе, который переходит в более узкий, почти параллельно край-
ний срединный отдел и далее — в дистальное расширение, имеющее вид
квадрата со сглаженными углами и отделенное снизу короткой узкой
выемкой; боковые края дистального отдела несут каждый по 2 длинных
щетинки. Фаллозома с очень короткой, узкой трубковидной текой и чрез-
вычайно массивным дистальным отделом, снабженным в вершинно-дор-
зальной части очень широкими плоскими, крупными зубцами. Передняя
часть парамер широкая, с 2 щетинками у основания, задняя заострена
к вершине; дорзальный отросток короткий, как и аподема. В остальном
внешность самца как у самки.
Ширина лба самки составляет 8 А ширины глаза, орбиты в 5 раз уже
широкой, выпуклой по краям бурой полосы; ширина скул не превышает
ширины 3-го членика усиков; щеки узкие, высота их в 4х/а—5 раз менее
вертикального диаметра глаза; передний верхний край дуговидно при-
поднят и заходит впереди за нижнюю границу глаза: затылок вогнутый
88
I. ТРИБА MUSCINI
в верхней и слабо выпуклый в нижней половине, выступает повади верх-
него края глаз. Усики коричневато-бурые, как и щупальцы; 3-й членик
их в 3 раза длиннее 2-го; щупальцы постепенно и сильно расширены
Рис. 97—103. Головы видов рода Orthellia.
97 — самца О. pacifica. sp. п., сверху. 98 — самки О. pa-
cifica, sp. п., сверху. 99 — самца О. pacifica, sp. п., сбоку.
100 — самки О. latipalpis,\ sp. in., сверху. 101 — самца
О. latipalpis, sp. п., сбоку. 102 — самки О. cornicina F.,
сверху. 103J— самца О. bimaculata St., сбоку.
к дистальному концу; наибольшая их ширина составляет s/t ширины
скулы в среднем ее отделе. Затылок, задняя треть орбит и задняя поло-
вина щек металлически зеленые; средина орбит и передняя часть щек
1. ORTHELLIA
89
блестяще черные; передняя треть орбит и скулы в сероватом светлом-
слабо блестящем налете. Затылки и щеки в черных коротких волосках,
орбиты с немногочисленными щетинистыми волосками, достигающие лишь
передних лобных щетинок, не заходя далее на скулы; орбитальных щети-
нок — 3; они короткие, толстые, расположены в задней трети орбит
и направлены вершинами вверх и вбок; лобная полоса с 2 щетинистыми
волосками в заднем отделе; наружные и внутренние теменные и затылоч-
ные щетинки и вибриссы крепкие, длинные; из лобных щетинок наиболь-
шей толщины достигает лишь одна их передняя пара. Глаза голые. Грудь
зеленая или сине-фиолетовая, блестящая, с 4 бронзовыми узкими впе-
реди шва и расширенными постепенно назад полосками; передний отдел
спинки в редком, но ясном сером налете; переднегрудь и супрасквамаль-
ное ребро в щетинистых волосках; гипоплевры с группой волосков впе-
Рис. 104. Фаллозома Orthellia pacifica, sp. п. (Ориг.).
Рис. 105. Фаллозома Orthellia latipalpis, sp. n. (Ориг.).
реди задних дыхалец и в нижнем углу. Переднее дыхальце рыжевато-
бурое. Крылья сероватые с желто-рыжими жилками; г4+5 в основной
половине первого отрезка со щетинками сверху и волосками с нижней
стороны, жужжальца сплошь палево-желтые; чешуйки рыжевато-пале-
вые; наружный отдел крыловой чешуйки белый. Ноги коричнево-бурые;
вентральная щетинка средних голеней крепкая, длинная; щетинки
по переднедорзальной поверхности средних голеней отсутствуют; задние
бедра по середине задне-вентральной поверхности с 1 щетинкой, по перед-
не-вентральной стороне с рядом щетинок более тонких и коротких
в основной половине бедра. Брюшко золотисто-зеленое, или синее, с фио-
летовым оттенком, блестящее, лишено налета, в черных щетинистых
волосках. Маргинальные щетинки короткие, тонкие, имеют характер
щетинистых волосков.
Вид описан по двум самцам и двум самкам из Китая и Японии.
Китай: Сычуань, Мао-Чжау, Матайги, 27 VIII 1893 (Потанин). Япония:
Комия бл. Суммо-Амами, 18 XII 1926 (Шмидт).
90
I. ТРИВА MUSCINI
Тип в Зоологическом институте Акад. Наук СССР в Ленинграде.
Вид весьма близок О. obscuripes Stein (1918) с острова Явы, от Кото-
рого отличается увеличенным количеством дорзоцентральных щетинок
— 2-4-3, j 2-(- 4 (5), против 2-|-2 у О. obscuripes], неразвитыми перед-
недорзальными щетинками на передних голенях и палевыми жужжаль-
цами. О. trispina Mall. (1928) с Суматры характеризуется крепкими, на-
правленными вперед орбитальными щетинками £ и количеством дорзоцен-
тральных щетинок, которых у О. trispina имеется 1-4-3 [вместо 2-|-4
(5)]. Очевидно систематически близким видом является также О. indica
R.-D. (1830), описанный из Индии, который нами не учтен в связи с отсут-
ствием сравнительного коллекционного материала и недостаточностью
оригинального описания.
2. Род MUSCA LINNE
Linne, 1758, Syst. Nat., X : 589; Robineau-Desvoidy, 1830, Essai Myod. : 394;
Schiner, 1862, Fauna Austriaca, I : 593; Schnabl u. Dzicdzicki, 1911 : 128; Bezzi u.
Stein, 1907 : 602; Stein, 1916 : 16; Awati, 1917 : 160—191, 484-195; Karl, 1928, Tierw.
Deutschl., 13, III : 6; Patton, 1932 : 347—405; он же, 1933 : 135—156; 397—430;
Ho, 1938 : 279—312; Townsend, 1937, Manual Myiology, V : 45—46; — Byomyia Robi-
neau-Desvoidy, 1830 : 392; — Plaxemyia Robineau-Desvoidy, 1830 : 392; — Philae-
matomyia Austen, 1909:295; — Pristirhynchomyia Brunetti, 1910, Rec. Ind. Mus.,
IV : 91; — Eumusca Townsend, 1911, Proc. Ent. Soc. Wash. XIII : 170; — Viviparo-
musca Townsend, 1915, Journ. Wash. Acad. Sci., V :435; — Ptilolepis Bezzi, 1921,
Ann. trop. Med. Parasitol., XIV : 335; — Awatia Townsend, 1921, Insec. Instic. Menst.,
IX : 132; — Lissosterna Bezzi, 1923, Bull. Soc. Roy. entom. Egypte : 108—118.
Тип рода — Musca domestica L. 1758.
Глаза голые или в волосках, жилка т с угловидным, более или
менее выраженным изгибом; жилка п — без щетинок; ариста с длинными
волосками с обеих сторон; ширина лба самца подвержена сильным коле-
баниям от вида к виду, занимая от Vio до 2/в ширины глаза, у самок —
составляет обычно около 1/з ширины головы; переднегрудь широкая,
в жестких волосках по краям. Средние голени с внутренней стороны
у середины без крепкой щетинки.
Средней величины и мелкие мухи обычно с темной окраской тела,
покрытого более или менее развитым светлым налетом, образующим
продольные полосы на спинке и переливчатый пятнистый рисунок на
брюшке. У некоторых видов грудь блестяще черно-оливковая, или
зеленая. Брюшко самцов у большинства видов с желтыми отметинами
в боковых отделах. Церки самцов в форме щитка, без длинных отростков
или крепких шипов и щетинок; кокситы 9-го тергостернита более или
менее квадратные; фаллозома с резко очерченной, обычно трубковидной
базальной склеротизованной частью и тонкой мембранозной вершиной.
6-й и 7-й тергиты в виде узких, асимметричных полуколец; 6-е и 7-е
дыхальцы брюшка у самцов имеются, у самок — отсутствуют. Хитиновые
части 6—9-го сегментов брюшка самок в виде продольно-вытянутых
узких тяжей.
В мировой фауне в роде Musca известно несколько десятков видов,
самостоятельность которых может быть с большей или меньшей степенью
точности установлена. Нельзя, однако, и сейчас с полной уверенностью
сказать, что видовой состав рода изучен с исчерпывающей полнотой.
Это в особенности относится к формам, обладающим большой индиви-
дуальной изменчивостью, как, например, группы видов — domestica
(vicina) — nebulo — cuthbertsoni — yerburyi, autumnalis — mesopotami-
2. MUSCA
91
ensis и им близкие виды, или — vitripennis (phasiaeiormis) — sugillat-
rix — osiris и некоторые другие.
Строение генитального аппарата самцов, исследованное Пэттоном
(1932) у многих видов, показывает, что эти морфологические признаки,
вопреки мнению Пэттона, сильно изменчивы, во всяком случае настолько,
что не могут служить критерием для разграничения всех видов.
И именно у видов, внешне сильно варьирующих, морфология половой
сферы самцов не дает надежных признаков. Более того, в роде Musca
у видов, хорошо отличающихся чертами внешней морфологии и окра-
ской, гениталии самцов дают мало ошутимые различия.
У М. domestica — vicina — nebulo — cuthbertsoni, кроме изменчивой
ширины лба самцов и еще менее постоянной окраски брюшка самок,
имеются лишь слабо выраженные различия в форме церок, дорзального
отростка фаллозомы и парамер, в связи с чем упомянутые формы едва ли
могут быть признаны хорошими видами.
Переходя к вопросу о систематическом единстве рода Musca нельзя
не признать наличия в нем хорошо очерченных группировок, которым
различные авторы придавали значение то родов, то подродов, то групп.
Не вдаваясь в подробности, укажем, что для видов нашей фауны Таун-
сенд (1915—1935) предусматривал вместо одного рода {Musca L.) 13 родов,
Мэллок (1919—1932) — 6 родов, Хо (1936) — 3 рода с 6 подродами;
Авати (1909—1916) различал в роде 3 типа видов, что близко подходит
к точке зрения Пэттона, наметившего в роде 3 группы видов, отказав-
шись от дробления рода на серию мелких родов. Учитывая морфологи-
ческие признаки взрослых и личинок, мы принимаем внутри рода Musca
4 группы видов, характеризующиеся следующими особенностями.
Группа I. Musca domestica L.
Проплевры в волосках впереди передних дыхалец; супрасквамаль-
ное ребро без щетинистых волосков; I стернит в волосках; лоб самцов
шире или равен ширине 3-го членика усиков; у самок лобная полоса
в 5—6 раз шире орбиты.
Конусовидные выросты на 8-м брюшном сегменте у зрелых личинок
отсутствуют, анальная пластинка очень коротка и широка; позади нее —
многочисленные шипики.
Мухи откладывают яйца.
В составе нашей фауны 1 вид — Musca domestica L.
Группа II. Musca sorbens Wd.
Проплевры, супрасквамальное ребро, как и I стернит брюшка,
лишены волосков; лоб самцов обычно уже 3-го членика усиков; орбиты
самок в два раза уже лобной полосы.
Зрелые личинки с коническими мелкими выростами вокруг задних
стигм, 8-й сегмент (брюшной) п мелких шипиках; анальная пластинка
узкая, ± длинная.
Мухи откладывают яйца.
К этой группе мы относим 2 вида нашей фауны: Musca sorbens 'Wi-
tt М- albina Wd.
Группа III. Musca vitripennis Mg.
Проплевры, супрасквамальное ребро и I стернит брюшка без воло-
сков; лоб самцов равен по ширине 3-му членику усиков или уже его;
орбиты самок шире или равны лобной полосе.
Зрелые личинки без щетинок и конических выростов на 8-м брюш-
ном сегменте вокруг задних дыхалец; анальная пластинка узкая, длин-
ная; позади нее шипиков нет или количество их невелико.
92
I. ТРИБА MUSCINI
Мухи откладывают яйца.
К этой группе относятся М. vitripennis Mg., М. osiris Wd., М. tem-
pestiva Fall., M. lucidula Lw.; вероятно сюда относится также М. era-
sirostris Stein.
Группа IV. Musca lusoriq Wd.
Проплевры голые; супрасквамальное ребро и I стернит брюшка
в волосках и щетиночках; лоб самцов обычно не шире 3-го членика уси-
ков; орбиты самок равны лобной полосе (или уже, но не более, чем
в 3 раза).
Зрелые личинки без конических мелких выростов и щетинок на 8-м
брюшном сегменте; анальная пластинка очень длинная, широкая, окру-
женная шипами.
Мухи яйценесущи и живородящи.
Из нашей фауны сюда относятся 4 вида: Musca autumnalis Deg.,
М. arnica, sp. nov., M. larvipara Portsch., M. convexifrons Thoms.
Из 74 достоверно хороших видов в Палеарктической области рас-
пространено 19 видов, в Неарктической — 21; в Ориентальной (включая
сюда Австралийскую) области — 32 вида, часть которых является общими
с Палеарктикой и Неарктической областью; из Эфиопской области
известно 25 видов, также частью общих с двумя предыдущими обла-
стями. Хотя это распределение является провизорным, оно все же
дает представление о распределении видов рода по областям.
Преобладающее число видов рода откладывают яйпа и лишь немно-
гим видам (группы lusoria) свойственно живорождение. Развитие личинок
протекает в экскрементах животных и фекалиях человека; многие
формы способны размножаться в гниюших веществах растительного
и животного происхождения. Зарегистрированы многочисленные случаи
тканевого и кишечного миаза (М. domestica, М. sorbens и др.).
Многие виды являются синантропами, ряд форм тесно связан
с животными. С медико-санитарной и ветеринарной точек зрения род
Musca представляет собою одну из наиболее опасных групп насекомых,
как распространителей возбудителей различных заболеваний.
ОПРЕДЕЛИТЕЛЬНАЯ ТАБЛИЦА ВИДОВ
Т (II). Глаза на темени сближены, ширина лба много меньше V2 ширины
глаза — SS.
II (I). Глаза наверху широко расставлены; ширина лба не менее п/г
ширины глаза — jj.
Самцы
Г (2). Подбородок сильно вздутый; ширина его превышает ширину
бедра; шупальны желтые..................9. М. crassirostris Stein
2 (1). Подбородок вытянутый; ширина его менее ширины переднего бедра.
3 (14). I стернит без волосков.
4 (5). Грудь металлически зеленая, в редком сероватом налете; глаза
голые; брюшко желтое, просвечивающее, на вершине металлически
зеленое, покрыто светлым налетом; щеки блестяще черные; лоб
уже или равен по ширине 3-му членику усиков; лобная полоса
в виде тонкой желтой линии; дорзоцентральные щетинки впереди
шва отсутствуют; длина тела 2.5—4 мм . . . *8. М. lucidula Lw.
5 (4). Грудь черно-бурая, с более или менее ясно развитым светлым
налетом, образующим на спинке пятна или продольные полосы;
2. MUSCA
аз
6
7
8
9
10
И
12
13
14
15
16
17
18
19
20
если грудь с металлически зеленым оттенком, то глаза в светлых
волосках и передние дорзоцентральные щетинки впереди шва
имеются.
(7). Стерноплевральные, а также дорзоцентральные щетинки впереди
поперечного шва спинки отсутствуют; грудь блестяще черная,
с пятнами светлосерого налета на плечах и верхней части бочков;
брюшко светложелтое; 5-й тергит и гипопигий черно-бурые; 5-й
тергит в густом сероватом налете, с двумя блестящими черными
округлыми пятнами; 4-й, 3-й и 1 -|-2 тергиты с маленьким пятном
посредине заднего края. Длина тела 3.8—6 мм. . *4. М. albina Wd.
(6). Стерноплевральные и дорзоцентральные щетинки впереди шва
явственно развиты.
(13). Среднеспинка без продольных резко выраженных чередующихся
темных и светлых полос, блестящая, черная или оливково-черная;
светлый налет образует пятна лишь на плечах и нотоплеврах.
(10). Брюшко черно-бурое, в желтовато-сером’налете, с суживаю-
щейся назад срединной бурой полоской и буроватыми узкими пере-
вязями по заднему краю тергитов; глаза голые; переднее дыхальце
груди темное: бурое, коричневое или темнорыжее. Длина тела
2.5—4-2 мм................................*7. М. tempestiva Fall.
(9). Брюшко с ясными желтыми пятнами по бокам; глаза в светлых
волосках; переднее дыхальце груди светлое; белое, палевое, желтое.
Длина тела 3—6.2 мм.
(12). Лобные щетинки располагаются в два ряда с каждой стороны
от лобной полосы, лоб шире 3-гб членика усиков; волоски в перед-
нем отделе щеки длинные, углом заходят вверх почти до нижнего
окончания лицевого шва. Длина тела 3—5.5 мм......................
.........................................*5. М. vitripennis Mg.
(11). Лобные щетинки располагаются в один ряд с каждой стороны
от лобной полосы; лоб уже или равен по ширине 3-му членику
усиков; волоски в переднем отделе щеки короткие, редкие, далеко
не доходят до окончания лицевого шва. Длина тела 3.2—6.2 мм
. . . .........................................*6. М. osiris Wd.
(8). Среднеспинка в густом сером налете, оставляющем за попереч-
ным швом две широких черных продольных полосы (основной
окраски груди), иногда раздвоенных перед поперечным швом.
Длина тела 7—9.5 мм..........................*3. М. sorbens Wd.
(3). I стернит по краю в волосках; если голый, то брюшко красно-
вато-желтое, без черных отметин.
(16) . Брюшко целиком красновато-желтое без черных отметин, реже
черный цвет развит в виде мелких пятен по средней линии тергитов
16. М. ventrosa Wd.
(15) . Брюшко черно-бурое, с желтыми пятнами по бокам, или желтое
с черной продольной средней полосой.
(18) . Все стерниты и вентральные края тергитов черные; глаза в ред-
ких волосках............................ 17. М. hervei Vill.
(17) . По крайней мере средние стерниты или края тергитов красно-
желтые; глаза голые.
(24) . Передние супрасквамальные щетинки отсутствуют; проплевры
в волосках.
(21) . Ширина лба составляет около Vs ширины глаза; верхняя часть
затылка сильно выпуклая; окраска брюшка темнее...................
.................................*1. М. domestica domestica L.
94
I. ТРИБА MUSCINI
21 (20). Ширина лба составляет не более Vs—'7s ширины глаза; затылок
в верхней половине слабо выпуклый; окраска брюшка светлее.
22 (23). Ширина лба равна ]/3 ширины глаза........................
................................*1- М. domestica vicina Mcq.
23 (22). Ширина лба равна х/5 ширины глаза.......2. М. nebulo F.
24 (19). Передние супрасквамальные щетинки имеются.
25 (30). Задние супрасквамальные волоски отсутствуют.
26 (27). Жилка г4+5 с нижней поверхности крыла по всей длине уса-
жена редкими щетинистыми волосками .... 14. М. lusoria Wd.
27 (26). Волоски нижней поверхности г44_5 не заходят далее 3Л длины
жилки, обычно располагаясь лишь у ее основания.
28 (29). Ширина скул на уровне 2-го членика усиков менее ширины
3-го членика усиков; 5-й тергит с резко выраженными темными
переливчатыми продольными полосками налета; верхняя граница
длинных тонких волосков в переднем отделе щеки заходит вверх
полукругом значительно далее нижнего края глаза. Длина тела
4.5—7.5 мм...............................*10. М. autumnalis Deg.
29 (28). Ширина скул на уровне 2-го членика усиков более ширины
3-го их членика или равна ей; 5-й тергит равномерно покрыт серо-
вато-желтым налетом, не образующим темных, резких продольных
полос; щеки с короткими волосками едва угловидно заходящими
за линию, соединяющую нижний край глаза с угловой вибриссой.
Длина тела 4.5—6.7 мм.....................*11. М. arnica, sp. nov.
30 (25). Задние супрасквамальные волоски имеются, равно как и перед-
ние супрасквамальные щетинки.
31 (32). Задняя половина тергита 1Д-2 сверху, исключая среднюю чер-
ную продольную полоску, красно-желтая; того же цвета боковые
отделы 3-го, 4-го и боковые части у основания 5-го тергитов; 5-й
тергит с тремя очень резкими темными узкими, проходящими
по всей длине тергита полосками, чередующимися с сильно бле-
стящими светлыми промежутками налета между ними; боковые
переливчатые темные продольные пятна на 3-м и 4-м тергитах
широкие, образуют по заднему краю последних поперечную полоску,
заходящую на вентральную сторону брюшка; задний край 4-го
и иногда 3-го тергитов с черной полосой, на 4-м тергите занимаю-
щей Vs—Vs длины тергита; жилка г4+5 снизу со щетинками,
в основной трети нередко заходящими далее гт поперечной жилки.
Крупнее: 7—9 мм........................*13. М. convexifrons Thoms.
32 (31). Верхняя сторона 1Д-2 тергита сплошь черно-бурая, реже
с узенькой полоской в боковых отделах заднего края, реже задний
край широко желтый; 5-й тергит без трех резких продольных тем-
ных полос, идущих по всей длине тергита; боковые темные пере-
ливчатые пятна на 3-м и 4-м тергитах узкие, не образуют длинной
полосы по заднему краю; задний край 4-го тергита с узкой черно-
бурой полоской, шириной не более г/«—x/s длины тергита, нередко
вовсе исчезающей; жилка г4+5 снизу со щетинками обычно лишь
у основания. Мельче: 4.8—8.3 мм. . . . *12. М. larvipara Portsch.
Самки
1 (2). Подбородок сильно вздутый, шире переднего бедра..........
.....................................9. М. crassirostris Stein
2 (1). Подбородок обычной конфигурации, не шире переднего бедра.
3 (4). Тело металлически зеленое в редком светлом налете; основание
2. MUSCA
95
4
5
6
7
8
9
10
И
12
13
14
15
16
17
18
19
20
21
22
брюшка часто медово-желтого цвета; лобная полоса черная с пря-
мыми краями, равномерно суженная к основанию усиков; ячейка
R в часто замкнута на краю крыла..........*8. М. lucidula Lav.
(3). Тело черно-бурое в сером налете, брюшко иногда с. более или
менее развитой желтой окраской, особенно с нижней стороны.
(6). Стерноплевральные, как и дорзоцентральные, щетинки впереди
шва отстуствуют; стерноплевры черные, блестящие; прочие части
тела в светло-сером густом налете; спинка с 2 черными ши-
рокими полосами раздвоенными впереди шва; брюшко с не-
сколькими черными блестящими округлыми пятнышкамй . . . .
..............................................*4. М. albina Wd.
(5). Имеются, по крайней мере, стерноплевральные щетинки.
(16). Первый стернит без волосков.
(9). Спинка с 2 широкими черными продольными полосами, раз-
двоенными впереди поперечного шва и разделенными здесь широ-
ким светлым промежутком налета..............*3. М. sorbens Wd.
(8). Спинка с 4 узкими продольными полосами.
(11). Брюшко сплошь красно-желтое без крупных черных отметин
............................................16. М. ventrosa Wd.
(10). Брюшко бурое или с красно-желтыми отметинами с нижней
стороны брюшка.
(13). Передние дыхальцы груди темнобурые, коричневые или рыжие;
глаза голые; лобная полоса в 2—2х/г раза шире орбиты; скулы
равны или чуть шире 3-го членика усиков........................
.......................................*7. М. tempestiva Fall.
(12). Передние дыхальцы груди светлые — белые или палево-желтые;
глаза в редких светлых волосках, иногда плохо заметных; лобная
полоса по ширине равна орбите или не более чем в 11/г раза шире
ее; скулы почти в 2 раза шире 3-го членика усиков.
(15). Орбиты, помимо покрывающих их коротких щетинистых воло-
сков, несут несколько длинных толстых волосков, обращенных
вершинами к лобной полосе и располагающихся в заднем или в заднем
и среднем отделах орбит около ряда лобных щетинок. Длина тела
3.2—6.2 мм................................*5. М. vitripennis Mg.
(14). Орбиты покрыты лишь короткими щетинистыми волосками,
направленными концами вперед или наружу. Длина тела 3.5—
5.5 мм. ......................................*6. М. osiris Wd.
(7). I стернит брюшка в черных волосках по краю или по краю
и в середине.
(28). Черные щетинки имеются, по крайней мере, в передней части
супрасквамального ребра (под основанием крыла); проплевры впе-
реди дыхалец посредине без волосков.
(19). Жилка г 4+5 снизу по всей длине до вершины крыла в воло-
сках ........................................14. М. lusoria Wd.
(18). Волоски на жилке г 4+5 снизу присутствуют лишь у основа-
ния первого отрезка или имеются и далее к вершине крыла,
не заходя за основную половину дистального отрезка этой жилки.
(21). Глаза в мелких редких волосках...........17. М. herirei Vill.
(20). Глаза лишены светлых волосков.
(25). Супрасквамальные щетинки есть лишь в передней части супра-
сквамального ребра, под основанием крыла; в задней части на вали-
кообразном возвышении ребра черные щетинистые волоски отсут-
ствуют.
08
I. ТРИБА MUSCINI
23 (24).. Лобные щетинки в среднем и заднем или только в заднем отделе
расположены с каждой стороны лба в 2 (реже 3) спутанных ряда,
из которых наружные неполные .... *10. М. autumnalis Deg.
24 (23). Лобные щетинки расположены в один ряд............. . . . .
......................................*11. М. arnica, sp. nov.
25 (22). Супрасквамальное ребро под основанием крыла и в заднем
отделе по всей его длине в щетинистых черных волосках.
26 (27). Ширина лба в наиболее узком его месте составляет 2/з—V2 ширины
глаза; орбита в 2 раза уже лобной полосы; боковые темные пере-
ливчатые пятна на 3-м и 4-м тергитах расширены сзади, образуя
поперечную темную боковую полоску по заднему краю тергита,
переходящую и на вентральную сторону брюшка; 5-й тергит с рез-
кими темными переливчатыми отметинами. Длина тела 7—9 мм
(Южное Приморье)......................*13. М. convexifrons Thoms.
27 (26). Ширина лба в наиболее узком месте составляет 3/< ширины глаза
или равна ей; темные боковые пятна 3-го и 4-го тергитов никогда
не заходят на вентральную сторону брюшка в виде полоски (на зад-
нем крае тергитов); ширина орбиты в П/а раза менее ширины лоб-
ной полосы или почти равна ей; переливчатые пятна 5-го тергита
не резко выражены, часто отсутствуют вовсе. Длина тела 4.8—8.3
(СССР на север до широты Ленинграда, кроме Сибири и Дальнего
Востока) ...............................*12. М. larvipara Portsch.
28 (17). Супрасквамальное ребро лишено щетинистых волосков; про-
. плевры впереди стигмы в середине с волосками.
29 (30). Брюшко красновато-желтое с неясно выделяющейся узкой бурой
продольной полоской посредине, не достигающей заднего конца
брюшка.............................................2. М. nebulo F.
30 (29). Бурая полоса брюшка ясная, образующая поперечные полосы
по заднему краю 3-го и 4-го тергитов.
31 (32). 3-й и 4-й тергиты с широкими красновато-желтыми пятнами
по бокам; часто брюшко, кроме средней узкой полосы, сплошь
красно-желтое; затылок сверху слабо выпуклый;.....................
.............................. *la. М. domestica vicina Mcq.
32 (31). 3-й и 5-й тергиты с маленькими иногда вовсе исчезающими
красновато-желтыми отметинами; затылок сверху более сильно
выпуклый.............................*1. М. domestica domestica L.
1 группа. Musca domestica L.
Лоб самца более или менее широкий, составляет х/«—2/в ширины глаза;
глаза голые; грудь в сером налете, с 4 продольными бурыми полосами
на спинке; проплевры впереди дыхальца с тонкими волосками; супра-
сквамальное ребро без щетинистых волосков; 1 стернит брюшка в воло-
сках по краю. . . .............
Сюда относятся: Musca domestica L. и М. nebulo F. 1
*1. Musca domestica L. Комнатная муха. (Рис. 41, 106—112, 118,
119, 123, 124, 128—131, 133—139, 140, 14.1).
Ыппё, 1758, Syst. Nat., X : 589; Linne, 1761, Fauna Suec., Ed. II : 453; Robi-
neau Desvoidy, 1830 : 398; Schiner, 1862,1 : 594; Bezzi u. Stein, 1907, III : 603; Stein,
1 Мима nebulo F. на территории СССР не обнаружена. Нахождение вида воз-
можно в южных частях наших среднеазиатских республик.
2. MUSCA
97
>1916 : 16; Awati, 1916, Ind. Journ. Med. Res., Ill : 510—529, pls. 35—50; Patton,
1923, Philipp. Journ. Sci., XXIII : 310—311; Malloch, 1925, Ann. Mag. Nat. Hist.,
XV, 85 : 131—142; Karl, 1928, Tierw. Deutschl., 13, III : 7; Patton, 1932, Ann.trop.
Med. Parasitol., XXVI : 347—405; Karl, 1935, Arb. morph, taxonom. Entomol., II :
29—49; Patton, 1937, Ann. trop. Med. Parasitol., XXXI, 1 : 128; Ho, 1938, там же,
XXXII : 287—312, figs. 2, 5, 7; Townsend, 1937, Manual Myiol., V : 45; —flavinervis
Thompson, 1868, Eugenies Resa, Dipt. : 547, 178; — lateralis Macquart, 1833, Recueil
Soc. Sc. Agric. Lille, 158, 5; — vicina Macquart, 1850, Dipt, exot., Suppl. IV : 253. —
vagatoria Robineau-Desvoidy, 1830 : 399.
Биология. Порчинский, 1885 :9—11, рис. 2—3; Howard, 1900, Proc.
Wash. Acad. Sci., II : 541—604; он же, 1911, The House Fly. Disease carrier, 2d Ed.;
Порчинский, 1910, 14—-22, рис. 10—14; он же, 1911; Hewitt, 1914; Кула-
гин, 1916, Любит, прир., XI, 3—4 : 93—100; Graham-Smith, 1913; он же, 1916, Para-
sitology, VIII, 4 : 440—544; Павловский, 1921; Кулагин, 1921; Квасникова, 1931 :
1—83; Петрищева, 1932 : 161—182; Смирнов и Кузина, 1933 : 96—109;. Nieschulz
u. du Toit, 1933 : 244—249 (vicina)', Roy a. Mukherjee, 1933 : 453—456 (vicina)', Смир-
нов, 1934, Мед. паразитолог., Ill, 4 : 330—336; Lorincz u. Makara, 1935, V, No 2 :
219—227; Kobayashi, 1935, Trans. Dynam. Develop., X : 385—395; Кузина, 1936:
329—339; Смирнов, 1937 : 872—879; Дербенева-Ухова, 1937 : 409—417; Кузина,
1938 : 244—257; Чеботаревич, 1937 :1—115, 9 рис.; Дербенева-Ухова и Кузина,
1938 : 399—405; М. Thomsen, 1938 : 1—352, 13 pl.; Кузина, 1940 : 340—349; Змеев,
1944 : 95—117; Зимин, 1944 : 133—192; Тетеровская, 1944 : 49—55; Семенов, 1945,
Мед. паразитолог., XIV, 6 : 50; Павловский, 1948 : 864—894, 20 рис.; Зимин, 1948 :
84—86, 1 рис. (vicina)', Дербенева-Ухова, 1949, Учебн. мед. энтомолог, под ред. Бекле-
мишева : 367—389, 30 рис.; Жовтый, 1950, О годовом цикле комнатной мухи. Ново-
сибирск : 1—25.
Ширина лба самца варьирует в пределах х/л—2/а ширины глаза;
орбиты в 6—8 раз уже лобной полосы; скулы равны, чуть уже или
несколько шире 3-го членика усиков, затылок слабо выпуклый в верх-
ней и значительно сильнее в нижней половине. Щупальцы бурые, слабо
утолщенные к вершине. 2-й членик усиков в 2—2х/г раза короче 3-го;
усики бурые, нередко с красноватым основанием 3-го и вершиной 2-го
члеников; голова черно-бурая, скулы и передний отдел щек, а иногда
и передний отдел лобной полосы красноватые. Передний отдел орбит
и скулы в блестящем белом или желтоватом налете; затылок за глазами
и щеки в сероватом более редком налете. Задняя поверхность затылка
в редком коричневато-желтом налете. Наружные и внутренние теменные
щетинки и вибриссы длинные, более или менее толстые; лобные щетинки
тонкие; орбиты с одиночными щетинистыми волосками, доходящими
до передних лобных щетинок. Затылок в более длинных снизу и коротких
вверху черных волосках; щеки в коротких тонких волосках того же
цвета. Грудь черно-бурая, нередко с коричневатыми отметинами на плев-
рах, покрыта сероватым или желтовато-серым налетом; спинка с 4 про-
дольными черными волосками, разделенными светлыми промежутками
налета; ширина средних черных полос в Р/г раза менее ширины разде-
ляющего их промежутка. Поверхность груди в черных волосках. Про-
плевры впереди дыхалец в тонких бурых волосках; переднегрудь в боковых
отделах в щетинистых, тонких волосках; гипоплевры в тонких волосках
впереди задних стигм и с группой волосков над основанием тазиков.
Щетинки груди: акростихальных — одна пара перед щитком (иногда
они не развиты); дорзоцентральных — 3 -4; интраалярных — 1 позади
шва; предкрыловых — 1, короткая; надкрыловых — 2, из которых зад-
няя короткая и тонкая; закрыловых — 3; плечевых — 3; заплечевых —1;
предшовных — 1; нотоплевральных — 2; стерноплевральных — 1 -f- 2;
на щитке — вершинных 2; основных — 2; предосновных — 2, короткие;
дискальных — 2 (1—3) пары впереди вершинных щетинок. Крылья серо-
ватые; жилки буровато-рыжие, в основной трети крыла на большем или
7 Фауна СССР, том XVIII, вып. 4.
98
I. ТРИБА MUSCINI
меньшем протяжении желтые; г 4+3 у основания снизу и сверху с оди-
ночными (1—3) волосками (на верхней стороне часто последние отсут-
ствуют). Основной ствол радиуса сверху с 1—2 (3) щетинистыми волос-
ками; чешуйки белые или желтоватые; жужжальца желтые. Ноги черно-
бурые, голени чаше коричневатые; эта окраска наиболее отчетливо выра-
жена на голенях средней пары ног. Средние голени по задне-дорзальной
Рис. 106. Самец Musca"domestica L. (Ориг.).
поверхности без крепких щетинок; последние есть в числе 3—5 на зад-
ней стороне тех же голеней. Количество щетинок на задних и средних
бедрах очень изменчиво. Брюшко узко яйцевидное, черно-бурое
с желтыми боковыми отделами, оставляющими по средней продольной
линии брюшка более или менее широкую черную полосу. Черный рисунок
крайне изменчив. У наиболее темных (северных) экземпляров красная
окраска занимает боковые трети переднего края 1 4-2 тергитов и боко-
вые трети 3-го тергита; снизу светлоокрашенными остаются стерниты
со 2-го по 4-й (и задняя половина 5-го), а также нижняя поверхность
1-j-2 и 3-го тергитов; прочие части брюшка в этом случае буро-черные.
У южных особей (Л/, domestica vicina) темная окраска занимает неболь-
2. MUSCA
99
шую часть поверхности и I стернит; от основания брюшка по средней
его продольной линии тянется узкая (ширина может достигать всего
1/i2 ширины тергитов) бурая полоска, образующая в срединном отделе
5-го тергита расширение в форме округлого бурого пятна. Между двумя
указанными типами окраски существуют постепенные переходы. Брюшко
Рис. 107—109. Musca domestica L.
107 — дорзальный отросток фаллозомы. 108 — V стер-
нит самца. 109 — церки самца. (По Пэттону).
Рис. 110—112. Musca domestica vicina Mcq.
110 — дорзальный отросток фаллозомы. Ill — V стер-
нит самца. 112 — церки самца. (По Пэттону).
Рис. ИЗ—115. Musca nebulo F.
113 — дорзальный отросток фаллозомы. 114 — V стер-
нит самца. 115 — церки самца. (По Пэттону).
Рис. 116. Церки и кокситы самца Musca osiris Wd.
(Ориг.). Рис. 117. Церки и правый коксит самца
Musca vitripennis Mg. (Ориг.).
покрыто сероватым налетом, приобретающим на дистальном конце более
или менее отчетливый желтый оттенок. Темные переливчатые пятна
по бокам 3-го и 4-го тергитов обычно узко-треугольные, нередко с рас-
ширением в форме полоски или клина на заднем крае тергитов. Форма
и размер весьма изменчивы. 5-й тергит с тремя узкими продольными
темными переливчатыми полосками, в некоторых случаях плохо выра-
7*
Рис. 118—127. Головы видов рода Musca.
118 — самца М. domestica vieina Mcq. (типичный экз.) сбоку. 119 — самца М. domes-
tica domestica L. (сбоку). 120 — самцаМ. larvipara Portsch. 121 — самки М. larvipara
Portsch. (сверху). 122 — самки М. autumnalis t)eg. (сверху).123 — самца М. domestica
vieina Mcq. (сверху) (схема). 124 — самца М. domestica domestica L. (сверху); 125-i-
передняя и боковая части головы самца М. autumnalis Deg. 126 — то же М.. arnica,
sp, n. 127 — голова самца М. autumnalis Deg. (сбоку). (Ориг,).
2. MUSCA
101
женными. Церки в виде щитка (рис. 109, 112), дорзальный отросток
изменчивой по форме фаллозомы (рис. 107, ПО) длинный, тонкий, имеет
широко, ланцетовидную форму. Передняя часть парамер крупнее задней,
овальная; задняя часть парамер представляется в виде маленького скле-
ротизованного придатка. V стернит расширен в заднем отделе (рис. 108,
111) с 2 тонкими, слабо согнутыми внутрь и зазубренными на вершине
отростками. Мембранозная часть фаллозомы длинная, равна, длиннее
или. короче основной склеротизованной части, по вентральному краю
с 3 угловидными выступами.
Ширина лба самки составляет 1/з ширины головы; лобная полоса
очень широкая, с сильно выпуклыми краями; орбиты узкие, ширина
орбиты в 6 раз менее ширины лобной полосы, в мелких щетинистых
волосках. Затылок сильно выпуклый, скулы в I1/*—IV2 раза шире 3-го
членика усиков. Окраска варьирует, как и у самцов. Брюшко бурое
с красноватой вентральной стороной 1 -|-2 тергита и краями стернитов,
или с желтоватыми отметинами на верхней и нижней стороне 1 -|-2
и 3-го тергитов и более развитой в основной половине брюшка красно-
ватой окраской снизу. Длина тела 7—9 мм; карликовые формы, раз-
вившиеся в ненормальных условиях, иногда не превышают в длину
4.5—5 мм.
Яйцо (рис. 41) цилиндрическое слабо дуговидно изогнутое, сужен-
ное от средины к обоим полюсам; верхний конец несколько острее ниж-
него; дорзальная сторона с двумя тонкими продольными бороздками,
проходящими от одного полюса к другому и расставленными на ширину,
равную 2/з поперечного диаметра яйца; хорион белый, слабо опалесци-
рующий, тонкий, с тонким рисунком из мелких шестигранных ячей.
Длина тела 1—1.2 мм.
Личинка I стадии метапнейстична (рис. 128, 131), задние дыхальцы
мелкие с округлым или сердцевидным просветом; тело удлиненное, валь-
коватое, постепенно и не сильно утонченное к переднему и обычно едва
заметно от средины к заднему концу; передние края брюшных сегмен-
тов, со 2-го по 7-й, снизу с валикообразными поперечными утолщениями,
покрытыми мелкими хитиновыми шипиками, собранными в поперечные
ряды; последний сегмент с шипиками впереди анального отверстия,
1-й и 2-й брюшные сегменты снизу с короткими узенькими полосками
из более мелких шипиков, число которых меньшее, чем на последующих.
Грудные сегменты без шипов, чем личинки отличаются от личинок I ста-
дии М. sorbens. Ротоглоточный аппарат сохраняет тип строения, свой-
ственный всем представителям рода Musca. Срединный дорзальный скле-
рит листовидный, широкий, по верхнему краю прямой или слабо вогну-
тый, на переднем конце с крючковидно или клювовидно загнутым вниз
зубцом, нижний край прямой, в переднем отделе широко закругленный
или несколько угловатый; гипостомальный склерит короткий с пере-
мычкой между боковыми частями, передние отростки фарингеального
склерита в виде двух тонких, длинных, хитиновых тяжей; сам склерит
в форме двух серповидных узких пластинок; дорзальные его отростки
равны по длине вентральным; дорзальная перемычка высокая, узкая.
Нижняя сторона головы впереди ротового отверстия с поперечно рас-
положенным рядом хитиновых зубчиков в числе 5—6 с каждой стороны
от продольной средней линии головы. Длина тела 1.2—2 мм.
Личинка II стадии (рис. 129, 130) амфипнейстична; передние
дыхальцы с длинными 6—7 пальцевидными выростами (лучами); задние
дыхальцы слабо склеротизованы, расставлены на расстояние диаметра
102
I. ТРИБА MUSCINI
дыхальца или несколько более того; каждое дыхальце с 2 короткими,
широкими и почти прямыми щелями. Тело без хитиновых шипов, с ниж-
ней стороны брюшных и грудных сегментов; брюшные сегменты, со 2-го
по 7-й, с поперечными короткими валикообразными утолщениями. Голов-
ной сегмент снизу впереди ротового отверстия с (2—4) крупными зуб-
цами, расположенными поперечным рядом в числе 1 (2) с каждой стороны
от средней продольной линии головы; внутренние зубцы длиннее и крепче
наружных, которые нередко отсутствуют; ротовые крючья срощены
в переднем отделе, неправильно дуговидно изогнуты, вершинная часть
Рис. 128—132.
128—ротоглоточный аппарат личинки Musca domestica L. I ста-
дии. 129 — то же II стадии. 130 — задние дыхальцы личинки
II стадии М. domestica L. 131 — срединный дорзальный склерит
личинки I стадии М. domestica L. при сильном увеличении. 132 —
задний конец тела личинки III стадии Haematobia stimulans L.
(сзади). (132 по Томсону, прочие—ориг.).
направлена вниз, задний отдел с направленными вверх и назад корот-
кими острыми отростками; угловидный склерит в виде пары узких,
согнутых склеритов, с концами обращенными назад; гипостомальный
склерит почти с прямыми верхними и слабо выпуклыми нижними кра-
ями; задний конец узкий, направлен назад и слегка вверх; эпистомаль-
ный склерит слабо выступает над верхней границей гипостомального.
Под основанием ротовых крючьев лежит пара широких с полукруглым
широким краем, суженных назад лопастей, сочлененных с узкими сильно
склеротизованными склеритами; от нижнепереднего угла гипостомаль-
ного склерита отходит вперед и вниз по узкому хитиновому тяжу, со-
единенному впереди с неправильно трапециевидным склеритом. Фарин-
геальный склерит с удлиненными и сравнительно тонкими передними
отростками, нижние углы которых оттянуты вперед и соединены снизу
вентральной перемычкой; верхний край склерита слабо выпуклый, ниж-
2. MUSCA
103
ний — почти прямой или слабо вогнутый в основной половине; дорзаль-
ные отростки крыловидные, слабо суженные к вершине; вентральные —
расширены к вершине, на конце косо срезаны и заострены; дорзальная
перемычка решетчатая, с узкими основными частями и расширенная
в дорзальной части; передний и задний края последней выемчатые посре-
дине; перемычка прикреплена к передним краям базального склерита,
соединяясь с последним лишь основаниями. Длина тела 2.5—3 мм.
Личинка III стадии (рис. 133—139) имеет постепенно суженное
кпереди тело; кутикула желтовато-белая, тонкая, гладкая. Головной
сегмент спереди двураздельный; ротовые бороздки покрывают всю ниж-
нюю его поверхность и переднюю половину боковых сторон, чуть заходя
на дорзальную сторону. Антенны короткие; 1-й их членик короткий,
кольцевидный, 2-й — короткий, усеченный на вершине. Нижнечелюст-
ные щупальцы в форме маленьких бугорочков, расположенных на непра-
вильно сферических выступах головного сегмента. Предротовые зубцы
плоские, широкие, располагаются дуговидно по бокам от ротового отвер-
стия. Передний край грудных и первого брюшного сегментов с много-
численными рядами мелких светлых шипиков, собранных в дуговидно
изогнутые короткие рядки. На 2-м грудном сегменте шипиками занята
1/в, на 3-м — х/в и на 1-м брюшном — х/в часть их длины. Брюшные сег-
менты, со 2-го по 7-й, вооружены шипиками лишь снизу, последние рас-
полагаются на поперечных валикообразных вздутиях переднего края.
Здесь шипики расставлены неправильными рядами; срединный ряд
состоит из более крупных шипиков. Передний и задний края вздутий
покрыты очень мелкими шипиками. Задние края сегментов у передней
границы вздутий с несколькими мелкими бугорочками, лежащими двумя
группами (по 8—10 бугорочков в каждой) по бокам от средней продоль-
ной линии сегмента. 8-й брюшной сегмент округло-выпуклый. Анальные
бугорки короткие, субанальные — сосцевидные, тупые, чуть длинее
анальных, от последних отставлены на расстояние, равное поперечнику
бугорка. Область позади анального отверстия слабо выпуклая, покры-
тая светлыми шипиками; часть шипиков (7—15) в средней части выпук-
лости с черной хитинизированной вершиной. Анальная пластинка корот-
кая, широкая, концы ее заходят далее Гранины субанальных бугорков.
Боковые отделы 9-го сегмента от краев анальной пластинки сплошь
покрыты мелкими светлыми шипиками. Передние стигмы обычно с шестью
короткими булавовидными выростами. Задние стигмы широко почко-
видные, или в форме долек апельсина. Перитрема очень узкая, в районе
стигмального диска несколько расширенная. Стигмальная пластинка
рыжая, у личинок готовых к окуклению — бурая. Дыхальцевые щели
широкие, змеевидно изогнутые, с концами, обращенными к центру
дыхальцевой пластинки. Дыхальцы расставлены на Vs ширины дыхаль-
цевой пластинки. Ротовой аппарат массивный; ротовые крючья неравной
длины: левый крючок короче правого; крючья серповидно изогнутые
в вершинной, узкой своей части; зубные склериты неправильно трапе-
циевидные. Гипостомальный склерит короткий, ширина его едва короче
длины. Передние отростки фарингеального склерита чуть длинее гипо-
стомального склерита, расширенные на конце. Вентральные отростки
фарингеального склерита короче и почти в два раза шире дорзальных;
по дорзальному краю с резким, большим угловидным выступом. Дор-
зальная перемычка решетчатая, широкая; передний край ее не достигает
переднего конца передних отростков фарингеального склерита. Фарин-
геальная мембрана широкая.
104
I. ТРИБА MUSCINI
Ложнококон рыжевато- или красновато-коричневый, эллиптический;
в передней половине чуть шире чем у задней; дыхальцевые рожки отсут-
ствуют, заменены мелкими светлыми пятнышками, лежащими с дорзаль-
ной стороны пупария в боковых отделах 2-го брюшного сегмента,
вблизи его границы с третьим. Длина тела 3.5—5 мм.
irrwwr
wr*’ **v*vx^T ’W»* 4>>v.»r»rr-
н ЧОТО*
»f>wwr
h’wwn' -ттгг *»«* *•»»»»
Ъиг««т
738
13?
/35
134
133
735
Рис. 133—139. Детали строения личинки III стадии Musca domestica vicina Mcq.
133 — ротоглоточный аппарат. 134 — задний конец тела сзади. 135 — он же сбоку.
136 — переднее дыхальце. 137 — задпее дыхальце. 138 — скульптура переднего
отдела верхней стороны 2-го грудного сегмента. 139 — вооружение снизу 5-го (2)
и 6-го (1) брюшных сегментов. (По Зимину).
Распространение. Musca domestica L. распространена все-
светно; она встречается от Арктики (Гренландия, Новая Земля, Шпиц-
берген, о-в Врангеля) до тропиков. В южных широтах М. domestica
domestica L. заменяется М. domestica vicina Mcq. Эта последняя известна
из тропических стран всех зоогеографических областей. Граница рас-
пространения той и другой может быть намечена лишь условно в связи
с существованием промежуточных форм, которые с одинаковым основа-
нием можно отнести и к северной и к южной форме М. domestica.
2. MUSCA
105
В СССР М. domestica domestica распространена на север от линии,
проходящей с запада на восток следующим образом: северное побережье
Черного моря и южное побережье Азовского моря, восточное побережье
Черного моря до Батуми. Между Черным и Каспийским морями граница
следует, примерно, по Главному Кавказскому Хребту, поднимаясь
на север по западному побережью Каспийского моря до его северной
части, откуда проходит к низовью Аму-дарьи, следуя далее несколько
севернее Муюн-кумов к дельте Или. В дальнейшем граница идет по тер-
ритории центральной Монголии, и, направляясь через Манчжурию
к северо-востоку, выходит вновь на территорию СССР в районе нижнего
течения Амура. К югу от намеченной линии встречается М. domestica
vicina Mcq., которая обитает, следовательно, в Закавказье, Средней
Азии, в Амурской области и южном Приморье.
Комнатная муха — синантроп, имеющая многосторонний и тесный
контакт с человеком; в отношении окружающей обстановки она крайне
неприхотлива.
Размножаться муха способна почти во всех отходах пищевого блока,
находящихся в состоянии разложения, независимо от того животного
они происхождения или растительного. С таким же успехом муха выпла-
живается в экскрементах животных, фекалиях человека, в любых нечи-
стотах. Она известна также как случайный паразит гусениц бабочек
и моллюсков. Личинки бывали находимы на ранах; зарегистрированы
также случаи нахождения личинок в полостях тела и кишечнике чело-
века и животных.
Таким образом, вблизи человека комнатная муха без труда находит
материал для пристраивания своего потомства и чем больше в населен-
ном пункте открытых нечистот, тем обильнее выплод насекомого.
Насколько многочислен бывает выплод мухи, даже из самых неболь-
ших скоплений нечистот, легко судить по тому, что для полного своего
развития личинка мухи, как минимум, может удовлетвориться 7 мили-
граммами питательного вещества. По данным венгерских исследователей
[Лоринч и Макара (Lorincz а. Makara), 1935] в 1 кг конского навоза может
развиться около 5000—8000 мух, тогда как то же весовое количество
свиного помета в состоянии продуцировать до 15000 экземпляров ком-
натной мухи.
Привлеченные запахом нечистот самки после краткого питания ими
приступают к откладке яиц. Как выяснено (Кузина, 1940 и др.), питание
субстратом, на который будет произведена откладка яиц, вызывает раз-
дражение у самки, имеющее значение стимула.
Гниющие вещества привлекают самок только на известной стадии
разложения. В силу этого обстоятельства некоторые из них в течение
первых дней их разложения или брожения крайне привлекательные,
с усилением в них процесса распада теряют для мух свои привлекательные
свойства. В этом отношении опыты Лоринча и Макара (1935) установили,
что ферментативные процессы в значительной степени влияют на дли-
тельность личиночной стадии, затягивая сроки развития; так, в све-
жем конском помете продолжительность личиночной стадии равня-
лась в опытах авторов 4—5 дням, тогда как в старом помете (при прочих
равных условиях) она затягивалась до 1—2 месяцев.
Кузина (1940) и другие авторы подтвердили, что предпочитаемые
для целей откладки яиц конский и свиной навоз теряют для самок свою
привлекательность через различные промежутки времени; если у первого
это наступает спустя 1—2 суток после его оставления животным, то для
108
I. ТРИБА MUSCINI
свиного навоза этот срок (при 25—40° С) колеблется между V2—1 меся-
цем. Порченое мясо утрачивает свои привлекательные свойства в зави-
симости от температуры через 3—5 суток после начала в нем процесса
гниения (Зимин, 1944).
Максимальное расстояние, на котором самки воспринимают запах
веществ, пригодных для кладки яиц, не определено; в природной обста-
новке значение имеет не только характер запаха, но и размер источника,
испускающего последний, направление движения воздуха, доносящего
запах до насекомого, и ряд других побочных факторов.
С наступлением теплых весенних дней самки, зимующие оплодотво-
ренными, покидают места зимовки и приступают обычно на 4—9 день
к откладке яиц. В редких случаях наблюдается непрекращающееся раз-
множение комнатной мухи и в зимние месяцы, когда этому способствуют
термические условия. В северных широтах эти условия комнатная муха
встречает в теплых помещениях; в экваториальных странах беспрерывное
размножение мухи на протяжении круглого года осуществляется и в при-
родной обстановке. Правда, в зимние месяцы численность яиц в кладках
М. domestica vieina Mcq. уменьшается против весеннего и летнего перио-
дов.
В соответствии с длительностью своей жизни, которая у комнатной
мухи варьирует в зависимости от характера пищи и окружающих усло-
вий от 2 до 112 (Кулагин, 1921) дней, в среднем равняясь, примерно,
14 дням, самки могут отложить различное число яиц. Яйца отклады-
ваются отдельными порциями через большие или меньшие промежутки
времени, что также связано с характером пищи самок. Количество яиц
в одной кладке и откладываемое комнатной мухой в течение всей ее жизни
определяется исследователями по-разному. Хьюитт (Hewitt, 1914) опре-
деляет число яиц в одной кладке в среднем в 120 яиц (от 100 до 150 штук),
считая, что одна самка в состоянии за свою жизнь дать 4—6 таких кладок,
что в сумме на одну самку составляет от 400 до 900 яиц. Лоринч и Макара
(1935) считают, что одна самка способна откладывать в среднем по 120 яиц
за раз, при этом число таких кладок у одной самки равно 3—5. В опытах
Кузиной (1936), среднее число яиц в кладке определилось в 93 (варьиро-
вание от 42 до 146). Большинство самок дали 1—2, реже 3 и в одном лишь
случае 4 кладки за свою жизнь.
Этих кратких данных достаточно, чтобы понять, насколько изменчи-
выми оказываются и длительность жизни комнатной мухи и ее плодо-
витость. Ранее мы уже указали на то, что все вышеизложенное в силь-
ной степени зависит от характера пищи. Известно, что при углеводистой,
лишенной белков пище самки до конца жизни остаются бесплодными.
При кормлении одним сахаром яичники не развиваются до конца, напо-
миная по степени развития яичники голодавших самок (Дербенева-Ухова,
1935). Сходную картину вызывает и кормление мух одним белком. Только
полноценная пища способна вызвать нормальный рост фолликулов
(Дербенева-Ухова, 1935). Согласно исследованиям того же автора, сахар
в пище мух способствует удлинению жизни насекомого.
Для продления жизни самкам необходимо включение в их пищевой
рацион сахаров и крахмала, тогда как для созревания яиц необходимы
протеины и пептоны. На основании этого Чеботаревич (1937) приходит
к выводу, что бытовая неряшливость в отношении хранения пищевых
продуктов и отбросов органического происхождения создает благопри-
ятную обстановку для размножения мух и способствует быстрому созре-
ванию яиц.
2. MUSCA
107
Вылет мух с мест зимовки начинается с первых теплых дней в марте—
апреле (в условиях Средней Азии) или апреле—мае (в Европейской части
СССР), когда самки, оплодотворенные с осени, приступают к откладке яиц.
Развитие яиц проходит в верхнем слое субстрата, на который были
отложены яйца. Продолжительность периода эмбрионального развития
колеблется в зависимости от температуры в пределах от 8 час. 5 мин.
до 51 час. 45 мин. (при относительной влажности воздуха равной 100%).
По данным Чеботаревича (1937), вылупление личинок из яиц
М. domestica vicina наступает спустя 8—12 часов после откладки.
По вылуплении из яиц личинки мигрируют вглубь субстрата, рас-
полагаясь в нем послойно в соответствии с температурой и влажностью.
Нужно заметить при этом, что отношение к различным температурам
у личинок различной степени зрелости неодинаково, в связи с чем и рас-
пределение в субстрате личинок различных стадий бывает неравномерным;
интерес в этом отношении представляют опыты Тетеровской (1944), Дер-
беневой-Уховой (1937—1949), а также Томсена (1938). Дербенева-Ухова
(1937), наблюдая за развитием личинок комнатной мухи в навозе разных
животных, установила, что распределение личинок в последнем всецело
зависит от температуры среды; температурные пределы для тех зон суб-
страта, где концентрировалась главная масса личинок, оказались весьма
широкими (20—41° С для свиного навоза, 23—49° С — для конского
навоза). По данным Тетеровской (1944), термический преферендум личи-
нок М. domestica vicina Mcq. заключен между 35.5—55° С. В субстрате
с температурой выше или ниже этой зоны личинки встречаются в значи-
тельно меньшем количестве.
При пониженной влажности субстрата личинки противостоят высо-
ким температурам дольше, нежели при высокой влажности среды. Так,
личинки в опытах автора выдерживали в сухой среде температуру от 44
до 45° С — около 3 часов, при 46—47° С — 21 минуту, при 48—49° С — 8
и при 50° С — 2х/2 минуты, тогда как увлажнение среды вызывало гибель
личинок при соответствующих температурах через 7, 2V2, 2 и 3/< минуты.
Автор считает поэтому, что температура выше 40° С является гибельной
для личинок.
Что касается реакции на низкие температуры, то, согласно данным
Петрищевой (1932), хорошо упитанные личинки III стадии переносят
без видимого для себя вреда температуры—1—2° С (давая с перенесением
их в теплую камеру пупарии) в продолжении двух недель. При воздей-
ствии более низких температур (—10° С) личинки той же стадии погибают
при экспозиции, превышающей 4 часа. Молодые личинки, обладающие
в этом отношении большей чувствительностью, не выдерживают более
З'/г часов пребывания в среде с температурой — 2° С. Температурный
минимум, при котором возможно развитие личинок, равен -Р7°С.'
Продолжительность личиночной фазы помимо характера пищи зави-
сит от условий температуры и влажности среды; в оптимальных усло-
виях она равна 3—4 дням, при неблагоприятной обстановке, затягиваясь
до 21 дня. Нужно заметить при этом, что особенно чувствительны к небла-
гоприятным условиям личинки в I и II стадиях.
Окукление личинок проходит в верхнем слое почвы, куда личинки
мигрируют из питавшего их субстрата, достигнув предельной зрелости.
В некоторых случаях личинки окукляются на поверхности или внутри
субстрата.
Длительность личиночной фазы может зависеть и от количества
пищи и, очевидно, от ее состава. Известно, что при недостатке питания
108
I. ТРИБА MUSCINI
личинки уходят раньше на окукление. Они дают мелких мух, плодови-
тость которых ниже, чем у насекомых нормального веса.
От момента образования ложнококона до вылета мухи проходит
в зависимости от режима среды, где был заключен пупарий, от 3 до 26
дней. Зимующие куколки находятся в субстрате значительно более про-
должительный срок, исчисляемый месяцами.
В местностях, где климатические условия позволяют мухам разви-
ваться беспрерывно в течение круглого года, если и бывает задержка
в развитии, то (экваториальные страны) она происходит под воздействием
слишком сухого, жаркого периода. Зимовка комнатной мухи может про-
текать в фазе личинки, куколки и взрослого насекомого (Порчинский,
Петрищева, Дербенева-Ухова и мн. др.).
При обследовании зимующих комнатных мух в г. Куйбышеве Петри-
щевой (1932) обнаружены в плохо отапливаемых помещениях только
самки, которые при перенесении в тепло дали потомство весной без нового
оплодотворения. Это указывает на перезимовку самок в оплодотворен-
ном состоянии. Установлено, что только у 26% собранных мух жировое
тело было развито, а среди самок всего 55% обладали развитыми яич-
никами .
В северных и средних широтах перезимовавшие мухи начинают
интенсивно размножаться, причем сроки отдельных генераций посте-
пенно сокращаются к лету и удлиняются к осени. Число генераций мухи
в природной обстановке зависит от длительности теплого сезона и от ком-
плекса условий, определяющих быстроту развития, как температура,
влажность, пища. Оптимальной термической зоной является темпера-
тура от 33 до 35° С; в обе стороны от нее продолжительность развития
возрастает. В опытах Петрищевой (1932) продолжительность развития
комнатной мухи (71/. domestica domestica) определилась в 20 дней при
16° С, в 15 дней при 20° С, 13.5 дней при 25° С, 10.5 дней при 28° С,
8 дней — при 30° С и 6—8 дней при 35° С. Чеботаревич (1937) для М.
domestica vieina получил длительность генерации в 8 дней и 20 час. при
24—32° С. Все эти данные сравнительно близки друг другу. Петрищева
допускает в природных условиях на широте г. Куйбышева наличие 6 по-
колений 71/. domestica domestica в течение времени с мая по сентябрь
включительно. Для 71/. domestica vieina Чеботаревич (в Ср. Азии) в сред-
нем предполагает существование 3 поколений в месяц. Следовательно,
в продолжение теплого времени года в этих широтах количество поколе-
ний комнатной мухи может достигнуть как минимум 15.
По теоретическим подсчетам потомство от одной пары мух
за летнее время определяется различными авторами от 5 000 000 000
до 604 000 000 000 000 000 особей, что с точки зрения некоторых иссле-
дователей оценивается как явное преуменьшение.
Перезимовавшие мухи с теплыми днями приступают к откладке
яиц, а при наличии самцов — к копуляции. Одной копуляции оказы-
вается достаточно для оплодотворения яиц за время всей жизни самки,
хотя в наблюдениях мы являемся свидетелями многочисленных случаев
повторного оплодотворения.
К спариванию (летние поколения) комнатная муха может приступать
в первые же сутки после вылупления из пупария, хотя чаще это наблю-
дается на 3—7 день. Петрищева (1932) имела случай проследить спари-
вание уже через 18 часов после вылета мух. Сроки копуляции в значи-
тельной степени зависят от характера пищи. При питании молоком мухи
в опытных условиях начинают спариваться на 4 день, тогда как при
2. MUSCA
109
растительной пище, копуляция наступает на 16 день. При голодании
мух спаривания не происходит.
Активность комнатной мухи (имагинес) связана ^температурными
условиями среды и освещением.
По Нишульцу и Дютуа (Nieschulz и du Toil) (1933), нормальная
активность мух наступает при температуре от 15 до 23.5° С. G повыше-
нием температуры до 35—43° С мухи проявляют беспокойство. ,
Значительные отклонения в температуре, при которой наступает
га или иная степень активности мух, указывают на присутствие и дру-
гих, одновременно действующих факторов, как, например, свет. Известно,
что при оптимальных температурах мухи могут пребывать в инактив-
ном состоянии.
В равных температурных условиях при рассеянном солнечном осве-
щении мухи менее деятельны, нежели в условиях прямой солнечной
радиации.
Отношение мух к различным частям солнечного спектра не одина-
ково. При синем, голубом освещении мухи ведут себя как в полной тем -
ноте, оказываясь крайне чувствительными к средним цветам спе-
ктра.
Дербенева-Ухова и Кузина (1938) в своих наблюдениях под Москвой
установили, что максимальный залет М. domestica domestica в жилища
происходит в период времени между 11 и 18 часами дня. Ночью залет
прекращается, если не считать слабой миграции, вызванной искусствен-
ным освещением. Наши многочисленные учетные работы в этом направле-
нии, проведенные в Гиссарской долине (Таджикистан) с М. domestica
vicina, за небольшими исключениями подтвердили данные вышеназван-
ных авторов.
Помимо действия температур и освещения, на миграцию мух в силь-
ной степени влияет наличие пищевых запасов в помещении, в силу чего
при равных метеорологических условиях «обмушение» таких помещений,
как кухни, столовые, магазины и т. д., происходит в наиболее сильной
степени.
Дальность перелетов мух из мест, где происходит их размножение,
большинством авторов определяется в 500—800 метров.
Имеются данные, свидетельствующие о миграциях мух на 21 км.
Чеботаревич (1937) указал на возможность залета мух от мест выплода
на расстояние 13 км. Судя по некоторым данным, муха в состоянии дер-
жаться в воздухе, не садясь, несколько часов подряд, причем скорость,
которая развивается при полете, может составлять около 20 км в час.
Большое значение в расселении мух имеет транспорт, при чем для
этой цели оказываются пригодными все его виды, начиная от гужевого
и кончая автомобилем и самолетом.
Наряду с неблагоприятными метеорологическими условиями, огра-
ничивающими размножение комнатной мухи, существенная роль в этом
отношении принадлежит паразитам и хищникам.
К числу первых относятся простейшие организмы, нематоды, клещи,
насекомые. Не малое значение имеет и деятельность паразитических
грибков.
Влияние на организм мух бактериальной флоры, почти постоянно
присутствующей в пищеварительном тракте и некоторых других вну-
тренних органах мух, окончательно не выяснено, хотя по отношению
некоторых видов {Bacterium delende-muscae Roub. Desc., 1923) установ-
лено патогенное для мух значение бактерий. В частности, упомянутый
110
I. ТРИБА MUSCINI
вид бактерии вызывает смертность куколок комнатной мухи и осенней
жигалки.
Взгляды исследователей на роль жгутиконосцев в жизни мух расхо-
дятся. Как известно из работ Хьюитта (Hewitt, 1914) и др., Herpetomonas
muscae domesticae встречается в пищеварительном тракте мух и является,
по убеждению некоторых авторов, паразитом имаго. Петрищева (1932)
в огромных количествах находила Herpetomonas muscarum в кишечнике
мух и личинок.
Автор на основе своих наблюдений считает возможным допустить,
что в отношении Н. muscarum имеет место не паразитизм, а мирное сожи-
тельство, не причиняющее вреда насекомому. Личинки заражаются
Н. muscarum через субстрат, который в свою очередь получает жгутико-
носцев от мух с их экскрементами.
Наибольшее заражение мух совпадает с летними месяцами, пони-
жаясь к осени. Пораженность мух может достигать 100%.
Рис. 140, 141.
140 — задний конец тела паразитической нематоды Allan-
tonema muscae Hoy. a. Makher. 141 — паразитическая пема-
тода A. striglandi Roy. ^a. Makher: n —папилла, я — яйца,
в — вульва, м —матка. (По Roy a. Makherey).
Еще в прошлом столетии были обнаружены в теле мух черви —
Habronema muscae.
Дальнейшие изыскания установили, что Н. muscae обитает в полости
тела как взрослых мух, так и куколок и сравнительно редко — личи-
нок. В теле насекомого Н. muscae проходит личиночную стадию. Имаги-
нальная фаза живет в желудке лошади. Нематоды обнаруживались
в полости головы, груди и в брюшке мух, причем в одной мухе удавалось
обнаружить до 8 паразитов. Численность личинок Н. muscae в одном насе-
комом может достигать 20 и более экземпляров. Развитие эмбрионов
нематоды проходит в жировом теле мух (Seguy, 1935).
В 1937 г. Рой и Мекери (Roy и Mukherey) описали два вида нематод
(рис. 140—141) Allantonema muscae и A. striglandi из Musca domestica.
Оба вида обнаружены (в брюшке мух) одновременно в стадии яйца, личи-
нок и самок, что в биологическом отношении резко отличает этих червей
от Habronema, для которой комнатная муха является лишь промежу-
точным хозяином.
В сравнении с вышеперечисленными паразитами, грибное заболева-
ние, вызываемое Empusa muscae, производит в популяции синантропных
мух более серьезные опустошения. Наибольшей силы болезнь достигает
2. MUSCA
111
во второй половине лета или осенью с наступлением дождливого вре-
мени года. Насекомое заражается спорами грибка через наружные по-
кровы. Росток споры проникает внутрь тела и, достигнув жирового тела,
дает многочисленные гифы, оплетающие жировую ткань и разрушающие
ее. Постепенно грибок заполняет всю полость тела насекомого и доводит
до полного разрушения все внутренние органы насекомого. Гибель мух
наступает летом через 4—15, осенью через 3—4 дня (Петрищева, 1932).
В качестве безусловных паразитов комнатной мухи известен наезд-
ник Muscidijurax brevicornis Asch. (=Мormoniella vitripennis Walk.),
заражающий пупарии комнатной мухи. Исследования Смирнова (1934)
и Смирнова и Кузиной (1933) показывают, что М. vitripennis является,
в сущности, наружным паразитом, т. к. личинки его высасывают куколку
мухи с поверхности, не проникая внутрь ее и располагаясь, следова-
тельно, между стенкой ложнококона и наружными покровами истинной
куколки. Самка паразита может заражать в среднем до 25 куколок ком-
натной мухи. В природных условиях зараженность паразитом пупариев
комнатной мухи, возможно, невелика, т. к. наездник не в состоянии
проникать в субстрат (гл. обр. почву), где окукливается муха.
По Хьюитту (1914) полный цикл развития паразита равен в сред-
нем 22х/г дням; продолжительность развития летом сокращается против
этого срока. Наездник зимует в пупарии мухи в стадии личинки; окукле-
ние весной. По свидетельству Смирнова и Кузиной (1933), М. brevicornis
в лабораторной обстановке развивается беспрерывно круглый год. Musci-
dijurax raptor Girault по биологии сходен с предыдущим.
Помимо вышеуказанных, известны в качестве паразитов домашней
мухи: Spalangia nigra Latr., заражающий пупарии М. domestica. Зара-
жение ложнококонов достигает местами 32%. Spalangia muscidarum
Rich., как и вышеназванные, паразитирует внутри пупариев мух. Томсен
(1938) приводит в списке паразитов, М. domestica Stenomalus muscarum,
St. muscae и несколько представителей сем. Cynipidae : Figites scutel-
larius, Fig. striolatus и Fig. anthomyidarum.
Данные по паразитизму на мухах красных клещей (Trombidium)
не достаточно полны. Что касается клещей, иногда весьма многочислен-
ных на двукрылых, в том числе и на комнатной мухе, то не лишено осно-
вания предположение, что мы имеем в данном случае перед собою
явление форезии, а не паразитизма.
Если в отношении паразитов список видов, связанных с комнатной
мухой, весьма ограничен, то в отношении хищников, уничтожающих
последнюю на различных фазах ее развития, он весьма обилен.
Опасность быть уничтоженной хищниками возникает для комнат-
ной мухи с момента откладки яиц. Особенную опасность для этой стадии
представляют муравьи (Pheidole, Cataglyphis, Camponotus и др.). Личинки
поедаются хищными жуками, принадлежащими нескольким семействам.
К ним должны быть отнесены жуки сем. Staphylinidae (Creophilus maxil-
losus, L., Ontholestes tessellatus Geoffr.J, Histeridae (Hister, Saprinus)
и Carabidae. В огромном количестве личинки комнатной мухи истребляются
хищными личинками мух (Myiospila meditabunda F., Hydrotaea dentipesF.,
H. armipesFaW., Ophyra leucostoma Wd., Polietes albolineata Fall., Mydaea
urbana Mg., Muscina stabulans Fall., M. pabulorum Fall., Mesembrina
mystacea L).
Наряду с мухами в большом количестве истребляют личинок, равно
как и пупарии комнатной мухи, птицы. В этом отношении большую
пользу приносит домашняя птица (Чеботаревич, 1937 и др.), использо-
112
I. ТРИБА MUSCINI
вание которой с этой целью имеет смысл лишь после предварительных
исследований червей, одновременно являющихся паразитами как неко-
торых личинок мух, так и птиц.
Пупарии мух, как и личинки, могут поедаться всеми вышеназван-
ными хищными насекомыми, равно как и птицами.
Взрослые мухи служат предметом случайной или специальной охоты
многих перепончатокрылых, главным образом ос, хищных мух (сем.
Asilidae), пауков, многоножек.
Комнатная муха в отношении большинства патогенных для человека
микроорганизмов является механическим переносчиком. Она, благодаря
попеременному контакту с источником заразы (больные люди, животные,
зараженные предметы, нечистоты и т. п.) и здоровыми людьми, осуще-
ствляет перенос возбудителя того или иного заболевания на наружных
частях своего тела. Другой способ передачи инфекции заключается в том,
что насекомое, питаясь зараженным материалом, вводит в свой кишеч-
ник и зоб культуру патогенных организмов, передавая их в дальнейшем
со своими экскрементами или с отрыгиваемой пищей. Таким путем мухи
благодаря своей колоссальной численности способствуют быстрой диссе-
минации болезнетворных организмов в населенных пунктах.
На поверхности тела мухи патогенные организмы, если не считать
особо устойчивых спор бактерий, остаются жизнеспособными в продол-
жение сравнительно небольшого промежутка времени, который всецело
зависит от окружающих насекомое условий (температура, солнечная
радиация и т. д.), и от резистентности самого организма по отношению
к тем или иным внешним факторам. Этот срок может в одних случаях
определяться минутами или часами, в других — днями.
В теле мухи многие бактерии оказываются вирулентными в продол-
жение нескольких суток. Bacillus prodigiosus в мухах, питавшихся
культурой бактерий, обнаруживалась на 17-й день. Лоринч и Макара
(1935) находили в мухах брюшнотифозную палочку на 2—4-е и на 18-е
сутки после питания мух на культуре бактерии.
До настоящего времени остается спорным вопрос о передаче инфек-
ции от одного поколения мух к другому через преимагинальные фазы
развития (яйцо, личинка, куколка). Так, Грегем-Смис (Graham-Smith,
1913) получил результат, доказывающий, что спорообразующие бактерии
(как, например, сибирской язвы) способны передаваться от личинок
к куколке и далее к имаго. Не образующие спор бактерии (В. typhosus,
В. enteritidis, В. prodigiosus), по убеждению автора, не способны пройти
через все стадии метаморфоза, и если подобного рода передача инфекции
возможна, то лишь как редкое исключение. При кормлении личинок
зараженной пищей Лоринч и Макара (1935) получили палочки брюш-
ного тифа у куколок и имаго в том случае, если пупарии оставались
нестерилизованными снаружи; при стерилизации пупариев были полу-
чены незараженные мухи. Это наводит на мысль о том, что в отношении
некоторых возбудителей инфекция может передаваться лишь наружными
покровами. Интересно отметить здесь, что авторы имели случай получить
брюшнотифозную палочку из мертвых мух, сохранявшихся при 8—12° С,
спустя 8 месяцев после кормления насекомых на культуре бактерий.
Комнатная муха известна как переносчик холеры (Павловский,
1948 и др.). Холерный вибрион обнаруживался как в кишечнике, так
и на наружных частях тела мухи. Многочисленные работы посвящены
вопросу передачи мухами возбудителей дизентерии и брюшного тифа.
Предполагается, что М. domestica участвует в диссеминации туберкулез-
2. MUSCA
113
ной палочки, которая регистрировалась в ее кишечнике и экскрементах'
Основываясь на ряде врачебных наблюдений, можно подозревать ком-
натную муху в переносе возбудителей глазных заболеваний (трахома,
конъюнктивит).
Как большинство мух, соприкасающихся с сибиреязвенными жи-
вотными, М. domestica, вероятно, участвует и в распространении сибир-
ской язвы.
Помимо распространения инфекций, мухи являются переносчиками
яиц глист. Известны следующие виды, яйца которых переносятся ком-
натной мухой: Taenia solium, Т. saginata, Т. serrata, Т. echinococcus,
Т. marginata, Dipilidium caninum (T. cucumerina), а также—Ascaris
lumbricoides, A. megacephala, Oxyuris vermicularis, Trichiuris trichiuris
(Trichocephalus dispar), Ancylostoma duodenale.
Принципы борьбы с комнатной мухой изложены в общей части
работы.
2. Musca nebulo (F.) Wd. (рис. 113—115).
Wiedemann, 1830, Aussereurop. zweiflug. Insekt., II : 416; Stein, 1917 : 104;
Patton, 1933, Ann. trop. Med. Parasitol., XXVII : 401—403, fig. 8; он же, 1936, там же,
XXX : 472; он же, 1937, там же, XXXI : 128.
Биология: Patton, 1933, цит. раб.; De la Paz, 1938, Mon. Bull. Bur. Hlth.
Philipp., I, 18 : 398—412, 515—519, 521—539 pl. 5.
Лоб самца узкий, равен Vs ширины глаза; передняя часть лба
и скулы кремовые; грудь серая с 4 черными продольными полосами,
ширина которых в два раза более ширины темени. Брюшко с преобла-
дающей оранжевой окраской; тергит 1 + 2-й с черной широкой срединной
полосой и таким же основанием; 3-й тергит светлооранжевый с заострен-
ной на вершине и часто незаконченной широкой черной полосой (иногда
неполно выраженной); по бокам от полосы располагаются области, покры-
тые белым налетом, а также краевые серебристые пятна; 4-й тергит той же
окраски, но с более узкой черной полоской и с сильнее развитыми пят-
нами налета; 5-й — в серебристом налете, нередко со срединным тем-
ным пятнышком, не образующим, однако, законченной продольной
полосы. Церки снизу с легкой закраинкой, с округленными наружными
и необразующими длинных конусовидных выступов внутренними углами.
Склеротизованная часть фаллозомы длинная, слабо утонченная у средины,
задний ее отросток маленький, не расширенный. Передний отдел пара-
мер длинный, с 4—5, реже 6 щетинками (одной средней по размеру, про-
чими мелкими); задняя часть парамер маленькая, невыступающая,
V стернит широкий, по заднему краю с сильными, загнутыми внутрь
выростами.
Лоб и скулы самки кремово-белые; ширина лба менее ширины глаза;
лобная полоса широкая, со слегка выпуклыми краями; ширина полосы
составляет 3/ь ширины темени. Среднеспинка как у самца; брюшко светло-
оранжевое; 1-{-2 тергит оранжевый, с маленьким круглым или оваль-
ным пятном у переднего края (размер пятна сильно варьирует); 3-й тер-
гит с узкой, темной срединной полоской, у некоторых особей вовсе исче-
зающей; налет серебристо белый или серый, резко выражен, как у самца;
4-й—почти целиком светлооранжевый, со слабо выраженным темным
штрихом; 5-й тергит на большем протяжении орнажевый. Длина тела
как у Musca domestica L.
Распространение. Широко распространен в Ориентальной
области, являясь обитателем жилищ и базаров.
8 Фауна СССР, том XVIII, вып. 4.
114
I. ТРИВА MUSCINI
Известен повсюду в Индии, Бирме, Ассаме, с Филиппинских остро-
вов и Малайского архипелага; нередок в восточной части Эфиопской
области; заходит в пределы Палеарктики (Египет). Пэттон (1936—1937)
высказывает сомнение, принадлежат ли африканские экземпляры дей-
ствительно к виду Musca nebulo. Обнаружение вида возможно и в пре-
делах Ирана и у южных границ Туркмении. М. nebulo систематически
близок М. domestica domestica и М. d. vicina, отличаясь от последних
деталями строения мужских гениталий, узким лбом самца и самки и более
светлой окраской тела (преимущественно брюшка).
Сходство названных видов наблюдается и со стороны биологических
их особенностей. Тот и другой являются формами синантропными, в раз-
множении своем связанными с нечистотами населенных пунктов (отбросы,
фекалии, конский навоз). На Филиппинах (Манила) М. nebulo вместе
с М. sorbens и М. domestica vicina является наиболее обычным двукры-
лым у нечистот, на базарах, в жилищах, лавках, ресторанах и т. п.
Исследование флоры бактерий поверхности тела названных видов мух
подтверждает предположение о распространении ими патогенных для
человека микроорганизмов.
Наши экземпляры Musca domestica vicina Mcq. из Южной Туркме-
нии по окраске являются близкими типичным Musca nebulo, разнясь
от последних деталями строения фаллозомы и чуть более широким лбом.
В этом отношении туркменские экземпляры занимают среднее положение
между типичными М. d. vicina и М. nebulo. Вероятнее всего, что Musca
domestica, domestica, М. d. vicina и M. nebulo являются одним видом Musca
domestica L., что по отношению двух первых представляется совершенно
несомненным.
Личинки, равно как и другие преимагинальные фазы М. nebulo,
подробно не известны. По Пэттону, личинки III стадии имеют 5—7-луче-
вые передние дыхальцы; задние дыхальцы сходны с таковыми М. dome-
stica vicina Mcq.
II. Группа Musca sorbens Wd.
Лоб самца обычно уже 3-го членика усиков; глаза голые; грудь
черная или в сером налете с 2 широкими черными полосами, раздвоен-
ными впереди шва; проплевры, супрасквамальное ребро и I стернит
брюшка без волосков.
В фауне СССР группа представлена 2 видами — Musca sorbens Wd.
и Musca albina Wd.
*3. Musca sorbens Wd. Базарная муха (рис. 142—154).
Wiedemann, 1830, Aussereurop. zweifliig. Insekt., II : 418, 58; Patton, 1923, Philipp.
Journ. Science, XXVII : 310—314; он же, 1932, Ann. trop. Med. Parasitol.. XXVI :
347—405, figs. 9b, 12, 13; он же, 1937, там же, XXXI : 133; Но, 1938, там же, XXXII :
287—312 fig. 10е. — humilis Wiedemann, 1830, 1. с. : 418, 59; Stein, 1918, Ann. Mus.
Nat. Hung., XXI : 490—491; Patton, 1922, Bull. ent. Research, XII : 412: Karl.
1935, Arb. morph, taxon. Entom., II : 34; — angustifrous Thomson, Stein (apud Beck-
er), Mitt. Zool. Mus. Berl., IV : 196, 111.
Экология и эпидемиологическое значение. Patton,
1922, 1923, 1. c.; Lamborn, 1933, Ann. Med. Rep. Nyasaland : 61—63; он же, 1934:
65—69; Austin a. Mayne, 1935, там же, 1934 : 73—82; Lamborn, 1936, Journ. trop.
Med. Hyg., XXXIX ; 235—239; он же, 1936, Ann. Rep. Med. Ent., 1935 : 50—52;
Чеботаревич, 1937, Tp. Сов. изуч. произв. сил, 9 : 39; Meng a. Winfield, 1938, Chin.
Med. Journ. suppl. : 463—486; De la Paz, 1938, Bull. Bur. Hlth. Philipp., I :
18:393—412; Зимин, 1944, Изв. Тадж. фил. АН СССР, 5:143—151; Зимин,
1944, Проблемы кишечн. инфекц. : 150—152; Зимин, 1947, Сб. работ клиники глазн.
бол. Сталинабад. Мед. инет. : 12—21, рис. 2; Зимин и Тетеровская, 1943, Мед. параавт.
2. MUSCA
115
и паразит, болезни : 44—53; Зимин, Парадоксов, Механикова, 1947, Эпидем. конъ-
юнктивит в Таджикистане и базарная муха как его переносчик, Сталинабад, 54 стр.
2 диагр.; Павловский, 1948 : 894—895; Зимин, 1948, Определ. личинок: 82; Дербе-
нева-Ухова, 1949.
Лоб самца равен или чуть шире 3-го членика усиков; скулы в два
раза уже щек; затылок плоский или слабо вогнутый в верхней половине;
3-й членик усиков в 3 раза длиннее 2-го, бурый; 2-й членик коричневато-
бурый; голова черно-бурая; орбиты, щеки и затылок между краем глаза
и заглазными щетинками в густом, шелковистом белом налете, имеющем
желтоватый оттенок на щеках и слабый серый оттенок на затылке; лицо
в темно сером матовом налете. Лобные щетинки, кроме передней креп-
кой пары, тонкие, волосовидные, короткие; орбиты с одиночными корот-
кими волосками. Внутренние теменные щетинки сравнительно длинные
и крепкие; затылочные щетинки в виде коротких волосков; щеки в корот-
ких, негустых бурых волосках. Щупальцы бурые, слабо и постепенно
утолщенные в вершине. Грудь черно-бурая в желтовато-сером светлом
налете; среднеспинка с 2 широкими бурыми или черными продольными
полосами позади шва, обычно раздвоенными перед ним; ширина полос
светлого налета уже темной полосы. Грудь в черных, негустых щети-
нистых и более коротких на спинке, тонких и длинных на плеврах —
бурых волосках; щиток в серовато-желтом налете; передние боковые
и средний отдел щитка затемнены. Щетинки груди: акростихальных —
1 пара перед щитком; дорзоцентральных t—2 впереди и 4 позади шва;
интраалярных — 1; закрыловых — 2—3; надкрыловых — 1—3; плече-
вых — 3; заплечевых — 2; нотоплевральных — 2; мезоплевральных —
5—6 по заднему краю склерита; стерноплевральных — 1-}-2; гипоплевры
впереди заднего дыхальца в многочисленных тонких волосках; на щитке —
2 основных, 2 пред основных, 2 вершинных, 2 боковых щетинки. Перед-
ние грудные дыхальцы желтовато-белые. Ноги черно-бурые, голени
коричневато-бурые; присоски серовато-желтые, немного короче 5-го чле-
ника лапки. Жужжальца желтые, чешуйки белые, грудная чешуйка
с желтоватым краем. Крылья прозрачные, светлосерые, иризирующие;
жилки в основной половине крыла бледножелтые, в вершинной — жел-
товато-серые; жилка т с мягким изгибом. Брюшко темножелтое, с бурым
основанием 1-]-2-го тергита, бурым гипопигием и бурой срединной сужен-
ной назад узкой полоской; 5-й тергит в середине с нечетким буроватым
пятном. Желтовато-серый налет образует квадратные пятна на 3-м, 4-м
и почти сплошь покрывает 5-й тергит. Пятна налета располагаются
в числе 4 на каждом тергите: по бокам от средней бурой полосы и по бо-
ковым краям сегментов. Нижняя сторона брюшка равномерно покрыта
редким желтовато-белым налетом. Церки типа М. domestica', свободный
край церок почти прямой, внутренние углы без выростов; склеротизо-
ванная часть фаллозомы узкая, короче мембранозного дистального отдела;
мембранозная часть фаллозомы шире основной, с 2 угловидными лопа-
стями на вентральном крае; длина основной склеротизованной части
равна длине мембранозного отдела фаллозомы; дорзальный отросток
длинный, расширен к дистальному концу, на вершине с прямым краем;
передняя часть парамер длиннее задней, с одной крепкой и несколькими
мелкими щетинками на дорзальной поверхности; задний отдел парамер
маленький, в форме короткого конуса; тека короткая, не всегда отчет-
лива в своих контурах. V стернит с двумя сравнительно длинными боко-
выми отростками, зазубренными на вершине; задняя половина V стер-
нита коричневая или буроватая.
8*
116
I. ТРИВА MUSCINI
q. Расстояние между глазами на темени составляет 2/з ширины глаза;
лобная полоса почти в 2 раза шире орбиты, ширина скулы на уровне
1-го членика усиков равна ширине щеки; сероватый налет на орбитах,
скулах, щеках и затылке более темный и матовый, чем у самцов. Лоб-
ные щетинки расположены в один ряд с каждой стороны от лобной
полосы; первая пара щетинок крепкая; теменные внутренние и темен-
ные наружные щетинки хорошо развиты: крепкие и длинные. Грудь
как у самца, но с более густым налетом; присоски на лапках в 2 раза
короче 5-го членика. Брюшко черно-бурое, снизу и в передних боко-
вых отделах 3-го и 4-го тергитов серо-желтое; окраска нижней
стороны брюшка темнее к его вершине. Верхняя поверхность брюшка
в густом желтовато-сером шелковистом налете, за исключением 1-|-2-го
тергита, где налет не всегда ясен. По середине 3-го и 4-го тергитов
проходит бурая полоса, более широкая на 3-м сегменте; по бокам от сред-
ней бурой полосы на 3-м и 4-м тергитах располагаются темные продоль-
ные пятна, по одному с каждой стороны; пятна расширены клиновидно
назад, нередко образуя по заднему краю узкую темную полоску. 5-й тер-
гит слабо затемнен посредине. Длина тела 4.5—7.3 мм.
Яйцо белое, удлиненное цилиндрическое с ясно выраженной ячеистой
скульптурой хориона, состоящей из шестигранных ячей, несколько
вытянутых по длинной оси яйпа; передний полюс чуть заострен. Дор-
зальная сторона со слабо заметными двумя продольными полосками.
Длина 3.0—3.6 мм, толщина, 0.3—0.5 мм.
Личинка I стадии (рис. 145, 147) белая, мелкая; начиная с 1-го
грудного сегмента передние края сегментов с нижней стороны с рядами
2. MUSCA
117
мелких рыжеватых тонких шипиков, расположенных в несколько (4—5
на грудных сегментах и 8—12 на брюшных сегментах) поперечных рядов;
передние и задние ряды составлены из мелких шипов, заходящих и на бо-
ковые отделы сегментов; средние ряды состоят из очень длинных крюч-
ковидно изогнутых шипиков, приуроченных лищь к валикообразным
поперечным выступам на вентральной поверхности тела личинки. Рото-
глоточный аппарат рыжевато-желтый; срединный дорзальный склерит
листовидный, узкий, с клювовидно изогнутой вершиной, ограниченной
на вентральном крае выступом; по дорзальному и вентральному краям
выпуклый, сзади раздвоен и сочленен с гипостомальным склеритом;
последний состоит из двух латерально лежащих, слабо дуговидно выгну-
тых хитиновых тяжей, соединенных с вентральной стороны тонкой, слабо
окрашенной перемычкой. Фарингеальный склерит тонкий, вместе с узкими
заостренными на конце вентральными и дорзальными отростками обра-
зует полукруг; передние отростки, соединенные дистальными концами
с гипостомальным склеритом, тонкие. Фарингеальные ребрышки выра-
женье слабо (на препаратах не всегда ясно видны). Боковые края рото-
вого отверстия укреплены хитиновыми пластинками. Антенны рыжева-
тые. Снизу головной сегмент впереди ротового отверстия снабжен попе-
речным рядом из частых желтоватых хитиновых пластинок. Задние
дыхальцы с одним отверстием.
Личинка II стадии (рис. 144, 146) белая или со слабым желтоватым
оттенком; передние дыхальцы с 5 пальцевидными выростами; задние
дыхальцы с 2 щелями каждое, дыхальцевая пластинка слабо хитинизи-
рована. Ротовые крючки тонкие, сильно склеротизованы, в переднем
отделе прямые, в заднем — резко загнуты на вентральную сторону
и слабо утолщены; вентрально от ротовых крючьев располагается пара
сагиттально вытянутых, загнутых на переднем конце и расширенных
в задней трети склеритов, по бокам и вниз от которых отходят широкие
неправильно листообразные лопасти, укрепляющие боковые стенки рото-
вого отверстия. Угловидный склерит парный, разделен на дорзальной
стороне и здесь загнут назад; гипостомальный склерит в форме перевер-
нутой широким основанием вверх трапеции, снизу соединен перемычкой;
базальный склерит высокий (широкий), с широкими крыловидными дор-
зальными и узкими, слабо расширенными на дистальном конце вентраль-
ными отростками; дистальный конец последних слабо скошен и слабо
выемчатый; передние отростки короткие, расширенные на конце, обра-
зуют здесь почти прямой верхний и оттянутый вниз и слегка вперед
нижний углы; дорзальная перемычка решетчатая, с заостренными отро-
стками на заднем крае; вентральная перемычка узкая, с полукруглыми
узкими, направленными назад отростками. Фарингс со слабо видимыми
ребрышками. Над ротовым отверстием располагается 4—6 коротких
хитиновых зубчиков; антенны и максиллярные пальпы хорошо заметны.
Длина тела 4—6 мм.
Личинка III стадии (рис. 148—154) с длинным постепенно утол-
щенным к заднему и заостренным кпереди телом;ксе сегменты, исключая
головной, по переднему краю с широкими полосками из шипиков; с ниж-
ней стороны по передним краям брюшных сегментов располагаются
поперечные валики, вооруженные наиболее крепкими и длинными зуб-
цами. Валикообразные утолщения на боковых отделах брюшных сегмен-
тов снабжены мелкими шипиками. Анальная пластинка узкая, бледно-
палевая, едва заходит на боковые отделы сегмента; латеральные отделы
пластинки едва расширены и на конце узко закруглены. Весь анальный
118
I. ТРИБА MUSCINI
сегмент, исключая заднюю поверхность вокруг дыхалец,’ усажен мел-
кими, густо расположенными светлыми шипиками. По бокам анального
сегмента расположено 8 маленьких бугорков (по 4 с каждой стороны).
Головной сегмент двулопастной; антенны и максиллярные щупальцы
ясные; передние дыхальцы с 6—7 лучами; задние дыхальцы с сильно
склеротизованной широкой черной перитремой; дыхальцевые щели
сравнительно широкие. Дыхальцевая пластинка слабо склеротизована,
Рис. 144—147. Детали строения личинок ран-
них стадий Musca sorbens Wd.
144—задние дыхальцы личинки II стадии. 145 —
как и стигмальный диск. Ды-
хальцы расставлены друг от
друга на расстояние, равное
—Vi их поперечника. Ро-
тоглоточный аппарат сильно
склеротизован; ротовые крю-
чья слабо согнутые,наподобие
косы, заостренные на верши-
не, в основании широкие,
скошены сверху вниз по' зад-
нему краю, снизу с зубцам;
один крючок уже и короче
другого; зубные склериты
округленные снизу, выемча-
тые по верхнему краю, не-
правильно треугольные; ги-
постомальный склерит корот-
кий, неправильно прямо-
угольный; базальный скле-
рит длинный; дорзальные от-
ростки узко крыловидные,
слабо выпуклые по дорзаль-
ному краю, вентральные от-
ростки почти в два раза ко-
роче дорзальных, к дисталь-
ному концу расширенные, на
вершине с широкой неглу-
бокой выемкой; фарингеаль-
ные бороздки хорошо видны;
дорзальная перемычка мелко
ротоглоточный аппарат личинки I стадий.
146 — то же II стадии. 147 — срединный дор-
зальный склерит личинки I стадии. (Ориг.),
решетчатая, с двумя острыми
выступами сзади; верхний пе-
редний край базального скле-
рита направлен вперед в ви-
де острого угла; передние отростки короткие, расширенные и выемча-
тые на вершине, снизу соединены вентральной перемычкой. Длина
тела 7—9 мм.
Пупарий рыжевато-коричневый, длина 3.7—4.5 мм; толщина
по середине 1.3—1.6 мм; поверхность слабо блестящая, сегмен-
тация ясная; рожки передних дыхалец тонкие, длинные, бурые. Аналь-
ный сегмент полушаровидно округлен; анальная пластинка короткая,
узкая, изогнута в виде скобки, анальный бугорок маленький,
по бокам его располагается по одному субанальному бугорку, отод-
винутому от анального бугорка на длину, равную тройному их диа-
метру. Впереди анальной пластинки и позади нее узкие шероховатые
области.
2. MUSCA
119
Распространение вида весьма широкое; им охватывается
Ориентальная, Эфиопская области и юг Палеарктики (крайний юг СССР,
Пран, Ирак, Китай). В СССР вид встречается по югу Закавказья,
в Туркмении, Узбекистане, южном Казахстане и Таджикистане.
/Л?
Рис. 148—154. Детали строения личинки III стадии Musca sorbens Wd.
148 — ротоглоточный аппарат. 149 — задний конец тела сзади. 150 — он же сбоку.
151 — заднее дыхальце. 152 — переднее дыхальце. 153 — вооружение на 5-м (2)
и 6-м (1) брюшных сегментах снизу. 154 — скульптура дорзальной поверхности
в средине 2-го грудного сегмента. (По Зимину).
Экология. М. sorbens является в Средней Азии обычным, в ста-
циальном отношении широко распространенным в населенных пунктах
синантропным видом. Базарная муха появляется поздно (конец июня—
начало июля), достигая наибольшей численности в августе и сентябре,
что совпадает с жарким и сравнительно сухим периодом лета. Задер-
живаются мухи в природе до октября, при теплой осени и дольше.
Базарная муха относится к категории теплолюбивых и гелиофиль-
ных форм. Именно этим обстоятельством определяется характер ее рас-
пределения в населенных пунктах. Мухи предпочитают открытые или
ярко освещаемые солнцем места, избегая затененных помещений и зале-
120
J. ТРИБА MUSCINI
тая в них лишь на короткий срок. Как правило, в противоположность
комнатной мухе, М. sorbens в помещениях не задерживается на ночь,
вылетая с наступлением сумерек. В Таджикистане, что в одинаковой
мере относится к Индии, Африке и Аравии, М. sorbens является обита-
телем базаров, открытых лавок, ларьков, свалочных мест и т. п.
В условиях Гиссарской долины (Таджикистан) в жилых помещениях
базарная муха значительно уступает по численности комнатной мухе,
в то время как на базарах в августе и сентябре она может количественно
преобладать над каждым из прочих синантропных видов, в том числе
и над М. domestica vicina. Наибольшая активность имаго М. sorbens
наблюдается в самое жаркое время дня, когда и осуществляется ее особо
интенсивный залет в жилища. Эта особенность резко отличает базарную
муху от комнатной, у которой при максимальных дневных температурах
замечается понижение активности. Максимум активности отмечен нами
при 26.0—34.0° С и относительной влажности воздуха 18—34%. В жар-
кие часы дня мухи становятся назойливыми, нападая на открытые части
тела человека и особенно на слизистые оболочки глаз, носа и на пора-
женные участки кожи.
В Средней Азии местами выплода базарной мухи служат фекалии
человека, оставленные на земле, фекальные подсыхающие массы в плохо
закрытых дворовых уборных, и в меньшей степени — свиной навоз.
На развитие мухи от яйца до вылета особей нового поколения
требуется при температуре от 25 до 28° С 9 суток (Зимин, 1944, 1945).
В тропических странах медико-санитарное значение базарной мухи
как распространителя различных патогенных микроорганизмов является
весьма существенным. Основываясь на данных бактериологического
анализа, некоторые исследователи считают базарную муху наравне
с Musca domestica vicina главнейшим распространителем патогенных
и гнилостных бактерий, опасных или вредных для человека. Пэттон
(1932, 1933) рассматривает М. sorbens как механического распространи-
теля острых кишечных инфекций, как дизентерия, холера, а также глаз-
ных заболеваний и, в частности, возбудителя конъюнктивита. Ламборн
(Lamborn, 1936) экспериментально показал возможность переноса мухами
этого вида Spirochaeta (Treponema) pertenve — возбудителя фрамбезии.
Правда, опыты того же автора в последующем (1937) году дади отрица-
тельный результат. Известно из работ того же автора (1936, 1937), что
М. sorbens подозревается в переносе возбудителей туберкулеза и про-
казы. До сего времени, однако, достаточно доказательных подтвержде-
ний этому мы не имеем. Подобно комнатной мухе, М. sorbens известна
как вид, вызывающий у человека явление тканевого миаза. Автор лично
наблюдал личинок этого вида на ранах человека (Таджикистан).
Не подлежит сомнению, что базарная муха в условиях Средней
Азии имеет большое значение как распространитель фекального загряз-
нения на продукты питания, хозяйственную утварь и самого человека.
Роль мухи как механического переносчика кишечных инфекций пока
не изучена. Доказано, что М. sorbens является в условиях Средней Азии
переносчиком эпидемического конъюнктивита, вызываемого палочкой
Кох-Викса (Зимин, 1947; Зимин Парадоксов, Механикова, 1947).
*4. Musca albina Wd. (рис. 159).
Wiedemann, 1830, Aussereurop. zweiflug. Insekt., II : 415; Stein (apud Becker),
1912, Ежегодн. Зоол. муз. Акад. Наук, XVII : 629: Patton, 1922, Bull. ent. Besearch,
XII : 423; он же, 1923, Philipp. Journ. Science, XXIII : 333; Bezzi, 1923, Bull. Soc. ent.
2. MUSCA
121
Egypte : 108—118 (Lissosterna)', Malloch, 1925, Ann. Mag. Nat. Hist. (9), XVI : 369
(Lissosterna); Patton, 1932, Atm. trop. Med. Parasitol., XXVI : 387—388, fig. 16 (Lis-
sosterna); он же, 1933, там же, XXVII : 408; он же, 1936, там же, XXX : 480; он же,
1937, там же, XXXI : 133; Чеботаревич, 1937, Тр. Сов. изуч. произв. сил, сер. турк-
менок., 9 : 36; Но, 1938, XXXII : 287—312, — leckeri Schn. & Dz. 1911 : 325 (о). —
speculifera Bezzi, 1911, Bull. Labor. Zool. VI : 96. — polita Malloch.
Лоб самца уже или почти равен по ширине 3-му членику усиков;
голова черная, передняя ее часть, орбиты и узкие полоски за глазами
в шелковисто-белом налете; лобная полоса черно-бурая; скулы узкие,
почти равны или чуть шире 3-го членика усиков; 3-й членик усиков
бурый, в два раза длиннее 2-го; 2-й членик буровато-коричневый. Щу-
пальцы бурые; лобные щетинки кроме 1-й пары волосовидные; орбиты
в нежных редких волосках; внутренние теменные щетинки крепкие корот-
кие; затылочные щетинки в виде тонких коротких волосков; щеки в 1х/2
раза шире скул, в тонких коротких бурых волосках. Грудь блестяще
черная, плечи и проплевры в серебристо-белом налете, спинка, плевры
и щиток в черных волосках, более длинных и густых на плеврах и спинке;
переднее дыхальце рыжеватое. Щетинки груди: дорзоцентральных —
1 пара перед щитком, надкрыловых — 2, закрыловых — 2 крепких
и 1 маленькая щетинка между ними; плечевых — 2—3; нотоплевраль-
ных — 2; мезоплевральных — 1, крепкая, прочие волосовидные; на
щитке — основных 2, предвершинных — 2; гипоплевры с многочислен-
ными тонкими волосками впереди заднего дыхальца; стерноплевральные
щетинки отсутствуют. Ноги черные, голени коричнево-бурые, присоски
палево-белые, немного короче 5-го членика лапки., Крылья прозрачные,
сильно блестящие и ирйзирующие; жилки крыла бледножелтые, слабо
затемненные у вершины; чешуйки белые. Брюшко воскового желтого
цвета, в редких, коротких волосках. 5-й тергит и 4-й в средней трети
в густом шелковистом желтовато-сером налете, в боковых отделах 5-го
тергита располагается по одному круглому бурому пятну; 3-й и 4-й тер-
гиты с бурым пятнышком у средины заднего края; на 3-м тергите пятно
мельче и светлее чем на 4-м; 1 -|-2-й тергит целиком желтый или с малень-
кой коричневатой отметиной у середины заднего края. Гипопигий бурый;
V стернит маленький, короткий и широкий; свободный край церок слабо
скошен к середине, без резких выступов на внутренних углах; склеро-
тизованная часть фаллозомы почти равна по длине мембранозной части,
длинная; дорзальный отросток фаллозомы сильно расширен к вершине,
у вершины по бокам широко закруглен, посередине вершинного края
с выемкой и округлыми выступами по бокам от нее; передняя часть пара-
мер маленькая, с 1 длинной и 4—5 мелкими щетинками; задний отдел
парамер конусовидный, с округленной вершиной.
Самка резко отличается по окраске от самца; тело черно-бурое,
в густом сером и голубоватом налете, образующем рисунок из полос
и пятен. Глаза на темени расставлены на ширину, равную 2/з ширины
глаза; лобная полоса в 2 раза уже орбиты, бурая; лобные щетинки, кроме
крепкой передней пары, короткие, волосовидные; орбиты в очень корот-
ких негустых бурых волосках; скулы в 2 раза шире 3-го членика усиков
и чуть уже щек. Окраска отдельных частей головы как у самца, за исклю-
чением затылка, который у самки покрыт редким налетом. Внутренние
и наружные теменные щетинки крепкие, короткие. Грудь буро-черная,
в голубовато сером налете; стерноплевры черные, сильно блестящие,
среднеспинка с 2 широкими бурыми полосами, раздвоенными перед попе-
речным швом и постепенно суживающимся назад; щетинки как у самца;
122
I. ТРИБА MUSCINI
передние бедра по задней поверхности в шелковистом сером налете;
в остальном окраска ног как у самцов. Брюшко черно-бурое; основная
половина 14-2-го тергита, боковые передние отделы 3-го, 4-го и 5-го тер-
гитов темные, красновато-желтые, вентральная поверхность брюшка
палево-желтая, с темными отметинами. Передняя половина 3-го тергита,
исключая средину его, которая занята черной полосой, весь 4-й тергит,
кроме узких боковых полос и тонкой средней линии, и весь 5-й тергит,
кроме двух черных круглых боковых пятен, — покрыты густым желто-
вато-серым налетом. Непокрытые налетом участки брюшка блестящие,
черно-бурые или темнокрасно-желтые (основание 14-2-го тергита). Длина
тела 3.8—6 мм.
Распространение. Вид известен из Эфиопской и Ориен-
тальной областей. В Палеарктику заходит лишь в западной части ареала
вдоль южной границы Средиземноморской подобласти; Северная Африка,
Сирия, Палестина, Ирак, Иран; найден в сев.-зап. Индии. В СССР вид
известен из Туркмении (Кушка, Мерв, Красноводск, северные склоны
Копет-дага, Гассан-кули, Репетек, Фараб, Имам-баба, Уч-аджи, Ахча-
куйма, Кара-кала, Ашхабад, Джебел), Узбекистана (Чангир Хатырчин-
ского р-на, Бага-абзал, Ходжа-давлет), юго-восточного Закавказья
(Ленкорань) и из юго-западной Украины (г. Алешки). Ксерофильный
вид, связанный с зоной песков и эфемеровых полупустынь.
Экология вида не изучена. В районе Ашхабада личинки
М. albino, были обнаружены 3 VII в норе черепахи (Testudo horsjieldi
Cray); через 5 дней личинки в лабораторных условиях дали мух;
об исключительной термофильности личинок можно судить по темпера-
туре почвы (на глубине 20 см), отмеченной в день нахождения этих личи-
нок: 7 ч.—31.5°, 13 ч. — 32.0°, 21 ч. — 36.2° С (данные Я. П. Власова).
Обособление М albina в отдельный род, как предложил Бецци (1923),
и даже подрод (Пэттон, Мэллок, Хо и др.) мы не считаем целесообраз-
ным, т. к. отсутствие етерноплевральных щетинок — признак «толь
изменчивый в пределах трибы Muscini и рода Musca — не представляется
нам для . этого достаточным, тем более, что в морфологии прочих частей
тела, включая сюда и строение мужских гениталий, не обнаруживается
-сколь-либо резких отличий от близких видов. III.
III. Группа Musca vitripennis Mg.
Лоб самцов по ширине равен 3 членику усиков или уже его, глаза
в волосках или голые. Проплевры, супрасквамальное ребро и 1 стернит
брюшка без волосков. Грудь с 4 темными полосами на спинке или без них.
Четыре вида этой группы обнаружены в СССР: М. vitripennis Mg.,
М. osiris ~Wd., М. tempestiva Fall., M. lucidula Bzz. Один вид (M. crassi-
rostris Stein.) вероятен у южных границ Туркмении, Узбекистана и Тад-
жикистана.
*5. Musca vitripennis Mg. (рис. 117, 155, 156, 206).
Meigen, 1826, System. Beschr., V : 78; S£guy, 1923 : 358, рис. 769—771; Seguy,
1933, Memor. estud. Mus. Zoolog. Univ. Coimbra, ser. I, n° 67 : 62; Patton, 1925, Phi-
lipp. Journ. Science, XXVII : 197.
Экология. Порчинский, 1910, Тр. Бюро энт., VIII, 8 : 34, 35; Дербенева-
Ухова, 1940, Мед. паразит, и паразит, болезни, IX : 337; Зимин, 1948, Проблемы
кишечных инфекций.
Глаза в светлых длинных волосках; ширина лба в Р/л—2 раза более
.ширины 3-го членика усиков или равна последней; лобная полоса в I1/»
2. MUSCA
123
раза шире орбиты или равна ей; скулы немного шире 3-го членика уси-
ков, высота щеки в 4—41/г раза менее вертикального диаметра глаза;
3-й членик усиков в 2Чг—3 раза длиннее 2-го, бурый; щупальцы слабо
утолщенные к дистальному концу; голова черно-бурая, затылок вверху
почти лишен налета, внизу сзади в редком сероватом, за глазами в густом
светлосером налете; скулы и орбиты в серовато-белом густом налете,
несколько более темном на орбитах; щеки в сером налете, более разви-
том в их передней части. Внутренние теменные щетинки и вибриссы
более или менее крепкие, длинные; лобные щетинки расположены
на каждой орбите в 2 — реже 3 ряда, близко стоящих друг к другу;
щетинки наружного ряда располагаются против интервала между щетин-
ками внутреннего ряда; лобные щетинки, кроме первой крепкой, корот-
кой пары, имеют характер длинных тонких волосков; затылочные ще-
тинки — волосовидные. Щеки в густых, тонких (сравнительно с М. osi-
ris) длинных черных волосках, передняя верхняя граница распростра-
нения которых несколько заходит за линию, проведенную от нижнего
края глаза к угловой вибриссе. Грудь синевато или оливково-черная,
блестящая, с очень слабо выраженным сероватым налетом на плевраль-
ных отделах; переднегрудь в мелких волосках, передние плевры, супрас-
квамальные ребра и гипоплевры впереди дыхальца без волосков; группа
щетинистых коротких волосков имеется лишь над тазиками задних ног.
Передние стигмы белые или светлопалевые. Щетинки груди тонкие,
длинные; акростихальных — 1 пара впереди щитка; дорзопентраль-
ных — 24-2 (передние пары позади шва отсутствуют); интраалярных —
1 позади шва; предкрыловых—1 (мелкая), надкрыловых — 1; закры-
ловых — 3; плечевых — 3; заплечевые не всегда явственны; предшов-
ная — 1; нотоплевральных — 2; стерноплевральных — 1 -|-2; на щитке
щетинки длинные: вершинных — 2, основных — 2; предосновная—1,
в виде тонкого короткого волоска; дискальных — 2, имеющих характер
тонких коротких щетиночек. Поверхность спинки в относительно тонких
бурых торчащих волосках. Крылья прозрачные, сильно блестящие,
иризирующие, с желтыми жилками; жилка т изогнута под тупым углом
(угол широко округлен); г4+5 с нижней стороны с несколькими мелкими
волосками при основании. Чешуйки белые или бледнопалевые по краю;
жужжальцы желтые. Ноги бурые; коготки почти такой же длины как
5-й членик лапки; присоски палевые. Переднедорзальные щетинки
на задних голенях развиты почти по всей длине голени; длина их почти
равна поперечнику голени. Брюшко широко яйцевидное, медово-жел-
тое, с темными отметинами. 1 4-2-й тергит черный или коричневый, кроме
нижней стороны и задне-боковых краев; 3-й тергит посредине с поло-
ской, ширина которой равна примерно Vs ширины тергита; 4-й тергит
с расширяющимися назад от середины переднего края треугольным
пятном и округлым пятном желтовато-белого налета; по середине зад-
него края — небольшое округлое блестящее черное пятно; 5-й тергит
с округлым бурым пятном, занимающим всю середину тергита, и пятном
желтоватого налета на том же месте; I стернит без волосков. Церки
с полукруглыми боковыми краями и более или менее резкими дисталь-
ными боковыми углами; внутренние отростки слабо выступающие, почти
прямоугольные; задние отростки V стернита тонкие, относительно длин-
ные; склеротизованная часть фаллозомы тонкая, длинная, суженная
у средины, дистальная — с 3 угловатыми лопастями по вентральному
краю; передняя часть парамер широкая, в виде лопасти, на верхнем
крае со щетинками (одной крупной и несколькими мелкими; число послед-
124
I. ТРИБА MUSCINI
них изменчиво; обычно их 2—3); задний отдел нарамер конусовидный
с склеротизованным основанием в виде колечка и мембранозной верши-
ной; дорзальный отросток фаллозомы расширен к концу, полукруглый
на вершине.
Рис. 155—162. Головы видов рода Musca.
155 — самца М. vitripennis Mg. сверху. 156—самки М. vitripe.nnis Mg. сверху.
157 — самца М. osiris Wd. сверху. 158 — самки М. osiris Wd. сверху. 159 —
самки М. albino. Wd. сверху. 160 — самки М. lucidula Lw. сверху. 161 —
самки М, tempestiva Fall, сверху. 162 — самка М. crassirostris Stein сбоку.
(Ориг.).
2. MUSCA
125
Лоб самки немного уже глаза; глаза в коротких очень редких волос"
ках; ширина выпуклой по бокам лобной полосы в Р/а—2 раза более
ширины орбиты; скулы в l1/* раза шире 3-го членика усиков; на скулах
и орбитах густой желтовато-серый налет; затылок в редком желтовато-
сером налете, как и щеки. Внутренние и наружные теменные щетинки
и вибриссы крепкие, относительно длинные; орбиты в коротких черных
волосках; по бокам от лобных щетинок располагается в среднем и зад-
нем или только в заднем отделе орбит несколько длинных щетинистых
волосков, длина которых почти равна длине лобных щетинок в этом
месте, направленных вершинами к продольной средней линии головы.
Грудь в густом сером, зеленовато или голубовато-сером налете, с 4 про-
дольными узкими, темными полосками на спинке, из которых средние
уже и короче боковых. Брюшко бурое, в сером или бронзово-сером налете,
блестящее, тергит 1 -|-2-й, исключая его задне-боковые отделы, бурый;
3-й тергит со срединной параллельно-крайней или слабо расширенной
назад бурой полоской; 4-й тергит с более узкой полоской того же цвета.
Длина тела 3.2—6.2 мм.
Распространение вида точно не может быть установлено
в связи с тем, что вид этот смешивался с М. osiris Wd., распространение
которого более широко, чем М. vitripennis. На территории СССР М. vitri-
pennis известен по коллекционному материалу из нижеследующих мест:
Курская область; Украина: Одесса, Аскания-Нова, Проскуров, Коро-
стышев, Ярески и Супоевка, Харьков, Бердянск, Мелитополь; Таганрог;
Крым (южный берег); Черноморское побережье Кавказа; Армения:
Ереван; Азербайджан: Ленкорань, Ордубад; Туркмения: Ашхабад. Вид
заходит в северный Иран (Тавриз).
Вид связан с пометом крупного рогатого скота. Личинки — копрофаги.
*6. Musca osiris Wd. (рис. 116, 157, 158, 163—169, 207).
Wiedemann, 1830, Aussereur. zweifliig. Insekt., II : 420; Patton, 1925, Philipp
Journ. Science, XXVII : 192; он же, 1932, Ann. trop. Med. Parasitol., XXVI : 347 —
405, рис. 23 (vitripennis Patt. nec Meig.); Seguy, 1933, Memor. Mus. Zool. Univ. Coimbra
(1) 67 : 62.
Экология. Чеботаревич, 1937, Тр. изуч. произв. сил, сер. туркменок.,
9 : 36 (М. vitripennis)', Вайнштейн и Родова, 1940, Мед. паразит, и паразит, болезни,
IX : 364—368 (М. vitripennis)', Зимин, 1948, Опред. лич. : 89, фиг. 49; он же, 1944,
Пробл. кишечн. инфекц., Сталинабад : 154.
Близок к предыдущему, от которого отличается следующими при-
знаками.
Ширина лба самца в два раза уже или равна ширине 3-го членика
усиков; лобные щетинки волосовидные, редкие, расположены на каждой
орбите в 1 ряд, иногда несколько передних пар сдвоены; волоски на щеках
короткие, редкие в переднем отделе и обычно не поднимаются за линию
нижнего края глаза. Налет на скулах и орбитах густой, белый, шелко-
висто-блестящий. Волоски на глазах короткие и редкие. Лобная полоса
линейная, никогда не бывает шире орбит, часто розоватая, или сплошь
покрыта серым налетом, реже черная. г4+5 «низу, как правило, без воло-
сков у основания. Церки сходны с таковыми М. vitripennis, отличаясь
мелкими деталями строения.
22 отличимы от самок М. vitripennis по отсутствию вблизи лобных
щетинок длинных, направленных внутрь черных волосков; орбиты несут
лишь короткие, почти равной длины черные волоски, длина которых
много менее таковой лобных щетинок. Щеки в редких, коротких черных
волосках, а не длинных и густых, как это имеет место у М. vitripennis.
126
I. ТРИБА MUSCINI
В остальном отличия от М. vitripennis сводятся к меньшей величине
тела и несколько более светлой окраске налета. Длина тела 3.0—5.5 мм.
Яйцо белое, сравнительно крупное, дл. 1.7—1.9 мм, шир. 0.4—
0.5 мм. Хорион матовый (скульптура на микроскопических препаратах
не видна), тонкий. Яйцо выпуклое по одной и слабо вогнутое по другой
стороне; к полюсам сужено, к переднему несколько сильнее, нежели
к заднему; две продольные бороздки очень нежные, не всегда хорошо
различимы на спиртовом материале.
Личинка I стадии белая; тело удлиненно цилиндрическое, слабо
сужено к переднему концу, где узко округлено; брюшные сегменты,
со 2-го по 7-й, снизу со слабо выступающими валикообразными попереч-
ными выпуклостями, по переднему краю сегментов вооруженными мно-
горядными мелкими шипиками; задние бледнорыжие дыхальцы с одним
отверстием. Ротовой аппарат сходен с таковым следующего вида. Средин-
ный дорзальный склерит под дистальным заостренным зубцом с малень-
ким добавочным зубчиком. Длина тела 1.5—1.8 мм.
Личинка II стадии белая с серым просвечивающим через наружные
покровы кишечником. Передние дыхальцы с 6—7 лучами; задние
дыхальцы бледнорыжие, каждое с двумя короткими щелями, более
длинными, чем у М. tempestiva-, нижняя щель длиннее верхней, в среднем
отделе выпуклая со стороны перитремы. Ротовой аппарат как у личи-
нок II стадии М. tempestiva, но с более тонкими ротовыми крючьями;
над ротовым отверстием расположено 6 зубцов, направленных верши-
нами назад; два внутренних зубца крупнее остальных. Длина тела 2.5—
3.0 мм.
Личинка III стадии (рис. 163—169) имеет полушаровидно округ-
ленный задний конец тела; шипики впереди и позади анального бугорка
есть; два конусовидных субанальных выроста; короткие полосы из шипи-
ков на средних брюшных сегментах достигают верхними концами средины
боковых отделов сегмента, в то время как у личинок М. tempestiva они
не распространяются далее вентральной стороны сегментов. Передние
дыхальцы с 7 лучами; задние дыхальцы с сильно склеротизованной,
широкой перитремой; щели сравнительно широкие, верхняя и нижняя
щели имеют по 4, средняя 3 изгиба; наружные концы верхней, нижней
и оба конца средней шели направлены наружу, прочие — к центру
дыхальцевой пластинки. Анальная пластинка и ротоглоточный аппарат
как у личинок М tempestiva, но с несколько удлиненными передними
отростками базального склерита; дистальные концы их выдвинуты далее
вперед, чем верхний угол базального склерита. Длина тела 6—8 мм.
Ложнококон красновато-коричневый или рыже-коричневый, глад-
кий, бочонкообразный, длиною 4—4.5 мм, диаметр 1.8—2 мм. По зад-
нему вентральному концу, сохраняющему признаки личинки III стадии,
легко отличается от пупариев М. tempestiva, более светлых и мелких.
Распространение. Вид распространен повсеместно в Эфи-
опской области, откуда заходит в южную Европу и проникает через
Аравию в Иран. Отсюда вид распространяется на восток. Северная его
граница проходит по территории СССР, южная — по Кашмиру и по
северной Индии, откуда поднимается к северным рубежам Китая
и смыкается здесь с северной границей ареала. В пределах СССР запад-
ная и северная границы ареала идут по восточному побережью Каспий-
ского моря, вверх по р. Уралу до г. Чкалова и далее на север до Уржума
(Кировской обл.). От этих границ ареал вида идет на восток, охватывая
всю Среднюю Азию и Казахстан на север до границы лесостепной звмм-
2. MUSCA
127
Из Сибири вид не известен, появляясь только в южном Приморье, куда
проникает вдоль Сихотэ-алиня из Манчжурии. Вид обычен в Монголии.
Рис. 163—170. Детали строения личинки III стадии Musca osiris Wd.
163 — задний конец тела сзади. 164 — ротоглоточный аппарат. 165 — он же сбоку.
166 — переднее дыхальце. 167 — заднее дыхальце. 168 — вооружение снизу 5-го (2}
и 6-го (1) брюшных сегментов. 169 — перианальная область: 1 — анальная пластинка;
2 — субанальные бугорки; 4 — анальные бугорки. -170—скульптура средне!!
части дорзальной поверхности 2-го грудного сегмента. (По Зимину).
Экология. Образ жизни М. osiris специально не изучался.
Известно (Чеботаревич, 1937; Вайнштейн и Родова, 1940; Зимин, 1944),
что развитие личинок происходит в помете животных; преимущество
отдается коровьему, свиному и ослиному помету. Вылупление личинок
в условиях Гиссарской долины (Зимин) наступает через сутки. При
128
I. ТРИБА MUSCINI
средней суточной температуре 25.7° С весь цикл развития от момента
откладки яйца до вылета мухи занимает (по нашим данным) 10 дней.
Мухи являются обитателями открытых городских и сельских биото-
пов. Обычны на базарах, во дворах, бойнях и других местах, где наблю-
дается скопление животных. Мухи относятся к группе кроволижущих
видов. Обычны на животных (вокруг глаз, носа и на спине), где питаются
слизистыми выделениями, потом и кровью с пораненных участков кожи.
Нередко нападают на человека. М. osiris — гелиофильный вид; в поме-
щения человека залетают редко и в единичных экземплярах.
Как многие другие виды рода, связанные с животными, М. osiris
в южных районах СССР, вероятно, имеет значение распространителя
возбудителей различных заболеваний; с этой стороны вид остается неизу-
ченным.
*7. Musca tempestiva Fall. (фиг. 161, 170—180).
Fallen, 1823, Muse. ; 53; Bezzi u. Stein, 1907, Catal. pal. Dipt., Ill : 604; Mal-
loch, 1925, Ann. Mag. Nat. Hist. (9), XVI : 372 (Biomyia); Patton, 1937, Ann. trop.
Med. Parasitol., XXXI' : 133; Ho, 1938, XXXII : 287—312, fig. 10; Tiensuu, 1935,
Acta Soc. Fauna Fl. Fenn., LVIII, n° 4 : 54.
Экология и стадии развития. Порчинский, 1910, Тр. Бюро
антом., VIII, 8 ; 34, фиг. 36; Чеботаревич, 1937, Тр. сов. изучен, произв. сил, сер.
туркменок., 9 : 37; Thomsen, 1938 : 309—310; Дербенева-Ухова, 1940, Мед. паразит,
и паразит, болезни, IX; 324, 337; Романов, 1940, там же: 356, 357, 361, 362, фиг. 2;
Зимин, 1948, Опред. лич. : 86, фиг. 46.
$. Глаза голые; лоб узкий, ширина его равна ч/з—Vz ширины 3-го
членика усиков; ширина скул в среднем отделе равна поперечнику
3-го членика усиков; высота щеки составляет Ve—1/ю вертикального
диаметра глаза; 2-й членик усиков в 2 раза короче 3-го; усики, как
и щупальцы, коричнево-бурые, первые в редком сероватом налете; ариста
утолщена в основной трети. Хоботок короткий. Голова черно-бурая;
затылок, исключая его блестящий верхний отдел, в редком сероватом,
орбиты и скулы в густом серовато-белом, щеки в редком сером налете,
как и лицо. Щеки в черных, коротких, затылок в нижнем отделе в более
длинных волосках того же цвета. Внутренние теменные щетинки и виб-
риссы крепкие, длинные; лобные щетинки тонкие, волосовидные в сред-
нем и заднем отделе лба, впереди слабо утолщенные, располагаются
в один ряд с каждой стороны лобной полосы; последняя в виде линии
или очень узкой черной полоски. Грудь черная, блестящая, в редком
сероватом налете, местами с коричневатым отблеском; переднегрудь
в мелких волосках; передние плевры и супрасквамальные ребрышки
без волосков: гипоплевры с несколькими волосками в нижнем отделе.
Передние дыхальцы бурые или коричневые. Щетинки груди: акрости-
хальных — 1—2 пары (в последнем случае передняя слабо развита);
дорзопентральных — 2 -[-4 (из этой группы 2 передние пары позади шва
короче остальных); интраалярных (за швом) — 1; предкрыловая — 1 мел-
кая; надкрыловая — 1; закрыловых — 3; плечевых — 3; заплечевая —
1; предшовная — 1; нотоплевральных — 2; стерноплевральных—l-f-2;
на щитке: вершинных — 2; основных — 2; предосновных — 2 и 1 пара
дискальных очень коротких; боковые щетинки щитка отсутствуют.
Крылья прозрачные, иризирующие; жилки бледножелтые, на вершине
крыла серовато-желтые; т изогнута под тупым углом (угол закруглен-
ный); г4+5 без волосков; чешуйки белые, с бледнопалевым краем или
сплошь белые; жужжальца желтые. Ноги черно-бурые или коричнево-
<бурые с обычной для рода хетотаксией. Коготки рыже-коричневые с тем-
2. MUSCA
129
ной вершиной, немного короче 5-го членика лапки. Брюшко черно-бурое,
коротко яйцевидное, в сравнительно густом сером или коричневато-
сером налете и черных волосках; 1 стернит голый; по дорзальной поверх-
ности брюшка проходит суженная назад бурая срединная полоска,
обычно не распространяющаяся далее заднего края 4-го тергита; задние
края 3-го и 4-го тергитов буроватые. Церки широкие; внутренние углы
вершинного их края очень слабо оттянутые, не образующие, однако,
кольцевидных отростков; боковые лопасти нередко с угловатыми высту-
пами; дистальный край очень слабо вогнутый или прямой, несколько
скошенный от середины к боковым отделам; основная часть фаллозомы
узкая, длинная, равна или несколько длиннее вытянутого, неровного
по вентральной стороне мембранозного отдела; дорзальный отросток
короткий, слабо расширенный к вершине, на дистальном крае прямой;
передняя часть парамер поперечно вытянутая, с 2 щетинками по краю;
задняя часть парамер имеет форму сахарной головы, вершинная треть
прозрачная, слабо склеротизованная; аподема утонченная к вершине,
длинная. V стернит с тонкими длинными, загнутыми внутрь отростками
ио заднему краю; наибольшая ширина стернита равна его длине.
j. Лоб немного уже глаза; лобная полоса черная или красновато-
желтая в переднем отделе, с выпуклыми боковыми краями, в 3—ЗДз раза
шире узких орбит; ширина скул равна ширине 3-го членика усиков;
щеки как у самца. Наружные и внутренние теменные щетинки и вибриссы
длинные; затылочные и лобные щетинки сравнительно длинные, тонкие;
орбитальные в виде ряда мелких, направленных вершинами вперед щети-
ночек, из которых задняя длиннее передних. Затылок за глазами в более
густом, чем у самца, налете. Налет на скулах и орбитах густой, серова-
тый или с легкой желтизной. Грудь в сером, более густом, чем у самца,
налете, на спинке с четырьмя ясно различимыми узкими продольными
темными полосами; средние полосы отстоят друг от друга примерно
на двойную свою ширину и заканчиваются посредине заднего отдела
спинки позади шва; боковые темные полосы шире и длиннее средних.
Щетинки — как у самца. Брюшко с темным треугольным пятном на 3-м
тергите; иногда этот тергит бурый, кроме небольших участков, покры-
тых серым налетом в боковых передних отделах; 1 ф-2-й тергит на боль-
шем протяжении бурый; 4-й тергит с бурым задним краем и узкой, рас-
ширенной кпереди срединной-полоской. Длина тела 2.5—4.2 мм.
Яйпо белое, удлиненное (длина в 5—6 раз более ширины), по вен-
тральной стороне более выпуклое, чем по дорзальной, с двумя слабо
выраженными, узко расставленными продольными бороздками; ячеистая
скульптура хориона не выражена. Длина 1.2—1.6 мм.
Личинка I стадии (рис. 170) белая с легкой желтизной; задние
дыхальцы рыжие, с одним округлым отверстием; шипики располагаются
по нижней и нижне-боковой поверхности брюшных сегментов; ротогло-
точный аппарат обычного для рода строения; срединный дорзальный
склерит удлиненно листовидный, прямой по верхнему и выпуклый по сере-
дине нижнего края, вершина его заострена; нижний край без зубчи-
ков; гипостомальные склериты в виде двух параллельных дуговидно
слабо изогнутых пластинок, соединенных тонкой перемычкой; фарин-
геальные склериты с широкорасставленными друг от друга дуговидными
дорзальным и вентральным (задними) отростками; дорзальная пере-
мычка высокая, слабо расширена в дорзальной части; передние отростки
фарингеальных склеритов тонкие, длинные, параллельные друг другу.
Длина тела 1.5—1.8 мм.
9 Фауна СССР, том XVIII, вып. 4.
130
I. TP ИВ A MUSCINI
Личинка II стадии (рис. 171, 173) желто-белая, с темносерым про-
свечивающим через наружные покровы кишечником; задний конец тела
широко закруглен, неправильно полущаровидный, с выступающими
дыхальцами; нижняя поверхность брюшных сегментов со слабо высту-
пающими поперечными валиками по переднему краю, покрыты мелкими
шипиками, не заходящими на дорзальную поверхность сегментов. Перед-
ние дыхальцы с 7 лучами; задние дыхальцы светлоокрашенные; равстоя.
Рис. 170а, 171. Ротоглоточный аппарат личинки Musca
temptstiva Fall.
170а — I стадии. 171 — II стадии.
Рис. 172. Общий вид взрослой личинки Musca
tempestiva Fall.
Рис. 173. Задние дыхальцы личинки II стадии Musca
tempestiva Fall. (Ориг.).
ние между ними равно поперечнику дыхальца; дыхальцевые щели широ-
кие, нижняя слабо изогнута в середине, длиннее верхней. Ротовые крючья
тонкие, почти прямые, слабо утончены к вершине и здесь заострены;
под ними располагается 2 хитиновых тяжа, отсылающих по бокам рото-
вого отверстия треугольные, заостренные книзу лопасти, не резко зазуб-
ренные по переднему краю. Угловидный склерит из двух S-образно согну-
тых тонких хитиновых тяжей; гипостомальные склериты удлиненные,
с оттянутым назад дорзальным верхним углом, сочленяющимся с перед-
ними отростками базального склерита. Последний длинный, дорзальные
задние его отростки немного тоньше вентральных, скошенных сверху
2. MUSCA
131
вниз на вершине; длина отростков почти равна длине основной части
склерита; дорзальная перемычка расширена в средине, с крупными
отверстиями; передние отростки фарингеальных склеритов тонкие, удли-
ненные, на вершине косо срезаны вниз и сзади наперед, внизу соединены
вентральной перемычкой, отсылающей назад несколько тонких выростов.
Длина тела 2.5—3 мм.
Личинка III стадии (рис. 172, 174—180) желтая с бурооливковым
просвечивающим кишечником; брюшные сегменты снизу с поперечными
валиками по переднему краю; 2-й и 3-й грудные сегменты по переднему
краю снизу с мелкими прозрачными плохо заметными шипиками; вали-
кообразные утолщения брюшных сегментов с крепкими многорядными
шипами; задний конец тела полушаровидный, со слабо выступающими
задними дыхальцами, расставленными на :/з—‘/д поперечника дыхальца;
анальная пластинка более или менее широкая, по середине переднего
края легко дуговидно вогнутая, по тому же краю в боковых отделах
слабо выпуклая; контур заднего края негативный по отношению к перед-
нему краю пластинки; впереди последней расположена узкая короткая
полоса шипиков; сосцевидные выросты по задней поверхности анального
сегмента, равно как и шипики позади анальных маленьких бугорков
отсутствуют. Боковые концы анальной пластинки более или менее широко
округлены и не достигают средины боковой поверхности тела личинки;
передние дыхальцы с 6—8 лучами; задние дыхальцы крупные, с рыже-
ватой пластинкой и узкой черной перитремой, стигмальный диск круп-
ный, с одним узеньким, лежащим в центре диска белым просветом;
щелей 3, глубоко изогнутые, толстые; каждая щель имеет по 4 изгиба;
наружные концы верхней и нижней и верхний конец средней щелей
направлены к перитреме, прочие — внутрь пластинки; периферия пла-
стинки с 6—7 периспиракулярными железами. Ротоглоточный аппарат
имеет ту же схему в строении, как и у других видов рода. Ротовые крючки
сильные, широкие, изогнутые, несколько притупленные на вершине;
один крючок короче другого; расширенное основание их с просветом
в вентральной части; зубные склериты глубоко выемчатые по верхнему
и заднему краю; гипостомальные склериты маленькие, базальные —
очень длинные и широкие; передние отростки очень короткие, широкие,
дуговидно вогнутые по переднему краю; дорзальная перемычка решет-
чатая, глубоко входит в стенки базальных склеритов, почти не выдаваясь
за их края, с 2 узкими длинными отростками сзади; длина базального
склерита немного более длины его задних отростков; последние широкие;
вентральные отростки перед срединой с высоким полукруглым выступом;
дистальные концы отростков постепенно сужены. Фарингеальные реб-
рышки резкие темные; вентральная перемычка базального склерита
с полукругло изогнутыми в вертикальной плоскости отростками. Длина
тела 4.5—8 мм.
Пупарий широко закругленный на заднем и широко конусовидно»
суженный на переднем конце, блестящий, гладкий, с хорошо различимой
сегментацией; задний конец тела сохраняет признаки личинки послед-
ней стадии. Длина 2.3—3.2, ширина 1.3—1.5 мм.
Распространение. Один из наиболее широкораспростра-
ненных в Палеарктике видов рода, заходящий и в Эфиопскую область.
В СССР встречается от широты Ленинграда до южной границы Европей-
ской части СССР и по всей Средней Азии и Казахстану; в Сибири изве-
стен от среднего и южного Урала до Забайкалья. В Зап. Европе известен
на север до средней Скандинавии; указан для палеарктической Африки,
q*
132
I. TPHBA MUSCINI
Ирана, Кашмира, северной Индии, северного и центрального Китая;
по Пэттону, широко распространен в Африке (1936).
Биология вида изучена недостаточно. Появляются мухи ран-
ним летом. В южных широтах (Таджикистан) наибольшая численность
180
Рис. 174—180. Детали строения личинки JII стадии Musca tempcstwa Fall.
174 — ротоглоточный аппарат. 175 — задний конец тела сзади. 176 —он же сбоку.
177 — заднее дыхальце. 178 —переднее дыхальце. 179 — шипики в переднем отделе
2-го грудного сегмента на дорзальной его стороне. 180 — вооружение снизу на 5-м (2)
и 6-м (7) брюшных сегментах. (По Зимину).
особей в природе наблюдается в июне; далее к осени количество мух
постепенно сокращается. По данным Романова (1940), зараженность
навоза личинками в 2.7 раза выше в июне, нежели в июле (южн. Тад-
жикистан). Откладка яиц происходит в коровий помет в утренние часы;
реже используется конский помет. Яйца откладываются группами
по нескольку штук в свежий субстрат на глубине 1—2 мм. Развитие
личинок в помете при 27—28° С заканчивается в 3 дня, после чего
2. MUSCA
133
личинки либо уходят для окукления в почву, либо остаются в помете,
где образуют уплотненный тонкий кокон из частичек помета. В этом
случае ложнококон оказывается заключенным в оболочку из сухих
частиц помета. Перед окуклением в почве личинки углубляются на 0.5—
2—3 см от ее поверхности под высыхающую лепешку помета, обычно
не расползаясь в стороны. Через 3—7 дней после образования пупариев
происходит вылет мух.
В условиях Средней Азии серьезную роль в размножении М- tem-
pestiva играет заготовка навоза на топливо (кизяки); заселение их личин-
ками достигает в июле весьма высоких цифр, измеряясь несколькими
сотнями особей на 1000 см3 массы субстрата.
М. tempestiva является кроволижущим видом, имеющим поперемен-
ный контакт то со здоровыми, то с больными животными, в связи с чем
можно предположить, что. М. tempestiva участвует в распространении
патогенных микробов. М. tempestiva на пастбищах крайне досаждает
животным. Ограничивающим фактором в размножении М- tempestiva
являются хищные личинки мух {Myiospila meditabunda F., Anaclysta
multipunctata Stein) (Зимин, 1944), муравьи и жуки-карапузики (Рома-
нов, 1940).
*8. Musca lucidula Lw. (рис. 160, 181—190).
Loew, 1856, Neue Beitr. Kenntn. Dipt., IV : 48 (Cyrtoneura)’, Bezzi, 1923, Bull.
Soc. ent. Egypte : 108—118; Patton, 1932, Ann. trop. lifed. Parasitol., XXVI : 391,
392, fig. 20; он же, 1933, XXVII : 414.
Биология. Чеботаревич, 1937, Тр. сов. изуч. произв. сил, сер. туркменск.,
9 : 37; Зимин, 1944, Пробл. кпшечн. инфекц. : 174—192; Зимин, 1948, Опред. личи-
нок : 89, фиг. 48.
<5. Лоб узкий, ширина его менее ширины 3-го членика усиков; лоб-
ная полоса в виде бороздки (представляет линию срощения орбит); скулы
шире лба и в 2 раза уже щек; 3-й членик в два раза длиннее 2-го, длина
его во столько же раз более ширины членика. Голова черная, орбиты,
скулы и узкая полоска за глазами в нижней половине головы — в густом
шелковисто-белом налете, лицо в темносером налете; щеки блестяще-
черные, в коротких черных волосках; затылок черный, с зеленоватым
металлическим отливом, щупальцы бурые; лобные щетинки, включая
и переднюю пару, очень короткие, редкие, волосовидные; вибриссальные
и внутренние теменные щетинки крепкие. Усики бурые, 2-й членик
на вершинной половине буровато-коричневый. Грудь блестящая, метал-
лически зеленая; проплевра и спинка перед поперечным швом в редком
светлосером налете; впереди спинка с 4 узенькими полосками, по бокам
попарно сближенными и широко разделенными посредине. Грудь в корот-
ких, редких, щетинистых волосках; щетинки груди: акростихальных —
1 пара перед щитком; дорзоцентральных — 3 пары в заднем отделе
спинки; длина их возрастает спереди назад; плечевых — 2; предшоь-
ных — 1; надкрыловых по 2 с каждой стороны; на мезоплевре развита
лишь одна щетинка у ее заднего края; стерноплевральных щетинок —
l-f-2; гипоплевры голые. Ноги смоляно-бурые, первые членики средних
и задних лапок темножелтые; присоски белые, в 1% раза короче 5-го чле-
ника лапки. Чешуйки белые, жужжальца светложелтые; крылья про-
зрачные, блестящие, иризирующие, жилки крыла светложелтые в основ-
ной и бледно серовато-желтые в дистальной половине; ячейка 7?в открыта
или замкнута на краю крыла. Брюшко широко яйцевидное, желтое,
в коротких редких черных волосках; 5-й тергит посредине широко затем-
134
I. ТРИБА MUSCINI
Глаза j на темени
нен, 4-й тергит посредине заднего края с маленьким коричневатым пят-
ном; гипопигий бурый, V стернит коричневатый в вершинной половине.
Брюшко блестящее, 5-й и 4-й тергиты сверху посредине в густом белом
налете. 1 стернит без волосков. Церки по боковым краям неправильно
округлены, внутренние углы их без ясных выростов. Хитиновая часть
фаллозомы по длине равна мембранозной части, в середине слабо сужена;
задний отросток фаллозомы расширенный; передний отдел парамер длин-
ный, изогнутый, с одной длинной и 4—5 мелкими щетинками; задняя
часть парамер с длинным узким пальцевидным выростом, как у М. cras-
sirostris St., V стернит маленький, полукруглый, с 2 тонкими длинными
зазубренными на вершине боковыми отростками по заднему краю.
Глаза J на темени расставлены на ширину, равную 2/з ширины глаза;
лобная полоса узкая, с прямы-
ми краями, суженная постепен-
но к основанию усиков; шири-
на лобной полосы в средине
равна или менее ширины орби-
ты; скулы шире, чем у самца,
равны ширине орбиты и в Р/г
раза уже щеки; окраска головы
как у самца, налет более тус-
клый; лобная полоса атласно-
черная; лобные щетинки ко-
роткие, редкие, волосовидные;
внутренние и наружные темен-
ные щетинки хорошо развиты.
Грудь блестяще зеленая; спин-
ка и проплевры в редком све-
тлосером налете, среднеспинка
с 4 узкими полосками основ-
ного цвета спинки; полоски не-
ясные, проходят назад далее
поперечного шва; присоски на
ногах в 2 раза короче 5-го
членика лапки. Брюшко метал-
лически зеленое или фиолето-
Рис. 181. Самец Musca lucidula Lw. (Ориг.). вое, с желтым основанием или
основной половиной и "нижней
поверхностью, в редком, не
всегда отчетливо выраженном, сероватом налете. Длина тела 2.5—4 мм.
Личинка III стадии (рис. 183—190). Близкий к Musca tempestiva
вид, отличающийся от него следующими морфологическими особенно-
стями. Шипики на переднем крае грудных сегментов располагаются,
как правило, в 6—7 рядов, менее тесно сближенных. 1-й брюшной сег-
мент сверху по переднему краю с 2—4 рядами шипиков, чего нет у М. tem-
pestiva. Валикообразные вздутия на переднем крае средних и задних брюш-
ных сегментов по всей длине передней стороны с 2—3 рядами редких
разбросанных шипиков; количество бугорков на заднем крае брюшных
сегментов в 2 раза больше, нежели у М. tempestiva и достигает 12—18 штук
(по 6—9 с каждой стороны от средней продольной линии сегмента). Ниж-
ний край 8-го брюшного сегмента (позади анальной пластинки) с 1—2
рядами из 5—10 шипиков. Передние дыхальцы с 5—6 округлыми выро-
стами; задние дыхальцы с короткими щелями, расположенными на свет-
2. MUSCA
135
лых полях стигмальной пластинки; щели своими концами направлены
как и у М. tempestiva. Ротоглоточный аппарат с более широкими фарин-
геальными склеритами и более широкими дорзальными отростками;
ротовые крючья с более короткой базальной частью. Длина тела 5—
6.7 мм; толщина в середине 1.2—1.5 мм.
Распространение. На территории СССР вид обнаружен
только в Средней Азии: Туркмении (Ахча-куйма, Ак-яйла, Баят-ходЖи
на р. Атреке, Кушка, Джебел), Узбекистане (Хатырчи, Чангир, Бага-
абзал) и различных пунктах южного и нейтрального Таджикистана (ст. Ха-
нака, Куляб, Модотовабад, Кировабад). За пределами СССР вид распро-
странен на крайнем юге Западной Европы (Сицилия), в северной и средней
Африке и в Передней Азии на восток до Иранского Белуджистана.
Вид развивается в помете
крупного рогатого скота. По
наблюдениям автора в южном
Таджикистане (Молотовабад),
М. lucidula появляется в при-
роде позже других калоядных
видов того же рода. Личинки
начинают встречаться единично
при анализе помета лишь со
средины июня до августа.
В этот же период численность
М tempestiva, обитающей в по-
мете вместе с М. lucidula, зна-
чительно превосходит таковую
последнего вида. В августе чис-
ленные соотношения названных
видов изменяются в противо-
положную сторону; одновре-
менно возрастает количество
М. lucidula в природе. Мухи от-
кладывают яйца на свежий ко-
ровий помет, предпочитая для
этой цели ярко освещенные и
хорошо прогреваемые выгоны и
его навоза, равно как и помет в
вида. В лабораторных условиях,
Рис. 182. Самка Musca lucidula Lw. (Ориг.).
проезжие дороги. Скопления коровь-
хлевах, не заселяются личинками этого
при 27—33°, С развитие мухи закан-
чивается в 8—9 дней, из которых 5—6 дней падает на фазу куколки.
При неблагоприятных условиях (пониженная влажность субстрата)
развитие куколок задерживается на срок до 10 дней. Весьма вероятно,
что в природной обстановке диапауза имеет, и более широкие пределы.
М. lucidula, по Пэттону (1933), кроволижущий вид, связанный
с позвоночными. В условиях Таджикистана М. lucidula встречается на ко-
ровах, верблюдах, реже на ослах, концентрируясь на слизистых оболоч-
ках носа, губ, глаз и на поранениях кожи, в связи с чем вид может быть
заподозрен в переносе заболеваний среди животных.
9. Musca crassirostris Stein, (рис. 4е, 162, 213).
Stein, 1903, Mitt. Zool. Mus. Berlin, II : 99; Austen, 1921, Bull, entom. Res.,
XXII : 120, 121; Patton, 1922, Bull, entom. Res., XIII : 411—426; он же, 1923, Phi-
lipp. Journ. Sci., XXIII : 311, 313, 320, 335; он же, там же, 1925, XXVII : 188, 189;
Malloch. 1925, Ann. Mag. Nat. Hist., XV : 132—135 (PhillaematomyiaY, Patton, 1932.
Vfmr
-wk *v ш
Рис. 183—190. Детали строения личинки III стадии Musca lucidula Wd.
183 — ротоглоточный аппарат. 184 — задний конец тела сзади. 185 —-он же сбоку.
186 — переднее дыхальце; 187 — заднее дыхальце. 188—перианальная область.
1 — анальная нластинка, 2 — субанальные бугорки; 4 — анальные бугорки. 189 —
шипики переднего отдела дорзальной стороны 2-го грудного сегмента. 196 — воору-
жение снизу на 5-м (2) и 6-м (7) брюшных сегментах. (По Зимину).
2. MUSCA
137
Ann. trop. .Mod. Parasitol., XXVI : 387, 391, figs. 18, 19; он же, там же, 1933, XXVII :
412, fig. 9; Malloch, 1932, Ann. Mag. Nat. Hist., (10), IX : 515 (PhillaematomyiaY, Pat-
ton, 1936, Ann. trop. Med. Parasitol., XXX : 480; он же, там же, 1937, XXXI : 134;
Но, 1938, ХХХ11 : 287—312. — insignis Austen 1909, Ann. Mag. Nat. Hist., Ill :
298 (Phillaematornyia).
Биология и медицинское значение. Mitzmain, 1914, Un. St.
Publ. Health., Hyg. Labor. Bull. 94 : 41; Nieschulz, u. Kronvaldd, 1929, Zentralbl.
Bakteriol., Orig., 113 : 403; Nieschulz, 1933, Ann. trop. Med. Parasitol., XXVII :
213—214; Nieschulz u. du Toit, 1933, Zentralbl. Bakteriol., LXXXIX, n° 8/12 : 244—
249; Du Toit u. Nieschulz, 1933, Journ. South Afric. Vet. Med. Ass., 4, n° 2 : 97—98.
S. Лоб узкий, ширина его равна или едва более ширины 3-го чле-
ника усиков; лобная полоса коричневато-бурая, посредине в 2 раза уже
орбиты; ширина скулы равна ширине лба и в 3 раза менее ширины щеки;
затылок слабо вогнутый; 3-й членик усиков в 3 раза длиннее 2-го; под-
бородок сильно вздутый, шире переднего бедра. Голова буроватая, перед-
ний край скул, щек и орбит красноватый; щупальцы желтые, подборо-
док красновато-коричневый; усики коричневато-бурые, 1-й и 2-й членики
их на вершине красноватые; скулы, орбиты и узкие полоски за глазами
в белом налете с легким желтоватым оттенком; лицо и щеки в сером мато-
вом налете, последние в редких коротких бурых щетинистых волосках;
затылок внизу с неясным красноватым оттенком. Лобные щетинки тон-
кие, волосовидные, короткие, но длиннее, чем у М. lucidula-, передняя
пара лобных щетинок несколько крепче остальных. Глазковые щетинки
тонкие, хорошо заметные. Спинка серо-бурая, в коротких щетинистых
черных волосках, плевры коричневатые, в тонких длинных волосках;
спинка в желтовато-сером или сером налете, с 4 продольными бурыми
полосами; средний промежуток светлого налета между внутренними
бурыми полосами шире каждой из полос; промежутки между внутрен-
ними и боковыми полосами уже бурых полос; щиток бурый в сером налете,
средина щитка и его переднебоковые отделы затемнены. Щетинки груди:
акростихальных — 1 пара перед щитком; дорзопентральных — 2 впереди
и 2—3 позади шва; плечевых — 3—4; заплечевых — 1—2; предшов-
ная — 1; интраалярных (позади шва) — 2; надкрыловых и закрыло-
вых — по 2 с каждой стороны; мезоплевры с одной щетинкой в передне-
верхнем отделе и с 6—7 щетинками по заднему краю; проплевры голые;
стерноплевральных щетинок 1-|—2; гипоплевры с несколькими волосками
около задних тазиков; волоски перед дыхальцем отсутствуют. На щитке
2 вершинных, 2 основных и 2 предшовных щетинки. Переднее дыхальце
белое. Ноги смоляно-бурые или рыжие; передние бедра в сером налете
по задней поверхности. Присоски белые, в Р/а раза короче 5-го членика
лапки. Жужжальца бледножелтые, чешуйки белые с бледно палевым
краем; крылья светлосерые, прозрачные; жилки в основной половине
крыла бледножелтые, в вершинной — серовато-желтые. Брюшко корич-
невато-бурое, в сером налете, основание 14 2 тергита, средние стерниты
и небольшие участки вентральной стороны тергитов — серо-желтые.
3-й тергит посредине переднего края с узкой бурой поперечной полоской,
не достигающей боковых краев сегмента, и коротким треугольным бурым
срединным пятном, отходящим назад от поперечной полосы; 4-й тергит
с треугольным пятном у переднего края; 5-й тергит с маленьким буро-
ватым клиновидным пятном у переднего края. I стернит без волосков.
Церки по наружным краям полукруглые, на внутренних углах без длин-
ных выступов; склеротизованная часть фаллозомы узкая, длинная, чуть
длиннее или равна мембранозной части; задний отросток фаллозомы
длинный, слабо расширен к вершине, на вершинном крае широко закруг-
138
I. ТРИБА MUSCINI
лен; передняя часть парамер длинная, дуговидная, с 1 длинной и 4—6
мелкими щетиночками; задний отдел парамер с узким пальцевидным
выростом, характерным для этого вида и М. lucidula. Аподема корот-
кая, узкая. V стернит маленький с длинными тонкими, на вершине зазуб-
ренными, боковыми выростами по заднему краю.
2- Отличается более широко расставленными глазами: ширина лба
почти равна ширине глаза; лобная полоса палево-коричневая, ширина
ее равна или чуть менее ширины орбиты. Лобные щетинки мелкие, тон-
кие; наружные, как и внутренние теменные щетинки, развиты. 3-й членик
усиков палево-коричневый, 1-й и 2-й — палево-красные. Серый налет
на груди гуще. Присоски короткие; темные отметины на брюшке слабс
выражены; брюшко палево-коричневое, в густом сером налете.
Распространение. Африканский вид, заходящий в Ориен-
тальную область и известный вдоль южной границы Палеарктики (Алжир,
Тунис, Египет, Аравия, Палестина, Ирак, Иран, Сев. Китай). В пре-
делах СССР М. crassirostris пока не обнаружен, хотя присутствие его пред-
ставляется вероятным в южной Туркмении.
Среди прочих палеарктических видов рода М. crassirostris представ-
ляется одной из наиболее своеобразных форм. По образу жизни муха
является кровососом. Она нападает на лошадей, буйволов, коров, ослов,
концентрируясь на таких частях тела, которые обладают наиболее тон-
кой кожей: ноги, живот, уши. Мухи гелиофильны и термофильны. Раз-
витие мух проходит исключительно в коровьем помете, куда отклады-
ваются сравнительно крупные яйца. Исходя из того, что вылупление
личинок из яиц происходит чрезвычайно быстро, допускается, что раз-
витие молодых личинок начинается еще в теле мухи.
М. crassirostris является переносчиком ряда заболеваний в эквато-
риальных странах. Механический перенос возбудителей с больных
на здоровых животных доказан экспериментально. В опытах с морскими
свинками установлена возможность прямой передачи возбудителей сибир-
ской язвы. Позитивный результат получен также в экспериментах с зара-
жением кроликов Bacillus bipolaris bubalisepticus, вызывающей острое
лихорадочное (инфекционное) заболевание среди буйволов, известное
под названием геморрагической септицемии буйволов; болезнь эта поль-
зуется широким распространением в местах разведения буйволов, как
на юге Европы и в Африке, так и в тропических странах востока. Способ-
ность к передаче заболевания сохраняется у мух до 24 часов. Подобно
многим кровососущим видам, М crassirostris способна осуществлять пере-
нос Trypanosoma evansi Steel, вызывающей среди однокопытных (лошадей,
ослов, мулов), а также среди крупного рогатого скота заболевание, извест-
ное под название «сурра».
М. crassirostris описан Штейном в 1903 г. из Египта. Спустя шесть
.чет (1909) Аустин вновь описал тот же вид под именем insignis Aust.,
поместив его во вновь установленный им род Phillaematomyia Austen
на основании, главным образом, строения хоботка (сильно развитый
подбородок). Той же точки зрения придерживались Мэллок, Пэттон,
Крэгг и Сеги. Ряд авторов склонен рассматривать Phillaematomyia как
один из подродов Musca L. (Хо, Карль и др.). Мы придерживаемся точки
зрения Штейна, сохраняя crassirostris в роде Musca L. на основании
оценки внешних морфологических признаков, лишь деталями отличаю-
щихся от таковых у прочих представителей рода. Это в значительной
мере относится к строению копулятивного аппарата самцов и гениталий
•самок. В систематическом отношении вид близок к Musca lucidula Lw.
2. MUSCA
139
IV. Группа Musca lusoria Wd.
Лоб самцов не шире 3-го членика усиков, глаза голые, реже в волос-
ках; проплевры голые, супрасквамальное ребро в щетиночках под
крылом или в волосках по всей длине; I стернит брюшка в волосках
и щетиночках. Грудь всегда с 4 темными полосами.
Фауна Советского Союза пока насчитывает 4 вида группы (Musca
autumnalis Deg., М. arnica, sp. n., M. larvipara Portsch., M. convexijrons
Thoms.). На Дальнем Востоке, в горных районах азиатской части СССР
и пустынях юга Средней Азии вероятно нахождение и других видов,
описание которых приводится ниже.
*10. Musca autumnalis Deg. (рис. 44, 122, 125, 127, 191—195,
198—205).
De Geer, 1776, Ins., VI : 83; Edwards, 1916; Entom. Month. Magaz., LII, 3, 15:
63; Patton, 1933, Ann. trop. Med. Parasitol,, XXVII : 428—430, figs.; on же, 1937,
XXXI : 207; Ho, 1938, Ann. trop. Med. Parasitol., XXXII : 287—312, figs.—
corvina Fabricius, Spec. Insect., II : 440; Karl, 1928 : 7; — prashadi Patton a. Se-
nior-White, 1924, Rec. Ind. Mus., XXVI : 553.
Биология и фазы развития. Порчинский, 1910 : 25—29, рис.
21—29; Keilin, 1916 : 393—415; Patton, 1933 : 430; Thomsen, 1938 : 304—309, figs.
78—81; Чеботаревич, 1937 : 37; Дербенева-Ухова, 1940 ; 337; Вайнштейн и Родова,
1940 : 364—368; Зимин, 1944, Проблемы кишечных инфекций : 146—153; Зимин,
1948 : 94, фиг. 51.
8. Лоб узкий, наименьшая ширина его у середины составляет около
3/з ширины 3-го членика усиков; ширина скулы в верхнем отделе
несколько менее ширины 3-го членика усика, книзу скулы заметно рас-
ширены и здесь шире 3-го членика усиков; щеки длинные, передний
верхний край широким полукругом заходит на верх за линию нижнего
уровня глаза; усики коричнево-бурые, 3-й их членик в 2‘/4 раза длиннее
2-го; ариста утолщена в основных 2/б своей длины; щупальцы бурые.
Голова черно-бурая; скулы, передний отдел орбит и узкие полоски за гла-
зами в густом серовато-белом блестящем налете; лицо, щеки и нередко
задний отдел орбит в слабо блестящем сером налете. Внутренние темен-
ные щетинки и вибриссы толстые, длинные; лобные щетинки помимо
передней крепкой пары и затылочные щетинки тонкие, имеющие харак-
тер щетинистых волосков; щеки с длинными, более или менее торчащими
и сравнительно густыми черными волосками, заходящими в переднем
отделе щеки выше горизонтальной линии, идущей вперед' от нижнего
края глаза; затылок в нижнем его отделе в черных, более густых и длин-
ных волосках. Грудь черно-бурая в сером или голубовато-сером налете;
спинка с 4 продольными черными полосами; средние разделены проме-
жутком светлого налета; боковые темные полосы длиннее и шире сред-
них; длина и ширина полос изменчивы. Переднегрудь сильно расширена
впереди, в длинных многочисленных черных щетинистых волосках; перед-
ние плевры лишены волосков; супрасквамальное ребрышко с группой
черных, мелких щетиночек лишь в переднем отделе (под основанием
крыла), далее назад — голое. Гипоплевры с группой черных волосков
тотчас над задними тазиками. Поверхность груди в черных сравнительно
толстых на спинке и тонких и длинных на плеврах волосах. Переднее
дыхальце белое, заднее—рыжее. Щетинки груди: акростихальных —
Г пара перед щитком; дорзоцентральных — 2 (3) + 4; интраалярных —
1; предкрыловых — 1; надкрыловых — 2 (задняя слабо развита); закры-
ловых — 3; плечевых — 3; заплечевых — 1; предшовных — 1; ното-
плевральных — 2; стерноплевральных — 1 -(- 2; на щитке вершинных —
140
I. ТРИБА MUSCINI
2 и основных — 2, длинные, толстые; предосновные — короткие, воло-
совидные; дискальных щетинок — 1—2 пары.
Крылья серые в основной трети, слегка желтоватые, жилки рыжие;
г4+5 сверху и снизу с несколькими волосками у основания; радиаль-
ный ствол у основания сверху с 3 щетинистыми волосками; чешуйки
белые, палевые или дымчатые в заднем отделе; жужжальца желтые.
Ноги черно-бурые с обычными для рода щетинками; коготки едва короче
5-го членика лапки. Брюшко коротко яйцевидное, темно или красножел-
тое. 1-й тергит сверху черно-бурый, 3-й — с черной срединной продольной
полоской, ширина которой составляет от до 1/в ширины тергита; перед-
ний конец полоски расширен, образуя в средине тергита поперечную
перевязь, постепенно суживающуюся к бокам; 4-й тергит с черной поло-
ской, узкой в его переднем отделе, расширяющейся назад и переходящей
на 5-й тергит; последний черно-бурый с желтыми переднебоковыми отде-
лами ; стерни ты в большей или меньшей степени затемненные, V стернит
черно-бурый на вершине; генитальные тергиты черно-бурые. V стернит
удлиненный, ширина его в середине в Iх/*—IV2 раза менее длины, по зад-
нему краю с двумя загнутыми внутрь (друг к другу) отростками, из кото-
рых правый толще левого; церки с полукруглыми боковыми краями,
по нижнему краю с глубокой выемкой по бокам от срединных (внутрен-
них) выростов; последние более или менее длинные, изогнутые вбок;
вершинные концы их достигают линии, соединяющей нижние края боко-
вых округлых выступов, или несколько не доходят до нее. Склеротизо-
ванная основная часть фаллозомы в Р/г—2 раза короче дистального
мембранозного отдела; последний по вентральному краю с двумя узкими
и одним широким базальным треугольными выступами;, дорзальный
отросток расширен к вершине, на конпе с глубокой выемкой посредине;
передние отделы парамер широкие, лопастевидные, с узким, лопасте-
видным отростком, отходящим в сторону аподемы от места соединения
с задней частью парамер; верхний край переднего отдела с 1 крепкой
и 1—2 мелкими щетинками; задняя часть парамер длинная, полукругло
изогнутая в дистальной трети, посредине с группой шипиков; аподема
длинная, слабо изогнута на вершине.
$. Лоб составляет около s/a ширины глаза; орбиты в Р/г (реже
в 2 раза) уже выпуклой по боковым краям черной лобной полосы, покры-
той редким шелковистым налетом; щеки в l3/^ раза шире 3-го членика
усиков, параллельнокрайние или несколько расширенные книзу; заты-
лок слабо выпуклый. Орбиты с многочисленными лобными щетинками,
расположенными по крайней мере в задней половине лба в 2—3 ряда;
один ряд располагается по наружному краю лобной полосы в непосред-
ственной близости к краю орбиты; 3-й ряд, если он имеется, распола-
гается кнаружи от настоящих лобных щетинок в числе нескольких тон-
ких щетинок; орбитальные щетинки направлены вперед и наружу в числе
2—3 с каждой стороны в задней части орбит; волоски на щеках далеко
заходят за линию нижнего уровня глаза, более или менее густые; скулы
в желтовато-серовато-белом, орбиты в желто-сером налете; орбиты, помимо
щетинок, несут несколько рядов коротких щетинистых волосков; внутрен-
ние и наружные теменные, затылочные и вибриссальные щетинки толстые,
но слабее развиты, чем у М. larvipara, что в равной степени относится
и к лобным щетинкам. Грудь в густом сером или сизом налете, с 4 про-
дольными черными полосами, достигающими переднего края щитка.
Брюшко черно-бурое в сером или желтовато-сером налете с темными
переливчатыми пятнами и с ± развитой желтой окраской на вентраль-
2. MV SC A
141
ных концах тергитов 1+2-го, 3-го всегда, 4-го и 5-го не у всех осо-
бей.
Дорзальная сторона брюшка с бурой продольной, суженной назад
и не распространяющейся обычно на 5-й тергит полоской; боковые пере-
ливчатые темные пятна узкие впереди, в заднем отделе — расширены,
образуя темную полоску по заднему боковому краю 3-го и 4-го тергитов.
Задние края 3-го и 4-го тергитов покрыты более редким налетом, отчего
кажутся несколько затемненными. Длина тела 4.5—7.5 мм.
Яйцо удлиненное, слабо суженное к обоим концам, на дорзальной
стороне выпуклое, на вентральной прямое или слабо вогнутое; на перед-
нем полюсе имеет длинный гладкий отросток (рис. 44), изогнутый на конце
и имеющий по вогнутой поверхности желобовидное углубление, про-
ходящее по всей его длине и продолжающееся далее между двумя тонкими
бороздками (отходящими от боковых отделов его основания) до заднего
полюса яйца; хорион с нежной скульптурой из слабо вытянутых вдоль
длинной оси яйца шестигранных ячеек; длина отростка в 5—-6 раз менее
Рис. 191. Общий вгд взрослой личинки Musca autumnalis
Deg. (Ориг.).
длины яйца; длина последнего 2.3—2.7 мм (без отростка), толщина
посредине 0.5—0.7 мм. Яйцо желтовато-белое, после пребывания в суб-
страте (навозе) рыже-желтое, с затемненной вершиной отростка.
Личинка I стадии (рис. 192, 195) белая или с легкой желтизной;
сегменты тела, начиная со 2-го грудного кольца, по переднему краю снаб-
жены полосами из шипиков, расположенными в несколько рядов и опоя-
сывающими сегменты целиком; шипики на вентральной стороне тела,
лежащие на легких валиковидных поперечных возвышениях, толще
и длиннее остальных; на грудных кольцах с дорзальной и боковой поверх-
ности шипики мелкие, с узким основанием. Задние дыхальцы с одним
сердцевидным отверстием, нередко разделенным поперечной тонкой поло-
ской на две части. Базальный склерит в виде двух узких полукруглых
полосок с заостренными дорзальными и косо срезанными на конце вен-
тральными отростками; дорзальная перемычка высокая, расширена
в верхней части; передние отростки тонкие, длинные; гипостомальный
склерит относительно широкий, с клювовидным, заостренным дисталь-
ным концом, по вентральному краю спереди с 2 шипиками; задний конец
коротко раздвоен и соединен с гипостомальным склеритом; впереди рото-
вого отверстия с продольными многочисленными склеротизованными
узкими полосками; боковые и вентральная стороны ротового отверстия
укреплены неправильно треугольными, зазубренными по наружному
(вентральному) краю пластинками. Длина тела 2.8—3.3 мм.
Личинка II стадии (рис. 193, 194) не имеет сильного вооружения
из темных шипов, лишь вентральные поперечные валики на брюшных
сегментах с плохо различимыми неровностями; переднее- дыхальце
с 7 лучами, задние дыхальцы имеют по 2 коротких щели. Фарингеальный
склерит длинный; дорзальные отростки крыловидные, постепенно сужены
142
I. ТРИБА MUSCINI
к дистальному концу, вентральные короче дорзальных, шире их и скошены
по дистальному краю. Дорзальная перемычка сильно склеротизована,
крупно-решетчатая, верхний край слегка выступает над верхним краем
стернита; передние отростки длинные, более или менее узкие, выемчатые
по переднему краю, снизу передние концы их далеко выдвинуты вперед
и по вентральному краю соединены перемычкой. Гипостомальный склерит
в профиль вытянуто треугольный, снизу боковые его части соединены
широкой перегородкой, так же как и вверху. Угловидные склериты
тонкие, не слиты своими концами; ротовые крючки сращены в переднем
отделе, позади раздвоены, узкие, слабо изогнутые и расширены сзади;
края ротового отверстия укреплены парой широких лопастей, обра-
зующих впереди поперечное расширение с зазубренным передне-вен-
Рис. 192—197. Детали строения личинок.
192 — ротоглоточный аппарат личинки I стадии М. autumnalis Deg. 193 — то же
личинки II стадии. 194 — задние дыхальцы личинки II стадии М. autumnalis Deg.
195 — срединный дорзальный склерит личинки I стадии того же вида, 196 — задние
дыхальцы личинки III стадии М. lusoria Wd. 197 — то же М. mrsopotamiensis Ptt.
(Рис. 196—197 по Пэттону, прочие—ориг.).
тральным краем. Впереди рта имеется по 4—5 зубцов, иногда двурядных,
симметрично расположенных по обе стороны от средней продольной
линии головы. Длина тела 3.2—4.3 мм. Личинка желтовато-белая; зад-
ний конец несколько утолщен.
Личинка III стадии (рис. 191, 198—204) ярко желтая с темным,
слабо просвечивающим кишечником; передние дыхальпы с 8 лучами;
задние, имея по 3 извитых щели, расставлены на 1/в—V? длины попе-
речника дыхальца; перитрема сильно склеротизована, не очень широкая;
щели широкие, каждая с 5 изгибами, тесно соприкасаются друг с дру-
гом; все концы щелей обращены внутрь к крупному светлому диску;
ечень редко верхний конец средней щели дает короткий отросток напра-
вленный наружу. Базальный склерит длинный, высокий; дорзальные
отростки широко крыловидные, суженные на конце, вентральные короче
дорзальных, шире их, косо срезаны на вершине, вследствие чего
2. MUSCA
143
на их верхнем крае образуется угловатый выступ; дорзальная сильно
склеротизованная с мелкими отверстиями; перемычка глубоко входит
в стенки склерита, по заднему краю с двумя тонкими отростками, слитыми
с верхним краем склерита; передние отростки относительно короткие,.
Рис. 198—205. Детали строения личинки III стадии Musca autumnalis Deg,
198 — ротоглоточный аппарат. 199 — задний конец тела сзади. 200— он же сбоку..
201 — переднее дыхальце. 202 — заднее дыхальце. 203 — вооружение снизу на 5-м
(2) и 6-м (Z) брюшных сегментах. 204— шипики на дорзальпой стороне и в переднем
отделе 2-го грудного сегмента. 205 — продольный разрез кутикулы в том же отделе
2-го грудного сегмента. (По Зимину).
иедалеко выступают за границу дорзальной перемычки, выемчатые
жо переднему краю, снизу соединены вентральной перемычкой, имеющей
то же строение, что и у личинок II стадии; фарингеальная область ясная,
темноокрашенная; гипостомальный склерит короткий широкий, непра-
вильных очертаний; ротовые крючки длинные, с расширенным основа-
нием, один крючок немного короче другого; зубные склериты мелкие,
треугольные; стенки ротового отверстия снаружи укреплены резко выра-
144
I. ТРИБА MUSCINI
женными, радиально расходящимися светлыми бороздками. Тело личинки
вытянуто, задний конец утолщен; брюшные сегменты по переднему краю
снизу с крупными поперечными валиками, покрытыми прозрачными
мелкими шипиками; на грудных сегментах эти неровности заходят и на
дорзальную поверхность, что не всегда наблюдается у сегментов брюш-
ных (исключая анальный). Анальная пластинка очень широкая, скле-
ротизованная, сильно и далеко заходящая на боковые отделы сег-
мента (до уровня верхней границы или средины задних дыхалец). Аналь-
ные бугорки маленькие, по бокам от них и несколько отступя назад
с каждой стороны расположено по маленькому субанальному бугорку;
вдоль всего переднего и заднего края до дистальных концов анальной
пластинки располагаются ряды прозрачных притупленных шипиков;
верхний край анальной пластинки тупой, слегка загнут вперед (при рас-
сматривании в профиль); задняя поверхность анального сегмента с неяс-
ными бугорчатыми выступами вокруг задних дыхалец. Верхняя сторона
с 3 бороздками, несущими по краям шипы. Длина тела 6—8 мм.
Пупарий белый или серый, имеет признаки личинки III стадии
(ясная анальная пластинка и шиповатость анального сегмента). Длина —
5—6 мм.
Распространение. Широко распространенный в палеаркти-
ческой области вид. В СССР распространен от Карельского перешейка
до южной границы Европейской части СССР; известен по всей Средней
Азии: на восток известен до линии Челябинск—Акмолинск и Алма-ата.
В Сибири к востоку от Челябинска замещен видом Musca arnica, sp. nov.
В Западной Европе распространен от Швеции и Норвегии до Испании,
Италии и Балканского полуострова.
Указания, относящиеся к Эфиопской области, как выяснил Пэттон
(1926), относятся к различным аберрантным формам того же вида, либо
к М. lusoria Wd. Данные Пэттона (1933) также не полностью могут быть
отцесены к М. autumnalis, в частности, северокитайские экземпляры
с несомненностью должны быть отнесены к М. arnica, sp. п., т. к. М. autum-
nalis в Азии не заходит далее восточных границ Казахстана и горных
областей Советской Средней Азии. Экземпляры, просмотренные нами
из северной, центральной и южной Монголии, принадлежат, как и восто-
чно-сибирские, к Musca arnica, sp. n.
В систематическом отношении наиболее близкими к М. autumnalis
видами являются М. arnica, М larvipara, М. convexijrons. От двух послед-
них видов М. autumnalis (по $$ и jj) отличается отсутствием щетинистых
волосков в заднем (приподнятом) отделе супрасквамального ребрышка.
Самцы М. autumnalis имеют, кроме того, высоко заходящий за нижний
уровень глаза край щеки, покрытый длинными волосками; самки обла-
дают 2—3 рядами лобных щетинок, у М. larvipara и М. convexijrons распо-
ложенных в один ряд.
От М. arnica самцы М. autumnalis легко отличимы по суженной
скуле, поперечник которой в верхнем отделе у первого вида более попе-
речника 3-го членика усиков, у второго — менее его; самки М. arnica
не имеют на орбитах вблизи лобной полосы добавочных многочисленных
лобных щетинок, составляющих у М.\ autumnalis дополнительные
ряды щетинок этой группы.
Вид относится к категории яйценесущих форм. Яйца откладываются
почти исключительно в коровий помет в поле. Самки предпочитают для
этой цели свежий субстрат. Яйца располагаются обычно тесной группой
вертикально: верхний отросток яйца остается над поверхностью помета.
2. MUSCA
145
Для кладки самки пользуются преимущественно утренними часами,
когда процесс протекает наиболее интенсивно. В жаркое время дня коли-
чество яйценесущих мух резко сокращается. По данным Порчинского,
число откладываемых самкой яиц достигает 24.
Через сутки из яйца вылупляется личинка. Личинки калоядны.
Развитие личинок протекает весьма быстро. По нашим наблюдениям
в Таджикистане, уже через сутки после вылупления из яйца личинки
достигают III стадии. По данным Дербеневой-Уховой для Кабарды,
на развитие личинки требуется 5—6 дней. Полный цикл развития мухи
при средней суточной температуре 25° С заканчивается в 9 дней. В усло-
виях Кабарды все развитие мухи завершается в 15—18 дней. Окукление
происходит в почве.
Взрослые мухи обычны как в степных, так и в лесостепных и лесных
районах, в особенности близ животных. Мухи являются постоянными
обитателями скотных дворов, боен, базаров. В помещения залетают
редко. На пастбищах и в открытых стойлах и загонах мухи (самки)
нередко массами и с болыцой настойчивостью нападают на глаза, нос
и кожные поранения животных, слизывая влажные выделения. М autum-
nalis относится к кроволижущим видам. Подобно другим кроволижущим
видам, М. autumnalis при помощи престомальных зубцов способна
наносить мелкие царапины заживающим ранкам и тонким слизистым
оболочкам.
В зоне степей, лесостепи и в хвойных лесах Европы вид весьма мно-
гочисленен и, несомненно, имеет значение как отрицательный фактор
в животноводстве. Как недавно установлено Клёсовым (1949) на Украине,
взрослые мухи этого вида являются промежуточным хозяином нематоды
Thelazia rhodesi Desm., вызывающей у крупного рогатого скота конъюнк-
тиво-кератит. Нематоды заглатываются мухами при слизывании слез
животных. Развитие личинок протекает в полости тела мух. Достигнув
инвазионной стадии, личинки нематоды передаются мухами при новом
контакте их с глазом животного. В южных широтах и особенно в полу-
пустынях Средней Азии вйд менее обилен; появление его там приурочено
к ранним летним месяцам.
Врагами личинок М- autumnalis являются различные хищные
личинки мух. Имагйнальная фаза подвержена грибковым заболеваниям
(Empusa muscae).
*11. Miisca arnica Zimin, sp. nov. (рис. 126, 208, 441, 442).
Близок к предыдущему, от которого отличается, меньшей величиной
и нижеперечисленными морфологическими особенностями.
<5. Лоб р’авен, шире или немного (3Д) уже 3-го членика усиков; скулы
широкие, в верхнем отделе ширина их более или равна ширине 3-го чле-
ника усиков, книзу слабее расширены; щеки в более густых волосках
и передний верхний край их не так далеко (и обычно угловидно) заходит
за уровень нижнего края глаза (рис. 126); передний внутренний край
глаза не так сильно вогнут. Основной ствол радиуса сверху с 1, реже
2 волосками, еще реже эти волоски не развиты вовсе. Темная продольная
полоса 3-го тергита обычно слабо расширена назад в среднем отделе
выемчатая; 5-й тергит чаще весь бурый; налет брюшка желтовато-серый.
V стернит с изогнутыми на вершине отростками; длина стернита в Р/граза
превышает его ширину в средней части. Церки уже, с менее широко
закругленными боковыми дистальными лопастями; фаллозома сходна
с таковой Musca autumnalis, отличаясь деталями строения парамер
Ю Фауна СССР, том XVIII, вып. 4.
146
I. ТРИБА MUSCINI
(передняя часть их короче и шире, задняя короче и уже), более длинным
мембранозным отделом фаллозомы и удлиненными вентральными лопа-
стями.
j. Лобные щетинки расположены в один ряд [реже присутствует
несколько (1—3) щетинистых волосков в заднем отделе]; переливчатые
темные боковые пятна на тергитах брюшка узкие, обычно не образуют
поперечного расширения на заднем крае по бокам 3-го и 4-го тергитов,
вытянутые параллельно продольной оси брюшка. Волоски в переднем
отделе щек реже и короче, чем у М. autumnalia Deg. Длина тела 4.5—6.6.
Вид описывается по многим экземплярам из различных мест Сибири
и Монголии. Крайним западным пунктом местонахождения вида является
Рис. 206—208. Фаллозома.
206 — Musca vitripennis Mg. 207 — М. osiris Wd. 208 — M. arnica, sp, n, (Ориг.).
г. Кустанай; вид найден в Омске, около Минусинска, близ Красноярска
(сел. Сорокине), Забайкалье (Зерентуй), р. Селенга, Ягодное на
оз. Щучьем, Нерчинск, Приамурье (Благовещенск), Приморском крае
(Камень-Рыболов, Ворошилов), в северной и центральной Монголии
(Урга, Хами, Эцзин-гол).
12. Musca mesopotamiensis Patt. (рис. 197, 230—234).
Patton, 1920, Ind. Journ. Medic. Research, VII; 4 : 751; он же, 1932, Ann. trop.
Med. Parasitol., XXVI : 389; он же, 1933, XXVII : 420—423, figs. : 16, 17, 18b; Ho,
1938, Ann. trop. Med. Parasitol., XXXII : 287—312 (Tiviparomusca).
3. Лоб равен Чв ширины глаза; скулы в сером налете, как и орбиты;
грудь в голубовато-сером налете с 4 черными продольными полосами.
Брюшко светло-оранжевое с широкими пятнами серебристого налета
на боках; тергит 1 + 2-й светло-оранжевый, с относительно узкой черной
срединной полосой и темной перевязью у основания; черная полоса 3-го
тергита, сохраняющего ту же окраску, что и предыдущий, расширена
вдоль переднего края; по бокам от черной полосы располагаются полосы
серебристого налета и по одному пятну той же окраски; 4-й тергит с узкой"
черной полоской и затемнением вдоль заднего своего края, серебристые
2. MUSCA 147
пятна налета выражены резче, чем на 3-м; 5-й тергит с неясной срединной
продольной полоской и серебристыми боковыми отделами; стерниты
1, II и дистальная часть V темнокоричневые. Наружные углы церок
широко закруглены, внутренние — с сосцевидными выступами. Склеро-
тизованная часть фаллозомы широкая, короче мембранозной; задний
ее отросток длинный, расширенный на вершине, оканчивающийся двумя
боковыми рожками; аподема маленькая. Передняя часть парамер длин-
ная, с 3 щетинками, из которых одна очень длинная; задний отдел пара-
мер широкий; 1 стернит слегка затемнен в вершинной половине, ширина
его равна длине; задние отростки короткие, широкие.
Орбиты в сером налете, ширина лба менее ширины глаза, лобная
полоса черная, относительно узкая с прямыми краями; ширина ее равна
1.3 ширины лба; лобные щетинки расположены в один ряд с каждой сто-
роны от лобной полосы. Среднеспинка как у самца. Брюшко черное
с отметинами серого налета; 1-|-2-й тергит с голубовато-серым пятном
с каждой стороны от средины, иногда расширяющимся вдоль заднего
края; 3-й тергит с узкой черной полоской и лежащими по бокам от нее
широкими полосами голубовато-серого налета; на 4-м тергите черная
полоска уже чем на 3-м; 5-й тергит с преобладающим голубовато-серым
налетом, в середине с темносерым затемнением. Стерниты от коричневой
до черной окраски.
Яйца М. mesopotamiensis не имеют отростка. Передние дыхальцы
взрослой личинки с 8—9 пальцевидными выростами. Задние дыхальцы
очень широкие, сильно склеротизованы; верхняя и нижняя дыхальцевые
щели длиннее средней. Расстояние между дыхальцами равно ширины
дыхальцевой пластинки.
Распространение. По Пэттону, вид является весьма обыч-
ным в среднем Ираке, особенно в долинах Тигра и Евфрата. В СССР
не обнаружен. Нахождение вида возможно ожидать в Закавказье и на юге
Туркмении.
М. mesopotamiensis развивается в лепешках коровьего помета
на полях. Самки яйценесущи. Мухи сильно досаждают крупному рогатому
скоту, лошадям, ослам и верблюдам на пастбищах, нападая на глаза,
нос и поранения кожи животных. Автор указывает, что мухи следуют
за стадами совместно со слепнями и жигалками, пользуясь для высасы-
вания крови ранками, наносимыми кровососами.
Вид представляет интерес как кроволижущая форма с сильно раз-
витыми престомальными зубцами. Эта особенность строения хоботка
позволяет мухе не только слизывать вытекающую из ранок кровь живот-
ных, но и самостоятельно вызывать кровотечение из заживающих ранок
и соскабливать засохшие эксудаты. По устройству престомальных зуб-
цов вид отнесен автором в группу М. lusoria.
*13. Musca larvipara Portsch. (рис. 120, 121, 209—212, 216—223).
Порчинский, 1910 : 30—35; Malloch, 1925, Ann. Mag. Nat. Hist., XV : 135 (Vivi-
paromusca)', Patton, 1932, Ann. trop. Med. Parasitol., XXVI : 387, 395; он же, 1933,
там же, XXVII : 416, 417, fig. 13; Awati, 1920, Ind. Journ. Med. Res., VIII : 89—92,
pl. XIII. — corvinoides Schabl u. Dziedzicki, 1911 : 128, figs. : 549, 550.
Биология стадии развития. Порчинский, 1910; Keilin, 1916:
393—415; Чеботаревич, 1937 : 37; Романов, 1940, Мед. паразит, и паразит, болезни,
IX : 355—363; Вайнштейн и Родова, 1940 : 364—368; Дербенева-Ухова, 1940 : 336;
Зимин, 1948 : 91 фиг. 50.
$. Лоб в наиболее узком месте составляет около 1/г ширины 3-го
членика усиков; скулы в IVa раза шире последнего; передний верхний
10*
148
I. ТРИБА MUSCINI
край щеки не заходит полукругом за нижний уровень глаза, что отли-
чает самцов этого вида от таковых М. autumnalis и М. arnica. Затылок
в верхней части вогнутый, внизу выпуклый; передний нижних край
глаза слабо вогнутый, фасетки верхней части глаз увеличенные; 3-й
членик коричнево-бурых усиков в 2—21/г раза длиннее 2-го; щупальцы
рыжевато-бурые, слабо утолщенные на конце. Голова черно-бурая. Заты-
лок почти лишен налета, за исключением нижней его части, покрытой
редким сероватым налетом, и полосок позади глаз, покрытых блестящим
серовато-белым густым налетом. Скулы в густом серовато-белом, белом
или желтовато-белом блестящем налете; орбиты в таком же налете только
в передней части или (у южных экземпляров) полностью. Внутренние
теменные щетинки и вибриссы длинные, крепкие; лобные щетинки,
исключая переднюю, сравнительно хорошо развитую пару, тонкие, почти
волосовидные, как и затылочные щетинки. Затылок в черных, более
длинных, щеки в коротких негустых волосках. Орбиты в переднем отделе
кнаружи от лобных щетинок иногда с 1—3 мелкими короткими черными
волосками. Грудь черно-бурая, в желтоватом или серовато-белом налете;
спинка с 4 продольными полосами, доходящими до щитка; промежутки
между ними в сером, слабо блестящем и сравнительно с плеврами, в более
темном томенте, благодаря чему светлый средний промежуток в передней
части спинки не резко выделяется. Ширина полос крайне изменчива,
поверхность их покрыта коричневатым налетом, не всегда отчетливо
заметным. Спинка и щиток в более толстых и коротких, чем на плеврах,
черных торчащих волосках. Переднегрудь широкая, с многочисленными
крепкими щетинистыми волосками в боковых отделах; проплевры впереди
дыхалец голые; гипоплевры с многочисленными тонкими черными воло-
сками лишь в нижнем своем отделе над основанием тазиков задних ног;
супрасквамальное ребро у основания крыла и по всему верхнему краю
почти до щитка в черных щетинистых волосках. Щетинки груди: акро-
стихальных — 1 пара перед щитком; дорзоцентральных — 2-J-4; интра-
алярных — 1 позади шва; предкрыловых (коротенькая) — 1; надкрыло-
вых — 2, задняя мелкая; закрыловых — 3; плечевых — 3; заплечевых —
1; предшовных — 1; стерноплевральных — 1 + 2; на щитке наибольшей
длины и толщины достигают вершинные (2) и основные (2) щетинки;
предосновные слабые; дискальных — 2—3 пары, они несколько крепче
предосновных щетинок. Переднее дыхальце белое; заднее округло-тре-
угольное, буровато-рыжее. Крылья сероватые, жилки крыла рыжевато-
бурые, при основании с более развитой желтой окраской (у южных экзем-
пляров в основной половине — бледножелтые). Основной ствол, как
правило, с 3 черными волосками; число волосков варьирует в пределах
1—5; г4+5 сверху при основании с 1—2 волосками, снизу с более длин-
ными и реже расставленными волосками, расположенными только у осно-
вания в числе 2-—3, или доходящими до г—т\ в редких случаях эти
волоски заходят даже в вершинный отрезок жилки (некоторые экзем-
пляры из Туркмении и Таджикистана). Чешуйки белые, верхняя по краю
палевая, нижняя обычно в задней половине дымчатая; жужжальца
желтые. Ноги черно-бурые; 5-й членик лапки едва длиннее коготков.
Передние голени, исключая предвершинную щетинку, без .крепких щети-
нок; передние бедра с обычными рядами заднедорзальных и задневен-
тральных щетинок; средние голени с 4—5 мелкими заднедорзальными
и задними щетинками; бедра средних ног в основной половине с перед-
невентральными и задневентральными щетинками; по задней поверх-
ности с рядом.щетинок длинных и толстых в вершинной части бедер.
2. MUSCA
149
Задние голени с 2—3 передневентральными щетинками у середины,
рядом коротких переднедорзальных и таким же рядом заднедорзальных
щетинок, из которых 2 развиты сильнее других. Задние бедра с полным
рядом переднедорзальных и прерванным у основания рядом передне-
вентральных щетинок; задневентральные щетинки крепкие в количестве
4—5. Брющко яйцевидное, красновато или (у среднеазиатских особей)
палево-желтое. 1-|-2-й тергит сверху черный, исключая узкую полоску
сзади в боковых отделах сегмента. У некоторых экземпляров из песков
Средней Азии боковые края 1 -|-2-го тергита по бокам широко желтые;
3-й тергит с бурой или коричнево-черной срединной полосой, образую-
щей узкое поперечное расширение вдоль переднего края сегмента; ширина
полосы варьирует в пределах от —1/в ширины тергита; 4-й тергит с более
узкой того же цвета полоской, переходящей на заднем крае тергита
в узкую поперечную перевязь, не заходящую на вентральную сторону
брюшка. У среднеазиатских экземпляров поперечная перевязь нередго
вовсе исчезает, отчего 4-й тергит имеет желтую окраску и тонкую сред-
нюю черную полоску. 5-й тергит черно-бурый, с более или менее раз-
витой желтой окраской в боковых частях. Стерниты, кроме буроватого
I, II и задней 1/з V, — желтые, как и вентральная поверхность тергитов
(кроме заднего края 4-го и задней половины 5-го). V стернит удлинен-
ный, со слабо вогнутыми в срединном отделе боковыми краями; задние
отростки короткие, широкие, с согнутыми по направлению друг к другу
вершинами. Церки с округленными боковыми и широко закругленными
дистальными наружными углами; срединные выступы короткие, верши-
ной направленные вниз и вбок, нередко слабо развиты. Склеротизован-
ная часть фаллозомы равна, чуть длийнее или явно короче мембранозного
отдела; передняя часть парамер поперечная, низкая, с несколькими
мелкими обычно тремя щетинками; задняя часть парамер узкая, длин-
ная, полукругло загнутая наверх, в середине с шипиками; дорзальный
отросток фаллозомы очень сильно расширен в заднем отделе и глубоко
выемчатый по заднему краю, вследствие чего дистальный его конец имеет
две узкие боковые, длинные лопасти. I стернит брюшка в волосках.
Шелковистый белый, или желтовато-белый налет брюшка хорошо развит;
боковые темные переливчатые пятна выражены не резко; на 3-м и 4-м
тергите они не образуют широкой резко выраженной поперечной полосы
по заднему краю; в передних 2/з ширина их обычно менее ширины пятна
светлого налета, расположенного от них к середине брюшка; 5-й тергит
сверху без резко выраженных, проходящих по всей длине тергита, про-
дольных боковых переливчатых пятен; если они есть, то имеют характер
коротких или неясно очерченных штрихов.
j. Лоб сужен назад, где ширина его составляет 2/з—3/< ширины глаза;
орбиты равны ширине лобной полосы или в Р/г раза уже ее; затылок
в светлосером налете, скулы и передний отдел орбит в шелковисто бле-
стящем белом или серовато белом, задняя часть и средина орбит в мато-
вом более темном (желто-сером или сером) налете. Скулы в 1х/з—2 раза
шире 3-го членика усиков. Лобные щетинки крепкие, расположены
в один ряд; лобная полоса черная, с серыми выпуклыми боковыми кра-
ями. Внутренние и наружные теменные щетинки, равно как и вибриссы,
крепкие. Грудь в густом желтовато-сером налете, на спинке с 4 бурыми
продольными полосами; ширина средних темных полос в 2—3 раза менее
ширины светлого промежутка между ними. Брюшко яйцевидное, заострен-
ное на вершине, черно-бурое. Нижняя поверхность (стерниты и края
тергитов) брюшка на большем или меньшем протяжении, вентральная
150
I. ТРИБА MUSCINI
сторона 1 + 2-го тергита обычно целиком палево-желтые; поверхность
брюшка в густом серо-желтом или желто-сером налете; бурая темная
продольная полоска сужена назад, достигает заднего края 4-го тергита
н иногда распространяется в виде узкого штриха на 5-й сегмент; боковые
переливчатые пятна на 3-м и 4-м тергитах узкие (в 2—3 раза уже свет-
лого пятна, расположенного от них к середине), слабо расширенные
назад с коротким темным поперечным отростком по заднему краю тер-
гитов, никогда, однако, не образующим поперечной темной полоски;
Рис. 209—215. Детали строения личинок.
209 — передний конец тела с ротоглоточным аппаратом личинки I стадии Musca
larvipara Portsch. 210 —• то же II стадии; 211 — срединный дорзальный склерит
личинки I стадии того же вида. 212 — задние дыхальцы личинки II стадии того же
вида. 213 — задние дыхальцы личинки III стадии М. crassirostris Stein. 214— тоже
М. hervei Vill. 215 — то же М. xanthomelas Wd. (рис. 213—215 по Пэттону, прочие —
ориг.).
г/г
-темные боковые пятна по бокам 5-го тергита отсутствуют, если есть,
то в виде коротких клиновидных штрихов лишь у основания сегментов.
Длина тела 4.8—8.3 мм.
М. larvipara относится к обширной группе видов, широко распро-
страненных в экваториальных странах Эфиопской и Ориентальной обла-
стей и на юго-востоке Палеарктики.
Отложенная личинка находится в стадии, лежащей на грани пере-
хода из I во II и совмещающей в себе признаки строения одновременно
I и частично II стадии. Это сказывается в строении стигм и ротового
аппарата. В некоторых случаях отложенная личинка относится к I ста-
дии; как исключение, наблюдающееся при воспитании насекомого в сад-
2. MUSCA
151
ках, откладывается яйцо, из которого, однако, личинка выбраться
не в состоянии.
Тело личинки I стадии на рубеже перехода во II стадию (рис. 209)
имеет хорошо развитый ротоглоточный аппарат (I стадии), на котором
располагаются части личинки II стадии. Срединный дорзальный склерит
(рис. 211) очень широкий, с узким дорзальным выступом на вершине;
вершина и передний широкий край склерита с зубчиками. Базальный
склерит и особенно дорзальная его перемычка шире, чем у соответствую-
щих по возрасту личинок М. autumnalis, с которыми имеет много общего.
Сегменты груди (исключая 1-й) и брюшка по переднему краю с полосками
из острых, мелких хитиновых шипиков; на 1-м грудном и часто на 8-м
брюшном сегментах шипики располагаются лишь на вентральной стороне
тела, на прочих сегментах груди и брюшка шипики кольцом опоясывают
весь сегмент. Впереди ротового отверстия расположен поперечный ряд
хитиновых узких пластинок; боковые стенки ротового отверстия укреп-
лены хитиновыми тяжами, зазубренными на вентральном крае. Перед-
ние дыхальцы (личинка II стадии) с 8 лучами; задние (I стадии) с сердце-
видным отверстием или (у личинок II стадии) с двумя тонкими изогну-
тыми щелями (нижняя с 5—6 изгибами, верхняя более короткая с 4—5).
Над рядом пластинок головного сегмента (рис. 210) 4—5 широких, креп-
ких, вершиной направленных назад зубцов.
Личинка III стадии сходна с таковой М. autumnalis, отличаясь
следующими признаками: узкими, длинными дыхальцевыми щелями,
широко треугольным гипостомальным и более широким и коротким фарин-
геальным склеритом.
Личинка III стадии (рис. 216—223) крупная, толстая, с булавовидно
утолщенным задним концом тела, тонкой блестящей желтовато-белой
кутикулой, морщинистой и часто с мелко ячеистой скульптурой на 8-м
брюшном сегменте. Головной сегмент крупный, двураздельный спереди;
антенны с коротким, широким, кольцевидным 1-м и маленьким конусо-
видным 2-м члеником; ротовые борозды заходят на боковые и частью
на дорзальную сторону сегмента. 1-й грудной сегмент сужен кпереди,
по переднему краю несет многочисленные мелкие шипики, занимающие
несколько менее 1/& длины сегмента. Шипики расположены на мелких
чешуйчатых возвышениях по их заднему приподнятому краю; шипики
очень мелкие, собраны по несколько штук и составляют как бы мелкие
гребешки. Посредине зона, покрытая шипиками, имеет сверху неболь-
шой полукруглый вырез, к границам которого примыкает гладкое, полу-
круглое возвышение, поделенное поперек узкой бороздкой. 2-й и 3-й
грудные, а также 1-й и 2-й брюшные сегменты вдоль всего переднего
края покрыты шипиками; ширина колец, занятых шипиками, составляет
около х/в длины сегмента. Шипики передних рядов расположены на чешуй-
чатых площадках и по существу являются приподнятыми задними их кра-
ями; задние шипы собраны в короткие полукруглые ряды; они мельче
передних и не всегда отчетливы. Брюшные сегменты с 1-го по 7-й снизу
по переднему краю с валикообразным вздутием, усаженным мелкими
и крупными зубчиками. Задний край брюшных сегментов с двумя неболь-
шими группами бугорочков вблизи средней продольной линии сегмен-
тов. 8-й брюшной сегмент несколько вздутый; на дорзальной поверх-
ности с тремя глубокими продольными бороздами. Задняя поверхность
сегмента почти плоская, со слабым вдавлением вокруг дыхалец, имеющим
тонко-морщинистую поверхность; вокруг дыхалец располагаются коль-
цом мелкие и не у всех особей хорошо выраженные пологие конусовид-
,152
I. ТРИБА MUSCINI
ные бугорки в количестве до 12; чаще всего отчетливо заметно лишь 6.
Анальная пластинка очень крупная; почти прямые ее концы заходят
Рис. 216—223. Детали строения личинки III стадии Musca larvipara Portsch.
216 — ротоглоточный аппарат; 217 — задний конец тела сзади. 218— он же сбоку.
219 — переднее дыхальце. 220 — заднее дыхальце. 221 — вооружение переднего
отдела на верхней стороне 2-го грудного сегмента. 222 —срез кутикулы в том же
отделе 2-го грудного сегмента. 223 — вооружение снизу 5-го (2) и 6-го (1) брюшных
сегментов. (По Зимину).
на верхнюю половину боковой стороны сегмента; пластинка широкая,
с изогнутыми передним и задним краями, позади с несколькими спу-
танными рядами шипов, далеко не достигающих концов анальной пла- •
2. MUSCA
153
стинки; такие же шипики покрывают узкую зону вдоль переднего края
анальной пластинки, где они доходят до угловидной выемки ее переднего
края. Субанальные и анальные бугорки маленькие. Передние дыхальцы
с поперечным атриумом, полукруглый край которого несет 10—12
коротко булавовидных выростов. Задние дыхальцы у вполне созревших
личинок крупные, с блестяще черными перитремой и дыхальцевой пла-
стинкой; стигмальный диск большой; дыхальцевые щели многократно
змеевидно изогнутые, концы их обращены внутрь пластинки. Дыхальцы
отставлены друг от друга на Ч2 поперечника дыхальца. Ротовой аппарат
с крепкими, не одинаковой длины широкими крючьями, слабо изогну-
тыми в вершинном отделе. Зубные склериты узко треугольные, гипо-
стомальные — неправильной формы, с округлым на дорзальной и угло-
видным выступом на вентральной сторонах. Фарингеальные склериты
с очень короткими, широкими передними отростками, передний край
которых простирается почти до уровня передней границы дорзальной
перемычки. Вентральная перемычка широкая, далеко выступающая;
-фарингеальная мембрана очень широкая. Вентральные отростки склерита
ниже дорзальных с угловидным выступом по верхнему краю, на концах
заострены; дорзальные отростки тонкие, суженные к вершине, на конце
закругленные. Дорзальная перемычка массивная.
Ложнококон рыжевато-коричневый, иногда с белесоватым или серо-
ватым оттенком. Ложнококон сохраняет признаки личинки последней
стадии (анальная пластинка, шипики позади нее и т. д.).
Распространение. Северная граница ареала вида прохо-
дит в лесной зоне по линии Луга, Ленинградской обл., — Гремячка,
Рязанской обл., — ст. Белебей, Башкирской АССР. Восточнее последнего
пункта в этих широтах вид не найден. Южная граница ареала совпадает
с южной границей Советского Союза от Молдавии до низовьев р. Или
(Казахстан), которая является крайней восточной точкой нахождения
вида в СССР. В Сибири вид отсутствует; северная и восточная границы
ареала между Уфой и р. Или идут на юг до Жигулей и далее — на Чкалов,
Чиили, Казалинск и Мерке, а оттуда — на о. Балхаш. Вне СССР вид
распространен в средней и южной Европе, в северной Африке и в северной
Монголии.
М. larvipara — живородящая муха. Она рождает живую личинку
(в искусственной лабораторной обстановке иногда — яйцо) I стадии,
реже II. Через 2—4 дня (при 28—38° С) личинка достигает предельной
зрелости и уходит в почву для окукления. Потомство муха пристраивает
на свежий коровий помет, который предпочитается всем остальным суб-
стратам. В редких случаях (Вайнштейн, Родова, Романов) личинки
встречаются в свином и ослином помете. При высоких температурах
субстрата, муха вылупляется из ложнококона через 6 (26—30° С),
4 (33° С), 7—8 (30—31° С) дней; по данным Дербеневой-Уховой, этот
период длится 15—20 дней, что, видимо, относится к случаям, связан-
ным с неблагоприятными условиями воспитания объекта. В южных
широтах (в Крыму, южном и центральном Таджикистане) личинки М. lar-
vipara являются одним из основных обитателей коровьего помета,
составляя в нем до 70—80%. Наибольшее обилие личинок в коровьем
помете наблюдается в июле или, на крайнем юге, — июне и с конца
августа по середину сентября.
Мухи относятся к числу кроволижущих форм, причем эта особен-
ность свойственна главнЬгм образом самкам; самцы обычны на помете,
навозе и растениях. Нападают мухи на животных преимущественно
154
I. ТРИБА MUSCINI
на пастбищах и в открытых загонах; в темные стойла залетают неохотно.
Как многие кроволижущие виды, М. laroipara часто следуют за истин-
ными кровососущими формами (слепнями, жигалками), пользуясь све-
жими кровоточащими ранками после их укусов.
Вид имеет несомненное значение в распространении патогенных
организмов среди животных, что подтверждается данными исследований
Украинского института экспериментальной ветеринарии (см. М autum-
nalis Deg.).
*14. Musca convexifrons Thoms, (рис. 224, 225).
Thomson, 1868, Eugenies Resa, Dipt. : 547; Stein, 1917 : 334; Patton, 1923,
Philipp. Journ. Science, XXIII : 333, 334; он же, 1925, там же, XXVII : 186, 198:
Patton, 1922, Bull. ent. Res., XII : 425—426; он же, 1933, Ann. trop. Med. Parasitol.,
XXVII : 417—420, figs. : 14, 15; Karl, 1925, Arb. morph, taxon. Entom., II : 33
(Eumusca); Ho, 1938, Ann. trop. Med. Parasitol., XXXII : 287—312, figs. 13b, 8c
(Viviparomusca).
о. Весьма близок к предыдущему, от которого отличается следую-
щим. 3-й членик усиков сампа длиннее (длина его в 3—Зт/а раза превы
шает таковую 2-го членика
усиков). Спинка груди с
очень резко выраженными
черными и блестящими бе-
лыми полосами налета, осо-
бенно в переднем отделе и
вдоль боковых границ; корич-
неватый налет ясно выра-
жен в основной и централь-
ной части щитка; жилка
г4+5 сверху с несколькими
(1—4) мелкими волосками у
основания, снизу с мелкими
щетинками в основной трети
или при основании и за г — т
поперечной; у некоторых осо-
бей щетинки располагаются
без перерыва от основания,
вплоть до вершинной четвер-
ти жилки. Брюшко красно-
вато-желтое с черным рисун-
ком; покрыто сильно бле-
стящим белым налетом, име-
ющим золотисто-желтый от-
тенок на последних сегмен-
тах; средняя черно-бурая по-
лоса на 3-м и 4-м тергитах в
коричневом налете. 1-|-2-й
Рис. 224. Самец Musca convexifrons Thoms, тергит сверху черный в основ-
(Ориг.). ной половине (редко в 2/з),
с узкой полоской посредине,
достигающей заднего края сегмента; задние боковые отделы и низ тер-
гита — желтые; 3-й и 4-й тергиты красно-желтые с продольной черно-
бурой полосой очень слабо суженной назад и образующей поперечную
перевязь (ширина ее Vs—Vs длины тергита) на 4-м и не всегда узкую
2. MV SC A
155
перевязь на 3-м тергите; 5-й тергит бурый, с желтыми боковыми краями
у основания; темные боковые переливчатые пятна отчетливо выражены,
с узкими границами (шире, чем у М. larnpara}, вдоль заднего края дают
расширение в виде поперечной полоски, переходящей и на вентральную
сторону; ширина темного бокового пятна на 4-м тергите почти равна
Рис, 225. Самка Musca convexifrons Thoms. (Ориг,).
светлому промежутку, лежащему сбоку срединной черной полосы
брюшка; 5-й тергит с проходящими по всей длине, суженными назад
тремя резкими, черно-бурыми полосками, которые придают сегменту
продольно-полосчатый вид. V стернит с короткими, резко изогнутыми
к середине задними отростками и выступающими далеко боковыми зад-
ними углами; церки с почти прямыми боковыми краями и широко закруг-
ленными наружными дистальными углами; внутренние выросты корот-
кие, ясные; передняя часть парамер с резким передне верхним углом;
в остальном гениталии самцов сходны с М. larvipara.
158
I. ТРИБА MUSCINI
9. Лоб сильнее сужен назад чем у М. larvipara (ширина его состав-
ляет 2/з—l/i ширины глаза); орбиты в два раза уже бархатисто-черной,
с серебристо-серыми краями, выпуклой по боковым краям лобной полосы;
лобные щетинки крепкие, расположенные в один ряд; щеки в Р/2—I1/*
раза шире 3-го членика усика. Спинка с резкими продольными полос-
ками. Брюшко черно-бурое, края тергитов и стернитов снизу на большем
или меньшем протяжении палево-желтые; 1 2-й тергит снизу целиком
светлый; боковые переливчатые пятна на 3-м и 4-м тергитах очень резкие,
образующие по заднему краю тергитов отчетливую темную полосу шири-
ной Vs—Ve длины тергита, переходящую и на задний край вентральной
стороны тергитов (у М. larvipara пятна не распространяются на вентраль-
ную сторону); 5-й тергит с более или менее резким всегда развитым рисун-
ком из темных и светлых переливчатых пятен. Крылья как у самца.
Длина тела 7—9 мм.
Распространение. Известен из Приморского края: Яков-
левка, Сучан, Седанка, Покровка, Тигровая, Евгеньевна, Каменушка,
ст. Пограничная, а также у Хабаровска. Северный, центральный и южный
Китай, включая Тайван.
М. convexifrons живородяща; личинка откладывается в I стадии,
подготовленной уже к линке на II. Развитие проходит в помете круп-
ного рогатого скота.
Как в биологическом, так и в морфологическом отношениях вид
весьма близок М. larvipara. По данным Крастина (1949), мухи являются
промежуточными хозяевами паразитической нематоды — Thelazia rho-
desi Desm.
15. Musca lusoria Wd. (рис. 196, 226—229).
Wiedemann, 1830, Ausser. zweifl. Insekt., II : 411; Patton, 1925, Philipp. Journ.
Science, XXVII : 185, 198, 199; Malloch, 1925, Ann. Mag. Nat. Hist., XV : 135 (Vivi-
paromusca); Seguy, 1933, Memor. estud. Mus. zool. Univ. Coimbra (I), 67 : 61, 63; Karl,
1935, Arb. morphol. taxon. Entomol., II : 29—49 (Eumusca); Patton, 1932, Ann. trop.
Med. Parasitol., XXVI : 347—405, figs. 3a, 7a, 8c, 14, 15; он же, 1936, там же,
XXX : 481, fig. 5а; Но, 1938, Ann. trop. Med. Parasitol., XXXII : 287—
312, fig. 13a fViviparomusca).
$. Глаза голые, лоб узкий, равен х/ю ширины глаза; скулы серебри-
стые, спинка серая с 4 широкими продольными черными полосами; перед-
ние и задние супрасквамальные щетинки имеются; жилка г4+5 снизу
с редкими волосками, развитыми по всей длине жилки до вершины крыла;
редко небольшой участок дистального отдела жилки лишен волосков.
Брюшко оранжевое, с темными отметинами и пятнами серебристого
налета. Тергит 1-|-2-й черный, 3-й — оранжевый с срединной черной
полосой, по бокам от которой располагаются пятна серебристого налета,
распространяющегося на задний край сегмента, часто затемненного;
4-й тергит с более узкой черной полоской, слабо выделяющимися пят-
нами светлого налета и яснее выраженной темной перевязью по зад-
нему краю. Средина 5-го тергита затемнена, поверхность его в серебри-
стом налете. Церки с сильно вытянутыми, пальцевидно изогнутыми
внутренними углами и лопастевидно расширенными наружными углами;
нижний край церок между наружными и внутренними углами выемча-
тый. Склеротизованная часть фаллозомы равна по длине мембранозной
части; задний отросток фаллозомы сильно расширен на конце и снабжен
выемкой на дистальном конце. Передний отдел парамер длинный, широ-
кий, с 3-5 щетинками, из коих 2 длиннее остальных; задняя часть пара-
2. MUSCA
157
мер длинная, изогнута и несет несколько шипиков. Г стернит относи-
тельно длинный, широкий, по бокам затемненный, по заднему краю
с широкими, загнутыми внутрь выступами, на конце зазубренными.
Лоб равен или почти равен ширине глаза; лобные щетинки
с каждой стороны от лобной полосы располагаются в два ряда. Спинка
как у самца. Брюшко черное в сером налете, образующем пятна, рас-
положенные в шахматном порядке; 1-^-2-й тергиты черные, с серыми
пятнами по бокам от средней
линии; 3-й тергит в сером на-
лете, с черной срединной по-
лоской; темные переливчатые
пятна расширены назад, обра-
зуя перевязь; задний край
сегмента серебристый; 4-й
тергит сходен с 3-м; 5-й тер-
гит в серебристо-сером томен-
те и с темной серединой.
По Пэттону, личинка
III стадии имеет 6—8-лу-
чевые передние дыхальцы.
Задние дыхальцы широкие,
крупные, сильно склеротизо-
ваны; щели их узкие, очень
длинные, сильно изогнутые.
Пупарий белый.
Распростране-
ние. Вид распространен в
Эфиопской области (цент-
ральная и южная Африка)
и на Цейлоне.
М. lusoria Wd. является
живородящей. Развитие ли-
чинок проходит в помете
Рис.. 226—229. Части генитального аппарата
самца Musca lusoria Wd.
226 — фаллозома с парамерами. 227— дорзаль-
ный отросток спереди. 228 — V стернит брюш-
ка. 229 — церки. (По Пэттону).
крупного рогатого скота.
Вид относится к кро-
волижущим формам, обыч-
ным на домашних животных.
Часто попадается на ранах
и у глаз животных.
16. Musca xanthomelas Wd. (рис. 215, 235—241).
Wiedemann, 1830,'Aussereurop. zweiflugel. Insekt., ll : 416; Stein, 1917 : 110
(Graphomyia)', Patton, 1923, Philipp. Journ. Science, XXIII : 333: он же, 1925,
там же, XXVII ; 184, 198; он же, 1932, Ann. trop. Med. Parasitol., XXVI : 387, 391—
392; он же, 1933, там же, XXVII : 423—426, figs. 19, 20а; он же,. 1937, там же,
XXXI ; 199; Но, 1938, Ann. trop. Med. Parasitol., XXXII : 287—312, fig. 12d
(Eumusca)', Patton, 1936, XXX : 481—482.
$. Лоб линейный, верхняя часть глаз с укрупненными фасетками;
нижняя часть скул в серебристом налёте. Грудь в голубоватом налете,
с 4 широкими продольными полосами, у темноокрашенных особей сли-
вающимися попарно. Брюшко оранжевое, с серебристыми пятнами налета
и узкими темными полосками; тергит 1-|-2-й от темно коричневой До чер-
ной окраски, реже задний край оранжевый; 3-й тергит оранжевый
Рис. 230—234. Части генитального аппарата самца
Musca mesopotamiensis Ptt.
230 — фаллозома с парамерами. 231 — дорзальный
ее отросток спереди. 232 — V стернит. 233 — церки.
234 — парамеры. (По Пэттону).
Рис. 235—241. Части генитального аппарата самца Musca xantho-
melas^NA.
235 — фаллозома. 236 — дорзальный ее отросток спереди. 237, 238 —
оп же в других положениях. 239 — V стернит. 240 — церки. 241 —
парамеры. (По Пэттону).
2. MUSCA
159
с широкой черной полосой, расширенной вдоль переднего края сегмента;
по бокам черной окраски располагаются серебристые полосы налета,
боковые края тергита серебристые; 4-й тергит с более узкой черной
полосой, расширяющейся вдоль заднего края; 5-й тергит темный в сере-
бристом налете. Наружные углы церок широко закругленные, внутрен-
ние — оттянутые вниз наподобие сосочков; склеротизованная часть фал-
лозомы составляет всего V2 длины мембранозной части; задний отросток
сильно капорообразно расширен; передняя часть парамер маленькая
с 2—3 сравнительно длинными щетинками; задний отдел парамер широ-
кий, изогнутый с одним выступающим у середины бугорком, несущим
сензорные образования в виде шипиков; V стернит вытянутый, темно-
бурый в дистальном своем отделе; отростки его заднего края длинные,
сильные, зазубренные.
Орбиты и скулы серебристые, ширина лба равна ширине глаза, или
уже его (3/<), ширина лобной полосы составляет V2 ширины лба; лобные
щетинки расположены с каждой стороны в 2 ряда; окраска спинки
у самки в общем светлее, нежели у самца. Брюшко с пятнами серебри-
стого налета и темными перевязями; тергит l-f-2-й от светлооранжевой
до темнокоричневой окраски. 3-й тергит темно или светлооранжевый,
с узкой темной срединной полосой и узкой ясно заметной темной пере-
вязью на заднем крае; 5-й тергит с такими же темными отметинами.
Помимо строения, гениталий вид отличается от соседних видов этой
группы темными перевязями по заднему краю 3-го и 4-го тергитов, сере-
бристо-белым теменем и щеками у обоих полов и голубовато-белым нале-
том на среднеспинке.
Распространение. М. xanthomelas распространен по всей
Африке, в Индии, где является обычной формой, в южном Китае, вплоть
до Кантона и острова Хайнан, а также и на Яве.
Муха откладывает яйца в коровий помет на выпасах; яйцо имеет
крючковидно загнутый отросток на верхнем полюсе, как у М. autumnalis.
Взрослая личинка серовато-белая с красновато-желтой дорзальной
поверхностью. Передние дыхальцы с 7—8 лучами; задние дыхальцы
крупные, расставленные друг от друга на */< поперечника дыхальца,
сильно склеротизованы. Пупарий белый.
Взрослые насекомые обычны на животных и вблизи них на паст-
бищах. Кроволижущий вид.
17. Musca ventrosa Wd. (рис. 242—245).
Wiedemann, 1830, Aussereur. zweifliig. Insekt., II : 657; Stein, 1917 : 105; Pat-
ton, 1922, Bull, entom. Research, XII : 419; он же, 1923, Philipp. Joum. Science,
XXIII : 310, 311, 315, 331; Malloch, 1925, Ann. Mag. Nat. Hist., XV : 133—136, fig.3;
Patton, 1932, Ann. trop. Med. Parasitol., XXVI : 387, 391, 392; он же, 1933, там же,
XXVII : 405—407, figs, :1с, 9b, 10; Seguy, 1933, Memor. estud. Mus. zool. Univ.
Coimbra (I), 67 : 63; Ho, 1938, Ann. trop. Med. Parasitol., XXXII : 287—312.
$. Темя линейное, нижняя часть темени и щеки серебристые, верх-
няя часть глаз с укрупненными фасетками; спинка в темно грифельно-
голубом налете и 4 широкими продольными черными полосами. Брюшко
оранжевое, лишено, как правило, темных отметин, реже имеет затемне-
ние по средней линии 3-го и 4-го тергитов. Церки типа М.. sorbens', наруж-
ные их углы округлены, внутренние — без сосцевидных выступов; фал-
лозома с тонкой склеротизованной частью, стянутой у средины, по длине
почти равной мембранозной; задний ее отросток сравнительно короткий,
на конце расширенный, с вильчатым раздвоением; парамеры с 1 длинной
160
I. ТРИБА MUSCINI
и 3—4 мелкими щетинками; задняя часть парамер выступает в форме
конического отростка, его вершина светлая. Длина I стернита более
ширины, задние его выступы ± короткие, широкие, на конце зазубрены.
Темя широкое, ширина его чуть более х/г ширины глаза; лобная
полоса красноватая, занимает 3/< ширины лба, боковые края слабо
выпуклые; лобные щетинки располагаются в один ряд с каждой стороны
полосы; спинка и брюшко как у самца.
Вид легко отличим от соседних видов красно-желтой окраской брюшка,
лишенной темных отметин, или в редких случаях имеющихся в форме
узких срединных штрихов на 3-м и 4-м тергитах.
245
Рис. 242—245. Части генитального аппарата самца
Musca ventrosa Wd.
242 —фаллозома. 243 — дорзальный ее отросток спе-
реди. 244 — И стернит. 245 — церки. (По Пэттону).
Распространение. М. ventrosa — один из обычных видов
рода в Ориентальной и Эфиопской областях. Он известен с островов
Малайского архипелага (Целебес, Суматра, Ява), с Тайвана, из конти-
нентального Китая, достигая северо-восточных его границ и проникая,
таким образом, в пределы Палеарктики.
Вид развивается в лепешках коровьего помета в поле. Муха отно-
сится к числу яйценесущих форм.
М. ventrosa является кроволижущим видом. Мухи слизывают выде-
ления с поранений кожи, слизистых оболочек глаз и носа животных.
18. Musca hervei Vill. (фиг. 214, 246—249).
Villeneuve, 1922, Ann. Sci. Nat., Zool., 10, 5 : 335; Patton, 1932, Ann. trop. Med.
Parasitol., XXVI : 389; он же, 1933, там же, XXVII : 426—428, figs. : 6,19, 21; он же,
1937, там же, XXXI : 199; Но, 1938, там же, XXXII : 287—312 (Viviparomusca).
i. Musca
lei
$. Лоб линейный, орбиты и скулы в серебристом налете, глаза слабо
волосистые, спинка в голубовато-сером налете с 4 широкими черными
полосами; задняя часть спинки и щиток в серовато-желтом налете.
Брюшко оранжевое в серебристом и желтовато-сером налете; тергит
l-f-2-й черный с маленьким округлым пятном серебристого налета
у середины заднего края, задний край часто коричневый; 3-й тергит
оранжевый, с широкой черной срединной полосой, по бокам от которой
располагается по серебристому или желтоватому пятну налета; 4-й тер-
гит сходен с 3-м, но с более узкой черной полосой и темной перевязью
вдоль заднего края, светлые пятна налета более желтого тона; 5-й тер-
Рис. 246—249. Части генитального аппарата самца
Musca hervei Vi 11.
246 — фаллозома. 247 — дорзальный ее отросток спе-
реди. 248 — V стернит. 249 — церки. (По Паттону).
гит почти полностью кремовый, с темным плохо выраженным продоль-
ным штрихом. Стерниты и края тергитов черные. Церки, как у М хап-
thomelas, но внутренние выступы длиннее и тоньше. Склеротизованная
часть фаллозомы широкая, короче мембранозной части; задний ее отро-
сток, длинный, сильно расширенный; передняя часть парамер длинная
с одной длинной и двумя короткими щетинками; задний отдел длинный,
несколько заостренный на вершине, изогнутый; V стернит маленький,
сплошь черный, с узкими боковыми выступами.
Орбиты в серебристом налете; ширина лба составляет 3/г ширины
лба; лобные щетинки иногда трехрядные. Глаза в реденьких мелких
волосках; спинка в сером или серовато-желтом налете, более темном
в ее переднем отделе, с 4 черными полосами. Тергит 1 -|-2-й как у самца,
3-й — черный, с широкими белыми или желтовато-белыми полосами
налета по бокам от средней чернбй полосы и серебристо белыми пятнами
11 Фаука СССР, том XVIII, вып. 4.
.162
I.- ТРИБА MUSCINI
у заднего края; 4-й — с более узкой черной полосой; в остальном как 3-й;
5-й тергит почти сплошь желтоватый или с темными боковыми пятнами;
стерниты и края тергитов желтые или кремовые.
Взрослая личинка имеет на передних стигмах 7—8 пальцевидных
выростов; задние дыхальцы с сильно хитинизированной пластинкой,
с очень узкими и длинными щелями. Расстояние между дыхальцами
почти равно 1/з поперечного диаметра пластинки. Ложнококон светлый
серовато-коричневый.
Распространение. Большая часть континентального
Китая, северная Индия.
Личинки мухи развиваются в коровьем помете на выпасах. Мухи
яйценесущи.
От близких видов М. hervei легко отличается по наличию волосков
на глазах (у самок мелких и редких). Самцы, помимо того, отличаются
от близких видов — М. autumnalis, М. convexifrons, М. larvipara —
желто-красной окраской брюшка и затемненными вентральными краями
тергитов и стернитами.
3. Род PYRELLIA R.-D.
Robineau-Desvoidy, 1830, Essai Myodaires ; 462; Rezzi u. Stein, 1907, Catal,
Palaarkt. Dipt., Ill : 606; Seguy, 1923: 360 (частью); Stein, 1916 : 17 (частью);
Schnabl u. Dziedzicki, 1911 : 177 (частью); Malloch, 1923, Ann. Mag. Nat. Hist. (9),
XII : 505—526 (частью); Karl, 1928 : 9 (частью).
Тип рода: Pyreilia aenea Zett. (1862).
Глаза голые, жилка гг, без щетинок, изгиб т в вершинной части
дуговидный; супрасквамальное ребро и переднегрудь без щетинистых
волосков.
Лоб самца узкий, орбитальные щетинки отсутствуют. Боковые
лопасти церок вытянутые, листовидные или треугольные, внутренние
выступы очень маленькие; фаллозома узкая, с небольшим количеством
шипиков в вершинной части, дорзальный отросток с мембранозной ниж-
ней поверхностью.
Фаллозома длинная, основная ее склеротизованная часть в Р/г—
2 раза короче мембранозного дистального отдела; последний расширен
к вершине, где заканчивается прозрачными, направленными вперед
лопастями, поверхность которых усажена плоскими крупными шипами;
дорзальный и вентральный края фаллозомы укреплены не всегда чет-
кими тяжами, несколько напоминающими по местоположению гипофалл
и парафалл Calliphoridae. Дорзальный отросток основного отдела фал-
лозомы короткий, реже удлиненный, с мембранозным вентральным краем
и склеротизованной дорзальной стороной, слабо изогнут на дистальном
конце; аподема тонкая, длинная, слабая склеротизованная на вершине,
соединяется основанием с фаллозомой и основанием передних отделов
парамер, которые длинные, треугольно расширенные у основания и снаб-
жены здесь несколькими длинными щетинками; дистальный конец фал-
лозомы мембранозный; задние отделы парамер короткие, слабо склеро-
тизованы в основной части и покрыты короткими шипами; вершинный
их отдел перепончатый и несет многочисленные тонкие щетинистые
волоски. Церки с сильно развитыми, треугольно или листовидно оттяну-
тыми боковыми лопастями и очень мелкими, коротенькими, обычно тре-
угольной формы внутренними выступами. Кокситы массивные, широкие,
с крупными лопастевидно выдающимся выступом по внутреннему краю.
3. PYHELLIA
16?
Род имеет много общих черт с родами Musca L., Orthellia R.-D. и
Dasyphora R.-D.
От Orthellia род отличается отсутствием супрасквамальных волосков
и узкой переднегрудью, обычно лишенной опушения. Только у двух видов
Рис. 250—258. Заднегрудное дыхальце, переднегрудь п вершина крыла.
250—252 —Pyreilia (in sp.) sp. 253—255 — РугеШа (Rypellia) senii-
lutea Mall. 256—258 — Orthellia cornicina F. (Ориг.). . .
переднегрудь широкая, а у подрода Dasyphoromima nov. — опушенная;
у последнего подрода заднегрудное дыхальце вытянутое. От рода Dasy-
phora род отличается голыми глазами и отсутствием щетинок на жилке п.
11*
164
I. ТРИБА MUSCINI
Род Pyreilia R.-D. распадается на 3 подрода. Центральным подро-
дом, охватывающим все известные нам палеарктические формы, является
подрод Pyreilia R.-D. s. str. с типом aenea Ztt. ПодродDasyphoromima
subgen. nov. «связывает» Pyreilia с родом Dasyphora на основе хетотакси-
ческих признаков настроения задних грудных стигм. Подрод Pyreilia
МаП. сближает род Pyreilia с родом Orthellia на основе строения
переднегруди, опушения ее и строения задних дыхалец груди. Род наи-
более богато представлен в Эфиопской и Неотропической областях: число
видов в первой достигает 23, во второй— 16; затем следует Ориенталь-
ная область, из которой известно 15 видов. Палеарктика, насчитываю-
щая 6, и Неарктическая область с 4 видами являются наиболее бедными.
Можно считать, что наибольшего расцвета род достигает в экваториальных
странах всех континентов.
Личинки всех известных представителей рода — копрофаги, раз-
виваются в помете различных позвоночных животных.
ОПРЕДЕЛИТЕЛЬНАЯ ТАБЛИЦА ПОДРОДОВ
1 (2). Переднегрудь широкая, в тонких волосках; надкрыловых щети-
нок — 2, предосновные на щитке ясно развиты, дорзоцентраль-
ных 3+4, стерноплевральных — 1-(-3...............................
................................3. Dasyphoromima, subg. nov.
2 (1). Переднегрудь без волосков; предкрыловых щетинок — 1 или 2;
предосновные щетинки щитка не развиты; дорзоцентральных — 2-|-
+4; стерноплевральных — 1+2, реже 1+3.
3 (4). Заднегрудное дыхальце длинное, крупное (рис. 253), стерноплев-
ральных щетинок 1-|-3, надкрыловых — 2; переднегрудь широкая
(рис. 254)....................................... 2. Rypellia Mall.
4 (3). Заднегрудное дыхальце треугольное, маленькое (рис. 250), его
вертикальный диаметр короче расстояния от его нижнего края до
нижнего края гипоплевры; стерноплевральных щетинок — 1+2
(или 0-4-2, реже 1+3); надкрыловых — 1; переднегрудь узкая
(фиг. 251)........................... . . . 1. Pyreilia R.-D. s. str.
1. Подрод Pyreilia R.-D. s. str.
Robineau-Desvoidy, 1830, Essai Myodaires : 462.
Тип подрода: Pyreilia aenea Ztt.
Заднегрудное дыхальце маленькое, треугольное; вертикальный диа-
метр его много меньше расстояния между нижним краем дыхальца и ниж-
ним краем гипоплевры. Переднегрудь узкая, желобовидная, реже (Р. са-
daoerina L.) расширена, всегда лишена волосков, как и супрасквамальное
ребро. Стерноплевральных щетинок — 1+2 (0+2 или 1+3), надкрыло-
вых — 1; щиток без предосновных щетинок; средние голени без передне-
дорзальных щетинок или они слабо развиты. Лоб самца узкий, орбиталь-
ные щетинки отсутствуют; у самки они имеются в числе двух с каждой
стороны; орбиты самки уже лобной полосы, скулы и щеки узкие. Жилка
т в вершинном отделе с дуговидным изгибом по направлению к перед-
нему краю; жилка г, без щетинок, г4+5 с волосками снизу и мелкими
щетинками сверху от основания до или почти до гт. Гениталии самцов:
церки широкие, с листовидно или треугольно вытянутыми боковыми лопа-
стями и мелкими внутренними отростками; кокситы массивные, с широ-
ким выступом по вентральному краю.
Подрод является центральным в роде и наиболее богатым видами.
3. PYRELLIA
165
ОПРЕДЕЛИТЕЛЬНАЯ ТАБЛИЦА ВИДОВ
1 (2). Переднее грудное дыхальце бурое или буровато-рыжее, тело ли-
шено налета; стерноплевральных щетинок — 1-J-3; переднегрудь
широкая, желобовидная; чешуйки крыла у самцов часто затемнен-
ные ...................................*1. Р. (in sp.) cadaverina L.
2 (1). Переднее грудное дыхальце белое или светлопалевое; тело без
налета или с развитой полосой светлого налета на спинке.
3 (4). Продольная полоса белого налета ясно развита в переднем отделе
среднеспинки..............................*4. Р. (in sp.) aenea Ztt.
4 (3). Среднеспинка блестяще зеленая, лишена налета.
5 (8). Лоб уже 3-го членика усиков (самцы).
6 (7). Скулы в нижнем своем отделе почти в 2 раза шире своего верх-
него отдела на уровне 1-го членика усиков, стерноплевральных
щетинок, как правило, 0-{-2; маленькая щетинка на переднедор-
зальной поверхности средних голеней имеется .......................
.................................*2. Р. (in sp.) secunda, sp. n.
7 (6). Ширина скулы в нижнем ее отделе превышает ширину ее верхнего
отдела не более чем в I1/*—IV2 раза; передне-верхняя стерноплев-
ральная щетинка имеется; переднедорзальная щетинка на сред-
них голенях отсутствует................*3. Р. (in. sp.) minuta, sp. n.
8 (5). Лоб в несколько раз шире 3-го членика усиков (самки).
9 (10). Ширина лба равна ширине глаза; орбиты в 2 раза уже лобной
полосы; 5-й тергит в 13/4—2 раза длиннее 4-го тергита..............
...................................*2. Р. (in sp.) secunda, sp. n.
10 (9). Ширина лба составляет 2/3 ширины глаза; орбиты в 3—4 раза
уже лобной полосы; 5-й тергит почти равен по длине 4-му тергиту
или едва превосходит его................*3. Р. (in sp.) minuta, sp. n.
*1. РугеШа (in sp.) cadaverina L. (рис. 36, 259, 264, 265, 267—273,
283).
Linn6, 1761, Fauna Suec., Ed. II : 451 (Musca)-, Schnabl u. Dziedzicki, 1911 :
177, figs. 634, 644; Stein, 1916 : 18; Malloch, 1923, Ann. Mag. Nat. Hist. (9), XII :
518; Karl, 1928 : 9; Karl, 1935, Arb. morph, taxon. Entom., II : 35.
Биология. Порчинский, 1910 : 35—37, фиг. 37; Keilin, 1917, Parasitology,
IX : 422, 423; Зимин, 1948, 79, фиг. 43.
<5. Лоб и скулы в верхнем отделе в 2 раза уже 3-го членика усиков;
лоб в средине сильно суженный; скулы в нижнем отделе в 2 раза шире,
чем в верхнем; лобная полоса в наиболее узком месте имеет характер
черной бороздки, слабо расширенной в передней и задней части; высота
щеки в 3Va—4 раза менее вертикального диаметра глаза; передний край
рта далее выступает вперед, нежели лоб; голова короткая, линия лба при
рассматривании сбоку сильно наклонена вниз; хоботок короткий; щу-
пальцы черно-бурые, слабо утолщенные к вершине. Затылок металли-
чески черно-синий или зеленый; щеки блестящие, черные; прочие части
головы черно-бурые; скулы и передняя часть орбит в серовато белом
шелковистом, лицо в матовом сером налете. Кроме внутренних теменных
щетинок и угловатых вибрисс, щетинки головы имеют характер тонких
черных волосков. Орбитальные щетинки отсутствуют. Грудь металлически
блестящая, зеленая или сине-зеленая, лишена налета; грудные дыхальцы
чернобурые; поверхность груди в тонких, коротких бурых волосках без
светлого налета. Щетинки груди: акростихальных — 1 пара, перед щит-
ком; дорзоцентральных — 24-4; интраалярных — 1, за швом; предкры-
186
I. ТРИБА MVSCINI
ловых— 1, короткая, тонкая; надкрыловых — 1; закрыловых — 2—3
(задняя в виде волоска); плечевых — 3; заплечевых — 1; предшовных —1;
нотоплевральных — 2; стерноплевральных — 1-|-3 (2, 4); гипоплевры
с группами мелких черных волосков перед задним дыхальцем и над зад-
ними тазиками; супрасквамальное ребро, передняя плевра и передне-
грудь лишены волосков. На щитке 3 пары щетинок: вершинных — 2,
боковых — 2, смещенных к диску щитка; основных — 2; предосновные
щетинки отсутствуют; расстояние дискальных щетинок от вершинных
в два раза меньше чем от основных щетинок. Ноги черно-бурые, бедра
с зеленым или синим металлическим блеском, в черных волосках; коготки
передних ног на Сз короче 5-го чле-
ника лапки. Средние голени в нижней
половине с одной передне-дорзальной
и двумя неполными рядами задне-
дорзальных и задневентральных ще-
тинок ; задние бедра снизу по передне-
вентральной и задне-вентральной по-
верхности с длинными тонкими ще-
тинками и длинными щетинистыми
волосками. Крылья сероватые, жилки
коричневато-рыжие; жилка п — го-
лая, г4+5 — снизу при основании
с 1—3 волосками сверху, до г—т
с редкими тонкими щетиночками;
т — в вершинной половине с мяг-
ким дуговидным изгибом по направ-
лению к переднему краю крыла; груд-
ная чешуйка белая, с желтым краем,
реже слабо дымчатая; жужжальца
желтые. Брюшко цвета груди, блестя-
щее, в черных волосках, более длин-
ных на 5-м тергите и на вентраль-
ной стороне брюшка. Края 3-го, 4-го
п 5-го тергитов с рядами щетини-
стых волосков. Церки широкие, с
Рис. 259. Самка Pyreilia cadaverina Mg. длинными ЛИСТОВИДНОЙ формы боко-
(Ориг.). выми лопастями; внутренние высту-
пы короткие, широкие с прямым
вершинным краем; кокситы неправильно треугольной формы с дуго-
видно округленным наружным краем, заостренным внутренним дисталь-
ным углом и широким, полукруглым свободным краем.
j. Лоб равен ширине глаза; блестяще черные орбиты в Ц/г-—2 раза уже
лобной полосы, имеющей выпуклые края; лобная полоса матово-черная
с одиночными волосками в заднем отделе с боков; орбиты в редких чер-
ных волосках; орбитальные щетинки в числе двух с каждой стороны,
из них задняя много короче передней; теменные наружные и внутренние
щетинки крепкие; ширина скулы чуть менее или равна ширине орбиты;
скулы в шелковистом серовато-белом, лицо в белом налете. Коготки
передних ног чуть короче 5-го членика лапки. Окраска как у самца; мар-
гинальные щетинки на верхней стороне тергитов мелкие; длина их едва
достигает 1/з длины тергитов. Длина тела 4—6.5 мм.
Личинка III стадии (рис. 267—273). Среднего размера, серовато-
елтые личинки с тонкой гладкой блестящей кутикулой. Головной сег-
3. PYRELLIA
167
мент глубоко раздвоен спереди; антенны с длинным цилиндрическим
1-м и коротким конусовидным 2-м члеником; нижняя поверхность сег-
мента вокруг ротового отверстия покрыта различными по форме мяг-
кими бугорками, кнаружи от которых радиально располагаются ротовые
бороздки, покрывающие часть нижней, боковые и среднюю область верх-
ние. 260—264. Головы видов рода Pyreilia.
260 — самца Р. secunda, sp. п., сбоку. 261 — самки того
же вида сверху. 262 — самца Р. minuta, sp. п., сбоку.
263 — самки того же вида сверху. 264 — самки Р. са-
daverina L. сверху. (Ориг.).
ней поверхности сегмента (на дорзальной поверхности бороздки правой
и левой стороны почти соприкасаются друг с другом). 1-й грудной сег-
мент по переднему краю с мелкими многорядными шипиками, занимаю-
щими около х/5 длины сегмента. В остальных частях 1-й грудной сегмент
гладкий, блестящий, как и последующие грудные и 1-й брюшной сег-
менты. Брюшные сегменты со 2-го по 7-й снизу по переднему краю с по-
перечными валикообразными вздутиями, покрытыми шипиками разного
размера. Средний ряд крупных шипов дуговидный, задние ряды не всегда
168
I. ТРИБА MUSCINI
правильные, состоят из более мягких шипиков, сзади которых (обычно
вблизи центра вздутия) имеется по 3—4 более крупных шипа, обращен-
ных вершинами назад и к средней продольной линии сегмента. Брюшные
сегменты со 2-го по 6-й снизу, по обе стороны от средней продольной линии
сегментов, у заднего края с группой конических бугорочков, стоящих
в 2(1) неправильных ряда. 8-й брюшной сегмент чуть короче 7-го; задняя
поверхность в центре вокруг дыхалец с легким вдавлением; анальная
пластинка относительно широкая, короткая, концы ее загнуты назад.
Б области анальной пластинки 7 бугорков: 2 крупных изогнутых суб-
анальных, 2 — маленьких экстраанальных, 2 — не всегда отчетливо
выраженных анальных и 1 постанальный, покрытый мелкими шипи-
ками. Вдоль обоих краев анальной пластинки расположены светлые шипы;
шипы вдоль заднего края пластинки стоят в 2 ряда и, подобно передне-
крайним, достигают концов анальной пластинки. Ротовой аппарат круп-
Рис. 265—266. Фаллозома.
265 — Pyreilia cadaverina L. 266-—Dasyphora pratorum. Mg. (Ориг.).
ный, с широкими ротовыми крючками. Дополнительные склериты у вен-
трального края ротовых крючков в виде двух хитиновых узких тяжей.
Базальный отдел ротовых крючьев с оттянутым вперед вентральным углом;
зубные склериты крупные, неправильно треугольных очертаний; гипо-
стомальные склериты короткие, широкие, в 2 раза короче ротовых
крючьев, неправильной формы. Фарингеальные склериты с очень корот-
кими и широкими передними отростками, едва выступающими вперед
за уровень дорзальной перемычки; фарингеальный склерит с широкими
вентральными отростками, снабженными на верхнем крае небольшим
угловидным выступом; дорзальные отростки почти в 2 раза уже вен-
тральных, заострены на конце и слабо дуговидно изогнуты. Фарингеаль-
ная мембрана очень широкая; вентральная перемычка широкая, с кольце-
видными отростками по заднему краю; дорзальная перемычка широкая,
с короткими задними отростками, срощенными с дорзальными краями
фарингеального склерита. Передние дыхальцы поперечные, с 7 сфериче-
скими отростками; задние дыхальцы крупные, расставленные на Ч*.—
г/з ширины дыхальца; перитрема очень широкая; дыхальцевые щели змее-
видно изогнутые, довольно широкие; внутренние концы верхней и ниж-
3. PYRELLIA
169
ней щелей и часто верхний конец средней щели обращены к крупному
стигмальному диску, прочие концы щелей направлены наружу. Длина
тела 10.5 мм; толщина у средины 1.3 мм.
Рис. 267—'273. Детали строения личинки III стадии Pyreilia cadaverina L.
267 — ротоглоточный аппарат. 268 — задний конец тела сзади. 269 — он же сбоку.
270 — перианальная область: 1 — анальная пластинка, 2— субанальные бугорки,
3 — экстраанальные бугорки, 4 — анальные бугорки, 5 — постанальный бугорок.
271—переднее дыхальце. 272 — заднее дыхальце. 273 — вооружение на нижней
стороне 5-го (2) и 6-го (1) брюшных сегментов. (По Зимину).
Распространение. В СССР вид встречается от Ленин-
градской обл. до Дальнего Востока и Закавказья и Средней Азии на юге.
Северная граница ареала проходит по линии Выборг—Ярославль-
170
I. ТРИБА MUSCINI
Калинин—Куйбышев; Чкалов, Троицк, Минусинск, Омск, Ачинск, Крас-
ноярск, Иркутск, Владивосток. Монголия, сев. Китай (Ганьсу).
Развитие мухи проходит в конском помете. Муха откладывает на
поверхность конского помета до 24—28 сравнительно крупных яиц. По
данным Порчинского, летом (наблюдения под Астраханью) развитие
мухи идет крайне быстро. В течение 1—2 суток заканчивается развитие
яиц и двух первых стадий личинок. Продолжительность фазы куколки
около 10 дней.
В конском помете, где протекает питание и развитие личинок Р. cada-
verina, последние подвергаются нападению личинок других хищных мух,
в частности личинок Polietes albolineata Fall. Этот факт ставится авто-
рами в связь с тем обстоятельством, чтоР. cadacerina сравнительно редка
в северных широтах, где встречается Polietes albolineata и, наоборот,
является обычным видом в средних и южных широтах, где этот хищник
отсутствует или редок. Порчинский указывает на частое нахождение
в теле мух филарий.
*2. Pyrellia (in sp.) secunda Zimin, sp. nov. (рис. 260,261, 278).
3. В наиболее узком месте ширина лба составляет х/з ширины 3-го
членика усиков; скулы заметно расширенные книзу, несколько уже 3-го
членика усиков; ширина их в нижнем отделе, примерно, в 2 раза шире
верхнего отдела на уровне 1-го членика усиков. Щеки узкие, высота их
не превышает Р»—г/в продольного диаметра глаза; щупальцы и усики
бурые; 3-й членик усиков в 2 раза длиннее 2-го; голова черно-бурая, заты-
лок, как и щеки, блестяще черные, с очень слабым металлическим синим
или зеленоватым отливом; желтовато-серый налет редкий, развит в ниж-
нем боковом отделе затылка; скулы в белом налете, как и передняя чет-
верть орбит, которые далее лишены его. Внутренние теменные щетинки
и вибриссы крепкие и длинные; глазковые и лобные щетинки, кроме
двух передних, короткие, тонкие, волосовидные. Грудь зеленая, сине-
зеленая или почти синяя, блестящая, лишена налета; переднее дыхальце
белое или палевое; щетинки груди: стерноплевральных — 0-J-3 или
0-[-2, очень редко 1 -|-3 (2); акростихальных — 1 пара перед щитком; дорзо-
центральных — 2-|-2 (3), позади шва наиболее сильно развита задняя
пара, впереди стоящие короткие, передние 2 пары или 1 за швом отсут-
ствуют; надкрыловых — 1; предкрыловыХ — 1; закрыловых — 2; интра-
алярных за швом — 1; плечевых — 3; заплечевых — 1; предшовных — 1;
на щитке: вершинных — 2, основных — 2 (длиннее остальных), боко-
вых — 2 (сдвинуты в верхней стороне щитка и могут быть приняты за
дискальные щетинки); предосновная в виде коротких волосков, едва
отличимых от прочих волосков щитка. Гипоплевры с 2—4 волосками
в заднем нижнем углу; переднегрудь и передние плевры голые. Заднее
дыхальце бурое.
Крылья бледносерые с желтыми жилками; жилка г4+5 с обычными
волосками с обеих сторон; чешуйки белые или палевые; жужжальца
желтые, с легким затемнением у середины. Ноги буро-черные, передняя
поверхность задних и задняя поверхность передних бедер с металлически
зеленым оттенком, блестящие. Щетинки на ногах имеют расположение,
свойственное роду Pyrellia. Средние голени с маленькой, но ясной передне-
дорзальной щетинкой в дистальной трети. Коготки и присоски короткие;
последние грязнобелые или бледножелтые. Брюшко коротко-яйцевидное,
зеленое или золотисто-зеленое, без налета, в черных коротких волосках.
Церки широкие, боковые их лопасти округло треугольные, длинные-
3. PYRELLIA
171
срединные выросты маленькие, треугольные, слабо скошенные вбок;
кокситы широко треугольные в дистальной половине и поперечные
в основной части, образующей широкий крупный выступ по наружному
своему краю.
Ширина лба равна ширине глаза; лобная полоса голая или
с несколькими волосками в задней трети боковых отделов, бархатисто-
черная, с выпуклыми краями; орбиты, кроме переднего их края, черные,
сильно блестящие, как и глазковый треугольник; орбиты в 2 раза уже
лобной полосы, скулы в блестящем белом налете. Наружные и внутрен-
ние теменные, глазковые щетинки и вибриссы толстые, хорошо развиты;
орбитальные щетинки присутствуют в виде 1—3 коротких щетинок в зад-
нем отделе орбит; задний отдел орбит, кроме того, с группой черных,
коротких, щетинистых волосков. 5-й тергит в РД—2 раза длиннее 4-го.
Окраска тела и хетотаксия как у самца. Длина тела <5 о 2—6 мм.
Распространение. Таджикистан: окр. Куляба, 26 VII—
14 VIII 1933 (Попов), Кабадиан, 19 VIII 1933 (Гуссаковский), Айвадж,
устье р. Кафирнигана, 1 VIII 1934 (Гуссаковский). Туркмения: Кара-
кала, 6 V—8 VIII 1932 (Петрищева), 1934 (Попов), Ахча-куйма,
5 VII 1934 (Попов), Чулли, 21 V 1914 (Гольбек), Арман-саад—Кзыл-
арват, 1896 (Ангер). Вост. Казахстан: Б. Алмаатинка, 21 VIII 1928 (Шнит-
ников). Крайние северные точки нахождения вида: Мугоджары: Джаксы-
тау, 26 VII 1904 (Дубянский), окрестности Уральска, 7 VII 1908 (Боро-
дин), Темир, 2 VI 1908 (Бородин и Уваров), с. Верхне-Днепровка,
Чкаловск, обл., на р. Урал, 11 VII 1934 (Зимин).
*3. Pyrellia (in sp.) minuta Zimin, sp. nov. (рис. 262, 263, 281).
<5. Лоб узкий, ширина его составляет от х/з до Чг ширины 3-го чле-
ника усиков; скулы посредине почти в два раза уже 3-го членика уси-
ков, книзу слабо и постепенно расширены; наибольшая ширина их в ниж-
нем отделе не более чем в Р/г—РД раза превосходит ширину верхнего их
отдела на уровне 1-го членика усиков. Передне-верхняя стерноплевраль-
ная щетинка всегда имеется; щетинка на передне-дорзальной поверх-
ности средних голеней отсутствует. Церки широкие, боковые лопасти
короче, чем у Р. secunda, с более широко закругленными дистальными
концами; внутренние выступы очень малы, округлотреугольные; ко-
кситы сравнительно широкие, с выступающим внутренним дистальным
углом, по дистальному наружному краю дуговидно округлен, по вну-
треннему краю с прямоугольным тупым выступом.
Ширина лба составляет 2/з — 3Д ширины глаза, в 3—4 раза уже
лобной полосы; ширина скул составляет 2/з ширины 3-го членика уси-
ков или (редко) почти равна ей. 5-й тергит в РД раза длиннее 4-го.
В остальном вид сходен с Р. secunda, но мельче. Длина тела 3—5 мм.
Распространение. Известен из долин в Туркмении: Чо-
люк 13 IX 1932 (Ушинский), Ак-яйла, 13 IX 1932 (Ушинский), Кара-
дегиш, 21 VIII 1932 (Ушинский).
*4. РугеШа (in sp.) aenea Zett. (рис. 274—277). •
Zetterstedt, 1845, Dipt. Scand., IV : 1.324: — suda Rond. Pandelle, 1898, Rev.
Entom., XVII : 37; — ignita Rob.-Desv., 1830 : 464; Stein, 1915 : 17, 18; S6guy,
1923 : 361; — serena Seg. (nec Meigen), Karl, 1928 : 9; — serena Karl (nec Meigen); —
serena Villeneuve (nec Meigen), 1919, Ann. Soc. ent. France, LXXXVIII : 259.
$. Лоб узкий, ширина его в наиболее узком месте равна х/г ширины
3-го членика усиков; скулы узкие, слабо расширенные книзу, в средин-
172
I. ТРИБА MUSCINI
ном отделе ширина скулы несколько превышает V2 ширины 3-го членика
усиков; усики коричневато-бурые или черно-бурые; 3-й их членик в 2—
13/4 раза длиннее 2-го; щупальцы бурые, слабо расширены к дисталь-
ному концу; хоботок сравнительно тонкий, удлиненный; голова черно-
бурая (у самцов с незатемненными крыльями — скулы красноватые).
Рис. 274—278.
274 — передняя половина крыла Pyrellia aenea Ztt. 275 — переднегрудь Р.
aenea Ztt. 276 — заднегрудное дыхальце Р. aenea Ztt. 277 — церки самца Р.
aen^a Ztt. 278 — то же Р. secunda, sp. п. (Ориг.).
затылок и щеки блестящие, черно-бурые; скулы и передний отдел орбит
в белом, шелковисто блестящем, густом налете; задний отдел орбит бурый.
Внутренние теменные щетинки и вибриссы крепкие, длинные; одна-две
передних пары лобных щетинок крепче прочих, имеющих вид щетини-
стых волосков, как и глазковые щетинки; в задней половине орбит лоб-
ные щетинки направлены вершинами вверх и вперед. Грудь, исключая
коричневый, задний отдел плевр, зеленая, металлически^' блестящая,
местами с бронзовым отливом; плевры в редком желтовато-сером налете,
Л. PYRELLIA
173
спинка в рыжевато-сером налете; посредине спинки проходит широкая,
далеко не достигающая щитка полоса сероватого налета, более отчет-
ливо выраженная впереди шва. Поверхность груди в редких черных
щетинистых волосках. Передние дыхальцы белые или светло-палевые.
Переднегрудь узкая, блестящая, лишена, как и передние плевры, воло-
сков; гипоплевры перед дыхальцем и в заднем нижнем углу с одиноч-
ными бурыми волосками; щетинки груди: акростихальных — 1 пара
перед щитком; дорзоцентральных — 3-|-4; интраалярных — 0-J-1
(реже 0-J-2, передняя в этом случае короткая); предкрыловых — 1, над-
крыловых — 1; закрыловых — 2, крепких передних и 1 позади воло-
совидная; плечевых — 3; заплечевых — 1; стерноплевральных — 1-|-
4-3; на щитке хорошо развиты вершинные, боковые (2), основные (2); ще-
тинки предосновные отсутствуют или имеются в виде щетинистых волос-
ков, почти такой же длины и толщины, как прочие волоски щитка. Крылья
дымчатые в передних 2/з или прозрачные, жилки рыжевато-желтые;
грудные чешуйки рыжевато-дымчатые или (у светлокрылых экземпляров)
бледнопалевые, реже белые с палевым краем. Ноги бурые; средние го-
лени с переднедорзальной щетинкой, расположенной на одном уровне
с вентральной щетинкой. Задние бедра с длинными щетинистыми волос-
ками и щетинками по передневентральной поверхности. Брюшко метал-
лически зеленое, местами с бронзовым реже пурпуровым отливом, в чер-
ных коротких сверху и длинных на вентральной поверхности волосках;
краевые щетинки на 3-м тергите равны или чуть длинее 1 /2 длины 4-го
тергита. Церки имеют листовидно вытянутые боковые лопасти, несколько
более короткие, чем у Р. secunda и более длинные, чем у Р. minuta-, вну-
тренние выросты треугольные, скошенные вбок; кокситы широкие, обыч-
ного строения, с небольшим выступом по внутреннему краю.
По окраске самка сходна с самцом. Лоб едва уже ширины глаза;
орбиты в 3 раза уже черно-бурой лобной полосы; последняя в заднем
отделе по бокам с одиночными щетинистыми волосками; скулы шире,
чем у самца; а в среднем отделе ширина их равна ширине 3-го членика
усиков; затылок металлически зеленый, оливковый или синеватый, равно-
мерно выпуклый; щетинки сравнительно крепкие; внутренние и наруж-
ные теменные щетинки, вибриссы и передняя орбитальная щетинка длин-
ные; задняя орбитальная щетинка в 2—4 раза короче передней. В осталь-
ном самка сходна с самцом. Брюшко заостренное к верщине, 5-й тергит
в Р/г раза длиннее предыдущего. Длина тела 4.5—6.5 мм.
Распространение. В пределах СССР вид встречается от
Ленинграда на восток до южного Приморья, в зоне тайги и лиственных
чесов. Северная граница распространения вида не вполне ясна. Южная
граница в СССР проходит по линии Киев—Конотоп—Ярески, Полтав-
ской обл., Харьков — горы Кавказа. В Азиатской части Советского
Союза южные точки нахождения вида следующие: Семипалатинск, Алтай,
Иркутск, Сидеми в южном Приморье. Вся Зап. Европа. Северная
Монголия.
2. Подрод Rypellia Mall.
Malloch, 1931, Ann. Mag. Nat. Hist., (10), VII : 190; — Orthellia Malloch,
1923, Ann. Mag. Nat. Hist., (9), XII : 515; — Pyrellia Enderlein, 1934, Sitzungsber.
Gesell. nat. Freunde Berlin : 422.
Тип подрода: Rypellia flavipes Malloch, 1931.
Глаза самца на лбу почти соприкасаются, голые; щеки узкие. Передне-
грудь и проплевра голые; щетинки груди: акростихальных — 1 пара
174
/. ТРИБА MUSCINI
перед щитком; дорзоцентральных — 24-4; интраалярных — 2; предкры-
ловых— 1; надкрыловых — 2 (задняя слабо развита); закрыловых—3
(задняя волосовидная); плечевых — 3; заплечевых — 2; предшовных —1;
стерноплевральных — 14~3; гипоплевры с волосками перед дыхальцем
и в нижнем углу; щетинки на щитке: вершинных — 2; боковых — 2,
основных — 2, все длинные, крепкие; дискальных — 2, волосовидные;
предосновные отсутствуют. Заднее дыхальце очень длинное, достига-
ющее передним концом переднего края вентрального отдела медио-
тергита; супрасквамальное ребро без щетинистых волосков. Жилка
т — с дуговидным изгибом; г4+5 лишь при основании (снизу) или
в основной половине Кго отрезка (сверху) в мелких щетинках и во-
лосках. Ноги длинные; переднедорзальная щетинка средних голеней
отсутствует, вентральная — сильно развита; нижняя поверхность зад-
них бедер в длинных щетинистых волосках; коготки и присоски короткие.
В род Pyrellia подрод Rypellia включен на основании следующих
признаков: отсутствие супрасквамальных волосков, голая переднегрудь,
строение головы (узкий лоб, узкие щеки), дуговидный изгиб т жилки.
С родом Orthellia данный подрод имеет общие черты в строении перед-
негруди (ширина) и заднегрудного дыхальца (удлиненность дыхальца).
Подрод насчитывает 4 вида, известных из Индо-Малайской области
и юго-восточной Палеарктики. В СССР представители подрода пока
не обнаружены.
5. Pyrellia (Rypellia) semilutea Mall. (рис. 253—255).
Malloch, 1923., Ann. Mag. Nat. Hist., (9), XII: 515 (Orthellia)', он же, 1932,
там же, (10), X : 306; —julvipes Malloch, 1932, там же, (10), X : 307 (syn.
nov.);—flavipes Enderlein, 1934, Sitzungber. Gesell. nat. Freunde Berlin : 422
(syn. nov.).
Лоб самца в 4 раза уже 3-го членика усиков, в профиль сильно накло-
нен вниз, со слабо выступающим передним углом, расположенным на од-
ной вертикальной линии с верхним краем рта; скулы слабо расширены
книзу, посредине ширина их не превышает 1/& ширины 3-го членика
усиков; высота щеки составляет Vs вертикального диаметра глаза; глаза
с сильно укрупненным фасетками в верхних 2/3. Усики длинные; 2-й чле-
ник в 21/я раза короче 3-го, красновато-коричневый; 3-й членик буро-
вато-коричневый; щупальцы коричневатые, постепенно и слабо расши-
рены к вершине; подбородок несколько удлиненный (длиннее 3-го чл.
ус.). Голова буровато-коричневая, скулы в светлосером налете. Затылок
и щеки в черных волосках; внутренние теменные щетинки и вибриссы
длинные, крепкие, прочие тонкие, короткие. Орбитальные щетинки отсут-
ствуют. Грудь коричневато-бурая, плевры желтовато-коричневые с не-
сколько более темными отдельными небольшими участками. Переднее
дыхальце палевое; заднее — сагиттальное вытянутое, длина его в 2 раза
превышает ширину в середине. Спинка и плевры в редком сероватом
налете. Переднегрудь широкая, желтовато-коричневая, без волосков.
Проплевры в центре голые, гипоплевры с группой волосков впереди зад-
него дыхальца и над задними тазиками. Супрасквамальное ребро лишено
щетинистых волосков. Щетинки груди охарактеризованы в описании
подрода. Грудь в черных редких волосках. Крылья бледносерые, с едва
заметной желтизной; г4+5 от основания до средины первого отрезка в редких
мелких щетинках сверху и с несколькими волосками при основании снизу.
Жилки крыла желтовато-рыжие; чешуйки темножелтые, жужжальца
желтые. Ноги длинные, тонкие. Передние тазики желтые, средние и зад-
3. PYRELLIA
175
ние коричневато-желтые, лапки рыже-желтые, прочие части ног желтые
с черными редкими волосками и щетинками. Задние голени слабо со-
гнуты. Переднедорзальная щетинка средних голеней отсутствует; зад-
ние бедра на 2/з от основания по задневентральной и по всей длине на
передневентральной стороне в редких длинных прямых щетинистых
волосках, переходящих у вершины бедер в тонкие, изогнутые ще-
тинки. Коготки темножелтые, с черной вершиной, желтые пульвиллы
короче 5-го членика лапки. Брюшко бурое, удлиненно яйцевидное,
с желтым основанием 1-го (1+2) тергита. Покрыто тонкими волосками
и щетиночками по краям тергитов (точнее окраску брюшка определить
не представилось возможным ввиду сильной деформации его). Длина
тела 7 мм.
В коллекции Зоологического института Академии Наук СССР
имеется 1 самец из Сычуаня: Тацзинлу, 9 VII 1893 (Потанин).
Описанный выше экземпляр весьма сходен с Pyrellia {Rypellia)
semilutea Mall., он отличается лишь деталями в окраске щитка и брюшка;
теми же признаками вид отличается от Pyrellia (Rypellia) /«Zpijpes Mall.
(1932), известного с о-ва Тайван и рассматриваемого нами как более
темный вариетет предыдущего. По морфологическим чертам Pyrellia
{Rypellia) semilutea занимает промежуточное положение между родами
Pyrellia и Orthellia.
3. Подрод Dasyphoromima Zimin, nov. subgen.
Тип подрода: Dasyphoromima pavlovskyi Zimin, sp. nov.
Новый подрод занимает промежуточное положение между родами Ру-
геШа и Dasyphora. С первым его сближают следующие признаки: жилка
п — без щетинок, глаза голые; с Dasyphora — развитие матового налета
на спинке, форма заднегрудного дыхальца (оно треугольное), наличие
хорошо развитой 2-й надкрыловой и предосновной щетинок, а также
переднедорзальной щетинки на средних голенях; переднегрудь широ-
кая, в волосках.
Щетинки груди, кроме вышеперечисленных: акростихальных — 1
пара перед щитком; дорзоцентральных 3+4; интраалярных — 2; пред-
крыловых — 1; плечевых — 3; заплечевых — 2; предшовных — 1; ното-
плевральных — 2; стерноплевральных — 1+3; супрасквамальное ребро
без щетинистых волосков; проплевры в центре голые; гипоплевры перед
дыхальцами и в нижней части с группой волосков. На щитке: вершин-
ных — 2, боковых — 2, основных — 2 и предосновных — 2, тонкие.
Сюда относится всего один вид с Дальнего Востока.
*6. РугеШа (Dasyphoromima) pavlovskyi Zimin, sp. n.
j. Лоб по ширине равен глазу, орбиты в 24i—2х/г раза уже черно-
бурой лобной полосы, несущей в задней половине сравнительно длинные
толстые щетинистые волоски; скулы посредине такой же ширины, как
3-й членик усиков; щеки в 2 раза шире орбиты; задняя поверхность
головы равномерно сильно выпуклая; усики бурые, 3-й членик в 2х/1
раза длиннее 2-го; щупальцы буровато-рыжие в основной и бурые в вер-
шинной половине, сравнительно широкие, у вершины шире, чем при
основании; голова черно-бурая, затылок и щеки слабо блестящие, с лег-
ким синеватым оттенком, в редком налете; орбиты и передняя половина
орбит в густом шелковисто-белом налете; задняя половина орбит с жирным
блеском, бурая. Наружные и внутренние теменные щетинки, вибриссы и
176
I. ТРЙБА MUSCINI
передняя из двух орбитальных щетинок крепкие, длинные. Грудь буро-
синяя, задние отделы плевр коричневые; в редком сероватом на плеврах и
густом, образующем продольные полосы, налете на спинке. Средняя светлая
полоса налета на спинке широкая (ширина ее в 237г раза более ширины
ограничивающих ее с боков темных полос); боковые полосы налета слабо
выражены, не шире внутренних темных полос; боковые темные полосы с
неясными границами, шире внутренних; щиток в сером налете, более
редком на вершине щитка; переднегрудь в тонких темных волосках. Перед-
ние плевры голые; гипоплевры с группой плохо заметных волосков впереди
дыхальца и в нижнем заднем углу; переднее дыхальце коричневато-
бурое; грудь в черных, щетинистых, коротких, редких волосках. Ще-
тинки груди длинные, крепкие. Крылья сероватые, жилки рыжие; m
с дуговидным изгибом; чешуйки белые, с бледнопалевым краем, жуж-
жальца желтые. Ноги смоляно бурые, коготки и присоски короткие, по-
следние палевые. Средние голени по переднедорзальной поверхности
с одной более крепкой щетинкой, расположенной на средине расстоя-
ния между вершиной и местом прикрепления вентральной щетинки,
2 мелкими щетинками и рядом коротких щетинистых волосков над нею.
Брюшко синее, металлически блестящее, в едва заметном сероватом
налете на дорзальной и сравнительно густом на вентральной его поверх-
ности; щетинки недлинные. Длина тела о — 6 мм.
Описан по 1 экз. (j) из с. Евсеевки, Приморского края 19—22IV1910
(Иконников). Тип в Зоологическом институте Академии Наук СССР
в Ленинграде.
4. Род DASYPHORA R.-D.
Robineau-Desvoidy, 1830, Essai Myod. : 409; Bezzi u. Stein, 1907, Catal. Pal.
Dipt., Ill : 605; Schnabl u. Dziedzicki, 1911 : 176; Stein, 1916 : 17; S6guy, 1923 : 361;
Karl, 1928 : 8; Ringdahl, 1929, Entom. Tidskr. : 8—13; Patton, 1934, Ann. trop.
Med. Parasitol., XXVIII : 571—578. —Pyrellia auct. nec Rob.-Desv.
Тип рода Dasyphora pratorum Meigen.
Средней величины мухи с металлически синей или зеленой окраской
брюшка и 4 темными продольными полосами на спинке, разделенными
светлыми промежутками налета. Глаза в светлых волосках или голые.
Лоб у самца узкий; ширина его варьирует в пределах от
Чз ширины 3-го членика усика до двойного поперечника последнего.
У самки ширина лба почти равна ширине глаза. Орбиты самок равны
х/з—1/з ширины лобной полосы, всегда матовые и снабжены 2 (реже
одной) орбитальными щетинками; орбитальные щетинки самца волосо-
видные, реже крепкие; как исключение, орбитальные щетинки могут
отсутствовать. Наружные теменные щетинки развиты лишь у самок.
Лобная полоса у самок всегда и у самцов редко со щетинистыми
волосками в боковых отделах. 3-й членик усиков в 2—З1/* раза длиннее
2-го, с длинноперистой аристой. Вибриссальные кили в мелких волосках,
поднимающихся вверх до 3/4 высоты килей.. Щупальцы длинные, слабо
утолщенные к вершине; хоботок короткий, толстый. Переднегрудь широ-
кая, желобовидно вдавленная, голая или в мелких волосках; проплевры
в центре голые; гипоплевры с группой волосков перед дыхальцем и в зад-
нем нижнем углу. Стерноплевральных щетинок — 1-}-3; акростихаль-
ные — постоянно присутствуют в числе 1 пары перед щитком, впереди же
шва отсутствуют или располагаются в числе 1—3 пар; интраалярная
щетинка впереди шва отсутствует; супрасквамальное ребрышко без
4. DASYPHORA
177
щетинистых волосков; дорзоцентральные весьма постоянны в числе 3-J-
+4 (позади шва); птероплевры по верхнему краю со щетинистыми воло-
сками; щиток с 2 вершинными, 2—4 боковыми, 2 основными, 2 пред-
основными и 0—2—4—6 ди скальными щетинками. Заднее дыхальце тре-
угольное; высота его равна или больше расстояния между его нижним
краем и нижним краем гипоплевры. У D. serena дыхальце маленькое.
Жилка т дуговидно или под тупым углом изогнута в направлении перед-
него края. Жилка п на верхней стороне в коротких щетинках, развитых
лишь у основания или почти по всей длине жилки; Г4+ев тонких щетини-
стых волосках как с верхней, так и с нижней поверхности крыла, более
длинных и гуще расположенных при основании жилки и коротких и ред-
ких в остальных ее отделах (почти до вершины крыла). Ноги короткие;
коготки и присоски короткие, длина их всегда менее длины 5-го чле-
ника лапки. Передневентралъная и задневентральная поверхность зад-
них бедер у самца всегда с рядами очень длинных прямых щетинистых
волосков; средние голени у обоих полов с крепкой вентральной щетин-
кой. Брюшко яйцевидное. Церки самца широкие с 2 короткими внутрен-
ними выступами по дистальному краю и полукругло или языковидно
оттянутыми вниз боковыми лопастями по дорзальному краю, несущему
зубчики по передней стороне снизу. Фаллозома с цилиндрической базаль-
ной частью, более короткой, чем бокаловидно расширенная к вершине
и снабженная мелкими тонкими шипиками дистальна-я ее часть; основная
часть фаллозомы с коротким дистальным выростом при основании. Перед-
няя часть парамер вытянутая с мембранозной вершиной и щетинками
по заднему краю при основании. Задний отдел парамер короткий, за-
остренный, с короткими шипиками по переднему краю и боковой наруж-
ной стороне. V стернит брюшка самца с короткими боковыми выступами.
Из известных нам 14 видов 13 ограничены в своем распространении
Палеарктикой и лишь один (D. serena) имеет циркумбореальпый ареал,
•обитая как в Евразии, так и в Северной Америке.
Насколько известно, представители рода связаны с широколиствен-
ными лесами, обитая в умеренном климате. В южных частях своего ареала
большинство форм придерживается горных областей, до зоны альпий-
ский лугов. Областями наибольшего видового многообразия следует
считать Средиземноморье (6 видов) и горные системы центральной Азии
(8 видов). * . . .
Большинство видов с биологической стороны не изучено. По отно-
шению к D. pratorum и D. asiatica выяснено лишь, что оба вида являются
живородящими. D. serena является яйценесущей формой. Развитие личи-
нок протекает в помете животных, главным образом жвачных. Данные
Шинера о размножении D. serena в разлагающихся веществах живот-
ного происхождения требуют проверки.
В настоящей работе границы рода несколько расширяются, по срав-
нению с общепринятыми, в связи со включением D. serena, D. суа-
nella и D. eriophthalma, имеющих много общих черт и с родом Pyrellia,
куда их ранее относили, и с родом Dasyphora R.-D. С родом Pyrellia
D. serena сближается по строению головы, отсутствию волосков на глазах,
строению половых придатков самцов; общие черты этого вида с родом
Dasyphora заключаются в наличии щетинок на п, хорошо развитой,
широкой переднегруди, величине заднего грудного дыхальца, хетотаксии
грудного отдела (надкрыловые, предосновные щетинки) и ног (переднедор-
зальные щетинки средних голеней, вентральные волоски задних
бедер).
12 Фауна СССР, том XVIII, вып. 4.
178
I. ТРИБА MUSCINI
Что касается D. cyanella и D. eriophthalma, то включение их в род
Pyrellia возможно лишь на основании строения конца брюшка самцов
(Patton a. Gibbins, 1934), в остальном внешняя морфология этих видов
всецело отвечает признакам рода Dasyphora, включая сюда и опушение
глаз.
ОПРЕДЕЛИТЕЛЬНАЯ ТАБЛИЦА ВИДОВ
1 (32). Ширина лба не превышает 1/з ширины глаза (самцы).
2 (3). Переднедорзальная щетинка средних голеней расположена на поло-
вине расстояния между местом прикрепления вентральной щетинки
и вершиной голени (не на одном уровне с вентральной щетинкой);
глаза голые, реже с едва заметными редкими волосками; средне-
спинка с широкой продольной полосой светлого налета...............
............................................*1- D. serena Mg.
3 (2). Вентральная и переднедорзальная щетинки средних голеней рас-
положены на одном уровне.
4 (9). 1-й членик задних лапок снизу при основании с пучком длинных
волосков, расположенных на бугорке.
5 (6). Переднегрудь голая; ширина лба в 3—4 раза менее ширины 3-го
членика усиков; средние голени на переднедорзальной поверх-
ности выше крепкой цереднедорзальной щетинки с несколькими
мелкими щетинками или щетинистыми волосками, длина которых
превышает длину прочих волосков голени. Крупнее 8.5—10.5 мм . .
.............................................*11. D. versicolor Mg.
6 (5). Переднегрудь в мелких щетинистых волосках; щетинки и длинные
волоски над переднедорзальной щетинкой на средних голенях
отсутствуют. Мельче 7—8 мм.
7 (8). Ширина лба равна или едва менее ширины 3-го членика усиков;
орбиты часто с ясными орбитальными щетинками или волосками.
Скулы бурые, ширина их равна или более ширины 3-го членика
усиков. Брюшко светло бронзовозеленое в густом сероватом налете.
Светлый промежуток между черными продольными полосами в перед-
ней части средйеспинки шире черной полосы или равен ей ... .
................................... . *10. D. paraversicolor, sp. n.
8 (7). Ширина лба в 3 раза менее ширины 3-го членика *усиков. Орбиты
без орбитальных щетинок или волосков. Скулы по переднему краю
желтые, ширина их чуть менее ширины 3-го членика усиков. Брюшко
синевато-зеленое, в густом сероватом налете; светлый промежуток
между черными полосами на спинке уже последних....................
.................................. *12. D. gussakovskii Zim.
9 (4). 1-й членик задних лапок обычного строения без пучка волосков.
10 (11). Передние голени по задней поверхности с одной или несколь-
кими крепкими короткими щетинками у средины; над переднедор-
зальной щетинкой средних голеней имеется несколько мелких щети-
нок. Грудь черно-синяя; промежуток светлого налета между чер-
ными полосами в переднем отделе спинки в 2 раза уже черной по-
лосы. Перед швом 1 пара крепких акростихальных щетинок. Щиток
с 2 боковыми щетинками. Брюшко зеленовато-синее в сероватом
налете.......................................*8. D. setitibia, sp. n.
И (10). Передние голени на задней стороне без щетинок у середины.
12 (25). Акростихальные щетинки впереди шва отсутствуют.
13 (16). Переднегрудь в коротких волосках.
4. DASYPHORA
179
14 (15). Ширина лба равна или более ширины 3-го членика усиков. Ор-
битальные щетинки или щетинистые волоски имеются; скулы шире
3-го членика...............................13. D. trichosterna, sp. n.
15 (14). Ширина лба в 2—3 раза менее ширины 3-го членика. Орбиталь-
ные щетинки отсутствуют. Ширина скулы не шире 3-го членика
усиков......................*14а. D. pratorum meridionalis, subsp. n.
16 (13). Переднегрудь голая.
17 (20). Между дорзоцентральными щетинками впереди шва распола-
гаются 2 ряда щетинистых волосков, следующих вдоль внутренних
границ продольных темных полос среднеспинки и резко отличаю-
щихся по длине и толщине от прочих волосков этого отдела спинки.
Тело блестящее зеленое или сине-зеленое, в редком налете.
18 (19). Ширина лба равна ширине 3-го членика усиков. Над передне-
дорзальной щетинкой средних голеней располагается 1—2 корот-
кие щетинки. Крылья серые, чешуйки белые или бледнопалевые . .
...........................................*3. D. eriophthalma Mcq.
19 (18). Лоб в 2—3 раза уже 3-го членика усиков. Щетинки над передне-
; дорзальной щетинкой средних голеней отсутствуют. Крылья по
переднему краю и у основания слабо дымчатые, чешуйки слабо дым-
чатые ...........................................2. D. cyanella Mg.
20 (17). Волоски в переднем отделе среднеспинки между щетинками
одинаковой длины и крепости.
21 (22). Ширина лба превышает ширину 3-го членика усиков или равна
ей; орбитальные щетинки или волоски имеются; щетинки над передне-
дорзальной щетинкой средних голеней отсутствуют.....................
...............................*14. D. pratorum pratorum Mg.
22 (21). Лоб в 2—3 раза уже 3-го членика усиков, часто без орбиталь-
ных щетинок.
23 (24). На средних голенях выше переднедорзальной щетинки есть доба-
вочная, более короткая щетинка; светлый промежуток между тем-
ными продольными полосами в передней части спинки шире тем-
ной полосы; брюшко синее.........................*9. D. similis, sp. n.
24 (23). Средние голени на переднедорзальной поверхности с одной
крепкой щетинкой; светлый промежуток между темными полосами
среднеспинки уже темной полосы или равен ей; брюшко бронзово-
зеленое....................*14а. D. pratorum meridionalis, subsp. n.
25 (12). Акростихальные щетинки впереди шва в числе 1—3 пар.
26 (27). Ширина лба в Р/г раза превышает ширину 3-го членика уси-
ков. Средние голени выше переднедорзальной щетинки с рядом
тонких щетинок и щетинистых волосков; латеральных щетинок
на щитке 4; грудь черно-синяя; брюшко синее. Дорзоцентральных
щетинок впереди шва 1 пара. Длина 10.3 мм...........................
•.........................................*7. D. latifrons. sp. n.
27 (26). Ширина лба в 2 раза уже 3-го членика усиков, реже равна ей;
средние голени с одной крепкой переднедорзальной щетинкой.
28 (29). Передняя половина скулы красновато-желтая; тело темносинее;
щиток с 4 боковыми щетинками, ширина лба почти в 3 раза менее
ширины 3-го членика усиков. Дорзоцентральных щетинок впереди
шва 3 пары (передняя слабее двух остальных). Длина тела 8 мм .
............................................6. D. quadrisetosa, sp. n.
29 (28). Скулы бурые, в светлом налете; щиток, как правило, с 1 парой
боковых щетинок. Тело зеленовато-бронзовое, в густом на-
лете.
12*
180
I. ТРИБА MUSCINI
30
31
32
33
34
35
36
37
38
39
40
41
42
(31). Дорзоцентральных щетинок впереди шва 2 пары (передняя
тогда слабо развита, но всегда ясная); длина краевых щетинок на
дорзальной стороне 3-го тергита почти равна длине 4-го тергита.
Тело более темное: грудь бронзово-оливковая, брюшко зеленовато-
темнобронзовое, в густом налете; ширина черных полос в переднем
отделе среднеспинки равна или чуть менее ширины светлого проме-
жутка между ними. Крупнее: 7—9 мм .... *4. D. saltuum Rd.
(30). Дорзоцентральных щетинок впереди шва 1 пара; длина крае-
вых щетинок на верхней стороне 3-го тергита не превышает 1/з—
г/г длины 4-го тергита; брюшко светлобронзово-зеленое или золотисто-
зеленое, в очень густом желтовато- или зеленовато-сером налете;
ширина темных продольных полос в переднем отделе среднеспинки
по крайней мере в 2 раза (обычно в 3) уже разделяющего их свет-
лого промежутка. Мельче: 6.5—8 мм...............................
...........................................*5. D. asiatica Zim.
(1). Ширина лба почти равна ширине глаза (самки).
(34). Переднедорзальная щетинка средних голеней расположена много
ниже места прикрепления вентральной щетинки, обычно на поло-
вине расстояния между местом прикрепления вентральной щетинки
и вершиной голени. Глаза голые или в редких едва различимых
волосках; тело блестящее синее или оливково-зеленое, с широкой
средней продольной полосой светлого налета на спинке ...........
............................................*1. D. serena Mg.
(33). Переднедорзальная и вентральная щетинки средних голеней
расположены почти на одном уровне от вершины голени.
(36). Передние голени вблизи средины задней поверхности с 1 или
несколькими щетинками, средние голени выше переднедорзальной
щетинки с 1—3 короткими щетинками; тело темносинее, брюшко
зеленовато-синее...........................*8. D. setitibia, sp. n.
(35). Передние голени на задней поверхности без щетинок у средины.
(40). Крепкие акростихальные щетинки впереди поперечного шва
спинки имеются в числе 1—3 пар.
(39). Длина краевых щетинок на верхней стороне 3-го тергита не
более 1/з длины 4-го тергита; акростихальных щетинок перед
швом 1 пара; ширина светлого промежутка между темными про-
дольными полосами в переднем отделе среднеспинки в 2 раза
более ширины темной средней полосы. Светлее, мельче: 6.5—8 мм .
........................................... *5. D. asiatica Zim.
(38). Длина краевых щетинок на верхней стороне 3-го тергита дости-
гает 3Д длины 4-го тергита; акростихальных щетинок впереди шва
2 пары, из них передняя слабее задней; средний светлый проме-
жуток между темными полосами спинки в 2 раза шире темной внут-
ренней полосы. Темнее, более крупный вид: 7—9 мм................
............................................*4. D. saltuum Rd.
(37) . Крепкие акростихальные щетинки впереди поперечного шва
спинки отсутствуют.
(44) . По внутренней границе внутренних темных продольных полос
спинки располагается по ряду толстых щетинистых волосков, выде-
ляющихся своей толщиной и длиной среди прочих волосков, нахо-
дящихся меджу дорзоцентральными щетинками впереди шва; брюшко
блестяще зеленое или синее, в очень редком сероватом налете.
(43) . Над переднедорзальной щетинкой средних голеней распола-
гается по 1—2 коротких щетинки . . . *3. D. eriophthalma Mctj.
4. DASYPHORA
181
43 (42). Добавочные мелкие щетинки над переднедорзальной щетинкой
на средних голенях отсутствуют....................2. D. cyanella Mg.
44 (41). Впереди шва среднеспинки между дорзоцентральными щетин-
ками волоски одной толщины и длины; брюшко в густом светлом
налете.
45 (48). Переднегрудь в мелких щетинистых волосках.
46 (47). 3-й и 4-й тергиты брюшка в густом желтовато-сером налете,
не образующем темных переливчатых пятен при основании сегмен-
тов; 3-й членик усиков буровато-коричневый....................... .
......................................13. D. trichosterna, sp. n.
47 (46). При основании в боковых отделах 3-го и 4-го сегментов брюшка
имеются полукруглые темные переливчатые пятна.........................
............................*12. D. gussakovskii, sp. n.
*15. D. pratorum meridionalis, subsp. n.
48 (45). Переднегрудь лишена волосков.
49 (50). Средние голени выше переднедорзальной щетинки с 1—3 корот-
кими щетинками или щетинистыми волосками, ясно выделяющимися
среди прочих волосков этой части голени; брюшко зеленовато-
синее; щиток, как правило, с 4 боковыми и 2 дискальными щетин-
ками ..........................................*11. D. versicolor Mg.
50 (49). Средние голени выше переднедорзальной щетинки без добавоч-
ных щетинок или длинных волосков; брюшко зеленое, реже сине-
зеленое; щиток обычно с 2 боковыми и 2 дискальными щетинками . .
...............................................*14. D. pratorum Mg.
*1. Dasyphora serena Mg. (рис. 282).
Meigen, 1826, System. Beschr., V : 59 (Musca)', — cyanicolor Zetterstedt, 1845,
Dipt. Scand., IV : 1323; Stein, 1915 : 18 (Pyrellia)-, Karl 1928 : 9.
Биология. Holmquist, 1928, Ann. Ent. Soc. Amer., I : 660—667 (Pyrellia).
8. Ширина лба составляет 1!з—Чг ширины 3-го членика усиков;
ширина скул в среднем отделе равна Чг ширины 3-го членика усиков;
усики бурые, 3-й членик в 2—24t раза длиннее 2-го; щупальцы бурые.
Голова черно-бурая; затылок и щеки блестящие, с очень слабым метал-
лически оливковым оттенком; скулы и передняя половина лба в густом
белом шелковистом налете; лицо в светлосером матовом налете; задняя
половина лба бурая, как и лобная полоса, в средней части имеющая вид
тонкой линии. Затылок и щеки в бурых волосках; из щетинок сильно раз-
виты лишь внутренние теменные и вибриссы; 3—4 передних пары лоб-
ных щетинок и глазковые щетинки развиты сильнее прочих лобных ще-
тинок. Орбитальные щетинки отсутствуют. Грудь черно-синяя, метал-
лически блестящая, на плеврах в редком желтовато-сером налете; перед-
ний отдел плеч, как и широкая полоса в переднем отделе по средине
спинки, в густом сероватом налете. По бокам от средины расположены
не четко ограниченные полосы слабого сероватого налета. Светлый
налет за поперечным швом спинки отсутствует. Грудь в чернобурых
волосках. Щетинки груди: акростихальных — 1 пара перед щитком;
дорзоцентральных — 2 (3)4-4; интраалярных — 2; надкрыловых — 2;
предкрыловых — 1; плечевых — 3; заплечевых — 1, реже 2, в этом
случае передняя слабая; предшовных — 1; нотоплевральных — 2; на
щитке: вершинных — 2; боковых — 2; основных — 2; предосновных — 2;
дискальных — 2, последние, как и предосновные, волосовидные. Стерно-
182
I. TP ИВ A MUSCINI
плевральные щетинки крепкие, в числе 14-3. Гипоплевры с группой
волосков впереди дыхальца; волоски над задними тазиками имеются.
Переднегрудь и проплевры лишены опушения. Крылья сероватые, жилки
Рис. 279—283. Церки самцов.
279 — Dasyphora cyanella Mg. 280 —D. eriophthalma Mcq. 281 —Pyrellia minuta,
sp. n. 282 — D. serena Mg. 283 — P. cadaverina L. (Ориг.).
рыжие, п и г4+5 с мелкими щетинками; чешуйки шелковисто белые, с па-
левым или бледнодымчатым краем; жужжальца рыжевато-желтые, с лег-
ким затемнением посредине. Передние дыхальцы темные; цвет их изме-
няется от рыжего до бурого (никогда не бывает белым или серовато-белым).
Ноги коричнево-бурые или бурые; бедра и лапки обычно окрашены тем-
нее голеней; присоски короткие, рыжевато-желтые; передние голени, за
4. DASYPHORA
183
исключением обычно присутствующих у всех видов щетинок, лишены
макрохет; средние голени с обычными для рода щетинками. Передне-
дорзальная щетинка средних голеней расположена обычно на средине
расстояния между вершиной голени и местом прикрепления вентраль-
ной щетинки. Над переднедорзальной щетинкой средних голеней у неко-
торых экземпляров развиты 1—2 мелкие щетиночки. В остальном ще-
тинки голени как у D. cyanella. Брюшко синее, лиловато-синее или темно-
зеленое, в черных волосках и щетинках. Дорзальная поверхность бле-
стящая, лишена налета; брюшко снизу в редком сероватом налете. Ще-
тинки брклпка тонкие. Церки в форме двух поперечных лопастей, с язы-
ковидно оттянутыми вниз наружными углами; внутренние углы, коротко
выступающие вниз и снабженные на дистальном конце 2 длинными ще-
тинками; внутренняя сторона боковых лопастей с многорядными мел-
кими зубчиками.
Самка сходна по окраске тела с самцом. Лоб едва уже или равен
по ширине глазу, в задней половине блестяще черный; впереди, как
и скулы, в густом блестящем белом налете; орбиты в 3 раза уже черно-
бурой лобной полосы. Последняя с многочисленными щетинистыми воло-
сками в боковых отделах своей задней трети; орбитальных щетинок — 2,
передняя, как правило, в 2—3 раза длиннее задней; внутренняя и наруж-
ная теменные и глазковые щетинки крепкие, длинные. Скулы почти равны
по ширине 3-му членику усиков, или едва уже последнего. Длина тела
<?$> 5.5—9 мм.
Распространение. В пределах СССР вид известен на север
до Мурманска, Архангельской обл.—Уржума—Забайкалья—устья р. Ал-
дана—Петропавловска на Камчатке; на юг вид идет до Киевской и Харь-
ковской областей, Предкавказья, Северного Казахстана и Алтая; на
Дальнем Востоке вид констатирован для южного Приморья. Данные по
Сибири ограничены. Большая часть Зап. Европы. Северная Америка.
Биология вида подробно не изучена. По Шинеру, личинки
развиваются в разлагающихся животных тканях. Зимовка мух, по дан-
ным Гольмквиста, проходит в дуплах деревьев, в трещинах коры и гни-
лых пнях. Перед зимовкой мухи собираются группами. Вылет мух с мест
зимовки происходит в апреле.
2. Dasyphora cyanella Mg. (рис. 279, 284—293).
Meigen, 1826, Syst. Beschr., V : 77 (Musca)', — lasiophthalma Mg. Ztt., 1845,
Dipt. Scand., IV : 1325, 1326; — eriophthalma Stein (nec Macquart); Patton, 1934,
Ann. trop. Med. Parasitol., XXVIII : 576, 577, figs. 6.
Биология. Banks, 1912, U.S. Bur. Ent. Techn. ser. n° 22; Thomson, 1937,
Parasitology, XXIX : 279—282, figs.
3. Лоб узкий, ширина его в наиболее узком месте составляет
от !/з до х/г ширины 3-го членика усиков; ширина скулы равна или
немного более Уг ширины 3-го членика усиков. Усики бурые, 3-й их
членик в 21/г раза длиннее 2-го; щупальцы бурые. Голова’ чернобурая;
затылок блестящий, с синеватым или зеленоватым металлическим от-
блеском, в сером налете, более развитом в верхних боковых и нижнем
его отделах. Скулы и передняя половина лба в белом, шелковисто бле-
стящем налете, несколько желтоватом в нижней трети скул. Внутрен-
ние теменные щетинки и вибриссы крепкие, длинные; лобные щетинки,
за исключением 2—3 передних пар, волосовидные, короткие; глазковые
такой же длины, как передние лобные; орбитальные не развиты. Грудь
металлически зеленая, синеватая или оливковая, местами с бронзовым
184
I. ТРИВА MUSCINI
отливом; плевры и спинка в редком сероватом налете; спинка с 4 узкими
буро-оливковыми продольными полосками; далеко не доходящими до
щитка; ширина средних темных полосок в 4, реже в 3 раза уже светлого,
разделяющего их промежутка. Поверхность груди в черных волосках.
Щетинки груди как у D. pratorum', впереди шва между дорзоцентраль-
ными щетинками имеется два ряда толстых и длинных щетинистых воло-
сков, расположенных вдоль внутреннего края средних темных полос
спинки; длина и толщина волосков возрастает от поперечного шва по
направлению к переднему краю спинки; передние дыхальцы палевые.
Крылья серые, с едва заметной буроватой дымкой в основной половине
и вдоль переднего края. Жилки буроватые в вершинной и рыжие
в основной половине крыла; чещуйки белые, с палевым краем или бледно-
дымчатые; жужжальца желтые. Ноги черно-бурые, наружная поверх-
ность бедер с зеленоватым или синеватым металлическим блеском; перед-
недорзальная щетинка средних голеней расположена несколько ближе
к вершине голени, чем вентральная щетинка. Короткие щетинки над
переднедорзальной щетинкой на средних голенях отсутствуют. Задние
бедра в длинных щетинистых волосках на вентральной, передне-и задне-
вентральной поверхностях; присоски палевые. Брюшко бронзово-зеле-
ное или синее, реже с фиолетовым оттенком, металлически блестящее,
в крайне редком сероватом налете. Церки с маленькими внутренними
выступами, из которых каждый снабжен 2 длинными щетинистыми волос-
ками по краю; наружные лопасти языковидно оттянуты вниз; по внут-
ренней поверхности нижнего края с многочисленными зубчиками. Кок-
ситы толстые, загнутые внутрь на вершине; при основании по внутрен-
нему краю с плоским прямоугольным коротеньким выступом.
2- Лоб равен по ширине или чуть уже глаза; черно-бурая лобная
полоса в 3—4 раза шире орбиты с немногочисленными щетинками
по бокам в задней половине; покрыта золотисто-бронзовым редким нале-
том, как и нижний отдел скул. Скулы в верхней 1/г и орбиты в передней
части в желтовато-белом или белом налете. Орбитальных щетинок — 2,
задняя обычно слабо развита. Крылья светлее, чем у самца.
Яйцо 1 сходно с таковым Orthellia, имеет в длину около 2 мм, однако
несколько тоньше его.
Почти по всей длине на слабо вогнутой поверхности яйца проходит
2 параллельных друг другу рубчика.
Личинка I стадии метапнейстична; задние дыхальцы слабо высту-
пающие назад. Снизу по переднему краю брюшных сегментов со 2-го
по 7-й располагаются ряды мелких шипиков, развитые слабее на I и VIII
брюшных кольцах. На нижней поверхности 8-го брюшного сегмента
отчетливо вырисовывается анальная пластинка, сходная с таковой Orthel-
lia. В строении ротового аппарата наблюдается большое сходство с личин-
ками I стадии Orthellia. Базальный склерит тонкий, с 4 направленными
назад отростками, из которых дорзальные снабжены дорзальной пере-
мычкой. Под срединным дорзальным склеритом помещен маленький
Т-образный склерит.
Личинка II стадии амфипнейстична; передние ее дыхальцы с 7 лучами;
задние дыхальцы имеют по 2 щели. Ряды шипиков имеются по переднему
краю 1—7 брюшных сегментов. Базальный склерит ротоглоточного аппа-
рата имеет решетчатую дорзальную перемычку; передние выросты соеди-
нены снизу вентральной перемычкой, как это имеет место у личинок
1 Описание фаз развития и биологии дано по Томсону.
4. DASYPHORA
185
той же стадии Orthellia-, сходство с последней обнаруживается и в устрой-
стве ротовых крючьев. Зубные склериты ясные. Длина тела 7—8 мм.
Рис. .284—293. Детали строения преимагинальных фаз Dasyphora cya-
nella Mg.
284 — ротоглоточный аппарат личинки I стадии. 285 — то же II стадии.
286 — то же III стадии. 287 — яйцо. 288 — переднее дыхальце личин-
ки. 289 — анальная пластинка. 290 —шипы на вентральной стороне
брюшных сегментов у личинки III стадии. 291 — ее задние дыхальцы.
292 — ложнококон. 293 — дыхальце куколки. (По Томсону).
Личинка III стадии амфипнейстична; передние дыхальцы с 7 лучами;
задние дыхальцы имеют по 3 изогнутых щели. Дыхательцевые пластинки
расставлены далее друг от друга, чем у Orthellia, и сильнее склероти-
зованы. Отверстия стигмальных желез, имеющиеся на каждой пластинке
186 I. ’ ТРИБА MUSCINI
в количестве 4, мелкие. Верхний край дыхальцевой щели направлен
наружу, вентральный — в сторону центра. Брюшные Сегменты со 2-го
по 7-й снабжены по переднему краю с нижней стороны рядами шипиков.
Анальная пластинка с выпуклым передним и вогнутым задним краем.
Позади анальной пластинки на границе вентральной поверхности
имеется 3 постанальных сосочка. Дорзальная перемычка базального
склерита имеет выемчатый, направленный назад отросток, с длинными,
широко расставленными тонкими концами. Под перемычкой базальный
склерит снабжен полукруглыми выступами, характерными для вида.
Вентральная поверхность базального склерита имеет 8 хорошо различ-
ных продольных полосок. Гипостомальный склерит массивный, обла-
дающий в боковых отделах по нижней стороне заостренными выступами.
Ротовые крючья длинные, не сильно согнутые. Левый крючок иногда
развит менее правого. Вентральная перемычка в боковых отделах пред-
ставлена треугольными зубными склеритами. Добавочные склериты,
имеющиеся в числе одной пары, крупные, удлиненно-треугольные.
У личинок Orthellia они мелкие, слабо развитые. Наибольшая длина
личинки 14 мм.
Пупарий типа Musca, длиной 6.5 мм., темнокоричневый, с широко
закругленным задним концом и' ясно выступающими задними дыхальце-
выми пластинками. Анальная пластинка плохо заметна.
Распространение. С достоверностью известно, что D. cya-
nella и D. eriophthalma обитают во Франции, Германии,1 Швеции. Мы рас-
полагаем, кроме того, материалом по D. cyanella из Сардинии и с.-в.
Ирана. В пределах СССР вид не обнаружен, но вероятен на Кавказе.
Биология. Мухи зимуют в фазе имаго в щелях и темных углах
помещений. В Англии появление мух в поле падает на первую половину
апреля. С мая начинается откладка яиц. Яйца помещаются кучками
в числе 25—30 штук на коровий помет. Через 2—3 суток происходит
вылупление личинок. При теплой погоде выход личинок из яиц уско-
ряется до 24—30 часов. Длительность развития личинки I стадии в сред-
нем равна, примерно, 25 часам. Обычно часть личинок несколько задер-
живается в развитии. При средней температуре 22° С продолжительность
жизни личинки во П стадии равна 30 часам. Полный цикл развития
охватывает, по Томсону, около 8 недель, из которых 4 падает на фазу
личинки; 3—4 недели насекомое находится в ложнококоне.
Личинки D. cyanella — копрофаги, развивающиеся в коровьем
помете.
* 3. Dasyphora eriophthalma Mcq. (рис. 280).
Macquart, 1833, Recueil Soc. Agric. Lille : 168 (Lucilia)', Hist. nat. Ins. Dipt.,
II : 258 (Lucilia)', — lasiophthalma Zetterstedt (nec Macquart) 1845, Dipt. Scand., IV :
1325; Patton, 1934, Ann. trop. Med. Parasitol., XXVIII : 576 (Pyrellia).
Сходна c D. cyanella, от которой отличается широким лбом самца,
достигающим ширины 3-го членика усиков или немного превышает тако-
вую; у обоих полов голени средних ног выще переднедорзальной щетинки
снабжены 1—2 (или более) короткими тонкими щетиночками. Окраска
тела синяя; бронзовый оттенок почти не заметен. Крылья серые, без
дымчатого тона по переднему краю. Церки того же строения, что у D. cya-
nella, но с более заостренными внутренними выростами.
1 В коллекции Зоологического института Академии Наук СССР имеется 2 экз.
D. cyanella, определенные Мейгеном.
4. DASYPHORA
187
Распространение вида не выяснено. В пределах СССР
с достоверностью можно отметить лишь два пункта, где вид найден: Крым
(без точного указания места) и Бакуриани (Грузия); коллекция ЗИН АН
СССР располагает, кроме того, 2 экз. из Нижней Австрии с этикеткой
Ковача (Kowarz) «.Dasyphora cyanella Mg.».
* 4. Dasyphora saltuum Rd. (рис. 297, 300, 301, 302, 313).
Rondani, 1862, Dipt. ital. Prodr., V : 207; Pandell£, 1898, Revue entom., XV1L :
40 (Pyrellia)-, Hough, 1900, Termesz Fiiz., XXIII : 249; —aeneomicans Portschinsky,
1882, Тр. Русс, энтом. общ., XVI : 143 (syn. nov. sec. typ.); Schnabl u.
Dziedzieki, 1911 : 228.
£. Лоб узкий, ширина его едва менее или почти равна ширине 3-го
членика усиков; скулы по ширине равны последнему, реже несколько
уже его; усики бурые, 3-й их членик в 2 х/г раза длиннее 2-го; щупальцы
бурые, слабо расширенные к дистальному концу. Голова черно-бурая,
затылок и щеки слабо блестящие, с легким металлически оливковым
отливом, в бурых волосках и очень редком сероватом налете; скулы
и орбиты в передней половине в густом шелковистом блестящем налете,
приобретающем в нижнем отделе скул более или менее отчетливо выра-
женный желтый оттенок; лицо в матовом сером налете. Из щетинок
хорошо развиты внутренние теменные щетинки и вибриссы. Прочие
щетинки тонкие; несколько передних пар лобных щетинок развиты силь-
нее прочих, расположенных за ними. Орбитальные щетинки в виде тон-
ких удлиненных волосков, согнутых вперед и расположенных почти
на одной продольной линии с лобными волосками. Грудь темная, буро-
оливковая или бронзово-зеленовато-бурая; плевры в редком сероватом
налете. Спинка и щиток в светлосером, сравнительно густом налете,
первая с 4 продольными, слабо расширенными назад и почти достигаю-
щими щитка темными полосами. Ширина средних полос в переднем отделе
спинки немного менее, равна или чуть более ширины разделяющего
их светлого промежутка. Грудь в черных волосках и крепких щетинках:
акростихальных щетинок — 2 (реже 3) пары впереди шва; передние пары
щетинок слабее развиты, нежели задняя; позади шва 1 пара крепких
акростихальных щетинок перед щитком; дорзоцентральных — 3 +4;
интраалярных — 2; надкрыловых — 2; предкрыловых — 1; закрыл fl-
вых — 3; плечевых — 3; заплечевых всегда по 2 с каждой стороны; пред-
шовных — 1; стерноплевральных — 1 4-3; переднегрудь и проплевры
голые; гипоплевры перед дыхальцем и в нижнем заднем углу с группой
волосков; на щитке вершинных щетинок — 2; основных — 2; пред-
основные тонкие и короткие; боковых — 2; иногда ясно развиты 2 тон-
ких дискальных щетинки. Крылья серые, с рыжими жилками; чешуйки
белые с широкой желтоватой или бледно дымчатой каймой по краю. Жуж-
жальца желтые. Ноги бурые, голени коричнево-бурые; присоски палевые.
Щетинки как у D. pratorum, но щетинистые волоски на вентральной
поверхности средних и особенно задних бедер более длинные и более
многочисленные. Брюшко оливково-бронзовое, в густом шелковистом
желтовато сером или сером налете, образующем темные переливчатые
полукруглые пятна на боковых отделах 3-го и 4-го тергитов у их осно-
вания; темная буроватая продольная полоса слабо заметна на 3-м и 4-м
тергитах. Щетинки брюшка длиннее, чем у D. asiatica. Длина краевых
щетинок на 3-м тергите сверху почти равна длине 4-го тергита или лишь
немного менее ее. Дискальные щетинки в задней трети 4-го тергита в боко-
вых отделах последнего достигает такой же длины, как и маргинальные
188
I. ТРИБА MUSCINI
его щетинки. Церки такого же строения, как у D. asiatica, но значительно
шире; шипы на нижней стороне боковых лопастей в числе 5 с каждой
стороны, крепки. Кокситы с более оттянутым внутренним углом своего
дистального конца.
Самка сходна по окраске с самцом. Ширина лба изменяется в пре-
делах 2/3—3Д ширины глаза, орбиты в 2—2г/а раза уже лобной полосы;
скулы равны по ширине 3-му членику усиков или едва шире последнего.
Лобная полоса в темнозолотистом налете; скулы и передняя половина
орбит местами в желтоватом налете. Орбитальных щетинок — 2, длин-
ные, хорошо развитые. Прочие щетинки головы как у D. asiatica. Сред-
ние темные полосы спинки в 2 раза уже светлого промежутка между
ними. Краевые щетинки на 3-м тергите немного короче (3Д длины) 4-го
тергита; ди скальный ряд щетинок на 4-м тергите выражен много слабее
маргинального его ряда. Щетинки тела крепче и длиннее, чем у D. asia-
tica. Длина тела 3 7—9 мм.
Распространение. В СССР обнаружен на южном берегу
Крыма (Алупка, Никитский сад), в Грузии (Тбилиси, Лагодехи), Арме-
нии (Ереван), Дагестане (Хочал-даг, гора Хили-хор, субальпийские
луга), Азербайджане (Билясар на р. Вашару-чай). На север заходит
по Черноморскому побережью Кавказа до Гагр, Туапсе и Майкопа.
Центральная и южная Европа. В пределах Средней Азии не обнаружен.
После просмотра типа D. aeneomicans Portsch. (1882) не остается сом-
нений, что он является синонимом вида D. saltuum Rd. (1862), описан-
ного из Италии.
* 5. Dasyphora asiatica- Zimin (рис. 294, 296, 314—321).
Зимин, 1945, Энтомолог, обозр., XXVIII, 3—4 : 116—120; он же, 1944 : 142—
156; он же, 1948 : 77, фиг. 42.
$. Ширина лба немного шире или равна ширине 3-го членика уси-
ков и почти равна ширине скулы. Усики коричнево-бурые, вершина
3-го членика с узким красновато-коричневым кольцом; длина 3-го чле-
ника в 2—21/г раза превышает длину 2-го членика. Щупальцы рыже-
бурые, с более темной вершиной. Голова черно-бурая, затылок слабо
блестящий, с не всегда ясным металлическим оттенком и в редком сером
или желтовато-сером, как и щеки, налете. Скулы и орбиты в серебристо-
белом густом налете. Внутренние теменные щетинки и вибриссы крепкие;
орбитальные щетинки волосовидные, в числе одной с каждой стороны;
передние 2—4 пары лобных щетинок крепче остальных. Грудь бронзово-
бурая, в очень густом светлосером налете; ширина темных средних полос
(из 4 присутствующих на спинке) среднеспинки в 2—3 раза менее ширины
разделяющего их промежутка; щетинки груди развиты слабее, нежели
у D. saltuum. Дорзоцентральных щетинок впереди шва — 1 пара;
ди скальные щетинки на щитке отсутствуют. В остальном хетотаксия
груди как у D- saltuum. Крылья светлосерые, с желтыми жилками.
Чешуйки белые, часто с бледнопалевым краем. Ноги как у D. saltuum,
но с менее длинными щетинками и волосками на нижней поверхности
средних и задних голеней. Брюшко золотисто-бронзовое или золотисто-
бледнозеленое, нередко с розовым или бледнофиолетовым оттенком.
Щетинки брюшка короче и тоньше, чем у D. saltuum. Длина краевых
щетинок 3-го тергита не превышает 1/з—х/г длины 4-го тергита. Дискаль-
ный ряд щетинок в вершинной трети 4-го тергита состоит из коротких
тонких щетинок, расположенных в боковых отделах тергита. Брюшко
4. DASYPHORA
189
сплошь покрыто очень густым желтовато- или зеленовато-серым светлым
налетом. Переливчатые пятна при основании боковых отделов 3-го и 4-го
тергита слабо выражены. Продольная темная полоса брюшка как
у D. saltuum.
$). Ширина лба составляет 2/з—3/а ширины глаза; лобная полоса
в 2—2х/2 раза шире орбиты, в сером, реже бледно желтовато-сером налете.
Скулы в блестящем белом, орбиты в светлосером более матовом и редком
налете; затылок и щеки в матовом сером налете. Орбитальных щети-
нок — 2, из которых задняя обычно в 2—3 раза тоньше и короче перед-
ней; вершина 2-го членика усиков со слабо развитым красновато-корич-
невым пятном. Лобная полоса с несколькими короткими тонкими волос-
ками в задней половине боко-
вых отделов. Грудь такой же
окраски, как у самца, но с бо-
лее густым светлосерым нале-
том и более узкими темными
продольными полосами спинки.
Ширина внутренних полос в 4
раза менее ширины светлого
промежутка между ними. Брю-
шко той же окраски, как у
самца, но с более ясными пе-
реливчатыми темными пятнами.
Краевые щетинки 3-го тергита
короткие, тонкие; длина их не
более Vs длины 4-го тергита.
Длина тела <?£ 6.5—8 мм.
Личинка III стадии. Круп-
ные толстые личинки с утол-
щением на заднем конце тела;
кутикула гладкая, блестящая,
светло-желтая. Головной сег-
мент в 2 раза короче 1-го
грудного сегмента, широкий,
спереди по бокам с двумя круп-
ными широкими коническими
выступами. Ротовые бороздки Рис. 294. Самец Dasyphora asiatica Zim.
покрывают нижнюю, боковую (Ориг.).
и часть верхней поверхности
сегмента. 1-й грудной сегмент по переднему краю с кольцевидным вали-
кообразным вздутием, покрытым мелкими шипиками, расположенными
короткими рядками на маленьких площадках, придающих поверхности
чешуйчатый вид; прочая поверхность сегмента гладкая; сегменты груди
2-й и 3-й гладкие без шипов; 1-й брюшной сегмент снизу с одним (иногда
двумя неполными) рядом крупных шипов; сегменты брюшка со 2-го
но 7-й снизу с валикообразными поперечными вздутиями по переднему
краю, покрытыми несколькими рядами разных по размеру шипов,
из числа которых 6 шипов в центре достигают наибольшей величины.
Задние края брюшных сегментов по бокам от продольной средней линии
несут по группе коротких бугорочков в количестве 4 с каждой стороны.
8-й брюшной сегмент в 2 раза короче 7-го, с гладкой блестящей поверх-
ностью, ниже дыхалец с 2 крупными коническими выступами. Анальная
нластинка широкая, короткая, слабо расширена, на концах вздутая,
190
I. ТРИБА MUSCINI
продольно-морщинистая. Анальные бугорки маленькие, субанальные
сосцевидные, крупнее анальных, направлены вершинами вниз и назад;
позади анальной пластинки в промежутке между субанальными бугор-
ками располагается группа светлых шипиков в количестве 4—7 штук.
Передние дыхальцы поперечные, с 14 выростами, округлыми и тесно
прижатыми друг к другу. Задние дыхальцы крупные, удалены друг
Рис. 295—299. Головы видов рода Dasyphora сверху.
295 — самца D. paraversicolor, sp. n. 296 — самца D. asiatica Zim.
297 — самки D. saltuum Rd. 298 — самца D. versicolor Mg. 299 — сам-
ца D. trichosterna, sp. n. (Ориг.).
от друга на Ut—-Vs поперечника дыхальца. Перитрема широкая,
у взрослых личинок сливающаяся со стигмальной пластинкой. Дыхаль-
цевые щели извилистые, тонкие. Наружные концы верхней, средней
и верхний конец средней щелей обращены к перитреме (наружу), концы
прочих щелей ориентированы к стигмальному диску. Ротоглоточный
аппарат крупный; ротовые крючья слабо изогнутые в вершинном отделе;
базальная их часть удлиненная, почти равна вершинному отделу. Зуб-
ные склериты крупные, неправильных очертаний. Гипостомальные скле-
риты почти в 3 раза короче ротовых крючьев, неправильной формы,
с выступающим округленным заднедорзальным углом. Фарингеальные
4. DASYPHORA
191
склериты с очень широкой мембраной; дорзальные задние отростки
значительно уже и немного длиннее вентральных, вентральные — с угло-
видным выростом по верхнему краю. Дорзальная перемычка решетча-
тая, впереди доходит до линии, пересекающей середину передних отрост-
ков. Вентральная перемычка далеко выступающая, с кольцевидными
образованиями по заднему краю. Длина тела И—13 мм, у средины
2—2.6 мм.
Распространение. Вид распространен по всей Средней
Азии, заходя на восток до Центрального Китая и внутренней Монголии;
известен из разных мест Туркмении, Узбекистана и Таджикистана, где
распространен как в равнинных местностях (долина Вахша), так
и в горах (Зап. Памир до 2000 м).
До сего времени вид смешивался с D. saltuum Rd., от которого от-
личается признаками, указанными в таблице, и по строению генитального
аппарата. Все данные, относящиеся к Средней Азии, должны быть отне-
сены к D. asiatica, а не к D. saltuum— типичному средиземноморскому
виду, известному у нас лишь с Кавказа и из Крыма. В соответствии
с этим сведения Порчинского, а также Вайнтшейна и Родовой (1940),
должны быть отнесены к D. asiatica. Муха живородяща. Личинки откла-
дываются на коровий помет. По данным Вайнштейна и Родовой (1940),
на южном склоне Гиссарского хребта (высота 1100 м) личинки D. asia-
tica составляли 2.1% всех калоядных личинок в коровьем помете; по тем
же сведениям, личинки этого вида в помете других животных не обна-
руживались. Мухи D. asiatica встречаются с конца марта по сентябрь.
Личинки развиваются в помете и редко в навозе коров и в свином
навозе. Личинки Dasyphora никогда не встречаются крупными скопле-
ниями: как правило, в пробах встречаются по 1—2 личинки, редко число
их достигает 5—7 штук; в Гиссарской долине (Таджикистан) частота
встречаемости личинок D. asiatica в субстрате повышается к весне и осени
и падает к средине лета.
6. Dasyphora quadrisetosa Zimin, sp. nov. (рис. 307),
Ширина лба в 3 раза менее ширины 3-го членика усиков; скулы
равны ширине последнего; 3-й членик усиков в 2x/t раза длиннее 2-го;
щупальцы бурые, слабо утолщенные к вершине. Голова черно-бурая,
затылок блестящий со слабым синеватым отливом, в редком сизоватом
налете. Лобная полоса в переднем отделе и передняя половина скул
красновато-желтые. Усики черно-бурые; орбиты и скулы в желтовато-
сером налете. Щеки, верхняя половина орбит и затылок в бурых волосках.
Внутренние теменные щетинки и вибриссы крепкие, длинные; 5 передних
пар лобных щетинок толще остальных; прочие лобные и глазковые
щетинки в виде тонких, коротких волосков. Грудь буро-синяя, с полосами
светлосерого налета на спинке и в редком равномерно распределенном
желтовато-сером налете на плеврах. Темные продольные полосы
спинки по ширине почти равны разделяющему их промежутку свет-
лого налета. Грудь в черных, негустых, коротких волосках. Переднее
дыхальце палевое. Щетинки груди: акростихальных — З-f-l, передняя
пара впереди шва слабее прочих; дорзоцентральных — 3-4-4; интра-
алярных — 3; плечевых — 3; заплечевых — 2; преДТповных — 1; надкры-
ловых — 2; предкрыловых — 1; закрыловых — 2; стерноплевральных —
1-4—3; на щитке: вершинных — 2; боковых — 4; основных — 2; предос-
новных — 2; слабо развитых дискальных — 2. Переднегрудь и про-
плевры без волосков; гипоплевры с группой волосков впереди дыхальца
192
I. ТРИБА MUSCINI
и в нижнем заднем углу. Крылья сероватые, более темные у основания;
чешуйки дымчато-желтые. Голени средних ног по переднедорзальной
поверхности с крепкой щетинкой, выше которой длинные волоски и ще-
тинки отсутствуют. Брюшко темносинее, в густом желтовато-сером на-
лете. Волоски брюшка щетинистые, сравнительно толстые. Церки несут
по 3 длинных зубца на нижнебоковом выступе; срединные выступы язы-
ковидные, слабо скошены вбок; кокситы обычные для этой группы видов.
Длина тела 7.3 мм.
Самка не известна.
Описан по 1 экземпляру (<5) из Сычуани: р. Фубянхо, Шиндянмардан,
6 VIII 1893 (Потанин). Тип в Зоологическом институте Академии Наук
СССР в Ленинграде.
*7. Dasyphora latifrons Zimin, sp. nov.
$. Лоб широкий, ширина его в Р/г раза превосходит ширину 3-го
членика усиков, бурая лобная полоса в 3 раза шире орбиты, скулы
почти в 2 раза шире 3-го членика усиков; 3-й членик усиков в 21/а раза
длиннее 2-го членика; усики черно-бурые; щупальцы рыжевато-бурые.
Голова черно-бурая, затылок и задний отдел щек со слабым синеватым
оттенком, в редком темносером налете, блестящие. Скулы и передняя поло-
вина лба в белом густом шелковистом налете; задняя половина лба
блестящая, бурая. Внутренние теменные щетинки и вибриссы крепкие,
длинные; глазковые лобные щетинки тонкие, в виде щетинистых волос-
ков; орбитальных щетинок имеется по одной с каждой стороны, такой
же толщины, как передние лобные; задний отдел лба в редких, щеки в гу-
стых тонких, затылок в жестких редких (вверху) и тонких длинных, густых
(в нижнем отделе) бурых волосках. Грудь черно-синяя, с 4 широкими
продольными полосами на спинке, разделенными промежутками сизого
налета; средний светлый промежуток равен по ширине темной средней
полосе спинки. Плевры в очень редком сероватом налете. Грудь и щиток
в черных редких волосках. Акростихальных щетинок — 14-1, хорошо
развитые; дорзоцентральных — 34-4; интраалярных — 2; надкрыловых
— 2; предкрыловая — 1, короткая; закрыловых — 3; плечевых — 3;
заплечевая — 1; предшовная— 1; нотоплевральных — 2; стерноплев-
ральных — 14-3; на щитке: вершинных— 2; боковых — 4; дискальных—
4; основных — 2; предосновных — 2, волосовидные. Переднегрудь и про-
плевры без волосков; гипоплевры с волосками перед дыхальцем и в ниж-
нем заднем углу. Передние дыхальцы рыжие. Крылья светло-серые,
жилки рыжие. Чешуйки шелковисто-белые; жужжальца палево-желтые
с буровато-рыжим стебельком. Ноги черно-бурые, голени смоляно-бурые;
средние голени над переднедорзальной щетинкой с гребнем коротких
щетинок и щетинистых волосков. Брюшко зеленовато-синее, в густом
шелковистом светлосером налете, более редком по заднему краю 3-го
и 4-го тергитов. Щетинки с обычным для группы pratorum расположением.
Длина тела 10.3 мм.
Описывается по 1 экземпляру (3) с верховьев реки М. Алмаатинки
в Семиречье, 25 VII (Шнитников). Тип в Зоологическом институте Ака-
демии Наук СССР в Ленинграде.
Вид наиболее близок D. quadrisetosa, D. similis и D. pratorum. С пер-
выми двумя видами его сближает строение средних голеней, хетотаксия
щитка и среднеспинки, общая окраска тела, с последним — ширина лба,
окраска головы.
4. DASYPHORA
193
*8. Dasyphora setitibia Zimin, sp. nov. (рис. 308).
$. Лоб в 2 раза уже 3-го членика усиков, скулы в Р/г—Р/л раза шире
последнего; 3-й членик усиков в 2 раза длиннее 2-го; усики и щупальцы
бурые. Голова черно-бурая, затылок и задние отделы щек с синеватым
оттенком; передняя половина орбит и скулы в шелковисто блестящем
белом, затылок и щеки в редком, тусклом темносером налете. Внутрен-
ние теменные щетинки и вибриссы крепкие, длинные, лобные щетинки,
исключая 4—5 передних пар, а также орбитальные и глазковые щетинки
тонкие, последние очень коротки. Задний отдел орбит в редких тонких,
щеки в густых, тонких, затылок вверху в щетинистых, внизу в тонких
длинных бурых волосках. Грудь черно-синяя, средние темные продоль-
ные полосы спинки в 2 раза шире промежутка светлого налета между
ними. Налет светло-серый, с легкой желтизной на плевральных отделах.
Щетинки груди: акростихальных — 1 крепкая-]-! слабо развитая пара
впереди шва и 1 пара перед щитком; дорзоцентральных — 3-]-4; интра-
алярных— 2; надкрыловых — 2; предкрыловых — 1; плечевых — 3; за-
плечевых — 2; предшовных — 1; стерноплевральных — 1 -|-3 5 на щитке
вершинных — 2; боковых — 4; основных — 2; предосновных — 2; ди-
скальных — 2, волосовидные. Переднегрудь и проплевры лишены волос-
ков; гипоплевры с группой волосков впереди заднего дыхальца и внизу
над тазиками задних ног. Переднее дыхальце палево-рыжее. Крылья се-
рые, жилки буровато-рыжие; чешуйки шелковисто-белые, жужжальца
палево-желтые, с рыжеватым основанием. Ноги черно-бурые; передние
голени на задней поверхности с 1—2 короткими, сравнительно крепкими
щетинками; выше передне-дорзальной щетинки на средних голенях
имеется 1—2 тонких щетинки и ряд щетинистых волосков. Брюшко зеле-
новато-синее, темное, блестящее, в сером негустом налете; 3-й и 4-й тер-
гиты сверху с узкой продольной буроватой полоской. Церки широкие,
с короткими внутренними выступами, скошенными по дистальному краю
за срединой. Боковые выступы церок несут по 3 коротких крепких зубца.
Кокситы обычного для рода строения.
$. Лоб широкий, в наиболее узком месте ширина его составляет 4/s
ширины глаза. Орбиты в 2—2г/г раза уже черно-бурой лобной полосы;
скулы почти в 2 раза шире 3-го членика усиков. Лобная полоса в зад-
ней половине боковых отделов с группой сравнительно длинных волос-
ков; орбиты в коротких редких волосках, доходящих до передних лоб-
ных щетинок; орбитальные щетинки крепкие, по 2—3 с каждой стороны;
внутренние и наружные теменные, глазковые, вибриссальные и 2—3 пе-
редних лобных щетинки — крепкие, длинные. Скулы и орбиты, исклю-
чая их задний отдел, в желтовато-белом густом, лобная полоса в слабо
блестящем светло сероватом или желтоватом налете. Темные продольные
полосы среднеспинки в 2—Р/г раза уже светлого промежутка между ними
в переднем отделе спинки; назад полосы сильно расширены и не дости-
гают основания щитка. Брюшко более блестящее, чем у самца. Длина
тела <5(5 9—9.2 мм, — 7.5—9.3 мм.
Распространение. Кызылджим, вост. Теньги, Бийск, окр.,
7 VII 1897 (Силантьев); Красноярск, IV 1891 (Вагнер), Красноярск,
Столбы, 29 VII 1928 (Левчук); Троицкосавск, Забайкалье, 4 VI 1928
^Лукьянович); р. Улахан-ан, лев. прит. р. Лены, 6 IX 1926 (Иванов)
(тип <5); Вестях, Якутск, окр., 30 V 1912 (Наумов). Монголия :ущ. Сюц-
зуктэ, ю.-з. Кентэй, 30 VIII 1924, 20 IV, 11 V 1925 (Козлов); р. Сугу-
нур, верх. Хара-гола, Кентэй, 6—10 VII 1924 (Козлов).
13 Фауна СССР, том XVIII, вып. 4.
194
I. ТРИБА MUSCINI
По внешним морфологическим признакам, равно как и по строению
генитального аппарата самца, вид относится к группе видов близких
D. pratorum, от которых он отличается признаком, редко повторяющимся
вообще у видов рода, — присутствием щетинок на задней поверхности
передних голеней. D. setitibia является в то же время наиболее темноокра-
шенным видом. Вид наиболее близок к D. quadrisetosa, D. latifrons,
D. similis. Вид описан по 2 <53 и 10 jj.
Тип в Зоологическом институте Академии Наук СССР в Ленин-
граде.
*9. Dasyphora similis Zimin, sp. nov. (рис. 309).
$. Ширина лба не превышает половины ширины 3-го членика уси-
ков; орбиты немного шире 3-го членика усиков; усики бурые, как и щу-
пальцы; 3-й их членик в 2х/г раза длиннее 2-го. Голова черно-бурая, перед-
няя часть скул красновато-желтая, затылок и щеки более или менее
блестящие, со слабым синеватым металлическим отливом, в сером светлом
.налете; скулы и передняя часть орбит в густом шелковисто белом налете.
Щетинки головы как у D. pratorum. Грудь темносиняя, с широкой поло-
сой сероватого налета в переднем отделе спинки, ширина которой более
ширины граничащих с нею темных полос; светлые промежутки между
боковыми и средними полосами спинки очень узки и плохо выражены.
Щетинки как у D. pratorum. Крылья серые, чешуйки белые, жужжальца
рыжеватые. Ноги смоляно-бурые; хетотаксия ног сходна с таковой у
D. versicolor. Брюшко темносинее, в густом сероватом налете. Церки
обычного для группы pratorum строения; боковые вершинные выступы
церок несут по 3 длинных крепких зубца. Кокситы как у D. pratorum.
Длина тела 9 мм.
Самка не известна.
Вид описан по 1 самцу из Чаган-обо, Саур, б. Семипалат, обл..
12 VII 1910 (Караваев). Тип в Зоологическом институте Академии Наук
СССР.
D. similis, sp. п., относится к группе видов, близких kD. pratorum.
По ширине лба, темносиней окраске тела и хетотаксии средних ног
вид весьма напоминает D. setitibia, sp. п., от которой отличается хето-
таксией передних голеней и по отсутствию акростихальных щетинок
перед швом. С D. pratorum вид сближает отсутствие акростихальных
щетинок. Отличительными признаками D. similis от D. pratorum являются
ширина лба самца, присутствие над переднедорзальной щетинкой сред-
них голеней добавочного вооружения и темно-синяя окраска.
*10. Dasyphora paraversicolor Zimin, sp. nov. (рис. 295, 304, 312).
<J. Лоб немного шире 3-го членика усиков или чуть уже его; скулы
в Р/г—13/а раза шире 3-го членика усиков; последний в 3 раза длиннее
2-го членика. Усики и щупальцы бурые, последние при основании жел-
товато-бурые. Скулы и лоб в серебристо белом налете; задний отдел лба
бурый; затылок, как и щеки, в редком светло сером томенте. Орбиталь-
ных щетинок одна пара (у 2 экземпляров отсутствуют вовсе). Прочее как
у Z>. pratorum. Грудь буровато-синяя, в сером, на спинке в голубовато-
сером налете. Темные полосы спинки в Р/г раза уже разделяющего их
светлого промежутка. Щетинки груди и щитка как у D. pratorum.
Переднегрудь в мелких черных щетинистых волосках; проплевры
голые; гипоплевры с группой волосков впереди дыхальца в нижней части
над тазиками. Крылья как у D. pratorum. Ноги черные, голени смоляно
300
300 — передняя половина крыла Dasyphora saltuum Rd. 301 —
заднегрудное дыхальце того же вгда. 302 — переднегрудь того же
вида. 303 — задняя лапка самца D. versicolor Mg. 304 — то же-
D. paraversiiotor, sp. n. 305 — Задние дыхальцы личинки III ста-
дии D. pratorum Mg. (Рис. 305 по Порчинскому с изменениями^
прочие — ориг.).
13’
196
I. ТРИБА MV SCI NI
бурые. Средние голени по переднедорзальной поверхности с крепкой
щетинкой; ряд мелких щетинок и длинных щетинистых волосков над нею
отсутствует. 1-й членик задних лапок снизу с конусовидным бугорком при
основании, несущим пучок золотистых щетинистых волосков.
Брюшко светлое, бронзово зеленое, в густом светлосером палете;
переливчатые пятна при основании 3-го и 4-го тергитов отсутствуют или
(у экземпляра из Средней Азии) слабо выражены. Щетинки с обычным
для этой группы мух расположением. Церки и внутренние их выступы
широкие, по нижнему краю скошены от средины кнаружи; боковые ло-
пасти церок несут по 4 крепких зубца. Кокситы с широко закругленным
краем дистальной части, в остальном сходны с таковыми у D. versicolor.
Длина тела 8.5—9 мм.
Самка не известна.
Распространение. Киргизия: Дмитриевское, Киргизский —
Ала-тау 16 V 1913 (Чернявин). Китай; Хуан-хэ, 6—20 X 1908, Хуан-чан,
вост, хребет Нань-шань (Козлов). Тип в Зоологическом институте Ака-
демии Наук СССР в Ленинграде.
*11. Dasyphora versicolor Mg. (рис. 298, 303, 310).
Meigen, 1826, System. Beschr., V : 77 (Musca); Pandelle, 1898 : 39 (Pyrellia); —
pratorum Brauer u. Bergenstamm (nec Meigen); Schnabl u. Dziedzicki, 1911 : 228;
Stein, 1915 : 17; Karl, 1928 : 8; Patton, 1934 : 571—578, figs. 3, 4.
. $. Лоб очень узкий, ширина его в 2—3 раза менее .ширины 3-го чле-
ника усиков; скулы едва шире 3-го членика усиков или равны его ширине;
усики бурые, 3-й их членик в три раза длиннее 2-го; щупальцы и усики
бурые. Голова черно-бурая, затылок с синеватым оттенком, блестящий,
в редком сероватом налете, в бурых волосках. Скулы и передняя поло-
вина орбит в шелковисто блестящем, белом густом налете; щеки
блестяще бурые; в матовом сером налете. Щетинки головы как у D. pra-
torum; волоски не длиннее и не толще лобных щетинок в этом отделе
лба; могут быть отличимы лишь по наклону вперед. Грудь оливково-бу-
рая, часто с синеватым оттенком. Среднеспинка с 4 темными полосами;
средние из них расширены назад; ширина полос в 1’/г раза уже разде-
ляющего их покрытого налетом светлого промежутка. Налет желтовато-
серый или серый. Щетинки груди как у D. pratorum, щиток с 4 боковыми
и 2—4 дискальными щетинками; последние, нередко не развиты вовсе.
Крылья серые, жилки буровато-рыжие; чешуйки шелковисто белые,
с палевым краем; жужжальца желтые, с. легким буровато-рыжим затем-
нением в середине. Ноги черно-бурые, бедра по наружной поверхности
нередко с металлическим оттенком. Выше переднедорзальной щетинки
средних голеней располагается ряд щетинистых волосков или мелких
щетинок, или тех и других одновременно. Длина щетинок и волосков не
менее 1/з длины переднедорзальной щетинки. 1-й членик задних лапок
снизу при основании с бугорком, на котором расположен пучок длинных
густых щетинистых золотистых волосков. Брюшко зеленовато-синее,
в сероватом налете, с узкой темной, не достигающей вершины брюшка
полоской. В боковых отделах 3-го, 4-го и 5-го, не всегда, тергитов рас-
полагается вблизи переднего края по темному переливчатому округ-
лому пятну. Щетинки брюшка как у D. pratorum. Церки сохраняют тип
строения таковых у D. pratorum; внутренние выступы слабо удлиненные,
угловатые; боковые углы несут по 2 крепких зубца; кокситы обычного
для рода строения.
4. DASYPHORA
197
j. Сходна с самкой D. pratorum, от которой может быть отличима
по наличию гребня из коротких щетинок и волосков над переднедорзаль-
ной щетинкой средних голеней. Самки D. versicolor имеют более синюю
окраску тела, резче выраженные боковые пятна на брюшке и более круп-
ные размеры тела.
Длина тела $ 8.5—10.5 мм, д — 8—9 мм.
Распространение. Горные области Кавказа (Армения,
Азербайджан, Дагестан), где заходит до зоны альпийских лугов. Сред-
няя и южная Европа.
* 12. Dasyphora gussakovskii Zimin.
Зимин, 1945, Энтомолог, обозр., XXVIII, 3—4 : 118—120.
(5. Лоб очень узкий, ширина его в наиболее узком месте составляет
Дз ширины 3-го членика усиков; скулы немного уже 3-гочленика усика;
усики и щупальцы бурые; голова черно-бурая, узкая лобная полоса в пе-
реднем отделе и скулы красновато-желтые; затылок слабо блестящий
с синеватым металлическим оттенком, в бурых волосках, в редком серова-
том налете; внутренние теменные щетинки и вибриссы длинные, толстые;
лобные и глазковые щетинки тонкие и короткие; орбитальные щетинки
не ясно выраженные. Грудь черно-синяя, блестящая, в сером на плеврах
и сизом на щитке и спинке налете; спинка с 4-мя широкими темными
полосами, доходящими до щитка; ширина средних темных полос немного
более ширины светлого промежутка между ними. Переднегрудь в корот-
ких, редких, тонких волосках; в остальном хетотаксия груди как
у D. pratorum. Крылья серые, жилки при основании и вдоль переднего
края бледные, серовато-желтые, в вершинной половине рыжевато-бурые;
чешуйки шелковистые, белые; жужжальца желтые с затемнением перед
булавой. Ноги черно-бурые, голени в вершинной половине смоляно-бу-
рые; 1-й членик задних лапок с конусовидным выростом снизу при осно-
вании, несущим пучок длинных бурых при основании и золотистых на
вершине щетинистых волосков; присоски палевые, короткие; выше перед-
недорзальной щетинки на средних голенях добавочные щетинки отсут-
ствуют. Брюшко синевато-оливковое, блестящее, в очень густом сером
налете; темные переливчатые пятна при основании 3-го и 4-го тергитов
маленькие, угловатые. Длина щетинок на 3-м тергите достигает J/z длины
4-го тергита; на этом тергите дискальные щетинки слабо выделяются
среди прочих щетинистых волосков тергита.
j. Лоб равен или немного уже глаза; орбиты в 2—Р/г раза уже лоб-
ной полосы; скулы в 2 раза шире 3-го членика усиков; скулы и орбиты
в белом шелковистом налете; орбитальных щетинок — 2, задняя из них
короче и тоньше передней. Грудь бурая со слабым синеватым или зеле-
новатым оттенком, в густом голубовато-сером или сером налете; ширина
средних продольных полос среднеспинки в 21/а—3 раза менее ширины
светлого промежутка между ними; переднегрудь всегда с мелкими щети-
нистыми волосками. Брюшко оливково-зеленое, блестящее, в густом се-
роватом или желтовато сером налете и буроватой узкой полоской, дости-
гаюшей передней половины 4-го тергита. Темные переливчатые пятна по
бокам 4-го тергита при его основании обычно в виде широких коротких
полосок.
Длина тела $$ 6 мм, 2$ — 7—9.2 мм.
Распространение. Таджикистан: Гиссарский хребет на
высотах 1000—2500 м.
198
I. ТРИБА MUSCINI
D. gussakovskii относится к группе видов pratorum — versicolor, систе-
матически чрезвычайно близких друг к другу. По опушению передне-
груди и наличию выроста на первом членике задних лапок самца D. gus-
sakovskii может быть смешан с D. paraversicolor, sp. п., от которого без
труда отличается узким лбом $ и широкими темными полосами средне-
спинки. Последний признак отличает вид от самок D. pratorum meridio-
nalis, иногда имеющих на переднегруди одиночные волоски.
13. Dasyphora trichosterna Zimin, sp. nov. (рис. 299, 311).
$. Лоб широкий, ширина его в Р/г—2 раза превышает ширину 3-го
членика усиков; скулы в 13/4 раза шире 3-го членика усиков; усики бурые,
их 3-й членик в 3—31/4 раза длиннее 2-го; щупальцы бурые. Голова черно-
бурая, затылок слабо блестящий, с легким синеватым отливом, в редком
голубовато-сером или в сером налете, как и щеки; скулы и орбиты,
исключая заднюю их треть, почти лишенную налета, в блестящем, густом,
шелковисто-белом налете; лобная полоса почти равна по ширине орбите.
Затылок, щеки и задний отдел лба в бурых волосках; орбитальных щети-
нок 2—3 пары, из них передняя имеет вид тонкой щетинки, задние —
тонких волосков. Прочие щетинки как у D. pratorum. Грудь буро-синяя,
в сером или голубовато-сером налете, с 4 темными продольными полосами
на спинке; ширина средних темных полос в 1г/г раза уже светлого проме-
жутка между ними или равна ему по ширине. Щетинки груди как у D. pra-
torum. Переднегрудь в мелких бурых волосках. Крылья серые, жилки
желтовато-рыжие; чешуйки белые, с очень легким желтоватым оттенком
по краю; жужжальца желтые. Ноги бурые, бедра со слабым блеском,
местами в сероватом налете. Хетотаксия как у D. versicolor. Выше перед-
недорзальной щетинки на средних голенях имеется ряд длинных щети-
нистых волосков, не имеющих характера щетинок, как это свойственно
D. versicolor. Присоски и коготки короткие, первые — палевого тона.
Брюшко бронзово-оливково-зеленое или синее, в сером налете, с узкой,
слабо выраженной буроватой продольной полоской на 3-м и 4-м тергитах.
Краевые щетинки длинные; на 3-м тергите длина их составляет 3/4 длины
4-го тергита; на 4-м тергите длина краевых щетинок равна длине тер-
гита; дискальные щетинки 4-го тергита много короче его краевых щети-
нок. На 5-м тергите щетинки имеют обычное для рода расположение.
Церки со слабо скошенными от середины вбок внутренними выступами,
сохраняющими тип строения таковых у D. pratorum. Боковые лопасти
церок несут по 3 длинных крепких зубца. Кокситы обычного для группы
pratorum строения.
Лоб сравнительно высоко приподнят над верхним уровнем глаз;
орбиты в Б2—21/з раза уже бурой лобной полосы; ширина лба равна
ширине глаза; скулы в 2 раза шире 3-го членика усиков, длина которого
в 3—ЗЦ4 раза превосходит длину 2-го членика; голова черно-бурая, скулы
и орбиты в густом, лобная полоса в редком светло-сером или почти белом
налете. Орбитальных щетинок — 2 с каждой стороны, крепкие; задняя
развита несколько слабее передней. Прочее как у D. pratorum. Грудь
в голубовато-сером налете; средние темные полосы спинки в 2—21/г раза
уже разделяющего их светлого промежутка. Брюшко бронзово-оливково-
зеленое, в густом желтовато-сером налете и узкой буроватой продольной
полоской, достигающей основания 5-го или вершины 4-го тергита. Длина
каревых щетинок на 3-м тергите не превышает ]/з—Дг длины4-го тергита;
на последнем дискальные щетинки почти не выделяются среди прочих
щетинистых волосков тергита.
4. DASYPHORA
199
Длина тела 7.2—9.3 мм.
Распространение. Монголия: оз. Орок-нор, сев. Гоби,
4—6 VI 1926 (Козлов); Ихе-богдо, сев. Гоби, 5—25 V 1926 (Козлов);
Кобдо, сев.-зап. Монголия, 1926 (Кулик); Мурэн-гол, Алашанский хребет,
Гоби, 12 V 1908 (Козлов); Курлык-баингол, вост. Цайдам, 31 V 1895 (Ро-
боровский и Козлов). Тип в Зоологическом институте Академии Наук
СССР в Ленинграде.
*14. Dasyphora pratorum Mg. (рис. 266, 305, 306).
Meigen, 1826, System. Beschr., V : 78 (Musca)-, Pandelle, 1898 : 39 (Pyrellia)',
Keilin, 1917, 55 : 397—398; Karl, 1928, III : 9; Thomson, 1937, Parasitology,
29 : 279—283; — viridescens Portschinsky, 1882 XVII: 4.
Биология. Порчинский, Tp. Гусск. энтомол. общ., VIII : 37—41, рис. 38—41.
$. Ширина лба в 1х/а—2 раза превышает ширину 3-го членика уси-
ков, реже равна последней; скулы в 1х/«—14а раза шире 3-го членика
усиков. 3-й членик усиков в 22/г—3 раза длиннее 2-го; усики, как
и щупальцы, бурые. Голова черно-бурая, со слабым металлически зеле-
ным или синеватым оттенком на затылке; скулы и передняя половина ор-
бит в блестящем белом, густом налете; затылок, задняя половина лба
и щеки в матовом сером или желтовато-сером налете. Опушение головы
обычное. Внутренние теменные щетинки и вибриссы крепкие, длинные;
лобные щетинки, за исключением нескольких передних пар, тонкие, воло-
совидные; такой же крепости глазковые и обычно орбитальные щетинки;
редко последние достигают толщины передних лобных щетинок. Грудь
оливково-бурая, реже с преобладанием синеватого тона; покрыта желто-
вато-серым налетом, более густым и распределенным в виде продольных
полос на спинке. Темные внутренние'продольные полосы спинки широ-
кие, ширина их равна или чаще — в 1Ча раза уже разделяющего их
светлого промежутка. Полосы сужены назад, перед основанием щитка
заострены и несколько загнуты концами наружу. Щетинки груди: акрости-
хальных — 1 пара перед щитком; дорзоцентральных — 3-|-4; интрааляр-
ных — 2; закрыловых — 3; надкрыловых — 2; предкрыловых — 1; пле-
чевых — 3; заплечевых — 1 (реже имеется вторая слабо развитая);
предшовных — 1; стерноплевральных—1-|-3; на щитке: вершинных — 2;
боковых — 4; основных — 2; предосновных волосовидных — 2; дискаль-
ные обычно отсутствуют, реже имеется 1 пара впереди вершинных щети-
нок. Переднегрудь и проплевры голые; гипоплевры с группой волосков
впереди дыхальца и в заднем нижнем углу. Переднее дыхальце палевое.
Крылья светлосерые, жилки в дистальной половине крыла буровато
рыжие, при основании желтоватые. Чешуйки шелковисто-белые, с бледно-
палевым краем; жужжальца желтовато-рыжие со слабо затемненным
стебельком. Ноги черно-бурые или смоляно бурые; присоски желтые;
короткие. Щетинки ног: передние бедра с гребнями дорзальных, задне-
дорзальных и задневентральных щетинок; передние голени, за исклю-
чением вершины, лишены вооружения; средние бедра посредине
с несколькими короткими крепкими передними щетинками в вершинной
трети заднедорзальными щетинками, переходящими у вершины голени
в дорзальные щетинки; нижняя поверхность бедра с крепкими задневен-
тральными щетинками, идущими от основания до средины бедра, и длин-
ными щетинистыми волосками, из которых на передневентральной поверх-
ности некоторые приобретают толщину щетинок. Средние голени с крепкой
вентральной и несколько более короткой переднедорзальной щетин-
200
I. ТРИВА MV SCI iNI
Рис. 306. Самец
кой, расположенными на одном уровне; голени несут б—8 коротких креп-
ких задних щетинок. Задние бедра с гребнями крепких переднедорзаль-
ных и передневентральных щетинок, развитых сильнее в дистальной
части бедра и длинных редких щетинистых задневентральных волосков
в основной половине. Голени задних ног вооружены гребневидно распо-
ложенными короткими переднедорзальными щетинками и волосками, из
которых 1 щетинка развита сильнее прочих этой же серии: дорзальная
щетинка расположена в дистальной четверти голени; передневентральные
щетинки крепкие, короткие, сосредоточены в нижней половине голени
в количестве 3—5; задневентральные волоски в числе 3—5, также рас-
положены в дистальной половине голени, короткие и тонкие. Брюшко
оливково-бронзовое или сине-
вато-зеленое, металлически бле-
стящее, в густом желтовато или
зеленовато-сером налете; в боко-
вых частях, при основании 3-го
и 4-го, а также (не всегда) 5-го
тергитов располагаются полу-
круглые, переливчатые темные
пятна, размер которых убывает
к вершине брюшка. 3-й, 4-й и
не всегда 5-й тергиты сверху
посредине с темной узкой полос-
кой. Тергит 3-й сверху с тон-
кими краевыми щетинками, дли-
на которых равна примерно д/г
длины 4-го тергита. 4-й тергит
снабжен рядом в два раза бо-
лее длинных (чем на 3-м тер-
гите) краевых и иногда по се-
редине тергита прерванным ря-
дом тонких коротких дискаль-
ных щетинок; 5-й тергит в ди-
стальной половине с 2 полными
и 1 прерванным посредине
Dasyphora pratorum Mg. {ЖД0М более крепких щетинок.
(Ориг.). Брюшко в черных волосках.
2- Лоб равен пли чуть уже
в 2 раза уже бурой лобной полосы; орбитальных щети-
равно как и глазковые
глаза; орбиты
нок — 2; наружные и внутренние теменные,
щетинки крепкие, длинные; скулы в IV2 раза шире 3-го членика уси-
ков. Окраска груди и брюшка сходна с таковой у самцов. Длина тела
$$ 8—9.2 мм, — 7.5—9.5 мм.
Яйцо сходно по форме с яйцом Musca larvipara, длиной в 2J/« мм.
Личинка I стадии метапнейстична; задние дыхальцы мелкие,
сердцевидной формы, без перегородок в середине. Тело личинки лишено
шипиков. G нижней стороны головного кольца сагиттально распола-
гаются две темные, тонкие склеротизованные полоски, передним, более
тонким своим концом достигающие ротового отверстия; длина тела 23Д мм.
Личинка II стадии имеет слабо выраженные передние дыхальцы;
задние дыхальцы широко эллиптические, снабженные каждое двумя
поперечно расположенными дыхальцевыми щелями. Посередине расстоя-
ния между ротовым отверстием и концом головного сегмента расположен
4. DASYPHORA
201.
короткий поперечный ряд мелких темных шипиков, направленных вер
шинами назад. По Порчинскому, эти шипики являются единственным
вооружением на теле личинки.
Рис. 307—314. Церки самцов.
307 — Dasyphora quadrisetosa, sp. n. 308 — D. setitibia, sp. n. 309 — D. similis,sp. n.
310—D. versicolor Mg. 311 —D. trichosterna, sp. n. 312 — D. paraversicolor, sp. n.
313 — D. saltuum Rd. 314 — D. asiatica Zim. (Ориг.).
Личинка II стадии, продолжая развиваться в теле мухи, достигает
в длину 6 мм. Только по достижении личинкой III стадии муха отклады-
вает ее на помет рогатого скота.
В III стадии личинка обладает крепкими ротовыми крючьями; тело
голое, с валиками поперек передних отделов брюшных сегментов; перед-
нее дыхальце с 12—13 лучами; задние дыхальцы крупные, разделены
202
/. ТР ИВ A MUSCINI
промежутком (V«—Vs поперечника дыхальца). Каждое дыхальце с 3 ще-
лями, сравнительно короткими и извилистыми. Наружные концы верхней,
средней щели и верхний конец средней щели обращены наружу; каждое
32?
Рис. 315—321, Детали строения личинки III стадии Dasyphora asiatica Zim.
315 — ротоглоточный аппарат. 316 — задний конец тела сзади. 317 — он же сбоку.
318 — анальная пластинка (7) с субанальными (2) и анальными (4) бугорками. 319—
переднее дыхальце. 320 — заднее дыхальце. 321 — вооружение снизу 5-го (2) и 6-го
(7) брюшпых сегментов. (По Зимину).
дыхальце имеет по 3 мелких округлых отверстия — стигмальных железы,
из. которых верхнее и нижнее отверстие лежат у средины соответствую-
щих дыхальцевых щелей, тогда как наружное располагается между кон-
цами верхней и средней щели.
5. MORELLIA
203
Распространение. Закавказье: Армения (Делижан),
Азербайджан (Акстафа). Средняя Европа на восток до Польши, южная
Европа.
Порчинский (1910) приводит вид для Крыма. Указание Порчинского
на нахождение D. pratorum в Средней Азии относится к другим видам.
Стадия развития вида и биологии описаны Порчинским (1910).
D. pratorum принадлежит к живородящим мухам, у которых развитие
I—II стадии личинок протекает в теле самки в специальном маткообразном
приемнике; личинка задерживается в теле самки до III стадии. Порчпн-
ский дает описание и двух первых стадий личинки, выделенных из мух.
Личинка развивается в помете рогатого скота, которым и питается.
Окукление происходит в почве, куда личинка переползает, достигнув
предельной зрелости. Развитие личинки заканчивается в 14—15 дней.
14а. Dasyphora pratorum meridionalis Zimin, subsp. nov.
От типичной формы подвид отличается узким лбом самца, который
равен всего г!з ширины 3-го членика усиков; передняя половина скул
красновато-желтая. Срединные темные полосы спинки шире или равны
светлому промежутку между ними. Переднегрудь иногда несет одиноч-
ные мелкие волоски, всегда отсутствующие у основной формы.
Самки описываемого подвида, кроме волосков на переднегруди, ко-
торые не всегда бывают развиты, отличаются от самок D. pratorum pra-
torum более узкими темными средними полосами спинки, ширина кото-
рых составляет Vs ширины светлого промежутка, в то время как у основ-
ной формы ширина темной полосы всего в 2 раза уже светлой средней
полосы налета.
Распространение. Туркмения: Кара-кала, май, август,
сентябрь 1931 (Петрищева); Фирюза, Копет-даг, 22 V 1934 (Попов). Тип
в Зоологическом институте Академии Наук СССР в Ленинграде.
5. Род MORELLIA R.-D.
Robineau-Desvoidy, 1830, Essai Myod. : 405; Schnabl a. Dziedzicki, 1911 : 173;
Stein, 1916 : 15; он же, 1918 : 108; Malloch, 1923, Ann. Mag. Nat. Hist. (9), XXII :
520, 523; — T richomorellia Stein, 1918 : 109; —• Dasymorellia Malloch, 1923 : 527.
Тип рода: Morellia hortorum Fallen, 1816.
Средней величины темно окрашенные мухи, с более пли менее раз-
витым светлым налетом на брюшке и груди. Спинка с 3 продольными
светлыми полосами налета. Лоб самцов но ширине равен, чуть уже или
шире 3-го членика усиков, у самки равен, примерно, Vs ширины головы.
Орбитальные щетинки развиты у обоих полов; глаза самцов снизу в мел-
ких волосках (у тропических видов иногда в длинных волосках у обоих
полов — Тrichomorellia Stein, Dasymorellia Mall.); у j волоски на глазах
не видны. Лобная полоса в заднем отделе с одиночными щетинистыми
полосками, m с дуговидным изгибом; п — голая; г4+6 сверху и снизу со
щетинками или волосками, не заходящими от основания далее г—m по-
перечной. Переднегрудь широкая, голая пли в волосках; проплевры
всегда голые; супрасквамальное ребро голое, гипоплевры с волосками
в нижнем отделе и в передне-верхнем углу (не всегда); заднегрудное
дыхальце округло-треугольное, крупное. Стерноплевральных щетинок —
1-(-2, реже 1 —3; акростихальные щетинки в числе 1—2—3 пар впереди
щитка; у некоторых видов они развиты п перед поперечным швом
спинки. Ноги самцов с добавочным вооружением из щетпнок и волосков.
204
I. ТРИБА MUSCINI
Церки крупные, сравнительно широкие, часто снабженные но дисталь-
ному краю шипиками или зубцами. Кокситы крупные, массивные,
сильно расширенные в основной половине и обычно суженные в дисталь-
ном отделе, имеющем лопастевидные или пальцевидные отростки
на вершине, где снабжены шипиками и часто щетиночками. Фаллозома с
бокаловидным расширенным дистальным концом; мебравозная часть фал-
лозомы в 2—3 раза длиннее склеротизованной базальной части; мембра-
нозный отдел фаллозомы склеротизован по дорзальному краю и в ди-
стальной расширенной своей части; вершинный край покрыт шипиками.
Дорзальный отросток фаллозомы более или менее длинный, слабо рас-
ширенный на дистальном конце, где слабо склеротизован и снабжен мел-
кими шипиками. Аподема толстая, длинная, утолщенная на конце. Перед-
ние отделы парамер длинные, при основании более или менее расширены
и снабжены щетинкой; дистальный их конец мембранозный. Задние от-
делы парамер короткие, покрыты многочисленными мелкими волосками
в вершинной половине и короткими толстыми шипиками у основания.
По строению ног самцов, переднегруди (у обоих полов) и по
устройству генитального аппарата самцов род распадается на два подрода.
В один из них (Morellia s. str.), по нашему мнению, должен быть
отнесен и род Dasymorellia Mall. (=Trichomorellia Stein), который отли-
чается от всех известных видов Morellia густыми волосками на глазах и
развитыми акростихальными щетинками впереди поперечного шва спинки.
Оба названных признака достигают слабого развития и у представителей
рода Morellia (волоски в нижнем отделе глаз у самцов и акростихальные
щетинки, имеющие характер щетинистых волосков, ясно развиты у боль-
шинства видов, что не является существенным).
Из числа хороших видов рода, известных в настоящее время, 14 от-
мечены для Эфиопской, 8 — для Индо-Малайской и Австралийской, 16 —
для Неотропической, 2 — для Неарктической областей (последние являются
общими с Палеарктикой). В Палеарктике известно 9 видов.
Личинки Morellia являются копрофагами, обитающими в помете
животных и в фекалиях человека.
Имеются указания (Англия) на вредоносность некоторых видов рода,
проявляющуюся по отношению к сельскохозяйственным животным.
В областях, где массовое появление Morellia наблюдается часто, они мо-
гут иметь известное значение в распространении патогенных организ-
мов среди животных.
ОПРЕДЕЛИТЕЛЬНАЯ ТАБЛИЦА ПОДРОДОВ
1 (2). Переднегрудь без волосков; средние голени самцов по передней
поверхности с продольным рядом мелких щетинистых волосков,
обращенных вершинами к дорзальной стороне голеней и с мелкими
шипиками у основания........................1. Morellia R.-D. s. str.
2 (1). Переднегрудь в тонких волосках по краям; средние голени сам-
цов без продольного ряда мелких волосков, направленных верши-
нами к дорзальной стороне голеней, и без шипиков у основания
.......................................2. Dasysterna, subgen. nov.
Подрод 1. Morellia R.-D. s. str.
Robineau-Desvoidy, 1830, Essai Myod. : 405; Schnabl u. Dziedzicki, 1911 : 122;
Stein, 1916 : 16; S<5guy, 1923 : 350; Malloch, 1923 : 321, 1925 : 87; Karl, 1928 : 13; —
Pyrellia Pandelle, 1898 : 35 (nec Robineau-resvoidy).
5. MORELLIA
205
Тип подрода Morellia hortorum Fallen, 1816.
Лоб самцов узкий, ширина его почти равна ширине 3-го членика
усиков; орбитальные щетинки у обоих полов в числе двух пар длинных
и нескольких пар более коротких щетинок, вершинами направленных
вперед; глаза самцов внизу в редких, мелких волосках; у самок глаза
голые; наружные теменные щетинки отсутствуют у обоих полов; щеки
узкие, высота их составляет Vi—1/з вертикального диаметра глаза. Грудь
черная или черная с синеватым отливом, с тремя продольными полосами
белого налета на спинке, из которых одна проходит по середине спинки и
2 на границе спинки и плевр от плечевых бугорков к основанию крыльев.
Щетинки груди: акростихальных — 2—3 пары перед щитком и иногда
несколько тонких щетиночек перед поперечным швом; дорзопентраль-
ных — З-pi; интраалярных — 1—2; предкрыловых — 1; надкрыло-
вых — 2; закрыловых — 3; плечевых — 4; заплечевых — 1; предшов-
ных— 1; на гцитке 3 пары крепких вершинных, боковых, основных и
одна пара более тонких и коротких предосновных щетинок. Проплевры
и переднегрудь, равно как и супрасквамальное ребро, без волосков; стер-
ноплевральных щетинок 1 -f-2 (реже 2-4-3); гипоплевры со сравнительно
длинными волосками в нижнем отделе; заднегрудное дыхальпе большое,
треугольное. Жилка т с дуговидным изгибом. Жилка п голая, г4+5 со
щетинками сверху и волосками с нижней стороны первого отрезка жилки.
Ноги самцов с добавочным вооружением из щетинок и волосков;средние
голени всегда с дорзальной стороны с рядом мелких волосков по всей
длине, обращенных вершинами к дорзальной стороне, и с шипиками у ос-
нования. Голени самцов (средние и задние) нередко изогнуты. Церки
самцов широкие или удлиненные с шипиками по дистальному краю. Кок-
ситы массивные.
ОПРЕДЕЛИТЕЛЬНАЯ ТАБЛИЦА ВИДОВ
1 (8). Ширина лба равна ширине 3-го членика усиков или едва шире
или уже его (самцы).
2 (3). Передние голени сзади вблизи средины с короткой крепкой щетин-
кой; средние голени изогнуты спереди, с рядом мелких коротких
волосков; средние бедра с одной крепкой толстой щетинкой снизу
в основной трети; первые 2—3 членика задних лапок снизу с длин-
ными толстыми щетинками, резко выделяющимися своей длиной
от прочих щетинок на лапках. Мельче: 4.5—6.5 см.................
...............................*4. М. (in sp.) aenescens R.-D.
3 (2). Передние голени сзади без крепкой щетинки; средние голени пря-
мые; средние бедра без крепкой щетинки у основания; задние лапки
снизу на всех члениках в мелких щетинках более или менее одина-
ковой длины.
4 (5). Передние бедра по задневентральной поверхности помимо ряда
щетинок снабжены многочисленными длинными, густыми щети-
нистыми волосками; средние голени без бугорка на переднедорзаль-
ной стороне у основания; передние голени сзади с несколькими тон-
кими длинными .щетинками у середины и 3—5 более короткими ще-
тинками ...........................*1. М. (in sp.) hortorum Fall.
5 (4). Передние бедра без густых длинных щетинистых волосков. Сред-
ние голени по переднедорзальной поверхности с бугорком, уса-
женным короткими толстыми шипиками.
206
I. ТРИВА MUSCINI
6 (7). Дорзоцентральные щетинки впереди шва отсутствуют..............
......................................3. М. (in sp.) nilotica Lw.
7 (6). Дорзоцентральные щетинки впереди шва в числе 3 пар............
................................. *2. М. (in sp.) podagrica Lw.
8 (1). Ширина лба равна примерно Vs ширины головы (самки).
9 (10). Дорзоцентральные щетинки впереди шва отсутствуют.............
......................................3. М. (in sp.) nilotica Lw.
10 (9). Дорзоцентральные щетинки впереди шва крепкие в числе 3 пар.
11 (12). Задняя щетинка на передних голенях хорошо развита . . . .
.................................*4. М. (in sp.) aenescens R.-D.
12 (И). Задняя щетинка на передних голенях отсутствует.
13 (14). Передние бедра снизу с короткими согнутыми щетинистыми
волосками; темные переливчатые пятна на боковых отделах 3-го
и 4-го тергитов плохо заметны; задние края 3-го и 4-го тергитов
темные................................*2. М. (in sp.) podagrica Lw.
14 (13). Передние бедра снизу помимо обычных мелких волосков несут
несколько щетинистых волосков почти такой же длины, как задне-
вентральные щетинки или немного короче их; темные пятна в бо-
ковых отделах 3-го if 4-го тергитов ясные; края тех же тергитов
не темнее прочей их поверхности ....................................
.................................*1. М. (in sp.) hortorum Fall.
*1» Morellia (in sp.) hortorum Fall. (рис. 322, 330, 332, 335, 340)»
FalUn, 1816, Acta Holm. : 252 (Musca)', Schnabl u. Dziedzicki, 1911 : 174,
figs. 533, 534; Stein, 1916 : 16; Malloch, 1923, Ann. Mag. Nat. Hist. (9), XII ; 521;
Karl, 1928 : 13; Babic u. Baranov, 1935, Veter, archiv, 5 : 163; Thomsen, 1938 : 310.
Биология. Порчинский, 1910 : 70—74 рис. : 65—74; Graham-Smith, 1919.
Parasitology, XI, 347—384.
$. Лоб в наиболее узком месте равен ширине 3-го членика усиков,
реже едва шире его; в том же месте орбиты немного шире лобной полосы;
скулы по ширине составляют около 2/з ширины 3-го членика!усиков;
глаза в нижнем отделе в очень редких, плохо заметных светлых волосках;
затылок в редком коричневатом, скулы, медиана и узкий край затылка
за глазами в нижней части головы в густом, шелковисто блестящем белом
или желтовато-белом, щеки в редком сероватом налете; затылок и щеки
в бурых волосках, щетинистых в переднем отделе щек и в верхнем отделе
затылка; усики и щупальцы бурые; 3-й членик усиков в 13/а,—П/г раза
длиннее 2-го; щупальцы слабо утолщены к вершине; внутренние темен-
ные и вибриссальные щетинки развиты сильнее прочих; лобные щетинки
длинные, сравнительно крепкие; орбитальных щетинок по 2 с каждой сто-
роны; они направлены вершинами вперед; за ними располагается ряд
тонких щетиночек. Голова черно-бурая, затылок блестящий, черный.
Грудь черная или синевато-черная, с двумя очень широкими черными
продольными полосами на спинке, разделенными каждая на две узким
штрихом сероватого налета; светлая полоса налета между полосами уз-
кая, наиболее ясно выражена в переднем отделе среднеспинки. Грудь
в черных волосках; переднегрудь без волосков; гипоплевры помимо во-
лосков в нижнем отделе, лишены опушения; переднее дыхальце бурое.
Щетинки груди: акростихальных — 1 пара, длинных крепких перед щит-
ком; впереди нее располагается 1—2 пары тонких щетинок; перед швом
несколько пар (наиболее развитых перед самым швом) имеют характер
длинных щетинистых волосков; дорзоцентральных — 3-|-4; интрааляр-
5. MORELLIA
207
ных — 1; предкрылспых — 1; надкрыловых — 2; закрыловых — 3-
(задняя слабая); плечевых — 4 или 3; заплечевая — 1 (реже имеется вто-
рая короткая щетинка вц< редп нее);' предшовная— 1; стерноплевраль-
ных — 1 2 (или 1-|-3; в этим случае передняя щетинка позади шва сла-
бая); на щитке длинные крепкие вершинные, боковые и основные; пред-
крыловая короткая тонкая; 2—4 пары дискальных щетинок; птероплевры
по верхнему краю с многочисленными щетинистыми волосками и мелкими
щетинками. Крылья сероватые, жилки желтовато-или буровато-рыжие,
на вершине темнее; жилка г4+5 сверху с мелкими щетинками от основа-
ния до г—т, снизу в волосках, не заходящих от основания жилки далее
Чз—Чг первого ее отрезка. Чешуйки белые, с палевым широким краем;
жужжальцы темножелтые. Ноги бурые, голени чаще смоляно-бурые,
в черных волосках; передние бедра снизу с рядом длинных задневентраль-
ных щетинок и располагающимися кнаружи от них многочисленными
длинными толстыми щетинистыми, густыми волосками; голени по зад-
ней стороне с 2 длинными, крепкими щетинками у середины и рядом из 3—
5 более коротких заднедорзальных щетинок в средней трети голени. Сред-
ние бедра с пучком длинных щетинок сверху перед вершиной; голени
средних ног по переднедорзальной поверхности в вершинной четверти
с несколькими рядами мелких коротких шипов, переходящих книзу сна-
чала в 3, 2 и далее в один ряд тонких мелких волосков, идущих вдоль всей
голени и обращенных вершинами к дорзальной стороне голени; сзади
голень несет несколько (4—5) длинных задних щетинок в средней трети
и 1—2 более крепких задневентральных щетинки. Задние голени с рядом
не длинных, но сравнительно толстых передневентральных щетинок,
из которых только средние достигают значительной величины; по задней
стороне голени снабжены 1 крепкой щетинкой в нижней трети и 2—3
мелкими щетинками в основной трети; задневентральные волоски корот-
кие, не длиннее поперечника голени и не всегда хорошо развиты.
Брюшко черное, с синеватым отливом, в более или менее густом сером,
местами желто-сером или голубовато-сером налете, с бурой продольной
полосой, ясно выраженной на 3-м и 4-м тергитах (ширина ее составляет
около г/в ширины тергита); по бокам тех же тергитов у заднего их края,
располагаются темные переливчатые пятна, идущие вперед и несколько
к средней линии брюшка; краевые и дискальные щетинки на 5-м тергите
длинные; длинными щетинками и волосками покрыты и стерниты. Церки
удлиненные, широко закругленные по боковым краям, с 2 шипиками
с каждой стороны на боковых дистальных краях. Кокситы широкие
в основной половине, далее почти в 2 раза сужены, с 2 щетинками и мно-
гочисленными мелкими шипиками на дистальном конце.
2- Лоб равен или едва уже глаза; орбиты в 2—24г раза уже лобной
полосы; последняя несет в заДнем отделе две тонких, направленных вер-
шинами друг к другу, щетинки; орбитальных щетинок — 2; из них перед-
няя длиннее if толще задней; наружные теменные щетинки не развиты;
спинка в более густом чем у <5 сероватом налете. Передние бедра снизу
в коротких прилегающих щетинистых волосках и с несколькими длин-
ными щетинистыми волосками, длиною достигающими 2/з или полной
длины задневенГральных щетинок (этот признак отличает самок
М. hortorum от самок М. podagrica). Передние голени с рядом выделяю-
щихся по длине переднедорзальных щетинистых волосков. Желтовато-
серый налет на брюшке хорошо развит, задние края тергитов без темных
поперечных полос, боковые косо расположенные переливчатые темные
пятна на 3-м и 4-м тергитах ясно выражены. Длина тела 6.5—9 мм.
Рис. 322—332. Ноги видов рода Morelliai
322 — передняя нога самца М. /zortowmfFalJ. сзади. 323 — то же М. podagrica Lw.
324 — переднее бедро самки М. podagrica Lw. спереди. 325 — задняя голень самца
М. simplex Lw. с внутренней и частью с задней стороны. 326 — передняя нога самца
М. simplex Lw. сзади. 327 — задняя голень и лапка самца М. aenescens R.-D. сбоку.
328 — бедро и голень средних ног самца М. aenescens R.-D. спереди. 329 — задняя
голень самца М. simplicissima, sp. п., с внутренней и частично с задней стороны.
-330 — переднее бедро самки М. hortorum Fall, спереди. 331 — средняя голень самца
М. podagrica Lw. спереди. 332 — то же самца М. hortorum Fall. (Ориг.).
5. MORELLIA
209
Личинка II стадии имеет пару крепких ротовых крючьев; тело по пе-
редним краям сегментов снабжено шипиками снизу, образующими здесь
сравнительно широкие поперечные полоски; каждый шипик заострен на
вершине, а не двураздельный, как это свойственно личинкам последующей
стадии. Задние дыхальцы имеют по две щели; верхняя шель прямая,
нижняя — изогнутая наподобие цифры 3.
Личинка Ill стадии. Задний конец плоский; задняя поверхность его
несколько выдается над прочей поверхностью сегмента. Головное кольцо
снабжено одним ротовым крючком, выше которого проходит полоска из
простых шипиков, расположенных в 3 неполных ряда. Все сегменты тела
по переднему краю имеют полоску шипиков. Шипики имеют вид сравни-
тельно широких пластинок, снабженных каждая 2, 3 и 4 зубчиками по
заднему краю. Поперечные валики снизу тела личинки почти не выра-
жены. Передние дыхальцы с 7—8 лучами; задние дыхальцевые пластинки
округлые с небольшим угловидным выступом снизу, расположены близко
одна от другой; ширина промежутка составляет всего около 1/ъ—Vi по-
перечника дыхальца. Дыхальцевые щели очень тонкие, длинные и сильно
извитые; каждое дыхальце имеет 3 щели, по строению напоминающие
щели личинок М esembrina meridiana. Щели хорошо заметны лишь у только
что слинявших личинок или в период перед линкой. После перехода в III
стадию дыхальцевые пластинки становятся почти черными, почему щелп
становятся различимы с трудом; в это время дыхальцы представляются
сплошными черными дисками.
Распространение. В СССР вид встречается на север до
Ленинградской обл., Петрозаводска, Усть-Цыльмы (на Печоре), Ки-
рова, Березова на Оби, Тобольска, Красноярска, Иркутска и Якутска;
в Европейской части СССР распространен до крайнего юга. В Средней
Азии вид не найден; далее Казахстана (низовья р. Или) в этой части
СССР вид на юг, повидимому, не заходит. Большая часть Западной
Европы; Монголия.
Личинки М. hortorum развиваются в коровьем помете и являются
чистыми копрофагами.
*2. Morellia (in sp.) podagrica Lw. (рис. 224, 331, 337).
Loew, 1852, Wien, entom. Monatschr,, I : 45 {Cyrtoneura); Pandelle, 1898 : 35
{Pyrellia)-, Malloch, 1923, Ann. Mag. Nat. Hist., (9), XII : 521; Karl, 1928 : 13.
3. Лоб в наиболее узком месте составляет ’-/г—2/з ширины 3-го чле-
ника усиков; скулы чуть уже лба, слабо и постепенно расширены книзу:
щеки узкие, как у следующего вида; усики бурые, 3-й членик в Р/г—
l3/i раза длиннее 2-го; щупальцы бурые, слабо расширены к дисталь-
ному концу. Внутренние теменные щетинки и вибриссы длинные, толстые;
лобные щетинки тонкие, длинные в переднем отделе; орбитальные щетинки
длинные, нетолстые в числе нескольких пар; позади орбитальных имеется
по несколько более коротких щетинок. Голова черная, затылок блестящий
в коричневом, слабо заметном налете, снизу позади глаз в шелковистом
густом налете, как и скулы и передние отделы орбит; щеки в редком се-
ром налете. Грудь блестящая черная, с легким синеватым оттенком:
плевры, исключая части их, расположенные под передним дыхальцем,
в редком желтовато сером налете. Задний отдел спинки в редком матово-
сером налете; щиток в более густом такого же цвета налете; вдоль
спинки проходят три узких полоски блестящего серовато-белого налета,
из которых боковые идут от плечевых бугорков к основанию крыла; сред-
14 Фауна СССР, том XVIII, вып. 4.
210
I. ТРИБА MUSCINI
няя полоса узкая, наиболее ясно выражена в переднем отделе спинки.
Переднегрудь, передние плевры и супрасквамальное ребро без волосков;
гипоплевры со сравнительно длинными волосками в нижнем отделе. Ще-
тинки груди: акростихальных позади шва —2 пары; дорзоцентральных—
интраалярных — 2; — предкрыловых — 1; закрыловых — 2; над-
крыловых — 2, иногда 3; плечевых — 4; заплечевых — 1; предшовная —
1; нотоплевральных — 2; стерноплевральных—14-2; на щитке: вер-
шинных — 2, боковых — 2, основных — 2, все хорошо развиты; пред-
основные и несколько пар дискальных щетинок тоньше остальных.
Крылья сероватые с легким затемнением при основании; жилки буровато-
коричневые; г4+- в основной половине первого отрезка сверху с мелкими
щетинками, снизу при основании с несколькими волосками; чешуйки
белые сочень слабо затемненным краем; жужжальца желтые. Ноги черно-
бурые, голени смоляно-бурые; передние бедра с рядом крепких заднедор-
зальных и задневентральных щетинок, ио вентральной поверхности с не-
многочисленными короткими и несколькими более длинными, торчащими
щетинистыми волосками; передние голени по переднедорзальной сто-
роне с рядом, идущим по всей длине голени, коротких крепких щетинок,
по задневентральной поверхности с 4—5 длинными и тонкими щетинками
по задней стороне. Средние голени без крепких щетинок, у вершины
с верхней стороны, с пучком толстых, тесно стоящих щетинок; средние
голени вблизи основания на дорзальной поверхности с бугорком, уса-
женным короткими толстыми шипиками, переходящими к дистальному
концу голени в ряд мелких тонких волосков, обращенных вершинами на-
зад; в средине голени между этими волосками имеются 2—3 щетинки;
задняя сторона голеней с рядом длинных коротких и толстых заднедор-
зальных и с несколькими короткими задневентральными щетинками.
Задние бедра с рядами переднедорзальных и передневентральных щети-
нок. Голени задней пары ноге очень длинными щетинистыми волосками
по всей длине переднедорзальной поверхности, рядом занимающим 4/s
или 3/* длины голени передневентральных щетинок и 3 щетинками на
заднедорзальной поверхности, из которых одна более длинная распо-
лагается в верхней, а две более коротких — в основной трети голени.
Задневентральная поверхность задних голеней с неполным рядом корот-
ких щетинистых, редко сидящих волосков, длиною не превышающих
поперечника голени. Присоски короче 5-го членика лапки. Брюшко сине-
вато-бурое, в черных волосках, в негустом сером налете, оставляющем
свободными задние края тергитов, кажущиеся в связи с этим более тем-
ными; средняя темная продольная полоска не имеет ясных контуров;
она достигает 5-го тергита. Щетинки тонкие, длинные. Церки удлиненные,
с полукруглым выступом посредине боковых краев; в боковых дистальных
отделах снабжены с каждой стороны 3 зубчиками. Кокситы очень широ-
кие при основании и узкие в дистальной половине, несущей несколько
мелких шипиков на верщине по внутреннему краю.
Самка сходна с таковой М. hortorum, от которой отличается более
крупными размерами, более темной окраской тела, почти не развитыми
темными переливчатыми косыми пятнами на 3-м и 4-м тергитах. Перед-
ние бедра снизу в редких, коротких, согнутых волосках (длинные щети-
нистые волоски отсутствуют); переднедорзальные щетинки развиты
сильнее, нежели у М. hortorum.
Отсутствие длинных щетинистых волосков на вентральной стороне
передних бедер служит наиболее удобным признаком для разграничения
самок М. hortorum и М. podagrica. Оба эти вида отличаются от М. sim-
5. MORELLIA
211
lex и M. simplicissima присутствием у последних волосков на передне-
груди.
Распространение. В СССР вид распространен в лесной
и лесостепной зонах, от Карельского перешейка на западе до Дальнего
Востока. На юге встречается в горных местностях. Наиболее южным ме-
стонахождением вида на территории СССР является пока Дагестан.
Большая часть Европы.
3. Morellia (in sp.) nilotica Lw.
Loew, 1856, NeueBeitr., IV : 48 (Cyrtoneura)-, Bezzi u. Stein, 1907, Katal. palaarkt-
Dipt., Ill : 601; Villeneuve, 1937, Bull. Ann. Soc. entom. Belg., LXXVII : 406; Mal-
loch, 1928, Ann. Mag. Nat. Hist., (10), I : 474;—? minor Mall.. Schnabl u. Dziedzicki,
1911 : 174.
Близок к M. podagrica, отличается меньшей величиной и отсутствием
дорзоцентральных щетинок впереди шва; передние щетинки той же
группы, расположенные позади поперечного шва спинки также отсут-
ствуют, или слабо развиты; лишь две дорзоцентральных щетинки перед
щитком достигают полного развития. Орбиты самки в белом налете
в передней половине, далее.— черные. Передние голени самца с наруж-
ной стороны от середины до вершины с 3—4 длинными нежными волос-
ками. Средние голени в общем как у М. podagrica, перед основанием утон-
чены, по дорзальной стороне после сужения — с бугорком, несущим гу-
стосидящие шипы, переходящие далее в ряд тонких коротких волосков,
следующих по всей длине голени до ее вершины; по задней стороне с 3>
щетинками. Задние голени снаружи на границе вершинной трети с 1 ще-
тинкой, с переднедорзальной поверхности по всей длине голени с рядом
щетинок, среди которых выделяются несколько более длинных; с передне-
вентральной стороны с 2—4 щетинками.
Распространение. В пределах СССР вид пока не обна-
ружен. Вид распространен в центральной и северной Африке и весьма
возможен у нас на южных границах Средней Азии.
*4. Morellia (in sp.) aenescens R.-D. (рис. 327, 328, 334).
Robinean-Desvoidy, 1830, Essai Myod.; 406; Pandelle, 1898 : 34 (Pyrellia); Stein,.
1916 : 15; Malloch, 1923, Ann. Mag. Nat. Hist (9), XII : 521;
Биология. Порчинский, 1910 : 74, фиг. 74.
<5. Ширина лба в IVi—1х/а раза превосходит ширину 3-го членика
усиков; орбиты в среднем отделе почти равны по ширине лобной полосе
в том же месте; ширина щеки в З1/*—4 раза менее вертикального диа-
метра глаза; усики и щупальцы бурые; 3-й членик усиков почти в 2 раза
длиннее 2-го; ширина скулы составляет около 3/i—2/з ширины 3-го членика
усиков; покрыты, как и передний отдел орбит и узкая полоска за глазами
в нижней части головы, шелковистым белым налетом; орбиты в остальной
части бурые; затылок блестяще черный в едва заметном коричнева-
том налете, щеки в редком сероватом налете. Затылок и щеки в черных
волосках. Внутренние теменные щетинки и вибриссы крепкие; лоб-
ные щетинки сравнительно тонкие; передние 2 орбитальные щетинки
длинные; позади них располагается по 3—5 коротких щетинистых волос-
ков. Грудь черная, блестящая, иногда с более или менее ясно выраженным
синеватым оттенком. Рисунок спинки, как у М. simplicissima, но с более
узкими полосками налета. Щетинки груди, как у М- simplex, исклю-
чая акростихальных щетинок впереди шва, которые у М- aenescens не раз-
виты в этом отделе спинки. Переднегрудь голая; гипоплевры с волосками
14*
212
I. ТРИБА MUSCINI
только в нижнем отделе. Крылья сероватые, с буроватыми жилками;
г4+5 сверху с мелкими щетинками в основной х/г или 2/з первого отрезка,
снизу с одиночными волосками лишь при основании. Чешуйки белые,
с палевым краем; жужжальца желтые. Передние ноги с обычным воору-
жением из щетинок. Передние голени имеют крепкую, короткую щетинку
на задней стороне. Средние бедра в основной трети снизу с 1 очень тол-
стой щетинкой; голени средних ног согнуты в основной половине на дор-
зальную сторону; по переднедорзальной стороне они снабжены по всей
длине рядом мелких тонких волосков, обращенных вершинами к дорзаль-
ному краю голени; по задней стороне голени вооружены 2—3 щетинками.
Задние голени, сильно изогнутые у середины, с 2—3 переднедорзаль-
ными щетинками, 1 длинной заднедорзальной в дистальной трети
(иногда имеется короткая щетинка в том же ряду в вершинной части
голени); у вершины голеней с передневентральной и задневентральной
сторон присутствует по 1—2 длинных, сильно изогнутых на вершине
щетинистых волоска. Два или три основных членика задних лапок снизу
по передневентральной и вентральной поверхности с 2—3 рядами креп-
ких, толстых щетинок, стоящих тесно друг к другу и по длине превышаю-
щих поперечник голени. Концевые щетинки члеников задних лапок длин-
ные. Коготки и присоски короче 5-го членика лапки; последние из них —
желтоватые. Церки широкие, закругленные на вершинном крае. Вблизи
задней границы с каждой стороны от средней линии располагается по 4
коротких шипика. Кокситы утонченные к вершине, при основании корот-
кие, по внутреннему краю вогнутые и здесь снабжены маленьким тре-
угольным выступом, на дистальном конце со щетинкой и в многочислен-
ных мелких шипиках.
2- Ширина лба составляет немного менее ширины глаза; орбиты
в 2—24г раза уже лобной полосы. Передние голени с задней щетинкой
у середины. Ноги обычного строения; голени задних ног слабо согнуты,
передние голени прямые, с 3—5 задними щетинками. Окраска тела как
у самца.
Длина тела 4.5—6.5 мм.
Распространение. В СССР вид найден во многих местах
как Европейской части: от Кольского п-ва и Архангельска до южного бе-
рега Крыма и до Кисловодска на Кавказе, так и в азиатской: северный
Казахстан, Семиречье, Алтай, Прибайкалье, Якутия, Приморский край.
Личинки обитают только в конском помете, встречаясь в природе
значительно реже других видов того же рода (Порчинский, 1910). Личинки
сходны с таковыми М. hortorum, отличаясь несколько приплюснутым
телом.
Подрод 2. Dasysterna Zimin, subgen. nov.
Cyrtoneura, Loew, 1857 : 45; —Pyrellia РапйеПё, 1898 : 35; —Morellia Schnabl
•u. Dziedzicki, 1911 ?122; Stein, 1916Ц- 16; S6guy, 1923 : 350; Malloch,1923 : 521;
1925Д 87; Karl, 1928 : 13.
Тип подрода: Morellia {Dasysterna') simplex Loew, 1857.
От n едыдущего подрода отличается наличием волосков на перед-
негруди (рис. 339). Самцы, помимо того, характеризуются отсутствием
вооружения из шипиков и торчащих волосков на средних голенях и
строением церок. Церки с длинными двумя зубцами по вершинному краю
около средины, по бокам от глубокой выемки. Кокситы массивнее, чем
в подроде Morellia.
5. MORELLIA
213
С достоверностью к этому подроду относятся 3 палеарктических
(один общий с Эфиопской и Ориентальной областями) и один южно-африкан-
ский вид. Для Союза ССР известны 2 вида, из которых М. simplexLw. рас-
пространен в Европейской его части и Средней Азии, не заходя в Сибирь.
Второй, описываемый в настоящей работе, свойствен Восточной Сибири
и Дальнему Востоку, не продвигаясь на запад далее Енисея.
Личинки М. (D.) simplex развиваются в коровьем помете. Мухи
обычны на животных; местами явно вредны для последних.
ОПРЕДЕЛИТЕЛЬНАЯ ТАБЛИЦА ВИДОВ
1 (2). Дорзопентральные щетинки впереди поперечного шва спинки и
2—3 пары позади него имеют характер тонких волосков................
....................................7. М. (D.) hortensia Wd.
2 (1). Дорзоцентральные щетинки шва крепкие, длинные.
3 (6). Лоб узкий; ширина его не более IVz ширины 3-го членика усиков
(самцы).
4 (5). Лоб уже 3-го членика усиков или по ширине почти равен ему; зад-
ние голени в вершинной половине с несколькими рядами длинных
щетинистых волосков, покрывающих заднюю, задневентральную и
частью вентральную поверхности; щетинки на верхней стороне
жилки г4+5 не распространяются далее основной трети 1-го отрезка
жилки; внутренние черные полосы спинки далеко не достигают
щитка....................................... *5. М. (D.) simplex Lw.
5 (4). Лоб шире 3-го членика усиков; задние голени в вершинной поло-
вине с одним рядом (или неполными двумя) редко стоящих волос-
ков по задневентральной стороне; первый отрезок жилки г4+5,
по крайней мере на половину своей длины от основания, усажен
щетинками; все 4 черные полосы спинки достигают щитка (иногда
слиты в задней части спинки попарно и в этом случае представлены
двумя широкими полосами). . . *6. М. (D.) simplicissima, sp. nov.
6 (3). Лоб широкий, ширина его по крайней мере в 3 раза более ширины
3-го членика усиков (самки).
7 (8). Первый отрезок жилки г4+5 сверху со щетинками в основных 2/з
или 3/ц отрезка; задние края тергитов без ясно выраженной бурой
окраски, в сером налете . . . *6. М. (D.) simplicissima, sp. nov.
8 (7). Первый отрезок жилки г4+5 сверху со щетинками, которые развиты
не более, чем на V2 (обычно 7з) длины отрезка; задние края тергитов,
как правило, буроватые, в очень редком налете......................
.......................................*5. М. (D.) simplex Lw.
*5. Morellia (Dasysterna) simplex Lw. (рис. 325, 326, 333, 338, 339,.
341—347, 351—361).
Loew, 1857, Wien, entomol. Monatschr., I : 45 (Cyrtoneura); Bezzi u. Stein, 1907,
Catal. palaarkt. Dipt., Ill : 601 {Morellia)', Malloch, 1923, Ann. Mag. Nat. Hist., (9),
XII : 523; он же, 1925, там же, (9), XVI; 87; Karl, 1928 : 13.
Биология. Thomson, 1937, Parasitology, XXIX : 290—294, figs. 25—31,
33—36, 44, 50, 51, 146, 156, 157; Thomsen, 1938 : 310; Дербенева-Ухова, 1940 : 324;
Зимин, 1948 : 75, фиг. 41;
$. Лоб уже, реже равен по ширине 3-му членику усиков, орбиты в наи-
более узком месте лба равны ширине лобной полосы или шире ее; послед-
няя слабо расширена к обоим концам; ширина скулы посредине менее
ширины 3-го членика усиков; 3-й членик в 2—21/* раза длиннее 2-го;
214
I. ТРИБА MUSCINI
ариста с длинными волосками по верхней стороне и более короткими по
нижнему краю; длина верхних волосков аристы в Р/г раза превышает
поперечник 3-го членика усиков; щупальцы черно-бурые. Голова черно-
бурая, как и усики. Орбиты в белом или желтоватом, скулы в шрлковисто
блестящем, серовато-белом густом, лицо в густом, темносером матовом,
щеки в затылок в редком сером матовом налете. Затылок и щеки в черно-
бурых коротких волосках. Лобные щетинки короткие, часто расставлен-
ные, в задней половине лба вершинами направлены вперед; глазковые
Рис. 333, Самец Morellia simplex Lw. (Ориг.).
щетинки короткие; над креп-
кими угловыми вибриссами ра-
сполагаются спутанными двумя
рядами короткие щетинистые
волоски, поднимающиеся до
половины высоты вибриссаль-
ных килей; внутренние, темен-
ные щетинки и вибриссы креп-
кие. Грудь черная, блестящая,
в сероватом налете, образую-
щем 3 продольных полоски на
спинке; боковые светлые поло-
сы располагаются вдоль гра-
ницы спинки и плевр. Средние
черные продольные полосы, ог-
раничивающие среднюю свет-
лую полосу налета разделены
вдоль серым тонким штрихом
налета на 2 узких, из коих вну-
тренние достигают лишь среди-
ны заднего отдела спинки за
швом, а наружные доходят до
щитка. Плевры в более редком
светлом налете, оставляющем
свободной косую полоску ос-
новного цвета груди в перед-
нем отделе мезоплевры. Поверх-
ность груди и щитка в чер-
'ных волосках; гипоплевры над
задними тазиками с группой
коротких волосков и с 1—3
волосками в верхнем переднем
углу; проплевры голые или с несколькими волосками в нижней
трети; переднегрудь с длинными щетинистыми бурыми волосками
по краям. Щетинки груди: акростихальные перед швом имеют вид
щетинистых волосков, из которых задняя пара наиболее крепкая;
за швом акростихальных — 1—3 пары, передние две пары щетийок
короче задней, часто вовсе отсутствуют; дорзоцентральных — 3-f-
4-4 (5, 6); интраалярных, за швом, — 1; предкрыловых — 1; надкрыло-
вых — 2; закрыл овых — 3; плечевых — 4; заплечевых — 1—2; пред-
шовных — 1; нотоплевральных — 2; стерноплевральных 1 4-2. На щитке
вершинных — 2, перекрещивающиеся, крепкие; основных — 2; пред-
основных — 2, в виде волосков; дискальных 2—3 пары, идущих от вер-
шины щитка вблизи боковых краев к его основанию. Супрасквамальное
ребро без волосков. Ноги черные, голени смоляно-бурые; передние бедра
5. MORELLIA
215
с гребнями щетинок на задневентральной и заднедорзальной стороне;
ковнутри от задневентрального ряда по нижней стороне бедра распо-
лагаются короткие волоски, длина которых в 2—3 раза менее длины ще-
тинок; передняя поверхность бедра в коротких прижатых, задняя —
в торчащих более длинных черных волосках. Голени передних ног без
крепкой щетинки на задней поверхности; коготки равны или чуть длин-
нее 5-го членика лапки; присоски чуть короче коготков. Голени средних
ног с неполными рядами щетинок по задневентральной и заднедорзальной
стороне. Средние бедра на вершине по дорзальной поверхности с тон-
кими короткими щетинками. Голени задних ног в вершинной половине
с длинными щетинистыми волосками, длина которых несколько превы-
шает поперечник голени, с задней, задневентральной и вентральной сто-
рон по заднедорзальной поверхности голень снабжена рядом щетинок,
более длинных в вершинной трети голени; переднедорзальная сторона
с рядом коротких щетинок почти равной длины; по передневентральной
поверхности голень несет в вершинной половине (или 2/з) тонкие
щетинки, длиной в Р/г раза превышающей диаметр голени; верхние ще-
тинки этого ряда короче. Крылья сероватые, 7'4+5 в основном отрезке
сверху с волосками, простирающимися от основания на 1/з расстояния
до г—т; снизу с несколькими волосками у основания. Жилки рыжева-
тые: грудная чешуйка слабо дымчатая, реже белая, с рыжеватым краем;
крыловая чешуйка обычно белая; жужжальца желтые. Брюшко яйцевид-
ное, оливково-бурое, с бурыми задними краями 1-ф2-го, 3-го и 4-го тер-
гитов и нерезко выраженной, суживающейся назад, тонкой, бурой, про-
дольной полоской, достигающей основания 5-го тергита. Брюшко в чер-
ных волосках, достигающих наибольшей длины в боковых отделах 4-го
тергита и на 5-м тергите. Переливчатый желтовато или слабо оливково-
серый светлый налет покрывает все брюшко, оставляя бурыми задние
края тергитов, кроме 5-го. Краевые щетинки сравнительно крепкие, но
короткие. Церки широкие, глубоко расщеплены по середине нижнего
края, на внутренних углах по дистальному краю снабжены 2 длинными
зубцами; по наружному дистальному краю округленные, лишены шипи-
ков; кокситы массивные, неправильной формы с мелкими шипиками и
1 длинной щетиночкой вблизи дистального конца.
2- Лоб равен 4/s ширины глаза, с параллельными краями; лобная
полоса голая, оливково-бурая или черная, в 3—4 раза шире орбиты,
почти не суживающаяся к лунке. Орбиты в редких коротких волосках.
Орбитальных щетинок — 2, расположенных в задней половине орбит;
наружные теменные отсутствуют, как у <5. Коготки короче 5-го членика
лапки. Брюшко шире, нежели у самца. Боковые черные полосы спинки
в заднем ее отделе за швом разбиты линией серого налета на 2 почти рав-
ной ширины полоски. Щетинки на верхней стороне 1-го отрезка жилки
?’4+- не занимают более половины его длины (обычно не более 1/з).
Длина тела 6—8.
Яйцо 1 по внешности напоминает яйцо Othellicr, продольные рубчики
нежные, не выступающие; длина 2 мм.
Личинка I стадии метапнейстична; задние дыхальцы выступают тем-
ными пластиночками, каждое из них снабжено одним почковидным отвер-
стием. По передним краям брюшных сегментов с 1-го по 7-й распола-
гаются хорошо заметные полоски мелких шипиков; такая же полоска
имеется и на 3-м грудном кольце, но с более мелкими шипиками; полоски
1 Описание фаз развития, кроме личинки III стадии, дано по Томсону.
216
I. TP ИВ A MUSCINI
из шипов окружают полностью края сегментов; последний брюшной
сегмент снабжен анальной пластинкой, позади и впереди которой рас-
полагается по полоске из шипиков; задний конец личинки плоский. Рото-
Рис. 334—340.
334 — церки и левый коксит Morellia aenescens R ,-D. 335 —
церки и правый коксит М. hortorum Fall. 336 — церки М. sim-
plicissima, sp. nov. 337 — церки и правый коксит М. podagrica.
Lw. 338 — то же М. simplex Lw. 339 — переднегрудь и основа-
ния ног М. simplex!™.340 — тоже М.hortorum Fall. (Ориг.).
глоточный аппарат устроен по типу Musca. Базальный склерит тонкий,
с 4 острыми, тонкими задними отростками, сильно расходящимися вверх
и вниз. Дорзальная перемычка имеет вид слабо склеротизованной по-
лоски; передние отростки длинные, соединены с гипостомальным скле-
5. MORELLIA
217
ритом, передний конец которого сочленен с длинным и несколько
S-образно изогнутым по краям срединным дорзальным склеритом. Вен-
трально от последнего расположены Т-образные склериты. Длина тела
(перед линькой на II стадии) 5 мм.
Личинка II стадии амфипнейстична; передние дыхальцы с 4—6 лу-
чами; задние дыхальцы имеют по 2 изогнутых щели. Полоски из пйшиков
развиты по переднему краю брюшных сегментов (с 1-го по 7-й); задняя
Рис. 341—347. Детали строения преимагинальных фаз
Morellia simplex Lw.
341 —задние дыхальцы личинки I стадии. 342 —то
же II стадии. 343 —яйцо. 344 — грудное дыхальце
куколки. 345 — головной конец тела личинки I стадии.
346 — тоже!! стадии. 347 — ложнококон. (По Томсону).
поверхность последнего сегмента округлена и несет полное кольцо ив
шипиков. Базальный склерит по дорзальному краю почти прямой, послед-
ний параллелен вентральному краю склерита; вентральные отростки
округлены на конце, дорзальные — острые. Дорзальные отростки соеди-
нены спереди грубосетчатой перемычкой; передние отростки базального
склерита снизу соединены крепким поперечным склеритом. При рас-
сматривании с дорзальной поверхности гипостомальный склерит имеет
строение в виде буквы Н; боковые лопасти треугольные; непосредственно
впереди гипостомального склерита причленены угловидные склериты,
разделенные снизу и тесно сближенные своими дорзальными отделами.
-218
I. ТРИБА MUSCINI
В этой части они не имеют направленного назад отростка. Ротовые
крючья слабо S-образно изогнуты и своим дистальным конном направ-
лены не к ротовому отверстию, а вверх и вперед. Вентральные и боковые
Рис. 348—350. Фаллозома.
348 — Morellia podagrica Lw. 349 — тоже Mesembrina mystacea L. 350—
то же Graphomyia maculata Scop. (Ориг.).
стенки ротового отверстия образованы двумя губными склеритами, сое-
диненными вместе наподобие желобка. Головной сегмент с парой антенн
и парой щупалец (максиллярных); вентральная поверхность головного
сегмента в бороздках, берущих свое начало у ротового отверстия и идущих
на боковые стороны сегмента. Личинки III стадии короткие толстые
с очень плотными покровами, плоским зачерненным задним конном тела;
5. MORELLIA
219
плевральные сегменты не выражены. Кутикула блестящая, желтовато-
рыжая, с грубой скульптурой, напоминающей тисненную кожу, 8-й сег-
мент в неглубоких крупных ямках. Передние края сегментов в редких
коротких черных шипиках. Голова впереди коротко-двураздельная; ро-
товые бороздки хорошо выражены; антенны короткие, двучленистые;
2-й членик конусовидный; максиллярные пальпы имеют вид слабо высту-
пающих бугорков. 1-й грудной сегмент с многорядными светлыми мел-
кими густостоящими шипиками по переднему краю. 2-й, 3-й грудные и
1-й и 2-й брюшные сегменты с черными редкими короткими двузубчатыми
передними маргинальными шипами; шипы расположены в 5—6 непра-
вильных рядов; 3-й брюшной сегмент снизу, 4—7-й сегменты вдоль всего
заднего края с 1—2 рядами шипиков и с 5 рядами более крупных, также
двузубчатых передних маргинальных; 8-й и 9-й сегменты с черными, длин-
ными тонкими, дуговидно изогнутыми черными шипами. 8-й сегмент сзади
почти плоский или вогнутый, с слабо утолщенным, резким краем. Аналь-
ная пластинка очень узкая, длинная; позади ее располагается валикооб-
разное возвышение, угловидно выдающееся назад от анального отверстия
л усаженное тремя (местами 4) рядами коротких двузубчатых передних
и длинных острых задних шипов. Угловидный выступ валикообразного
возвышения, сзади упирается в полулунную выпуклость, вооруженную
черными, изогнутыми тонкими шипами. Передние дыхальцы с 5 (реже
с 4) короткими овальными выростами. Задние дыхальцы у взрослой
личинки с сильно склеротизованной пластинкой, черные; дыхальце-
вые щели многократно изогнутые, тонкие; расстояние между дыхаль-
цами равно двойному-тройному поперечнику стигмы. Центральный диск
приближен к внутреннему краю дыхальца; перитрема узкая; стигмальные
железы в форме маленьких светлых пятнышек, лежащих по одному
кнаружи от каждой щели. Ротовой аппарат массивный, с очень крупным
склеритом. Ротовые крючья вытянутые, слабо изогнутые, с узкой базаль-
ной частью; зубные склериты неправильно треугольные, с оттянутым
нижним углом; гипостомальные — неправильно прямоугольные, длина
их почти в IVa раза превышает ширину склерита посредине; вентральный
край с коротким выступом у середины. Фарингеальные склериты широ-
кие, крупные, сильно склеротизованные, длина их равна или чуть менее
удвоенной длины гипостомальных склеритов и ротовых крючьев, вместе
взятых; поперечник склеритов без фарингса, более чем в два раза короче
его длины; передние отростки несколько короче гипостомальных скле-
ритов, широкие; дорзальная перемычка решетчатая с округлыми перед-
ними и узкими заостренными задними отростками; задние верхние от-
ростки в 2—21/г раза уже нижних и почти равны по длине, на конце ок-
руглены и в задней половине снабжены хитиновой широкой кромкой,
имеющей зазубренный край; нижняя фарингеальная перемычка узкая
с мелкими кольцевидными выростами по заднему краю; фарингеальная
мембрана хорошо выражена, широкая, хитинизированная.
Длина тела личинки 7—9 мм (средняя 7.6 мм): толщина у средины —
3—-3.3 мм.
Ложнококон бочонкообразный с ячеистой скульптурой на поверх-
ности, как у личинки. Дыхальцы хорошо развиты, имеют вид рожек,
внутри с многочисленными дыхальцевыми сосочками.
Распространение. В Европейской части СССР на север
доходит до Карельского перешейка, Ярославля, Кирова и среднего Урала;
известен с Кавказа, из Средней Азии и Казахстана (до северной его гра-
ницы с Сибирью). В Сибири отсутствует. Большая часть Зап. Европы.
Рис. 351—361. Детали строения личинки III стадии Morellia simplex Lw.
351 — ротоглоточный аппарат. 352 — задний конец тела сзади. 353 — он же
снизу. 354 — он же сбоку. 355 — переднее дыхальце. 356 — заднее дыхальце.
357 — вооружение снизу 5-го (2) и 6-го (1) брюшных сегментов. 358 —
скульптура среднего отдела дорзальной поверхности 2-го грудного сегмента.
359 — сильно увеличенные шипы 2-го грудного сегмента. 360 — шипы
позади анальной пластинки при сильном увеличении. 361—шипы на
1-м грудном сегменте. (По Зимину).
5. MORELLIA
221
Томсон (1937) указывает, что развитие личинок проходит в помете
крупного рогатого скота. В Кабарде, по данным Дербеневой-Уховой
(1940), мухи развиваются и в коровьем навозе в хлеву. Самки отклады-
вают яйца группами по 25 штук под верхнюю корочку помета. Откладка
яиц предшествует питанию самки жидким содержимым помета. Про-
должительность фазы яйца равна, примерно, 28 часам (при 20° С); в иных
случаях она затягивается до 36—40 часов. Личинки I стадии, как и все
прочие стадии, — копрофаги. Продолжительность I стадии равна почти
18 часам. Личинок II стадии автор находил через 42 часа после вылупле-
ния личинок из яиц.
Личинки мало поворотливы; вся их жизнь протекает в коровьем
помете, в нем же проходит и окукление личинок. В пупарии муха пре-
бывает 17—18 дней. Зимует М. (D.) simplex в фазе личинки. В Англии
вылет мух происходит в средине мая. В природных условиях они задер-
живаются до октября.
В средней Европе мухи имеют некоторое отрицательное значение
в животноводстве. По утверждению Томсона, они досаждают скоту, в осо-
бенности ранним летом (лошадям, коровам), в массах пристраиваясь
вокруг глаз и на пораженных местах кожи, слизывая выделения.
В Таджикистане, по нашим данным, М. simplex является обычным
синантропным видом, встречаясь на скотных дворях, у помойных и мусор-
ных ям, изредка посещая и жилище человека. Самки иногда бывают очень
назойливы обычно в предвечерние часы (Таджикистан).
6. Morellia (Dasysterna) simplicissima Zimin, sp. nov. (рис. 329, 336).
Близок к предыдущему виду. Несколько крупнее D. simplex-, налет
на брюшке развит сильнее, последнее обычно не имеет темных полос по
задним краям тергитов. У j лоб много уже, чем у j предыдущего вида.
Прочие отличительные признаки и указаны в определительной
таблице. Длина 6.2—8.3 мм.
Распространение. Сибирь от Челябинска на восток до
Приморского края; Северный Казахстан. Известен из следующих мест:
окрестности Минусинска, 11 V 1922 (Сушкин и Редикорцев), Минусинск
3—7 VI (Вагнер); с. Новоселово, Минусинского района, 10 VIII 1924
(Виноградов); с. Сорокине бл. Красноярска, 22 VI 1897 (Вагнер); раз.
Смолино бл. Челябинска 16 VI 1916 (Кузнецов); Боровое Кокчетавского
р-на, Сев.-казахст. обл., 22 VII 1932 (Попов), 15 VII 1916 (Родионов),
ст. Лиственичное, Иркутского окр., 29 VII 1912 (Крюкова); Якутия:
Якутск, 16 VI—21 VII 1927 (Москвин); с. Намское, Якутского р-на,
20 VII 1926 (Москвин) Хамурган бл. устья р. Алдана, 29 VI 1926
(Бианки); Приморский край: Ворошилов 25 VIII 1933 (Кривенко); Шко-
товский р-н, Владивостокского окр. 15 IX 1929, полуостров Янковского
26—29 VII 1905 (Гудзенко).
Вид близок также к М. hortensia Wd. (см. ниже), от которой отли-
чается наличием крепких предшовных щетинок.
7. Morellia (Dasysterna) hortensia Wd.
Wiedemann, 1830, Aussereur. zweifliig. Ins., II : 413 (Musca); Stein, 1917 : 108;
Karl, 1935, Arb. morph, taxon. Entomol., II : 36; Malloch, 1925, Ann. Mag. Nat. Hist.,
(9), XVI : 86.
Вид является весьма близким к М. simplicissima и М. simplex. От
названных двух видов он отличается слабо развитыми, тонкими, коротень-
кими de впереди шва (и первыми парами позади него). От М. simplex по-
222
I. ТРИБА MUSCINI
мимо того М. hortensia отличается наличием лишь одного ряда задневен-
тральных щетинистых волосков, которые у М. simplex располагаются
в несколько неправильных рядов.
Вид широко распространен в Ориентальной области: Индия, Цейлон,
Индокитай, Ява, Тайван.
Приводимый Мэллоком (1923 : 523) под именем М. hortensis Wd.
вид, а также упомянутый им в последующей его работе (1925 : 86, 87)
М. hortensis с о. Сокотры (вост. Африка) по всей вероятности представ-
ляют собою самостоятельный вид, отличающийся от ориентальных экзем-
пляров М. hortensis волосками на переднегруди.
6. Род MESEMBRINA MG.
Meigen, 1826, Syst. Beschr., V : 10;—Neomesembrina Schnabl u. Dziedzicki,
1911 : 226; — Hypodermodes Townsend, 1912, Proc. Ent. Soc. Wash., XIV : 46; S<5gin .
1923 : 363; — Metamesembrina Townsend, 1912 : 45—53.
Тип рода Mesembrina meridiana Linne (1761).
Крупные мухи с черным, обычно блестящим телом; грудь покрыта
то черными короткими, то густыми, темными и светлыми волосками,
делающими этих мух похожими на шмелей.
Глаза голые. Щетинка усиков длинноперистая; жилка т в вершин-
ном отделе всегда изогнута дуговидно. Гипоплевры, метаплевры, супра-
сквамальное ребро и слабо расширенная впереди переднегрудь лишены
волосков. Щетинки среднеспинки, исключая краевых щетинки щитка,
развиты сравнительно слабо; акростихальные щетинки волосовидные,
присутствуют лишь в заднем отделе спинки; дорзоцентральных щетинок
за поперечным швом — 4; щиток по краю с 10—16 щетинками; стерноплев-
ральных щетинок 1—1, 1—2, 1—3.
Лоб самцов уже глаза, у самок равен, уже или шире глаза; скулы
широкие; лобная полоса со щетинистыми волосками в заднем отделе,
достигающими у некоторых видов крепости щетинок. Орбитальные ще-
тинки слабо развиты; у SS отсутствуют, реже имеются; у самки они сог-
нуты вершинами или вперед и вниз или назад. Скулы голые или с волос-
ками в верхнем отделе. Ноги длинные, у самцов некоторых видов с очень
сильно развитыми волосками, щетинками и шипиками на внутренней сто-
роне средних голеней. 5-й тергит самок нередко с пятнами из густых
коротких волосков в основной половине. Фаллозома с очень короткой
склеротизованной базальной частью, длина которой в 4—5 раз менее
таковой мембранозной апикальной части; последняя в вершинном отделе
в многочисленных длинных тонких светлых шипиках. Базальный отро-
сток фаллозомы длинный, крепкий, изогнутый на дистальном конце.
Относящиеся к этому роду 9 видов известны из Палеарктической
области; один вид является общим с Северной Америкой. Два вида срав-
нительно недавно описаны из южного Китая.
Большинство видов рода свойственно таежно-лесной зоне. В южных
провинциях, бедных видами, представители рода встречаются лишь
в горных областях в поясе лесов и на альпийских лугах на сравнительно
большой высоте (до 4000 м).
Данные по биологии, относящиеся к двум широко распространенным
в Европе видам, указывают, что развитие личинок проходит в помете круп-
ного рогатого скота, где личинки ведут себя то как обычные калоядные
формы, то как хищники, истребляющие личинок других мух.
6. MESEMBRINA
223
В морфологическом отношении род является весьма хорошо отгра-
ниченным от соседних родов, составляя монолитную группу видов. Незна-
чительные уклонения от общей схемы строения наблюдаются лишь у трех
видов, обладающих развитым волосяным покровом груди и брюшка и
более выраженным половым диморфизмом (волосяной покров и щетинки
средних голеней <5<5). Признаки эти, столь обычные для многих родов
семейства, не дают оснований для дробления рода, как это совершенно
формально сделали Шнабль й Дзедзицкий (1911) и другие.
ОПРЕДЕЛИТЕЛЬНАЯ ТАБЛИЦА ВИДОВ
1 (4). Среднеспинка и дистальная половина брюшка в светлых (желтых,
серовато-желтых) длинных и густых волосках.
2 (3). Подкрыловые чешуйки коричнево-бурые. Задний отдел средне-
спинки перед щитком и щиток в черных волосках, реже вся сред-
неспинка (включая щиток) в светлых волосках........................
..........................................*1. М. mystacea L.
3 (2). Подкрыловые чешуйки желтые. Вся среднеспинка в светложел-
тых волосках...................................*2. М. decipiens Lw.
4 (1). Среднеспинка в чернобурых коротких волосках.
5 (8). Отрезок костальной жилки между г4+- и т равен или едва короче
предыдущего отрезка костальной жилки, между жилками г2+3
и Г4+5-
6 (7). Продольная срединная полоса светлосерого налета на средне-
спинке широкая; ширина ее равна расстоянию между наружным ее
краем и внутренней границей плечевого бугорка. Пятна из густых
мелких волосков в передней половине 5-го тергита у j ясно выра-
жены (рис. 378—379), Длина тела 11—13 мм..........................
...................................*3. М. resplendens Wahlb.
7 (6). Полоса светлого налета на среднеспинке узкая, ширина ее в Р/г—
2 раза менее расстояния между наружной ее границей и внутрен-
ним краем плечевого бугорка. Пятна из волосков в основной поло-
вине 5-го тергита $> не выражены (рис. 377). Длина тела 7—11мм. .
..........................................*6. М. gracilior, sp. nov.
8 (5). Отрезок костальной жилки между жилками г4+5 и т в 2—4 раза
короче предыдущего отрезка костальной жилки.
9 (14). Лоб почти равен ширине глаза или шире его (самки).
10 (И)- Скулы со щетинистыми волосками в верхнем отделе (под лоб-
ными щетинками). Среднеспинка с 3 узкими, часто исчезающими
полосками сероватого налета. Лобная полоса в 1х/г раза шире ор-
биты ..........................................*4. М. meridiana L.
11 (10). Скулы под лобными щетинками без черных щетинистых во-
лосков.
12 (13). Высота щеки равна 4/s вертикального диаметра глаза; ширина
орбиты почти равна ширине лобной полосы. Полосы светлого налета
на среднеспинке отсутствуют...............7. М. montana, sp. nov.
13 (12). Высота щеки не превышает а/г вертикального диаметра глаза;
орбита в 2 раза уже лобной полосы. Среднеспинка с продольной
полоской светлосерого налета..............*5. М. intermedia Zett.
14 (9). Лоб в 2—5 раз уже глаза (самцы).
15 (16). Скулы в верхнем отделе с немногочисленными щетинистыми
волосками; орбитальные щетинки отсутствуют; среднеспинка беи
продольных полос светлого налета, реже с 3 нерезко выраженными
224
I. ТРИБА MUSCINI
узкими сероватыми полосками в переднем отделе. Скулы в 2 раза
шире 3-го членика усиков; лоб в 4—5 раз уже глаза...............
• • ..................................... *4. М. meridiana L.
16 (15). Скулы в верхнем отдела без черных щетинистых волосков; орби-
тальные щетинки имеются и направлены вершинами назад и вбок;
среднеспинка в переднем отделе с ясной узкой полосой светлосерого
налета; скулы не шире 3-го членика усиков; лоб в 2*/г раза уже
глаза.........................................*5. М. intermedia Zett.
*1. Mesembrina mystacea L. — Шмелевидная навозница (рис. 349,
369, 371, 373, 383, 401).
Linn6, 1761, Fauna Suec., Ed. II : 442 (Musca); Stein, 1916, Arch. Naturg.,
A, 10 : 19; — matutina Rondani, 1862 : 210; — vespertina Rondani, 1862 : 211.
Биология и экология. Порчинский, 1910: 45—49, фиг. 46—50;
Handschin, 1934, Rev. Suiss. Zool., VI : 8.
5- Лоб равен, чуть уже, пли немного шире 3-го членика усиков,
коричневой или бурой окраски, лобная полоса сужена посредине; орбиты
в 2^2—3 раза уже скул, последние на уровне прикрепления усиков равны
по ширине 3-му их членику, далее вниз суживаются; высота щеки в 3
раза менее вертикального диаметра глаза; нижний край лица выступает
вперед едва далее лба; 3-й членик усиков с параллельными краями, вы-
тянутый, в 21/г раза длиннее 2-го, хоботок с широкими сосательными ло-
пастями, толстый, пальпы сплюснуты с боков, широкие, булавовидно
расширены к вершине, в длинных, густых черных волосках по краям
и в коротких по боковым поверхностям. Голова черно-бурая, скулы и ме-
диана рыжие или коричнево-рыжие, 2-й членик усиков желто-рыжий,
3-й — такого же цвета при основании, далее к вершине слабо затемнен-
ный. Щетинка усиков с рядами длинных и коротеньких волосков. Лицо
в сером, орбиты и наружные края затылка в желтовато-сером, скулы
в шелковистом, бледножелтом, медиана в темножелтом налете. Вибриссы,
передние стомальные и внутренние теменные щетинки длиннее и крепче
щетинок головы, имеющих характер щетинистых волосков. Затылок
и щеки в длинных густых бурых волосках, задняя половина орбит в ко-
ротких тонких волосках того же цвета. Вибриссальные кили с рядом
коротких щетиночек, развитых до половины высоты килей. Грудь бле-
стяще бурая, покрытая густыми черными волосками, за исключением
спинки, покрытой желтыми волосками. Желтое опушение несколько не
достигает задней границы спинки, на бочках груди не распространяясь
ниже нотоплевры. Щиток в черных волосках. Гипоплевра, проплевра,
переднегрудь и супрасквамальное ребрышко лишены волосков; дыхальцы
коричнево-бурые, заднегрудное дыхальце длинное (высота менее длины).
Щетинки груди: акростихальные в виде тонких волосков, 1 пара
перед шитком может отсутствовать; дорзопентральные — 1—2 пары за
швом, очень тонкие, длиннее волосков среднеспинки; плечевые — 1—
2; предшовная — 1 (желтая или черная); предкрыловые — 1; надкры-
ловые— 3—4; закрыловые—2 крепких, между которыми имеется 1—2
более коротких, тонких, щетинистых волоска; нотоплевральная — 1
впереди и 3—6 щетинок сзади, нередко имеющих характер толстых волос-
ков; передняя верхняя щетинка на мезоплеврах отсутствует; стерноплев-
ральных— 0 (реже 1 в виде волоска) 4-1. На щитке: вершинных—2,
боковых—-4 пары, предосновных — 1, основных—1. Ноги черно-бу-
рые, голени смоляно-бурые, присоски грязно-желто-серые, вершинные
членики лапок часто рыжеватые. Голени передних ног по задней'стороне
6. MESEMBRINA
225
co щетинистыми^олосками по всей длине (длина в средине равна диа-
метру голени), стоящими в несколько неправильных рядов; задневен-
тральная сторона с рядом Длинных щетинистых волосков (длина их в 1’/2—
Рис. 362—370.
362 — голова самца Mesembrina meridiana L. сверху. 363 — голова сам-
ки того же вида сверху. 364 — то же самца М. resplendens Wahlb.
сверху. 365 — то же самки М. resplendens Wahlb. 366— голова самца
М. intermedia Ztt. сверху. 367 — то же самки М. intermedia Ztt. 368 —
то же самки М. gracilior, sp. 11. 369 — средняя голень самца М. mystacea
L. сзади. 370 — то же самца М. meridiana 1.. (Ориг.).
15 Фауна СССР том XVIII, вып. 4.
226
I. ТРИБА MUSCINI
3 раза превышает диаметр голени), переднедорзальная поверхность с ко-
роткими щетинками, составляющими гребень по длине голени; бедра пе-
редних ног в черных длинных волосках; коготки передних ног по длине
составляют 4/s длины 5-го членика лапки. Бедра средних ног в волосках
длинных и щетинистых с нижней и более коротких с прочих сторон бедра;
Риг. 371. Самец Mesembrina mystacea L. (Ориг.).
голени средних ног с гребнем коротких крепких щетинок ио заднедор-
зальной поверхности, с задневентральной стороны с рядом щетинистых
рыжих волосков (длина их у вершины голени в 3*/2—4 раза более по-
перечника голени), длина которых возрастает к вершине голени; вен-
тральная поверхность в очень коротких, крючковидно загнутых на вер-
шине волосках; и с рядом длинных щетинистых волосков; заднедорзаль-
ная сторона в щетинистых волосках (в 2—2х/г раза более длинных, чем
поперечник голени); задние бедра в бурых, тонких волосках; задние го-
лени на заднедорзальной стороне с 1 длинной, тонкой щетинкой
6. MESEMBRINA
227
у средины и рядом коротких тонких щетиночек по всей длине; переднедор-
зальная поверхность с рядом более крепких, коротких щетинок по всей
длине; передневентральная сторона в длинных и коротких щетинистых
волосках разной толщины. Коготки передних лапок немного короче 5-го
членика лапки. Крылья серые, с легким затемнением вдоль жилок, при
основании бледножелтые, с такого же цвета жилками в этой части крыла;
жилка 7'4+5 лишена волосков; т — в вершинной части мягко дуговидно
изогнута, ячейка R& широко открыта, отрезок костальной жилки, заклю-
ченный между г4+6 и т, в 2 раза короче предыдущего ее отрезка, распо-
ложенного между г2+3 и г4+5; чешуйки коричневые, жужжальца буро-
ватые. Брюшко коротко яйцевидное, блестяще черное в очень густых
тонких торчащих волосках на тергитах 1 -|~2-м, по переднему краю 4-го
тергита и одноименных стернитах; тергиты 4-й и 5-й в более длинных
тонких густых серовато-желтых волосках. Церки расширенные в основ-
ной половине, резко суженные в дистальной части, оканчивающейся двумя
средними короткими и двумя узкими пальцевидными боковыми отрост-
ками. Кокситы с коротким округлым выступом посередине внутреннего
края (рис. 383).
j. Лоб шире глаза, лобная полоса широкая, суженная кпереди,
коричневая, в волосках в боковых отделах задней своей половины и
с парой хорошо развитых перекрещивающихся вершинами щетинок впе-
реди глазкового треугольника; орбиты сужены назад, в 3 раза (в среднем
отделе) уже лобной полосы; внутренние и наружные теменные щетинки
сильные, длинные, орбиты в полосках; орбитальные щетинки не выра-
жены; щетинки, расположенные на затылке за внутренними теменными,
лишь немного короче их, часто двойные. Щетинки на средних голенях:
нереднедорзальные—1, заднедорзальных — ряд из нескольких щети-
нок; задневентралъных — 3.
Длина тела 12—16 мм.
Среди обширного материала в коллекциях Зоологического института
Академии Наук СССР мы имели возможность отметить ряд экземпляров
с сильно уклоняющейся окраской волосков. Особенно изменчивой оказы-
вается расцветка волосяного покрова на плевральных отделах груди и
верхней поверхности щитка (от черных до светложелтых), что дало повод
Рондани к описанию этих цветковых аберраций как самостоятельных
видов (М malutina, М. oesperti.no). Весьма близким к М- туstacea пред-
ставляется и М. tristis Aldr., описанный из Китая (Сычуань).
Яйцо длиной до 5 мм; удлиненное с глубокой продольной бороздкой
на верхней стороне, ограниченной по краям закраинкой, образующей
на верхнем полюсе подобие ушек.
Личинка I стадии толстая, короткая; задние дыхальцы мелкие, серд-
цевидной формы; тело голое сверху, снизу, начиная с 5-го сегмента до по-
следнего, по переднему краю с полоской из мелких, темных, густо рас-
положенных шипиков; полоска из шипиков достигает наибольшей ширины
в своем среднем отделе; к боковым концам сужена.
У личинки II стадии задние дыхальцы имеют по 2 коротких щели,
из которых верхняя щель расположена наискось, а нижняя дуговидная,
обращенная выпуклой стороной книзу; концы нижней щели обращены
кверху.
Личинка III стадии, начиная с 7-го сегмента, снизу несет попереч-
ные валикообразные утолщения. Передний край второго сегмента снабжен
многочисленными, мелкими темными шипиками, заходящими и на верх-
нюю сторону сегмента. На боковых отделах сегмента шипики располо-
15*
228
I. ТРИБА MUSCINI
жены группами по 2—3 шипа. Задние дыхальцевые пластинки округло
треугольных очертаний, с 3 S-образно изогнутыми, сравнительно корот-
кими щелями. Передние дыхальцы с 10—13 лучами. Задний конец тела
с 7 сосочками, из которых 5 располагаются вблизи анального отверстия,
а 2 удалены от последнего, возвышаясь по бокам у основания анальной
площадки.
Распространение. Широко распространенный в Пале-
арктике вид; на юге редок и встречается только в горах. Р> СССР известен
на север до Мурманска, Соловецких островов, Усть-Цыльмы (Печора),
в Сибири — до р. Нижней Тунгуски и Якутска, на юг — до Киева,
Сухуми и далее на восток — до о. Зайсана. Вид наиболее обычен в зоне
хвойных лесов. За пределами СССР М. mystacea известен от Швеции
и Норвегии до Пиренеев и Атласских гор (Алжир). Монголия.
Личинки проводят жизнь в помете крупного рогатого скота. Муха
откладывает крупные яйца обычно поодиночке на кучки совсем свежего
помета, погружая их почти целиком в массу последнего. К кладке яиц
в северных широтах муха приступает в июле—августе. Развитие личинок
совершается крайне быстро. По выходе из яйца молодые личинки пи-
таются навозом, переходя со второй стадии на хищный образ жизни,
нападая и быстро уничтожая личинок не только других видов, находя-
щихся с ними по соседству, но и личинок того же вида.
Хищничество личинок шмелевидной навозницы дало повод [Сим-
мондс (Simmonds)] испытать их в борьбе с вредными синантропными
видами мух. Однако, этот опыт, равно как и попытка акклиматизации
вида для целой борьбы, не дал положительных результатов [Гандшин
(Handschin)].
*2. Mesembrina decipiens Loew. (рис. 382).
Loew, 1873, Bcschr. europ. Dipt., Ill : 239; — putziloi .Portschinsky, 1874, Tp.
Русс, энтомолог, общ., VII : 60; 1875, Horae Soc. ent. Ross, XI : 29.
Сходен с M. mystacea, от которого отличается желтыми крыловыми
чешуйками и желтым опушением щитка. Черные волоски, если они при-
сутствуют, развиты слабо, никогда не образуя на нем сплошного черного
покрова. Церки в дистальном отделе уже, чем у М. mystacea, и несут
более вытянутые и заостренные выступы (внутренние) на вершине: выступ
на внутреннем крае кокситов короче и несколько шире.
Распространение. Вид широко распространен в зоне хвой-
ных лесов Сибири и Камчатки, на юг спускаясь до Монголии и Кореи.
Крайней западной точкой нахождения вида пока является Байкпт на Под-
каменной Тунгуске в Красноярском крае.
*3. Mesembrina resplendent Wahlb. (рис. 364, 365, 375, 378, 384).
Wahlberg, 1844, Ofv. К. Vet. Akad. Forhandl., I: 66; Порчинский, 1874, Tp.
Русск. энтомолог, общ., VIII : 60; Frey, 1909, Acta Soc. Fauna Flora Fenn., 31, 9 : 10:
Stein, 1916 : 19.
S. Ширина лба равна ширине 3-го членика усиков; лобная полоса
в середине уже орбит, имеет характер черной бороздки, сильно расши-
ренной вперед п слабо, постепенно, — назад; скулы равны ширине
3-го членика усиков или слегка уже его; щеки равны -'/г вертикального
диаметра глаза. 3-й членик усиков узкий, слабо вогнутый по переднему
краю, в 3 раза длиннее 2-го. Щупальцы бурые. Усики буровато-коричне-
вые, вершинный край 2-го и основание 3-го членика снизу красновато-
Рпе. 3.2—379.
372 — вершинная половина крыла Mesembrina intermedia Ztt, 373 — то же М. mysla-
сеа L. Зг4 — то же М. meridiana L. 375 — то же М. resplendens Wahlb. 376 — то же
М. gracilior, sp. n. 377 — пятый тергит брюшка самки М. gracilior, sp. n. 378 — то же
М. resplendens Wahlb. 379 — то же М. meridiana L. (Ориг.),
230
I. ТРИБА MUSCINI
коричневые; утолщенная основная треть аристы красновато-желтая.
Голова черно-бурая; орбиты в тонких коротких волосках; скулы без
щетинистых волосков в верхней трети, в сероватом налете, имеющем
желтоватый оттенок в нижнем их отделе; затылок черный, в бурых воло-
сках, коротких и щетинистых вверху и тонких длинных в нижней поло-
вине; щеки в длинных бурых волосках. Щетинки головы тонкие, длин-
ные; внутренние теменные щетинки и вибриссы развиты сильнее прочих
щетинок головы; наружные теменные явственные. Грудь черная, блестя-
щая; серый налет покрывает плечевые бугорки и образует широкую
продольную полосу по середине спинки, доходящую до поперечного шва.
Поверхность груди в тонких буро-черных торчащих волосках. Гипо-
плевры, проплевры и переднегрудь без волосков. Заднее дыхальце с оття-
нутым вперед передним углом. Щетинки груди: акростихальных — 1 пара
перед щитком; дорзоцентральных — 3 -|-3 (передняя слабо развита);
стерноплевральных —1 -J-2; прочие щетинки как у М. gracilior, sp. п. Ноги
черно-бурые; 4-й и 5-й членики передних лапок рыжевато-бурые; чле-
ники лапок тонкие; 5-й членик передних лапок равен по длине когот-
кам; длина 4-го членика в Р/а раза более его ширины. Средние голени
с 1 длинной переднедорзальной щетинкой, рядом коротких задних и 2—3
задневентральными щетинками. Крылья сероватые, с желтым основа-
нием; жилка т в вершинной части с S-образным слабым изгибом; отре-
зок костальной жилки между г4+5 и т едва короче предыдущего отрезка
косты. Ячейка Rs сильно расширена к вершине. Чешуйки яркожелтые,
жужжальца рыжие, с бурой вершиной. Брюшко блестящее, черное,
с слабым оливковым отливом, в тонких, торчащих, бурых волосках,
более длинных и редких на 5-м тергйте и стернитах. Церки самца уже,
чем у прочих видов рода, вершинные наружные выступы направлены
дистальными концами вбок; внутренние дистальные выступы удлиненно
треугольные; кокситы очень узкие, длинные с широким коротким высту-
пом на внутреннем крае; наружный край кокситов почти прямой.
Самка отличается от М. gracilior более узким лбом (уже глаза);
полоса серого налета спинки шире (ширина равна половине ширины
темени) и длиннее (достигает поперечного шва). Брюшко шире, с ясно
выраженными небольшими пятнами в передней половине 5-го тергита,
состоящими из мелких тонких волосков. В остальном сходна с М. gra-
cilior.
Распространение. Известен из лесной и таежной зоны
от Швеции, Норвегии до Венгрии и в СССР от Ленинградской области
и Могилева на западе до Приморского края на востоке. Указания (Hough,
Aldrich) на нахождение вида в Северной Америке (М. latreillei R.-D.)
требуют проверки.
*4. Mesembrina meridiana Linne (рис- 362, 363, 370, 374, 379, 380,
385—400).
Linn6, 1761, Faune Suec., Ed. II : 451 (Musca}'. 'Ftey, 1909, Acta Soc. Fauna
Flora Fenn., 31, 9 : 10; Schnabl u. Dziedzicki, 1911 : 226—227 (Neomesembrina); Town-
send, 1912, Proc. Ent. Soc. Wash. XIV : 45—53 (Metamesembrina); Stein, 1916 : 18; —
latreillei Robineau-Desvoidy, 1830 : 401; — ingrica Portschinsky, 1875, VII : 59 (syn.
nov.).
Биология и экология. Порчинский, 1910:41—45; Keilin, 1915,
Bull. Sci. Fr.-Belg. 7, 49 : 15-—198; Keilin a. Fate, 1930, Parasitology, 22 : 168—181;
Thomson, 1937, Parasitology, 29 : 298—303, figs.
<5. Лоб в 2 раза шире 3-го членика усиков, орбиты в наиболее узком
месте в 3 раза уже коричневато-бурой лобной полосы; последняя посте-
6. MESEMBRINA
231
пенно расширена кпереди, покрыта бурыми волосками, наиболее густыми
в задней половине лба; скулы в РА раза шире 3-го членика усиков,
с несколькими щетинистыми волосками в верхней трети; высота щеки
в 2 раза превышает вертикальный диаметр глаза; щупальцы бурые,
в длинных и коротких многочисленных волосках; 3-й членик усиков
постепенно и слабо сужен к дистальному концу, по верхнему краю слабо
вогнутый, в 2 раза длиннее 2-го членика усиков; щетинка усиков жел-
товато-рыжая, утолщена в основной половине; жилки буро-коричневые;
голова черно-бурая, вибриссальные кили коричневые; узкая полоска
позади глаз в бледножелтом, скулы в золотисто-желтом налете, передние
края орбит (не всегда) в редком желтовато-сером, затылок и лицо в сером
редком налете. Щеки блестящие, бурые. Орбиты в тонких бурых, заты-
лок вверху в щетинистых, коротких бурых, внизу в длинных тонких
бурых волосках, как и щеки. Щетинки головы, кроме крепких вибрисс
и Внутренних теменных щетинок, тонкие недлинные; наружные темен-
ные ясные; вибриссальные кили с несколькими неправильными рядами
коротких щетинистых волосков. Грудь черная, блестящая, лишена свет-
лого Налета, за исключением переднего отдела плечевых бугорков; грудь
как и щиток, в коротких тонких (несколько более редких и толстых
на щитке) черно-бурых волосках. Гипоплевры проплевры и переднегрудь
лишены волосков. Щетинки груди: акростихальных — 2 пары перед
щитком, имеющих характер длинных щетинистых волосков или тонких
Щетиночек; дорзоцентральных (лишь в заднем отделе спинки) — 2,
если бывают развиты перед швом, то имеют вид волосков (1 пара); интра-
алярных — 2 позади шва; предкрыловых — 1; надкрыловых — 3; закры-
ловых — 2; плечевых — 3; заплечевых — 1; предшовных — 1; ното-
плевральных — 1 впереди и 2—3 сближенных щетинки в задней части
склерита; стерноплевральных — O-f-1; передневерхняя щетинка на мезо-
плевре отсутствует. Щиток короткий, широкий, слабо выпуклый; длина
его в 2 раза менее ширины; несет следующие щетинки: вершинные — 2,
перекрещивающихся; боковые — 4 пары; предосновные — 2; основ-
ные — 2, вдоль краев щитка, кроме того, располагается по 3—5 тонких
щетинок. Щиток буро-коричневый. Заднегрудное дыхальце высокое,
длина его не превышает высоты. Ноги черно-бурые, 2—4 конечных чле-
ника передних и 5-й или 5-й и 4-й членики средних лапок желтовато-
рыжие; присоски желтоватые. Коготки передних ног равны или чуть
длиннее 5-го членика лапки, 4-й членик передних лапок шире своей
длины. Голени средних ног с крепкой переднедорзальной щетинкой
в нижней половине, рядом коротких крепких задних щетинок и 2 креп-
кими щетинками на задневентральной поверхности. Крылья яркожелтые
в основной половине, в вершинной — сероватые, с легким затемнением
вдоль жилок; т в вершинной части более или менее резко дуговидно
изогнута; ячейка Rs сильно сужена на вершине; отрезок костальной
жилки между г4+5 и т жилками в Р/г—2 раза короче предыдущего ее
отрезка (между г2+3 и г4+5). Чешуйки яркожелтые, жужжальца бурые,
с буровато тонким основанием. Брюшко коротко-яйцевидное, блестяще
черное, с коричневыми боковыми отделами первых 3 или 4 тергитов,
покрыто черными, тонкими, густыми волосками, длина которых возра-
стает к вершине брюшка. Задние края 4-го, 5-го тергитов и стернитов
покрыты превосходящими прочее опушение своей длиною тонкими щети-
нистыми волосками. Церки широкие, на дистальном конце заканчиваются
4 длинными отростками: 2 более тонкими и изогнутыми наружными
и 2 тупыми, широкими внутренними. Кокситы широкие, с не-
232
/. ТР ИВ A MUSCINI
большим коротким выступом по внутреннему краю ниже середины
(рис. 380).
Ри'. 380- -384. Церки и кокситы самцов.
380 — - Mesembrina meridiana L. 381 —М. intermedia Zett. 382 -- M. decipiens Lxv.
383 — M. mystacea L. 384 — M. resplendens Wahlb. (Ориг.).
j. Лоб шире глаза, лобная полоса в Р/г раза шире орбиты, покрыта
волосками, более густыми в задней ее половине; крепкие щетинки лоб-
ной полосы отсутствуют. Скулы широкие, ширина их в 3 раза превышает
ширину 3-го членика усиков, в верхнем отделе с несколькими черными
6. MESEMBRINA
233
щетинистыми волосками, спускающимися до уровня основания 3-го чле-
ника усиков. Орбиты бурые, в редком желтоватом налете в переднем
отделе; скулы в золотисто-желтом ярком налете; затылок в сравнительно
густом желтовато-сером налете. Щетинки головы крепче нежели у самца.
Спинка черная, в передней четверти с намечающимися узкими тремя
полосками желтовато-серого палета; ширина средней светлой полосы
Рис. 3S5. Самец Mesembrina meridiana L. (Ориг.).
налета менее ширины черных боковых полос. Такого же цвета налет
покрывает плечевые бугорки. 5-й тергит брюшка сверху, по обе стороны
от продольной средней линии тергита, в передней половине имеет по пятну
из очень густо расположенных коротеньких тонких волосков. Членики
передних лапок широкие; 4-й сердцевидный, ширина его почти равна
длине, 5-й — квадратный или слегка вытянут.
Длина тела 11—13 мм.
Вид мало изменчив в окраске, как и в величине. Исключение пред-
ставляют полоски светлого налета в переднем отделе спинки. В боль-
шинстве случаев они выражены весьма слабо, причем: у самцов слабее,
234
I. ТРИБА MUSCINI
чем у самок. Встречается, однако, особи с резко очерченными светлыми
полосками (М. ingrica Portsch.), иди, наоборот, совершенно их лишен-
ные.
Яйцо крупное — 4.5 мм длины, с узкой продольной бороздкой
по дорзальной стороне, ограниченной тонкими параллельными ребрыш-
ками; вентральная поверхность выпуклая или слабо уплощена. Верхний
полюс яйца заострен несколько более нижнего.
Личинка I стадии метапнейстична. Задние дыхальцы мелкие, коротко-
почковидные, иногда разделенные поперек тонкой перегородкой. Брюш-
ные сегменты со 2-го по 7-й снизу по переднему краю снабжены широ-
кими полосками шипиков; полоски шипов заходят на боковые отделы
дорзальной поверхности сегментов. Базальный склерит ротового аппа-
рата с постепенно суженными назад дорзальными и расширенными,
скошенными на конце вентральными отростками. Дорзальная перемычка
базального склерита большая, высоко приподнятая, расширенная по дор-
зальной стороне. Передние отростки базального склерита тонкие, длин-
ные, соединены с гипостомальным склеритом. Средний дорзальный скле-
рит широко-листовидный, соединен с верхней стенкой ротового отвер-
стия; по заднему краю с двумя отросточками, сочленяющимися с перед-
ним отделом гипостомального склерита.
Личинки II стадии до 7 мм длины; амфипнейстичны. Передние
дыхальцы с 16—18 лучами; задние стигмы светлоокрашенные, с двумя
подковообразно изогнутыми щелями. Дорзальная перемычка базального
склерита неполная; ее верхний отдел, соединяющий боковые части,
слабы и представлен тонкой полоской. Передние отростки базального
склерита снизу соединены поперечной перемычкой. Ротовые крючки
парные, тонкие, близко сходящиеся передними своими концами и широко
разделенные позади в месте своего прикрепления с гипостомальным
склеритом. Вентрально от ротовых крючьев помещается две пары хити-
новых образований; первая из них сближена и располагается непосред-
ственно под вершиной ротовых крючков; Вторая, лежащая непосред-
ственно за первой, составлена из крупных обособленных склеритов,
отвечающих губным склеритам Orthellia.
Личинки III стадии амфипнейстичны; передние стигмы с 16—18
лучами и резко выраженным внутренним черным рубцом. По количеству
и расположению лучей передние дыхальцы отличают личинку этого вида
от всех прочих Muscini. Наружный рубчик и перитрема со слабо выра-
женными границами. Анальная пластинка хорошо развита. Базальный
склерит вытянутый; его вентральные задние отростки с характерными
темными, сильно хитинизированными полосками. Дорзальная перемычка
нависает над передними отростками базального склерита. Гипостомаль-
ный склерит массивный, по передне-нижнему краю снабжен пальцевид-
ным выростом с каждой стороны. Ротовые крючки с широкой базаль-
ной частью, изогнутые, заостренные на вершине. Под крючьями распо-
лагается пара вытянутых тяжей, соединенных снаружи с зазубренными
изнутри у вершины второй парой хитиновых, сагиттально вытянутых
образований. Зубные склериты треугольные, нижние концы их соеди-
нены, благодаря чему образуется срединная вентральная перемычка,
наличие которой характерно для плотоядных и полуплотоядных личинок
мух, как Muscina, Myiospila, Mydaea и др. Личинка III стадии дости-
гает в длину 20 мм. Окраска тела желтая.
Ложнококон длиной 14 мм, в поперечном сечении несколько сплюс-
нут и снабжен боковыми выступами в виде рубчиков. Передние дыхальцы
Рис. 386—401. Детали строения преимагинальных фаз Mesembrina
meridiana L.
386 — ротоглоточный аппарат личинки III стадии сбоку. 387 — то же
II стадии. 388 — то же III стадии сверху. 389 — головной конец
личинки III стадии снизу. 390 — задние стигмы личинки II стадии.
391 — анальная пластинка личинки III стадии. 392 — ротоглоточный
аппарат личинки I стадии сбоку. 393 — языковидный склерит личинки
I стадии. 394 — шипы на вентральной стороне тела личинки I стадии.
395 — задние стигмы личинки I стадии. 396 — грудное дыхальце куколки.
397 —’ложнококон. 398 — яйцо. 399 — переднее дыхальце личинки III
стадии. 400 — задние дыхальцы личинки III стадии. 401 — заднее ды-
хальце (правая пластинка) личинки III стадии Mesembrina mystacea L.
(Рис.'401 по Порчипскому, прочие — по Томсону).
236
I. ТРИБА MUSCINI
выступают в форме рожков. Задний конец пупария уплощен и имеет
ребрышко, окружающее его. Анальная пластинка сильно склеротизована.
Распространение. Наиболее обычный и широко распро-
страненный в Палеарктике вид рода; как и все виды этого рода, М. meri-
diana связан с лесным ландшафтом. На юг проникает далее прочих
видов рода, достигая восточного Нань-шаня в Монголии. Является,
если не считать М. resplendens, единственным представителем рода, захо-
дящим в Северную Америку. Северная граница вида проходит от Вели-
кобритании через Скандинавию по СССР, где через Петрозаводск — Усть-
Цыльму (Печора)—Тобольск—Томск—Иркутск—Жиганск (на Лене) дохо-
дит до берега Охотского моря (Алдан). Южной границей на западе нужно
считать крайний юг Зап. Европы. В СССР южная граница проходит по
линии Полтава—южный берег Крыма—Сухуми—Тбилиси. Южная гра-
ница в Азии не достаточно выяснена; здесь наиболее южными точками
являются пока оз. Зайсан, Абаканский хребет (Кузнецкий р-н), Амур-
ская обл. Под именем М. latreillei вид приводится для Японии.
Муха откладывает яйца на свежий коровий помет. Яйца отклады-
ваются по-одному. Созревание яиц в теле самки идет не одновременно,
а первоначально заканчивается в одном яичнике. В последнем полной
зрелости достигает только одно из трех находящихся в яичнике яиц,
которое и откладывается. В момент откладки первого яйца второе яйцо
имеет крупные размеры, в то время как 3-е яйцо в том же яичнике, равно
как и яйца второго яичцика, остаются еще недоразвитыми.
Вылупление личинки из яйца наступает в момент откладки его сам-
кой, или вскоре после этого. Из яйца выходит обычно личинка I стадии,
хотя в некоторых случаях установлен факт отрождения личинки II ста-
дии. При вылуплении личинки I стадии линка на следующую стадию
наступает (по Томсону) через 3—5 часов, реже развитие задерживается
до 12г/2 часов. Во II стадии личинка находится около 35 часов. Все раз-
витие заканчивается в 15—17 дней.
По наблюдениям Порчинского, личинки черной навозницы — копро-
фаги. Томсон на основании своих опытов, в которых личинки этого вида
вели себя то как копрофаги, то как хищники, считает М meridiana
факультативно плотоядной формой. В экспериментах этого автора ли-
чинки М. meridiana весьма активно уничтожали личинок Orthellia. Dasy-
phora, Haematobia.
*5. Mesembrina intermedia Zett. (рис. 366, 367, 372, 381).
Zetterstedt, 1849, Dipt. Scandin., VIII : 3274; Порчинский, 1874, Tp. Русск.
энтомолог, общ., VII : 59; Brauer, 1895, Sitzungsber. Kais. Ak. Wiss. Wien, Math.-
nat. Klasse, CIV, Abt. I : 596; Frey, 1909, Acta Soc. Fauna Flora Fenn., 31, 9 : 10.
Лоб весьма широкий, в наиболее узком месте ширина его в 3 раза
превосходит ширину 3-го членика усиков, орбиты в 2 раза уже лобной
полосы, скулы немного уже 3-го членика усиков, высота щеки в 23/а раза
короче вертикального диаметра глаза; щупальцы бурые; усики корич-
нево-бурые, 3-й членик в 21/*—24% раза длиннее 2-го;_ щетинка в основ-
ной утолщенной половине желто-рыжая. Голова черно-бурая, передний
край рта и вибриссальные кили — красноватые, щеки блестящие, буро-
коричневые. Орбиты в коротких, нижняя часть затылка и щеки в длин-
ных тонких черно-бурых волосках, верхняя часть затылка в коротких,
более редких щетинистых волосках. Скулы в верхней трети голые.
Щетинки головы хорошо развиты; наружные теменные щетинки в P/s
раза короче внутренних теменных; орбитальных щетинок — 2, из кото-
6. MESEMBRINA 237
рых передняя крепче задней, направлены они вершинами назад. Скулы
и передняя часть орбит в желтом, затылок позади глаз в светлом жел-
товато сером, далее в очень реденьком сероватом налете. Грудь буро-
черная, блестящая, плечи и щиток буро-коричневые; плечевые бугорки
в переднем отделе и спинка посредине с полосой светлосерого налета,
достигающей поперечного шва; ширина полосы налета в 2 раза менее
ширины темени. Поверхность груди в бурых тонких торчащих волосках,
более длинных на плеврах. Дыхальце заднегруди с оттянутым вперед
передним углом; длина его явно превышает высоту. Щетинки груди:
акростихальных—2 пары перед щитком (впереди шва 1 пара очень тонких
щетинистых волосков); дорзоцентральных — 1 пара впереди шва, тон-
ких, длинных, позади шва — 4 пары, из которых 2 передние короче
и тоньше задних; интраалярных — 2 позади шва; предкрыловых — 1;
надкрыловых — 3 (средняя крепче и длиннее остальных); закрыловых —
2; плечевых — 3; заплечевых — 1; предшовных — 1; нотоплевраль-
ных — 1 4-2 (сзади); стерноплевральных — 1 4-1гипоплевры, проплевры
и переднегрудь — голые; на щитке: вершинных щетинок — 2; боко-
вых — 3; основных — 1; предосновных — 1; дискальных — 2—3 пары
тонких у боковых краев. Ноги смоляно-бурые. Задняя половина 5-го,
4-й, 3-й и 2-й и передние 2/з или х/а 1-го члеников передних лапок жел-
тые, коготки передней пары почти в 2 раза короче 5-го членика лапки.
Задневентральная поверхность передних голеней в длинных тонких
волосках; средние голени с 1 переднедорзальной щетинкой, рядом креп-
ких щетинок и длинными, тонкими, редкими по задней поверхности,
такими же, но более густыми волосками и 1 крепкой щетинкой по задне-
вентральной стороне. Задние голени по переднедорзальной поверхности
с рядом толстых щетинок (3—5 штук), рядом щетинистых волосков
по передней поверхности, рядом длинных тонких щетинок вдоль всей
передневентральной стороны (длина щетинок почти равна двойному
поперечнику голени), с 2—3 длинными заднедорзальнымй и рядом тон-
ких длинных щетинистых волосков по задневентральной стороне. Крылья
светлосерые, с бледножелтым основанием; жилка т с равномерно дуго-
видным изгибом; ячейка R-a сильно сужена; отрезок косты, располо-
женный между r4+s и т, в 2 раза короче отрезка ее между г2+3 и г4+.;
чешуйки яркожелтые, жужжальца рыже-желтые, с бурой булавой.
Брюшко короткояйцевидное, сине-черное, блестящее, с темнокоричне-
выми пятнами на боках, сверху покрыто чернобурыми тонкими корот-
кими волосками; 5-й тергит сверху, задний край 4-го тергита и нижняя
сторона брюшка в длинных волосках того же цвета. Церки с расширен-
ной базальной и суженной вершинной частью; вершина церок с 4 отро-
стками — пальцевидным и, направленными вбок, заостренными наруж-,
ными и сближенными, более широкими, тупыми внутренними. Кокситы
в виде широких, округлых на вершине лопастей, несущих по внутрен-
нему краю округлый пологий выступ.
Лоб самки едва шире глаза, орбиты в 2 раза уже лобной полосы,
последняя в коротких черных волосках в боковых отделах задней ее
половины, лишена крепких щетинок; орбитальные щетинки короткие,
направлены назад; скулы в золотисто-желтом налете, ширина их в наи-
более узким месте (внизу) в Р/г раза превышает ширину 3-го членика
усиков, высота щеки в 2 раза менее вертикального диаметра глаза; ноги
сплошь бурые, голени лишены тонких длинных волосков; брюшко чер-
ное, со слабым синеватым оттенком; пятна из коротких тонких волосков
на 5-м тергите отсутствуют.
238
I. ТРИБА MUSCINI
Длина тела 9—12.5 мм.
Вид близок М. meridiana, от которого легко отличается резко выра-
женной широкой полосой светлого налета на спинке, более выпуклым
щитком и строением генитальных придатков самца.
Распространение. Вид известен из зоны хвойных лесов
от Ленинградской обл. до Якутии; южнее Алтая, где встречается в аль-
пийском поясе, не известен. За пределами Советского Союза отмечен
для Скандинавии. Биология и фазы развития не известны.
*6. Mesembrina gracilior Zimin, sp. nov. (рис. 368, 376, 377).
Лоб самки уже глаза, орбиты узкие, ширина их в l3/^—IV2 раза
менее ширины лобной полосы; последняя лишена крепких щетинок
и покрыта тонкими волосками, особенно многочисленными в боковых отде-
лах задней ее половины. Скулы в 1]/2 раза шире 3-го членика усиков,
заметно суженные сверху вниз, без волосков в верхней своей трети,
в бледно-желтовато-сером налете; лоб несколько далее выступает вперед,
нежели верхний край рта, высота щеки равна половинному диаметру
глаза; 3-й членик усиков в 21/г раза длиннее 2-го; голова черно-бурая,
вибриссальные кили красновато-коричневые, щеки блестящие, черно-
бурые; вершина 2-го членика усиков красновато-коричневая, прочие
членики буро-коричневые, основание аристы рыже-желтое; щупальцы
черно-бурые. Щетинки головы недлинные, сравнительно хорошо развиты;
орбиты в черных коротких волосках с двумя недлинными орбитальными
щетинками с каждой стороны, нижняя часть затылка и щеки в длинных
черно-бурых тонких, верхний отдел затылка в коротких щетинистых
волосках. Грудь блестяще-черная, как и щиток, с короткой, недостигаю-
щей поперечного шва полосой светлосерого налета на спинке; ширина
полосы в 4 раза менее ширины темени; плечи в светлосером налете. Щиток
сильно выпуклый. Грудь в черных тонких волосках, на спинке более ред-
ких, чем у М. meridiana. Гипоплевры, проплевры и переднегрудь без
волосков. Щетинки груди: акростихальных — 1-|-3; дорзоцентраль-
ных— ЗЦ-4; интраалярных — 2 за швом; предкрыловых — 1; надкры-
ловых— 3; закрыловых — 2; плечевых — 3 (2); заплечевых — 1; пред-
шовных — 1; нотоплевральных — 1-(-2 (сзади); стерноплевральных —
1-J-2; передне-верхняя мезоплевральная щетинка отсутствует. На щитке:
вершинных щетинок — 2; боковых — 2; основных — 2; предосновных —
2; вблизи края щитка по обе стороны выделяется 2—3 щетинистых
волоска. Ноги черные, членики лапок узкие; коготки передних ног
в IV2—2 раза короче 5-го членика лапки; 4-й членик удлиненный: длина
его в 1х/г раза превышает ширину. Крылья яркожелтые при основании.
Жилка т слабо S-образно изогнута; ячейка Нъ широко открытая; отре-
зок костальной жилки между F4+s и т едва короче предыдущего отрезка
косты между г2+3 и г4+Б. Чешуйки темножелтые, жужжальца желтоватые
с бурой головкой. Брюшко яйцевидное, заостренное на дистальном конце;
5-й тергит несколько сдавлен с боков; брюшко черное с оливковым оттен-
ком, в черных редких торчащих волосках. Пятна из мелких волосков
на 5-м тергите не выражены.
Длина тела 7.5—11 мм.
Тело уже, чем у М. resplendens, более стройное. Самец не известен.
Вид наиболее близок к М- resplendens, отличаясь от нее меньшей вели-
чинной, отсутствием пятен из густых волосков при основании 5-го тер-
гита и суженной срединной полоской светлого налета в переднем отделе
среднеспинки.
6. MESEMBRINA
239
Распространение. Камчатка: с. Камаки, с. Коряки,
с. Ключевское, Козыревск, Петропавловск (Державин, Новограбленов);
о-в Шантар, Якшина губа (Дулькейт); Сахалин (Супруненко); Хабаров-
ский край: Андома бл. Аяна (Попов); р. Чхиль, лиман р. Амура (Сол-
датов); Сихотэ-алинь (Емельянов); Якутия: Верхоянский округ (Тка-
ченко); р. Амга (Иванов); Байкит (Тунгуска, Красноярский край); Алтай,
слияние рек Балыксы и Томи (Хворов); Малый Урал, бассейн р. Маньи
(Флеров); г. Вой-сабля-из, б. Б. Сыни, Печора (Журавский). Вид опи-
сывается по 25 экземплярам (jj).
Тип в Зоологическом институте Академии Наук СССР в Ленинграде.
7. Mesembrina montana Zimin, sp. nov.
j. Лоб шире глаза (3/<); орбиты почти равны ширине лобной полосы;
скулы~на уровне основания усиков равны двойной ширине 3-го членика
усрков, книзу сужены (здесь ширина скулы не превышает полуторного
диаметра 3-го членика усиков); высота щеки немногим меньше верти-
кального диаметра глаза; бурые с буро-рыжим основанием щупальцы
слабо и.постепенно расширены к вершине; 3-й членик усиков в 2х/г раза
длиннее 2-го, буровато-коричневый, 2-й членик и основание аристы
коричневато-красные. Голова черно-бурая, вибриссальные кили и ме-
диана красноватые, лобная полоса рыжевато-буро-коричневая; затылок,
скулы и передняя часть орбит в бледном серовато-желтом, лицо в сером
более редком налете, щеки блестящие. Орбиты в редких бурых волосках,
с тонкими орбитальными щетинками; лобная полоса в задней половине
с редкими сравнительно длинными волосками и тонкими (с одной сто-
роны сдвоенными) перекрещивающимися щетинками; вибриссы крепкие;
над угловыми вибриссами располагаются короткие щетинистые волоски,
ковнутри от которых по внутреннему краю вибриссальных килей имеется
несколько крепких коротких щетинок. Орбиты в верхней трети без
группы щетинистых волосков. Внутренние теменные щетинки длинные,
тонкие, наружные теменные — короткие; заглазные реснички длинные,
очень тонкие; щеки и затылок; исключая его верхнюю половину, покры-
тую короткими волосками, в длинных бурых волосках. Грудь черно-
бурая, плечи и задняя поверхность в слабо заметном рыжевато-сером
налете; спинка со слабым блеском, в передней половине почти вовсе
лишена его. Поверхность груди в тонких сравнительно длинных, бурых
волосках и тонких щетинках, местами имеющих характер щетинистых
волосков. Щетинки груди: акростихальных — 1 пара тонких перед
щитком;’дорзоцентральных — 4 позади шва, из которых две передние
имеют характер тонких волосков; впереди шва по одному волоску, с тру-
дом отличимому от прочего опушения спинки; интраалярных — 2
за швом; цредкрыловых — по 2 с каждой стороны, сближенные своими
основаниями; надкрыловых—ч4, из которых 1 длиннее и крепче про-
чих щетинок этой группы; закрыловых — 2; нотоплевральных — 1 4-2
(сзади); плечевых — 3; заплечевые — не развиты; предшовная — 1;
стерноплевральных 0-J-1, гипоплевры, проплевры, переднегрудь и супра-
сквамальное ребро — голые; длина заднего’ Дыхальца несколько превос-
ходит вертикальный его диаметр. На щитке: вершинных щетинок — 2;
боковых — 3; основных — 2; предосновных — 2; вблизи боковых краев
щитка несколько щетинистых волосков, расположенных в ряд. Ноги
бурые, голени и лапки смоляно-бурые. Передние лапки с длинными
узкими члениками; коготки передних ног в 1х/г раза короче 5-го членика
лапки, 4-й членик в длину больше, чем в ширину. Передние голени по задне -
240 ____ II. ТРЙБA STOMOXYDINI
вентральной и частично по задней поверхности в длинных, тонких воло-
сках; длина их в IV2 раза превосходит поперечник голени; по передне-
дорзальной стороне с обычным для рода рядом щетиночек; прочие
щетинки ног как у предыдущего вида. Крылья с желтым основанием.
Жилка m в вершинной части с правильным дуговидным изгибом, ячейка
/?. сально сужена; отрезок костальной жилки между г4+5 и m в 4 раза
короче предшествующего (между г 2+3 и г4+5). Чешуйки желтые, жуж-
жальца с коричневато-рыжим основанием и бурой вершиной. Брюшко
яйцевидное, черное, блестящее в черных, торчащих волосках, более
длинных на' 5-м тергите, стернитах и по заднему краю 4-го сегмента.
Пятна из мелких волосков на верхней стороне 5-го тергита отсутствуют.
Длина тела 12.5—14.5 мм.
Самец не известен. Описывается по 3 самкам из Китая: 1 о — верх,
р. Хи-чю, бассейн р. Голубой, 4000 м, июль 1900 г. (Козлов); 2 <уд—
бас. р? Джагын-гол, восточный Тибет, 4000 м, 1—6 VII 1900 (Козлов).
Тип в Зоологическом институте Академии Наук СССР в. Ленин-
раде.
Триба Stomoxydini
Задний край глаз вогнутый в нижнем отделе, реже (Acridomyia) выпу-
клый; нижний край лица высоко приподнят, затылок выпуклый; нижние
чешуйки (грудные) узкие, широкие или едва развиты; темный рисунок
среднеспинки представлен 5 полосами, часто разорванными на изоли-
рованные пятна; хоботок удлиненный, тонкий; жилка m изогнутая или
(Acridomyia) прямая; мембрана брюшка развита, часто в волосках.
Триба содержит по преимуществу кровососущие реже (Graphomyia)
растительноядные формы. Личинки — копрофаги, хищники, либо пара-
зиты (Acridomyia).
В трибе мы различаем 3 подтрибы: 1 — Graphoyina, 2 — Stomoxy-
dina и 3 — Acridomyina.
ОПРЕДЕЛИТЕЛЬНАЯ ТАБЛИЦА РОДОВ
1 (2). Жилка m в вершинном отрезке прямая; ариста без длинных волос-
ков; затылок в нижней половине сильно вздутый; вибриссальные
щетинки отсутсвуют. (Паразит азиатской саранчи) ........
....................................5. Acridomyia Stack.
2 (1). Жилка m в вершинном отрезке дуговидно или угловидно изогнута
по направлению к вершине крыла, отчего ячейка Ri сужена
в дистальной части. АрисДа с длинными волосками Сверху йли
4 сверху и снизу одновременно. (Главным образом кровососущие
формы).
3 (8). Жилка m изогнута дуговидно.
4 (7). Ариста в длинных волосках только по_ верхней стороне.
5 (6). Щупальцы цилиндрические, тонкие, в 3.5—5 раз короче хоботка;
пефеднегрудь очень широкая, в щетинистых волосках; проплевры
и гипоплевры впереди дыхалец с группой тонких волосков; глаза
узкие, с глубокой выемкой по заднему краю: затылок внизу слабо
выпуклый.....................................2. Stomoxys Geoffr
6 (5). Щупальцы сплющены с боков, сильно расширены в вершинной
половине (или 2/з); длина их не более чем в 2 раза короче хоботка
1. GRAPHOMYIA
241
(обычно длиннее его половины); проплевры и гипоплевры впереди
дыхалец без волосков; переднегрудь узкая, без щетинистых волос-
ков; глаза широкие со слабой выемкой по заднему краю.; затылок
сильно вздутый в нижней половине .... 4. Lyperosia Rond.
7 (4). Ариста в длинных волосках по всей верхней стороне и с несколь-
кими такими же волосками по нижней в ее дистальной части; передне-
грудь широкая с щетинистыми волосками; щупальцы сильно рас-
ширены в дистальной половине, длиною почти равны хоботку . .
......................................3. Haematobia R.-D.
8 (3). Жилка т с угловидным изгибом, грудная чешуйка широкая . . .
........................................1. Graphomyia R.-D.
1. Подтриба GRAPHOMYINA
Глаза с вогнутым в нижней части задним краем; щеки очень узкие;
, передне-верхний край рта высоко Приподнят над нижним уровнем глаза;
жилка т с мягким угловидным изгибом в вершинном отделе; Тека мас-
сивная, в несколько раз шире дистального отдела фаллозомы, передние
отделы парамер не четко дифференцированы; 6—9-й сегменты брюшка
самок' с развитыми в виде широких хитиновых пластинок тергитамй
и стернитами. Брюшко с пятнистым рисунком.
Личинки, с суженным к обоим концам телом; факультативные хищ-
ники.
1. Род GRAPHOMYIA R.-D.
Robineau-Desvoidy, 1830, Essai Myod : 403; Pandelle, 1898,; Rev. ent., XVII i
46; Bezzi u. Stein, 1907, Catal. pal. Dipt., HI : 599; Schnabl u. Dziedzicki, 1911 : 173;
Stein, 1916 : 15; Stein, 1917 : 110; Karl, 1928 : 10; Ringdahl, 1929, Ent. Tidskr,
(sep.) : 10.
Тип рода Graphomyia maculata Scop. (1763).
Средней величины мухи, с тремя резкими широкими бархатисто
черными продольными полосами на спинке и большими черными пят-
нами на брюшке.
Глаза в волосках, более густых и длинных у самцов; у некоторых
экзотических видов опушение глаз не явственно. Лоб самцов узкий,
ширина его не более тройной ширины 3-го членика усиков, у самок равна
или чуть уже глаза; лобная полоса в сравнительно многочисленных
и длинных волосках, наиболее густых в переднем отделе полосы; у д лоб-
ная полоса с выпуклыми краями, в 4—5 раз шире орбиты; последние
у обоих полов в щетинистых редких волосках, без орбитальных щетинок;
SS — без наружных теменных щетинок; внутренние теменные и затылоч-
ные щетинки £ слабые, часто имеют характер волосков, у j — крепкие,
длинные; скулы и щеки очень узкие, затылок в верхней части вогнутый;
глаза длинные, с вогнутым задним краем в нижней */г или у середины;
щупальцы слабо, но явственно расширены к вершине, сравнительно
толстые; подбородок удлиненный. 3-й членик усиков параллельно край-
ний, удлиненный, в 2—4 раза длиннее 2-го; ариста в длинных волосках,
более густых на верхней стороне. Налет на груди развит; переднегрудь
в переднем отделе голая, в задних боковых отделах (на боковых утол-
щенных краях) с очень мелкими густыми светлыми волосками; передние
плевры голые в центре; гипоплевры в длинных щетинистых волосках
в передневерхнем углу, впереди дыхальца и с несколькими волосками
16 Фауна СССР, том XVIII, вып. 4.
242 II. ТРИБА STOMOXYDINI
выше- задних тазиков, Супрасквамальное ребро без щетинистых темных
волосков. Щетинки груди: акростихальных — 1 пара перед щитком,
дорзоцентральных—‘2 + 4; интраалярных — 1 позади шва; предкры-
лоцая короткая, крепкая; надкрыловых — 1; закрыловых — 3; пле-
чевых — 2; предшовных — 1; нотоплевральных — 2; стерноплевральных
— О -|- 2 (1 -}-1; 0+1); на Щитке: вершинных — 2; боковых — 2;
основных — 2; пребазальные. отсутствуют; дискальных — 2, крепкие;
задние дыхальцы округло треугольные, высота их почти равна высоте
гипоцлевры от нижнего уровня дыхальца до нижнего края гипоплевры.
Жилца п голая, г«+5 в первом своем отрезке при основании или почти
по всей длине со щетиночками сверху и волосками снизу; т — с мягким
угловидным, иногда с полукруглым изгибом. Ноги бурые, с более свет-
лыми, часто желто-красными голенями; щетинки малочисленны. Перед-
ние бедра с гребнями заднедорзальных и задневентральных щетинок;
передние голени у палеарктических видов без щетинок помимо вершины;
средние бедра с щетинистыми волосками или тонкими щетиночками
по задневентральной стороне в основной половине и щетинками по перед-
ней поверхности бедра; средние голени с 2—3 щетинками на задней
поверхности и 1—2 (последние могут отсутствовать) переднедорзальными
щетинками; задние бедра с переднедорзальными и передневентральными
щетинками; последние доходят до */з или длины бедра, реже развиты
по всей его длине; задние голени с 1 (2) переднедорзальной, 1 (2) перед-
невентральной и 1 предвершинной дорзальной щетинками; задняя
поверхность без толстых щетинок. Брюшко широко яйцевидное, более
или менее сильно выпуклое, с пятнистым рисунком, в черных щетинистых
волосках и тонкими, но длинными на 4 и 5 тергитах маргинальными
щетинками; стерниты в длинных и коротких волосках (начиная от I
и кончая V стернитом). V стернит с маленькими боковыми выступами,
сравнительно широкий. Церки глубоко вырезаны сверху и с треуголь-
ной короткой срединной выемкой снизу, образующей два дистальных
треугольных выступа. Кокситы широкие, полукруглые на дистальном
конце, Фаллозома с очень широкой основной частью, ширина которой
почти в 4 раза превосходит дистальный мембранозный отдел фаллозомы.
Дистальная часть прямая, удлиненная, цилиндрическая, при основании
слабо склеротизована. Задний отросток фаллозомы очень широкий,
округлый на вершине или слабо выемчатый; задний отдел парамер широ-
кий, на вершине сужен и закруглен, при основании снабжен 2 щетиноч-
ками. Передние части парамер широкие. Аподема длинная, тонкая, почти
прямая; ампула с тонким длинным проксимальным концом и грушевидно
расширенным дистальным, имеющим многочисленные мелкие просветы,
концом. Дистальная часть фаллозомы без шипиков, свойственных мно-
гим родам трибы. ’
Виды рода встречаются на всех континентах от самых северных гра-
ниц до экватора. Из известных до сего времени, примерно, 35 видов рода
только один заходит в Палеарктику, обитая одновременно в Неарктичес-
кой, Неотропической и Эфиопской областях. Наиболее богаты видами
Ориентальная и Эфиопская области, в первой из названных насчиты-
вается 16, во второй — 11 видов; Неарктическая область имеет 6 видов
рода, включая "сюда и один палеарктический вид.
ч
*1. Graphomyia maculata Scop. (рис. 350, 402—415).
Scopoli, 1763, Entom. earn. : 326 (Musca)\ Bezzi u. Stein, 1907, Catal. pal.
HI : 599; Stein, 1916 : 15; Malloch, 1925, Ann. Mag. Nat. Hist., (9), XVI : 85; Karl:
1. GRAPHOMYIA 24?
1928 ' 10; Ringdahl, 1930, Arkiv zool., 21, A, 20 : 3; Tiensuu, 1935, Acta.Sop. fauna
Flora Fenn., 58, n° 4 : 53; Oucbi, 1933, Journ. Shanghai Sc. Inst;, III, IV ; 236.
Биология и фазы развития. Keilin, 1917, Parasitology, IX : 354,
fig. 5, VIII—XII; Graham-Smith, 1919, Parasitology, XI : 350; Зимин, 1948 : 70.
Голова сильно выпуклая, лоб впереди глазкового треугольника
составляет от 1/в до 1/а ширины глаза; орбиты в наиболее узком месте
равны х/2 ширины лобной полосы, резко расширенной вперед. Скулы
посредине в 1/« — IW раза шире 3-го членика усиков; щеки в 2 раза
шире скул. Затылок в верхней половине вдавлен, в нижней слабо выпу-
клый. 1-й членик усиков кольцеобразный, 2-й — расширенный к вершине,
3-й — слабо и постепенно суженный к дистальному концу в — 2 раза
превышает по длине 2-й членик. Волоски аристы (снизу) почти равны,
сверху в Р/г—2 раза длиннее поперечника 3-го членика усиков. Глаза
большие с вогнутым задним краем в нижних 2/з, в желтоватых волосках.
Щупальцы дуговидно слабо изогнутые, слабо утолщенные на вершине.
Голова чернобурая, вершина 2-го членика усиков коричневатая, ариста
и основная половина щупалец рыжие. Орбиты в светлосером, щеки
в сером или желтоватом налете, лицо в сером налете, постепенно пере-
ходящем в нижней х/г лица в желтовато-коричневый; лобная полоса
черная; затылок по краю глаз, в нижней половине и щеки — в сером
светлом налете. Глазковый треугольник, края лобной полосы и внутрен-
ние края орбит в длинных достаточно густых черных волосках, щеки
и затылок в нижней части в более коротких волосках. Верхний отдел
затылка в коротких щетинистых волосках. Грудь коричневато-бурая;
спинка с тремя черными широкими продольными полосами, разделен-
ными промежутками светло серого (с голубоватым или желтоватым оттен-
ком) налета. Бочки в матовом сером налете. Срединная черная полоса
начинается от переднего края спинки, она состоит из двух узких полос,
слабо расходящихся назад и достигающих средины заднего отдела спинки.
Сзади между срединными полосками вклинивается узкое, длинное чер-
ное треугольное пятно, основанием соприкасающиеся со щитком. Свет-
лые волосы налета между черными полосами спинки уже последних.’
Боковые черные полосы спинки слабо выпуклые по краям; над основа-
нием крыльев располагаются сагиттально вытянутые узкие черные пятна.
Щиток рыжий, розовато-коричневый или коричневый с широким тре-
угольным черным пятном на верхней поверхности и черными треуголь-
ными пятнами на боках при основании. Щетинки груди — см. диагноз
рода. Переднее дыхальце желтоватое. Поверхность груди, плевр и щитка
в черных, тонких торчащих волосках. Ноги смоляно-бурые, голени часто
рыжеватые, лапки черные; коготки короче 5-го членика лапки; пуль-
виллы равны длине коготков. Переднедорзальная щетинка средних голе-
ней, как правило, отсутствует. Крылья сероватые, иногда с дымчатыми
слабыми полосками вдоль жилок; костальный шип отсутствует; г1+5
снизу и сверху с несколькими короткими щетиночками, снизу заходя-
щими на 1/з расстояния от основания. Б'рюпко широко яйцевидное,
сильно выпуклое, чуть шире груди, рыжеватое; с бурым основанием
1+2-го тергита, вершиной, стернитами и примыкающими к ним концами
тергитов. Верхняя поверхность брюшка с черными пятнами: 3-й тер-
гит с крупным листовидной или округлой формы срединным пятном,
лежащим на средней продольной линии тергита, в передних 2/з с двумя
расположенными по бокам от последнего неправильно-треугольными
большими бурыми с черным, как правило, задними краями и двумя
маленькими округлыми пятнами на боках. На 4-м тергите порядок
16*
244
II. ТРИБА STOMOXYDINI
расположения пятен остается тем же, что на 3-м тергите, но сред-
нее пятно эллипсовидное и все пятна меньшего размера. 5-й тергит
с двумя округлыми мелкими пятнышками по бокам от средней линии.
Брюшко в желтовато-сером налете и черных негустых тонких волосках,
более длинных и щетинистых на конце брюшка. Маргинальные щетинки
на 3-м, 4-м и 5-м тергитах тонкие, длинные, на 3-м тергите короче 2/г
1’ис. 402. Самец Graphomyia maculata Scop. (Ориг.).
длины тергита. Стерниты в торчащих тонких волосках и тонких длин-
ных щетинках. Генитальные'придатки — см. диагноз рода.
По внешности личинка G. maculata резко отличается от личинок
других Muscidae. Она имеет сигаровидную форму тела; его наружные
покровы плотные, эластичные, продольно-морщинистые, желтовато-
розовой окраски. Головной сегмент личинки несколько втянут в 1-й
грудной сегмент, снабжен несколькими парами сензорных образований:
антенны с вытянутым вершинным цилиндрической формы члеником,
и втянутым внутрь базальным сегментом, склеротизованным и имею-
щим вид небольшого тяжа. Максиллярные щупальцы крупные, высту-
1. GRAPHOMYIA
245
лающие, на вершине имеют широкую чувствующую область, обладающую
несколькими сензорными сосочками, помимо которых имеется несколько
таких же сосочков в периферических частях щупалец. Нижнегубные
щупальцы сравнительно крупные, на конце снабженные волоском, рас-
положенные с нижней стороны псевдоцефала позади ротового отверстия
в непосредственной к нему близости. Впереди ротового отверстия также
снизу головного сегмента имеется пара мелких сензорных образований.
Ротоглоточный аппарат сложного устройства. Базальный склерит длин-
ный, щирокий, по переднему краю с глубокой угловидной выемкой.
Гипостомальный склерит короткий, массивный, неправильно прямо-
угольный. Ротовые крючья серповидно изогнутые, узкие, длинные,
заостренные на вершине. Зубные склериты треугольные, снизу соеди-
нены друг с другом зазубренной по переднему краю перемычкой. Впереди
ротовых крючьев располагается несколько хитиновых тяжей, имеющих
гид полос и пластинок: ротовой тяж, передняя и задняя хитиновые
полоски и срединная полоска.
Личинка III стадии амфипнейстична; передние стигмы с 4 корот-
кими, округленными на вершине, сосочками; впереди каждого сосочка
располагается по одной маленькой блестящей точке, являющейся
выходным отверстием стигмальной железы. Задние дыхальцы распо-
ложены на бугорках, сильно склеротизованных и снабженных каждое
тремя широкими, короткими щелями, один конец которых сужен
и несколько изогнут, другой — более широко округленный, прямой.
Каждый брюшной сегмент тела личинки снизу, считая со 2-го по 4-й,
снабжен двумя втягивающимися мясистыми бугорками, вооруженными
изогнутыми назад шипами, более длинными на вершине бугорков. По-
следний брюшной сегмент вытянутый, с мясистыми выростами, несу-
щими хитиновые шипики; один из выростов расположен по средине
сегмента с нижней его стороны впереди анального отверстия; он окру-
гленный и соответствует вентральному выросту предыдущего сегмента;
позади анального отверстия сегмент имеет пару выростов, несущих
кисточку волосков, среди которых скрыты сензорные сосочки; по бокам
от анального отверстия имеется по одному более вытянутому выросту,
несущему два чувствительных сосочка; вблизи задних стигм располо-
жено, кроме того, 3 пары мягких выростов.
Пупарий сужен к заднему концу, снабженному коротким согнутым
набок отростком (последний сегмент тела личинки); дорзовентрально
пупарий слабо сплюснут; на верхней стороне пупария на границе 1-го
и 2-го брюшных сегментов выступают темноокрашенные рожки, под
которыми расположены на небольшом выросте дыхательные сосочки.
Распространение. Вид широко распространен в Палеарк-
тике от ее северной океанической границы до южных пределов области.
В Сибири границы распространения вида пока не ясны. Вид известен
также из Африки, Северной и Южной Америки.
На протяжении этого обширного ареала вид дает серию форм, укло-
няющихся от типичной формы, за которую нужно принять средиземно-
морские экземпляры вида. Отличия между отмеченными нами вариаци-
ями сводятся к мелким морфологическим и цветовым признакам, не
всегда постоянным и связанным рядом постепенных переходов. Наши
попытки отыскать различия в строении генитального аппарата самцов
оказались неудачными; у всех форм, помещенных нами ниже в опреде-
лительную таблицу, строение генитальных частей оказалось тожде-
ственным .
246
II. ТРИВА STOMOXYDINI
Для распознавания этих форм приводим определительную таблицу
й краткое описание каждого вариетета с указанием его распростране-
ния на территории СССР.
ОПРЕДЕЛИТЕЛЬНАЯ ТАБЛИЦА ВАРИАЦИИ (ПО САМЦАМ)
1 (4). Ширина лба в 21/э—3 раза превышает ширину 3-го членика уси-
ков; край затылка выступает в виде полоски позади верхней гра-
ницы глаз, скулы в I1/*—P/а раза шире 3-го членика усиков; перед-
недорзальная щетинка на средних голенях имеется; в средней части
3-го и 4-го тергитов развиты длинные щетинистые волоски. Мельче;
средние пятна на 3 тергите узкие, клиновидные.
2 (3). Среднее пятно на 4-м тергите в форме узкого штриха; ширина
боковых пятен на том же тергите в 2 раза уже светлого промежутка
, между ними и срединным пятном................. maculata var. d
3 (2). Среднее пятно.на 4-м тергите в виде клина; боковые пятна круп-
нее; ширина их равна или превышает ширину светлого промежутка
• ......................................maculata var. е
4 (1). Ширина лба равна или шире 3-го членика усиков в 1т/2—2 раза;
затылок позади верхнего края глаз не образует выступающей
полоски; скулы равны или уже 3-го членика усиков (реже чуть
шире его); длинные щетинистые волоски в центре 4-го тергита отсут-
ствуют. Крупнее; средние пятна на 3-м тергите более или менее
ркруглые.
5 (8). Переднедорзальная щетинка средних голеней отсутствует.
6 . (7). Средняя черная полоса спинки впереди шва не разделена вдоль
двумя серыми тонкими линиями; сплошная; лоб равен ширине
3-го членика усиков или едва шире его; скулы в середине не шире
3-го членика усиков; брюшко бурое с едва заметными рыже-жел-
тыми пятнами по бокам; пятна на брюшке сплошь черно-бурые . .
• . .......................................maculata var. с
7 (6). Средняя черная полоса спинки впереди шва разделена.вдоль двумя
тонкими серыми линиями; ширина лба в IVa—2 раза шире 3-го чле-
ника усиков; брюшко с преобладающей красно-желтой окраской;
темные пятна брюшка красновато-коричневые в переднем отделе
3-го и 4-го тергитов.............................maculata var. а
8 (5). Переднедорзальная щетинка на средних голенях имеется . . .
...........................................maculata var. Ь
Самки перечисленных вариететов в большинстве случаев очень
сходны. Исключением могут служить лишь самки вариаций b, d, и е,
имеющие передне дор зальную щетинку на средних голенях. Что касается
цветовых признаков и размеров, то они настолько изменчивы у назван-
ных вариететов, что установить здесь границы не представилось воз-
можным.
Приводим данные по местам нахождения различных вариететов в
в пределах СССР.
Gr. maculata var. а. (Длина тела 6.5—8 мм). Северная граница аре-
ала идет от Ленинградской обл. на юго-восток через Ярославль, Рязань,
Куйбышев, Таганрог. Южная граница в Европейской части СССР не ясна.
На западе наиболее южными точками являются Проскуров (Подольский),
Киев и Харьков. В азиатской части СССР этот вариетет обнаружен на
Алтае и в Семиречье, каковые территории, видимо, и являются восточ-
ной гр'аницей ареала вариации а в этих широтах. В пределах Средней
1. GRAPHOMYIA
Азии известны по коллекционному материалу лишь 3 местонахождения:
Сталинабад (Варзобское ущелье), Катта-курган, Копет-даг. .
Рис. 403—413.
-103 — голова самца Graphomyia maculata Scop. var. d сбоку.
404 — то же самца G. maculata Scop. var. с. 405 — боковой край
затылка самки G. maculata Scop. var. d сзади. 406 — то же самки
G. maculata Scop. var. d. 407 — голова самца G. maculata Scop. var.
d сверху. 408 — то же самца G. maculata Scop. var. с сверху. 409 —
то же самки G. maculata Scop. var. а сверху. 410 — то Же самки G.
maculata Scop. var. d сверху. 411 — задняя стигма личинки III ста-
дии G. maculata Scop. 412 — переднее дыхальце личинки III ста-
дии того же вида. 413 — личинка G. maculata Scop, вид сбоку.
(Рис. 411—413 по Кейлину, прочие — ориг.).
6r. maculata var. Ь. (Длина тела 7.5—10 мм). Эта форма' связана
с зоной хвойных лесов,- начиная от Хибин и Ленинграда, на восток
248
II. ТРИВА STOMOXYDINI
до Камчатки. Вариетет зарегистрирован в нижеследующих пунктах:
Хибины (Кояфский п-ов.), Ленинград, Рязань, Казань, Кыштым, Малый
Урал, ср. течение р. Енисея, Иркутск, Якутский окр., Ключевское на
Камчатке; северная Монголия.
Gr. maculata var. с. (Длина тела 7.5—9.2 мм). Известна с Дальнего
Востока: Славянский залив, Яковлевка, Спасского р-на, Шкотовский
р-н, Гармахта бл. Хабаровска.
Gr. maculata var. d. (Длина тела 6—8 мм). В коллекции Зоологичес-
кого института Академии Наук СССР имеются экземпляры из северо-
восточного Цайдама и южного Нань-шаня.
Gr. maculata var. е. (Длина тела 6—8 мм). Как и вариетет с распро-
странен по преимуществу в зоне хвойных и лиственных лесов, не про-
двигаясь далее на юг в западных частях ареала.
414 — вершина крыла Graphomyia maculata Scop. 415—церки и кокситы
самца G. maculata Scop. 416 — крыло Acridomyia sacharovi Stack. (Ориг,).
В пределах СССР известен из следующих мест: окр. Ленинграда,
Витебская обл., Ярославль, Рязань, Проскуров (Подольский), Киев-
ская обл., Харьков, Полтава, Орловская обл., Жигули, Кокчетав, Севе-
роказахстанской обл., Иркутская обл., Якутия.
Личинки G. maculata встречаются в экскрементах животных или
в разлагающихся веществах растительного происхождения, где ведут
хищный образ жизни, охотясь за личинками мух, обитающих в тех же
веществах. В лабораторных условиях (Keilin) личинки G. maculata про-
являют себя как хищники, нападая на личинок Eristalis или (Зимин)
как сапрофаги.
2. Подтриба STOMOXYDINA
Задний край глаз вогнутый, жилка m дуговидно изогнута на вер-
шине; щупальцы нередко расширены; фаллозома с поперечной лопастью
на вершине, оканчивающейся зубцами, дорзальный отросток фаллозомы
2. STOMOXYS 24»
отсутствует. Мембрана брюшка хорошо развита. Грудные чешуйки узкие,
языковидные ± длинные.
В имагинальной фазе мухи кровососущи. Личинки развиваются
в помете животных и влажных растительных остатках, как исключение —
в почве. Задние стигмы взрослых личинок с S-образными щелями. Вали-
кообразные вздутия на брюшных сегментах с косыми ребрами.
2. Род STOMOXYS GEOFFR.
Geoffroy, 1764, Hist. Ins., II : 538; Bezzi u. Stein, 1907, Cat. Pal. Dipt., Ill -
611; Schnabl u. Dziedzicki, 1911 : 126; Townsend, 1914, Ann. Entom. Soc. Amer.,
VII : 160—167; Stein, 1916 : 19; Stein, 1917 : 102; Karl, 1928 : 14; Malloch, 1932,
Ann. Mag. Nat. Hist. (10), IX : 381; Pinto et Souza Lopes, 1933, X, n° 1 : 87; Patton,
1933, Ann. trop. Med. Parasitol., XXVII : 501—537; Ho, 1936, Bull. Fan Mem. Inst.
Biol., VI, n° 5 : 185—206; Zuinpt, 1938, Zeitschr. ang. Entom., XXV : 337—353.
Тип рода Stomoxys calcitrans Linne (1761).
Глаза без волосков, длинные, с выемкой по заднему краю; щеки
очень узкие; затылок слабо выпуклый в нижнем отделе, передний верх-
ний край рта высоко приподнят над нижним уровнем глаз; ариста с длин-
ными волосками по верхней стороне; хоботок тонкий, длинный, высту-
пающий далеко вперед из ротового отверстия; щупальцы цилиндричес-
кие, почти в 3 раза короче хоботка. Переднегрудь очень сильно расши-
рена впереди, со щетинистыми волосками в боковых отделах; передние
плевры в тонких волосках; гипоплевры с группой волосков впереди
стигм и с многочисленными волосками в передне-верхнем углу; заднее
Дыхальце очень маленькое, округло-треугольное. Щетинки груди
и щитка — см. описание вида, т с легким дуговидным изгибом;
с волосками, по обеим сторонам крыла, развитыми на 1/s—]/а или на всем
протяжении (у некоторых тропических форм) основного отрезка жилки.
Чешуйки суженные к вершине. Ноги у самцов нередко (некоторые тро-
пические виды) со своеобразным вооружением из щетиночек или волос-
ков. Хетотаксия ног слабее развита, нежели у трибы Muscini. Грудь
с полосчатым, брюшко с пятнистым или (у тропических видов) полосчатым
рисунком.
Фаллозома без дорзального отростка, вершинная ее часть с попе-
речной, склеротизованнрй лопастью, несущей зубчики на концах. Пара-
меры в переднем и заднем отделах хорошо развиты; задние части пара-
мер серповидно заостренные, передние — тупые, широкие. Тека корот-
кая. V стернит с 2 выростами по бокам заднего края; церки поперечные,
с 3 округлыми выступами по свободному вершинному краю. 6-е и 7-е
дыхальцы имеются; 6-й и 7-й тергиты узкие, асимметричные, как у Musca.-
Средней величины мухи.
Из 36 видов, самостоятельность которых установлена с достаточной
достоверностью (Мэллок, Пэттон, Сеги, Цумп), лишь один вид, распро
страненный всесветно, известен из Палеарктики. Наибольшего видового
многообразия род достигает в Эфиопской и Ориентальной областях;
в первой из них насчитывается 23, во второй—.11 видов. Общими для
них можно с достоверностью считать лишь 2 формы, прочие являются
эндемиками. Неарктическая и Неотропическая области бедны видами;
так для последней нам известен лишь 1 эндемичный вид (St. concolor
Walk.), для Неарктики — 3, в число которых входит и Stomoxys calci-
trans.
В биологическом отношении виды рода заслуживают интереса, как
формы кровососущие, тесно связанные с млекопитающими и птицами_
'250
II. ТР ИВ A STOMOXYDINI
Развитие личинок протекает либо в экскрементах животных, либо
в гниющих растительных остатках. Известны виды этого рода, отклады-
вающие яйца во-влажный песок вблизи засоленных водоемов.
С хозяйственной стороны виды этой группы мух имеет большое значе-
ние как распространители среди животных и людей возбудителей различных
заболеваний, как сибирская язва, трипанозомозы. Вместе с тем жигалки
имеют значение как промежуточные хозяева для некоторых глист
обычных у домашних птиц (Hymenolepis carioca Megalhaes, Habronema
microstoma Schneid.) и других. Бразильский вид рода зарегистрирован
как случайный транспортер яиц мухи Dermatobia hominis (Neiva et Gomes,
1917; Pinto da Fonseca, 1930).
Экспериментально доказана возможность передачи мухами болот-
ной лихорадки, анемии лошадей (Seguy) и желтой лихорадки.
*1. Stomoxys calcitrans Linne — Осенняя жигалка (рис. 417—428,
-431, 434, 440).
Linne, 1758, Fauna Suec., Ed., II : 467 (Conops); Schnabl u. Dziedzicki, 1911 :
539—540; Bezzi u. Stein, 1907, Cat. pal. Dipt., Ill : 611; Stein, 1916 : 19; Stein, 1917 :
102; •Karl, 1928 : 14; Patton, 1933, Ann. trop. Med, Parasitol., XXVII : 501—537;
Eidemann, 1935, Arb. morph, taxon. Ent., II : 99; Ho, 1936, Bull. Fan Mem. Inst.
Biol., VI, n° 5 : 186; Malloch, 1932, Ann. Mag. Nat. Hist., 10, IX : 402; Zumpt, 1938,
Zeitschr. ang. Entom., XXV, 2 : 339; Dinulescu, 1930, Ann. Parasitol., VIII : 61.
Биология и фазы развития. Порчинский, 1910 : 1—90; Mitzmain,
1913, Philipp. Journ. Sci. В. Trop. Med., VIII; Newstead, 1915, Journ. econom.
Biol., I : 157—166; Thomsen a. Hammer, 1938, Bull. ent. Res., XXVII : 559—587;
Thomsen, 1938 : 253—258; Mesnard et Toumanoff, 1939, Rev. m6d. franQ. Extr. orient.,
XVII, 8 : 995—999; Mohler, 1937 (38), Rep. Chief Bur. Anim. Industry : 85; Дербе-
нева-Ухова, 1940, Мед. паразит, и паразит, болезни, IX : 334—335; Зимин, 1944:
-156—159, 167—176; Зимин, 1948 : 78, фиг. 40; Павловский, 1948 : 860—863; Дербе-
.нева-Ухова, 1949 : 392—394.
$. Ширина лба составляет около 2/з глаза; лобная полоса шире
орбит в 4—5 раз; на уровне основания усиков ширина скулы превышает
ширину 3-го членика усиков, в среднем своем отделе немного менее послед-
ней; щеки узкие, высота их чуть более ширины скулы (посредине) и, при-
мерно, в 9 раз менее вертикального диаметра глаза. Усики бурые, вер-
шина 2-го членика их красноватая, ариста оперенная с верхней стороны,
рыжая; 3-й членик усиков в 2—2х/< раза длиннее 2-го; хоботок длинный,
тонкий, блестящий, буро-коричневый с маленькими сосательными лопа-
стями; щупальцы в 3 раза короче хоботка, цилиндрические, желтые.
Лобная полоса черно-бурая, в шелковистом, желтоватом редком, орбиты
и скулы — в густом желтовато-белом налете; затылок, исключая узкие
полосы сероватого густого налета за краем глаз, в редком налете; щеки
в редком сероватом налете и редких буроватых волосках. Орбиты в зад-
ней Va или 1/з с несколькими щетинистыми волосками, направленными
вперед и вверх; передние лобные тонкие, немного не доходят до перед-
него края лба; наружные и внутренние теменные щетинки тонкие,
но ясные; вибриссы — более крепкие. Грудь бурая, с 4 бурыми полосами
на спинке, разделенными узкими промежутками желтовато-серого свет-
лого налета. Переднегрудь очень широкая, трапециевидно расширенная
впереди, с щетинистыми крепкими волосками в боковых отделах; про-
плевры впереди дыхалец в тонких волосках, супрасквамальное ребро
голое; гипоплевры с тонкими волосками перед задним дыхальцем
и многочисленными тонкими волосками в передне-верхнем углу;
заднее дыхальце маленькое, округло-треугольное, передний его
.край достигает лишь середины вентрального отдела медпотергита.
2. STOMOXYS
251
Щетинки груди: акростихальных — 1 пара перед щитком; дорзо-
центральные тонкие, крепости щетинок достигают лишь 1—2 пары впе-
реди щитка и 1 пара перед поперечным швом, прочие в числе 3—4 впе-
реди шва и 5—6 пар позади него, имеют характер волосков; интрааляр-
ных — 1 позади шва, предкрыловые отсутствуют; надкрыловых — 1; закры-
ловых — 2; плечевых — 3; заплечевых — 1; предшовных — 1; нотоплев-
ральных— 2; стерноплевральных — 0-|-1. На щитке — вершинных 2;
основных — 2; предосновных — 2 (слабые); дискальных — 1—2 пары.
Нижняя сторона щитка и боковые края его без волосков в основном
отделе. Крылья сероватые, жилки рыжие; жилка т с дуговидным или
S-образным изгибом в вершинной части; г4+5 с волосками снизу и сверху,
Рис. 417. Самец Stomoxys calcitrans L. (Ориг.)
доходящими до Чг длины основного отрезка жилки; базальный участок
радиуса около места разветвления п и г4+5 ветвей сверху с группой из
2—4 волосков. Чешуйки на большем или меньшем протяжении пале-
вые по краю; нижняя из них суженная к вершине; жужжальца желтые.
Ноги бурые, голени, исключая желтоокрашенную основную треть, буро-
вато-коричневые; коготки несколько короче 5-г.о членика лапки; пуль-
виллы сероватые. Брюшко широко яйцевидное или яйцевидное, бурое,
в желтовато-сером шелковистом налете. 1 4-2-й тергит затемнен в основ-
ной половине, 3-й, 4-й и 5-й — с коричнево-бурыми и округлыми или
треугольными пятнами посредине, нередко сливающимися и образую-
щими в этом случае продольную полоску на брюшке. Передние края
тех же тергитов и коричневатой узкой поперечной полоской, образую-
щей в боковых отделах округлые выступы; задние края тергитов (кроме
1-1-2-го п 5-го) по бокам имеют по округлому пятну, вблизи заднего края,
нередко затемненному. Стерниты III и IV узкие, удлиненно эллипсоид-
ные. V стернит с двумя узкими выростами по заднему краю; I стернит
252
II. ТР ИВ A STOMOXYDINI
в волосках. Церки поперечные, нижний край с 3 выступами, из которых
средний короче боковых и более широко округлен. Фаллозома без дор-
зального отростка, тека короче penis, последний с хитинизированным
и имеющим вид трубки основанием; на вершине с передней поверхности
несет поперечно расположенную склеротизованную полоску, снабжен-
ную на концах зубцом (иногда раздвоенным). Передняя часть парамер
широкая, задняя серповидная, заостренная на вершине.
$. Лоб равен или немного шире глаза; орбиты в 3^2—4 раза уже
выпуклой по краю лобной полосы, в заднем отделе с 1—2 орбитальными
и группой коротких щетиночек, направленных вершинами вперед. Лоб-
ная полоса с несколькими щетинистыми волосками в боковых частях
заднего отдела, направленными вершинами к средней линии головы.
Длина тела 5.5—7 мм.
Яйцо желтовато-белое, цилиндрическое, к верхнему полюсу сужен-
ное несколько сильнее, чем к нижнему; с дорзальной стороны с 2 почти
параллельными продольными бороздками, расставленными на расстоя-
ние несколько меньшее чем Vs поперечника яйца; поверхность хориона
в мелких шестигранных ячейках, образованных тонкими ребрышками;
этот скульптурный рисунок виден при увеличении в 40—60 раз.
Длина яйца 0.9—1.0 мм.
Личинка I стадии метапнейстична; задние стигмы с сердцевидным
или округлым отверстием каждая. Тело без склеротизованных шипов;
с нижней стороны брюшных сегментов с короткими, поперечными вали-
кообразными утолщениями по переднему краю. Срединный дорзальным
склерит с узким удлиненно-клиновидной формы передним отделом, окан-
чивающимся согнутым вниз зубцом; основные 2/з склерита желобовидно
согнутые, в 3 раза шире передней трети; от основания передней узкой
части края склерита направлены вентрально, образуя почти прямой
угол, откуда направлены назад, где соединяются с гипостомальным скле-
ритом. Снизу под дистальной частью и несколько впереди от ее вершины
располагается 3 мелких добавочных склерита средний, в виде узкой
маленькой полоски, и 2 боковых, неправильно клиновидных. Гипосто-
мальный склерит в виде двух узких коротких тяжей, соединенных в зад-
нем отделе поперечной перемычкой; базальный склерит крупный, длин-
ный, длина его средней части почти равна длине узких тонких передних
его отростков; дорзальные отростки почти в 3 раза короче среднего
отдела склерита, заостренные на вершине, очень узкие в верхнем отделе,
вентральные отростки равны или чуть длиннее срединной части склерита,
много длиннее дорзальных отростков; склеротизация вентральных отро-
стков неравномерная; средняя продольная их часть почти прозрачная,
слабо склеротизована. Дорзальная перемычка высокая, узкая. Длина
тела 1.1—1.3.
Личинка П стадии амфипнейстична; передниедыхальцыс5—6 лучами;
задние дыхальцы со слабо склеротизованной перитремой; каждое имеет
по 2 наискось направленных коротких, почти прямых щели, из которых
внутренняя (верхняя) в Р/г раза короче наружной; дыхальцы малень-
кие, далеко расставленные друг от друга (на 5—6 диаметров дыхальца).
Тело лишено хитиновых шипов, головной сегмент двураздельный,
каждая из боковых его частей имеет вид полушарий; антенны и максил-
лярные щупальцы хорошо развиты; передние края брюшных сегментов
с валикообразными поперечными утолщениями, снабженными каждый
мелкими мягкими заостренными выростами, собранными в несколько
рядов; анальный сегмент сзади в верхнем отделе (вокруг дыхалец) в мне-
2. STOMOXYS
253
гочисленных пологих ямках, придающих кутикуле шероховатость; сред-
ний отдел ниже дыхалец слабо выпуклый, блестящий, лишен пункти-
ровки ; нижний край задней поверхности в мелких конических или округ-
лых выпуклостях, из которых 2, лежащие посредине, крупнее остальных,
вытянутые в вертикальном направлении. Ротовые крючья очень малень-
кие, удлиненно треугольные; тонко заостренные на вершине, в дисталь-
ном отделе сращены. По бокам от ротовых крючьев расположено
по одному неправильно треугольному, более широкому, чем ротовые
крючки, губному склериту. Угловидный склерит в виде пары верти-
Рис. -418—421. Детали строения личинок ранних стадий
Stomoxys calcitrans L.
418 — головной конец личинки I стадии. 419 — то же II ста-
дии. 420 — задние стигмы личинки II стадии. 421 — средин-
ный дорзальный склерит личинки I стадии. (Ориг.).
кально лежащих, слабо S-образно изогнутых хитиновых тяжей, сходя-
щихся своими верхними концами. Гипостомальный склерит короткий,
С глубокой вырезкой по заднему краю, по бокам от которой отходят назад
слабо разведенные вершинами друг от друга два длинных отростка,
сочленяющихся с передними отростками фарингеального склерита;
от верхнего вогнутого по середине края гипостомального склерита отхо-
дят вперед и несколько вверх два тонких отростка, сочленяющихся с вер-
шинами угловидных склеритов; эпистомальный склерит желобовидный,
выпуклой стороной обращен вверх; боковые лопасти его загнуты вниз
и соединены с передними отростками гипостомального склерита. Фарин-
геальный склерит сравнительно с расположенными впереди от него
частями ротоглоточного аппарата очень длинный и крупный. Передние
254-
II. ТР ИВ A STOMOXYDINI
отростки его немного короче гипостомального склерита, расширены
на переднем конце, где образуют заостренные нижний, более оттянутый
клювовидно вниз, и верхний короткий углы, передний край отростков
слабо выемчатый у средины; нижние края отростков соединены массив-
ной вентральной перемычкой, состоящей из передней поперечной, узкой
лопасти и отходящих от ее заднего края в кольцеобразных, лежащих
в вертикальной плоскости отростков; дорзальная перемычка узкая,
решетчатая, слегка выдается своим верхним краем над краями базаль-
ного склерита при рассматривании в профиль ее верхний край угло-
видно выдается вперед, нависая над передними отростками; задние
отростки различны по длине и ширине: вентральные очень массивные,
несколько сужены в основной половине, далее к вершине слабо расши-
рены; их нижний край почти прямой, верхний, вогнутый в основной
и выпуклый в дистальной частях; на конце заострены. Дорзальные
отростки в 2 раза короче и почти в 5 раз уже вентральных, слабо
выпуклые по верхнему и вогнуты по нижнему краям, на вершине
заостренные. Длина тела 2.2—4.3 мм.
Личинка III стадии амфипнейстична; передние дыхальцы с 6 (реже
с 5—7) лучами; задние — расставлены на длину двойного или полутор-
ного диаметра дыхальца, округло 5-угольные или треугольные; дыхаль-
цевая пластинка коричневато-рыжая, с 3 овальными или неправильной
почковидной формы просветами, заключающими в себе по одной S
образно изогнутой дыхальцевой щели. Тело личинки вытянутое, тоньше,
нежели у личинок рода Musca или Orthellia. Длина тела 6—10 мм.
Ложнококон того же типа, что и у Musca коричневато-краснова-
тый, в переднем отделе несколько более расширен, чем в заднем; сегмен-
тация ясная; дыхательные рожки отсутствуют, заменены маленькими
округлыми слабо выступающими светлыми выпуклостями. Задние стигмы
широко расставлены, как у личинок, округло треугольные; анальный
бугорок спереди и сзади опоясан широкой шероховатой полосой. Длина
4—4.5 мм; толщина в середине 1.8—2.1 мм.
Распространение. Вид обладает всесветным распростра-
нением. В СССР известен из многих мест Европейской части СССР, где
доходит на севере до Хибин и Архангельска, на восток известен до Баш-
кирии; повсеместно распространен в степной и полупустынной частях
Казахстана, а также в Средней Азии, откуда известен как из горных,
так и из равнинных местностей. Материала по Сибири в коллекции
ЗИН нет.
Появляются мухи в природе в начале лета (IV—V), достигая макси-
мума в конце его (VII) или осенью (VIII—IX). На юге развитие мух
протекает беспрерывно круглый год. Откладка яиц производится в навоз
животных или в перегнивающие растительные остатки. По данным ряда
исследователей, конский навоз является предпочитаемым материалом
для кладки яиц, хотя при отсутствии такового мухи охотно отклады-
вают яйца на свиной, телячий и коровий помет. Известны случаи мас-
сового размножения осенней жигалки во влажных, гниющих раститель-
ных остатках в полях. Нередко жигалка размножается в отбросах
на свалках. Известны, наконец, случаи нахождения личинок в ранах
животных и людей. Яйца откладываются в утренние часы, причем длй
этой цели самки предпочитают затененные помещения и теневые части
двора. Количество откладываемых яиц весьма различно: от 48 до 1.04штук.
Муха дает несколько поколений в течение теплого периода года.
Количество поколений зависит от гигротермических условий среды,.
2. STOMOXYS
255-
где происходит развитие преимагинальных стадий. По отношению к тем-
пературе и влажности среды осенняя жигалка крайне чувствительна.
Так, например, под влиянием излишней влажности (и освещения) раз-
427
Рис. 422—427. Детали строения личинки III стадии Stomoxys calcitrans L.
422 — ротоглоточный аппарат. 423 — задний конец тела сзади. 424 — он же сбоку.
425 — переднее дыхальце. 426 — заднее дыхальце.. 427 — вооружение снизу 5-го (2}
и 6-го (7) брюшных сегментов. (По Зимину).
питие личинок сильно задерживается. Обычно стадия личинки длится
от 14 до 21 дня.
На стадию куколки требуется от 9 до 17 дней. Полный цикл раз-*
вития мухи продолжается 25—37 (Ньюстид), 32—38 (Порчинский) дней.
По нашим наблюдениям в Таджикистане, развитие личинок при темпе-
256
II. ТРИБА STOMOXYDINI
ратуре 28.6° С заканчивается в 17 дней; продолжительность стадии
куколки при той же температуре равна 5—8 дням.
По данным Ньюстида, длительность личиночной фазы при условиях
повышенной влажности субстрата может достигнуть весьма высоких
пределов (нескольких месяцев). Предельные сроки диапаузирования
для личинок не установлены.
В отличие от других жигалок {Lyperosia, Haematobia'), у обыкно-
венной жигалки наблюдается лишь кратковременный контакт с жи-
вотными во время ее питания. В остальное время жигалки оставляют
животное, располагаясь на стенах стойл, домов, на заборах, ство-
лах деревьев. В полях и открытых не защищенных от солнца и ветра
местах жигалка не встречается в больших количествах, предпочитая
защищенные, тенистые скотные дворы, помещения конюшен, свинарни-
ков и т. д.
Рис. 428—430. Фаллозома.
428 — Stomoxys calcitrans L. 429 — то же Lyperosia irritans L. 430 — то же
Haematobia atripalpis Bzz. (Ориг.).
Осенняя жигалка — кровосос, питающийся кровью различных млеко-
питающих или (реже) птиц. Предпочтение отдает крови лошадей и коров,
хотя нападает также на верблюдов, буйволов, ослов, собак. Она пресле-
дует и человека как в природе, так и в его жилище. Жигалка неодно-
кратно зарегистрирована и на свежих трупах животных во время пита-
ния их кровью.
В силу попеременного контакта мух то с больными (или даже пав-
шими), то со здоровыми животными и человеком, осенняя жигалка спо-
собствует распространению возбудителей различных заболеваний.
Экспериментально доказана возможность переноса жигалками сибир-
ской язвы [Сеги (Seguy), 1935]. Жигалке принадлежит существенная
роль в переносе различных трипанозом. На о-ве Яве жигалка совместно
со слепнями является основным агентом в распространении Trypanosoma
^evansi — возбудителя болезни «сурра».
Жигалки участвуют и в переносе сходного заболевания, вызывае-
мого Trypanosoma evansi var. mbori Laveran, известного под наименова-
3. HAEMATOBIA
25T
нием «мушиной болезни». По свидетельству Буфара и Пэко (Bouffard,
Pecaud),* 1 Stomoxys подозревается в распространении Trypanosoma vivdx
(=cazolboni) Laveran, вызывающей сильную смертность лошадей и ослов
в Судане. Мартен, Лебёф и Рубо (Martin, Leboeuf et Roubaud)1 пока-
зали, что жигалка может распространять Trypanosoma brucei Pl. et Вг. —
возбудителя болезни «нагана», причем передача происходит механически
в момент укола животного загрязненным хоботком мухи. Осенняя
жигалка является также агентом в перенесении Trypanosoma equina-
voger среди лошадей в Южной Америке [Сеги (Seguy), 1935], Trypano-
soma soudanense Laver. — среди верблюдов, лошадей в Северной Африке,
Trypanosoma berberum—среди дромадеров, собак [Сержан, Донатиен
и Лестокар (Sergent, Donation et Lestoquard)]1, Trypanosoma annamense—
в Индокитае среди буйволов и быков [Менар и Туманов (Mesnard и. Тои-
manoff)]. Молер (Mohler) подозревает осеннюю жигалку в распростра-
нении энцефаломиелита среди лошадей в Северной Америке. В опыте
с обезьянами доказана возможность переноса жигалкой вируса желтой
.лихорадки от больных к здоровым животным.
Также опытным путем установлена способность мух-жигалок осу-
ществлять перенос Treponema recurrentis и ictero-haemorrhagiae, Treponema
galliarum (Seguy, 1935). Тот же автор указывает на распространение
жигалкой анемии лошадей и туляремии. Барбериэн (Berberian, 1939)
экспериментально доказал возможность переноса жигалкой пендинской
язвы. Stomoxys подозреваются в распространении ящура и проказы,
что, правда, остается пока недоказанным.
Жигалка является промежуточным хозяином нематоды Наргопета
microstoma Schneid., во взрослой стадии живущей в желудке лощади; раз-
витие этой нематоды происходит исключительно в жировом теле нимф
(наподобие Habronema muscae домашней мухи) [Сеги (Seguy), 1935].
Жигалка является также промежуточным хозяином и для паразитиче-
ской нематоды Hymenolpis carioca (Megalhaeus), обычной у домашних
птиц.
Помимо распространения патогенных организмов осенняя жигалКа,
правда сравнительно редко, встречается в фазе личинки в ранах живот-
ных и человека.
Борьба с жигалкой сводится главным образом к уничтожению личи-
нок в местах размножения (навоз, нечистоты и т. д.), а также к тому,
чтобы воспрепятствовать контакту мух как с животными, так и с тем
материалом, где возможна откладка яиц и развитие личинок. Для осен-
ней жигалки, как и других синантропных мух, высокотоксичным ока-
зывается ДДТ.
3. Род HAEMATOBIA R.-D.
Robineau-Desvoidy, 1830, Essai Myod.: 388; Bezzi u. Stein, 1907, Catal.pal.
Drpt. : 612; Stein, 1916 : 19; Seguy, 1923, Diptera : 343; Zumpt, 1938, Zeitschr. ang.
Entom., XXV : 350; — Haematobosca Bezzi, 1907, Zeitschr. Hym. Dipt., VII : 414;
Malloch, 1932 : 501; — Bdellolarynx Austen, 1909, Ann. Mag. Nat. Hist., 8, III : 290;
Malloch, 1932, 10, IX : 434; — Lyperosiops Townsend, 1912, Proc. Ent. Soc. Wash.,
XIV : 47; — Bdellia Enderlein, 1928, Zeitschr. ang. Ent;, XIV : 359; — Haema-
tobina Malloch, 1932, Ann. Mag. Nat. Hist., (10), IX : 442; — Neivamyia Pinto da
Fonseca, 1930, Rev. Med. Cir. Bras., XXXVI : 6.
Тип рода Haematobia stimulans Mg. (1824).
Глаза голые, сравнительно широкие (по сравнению с Stomoxys)
и длинные (по сравнению с Lyperosia) с выемкой по заднему краю; хобо-
1 Цитировано по Сеги (S6guy, 1935).
1 7 Фауна СССР, том XVIII, вып. 4.
258
II. ТРИВА STOMOXYDINI
ток с тонким длинным сосальцем и маленькими сосательными лопастями;
щупальцы длинные, сплюснутые с боков и сильно расширенные к вер-
шине, наружная их поверхность в коротких и длинных щетиночках;
верхняя часть затылка плоская, нижняя выпуклая, щеки очень узкие;
передний верхний край рта сильно выступает вперед и высоко припод-
нят над нижним уровнем глаз; с верхней и нижней стороны с длинными
волосками, переднегрудь у. палеарктических видов сильно расширена
впереди стигм и гипоплевры перед дыхальцами в волосках или без них;
передне-верхний угол гипоплевры голый или с одиночными волосками.
Рис. 431—436.
431 — вершинная половина крыла Stomoxys calcitrans L. 432 — то же Haematobia
stimulans Mg. 433 — то же Lyperosia irritans L. 434 — голова самца Stomoxys calci-
trans L. в профиль. 435 — то же Haematobia stimulans Mg. 436 — то же Lyperosia
irritans L. (Ориг.).
Акростихальные щетинки в числе 1 пары впереди щитка, дорзоцентраль-
ных — 1 пара перед швом и несколько пар позади него, из которых 1—2
пары перед щитком достигают более сильного развития; стерноплевраль-
ных щетинок Жилка т в дистальном отрезке дуговидно изогнута
по направлению к вершине крыла; п с волосками в основной половине
или голая. Гениталии самцов сходны с таковыми Stomoxys-, церки самца
обычно не имеют сильно оттянутых внутренних углов. Спинка груди
с продольными полосами; брюшко с 3 рядами пятен.
Из 23 видов, относимых нами к данному роду, 2 свойственны Пале-
арктике (один из них встречается и в Северной Америке). Н. crassipalpis
Ringd. по всей вероятности должен быть, как это полагает и Цумпт (Zumpt),
отнесен в род L yperosia как вид, у которого волоски располагаются лишь
с верхней стороны аристы. Кроме вышеотмеченного одного вида из Север-
3. HAEMATOBIA
259
ной Америки, известен еще 1 вид, свойственный, вероятно, лишь Неарк-
тике. Неотропическая область имеет 3 эндемичные вида. Значительно
многообразнее род представлен в Ориентальной (9 видов) и Эфиопской
(б видов) областях, причем, насколько нам известно, виды эти являются
для них эндемичными.
Все виды рода в фазе, имаго являются кровососами. Личинки раз-
виваются в помете животных.
ОПРЕДЕЛИТЕЛЬНАЯ ТАБЛИЦА ВИДОВ
1 (2). Проплевры впереди дыхалец и центр гипоплевр без волосков;
жилка п с волосками в основной 1/г; щупальцы красно-жел-
тые ...........................................*1. Н. stimulans Mg.
2 (1). Проплевры и гипоплевры впереди дыхалец в тонких волосках;
жилка п без волосков; щупальцы бурые или буровато-ры-
жие ...........................................*2. Н. atripalpis Bzz.
*1. Haematobia stimulans Meig. (рис. 132,432, 435, 438,444,445—452).
Meigen, 1824, Syst. Beschr., IV : 161 (Musca)-, Bezzi u. Stein, 1907, Catal. pal.
Dipt., Ill : 612; Schnabl u. Dziedzicki, 1911 : 126, figs. 535, 536; Townsend, 1912,
Proc. Ent. Soc. Wash., XIV : 47 (Lyperosiops)', Malloch, 1932, Ann. Mag. Nat. Hist.,
(9), X : 436 (Bdellolarynx)-, Patton, 1933, Ann. trop. Med. Parasitol., XXVII : 501—
537, figs. 3a, 4c, 5b, 8, 10, 14b; Se'guy, 1935, Diptera VIII : 47; Zumpt, 1938,
Zeitschr. angew. Entom., XXV : 351.
Биология и стадии развития. Порчинский, 1910:11—13,
фиг. 7—9; Hewitt, 1912 : 267; Thomsen, 1936, Bull. Entomol. Research, XXVII :
380; он же, 1935, Proc. Zool. Soc. London; Thomsen, 1937, Zeitschr. Physiol., XXIV,
n° 3 : 343—380, figs.; Thomson, 1937, Parasitology, XXIX : 282—290, figs. 13—18,
20, 54 , 55, 147; Thomsen, 1938 : 295—304, figs. 74—77.
<5. Ширина лба в наиболее узком месте равна или чуть превышает
х/г ширины 3-го членика усиков, скулы по середине менее Va ширины 3-го
членика усиков, слабо суженные книзу; щеки едва шире скул; затылок
в верхней трети плоский, далее в нижней части сильно выпуклый; лоб-
ная полоса черная, в середине суженная и в этом месте равна или шире
(не более чем в 2 раза) орбиты; верхний передний край рта сильно угло-
видно выставлен вперед; усики черно-бурые, 3-й их членик в Р/г раза
длинее 2-го; ариста утолщена в основной трети с 8—10 длинными волос-
ками на верхней и 2—5 короткими волосками на нижней стороне;
щупальцы сильно и резко утолщены в вершинной половине, красно-
желтые, длина их немного менее длины сосальца; наибольшая ширина
их в дистальном отделе составляет 3/а ширины 3-го членика усиков.
Щетинки головы тонкие, короткие; наружные теменные не развиты;
вибриссы и внутренние теменные развиты сильнее прочих. Голова черно-
бурая; скулы внизу и часто орбиты в густом серовато-белом налете;
в верхнем отделе скул и передней части орбит окраска налета нередко
желтовато-серая. Затылок в матовом сероватом налете, как и щеки,
покрыт бурыми сравнительно редкими волосками. Грудь бурая, средне-
спинка с 4 ясными и 1 иногда исчезающей продольными полосами, начи-
нающимися от переднего края среднеспинки, и короткой срединной
Полоской, направленной от основания шитка впереди имеющей вид удли-
ненного треугольника: боковые полосы среднеспинки шире срединных;
впереди шва они имеют характер вытянутых треугольников. Задние
конпы боковых бурых полос немного не достигают края щитка, средние
узкие полосы развиты до средины заднего отдела спинки. Спинка Н корич-
17*
260
II. ТРИБА STOMOXYDINI
невато-желтом или желтовато-сером налете, местами переходящем в сизо-
голубой; плевры в более светлом налете. Поверхность груди в тонких
бурых волосках. Щетинки груди: акростихальных — 1 пара перед щит-
ком, дорзоцентральных — 1 перед поперечным швом и 2—3 пары впереди
Рис. 437—443.
437 — церки и кокситьТсамца Lyperosia irritans L. 438 — то
же Haematobia stimulans Mg. 439 — то же Haematobia atripal-
pis Bzz. 440 — церки самца Stomoxys calcitrans L. 441 — F
стернит самца Musca arnica, sp. nov. 442 — церки и кокситы
самца Musca arnica, sp. n. 443 — церки и кокситы самца
Acridomyia sacharovi Stack. (Ориг.).
щитка, из которых лишь 2 задние пары достигают нормальной толщины;
мнтраалярных — 1 позади шва; надкрыловых — 1; закрыловых — 2;
предкрыловая щетинка отсутствует; плечевых щетинок 3 (2); заплече-
вых — 1; предшовных — 1; нотоплевральных — 1 + 1; на щитке: вер-
шинных — 2 и боковых — 2 (развиты сильнее других); основных — 2,
предосновных — 2. Переднегрудь широкая, с многочисленными жест-
3. HAEMATOBIA
261
кими волосками по краю; передние плевры голые, гипоплевры без волос-
ков. Крылья слабо дымчатые, жилки коричневато-рыжие; п в основной
половине сверху со щетинками, r4+s при основании сверху с 2—5
щетинистыми волосками; т с дуговидным изгибом в дистальном отделе.
Чешуйки узкие, с полукруглым свободным краем, на вершине палевые,
в средней части белые; жужжальцы палево-желтые, ноги черно-бурые,
вершины бедер коричневатые, основания голеней бледножелтые. Сред-
ние голени с одной щетинкой у средины задней поверхности; задние
бедра, перед вершиной на передневентральной поверхности с 2 (1) щетин-
ками и рядом переднедорзальных щетинок; бедра снизу у основания с 1—2
длинными тонкими щетинистыми волосками. Задние голени с 1 передне-
дорзальной щетинкой у средины и 1 предвершинной на дорзальной стороне;
коготки в 1х/г раза короче 5 членика лапки, присоски грязножелтовато-
серые. Брюшко широко-яйцевидное, бурое, мембрана палевая в редких
черных, коротких щетинистых волосках. 1 стернит короткий, полулун-
ной формы, II, III и IV стерниты узкие, эллиптической формы, V — широ-
кий с двумя тонкими, длинными отростками по заднему краю, между
которыми ясно обозначается хитиновая двураздельная полоска (= VI стер-
ниту). Верхняя сторона брюшка в серовато-желтом (реже голубоватом)
налете, переходящем на местах расположения пятнистого рисунка
в коричневатый. Бурые пятна расположены в 3 ряда. Средний ряд состоит
из узких клиновидно расширенных к основанию каждого сегмента пятен,
не достигающих задних краев тергитов; боковые ряды образуются
262
II. ТРИБА STOMOXYDINI
округлыми пятнами, размер которых убывает вперед и назад от 3-го
сегмента. Поверхность брюшка в щетинистых волосках, приобретающих
на задних краях сегментов характер тонких щетиночек. Задние отделы
парамер заостренные, серповидные, передние — широкие, тупые на конец.
Дистальная часть фаллозомы с поперечной лопастью, несущей на концах
по шипу.
2- Отличается широким лбом (поперечник его в заднем отделе равен
ширине глаза); лобная полоса слабо расширена впереди, в 3—З1^ раза
шире орбиты; щупальцы чуть шире, чем у самцов. Задний отдел орбит
с каждой стороны с 2—3 короткими, направленными вперед и несколько
вбок орбитальными щетинками, позади от которых располагается группа
более мелких щетинистых волосков. Налет на груди светлее, чем у <5
(светложелтовато-серый), волоски на среднеспинке короче; крылья
серые. Средние пятна брюшка нередко образуют сплошную узкую про-
дольную полоску, суженную назад и не достигающую вершины брюшка.
Бедра средних и задних ног красновато-желтые, с более или менее раз-
витым широким кольцевидным затемнением на вершине.
Длина тела 4.8—6.2 мм.
Яйцо овальное, по дорзальной поверхности с 2 тонкими бороздками,
расставленными на х/з ширины яйца; хорион в мелких ячейках, белый.1
Длина около 1 мм.
Личинка I стадии метапнейстична; задние дыхальцы выступают
в виде округлых бугорков. По переднему краю на вентральной стороне
брюшных сегментов со 2-го по 7-й проходят полоски из шипиков. Усики
и максиллярные щупальцы явственны. Срединный дорзальный склерит
ротоглоточного аппарата узкий, вытянутый. Вентрально от него рас-
положена 1 пара Т-образных склеритов, узких, вытянутых и слабо утол-
щенных на вершине. Гипостомальный склерит короткий, с узкими боко-
выми дуговидно изогнутыми частями и широкой перемычкой между ними
Фарингеальный склерит с узкой срединой, очень узкими дуговидными
дорзальными и значительно превышающими их по длине вентральными
отростками. Те и другие заострены на концах. Дорзальная перемычка
высоко приподнята над краями фарингеального склерита, узкая. Перед-
ние дыхальцы с 6 лучами; задние дыхальцы снабжены каждый двумя
дыхальцевыми щелями. Ротовые крючки короткие, узко клиновидные,
заостренные на конце, тесно сближены; латерально от них располагается
по добавочному склериту, клювовидно заостренному внизу (по Томсону
последний соответствует лабиальному склериту). Ангулярные склериты,
лежащие между ротовыми крючьями и дистальными концами гипосто-
мального склерита, представлены двумя маленькими прямоугольными
хитиновыми склеритами. Гипостомальный склерит вытянутый, продол-
жающийся впереди в тонкий тяж, сзади слабо расширен и образует два
угловидных узких выступа, сочлененных с фарингеальным склеритом.
Базальный склерит очень длинный, с острыми, тонкими и короткими
Дорзальными и превосходящими их по длине коротко заостренными на вер-
шине, широкими вентральными отростками; дорзальная перемычка
маленькая, расширена в дорзальном отделе, с крупными отверстиями;
передние отростки, соединенные снизу узкой вентральной перемычкой,
узкие, почти такой же длины, как гипостомальный склерит, с которым
они соединены.
1 По Порчинскому, цвет яиц кирпично-красный. По всей вероятности, автор
смешивал яйца Haematobia. с яйцами Lyperosia irritans L.
Рис. 445—452. Детали строения преимагинальных фаз Haematobia
stimulans Mg.
445 — ротоглоточный аппарат личинки 1'стадии. 446 — то же II ста-
дии. 447 — то же III стадии. 448 — анальная пластинка личинки III
стадии. 449 — дыхальце куколки. 450 яйцо. 451 — ложнококон.
452 — задние дыхальцы личинки III стадии. (По Томсону).
264
II. ТРИБА STOMOXYDINI
Личинка III стадии амфипнейстична; головной сегмент двураздель-
ный спереди, нижняя его сторона покрыта радиально расходящимися
от ротового отверстия бороздками. Передние дыхальцы с 7 лучами; зад-
ние, расставленные на расстояние, — в Р/г—2 раза превышающие попе-
речный диаметр дыхальца. Каждое дыхальце с 3 короткими, S-образно
изогнутыми щелями; перитрема слабо склеротизована. Снизу по перед-
нему краю брюшных сегментов со 2-го по 7-й располагаются полоски
мелких шипиков. Анальная пластинка короткая, широкая, с дуговидным
задним и 3-лопастным передним краем. Ротовые крючья неравный
по длине: левый крючок много короче правого; тот и другой массивные,
суженные к концу и притупленные на вершине; зубные склериты окру-
гленно-треугольной формы, маленькие. Гипостомальный склерит почти
с прямым дорзальным и закругленным вентральным краями; фарингеаль-
ный — длинный; длина вентрального заднего отростка равна или менее
длины склерита. Дорзальные отростки очень тонкие (в 5—6 раз уже
вентральных), острые; вентральные — широкие, много длиннее дорзаль-
ных отростков, узко округленные на конце. Дорзальная перемычка
не выступает над дорзальным краем склерита, с крупными отверстиями;
передний край дорзальной ее стороны, при рассматривании в профиль,,
угловидно направлен вперед. Передние отростки умеренной длины, рас-
ширенные на конце, где имеются дорзальный и вентральные угловидные-
выступы. Вентральная перемычка узкая. Фарингеальные ребрышки ясно'
выражены, присутствуют в числе 6.
Длина тела около 9 мм.
Пупарий коричневый, длиной 4.5 мм, как у Musca. Дыхальцы в виде-
мелких темных рожек на границе 1-го и 2-го брюшных сегментов, явствен-
ные.
Распространение. Данные по СССР отрывочны; в коллек-
ции ЗИН АН СССР имеется материал из нижеследующих мест: Карель-
ский перешеек (Выборг, Зеленогорск, Ленинградская обл., южный берег
Крыма, Алтай (Онгудай). Вид широко распространен в Зап. Европе от
южной Швеции до Италии; указан для северной Монголии.
Развитие протекает в коровьем помете. Откладка яиц происходит
на совершенно свежий, только что оставленный животным помет. Через
43—48 часов из яиц вылупляется личинка I стадии. Личинки Haema-
tobia — копрофаги, проводящие жизнь в помете рогатого скота. Длитель-
ность личиночных стадий, по Томсону, следующая: I стадия — около
25 часов, II — от 69 до 80 часов, III — около 8 дней. Исключение пред-
ставляют личинки III стадии, остающиеся на зимовку. В стадии куколки
насекомое пребывает около 14 дней, следовательно, полный цикл разви-
тия от момента яйцекладки до вылупления мух следующего поколения
в средние лета заканчивается в 34 дня.
По данным М. и Е. Томсен (1937) предпочитаемые температуры
для личинок равны 15—26° С; их пределы, следовательно, несколько
ниже чем у L. irritans (27—33° С) и S. calcitrans (23—26° С).
Haematobia — полевые мухи, редко встречающиеся в стойлах. Пита-
ние кровью животных наблюдается в течение светлого времени суток,
причем нападению подвергаются преимущественно передняя часть тела
животного (голова, шея, грудь); мухи предпочитают животных темной
окраски (как и Lyperosia), сосредоточиваясь при пятнистой расцветке
животного на темных участках шерсти. За один раз на одну порцию
помета самка откладывает 1—2 яйца, перелетая потом на следующую.
Перед откладкой яиц самка питается навозной жидкостью.
5. HAEMATOBIA
265
Хозяйственное значение H. stimulans в северных широтах, невиди-
мому, невелико.
*2. Haematobia atripalpis Bzz. (рис. 430, 439).
Bezzi, 1895, Bull. Soc. entom., Ital., XXVII : 50; Bezzi u. Stein, 1907, Catal.
pal. Dipt., Ill : 612; Schnabl u. Dziedzicki, 1911 : 126; Malloch, 1932, Ann. Mag.
Nat. Hist., 10, IX : 501 (Jlaematoboscay. Patton, 1933, Ann. trop. Med. Parasitol.,
XXVII : 501—537 (Stomoxys Patt. nec Geoffroy); Seguy, 1935, Diptera : 47 (Haema-
tobosca); Zumpt, 1938, Zeitsch. angew. Entom., XXV : 350; — irritans Haugh, 1900,
Term. Eiizet., XXIII : 250.
$. Ширина лба варьирует в пределах 1/л—х/е ширины глаза; орбиты
в наиболее узком месте лба чуть шире или равны ширине лобной полосы;
скулы в среднем своем отделе равны 3/а ширины 3-го членика усиков,
щеки едва шире скул. Затылок в верхней 1/i вогнутый или плоский,
далее книзу резко и сильно вздутый, ширина его в профиль составляет
чуть более х/а ширины глаза; усики бурые, их 1-й и 2-й членики
на дистальном конце на большем или меньшем протяжении красновато-
желтые, 2-й членик на А'а длины короче 3-го; ариста сверху на всем про-
тяжении в длинных волосках, снизу с 1—3 ресничками. Хоботок длин-
ный, подбородок утолщен в основной половине, щупальцы сплошь черно-
бурые, или с рыжеватой основной половиной; поперечник щупалец
на вершине почти равен диаметру 3-го членика усиков; щупальцы пло-
ские, снаружи в коротких темных щетиночках; голова черно-бурая;
лобная полоса черная, у свежих экземпляров — красновато-желтая;
орбиты, скулы и вибриссальные кили в густом блестящем желтовато-
сером налете, местами с коричневато-желтым отливом; затылок и щеки
в матовом, более темном желтовато-сером или сером налете, лицо в темно-
сером налете. Щетинки головы черные; наружные теменные щетинки
слабо развиты, внутренние теменные короткие, вибриссы несколько
толще лобных щетинок; задняя треть орбит с группой черных коротких
щетинистых волосков; щеки в редких, как и затылок, бурых волосках.
Грудь бурая, в густом серовато-желтом налете, местами с голубовато-
серым тоном. Спинка с продольными буровато-коричневыми полосками,
начинающимися от переднего края спинки. Внутренние полоски узкие,
достигают лишь средины задней части спинки, наружные впереди шва
имеют вид косо расположенных неправильно-треугольных штрихов, за
швом имеют, характер суженных назад клиновидных, более длинных поло-
сок, несколько не достигающих заднего края спинки. От основания щитка
вперед отходит темное узко-треугольное пятно, не всегда хорошо выра-
женное; у темных экземпляров оно заходит на поверхность щитка.
Поверхность груди в бурых, негустых и более длинных на плеврах
волосках. Переднегрудь широкая, трапециевидная, в сером налете и более
или менее густых тонких волосках; проплевры впереди дыхалец с груп-
пой тонких, бурых волосков; гипоплевры с такими же волосками впереди
заднего дыхальца и в передне-верхнем углу. Крылья с бледно-желтовато-
серой мембраной и рыжевато-желтыми жилками; т с мягким дуговидным
изгибом; жужжальца бледножелтые; чешуйки палево-желтые, грудные —
с прямым или слабо вогнутым внутренним краем, суженные к вершине.
Ноги бурые, вершины бедер коричневато-красные или желтые, основа-
ния голеней желтые; коготки немного короче 5-го членика лапки; при-
соски желтрвато-белые. Передние голени (исключая концевые щетинки)
лишены вооружения, как и средние голени; задние голени по внутрен-
ней поверхности с несколькими неправильными рядами щетинистых
266 II. ТР ИВ A STOMOXYDINI
волосков, длина которых почти в 2 раза превышает поперечник голени
(при осмотре с дорзальной или вентральной стороны); переднедорзаль-
ный ряд волосков хорошо выражен; предвершинная щетинка тонкая,
почти прямая; задние бедра перед вершиной по передневентральной
стороне с 1—2 длинными щетинками и с 1 сравнительно крепкой щетин-
кой с вентральной стороны вблизи середины.
Щетинки груди: акростихальные 1—2 пары впереди щитка; дорзо-
центральные — в заднем отделе спинки в числе 3 (4-4) пар, из которых
наибольшего развития достигает лишь задняя пара; интраалярных — 1;
надкрыловых — 1; закрыловых — 2; плечевых — 1—2; заплечевые —
не ясны; предшовная — 1; нотоплевральных — 2; стерноплевральных —
1 4-1; вблизи задней щетинки нередко располагается очень близко от нее
2-я щетинка; мезоплевры в передневерхнем углу со щетинкой, обращен-
ной вершиной вверх. Брюшко широко-овальное, черно-бурое, в густом
серовато-желтом или золотистом налете и черных негустых щетинистых
волосках. Посредине дорзальной поверхности 3-го, 4-го и часто 5-го
тергитов располагаются буровато-коричневые пятна, образующие пре-
рванную у задних краев тергитов продольную полоску; ширина пятен
убывает от 3-го к 5-му тергиту. I стернит в щетинистых волосках. Мем-
брана брюшка с редкими черными короткими волосками, хорошо замет-
ными на задней половине брюшка; 2-й, 3-й и 4-й тергиты эллипсовидные;
5-й широкий, сильно расширен к заднему концу, по заднему краю с неглу-
бокой выемкой, по бокам от которой располагаются 2 тонких сравни-
тельно длинных выступа. Церки щитовидные; дистальный край почти
ровный, иногда со слабо выступающими внутренними углами; наружные
вершинные углы широко закругленные, реже прямоугольные; кокситы
"9-го тергостернита языковидные, суженные к вершине и с узкой, длин-
ной базальной частью, внутренний их край с незначительно выдающимся
выступом. Фаллозома имеет тот же тип строения, что и у прочих видов
подтрибы. Передние части парамер широкие, короткие, задние — длин-
ные, саблевидно согнутые, суженные к вершине.
j. Несколько крупнее самца; ширина лба равна ширине глаза;
лобная полоса равна или немного шире орбиты; последние с группой
щетинистых черных волосков и 2—3 направленными вверх и вбок корот-
кими орбитальными щетинками; ширина скулы составляет около 2/з
ширины 3-го членика усиков; голова в светлом желтовато-сером налете.
Темные продольные полосы выражены несколько слабее, чем у самцов;
пятна посредине брюшка на 4-м и 5-м тергитах иногда не развиты. Длина
тела 4—5.2 мм.
Распространение. Украина (Харьков: Куряж), Крым,
южное Поволжье (Сарепта), окрестности Чкалова (В. Днепровка, Маяч-
ная), Темир, Уральск, обл., Закавказье (Ордубад, Арарат); Туркме-
ния (Копет-даг: Кара-кала, Атрек: Баят-ходжи); Киргизия: Фрунзе;
Казахстан: Алмаатинская и Восточноказахстанская обл. Южная
Европа.
Экземпляр (<J) с Дальнего Востока (Владивосток) несколько отли-
чается от типичных экземпляров более узким лбом, линейной лобной
полосой и деталями в окраске тела.
Точно также незначительные отличия наблюдаются между крым-
скими, кавказскими и киргизскими экземплярами. Различия сказываются
в деталях строения церок, более темной окраске и несколько большей
ширине щупалец, т. е. в тех признаках, которые среди жигалок подвер-
жены особенно сильной изменчивости. Именно поэтому мы не считали
4. LU PE ROS IA
26T
себя в праве и выделять уклоняющиеся экземпляры в самостоятельные
виды или даже подвиды.
4. Род LYPEROSIA R.-D.
Rondani, 1856, Dipt. ital. Prodr., I : 93; Rezzi u. Stein, 1907, Catal. pal. Dipt.,
Ill : 612; Stein, 1916 : 19; Zumpt, 1938, Zeitschr. ang. Entom., XXV : 351; —Styge-
romyia Austen, 1907, Ann. Mag. Nat. Hist. (7), XIX ; 445; — Haphospatha Enderlein,
1924, Konowia, III : 51; — Brucemyia Malloch, 1932, Ann, Mag. Nat. Hist. (10), IX :
43; —Haematobia Malloch (nec Rob.-Desv.), 1932, Ann. Mag. Nat. Hist., 10, IX : 501.
Тип рода Lyperosia irritans Linne (1758).
Глаза голые, сравнительно широкие, слегка вогнутые в нижней
половине их заднего края; хоботок тонкий, с длинным сосальцем, несу-
щим на конце маленькие сосательные лопасти; щупальцы длинные, зна-
чительно расширенные к вершине и сплюснутые с боков; наружная
поверхность их в недлинных щетиночках; верхняя часть затылка плос-
кая, нижняя сильно выпуклая; щеки очень узкие; ариста с волосками
по верхней стороне; передний нижний край лица высоко приподнят над
нижним уровнем глаз. Переднегрудь уже, чем у предыдущего рода, слабо
укороченная, голая; проплевры и гипоплевры лишены волосков; стерно-
плевральные щетинки 1 4-1; акростихальные в числе 1, реже 2 пир
в заднем отделе; дорзоцентральные в числе нескольких пар, развитых
по всей длине среднеспинки, из них только 1 пара перед швом и 1—3 пары
впереди щитка имеют характер щетинок. Жилка т с легким дуговидным
изгибом в вершинном отрезке. Среднеспинка с продольными полосами,
из которых боковые более широкие и впереди поперечного шва имеют
треугольные очертания. Брюшко с вытянутыми пятнами по средней про-
дольной линии. V стернит <3 с длинными боковыми выростами по заднему
краю и ясно развитым склеритом между ними. Церки поперечные, с ясно
оттянутыми внутренними углами. Фаллозома без дистального отростка.
В состав рода входит 14 видов. Четыре из них (включая Haematobia
crassipalpis Ringd.) свойственны Палеарктике, причем один (L. irritans)
является общим с Северной Америкой. Наибольшего расцвета род дости-
гает в Эфиопской области, откуда известно 9 видов. Два из них обнару-
жены также в Ориентальной области; помимо них эта область имеет еще
один эндемичный вид.
До настоящего времени на территории СССР зарегистрировано
лишь 2 вида. Весьма вероятно ожидать в южных частях СССР L. minuta
Bzz., а на северо-западе Советского Союза L. {Haematobia) crassipalpis
Ringd., известного из Швеции.
Мухи кровососущи, тесно связаны с телом хозяина (млекопитающих)
и оставляют его лишь на короткие промежутки времени.
Как кровососы мухи участвуют в распространении ряда патоген-
ных организмов, в частности трипанозом (Рубо, Сеги).
Развитие личинок протекает в помете животных.
ОПРЕДЕЛИТЕЛЬНАЯ ТАБЛИЦА ВИДОВ
1 (4). Анальная жилка толстая, короткая, не утончаясь обрывается
в основной трети крыла; задние лапки самцов обычного строения.
2 (3). Средние голени с маленькой передневентральной щетинкой вблизи
середины; мезоплевры в передневерхнем углу с направленной вверх
щетинкой. Щупальцы резко расширены к вершине, относительно
2(58
II. ТРИБА STOMOXYDINI
короткие: длина их чуть более Vz длины сосальца обычно в вершин-
ной половине или сплошь коричневые ... *1. L. titillans Bzz.
3 (2). Средние голени на передневентральной поверхности и мезоплевры
в передне-верхнем углу без щетинок; щупальцы постепенно утол-
Рис, 453—458.
453 — голова самки Acridomyia sacharovi Stack, сбоку. 454 — голова
самцаLyperosia titillans Bzz. сбоку. 455 — то Же Lyperosia equina End.
(L. titillans Bzz.). 456 — V и VI стерниты самца Lyperosia irritans L.
457 — задняя лапка самца L. titillans Bzz. сбоку. 458 —= задняя лапка
самца L. irritans L. (рис. 455 по Пеусу, прочие — ориг.).
щаются к вершине; длина их составляет почти 3/а длины сосальца;
тело светлоокрашенное, желтое................2. L. minuta Bzz.
4 (1). Анальная жилка длинная, постепенно утончаясь к концу, пере-
ходит в складку, вершина которой близко (на Vs расстояния от осно-
вания анальной жилки до края крыла) подходит к задйему краю
крыла. Задние лапки самцов (3 первых членика) сверху с длинными
волосками, сплюснутые с боков и расширенные ......................
*3. L. irritans L.
4. LU PEROS I A
289
*1. Lyperosia titillans Bzz. (рис. 454, 455, 457, 459—464).
Bezzi, 1907, Rend. 1st. Lombard, (2), XL : 454; Stein, 1916 ; 19; Karl, 1930, Zool.
Anz., 86 : 162 (Haphospatha)', Malloch, 1932, Ann. Mag. Nat. Hist., (9), X : 511, fig. 26
{Haematobia}', Patton, 1933, Ann. trop. Med. Parasitol., XXVII ; 501—537 (Stomoxys)',
Seguy, 1935, Diptera VIII; Zumpt, 1938, Zeitschr. angew. Ent., XXV : 347; —
irritans Rondani (nec Linne), 1862, Prodr. : 231; — equina Enderlein, 1929,
Zeitschr. angew. Ent., XIV : 364; Pens, 1937, Zeitschr. angew. Entom., XXIV :
152; — bovina Pens, 1937 : 152; Zumpt, 1938, Zeitschr. angew. Entom. : 352; — scolopax
Peus, 1937 : 152; Zumpt, 1938 ; 352; — latirostris Enderlein, 1929, Zeitschr. angew.
Ent., XIV : 364.
Биология и фазы развития. Зимин, 1948 : фиг. 38, 71.
Ширина лба составляет от 'А до 3/< ширины 3-го членика усиков;
лобная полоса бурая, реже коричнево-красная в переднем отделе, ширина
ее посредине менее ширины орбит, реже равна ей; скулы очень узкие,
равные по ширине орбите в своем среднем отделе; высота щеки составляет
не более 2/з ширины 3-го членика усиков; нижний край лица высоко
приподнят над уровнем нижнего края глаз, едва выступающий над перед-
ней линией скулы; задняя линия глаза слабо вогнута в нижней поло-
вине; усики бурые, 1-й и 2-й их членики красновато-желтые, реже свет-
лая окраска сохраняется только по вершинному краю и на внутренней
стороне 2-го членика; ариста утолщенная в основной трети, с волосками
по верхней стороне; щупальцы длинные, плоские, расширенные в вер-
шинной половине, где поперечник их составляет 2/з ширины 3-го членика
усиков, буровато-коричневые, у светлых экземпляров рыжевато-желтые;
наружная поверхность в мелких светлых, частью темных щетинках;
длина щетинок, расположенных по нижнему краю, равна, чуть меньше
или несколько превышает диаметр щупалец; хоботок с тонким, длинным
сосальцем, утолщенным у основания, сосательные лопасти узкие; хобо-
ток коричневый или желтовато-коричневый. Щетинки головы буроватые,
частью желтые; наружные и внутренние теменные короткие, но ясные.
Голова черно-бурая, орбиты и скулы в светлом серовато-белом налете,
внизу скул более светлом; затылок в матовом светлосером налете. Грудь
бурая, плевры местами с рыжевато-коричневыми участками; налет,
покрывающий грудь, желтовато-серый; спинка с 4 плохо выраженными
продольными полосами, узкими и не всегда отчетливо различными
внутренними короткими и широкими боковыми, несколько не достигаю-
щими переднего края щитка; переднегрудь уже, чем у Stomoxys и Hae-
matobia, слабо расширенная впереди или с параллельными краями,
лишена волосков, как передние плевры и гипоплевры. Щетинки груди:
акростихальных — 1 пара, дорзоцентральных — 1—2 пары впереди
щитка; интраалярных — 1; надкрыловых — 1; закрыловых — 2; пле-
чевых — 3, заплечевая — неясная, предшовных — 1; нотоплевральных —
2; стерноплевральных— 1 4-1; мезоплевры со щетинкой в передне-
верхнем углу и с рядом щетинок вдоль заднего края. Помимо перечислен-
ных щетинок впереди акростихальных и дорзоцентральных щетинок
иногда имеются впереди поперечного шва и позади него несколько мел-
ких щетинок, слабо отличающихся по длине от щетинистых волосков
спинки. Щиток с длинными вершинными и основными короткими, воло-
совидными предосновными щетинками. Щетинки на спинке бурые, реже
с желтоватой вершиной, на плеврах и щитке чаще светлые. Крылья
с бледно желтовато-серой мембраной и рыжевато-желтыми жилками;
:т очень слабо изогнута; чешуйки с палевым краем, грудные чешуйки
длинные, языковидные, с вогнутым внутренним краем у его основания.
Жужжальца рыжеватые, иногда с легким затемнением на вершине. Ноги
270
II. ТРИВА STOMOXYDINI
короткие, рыжевато-бурые, бедра нередко бурые, вершины бедер и осно-
вания голеней всегда желтые или рыжевато-желтые, у светлых особей
передние голени, а иногда и средние — сплошь желтые. Средние голени
с маленькой, но явственной переднедорзальной щетинкой у середины.
Коготки черные в вершинных 3/«, сильно изогнутые, не короче 5-го чле-
ника лапки; присоски желтовато-белые; членики задних лапок цилин-
дрические, без длинных щетинистых волосков на верхней и внутренней
поверхности. Брюшко рыжевато-бурое или бурое, в густом желтовато-
сером налете, с широким округлым, квадратным или широко-треуголь-
ным пятном в передней V2 или 2/з посредине 3-го тергита и более узким
буроватым пятном в переднем отделе 4-го тергита; иногда затемнение
наблюдается и при основании 5-го тергита; поверхность брюшка в сравни-
тельно длинных желтых (или с затемненным основанием) волосках
и краевых щетинках. II, III и IV стерниты широко овальные, V—широкий,
с 2 тонкими длинными выростами по бокам заднего края, I I стернит-
в виде ясной хитиновой поперечной полоски, расположенной между
внутренними краями боковых отростков V стернита. Церки в форме
щитка, с глубоким треугольным вырезом сверху; внутренние углы их
слабо выступают над дистальным краем церок, боковые углы (наруж-
ные) неправильно закругленные. Фаллозома сохраняет тип строения,
свойственный представителям подтрибы.
Самка несколько крупнее самца. Ширина лба равна 3Д—2/з ширины
глаза; черная лобная полоса слабо сужена в среднем отделе, равна
ширине (или уже) светлых желтовато-серых орбит; щетинки головы пре-
имущественно желтые с бурыми основаниями; орбитальных щетинок —2;
они далеко отставлены друг от друга, длинные; орбиты с одиночными
короткими волосками в заднем отделе; реже они отсутствуют. Окраска
щупалец, бедер и голеней обычно светлее, чем у самцов. Пятна посре-
дине 3-го и 4-го тергитов уже, нежели у самцов.
Длина тела 2.3—3.7 мм.
Личинки III стадии мелкие, желтовато-белые, обычно с затемнен-
ными участками в области кишечника. Кутикула тонкая, блестящая,
неправильно редко-морщинистая. Голова спереди с коническими боко-
выми выростами, почему передний отдел при осмотре сверху представ-
ляется расщепленным вдоль на две части. Грудные сегменты без шипов
по переднему краю, равно как и 1-й брюшной сегмент. Брюшные сег-
менты со 2-го по 7-й снизу по переднему краю с валикообразными низ-
кими возвышениями, покрытыми косыми ребрышками, составляющими
передний ряд, который у середины прерван, и бугорками позади них,
расположенными в 1—2 изогнутых ряда. 8-й брюшной сегмент округлый,
короче 7-го, задняя поверхность вокруг дыхалец слабо вдавлена, с кон-
центрически расходящимися от дыхалец мелкими морщинками, придаю-
щими поверхности некоторую матовость. Дорзальная поверхность сег-
мента с 3 продольными бороздками. Анальная пластинка относительно
широкая; в области анального отверстия 8 бугорков; 2 крупных, округ-
лых анальных, 2 слабо выступающих преанальных, 2 параанальных,
лежащих на концах анальной пластинки, 2 маленьких сосцевидных суб-
анальных. Параанальные бугорки выступают сильнее, чем у L. irritans,
благодаря чему образуется как бы сглаженный угол, отчетливо видимый
при осмотре объекта сзади. Вокруг анальной пластинки расположены
длинные, прижатые, притупленные зубцы, часть которых заменена
ребрышками. Позади анальной пластинки зубцы располагаются в один
ряд. Передние дыхальцы короткие, маленькие, с 4—5 сферическими
4. LYPEROSIA
271
выростами по краю. Задние дыхальцы маленькие, расставлены на Чъ
поперечника дыхальца, округленно треугольные, с узкой перитремой.
Дыхальцевые щели лежат на светлых участках стигмальной пластинки,,
сильно S-образно изогнутые, короткие; стигмальный диск очень велик,.
Рис. 459—464. Детали строения личинки III стадии Lyperosia titillans Bzz.
459 — ротоглоточный аппарат. 460 — задний конец тела сзади. 461 — он же сбоку.
462 — переднее дыхальце. 463 — заднее дыхальце. 464 — вооружение снизу
5-го (2) и 6-го (1) брюшных сегментов. (По Зимину).
темный. Ротоглоточный аппарат с очень крупными фарингеальными
склеритами. Ротовые крючья маленькие, с сагиттально вытянутой
базальной частью и сильно выдающимся остроугольным выростом по вен-
тральному краю. Зубные и лабиальные склериты крупные, первые длин-
ные, вторые — треугольные, с оттянутыми назад задними углами, вогну-
тые по верхней и выпуклые по нижней границе. Передние отростки фарин-
.272
II. ТРИБА STOMOXYDINI
геальных склеритов длинные, с торчащими углами; вентральные отростки
широко ланцетовидные, слабо склеротизованные в середине, в 2 раза шире
дорзальных и примерно на К? длиннее их; дорзальные отростки тонкие,
заостренные на вершине. Дорзальная перемычка большая, крупнорешет-
чатая, далеко заходит вперед; фарингеальная мембрана очень широкая;
вентральная перемычка склеротизованная, далеко выступающая. Длина
тела 5—6.5 мм, толщина у середины 0.5—0.6 мм.
Распространение. В Европейской части СССР известен
с юга Украины и из Крыма. Местами распространен в Закавказье.
Повсеместно распространен в Средней Азии и южном Казахстане.
По данным Пеуса, распространен в Латвии. Вне СССР известен из юж-
ной Европы и Монголии.
В окраске тела, как и Lyperosia irritans, крайне непостоянен, что
отмечается у особей даже из одного и того же места, пойманных одно-
временно. Особенно сильно варьируют окраска ног, щупалец, и даже
такие мало изменчивые признаки, как ширина лба, лобной полосы и 3-го
членика усиков.
L- minuta Bzz. отличается от L. titillans Bzz. по отсутствию перед-
невентрапьной щетинки на средних голенях и передне-верхней щетинки
на мезоплеврах. По указанным признакам L. minuta должен быть сбли-
жен с L. irritans, от которого без труда отличается по отсутствию у <5<5
длинных волосков на задних лапках и нерасширенными члениками
последних, а также короткой ап.
Биология вида не изучалась. Как кровосос, весьма много-
численный в южных районах Казахстана и Средней Азии, вид должен
иметь существенное отрицательное значение в животноводстве, в част-
ности, как механический переносчик патогенных микроорганизмов среди
животных.
Вид этот способен нападать не только на животных, но и на человека,
причем известны случаи укусов в ночное время в освещенных помеще-
ниях. Развитие личинок протекает в коровьем помете.
2. Lyperosia minuta Bzz.
Bezzi, 1892, Ann. Mus. Civ. Genova, XXXII : 192; Stein (apud Becker), 1912,
Ежегодник Зоол. муз. Акад. Наук, XVII : 630; Austen, 1921, Bull, entom. Res.,
XII : 121; Malloch, 1932, Ann. Mag. Nat. Hist. (9), X : 510 (Haematobosca); Patton,
1933, Ann. trop. Med. Parasitol., XXVII : 501—537; Zumpt, 1938, Zeitschr. angew.
Entom., XXV : 346, 352; — longipalpis Roubaud, 1906 : 896; — potrix Enderlein,
1929 : 362; Zumpt, 1938, цит. раб. ; 346, 352.
Биология. Gibbins, 1937, Not. Breed. Hab. House-frequ. Fl. Uganda; du
Toit a. Smith, 1939, Horn fly probl. South Afr. : 2.
Лоб в сероватом налете, лобная полоса черная, суженная у середины;
лобные щетинки желтые, того же цвета тонкие волоски на щеках;
щупальцы длиною с хоботок, у основания светлые, реже сплошь свет-
лые, в редких коротких волосках; лоб самца в 2 раза уже, чем у самки.
Затылок в сером налете, как и антенны; 2-й членик усиков красноватый;
ариста с редкими волосками по верхней стороне. Грудь, включая плевры
и щиток, в сером налете; поверхность груди в слабых желтых щетинках,
и волосках; чешуйки сероватые, по краю узко-белые, жужжальца белые.
Брюшко одноцветно серое, в коротких желтых волосках, у самки более
широкое, снизу бледножелтоватое. Ноги преобладающей желтой окраски,
в желтых волосках; задние бедра редко затемнены. Крылья желтова-
тые со светлыми жилками; жилка т слабо изогнута на вершине.
Длина тела 5 1.8—2.3 мм, д — 3—3.8 мм.
4. LYPEROSIA
273
Распространение. Бецци описал вид из восточной Африки
?(Овво). Под именем L. (filossinella) schillings вид оттуда же указывается
Грюнбергом. Аустен приводит L. minuta из Палестины, Штейн — из пер-
сидского Белуджистана. Вид распространен в южной Африке и ио всему
африканскому побережью от юго-западного до северо-восточного его
отделов. Расселение вида вдоль побережья вследствие усиленной транс-
портизации животных произошло очень быстро (около 10 лет). В пре-
делах Союза ССР вид пока не встречен, но весьма вероятен вдоль южной
границы Средней Азии.
Биология вида не изучена. Личинки развиваются в помете
коров. Взрослые кровососущи, нападают на коров и овец. Отмечены
частые случаи нападения мухи и на человека, причем зарегистрированы
укусы мух и в ночное время.
В южной Африке является вредным в животноводстве видом; вред
заключается в снижении удоя молока у коров, которые подвергаются
нападению мух; кроме того, животные теряют в весе.
*3. Lyperosia irritans Linne (рис. 429, 433, 436, 437, 456, 453, 465,
466—470).
Linne, 1758, Syst. Naturae, 10 : 604 (Conops); Pandelle, 1898, Rev. Entom.,
XVII : 43 (Stomoxys); Malloch, 1932, Ann. Mag. Nat. Hist. (9) X, 504; Seguy, 1935,
Diptera, VIII; Peus, 1937, Zeitschr. angew. Entom., XXIV : 150—154; Zumpt, 1938,
Zeitschr. angew. Entom., XXV : 352; — meridionalis Bezzi, 1911, Archiv Parasitol.,
XV : 131; — weissi Bezzi, 1911 : 131.
Биология. Порчинский, 1910:4—11; Melvin, 1934, Ann. Entom. Soc.
Amer., XXVII : 406—410; M. Thomsen a. E. Thomsen, 1937, Zeitschr. vergl. Physiol.,
XXIV, n° 3 : 343—380; Meyers, 1938, Journ. Conn. Sci. industr. Res. Aust., XI : 35—
46; Kuwayama a. Kato, 1939, Oyo Kontyu, I : 151—159; Романов, 1940, Мед. пара-
зит. и паразит, болезни, IX 4 : 353—363; Зимин, 1944 : 159; Зимин, 1948 : 72,
•фиг. 39.
3. Лоб немного уже 3-го членика усиков или равен ему по ширине;
лобная полоса черная или красновато-коричневая, уже, равна или
несколько шире орбиты; скулы на уровне 1-го членика усиков составляют
3/и ширины 3-го членика усиков, далее книзу постепенно сужены; высота
щеки равна наибольшей ширине скулы; передне-верхний край рта высту-
пает за переднюю линию скулы маленьким уголком. Усики короткие.
3-й членик едва длиннее 2-го, сплошь или только в вершинной ]/г бурый;
1-й и 2-й членики усиков красновато-желтые, иногда более или менее
затемненные; ариста утолщена в основной трети с длинными волосками
но верхней стороне; хоботок как у L. titillans', щупальцы длиннее, чем
у названного вида, плоские, расширяются к вершине от основной чет-
верти, а не от средины, как у L. titillans, бурые или рыжевато-бурые,
реже рыжие; наибольшая их ширина равна ®/< ширины 3-го членика уси-
ков; наружная поверхность щупалец в коротких черных и светлых щетин-
ках. Голова черно-бурая, в густом налете на орбитах и скулах; окраска
налета изменяется от голубовато или серовато-белого до.желтовато-серого;
затылок в матовом желтовато-сером налете, сильнее развитом на выпук-
лой части затылка. Щетинки, сравнительно с L. titillans, более длинные,
тонкие, желтые, рыжие или буроватые; внутренние и наружные теменные
развиты; щеки в длинных редких светлых волосках. Грудь бурая,
на плеврах часто с неясными рыжеватыми отметинами, покрыта на спинке
коричневато-желто-серым, а на плеврах, плечах и нотоплевре желтовато-
серым налетом. Спинка с двумя неявственно выраженными короткими,
узкими срединными и длинными, широкими боковыми коричнево-бурыми
18 Фауна СССР, том XVIII, вып. 4.
274
II. ТРИБА STOMOXYDINI
продольными полосами. Спинка в коротких черных жестких, плевры
в желтых более тонких волосках, иногда с примесью бурых. Передне-
грудь как у L. titillans, но несколько укороченная, лишена волосков, как
проплевры и гипоплевры. Щетинки груди на спинке черные, на плеврах
желтые; акростихальных — 2—3 пары впереди щитка, из которых 1—2
достигают большей длины и толщины, дорзоцентральных — 3 -f-3(4);
длина щетинок убывает спереди назад; интраалярных — 2, расположены
в один ряд; надкрыловых — 1; закрыловых — 2; плечевых — 3 (4);
заплечевых — 1; предшовных — 1; стерноплевральных — 1 + 1; ното-
Рис. 465. Самец Lyperosia irritans L. (Ориг.).
плевральных — 2; в передне-верхнем углу мезоплевры щетинка отсут-
ствует; на щитке развиты вершинные, боковые, основные и укороченные
предосновные щетинки. Крылья от бледно рыжевато-серых до темных —
дымчатых; жилки рыжевато-бурые или рыжие; т с легким дуговидным
изгибом; чешуйки шире, чем у L. titillans, с прямыми или слабо выпуклым
внутренним краем; окраска их варьирует от белых с палевым краем
до дымчатых с бурым краем и бурой бахромкой. Жужжальца желтые,
у темных экземпляров с буроватой вершиной. Ноги обычно с бурыми
или коричневыми бедрами и лапками. Расцветка их варьирует от почти
черного до рыжеватого тона; в последнем случае основание или задняя
сторона передних и вершинные отделы средних и задних бедер остаются
на большем или меньшем протяжении затемненными. Голени всегда
4. LYPEROSIA
275
с желтым основанием, более широким на передней паре ног; у светлых
особей передние, а иногда средние и задние голени могут быть рыжевато-
желтыми. Средние голени без передне-вентральной щетинки у средины.
Задние голени с длинной тонкой предвершинной щетинкой; нижняя
поверхность задних бедер при основании с толстой щетинкой, длина кото-
рой едва превышает поперечник бедра в этом месте. Первые 3 членика
задних лапок сплюснуты с боков, вытянуто-треугольные с длинными
волосками в дистальных отделах и с закрученными вниз волосками
на внутренней стороне. Брюшко удлиненно-яйцевидное, бурое, в густом
желтовато-коричневом налете, длинных волосках и с маргинальными
щетинками; 3-й и 4-й тергиты с удлиненными срединными пятнами.
V стернит сильно расширен назад, с 2 очень длинными узкими боковыми
выростами и хитиновой поперечной перемычкой (VI стернит) между ними.
Церки с пальцевидно оттянутыми вниз и слабо изогнутыми вбок внутрен-
ними углами, наружные углы слабо оттянуты вниз, благодаря чему ниж-
ний (вершинный) край церок с глубокими выемками по бокам от средины.
Фаллозома сохраняет тип строения, свойственный всей подтрибе Stomoxy-
dina.
Ширина лба изменяется в пределах 2/з—3/4 ширины глаза; лоб-
ная полоса равна орбите (или слабо изменяется в том и другом направле-
нии). Щупальцы расширены от основной 1/i до дистального конца. Ноги
более светлой окраски чем у <5; бедра средних и задних ног чаще рыже-
вато-желтые, с затемненной вершиной; передние бедра с буроватой или
рыже-бурой задней поверхностью.
Длина тела 2.7—4.5 мм.
Яйцо удлиненно цилиндрическое, коричневато-оранжевое, по дор-
зальной стороне с 2 слабо расходящимися к нижнему полюсу бороздками,
расстояние между которыми равно (в средине) 1/з ширины яйца; хорион
в мелких шестигранных ячейках; длина около 1 мм.
Личинки I и II стадии с достоверностью не известны. Личинка III
стадии имеет передние дыхальцы с 6—7 лучами. Тело вытянутое, сильно
суженное в передней части; задний конец в профиль полукруглый; перед-
ние края брюшных сегментов с 4-го по 7-й с нижней стороны с валико-
образными поперечными утолщениями, снабженными косыми ребрыш-
ками, часть которых заострена и имеет характер шипиков. Тело желтое,
с просвечивающим оливково-бурым кишечником. Задний конец тела снизу
посредине с 2 крупными и несколькими мелкими бугорками. Задние
дыхальцы расположены вблизи верхнего края сегмента, расставлены
на небольшое расстояние, равное 1/з—1/5 ширины дыхальца; последнее
имеет 3 тонких извитых щели. Верхний конец верхней, аналогичный
конец средней и наружный конец нижней щели обращены к перитреме,
прочие — к центру дыхальцевой пластинки. Направление концов щелей
сильно варьирует. Ротоглоточный аппарат сходен с таковым Haematobia,
отличаясь в два раза более широкими дорзальными и короткими перед-
ними отростками фарингеального склерита. Последние со слабо оттяну-
тыми короткими верхними дистальными углами. Левый ротовой крючок,
как у всех видов подтрибы Stomoxydina, значительно короче правого.
Длина тела личинки 8—9 мм.
Пупарий коричнево-бурый.
Распространение. Европейская часть СССР на север
до Карельского перешейка; Закавказье; повсеместно в Средней Азии
и в Казахстане на север до северной границы полынно-злаковых степей.
Данные по Сибири отсутствуют. Вся Зап. Европа от Скандинавии и
<8*
276
И. ТРИБА STOMOXYDINI
южной Финляндии до средиземноморского побережья; северная Африка;
Северная Америка, острова Гавайские, Гаити.
Экземпляр самца с Дальнего Востока отличается от прочих отсут-
ствием темного рисунка на спинке и брюшке. Генитальный аппарат
не дает надежных отличий.
Личинки L. irritans развиваются в коровьем помете. Самки на корот-
кое время (х/2—1 мин.) оставляют животное, отлетая для откладки яиц.
Рис. 466—470. Детали строения личинки III стадии Lyperosia irritans L.
466 — ротоглоточный аппарат. 467 — задний конец тела сзади. 468 —
он же сбоку. 469 — переднее дыхальце. 470 — заднее дыхальце. (По
Зимину).
Помещаются яйца в только что оставленный коровий помет на его поверх-
ности. Акт яйцекладки протекает быстро, после чего самки спешат вер-
нуться на тело хозяина. Размножение мух идет быстро; количество
их на животных сильно возрастает к осени, достигая 1000 экземпляров
на 1 животное. Своими уколами и высасыванием крови мухи сильно доса-
ждают скоту. Животные теряют в весе, удой молока понижается. Как
и многие другие виды этой группы, L. irritans предпочитает темные масти
животных светлым. Вместе с тем мухи предпочитают места с тонкой кожей:
вокруг рогов, на ушах, морде, животе. По свидетельству Порчинского,
процесс яйцекладки проходит в наиболее жаркое время дня. Спустя
5. ACRIDOMYIA
277
сутки после откладки яиц из них вылупляются личинки, проводящие
всю свою жизнь в помете животного. Термический преферендум личинок
лежит (Thomsen, 1938) между 27.0—33° С, т. е. значительно выше, чем
у Haematobia stimulans, и несколько выше, чем у Stomoxys calcitrans.
Достигнув предельной зрелости, личинка уходит для окукления в землю
или окукляется в нижнем слое лепешки коровьего помета, опускаясь
на глубину 1—2 см.
Использование паразитов в борьбе с Lyperosia, в частности ввоз халь-
цида Spalangia в Австралию, хищных жуков Sphaeridium scarabaeoides L.
на Гавайские и Антильские о-ва из Германии и США (Техаса), не оправ-
дало возлагавшихся на них надежд. Повидимому, лучшие результаты
в борьбе с Lyperosia дает деятельность хищных личинок жуков Histeridae,
хищных личинок мух Limnophora, а также навозных жуков Aphodius-,
последние, разрушая навоз, косвенным образом препятствуют размноже-
нию в нем личинок вредных мух, в том числе и личинок Lyperosia. Рядом
исследователей отмечается уничтожение личинок муравьями родов Phei-
dole, Cataglyphis, Camponotus и других.
2. Подтриба ACRIDOMYINA
Задний край глаз выпуклый; жилка т прямая на всем протяжении;
затылок за глазами сильно вздутый; щупальцы отсутствуют. Фаллозома
с 2 острыми дистальными выступами, без поперечной лопасти. Грудная
чешуйка крайне мала.
Паразиты саранчевых.
5. Род ACRIDOMYIA STACK.
Штакельберг, 1929, Изв. прикл. энтомолог., IV, 1 : 121—123.
Тип рода: Acridomyia sacharovi Stack. (1929).
Голова удлиненная, затылок плоский в верхнем и сильно выпуклый
в нижнем отделе; глаза голые широкие с выпуклым передним и задним
краями; скулы и щеки узкие, последние равны 1/s—1/у вертикального
диаметра глаза; хоботок сильно склеротизован, у. $ с крепкими престо-
мальными зубцами; сосательные лопасти маленькие; верхний передний
край рта высоко приподнят; вибриссальные щетинки отсутствуют; орби-
тальных щетинок у j имеется по 1 с каждой стороны, у <3 они отсут-
ствуют; усики очень короткие, 3-й членик почти равен 2-му; ариста
в микроскопических волосках, сильно утолщенная в основной трети;
щетинки головы короткие. Чешуйки крыла маленькие, грудная — в виде
полоски; заднее грудное дыхальце очень маленькое, округлое; передне-
грудь очень широкая в переднем отделе, с группой светлых волосков
по краю; передние плевры в центре и супрасквамальное ребро лишены
волосков; стерноплевральных щетинок — 1 (2) пли они отсутствуют;
крылья узкие; г4+5 и п лишены щетинок, шип на костальной жилке
отсутствует; предкрыловая щетинка груди, боковые и основные щетинки
щитка отсутствуют. Ноги со слабо развитыми щетинками, средние голени
ниже середины с 1 передневентральной щетинкой. Брюшко удлиненно
яйцевидное, у g длиннее, чем у <3, заостренное на вершине; I стернит
удлиненный без волосков.; V стернит <5 без отростков по заднему краю;
церки короткие; кокситы длинные с зубном по внутренней стороне у вер-
шины. Фаллозома с двумя выступающими на вершине острыми склеро-
278
II. ТРИБА STOMOXYDINI
тизованными отростками. Основной отдел фаллозомы тонкий, длинный,
по дорзальной стороне склеротизованный. Дорзальный отросток фал
лозомы длинный ± тонкий; аподема вытянутая, изогнутая, утолщенная
на вершине. Передняя часть парамер широкая, короткая, задняя — тон-
кая, длинная. У обоих полов мембрана на вентральной стороне брюшка
широко разделяет стерниты от тергитов.
К этому роду относится A. sacharovi, паразитирующая в личинках
и взрослых особях азиатской саранчи (Locusta migratoria L.). До сих пор
вид известен только из пределов СССР. Из Северной Америки описано
еще 2 вида этого рода.
Род является несомненным представителем трибы Stomoxydini,
составляя в ней самостоятельную подтрибу. Принадлежность Acridomyia
к Stomoxydini определяется на основании следующих морфологических
черт. Треугольно сильно расширенная переднегрудь; строение головы:
вздутый в нижних 2/з затылок, узкие скулы, высоко приподнятый ниж-
ний край лица. Существенными признаками, свойственными всем Stomoxy-
dini, являются строение заднегрудного дыхальца и сильное развитие
мембраны брюшка. В устройстве генитального аппарата самцов наблю-
дается известное своеобразие, однако и по этим признакам с видами
трибы Stomoxydini наблюдается большее сходство, нежели с представите-
лями других триб Muscini. В частности склеротизованные выросты фал-
лозомы, направленные у Acridomyia дистально, а не латерально, как
это имеет место у прочих Stomoxydini, являются весьма характерными
для трибы. Полосовидная лопасть фаллозомы располагается у жигалок
поперечно, тогда как у Acridomyia она несколько сдвинута вентрально,
и развита значительно слабее. Чтобы представить вышеуказанное сход-
ство, достаточно вывести зубцы фаллозомы жигалок из бокового поло-
жения в сагиттальное, расположив их рядом на вершине penis. Тогда
и поперечная лопасть при осмотре фаллозомы в профиль займет такое же
положение, как у Acridomyia.
Отличительной особенностью Acridomyia от жигалок, на основании
которой мы считаем целесообразным выделить род в самостоятельную
подтрибу, служит строение глаз, не имеющих у Acridomyia вогнутости
по заднему краю, наличие длинного дорзального отростка фаллозомы,
слабо развитого и направленного у'жигалок сагиттально, и прямая меди-
альная жилка.
На близость рода к трибе Stomoxydini указал Штакельберг (1929),
исходивший из данных сравнительного анализа морфологии головы
(лицевой шов, затылок и др.). К тому же выводу на основании изучения
хоботка приходит и Рукавишников (1930), сближающий Acridomyia
с родом Haematobia.
*1. Acridomyia sacharovi Stack, (рис. 416, 443, 453, 471, 472).
Штакельберг, 1929, Изв. прикл. энтомолог., IV, 1 : 123.
Биология. Сахаров, 1913, Отчет о деят. Энтом. стан, за 1912 г., Астрахань;
Предтеченский, 1928, Зап. Астрахан. станции защ. раст.; Олсуфьев, 1929, Изв. Отд.
прикл. энтомолог., IV : 90—94; Рукавишников, 1930, Тр. защ. раст., сер. энтомо-
лог., 1, 1 : 205—225, фиг. : 5—23, табл. фиг. : 1, 2.
$>. Голова удлиненная, нижний край лица выступает вперед
несколько далее лба, закругленный или угловидный (в профиль); глаза
округлых очертаний, широкие; лоб с ровными краями, ширина его равна
ширине глаза; лобная полоса параллельнокрайняя, или слабо вогнутая
в среднем отделе, темножелтая, красноватая в задней половине; орбита
5. ACRIDOMYIA
279
почти в 2 раза уже лобной полосы; скулы уже 3-го членика усиков; щеки
в 5i/2—7 раз уже вертикального диаметра глаза; затылок в верхнем отделе
плоский, далее книзу выпуклый, боковые его нижние отделы сильно
вздуты. Нижний край лица (fronto—clypeus) приподнят и расположен
почти на уровне средней горизонтальной линии глаза. Усики короткие,
3-й членик буро-коричневый, с красноватым основанием, ширина его
несколько превосходит его длину или равна ей; 2-й членик усиков едва
короче 3-го, как и 1-й члений темножелтый; ариста красно-желтая, с тем-
ным концом, утолщена в основной трети, далее резко утончена, в мелких
Рис. 471. Самец Acridomyia sacharovi Stack. (Ориг.).
волосках, заметных лишь при очень сильном увеличении. Хоботок красно-
желтый, с длинным подбородком. Голова бледножелтая, верхняя часть
затылка и глазковый треугольник буроватые. Скулы в белом блестящем,
щеки в бледно желтовато-белом, скулы и затылок в матовом желтовато-
белом налете, несколько затемненном на затылке. Щетинки головы жел-
тые; все слабо развиты, имеют характер коротеньких щетинистых волос-
ков; орбитальных щетинок по 1 с каждой стороны, лобных — 4—5;
вибриссы отсутствуют. Грудь светло красновато-желтая; спинка, верхняя
сторона щитка и задняя треть плевральных отделов — бурые; налет
густой, светло серовато-белый на плеврах и золотисто-желтый в среднем
отделе спинки. Переднегрудь очень сильно расширена впереди, с округ-
ленными боковыми отделами в этом отделе. Заднее дыхальце очень
280
II. ТРИБА STOMOXYDINI
маленькое, округлое, диаметр его в 4 раза короче вертикального диа-
метра гипоплевры. Переднее дыхальце бледно палево-белое; передние
плевры широкие, голые; переднегрудь с несколькими светлыми волос-
ками. Щетинки груди: акростихальных щетинок в переднем отделе спинки
1 — реже 2 (асимметрично расположенные) пары; дорзоцентральных —
1 4- 4 (3); интраалярных позади шва — 2 (передняя приближена к попе-
речному шву); плечевых — 2; заплечевых — 2; предшовных — 1; над-
крыловых— 1; закрыловых'—2; предкрыловая — отсутствует; стерно-
плевральных — 0 -|- 1 (2) или 0; мезоплевральных — 2—4; птероплевра
и гипоплевра — голые; на щитке: вершинных щетинки — 2; основ-
ных — 2; поверхность груди в очень редких светлых волосках. Крылья
узкие, с бледно желтовато-серой мембраной и бледножелтыми жилками;,
чешуйки белые, маленькие; жужжальца желтые, с желтовато-белой вер-
шиной, короткие. Ноги желтые, в коротких черных щетиночках; средние
голени с передневентральной щетинкой ниже средины. Коготки коричне-
вые с бурой вершиной, присоски белые, короче 5-го членика лапки.
Брюшко удлиненное, буроватое, стерниты красновато-желтые, первый
из них широкий палево-белый, без волосков, с выемкой у середины зад-
него края. Поверхность брюшка в желтовато-сером светлом налете и вытя-
нуто-треугольными коричневыми пятнами по средней линии каждого
тергита, начинающимися от переднего края последних и прекращаю-
щихся в задней их трети; по бокам тергитов у переднего их края распо-
лагаются полукруглые поперечно вытянутые пятна коричневого цвета.
Брюшко в редких желтых и частью (на дистальном конце брюшка}
черных волосках.
Яйцеклад состоит из двух ясно различимых склеротизированных
сегментов.
S. Отличается узким лбом, ширина которого составляет около 1/?-
части ширины головы. Щеки уже, хоботок тоньше, лишен престомаль-
ных зубцов; орбитальные щетинки отсутствуют. V стернит на заднем'
конце двураздельный, с глубокой срединной выемкой, по заднему краю
без выростов. Церки короткие, на дистальном конце закругленные,
с неглубокой выемкой посредине; кокситы удлиненно треугольные, слабо-
изогнутые, вытянутые пальцевидно, дистальные отделы их снабжены
по внутренней стороне изогнутым коротким зубцом близ вершины. Фал-
лозома длинная; ее вершинная часть широкая, выпуклая по дорзальной
и вентральной сторонам; по дорзальной стороне проходят две склероти-
зованные полоски, образующие на дистальном конце фаллозомы два за-
остренных выступа; срединные боковые отделы широко склеротизованы,
вентральный вершинный отдел выступает в виде лоскута; основной отдел
фаллозомы тонкий, почти равен по длине дистальной части, с узкой скле-
ротизованной полоской по дорзальной стороне; дорзальный выступ длин-
ный, тонкий, слабо загнут кверху; задний отдел парамер расширенный
при основании, на вершине закруглен; передняя часть парамер широкая,
тесно срощена с IX тергостернитом. Аподема очень длинная, дуговидно-
изогнутая, булавовидно утолщенная на дистальном конце. Длина тела
2.5—3 мм.
Яйцо1 белое веретеновидной формы, более заостренное к переднему
полюсу; структура хориона неразличима; длина 0.46—0.48 мм.
Личинка I стадии не резко сегментирована, гладкая, без шипиков;
дыхальцы неразличимы. По бокам анального отверстия имеется
1 Данные о строении яйца и личинки приводятся по Олсуфьеву.
5. ACRIDOMYIA
28 i'
с 2 небольшими выступами
Рис. 472. Фаллозома Acridomyia
sacharovi Stack. (Ориг,).
по одному полушаровидному бугорку. Срединный дорзальный склерит
ротоглоточного аппарата удлиненный, заостренный на вершине и согнут
клювовидно на вентральную сторону. Фарингеальный склерит тонкий,
с широко расставленными друг от друга дорзальными и вентральными
отростками; последние короче первых, косо срезаны на дистальном конце.
Передние отростки фарингеального склерита короткие. Длина тела 0.85—
1.17 мм.
Во II стадии личинка; как и в предыдущей стадии, неясно сегмен-
тирована. Передние дыхальцы имеются, задние редуцированы. Дыхальце-
вых отверстий 7—9. Анальное отверстие
по краям. Ротовые крючки массивные,
короткие, загнутые крючком. Гипосто-
мальный склерит вытянутый, с длинным
направленным вниз и вперед тонким
изогнутым отростком. Фарингеальный
склерит с широкими задними отростками.
Длина тела 2.4—4.4 мм.
В третьей стадии личинка имеет
желтовато-белую окраску. Головной сег-
мент с ясно развитыми усиками и мак-
силлярными щупальцами. Передние ды-
хальцы расположены в кутикуле 2-го
видимого сегмента; они имеют вид пло-
щадки с 7—12 отверстиями, расположен-
ными веерообразно по переднему краю
площадки. Последний сегмент тела ли-
чинки округленный, скошен по заднему
краю сверху вниз; задняя поверхность
притупленная, снабжена 2 дыхальцами;
последние имеют вид розетки с 8—12
отверстиями. Ротовые крючки сильные,
выпуклые по дорзальной и вогнутые по
вентральной стороне: у основания с вен-
тральной стороны располагаются длин-
ные треугольные зубные склериты. Гипо-
стомальный склерит неправильно ромбовидный, с тонким изогнутым и
направленным вниз и вперед отростком. Фарингеальный склерит с ши-
рокими дорзальными и вентральными и очень короткими заостренными
передними отростками. Дорзальные и вентральные отростки сильно
пигментированы лишь по внутреннему краю. Первые из них на дисталь-
ном конце с грушевидным резко очерченным просветом. Вентральные
отростки в верхнем отделе дистального конца с отвесным краем, внизу
переходящим в фарингс. Длина тела 6.0—7.5 мм, толщина 1.5—1.8 мм.
Пупарий от светло- до темнокрасно-коричневой окраски, в переднем
отделе слабо дорзовентрально сдавлен. Задние дыхальцы расположены на
коротких выступах. Сегментация не ясная. Длина 2.8—4.6 мм, ширина
1.1—1.8 мм .
Распространение. Вид известен из зоны полупустынь, где
приурочен к поймам рек, что находится в связи с условиями обитания
азиатской саранчи. До сего времени встречен в дельтах рек Урала,
Волги, Сыр-дарьи, а также Терека и Кумы.
A. sacharovi представляет собою своеобразно специализированного
паразита азиатской саранчи, в теле которой протекает развитие ее-
-282
II. ТРИБА STOMOXYDINI
преимагинальных стадий. Муха откладывает яйца в тело саранчи, обычно
в брюшную полость, реже в область груди. Чтобы пристроить свое потом-
ство, самки при помощи крепких престомальных зубцов хоботка делают
прокол межсегментной перепонки на брюшке саранчи или ее личинки
и, подведя к образовавшейся ране яйцеклад, просовывает его в тело
жертвы. Таким образом, яйца мухи оказываются внутри тела саранчи.
За один раз самка в состоянии отложить от 25 до 46 яиц (лабораторные
данные). Судя, однако, по числу личинок в саранчуках, обнаруживаю-
щихся при вскрытии последних (до 157 штук), размер яйцекладки может
-быть много выше. Сам процесс заражения саранчи протекает очень быстро
и занимает менее 1—Р/г минут. Мухи термофильны; развитие мух и наивыс-
дпая их активность проходит при высоких температурах. Рукавишников
(1930) отмечает, например, что при 20° С развитие куколок приостанав-
ливается, в то время как у других мух, паразитирующих в саранче
и, в частности, у видов Blaesoxipha, развитие в этих условиях протекает
нормально. A. sacharovi принадлежит к числу гелиофильных насекомых,
-что сказывается и на общем поведении мух и на условиях яйцекладки,
которая осуществляется при ярком солнечном освещении. В лаборатор-
ной обстановке попытка заставить самок откладывать яйца удавалась
лишь после сильного облучения последних сильными электролампами.
По данным Рукавишникова, самки питаются гемолимфой саранчи, после
нанесения ей поранений при откладке яиц. Остается, однако, неизвест-
ным, прибегают ли самки к названной форме питания в промежутках
между яйцекладками. Рукавишников склоняется к убеждению, что
самцы этого вида вообще не питаются.
Что касается питания личинок, то, по свидетельству того же автора,
оно проходит преимущественно за счет жирового тела, в котором, как
правило, происходит концентрация паразитов. В теле хозяина развитие
личинок заканчивается через 11—14 дней. За этот период личинка про-
ходит три стадии. Достигнув предельной зрелости личинки, собравшись
группами, пробуравливают межсегментную перепонку на брюшке или
у основания ног саранчи и, выбравшись наружу, уходят в почву для
окукления. Последнее проходит на сравнительно небольшой глубине
(2—3 см). Продолжительность стадии куколки определена Рукавишни-
ковым в 22—25 дней. Полный цикл развития мухи охватывает около
33—38 дней. На созревание яиц требуется 3—5 дней, считая со дня вылуп-
ления самки из ложнококона. Зимуют ложнококоны. В Сырдарьинских
плавнях, по подсчетам Рукавишникова, зараженность саранчуков не пре-
вышала 3.2%, что имело место в конце августа и начале сентября; в дру-
гое время саранча была заражена значительно слабее. Сходные данные
по той же местности получил и Олсуфьев. Что касается продукции яиц
у A. sacharovi, то, по Рукавишникову, она составляет в среднем около
200 штук при колебании от 141 до 261.
АЛФАВИТНЫМ УКАЗАТЕЛЬ НАЗВАНИИ НАСЕКОМЫХ i
Acridomyia 6, 12, 13, 14, 20, 28, 31, 36,
46, 49, 50, 53, 54, 57, 60, 71, 240, 277,
278
Acridomyina 6, 48, 240, 277
aenea, Pyrellia 5, 47, 52, 162, 164, 165,
171, 172
aeneomicans, Dasyphora 187, 188
aenescens, Morellia 6, 22, 23, 205, 206,
208, 211
Aethiopomyia 47, 49
Agalmia 73
albina, Musca 5, 18, 51, 91, 93, 114, 120,
122, 124
Alluaudinella 47, 50, 53
arnica, Musca 5, 51, 92, 96, 139, 144, 145,
146, 148, 260
Anacrostychia 79
angustifrons, Musca 114
argentigena, Orthellia 86
asetosa, Morellia 52
asiatica, Cryptolucilia 75
asiatica, Dasyphora 6, 31, 54, 55, 70, 177,
180, 187, 188, 189, 190, 201, 202
atripalpis, Haematobia 256, 259, 260, 265
autumnalis Musca 5, 33, 34, 40, 54, 70, 90,
92, 94, 96, 100, 139, 140, 142, 143, 144,
145, 146, 148, 151, 154, 159, 162
Bdellia 257
Bdellolarynx 257, 259
beckeri, Musca 121
bimaculata, Orthellia 86, 88
Biomyia 90, 128
booina, Haphospatha 269
Brucemyia 267
cadaverina, Pyrellia 5, 28, 52, 70, 164,
165, 166, 167, 168, 169, 170, 182
caesarion, Orthellia 5, 51, 73, 75, 76, 77,
78, 79, 86
calcitrans, Stomoxys 6, 29, 53, 67, 68, 69,
71, 249, 250, 251, 253, 255, 256, 258,
260, 264, 277
chalybaea, Orthellia 86
Commo sia 73
concolor, Stomoxys 249
convexifrons, Musca 5, 31, 51, 54, 92, 94,
96, 139, 144, 154, 155, 156, 162
cornicina, Lucilia 75
cornicina, Orthellia 5, 29, 51, 70, 71, 73,
75, 81, 83, 84, 86, 88
coroina, Musca 139
coroinoides, Musca 147
crassipalpis, Haematobia 258, 267
crassirostris, Musca, 5, 67, 92, 94, 122,
124, 135, 138, 150
Cryptolucilia 73, 81
cuthbertsoni, Musca 90, 91
cyanella, Dasyphora 6, 47, 52, 178, 179,
181, 182, 183, 185, 186, 187
cyanicolor, Pyrellia 181
Cyrtoneura 133, 209, 211, 212, 213
Dasymorellia 47, 203, 204
Dasyphora 6, 8, 13, 18, 19, 20, 21, 22,
23, 25, 26, 27, 30, 31, 36, 41, 42, 44,
45, 47, 49, 50, 52, 53, 54, 55, 72,
163, 164, 175, 176, 177, 178, 190, 236
Dasyphoromima 6, 52, 163, 164, 175
Dasysterna 6, 204, 212
decipiens, Mesembrina 6, 22, 223, 228, 232
diadema, Orthellia 86
domestica, Musca 5, 12, 33, 34, 51, 55,
64, 65, 66, 67, 68, 70, 90, 91, 92, 96,
98, 99, 102, 104, 111, 112, 113, 115
domestica, Musca domestica 94, 96, 100,
104, 105, 108, 109, 114
domestica, Musca vicina 94, 96, 98, 99,
100, 104, 105, 106, 107, 108, 114, 120
dubia, Orthellia 86
equina, Lyperosia 268, 269
eriophthalma, Dasyphora 6, 47, 177, 178
179, 180, 182, 186
eriophthalma, Pyrellia 183, 186
Eumusca 90, 154, 156, 157
fennica, Cryptolucilia 75
flavinervis, Musca 97
flavipes, Pyrellia 173
Fraseria 47, 50, 52
fulvipes, Orthellia 175
1 Названия семейств, подсемейств, триб и подтриб выделены жирным шрифтом.
Синонимы и прежние родовые названия выделены курсивом. Все видовые названия
начинаются со строчной буквы. Цифрами отмечены страницы, где цитируется дан-
ное название, причем жирным шрифтом показана страница с описанием видов,
родов, триб и подтриб.
284
АЛФАВИТНЫЙ УКАЗАТЕЛЬ НАЗВАНИЙ НАСЕКОМЫХ
Glossinella 273
Gordonia 50, 53
gracilior, Mesembrina 6, 223, 225, 229,
230, 238
Graphomyia 6, 9, 12, 13, 18, 22, 24, 27,
28, 30, 31, 33, 39, 42, 43, 46, 48, 49,
50, 53, 54, 60, 71, 157, 240, 241
Graphomyina 6, 48, 240, 241
Graphomyini 45
gussakovskii, Dasyphora 6, 178, 180, 197,
198
Haematobia 6, 19, 25, 27, 29, 31, 41, 42,
50, 53, 54, 236, 241, 256, 257, 262,
264, 269, 275, 278
Haematobia 267
Haematobina 257
Haematol osca 19, 257, 265, 272
Haematostoma 50, 53
Haphospatha 68, 267, 269
hervei, Musca 5, 93, 95, 150, 160, 161,
162
hortensia, Morellia 52, 213, 221, 222
hortensis, Morellia 222
hortorum, Morellia 6, 22, 29, 205, 206,
207, 208, 210, 211, 216
humilis, Musca 114
Hypodermodes 222
ignita, Pyrellia 171
indica, Orthellia 87, 90
inferior, Musca 51
ingrica, Mesembrina 230, 234
insignis, Phillaematomyia 137, 138
intermedia, Mesembrina 6, 223, 224, 225,
229, 232, 236
inventrix, Pyrellia 47
irritans, Lyperosia 6, 34, 53, 68, 256,
258, 260, 262, 264, 267, 268, 270, 272,
273, 274, 276
irritans, Lyperosia 269
larvipara, Musca 5, 31, 40, 51, 54, 55,
70, 92, 94, 96, 100, 139, 140, 144,
147, 150, 152, 153, 155, 156, 162, 200
lasiopa, Musca 51
lasiophthalma, Pyrellia 183
lateralis, Musca 97
latifrons, Dasyphora 6, 179, 192, 194
latipalpis, Lyperosia 272
latipalpis, Orthellia 5, 77, 86, 87, 88, 89
latirostris, Haphospatha 269
latreillei, Mesembrina 230, 236
lauta, Orthellia 86
Lissosterna 90, 121
lucidula, Musca 5, 16, 20, 51, 70, 92, 95,
122, 124, 133, 134, 135, 136, 137,-138
lusoria, Musca 5, 54, 92, 94, 95, 139, 142,
144, 156, 157
Lyperosia 6, 20, 22, 50, 53, 54, 241, 256,
257, 258, 264, 267, 277
Lyperosiops 257, 259
maculata, Graphomyia 6, 28, 29, 43, 53,
69, 218, 241, 242, 244, 246, 247, 248
matutina, Mesembrina 224, 227
meridiana, Mesembrina 6, 14, 29, 31, 34,.
52, 69, 71, 209, 222, 223, 224, 225,-
229, 230, 232, 233, 235, 236,. 238
meridionalis, Lyperosia 273
Mesembrina 6, 8, 12, 13, 16, 17, 18, 19r
20, 22, 25, 27, 28, 29, 30, 35, 41, 42,
45, 47, 48, 49, 50, 52, 53, 54, 55, 60,.
72, 222
Mesembrinini 45
mesopot.amiensis, Musca 5, 90, 142, 146,
147, 158
Metamesembrina 222, 230
minor, Morellia 211
minuta, Lyperosia 6, 267, 268, 272
minuta, Pyrellia 5, 52, 165, 167, 171,
173, 182
montana, Mesembrina 6, 223, 239
Morellia 6, 8, 13, 19, 20, 22, 23, 27, 30,.
31, 40, 45, 47, 50, 52, 73, 203, 204,.
208, 212, 213.
Musca 5, 8, 10, 12, 13, 17, 18, 19, 20,.
21, 22, 25, 26, 27, 28, 30, 31, 33, 35,
36, 41, 42, 44, 45, 48, 49, 50, 54, 55,
66, 67, 68, 73, 78, 80, 81, 83, 90, 91,
101, 122, 138, 163, 165, 216, 254, 264
Muscidae 5, 35, 43, 44, 46, 48, 66, 67,.
68, 244
Muscinae 44, 45, 46,‘ 47, 48, 67, 68
Muscini 5, 8, 9, 13, 14, 17, 18, 21, 22,.
23, 24, 26, ,27, 28, 31, 33, 35, 36, 38,.
40, 42, 43, 44, 45, 46, 47, 48, 49, 50,
53, 54, 55, 57, 60, 71, 72, 122, 234,
249
mystacea, Mesembrina6, 22, 54, 218, 223,
224, 225, 226, 227, 228, 229, 232, 235
nebulo, Musca 5, 90, 91, 94, 96, 99,.
113, 114
Л’eivamyia 257
TVeomesembrina 222, 230
Neomusca 47, 50, 53
Neomyia 73, 86
nilotica, Morellia 6, 52, 206, 211
obscuripes, Orthellia 90
Ochromusca 47, 49
Orthellia 5, 13, 16, 17, 19, 20, 22, 25,.
27, 28, 29,'30, 31, 36, 41, 42, 44,
48, 49, 50, 51, 54, 66, 72, 73, 74, 75,
81, 86, 87, 88, 163, 164, 173, 174,
175, 184, 185, 186, 215, 234, 236, 254
osiris, Musca 5, 51, 57, 70, 92, 93, 95,
99, 122, 124, 125, 127, 146
pacifica, Orthellia 5, 51, 75, 77, 85, 86,
88, 89
paraversicolor, Dasyphora 6, 178, 190,
194, 195, 198,- 201
pavlovskyi, Pyrellia 6, 52, 175
phasiaeformis, Musca 91
Philaematomyia 90, 135, 137, 138
Plaxemyia 90
podagrica, Morellia 6, 22, 206, 207, 208,.
209, 210, 211, 218
polita, Musca 121
potrix, Lyperosia 272
АЛФАВИТНЫЙ УКАЗАТЕЛЬ НАЗВАНИЙ НАСЕКОМЫХ
285
prashadi, Musca 139
pratorum, Dasyphora 6, 54, 55, 168,
176, 177, 180, 184, 187, 192, 194, 195,
196, 197, 198, 199, 200
pratorum, Dasyphora 52, 196
pratorum, Dasyphora meridionalis 6, 179,
180, 198, 203
pratorum, Dasyphora pratorum 179, 199,
203
Pristirhynchomyia 90
Pseudopyrellia 73, 75, 81
Ptilolepis 90
putziloi, Mesembrina 228
Pyrellia, 5, 13, 18, 19, 20, 23, 25, 26,
27, 29, 30, 31, 36, 44, 47, 48, 49, 50,
52, 73, 87, 162, 163, 164, 167, 169,
170, 173, 174, 1,75, 176, 177, 178, 180,
186, 199, 203, 204, 209, 211, 212
Pyrellina 47, 49
Pyrelliomima 5, 51, 73, 74, 77, 86, 87
quadrisetosa, Dasyphora 6, 179, 191, 192,
194, 201
Heinwardtia 47, 49
lesplendens, Mesembrina 6, 223, 225, 228,
229, 232, 236, 238
jacharovi, Acridomyia 6, 9, 18, 268, 277,
278, 279, 281, 282, 283
saltuum, Dasyphora 6, 29, 31, 52, 54,
180, 187, 188, 190, 191, 195, 201
schillingi, Glossinella 273
scolopax, Haphospatha 269
Scutellorthellia 73
secunda, Pyrellia 5, 52, 165, 167, 170,
171, 172, 173
semilutea, Pyrellia 5, 163, 174, 175
serena, Dasyphora 6, 29, 47, 52, 177, 178,
180, 181
serena, Pyrellia 171
•setitibia, Dasyphora 6, 23, 178, 180, 193,
194, 201
similis, Dasyphora 6, 179, 192, 194, 201
simplex, Morellia 6, 69, 208, 211, 212,
213, 214, 216, 217, 220, 221, 222
simplicissima, Morellia 6, 52, 208, 211,
213, 216, 221
sorbens, Musca 5, 51, 57, 59, 64, 65, 70,
91, 92, 93, 95, 101, 114, 116, 118, 119,
120, 159
speculifera, Musca 121
Stenomitra 73
stimulans, Haematobia 6, 53, 69, 102,
257, 258, 259, 261, 263, 265, 277
Stomoxydidae 68
Stomoxydinae 48
Stomoxydina 6, 48, 240, 248, 275
Stomoxydini 6, 9, 13, 14, 17, 18, 20, 21,
22, 23, 24, 26, 27, 31, 44, 45, 46, 48,
49, 50, 53, 54, 57, 60, 71, 240, 278
Stomoxynae 67
Stomoxys 6, 19, 20, 22, 25, 27, 30, 31,
33, 35, 36, 38, 40, 41, 42, 43, 44, 48,
50, 53, 54, 56, 58, 240, 249, 257, 269,
278
Stygeromyia 267
suda, Pyrellia 171
sugillatrix, Musca 91
superba, Orthellia 74
Synthesiomyia 47, 49
tempestiva, Musca 5, 54, 56, 70, 92, 93,
95, 122, 124, 126, 128, 130,- 132, 133,
134, 135
titillans, Lyperosia 6, 70, 268, 269, 271,
272 273 274
Trichomorellia 203, 204
trichosterna, Dasyphora 6, 179, 180, 190,
198, 201
trispina, Orthellia 87, 90
tristis, Mesembrina 227
vagatoria, Musca 97
ventrosa, Musca 5, 93, 95, 159, 160
versicolor, Dasyphora 6, 52, 178, 180,
190, 194, 195, 196, 197, 198, 201
vesper tina, Mesembrina 224, 227
vieina, Musca 65, 67, 90, 91, 971
viridescens, Dasyphora 199
vitripennis, Musca 5, 51,' 91, 93, 95, 99,
122, 124, 126, 146
vitripennis, Musca 125
Viviparomusca 90, 146, 147, 154, 156, 160
iveissi, Lyperosia 273
xanthomelas, Musca 5, 150, 157, 158,159,
160
Xenomorellia 47, 50, 52
yerburyi, Musca 90
1 См. также domestica vieina, Musca — стр. 283.
СОДЕРЖАНИЕ
Стр~
Предисловие............................................................... Л
Систематический указатель видов........................................... 5
Введение
Морфология взрослого насекомого ..................................... 7
Морфология фаз развития ........................................... 33-
Систематическое положение группы.................................. 43.
Географическое распространение ..................................... 49
Экология группы ................................................... 53-
Эпидемиологическое и хозяйственное значение группы ................. 58
Методы борьбы с мухами............................................. 60>
Литература.......................................................... 64
Специальная часть
Определительная таблица личинок III стадии ...................... 69
Определительная таблица триб по взрослой фазе .................... 71
I. Триба Muscini.................................................. 72
2. Триба Stomoxydini .......................................... 240
Алфавитный указатель названий насекомых......................... 283.
Печатается по постановлению
Редакционно-издательского Совета
Академии Наук СССР
*
Редактор издательства К. А. Борисов
Техн, редактор Д. М. Крол
Корректор Л. С. Халиманович
РИСО АН СССР № 4300. Подписано к печати
1/Ш 1951 г. М-18842. Бумага 70 X 108V16-
Бум. л. 9. Печ. л. 24.66. Уч.-изд. л. 23.6.
Тираж 2000 экз. Зак. 1805. Цена в пере-
плете 22 руб.
1-я типография Издательства Академии Наук
СССР. Ленинград, В. О., 9 линия, дом 12.
ИСПРАВЛЕНИЯ И ОПЕЧАТКИ
Страница Строка Напечатано Должно быть
18 21 сверху НИХ нее
22 21 » и неподвижно — о вертлугом и с вертлугом
26 22 внизу (в частности, Dasyphora, (в частности, Dasyphora),
26 20 » V стернита). V стернита.
63 18 сверху пиретрум пиретрин
164 5 » Pyrellia Rypellia
257 &— 9 » eguina-voger equina Voger
Л. С. Зимин. Сем. Muscidae. Настоящие мухи.