/











































































































































































































































































































































































































































































































































































































































































































































































Similar




Text
Л1Т0ИИЯ
ГПдручник
За редакщею
професора М.Н. Зайка,
професора К).В. Биця,
професора М.В. Кришталя
АВГОРСЬКИЙ колектив
Биць Юрш Викторович — член-кореспондент НАПН Украши, доктор медичних наук,
профссор
Бутенко Геннадш Михайлович — академтк НАМИ Украши. член-кореспондент
НАН Украши, член-кореспондент АМН Роси, доктор медичних наук, професор, директор
ДУ ‘Институт генетичнот та регенеративно! медицини НАМИ Украши”
Гоженко Анатолий Званович — доктор медичних наук, професор директор ДУ
“Украшський НД1 медицини транспорту МОЗ Украши”, до 2011 р. — завтдувач кафедри
патофтзюлоп! Одеського державного медичного университету
Горбань Владислав Олександрович — доктор медичних наук, професор, завтдувач
дисциплши патофтзюлогп кафедри фармакологи Медичного университету Украшсько!
асошацт! народно! медицини
Данилова Людмила Яплвна — доктор медичних наук, професор
Досенко Виктор Свгенович — доктор медичних наук, професор. провтдний науковий
сшвроб'тник втддтлу загально! т молекулярно! патоф! полон! Гнети гуту фтзюлоп! тмент
О О Богомольца НАН Украши
Гльський В/ктор Миколайович — член-кореспондент НАМИ Украши, доктор медич-
них наук, професор, завтдувач кафедри патофтзюлогп Донецького нацюнального медич-
пого ушверситету
Зайко Микола Никифорович — член-кореспондент АМН СРСР, доктор медичних
наук, професор
Заярна Лариса Павлгвна — кандидат медичних наук, доцент кафедри патофтзтолоп!
Вацюнадьного медичного университету !меш О О. Богомольца
Кришталь Микола Васильевич — доктор медичних наук, професор, завтдувач ка-
федри патофтзюлогп Национального медичното университету тмент О.О. Богомольца
Кубишк1н Анатолш Володимирович — доктор медичних наук, професор. завтдувач
кафедри патофтзтолоп! Криме ького державного медичного ушверенгету .мент С.1. Георп-
евського
Мислицький Валентин Франиович — доктор медичних наук, професор кафедри
патофтзюлогп Буковине ькс-т о державного медичного ушверситету
Мгхньое Володимир Анатолшович — член-кореспондент НАМИ Украши, доктор
медичних наук, професор кафедри патофтзюлоп! Напюнатьного медичного ушверситету
тмен! О.О. Богомольца
Попова Лариса ОлександрЗвна — доктор медичних наук, професор кафедри ватео-
лоп! Нацюнально! медично! академт! шеладипломно! освтти тмен; П.Л Шулика
Потоцька 1рина Зватвна — кандидат медичних наук, доцент кафедри патофтзтолоп!
Нацюнального медичного ушверситету тмент 0.0. Бот-омольця
Резшков Олександр Григорович — академик НАМ11 Украши, член-кореспондент
НАН Украши, доктор медичних наук, професор, завтдувач втддтлу ендокринолоп! репро-
дукцп та адаптацп ДУ 'Институт ендокринолоп! та обмтну речовин тмент В.П. Комтсаренка
НАМИ Украши”
Репецька Ганна Григоргвна — кандидат медичних наук, доцент кафедри патофтзю-
логц Нацюнального медичного ушверситету тмен! 0.0 Богомольца
Омеонова Наталия Костянтингвна — кандидат медичних наук, доцент кафедри
патофтзтологт! Нацюнального медичного ушверситету тмент 0.0. Богомольца
Сукманський Олег Званович — доктор медичних наук, професор, кертвннк курсу
патофтзюлогп Одеського державного аграрного ушверситету
Хара Мар1я Романовна — доктор медичних наук, професор, завтдувач кафедри
патофтзюлогп Терногпльського державного медичного ушверситету тмеш 1.Я. Горбачев-
ського
1ЙТ0ФШ0ГШ
Четверте видання, перероблене I доповнене
За редакцгею
професора М.Н. Зайка,
професора Ю.В. Биця,
професора М.В. Криштпаля
Затверджено
Министерством охорони здоров'я УкраГни
як базовий шдручник для студенив
вищих медичних навчальних закладйв
III—IV р1вшв акредитацн
Затверджено
Мшютерством осв1ти 1 науки Украши
як шдручник для студентов
вищих медичних нанчальних-дакляд1в
Фонд пщручниюв
паукова оюлютека
I
Ки1В
ВСВ “Медицина'
2014
9000000005
НМУ1М. О.О. Богомольця
УДК 616-092(075.8)
ББК 52.5я73
П20
А в т о р и .
Ю В. Биць, Г.М. Бутенко, А.1 Гоженко, В.О. Горбань, Л.Я. Данилова,
В.€. Досснко, В.М. €льський, М.Н Зайко |, Л.П. Заярна, М.В. Кришталь,
А.Б. Кубишкш, В.Ф. Мислицький, В А. М1хнъов, Л.О Попова, 1.1. Потоць-
ка, О.Г. Резников, Г.Г. Репецька, Н.К. Симеонова. 0.1. Сукманський,
М Р. Хара
Рецензенти:
О.О. Мойбенко — академ!к НАН Украши, доктор медичних наук, про-
фесор, завщувач вщдьту загально! та молекулярно! патофЫологй 1нституту
ф!з!ологц !мен! О О Богомольця Н АН Украши;
Ю.М. Колесник — заслужений дзяч науки I техюки Украши, доктор ме-
дичних наук, професор. завиувач кафедри патофззюлоги Запор!зького дер-
жавного медичного ун!верситсту;
М.С. Регеда — заслужений прашвник освгти Украши, доктор медичних
наук, професор, зав!дувач кафедри патофгзюлогп Л ьвзвського на зонально-
го медичного университету зм. Данила Галицького
Патоф!зюлопя: нщручник / Ю.В. Биць, Г.М Бутенко, А.1. Гоженко та ш.;
П20 за ред. М.Н. Зайка, Ю.В. Биця, М.В Кришталя. — 4-е вид., переробл. 1
допов. — К.: ВСВ '‘Медицина”, 2014. — 752 с. + 4 с. кольор. вкл.
15ВК 978 -617-505-295-2
Пщручник пщготовлений авторським колекгивом сп!вроб1тниюв виших навчальних за-
кладдв 1 академ!чних науково-дослфиих заклад!в Украши, яю е визнаними фахшцями з р1зних
питань пагофгзюлогн, шо дае змогу вважати цей базовий гцдручник нацюнальним.
Пор!вняно з останшми виданнями суттево перероблеш таю роздши. як “Паголопчна
фЬюлопя шунноТ системи”, “Алерпя”, “Запалення”, “Пухлина”, “Порушення вутлеводного
обмшу”, “Порушення кислотно-основного стану”, “Порушення водно-електролггного обм1
ну”, “Роздали кровооб1гу, повязан! з порушенням функцн судин” та ш. Заново написан!
роза ши "Патоф^зюлопя обмшу м! крое л сменив”, “Патоф!зюлопя обкину впамйпв”, “Вибухова
шахтна травма”.
Матер!ал пщручника викладено вадповшно до ново! навчально! программ з патолопчно!
ф13юлогц, складено! на основ! кредитно-модульно! системи оргашзацн навчяльного пронесу.
Для студенпв виших медичних навчальних заклад!в IV ршня акредитанн. ГПдручник може
бути корисним для викладач!в I л!кар!в ус!х спешачьностей.
УДК 616-092(075.8)
ББК 52.5я73
I8В^ 978-617-505-295-2
© Ю В. Биць, Г.М. Бутенко, А.1. Гоженко, В О. Горбань.
Л.Я. Данилова, В.€. Досенко. В М Ельський. М.Н. Зайко,
Л.П. Заярна, М.В. Кришталь, А В Кубишкш, В.Ф. Мисли-
цький, В.А. Мзхньов, Л.О. Попова, 1.1. Потоцька, О.Г. Рез-
шков, Г.Г. Репецька, Н.К. С'меонова. О I Сукманський,
М.Р Хара, 2008. 2014
© ВСВ “Мединина”, оформления, 2014
Зм1ст
Список скорочень.....................................-.......................11
Передмова....................................................................14
Вступ (д-р мед. наук, проф. М.Н. Зайко)......................................15
Предмет 1 завдання патолопчно’1 фтзюлогп.....................................15
Зв’язок патолопчно! фхзюлоги з шшими медичними науками,
П значения для клшпои........................................................16
Експеримент — основний метод патолопчно! фхзюлоги............................17
[сторичний нарис розвитку патолопчно! фхзюлоги
(д-р мед. наук, проф. М.Н. Зайко, д-р мед. наук, проф. Ю.В. Биць,
д-р мед. наук, проф. В. Ф. Мислицъкий).......................................20
Частина перша
3 АГ АЛЕНА НОЗОЛОГ1Я
Роздал I. Загальне вчення про хворобу (д-р мед. наук, проф. М.Н. Зайко)......29
Здоров’я.....................................................................29
Хвороба......................................................................30
Принципи класифхкацц хвороб...............................................33
Патолопчна реакигя, патолопчний процес, патолопчний стан.....................34
Типов! патолопчш процеси..................................................34
Основш перюди (стадй) розвитку хвороби.......................................35
Роздал II. Вчення про етюлогло 1 патогенез (д-р мед. наук, проф. М.Н. Зайко).40
Етюлопя.................................................................... 40
Патогенез....................................................................44
Причинно-наслщковий зв’язок...............................................46
Загальне 1 мюцеве.........................................................47
Структура 1 функция.......................................................47
Загальне (неспецифгчне) 1 специфхчне......................................48
Адаптащя 1 компенсащя.....................................................49
Роздал Ш. Патогенна дая на оргашзм фактор1в навколипшього середовища
(канд. мед. наук, доц. НХ. Омеонова).................................51
Мехашчна травма..............................................................51
Термтчна травма....................•.........................................51
Баротравма...................................................................55
Променева травма.............................................................57
Етюлопя...................................................................57
Патогенез.................................................................58
Компенсаторно-пристосувальш реакцн........................................64
Внутрппне (шкорпороване) опромшення.......................................65
Г остра променева хвороба.................................................65
ГТ
Хроючна променева хвороба .............................. . ...........66
Корекщя променевого ураження......................................... .67
Електрична травма................................................. 67
Патогенна д!я факторов космочного польоту .......................... . ____68
Патогенна доя хомочних факторов (канд. мед. наук, доц Г.Г. Репецька)....... .69
Патогенна доя боологочних (1нфекщйних) факторов
(д-р мед. наук, проф. В.€. Досенко)........................................ 77
Системно прояви шфекшйного процесу................................... 86
Роздал IV. Патолопчна фозюлопя кптини (д-р мед. наук, проф. Г.М Бутенко)......88
Порушення функций клггин ................................................ 88
Порушення процесов обману та взаемодц з навколишшм середовищем...............88
Порушення енергетичного забезпечення клштни...............................90
Порушення зберсження 1 передавания генетичноо шформапоо...................91
Порушення процессе синтезу............................................... 93
Порушення процесов розпаду........................................... 94
Ушкодження клотини та н загибель.............................................95
Роздал V. Роль спадковоеп, конституцп та вокових змш у патологп
(канд. мед. наук. доц. Н.К. Омеонова).................................... 98
Генетично детермшоваж хвороби ............................................. 98
Класифокацоя..................................................... 98
Етюлопя............................................................... 99
Патогенез.............................................................. 100
Молекулярно-генно (спадково) хвороби.....................................102
Хромосомно хвороби ................................................... 107
Спадкова схильность до хвороб....... ....................................109
Генетичне обстеження хворого.............................................110
Принципи локування о профилактики........................................113
Роль конституцп в патолога..................................................114
Патолопчна фозюлопя внутршньо'дробного розвитку
(д-р мед. наук, проф. Л. О. Попова).........................................119
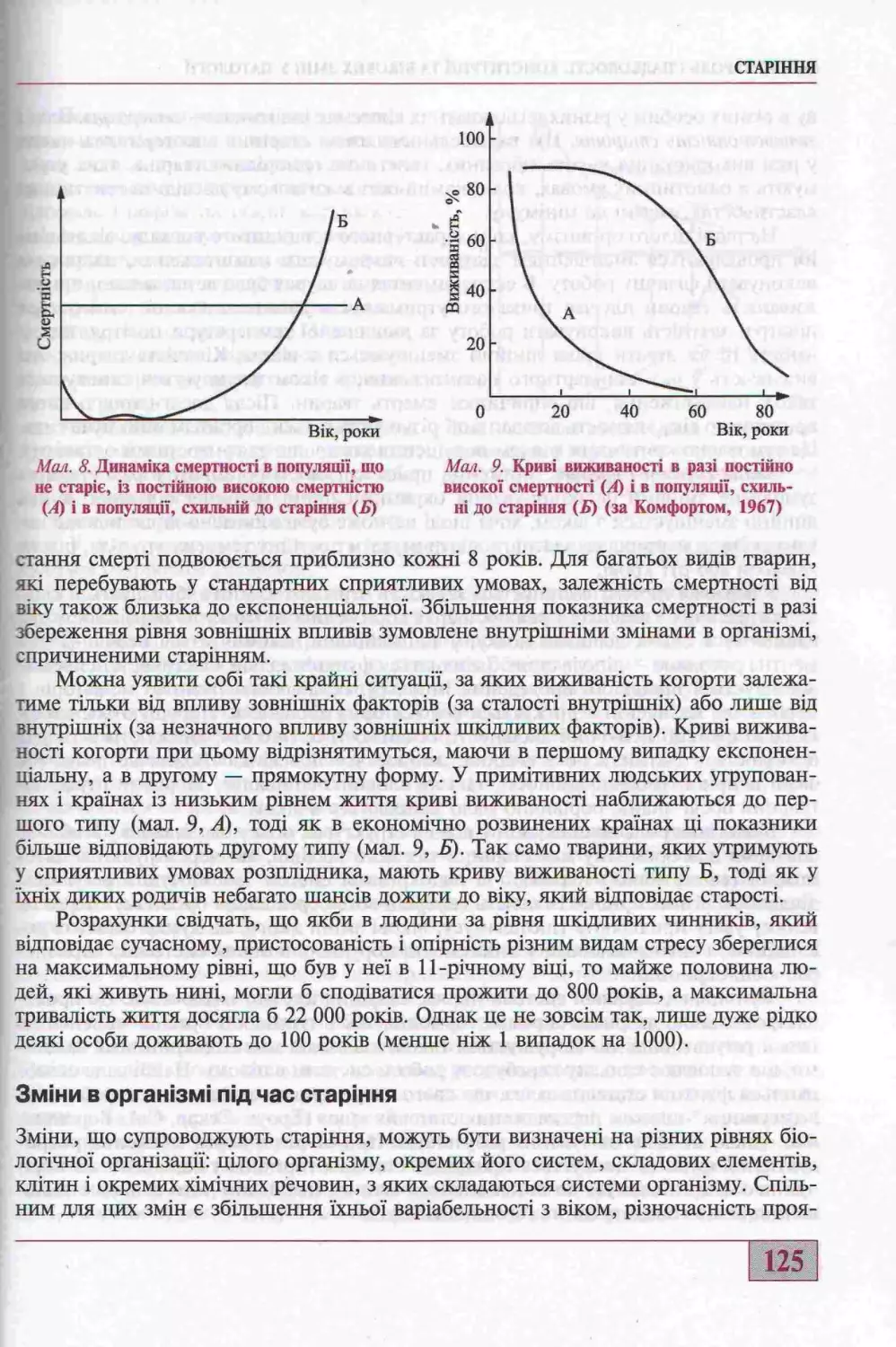
Старения (д-р мед. наук, проф. Г.М. Бутенко)............................... 123
Змони в органозмо под час старшня...................................... 125
Старшня о хвороби...................................................... 132
Причини I механозми старшня......................-.......................133
Шляхи впливу на старшня. .............................................. 135
Роздал VI. Патолопчна фозюлопя сполучно! тканини
(д-р мед. наук, проф. М.Н. Зайко, д-р мед. наук, проф. М. Р Хара).................. 137
Сполучна тканина як едина система.............................................137
Порушення структури 1 функцш сполучно! тканини..............................139
Патология костковоо тканини................................................ 152
Роздал VII. Реактивность о П роль у патологп
(д-р мед. наук, проф. Л.Я. Данилова).................................................................. 157
Резистентность. взаемозв'язок з реактивнктю.................................159
Реактивность на розних ршнях оргашзацп живих систем.........................160
Еволюцоя реактивност!..................................................... 161
Механозми реактивност!. Боолопчн! бар’ери.................................. 162
Р злй УШ. Патолопчна фзззологзя змунноз сисгеми
(д-р мед. наук, проф В О. Горбань)......................................, 165
Основы функцп та структура шунноз системы....................................165
Загальш закояомзрностз порушень змунноз системы...................... .. ____172
Гмунодефзпитнз хворобы ............................................... 173
Тмунодепресывн! станы............................................. .. .. 179
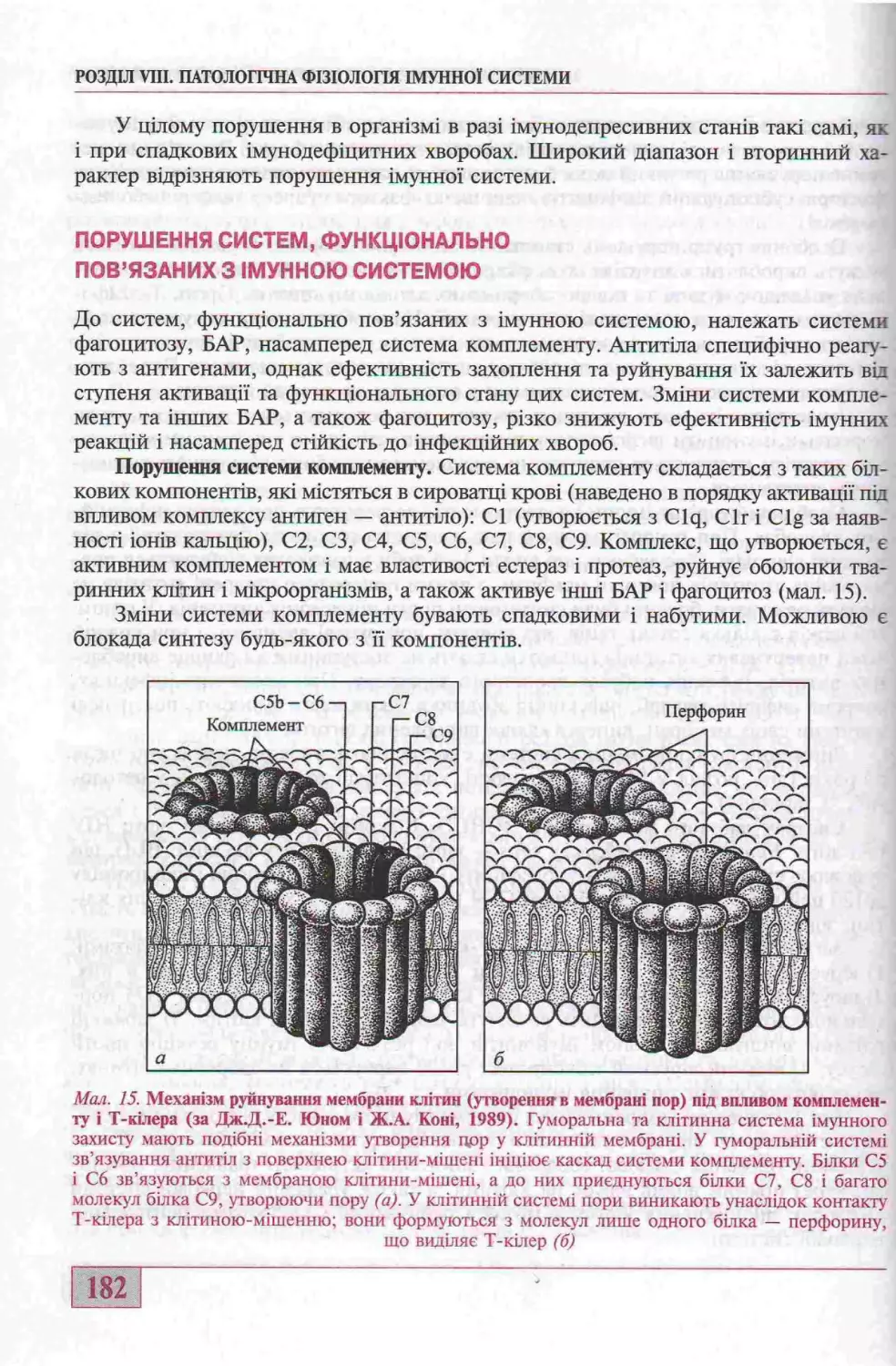
Порушення систем, функщонально пов’язаних з ззиунною системою.............. 182
^нтигенасоцшовазп хвороби....................................................184
Частина друга
ТИПОВ1 ПАТОЛОПЧН1 ПРОЦЕСИ
Роздал IX. Патолопчна фзззолопя периферийного кровообпу
(д-р мед. наук, проф. Ю В. Бицъ, д-р мед. наук, проф. А.В. Кубишкш).......186
^ртерзал ьна гшеремзя........................................................190
Венозна гшеремзя.............................................................196
1шем1я.............................................................. . .197
Стаз....................................................................... .202
Тромбоз...................................................................... 203
Емболзя.................................................................. .206
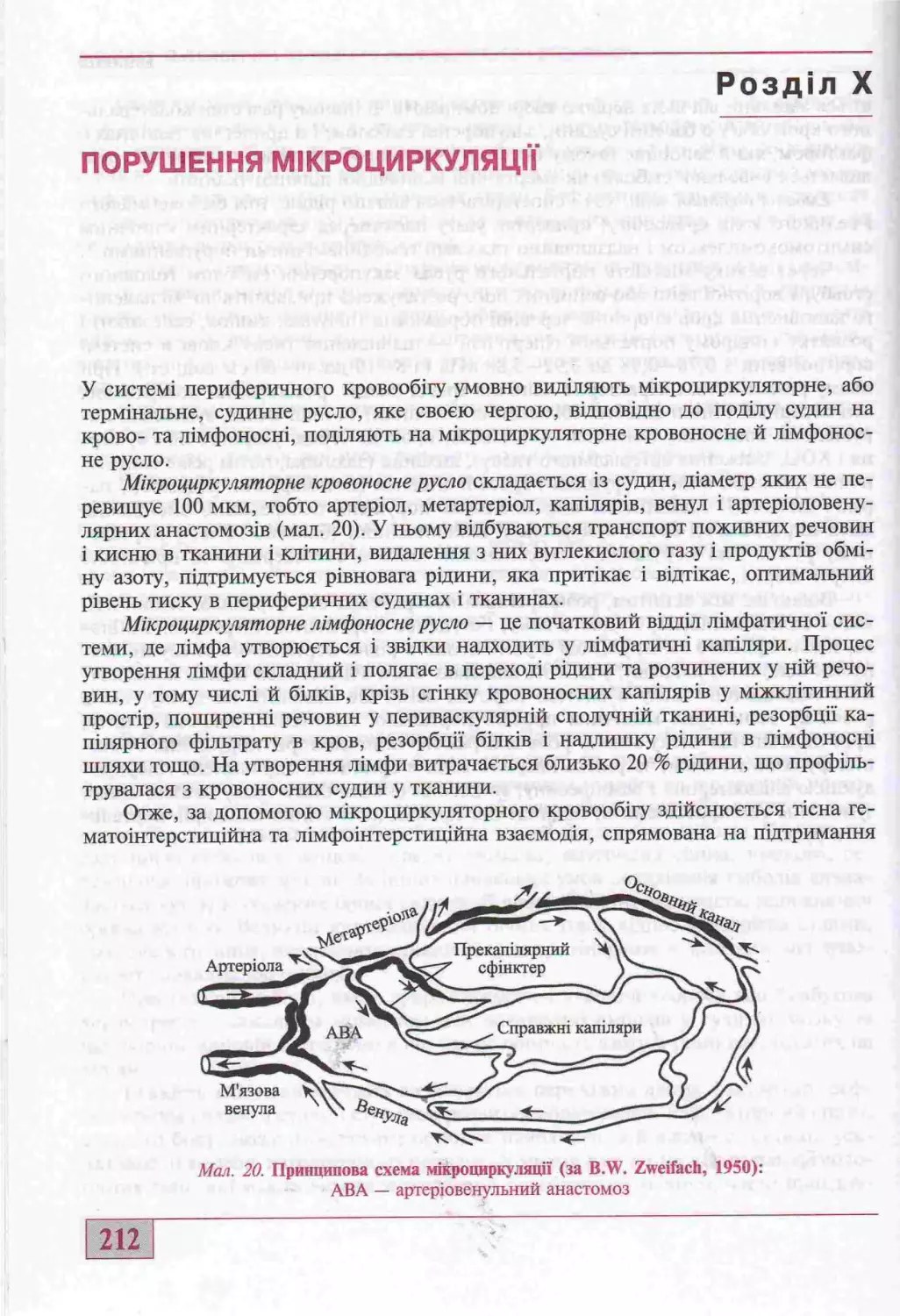
отдал X. Порушення мзкрозшркуляцн
(д-р мед. наук. проф. Ю.В. Бицъ, д-р мел. наук, проф. А.В Кубишкш)........212
Внугрз шн ьосуди н нз поруш енн я мзкропиркул яцн............................213
Позасудинн! порушення мзкроциркулянзз........................................216
Роздал XI. Запатення
(д-р мед. наук, проф М.Н Зайко, д-р мед. наук, проф. М.В. Кришталь).......219
Експериментальне выдворения запатення ...................................... 220
Етзологзя................................................................... 221
Патогенез................................................................ 221
Альтсрацзя............................................................. 224
Ексудацзя та емзграцзя ............................................. ..234
Пролзферацзя з рстзарацзя............................................. 244
Запалення та змунологзчна реактивнзеть органззму.............................249
Класифзкацзя.............................................................. . 250
Вззлив на запалення нервових з гормональных факторы .........................251
Формування загальноз теорн розвитку запалення........................... 252
Значения запалення для органззму....................................... 253
Роздал XII. Алерзтя (д-р мед. наук, проф. В. О Горбань)........................................... 254
Етзологзя........................................................... 254
Патогенез............................................................. 256
1мунолопчна стадзя алергзчних реакшй з'зх класифзказтзя...................256
Патохзмзчна стадзя алерпчних реакшй ................................... 261
Патоф’ззологзчна стадзя алергзчних реакцзи.......................... 266
Механззми формуванззя алергзчних реакцзй................................. 267
Автоалергзчнз хвороби................................................... 269
Параалергзя, гетероалергзя...................................................271
Зап об дан ня алерзи. Десенсибзлззаизя..................................... 272
Алерпя 1 спадковгсть............................................................273
Алерпя 1 запалення..............................................................273
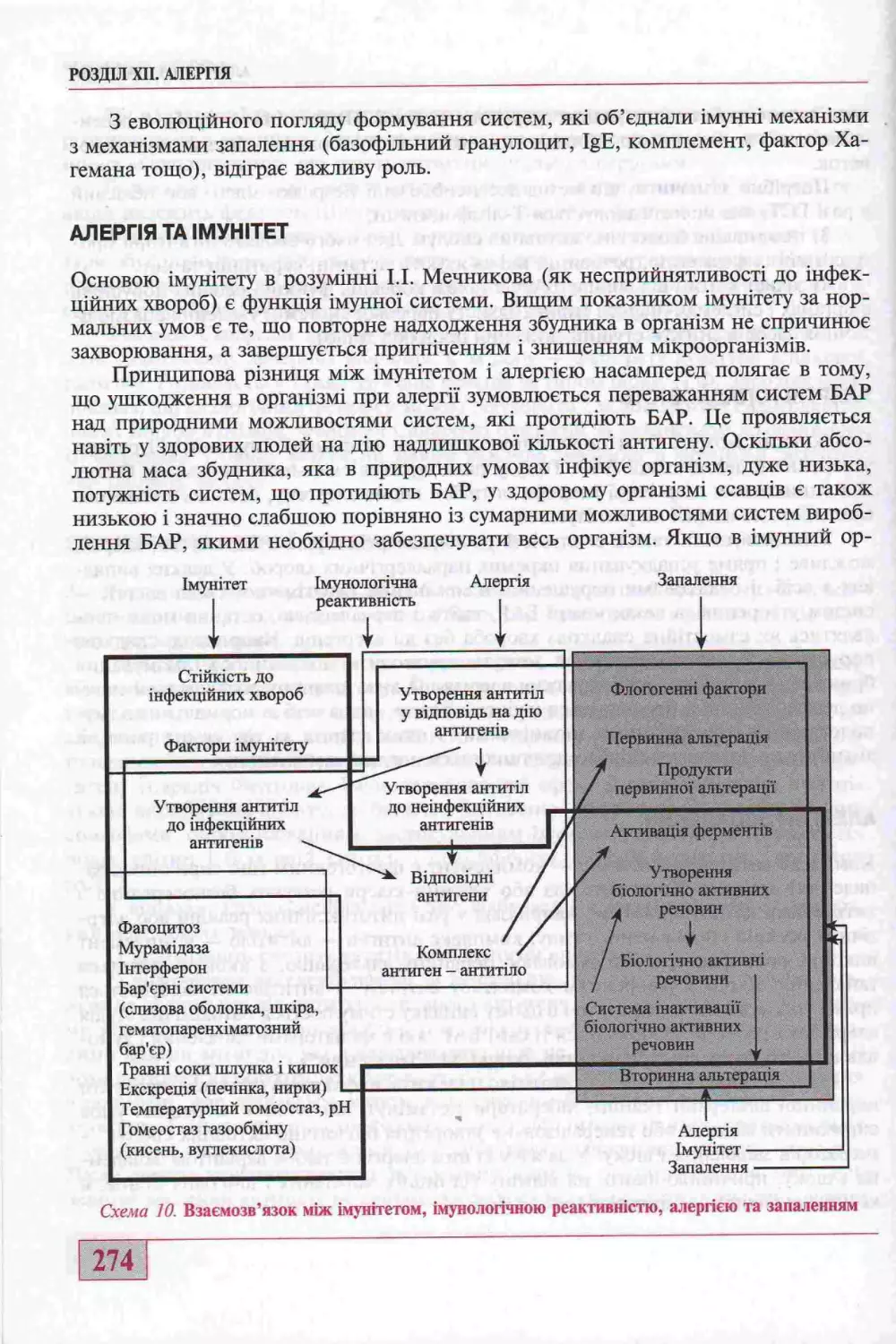
Алерпя та шунггет...............................................................274
Роздал XIII. Гарячка (д-р мед. наук, проф. М.Н. Зайко).......................................... 276
Експериментальне вивчення гарячки...............................................276
Етюлопя.........................................................................277
Патогенез.......................................................................278
Роль центр1в терморегуляцп у розвитку гарячки...................................280
Стадп гарячки. Сшввгдношення мгж теплопродукщею 1 тепловщдачею..................282
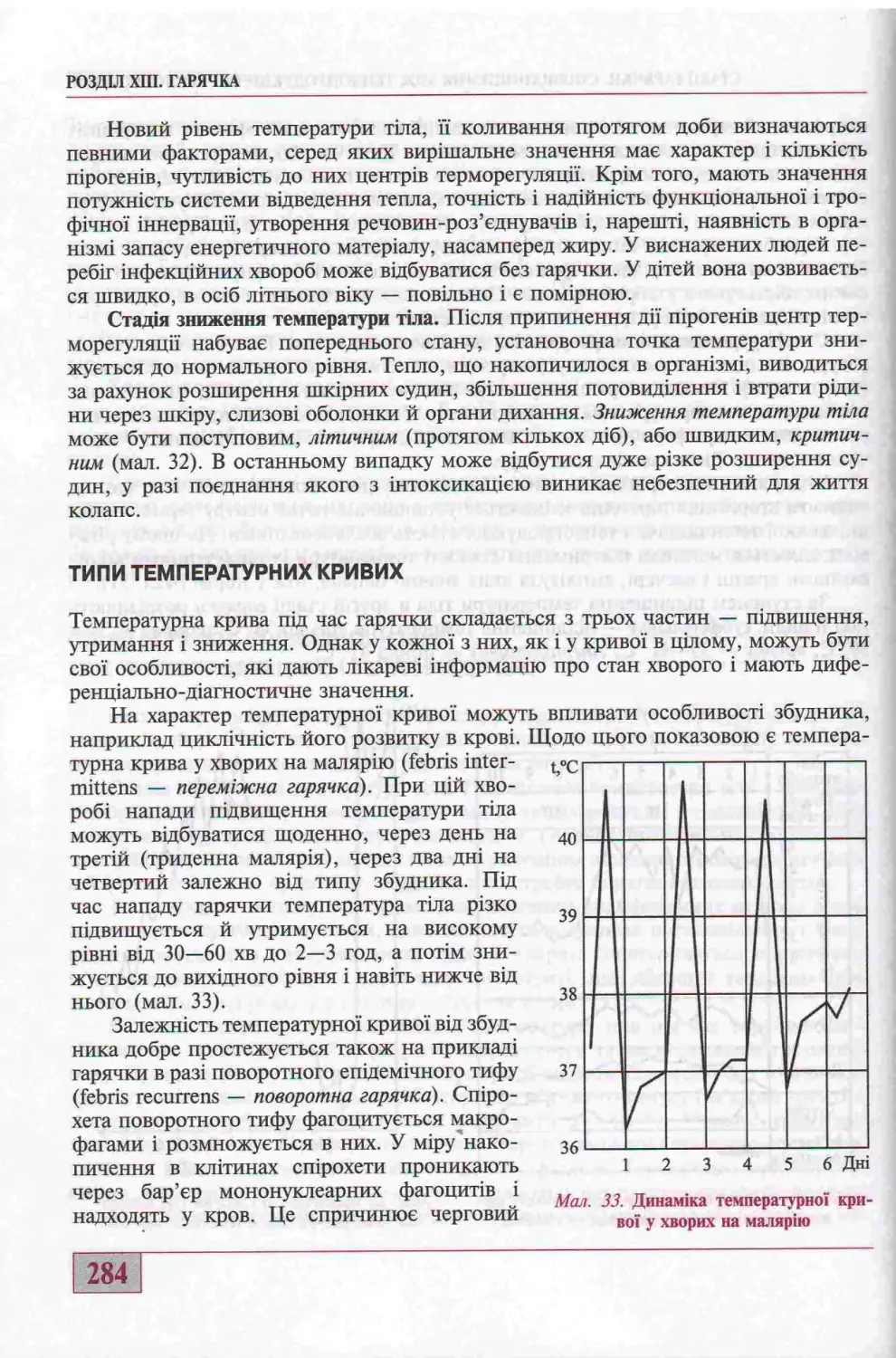
Типи температурних крив их......................................................284
Гшертерм1я та и вщмпппсть в!д гарячки...........................................286
Змзни в органах 1 системах у раз! гарячки.......................................287
Значения гарячки................................................................288
Роздал XIV. Пухлина (д-р мед. наук, проф. В. О. Горбань)........................290
Експериментальне в!дтворення пухлини............................................290
Етюлопя.........................................................................292
Патогенез...................................................;...................296
Особливост! пухлинно! тканини...................................................305
Вплив пухлини на оргашзм........................................................310
Роль оргашзму в пухлинному процес!..............................................311
Принципи лгкування пухлини......................................................312
Роздал XV. Г1ПОКС1Я (канд. мед. наук, доц. Н.К. Сгмеонова)......................314
Етюлопя.........................................................................314
Патогенез.......................................................................315
Компенсаторно-пристосувальн! реакцп.............................................316
Негайн! компенсаторно-пристосувальн! реакцп.................................316
Тривал! компенсаторно-пристосувальн! реакцп.................................318
Патолопчн! порушення при ппоксп.................................................320
Принципи л!кування та профилактики ппоксп.......................................323
Роздал XVI. Порушення енергетичного обмшу
(д-р мед. наук, проф. Л.Я. Данилова)........................................325
Роздал XVII. Голодування (д-р мед. наук, проф. Л.Я. Данилова).............................. 329
Повне голодування...............................................................329
Неповне голодування.............................................................335
Часткове голодування............................................................335
Роздал XVIII. Порушення вутлеводного обмшу
(д-р мед. наук, проф. Л.Я. Данилова)................................................................ 338
Цукровий д!абет.................................................................344
Цукровий даабет типу 1......................................................346
Цукровий даабет типу 2......................................................351
Роздал XIX. Порушення жирового обмшу (д-р мед. наук, проф. Л.Я. Данилова).......355
Розлад усмокгування 1 видалення жир!в...........................................355
Порушення транспорту 1 депонування жир! в.......................................356
Порушення промгжного жирового обмшу.............................................357
Порушення обмшу жир!в у жировш тканин!................................358
Лпндоз................................................................365
Роздал XX. Порушення бокового обмшу
(д-р мед. наук, проф. Л.О. Попова, д-р мед. наук. проф. В.С. Досенко)...367
Порушення перетравлювання I синтезу б!лк!в...................7........367
Порушення руйнування бы к!в...........................................368
Порушення обмшу амшокислот..........................7.................369
Порушення кшцевих еташв бшкового обмшу................................372
Порушення бшкового складу кров!.......................................373
Порушення метабол!зму нуклешових основ................................374
Роздал XXI. Порушення кислотно-основного стану
(д-р мед. наук, проф. М.В. Кришталь, д-р мед. наук, проф. Л. О. Попова).376
Етюлопя та форми порушень КОС.........................................378
Мехашзми компенсацп порушень КОС......................................381
Патогенез порушень КОС................................................384
Роздал XXII. Порушення водно-електролп'ного обмшу (д-р мед.
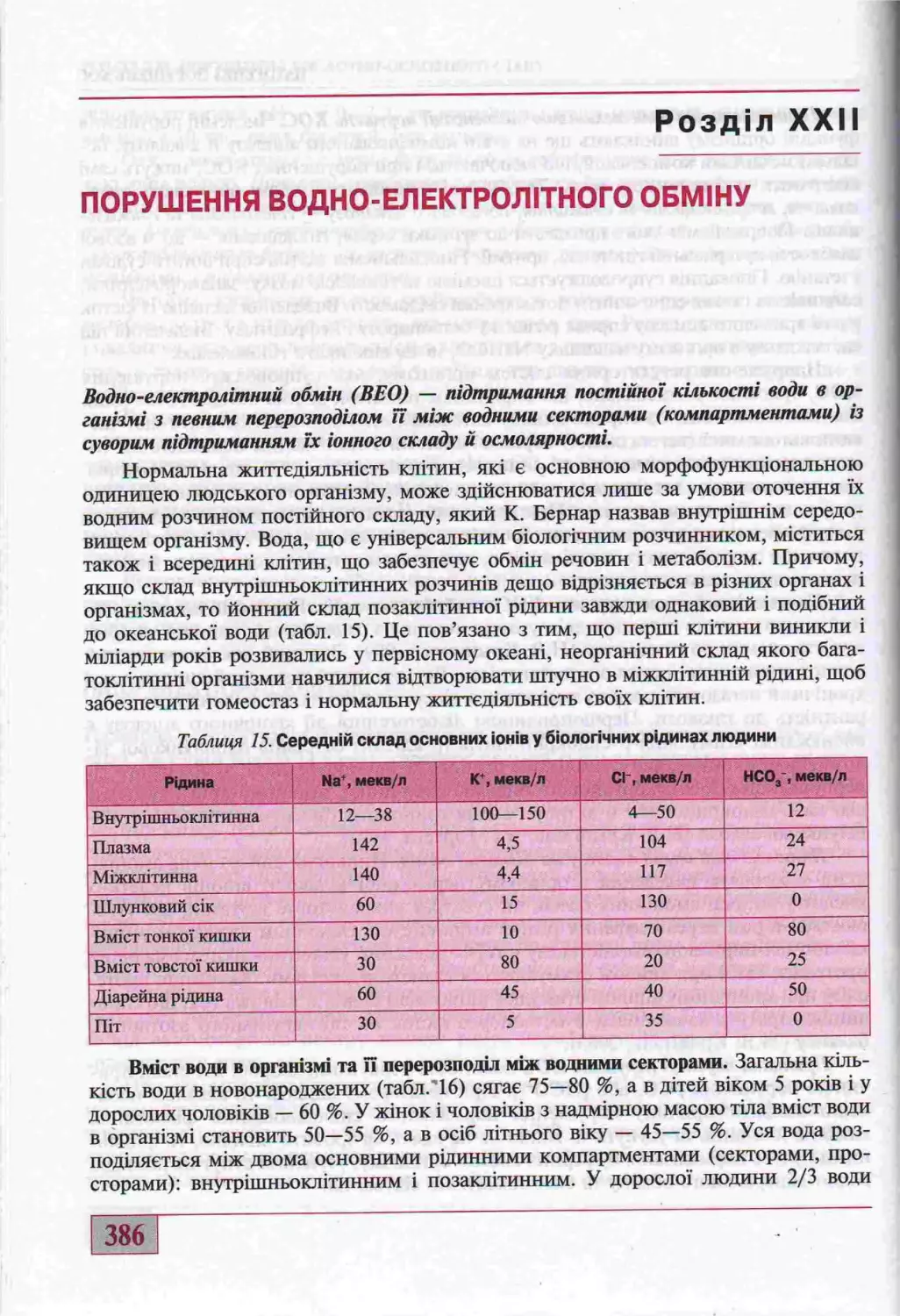
наук, проф. 0.1. Сукманський, д-р мед. наук, проф. М.В. Кришталь)...386
Р1зновиди, причини та насладки порушень водно-електролпного обмшу..........390
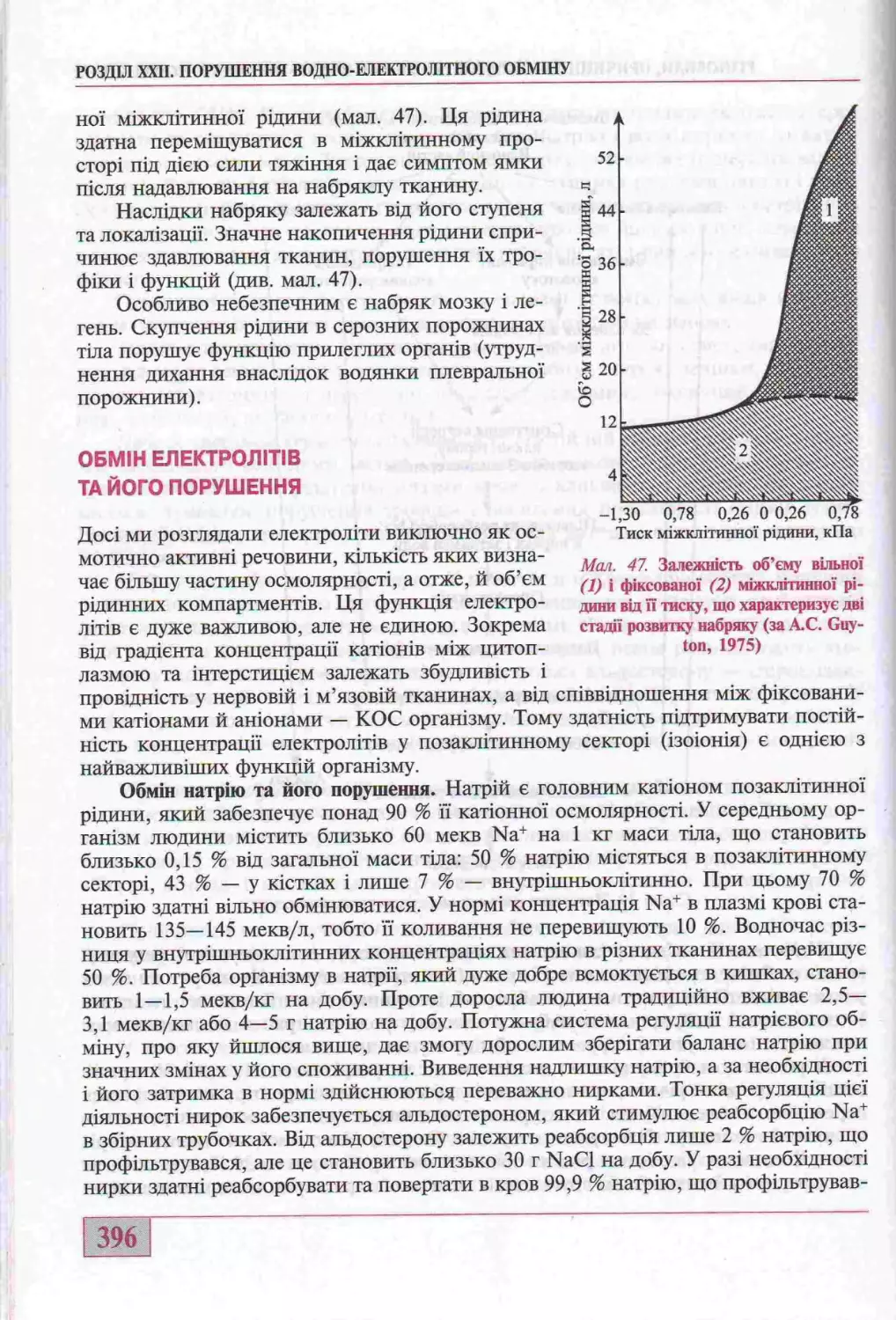
Обмш електролтв та його порушення.....................................396
Роздал XXIII. Патолопчна ф!зюлопя обмшу м!кроелеменпв
(д-р мед. наук, проф. М.В. Кришталь)...............................405
Обмш залгза та його порушення.........................................406
Обмш мщ! та його порушення............................................410
Обмш цинку та його порушення..........................................413
Обмш марганцю та його порушення.......................................415
Обмш хрому та його роль в оргашзм!....................................417
Обмш ванадпо та його роль в оргашзм!..................................418
Патолопчна фхзюлопя обмшу молгбдену...................................419
Патолопчна фхзюлопя обмшу селену......................................419
Патолопчна ф!зюлог!я обмшу йоду.......................................424
Патолопчна ф!зюлог1я обмшу фтору......................................425
Роздал XXIV. Патолопчна ф!зюлопя обмшу вггамппв
(д-р мед. наук, проф. М.В. Кришталь)................................428
Класиф!кац!я та загальна характеристика в!там!н!в людини..............428
Джерела надходження вгтампйв та !х метабол!зм в оргашзм! ............ 432
Основы! причини виникнення вггамшно! недостатносп.....................434
Недостатшсть антиневритного впамшу В,..............................436
Недостатшсть рибофлавшу............................................437
Недостатшсть антипелагричного в!тамшу РР......................... 438
Недостатшсть антидерматитного вггамшу В6...........................438
Недостатшсть фолгево! кислота......................................439
Недостатшсть антианем!чного в!тамшу В12............................440
Недостатшсть антицинготного впамшу С...............................442
Недостатшсть бютину................................................442
Недостатшсть рутину................................................443
Недостатшсть антиксерофтальмгчного вггамшу А........................443
Недостатшсть антирахггичного в!тамшу Б..............................444
Недостатшсть вггамшу Е..............................................446
Недостатшсть вггамшу Р..............................................447
Недостатшсть антигеморапчного вггамшу К.............................447
Гшервпамшози...........................................................448
Частина трети
ПАТОЛОПЧНА Ф13ЮЛОПЯ ОРГАН1В I СИСТЕМ
Роздал XXV. Патолопчна ф!з!олопя системи кров!
(канд. мед. наук, доц. 1.1. Потоцъка)..............................450
Загальна характеристика патолопчних змш у систем! кров!............,...450
Порушення загального об’ему кров!......................................451
Крововтрата.........................................................452
Патолопчш змши еритроцита..............................................454
Еритроцитоз.........................................................456
Анемгя..............................................................457
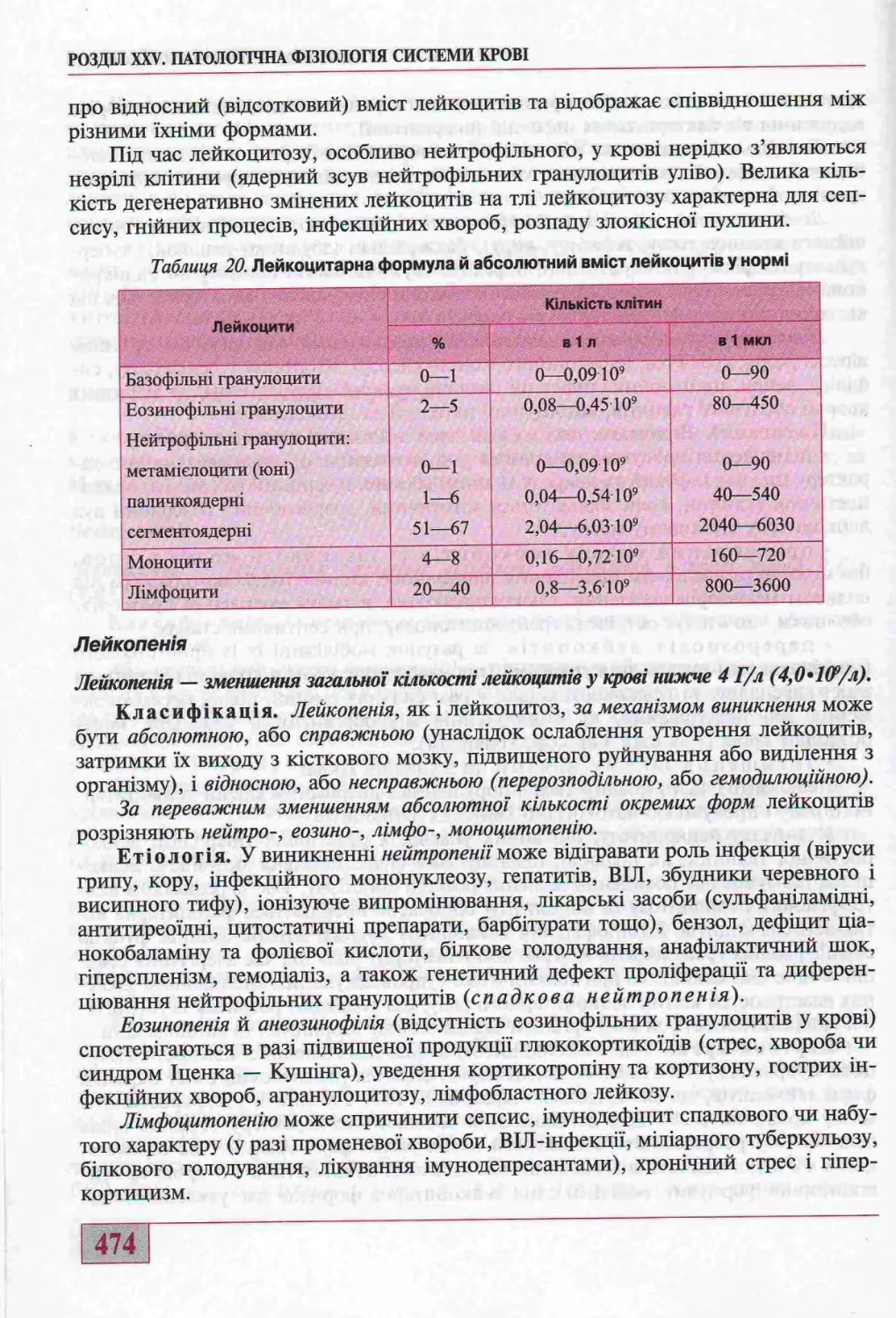
Патолопчш змши лейкоцита............................................. 470
Порушення лейкопоезу................................................470
К!лыасн! змши лейкоцита у кров!.....................................472
Якгсш змши лейкоцита у кров!........................................477
Гемобластоз. Лейкоз.................................................481
Порушення гемостазу....................................................488
Зниження задания кров! (ппокоагулящя)...............................489
Пщвишення зсщання кров! (гшеркоагулящя).............................495
Роздал XXVI. Патолопчна фгзюлопя системного кровообпу
(д-р мед. наук, проф. Г.М. Бутенко)................................498
Розлади кровообпу, зумовлеш порушенням функцп серия....................499
Недостатшсть серця..................................................499
Порушення серцевого ритму...........................................516
Недостатшсть кровообггу в раз! порушення надходження кров! до серця.522
Розлади кровообггу, пов’язаш з порушенням функци судин
(д-р мед. наук, проф. Ю.В. Бицъ, д-р мед. наук, проф. М.В. Кришталь)...523
Патолопчш змши в судинах компенсацшного типу. Атеросклероз.........525
Патолопчш змши в судинах резистивного типу.........................537
Роздал XXVII. Патолопчна фгзюлопя зовнппнього дихання
(д-р мед. наук, проф. В.А. Мёкньов)................................550
Порушення вентиляци альвеол............................................550
Дисрегуляторна вентиляцшна недостатшсть дихання.....................551
Обструктивна вентилящйна недостатшсть дихання.......................556
Рестриктивна вентилящйна недостатшсть дихання.......................559
Змши показникгв зовнппнього дихання щд час обструктивних
1 рестриктивних процеов.............................................561
Порушення дифузц газ!в у легенях.......................................563
Зменшення загального току кров! в легенях..............................565
?рушення загального 1 регюнарного вентилящйно-перфузшного
ешввщношення у легенях................................................ 565
Валив недостатност! дихання на оргашзм.............................. .567
Порушення нересшраторних функцш легень.................................. 570
Раздал ХХУТП. Патолопчна ф!зюлопя травления
(д-р мед. наук, проф 0.1. Сукмансъкий)........................................................... 572
Порушення апетиту.................................................... 572
Порушення гравлення в порожниш рота...................................... 573
Кариес зуб1в......................................................... 573
Пародонгит..................................... ... ............574
Порушення функцн слинних залоз.........................................575
Порушення ковтання................................................. 576
Порушення травления в шлунку..................................... ,...,$76
Порушення секреторно! функцн шлунка....................................576
Розлад рухово! функцн шлунка...........................................578
Пептична виразка.......................................................580
Порушення кишкового травления........................................... 583
Порушення травления, пов’язане з розладом видыення жовч)
. панкреатичного соку................................................... 583
Панкреатит....................................................... 584
Порушення мембранного травления .................................... 585
Порушення всмоктувально! та видел ьно! функций кишок...................586
Розлад рухово! функцц кишок........................................... 587
Непрохцпйсть кишок................................................ 588
Кишкова автопггоксикащя............................................. 589
Порушення синтезу гормошв системи травления.................... . .. . . 590
Роздал XXIX. Патолопчна ф!з1олопя печшки
(канд. мед. наук, доц. /./. Потоцъка)................................. 593
Загальна характеристика патолопчних процеств у печшцЬ.....................593
Недостатнкть печшки.......................................................595
Порушення метабол1чно! функцн печшки................................ 597
Порушення захисно! функцн печшки.......................................601
Порушення жовчоутворювально! та жовчовидшьно! функшй печшки.............603
Порушення гемодинам1чно! функцн печшки.......................... 613
Порушення кроветворно! функцц печшки...................................614
Роздал XXX. Патолопчна ф1зюлопя нирок (д-р мед. наук, проф. Ю.В. Бицъ,
д-р мед. наук, проф. А.1. Гоженко, д-р мед. наук, проф. М.В. Кришталь)..615
Порушення основних ниркових функшй та процесш ............................616
Порушення процесу клубочково! фшьтрацц..................................616
Порушення канальцевих процешв......................................... 618
Патогенез ниркових сицдромтв.............................. . ...........622
Нсфротичний синдром........................................ . .....624
Гостра ниркова недостатшсть.......................................... 625
Хрошчна ниркова недостагшсть........................................ 627
Тубулоштерстицшний синдром.............................................629
Етюлопя 1 патогенез основних хвороб нирок........ ... ......... 629
Гломерулонефрит.........................................................629
Пгелонефрит.............................................................633
Роздал XXXI. Патолопчна фгзюлопя евдокринног системи
(д-р мед. наук, проф. О.Г. Резников)....................................634
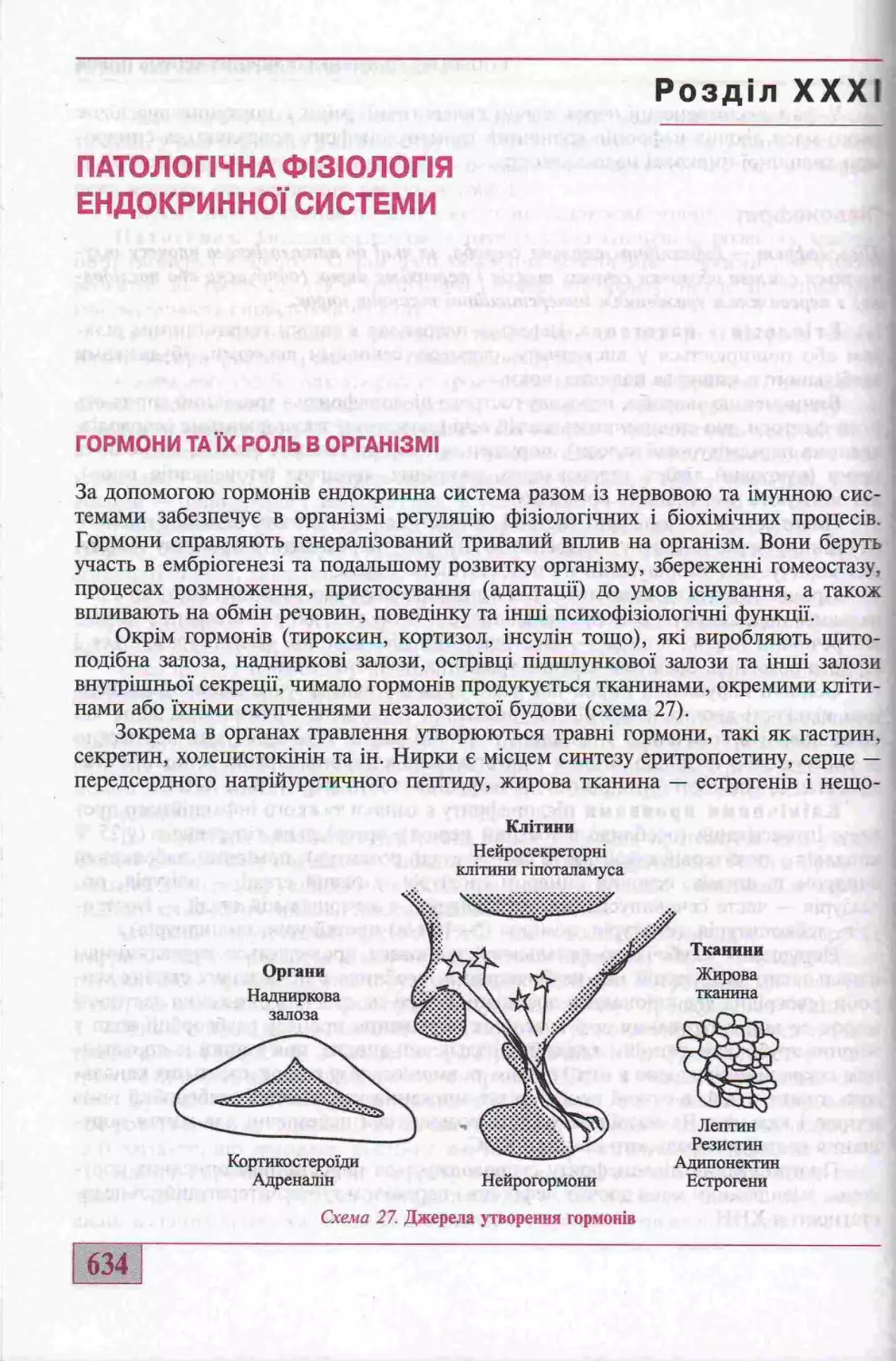
Гормони та гх роль в оргашзм!..............................................634
Етюлопя г патогенез ендокринних розладхв...................................636
Порушення функцгй ппоталамо-ппофгзарног системи............................646
Порушення функцгй ппофгза...............................................646
Нейроендокриннг хвороби.................................................649
Порушення функщй гнших ендокринних органгв.................................650
Порушення функцгй щитоподгбног залози...................................650
Порушення функцгй прищитоподгбних зал оз................................653
Порушення функцгй надниркових залоз.....................................655
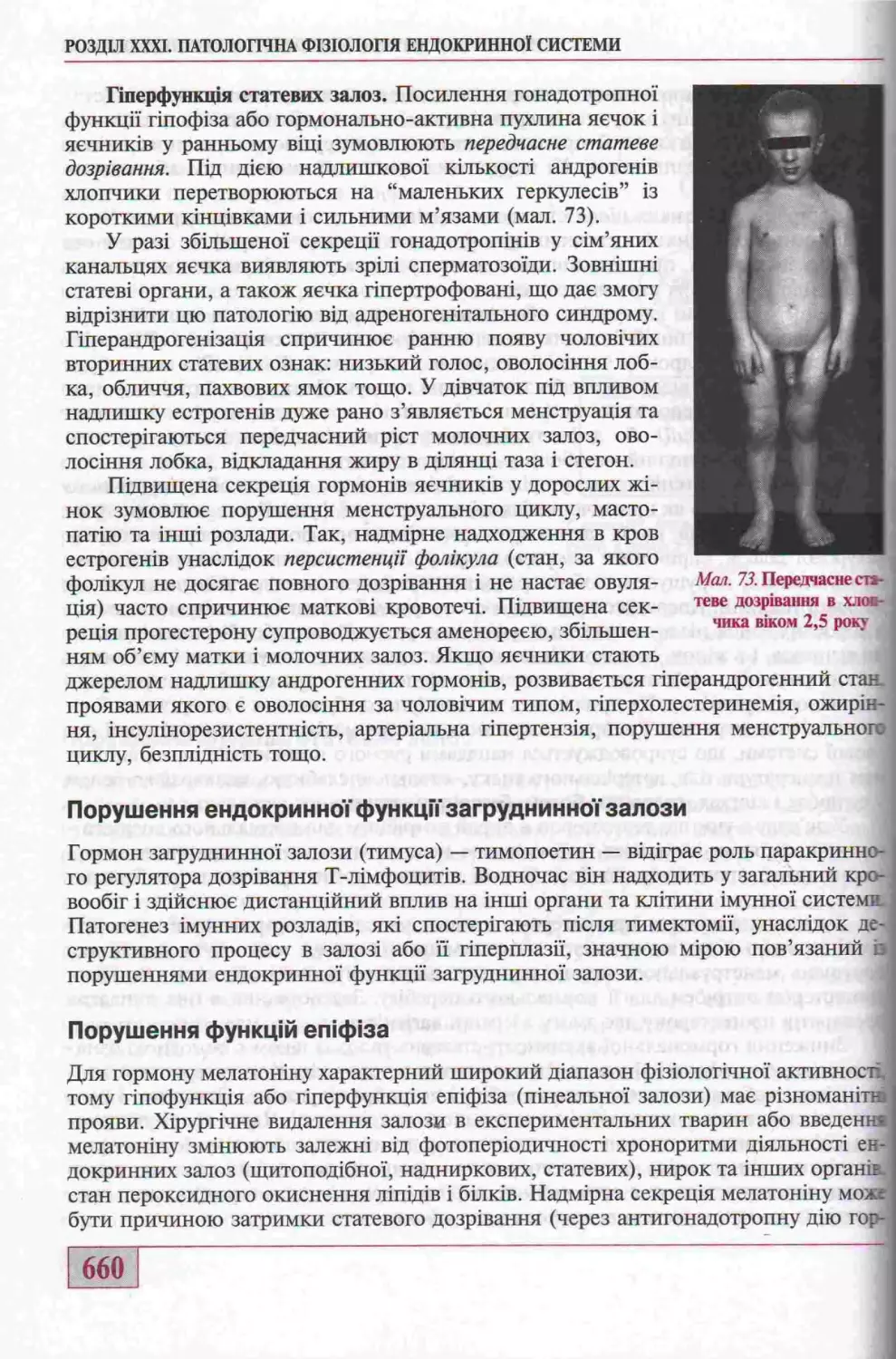
Порушення функцгй статевих залоз........................................658
Порушення ендокринног функцц загруднинног залози........................660
Порушення функцгй епгфгза...............................................660
Стрес г загальний адаптацгйний синдром.....................................661
Роздал XXXII. Патолопчна фгзголопя нервовог системи
(д-р мед. наук, проф. В.А. М1хнъов).....................................664
Етюлопя г патогенез порушень функцгй нервовог системи......................664
Порушення функщй нервовог системи..........................................667
Порушення функцгй нервових клггин г провгдникгв.........................667
Порушення функцгй синапсгв..............................................669
Порушення чутливосп.....................................................672
Бгль....................................................................674
Порушення руховог функцп нервовог системи...............................679
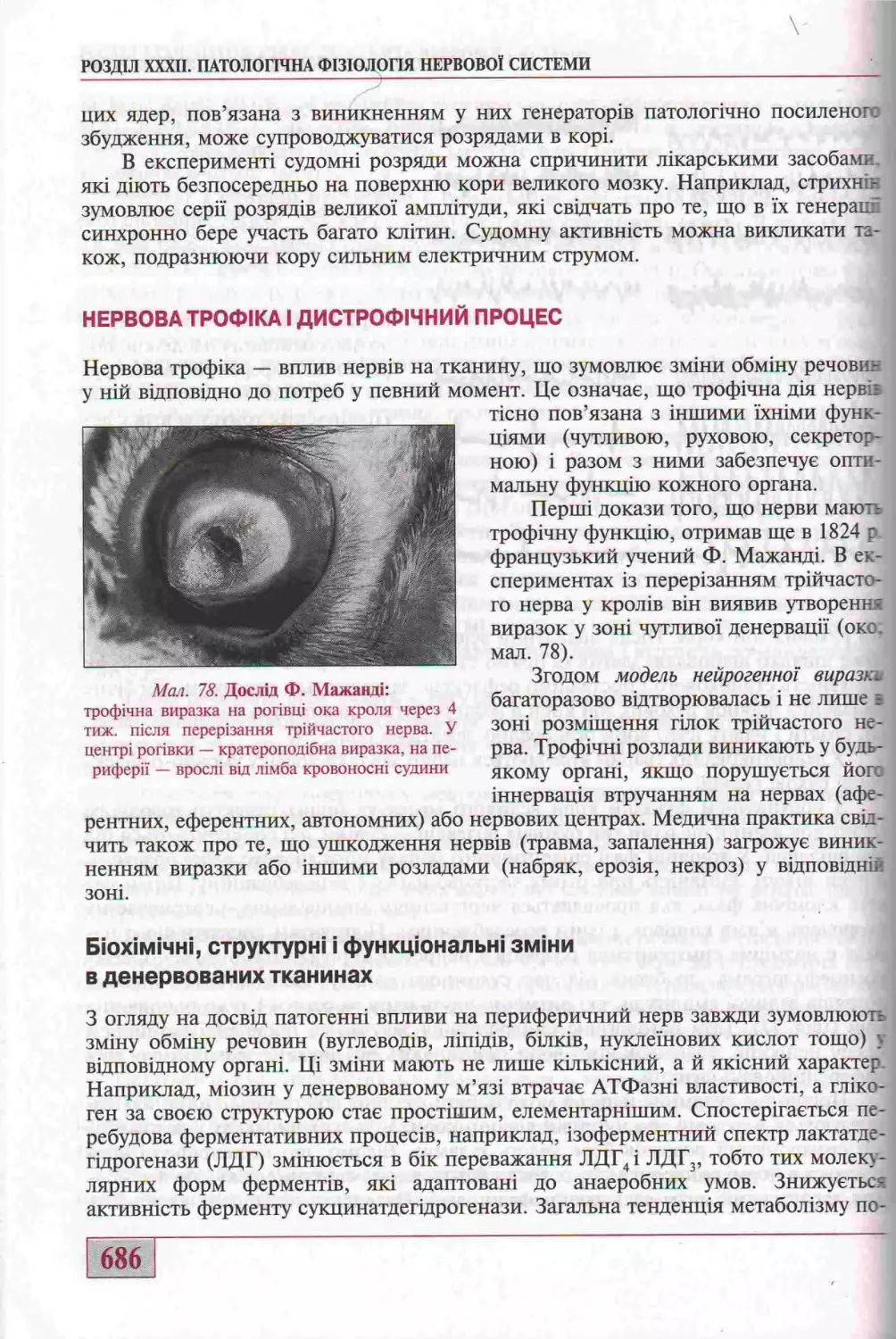
Нервова трофгка г дистрофгчний процес
(д-р мед. наук, проф. М.Н. Зайко)..........................................686
Бгохгмгчнг, структурнг г функцгональш змгни в денервованих тканинах......686
Мехашзми трофгчного впливу нервгв.......................................688
Порушення функцгй автономно! нервовог системи
(д-р мед. наук, проф. В.А. Мйсньов)........................................690
Експериментальне вивчення емоцгйних реакцгй г гх порушень..................692
Роздал XXXIII. Патолопчна фгзголопя екстремальних стангв
(канд. мед. наук, доц. Л.П. Заярна)................................................................... 697
Шок........................................................................697
Особливостг розвитку ргзних видгв шоку..................................711
Колапс.....................................................................719
Кома..................................................................... 723
Вибухова шахтна травма (д-р мед. наук, проф., В.М. бльський)...............730
Предметний покажчик........................................................732
Список скорочень
АА — апластична анемзя
Ай — альвеолярна вентиляция
ДГ — антидзуретичний гормон
ДФ — аленозинфосфат
ЗКП — антитзлозалежна клзтинна цитоток-
сичнзсть
)Л — аквапорин
сКР — активний кисневий радикал
ЧК1Г — адренокортикотропний гормон
ЧлАт — аланзнамзнотрансфераза
- ЛК — амшолевулшова кислота
Л С — антилзмфоцитна сироватка
А'ЛФ — аденозинмонофосфат
- ПК — антиген пре зентуточа клзтина
ПФ — ангюте нзинперетворзоючий фер-
мент
сАТ — аспартатамшотрансфераза
— артерзальний тиск
ТФ — аденозинтрифосфат, аденозингри-
фосфорна кислота
лФК — активна форма кисню
АХ — ацетилхолш
ЩС — антилзмфоцитарна цитотоксична
сироватка
БАР — бюлопчно активна речовина
ВГЛ — великий грану лярний лзмфоцит
- ЕБ — взрус Епстайна—Барр
ВЕС — водно-електролзтний обмзн
ВЫ — взрус шунодефзпиту людини
' ГП — вазоактивний знтестинальний пеп-
тид
*ЮЗК — внутрзшньосудинне задания кровз
" зПТ — вибухова шахтна травма
Г-6-Ф — глюкозо-6-фосфат
ГБ-ФДГ — глюкозо-6-фосфатдегздрогеназа
ГАМК - у-амзномасляна кислота
ГДФ — гуанозиндифосфат
ГЗ-ЕФР (НВ-ЕОЕ) — гепапинзв’язуючий
епзцермалъний фактор росту
111 М — гшерпродукцзя 1₽М
ПФ (Н1Е) — гшоксзе-зндуцчбельний фактор
’ГКГС (МНС, НЬА) — головний комплекс
гзсгосумзсносп
ГДЮТ — тчюкозний транспортер
ГМФ — гуанозинмонофосфат
ГНН — гостра ниркова недостатшсть
ГП — глзкозтротезн
ГРДСД (аКЛ8) — гострий респзраторний
дистрес-синдром дорослих
ГСТ — гшерчутливзсть сповпьненого типу
ГТК — 1 щростатичний тиск кровз
ГТФ — гуанозинтрифосфат
ГФРТ — ппоксантиигуанзнфосфорибозил-
трансфераза
ДАБ — диметиламзззоазобензол
ДАТ — дзастолзчний артерзальний тиск
ДБА — дибензантрацен
ДВЗ — дисемзноване внутрпиньосудинне
зсздаззня кровз
ДМБА — циметилбензантрацен
ДНК — дезоксирибонузслезнова кислота
ДФГ — дифосфоглщерат
ЕКЛ — ендозелзальна клзти на-лопереди ик
ЕП (ЕрО) — еригропоетин
ЕР — ендоплазматичний ретикулум
ЕФР (ЕОГ) — епздермальний фактор росту
ЕЦОК — ефектзгвно циркулюючий обмш
кровз
ЗВ1Д — загальний варзабельний змунодефь
цит
ЗДА — залззодефщитна анемзя
ЗЛО — загальний периферичний ошр
13ЦД — шсул1нозалежний цукровий дзабет
1Л — штерлейкш
1НЦД — знсулзнонезалежний цукровий дза-
бет
ШФР — знсулзноподзбний фазстор росту
зРНК — шформацзйна РНК
РНК! — знтерферуючэ РНК
1Ф — знтерферон
1ФС — штенсивнзсть функцзозлування
структури
1ХС — зшемзчна хвороба серия
Ко А — коензим А
КОС — кислотно-основний стан
КП — колзрний показник
КСФ — колошестимулюючий фактор
КФ — креатинфосфат
ЛАК — лзмфокзнактивованпй юл ер
ЛДГ — лактатдегщрогеназа
ЛП — лейкоцитний троген
ЛПВЩ — лшспротезд високоз зшльностз
ЛПДНЩ — лшопротезд дуже иизькоз зцсль-
ностз
ЛПНЩ — лшопротещ низько! пцльносп
ЛППЩ — лшопротещ помтрнот щйгьносп
ЛПС (БР8) — лшополюахарид
ЛРТ — лппдний радютоксин
ЛТ — лшфощний токсин
МД С — м!елодиспластичннй синдром
МЗБ (МЕР) — макрофагальний запальний 61-
лок
МКСФ — макрофаго колсшестимулюючий
фактор
МОД (МОЙ8) — синдром мультиорганно'1
дисфункцц
мРНК — магрична РНК
МСГ — меланоцигостимулюва-тьний гор-
мон
МТФ (МТР) — метал отранскритпцйний фак-
тор
МХБ (МСР) — моноцитний хемоатрактант-
ний быок
НАБ (МАР) — нейтрофшьний акгивуючий
61ЛОК
НАД — н1котинам1дадеи1ндинуклеотид
НАДФ — шкотинамщадешндинуклеотидфос-
фат
НАЛ — недостатшсть адгезц лейкоцита
НЕЖК — неестериф^кована жирна кислота
НК (МК) — клггини — натуральн! (природа!)
кыери
НПЗЗ — нестерощний протизапальний заей
ОБО — основна багатокдггинна одинипя
ОО — основний обмпт
ОТ К — онкотичний тиск кров!
ОТТР — онкотичний тиск тканинно! рщини
ОФВ — об’ем форсованого видиху
ОЦК — об’ем пиркулюючо! кровт
ПАВ — ПОЛ1КЦИКЛ1ЧНИЙ аромата чний вуг-
леводень
ПГ — простагландин
ПЛР — пол!меразна ланцюгова реакция
ПНЖК — полшенасичена жирна кислота
ПНП (АНП) — нередсердний (атртальний)
натртйуретичний пептид
ПО — периферичний оп!р
ПОЛ — пероксидне окиснення лпцдтв
ПОН — полюрганна недостатшсть
ПРОА — повптьно рсагуюча субсганшя ак-
афттаксп
ПФШ — пентозо-фосфатний шунт
РААС — решн-анпотензин-альдостеронова
система
РАС — решн-альдостеронова система
РНК — рибонукле'!нова кислота
САА — сироватковий амйкмд А
САТ — систол!чний артер!альний тиск
сАТ (Р) — системный артср!алъний тиск
СГЛУ — синдром гострого легене вого уш-
кодження
СЕФР (УЕ6Р) — судинний ендотел!альний
фактор росту
СМФ — система мононуклеарних фаго-
цита
СН1Д — синдром набутого !мунолефщиту
СОД — супероксидашондис мутаза
СПА — сироватковий попередиик амиоТ-
дозу
СРП — С-реактивний протеш
ССЗВ — синдром системно? запально!' вщ-
ПОВ1Д1
Т3 — трийодтиронш
Т4 — тироксин
Тте§ — Т-л!мфоцит-хелпер регуляторний
ТГФК — тетрапдрофолква кислота
ТЗКЛА — тиск заклинивания кашлярга леге-
нево! артерп
ТК1Д — тяжкий комбшовдаий (мунодефт-
цит
Тл — Т--|Пмфопит
Тл-сф — Т-л!мфоцит-альфа-бета
Тл-у8 — Т-лтмфоцит-гамма-дельта
тРНК — транспортов РНК
ТФ (РЕ) — тромбоцитний фактор
ТФР (ТСР) — трансформуючий фактор росту
Тх СПт) — Т-л!мфопит-хелпер
УДФ — урилинфосфат
УЗД — ультразвукове дослщження
УОС — ударный об’ем серця
ФАМ — фактор агрегацп макрофапв
ФАТ (РАЕ) — фактор актавацп тромбоцита
ФГД — фосфоглюконатдетщрогеназа
ФЖСЛ — форсована житгева емшеть легень
Ф1М — фактор (нпбування макрофапв
Ф1ММ — фактор, що шпбуе мпрацпо мак-
рофапв
ФНП — фактор некрозу пухлин
ФРК (КОР) — фактор росту кератинонита
ФРН (МОЕ) — фактор росту нерв!в
ФРПС — фосфорибозилтроФосфатспнтетаза
ФРТ (РОСЕ) — фактор росту тромбоцита
ФРФ (РОР) — фактор росту ф’бробласта
ФТ — фщьгращиний тиск
ФХЕ — фактор хемотаксису еозиноф!л!в
ФХМ — фактор хемотаксису макрофапв
ФХН — фактор хемотаксису нейтрофпнв
Х-АГ — Х-зчеплена ага.ммаглобул!нем!я
ХНН — хрошчна ниркова недостатшсть
ХОС — хвилинний об’ем серця
ХРТ — хшоновий радютоксин
цАМФ — цитепчний аденозинмонофосфат
цГМФ — ЦИКЛ1ЧНИЙ гуанозинмонофосфаз
ЦЗК — циклшзалежна кшаза
дк. — циркулюючий 1мунний комплекс ЛЧО8 - щдуцибельна КО-синтетаза
дне — центральна нервова система 1КЕ - залгзорегулюючий елемент
ТУК — цикл трикарбонових кислот ПЕР - зал1зочутлиьий биток
ЗФНК — цитотоксичний фактор НК 1г-ген — шунореакшвний ген
чес — частота серцевих скорочень ЕВР - лцтополгеахаридзв’язувальний 61-
Т)КФ — птвидкють клуб очков о! фшьтрацп лок кров) ш
НОЕ — швидккть осщання еритроципв МТГ - металрегульований фактор транс-
-М — а-макроглобулш крипип
с — компонент комплементу МТР - биток — переносник металу
СТ) — кластер диференщацп ОТ-кВ - нуклеарний фактор кап па-В
СЧО8 — конститутивна ЫО-синтетаза п18О8 - нейрональна ЪЮ-синтетаза
СК — рецептор комплементу ЫО8 - МО-синтетаза
От — переносник мцц рО, (РаО2) — парщальний тиск кисню в ар-
ЗХЕ — бйюк-сироватковий шпбгтор ком- плементу рСО2 - тер1альшй кров, паршальний тиск вуглекислого
4АТР — дезоксиаденозинтрифосфат газу
Х5ТР — дезоксигуанозинтрифосфат НаОТ - рецегпори на остеокластах, через
О МТ — переносник двовалентних метал1в як1 активуеться ОТ-кВ
пет — переносник двовалентних катюшв КАМК1- лиганд ВЛМК
е\О8 — ештел1альна ЫО-синтетаза ВАОТЕ8 - регулятор активапй, експресп та
НО- — концентрация кисню у видихувано- МУ ПОВ1Тр1 ТЕК - секрецп нормальних Т-кл1гин То11-под1бнии рецептор
ШЕ — транскрипщйний фактор УСАМ - молекула адгезп судинного ендо-
Н5Р — протеш теплового шоку телпо
1САМ — молекула мЬкклггинно! адгезп 2г.1Р - биок — хмпортер цинку
12 — 1муноглобулш 2пТ - переносник цинку
Передмова
Цей шдручник мае 36-рхчну хсторпо видання. За зазначений перюд в!н неод-
норазово виходив у св1т рхзними мовами: росшською (1977, 1985, 1996), румунсь-
кою (1991), украшською (1995, 2008, 2010). Варто вщзначити, що стереотипне
перевидання третьего видання пщручника (1996) росшською мовою (2006, 2007,
2008) отримало рекомендащйний гриф МОЗ РФ щодо використання його як пщ-
ручника для студенпв вищих медичних навчальних закладхв уехх спещальностей.
Для пщручника завжди характерними були достатня повнота 1 доступнхсть
викладу фактичного матерхалу з питань загально! та специально! патолопчно!
фхзюлогп вщповщно до стану й розвитку в евгп на момент виходу друком. Завдя-
ки цьому перше видання (1977) росшською мовою було вщзначено Державною
премхею Укра'ши в галуз! науки х техшки. 1нпй видання також отримували пози-
тивы вщгуки користувачхв.
Вщ моменту виходу в евхт першого видання “Патолопчно! фхзюлогп” ук-
рашською мовою минуло 18 рокхв. За цей час у цш галузх вщкрито нов! факти х
закономхрностх шодо етхологй, патогенезу, проявгв найпоширенххххих патолопч-
них станхв, патофхзюлопчних синдромхв, хвороб, з’явилися нов! науковх узагаль-
нення х напрями. Через це у подалыпих виданнях навчальний матерхал постшно
оновлювався, доповнювався й осучаснювався.
У четвертому виданы оновлено змхет майже вехх роздхлхв хз загально! патоло-
ги та патолопчно! фхзюлогп оргаыв ! систем. У викладх питань спецхально! пато-
лопчно! фхзюлогп основну увагу придыено загалъним х молекулярним мехаызмам
порушень х компенсацп функцш оргаыв ! систем. У бшьшосп випадкхв як пато-
лопчний стан розглядаеться недостатнхсть того чи того органа (системи оргаыв).
Порхвняно з останнхми виданнями в четвертому виданы суттево перероблено
тага роздхли, як “Патолопчна фхзюлопя хмунно! системи”, “Алерпя”, “Запален-
ня”, “Пухлина”, “Порушення вуглеводного обмхну”, “Порушення кислотно-ос-
новного стану”, “Порушення водно-електролхтного обмхну”, “Розлади кровообх-
гу, пов’язаы з порушенням функцн судин” та !н. Заново написано роздхли “Па-
тофхзюлогхя обмхну мхкроелеменпв”, “Патофхзюлопя обмхну вггамхшв”,
“Вибухова шахтна травма”.
Пщручник написано авторським колективом провщних фаххвцхв рхзних ви-
щих навчальних закладхв та академхчних науково-дослщних шетитутхв Украши,
якх е визнаними фаххвцями з рхзних питань патофхзюлогп, що дае змогу вважати
цей базовий пщручник нацюнальним.
Матерхал пщручника викладено вщповщно до нових навчальних програм
(2006—2010) з патолопчно! фхзюлогп, складених на основ! кредитно-модульно!
системи оргаызацп навчального процесу.
Пщручник призначений для студенпв вищих медичних навчальних закладхв
III—IV рхвнхв акредитацп. Вш може бути корисним ! для викладачхв цих закладхв,
а також лхкархв уехх спещальностей.
Доктор медичних наук,
професор М.В. Кришталь
Вступ
... Коли починаеться хвороба — тут
кьнчаеоься талузь веления нормально!
фозюлогц, починаеться фозоологоя хворого,
патологичного оргашзму.
В. В. ГВдвисоиъкий
- РЕДМЕТIЗАВДАННЯ ПАТОЛОПЧНО1Ф13ЮЛОГН
Тх^о.юг1чна ф1з1олог1я — це наука, що вивчас життсд1я.1ьн1сть хворого оргашзму.
~ вчальна програма, за якою здшснюеться подготовка майбутнох л (каров о прово-
-»!13 у вищих медичних о фармацевтичних навчалъних закладах Укра'Гни, побудо-
ша так, що опечатку вивчаюоъся предмета, яко розглялають структуру, функцй
метаболозм нормального оргашзму (нормальна анатомоя, псоологоя. бюхомоя.
рмальна фозоологоя), а потом — предмета, завдання яких — донести до студента
з плоя про змови струк тури о функцо!, метаболой но порушення хворого оргашзму,
с там самим подвести його безпосередньо до клинки. Тако завдання виконують
юлопчна анатомоя о патолопчна фозоологоя.
Отже, патологична ф!зюлог1я — це наука, що вивчае загальн! закономерности
ялникнення, розвитк) та завершения хвороби.
Ниш налочуютъ близько 4000 нозолопчних форм хвороб. Практичний локар
Чрагне визначити хворобу, водрознивши и вод !шпоо, гобто встановити правилъний
гноз о призначити рацоональне локування згщно оз загальноприйнятими стан-
зоами надання медичноо допомоги. Патофо зоолог абстрагуеться вод частко-
осгей, намагаючись знайти те загальне о головне, шо характеризуе велико грули
.|роб або навоть одну конкретну хворобу. Концевою метою патолопчноо фозооло-
гм е розкриття законов, за якими розвивасться хвороба, 1 тут патолопчна фозооло-
~ вступае в тосний зв’язок оз фитософоею.
Водповодно до нових навчалъних програм для студентов розних факультетов
иих медичних навчалъних закладов III—IV ровное акредитацоо (Кшв, 2006—2010),
г.тадених на основ! кредитно-модульно! системи орган! зашо навчатьного проце-
у, курс патолопчноо фозюлоон подоляеться на два модул!.
Модуль I "Загальна патология ''мостить три змостових модуль Перший змосто-
ни модуль — це “Загальна нозологгя”, або ‘Загальне вчення про хворобу”. Под час
ьч алозу хвороби локар повинен дата водповодь на два зэпитання: чому виниюта
о роба? який механозм н розвитку? Обощва запитання (етоолопя ! патогенез) е
п зовними в паоологочной фозюлоги. Виргшення !х передбачас аналоз складних
э демозв язков мож навколишном середовищем о организмом. Нэвколишне середо-
шце розглядаеться як джерело розних, у тому число й патогенних, вооливов, а ор-
анозм — як боолопчна система з розним и ровнями реоуляпо!, йооо спааковостю о
0КТИВН0С1Ю.
Другий змгстовий модуль “ Типов! пато.юггчш процеси ” мктить дан! про про-
неси, що лежать в основ! багатьох хвороб: ушкодження клгтини, алерпю, пору-
шення периферийного кровообпу та мгкроциркуляцв, запалення. гарячку, пухли-
ни, голодування. гшокспо.
Трепй змктовий модуль “ Типов! порушення обмшу речовин" передбачае послг-
довне вивчення поргтиень енергетичного, вутлеводного. жирового та бокового об-
ману. порушень обмшу пуринових 1 шримщинових основ, обмшу в!там!н!в. м!кро-
елементгв, водно-електролггного обмшу та кислотно-основного стану (КОС).
Модуль II “Патологлчна ф!31олог!я оргашв ? систем ” передбачае вивчення чо-
тирьох зм1стових модулгв.
Четвертой змктовий модуль вивчае патолопчну фгзюлогйо системи кров!,
п’ятий — патолопчну ф!з!ологпо системного кровообпу та зовнппнього дихання,
шостий — патолопчну ф!з!олог!ю травления, печшки, нирок, сьомий — патоло-
пчну ф!з!оло1!ю регуляторних систем (ендокринно!, нервово!) та екстремальних
стан!в. I хоча ня частона називаеться спещальною патолопчною фгзюлопею, тут
знову-таки основними е загальнопатолопчн! аспекта. Патоф!з!олог прагне з’ясу-
вати загальш закономгрност! порушень функцгй вцшовшгих орган!в 1 систем,
компенсаторш можливост! оргашзму. ГПд час патофгзюлопчного анал!зу прояви
хвороб лгкар вщдае перевагу не стгтьки характерним симптомам, скьтьки синдро-
мам г патолончним станам, насам перед недостатност! оргашв ! систем на еташ
компенсованого переб!гу г в раз! розвитку деком пенсацц. П!д ним кутом подаеть-
ся потр!бний материал г з найпоширеншгих нозолопчних форм. Природно, шо
проблеми гх етюлогн га патогенезу е ключовими.
Отже, патолопчна ф!з!олопя е штегральною професпшо зоргентованою ме-
ди ч ною дисциплшою, своергдним мктком мгж базисними медико-бюлопчними
дисшптлшами та клшгкою.
ЗВ'ЯЗОК ПАТОЛОПЧНО1 ФЕЗЮЛОГП З 1НШИМИ
МЕДИЧНИМИ НАУКАМИ, ТГЗНАЧЕНИЯ ДЛЯ КЛ1Н1КИ
Патолопчна ф!з!олопя спираетъся на науки, що передують !й, насамперед на и
дан!, як! дають нормальна ф!з!олопя I бюхгмгя. Дуже багато сшльного у патоло-
пчно! фгзюлоп! з нормальною фтзюлопею (патолопчну фгзюлопю шод! назива-
ють фтзюлопею хворого оргашзму). Обидв! прагнуть в!дкрити закони, за якими
живе г функшонуе оргашзм. П! закони мають велике значения для розумшня
хвороби. Однак було б помилкою думати, що хвороба е лише якоюсь шитою ком-
бшашгю нормальних пронесгв. Патолопчний процес зумовлюе якгсно новий стан
оргашзму. Звщси й вщмшшсть патолопчно! ф!з!слогп вщ нормально’!, а основне
п завдання — вивчення вс!х прояв!в хвороби для того, шоб вгцкрити закони, за
якими вона розвиваеться.
Патолопчна ф!з!олог!я псно пов’язана г з патолопчною анатомгсю. Понад
140 рок!в тому це була одна наука. Под!л ц був зумовлений тим, що самого мор-
фолопчного анализу для розумшня динамгки паголопчного процесу було недо-
статньо. Функтцональш змпги псно пов’язаш з морфолопчними, проте м!ж ними
не завжди спостерггаеться сувора в!дпов!дн!сть. Пе вгдбуваегься зокрема тому, що
кожен орган мае пристосувадаш та компенсаторш можливост! г його функп!я в
---—---------------------------------------------------------------
раз! змшено! струкгури може бути не порушеною. Тому деяк! питания клшхки не
могли бути вирпнеш тд час розтину трупа з подальшим мхкроскопхчним дослщ-
женням секщйного матерхалу. Таку можливхсть надавало лише спостереження за
розвитком патолопчного процесу в живому органхзмх, передусхм в експеримент!.
Широке використання експерименту е вщмхнною ознакою патолопчно! фхзюлогп
вщ патолопчно! анатомп.
Патолопчна фхзюлопя пов’язана з клгнхчними науками. Кхнцева мета в них
спхльна. I клхнхчнх науки, х патолопчна фхзюлопя вивчають хворобу для того, щоб
ефективно запобхгти хй х лхкувати хворих. Проте найближчх завдання, а також
методи й об’екти у них рхзнх. Об’ектом вивчення клхнхчних наук е хвора людина
з конкретними проявами хвороби. Наприклад, клхнхка внутрххххнхх хвороб вивчае
причини виникнення, симптоми, особливост! перебхгу, метода лхкування х про-
гноз таких захворювань, як хронхчний гломерулонефрит або системний червоний
вовчак. Невролопчна клхнхка в тих самих напрямах вивчае енцефаломхелхт, що
розвиваеться, наприклад, теля антирабхчного щеплення. Клшхка хвороб очей
здавна стикаеться з тим, що коли уражене одне око, то з часом може захворхти
шше, хнтактне. Усе не — рхзнх хвороби з рхзними симптомами, локалхзацхею, про-
гнозом. Та водночас мхж ними е щось спхльне. Виявляеться, усх щ захворювання
мають стльний автохмунний мехашзм. У розкритп зазначено! закономхрностх ве-
лике значения мае патолопчна фхзюлопя.
Ще приклад. Клхнхчнх дисциплши вивчають конкретнх форми прояву добро-
якхсних 1 злоякхсних пухлин. Щоб установи™ дхагноз раку шлунка, саркоми стег-
на, мюми матки, потрхбно знати, що !х вщрхзняе. Перебхг ! прогноз 1х рхзнх. Однак
поглиблене розумхння цих хвороб потребуе визначення спхльних ознак: як нор-
мальна клхтхша перетворюеться на пухлинну, як змхнюеться п спадковхсть, звщки
бере вона енергио для свого розмноження, як! ланки ц метаболхзму найчутливппх
до впливу лхкарських засобхв тощо. Усе не — сфера патолопчно! фхзюлогп.
Отже, патолопчна фхзюлопя з погляду спецхальних клхнхчних дасциплхн — це
наука, що вивчае пронеси, спхльн! для всхх хвороб або для великих ххнхх труп.
Зрозумхло, що клпххчнх науки х самостхйно можуть отримати важдав! дан! про
сутнхсть хвороби, механхзм хх розвитку, особливо тепер, коли вони “озброенх”
методиками тонких бюххмхчних х фхзхолопчних дослщжень. I все-таки клхнхка
потребуе фундаментальних даних про хвороби х закони ххнього розвитку. Цх дан!
вона може отримата в!д патолопчно! фхзюлогп, що мае метод, якого немае х не
може буш в клхнщх, — метод патоф!зюлог1чного експерименту на тваринах.
ЕКСПЕРИМЕНТ - ОСНОВНИЙ МЕТОД ПАТОЛОПЧНО!Ф13ЮЛ0ГН
Об’ектом вивчення патолопчно! фхзюлогп е хвороба, методом — патофхзюлопч-
ний експеримент. Експериментом користуються багато наук (нормальна фхзюло-
пя, фармаколопя тошо). Сутахсть експерименту в патолопчнхй фхзюлогп полягае
в тому, щоб спричини™ хворобу у тварини, вивчити н й отриманх данх перенести
в клхнхку. Гнодх заперечують, що завдяки дослщам на тваринах неможливо роз-
крхгш сутнхсть хвороб людани та вщтворити вех наявнх хвороби. Тодх на допомогу
приходить клшхко-патофхзюлопчнх метода дослщження з використанням сучас-
но! хнетрументально! технхки, тонких цитолопчних, бюххмхчних, хмунологхчних та
1нших методы. При иъому важливо вщзначити, щс проведения експерименталь-
них дослщженъ у сучасних умовах потребуй дотри мания принцшпв “Свропейсь-
ко! коквешги про захист хребетних тварии, яких використовують для експери-
ментальних ) наукових гилей” (Страсбург, 1985) 1 вимог (морально-етичиих норм)
Нацюнального комитету з бюетики та комкай з бюетики навчалъних закладов 1
наукових устаиов
Хвороба людини настшьки складна, а мехаызми и шод) так глибоко прихо-
ваш, що розкрити тх складно або нав’тъ неможливо попри застосування найсучас-
ыших метод1в дослщження. Експериментальна модель хвороби завжди простила
за хворобу людини. Це евщоме спрощення дае змогу експериментаторовт роз-
класти хворобу на основы елементи для того, щоб падай реконструювати и сто-
совно людини. Таке спрощення е не минусом, а плюсом на певному еташ науко-
вого анализу. Доказав цього багато, та до найперекоиливппих. мабуть, можна вщ-
нести дослыи 1.1. Мечникова щд час вивчення запалення, коли не на людны, не
на кроль а на прозорш особиш морсько'1 зтрки в)и вщкрив явише фагоцитозу. У
дослщах на мишах злобу го фундаменталый даы про рак, а на собаках — валет ив 1
даы про цукровий дтабет Тепер технтчнт засоби дають змогу проводили експери-
менти на окремих кл) тинах 1 навггь на тхнтх частинах.
Експерименти подътяють на гостр! й хроычы, залежно вщ мети, яку ставить
перед собою дослщник. Для вивчення наслщктв втрати кровт застосовуютъ го-
стрий експеримент. Пришеплюваысть пухлини можна дослщити лише за лопо-
могою хроычного експерименту.
Для вивчення патолопчних процесс в застосовуютъ так! основы експеримен-
тальы методики: вилучення; подразнення; уведення в оргаызм ртзних речовин;
трансплантата оргаыв 1 тканин; методику тзольованих орган)в т тканинних куль-
тур; моделювання генетично зумовленот патологп.
Методика вилучення пол я гае у видал ей ы того чи того органа йрурпчним шля-
хом або фармаколопчними засобами, високими дозами спепиф|чних антипл
тощо. Цю методику застосовуютъ давно. Вона особливо шформатавна пщ час
вивчення еидокринних порушень. Саме видалення ыдылунковот залози дало
змогу встановити, що цукровий дтабет пов’язаний з гшофункщею и остршцевого
апарату. Завдяки видаленню одного з ларних оргаыв (наприклад нирки) було
визначено компенсаторы та пластичы можливост) органа, що залишився. Вилу-
чення частини непарних оргаыв дало змогу визначити ту мтру вграти паренхтма-
тозних елеменйв, за я кою настав латолопчний етап 1х недостатностт. Методику
вилучення широко застосовуютъ щд час вивчення патологи нервово) системы
Так, Ф. Мажанд! розробив спошб вщтворення рухових 1 чутливих порушень шля-
хом перерззання передых або задних коршщв спинного мозку.
Методику подразнення застосовуютъ толь коли пстр)бно зм)нити функцц того
чи того органа. Так, О.П. Вальтер, подразнюючи симпатичы нерви в експеримен-
т), вия ви в звуження артерш, що стало щдставою для подал много вивчення пато-
генезу спонтанно)' гангрени в людини.
Уведення в организм ргзних речовин (гормошв, гнших бюлопчно активних речо-
вин, ферменпв та 1х 1нпбггор)в, антигены тошо) також широко застосовуютъ у
патологичны фхзюлогп. Експерименти в галуз! пересадки органгв I тканин мають
велике значения для х)рурги.
4ям ввсс .-зольованих органов / тканинних культур Патофтзтологи давно вико-
г жяь аю методику, осклльки вона дае важливу ш формацию про сутшсть
- и процеств. Наприклад, киггинна теор!я анафщаксп грунтуеться на
гких було доведено, що анафйтактичну реакшю можна спричинити .
. нп~1 киппп, коли подхяти на не! антигеном, яким було сенсибьнзовано
^ерез акту альшсть проблеми спадкових хвороб або спадково! схильносп до
И * . часних наукових дослщженнях широко застосовують полшеразну лан-
[й, - -г лачяо з метою з'ясувати генетичш варташ! (пол!морф1зм) певних ген!в у
_ з "вкож методику моделювання генетично зумовлено! патологи на гпддо-
г .аринах (нокаутних, трансгенних, генетично модифЮкованих).
^емэе потреби дал! перераховувати спещалып методики, як! застосовуе пато-
ф югу свой робот!, однак слщ зтадаги тт положения, як! мають загальне мето-
14, йог ше значения. Бажливу роль в1дтграе пор^вняльний. або еволюшйний, ме-
I 0 ' злопчний пронес, який спостерггаеться в людини ) тварин, е наслщком
с- - чня вщповздних реакцш в еволющ! тваринного св!ту. Так! патолопчн]
Кр ккзг, як запалення, гарячка. вивикали й ускладнювалися в еволюпп у зв’язку
пяенням 1 вдосконаленням захисних 1 пристосувальних реакцш орган!зм!в
.тав! умови життя. Тому правильний науковий анализ реакцш людини на
ге лний валив погребуе якомога повинного знания шлях!в1 форм 1'хнього ста-
] . -ля. А це можливо .лише в раз! застосування 1сторичного методу — методу
а к мп патологи. Виняткове значения нього методу' довтв 1.1. Мечников на
. л: вивчення запалення 1 несприйнятливост! до шфекщйних хвороб
Як; будь-який експеримент, патофтзюлопчний дослщ складаеться з чотирьох
Перший етап — тдготовка питания, на яке експериментатор хоче одержали
1 дь. Кожному експерименту мае переду вати лпотеза. Не можна проводили
1. з дження. сподтваючись на ле, шо може вийли шось шкаве. “Якшо немас в
1 пей, ло не побачиш ! фактлв”, — говорив 1.П. Павлов.
Друтий елап — вибгр спещально/ методики, яка найбптьше вщповщае постав-
- -: му ?авданию, т забезпечення надежного контролю. Контролем може бути
стяй” дослщ, який ставиться паралельно з основним. Наприклад, однш тва-
। к о зять пстамш, а тнппй — !зотон!чний розчин натрно хлориду. Якщо йдеть-
зо експеримент на людях, то пл я отримання психотерапевтичного ефекту
। ш людин) вводять лткарський застб, а 1ншш — лщиферентну речовину, причо-
I про це не знае не тшьки шддослщний, а й медичний персонал. Така шлифе -
’та речовина, яку вводять як контроль пщ час клппчного дослщження лт-
кдкьких засоб1в, називаеться плацебо (вщ лат. р1асеге — подобаеться). Важливо,
_ эб п!д час проведения дослшу не було жодних суб’ективних фактор!в, яю неза-
е <но вщ вол! експериментатора могли б позиачитися на результатах.
Третш етап — планування експерименту. Дослщ .лише тодт дасть вшповщь на
-.тавлене запитання, коли вш правильно спланований. Насампсред слщ
зм'ятати, шо живий об’ект вщртзняетъся вщ об’екпв, яю вивчають точн! науки.
-~ан!зм складаеться з орган 1в, орган и — з клггин, кФтини — !з субюптинних
— }ктур (ядер, мттохондрш. рибосом), якт е ансамблями молекуляриих систем.
кна гака система — це окрема функшонуюча одиниця, шо мтстить системи
- джчого порядку т водночас входить до складу систем вишого порядку. Так в
оргашзм! формуються склада! динамтчн! взаемозв’язки регуляцл та пщпорядку-
вання.
Така складна система може неадекватно реагувати на вплив стандартного
подразника. Кр!м того, вона е вщкритою 1 шяк не може бути захигцена в!д випад-
кових вплив!в середовища, що можуть спотворити результати експерименту. Сл1д
також мати на уваз!, що однаков! на перший погляд тварини в!др!зняються спад-
ковими ознаками й тндивщуалъним розвитком. Тому експериментатор часто от-
римуе вщповшь, перекручену внаслщок впливу додаткових, випадкових фактор]в.
Закономтрнтсть мае пробитися кртзь безл!ч випадковостей.
Розумтючи це, вчен! стали не титьки враховувати випадковосп, а й планувати
!х. При цьому використовуеться математичний апарат теорн втрогщност!. Щоб
внести в експеримент випадковгсть як важливий елемент, застосовують спещаль-
ний прийом — рандомтзацпо (в!д англ. гапс1от — випадок). € також шпп методи
планування, як! дають змогу проводили експеримент у мшливих умовах або до-
слщжувати одаочасну д!ю багатьох факторгв.
Четвертий етап — обробка отриманих даних. П!сля закшчення експерименту
на твариш дослщження тривае: ощнюють результати реестрацп житгево важливих
функшй оргашзму, проводить морфолопчнт, бтохтмтчн! та !нш! дослщження тка-
нин, узятих за життя 1 п!сля загибел! тварини. Увесь цифровий материал шддаеть-
ся статистичшй обробц!, шоб визначити середа! величини дослщжуваних показ-
ник!в, вщхилення вщ середнтх значень, вщмшност! м!ж середнтми величинами,
!хню втрогщшеть.
1СТОРИЧНИЙ НАРИС РОЗВИТКУ ПАТОЛОПЧНОТ Ф13ЮЛ0П1
Основоположником експериментального методу в бюлопчних 1 медичних дослщ-
женнях вважають видатного давньоримського лжаря Клавдтя Галена (130—200 рр.
н. е.). В!н створив учения про хворобу, яким користувалися лжарт впродовж шв-
тори тисяч! роюв ! яке було замшено лише у зв’язку з подальшим розвитком
медико-бюлопчних наук 1 накопиченням нових клштчних ! експериментальних
даних.
У свотх працях К. Гален пропагував необхщшеть зв’язку практично! медици-
ни з анатомтею т ф131олопею. В!н дуже точно описав низку анатомтчних структур
1 першим розпочав дослщи на тваринах. 3 ртзних причин експерименти на твари-
нах вщродилися титьки на початку XVII ст., коли англшський хтрург, особистий
лткар короля Якова I, Ушьям Гарвей (1578—1657) описав велике 1 мале коло кро-
вообпу 1 цим спростував хибн! уявлення, як! панували з чаФв К. Галена. У на-
ступи! столптя експерименти мали важливе значения в бюлоги т медицин!. 1х
називали фгзюлопчними, аде здебшьшого по сутт 1 за методичними прийомами
вони були патофтзюлопяними.
Формування термпнв “патолопчна фтзтолопя”, “загальна патолопя” мае три-
валу тсторпо 1 !х поява припадае на ктнець XVI — початок XVII ст. Праця Ж. Фер-
неля (1497—1558) “Про природа! частини медицини” (“Бе паШгаИ рапе
шеФсшае”), яка вийшла у евгг 1542 р., постулюе ц!лком стаде переконання, що
виникнення хвороби супроводжуеться переходом орган!зму до нових 1 своерщних
законтв. Цт закони не щентичш тим, яким пщпорядковуеться дтяльшеть здорово-
- тргашзму. Ту галузь медицини, яка вивчае життед!яльшсть хворого организму,
кгггр позначив поняттям “патолопя”, тобто “патолопчна фззюлопя”.
Праця А.Ф. Геккера “Основи патолопчно! фгзюлоп!” (“ЬшпаНкк бег РЫ8ю1о§1е
раёюй^цса”) побачила свгг у 1791 р., а книга Л. Гайо “Загальна патолопя 1 пато-
л "чна ф1зюлопя“ (“РаШо1оё!е §епега1е е! рИумо1о§1е раЙ1о1о§!дие”) була надруку-
пна у 1819 р.
Отже, термш 1 зм!ст основ патофтзюлогп сформулювалися впродовж XVIII ст.
Здсдовжував розвиватися 1 вдосконалюватися експеримент. Перший вторично
□ теестрований експеримент з метою моделювання патологичного процесу здшс-
мв у 1823 р. французький фгзюлог Франсуа Мажавдц (1783—1855). У кроля од-
иобгано перерхзали тршчастий нерв, 1 через юлька дшв в ощ, розташованому на
тот ушкодженого нерва, з’явилися виразки. Так були вщкрип троф!чш функцп
г.’рвово! системи через вплив на обмш речовин у тканинах.
Основоположником експериментального методу у фгзюлоп! 1 патологп вва-
жають учня Ф. Мажанд! великого французького ф1зюлога Клода Бернара (1813—
1878), який завдяки створенню експериментально! школи посилив значения екс-
тезименту 1 дав потужний поштовх до вивчення динам 1ки патолопчних про-
таяв.
Наукова д!яльшсть К. Бернара охоплювала майже всю тодппню фхзюлопю
-ормальну 1 патолопчну), 1 в кожнш ц галуз! вгн робив блискуч! вщкриття. К. Бер-
нар надавав експерименту наспльки важливого значения, що навпъ сформулю-
вав таке афористичне положения: “Л!кар-експериментатор — це лгкар майбут-
нього”.
Значних усшх!в досягла патолопя в особ! засновника целюлярно! патологп
Адольфа Ейхрова (1821—1902). Целюлярна патолопя ознаменувала поворотний
’.нкг у розвитку медицини, нову епоху в п гсторп, 1 мала колосальне значения в
розвитку патологп як науки. Величезну роль у розвитку патолопчно! ф1зюлогп
в играли надзвичайш успехи бактерюлогп, основу яко! як експериментально! на-
ки заклали Лу! Пастер (1822—1895) 1 його сучасники Р. Кох, Д. Лютер, 1.1. Меч-
ников та гн.
Видатний В1ТЧИЗНЯНИЙ учений, лауреат Нобел 1всь ко! премп з ф1зюлоп! та
медицини за 1908 р. 1.1. Мечников (1845—1916) започаткував пор!вняльне експе-
риментальне вивчення патолопчних процесхв 1 сво!ми дослщженнями значно зба-
татив низку провщних роздшв патологп (наприклад запалення, !мун!тет, реактив-
шсть тощо). 1.1. Мечников заснував новий напрям у медицин! — геронтологпо —
I запропонував його назву.
Створення ! розвиток кафедр пагофЫологн. У 40-1 роки XIX ст. в Росшськш
шперп розпочалася шдготовка до чертово! реформи системи медично! освтти.
Передбачалася орган!зац!я медичного факультету в ушверситет! Св. Володи-
мира в Киев! (для потреб швденно! частили Росшсько! !мперн) 1 реоргашзацхя
медичного факультету Московського ушверситету (для подготовки л!кар!в у шв-
шчному репон! Росшсько! !мперй).
В оргашзацп медичного факультету в ушверситет! Св. Володимира, вщкрито-
го у 1841 р., акгивну участь брав видатний х!рург Микола 1ванович Пирогов, який
наполи на необхцшост! у створенш на цьому факультет! окремо! спешально! ка-
федри патолопчно! анатомн 1 патолопчно! ф!з!олош.
Н.А, Хржоншевський
(1836-1906)
Професор М.1. Пирогов вважав. що ця кафедра
мае навчити майбутннх л!кар!в патологи в повному
розумшн! цього слова, тобто патолопчнш анатоми 1
патолопчнш ф1зюлоги одночасно.
Вщповщно до прийнятого у 1842 р. уставу уншер-
ситету в 1845 р. в Киев1 розпочала роботу перша в
Росшськш 1м пери кафедра патологично! ф1з1ологи з па-
толог!чною анатомию.
Очолив и 1 завщував нею з 1841 по 1853 р. учень
М.1. Пирогова М.1. Козлов. 1з 1854 по 1869 р. завщу-
вачем кафедри був вщомий патолог 1 терапевт Ю.1. Ма-
нон.
Зпдно з уннверситегським статутом, прийнятим у
1863 р., у 1869 р. було засновано самостшну кафедру
загально) патологи. Г! першим професором т завщува-
чем, за рекомендашею Р. Внрхова, було обрано
Н.А. Хржонщевського, вихованця Казанського уни-
верситету, доцента курсу плени та загально'! патоло-
ги, засновника кафедри пстологи, ембрюлоп! та
поршняльно! анатоми Харкнр.ського университету. Вш був широко вщомий сво!ми
експериментальними науковими дослщженнями з використанням методу ф1з!о-
лопчних ш'екщй впал ьн их барвникгв, який отримав високу ошнку його науково-
го куратора Р Внрхова
За допомогою цього методу було вирппено стрне питания про наявшсть ет-
телто в легеневих альвеолах, проведено фундаментальна дослщження мйфоскошч-
но! будови нирок, печшки, селезенки, л!мфатичних судин як у норм!, так 1 при
патологи. До приоритета их слщ зарахувати роботу Н.А. Хржонщевського “Про
вазомоторн! нерви”.
Попри штенсивну наукову днялыпсть на царинп пстологп вш розвивав ф!з!о-
лопю та патололю, а себе завжди називав патоф!зюлогом.
Н А. Хржонщевський був деканом медичного факультету (1870—1872), трива-
лий час очолював Товариство л!кар!в (оргашзував при пьому товариств! першу в
Киев! безкоштовну л!карню), головою комки !з загально доступ них народних чи-
тан ь.
На медичному' факультет! Московського университету кафедра патолопчно!
анатоми 1 патолопчно! ф!з!ологй запрацювала в 1848—1849 рр., коли и очолив
професор 01. Полунин, якого вважають одним нз основоположников педагопчно!
та науково! школи патоф!зюлопв. За 20 рок!в завсршилося становления патоло-
пчно'! анатоми як самоспйно! навчально! дисциплши в систем! подготовки л!-
каров. У зв’язку оз цим було организовано окрему кафедру патолопчно! анатом!!,
яку очолив учень 0.1. Полутона 1.Ф. Клейн. Самого 01. Полунина було обрано
завщувачем кафедри загально! патологи.
Надал! окрем! вщ патолопчно'! анатоми кафедри загально! патологи було вщ-
крито 1 в ннших университетах Росшськон 1мпери: у 1872 р. окрему кафедру пато-
логи було организовано на медичному факультет! Харювського университету; у
1874 р. — кафедру загально! та експериментально! патологи (завщувач — профе-
сор В В. Пашутш) у Казанському университет!; у 1879 р. у Пегербурзькнй вшсько-
В.В. Пашупн
(1845—1901)
: ~-д< академй (завыувач — професор В В.
[”_1- у 1891 р. — кафедру загально! патологи
а зач — професор П.М Альбщький) у Томсь-
- -ерсите’п; у 1902 р. — кафедру загально’] па-
. вцувач — професор В В. Пщвисоцький)
I в эму факультет! Новоросшського ушверси-
-- Одеса).
к ремо необхщно сказати про кафедру Львзвсь-
I - ;ерситету. На той час (кшець XIX — поча-
। не XXст. до 1918 р.) Льв1в пщ назвою Лемберг вхо-
рв 2 2 складу Австро-Угорсько! 1мперп. Кафедру
щлювано у 1896—1897 рр. (завщувач — профе-
I яр Яь Прус) пщ назвою “Кафедра загально! та екс-
ентально! патологи”.
3 'вторично! точки зору важливо констатувати,
. юха оргаюзацй кафедр патоф!з!ологй в Роси
т ваяа на кшець XIX ст., навпъ дещо ранние, нок
фситетах Захщно! Европи.
Уважают ь, що трьома китами, на яких грун-
як* сучасна патоф!з!олопчна наука, е Казансько-Петербурзька школа осново-
. гника вггчизняно! патоф!з!ологй В.В. Пашутша, Московська школа О Б. Фох-
. - । украшська школа В.В. Пщвисоцького.
Особлива заслуга В В. Пашутша (1845—1901) полягас в тому, що вш як експе-
,;нтагор-ф1з!олог — безлосередшй учень 1.М. Сеченова — сформулював за-
ня ново! науки й реал!зував !х на практиш. У 1874 р. вш оргашзував кафедру
ально! патологи в Казанському ушверситеть а через 5 роюв — у Вшськово-ме-
чш академй в Санкт-Пегербурз!, де повною м!рою прояви вся його талант як
чего й педагога. У патолопчнш ф1зюлоп! тод! щс не було ш традинш, ш пщруч-
-ш»лв 1 навчальних програм, вщсутнш бу в < досвщ науково! роботи. В. В. Пашутш
.. це створив. Видаю ним "Лекции по общей патологии (патологической физио-
ми)” свшчать про високий р!вень викладання дисциплши. В.В. Пашутш з! сво!-
чм учнями розробляв насамперед питания обмшу речовин у здоровому ! хворому
’зшзмг. На кафедр! у великих масштабах було проведено дослщження з голоду-
лдння. В В. Пашутш уперше в свт сконструював калориметр, який лав змогу з
е гикою точюстю визначати теплообмш у тварин 1 людини пщ час хвороби.
В В. Пашутш створив вщому наукову школу патоф!з!олог!в. Його учн! надал1
чолювали кафедри патолопчно! ф!з!ологй в Санкт-Петербурз! (П.М. Альбиць-
кий), Томську 1 Харков! (О.В. Репръов), у Варшавк Одес! й Баку (М.Г. Ушинсь-
кий). В В Пашутш був талановитим организатором, що особливо проявилося,
’ коли вш став начальником Ыйськово-медично! академй. Це був перюд розкв!гу
_ього навчального закладу. Багато блискучих учених, зокрема 1.П. Павлов, були
залучен! ним до роботи в академй.
Кафедру загально! патологи на медичному факультет! Московського уншер-
.птету було засновано водночас з кшвською у 1869 р. Першим завщувачем ’п був
професор 0.1. Полунгн — вщомий патологоанатом. Засновником Московсько!
школи став професор О.Б Фохт (1848—1930) — видатний експериментатор-пато-
Ф131ОЛОГ,
О.Б. Фохт
(1848-1930)
Особливктю ше! школи був пений зв’язок екс-
периментальних дослщжень 13 запитами клшгки.
Науков! дослщження О.Б. Фохта 1 його учшв стосу-
валися переважно патолопчно! фгзюлогп серця 1 су-
дин. Йому належать класичш експериментальш до-
слщження з емболп впщевих 1 легенево! артерш.
Пщ кер1вництвом О.Б. Фохта отримали пщго-
товку видатн! вчеш, як! попм очолювали кафедри в
Киев! (В.К. Лйщеман), Одес!, Тбшс! (В.В. Воронш),
Москв! (Г.П. Сахаров), Мгнську (Ф.А. Андреев),
Катерин ослав! (ниш — Дншропетровськ) (А.1. Таль-
янцев).
У 30-х роках XX ст. у Москв! було створено На-
уково-дослщний гнетитут загально! патологп та па-
толопчно! ф1зюлоп1. Його очолювали видатн! пато-
ф!зюлоги: академик О.Д. Сперанський, учень 1.П. Пав-
лова, згодом його учн!: до 1982 р. — академгк АМН
СРСР О.М. Чернух, а з 1982 по 2002 р. — академик
АМН СРСР Г.М. Крижашвський. Як головна науко-
ва установа шетитут вццгравав важливу роль координатора наукових дослщжень з
проблем патолопчно! ф!зюлогп до 1991 р.
Найпотужнгшими патофгзюлопчними школами в Укра'йн стали Кшвська та
Харкгвська. Кафедру Кшвського ушверситету в 1888 р. очолив В.В. Шдвисоцький
(1857—1913), якого вважають фундатором укра!нсько! школи патоф!зюлог!в. За
нетривалий час (2 роки в Киев! 1 7 рок!в в Одес!) вш зумгв зробити так багато, що
пльки за це його !м’я мае бути вписане золотими ливрами в кторйо украшсько!
науки. Коло наукових !нтерес!в Володимира Валер’яновича було надзвичайно
широким. Класичними лишаються його дослщження з регенерац!! печшки, ни-
рок, слинних залоз. Програмними на багато роив стали порушен! ним проблеми
шфекцшно! патологп, епщемюлогп, реактивност! ор-
гашзму, !мун!тету, ендокринологц та !н.
Питания епологй пухлин та !х хгмютерапп, запо-
чатковаш на медичному факультет! Новоросшського
ушверситету в Одес!, який В.В. Пщвисоцький очолив
у 1900 р., залишалися актуальними 1 в 1нституп екс-
периментатьно! медицини у Санкт-Петербург!, ди-
ректором якого його було призначено (1905—1913).
Одеський перюд життя ! д!яльносп В.В. Пщвисопь-
кого ознаменувся створенням прекрасного архпек-
турного комплексу примпцень нового медичного фа-
культету. Вершиною науково-педагопчно! дгяльносп
В.В. Пщвисоцького стала його класична, всесвггнъо
визнана праця “Основы общей и экспериментальной
патологии”(1891). Ця книга отримала не пльки все-
росшське, але й евггове визнання. Г! було перекладе-
но французькою, шмецькою, грецькою 1 японською
мовами.
В.В. Пщвисоцький
(1857-1913)
3 1900 по 1922 р. кафедру загально! ф1зюлоги в Кшвському университет! очо-
в професор В.К. Лшдеман — учень О.Б. Фохта. В.К. Лщдеман як в!рний учень
_ лп О.Б. Фохта спрямував наукову роботу кафедри на вивчення конкретних
гань 1 проблем патоф!з!олоп! орган!в 1 систем. Вщом! його науков! пращ з вив-
патогенезу емболи легенево! артерп (разом 13 професором О.Б. Фохтом), 31
~ення експериментально! модел! цитотоксичного гломерулонефриту (1901),
и. через 32 роки вдосконалив японський дослщник М. Масуп. У 1910—1912 рр.
1 К Лщдеман видав “Учебник общей патологии”, який був дуже популярним
. р ед студенпв, викладачгв, лгкар!в, наукових сп!вроб!тник!в. Одночасно !з кер!в-
пшом кафедрою В.К. Лшдемана з 1910 р. було обрано директором новостворе-
Кшвського бактерюлопчного шституту, а в часи Першо! свггово! вшни вш
е й очолював ком!с!ю з протих1м!чного захисту.
Професор М.П. Вашетко, учень В. К. Лшдемана, за перюд зав!дування кафед-
рою (1922—1931) займався вивченням проблеми патоф!зюлогп сечово! системи.
Починаючи з 1930 р. в украшськш патоф!зюлогп розпочинаеться ера Олек-
- тра Олександровича Богомольця, який, до речц 1 запропонував перейменувати
офедри загально! патологи в кафедри патолопчно! ф!з!ологи.
О.О. Богомолець (1881—1946) — один !з найвидатнппих патоф!зюлопв пер-
половини XX ст., а започаткована ним наукова школа — потужна плка ук-
ра нсько! патоф!зюлог!!'.
Медичну освпу О.О. Богомолець здобув у Новоросшському уншерситеп. У
>)9 р. вш захистив докторську дисертащю, присвячену патологи надниркових
«оз, а в 1911 р. у 29-р!чному вщ! очолив кафедру загально! патологи Саратовсь-
пвго ушверситету. 3 1924 по 1930 р. керував кафедрою патолопчно! ф!з!олоп!
11 Московського медичного шституту. У 1930 р. О.О. Богомольця обирають пре-
зидентом Академи наук Украши. Цей видатний д!яч залишив шсля себе прекрас-
н) наукову спадщину, а створений ним Тнститут фгзюлоги НАН Украши в Киев!
носить його !м’я. Учн! академ!ка О.О. Богомольця стали оргашзаторами 1 кер!в-
-пками самоспйних науково-дослщних установ: геронтологи (академгк М.М. Го-
в), ендокринологП та обмшу речовин (академик
В.П. Комкаренко), патологи, онкологи та радюбю-
тоги (академж Р.С. Кавецький), Ельбрусько! медич-
во-бюлопчно! станщ! (академик М.М. Сиротинш).
3 1931 по 1986 р. кафедрою патолопчно! ф!зюлоги
Киев! керували учн! О.О. Богомольця — член-ко-
песпондент АН Украши €.0. Татаринов (1931—1950),
кадемгк АН Украши В.П. Ко.шсаренко (1950—1954),
зкадемгк АМН СРСР М.М. Сиротинт (1955—1961),
ялен-кореспондент АМН СРСР М.Н. Зайко (1961—
1986). Ус! вони розробляли окрем! напрями науково!
гпадщини О.О. Богомольця: €.0. Татаринов — роль
люлучно! тканини в реактивност! оргашзму, В.П. Ко-
псаренко — патоф!з!олопчн! проблеми ендокрино-
логп, М.М. Сиротинш — проблеми алерги, !мунпе-
ту, ппокси, гшотерми, медико-бюлопчш аспекта
космхчного польоту тощо. Основний напрям науко-
во! д!яльност! професора М.Н. Зайка — проблема
О.О. Богомолець
(1881-1946)
М.М. Сиротишн
(1896—1977)
В. В. Вороши
(1870-1960)
М.Н. Зайко
(1908-1991)
нервово) троф1ки, нейрогенних дистрофш 1 проникносте судинно! стенки. За ре-
дакщею М.Н. Зайка в 1977 р було видано третей в кторц кафедри пщручник з
патолопчно! ф1з1ологц. За час роботи на кафедр! в!н пщготував 13 кандидатев 1
7 докторгв наук.
Оз же. якшо на початковому еташ кнування кафедри загально! патологи (па-
толопчно! фнюлогп) в Киев! бгля Г! витоюв як засновники стояли так1 видатеп
патологи, як Н.А. Хржонщевський, В.В. Пщвисоцький 1 В.К Л1ндеман, то шзш-
ше стан Г! значною лирою визначався д!яльнктю учшв О.О. Богомольца.
3 1986 по 2(108 р. кафедрою керував учень М.Н. Зайка — член-кореспондент
АМН УкраТни, професор Юрш Викторович Биць. 3 2009 р. кафедру очолюе профе-
сор Миком Васильевич Кришталь.
Пщручник "Патологическая физиология’’ (1977),
створений авторським колективом за редакщею чле-
на-корсспондента АМН СРСР, професора М Н. Зай-
ка, удостоений Державно! премп УРСР.
Заслутовуе право бути серед фундаторхв укра!нсь-
ког патофпюлогй кафедра медичного факультету Хар-
кУвського унУверситету (1872). Засновником 1 першим
завщувачем ц був професор 1.М. Оболенський— вихо-
ванець Санкт-ПетербурзькоУ медико-хУрурпчно! ака-
демп. У 1886 р. кафедру очолив учень засновника
Росгйсько! патоф131олопчно! школи В.В. Пашупна
професор С.Д. Костюрт.
У 1895 р. завщувачем кафедри було обрано профе-
сора О.В. Репрьова — учня В.В. Пашупна. За 30 роив
кер1вництва кафедрою О.В. Репрьов пщготував 20 про-
Ю.В. Биць фесорлв, серед яких були оргашзатори 1 засновники
(1938—2008) власних наукових шюл. Це професори М.М. Павлов,
:Пзппо, Д.О. Альперн, С.Г. Генес, С.М. Лей-
и Ф 1 Бр!ккер та ш. Його пщручник “Основы об-
Е экспериментальной патологии” (1908) ко-
- загальним визнанням. О.В. Репрьов був
. засновниюв украшсько! ендокринологц.
Найбътыпий внесок у подалыпий розвиток нау-
вяк досягнень Харювсько! школи патофхзюлопв
стю ?7. О. Альперн, який очолював кафедру протя-
- роюв, до 1968 р. У цей период було видано
^ручник “Патолопчна ф1зюлопя” (одне з видань
чською мовою), а основним напрямом науко-
й щяльносп кафедри були проблеми запалення,
- -гуморально! регуляцп в патологи.
У 1902 р. в Одес! на медичному факультет! Но-
х>|т.<шського унхверситету було створено кафедру
ягатьшл патологп, якш судилося стати останньою
;оревалющйною. Благословив Г! на життя 1 дгяль-
- <ть В.В. Шдвисоцький. По його в!д’!зд! до Санкт-
г ербургу п очолювали таю видатн! вчеш, як про-
вес эр М.Г. Ушинсысий (1907—1908), професор В.В. Вороши (1908—1923), про-
десор БА. Шацило (1923—1947), професор М.Н. Зайко (1950—1960). Кожей !з
па ччених зробив вагомий внесок у розвиток патолопчно! ф!з!олоп!.
П!сля 1918 р. в Украш! створюються нов! медичш шститути, вщкриваються
-янп кафедри патолопчно! ф!з!ологп. Зокрема в 1918 р. у Катеринославському (зго-
юм — Дншропетровському) ушверситет! (перший завщувач — професор А.1. Та-
пднцев, орипнальний 1 обдарований експериментатор, учень професора О.Б. Фох-
га , у 1920 р. — у Кримському медичному шституп (оргашзатор кафедри — про-
*>есор О.В. Репрьов), у 1933 р. — у Донецькому медичному шституп (перший
члувач — професор М.П. Вашетко), у 1936 р. — у Вшницькому медичному шс-
гул (оргашзатор кафедри — професор М.М. Смик), у 1946 р. — у Чершвецькому
медичному шституп (кафедру очолив професор 1.Г. Федоров), у 1948 р. — в 1вано-
Франювському медичному шституп (перший завщувач — професор М.М. Смик),
> 1957 р. — у Луганському (оргашзатор кафедри — професор М.М. Смик) 1 Тер-
оп!льському медичних шститутах (кафедру оргашзував 1 протягом 20 роюв очо-
тював професор Е.Н. Бергер, учень Д.О. Альперна), у 1965 р. — у Запор!зькому
медичному шституп (професор Ю.М. Орестенко). Кафедри було створено також
на медичних факультетах ушверситет! в у повоенний час: у 1947 р. — в Ужгородсь-
кому державному ушверситет! (доцент 1.М. Куд!енко), а в 1993 р. — у Сумському
державному ушверситет! (професор О.В. Атаман).
Цей нарис !сторп украшсько! патолопчно! ф!з!ологп, особливо на початково-
му еташ виникнення 1 становления Г! як навчально! дисциплши 1 науки, був би
неповним без розкритгя н природних зв’язюв з шшими визнаними патоф!зюло-
пчними школами, у першу черту Петербурзькою 1 Московською. Про те, що щ
зв’язки трьох наукових шк!л, яю виникли майже одночасно, були плщними, св!д-
чить поспйний взаемообмш м!ж найвидатшшими !х представниками. Природно,
що таким чином вщбувалось 1 взаемозбагачення наукових напрям!в.
Як уже згадувалося, в Украш! працювали так! видатн! представники Петер-
бурзько! школи патоф!зюлопв, як О.В. Репрьов (Харюв), М.Г. Ушинський (Оде-
са) та Московсько! школи — В.К. Лшдеман (Кшв), В.В. Воронш (Одеса), А.1. Та-
льянцев (Катер инослав — Дншропетровськ). У той самий час у росшських уни-
верситетах та шших наукових закладах тривалий час працювали не менш видатн!
представники Украшсько! школи патоф1зюлопв: В.В. Пщвисоцький (Санкт-Пе-
тербург), О.О. Богомолець (Саратов, Москва), Л.О. Тарасевич (Москва), 1.Г. Сав-
ченко (Казань, Краснодар) та ш.
Велику роль у злагодженосп дгяльност! наукових 1 педагопчних колектив!в
вщгграе Наукове товариство патоф!зюлопв Украши, яке очолювали й очолюють
видатн! патоф!зюлоги: академик АН УРСР Р.С. Кавецький (1954—1958), академгк
АМН СРСР М.М. Сиротин!н (1958—1966), член-кореспондент АМН СРСР
М.Н. Зайко (1966—1991), академик НАН Украши 0.0. Мойбенко (з 1992 р.).
Дуже важливим для розвитку патолопчно! ф!зюлоп! на сучасному етап! та для
координаций наукових дослщжень у р!зних п напрямах, а також для викладання
ще! дисциплши в унгверситетах свпу було створення у 1991 р. на Установчому
мгжнародному конгрес! М!жнародного товариства з патоф!зюлоп! з! штаб-квар-
тирою у Москв! (фундатором став академик Г.М. Крижангвський) 1 видання в
Японп (Кюто) наукового журналу “Ра1ИорЬумо1о§у”.
Вщтод! проведено пйсть мгжнародних конгрес!в з патоф!зюлогп: у 1991 р. в
Москв! (Рос!я), у 1994 р. в Кюто (Япон!я), у 1998 р. в Лахт! (Фшлящця), у 2002 р.
в Будапешт! (Угорщина), у 2006 р. в Пекш! (Китай), у 2010 р. в Монреал! (Кана-
да). Це дало змогу об’еднати зусилля патоф!зюлопв р!зних кра'ш свпу, фаххвщв
шшого профыю для вир!шення актуальних наукових питань у галуз! патолопчно!
ф!з!олоп!. На них конгресах широко обговорювалися й актуальш питания викла-
дання патолопчно! ф!з!олоп! як фундаментально! професшно зор!ентовано! ме-
дично! дисциплши на медичних факультетах унгверситепв свпу.
Частина перша
ЗАГАЛЬНА НОЗОЛОПЯ
г®
Р03Д1Л I
ЗАГАЛЬНЕ ВЧЕННЯ ПРО ХВОРОБУ
В-Агяня про хворобу, або загальна нозолопя (вщ грец. удаод— хвороба), наложить
I - й завнппих проблем медицини. Стан здоров’я 1 хвороби, як правило, чергу-
шся 1 переходять один в другий часто без помггних меж. Для л псаря важливо
и загальн! критерп, яю б давали йому змогу безпомилково вщргзняти здоров’я
и хвороби.
ЗДОРОВ’Я
У практичной Д1яльносп л!каря часто застосовуються таю поняття, як нормальна
-емпература тша, нормальний тиск кров! тощо. При цьому йдеться про усеред-
веш показники тих чи шших вим!рювань у певнш популяци. Показники, яю
гпначаються найчастпие, приймаються за нормалью, а людина, у яко! !х визна-
-лють, вважаеться здоровою. Однак щ показники можуть значно коливатися за-
-ежно вщ умов навколишнього середовища, сощально-побутових умов, вгку лю-
лини, 1 тому !х не завжди можна використовувати для визначення стану здоров’я.
Щоб мати пщстави вважати себе здоровим, потребно щоб були стабшьними, оп-
пгмальними показники норми. Такий пщхщ дае чггю оргентири для профщакти-
ки, диспансеризацп 1 визначення мгри здоров’я.
Всесв!тня оргашзащя охорони здоров’я (ВООЗ) в 1946 р. у преамбул! свого
статуту ухвалила таке визначення здоров’я: “Здоров’я — це стан повного ф!зично-
го, душевного й социального благополучия, а не пльки вщсутшсть хвороб 1 ф!зич-
них дефектов”. Звичайно, це визначення надто загальне 1 швидше харакгеризуе
те, до чого слщ прагнути.
Здоров’я — це насамперед стан шчим не обмеженоТ реалгзаци генетично за-
нрограмованих процессе саморегуляци, самовгдновлення та самовгдтворення организ-
му, за якого спостерггаються в1дпов1дн1сть структуры / функци, а також здат-
шсть регуляторных систем п1дтримувати стал1сть внутрйинього середовища (го-
меостаз).
I 29
Ця здатшсть забезпечуеться можливтстю орташзму у вщповтдь на вплив чис-
ленних подразниктв навколишнього середовиша вщповшаги пристосувалъними
реакщями, адекватними за характером т силою, часом 1 тривалтстю. Висновок про
здоровий чи нездоровий, нормальний чи ненормальний стан оргашзму роблять
на лщставт антропометричних. фтзюлогтчних т бюх1м1чних дослщжень. Для ощн-
ки стану здоров’я людини мають значения 1 сошалын критерп, насамперед мож-
ливтсть здшснювати трудову дтяльшстъ.
Птдсумовуючи викладене, варто навести визначення поняття здоров’я, яке
належить видатному росшському патологу професору В.В, Пангупну, сформульо-
ване ним наприкшщ XIX ст.: “Здоровий стан оргашзму — це максимум гармони
всередиш та максимум його сили в боротьбт за кнування”. Воно I сьогодю не
втратило свого значения, оскитьки в потому правильно вщображае загальнофтло-
софську та медико-бюлопчну сутнтсть цього складного поняття.
ХВОРОБА
Хвороба — це порушення нормально! життедгяльностг оргашзму внасмдок впли-
ву на нього ииадливих факторов, шо спричинюе зниження його пристосувальних мож-
ливостей, працездатност!I збйьшус 1мов1ршсть смерть
Це визначення Грунтуеться на тому положе них, що нормальне житгя е наел та-
ком посттйного присгосування оргашзму до умов навколишнього середовиша, яка
безперервно змшюються. У разт зниження цих можливостей хвороботворний
фактор (надзвичайиий подразник) може спричинити змши, якт спроможвд приз-
вести до порушення гомеостазу т тако'! важливот функци людини, як здатшеть до
пращ.
У розвитку хвороби завжди выбуваюгься два протилежних процеси. Один тз
них 1.П. Павлов назвав фтзюлопчним заходом проти хвороби, шший — власне
патолопчним. Вони не кнують портзно, а можливт ильки в цталектичнш едностт
й боротьбт. Наприклад, у разт абсцесу виявляють патолопчш ознаки (загибель
обмеженот дшянки тканини, бхлъ), однак одночасно визначають т заходи проти
хвороби, якт мобппзують оргашзм (емйращя лейкоцитов, фагоцитоз, утворення
бар’ера навколо вогниша запалення тошо). П1д час гарячки одночасно з високою
температурою тгла, головним болем та тншими лроявами спостертгаються бтльш
активе вироблення антитез, енергтйнштий фагоцитоз тощо, гобто заходи проти
хвороби.
Навхть у випадку такот хвороби, як рак, коли, здавалося б, немае жодного
захисту, уважний дослщник знайде його. Патологтя т захцт проти хвороби кнують
нерозривно, т немае хвороби, якшо немае такот едностт. Повна втдеутшеть захис-
ного пронесу означав смерть. Повна вщеутшеть патолопчного пронесу' означав
здоров’я.
Викладене вище евщчить, що хвороба — це едтнеть двох протилежних тен-
денций: руйнтвнот га захиснот, що перебувають у постнппй боротьбт. Ця боротьба
е головним, шо “створюе” хворобу, надае тй певнот спрямованосп, перебтгу.
Складнтсть полягае в тому, що в процест хвороби шощ складно втдртзнити власне
хворобу вц] захисту. Умтння розртзнити тх е чи не найголовнтшим заеданиям лтка-
ря. Чи застосовувати жарознижувалып засоби пцт час гарячки? Чи видаляти миг-
I
. _тики в раз! хрошчного тонзилггу? Чи знижувати артергальний тиск при вираже-
му атеросклероз!? Правильну тактику лжарев! щдкаже розумшня хвороби як
тност! та боротьби протилежностей. Потребно прагнути 1 до того, щоб усунути
зсне патолопчне 1 водночас стимулювати захисне,
Однак досягти цього вдаеться не завжди. Часто лжувальш заходи, спрямоваш
за пригшчення хвороботворного фактора, пею чи ышою м!рою пригтйчують 1
захисш сили оргашзму (наприклад, променева терашя п!д час раку, деяю лжарсью
засоби, що одночасно з користю завдають шкоди).
Формування ново! якост! у хвороб!. 1снус думка, що хвороба не е чимось при-
"Ципово новим поршняно з нормою. Свого часу Р. В!рхов писав, що щд час хво-
роби маемо все таке саме, що й у норм!, тыьки в быышй або меншш юлькосп
быьше кл1тин при пухлинах, менше — при анеми). В!н назвав це гетерометр1сю.
П!д час лейкозу кроветворения здшснюеться не тыьки в юстковому мозку, а й на
перифери — екстрамедулярно, чого не бувае в норм!, — гетерототя. Р. В!рхов
розр!зняв ще гетерохронию, тобто появу кгптин не у свш час. Наприклад, мегало-
бласта в норм! виявляють -пльки в ембрюна, а в раз! В12-дефщитно! анеми вони
з:являються ! в дорослих.
Кыьюсна характеристика хвороби, представлена в тр!ад! Вхрхова, е спроще-
ною. Насправд! хвороба несе в соб! нову яюсть, що е наслщком ! юльюсних змш.
В оргашзм! можуть з’явитися быки (ам!лощ А, раково-ембрюнальш быки, анти-
тыа), яких немае в норм!. Надмгрна юльюсть, як 1 дефщит ферменпв, можуть
спричинити таю метабол!чш ! структурш порушення, яю виявляють пльки шд
час хвороби. 1.П. Павлов зазначав, що “св!т патолопчних явшц — це несюнчен-
ний ряд всыяких особливостей, тобто комбшащй ф!зюлопчних явищ, яю не ви-
никають протягом нормального життя”.
Анал!з хвороби за р!внями абстракщ!. У патолопчшй ф!зюлоп! прийнято роз-
глядати хворобу на чотарьох ргвнях (мал. 1). На найвищому р!вш хвороба уяв-
ляеться ! визначаеться як абстракц!я, фиюсофсъке узагалънення.
Нижчим ргвнем узагальнення е поширене в патологи понятая про типов! па-
толог1чн1 процеси. Типовими е патолопчш процеси, як! розвиваються за однако-
вими законами, незалежно в!д характеру причини, що зумовила !х, локал!зацй 1
виду тварини. Яскравим прикладом цього е запалення. Воно розвиваеться 1 в
жаби, 1 в кроля, 1 в людини, причому
основн! його законом!рносп однаков!.
Тому так щнуються дан!, здобут! в екс-
перимент! на тваринах, яю перебувають
на рхзних ступенях еволюцшного роз-
витку. Так формувалося загальне уяв-
лення про запалення, гарячку, гшокыю
тощо.
Поняття про типов! патолопчш про-
цеси важливе для лжаря в його м!рку-
вашй вщ загального до часткового, кон-
кретного. Як зазначалося ранние, тапо-
вий патолопчний процес не пов’язують
з якоюсь одшею причиною, там чи ш-
шим видом тварин або органом. Конк-
Мал. 1. Р1ВШ хвороби (ГУ—I)! сшввцшошення
абстрактного та конкретного на кожному з них
ретизацёя вщбуваеться в процесё дедуктивного мислення, коли лёкар точно вста-
новлюе причину хвороби (наприклад, мёкобактерп туберкульозу), п патогене.'
(залежно вщ переважаючого патолопчного пронесу — запальное чи ггухлиннс
природа, зумовленого порушенням обмёну речовин або периферийного кровоос
гу та ён.), визначае ушкоджений орган. Тоде здёйснюеться перехед вщ абстрактнс
го третього рёвня до бёльш конкретного уявлення про хворобу, наприклад, пр
туберкульоз легень, рак шлунка, цукровий деабет, ёшемечну хворобу серия. Це то»
ревень анализу за ступенем узагальнення, коли формуеться уявлення про нозоло-
г1чну форму хвороби.
Нарештё, у своей практичней дёяльностё лекар пед час аналёзу хвороби повине
перейти до максимально! конкретизацёё не лише причин, патогенезу ё локалёзац
хвороби, а й ёндивёдуальних ёё проявёв. Так виникае уявлення про диагноз захворк
вання у конкретно! людини, наприклад: запалення — туберкульозне — легень —
хворого Н. (стать, вёк та ён.).
Хвороба людини ё соцёальнё фактори. Людина — ёстота соцёальна, через те ~
хвороба характеризуеться багатьма особдавостями. Насамперед це стосуетьс:
причин хвороби. Для тварини середовище складаеться з фёзичних, хёмёчних ё бес
лопчних факторёв, для людини воно збагачуеться, а разом з тим ё ускладнюетьсй
сощальними факторами, причому ехня роль зростае.
Про роль соцёальних факторёв свёдчать, наприклад, ёстотнё змёни структур
захворюваностё у зв’язку з урбанезацеею, збёльшенням тривалостё житгя населен-
ия. Тривожнё данё про пёдвищення в усьому свётё поширеностё серцево-судинних
хвороб, хвороб пухлинноё, алергёчноё природи, неврозёв свёдчать про важлив
роль забруднення навколишнього середовища, мёнливих умов працё й побуту (на-
приклад, напружений ритм мёського житгя, сучасного виробництва).
Соцёальний бёк хвороби визначаеться також тим, що хвороба порушуе труде-
ву дёяльнёсть людини або колективу людей, завдаючи матерёальних ё моральней
збиткёв як конкретней люданё, так ё суспёлъству в целому.
Отже, у хворобе людини е тварин е спельне (зведси значения експерименталь-
його моделювання хвороб) е водночас резне, бднесть соцёального е бёологёчног
створюе ту особливу картиегу, яка характеризуе хворобу людини.
Людина зал ежить вщ природа, але ё природа не меншою мёрою зал ежить вт
людини. Ранёше екологёя вивчала процеси ревноваги, яке еволюцёйно склалися и
природё (виживання, розвиток, розмноження). Тепер поняття екологн без люди-
ни втрачае свей змёст — настельки велике втручання людини в природу. Негатив-
нё наслёдки цього втручання (забруднення навколишнього середовища, дефецег
кисню, пресное води) входять у поняття причини хвороби. Крем того, понятт'
екологёё людини мае й ёнший, не менш важливий змёст. Воно охоплюе найваж-
ливёшё проблеми ё тенденцп сучасностё, насамперед захист природи, охорону нав-
колишнього середовища заради здоров’я людини.
Даючи визначення хворобе, ми визначаемо критерёё, яке були б найбельш по-
стёйними. Щодо хвороби людини таким критерёем е обмеження працездатност
або ёгвтрата, тобто порушення найважливёшоё сощальноё функцц людини — пра
цездатностё.
У хворое людини складаються новё вщносини з навкодашнём середовище^
Житгя тривае, але воно “стиснене”, не вёльне. “Що таке хвороба, як не обмежеш
у своей свободе життя?” (К. Маркс). 1з цим фёлософським визначенням хвороб;
ЗГ1
ерегукуються визначення Р. Вгрхова: “Хвороба — це житгя в ненормальних умо-
вах”, О.О. Богомольця: “Хвороба — це порушення р1вноваги оргашзму 1 середо-
вища”.
Ус! Ц1 визначення розкривають лише один бгк захворювання — руйнгвний.
Однак завжди слщ пам’ятати, що хвороба — це не пасивне порушення функций
шкоджених оргашв г систем, а боротьба оргашзму з хвороботворним фактором.
Цей бгк хвороби вдало розкривае визначення С.П. Боткгна: “Реакщя оргашзму на
шкщливий вплив навколишнього середовища 1 становить суть хвороби”.
Принципи класиф1кацп хвороб
Ниш налгчуеться близько чотирьох тисяч хвороб (нозолопчних форм), 1з яких
майже 2500 мають спадкову основу. Кшькгсть гх увесь час змгнюеться. Деякг но-
юлопчш форми розчленовуються. Однг хвороби зникають, ппш з’являються. На-
приклад, променева хвороба не гснувала, поки не було застосовано рентген 1вське
випромшювання. Не 1снувала й космгчна медицина, поки не з’явилися космгчш
польоти.
В основу дпочог М1жнародно1 класифгкацц хвороб (перегляд н здшснюе ВООЗ
кожш 10 рок!в) покладено илька критерив (принцишв).
1. Ет1олог1чний; грунтуеться на сшльносп причини для групи хвороб (напри-
клад, хвороби набуп та спадковц гнфекцгйш та негнфекщйш).
2. Топографоанатомлчний; дае змогу розглядати хвороби на р!вш орган1в 1 сис-
тем (хвороби серця, нирок, печшки та ш.). Ця класифгкащя не раз зазнавала
серйозно! критики на тгй пщставц що хвороби органа взагал! не бувае, будь-яке
.псце ушкодження неминуче залучае до реакцп весь оргашзм. Однак ця класифг-
кащя поширена, осюльки вона зручна на практищ. Кргм того, вона вщповщае
сучаснгй тенденцп до спещалгзацн лгкарсько! допомоги. Ця класифгкащя поед-
нуеться з класифгкащею за функцюнальними системами: хвороби системи кров!,
нервово!, ендокринног, 1мунно'1 систем тошо.
3. За ежом I статтю; розргзняють хвороби дитячого (у тому числ! перина-
тального) та старечого вгку. Останшм часом геронтолопя 1 гер1атр!я виокремили-
ся в самостгйш науки, значения яких зростае у зв’язку з! збгльшенням тривалост!
життя населения 1 кглькосп осгб похилого вшу серед пащенпв. Спещальним роз-
дглом медицини е пнеколопя. Так само гснують хвороби чолов1чо‘1 стаи, як! вив-
чае андролопя.
4. Еколог1чний; враховуе роль умов гснування людини у виникненш хвороб.
Температура повгтря, атмосферний тиск, сонячне освгглення, чергування дня 1
ноч1, вмгст макро- та мгкроелеменпв у питнгй вод!, грунп, продуктах харчування
позначаються на сташ здоров’я населения певних репошв. У зв’язку 1з цим вщц-
ляють хвороби Крайньо! ГПвночг, помгрного кшмату, тропгчш хвороби. Це Край-
ова, або географгчна, патолопя (маляргя, серпопод1бно-кл1тинна анем1я, ендем1ч-
ний зоб, подагра, флюороз та ш.).
5. За стлыистю патогенезу — хвороби алерпчио!, запальноц пухлинно! та
шшо! природи.
6. Социальный; розргзняють хвороби професшнг, военного часу, хвороби
цившзацп.
7. За характером перебггу — гострг та хрошчш хвороби.
8. Залежно в1д метод1в л1кування — хвороби хёрургёчного ё терапевтичного про-
филю.
В останнёй (10-го перегляду) Мёжнароднёй класифёкаци хвороб (МКХ-10)
вшил ено 21 клас хвороб переважно за топографоанатомёчним принципом (лише
частково за етёологёею та патогенезом). Детальнёшу розробку класифёкацёй хвороб
вёдповёдних класёв здёйснюють експерти ВООЗ, враховуючм й ёнпп наведен! при-
нципи.
ПАТОЛОПЧНА РЕАКЦ1Я, ПАТОЛОПЧНИЙ ПРОЦЕС,
ПАТОЛОПЧНИЙ СТАН
Патолопчна реакщя — це неадекватна за силою чи спрямованёстю, шкёдлива або
недощльна реакцёя оргашзму на дно як патолопчних, так ё нормальних факторгв.
Патолопчний пронес — це поеднання мёсцевих ё загальних реакцёй, якё вини-
кають в оргашзм! у вёдповёдь на ушкоджувальш дп хвороботворного фактора.
Розвиток патолопчного процесу залежить як в!д етёолопчного фактора, так ё в!д
реактивних властивостей оргашзму. Ушкоджувальний фактор може припинити
свою дёю, а патолопчний процес продовжуе розвиватися вщповёдно до програми,
вироблено! та закршлено! в процес! еволюцёйного розвитку (наприклад, гостре
запалення).
Патолопчний стан — це патолопчний процес, який розвиваеться пов!льно.
Хворобливё порушення, що при цьому спостерцаються, малодинамёчнё. Вони зали-
шаються майже незмённими протягом тривалого часу (роки, десятирёччя). Патоло-
пчний стан часто е наслёдком патолопчного процесу. Так, запалення ропвки може
закёнчитись утворенням бельма, яке зберёгаеться на все життя. Виразка шлунка
(патолопчний процес) може закёнчитися рубцюванням ё звуженням воротаря (па-
толопчним станом). Надал! це може стати умовою для переходу вказаного патоло-
пчного стану в ёнший патолопчний процес. Наприклад, на мёсщ шеляопёкового
рубця гцц впливом канцерогенних фактор!в може утворитися ракова пухлина.
Наявюсть патолопчного процесу або патолопчного стану ще не означае, що
е хвороба в повному п обсязё. Однак це може статися, якщо оргашзм ослаблений
або процес набувае значного поширення й особливо!’ агресивностё. Наприклад,
запалення волосяного фолёкула — фурункул — е патологёчним процесом на шкёрё,
а не хворобою. Виникнення множинних фурункулёв може дати розгорнуту клёнёч-
ну картину хвороби з гарячкою, ёнтоксикацёею тощо.
Особливу категорёю патолопчних станёв становить недостатшсть органёв ё
систем (ЦНС, ендокринна, ниркова, печёнкова, дихальна тощо). 1х особливёстю е
гострий чи хронёчний, компенсований чи некомпенсований характер.
Типов! патолопчн! процеси
Типовим» патологёчними процесами називають такё, що розвиваються за однако-
вими основними закономерностями, незалежно в!д причини, локалёзацп, виду
тварини. Гнакше кажучи, це процеси, якё вщбуваються в основному однаково за
рёзних патогенних впливгв, у рёзних органах ё в рёзних тварин. Це запалення, га-
рячка, пухлина, ппоксёя, голодування, гёперемёя, тромбоз, емболёя, ёшемёя, ти-
' мм порушення обмшу речовин. Той факт, шо вс! тварини 1 людина однаково (в
1 иному) голодують, однаково реагують на нестачу кисню, що в ус!х тварин 1
лиини за однаковими законами розвиваеться запалення, евщчить про те, що вс!
~опеси сформувалися еволюцшно 1 можлив1сть !хнього розвитку успадко-
т пъся. Звичайно, запалення або пухлини у рззних бюлопчних вщцв мають сво!
соб 'ивост1, однак патолопчна ф!зюлопя абстрагуеться вщ деталей, а бачить свое
иссиня в тому, щоб розкривати в них загальн! ознаки 1 тим самим наблизитися
р розумшня закономзрностей !х розвитку в людини.
:-:новн1 перюди (стадн) розвитку хвороби
У - авитку хвороби розргзняють чотири перюди (стадц): латентний, продромаль-
перюд розпалу хвороби 1 перюд завершения хвороби. Така периодизация
р чувалася на пщетав! клппчного анал1зу гострих шфекцшних хвороб (черев-
>юто тифу, скарлатини тощо). Перебц шших хвороб (серцево-судинних, ендок-
- -них, онколопчних) вщбуваеться за шшими законом!рностями, 1 тому наведе-
лерюдизацпо не можна застосовувати до них. А.Д. Адо видщяв три стадп роз-
етку хвороби: початок, власне хворобу, кшець.
Латентний перюд (щодо шфекцшних хвороб — шкубацшний перюд) тривае
моменту впливу на оргашзм причини до появи перших юпшчних ознак хво-
роби. Цей перюд може бути коротким, як пщ час дп силънодпочих отруйних ре-
. . вин, 1 луже тривалим, як у випадку прокази (кыька рокгв). У цей перюд вщбува-
- ”ься мобйпзащя захисних сил оргашзму, спрямованих на компенсацию можливих
-орушень, на знищення хвороботворних факторгв або на видалення !х з оргашз-
му. Латентний перюд слщ ураховувати пщ час проведения ргзних профшактичних
цаод!в (130ЛЯЦП в раз! шфекцп), а також у раз! лпсування, яке нерщко бувае ефек-
тивним тыьки в цей перюд (сказ).
Продромальный перюд — промгжок часу вщ появи перших ознак хвороби до
зевного ц прояву. 1нод! цей перюд перебцае виражено (крупозна пневмония, ди-
зентергя), в шших випадках характеризуемся слабким, але чггким проявом хво-
роби. У раз! прсько! хвороби, наприклад, це безпричинна веселють (ейфор!я),
ри кору — плями Бельського — Филатова — Коплгка та ш. Усе це важливо для
шференщально! д!агностики. Водночас видшення продромального перюду при
"агатьох хрошчних хворобах часто утруднене.
Перюд виражених проявив, або розпалу, хвороби характеризуемся повним роз-
витком клппчно! картини: судоми в раз! недостатност! прищитопод1бних залоз,
лейкопензя — при променевш хвороб!; типова тр!ада (гшергл!кем!я, глюкозур!я,
пол!ур!я) у хворих на цукровий д!абет. Тривалють перюду розпалу деяких хвороб
крупозно! пневмонп, кору) пор!вняно легко визначити. Пщ час хрошчних хвороб
з !х повшьним перебцом змша перюдгв невловима. Якщо це таю хвороби, як ту-
беркульоз, сифшс, перебц процесу характеризуемся наявшетю загострень, при-
чому нов! спалахи шод! пом!тно в!др!зняюмся вщ первинного прояву хвороби.
Кшець (завершения) хвороби може бути рззним: видужання (повне й неповне),
рецидив, перехщ у хрошчну форму, смерть.
Видужання — це процес, який характеризуемся лгквщашею порушень жит-
тед!яльност!, спричинених хворобою, вщновленням нормальних зв’язюв оргашз-
му 1з середовищем, для людини — насамперед це вщновлення ц працездатносп.
Видужання може бути повним 1 неповним. Повне видужання — це стан, за
якого зникають ус! прояви хвороби, а оргашзм повшстю вщновлюе своТ присто-
сувальш можливост!. Видужання, однак, не завжди означав повернення до почат-
кового стану. Унаслщок хвороби можуть виникати 1 зберггатися надал! змши з
боку резних систем.
У раз! неповного видужання спостерйаються значш наслвдки хвороби. Вони
залишаються надовго ! навив назавжди (зрощення листк!в плеври, звуження мгг-
рального отвору). Изниця мгж повним 1 неповним видужанням вщносна. Виду-
жання може бути практично повним, незважаючи на стшкий анатомгчний дефект
(наприклад, вщсутшсть одше! нирки, якщо шша повшстю компенсуе н функцию).
Не слщ думати, що видужання починаеться шсля того, як минули попередн! стада
хвороби. Процеси видужання починаються вщ моменту виникнення хвороби.
Уявлення про мехашзми видужання грунтуеться на тому загальному положен-
ш, що хвороба — це едшсть двох протилежних явищ: власне патолопчного 1 за-
хисно-компенсаторного. Переважання одного з них визначае кшець хвороби. Ви-
дужання настав тод1, коли комплекс пристосувальних реакщй виявляеться до-
статньо сильним, щоб компенсувати можлив! порушення.
Серед мехашзмгв видужання розр!зняютъ термшов! (аваршш) ! триваль До
термшових механхзмав належать так! рефлекторш захисш реакцп, як зм!ни
дихання 1 кровообггу, видшення адреналшу ! глюкокортикоида пщ час стресових
ситуацш, а також ус! мехашзми, спрямоваш на збереження сталост! внутргшнього
середовища (рН, ргвень глюкози в кров!, тиск кров! тощо). Тривал! реакц!!
виявляються дещо шзшше ! дпоть протягом усього перюду захворювання. Вони
вщбуваються насамперед за рахунок резервних можливостей функцюнальних
систем. Цукровий д!абет не виникае в раз! втрати навпъ 3/4 остр!вц!в пццштунко-
во! залози. Людина може жити з одшею легенею чи одшею ниркою. Здорове
серце щд час навантаження може виконувати роботу у 5 разгв б!льшу, шж у стан!
спокою.
Наростання функцп вщбуваеться не лише за рахунок уведення в д!ю струк-
турно-функщональних одиниць, як! рашше не працювали (наприклад нефрошв),
а й збшыиення штенсивност! !хньо! робота, що в свою черту зумовлюе активацйо
пластичних процеав 1 наростання маси органа (гшертрофхя) до р!вня, коли на-
вантаження на кожну функщонуючу одиницю маси органа не перевищуе нор-
мальш показники.
Уведення компенсаторних мехашзмгв так само, як 1 припинення !хньо! дгяль-
ност!, може залежати в!д нервово! системи. П.К. Анохш сформулював уявлення
про функщональш системи, як! специф!чно компенсують функцюнальний де-
фект, спричинений ушкодженням. Ц! функщональш системи утворюються 1 пра-
цюють у певнш послгдовност!.
1. Сигналгзашя про порушення, завдяки чому починають функшонувати в!д-
повщю компенсаторш мехашзми.
2. Прогресуюча мобйпзащя запасних компенсаторних мехашзмгв.
3. Зворотна аферентащя про послгдовш етапи вцщовлення порушених функщй.
4. Формування в центральшй нервовш систем! тако! комбшацп збуджень, яка
визначае ефективне вцщовлення функцш у периферичному орган!.
5. Оцшка адекватност! 1 надгйносп кшцево! компенсацп в динампц.
6. Розпад системи через непотргбшсть.
киа вшсть етап1в компенсаций можна простежита на прикладо кульгання
। дження ноги: 1) сигнал!защя про порушення ровноваги з присшково-
твою органа; 2) перебудова робота моторных центрш 1 м’язових груп для
рок .я ргвноваги 1 можливостх рухагися; 3) постшна комбшащя аферентацш
(к стабильного анатом!чного дефекту), що надхоцять у випц вщтпли цент-
ре ово! системи, ! утворення тимчасових зв’язюв, яю забезпечуютъ оп-
____’ компенсацпо, гобто можливкть ходили з мхшмалыюю кульгавхстю.
1 -ггнсаторш мехашзми можуть активгзуватася також гормональними або
тшгми стимулами.
ига — це новый лрояв хвороби теля удаваного або неповного Г! припи-
наггиклад, вщновлення нападгв маляры теля бшьш-менш три вал ого без-
го периоду.
- -сллчно може виникаги рецидив запалення легень, коллу тощо.
БЬерехц у хрошчну форму означае, що перебхг хвороби вщбуваеться повшьно з
гм и перюдами ре моей (м!сящ! навпь роки). Такий переб!г хвороби зумов-
г« I’ в;рулентшстю збудника 1 здебгльшого реактившетю оргашзму. Так, в ос!б
жкжтого 1 старечого в!ку деяю хвороби (пневмония, коли) набувають хрошчного
^актеру.
Терли палый стани. Припинення життя вщбуваеться поступово, навпь у разг,
.иъалося б, митте во! смерт!. Термшальний стан — це оборотний стан згасання
| чкшй оргашзму, який передуе бюлопчшй смерт! 1 в якому можна видиити
«_ -ка стангв: преагональний, агонгю, клпйчну смерть.
Преагоналъний стан (преагон1я) — термшальний стан, що передуе агонп, ха-
жкгеризуеться р!зною тривалктю (години, дш), задишкою, зниженням артер!аль-
т го таску до 60 мм рт. ст. 1 нижче, гах!кард!ею, розвитком гальмування у вищих
эьдитах центрально! нервово! системи. У людини спостерггаеться потьмарення
нэмосп. Преагошя переходить в агошю.
Агон1я (вщ грец. ауосогга — боротьба) — термшальний стан, що передуе на-
—линю смерт!; харакгеризуеться поступовим глибоким порушенным функцш ор-
лшзму. особливо кори пшкуль великого мозку, з одночас ним збудженням дов-
гэсгого мозку 1 надзвичайним напруженным захисних функцш, що вже втрачають
зою цоцшьшеть (судоми, термшальне дихання). Тривалхсть агонп становить 2—
а, 1нколи бгльше.
Клййчна смерть — термшальний стан, що настае теля припинення дихання 1
гобоги серця 1 призводить до незворотних змш у вищих вылетах центрально! не-
зврво! системи. На цьому етат ще тривае обмш речовин, 1 житгя може бута вщ-
жжлене. Саме тому стад!я клппчно! смерт! привертае особливу увагу клшщиспв
• экспериментаторов.
Ыологгчна смерть харакгеризуеться незворотним припиненням життедгяль-
аосп оргашзму, е неминучою завершальною стадхею його шдивщуального гсну-
ьышя.
Дослщи на тваринах, насамперед на собаках, дали змогу детально вивчити
чкцюналып, бюхгм1чш та морфолопчю змши на вехх етэпах умирания.
Умирания — процес припинення житгедгяльност! оргашзму, що послщовно
зозвтгваеться в термшальному стан!, розпад його щлост!. Оргашзм перестае бута
саморегулюючою системою. При цьому спочэтку руйнуються т! системи, яю
» еднують оргашзм у едине щле. Це насамперед нервова система. Водночас ни-
жч! ргвнз регуляш! псвною игрою зберггаються. У нервовш систем! спостерггаеть-
ся певна послгдовшсть умирания ргзних п вщдьвв. Найчутливтша до ппоксп кор
великого мозку. Пщ час асфгксп або гостро! крововтрати спочатку вщбуваетьс^
активацтя нейронгв. У зв’язку гз цим виникаютъ рухове збудження. прискоренн'
дпхання г пульсу, щдвищення артергального гиску. Потгм настае гальмування в
кор!, що мае захисне значения, оскгльки на деякий час може зберегти клггини вь
за гибель У разг подальпюго вмирання пронес збудження, а поим гальмування 1
виснаження погиирюеться нггжче, на стовбурову частину головного мозку . на
сгтчасту речовину (ретикулярну формаппо). Цг фглогенетично найдавнгшг вгддьт;
мозку бгльш стгйкг до кисневого голодування (цснтри довгастого мозку7 можул
витримувати нестатгу кисню протягом 40 хв).
У такгй самгй послщовност! вщбуваються змгни в шших органах 1 системах
Пщ час смертельно! крововтрати, наприклад, протягом першо! хвилини, диханн
ргзко поглиблюеться г прискорюеться, Потгм поругпуеться його ритм, вдихи ста-
ють го дуже глибокими, то поверхневими. Нарепгп, збудження дихального центр
сягае максимуму, що проявляться особливо глибоким диханням, яке мае знач-
ний шсшраторний характер. Пгсля цього дихання ослаблюсться або навгть при-
пиггяеться. Це термгнальна пауза, яка тривае 30—60 с. Потгм дихання тимчасовс
вгдновлюеться, набуваючи характеру ргдких. спочатку глибоких, а потгм деда.7,
бгльш поверхневих вдихгв. Разом з дихальним активуеться судиноруховий центр
Тонус судин пцгвишуеться, скорочення серия посилюються, однак незабароу
припиняються 1 тонус судин ргзко знижуеться.
Важливо зазначити, шо пгсля припинення роботи серця система, що генерус
та проводить збудження, процовжуе функцгонувати ще досить довго. Зггдно з да-
ними ЕКГ, бюструми зберггаються протягом 30—60 хв пгсля зникнення пульсу
Пщ час умирания вщбуваються характернг змгни обмшу речовин, зумовлен
переважно кисневим голодуванням, яке дедалг поглиблюеться. Окиси! шляхи ме-
таболгзму блокуються, г оргашзм одержуе енергпо за рахунок глпсолгзу. I Усилен-
ия цього типу обмшу речовин мае компенсаторне значения, але його низыа
ефективнгсть неминуче призводить до декомпенсацп, що поглиблюеться внаслг-
док ацидозу. Настае клгнгчна смерть. Припиняються дихання, кровообтг, зника-
ють рефлекси, однак обмш речовин, хоча ! на дуже низькому ргвнг, ще тривае.
Цього достатньо для пгдтримання "мгнгмального життя” нервових клгтин. Саме
ним пояснюеться оборотнгсть пронесу клппчно! смертг, тобто в цей перюд мож-
ливе оживления.
Реашмащя, або оживления, оргашзму охоплюе заходи, спрямованг насамперед
на вщновлення кровообггу г .дихання: масаж серия, штучна вентилящя легенг
дефгбриляцгя серця. Для проведения останнього заходу потргбна вщповгдна апа-
ратура, тому його необхгдно здгйснювати в спешальних умовах
Дуже важливгш. е питания про термгни. протягом яких можлива г доцгльш.
реангмащя. Адже оживления виправдане лише в разг вщновлення нормальнг
психгчног гцяльностг. Завдякп дослщженням академгка АМН СРСР В О. Неговсь-
кого та його послгдовни кгв прийнято вважати, що позитивний результат реагггма-
цц можливий не пгзнтше шж через 5—6 хв вщ початку кдгнгчно! смертг. Ятощ
процес умирания вщбуваеться на тл! ргзкого та швидкого виснаження резервп
креатинфосфату й аденозинтрифосфату (АТФ), то перюд клппчно! смертг ще ко-
роли ий. Тим часом пщ час ппотерми оживления можливе навгть через годину
шсля настання клпнчно! смерть У лабораторп академжа АМН СРСР М.М. Сиро-
тишна було встановлено, що оживили собаку можна через 20 хв шсля смерт! вщ
крововтрати з подалыпим повним вщновленням Д1яльност1 головного мозку, якщо
реашмащйш заходи здшснювати за методом професора С.С. Ьрюхоненка. Вико-
ристовуючи створен ий ним (одним 13 перших у свт) апарат штучного кровообпу
орипнально! конструкцп, можна своечасно видаляти з оргашзму так зваш ппемзч-
ш токсини. Слщ однак мати на увазх, шо гшокс1я мозку в людини спричинюе
сутгевпш змши, н!ж у тварин.
Р03Д1Л II
ВЧЕННЯ ПРО ЕТЮЛОПЮ I ПАТОГЕНЕЗ
ЕТЮЛОПЯ ,
Етюлог1я (вщ грец. ата — причина, Хдуо<; — вчення) — вчення про причини й умо-
ви виникнення хвороби.
Перше запитання, що виникае в лжаря пщ час зггкнення його з хворобою,
стосуеться и причини. Виявити причину хвороби означало б знайти шлях до й
лжування 1 профилактики. Однак це часто виявлясться дуже нелегким завданням.
За р1зних часгв у медицин! його вирппували по-р!зному. Це визначалося загаль-
ним р!внем розвитку науки, а також свггоглядом л!кар!в, тобто пею методолопч-
ною позищею, на якш вони стояли. Теор!я причинност! в патологп — завжди
“дитя свого часу”. Вона е суто кторичною, хоча сам принцип причинност! непо-
рушний, абсолютний, осюльки в його основ! лежить ф!лософське вчення про
детермтзм (вщ лат. (1е1егт1по — обмежую, визначаю) — вчення про всеохоплюю-
чий об’ективний ун!версальний взаемозв’язок I причинну зумовленкть процес!в 1
явищ природи, сусшльства й мислення. Принцип причинност! в патологи озна-
чав, що будь-як! змши в стан! живо! системи детермшоваш взаемод!ею ще! сис-
теми з факторами навколишнього та внутршгнього середовищ.
У пер!од, коли медицини як науки ще не кнувало, причиною хвороб вважали
злий дух, що вселявся в людину ! завдавав !й страждань. Рел!г!я сприяла цим уяв-
ленням, проповщувала, що позбавитися хвороби можна, вигнавши злого духа за
допомогою божественного начала. кнувало й шше уявлення, зпдно з яким хво-
роба е наслщком гн!ву Божого, якого треба умилостивити молитвою 1 пожер-
твою.
Становище ктотно змшилося наприкпщ! XIX ст., коли було виявлено збуд-
никш багатьох шфскщйних хвороб. Вщкритгя Л. Пастера, Р. Коха та шших уче-
них того часу мали важливе значения для матергалктичного розумшня етюлопТ
хвороб. П!д впливом цих вщкритпв значно посилилися подалыш пошуки причин
хвороб, причому не титьки шфекщйно!, а й нешфекцшно! природи. У зв’язку !з
цим виник 1 деякий час кнував погляд на причиннкть у медицин!, вщомий як
монокаузал!зм.
Монокаузал1зм (вщ грец. — один I лат. саиза — причина) — напрям в
етюлогп, зпдно з яким будь-яка хвороба мае одну-едину причину 1 зггкнення ор-
гаюзму з нею мае неодмшно призвести до виникнення хвороби. На перший по-
гляд це здаеться переконливим, однак, як з’ясувалося, така переконливкть лише
, ак. По-перше, не кожна хвороба спричинюстьея бактериями. Кргм того, на-
. гх в оргашзм! ше не означав наявшсть хвороби. В оргашзм! протягом
со часу можуть перебувати гнфекщйш збудники, а людина залит аеться
зою (бактерюносшство).
Пщ час розтнну труп!в людей, як! померли не в!д туберкульозу, у 95 % випад-
ж> виявлено первинний афект. Це пояснюеться тим, шо вони були шфгко-
;обакгер!ями туберкульозу, але за життя виражених ознак хвороби в них не
пялено
Дат! з’яс^валося, що в раз! заражения групи людей одним ! тим самим видом
-нних мгкрооргашзмгв реакщя гх нерщко р!зна: в одних розвиваеться тяжка
в шших— слабка, грет! — зобс!м не хворпоть. Ц! спостереження похитну-
анепшю монокаузалгзму.
Монокаузалгзм не зм!г визначити правильний шлях виргшення проблеми. В!н
ахов звав взаемодгю хвороботворного фактора й оргашзму, мгнливгсть пер-
великт захисш можливост! другого, а також того, що причина хвороби дге
дча. Разом з нею оргашзм вёдчувае. вплив 1 багатьох шших факторгв, як! мо-
-5 сприяти або запоб!гати й ди.
Монокаузалгзм як етап у вченш про причини хвороб був закономгрним 1 пот-
м. оскыьки выображав прогресившш! погляди на причини хвороб.
На початку XX ст. у зв’язку з кризою в природознавств! набула поширення
_ы доктрина, в! дома як кондишон&йзм
Кондицюнатзм (в!д лат. сопсНиопакз — умовний) — напрям у патологи, основ-
я зложення якого полягають у тому, що поняття про мехашчну причиншсть, як
дальне мислення взагал!, застарыи. Науковий пщх!д мае зводитися не до по-
те причин хвороби, а до розгляду та опшки вс!е! сукутшост! умов, за яких цю
! робу виявили. Ус! умови принципово рзвнозначш, оскыьки вони нотргбш.
Ферворн (1912) сгверджував, що причин хвороб не !снуе г пошуки !х марш,
хто шукае причину, виявляе лише недостатшсть свого мислення, вш уподгб-
с гься дикуну, який думас, що “п!сля цього” означае “внаелгдок цього”, на-
- и стад, причиною дня е нгч. Насправд! хвороба е наслыком багатьох факторгв,
>оден з них не може бути виокремлений, видыений нал !ншими 1 названий
чиною. Ус! п! фактори — сутшсть умови, г повного аналгзу може бути досяг-
то лише тод!, коли вс!х !х буде вивчено Видыення одного з фактор!в шчого не
г-юе, оскыьки е суб'ективною думкою лгкаря.
Удавана переконлиюсть цих доказгв привернула свого часу увагу багатьох до-
дникгв. Кондищонал!зм 1 нин! досить поширений.
Фыософською базою виникнення кондишонап’зму е махизм. Як мах!зм. так г
чдицюналгзм мають суб’ективно-щеалгстичний характер. Заперечуеться об’ек-
. не !снування закошв природи ! стверджуеться, шо закономгршеть породжуеть-
суб’ективною д!яльшстю людини. Мах писав, що матергя як об’ективна реаль-
1сть не !снуе, вона генуе лише в наших вщчуттях як суб’ективна реальнгсть. Тим
сом деяк! представники вказаного напряму (Шор, А. Опель) формально не щд-
(мували позицп мах!зму, вважаючи умови об’ективно генуючими поза нашою
.ДОМ1СТЮ.
У цыому кондищоналгзм обеззброюс л!каря. Оскыьки немае причини, немае
потреби шукати п 1 боротися проти не!. Водночас вивчити вс! умови хвороби,
клиги найважливпиу в шдивщуальному варганп переб!гу практично неможли-
во. 1з цих причин кондицйоналйзм як науковий напрям не е переконливим й про-
дуктивним.
Конституц1онал1зм — ще один важливий напрям у загальнйй етйолопй, яки?
наголошуе на вирйшальному значений у виникненни та розвитку хвороб спадковс;
патологи метаболизму, функцц та структури вйдповйдних органйв й систем. Тобт<
неповноцйннйсть генотипу е вйдхиленням вйд нормального типу його конституций.
Такй аномалий конституций водночас йстотно впливають на реактивнйсть организм)
його здатнйсть вйдповйдати певним чином на змйни навколишнього та внутрйшньо-
го середовищ. Природно, шо пристосувальнй можливостй оргашзму до таких змйв
е обмеженими.
На сучасному етапй цей напрям належить до кола проблем так званой генетыч-
ноИ модели медицины (Л. Людвиг, 1980).
Сучасна медицина не може обййтися без знания роли порушень з боку психйч-
ной сфери организму у виникненни соматичних хвороб. Це так званий психосома-
тичний напрям у загальнйй етйолопй. Для його обгрунтування психосоматики ант-
лйець Холлидей (1949) та американець Александер (1950) використали давни спос-
тереження щодо можливостй розвитку ятрогенних (вйд грец. гатроз — лйкар) хвороб,
про значения пережитих страждань у виникненни цукрового диабету, гипертонич-
ной, виразковой хвороб й навить раку, а також сучаснйшй данй про так званй хвороб;
цивилизаций як своерйдну “розплату за прогрес”, про здатнйсть центральной нерво-
вой системи “зберйгати слйди” вйд отриманих протягом житгя негативних впливйи
Особливого значения розробники психосоматычно! моделг медицины надают е
конфликтам раннього дитячого вйку (несвоечасне та неповне задоволення харчо-
вого инстинкту — до 6 мйс., емоцййна атмосфера в родинй в период наййнтенсивип-
шой реализаций природной потреби дитини аналйзувати речй та оточення — вйд 6 мйс.
до 3 рокйв, а також пйд час формування почуггя власной гйдностй, власного “Я” та
свого мнения в родинй, колективй — вйд 3 до 6 рокйв) як причинам розвитку сома-
тичних хвороб. Такй конфликта мйж инстинктами дитини та можливйстю их задо-
вольнита спричинюють формування загальмованой чи явной агресивной поведин-
ки, йстерйй й, як наелйдок, порушення нервовой регуляций внутрйшнйх органйв й роз-
виток хвороб.
У зв’язку з прогресуючим зростанням захворюваностй на так званй основш
нейнфекцййнй хвороби (артерйальна ппертензйя, йшемйчна хвороба серця, пептидаа
виразка шлунка та дванадцятапалон кишки, цукровий диабет, ожирйння, атеро-
склероз, рак), на частку яких припадае 80—85 % захворюваностй на патологи
нейнфекцййной природи, в останнйй чвертй XX ст. запропоновано ще двй концепп
загальной етйолопй.
Онтогенетычна модель медицины (Ф. Людвиг, 1980), згйдно з я кою великого
значения у виникненни основних нейнфекцййних хвороб надають закономйрнс
зростаючим вйдхиленням найважливйших гомеостатичних констант (артериального
таску, вмнету глюКози, холестерину в крови тощо) не тйльки у зв’язку з ростом
розвитком у дитячому й пйдлйтковому вйцй, а й у подалыш вйковй периоди, коли рйез
й розвиток припиняються, а вйдхилення гомеостатичних констант посилюються.
Вйдомий росййський геронтолог професор В.М. Дйльман (1987) до онтогенетич-
ной концепций етйолопй пропонуе додати акумуляцшну. На його думку, розвито-
основних нейнфекцййних хвороб (за проявами вони е дегенеративними, йнволюцйй-
ними, регресивними) зумовлений накопиченням помилок обмину речовин унаелй-
д 2.11 як зовшшшх, так ! внутргшшх факгоргв зг стгйкими порушенными гомеоста-
тропонова ну модель медицина було названо тволющйною, або метаболгчною.
Завершуючи цей стислий гсторичний огляд основних напрямгв загальног етю-
V’ । сл!д навести основы! положения материалистично! д1алектики про причин-
.ч у патологп: 1) ус! я вита в природ! мають свою причину; 2) причина ма-
| иьна. вона генуе незалежно в!д нас; 3) причина хвороби взаемодге з оргашз-
4) причина надае процесу (хвороб!) специфгчносп; 5) причина дге за певних
I !. як! можузъ вплинути на остаточний результат.
Найважливппим у цьому уявленн! е визнання об'ективно! реальност! причини
— взаемодгя причини й оргашзму). На цьому потргбно наголоепти, осюльки
:ч матергальний фактор (мгкрооргашзм, онкоген, отрута) не е причиною хво-
Би (фурункульоз, рак, огруення) доти, доки вш не подгяв на оргашзм ё останшй
?еагував. При цьому змппоються обидв! сторони: в оргашзм!, наприклад, ви-
яюгься антитеза, в мткробг вщбуваються мутацп. змшюються його спадковг
к гвост!. Тобто причини поза взаемолгею немас. Правильною вщповйщю на
в пання: “Що е причиною — р!ч чи взаемодгя?” — була б. "Взаемодгя речей”.
Важливо зазначити, що причина впливае на оргашзм неодмшно в конкретных
зах: або в тих, шо сприяють дп причини, або в тих, шо перешкоджають ш.
Нареши, причиною можна назвати лише те, що надае патолопчному процесу
ыфгчносп, тобто серед багатьох факторов, якт впливають на оргашзм, е один,
кий надае ггроцесу своерщних, неповторних ознак. Усе 1нше — це умови. 3 ог-
на це дослщник шукае причину 1, знайшовши п, визначае шляхи усунення
ч ише хвороби, а й само! можливост! ц виникнення. I якими б важливими не
л умови виникнення хвороби, вони не можуть створиги нозололчну одиницю.
9 туберкульозу луже важливими е 1 харчування, т умови працг та побуту, проте
вороба в!к може виникнути лише пш впливом мткобактерн туберкульозу.
на г е причиною ц!е! хвороби.
У тому раз!, коли причина хвороби ниш невщома г якщо ще не видтлений з
_"агьох вщомих факторов один, що вщповшае вищезазначешй вимозг, тод! в бо-
гьбт з хворобою може бути ефективним усунення яког'сь умови (наприклад,
плен ня болтт, де розмножуеться малярийный комар). Можна вилшувати хворо-
лтдвищивши неспецифгчну резистентшсть його ошашзму. Однак не можна
гзати, що причиною пневмон!! е ослабления оргашзму, а не пневмокок.
У медицин! поширена точка зору, зпдно з якою одна хвороба може виникну-
з кглькох причин. Така полтетюлопчна точка зору не вщновщае правильному
вленню про епологно. Вона лише евщчить про труднощ!, проблеми т нелостаг-
ть наших знань. Ми не завжди можемо вщокремити причину вщ шов, г все,
К впливае на розвиток процесу, называемо причиною. Те. шо причину в багатьох
.гпадках визначити складно, зовегм не дае шдстави вважати, шо !х багато. 3 ме-
одолопчно! точки зору це уявлення близьке до кондишоналтзму з його запере-
: дням причин хвороб узагал!. 1з розширенням знань про сутшсть хвороби засто-
зують нов!, досконалши диференшальш дгагностичн! метода. Багато нозолопч-
гих форм, що рашше здавалися единими, подшяються на к!лька ргзних хвороб,
“зк. геморапчний д!атез, яким фггурував у старей класифжацп як эб’е.днуюче по-
гтя для всех варганив пгтвишено! схильносп до кровогочивоегг, подыений ниш
а багато ргзних хвороб спадковог (гемофглгя) чи набутот (цинга) природи. Те
саме стосуеться г пухлин. 1х ргзн! види характеризуються не титьки сшльними, а й
вщмгнними властивостями, що дае змогу передбачити ргзш причини.
Останнтм часом у медичнш практищ широко застосовують термш “фактори
ризику” щодо так званих основних нешфекщйних хвороб (виразка шлунка, ар-
тергальна ппертенз!я, цукровий д!абет, ожиршня, атеросклероз тощо), на яю при-
падае левова частка захворюваносп населения. Так, серед факторов, яю зумовлю-
ють атеросклероз, видшяють ожиршня, куршня, гшодинампо, порушення обмшу
лшопротещгв, спадков! дефекти фермента, стрес, цукровий дгабет. Це дае змогу
з велико! кшькост! внутршгшх г зовшшюх фактор! в у кожному шдивщуальному
чи груповому вар!ант! перебггу видшити таю, шо якнайбшыпе стосуються форму-
вання конкретного патолопчного пронесу. Складюсть проблеми в тому, що до
факторгв ризику належать таю, що е причинами, умовами чи навпь ланками па-
тогенезу вщповщно! хвороби, а це потребуе окремого аналгзу.
Умови виникнення хвороби. Взаемодгя причини хвороби з организмом завжди
вщбуваеться за певних умов. Вщмшюсть умов вщ причини полягае в тому, що
причина одна, а умов багато г що бптышсть гз них не обов’язкова для виникнення
хвороби. Умови не надають хвороб! специфичность I хоч якими важливими для
виникнення туберкульозу е таю фактори, як харчування, умови пращ й побуту,
однак не вони визначають розвиток ще! хвороби. Порушення харчування, вплива-
ючи на гмунолопчну реактившсть оргашзму, може мати значения у виникненн!
ргзних шфекцшних хвороб, але лише мгкобактергя туберкульозу е його причиною.
Розргзняють умови, як1 сприяють ди причини / запоб1гають ш. Наприклад,
сприяють виникненню хвороби перенапруження нервово! системи, порушення
режиму пращ й вщпочинку, неращональне харчування, алкоголизм, спадкова об-
тяжешсть тошо, запобпають — правильний режим пращ й вщпочинку, ращо-
нальне харчування, загартовування оргашзму, занятгя фгзкультурою тощо.
Головною умовою для виникнення хвороби е здатшсть оргашзму вщповщати
на дно патогенного фактора певними захисними реакцгями, быышсть з яких не е
досконалими ! можуть сам! ушкоджувати органи г тканини.
ПАТОГЕНЕЗ
Патогенез (вщ грец. лавод — страждання, уесяд — походження) — це вчення про
механизмы розвитку I к1нця хвороби.
Тод! як етюлопя трактуе питания, що стосуються причин г умов виникнення
хвороби, патогенез вивчае все те, що вщбуваеться шсля да! причини. 1нод! причи-
на (травма, ошк, юшзуюче випромшювання), здшснивши акт агресп, зникае.
Тод! чпко видно ргзницю м!ж етюлопею г патогенезом у часовому аспект!: спо-
чатку д1я еполопчного фактора, попм патогенез, спочатку “чому?”, а поим “як?”.
В шшому варгант! причинний фактор дае протягом усього процесу ! вщгграе вирг-
шальну роль у його розвитку г перебггу (електротравма ! гостр! отруення зг смер-
тельним наслщком). Власне мехашзмом да! причинного фактора можна пояснити
весь перебй патолопчного процесу. У цьому варганп причина доргвнюе дп. Та
найчастппе д!я етюлопчного фактора зберггаеться протягом усього перебггу хво-
роби, однак його роль на рхзних етапах неоднакова. Така картина притаманна
шфекщйним хворобам, коли збудник, спричинивши хворобу, продовжуе дгяти, а
? разг хрошчного перебпу характеризуеться наявшстю решен та загострення.
Гэбто в цьому випадку етюлопчний фактор завдяки власному механизму ди про-
эгсае в патогенез, впливае на нього.
Отже, патогенез е ун!версальним посередником етюлопчних факторгв, ос-
ь_ ъки “будь-яка зовшшня причина дге т!льки завдяки патогенезу г на його ос-
юа”. Патогенез запрограмований {вторично, еволюцгйно завдяки саморозвитку,
-ичоруху та саморегуляцп. “Природа навив у стан! хаосу може д!ята титьки пра-
мтьно 1 злагоджено,” — висл!в видатного росшського патолога академика 1.В. Да-
тиловського, що якнайкраще характеризуе цю ушкалъну здатнгсть оргашзму про-
грамувати навгть патолопчний процес, хворобу.
Характеризуючи в гсторичному аспект! основы! науков! напрями вчення про
-_этогенез, у загальному вченш про хворобу слщ назвати найважливши, як! збе-
регли свое значения дошил.
1. Гшпократом (V—IV ст. до н. е.) — засновником медицини як науки — було
„точатковано гуморальну (в!д лат. китог — рщина) теории патологи. Зпдно з щею
тезргею здоровий стан оргашзму характеризуеться правильным стввгднишенням
сразив), гармонгею та постштстю складу ргдинних середовищ, тод! як у випадку
свороби спостерггаеться явище дискрази — неправильного змгшування рщин 1
~орушення !хнього складу. Наведена теоргя виявилася дуже продуктивною. По-
альший розвиток ендокринологи, вчення про бюлоггчно активш речовини, у
тщому про систему гуморально! регуляцн фгзюлоггчних процесгв г функцгй, вчен-
ня про так зване внутргшне середовище (кров та штерстицгйна р!дина) оргашзму,
торушення !хнього складу за основными параметрами гомеостатичних констант
вшсту субстрапв, ферментгв, виампггв, гмуноглобулшгв тощо, водно-мгнераль-
чого обмшу та КОС) дае змогу сучасному лгкаревг правильно визначати як основ-
ну, так т допомгжш ланки патогенезу, своечасно та адекватно коригувати вщ-
повщнг змши при багатьох хворобах.
Найвщомппими представниками гуморального напряму в патологи були про-
фесори К. Рокитянський (Австргя, XIX ст.) 1 Г. Селье (Канада, XX ст.).
2. Солидарна (в!д лат. зоПдиз — твердый, мщний) теория патологи — система
теоретичных поглядгв Демокрита (II ст. до н. е.) на патогенез як наслздок пору-
шення сшввщношення мгж елементарними складниками твердых частин оргашз-
му. Досягнення сучасно! молекулярно! бюлоги та молекулярно! патологи пщтвер-
джують зазначену теоргю.
3. Теории целюлярно! патологи запропонував гешальний шмецький учений па-
тологоанатом Р. Вгрхов у середин! XIX ст. (1855—1858). Уже 13 само! назви вказа-
ного напряму патогенезу зрозумшо, що матергальним субстратом хвороби ор-
гашзму е “хвороба” певного типу клгтин за природою, походженням ! здгйснюва-
ними функциями. Це дало змогу автору та його послщовникам установи™ при
ргзних хворобах локалгзащю патологи в тому чи тому орган!, юлыасш та якгсш
морфолопчш змши клггин у вигляд! гетерометрп (не в т!й кщькостг), гетерохрони
(не в той час) г гетеротош! (не в тому м!сщ).
Автору теорн целюлярно! патологи дор!кали за ггнорування принципу цглгс-
ностг оргашзму, едност! оргашзму та навколишнього середовиша, за надмгрне
використання анатомгчного напряму в патологи (1.М. Сеченов, В.В. Пашутш).
Проте слщ зазначити, що в XX ст. з упровадженням електронно-мжроскошчного
та шших методов цитолопчного дослщження “целюлярна патолопя” як напрям
збагатилася новими фундаментальними вщкритгями, яю дали додатковий пош-
товх розвитку ще! теори. Зокрема це дало змогу описати низку хвороб, пов’язаних
з хзольованою патолопею субклггинних структур (мембран, лхзосом, мпохондрш
тощо).
4. Функцюналъний напрям у патологи витгкае, з одного боку, 13 фшософського
принципу переважання зм1сту над формою, функцп над структурою, з шшого — з
1хньо! нерозривност! та взаемозумовленосп. Представники цього напряму розви-
ток хвороби пов’язували з порушеннями функцюнальних зв’язюв мгж окремими
елементами та системами оргашзму. Осюльки таю зв’язки забезпечуються нерво-
вою системою, то концепция нерв1зму, започаткована ще в XIX ст. видатними ф!зю-
логами Ф. Мажандц К. Бернаром, 1.М. Сеченовим, 1.П. Павловим та ш., як 1
сучасю досягнення нейрофхзюлогп, у тому числ! всесвзтньо вщомо! науково! шко-
ли видатного украшського вченого академ пса П.Г. Костюка, суттево розширили
можливосп розвитку функцюналъного напряму в патологп.
Що стосуеться термшхв “функщональна патолопя” та “функщональш хворо-
би” (наприклад, дисюнезхя кишок, жовчних шлях!в, стравоходу, функщональна
диспепсия, вегетативно-судинна дистон!я тощо), то !х використання вступае в
протирхччя з уже згадуваним фшософським постулатом про нерозрившсть 1 взаемо-
зумовлешсть змзсту 1 форми, функци та структури.
У природ! не 1снуе ш суто функцюнальних (без вщповщних структурних
зм!н), н! суто оргашчних (без вщповщних функцюнальних проявив) хвороб. Може
йтися про р1зне спхввщношення тих чи мних змш у динампц хвороби на р!зних
!Т етапах.
5. Сучасний етап медицини (починаючи з друго! половини XX ст.) пов’язаний
з бурхливим розвитком молекулярно! бюлоги, упровадженням вщповщних ме-
тод1в у теорпо та практику медицини. У вченш про хворобу сформувався напрям,
який отримав назву молекулярной патологи та молекулярно! медицини. За прогноза-
ми нобелхвських лауреапв, в!н буде домшуючим 1 у XXI ст.
Зусилля представниюв теоретично! медицини спрямоваш на з’ясування мо-
лекулярних мехашзм1в виникнення та розвитку хвороб, а представниюв юпшчно!
медицини — на використання вщповщних метод!в молекулярно! бюлоги з д!агнос-
тичною та лткувальною метою. У першу черту це стосуеться спадкових хвороб 1
хвороб 31 спадковою схильшстю. Природно, що метода генно! шженери вщповщ-
них порушень, яю спостер!гають пщ час найпоширенппих хвороб спадково! при-
роди, мають упроваджуватися в практику медицини.
Причинно-наслщковий зв’язок
У патогенез! хвороби можна видыити юлька еталгв, або ланок, пов’язаних м!ж
собою причинно-наслщковими зв’язками. Це означав, що змши, яю виникли пщ
час хвороби, стають причинами нових порушень. Наприклад, перша ланка в па-
тогенез! травматичного шоку — б!ль. Сильний быь зумовлюе пригшчення житге-
во важливих функцш, зокрема зниження артер!ального тиску, що е причиною
кисневого голодування. Гшоксхя головного мозку спричишое ще бшыпе пригш-
чення судинорухового центру 1 зниження артер!ального тиску. Як видно, змша
причин 1 наслщюв призвела до утворення порочного кола (сггсиШз уШобиз). Зна-
ення порочного кола можна прошюструвати також на приклад! метеоризму,
'лупя кишок галъмуе !х моторну й секреторну функцц, а це зумовлюе бродшня,
зорення газ!в 1 ще бгтьший метеоризм.
Поняття про причинно-насл йд ковй зв'язки у патогенез! становить великий
рзктичний штерес, оскьтьки дае змогу лткаревт ишеспрямовано втручатися в
зозвиток хвороби. При цьому потрйбно враховувати, що не вс! ланки патогенезу
знаково важлив!; серед них е головн! 1 другорядн!. Головною ланкою патогенезу
азиваютъ той процес, шо потргбний для розгортання вс1х гнших. Своечасна лйквт-
гц!я головнот ланки сприяе усуненню процесу в щлому. Так, при шсулшозалеж-
юму цукровому дтабеп 1-го типу головною ланкою патогенезу е нестача шсулшу.
Усунення п (увеаення гормону) сприяе зникненню шших проявив хвороби (ппер-
йкемп, кетоацидозу, коми). У раз! гострого панкреатиту виникае багато пору-
тень з боку всйх систем, однак гол овна ланка — це актива щя протеолйтичних
^ерменпв шдшлунково! завози, як! ушкоджують тканину й га !нших орган!в.
Найефективнишим заходом протидц цьому е пригючення активност! протеиназ.
Загальне! мкщеве
Анализ патогенезу потребуе чёткого розумйння сшввщношення миж загальним 1
йсцевим, вщ чого певною м!рою залежить ефективнйсть лйкувальних заходы. У
едност! загального й мйсцевого потрйбно знайти провщну ланку, яка визначае
тенденцию процесу. Якшо, скажимо, встановлено, шо кар!ес зуба пйддаеться м!с-
цевому лйкуванню, то достатньо поставили пломбу, а коли каркс е наслщком за-
гатьних порушень минерального 1 бокового обмшу, то й лйкування мае бути на-
самперед загагьним.
Сшввщношення м!ж загальним 1 мкцевим зминюегься в час!. Мкцеве (фу-
пункул) може поширитися 1 стати загальним (сепсис)!, навпаки, загальний пато-
лопчний процес завдяки захисним функциям оргашзму може обмежитися, локалй-
иуватись и зникнути.
Структура । функция
Хвороба завжди мае той чи той морфолопчний прояв. Як уже зазначалось, кну-
лання поняття функшонально! хвороби можна пояснити тим, що морфолопчн!
ознаки ц поки не встановлено (наприклад деяк! розлади психики). Про це свщ-
чить и те, що кйльккть функшэнальних хвороб з часом зменшуеться. Змгнюеться
1 поняття про морфолопчний субстрат хвороб Засновник патолопчно! анатом!!
Р. Вирхов починав дослщження трупа, розтинаючи його ножем, а пот!м застосо-
вував микроскоп. Тепер морфологи користуються складними онтичними й елект-
ронними микроскопами, фиксу ють зм!ни не тйльки на клйтинному, а й на субклй-
тинному ривни.
Практичний л!кар вивчае насамперед функцию. Це дае йому информацию про
пристосувальнй можливостй оргашзму. Однак поеднання функцйональних и мор-
фолопчних дослщжень дае повшше уявлення про це, оскмьки фйзюлогйчнй га
морфолопчн! явища, функшя й форма взаемно зумовлюють одне одного
Загальне (неспецифЕчне) Е специфике
У хвороб! завжли можна розр!знити ознаки (симпгоми), характерш лише для не!
(наприклад, поширення болю при стенокардн), та ознаки, характерш для багать-
ох або навпъ для вс!х хвороб. Це загальна, неспеиифзчна ознака патогенезу, що
визначаеться реакциями оргашзму, як! виникли в процес! еволюци та успадкову-
ються. Значения них реакцш полягас в захист! оргашзму, який використовуе !х
завжли в тих випадках, коли виникае патолопчна ситуащя. Таких неспецифгчних
реакцш (факторов патогенезу) принайми! п’ять, 1 вез вони визначаються нервовою
або нервовою та ендокринною системами: патолопчний парабиоз, патолопчна
домшанта, порушення кордико-вгенерально! динамита. нервоьо-троф!чних функ-
щй, стрес.
Парабюз — це застшне збудження, яке не поширюеться т виникае в раз) уш-
кодження збудливо’1 тканини. Мас значения, наприклад, у патогенез! деяких форм
блокади серця.
Домшанта. тобто наявшеть у центральнш нервовтй систем! пан!вного вогни-
ща збудження, яке шбито шдпорядковуе собт вс! !нш! центри, може зумовлювати
багато патолопчних проявив у раз! психтчних хвороб (наприклад, марення пере-
слщування). Дом!нантою можна поясните загострення болю в ус!х випадках, коли
на оргашзм впливае нав!ть шдиферентний подразник (наприклад, свило, звук).
При гшертошчшй хвороб! в головному мозку виникае заст!йне вопгище збуджен-
ня, що реагус на будь-яке по дразнения спазмом ! звуженням судин.
Наступнии фактор патогенезу — порушення кортико-вгсцеральноТ тинам! км —
зумовлений наявшстю т!сного зв'язку м!ж корою великого мозку ! внутршгшми
органами (К.М. Биков). Зв’язок цей насамперед регуляторний, позитивний, однак
в!н може виконувати роль 1 патогенного фактора. Не можна зрозумгте, як розви-
ваеться виразка шлунка ! дванадцятипало! кишки або, скаж!мо, тиреотоксикоз без
ураху вання того, шо шлунок! шитопощбна залоза якнайпеншге пов’язаш з нерво-
вою системою. У раз! порушення функпп нервовог системи (насамперед кори ве-
ликого мозку) саме ц! органи можуть стати мштенями патолопчно!' тмпульсащ!'.
Патогенез охоплюс також троф!чну функщю нервов»? системи О.Д. Сперансь-
кий вважав, що немае жодног хвороби, у якгй би не вшграваь рол! мехашзм по-
рушеного троф!чного впливу нервово? системи на вгдповщний орган. Цей вплив,
за сучасними даними, мае бсз!мпульсну природу 1 здшснюегься у форм! двоич-
ного аксоплазм атичного току р!зних речовин, як! виконують функшю трофогешв —
регуляторгв структурно-метабол!чних процесгв 1 м!жкт!тинно! взаемодГ? м!ж ней-
ронами ! структурами-мипеням и. Припускаюсь можливгеть утворення паготрофо-
гешв (Г.М. Крижашвський, 1997).
Уявлення про неспециф!чне у хвороб! особливо збагатилося вченням Г. Селье
про стрес. Автор показав, шо р!зш подразники (тепло, холод, токсини, б!ль) за-
вжди спричинюють стандартну, неспециф!чну реакцию. В ус!х випадках це акти-
вапгя симпатико-адреналово! системи та викид корта котрогйну — гормону ппо-
ф!за, на який юркова речовина надниркових’ залоз вгдповшае пгдвшценим синте-
зом глюкокортикощних гормошв. Якшо вплив подразник!в не дуже гривалий 1 не
дуже значний, гормони надниркових залоз лише допомагають оргашзму адапту-
ватися до сбставин. Якщо вплив ушкоджчвального фактора надмфний. розви-
ваеться хвороба або смерть
Щодо специфичного в патогенез!, тобто тих ознак, за якими встановлюють
хапюз, то тут основну роль в!д!грае етюлопчний фактор. На загальне, однакове
вшаровуеться особливе, окреме. Причина н!би вишивае св!й в!зерунок на за-
-^ъг эму ТЛ1. Кр!м того, сл!д мати на уваз! т! п’ять фактор!в патогенезу (парабюз,
. . -анту, порушення кортико-в!сцеральних зв’язюв, нервово-троф!чно!.функ-
стрес), яю сам! по соб! неспецифзчш, але в складшй комбшаш! м!ж собою
- к можуть давати особливе саме для ще! хвороби.
Огже, кожна хвороба е “сплавом” специф!чного 1 неспецифзчного, особливо-
г зкремого) та загального. 1х послщовшсть така, що на самому початку хвороба
являеться быьш загальними, неспециф!чними ознаками, властивими р!зним
за своею природою хворобам, тобто проявами “хвороби взагал!”. До них належать
_-2Льна слабгсть, нездужання, втрата апетиту, порушення сну, розбипсть, стан
гривоги, депрес!я, драт!влив!сть, зниження працездатност! тощо. У раз! виник-
• г <ня хвороби запально! природи до перерахованих неспециф!чних ознак “ста-
- в тення хвороби” (“хвороби взагал!”) приеднуються ще й загальш прояви м!с-
_ в вого запального процесу (див. роздал XI “Запалення”). Згодом до цих неспе-
_.ф!чних прояв!в додаються специфхчш, !з характерною машфестащею залежно
вщ етюлопчного фактора, його механизму ди та патогенезу в целому.
Ддаптащя I компенсащя
Ущптащя — пристосування оргашзму до умов (снування шляхом адекватних зм!н
?ункцн, метабол!зму та структури його орган!в ! систем. У кожного бюлопчного
иду щ змёни яюсно та юльюсно вщр!зняються. У людини адаптация пов’язана зё
береженням за певних умов не тыьки и бёологёчних, а й сощальних функцёй —
щатностё брати участь у суспыьно-корисшй робот!, здшенювати творчу дёяль-
шеть. Мехашзми адаптацп вироблеш еволюцёйно ! спрямован! на збереження
гомеостазу. Так, пщ час пщнёмання на висоту спостерёгаеться посилення легене-
во! вентиляцп та кровообёгу, збшьшення вмёсту гемоглобшу та кёлькостё еритро-
цитёв у кров!, при посилешй робот! — ппертрофёя серця та ён. Може йтися про
адаптацию до темряви, до тепла чи холоду тощо. Адаптацёйн! мехашзми завжди
напоготов! ё вводиться в дно у вщповщь на певний сигнал. У цьому й полягае !х
гомеостатична роль.
1нша ситуацёя виникае п!д час хвороби. Тода вплив патогенного фактора такий
потужний, що звичайних адаптащйних мехашзмгв стае недостатньо, ё гомеостаз
порушуеться. Змёнюються температура тыа, р!вень глюкози в кров!, рН. Почина-
ються руйнёвш процеси в тканинах. У цей момент у реакцйо оргашзму вводиться
крём адаптащйних шип мехашзми, якё називають компенсаторними. Останш також
виникають у процес! еволюцёйного розвитку, однак вщр!зняються вщ адаптацёй-
зих як юльюсно, так 1 яюсно. Завдання в них теж рёзш. Адаптацгя забезпечуе
лалёсть основних фёзюлогёчних параметр!в, тобто гомеостаз у норм!, у вузькому
цапазош змш зовнёшнього та внутрёшнього середовищ, а компенсация — це бо-
зотьба за гомеостаз в умовах, коли вён уже порушений ё можливе подальше по-
'либлення порушень.
Точнёше, коли хвороба вже почалася, розмежувати адаптацёйн! та компенса-
орш мехашзми неможливо, вони ёснують 1 дёють разом, проте вирппальну роль
вдпрають компенсаторш. Ю. Конгейм наголошував, що хвороба — це життя в
Зовшппп фактори
Гомеостаз збережений
Боротьба за гомеостаз
Адаптащя
Компенсащя
Мал. 2. Сшввщношення адапташ! 1 компснсапп в норм! та шд час хвороби
ненормальних умовах, яке можливе завдяки компенсаторним пристосуванням.
Сшввщношення мЬк адаптащею 1 компенсащею схематично наведено на мал. 2.
Як узагальнення наводимо передне найважливниих принцишв, на яких Грун-
туються адаптацхя та компенсация незалежно вщ того, на якому р!вн! вони здше-
нюються.
Л!воруч у кол! наведено адаптащйн! мехашзми, що дпоть у межах норми. При
хвороб! & бракуе, ! тод! до них додаються компенсаторю (праворуч у кол!). Стрел-
ка показуе, що адаптацию мехашзми продовжують д!яти 1 пщ час хвороби:
1) “запас мщност!” (В. Кеннон) — характерна для оргашзму вщеутшеть обме-
женого л!мпу та жорстко! економп. Наприклад, наявшеть в артер!альшй кров! в
3,5 раза бшьше кисню пор!вняно з потребою в ньому тканин. Винятком е мю-
кард, у якому використовуеться 75 % кисню, що надходить по систем! вшцевих
артерш;
2) використання резервних можливостей за рахунок посилення обм!ну речо-
вин 1 функцш оргашв;
3) посилення фхзюлопчно! регенераци клггин 1 клхтинно! репарацп для вщ-
новлення;
4) принцип економи: кшыасть кров! менша н!ж об’ем судинного русла, що
дае змогу за рахунок адекватних змш того чи шшого параметра забезпечувати
необхщний рхвень кровопостачання органов 1 тканин;
5) принцип парност! оргашв; у раз! ушкодження одного з парних оргашв ви-
користовуються резервн! можливост! шшого;
6) дублювання функцш рхзними органами ! системами, навпь органелами
кл!тин.
7) Принцип системност! вщповцц, залучения одночасно чи в певнш послщов-
ност! юлькох систем оргашзму з метою лпевщувати наслщки того чи того ушкод-
жувального фактора: наприклад, системна вщповщь оргашзму в умовах киснево!
недостатносп.
Р03Д1Л III
ПАТОГЕННА Д1Я НА ОРГАНОМ
ОДКТОР1В НАВКОЛИШНЬОГО СЕРЕДОВИЩА
:-ня1 фактори навколишнього середовиша (ф!зичш, х!м!чш, бюлопчш), з
г "чи постшно стикаеться людина, можуть стати причиною багатьох травм 1 хво-
Б ’ . раз! надмтрно! Гх ди, за наявносп умов, як! сприяють ушкодженню, а також
—тк зшш реактивност! оргашзму. 1х вплив стае дсдал! штенсившшим, оскГльки
Е зднюеться виробнича дГяльшсть людини. Природы фактори середовиша за
I нрно! ди в екстремальних умовах, а також новГтнГ техногенш фактори, з яки-
м впродовж еволюци оргашзм не стикався 1 до яких пристосування майже не
инулося (нов! види ви проми новання, фактори косхпчного польоту тощо),
гчинюють значке фГзичне ! психГчне напруження, формують екстремалън!
жыи га стрес.
На приклад! патогенно! ди фтзичних факторов навколишнього середовиша на
. -ровий оргашзм можна легко засвогги основы патофГзюлопчш поняття — ет'ю-
) (етюлопчн! фактори 1 значения умови !хньо! ди) та патогенез (патолопчн!
рушения та компенсаторно-пристосувальн! реакци на р!зних р!внях оргашзацй
р во! матери, причин но-насл!дков! зв'язки, стад!! комленсаци та декомпенсацп
штатного процесу тощо).
4ЕХАН1ЧНА ТРАВМА
полопя. Етюлопчними факторами мехашчно! травми е тверд! ила або ударна
лнля, як! д!ють мгсцево 1 спричинюють розриви, заб!й, переломи, розчавлювання
-тин. Важливою умовою. яка визначае тяжюсть мехашчно! травми, е и локал!-
. шя, а також наявшсть супутнього ушкодження (крововтрати, ушкодження не-
т-эвих стовбурш тошо).
Патогенез. Першою ланкою патогенезу загатьних 1 мюцевих порушень часто
Е быь. На М1сщ травми виникае запалення. Загальш порушення за сво!м значен-
ии можуть переважати мгсдев!! проявлятися крововтратою, травматичным ию-
тч, синдромом тривалого розчавлювання.
ТЕРМИНА ТРАВМА
.полопя. Термтчна травма зумовтена д!ею на оргашзм зовшшнього середовиша
: змшеними температурними параметрами (комфортна температура пов.тря для
"юдини — 18 °С). Ця травма проявляеться низкою патолопчних стан!в, етюлопя
патогенез яких в!др!зняеться.
ТермЁчнЁ фактори (холод, тепло) у раз! надмЁрно! ди (стихшие лихо, вщеут-
шеть штучно! термоЁзоляци тощо) можуть стати етюлопчним фактором ушкод-
ження ё спричинити низку патолопчних процесгв: охолодження, перегрЁвання,
тепловий удар, вЁдмороження, опёк, шок. При цьому важливою умовою, що ви-
значае перебЁг термЁчноЁ травми, е загальний чи мЁсцевий характер ди етЁолопч-
ного фактора.
Патогенез термЁчно! травми проявляеться комплексом патолопчних порушень
ё компенсаторних реакцЁй, якё полягають у напруженш фЁзично‘1 ё хёмёчноё термо-
регуляцп (тепловЁддачЁ ё теплопродукци). У патогенезЁ видёляють двё стадй — стадЁю
компенсаци Ё стадЁю декомпенсацп. У стадп компенсаци пристосувальш реакцЁЁ
переважають, ё гомеостаз не порушуеться (температура тЁла пщтримуеться на
вихЁдному рЁвнЁ). СтадЁя декомпенсаш'1 настае тодё, коли захисш реакцЁЁ виснажу-
ються, гомеостаз порушуеться Ё температура тЁла змЁнюеться (знижуеться чи
пЁдвищуеться).
Загальна дёя низькоё температуря може зумовити зниження температури тЁла ё
розвиток гшотермп (охолодження).
У стадп компенсаци температура тЁла пЁдтримуеться на вихЁдному рЁвнЁ. При-
стосування полягае в напруженш фЁзЁолопчних мехашзмЁв фЁзичноЁ терморегуля-
цц, що спричинюе обмеження теплов1ддач1 шляхом зменшення випаровування
(потовидЁлення), зниження теплопроведення Ё тепловипромЁнення (за рахунок
звуження судин шкЁри) ё додатковоЁ теплоЁзолягщ (за допомогою шерстЁ у тварин,
одягу — у людини). У разЁ ЁнтенсивнЁшоЁ дп холоду в тканинах, на якё вён впливае
безпосередньо, вЁдбуваеться своерЁдна судинна реакцЁя (“гра судин”), коли вод-
ночас спостерЁгаються охолодження шкЁри та и почервонЁння (артерЁальна гЁпере-
мёя запобЁгае вЁдмороженню).
ХЁмЁчна терморегуляцЁя спрямована на збыъшення теплопродукци. При цьому
посилюеться обмЁн речовин, збЁльшуються розпад глЁкогену в печЁнцЁ ё м’язах, а
також вмёст глюкози в кровЁ ё споживання кисню, ЁнтенсивнЁше функцЁонують
системи транспорту кисню до тканин. ОбмЁн речовин не тёльки пЁдвищуеться, а
й перебудовуеться. Додаткова теплова енергЁя утворюеться за рахунок роз’еднання
окиснення Ё фосфорилювання. В оргашзмЁ, адаптованому до холоду, збЁльшуеться
потужнЁсть мЁтохондрЁальноЁ системи, пЁдвищуеться актившеть ферментЁв циклу
трикарбонових кислот Ё дихального апарату.
Ця перебудова терморегуляци контролюеться нейрогуморальними механизма-
ми. Терморецептори шкЁри першими сприймають Холодове подразнення Ё чутли-
вими шляхами посилають Ёмпульси в центр терморегуляци ппоталамуса, звёдки у
зворотному напрямку надходять сигнали до оргашв Ё систем, якё пЁдтримують
температуру тЁла. У терморегуляцЁЁ бере участь гшофЁз, який за допомогою тропних
гормошв залучае ендокринш залози — щитоподЁбну (тироксин пЁдвищуе мётохон-
дрЁальне окиснення глюкози Ё жирних кислот) та кЁркову речовину надниркових
залоз (глюкокортикощи стимулюють новоутворення вуглеводЁв ё викликають
стрес). Енерпя, що вивЁлънюеться при окисненнЁ, частково накопичуеться у ви-
глядё ДТФ, а частково розсЁюеться у виглядё тепла (первинно розсЁяне тепло).
Через симпатичнЁ нерви збудження досягае мозково! речовини надниркових за-
лоз, де посилюеться секрецЁя адреналЁну, який сприяе розвитку судинноЁ реакцЁЁ,
мобЁлЁзуе глюкозу ёз глЁкогену печшки та стимулюе роз’еднання окиснення ё фос-
форилювання в мЁтохондрЁях, завдяки чому частка енергп, що йде на вироблення
т на, збтльшуеться. Через руховт нерви тмпульси надходять у м'язи, у яких тпдви-
_ еться терморегуляторний тонус, 1 починаеться тремттння, ню спричинюе роз-
: \ТФ 1 утворення тепла (вторично розстяне тепло).
За умови тривалого або пгтенсивного впливу холоду мехашзми терморегуляцн
-чажуються, температура тша знижуеться. т настае друга стадтя гшотермп —
-шаШя дексмпенсаци. У пей пертод спостерпасться зниження не .лише температу-
пла, а й метаболгчних процес!в 1 споживання кисню; життево важливт функцн
- итпчуються. Порушення дихання та гемодинамики зумовлюють кисневе голо-
. зання, гальмування центрально’] нервовоТ системи, зниження тмунолопчно’т ре-
активност!.
У друпй стади лпотермй пристосувальнт й патолопчш явиша ясно пов’язаю.
- 1ыпе того, одш й тт самт порушення, шо, з одного боку, е патологтчними, з
того — можна оцтнити як пристосувальнт. Так, лригшчення функцш централь-
, нервово! системи можна назвали охоронним, осюльки знижуеться чутливтсть
.-рвових КЛ1ТИН до нестач! кисню 1 подальшого зниження температури тша. Ос-
лабления обмтну речовин у свою чергу зменшуе потребу оргашзму в кисит. У стан!
гг.отермп оргашзм стае менш чутливим до ди екстремальних факторов т неспри-
- швих умов навколишнього середовиша (нестачт Ькт, штоксикапп. тнфекцн, уш-
лжувальноУ ди юшзуючого випромтнювання. електричного струму тощо). Штуч-
з. ппогермпо (пбернацно) засгосовують для вцттворення пасивноУ резистентностУ
ргашзму.
Мкцева щя низькоУ температури спричинюе в^дмороження, у патогенез! якого
зовщну роль вшграе порушення мтсцевого кровообпу внаслщок спазму та паре-
периферичних судин, тромбоутворення, а також прямот ушкоджувальноТ ди
изькот температури на колотднт системи цитоплазми. Якшо втдмороження суп-
оводжуеться птфткуванням, перебп остан нього на тлт загальних порушень кро-
ообтгу та зниження тмунолопчноУ реактивнослт тяжкий, як сепсис.
Загальна дтя високот температури спостерйасться в умовах виробництва або
жаркого клтмату. Якщо порушуеться ртвновага мтж утворенням тепла в органтзм! т
йддачею його в навколишне середовище, розвиваеться гтертермгя (перегргвання).
Пристосування полягае в зб'шыиенш теплов1ддач1 шляхом розширення судин
шктри (пщвишення теплопроведення т тепловипромтнення) т збйтьшення пото-
идтлення (щдсилення випаровування). Це забезпечуе стадно компенсаци. У раз!
значного тпдвишення температури повттря, теплопроведення 1 тепловипромтнен-
ня утруднюються, а за температури повттря 33 °С (тцо дортвнюе температур] шкт-
ри) блокуються. Можливостт судинноУ реакцтУ вичерпуються, т вона стае неефек-
гивною. €диною реакшею терморегуляцн, спрямованою на шдсилення теплов’щ-
тачт, залишаеться лотовиддлення (випаровування 1 мл води вичерпуе 0,58 ккат/год
тепла) . випаровування з поверхн! повттропровщних шляхтв. В умовах шдвишеноУ
волотостт повттря пронес вцтдачт тепла утруднюеться г цим останшм механтзмом.
Якщо температура навколишнього середовища значно виша за температуру ши-
ри (в саую вона сягае близько 100 °С), остання охолоджуеться кров’ю гз розши-
рених периферичних судин, шо запобпае олтку.
Перенапруження функпп терморегуляцн переводить процес перегртвання в
стадно декомпенеацц. Найбшьше навантаження припадае на серцево судинну
систему Унаслтдок розширення судин можливе зниження артертального тиску г
навить колапс. Тахткардтя — перший симптом перегргвання. Перегр)вання супро-
воджуеться збудженням центрально! нервово! системи, задишкою. Надапрне но-
товидшення як компенсаторна реакщя мае 1 негативн! насл1дки: зменшення
об ему пиркулюючо! кров! (ОПК), зневоднсння (втрата рщини може сятати 3,5 л
за 1 год), згущення кров! й шдвншеняя ц в’язкост!. що створюе додаткове наван-
таження на систему кровообпу 1 зумовлюе розвиток серпе во! недостатносп. У шй
стали спостернають порушення електролттного балансу, зумовлене втратою на-
тр! ю т хлорид!в !з потом, пперкалЁемтею. Пщвишуються розпад б!лив г виведення
азотистих сполук хз потом. Пщвищення температуря ттда посилюе обм!нн! проце-
си, що призводить до збшыпення споживання кисню 1 пщвищення вироблення
ендогенного тепла, що за механизмом порочного кола збыъшуе нагртвання тиа.
На тлх зростання проявив кисневого голодування розввваються судоми. Смерть
настае вщ зупинки дихання. Гостре перед ртвання з! швидким пщвищенням тем-
пературя т!ла називаеться тепловим ударом
Сонячний удар за своею клппчною картиною нагадуе тепловий удар (пщви-
щення температури тгла, збудження 1 пригшчення нервово! системи, порушення
кровообпу та дихання), проте !х етюлопя 1 патогенез вщрпняються. У раз, соняч-
ного удару етюлопчним фактором е сонячне випромпповання за умови д!! на
непокриту голову. При цьому, на вщм!ну вщ теплового удару, патолопчний про-
нес не завершуеться, а починаеться порушення нервово! д!яльност! (в оболонах
мозку розвиваються гшеремгя та набряк), а температура тша пщвищуеться вже
унаслщок цього.
Мкнева д!я высоко! температури с причин юс отк ! проявляеться мтсцевими де-
структивними 1 реакгивними змшами. За тяжнстю розр!зняють так! ступен! опгку:
I — почервоншня шири (еритема), слабка запальна реакц!я без порушення пслосп
шири; II — гостре ексудативне запалення шири, утворсння пухир!в з вщшаруван-
ням епщерм^су; III — частковий некроз шири й утворення виразок; IV — обвуг-
лювання тканин, некроз, шо поширюетъся за меж! шири.
Однак було б неправильно розглядати ошк лише як мюцеве явище. Нерщко
небезпека затальних порушень перевишуе значения мтсневих. 1деться про опйсову
хворобу.
У клппчному перебну отково! хвороби розр!зняють таи стали: ошковий шок,
ошкова токсемия, ошкова !нфекн!я. откове виснаження, видужання (або
смерть).
У розвитку оптового шоку головну роль вщпрають больовий фактор ! надмтр-
на аферентна 1мпульсан!я в централину нервову систему, унаслщок чого теля
короткочасного збудження вщбуваетъея виснаження нервових центров, що пору-
шус регулярно судинного тонусу, дихання 1 функш! серця. Зменшуеться ОЦК,
розвиваеться гемоконпентрац!я, гостра недостатшсть нирок (ол!гур!я).
Надмтрна 1нтоксикац1я, що розвиваеться пщ час огпку, поглиблюс шок. Ток-
сичш речовини утворюються в оргашзм! внаслщок порушення обм!ну речовин,
але найбьтьше !х утворюеться в зон! подразнення, звщки вони 1 всмоктуються в
кров. Серйозних порушень зазнае быковий обм!н. Кр!м значно. втрати б!лив
через ушкоджену поверхню спостернаються генерал! зований !х розпад 1 тяжи
порушення азотного обм!ну. У кров надходять денатурований биток 1 токсичш
продукта його ферментативного гщрол!зу.
Тяжким усютадненням опгково! хвороби е зневоднення. Втрата б!лиз ! рщини
слостер!гаеться переважно в м!сш ураження як наслщок пщвищення проникност!
"м-’
. т судин. Згущення кров! Ё щцвищення п в’язкостё утруднюють кровообЁг Ё
лу серця.
Порушуеться водно-линералъний обмш. У перш! години шсля ошку ушкодженЁ
ини затримують натр!й, який разом з водою переходить у кштинний простЁр,
- аиваеться клЁтинна гшерпдратащя. ПерерозподЁл калЁю полягае у виход! його
. .. тинного простору в плазму (гтперкалЁемЁя) Ё проявляеться порушенням ритму
хгся Спостер!гаеться зменшення активност! дихальних ферментов, що нар1вш з
ч^шенням дихання ! кровообЁгу пояснюе причину кисневого голодування. Важ-
им механизмом ушкодження оргашзму е доя протеолЁтичних ферменпв, як!
апляють у кров з уражених клётин. Якщо хворий не загинув у раншх стад!ях
.- ково! хвороби, то згодом може приеднатися автоЁмунний процес: ошков! анти-
:и можуть бути причиною автоЁмушзацп'.
1нфекц1я — ПОСТ1ЙНИЙ супутник ошково! хвороби, що посилюе штоксикацпо
гдопзму. Джерелом Ёнфекцп е ушкоджен! тканини Ё вмёст кишок.
При оптовому виснаженш прогресуе кахексия, розвиваються набряки, анем!я,
2уггроф1чн! зм1ни у внутр!шн!х органах, пневмон!я, гломерулонефрит, недостат-
.:ть функцц надниркових залоз. Саме щ наслЁдки опеку можуть стати причиною
гмертЁ.
За сприятливого перебЁгу ошково! хвороби настае видужання. Некротичш
-канини вЁдторгаються, дефект виповнюеться грануляцЁями, рана рубцюеться й
ештелЁзуеться. Успехи в клонувашп органЁв людини, можливо, наблизять той час,
коли трансплантацЁя шк!ри хворим на ошкову хворобу стане звичайною опера-
шею.
БАРОТРАВМА
чаротравма зумовлена патогенною дЁею на оргашзм змЁненого атмосферного тиску.
Етюлопя ! патогенез. Якщо атмосферний тиск змЁнюеться вёдносно норми,
зён може стати етюлопчним фактором баротравми, тяжкёсть яко! залежить вщ
ступени 1 швидкостё компресн або декомпресГ!. Вщ ртвня атмосферного тиску, як
ведомо, залежать деякЁ фЁзичнЁ властивост! газЁв ё редин оргашзму (об’ем газЁв у
порожнинах оргашзму, !х розчиншсть у кров!, точка кишння рщини тощо). Па-
-огенез баротравми залежить вщ напрямку змёни атмосферного тиску (зниження
або пЁдвшцення). Видёляють стадЁю компенсаци ё стадЁю декомпенсацп.
Дно зниженого атмосферного тиску людина вЁдчувае на висотё (у горах, у разЁ
розгерметизацЁЁ салону лггака). У лабораторних умовах такий стан (гтобар1я) мо-
делюеться в барокамерЁ шляхом штучного розрЁдження повЁтря. ПатологЁчш змё-
ни, що виникають при цьому, зумовленЁ двома факторами: зниженням паршалъ-
ного тиску кисню (рО2) у вдихуваному повЁтрЁ, що зумовлюе гтоксичну гтоксйо
(про компенсаторнЁ реакцп ё патологЁчш порушення читайте в розд!лЁ “Гёпоксёя”),
та зниженням атмосферного тиску, яке супроводжуеться комплексом проявЁв —
синдромом декомпреси. Гази, шо мёстяться в порожнинах тЁла, розширюються, Ё
людина вЁдчувае бЁль у вухах ё лобових пазухах, суглобах, кишках (висотний мете-
оризм). Можлива кровотеча з носа внаслЁдок розшпрения та розриву дрЁбних су-
дин. У рЁдинному середовищЁ розчиншсть газЁв зменшуеться (десатурафя), Ё воеш
у виглядо бульбашок виходять у тканини Ё кров (газова емболгя). На висоту 19 000 м
не можна пщшматися без надшноТ герметизацп кабгни, оскиьки саме на шй ви-
сот! рщини оргашзму закипають за температури пла.
ГПл час швидкого перепаду (зниження) атмосферного тиску (у раз! порушен-
ня герметизаци косшчного корабля або висотного л! така) розвиваеться синдром
вибуховоИ декомпреси. Смерть настае миттево Причинами цього е:
• закипания кров! та гнших рщин оргашзму за температури пла;
• травма легеиь (розрив), серия 1 великих судин середостшня через р!зке
шдвищення внутргшньолегеневого тиску ! р!зке розширення легень:
• газова емболхя внаслщок зниження розчинност! газ!в у кров: 1 р! динах ор-
гашзму. Бульбашки газу вившьнюються в кров, газова ембол1я посилюеться вна-
слцюк розриву альвеол 1 судин легень;
• I остра форма ппокси.
Д1Ю шдвищеного атмосферного тиску людина вщчувае в раз! занурення у воду
на значну глибину пщ час водолазних ! кесонних робп. Унаслщок вдавлювання
барабанних перетинок виникае 61ль у вухах. а при резкому . швидкому заглиблен-
ш можлива травма легеневих альвеол. Однак набагато бшьше значения мае той
факт, шо в умовах ппербард людина мае дихати повггрям або газовими сумштами
шд пщвищеним тиском, унаслщок чого в кров! 1 зканинах розчиняеться додатко-
ва к!льк!сть газш (ситурищя).
1з складниюв пов!тря найб!лыне значения мають азот I кисень. Через легку
розчиншсть азоту в лшщах найбъзыпе потерпае нервова система. Спочатку вини-
кае збудження, що нагадуе ейфорпо, дал! — штоксикащя, що нагадуе наркоз
(табл, 1). Надлишок кисню (гтерокая) спочатку лолшшуе тканинне дихання, а
поим Д1е токсично (под!бно до радюактивного опромшення; в обох випадках
утворюються в!льн1 радикали !з сильными окисними властивостями).
Таблиця 1. Симптоми порушення функшй центрально!нервовоТсистеми
у нетренованих людей зале» но вщ глибини занурення у воду
Глибиназанурення уводу, м Симптоми порушення функцш центрально'!' нервовоТ системи
30- -60 Ейфорт
60—75 Безлричинний см!х ! перш! ознаки !стерп. Ослабления концентрацп у ваги. Помилки п!д час виконання профес!йних 1 розумових !авдань. Недооцшка особист о! безпеки
100 Депресгя, втрата здатност! до четкого мислення. Порушення нервово- м’язово’Г координацп
115 Можливе знепритом нения
У стада компенсаци розвиваються реакцй, спрямован! на обмеження п!дви-
щення концентрацп кисню в тканин! головного мозку, — знижуеться збудлив!сть
хеморецепторш, унаслщок чого сповьтьнюються дихання ! пульс, зменшуеться
О ЦК. а також ршень ери тропит! в 1 гемоглобшу (за рахунок депонування кров! та
розпаду еритроциттв), звужуюзься судини головного мозку. У стада декомпенсаца
виникае своерщна “асфгксзя тканин”, пов'язана з зим, що молекула гемоглобшу
блокуеться киснем (гемоглобш перебсвае в стан! оксигемоглобшу не тыьки в
артер!альн!й, а й у венознш кров1) ! втрачае здатшсть виводити вуглекислий газ.
:иметься це тим, шо тканини насамперед використовують той кисень, який
чно розчинений у плазм!, а в умовах ппербаро вмтст розчиненого в плазм!
'.л кисню збптьшуеться (пщ тиском 5 атм. у кровх додатково розчиняеться ки-
що забезпечуе потреби тканин у ньому). При ньому оксигемоглобш маиже
дисоцпое ! гемоглобш не може зв’язувати 1 виводити з оргашзму вуглекислий
. Накопичення в кров! вугщьно! кислота спричинюе розвиток газового ацидо-
Щоб запоб!гги токсичнш ди кисню пщ час дихання в умовах пщвищеного
_._<у, у пщводн! пристро! поцають газов! сумйш з низьким вм!стом кисню. а азот
аюють на гелш. Наприклад, при зануренш на глибину 100 м концентрашя
. по в газовгй сумхш! мае становити не бътьше 2 %.
Пщ час повернення людини з глибини в умови нормального атмосферного
.ку виникае ще один вархант синдрому декомпресп, пщ час якого теж спостер!-
ться десатурация — виведення надмхрно! юлькосп розчинених газгв через кров
егеш. Декомпрес!ю слщ проводите пов!льно, шоб швидюсть утворення газ!в не
евищувала можливост! легень виводити гх, гнакше бульбашки повгтря затри-
лимуться в кров! 1 тканинах (га зов а ембол1я). Клппчна картина при ньому
1 значаеться локалгзацгею газових бульбашок (б!ль у суглобах, свербхж шк!ри, у
кких випадках — порушення зору, парал!ч, потьмарення свщомост/ та шш!
таки уряжен ня головного г спинного мозку).
“РОМЕНЕВА ТРАВМА
\юменева гравма зумовлена патогенною д!ею на оргашзм юшзуючого випром!-
зювання, яке мае найбшьше значения в медицин! пор!вняно з оптичним — св!т-
ом, улътрафюлетовим та шфрачервоним, а також улыразвуковим випромшю-
-нням 1 радюхвитями гощо).
Вплив юшзуючого випромшювання людина зазнае через застосування атом-
• 7 енергп у виробництв! (атомш електростанцп), у раз! авар!й на виробництв!,
.тосування джерел юшзуючого випром!нювання з дгагностичною ! лгкувальною
етою (радюфармацевтичн! препарата, рентген!вськ! трубки тощо), а також при
икористанш атомно! енергп у вигляд! ядерно! збро!. Жоден вид енергп, поглину-
им в однаковш кглькост!, не супроводжуегься такою тяжкою травмою, яко! завдае
зшзуюче випромшювання.
*
Етюлопя
Етюлопчним фактором променевого ураження е юшзуюче випромшювання, яке
л своею природою подйгяеться на хвильове (рентгенгвське та у-випромшювання)
*а корпускулярне (а-, 0-частанки, нейтронне випромшювання тощо). 1х загаль-
ною властивхстю е здатшеть проникати в середовище, поглинатася ним г спричи-
нювата юшзаппо атомгв ! молекул.
Характер ! стугпнь променевого ураження залежать вщ виду юшзуючого випро-
гнювання. дози й умов опромшення, його експозицп, а також вщ вгрогщност! пот-
рапляння в ту або ту (молекулярну чи надмолекулярну) важливу для життя мппень.
Здатшеть до проникнення в оргашзм залежить в!д наявност! чи вщсутност]
заряду юшзуючого випромшювання. Так, найб!лыиу проникшеть мають нейтро-
ни ё рентгенЁвськЁ та у-промеш, що не мають заряду, а найменшу — а- Ё р-про-
меж, кванти яких мають заряд ё гальмуються в раз! взаемодн Ёз зарядженими
молекулами бЁлкЁв.
Ёошзуюча здатшсть випромЁнювання залежить вщ маси корпускулЁв. Вщ-
повщно, найбшыпу Ёошзуючу силу мають нейтрони ! а-промен!, а найменшу —
рентгенЁвськЁ ё у-промен!.
Щодо дози опромЁнення, то пряма залежшсть вщ не! спостерггаеться лише в
раз! високих Ё середнгх доз. ЗакономЁрностЁ дп низьких доз випромпповання дещо
ёншё. СпостерЁгаеться так званий рад1об1олог1чний парадокс, який полягае в тому,
що клёнёчний ефекг опромЁнення може досягнути катастрофЁчних масштабЁв на-
вёть при незначнЁй кёлъкостё поглинуто! енергп, оскёльки первинне радЁацЁйне
опромЁнення пщсилюеться бЁолопчними механЁзмами. КрЁм того, доза опромЁню-
вання оцЁнюеться по-рЁзному щодо Ёндивщуума та популяцп. € мЁнЁмальн! рЁвнЁ
Ёошзуючого випромЁнювання, якё Ёстотно не позначаються на захворюваностЁ по-
пуляпп, але навЁть одного кванта енергп достатньо для того, щоб вщбулася мута-
цёя, наслЁдки яко! можуть бути дуже серйозними для конкретно! людини. Зважа-
ючи на це, жодну дозу опромЁнення не можна вважати абсолютно безпечною для
людини.
Важливою умовою, яка суттево впливае на характер Ё ступЁнь променевого
ураження, е шлях впливу Ёошзуючого випромЁнювання (дистанцЁйний, контакт-
ний, ЁнгаляцЁйний чи пероральний). РозрЁзняють зовнйине опромшення, коли дже-
рело мёститься поза органЁзмом, Ё внутриине (шкорпорованё) — у разЁ потрапляння
радЁоактивних речовин усередину оргашзму.
Патогенез
Як ё будь-який Ёнший патолопчний процес, променеве ураження е комплексом
патолопчних змён ё компенсаторно-пристосувальних реакщй, якё розгортаються
на рЁзних рЁвнях оргашзацн живо! матери: молекулярному, клЁтинному, тканин-
ному, а також на рЁвнЁ оргашзму в цЁлому.
ПервиннЁ фЁзико-хЁмЁчнЁ процеси е початковою ланкою патогенезу Ё полягають
у радЁохЁмЁчному перетвореннЁ атомЁв ё молекул, !х Ёошзацп та збудженш, тому що
кЁнетична енергЁя Ёошзуючого випромЁнювання перевишуе енергЁю внутрЁшньо-
молекулярних та внутрЁшньоатомних зв’язкёв. ХЁмЁчне перетворення речовин пёд
дЁею Ёошзуючого випромЁнювання з утворенням активних промЁжних продуктов
називають радйшзом.
1з первинних радЁохЁмЁчних перетворень найбЁльше значения мае радюмз
води, що е основним розчинником у бЁолопчних середовищах ё вмёст яко! най-
бЁльший. УнаслЁдок ЁошзацЁ! молекули води утворюються вёльнё водневЁ та гЁдрок-
сильнё радикали (Н‘, 'ОН), якё вступають у взаемодЁю з вёльним електроном, збуд-
женою молекулою води, киснем тканин. За наявностЁ кисню утворюються нов1
продукта: водню пероксид (Н2О2), гЁдроксильний радикал ('ОН), атомарний ки-
сель (О,), супероксидний ашон-радикал СО2“), а також так званий синглетний
кисень (*О2).
Продукта радЁолЁзу води 6ёохёмёчно дуже актавнЁ Ё спричинюють нефермента-
тавне окиснення через будь-якЁ зв’язки, у тому числё ё через стёйкё при звичайних
окисно-вёдновних перетвореннях (непряма д1я рад1аци). Накопичення цих продук-
сгивних форм кисню) залежить в!д наявностг води г концентрацп кисню в
;нах: за наявностг кисню !х кглькгсть бшьша й ушкодження посилюеться, г
швпаки, якщо концентрацгя кисню в тканинах зменшена, ступшь променевого
тгж-ння знижуеться (кисневий ефект).
Щодо великих бюлопчних молекул, то вони можуть бути мошенню прямо!
д_ходжувально! дп юшзуючого випромшювання (пряма д1я рад/аци). Ёнерпя,
наючись макромолекулою, мггруе в нш, ушкоджуючи найуразлившп мгсця,
.«.риваючи слабкг зв’язки г спричинюючи внутргшньомолекулярну перебудову,
Ж -хтурац1Йн1 змши (в!дрив вгльних радикалгв, деполгмеризацпо тощо). Мппенню
* с стати будь-який компонент живо! системи — оргашчю макромолекули (ДНК,
д^щи, ферменти, быки, впамши, гемопротещ, полгмери), а також будь-яка
<т-ктура клгтини (теор1я мииеш).
Таким чином, початков! ланки патогенезу променевого ураження поеднують
гьму д!ю юшзуючого випромшювання на макромолекулярш мгшеш г непряму
I срез продукта радюлгзу води). Доказом того, що п!д час опромшення бгльппсть
мгчних змш опосередкована продуктами радюлгзу води, е радгостгйкгсть су-
I г порошкопод!бних ферментов поргвняно з гх водними розчинами.
Бюхгмгчш порушення полягають в ушкодженш бюлопчних макромолекул як
шляхом прямого ураження енерпею юшзуючого випромшювання, так г гх атаки
* тивними окисниками. При цьому також виникають рад икал и, як! мають вшьнг
е ?ктрони г тому дуже реакцшноздатш. Хгмгчнг й бюхгмгчш реакцп, як! вщбува-
-.~ься одна за одною, можуть швидко наростати, набуваючи характеру ланцюгових
. згалужених реакцш. У лпцдному середовшщ щ реакцп найнебезпечншл, бо
- 1ьк!сть активних продуктов швидко г значно збшьшуеться, шщгюеться процес
роксидного окиснення лшццв (ПОЛ). Кр!м того, утворюються радютоксини —
попчно активн! речовини ргзно! природи. Унаслщок окиснення ненасичених
гчрних кислот г фенолгв утворюються так! аномальш метаболии: лппдш радготок-
:.гни — ЛРТ (пероксиди, епоксиди, альдегщи, кетони) г хшоновг радютоксини —
ХРТ (попередниками е тирозин, триптофан, серотонш, катехоламши). До радю-
- ксингв належать г шш! токсичн! метаболии, продукта клпинного розпаду.
П!д впливом юшзуючого випромшювання прямо чи через продукта радюлгзу
щ г радютоксини ушкоджуютъся ферменти. Значна радгочутливгсть властава тю-
вим ферментам, якг мгстять сульфгщрильш групи (8Н), що легко окиснюються
втратою активностг. Деякг ферменти, особливо тг, як! виходять у цитоплазму з
шкоджених лгзосом, навпаки, активуються. Надалг зростае штенсивнгсть фермен-
тативних реакцш: посилюеться ферментативний розпад бглкгв г нуклегнових кис-
лот, ушкоджуються окисне фосфорилювання та антиоксидантна активность. Висо-
кополгмернг полгсахариди (наприклад палуронова кислота) деполгмеризуються.
Варто наголосити на ушкодженш унгкалъно! макромолекули ДНК (прямо, а
гкож через продукта радюлгзу води, радютоксини г ферментативну атаку). П!д час
|промшення розчишв нуклегнових кислот спостерггаеться радгацгйно-хгмгчне окис-
нення шримщинових г дезамшування пуринових основ. На схем! 1 вщображено
гехангзми ушкодження ДНК юнгзуючим випром!нюванням, фактори, якг модифгку-
эть г вщновлюють це ушкодження, клгтинш ефекти радгагцйного ураження ДНК.
Пригшчуеться бглоксинтетична функцгя. Масова загибель клгтин пщ час мг-
тозу призводить до збшьшення концентрацп азотовмгсних речовин у кровг. Азо-
тистий баланс стае негативним, пщвищуеться виведення амшокислот гз сечею.
Схема 1. Мехашзми ушкодження 1 вцновлення ДНК пщ час опромшегшя. Фактори, шо мс-
дифпсують । вцшовлюють ушкодження
Порушення бюлопчних проиесш у кл(тинах. Будь-яка кттинна структура може
стати миненню для променево! енергп, а також об'екгом атаки активными про-
дуктами окиснсння, радют оксинами 1 ферментами.
За допомогою мпсроскопп виявляють ознаки рад1ацтйного ураження ядра (на-
бухания, шкноз, Л1зис), а гакож хромосомы' мутацп (поломки, перебудови, фраг-
яю разом 13 генними мутац1ями спричинюють порушення спадкових
е~д-~и воете й клётини, пригшчення синтезу ДНК ё бЁлкЁв. Ушкодження ядра
И№ чзане не лише з безпосередньою дЁею Ёошзуючого випромЁнювання на моле-
< ДНК ё структуру хромосом, а й ёз порушеннями в Ёнших органо'Ёдах, якё
1 иваються за типом порочного кола.
ВЁльнорадикальнЁ реакцп активно розвиваються у фосфолЁпЁдному бЁшарЁ
I нембран, ре вщбуваеться постёйний контакт молекулярного кисню з лЁшдами.
12г черетворюе мембрани на мЁшень ё “критичну структуру”. ПЁдвищуеться про-
' енёсть клётинноё мембрани для електролЁтЁв ё води, калЁй виходить ёз клётин. У
ушкодження л1зосом у цитоплазму вивёльнюються лётичнё ферменти (рибо-
зе кзеаза, дезоксирибонуклеаза, катепсини). Порушення окисного фосфорилю-
и^.чя Ё транспорту електронЁв у м1тохондр1ях зумовлюе порушення енергетично-
обмЁну, гальмування бЁосинтетичних процесЁв Ё розлад багатьох клётинних
КлЁтина як бЁолопчний об’ект е динамЁчною системою, що постёйно перебу-
ы в стадЁЁ переходу з одного стану в Ёнший. в двё форми клётинноё загибелЁ.
Верша — репродуктивна, або мЁтотична (год час подЁлу), друга — Ёнтерфазна.
' . одки ушкодження ДНК можуть не виявлятися в диференцЁйованих клЁтинах,
(кё не дЁляться. Якщо ж це камбЁальна клЁтина, здатна до мётотичного подЁлу, то
-щщцЁйне порушення структури ДНК, як правило, призводить до загибелЁ клёти-
вм шд час мЁтозу. Якщо клЁтина з генетичними порушеннями не гине, то вона
може стати носЁем змЁнених спадкових властивостей. Соматична клЁтина може
дретворитися на злоякЁсну, а хромосомы аберацЁ! у статевих клЁтинах можуть
-ричинити розвиток спадкових хвороб у нащадкЁв.
Порушення на тканинному рЁвнЁ. Хоча радЁочутливЁсть ядра не вища, нёж в ён-
ших органощах, порушення саме ядерних структур бЁтыпе позначаеться на жит-
ЦДЁЯЛЬНОСТЁ клётини. Тому легко зрозумЁти ТЁ ЗакономЁрностЁ, ЯКЁ виявляють ПЁД
с вивчення порЁвняльноЁ радЁочутливостЁ тканин: вона прямо пропоршйна про-
-1феративнЁй активностЁ та обернено пропорцЁйна ступеню диференцЁацп клётин.
Тэбто найвищу радЁочутливЁсть мають тканини, яким властива велика швидкёсть
А-йтинного оновлення. Це насамперед кроветворна Ё лЁмфощна тканини, де про-
ткс подЁлу клётин вЁдбуваеться постёйно. Наступною в цьому ряду е ештелЁальна
тканина (особливо залозистий ештелЁй травних ё статевих зал оз), потём покривний
цптелЁй шкЁри та ендотелЁй судин. А завершують перелЁк фЁброзна, хрящова, кёс-
ткова, м’язова Ё нервова тканини. НервовЁ клётини позбавлеш здатностЁ до подЁлу
тому в разЁ прямого опромЁнення гинуть останнЁми.
Порушення функцш оргашзму та основш патофЁзЁолопчнЁ с и ядром и. При опро-
мЁненнЁ летальними дозами радЁацп переважае Ёнтерфазна загибель клётин ё
?мерть настае протягом кёлькох годин теля опромЁнення. У разЁ опромЁнення
середнЁми дозами життя можливе, але у всёх Фёзёологёчних системах вЁдбуваються
татолопчш порушення вёдповёдно до порЁвняльноЁ радЁочутливостЁ тканин. У
тазЁ опромЁнення всього оргашзму клётини ушкоджуються бЁлыпе, нёж пёсля оп-
ромЁнення Ёх у тканиншй культурЁ. Це означае, що променева хвороба — це не
сукупнЁсть клётинних ё тканинних змён, а наслЁдок значно складнЁших бЁолопч-
них явищ — порушення регуляторних механЁзмЁв ё взаемодп рЁзних функщональ-
них систем.
й
На ровно Ц1лого оргашзму патогенез радоацшного ушкодження складаеться з
низки патофозоолопчних синдромов (синдром — це комплекс симптомов, поедна-
них спольшстю патогенезу).
Гематолог1чний синдром найбшьш характерный для всох форм променевого
ураження. Вш полятае в порушены! структури гемопоетично! тканини, прогресу-
ючш атрофп косткового мозку, ломфатичних залоз о селезшки та супроводжуеться
змонами периферично! кров!. Спостерггаються зменшення кыькосто вс!х клггин
кров!, а також ох функциональна неповноцшность. Спочатку розвиваеться лгмфо-
пешя, бо саме л!мфоцити найчутлившп з ус!х клггин кров!: вони гинуть одразу
посля опромшення не лише в ломфатичних вузлах о костковому мозку, а й у кров!
(механозм загибел! штерфазний). Позноше розвиваються гранулоцитопетя, тром-
боцитопения, анем1я.
Гемораггчний синдром е типовим проявом променевого ураження у виглядо
малых о великих крововилив!в у покору, слизов! оболонки, внутрппш органи, кро-
вотеч! з носа, кишок, сеч!вника. У патогенез! цього синдрому мають значения
нижченаведен! мехашзми.
1. Зменшення к!лькост! тромбоцита у периферичнш кров! (тромбоцитопе-
н!я), що призводить до зниження вмосту факторов зсщання кров!. Причинами
тромбоцитопен!! е:
— пригшчення подшу попередников тромбоцит!в у костковому мозку;
— затримання дозргвання тромбоцитов;
— руйнування тромбоцита у кров!;
— втрата тромбоцитов при кровотечо.
2. Зниження здатносто тромбоцита до агрегаци через змшу ультраструктури
охнгх мембран (бюлопчш фактори зсщання крово виволънюються саме при агрега-
ци тромбоцитов).
3. Зниження синтезу прокоагулянтов у печонцо.
4 Змша молекулярно! структури фобриногену о фобрину зумовлюе ослабления
здатносто волокон фобрину до скорочення, а кров’яного згустка — до ретракцп.
5. Активацоя протизсщально! системи крово завдяки появо в кров! антикоагу-
лянта (гепарин вившьнюеться в разо дегрануляцо! базоф!льних гранулоцитов).
6. Активацоя фобринолозу.
7. Порушення трофочноо функци тромбоцитов щодо ендотелпо.
Судинний синдром. Судини (особливо дробно) зазнають патолопчних змш. Ме-
хашзми ушкодження тако:
— судинний ендотелш стае мошенню для прямо! доо радоаци;
— ендотелш ушкоджуеться окисниками (продуктами радюлозу води) о радоо-
токсинами;
— трофока судин порушуеться внаслщок дефщиту тромбоцитов (тромбоциты
беруть участь у пщтриманно цшосто судинноо стшки, п еластичносто та резистент-
ност!);
— ушкоджений ендотелш втрачае здатшеть виробляти полосахаридно-бшков!
комплекси для побудови базальноо мембрани;
— ушкоджений ендотелш злущуеться;
— периваскулярна тканина, яка е мехашчною опорою судини, зазнае де-
структивных змш: сполучнотканинно волокна руйнуються, основна речовина де-
поломеризуеться, колаген якосно змшюеться;
— ушкоджен! тканини вивпьнюють бюлопчно активы! речовини (БАР), до
г к належать протеол!тичю ферменти з л!зосом, юн!ни, палурощдаза, як! пог-
'.поюгь ушкодження судинно! ст!нки;
— порушуються тонус ! резистентшсть судин, а також проникнуть 1 обмтн
(вин М1ж кров'ю ! тканинами
Синдром зниження 1мунолог1чно'[ реактивност! проявляеться зниженням ак-
двност! фагоцитозу, ггригшченням утворення антитьт, тому шфекщя — найбыьш
-ачне 1 тяжке ускладнення опромшення. Не тьтьки зовнпине, а й внутршше се-
довище организму стае джерелом шфекцп (автошфекнш). У ротовш порожний!
розвиваегься тяжка форма запалення. Перебп тонзил!ту мае некротичний харак-
. Пневмоюя стае частою причиною смерт! хворого. Тнфекщя розвиваеться в
«а пиках.
Шлунково-кишковий синдром — це комплекс симптомов з боку травного кана-
ту, що призводить до порушення травления й штоксикацп. Це одна з причин
мери хворого. У його патогенез! мають значения так! мехашзми:
• пряме ! непряме ушкодження ештел!ю слизово! оболонки травною тракту;
• пряме 1 непряме ушкодження ештелпо кишкових залоз;
• штерфазна загибель юптин ештелпо;
• зупинка мгготичного под!лу ! регенераци юптин егател!ю;
• оголення ворсинок, слустошення крипт;
• зм!на нормально! та патогенно! мгкрофлори кишок (авто!нфекц!я);
• втрага захисних властивостей слизово! оболонки кишок (бар’ерно! функцц);
• усмоктування в кров токсичных продукта бактер!ального 1 кишкового по-
ходже] шя;
• вих!д плазм и кровх в кишки;
• тяжке некротичне запалення слизово! оболонки ротово! порожнини;
• метеоризм, д!арея;
• б1ль;
• неможливУть травления, як наслщок — виснаження оргашзму.
Нервово-церебрильний синдром пщтверджуе високу чутливУть нервово! систе-
ми до будь-якого впливу, зокрема до юшзуючого випромшювання.
Вище йшлося про поршняно високу радюстшкдсть нервових кл!тин до прямо!
и! радхацп, осюльки !х загибель настав в!д виших доз опромшення. Однак струк-
турн! змши не завжди в!дпов!дають функшональним. За юлька секунд шсля оп-
ромшення нервов) репептори зазнають подразнення продуктами радюл!зу, пер-
зинними 1 вторинними впшними радикалами, радютоксинами. Патолопчш 1м-
пульси надходять у нервов! пентри, порушуючи функцюнальний стан центрально!
та автономно! (вегетативно!) нервово! системи. Змши бюелектрично! активност!
юловного мозку можна зарееструвати в перш! хвилини шсля опромшення. Спо-
чатку спостерпаеться збудження, потш гальм^ъання ! пригшчення. Огже, нерво-
во-рефлекторна д!яльнУть, а також кортико-вУцеральш взаемозв’язки порушу-
ються ранппе, н!ж з’являються шш! типов! симптоми промене во! хвороби. 1з цим
пов’язаш спочагку функшональш, а тпзшше й вираженшп порушення оргашв 1
систем. Летальн! дози юшзуючого опромшення спричинюють церебральну форму
гостро! променево! хвороби.
Синдром екдокринно? недостатноспй розвиваеться внаслщок прямого ! непря-
мого ушкодження ештелпо ендокринних залоз. Кр1М того, ураження ендокринно!
системи вщбуваеться внаслщок стресу (гшерсекрец!я кЗрково! речовини наднир-
кових залоз, активащя гшоф!за, зменшення розмГрш загруднинно! залози, л!мфо-
пен!я). За патогенезом це здебшыпого опосередковаш й реактивш ефекти опро-
мшення. Початкове пщвищення активносп ендокринних залоз змшюеться на при-
гшчення з подалыпим виснаженням !'х функцн. Додатково сл!д зазначити дпо
радю1зотошв (радюактивний йод ушкоджуе щитопод!бну залозу).
Компенсаторно-пристосувальж реакцп
На молекулярному р!вш первинш ф!зико-х!м!чю змши компенсуються природ-
ними антиоксидантными системами (перехоплювачами вйьних радикалш, шак-
тиваторами пероксидов, донаторами сульфгщрильних труп). Це металоферменти
(каталаза, глутапонредуктаза, супероксиддисмутаза, глутапонпероксидаза), в!та-
мши (аскорбшова кислота — вггамгн С, нгкотинова кислота — вггамш РР, токо-
феролу ацетат — впамш Е, рибофлавш — впамш В2), полов! сполуки (глутатюн).
Здатшсть клггин вщновлювати ушкодження ДНК (ферментами репарацп; мал. 3)
— один з основних фактори, як! визначають стшктсть оргашзму до впливу !ош-
зуючого випромшювання. Генуе система контролю за утворенням та шактивашею
вс!х бюлопчно активних речовин. Ушкоджен! клггини видаляються завдяки ме-
хашзмам апоптозу, ципшзу та фагоцитозу. Оскшьки быышеть тканин — асин-
хронна популяцщ кл1тин, як! перебувають на р!зних стадиях життевого циклу,
майже завжди е клпини, здатш до репаративно! регенерацц. Отже, перебп проме-
невого ураження визначаеться не лише дозою опромшення, а й реактившетю орга-
шзму.
При реадозацп радюбюлопчного ураження виникае безл!ч випадкових обста-
вин, зокрема в!ропдшсть випадкового потрапляння юшзуючо! рад!ацп в той чи
той життево важливий м!крооб’ем (мппень). Це зумовлюе ще один напрям у тео-
ретичних поглядах на патогенез променевого ураження. Як доповнення до теори
мпиет (детермшованост! променевого ураження первинними ф!зико-х!м!чними
явищами) пропонуеться 1мов1рн1сна (стохастична) теория, яка враховуе певну ви-
падковГсть причинно-наслщкових зв’язюв (залежно в!д стану того чи того бюло-
пчного об’екта, особливостей його клпинних реакцш, здатност! саме його до
вщновлення, а оргашзму в ц!лому — до компенсаци).
1. ПЬнавання
4. Подальше
розщеплення
Екзонуклеаза
Ендонуклеаза
2. Перер!зування ______▼ ____________
дефектно! ДНК ------------------------
5. Синтез
нормально! ДНК
Полимераза
3. Розвцеплення
Екзонуклеаза
——
6. З'еднання нового
фрагмента ДНК
Л!газа
Мал. 3. Вщновлення ушкодженого ланцюга ДНК за допомогою репарувальних ферменттв
Внутр) ш не (1нкорпороване) опром!нення
Внутршше (шкорпороване) опромшення виникае внаслщок шгаляцшного або
перорального надходження радюнуклццв усередину оргашзму. Перебування
рад!онуклщ!в в оргашзм! пов’язане з розм!рами 1'хшх частинок, розчиншстю, пе-
рюдом пгврозпаду, особливостями метаболозму, здатшстю до виведення з оргашз-
му. Для вс!х радюактивних речовин характерна нергвноморшсть (органотропшсть)
розподщу в оргашзм!. Так, радюнуклщи йоду накопичуються в щитопод!бн!й за-
лоз!, радюнуклщи стронцию зам!щують кальцш у юстках, радюнуклщи цезпо е
конкурентами калйо 1, зампцуючи його, спричинюють гшокалоемпо. Залежно вщ
переважного тканинного накопичення розрозняють остеотропш, гепатотропн!,
тиреотропн! радюнуклщи. Саме цим визначаеться особливють !хнъо! бюлопчно!
ди. Як джерело високоюшзуючих а- 1 р-частинок радюнуклщи локально створю-
ють високу дозу опромшення, 1 таким чином у мгсцях !х депонування, а також у
тканинах, як! безпосередньо прилягають до цих мосць, гмоворшсть ! стушнь рад!а-
цшного ураження дуже висок! (при а-опромшюванш густина юшзацп в тканин!
становить калька тисяч пар юн!в на 1 мкм шляху, при р-опромшюванн! — та сама
юлькость пар юшв на 0,16 мм шляху). Вважають, що вщновлення кл!тин п!сля
ураження юшзуючим випромшюванням з високою густиною юшзацп майже не-
можливе.
Гостра променева хвороба
Вид!ляютъ три форми гостро! променево! хвороби: юстковомозкову, кишкову 1
церебральну.
Юстковомозкова форма мае чотири клпйчних перюди.
Початковий перюд (перюд первинно! загально! реакцп), тривалють якого ста-
новить вщ колькох годин до 1—3 д!б, характеризуеться реакщею нервових 1 гор-
мональних механизмов на опромшення, що проявляеться нервовим збудженням,
головним болем, порушенням вегетативних функций, лабольшстю вазомоторних
реакцш, артериального тиску о пульсу, функцюнальними розладами внутрошнох
органов, нудотою, блюванням ! проносом (“рентгешвське похмшля”). Температу-
ра тгла може пщвишуватися внаслщок розладу терморегуляций Спостерогаеться
короткочасний перерозподшьний нейтрофольний лейкоцитоз з ломфопешею. Ак-
тивуеться гшофозарно-наднирникова о симпатико-адреналова системи. У тяжких
випадках можливий променевий шок.
Прихований перюд — це перюд удаваного благополучия (тривалйсть — вщ 10
доб до 1 мос.), коли прояви, пов’язаш з надм!рним збудженням нервово! системи,
зникають, а типов! симптоми променево! хвороби ще не з’явилися. Однак про-
гресуе ломфопеноя, також зменшуеться юлыасть гранулоцитов о тромбоцитов.
У перюд розпалу (тривалость — 2—5 тиж.) розгортаються основш синдроми
променевого ураження, прогресують лейкопешя о тромбоцитопеноя, розвивають-
ся геморапчно прояви, анем!я, шфекцшш ускладнення. Стан ротово! порожнини:
автошфекщя спричинюе некротичне запалення язика, ясен, мигдаликов; спожи-
вання ожо утруднене.
К1нецъ хвороби. Максимум летальноси припадае на перюд значно! гранулоци-
топенп о тромбоцитопени. Основними причинами смерт! е кровотеч! та шфекцоо
(найчастппе це пневмония, перебй яко! на тлг зниження хмунолопчно! реактив
носи луже тяжкий).
Ознаками видужання е полшшення самопочуття, нормал!зац!я картини крой
(пул юптин кров! поповнюеться за рахунок пролгферацп неушкоджених стовбур -
вих юптин). Однак ще тривалий час (до 6 м!с.) зберпаються таю прояви, як I
тен1я, стомлювашсть, загальна слабить, неспйюсть гемопоезу, ендокриню поч-
тения, розлади статево! функцп, ослабления гмунггету.
В1ддален1 насл1дки хвороби подшяють на соматичш (стерильшсть, ослаблен- к
гмунпету, катаракта, нефросклероз, нейродистроф!чн! порушення, зниження
адаптащйних можливостей, що зумовлюе передчасне старшня ! скорочення 'туям
валост! життя) ! генетичн! (нестабшьшсть спадкового апарату, накопичення < --
коджень у генетичному апарап соматичних юптин, лейкози, пухлини, спадкяч
хвороби).
Каткова форма гостро! променево! хвороби розвиваеться внаслщок мае о I
загибел! юптин кишкового ештелпо пщ час !х мпотичного подщу. Слизова обо-а
лонка кишок, позбавлена ештелпо, стае вщкритими воротами для шфекцп. М н н
ливий розвиток колапсу внаслщок всмоктування в кров токсичних речовин "и-!
термального 1 тканинного походження, втрати води та електролтв. Клппчна им
тина доповнюеться блюванням, анорексгею, млявютю, пщвищенням темпера-
тща, болем у р!зних дыянках кишок, домппкою кров! в кал!. Можливими е .4
ралхтична непрохщшеть кишок, перитонп. Смерть настае через 8—12 д!б.
Церебральна форма розвиваеться внаслщок прямо! ушкоджувально! дц ва Я
ких доз юшзуючого опромшення на нервову тканину, що спричинюе струю
змши 1 нав!ть загибель нервових юптин, порушення терморегуляцп, тонусу суя -I
розвиток судомно-паралггичного синдрому. Можливою е токсична (суди->^|
форма променевого ураження, коли смерть настае на 4—7-у добу з розвиткчи
меншгеально! симптоматики або в 1—3-й день — з колапсом 1 судомами.
Найгострппа форма гостро! променево! хвороби (смерть “пщ промен: I
спостерпаеться через юлька хвилин (годин) унаслщок денатурацшно! шактиьли
юптин 1 штерфазно! !хнъо! загибел!.
Хрожчна променева хвороба
Хрошчна променева хвороба е наслщком повторних опромшень невисокимг _ I
зами. Патогенез 1 клппчна картина по суп таю сам!, як 1 при гострш промен г
хвороб!, проте динамгка ! ступшь тяжкост! окремих прояви мають вщмшност |
Видшяють три ступеш тяжкост! хрошчно! променево! хвороби.
У раз! першого ступеня порушення мають характер оборотних функции -I
них розладгв найчутлившшх систем оргашзму. 1нод! самопочуття хворог- яй
довшьне, однак пщ час дослщження кров! виявляють помхрну нестшку лейк
нпо 1 тромбоцитопенпо.
Хвороба другого ступеня тяжкост! характеризуеться виражешшими змша ья
боку нервово! та кроветворно! систем, наявшетю геморапчного синдрому 1 ЗОВ
же пням тмуштету. Лейкопешя 1 л!мфопен!я набувають спйкого характеру, ' |
юсть тромбоцит!в також зменшена.
Для хвороби третьего ступеня характерними е тяжю необоротю змши в -
ганах, глибока дистрофхя тканин, зокрема в нервовш систем!. Функцгя гшек, ।
надниркових залоз виснажена. Кроветворения рёзко пригшчене, тонус судин зни-
кений, а проникшеть !хньо1 стшки пёдвшцена. Спостерёгаються виразково-не-
кротачш ураження слизових оболонок. 1нфекцёйш та запальш процеси, що при-
еднуються, також набувають некротичного характеру.
Хрошчна променева хвороба будь-якого ступеня може зумовити патологёчш
.мши у вёддалеш термши. Оскёльки в даференцёйованих тканинах при хрошчнёй
троменевёй хвороб! ёнтерфазна, загибель клётин не вёдбуваетъея, у них можуть
берёгатися ушкодження ДНК ё пёдвищуеться схильнёсть до виникнення пухлин,
раннёх дистрофёчних змён. Органёзм, який зазнав опромёнення низькими дозами
ёошзуючо! радёацп, передчасно старёе, ё вважають, що ця властивёсть успадко-
вуеться.
Корекщя променевого ураження
Хёмёчнё засоби протипроменевого захисту мають назву радюпротекторгв. Це пере-
хоплювачё активних радикалёв, антиоксиданта, речовини, що зв’язують радёоёзо-
топи важких метадёв, а також речовини, якё створюють тканинну гёпоксёю (нап-
риклад метгемоглобёнутворювачё). Вони захищають сульфгёдрильнё групи бёлкёв,
стимулюють клётиннё вёдновш системи, збёльшують вмёст ендогенних амёнёв (се-
ротонёну, дофамёну, пстамёну), якё, активуючи систему циклёчних нукле,отидёв,
впливають на клётинний подёл, синтез нуклеёнових кислот, глёколёз, проникнёсть
мембран та ёншё клётиннё процеси. Якщо радёопротектори ввести перед опромё-
ненням, вони зменшують ураження, блокують розвиток ланцюгових радёацёйно-
хёмёчних реакцёй ё переводять тканини в стан радёорезистентного метаболёзму.
ЕЛЕКТРИЧНА ТРАВМА
Етюлопя. Ушкодження оргашзму електричним струмом залежить вёд його фёзич-
них параметрёв: сили (струм понад 100 мА для людини смертельний), напруги (до
100 В спричинюе низьковольтну електротравму, понад 1000 В — високовольтну;
ураження блискавкою зумовлюе надвисоковольтну електротравму), характеру
(постёйний струм напругою близько 500 В ушкоджуе менше, нёж змённий, ё нав-
паки), а також вёд опору середовища. За електричними властивостями органёзм
неоднорёдний: рёдини — добрё провёдники, а епёдермёс мае дуже високу природну
опёршеть. Важливою умовою, що визначае ушкоджуючу дёю електричного стру-
му, е шлях проходження його в оргашзмё.
Патогенез. Пёдгрунтям реакцёй оргашзму на елекгричний струм е первинш
змёни в тканинах, через якё пройшов струм, ё якё е наслёдком переходу електрич-
но'1 енергп' в ёншё вида енергп — теплову, механёчну, хёмёчну. У бёолопчному се-
редовипц електричний струм поляризуе атоми ё молекули, змёнюе просторове
орёентування заряджених частинок ё посилюе ё'хнёй рух: електрична енергёя пере-
ходить у теплову. Порушення цёлостё тканин (розриви ё навёть переломи кёсток) —
прояв мехашчно! дп струму.
Для електропатологп найбёлыпе значения мае так звана специфёчна дёя стру-
му, зумовлена перерозподёлом ёошв (електролгзо.м) бёля клётинних мембран, що
порушуе бёострум дп ё створюе бёопотенцёали ушкодження (збуджуваш структури —
нервов!, найчутлившп — м’язов!). Електричний струм змшюс також стан колош! в
(завислих заряджених частинок).
Отже, патогенез електротравми визначаеться електрох1.хичним, електротер-
мгчним 1 електромехашчним впливом на оргашзм.
Електротравма зумовлюе м!сцев! (ошки) 1 загальш змши в оргашзм].
Збудження нервово! системи спричинюе гшерсекрецпо катехоламппв з по-
далыпими соматичними ! в!сцеральними змшами. М’язов! спазми 1 судоми мо-
жуть зумовити переломи исток, порушення дихання, пщвищення артер!ального
тиску, мимовшьш дефекацпо 1 сечовипускання.
У людини найнебезпечшшим е проходження електричного струму кр!зь сер-
цевий м ’яз, який найчутливппий до нього в сташ рефрактерност!. Електрол!з у
м!окард!альному синтицй скорочуе рефрактерну фазу 1 змшюе серцевий ритм.
Шлуночкова ф!брилящя, що при цьому розвиваеться, смертельна 1 спонтанно не
зникае (у деяких тварин вона оборотна). Порушення ритму серия (тах!кард!я, бра-
дикард!я, екстрасистол!я, блокада) можуть виникати 1 в тих випадках, коли елек-
тричний струм кр!зь серпе не проходить ! е наслщком рефлекторного порушення
вшцевого кровооб!гу або пщвищення тонусу блукаючого нерва. 3 цих самих при-
чин може настати зупинка серця.
Р!зке порушення дихання або його зупинка при елекгротравм! мае централь-
ний (парал!ч дихального центру, зупинка дихання теля проходження струму че-
рез трансбульбарну петлю)! периферичний (рефлекторне порушення роботи ди-
хального центру внаслщок подразнення чутливих нерв!в та штерорецептор!в) ме-
хашзми. Спазм голосово! пилили 1 вертебральних судин, як! постачають кров до
дихального центру, або теташчне скорочення дихальних м’язгв також значно ут-
руднюють дихання (гостра електрична асф!кс1я).
Шок, який виникае внаслщок електротравми, належить до больового.
Тяжюсть електрично! травми залежить вщ вихщного стану оргашзму. Електро-
травма менш тяжка у сташ наркозу або пщ час сну внаслщок гальмування кори
великого мозку, а також у раз! загального охолодження 1 ппотиреозу. Резистент-
шеть до електротравми знижуеться на тл! виснаження, втоми, крововтрати, сер-
цево-судинних хвороб, туберкульозу, алкогольного сп’яншня. Навгть ом!чний
ошр шюри змшюеться залежно вщ стану нервово! системи — бшь знижуе його.
ПАТОГЕННА Д1Я ФАКТОР1В КОСМ1ЧНОГО ПОЛЬОТУ
Фактори косм1чного польоту ц!кав! тим, що з ними (особливо з невагомктю)
оргашзм в еволюцп не стикався 1 пристосування до них обмежене. Тяжюсть по-
рушень залежить вщ тривалост! перебування в умовах невагомост!, ступеня пере-
вантаження на динам!чних етапах польоту, а також тренованост! космонавта.
Перевантаження — це сила, що д!е на оргашзм пщ час руху з прискоренням
на динамгчних дщянках польоту (на старт! 1 пщ час приземления). Порушення,
як! виникають, належать до к1нетоз1в. Провщною ланкою в патогенез! порушень
е змпцення органов 1 рщинних середовищ у напрямку, зворотному рухов!. Тому
пщ час польоту т!ло космонавта зор!ентоване так, щоб д!я перевантаження не
збиалася з поздовжньою в!ссю тща (тому що в цьому випадку р!зко порушуеться
мозковий кровообГг), а була спрямована впоперек. При цьому патолопчн! змши
«чнкають переважно в легенях У полягають у порушеннУ зовнУшнього дихання,
легЕневого кровообпу та газообмУну (розлад перфузУйно-дифузУйних спУввщно-
лень у вентральных У дорзальных вУддУлах легень). Не менш важливими е змпцен-
.нутршппх орган ГВ, надмУрне подразнення Унтерорецепторгв У значна аферент-
I чпульсУя в центральну нервову систему.
НевагомУсть — це стан, що виникае на основному етагп польоту в умовах
икнення гравУтацп. ЗначнУ змши розвиваються в опорно-руховому апарапи
ас идок гшокшезп У зниження мехаючно’1 компресй кУсток. Знижусться м’язовий
Т“ус (у норм} певнУ зусилля м’язУв витрачаються на подолання сили земного
техуння) У пов’язаш з ним енерговитрати. Зменшуеться маса скелетних м’язУв У
। . 'ок, яю втрачають кальцш 1 фосфор.
ЗначнУ змУни розвиваються в систем! кровообггу. Вщбуваеться иерерозподУл
св1 зУ збУльшенням кровонаповнення судин верхньоУ половили тулуба. Розтяг-
—ня передсердь У подразнення волюморенепторУв призводять до збУльшення
крещУ кардУомУопитами передсердною нагрУйуретичного пептиду та гальмуван-
видшення вазопресину й альдостерону, шо спричинюе вграту нагрУю У води
тасками, зменшення об’сму циркулюючоУ кровУ та навантаження на серпе (див.
*кУл XXII “Порушення водно-електролУтного обмУну”). Негативный азотный ба-
1_'чс 1 втрата води зумовлюють зменшення маси тала.
У патогенез! змш пщ час невагомостУ певне значения мае розлад нервово! тро-
ки. Достатня аферентацУя е необхщною ланкою трофУчного рефлексу, а в нева-
(ииостУ створюеться дефУцит аферентно’У Умпульсацп з усгх органУв (опорно-рухо-
- х, мозочка, судинно/ стУнки). функпУя яких залежить вщ сили гравУтацП. У цих
гганах виникае стан функшонально'У деаферентацп. А не означав, що при три-
>.-тш невагомостУ в зазначених органах виникають не лише атрофУчнУ, а й нейро-
лрофУчш змУни (Н.К. СУмеонова, 1977).
ЗначнУ проблеми виникають пщ час адаптапй до умов нормально!' гравУтацп
зля приземления.
ЕкстремальнУ фактори космУчного польоту суттево змУнюють загальну реак-
внУсть оргашзму, У на цьому тлУ рУзнУ додатковУ вплыви (гУпоксУя. перегртвання,
элодження, УнтоксикацУя тошо), а також дгя лгкарських засобгв можуть бути
. передбачуваними (Н.К СУмеонова. 1965).
"ДТОГЕННА Д1Я Х1М1ЧНИХ ФАКТОР1В
.1 мУльйонтв хУмУчних сполук десятки тысяч використовують у побутУ, медицин!,
"ромисловостУ, сшьському господарствУ. ЬУл вплоть Уз них може зашкодити здо-
"ов'ю людини. ТоксичкУ власти вост, мають хУмУчнУ речовини як екзогенного по-
дження, що потрапляюгь в органУзм рщними шляхами, так У речовини ендоген-
ного походження. Будь-яка неорганична га органУчна хУмгчна речовина залежно
щ дУючо! кУлькоста може бути корисною, нейтральною або токсичною (отруй-
аю) для оргашзму
За походженням розрУзняють екзогенн! токсичн! речовини природного поход-
ження та синтетичн! речовини До природных належать токсичнУ речовини бУолс-
йчного походження (бакгерУальнУ токсини, струги рослинного 1 тваринного по-
। чодження), неорганУчнУ сполуки та органУчнУ сполуки небУологУчного походження.
Серед екзогенних токсичних речовин видтляють окрему трупу ксенобютикш —
стороннтх для людини, штучно синтезованих хтмтчних речовин, якг не поширея!
в природ! (сполуки важких металтв, пестицида, радтонуклщи тощо).
За способом використання людиною видтляють продукт хтмтчного синтезу
спецтальних видтв виробництв, пестициди, лткарськт засоби т харчов! добавки
паливо т мастила, розчинники т барвники, косметику, стороннт продукта хтмтчн -
го виробництва, домтшки, вщходи.
За умовами впливу на органтзм розртзняють забруднювач! навколишнього -
редовища, виробнич! та побутовт токсичнт речовини, наркотичнт речовини, яю
формують у людини ШК1ДЛИВ1 звички т залежнтсть, бойов! отруйнт речовини.
Токсичшсть х1м1чно'1 речовини визначаеться низкою факторов, а саме: фтзичтш-
ми т хтмтчними властивостями, дозою, концентрацтею, шляхом т швидктстю про-
никнем ня в оргашзм, втком, статно, масою пла, реактившспо оргашзму, режи-
мом харчування, наявнтстю хвороб.
Вщповщь оргашзму на токсичну дтю хтмтчно! речовини характеризуемся мо-
таболтчними, функцюнальними, морфологтчними та клппчними проявами к
наслщок ушкодження), ттсно пов’язаними з патогенетичними мехашзмами да от-ч
рута. Токсичнтсм хтмтчних сполук залежить в!д наявносп реактивних хтмтчв »
труп т проявляв себе на ртвнт оргашзму в цтлому, органтв т систем, тканиннокг.
клттинному та субклггинному ртвнях.
Основними фтзичними властивостями хтмтчних речовин, якг визначаюп
стутшть ураження оргашзму, вважають стан агрегацп речовини, ртвень п дисперс*
ностт, кристалтчний полтморфтзм, летктсм, розчиншсм, здатшсть до тонтзаь 1
Токсичний потеншал речовини, що перебувае в газоподтбному стант, значно ьл-
щий, н!ж речовини, що мае рщкий або твердий стан. Газоподтбш речовини дуж
швидко потрапляють в органтзм через дихалъш шляхи. Що б стыл а летктсть реч >
вини, то вищий ризик отруення людини за рахунок абсорбцп хтмтчнот речовин®
дихальними шляхами. Що вища дисперснтсть твердих токсичних речовин, ъ
виша тх здатнтсть до проникнення в органтзм. особливо через травний канал.
Водо- т жиророзчиннтсть хтмтчних речовин — властивосп, в!д як.®
залежить ступтнь ураження тпд час отруення. Висока жиророзчиншсть сприяи
значшй абсорбцп токсичних речовин шктрою т травним каналом, тх дифузй
внутрштньому клттинному секгорт.
Токсичшсть водорозчинних речовин прямо пропорщйна ступеню ’тхньо! рам
чинностт у водт, мае важливе значения в абсорбцп дихальними шляхами п!д ч Ц
шгаляцп легких речовин. Речовини з большою розчиннтстю абсорбуються швидтиг
! в бтлышй кшькостт за одиницю часу. Наприклад, розчинтп у водт солт бартю нал*
звичайно токсичнт, а нерозчинний бартю сульфат не мае токсичних властивосп :
Ступтнь тонтзацтт може бути виртшальним для токсичность Токсичнкт!1
сполук важких металтв залежить вщ тх здатносп вившьняти тони металу. Оптичж.
та геометричш властивостт хтмтчних речовин важливт пщ час взаемодй з рецептом
рами на клттинному ртвнт.
Доза е найголовшшим фактором^ що визначае токсичшсть речовини. Розр -
няють поглинуту й абсорбовану дозу, тобто ту юльктсть речовини, що потрапк.-.
в кров, мтжклттинну рщину та клттини. Залежно вщ величини смертельно! (леталь-
но!) дози (ктлькостт речовини, яка спричинюе смерть дорослого оргашзму) розри-
няють штсть ступешв токсичностт речовин екзогенного походження, що надходог-.
7Р 1'
з оргашзм перорально: надзвичайно токсичш (5 мг на 1 кг маси тша); вкрай ток-
. ичш (5—50 мг/кг); високотоксичш (50—500 мг/кг); середньотоксичн! (500—
’’ XX) мг/кг); малотоксичш (5000—15 000 мг/кг); практично нетоксичш (15 000 мг/кг)
Г. Могош, 1984). Щодо лгкарських препарата, то !снують три основш типи доз:
: ^апевтичш, токсичш, смертельш. Токсична доза лжарського препарату зумо-
ьтюе в оргашзм! патолопчш змши, як! не призводять до летального наслщку.
Концентрагця х!м!чних речовин посщае одне з найголовшших м!сць у
зетермгшзм! токсичност!. Доза в 5 мл концентровано! сульфатно! (сгрчано!) кис-
ета може бути летальною, а та сама к!льк!сть кислота у вигляд! 0,002—0,003 %
тозчину не завдасть шкоди оргашзму. Токсичш гази 1 летк! речовини зашкодять
оргашзму большою м!рою в раз! бьчьшо! !х концентрацп в пов!тр!.
Токсичшсть речовин може бути значно бшыиою, яюцо вони надходять в ор-
гашзм парентеральним шляхом або через дихальш шляхи пор!вняно з уведенням
через травний канал. Токсичшсть змпно! отрути, гепарину в раз! потрапляння
ерез травний канал може бути мгшмальною. У травному канал! токсичшсть
х!м!чних речовин може модифжуватася завдяки розвеценню, зменшенню або
збшьшенню Тхньо! розчинност!, утворенню окремих сполук з незначною токсич-
шстю. Жири молока та шших продуктов харчування можуть щцвищувата токсич-
псть жиророзчинних речовин або сповпгьнювати !з абсорбцпо (наприклад солей
ииш’яку).
Токсичшсть х!м!чних речовин для дней вища, шж для дорослих. Наприклад,
барбггурати токсичнпш для дхтей за рахунок пропорщйно бгльшого розм!ру голо-
вного 1 спинного мозку, Н1Ж таа. Ж!нки менш резистентш до х!м!чно! агресй.
1хня чутливють до токсичних речовин вища у перюд ваптност! та лактацп.
Ступшъ чутливосп до окремих х!м!чних речовин може бути вродженим або
набутим. Найчастние набута модификация чутливост! пов’язана з наявшстю в ор-
гашзм! шших патолопчних процесхв. Пщ час гарячки пщвищуеться токсичшсть
антишретачних х знижуеться токсичшсть дипталхсних препарата. Токсичшсть
барбитурата нижча у хворих на епхлепсхю, як! тривалий час отримували ц! препа-
рата 1 мають попередньо шдуковаш посилеш процеси детоксикацп.
Токсичный вплив х1м1чних речовин на р1вн1 людсько'1 популяци проявляется зрос-
танням захворюваностх, смертност!, збхлыпенням кхлькосп вроджених вад розвит-
ку, зменшенням народжуваност!, порушенням демографхчних характеристик,
зниженням рхвня середньо! тривалостх життя, культурною деградащею.
Токсичш процеси на р1вн1 цшсного оргашзму характеризуються хворобами ххм!ч-
нох етюлогп, транзиторними токсичними реакщями (тимчасова втрата дхездат-
ностх, що швидко 1 самостшно минае, наприклад, подразнення дихальних шляххв,
психодиспептичний стан), стшкими змшами реактивност! оргашзму до дп фхзич-
них, ххмхчних, бюлопчних факторгв навколишнього середовища, а також до
психхчних х фхзичних навантажень (алерпя, !му посул ресхя, швидка стомлю-
вашсть), спещальними токсичними реакщями з тривалим прихованим перюдом,
що розвиваються лише в частики популяцп (онкогенез, ембрютоксичшсть, пору-
шення репродуктивно! функцп).
На р!вш оргашзму розр!зняють процеси, що формуються за порого-
вим принципом. Пщ д!ею хгмхчних речовин у дозах, нижчих певного ргвня,
токсичний процес не розвиваеться. За умови досягнення певно! дози процес роз-
виваеться обов’язково. Простежуеться залежшсть доза — ефект у конкретного
оргашзму. Що вшца доза, то значншп прояви токсичного ефекту. До цтет гр>
належать пттоксикаци, транзиторн! токсичш реакцт!. Упроцесах, що фор-- Ч
ються за безпороговим принципом, тмовтрнтсть розвитку ефекту зберД
гаеться за наявностт навпь одшет молекули токсично! речовини. Але процес х &
не розвинутися за умови ди дози, навпь наближено! до летально!. Дозова зале»
шстъ простежуеться титьки на ртвнт популяцн. До таких процеств належать а:е>
пя, онкогенез, тератогенез.
Упгкоджувальна д!я хйтчних речовин на окрем! органи ! системи проявлять»
нейро-, гепато-, гемато- 1 нефротоксичшстю. Для цих речовин характернина |
хвороби оргашв, патолопчш функщональш реакцп (короткочасне зниження । >
тертального тиску, збйтьшення частоти дихання, посилення дтурезу), неопластжм
н! процеси.
Цитотоксичность на р1вн1 клипини проявляеться оборотними структурно-ф;
щональними змшами клпини (змша форми, рухомостт), загибеллю клтгин (петгн
дчасний апоптоз, некроз), мутацтями. Детализация порушень функщональннх 1
бтохтмтчних процеств за умов ди хтмтчних факторов можлива титьки на субклтык
ному р1вн1.
Найпоширенипим проявом токсичного процесу е штоксикац!я. За тривал?с~в
взаемоди хтмтчно! речовини й оргашзму розртзняють: гостру тнтоксикатпиу
що розвиваеться внаслщок одноразово! або повторно! ди речовини впродовж он
шет доби; птдгостру тнтоксикацтю — птсля безперервно! або переривчасти
ди хтмтчнот речовини тривалтстю до 90 дтб; хронтчну тнтоксикащю, що 1-4
никае через тривалу дйо токсично! речовини (тнколи навпь роками).
У разт ди шкщливих речовин у пщпорогових концентрациях важливе значемя
мае накопичення цих речовин або тх ефекгтв в оргашзм! (кумулящя матерталъ-^1
функциональна). Унаслщок матертально! кумуляци отрути можуть накопичувзпи
ся, наприклад, у ктстках, паренхтматозних органах, щдшктрнтй жировгй клипе
винт. Особливо це характерно для важких металтв (ртутт, свинцю). Пщ вплг’ в
додаткових фактортв (тнфекци, значних фтзичних навантажень) може вщбува тя
вихщ накопичених речовин з депо, що зумовлюе появу тяжких клтнтчних озня
тнтоксикаци.
Видаляють так! пергоди !нтоксикацП:
— контакту з хтмтчною речовиною;
— прихований;
— розпалу захворювання;
— видужання.
Ступшь тяжкостт т тривалтсть кожного з пертодтв залежать вщ властивоскЙ
токсично! речовини, и дози, умов взаемоди з органтзмом, його реактивност »
резистентности
За локашзащею патолопчного процесу токсична дтя може бути загальнок 1
мтсцевою. У раз! загально! токсично! ди в патолопчному процест беруть учаи’’
зазвичай калька оргашв т систем, вщдалених вщ мтсця ди (аплткацй) токсичая
речовини. Загальна дтя визначаеться,резорбшею (усмоктуванням) хтмтчних речи
вин, продукт!в розпаду тканин у внутрттттне середовище, рефлекторними механлм
мами. У разт м!сцево! токсично! дП патолопчний процес розвиваеться безпосер=_ц
ньо на мтсщ аплйкацп токсично! речовини. Можливим е мтсцеве ураження шк I
дихалытих шляхтв т легень, травного каналу. Мтсцева альтеращя у виглядт запая^
но-некротичних змш характерна для уражень шири, слизово! оболонки, оргашв
травления, легень кислотами, лугами, шритом, лкизитом.
У раз! низького порогу чутливосп будь-якого органа або системи до токсич-
но! речовини можливим е виб1ркове ушкодження. Речовини, порш чутливосп до
яких ргзний, називають виб!рково дпочими. Розргзняють нейро-, нефро-, гепато-
, гематотоксичш речовини. Надзвичайно токсичш речовини (ботулотоксин, тет-
родотоксин, амаштин) можуть слричинювати виб!ркове ушкодження. Кр!м того,
можливим е виб!ркове ушкодження субклггинних структур мггохондр1альними,
лгзосомними, цитоплазматичними отрутами, мембрано-, генотоксичними речо-
винами. Бшышсть ксенобютик!в спричинюе розвиток патолопчних процесгв у
кшькох органах 1 системах. Гнтоксикацц здебшьшого мають змпианий характер з
компонентами загальних ! мгсцевих ушкоджень.
Залежно вщ штенсивносп ди токсично! речовини розр!зняють тяжку !н-
токсикащю 13 загрозою для життя, !нтоксикац!ю середнього ступеня
тяжкост! з тривалим перебйом, розвитком ускладнень, незворотними ушко-
дженнями оргашв ! систем 1легку гнтоксикащюз повним одужанням протя-
гом кшькох Д1б.
Кшетика х1м!чних речовин складаеться з абсорбцп, розподшу !х в оргашзм!,
метаболтчних перетворень та екскрецц. На бюдоступшсть — абсорбщю х!м!чних
речовин — впливаютъ наявн! захворювання !ншо! етюлогп, одночасна д!я к!лькох
х!м!чних речовин ! низка додаткових факторгв (мюце проникнення, дхетичн! упо-
добання, голодування). Взаемодья кшькох хш!чних речовин може детермшувати
токсичшсть. Ефекти, що виникають в оргашзм! у вщповщь на токсичну речовину,
можуть бути шдиферентними, адитивними, синерпчними або антагошстичними.
На розподш х!м!чно! речовини — процес ц перемпцення по р!зних компарт-
ментах (секторах) оргашзму — впливае багато факторов. Речовини, що псно
зв’язаш з бшками плазми, повшьно вившьнюються !з кров!. Для добре розчинних
у жирах токсичних речовин характерний високий стушнь розподшу. Для подо-
лання лшщного бгшару клпинно! мембрани х!м!чна речовина мае бути в неюш-
зованш форм!. Гошзоваш речовини здебшьшого залишаються в компартментах
первинного проникнення.
Мехашзми токсично! дн бшьшост! х!м!чних речовин вщом! частково. Основу
токсично! дГ! х!м!чних речовин становить !х взаемод!я з певними структурними
елементами (м!шенями) оргашзму. Роль мнпеней можуть виконувати структурш
елементи клггин, м!жкл!тинного простору, систем регуляцп клпинно! актив-
ност!.
У М1ЖКЛ1ТИННШ р!дин! отрути, що зв’язують юни (комплексоутворювач! або
етиленглжоль, який метабол !зуеться до щавлево! кислота), спричинюють ппо-
кальщсмпо. Гостра гшокальщемгя порушуе д!яльшсть нервово! системи, м’язовий
тонус, гемостаз. Порушення КОС внутргшнього середовиша зумовлюе розвиток
газового та метаболхчного ацидозу й алкалозу. У плазм! кров! ушкоджуються фак-
тори зсщання кров!, гщролггичш фермента (естерази). Пригшчення активност!
карбоксилестерази пщвишуе токсичшсть фосфороргашчних сполук. Порушення
осмотичного таску кров! та мгжклггинного простору здебшьшого мають вторин-
ний характер за рахунок ушкодження печшки, нирок, легень.
У клгтинах структури-мппеш представлен! бшками, нуклешовими кислотами,
л!пщами, бюлопчними мембранами, рецепторами гормонгв ! нейромед!атор!в.
Токсичнт ефекти хтмтчних речовин проявляються порушенням транспортних,
структурних, ензиматичних функцтй бтлктв. Незалежно вщ функционального при-
значення бтлктв ушкодження внутр! шньобшкових зв’язктв (п!д дтею концентрова-
них кислот, лугтв, окиснювачгв, юнтв важких металтв) спричинюе денатурацтю.
Ртуть, миш’як, сурма, талтй приеднуються до сульфпдрилъних труп бтлктв, а сви-
нець, кадмгй, мщь, кобальт — до карбоксилъних труп.
Каталттична актившсть ферменттв (ензимтв) птд дтею токсичних речовин може
посилюватися за рахунок гхнъот активацп, пщвищення синтезу та блокади тх руй-
нування. Активацтя лтзосомальних ферменттв т вихщ тх у плазму пщ час отруення
шритом, чотирихлористим вуглецем гнщпое автолтз юптин. Пригнтчення каташтич-
нот активностт можливе шляхом пригнтчення специфтчнот активност! та синтезу
ферменттв, прискорення тх руйнування. Пригнтчення каталттичнот активностт бгл-
ктв за мехашзмом конкурентного пригнтчення передбачае зампцення токсичною
речовиною субстрату (фторлимонна кислота) або приму и взаемодтю з активним
центром (карбамата). Неконкурентне пригнтчення каталттичнот активностт бтлктв
щд час взаемодп токсично!' речовини з додатковим центром ферменту знижуе
споргднентсть субстрату до активного центру (стрководень, монооксид вуглецю).
Комплексоутворювачт, салщилова кислота зв’язують метали, наявнт в реакцтйно-
му середовищт, а цтантди, монооксид вуглецю (СО, чадний газ), сульфщи взае-
модтють з металами, що входить до струкгури складнштих простетичних груп фер-
ментгв.
Ензиматичне пригнтчення може бути зворотним т незворотним. Пригнтчення
ферменту може вщбуватись одночасно за кшькома механтзмами.
Токсичнт речовини взаемодгють тз селективними рецепторами—протетнами
мембран, якг формують тоннт канали та беруть участь у проведенш нервових гм-
пульств у нервовш систем!, з тншими регуляторними протетнами (О-протетнами,
рецепторами з тирозинктназною активнтстю).
Речовини, що змтнюють структуру лшщгв т гщрофобш зв’язки мтж гхнтми мо-
лекулами, вважаються мембранотоксичними (спирта, детергенти, змпнт отрути з
фосфолшазною активнтстю).
Ртст т диференцповання клттин нерозривно пов’язант з обмтном нуклегнових
кислот (ДНК, РНК) г синтезом бтлка. Ушкодження виникають на етапах синтезу
ДНК, РНК або бтлка. Порушення реплгкацп може бута наслщком хтмтчнот взаемо-
дп токсичних речовин з ДНК. Втдомг й ппш мехашзми ушкодження. 1прит, цик-
лофосфан зв’язуються ковалентно з ДНК, актиномщин — нековалентно. Хтмтчн!
речовини пригнтчують синтез нуклегнових кислот ! бтлктв шляхом взаемодп з
ферментами (полтмеразами, ревертазами), порушення синтезу нуклеотадтв, обм!-
ну фолгево! кислота, шримщину, пурину, тндукцп однониткових розривтв ДНК.
Бтолопчне значения хтмтчних речовин, як! “втручаються” у процес синтезу бтлка
г юптинний подтл, зумовлене 1'хньою цитостатичною, гмуносупресорною, мута-
генною, тератогенною т канцерогенною дтею.
Ртзнт типи бюлопчних мембран мають високу чутливтсть до дп хтмтчних ре-
човин, яка виявляеться численними транспортними, метаболтчними, структур-
ними ! функцюнальними порушеннями. Токсичнт речовини пщвишують про-
никнтсть мембран, змппоють процеси бюсинтезу, знижують ртвень життево важ-
ливих енергозалежних процеств (активного транспорту юшв, процеств
енергозалежного транспорту, функцюнування скоротливих систем). Ушкоджен-
ня ультраструктурно! оргашзацт! мембран, рецепторов, порушення активного т
пасивного транспорту речовин через мембрани змтнюють внутрштньоклттинний
пул тонтв, бюелектричш потен шали, проведения нервових тмпульств, енергетичш
процеси. Ушкоджувальна дтя хтмтчних речовин (бензол, толуол, хлороформ,
мило, сапонши, важкт метали) на мембрани може бути прямою неспецифтчною
з порушенням структурно!' цтлостт клггйни, и лтзосом 1 може спричинити заги-
бель клттини. Специфтчна пряма ушкоджувальна д!я е наслщком дц токсичних
речовин на ферменти та рецепторн! бшковт комплекси. Опосередковане ушкод-
ження бюлопчних мембран пов’язане з механтзмами активацп пероксидного
окиснення лппдтв т фосфолшазно! активность Вивтльнена з бюлопчних мембран
щд дтею фосфолшази арахщонова кислота метабол (зуеться циклооксигена-
зним або лшоксигеназним шляхом з утворенням БАР. Для хтмтчних речовин з
прооксидантними й антиоксидантними властивостями мае значения ступить на-
сиченостт киснем тканин.
Залежно вщ внутртшньоклттинних мембран-мгшеней розртзняють отрути, що
порушують енергетичний обмгн (мттохондрй), процеси синтезу бтлка (шорстка
ендоплазматична сттка), тндукують або пригнтчують метаболтзм ксенобтотиктв
(гладка ендоплазматична сттка), зумовлюють автолтз клттин (лтзосоми).
1стотш порушення енергетичного обмпту в органтзмт пщ дтею токсичних ре-
човин дослтджент на механтзмах бюлопчного окиснення (ферменти циклу три-
карбонових кислот I дихального ланцюга), поеднаного бюлопчного окиснення т
фосфорилювання (утворення АТФ з аденозинфосфату (АДФ) т неоргантчного
фосфату), доставки кисню до клттин кров’ю. Пригнтчення активностт ферменттв
глтколтзу рщко призводить до значних порушень енергетичного обмпту, оскшьки
метаболтзм жиртв т бтлктв може компенсувати цей вид порушень.
Токсичнт речовини впливають на енергетичн! процеси по-ртзному. Миш’як,
ртуть, тхнт оргаючш та неоргашчн! сполуки, тнпп важкт метали можуть взаемодтя-
ти тз сульфпдрильними трупами ферменттв глтколтзу т циклу трикарбонових кис-
лот т пригнтчувати тх активнтсть (сульфгщрильнт отрути). Токсичнт речовини, що
блокують елементи ланцюга дихальних ферменттв (цтантди, сульфщи), можуть
протягом кшькох хвилин зумовити загибель оргашзму. Механтзм дц роз’еднувачтв
процеств бюлопчного окиснення т фосфорилювання (2,4-динттрофенол, хлорфе-
ноли, дикумарол, салтцилати, арсенати, тироксин) до ктнця не з’ясовано. Вважа-
ють, що вони полегшують перехщ протошв безпосередньо через мембрану з мтто-
хондртй у цитоплазму. Порушення енергозабезпечення клттини можливе також
унаслщок пригнтчення мттохондртальнот транслокази, яка забезпечуе транспорту-
вання синтезованот АТФ з мттохондргй у цитоплазму.
Зниження токсичними речовинами парцтального тиску кисню в тканинах
вважають непрямою цитотоксичною дтею. Чутливими до ггпокст! та аноксп е клт-
тини з штенсивним енергообмтном. Необоротнт змши в центральшй нервовтй
систем! виникають через 4—5 хв шсля повного припинення постачання нейрошв
киснем. Надзвичайно токсичнт хтмтчнт речовини порушують транспорту кисню в
органтзмт (оксид вуглецю (II), штро-, амшосполуки). Оксид вуглецю зв’язуеться з
двовалентним залтзом гема з утворенням карбоксигемоглобшу, метгемоглобшут-
ворювачт переводить залтзо гема з двовалентнот в тривалентну форму. В обох ви-
падках гемоглобтн втрачае здатнтсть зв’язувати кисень т транспортувати його в
тканини.
Цитотоксичний ефект шдвищеного вм!сту кальцпо в цитоплазм! пов’язаний з
ушкодженням цитоскелета 1 неконтрольованою акти вашею катабол!чних фермен-
Т1в. Кальцш 13 мттохондрш вившьнюють динпрофенол, диштрокрезол, хшони,
пероксиди, загпзо, кадмий, а з ендоплазматично! сггки — чотирихлористий вуг-
лець, бромбензол, х!нони, пероксиди, альдегщи. Надходження кальцпо через
плазматичну мембрану посилюють чотирихлористий вуглець, диметилштрозамш,
парацетамол, 2, 3, 7, 8-дюксин. Кальцш бере участь в актавацп ендонуклеаз.
Спричинена глюкокортикощами фрагментацгя ДНК тимоципв 1 загибель цих кл!-
тин пов’язаш 31 стшким зб1лыпенням вм!сту кальщю в цитоплазм!.
Активацгя вшьнорадикальних пропес!в з утворенням реактивних метабол!т!в
або вторинних продукпв гхнього перетворення, взаемод!я з молекулами-мппеня-
ми спричинюють порушення клггин них функщй. Змши в кл!тинах можуть бути
наслщком не тшьки виб!ркового ушкодження одше! структури, а й результатом
поеднаного ушкодження юлькох структурно-функцюнальних елеменпв. Поси-
люеться утворення вшьних радикал!в пщ час метабол!зму цитостатиюв антрацик-
лшово! групи, штрофурантюну, параквату, феншпдразину, чотирихлористого вуг-
лецю, бензшрену.
Ксенобютики в оргашзм! зазнають бютрансформацн та вид!ляються у вигляд!
метабол!™. П!д час бютрансформацн в перипй фаз! метаболизму завдяки окисно-
вщновним або пдролггичним реакцгям х!м!чн! речовини стають бшьш полярни-
ми. Печшка е основним органом бютрансформацн ксенобютиюв. Нирки ! леген!
також мютять фермента першо! та друго! фаз метабол!зму. У першш фаз! беруть
участь оксидази змнпано! функцн: цитохром Р450 1 флавшумкш монооксигенази,
алкогольдегщрогенази й альдегщдепдрогенази, флавопротеТнредуктази, епоксид-
пдролази, естерази й ам!дази. Утворюеться пром!жний реактивний продукт, який
у реакщях друго! фази метаболизму може бута кон’югований з глюкуроновою кис-
лотою, глюкозою, !з залишками оцтово! кислота (ацетилювання), неоргашчними
юнами (сульфатащя), глутатюном, амшокислотами. Незалежно вщ типу реакцп
утворюються полярниц!, бшьш розчинн! сполуки, 13 кращою здатшстю до екс-
крецц оргашзмом через нирки 1 травний канал. Наслщком бютрансформацн ксе-
нобютикзв може стати змша характеру токсично! дн й шщ!ац!я токсичного про-
несу. Метабол!зм ксенобютиюв не завжди е детоксикащею та бютрансформащею.
У раз! метабол!зацн токсичш властивосп х!м!чних речовин зазвичай знижуються.
Але деяк! х!м!чш речовини утворюють токсичншп сполуки метабол!зму.
У раз! потрапляння х!м!чних речовин через травний канал можливе !х мета-
болгчне перетворення за перше проходження (абсорбц!я з кишок у кровообп ворггних
вен, перфуз!я печшки), тобто до надходження отрута в системну циркуляцно.
Здатшеть до метабол!чно! трансформацн х!м!чних речовин у людини мае ге-
нетичш вщмшност!. Вщомо генетично зумовлеш типи ! кшькзен! вщмшност! мо-
нооксигеназно! системи печшки, а також вар!анти здатност! до ацетилювання
х!м!чних речовин. Метабол!зац!я х!м!чних речовин попршуеться при хворобах пе-
чшки та в раз! дотримання гшопротешових д!ет. У деяких випадках, наприклад
шеля отруення гептахлором, гшоцротешовий режим харчування скорочуе пул
мщросомних фермент!в, що зменшуе трансформащю в токсичшш! метаболгги. У
немовлят метабол!чна актившеть реакцш кон’югацп та окиснення невисока. В
ос!б похилого внеу здатшеть до метабол!чних перетворень х!м!чних речовин також
знижена.
Основний шлях екскреци хёмёчних речовин з оргашзму — через нирки. Хво-
роби нирок посилюють токсичшсть хёмёчних речовин через зменшення виведен-
ня ёхнёх метаболёпв. Виведення через кишки вщбуваеться за участё бёлёарноё сек-
рецп речовини. Частина хёмёчноё речовини виводиться з калом, а частина повтор-
но реабсорбуеться з утворенням кишково-печёнковоё циркуляцн. Деякё хёмёчш
речовини виводяться з оргашзму через легенё (азоту II оксид), з потом, единою,
молоком, сльозами.
Швидкёсть виведення одних хёмёчних речовин з оргашзму за умови насинен-
ня ферментних систем може бути постшною, шших — зростаючою в раз! шдви-
щення !х концентраци у внутрёшньому середовищё. Елёмшацёя хёмёчних речовин
за обома варёантами одночасно характерна для отруень етиловим спиртом, аце-
тилсалшиловою кислотою.
До деяких хёмёчних речовин може виникати звикання в раз! повторного ёх
вживания та формування психёчноё залежност! найчастёше без тенденцп до збёль-
шення дози. Випадки зё збёлыпенням дози характеризуються поступовою адапта-
цёею оргашв, тканин, клётин до токсичних ефекпв х!м!чних речовин ё можливёстю
переносити так! дози, як! у незвичних до цього ос!б можуть бути летальними. Зви-
кання виникае до барбггуратёв, амфетамёну, проносних, знеболювальних засобёв.
На в!дм!ну в!д звикання, залежшеть ё токсикомашя не супроводжуються пос-
туповим зростанням толерантност! до хёмёчних речовин. Фармакозалежшсть виз-
начаеться психёчними, а исколи й ф!зичними станами, як! розвиваються як насл!-
док взаемоди живого оргашзму та препарату з модифшашею поведении та пешими
рсакцёями, з намаганням приймати препарат для закрепления його психёчних
ефекпв, нередко для уникнення розладёв, яке виникають у разе його вщмёни. Од-
ночасно може сформуватися залежшеть вщ кшькох препаратов або токсичних ре-
човин. Найчаспеешми видами е залежшеть вщ етилового спирту, амфетамшу,
барбггурапв, седативних засобёв, деяких сполук СапаЬёз кабуа (марихуана), кокаё-
ну, галюциногенёв (ЛСД, мескалён, псилоцибён), ошатёв (морфёй, кодеён), синте-
тичних похщних ошат!в (метадон), деяких оргашчних летких розчинникёв (толу-
ол, етиловий ефёр, ацетон, чотирихлористий вуглець).
У разе розвитку патолопчних процесёв в органёзм! з’являються продукт пору-
шеного метаболёзму. Деяке з них мають токсичш властивостё. Це р!зш за хёмёчною
природою продукт змёненого метаболезму клетин ё структур оргашзму, що можуть
утворюватися за рёзних видёв патологи та робити значний внесок у розвиток пато-
лопчних процесёв. Знешкодження цих речовин та ёх виведення може бути недо-
статнём у раз! ураження печёнки, нирок, порушення обмену речовин. Надлишкове
накопичення в органёзмё фенольних, азотистих, амошйних сполук, БАР проявля-
ться клёшчно та ускладнюе основне захворювання.
Характеристики дп хёмёчних речовин ендогенного походження на органёзм
наведено у вщповёдних роздёлах.
ПАТОГЕННА Д1Я БЮЛОПЧНИХ (1НФЕКЦ1ЙНИХ) ФАКТОР1В
1нфекцшний процес — це комплекс захисно-пристосувалъних компенсаторних / пато-
логичных реакцш, що виникае при взаемоди микроорганизму (людини чи тварин) та
мисрооргашзму (збудника шфекцшноТ хвороби) за певних умов навколишнього середо-
вища.
1нфекцшна хвороба — це один гз проявив шфекщйного процесу, наслщок заго-
стрення мгжвидовог боротьби мгж оргашзмом-хазяшом та оргашзмом-паразитом.
Гострота цього конфлгкту може бути ргзною г визначаеться як властивостями
макрооргашзму (станом неспецифгчно! та специфгчно! реактивности), так ! пато-
геншстю збудника. Залежно вщ цього усг шфекщйш хвороби подыяють на гостр!,
пщгострг, хрошчш та повыьш. Останн! характеризуются досить тривалим пере-
бггом, але невпинно призводять до летального наслщку. Кр!м того, видшяють ще
одну форму шфекцн — латентну (персистентна, носгйство), за яко! не спостериа-
ють жодних юпшчних симптом1в, але може вщбуватися репродукция мгкроор-
гашзму та видыення його в навколишне середовище.
Патогеншстъ визначаеться як здатшсть мгкрооргашзму спричинювати розви-
ток шфекцгйного процесу. В1рулентн1сть — це ступшь патогенное!! мгкрооргангз-
му, комплекс фенотипових ознак, яю дають змогу шфекщйному агенту прикрш-
люватися до вщповщних юптин (адгезивность), проникати кр!зь слизовг та спо-
лучнотканинш бар’ери (твазивность), колошзувати й ушкоджувати тканини про-
дуктами свое! життедгяльност! (токсиногеншсть). Однак не варто розглядати мак-
рооргашзм як пасивного спостерггача, поживне середовище для мгкрооргашзму.
Для виникнення власне шфекцгйного процесу потр!бш не тыьки патогенний мгк-
рооргашзм, а й реакцгя з боку чутливого до цього шфекцгйного агента макроор-
гашзму, здатного реагувати на вторгнення збудника. Так, виникнення деяких ди-
тячих шфекщй (юр, краснуха, впряна вгепа) тыьки в певному впц евщчить про
те, що не фактична наявшеть гнфекга, а властивосп оргашзму, що розвиваеться,
зумовлюють розвиток шфекщйно! хвороби (1.В. Давидовський). 1мовгршсть роз-
витку г тяжюсть гнфекщйного процесу залежать не тыьки вщ шдивщуальног, а й
групово!, популяцшног реактивност!. Не виключено, що популяц!йна реактив-
шеть може визначити г власне вгрулентшеть мгкрооргашзму, впливаючи на р!вень
фенотипових проявгв генетично детермшованих факторгв патогенност!.
Серед фактор!в неспецифгчно! реактивност! оргашзму у протигнфекцгйному
захистг мають значения бактерицидн! компоненти секретгв слизових оболонок г
шюри (зокрема лгзоцим), фагоцитоз, быки системи комплементу, гнтерферони.
Проте важливу роль у боротьб! з гнфекцгйними агентами у ссавцгв вщгграють фак-
тори специфгчног реактивност! — гуморальш та клгтинш фактори гмунно! систе-
ми. Залежно вщ властивостей мшрооргангзму щодо паразитування у позаюптин-
ному простор! або внутргшнъоюптинно дгють принципово ргзш мехашзми гмун-
ного захисту (табл. 2).
У раз! першого вар!анта антигени збудника презентуються макрофагами та
шшими антигенпрезентуючими клгтинами (АПК) у комплекс! з молекулами голо-
вного комплексу пстосумгсносп (ГКГС; та)ог ЫзГосотраПЫШу сотр1ех; МНС) II
класу. При цьому обов’язковим моментом е видыення певних штерлейкгнгв (1Л-4,
1Л-5), необхщних для залучения до взаемодп з АПК В-л!мфоципв. Останн! теля
антигенно! стимуляци перетворюються на плазмобласти, згодом на плазмоцити,
як! й синтезують антитыа, специфгчнг до антигеюв збудника. Ргвень (титр) цих
антиты у кров! 1 тканинах макрооргашзму г визначае протимгкробний гмунгтет.
Якщо ж збудник здатний до внутрппньоклгтинного паразитування, спрацьо-
вуе гнший механгзм. Презентацгю антигену забезпечують як гмунш юптини, так г
будь-як! ядерн! юптини оргашзму, здатн! до утворення комплексгв антиген збуд-
ника — молекула ГКГС I класу. Видыення при цьому гнших цитоюшв (1Л-2, у-
Таблиця 2. Найпоширежил патогены м!крооргажзми людини
Позаклпинн! М1кроорган!зми Внутр1шньокл(тинн1 М1кроорган1зми
обл1гатн1 факультативы
Бакгерй: • стафшококи •стрептококи • пневмококи • меншгококи* • гонококи* • шигели • клостридп та ш. Гриби: • кандида* • криптококи* та ш. В1руси Рикетсп Хлам!дп Протозоа: токсоид азми лейшмашГ та ш. Бактери: • М1кобактерн • сальмонели • бруцели та ш. Гриби: • кандида • криптококи • кокцидомщети • пстоплазми та ш. Протозоа: • плазмодп • трипаносоми • пневмоцисти та ш.
*Можуть бути також факулътативними внутрииньоклгтинними паразитами.
штерферону) зумовлюе залучения Т-л!мфоцит!в до взаемоди з АПК. Пгсля бласт-
трансформаип антигенактивованих Т-л!мфоцит!в вщбуваеться !х прол!феращя з
утворенням певно! юлькост! !муноцит!в, здатних розшзнавати антигени збудника,
представлен! на поверхш клггин у поеднанн! з власними антигенами ГКГС I кла-
су. Важливо пам’ятати, що !з цих двох р!зних “сценарив” ьмунно! вщповцц синтез
антиты плазмоцитами контролюеться специф!чними Т-л!мфоцитами — Т-хелпе-
рами 2-го типу, а в раз! кл!тинного типу кр!м пулу цитотоксичних л!мфоципв
утворюються антипла (1§М), що також мають значения в знешкодженш внутрпп-
ньоклггинного паразита. Тобто клггинна та гуморальна ланки !мунно! системи
активно взаемодпоть, забезпечуючи протимжробний тмуштет.
Антипла формують к!лька еташв (“ешелон!в”) захисту вщ збудника шфекщй-
но'1 хвороби. На першому еташ м!крооргашзм мае подолати дпо секреторних 1&А,
яю, зв’язуючись з антигенами шфекщйного агента, блокують його прикргплення
до еп!тел!альних клггин. На другому еташ мгкрооргашзм, що проник у пщслизо-
вий прошарок, зв’язуеться з 1§Е та 1§О4, як! приеднаш до поверхш базофщьних
гранулоцита (тканинних базофшв) своТми Рс-фрагментами. Утворення комплек-
су антиген — антитшо на поверхш базофщьних гранулоцита зумовлюе !х дегра-
нуляцпо, а мед!атори, що видщяються, сприяють хемотаксису нейтрофщьних та
еозинофщьних гранулоцит! в, а також макрофата у м!сце проникнем ня шфекщй-
ного агента. Якщо ж останнъому вдалось якимось чином уникнути описания пе-
репгкод, то в кров! його “зустр!чають” 1§М 11§С12, як! найбшыпе здатш до акти-
вацп системи комплементу. Мембранатакуючий комплекс, шо утворюеться на
поверхш розшзнано! антиплами мгкробно! клпини, зумовлюе цитол!з 1 ел!м!на-
щю мжрооргашзму.
У КЛ1ТИНШЙ (Т-л!мфоцитшй) !муншй вщповщ! також спостерггаеться певна
етапшсть. В ештели дихальних шлях!в, травно!, сечово! 1 статево! систем велике
значения мають Т-л!мфоцити-гамма-дельта (Тл-у§), як! розшзнають представлен!
в поеднанн! з антигенами ГКГС антигени збудника або реагують !з протешами
теплового шоку (ЬеаГ айоск ргоГеш, Н8Р), що експресуються на поверхш шф1ко-
ваних клггин. Розташовуючись м!ж сусщшми ештелкщитами, Тл-у<5 мають мож-
ливгсть шшьно контактувати з юптинами слизово! оболонки, що дае !м змогу
щдтримувати антигенний гомеостаз ! негайно знищувати юпгини, у яю проник
шфекцшний агент Т-лшфоцити-альфа-бета (Тл-оф) також мктягься в слизовгй
оболонц! бар'ерних оргашв, але бшьше значения в 1мунному захисп вони мають
у раз! проникнення збуцника в глибпп середовища оргашзму, Мехашзми цито-
токсично! дп Л1мфоцит!в рззних популяцш ушверсальш. Це або утворення отвору
в мембпаш шф]ковано! юнтини за допомогою перфоришв та ждукшя апоптозу
клгтини внаслщок уведення гранзим!в (специфЁчних фермента, як! моститься в
гранулах Т-лимфоцита), або запуск апоптичного пронесу через взаемодпо Раь-
лхганд / Раь-рецептор. Обидва мехашзми зумовлюють загибель шфгковано! клни-
ни разом 13 внутргшньоюътинним паразитом.
Видшено в1с!м гишв шфекцшних агента (розташоваш в порядку зменшення
геометричних розм!р1в): гельмшти, найпростшп, гриби, бактерп, хламщп, рикет-
сп, в!руси га прюни. Кожей !з них використовуе специф!чш засоби, що дають
змогу вести паразитичне кнування за умови протидп систем антимгкробного за-
хисту макрооргашзму.
Паразитами! та протозойн! шфекцп. Найпростшп та гельмшти е причиною
дуже поширених хвороб в усьому свгп. Наприклад, вщ малярп, за даними ВООЗ,
шоргчно помирають вщ 1 до 2 млн ощб. Кишковими гельмштами заражена трели-
на всього населения Земл!, що мае особливо небезпечш наслщки для дней.
Быышсть паразита мае складний життевий цикл. Причому в р!зних стад!ях
розвитку вони синтезують ргзн! антигени, що значно утруднюе роботу !мунних
клггин оргашзму. Властивкть знищувати як найпростшпи. так 1 гельмшпв мають
макрофаги, гранулоцита (нейтрофьтьш, еозинофыьн!), Т-л!мфоцити 1 навпь
тромбоцита (ефекторш клпини).
Гуморальш мехашзми Гмушгету (антитша, протеши системи комплементу)
гакож мають значения у забезпечешп захисту вщ позаюптинних паразита, що
локшпзуються в кров!, л!мфатичних судинах, кишках. Однак !хня дш е найефек-
тивнппою тод1, коли вони функцюнують одночасно з ефекгорними юптинами.
На першому еташ протапаразитно! реакцп оргашзму макрофаги поглинають па-
разита та секретують речовини (ЕЛ-1,1Л-2, фактор некрозу пухлин альфа (ФНП-
а) тошо), як! регу.поють розвиток запалення. Антигени паразита презентуюлься
на поверхш макрофапв у поеднанш з молекулами ГКГС, 1 починаеться власне
специфхчна Ёмунна вщповщь — пщбираються та клонуються Т- 1 В-л!мфоцита.
Плазмоцита, шо утворюються з В-л!мфоцита, виробляють антиила, як!. взае-
модпочи з поверхневими антигенами паразита, “навантажують” !х 1 полегшують
!х розшзнавання та поглинання макрофагами, гранулоцитами (нейтрофшьними.
еозинофщьними) та шшими юптинами, що мають репептори до Рс-фрагмента
антит!л. Пщ час фагоцитозу у макрофагах 1 нейтрофшьних гранулоцитах утворю-
ються високоэктавш метаболии кисню (супероксид-ашонрадикал), пероксид
водню, пероксиштрит (О1\ЮО') та шип потужн! окисники. Вони забезпечують
руйнування бшшщного шару юптанно! стшки паразита та його дерет равлювання
в л!зосомах.
Принципове значения у формуванш !мун!тету та елхмшаци паразита мае ак-
тивашя Т-л)мфоцит!в-хелпер]в (С04) 1 цитотоксичних клггин (С08) Пщ час па-
разитично! шфекцй ргзнт субпопуляцп Т-хелпер гв (Тх) 1-го та 2-го тишв активу-
ються ргзною лирою. 1 !х сп!вв!дношення може змшюватися протягом шфекщй-
ного процесу. Так, Тх 1-го типудгють, з одного боку, на печшковш стадп маляр!!
г вид!ляючи у-штерферони, сприяють елгмгнацц паразит!в через активащю фаго-
: тпв. 3 шшого боку, знищеыня паразита на еритроцитнш стадп вщбуваеться
.авдяки активацп Тх 2-го типу, шо стимулюють вид!лення специфгчних антиттл.
У разг шших паразитичних хвороб спостерйають антагошстичн! взаемовщно-
шення М1Ж. цими субпопуляпдями Т-лшфощгпв. На лш!ях мишей, резистентних
до лейшманюзу та чут.тивтгх до нього, доведено, що в одних домшують Тх 1-го
типу, яю виробляють у-штерферон, а в шших переважають Тх 2-го типу, здатш до
еекрепп 1Л-4, який пригшчуе утворення у-штерферону Тх 1-го типу. Уведення
антитЬ] до 1Л-4 сприяло знищенню шфекцп через зменшення активност! Тх 2-го
типу 1 зб!лыпення чисельност! популяцп Тх 1-го типу. У людини в раз! дифузного
лейшманюзу шюри та прогресуючого вюцерального лейшманюзу спостерйэсться
недостатюсть синтезу у-штерферону та надмтрне видшення 1Л-10 — питокшу, що
Биробляеться Тх 2-го типу 1 пригшчуе функшональну актившсть Тх 1-го типу.
У виладку кишкових гельмштозгв, навпаки, для ел!мшапп паразитов необхш-
ною е активашя Тх 2-го типу. Саме ц! юптини вщповщають на паразитичн! анти-
гени видшенням 1Л-4 та 1Л-5, яю стимулюють лрол!феран!ю В-л!мфоцит!в у
кишкевш стшщ та синтез ними 1§Е, що опсошзують гельмшти 1 тим самим спри-
яють прилипанию до них еозиноф!л!в. Кр!м того, 1Л-5, який називають еозино-
фыьним фактором, посилюе прол!ферашю 1 хемотаксис еозинофкпв та акгивашю
секрецн ними цитотоксичних б!лк!в. 1Л-3, 1Л-4, 1Л-9 та 1Л-10, що видыяються Тх
2-го типу, стимулюють пролгферацпо базоф!льних гранулоцита. На !хн!й поверх-
ю вщбуваеться утворення комплекств 1§Е та паразитичних ангигешв, наслщком
чого е дегранулящя гранулоцита 1 видщення велико! юлькост! БАР. При цьому
макрофаги секретують питоюни, що сприяють прол!ферацй келихоподтбних ск-
зокриноцита кишок 1 спричинюють шдвищений синтез слизу. Блокований анти-
ттлами, ушкоджений еозинофшами та оброблений слизом гельм!нт може бути
легко видалений з просвтту ют шок.
У деяких випадках тмунна система не здатна повшетю позбутися паразита,
проте 1золюе !х в!д здорових тканин зап альним бар’сром. Клттини, що шфыьтру-
ють тканини й оточують паразита, переважно представлен! Тх Его типу.
Однак у процес! еволюцп паразита винайшли багато способов уникати дп
майже досконалих систем захисту макрооргашзм!в. Наприклад. токсоплазми про-
никають у макрофаги не шляхом фагоцитозу ! тому не спричинюють '‘ресшратор-
ний вибух” та утворення вьтьних радикал!в кисню. Лейшманп проникають у мак-
рофаги. використовуючи рецептори комплементу. Кр!м того, вони здатш синте-
зувати супероксиданюндисмутазу (СОД) — фермент, що швидко шактивуе
активш форми кисню. Блокуеться також актившсть кисненезалежних мехашзлнв
фагоцитозу. Лейшманп' синтезують глткопротеш. шо д!е як шг!б!тор протешаз
лтзосомного походження. Серед шших мехашзм!в захисту паразита вщ дп юптин
1мунно! системи варто назвата високу антигенну м)нлив!сть (Р1а&то(Нит, Тгурапо-
зота), видщення розчинних анти ген! в. яю блокують антипла ще на “шдс гулах”
до юптини-паразита (ЗсН/'зГозота), здатшеть до утворення кутикули (1Уис1гегег1а) та
перехщ у цистну форму (ТпсЫпе11а).
Ус! зазначеш пристосувальш мехашзми м!кроорган!зм!в зумовлюють пр]
нання до шфекцшного процесу 1мунопатолопчних реакцш. Наприклад, це вт
ваеться тод!, коли замтсть юптин паразита уражуються власш юптини макр
гашзму, як! несуть на свош поверхш антигени, схож! на паразитичш, або ги
унаслщок дп надлишку мещаторгв запалення, оксидант) в ! л!зосомних фермен~в|
як! видгляють 1 мунш юптини у вогнищ! локалгзацп паразита.
Бактер!алып, грибков!, хламщшш та рикетсинп шфекцп. Тмуштет, що
муеться до цих шфекцшних агенпв, може бути власне антамткробним 1 ант
сичним. В останньому випадку специф!чш антитыа блокують токсини, що
ляються мгкрооргашзмами. Як правило, ел!мшац!я шфекцшного збудника з
гашзму потребуе поеднання кщькох популяцш гмунних юптин. Наприклад, 1
стимулюють фагоцитну актившсть макрофапв за рахунок активацц шдуцибел
ЫО-синтетази, г!перекспрес!я яко! зумовлюе утворення велико! к!лькост! перс
штриту — одного з найпотужшших оксиданпв. У випадку грибково! шфекцп,
рема при кандидоз!, зниження кыькост! Тл-уЗ спричинюе незавершений фаг
тоз унаслщок недостатньо! активацц фагоцит) в. 1нш! мехашзми гмунного заз
реалгзуються за описаною схемою залежно вщ типу паразитування бактерн
заюптинний чи внутршшьоклгтанний) або за рахунок роботи юптинного !му
ту в раз! грибкових (факультативш внутршшъоюптинш паразита), хламщп
рикетсшних (облйатн! внутршшьоюптинш паразита) шфекщй.
Бактертальш та грибков! мткрооргашзми для уникнення дп описаних г
мжробних систем застосовуютъ численш мехашзми, шо дають !м змогу пара
вата та спричинювата шфекцшш хвороби (табл. 3).
Таблица 3. Мехашзми уникнення тфекцшними агентами
дизахисних систем макрооргашзму
Фактор захисту макрооргашзму -1 =^=—~ '— —' 7 ' Пристосувальш мехашзми м*1крооргажзм1в Приклад
Хемотаксис Видшення речовин, що пригшчують хемотаксис Мпрастатин 31гер1отусез, металопротетнази ЬеуюпеНи !
Фагоцитоз Пригшчення злиття лхзосом 13 фагосо- мами Амоншш сполуки МусоЬас1егшт 1иЬегси1от. Ье^Итата
Видыення токсичних для фагоцитов речовин Лейкоцидини 8(арИу1осоаж 1 С1оМг1сНит
1нпбування протонового насоса в лтзо- сомах, порушення ацидифткацп л!зосом Тнпбтгори Н+-АТФази Уегхйиа, МусоЪас1еггит 1иЬегси1оз15
1нактивац1я водню пероксиду, що вид)- ляеться фагоцитами Каталаза 81арку1ососси$, Еп1егососси5 та !нших мткроеа ГЭШЗМ1В
Презентация анти- гешв у поеднанн! з ГКГС Порушення презентацм антигетв шф1- кованою кл1тиною Токсини МусоЪасСепит 1иЬегси1омз ЕзскепМа сок
Розщеплення фактора транскрипцп, необх1дного для експресн ГКГС Протеаза СМатуЛа
Закшчеъня табл. 3
Фактор захисту макрооргашзму Пристосувальн! мехажзми _ м1крооргажзм1в Приклад
Антитела Антигенна мтнливтсть СатруЫасгег, (Зопососсиз, Мусор!аята та переважна б!ль- штсть мткрооргангзмтв
Система комплементу С!ал!зац!я юптинно! поверх ю, що лере- шкоджае актпвацп компоненпв компле- менту (СЗЬ) Залишки сталових кислот Метп^осоесия та шших мткро- органтпв
Адсорбт» на поверхн! шпбггор1В СЗЬ
Експресгя мембранасоцшованих фер- ментгв, що пдролхзують компонента комплементу Протсази Уегят!а, Рзеи^ото/гая, ВоггеНа
Гнактиващя хемотаксичних властивос- тей 61 нов комплементу (СЗ, С5)
Цитокши Блокування В1ДПОВ1Д1 макрофапв на у- штерферон Л йтоарабшомана н МусоЬааегтт шЬегси1о818
Хндукщя синтезу 1Л-10 11Л-12, що при- гючують апоптоз монощгпв СЫатуЛа
Кр1м резистентност! до дп факторгв природного захисту макрооргашзму ш-
фекцшш агенти можуть набути стшкосп 1 до ди антибактергальних препарапв.
Так, деяк! шгами стрептококтв мають ген, що кодуе додатковий пенщилшзв’язу-
вальний биток. Ентеробактерц набувають резистентност! до р-лактам!в завдяки
мутащям у генах лактамаз, як! розширюють спектр ди цих ферментгв. А МусоЬас-
епит шЬегсиЬж стае непугливою до ди гзошазиду та рифамппшну в раз! мутаци
в ген! каталазпероксидази г РНК-полгмсрази вгтповтдно.
Досить часто тмунна вщповгдь на бактертальш (грибков!, хламщшн!) антигени
супроводжусться ушкодженням власних кл!тин за рахунок видитення надлишку
цитокгшв, перехресного розшзнавання антигенгв власного оргашзму та шших ме-
зашзмтв. Уст вони е щдГрунтям для формування алерпчних реакцш за цитоток-
спчним, тмунокомплексним г клггинним типом гз розвитком так званих шфекщй-
но-алерпчних хвороб. Не виключають роль шфекцшних агенпв г в патогенез!
алерпчних реакцш стимулюючого типу. Наприклад. гнфекшю, спричинену клебс-
гелами, розглядають як один гз пускових мехашзмгв хвороби Грейвса (тиреоток-
сикоз).
В!русш шфекцп. Л1кв!дац!я в1рус!в вщбуваеться за упаси тих самих ефекторних
клггин, шо й при внутр! шньоюптинних бакгергальних (грибкових, хламщшних)
(нфекцтях. Дещо битьше значения мають нагуральш китери (НК) — клгтини, здатнг
руйнувати !нф!кован! клгтин и без попередньо! антигенно! стимуляци. НК розшзна-
ють в!русшф!коваш клиини або за появою на поверхш Н8Р, або за зменшенням
китькост! експресованпх молекул ГКГС. Проте вгрусн! гнфекшйнг агенти здатн!
спршгинювати в оргашзм! значно гяжч! порушення поргвняно з шшими збудника-
ми. Перше завдання, яке “виртшують” вгруси, — специф!чне проникнення б клг-
тини оргашзму певного типу'. У кожною вгрусу, здатного паразитувати в оргашзм!
людини чи тварин, е битки, аналопчш проте'тнам макрооргашзму. Взагал! еволюцтя
та добтр мгкрооргантзмтв т насамперед втруств вщбуваеться за наявностг бтлктв,
подтбних до протетнтв оргашзму людини як найбтлып “привабливого” об’екта па-
разитування. Таким чином, мгкрооргашзм виртшуе одразу ктлька проблем, основна
з яких — проникнення в клттину. Класичним прикладом цього е взаемощя глт-
копротетну (§р120) втрусу тмунодефтциту людини (В1Л, втрус СН1Д) та СВ4-рецеп-
тора Т-хелпер!в. Втрус Епстайна — Барр використовуе для цього рецептори комп-
лементу 2-го типу (СК2), риновтруси — молекули клттинно! адгезй (1САМ-1).
Головним питаниям для втрусу шсля проникнення в клттину с налагодження
редуплткацп власного генетичного матерталу й утворення нових втртошв. Для цьо-
го втруси використовують ферментш системи клттини-хазяша. Зрозумтло, що вт-
рус “не зацткавлений” у руйнуваннт клттини, яку вш хоче використати як “фаб-
рику” для побудови втрусних частинок. Деяк! втруси майже позбулися в процес!
еволюцп цитопатичностт т, навпаки, дозволяютъ клттинт уникнути апоптозу. 3
шею метою втруси використовують быки — супресори апоптозу. Так, втрус кон-
тагтозного молюска мае биток, подтбний до внутрпиньоклттинно! частили Газ-ре-
цептора, який порушуе передачу сигналу в!д активованого Раз-лтгандом рецепто-
ра до ферменттв (каспаз), залучених до реалтзацп программ апоптозу. Аденовтруси
здатн! до видглення аналогтв клттин них битктв (наприклад Ьс1-2), що запобтгае
активацп каспаз т розвитку апоптозу. Особливого значения цей процес набувае в
раз! онкогенност! втрусу. Тодт втн одночасно спричинюе трансформацию клттини
та зумовлюе п уникнення апоптозу. Деяк! онкогеннт втруси (НТЬУ-1) здатнт ви-
користовувати клттини тмунно! системи для стимуляцп подглу тнфткованот клтти-
ни. Так, гранулоцитно-моноцитний колошестимулюючий фактор, що видптяеть-
ся втрусшфткованими Т-л!мфоцитами, стимулюе макрофаги до продукци 1Л-2,
що е фактором росту для Т-лтмфоцитгв.
Втруси також здатнт щдтримувати життедгяльнтсть тнфткованот клттини, ак-
тивно протидпочи тмуннш систем!, що намагаеться елтмгнувати клттину зт сторон-
ньою генетичною тнформацтею з оргашзму (табл. 4).
Таблица 4. Мехашзми протидп втруств захисним системам макрооргашзму
Захисний фактор В!рус (и) Втрусний продукт Мехажзм дп
Експрестя моле- кул ГКГС Аденовтруси Бтлок ЕЗ Блокада транспорту молекул ГКГС на поверхню клттини
Герпесвтруси Убтквттинлтгаза Протеасомна деградацтя молекул ГКГС
Цитотани Втрус мтксоми Гомолог рецепто- ра у-штерферону Зв’язування у-штерферону та запобь гання його дп на клггини
Втрус Епстайна — Барр Гомолог 1Л-10 Пригнтчення активностт клттинного тмуштету
Комплемент Втрус простого герпесу 1 -го типу Глткопротетни Зв’язування 61 лгав системи комплемен- ту
Антиттла Цитомегаловтрус ГлткопротеТни з властивостями Гс-рецептор!в Блокада противтрусних антиттл через зв’язування Е^-фрагменпв 1§Сг
1мунопатолопчш реакцп досить характерш для багатьох в!русних шфекцш.
Кргм перехресного реагування против!русних антитщ 13 власними антигенами в!-
руси здатш змшювати протеши макрооргашзму та порушувати гмунолопчну то-
лерантшсть. У деяких випадках, наприклад у раз! шфгкування в!русом гепатиту В,
авто!мунний мехашзм руйнування власних юптин стае визначальним 1 набувае
неконтрольованого характеру. Це призводить до руйнування одночасно значно!
кшькосп навпь нешфгкованих юптин 1 зумовлюе розвиток хрошчного активного
гепатиту.
Пр1онов! шфекцп. Особливий характер взаемоди оргашзму 31 збудником шфек-
шйно! хвороби виникае, коли в рол! шфекцшного агента виступае протеин прюн
(протеш шфекцшний). В!н власне не е мжрооргашзмом, але спричинюе розви-
ток шфекцшного процесу. Перш! сенсацшш повщомлення про патогеншсть про-
теппв з’явилися ще у 1975 р., коли Оазбизек (Нобел!вський лауреат, 1976) дов!в,
що збудник, вщповщальний за розвиток невролопчно! хвороби в кашбал!в Ново!
Гвше! (хвороба куру), не належить аш до бактерш, аш до в!рус!в.
Нин! встановлено етюлопчну роль протеппв — збудникгв хвороб у людини
(хвороби Крейтцфельдта — Якоба та Герстманна — Штрауслера), овець (скреп!) 1
кор!в (коров’ячий сказ).
Патогенез прюнових хвороб е досить цпсавим 1 незвичайним. Установлено,
що хвороба виникае внаслщок потрапляння в оргашзм прюнового протешу (РгР5с;
молекулярна маса — 33—35 кДа), який е аналогом нормального юитинного про-
тешу (РгР). Основними шляхами шфгкування варто вважати проникнення прюшв
кр!зь кишкову стшку п!д час споживання м’яса шфгкованих тварин (доведено, що
прюнов! протеши людини аналопчш прюнам тварин), а також трансплантацп
(ятрогенний шлях). Р!зняться щ протеши (РгР8с та РгРс) переважно конформа-
щею: певним сшввщношенням а-сшральних 1 р-плоских структур. РгРс е звичай-
ним компонентом мембран клггин нервово! та гмунно! систем. Взаемод!я транс-
формованого екзогенного прюнового протешу спричинюе перетворення нор-
мального ендогенного протегну, внаслщок чого вш набувае властивостей
екзогенного шфекцшного протешу, а також здатност! до трансформацп шших
прюнових протешгв шфгкованого оргашзму. Накопичення трансформованих б!л-
к!в у юптин! створюе певш проблеми щодо !х утюпзацп (прюнов! протеши досить
резистентш до дц протешаз), 1 поступово в клггин! утворюються бшков! вщкла-
дення, що складаються з фрагмента прюнових протеппв. Ц! вщкладення е одним
13 варианта амшощу.
Отже, саме амыо'щоз спричинюе розвиток характерного нейродегенеративно-
го процесу, що проявляеться спонгюформними губчастими енцефалопатгями
(хвороба Крейтцфельдта — Якоба). Зрозум!ло, що цей процес може тривати де-
сятки рокгв (пщтримання протягом певного часу балансу м!ж накопиченням
прюнових протеппв та !х деградащею). Тобто прюнов! шфекцп належать до
повшьних. 1мунну вщповщь п!д час прюнових шфекцш майже не спостерйають,
шо зумовлено, з одного боку, толерантшстю до власного прюнового протешу
(амшокислотна послщовшсть, а отже, антигени! властивост! трансформованих
протеппв не в!др!зняються в!д нативних), а з шшого — вщокремлешстю патоло-
пчного процесу вщ гмунно! системи гематоенцефал!чним бар’ером.
Засоби профилактики прюнових гнфекцш можуть бути пов’язан! з ранньок
диагностикою та вибраковуванням гнфгкованих тварин або з обмеженням спожи-
вання м’яса ! мясопродуктов. Лгкування прюнових гнфекцш майже не розробле-
но, прогноз дуже несприятливий — нейродегенеративний процес неминуче при-
зводить до загибелг оргашзму.
Системы прояви <нфекц1йного процесу
1нфекщйний процес нерозривно пов’язаний з такими патолопчними процесами.
як запалення та гарячка (див. роздгл XI “Запалення”, роздгл XIII “Гарячка”).
Саме запалення як типовий патолопчний процес еволюцшно сформувалося в
процес! налагодження мехашзмгв захисту вщ шфекцшних агентов. Удосконалення
цих механ1зм!в сприяло формуванню !мунно! системи, яка дае змогу специфично,
з мппмальною шкодою для клгтин власного органгзму елгмгнувати збудника пй
час шфекцгйного процесу г запобггти в бглыпостг випадкгв розвитку гнфекцшнс
хвороби. Якщо шфекцгйний агент виходить за меж! певно! тканини, мгсцева за-
пальна реакцгя доповнюеться акгиващею систем антимикробного захисту всьогс
органгзму. Насамперед це проявляеться гарячкою, що виникае у вщповгдь на д!ю
певних речовин мгкрооргангзмгв — екзогенних шрогешв. Розвиток шфекцгйного
процесу зумовлюе активацпо шших фгзюлопчних систем органгзму: нейроендок-
ринног системи, мгкроциркуляцц, сполучно! тканини, зовнгшнього дихання, ге-
мостазу та гн. Активне функцюнування зазначених систем пщ час шфекцгйного
процесу, з одного боку, дае змогу оргашзму мобглгзувати додатковг мехашзми для
боротьби з! збудником шфекщйно! хвороби, а з шшого — може призвести до не-
бажаних патолопчних наслгдкгв (шок, шсульт, синдром дисемгнованого внутргш-
ньосудинного зсщання кров!, синдром гостро! недостатносто легень г нирок тощо).
Наприклад, активацгя ппоталамо-ггпофгзарно-наднирково! та симпатико-адрена-
лово! систем, що супроводжуе розвиток шфекцгйного процесу, спричинюе поси-
лення глгкогенолгзу та лгполгзу зг збглыпенням концентрацп глюкози та вгльних
жирних кислот у кровь Цг речовини е енергетичним субстратом для клггин гмун-
но! системи. Кргм того, глюкокортикощи безпосередньо регулюють гмуннг проце-
си, впливаючи на актившсть тимоцитгв 1 лгмфоцитов. Глюкокортикощи е одним
гз найпотужнгших фактор!в шдукцп апоптозу в гмунних клгтинах. Це певною
м!рою стримуе гмунну вщповщь оргашзму 1 дае змогу запобпти автогмунному ушко-
дженню власних клггин. Проте надмгрна активацгя зазначених нейроендокринних
систем зумовлюе виснаження запашв глгкогену з розвитком недостатносто деяких
оргашв, наприклад печшки. Надлишок глюкокортикощг'в, безумовно, спричинить
розвиток гмунодепресивного стану.
Активацгя системи кровообггу, що, як правило, виникае пщ час шфекцгйного
процесу, з одного боку, сприяе актившшому перебггу метабол!чних процесгв у тка-
нинах, полшшенню гх оксигенаци, прискоренню “доставки” клггин, залучених до
гмунно! вгдповщ!, до вогнища запалення. Однак, з шшого боку, пщвищення ар-
тер!ального тиску та штенсифгкащя роботи серця пщ впливом симпатико-адрена-
лово! системи зумовлюе розвиток вщносно! недостатносто коронарного кровообпу
тахгкардп, екстрасистолп та шших аритмш, недостатносто зовшшнього дихання.
Завдяки вдосконаленню методгв мгкробюлопчного дослщження вдалося до-
вести патогенетичне значения бактерш 1 в!рус!в у розвитку найпоширенгших не-
чфекщйних соматичних хвороб. Наприклад. активно вивчаеться роль в!рус1в гер-
есу, цитомегалов]русу, НеНсоЬастегру1оп та СЫату<Иа рпеитопла в патогенез! ате-
тосклерозу й 1шсм1чно1 хвороби серця. вирусу Епстайна — Барр, НТЬУ-1, НТЬУ-2
Рароу-а В1рус1в — у патогенез] пухлин, НеНсоЬааег ру1оп — хрошчного гастриту
та виразки шлунка. Це значно збшыпуе медико-сощальне значения шфекщйних
хвороб, вису вас низку пробл емких питань, що стосуються демарка цп соматичних
власне шфекщйних хвороб.
Роздал IV
ПАТОЛОПЧНА Ф13ЮЛ0ПЯ КЛ1ТИНИ
Клпина е найменшою структурно-фхзхолопчною единицею тша багатокштинного
оргашзму. 1й притаманн! вех основш процеси його життезабезпечення. Тому ко-
жей патолопчний процес виникае та розвиваеться за обов’язковою участю юп-
тин, а будь-якх патолопчш змши, що виникають на рхвш органа або системи,
неодмшно грунтуються на патолопчних змшах у клпинах, спотворенш або втрап
ххнхх функшй, а нерхдко х втрап самих клггин.
Отже, можна стверджувати, що патолопя юптини супроводжуе вех патолопч-
ш процеси, але не вичерпуе повшетю тих змш, що при цьому спостерхгаються.
ПОРУШЕННЯ ФУНКЦ1Й КЛ1ТИН
Усх численнх процеси, що вщбуваються в юптиш, можна умовно об’еднати в двх
основн! групи: процеси, необхщш для забезпечення життедхяльносп й самовхд-
творення клпини, та процеси, пов’язаш з виконанням спецхалхзованих функцш
диференцшованих юптин у багатоклпинному оргашзмх. До першоТ групи нале-
жать: процеси обмшу речовин х зв’язку клхтини з й оточенням; збереження, пере-
давания та реалхзацхя генетично'х шформацп; синтез х внутрхшньоюптинний роз-
пад речовин, забезпечення енерпею; подш х пролхферацхя юптин, загибель клпи-
ни. Друга трупа охоплюе процеси генерацй та передавания сигналу збудження,
скорочення, вироблення х видщення рхзних бнткхв, а саме: бнпов кровь ферменпв
травнох системи, гормошв, медхаторхв тощо; утворення спещалхзованих юптин х
структур — еритроцитхв, кришталика, волосся, м’язхв, исток. Безумовно, базов!
та спецхалхзоваш функцн клпини взаемозалежнх. Але порушення спещалхзованих
функшй буде розглянуто в роздшах, присвячених патологхх орган! в х систем. У
цьому ж роздш розглянемо лише найбшьш загальш ознаки розвитку патолопч-
ного процесу, характерш для вехх клхтин.
ПОРУШЕННЯ ПРОЦЕС1В ОБМ1НУ
ТА ВЗАеМОДПЗ НАВКОЛИШН1М СЕРЕДОВИЩЕМ
Обмш речовинами, що надходять урередину юптини або видшяються нею назов-
ш, а також обмш рхзними сигналами з мхкро- та макрооточенням вщбуваеться
через зовшшню мембрану клпини. Як вщомо, юптинна мембрана е лпххдним бх-
шаром, у який вбудоваш рхзнх бшковх молекули, що виконують роль спещалхзо-
ваних рецепторхв, йонних каналхв, пристро’хв, якх активно переносить або видаля-
мъ ртзш хтмтчнт речовини, можклгтинних контактов тощо. У здорових юптинах
евкарютов мембрани е асимегричними за розподном фосфолншпв: зовншппй
шар складаеться зт сфптгомтелгну та фосфатидилхолтну, внутрштюй — тз фосфати-
дилсерину та фосфатидилетаыоламону. Пщтримання цте! асиметрп потребуе за-
трат енергй. Тому в раз! ушкодження клотини, и шфткування, енергетичного го-
тодування вщбуваеться збагачення зовшшнього шару мембрани невластивими !й
фосфолшщами, що стае сигналом для шших клттин 1 ферменттв про ушкодження
клттини з вщповодною реакшею на це. Найбгтьше значения при цьому мае роз-
чинна форма фосфолшази А2, яка вщщеплюе арахыонову кислоту та створюе
лтзоформи з витценазваних фосфолштдтв. Арахщонова кислота е лтмттуючою лан-
кою для створення таких медтатортв запалення, як ейкозанотди, а до лтзоформ у
мембран! приеднуються такт захпсн! молекули, як пентраксини (С-реактивний
лротеон, попередники амйтотдних бтлктв) з наступною активацтею системи комп-
лементу за класичним шляхом । руйнуванням клттини.
Будова мембрани сприяе збереженню особливостей внутрштнього середовища
клттини, його вщмонностей в!д середовиша, що п оточуе. Це забезпечуеться вибтр-
ковою проникнтспо юптиннот мембрани, тснуванням у нш механтзмтв активного
транспорту. 1х порушення внаслщок прямого ушкодження, наприклад, тетродо-
токсином, уабатном, тетраетиламонтем, або в разт недостатнього енергетичного за-
безпечення вщповщних “насоств” призводить до порушення електролттного складу
клттини, змтн у н!й метаболтзму, порушення специфтчних функцтй — скорочення,
проведения тмпульсу збудження тощо. Порушення клттин них йонних каналов
(кальптевих, натртсвих ! калтевих) у людини може бути також генетично зумовле-
ним мутацтею гентв, якг втдповщають за структуру канал!в. Цт так зван! каналопатп
спричинюють спадково хвороби нервово! та м’язовот систем. Надмтрне надходжен-
ня всередину клттини води може призвести до П розриву — цитолтзу, що вщбува-
тиметься за рахунок перфорацц мембрани птд дтею специфтчних антиттл, активацц
комплементу або атаки шгготоксичних лтмфоциттв т натуральных юл ер! в.
У клотинну мембрану вбудовано безлтч рецепторов — структур, як! в раз! поед-
нання з вщповщними спенифтчиими сигнальиими молекулами, лиан дам и, пере-
дають сигнал усередину клттини. Це вщбуваеться за допомогою ртзних сигнальных
каскадтв, якт складаються з низки ферментно активных молекул, що послщовно
активуються т врештт-решт призводять до реалтзацп ртзних клттинних програм,
таких як ртст т иро.пферацтя, диференшювання, рухомтсть, стартння, загибель клт-
тини. Ктльюсть таютх регуляторних каскадтв досить велика, хоча до конпя ц ще не
визначено. Система рецепторов т поьднэних тз ними регуляторних каскадтв тснуе
також усередитп клттини. Вони сгворюють усерединт клттини певну регуляторну
мережу' з пунктами концентрацп, розподйту та вибору податьшого шляху сигналу
залежно вщ фунгапонального стану ктттини, етапу и розвитку, одночасно! дп сиг-
налтв з шших рецепторов. Наслщком цього може бути гальмування або посилення
сигналу, спрямування його по оншому регуляторному1 шляху. Як рецепторный
апарат, так т шляхи передавания сигналу через сигналил каскады, наприклад до
ядра, можуть порушу ватися внаслщок генетичного дефекту, який виникае як
вроджена вада на ртвш оргашзму або через соматичну мутацпо в певному титл
клттин. Цт мехашзми можуть ушкоджуватись тнфекцтиними агентами, токсинами,
а також змтнюватися в процест стартння. Завершальним етапом цього може бути
порушення функшй юптини, процесов пролофецацп та диференцповання.
На поверхш клггин розташовано також молекули, яко вщшрають важливу роль
у процесах можклотинно! взаемодп. До них можуть належати антигени тканиннс
сумосносто, тканинно-специфочш, диференцповальш антигени тощо. Змоош в скла
щ таких молекул зумовлюють порушення можклггинних взаемодш о можуть стати
причиною включения водповщних мехашзмгв еломшацп таких клггин, адже вони
становлять певну небезпеку для цолосност! оргашзму як резервуар шфекцп, особ-
ливо ворусно!, або як потенщйш гшцоатори пухлинного росту.
Порушення енергетичного забезпечення клггини
Джерелом енергп в клотин! е харчово продукта, шсля розпаду яких до кшцевих
речовин видгляеться енерпя. Головним мгсцем енергоутворення е мггохондрп, у
яких речовини окиснюються за допомогою ферментов дихального ланцюга. Окис-
нення — основний постачальник енергп, тому що за рахунок глгколозу з тако1,
само! колькосто субстратов окиснення (глюкози) вивыьнюеться поровняно з окис-
ненням не больше 5 % енергп. Близько 60 % енергп окиснення акумулюеться
шляхом окисного фосфорилювання в макроерпчних фосфатах (АТФ, КФ), решта
розсоюеться у виглядо тепла. Надало макроергочш фосфата використовуються кло-
тиною для таких процесов, як синтез, подол, рух, секрецоя тощо Видоляють три
мехашзми, ушкодження яких може спричинити порушення забезпечення клотинп
енерпею: перший — фермента енергетичного обмшу, другий — мехашзм окисно-
го фосфорилювання, третш — мехашзми використання енергп.
Об’ем, форма та кглькость мотохондрш залежать вод функцоонального стану .
типу клотин. У раз! надлишку АТФ вони скорочуються, за до! токсинов, Йонов
кальцою, жирових кислот, тироксину, онсулшу, диштрофенолу, навпаки, набряка-
ють. При цьому збольшуеться водстань мож внутрошньою 1 зовнппньою мембрана-
ми мотохондрой. Це зумовлюе ослабления окисного фосфорилювання так, що
болына частка енергп розсоюеться як тепло 1 зменшуеться утворення макроерпч-
них сполук. Цей мехашзм роз’еднання окиснення та фосфорилювання викорис-
товуеться клпиною для регулювання теплопродукцп в гомойотермних орган озмов
для шдтримання постшноо температури тола под час охолодження або И шдвищен-
ня под час гарячки. Значно змони в структур! мотохондрш та енергетичному мета-
бол озмо спостерогаються в разо тиреотоксикозу. Цо змони спочатку оборота!, але
шсля певно! меж! стають необоротними. Мотохондрц при цьому фрагментуються
розпадаються або набрякають, втрачають криста, перетворюючись на вакуоло, та
зрештою накопичують таю речовини, як палон, феритан, кальцой, лшофусцин. 3
хворих на цингу вщбуваеться злиття мотохондрш з утворенням хондрюсфер, мож-
ливо, за рахунок ушкодження мембран перекисними сполуками. Значно ушкод-
ження мотохондрш виникають под доею юшзуючого випромшювання, под час пе-
ретворення нормально! клгтани на злояюсну.
Мггохондрп е потужним депо кальцою, де його концентрацоя на колька поряд-
ков перевшцуе таку в цитоплазм!. У раз! ушкодження мотохондрш кальцой вихо-
дить у цитоплазму, зумовлюючи порушення функцш вщповщноо клгтани, напри-
клад розслаблення м’язов — кальцоево контрактури або навоть “кальцоеву смерть”
у нейронах. Унаслщок порушення функционально! здатносто мотохондрш розко
збольшуеться утворення вольнорадикальних перекисних сполук, яю мають дуже
високу реакцшну здатность о тому ушкоджують важлив! компонента клггини —
-уклешов! кислоти, б1лки та лхпщи. Це явите споете рйаеться в раз! так званого
ксидащйного стресу 1 може мати негативн! наслщки для кнування клпини. Так,
пкодження зовнниньо! мембрани: мпохондрп призводить до виходу в цитоплаз-
му речовин, яю мктяться в мокмембранному простор!, насамперед цитохрому С
та низки шших бюлопчно активних сполук, як! започатковують ланцюгов) реак-
дп, що спричинюе запрограмовану загибель клпини (апоптоз'). Ушкоджуючи
таен! ДНК мпохондрш, вЕшнорадикальш реакцп спотворюють генетичну шфор-
..ашю на вироблення деяких фермент!в дихального ланцюга, що продукуються
_аме в мпохондргях. Це призводить до ще большого порушення окисних провесов
загал! власний генез ичний апарат мпохондрш поршняно з генетичним апаратом
дра г!рше захищений вщ шкщливих впливгв, здатних порушувати закодовану в
льому генетичну шформашю. Як наслщок виникае порушення функцш мпохон-
дрш протягом життя, наприклад у раз! стар!ння, за злояккно! трансформацп юп-
юни, а також на тл! спадкових мпохондр!альних хвороб, пов'язаних з мутащею
ч!тохондр!ально! ДНК у яйцеклиинь Нин! описано понад 50 м!тохондр!алъних
утапш, як! спричинюють спадковт дегенеративнт хвороби нервово! та м’язово!
систем. Вони передаються до дитини винятково вщ матер!, тому що мпохондрп
сперматозоида не входять до складу зиготи ц вщповщно, нового оргашзму.
Порушення збереження! передавания генетичнойнформаци
Ядро юптини мтстить быыпу частину генетично! шформацп 1 тим самим забезпе-
туе ц нормальне функцюнування. За допомогою вибтрково! експресГ! ген!в воно
координуе роботу юптини в штерфазц збериае генетичну гнформаппо, вщтворюе
1 передае генетичний матертал у раз! подгду юптин. У ядр! вщбуваеться регопкащя
ДНК 1 транскрипция РНК. Ртзн! патогенн! фактори, таю як ультрафюлетове та
мипзуюче випромпповання, вьтьнорадикальне окиснення, х!м!чн! речовини, в!ру-
си, здатн! ушкоджувати ДНК. Пщраховано, що кожна клпина теплокровно! тва-
рини втрачае таким чином за добу бшьше юж 10 000 основ. Сюди варто додати
порушення в раз! копповання пщ час под!лу. За збереження цих ушкоджень юп-
тина не здатна була б вижити. Захист полягае в кнуваннх потужних систем репа-
рацп, таких як ультрафюлетова ендонуклеаза, система реларативно! реплгкацп та
рекомбшащйного вщновлення, як! зампцуюгь порушення ДНК. Генетичн! дефек-
ти в репаративних системах спричинюють виникнення хвороб, яю залежать вщ
пщвшцено! чутливост! до фактор)в. що ушкоджуюгь ДНК. Це пкментна ксеро-
дерма, деяю синдроми прискореного старшня, шо зазвичай мають пщвищену
схилыпсть до розвитку злояюсних пухлин.
Система регуляцп процесгв репл!кацп ДНК, транскрипцй 1РНК, трансляцп
генетично!' шформацп з нуклешових кислот у структуру б!люв е досить складною
та багаторГвневою. Окр!м регуляторпих каскадов, яю запускаюгь дпо факторзв
транскрипцп загалъною юльюстю понад 3000, що активують певш гени, кнуе ще
багаторшнева регуляторна система, шо опосередковуеться малими молекулами
РНК (штерферуюч! РНК; РНК!). Геном людини, що скчадаеться приблизно з 3
млрд пуринових 1 шримщинових основ, лише на 2 % зайнятий структурнимп ге-
нами, що вщповщаюзъ за синтез б!лк!в. Решта зайняга синтезом регуляторних
РНК, яю одночасно з факторами транскрипш! активують або блокують роботу
структурних генгв на ргвн! ДНК у хромосомах або впливають на процеси транс-
ляцй з мРНК при утворенш молекули полшептиду в цитоплазм!. Порушення ге-
нетично! шформацг! може в!дбуватися як на р!вн! структурних ген!в, так г регуля-
торно! частини ДНК з вщповщним вщображенням у вигляд! ргзномаштних спад-
кових захворювань.
Останнш часом велику увагу привертають змши в генетичному материал!,
як! вщбуваються пщ час гндивщуального розвитку оргашзму 1 пов’язанг з гальму-
ванням або активащею певних дшянок ДНК 1 хромосом за рахунок !х метилюван-
ня, ацетилювання та фосфорилювання. Цг змши зберггаються надовго, часом
протягом усього житгя оргашзму вщ ембрюгенезу до старост!, г отримали назву
еплгеномно! спадковоспй.
Розмноженню клиин зг змгненою генетичною гнформацгею перешкоджають
також системи (фактори) контролю проходження мпотичного циклу. Вони взае-
модгють 1з циклозалежними протешкшазами та !х каталиичними субодиницями —
циклшами — г блокують проходження клпиною повного мистичного циклу, зу-
пиняючи подш на меж! м!ж пресинтетичною та синтетичною фазами (блок 01/8)
аж до завершения репарацй ДНК, а в раз! ц неможливост! — гнщгюючи запрогра-
мовану смерть клггини. До таких факторгв належить ген р53, мутацгя якого спри-
чинюе втрату контролю над пролгферацгею трансформованих клггин; ц спостерг-
гають майже в половин! випадюв раку в людини. Другий контрольний пункт
проходження мистичного циклу мютиться на меж! 02 / М. Тут контролюеться
правильшсть розподшу хромосомного матер!алу м!ж дочгрнгми клиинами в миоз!
або мейозг за допомогою комплексу контролюючих механгзмгв за клгтинними
веретеном, центром 1 центром!рами (кшетохорами). Неефектившсть цих механгз-
м!в призводить до порушення в розподыг хромосом або гхнгх частин, що прояв-
ляться вщсутнгстю яког-небудь хромосоми в однш гз дочгрюх юптин (анев-
плогдгя), наявшстю зайвог хромосоми (полгплощгя), вщривом частини хромосоми
(делецгя) та перенесениям Г! на гншу хромосому (транслокац!я). Так! явища дуже
часто супроводжують розмноження злоякгсно перероджених 1 трансформованих
клиин. Якщо ж це вщбуваеться пщ час мейозу зг статевими юптинами, то приз-
водить або до загибелг плода на ранньому етап! ембрюнального розвитку, або до
появи органгзму з хромосомною хворобою (див. вщповщний роздгл).
Неконтрольоване розмноження клиин пщ час росту пухлин виникае як на-
слщок мутацп в генах, що контролюють пролгферацпо клиин г одержали назву
онкогешв. Серед понад 70 вщомих ниш онкогешв б!лыпа частина належить до
компоненпв регуляцг! росту юптини, частина е факторами транскрипцй, як! ре-
гулюють актившсть генгв, а також факторами, що гальмують подгл г р!ст клиин.
Ще одним фактором, що обмежуе надмгрну експанс!ю (поширення) пролгферую-
чих клиин, е вкорочення кшщв хромосом — теломер!в, як! не здатн! внаслщок
суто стеричних взаемовщношень повшстю реплгкуватися, тому гпсля кожного
подшу юптини теломери вкорочуються на певну частину основ. Таким чином,
пролгферуючг юптини дорослого органгзму шсля певно! кшькостг подпив (зазви-
чай вщ 20 до 100 залежно вщ виду органгзму та його вгку) вичерпують довжину
теломера 1 подальша реплгкацгя хромосом припиняеться. Це явище не виникае в
статевих та ембргональних клгтинах завдяки наявносп ферменту теломерази, який
вщновлюе довжину теломергв пгсля кожного подолу. У бгльшосп клггин дорослих
ГйП
рган!зм!в теломераза заблокована, але, на жаль, вона здатна вщновлюватися в
юптинах злояюсних пухлин.
Зв’язок мгж ядром т цитоплазмою, транспорту речовин в обох напрямках вщ-
'уваються через пори в ядернш мембран! за участю спещальних транспортних
систем з! споживанням енергп. Таким чином транспортуються до ядра енергетич-
ш та пластичн! речовини, сигнальн! молекули, так! як фактори транскрипцп.
Зворотний пот!к виносить до цитоплазми молекули шформацтйнот 1 транспортно!
РНК, рибосоми, необхщн! для синтезу б!лка в юптин!. Цей самий шлях перене-
сения речовин властивий 1 в!русам, зокрема таким, як В1Л. Вони переносять св!й
генетичний матер!ал у ядро юптини-хазяша з подалыпим включениям його в ге-
ном хазяша та перенесениям новостворено! в!русно! РНК у цитоплазму для по-
дальшого синтезу бтлктв нових в!русних частинок.
Порушення процес!в синтезу
Процеси бтлкового синтезу вщбуваються в цистернах ендоплазматично! стгки, ттс-
но пов’язаних !з порами в ядернш мембран!, куди надходять рибосоми, транспор-
та! та шформацшн! РНК. Тут здшснюеться синтез полшептидних ланщожюв, яю
згодом набувають свого юнцевого стану в гладеньюй ендоплазматичнш с!тщ та
пластинчастому комплекс! (комплекс! Гольдою), де зазнають посттранслящйнот
модифткацп в поеднанн! з молекулами вуглеводав 1 лш!д!в. Новостворен! битков!
молекули не лишаються на мтсцт синтезу, а за допомогою складного регульовано-
го процесу, що мае назву протешкшезису, активно переноситься до т!е! !зольова-
но! частини юптини, де вони виконуватимуть призначену !м функцпо. При цьому
дуже важливим етапом е складання перенесено! молекули у вщповщну просторо-
ву конфтгурацпо, здатну виконувати властиву !й функцпо. Таке складання вщбу-
ваеться за допомогою спещальних ферменттв або на матриц! спещалтзованих б!л-
кових молекул — шаперошв, яю допомагають бтлковш молекул!, новостворентй
або змтнентй пщ дтею зовшшнього впливу, набути правильно! тривимтрно! струк-
тури. У раз! несприятливого впливу на клттину, коли виникае ймовтршсть пору-
шення структури бшкових молекул (наприклад, пщ час пщвищення температури
ттла, при шфекцп, штоксикацп) концентращя шаперошв у клттин! ртзко зб!ль-
шуеться. Тому так! молекули отримали ще назву стрес-бииав, або б1лк1в теплового
шоку. Порушення складання бшковот молекули призводить до появи хтмтчно
шертних контломераттв, яю вщкладаються в клттиш або поза нею при амшощоз!,
хвороб! Альцгеймера тощо. Школи матрицею може слугувати попередньо складе-
на аналопчна молекула, 1 в цьому раз!, якщо первинне складання вщбулося не-
правильно, вст наступи! молекули також складатимуться дефектно. Така ситуация
виникае при так званих прюнових хворобах (скреп! в овець, коров’ячий сказ,
Куру, хвороба Крейтцфельдта — Якоба в людини), коли дефектно складаеться
один !з мембранних бщюв нервово! юптини з наступним накопиченням шертних
мае усередин! клттини, яю порушують п життедтяльшеть.
Порушення процеств синтезу в клттиш можуть вщбуватися на р!зних його
етапах: транскрипцтя РНК у ядр!, транслящя полптептщцв у рибосомах, шс-
лятрансляцшна модифткащя, пперметилювання та гл!козування бптково! молеку-
ли, транспорт 1 розподцт бщюв у юптинт та виведення !х назови!. За цим можна
спостерйгати збшьшення або зменшення кйлькостй рибосом, розпад полирибосом,
розширення цистерн грубо! ендоплазматично! спки, втрату нею рибосом, виник-
нення везикул й вакуоль. Так, у раз! отруення блйдою поганкою ушкоджуетьс.
фермент РНК-полймераза, що порушуе транскрипцию. Дифтерййний токсин, йна-
ктивуючи фактор елонгацп, порушуе процеси трансляции, що зумовлюе ушкод
ження миокарда. Порушення деяких специфичних бйлкових синтезов можуть спри-
чинювати йнфекцййн! агенти. Наприклад, вйруси герпесно! групи зменшують син-
тез и експреспо молекул антиген!в головного комплексу пстосумйсностй, що дае !?
змогу частково уникнути ймунного контролю, бацили чуми гальмують синтез
медиаторйв гострого запалення. Поява незвичайних бйлкйв може призупиняти Ь
подалыпий розпад ! призводити до накопичення инертного або навпь токсичного
матер!алу. Цьому певною мирою можуть сприяти ! порушення процесйв розпаду.
Порушення процес1в розпаду
Одночасно из синтезом быка в клггин! безперервно вщбуваеться його розпад. У
нормальних умовах це мае велике регуляторне та формоутворювальне значения,
як, наприклад, пщ час активации неактивних форм фермента, бйлкових гормошв
бйлкйв мистичного циклу. Нормалью р!ст й розвиток клпини потребують тонко
контрольованого балансу мйж синтезом й деградацйею бйлкйв та органел. Однак
процес! синтезу б!лк!в унаслщок помилок у робот! синтезуючого апарату, ано-
мального складання бйлково! молекули, и ушкодження хймйчними та бактерйаль-
ними агентами постййно утворюеться досить велика килькйсть дефектних моле-
кул. За деякими оценками их частка становить близько третини вс!х синтезованих
61ЛК1В.
Клпини ссавцйв мають к!лька головних шлях1в руйнування бык1в\ через лйзо-
сомнй протеази, через кальцййзалежнй протешази й через систему протеасом. Кр!м
того, е ще й спец!ал!зован! протешази, наприклад каспази. Основною органелою
де вщбуваеться деградация речовин в евкарйотичних клйтинах, е л!зосома, що мйс-
тить числений гщролйтичн! ферменти. За рахунок процесйв ендоцитозу та р!зних
типйв автофаги в л!зосомах и фаголйзосомах руйнуються як дефекта! бйлковй моле-
кули, так й цйлй органели: ушкоджен! мйтохондри, дйлянки плазматичнои мембра-
ни, деяк! екстрацелюлярнй бйлки, вмйст секреторних гранул.
Важливим механизмом деградации бйлкйв е протеасома — мультикаталпична
протешазна структура складно! будови, що миститься в цитозол!, ядри, на ендо-
плазматичний спщ та мембран! клпини. Ця ферментна система вщповщае за руй-
нування ушкоджених б!лкйв, а також здорових бшкйв, якй мають бути вилученй для
нормального функцйонування клпини. При цьому бйлки, приреченй на руйнуван-
ня, попередньо намйчаються, поеднуючись зй спеиифйчним полипептидом убйкви-
тином. Але частково в протеасомах можуть руйнуватись й неубйквйтованй бйлки.
Процесування бйлково! молекули в протеасомах до коротких полйпептидйв йз на-
ступною их презентацйею в склад! молекули тканиннои сумйсностй I типу е важли-
вою ланкою в здййсненнй !мунного контролю за антигенним гомеостазом организ-
му. У раз! послабления функций протеасом вйдбуваеться акумуляцйя ушкоджених и
непотрибних бшкйв, що супроводжують стариння клйтини. Порушення деградации
циклйнзалежних бйлкйв призводить до порушення клйтинного подйлу, деградация
, екреторних бУлкУв — до розвитку цистофУброзу. I навпаки, пщвищення функгш
.ротеасом супроводжуе виснаження оргашзму (СН1Д, рак).
У випадку генетично зумовлених порушень деградацй бУлкУв оргашзм е не-
мптездатним 1 гине в рашпх стадУях ембрУогенезу. Якщо ж порушення належать
го розпаду жирУв або вуглеводУв, то виникають хвороби накопичення (тезаурисмо-
зи). За ним усерединУ юптини накопичуються незвичш юлькостУ певних речовин
або продуктов гхнього неповного розпаду — лптУшв, полУсахаридУв, що суттево
пкоджуе функции юптини. Найчастпне не спостерУгаеться в ештелУоцитах печш-
ки (гепатоцитах), нейронах, фибробластах У макрофагопитах.
Набуп порушення процесУв розпаду речовин можуть виникати як наслщок
патолопчних процесУв (наприклад, бУлково), жирово’1, вутлеводно‘1 1 пУгментноУ
дисгрофп) та супроводжуватися поя вою незвичних речовин. Порушення в сис-
тем! лУзосомного протеолУзу призводить до зменшення адаптанп гид час голоду-
вання або пщвищеного навантаження, до виникнення деяких ендокринних дис-
функций — зниження рУвня шсулшу, тиреоглобулшу, цитокУнгв та ЁхнУх рецепторов.
Порушення деградаш"1 бУлкУв сповЕгьнюють швидкУсть загоення ран, зумовлюють
виникнення атеросклерозу, впливають на Умунну вУдповщь. При ппоксп, змий
внутрУшньоклУтинного рН, променевому ураженнУ, коли виникае посилена пер-
0КСИДЗЦ1Я мембранних лУшдУв, а також пщ впливом лУзосомотропних речовин —
ендотоксинУв бактерш, метаболтв токсичних грибУв (спорофузарин), криста.пв
.•ксиду кремнУю — змУнюеться стабУльшсть лУзосомноУ мембрани, виыльнюються
в цитоплазму активованУ лгзосомнУ фермента, що с причин юе руйнування струк-
тур юптини та п загибель.
УШКОДЖЕННЯ КЛПННИ ТАН ЗАГИБЕЛЬ
Змши в робоп будь-якоУ органели або функцУУ клУтини не залишаються вщокрем-
леними, вони зазвичай зумовлюють низку пос.тщовних явищ, яю спричинюють
розлад у функцУонуваннУ всУе! юптини. Протс ш змУни не обмежуються лише ви-
никненням порушень. На ргвнУ клУтини залучаються мехашзми 'и захисту вш па-
тогенного впливу, компе нсацп та вщновлення ушкоджень, пристосування до змУ-
нених умов Уснування. Так, у вщповщь на гшоксУю вщбуваються адаптацУйш змУ-
ни в метаболУзмУ, перемикання з окиснення на глУколУз. Проникнення у клУтину
вУрусгв спричинюе вироблення УнтерферонУв та шших речовин, яю гальмуюгь
розмноження ырусних частинок, руйнують чужорщнУ нуклешов) кислота. Збшь-
шення навантаження зумовлюе гшерплазУю та ппертрофУю певних органел, заги-
бель дУлянки цитоплазми супроводжуеться У1 ендоцитозом У перетравленням або
вщгоргненням Уз наступною регенерате ю втрачених структур. Тобто таю основш
ознаки патолопчного процесу, як ушкодження У захист, порушення У компенса-
цУя, можуть проявлятися вже на кл У тинному ргвнУ. Але якщо патогенний вплив
перевищуе компенсаторнУ можливост) ушкодженоТ клУтини або якщо за деяких
спепифУчних обставил подальше в Уснування недодУльне, або не загрожуе цУлУс-
ностУ органгзму, клУтина гине.
РозрУзняють два типи загибелУ юптини. 1) некроз, який виникае внаслщок уш-
кодження клУтини пщ дУею фгзичних, хУмУчних або бУолопчних факторУв, Унтен-
сившсть яких перевищуе захисну спроможшсть клУтини; 2) запрограмована кл1-
тинна загибель у виглщи апоптозу (вщ треп арорюж — опадания листа) або авто-
фагп, коли юггтина тине активно шсля надходження специфичного сигналу та
включения передгснуючих виконуючих мехашзмгв.
Некротична загибель юптини виникае пщ впливом зовнгшнгх ушкоджуваль-
них факторгв. як! можуть специфгчно взаемодгяти з певними структурами абс
процесами в клгтин!. Так. мембрани ушкоджуються шд дтею лшотропних фак-
торгв, токсин макрооргашзму, зминог отрути, активоваяого комплементу, оксида-
пгеного стресу. Ушкодження мембрани зумовлюе зникнення вибгрково! про-
никностг, втрату внутргшньоюп тинного калпо, збагачення юптини натргем, каль-
ц1€м, хлоридами. Це супроводжусться розладом метаболгчних процесгв. накопи-
ченням у клгтинг води (набряк), збътыпенням п об’ему, розривом юптинно! обо-
лонки, ЦИТОЛ13ОМ. Функшя генетичного апарату ушкоджуеться внаслщок юшзую-
чо1 дп випромгнювання. деяких антибютикгв, аналопв нуклегнових основ. вгрусгв
Специфгчного ушкодження також можуть зазнавати й !нш! органощи — л!зосоми,
мгтохондрп, про що йшлося ранппе. При цьому спостерггаеться пщвишення ПОЛ
унаслщок надмгрного утворення первинних выьних радикал гв або порушення
функшонування антиоксидантних систем, активацп мембранних фосфолшаз.
Порушуеться дгяльшсть йонних канапв, зокрема кальшевих, 1 кальщевих насоав,
що призводить до пщвишення концентрацп йонгв кальцгю в цитоплазм!, розладу
багатьох функцгй. Наколичення йонгв водню спричинюе розвиток внутршшь-
оклгтинного ацидозу. Порушуеться вироблення енергп мпохондргями, п ьико-
ристання для специфтчних функщй. Руйнування оболонки лгзосом гз вивгльнен-
ням активованих лгтичних ферменпв призводить до автолгзу юптини. За тако!
“насильницьког” загибел! юптини в середовище, що !Т оточуе. вивгльнюеться без-
лгч БАР (ейкозаногдгв. цитокгнгв, кшшгв), як! сигналгзують про небезпеку в ор-
гашзм! й активують дгяльшсть захисних мехашзмгв (наприклад фагоштпв), як!
спричипюють розвиток запалення.
€ й 1НШИЙ механизм юптинно! смерт! — це запрограмована загибель юптини.
або апоптоз та автофапя. В усгх юптин, що мають ядро, наявний г мехашзм само-
губства. На ещмгну вщ некрозу юптини, у раз! апоптозу не вщбуваеться така акти-
вапгя фагоцитгв г юптин гмунног системи. яка була б здатна спричинити за пальну
реакции або !мунну вщповгль. .Апоптоз вщ!грае важливу роль у нормальнш жит-
тед!яльност! оргашзму, бере участь у формоутворенш оргашзму, регулюючи к!ль-
кгсний 1 як’.сний склад його юптин, вилучаючи нежиттездатш або потенцшно
небезпечш клттини. Його можна згншгювати за допомогою кьтькох специфтчних
сигналгв. як! надходять ззовнг у вигляд! таких молекул, як фактор некрозу пухлин
та лея кг пыл питою ни, шо сполучаються зг спещальними рецепторами на повер-
хнг юптини. Примусити юптину до самогубсзва можуть також спецгалгзоваш лгм-
фоцити — натуральш кьгери або нгунн цитотоксичш лгмфоцити. Вони влорску-
ють у юйтину-мгшень гранули з ферментами, що розпочинають апоптоз.
Усередин! юптини розпочати апоптоз здатн! фактори, що м!стяться м!ж зов
ншшьою та внутргшньою мембранами мгтохондрш, зокрема питохром С, пши
фактори, якг шдукують апоптоз г видгляються в цитоплазму шсля ушкодження
мгтохондрп, а також порушення юйтинного циклу, яке спостерггаеться пщ час
ушкодження молекули ДНК ! включения системи контролю за проходженням
юйтинного циклу (наприклад б!лок р53). При цьому послщовно активуеться
складна ферментна система, каспазний каскад, який зрештою сприяе активаш"!
ядерних ендонуклеаз, швидкёй фрагментацёё ДНК, руйнуванню багатьох важли-
аих ферментёв синтезу, активащё лётичних ферментёв, перемещению назови! мем-
брани фосфатидилсерину, що зумовлюе не тёльки майже миттеве припинення
всёх процесёв життедёяльностё клётини, а й швидке и поглинання макрофагами з
-аступним перетравлюванням усерединё цих клётин.
Одним ёз рёзновидёв запрограмовано'1 клётинноё загибелё е автофапя — конт-
рольована деградацёя цитоплазматачних бёлкёв, органел ё самих клётин за участю
активацп лёзосомального апарату. Вона виникае за умов голодування (харчовоё
депривацц) як форма компенсаци, що спрямована на виживання клётини та ор-
ганёзму в целому, а також унаслёдок дн рёзноманётних патогенних факторёв (анта-
бёотик рапамёцин, вёруси герпесу, внутрёшньоклётиннё паразита), а також при
реперфузёё пёсля ёшемп мёокарда.
Отже, клётана, “приречена” на загибелъ, досить швидко й ефективно вилу-
чаеться з кругообёгу в оргашзмё. Це мае велике значения в разё п злоякёсноё
трансформацп або ёнфёкування вёрусами, що в бёльшостё випадкёв запобёгае реп-
лёкацп вёрусёв пёд час ёнфекцн. Запрограмована загибелъ клётини е тонко вёдрегу-
льованим процесом, який окрём проапоптозних сигналёв мае ще й потужну анта-
апоптозну систему, ё тому остаточне “вирёшення долё” клётини залежить вёд бага-
тьох сукупно дёючих факторёв.
Акгивацёя апоптозу спостерёгаеться пёсля ёошзуючого опромёнення, гёпоксёё,
пёд дёею продуктёв життедёяльностё бактерёй (наприклад суперантигенёв стафёло-
кокёв або В1Л, якё спричинюють апоптоз в ёмунокомпетентних клётинах ё тим
самим обеззброюють органёзм) тощо. Проте в деяких випадках апоптоз може
гальмуватася Наприклад, деякё вёруси здатнё блокувати апоптоз в ёнфёкованих
клётинах, забезпечуючи тим самим умови для свое! реплёкащё. ёнактивашя про-
апоптозного сигналу внаслёдок соматачноё мутацц гена бёлка р53 або нерегульо-
вана активацёя антиапоптозного сигналу (гена Вс1-2) спостерёгаеться в бёльшостё
злоякёсних клётин.
Варто зазначита, що тривале невикористання якоё-небудь групи клётин зумо-
влюе зменшення ёх кёлькостё шляхом апоптозу (“смерть вёд зневаги”) ё, навпаки,
функцёональне завантаження сприяе ёх збереженню. Це правило “вживай, бо
втратиш” стосуеться багатьох функцёонуючих клётинних систем, у тому числё й
клётин ёмунноё системи та головного мозку.
Розд|д V
РОЛЬ СПАДК0В0СТ1, КОНСТИТУЦП
ТА В1КОВИХ ЗМ1Н У ПАТОЛОГИ
Окргм патогенних факторгв навколишнього серсдовища у розвитку хвороб люди-
ни значну роль вщ1грають генетичнг фактори (спадковгсть г конституция).
У мгру того як удосконалюються метода лгкування та профилактики шфекшй-
них хвороб, структура захворюваносп населения змшюеться, г все бгльшого зна-
чения набувають хвороби, яю виникають пщ впливом генетичних факторгв. Ниш
вщ спадково! патологи помирають бгльше людей, шж вщ шфекщй; нею зумовле-
но 40 % ранньо! смертностг та гнвалщностг з дитинства, у 4 % дгтей виявляють
генетичнг дефекта.
Хвороби, причиною яких е генетичнг фактори, називають генетично детерлй-
нованими, Часто гх помилково називають спадковими, але такими е тгльки части-
ца з них. Це залежить вщ розмгру ушкодження генетичного апарату, типу успад-
кувашгя, а також вщ здатностг носгя генетично! аномали вщтворювати потомство.
Багато генетичних аномалгй не успадковуеться, оскгльки супроводжуеться без-
плщнгстю носгя. Кргм того, носгй патолопчного гена не завжда хворге завдяки
механгзмам компенсацй генетично! аномали. Якшо хвороби, зумовлеш порушен -
ням генетичного апарату, передаються в поколгннях, вони отримують назву спад-
кових хвороб. Близьким поняттям е вродженг хвороби, до яких належать хвороби,
що проявлятоться одразу пгсля народження. Своею чертою останнг можуть бута
генетично детермгнованими або не пов’язаними з генетичними порушеннями
(наприклад акушерська патолопя, а також токсоплазмоз або сифшс у раз! внут-
ргшньоутробного гнфгкування плода). Щодо часу виявлення суто спадкових хво-
роб, то вони можуть бута чи не бути вродженими, тобто гнодг виявляються одра-
зу шсля народження (дефекта фгзичного розвитку, скелетнг аномали, наприклад
полщактилгя, муковгсцидоз), а гнодг пгзниие (хорея Гентгнгтона проявляеться у 30—
35 рокгв, подагра — у похилому вщг).
Хвороби, розвиток яких визначаеться факторами навколишнього середови-
ща, але клгнгчна картина яких гдентична вщомим спадковим хворобам, називають
фенокошями.
ГЕНЕТИЧНО ДЕТЕРМ1НОВАН1 ХВОРОБИ
Класификащя
Класификацгя генетично детермгнованих хвороб Грунтуеться на кглькох принципах.
За розмграми ушкодження генетичного апарату статевих клггин хвороби подг-
ляють на молекулярно-генш г хромосомы. Своею чертою пергш подгляють на моно-
гены та пол1генн1 (зумовлеш вщповгдно одним або кглькома генами). Моногенш
хвороби класифгкують залежно вщ тапу успадкування на домтантш, рецесивш та
>чеплет з1 статтю. Полпенн! захворювання називають мультифакторгальними.
Кр!м того, е анатомо-топографгчна (органно-системна) класифгкашя спадкових
захворювань (хвороби кров!, нирок, опорно-рухового апарату тощо). Класифпса-
шя, заснована на типових порушеннях обмшу речовин, корисна для медично!
практики: за субстратним принципом розрхзняють хвороби вугл вводного, жирово-
го, бшкового обмшу, амшокислотного, пуринового метаболгзму, а також в!там!-
гюзалежш захворювання 1 хвороби, зумовлеш порушенням обмшу металзв, тощо.
Етюлопя
Матер!альною основою генетично детермшованих хвороб е мутац1я в статевих
юптинах. Однак мутащю не можна назвати етюлопею або етюлопчним факто-
ром, виходячи 1з загальноприйнято'1 концепцй цих понять. Особливгстю спадко-
вих хвороб е те, що етюлопя, патогенез 1 юпшчш прояви роздшеш в поколшнях.
Тобто д!я етюлопчного фактора 1 первкже ушкодження генетичного апарату (му-
тащя) вщбуваються в одше! людини, а хворпоть п нащадки за умови, якщо вони
отримали мутацпо у спадок.
Етюлопчними факторами, яю зумовлюють мутаццо, е екзогенн! ф!зичн!,
Х1М1ЧН1 та бюлопчш фактори за умови, що вони здатш змшити генетичний апарат
статевих клггин. 1х називають мутагенами. Х!м!чш мутагени можуть бути ендо-
генними, наприклад активн! киснев! радикали.
Серед фгзичних мутагенов найважлившшм е юшзуюче випромшювання, яке
безпосередньо або через продукт радюл!зу ушкоджуе генетичний апарат. 1нод!
мутащю зумовлюе випромпповання в таюй мппмальюй доз!, яка не спричинюе
променеве ураження оргашзму.
Серед х1М1чних мутагенов найсильншгими е аналоги пуринових 1 шримщино-
вих основ, помилкове включения яких у полшуклеотидний ланцюг призводить до
спотворення триплетного генетичного коду, а також алюлуюч! (шрит, що мети-
люе азотист! основи) та дезамгнуюч! (нггритна (азотиста) кислота, речовини, що
в процес! метаболизму можуть перетворюватися на штрити, шгрозамши) агенти.
Серед х!м!чних шюдливостей, яю постхйно накопичуються в навколишньому се-
редовипц, е багато х!м!чних мутагешв (сшъськогосподарських — пестициди, гер-
бщиди; виробничих — епоксидю смоли, фенол, формальдегщ; харчових — аце-
тальдепд, який використовують у виробництв! консерванпв; побутових тощо). 1з
вихлопними газами автомобшв у повпря надходять ароматичш вуглеводш, сви-
нець, оксид азоту. Роль хзмхчних мутагенгв можуть вщиравати лгкарськ! засоби,
особливо цитостатики (шпбгтори синтезу ДНК, похщш фол!ево! кислота, анало-
ги пуринових 1 шримщинових основ та !х похщш — бромурацил, амшопурин).
Небезпеку становлять х!м!чн! сполуки, як!, не будучи мутагенами, п!д д!ею фак-
тор!в навколишнього середовища або включившись у метабол!зм перетворюються
на них. Наприклад, ртуть пщ впливом мгкрооргашзмгв може перетворюватися на
мутаген метил- чи диметилртуть; немутагенш виробнич! в!дходи внаслщок взае-
модп 13 солями важких меташв можуть утворювата мутагенш комплекси; цикло-
фосфамщ, який використовують як цитостатик, не е мутагеном, але в оргашзм!
перетворюеться на високомутагенну речовину.
До биэлоггчних мутагешв належать в!руси шфекщйного мононуклеозу, в!спи,
гепатиту, кору, краснухи, еп!дем!чного паротиту. Так, у ваптних, яю перехворши
на краснуху чи вёрусний гепатит, трапляються мимовёльш викиднё, а в клггинах
плода виявляють хромосомш аберацп. У потомства цих жёнок частёше вщ шших
спостерёгають хромосомш хвороби. Продукти житгедёялъностё деяких патогенних
грибёв (афлотоксин) можуть також бути мутагенами.
Изю мутагени спроможнё потенцповати дёю один одного, тобто доли синерпчно
Екзогенш мутагени здатш шдукувати утворення ендогенних мутагенёв — актив-
них форм кисню, вшьних радикалёв, радёотоксинёв тощо.
Патогенез
Первинною та головною ланкою патогенезу генетичноё' патологи е мутащя —
стёйке стрипкоподёбне порушення генетичного апарату, не пов’язане з подёлом
клётини або звичайною рекомбёнаиёею хромосом. Близьким до мутацп поняттям
е однонуклеотидш полёморфёзми (алелью варианта генёв). Ниш описано десятки
алельних варёантёв певних гешв.
Особливёсть патогенезу полягае в тому, що мутацёя вщбуваеться в однёе'ё лю-
дини (у батькёв), яка залишаеться здоровою, а наступи! ланки патогенезу та
клёнёчнё прояви розгортаються в нащадкёв.
Розрёзняють калька видёв мутацп, як! виникають пщ впливом мутагенёв.
Соматична мутащя виникае в соматичних клётинах ё зникае з популяцп зё
смертю носёя, не передаючись у поколениях (цю мутацёю в цьому роздёлё пщруч-
ника не розглянуто).
Гермшативна (гаметична) мутащя виникае в статевих клётинах ё впливае на
спадковёсть потомства. Мутащя може бути корисною (як фактор еволюци) та шклд-
ливою. За причинами виникнення розрезняють спонтанну (спричинену певними
природними факторами) та тдуковану (зумовлену мутагенами) мутацию. Залежно
вщ розмёрёв ушкодження генетичного апарату розрёзняють генну та хромосомну
мутацёю.
Генна мутащя — це порушення триплетного генетичного коду молекули ДНК.
В основ! н — порушення специфёчноё послёдовностё пуринових ё пёримёдинових
основ (точкова генна мутацёя, що серед генних мутацёй становить 95 %). Рёдше
спостерёгаються структурнё змёни молекули ДНК — внутрёшньогенна делещя
(вёдрив ё втрата частили гена), ёнверсёя, ёнсерцёя (вставки), транслокацёя, дуплёка-
цёя. Спонтанш мутацёё' виникають унаслёдок дп ще недостатньо вивчених фак-
торёв ендогенного характеру (похибок у роботё генетичного апарату гцд час реалё-
зацёё генетичноё ёнформацёё). Частота ёх низька порёвняно з ёндукованими мута-
цёями. Мутацёя може стосуватися як структурних, так ё регуляторних генёв. Генна
мутацёя може бути притаманна будь-якому гену (домёнантному, рецесивному,
зчепленому зё статевою хромосомою).
Мётохондрёальш тени теж можуть мутувати, причому частота мутацёй у них
вища, нёж у ядерних генёв, якё також кодують ферменти окисного фосфорилю-
вання.
Хромосомна мутащя — тяжче порушення спадкового апарату. Вона полягае в
змёнё або структури хромосом (структурна аномалёя або хромосомна аберацёя —
делецёя, транслокацёя, транспозицёя, ёнверсёя, дуплёкацёя; хромосомна нитка може
розёрватися в одному чи кёлькох мёсцях, н сегмента можуть втратитися чи непра-
вильно з’еднатися), або ёх кёлькостё (як наслёдок — нерозходження гомологёчних
хромосом у гаметогенез!). Делены призводить до втрати хромосомного матер!алу
та спричинюе загрозлившй наслщки, нгж транслокашя й шверстя. У виникненш
деяких хромосомних аберащй мають значения не одна, а илька первинних подгй
(для формування деяких транслокацгй необхщне майже одночасне виникнення
двох делецш з наступним об’еднанням розгрваних частин).
Завдяки вдосконаленню методов диференщального забарвлення спадкового
матер!алу меж! м!ж генними 1 структурними хромосомними мутащями почали
звужуватися, однак поды мутащй на гешл 1 хромосоми! все-таки залишаеться
корисним для практики.
Антимутащйн! мехашзми. В оргашзм! е багато мехашзмгв, як! запобггають по-
яв! мутацп, вщновлюють мутантний ген, перешкоджають його реалгзацц або ком-
пенсують спричинеш ним порушення.
На молекулярному ргвш пристосування полягае в шактивацц ендогенних му-
тагешв (наприклад активних форм кисню) за допомогою природних антиокси-
дантных систем.
Генетичному апарату притаманна певна надгйшсть. Не кожна замша азотис-
то! основи в молекул! ДНК зумовить помилку в раз! ц редушпкапп. Подвгйний
характер сшрал! ДНК — один 1з фактор! в ще! надшност!, осюльки в раз! однонит-
кового ушкодження молекули ДНК вщновлення вщбуваеться за матрицею пшюго
ланцюга. Кр!м того, у клиин! е система ферменпйв репарацп ушкоджено! ДНК
(див. мал. 3), як! розшзнають дефект, вир!зають цей фрагмент (ендонуклеазою),
розщеплюють його (екзонуклеазою), синтезують нормальний фрагмент (пол!ме-
разою) та вшивають його (лиазою). Цей захисний мехашзм вщновлюе близько 95 %
спонтанних мутащй.
Оск!льки кодуюч! послщовност! ДНК становлять не быьше шж 10 % геному
(вважають, що навпь 2—5 %), а основна частина ДНК не несе шформащю про
амшокислотну послщовшсть б!лк!в 1 не кодуе структури рибосомних та шших
типгв РНК, мутацц в цих частиках геному, вочевидь, “нейтральш”.
Так! груб! порушення генетичного апарату, як хромосоми! аберацп, не репа-
руються, тобто морфолопчно не вщновлюються. Однак шод! !х юнування яки-
мось чином компенсуеться. Наприклад, збалансованими шод! бувають хромосом-
на транслокацц, можливо, тому що наб!р генетичного материалу все-таки повний
(а не дефщитний).
На вщмгну вщ мутантних соматичних клггин, проти яких е !мушп мехашзми
захисту (елгмшащя аномальних быкгв за допомогою антиты, а мутантних — шля-
хом цитол!зу 1 фагоцитозу), мутантш статев! клггини не пщпадають пщ дгю цих
захисних мехашзмгв. Але, якщо вони виживають, то часто втрачають здатшеть до
заплщнення. Кр!м того, з вгком репродуктивна функщя людини ослаблюеться,
статев! юптини рщше заплщнюються г чимало хромосомних аберащй у статевих
клггинах, частота яких з в!ком збшыпуеться, залишаеться без наслщкгв.
Наступи! захисш мехашзми розгортаються вже у нащадкгв, як! отримали му-
гащю у спадок.
кнують генетичш мехашзми компенсаци. Так, якщо мутантний ген у статевш
клггин! е рецесивним, то в нащадкгв у гетерозигот! за наявност! нормального
алельного гена мутащя фенотипово не проявляется (нормальний ген домгнуе
над патолопчним алелем). 1нод! гетерозиготи навгть отримують селективну пере-
вагу (носп гена серпоподгбностг еритроцитгв стшкг проти маляр!!). Якщо мутант-
ний ген локализований у Х-хромосомо, яка в людини вщповщае за багато патоло-
пчних генетичних ознак, болышсть оз яких рецесивно (наприклад ген гемофол!!'),
то прояви залежатимуть вщ стат! носоя: жонки будуть здоровими, у них наявшсть
Х-хромосоми з ушкодженим геном компенсуватиметься шшою нормальною X-
хромосомою.
Метаболизм теж мае певний запас мщносто. Не кожне порушення первинноо
структури болкових молекул призводить до порушення Гх конформаип, тому що
вони пластично (нимало мутацш генов гемоглобшу зовсом не мае шкщливих на-
слщков). Функцоя быка може зберйатися ! в раз! зниження його концентрацп
(шодо достатньо 50 % активност! ферменту). Нимало функцш в оргашзм! забезпе-
чуеться розними метаболочними шляхами, тому при ушкодженш одного з них
компенсащя може вщбуватися за рахунок шших.
На р!вн1 цолого оргашзму оснують так! пристосувальш феномени, як пене-
трантшстъ (омоворшсть фенотипового прояву гена) та експресившстъ (ступшь
клшочного прояву) гена Вони залежать як в!д генетичних фактор!в, так о вщ фак-
торов навколишнього середовища. Пенетрантность ноколи не бувае 100 %. Навоть
домшантним ознакам властив! неповна пенетрантность о вароабельна експресив-
ность. Тнодо експресоя обмежена ровнем статевих гормонов (наприклад, реалозацоя
генов, водповщальних за облисшня, залежить вщ ровня андрогенов, тому вона об-
межена в жшок). Тривае дискусоя щодо видолення класу так званих диналйчних
мутацш, яким властива вароабельна пенетрантность у поеднанн! з неповним дом!-
нуванням. Для деяких хвороб, спричинених саме такими мутацоями, характерне
наростаногя тяжкосто хвороби в поколшнях (наприклад синдром ламкоо Х-хромо-
соми, моотоночна дистрофоя, нейродегенеративш хвороби, хорея Гентонгтона).
Популяцоя не пщтримуе носив шкщливих мутацш. Не виживають або не да-
ють потомства болышсть носив хромосомних аберацш о шкщливих домшантних
генов (природний в1дб1р). На цьому грунтуеться о традицоя обмеження шлюбов серед
кровних родич!в.
Подалыш ланки патогенезу залежать вщ виду мутаци — генна вона чи хромо-
сомна. На цш подстав! розрозняють дво групи хвороб — молекулярно-генно (власне
спадково) о хромосомно.
Молекулярно-генн! (спадков!) хвороби
Оскольки генна мутацоя поровняно з хромосомною стосуеться невелико! частили
генетичного матероалу, то и носой найчастгше зберогае репродуктивну функцою.
Тому молекулярно-генно хвороби можуть передаватися в поколшнях о е спадко-
вими.
Спадкова хвороба — це хвороба, зумовлена мутаоиею (частице генною) у ста-
тевш клггин! о може передаватися в поколшнях.
Термгни, яко означають таку патологпо, зазвичай мають закшчення -пагшя:
ферментопатоя, гемоглобшопатоя, мембранопатоя, тромбоцитопатоя тощо.
У носоя, який отримав генну мутащю у спадок, розвиваеться ланцюг молекуляр-
них, клотинних г системних порушень, що становлять патогенез спадковоо хвороби.
Порушення на молекулярному (бюхгмгчному) р1вш. Мутацоя структурного гена
призводить до порушення структури болковоо молекули, тобто послщовносто ам!-
юкислот у полшептидних ланцюгах бтлка без змши його юлькостт. Мутащя регу-
ляторного гена спричинюе змгни юлькостт бика без змши його структури.
Бюхтмтчю наслщки мутацп залежать вщ функцп того битка, синтез якого по-
пущений.
Якщо це ферментопапйя (ензимопапия), то порушення навтть одного ферменту
може клтшчно проявится недоумством, хворобами системи кровт, ендокринно!
та шших систем. Ферментопатй лежать в основ! багатьох хвороб, але для тх
гцагностики завжди необхщне бтохтмтчне пщтвердження. Так, якщо синтез фер-
менту повнтстю вщсутшй, то за допомогою тмунологтчних метод! в цей биюк (фер-
мент) не виявляють. Може порушуватися швидктсть синтезу ферменту, т тодт
зменшиться його юльюсть. Осюльки синтез битковот молекули часттше кодуеться
не одним, а китькома генами, то порушення структури певного ферменту часттше
детермшоване мутащею ртзних гентв.
Наслщком порушення структури молекули ферменту можуть бути зниження
и стабшьностт, протеолтз т руйнування змшеного мутащею ферменту.
Каталттична актившсть ферменту може бути зниженою чи вщсутньою навтть
за нормально!' його юлькостт. Це може бути наслщком вщсутностт активатортв або
надлишку тнпбттортв ферменту, вщсутностт рецептортв до нього, порушення то-
пографп розподиту ферменту в клттин!, порушення його каталггичного центру. В
останньому випадку, якщо порушена дитянка ферменту, призначена для його
взаемодп з шшими молекулами, то порушуеться вшзнавання ферменту або його
спорщнентсть до субстрату.
У гомозиготного ностя мутантного рецесивного гена актившсть ферменту ртз-
ко знижена, гнодт повнтстю вщсутня, у гетерозиготного — зниження активностт
менше. В останьому випадку ферментопаття виявляеться титьки пщ час наванта-
ження харчовими, виробничими, середовищними факторами чи лткарськими за-
собами т проявляеться як спадкова схильшсть до хвороби (див. нижче).
Окртм зниження активностт ферменту може спостертгатися тт пщвищення (у
разт пригнтчення шпбування), ! тодт може виникнути неконтрольований синтез
кшцевих продукттв (наприклад при подагр!).
Кртм того що клштчш прояви ензимопатн залежать вщ зниження або пщви-
щення активностт ферменту, вирппальне значения мае також приналежнтсть ен-
зимопатп до метаболтчних шляхтв: анаболтзму чи катаболтзму.
У разт пщвищення активностт ферменттв анаболизму збитыпуеться синтез ктн-
цевих продукттв. I навпаки, при блокуваннт анаболтчних шляхтв порушуеться бт-
локсинтетичний процес, що зумовлюе дефщит ктнцевого продукту т накопичення
субстрату-попередника (так, дефщит глткогенсинтетази в печшцт спричинюе
зменшення вмтсту глткогену в печшцт т накопичення глюкози в кровт).
У разт дефициту ферменттв катаботзму вихщний субстрат накопичуеться, що
спричинюе розвиток так званих хвороб накопичення. До них належать гшерхолес-
теринемтя, гшертриглщеридемтя, мукополтсахаридоз, муковтсцидоз. Наприклад,
при дефщит! в печшш фосфорилази розвиваеться хвороба Прке (глткогеноз), що
проявляеться збшьшенням розмтртв печшки т вмтсту в шй глткогену. До цте! само!
категорп хвороб належить накопичення металовмтсних сполук унаслщок пору-
шення тх транспорту за допомогою бшкових переносниюв (наприклад, вщкладен-
ня гемосидерину в тканинах).
Якщо взяти до уваги, шо в оргашзм! людини мктиться близько 10 000 фер-
ментов, то 200 ферментопатш, як! вже описано, становлять 2 %. Це означае, що
напп знания про спадков! порушення метабол!зму недостатш. Краше вивчено де-
фекти катабол1чних шлях!в, шж анабол!чних. Можливо, дефщит фермента ос-
новних анаболгчних шлямв зумовлюе передчасну смерть.
Патогенетичною основою мггохондрхальних хвороб е порушення в систем!
окисного фосфорилювання. Оскшьки зигота отримуе мпохондрп в!д яйцеклгги-
ни, то цим хворобам притаманний материнський тип успадкування, а також гпз-
нш початок 1 кумулятивний ефект.
Актуальним аспектом зазначено! проблеми е порушення ферментно! системи
репарацп ушкоджено! ДНК, яка зумовлюе чутливкть оргашзму до мутагешв. До-
ведено, що патолопя саме ще! ферментно! системи е важливою ланкою патогене-
зу злояюсного росту 1 передчасного старшня. До ще! патологи належить синдром
спонтанно! хромосомно! нестабьтьност!, який проявляеться грубими хромосом-
ними аберащями 1 добре вивчений у препаратах метафазних пластинок культури
л1мфоцит1в пащента. I хоча щ порушення рееструють пщ час вивчення культури
клггин пащента в раз! !хньо! фпогемаглютиншово! стимуляцп т уНго, вважаютъ,
що нестшюсть хромосом спостерйаеться т у1уо. Такий хворий бшьш схильний до
раку.
У контекст! теми, що розглядаеться, варто зауважити, що метабол!зм бшь-
шост! стороншх речовин, як! потрапляють в оргашзм (продукта харчування, рос-
лин, шгаляцшних речовин тощо), перебувае пщ генетичним контролем актив-
ност! фермента. Розщеплення 1 перетворення цих речовин в оргашзм! може бути
аномальним з утворенням токсичних пром!жних продукта (така непереносимкть
речовин шод! помилково розглядаеться як алерпя, але в основ! лежать не гмунн!
мехашзми, а ферментопатп). Ця група порушень вивчаеться екогенетикою. Особ-
ливо це актуально щодо л!карських препарата, перюд шврозпаду яких в оргашз-
М1 також визначаеться генетичними факторами 1 в 2,5 % населения в!н утрич!
менший вщ середнього. Перюд п!врозпаду настшьки шдивщуальний, що у вщ-
повщь на однакову дозу лгкарсько! речовини в одних пащента терапевтичний
ефект може не виявлятися, в шших ос!б виникають побгчш д!1 1 розвиваються
токсичш ускладнення (фенацетин спричинюе метгемоглобшем!ю, каптоприл —
агранулоцитоз). Ус! щ проблеми визначили виникнення окремого наукового на-
пряму — фармакогенетики.
Гемоглобгнопапйя — наслщок мутацп гена, вщповщального за синтез гемоглобг-
ну, коагулопатия — генетично детермшований дефщит факторов гемостазу (див.
“Патолопчна ф!зюлопя системи кров!”), 1мунодефщити — генетично детермшо-
ваш порушення гмунно! вщповцц (див. “Патолопчна фгзюлопя гмунно! системи”),
мембранопаття — генетичний дефект б!лкових молекул клпинних мембран (на-
приклад, монотонна гшерхолестеринемгя, що виникае внаслщок порушення робо-
ти рецепторгв лшопротещгв з низькою щшьшстю; муковйцидоз, при якому пору-
шене утворення трансмембранного регулятора хлорних канал!в, що регулюють
транспорт юшв хлору через ашкальн! мембрани ештелгальних кл!тин).
Втам1нозалежн1 спадковг хвороби — генетично зумовлеш порушення обм!ну
того чи того вггамшу. Так, у раз! порушення обмшу памшу, шридоксину, ща-
нокобаламшу (вггамш В12), бютину (вггамш Н), фолгево! кислоти (вггамш В9)
эиникають порушення функщй нервово! системи, при дефщит! кальциферолов
зотамш В) — костково! тканини; фыохшонов (вггамши групи К), щанокобаламо-
ну — розвиваеться патолопя кров!; при ав!тамшозах А та Е виникають мембра-
нопатп.
Можливий генетично детермшованцй дефщит транспортних бшков (при де-
фщит! трансферину — переносника залоза — розвиваеться залозодефщитна
1нем!я), а також молекул пептидних гормонов (наприклад гшотиреоз як насл!док
недостатнього синтезу гормону шитоподобно'! залози).
Порушення на клгтинному р1вн1 при спадкових хворобах проявляються змшою
форми ! функщй клотини. Прикладом можуть бути серпо-, сферо- та мппене-
подобга еритроцити, що утворюються в раз! спадково! анеми. Тромбоцитопатия —
це порушення функцн тромбоцитов без змши охю>оо осолькосто. Описано спадков!
дефекти р!зних частин о органощов юптин — мембран (дефщит спектрину в мем-
бран! еритроощтов спричинюе порушення Г! еластичносп о перетворення еритро-
цитов у мгкросфероцити при мокро сфероцитарной гемолотичнш анемп), специфоч-
них гранул (у нейтрофшов), внутршшьоклотинних месенджергв (при псевдогшопа-
ратиреозо), рецептор1в (рецепторно хвороби, наприклад моногенна гшерхолестери-
немоя, що виникае внаслщок патологп рецепторов ендотелпо судин для лшопро-
тещов низькоо щщьносто, в результат! чого порушуеться клотинний ендоцитоз о не
досягаеться нормальна швидкость виведення холестерину з кров!).
Порушення на р1вш всього оргашзму (тобто фенотипочга прояви мутацп) зале-
жать вщ характеру метаболочних порушень, а також типу клотин, структура о функ-
ция яких порушено. Грубо генетично аномалн настольки сутгево змонюють фенотип,
що репродуктивна здатгасть носпв генетично! аномалп зменшуеться або припи-
няеться. Загибель носоя може статися внутрошньоутробно (призведе до мимоволь-
ного викидня), а загибель зигот до настання ох омплантацп частппе вщбуваеться
непомотно. Больошсть емброонов з трисомоями за 13-ю о 18-ю парами хромосом ги-
нуть у рангах стадоях розвитку. Якщо ж мутацоя сумосна з постнатальним оснуван-
ням, розвиваються хвороби як у носоя генетично! аномал!!, так о у його потомства
за збереження репродуктивно! функцп.
На популяцшному ргвш мутантно гени накопичуються (генетичний вантаж по-
пуляцп), якщо хворо зо спадковими хворобами виживають о вщтворюють потомс-
тво. Це стосуеться в основному молекулярно-генних скелетних о стоматолопчних
аномалой, а також розумово! недостатносто. Вважають, що майже кожна людина е
гетерозиготною за колькома генами, летальооими в гомозиготному стага гена. У
популяцп виникають ново шкодливо мутацп пщ впливом юшзуючого випромшю-
вання, шших мутагенних факторов навколиооонього середовища. Теоретично оснуе
критичний ргвень генетичного вантажу як мехашзм виродження популяцп. Вели-
ке значения в накопиченно мутантних генов мае кровне спорщнеогня батьков (вони
мають пщвищену ймоворшсть наявносп однакових рецесивних патолопчних генов
у гетерозиготному стано). Деяко спадково хвороби описано тольки в популяцоях
невеликого розмору (озолятах).
Розвиток спадкових хвороб залежить вщ типу успадкуваогня, тобто мае зна-
чения, який вид гена (домшантний або рецесивний) зазнав мутацп, чи е мутацоя
23 пари (статевих) хромосом, а також те, одним чи групою генов детермоновано
патологпо.
Мал. 4. Пол шакти Л1я
Хвороби з домшантним типом ус-
падкування. Мутащя домшантних ге-
Н1в виявляеться як у гомо-, так 1 в
гетерозиготному стаж. Виживають 1
успадковують патолопю носи тих
домшантних мутантних гешв, яю
сутгево не порушують житгездат-
н1сть, не перешкоджають розмно-
женню, а отже, мало пщпадають п!д
природний в1дб1р. Домшантш генн!
мутацп призводять до добре вщомих
фенотишв. Це найчастппе скелетш
аномалп: багатопалгсть (мал. 4), ко-
роткопал1сть, зротцення чи викрив-
лення пальц!в, аномал п зуб!в, нап-
риклад, в!дсутн!сть б1чних р!зщв, а
також далекозорють, короткозорють,
астигматизм, отосклероз, ахондро-
плазия (карликовий зр!ст 1 непропор-
щйна будова тша).
За автосомм о-домшантним ти-
пом успадковуються вроджена ката-
ракта, ПОЛ1ЮСТОЗ нирки, деяю фор-
ми м’язово! атрофи, прогресуюча хорея Гентшгтона (дегенеративш зм!ни нерво-
вих кл1тин у базальних ганппях), множинний полшоз товсто! кишки (мае
тенденцпо до малптазаци), нейроф1броматоз (хвороба Реклштгаузена). Быышсть
13 цих тяжких хвороб е наслщком мутацп де поуо у батьюв, у яких немае ц!е! па-
тологи. Моногенн! хвороби з частковим (неповним) домшуванням виявляються у
гомо- 1 гетерозигот, але тяжч! в гомозигот.
Хвороби з рецесивним типом успадкування. Якщо мутантний ген е репесивним,
то хвороба розвиваеться тйгьки в гомозиготному стан! гена, тобто тод!, коли ди-
тина отримуе патолопчний ген в!д обох батьюв, яю, будучи гетерозиготними
носхями ознаки, лишаються здоровими. Так успадковуються ферментопатп. До
ще! ж групи хвороб належать вроджена глухошмота, пггментний ретишт, микро-
цефалия.
Хвороби з успадкуванням, зчепленим 31 статевою хромосомою. Захворювашсть
носив мутантних гешв в X- або У-хромосом! залежить в!д стаи пащента. Мутащя
в У-хромосом! передаеться по чолов!чш лши незалежно вщ того, домшантний ген
чи рецесивний. Якщо мутац!я вщбулася в ген!, локал!зованому в Х-хромосом!, то
мае значения, домшантний вш чи рецесивний. Якщо вш домшантний (напри-
клад ген гшофосфатем!чного рахпу, який не пщдаеться л!куванню в!тамшом О2),
то проявляеться патолопею як у чоловжгв, так 1 в жшок у гетерозиготному стан!
гена. Якщо вш рецесивний (при гемофш! А, дальтошзм!, юнацьюй глауком!, ге-
мералопп — вщсутност! сутшкового зору), то можуть хворгти пльки чолов!ки,
осюльки мають тшьки одну Х-хромосому. Щодо ж! но к (носи двох Х-хромосом),
то гетерозиготю жшки залишаються здоровими 1 водночас е джерелом подальшо’1
передач! гена потомству чоловзчо! стат! (жшки можуть хворгги ильки в гомозигот-
ному сташ мутантного гена, що бувае рщко).
Хвороби з аолшенним зилом успадкування. Оскгльки бглышсть ознак в оргашз-
М1 детермшована не одним, а кьтькома генами, серед яких можуть бути домшан-
тн!, рецесивш. автосомш та зчеплеш зг стапю, то це зумовлюе розвиток хвороб 13
полпенним типом успадкування. Для деяких випадкгв групи них генгв точно виз-
начеш (так, розвиток подагри детермшуеться двома генами, як! розташоваш в
Х-хромосом1; схильшсть до алкоголизму детермшована генами ферменту алко-
гольдегщрогенази та опю!д!в).
Успадкування найпоширешших нешфекцшних хвороб (артергально! гшертен-
зп, бронхгально! астми, пептидно! виразки шлунка, ожиршня, цукрового д!абету,
атеросклерозу, алерпчних та онколопчних хвороб), загальномедичне та сощатьне
значения яких дуже важливе, не може бути пояснене законами Менделя. 3 одного
боку, участь генетичних фактор!в у патогенез! цих хвороб виявляеться статистично
захворювашсть у родин! чи у близнюкгв перевищуе таку в цопуляцц), з шшого —
не можна встановити н! класичний тип успадкування. ш хромосомну аномалдю.
хоча е певн! досягнення молекулярно! генетики в розшифруванн! генетичних ме-
хан!зм!в багатьох !з них. Для пояснения ще! категорп хвороб запропоновано кон-
цепшю полкенно-адштшвного успадкування з пороговом ефектом. Феномен алелышх
пол1морф!зм1в ген1в гакож залучають для пояснения патогенезу цих хвороб. При
таких хворобах показники гомеостазу визначаються не тьгьки генетичними факто-
рами, а й факторахчи навколишнього середовиша, причому !снуе порш, за яким
гомеостаз легко порушуеться. Пороговим е певний лгмгтуючий фактор навколиш-
нього середовища (сольове чи цукрове навантаження, вм!ст холестерину в д!ет!,
гшокс1я), адаптащя до якого контролюеться певною групою гешв. Щ хвороби от-
римали додаткову назву багатофакторних (мультифакторних). Характерними для
них е значний пол!морф1зм симптомгв, велике розмаптя клппчних форм та шдив!-
дуальних прояв1в, значна поширешсть у популяцп. незалежно вщ географ!чних,
етшчних । сошальних факторгв, ргзний вгк хворих. Клш!чний перебгг може бути
явним або прихованим. ревень пснетрантност! — вщносно низьким. Терапевгичн!
усп!хи сучасно! медицин и сприяють виживанню таких хворих.
Хромосомы хвороби
Хромосомна мутан!я ушкоджуе бггьшу частину генетичного апарату 1 тому спри-
чинюе серйозншй порушення. шж генна мутащя. Проте наявносп тако! мутацп в
статевш клгтин! ще недостатньо для розвитку хромосомно! хвороби, необхщною
е низка умов. Для реалгзацц ще! мутацп та розвитку хромосомно! хвороби мають
значения так! обставили:
• виникла хромосомна мутащя ае похо в статев!й клггиш здорово! людини чи
отримана у спадок;
• чи життездатна мутантна статева юптина (дуже часто вона гине; б!льш жит-
тсздатною е мутантна кштина з К1льк!сною, а не структурною змшою хромосом;
причиною кгльюсних порушень е нерозходжсння хромосом у мейоз!);
• чи заплщнена мутантна статева юптина. Якшо вона з р1зних причин не за-
плщнена. то хромосомна муташя не мае жодних наслщкгв (часто така стагева
кл!тина втрачае здатшеть до заплщненняц
• чи збережена житгездатшсть ембрёона ё плода в раз! заплщнення мутантно!
статево! юптини плёд може загинути на рёзних етапах свого розвитку; больше 40 %
мимовёльних викиднёв зумовленё хромосомними аномалиями);
• чи збережена постнатальна житгездатшсть дитини (хромосомш аномал!! зу-
мовлюють близько 6 % мертвонародженосп).
Етёолопчш фактори, що зумовлюють хромосомну мутацёю де пог о, е мутаге-
нами фёзично!, хёмёчно! та бёологёчно! природи. Етюлопя хромосомних хвороб
мае таю особливостё:
• етюлопчш фактори зазнають впливу на батькёв дитини, яка хворёе;
• порушення гаметогенезу ё нерозходження хромосом частппе спостерёгають
у людей старшого вёку, але !х статевё юптини рёдше заплёднюються;
• сёмейна схильшсть до нерозходження хромосом.
Якщо хромосомний дисбаланс сумёсний ёз постнатальним ёснуванням, то в
тако! дитини порушений соматичний ё психёчний розвиток. Якщо хвора дитина
досягае певного вшу, то така людина найчастёше стерильна ё тому не передав
свою хворобу потомству (зё смертю хворого не лишаеться ё слёду вщ ще! мутацп).
Хромосомш аномал!! не накопичуються в популяцп. Тобто хромосомш хвороби
належать до генетично детермёнованих, але не спадкових. У тих виняткових ви-
падках, коли репродуктивна функщя все ж зберёгаеться (3—5 %), потомство ус-
падковуе ту саму хромосомну аномалёю.
Хромосомна хвороба — це хвороба, зумовлена хромосомною мутащею в гермёна-
тившй юптин! одного з батыов, що виявляеться у потомства за умов: а) збереження
життездатностё мутантно! статево! юптини; б) н заплщнення; в) збереження жигге-
здатност! ембрюна 1 плода; г) можливосп постнатального кнування.
У лётературё описано близько 300 хромосомних аномалёй. Найменше пору-
шуеться житгездатшсть носИв кёлькёсних мутацёй 23-1 пари хромосом (статевих).
Завдяки виживанню таких хворих так зван! хромосомна синдроми добре вивчено й
описано.
Трисолйя за Х-хромосомою (карёотип 47,XXX) у ж!нок ёнодё не мае очевидних
клёнёчних наслёдкёв. Деяк! з них нормально розвинуп, мають дней, але зазвичай
цей синдром супроводжуеться гшогонадизмом, зниженням фертильност! (плёд-
ностё), ёнтелекту.
Трисо.шя типу XXV у хворих чоловёчо! стат! (синдром Клайнфелтера) супро-
воджуеться первинним гшогонадизмом, евнухощшстю, як правило, безплщнёстю,
зниженням штелекту; хвор! висок! на зр!ст, мають астешчну будову пла. При
трисомп типу ХУУ (можлив! вар!анти ХХХУУ) чоловёки дуже висок! на зр!ст, мо-
жуть мати нормальний статевий розвиток 1 навёть пёдвшцену плёднёсть; рёвень
1нтелекту нормальний або середшй; спостерёгають схильшсть до агресивно! по-
ведёнки.
Синдром Тернера (Шерешевського — Тернера) характеризуеться карёотипом 45,ХО
(вщсутня одна Х-хромосома в жёнок). Плоди часто абортують, а якщо виживають,
то дёвчата — низького зросту ё часто розумово вёдсталё, безплёднё (унаслёдок пер-
винно! аменорег), схильш до лёмфостазу ё тирео'щиту Гашёмото. У гхнёх родичёв
спостерёгаеться висока частота розвитку цукрового дёабету.
Носёйство карёотипу УО не сум!сне ёз життям. Такё ембрёони гинуть у раннёх
стадёях внутрёшньоутробного розвитку.
Щодо мутацп автосом, то вона мае тяжч! наслщки. юж мутацш статевих хро-
мосом. Своею чертою, дефщит автосом ппадтившшй, н!ж !х надлишок (випадки
моносомп Клашино не описано).
Трисомйо автосом за найб1льшими автосомами не описано Якщо вона сумтс-
на 1з життям, то призводить до розвитку численних фгзичних вад 1 дуже високоТ
тегальност! в ранньому дигячому вцц.
3 ус!х носив авгосомних трисомш найжитте здатн!шими е носи трисоми 21-1
пари кромосом — синдром Дауна, Чоловжи безтпдн!, жшки 1нод1 мають дней.
Осюльки це домшантна ознака, половина датей, народжених такими жшками,
здоров!, а половина мають таку саму' хворобу, Вона яроявляеться розумовою в!д-
сталютю, передчасним старениям, невролопчними порушеннями, катарактою,
пщвищеним синтезом пуритйв, амйтощозом, продукшею б1лк1в-онкоген!в, щдви-
щеною частотою розвитку лейкозгв, активашею вьльнорадикдтьного окиснення,
Антитиреощш антипла тдвищують вфоидшеть нерозходжеиня 21-! пари хромо-
сом у гаметогенез!, через шо ппофунки!я шитопод1бно'1 задози е фактором ризи-
ку. У Ж1нок В1ком понад 35—39 рокйв, а у чоловпов веком понад 55 роктв збшь-
шусться частота нерозходжеиня хромосом < ризик мати д1тей 1з синдромом Дауна
Описано родинну схильшсть до нерозходжеиня хромосом
Частина хромосомних хвороб зумовлена структурними аномал!ями хромосом.
1х виявляють рщше, але вони мають тяжч! наслщки поршняно з кшыасними зм!-
нами хромосом. Небагато дней вижквають теля народження, мають тяжю ф1зич-
ш вади, розумово вщетал! 1 потребують постшного медичного догляду, але бгль-
цйсть невдовз! помирають.
Спадкова схильшсть до хвороб
В одних випадках (гостра променева хвороба) розвиток хвороби зумовлюеться
факторами навколишнього середовища, в шших — ильки тенетичними фактора-
ми (хвороба Дауна). Проте с хвороби, розвиток яких детермшований як генетич-
ними, так 1 екзогенними факторами. У таких випадках говорить про спадкову
схильшсть. I! основою е: а) гетерозиготне восшство рецесивного патолопчного
гена; б) полпенний тип успадкування; в) алельний полшорф!зм гешв.
Один 1 той самий зовншппй фактор у рхзних людей залежно вщ спадково)'
схильноси може спричинити р!зш хвороби. Наприклад, псюячне перевантаження
1 негативн! емоцп в одних людей зумовлюють розвиток ппертошчно! хвороби, в
шших — виникнення нервових 1 псих!чних захворювань, у деяких — цукрового
д!абету або тиреотоксикозу.
Для кл1н1чно] практики важливим е знания факторыризику, про як! потр!бно
пошформувати хворого (фактори ризику сл!д уникати). Так, навантаження на
певний ланцюг обмшу речовин у гетерозиготного носи ферментопатп може вия-
вити функцюнадьиу недостатшсть! призвести до розвитку хвороби. Переважання
в ращон! тваринних жирй е фактором ризику для хворих, схильних до атероскле-
розу, зловживання кухонною стилю — для оехб, схильних до гшертон!чно'1 хворо-
би. Для розвитку 1мунодеф!циту фактором ризику е шфекцгя. Вплив юшзуючого
випромшювання ! мутаген] в с фактором ризику при схильноси до онколопчних
здхворювань (в основ! — або генетично детермшована недостатшсть !мунного
нагляду над мутащями, або аномалы обмшу онкогенов, або наявшсть фермента,
що перетворюють проонкогени на онкогени). Змшюючи умови життя та врахову-
ючи фактори ризику, можна змшити перебгг быьшосп багатофакторних хвороб.
Генетичне обстеження хворого
Значения. Для вибору методу лгкування будь-якого хворого важливо знати причину
1 визначити вплив фактор!в навколишнього середовища 1 генетичних факторгв.
Деяк! хвороби КЛ1Н1ЧН0 подтбш до спадкових, але !х розвиток зумовлений
факторами навколишнього середовища (фенокопи). Наприклад, серед гормональ-
них порушень у людини частою фенокошею е ппотиреоз, зумовлений дефицитом
йоду в об’ектах навколишнього середовища. Хоча цей випадок клппчно под1бний
до генетично детермшованого порушення синтезу тирео'щного гормону, лпсуван-
ня 1 прогнози р!ЗН1, оскыьки фенокопп не успадковуються.
Лткарям загально! практики варто розум!ти, що коли обидва батьки хвор! на
одну й ту саму хворобу (наприклад, на гшертошчну хворобу або цукровий дхабет,
що бувае дуже часто), то у !хн!х дней пщвищена ймов1рн!сть захворгги на ту саму
хворобу. 1з феноменом спадково! схильносп стикаються лжар! будь-яко! спе-
щалхзацп, але (иод! потр!бно звернутися до спещалзста-генетика за генетичною
консул ьтацтею
Гетерозиготн! нос!! патолопчного гена, як правило, клппчно здоров!, але для
прогнозування потомства в таких хворих потр!бно провести генетичний аналтз.
Наприклад, у ж!нки — нос!я гена гемофып А у шлюб! з! здоровим чоловгком по-
ловина син!в хворггимуть на гемофыйо, а вс! дочки будуть здоровими (половина
з них стануть нос!ями патолопчного гена). Тому щй жпщ! можна порадити зберг-
гати ту ваптшсть, за яко! встановлено (методом амшоцентезу) жшочу стать плода.
У людини з автосомно-домшантною скелетною аномал!ею (брахщактшпею) в!ро-
гщшсть появи ще! вади у дней становить 50 %, а в гомозиготного нос!я цього
гена вс! дгги будуть народжуватися з такою самою аномал!ею.
Якщо приводом звернення за генетичною консультащею е народження хво-
ро! дитини ! кртм того батькгв шкавить прогноз для подалъшого потомства, то
роль генетичних фактор!в ще потребно довести. Так, якщо мутац!я вщбулася в
ембрюна чи плода пщ час ваптносп, то повторения тако! само! мутацц при на-
ступил! ваптносп малоймов!рне. Якщо встановлено, що причина народження
хворо! дитини полягае в акушерськш патологи, то цьому можна запоб!гти пщ час
наступно! ваптносп. Якщо в новонародженого в!д жшки, хворо! на сифппс, ви-
явлено сифшс (вроджений сифшс), то це не спадкова хвороба, оскыьки вона
спричинена (нфжуванням пщ час полота.
Для виргшення ус!х цих питань е калька методов, як! поспйно вдосконалю-
ються.
Генний метод. Генний р!вень анал!зу можливий, 1 при моногенних спадкових
хворобах вдаеться встановити мутащйну перебудову або вар!ац!1 гешв на р!вш
ДНК. Вщкриття рол! пол!морф!зму вщповщних гешв дае змогу на сучасному
ртвш оцшити ’!х клппчне значения, використовуючи генетичш маркери. € дв!
можливост!. Пряма ДНК-д1агностика полягае у визначешн генно! мутацп на р1в-
н! нуклеотидно! поел (довноси змшеного гена (секвенування) за допомогою пол!-
меразно! ланцюгово! реакцп (ПЛР), специф!чних зондтв. Змшеш ДНК назива-
югься ДНК-маркерами, 1 бшышсть 13 них уже визначено. Якшо нуклеотидш по-
слщовност! зм!неного гена виявляються б!ох!м!чними тестами (наявшстю бю-
х1М1чних продукт цього гена) та за юшичними проявами, то це непряма ДНК-
дшгностика
Бюх1лйчний метод. Ферментопатйо щагносгують за активнктю ферменту або
продукт реакцп, що каталгзуються ним ферментом. Шоб дефект ферменту мож-
на було виявити, в!н мае призводити до ч!тких лабораторних змш. Ате, якшо
порушення не с^проводжуеться б!ох!м!чними вщхиленнями, то втрогщно д!агносту-
вати ферментопатйо неможливо.
У М1ру вивчення спадкових хвороб з'являються нов! методи б!ох!м!чно1 щен-
тиф!кацп. Розроблено нимало експрес-метошв диагностики спадкових порушень об-
ману речовин шляхом дослщження сеч! або краплин кров! новонародженого. За до-
помогою 610ХШ1ЧН0Г0 методу можна дослщити фактори зсщання кров!, БАР,
бюхЬмпчну д!ю гормон!® тощо.
ЦитологЫний метод. Дослгдження карютипу. Пщ час под!лу юптини в
стадГ! профази хромое оми можна побачити пщ мщроскопом, а в стад!! метафази
Ч1гко визначають !х к!льк!сть 1 морфолопчн! особливост! Карютип можна дослщ-
жувати у юптинах юсткового мозку, але придатн!ип ятя цього культури лейко-
цит. На мал. 5 наведено к!льк!сн! патолопчн! карютипи та структури! змши
генетичного апарату, шо зумовлен! хромосомною мутащею 1 проявляються розри-
вами, дицентричними та к!льцепод!бними хромосомами, !х фрагментами, хромо-
сом ними асощатами. Вони виникли в дослщжуваних лимфоцитах уже в культур!!
евщчать про хромосомну нестабшьшсть генетичного апарату пащента.
Доайдження статевого хроматину. Статевий хроматин в штерфазних ядрах
(тшьця Барра) мае вигляд сгорал (зовано! Х-хромосоми тол!, коли в хромосомному
набор! !х дв!. Природно, шо в норм! статевий хроматин можна виявити лише в
ос!б Ж1ночо1 стать За наявносп. в юптин! юлькох Х-хромосом юльюсть статевого
хроматину дорщнюе гхшй юлькост! м!нус 1. Не кожна соматична юптина ж!нки
м!стить статевий хроматин. Кетьк!сть хроматинпозитивних ядер у вщеотках вщоб-
ражуе функцюнальний стан оргашзму жшки.
Прикладом використання 1мунолог1чного метооу може бути виявлення гетеро-
зиготност! при гемофшп А за визначенням антипл до антигемофыьного глобулину.
Популящйно-статистичний метод засгосовують для встановлення спадково!
природи захворювання в конкретшй родин!. В!н полягае в пор!внянш частоти
виникнешгя хвороби в родин! хворого з частотою пього захворювання в популяцп
(за даними медично! статистики). Р!зниця мае бути статистично в!рогщною. У
раз! мало! юлькост! членгв родини застосування пього методу утруднене. Однак
год час пщрахунюв умовно можна вважати хворими тих членгв с!м’1, як! юпшчно
здоров!, аде результата лабораторних дослщжень або функцюнальних проб не-
сприятлив!. Велике значения для практично! охорони здоров’я мае вивчення по-
ведшки мутантних гешв у популяцп, прогнозування !х поширення в популяцп,
розроблення простих метод!в виявлення носив таких ген!в.
Генеалоггчний метод, тобто складання родоводу, дае змогу визначита тип ус-
падкування 1 стушнь ризику повторения спадково! хвороби в родин! хворого.
За допомогою близнюкового методу можна розргзнити роль спадкових фак-
тор!в 1 фактор!в навколишнього середовища. Однояйцев! близнюки генетично
абсолютно однаков1, 1 вщмшшеть м!ж ними визначаеться тшьки факторами нав-
I) л и п
1 2 3 4 5
п п и и и и я
6-12
А4 Н
13-15
XX «X <*
16 17 18
«X «Я
19 20
4 А 44
21 22
а
ХХХТУ
К в в
1 2 3
ни
4-5
|> н и
6 7 8
13-15
и *л ад
16 17 18
ллл*
19-20 21-22
пик»
9 10 11 12
Н1
XXX
АА
А
Мал. 5. Хромосомн! мутаюцк
А — юлыасш: а — карютип (ХХХУУ) хворого
чолов!ка (синдром Клайнфелтера); б — карю-
тип (XXX) хворо! Ж1нки (трисом1я за Х-хромо-
сомою); Б — структури!: 1 — делещя; 2 — хро-
мосомний асощат; 3 — К1льцепод1бна хромо-
сома
колишнього середовища. Двояйцев)
близиюки генетично схож! не бглыпе,
н!ж узагал! брати 1 сестри, тому !х за-
хворювашсть вщображуе вплив подгб-
них умов на р!зш генотипи. Напри-
клад, у розвитку недоумства 1 деяких
псих 1ч них хвороб генетичш фактори
вгшграють важлив1шу роль, шж при
таких хворобах, як ендем1чний зоб або
туберкульоз.
Оскглъки КЛ1НЩИСТ1В не задовольняе д!агностика спадково! патологи теля
народження, розробляються методи допологовоТ (пренатальног) диагностики. Для
цього за допомогою М1крох1рургй вщбирають проби тканин плодового походжен-
ня. Зразки ДНК плода видыяють з бюптаттв плаценти, кмтин амнютично! рщини
або з лгмфоцитгв пуповинно! кровь У навколоплщшй рщиш можна визначити
актившсть ферменпв, вм!ст продукпв метаболхзму, а також отримати (а пот1м
культивувати) клпини плода, цитолопчне дослщження яких полягае у визначенш
статевого хроматину (стат! плода) 1 карютипу. Можливо провести бюпегю печшки
чи м’яза (наприклад для даагностики мюпати). За допомогою методу ультразвуко-
вой ехографи виявляють дефекти скелета (дисплазио исток, дефекти тулуба або
к1нц1вок), нервово! трубки, мгкроцефалгю, пдроцефалпо. У раз1 виявлення не-
сприятливо! спалковосп ваптшеть можна перервати.
11Г
Достов1рн1 даю про роль генетичних факторгв у розвитку спадкових хвороб
юдини може надати експериментальне моделювання цих хвороб у тварин. Шля-
хом гмбридингу або оброблення тварин сильними мутагенами отримують лшн
хтантних тварин. Так виведено високо- та низькораков! лпш тварин з високою
частотою розвитку ппертошчно! хвороби й атеросклерозу, собак гз гемофшею,
:ш!г мишей з! спонтанною розколиною щцнебшня, хом’якгв г курей 1з м’язовою
дистрофгею. На мишах змодельовано також альфа-таласемпо, полщитемпо, нир-
ковий ацидоз. Сучасними е моделг спадкових хвороб людини на трансгенних
твари нах, яких одержують шляхом спрямованого перенесения чужоргдного гене-
тичного матергалу, фрагмента гена або гншо! послщовносп ДНК у заплщнену
яйцеклгтину або зародок на ранн1х етапах розвитку.
Принципи лЕкування 1 профшактики
Етюлопчна тератя г профшактика генетично детермшованих хвороб полягають у
запобианнг надходженню або знешкодженш мутагенних факторов. Нейгралгзащя
ендогенних мутагенгв вщбуваеться за допомогою штучних антиоксиданта.
Патогенетичиа тератя передбачае виправлення гена, ушкодженого мутацгею,
або замшу дефектного гена на нормальний, тобто генетична тератя — це спря-
мована маншуляшя генетичним матергалом (генна 1нженер1я). Бютехнолопю гене-
тичного принципу лгкування поспйно вдосконалюють. Будь-який ген може бути
видгленим з оргашзму г проклонованим. Новий генетичний матергал можна вне-
сти в геном у склад! вгрусних частинок (попередньо позбавлених здатност! до
реплгкацп) або в комплекс! з лшосомами. Можливими е видыення г культивуван-
ня кл1тин пащента, уведення в них чужорщних генгв г решфузгя цих клгтин тому
самому пащенту.
Замгсна тератя. Уведення антигемофгльного глобулшу (фактора VIII) хворим
на гемофглгю А збйгыггуе тривалгсть життя, дае змогу деяким з них вести майже
нормальний спосгб життя, мати дней. Такого самого застосування набули у-гло-
булш, гормони (шсулш, тироксин), фермента, метаболии тощо. За допомогою
методу генног гнженерп створюють химерш форми мгкрооргангзмгв, здатних син-
тезувати кориснг для людини б!лки (штерферон, гормони, фермента тощо). Од-
нак екзогенне введения бйгка може супроводжуватась гмунною реакщею вгдторг-
нення (розробляеться метод лгпосомного захисту екзогенного продукту в склад!
напгвпроникних капсул).
Бюхгмгчна корекщя або зменшенггя порушень обмгну речовин, мае чимало
можливостей. Дгетичш поради полягають у виключеннг з рашону небезпечних
речовин, обмш яких порушений. Так, при фенглкетонурц переведения дитани на
дгету, майже позбавлену фенглалангну, запобггае розвитку недоумства. Можливою
е шактивацгя токсичних продукта, як! утворюються в процесг порушення обмгну
речовин. Додавання ферменту або нормального метаболиту вже було названо як
метод замгсно! терапп.
Симптоматична терагпя. За наявносп психгчних порушень при спадковгй хво-
роб! важливим е застосування психотропних засобгв. Так, седатавна терапгя пок-
ращуе якгсть життя хворих з емощйними порушеннями, симптомами збудження
й агресн. Застосування статевих горможв у дгвчаток гз синдромом Тернера г хлоп-
чик!в гз синдромом Клайнфелтера сприяе полгпшенню розвитку вторинних ста-
тевих ознак, поя во менструащй у довчаток, у деяких випадках шдвищуе потенщю
в юнаков о позитивно впливае на психгку. Для стамуляцп мехашзмов протимута-
цшного омунотету корисними е омуномодулятори. Хорурпчне локування з приводу
аномалш розвитку (розколини губи о пщнебоння, вовчо! пащо) дуже ефективне.
Видаленню подлягае ободова кишка зо спадковим полшозом, який мае ризик
малогшзацп.
Профьпактищ слщ надавати перевагу, оскольки локування спадкових хвороб
мае низку обмежень.
Велике значения мае еколопя. Гошзуюча радоашя завдае особливо! шкоди.
Будь-яка нова речовина (локарськ! та косметачно засоби, пестицида, консерван-
та), що використовуеться людьми, мае прохода™ переворку на мутагенность.
У разо мультафакторних полггенних хвороб профилактика полягае в запобо-
ганно впливу факторов ризику.
Попередження особ з обтяженою спадковостю про можливость народження у
них хворих дотей о визначення ступени цього ризику е подставою для рацюналь-
ного вирппення питания про дотонародження. У розвитку деяких спадкових хво-
роб е критично воково перюди, коли ох омоворшсть найвища. Тому питания про
дотонародження можна вирошувати теля того, як людина досягда цього воку.
У запобогашп спадковим хворобам мають значения деяко сощально фактори
(обмеження шлюбов мож близькими родичами, боротьба оз забобонами, що приз-
водять до озоляцп за расовими 1 релопйними ознаками). Не позбавлена сенсу по-
рада локаря генетика народжувати дней у молодшому виц.
Ново перспективи у вивченно спадкових хвороб людини вщкриються оз вста-
новленням функцш геному, що лежать за межами законов класично! генетики.
Належить ще розкрита таемницю “надлишковосто” ДНК. Вважають, що в неко-
дуючш частит геному е особливий, поки ще не вивчений код. Можливо, що в
геномо людини наявш структури вищого оерархочного порядку. Поняття про хви-
лъовий геном уже виникло.
РОЛЬ КОНСТИТУЦЙ В ПАТОЛОГИ
Конститущя — це ушкальний комплекс морфолог1чних, функциональных I психгчних,
досить спййких особливостей оргашзму, що визначають його реактившеть I сфор-
мувалися (виникли) на генетичнш основ! п1д впливом факторов навколишнього сере-
довища.
Пщ генетичною основою конституцп елш розумгги насамперед поломорфозм
гешв, як! зумовлюють вароативность морфолопчних структур, рецепторов, фермен-
тов тощо. Це своею чертою обумовлюе вароатившеть видових та ондавщуальних
особливостей оргашзму, типи конституцп, нормальш та патолопчно схильноси.
У тлумаченш конституцп неправильним е перебшыпення роло генетичних
факторов 1 виведення з цього неминучост! виникнення хвороби в разо наявносто
генетично! схильноси (конституцюнашм}. Проте не варто переошнювати о роль
зовнпиюх факторов о вважата, що конституцгю оргашзму можна довольно змоню-
вати. Навколишне середовище е умовою реалозацп спадкових ознак о водночас
може сприята змоненню реактивносто о резистентносто о навоть зовншшього виг-
ляду, формуванню нових ознак, яко мають конституцоональне значения. Для лю-
дини також безперечно важливими е сощально-гопеночш фактори — умови побу-
ту, пращ, харчування тощо.
Кожна хвороба мае свою причину (етоолопю). Конститущя не е етюлогоею.
Вона визначае ондиводуальну реактивность оргашзму, його адаптацойш особли-
восто, своерщность перебпу фозюлопчних_Д патолопчних процесов, патолопчну
схильшсть.
Значения конституцп розумоли ще давно лгкаро о вказували на зв’язок мож бу-
довою тола о схильшстю до певних хвороб. Таких зв’язков описано чимало. За-
вдання науковцов полягае у вивченш природи встановлених зв’язков, метаболоч-
них о функцюнальних особливостей розних типов конституцп.
Класифокацоя конституцюнальних гиопв. За велико! розномаштносто ондиводу-
альних особливостей оргашзму практична медицина прагне класифокувати ох, бе-
ручи за основу розно ознаки, — морфолопчно, боохомочно, серолопчш, темпера-
мент, типи психочноо (вищо'о нервово!) дояльносто тощо. Водповодно до них видо-
ляють колька конституцюнальних типов. Розни автори под розними назвами та за
розними критероями описуооть одно й то само типи конституцп.
Першу класифокащю запропонував видатний локар античносто Пппократ. Вон
звернув увагу на водмонность мож людьми, шо полягае в особливостях темперамен-
ту о поведшки в сусшльство. Саме цо спостереження Гшпократ поклав в основу
свое! класифокацп. Холерик, сангвоюк, флегматик о меланхолок — це термшолопя
Гшпократа, яка оснуе й досо.
Холерик — поривчастий, запальний, онодо нестримний, дратовливий, з висо-
кою, але непостшною працездатностю. Флегматик — спокшний о повольний, але
стойкий. Меланхолйс — замкнений, онодо пригшчений, нершгучий. Сангвтк —
жвавий, рухливий, емоцойний, схильний до повнокров’я, апоплексп, цукрового
доабету (таким людям частоте, шж пилим, призначали такий популярний у дав-
нину метод локування, як кровопускания). Гшпократ ще видоляв фтизичний тип
людей (видовжений тип, схильний до туберкульозу) та апоплексичний — схильний
до порушення мозкового кровообогу.
Пояснения конституцюнальних вщмонностей Гшпократ вбачав у розшй про-
порцп змппування основних “соков” (“гуморов”) оргашзму (крово, слизу, жовтоо о
так звано! чорноо жовчо). Правильне змошування цих “соков” (кразис) оз перева-
жанням одного з них формуе той чи той конституцюнальний тип. Неправильно
ох змошування (дискразоя), на його думку, е головною причиною хворобливих
станов. На'овшсть цих суджень е тольки уявною. Гуморальна патологоя Гшпократа
сповзвучна оз сучасним боохомочним уявленням про роль гормонов о БАР у розних
фозоолопчних о патолопчних процесах.
В основу класифокацп шзнппого перооду було покладено морфолопчний при-
нцип. Вщмонними ознаками е пропорцп скелета, сшвводношення мож вертикаль-
ними та горизонтальними розморами тола, розвиток тоео чи тое'о фозюлопчноо сис-
теми. Так, Сото за переважним розвитком тое'о чи тое'о фозюлопчноо системи розроз-
оояв чотири конституцюнальних типи: дихалъний, травний, м’язовий, мозковий
(мал. 6). Сото належав до тих дослодников, яко вважали, що конституцоя людини
формуеться протоном життя, переважно в дитинство, шд впливом навколишнього
середовища о тренувань.
Цоншсть будь-якоо морфолопчно! класифокацп визначаеться ступеней коре-
лятивних зв’язков мож морфологочними та функцюнальними ознаками. Форма —
Мал. 6. Класифжащя конститущональних ги-
те за Сто:
а — дихальний; б — травний; в — м’язовий; г —
мозковий
це пластичне вираження функцп. За ।
словами Е. Кречмера, обличчя — в!-
зипвка загально! конституцп людини
Вчення про конституцпо мае дати змо-
гу практичному лгкарю з першого пог-
ляду на хворого зробити припущення
про його функщональш особливост! та
патолопчш схильностг
Е. Кречмер за морфолопчними особ-
ливостями розр!зняв три конститущо-
нальних типи: астешчний, атлетичный.
пйаачний (мал. 7). Псих!атр за фахом.
вш намагався пов’язати морфолопчш
особливосп людини не тальки з особли-
востями характеру, псих!ки й темпера-
менту, а й з! схильшстю до певних
ПСИХ1ЧНИХ хвороб. Серед хворих на шизофрению ос!б астеючного типу спостерпа-
ють частппе, юж шли конститушоналып типи, хвор! на ешлепспо — переважш
атлетичного типу, на машакально-депресивний психоз — шкшчного типу. Харак-
тер симптомгв при неврозах теж пов’язаний з будовою т!ла. Так, !стер!я 1 депрес!я
б!льш властив! особам атлетичного 1 шкшчного типгв, страх — астеючного типу.
У клшщ! досить поширеною е класифгкацгя М.В. Чорноруцького. Кожному з
конститущональних тигпв — гтостетчному, гтерстешчному, нормостешчному —
вш дав характеристику з огляду на основю функцп та обмш речовин. Так, в ос!б
гшостеючного типу знижею артер!альний тиск 1 всмоктувальна здатшеть кишок,
пщвищений обмш речовин. Для гшерстеючного типу характерними е високий
артергальний тиск, уповптънею обмшш процеси, зниження толерантност! до вуг-
левод!в, схильшсть до ожир!ння, цукрового д!абету типу 2, плем!чно! хвороби
серця, шфаркту мюкарда, а також гшертроф!чного ришту та синуситу, кар!есу.
У. Шелдон поклав в основу свое! класифжацп переважний розвиток похщних
певного зародкового листка: ектодерми (долихоморфный 1 гтотроф1чнии), ентодер-
ми (брахшорфний 1 гйгертроф1чний), мезодерми (нормотроф1чний). Його класиф!-
кащя Трунтуеться на антропометричних индексах 1 оцппц габпусу (зовншшього
вигляду) шдивща в балах.
Мал. 7. Класифшащя конститущональних тшпв за Кречмером:
а — астеичний; б — атлетичний; в — гпкшчний
Т.П. Павлов керувався тим, що внутрппня едшсть ус!х частин оргашзму, його
е актившсть 1 вртвноважешсть !з навколишшм середовищем забезпечуються цен-
гзльною нервовою системою. Спираючись на те, що психгчна (вища нервова)
ыльшсть харакгеризуеться такими властивостями, як сила процес!в збудження 1
-дтьмування. Тх рухлив!сть ! вр!вноважендсть, вш видении чотири конститушо-
альних типи, що збггалися з типами Гппюкрата: сильный рухливий невргвноваже-
ай (13 сильними процесами збудження 1 гальмування, але з деяким переважан-
ям першого — холерик); сильный рухливий урмноважений (сангвгшк); сильный
твноважений повиьний (шертшсть основних нервових процесш — флегматик);
аабкий (слабюсть обох процессе !з дсяким переважанням гальмування — мелан-
.олгк). Для людини Т.П. Павлов запропонував ще одну класифгкашю, шо грун-
Т; еться на переважанш першоТ або цруго! сигнатьно! системи. Залежно вщ цього
1Н розрТзняв розумовий 1 художнш типи.
О.О. Богомолець вважав конституцию властивгстю динамТчною, а не морфо-
топчною, яка проявляеться в реакцп, а не в шдексах. Вш, як 1 Т.П. Павлов, в
основу свое! класифжацп поклав характеристику системи, шо об’еднуе сукуп-
шсть оргашв у единий ф1з!олог!чний тип. На думку О.О. Богомольця, це ф!з!оло-
пчна система сполучно! тканини, 1 тому вш подшив людей за особливостями
активно! мезенхТми. Сполучна тканина в астетчного типу гонка 1 н!жна, у фиброз-
ного — щ!льна й волокниста, у л'томатозного — здатна до жировоТ шфыьтрацн та
декомпозиш! лшощного характеру, у пастозного — набрякла ! пухка.
Видщення чотирьох конституцюнальних тишв залежно вщ групи кров! теж
грунтуеться на вщмшностях реактивност! оргашзму, кореляци м!ж хворобами та
еритроцитними аглютиногенами, яю визначають не тельки групи кров!, а й особ-
ливост! (мунно! вщповш!. Так, носи I (0) групи кров!, у яких немае еритроцитних
аглютиногсшв, характеризуються бшып активним протшнфекщйним ! протипух-
линним !мун!тетом. У них виявлено тенденцйо до нижчого р!вня холестерину в
плазм! кров! г меншу схильшсть до атеросклерозу; серед них бщыпе довгожителхв.
1м власти в! нижчий потенщал гшоталамо-ппоф!зарно-наднирниково! системи
(знижена продукщя стерощних гормошв) г ослаблена сттйкзсть проти стресу (част!-
ше розвиваеться пептидна виразка дванадцятипало! кишки). Носи П (А) групи
кров) схильш до гшерхолестеринемп та атеросклерозу, артергально! гшертензи, ш-
фаркту мюкарда, раку шлунка. Носи III (В) групи хворпоть мало, мають резистен-
тну систему травления ! спйку !мунну систему, У носив IV (АВ) групи вщзначають
низьку кислотшсть шлункового соку 1 гальм!вну реакцпо шеулярного апарату.
Пщ час вивчення конституцюнальних тишв стае очевидним, шо до чистих
тип!в належать менписть людей, а бшышеть е про.ьйжними типами. I хоча раню
нально! класифгкацп пром!жних тишв констигуцй дос! ше не створено, !х постш-
но намагаються охарактеризувати. Так, завдяки устпхам у молекулярнш генетиш
та (мувологп виявлено ргзномашття гешв ГКГС, яю становлять основу антиген-
но! шдивщуальност! й !мунолопчно! реактивност!, беруть участь у маркуванн!
власних кл!тин, розшзнавашп “свого” пщ час !мучолопчних м!жкт!тинних взае-
М0Д1И.
Корисним для практики виявився подш людей на конституцюнальш типи
залежно вщ рхзно! чутливост! до лгкарських засоб!в, запропонований шмецьким
гомеопатом Ганеманом. Це стало основою для використання лжарських засоб!в у
низьких дозах.
Вважають, що бюлог1чн1 ритми, пов’язанё з планетарными ё так званими зодёа-
кальними ритмами, також визначають реактившсть оргашзму ё мають бути врахо-
ванё при вивченнё хвороб.
Вивчення слабких, найуразливёших сторён конституцп дае змогу вчасно вияв-
ляти схильшсть до хвороби, передбачати и перебёг, ёндивёдуалёзувати лёкування
Профёлактична медицина, враховуючи це, може рекомендувати найсприятлившп
умови життя ё дёяльностё людям будь-якого типу конституцп. Аналёз переваг кож-
ного фёзёологёчного типу мае сприяти повинному розкритгю можливостей ёндивё-
да, правильней професёйнёй орёентацп ё повнёшёй реалёзацп здёбностей усього сус-
пёльства.
Для практичных цёлей медицини важливо встановити тип конституцп з ран-
нього вгку людини. I це не складно, осюльки саме в дитинствё ще недосконалё
мехашзми гомеостазу.
Дёатез. 3 учениям про спадковёсть ё конституцёю тесно пов’язане уявлення
про дёатез, який виявляеться у дитячому вщЁ Дёатез — не хвороба, а вроджений
стан, зумовлений спадковёстю (здебёльшого полёморфёзмом певних генёв). Це
аномалёя конституцп, при якёй дитина бёльш схильна до розвитку певних захво-
рювань. В основе дёатезу лежать ёндивёдуальнё особливостё реакцёй оргашзму, дл=
яких характерна неадекватнёсть на звичайш зовнёшнё та внутрёшнё подразники.
Саме це зумовлюе легше виникнення ё тяжчий перебёг захворювань. Дёатез може
трансформуватися в хворобу лише за несприятливого збёгу обставин.
Д1атез — це своер1дна аномамя конституцп, що характеризуешься неадекват-
ною реакщею оргашзму на ф1зюлог1чш та патологгчш подразники.
Дёатез — це типи реакгивностё оргашзму на межё з патологёею. Цё типи реак-
тивностё лежать в основё подёлу дёатезу на види.
Ексудативно-катаральний дзатез. Зовнёшшй вигляд дитини нормальний або
пастозний; вёдзначають високу схильшсть до запальных процесёв з утворенням
ексудату, затяжным перебёгом та алергёчними реакцёями. Спостерёгаються екзема-
тознё прояви на шкёрё, атопёчнё особливостё ёмунноё вёдповёдё, тенденцёя до над-
лишково'ё продукцп ёмуноглобулёшв Е та О, щдвищеноё активностё Т-хелперёв 1
кёнёново'ё системи. Легко виникають алерпчнё реакцп анафёлактичного типу з
вивёльненням надлишку БАР (серотонёну, гёстамёну), чутливёсть до яких щдвище-
на (у клётинах переважають Н^гёстамёновё рецептори при вёдносному дефёцип
Н2-рецепторёв). Сукупнёсть цих факторёв зумовлюе в таких дётей пёдвищений ри-
зик гёперерпчного перебёгу запалення. Клёнёчнё прояви — бронхёальна астма, кро-
пив’янка, набряк Квёнке, круп, анафёлактичний шок. У кровё виявляеться еози-
нофёлёя (проте цё дёти бёльше захищеш вёд гельмёнтозев).
Л1мфатико-г1попластичний д1атез. Друга назва цього стану — зёаШз ёЬутёсоёут-
рйаёёспз. В основё його лежить затримка ёнволюцёё загруднинноё залози та пору-
шення регуляторних впливёв гормошв кёрковоё речовини надниркових залоз.
Клёшчна симптоматика рёзномаштна. 3 одного боку — це явища пперплазп (ё вод-
ночас недостатностё) загруднинно-лёмфатичного комплексу, збёльшення загруд-
нинно! залози, мигдаликёв, лёмфатичних вузлёв, селезёнки. У кровё — лёмфоцитоз.
3 ёншого — ознаки гёпопластичного характеру: гёпоплазёя надниркових залоз, хро-
мафённо’1 тканини, щитоподёбноё залози, статевих оргашв, ранне виснаження ре-
паративных властивостей мезенхёми. Зовнёшшй вигляд хворого характеризуешься
соматичним ё психёчним ёнфантилёзмом, пастозшстю ё блёдёстю, слабким розвит-
ком м’язово! тканини, ранньою тшоплазхею органов, передчасним старшням. Час-
то розвиваються ангша й фаринпт, спостерйаеться схилыйсть до автоалерпчних
вороб, раннього атеросклерозу. Дхатез характеризуеться зниженими функцю-
аальними можливостями юрково! речовини надниркових залоз 1 адаптацшними
ожливостями, малою стресорезистентшстю, легким розвитком фази виснаження
в раз! сгресу. Оргашзм дуже вразливий, можлива раптова смерть з незначних при-
еин. Наприклад. такими причинами можуть бути стрхмкий розвиток иитотоксич-
них та автохмунних реакцш, токсико-алерпчне ураження центрально! нервово!
системи I мю карда.
Нервово-артритичний д/атез характеризуеться штенсивним пуриновим обм!-
ном, тдвищеним утворенням ендогенних ураттв з нуклезнових кислот. Кофешо-
подхбна Д1Я урат!в як ендогенний доп!нг формуе силъний невр!вноважений тип
психзчно! нервово! дхяльност!, пщвишуе нервову збудливкть, нейровегетативну та
емощйну лабпьшсть. € схильшсть до порушення психтки, не врали!, мггреш, де-
^ормуючих хвороб суглобгв нешфскшйного походження, сечокам’яно! або жовч-
нокам’яно! хвороби, лодагри, цукрового дхабету, ревматизму, а також до перю-
дичногс пщвищення в кров! р!вня кетонових т!л 13 розвитком метабол!чного аци-
дозу, що супроводжуеться блюванням.
Для астеючного дйнпезу характерними е адинам!я, лабшьшстъ судинних резк-
ий, схильшсть до виразок шлунка ! дванадпятипало! кишки, гастриту, артерхаль-
но! гшотензц, хрошчно! недостатност! надниркових залоз, хрошчних бронхолеге-
невих хвороб. Так, у раз! туберкульозу первинне шфгкування не залежить в!д бу-
дови т!ла, але переб!г хвороби тяжчий в осхб астешчного типу.
Рашше розр!зняли спазмоф!льний, геморапчний та шип види дхатезу. Але
поступово з них почали видьтяти певш хвороби та синдроми.
Можливх змхшаш прояви р1зних ВИД1В д!атезу.
Таким чином, при дхатезх дхти не хвор!, але перебувають у стан! мппмально!
резистентност! та максимального ризику розвитку тих чи тих хвороб. Умови нав-
колишнього середовиша можуть сприяти або запобйати !хшм проявам. На жаль,
на практиц! л!атез часто вважають хворобою.
ПАТОЛОПЧНА Ф131ОЛОПЯ
ВНУТР1ШНБОУТРОБНОГО РОЗВИТКУ
Внутршшъоутробний перюд .життя — це насамперед перюд реалхзацн генетично!
програми розвитку. Перебц' його вщбуваеться в умовах особливо! захищеностх.
Однак вш не захишае в!д впливу патогенних фактор!в оргашзм, шо розвиваеться
1х ушкоджувальна дхя може проявитися на будь-якому егап! раннього онтогенезу,
а особливо в так зван! критичш перюди.
Критичный перюд — це такий пром!жок часу в розвитку оргашзму, коли шсля
чертово! стали визначаються подалыш шляхи формування всього зародка або ок-
ремих його зачаткхв, перехщ !х до нового етапу морфогенезу. Для такого перюду
характерними е висока метаболхчна активность у певних зачатках, пщвишена чут-
ливхсть !х до дц ушкоджувальних фактори.
Розрозххяють два основних кригичних перюди: перший — перюд переходу за-
родка в!д стадп дробления до диференшювання на три зародков! листки, який за-
вершуеться гмплантащею (1-й тиждень); другий — перюд закладання оргашв, зок-
рема й плаценти, — перюд плацентаци 1 великого органогенезу (3—8-й тиждень).
Класиф1кац1Я. Усг порушення внутршшьоутробного розвитку класифжуют
за часовою ознакою, тобто залежно вщ того, у який перюд розвитку вони вщбу-
лися. До заплщнення, щд час гаметогенезу, можуть виникати гаметопати, яа
впливають поим на формування утвореного оргашзму.
1з порушень подальшого внутршгнъоутробного розвитку основними е бласто-
пат!я, ембрюпапя 1 фетопапя.
Бластопат1я формуеться у перш! 15 д!б розвитку зародка (бластоцисти), колг
в1н шсля заплщнення через маткову трубу проникав в матку, перебувае в нт
близько 3 д!б 1 на початку другого тижня !мплантуеться. У цей пер! од вщбуваеть-
ся дробления зародка, тобто утворення ембрюбласта, трофобласта ! диференцпо-
вання на три зародков! листки.
Ембрюпапйя об’еднуе порушення, що виникають теля диференцтювання ем-
брюбласта до завершения закладання оргашв (!з 16-! доби до 12-го тижня).
Фетопапйя — порушення розвитку плода; охоплюе патологпо раннього фето-
генезу, протягом якого вщбуваеться утворення тонких структур 1 досягаеться жит-
тездатшеть плода (з 12-го тижня до 7-го мшяця); порушення шзнього фетогенезд
коли вщбуваеться становления функцш плода та одночасне стар!ння плаценти (11
7-ГО М1СЯЦЯ до полопв).
Еполопя. Причини порушення внутршшьоутробного розвитку можна об’ед-
нати в три основн! групи.
1. Неповнощншсть статевих кл1тин (гаметопат!я), що може бути наелщко»
спадкових дефекпв, впливу патогенних фактор!в навколишнього середовища
порушень в оргашзм! матер!. За учасп в заплщненн! неповноцшних статевих кл!-
тин утворюеться зигота, яка зазвичай тине або дае аномальний плщ.
2. Патогенн! фактори навколишнього середовища, що зумовлюють розвитем
бласто-, ембрю-1 фетопатш: дефщит кисню, юшзуюче випромшювання, неращо-
нальне харчування ваптно!, х!м!чн!, термхчн!, бюлопчш фактори, а також психо-
емоцшш впливи, порушення адаптацц жшки до ваптност!. Х!м!чними факторами
е: 1) шкотин, який зумовлюе спазм судин матки ! плода, що призводить до гшок-
си ! гшотрофп плода, пригшчення гмуногенезу; 2) алкоголь, який спричинюе !н-
токсикацпо аж до алкогольного синдрому плода (затримка росту ! розвитку, енце-
фалопат!я, множинний дисморфоз тощо); 3) пестициди 1 деяю лгкарськ! засоби.
1з бюлопчних факторхв найнебезпечншшми е в!руси, особливо краснухи,
кору, грипу, що проникають у плщ кр!зь неушкоджену плаценту 1 зумовлюють
тератогенш та мутагени! ефекти. Серед хвороб плода, спричинених найпрост! ши-
ми, найвщомшюю е токсоплазмоз.
3. Хвороби вагтноТ. На перших етапах внутрццньоутробного розвитку вирь
шальним фактором може бути патолопя внутрхшшх статевих оргашв ж!нки, а
саме: недостатшсть яечник!в, ушкодження ендометргя. В останньому випадю
особливо небезпечним е травмування чутливих нервових заюнченъ матки, щс
призводить до розвитку рефлекторних зв’язктв М1ж маткою 1 яечниками 1 до вто-
ринного зниження синтезу естрогешв. У раз! зниження впливу гормошв яечникш
на матку порушуеться пщготовка трофхчного середовища для зародка, сам процес
гмплантац!!, а гпзнцце — плацентация (Л.О. Попова). Це зумовлюе формування
первинно! недостатност! плаценти.
У фсгальний перюд (период плода) найнебезпечшшими е хвороби ваптно!,
до зумовлюють зменшення плацентарного кровооб1гу 1 розвиток вторинно! недо-
гатносп плацеяти (гшертошчна хвороба, анемзя, шзшй токсикоз ваптних). При
раженн! окремих орган !в 1 систем материнського оргашзму можуть виявмтися
таен и ми функцц тих самих структур у плода. Причиною внутршлгьоутробноТ
атологп може бути гакож гмунна агрес!я оргашзму ваптно! щодо плода.
Сучасне уявлення про порушення внутрппньоутробною розвитку може бути
эглибленим, якшо розглядати людину з цйпсних позиций, гобто як бюенергош-
юрмащйну систему. Людина, вщповщно до цього уявлення. — триедина система
пло, психтка 1 вищ1 аспекты свщомосп) вщкритого типу, що самооргашзуеться,
саморозвиваеться, самовщтворюеться. Кргм ф!зичного тша вона мае хвильову
тольову) структуру, що керуе процесами формоутворення (Р Шелдрейк, 1981).
Трограму розвитку пла закладено в хромосомному апарат! клггин, а розкриття
СИХ1ЧНОГО потеншалу людини пов’язане !з хвильовим компонентом цього апара-
ту (П. Гаряев. 1998).
У внутрцнньоутробний перюд пл!д одночасно з! сгворенням структур ф!зич-
ного т!ла набувае базового психгчного досв!ду, що останшм часом вважають важ-
-ивим для подалыного формування характеру людини (С. Гроф, 1994).
Загальн/ законом1рност) розвитку патолопчних проце&в
у внутр1шньоутробний перюд
1. Змши характеру реакц1й оргашзму на дно патогенных факторов затежно вщ сзади
внутр! шнъоутробного розвитку. Для равшх стадш розвитку властива лише альте-
ращя. Тому будъ-яке ушкодження супроводжуеться зниженням дп ембрюнальних
ндуктор!в. Це своею чертою призводить до порушення процес!в формоутворен-
ня. Останне лолягае в затримц! розвитку оргашзму або окремих його орган !в, а
також у дискоординацп темшв росту та диференц!ювання р!зних тканинних ком-
поненпв. формуванш аномалгй.
У плода вже спостериаеться прол!ферац!я, однак судинний компонент за-
пально! реакцп виявляеться ще слабко. Мехашзми запалення та гмунолопчна ре-
актившеть не сформован! навпь до народження ! становления !х три вас в постна-
тальний перюд. У шзньому фетогенез! перебгг патологи плода переважно гакий,
як у доросло! людини, але з деякими особливостями: 1) будь-який патолопчний
процес у будь-якому орган! зумовлюе порушення ще не завершеного диферен-
цповання його тканин; 2) шфекщйний процес завжди мае генерал! зова нии харак-
тер; 3) процес регенерацп починаючи з раннього фетогенезу вщбуваеться з! злач-
ною пперпродукшею мезенхгмних елементгв, що спричинюе ф!броз оргашв
2. Зниження чутливоспй до ушкоджуваяъних факторов у процесг внутриинъоут-
робного розвитку з наближенням п в к!нш ваптност! до тако! в дорослого орган 13-
му. Характер ушкодження при цьому визначаеться не специфгчшстю фактора, а
сгащею розвитку оргашзму в момент його дп.
Будь-який пагогенний фактор у конкретнш стад!! розвитку оргашзму зумо-
влюе однотипн! порушення — аномалп тих структур, як! в цей момент найчут-
ливпш до ушкодження, тобто проходять свш Критичний перюд. Водночас одна й
та сама д!я фактора в р!зш стад!! розвитку спричинюе аномалп ргзних оргашв:
саме тих, критичний перюд яких припадас на цю стадно. Наприклад. опром!нен-
ня щургв однаковою дозою рентгешвського випромшювання на 10-й день вапт-
ност! зумовлюе аненцефалпо, на 11-й — м!кро- й анофтальмйо, на 14-й — ано-
малп к1нц1вок. Знания цих критичних перюд1в дае змогу диференщювати врод-
жеш та спадков! вади розвитку.
3. Змши реакцп плода залежно в1д силы вплыву патогенного фактора. У раз
надмхрного впливу фактора вщбуваеться загальне гальмування розвитку функцю-
нальних систем, що спричинюе загибель плода або появу тяжких форм патолоп
Вплив середньо! сили зумовлюе перезрхвання функшоналытих систем плода, як>
беруть участь у компенсапп навантаження, 1 недостатшй розвиток шших систем
(ф!з!олопчна незрйпсть). При слабкому вплив! виникае ф!зюлог!чний стрес, яки
тренуе адаптащйш можливост! плода.
Бластопатш. Шсля заплщнення починаеться реашзащя генетично! програм:
розвитку зародка, результат яко! залежить як вщ повноцшност! гамет, так 1 в!
умов реалззацп. Змша умов вщбуваеться з двох причин: по-перше, унаслщок па-
тогенного впливу фактор!в середовища, який перешкоджае розходженню хромо-
сом у клггинах, що проявляеться порушенням формування осей 1 загального пла-
ну будови зародка. Не маючи мехашзм!в регенерацп, зародок вщповщае на уш-
кодження утворенням додаткового цшого оргашзму або його частини у вигляд
вшьних або зрослих близнююв, циклотею, тератомою тощо. По-друге, факто-
ром, який перешкоджае розвитку зародка ! часто спричинюе його загибель, е за-
тримка процесу !мплантацп.
Ембрюпапя. Особливктю перюду ембрюгенезу е формування плаценти 1 б!ль-
шост1 оргашв оргашзму, що розвиваються. Порушення плацентацп !, отже, ста-
новления системи мати — плщ зумовлюе загибель або ргзку затримку завального
розвитку ембрюна ! зниження його життездатност!. Порушення органогенез)
проявляеться вадами розвитку окремих оргашв (головного мозку, очей, серия
тощо).
Фетопапя, У фетальний перюд, коли органогенез в основному вже заверше
ний, патогенш фактори спричинюють порушення розвитку тих оргашв 1 систем
формування яких ще тривае, — центрально! нервово!, ендокринно!, статево!, сер-
цево-судинно! систем 1 системи сполучно! тканини. Окрему трупу становлять по-
рушення, зумовлеш персистуванням (сповшьненням зворотного розвитку) емб-
рюнальних структур (наприклад сечово! протоки — урахуса) або збереженням
початкового розмйцення деяких оргашв (наприклад крипторх!зм).
Патолопя фетального перюду значною мхрою пов’язана з порушенням систе-
ми мати — плщ. Якщо порушуються кровообш у плацент! та вс! и функцп (ди-
хальна, троф!чна, захисна, вщцльна, гормоноутворювальна), то розвиваеться уш-
версальний синдром вторинно! недостатносп плаценти. Як ! у випадку порушен-
ня плацентацй, це зумовлюе зниження трофгки плода, його ф!зюлопчну незршсть
(неспецифгчна фетопапйя).
Якщо внаслщок соматичних захворювань у ваптно! порушена функшя лише
якогось певного органа, у плода через системну оргашзацпо його взаемодп з ор-
гашзмом матер! змшюеться функц!я гомолопчного органа — специф1чна фето-
папйя. Так, Ж1нки !з хворобами серця частице народжують дггей з вадами серця.
Припускають !мунн! мехашзми цього ураження. Зниження функцп окремих ен-
докринних залоз в оргашзм! ваптно! зумовлюе передчасне посилення активност!
“к самих залоз у плода, шо може порушили нормальний сисгемогенез, унаслщок
эго новонароджений одночасно матиме ознаки недостатньоТ функцп одних ор-
1Н1В 1 систем 1 пперфункий !нших. Така дострокова актива цы згодом зумовлюе
^искажения функцп 1 п недостатшсть (особливо в пубертатний перюд). Прикла-
1эм таких порушень може бути гшерплазгя остршцгв шдшлунково! залози в дити-
и. мати яко! хвора на цукровий д!абет. Наслщками цього е ппотлшемгя, збшь-
.ення маси тьла цилини, р!зш тератогенш ознаки у вигляд! ураження кзстково-
язово!, серцево-судинно!, центрально! нервово! систем, раншй розвиток цук-
тэвого дгабету.
СТАРШНЯ
Тривайсть шдивщуального життя багатоклгтинних оргашзмгв обмежена певною
характерною для кожного виду часовою межею, теля яко! настае смерть, У б!ль-
шостг випадкгв цьому перепуг прогресуюче незворотне порушення функщй, на-
слщком якого е зменшення здатностг оргашзму адаптуватися до умов навколиш-
нього середовища та збшьшення ризику захворпи та вмерпи.
Отже, старшня — це процес прогрееуючого зниження функцшналъних можливос-
тей оргашзму шсля досягнення ним зриостг.
Будь-який живий оргашзм в!д моменту зародження 1 протягом усього життя
зазнае певних змш у структур!, метаболизм!, функцгях 1 поведшш, поелшовно
проходячи етапи ембрюнального 1 постембрюнального розвитку, зрьлосл! та ста-
рост!, що закшчуеться смертю. Стар!сть 1 смерть неминуче настають нав!ль тодг,
коли оргашзм перебувае в максимально сприятливих умовах середовища життя !
харчування. Незважаючи на те що середовище може вп лива ли на темп 1 характер
вгкових змш, поки що шкому не вдалося значно загальмувати старшня 1 тим боль-
ше зумовити зворотний розвиток. Це евщчитъ насамперед про внутр!шню приро-
ду процесу старшня
Неминучють старшня 1 фатальшеть смерт!, бажання затримати ! повернута
молодгсть. шо минае, здавна хвилювали людський розум. Безл!ч легенд, фантас-
личних намагань 1 необтрунтованих рекомендащй, як! припиши з г.ли би ни вгк!в 1
поширеш в наш! дш, общяють швидке досягнення тривало! молодост!, довгойття
г позбавлення вщ ус!х хвороб Найчастше ц! методи “омолодження” на!вит 1 в
кращому випадку вари уваги. Однак вони можуть виявитися потешпйно небез-
печними для здоров’я. Так! рекомендащ! мають Грунтуватися лише на науковому
розумшн! мехашзмгв старшня ! всебгчнш експерименлальшй леревгрц!.
Науковий п!дх!д до проблеми старшня почав розвиватися поргвняно недавно.
Одним !з перших доелцщиюв, який серйозно вивчав ню проблему, був 1.1. Мечни-
ков Багато пробили для вивчення старшня Броун-Секар, О.О. Богомолеиь, О Б. На-
горний, Ф. Вернар. В.В. Фролькю. Д.Ф Чеботарьов.
На особливу увагу заслуговуе вивчення процесу старшня тепер. Одшею з
причин цього е бурхливий розвиток бюлопчно! науки, поява нових методичних
п!дход!в, як! дають змогу проникали в таемнищ живого оргашзму, розумЁги ос-
новн! закони його розвитку та життед!яльност1 1, отже, вивчати причини 1 ме-
хашзми старшня на експериментальшй основ!,
Друга причина пов’язана з тим, що вперше за всю тсторпо людства, незважа-
ючи на величезн! досягнення медицини в розумшш, розшзнаванш та лпсувань
хвороб, середня тривалгсть житгя людини, перейшовши в економ!чно розвинени!
крашах руб!ж 70 роюв, або перестала збглыпуватися, або збшьшуеться дул.
пов!льно. Дшсно, починаючи з кам’яного впсу, коли середня тривалгсть житгя
ледь сягала 18 рокгв, протягом усге! юторп розвитку людсько! цившзацп вон,-
збгдыпувалася, особливо швидко з юнця XIX — до середини XX ст. Цьому спри-
яли полшшення ппешчних умов життя, рап!онал!защя харчування г значит усшхн
в медицин!, здебЬтыпого в галуз! боротьби з шфекшйними хворобами, що супро-
воджувалося значним зменшенням дитячо! смертности зниженням смертносп в
шших впсових трупах 1 зб!льшенням китькостт людей лггнього ! старечого втку в
сустпльств!. Своею чертою така змша демограф1чно! структури популяцй зумови-
ла певш медичш та сощальш проблеми. Так, у виц понад 65 роюв кожн! плеть 1
семи осгб страждають на одну, а частппе — на киька хрошчних хвороб. Надаг
кыьюсть здорових осгб зменшуеться. Характерною ознакою старост! е наявшег
одночасно юлькох загрозливих хвороб — мультиморб!дн!сть. Хвор! старшого вне-
становлять значну частку контингенту лткувальних закладхв, маючи в середньом
три з половиною д!агнози на одну людину, хоча е пащенти з 10—12 рхзними за-
хворюваннями одночасно.
Старпоча людина прше пристосовуеться до мшливих умов житгя 1 виробни-
чого процесу, у не! зменшуеться продуктившеть пращ. Збитылуеться чисельшстъ
непрацездатних члешв суспшьства. У багатьох крашах к!льк!сть людей лггньогс
впсу перевищуе 13 %, а в деяких крашах — нав!ть 20 %. СпостерФають тенденцпе
до подалыпого зростання нього показника. За прогнозом ВООЗ, у першш поло-
вин! XXI ст. постарФпання охопить населения битылост! краю евггу. Особливо
гострою ця проблема е для сучасно! Украши, де частка людей л!тнього втку набли-
жаеться до 25 % на тл! одночасного катастрофхчного зменшення народжуваносп.
Загострюються питания життевлаштування, лгкування, догляду за особами старе-
чого впсу, непрацездатними, часто хворими членами суспшьства. Усе це зумовлюе
вирштення невщкладного завдання з вивчення старшня в ус!х його аспектах: бю-
лопчному, медичному та сошальному. Ось чому таким важливим е високий рь
вень розвитку геронтологи — науки про стар!ння — г гер!атри — науки про хворо-
би ос!б лггнього 1 старечого втку.
Для старшня характерш дв! ключов!, пов’язаш м!ж собою ознаки: зниження
пристосованост! до впливтв навколишнього середовища г збшьшення вхропдност:
смерт! з впеом. Як правило, оргашзм гине не в!д старост! як тако!, а в!д рхзних
хвороб г шкщливостей. Тому виживашеть оргашзму е наслщком взаемоди двох
груп факторов — зовшшшх х внутрхшнхх: характеру й пттенсивностх несприятливих
впливхв навколишнього середовища та зниження адаптивних 1 компенсаторних
властивостей фхзюлогхчних систем.
Згщно з аналхзом смертност! в людськш популяцй для криво! вимирання ко-
горта (групи оргашзм!в, як! народились одночасно) характерю певш фази. Перша
фаза вщображае швидке зниження смертност! в молодших вгкових трупах, яке
сягае найменшо! величини в перюд, що передуе статевому дозргванню. За нею
йде фаза битьш-менш постхйно! смертност! або незначного повитьного п шдви-
щення. Третя фаза, що охоплюе впе вщ 35 до 90 рок!в, мае характер експонен-
щального збыыпення смертност! (мал. 8). У людини в ц!й фаз! ймовтрнхеть на-
Мал. 8. Динамита смертности в популяцп, що
не стар!е, !з поспйною высокою смертшстю
(Л) 1 в популяшТ. ехилылй до старшчя (Б)
Мал. 9. Крив! виживаносН в раз! поспйно
внсоко! смертност! (А) 1 в популяцп, схиль-
н! до старшня (Б) (за Комфортом, 1967)
:гання смертг подвоюеться приблизно кожи! 8 роюв Для багатьох видгв тварин,
яю перебувають у стандартних сприятливих умовах, залежшсть смертносп вщ
вгку також близька до експоненщально]. Збшьшення показника смертносп в разг
збереження ргвкя зовншгшх впливш зумовлене внутргшшми змшами в оргашзм!,
спричиненими старениям.
Можна у явит и соб! так! крайн! ситуанй, за яких виживашсть когорта затежа-
тиме титьки вщ впливу зовншппх факгортв (за сталостг внугршшгх) або лише вш
внутргшнгх (за незначного впливу зовншгшх шюдливих факторгв). Крив! вижива-
ностг когорта при цьому вщргзнятимуться, маючи в перш ому випадку експонен-
шальну, а в другому — прямокутну ферму. У прим!тивних людських угрупован-
нях г крашах гз низьким ргвнем життя крив! виживаносп наближаються до пер-
гпого типу (мал. 9. А), тод! як в економгчно розвинених крагвах цг показники
бшьше вщповщають другому типу (мал. 9. Б). Так само тварини, яких утримують
у сприятливих умовах розплгдника, мають криву виживаност! типу Б, тод! як у
гхнгх диких родичгв небагато шанс!в дожита до вгку, який вщповщае старост!.
Розрахунки евгдчать, що якби в людини за ргвня шюдливик чинвикгв, який
вщповщае сучасному, пристосовашсть 1 отрнгсть р!зним видам стресу збереглися
на максимальному ргвш, що був у не! в 11-р!чному вгц!. то майже половина лю-
дей, як! живуть нин!, могли б сподгватися прожиги до 800 рокгв, а максимальна
тривапсть життя досягла б 22 000 рокгв. Однак це не зовегм так, лише дуже рщко
деяк! особи доживають до 100 роюв (менше н!ж 1 випадок на 1000).
Зм!ни в орган1зм! пщ час старшня
Зм!ни, що супроводжують старгнкя, можуть бути визначеш на ргзних р!внях бю-
лопчно! органгзацн: цйюго органгзму, окремих його систем, складових елементгв,
кл1тин г окремих хгмгчних речовин, з яких складаються системи оргашзму. Спгль-
ним для цих змгн е збгльшення гхньог варгабельностг з в! ком, ргзночасшсть проя-
ву в розних особин у розних фозоолопчшгх системах о клотинах — гетерохроннгсть
гетеротоптстъ старшня. Цю вароабельность ознак старшня спостерогають нави
у разо використання високошбредних, генетично однорщних тварин, яких утри
мують в однотипних умовах, коли вщмшносто в життевому досвщо та генетичних
властивостях зведено до мономуму.
На ровно цшого оргашзму, кром характерного зовншшього вигляду, воково змо-
ни проявляються зменшенням здатносто витримувати навантаження, наприкла_
виконувати фозичну роботу. В експериментах на щурах було встановлено, що ви-
живашсть тварин пщ час тривалого утримання в умовах знижено! температур:
повотря, здатшсть виконувати роботу за пщвищеноо температури повотря, вижи
ваность теля втрати кров! лшйно зменшуються з вгком. Кольосость тварин, як
виживають у раз! стандартного навантаження, з в!ком зменшуеться, знижуеться
також навантаження, що спричинюе смерть тварин. Шсля досягнення певног.1
критичного воку здатшсть до адаптацп розко знижуеться о оргашзм неминуче гинс
Це зумовлено критичним ровней порушення контролю за гомеостазом оргашзм)
Змши систем I оргашв. Зниження пристосованосто оргашзму в раз! староош-
зумовлене змшами функшонування окремих систем. Функция большосто з них
лшшно зменшуеться з вгком, хоча шодо це може бути виявлено лише пщ час на-
вантаження, наприклад, здатшсть пщтримувати постшну температуру т!ла, р!венъ
глюкози або рН кров!.
У нерповой систем! зменшуеться колькость нервових клотин о зб!льшуеться коло
кость глоальних елементов у деяких шарах кори великого мозку за незмшносто шг
елемент!в в шших дшянках мозку, у т!л! нейронов накопичуеться особлива шг-
ментна речовина — лшофусцин. Змшюються функщональш властивост! нейронп
зменшуеться швидкость проведения 1мпульсу, ослаблюеться синтез мед!атор!в у
нервовому законченно о знижуеться рефлекторна лояльность. Порушуються такол
складно функцп псих!чно! дояльносто: ослаблюеться пам’ять, знижуються творч.
актившсть о здатшсть до навчання, змшюеться поведшка. Водночас деяко та-
зван! викристал!зован! зд!бност!, так! як словниковий запас, здоровий глузд, за-
гальний обсяг знань, поровогяно мало змшюються з в!ком.
Боолопчна роль вокових змш у розних структурах мозку неоднакова. Особливе
значения для оргашзму мае старшня тих його вщдшов, як! через функшю авто-
номно! (вегетативно!) нервово! та ендокринно! систем забезпечують регулящк
дояльносто розних оргашв о сталость внутрппнього середовища оргашзму. Через ш
велику увагу придщяють гшоталамусу, воково змши якого, на думку багатьох дс-
слщников, е визначальними у виникненш порушень в шших системах, насампе-
ред в ендокриншй.
Функц!я ендокринно! системи пщ час старшня остотно змшюеться. Це прояв-
ляеться змшою як ровня окремих гормонов, так о чутливост! оргашв-мппеней дс
1'хньоо регуляторно! дп. Порушуеться також взаемодоя м!ж ендокринними залозг-
ми, що зумовлюе складну перебудову роботи системи в щлому. Найбшыие ослаб-
люеться функцоя статевих залоз, що свого часу сприяло здойсненню спроб “омо-
лоджування” шляхом пересадження статевих залоз (Броун-Секар, С.А. Воронов1
яко, однак, не дали очокуваних результатов. При старшно окром зниження репро-
дуктивно! функцп знижуеться продукция статевих гормонов о пщвищуеться пори
чутливосп гшоталамуса до ох гальмовного впливу, наслщком чого е значне збшь-
шення ровня гонадотропних гормонов гшофоза.
Порушуються взаемозв’язки в ппоф!-
трно-напнирниковш систем], зменшу-
.гься вцшовщь надниркових залоз на
"ропш гормони ппофгза, так само як 1
дпов1дь ппоф!за на ргвень кортикосге-
31 цв у кровг, знижуеться вироблення 17-
кетостероццв, змшюсться якгсть 1 змен-
луеться к!льк!сть репеп горгв до корта-
костероТд1в на кмтинах-мшгенях (л!м-
фоцитах, гепатоцитах тошо). Очевидно,
саме ним значною мхрою можна пояс-
нили зниження резистентноеп оргашз-
му до стресгв.
Знижуеться тиреощна регуляция
чроцеслв обмшу за рахунок зниження
тиреотропно! функцц гшофгза, ослаб-
ления дгяльносп щитоподгбко! залози
Мал. 10. Швов! змши ыунних реакшй: зни-
ження 1мунноТ ьццтовщ! на сторонн) антигени
(“не свое”): шдвишення автшмгнних реакшй
(“свое”)
та зниження реакщйно! здатносп тка-
нин. У цьому можуть брата участь гакож неведом! гальм!вш речовини, як! проду-
куються стар!ючим г!поф!зом.
Пщ час старшня порушуеться забезпечення оргашзму шеулшом унаелшок
зниження функщоналъних можливостей тдшлунково! залози. пригшчення ш-
сулшу в кров! та зниження реакцшно! здатносп тканин, шо може спричинити
порушення регуляцн вуглеводного та лшщною обмшу в р!зних тканинах.
Значних вгкових змш зазнае загруднинна залоза (тимус)- зменшуються (почи-
наючи в!д перюду статевого дозргвання) ц розмгри 1 функщональна актившсть.
При пьому спостерйаються гстотш змши структури залози: зменшуеться юльшеть
лтмфоципв (особливо в кзрковш речовин!), змшюються к!льк1сть 1 характер еш-
телторетикулоципв, зникають пльця тимуса (пльця Гассалля). Ц! зм!ни суттево
позначаються на функцн гмунно! системи.
Змши гмунно! системи в раз! старшня можна схарактеризувати ознаками, що
об’еднан! в три основн! групи: зниженням реактнвност! на сторонн! антигени
(!мунна недостатшсть), збптьшенням частота 1 ступени (мунних реакшй проти
антигешв власного оргашзму (автотмуншеть), схилыпстю до появи лгмфопрол!-
феративних хвороб (мал. 10).
При цьому змшюеться спектр питокппв. як! видьтяються клггинами !мунно!
системи, з появою так званого прошфламаторного цитокшовою профппо, що
зумовлюе схильшсть оргашзму людини старечого в!ку до певних хрошчних за-
пальних процеств Так! змши вщбуваються на тл! незначних вщхилень вмгсту ос-
новних класгв лтмфоцитгв, плазмоцита г за гпдвищення концентрата! !муногло-
булппв у кров!. Вони, очевидно, зумовлеш регуляторними порушеннями в !мун-
н!й систем!, призводять до обмеження можливосп пщтримувати сташсть анти-
генного складу оргашзму, авгоагресп проти компонентов власного оргашзму 1
псно пов язан! з патолопею, що виникае в ос!б л!тнього та старечого в!ку На-
справд!, так характерш для старшня патолопчш процеси, як тадвищена чут-
лив!сть до шфекцш, схильшсть до утворення злоякгених пухлин, старений амгло-
щоз, хвороби судин, у тому чиелг й атеросклеротичного типу, деяк! дегенерагивн!
хвороби мозку, опорно-рухового апарату, порушення функцгй пгдшлунковог
щитоподгбног залоз, деякг вида В12-фолгеводефщитног анемп бгльшою чи мен-
шою игрою пов’язанг з впсовими змшами гмунгтету.
Особливе мгсце в цьому ряду змш посгдають активацгя хронгчног вгрусног ш-
фекцп, розгальмування геному вгрусгв, штегрованого в геном органгзму-хазя'ша. 3
одного боку, це сприяе перебпу шфекцгйного процесу за типом повгльних шфек-
цш, з чим пов’язують деяк! види старечо! патологи мозку, автогмуннг процеси, з
шшого — може призводати до пухлинног трансформацп клиин. Захист оргашзм -
ос!б старечого вгку гстотно ослаблюеться також через порушення мехашзмгв де-
токсикацп в систем! мгкросомального окиснення, зниження активностг та здат-
ностг до шдукцп ферментгв системи цитохрому Р450, здебшьшого печшки. Це
зумовлюе уразливгсть старечого органгзму не лише до токсичних, а й до ргзни .
лгкарських речовин.
Значнг змгни розвиваються в процесг старшня в систем! сполучног тканини
Цим змшам свого часу великого значения надавав О.О. Богомолець, вважаючи
що вгк людини визначаеться станом и сполучног тканини. При цьому змшюютьс
клгтинний склад г залежне вщ нього спгввщношення основних структурних мак-
ромолекул мгжклгтинного матриксу. Збшьшуеться кглькгсть сполучног тканини в
ргзних органах (печгнцг, серцг, нирках), зменшуеться вмгст структурних глгкопро-
тещгв г протеоглгкангв, кглькгсть еластичних волокон на тлг збгльшення вмгст
колагену. Цей “старечий” колаген погано розчиняеться, його молекула стабглгзо-
вана внутрппньо- г мгжмолекулярними зв’язками-зшивками, що спричинюе по-
пршення механгчних властивостей сполучног тканини, склероз органгв. Справля-
ючи трофгчний г регулюючий вплив на функцгю спецгалгзованих клггин, сполуч-
нотканинна строма органа може впливати на його роботу. 3 порушенням стану
сполучног тканини пов’язанг вгков! змши судин г органгв опорно-рухового апара-
ту. Порушуються структура та мгнералгзацгя кгстковог тканини, суглобових г мгж-
хребцевих хрящгв, що зумовлюе порушення мщностг скелета, його деформацгю.
яка значною игрою визначае характерну старечу поставу та схильнгсть до перело-
мгв кгсток.
У процесг старшня знижуються також репаративнг функцп сполучног ткани-
ни. Вгдомо, що загоення ран, зростання кгсток шсля перелому та шшг вгдновнг
процеси в старечому вщг вщбуваються ггрше, юж у молодому.
Закономгрнг вгковг змши виникають г в систем! кровообггу. Вони стосуються
структури г функцгй як серця, так г судин. Зменшуються хвилинний об’ем серця
(ХОС) г сила скорочення шлуночкгв (приблизно на 1 % за ргк), зростае загальний
периферичний ошр (на 1,7 % за ргк). Зменшуеться щгльнгсть кагпляргв у тканинах
унаслщок стовщення базально'г мембрани стгнки капглярних судин, особливо в
легенях, зменшуеться здатшеть до дифузп речовин гз кров! в тканини г в зворот-
ному напрямку. Усе це попршуе перфузгю тканин кров’ю, забезпечення гх кис-
нем. Пщвищуеться артергалъний тиск. Серце прше компенсуе шдвищене наван-
таження г менше здатне до гшертрофц. Судини стають менш еластичними, в них
розвиваються склерозуючг процеси. Зростае частота ураження судин атеросклеро-
зом. 3 вгком порушуеться також нервово-гуморальна регуляцгя функцп серця г
судин, що може легко призвести до порушення судинного тонусу (артергальна
гшертензгя), дгяльностг серця.
Спостерпаеться ослабления функцш шших систем: попршуеться регенера-
орна здатшсть кровотворних оргашв, знижуеться функцтя нирок ! звужуються меж!
< пристосованост!, зменшуються секреторна актившсть травних залоз, сила м’язтв,
^нижуеться функция аналтзатортв.
Однак процес старшня — це не лише ослабления встх функцш оргашзму,
сюльки паралельно вщбуваеться в певних межах пристосування до нових умов.
~обто вищезазначеш процеси не е простим воображениям регресивних змш, а
.тановлять складний комплекс внутр! шньо суперечливих явищ порушення функ-
ций та тхньот компенсацц.
Змши кмтин. Оргашзм людини, так само як т шших хребетних, складаеться з
лттин, яю мають ртзнт здатшсть до подшу т тривалтсть життя. Одш з них посттйно
або пертодично дшяться, даючи початок мтотичним (спецталтзованим) кмтинам
з коротким периодом життя. До цих клттин можна зараховувати стовбуров! клтти-
ни юсткового мозку, сперматогонп, базальн! клттини ештелпо, примттивш клтти-
ни крипт кишок, а також клттини, що диференцпоють, такт як еритробласти,
мтелобласти, сперматоцити. Другий вид клттин — це зршт високоспецталтзовант
клттини, у норм! не дтляться або дйтяться рщко, але за певних умов здатнт до
швидкого подтлу. Це оборотно постмйпотичш. клйпини, до яких належать фтброци-
ти, хондроцити, остеоцити, клттини ендотелпо, печтнки, нирок та шших оргашв.
I, нарештт, третш вид клтин — необоротно постмтотичш високодиференцшо-
вант клттини, якт не здатнт дыитися за жодних умов. 1х можна роздшити на клтти-
ни з коротким перюдом життя (еритроцити, гранулоцита, епттелтальш клттини,
сперматозоони) т клттини з тривалим пертодом життя (клттини нервово! та м’язово!
тканин, овоцити).
Старшня мттотичних клттин може проявляйся зменшенням тх юлькостт та зм!-
ною здатностт до подтлу. Щоправда, встановлено помтрне зменшення юлькостт
стовбурових клттин юсткового мозку т крипт кишок з втком. Змшюеться також
поды цих клттин, що проявляеться подовженням мистичного циклу т змшою дина-
мтки росту тхньот популяцп. При цьому утруднюеться вступ цих клттин у мттотач-
ний цикл (блокада О0/ (3,), перехщ вщ пресинтетично! фази до синтезу ДНК (бло-
када О] / 8), сповыьнюються синтез ДНК 1 перехщ до мггозу (блокада О2/ М).
Цпсаво, що перенесения стовбурових клттин старо! тварини молодому реци-
птенту в деяких випадках сприяе зникненню ознак старшня т продующ нормаль-
них постмттотичних клттин. I навпаки, трансплантацтя стовбурових клттин моло-
дого оргашзму старому рецишенту спричинюе тх швидке старшня, що свщчить
про роль середовища, яке оточуе клттину. Зокрема доведено, що плазма т сироват-
ка кровт, взятт у старих тварин, гальмують пролтферацпо клттин у культур!.
Ще один феномен, який супроводжуе старшня клттин, шо дшяться, полягае
в порушенш регуляторних впливтв на ртвн! клттиннот популятттт. Кшьюсний склад
клттин певного типу пщтримуеться на постшному ртвш двома протилежно спря-
мованими трупами впливтв: стимулюючими (мттогени, фактори росту) т гальмтв-
ними, у тому числт й тими, яю походять вщ клттин, що розмножуються (кейлони).
У процест старшня не лише знижуеться утворення як перших, так т других речо-
вин, а й значно зменшуються юльюсть рецептортв т чутливтсть клттин, що дтлять-
ся, унаслщок порушення передач! сигналу всерединт клттини вторинними месен-
джерами. Це супроводжуеться попршенням контролю за розмноженням клттин,
тому одночасно зг зниженням пролгфсрацй можна нерщко спостершати схилд
н!сть до некоординованого надмгрного розмноження клггин. Можливо, це явить
лежить в основ! схильносп старого оргашзму до виникнення пухлин. зокрема
за рахунок порушення регуля гш апоптозу.
Ц!кав! результата одержан! пщ час тривалого вивчення розмноження кип
у культур! (Хейфлж). Якщо фибробласта новонародженого, перенесен! в певт-
культуральне середовище, у М1ру росту клггинного пласта переносити в нов! чаи
ки, то вони пгсля майже 50 подвоень клгтин но! маси неминуче гинуть. У ра
культивутапня фгбробласпв, узятих в ос!б разного впсу, було встановлено. ш
кглькгсть подвоень, яке вони можуть дата в культур!, зменшуеться з! збшьшеннз
в!ку донора клгтин. Це дало змогу дшти висновку, що в ядр! кожно! клгтани за-
кл адено мехашзм старшня, який обмежуе кглькгсть подътгв клгтин, однак вш мол
бута втрачений у разг грансформацП клгтин у пухлин!. Осташпм часом одержав. I
факта, що не тгльки пщтвсрджують, а й спроса овують цю точку зору. Зокрема
деяких видгв тварин з короткою видовою тривалгстю життя кглькгсть подвоем
як! можуть дата гхнг клгтани в культур!, значно бьтьша, нгж у вид!в з довшот
тривалютю життя (щури (2—2,5 року), подвоень 100; людина (70—85 рокгв), под
воень 50). У деяких експериментальних моделей вдалося отримати нетрансфт
мован! безсмертш клггиннг клони.
Морфолопчн! та бгохзмтчш змши, шо !х спостернають у кл!тинах, як! старея
в оргашзм! 1 в культур!, дсшо вщргзняються. Усе це дае пщстави розглядати л!м •
клгтинних лодгл!в як цгнний феномен диференц!ювання фгбробластгв у культур
який безпосередньо стар!ння як такого не стосуеться. Цей феномен змогли пог.
нити, коли вщкрили явище вкорочення теломертв у хромосомах клгтин пщ чж
кожного подгту клгтани.
Ст аршня обороти о постмгтотичних кл!тин вивчено набагато менше. В!до\
шо у втдповгдь на ушкодження (поранения, резекцгя частини печшки) або збгль
шення навантаження (однобгчна нефректомгя) здатшеть цих клгтин до лролщ
рацп в процес! старшня зменшуеться. При цьому про порш йно вшу подовжуетг.
час м1ж надходженням стимулюючого сигналу 1 початком клгтанно/ пролгфера.
(лаг-пертод). Змшюеться також функшя цих клгтин, про що евщчить змша фунт,
щй тих органгв. до складу яких вони входять. Та, оекзльки функшя органа е на
слщком взаемодн багатьох типгв клгтин, як! входять до його складу (зокрема стр
ми га паренхгми), а також впливу нервово! та гуморально! регуляцп, однознач-ч
трактування виявлених змш нерщко бувае утрудненим.
У необоротно постмгтотичних клгтин з коротким пергодом життя старшие
часто називають гх диференщювання г подальш! змши, як! зумовлюють загиба
клгтин. Таке старшня, очевидно, вщбуваеться багаторазово протягом життя ор-
гашзму г сносгерггаеться як у молодому, так 1 в будъ-якому шшому в!щ, г йог
потргбно вщргзняти вщ зм!н у цих клгтинах в оргашзм! людини старечого вщ
При ньому пор!вняно невелик! змши виявляють у таких клгтинах, як еритроци
знижуеться фагопитна ! бактерицидна активнтсть нейтрофшьних гранулопи"
Зменшуеться здатшеть до заплщнення г шдвищуеться частота хромосомних по;-
шень у сперматозоонах.
Необоротно постмгтотичн! клгтани з тривалим периодом життя можзпь В1 I
витися дуже важливим елементом у мехашзм! старшня- вони зберпаються прол
гом усього життя оргашзму, г старгння значною мгрою може бута зумовлене вахт
вими змёнами саме цих клггин. Оскёльки в постнатальний перюд вони не делить-
ся ё мало здатш до замещения, то навёть випадкова ёх загибелъ зумовлюе необоротну
втрату функщонуючих одиниць. Така втрата клётин визначаеться в корё лобовоё
частки (пёрамёднё клётини) ё в мозочку (грушоподёбнё нейрони; клётини Пуркёнье),
• мёокардё ё скелетшй м’язовёй тканинё. 1ншё клётини змушенё нести пёдвищене
навантаження, тому в цих органах окрём атрофёчних можна виявити ппертрофёч-
ш клётини. Клётини, що старёють, гёрше вёдповёдають на навантаження, у них
тривалёший латентний перёод, тривалёший ё менш досконалий перёод вёднов-
лення.
Для клётин старих тварин характерними е морфологёчнё та функцёональнё
змёни: змёни структури ядра, збёлыпення розмёрёв ё порушення будови мётохонд-
рёй, зменшення кёлькостё рибосом (апарату, де синтезуеться бёлок), збёлыпення
кёлькостё ё розмёрёв лёзосом, стовщення плазматичноё мембрани.
Дуже часто в клётинах накопичуються вакуолё з неперетравленими залишка-
ми мембран клётинних органоёдёв, лёпофусцин. Знижуеться енергетичний обмён,
зменшуеться внутрёшньоклётинний вмёст калёю, що зумовлюе зниження мемб-
ранного потенцёалу ё пов’язаних ёз ним функцёй збудливостё та провёдностё. Змё-
нюються чутливёсть ё вёдповёдь клётин на регуляторш впливи, взаемодёя клётин.
Дёапазон цих змён у рёзних клётинах досить широкий.
Змши молекул. Старёння супроводжуеться накопиченням речовин, якё в моло-
дому вёцё виникають лише за патолопчних станёв. Так, внутрёшньоклётинно нако-
пичуеться лёпофусцин, у клётинах макрофагоцитноё системи — гемосидерин. Мёж
клётинами з’являються скупчення амёлоёду, який можна виявити в усёх людей вё-
ком понад 80 рокёв. Для старечого амёлоёдозу характерною е трёада: амёлоцюз
мозку, серця й острёвцёв щдшлунковоё залози; часто приеднуеться також амёлоё-
доз судинноё стёнки та гонад. Молекули колагену стають стабёльнёшими, менш
розчинними, легко пёдцаються тепловому скороченню за рахунок збёльшення
кёлькостё мёжмолекулярних зшивок. Накопичуються позбавленё специфёчноё ак-
тивностё молекули ферментёв, хоча сумарна ферментна актившсть змёнюеться не-
однозначно: одночасно зё зниженням активностё одних ферментёв вёдбуваеться ёё
збёльшення в ёнших, а в деяких вона залишаеться незмёненою. Однак, мабуть,
найцёкавёшими ё бёологёчно значущими могли б бути змши, що вёдбуваються в
процесё старёння в молекулах ДНК ё РНК — носёёв генетичноё ёнформацп. Проте й
досё однозначнё змёни в цих молекулах встановити не вдалося. Виявлено збёль-
шення мёцностё зв’язку ДНК з пстонами й металами, легшу фрагментацёю ДНК
за деяких хёмёчних впливёв, збёльшення частота хромосомних аберацёй, змёну
синтезу деяких видёв РНК, порушення системи ферментёв, якё репарують ДНК
пёсля ушкодження. Значно змёнюеться ДНК мётохондрёй.
Пёд час аналёзу вёкових змён, якё рееструються в системах рёзноё складностё,
привертають увагу високий ёхнёй ступёнь ё визначенёсть на вищих ргвнях органё-
зацёё порёвняно з нижчими. I хоча основш процеси, що характеризують старёння,
вёдбуваються, очевидно, на нижчих рёвнях органёзацёё — молекулярному ё клётин-
ному, вёковё змёни бёльше проявляються там, де потрёбна ёнтегральна актившсть
багатьох елементёв, тобто на рёвнё фёзёолопчних систем ё органёзму в цёлому. При
цьому ефектившсть багатьох процесёв бёлыпою мёрою залежить вёд збереженостё
мехашзмёв регуляцп, якё сприяють пёдтриманню гомеостазу.
I
Старшня । хвороби
Отже, старшня призводить до зниження практично всхх функцш оргашзму. Це
значно обмежуе здатшсть адаптуватися до змш, компенсувати порушен! функцих
може спричинити виххд життево важливих показникгв за меж!, як! забезпечують
збереження сталостх внутрхшнього середовища оргашзму, 1 виникнення патолог!!.
ГЦдвищена частота розвитку рхзних патолопчних процес!в, що спостерхгаеться в
старост!, е не лише наслщком несприятливих впливхв на оргашзм протягом жит-
тя. Пщтрунтя пщвищено! захворюваност! в осхб лхтнього х старечого вжу сл!д
шукати у вхкових змшах юптин, оргашв ! систем, а хвороби старост! внутршшьо
властив! процесу старшня, хоча глибинш мехашзми, як! пов’язують впс 1 патоло-
пю, багато в чому залишаються незрозумхлими.
Перебп патолопчних процес!в у старечому вщ! значно в!др!зняеться в!д пере-
бпу в молодшому виц, ступшь компенсаторних процесхв, ппертрофц та регенера-
цц нижчий через значке зменшення резервних можливостей старого оргашзму.
Це робить оргашзм у старост! уразлившшм, менш здатним протид!яти хворобам
Порушення секрецп та рецепцп гормонхв х мед!атор!в, неправилен! реакцп на ни-,
зумовлюють, наприклад, менш чггку гарячкову реакцпо, запальний процес, за-
гальний адаптацшний синдром хз млявим перебйом. Часто переважають процесс
ушкодження, а не захисту, вхдновлення.
Характерним для старшня е певний спектр хвороб: серцево-судинш, шфек-
ц!йш, злояк!сн1 пухлини, дегенеративш хвороби мозку х внутрхшнхх оргашв, цук-
ровий дхабет типу 2. Зх збхлъшенням вхку ймовхршсть тяжких хвороб, як! спричи-
нхохоть смерть, зростае. Так, смертшсть вхд серцево-судинних хвороб протягом
кожного десятирхччя збхльшуеться в 2—3 рази. Так само закономхрно збхльшуеть-
ся смертшсть в!д шфекцшних хвороб х раку. Така сама картина смертност! спос-
терхгаеться х в пхддослхдних тварин. Атеросклероз не е характерною хворобою для
бхльшостх тварин (хоча спонтанний атеросклероз спостерхгаеться при старших ..
слонхв, китхв, свиней, людиноподхбних мавп, курей х голубхв), але збхльшуеться
частота розвитку хнших видхв ураження судин, наприклад, вузликового перхар-
терпту х склерозу гломерулярного апарату нирок у гризуюв х собак. Щодо частота
розвитку пухлин, дистрофхчних змш у внутрхшнхх органах, остеопорозу х хвороб
суглобхв, то вона, як х в людей, збхльшуеться з вхком.
За найоптимхстичшшими прогнозами, повна перемога над серцево-судинни-
ми хворобами х пухли нами сприятиме збшьшенню середньо! тривалостх житгя
людини на 10—12 рокхв. Ураховуючи велику залежшеть цих хвороб вщ змш, ш.
вхдбуваються в оргашзм! в процес! старшня, очевидно, ефективнппими будут*
заходи, спрямоваш на сповхлънення вхкових змш.
3 цього погляду щкаво розглянути питания про швидкхсть перебхгу стархння
в рхзних шдивхдуумгв одного виду. Тривалхсть житгя, як уже зазначалося, е ре-
зультатом взаемоди зовшшшх 1 внутрхшнхх факторгв. Своею чертою внутрхшг.
фактори зумовлеш спадковхстю х представлен! двома компонентами. Перший й
них — життездатшеть, яка забезпечуе пристосувальш можливостх оргашзму ! за-
лежить вхд багатьох гешв. Другим компонентом е темп старшня, тобто швидкхст
вгкового зниження життездатност!, що пов’язано з одним або кхлькома генами. У
популяцй е певний дхапазон коливань життездатностх х темпу старшня, який зу-
мовлюе хстотш вархацп тривалостх життя рхзних людей.
Одним 13 методов оцшки загального стану здоров’я оргашзму 1 швидкост! його
старшня е визначення так званого бюлопчного вису. Бюлопчний в!к людини часто
не вщповщае календарному. Його ощнка грунтуеться на всеб!чному аналгз! деяких
ключових ф131олопчних параметр1в (стану серцево-судинно!, дихально! та нервово!
систем, здатносп переносити ф!зичне навантаження тощо) 1 дае змогу скласти уяв-
тення про р!вень здоров’я людини, шдивщуальну швидгасть старшня, вщповщшсть
його вшовим стандартам, що може виявитися прогностично цшним.
Одш люди старпоть швидко, шип — повыъно. 1нколи спостерйають патоло-
пчш стани, для яких характерний швидкий розвиток змш, як! нагадують старечь
Прикладом такого прискореного старшня е деяк! рщюсш спадков! синдроми, що
отри мали назву прогерш.
Синдром Гетчшсона — Плфорда е рщюсною автосомно-рецесивною хворобою
дитячого в!ку. Симптоми хвороби починають проявлятися на першому рощ жит-
тя, швидко прогресують ! закшчуються смертю протягом першого — на початку
другого десятир!ччя, найчастиие вщ шфаркту мюкарда. Характерними ознаками е
затримка росту, посившня волосся, облисшня, стоншешсть 1 в’ялють ипари, ка-
таракта, глаукома, емф!зема легень, гиперхолестеринемия, атеросклероз, порушен-
ня у м’язовш 1 юстковш тканинах. Причиною цього е мутацхя в геш, що вщпов!-
дас за синтез быка мембрани ядра — ламшшу.
Синдром Вернера також автосомно-рецесивна хвороба, шо виникае у вщ! 15—
25 рокгв. Характерними ознаками е затримка росту в пубертатний перюд, поси-
вшня волосся, облисшня, атрофтя шюри й гшеркератоз, катаракта, зниження
гостроти слуху, розвиток цукрового д!абету, атеросклерозу, що спричинюють ш-
фаркт мюкарда, 1 висока частота виникнення пухлин мезенхгмного походження.
У раз! цього синдрому виявлено мутацпо в короткому плеч! хромосоми 8, у ло-
кус!, що вщповщае за синтез ферменту гелгкази, який розплггае парку сшраль
ДНК, що необхщно для репарацп, репл!кацц й експреси гешв.
Характерними для обох синдромов е патолопчш змши сполучно! тканини. Ц!
змши надають !м под!бност! до природного старшня. Ф!бробласти вщ хворих на
прогерпо погано ростуть у культур!, на них мало виражеш антигени пстосумю-
ност!, порушений синтез ДНК.
Деяк! ознаки, що нагадують старшня, можуть бути виявлеш також у людей з
аномал!ями статевих хромосом, наприклад при синдром! Шерешевського — Тер-
нера, а також при хвороб! Дауна.
Причини мехашзми старшня
Нин! не можна дшти остаточного висновку про причини старшня. Численш ппо-
тези щодо цього можна подшити на дв! основш групи. Ппотези першо! групи
передбачають, що старшня е наслщком накопичення невщновлюваних ушкод-
жень, яких оргашзм зазнае протягом життя. Ушкоджувальними факторами мо-
жуть бути юшзуюче випромшювання, у тому числ! й фонове, а також випадков!
флюктуацц рН ! температури в м!крооб’емах середовища. Кр!м того, велику уш-
коджувальну дпо мають в!льш радикал и та пероксиди, як! можуть утворюватися
внаслщок окиснення речовин в оргашзм!. Як вщомо, щ сполуки х!м!чно надзви-
чайно активш 1 можуть спричинювати ушкодження молекул, розриваючи !х або
утворюючи внутршшьомолекулярш та мгжмолекулярн! зшивки. Виявлене нако-
пичення год час старшня погано розчинного колагену й лгпофусцину, можливо,
е наслщком такого вгльнорадикального окиснення.
Найзагрозлившп наслщки при старшнг можуть мати ушкодження молекули
ДНК або ферментних систем, яю беруть участь у синтез! нукле'шових кислот,
осюльки це мае супроводжуватися накопиченням помилок г суттевих змш у гене-
тичному апаратг, виникненням мутащй. € припущення, що накопичення одних
помилок згодом призводить до появи нових, зумовлюючи лавиноподгбне нарос-
тання процесу, г закгнчуеться “катастрофою помилок”. Дшсно, пщ час старшня
збгльшуеться частота розвитку хромосомних аберащй у клгтинах, однак немае
доказгв почастппання мутацгй г появи “помилкових” бглкгв. Кргм того, в оргашз-
М1 е могутш захиснг системи — антиоксиданта!, що репарують ушкоджену ДНК,
контролюють появу аномальних бпгкгв. Однак швидюсть змш, яю з’являються в
раз! старшня, вища за ту, яка могла б бути зумовлена згаданими випадковими
процесами.
Згщно з гипотезами другог групи старшня е генетично запрограмованим про-
цесом, що виробляеться в еволюци як шструмент, який обмежуе тривалгсть ш-
дивщуального життя. 3 огляду на це швидка змша поколгнь сприяе кращш при-
стосованост! до умов гснування г захищае вид вщ вимирання бгльшою игрою, нгж
потенцшне безсмертя окремог особи. Яскравим доказом цього е велика вщмш-
нгсть у максимальнгй тривалостг життя тварин ргзних видгв. У такому разг старш-
ня слщ розглядати як один з етапгв онтогенезу, коли досягнення однгег фази
розвитку супроводжуеться змшою г виробленням гндукгоргв для наступно! фази.
Данг, отриман! при пересадженнг тканин молодо! тварини старш г навпаки свщ-
чать про активний вплив тканин старого органгзму у бгк старшня. Деякг дослщ-
ники припускають, що генетична запрограмовашсть виникае як наслщок про-
цесгв диференцгацп органгзму, що автоматично продовжуються в пострепродук-
тивний пергод, який перебувае поза дгею природного вщбору.
€ кглька варгантгв мехашзму розвитку вгкових змш (мал. 11).
Варгант перший передбачае, що причини старшня приблизно однаковою
игрою й одночасно дгють на ргзнг елементи органгзму (позначен! А, В г С), зумо-
Мал. 11. Мехашзм розвитку старшня:
а — “р1вном1цний вГзок”; б — “водш” старшня; в — “годинник”. Строками позначено зовншш! та
внугрГшш впливи, що спричинюють старшня окремих систем (А, В, С), ) взаемш впливи мЬк ними.
Заштрихован! дглянки позначають р1вш зниження функцп з вжом
влюючи ргвнокпрне !х порушення. Вариант другий видные одну ланку в оргашзм!,
яка через свою слабюсть або шдвищене навантаження на не! першою виходить з
ладу, стаючи надал! своерщним “вод!ем” (пейсмекером) в!кових змш 1 зумовлю-
ючи вторинш змши в шших, спйюших ланках. На роль такого “вод!я” старшня
могли б претендувати гшоталамус, етф!з, сполучна тканина, Гмунна система.
Третш варгант передбачае, що старшня виникае як наслщок дхяльносп певних
мехашзм1в, для яких генеращя вжових змш в шших тканинах е гхньою нормаль-
ною функщею (“годинник”), а не наслщком якихось ушкоджень. Така функцю-
нальна актившсть механизму може бути неоднаково сприйнятою ргзними органа-
ми або диференпшовано адресуватися рхзним тканинам. При цьому не обов’язково,
щоб роль “годинника” протягом усього процесу старшня належала одним 1 тим
самим утворам, вона послщовно може переходити вщ одних до шших.
Накопичеш експериментальш даш не пщтверджують перший 1з пропонова-
них мехашзмгв старшня, а подальнп дослщження мають дата пщстави для вибору
одного з двох, що залишилися.
Шляхи впливу на старшня
Можливгсть сповыьнення процесу старшня пов’язана з! створенням експеримен-
тально! модел!, на як!й можна було б регулювати темп впсових змш. € илька
методгв, за допомогою яких можна вщтворити прискорення окремих процесгв, яю
спостерп'ають пщ час старшня тварин, однак поки що не вдаеться вщтворити весь
комплекс В1КОВИХ змш, яю при цьому розвиваються. Вщомц наприклад, деяю
лшц експериментальних тварин, у яких в!ков! змши окремих систем вщбуваються
шби в прискореному темп!. До них належать атимусш, гол!, карликов! ппошту!-
тарш мини, деяю лшГ! мишей 1 щур!в з! схильшстю до автсймунно! патологп,
спонтанних злояюсних пухлин, цукрового дгабету або артер!ально! гшертензц.
Кр!м того, процес старшня намагаються прискорити у тварин шляхом трива-
лого впливу низьких доз юшзуючого випромшювання, призначення дхети, бщно!
на антиоксиданта (речовини, яю перешкоджають утворенню пероксщцв в ор-
гашзм!), деяких препарат!в кальциферолу — в!там!н!в групи В (дигщротахюте-
рол), тривало! гшодинами, створення в оргашзм! хмунного конфлгкту (наприклад
у раз! парабюзу партнеров, як! вщр!зняються один вщ одного слабким антигеном
тканинно! сумюност!), парабюзу тварин протилежного (молодого — старечого)
в!ку тощо.
Генуе також юлька шляхзв подовження життя експериментальних тварин.
Один 13 них полягае у сповшъненга вйсових змш. У теплокровних тварин найзнач-
нши результата отримують у випадку перюдичного або тривалого утримання тва-
рин на д!ет!, яка стримуе рют, яюсно повноцшюй, юльюсно обмежешй. Таку
дгету вперше застосував Мак-Кей 1 досягнув значного (на 1/3) збыьшення трива-
лост! життя щур!в. Под!бш результата було одержано в шших видгв ссавщв. Об-
меження д!ети сповЬгьнюе в!ков! змши з боку нейроендокринно! та гмунно! сис-
тем, зменшуе частоту розвитку хвороб судин 1 пухлин. Однак у молодому вщ!
обмеження д!ети призводить до зменшення резистентност! та пщвищення р!вня
смертност! (мал. 12). Це примушуе утримуватися вщ рекомендацш щодо обме-
ження ддети в людини. Установлено, що надм!рне харчування, недостатня витра-
та енергп та ожирпшя е несприятливими факторами, яю пщвищують захворю-
Мал. 12. Смертшсть тварин, яких угримували на
обмежувалыпй (Л) 1 стримуючш р!ст (Б) даеп
вашсть 1 зменшують тривалють житгя.
Тому рацюнальне збалансоване харчу-
вання без пере'щання 1 помгрш фгзичш
навантаження (вщповщно до можли-
востей оргашзму) сприятливо вплива-
ють як на загальний стан здоров’я, так
1 на тривалють житгя.
Поспйне зниження температури
пла у пойюлотермних (з непостшною
температурою пла) тварин також спри-
яе значному збщыпенню тривалосп
житгя.
Певного успеху вдалося досягги
шляхом тривалого застосування речо-
вин, яю блокують синтез нуклешових
кислот.
Другий шлях полягае у застосуван-
ш метод1В, спрямованих на подолання висових зм1н, що еже виникли, або 1х насл1д-
к1в. Тут можливими е замша органа або юптин, яю постарши, втручання в гене-
тичний апарат юптини для вщновлення або розгальмування певних дшянок гено-
му, активащя певних ферменпв. Слщ однак зазначити, що клпини й органи
молодих тварин, уведен) в старий оргашзм, швидко старпоть, а для цшеспрямова-
но! змши геному ще не розроблено належних методтв.
Як згадувалося, у виникнешп ушкоджень на молекулярному р)вш велику роль
вщгграе вгльнорадикальне окиснення речовин. Тому деяю дослщники для подов-
ження життя застосовують антиоксиданта (токоферол, аскорбшову кислоту, ру-
тин, штучно синтезоваш речовини, що дають такий самий ефект) з метою за-
поб1гта цим ушкодженням. Щоб запобпти виникненню, а також для руйнування
поперечних водневих зв’язюв у молекулах пропонують застосовувати невелику
юльюсть хелатоутворювальних сполук 1 латирогенгв — речовин, яю руйнують
м1жмолекулярн1 поперечш зв’язки. Кр1м того, застосовують метода 1 засоби (д!е-
тичш, стимулюючг, лгкувальш), яю полшшують д)яльшсть ргзних оргашв 1 систем
в оргашзм) людини, що стар!е, а також спрямован) на боротьбу з наявними пато-
лопчними процесами. Останн1м часом у свгп поширюеться рух за здорове “ус-
пшше” старшня, який грунтуеться на використанш досягнень науки з ращональ-
ного створення активного способу житгя, уникнення шкщливих звичок, шоб вщ-
строчита появу характерних для старшня патолопчних змш.
Роздш VI
ПАТОЛОПЧНА Ф13ЮЛ0ПЯ СПОЛУЧНОГТКАНИНИ
СПОЛУЧНАТКАНИНА ЯК ЕДИНА СИСТЕМА
Сполучна тканина — це комплекс клйпин, волокон / основно!речовини, яю об'сднуютъся
спиътстю походження I виконуваних функцш.
У вивчення функщй 1 структури сполучноУ тканини значний внесок зробили
1.1. Мечников, О О. Максимов, О.О. Заварзён, Л. Ашоф. Особлива заслуга в цьому
належить 0,0. Богомолы] ю, який першим запропонував назвати сполучну ткани-
ну фнзюлоглчною системою сполучно! тканини. Найхарактернгшими ознаками цгег
системи е унУверсалыпсть, гетерогеннгсть У висока пристосовашсть (А.В. Шехтер).
Ушверсалыйстъ сполучноУ тканини визначаеться значним Ут поширенням в
органёзмг; строма внутрУшнУх оргашв, основа шкУри, серозног ё синовёальноТ обо-
лонок, зв’язок, сухожилкУв, апоневрозУв, оболонок м’язУв г нервУв, утворення су-
ДИНН01 стУнки.
Гетерогенность зумовлена особливУстю клУтин У мУжклУтинних компонент^,
якУ представляюгь сполучну тканину. Три типи клУтин У волокнистих структур,
кУлька типёв глУкозамУноглУканУв — усе це робить нескёнченною кёлькёсть комбг-
нацУй, якУ можуть виникнути год час патолош сполучноУ тканини. Усг клУтини
сполучног тканини здатнУ до розмноження та дисререншювання, шо зумовлене
наявшстю клУтин рУзного ступени зрёлостё У функщонально’У активноси. Усе це
створюе всличезний резерв, здатний мобййзовуватися органУзмом для пластичних
та шших цУлей, наприклад тд час загоення ран.
1з гетсрогеннёстю сполучноУ тканини пов’язана г незвичайна рУзноманУтнУсть
ц функцгй. 0.0. Богомолець розргзняв пластичну, захисну, трофгчну та опорну
функцп. До них згодом додали ще й морфогенетичну. Сполучна тканина е не
лише субстратом для прикрепления клУтин (“просторовий оргентир”), а й впливае
на процес гх диференцгювання та морфогенез. Припускають, шо в колагенових
золокнах е полёпептиднё дУлянки, якё передають клгтинам, шо з ними контакту-
ють, сигналя, що сиецифУчно впливають на гхнгй метаболёзм г рухову актившеть.
Регулящя функщй сполучноУ тканини здУйснюеться на вегх ргвнях орган гзацп:
клУтинному, органному, цглого оргашзму. На клгтинному рУвнё мають значения
мгжклУтиннё контакта, ефекторш речовини, тгено пов'язаш з мембраною клгтани,
а також медгатори, якУ видьгяються у мёжклгтинний простёр лимфоцитами (лУм-
фокёни), моноцитами (монокгни), фУбробластами (фёброкёни), тканинними базо-
фглами, у тому чиелг базофёлъними гранулоцитами (лаброкёки). Кргм специфгч-
"П7
них мегпатор!в. для яких на юптинних мембранах е вщповТдн! рецеитори, таку
саму роль виконують просгагландини, мурам!даза, фибронектин, протешази.
ВзаемодТя мок елементами сполучноТ тканини здшснюеться за принципом
зворотного зв’язку, що за нормачьних умов забезпечуе адекватшсть вщповцц, а
при патолопТ — високу пристосовашсть 1 надтйшстъ. Авторегулящя, шо грун-
туеться на кооперативнш взаемоди м!ж юптинами, доповнюеться ендокринною
та нервовою регулящею. побудованою за 1ерарх!чним принципом (згори донизу).
У цьому важливу роль вщшрае соматотропш (гормон аденоппофзза), який сти-
мулюе розмноження клпин сполучноТ тканини та синтетичш процеси в них. Кор-
тикотрошн 1 глюкокортикоТди пригшчують прол1ферацпо, спричинюють перед-
часне диференщювання та дозртвання ф’бробласпв. що супроводжуеться пору-
шенням колагеногенезу. Роль шсулшу в регуляпп структури 1 функцш сполучноТ
тканини поля гае в прискоренш обм)ну палуроновоТ кислот и й хондроТтинсульфа-
ту, що пояснюе серйозш порушення сполучноТ тканини взагалт та судинноТ стш-
ки, зокрема при цукровому д!абеи (дТабетична ангТопатТя).
Порушення регуляцп системи сполучноТ тканини можуть вщбуватися на будь-
якому р)вш. У самгй сполучнш тканин) можуть порушуватись авторегулящя та п
зв’язки з юптинами паренхТми. Порушення може виявлятися в тому або шшому
орган) (у суглобах при ревматизм 1, шюр! при системному червоному вовчаку), в
оргашзм) в щлому (розростання юсток при акромегалп, карликовхстъ 1 слизовий
набряк при ппофункцц щитоподТбноТ залози).
Стан сполучноТ тканини вщТграе важлтгву роль у процес! стар)ння. Свого часу
О.О. Богомолець зазначав, що “старшня починаеться саме з! сполучноТ тканини”.
Причину цього вш вбачав у ф!зико-х1м!чних змшах п макромолекулярних компо-
нентов — “дозрхванш юптинних колошТв 1 мщелоцпв, перетворенш Тх на преци-
штати та флокуляти, що утворюють бюлопчно шерил включения, як! гальмують
житгед!яльн!сть юптин”.
Зпдно з результатами сучасних дослщжень, з в)ком у сполучнш тканин) в)д-
буваються змши, яю порушують и трофочну, захисну та шип функцн. У шюр!, су-
хожилках. хрящовш тканин!, аорт) зменшуються кьтыпсть юптин, Тх розмгр, а та-
кож розмор ядер. У людей лггнього вгку розмноження ф!бробласт)в при запаленн)
словшьнене. У волокнистих структурах в!ков) змши лроявляються зб!льшенням
юлькоси поперечних зв’язюв м!ж волокнами колагену. Колаген !з надм!рною
юльюсио макромолекулярних зшивок набувае нових властивостей. Вш стае стш-
ю шим до температурних вплив)в, знижуеться його розчиншсть, здатшсть зв’язувати
воду. Метаболзчно стабшъний у норм!, колаген стае дедал! шертншшм. Сповьть-
нюеться його самовщновлення, що неминуче призводить до накопичення в моле-
кулах нових помилок. В основнш речовин! зменшуеться вмтст палуроново! кисло-
ти, шо знижуе з вгком здатшсть тканин зв’язувати воду. Водночас збьтыпуеться
юльюсть хондроТтинсульфату в судиннш стптщ. Останне зумовлюе кальцифйсацпо
судин, осюльки сульфатоваш гл!козамшогл!кани спорщнеш до ююв кальцпо.
Захисна функщя сполучноТ тканини проявляв гься в тому, шо як структурний
елемент шюри, слизовоТ га волоки исто!. (ф)брозноТ) оболонок, а також спещал!-
зованих бар’ерТв вона виконуе роль мехашчно! переткоди. Деяю клпини сполуч-
ноТ тканини здатш фаплштувати (макрофаги), а також вшокремлювати ушкодже-
ну тканину7 в!д нормально! (грануляцшний вал). Нареши, юптини сполучноТ тка-
нини, вступаючи у взаемодйо з лТмфоцитами, берулъ участь в гмуншй вщповщ!.
0,0. Богомол едь був першим, хто повною м)рою ошнив захмсну роль сполуч-
но] тканини. Вш та його учш довели, що низька реактивность сполучно! тканини,
як правило, поеднуеться з тяжчим перебпом хвороб (повыьно загоюються рани,
прше зростаються к)стки теля перелому). Усе це дало змогу зробити припушення
про те, шо, впливаючи на сполучну тканину, можна попршити або покращити
переб1г хвороби, потртбно лише винайти способ стимулювання Г! функцш. Певних
результатов можна було б досягти завдяки застосуванню таких методов, як локу-
валъна ф)з культура, масаж, шеоляшя, д)ета. Однак 0.0 Богомолець мав на у ваз)
специф)чну вибфкову стимуляцию вегх елемент’ш сполучно! тканини. де б вони в
оргашзм) не м)стилися Цього було досягнуго гмунолопчним шляхом, тобто вве-
дениям людин 1 сироватки, добуто! теля )мушзацп тварин (коней) органами, ба-
гатими на сполучну тканину (юстковий мозок, селезенка). Ця сироватка отримала
назву антирепшкулярна цитотоксична сироватка (АЦС) ) була детально вивчена
0.0. Богомольцем та його учнями, а пойм застосована в клшщ).
Встановлено, що ефект АЦС залежить в!д й дози. Висок) дози сироватки справ-
ляли питотоксичну Д1ю, тобто руйнували клггини сполучно! тканини, низьк) —
давали стимулювальний ефект. Це проявлялося шдвищенням обм)ну речовин у
макрофагах тканини га активацц фагоцитозу, збыьшенням титру прочим,кробних
антит)л, нормал )зашсю волно-мгнерального й лппдного обм)ну. Активапы фер-
ментних процесгв дае змогу сполучнш тканин) звыьнятися вш баластних речовин,
як) накопичуються в шй пщ час р)зних хвороб обм)ну речовин, а також у процес)
старшня. При цьому можуть утворюватися речовини, яю справляють неспециф1Ч-
ну Д1Ю на клттини оргашзму 1 дають загальний стимулювальний ефект.
ПОРУШЕННЯ СТРУКТУРИ I ФУНКЦ1Й СПОЛУЧНО! ТКАНИНИ
До клйнин сполучно! тканини належать ф)бробласти, макрофаги (пейоцити), базо-
фыьн) гранул оцити (тканинн) базофыи, лаброцити).
Ф1бробласти — основн) клггини сполучно! тканини. 3 !х життедгяльшетю
пов’язан) утворення ) видыення в середовише основно! м)жкл)тинно! речовини
(глжозамшоглйсашв), а також синтез ) секрецтя волокнистих структур (колагену,
еластцну, ретикулшу). Спадков) та набут) порушення синтетично! функцп фибро-
бласты € основою мукопол)сахаридоз)в ) дифузних уражень сполучно! тканини.
Макрофаги поход ять вы стовбурово! кроветворно! клггини юсткового мозку.
Як мононити вони заносяться з током кров] в р)зш органи, де залежно вщ кл)тин-
ного оточення, набувають фенотипових особливостей. Головною функщею цих
юптин е фагоцитоз. У печ)нш !х називають з)рчастими ретикулоендотелюцитами
(клггини Купфера), у легенях — альвеолярними макрофагами, у серозних порож-
нинах — плевратьними та перитонеальними макрофагами, у ктстковш тканин) —
остеокластами, у нервовш тканин) — клггинами мгкроглн. Завдяки структурним
особливостям цим клггинам властива висока фагоцитна ) метабол)чна актившсть.
Цитоплазма макрофапв м:стить велику кшьюсть вакуоль ) фагосом з) значним
вмютом ферменты. На поверхневих мембранах цих клггин е рецептори, контакт з
якими р)зних речовин, у тому числ) й антигешв, зумовлюе збудження. При цьому
вони можуть викидати в середовише р)зн) ферменти (колагеназу, еластазу), а та-
кож антибактерталъш речовини (л)зоцим). Макрофаги виробляють спещальн) ре-
човини, як! за аналойею до лшфокшш отримали назву монокпнв. Моною
вптивають на шгш юптини, як! перебувають поблизу (мшрооточення): активп.
одних стимулюють (колошестимулювалъяий, л1мфостимулювальний факте р
шших — пригшчуютъ (шпбггор росту КЛ1ТИН тощо). У цьому полягае важлг*.
роль макрофапв гад час запалення.
Ус1 макрофаги виконують едину функшю — очищения оргашзму вщ ипо.т
вих 1 стороншх речовин, мертвих або живих клггин, у тому числ! пухлинн.
бакгерш, в1рус1в, продукты вщпрапьовано! автолопчно! тканини, антигены, лг
карсысих речовин, мжроагрегатгв ф<брину тощо. Усе, шо потрапляе в клгпп |
лщдаеться бютрансформацн, а поим виводиться, у тому числ!1з секретами (же
чю, единою, потом). Завдяки радюнуюпдному дослщженню фагоцитного кд!рс
су, тобто швидкосп очищения оргашзму вы будь яко! речовини (наприклад оке -
ду торпо у вигляд! колошу), доведено, що в норм! очищения вщбуваеться за пе
ний час (фагоцитний шдекс). Фагоцитний !ндекс використовують для вивчен-
потужност! системи мононуклеарних фагоциты (СМФ) за р!зних ушкодже>д|
(травма, оп!к, хфурпчне втручання). При гшокси печшки (у норм! ц фагощп>-
актившеть сгановить близько 80 % в!д загально!) фагоцитний юпренс р!зко зе
жений. Выомо, шо блокада ц!е! системи гндиферентним колощом, напршс
тушшю, знижуе резистентшсть оргашзму до травми, а и стимулящя (естрогенами. I
холгном, зимозаном, АЦС) гадвищуе огаршеть оргашзму.
У раз! патологи системи макрофапв порушуеться основна и функшя — здш
шеть захопдювати 1 перетравлювати сторонн! та шкщлив! для оргашзму речовинт
У юпшга вшома група хвороб, як! отримали назву хвороб накопичення.
Базоф1льш гранулоциты м!стять у цитоплазм! гранул и, яю мстахромагично за-
барвлюються ашлшовими барвниками. У гранулах е високоактивн! речовинт
пстамш, гепарин, серотонш (у щур’ы). Пстамш у юйтинах представлений не?
тивною формою, осюльки зв’язаний з быково-гепариновим комплексом. Вык-
ипеть пстам1ну мютиться в базофыьних гранулоцитах сполучно! тканини 1 крс I
(на 10° клггин кров! припадае 20—30 мкг пстамшу). Вивыьнення пстамшу з гра
нуд (дегрануляиы) вщбуваеться пщ шею л!бератор!в — речовин низькомолеку-
лярно! (моноамши, замипен! ароматичн! ам!ни, тубокурарин, морфш) 1 високо-
молекулярно! (декстрани, пептони, протеолгтичш ферменти, отрути, комплекс!
антиген — антитыо) природи. Провес дегрануляцГ! вщбуваеться з використанняз
енергп АТ Ф.
Пстамш, що вивыьнився з гранул тканинних базофыьних гранулоциты, з)
мовлюс характерш змши в тканинах, а саме бронхоспазм, розширення судин
збшьшення проникносп шлю! стшки (див. “Запалення”).
Основна речовина сполучно! тканини. Найважливппим компонентом сполучнт
тканини е основна мгжклггинна речовина, або матрикс, яка складаеться переваж-
но з глгкозамшоглгкашв (мукополисахариды). Останш е високомолекулярним!
полгеахаридами, до складу яких входять амшоцукри, залишки глюкуроновен та
сгалових кислот, ашони сгрчано! й оцтово! кислот.
У сподучшй тканин1 людини е 9 типы глгкозамшоппкашв, як! вщргзняються
один в!д одного набором амшоцукры (галактозамш, глюкозамш), наявшстю абс
вщеутшетю сульфатних груп (сульфатоваш та несульфатоваш гл!козамшогл!ка-
ни), типом зампцення в амшнш груп! тощо. Сшлъним для них е те, що полгеаха-
ридн! ланцюги з приблизно однаковою молекулярною масою прикршлюються до
одного бокового стрижня, формуючи таким чином велим полтмернт молекули —
протеоглткани. Структура глткозамтноглткантв змтнюеться залежно вщ виду ткани-
ни (пиара, хряш, ропвка, склисте тыо тощо).
Бюлопчне значения глткозамтноглткантв не обмсжуеться тим, шо вони е “мас-
тильним”, “склеювальним” т ‘‘опорним” материалом Цт бтополтмери, легко утво-
рюючи комплекси з гншими молекулами, можуть затримувати та вившьнювати
ртзнт речовини. у тому числт сторонн! г токсичнт. Структурна г хтмтчна органтзацгя
глткозамтноглткантв така, що вони можуть утворювати впорядкован! сггки з пора-
ми певних розмлртв, як! забезпечують селективну проникшсть для ртзних речовин
(молекулярне сито). Саме в цьому полягае !х бар’ерна роль. Глткозамтноглткани
можуть зв’язувати велику ктльктсть води (близько 500 молекул на одну макромо-
лекулу). Порушення шет властивостт може призвести до зневоднення або, навпа-
ки, до надапрного накопичення води (мткседема).
Синтезуються глткозамтноглткани фтбробластами т макрофагами. Цей процес
каталтзуеться специфтчними ферментами — глткозилтрансферазами, вит актив-
ностт яких залежать швидктсть складання полтсахаридних ланцюгтв т чергування в
них глткозильних залишктв Для глткозамтноглткантв характерним е тнгенсивний
метаболтзм. Наприклад, пертод птвжиття палуроновот кислоти (одного з глткоз-
амшоглжантв) становить 2—4 дш. Деградащя тлткозамтноглткаюв вщбуваеться пе-
реважно в клпинах (макрофагах). Гол овну роль у цьому вщтграють лтзосомш фер-
менти: екзо- й ендоглткозидази, сульфотщролази т, можливо. протетнази.
Патолопю глткозамтноглткантв зумовлюе порушення тх синтезу або розпаду
Описано деяк! .хвороби (синдроми), об’еднант пщ назвою мукопол1сахаридоз. Най-
характернтшим для цих хвороб е накопичення глткозамтноглткантв у клпинах т
водночас збиьшене видыення тх з фекалтями. Клттини мезенхтми та паренхтми
виявляються переповненими глткозамтноглтканами, якт не катаболтзуються, що
може зактнчитись загибеллю клттин. Причиною цих хвороб е дефщит ферменттв,
потртбних для деградацп глткозамтноглткантв.
Зтдцно з результатами дослщження т уИго фтбробласттв людей, якт хворттоть
на мукополтсахаридоз, мтчеш радтоактивними речовинами попередники глткоз-
амтноглткашв засвоюються ктггинами, але повыьно виводяться з них. Навтть птс-
ля триденного перебування в культуральнш рщинт 70 % радюактивнот мттки ли-
шаеться всерединт клттин. Мтсдем накопичення глткозамтноглткантв у клттинах е
лтзосоми. Якщо в середовище, де культивуються цт фтбробласти, внести нормаль-
нт клттини, то швидктсть катаболтзму глткозамтноглткантв збтльшуеться. Очевидно,
нормальш клттини видтляють яктсь речовини, здатнт проникати в клттину хворого
га усувати наявний дефект. Такими коригувальними речовинами е .лтзосомш фер-
менти, дефщит яких при мукополтсахаридоз! мае спадковий характер.
У тканинах людини е кгтька тип!в глткозамтноглткантв: хондропинсульфати
(найпоширентшт глткозамтноглткани, мтстяться в хрящт, шктрт, сухожилках, зв’яз-
ках, артертях, ропвцт ока); гепарин; гепарансульфаз (входить до складу протеоглт-
кашв базальних мембран, е поспйним компонентом ктттинно! поверхнт); кера-
тансульфати (найбтльш гетерогенит глткозамтноглткани, вщртзняються один вщ
одного сумарним вмтстом вуглеводтв, мтстяться в ктстковтй т хрящовтй тканинах,
ропвцт); палуронова кислота (мтститься в багатьох органах, у хрящт зв’язана з
битком т бере участь в утворенн. протеоглтканових агрегаттв, у скли стому ты! ока,
пупковот»гу канатику та суглобовш рщинт перебхъае також у выьному станг); де-
рматансульфат (значно поширений у тканинах тварин, особливо характерний д_.
шктри, кровоносних судин, клапашв серия, у склад! малих протеоглшашв — бй -
кан 1 декорин — мктиться в мГжклггиншй речовин! хрятшв, мтжхребцевих диски
менкюв), Катаболхзм кожного з них визначаегься спениф!чним ферментом, ш
пояснюе розмаптя нозолопчних одиниць, якг вщображають хвороби накопичен-
ня. При накопиченн! в кл!тинах ! пперекскрецй дерматансульфату 1 гепарансулъ-
фату (низька або нульова актившсть глюкурошдази та палуроншази) виникае сис
дром Гурлера, хондро!тинсульфату 1 кератансульфату (дефект ферменту галактеза-
мш-6-сульфат-сульфатази) — синдром Маркхо, гепарансульфату (дефект фермен’
гепарансульфата зи, М-ацетил-а-В-глюкозамшщази чи ацетилтрансферази) —
хвороба Санфьпппо, дерматансульфату (дефект ферменту щуронатсульфатази) —
хвороба Гантера, хондроТтинсульфату (дефект ферменту 0-глюкуронщази) — хво-
роба Слая. Для вс!х них характерним е генерал !зоване ураження сполучноТ ткаш-
ни. Наприклад, Гурлер-подТбний фенотип визначаеться грубими змшами рш
обличчя, деформащею скелета, множинним дизостозом, гепатоспленомегалТе к
При синдром! Маркю спостер]гаються ураження скелета, карликовость, малор)
хомкть суглобтв, недостатшсть клапана аорта, помутнгння ропвки.
Другим нефтбрилярним компонентом основной речовини е гл!копроте'Тни —
пол (мери, як! мктять бьток 1 ковачентно зв’язаш з ним вуглеводш компонент!
Моносахаридними складниками глТкопротепмв е гексози, пентози й амшоцукр!
КТльккть вутлеводних ланцюпв ргзна — вщ одиниць до сотень. Важливим компо-
нентом гл!копротеТн!в е аалов! кислота, при ферментному вщщепленн! яких зма-
нюються властивост! гл!копротеТн!в, р!зко зменшуеться тривалкть Тх П1врознад>
Ус! глгкопротеТни под!ляються на три групи: функцюнатьш (ферменти, гар-
мони), штегроваш (у мембранах) ! структури!. Кожна !з зазначених труп виконуд
специфТчш функцн. Структури! гл!копротеТни утворюються у фхбробластах, зв!д-
ки надходять у навколишпе середовище. Туди ж виходять еластинов! та колаге-
нов1 бшки (проеластин, проколаген). Подальше дозр!вання цих фТбрилярних 61л-
к!в значною м1рою визначаеться наявнктю в середовипц глшопрогенов. Так
морфолопчн! особливост!, як д 1аметр фкрил, Тх розм!щення, еркнтування в про-
стор!, СП1ВВ1Дношения м!ж еластиновими та колагеновими нитками, пов’язаш
юлькктю та яккгю структур них глткопротеТнш. Змша синтезу структурних гль
копротеТшв позначаеться на регенераил, зокрема на загоенш ран, мае значения
при розвитку атеросклерозу, гломерулонефриту та шших хвороб
Структурна особливктъ гл!ко протеин в визначае Тх Тмуногеншстъ. В екс пери-
мент! на тваринах доведено, шо парентеральне введения кролям очишених струк-
гурних глГкопротеТнтв сприяе появт в кров! специфтчних антитш. Цими речовина-
ми можна шдукувати атерпчш реакцп у шур!в та морських свинок. 1нод! антит!"
до структурних глткопрогеТшв ниявлятсть 1 в людини. Пе пояснюеться тим, щс
глткопротеТни м!стятъся в мембранах мжрооргашзмГв ) пкля перенесено! шфекш
антипла, як! утворилися при цьому, можуть взаемодшти з гл!копротешами спо-
лучноТ тканини людини. Тому дуже важливо встановити, як! саме компоненте
сполучно! тканини можуть бута джерелом антигешв при автотмунних захворю-
ваннях, у тому числ! й при дифузних й ураженнях.
Порушення ВО.ЮКНИСТИХ структур сполучноТ тканини. Головною волокнистою
структурою сполучноТ тканини е колагеновг волокна. Основна функцш колагену —
пштримувати специф!чну структуру органш 1 тканин у продеа розвитку оргашз-
1. З’еднання трьох полтеп-
тидних ланцюпв
2. Присднання пдроксилш
3. Приеднання ГЛК1 ГАЛ
до гщрокситзину
4. Утворення дисульфщних
зв’язюв
5. Формування потрыно!
страт
7. Утвооення поперечник
зв’язюв кож молекулами
проколагену
Мал. 13. Основы етапи бюсинтезу колагену
му, що вщбуваеться завдяки здатност! впорядковувати 1 стабплзуваш юитинш
комплекси, з якими вш контактуе. Ця властивгсть забезиечуеться су во рою впо-
рядковаюстю 1 стабшьшстю самого колагену.
Колатен (його молекулу шод! називають тропоколагеном) сгановить 25—33 %
загально! юлькост! вс!х быюв оргашзму людини. Амшокислотний склад речовини
характеризуеться високим вмютом глщину (33 %). прол1ну та гщроксипролшу (21 %),
аланшу (10 %), оксилгзину, вщсутшстю триптофану та цистешу, дуже низьким
вм1стом тирозину та метюшну. И ото бюсинтез здшснюеться фгбробластами 1 не-
лосмугованими м’язовими волокнами вцшовщно до генетичного коду. 1нформа-
Ц1я, що передаеться через мРНК, реал!зуеться на рибосомах у вигляд! полшеп-
тидних ланцюпв, яю поим у цитоплазм! зазнають певних перетворснь, теля чого
вже позаклпинно утворюетъея зрший нерозчинний кола ген (мал. 13).
Розчинний колатен розглядають лише як пром!жну стадию мок мономерними
макромолекулами тропоколагену та ф!брилярними нерозчинними агрегатами цих
молекул. Розр!зняють нейтральмосолерозчинний та кислоторозчинний колатен. У
них фракц!ях колагену слабко розвинен! внутрштньомолекулярн! ковалентн! по-
перечн! зв’язки.
Порушення кожного з етатв синтезу може проявится хворобою сполучно!
тканини спадкового та неспадкового характеру. Мутаи!я гена, який кодуе утво-
рення колагену, може змгнити амтнокислотний профиь первинного полшептх
ного ланцюга, а це своею чертою позначиться на мщност! його фтбрили. О
зазначити, що колаген виртзняеться особливим сшввщношенням амшокислог 1
!х розмштеиням у ланцюгч. 3 усгх амшокислот трстина ирипадае на глщин, я1.
займае кожне трете мтсце в полшептидному ланпюгу. Порушення генетичн
коду, дефщит глщину або вщповщного ферменту можуть зумовити порушень
синтезу всте! молекули колагену.
Первиннт полшептидн! ланцюги (а-ланцюги), що утворилися в рибосомд
далт проходять юлька етагнв, чотири з яких можна визначити як критичш, оскг
ки саме тх порушення найчаслше призводить до патолопчних стантв.
Перший етап — ггдроксилювання мзину та промну (приеднання до заэначенв
амшокислотних залишктв ОН-груп). В!н потртбний для того, щоб мтж трьо*
первинними амптокислотними ланцюгами, об’еднаними мтж собою, виникли к
валентнт зв’язки г розпочалася сшралтзацтя потртйноТ молекули. На цьому еташ
потреба ь активних лтзин- або пролшгщроксилаз! та кофактор! — аскорбшовш
кислой.
Другий етап — утворення альдег1дних груп унаслщок дезам1нування лтзину
оксилтзину. Альдепдш групи двох сусщнтх а-ланнюпв, взаемодгючи м!ж собой
угворюють поперечнг ковалентн! зв’язки, погртбнт для подальше! стабйпзацп мс
лекул колагену. особливо там, де одна фтбрила закшчуеться 1 починаеться друг?
де вони на вщртзку 1/4 довжини накладаються одна на одну. На другому етаг
синтезу колагену необхщною е наявнтсть лтзиноксидази та кофактора, роль якого
виконуе мщь.
Трети етап — гткозилювання колагену. Молекула колагену завжди мтстить пер-
ну кйьктсть вуглеводтв (глюкоза або пикозилгалактоза), лрисднаних до гщроксил--
зину. Ця реакцтя каталтзуеться двома ферментами — галактозилтрансферазою
глткозилтрансферазою Значения приеднання вуглеводтв ще не з’ясовано. Очевид-
но, воно вщгграе роль у транспорт! готових молекул колагену з фтбробласттв у мтж-
клттинний просттр, в тх укчаданнт та подальших конформантйних змпгах.
Четвертей етап — секретя колагену в м1жклтинпий матрикс. Птсля того я-
колаген пройшов лерпп етапи синтезу й утворилися потршш сптралт, останн! за-
лишають клттину. Цей пронес утруднюеться при порушеннт гщроксилювання
пролшу га лтзину.
Контроль за синтезом колагену7 здгйснюеться на встх його етапах. У рибосо-
мах цей процес обмежуеться кйьктстю втлытого окси прол! ну. На поцальших ета-
пах втн залежить вщ специфтчних ферменттв, як! забезпечують гщроксилювання
та глткозилювання. На ступтнь внутртшньоклгги иного синтезу колагену впливае
також та його кшьктсть, яка м ютиться поза клггиною. У цьому певне значения
мають полшептиди лроколагену, як! вщщеплюються при його дозртванн!. 1х вплив
на синтез колагену в клттиш мае зворотну залежктсть, тобто що бтльше полшеп-
тицтв поза юттшою, то менший синтез.
1з неспецифтчних факгортв, якт впливають на синтез колагену, слщ видйити
тони кальцтю, залтза, а-кетоглутарат, аскорбшову кислоту. Цинга (скорбут) завж-
ди асошюеться з порушенням колагенового метаболтзму. У клттинних культурах
тварин, хворих на штату, накопичуеться незрглий колаген. Той факт, що для ре-
акцп гщроксилювання потртбен а-кетоглутарат, евщчить про зв’язок мгж синте-
зом колагену та енергетичними процесами в клттиш.
144~] 1
Колатен — невщ’емна складова м!жкл!тинно! речовини. Хоча його взаемо-
зв'язок з шшими речовинами, що шстяться в н!й, ще повшстю не вивчено, вщо-
мо, шо струкгурно-функшональна роль колагену, його участь у метабол!зм! зале-
жать вщ середовища, у якому вш перебувае, тобто вщ взаемозв’язку з гл!козам!-
ноглжанами та глжопротещами. У регуляцп синтезу колагену беруть участь також
гормони. В експериментах 13 застосуванням радюнуклццв виявлено, що в гшоф1з-
ектомованих тварин синтез колагену знижусться. Кр!м того, знижуеться ощр су-
уожмлктв до розтягнення.
Дан! про хвороби (патолопчн! стали), в основ! яких — порушення бюсинтезу
колагену, наведено в табл. 5. Деяк! !з цих хвороб успадковуються, !нш! розвива-
ються внаслщок дефщиту кофактор!в ферменпв, як! беруть участь у модиф!кацп
колагену.
Таблиц# 5 Порушення основных ет ап)в синтезу колагену I хвороби
(патолопчн! стани), як! вони зумовлюють
Етап утворення колагенових структур Значения етапу Характер порушень сии тезу Хвороба (патолопчний стан)
Синтез молекула колагену
Транскрипщя 1 транс- ляция Утворення первин- них структур Дефект структурних генгв, вцщоводальних за синтез колагену. Спадко- ва зм1на первинноо струк- тури. Прискорений син- тез Синдром Елерса — Данлоса Недосконалий остеоге- нез. Келощ Дироз печшки Ф!броз легень
Посттрансляцшш модифшщН
Пдроксилювання про- Л1Ну Утворення потрш- ноТ сшраш Дефщит аскорбшова кислота (кофактора про- лй птдроксплази) Циша
Пдроксилювання ло- зину Ппкозилювання й утворення попереч- ных зв’язков Зниження активноеп лт- зинпдроксилази Синдром Елерса — Данлоса
Приедыання глюкоз и та гадактози до г!дроксил!зину У творения кслаге нових ф’.брил Пщвищення активност! ппкозилтрансфераз Цукровий д!абег
Перетворення проко- лагену на колаген Утворення колаге- нових ф1бри.т Недостатня активность проколагенпептидази Дерматоспараксис овець 1 велико! рогато! худоби
Окисне дезамонування лизину й оксилозину Утворення попе- речных зв’язков Недостатня активность лозиноксидази Латиризм Синдром Марфана
Прискорений розпад колагену Утворення попе- речных зв’язков Пщвищення активност! кочагешптачних фермен- 11В Деструкщя хряща при ревматогдному артрит!
На сьогодн! видшено 12 тип!в колагену, якг роняться первинною структурою,
набором ланцюпв у молекул!, вмзстом вуглевод!в, органною 1 тканинною локалз-
защею. Найпоширенплими с 4 типи (табл. 6), шли — менше поширен! 1 ще недо-
статньо вивчен!. У норм! ф!бробласти синтезуюь п’ять первинних пол!пептидн>_
лантгюпв: а/1), АДП), а/Ш), 6, (IV) 1 а2, як! вщр!зняються один вщ одного набо-
ром амшокислот та пшгими ознаками. Так. якшо в а/Ч-ланиюгу гщроксильов.
лише 5 13 30 залишюв л!зину, то в а,(П)-ланцюгу — 23 13 26. Вщ цього залежт
юльккгь поперечник зв’язюв м!ж ланшогами 1 юльюсть зв’язаних вуглевощв. Е
на комбшащя п’яти первинних ланцюпв у трисшральних молекулах 1 зумовг- и
виникнення чотирьох згаданих тишв колагену.
Таблица 6. Генетичж типи колагену
Тип Комб1нац1я ланцюпв Локал!зац1я Властивост!
I а2 Шк!ра, сухожилки, метки, ропвка. склера Ошр розтягненню
П [а1(П)]3 Хрятц, склисте тою Огар стисненню
Ш [й1(Ш)]3 Шк1ра плода, стшка великих кровоносних судин, пухлина, ретикулшов! волокна Гнучкхсть, розтяжшси в устх напрямках
IV [6,(1У)]3 Батальна мембрана Проникшсть
Молекулярна орган 1защя колагену вщповщае його функциональному наван'.
женню в певному оргаш. Так, сухожилок зазнае навантаження розтягнення1»
тому його складниками с парад ельш товсп пучки колагену 1 типу; хрящова ткз-
нина стискаеться й архггекгонйса колагену в шй шша: тонк! нитки колагену
типу мають тривимхрну просторову конфпурашю ! псно зв’язаш з глисонротеь-
ми. Шк1ра I кровоносш судини мають добре розтягуватися в ус!х напрямках, ток
комбшащя колагену I типу I III типу тут оптимальна. Молекулярна струки г.
колагену IV типу гака, що дае змогу забезпечити головну функшю базально! меь
брани — проникшсть.
Розподы тишв колагену в органах може змшюватися при патологи. Ф1бро-
бласти або непосмуговаш м’язов! юптини внаслщок мутацп можуть синтезува г
не властивий для них колаген. Синдром Елерса — Данлоса (спадкова хворое,
слолучно! тканини, молекулярно-генетично щентифпсовано 10 тип1в синдром]
харакгеризуеться вщсутшстю колагену III типу Головна ознака патологи — гшер-
еластичшсгь шк!ри, яка дуже легко ушкоджуеться; на нгй неправильно загоюклт-
ся рани, формуються рубц!. При шй хвороб! ураження зазнають . суглоби, щг
проявляеться гшеррухом!стю (перерозгинання суглобтв). Так! люди страждак-
вщ рхзних травм (вивих, щдвивих, розтягнення зв’язок, деформац1я, шош розрщ
м’яких тканин). Характсрними е птоз внутришпх оргашв (шлунка, нирок, матку
гриж!, можлива спонтанна перфоращя кишки, аневризма судин. При шппй спая
ковш патологи — недосконалому остеогенез! (озтео^епезга ипрегГеси) — у юст-
ковш тканиш не синтезуеться колаген I типу, а виявляеться колаген III типу, тс
як у норм! його не мае бути. Хвороба харакгеризуеться ламюсгю исток. Перело-
ми виникають спонтанно або при незначних наван гаженнях, особливо небезпеч-
ними е компрестйш переломи хребщв.
Ще одним прикладом колагенопатн може бути синдром Марфана (наслщс-
мутаци гена ф1брил!ну-1), характерними проявами якого е юстков! аномал!! (вя-
довження ктсток скелета, тх деформацп — ктфосколтоз, пперрухомтсть суглобтв),
патология серцево-судинно! системи (аневризма аорти, пролапс морального кла-
пана) та очей (пщвивихи кришталика, вщшарування ставки, мюптя), рщко — ге-
морапчний синдром, спонтанний пневмоторакс. Атиповт форми синдрому вини-
кають при мутацп гена трансформуючого фактора росту.
Поява в тканинах невластивих для них титв колагену зумовлена дтею ртзних
патогенних фактортв на постгрансляцтйш етапи бюсинтезу колагену. При атеро-
склероз!, наприклад, у стшш судин ртзко зменшуеться ктльктсть властивого для
не! колагену III типу. У раз! запальних процес!в у тканинах, особливо в стад!!
пролтферацп, збыьшуеться ктльктсть фетального колагену III типу. Можливо, у
цьому виявляються диферентцювання клттин гранулящйнот тканини ! наближен-
ня тх за бюсинтетичними можливостями до ембрюнально! тканини. Нарешт!, па-
тологпо колагену може спричинити порушення сшввщношення мтж ним та шши-
ми елементами сполучно! тканини (кислими глткозамшоглтканами, глткопротет-
нами), якт справляють модулюючий вплив на конформацпо колагену.
Ураження сполучно! тканини в умовах експерименту вдалося вщтворити при
вигодовуванш тварин насшням горошку запашного (ЬаШугиз одогаТиз). Це прояв-
лялося порушенням росту исток ! хрящово! тканини. Трудна клттка у таких тва-
рин деформована, хребет викривлений, м!жхребцев! диски сплощен!, суглоби роз-
хитан!. Судини втрачають еластичшсть, утворюються аневризми. При цьому спос-
тертгаеться пщвищена пролтферацтя фтбробласпв, однак колаген, який вони про-
дукують, мае неправильне невпорядковане розмтщення. Описана експерименталь-
на модель мае прояви, подтбн! до таких при хвороб! сполучно! тканини в людини.
Аналопчн! змши виникають ! при введенш деяких синтетичних речовин: латиро-
ген!в (амшопроптоштрил, семикарбазид, Д-пентциламтн), механтзм дп яких, оче-
видно, полягае у зв’язуванн! карбоншьних груп амшокислот (лтзину) т блокуванн!
зв’язку м!ж ними й амшною групою лтзину сусщнього а-ланцюга. Це ослаблюе
внутртшньомолекулярн! поперечн! зв’язки, як! е дуже важливими для сттйкосп
колагену до впливу ферменттв та шших патогенних фактортв (коливання рН, тем-
ператури навколишнього середовища, юшзуючого випромшювання).
У норм! для колагену характерною е вщносно низька тнтенсившсть обмпту.
Перюд птврозпаду ртзних титв колагену коливаеться вщ ктлькох днтв до року. У
розпадт колагенових бтлктв головну роль вщтграе спецтальний фермент колагена-
за, який розщеплюе молекулу колагену на дат неоднаковт частини. Подалыпий
розпад вщбуваеться птд дтею звичайних протеолттичних ферменттв, якт не вплива-
ють на нативний колаген.
Зрглий колаген не справляе виражену антигенну дтю. Для того шоб одержати
тмунну вщповщь, потртбно разом з колагеном увести тварин! ад’ювант Фрейнда
(препарат, що мтстить мткобактерт! та мшеральш масла мае здатшсть неспецифтч-
но посилювати тмуннт реакцп). Проколаген щодо цього активнтший. Це пояс-
нюеться тим, що молекули проколагену на обох свотх кшцях мають несшралтзо-
ваш дтлянки (16 амшокислотних залишктв на амшному к!нцт! 26 — на карбоксиль-
ному) — телопептиди. € припущення, що саме вони несуть на соб! детермтнантнт
групи. Цих груп у зртлому колаген! немае, осктльки його дозртвання поза клтти-
ною обов’язково поеднуеться з ферментним вщщепленням телопептидтв. Можли-
во, птд час денатурацп колагену, наприклад, лтзосомними ферментами, вщкрива-
ються тншт детермшанти а-ланцюгтв, т колаген набувае високот антигенностт.
Дифузш хвороби сполучно! тканини (колагенози) — особлива трупа хвороб, я
характеризуються генерал !зованим и ураженням. До головних клёшчних прож 1
належать ревматоТдний артрит (ураження переважно суглобёв), ревматизм (ура-
ження серця), склеродерм!я (ураження шкёри), дерматомюзит (системне ураже-
ня скелетних ё непосмутованих м’язёв 1 шкёри), вузликовий пер1артерпт (ураження 1
судин), системний червоний вовчак (ураження всёе! сполучно! тканини). Причи- I
нами можуть стати ёнфекщя, особливо стрептококова, переохолодження, надое:- I
на ёнсоляцёя, непереносим!сть деяких лгкарських засобев. Певну роль при пьо'
вёдпрае спадкова схильшсть.
Дифузнё хвороби сполучно! тканини е типовим прикладом авто!мунних х&"
роб, що шдтверджуеться наявшстю в кров! хворих антитёл, якё реагують на влас-А
тканини оргашзму, скупченням в уражешй тканин! макрофапв, лёмфоцитёв ё плаз
моцидёв, наявшстю у вогнипп ураження комплексов антиген — антитело, ефектив- ।
шетю ёмунодепресантёв.
Патолопчний процес починаеться з утворення автоантигену з наступною акт
вацгею ёмунокомпетентних клгтин, як! починають виробляти антитёл а до влась;
тканини. П!д впливом етюлопчного фактора лёмфоцити можуть змёнювата св
властивост! та починають реагувати на свое, як на чуже (заборонен! клони). !_ I
стосуеться як В-, так ё Т-л!мфоцит!в. При цьому утворюються антитела до вс!х еле- I
ментёв сполучно! тканини, у тому числ! до ядерно! ДНК ё цёлих ядер клггин. Ъ
таких умов можливою е специф!чна автогмунна реакц!я проти колагену, в!ропд- |
шеть яко! незначна. Ушкодження колагену найчастёше е результатом проходженн*
крёзь судинну стёнку циркулюючих гмунних комплексёв (Ц1К), як! затримуються нв
базалыпй мембран! (до и складу входить колаген). До комплексёв антиген — анти-
тело приеднуеться комплемент, який спричинюе додаткове ушкодження колагещ
Процес ускладнюеться накопиченням у щй дёлянщ нейтрофёльних гранулоцита п
1Н1ИИХ клпин запалення, якё, фагоцитуючи комплекси, сам! при цьому часто руй-
нуються ё вивёльнюють лёзосомш фермента. Останш г!дрол!зують белки, нуклешо»
кислоти 1 тим самим створюють умови для появи нових автоантигешв. Формуетъсж
своерщне порочне коло, чим пояснюють системшсть ураження ! тривалють перебе-
гу хвороби. При цьому ёмунш роздали завжди поеднуються з порушеннями м!кр.
циркуляр!!. ёмунш комплекси, осщаючи на мембранах кап!ляр!в, збельшують д
проникшеть. Контакт з ушкодженою судинною стойкою активуе фактор Хагеман,
й кёнёнову систему. Вихёд фёбрину за меж! судини й вщкладання його мёж кола:г
новими волокнами та всередин! них зумовлюють фобринощне набухания, що <
одшею з найхарактернппих ознак дифузних хвороб сполучно! тканини.
Ревмапиндний артрит — хрошчне системне захворювання сполучно! тканин* 1
авто!мунного генезу, для якого характерним е ураження переважно дрёбних суг-
лобёв, зокрема ерозивно-деструктивний полёартрит з ушкодженням хряща, ис-
ток, сухожилкёв ! зв’язок, шо утворюють суглоб. Етюлопя ревматоёцного артрит
неведома. На сьогодш видетяють три групи факгор!в, як! зумовлюють розвито-
захворювання (так звана ревматолопчна трёада):
1) спадкова схильшсть до автоёмунних реакцш;
2) ёнфекцёйний фактор (найёмовёрнёше, парамёксовёруси — вёруси паротит,
кору, ресшраторно-синципально! шфекцщ в!рус гепатиту В; герпесвгруси — вгру-
си простого герпесу, опервувального герпесу, питомегаловёрус, вёрус Епстайна —
Барр, ретровёруси);
3) пусковым фактор (переохолодження, гшергнсолящя, онтокси кацоя, локарсыа
препарати мутагенноо дп, ендокринопатп, стреси).
Серед основних механозмов ураження суглобов видоляють активацпо Т-л!мфо-
цит!в 1 макрофагов з наступним утворенням 1 вивольненням прозапальних ци-
токппв (ФНП-а, 1Л-1, 1Л-6, 1Л-8, 1Л-12). Останно вщповщають за стимуляцйо
клотан, як! спричинюють деструкцйо суглобового хряща (моноцита, синовооцити,
хондроцита, фобробласти, нейтрофоли), утворення простагландинов, активацою
металопротешаз, системи комплементу, лозосомних ферментов, утворення актив-
них форм кисню. У початковгй стадп хвороби домонують припухлость суглобов о
боль. Дало внаслщок стрГмкоо пролофераци клотин вщбуваеться ущольнення си-
новоально! оболонки. Активне видщення юптинами запалення лггичних фермен-
тов спричинюе ураження истки та хряща, що призводить до деформацп суглоба,
посилення болю о втрата рухово! функцц.
Ревматизм — системне захворювання сполучно! тканини запального генезу,
при якому домонуе ураження серия (моокардит, ендокардит, ендомюкардит, пери-
кардит, панкардит). Хвороба найчастоше виникае в дитячому вщо (7—15 рок») шсля
перенесено! стрептококово! онфекцо! (81гер1ососсиз руо^епез — р-гемолотичний стреп-
токок групи А) в особ зо спадковою схильностю до автоомунних захворювань. У пато-
генез! ураження серця провщну роль вщограе сенсиболозацоя оргашзму автоантигена-
ми. Доведено, що стрептококи виробляють речовини, яко справляють кардоотоксич-
ний вплив, ушкоджують мембрани лозосом, основну речовину сполучно! тканини.
Наивность антигенно! спольносто мож стрептококами 1 кардюмюпитами спричинюе
активацпо автоомунного мехашзму з появою автоантапл до кардоомооцитов о розвит-
ком омунного запалення. Антигенным компонентом сполучно! тканини за таких
умов е глокопротеши та протеоглокани. Окром серця при ревматизм! уражуються
судини (артерит), велико суглоби (полоартрит), шкора (вузлувата еритема), централь-
на нервова система (мала хорея). Головною причиною онвалщносто таких хворих е
ускладнення ревмокардиту, зокрема вади серця, аритмп.
Склеродермы (вогнищева, системна — системний склероз) — захворювання
сполучно! тканини, що харакгеризуеться склеротичними змонами шкори, опорно-
рухового апарату, а при системному характер! — ураженням о внутрошнох органов.
Причина захворювання на сьогодношной день залишаеться невщомою; частоше на
цю недугу хворпоть жонки; головну роль вщводять патолого! омунно! системи (на-
бутой чи вродженш) та розвитку автоомунних реакцш. Серед сприятливих фак-
торов слщ видшити переохолодження, бактероально та в!русн! онфекцц, вобрацйо,
травму, контакт оз деякими хомочними речовинами, нейроендокринно порушення,
сенсиболозацпо. Домонуючими в патогенез! склеродерм!! е омунно порушення, зок-
рема зменшення супресорно! функцц Т-ломфоцитов, зболылення в пероод загос-
трення вмосту В-л!мфоцит!в, що сприяе поя во антануклеарних антитол о антитш до
колагену. При склеродерм!! збольшуеться бюсинтез колагену фобробластами та
шшими колагенсинтезуючими клотанами. Надморие утворення при цьому роз-
чинних форм колагену й ураження ендотелпо судин спричинюе розлади мпсро-
циркуляцп. 1х посилення зумовлюють цитотоксично ломфоцита. Вони ушкоджу-
ють ендотелюцита, що своею чертою актавуе адгезпо й агрегащю тромбоцитов,
.быьшуе коагуляцшний потенщал крово. Вившьнення медоаторгв запалення зболь-
доуе проникнкть кашляров, !х стонки просочуються плазмою, що одночасно !з
-щкладанням фобрину спричинюе звуження !х просвоту.
Клппчна машфестащя склеродерм!! починаеться з! шк!ри (вогнищева скле~ч>
дермтя), яка поступово втрачае еластичшсть, чутлив!сть та зрештою атрофуенв
Найтяжч! КЛ1ШЧН1 прояви властив! захворюванню !з системним характером череа
залучения до патолопчного процесу внутршш1х оргашв. Унаслщок уражев»
м’язтв порушуються ковтання та функцп стравоходу, що часто ускладнюеться жн
шрацшною пневмошею, розвиваеться дилатация шлунка, атошя тонко! кие.
що ускладнюеться кишковою непрохщшстю. Патолопя легень при склеродс р I
проявляеться ф!брозуючим алъвеол!том, дифузним пневмосклерозом, пато.
серця — кардюсклерозом. Найтяжч! ускладнення виникають у раз! ушкоджен
судин нирок 1 розвитку нефропатп.
Дерматомюзит — дифузне прогресуюче захворювання сполучно! тканини .
пального генезу, що характеризуется переважним ураженням м’язхв 1 шк!ри. Ет -
лопя ще! хвороби невщома, важливу роль у п виникненш вццграе спадков 1
Серед сприятливих (пускових) фактор) в важливе м!сце посщають загострення в. а
нищево! шфекцп, переохолодження, перегр!вання, надм!рна шсолящя, вакш -
ц!я, алерпя на лхкарсью препарата. У патогенез! розвитку дерматомюзиту важное
значения мае хрошчне персистування в!рус!в у м’язах, що спричинюе актива г
!мунно! в!дпов!д! та розвиток полгмюзиту. Антигенна пощбшсть збудниюв ! м’я
е щдГрунтям для появи автоантапл до цитоплазматичних б!лк!в 1 рибонуклеп.
вих кислот м’язових волокон. За таких умов реалззаци авто!мунних мехашзч к
сприяють дисбаланс у сшввщношенш Т- 1 В-л!мфоцит!в, зниження супресорнс»
функцп Т-л!мфоцит!в. Головними проявами прогресуючого ушкодження ске. е
них м’яз!в при дерматомюзит! е м!алпя (при рухах! навпъ у стан! спокою), м’язс ш
слабк!сть. Ураження дхафрагми та м!жреберних м’яз!в спричинюе гшовентиляц
част! пневмонп, дихальну недостатшсть. Поширення патолопчного процесу _
мюкард зумовлюе розвиток серцево! недостатносп. При ушкодженш травнсте
тракту виникають перфорацп, кровотеч!, кишкова непрохщшсть. Дистроф!я т
руйнування м’язових волокон спричинюють розвиток мюфгброзу, атрофП, кои!
трактур, ступшь яких можна оцшити за штенсившстю креатинурп та ферменте? I
Ураження ппари характеризуется щщьним I! набряком, появою еритеми, зр< -
говшням, порушенням шгментацп. У людей молодого вгку досить часто в ушксл
жених дшянках шк!ри та шдппарно! жирово! клпковини вщкладаеться калы а
Так! хвор! помирають найчастпие вщ злоягасних пухлин, асшрацшно! пневмок I
серцево-легенево! недостатносп, ускладнень кортикостерощно! терапп.
Вузликовий пергартерит е прикладом гшерерпчно! вщповщ! гмунно! систе
на компонента судинно! спнки. Цю хворобу зараховують до групи так зван»
великих колагеноз!в. На вузликовий пер!артерпт частице хворйоть хлопчики та
ЧОЛОВ1КИ В1ком 30—50 рок!в. Етюлопя хвороби достеменно невщома, хоча нале-
теть у кров! таких хворих антигешв в!русу гепатиту В (НВзА§) та антипл до не
го в склад! Ц1К дае змогу припустите роль збудника у виникненш ц!е! патолоп
Для цього захворювання характерним е системне !мунне запалення артера
м’язового типу (др!бних 1 середнгх) !з деструктивно-прол!феративними змшач»
ус!х шар!в судини, унаслщок чого розвиваються периферична та висцеральна шге-
м!я 1 некроз оргашв. До провокуючих факторов належать перенесен! шфекг.
!нтоксикацп, уведення вакцин, сироваток, уживання певних лгкарських засобт
переохолодження, шеолящя.
Ушкодження судин при вузликовому пергартерптт е наслщком вщкладання в
них тмунних комплекс1в г активаци комплементу. Важшву роль при цьому вцц-
грають ендотелюцити, як! мають рецептори до Гс-фрагмента ]§6 г першо! фракш!
комплементу С1р, що значно полегшуе взаемодйо тмунних комплексов !з судин-
ною стшкою Активоваш нейтрофтли при захоплешп тмунних комплекств вилшя-
ють велику кшьккгь лтзосомних протешаз, активних форм кисню, що поглиблюе
прояви руйнування судин. За таких умов ендотелш активно видгляе фактори за-
дания кров!. гншгюе тромбоз, що носил юс негативно наслшки васкулггу, серед
яких — облттерацтя судин, аневризма, шфаркг внугршппх органгв, крово геча.
Системный червоний вовчак — найтяжчий вариант патологи сполучно! ткани-
ни, шо характеризуеться полкиндромнктю г виникае внаслщок генетично детер-
мптованих порушень тмунгтету (пперреактивносп гуморально! ланки). Етюлопю
хвороби остаточно не встановлено, проте отримано ваг ом) докази рол! РНК
вгруств 1 ретровгрусгв (кору). Так! в!руси здатн! утвооювати пбрид свого геному г
ДНК клгтин людини. Серед провокуючих факторгв розвитку системного червоно-
го вовчака вцщляють гнсоляшю, перенесен! шфекцп, медикаментозн! чи поствак-
цинальн! реакцп. Важливе значения мае й гшерестрогенемш, адже хворпоть на
цю недугу переважно молод! жшки та д!вчатка-п!дл!тки (факторами ризику виз-
нано аборта, ваптнкть, пологи, початок менархе). Основою патогенезу систем-
ного червоного вовчака е сенсибыгзацтя оргашзму до нуклешових з’еднань втрус-
ного або ядерного походження, утворення антинуклешових г антинуклеарних
штапл унаслщок дисфункцц Т-кл!тинно! ланки !мунно! вщповщ! та гшерактив-
ностг В-лтмфоциттв Порушення Т-кл!тинно! вщповщ! характеризуються недо-
статнктю Т-хелпертв типу 1, гшерактивнктю Т-хелпертв тапу 2 з насту иною ак-
гиващею В-л!мфоцит!в. Серед В-клпинних дефект!в основними за ц!е! патолог!!
вважають над м гр ну вщповщь цих клгтин на гмуностпмулюючг впливи та надлиш-
кове продукування ними таких сигнал!в, знпжену чутливкть до супресорних чин-
никгв. Втрата гмунно! толерантносп до власних антигенов спричинюе надлишкове
хтворення антитгл, автоантитгл та тмунних комплексов, як! вщкладаються на ба-
зальшй мембран! судин г спричинюють розвиток васкулгту. Активацгя системи
комплементу, нейтрофглш, вивгльнення кгнгнгв, простагландишв, лтзосомних
ферментов та шших БАР, активацгя зсщально! системи кров! поглиблюють розла-
ди мгкроциркуляци, ушкодження судин г тканин Виражену цитотоксичну дно за
таких умов справляють г сенсибгл!зован! Т-ктлери. Системний характер ураження
сполучног тканини г паренхгмат озних органгв пояснюеться розвитком автогмун-
ного та гмунокомплексного хронгчного запалення. Дезоргангзацгя сполучно! тка-
нини й ушкодження судин у нирках спричинюе розвиток нефропатт! (тмуноком-
плексний нефрит), у шктрт — агрофгю епщермгсу, плеркератоз, дезорган! зато ю
дерми, у синовталыпй оболонцг — синовцт (при цьому вщбуваеться розволокнен-
ня тканини синовтально! оболонки, ушкоджуються суглобовий хрящ, ккткова
тканина епгфгзтв). Ушкодження серия характеризуеться розвитком панкардиту,
легень — специфгчним потовщенням альвеолярних перегородок ! стшок судин на
ргвнг м!кроциркуляторного русла з розвитком дихалъно! недостатностт, централь-
но! । периферично! нервово! системи — мешнгоенцефа.томтелпом, радикулттом,
невритом, плекситом. Про генералгзований характер цього захворювання евщчать
також л!мфаденопаття, силе но- г гепатомегалгя.
ПАТОЛОПЯ К1СТКОВО1 ТКАНИНИ
Одним 31 специф1чних р!зновид!в сполучно! гканини е мсткова тканина, шо в
конус функцн мехашчно! опори, а також депо фосфору та кальшю. Щ!льна
губчаста (трабекулярна) речовина метки утворюе каркас, де мктяться клпт-
мсткового мозку, з яким метка ткно поеднана не лише анатошчно, а й фунм
нально. Розвиток меток в ембрюнальний! постембрюнальний перюди ! функз
нування !х у дорослому оргашзм! е ф!зюлог!чно контрольованим процесс-1
пов’язаним хз синтезом мсткового матриксу остеобластами та координовав
резорбшею меток остеокластами. Обидва типи клпин походить з незр!лих клпин
попереднимв мсткового мозку: остеобласта з мультипотентно! мезенхгмно! ст
бурово! клпини, що входить до складу строми мсткового мозку, остеокласта — 1
гемопоетичних клпин — попереднимв макрофагового ряду.
Протягом усього житгя вщбуваеться постшне руйнування та вщновлення к
ково! тканини — ремоделювання, завдяки чому в доросло'! людини виникаюп
кодження 1 деформацц, пов’язаш з мехашчним перевантаженням меток. У де-
ло! людини завдяки ремоделюванню в середньому один раз на 10 роив вщбув эея
ся повна замша век! кктково! тканини, хоча певш дшянки меток моя-^,
ремоделюватися швидше або повмьнппе. Ремоделювання вщбуваеться за уч -
клпин, ям становлять основну багатоклпинну одиницю (ОБО) завдовжки 1—2
1 завширшки 0,2—0,4 мм, що складаеться з остеокластов спереду та остеобла
позаду. При вщновленн! структури метки в оргашзм! працюе майже 1 млн С
середня гривалкть жиггя кожно! !з них становить вщ 6 до 9 М1С. Проте трива.’
жигтя кожно! юптини, що входить до складу' ОБО, набагаю коротша: остеокласт : -
до 2 таж., остеобласт — приблизно 3 м!с. Загибелъ юптин вщбуваеться за рагу
апоптозу, а поповнення — шляхом диференцповання з юптин-попередниюв.
Ремоделювання метки починаеться з активацн остеокласта. Це багатояде
велика повнктю диференц!йована клпина. яка формуеться шляхом злитгя мп
нуклеарних клггин 1 мае велику мльмсть мпохондрш, л!зосом, вьтьних рибос
складну систему пальпепод!бних вшросгмв. Останш оснашеш протоновою п<
пою, що сприяе закисненню середовища та розчиненню мшерального компов
та метки — гщроксиапатиту [Са10(РО4)6(ОН)2], а також ферментами — кис.:.
фосфатазою, металопротешазою та катепсинами К, В, Ь, ям руйнують органг
настану матриксу. Остеокласта активуються сигналами з остеопипв 1 клпин вв.
тилки (своерщна рецепторна система мстм]), паратгормоном, гормонально-ас
тивним метабо.птом впам!ну Э (1,25-дипдроксихолекальциферол), тироксине-
проза!гальними цитомнами (1Л-1, 1Л-6, 1Л-11, ФНП-а, ФНП-0, простагланди-
ни). Велике значения для морфогенезу й активацп остеокласт мае !х безпосеред-
ня взаемоди з клпинами с громи та остеобластами. На поверхн! остеокласт та сх
попереднимв е спещальш рецептори, через як! активуеться нуклеарний факте'
каппа-В (ВА1ЧК), що е ключовим елементом активацп клпин. На поверхн! осте-
областа мктиться вщповщний лпанд (ВАМК.Б), лише у процес! взаемодп з яки:
може вщбутися активашя розвитку та функщонування остеокласта. Тамгй ме-
хан!зм зап об, гас !зольовашй активацп остеокласт без в!дповщно! пщгримки ос-
теобластами та забезпечуе гармошйне ремоделювання метки.
Остеобласта, як! з надкктя рухаються за остеокластами й активуються про-
дуктами розпаду мелей, синтезують оргашчний матрикс, до сктаду якого входя?-
Остеоктаст
Мал. 14. Регуляшя взаемодп остеокластов та остеобластов при ремоделюванш истки:
1 — КАИК — реиептори на остеокластах, через як! активуетъся МР-кВ); 2 — КаМСЬ —
л!ганди для ВА1ЧК на остеобластах о аюивованих Т-ломфоцоттах; 3 — походження; 4 —
вплив (+ позитивний. — негативний)
колаген I типу, остеокальцин, остеонектин, глокозамшоглокани. остеопонтин,
фобронектин, витронектин тошо Матрикс пщ впливом цих самих клттин надало
монера-йзуеться з утворенням кристалов гьцроксиапатиту, У со регуляторш фактори
витого порядку накладаютъся на щ базово мехашзми, забезпечуючи ровновагу
мож активнтстю остеокластов о остеобластов (мал, 14).
Правильно функщонування костковоТ тканини забезпечуеться балансом мож
процесами руйнування й утворення костки, тобто мож только стю й актившстю ос-
теокластов о остеобластов. Адекватне регулювання утворення та загибел! водповод-
них КЛ1ТИННИХ погоуляпой. а також регулювання ох активносто контролюеться сис-
темними гормонами (паратгормон, вотамон В, калыоитонш, естрогени, андрогени,
тироксин, соматотротпн, глюкокортикооди, тнсулон, глюкагон, лептин), регуля-
торн ими факторами мокрооточення (цитокгни та фактори росту), а також прямою
взаемодоею (клотина — клотина та клотина — матрикс). Поруоиення балансу мож
утворснням юстково! тканини та ц руйнуванням призводить до патологи юсп
що може проявляйся остеопорозом, остеопорозом, остеомаляшею / раитом.
Остеопетроз (си нош ми — остеосклероз, мармурова х вороба) — спадкове
хворювання. зумовлене функшональною неспроможшстю остеокластов, для як.
характерне зб!лъшення частки юстково! тканини в склад] юстки. € рання та г
фсрми ще! хвороби. Рання форма остеопетрозу успадковуеться за автосомно-реь
сивним типом, проявляеться в ранньому дитинствх 1 мае злоякюний перебгг, нерд
ко завершуеться летально (причина смерп — анемзя, пневмошя, сепсис). Об’
юстково-мозкових порожнин за таких умов зменшуеться. Порушення кровог
рення, основними проявами якого е тяжка анем!я 1 тромбоцитопешя з подалыд I
розвитком геморапчного синдрому, спричинюють появу екстрамедулярних в. -
нищ кровотворення в печ!нш, селезшц! та л!мфатичних вузлах, шо зумовлкх.
збьлынення (мае компенсаторний характер). Недосконала архпектошка та нев"
рядковане утворення юсткових структур провокуе множинш переломи. Гншн 1
остеомоелп, що виникае при цьому, може стати причиною сепсису та смерп. П
остеопетроз! найбшъше ушкоджуються трубчасп юстки, юстки основи череп
таза, хребет, ребра. Кзсткова маса при цш хвороб! займае вс! порожнини юсток, -
м!сц1 губчасто! речовини виявляють шьльну однорщну юсгкову тканину, яка .
розпил! юстки нагадуе шл!фований мармур, звщси ! походить !нша назва недуги -
мармурова хвороба. Шзня форма остеопетрозу передаеться за автосомно-доми
тним типом, мае б!льш сприятливий перебш. Ураження юсток — обмежене.
Остеопороз — захворювання, зумовлене порушенням метабол!чних проце
у юстках, що призводить до вграти юстково’! маси при збереженн! вм!сту калы
! фосфору на одинипю маси юстки (синошм — атроф!я юстки). Локальний оспи
пороз спостернають при запаленш (наприклад. ревматощний артрит, пародонтгг
Мехашзм остеопорозу за таких умов передбачае появу у вогнищ! запалення аю
вованих Т-л!мфоципв, як! несуть на сво!й поверхш КАМКЕ! здатн! таким чин
активувати остеокласти без учасп остеобластов. Активоваш остеокласти розчиз
ють мшеральний компонент юстки (пдроксиапатит) за допомогою оргашчн'-
кислот та оргашчну и частину шляхом вившьнення л!зосомних ферменпв. Зр< .
танню активност! остеокластов сприяють також прозапальш питоюни. При тр; -
матичних ушкодженнях юсток розвиток остеопорозу може бути наслщком вш
ристання неушкоджено’! юстки як депо кальцпо для ремоделювання ушкодже] ।
частини, особливо при масивних переломах.
Загрозлив!шою медичною проблемою сьогодення е генерализован ий остеа
роз, який характеризуеться ураження м ус!х юсток. Головними його проявам!
стоншення 1 деформаптя юсткових балок, збгтъшення поротичносп юсток, по;
шення юстково! м!кроэрх!тектон!ки. Так! юстки стають крихкими, немщнш
легко ламаються навпъ при незначних травмах. Небезпечними е комлресгйн! пе
реломи т!л хребшв, переломи шийки стегна. Причиною генералтзованого остеог
розу може бути старшня. Це зумовлено зменшенням утворення естрогенш у х
нок, як! контролюють екскрешю кальшю 13 сечею, синтез гормонально-активо
форми вггамш у В в нирках (особливо пщ час лактац!! та ваптносп)! андрогенн
чолов1юв (збшьшують ф!ксац!ю кальцпо 1 Фосфору юстковою тканиною), горз
нальними порушеннями (гонадектом!я, пперпаратиреоз, гшертиреоз, синдр
1ценка — Кушшга, цукровий шабет), пподинамтею (тривала (ммобйпзашя гт
травмах, тяжю хвороби, зниження ф!зично! активносп здорово! людини), нева
•□стю (у космонавта), недостаппстю вттамшу В (контролюе засвоення кальщю в
кишечнику га реабсорбщю кальщю т фосфата у нирках, щдсилюе мшералтзацпо
исток; див. роздЕт XXIV “Патофтзюлопя обмшу впампйв'’), хрошчною патоло-
пею кишок (синдром матьабсорбцп, резекптя — зменшуеться засвоення кальшю
та фосфата), хронтчними захворюваннями нирок (втрата кальшю тз сечею), не-
досконалим остеогенезом (у тому числт при хвороб) Педжета), тривалим застосу-
занням глюкокортикоццв (пригшчуюгь бюсинтез колагену та такозамшоглёкантв),
дтуретиктв, антиконвульсанта. Серед провокуючих фактортв слщ видтлити незба-
лансоване за кальщем харчування та вегстартанство (основним джерелом кальшю
е молочн) продукта, риба). Усе це зумовлюе ризик виникнення переломи та ш-
ших ускладнень т потребуе вщповщних профтлактичних т лжувальних заходов.
Остеомаляция (рахт — у дйпей) — розм’якшення исток унаслщок порушен-
ня тх мшералтзацп. Причиною остеомаляцп може бути дефшит кальцпо та / або
фосфору в рацюнц порушення засвоення них елемента, !х надмтрна втрата. Най-
частште остеомаляция виникае при дефшит) визиту О в ращош (сувора веге-
тар)анська дзета), порушенн) його синтезу в печптш (кальциферол-25-ОН впамш
О), незасвоснн) у раз) розладу жовчовидшення, тривалому застосуваннт антикон-
вульсанта (фенобарбитал, фениотн), як) активують печшкову пдроксилазу) поси-
люють деградацпо гормонально-активно) форми впамшу О кальвитрюлу [(1,25-
(ОН)2 впамш 13]. Кальц)й 1 фосфор погано засвоюються при патологи травления
сгастректомтя. панкреатит, синдром мальабсорбцн). При хрошчнш нирковш не-
досгатност) (тубулопатп) вщбуваеться не лише втрата фосфата )з сечею, а й зни-
жуеться синтез кальцитрюлу (цей процес вщбуваеться в нирках т контролюеться
паратгормоном). Остеомалящя е частим супутником гшерпаратиреозу як компен-
саторно! реакцп на гшокальщемно. Надлишок паратгормону за таких умов спри-
чинюе некойгрольовану резорбшю мтнерально! частини клетки, шо е зрозумтлим
з огляду на головну функшю паратгормону — пщвищення ртвня кальщю в кров).
Клпнчно остеомалящя проявляется болем у ктстках, тх деформашею, м’язовою
слабктстю, порушенням тогнки органтв.
Небезпечним е дефицит кальцпо та фосфору для д]тей, адже швидкий р)ст
ктсток вимагае значно! кшькост) цих елемента. 1х нестача спричинюе розвиток
рахту, що е пед'атричним еквтвалентом остеомаляш! 1 зазвичай виникае в дней
В1ком 6 м)с — 3 роки. На сьогодш домшують зроджен) йот о форми: впамш-Л-
залежн) та фосфопешчн). Впамш-П-залежний рах)т 1-го типу зумовлений спад-
ковим дефектом його синтезу в нирках, легко шддаеться лткуванню Вттамтн- О-
залежний рахп 2-го типу (або впамш-Э-резистентний рахтт) е наслщком нечут-
ливост! оргашв-мштеней до кальцитрюлу т мае тяжкий перебгг. Головна ознака
устх фосфопентчних форм рахтту — спадковий дефект синтезу фермента, якт вщ-
ловщають за транспорт фосфата у нирках. Серед них — зчеплена з Х-хромосо-
мою гшофосфатемтя, автосомно-домшантне гшофосфатемтчне ураження к)сток,
автосом но-домшантн ий ппофосфатемтчний рахтт, синдроми Фанкою (трупа за-
хворювань, як) проявляються генералтзованим порушенням функцп ниркових ка-
нальцтв т втратою фосфата). Набут) форми рахтту в дней найчаст)ше спричинент
дефшитом виамгну О в рашон), недостатньою шеолящею чи латолопею органтв,
вщповтдальних за його засвоення.
При остеомаляцп / рахит ктстка мтстить менше калъппо т фосфору на одини-
цю маси, дем)нерал)зована тверда субстанцтя зам)щуеться м’якою остеощною тка-
ниною. Оргашчна матрица погано мшералхзуеться, що в дггей проявляеться за-
тримкою росту, деформащею исток, особливо нижнгх к!нц!вок (варусна О-подаб-
на та вальгусна Х-под!бна), затримкою розвитку зуб1в, зниженою пГгменташею
шири, у дорослих — деформащею исток.
Окремо слщ ВИД1ЛИТИ розвиток остеодистрофп при гшеркальщемп, що най-
частпие супроводжують пперпаратиреоз. Гострий декальциноз скелета за таких
умов зумовлюе появу множинних вогнищ истозно! дегенерацп. Порушення фор-
мування скелета при пперпродукцп паратгормону е результатом розсмоктування
ус!х компонентов истки, а не лише и демгнералззаци. Пщ впливом паратгормону
збыьшуеться ильисть активних остеокластгв, шдвищуеться !х метабол!чна актив-
шсть. Мехашзм декальцинування истки полягае в накопиченн! оргашчних кис-
лот. Найбьтыпий вплив на розчиншсть пдроксиапатиту справляе лимонна кисло-
та, що утворюе з калыцем легкорозчинш сполуки. .Шзосомш ферменти остеоклас-
тов розщеплюють колаген. Унаслщок втрати истково! матриц! истка стае м’якою,
дшянки розсмоктування зампцуються сполучною тканиною, утворюються зони
остеощно! речовини. Процес м!нерал!зацн исток за таких умов порушений 1 пе-
ребггае дуже повшьно, истка залишаеться незршою, розвиваеться остеоф!броз.
Окремо видыяють остеопент, що не е самостойним захворюванням, а твид-
па е характеризуе втрату истково! тканини, яку виявляють за допомогою спещаль-
них методов дослщження (рентгенодхагностика, денситометрхя). Основними при-
чинами остеопени е остеопороз, остеомаляция, множинна м!елома, ендокрино-
патй.
Розд1л VII
РЕАКТИВШСТЬ IТГ РОЛЬ У ПАТОЛОГИ
Реактивность — це здатшсть оргашзму змшювати життсдояльшсть у водповодь на
р1зш впливи навколишнього середовища
Реактившсть е такою самою властивктю оргашзму, як р!ст, розмноження,
харчування, обмш речовин (А Д Адо). Вона формуегься в процес! фйо- та онто-
генезу й вщображае видов!, групов1 та шдивщуальш особливост! реагування. Ре-
актившсть — одна з основних форм зв'язку та взаемодп оргашзму з навколишшм
середовищем переважно захисного та пристосувального характеру.
Перш! згадки про реактившсть знаходимо ще в лгкаргв давнгх час!в, яю зазна-
чали, шо р!зн1 люди хвориоть неоднаково. Проте поняття реактивност! виникло
лише на початку XX ст., коли патологи почали видьтяти р!зн! типи реагування
оргашзму. Вчення про реактившсть з позицп загально/ патологп першими почали
розробляти росшськ! та украУнськ! патологи, як! мають значн! досягнення в ц!й
галузь Важливу роль у вирппенш ще! проблеми вздирали поргвняльно-патолопч-
ш дослщження 1.1. Мечникова про фагоцитоз 1 його участь у запаленш та шуш-
тет!; прац! М.М Сиротишна, що встановив загальш закономтрност! еволюцп ре-
активност! у фйю- та онтогенез!; уявлення О.О. Ьогомольця, який пов’язав кон-
ституцпо 1 реактившсть !з властивостями сполучно! тканини.
Поняття реакгивност! надшно ьв!йшло в практичну медицину. Це дае змогу
об’ективно ошнювати стан хворого. Реактившсть оргашзму е одним !з найваж-
лив!ших факторы патогенезу хвороб, оск!льки патолопчна реактившсть харакге-
ризуеться зниженням пристосувальних можливосгей оргашзму.
Будь-який патолопчний процес пею чи нею м!рою змшюс реактившсть ор-
гашзму Водночас змйза реактивност!, що псревишила ф!з!олопчш меж!, може ста-
ти основою розвитку хвороби. Тому вивчення реакгивност! та л мехашзмгв мае
важливе значения для розумзння патогенезу хвороб ! пглесирямованого лгкування.
Види реактивност!. Найзагалыпшою формою реактивност! е бюлогочна, або
видова, реактившсть, яка зумовлсна насамперед спадковими факторами : вщоб-
ражае здатшсть оргашзму реагувати на ргзномаштш впливи навколишнього сере-
довища (токсини, 1шокс!ю, радгальне прискорення тошо) зм!ною життедгяльност!
захисно-пристосувального характеру I! називають щс первинною.
Видовг особливост! реактивност! визначають видовий !мун!тет проги щфскшй
них хвороб. Так, видовим (муштетом пояснюеться несприйнятливкть людини до
чуми велико/ рогато/ худоби. Прикладами видових змш реактивност! е зимова спляч-
ка тварин, ссзонна мпращя риб г птах!в. Пш час зимовог сплячки, що характери-
зуеться глибоким пригшченням активност! нервсво/ та ендокринно! систем, обмгну
речовин г зниженням у зв'язку гз цим температури тьта (до 3—4 °С у прямы киты),
значно знижуеться реакгившсть до багатьох факторгв. Ховрахи, заражен! в перю.
сплячки чумою, туберкульозом, не уразливг до них, а шсля пробудження — захво-
рюють г гинуть. Сплячка пщвищуе стшкгсть до стрихншу та шших отрут. Водноча;
запалення, фагоцитоз, утворення антипл, загоювання ран пригшчуються. Видов
реакгившсть спрямована на збереження виду в щлому 1 кожно! особини зокрема.
На основ! видово! реактивносп формуеться групова та шдивщуальна. Групова
реактивн1стъ характерна для певно! категорп людей або тварин, як! об’еднан! деяки-
ми спглышми ознаками, наприклад типами конституцг! або трупами кров!, пщвище-
ною чи, навпаки, зниженою сприйнятливктю до дп тих чи тих факторгв. схильшсп
до розвитку тих чи тих хвороб. Так, за Кречмером, астенгки схильнг до шизофренП.
люди з I групою кров! частгше хворгють на виразку шлунка. Описано лшп тварин з
пщвищеною схилъшстю до розвитку артергально!' гшертензн, атеросклерозу.
1ндив1дуальна реактившстъ зумовлена спадковими та набутими факторами
Вона залежить як в!д генетичного полгморфгзму, так г вщ тих умов навколишньо-
го середовища, у яких оргашзм розвиваеться, — характеру харчування, клгматич-
ного поясу, вмгсту кисню в атмосферному повггрг тошо.
Реакгившсть залежить вщ стат!. Так, реакгившсть жшочого органгзму змг-
нюеться у зв’язку з менструальним циклом, ваптшстю. Жшочий оргашзм спйкг-
ший до ппоксп, крововтрати, рад!ального прискорення, голодування.
Вщома роль вгку в реактивност!. Для раннього дитячого вгку характерною е
низька реакгившсть. Це визначаеться незавершеним розвитком нервово!, ендо-
кринног та гмунно! систем, недосконалктю зовшшшх г внутргшнгх бар’ергв. Най-
вища реакгившсть спостерггаеться у зрглому вщ!, поступово знижуючись зг старен-
иям оргашзму. Особи лгтнього та старечого в!ку дуже сприйнятлив! до шфекцп, у
них часто розвиваються запальш процеси в легенях, шодермгя, ураження слизово!
оболонки. Причина цього полягае в ослабленш гмунних реакцш та зниженн! бар’ер-
них функцгй у процес! старшня органгзму.
Хндивщуальна реакгившсть може бути специфгчною г неспецифгчною. Спе-
цифична реактившстъ проявляеться здатнгстю органгзму вщповщати на певнг по-
дразнення. Таким вимогам вщповщае й 1мунолог1чна реактившстъ. Вона забезпе-
чуе стгйкгсть (гмуштет) до шфекцшних збудникгв, реакцп ггстолопчног несумгс-
ностг тканин (вщторгнення трансплантата), пщвищеног чутливостг (алерпя).
Неспецифгчна реактившстъ проявляеться пщ час дп на оргашзм ргзних не-
сприятливих факторгв навколишнього середовища. Вона реалгзуеться за допомо-
гою таких механгзмгв, як стрес, змгна функцгонального стану нервово! системи,
парабюз, фагоцитоз, бюлопчн! бар’ери.
Специфгчна та неспецифгчна реакгившсть може бути як фгзголопчною, так г
патолопчною.
Ф1з1олог1чна реактившстъ охоплюе реакцп здорового органгзму у сприятливих
умовах гснування. Прикладом може бути гмунгтет (специфгчна реакгившсть), ре-
акцгя органгзму на дгю ргзних факторгв навколишнього середовища в межах, якг
не порушують гомеостаз (неспецифгчна реактившстъ).
Патолог1чна реактившстъ проявляеться в разг дп на оргашзм як нормальних,
так г хвороботворних факторгв. Вона якгсно або кглькгсно вщргзняеться вщ фгзго-
лопчно! реактивносп г характеризуеться незвичайною формою реагування. Зага-
лом патолопчна реакгившсть проявляеться обмеженням пристосувальних мож-
ливостей органгзму.
Прикладами специф!чно! патолопчно! реактивност! е алерпя, хмунодефщитш
та гмунодепресивш стани. Прояви неспециф!чно! патолопчно! реактивност! спо-
стершають год час травматичного шоку, наркозу. Пщ час шоку приппчуеться ре-
актившсть щодо шфекшйних та !нших хвороботворних фактор!в. Кр!м того, при-
ппчуеться фагоцитоз, змшюеться чутливгсть до лгкарських засоб!в.
За формами прояву розр!зняють реактившсть пщвигцену — гтерергио, зниже-
ну — гтергио та спотворену — дизергио.
У кл1н!чн!й картин! внутршгшх та шфекцшних хвороб розр!зняють гшер-
ерпчний 1 гшерпчний перебгг пневмонп, туберкульозу, дизентери та шших хво-
роб. Гшерерпчними називають хвороби з! швидким та штенсивним перебггом,
начними змшами функцш оргашв 1 систем. П!д гшерпчними розумпоть хвороби
з млявим перебггом, стертими симптомами, низьким р!внем антиплоутворення та
фагоцитозу.
Показниками неспециф!чно! реактивност! е драпивлив1сть, збудливкть 1 чут-
ив1сть (поняття, аналопчне збудливосп, але застосовуеться щодо складшших
процесхв в оргашзм!). Можлив! змши чутливост! до больового 1 температурного
подразнишь, порушення чутливост! органтв чуття (зору, слуху, нюху тощо). По-
казниками неспециф!чно! реактивност! можуть бути також швидюсть та штен-
сившсть розвитку загального адаптацшного синдрому, здатшсть вщповщати на
подразник посиленням секрецц адреналшу.
Для оцшки специфично! реактивносп визначають р!вень !мунно! вщповцц
штенсившсть утворення антипл, актившсть фагоцитозу тощо).
РЕЗИСТЕНТНЮТЬ. ВЗАбМОЗВ’ЯЗОК 3 РЕАКТИВН1СТЮ
Резистентшсть — це стшклсть оргашзму до ди патогенних факторов.
У процес! еволюцп оргашзм виробив певш пристосувальш мехашзми для за-
безпечення свого !снування в умовах постёйно! взаемоди з навколишшм середо-
вищем, р!зномаштш фактори якого могли б спричинити не ильки порушення
життед!яльносп, а й його загибель у раз! вщсутносп або недосконалосп цих ме-
хан!зм!в.
Резистентшсть псно пов’язана з реактившстю. Здатшсть оргашзму протисто-
яти ушкоджувальним впливам визначаеться його реакшею як единого ц!лого, !
тому резистентшсть е одним з основних наслщкгв реактивност! оргашзму.
Розр!зняють резистентшсть первинну, пов’язану з! спадковими факторами, 1
вторинну — набуту. Остання може бути активною та пасивною. Пасивна резистен-
тшсть не пов’язана з акгивними реакщями на вплив, а зумовлена анатомо-ф!зю-
лопчними особливостями оргашзму — будовою шири, слизово! оболонки, к!ст-
ково! тканини, пцльних покривгв комах, черепах. Пасивну резистентшсть спо-
стерйають ! тод!, коли мехашзми реагування вщсутш або “зайнят!”.
У раз! дп двох або б!льшо! шлькост! надзвичайних (екстремальних) подразни-
к!в оргашзм нер!дко реагуе лише на один, лишаючись “глухим” до ди шших
подразник!в. Наприклад, якщо тварин! пщ час судоми нанести на шк!ру краплю
лю!зиту, ураження буде пор!вняно слабким. Тварини, як! зазнали рад!ального
прискорення, витримують смертельну дозу стрихншу, виявляють високу вижи-
вашсть в умовах гшокси, перегргвання. Така форма реагування може бути зарахо-
вана до пасивно! резистентность Але е й шший погляд. На думку 1.А. Варшаь
кого, це явище не е резистентнтстю, осюльки в цих умовах оргашзм не може
тивно лротпстояти впливу навколишнього середовища, збертгаючи гомеостаз 1
лише генуе п!д шею пього впливу в стан) глибокого пригнтчення життедтяльнс.
Учений запропонував назвати таку форму резистентное^ витривалгегю.
Витривалктъ зазвичай спостерегають у менш досконалих органтзмтв, на^;
клад у холоднокровних або в ранш перюди постнатального розвитку, особлит
недоношених тварин (шургв, мишей). Одним тз пристосувальних мехашзми
цих умов с перехщ на белый давнш г еггйкий, хоча 1 менш економ)чний, ш.
утворення енергп — глтколгз.
Резистентш’сть, на противагу витривалостц забезпечуе активну протидпо оМ
гашзму патогенним факторам, унаслщок чого збер!гаються сталгсть внутрпшгы
середовища та цостатнш ртвень життсдтяльносп оргашзму
Активна резистентнгсть зумовлена захисно-пристосуватьними механтзма’
Так, сийкгсть до гшоксгё пов’язана зт збшыиенням вентиляш) легень, пр искор
ням току крове, пщвищення м р)вня еритроштттв 1 гемоглобшу в кров! тощо, ст
ктсть до шфекщй бмуштет) — з утворенням антител ) акти вашею фагоцитозу
Активна набута резистентнгсть виникае також теля вакцинацй. пасивна тд
бута резистентнгсть — шсля серотерапй.
Неспецифгчна резистентнгсть — це еттйкгеть до багатьох факторгв, специф
на — до одного фактора.
Зазвичай реактившсть ) резистенттпсть змшюються односпрямовано. Одт
можливим е варгант, коли реактившсть знижуеться, а резистентнгсть пщвишуе
ся, як це бувае в умовах зимовоТ сплячки, пщ час глибокоё ппотермп, наркоз?
(М.М. Сиротинш).
Стан реактивносп ногртбно розглядати не взагалг, а конкретно, шодо од и.
або ктлькох однорщних факторгв. 1нод1 пщвищена реактившсть до одною подр
ника поеднуеться зт зниженням реактивностт до тншого.
РЕАКТИВШСТЬ НА Р13НИХ Р1ВНЯХ
□РГАН13АЦН ЖИВИХ СИСТЕМ
Реактившсть, формуючись на основ) с падковост), конституцп, певного ргвня р<
витку нервовог, ендокринно) га )мунно) систем, е властивтстю оргашзму як щл ,
ноТ системи. Водночас про реактившсть можна говорит на ртзних рёвнях орган
зацп ще! системи, починаючи тз субклттинного. Приклад реактивности на моле,
лярному р!вн1 — збтльшення спорщненостт гемоглоб)ну до кисню при ппокс.
насл)дком якот е змпцення кривот дисошадп влево та покращання киснево-тра;
портноё функцп еритроцитёв.
Клтинна реактившсть пов’язана з мембранними процесами, шо забезпеч>
ють взаемодпо клггини 1з середовищем. яке п оточуе, за допомогою вбудовашг-
нет бтлкових структур, що виконують функцпо клттинних рецептортв та юння
каналтв. Реактившсть на клттинному ртвш спостериають пщ час здтйснення ле
коцитами фагоцитозу, дегрануляцп базофтльних гранулоцитгв (лаброциттв. ткя
нин них базофппв) пщ дтею комплекств антиген — антитшо.
Реактившстъ на р'шш органа проявляеться, наприклад, спазмом його непо-
смугованих м’язових волокон у вгдповщь на повторне надходження в оргашзм
алергену (див. “Алерпя”).
Прикладом реактивностт системи оргашв I оргашзму в щлому е перебудова
терморегуляцп та основних життезабезпечувальних систем у вщповгдь на дпо
шрогену (див. “Гарячка”). У розвитку багатьох патолопчних процесгв (алерги,
запалення тощо) можна простежити змши реактивносп на ргзних ргвнях.
ЕВОЛЮЦ1Я РЕАКТИВНОСП
ФЬюгенез. Реакгившсть ргзного ступеня мають усг оргашзми. Що вище столь
тварина фыогенетично, то складншп, досконалпш и реакцп на ргзш ди. У най-
проспших ! багатьох безхребетних первинна реактившстъ розвинена слабко, а
гмунолопчног — немае. Мехашзм реактивносп в них обмежуеться змшами обмгну
речовин, що дае змогу витримувати значне зниження температури та зменшення
вмгсту кисню в навколишньому середовшщ. Досконалпш та ргзномангтшшг ме-
хашзми реактивносп спостерггають у хребетних.
Становления реактивносп поеднуеться з виникненням мехашзм! в активного
пристосування до дп шкгдливих факторгв. У риб з’являються комплемент г анти-
пла, однак останш не наспльки специфгчш, як у теплокровних; алерпчну реак-
цию спричинити неможливо. У земноводних ! рептилш алерпя виражена слабко,
при цьому добре розвинута запальна реакшя, що проявляеться не пльки фагоци-
тозом, а й судинною реакщею.
Теплокровш мають сильшшу реакгившсть, що пов’язано з посиленням мета-
болгчних процесгв г щдвищенням ргвня розвитку ендокринно! та нервово! систем.
У них бгльша реакгившсть до ди ргзних факторгв г водночас краше розвинеш при-
стосувальнг реакцг! на нестачу кисню, пгдвищення г зниження температури навко-
лишнього середовища через змгну теплопродукци та тепловгддачг. В усгх теплокров-
них чгтко проявляеться гмунолопчна реактившстъ. Лише теплокровним, г особливо
ссавцям, властива автоалерпя. Яскраво вираженими е вег елементи запалення.
Отже, у процесг еволюци вдосконалюються мехашзми, за допомогою яких
оргашзм реагуе на ргзш фактори навколишнього середовища. Водночас з’являють-
ся мехашзми, як! дають змогу органгзму активно пристосовуватися до мшливих
умов середовища, що забезпечуе можливгеть збереження його гомеостазу й актив-
но! життедгяльносп.
Мехашзми пасивно! резистентностг у холоднокровиях ! безхребетних мають
шше значения. Вони не завжди спрямованг на пгдтримання гомеостазу. Гомеостаз
може змгнюватися (знижуються обмш речовин, температура тгла, потреба в кис-
ш), але виживашеть при цьому навгть пгдвишуеться. Холоднокровнг в раз! зни-
ження температури навколишнього середовища переходять у стан анабгозу.
Онтогенез. Особливу увагу привертае становления реактивносп в процесг онтоге-
незу. За зршетю до моменту народження тварин подгляють на незршонароджених
(гцури, гфолг) г зрглонароджених (гвшейськг свинки). Перш! народжуються елшими г
не покритими шерстю. 1х здатшеть пгдтримувати гомойотермпо занадто низька,! вони
практично пойкзлотермш. До моменту прозргвання, тобто до 13—14-го дня, у незргло-
народжених закшчуеться морфолопчний розвиток кори великого мозку 1 встанов-
люеться нормальна енцефатограма. Температура тата стабсвзуеться на 2—4-й тиждень
житгя. Пе забезпечуеться розвитком х»йчно1, а нотам фгзично! терморегуляци.
Перюд прозргвання с важдивим егапом формування реактивноста в незрьто-
народжених. До прозр!вання новонароджен! тварини можуть ви гриму вати глибо-
ку ггпотермпо, гшоксгю, вони легше переносять радгатьне прискорення. 3 удоско-
наленкям механгзмгв реактивноста, розвитком нервово! та ендокринног систем,
установлениям гомойотерми та можливоста активного пристосування до мгнливьх
умов навколишнього середовища у них знижуеться стайкгсть до впливу ексгре-
мальних факторгв (повторения закономгрностей филогенезу). Реактившсть у зрг-
лонароджених тварин наближаеться до реактивноста дорослих особин.
У людини п!д час внутргшньоутробногс розвитку первинна реактившсть зни-
жена, гмунолопчна забезпечуеться природними антиталами, алерпчна — вщсутня
Стан реактивноста новонароджених визначаеться як спадковими факторами
так ! особливостями внутршгньоутробного розвитку, а також впливом навколиш-
нього середовища. Зокрема низькою е збудливгсть кори великого мозку, перева-
жають шдкгрков! впливи. Больовг подразнення не локал! зуються. У зв’язку з не-
зршгстю гшоталамо-гшофгзарно-наднирникова система в перил 2—4 дю теля
народження не реагуе на стресори. У новонародженого недостатньо розвинен!
бюлопчш бар’ери. Епгдермгс складасться з двох-трьох шарге клпин замгсть семи,
спостсрггаються висок! значения рН на поверхн! шкгри, що зумовлюе легше про-
никнення гнфекцп. У перш! дш теля народження не мае лгзоциму, титр компле-
менту в кров! низький, фагоцитоз, як правило, не завершений До 2 роюв син-
тезуеться незначна кглькгеть интерферону.
Становления гмунолопчно! реактивноста почииаеться у внутргшньоутробний
перюд г пов’язане з функшею загруднинног залози та лгмфощно!' тканини. Дитииа
народжуеться з гмуноглобулгнами (антителами), одержаними в!д матер!. Це гму-
ноглобулши класу С (1§С), як! збернаються протягом 3 м!с. Першим гмуногло-
бул!ном, шо синтезуеться в оргашзм! новонародженого, е 1§М. Його вмгст пгдви-
щуеться протягом 1-го тижня 1 до 1 року сягае ргвня дорослих. Г&А синтезуеться
з 2—3-го тижня життя. Пщвищення синтезу !§С визначаеться лише з 2—3-го м!-
сяця. Антиталоутворення пщвищуеться з розвитком лгмфощног тканини, що вгд-
буваегься з першого року життя ! тривае лише до перюду статево!’ зрглоста.
Пш час внутргшньоутробного розвитку не проявляеться патогенний вшив де-
яких бактергальних гоксишв (черевного, висипного тифу), очевидно, завдяки в!дсут-
носта специфгчних рецепторгв на поверхш кл!тин. Так пояснюютъ ниш клггинну ре-
актившсть у разг вродженого тмунгтету до гоксишв г вгрусгв. Водночас до шших ток-
сингв (гноетворних мгкрооргашзмгв, збудника дифгерй тощо) резистентшсть низька.
Мехашзми резистентност! формуються протягом перших рокгв життя дитини.
МЕХАШЗМИ РЕАКТИВНОСП. Б10Л0Г1ЧН1 БАР’СРИ
У формуванш механгзмгв реактивноста_важливу роль вцпграють нервова й ендо-
кринна системи. Даш онто- й фшогенезу евщчать, що становления реактивноста
пов’язане з удоеконаленням цих систем. У дослщженнях 1.П. Павлова та пред-
ставниюв його школи встановлено значения ргзних вщдглгв центрально! та авто-
номно! (вегетативно!) нервсво! система в реактивности Наприклад, вгломо. шо шд
’162 ! I
час глибокого сну т в стант наркозу уражувантсть електричним струмом менша,
нтж у стан! неспання. Зимова сплячка тлюструс роль нервово!' системи тварин в
тмунолопчнш реактивносп та резистентностт до шфекщй.
Кеннон 1 Селье важливу роль у реактивносп та резистентносп вщводять ен-
докриншй систем!. В умовах. що вимагаютъ вщ оргашзму напруження та мобшза-
цц пристосувальних мехашзмгв, Кеннон вбачае провтдну роль адреналтну (“авартй-
ного гормону”), Селье — гормонтв аденогптофУза (передньот частки ппофтза) т
кУрково!' речовини надниркових залоз. Особливо показовою щодо цього с участь
кортикостерощтв у реалтзацп запалення, коли глюкокортикощи виконутоть роль
протизапальних, а мУнералокортико'тди — прозапальних фактортв
Вщомо також, шо в раз! гтперфункцц щитоподтбноз залози перебтг запалення
тнтенсившший, а в раз! гшофункцй — млявий. Значно знижуеться реактившсть
при цукровому дтабетт (тривале загоювання ран, гподермУя, часто приеднуеться
туберкульоз).
Важливу роль у реактивностУ вщУграе сполучно тканина, елементи якот беруть
участь в Умунних реакцУях. фагоцитоз!, забезпечують загоювання ран, виконують
бар’ерну функцпо.
Фактори навколишнього середовища також впливають на процес реактивностУ.
Так. з пУдвищенням температури повттря (води) реактившсть гпдвищуетъся навУть у
холоднокровиях. У цих умовах у рептилУй (варантв) виникае анафУлаксУя, у жаб —
правець У “камфорнт судоми”. У геплокровних пщ час гарячки пУдвишуеться титр
антиттл, посилюеться фагоцитоз. СпоетерУгамль тяжкий перебтг УнфекцУйних хвороб
без гарячки (“холодна дифтерУя”). Зниження температури тыа гцдвищуе стУйкУсть до
ппоксц, дп механУчних факторУв. Реактившсть знижуеться пщ час повного У, особ-
ливо, часткового голодування. Цпсаво, що окремг види реактивностУ пригнтчуються
в послщовностУ, оберненш тх розвитку в еволюцУ!'. Наприклад, при голодуваннУ пер-
шими зникають алерпчш реакцп. а поттм пригнУчуеться тмунттет. Ртзка змУна пого-
ди, пора року У юпмат також визначають стан реакптвностУ та резистентностт.
До факторш навколишнього середовиша. якУ впливають на людину, належать
також сошальнУ. Прикладом остального опосередкування реактивностУ в людини
е залучения людини до виробничого процесу. Порушення роботи цУе! соцУальнот
системи, наприктад вУдставання вщ темшв виробничого процесу. може стати
причиною лсихУчних розладтв Порушення мУкросошатьних вщносин на робой, у
родин! може зумовити розвиток неврогичних стантв, виникнення неадекватно!'
реакцп на соцУатьне та бюлопчне середовише.
Важливим мехашзмом реактивностУ, шо склався еволюцшно, е бУолопчний
бар’ер — спецУалУзованУ тканиннУ структури, якт захищаютъ органтзм або певнт
йот о части ни втд патогенних фактортв навколишнього середовища У забезпечують
збереження гомеостазу.
Розртзняють зовнтшнУ та внутрУшнУ бар’ери. До зовншшх бар’ерю належать
шкра, слизова оболонка. якУ захишають органтзм вщ впливу фтзичних, хтмтчних У
бтологУчних фактортв навколишнього середовища: органи дихання, якт затриму-
ють шкщливУ речовини атмосфсри; травний канал (бактерицидна дтя шлункового
соку, позбавлення харчових елементУв антигенних властивостей); печтнка. яка
знезаражуе токсичнт речовини, що надходять з тжею або утворюються в кишках:
селезтнка, лУмфатичнт вузли та решта ортантв, у яких мтстяться клУтини системи
мононуклеарних фагоцит Ув.
Внутриин! бар 'ери регулюють надходження з кров! в органи 1 тканини потргб-
них поживних речовин, води, електролгтгв, БАР г перешкоджають проникненню
стороншх 1 токсичних речовин.
Основоположник учения про бар’ерн! функцп Л.С. Штерн у 1921 р. обгрун-
тувала наявшсть мгж кров’ю 1 тканинною рщиною диференцшованих захисно-
регуляторних пристосувань, як! вона назвала пстогематичними бар’ерами. Кож-
ний орган, на думку Л.С. Штерн, мае свое середовище, тому що кров не контак-
туе з юптинами оргашв.
Функцюнальна характеристика бар’ер!в залежить вщ фгзюлопчних 1 морфоло-
пчних особливостей окремих оргашв 1 тканин. Особливютю кожного внутргшнього
бар’ера е його вибфкова (селективна) проникшсть. В окрему трупу видшено спе-
ц!ал!зоваш бар’ери: гематоенцефалхчний, гематоофгальм1чний, гематолаб!ринтний.
Основним структурним елементом внутргшшх бар’ергв е кровоносш капигяр-
н! судини, стажа яких складаеться з ендотелпо г базально! мембрани. Ендотелш
кашляргв, як 1 базальна мембрана, мае характерш для кожного органа особли-
вост!. Вони е морфолопчною основою виб!рково! проникност! бар’ер!в.
Гематоенцефал^чний бар’ер мае найскладшшу организацию. Кр!м ендотел!ю !
базально! мембрани до його складу входять основна арпрофгльна речовина, обо-
лони мозку (м’яка та павутинна), а також астроцити нейроглп. Перш! дан! про
гематоенцефал!чний бар’ер було отримано П. Ерлгхом (1885), який виявив, що
барвник трипановий сишй у раз! введения в кров щура забарвлюе вс! органи,
кргм мозку. Тепер вщомо, що в мозок не проникают антитгла, л!мфоцити, деяк!
лжарсыа препарата. Щодо метаболгпв, гормонгв, БАР бар’ер д!е вибгрково, ре-
гулюючи проникнення цих речовин безпосередньо в юптини мозку. Ця влас-
тив!сть пов’язана з локалгзованими в бар’ер! ферментними системами, як! руйну-
ють одш речовини та сприяють проникненню шших. Такий ферментний бар’ер
генуе, наприклад, для у-амшомасляно! кислота г дгоксифенглаланшу. Отже, бар’ер
може активно вщбирати з кров! речовини, потргбш для функцп органа.
В основ! функцп бюлопчних бар’ер!в лежать мехашзми д!ал!зу, ультрафшьтрацп,
осмосу, а також метаболгчна актившеть клгтин, яю входять у структуру бар’ера. 1нтен-
сившсть транспорту через бар’ер залежить вщ потреб органа, гемодинамгки, нервових
1 гуморальних впливгв, наявносп морфолопчних ! функщональних порушень.
Виконуючи захисну г регулюючу функцп, бюлопчш бар’ери пщтримують оп-
тимальний склад живильного середовища для органа г сприяють збереженню клЬ
тинного гомеостазу.
Бар’ерна функцгя змгнюеться залежно вщ стат!, вгку, нервових г гуморальних
впливгв, стану зовнппнього та внутргшнього середовищ. На функцп бар’ергв мо-
жуть впливати змгна сну ! неспання, голодування, втома, травма, д!я шфрачерво-
ного, ультрафюлетового та рентгенгвського випромшювання, ультракоротких ।
високочастотних хвиль, ультразвуку. Алкоголь, ацетилхолгн, пстамш, кгнгни, па-
луронщаза — засоби, що збуджують центральну нервову систему, шдвигцують про-
никшсть бар’ергв, а катехоламгни, сол! кальцпо, флавонощи (вгтамгн Р), снодшш
засоби справляють проталежний ефект.
Проникшсть бюлопчних бар’ергв змгнюеться при патолопчних процесах.
Пщвищення проникносп робить органи чутливппими до гнтоксикацп, посилюе
пухлинний ргст. 3 порушенням бар’ерних функцш пов’язують можлив!сть авто-
гмунного ушкодження органгв.
Розд|л VIII
ПАТОЛОПЧНА Ф13ЮЛ0ПЯ 1МУННО1 СИСТЕМИ
ОСНОВН1 ФУНКЦГГТА СТРУКТУРА 1МУННО1 СИСТЕМИ
Головною функцхею хмуннох системи е тдтримання антигенного гомеостазу в ор-
гашзм!. При цьому хмунна система забезпечуе зв’язування х руйнування як хнфек-
щйних, так 1 нешфекщйних антитешв, виконуючи тим самим захисну функцйо.
Захист (спишешь, резистентшсть) оргашзму проти чужоргдних шфекцшних /
нетфекцшних, наприклад пухлинних, антигешв визначаеться як гмуштет I бувае
вродженим (природном) 1 набутим (адаптивном).
Мехашзми вродженого хмуштету неспецифхчш х спрямоваш проти будь-якого
збудника хвороб. Цх мехашзми включаються швидко, але хх недолгком е те, що
дхють вони хнколи неадекватно та позбавлеш хмунологхчно! пам’ятх. Вони подхля-
ються на клхтинш, гуморальш х додатковх.
• До клггинних мехашзмхв вродженого хмунхтету належать моноцити х тканин-
н1 макрофаги, нейтрофйш, еозинофели та натуральна (природы) кглери (НК, пайхга!
кШег, ТЧК).
• До гуморальних механгзмгв вродженого хмунхтету належать комплемент, 61-
лок пропердин, що активуе систему комплементу по альтернативному шляху, ан-
тибактерхальний быок — бета-л(зин, лактоферин, що забирае у мхкробхв залхзо, а
також антивхрусш а- х ф-штерферони (1Ф).
• До додаткових мехашзмхв вродженого хмуштету вщносять зовшшш I внут-
рйиш бар ’ери (неушкоджену шкхру та слизовх оболонки), соляну кислоту шлунково-
го соку, жирш кислоти сальних залоз, молочну кислоту вагшального секрету та
потових залоз, л1зоцим сльозовох рхдини х слини, хншх секрети, що видаляють мхк-
рооргашзми, кисень у тканинах (проти анаеробних мхкробхв), температуру тхла.
Набутий хмуштет формуеться пхсля першого потрапляння збудника в органхзм
та його фагоцитозу АПК. Цей хмуштет е специфхчним до збудника, зберхгае хму-
нолопчну пам’ять про антиген, а тому швидкхсть х сила реакцп хмунно'Т системи
на антиген значно зростае при повторному контакт! з ним.
Мехашзми набутого (адаптивного) хмунхтету теж подхляють на клхтинш та
гуморальш.
• Клхтинш мехашзми набутого хмуштету реалхзуються Т-лхмфоцитами за
участх АПК (макрофагхв х дендритних клггин).
• Гуморальш мехашзми набутого хмунхтету представлен! хмуноглобулшами,
що продукуються В-лхмфоцитами, та хщтокхнами, що синтезуються активовани-
ми Т-лхмфоцитами ! моноцитами-макрофагами.
Залежно вщ того де мгстяться чужорщн! антигени, !мун!тет у функцюнально-
му аспект! також можна под!лити (схема 2) на гуморальний (позаклгтинний) )
КЛ1ТИННИЙ (протиклггинний).
• Гуморальний гмуштет (не слщ плутати з гуморальними мехашзмами гмун!-
тету) забезпечуе резистентшсть до позаклггинних антиген!в (гноетв!рн! бактерп,
гельмгнти), як! мютяться в плазм! кров! ! тканиншй рщин! поза клггинами ор-
гашзму. Такий гмуштет забезпечуеться узгодженою д!ею комплементу, нейтро-
фгл!в, еозинофщгв (неспециф!чн! вроджен! мехашзми) та гмуноглобулшгв (спе-
циф!чш набут! мехашзми).
• Клгтинний гмуштет (не слщ плутати з клггинними мехашзмами гмуштету)
забезпечуе знешкодження пухлинних клггин 1 клттин, у яких паразитують в!руси,
гриби, найпростшп та деяк! бактерп (мжобактерп туберкульозу, збудники лепри,
сифппсу, гоноре!, бруцельозу тощо). Такий гмуштет забезпечуеться НК (неспе-
циф!чний вроджений мехашзм) та макрофагами, Т-лимфоцитами, монокшами 1
лгмфокгнами (специф!чш набут! мехашзми).
3 викладеного вище видно, що неспецифгчний вроджений ! специф!чний на-
бутий !мун!тети дуже псно взаемодпоть м!ж собою, пщтримують 1 доповнюють
один одного.
1мунна система складаеться з центральных оргашв (кгстковий мозок, загруд-
нинна залоза (тимус), фабрициева сумка птах!в та и аналог у людини) 1 периферий-
ных (селезшка, лгмфатичн! вузли, лгмфощна тканина травно! системи, мигдали-
ки). Кр!м того, у систему входять рухлив! !муноцити — лгмфоцити, як! перено-
ситься кров’ю 1 лгмфою.
Антигенами е р!зш за структурою 1 походженням речовини, що зумовлюють
гмунш реакцц. Розр!зняють антигени повн! й неповш (гаптен). На вщмшу вщ
повних антигешв, гаптени можуть спричинювати гмунну реакцпо разом !з вели-
комолекулярним нос!ем.
Схема 2. Дш тр!ади гуморальних > кзптинних мехашзмш вродженого та набутого виуштету, що забез-
печують гуморальний гмуштет (захист вщ позаклггинних агентов) I кл1тинний 1муштет (захист вщ
пухлинних юптин । внутрппньоюптинних чужорщних агенпв), за А. Ройтом (1991)
Генез г функшя Т- 1 В-л1мфоцитгв. Основними ефекторами гмунно! вщповщ! е
два види гмуноцитгв: Т-лгмфоцити (тимусзалежш) 1 В-лгмфоцити (залежи! вщ
фабрицгево! сумки у птах!в та и аналога в людини). Т-л!мфоцити здгйснюють клг-
тиннг гмуннг реакцп. В-лгмфоцити, що виробляють гмуноглобулши (антитгла),
забезпечують гуморальш гмунш реакцп.
Обидв! лгнп лгмфоцитгв розвиваються з! спглъног кроветворно! частково ди-
ференцйованог мультипотентног стовбуровог клггини. Т-л!мфоцити утворюються
з клгтини-попередника в тимус!, В-л!мфоцити — у птахгв у фабрицгевш сумш, ана-
логом яко! в людини, очевидно, е ембргональна печгнка, а шсля народження —
кгетковий мозок.
Види Т-л!мфоцит!в. СубпопуляцГ! лгмфоцитгв ргзняться як специфгчними до
антигену рецепторами, так г свогми функц!ями. Кргм того, згщно з мгжнародною
класифгкацгею лгмфоцити розргзняють за наявшстю певних трансмембранних
глгкопротегнгв — маркерних антигенов клйпин, як! також називають кластерами
диференшацп (с!а8!ег оГ (ИГГегепГгаИоп, СБ). Т-л!мфоцити, частка яких у кров! ста-
новии, 55—75 %, подшяють на дв! велик! групи.
1. Т-лшфоцити-хелпери, що становлять 55—60 % ус!х Т-лгмфоцитгв кров!.
Вони мають на свой поверхш СБ4 г розшзнають чужорщш антигени ильки пгсля
гх обмеженого протеолгзу (процесингу) та експресп на свой поверхш макрофага-
ми й гншими АПК у комплекс! з антигенами ГКГС II класу. Основна роль Тх
полягае в активацн В-лгмфоцитгв, л!мфоципв-к!лер!в ! макрофапв.
2. Т-л1мфоцити-к1лери (Тк; вщ англ. клИег — убивця). Вони становлять 20—
30 %, несуть на соб! СП8 г розшзнають чужорщш антигени на клгтиш, що мае
ядро, у комплекс! з антигенами ГКГС I класу. Основна !х функцгя — запуск ци-
толггичног реакцп або апоптозу в пухлинних чи гнфгкованих клгтинах.
Т-хелпери своею чертою подшяють на Тх 0-го, 1-го, 2-го г 17-го типу (ТхО,
Тх1, Тх2, Тх17):
• лгмфоцити ТхО (“нагвш”) — це попередники шших видгв Т-хелпергв. Зок-
рема пщ впливом 1Л-12, який продукуеться активованими АПК, ТхО диферен-
цпоються на Тх1, пщ впливом 1Л-4, що виробляеться тучними клгтинами, — на
Тх2, а в разг послщовног дп трансформуючого фактора росту бета (ТФР-0), 1Л-1,
1Л-6, 1Л-21 г особливо 1Л-23 — на Тх17;
• Тх 1-го типу продукують 1Л-2, гамма-штерферон (1Ф-у) г ФНП-а, як! акти-
вують макрофаги, Т-кглери г НК, що забезпечуе посилення клгтинного гмунгтету,
у тому чиелг захист вщ внутргшнъоклгтинног гнфекцп;
• Тх 2-го типу продукують 1Л-4, 1Л-5, 1Л-10 г 1Л-13, якг сприяють перетво-
ренню В-лгмфоцитгв на плазматичнг клггини, збглыпують продукцию гмуногло-
булгнгв г тим самим пщеилюють гуморальний гмуштет;
• Тх 17-го типу виробляють переважно 1Л-17, який об’еднуе низку цитокгнгв
(1Л-17А, 1Л-171, 1Л-17С, 1Л-17В, 1Л-17Е г 1Л-17Р, ФНП-а, 1Л-6, 1Л-8, 1Л-23
тощо) г хемокгшв, основне призначення яких полягае в посиленш гуморального
гмунгтету шляхом активацн нейтрофглгв для боротьби з грамнегативними бак-
тергями г деякими видами грибгв. При гнфгкуванш мгкобактергями туберкульозу
Тх17 продукують хемокгни СХСЬ9, СХСЫО, СХСЫ1, якг стимулюють хемотак-
сис Тх 1-го типу в легеневу тканину для боротьби гз ними внутргшньоклгтинними
бактергями, тобто посилюють г клгтинний гмуштет.
Супресорна функшя лтмфоципв. Рашше вважатося, шо !снуе окрема популящя
Т-л1мфоцит]в-супресор1Б. Нин! з’ясувалося, що таких юптин немае, а супресорш
функшТ виконують 1 Т-хелпери, 1 Т-кшери. Так, Тх2 продукують 1Л-10, який при-
гшчуе актившсть Тх1. Своею чертою Тх1 продукують )Ф-у, який гальмуе актив-
шсть Тх2 1 тим самим пригшчуе перетворення В-л!мфоципв на плазматичш юн-
тини 1 зменшуе продукцию 1ёЕ.
З’ясувалося. що СОЗ Т-юлери е двох вщпв. якз разняться наявшстю рецепто-
ра СО28 I вщповщно функцию: СО8* СО28Т Т-кшери (що мають одночасно СОЕ
1 СО28) е юлерами, а другий вид СО8" СО28’ Т-л!мфоцити (у яких немае СО28)
у дшсносп е супресорами, що виробляють гальмшш цитоюни 1Л-10, 1Л-6. 1Ф-у,
яю пригшчують актившсть АЛК > Т-кыерш. Накопичення СО8’ СО28- Т-Л1мфо-
цитщ виявлено в пухлинах, що пояснюс гальмування !х ьмунного знищення. Та-
кож було встановлено, що в раз! збтьшення юлькосп цих супресорш в!русна ш-
фекшя може набути хрошчного переб!гу.
Кр1м того, виявлено Т-хелпери, на яких кргм антигену СО4 екпресуеться
СО25, а також е ген РохрЗ, на якому синтезуеться бьток ГохрЗ, який е репресо-
ром транскришш ДНК, пщ д!ею якого гальмуеться активаця Т-Л1мфоцит1в. Ц!
СО4+ СО25Т Т-хелпери назвали Тге^ (регуляторними). Вони не продукують сти-
мулювальний 1Л-2, але здатш синтезувати гальм!вний для Тх1 1Л-10 ! ТФР-р. Усе
це супресуе не ильки Т-л!мфопити, а й АЛК.
Натуралып юлери — це велик! грануловмюш лгмфопити, що не мають аш
поверхневих !мунотлобул!нових рецептортв, аш специфтчного Т-юптинного ре-
цептора Проте НК здатш швидко розшзнавати та руйнувагп деяю пухлин ш •
втрусшф]кован! юптини за допомогою лекгинових га шших репеиторгв, як! реагу-
ють на неспецифхчш змши антигенш юптин.
Генез । види В-л!мфоцит!в. В антигензалежний перюд В-л!мфоцити кров) та
периферичних оргашв !мунно! системи стимулюються антигеном 1 осщають у В-
зонах селезшки та лшфатичних вузл!в (у фолгкулах ! центрах розмноження), де
зазнають бласттрансформацп: з малих л!мфоцит!в перетворюються на велике, що
розмножуються, а поттм — на плазматичш юптини. У них вщбуваеться синтез
хмуногл обули ив, ям надходять у кров. У людини вщомо пять клас!в !муногло-
бул!н!в: !§М, 1§Сг, [§Е, 1?А, (див. схему 4).
Будова (муноглобулшв 1муноглобулши класу О, Л 1 Е складаються з двох
легких (Ь) 1 двох важких (Н) полшептидних ланцюпв, зв’язаних дисульфщними
милками. Вшьн! ЕШ2-кшщ амшокислотних зазишгав легкого 1 важкого ланцюпв
(муноглобулппв збиаються Саме тут моститься активний центр антипла, за допо-
могою якого зоно реагуе з детермтантою антигену (ештопом). 1&А под1бний до
1$О. однак у раз! секрет! слизовою оболонкою перетворюеться на здвоену моле-
кулу7 — димер. 1§М е пенгамером, до складу7 якого входять 5 пар легких 1 важких
ланцюпв. УФ шуноглобулши мають лише два типи легких ланиюпв — к 1 X Важ-
ю ланнюги ь кожного класу 1муноглобулЁшв власн): ц, §, е, а, у.
Функщональт особаивосп 1муноглобулш1в. мають великий молекулярний
розм!р, через що мало проникають у тканини 1 слизову оболонку, дпоть в основ
ному в кров!, максимально прециштують 1 аглютинують антиген, сильно активу-
ють комплемент по класичному шляху, справляють цитотоксичну д!ю. Вони пер-
шими синтезуються в новонароджених, е незалежними вщ Т-л!мфопит1в ! акти-
Лб8
вують хемотаксис фагоцита. 1§М беруть участь у цитотоксичних та тмунокомп-
лексних алерпчних реакцтях.
1§А — секретарш тмуноглобулши, що мктяться в основному в слизу на сли-
зовш оболонцт та захищають и вщ мткробтв. У кров! тх значно менше, але вони
здатш активувати комплемент по альтернативному шляху т знешкоджувати мткро-
би 1 токсини, що циркулюють у кровь Беруть участь в утворенш комплекта з
антигенами в патогенез! алерпчних реакцш III типу (тмунокомплексних).
1§Е — !муноглобул!ни мал их розмтртв, у норм! в кров! вони мктяться в дуже
малш ктлькостт, легко проникають через судинну сттнку ! призначеш для клттин,
на яких е спецтальш рецептори для цих тмуноглобулштв. Вони не прециштують
антиген 1 не активують комплемент. 1§Е опсошзують гельмшта й активують еози-
нофйш, а разом з 1§А захищають слизов! оболонки. У раз! збтльшення тх синтезу
в десятки 1 сотш разтв розвиваеться анафтлактичний тип алерпчних реакцш.
1§(л — тимусзалежнт тмуноглобулши, що виробляються при повторит тмун-
нш вщповщт за обов’язковот участт Т-л!мфоцита, мають властивост! вс!х титв
тмуноглобулштв, але нижчого ступеня: прециштують антиген 1 активують компле-
мент, як 1§М; 1§О4 проникають у тканини т сорбуютъся на мембранах клттин, як
1§Е; транспортуються в слиз т секрета, як 1§А. Отже, 1§О беруть участь у встх
алерпчних реакцгях негайного типу, зокрема стимулюючих т гальмуючих, але на-
самперед цитотоксичних реакшях.
Функцп тмунно! системи. 1мунна система в раз! надходження антигенних речо-
вин в органтзм вщповщае за: 1) розптзнавання (процесинг) антигену; 2) розмно-
ження Т- г В-лтмфоцита клона, що несе рецептори або антипла до цього антиге-
ну, яке завершуеться утворенням субпопуляцтй лтмфоциттв г гуморальних антитш;
3) специфтчну взаемодтю субпопуляцтй Т- т В-лтмфоциттв т гуморальних антиттл з
антигеном; 4) утворення комплекта антиген — антиттло, як! активують лейкоци-
та кровт, БАР, якт прискорюють шакгивацтю антигену в органтзмт; 5) формування
тмунолопчнот пам’ятт; 6) запобтгання виробленню антиттл до структур власного
оргашзму ! пригнтчення цього вироблення, тобто шдукцтю т пщтримання тмуно-
лопчно'т толерантностт до свотх антигентв.
Специф1чн1стъ взаемодп з антигеном досягаеться завдяки вщповщностт актив-
ного центру антиттла детермшантт антигену. Ця вщповщшсть зумовлена послщов-
нтстю амтнокислотних залишктв М Н2-ктнпевих частан важкого т легкого ланцюпв
тмуноглобултну. Мтнливкть послщовностт спостертгаеться серед 100 амтнокислот-
них залишктв кожного ланцюга (вартабельних дтлянок), якт позначають вщповщ-
но Ун т Уь, а сталт (константнт) — Сн т Сь.
1з 200 вартабельних амтнокислотних залишктв з детермшантою антигену од-
ночасно реагують близько 200 амшокислот. Послщовнтсть амшокислот визначае
форму вигину ланцюга тмуноглобултну т наявнтсть радикалтв для взаемодп з анти-
геном. Вартабельнт дшянки важкого т легкого ланцюггв розташовуються в про-
стор! так, що певш радикали амшокислот точно пщходять до радикалтв антигену.
Цим т пояснюеться висока специфтчнтсть тмунних реакцш. Навтть портвняно не-
значн! змши в структур! антигену, його просторовот форми послаблюють або
усувають цю вщповщшсть т взаемодтю.
Ун1версалън1стъ взаемодп антипйл т рецепторгв з антигенами ртзнот хтмтчнот
природи пояснюеться здатшстю амтнокислотних залишктв тмуноглобултну утво-
рювати ртзш види зв’язктв: тонний, водень-водневий т зв’язок на основ! сил Ван-
дер-Ваальса. Через зв’язки Ван-дер-Ваальса антитгла реагують навпъ з пластмаса-
ми, стгйкими до дп кислот 1 лупв.
Оргашзм здатний давати гмунну вщповщь на велику кглькгсть ргзних речовин
1 навпъ на штучно створюваш речовини, яких рашше в природа не було. Ф.М. Бер-
нет показав, що ушверсальна гмунна вщповщь (вироблення антипл до будь-якого
антигену) можлива вже в раз! наявностг 100 000 клонгв гмуноцитгв, що для люди-
ни становить мгзерну частку вщ загально! кглькостг клгтин. Ргзномангтнгсть гмун-
ног вщповщг зумовлюеться величезною кглькгстю клонгв гмуноцитгв. Утворення
клонгв гмуноцитгв, якг несуть антитгла до ргзних антигенгв, досягаеться кгпъкома
шляхами.
1. Деякг гени, якг кодують варгабелъш дшянки гмуноглобулгнгв, успадкову-
ються.
2. кнують системи ферменпв, якг переставляють гени в геномг, що перебува-
ють навгть у ргзних хромосомах. При цьому ферменти розргзують г зшивають да-
лянки ДНК. За допомогою мехашзмгв перестановки гени варгабельних г констан-
тних дглянок гмуноглобулгнгв перемпцуються, або рекомбгнуються, у геномг г
об’еднуються в складнг гени-транслокони, на яких вщбуваеться синтез остаточ-
них полшептидгв гмуноглобулгшв. За допомогою рекомбгнацш лише 1000 кодую-
чих дглянок ДНК можуть забезпечити синтез мшгардгв гмуноглобулгнгв ргзно! спе-
цифгчностг.
3. Завдяки унгкальному мехаюзму мутацгй, який здгйснюеться в клпинах-по-
передниках за допомогою ферменту кгнцевог нуклеотидтрансферази, оргашзм
протягом гндивщуального розвитку набувае додатковог юлькосп гмуноглобулгнгв.
Доведено, що кглькгсть клонгв у новонародженого в 1000 разгв бгльша, шж у пло-
да. Навгть в шбредних тварин, щентичних, як однояйцевг близнюки, виявляють
гмуноглобулши. характерш для кожног особини.
4. Як вщомо, варгабелъна дшянка гмуноглобулгну утворена 200 амгнокислот-
ними залишками, а реагують з одним антигеном лише 20 амгнокислот. На тгй
самш дглянцг потенцгйно реактивними е й пшп комбгнацп амшокислотних за-
лишкгв. Встановлено, що на варгабелыпй дглянцг гмуноглобулгну одночасно мож-
ливг 100 комбшацгй по 20 амшокислотних залишкгв гз 200. Це означав, що один
клон гмуноцитгв потенцгйно може реагувати зг 100 ргзними антигенами з уси
можливих у природг.
Отже, завдяки сукупностг механгзмгв, якг забезпечують мутацпо г ргзноманп-
нгсть варгабельних дглянок, гх рекомбшацпо г потенцгйну полгвалентнгсть гмуно-
глобулшгв, оргашзм мае властивгсть реагувати на велику кглькгсть ргзних анти-
гешв.
Ефективн1сть руйнування антигену в процесг гмунних реакцш зумовлена фак-
торами, здатними вбивати клпини, що несуть антигени, г знешкоджувати поза-
клгтинш гнфекцгйнг агенти. Проти антигену в бглыпостг випадкгв активуються
клгтиннг та гуморальш гмуннг реакцп. Далг гмуноцити та комплекси антиген —
антитгло активують комплемент, протеолгтичнг ферменти та шшг БАР, якг руйну-
ють антиген. Лгмфокши залучають до процесу захоплення г руйнування антигенч
моноцити-макрофаги та шгш клгтини кровг й тканин.
Над1йн1сть в1дпов1д! оргашзму на антиген забезпечуеться розвитком клгтинних
г гуморальних гмунних реакцгй ргзних тишв одночасно, а також тим, що на одну
молекулу антигену може вироблятися 100 000 молекул антитгл. Цей ефект пояс-
170
нюеться кооперацоею макрофапв, Т-о В-тпмфоципв, розмноженням клггин клошв
унаслщок стимуляцо! антигеном.
У сповпрацо з омуноцитами беруть участь клгтини групи А (допоможш клпини
омуногенезу): клгтини ендотелпо судин, фобробласти, макрофаги, нейтрофоли,
еозинофоли. Ральф Стейнмен (лауреат Нобеловськоо премп 2011 р.) вщкрив ден-
дритно клпини та ох роль в активацп Т-ломфоцитов ! стимуляцн омунно! вщповщо.
Деоодритш юптини (вщ англ. бепбпИс — деревоподобний) — це юптини з вщрост-
ками, яю збольшують поверхню клпини, на яюй вони виставляють для Т-лгмфо-
цитов-хелперов чужорщн! антигени в комплекс! з антигенами ГКГС II класу, що
активуе клпинно о гуморальш реакцп проти цих антигенов. Дендритними клпина-
ми е грануляции! дендроцити епщермосу — юптини Лангерганса (отже, шюра е
своерщним органом омуногенезу), гнтердиптатш клпини центр!в розмноження
ломфатично! тканини та загруднинно! залози (тимусу).
Активно клпини омунно! системи в раз! кооперативно! взаемоди виробляють-
багато мед!атор!в, зокрема 1Л-1 (макрофапв о дендритних клпин), против!русн! та
протипухлинш 1Ф-а (лейкоцитов) 11Ф-0 (фобробластов), активувально юитиню та
блокувальш гуморальш омунно реакцп 1Ф-у, ФНП-а, 1Л-2 (Тх1), 1Л-4, 1Л-5,1Л-13 —
фактори диференцоацн, пролоферацп й активацп В-л!мфоцит!в, еозинофйпв, туч-
них клпин (Т-хелпер!в 2-го типу), макрофагальний 1Л-8 — стимулятор хемотак-
сису й активацп нейтрофшв, ГЛ-10, що виробляеться Тх2 1 пригшчуе Тх1, а також
шшо штерлейюни, колошестимулювальн! фактори о фактори росту епщермосу,
гранулоцитов, макрофапв, тромбоцит!в, нерв!в.
1мунолог1чна толерантшсть (або специфгчна витривал1стъ, ареактившсть) —
вгдсутшсть 1мунолог1чно1 реактивности до певних антигешв. Толерантность до
власних антигешв називають ф1зюлог1чною, а до чужих — патологичною. За кло-
нально-селекцойною гшотезою Ф.М. Бернета передбачалося, що функционально
незрыо гмуноцити на раншх етапах онтогенезу зустр!чаються в оргашзм! плода зо
сво!ми антигенами та блокуються ними. Згодом було встановлено, що надлишок
антигену справд! спричинюе блокаду свого клона омуноципв. Набута толерант-
ность такого типу називаеться високодозовою, а толерантшсть, зумовлена невисо-
кими дозами антигену, як! можуть спричинювати випереджальну стимуляцою Т-
ломфоцитов, що мають супресорну дно, — низъкодозовою. Доза антигену, достатки
для стимуляцн супресорно! вщповщо, менша за ту, яка потробна для стимуляцо!
хелперно! дп.
Формування толерантност! вщбуваеться протягом усього житгя, оскшьки це
пов’язано з утворенням клонов Т- о В-ломфоцитов зо стовбурових клпин, яке вщ-
буваеться також протягом усього житгя. Пгсля цього вщбуваеться мутацоя генов з
утворенням нових клонов Т- о В-ломфоцопов, зокрема здатних реагувати зо свооми
антигенами. У загрудниншй залозо тако клони еломонуються. Водночас В-ломфо-
цити, що утворюються в костковому мозку або в периферичних органах омунно!
системи, не мають таких умов, як Т-ломфоцити в загрудниншй залозо. Заборонен!
клони В-ломфоцитов, нетолерантш до антигенов свого оргашзму, або гальмуються
за типом високодозовоо толерантносто, або лишаються незагальмованими.
З’ясувалось, що у здорових людей частина В-ломфоцитов нетолерантна до анти-
генов свого оргашзму. Однак автоомунний процес при цьому не виникае, осюльки
Т-ломфоцити в норм! толерантно до сво'ох антигенов, а без Т-хелперов В-ломфоцити
"ТтГ
не стимулюються антигеном. Очевидно, у частиш випадктв до антигешв свого
оргашзму встановлюеться низькодозова толерантность, а коли все-таки з’являються
заборонен! клони Т-ломфонипв, в оргашзм! подтримуеться невисока концентра-
Ц1я антигену, достатня для стимуляцп супресорно! функцп Т-ломфоцитов.
Тмунолопчна толерантность принципово в!др!зняеться в!д омунодепресп своею
специфочшстю: при толерантност! до певного антигену антит!ла не виробляються
пльки до нього, а до гнших антигешв вироблення антитол повноцшне; у раз! ому-
нодепресп гальмуеться синтез антитол до болыпосто антиген!в.
ЗАГАЛЬН! ЗАК0Н0М1РН0СТ1 ПОРУШЕНЬ 1МУННОТСИСТЕМИ
Тмунна система мае певш особливост!, як! впливають на розвиток патолопчних
процес!в у шй.
Клггини омунноо системи, що поспи но розмножуються, високочутлив! до дп
мутагенних, канцерогенних 1 цитостатичних факторов, зокрема! певних л!карських
засобов. 3 ще! причини омунна система вразлива до дефициту вотамшов 1 болков.
Особливе значения мае вразливость до канцерогенних факторов, оскольки навоть
лгмфоцити периферично! кров! здатш до розмноження й онкогенезу.
Порушення диференцповання та функцш омуноцитов можуть спричинювати-
ся патологоею органогенезу ще! системи та дефектами генов у стовбурових копти-
нах, а також розладом взаемодп мож субпопуляцоями ломфоципв, недостатностю
гормонов загруднинно! залози, змшою функщй шших ендокринних залоз. Особ-
ливого значения набувають колыасно змши гормонов при стресо, надморность яких
зумовлюе омунодепресою.
Оскольки клотини омунно! системи при взаемодп з шфекцшними агентами та
!х токсинами можуть змшювати свою антигенность, в оргашзм! можуть виникати
омунно й автоомунно ушкодження цих клттин.
Диференцоацоя та функцп омунноо системи, зокрема й формування толерант-
носто, залежать вод контакту з антигеном, а тому вплив антигену може порушити
цо процеси, наприклад шдукувати толерантность до шфекцшного антигену.
Розлад функцш омунно! системи може проявлятися гопер-, дис- та гшофунк-
цоею, змшою голерантносто до антигенов.
Пперфункц1я шунног системи розвиваеться в раз! перенапруження цоео системи
антигеном, зокрема внаслщок надходження в органозм стимуляторов омунноо вщ-
поводо. Гшерфункцно можуть спричинювати спадково змши синтезу омуногло-
булшов, наприклад, 1г-генами (омунореактивними генами), що зумовлюють поси-
лену омунну водповодь на будь-який антиген. До гшерфункцо! може призвести
зменшення регуляторного гальмування всередиш омунно! системи: зниження п
супресорно! функци, а також оззовно — недостатшсть функцп гшоталамо-ппофоз-
но-наднирниковоо системи.
Особливе мосце посодае пперфункцоя под час формування пухлин з клттин
тмунокомпетентно! тканини. При цьому спостерпаеться збольшення колькосп
клттин та омуноглобулшов одного типу, шо вщображае втрату пухлинними омуно-
цитами контролю над процесами синтезу о розмноження.
У раз! пперфункщ! омунно! системи в органозм! створюються умови для роз-
витку алергн.
Дисфункщя шунног системи може розвиватися, наприклад, у разё зниження
функцп Т-лёмфоцитёв, що зумовлюе недостатню стёйкёсть оргашзму до шфекцп,
особливо вёрусёв ё грибёв. У таких випадках через дефщит супресорних впливёв
можуть збёльшуватася реакцёя В-лёмфоцитёв ё вироблення антитёл, зокрема 1§Е,
що спричинюе алергёчнё реакцп на ёнфекцёёёш антигени (наприклад при брон-
хёальнёй астмё). Уведення хворому засобёв, якё стимулюють Т-лёмфоцити (напри-
клад левамёзолу), може призупинити розвиток шфекцгйного захворювання й од-
ночасно напади бронхёальноё астми. Дисфункщя ёмунноё системи часто поед-
нуеться з п гёпофункцёею.
Гтофункщя гмунно! системи е дуже поширеним порушенням. Хвороби, що
супроводжуються гёпофункцёею гмунноё системи, подёляють на ёмунодефёцитнё
(спадковё, первинш) та ёмунодепресивш (набутё, вториннё).
Експериментальне вёдтворення ппофункцп ёмунноё системи. Спадковё форми
недостатностё системи Т-лёмфоцитёв вёдтворюються у тварин ёнбредних (одержа-
них унаслёдок близькоспорёдненого схрещування) лёнёй. Виведено лёнёю мишей
писёе (голё), для яких характерними е спадково зумовлений недостатнёй розвиток
загруднинноё залози, вёдсутнёсть шерстё; клётиннё реакцп сповёльненого типу в
них не вёдбуваються.
Недостатшсть системи вёдтворюеться видаленням загруднинноё залози в
плодёв або новонароджених тварин. При цьому на тлё збережених гуморальних
реакцёй у тварин розвиваеться вастинг-хвороба (виснажлива хвороба), характер-
ними для якоё е рёзке зменшення кёлькостё лёмфоцитёв у кровё, зниження клётин-
них ёмунних реакцёй, спленомегалёя, ёнфекцёйш хвороби, зокрема вёрусний гепа-
тит, кишковё ёнфекцёё, припинення росту ё загибель. ёнфекцёйнё хвороби набува-
ють особливо тяжкого перебёгу.
Недостатшсть системи В-лёмфоцитёв вёдтворюеться у птахёв в ембрёональний
перёод або в курчат шляхом видалення фабрицёевоё сумки. Розвиток Т-лёмфо-
цитёв при цьому не порушуеться.
Зниження функцп Т- ё В-систем або окремих субпопуляцёй лёмфоцитёв мож-
на досягти за допомогою антилёмфоцитарних сироваток проти антигешв, якё ма-
ють рёзнё лёнёё та субпопуляцп лёмфоцитёв. Пёд час експерименту ёмунодепресив-
ного ефекту досягають завдяки використанню ёонёзуючого випромёнювання, хё-
мёчних ёмунодепресантёв, глюкокортакоёдёв.
Порушення рёзних функцёй ёмуноцитёв можна вёдтворювати пёд час ёх культи-
вування. Так, усуваючи Т-хелпери, пригнёчують вироблення антитёл В-лёмфоци-
тами.
1мунодефщитж хвороби
ёмунодефёцитнё, або спадковё, хвороби називають ще первинними, на вёдмёну вёд
вторинних порушень ёмунноё системи, якё виникають унаслёдок ёнших хвороб.
Отже, можуть розвиватися ёмунодефёцитнё хвороби окремо системи Т-лёмфоцитёв
ё системи В-лёмфоцитёв, а також ёхнё поеднання. Оскёльки спадково зумовлена
блокада можлива на всёх етапах розмноження та диференщювання обох лёнёй
ёмуноцитёв та ёнших клётин ёмунноё системи, варто розрёзняти спадковё порушен-
ня: 1) органогенезу ёмунноё системи; 2) генетичного апарату ёмуноцитёв; 3) 1г-
генёв ё, як наслёдок, спадково зумовлеш дефекта вироблення антитёл.
Схема 3. Генез Т-тмфоцитш (за Б.Д. Бройлзом ] О.В Рохлшим, 1978)
На пгцсгав! анайзу виявлених у людини 70 вид1в первинного !мунодефщиту в
2006 р. трупа експерпв УИжнародного товариства з вивчення гмунодефшитгв зап-
ропонувала класифпгащю ]мунодефщитних хвороб. В п основу було покладено
таю ознаки: кл!н!чн! прояви захворювання, зокрема 1 зчеплення з не!мунною па-
толопею, вид порушення функип Т- 1 В-л!мфоцит!в окремо або в поеднанн!; р!-
венъ генетичноТ блокади в генез! шунопипв; тип успадкування.
1мунодефщитт хвороби, пов’язаш з1 змтами в систем Т-ммфоццпйв. Генетич-
на блокада розмноження та диферешнювання Т-л!мфоцит!в можлива на будь-
якому етап! генезу. У генез! Т-л!мфоцит!в можна вид!лити три перюди: дотимус-
ний, тимусний, шслятимусний (схема 3). За схемою можна простежиги, генез
яких субпопулянш 1 тип!в гмуноципв порушуеться шсля блокади.
У дотимусний перюд на I етап! стовбурова кроветворна кл!тина псретворюеть-
ся на нашвсговбурову л!мфо!дну юптину-попередника. сшльну для Т- 1 В-лгмфо-
щгпв. Блокада на пьому р!вн! призводить до полною розладу утворення л!мфоцит!в
обох лгн!й 31 збереженням генезу шших кл!тин кров! — гранулоцита, еритроцита
тощо. За такого ступени гмунно! незахищепосп можлива смерть у ранньому виц.
На II етап! утворюстъся нашвстовбурова юптина — попередник Т-л1мфоципв.
Блокада на цьому ргвш генезу зумовлюе вщсутшсть Т-.пмфошгпв за наявносп В-
лтмфоципв, розвиток тяжко! 1мунодефщитно! хвороби через вщсутшсть тмунних
реакшй клпинного типу 1 випадання шших функцш Т-л!мфоцита.
Генетична блокада в тимусний перюд диференцповання клпин (III—IV ета-
пи) спричинюе порушення утворення Т-л!мфоцит!в. однак не завжди призводить
до !х повно! вщсутност! в кровь
У раз! блокади шслятимусного диференц! ювання юптин може порушились
утворення окремих субпопуляцш Т-лгмфоцита (V 1 VI етапи).
Наведемо передне хвороб (синдром !в), пов’язаних 13 розладом утворення Т-
лтмфоциттв (ВООЗ, 1978).
Первинна недостатшсть Т-л!мфоцит!в виникае у раз! блокади на раншх етапах
утворення Т-л!мфоцит!в або розладу утворення Т-хелпер!в. Установлено спадко-
вий характер хвороби.
Плоплаз^я загруднинно! залози (синдром Д1 Джордж!) розвиваеться внаслщок
порушеното формування третьо! 1 четверто! приглотково! зяброво! кишен! в емб-
рюнальний перюд. Унаслшок цього кр!м вщсутност! загруднинно! залози 1 Т-л!м-
фоцита виникають дефекти обличия, щитоподтбно! та пришитопол!бних залоз,
вади оргашв кровообпу. Не розвиваються !мунш реакцп клпинного типу, не вщ-
буваеться або затримуеться вщторгнення трансплантата.
1мунодефшит Т-л1мфоцита, зумовлений генетично спричиненим дефектом адсно-
зиндезамшази або пуриннуклеотидфосфорилази, харакгеризуеться зниженням фун-
кцп Т-л!мфоцита. Деф)цит аденозиндезамёнази або пуриннуклеотидфосфорила-
зи, шо розшеплюють пурини, призводить до накопичення метаболшв — дез-
оксиаденозинтрифосфату (6АТР) 1 дезоксигуанозинтрифосфату (6ОТР), токсичних
для стовбурових Л1мфо!цних клггин. Ц1 метаболии шпбують фермент рибонуклео-
тидредуктазу, необхщну для синтезу ДНК I, вщповщно, для розмноження кл!тин.
Кр1м того, у Л1мфо!дних кптинах визначаеться низька актившсть 5’-нуклеотида-
зи, яка в !нших ют пи нах оргашзму компенсуе недостатшсть аденозиндезаменази 1
пуриннуклеотидфосфорилази шляхом запобпання накопиченню б АМР ! 6ОМР
Недостатшсть експресп антигешв ГКГС II класу спричинсна дефектом промо-
торних б!лк!в, що зв’язуютъся з 5’-д1лянкою гешв II класу. Без молекул ГКГС II
класу в загрудниншй залоз! не утворюються Т-хелпери СО4+. Ней синдром успад-
ковуеться за автосомно-рецесивним типом. У хворих тз недостатшстю експресп
антигешв ГКГС II класу знижуеться продукция антитш, спостер!гаю1ься повтори!
шфекш! травного каналу.
1мунодефщитн1 хвороби, нов ’язан! з/ злйнами в систем! В-лЫфоципйв. Генетич-
но зумовлеш порушення формування В-лтмфоцита можлив! на вс!х етапах !х ут-
ворення (схема 4). Якщо генетична блокада розвиваеться на р!вн! утворення стиль-
ного попередника Т- ! В-л!мфопит!в (I етап), то може виникнути дефщит обох
вид1в л!мфоцит!в. Якщо дефект перешкоджатиме диференцпованню В-л!мфоцит!в
на II, III, IV етапах. розвинеться недостатшсть В-лтмфодита р!зних субпопуляшй.
а формування системи Т-лтмфошптв може здшснюватися звичайним шляхом.
У раз! порушення остаточного диференцповання В-л!мфоцит!в 1 клпин
пам’ят! на V, VI, VII, VIII етапах може припинятися синтез окремих кластв тму-
Схема 4. Генез В-лзмфоцитш (ВООЗ, 1981)
ноглобулпнв, що призводить до розвитку 1мунодефщиту системи В-л!мфо-
ЦИТ1В.
Ф!з!олопчна ппогаммаглобулшемы виникае в дней в!ком вщ 4 до 12 тиж. У цей
перюд знижуеться рзвень 1муноглобулпнв, яю дитина успадкувала вщ матер!, 1
починаеться синтез власних змуноглобулппв.
Транзиторна гшогаммаглобулшем!я дитячого вису характеризуеться затримкою
синтезу що може продовжуватися до трир!чного в!ку. В-клггини таких дней
не мають дефектов, проте, очевидно, не одержують вщ Т-юптин СО4+ достатньо!
допомоги для продукцп антит!л.
Гшогаммаглобулшемш Брутона (Х-зчеплена агаммаглобулшемгя — Х-АГ) —
класична форма В-1мунодефщиту. При цьому утворення всхх тип1в плазматичних
юптин блокуеться так, що кыыасть 1§С знижуеться в 10, а 1§А 1 1§М — у 100
раз1в. Генетична блокада вщбуваеться на ргвш юптини — попередника В-Л1мфо-
цит1в. У цих юптинах порушений ген цитоплазматично! тирозинкшази, що нале-
жить до родини онкогену зге; попередники В-л1мфоцит1в не можуть перетворю-
ватися на зрпп форми. Хвороба передаеться за зчепленим 31 статтю типом 1 про-
являеться у хлопчикгв. Реакцп Т-л1мфоципв збернаються.
Загальний вар1абелышй 1мунодефщит (ЗВ1Д), пов’язаний з порушенням переда-
вания сигнал1в вщ Г- до В-юптин (шзшй 1мунодефщит), розвиваеться в ос1б вжом
вщ 10 до 30 роюв 1 старше як агаммаглобулшемхя теля вгрусно! гнфекцп, зокрема
спричинено! в!русом Епстайна — Барр (ВЕБ). Хвор! вразлив! до гншно! шфекцп
та кишкового паразита 61агсНа 1атЬНа, що зумовлюе тяжку диарею, а також злоя-
к!сну анем1ю. У бшьшосп хворих на ЗВЩ В-юптини незрш, бо не одержують
необхщних для активацп сигналш вщ Т-л1мфощгпв, можливо, через ушкодження
останшх. ЗВ1Д не е спадковою хворобою, проте вш асоцгйований з ГКГС-гапло-
типами НГА-В8 1 НЬА-ПКЗ.
1мунодефщит, пов’язаний з порушенням синтезу 1§А, розвиваеться в раз! гене-
тично! блокади на завершальному еташ диференщювання В-лгмфоципв у плаз-
матичнт юптини, що синтезують 1§А. Передаеться як автосомна, рецесивна, а
школи як доминантна ознака. У деяких випадках у хворих виявляеться делец!я
хромосоми 18. Синтез хмуноглобулппв шших класгв зазвичай зберпаеться.
Кр1м блокади диференщювання В-лгмфоципв дефщит 1§А спричинюеться
генетично зумовленим порушенням синтезу секреторного компонента (8) в егп-
телп залозистих оргашв. Полшептид 8 потргбний для захисту 1§А вщ протеолггич-
них ферменттв.
1мунодефшит унаслщок генетично! блокади синтезу легких ланцюпв 1муногло-
булппв, тобто появи неповноланцюгових хмуноглобулппв.
1мунодефщит унаслщок дисфункщ! гмуноципв характеризуеться одночасним зни-
женням продукцп одних хмуноглобулппв 1 зростанням синтезу шших. До таких
хвороб належить 1мунодеф1цит, пов’язаний з блокадою синтезу та 1§А внаелг-
док гшерпродукцп 1§М (ГТГМ). При цьому у В-юптинах не вщбуваеться переклю-
чения синтезу 1§М на синтез 1§О, 1§А 11§Е. Генетична блокада може зумовлюва-
тися мутащями в геш, що кодуе лгганд (радикал зв’язування) СВ40, який у норм!
забезпечуе зв’язування В-лгмфоципв з Т-лгмфоцитами 1 переключения синтезу
1§М на синтез 1§С 11§А. Втрата СО40 робить неможливим цей перехщ. Кр1м того,
може бути порушений шший мехаюзм переключения синтезу 1§М — зв’язування
1Л-4 з рецептором до 1Л-4 на поверхш В-юйтин у раз! мутацп гена рецептора 1Л-4.
У разг блокади переключения вех плазматичш юптини можуть синтезувати пльки
1§М, кшьюсть яких стае надапрною на тл1 вщсутносп шших 1муноглобулш!в. Цей
1мунодефщит спричинюе гшйш шфекцп та автохмунш хвороби. Вш успадковуеть-
ся за зчепленим з! статтю (Х-хромосомою) типом.
Комбшоват {мунодефщитш хвороби. Найбшьша кшькгеть виявлених 1муноде-
фщитних хвороб е комбшованими з одночасним дефектом Т- 1 В-лгмфоцитгв.
Серед них видшяють так!.
Тяжкий комбшований 1мунодефщит (ТК1Д), Хвор! на ТК1Д д1ти гинуть зазви-
чай у перпп два роки життя вщ шфекцп, спричинено! умовно-патогенними г
сапрофгтними збудниками — ротавгрусом або кишковою бактергею (дгарея), най-
проспшими Рпеитосузпз саппи (пневмонш), др!жджоподгбними грибами Сап<И<1а
а1Ысапз (стоматит, дерматит). Навпъ гмунгзацгя живи ми вакцинами проти полго-
мгелгту г туберкульозу (БНЖ) спричинюе нрогресуючу гнфекшю г смерть. Проте
шсля транс пл анташ । кгсткового мозку дитина може нормально розвиватися. Кров
хворих на ТК1Д дней мгстить дуже мачо лгмфоцитгв (< 3000 в 1 мл) У затрудни н-
нш залозг вщсутнг стовбуровг лгмфогдю клггини, через шо вона втрачае значения
л!мфо(цного органа.
ТК1Д часпше розвиваеться у хлопчикгв, юж у дтвчаток (3 : 1). У половини ви-
падкгв хвороба спричинюеться дефектом гена, розташованого в Х-хромосомГ Цей
ген кодуе у-ланнюг рецептора до 1Л-2, шо входить також до складу рецепторгв для
шших штерлейкпйв: 1Л-4, 1Л-7, 1Л-И г 1Л-15. Для нормально! функш! Т-клгтин
важливе значения мае вплив 1Л-7 на сво1 реп ел тори, оскгльки цей штерлейкш
шдукуе диференгпацгю стовбурових клгтин у попередники Т- 1 В-лгмфоцитгв, по-
сидите пролгферашю В- г Т-лгмфоцитгв г активуе зрит Т-лгмфоцити.
Отже, при ТК1Д стовбуроы лгмфощш клгтани не можуть одержувати сигнали,
необхщнг для росту та диференцповання. В шших випадках ТК.1Д пов’язаний з
дефектами в рецесивних генах шших хромосом.
Як близький до ТК1Д видьтяють гмунодефщит швейпарського типу, що успад-
ковуеться за автосомно-рецесивним типом 1 проявляеться лгмфопенгею та пло-
там маглобулгнемгею (деф! питом Т- 1 В-лппй лгмфоцитгв одночасно) у перш! тиж-
н! життя. При цьому загруднинна затоза мае зачаткову форму; кгркова г мозкова
речовини п не диференцноються. У периферичних лгмфощних органах (селезшцг,
лгмфатичних вузлах) спостерйаеться р!зке зменшення клтькост) лгмфоцитгв г плаз-
матичних клгтин. Трансплантата тканин не вгдгоргаються. реакцп сповй.ьнено!
гшерчутливосп немае. Спостерпають слгди 1§С, 1§М та 1§А вщсутнг. До цих по-
рушень призводить дефщит ферменпв аденозиндезамгнази або пуришгуклео-
тидфосфорилази.
Лмунодсфщит з телеанпектазгею та атаксгею (синдром Луг-Бар) успадковуеться
як автосомно-репесивпа хвороба. Порушен) функцгя систем Т- г В-лгмфоцитгв,
диференцповання Т-лгмфоципв на раннгх етапах генезу. Загруднинна залоза пе-
ребувае в зачатковому сташ, кглькгсть Т-лгмфоципв знижена, вгдеутнгй, зни-
жений або залишаеться нормальним ргвень за нормального ргвня 1§М. Пору-
шен! кгнцев! етапи диференшювання В-лгмфонипв. П!д час цитогенетичного до-
елгдження в клгтинах виявляготъ розриви хромосом 7 г 14 у дглянцг генгв
Т-клпинного рецептора г генгв, якг кодують важкг ланцюги гмуноглобулппв. Ген.
дефектом якого зумовлений цей гмунодефгцит, кодуе биток, що бере участь у ре-
парацп розрив!в д волан цюговог ДНК.
Тмунодефщит з громбоцитопешею та екземою (синдром Вгскотта — Олдргча) успад-
ковуеться за ознакою, зчепленою зг статно, виявляеться у хлопчикгв у вгцг 10 ро-
кгв. Прогресуе порушення функцгй системи Т-лгмфоципв, спустошуються Т-зони
в лгмфатачних вузлах. Одночасно порушуються гуморальн! !муннг реакцп через
ураження В-лшш знижуеться ргвень 1§М за нормального вмгсту 1§А г 1§6. Вста-
новлено, шо за цгег хвороби порушуеться цитоскелет Т-кдгтин. зменшуеться кгль-
к!сть мшроворсинок на поверхнг цих клгтин поргвняно з нормальними Т-лгмфо-
цитами. Вгдомо, що в ход! кооперацгг Т- г В-клгтан пщ час антиплоутворення
цитоскелет Т-клгтини зм!нюе свою оргенташю або по.тяризуеться в напрямку В-
юптини. У випадку синдрому Взскотта — Отдртча це не вздбуваеться !, як наслз-
цок, порушуеться взаем одзя юптин зм^ттно! системи.
Гмунодефщити часто мають тяжкт наслздки. Спостерзгаюгь рззке зниження
змунзтету. Навпь ослаблен! вакцини, застосовуваш для щеплень, можуть спричи-
нити шфекшйну хворобу 1 смерть.
У раз! змунодефтциту спостерзгають значке збзлыпення частоти пухлинних
хвороб. За дефщиту Т-лзмфоцитзв частота утворення пухлин збхльшуеться у 1000
раз!в. Недостатшсть змунноз системи харакгеризуеться збзльшенням кзлькостз ви-
палюв автозмунних хвороб, пов’язаних з порушенням п регулящУ й ослаблениям
змунного захисту проти мутацш.
€ певш вздмзнностз мзж наслцзками дефншту Т- ! В-лзмфопитзв, а також м!ж
окремими хворобами всерединз кожно! групи. Порушення в разз дефшиту Т-лзм-
фоцитзв проявляеться вздсутнзстю або зниженням юптинних змунних реакпш, ре-
гуляторноз та кооперативно! фунзсцш Т-лзмфоцитзв. Значно знижена стзйкзсть до
ураження втрусами з грибами, меншою мзрою — до гноетвзрноз та пневмококово!
шфекцзт, знижуеться здатшсть органззму вздторгати трансплантат. Дефшит В-л1м-
фоцитзв проявляеться вздсутнзстю або зниженням гуморальних змунних реакцш
через дефшит змуногл обул пив одного або кзлькох класзв. Спостерхгаеться значне
зниження стзйкостз до стрептококово!, пневмококово!, кишковоз знфекцзз однак
зберзгаеться стзйкзсть до втруств 1 гриб! в.
Тип дсфзцитних змунотлобулзшв визначае характер хвороби: при дефзцитз
]§М знижуеться стзйкзсть до з'рамнегативних бакгерзй, у раз! дефщиту 1§А ослаб-
люеться захист слизово! оболонки травного каналу, очей тошо.
Перебзг комбшованого змунодефшиту найтяжчий. Так, при гшогаммаглобулз-
немзз швейпарського типу дуже швидко розвиваеться пневмо Кокова знфекшя, шо
рззко прогресуг 1 часто призводить до смерть
1мунодепресивж стани
Набутими можуть бути порушення генезу центральных г периферичних оргазтзв
змунноз системи, змунолопчноз толерантност! та найчастште — порушення генезу
1 функцз! змуноцитзв.
Порушення органогенезу змунно! системи, наприклад порушення розвитку за-
груднинно! залози з глоткових кишень, можуть вздбуватися пзд впливом еколопч-
них факгорзв у перзоди ембрзогенезу га формування органа. До таких факторзв
належать зошзуюче випромштовання, питостатичю речовини (знпбзтори синтезу
нуклешових кислот I бзлкзв — антибзотики, аналоги пуринових з пзримздззнових
основ, антиметабол зги фолзево! кислота), токсини бакгерзй, взруси (наприклад
взрус краснухи). ПздГрунтям змзн можуть бути порушення синтезу та рецепт!
зндукторзв органогенезу.
Рохтад формування; тпдтримання змунолопчноз толерантностз може стосуватися
Т- 1 В-лппй лзмфопитзв окремо чи в поеднанш. Проявляеться втратою толерант-
ностз до антигензв свого оргашзму' (див. роздзл ХП “Алергзя”) або набуття толе-
рантностз до чужих антигешв. У другому випадку причинами розвитку толерант-
ностз можуть бута надходження антигензв в оргашзм плода вза хворо! матерт.
порушення плацентарного бар’ера, у дорослого — дзя розничного антигену у
пщвищешй доз! (парад 1ч Фелтона); 1ндукц!я Т-супресорно! функцп невисокими
дозами антигену (тимусзалежна низькодозова голерантшсть).
Порушення генезу та функцш 1муноцит!в вщбуваеться внаслщок дп р!зних
етюлопчних факторов. Унаслщок цього можуть розвиватися порушення на етапах
розмноження, диференщювання л!мфопит!в ! продукпп !муноглобул!н1в. Ц! пору-
шення стосуютъся кожно! л!нп Л1мфоцит!в окремо або в поеднанш, одного клона
!мунопит!в, шо реагують на один антиген, або юлькох клошв.
Можлив!сть розвитку ртзних патолопчних процеФв в !муннш систем! визна-
чаеться тим, що 1мунокомпетентна тканина е мгготичною 1 чутливою до дц бага-
тьох етюлопчних факторов — канцерогенних, мутагенних, цитостатичних, як! зу-
мовлюють порушення подиу юптин 1 синтезу б!лк!в у них.
Пщ впливом канцерогенних факторгв !мунокомпетентн! кл1тини дають поча-
ток розвитку пухлини, лейкозу. Онкогенез можливий не лише в стовбурових кро-
вотворних кл!тинах, а й у б!льш диференцшованих Т- 1 В-л!мфоцитах, здатних до
бласттрансформацп га розмноження. а також у плазматичних юптинах.
Мутагеннг фактори спричинюють соматичю мутацп юптин !мунно! системи,
що призводить до змши функционально'] знатноеп !муноцит1в ! ступеня реакцп на
антиген. Соматичн! мутацп гмуношгнв також сприяють почв) заборонених клошв,
здатних реагувати з антигенами свого оргашзму 1 спричинювати розвиток автс-
!мунних хвороб.
У раз! дп цитостатичних факторов вщбуваеться пригшчення ёмунно! вщповщ!
внаслщок порушення розмноження, диференцповання кл!тин 1 ослабления синте-
зу антиттл. Цитостатичний ефект дають х!м!чн! шпбпори синтезу ДНК, РНК, нук
лешових кислот, юшзуюче випромшювання, лжарсъю препарата, зокрема циюто-
фосфамщ. азатюприн, мткофенофетат безпосередньо д1ють на ДНК або н синтез.
Однак варто видшити деяк! особливост! редкий тмунно' системи на Д1ю щет
групи ьмунодепресанпв. Так, в !мунн!й систем! е досить радюрезистентн! тмуно-
цити. Вщ тмунодепресанпв найбшыпе потерпають юптини клон!в, шо розмножу-
ються т стимульован! в цей момент антигеном. На практиц! не використовують
для виб!рково! (мунодепресп щодо будь-якого антигену. 1мунодепреси може роз-
виватися внаслщок штоксикацп при тяжких хворобах ! отруеннях.
Потреба в постшному розмноженн! гмунокомпетентних юптин зумовлюе враз-
ливтсть !мунно! системи до дефщиту бйгкгв I вгтамгнгв. За вщсутност! аскорб! ново!
кислота може пригшчуватися (мунна вщповщь, що перешкоджатиме розвитку ав-
то!мунного захворювання — експериментального алерпчного енцефал!ту, який
виникае в раз! !мун!зацп тканиною мозку. Недостатшсть ретинолу (вггамшу А),
шридоксину (в!там]ну В6) ! фол!ево! кислота, а також мгкроелементгв, таких як
цинк, селен, м!дь або залхзо, призводить до ослабления функцш !мунно! системи.
Стрес, стерогдш гормона спричинюють лейкоцитопешю, галъмують вироб-
лення цигокппв, впливають на мйрацйо юптин. 3 д!ею глюкокортикощгв, як!
ослаблюють розмноження ! синтез б спав, пов’язаний розвиток тмунодепресивно-
го стану тип, час стресу. Це може зумовити виникнення шфекшйних хвороб, таких
як грип, туберкульоз тощо. Хзрурпчне втручання може сиричинити стрес 1 !муно-
депреФю Однак поряд з кортизончутливими е й кортизоннечутлив! !муноцити.
Розлад внутрйяньосис темно! гормонально! регуляцй гмунно!' системи спостерйа-
ють при хворобах затрудни нно! залози, шсля п видалення, у раз! порушення сек-
рецп тирозину та шших гормошв, шо призводить до змши стану та генезу гмуно-
цитгв 1 недостатност! гмунно! вщповщ-, шод! в прихованш форм! Внутршшьосис-
темне порушення регуляцп може бути зумовлене розладом утворення гуморальних
факторгв субпопуляцш лгмфоцитгв, наприклад фактора супресп хелпер!в або лгм-
фокшгв.
Особливу трупу порушень становлять автамушп хвороби. В умовах патологи
можуть вироблятися анти тгла до маркерних антигешв Т- г В-лгмфоципв, напри-
клад унаслщок мугаци та появи заборонених клонгв гмуноциггв. Проти Т-лгмфо-
цит1в розвиваються гуморальш гмунш реакцп. На особливу увагу заслуговуе мож-
ливгсть вироблення антиты до варгабельних дглянок гмуноглобулппв (антиантиты).
Варгабсльш дыянки сам! е щютиповими антигенними детерминантами. Вироблен-
ня антиантиты може пригшчуваги 1 му ян у вщповщь на певний антиген.
1мунодепрес!ю може зумовити пухлина. шо розвиваеться в оргашзм!, коли
нормальш гмуноцити витгсняються пухлинними юптинами г юптинами метаста-
31В. оргашзм насичуеться продуктами порушеного метаболизму, а також пухлин-
ними антигенами.
Особливий варган г гмунно! недостатност! спостершаютъ при деяких шфекщй-
них хворобах. При гоноре! це зумовлено здатшстю гонококк “вислизати” з-п!д
!мунно! вщповщ!. З’ясувалося, що кожи! 3—4 доби в гонококах вщбуваеться пов-
на замша антигешв поверхн! мембран, з якими попередньо утворен! антитыа не
можуть реагувати, бо вони були синтезоваш проти попередшх антигешв. У геном!
гонококзв е кыька сотень гешв, якг кодують поверхнев! антигени, г при кожнш
змш! поверхневих антигешв гонококи стають не доступними дн ранппе виробле-
них антиты, шфекцгя набувае хрон!чного характеру. При хрошчних шфекцгях,
зокрема сифшсг, маляр!!, шфекшйш збудники також часто змшюють поверхнев)
антигени сво!х мембран, випереджаючи вироблення антит!л.
Зниження функшй гмунно! системи спостерхгають з вгком, переважно теля
50 рок!в (див. розды V “Роль спадковосп, конституцп та вгкових змш у патоло-
ги”; “Старшня”).
Синдром набутого гмунодефщиту (СН1Д). Вщкрито РНК-вмтсний в!рус Н1У
(В1Д англ. Нитап иптипоНераепсу уйиз — в!рус гмунодефщиту людини, В1Л), що
зумовлюе синдром набутого гмунолефшиту — СШД. За допомогою глгкопроге'щу
ёр! 20 цей и!рус зв’язуеться з быком СЭ4 на мембранах Т-хелпертв та шших юп-
тин, зокрема клггин мозку.
Загибель Т-хелпер!в та 1нших СВ4’-юптин вщбуваеться кыькома шляхами:
1) в!рус безпосередньо руйнуе заражен! клггини, коли розмножуеться в них;
2) вгрусний §р!20 у мембран! заражено! юптини з’еднусться з быком СВ4 нор-
мально! юптини, що призводить до злигтя здорово! та хворо! клгтин; 3) заражеш
юптини знищуються клоном !муноцит!в, яю реал!зують тмунну реакцйо проти
в!русу; 4) выьний вгрусний глгкопротещ ®р!20 сорбуеться на здорових юптинах,
яю стають об’ектом знищення гмуноцитами.
Без Т-хелперних л!мфопит!в уся система !мунного захисту ргзко слабне, особ-
ливо стгйкгсть до в!русгв, гриб!в, патогенних найпростгших, мгкобактерш. Спос-
тернають ураження слизово) оболонки ! зниження активност! травления, можли-
во, через прямий вплив вгрусу на юптини, а також ураження нервовог системи
внаслщок проникнення вгрусу в мозок 1 руйнування СЕ>4+-клгтин центра.тьно!
нервово! сисгеми.
У щлому порушення в оргашзм] в раз! гмуноделресивних сташв такт самт, як
1 при спадкових гмунодефшитних хворобах. Широкий .шапазон 1 вторинний ха-
рактер в(др]зняють порушення тмунноТ системи.
ПОРУШЕННЯ СИСТЕМ, ФУНКЦЮИАЛЬНО
ПОВ’ЯЗАНИХ 3 1МУННОЮ СИСТЕМОЮ
До систем, функгуонально пов’язаних з гмунною системою, належать систем»
фагоцитозу, БАР, насамперед система комплементу. Антипла специфично реагу-
ють з антигенами, однак ефектившсть захоплення та руйнування гх залежить вщ
ступени активацп та функцюнального стану цих систем. Змгни системи компле-
менту та шших БАР. а також фагоцитозу. р!зко знижують ефектившсть хмунних
реакцдй 1 насамперед спйкгсть до шфекцш них хвороб.
Порушення системи комплементу. Система комплементу складаеться з таких бгт-
кових компонента, яю мгстяться в сироватц! кров! (наведено в порядку активацп год
вшивом комплексу антиген — антитйо): С1 (ушорюеться з С1д, С1г I С1§ за наяв-
носп юив кальшю), С2, СЗ, С4, С5, С6, С7, С8, С9 Комплекс, що утворюеться, е
активним комплементом 1 мае властивосп естераз г протеаз, руйнуе оболонки тва-
ринних клггин 1 мпсрооргашзмтв. а також активуе шип БАР 1 фагоцитоз (мал. 15).
Змши системи комплементу бувають спадковими т набутими Можливою е
блокада синтезу будь-якого з п компонента.
Мал. 15. Мехашзм руйнування мембран» клггин (утворення в мембран! пор) шд вшивом комплемен-
ту 1 Т-йлера (за Дж.Д.-Е. Юном 1 Ж.А. Кот, 1989). Гуморелъна га клггинна система !мунного
захисту мають полдбш мехашзми утворення пор у кл!тинн!й мембран!. У гуморальнтй систем!
зв’язування антиттл з поверхнею юптиии-мштен! тнпцюе каскад системи комплементу. Б!лки С5
! С6 зв’язуються з мембраною кл!тини-м!шент, а до них приеднуються битки С7, С8 1 багато
молекул бглка С9. утворюючи пору (а). У клпиннш систем! пори виникають унаслщок контакту
Т-ктлера з юптиною-мштенню: вони формуються з молекул лише одного бгтка — перфорину,
шо видшяс Т-юлер (б)
У раз! дефщиту С1 сироватка кров! втрачае бактерицидшсть. Розвиваються
повтори! шфекцп верхшх дихальних шляхтв, отит, ураження суглобгв, хрошчний
гломерулонефрит. Нерщко спостерпають летальний наслщок. Цей дефщит успад-
ковуеться як автосомна ознака.
Дефщит С2 зумовлюе зниження бактерицидност! сироватки кров!, розвиток
в!русних шфекщй, дифузних хвороб сполучно! тканини, гломерулонефриту! тром-
бопенИ.
Компонент СЗ е ключовим у формуванш ферментних ! регуляторних власти-
востей комплементу. Синтез СЗ визначаеться двома алельними генами як кодо-
мшантна ознака; якщо в кров! немае одного з гешв, кыыасть СЗ у шй змен-
шуеться вдв!ч!. Дефщит СЗ спричинюе високу смертшсть.
Дефщит С5 зумовлюе тяжю кишков! шфекцп, збудниками яких е грамнега-
тивш бактерп. Характерним е також ураження шк!ри. Спадкова недостатшсть С5
уже в ранньому дитячому вщ! проявляеться дерматитом, проносом, порушеннями
розвитку дитини.
У раз! дефщиту С6 спостер!гають хворобу суглобгв — специф!чний артрит.
Порушуеться здатшсть кров! до зсщання. Дефщит С7 зумовлюе розвиток дифуз-
них хвороб сполучно! тканини. Недостатшсть термшальних компонент (С5, С6,
С7 1 С8), а також компонент альтернативного шляху активацп комплементу
(фактора Ц 1 пропердину) зумовлюе особливу схильшсть до шфекщй, збудниками
яких е два види роду бактерш №1ззег1а — № ^опоггдоеае ! У. тетп&НйНз.
У раз! спадково зумовленого дефщиту шпбпора компонента С1 полегшуеть-
ся активац!я комплементу, який е патогенетичною основою анпоневротичного
набряку Квшке.
Набуту недостатшсть комплементу спостерйають у раз! розвитку в людини
ендокардиту, сепсису, малярп, деяких в!русних шфекщй. Ц! хвороби можуть
спричинювати виникнення гломерулонефриту, можливо, внаслщок нагрома-
дження в оргашзм! незруйнованих за вщсутност! комплементу комплект анти-
ген — антитшо. Актившсть комплементу знижуеться також при червоному вовча-
ку, ревматощному артрит! тощо.
Порушення фагоцитозу. Актившсть фагоцитозу може знижуватися внаслщок
розладу утворення фагоцит 1 пригшчення !х активност!. Ц! порушення можуть
бути спадковими 1 набутими.
Зниження активности фагоцитозу вщбуваеться щд впливом гормошв — глюко-
кортикоида, мед!атора нервово! системи — ацетилхолшу, у раз! нестач! в оргашзм!
певних гормонгв, впампйв, порушення водно-мшерального балансу. Актившсть
фагоцитозу може знижуватися пщ впливом гл!кол!тично! отрути — монойодацета-
ту, який порушуе вироблення енергп в лейкоцитах, бакгер!альних токсишв, що
призводить до незавершеного фагоцитозу. Фагоцитарна актившсть деяких фер-
мент у лейкоцитах р!зко знижена при лейкоз!, тмуншй недостатност! та дефщит!
комплементу, осюльки в норм! антитша й комплемент стимулюють фагоцитоз.
У раз! синдрому Чед!ака — Х!гас! виникае спадковий дефект утворення л!зо-
сом, порушуеться фагоцитарна функция лейкоцит, що супроводжуеться накопи-
ченням у них фагоцитованих незруйнованих м!кроорган!зм!в. Успадковуеться за
автосомно-рецесивним типом.
При хрошчному гранульоматоз! виявлено спадков! дефекта фермент лейко-
цит, потргбних для внутрппньоклгтинного руйнування бактерш, зокрема деф1-
~ 183
цит НАДФ-оксидази, який спричинюе недостатшсть утворення пероксиду водит
та порушення фагоцитозу. Фагоцитоз стае незавершеним. Хвор! на хрошчни
гранульоматоз резистентш до пневмо-, стрепто-, мешнгококово! шфекцп, тодг я
стафглококи, кишкова паличка спричинюють у них розвиток тяжких хвороб. Д
подгбних порушень призводить дефект лпелопероксидази. Це дае пщстави стверд-
жувати, що, можливо, основну функцгю захисту оргашзму вщ сапрофггних шфек-
цгй виконуе фагоцитоз.
Установлено також спадковг дефекта быка м’язових волокон — актину, за вщсуг-
ностг якого порушуеться змикання цитоплазматичног мембрани над фагосомою
Спадковий дефект фермент! в вутлеводного й енергетичного обмшу знижу
загальний функцюнальний потенцгал фагоцитгв. Прикладом е хвороба Альдера —
спадкове порушення метаболгзму полгсахаридгв у лейкоцитах.
Недостатшсть адгезп лейкоцитгв (НАЛ) у раз! порушення складу штегришв. У фа-
гоцитах змгнюеться вмгст одразу трьох штегришв: штегрину СК.З, штегрину ЬРА-
1 (лейкоцитний функцюнальний антиген) г р150,95, оскгльки гх структура пред-
ставлена одним г тим самим р-ланцюгом. Встановлено дефект гена р-ланцюга.
який мгстаться в хромосом! 21.
Тнтегрин СКЗ необхщний для поглинання мгкрооргашзмгв фагоцитами, ос-
кгльки вгн е мембранним рецептором 3 для зв’язку з активованою фракщею ком-
плементу СЗЫ на поверхш опсошзованих мгкробних клгтин. У разг вщсутностг гн-
тегрину СКЗ розвиваються тяжкг бактергальш шфекци, зокрема в порожниш рота
г травному канал!. Молекула штегрину СКЗ побудована з а-ланцюга з молекуляр-
ною масою 165 кДа (СИНЬ) г р-ланцюга з молекулярною масою 95 кДа (СВ 18).
У структуру гнтегрингв ЬРА-1 ! р150,95 входить також р-ланцюг. Незважаючи
на те що кожен гз цих бглкгв мае свтй а-ланцюг (СВПа г СВПс вщповщно), оби-
два вони пщ час НАЛ дефекта!. Молекула штегрину ЬРА-1 взаемодге з молеку-
лою мгжклгтинног адгезп (1САМ-1) на поверхш ендотелюцитгв та шших клгтин-
них мембранах. Унаслщок дефекту ЬРА-1 фагоцита хворих на НАЛ не здатн!
прикршлятися до ендотелгю судин г тому не можуть мггрувати з кровоносних су-
дин у дглянку проникнення шфекцгйного агента. Унаслщок цього гнфекцгя по-
ширюеться по всьому органгзму.
Разлад утворення I диференцйовання фагоцитов вщбуваеться пщ впливом гнпбг-
торгв синтезу нуклегнових кислот, у разг пухлин кгсткового мозку та лейкозу, гн-
ших факторгв, як! порушують подш клгтин. За наявностг деяких спадкових хвороб
(наприклад спадково!' нейтропени) може вщбуватася блокування подогу клгтин, а
також спостерггаються функцгональна неповноцгннгсть дозргваючих лейкоцитгв,
автогмушп ураження ргзних видгв лейкоцитгв г на цш основ! — лейкопенгя.
АНТИГЕНАСОЦ1ЙОВАН1 ХВОРОБИ
Роль антигешв можуть виконувата ферменти, структури! быки, полгсахариди, глг-
копротещи та гншг речовини, як! виробляються в самому оргашзм!. Наявшсть цих
речовин, гхня структура генетично зумовлеш г визначають властивостг тканин
органгзму, зокрема спадкову схильшсть до хвороб.
Зв’язок мгж антигенами оргашзму г хворобами можна встановити ргзними
шляхами, адже антигени! властивостг якого-небудь компонента органгзму можуть
зумовлювати ймов!ршсть розвитку хвороби. Якщо, наприклад, у тканинах е анти-
ген, близький до антигену збудника, то такий збудник може уражувати цей ор-
гашзм, осюльки 1х антигенна сшльнУсть знижуе ймовУрнУсть 1мунноУ вщповщУ.
Вщомо, що вУруси грипу вступають у генетичнУ рекомбшацп з тканинами люди-
ни, тварин У птахУв У набувають антигешв цих тканин. Так утворюються нов! шта-
ми в1русу грипу, що спричинюе виникнення повторних епщемУй, осюльки цей
В1рус грипу в новш быковш оболонш не розшзнають антипла, яю виробилися до
попереднього штаму в!русу.
За даними Альберта 1 Итце (1977), у людей, яю мають трансплантацшний
антиген НБА-В8 ГКГС (кодуеться трьома генами, розмпценими в людини в хро-
мосом! 6-1 пари), Устотно збшыиена частота розвитку автоУмунних ушкоджень
м’яз!в, надниркових залоз, печшки, шюри, щитоподУбно! залози, кишок. У лю-
дей, яю мають трансплантащйш антигени НЬА-В8 1 НЬА-В\у15, найчастУше спос-
терУгають юнацький д!абет, у носив Н1А-В27 — ураження сечово! У статевоУ сис-
тем (хвороба Рейтера) 1 юнацький ревматощний полУартрит, у нос!Ув Н1А-В13,
НЬА-17 У НГА-Вау38 — псор!аз. Унсулшозалежний цукровий дУабет чпко асоцшо-
ваний з успадкуванням антигешв ГКГС НЬА-Р9>уЗ,2 У НГА-ЭК4, у яких у поло-
женш 57 р-ланцюга стоУть валш або серин (якщо сто'Уть аспарапн, то цукровий
дУабет не розвиваеться).
Для розумшня антигенасоцшованих хвороб важливим е вщкриття прямо!
участУ антигешв ГКГС у специфУчних рецепторах Т-лУмфоцитУв, яки ми цУ лУмфо-
цити розшзнають антигени. Отже, наявнУсть ГКГС визначае, вщ яких антигешв
лУмфоцити не можуть захищатися або, навпаки, на яю антигени лгмфоцити схиль-
ш нападати. Тому деяю антигенасоцшованУ хвороби е наслщком порушення нор-
мально! регуляцн Умунно! вщповщУ антигенами ГКГС.
Другу трупу антигенасоцУйованих хвороб становлять тУ хвороби, при яких
роль певно! речовини в мехашзм! захворювання визначаеться не антигенними, а
п структурними або функцюнальними особливостями. Так, при спадковш фер-
ментопатп дефщит ферменту може бути виявлений не лише бУохУмгчно, а й за
допомогою антитш, одержуваних теля Умушзацп ферментом.
Завдяки Умунолопчним методам в оргашзм! виявлено рУзш антигени, функция
якихУ дос! лишаеться невщомою. Однак встановлено зв’язок м!ж ними антигена-
ми У хворобами. Наприклад, в осУб, гомозиготних за антигенами групи кров! 0(1),
утрич! частпие розвиваеться виразка шлунка. Можливо, антигени А- 1 В-груп
кров! потр!бш для мембран юптин слизово! оболонки шлунка, щоб пщвищити
Ухню стгйюсть до дп соляноУ кислота, а саме цих антигешв немае в ос!б з групою
кров! 0(1).
Антигенний склад оргашзму може зумовлювати також щдвищення !муштету.
Так, в ос!б з групою кров! В(Ш) захворювашеть на чуму значно нижча, юж в ос!б
з шшими трупами кров!.
Антигени труп кров! як складова частина мембран та шших структурних еле-
ментгв юптин, а також трансплантацУйш антигени не ильки визначають антиген-
ну сумюшсть тканин, а й, очевидно, можуть впливата на рецепцгю юптинами
гормонУв, мед!атор!в, БАР (мембранний транспорт). Це, можливо, позначаеться
на УндивУдуальних особливостях реактивностУ, зокрема вщповщУ на дУю лпсарських
засобУв.
Частина друга
ТИП0В1 ПАТОЛОПЧШ ПРОЦЕСИ
Розд1л IX
ПАТОЛОПЧНА Ф13ЮЛ0ПЯ ПЕРИФЕРИЧНОГО
КРОВООБ1ГУ
Перш юж обговорювати основы! мехашзми порушень периферийного кровообпу,
слщ нагадати, як функцюнуе система кровообпу в цьлому 1 яке мкце в шй посщае
периферичний кровообгг. Гемодинамгка, або рух кров! по судинах, виникае
внаслщок р!зниц! гщростатичного тиску в р!зних дшянках судинно! системи (кров
рухаеться з дьтянки високого тиску в д!лянку низького). Р!вень тиску в р!зних
вщдщах судинного русла залежить вщ трьох основних мехашзм!в: капитально!
функцп серця, ОЦК 1 периферичного опору.
Нагшталъна функцгя серця забезпечуеться викидом у судинну систему 60—
70 мл кров! пщ час кожного скорочення. Основним гемодинамтчним показником
е об’емна швидкгсть кровотоку (0), або ХОС — кглькють кров!, що викидаеться
серцем в аорту за 1 хв. ХОС дор!внюе добутку об’ему кров!, що викидаеться при
кожному скороченш (ударний об’ем), на частоту скорочень серця 1 в доросло!
людини становить 4—6 л/хв.
ОЦК е сумарним об’емом кров!, що м!ститься у функцюнуючих кровоносних
судинах. У доросло! людини вш становить приблизно 6—8 % маси т!ла або 4,5—
5,5 л. Приблизно 20 % ОЦК мгститься в артериях, що становлять систему судин
високого тиску. Близько 80 % ОЦК мгститься у венах великого кола кровооб!гу !
в малому кол! кровообхгу, що е системою судин низького тиску.
Периферичний оп1р (ПО) виникае внаслщок тертя кров! об судинну стшку та
в’язкост! само! кров!. Величина периферичного опору в основному визначаеться
тонусом артерюл, що характеризуеться м!рою постгйного скорочення непосмуго-
ваних м’яз!в стшок цих судин. Це створюе перешкоду для вщтоку кров! з ар-
тер!ально! системи 1 забезпечуе пщтримку певного р!вня артер!ального тиску. 3
шшого боку, змгна просвпу артерюл 1 опору судин регулюе величину кровотоку
через окрем! судинш дщянки, приводячи його у вщповщшсть до штенсивносп
житгед!яльност! тканини та и потреб у кисш 1 поживних речовинах. В штенсивно
пращоючих тканинах, наприклад у м’язц що скорочуеться, кровотж може збшь-
шуватися в 100 раззв 1 бзлыпе, причоиу величина загалъного артерзального тиску
1 ХОС можуть зстотно не змгнюватася.
Завдяки сицгьнгй роботз вказаних трьох мехашзмзв в аорт! й артерзях кров пе-
ребувае щд високим тиском (для людини в норм! приблизно 120/70 мм рт. ст.).
Рзвень його визначаеться стввздношенням мзж ХОС 1 опером периферичних су-
дин (у першу черту7 артерюл). Лшзйна швидкзеть руху кров! при постзйному ХОС
залежить вщ сумарноз площ! перер!зу судин. При розгалуженн! артерш спостерз-
гаеться розширення сумарного судинного русла, що досягае максимальных значень
у капшярнзй с!тц! (сумарний просв!т кап!ляр!в на 2—3 порядки перевищуе проевзт
аорти). Тому швидкзеть кровотоку' висока в артерзях (у людини близько 50 см/с) !
артсрюлах 1 низька в капзлярах (близько 0,5 мм/с) На посткапзлярних дзл янках
тиск кровз продовжуе знижуватися, досягаючи в передсерд! нульових ! навпъ нега-
тивных значень, а швидкзеть кровотоку збзлыцуЕться через звуження кров’яного
русла. Рух кров! по венах здшснюеться головним чином за рахунок енергп, що
забезпечуеться роботою серця; гх он!р незначний, унаслщок чого повернення кровз
до серця вщбуваеться при невеликому градзенп тиску у венознш систем!.
Кровообзг на дзлянш периферичного судинного русла включав таю структуры!
елементи: дрзбн! артерп, артерюли, метартерюли, прскапзлярн! сфзнкгери, капзля-
ри, посткапщярш сфгнктери. аргерюловенулярнз анастомози, венули 1 др!бн! вени.
У систем! периферичного кровообпу умовно видзляють м!кроциркуляторне русло,
до складу якого входять терм шалънз судини, д!аметр яких не перевищуе 100 мкм.
тобто артерюли, метартерюли, капзляри, венули. артерзовенулярзп анастомози.
Окрему трупу становлять судини лзмфатично! системи. Але у фзззологПз пато-
фшологп част!ше використовують функцюнальну класифзкацзю судин мзкроцир-
куляторного русла, яка включае три групи судин: резиставн!, обмш из та емнзенз.
Вздповщнзсть анатомзчноз та функшональноз класифгкащй наведено в табл 7.
Таблица 7. Анатом!чна! функциональна класиф!кац!я
периферичного судинного русла
Анатомзчна класифзкащя Функциональна класифзкафя
Артерюли Прекапзляпи Мез артерюли Прекапзлярнз сфзнкгери Резистивнз судини
Капзляри Обмзннз судини
По ст капзляри Посткаи! зярн! ефзнктери Венули Дрзбш вени €мнзст судини
Артерзовеьулярнз анастомози Резервнз судини
Лзмфатзгчна суаззнна система Резорбтивнз судини
Вщззовгдно до функщонально! класифзкацп. перша трупа судин передуезм за-
безпечуе функцзю периферичного опору, капьтярам властива функи!я обмшу, а
вщ третьоУ групи судин залежать ефект депонування кров! та регуляцзя венозного
повернення. Судин и л!мфатично! системи забезпечують функцпо резорбцп або
збирання тканинно! рщини 1 и повернення в судинне русло.
Зулин имося на двох перших функцхях деталънпне. Як уже зазначалося, пери-
феричний ошр, який залежить вщ стану резистивних судин, вил грае ключову
роль як у регуляцн системного артерхального тиску, так 1 в регуляш! кровотоку в
систем! мйфоциркуляцП, Причому системна змша тонусу периферичних судин
часише спричинюе змгни ргвня системного артергального тиску (спазм забезпе-
чуе пшвишення тиску. дилаташя — його зниження), а локальна змша тонусу (в
окремому оргаш або дшянщ тканини) зумовлюе розвиток порушень кровотоку на
р1вн1 мгкроциркуляцц (табл. 8).
Таблица 8. Развитой патолопчних стаже,
пг-Уязаних 31 зм!ною тонусу резистивних судин
Стан резистивнихсудин Системна реакщя Локапьн а реакцкя
Вазоконстрцкщя Гшертензхя 1шем1я
Вазодилатация Гшотензи Артергальна пперем.’я
Мехашзми регуляш! регюнариого кровообггу включаютъ, з одного боку, внлив
судинозвужувально! 1 судинорозширювально! шнервацп, з шшого — лпо на су-
динну стжку неспецифгчних метаболгпв, неоргашчних юн!в, мгсцевих БАР 1 гор-
М0Н1В, як. надходять з кров’ю. Вважають, що з< зменшенням дгаметра судин зна-
чения нервово" регуляцй зменшуеться, а метаболгчно!, навпаки, збшыцуеться.
Рёзномаштш регуляторн! чинники забезпечують стан тонусу резистивних судин.
Ефекти ключових компоненпв наведено в табл. 9.
Таблица 9. Мехашзми регуляцн тонусу резистивних судин
Ргвень регуляцг Вазоконстрикторы Вазодилататор^
Компоненте су- динноТ стшки Скоротливий автоматизм гладком ’язових клггин судин
Нейрогенна регу- ляция Симпатичш вазоконстриктори (мед!атори — адреналш, норадре- налш — Д1я на а-адренорецспто- ри) Симпатичн! вазодилататор и (медиатор — адреналш — д(я на [3-адренорецептори); нараспмпатичш вазодилататори (медиа- тор— ацетилхотн)
Гуморальна регу- ЛЯЦ1Я бюЛОПЧНО- активними речо- винами 1 гормона- ми Катехолампги; ангютензин П; вазопресин; ендотелтни; деяю лейкотркни Кшши; пстамш; простагландини; лейкотр.сни
Гуморальна регу- ляция вазоактив- ними продуктами тканинного мета- болизму я Вуглекислий газ; мтсцева ппоюля; молочна кислота; !они кал1Ю 1 водню; продукты пдролву АТФ, у першу черту аденозин; азоту оксид ЫО
Обмшн! судаки (кашляри) окрём руху кров! забезпечують обмён боди, елект-
ролётёв, газ1в, необхёдних доживши речовин ё метаболтпв по систем! кров — тка-
нина — кров
Морфологёчними основами проникностё какёлярних судин ё венул е ендо-
телш 1 базальна мембрана. Р!зна за ёнгенсивнёстю та мёрою вибфковосп проник-
нуть кал1ляр!ь ё венул рёзних судинних дёлянок пояснюеться особливостями !х
будови (товщиною ендотел1оцит1в. базатьно! мембрани, наявнёстю або вёдсутшс-
тю пор, П1НОЦИТОЗНИХ шлях!в) 1 функцюнального стану вказаних структур.
Механёзм переходу речовини через судинну’ стенку може бути активним I па-
сивним. Якщо сили, яю забезпечують транспорту речовин, перебувають за межа-
ми судинно! сгёнки, а транспорт здшснюеться вщповщно до осмотичних, концен-
трашйних 1 електрохёмёчних градУнлв. такий вид транспорту називають пасив-
ним. 1снуе вён головним чином для перенесения води, розчинених газёв 1
низькомолекулярних речовин, тобто таких речовин, як! вьтьно проникають через
стёнку судин обмёну.
Активним характер транспорту речовин е тодё, коли вш здшснюеться проти
кониентрацёйного або електрохёмёчного градкнта (транспорт “угору”), для чого
потрёбнё наявшеть спешальних механёзмёв та певна кёлькёсть енерги, що забезне-
чуе ёх роботу. Особливо важлива роль цього механёзму в транспорт! бхтюв та ён-
ших, у тому числё стороннёх, макромолекул.
Пасивнии мехашзм переходу речовин через стёнку мёкросудин може бути роз-
дьтений на два види: ультрафшьтращю та дафуз!ю. Для позначення процесёв пе-
реходу рёдини через капёлярну мембрану використовують термин “фёльтрацёя”,
процесш обмёну розчинених у водё речовин ё газёв — термён “дафузёя”.
Таким чином, обмина функцёя в кашлярах забезпечуеться спёльною роботою
трьох процесёв: двох пасивних — фьчьтрацёя ё дифузёя та одного активного — транс-
порт мёкровезикул.
Ф1лътрац1я води I дифуз1я речовин У нормё фёльтрацёя води ё розчинених у
плазм! низькомолекулярних речовин здёйснюеться в капёлярах через функцёо-
нальнё м!кропори в стёнц! судин, дёаметр яких становить близько 6—8 нм. На-
справдё, цё пори е мёжклёгинними промёжками мёж сусёдшми ендотелёоцитами У
капётярах головного мозку вони луже щёльш ё пропускають тёльки воду, кисень ё
вуглекислий газ. У кашлярах печшки пори велию ё здатш пропускали всё компо-
нента плазми У бёльшост! органёв пори мають середнё розмёри. Шодня через цё
капшярш пори фёльтруеться 1 потраптяе в тканини близько 20 л рёдини. Близько
18 л повертаються з тканин у каньтяри шляхом резорбцв, а ще приблизно 2 л
повертаються в систему кровсобёгу через лёмфатичну систему.
Вёдповёдно до класично! теорй Старлёнга (1909), перехщ рёдини ё розчинених
у ши низькомолекулярних речовин ёз кров! в тканини здёйснюеться через нашв-
проникну мембрану кап!лярних судин шд впливом фёлътрацёйного тиску:
ФТ = (ГТК + ОТТ) - (ПТ + ОТК),
де ФТ— фёльтрашйний тиск; ГТК— гёдростатичний тиск кров! на стёнку кашляр-
них судин (в артерёальнёй дёлянцё капшярно! судини ПК становить близько 30—
35 мм рт. ст.); ОТТ — онкотичний тиск тканини — 4—7 мм рт. ст.; ГТТ — гёдро-
статичний тиск тканини + 6 мм рт. ст., тобто ~ 0 мм рт. ст.; ОТК — онкотичний
тиск кров.----28 мм рт. ст. Онкотичний тиск практично не змёнюеться в ар-
тер!альнш 1 венознш дшянках кашляра 1 залежить вщ концентрат! б!лк!в у сиро-
ватщ кров!, яка в норм! становить 60—70 г/л.
Вщповщно до наведено! формули, в артергальнш частит катляргв ефектив-
ний ФТ становить близько 10 мм рт. ст., який ! визначае перехщ рщини !з кров]
в тканини.
У венозному вщртзку кашляргв 1 у венулах пдродинам!чний тиск кров! зни-
жуеться до 15 мм рт. ст. Унаслщок цього ФТ стае негативним 1 становить близько
—7 мм рт. ст., що 1 зумовлюе часткову резорбцпо рщини !з тканини в кров. Кр!м
того, резорбц!я друго! частини штерстищально! рщини здшснюеться л!мфатични-
ми судинами 1 по них рщина повертаеться в судинне русло.
Дифуз1я газ1в у капщярах вщбуваеться за рахунок р!знищ в ргвн! парщального
тиску в кров! 1 тканинах. Парщальний тиск кисню в артер!альнш кров! становить
близько 85—100 мм рт. ст., а у тканинах — близько 10—20 мм рт. ст. У зв’язку !з
цим кисень дуже активно переходить у тканини 1 у венознш д!лянщ капшяра його
парщальний тиск знижуеться до 40—50 мм рт. ст. На вщмшу вщ кисню, вугле-
кислий газ, що утворюеться в тканинах, дифундуе в кров, унаслщок чого тканини
звшьняються вщ його надлишку, а парщальний тиск вуглекислого газу !з 40 мм
рт. ст. в артер!альнш д!лянщ кашляра пщвищуеться до 46—48 мм рт. ст. у ве-
нознш частит.
М1кровезикулярний транспорт — активний транспорт макромолекул через ци-
топлазму ендотелюципв, що потребуе витрат енергп. Вщкриття елеменпв везикул
у цитоплазм! ендотел!альних клггин, з’ясування механ!зм!в !х утворення з повер-
хнево! оболонки, доведения учасп !х у трансендотел!альному перенесенн! речо-
вин пов’язаш з !м’ям американського дослщника С. Ра1абе. У 1953 р. вш першим
описав ультраструктуру кровоносних капшяргв ! наявшсть в ендотелюцитах вези-
кул, функцгя яких полягала в трансмембранному перенесенн! речовин. Щ мгкрош-
ноцитозш везикули здатн! захоплювати рщину з розчиненими в н!й речовинами
на одному боц! клггини I перемйцуватися до пппо!. Калька таких везикул можуть
утворювати везикулярний канал. Проте нит вважають (А.К. Гайтон, Дж.Е. Холл,
2008), що в кшьккному вщношенш везикулярний транспорт за нормальних умов
не мае важливого значения.
У раз! розвитку функщональних, структурних або патолопчних зм!н в органах
чи тканинах можуть виникати м!сцев! порушення кровообйу. До найпоширен!-
ших форм мкщевого порушення кровообпу належать гшеремгя (збщьшення кро-
вонаповнення органгв 1 тканин), яку пщроздшяють на артер!альну 1 венозну, а
також ппем!я, стаз, тромбоз 1 ембол!я.
АРТЕР1АЛБНА Г1ПЕРЕМ1Я
Артергальна гтеремгя (активна гтерем1я) (вш грец. опер — бшыпе 1 аща — кров) —
це збиьшення кровонаповнення оргашв унасйдок надлишкового надходження кров!
артериальными судинами.
Збщьшення надходження кров! в мгкроциркуляторне русло пов’язане з ло-
кальним розширенням резистивних судин. У зв’язку з! збгльшенням припливу
кров! артерхальну гшеремпо часто називають активною. Г! розвиток характеризуе
низка функщональних змш 1 клгшчних ознак. До основних характеристик ар-
Мал. 16. Динамека тиску кров! в резнях вщщлах кро-
воносного русла в норм! ё при артерГалыйй пперемп
терёальноё гёперемёё можна зарахува-
ти: розлите почервонёння, розши-
рення дрёбних артерёй, артерёол г
капшярёв, пульсацёю дргбних ар-
терит ё капёлярёв, збёльшення кёль-
костё функцёонуючих капёлярёв, мёс-
цеве гцдвигцення температуря, збёль-
шення об’ему гёперемёйованоё делян-
ки, пщвищення тургору тканини,
збёльшення тиску в артерёолах, капё-
лярах ё венах (мал. 16), прискорення
кровотоку, пёдвищення обмшу та по-
силення функцц органа. Причина-
ми артерёальноё гшеремп можуть
бути: впливи рхзних факторов навколишнього середовища, включаючи бёолопчнё,
фёзичнё, хёмёчнё; збёльшення навантаження на орган або делянку тканини, а також
психогенний вплив. Осюльки частина цих факторев е звичайними фезюлопчними
подразниками (збельшення навантаження на орган, психогенне дп), артерёальну
пперемею, що виникае пщ ёх впливом, слщ вважати фезеолотечною. Основним
резновидом фёзёологёчноё артерёальноё гшеремп е робоча, або функцёональна, а
також реактивна гёперемёя.
Робоча гтеремгя — це збёльшення кровотоку в орган!, що супроводжуе поси-
лення його функцш (гёперемёя пёдшлунковоё залози пщ час травления, скелетно-
го м’яза пщ час його скорочення, збельшення коронарного кровотоку при поси-
ленне робота серця, приплив крове до головного мозку в разе психёчного наванта-
ження).
Реактивна гтерем1я е збёльшенням кровотоку песля його короткочасного об-
меження. Розвиваеться зазвичай у нирках, головному мозку, шкёрё, кишках,
м’язах. Максимум реакцп спостерёгають через келька секунд пёсля вщновлення
перфузп. I! тривалёсть визначаеться тривалёстю оклюзй. За рахунок реактивное
гёперемёё усуваеться “заборгованесть” по кровотоку, що виникла пёд час оклюзн.
Патолопчна артерёальна гёперемёя розвиваеться пёд дёею незвичайних (пато-
логёчних) подразникёв (хёмёчнё речовини, токсини, механёчнё фактори, продукта
порушеного обмену, медёатори, що утворюються в разё запалення, опеку та деяких
ёшпих станёв). У поодиноких випадках умовою виникнення патологёчноё артерё-
альноё гёперемёё е педвищення чутливостё судин до подразникёв, що спостерёгаеть-
ся, наприклад, при алергёё.
1нфекцёйний висип, почервонёння обличчя при багатьох ёнфекцёйних захво-
рюваннях (кёр, висипний тиф, скарлатина), вазомоторнё розлади при системному
червоному вовчаку, почервонёння шкёри кенцёвок при уееекодженнё певних нерво-
вих схелетень, почервонёння половини обличчя при невралгп, пов’язанёй з подраз-
ненням трёйчастого нерва, е клёнёчними ознаками патологёчноё артерёальноё гёпе-
ремёё. Залежно вед фактора, що спричинив патологёчну артереальну геперемею,
можна виделити запальну, теплову пперемёю, ультрафёолетову еритему та ён.
За патогенезом розрёзняють два вида патологёчноё артерёальноё гёперемёё —
нейрогенну (нейротонёчного ё нейропаралётичного типу) ё зумовлену дёею мёсце-
вих БАР ё метаболёчних факторев.
Нейрогенна артер!альна гтеремзя розвиваеться внаслщок змши тонусу ргзнн
вщццпв вегетативно! нервово! системи. Як вщомо, активашя симпатичного вугц-
лу нервово! системи супроводжуеться спазмом резистивних судин шк!ри (заучаст
а-адренорецептор!в), а активашя парасимпа точного вщдшу — !х розширенням. >
зв’язку 13 цим для розвитку пперем!! необхщним е або пщвищення тонусу парт-
симпатичного вщдшу нервово'! системи (нейротошчна гтерелия), або пригшченн
тонусу симпатичного вщдшу нервово! системи (нейропарамтична гтеремгя).
Арте реальна гтеремгя нейротошчного типу може виникати рефлекторно у
зв’язку з подразненням екстеро 1 штерорецептор!в, а також при подразненш су-
динорозширювальних нерВ1В ! ценгр!в. Подразниками можуть бути псюачш, ме-
хашчш, температурю (тепло). х!м!чн! (скипидар, прчична ол!я та ш.) ! б!олопчн:
фактори.
Типовим прикладом нейрогенно! артерьально! гшеремп е почервоншня об-
личия та ши! при патолопчних пропесах у вштрпшпх органах (яечниках, серш
печшщ, легенях)
Б експеримент! на тваринах (кролики 1 собаки) нейроген ну артер!атьну ппе-
ремпо уперше вщтворив К. Бернар шляхом подразнення сйсгйа 1ушраш — г!лк
п. Гас!а1!з, що складаеться з парасимпатичних судинорозширювальних волокон
Як реакщя у вщповшь вин икали г!перем!я 1 посилення секреци лщнижньощеле-
пно! слинно! залози.
Аргергальна пперем!я, зумовлена холшерпчним мехашзмом (вплив ацетил-
холшу), можлива ! в шших органах 1 тканинах (язик, зовшшн! статев! органи та
ш.), судини яких шнервуються парас импатични ми нсрвовими волокнами.
У деяких випадках нейротонгчна гшерем!я може бути зумовлена активапн»
симпатичного вщд!лу нервово! системи (холшерпчною. пстамшерпчною 1 р-ад-
ренерпчною), представлении на перифер!! вщповщними волокнами, медиаторами
1 рецепторами (Н,-рецептори для пстам!ну, р2-адренорецептори для адреналин
мускаринов! рецептори для ацеталхолшу).
Сим пати чн! холшерпчн! нерви розширюють др1бш артерп й артерюли ске-
летних м’яз!в. м’яз!в обличчя. слизово! оболонки шж, кишок. 1х мед!атором також
е ацетилхолш, Вважаюгь, що в частин! ьиладк!в гшеремхя е наслщком утвореннт
в нервових заюнченнях або в шнервовашй тканин! простат ландин!в — БАР —
похщних полшенасичених жирних кислот.
Простагландини Е 1 А (ПГЕ, ПГА) справляють судинорозширювальну дно на
артерюли, метартерюли, прекатляри 1 венули. 3 м’язово! стшки судин видшенс
простагландин 12 (ПП2 — простациклш), який окр!м розслаблюючо! ди на артерй
р!зного д!аметра чинить потужний антиагрегуючий вплив на тромбоцита.
Нейроген ну артергальну гтеремйо нейропараттичного типу можна спостерпати
в клшщ1 та експеримент! на тваринах при перергзанн! симпатичних а-адренерпч-
них волокон । нерв!в, як! справляють судинозвужувальну дио. У 1842 р. О.П. Валь-
тер уперше вщзначив розширення судин лапки жаби теля перетзування симпа-
тичних волокон ещничного нерва. Пщнипе К. Бернар (1851) спостершав почер-
воншня 1 пщвищення температури шкзри голови кроля на бош видалення
шийного вузла симпатичного стовбура. Особливо це помпно на вус! (мал. 17).
Аналопчш ефекти спостернаються також у раз! порушення шлосл зм!шаних не-
рв!в, як! м!стять у своему склад! а-адренерпчн1 судинозвужувальн! волокна.
Мал. 17. Дослад К. Бернара (1851): пра-
воруч вилучено вузол симпатичного
стовбура
симпатичних нервових заюнчень
Симпатичш судинозвужувальн! нерви
тошчно активы) 1 в звичайних умовах постш-
но несуть [мггульси центрального походження
(1—3 гмпульси за 1 с у стан! спокою), яю 1
визначають нейрогенний (вазомоторний) ком-
понент судинного тонусу. 1х медгатором е но-
радреналш.
У людини 1 тварин тошчна !мпульсац!я
властива симпатичним нервам, яю прямують
до судин шюри верхних юнщвок, вух, скелет-
них м’язгв, травного каналу та ш. Перер!зуван-
ня цих нерв1в у кожному !з зазначених органгв
зумовлюе збшьшення току кров! в артер!аль-
них судинах. На цьому ефект! Грунтуеться за-
стосування периартер!ально! 1 ганглюнарно!
симлатектомн при ендартерпп, що супровод-
жуеться тривалими спазмами судин.
Артергальну пперемпо нейропаралпично-
ГО ТИПУ МОЖНа СПрИЧИНИТИ Й Х1М1ЧНИМ шля-
хом, блокуючи передачу центральних нерво-
вих 1мпульс1в у д!лянщ симпатичних вузл!в (за
допомогою ганглюблокаторгв) або на р!вш
(за допомогою симпатолгтичних або а-адре-
ноблокуючих засобгв). У цих умовах блокуються потенщалзалежш повшьш каль-
щев! канали, порушуються надходження в гладком’язов! юптини позаюптинного
Са2+ за електрох!м!чним градиентом, а також вившьнення Са2+ !з саркоплазматич-
ного ретикулума. Унаслщок цього юптини непосмуговано! м’язовоТ тканини роз-
слаблюються.
Нейропаралггичний мехашзм артер!ально! гшеремп частково е основою за-
пально! гшеремп, ультрафюлетово! еритеми та ш.
Уявлення про артер!альну пперемпо (ф!зюлопчну та патолопчну), зумовлену
мгсцевими метаболгчними (хгмгчними) факторами та БАР, яю утворюються на
м!сш або надходять з кров’ю, грунтуеться на тому, що вказаш фактори спричи-
нюють розширення судин, дпочи безпосередньо на непосмуговаш м’язов! еле-
менти !хньо! стпгки, незалежно вщ гннервацгйних вплив!в. Це шдтверджуеться
також тим, що повна денерващя не запобггае розвитку н! робочо!, ш реактивно!,
ш запально! артер!ально! гшеремп.
Розширення судин виникае внаслщок зниження в кров! 1 тканинах стгнки
судин напруження кисню (рО2), що спостерйаеться, коли юптини споживають
кисень швидше, н!ж вш надходить у тканини з кров’ю.
Мехашзм вазодилататорного впливу ппоксп полягае в тому, що зниження
рО2 зумовлюе зменшення продукцп АТФ. Це призводить до вщкритгя АТФ-за-
лежних кал!евих канал!в у мембран! гладком’язових юптин. Вившьнення калпо !з
клиин спричинюе пперполяризацпо юйтинно! мембрани, що зумовлюе закриття
електрокерованих кальшевих каналгв. Унаслщок цього ргвень вшьного кальцпо в
цитозол! знижуеться, що розслаблюе непосмуговану м’язову тканину 1 спричи-
нюе розширення судин.
АртерУ ал ьна гшеремУя виникае також унаслщок пщвищення рСО2. надлишк
неспецифУчних метаболт в 1 неоргашчних УонУв (молочна кислота, органУчнУ кис-
лоти циклу Кребса, АДФ, АМФ, аденозин, юни кал1ю). мУсцевих БАР (брадп-
кУнгн, калщин, пстамУн, ПГЕ, ПГА, ПП2, немедУагорний ацетилхолш, субстан
Р, у-амшомасляна кислота) У гормонУв. якУ надхолять в органи з кров’ю (атрУаль-
ний натргйуретичний пептид, адреномедулш, вазоактивний Уггтестинальний пеп-
тид). Значения кожного з наведених факторов у розвитку артергальноУ гУпере
разного генезу неоднакове У виникненш реактивно! та робочоУ артергальноУ гша-
ремн внаслщок гшоксй особливого значения набувае аденозин, що утворюетьс
внаслщок ди на АМФ екто-5’-нуклеотидази. Джерелом аденозину е гладком’яз
й ендотелУалып клггини судин. кштини паренхУми органа, з яких певна його час-
тина потрапляе в позаклУтинний проспр, спричинюючи розширення судин. Аде-
нозиндезамшаза забезпечуе руйнування аденозину. Тим самим пщтримуеты.
його баланс усерединУ клггини У поза нею. Аденозин, пстамУн У атрУальний на-
грУйуретичний пептид дУють на гладком’язовУ клУтини через Н2-рецеггтори.
Ще одним фУзУологУчно важливим, дуже активним У унУверсальним факторе-
вазорелаксацУУ е азоту оксид 1МО В1н утворюеться юптинами Унтактного ендо-
телпо з Ь-арггнУну за наявноси НАДФН У кисню.
Синтез оксиду азоту може вщбуватися кУлькома шляхами. У людини синте
здУйснююе родина М0-синтетаз (МО8). МО8-ферменти — члени гемов мУсноУ су-
перродини ферментУв, якУ отримали назву монооксигеназ. Залежно вш структур
1 функцш 1408 можуть бути роздшенУ на три групи: ендотелгальнУ (еМО8). нейро-
налью (пМ08) та УндуцибельнУ (1МО8). В активний центр будь-яког з МО8 вхо-
дить залУзопорфУриновий комплекс, який мУстить аксУальний координовани
цисте’Ун або метУонш. Хоча вег Узоформи 1408 каталУзують утворення МО, вони €
продуктами рУзних генУв, кожна з них мае певнУ особливостУ як у механУзмах ди
локалУзацУУ, так У в бУологУчному значеннУ для оргашзму. Тому вказаш Узоформи
прийнято також тдроздйгяти на конститутивну 1408 (сЫОЗ) та 1М08. ВщмУнност
полягають у тому, що конститутивний фермент синтезуеться безперервно, а Унда-
цибельний (адаптивний) — тУльки за наявностУ в середовищУ вщповщних фак-
торУв.
Установлено, шо МО е лэбУльною, короткоживучою (юлька секунд) молеку-
лою. У процесг взаемоди з киснем 1 водою МО швидко перетворюеться на ютрата
У нУтрити. Судини малого калУбра синтезують його в бУлышй кыькостУ, нгж судини
великого калгбра. Конститутивна (шгредУентна) М08 реагуе збшьшеною продук-
цУею та видшенням протягом кьтькох хвилин МО у вУаповУдь на пщвищення
ендотелУоцитах рУвня Са2+ пУд впливом низки вазодилататор!в (ацетилхолУну, бра-
дикУнУну, калшину, адреномедулУну, субстанпп Р, вазоактивного Унтестинального
пептиду), ппоксп, естрогенУв 1 посилення напруження змйцення кров! щодо ен-
дотелУальних клУтин. МУж утворенням МО та активнУстю сМ08 е негативний зво-
ротний зв’язок. Дифундуючи з ендотелУоцитУв до непосмугованих м’язових клУ-
тин, МО стимул юс в них гуаншатциктазу. утворення (кГМФ) з подальшою акти-
ващею цГМФ-залежних протеУнкУназ. кальшевих АТФаз, Унозитол-1,4,5-трифос-
фату га калъцУйзалежних калУевих канате. Як наслщок знижуеться концентращя
Са2+у непосмугованих м’язових клУтинах, виникають Ух розслаблення та вазоди-
латашя. Цього ефекту, крУм того, можна досятти за рахунок пригнУчувального
впливу МО на кальцУйчутдивУсть скоротливих бУлкУв.
ИИ _ 4
У патолопчних умовах (ошк, травма, ушкоджувальна д!я ультрафюлетового
або юшзуючого випромшювання, шфекщйних збудниюв, розвиток унаслщок
цього запалення — асептичного або септичного — !з залучениям гмунних мехашз-
М1в ушкодження або без них) великого значения в розвитку артер!ально! гшеремп
набувае шдукована кальшйзалежна 1МО8. I! синтез здшснюеться за рахунок екс-
пресп вщповщного гена (розмйцений у хромосом! 7), щд д!ею прозапальних та
гмуногенних фактор!в (ендотоксини, штерферони, 1Л-1, ФНП-а тощо). При ць-
ому В1ДПОВ1ДШ клттини (ендотелюцити, нейтрофши, ф!бробласти, але в першу
черту моноцити 1 макрофаги) продукують 1 видшяють велику кыыасть МО протя-
гом багатьох годин. Надлишок МО в цьому випадку спричинюе не пльки значну
та пролонговану артерхальну гшеремпо, а й цитолхз клпин.
Крхм того, неушкоджеш ендотелюцити продукують ендотелшзалежний гшер-
поляризуючий фактор, який також е модулятором вазодилатацп. На вщмшу вщ
МО його вивчено меншою мхрою.
Важливу роль у збшьшенш кровотоку при мюцевих судинних реакцхях вццг-
рають змши рН тканинного середовища — зрушення реакцп середовища в б!к
ацидозу сприяе розширенню судин завдяки пщвшценню чутливосп ендотелю-
цитгв до вщповщних факторхв 1 клпин непосмуговано! м’язово! тканини щодо
МО.
При артерхальшй гшеремп в мгкроциркуляторному русл! вщбуваються змши
в процесах ф!льтрацц та дифузп. У зв’язку з! збшьшенням припливу кров! в мпсро-
циркуляторне русло шдвищуеться р!вень гщростатичного тиску як в артерхальнш,
так 1 венознш делянках кашляргв. Причому пщвищення гщростатичного тиску на
10 мм рт. ст. призводить до пщвищення ФТ в артер!альн!й дшяшц до 20 мм рт. ст.,
тобто рщина активнппе виходить !з судин у тканини. При цьому у венознш дшян-
Ц1 гщростатичний 1 онкотичний тиск вир!внюються ! резорбцгя рщини не вщбу-
ваеться. Але слщ зазначити, що гстотне накопичення рщини в тканинах не спос-
терёгаеться. Це пов’язано з штенсивнгшим поверненням рщини в л!мфатичну
систему, резорбтивна актившсть яко! значно збшьшуеться. У зв’язку 13 цим вщбу-
ваеться збшьшення дренажу тканин, шдвищуеться штенсившсть транспорту до
них поживних речовин 1 виведення метаболтв.
Одночасно зё збшьшенням дренажу тканин шдвищуеться штенсившсть про-
цес!в дифузп. При артер!альнш гшеремп приплив артер!ально! кров!, насичено!
киснем, до тканин збёльшуеться, що ютотно покращуе !х оксигенащю. Водночас
швидккть лшшного й об'емного кровотоку може так збшьшуватися, що весь ки-
сень не утшпзуеться тканиною 1 вщбуваеться оксигенац!я венозно! кров!. При
цьому парщальний тиск О2 у венознш кров! може сягати 60—70 мм рт. ст. Вугле-
кислий газ при артер!альнш пперем!! повшстю виводиться з тканин, що можна
вважати позитивним ефектом артер!ально! гшеремп. М!кровезикулярний транс-
порт при артер!альнш гшеремп ютотно не змшюеться.
Результат артер!ально! гшеремп може бути рхзним. Розр!зняють позитивш
(адаптивш) ! негативш (патолопчш) змши при артер!альн!й пперем!!, причому
вщзначаеться переважання позитивних ефекпв, пов’язаних !з посиленням дрена-
жу тканин, пол!шпенням газообмшу, посиленням обмшу речовин 1 функцш ор-
гана. Ц! змши можна розцшити як пристосовш або адаптивш. Вони сприяють
стимуляци вцщовлення тканин шсля !х ушкодження. Штучне створення ар-
тер!ально! гшеремп часто використовують як лгкувальний фактор для стимуляцп
процес1в вщновлення тканин (масаж, ф!з!отерашя та ш.). Проте можлив! 1 не-
сприятлив! наслщки. Наприклад, при атеросклероз! р!зке розширення судини
може супроводжуватися розривом п стшки ! крововиливом у тканину. Особливо
небезпечними е под!бн! явища в головному мозку.
ВЕНОЗНА ППЕРЕМ1Я
Венозна гтеремгя (пасивна гтерелая, застш) розвиваеться внасл1док збмыиення
кровонаповнення органа або дтянки тканини внасмдок утрудненого вгдтоку кров1 по
венах.
На вщмшу вщ артер!ально! гшеремп розвиток венозно! гшеремп пов’язаний
13 порушенням вщтоку кров! з мгкроциркуляцп, у зв’язку 13 чим розвиваеться за-
стш кров!, тому венозну пперемпо ще називають пасивною. Причинами Г! роз-
витку е закупорка вен тромбом або емболом, здавлення пухлиною, рубнем, зб!ль-
шеною маткою. Тонкостгнн! вени можуть здавлюватися також у дшянках р!зкого
пщвищення тканинного 1 гщростатичного тиску (у вогнищ! запалення, у нирках
при пдронефроз!).
У деяких випадках фактором, що спричинюе розвиток венозно! гшеремп, е
конститущональна слабюсть еластичного апарату вен, недостатнш розвиток 1
знижений тонус гладком’язових елемент!в !хшх стшок. Нерщко така схильшсть
мае с1мейний характер.
Професп, що вимагають щоденного тривалого (протягом багатьох годин) пе-
ребування у вертикальному положенн!, е факторами ризику, сприяють розвитку
венозно! гшеремп в дистальних вщдщах нижшх кшц!вок в ос! б з конститущо-
нально зумовленою неповнощншстю еластичних 1 гладком’язових елеменпв стш-
ки венозних судин.
Вени, як 1 артерп, хоча 1 меншою м!рою, е рефлексогенними зонами, що дае
змогу припустити нервово-рефлекторну природу венозно! гшеремп. Морфолопч-
ною основою вазомоторно! функцп вен е нервово-м’язовий апарат, що включае
гладком’язов! елементи й ефекгорш нервов! закшчення.
Венозна гшерем!я розвиваеться також при ослабленш функщй правого шлу-
ночка серця ! зменшенн! присмоктуючо! дп трудно! клики (ексудативний плев-
рит, гемоторакс). При ц!й патологи застш кров! розвиваеться у венознш систем!
великого кола кровообпу, 1 можна говорите про системну венозну пперемпо.
Утруднення кровотоку в малому кол! кровообпу при пневмосклероз!, емф!зем!
легень, ослабленш функцп л!вого шлуночка призводить до застою кров! у венах
малого кола кровообггу.
Клппчно венозна гшерем!я проявляеться збшыпенням органа або дшянки
тканини, щанозом, мкщевим зниженням температури, набряком, пщвищенням
тиску у венах 1 кашлярах застшно! д!лянки, спов!льненням кровотоку, д!апедезом
еритроципв. На завершальному етап!. гшеремп можлив! маятникопод!бний рух
кров! 1 стаз.
Тривале розширення вен призводить до розтягування !хньо! стшки, що може
супроводжуватися ппертроф!ею Г! м’язово! оболонки та проявами флебосклерозу
1 варикозного розширення вен.
Тривалий венозний застш супроводжуеться значними змшами функцгональ-
них елементгв стшки вени, Тх атрофгею 1 загибеллю. Разом гз цим у дщянцг веноз-
ноТ гшеремп вщбуваеться замгсне розростання сполучноТ тканини. Класичним
прикладом е цироз печгнки при недостатностг функцй серця, спричинений ве-
нозним застоем.
При венознш гшеремп значно змшюються фыьтрацгя рщини 1 дренаж тка-
нин. Порушення вщтоку з мТкроциркуляторного русла супроводжуеться пщви-
щенням гщростатичного тиску у венознш дГлянц! кашляра. Причому пщвищен-
ня тиску всього на 10 мм рт. ст. призводить до вирТвнювання гщростатичного й
онкотичного тиску у венознш дьтянц! кашляра. В артерТальнш дглянщ рщина
продовжуе фшьтруватися в тканини, а у венознш дглянщ резорбщя не вщбу-
ваеться. Рщина накопичуеться в тканинах, що може призвести до значних на-
бряюв. Це пов’язано з тим, що в раз! порушення венозного вщтоку вщбуваеться
рефлекторна блокада лгмфовщтоку ! виникае одночасний венозний 1 лгмфатич-
ний застш.
1ншим ключовим фактором, що зумовлюе мгсцев! змгни при венознш гшере-
мп, е кисневе голодування (гшоксгя) тканини, розвиток яко! пов’язаний з! зм!-
ною дифузн. Ппоксгя при цьому спочатку спричинена обмеженням припливу
артергальног кров!, пот!м д!ею на тканинн! ферментнг системи продуктов пору-
шення обмшу, наслщком чого е порушення утилгзацИ кисню. Кисневе голодуван-
ня при венознш гшеремп зумовлюе порушення тканинного обмшу, атрофгчн! г
дистроф!чнг змши та надмгрне розростання сполучноТ тканини.
Одночасно з мгсцевими змшами при венознш гшеремп, особливо якщо вона
зумовлена загальними причинами 1 мае генерал(зований характер, можлив! г за-
гадил гемодинамгчн! порушення з дуже тяжкими наслщками. Найчастгше вони
виникають при закупорщ великих венозних колекгоргв: ворггноТ та нижньог по-
рожнистог вен. Скупчення кров! у вказаних судинних резервуарах (близько 90 %
усге! кров!) супроводжуеться ргзким зниженням артергального тиску, порушенням
живлення житгево важливих органгв (серця, мозку). Унаслщок недостатност! сер-
ця або паралгчу дихання можливий летальний наслщок.
1ШЕМ1Я
Порушення периферийного кровообйу, тдгрунтям якого е обмеження або повне при-
пинення припливу артергально! кров!, називають йиелйею (вщ грец. 10%ауаа> — за-
тримувати, зупиняти; агра — кров), або мюцевим недокр1в’ям.
Мехашзм розвитку гшемп полягае в зменшенш притоку кров! в систему
мгкроциркуляцп. У зв’язку !з цим йпемгя харакгеризуеться такими ознаками:
зблщненням гшемгзованог дщянки органа, зниженням мгсцево! температури, по-
рушенням чутливост! у виглядг парестезп (вщчуття онгмшня, поколювання, пов-
зання мурах), больовим синдромом, зменшенням швидкост! кровотоку й об’ему
органа, зниженням артергального тиску в дглянщ артерп, розташованоТ нижче за
перешкоду, напруження кисню в ппемгзованш дглянщ органа або тканини, змен-
шенням утворення м!жтканинноТ рщини та зниженням тургору тканини, пору-
шенням функцц органа або тканини, дистрофгчними змгнами.
Причиною ёшемё! можуть бути рёзнё фактори: здавлення артерн, обтурашя и
просвёту, Д1я на нервово-м’язовий апарат артерёально'ё стёнки. Вёдповёдно до цьо-
го розрёзняють компресёйний, обтурацёйний ё анпоспастичний типи ппеми.
Компресёйна ёшемёя виникае внаслщок здавлення привёдноё артерн лигатурою,
рубцем, пухлиною, стороннём тёлом та ён.
Обтурацшна ёшемёя е наслщком часткового звуження або повного закриття
просвету артерн тромбом чи емболом. Продуктивно-ёнфёльтратавш та запальш
змёни стёнки артерн, що виникають при атеросклероз!, облётеруючому ендар-
терптё, вузликовому периартерётп, також спричинюють обмеження мёсцевого кро-
вотоку за типом обтурацёйноё ёшемё!.
Анпоспастична йиемёя виникае внаслщок подразнення судинозвужувального
апарату судин ё ёх рефлекторного спазму, спричиненого емоцёйною дёею (страх,
б!ль, гнёв), фёзичними (холод, травма, механёчне подразнення) ё хёмёчними факто-
рами, бёологёчними подразниками (бактерёйнё токсини) та ён. В умовах патологи
анпоспазм характеризуеться вщносною тривалёстю ё значною вираженёстю, що
може бути причиною рёзкого сповёльнення кровотоку, аж до повноё його зупинки.
Найчастёше анпоспазм развиваеться в артерёях вёдносно великого дёаметра всере-
динё органа за типом судинних безумовних рефлексёв з вщповёдних ёнтерорецеп-
торёв. Цё рефлекси характеризуються значною ёнертнёстю й автономнёстю. При-
кладом такого типу судинних реакцёй можуть слугувати спазм вённевих артерёй
серця при подразненнё рецепторёв внутрёшнёх органёв (кишок, жовчних шляхёв,
сечоводёв, сечового мёхура, легенёв, матки), рефлекторний спазм судин парного
органа (нирки, кёнцёвки) при подразненнё протилежно розташованого. Анпоспас-
тична ёшемёя може мати також умовно-рефлекторний характер. Нарештё, пряме
подразнення розташованого в пёдкёрковёй дёлянцё судинорухового центру токсич-
ними речовинами, якё мёстяться в кровё, що омивае його, механёчне подразнення
пёдкёркових утворень, якё регулюють судинний тонус (при пухлинному процесё в
головному мозку, крововиливах у мозок, пёдвищеннё внутрёшньочерепного тис-
ку), наявнёсть патолопчного, зокрема запального, процесу в дёлянцё промёжного
мозку також часто призводять до виражених анпоспастичних проявёв.
Таким чином, анпоспазм розвиваеться передусём унаслёдок активацп нейро-
генних а-адренерпчних, Н^пстамёнерпчних, серотонёнерпчних, дофамёнергёчних
механёзмёв. Джерелом бёогенних амёнёв немедёаторно! природи е наднирковё зало-
зи (катехоламши) ё клётини дифузно! нейроендокринноё системи (АРЕЮ — систе-
ма захоплення попередникёв амёнёв ё !х декарбоксилювання), що розкиданё по
рёзних органах ё тканинах, мають здатшсть виробляти 5-гёдрокситриптамён (серо-
тонён), гёстамён ё дофамён. У пухкёй сполучнёй тканинё джерелом пстамёну, крём
того, е тканиннё базофёли, у кровё — базофёли (гёстамён) ё тромбоцита (гёстамён,
серотонён). У розвитку анпоспазму певну роль вёдёграють простагландини (ПГР2а
ё тромбоксан А2). Останшй утворюеться в тромбоцитах, його видёлення особливо
рёзко зростае при !х адгезё! та агрегаци на ушкодженёй стёнцё судин. Нарештё, ан-
гёоспастичну дёю мають вазоактивш пептиди — вазопресин ё ангиотензин II.
Важливого значения в розвитку анпрспастичних реакцёй надають ендотелёну-1,
який виробляеться ендотелёем судин. Ендотелёй спочатку утворюе прогормон —
великий ендотелён-1, який складаеться ёз 39 амёнокислотних залишкёв. Пщ впли-
вом ендотелёнперетворюючого ферменту з великого ендотелёну-1 утворюеться ендо-
телш-1, який мёстать 21 амёнокислотний залишок. Вён е найпотужнёшим ёз вёдо-
мих судинозвужувальних факторгв, причому його актившсть бгльше нгж у 10 разгв
перевищуе актившсть анпотензину II.
Основний мехашзм дп ендотелшу-1 полягае в рецепторопосередкованому вщ-
криттг потенщалзалежних кальцгевих каналгв г вивгтьненнг кальцпо в цитозоль
гладком’язових клггин гз внугргшньоклпинних депо, шо спричинюе гх скорочен-
ня та пролгферацгю з потовщенням стшки судин г зменшенням гх дгаметра.
Синтез ендотелшу-1 посилюють ангготензин II, катехоламши, тромбш, ггпок-
сгя, фактори росту, ушкодження ендотелгю окисненими лшопроте'щами або пщви-
щеним артергальним тиском. Усг цг фактори активують транскрипцгю гена ендо-
телшу-1. КО, ПГЕ2, ПП2, атргальний натргйуретичний пептид, навпаки, пригнг-
чують актившсть цього гена г зменшують секрецгю ендотелгну-1. Ендотелш-1
причетний до виникнення низки патолопчних процесгв (шфаркту мюкарда, по-
рушення ритму серця, легеневог та системно! гшертензп, атеросклерозу та ш.).
Мехашзм активацн скоротливих бглкгв гладком’язових клгтин залежить вщ
стану специфгчних рецепторгв. Щодо катехоламгнгв це зокрема а-адренорецепто-
ри. У разг гх збудження, певно, передусгм змгнюються електричнг властивостг мем-
брани гладком’язових клгтин унаслщок пщвищення проникностг клгтинног мемб-
рани для №+, Са2+, К+, СГ. Особливу роль вщгграе Са2+, який надходить через
клгтинну мембрану з мгжклгтинног рщини та вивгльнюеться гз саркоплазматично-
го ретикулума. Наслщками цього е активацгя скоротливих бщкгв гладеньком’язових
клгтин г розвиток гх скорочення. Використовуючи блокатори повгльних кальцге-
вих каналгв (антагонгсти Са2+ — верапамгл, нгфедипш, бефщил), можна запобггти
(у середньому на 50 %) скороченню, спричиненому норадреналгном. Ангготензин
II — одна з найпотужнгших судинозвужувальних речовин — справляе свою пряму
дно на гладком’язовг клггини шляхом гх деполяризацц внаслщок збглыггення про-
вгдностг каналгв для Ка+ г Са2+.
Велику роль у виникненш анггоспастичног гшемп вщгграе змгна чутливостг
м’язових елеменпв стшки судин щодо норадреналшу г вазоактивних пептидгв.
Наприклад, гони натргю, накопичуючись у м’язових волокнах судини, пгдвищу-
ють його чутливгсть до пресорних речовин — катехоламгнгв, вазопресину й ангго-
тензину И.
Останнгм часом вщзначають участь ендотелгю в нормальному функцгонуваннг
механгзму скорочення — розслабленнг непосмугованих м’язгв судин. Ушкоджен-
ня ендотелгю позбавляе його здатностг видгляти фактор релаксацп, унаслщок чого
посилюються спастичнг реакцп. Останнг можуть виникати навгть на ацетилхолгн
г брадикшш, якг в нормг спричинюють розширення судин.
Характер обмшних, функшональних г структурних змш в гшемгзовангй дг-
лянцг тканини або органа визначаеться ступеней кисневого голодування, тяж-
кгсть якого залежить вщ швидкостг розвитку г типу гшемп, п тривалостг, локалг-
зацп, характеру колатерального кровообггу, функцгонального стану органа або
тканини.
1шемгя, що виникае в дглянцг повног обтураци або компресп артерш, за ш-
ших ргвних умов спричинюе суттевшп змши, шж при спазмг. 1шемгя, що швидко
розвиваеться, як г тривала, мае тяжчий перебгг, шж така, що розвиваеться повгль-
но або е нетривалою. Особливо велике значения в розвитку гшемп мае раптова
обтурацгя артергй, оскгльки при цьому може приеднатися рефлекгорний спазм
системи розгалужень цгег артерп.
1шсм)я життево важливих оргашв (мозку, серия) мае тяжч) наслщки, н!ж
)шем)я нирок, селезенки, легень, а пдемтя останшх — тяжч! поршняно з ппемтсю
скелетнс'1, м’язовот, юстковот або хряшовот тканини Це пов’язано з тим, що чут-
ливтсть тканин до гшоксц фактично визначаеться двома факторами: метабол 14-
ною актившстю тканини т ртвнем розвитку колатерального кровообпу. Тканини з
вищим ошнем метабол тзму та специфично) активност) чутлившп до ппоксичного
ушкодження. Так. нейрони зазнають незворотно! деструкш! вже через 3—5 хв
повноТ ппемп, кардюмюцити — протягом 20—30 хв. 3 шшого боку, фтбробласти в
мюкарц) здаттн збертгати житте здатшсть упродовж кшькох годин.
Ревень колатерального кровообтгу е другим лтмпуючим фактором стшкост)
тканин до ппемп. За ступенем розвитку колатерального кровообтгу вс) тканини
можна пщроздптити на три типи: абсолютно достатн), вщносно достатн) й абсо-
лютно недостатнт Якшо колатералып судини функционально абсолютно або вщ-
носно не здатн! компенсувати порушення кровооб)гу. то тканина бшьше схильна
до йпемтчного ушкодження. Класичним прикладом абсолютно недостатнього ко-
латерального кровообпу е тканини головного мозку 1 мюкард. До тканин з вщ-
носно достатнтм розвитком колатералей можна зарахувати кишки, а абсолютно
достатнш колатеральний кровооб)г спостертгають у м’язовш тканин) та шктрт
Нарепгп. велике значения в розвитку ппемп мае попереднш функшональний
стан органа або тканини. Утруднення припливу артертальноТ кров) в умовах пшви-
шено! функщонально) активност] органа або тканини небезпечшше, шж у стан)
спокою Особливо важливою е роль невщповщност) функцц органа 1 його крово-
постачання за наявностт органтчних змш в артертях. Це пов’язано з тим, що ор-
гашчнт змши судиннот стгнки, з одного боку, обмежують и здатшсть до розши-
рення в раз) тцдвищеного навантаження, а з шшого - пщвищують п чутливтсть
по р)зних вазоконстрикторних фактор)в. Кртм того, можливтсгь посилення кола-
терального кровообтгу в склеротично змшених судинах також дуже обмсжена.
Сл)д зазначити, що при ппемп вщзначакпъ змши процеств фтлырацп, дифузн т
транспорту мткровезикули. Зниження тиску в систем) мпфоциркуля цп на 10 мм рт. ст.
призводить до практично повного припинення процесу фшьтрац)! та дренажу
тканин. У тканиш ртзко скорочуеться або припиняеться надходження кисню, що
зумовлюе розвиток гптоксп в тканинах т перебудову метабол)зму на анаеробний
глтколтз. Мала кйтьктсть енергп позначаеться на порушенн) пропеав транспорту
мткровезикул.
Схематично змти в тканинах при 1шемн можна роздцтити на кшька послщов-
них стадий.
1. Зниження ефекгивностт циклу Кребса, пщвищення штенсивностт глтколтзу
т пентозного циклу, зниження гнтенсивност) енергетичного обмшу в цыому. По-
рушення утворення енергн в дглянщ ппемп патогенстично пов’язане з недостат-
н)м надходженням кисню ) необхщних для окиснення субстрат)в. зниженням ак-
тивност) та синтезу фермента, виходом тх з ушкоджених юптин у зв’язку з пщви-
щенням проникностт юптинних мембран, вщо кремле нням процеств окиснення )
фосфсрилюван ня
2. Виснаження запас) в макроерпчних фосфата супроводжуеться сумарним зрос-
танням вмтсту Са2+у юптин). особливо шсля тойвал их перюдш ппемп (схема 5).
На схем) 5 показано джерела га мехашзми пщвищення цигоплазматично)’ кон-
центрацп Са2’т послщовшсть подш, зумовлених (х надлишковим вмтстом. Вщбу-
Г1ПОКС1Я
Ьпемгя
Вившьнення ендогенних
катехоламш!в
Зменшення тианинних
--- запасов АТФ
Активация
повшьних
канал 1в
Пригшчення
сарколемно!
Са -АТФази
Перерозподы
Са2, всередиш
клггини
Пригшчення
МаТК’-насоса
Надходження
Са2+
Прирост
тканинного
вмюту №+
Збшьшення концентрацп №+_ Са2-чзбм1н
Са2 в цитозол! -<---------------------
Ушкодження мембрани
Активащя Са2+-чутливих
протеаз 1 фосфолшаз
Втрата аден1ннуклеотид1в Масивне
(аденозину, шозину) надходження Са2+
Схема 5. Послщовшсть змш у тканинах, що запускаеться ппокскю або
(шемгсю (за В.Г. Нейлером 1 М.Дж- Дейшем, 1988)
ваеться активащя Са2+-чутливих протешаз 1 фосфолшаз, виникають незворотш
змши структури та щлост! юптинно! мембрани, втрачаються внутршшьоюптишп
адешлов! основи (попередники синтезу АТФ).
У зв’язку 13 зазначеними змшами порушуються специф1чю функцп юптин
(скорочення, секрещя та ш.), робота юнних насоов, розвиваеться некробиоз.
Перпп ознаки ультраструктурних змш з’являються через илька хвилин вщ
моменту виникнення 1шемп 1 характеризуються змшами внутр! шньо'Г структури
мгтохондрш. Спостериають !х набухания, поступове накопичення жирових кра-
пель, наступне зникнення крист 1 зампцення !х гранулярною субстанщею. По-
дальший розпад мгтохондрш, а також ендоплазматичного ретикулума 1 юптинних
ядер може закшчитися утворенням вогнища некрозу — шфаркгу. Це, як правило,
вщбуваеться в пареюпматозних органах, як! виртзняються шдвищеною чутливю-
тю до кисневого голодування й особливостями анпоархггектонпси, що не дають
змоги швидко й ефективно усунути порушення кровообпу внаслщок розвитку
колатералей. Тривала 1шем1я ппари, скелетних м’яз1в 1 истково) тканини не спри-
чинюе таю тяжю функщональш 1 морфолопчш наслщки. Спроби запобхгти роз-
витку постппемтчних некробютичних змш за допомогою реперфузп не дали пози-
тивних результата, осильки в умовах реперфузп вщбуваються подальше гцдви-
щення тканинного вмгсту калыцю та посилення ушкодження мембран г
мгтохондргй. Уперше це було встановлено при гшемп мюкарда. Таку реперфузгю
проводить пщ захистом антагоыста кальцгевих каналгв та антиоксиданта.
3. Стадия посиленого бюсинтезу основних бюхгмгчних компонента сполучно/
тканини — колагену, кислих г нейтральних глгкозамгноглгкангв, що е основою для
наступного склерозування гшемгзовано! дыянки тканини або органа. Необхщною
умовою для його здшснення е посилення синтезу нуклешових кислот.
Реперфузшний синдром. Реперфузгйш ушкодження тканин виникають у ситу-
ацгях, коли вщновлюеться кровопостачання гшемгзовано! дыянки тканини або
органа. Ступгнь вираженостг реперфузгйного синдрому залежить вщ об’ему гшемг-
зованог тканини г тривалостг гшемп. У раз! нетривало! гшемп можливим е вщнов-
лення функцп гшемгзовано'г тканини. При тривалгй гшемп, особливо такгй, що
розвиваеться в тканинах з вгдносною або абсолютною недостатнгстю колатераль-
ного кровообггу, функщональне вщновлення тканини не вщбуваеться, навпаки,
посилюеться п ушкодження. Вщсутысть кисню г поживних речовин протягом
перюду гшемп створюе в тканинах умови, за яких виснажуються г не вщновлю-
ються антиоксиданты системи. Тому вщновлення циркуляцп зумовлюе розвиток
у зонг гшемп выьнорадикального ушкодження тканин унаслщок утворення гз кис-
ню, що починае надходити, активних кисневих радикалгв на тлг вщсутносп анти-
оксидантного захисту.
Слщ зазначити, якщо дглянка ушкодження тканини при ппемп-реперфузп
велика, що е можливим, наприклад, при синдром! тривалого здавлення, то пот-
рапляння велико! кыькостг БАР ! метаболгтгв гз вогнища гшемп при реперфузп в
системний кровопк може спричинити системы ефекти, аж до розвитку шоку.
СТАЗ
Стаз (вщ грец. — стояния, нерухомгсть, застш) — уповиьнення I зупинка
току кров1 в капиярах, др1бних артергях / венах.
Розргзняють кыька варгантгв стазу: а) справжнгй (кашлярний) — виникае
внаслщок патолопчних змгн у капыярах або порушення реологгчних властивостей
кровг; б) гшемгчний — розвиваеться внаслщок повного припинення припливу
кров! з вщповщних артергй у капыярну сгтку; в) венозний — пов’язаний !з блока-
дою вщтоку кровг з капыярног сггки.
Венозний 1 гшемгчний стаз е наслщком простих сповыьнення г зупинки крово-
току. Цг стани виникають з тих самих причин, що й венозна ггперемгя та гшемгя.
Венозний стаз може бути результатом здавлення вен, закупорки гх тромбом або
емболом, а гшемгчний — наслщком спазму, здавлення або закупорки артергй.
Усунення причин стазу сприяе вщновленню нормального кровотоку. Навпаки,
прогрес гшемгчного г венозного стазу зумовлюе розвиток справжнього.
При справжньому стазг рух кровг в капыярах г дргбних венах припиняеться,
кров гомогеызуеться, еритроцити набухають г втрачають значку частину гемог-
лобгну. Плазма разом гз гемоглобгном, що вивыьнився, виходить за меж! судинно!
стшки. У тканинах вогнища кагйлярного стазу спостерггають ознаки ргзкого по-
рушення живлення, змертвгння.
Причинами справжнъого стазу можуть бути ф!зичн! (холод, тепло), х!м!чнг (от-
рути, концентрований розчин натрпо хлориду та шших солей, скипидар, прчична
1 кротонова олгя) 1 бюлопчш (токсини мгкрооргашзмгв) фактори.
Мехашзм розвитку справжнъого стазу пояснюють внутрппньокашлярною аг-
регащею еритроципв, тобто Гх склеюванцям 1 утворенням конгломератгв, як! ут-
руднюють рух кровт При цьому шдвищуеться периферичний ошр.
Агрегация виникае внаслщок змши фГзичних властивостей плазмолеми еритро-
ципв щд д!ею факторов, як! проникають усередину катплярно! судини. При елект-
ронно-мгкроскошчному вивченш феномену агрегацп еритроципв з’ясували, що Гх
поверхня, гладка в нормальних умовах, при посилешй агрегацп стае неравною,
“шорсткою”. При цьому змшюються сорбщйш властивосп еритроципв вщносно
деяких барвникгв, що свщчить про порушення Гх фгзико-хгмгчного стану.
У патогенез! справжнъого стазу важливе значения мае вповшьнення току кро-
в! в кашлярних судинах унаслщок ц згущування. Провщну роль при цьому вщгграе
пщвищена проникшсть стшки кашлярних судин, розташованих у зон! стазу. Цьо-
му сприяють етюлопчш фактори, що спричинюють стаз, ! метаболтги, що утво-
рюються в тканинах. Особливе значения в мехашзм! стазу належить БАР (серо-
тонш, брадикшш, пстамш), а також ацидотичному зрушенню тканинноГ реакцн
середовища та його колощному стану. Як наслщок, вщзначають пщвищення про-
никност! судинно! стшки 1 розширення судин, що зумовлюють згущування та
сповГльнення току кров!, агрегацно еритроципв !, зрештою, стаз.
Особливо важливим е вихщ у тканини плазмових альбумш!в, як! зумовлюють
зниження негативного заряду еритроципв, що може супроводжуватися випадан-
ням Гх 13 завислого стану.
ТРОМБОЗ
Тромбоз (вщ грец. троц^оС — грудка) — це процес прижиттевого утворення на
внутршшш поверхш стшки судин згустюв кровг, якг складаютъся з п елементгв.
Згустки кров! можуть бути пристшковими (частково зменшують просвп су-
дин) 1 закупорювальними. Перш! з’являються переважно в серщ 1 стовбурах ма-
пстральних судин, друг! — у дргбних артер!ях 1 венах.
Залежно вщ того як! компоненти переважають у структур! тромбу, розргзня-
ють бш, червою 1 змниаш тромби. Перш! утворюються тромбоцитами, лейкоцита-
ми, а також невеликою кглызстю б!люв плазми, друг! — еритроцитами, скршле-
ними нитками фхбрину. Змгшан! тромби — це б!л! й червош кул!, як! чергуються
м!ж собою.
Ще в XIX ст. сформувалося чихе уявлення про основш фактори тромбоутво-
рення у вигляд! тргади В1рхова.
1. Ушкодження судинно! спнки, що виникае пщ впливом ф!зичних (механич-
на травма, електричний струм), хгмгчних (№С1, РеС13, Н§С12, Н§ЫО3) г бюлопч-
них (ендо- й екзотоксини мжрооргашзмгв) факторгв унаслщок порушення н жив-
лення 1 метаболгзму. Зазначеними порушеннями, кргм того, супроводжуються
атеросклероз, артергальна гшертенз!я, алерпчш процеси.
Пристшковий тромб утворюеться насамперед у дшянц! ушкодження стшки
судини. Це пояснюють, з одного боку, тим, що з ушкоджено! внутршшьо! обо-
лонки судини видыяються фактори зсщання кров!, яю активують процес тромбо-
утворення, а з шшого — локальним пригшчеошям фобринолозу, зменшенням ут-
ворення в ендотелоо кровоносних судин ПП2 (простациклш) та його ендоперок-
сидов, МО, яю в норм! справляють значну антиагрегащйну дно на тромбоцити.
У разо ушкодження ендотелйо збольшуеться його здатшеть до синтезу алюл-
умосних глщерофосфохолшов, зокрема фактора активагш тромбоцитов (ФАТ). 3 його
участю пов’язують агрегащю о дегранулящю тромбоцитов, вившьнення з них адре-
налину та вазоактивних амшов (пстамшу, серотоншу), АДФ, активацйо фосфоло-
пази А^, посилення бюсинтезу тромбоксану експресп адгезивних болков на
поверхш лейкоцитов о ох прилипания до ендотелио. Кром того, ФАТ, як о тромбок-
сан А2, е потужним агрегантом оз сильною судинозвужувальною здатшетю, що
зменшуе вникания крово в ушкоджешй долянцо. Впливаючи на венули, ФАТ розко
збшыпуе ох проникшсть (у 100 разов сильшше, нож гостамон).
2. Порушення активносто систем зсщання о протизещання крово та судигоноо
стшки. Пщвищення активносто системи зсщання осрово за рахунок збольшення в
нш концентрацп прокоагулянтов (тромбопластину, тромбону), як о зниження ак-
тивносто системи протизещання крово (зменшення вмосту в крово антикоагулянтов
або збольшення активносто ох шпботоров), у тому число о фобринолотичноо, як пра-
вило, призводять до розвитку внутрошньосудинного зещаошя крово (ВСЗК) о
тромбозу.
ВСЗК зумовлене онвидким о значним надходженням у судинне русло факторов
зсщання крово, зокрема тканинного тромбопластину, що е мембранним фосфоло-
поглокопротещом, який утворюеться при руйнуванш клотин у разо передчасного
вщшарування плаценти, емболп навколоплщними водами, масивного травмуван-
ня, гострого гемолозу еритроцитов. В експерименто на собаках або кролях ВСЗК
можна вщтворити шляхом уведення в систему загальноо циркуляцп крово актив-
ного тромбону або тромбопластину.
ВСЗК може бути генералозованим (дисемшоваогим — ДВЗ-синдром) о локаль-
ним. Цей процес оборотний, особливо в разо своечасноо терапц аоятикоагулянтами.
Переход ВСЗК у тромбоз вщбуваеться пщ впливом факторов, яю вивыьню-
ються в разо ушкодження судинноо стшки, 1 тромбоцитов.
Унаслщок посиленого використання факторов зсщання крово о тромбоцитов,
вторинноо активацн системи протизещання о фобринолозу у другой фазо дисемшо-
ваного внутрошньосудинного зсщання крово розвиваеться тромбогеморагочошй
або гопергшокоагуляцойогий сиощром.
3. Сповшьнення о порушеошя (завихрения в долянцо аневризми) кровотоку.
Цей фактор, очевидно, мае меоппе значения, однак вон дае змогу пояснили, чому
у венах тромби утворюються у 5 разов частоте, нож в артероях, у венах нижнох кон-
цовок — у 3 рази частице, нож у венах верхнох концовок, а також високу частоту
тромбоутворення в разо декомпенсацп кровообогу, тривалого посильного режиму.
Процес тромбоутворення умовно можна подолити на дво фази: клотинну —
фазу адгезп, агрегацц та аглютинацн тромбоцитов о плазматичну — фазу коагуляцп
крово. »
Фозико-хомочно властивосто клтинно! фази (первинний, або судинно-тромбо-
цитний гемостаз) тромбоутворення полягають у змпп елеостричного потенцоалу
судинноо стонки, заряду тромбоцитов та онопих клотин крово, пщвищенно адгезив-
но-агрегацойноо здатносто тромбоцитов, що спричинюе осщання ох на уопкоджешй
(“чужорщнУй” з оголеними детерминантами молекул колагену субендотелоального
позаклотивного матриксу) поверхн! внутргшньо! оболонки судин (адгезоя) У при-
липания один до одного (агрегацоя).
За сучасними у'явлениями, основним механизмом адгезп та агрегацп тромбо-
цитов е експрес!я на ох поверхн] та на мембран] сндотелюпитов У лейкоцит,в бтлков
клггинно‘1 адгезп (див. роздол XI “Запалення”), але певне значения в цьому про-
нес) мае о зниження ох негативного заряду внасл!док зменшення вм!сту АТФ У
збьтьшення р!вня АДФ в ушкоджених дУлянках судин 1 тканин, а також у тромбо-
цитах, гальмування антиагрегашйних властивостей судинно/ стонки в раз1 и уш-
кодження внаслщок зниження синтезу ПП2 (простациклшу) на тло пщвищення
синтезу в тромбоцитах ШТ>2, ПГН2, тромбоксану ФАТ. ОстаннУ, особливо
тромбоксан А^ на противагу ПГ12 е активними Унгшаторами агрегат"] тромбо-
цитов. Така доя цих речовин, очевидно, реалхзуеться через вивыьнення тромбоци-
тами АДФ, що й призводить до ох агрегащУ. Пщвищений синтез простагландинУв
зумовлений активуточим влливом ртзних стимуляторов агрегацп (тромбону, кола-
гену, серотоншу, адреналУну. норадреналону тощо) на фосфолшазу А, (яка вивУль-
няе з фосфолшщов мембран тромбоцигш арахУдонову кислоту' — вихУдний суб-
страт для синтезу простагландинов), циклооксигеназу та УншУ фермента. Певну
участь у процесах адгезп та агрегацп тромбоцитов беруть 1 плазматичш фактори
зсщання крово (фтбриноген, фактори VIII, XIII).
Важливо зазначити, шо на етат необоротноУ агрегащУ з тромбоцитов у кров
викидаеться додаткова колькУсть АДФ, серотоншу. адреналгну, норадреналУну, Ун-
ших БАР. Отже, процес набувае характеру порочного кола.
Необоротна змша тромбоцитов починаеться через 2—3 хв вщ моменту ушко-
дження внутрУшньоУ оболонки судин. При цьому спостертгають розширення Ухнь-
оУ цитоплазми, появу множинних псевдоподгй, втрату тромбоцитних гранул по
краях агрегатУв тромбоцитУв, прилипания лейкоциттв о утворення на Ухной повер-
хю фУбриногенових мУсткУв, як1 сприяють коясолУдащУ первинноУ тромбопитно1
пробки. Подальша дезУнтеградУя поширюеться вглиб маси, чому сприяють акта-
вашя автолУтичних ферментов, пУдвищення проникностУ та розчинення плазма-
тичних мембран. Це сшорюе умови для пУдвищеного проникнення Уз сироватки
кров! в тромбоцита кальцою, активацУУ в них Са2+-залежноУ АТФази, порушення
сшввУдношення АТФ / АДФ у бУк збУльшення .АДФ У, як наслУдок, подальшого та
лрогресуючого посилення алгезУУ й агрегаш’1.
Вщ моменту розпаду тромбоцитУв, вивУльнення тромбоцитних У активацУУ
плазмових факторов системи зсУдання кров! починаеться наступили етап тромбо-
утворення — плазматтна фаза (фаза коагуляшо крово}. Ф’зико-хомтчна та бУохомоч-
на сутность цоео фази полятае в кУлъкох послодовних перетвореногях за типом про-
фермент — фермент. Бшышсть Уз цих перетворень мають справжню ферментну
природу. Зазначено боохтмочнУ реакщ] розгортаються на оголеному колагено субен-
дотелоального шару га фосфолУпщних матриц ях агрегованих тромбоцитов. Резуль-
татом е утворення фобрину, який перетворюе нестабУльну тромбоцитну пробку
(первинну) на стабольну гемостатичну.
На першому етат вщбуваеться утворення активно) протромбУнази Це мем-
бранозв’язаний комплекс протешаз о бшкових кофакторов, що мае здатопсть фер-
ментативно впливати на протромбш. Вш може утворюватася зовшшнтм У внут-
рошном шляхом. ЗовнштнУй шлях (основний) забезпечуеться взаемодУею ткаошн-
них (тромбопластин) о плазматичних (фактори зсщанооя крово VII, X, V, Са2+)
компонентов зсщально! системи кровь Внутршшш шлях активацп зсщання кров!
забезпечуеться взаемодоею лише факторов зсщання, яю моститься в крово (XII, XI,
IX, VIII, X, V, II, фосфолшщ о Са2+). Час, протягом якого вщбуваеться зовншшш
ооолях, становить колька секунд, тодо як для внутршшього потробно хвилини.
Другий етап — утворення активного тромбону. Пщ впливом активно! протром-
бонази, яка складаеться оз Ха, Vа, Са2+ та фосфолшщу, вщбуваеться розщеплення
в двох мосцях молекули протромбону, оз фрагментов якого утворюеться тромбон —
високоспецифочний протеолотичний фермент. Взаемодоя протромбшазного комп-
лексу з протромбшом вщбуваеться в долянцо плазматично! мембрани оз залучен-
иям тромбоцитного скелета.
На третьему етат пщ впливом тромбону здшснюеться перетворення фобри-
ногену на фобрин з утворенням згустка. Потом можмолекулярно воднево зв’язки у
фобриш-поломеро стають ще моцнппими пщ впливом фобринстаболозуючого факто-
ра плазми крово.
Фобрин мае вигляд ниток, яко розташоваш пухко або компактно, о становить
основну масу тромбу. У коморках сотки, що утворилася, розмпцуються клггини
крово (агреговано тромбоцита, скупчення лейкоцитов та еритроцитов).
На завершальному етат зсщання крово пщ доею тромбастеншу (ретрактози-
му), який видшяеться з онтактних тромбоцитов, вщбуваеться скорочення (за ти-
пом скорочення актомоозину) фобринових волокон о волоконець тромбоцитов.
Наслщком цього е ретракция (стиснення) й ущольнення згустка. Для нормально!
ретракцп потробно оони кальцпо, глюкоза, АТФ, фозоолопчний перебог глоколозу,
певно споввщношення мож концентрацоею тромбону о фобриногену, а також фобри-
ногену о тромбоцитов.
Можливо розно наамдки тромбозу. Враховуючи його значения як кровоспин-
ного механозму пщ час гостро! травми, що супроводжуеться кровотечею, тромбоз
слщ розглядати як пристосувальне явище.
Водночас тромбоз при розних хворобах (атеросклероз, облотеруючий ендар-
терпт, цукровий доабет тощо) може супроводжуватися тяжкими наслщками, зумо-
вленими гострим порушенням кровообогу в зош тромбованоо судини (ппемоя в
разо тромбозу артерой, застой крово при тромбозо вен).
Розвиток некрозу (шфаркту, гангреоги) в басейно тромбованоо та позбавленоо
колатералей судини — завершальний етап тромбозу. Особливо великою е роль
тромбозу вонцевих артерш у розвитку шфаркту мюкарда.
Завершениям тромбозу можуть бути асептичне (ферментне, автолотичне) або
септичне (гнойне) розплавлення, органозащя (розсмоктуванооя оз замощениям спо-
лучною тканиною), реканалозацоя. Септичне розплавлення дуже небезпечне, осколь-
ки спричинюе септикошемою й утворення множинних абсцесов у розних органах.
ЕМБОЛ1Я
Ембол1я (вщ грец. ер^оДкоз — кинута всередину) — це закупорювання судин пилами
(емболами), як1 надходять з током кровг або л1мфи.
Залежно вод характеру емболов розрозняють емболпо екзогенну (повотряну,
ощльними сторонноми толами, бактероальну, паразитичну) та ендогенну, спричи-
Мал. 18. Джерела Г шляхи проходження емболш у
динах великого га малого кола кровообпу
1 — права легенд; 2 — .ива легенд; 3 — головний мо-
зок; 4 — сонна артерш; 5 — аорта; 6 — права пипка,
7 — кишки; 8 — печшка; 9 — нижня порожписта
ш вена; 10 — легеневий стовбур
йену тромбом, краплями жиру, ргзними
тканинами, навколоплщними водами,
бульбашками газу при десагурацп.
За локалгзацгею розргзняють емболпо
судин великого, малого кола кровообггу га
системи воргтног вени.
В усгх цих випадках рух емболгв зазви-
чай може вщбуватися вщповщно до при-
родного руху крови Звщси зрозумпо, шо
джерелом емболп судин великого кола кро-
ьообггу е патологгчнг процеси в легеневих
венах, порожнинах лгвог половини серия,
артергях великого кола кровообпу; джере-
лом емболп судин малого кола кровообггу —
патолопчнг змши у венах великого кола
кровообггу г право! половини серця. Ви-
никнення емболп воргтно! вени зумовлю-
ють патологгчн! змши в басейнг цгег вени. Винятком е ретроградна емболгя, коли
рух ембола вщбуваеться не за гемодинамгчними законами, а вщповщно до сили
тяж1ння самого ембола. Така емболгя розвиваеться у великих венозних стовбурах
у раз! сповгтьнення току кров! га зменшення присмоктувально! дц трудно! клгтки.
Спостернаеться також парадокс альни емболгя в раз! незарощення мгжпередсердно!'
або мгжшлуночковог перегородки, унаслщок чого емболи з вен великого кола
кровооб!гу та право! половини серця переходить у лгву, оминаючи мале коло кро-
вооб!гу (мал. 18).
Емболы екзогенного походжепня. Повйпряна ембол1я виникае пгсля поранения
великих вен (яремно‘1. пщключичног, синусгв твердо! оболони головного мозку),
як! погано спадаються г тиск у яких близький до нуля або негативний. Це може
бути причиною повгтряно! емболп пщ час виконання лгкувальних маншулящй,
наприклад, гнфузп розчишв у велик! вени. Унаслщок цього у вени всмоктуеться
повгтря, особливо на висотг вдиху, 1 виникае емболгя судин малого кола кровообг-
гу. Такт сам! умови створюються в раз! поранения легеш або деструктивних про-
цесгв у нш, а також ищ час накладання пневмотораксу. У таких випадках, однак,
спостерггаеться емболгя: судин великого кола кровообггу. Аналоггчн! наслщки мае
надходження велико! кыькостг пов!тря з легенгв у кров у раз! дп на людину вибу-
ховог ударно! хвил! (повггряног, водяног), а також год час “вибуховог декомпреегг”
г швидкого гшгняття на значну висогу. Ргзке розширення легеневих альвеол, що
при цьому виникае, розрив гхньог стгнки г надходження пов!тря в капътяри зумо-
влюготь неминучу емболпо судин великого кола кровообггу. Чутливгстъ ргзних
тварин । людини до повггряно! емболп неоднакова. Кргль гине шсля внутршшьо-
венного введения 2—3 мл повзтря, тодз як собака переносить об’ем уведеного
повзтря 50—70 мл/кг. Людина щодо цього поезд ае промзжне мзеце.
Емболзя ендогенного походження. Причиною тромбоемболи е частника тромбу,
що вщзрвалася. Вщрив частинок тромбу е ознакою його неповноцзнносп (“хво-
рий” тромб). У бзльшостз випадкзв “хворз” тромби утворюються у венах великого
кола кровообпу (вени нижнзх кшцзвок, таза, печшки), чим з пояснюеться бьтьша
частота виникнення тромбоемболи малого кола зфовообзгу. Запальнз змши в за-
елшцз клапана легеневого стовбура з в правому передсердно-шлуночковому (три-
стулковому) клапан!, як! е причиною тромбоендокардиту, нерщко супроводжу-
ються тромбоемболзею легеневих артерш. Лише в тому раз!, коли тромби утворю-
ються в лзвзй половинз серця (ендокардит, аневризма) або в артерзях (атеросклероз),
розвиваеться емболзя судин великого кола кровообпу.
Причинами неповноцшностз тромбу, вщриву його частинок ! тромбоемболи
е асептичне або гншне розплавлення тромбу, порушення фази ретракци тромбо-
утворення, а також зсщання кровз.
Жирова ембол1я виникае в разз надходження в кров крапель жиру, переважно
ендогенного походження. Причинами потрапляння жирових крапель у кров е
роздроблення трубчастих кзеток, ушкодження пздшкзрно’з або тазово’з клзтковини
та жирових скупчень, печшки в станз жирового переродження.
3 взком у зв’язку зз замиценням червоного кзеткового мозку трубчастих кзеток
жовтим з внаслщок збзлыпення в ньому вмзету жирзв з низькою температурою
плавления небезпека виникнення жирово'з емболи зростае.
Оскзльки джерело емболи розмззззуеться переважно в басейн! вен великого
кола кровообзгу, жирова емболзя виникае насамперед у судинах малого кола кро-
вообзгу. Лише згодом можливе проникнення жирових крапель через легеневз ка-
пзляри (або артерзоловенознз анастомози малого кола кровообзгу) в лзву половину
серця та артерзз великого кола кровообзгу.
Кзлькзсть жиру, яка може спричинити смертельну жирову емболзю, у рззних
тварин становить вщ 0,9 до 3 см3/кг.
Тканинна (клипинна) емболгя. У разз травми можуть заноситися частники рзз-
них тканин органззму (кзеткового мозку, м’яззв, головного мозку, печзнки, тро-
фобласта), особливо багатих на воду, у систему циркуляцп кровз, передуезм мало-
го кола кровообзгу. Вщрив кашкоподзбних жирових мае атером вщ атеросклеро-
тично змзненоз артерзальноз стшки з потрапляння зх у кров супроводжуються
емболзею артерш великого кола кровообзгу.
Особливе значения мае емболзя судин клзтинами (часто у виглядз агрегатзв)
злоякзених пухлин, оскыьки вона е основним механззмом утворення метастаззв.
Емболгя навколоплгдними водами — надходження навколоплщних вод пщ час
полопв в ушкодженз судини матки в дзлянцз плаценти, що вщокремилася. В ар-
терзолах з капыярах легень затримуються щзльн! частники амнзотично’з рщини
(меконпо, уегпзх сазеоза), що клшзчно проявляеться клппчною картиною емболи
малого кола кровообзгу. Цей вид емболи вщрззняеться вщ тканинно’з емболи
пщвищенням активностз фзбринолзтично! системи кровз, рззким зниженням вмзе-
ту фзбриногену в кровз (гшо- та афзбриногенемзя), порушенням зсщання кровз
(вторинним) з тривалою кровотечею з матки.
Клзнзчю прояви емболи визначаються и локалззацзею (мале або велике коло
кровообзгу), особливостями ангзоархзтектошки, зокрема станом колатерального
Мал. 19. Реакцкя дихання та кровообпу соба-
ки на внутршньовснне введения повггря (за
II.С. Смирновой», 1972).
а — дихання: б — центральний венозний тиск;
в — аргер1альний тиск; г — тиск у легеневих
ы артертях: г — позначна введения повотря; д —
позначка часу (5 с)
кровообпу та його нейрогуморально!
регуляцц. розм!ром 1 складом ем-
болтв, !х загальною масою, швидюс-
тю надходження в кровот!к, реактив-
нтстю оргашзму.
Газова емболзя е головною пато-
генетичною ланкою стану декомп-
респ, зокрема кесонно! хвороби. Пе-
репад атмосферного тиску вщ пщви-
щеного до нормального (у робггниюв
кесон 1 в 1 водолаз!в) або вщ нормаль-
ного до р>зко зниженого (пщ час
швидкого Л1ДНЯТТЯ на висоту або роз-
герметизаш! кабши висотного леталь-
ного апарата) зумовлюе зниження
розчинност! газ1в (азоту, вуглекисло-
го газу, кисню) у тканинах 1 кроы
(десатурап!я) та закупорюваиня буль-
башками цих газ!в (насамперед азоту) кап!ляр!в, розм!шених переважно в басейш
судин великого кола кровообпу.
Газова ембол!я можлива також при анаеробнш (газовгй) гангрен!.
Ембол!я судин малого кола кровообпу. Наиважливплим функщональним пору-
шенням у раз! емболп судин малого кола кровообпу е ргзке зниження артергаль-
ного тиску в систем! великого кола кровообзгу (мал. 19). € к!лька гшотез, яю пояс-
нюють мехашзм гшотензивного ефекту емболп легенево! артерп.
1. Дехто з дослщниюв пов’язуе р!зке зниження артертального тиску в систем!
великого кола кровообпу з! зменшенням ХОС, зумовленим мехашчним закупо-
рюванням легенево! ар герц та правошлуночковою недостатшстю серця. Однак
подалыш дослщження довели, що мехашчне закриття наветь б!льшо! частини ле-
геневих судин не спричинюе таю порушення, як при емболп.
2. Поширеною е думка, зпдно з якою р!зке зниження артер1альното тиску
розглядаюзь як рефлекторну гшотензпо (розвантажувальний рефлекс Швггка —
Парша) Вважають, що депресорний рефлекс при цьому спричинюеться подраз-
ненням рецептор)в, розм!щених у русл! легенево! артерп. Зпдно з даними О.
Б. Фохт 1 В.К. Л!ндемана (1903), ваготомзя, а також атропш!зац!я тварин ослаб-
люють цепресивну реакнпо, що шдтверджуе п рефлекторний мехашзм.
3. Певного значения у зниженю артериального тиску пщ час емболп легенево!
артерн надають ослаблению функцп серця внаслщок гшоксй мюкарда, що е на-
слщком збщыпення навантаження на праву половину серия та резкого зниження
артерзального тиску.
Обов’язковим гемодинамгчним ефектом емболп судин малого кола кровообг-
гу е гндвищення тиску кров! в легеневш артери (у норм! систолгчний артер!альний
тиск становить 20 мм рт. ст., д!астол!чний — 8 мм рт. ст.) г р!зке збгльшення
градгента тиску на дгтянщ легенева артергя — кап!ляри, що розглядаеться як на-
слщок рефлекторного спазму легеневих судин. Рефлекторне звуження легеневих
артерюл необхщне для пщтримання тиску в легеневих каггглярах нижче ргвня, за
якого може розвинутися набряк легень.
До такого самого ефекту — подразнення рецепторов легеневих судин г внаслг-
док цього виникнення спазму — можуть призвести пщвищення тиску в артерюлах
легень, механгчне подразнення судин емболами, зменшення току кров! в судию
нижче ембола, видшення в мгсцг закупорювання речовин (серотонгну, пстамгну,
ендотел!н!в), як! здатн! зумовлювати скорочення непосмугованих м’язових воло-
кон судин.
Унаслщок зазначених гемодинамгчних порушень ргзко пщвигцуеться цент-
ральний венозний тиск, розвиваеться синдром гострого легеневого серця (синдром
гостро! правошлуночковог недостатностг), який часто е причиною смерть
Порушення гемодинамгки в малому г великому кол! кровообпу п!д час ем-
болп легенево! артери призводить до зм!н вентиляцгйно-перфузгйного спгввщно-
шення в легенях !, як наел (док, до вторинних змш газового складу кров! — збгль-
шення рСО2, зменшення рО2 (див. роздгл XXVII “Патолопчна фЫолопя зовншг-
нього дихання”). Як пристосувальна реакция, спрямована на нормалгзацгю
газового складу кров!, розвиваеться задишка. Вважають, що порушення зовн!ш-
нього дихання внаслщок емболп легенево! артерп е рефлекторною реакшею, яка
виникае як з рецепторного поля малого кола кровообггу, так г внаслщок подраз-
нення рефлексогенних зон великого кола кровообггу кров’ю зг зниженим вмгстом
кисню. Експериментально доведено, що шляхом перергзання блукаючих нервгв
можна значно зменшити порушення дихання.
Ембо.пя судин великого кола кровообпу. Причинами емболп судин великого
кола кровообпу, як зазначалося, е: патолопчнг процеси (тромбоендокардит, ин-
фаркт мюкарда), що супроводжуються утворенням тромбгв на внутргшнш повер-
хнг порожнин лгвог половини серця; тромбоутворення в артергях великого кола
кровообпу з подалыпою тромбоембол!ею; газова або жирова емболгя. Частою ло-
калгзашею емболгв е вгнцевг, середня мозкова, внутрппня сонна, ниркова, се-
лезшкова, брижова артерп. За гнших однакових умов локалгзацгя емболгв визна-
чаеться кутом вщхилення бгчног судини, п дгаметром, гнтенсивнгстю наповнення
органа кров’ю. Великий кут вщхилення бгчних гглок вщносно вщргзка судини,
розмпценого вище, поргвняно великий гх дгаметр, ггперемгя — фактори, якг зумо-
влюють локалгзацгю емболгв.
При газовгй емболп, якою супроводжуеться кесонна хвороба або “вибухова
декомпреегя”, важливим моментом для локалгзацп емболгв у судинах мозку та
пщшкгрнгй жировгй клгтковинг е висока розчиннгсть азоту в тканинах, багатих на
лппди.
Тяжкгсть клппчно! картини визначаеться переважно двома факторами: реф-
лекгорним спазмом судин г ступенем розвитку колатералей. Рефлекторний спазм,
з одного боку, може охоплювати не лише найближчг, а й вщцален! судини, уск-
ладнюючи перебгг патолопчного процесу. У цьому разг до мгецевих патофгзголо-
пчних змгн, якг мають вигляд ушкоджено! гшемгзовано! дглянки, часто приедну-
ються загальш, вщ яких нерщко хвор! помирають. В !ншому разг стан колатераль-
ного кровообпу в ба сейш судини, закупорено! емболом, г в прилеглих тканинах е
фактором, яким запобпае такому тяжкому ! часто необоротному процесу, що роз-
виваеться внаслщок емболи, як змертвгння вщповщно!' дщянки тканини.
Емботя ворипно!вени, хоч ! слостерйаеться значно рщше, н!ж емболш малого
1 великого кола кровооб!гу, привертас увагу насамперед характерним клш!чним
симптомокомплексом г надзвичайно тяжкими гемодинам!чними порушеннями.
Через велику мгсткйть портального русла закупорення емболом головного
стовбура ворггно! вени або основних його розгатужень призводить до збшьшено-
го наповнення кров’ю оргашв черевно! порожними (шлунка. кишок, селезшки) !
розвитку синдрому портально! гшертензп — пщвищення тиску кров! в систем!
ворйно! вени з 0.78—0.98 до 3,92—5,88 кПа (з 8—10 до 40—60 см вод. ст.). При
цьому розвиваються характерна клппчна тргада (асцит, розширення повсрхневих
вен передньо! стшки живота, збёльшення селезёнки) та пшп змгни, зумовлен! по-
рушенням кровообпу (зменшення припливу кров! до серця, ударного общему сер-
ця > ХОС, зниження артергального тиску), дихання (задишка, поттм ргзке сповгль-
нення дихання, апное) ! функцц нервово! системи (потьмарення свщомосп, па-
рал!ч дихання). Мехашзмом цих загальних порушень е пере важно зменшення
маси циркулюючо! кров?', зумовлене п накопиченням (близько 90 %) у порталь-
ному русла. Такт порушення гемодинамгки часто е безпосередньою причиною
смерт! хворих
Водночас мгж асцитом, розширенням поверхневих вен передньо! стшки жи-
вота 1 спленомегалйю. з одного боку, । ступенем портально! гшертензп, з гншо-
го, немае прямого зв’язку'. 1нод! в раз! високого р!вня портального тиску щ сим-
птоми вщсутн! 1, навпаки, у деяких випадках вони з’являються нав!ть за незнач-
ного пщвишення тиску в систем! ворйно!' вени. Це наводить на думку, що в
розвитку зазначених клгшчних проявив портально! гшертензп, зокрема асциту,
кргм пщвищення тиску велику роль вщ!грають ! шш! фактори: порушення обмгн-
них функшй печшки; зазримка натр!ю 1 води в оргашзм!, спричинена гшерпро-
дукцгею альдостерону 1 вазопресину; зниження онкотичного тиску плазми кров!,
зумовлене гшопротешемгею; пщвишення проникност! стшки капгляргв порталь-
ного русла.
ПОРУШЕННЯ М1КРОЦИРКУЛЯЦП
У систем! периферичного кровооб!гу умовно видшяють мпсроциркуляторне, або
термшальне, судинне русло, яке своею чертою, вщповщно до под!лу судин на
крово- та лгмфоносн!, подшяють на мпсроциркуляторне кровоносне й л!мфонос-
не русло.
Мйсроциркуляторне кровоносне русло складаеться 13 судин, д!аметр яких не пе-
ревищуе 100 мкм, тобто артерюл, метартерюл, кап!ляр!в, венул 1 артерюловену-
лярних анастомозгв (мал. 20). У ньому вщбуваються транспорт поживних речовин
1 кисню в тканини 1 клггини, видалення з них вуглекислого газу ! продуктов обма-
ну азоту, пщтримуеться р!вновага рщини, яка притгкае 1 вщтакае, оптимальний
ргвень тиску в периферичних судинах 1 тканинах.
Мгкроциркуляторне лгмфоносне русло — це початковий вщдш лгмфатично! сис-
теми, де л1мфа утворюеться 1 звщки надходить у л!мфатичн! катляри. Процес
утворення л1мфи складний 1 полягае в переход! рщини та розчинених у шй речо-
вин, у тому числ! й б!лк!в, кр!зь станку кровоносних капшярхв у мЬкюптинний
простар, поширенн! речовин у периваскуляршй сполучшй тканин!, резорбцп ка-
пщярного фыьтрату в кров, резорбцп бйпов 1 надлишку рщини в лгмфоносш
шляхи тощо. На утворення л!мфи витрачаеться близько 20 % рщини, що профщь-
трувалася з кровоносних судин у тканини.
Отже, за допомогою мгкроциркуляторного кровообпу здшснюеться таена ге-
матоштерстицшна та лшфоштерстищйна взаемодгя, спрямована на пщтримання
Мал. 20. Принципова схема мжроциркуляцп (за В.XV. 2«еИасК, 1950):
АВА — артерювенульний анастомоз
Р1вня метаболизму в органах 1 тканинах вщповщно до тх власних потреб, а також
потреб оргашзму в цйюму.
Вщповщно до загальноприйнято! класифгкацп, зэпропоновано! Е. Ма^ёю
(1965), роздали мгкроциркуляцП подпятотъ на внутрппньосудинш порушення,
пов’язаш 31 змшою самих судин, ! позасудинн! порушення.
ВНУТР1ШНЬОСУДИНН1 ПОРУШЕННЯ М1КРОЦИРКУЛЯЦП
Порушення реолопчннх властивостей крови Найважливппими внутр,шньосудинни-
ми порушеннями е роздали особливостей реологп кров! у зв’язку з! змшою сус-
пензшно! стабыьност, клпин кров) та ц в’язкостт. 3 ними частково пов’язаш по-
рушення зсщання кров! та утворення гемокоагулящйних мшротромбтв, а також
порушення перфузп кров! через м!кроциркуляторне русло у зв’язку зт змшою
швидкосп току крови
У нормальних умовах кров мае характер стабильно! суспензн клпин кров! в
рщк!й Г! частив и Збереження суспензшно! стабйгьносп кровт забезпечуеться вели-
чиною негативного заряду еритроципв ! тромбоцитгв, певним сшввщношениям
бнткових фракцш плазми (альбумшу, з одного боку, глобулгшв ! фтбриногену, з
шшого), а також досгатньою швидктстю кровотоку. Антиадгезивш й антиагрегат-
ш властивост! тромбопипв визначаються спещальними механизмами, розтлянути-
ми в роздш ЕХ “Патолопчна фтзюлопя периферичного кровообзгу”.
Зменшення величини негативного заряду еритроципв, причиною якого най-
частште е абсолютно або вщносне збыьшення вмтсту позитивно заряджених мак-
ромолекул глобушшв 1 / або ф1бриногену та !х адсорбшя на поверхн! еритроципв,
зумовлюе зниження суспензшно! стабыьност! кров!, а1регащю еритроципв та ш-
ших КЛ1ТИН кров!. Зниження швидкосп кровотоку посилюе цей процес. Описа-
ний феномен отримав назву “сладж” (вщ англ. — густий бруд, твань. мул).
Основними особливостями сладжовано! кров! е прилипания один до одного ерит-
роципв, лейкоципв 1 тромбоиипв ! пщвищення в'язкосп кров!, що утруднюе и
перфузпо через м!кросудини. Внутршшьосудинне утворення агрегапв еритроципв
та 1нших клпин кров) спостерггаеться при: перев’язщ судин; ушкодженн! тканин;
надлишку в кров! брадикшшу, серотоншу, тромбину, норадреналину; внутр! шньо-
венному введен н! високомолекулярних речовин (декстрану, метилцелюлози); от-
руенш миш'яком, кадм!ем, бензолом, толуолом, ашлшом, при р!зних вицах шоку,
олпурП, гострш судин шй недостатносп; екстракорпоральному кровооб!гу; гшо-
термп; захворюваннях, що супроводжуються збшьшенням у кров! вм!сту ф!брино-
гену I глобул! н!в 1 зниженням вмюту альбумину (множинна м!елома, макрогло-
булшемзя та ш.).
Залежно в!д характеру дп сладж може бути оборотним (за наявност! пльки аг-
регаци еритропипв) 1 необоротним. В останньому випадку мае м!сце аглютинашя
еритроципв. Розмгри агрегапв при сладж! коливаються в(д 10x10 до 100x200 мкм 1
больше.
Процес формування агрегапв кл!тин кров! мае певну послщовшсть. У перш!
хвилини теля ушкодження. переважно в супинах кап!ляр!в 1 венулах, утворюють-
ся агрегата з тромбоципв 1 хиомшрошв (велик! лшйпй частники розм!ром 0.1—
1 мкм, як! м!стятъ в основному триглшериди; надходять у кров з лхмфи кишок 1
циркулюють у формЁ стабЁльно! емульсп). Вони щёльно фЁксуються до стёнки
мЁкросудин, утворюючи “61ЛИЙ” тромб, або ВЁДНОСЯТЬСЯ В ЁНШЁ ВЁДДЁЛИ судинно!
системи до нових вогнищ тромбоутворення.
ЕритроцитнЁ агрегати утворюються в першЁ години шсля ушкодження, спо-
чатку у венулах, а потём в артерЁолах. Це зумовлено зниженням швидкостё крово-
току. Через 12—18 год зазначеш порушення прогресують як за вираженЁстю про-
явёв, так 1 за поширенЁстю. Можливий Ё зворотний розвиток процесу в напрямк)
дезагрегацЁЁ.
Патоф1з1олог1чн1 насл1дки агрегаци еритроцитЁв проявляються порушенням
мЁкроциркуляцц ё внаслЁдок цього порушеннями метаболЁзму та функщй оргашв
ё систем.
Порушення мЁкроциркуляцц зумовлеш:
1) парцЁальною обтурацЁею мЁкросудин унаслщок осЁдання на ’ёхнёй внутрЁш-
нёй оболонцЁ еритроцитних агрегатЁв, що мають бЁльшу масу, нёж окремЁ еритро-
цити. Зниження швидкостё кровотоку, збЁльшення розмЁрЁв агрегатЁв, прилипания
еритроцитЁв до стёнки судин, пЁдвищення в’язкостё кровЁ — фактори, шо приско-
рюють процес осЁдання агрегатних комплексЁв на внутрЁшнЁй оболонцЁ мЁкро-
судин;
2) повною обтурацЁею мЁкросудин агрегатами тромбоцитЁв ё еритроцитЁв. При
цьому великЁ агрегати, що складаються ёз кёлькох десяткЁв ё сотень еритроцитЁв,
можуть повнёстю перекривати просвЁт артерЁол ё венул. Агрегати менших розмЁрЁв
досягають дрЁбнЁших судин, аж до капЁлярних, спричинюючи ’!х емболЁю;
3) рЁзким уповЁльненням кровотоку, сепарацЁею (вЁддЁленням) плазми вёд ерит-
роцитЁв, маятникоподЁбним рухом плазми ёз завислими в шй агрегатами, стазом.
У зв’язку ёз закупоркою термЁнальних артерЁол великою кёлькёстю еритроцитних
агрегатЁв капЁлярю судини пропускають тёльки плазму. При цьому ушкоджуеться
стЁнка мЁкросудин (набухания Ё десквамацЁя ендотелЁю). Посилюють цей процес
кисла реакцЁя середовища, мЁсцевЁ метаболЁти, БАР (серотонЁн, гЁстамЁн, гепа-
рин), що надходять у кров у зв’язку з масовою дегранулящею тканинних базо-
фЁлЁв прилегло! сполучно! тканини. ПЁдвищення проникностЁ венул ё капЁлярних
судин, яке виникае внаслЁдок цього, сприяе виходу рщини за !х межЁ, згущуванню
кровЁ, шдвищенню н в’язкостё. Створюються умови (ушкодження судинно! стён-
ки, агрегацЁя тромбоцитЁв та !х ушкодження, сповЁльнення кровотоку) для утво-
рення множинних гемокоагуляцЁйних мЁкротромбЁв з подальшим наростанням
тяжкостё мЁкроциркуляторних розладЁв.
Комплекс описаних вище патофЁзЁолопчних порушень мЁкроциркуляцц на
завершальному етапЁ розвитку сладжа, що характеризуеться вираженими пору-
шеннями метаболЁзму Ё функцЁй оргашв Ё тканин, а також недостатшм рЁвнем
трофЁчного забезпечення, називають капЁлярно-трофЁчною недостатнЁстю.
Таким чином, сладж-феномен, що виникае спочатку як мЁсцева реакцЁя тка-
нини на ушкодження, у подальшому своему розвитку може набути характеру сис-
темно!’ реакщ!, генералЁзовано! вёдповёдё оргашзму. У цьому полягае його загаль-
нопатологЁчне значения.
Порушення проникностЁ судин обмшу. Судини обману (капЁляри) характеризу-
ються двома основними функцЁями: здЁйсненням руху кровЁ Ё здатшстю пропус-
кали в напрямку кров — тканина Ё назад воду, розчинею гази, кристалогЁдрати ё
великомолекулярнЁ (бЁлковЁ) речовини.
Мал 21. Шляхи транспорт} речовин кртзь стшку капалярних судин
(за Р.П. Кптгйе, 1971)
Ре гуляша кровообёгу в систем! судин обману шлком падпорядковуеться зако-
номерностям пре- ё посткапалярного кровотоку, а також мёсцевим ауморальним
впливам.
При патологп часто спостерёгаеться збёльшення або зменшення ёнтенсивноста
переходу речовин через судинну стшку не лише внаслщок змши ёнтенсивностё
кровотоку, а й у результата справжнього порушення судинноа проникностё, що
супроводжуеться змшою структури стшки судин обману, У морфол отёчному вёд-
ношеаш! падвищення судинноё проникносп характеризуеться збёльшенням про-
мажкёв мёж ендотелёоцитами внаслщок ёх скорочення та посиленням утворення
транслортних везикул (мал. 21), у функцёональному — посиленим переходом ви-
сокомолекулярних речовин (бёлкёв) через капёлярну стшку.
У мехашзм! пщвищення судинноа ироникноста при травма, опаку, запаленнё,
алергп великого значения надають кисневому голодуванню тканин, ашшотично-
му зрушенню реакцёё середовища, накопиченню маспевих метаболпёв, утворенню
БАР, активним глобулёнам плазми крова (а-, 0-глобулёнам), катаонним балкам ё
нейтрофалъним лазосомним ферментам гранулоцитёв. При шоц! разного похо-
дження можливе ! генералазоване порушення судинноё проникносп.
За сучасними уявленнями, бёолопчно активна амани (гёстамён, серотонён) та
ёх природна лабератори, а також брадикёнш, фактори комплементу й ейкозаноёди
(простагландини 1 лейкотрёени) чинягь короткочасну даю на проникнасзъ судин-
ноа спнки за допомогою впливу на конграктильна елементи ендотелёю судин,
переважно венул, що спричинюе ёх заокруглення ! збальшення мёжклатинних про-
мажкёв мёж ними. При разних патолопчних процесах, особливо при запаленна,
спричиненому слабкими факторами (тепло, ультрафёолетова променё, деяк! хёмач-
н! речовини), ц! чинники вёдтворюють ранню фазу падвищення судинноё про-
никносп (!0—60 хв). Пазнаша порушення проникносп судинноё станки (вёд 60 хв
до калькох дёб) зумовлюютъся а посиленням трансцигозу, ё протеёназами, лазосом-
ними гщролазами, капонними балками нейтрофальних гранулоцитёв, дая яких
спрямована на стшку капалярних судин (мёжклатиннё зв’язки ендотелпо ё базальну
мембрану) ё полягае у фёзико-хёмёчних змёнах (зокрема деполёмеризацаё) складних
бёлково-полёсахаридних комплексёв. При сильному ушкодженнё тканин падвп-
щення проникност! судинно! стшки мае монофазний характер 1 зумовлене впли-
вом протешаз 1 юшюв.
При деяких патолопчних процесах (феномени Шварцмана, Артюса) ! захво-
рюваннях шфекщйно! природи (к!р, скарлатина, грип та !н.), у раз! ди силъних
ушкоджувальних фактор!в (терм!чних, юшзуючого випромшювання та ш.) разом
з ознаками щдвищено! проникност! судин у вигляд! збыьшеного виходу макромо-
лекулярних речовин можна спостерпати д!апедез еритроципв! навив мгкрокрово-
виливи. Передбачаеться, що д!апедез еритроципв у навколосудинну тканину здш-
снюеться пасивно крхзь м!жендотел!альн! промгжки пщ тиском кров!. Мгкрокро-
вовиливи е наслщком виражених структурних порушень цыосп судинно! стшки,
що пщвищують и ламгасть.
П03АСУДИНН1 ПОРУШЕННЯ М1КРОЦИРКУЛЯЦ1Т
Найважливйпими е два типи позасудинних порушень. Один з них значно впливае
на стан мжроциркуляци, слугуе додатковим патогенетичним мехашзмом Г! пору-
шень в умовах патологи. Передус!м це реакц!я тканинних базофшв сполучно!
тканини, що оточуе судини, на ушкоджувальш фактори.
При деяких патолопчних процесах (запалення, алерпчне ушкодження тканин
та !н.) 13 тканинних базофцпв у раз! 1х дегрануляци в штерстищальний проспр,
який оточуе мпсросудини, викидаються БАР 1 ферменти. Доведено важливу роль
деяких 13 них у пщвищенн! протизсщально! активност! кров! та змш! и реолопч-
них властивостей (гепарин), у явшцах вазоконстрикци (серотонш) 1 вазодилатаци
(пстамш), змш! швидкост! кровотоку та судинно! проникност! (пстамш, серо-
тонш). Як було зазначено вище, при ди пстамшу та серотоншу вщбуваеться агре-
гац!я еритроципв 1 тромбоципв.
1нший тип порушень прилегло! сполучно! тканини включае змши периваску-
лярного перем!щення !нтерстиц!ально! рщини разом з розчиненими в шй речови-
нами, утворення 1 транспорту л!мфи. Ц! порушення пов’язаш передусгм з розла-
дами ф!льтрацй рщини через нап!впроникну мембрану вщповщно до теори Стар-
лшга 1 31 змшами р!вня ФТ.
Посилення транссудаци рщини спостерпають при: зб!льшенш г!дродинам!ч-
ного тиску кров! на стшки м!кросудин (найчаспшою причиною цього е застш
кров! м!сцевого характеру або спричинений загальною недостатшстю кровооб!гу);
зменшенш онкотичного тиску кров! (основними причинами е зниження продук-
цц быкгв плазми, передусгм альбумшу, наприклад, при голодуванш, запальних 1
дистрофтчних змшах у паренх!м! печ!нки, розладах травления ! кишкового всмок-
тування); значка втрата б!лк!в при великих ошках, ентероколт, гемораги, л!мфо-
раги, а також при захворюваннях нирок запально! 1 дистроф!чно! природи. У раз!
затримки юшв натрйо в оргашзм! у зв’язку !з вторинним пперальдостерошзмом
(див. роздщ XXII “Порушення водно-електролпного обмшу”) накопичення рщи-
ни в штерстищальному простор! зумовлене не лише шдвищеною транссудащею,
а й перемпценям и !з внутрппньоюптинного простору з метою виргвнювання ос-
мотично! концентрацп електролтв. Збщьшення транссудаци быюв 1 м!жтканин-
но! рщини через спнку мЁкросудин бувае пов’язане з р!зким пщвищенням судин-
но! проникност! пщ д1ею запальних фактор!в (мембраногенний мехашзм). Особ-
Рщина 20 л - 16-18 л - 2-4 л
Емок 80 200 г-5 -75-195 г
Мал. 22. Взаемозв’язок шж фозырашею, ре-
зорбщею та шмфоутвореяням у систем) м1кро-
ииркуляцц (за Б. Фолковим 1 Е. Нелом, 1976)
ливостю останнього е виша концентрация
бйтк!в в (нтерстишальнш рщиш (1,5—
2 %) пор1вняно з нормою (0,3—1,5 %).
Як показано на мал. 22, з 20 л фшь-
тр^ту. що утворюеться з плазми за добу,
16—18 л резорбуеться у венозних кап!-
лярних супинах 1 венулах, решта фйтьтра-
ту (2—4 л) 1 практично вся маса бйпав
(80—200 г) виорачаеться на утворення
Л1мфи та повертасться в кров по ломфа-
тичнш систем! Завдяки пьому не вщбу-
ваеться накопичення рщини в тканинах.
В отловах патологи внаслщок недо-
статносп механизмов резорбци М1жтка-
нинноТ рщини в кров або зменшення ц
притоку в ломфатичн! капйояри певна час-
тина рщини затримуеться в тканинах, зу-
мовлюючи розвиток набряку (див. розд!л
XXII “Порушення водно-електролггного
обмшу”). Порушення резорбци рщини у
венознш частиш кашляра 1 посткаппяр-
Н1И венулх наичастпне е наслщком пщви
щення тканинного колощно-осмотично
го тиску 1 меншою морою зумовлене зм!
нами 1нших параметрш р!вняння Стар.
лшга. Окр1м розглянутого вище мембраногенного механизму пщвишення тканин-
ного колощно-осмотичного (онкотичносо) тиску можуть спричинити затримка
бйпав в шгерстици внаслщок порушення виведення ох у лгмфатичну систему, а
також розщеплення великих бшкових молекул на дртбниш пщ д!ею протеолотич-
них ферментов макрофагального о нейтрофшьного походження, що вивщьнюють-
ся з лейкоцитов при запаленн!.
Нареоогп. репонарш особливост! булови периваскулярно! сполучно! тканини,
розн! просторов! в!цношення мож спещал!зованими клотинами о мокросудинами
можуть значно впливати на транспорт речовин, у тому число 1 битков, через штер-
стицой. Допускають, що колагенов! волокна вщшрають роль свогрщних фобропро-
вод1в у здшененно переважного або виборкового транспорту речовин через штер-
стицой у напрямку до спецоалозованих клотин 1 л!мфатичних кап!ляр!в, з одного
боку, о до посткапйоярних венул, з шшого.
Очевидно, що в умовах патологи при ушкодженш тканин, для якого харак-
терно дезоргашзащя волокнистих структур о нерегульоване ох новоутворення. по-
рушення нормальних шляхов перикапшярного й !нтерстишального транспорту
речовин 1 ровня резорбци !нтерстицоатьно! рщини в цйюму може набуваги вира-
женшюго характеру.
Описано вище мехашзми порушення транссудацп о резорбци можтканинноо
рщини було покладено в основу сучасних уявлень про недостатшсть ломфатично!
системи та и класифшашю. Недостатшсть лЕмфатично! системи (л!мфообпу) — це
стан, при якому ламфатично судини не виконують свою основну функцию: здойс-
нення постГйного 1 ефективного дренажу штерстицпо. Вщповщно до класифЁкацп
I. Русняк, М. Фелд! 1Д. Сабо (1957) розр!зняють так! форми недостатносп лшфо-
обйу:
1) мехашчна недостатшсть, за яко! рух л!мфи утруднений у зв’язку з наявшс-
тю оргашчних (здавлення пухлиною, рубцем, екстирпащя шмфатичних вузл!в 1
судин, обл!терац!я лшфатичних судин при !х запаленш, тромбоз! та ш.) або фун-
кщональних (пщвищення тиску в мапстральних венозних судинах, спазм л!мфа-
тичних судин, недостатшсть клапан!в л!мфатичних судин, припинення м’язових
скорочень — “акшетична недостатшсть”) причин;
2) диналйчна недостатшсть: об’ем транссудацп м!жтканинно! рщини переви-
щуе можливгсть л!мфатично! системи забезпечувати ефективний дренаж штер-
стищально! тканини;
3) резорбцшна недостатшсть, зумовлена структурними змшами основно! ре-
човини, накопиченням б!лк!в ! вщкладенням !хшх патолопчних вид!в в штерсти-
щальшй тканин! та ш.
Основними клтшчними та патофгзюлопчними проявами недостатносп л!м-
фообпу в гостр!й стада е набряк, накопичення бнтктв 1 продукпв !х розпаду в ш-
терстшцальшй тканин! 1 наступний розвиток ф!брозу 1 склерозу — у хрошчнш
стадп.
218
Роздал XI
ЗАПАЛЕННЯ
Запалення — це пгиповий патолог/чний процес, що виникае в органах / тканинах у
вгдпов/Оъ на Тх ушкодження I слугу с для Тзоляци, знешкодження та видалення як па-
толог/ чного фактора, так I некротизованих клипин / тканин, з подальшим замйцен-
ням сполучною тканиною дефекту, що утворився.
Це переважно мкщевий процес, основне завдання якого — створити бар’ер
м!ж ушкоджувальним фактором разом з медиаторами западения, що утворюються.
та загальним кровообпом. Порушення цього бар’ера може спричинити сетич-
ний або асептичний шок.
Мюцевий характер запального процесу було тдкреслено шотландським хирур-
гом Д. Хантером ще в 1793 р. Цей “класик’’ хгрурп! цьлком слушно стверджував,
що запалення — не хвороба, а неспецифгчна захисна реакция, яка може бути тшь-
ки “тЮ /Не огран”, а не ‘‘о/ап ог%ап”, тобто неможливе запалення всього органа.
Тим бглыпе, “не генуе запалення всього оргашзму’ (Ф.Ш. Зайчик, Л.П. Чурилов,
2001).
М1сцев1 ознаки запалення:
— почервоншня (гиЬог);
— збшьшення м1сцево1 температури (са!ог);
— виникнення болю (сЫог);
— прилухлгеть (Штог);
— порушення функцп органа (Шпсйо 1аеза).
Вважаютъ, шо перни чотири кардиналый ознаки було описано римським
письменником 1 науковцем К. Цельсом (30 р. до н. е. — 38 р. н. е ), а остання
п'ята — римським л жаре м К. Галеном (130—200 рр. н. е.; мал. 23).
За нормального перебпу процесу !з запального вогнища у кров потрапляе незнач-
на частина медаатортв, що проявляеться такими системными реакциями, як нейгроф!-
льоз, гарячка, збщьшення швидкосп осщання еритроципв (ШОЕ), синтез печшкою
гострофазових б!лк!в. При цьому реакщя органгзму на мгспеве ушкодження не спри-
чинюе значит системи! порушення, осюльки дгя мед!атор!ь запалення на системному
р!вш блокуеться п Цви щепою продукщею глюко кортикоццв ! адреналшу.
При великих, множинних, поширених та ппенсивних запальних процесах
загалып прояви запалення можуть бути досить вираженими 1 супроводжуватися
виснаженням оргашзму. що дае пщетави називати таке явише синдромом систем-
но! дй медтатортв запалення, або синдромом системно! запально! вцповЫ (ССЗВ,
зузгегше шЛаттаюгу гекропье зупбготе, 81К8), але не “системним запаленням”.
У ньому випадку формуеться шший патолопчний процес — шок.
Запалення е фьюгенетично давшм процесом, який упродовж еволюци уск-
ладнювався одночасно з ускладненням оргашзаш! живих систем, 1 у виших ссав-
Мал. 23. Кардиналып ознаки запалення за Цельсом I Галеном
щв 1 людини набув ознак багатогранного та комплексного явища. Однак завжди
можна виявити дв1 складов! патолопчного процесу: перша — це екзогенне й ен-
догенне ушкодження юптин, тканин або оргашв, тобто власне патолог1чне, друга —
мобшзацгя захисних мехашзмгв, спрямованих на боротьбу як з патогенним фак-
тором, так г з руйнгвними наслщками захисних реакцш, тобто ф1зюлог1чне.
Запалення е одним !з найпоширешших патолопчних процесгв. Патолопчним
пщгрунтям понад 80 % усгх хвороб, з якими доводиться стикатися лгкарям ртзних
спещальностей, е запалення. Тому дуже важливо, щоб знания, досвщ г вмшня
лжаря дали йому змогу розшзнати серед ргзномаштних проявгв, яю спостергга-
ються пщ час запалення, власне патолопчне та вщргзнити його вщ захисного, щоб
вщповщним чином 1 своечасно втрутитися в перебгг хвороби.
ЕКСПЕРИМЕНТАЛЬНЕ В1ДТВОРЕННЯ ЗАПАЛЕННЯ
Запалення е важливою проблемою 1 предметом вивчення вс!х галузей медицини.
Вщмшшсть полягае лише в методах дослщження. Наприклад, лгкар терапевт
спостерггае переб!г запалення легень (пневмошя) бщя лгжка хворого, патолого-
анатом — пщ час розтину трупа, а патофгзюлог — в експерименп на тваринах.
Учень Р. Вгрхова Ю. Конгейм (1867) уперше вивчив в експеримент! на жаб!
кровообгг у брижг пщ час запалення, встановивши вег його стад!! вщ гшеремп до
стазу. Вш також описав процес емгграцй лейкоцит кргзь судинну стшку. Його
експериментапьну модель широко використовують! ниш на практичних заняттях
зт студентами (дослщ Конгейма) та в наукових дослщженнях.
Подальше вивчення рол! судинно! реакцп на запалення було здшснено Е.Л. Клар-
ком х Е.Р. Кларком (на вусх кроля, 1920), Г. Селье (на защхчних милках хом’яка),
О.М. Чернухом (на бриж! щурхв, 1975); В. Менкш у 1948 р. визначив бхоххмхчш
змхни у ВОГНИЩ1 запалення.
Особливе значения для розумшня бхолопчнох ролх запалення мали експери-
менти 1.1. Мечникова на личишц морсько! зхрки (1883). У цих дослщженнях, а
також пщ час подальшого вивчення запалення у тварин, як! перебувають на рхз-
них рхвнях еволюцшного розвитку, було доведено роль фагоцитгв як одного з
основних елементгв захиснох реакцп оргашзму у вщповщь на вторгнення флого-
генних запальних факторхв На основ! цих спостережень було створено теорхю
запалення, яка увшшла в науку пщ назвою порхвняльно-патолопчнох, або еволю-
цгйно!. Згодом патологи стали широко використовувати еволюцшний принцип в
експериментальному моделюваннх, виходячи з того, що патолопчш прояви в ни-
жчих тварин, являючи собою умови найпростпш та первхсш, вщкривають шлях до
розумшня складних патолопчних процесгв.
Вщтод! отримано багато нових даних, яю дають змогу глибше зрозумхти про-
цеси, що вщбуваються п!д час виникнення та розвитку запалення. Майже щороку
з’являються нов! факти, яю уточнюють 1 доповнюють цю складну й рхзноманггну
картину.
ЕТЮЛОПЯ
Фактори, що спричинюють ушкодження та запалення, називають флогогенними
(вщ грец. (р/.ос, — полум’я х уеууаа> — народжуючий). Такими можуть бути будь-
яю фактори, яю за своею силою х тривалхстю дп переважають адаптащйш мож-
ливостх тканин ! можуть спричинити тх ушкодження. Ус! флогогешп фактори
прийнято подшяти на зовхшшп (екзогенш) та внутрхшш (ендогенн!). До зовнпп-
н1х належать бюлопчш фактори: мгкроорганхзми (бактерп, вхруси, найпростпш,
сшрохети, рикетсп, гриби), тваринш оргаюзми (черви, клпщ, комахи), ендо- й
екзотоксини (отрута зм!й ! комах); ххмтчнх речовини (кислоти, дуги, бойов! отруй-
н! речовини, ксенобютики тощо); фхзичш фактори (мехашчн!, термхчш, елект-
ричш впливи, хошзуюче та ультрафюлетове випромпповання). Причиною ушко-
дження юптин може бути х г!покс!я, яка через бхлок Н1Р-1а хндукуе запальну
вщповщь.
До внутрхшнхх факторхв запалення належать тх, що виникають у самому ор-
гашзм! внаслщок шшого захворювання. Наприклад, запалення може розвинутися
як реакщя на пухлину, жовчш або сечов! камент, тромб, що утворився в судинах.
Причиною запалення можуть стати комплекси антиген — антитхло — компле-
мент, якщо вони фхксуються в якомусь орган!.
ПАТОГЕНЕЗ
Мехашзми запалення включають альтерацпо, вироблення, вивхльнення й активацпо
БАР — медхаторхв запалення, змши мхсцевого кровообпу, пщвищення проникносп
стшки судин, ексудацпо, емхграцпо лейкоцитхв, пролхферацпо та репарацию.
Основн! компонента, що беруть участь у запальному процес!:
Дгя ----Первинна (Г) та втпринна (1Г) альтерап!я
— кровоносн! судини (вщсутнтсть судин у ропвщ, склистому пл!. клапанах
серия, хрящ! уиеможливлюе виникнення там запалення доти, доки туди не
вростуть судини);
— лейкоиити;
— медтатори.
Класичш стали запалення
]. Альтеращя:
а) первинна — спричинюеться ушкоджуватьним фактором;
б) вгоринна — зумовлюеться медтаторами запалення.
II. Ексудаи1я та ем)грац1я.
III. Про.йферацт / репарация.
Слад зазначиги, що щ стади запалення, прийнятт в класичшй патологи, с
дещо спрощеними, осюльки ексудащя та емпрашя починаються в перш! хвилини
первинно! альтерацй, а вторинна альтеращя набирас потужност! лише шсля про-
никнення у вогнище запалення лейкоципв. Кртм того, пщ час запалення одно-
часно з альтеращею, ексудашею та емтграшею змшюються судиннт реакцп мткро-
циркуляторного русла т посладовно виникають т зникають про- ! протизапальн!
медтатори (мал. 24).
Повну послщовшсть розвитку оеновних подш при запаленн! наведено на схем! 6:
• первинна альтеращя, тобто ушкодження тканин, у першу черту мембран
юптин, при ди на них флогогенного агента, з одночасним викидом мед!атор!в
западения (метабол!ти та БАР) ушкоцженими клттинами. макрофагами ! тканин-
ними базофтлами;
• коротко1 тасний спазм артерюл, актива цтя глтколтзу, розвиток внутршшьо- т
мтжклггинного мтсцевого ацидозу, активащя лтзосомних ферменттв — початок вто-
ринно! атьтераци; утворення нових медтатортв запалення, артертатьна пперемтя;
• подалыиий розвиток порушень мткроциркуляип (венозна пперемтя, стаз),
пщвищення проникносп судинно! стшки. ексудацтя та емтграшя нейтрофшв;
• иосилення вториннот альтераип за рахунок руйнування клттин ! макромоле-
кул актив ни ми кисневими радикатами, лтзосомними ферментами, каттонними
боками, лактоферином ! лтзоцимом, що виробляються нейлрофшами;
Схема 6. Патогенез запалення. Послщовшсть змш, що вщ-
буваються у вогнищ! запалення
• розвиток пперонки у вогнищ! запалення з посиленням ексудацп та нор-
малгзащею рН;
• ем1грац1я моноципв-макрофапв, а потш 1 лшфоцитзв; очищения вогнища
запалення, нейтрал!защя 1 знешкодження БАР, що стимулюють запальний про-
цес; посилення вироблення протизапальних мед1атор1в;
• пролгферацгя фхбробластгв, ендотелзальних, гладком’язових, епщермальних
клггин унаслщок зменшення кейлонхв, усунення контактного галъмування, збщь-
шення продукцп фактор1в росту макрофагами;
• зампцення дефекту тканин (репарация) шляхом регенерацп 1 ф1броплазц.
Альтерацёя
Первична альтеращя. Запалення завжди починаеться з ушкодження тканини. Пщ
дгею ушкоджувального фактора в гептанах виникають оборотит та необоротнг
функщональш, метаболтчш г структури! змши, прояви яких залежать в!д характе-
ру ! сили ушкодження, виду клгтин, ступени ёхньот зр!лост! тощо. За власними
закономерностями розвиваеться первинне ушкодження позаклттинних структур у
вогнищ! запалення.
Отже, унаслщок первинно! альтерацп в тканин! з’являються продукта ушкод-
ження та руйнування клггин ё мёжклётинних структур. Це продукта первинно!'
альтерацп, як! активують наявнё БАР та утворення нових.
Вторинна альтеращя. Важливе значения в розвитку вторинно! альтерацп мае
активацёя лёзосомних ферментёв усёх клётин в умовах мёсцевого ацидозу, який е
наслщком активацн глёколёзу. Проте головними ефекгорами вторинно! альтерацп
е лёзосомнё фермента й активнё кисневё радикали (АКР) лейкоцитгв, якё емёгру-
ють у вогнище запалення. Вони руйнують збудника запалення бюлопчно! приро-
ди та продукта первинно!' альтерацп. Саме в цьому полягае !х захисне значения.
Проте лёзосомнё фермента та АКР руйнують ё неушкодженё клгтани та структури
власного оргашзму. Тобто первинна альтерацёя — це ушкодження, завдане ззовнё,
а вторинна альтерацёя — це самоушкодження.
Ушкодження клётин передусём спричинюе порушення структури та функцп
клётинно! мембрани, мётохондрёй ё структур, що стосуються синтезу та упакову-
вання протегнёв ё ДНК.
Молекулярш мехашзми альтерацп клётин. Ушкодження клётинно!' мембрани
зумовлюе вихёд ёз клётин ёонёв калёю ё надходження в них ёонёв натрпо ё кальцпо,
що вщбуваеться за градёентом гх концентрацё!. При цьому одна молекула К+ гёд-
ратуеться 10 молекулами води, а одна молекула Ка+— 14 молекулами води. Збёль-
шення концентрацп Иа+ у клгтинах зумовлюе накопичення внутрёшньоклётинно!
води, що призводить до !х набухання. Крём того, в1лыа 1они кальцт активують
мембрашй фосфолтази та цитозолый проте тази, що руйнуе мембрани, ферменти
та структури! быки.
Викачування ёонёв натргю ё кальцёю ёз клгтин з одночасним закачуванням
ёонёв калгю залежить вщ роботи №+-, КГ-АТФази ! Са2+-АТФази, що потребують
енергп АТФ, синтез яко! забезпечуеться окисним фосфорилюванням у мгтохонд-
ргях або анаеробним глгколёзом. У разг ушкодження мгтохондргй порушуеться мг-
тохондргальний синтез АТФ, який залежить вёд створення в процесг аеробного
мгтохондргальго окиснення високого градёента концентрацп гонгв водню в мгжмем-
бранному простор! мгтохондргй. Цей градгент е рушгйною силою активацн АТФ-
синтетази при перемгщенш через не!' у матрикс мгтохондргй гонгв водню. Дёя
ушкоджувальних факторгв на мембрани мгтохондргй спричинюе формування ви-
сокопровгдних регульованих бшкових каналгв^ якг називають м!тохондр1алъними
порами (пШосИопбпа! реппеаЬййу (гапзпгоп роге). Вгдкриття цих каналгв зумовлюе
вихёд Н+ гз мгжмембранного простору повз АТФ-синтетази, втрату водневого
градгента, роз’еднання окиснення та фосфорилювання г пригнгчення синтезу
АТФ у мгтохондрёях. Унаслщок цього активуеться гл!кол1з, який повшстю не за-
безпечуе потребу клгтани в АТФ. але сприяе накопиченню молочно!' кислота в
цитозоль Розвиваеться мюцевий ацидоз, ёнактивуеться при зменшенн! рН бгль-
штсть юптинних ферменттв, але активуються г1дролтичн1 ферменты, як! виттка-
ють 13 Л1зосом при ушкодженш !хн!х мембран, що спричинюе автолиз 1 некроз
юптин.
Порушення Ц1ЛОСТ1 юйтинно! мембрани, зниження активност! Са2+-АТФази й
ушкодження мггохондрш т ендоплазматичного ретикулума (ЕР), яю е внутрппньо-
юптинними депо кальцпо, спричинюють значне збглыиення концентраци виьного
Са2+у цитозолг клтин. Це змушуе витрачати енергпо мттохондртального окиснен-
ня на транспорт кальцпо, а не на створення градтента Н+, що також порушуе
поеднання окиснення т фосфорилювання та, як наслщок, ще бшьше зменшуе
утворення АТФ.
Надлишок цитозольного Са2+ активуе ензими: мембрани! фосфолшази, що
спричинюе ушкодження мембран т збшьшення продукцп ейкозанощтв; протеша-
зи, у тому числ!1 каспази, яю руйнують бшки цитоскелета, мембран 1 ядра; ендо-
нуклеази, яю фрагментують ДНК 1 запускають апоптоз. Кр!м того, вщкриття “м!-
тохондртально! пори” зумовлюе вихщ !з мггохондрш у цитозоль цитохрому С т
деяких шших мггохондртальних бтлюв, яю разом з цитоплазматичними битками,
зокрема активуючим апоптоз фактором 1 (арорФзтз аейуайоп ГасГог 1, АРАЕ-1),
активують каспазу-9. Це через активацпо шших прокаспаз призводить зрештою
до розщеплення ДНК ендонуклеазами й апоптотично! загибелт юптини.
Тривалий дефщит кисню, глюкози й АТФ спричинюе вщшарування рибосом
в!д ЕР з порушенням синтезу протештв. Бшки при цьому можуть неправильно
упаковуватися, що ушкоджуе юптину (ипГоМеб рго1еш гекропзе) т може спричини-
ти п загибелъ.
АКР продукуються в нормальних юптинах п!д час мттохондртального дихання
! знешкоджуються компонентами антиоксидантно!' системи. Коли тх продукшя
значно збшьшуеться при радтацтйних т деяких х!мтчних ураженнях або внаслщок
недостатньо! активност! антиоксидантних систем в умовах дефщиту АТФ при тше-
мп та ушкодженш, а також при синдром! штемп-реперфузп, вщбуваеться накопи-
чення втльних радикалтв т реалтзуеться тх руйшвний вплив на юптини. Особливо
велик! юлькосп найагресивнппих АКР утворюються активованими при фагоци-
тоз! нейтрофтлами т макрофагами.
При фагоцитоз! вщбуваються швидке формування в мембран! фагосоми бага-
токомпонентно! НАДФ-Н-оксидази тз мембранних 1 цитозольн их б!люв та п ак-
тивацтя. Активована об’ектом фагоцитозу фагоцитарна оксидаза переносить елек-
трони на кисень, вщбираючи тх у вщновлених молекул. Такими молекулами е
насамперед НАДФ-Н, яю окиснюються до НАДФ, а дефтцит НАДФ-Н компен-
суеться за рахунок окиснення глюкози в пентозному шунт!. При цьому О2 вщнов-
люеться до першого з АКР — супероксид-ашона (‘О2”). Процес утворення АКР
збтлыпуе споживання лейкоцитами кисню майже в 10 разтв. Цей процес отримав
назву “ресшраторний вибух”.
За рахунок спонтанно! дисмутацп супероксид-анюни радикали перетворю-
ються на пероксид водню (Н2О2), який не справляе значну юлерну дпо. Проте за
наявностт вщновлених форм металтв з перемшною валентшетю, наприклад Ре2+
або Сп2+, пероксид водню перетворюеться на дуже агресивний гщроксилышй ра-
дикал (реакцтя Фентона: Н2О2 + Ре2+ —> Ре3+ + 'ОН + ОН-). Причому вщновлення
окиснених форм залтза чи мщ! до Ре2+ / Сп2+ прискорюеться за наявност! суперок-
сид-ашошв радикалтв.
Кргм того, нейтрофши мгстять фермент миелопероксидазу, яка за наявностг
галощгв, наприклад СГ, перетворюе пероксид водню на гшохлорит (Н2О2 + СГ —>
ОСГ + Н2О), що мае дуже потужну бактерицидну щю. Агресившсть гшохлориту,
який руйнуе сульфпдрильш групи мгкробних ферментгв г пептидш елементи клг-
тинних мембран, настшьки велика, що при його утвореннг вщбуваеться загибель
1 самого нейтрофиа.
Оксид азоту, що утворюеться у великш кглькостг в активованих макрофагах за
допомогою 1МО8 г мае помгрну бактерицидну дпо, пщ впливом супероксид-ашона
перетворюеться на високоактивний вгльний радикал — пероксингтрит (О1ЧОО).
Усг щ високоактивш сполуки зумовлюють пероксидне окиснення лшщгв, бш-
кгв, нуклегнових кислот г ушкоджують вщповщш структури мгкрооргангзмгв г влас-
них клгтан, побудоваш гз цих речовин. Дуже легко окиснюються ненасичею жирш
кислота, яю входять до складу клгтинних мембран. Пщ впливом АКР лшщи (ЬН)
утворюють лшцш радикал и (Ь’), якг за наявностг кисню перетворюються на лйто-
пероксиди (ЬО2’), пдропероксиди лшщгв (ЬООН), новг лгпщю радикали (Ь’), а за
наявностг Ре2+ або Си2+ — на АКР лшщгв (Ь0‘). Тобто ця реакцгя набувае розгалу-
женого ланцюгового характеру. У такгй ситуагш фагоцит г сам зазнае агресивного
впливу зазначених речовин. Нейтрофши при цьому гинуть, але моноцита-макрофа-
ги виживаютй, тому що мають потужний антиоксидантний мехашзм, завдяки якому
не вщбуваеться надмгрне накопичення активних форм кисню поза л!зофагосомою.
Опосередковане лейкоцитами ушкодження нормальних клтгин та мгжклггин-
ного матриксу, яке становить основу вторинно! альтерацп, може значно переви-
щувати ушкодження тканин, спричинене флогогенним фактором. Мехашзми
вторинно! альтерацп створювались у фшогенезг для знешкодження шфекцшних
факторов. Проте бшышсть гз цих мехашзмгв не е селективними, а тому вони час-
то зумовлюють значне руйнування власних тканин, навгть тодг, коли первинне
ушкодження було асептичним.
Клггини запалення. Клгтини, функцгг яких активуються пщ час запалення,
роздшяють на двг групи за мгсцем гх перебування в тканинах: перша — дендритш
клгтини, макрофаги й лаброцити, що мгстяться в тканинах постгйно (клгтини-ре-
зиденти), друга — гншг клгтини, що надходять у зону запалення ззовнг (клгтани-
емггранти). До них належать гранулоцита (нейтрофгльш, еозинофгльш), моноци-
та г лгмфоцити.
Дендритш к/шпини разом з моноцитами-макрофагами належать до АПК, яю
здатш презентувата Т-хелперам на свогй цитоплазматичшй мембранг чужорщнг
процесованг антигени разом з молекулами ГКГС II класу. Завдяки наявностг роз-
галужених вгдростюв вони утворюють важливу систему захисту в поверхневих
структурах оргашзму (шкгрг та слизовгй оболошц) та в органах (лгмфатичш вузли,
селезгнка, легеш), розташованих на шляху найбгльш гмовгрного проникнення
флогогенгв. Кргм того, активованг АПК продукують специфгчнг си гнали — “за-
грози”. До ще! групи належать прошфламаторн! цитокши, таю як 1Л-1, 1Л-6,
ФНП-а, -0 та у-1Ф, ргзш хемокгни та ш. Роль 1Л-1 особливо важлива. Пщ його
впливом Т-лгмфоцити-хелпери 1-го типу в момент презентацп антигену почина-
ють продукувати 1Л-2, який активуе пролгферашю г диференщацпо Т-лгмфоцитгв-
кглергв та активуе функщю моноципв-макрофаггв 1 натуральних кглергв.
Макрофаги. “Класично активованг” на початку запального процесу гнфекцгй-
ними факторами г Т-хелперами 1-го типу макрофаги синтезують 1Л-1. Вш знахо-
5К
Мжробний
токсин
Мал. 25. 1Л-1 та його мшгеш. У центр! ма-
люнка розм!щено макрофаг, який шсля
активаш! (наприклад мгкробним токси-
ном) починае виробляти 1Л-1. Останнш
впливае на сво! мтшен!, якими е юптини
юсткового мозку, печгнки, ппоталамуса,
лгмфоцити, ф!бробласти тощо
дить свог мгшен!, якими е юптини
м’язово!, юстково!, нервово! тканин,
синовгоцити, гепатоцита, лгмфоци-
ти. На мембранах цих клгтин е спе-
цифгчнг рецептори дня 1Л-1. Дгя 1Л-1
стимулююча й универсальна, тобто
проявляеться при будь-якгй хвороб!
запальног природи (шфекцгйного чи
негнфекцгйного походження), при-
чому на рангах п етапах. € пгдстави
вважата, що ранга симптоми хворо-
би (головний бгль, б!ль у м’язах г
суглобах, сонливгсть, гарячка, лей-
коцитоз, збщьшення вмгсту бглкгв, у
тому числг й гмуноглобулгнгв) пояс-
нюються саме дгею 1Л-1 (мал. 25).
Роль зазначених макрофапв не обмежуеться секрецгею 1Л-1. У цих клгтинах
синтезуються понад 100 бгологгчно активних прозапалъних цитокшгв, фосфол! -
паз, протешаз, лгзосомних пдролаз, факторгв комплементу, ейкозаногдгв, актав-
них кисневих радикалгв, у тому числг МО, тощо.
Основна функцгя цих макрофапв — фагоцитоз власних клгтин, гнфгкованих
бактергями, вгрусами, простгшими, грибами, та загиблих клгтин, у тому числг ней-
трофглгв.
“Альтернативно активоваш” Т-хелперами 2-го тапу макрофаги синтезують
протазапальнг цитокгни, гнггбгтори протешаз, антифосфолшази, антиоксиданта,
гнпбгтори комплементу, бгогенних амгнгв, кгнгнгв, фгбринолгзу, фактори росту та
компонента позаклгтинного матриксу. Така змша функцп моноцитгв-макрофапв
вщбуваеться лише пгсля знешкодження флогогенного фактора, його перетравлю-
вання г перетворення антигену на розчинну форму.
Опасистгй клйпини (тканинш базофгли, мастоцити, лаброцити, тучнг клгтани)
шсля ушкодження викидають пстамш, гепарин г лейкотрген Г),. фактори хемо-
таксису нейтрофглгв (ФХН) г еозинофглгв (ФХЕ), фактор активацп тромбоцитгв
(ФАТ), що мгстяться в гхнгх гранулах. Дгя пстамгну та гепарину позначаеться пе-
редусгм на судинах (гшеремгя). Лейкотрген Э4 е хемоатрактантом, а також спри-
чинюе спазм непосмугованих м’язгв бронхгол г кишечнику.
Нейтрофглый гранулоцити. Основна функцгя цих клгтин — фагоцитоз грамне-
гативних бактерш г деяких грибгв (гноетвгрна мгкрофлора). Подранивши у кров гз
кгсткового мозку, вони пщ впливом хемоатрактантгв мггрують гз судин г у великгй
кглъкостг скупчуються у вогнищг запалення. Розмноження клгтин, мгграцгя, участь
у фагоцитоз! регулюються БАР, для яких на поверхневш мембран! нейтроф!л!в
оснують специфично рецептори.
У цитоплазм! нейтрофыьних гранулоцит е два типи гранул: первинн! азуро-
ф!льн! (быып! за розм!рами) — звичайн! лозосоми, у яких мгстяться кисл! пдрола-
зи, Л1зоцим, миелопероксидаза. Вторинн! (специфочн!) гранули дробный, а голо-
вне — вони мостять 1нший наб!р речовин (ферментних 1 неферментних), лужну
фосфатазу, л!зоцим о неферментн! (катюнш) быки. Гл!копроте'щ лактоферин е
хелатором зал!за (зв’язуе два моля залоза на один моль б!лка), що вщбирае важли-
ве для життедгяльност! залозо у бактерш. Водночас зниження завдяки лактофери-
ну коошентрацп Ре2+ зменшуе утворення пдроксильного радикала в реакцп Фен-
тона, що зменшуе ушкодження власних тканин. Найпотужношим бактерицидним
фактором нейтрофыгв е активол киснев! радикали, особливо гшохлорит.
Еозиноф1льн1 гранулоциты. 1х роль у запаленн! визначаеться здатшстю адсор-
буватио руйнувати выьний пстамш за допомогою пдролггичного ензиму пстамш-
ази та знешкоджувати лейкотроени арилсульфатазою-В. Антихомотрипсин-а, !нп-
буе серинов! протешази. Важливу роль вщгграють еозинофыьш цитотоксичш бы-
ки (основний быок еозинофшв о катоонний быок еозинофшв), як! здатн!
ушкоджувати гельмшти та охш личинки о нейтралозувати гепарин. Мограцоя й ак-
тивацоя еозинофшьних гранулоцогтов вщбуваються пщ д!ею фрагмент компле-
менту (С5а о С5, С6, С7), пептидов тканинних базофщьних гранулоцит, лей-
котроенов о речовин, як! виробляються гельмштами.
Тромбоциты. Роль тромбоцит у процес! запалення полягае здебыьшого в
тому, що вони беруть безпосередню участь у процесах мгкроциркуляцп. Очевид-
но, це найбыып постойно й уноверсальш клггини запалення. У них мостяться речо-
вини, як! впливають на проникшсть стшки судин, охнш тонус та адгезивно влас-
тивост!, на р!ст о розмноження клпин, а головне — на здатшсть кров! зсщатися.
Лимфоциты беруть участь у будь-якому запаленн!, особливо омунному. 1мунна
вщповщь, як о ефекторна фаза омунних реакцш, регулюеться розчинними медоа-
торами (штерлейкшами). Основними продуцентами штерлейкшов е Т-ломфоци-
ти, моноцити о тканини! макрофаги.
Ф^бробласти прол!ферують о активуються в останнш стадп запалення, нако-
пичуючись у вогнищо запалення о збыьшуючи синтез колагену, еластину та глокоз-
амшоппканов. Узагальнеш дан! про юптини запалення наведено в табл. 10.
Таблиця 10. Клпини запалення
Клггини Синтезован! речовини Участь у запаленн!
Дендритш юйти- ни 1Л-1,1Л-6,1Л-18, ФНП-а о -0, у-1Ф, хемокши тощо Презентують антигени. Активують адгезгю лейкоцитов. Блокують дно ворусов. При сис- темной дп спричинюють гарячку, сонливость, анорекаю, продукщю гепатоцитами быков гостроо фази
“Класично акги- воваш” макрофа- ги 1Л-1,1Л-6,1Л-8, ФНП-а, ФНП-0, ФАТ, АКР, МО, а4Ф, фосфолшази, компонента комплементу, протешази, ад- гезивш быки тощо Презентують антигени. Стимулюють могра- цпо о хемотаксис лейкоцитов. Фагоцитують клггини, заражено ворусами, грибами, бак- тероями, та мертво клггини, у тому число ней- трофши. Блокують дою ворусов. Забезпечують поеднання з оншими клотинами запалення
Зактчення табл. 10
Клгтини Синтезован! речовини Участь у запаленн)
“Альтернативно активоваш” мак- рофаги 1Л-4,1Л-10,1Л-13, шпбпюри протешаз (а2-макроптобулш та ш.), полтамши, хондроТ- тин-сульфат, антиоксиданта, антифосфолшази, фактори росту, гепарин, колаген, фтбронектин Пригшчують продукцию прозапальних ци- токнив, 1нпбують актившсть кшппв, комп- лементу, протеТназ, бюгенних ампйв, АКР, факторТв коагуляцн, ф1бринолтзу. Зменшу- ють проникшсть судинноТ стшки т гальму- ють ексудацпо та емпрацно. Очищують вог- нище запалення вщ юптинного детриту. Ак- тивують фибробласта 1 стамулюють пролТферацпо , репарацию
Лаброцити Пстамш, гепарин, ФХН, ФХЕ, ФАТ, повыьно реагую- ча субстанщя анафыаксй (лейкотртени) Спричинюють артер^альну та венозну гше- ремпо. Збыьшують проникшсть судинноТ стшки й ексудащю. Активують хемотаксис 1 емпращю лейкоциты
Нейтрофыып гра- нулоцита Лтзосомн! ферменти, катюнш быки, АКР, ФАТ, лтзоцим, лактоферин, ейкозано'щи Фагоцитоз гноствТрноТ мйсрофлори. Потужна бактерицидна 1 цитотоксична Д1я
Еозинофыьш гра- нулоцитй Пстамшаза, арилсульфатаза, сериновт протешази, еозино- фыьнт ЦИТОТОКСИЧШ быки Руйнування пстамшу, лейкотрТешв та шших мед1атор!в запалення. Знешкодження гель- мпгпв та Тхшх личинок
Тромбоцита Тромбоксани, лейкотр!ени, простагландини, серотонш, адреналш, тромбоцитний фактор росту Адгезия й агрегащя тромбоципв, активация зсщання кровь Стимуляция прол^ферацп та репарацн
Ллмфоцити: Тх1 Тх2 В у-1Ф, 1Л-2, ФНП-а 1Л-4,1Л-5,1Л-6,1Л-10,Ш-13 Тмуноглобулши Активащя макрофапв, Тк 1 НК, реакцп кль тинного хмуштету. Блокада Тх2 Активащя В-Л1мфоцит1в, пригшчення Тх1 Реакцп гуморального 1мунггету
Фтбробласти Колаген, фибронектин, ппкоз- амшоглткани, фактори росту Створення М1ЖКЛ1ТИНН0Г0 сполучнотканин- ного матриксу
Медаатори запалення — це БАР, яю е центральною ланкою патогенезу запа-
лення, осюльки вони справляють безпосереднш регуляторний вплив на запаль-
ний процес, як правило, у межах вогнища запалення. Розрззняють гуморальш та
кл1тинн1 мед1атори запалення.
Гуморальш медтатори запалення становлять систему проте'Гназ плазми кров1, яю
синтезуються переважно в макрофагах 1 гепатоцитах 1 потрапляють у кров у неактив-
ному стань Кл1тинн1 мед1атори або синтезуються заново при запаленш в активному
стаю (ейкозано'щи 1 АКР), або депонуються в гранулах 1 везикулах юптин з подаль-
шими секрещею 1 активащею. Основною причиною утворення 1 / або вивыьнення
мед1атор1в юптинного походження е альтеращя. Саме внаслщок ушкодження клттин
видыяються й активуються лтзосомш ферменти, яю своею чертою активують ппш
ферменти, у тому числт й протепгази плазми кровь При цьому протеолпичш фер-
менти розщеплюють быки не до юнця (обмежений протеолтз), унаслщок чого утво-
рюються специфтчш активш речовини, яю дають певний патофтзюлопчний ефект.
З’ясувалося, що одш з них дпоть переважно на судини, посилюючи проникшсть
тхньот стшки, ппш — на мтграцтю лейкоциттв, а деяю — на розмноження клттин.
Першим, хто побачив у запаленнт певний “порядок”, закономтрнкть, був
В. Менкш (1948) У запальному ексудат, взн выявив х!м1чн1 речовини 1 поржняв !х
з деякими характерними для запалення реакщями: гшеремтею, ексудащею, лей-
коцитозом, хемотаксисом. Вйггол! у фактичному магерхал! багато шо змшилося,
однак цей напрям (х!м1я запалення) ще юнуе. У табл. 11 наведено найважлившй
медгатори запалення (юптинного та гуморального походження).
Таблиця 11 МедЕатори запалення
Медзтори Д1я Основне походження
Клгтинш
Бюгенн! амти
Пстамш Мюцеве розширення артерюл через 180. Пщвищення проникност! стшки судин. СвербЬк 1 быв У великш кон- центрацп звужуе венули Гранули лаброципв 1 базофшьвпх грануло- ЦИТ1В
Серотонш Розширення артерюл через ЫО та зву- ження великих судин. Активашя адгезп й агрегащ! тромбоцит! в 1 моноципв Тромбоцита, хрома- фшЩ КЛ1ТИНИ слизово’Г оболонки травного канату
Пол1ам1ни: спермш, спермь дин, кадаверин, путресцин Зменшення проникност! судин. С'тиму- ляц1я прол1ферацп та репарацп УС! КЛ1ТИНИ
Адреналш, норадрензлш Короткочасний спазм артерюл 1 змен- шення проникност! судин, активашя агрегаш! тромбоцитЕв Тромбоцита
Фактори вторично! альтерацп
Пдролггичш ферменти лизо- сом Псретравлення в кислому ссредовипп вбитих бактерш 1 клиин Руйнування колагену, еластину. пикетам Еноппкашв. Активацгя ганшовоУ системи Гранулоцити, макро- фаги
Катюш п бики’ дефензнни, катепсин О, азуроцидин А Вбудов}вання в мембрану бактерш 1 власних клттин 13 формуванням юнних каналгв, Руйнування кл!гин. Еластолтз 1 пщвищення проникност! стшки судин Активашя комплементу Вейтроф1льн1 грануло- цити, макрофаги
Еозинофптьш цитотоксичгп бщки (головний ОСНОВНПИ 61- лок еозиноф1Л1в Е катюнний бпюк еозинофйнв) Ушкодження зовшшньо! оболонки га за1ибель гельмЕнпв та шпх личинок Еозинофыи
Мембраноагакуючий комп- лекс С5Ь — С9 комплемент} Утворення пори в мембран! клпин-м1- шеней. шо зумовлюе й лтзис та / або апоптоз Сироватков! бшки, що утворюються макро- фагами та гепатоцита- ми
Активш киснев! радикали (АКР: ‘О", 'ОН, Н2О2, ОСТ) Внутр1шньокл1тинне 1 позаклггинне знешкодження бактерш 1 паразит1в та ушкодження власних кл1тин —— —< — Нейтрофыи. макрофа- ги
Продовження табл. 11
Мед!атори Д!я Основне походження
Оксид азоту, пероксиштрит Дёють як ёншё АКР Макрофаги
Лт1дн1 продукта розщеплення арюадоново'! кислоту (ейкозаногди)
ПГЕ , ПГ Е„ ПГА , ШТ, 1’ 2’ 2’ 2а Розширення судин ё посилення про- никностё гхньоГ спнки, набряк. Бёль. Адгезёя, хемотаксис ё фагоцитоз лейко- цитёв. ПГЕ опосередковують дёю пёро- генёв на гёпоталамус Арахёдонова кислота з фосфолёпёдёв мембран ушкоджених клётин пщ дёею циклооксиге- нази
Тромбоксан А2 Агрегация тромбоципв. Звуження су- дин. Акливащя системи гемостазу Тромбоцита пёд впли- вом тромбоксансин- тетази
Простациклён (ПП2) Дезагрегация тромбоцитов. Розширення судин Ендотелооцити под досю простациклш- синтетази
Лейкотрёени Хемотаксис. Скорочення непосмугова- них м’язових волокон. Набряк Лейкоцити, лаброцити под впливом лопокси- генази
Лтдш продукты розщеплення 1-алк1л-2-ацетил-фосфатидилхол1ну
ФАТ Активус фосфолёпазу С, що зумовлюе утворення дёацилглёцеролу, який вивёльнюе гранули тромбоципв ё з Са2+ активус фосфолшазу шо вивёльнюе з мембран арахёдонову кислоту Нейтрофёли, моноци- та, тромбоцита
Цитоклни — низькомолекулярш быки, що продукуютъся шуноцитами кров! та строми /забезпечують лижклипиннг взаемоди
ФНП Посилюе утворення АКР 1 ИО, ушко- джус клггини Макрофаги, Т-ломфо- цити
1Л-1 — 1Л-21 Активацоя або гальмування всох реакций запального процесу Макрофаги, ломфоци- ти та оншо клотини омунноо системи
Хемокши Активацоя мпрацп лейкоцитов о фагоци- тозу Моноцити, макрофаги, лёмфоцити, фёброблас- ти та ён.
1Ф Збёльшення резистентност! клггин до вёрусёв. Хемотаксис. Тмунна вщповщь а-1Ф — лейкоцитами, Р-1Ф — фёбробласта- ми, у-1Ф — Т-хелперами 1-го типу
Фактори росту Регуляция росту ё диференцёацп юптин У багатьох клётинах, але найбёльше — в макрофагах
Гуморалый
Кононова система: брадиконш, КЯЛОДИН Розширення артерёол, збёльшення про- никностё судинноГ синки. Бёль. Свербёж а2-Глобулёни кровё
Зактчення табл. 11
Мед<атори Д1я Основне походження
Система комплементу Опсошзащя м!кроорган!зм!в. Активащя хемотаксису 1 фагоцитозу. Цитолтз чу- жорщних клттин Плазма кров!
Зс1дальна 1 ф1бринолггична системи Тромбш активуе хемотаксис лейко- цит! в. Плазмш 1 тромбш активують комплемент Плазма кров!
Гуморальш мед1атори запалення представлен! в основному кшшами 1 систе-
мою комплементу.
Кийни — трупа вазоактивных полшептщцв, як! утворюються внаслщок каскаду
бюх!м!чних реакцш, що починаються з активацп сериново'1 протешази — фактора
Хагемана (XII фактор зсщально! системи кров!). Зггкнення з ушкодженою повер-
хнею або змша внутршшього середовища (температури, рН) призводить до того,
що цей фактор стае активним ! д!е на прекал!кре!н, який е неактивною протеш-
азою, що мютиться в плазм! кров!, перетворюючи його на активний фермент
калпсреш. Останнш своею чертою д!е на а2-глобулши (кппноген), вщщеплюючи
в!д них полгпептидний ланцюг, який складаеться з 9 (брадикппн) або 10 (калщин)
амтнокислотних залишктв. Кшши плазми кров! безпосередньо впливають на тонус
1 проникшсть судинно! стшки, зумовлюючи розширення артерюл 1 зб!льшуючи
проникшсть стшки капшяргв. Кргм того, вони викликають типов! для запалення
свербЬк 1 б!ль. Медтатори калткре'ш-кшшово'т системи пщ час запалення впливають
на реолопчнт властивосп кровт, п здатшсть до зсщання та фтбринолтзу. На схем! 7
показано, як акгивований фактор Хагемана шщное процеси ктшноутворення, ге-
мокоагуляцц та фтбринолтзу. Випадання ниток фтбрину й утворення тромбтв у зонт
запалення певним чином пов’язаш зт станом калткреш-кшшовот системи.
Фактор Хагемана (ХП фактор)
Контакт (скло, колаген)
Трипсин
Кал1кре!н
Ф!бринол!зин
Включения
системи
коагуляцп
Активащя
прекал1кре'1ну
Ф1брин
Прекал!креш
Включения
ф!бринол1тично!
системи
Ф1бринол!зин
-> Кал1кре'!н
Кппноген
> Юган (брадикшш)
Кнпназа
Тнакгивоваш петиди
Схема 7. Система плазмових кшнпв
Ще одним важливим медгатором запалення гуморально! природи е каскадна
система протешаз — комплемент. Вщомо, що комплемент е захисним фактором
оргашзму, однак разом з тим в, и може спричинювати ушкодження власних тка-
нин, що вщбуваеться пщ час запалення, особливо гмунного. Це пояснюеться тим,
що компоненти комплементу СЗа ! С5а активують викид медгаторгв з лаброцитгв
та утворення лейкоцитами АКР, як! ушкоджують як мгкрооргангзми, так г власнг
клгтини. Компонент С5Ь мае властивкть фгксуватися на сенсибшгзованих г не-
сенсибглгзованих антитглами клгтинах, утворювати мембраноатакуючий комплекс
С5Ь6789, що руйнуе гх мембрани та здгйснюе лгзис ушкоджених клггин. Фрагмент
С5а посилюе хемотаксис нейтрофглгв г збщьшуе проникшсть судин. Наре шт!,
мгкрооргангзми, мембрана яких покрита (опсошзована) фрагментом комплементу
СЗЬ, екпресують на фагоцитах рецептор комплементу 1-го типу (СК1) г стають
об’ектом активного фагоцитозу. Опсошзацй бактерш фрагментом СЗЬ сприяе по-
передне зв’язування фосфатидилхолшу гх мембран з бшком гостро! фази — С-
реактивним протегном (СРП).
Медгатори запалення здшснюють свш вплив переважно мгсцево у вогнипц
запалення, що описуеться при характеристицг перебггу запального процесу. У
норм! незначна кглькгсть медгаторгв, а за несприятливого перебггу запалення —
бшьша, потрапляе в загальний кровообгг, що проявляеться загальними ознаками
мгсцевого процесу.
Вплив мед<атор1в запалення на р1вн! орган'/зму
1. Вплив на гтоталамус. Прошфламаторнг медгатори, таю як 1Л-1, 1Л-6, ФНП-а,
через загальну циркуляцию дгють на ппоталамус. Це мае принаймш два наслщки:
по-перше, сприяе пщвищенню локально! продукцг! ПГЕ2, який своею чертою,
змгнюючи ргвень терморегуляцгг, зумовлюе пщвищення температури тгла (див.
роздш XIII “Гарячка”); по-друге, пщвищуючи викид кортикотрошн-рилгзинг-
гормону, вони збшыпують продукцпо кортикотропшу (АКТЕ) аденогшофгзом, а
також видглення глюкокортикощгв кгрковою речовиною надниркових залоз. Це
пщвищення ргвня глюкокортикощгв у кров! д!е як негативний зворотний зв’язок,
який гальмуе необмежене наростання проявгв запалення.
2. Вплив мед1атор1в запалення на печшку, яка у вщповщь на гх д!ю р!зко пщви-
щуе продукцпо так званих бшюв гостро! фази. Нин! вщомо калька десятюв таких
бшюв, що в гострий перюд запалення вщгграють важливу загальну захисну функ-
цгю вщ бактерш г речовин, якг утворюються внаслщок деструкцг! власних тканин.
До таких бшюв насамперед належить СРП г сироватковий попередник амшощу
(СПА). За хгмгчною будовою вони складаються з п’яти полшептадних юлець (у
СПА — в подвосному набор!) г тому мають назву пентраксиюв. 1х ргвень у кров!
при запаленш може пщвищуватися у 100—1000 разгв. Тому вони е дуже чутливи-
ми шдикаторами, яю сигнал гзують про виникнення запалення в оргашзм!, що
широко використовуеться в клгнщг.
Серед шших бшюв гостро! фази варто назвати сироватковий амглощ А (САА),
гаптоглобш, гемопексин, церулоплазмш, трансферин, лактоферин, гнггбгтори про-
теаз, активоваш компоненти комплементу, ргзн! фактори росту. 1х ргвень пщ час
запалення пщвишусться не так виражено, лише в юлька разгв. Бглки гостро! фази
вщгграють ргзномангтну роль. Вони зв’язують шкщливг продукти, роблять недо-
ступ ними для бактергй важлив! нутр!енти (залтзо, цинк, м!дь, щанокобаламш),
ослаблюють генерат1зац1Ю реакцп та стимулюють репаративн! процеси. 11роте в
раз! надто потужного впливу патогенних прошфламаторних фактор!в може спос-
терггатися загрозлива для життя генерал! зацш запального процесу, шо отримала
назву синдрому системно! дп медгаторгв запалення. В шитому випадку пщ час
хрон!чного запалення тривала продукцтя САА та СПА призводить до !х накопи-
чення як у вогниш! западения, так : в шших д!лянках оргашзму (селез!нщ, нир-
ках) у вигляд! аморфно! нерозчинно! бглково! маси — амио'щу, що порушуе
структуру 1 функшю клетин 1 оргашв.
Ексудацся та ем!грац)я
У вщповщь на дпо БАР, як! отри мал и назву мед!агор!в запалення. вщбуваються
порушення мгсцевого крово- 1 лгмфообпу, передусгм мжроциркуляцп. Мшроцир-
кулящею прийнято називати рух кров! в терминальному судинному русл! (в ар-
терюлах, метартерюлах, кашлярах г венулах), а також транспорту р!зних речовин
кр!зь стшку цих судин.
Судини! реакцп при запаленн! виникають у послшовност!, описаюй Ю. Конгей-
мом: короткочасний спазм артерюл змшюеться артериальною гшерем!ею, яка пе-
реходить у венозну гшеремпо, престаз 1 стаз кров!. Так! судинн! редкий забезпе-
чують головну мету запального процесу: вщокремлення вогнища ушкодження вщ
загального кровообпу та локальне знешкодження фактора, що спричинив ушкод-
ження КЛ1ТИН.
Ус! реакдп, як! супроводжують запальний процес, здшснюються пщ регуля-
торним вшивом мед!атортв запалення.
Мгкроциркуляпйо зручно вивчати за допомогою дослщу Ю. Конгейма (мал. 26).
При цьому пщ мгкроскопом можна побачити, як одразу июля дц подразника
(наприклад травми пщ час витягання кишки) виникае спазм артерюл, який мае
рефлекторний характер ! швидко
минае. У його розвитку. кр!м того.
беруть участь енд отел гни. як! вивыь-
нюються з ушкоджених енд отел юци-
тгв, та катехоламгни г лейкотр!ени —
1з тромбоципв. У подалыпому вини-
кае артергальна гтеремгя. Вона е на-
слщком утворення у вогниш! запа-
лення велико! киькост! вазоактив-
них речовин — метабол! т!в з ушко-
джених клггин (К+, Н+, СО,, АДФ,
АМФ. аденозину, неоргашчного фос-
фору) та БАР (г!стам!ну, к!шн!в,
ПГЕ2, ПП2). Ц! мед!атори запален-
ня розслаблюють м'язов! елементи
ст!нки артерюл 1 прекашлярнз. що
зумовлюе збыьшення припливу ар- Мал 26 Запалення бриж1
тергально! кров!, прискорення П ру- у _ лейкоцита, що емзгрували; 2 — емцращя лейко-
ху, вщкриття кап!ляр!в, як! рашше > цитш; 3 — еритроиити в тканиш
не функцоонували, пщвищення в них тиску. Кргм того, розширення привщних
судин виникае внаслщок паралочу вазоконстрикторов о зниження еластичносп
сполучно! тканини, що оточуе судини.
Через 30—60 хв теля початку експерименту перебн запалення поступово змо-
нюеться: артергалъна гтерем1я переходить у венозну. При цьому швищкость руху
кров! зменшуеться, змшюеться характер току кров!. Зокрема, якщо пщ час ар-
тероально! гшеремп клггини кров! розмгщувалися переважно в центр! судини
(осьова течоя), а бшя стшок були плазма о невелика к!льк!сть лейкоцитов (плазма-
тична течоя), то при венозной гшеремп такий розподщ поруоооуеться. Змшюються
реолопчш властивосп крово. Вона стае густотою о в’язкошою, еритроцити набуха-
ють, утворюючи агрегата — хаотично з’еднано мож собою скупчення еритроцитов,
яю повшьно рухаються або зовсом зупиняються в судинах з малим доаметром.
Переход артероальноо гшеремп у венозну пояснюеться доею кшькох факторов,
яю можна подолити на три групи: фаосгори крово, судинно! стшки о прилеглих
тканин. До факторов крово належать крайове розмощення лейкоцит! в, набухания
еритроцитов, виход рщко! частани крово у запалену тканину та згущення крово,
утворення тромбов унаслщок активацп фактора Хагемана, зменшення вмосту гепа-
рину. Вплив факторов судинно! стшки на венозну пперемою пояснюеться як на-
буханиям ендотелпо, так о спазмом вен пщ впливом серотоогшу, тромбоксану
ПГР2а о великих концентрацой пстамону. Змшено венули втрачають еластичшсть о
стають уразливошими до стиснення шфольтратом. Прояв тканинного фактора по-
лягае в тому, що набрякла тканина, стискуючи вени та ломфатичн! судини, сприяе
розвитку венозноо гшеремп.
3 розвитком престатичного стаооу спостерогаеться маятникоподобний рух
крово: пщ час систоли вона рухаеться вщ артерой до вен, пщ час доастоли — у про-
талежному напрямку. Зрештою рух крово може повшстю припинитися, розви-
ваеться стаз. Наслщком стазу можуть бута необоротно змши клотин крово о тка-
нин.
Ексудащя — вихщ рщко! частани крово оз судин у тканини — зпдно з класич-
ною теороею Старлонга може збшьшуватися або при пщвищенш гщростатичного
тиску крово (ГТК), або при зменшенш онкотичного тиску крово (ОТК), або при
збольшенно онкотичного тиску тканинно! рщини (ОТТР). При запаленно ГТК по-
силюеться внаслщок розвитку артероальноо та венозноо гшеремп. ОТК зменшуеть-
ся, а ОТТР збольшуеться завдяки посиленню проникносп судинно! стшки, через
яку оз крово в тканини надходять бшки плазми кров!. Кром того, збольшенню
ОТТР сприяе гщролотичне розшеплення лгзосомними ферментами нейтрофглов
макромолекул, осюльки онкотичний тиск визначаеться не розморами, а кшькостю
молекул.
Основною причиною змони онкотичного тиску о розвитку ексудацп при запа-
ленно е пщвищення проникносп пстогематичного бар’ера, тобто стшюо судин,
насамперед каполяров о вен, для бпгкових молекул. У дослщженнях доведено, що
пщвищення проникносп судинно! стшки при запаленш мае дво фази: ранню о
позню. У ранню фазу, яка троовае вщ 5 до 30 хв шсля уооокодження, пщвищена про-
никность стгнки судин зумовлена заокругленням ендотелооощпв пщ час скорочен-
ня ох контрактильних структур о збольшенням проможков мож ними пщ впливом
пстамшу, серотоншу, компонентов комплементу, простагландинов, лейкотроенов,
брадикшшу, калщину. Пщвищення проникносп стшки судин у п1зню фазу, мак-
розди XI. запалення
симум яко! можливий меж 5-ю годиною 1 7-ю добою пёсля ушкодження, пов’язане
з впливом Ш-1, ФНП, у-1Ф, як! посилюють транснитоз (.транспорт речовин крёзь
цитоплазму ендотелюципв шляхом пёнонитозу), коли клггини ендотелёю здатн!
‘‘ковтати” найдрёбнётш краплё рщини (мткропёноцптоз), переправляти !х на про-
тилсжний кшець кштини та викидати назови! (екструз!я). Електронний мёкроскоп
дае змогу не титьки побачити ш мёкровезикули, а й вимёряти ё пёдрахувати тх кёль-
кёсть. З’ясувалося, шо шд час запалення вщбуваеться активашя енергозалежного
мёкровезикулярного транспорту. Про пе свёдчить його припинення п!д д!ею ён-
пбтторёв утворення макроерпчних сполук.
Водночас лёзосомнё ферменти нейтрофёлёв руйнують базальну мембрану су-
дин. що також збёльшуе тх проникшсть.
Ексудат вщрёзняеться вщ транссудату тим, шо мтстить бё.тыпе бшюв (понад
2 %). Якщо проникшсть стшки судин порушена незначно, то в ексудат, як пра-
вило, проникають альбумёни ё глобултни. У раз! ц значного порушення з плазми
в тканину виходить бьток з великою молекулярною масою (фибриноген). Пщ час
первинно!'. а пот!м ё вторинно! альтерацп, яка охоптюе е структури судинно! стш-
ки, крёзь останню починають проникати не лише бёлки, а й клггини. Цьому сприяе
те, що в раз! венозноз гшеремп лейкоцити розмпцуються вздовж внутртшньо! обо-
лонки дрёбних судин, бёлын-менш мёцно прикрёплюючись до ендотелёю (фено-
мен крайового стояния лейкоцитгв).
Емтграцтя лейкоцитов (трансмёграшя. д!апедез) з просвету судин (мал. 27) кр!зь
судинну стёнку в мёжклттинний простёр вогнища запалення вщбуваеться в певшй
послщовностё, яка значною мтрою залежить вёд етёологёчного фактора: у бёльшостё
С залогпзкоз .роз ез ь
лейкоцита
I Р-селектин
1нтегрив
V 1САМ-1,-2;
УСАМ-1
| Ь-селектин
иСзалогазкопротеТк
ендотелно
Е-селектин
Мал. 27 Маргша.озащя, емпразця, хемотаксис лейкоцитгв । фагоцитоз у вогннпй запалення:
1 — ролзш (катания) лейкоцита; 2 — адгезёя (прилипания) лейкоципв; 3 — емёграци лейкоципв;
4 — хемотаксис; 5 — адгеззя до об’екта фагоцитозу; б — поглинання об’екта й утворення фатосомп;
7— утворення фагол1зосоми з переправления об’екта
випадюв гострого запалення, особливо коли воно зумовлене гноетв!рними бак-
терзями (стафыококами, стрептококами, пневмококами тощо), спочатку емйру-
ють у великш кшькост! нейтрофгльш гранулоцита, потгм моноцита 1, нарешп,
лгмфоцити. Проте при вхрусних гнфекщях ! залаленш, спричиненому бактер!ями,
що паразитують усередин! кл!тин (туберкульоз, сифшс, тулярем!я, бруцельоз
тощо), першими у вогнипц запалення можуть з’явитися лимфоцита, а в ексудат!
будуть переважата лхмфоцити та моноцита. У випадках, коли перебгг запалення
мае алерпчне походження або зумовлений паразитами (гельмштами), спостер!-
гаеться шфиьтращя еозинофйами.
За сучасними уявленнями, головний мехашзм емпрацп лейкоцита полягае в
комплементарнгй лгганд-рецепторшй взаемодп м!ж лейкоцитами та компонента-
ми судинно! стшки. При цьому роль рецептор!в та !хн!х лйащцв виконують моле-
кули клтиншн адгези, що експресуються на поверхш лейкоцита 1 ендотелюцита
щд впливом певних мед!атор!в запалення. Таких адгезивних молекул !снуе вели-
чезна кшыастъ. У табл. 12 наведено лише основш. За нормальних умов быышсть
13 них не функцюнуе. Вони або депоноваш у внутршгньоклггинн их гранулах, або
активуються шляхом конформацй, або синтезуються де поуо. Ц! молекули подшя-
ють на юлька родин.
Таблиця 12. Основы адгезивн! молекули лейкоципв
та ендотелюципв таТх взаемод!я
Адгезивн! молекули М1сце експреси 1хж Л(ганди (комплементарш молекули)
Селектини: Р-селектин Е-селектин Ендотелюцити Модиф1кован! с!алогл!копротеши плазматично! мембрани нейтроф1л!в 1 моноцитов (51а1у1-Ьеу/13 X — тоШбеб §1усорго1етз)
Ь-селектин Лейкоцита С!алогл!копротеТни ендотел1ю (8ср- 200) та вуглеводи у склад! МА<1САМ-1
Суперродина шуноглобултгв: Молекула м!жкл!тинноТ адгези 1-го (ш1егсе11и1аг айЬезюп то1еси1а-1,1САМ-1) 1 2-го (1САМ-2) титв Переважно ендотелш Тнтегрини нейтроф!л!в 1 моноцитов
Молекула адгези судинного ендо- тел!ю (уазси1аг се11и1аг адйезюп пзо1еси1а-1, УСАМ-1) Ендотелш, (фибро- бласта, макрофаги 1нтегрини лейкоципв
Молекула адгези лшфоципв або адресин 1-го типу (тисоза! абтеззт-1, МАССАМ-1) Високий ендотелш венул 1нтегрини л!мфоцит!в
1нтегрини: Р^штегрини (дуже шзнш антиген, уегу 1а1е апн§еп, УЬА) Лейкоцита та шип клгтини Колаген, ламшш, ф!бронектин, тромбоспондин, УСАМ-1
Законченна табл. 12
Адгезивн) молекули - - - М>сце експресп |ХН1 Л1ГЭНДИ (комплементарн) молекули)
р,-1нтегрини (антиген, що асошйо- ваний з функщею л!мфоцит!в-1, 1утрЬосу1:е Гипсйоп-а58ос!а1е<1 апй§еп-1, ЬРА-1; СКЗ, СК4) Лейкоцита 1САМ-1,1САМ-2, СЗЬ
Р3-!нтегрини Нейтрофши 1САМ-1,1САМ-2
р7-штегрини (ЕРАМ) Т-тмфоцита МАйСАМ-1
Селектини — трансмембранн! бшки-рецептори, зовншппй (позаюптинний)
кшцевий домен яких здатен зв’язувати олйосахариди, тобто е битком-лектином.
Пщ впливом цитокппв, як! утворюються лаброцитами, тканинними макрофагами
й ендотелюцитами у вщповщь на контакт з мгкрооргашзмами й ушкодження юй-
тин, на мембран! ендотелюцит!в експресуються Е-селектини, на мембран! ендо-
тел!оцит!в 1 тромбоципв — Р-селектини, а на поверхн! лейкоцита — Е-селекти-
ни. Л п андами для Е-селектишв е с!аловм!сн! олйосахаридн! молекули глпсопро-
теййв на мембран! нейтроф!л!в, для Р-селектингв — на вс!х лейкоцитах, а для
Е-селектин!в — на ендотелтальних юйтинах.
1нтегрини — гетеродимерн! (складаються з двох нековалентно зв’язаних суб-
одиниць: а 1 р) трансмембранн! глткопротешов! рецептори, що еспресуються гцд впли-
вом мед!атор!в запалення на мембран! лейкоципв, ф!бробласт!в, тромбоцита! юй-
тинах деяких внутргшшх оргашв! зв’язують кл!тини з компонентами позаюйтинного
матриксу: фтбронектином, колагеном 1 ламшшом, тромбоспондином, впронекти-
ном, а також з ендотелюцитами. Кр!м адгезп лейкоципв 1 тромбоципв вони забез-
печують д!апедез лейкоципв через судинну стшку, хоумшг л!мфоцит!в ! прикрш-
лення юптин до мхжюйтинно! речовини, що сприяе самоформуванню тканин.
Ылки суперродини 1муноглобул1н1в — трансмембранн! протеши з п’ятьма-пйсть-
ма позаюйтинними доменами, под!бними до !муноглобулш!в, що експресуються
тпд впливом Ш-1, ФНП 1 у-1Ф на мембран! переважно ендотелпо. Ц! бшки ком-
плементарн! р!зним штегринам, а тому забезпечують адгез!ю 1 проходження лей-
коцита через судинну спнку.
Адресини — бшки високого ендотелпо венул, як! забезпечують емтграцпо пе-
реважно лхмфоципв.
Певна послщовнють емгграцп лейкоцита пов’язана з тим, що експрестя рхз-
них адгезивних молекул вщбуваеться не одночасно. Першими пщ впливом мед!а-
торгв запалення експресуються селектини. Уже в перш! хвилини дп на судинну
стшку пстамшу, тромбшу, ендотоксин!в бактерш (лшополюахарщцв; ЛПС, Иро-
ро1у8асс11ап(1е, ЬР8), фосфолшщного фактора активацп тромбоцита (ФАТ, р1а!е1е!
асйуайоп Гас!ог, РАЕ) вщбуваеться перерозподш Р-селектину тз його внутршшьо-
юйтинного депо — гранул ендотелтальних юптин (тшьця У/ейэе! — Ра!айе) на по-
верхню плазматично! мембрани. Через, 1—2 год пщ впливом фрагмента компле-
менту (С5а, ВЬ), лейкотрхену В4, ФНП-а експресуються Ь-селектини лейкоципв,
а внаслщок дп ЛПС бактерш, Ш-1, ФНП-а, ФНП-р, Ш-8 та шших цитокпйв —
Е-селектини, а також !хн! лйанди. Тнтегрини, бшки суперродини гмуноглобулпйв
1 адресини з’являються на мембранах лейкоцита 1 ендотелюципв значно гйзнппе.
”238
Тому максимальна швидкзеть виходу нейтрофолов припадае на перил 2 год о знач-
но знижуеться через 4—6 год. Емограцоя моноцитов починаеться разом з нейтро-
фолами, але сягае максимуму за 16—24 год.
Взаемодоя селектишв з охшми ологосахаридогими логандами не дуже мщна
(низькоафшна) та легко руйнуеться токрм кров! (оборотпна адгезгя). Селектини
притягують лейкоцита до судинно! стшки о на деякий час утримують !х, вщпуска-
ють 1 знову прикршлюють, що створюе ефект катания вздовж судинно! стшки
(гоШп§).
Пщ впливом медоаторов, як! вивольнюються при ушкодженш лаброцит!в, а
потом зв’язуютъея з поверхнею ендотелпо (ФАТ, ФХЕ тощо), або хемокшгв —
низькомолекулярних цитокошв, як! синтезуються активованими моноцитами-
макрофагами, лимфоцитами, ф!бробластами й ендотелооцитами та стимулюють
мйрашю й активацйо лейкоцитов {(1Л-8, нейтроф!льний активуючий бшок-2
(НАБ-2, пеШгоркП аейуабоп ргоГет-2, МАР-2), моноцитарний хемоатрактантний
бшок-1 (МХБ-1, топосуСе сйетоайгасСаШ ргоГет-1, МСР-1), макрофагальний за-
пальний болок-1а (МЗБ-1а, тасгор!1а§е тйаттаоогу рго1еоп-1а, М1Р-1а), МЗБ-
1р тощо)} вщбуваеться конформащйна змша й актавац!я штегришв на поверхш
лейкоцитов або ох перемощения !з внутрппньоклотинних депо. При збольшенн! ло-
кально! внутрошньооаоггинноо концентрацп Са2+о М§2+ онтегрини ще й перерозподо-
ляються на поверхш лейкоцитов о утворюють кластери (плями), що розко збольшуе
афшность (мощность) ох зв’язку з логандами на поверхш ендотелпо.
Активацоя штегришв супроводжуеться експресоею на поверхш ендотелооцитов
пщ впливом ФНП, 1Л-1, 1Л-2, у-1Ф, ЛПС тощо адгезивних молекул суперродини
омуноглобулошв. Усе це забезпечуе мщний зв’язок лейкоцитов зо стойкою судини,
розпластування ох на поверхш ендотелпо (необоротна адгезгя), унаслщок чого вони
починають проникати крозь розширено проможки мож ендотелоальними клотинами
каполяров о венул (елигращя лейкоцитгв). При цьому нейтрофоли о моноцита про-
тискаються мож ендотелоальними клотинами амебоподобно, випускаючи псевдо-
подп та видшяючи колагеназу й еластазу, яко роблять отвори в базальной мемб-
ран!. Ломфоцита, кром того, мають здатшеть проникати прямо крозь цитоплазму
ендотелоальних клггин, не ушкоджуючи !х. Цей способ мае назву “емпериполез ”.
Емограцоя лейкоцитов — це нормалышй процес, яосий зазвичай здшснюеться
через високий ендотелой венул усього оргашзму. При запаленно посилена о при-
скорбна емограцоя вщбуваеться через стшку каполяров о венул обмеженоо дшянки
судинного русла, що безпосередньо контаостуе з мосцем ушкодження. Шсля пот-
рапляння в позасудинний простор унаслщок ферментативного розщеплення лей-
коцита втрачають зо свое! поверхно Ь-селектини о починають цолеспрямовано о
швидко могрувата (рухатись) у напрямку флогогенного фактора о / або зруйнова-
них клггин за допомогою амебоподобних рухов.
Цей процес е першою стадоею фагоцитозу г мае назву “хемотаксис ”. Це яви-
ще описав о дав йому назву 1.1. Мечников. Як о емограцоя, хемотаксис значною
морою визначаеться градоентом концентрацп хемоатрактанпйв — молекул, яко
зв’язуються з вщповщними рецепторами на поверхно лейкоцитов, активують ох о
зумовлюють спрямовану мйрашю. Сигнал вод активованогх рецепторов через сис-
тему вторинних месенджеров, у тому число Йонов кальцою, спричинюе поломериза-
цпо актину на передньому кшцо лейкоцита о сосупчення моозинових фоламентов у
задной частино клггини, що пщштовхуе лейкоцит у напрямку джерела хемоатрак-
тантгв. Кром того, завдяки взаемодп Р]-1нтегрин1в з можюптинним матриксом лей-
коцита пщтягуються в напрямку джерела хемоатраосгантгв.
Якщо запалення е наслщком шфокування тканин, то головними хемоатрак-
тантами виступають компонента мокроорганозмгв, продукта гх життедояльносто
або опсонши, що покривають ох поверхню. Вщповщно, на поверхш лейкоцитов
розмгщено спецоальш рецептори, що реагують на наивность мокрооргашзмов г
включають мехашзми ох знешкодження, у першу черту — наближення мокро- о
макрофапв до об’екпв фагоцитозу.
Зараз водомо близько 20 мембранних глокопротеонових рецепторов лейкоцитов,
подобной до ОоИ-родини болков дрозофыи (1о11-Нке гесерЮгз, ТЬК.), яко у ссавцов ви-
борково зв’язуються й активуються певними трупами бактерш, ворусами, грибами,
найпростошими. Так, ЛПС грамнегативних бактергй активують ТЬК4, лшопротео-
ди грампозитивних бактерш е лггандами для ТЫ^, ТЫЦ, ТЬК5, одно- та дволан-
цюгова РНК ворусов зв’язуються з ТЬК3, ТЬК4, ТЬК,, ТЬК8, триби — з ТЬК6.
Розшзнавання нейтрофольними о моноцитно-макрофагальними ТЬК онфекцшних
факторов призводить до активацп транскрипцшного фактора МР-кВ, що через
активацою вщповщних генов стимулюе продувдпо прозапальних цитокгнов (ФНП-
а, 1Л-1, 1Л-6, 1Л-8, 1Л-12, 1Л-18, а-1Ф, р-1Ф), яко надало впливають на ендотелш,
фобробласти та оннп клггини, у тому число лейкоцита.
(З-протетасоцшоват рецептори лейкоцитов можуть активуватися П-форм1лме-
тюнтом — амшокислотою, що моститься в бшках бактерш о вщсутня у людини.
Лейкоцитарно рецептори до опсонйав, тобто болков, що покривають мокроор-
гашзми для полегшення ох фагоцитозу, експресуються за наявностг певних оп-
сононов. Так, гмуноглобулиш О, що покривають мокроби, розшзнаються о зв’язуються
Рсу-рецепторами фагоцитов. Кргм того, лейкопептидаза фагоцитов вившьнюе оз
у-глобулшгв тетрапептид тафтсин, який мае виражену хемотаксичну й опсонозу-
ючу дою, активуе лейкоцита та прискорюе фагоцитоз. Фрагмент комплементу СЗЪ
тел я фоксацп на мгкрооргашзмах експресуе на фагоцитах рецептор комплементу
1-го тапу (СВ.1). У глоколшщах о глгкопротегнах клотин бактерш, на вщмону вщ
тварин о людини, багато таких вуглеводов, як маноза о фукоза. Манозозв’язуючий
лектин плазми крово здатен розпознавати бактерп за великим вмостом у вуглевод-
ному компонент! охньоо клотинноо стонки манози або фукози, фоксуватися на ной,
активувати комплемент класичним шляхом о виконувати функцпо опсоншу для
макрофагального манозного рецептора. СРП, що зв’язуеться з фосфатидилхоло-
ном клотинноо стонки деяких мгкробов, також може активувати ох атаку фагоцита-
ми та комплементом.
1нпп хемоатрактанта е продуктами ушкодження власних тканин як боолопч-
ними, так о фозико-хгмочними факторами. У раз! ушкодження або подразнення
лаброципв, макрофагов, ендотелооцитгв, ломфоцитгв та шших клотин утворюються
або активуються о видшяються простагландини, лейкотроени (особливо В4), тром-
боксан фрагмента комплементу (особливо С5а), ФАТ, ФХЕ, Н8Р, деяко фак-
тори росту, цитокши (ФНП-а, 1Л-1, у-йФ), хемокони (НАБ-2, МХБ-1, МЗБ-1а,
МЗБ-10, 1Л-8) тощо. На лейкоцитах пщ впливом хемоатрактантгв експресуються
комплементарно для кожного з них рецептори.
У разо ушкодження клотин у мгжклотинний простор потрапляють ДНК й АТФ
о продукта ох розщеплення, як! впливають на пуринов! рецептори лейкоощтов. Руй-
нування м!жкл!тинного матриксу своею чертою призводить до утворення тромбшу
та продуктов деградацн колагену, ф!брину, фдбронектину тощо. Вони можуть акти-
вувата на лейкоцитах як ашттевг рецепторы, так 1 ТЬК.
Якщо завдання нейтрофйпв як одного з головних фактор!в гуморального
хмуштету полягае у знешкодженш патолопчного шфекцшного фактора, у першу
черту грамнегативних бактерш, то не дивно, що !х хемотаксис акгивуеться ендо-
токсинами бактерш (ЛПС), К-формшметооншом, гмунними комплексами, 1Л-8
або НАБ-1, НАБ-2, тромбоцитним фактором-4 (ТФ-4, р1а1е1е! ГасДог-4, РР-4),
фрагментами комплементу, лейкотриеном В4, ПГЕ2, пстамшом, калгкрешом, про-
дуктами розпаду колагену та ф!брину, тонами водню.
Роль макрофапв полягае головним чином у знешкодженш юптин, у яких
паразитують втруси, деяю бактерц та гриби, а також в очищены вогнища запален-
ня вщ загиблих юптин, у тому числ! нейтрофшгв, 1 утворенш протизапальних
мед!атор!в, що руйнують (арилсульфатаза, пстамшаза, юшназа тощо) або ней-
тралтзують (гепарин, хондроггин-сульфат, шпбггори протешаз, антифосфолшази,
антиоксиданти, шшамши, лшоксини, ГЛ-10, пстамш через Н2-рецептори) мед!а-
тори запалення. Хемотаксис моноцитов ! Т-л!мфоцит!в як основних ефекторгв
клпинного хмуштету та регуляторхв завершально! стадп запалення активуеться
вхрусними РНК, екзотоксинами грампозитивних бактерш, продуктами розпаду
нейтрофыхв, лхмфо- 1 монохемоюнами: МХБ-1, МХБ-2, МХБ-3, МЗБ-1а, МЗБ-
10, регулятором активацп, експресп та секрецп нормальних Т-клпин (ге§и1а1ес! оп
асйуайоп поппа! Т-се11 ехргеззей ап<1 зесге1ес1, КАЫТЕ8), але блокуеться Н+.
Фагоцитоз. У вогнигщ запалення лейкоцити виконують рхзн! функцн та голо-
вна з них — поглинання 1 перетравлювання стороншх частинок (фагоцитоз). Для
одноклпинних оргашзмгв фагоцитоз е способом живлення, а у вищих оргашзмхв
ця функция збереглася тольки в деяких юптинах х набула захисного характеру. Уст
фагоцитуючт клггини 1.1. Мечников подшив на мхкро- й макрофаги (макрофаго-
цити). Перш! (полхморфноядернх лейкоцити) фагоцитують мхкрооргашзми, друг!
(моноцити, тканинн! макрофаги, пстооцити) поглинають х быыш частники, у
тому числт шфхковаш й ушкоджен! клпини та ххш фрагмента.
Розрхзняють чотири стадп фагоцитозу: наближення (хемотаксис), прилипан-
ия, поглинання, перетравлювання. Першу стадно — хемотаксис (спрямований
рух лейкоцитов) — було розглянуто вище.
Друга стадия — прилипания (адгезия) — пояснюеться здатшстю лейкоцитних
рецепторгв та штегринхв, яю беруть участь у хемотаксис! та були писан! вище,
розшзнавати поверхнев! лхганди м!кроорган!зм!в та пгфхкованих х ушкоджених
власних клпин ! модифгкованих компонентов тканин х мщно приеднуватися до
них. Певне значения при цьому мае поверхневий заряд лейкоцитов. Негативно
заряджен! лейкоцита краще прилипають до об’екта з позитивним зарядом.
Прилипания фагоцита до об’екта фагоцитозу започатковуе метаболхчш змгни
в мхкро- 1 макрофагах, яю мають назву “метаболхчний вибух”. Активуються гек-
созомонофосфатний шунт х глгколгз, накопичуеться НАДФ-Н, необхщний для
подальшого утворення активних форм кисню (АФК) або АКР, синтезуються ей-
козанощи, макрофагами видшяються ГЛ-1, 1Л-6, ФНП-а.
Поглинання об’екта лейкоцитами — третя стадхя — може вщбуватися в два
способи. Дшянка цитоплазми, що контактуе з об’ектом, утягуеться всередину
юптини, а разом хз нею — х об’ект. Другий спос!б — фагоцит торкаеться об’екта
Фосфолшаза А2,
колагеназа, еластаза,
катюнш битки, лтзоцим
Мал. 28. Речовини, що видтляються активними полтнуклеарами шд час фаго-
цитозу
сво'1ми довгими тонкими псевдоподтями, а потгм усш плом щдтягуеться в б!к
об’екта й обволжае його. I в першому, 1 в другому випадку стороння частника
виявляеться оточеною цитоплазматичною мембраною 1 затягнутою всередину
юптини. Вщбуваеться взаемодгя опсонпйв з опсоншовими рецепторами фаго-
цит1в, а також пол!меризац!я актинових фыаменпв. Як наслщок, утворюеться
своерщний мппечок з! стороншм тйтом (фагосома).
Четверга стадтя фагоцитозу — перетравлювання (знешкодження I деградащя).
Лтзосома наближаеться до фагосоми, поп мембрани зливаються, утворюючи едину
вакуолю, у якш мктяться поглинута частника та лтзосомн! ферменти (фагол!зосо-
ма). У фагол!зосомах встановлюеться оптимальна для дп ферменпв реакщя (рН
близько 5,0) 1 починаеться знешкодження 1 перетравлювання об’екта фагоцитозу.
На мал. 28 показано весь “арсенал”, який е в розпорядженш активного мгкро-
фага 1 який вш застосовуе на р!зних етапах фагоцитозу. Варто зазначити, що сам!
лише активоваш л!зосомн! ферменти не можуть забезпечити достатню юлерну
дпо. Ефектившсть фагоцитозу зростае, коли до процесу залучаються АКР.
Лейкоцита утворюють ! при фагоцитоз! активують 1 шпй бактерицидна речо-
вини. Це фосфолшаза еластаза, колагеназа, л!зоцим, капонн! бпгки (дефензи-
ни, катепсини), еозинофшып цитотоксичш б!лки тощо, як! порушують структуру
мембран мжрооргантзмтв. Спыьна д!я бактерицидних речовин призводить до руй-
нування об’екта фагоцитозу. Кр!м того, жив! фагоцита п!д час фагоцитозу та за-
гибл! нейтроф!ли вид!ляють у м!жкл!танний проспр властив! !м ферменти! та
неферментн! БАР, як! дпоть екстрацелюлярно як на бактерп, так 1 на власш юп-
тини. Контрольована секрещя лейкоцитами лтзосомних ферменпв ! АКР виникае
в раз! неможливосп поглинута патолопчний об’ект 1 е нормальною реакщею ак-
тивованого фагоцита. АКР шдукують некробюз або апоптоз як фагоцитованих,
так 1 прилеглих юптин, а макрофаги фагоцитують мертв! юптини та шп уламки !
перетравлюють !х.
Порушення обмшу речовин у вогниир запалення. Гнтенсившсть обмгну речовин
гцд час запалення, особливо в центр! вогнища, посилюеться. Фермента, що вив!ль-
нилися з ушкоджених лхзосом, г!дрол!зують вуглеводи, б!лки, нуклешов! кислота,
жири, яю мктяться у вогшпцт запалення. Продукта гщролтзу зазнають дп фермен-
та глтколтзу, актившсть яких також шдвищуеться. Це певною мтрою стосуеться т
ферментов аеробного окиснення. При експериментальному вивченш ди запально-
го фактора (кротоново! олп) на шюру виявлено, що споживання кисню при цьому
зростае на 30—35 %, можливо, в основному за рахунок утворення АКР, осюльки
воно не пригшчуеться щашдами. Однак це тривае недовго — 2—3 год. Подальша
альтерацтя клттин супроводжуеться ушкодженням мгтохондрш — морфолопчного
субстрату, на якому локалтзуються фермента циклу трикарбонових кислот т де
вщбуваеться аеробне окиснення т поеднане з ним окисне фосфорилювання. Унас-
лщок цього окиснення ще бтльше порушуеться за умови майже незмшеного глт-
колтзу, що призводить до збтльшення вмтсту молочно! та трикарбонових кислот
(а-кетоглутаровот, яблучно!, бурштиновот). Окиснення при цьому не завершуеться
в цикл! Кребса. Зменшуеться утворення вуглекислота, знижуеться дихальний ко-
ефщтент.
Для характеристики метаболтзму у вогнищ! запалення здавна застосовують
термш “пожежа обмтну": обмтн речовин ртзко посилений, “горшня” вщбуваеться
не до юнця, а з утворенням недоокиснених продукттв (полтпептидтв, жирових
кислот, кетонових т!л).
Отже, запалення завжди починаеться з посилення обмгну речовин. Цим знач-
ною мтрою пояснюеться одна з кардинальних ознак процесу — мгсцеве пщвищен-
ня температури. Згодом штенсившсть метаболтзму ослаблюеться, а разом тз цим
змшюеться т його спрямовашсть. Якщо спочатку, тобто в гострий перюд запален-
ня, переважають процеси розпаду, то надалт — процеси синтезу. Розмежувати тх
за часом майже неможливо. Коли переважають катаболтчнт процеси, спостертга-
ються деполтмеризацтя бшково-глткозамшоглтканових комплекств, розпад бтлктв,
жиртв т вуглеводтв, виявляються втльш амшокислота, полшептиди, амтноцукри,
уроновт кислота. Деяю речовини (кшши, простагландини), що утворюються, бе-
руть участь у регуляцп процесу запалення.
Анаболтчн! процеси проявляються вже в раннтх стадтях запалення, однак пе-
реважають у шзшших стадтях, коли виникають вщновш (репаративнт) тенденцц.
Унаслщок акгивування певних ферменттв посилюеться синтез ДНК т РНК. Зростае
актившсть пстюциттв т фтбробласттв. У них активуються процеси окиснення й
окисного фосфорилювання, посилюеться вихщ макроерпв.
Ф1зико-х1м1чн1 змши у вогнищ! запалення. Унаслщок порушення тканинного
окиснення та накопичення в тканинах недоокиснених продукттв розвиваеться
мзсцевий метаболтчний ацидоз. Спочатку вш компенсуеться буферними мехашз-
мами, а поттм стае декомпенсованим, унаслщок чого рН ексудату знижуеться.
Концентрацтя тошв водню там вища, чим штенсившшим е запалення. У разт во-
строго абсцесу рН гною може знизитися до 5,3. Водночас тз пщвищеною кислот-
нтстю у запалешй тканинт шдвищуеться осмотичний тиск за рахунок зростання
онкотичного таску. Це е наслщком як пщвищення проникностт судинно! стшки
та виходу альбумштв з плазми кровт в тканини, так т посилення катаболтчних про-
цеств: велик, молекули розщеплюються активованими ацидозом лтзосомними
ферментами на дртбшшт, гх концентрацтя, а отже, й онкотичний тиск зростають.
Змшюеться також вмтст електролтттв (Ма+, К+, Са2+). В ушкоджених юптинах
зб!лыпуеться концентрацтя натрпо т кальцтю, а в мтжклгтаншй рщин! — вмтст
юшв калтю, якт вившьнюються тз загиблих клттин. У гшйному ексудатт концент-
ращя калпо може сягати 25,6—51,1 ммоль/л, тодо як у нормальной можклггиннш
рщино вона не перевищуе 5,1 ммоль/л.
Пщвищення онкотичного тиску збольшуе ексудацйо о м!сцевий набряк. Цим
пояснюються таю ознаки запалення, як припухлость 1 боль, який з’являеться
внаслщок натягу тканини та ди мед!атор!в запалення на больово рецептори.
Ексудацоя, яка кр!м пщвищення ОТТР зумовлена зболыпенням ГТК в умовах
артероально'о та венозно! гшеремп, поступово знижуе в мост ушкодження концен-
трацпо оонов водню й осмотично-активних речовин, нормалозуе рН о осмотичний
тиск, що сприяе емограцп моноцитов-макрофагов о ломфоцитов, завершению фаго-
цитозу й очищению вогнища запалення вщ патолопчних факторов о ушкоджених
тканин.
Тыьки шсля знешкодження патолопчного фактора, затухания процесов аль-
терацп та ексудацп, очищения вогнища запалення вщ уламгав клотин о прозапаль-
них медоаторов можлива репарацоя дефектов ушкоджених тканин.
Проя1ферац1я । репаращя
Руйшвш процеси поступово припиняються 1 починаються вщновш. Передусом це
розмноження клпин о компенсування дефекту. Головною умовою для Гх здшс-
нення е припинення запального процесу: знешкодження, деградац!я 1 видалення
шфекцшних факторов о некротизованих тканин; шпбпця гщролотичних фермен-
тов; дезактивация медиаторов запалення; розщеплення та виведення токсичних ре-
човин.
Головними регуляторами переведения запалення з рейок катаболозму на шлях
загоення е “альтернативно активоваш” макрофаги, що утворюються оз моноцитов-
омогрантов.
“Класична активацоя” макрофагов здшснюеться шфекцгйними факторами о
цитоганами, яга продукуються Т-хелперами 1-го типу (у-ТФ, 1Л-2, ФНП-а) шсля
Гх активацп АПК. Тага макрофаги продукують прозапальш цитокони (1Л-1, ГЛ-6,
ГЛ-8, ФНП-а, КАЯТЕ8), Т>0, протеолотично фермента, адгезивно быки тощо.
“Альтернативна активацоя” макрофагов вщбуваеться завдяки 1Л-4 о ГЛ-13, яко
синтезуються Т-хелперами 2-го типу. Цо макрофаги продукують протизапально
цитокши (ГЛ-4, ГЛ-10, ГЛ-13), шпботори протеГназ, хондроотинсульфат, гепарин,
фобронектин, полоамши, антиоксиданта, антифосфолшази, фактори росту.
Протизапальна доя ГЛ-10 зумовлена шпбуючим впливом цього онтерлейкона на
функцпо Т-хелперов 1-го типу. ГП-4 о ГЛ-13 пригночують продукцою прозапальних
1Л-1, ГЛ-6, ГЛ-8, ФНП-а, зменшують активность Т-хелперов 1-го тапу за рахунок
гальмування доо у-1Ф, а також знижують цитотоксичность макрофагов о Т-галеров.
Серед 1нг1б1тор1в протешаз найважливпиу роль вщограе а2-макроглобулш (а2-М).
Цей быок мае широкий спектр доо, е основним шгоботором кшоооутворювальних
ферментов крово о, отже, усувае ефект юнонов: розширення судин о збольшення
проникносто Гхньоо стшки. Кром того, вон пригшчуе болышсть протеГназ лейко-
цитов, у тому число й колагеназу та елэстазу, захищаючи вщ руйнування елемента
сполучно! тканини. Нарешто, а2-М може зв’язуватися з мембранами нейтрофщь-
них гранулоцитов 1 таким чином гальмувати Гх реакцпо на СЗа о С5а (хемотаксис).
До контролю за западениям залучаються и шшо шпботори, у тому число а,-анти-
хомотрипсин, який гальмуе катепсин О о хомотрипсин. Антитромбон ГП о а2-анта-
1Н
фгбринолгзин нригшчують серинов! ферменти 1 е основными шпбгторами систем
коагуляцп. фтбриношзу та комплементу.
Хондротинсульфат стабытзуе судинну стгнку, що зменшуе и проникшсть «
гальмуе ексудацтю.
Гепарин шпбуе актившсть комплементу, кпйшв, бюгенних амштв, разом з
фгбронектином входить до складу мтжклггинного матриксу.
Полшмши (спермш, спермедин, кадаверин, путресцин) зменшують проник-
шсть судинно! стшки 1 значно прискорюють пролгферацгю клгтин.
Киснев) радикалы, що виробляються лейкоцитами на початку запального
процесу для знешкодження шфекцшних факторгв, руйнують г власнт клггини.
Потужним джерелом ферментативных I неферментативних антиоксидант 1в лакож
е макрофаги. Головну захисну роль при цьому вщгграють насамперед два фермен-
ти: глутатюнпероксидаза та глутатгонредуктаза. Перша переносить водень на
окиснений глутатюн, а друга — выбирае цей водень г переносить на Н2О2, унас-
лшок чого ^творюютъся двт молекулы води. Знешкодження супероксидною аню-
на забезпечус фермент супероксиддисмутаза (2"О2‘ + 2Н’ —> О2 + Н2О2). Певну
роль вщгграе каталаза, яка звглъняе клггини вщ надлишку пероксиду водню (2Н2О2 -»
—> 2Н2О + О2). Зниження ршня залгза । мщ) за допомогою !х зв'язування лактофе-
рином лейкоцитгв, СРП, феритином, трансферином мшгмгзуе формування агрес-
сивного пдроксильного радикала. Церулоплазмш, який е ферооксидазою, окис-
нюе Ге2+ до Ре3+, що також блокуе реакшю Фентона й утворення ОН.
В шактивацц клгтин запалення кргм мгсдевих велику роль вщгграють загальнг
фактори, у тому числг й ендокринш. Гормон кгрковог речовини надниркових за-
лоз пдрокортизон гальмуе синтез вазоактивных речовин у клгтинах, спричинюе
лгмфопенгю, зменшуе кглькгсть базофыьних та еозинофгльних гранулоципв. Кргм
того, вш стабглгзуе мембрани лгзосом г гальмуе синтез ейкозанощгв завдяки сти-
муляцп синтезу в макрофагах антифосфолтаз, якг пригнгчують фосфолгпазу
Кргм того, що пгсля знешкодження шфекцгйного фактора припиняють ви-
роблятися однг медгатори г починають синтезуватыся гнил, на той самый медгатор
клгтина може дати зовсгм шшу вщповщь, оскыьки на и поверхш виникають гншг
рецепторы, а попсреднг йдуть усередину (штерналгзащя). Це, гмовгрно, пов’язано
з дгею на юптини-мппеш макрофагальных цитокгшв.
Так, пстамш — типовий фактор запалення, однак його ефект у кшцг запален-
ня може вшргзнятися вщ того, що був на початку. Якщо на ефекторних клгтинах
у цей момент експресованг (выставлен)) Н^ренептори, то ефект буде прозапаль-
ним, а якшо Н2 — протизапальним. Пстамш, дгючи на Н,-рецепторы нейтрофгль-
них гранулоцитгв, гальмуе гхнгй хемотаксис, фагоцитну актившсть, вивгльнення
лтзосомних протешаз.
Про склада) вщношення мгж клпинами запалення евщчить такий факт. Базо-
фтльш гранулоцита пщ час дегрануляцп видгляють два медгатори запалення — пс-
тамш ) лейкотрген, а в еозинофгльних гранулоцитах мгстяться два ферменти. якг
руйнують гц медгатори, — пстамшаза й арил сульфатаза. Кргм того, у базофыьних
гранулоцитах е ще одна речовина, яка “приваблюе” еозинофыьш гранулоциты.
Це фактор емгграцп еозинофыгв. Урештг-решт еозинофыьш гранулоцита, якг
емггрують у зону запалення, руйнують медгатори, вшгглеш базофыьними грануло-
цитами, г тим самим “гасять” запалення.
Гол обними регуляторами пролёферацё!’ клётин е полшептиди, що мають назву
факторгв росту. Вони е лшандами, що взаемодёють зё специфёчними поверхневи-
ми рецепторами клггин, цитоплазматичний домен яких найчастёше мёстить фер-
мент тирозинкёназу. Зв’язування фактора росту з вёдповёдним рецептором приз-
водить до його димеризаци й активацп шляхом самофосфорилювання рецептор-
ного тирозину. Активований рецептор фосфорилюе й активуе каскад ферментёв ё
структурних бёлкёв. Основним ферментом тирозинкёназного каскаду е Вак-биток —
мембранна ГТФаза. Активацёя Вак зумовлюе низку активащй внутрёшньоклётин-
них кёназ, з яких першою е ВаГ-1 — серин-треошнкёназа, а дал! — тирозинов! та
треонёновё кёнази, що активують циклёни, циклёнзалежнё кёнази (ЦЗК), ядерн!
транскрипцёйш фактори та ёншё регуляторн! быки, необхёднё для стимуляцГё
транскритщп генёв, у тому числ! протоонкогешв, якг контролюють запуск клётин-
ного циклу, синтез рибосом ё трансляцию бёлкёв. Для активащ!' клётинного циклу
передусём необхёднё б!лки циклёни та ЦЗК. Комплекси активованих ЦЗК ё цик-
лёнёв фосфорилюють быки, що регулюють фазов! переходи клётинного циклу.
Так, для переходу ёз фази О] (пресинтетична) у фазу 8 (синтез ДНК) необхщнё
циклёни В ё ЦЗК 2, ЦЗК 4, ЦЗК 5, а для переходу в!д С2до М (мётоз) — циклён
В ! ЦЗК 1. Актившсть комплексёв ЦЗК ё циклёнёв своею чертою контролюеться
бёлковими ёнпбёторами ЦЗК (р!5, рё6, р21, р27 ё р57). Деякё фактори росту мо-
жуть вёдключати синтез цих ёнгёбёторёв.
Хоча фактори росту утворюються рёзними клётинами (лёмфоцитами, ендо-
телёоцитами, кератиноцитами, гладкими мёоцитами, лаброцитами, фёбробласта-
ми, тромбоцитами тощо), головним ёх джерелом е “альтернативно активованё”
макрофаги. Крём подёлу клётин фактори росту можуть впливати на диференцёацёю
й акгивацёю клётин, ’ёх хемотаксис, синтез ними елементёв мёжклётинного матрик-
су й ангёогенез. Фактори росту можуть мати загальну дёю або впливати на обме-
жений рёзновид клётин.
До найважливёших стимуляторёв регенерацёё та репарацёё', що мають широ-
кий спектр дёг, належать: епёдермальний фактор росту (ЕФР, ЕСЕ), фактор рос-
ту тромбоцитёв (ФРГ, РОСЕ), фактор росту фёбробластёв (ФРФ, ГОЕ), ТФР-а
(ТОЕ-а), ТФР-р (ТОГ-р), ёнсулёноподёбний фактор росту 1 (соматомедин С).
Вважають, що соматомедини опосередковують мётогенну актившсть та анаболёч-
нё ефекти соматотропёну — гормону аденогёпофёза, особливо в першё два деся-
тилёття життя. Своею чертою мётогенний вплив соматомединёв на хондроцити,
остеобласти, мёоцити опосередковуеться виробленням у тканинах полёамёнёв.
Тим часом, соматотропён, ёнсулён ё анпотензин II, а також 1Л-1 — 1Л-6 ё ФНП
за рахунок прямого стимулюючого впливу на пролёферацёю й анаболёзм клётин
рёзного типу теж можна умовно зарахувати до факторёв росту або, точнёше, до-
помёжних факторёв пролёферацп унёверсального призначення. Значний неспе-
цифёчний вплив на процеси репарацп чинять також гормони щитоподёбноё зало-
зи й андрогени. Так, у разё дефёциту тиреощних гормонёв ефекти соматотропёну
ё соматомединёв значно пригнёчуються.
До факторёв росту, що мають виражену специфёчнёсть впливу, можна зараху-
вати: судинний ендотелёальний фактор росту (СЕФР, ангёогенён, УЕСгЕ), фактор
росту нервёв (ФРН, А1СЕ), еритропоетин (ЕП, ЕРО), фактор росту кератиноцитёв
(ФРК, КОЕ), гепаринзв’язуючий епёдермальний фактор росту (ГЗ-ЕФР, НВ-
ЕСгЕ) тощо.
1нколи рецсптори факторов росту замгстъ тирозинкгназного мУстять серин-
треонУнкшазний домен, наприклад рецептор ТФР-|< Цей фактор росту 1 його
зоформи беруть участь у процесах ембрюгенезу, морфогенезу хряппв. кзеток У
статевих оргашв. Пщ час усього фглогенезу ТФР (3 трансформуе мультилотентнУ
та соматичю (дороелг) стовбуровУ юптини в рУзгп клггини-попередники та забез-
печуе тим самим тканинне рУзноманУтгя При запаленн! ТФР-р стимулюе синтез
быюв позаклгти иного матриксу та зламаних кгеток, справляв хемотаксичний
вплив на нейтрофши, моноцити, фУбробласти । гладком’язовг юптини. ОсобливУс-
тю цього фактора е те, шо вш затримуе клгтинний цикл У пригшчуе пролУферашю
та функшю лгмфоцитУв, ендотелУонитУв У епгтелУальних юптин.
Для запобггання угворенню надлишковоУ кшькостУ юптин Уснують й гншУ фак-
тори, що гальмуютъ клгтинний подУл. До них зокрема належать тканинепепифш-
нг глУконротегди — кейлона. Ц1 речовини синтезуюгься та депонуються в зрУлих
клгтинах. Коли останнУх стае мало, зменшуеться У кУлькУсть кейлонУв, а отже, ос-
лаблюеться ефект гальмування — розмноження юптин прискорюеться. Для вог-
нища запалення шсля альтерацп характерный е мала кУлькгсть зрУлих клУтин, а
отже, низький вмгст кейлонУв — депресоргв подглу. Коли внаслщок пролУферацп
кУлькУсть клУтин вщновлюеться, збпгьшуеться г концентрацпг кейлонУв, а отже,
посилюеться гх паракринний гальмУвний вплив на поды клУтин, здатних до про-
лУферашУ.
КрУм того, клУтинний подУл може лригнУчуватися внаслщок контактного галь-
мування. шо здУйснюеться шляхом передачу сигналУв вщ юптин або позаюгУтинно-
го матриксу через УнтегриновУ рецептори, описаю вище. ЗбУльшення кыькостУ
ютУтин призводить до контакту Унтегринових рецепторы з Ухнгми лУгандами, що
включае механУзми гальмування клпинного подглу.
1з гормональних УнпбУторУв росту варто вщмУтити глюкокортико'щи. КрУм
того, важливу роль у припиненнУ подшу клУтин У подальшому гх диференшюванн!
впшрае збУльшення в них сшввщношення цАМФ / пГМФ Зрештою, пригнУчу-
вальний вплив на клУтинну пролУферашю виявляють соматостатини через змен-
шення секрецп соматогрошну та соматомедингв.
Коли запальний фактор знешкоджено та / або локализовано, подальшУ проне-
си спрямоваш на те, щоб зону запалення, вщмежовану вщ решти тканин, очисти-
ти, а потУм замУнити п новою, здоровою тканиною, шо досягаеться шляхом реге-
нерашТ та репарацп.
Регенерация — пе про.йферащя (розмноження) попередникУв паренхУматозних
клУтин, що залишилися. У цьому випадку вщбуваеться замУна втрачених клУтин
клгтинами того самого типу. РегенерацУя характерна для тканин, юптини яких
посгУйно пролгферують (ештелгальна У кровотворна) У тканин, що в норм! мають
низький рУвень реплгкацп, але за необхщностУ пщ впливом вУпповшних факторУв
росту можуть розпочати компенсаторну гшерплазУю (гладком’язовг югУтини та па-
ренхгматознУ юптини печУнки, нирок г пщшлунковог залози).
У разг ушкодження тканин, юптини яких у постнатальний перУод не дУляться
або регенерують у дуже обмеженУй юлькостУ (нерьова г поперечно посмугована
м’язова тканини) та при деструкцп великих масивУв юптин з ушкодженням ба-
зальних мембран У позаклУтинного матриксу замппення дефекту вщбуваеться
шляхом репарацп, яка е комбУнашею регенерат) та фгброплазН — розмноження
мезенхгмних клгтин строми з ггодальшим диференипованням 1'х у фгбробласти.
продукцию фгбробластами колагену та гдгкозамшоглгкангв, з яких формуються
фгбрилярнг структури г грануляцгйна тканина, шо перетворюються на сполучну
тканину. Регенерацгя здшснюеться за рахунок активаш! дорослих (соматичних,
уншотентних) стовбурових клггин, як! мають обмежену здатшеть до диференща-
цц, але високу здатшеть до пролгферацп (наприклад, гемопоетичш сговбуров!
клгтини, печгнковг стовбуровг юптини тощо).
Вщновлення 1 новоутворення сполучно! тканини (фгброплазгя) здшснюються
за рахунок активацл прол1ферапп фгбробласпв, ендотелюцилв, гладком’язових
юптин пщ впливом ЕФР, ФРТ, ФРФ. ТФР-а, СЕФР тощо. Фгброплазгя, навпъ
при повнш загибелг тканини, може вщбутися в разг потрапляння в ушкоджену
тканину стромальних (мезенхгмальних) стовбурових клггин червоного ккткового
мозку, яю е мультипотентними. Таю клгтини пщ впливом ТФР-р можуть пере-
творюватися на фгбробласти, адипоцити, хондроцити, остеобласта, мюбласти й
енлотелюцити.
Головними ефекторами фгброплази е фгбробласти, яю активно прол1ферують
г синтезуютъ колаген, еластан, колагенасоцгйоваш битки та протеоглткани, шо
становлять основу мгжюптинног речовини сполучно! тканини. Нитки колагену в
мгжклгтинному простор! розмпцуються в певному порядку г виконують не лише
мехаючну (розтягнення, стиснення), а й структурну (порядок у розмгщеннг фгбро-
бластгв, ештелгальних та шших юптин) функцгю.
Фундаментальним процесом, який забезпечуе репарацгю ушкоджених тка-
нин, е англогенез, що може бути наслщком розгалуження г розростання наявних
судин або утворення нових судин зг стовбурових клиин кгеткового мозку. Вони
отримали назву “ендотелгальнг клгтини-попередники” (ЕКП). Пролгфсрацгя г мгг-
ращя ендотелюцилв 1 ЕКП та диференцгацгя ЕКП в ендотелгоцити стимулюють-
ся СЕФР (анпогешном). Своею чертою транскрипшя СЕФР та екепреегя його
редепгоргв стимулюсться деякими цитоюнами г такими факторами росту, як
ТФР-р, ТФР-а, ФРТ, а також транскрипцгйним фактором Н1Р, що шдукуеться
гшоксгею. Осганне пояснюе той факт, шо напрямок руху (таксис) ендотелюцилв
г ЕКП здшснюеться за градгентом кисню (у бгк, де його менше). Кргм того, ггпок-
егя гакож стимулюе екепреейо рз-гнтегришв на поверхш ендотелюцилв, що спри-
яе взаемоцГ! ендотелгоцити при утвореннг кашлярних цилгндргв.
Новоутворен! судини е ламкими г мають бута стабглгзоваш. Для цього мобщгзу-
ються перицита, гладком’язовг юптини та бглки позаюйтинного матриксу. Залучен-
ия периципв стамулюеться аьпопоетинами. гладком’язових юптин — ФРТ, а пщви-
щення синтезу бглкгв сполучно! тканини — ТФР-р. Кргм ф1бробластгв бглки позак-
лпинного матриксу можуть продукуватися гладком’язовими юптинами,
макрофагами, а деяю й ендотелюпитами. У разг ушкодження базагьно! мембрани
меди судив гладком ’язов/ клгтини пщ впливом ФРТ 1 ФРФ втрачаюгь мюзиновг фаб-
рили, пролгферують, мггрують г за участг ТФР-р, як г фгбробласти, виробляють ко-
лаген, еластан 1 глгкозамгноглгкани. Коли васкуляризацгя сягае максимального пгв-
ня, подальше утворення судин припиняеться пщ контролем загалъних г мгецевих
фактор!В, яю можуть як стимулювата, так г гатьмувата пролгферащю.
Мгграшя. пролгферапгя, поляржсть 1 функшональна актившегь клгтин зумо-
влеш гх взаемодгею з компонентами мгжюптанно! речовини. Угворення структу-
зж
рованих, функцюнально активних тканин можливе лише завдяки розшзнаванню
яюрних молекул колагенових 1 еластичних волокон 1 гл!козам!ногл!кашв базаль-
них мембран поверхневими структурами юптин. Зв’язок штегринових рецептор!в
юптин 1з позаюптинним матриксом здшснюють клейю колагенасоцшоваш гл!-
копротещи та протеоглткани, до яких належать глжопротещи — фибронектин,
ламшш, тромбоспондин та протеоглпсан — синдекан. 1нтегринов! рецептори, яю
через быки талш 1 вшкулш взаемодпоть 1з внутр!шньокл!тинним цитоскелетом,
зв’язують волокна м!жкл!тинно! речовини з актиновими мГкрофыаментами юп-
тин. Усе це забезпечуе вцщовлення щлосп, структурованостг та стабыъност! тка-
нин шсля ушкодження. Якщо цйпсть базальних мембран, головним адгезивним
быком яких е ламшш, не вщновлена або порушена взаемодгя цитоскелета юптин
1 позаюптинного матриксу, виникае порушення структури ! функцп органа з ут-
воренням остр1вщв паренхзматозних юптин м!ж надлишковими скупченнями
сполучно! тканини (як приклад — цироз печшки).
ЗАПАЛЕННЯ ТА 1МУНОЛОПЧНА РЕАКТИВШСТЬ ОРГАШЗМУ
У раз! дп на оргашзм бюлопчних факторгв великого значения набувае хмунний
стан оргашзму щодо збудника запалення.
Шсля попередньо! Гмушзацп збудником оргашзм перебувае в сташ гмуштету.
У раз! повторно! дп збудника в шфгкуючш доз! починають д!яги гуморальш та
юптинш 1мунн! реакцп, вщповщш БАР, яю знищують збудника ще до розвитку
первинно! альтерацп. Сам! БАР дезактивуються вщповщними системами оргашз-
му. Запалення в цьому випадку не розвиваеться.
У раз! проникнення збудника або стороннгх антигешв у не!мун!зований ор-
гашзм також спрацьовують системи гуморальних / кмтинних факторгв неспецифгч-
ного (вродженого) захисту. Ушкоджеш юптини, з одного боку, стають джерелом
р!зних БАР ! сигналов, яю розшзнаються цими неспециф!чними захисними факто-
рами. 3 1ншого боку, мжрооргашзми мають сво! постшш характерш ознаки (па-
терни), таю як переважання певних сахарщцв (маноза, фукоза) у вуглеводних спо-
луках, вщмшност! у структур! пептид!в, нуклешових кислот, фосфолшдав зовшш-
нього шару юптинних мембран, у будов! лшопол!сахарид!в та ппш ознаки, прита-
манш юптинам багатоюптинних оргашзм!в. Уроджен! мехашзми багатоюптинних
оргашзм!в (медгатори: л!зоцим, р-л!зин, фосфолшаза А^,, система комплементу,
колекгини, пентраксини, катюши быки (дефензини); юптини: дендритш юптини,
макрофаги, тканинш базофыьш гранулоцита, натуральна юлери, нейтрофыьш
гранулоцити) розр!зняють щ ознаки чужорщносп так само, як 1 продукта деструк-
цп власних юптин 1 тканин, 1 започатковують активащйш каскади, год час яких
вивыьнюеться велика юльюсть мед!атор!в запалення 1 вщбуваються ушкодження
та локал!зац!я патогенного фактора. Наприклад, манозозв’язувальш быки, про яю
вже згадувалося, розшзнають бактерп за високим вм!стом у вуглеводному компо-
нент! !хньо! юптинно! стшки манози, фгксуються на н!й, змшюють свою молеку-
лярну конформацпо й започатковують активацпо системи комплементу за класич-
ним шляхом, а також виконують роль опсошшв для фагоцитуючих юптин. Част-
ково активащя комплементу може вщбуватися 1 безпосередньо бактер!альними
речовинами за рахунок альтернативного шляху. Наслщком активацп комплементу
е вироблення низки БАР з опсонёзуючою та хемоатрактавною актившстю. На за-
вершальному еташ активацп комплементу утворюеться мембран оатакуючий комп-
лекс (С5Ь — С9). який пробивае клётинну спнку бактерп ё призводить до п заги-
бел±.
Пробивания бактерёально! стшки може вёдбуватжся також шд дёею досить
невеликих полёпептадёв — дефензишв. Додатково дефензини златнё активувати
систему комплементу.
Дендритнё клётини вёдгграють важливу роль у захисп оргашзму вёд чужорёдно-
го впливу. У поверхневих структурах органёзму (шкгрё. слизових оболонках) та
мёсцях найбёлып можливого проникнення патогешв (у лёмфатачних вузлах, се-
лезёнцё, легенях тощо) вони виконують функшю сьоерёдних “вартових”, якё пер-
шими зустрёчаютъ агресора.
Сигнали '‘загрози” мають юлька шляхёв подальше! реалёзацёё; 1) мгспевий ёз
виникненням ё розвитком характерних ознак запалення; 2) через печёнку з ёшша-
цёею та розвитком гостроё фази запалення; 3) через гёпоталамёчну дёлянку з по-
далыпим розвитком гарячковоё реакцп. змшами функцш ендокриннот та нервово!
систем, 4) мёграцёйний, яким активованё дендритнё клпини, шо поглинули сто-
роннё речовини (антигени), набувають здатносп до мгграцп лёмфатичними суди-
нами до репонарних лёмфатачних вузлёв. За час своёх “мандрёв” вони змёнюють
поверхневё рецепторы, перетворюються на АПК, якё осёдають у Т-залежних зонах
лёмфатачних вузлёв ё там активують антигенспецифёчнё Т-лёмфоиити, розпочина-
ючи таким чином специфёчну набуту ёмунну вщповщь. На первинну агресёю окрём
дендритних клётин реа1ують також тканинш макрофаги, моноцита кровз ё тка-
ниннё базофёльнё гранулоцита (мастоцити), якё також вивёльнюють у навколишнс
середовище багато БАР.
Флогогеном може стати ё комплекс антиген — ангипло. У такому випадку
розвиваеться алергёя (див. роздёл XII ‘'Алергёя”).
КЛАСИФ1КАЦ1Я
Ступёнь проявзв запалення залежить не ильки вщ причинного фактора, а й в!д
реактивносп органззму. Якшо реакцёя оргашзму на флогогенний фактор не вихо-
дить за межё пег, яка спостерегаеться найчастёше, таке запалення називають
нормерг1чним (Рессле). Коли за дальний фактор зумовлюе лише слабку затяжну
реакшю з переважанням альтерацп, таке запалення називаеться гтерг!чним. Це
спостерёгаеться, наприклад, пщ час голодування. Проте в деяких випадках перебёг
запалення наспльки ёнтенсивний, шо виникае невёдповёднёсть м!ж силою подраз-
яика ё реакпкю — вёдповёддю «мёсцевою ё загатьною) оргашзму. Таке запалення
отримало назву гтерерггчного. Його особливгсть полягае в тому, що найчастпие
воно розвиваеться на гмуннгй (алерпчшй) основ! (А.1. Струков).
Залежно вщ характеру домёнуючого месцевого процесу (альтерация, ексудашя
або пролзферащя) розргзняють три вили запалення.
При алыператпивному запаленн! переважають ушкодження, дистрофгя, некроз.
Воно спостерёгаеться здебёльшого в паренхёматозних органах при шфекшйних
хворобах, якг перебёгають зё значною ёнгоксикащею (казеозний розпад наднирко-
вих залоз або легень у хворих на туберкульоз).
Для ексудативного запалення характерне значке порушення кровообпу з яви-
щами ексудацп й емпрапп лейкоштпв За характером ексудату розрпняють серо-
зне, гншне, гемораг!чне, фюршюзне, злашане запалення. Якщо запалення охоппое
слизову оболонку, наприклад дихатьних шляхи або травного каналу. 1 ексудат
мютить багато слизу. це евщчить про катарапьне запалення.
Прол1феративне, або продук пшене, запалення визначаеться переважним роз-
множенням юптин гематогенного та пстюгенного походження. У запальнш зон)
виникають кгптинш шфыьтрати, як) залежно вы характеру накопичених юптин
подшяють на круглоюптинн! (л)мфопити, пстюпити), плазмоюйгинш, еозино
фыьно-юйтинн), ештелющно-юйтинн), макрофага.!ый. Пщ час запалення юйти-
ни 1з закшченим никлом розвитку (зрйй) гинуть, а мезенхгмн) юптини зазнають
трансформацп та диференцповання, унаслщок чого утворюетъея нова сполучна
тканина. Орган або його частина пронизуеться сполучнотканинними тяжами. У
ШЗН1Х стадых запалення це може призвести до пирозу.
За характером перебггу запалення полгляють на гостре та хрошчне. Гостре
запалення завершуеться за умови усунення причинного фактора. Однак, якщо
бюлопчний збудник мтсгиться в тканинах або флогогенний фактор е таким, що
не пщдаеться метаболтчним перетворенням (наприклад, кристали кремнпо при
силгкоз!, кристали урат) в при подагр), сторонн) тыа тощо), то запалення набувае
хрошчного перебггу. Для такого запалення характерними е ознаки альтерацп й
ексудацп, а також розгорнуто! пролтферацп. Саме таке поеднання загострень аль-
теративно-ексудативних ) прол)феративних проявив переважно мононуклеарно!
шфшьтрацп лгмфопитами ) макрофагами ушкоджених тканин визначае характер
хрошчного запалення.
ВПЛИВ НА ЗАПАЛЕННЯ НЕРВОВИХ I ГОРМОНАЛЬНИХ ФАКТОР1В
У явлен ня про роль нервово'1 системи в розвитку запального процесу залишаються
суперечливими. Так, теля перергзування тр)йчастого нерва у кроля розвиваеться
кератит У раз) ушкодження верхшх вузл)в симпатичного стовбура може виникну-
ти пневмошя (А.В. Тонких). Ушкодження дыянки сфого горба спричинюе поши-
ре нг запальн) ураження шюри ) слизово! оболонки, шо пояснюеться змшою гро-
ф)ки тканин ) ослаблениям ТхньоТ стшкост) до дп ушкоджувалытих факторгв
(О.Д. Сперанський). Наре шт), вщом) випадки, коли явн) ознаки запалення вини-
кали в людей, яким пщ ппнозом навтювали, що до шюри прикладають розпече-
ний предмет.
При цьому новокаТнова блокада або повна денерващя органа не гальмуе роз-
виток запалення при ушкоджених тканин Вплив центрально! нервово! системи на
запалення заперечують дат про розвиток запального процесу пш наркозом г в
гзольованих органах.
Про вплив ендокринно! системи на запальний пронес евщчать численн) юйнгч-
н! спостереження. Наприклад, при гйтертиреоз! перебгг запалення вщбуваеться
)нтенсивн1ше. при гшотиреоз) (мгкседема) — мляво. У хворих на цукровий дгабет
часто спостерггаеться фурункульоз. Особливпй вплив на перебй запалення справ-
ляють гормони гшоф)за 1 надниркових залоз. При цьому одш гормони юрково!
речовини надниркових затоз чинять дно, протилежну западению, тому гх було
названо протизапальними (глюкокортако'Уди), Уншг отри мал и назву прозапальних
(мгнералокортикощи). Гщрокортизон та гнгш глюкокортикощи ослаблюють за-
пальну реакцгю У тому набули значного поширення в клнпцУ. Така дгя глюкокор-
тикощгв пояснюеться там, що вони зменшують кглькУсть базофшьних грануло-
цит1в, зменшують актившсть пстидиндекарбоксилази й одночасно пщвигцують
актившсть ферменту, який руйнуе ггстамгн (ггстамгназа). Зменшуеться також ут-
ворення серотонгну за рахунок пригшчення гщрокортазоном 5-окситриптамгнде-
карбоксилази. Отже, год впливом гщрокортизону зменшуеться утворення двох
важливих медУаторУв запалення. Протизапальна дУя глюкокортикоццв пояснюеть-
ся також тим, що вони стабУлУзують мембрани лУзосом й шактивують кислу фос-
фатазу, рибонуклеазу та шип ферменти, якУ мУстяться в них. Пригшчення фер-
ментУв проявляеться також зниженням енергетичного потенцУалу клггин, змен-
шенням гх подУлу та рухомостУ (емгграцгя, фагоцитоз).
Глюкокортикощи Ундукують синтез специфУчних бонов (лУпомодулУну, макро-
кортину), якУ е УнпбУторами фосфолУпази тобто блокують синтез усге’г групи
похщних арахщоновог кислота (простагландишв г лейкотргешв).
Прозапальна дУя мгнералокортикощгв (альдостерону, дезоксикортикостерону)
пов’язана з там, що вони збУльшують ексудацпо. КрУм того, пщ гх дУею прискорю-
ються розмноження клгтин, утворення нових капУляргв, синтез основной речови-
ни сполучно! тканини.
ФОРМУВАННЯ ЗАГАЛЬНОТТЕОРП РОЗВИТКУ ЗАПАЛЕННЯ
Першу теорпо запалення сформулював Ю. Конгейм (1887). ВУн вважав, що всУ
клУнУчнУ ознаки запалення пояснюються порушенням мУкроциркуляци. Розши-
рення артерУол У збУльшений приплив артерУальног кровг у вогнище запалення
зумовлюють жар г почервонУння, пщвищення проникностг капУлярУв — припух-
лость, утворення гнфУльтрату — стиснення нервгв г бшь, а в поеднаннУ — порушен-
ня функцп. Судинна теор1я запалення Ю. Конгейма завдяки своУи зрозумшостг У
простотУ набула значного поширення. Сучасш електронно- та бУомгкроскошчнУ
дослщження дають новг пщтвердження основних положень цгег теорц.
Друга теория запалення, бюлоггчна, належить 1.1. Мечникову (1892). Для трак-
тування запалення вУн використав широкий бУологУчний пщхщ: запалення розгля-
даеться як реакцгя пристосування У захисту вщ шкУдливих факторгв. Великого
значения в механгзмг боротьби з “агресором” 1.1. Мечников надавав фагоцитозу.
УсУ попередники вченого розглядали запалення як процес мгсцевий. 1.1. Мечни-
ков уявляв запалення як процес, що розвиваеться в оргашзмг на всУх його ргвнях:
клгтинному (фагоцитоз), системному (Умунна система), цУлгсному (еволюцгя запа-
лення з еволюцгею органгзмгв).
Наступна узагалыгююча теоргя запалення виникла у зв’язку з глибокими фг-
зико-хУмгчними й патохгмгчними дослщженнями цього процесу. ДовУвши, що пщ
час запалення спостерггаються тканинний ацидоз У пщвищення онкотичного тас-
ку, Шаде (1923) запропонував ф1зико-х1м1чну теорио запалення. Саме з цих пози-
цш вгн пояснював вщому мгсцеву клУнгчну пентаду симптомгв запалення, визна-
чених ще лгкарями давнини.
В. Менкш (1948), використавши у власних дослщах передов! для свого часу
метода б!олог1чно1 х!м!1 ! техшки препарування, вид!лив специф!чн! для розвитку
запалення речовини: лейкотаксин, ексудин, шрексин, некрозин, фактор лейко-
цитозу та ш. Не все з того, що передбачав В. Менкш, пздтвердилося, однак вш
заклав пщтрунтя 61ох1м1чно1 теорп запаленця.
Нин! патогенез запалення розглядають значно ширше. в спроби узагальнити
накопляет дан! з цього питания 1 побудувати сучасну теорпо запалення, викорис-
товуючи дослщження молекулярно! бюлогп та медицини.
ЗНАЧЕНИЯ ЗАПАЛЕННЯ ДЛЯ ОРГАШЗМУ
Процес запалення, як 1 будь-який патолопчний процес, за своею суппстю су-
перечливий. Вш поеднуе 1 мобшзацпо захисних сил оргашзму, 1 ушкодження
(поломку). Виникнувши у филогенез! як явище пристосувальне, запалення зберег-
ло цю властив!сть 1 у вищих тварин. Оргашзм захищаеться вщ впдаву стороннгх 1
шкщливих для нього факторов вщмежуванням вогнища запалення вщ усього ор-
гашзму. Це запобшае поширенню 1 генерал!зацц запального процесу, зосереджу-
ючи боротьбу з! шкщливим фактором в одному м!сц!. Вогнище запалення не
т!льки ф!ксуе все, що мютиться в ньому, а й поглинае токсичш речовини, як!
циркулюють у кров!. Це пояснюеться тим, що навколо вогнища формуеться
своерщний бар’ер з одноб!чною проникшстю. Спочатку вш створюеться внасл!-
док закупорювання вщвщних лхмфатичних 1 кровоносних судин ! за рахунок бло-
кади позасудинного тканинного транспорту. Згодом цей бар’ер формуеться за
рахунок розмноження сполучнотканинних клггин 1 формування грануляцшного
валу на меж! нормально! та уражено! тканин. Захисна роль запального бар’ера
наочно продемонстрована в експеримент! з! стрихншом, смертельна доза якого,
уведена у вогнище запалення, не спричинюе загибелъ тварини.
У вогнипц запалення створюються несприятлив! умови для м!крооргашзм1в.
У цьому основну роль вщирають фагоцити 1 специф!чн! антипла, а також фер-
мента та основн! б!лки. Цшюпц властивосп запалення особливо чггко проявля-
ються в стадй прол!ферац!! та регенерацц.
Проте все викладене вище вщображае лише один (позитивний) б!к запален-
ня. Тнший, протилежний, полягае в тому, що запалення завжди несе в соб! еле-
мент руйнування. Боротьба з “агресором” у зош запалення неминуче поеднуеться
13 загибеллю власних юптин. У деяких випадках починае переважати альтеращя,
що призводить до загибел! тканини або навпь цщого органа. Ексудац!я може зу-
мовити розлад живлення тканини, ферментне п розплавлення, гшокс!ю та загаль-
ну штоксикащю. 1.1. Мечников писав: “Цшюща сила природи, головний елемент
яко! становии» запальна реакщя, ще не е пристосуванням, яке досягло доскона-
лост!”.
Уявлення про запалення як патолопчний процес, у якому захисне 1 власне
патолопчне перебувають у едносп та боротьб!, дае л 1 карев! стимул до дп.
Розд1л XII
АЛЕРПЯ
1сторично алерпю (вщ грец. — шший, ёрусуу — робота, д!я) було вщкри-
то як змшену реакцию оргашзму на дно речовин антигенно! природи, що призво-
дить до ушкодження тканин 1 р1зних порушень в оргашзм] — запалення, спазм',
бронхгв, некрозу, шоку тощо. Отже, алерпя — це порушення, спричинеш !мун-
ними реакщями. Осюльки мехашзми, що лежать в основ! таких змшених реакцш.
принципово не вщрхзняються вщ тих, яю використовуються оргашзмом для за-
безпечення гмуштету вщ шфекцшних ! нешфеюцйних антигешв, слщ визнати, що
алерпчш реакцп мають переважно юльюсш, а не яюсш вщмшност!.
Отже, алерг1я — це неадекватно сильна (гтерерг1я, гтерчутливгсть) або змшена
реакция оргашзму на дт речовин антигенной природи, що може спричинити ушкод-
ження тканин.
ЕТЮЛОПЯ
Причиною алергц е алергени, а також алерпзуюч! фактори, яю сам! не е алерге-
нами, але сприяють появ! в оргашзм! алергенгв.
Алергени — це антигени, як! називаються алергенами в тому випадку, якщо
вони спричинюють таку !мунну вщповщь, що призводить до ушкоджень в ор-
гашзм!. Алергенами можуть бути повш ! неповш антигени (гаптени). Неповш
антигени зумовлюють алерпю юлькома шляхами: 1) з’еднуючись !з макромолеку-
лами оргашзму та шдукуючи вироблення антиттл, специф!чшсть яких спрямована
проти гаптена, а не проти його нос!я; 2) формуючи антигенн! комплекси з моле-
кулами оргашзму. Антипла, що утворюються при цьому, реагують лише з комп-
лексом, а не з його компонентами.
Алерг1зуюч1 фактори — це фгзичш, х!м!чш та бюлопчю фактори, яю не е ан-
тигенами, а лише зумовлюють !х появу. У цьому випадку ф!зичш (тепло, холод,
юшзуюче випромшювання), хгмгчш (галогени, метали) та бюлопчн! (в!руси, ток-
сини) фактори спричинюють утворення алергешв з молекул оргашзму шляхом
демаскування прихованих антигенних детермшант або утворення нових антиген-
них детермшант унаслщок денатурацц цих молекул. 3 утвореними антиплами
демаскуючий або денатуруючий фактор не реагуе.
За повторно! д!! алерпзуючого фактора процес демаскування антигенних де-
термшант або денатурацц власних молекул повторюеться, виробляються антипла,
яю вступають у реаюцю з утвореними антигенами, г розвиваеться алерпчна реак-
ц!я. Наприклад, у деяких людей охолодження т!ла чи органа призводить до змши
Алергени
Екзогекш
Ендогешп
Бактериальна
— РоСЛИНН!
Природа
Набут:
Бактерп
— Л1карсыа
Мозок
Нешфекщйга
1нфекцпйп
Гриби
— Харчовг
Криыгалик
ХОЛОДОВ1
Прост) Комплексы)
В1руси
— Побутов!
Прост!
— Х1М1ЧН1
речовини
Статев! залози
Щитопод1бна залоза
Ошков!
Променев»
Тканина — микроорганизм
Тканина — токсин
Схема 8. Класиф1кашя алергешв (за А.Д. Адо)
конфшурацп крюглобул!н1в. унаслщок чого в гх молекулах з'являються антигеннг
детсрмшанта. до яких виробляються антипла. Шсля повторного охолодження
кр1оглобул1ни тим самим способом змшюють свою третинну структуру, у них
з’являються невластивг за нормально! температури антигенш детермгнанта, до
яких в орган!змг ьже е антипла. Це спричинюе виникнення реакцп антиген —
антител о г холодово! алергп. За подгбним мехашзмом може розвиватися теплова 1
променева алерпя.
Класифжащя алергешв. Алергени можуть бути класифгкованг за походженням
та за шляхом дц на оргашзм.
Класиф1кац1я алергешв за походженням. Розргзняютъ алергени зовнгшш (екзо-
алергени) та внутргшнг (ендоалергени; схема 8).
Зовнииш алергени своею чертою подгляють на шфекцгйнг, паразитичш та не-
шфекщйш. Надлишок шфекщй них антигешв може спричинити алерпчну реак-
шю. наприклад туберкулшовий шок у раз! парентерального введения туберкул гну.
Прикладом паразитичних алергешв е гнвазп гельмштами, шо завжди супроводжу-
ються алерпчними реакциями на гхш антигени. Нешфекщйнг алергени: хгмгчн!
(промислов!. побутов!, лгкарськг засоби, зокрема антибготики, знеболювальш пре-
парата г нав1ть деяю вгтамши, косметачнг засоби); росли нш (пилок кв!тгв, пил
перетертих рослин, борошно); тваринш (ешдермгс, видшення тварин, шр’я 1 пух
пгахгв), алергени комах (отрута бдж!л та ос, речовини, як! видшяють клопи, бло-
хи в рану щд час укус!в, видьтення гаргашв, токсини комах). З’ясувалося, що для
80 % хворих на бронхгальну астму алергеном е домашни пил. Головна роль при
цьому належить мгкроскошчним клипам Леппа1орка%рй1ех ргегопуззтиз г продуктам
гх жнттед!яльност1. як! мктяться в домашньому пилу. Часто бронхгальну астму
зумовлюе компонент кхмнатного пилу, що е вутлеводом — продуктом природного
або бактер1ального розпаду целюлози з бавовни. Цей атерген виявляють лише в
житлових прим!шеннях. У реши випадк!в анафыактичн! реакцп, у тому числх й
бронхгальну астму, спричинюють !шш алергени (пилок рослин, злущений еп!-
дерм!с, шерсть тварин), речовини, що потрапляюгь в оргашзм парентерально, а
також ентерально, зокрема й деяк! лпсарсый засоби — ацетилсалщилова кислота,
антитрин, морфш тощо. Серед пилкових алерген!в, як! е найчисленнппими
(близько 100), найбшыпу алерпзуючу актившсть виявляють пилок амброзп та
злакових трав.
Внутришй алергени: природа! автоалергени (автоантигени) та набут! анти-
гени. Природш автоалергени формуються в норм! у самому оргашзм!, наприклад
автоантигени головного мозку, кришталика ока, яечка, щитопод!бно! залози, к!р-
ково! речовини надниркових залоз, як! утворюються за пстогематичним бар’ером
! лишаються гзольованими цим бар’ером вщ !муноцит!в як у ембрюна пщ час
формування !мунолопчно! толерантност!, так 1 у здорових дорослих людей.
Набут! (внутршгш) алергени утворюються в самому оргашзм! пщ д!ею алерп-
зуючих фактор!в, наприклад юшзуючого випромпповання або йоду.
Класифгкацгя алергешв за шляхом дина оргашзм. Розр!зняють алергени ш’екщйш.
як! потрапляють в оргашзм парентерально, наприклад протиправцева сироватка
або новокаш; контакта! — речовини, що дпоть безпосередньо на шк!ру та слизову
оболонку, наприклад фарби, пол!мери, метали, косметичю засоби; рестраторш —
речовини, що надходять через дихальш шляхи, наприклад пилок квтв; харчов! —
дпоть через органи травления, наприклад антигени полунищ» 1 суниць.
ПАТОГЕНЕЗ
Р!зш за юйшчними проявами алерпчш реакцп мають сшльш патогенетичш ме-
хашзми. Розр!зняють три стадп алерпчних реакцгй: !мунолопчну, патох!м!чну
(бюх!м!чну) 1 патофьзюлопчну (стадпо функцюнальних 1 структурних порушень).
1мунолопчна стад!я алерпчних реакцш
нх класиф|кац1я
1мунолог!чна стад!я починаеться теля першого зггкнення оргашзму з алергеном
1 завершуеться його взаемод!ею з антиплами або Т-л!мфоцитами шсля повторно!
дп антигену. У цей перюд вщбуваеться сенсибппзащя оргашзму, тобто пщвищення
чутливост! та набуття здатносп бурхливо реагувати на повторне введения антиге-
ну. Перше введения алергену називаеться сенсибппзуючим, а повторне, яке без-
посередньо спричинюе прояви алергп, — завершальним. Установлено, що напря-
мок !мунно! реакцп, який може призвести до розвитку алергп анафшактичного
типу, визначаеться на початку гмушзацн сшввщношенням Т-хелпер!в 1-го типу
(ТЫ), як! гальмують синтез 1§Е 1 запобггають анафыаксп, та Т-хелпергв 2-го типу
(ТЪ2), як! активують синтез 1§Е 1 розвиток анафглаксп.
Сенсибшзащя бувае активною 1 пасивною. Активна сенсибййзацгя вщбуваеть-
ся в раз! гмушзац!! антигеном, коли у вщповщь вмикаеться власна гмунна система.
Мехашзми активно! сенсибйпзацп так!:
1) розшзнавання антигену, утворення на кожен антиген специф!чних гмуно-
глобулппв (антипл) 1 субпопуляцш Т-лгмфоципв, хоча кглыйсть !х може бути
разною, що залежить вщ регуляцн !мунно! вщповщ!;
2) розподщ антитш в оргашзм!, !х проникнення в тканини 1 фжсащя на тка-
нинних базофщьних гранулоцитах або циркуляц!я в кров! зумовлею тим, що ко-
жен клас гмуноглобулгнгв мае певнг можливост! ё пристосований до виконання
певних функцгй у певному простор! оргашзму: 1§М та 1§6 — у кров! та мгжклг-
тинному простор!; 1§Е та 1§О4 — у тканинах; 1§А г 1§Е транспортуються в слиз г
секрета; к!льк!сть 1§Ц збёльшуеться шд час ваптностг.
На характер алерпчноё реакцп впливають так! особливост! гмуноглобулгнгв:
1) здатшеть зв’язувата комплемент, яка максимально виражена в 1§М, помгрно —
в г майже вёдсутня в 1§Е; 2) здатшеть проникати в тканини, обмежена у ви-
сокомолекулярних 1§М, пом!рна — в 1§О, значна — в 1§Е та 1§В; 3) здатшеть до
сорбцёё на тканинних базофтлах значна в 1§Е ё властива 1§О4; 4) здатшеть до пре-
ципгтацп найвираженппа в 1§М та 1§Сг; 1§Е зазвичай е непрециштуючими анти-
телами (однак реакцп прециштацц, аглютинацн та флокуляцп залежать не ильки
в!д класу гмуноглобулгнгв, а й в!д властивостей антигену, тому з деякими антиге-
нами навгть 1§А можуть давати реакцпо прециштацц; 5) проникнення в секрета г
слиз (основним секреторним типом антитёл е 1§А, значно менше це властиво 1§С
г1§Е); 6) здатшеть проникати крезь плаценту, яка, з одного боку, вёдёграе важливу
роль в гндукцц гмунгтету в плода, з шшого — у виникненне емунного конфлёкту
меж матёр’ю ё плодом та розвитку алергёё у плода ё новонародженого. Таку здат-
шеть у людини мають переважно 1§О.
У разе повторного введения антиген реагуе з усёма специфечними антителами,
зокрема й на зазначених клгтинах. У гвёнейськоё свинки алергечну реакцгю можна
спричинита на 6—7-й день шсля введения сенсибелезуючоё дози алергену.
Пасивна сенсибиизацгя ведбуваеться в неёмунёзованому органёзмё шсля введен-
ия гуморальних антитёл або емуноцитев, одержаних з оргашзму, гмунёзованого
цим антигеном. Здатнесть вёдповёдата алерггчною реакцеею проявляеться зазвичай
за кёлька годин. Цей час потребен для розподёлу антитёл в органёзмё та фёксацп гх
на клгтинах.
Класиф1кац1я алерпчних реакцш
Класиф1кац1я алерг1чних реакцш за часом ёх розвитку. В емунолопчней стадп виде-
ляють негайний г сповельнений типи алерпчноё реакцп. За алерпчноё реакцёё не-
гайного типу гмуноглобулши — гуморальш антитела — поширюються в органёзмё,
виходять у тканини й секрета г в разё повторного введения антигену, ведразу ста-
каючись ёз ним, негайно залучаються до реакцёё антиген — антитело.
Алерггчна реакцгя сповёльненого, або клгтанного, типу розвиваеться пёсля
повторного надходження антигену в тканини, коли при сенсибёлгзацп утворю-
ються переважно Т-лгмфоцити. Гё ознаки проявляються протягом кёлькох годин г
посилюються впродовж перших двох дгб.
Алерггчна реакцгя обох тапёв може вёдбуватися одночасно проти багатьох ан-
тигенгв. Така алерггчна реакцёя мае змёшаний характер.
Класиф1кац1я алерггчних реакцш за Кумбсом I Джеллом (зг змёнами). Кумбс ё
Джелл (1968) в основу класифгкацп поклали мгеце перебування в тканинах анти-
гене в г антитёл г мгеце реакцёё антигешв з антителами ведносно клётин у тканинах.
За цими критергями ёмунолопчноё стадп автори видёлили типи алерпчних реак-
цш, наведенё нижче.
Тип I — анафёлактичш реакеш (мал. 29). Антитела зв’язаш зё свогми рецепторами
на тучней клётинё, а дребнодисперсний антиген надходить у тканини через судинну
Мал. 29. Загалып мехашзми розвитку алерпйноТ реакш! анафглактичного шоку
(за В.1. Пицьким, 1984):
1 — базофцтьний гранулоцит; 2 — еозинофгаьний гранулоцит; 3 — нейтрофцгьний
гранулоцит; 4 — тромбоцита; ЕХФА — еозинофыьний хемотаксичний фактор ана-
фшаксй; ЕХФПММ — еозинофшьний хемотаксичний фактор промЬкно! молекуляр-
но! маси; ПРСА — повально реагуюча субстанц1я анафшакси; ДАО — диамшоксидаза
(пстамшаза); ВНХФ — високомолекулярний нейтрофшьний хемотаксичний фактор;
ЛХФ1 ХФ — лшщний хемотаксичний 1 хемокшетичний фактори; ТАФ — тромбоцит-
активуючий фактор; ПГ — простагландини; Аг — антиген; Ат — антапло
стшку 1з кров! та через слизов! оболонки. Комплекс антиген — антипло утворюеть-
ся на юптинах, яю несуть антипла. Антителами е 1§Е та 1§С4, фхксоваш на лабро-
цитах (тканинних базофшьних гранулоцитах). Взаемод1я антигену з 1§Е та 1§С4
призводить до дегрануляцп цих юптин з вивгльненням мед1атор1в, яю збшьшують
проникшсть спнки судин, спричинюють М1грашю нейтрофыьних гранулоципв 1
еозинофцпв, бронхоспазм, секрецпо слизу тощо. Осюльки гомоцитотропш 1§Е та
1§С4 у людини здшснюють анафшактичш реакцп, вони отримали назву реагппв.
До цього типу реакцш належить анаф1лакс1я загальна та мюцева. Загальна
анафйгаксхя — це анафшактичний шок. Мтсцеву анафшакспо подшяють на шюр-
ну (кропив’янка, феномен О вер!) й анафшакспо шших оргашв (бронхзальна аст-
ма, сшна лихоманка).
Реакцп цього типу рашше називали атошчними. Ниш частина вчених нази-
вають атошчними и анафшактичш реакцп, до яких е спадкова схильшсть. Нап-
риклад, частина вар1анттв бронхеальноё астми, основою яких е анафыактична ре-
акция, може бути названа ще й атотчною бронххальною астмою в раз! виявлення
у хворого успадкування мехашзм1в, яю полегшують розвиток ще! хвороби.
Тал II — реакция питолву, або цитотоксично! ли. Антиген е компонентом юп-
тини або сорбований на иёй, а 1§О I1?М вёльно ниркулюють у кров!. Алерпчна
реакщя починаеться внаслщок прямо! взаемоди антитёл з антигенами юптинних
мембран ! активацп комплементу, що призводить до ушкодження клггин.
1ншим мехашзмом шггошзу е антитётозалежний фагоцитоз кл!тин-м!шеней
нейтрофилами ! макрофагами завдяки хемотаксичнш ! опсошчнёй дп ёмуногло-
булппв । фрагменте активованого комплементу, зокрема СЗЬ.
Кр!м того, юптини-мппен!, що покриваюгься малою юльюстю 1§С, можуть зне-
шкоджуватися за механтзмом антиплозалежно!’ клпинно! цитотоксичност! (АЗКЦ).
При цьому клттини-мшен! не фагошттуються, а знешкоджуються позаюптинними
механизмами, пригаманними нейтрофёлам, макрофагам 1 натуральним кёлерам (див.
оозды XI “Запалення”), яю зв’язуються сво'1'ми рецепторами з Ес-фрагментом 1§С.
За II типом вщбуваються гемолёз еритроципв при передиванн! несумёсноУ
кров!, 1мунне знешкодження пухлинних клпин, реакц!я вщторгнення трансплан-
тата та численш автоалерпчн! реакцп, зокрема аьго!мунн! анемп, автошунне руй-
нування тромбоципв при хвороб! Верльгофа, цитолтгична форма гломеруло-
нефриту, синдром Гудпасчера, за якого утворюються антитша до базальноё мемб-
рани ниркових юлубочкёв 1 легеневих альвеол. Цитотоксичну алерпчну реакцёю
зумовлюють 1 висок! дози АЦС.
Тип III — гмунокомллексш реакцп. Н! антиген, н! антипло при цьому не е
компонентами кл!тин, 1 утворення комплексу антиген — антитёло — комплемент
вщбуваеться в кров1 та мёжютётиннёй рщин!. Роль прециштуючих антител викону-
ють 1§М 1 1^гС! •
У раз! надлишку антитш швидко утворюються нерозчиню комплекси, шо
локал!зуються периваскулярно ! в судиннш стёнщ, у м!сш введения антигену I
спричинюють гостру запальну реакцпо. ГИсля активацп комплементу анафёлаток-
сини СЗа 1 С5а зумовлюють дегрануляшю лаброципв. артер!альну пперемпо ।
збшыпення проникносп судинно! стшки, птщпоють хемотаксис 1 активацпо ней-
троф!л!в Захоплення нерозчинних комплекс!в макрофагами стимулюе секрецёю
1Л-1, ФНП-а 1 активних форм кисню та ГМО що збшьшуе ушкодження тканин.
Таку реакцпо (реакция Артюса) продемонстрував Морис Артюс шсля внут-
ршшьошкёрного введения розчинною антигену пперёмушзованому кролю. За го-
дину з’являеться набряк 1 почервошння Попм виникае ёнф!льтрац!я нейтрофёла-
ми, яка за 8—12 год може розсмоктатися, а може спричинити некроз. Такий са-
мий мехашзм мають алерпчн! ушкодження легень у разг повторного вдихання
алергешв, зокрема й мёкозного походження; легеня фермера на антигени м!кро-
полтспори з шпснявого сёна, на антигени шших паразитичних грибтв. зокрема
роду Азрег&Пиг, професшн! алерпчн! ушкодження оргашв дихання антигенами
шкёри коней у конюх1в, вщповщними антигенами птахёв у пташниюв, шшими
антигенами бюлопчного середовища людини.
У раз! надлишку антигену утворюються др!бш розчинн! комплекси, яю мтс-
тять СЗЬ-фрагмент комплементу, яким вони зв язуються з рецептором СК1 на
еригроцитах людини, шо дае змогу транспортуваги Ух у печён ку. де комплекси
фагопитуються ! знешкоджуються макрофагами. Проте в раз! недостатносп ком-
понентов або дефектност! мехашзм!в активацп комплементу за класичним шля-
хом, порушення функпш фагоцит.в або зменшення Ух кёлькосп. порушення будо-
ви або блокади антиплами СЯ1 -рецептора еритропитёв та значного переванта-
ження ще! системи антигеном комплекси не видаляються, а циркулюють у кровг
та осщають у стшках судин у мгсцях зг зниженою швидюстю кровотоку (нирки,
суглоби, шкгра, судинне сплетения мозку тощо). Це спричинюе порушення мгкро-
циркуляцп г вторинну альтерацгю тканин активованими комплементом, нейтро-
фыами та макрофагами.
До реакцш такого типу належать сироваткова хвороба, системний червоний
вовчак, ревматощний артрит, вузликовий пергартернт, гмунокомплексний гломе-
рулонефрит, склерозуючий паненцефаломгелгг тощо. Особливгстю сироватковог
хвороби е те, що вона може розвинутися не лише шсля повторного, а й шсля
разового введения велико! юлькосп сироватки, коли П быки зберггаються в тка-
нинах до появи антиты до них.
Тип IV — реакцш гшерчутливостг сповыьненого типу (ГСТ). Цей тип гшерчутли-
востг являе собою неадекватно сильнг реакцп, що забезпечують клгтинний гмуштет.
Основна особливгсть тако! реакцп' полягае в тому, що з антигеном, який е внутргш-
нгм або зовнгшнгм компонентом юптин, взаемодпоть сенсибглгзованг Т-хелпери, що
пгсля стимуляцп АПК активують макрофаги, Т-юлери та шил юлернг юптини. Ре-
акцгя ГСТ не менш специфгчна щодо антигену, нгж реакцгя з гмуноглобулгнами.
Однак у тканин!, де вщбуваеться ця реакцгя, серед безлгчг юптин, яю руйнують ан-
тиген г тканину, виявляються лише юлька вщсоткгв Т-лгмфоцит!в, що можуть спе-
циф!чно реагувати з антигеном. Цей факт став зрозумыим шсля вщкриття лгм-
фоююв — особливих речовин, яю видыяються Т-лгмфоцитами г стимулюють шшг
лейкоцита до боротьби з антигеном. Завдяки гм навгть невелика кглькгсть тмунних
Т-лгмфоципв може стати факторами руйнування антигену нейтрофыами г макро-
фагами. Розвиваеться така реакщя протягом 24—48 год 1 проявляеться еритемою й
ущыьненням тканин за рахунок гх гнфыьтрацп, переважно мононуклеарами.
Прикладами реакцш такого типу е реакщя Манту на туберкулш, контактний
дерматит, псор!аз, укуси комах, гранульоми при туберкульоз!, лепр!, вгспг, кору,
герпес!, грибкових ураженнях, лейшмангозг тощо. Разом з реакцгею цитолву вона
бере участь у вщторгненнг трансплантата.
Тип V — стимулююч! та галышвш алерпчш реакцп, якг також мають назву па-
торегул яторн!, або рецепторзалежш. Унаслщок дп антиты на юптини, що несуть
антиген, функцп цих юптин або стимулюються, або гальмуються. Мехашзм таких
реакцш пояснюеться тим, що утворюються антитгла (насамперед 1§6), специфгч-
ш до юптинних рецепторгв, призначених для гормонгв або мед!атор!в. У випадку
подразнення рецепторгв виникае ефект стимулювання, а при блокад! антитыами
чи автогмунному комплементзалежному руйнуванн! рецептор!в дгя гормонгв г
медгаторгв гальмуеться.
Стимулююч! алерпчш реакш!. Стимулююч! антитыа фактично гмгтують дпо
стимулюючого гормону, але продукцгя гх не шддаеться нормальней регуляцн. У
норм! вироблення гормону вчасно зменшуеться або припиняеться за рахунок не-
гативного зворотного зв’язку при посиленнг функцп стимульованого органа чи
тканини. Однак лгмфоцити не мають такого зворотного зв’язку з функцгею орга-
на, який вони ушкоджують. 1муноцити продовжують синтез сво'!х стимулюючих
антиты, що спричинюе розвиток хвороби, тяжюсть яко! визначаеться тим, на-
скглъки важливий регуляторний мехашзм ушкоджуеться.
Саме цей мехашзм зумовлюе патогенез автогмунного тиреотоксикозу при ди-
фузному токсичному зобг (базедова хвороба), за якого антитгла реагують з глг-
копроте'/дною частиною репепторёв фолёкулярних еещокриноцитёв (тироцитев),
призначених для тиреотропёну (тиреотропного гормону гёпофёза), а ревень самого
тиреотропену в кров! дуже низький. При цьому до рецептора можуть вироблятися
одночасно два типи автоантит!л. Автоантитела до ганглёозидноё частики рецепто-
ра стимулюють вироблення ! секрецёю тиреоедних гормоны, а автоантипла до
його бшковоё частини спричинюють гшерплазйо залози. Ведоме випадки, коли у
хворого виробляеться тыьки один вид цих автоантитёл ё розвиваеться або тирео-
токсикоз без гёпергыазёё щитоподёбноё залози, або дифузна гёперплазёя залози без
геперфункцёё, що ведома як спорадичний зоб.
Ще одним прикладом стимулюючоё алергёчноё реакцп е едеопатичний прсу-
тизм, коли в жёнок виробляються антипла до тестостеронових рецепторёв, розта-
шованих на волосяних фолёкулах. Це спричинюе оволосшня за чоловечим типом.
При подразненн! автоантитыами енсулёнових рецепторгв може виникнути п-
поглёкемы.
Гальмёвнё алерпчнё реакцп теж досить поширеш. Доведено, що мехашзм алер-
пчних гальмёвних рецепторзалежних реакцш лежить в основ! багатьох хвороб,
патогенез яких ранипе був неведомий. Так, мёастенея зумовлена утворенням авто-
антипл до ацетилхолёнових рецепторы на мембран! м’язового волокна. Автоан-
типла блокують ё руйнують цё рецептори, що спричинюе м’язову слабкесть. Бло-
када автоантитыами р2-адренорецепторш у бронхёолах зумовлюе одну з форм
бронхёальноё астми. Руйнування або блокада автоантитыами ёнсулёнових рецеп-
торы на органах-мгшенях спричинюе виникнення однёеё з форм енсулённезалеж-
ного цукрового д!абету.
Тмушп реакцп проти трансплантата. Ведторгнення трансплантата вщбуваеться
через ведмееешсть антигешв пересадженоё тканини вщ антигешв, до яких в ор-
гашзм! рецишента сформувалася толерантнёсть.
За ступеней еволюцёйноё спорёдненостё розр!зняють: алоантигени (езоантиге-
ни) — антигени в осеб одного виду, гетероантигени — антигени в осёб р!зних
видев, ксеногенн! антигени — антигени чужорщних, зокрема ё штучних речовин.
Антигенш ведменностё у двох неёнбредних ёндивёдёв одного виду можуть ёсну-
вати щодо тисяч антигены одночасно. Проте не вс! вони мають однакову силу
(ёмуногеншсть). Найсильнёшё трансплантацёйнё антигени кодуються в локус! хро-
мосоми 6 у людини ё е антигенами ГКГС. Патогенез вщторгнення трансплантата
вщповщае загалъним закономерностям патогенезу алерпчних реакцш змёшаного
типу. Переважне значения мае клётинна реакцш сповыьненого типу. Гуморальш
антитела деють за цитотоксичним типом (класифёкацёё Джелла — Кумбса).
Пёсля пересадження опромёнешй тваринё алогенного кёсткового мозку здоро-
вого донора його Т- ё В-лёмфоцити починають ёмунну реакцёю проти тканин оп-
ромёненого реципёента. Цю реакцёю називають реакц1ею трансплантата проти
хазяша.
Патохёмёчна стадёя алерпчних реакцёй
Сутнёсть патохёмёчноё (бёохёмёчноё) стадц полягае в утвореннё або активацц БАР,
що починаються вёд моменту поеднанеы антигену з антителом (схема 9).
Водночас активуються також механёзми ! системи дезактивацёё БАР. Отже,
результат патохёмёчноё стадп залежить вщ балансу обох процесёв — утворення ё
Схема 9. Утворення БАР шд час алерпчних реакцш
(ПРСА — повольно реагиоча субстанция анафолаксп)
дезактивацп БАР. Цим результатом може бути алергоя, коли надлишков! колькосто
БАР спричинюють третю сталою алергочноо реакш! — стадию структурних о функ-
цюнальних порушень, або омуштет, коли руйнуеться збудник хвороби, а системи
дезактивацп БАР всзигають вчасно загальмувати щ речовини о не допустити уш-
кодження власних тканин оргашзму.
Патохомочна стадая алерпчних реакиш негайного типу. За алерпчних реакцш
негайного типу вщбуваються процеси, описан! нижче
1. Активацгя системи комплементу Акшвний комплемент мае ферментну ак-
тившсть. здатшеть руйнувати мембрани бактероальних I тканинних клгтин. зумо-
влюючи вившьнення нових БАР, активувати фагоцитоз, протеолггичш ферменти
кровь фактор Хагемана, дегранулящю базофыьних гранулоцитов.
Частина фракпш комплементу СЗа о С5а не входить до загального комплексу,
а дое як самостшн! БАР. зумовлюючи реакцию клгтин: дегранулящю базофольних
гранулопипв; адгезою до ендотелюцилв судин лейкоцитов, ок хемотаксис о актива-
щю; утворення та вивольнення АКР. ейкозанопив, катоонних болю в нейтрофолъ-
ними гранулоцитами о макрофагами; продукцию о секрешю макрофагами 1Л-1 та
МО; селективний вплив на розно субпопуляшо ломфонитов. модулювання рецирку-
ля цц, пролоферацн, диференпповання та обробку антигенов ломфонитами Компо-
ненти СЗа 1 С5а е анафолатоксином, оснування якого як мешатора анафолаксп
передбачали раноше. СЗа о С5а спричинюють збольшення проникност! стонки су-
дин о скорочення непосмугованоо м'язовоо тканини.
2. Активация фактора Хагемана — XII фактора зсщання кров!, який активус
систему зсщання крови, систему комплементу7 та протеол!тичш ферменти
кровь
3. Активации протео.йтичних ферментов кров! — трипсиногену, клазмшогену
(проф!бринол!зину), калгкрешогену. Ыолопчна актившсть цих фермента прояв-
ляеться розщепленням б!лк!в на полшептиди, взаемоактиващею один одного, ак-
тиващею фактора Хагемана, системи комплементу, базофщьних гранулоцита,
здатшстю ушкоджувата клпини тканин, руйнувати фтбрин за допомогою плазмг-
ну (ф1бринол1зину).
КалгкреУн як провщний фермент у кшшовёй систем] кров! вщщеплюе вщ гло-
булину кров. — кппногену — полшептиди. що отримали назву кппюв. До них
належать брадикшш г калщин, який пщ впливом трипсину перетворюеться на
брадикшш Брадикшш шдвищуе проникнгсть стшки судин, знижуе тонус 1 роз-
ширюс !х, зумовлюе спазм непосмуговано! м’язово! тканини деяких оргашв. е
медиатором болю. Калщин менш активный, н!ж брадикшш.
4. Збудження нервових рецептора болю. 1з репептор!в болю пщ час !х збуджен-
ня в тканину видыяеться полшептид Р — один тз найактивншгих медгаторгв запа-
лення. Полшептид Р ртзко зб!льшуе проникшсть стшки судин, активуе шип БАР;
зумовлюе дегрануляцпо базофщьних гранулоцита, активуе кшшову систему.
5. Активащя I вившьнення протеолтичних ферменгтв тканин — колагенази,
еластази, палурошдази.
6. Дегрануляцы базофиъних гранулоцштв, на яких сорбоваш 1§Е, унаслщок
приеднання до !муноглобулш!в антигену. При цьому базофшый гранулоцита
вившьнюють дв1 ]рупи БАР: 1) синтезоваш попередньо 1 зв язан< з гранулами —
пстамш, гепарин, брадикшш, серотонгн, фактор хемотаксису нейтроф!льних гра-
нулоцита, запальний фактор анафшакси, ферменти; 2) утвореш у пропес! дегра-
нуляцп з компонента мембрани — лейкотргени ЬТС4, БТЕ4, ЬТО4 (сумпл цих
трьох лейкотр!ешв — це ПРСА. ЫВ4 (ФХЕ), р)зн! за дгею (про- та протизапальш)
простагландины; ФАТ. Пстамш через рецептори Н, 1 серотошн, под1бно до бра-
дикш1ну, пщвищуе проникшсть стшки судин, скорочуе м’язи бронхгв, кишок,
матки, спричинюе бшь, свербгж., печшня, шок 1 некроз, впливае на шш! нервов]
рецептори. Бодночас вплив пстамшу на рецептори типу Н2 зумовлюе проталеж-
ний ефект. Гепарин перешкоджае зсщанню кровг, гальмуе вироблення антитш,
хемотаксис, ПРСА зумовлюе поступове, але тривале скорочення м'язгв бронх!в,
що мае важливе значения в патогенез! бронхгально! астми. Вившьнення пстамшу
й серотоншу вщбуваеться в процес! розпаду тромбоцита ! базофщьних грануло-
цита.
7. Пероксидне окиснення л1тд1в ! синтез похгдних фосфолтд1в. Пероксидне
окиснення лшщ!в клетинних мембран разом з фосфолшазою А, сприяе !х руйну-
ванню 1 вившьненню з них фосфолшшгв 1 жирних кислот, з яких можуть утворю-
ватися БАР. Вщомг дв1 групи лшщних БАР, якг беруть участь в алергп: 1) похщн!
фосфолш!д!в (похшн! 1-алк1т-2-апетил-фосфатидилхол!ну), зокрема ФАТ, який
спричинюе бронхоспазм ! водночас розширюе судини з! зниженням артергально-
го тиску, зменшуе в!нцевии кровоток; 2) похщш арахщоново! (ейкозатет рае ново!)
кислота — ейкозанощи. Метаболгзм арахщоново! кислота може вщбуватися дво-
ма шляхами. Перший шлях оксигенування — лшоксигеназний, при якому пщ
впливом ферменту лшоксигенази утворюються лейкотртени — представники двох
груп: лейкотртени В т лейкотртени С, В. Е (сульфолейкотртени). Останн! утворю-
ються в раз) перенесения цисте'шово! сульфогрупи вщ глугатюну. Спазм бронххв
при бронхтальнтй астмт зумовлюеться ПРСа (див. вище).
Другий шлях перетворення арахщоновот кислота — циклооксигеназний: пщ
впливом ферментов пиклооксигенази та пероксидази утворюються простэгланди-
ни т тромбоксани. Тромбоксан А?, що утворюеться в тромбоцитах, спричинюе
скорочення непосмугованих м’язтв г зумовлюе спазм судин т агрегацгю тромбо-
цита. Простациклин (ПП2), шо синтезуеться в енд отел юнитах. навпаки. тнпбуе
агрегацтю тромбоцита т розширюе судини. ПГЕ, лаброиипв т лейконита сприяе
нагромадженню цАМФ у клттин ах, зумовлюючи розел аблення бронхтв, розши-
рення судин I зменшеиня видшень тз базофыьних гранулоцита БАР. ПГЕ2а, нав-
паки, зумовлюе значний бронхоспазм, пщвищення тонусу судин, пщеилюе агре-
гацпо тромбоцита.
8. Нагромадження продукттв руинування клтшн кров! г тканин, яю як продукта
альтерацй додагково активують мехашзми запалення.
9 Розпад лейкоцигтв I вив1лънення л1зосомних факторов (див. розды XI “Запа-
лення”).
10. Змта активност! холшестерази / збглъшення вив1лънення аиетилхолгну.
11. Зм1на вмгсту електролитв. Пщвищення концентрацп юшв калпо т кальцтю
зумовлюе змтну збудливостт тканин.
Уст системи БАР пов’язант мтж собою насамперед мехашзмами взаемоактива-
цц, однак дпоть також мехашзми гальмування. У визначенш характеру алерпчних
реакций особливого значения можуть набувати окремт системи БАР: 1) залежш вщ
базофшьних гранулоцита (анафыакстя, бронхтальна астма, кропив’янка); 2) за-
лежш вщ комплементу (васкултт. пневмошт); 3) цитотоксичного типу (васкул!т.
пурпура).
Патохтмтчна стадтя алерпчних реакцш сповшьненого типу, Алерпчш реакцп
сповшьненого юптинного типу мають важливт особливостё реакщя пперчутли-
востт сповшьненого типу вщбуваеться в раз! безпосереднього контакту лтмфоцита
з антигеном. Якщо антиген е компонентом клттини, то до цте’т клттини прикршлю-
ються Т-юлери т спричинюють загибель клттини-мштент. Кшер видыяе перфорин,
який, подтбно до компонента комплементу, активно занурюеться в мембрану
клттини або бактерп т полтмеризуеться, утворюючи в мембран! наскртзш канали,
через якт в клттину некойтрольовано надходять вода, електролтти, що зумовлюе
швидку загибель юптини-мштеш. До того ж Т-ктлер видшяе гранзими — гранули
тз сериновими протеТназами, що через перфоринов, пори в мембрана входять у
цитоплазму юптини й активують групу каспаз (10 тишв серинових протетназ), з
яких каспаза 7 активуе в ядрт ендонуклеази. що руйнують нуктешов! кислота
ядра клттини. Т-юлер свотм рецептором апоптозу ЕаьйёапО активуе рецептор клт-
тини Раз, який також активуе каспази цитоплазми: ендонуклеази ядра, що приз-
водить до апоптотичнот смертт клттини.
11щ час контакту з антигеном Т-лтмфопити впробляють лтмфокгни, якт е БАР.
За допомогою лтмфоюнтв Т-лтмфоцити керують функцгею тнших лейкопита. Лтм-
фоктни бувають стимулюючими та гатьмтвними. Залежно вщ того на яю клттини
вони дпоть. видшяють п’ять груп .пмфоюшв.
1. Трупа А — лгмфокгни, що впливають на макрофаги: фактор шпбування мгг-
рацй макрофапв (Ф1М); фактор агрегацй макрофапв (ФАМ); фактор хемотаксису
макрофапв (ФХМ); фактор резистентностз макрофагов. ФХМ спричинюе, а Ф1М
пригнгчуе мгграцпо макрофапв з тканини, що зумовлюе гх нагромадження у вог-
нищг реакцп.
2. Трупа Б — лгмфокгни, що впливають на лгмфоцити: фактор бласттрансфор-
мацй; фактор допомоги (фактор хелпергв); фактор посилення; фактор супресй;
фактор перенесения (Лоуренса) та ш. Фактор хелпергв сприяе залучению до гмун-
ног вгдповгдг В- г Т-лгмфоцитгв шших субпопуляцгй, фактор супресй, навпаки,
гальмуе гмунну вгдповгдь або запобггае гй. Фактор бласттрансформацй зумовлюе
трансформацгю шших лгмфоцитгв, акгивуючи синтез нуклешових кислот. Особ-
ливу зацгкавлешсть викликае фактор перенесения; його видгляють з лгмфоцитгв
сенсибшзованого антигеном органгзму. Пгсля введения його в десенсибглгзований
органгзм з’являються лгмфоцити, здатнг специфгчно реагувати з антигеном. Фак-
тор перенесения може набути широкого застосування в лгкуванш гмунодефщит-
них хвороб.
3. Трупа В — лгмфокгни, що впливають на гранулоцита: хемотаксичний фактор;
фактор гнггбщй. Хемотаксичний фактор зумовлюе, а фактор гнпбгцй пригнгчуе
емгграцгю лейкоцитгв, що, як г за дй Ф1М, може сприяти накопиченню лейко-
цитгв у вогнищг реакцй.
4. Трупа Г — лгмфокгни, якг впливають на клгтиннг культури: гнтерферон, що
пригнгчуе синтез нуклешових кислот г захищае клгтину вщ вгрусних шфекцгй;
фактор, який пригнгчуе пролгферацпо клгтин культури тканин, та ш.
5. Трупа Д — лгмфокгни, що дгють у цшсному органгзмг: фактори, що спричи-
нюють шкгрну реакцгю г сприяють пгдвищенню проникностг стшки судин, роз-
витку набряку, проникненню лейкоцитгв у тканину. Пгд впливом лгмфокгнгв нав-
коло антигену протягом кглькох годин накопичуються лейкоцита — макрофаги,
лгмфоцити, гранулощгти, змгнюеться проникшсть стшки судин г розвиваеться
запальний процес. В гмунокомпетентшй тканинг (лгмфатичнг вузли, селезёнка та
ш.) спостерггаються бласггрансформацгя, активацгя синтезу антатгл г утворення
Т-лгмфоцитгв. 1з факторгв, якг утворюються лгмфоцитами, видгляють трупу штер-
лейкшгв (1Л-1 — 1Л-6 та ш.).
В органгзмг гснують мехашзми дезактивацп' БАР г захисту оргашв-мшгеней вщ
гх впливу.
1. Припинення секрецп БАР: цАМФ гальмуе дегрануляцпо базофгльних грану-
лоцитгв, ггдрокортизон пригнгчуе активацгю лгзосом г утворення лтзосомних фер-
ментгв.
2. Пригшчення БАР (протеолгтичних ферменпв кровг — трипсину, плазмшу,
калгкрегну, а також комплементу). а2-М — шггбгтор протеолгтичних ферменпв
лгзосом лейкоцитгв г кшшовог системи; а,-антитрипсин — гнггбгтор трипсину та
хгмотрипсину; антитромбгн III г а2-антиплазмгн пригнгчують протеолгтичнг фер-
менти кровг, гальмугочи системи коагуляцй, фгбринолгзу г комплементу; ггстамш
через рецептори Н2 гальмуе актившсть Т-кглергв, секрецгю лгмфоцитами лгм-
фокгшв. 1нпбгтор продукцй ейкозанощгв — лшомодулш, який е посередником
дй глюкокортикогдгв. пригнгчуе фосфолшазу А^, що вивгльняе арахщонову кис-
лоту з лггпдгв мембран. 1нпбгтором широкого спектра дй е гепарин.
3. Руйнування БАР. 1снують системи руйнування вс!х БАР. Цю функцпо вико-
нують ферменти з вщповщною специфгчшстю: пстамшаза. карбоксипептидази,
протешази, холшестерази; ферменти руйнування вс!х ейкозанощш, наприклад
арилсульфатази А 1 В, руйнують за тюеф!рпим зв’язком лейкотрхен О4 у склад!
ПРСА; супероксиддисмутаза (внуцлпшьоклпинний фермент), церулоплазмш (у
кров! й мЬюсптиннш рщин!) руйнують супероксидний ашон О;, що е небезпеч-
ним окисни ком пщ час выьнорадикалъного окиснення.
Особливе М1сне в системах припйчення та руйнування БАР поедають еози-
нофетьн! гранулопити. Вони мйтять пстамшазу, арил сульфатазу та шпп системи
руйнування ейкозанощш, особливий "великий биток еозинофьшв”, за допомогою
якого дезактивуюгь найр!зноманггншп речовини.
4. Захист кл1тин-м1шеней вЮ впливу БаР за допомогою контррегулятор них
гормошв — антагошспв БАР (адреналшу. пдрокортизону) або шляхом змши
функционального стану клггин (наркозу).
Алерпчна реаки!я розвиваеться тодт, коли секретя БАР пщ впливом комп-
лексу антиген — анттгпло перевищуе можливост! систем дезактиваци БАР 1 захис-
ту кл1тин або коли комплемент 1 Т-кшери безпосередньо ушкоджують клггину.
Якщо БАР утворюеться бьтьше. юж може бути дезактивовано власними систе-
мами оргашзму, спостерйаеться лавинопод!бне наростання алерпчного процесу тз
розвитком шоку. Це пов’язано 31 здатшстю одних БАР активувати утворення шших
без учасп комплексу антиген — антитгто. Цим. очевидно, можна пояснити розви-
ток тяжких алерпчн их реакшй на поршняно низьк! завершальн! дози антигену.
1нтенсившсть утворення БаР залежить також вщ стану 1 спадково зумовлених
можливостей систем, як! утворюють БАР. Щ системи за функщональними особ-
ливостями, а шодт 1 яюсно вщрвняються в р!зних оргашзмах. Отже, у двох ш-
дивш1в з однаковими характеристиками !мунолопчно! стадп алерпчна реакция в
патох1м1чшй стадп може бути неоднаковою.
Г1этоф!з!олопчна стадия алерпчних резкий
Функшоналып 1 структури! роздали в органах пщ час алергп' можуть розвиватися
внаслщок прямого ушкодження клггин шмфонитами-кьтерами й активованим
комплементом, впливу БАР, шдукованих комплексом антиген — антитшо, а та-
кож повторно як реакшя на первинш атерпчш змши в будь-якому орган!.
Система кровообзгу. Пщ час алергп змшюсться робота серця, знижуеться ар-
тер!альний тиск, р!зко пшвищуетъея проникшсть судин. Може спостерйатися
раптова асистол!я. яку в експеримент! вдаеться спричинити введениям брадикш!-
ну Зниження артер1ального тиску зумовлене переважно д!ею брадикшшу й аце-
тилхолшу, а також пстам!ну, серотошну, деяких простагландишв. Бюгенн! амши
та брадикшш ищвищують проникшсть стшки судин, а тому пщ час алергп в ба-
гатьох випадках розвиваеться набряк. Окр!м розширення судин спостерйаеться !х
спазм у деяких органах. Так. у кролгв алерпчна реакцш проявляеться як спазм
судин легень.
Органи дихання. Кшши, серотошн, пстамш, ПРСА, ПГР2а спричинюють ско-
рочення непосмуговано! м’язово! тканини бронхш. Спазм бронхш призводить до
порушення вентиляцп легень, кисневого голодуваиня.
Система кровь Алерпя активуе систему зсщання кров! через активацию фак-
тора Хагемана, протизсщальну — завдяки вившьненню гепарину, фтбринолттичну —
шляхом перетворення плазминогену на плазмтн. Сумарний ефект порушення за-
дания кров! неоднаковий на ртзних ртвнях кровоносного русла. Пщ час анафшак-
тичного шоку кров в аорт!! великих судинах мае знижену здатшсть до зсщання,
тодт як у кашлярах спостертгаеться тромбоз.
Нервова система. Бтолопчно активш ам!ни й кшши за нормальних умов е
медиаторами больово! чутливост!. Навггь у дуже малих кшькостях вони спричиню-
ють бтль, печтння, свербтж, можуть впливати на тншт нервов! рецептори в крово-
носних судинах! тканинах.
У патофгзюлоггчнш стада алерпчнт реакцп посилюються в остб з дефщитом
гормонтв т речовин, як! е контррегуляторними щодо флогогенних (що зумовлю-
ють запальну реакцтю) БАР — катехоламштв, глюкокортико'щтв, кортикотрошну,
а також в остб з пщвищеною чутливтстю клттин-мштеней до дп БАР.
Мехашзми формування алерпчних реакцш
Кр!м алергентв у виникненш алерпчних реакцш мае значения стан оргашзму. У
зв’язку тз цим розртзняють два основних види алергп: у людей з первинно нор-
мальною тмунною системою ! в остб з первинним дисбалансом тмуннот системи.
Алерпя в людей тз нормальною тмунною системою за нормального утворення
антипл т БАР спричинюеться надмтрною кптьктстю антигену; и механтзм полягае
в перенапруженш тмуннот системи т систем утворення БАР. Установлено, що
ктльктсть антигешв, якт надходять в органтзм у природних умовах, дуже мала, т
надмтрним для нормально! здорово! людини може бути 0,1 мл сироватки коня.
Можливтсть розвитку алергп в здорових людей зумовлена, очевидно, перева-
жанням сумарно! потужностт систем утворення БАР пщ впливом велико! ктль-
костт комплекств антиген — антитшо над системами дезактивацп БАР. Причиною
тако! нертвноваги е те. що системи продукцт! БАР т протилежш !м системи дезак-
тивацп БАР формувались як ртзнт фактори природного вщбору. Еволюцтя зумо-
вила формування таких систем, якт можуть реагувати викидом БАР на повторне
потрапляння в органтзм навтть единичного мткрооргашзму. Осктльки збудник
(антиген) може проникнута в органтзм у будь-якому мтец!, то й антитша та
пов’язант з тмунними мехашзмами системи продукцт! БАР (наприклад базофшьш
гранулоцита) мали б бути наявними в устх тканинах оргашзму. Вщомо, що для
забезпечення встх тканин антиттлами органтзм виробляе в середньому 100 000
молекул антиттл на одну молекулу антигену. Кртм того, вст дтлянки оргашзму
забезпечент системами продукцп й активацц БАР. Сумарна здатшсть устх базо-
фтльних гранулоциттв оргашзму до викиду бтогенних амтнтв у разт одномоментно!
дегрануляци висока.
Отже, генералтзацтя активацп БАР завжди смертельно небезпечна т призво-
дить зокрема до розвитку шоку.
Однак природний вщбтр зумовив низыа функцтональнт можливостт систем
дезактивацп, здатних знешкоджувати портвняно невисокт дози БАР, осктльки в
органтзм у природних умовах потрапляе невелика кптъктсть (за масою) тнфекцш-
ного антигену. 3 точки зору шфекцшност!, 1000 мткрооргаюзмтв можуть вияви-
тися пгантською дозою у випадку особливо небезпечних шфекцшних хвороб.
Але маса 1000 мгкрооргашзмгв дуже мала як кглькгсть антигену. Цим г пояснюеть-
ся переважання потенцгйног потужностг систем утворення БАР над системами гх
дезактивацп.
Постае практичне запитання: яка найменша кглькгсть антигену вже спричи-
нить алерпю, оскыьки ця доза антигену доргвггюе кыькосп БАР, що перевищить
потужшсть систем дезактивацп БАР. Вщомо, що завершальна доза, здатна спричи-
нити в здоровому оргашзм! смертельний анафыактичний шок, приблизно в 10 ра-
згв перевищуе сенсибыгзуючу дозу, яка становить 10-6 мл (0,07 мкг б1лк!в) сиро-
ватки коня. Ц1 фактично дуже низыл дози 1 е надлишком антигену для здорово!
людини. А в сучасних умовах можливе значно быьше навантаження антигеном,
зокрема в раз! введения знеболювальних речовин, антибютикгв тощо. Так, парен-
терально введения 2 мл 2 % розчину новокашу чи лщокашу у сенсибыгзовано!
людини може спричинити смертельний анафыактичний шок.
Потрапляння антигену в оргашзм у високих дозах, здатних зумовити розви-
ток алергп, можливе при парентеральному введенш вакцин г лгкарських засобгв.
Сприяе цьому 1 недостатшсть вироблення 1&А або його секреторного компонента
8, а також пщвищення проникност! стшки судин або слизово! оболонки бронхгв
г кишок. Так, вщомо, що у дней раннього в!ку, на вщмшу вщ дорослих, через
кишки можуть потрапляти в кров високомолекулярш нативш б!лки. Це спричи-
нюе збыьшення частоти виникнення харчових алерпй у дней цього вису.
Навгть у раз! потрапляння незначних кыькостей антигену в оргашзм через
шк!ру 1 слизов! оболонки вш може накопичуватися в кров! г тканинах на тлг
ушкодження систем, як! забезпечують його знешкодження г виведення. Оскыь-
ки знешкоджують ксенобютики, головним чином, макрофаги печшки, а виво-
дять з оргашзму нирки, частою причиною розвитку алерпчних реакцш е хвор!
печшка або нирки.
Алерпя в людей з первинним дисбалансом гмунно! системи може розвинутися пщ
впливом звичайних г навгть зменшених доз антигену, отримавши як! здоров! осо-
би не захворгють. Основою цього е набутг або спадковг порушення регуляцп гмун-
но! системи.
Оскыьки для розвитку анафыактичних алерпчних реакцгй велике значения
мае кглькгсть 1§Е, то сшввщношення мгж ТЫ г Т1г2 може визначити напрямок
гмунно! вщповщ!. Так, з одного боку, синтез 1§Е активують Т-хелпери 2-го типу,
якг виробляють гнтерлейкши 1Л-4, 1Л-5 та 1Л-13, що стимулюють вироблення
гмуноглобулгнгв, а також 1Л-10, що гальмуе ТЫ. 3 шшого боку, ТЫ е контррегу-
ляторними вщносно Т112, оскгльки спрямовують гмуногенез у напрямку активацг!
клгтинного гмунгтету. Вони виробляють 1Л-2 г у-1Ф, як! стимулюють макрофаги,
Т-лгмфоцити-кглери г НК г гальмують ТТг2, а отже, ! синтез гмуноглобулгнгв. Мак-
рофаги виробляють 1Л-12, який збгльшуе кглькгсть Т-хелпер!в 1-го типу г тим
самим зменгггуе актившсть Тй2, гальмуючи вироблення 1§Е г розвиток анафг-
лаксгг.
Отже, у разг переважання активуючог функцп ТИ2 збглыцуеться вироблення
гомоцитотропних антитгл 1§Е та 1§О4, а алерггчна реакцгя стае анафыактичною.
Навпаки, при переважаннг активностг макрофапв г Т-хелпергв 1-го типу ан-
афглактична алерггчна реакцгя може не розвинутися. Така дисфункция гмунно!
системи може бути наслщком дц як спадкових чи вроджених, гак о набутих чин-
ниюв. Так, у жшок естрогени активують ТЬ2, що збзльшуе ’охню стойкость до по-
заклггинних збудников шфекцп, але одночасно пщвишус й моворшсть розвитку
алерпчних реакшй анафшактичного типу. Так само активность ТЪ2 збольпгена о в
людей, яю хворйоть або колись хворый на гельмштоз, осюльки пщвищення вп-
роблення необхщне для знешкодження ох еозинофыами.
Така ситуашя може виникати о в разо дефщиту Т-ломфоципв при шволюцП
загруднинноТ залози шсля сгресу або в сгаречому вщо, але за збереження здат-
носп виробляти гуморальш антипла. Стимуляция гмунокомпетентних клотин у
цьому разо сприяе локвщацп онфекцойнооо процесу 1 водночас — припиненню
алергочноо реакш'о.
АВТОАЛЕРПЧН1 ХВОРОБИ
Автоалерг/чш (автогмунш) хвороби розвиваються внаслгдок утворення антштл, як!
можуть взаемодгяти з антигенами власного оргашзму. Це може в спохватись у раз!
демаскування антигешв, усунення омуооолопчноо голерантносп та еоматичних му-
тацой Т-ломфощгпв.
Демаскування антигешв спостерогають у високодиференцойованих органах
(головний мозок, кришталик ока, яечко. шитоподобна затоза тошо) у раз! пору-
шення гостогематичних бар’еров. У цих органах моститься речовини, яю е авто-
антигенами, осюльки в перюд формування омунолопчноо толера нтност! ! падало
вони були озольовано вщ омунноо тканини псгогематичними бар’ерами. Пору-
шення бар’еров зумовлюе демаскування цих автоантигенов о вироблення до них
автоантиты та / або Т-юлеров. За таким типом розвиваеться руйнування Т-коле-
рами й автоантитыами тканини щитоподобноо затози при тиреощит! Хашомото.
Демаскування автоантигешв, прихованих усередино клотин о бюлопчних мак-
ромолекул, виявляеться в органах, де спешалозованих бар’еров немае. Под впли-
вом фозичних. хомочних 1 бюлопчних факторов можливими е виявлення потенцш-
но автоантигенних детермонант о виникнення авгоалерпчно! реакцп. Так, при
розе одному склероз! виявляються Т-колери й антитша, специфочно до компонентов
шелону нервово! тканини. При синдром! Гудпасчера виникають автоантитша до
компонента колагену IV типу, шо входить до базальноо мембрани легеневих аль-
веол о ниркових клубочков. При первинному болоарному цирозо виявляються анти-
тола до антигенов мотохондрой. При щоопатичной хворобо Аддосона в кров! спос-
терогаються автоантитша до антигенов надниркових залоз. При щюпатичному п-
попаратиреоз! утворюються автоанпотыа до питоплазматичних антигенов клотин
паратиреощних залоз.
Усунення омунолопчноо толерантиосто до нормально» компонентов тканин. Мож-
ливе, наприклад, усунення толерантное п до гаптенов свого оргашзму шсля зам они
носоя гаптенов. Вщомо, що до велико! колькосто антигенов свого оргашзму В-лом-
фоцити нетолерантно. Це пояснюеться вщмшностями в умовах формування ому-
нолопчноо голерантносп у Т- I В-ломфоцитов: у загрудниншй залоз! формуеться
толерантность толъки Т-ломфоципв, а В-ломфоцити нетолерантн!. Однак у норм!
В-лгмфоцити не виробляють автоантитша до компонентов власного оргашзму.
осюльки до них толерантно Т-ломфоцити. Пщ час зустроч! з цими антигенами Т-
хелпери не активуються т не всгупають у взаемод!ю з В-лтмфоцигами, без чого
нетолерантш В-лтмфопити не залучаються до шуннот вщповщ!.
Про те можливою е акгивацтя лшфоциттв на антиген власного оргашзму шсля
потрапляння в нього чужих макро молекул ярних ан тигешв. до яких приеднуються
власш. У пьому випацку Т-лтмфопити починають реагувати на чужт носп анти-
гешв 1 посднуватися з В-лтмфоцитами, а В-лтмфоцити своею чертою — на гапте-
ни власного оргашзму, що увтйшли в антигенний комплекс тз чужорщним ностем.
Очевидно, за таким механизмом мжрооргашзми 1 макромолекулярш речовини,
як! виконують в оргашзм] функцй носив гаптетнв, здатш шдукувати авгошунш
хвороби.
В обхщ Т-хелпертв В-л!мфоцити можуть безпосередньо активуватися високо-
молекулярними антигенами типу пневмококових пол1сахарид1в (полтклональш
або суперантигени) або огримати стимуляппо шсля аномально! експресп власних
антигешв ГКГС II класу кчттинами, як! в нормт не презентують антиген, напри-
клад ешгелюцитами печшки (гепатоцита).
Шодо багатьох речовин в органтзмт гпдтримуеться тмунолопчна голерангшсть,
в основ] яко’ — низькодозова активащя антигенами Т-супресорних функшй л!м-
фоциттв. Несприятливт вплпви на оргашзм, а також спадковт порушення можуть
спричиниги зниження супресорно! функцп Т-лтмфоципв та !мунну вщповщь
проти нормального компонента оргашзму.
Усунення 1мунолог1чно’| толерантностг можливе в раз! !мун!заш! близько-
спорщненими антигенами. Цей феномен може розвинутися за рахунок мткрогете-
роген носи клона !муноциттв т згасання супресорно! активность
Соматичн! мутатш в р!зних органах можуть призвести до появи клттин або
молекул, яю мають незвичайш антигенш властивость У бьтьшост! випадюв це
завершуегься швидкою елгмшашею мутованого клона соматичних клпин навпь
без розвитку алерпчно! хвороби. Проте, якщо наслщки мутацп збережуться, може
виникнути алерпчна реакщя на эмитент клпини чи молекули. Так, характерною
ознакою ревматощного артриту е наявшсть 1йМ, специфтчних до Ес-дтлянки 1§О,
що вщртзняеться аномалытим складом олтгосахаридното компонента Можна
припустили, що причиною змши будови Ес-дьтянки ]§С може бути соматична
мутащя В-тмфоцита.
Мутаци Т-л1мфоцит1в мають особливе значения, оскьтьки вони можуть стати
причиною розвитку автотмунних захворювань у зв’язку з тим, шо призводять до
появи заборонених клонтв Т-лтмфопиттв, яю сприймають нормальш компонента
оргашзму як чужорщн! антигени т можуть атакувати тх. За таким типом може роз-
виватися руйнування Т-кшерами 0-клпин остр!вптв Лангерганса при жсулшза-
лежному цукровому дтабетт типу 1.
Мутацп Т-л!мфоцит!в первинно можуть зумовити ослабления ’!х супресорно!
функцц шодо В-л1мфоциттв, яю не мають толерантност! до власних антигешв.
Унаслщок цього нетолерантш В-л!мфоцити починають реатувати на нормальш
компонента тканин свого оргашзму. До авпимунних захворювань, яю можуть
виникати за цим типом, належать системний червоний вовчак, автотмунна ге-
молттична анемтя хвороба Верльгофа т лейкопентя. У патогенез! системного чер-
воного вовчака основне значения мають протиядерш автоантитша до дволанцю-
говот ДНК, яю реагують з компонентами ядер клпин кровз т тканин.
ПАРААПЕРГ1Я, ГЕТЕРОАПЕРПЯ
Рашше термоном “параатергоя” позначали ачерпчний стан, зумовлений одним
алергеном щодо шшого. Майже такий самий змост мало поняття ’Тетероатер-
г!я” як реакцп алерпчного типу в органозмо, сенсиболозованому одними речови-
нами, на завершальне введения шших речовин, частице бактерш або охнгх ток-
син ОВ.
Доведения того факту, що алерпчш реакцп обов’язково лочинаються з омуно-
лопчно! стадп, дало змогу А.Д. Адо видьлити справжш та несправжн! алерпчш
реакцп. Справжш агерг'ьчш реакцп починаються омунними механизмами 1 взае-
модоею антигенов з анти илами. У мехашзм! несправжшх алерг1чних реакцш не вщ-
буваеться реакпоя алергену з ангитшом.
Отже, за еучасними уявленнями, параалерпя (алергощн! реакцв) ! гетероалер-
|тя — це трупа реакцш, за проявами о способами вщ творения подобних до алергп, що
однак вщрозняються вщ не! вщсутюстю омунолопчно! стадп о реакцп антиген —
антитшо. Алергощн! реакцп виникають досить часто Так, близько 40 % випадюв
бронхоалъно! астми в д!тей, як! проживають у мост!, зумовлеш несправжньою
алерпчною реакшею.
Виникнення параалерпчних реакцш пояснюеться тим, що речовини, як!
спричинюють параалергпо, здатн! активувати вироблення БАР без взаемодп з ан-
титщами, даючи кл!шчн! прояви, подобно до алергп. Зокрема параалергпо здатн!
спричинювати лобератори пстамону — фактори, под прямим впливом яких водбу-
ваеться деорануляцоя лабропипв 1 базофольних гранулоцитов кров! без участо 1§Е.
До таких факторов належать ацетилсалщилова кислота, ошати, цитостатичш пре-
парата, мюрелаксанти, радооконтрастно речовини, нестерощш протизапальш пре-
парата, холод, оошзуюче й ультрафюлетове випромшювання. фозичне наванта-
ження, компоненти комплементу СЗа о С5а. автоантитша до 1§Е, автоанттгпла до
фрагмента Есе-Т рецептора до омуноглобулшу Е на лаброцитах, 1Л-3, 1Л-8, деяк!
хемокши та окрем! компонента продуктов харчування, наприклад полуничний
лектин.
БАР можуть утворюватись о без дегрануляцл лабропипв. Так, алергошний
астматичннй синдром, шо досить часчо виникае при вживанш ацетилсалщиловоо
кислота, зокрема зумовлений тим, що блокада циклооксигенази призводить до
зменшення продукцй протазапальних простагландишв о неадекватного збщьшен-
ня синтезу лейкотроенов з арахщоново! кислота.
Алергощш реакцп можуть виникати о при порушенш балансу систем актива-
ц!! о дезактивашо БАР. Можуть посилюватися утворення й активапоя БАР, вившь-
нення БАР, наприклад за зростаючоо дегрануляпп базофшьних гранулоцитов. По-
рушення систем дезактивацп та пригшчення БАР спостерпаються в раз!: недо-
статнього утворення шпботоров БАР, наприклад, у випадку набряку Квшке, який
розвиваеться в раз! спадкового деф!циту шпботоров калокреону! деяких фрагментов
комплементу; дефщиту ферментов, як! руйнують БАР: порушення функцгй ор-
гашв. шо дезактивують Б/тР.
Параалерпею варто називати несправжн! алерпчш реакцп, зумовлеш введен-
иям в оргашзм одшео! тосо само! речовини, гетероатерпею — реакцп оргашзму,
“подготовленого" од шею речовиною, на провокуюче введения оншо! речовини.
Вщ параалергп та гетероалергп варто вщр!зняти перехресн! алерг1чн1 реакцп, як!
розвиваються в оргашзм!, !мушзованому одними мжрооргашзмами, на введения
шших м!крооргашзм!в, що мають антигени, спшъш з першими.
Параалерпчн! реакцп можуть бути мюцевими 1 загальними. До мгсцевих ре-
акцш належить феномен Швартцмана, до загальних — феномен Санарелл!.
Феномен Швартцмана виникае в результат! уведення тварин! фшьтрату куль-
тури збудниктв черевного тифу в пиару, а за добу — внутрппньовенно. На м!сщ
першого введения виникае геморапчно-некротичне запалення. Основою цього
ефекту е сумарна токсична д!я ендотоксину збудника черевного тифу.
Феномен Санарелл! вщтворюють внутргшнъовенним уведенням нелетально!
дози ендотоксину холерних в!брюн1в, а за добу — фшьтрату культури кишково!
палички. Розвиваеться тяжка загальна реакц!я за типом шоку. П.Ф. Здродовський
показав, що аналопчний феномен можна вщтворити 1 за допомогою ендотоксишв
шших м!кроорган!зм!в. Феномен Санарелл! близький за мехашзмом до феномену
Швартцмана. Обидва феномени мають важливе значения в мехашзм! розвитку
шфекцшних хвороб.
ЗАП0Б1ГАННЯ АЛЕРГП. ДЕСЕНСИБ1Л13АЦ1Я
Запобпти розвитку алерпчних хвороб можна завдяки !золяцп оргашзму вщ потен-
щйного антигену, вщтворенню специфично! гмунолопчно! толерантност! або !му-
нодепресивних сташв.
Тмунолопчну толерантшсть спричинюють уведенням новонародженому
або ембрюну певного антигену. Г! вщтворення становить штерес для галуз!
трансплантологи, а також щодо запобйання реал!зацп спадково! схильност! до
алерпчних реакцш на якийсь антиген. У дорослих специф!чну !мунолопчну
толерантшсть можна забезпечити введениям дуже високих доз розчинного ан-
тигену (паралич Фелтона). Тмунодепресивний ефект можна зумовити, пригш-
чуючи вироблення антитш до багатьох антигешв. Цього можна досягги трьома
способами: опромшюванням, застосуванням !мунодепресант!в, як! гальмують
поды клггин 1 бшковий синтез, 1 специфтчних антшпмфоцитних сироваток
(АЛ С).
У випадку, коли сенсибшзац!я вже вщбулася ! почалася алерпчна хвороба,
уживають таких заход!в:
1) пригшчення синтезу антитш зазначеними способами;
2) специфична десенсибшзацгя за Безредкою, що дае змогу звшьнити орга-
шзм на деякий час вщ антитш до певного антигену, хоча 1 не повшстю. Мехашз-
ми зазначеного методу полягають у тому, шо десенсибшгзацно проводять невисс-
кими дозами антигену, як! добирають так, щоб не спричинити тяжк! реакцп. Ц!
дози антигену активують таку кшьисть БАР, яка не зумовлюе тяжк! ушкодження
в оргашзм!. Але лишаеться загроза накопичення БАР у раз! повторних уведень
антигену. Тому повторно дози вводить через певш штервали часу, протягом яких
в оргашзм! системи дезактивацп БАР шактивують бюлопчно акгивн! сполуки.
Шсля такого неушкоджуючого зв’язування антитш, достатнього для того щоб
чергове введения антигену не супроводжувалося реакщею, можна ввести основну
дозу антигену без виникнення алерпчно! реакцп. Найчастппе потреба в десен-
сиб!л!зацц за Безредкою виникае на введения чужорщних лжувальних сиро-
ваток.
Потргбно зазначити, що метод десенсибшзацп Безредки мало- або недтевий
у раз! ГСТ, яка опосередковуеться Т-л1мфоцитами;
3) шактиващя бюлопчно активних сполук. Для цього вводить !нпбпори про-
теолтгичних фермента, речовини, як! зв’язують пстамш, серотошн та ш.;
4) захист клпин вщ впливу БАР, а також корекцхя функцюнальних порушень
в органах 1 системах (наркоз сприяе захисту нервово! системи, уведення спазмол!-
тичних засобхв знижуе ступшь звуження бронхюл тощо).
АЛЕРПЯ I СПАДКОВЮТЬ
Спадковими можуть бути порушення структури та функцп хмуноцита 1 систем
утворення й дезактивацп БАР. Тому успадковуеться схильшсть до алергп. Однак
для виникнення алерпчно! хвороби потр!бен вплив алергену, без якого спадкова
схильшсть до алергп не реалгзуеться.
Успадковуеться також схильшсть до параалергп, але, на вщмшу вщ алергп,
можливе 1 пряме успадкування окремих параалерпчних хвороб. У деяких випад-
ках в осхб з! спадковими порушеннями мехаюзм!в патох!м!чно! стадп алергп —
систем утворення та дезактивацп БАР, тобто з параалерпею, остання може про-
являтись як самостшна спадкова хвороба без дп алерген!в. Наприклад, спадков!
порушення р2-адренорецептор1в можуть призводити до значного спазмування
бронх!ол. Але й у цих ос!б справжня алерпя пщ д!ею алергену розвиваеться знач-
но легше ! юпшчно проявляеться набагато тяжче, шж в ос!б 13 нормальними !му-
нолопчними ! б!ох!м!чними мехашзмами, у яких алерпя за тих самих умов дп
антигену не розвиваеться або спричинюеться надлишком антигену.
АЛЕРПЯ I ЗАПАЛЕННЯ
Комплекс антиген — антипло — комплемент е флогогенним (що спричинюе за-
палення) агентом. Коли антит!ла або клггини-к!лери реагують безпосередньо з
антигенами клпин оргашзму, наприклад у раз! цитотоксичних реакщй або алер-
пчних реакщй сповыьненого типу, комплекс антиген — антипло — комплемент
виконуе роль фактора, що зумовлюе первинну альтерацпо, з яко! починаеться
запалення. Якщо з утворенням комплексу антиген — антипло не вщбуваеться
пряме ушкодження клпини, то 1 в цьому випадку стимулюеться патох!м!чна стад!я
алерпчних реакцш, утворюються т! сам! БАР, яю е мед!аторами запалення 1 зумо-
влюють вторинну альтерацпо (див. розды XI “Запалення”).
Отже, комплекс антиген — антипло нал ежить до факторгв (гшокс!я, продукти
первинно! альтеращ! тканин, лхбератори пстам!ну), яю можуть за певних умов
спричинити мкцеве або генерал!зоване утворення бюлопчно активних сполук —
мед!атор!в запалення 1 шоку. У зв’язку 13 цим алерпя е таким вар!антом запален-
ня ! шоку, причиною якого, на вщмшу вщ шших запальних 1 шокових сташв, е
комплекс антиген — антипло.
3 еволюцшного погляду формування систем, як! об’еднали !мунш мехашзми
з мехашзмами запалення (базофйгьний гранулоцит, [§Е, комплемент, фактор Ха-
гемана тощо), вццграе важливу роль.
АЛЕРПЯ ТА 1МУН1ТЕТ
Основою !муштету в розумшш 1.1. Мечникова (як несприйнятливосп до шфек-
цшних хвороб) е функцгя !мунно! системи. Вшцим показником гмунгтету за нор-
мал ьних умов е те, що повторне надходження збудника в оргашзм не спричинюе
захворювання, а завершуеться пригшченням ! знищенням м!крооргангзм!в.
Принципова ргзниця м!ж гмуштетом 1 алерпею насамперед полягае в тому,
що ушкодження в оргашзм! при алергп зумовлюеться переважанням систем БАР
над природними можливостями систем, яю протидпоть БАР. Це проявляеться
навпъ у здорових людей на дгю надлишково! юлькосп антигену. Осюльки абсо-
лютна маса збудника, яка в природних умовах шфгкуе оргашзм, дуже низька,
потужшсть систем, що протидпоть БАР, у здоровому оргашзм! ссавщв е також
низъкою 1 значно слабшою поргвняно гз сумарними можливостями систем вироб-
лення БАР, якими необхщно забезпечувати весь оргашзм. Якщо в гмунний ор-
1мун1тет
V
1мунолопчна
реактившстъ
Алерпя
V
Запалення
Спйюсть до
шфекщйних хвороб
Фактори 1му штету
Утворення антиты
у В1дпов1дь на д!ю
антигешв
Флогогенш фактори
Утворення антиты
до шфекцшних
антигешв
Утворення антиты
до нешфекцшних
антигешв
Первинна альтеращя
Продукта
первинно! альтераш!
Фагоцитоз
Мурамщаза
1нтерферон
Бар'ерш системи —
(слизова оболонка, ппара,
гематопаренх1матозний
бар'ер)
Травш соки шлунка 1 кишок
Екскрещя (печшка, нирки) 1—
Температурний гомеостаз, рН
Гомеостаз газообмену
(кисень, вутлекислота)
Комплекс
антиген - антитшо
В1ДП0В1ДШ
антигени
Активацгя ферментов
Утворення
бюлопчно акгивних
* речовин
Бюлопчно активш
речовини
Система шактиваш!
бюлопчно акгивних
речовин г
Вторинна альтеращя
Алерпя
1муштет
Запалення
Схема 10. Взаемозв’язок мёж ёмуштетом, ёмунолопчною реактившстю, алерпею та запаленням
ганёзм уводити ёнфекщйний антиген у дозах, близьких до завершалъноё, шо зумо-
влюють алергёчш реакцп, то заметь ёмунётету можна спричинити ёнфекцёйну
алергёю. Отже, алерпя охоплюе й шфекщйну алергёю, перебёг я ко е вёдбуваеться за
загальними закономерностями алерпчних реакцёй.
ёндукцёя патохёмёчноё стали алерпчних реакцёй комплексами антител з тнфек-
пёйними антигенами залежить вёд кёлькзених спёввёцношень ё якёсних особливос-
тей антигенёв ё антитёл, а також вщ систем оргашзму, якё беруть участь у вироб-
леннё й ёнактивацп БАР. На цёй пёдставё можна припустити, шо стан специфично-
го ёмунётету визначаеться таким рёвнем процесёь в емунологёчнёй ё патохёмёчшй
стадёях. за якого не розвиваеться стадёя функиёональних ё сгруктурних порушень,
однак досягаеться знищення ёнфекцёйного збудника (схема 10).
Цей рёвень пропесёв ё стан ёмунётету встановлюються, коли системи дезакти-
вацёГ повнёстю ёнактивують БАР шсля того, як за ёх допомоги було знищено
збудника ёнфекцп. Алерпя розвиваеться тодё, коли шд впливом комплексу анти-
ген — антитело утворюеться така кёлькёсть БАР, що системи ёнактивацее БАР не
можуть ёх пригнёчувати ё тё починають дёяти не лише на збудника ёнфекцёё, а й на
тканини власного оргашзму. Розвиваеться вторинна атьтерацёя, яка ё е проявом
алергп.
Розд1Л XIII
ГАРЯЧКА
Гарячка (вщ грец. ттр — вогонь, жар) — типовий патологичный процес, який прояв-
ляешься тдвищенням температуря тиа вна&йдок перебудови терморегуляци у вгд-
повгдь на дт трогенних факторов.
В еволюцп гарячка виникла як реакщя оргашзму на шфекцпо, о тому кром
жару спостерогають шш1 ознаки, характерш для шфекцшного процесу. 1нтокси-
кацоя о самонагровання оргашзму створюють складну картину, у якш явища уш-
кодження та захисту поеднуються. Проте можливий розвиток гарячки внаслщок
ушкодження агентами нешфекцшно! природи (асептична гарячка).
У норм! терморегуляцоя здойснюеться за участю складно! системи нейрогумо-
ральних впливов. На периферп оргашзму (шкора, внутрппш органи) е холодов! та
теплов! рецептори, як! сприймають температурн! коливання зовношнього о внут-
рппнього середовищ о з яких надходить шформацоя в центр терморегуляци, роз-
мпцений у гшоталамусо. Нейрони, що мостяться тут, е чутливими як до тепла, так
о до холоду. Хнтеграцоя температурних сигнал!в з периферп о температура самого
ппоталамуса формуе так звану установочну точку, яка за нормальних умов доров-
нюе 37,2 °С — температур! ядра (внутрппш органи), а вже ефекторш мехашзми
визначають р!вень теплопродукци за рахунок змш обмшу речовин о м’язово! ак-
тивност! та регулюють штенсивность тепловщдач!, змшюючи интенсивность пери-
ферийного кровооб!гу, потовидолення (персшрашя) та втрати води з поверхн! ле-
гень, слизових оболонок.
Отже, гарячка починаеться з перебудови терморегуляци на вищий р!вень,
тобто пщвищення температури т!ла досягаеться за рахунок змши теплового ба-
лансу в бок накопичення додатково! кшькосто тепла.
ЕКСПЕРИМЕНТАЛЬНЕ ВИВЧЕННЯ ГАРЯЧКИ
Здатшсть регулювати сталкть температури т!ла, як о здатшсть до розвитку гаряч-
ки, у фологенезо виникли досить шзно. Вони характерш тольки для вищих гомойо-
термних тварин — тварин з постойною температурою тола, що не залежить вщ
температури навколишнього середовища. Гарячка чотко проявляеться у приматов
о особливо в людини. Органозм тварин, яко не мають стойко! температури тола
(гомойотерми), на дою патогенних факторов вщповщае лише слабкою о нетиповою
гарячковою реакцоею.
В онтогенез! того чи шшого виду тварин здатшсть вщповщати гарячковою ре-
акюоею формуеться по-розному залежно вщ сту пеня розвитку центрально! нервово!
системи до моменту народження, Тварини. як! народжуються зрйшми (копитн!,
гвкнейськг свинки, деяк! види птах!в), здатн! утримувати на гюстшному ргвн! темпе-
ратуру тгла вже з перших годин самостшного життя. Новонароджен! в людейг щур!в
не вщразу набувають цгег здатностг, терморегулящя розвиваеться у них поступово
Недоношен! дгти не мають гомойогермй 1 залежно в!д температури навколишнього
середовища або легко перегршаються, або персохолоджуються. Оргашзм новона-
роджених цупенят г кроленят у нерш! два м!сяц! життя реагуе слабкою й атиповою
гарячкою. У немовлят у перил м!сящ життя переб!г пневмон!! вщбуваеться за суб-
фебрилъног температури ила або взагал! без п пщвищення. Вщсутнгсть гарячки в
немовлят на ранньому етапг постнатального перюду пояснюегься насампеоед тим,
що в них ше недозргта ф!зична терморегулящя, тобто здатшеть швидко й ефекгив-
но обмежувати тепловщдачу. Гарячка в них не виникае г в раз! пщвищеного тепло-
утворення, оскгльки ще не сформувалися судинозвужувальнг реакцй шк!ри.
ЕТЮЛОПЯ
Розргзняють шфекцшю та негнфекцгйнг причини гарячки. У процес! еволюцшно-
го розвитку гарячкова реакщя формувалася насамперед як вщповщь органгзму на
проникнення м1крооргангзм!в та гхнгх токсишв. Водночас вщомо, що вона може
виникнути г в раз! потрапляння в оргашзм або утворення в ньому речовин, як! не
мають вщношення до шфекцп, наприклад, пщ час переливания кров!, уведення
61ДК1В для парентерального харчування, асептичного ушкодження тошо.
ГПрогенними (жароутворювальними) речовинами називають гак!, як!, потрап-
ляючи ь оргашзм або утворюючись усередию нього, спричинюють гарячку За
походженням гпрогенш речовини подшяють на екзогенш (бакгергальн!, небак-
тергальн!) й ендогенш (з клгтин-продуценпв макрофагово-моноцитного ряду, ней-
грофшьних гранулоцит!в тошо), а за мехаюзмом дй — на первинн! (гндукторнг) та
вторинш (справжнг). Первиню юрогени, проникаючи в оргашзм, ще не спричи-
нюють гарячку, а лише шщгюють цей пронес, спонукаючи власш юптини до ут-
ворення бьткових речовин (вторинн! шрогени), як! своею чертою дпоть на ме-
хашзми терморе гуля цп та викликають гарячку.
Отже. первинн! шрогени — це етголопчн! фактори, вториннг — патогенетичю.
Первиню шрогени проникають в оргашзм разом з шфекцшними факторами.
Пгрогенну актившсть мають практично вс! бактерй (як грампозитивнг, так г грам-
негативш), а також деяю вгруси, епгрохети, рикетей та плгенявг гриби. Найкраше
вивчено роль бактергальних токсин!в (екзотоксингв, ендотоксишв) як джерела
первинних пгрогеюв. З’ясовано зокрема, що ендотоксини грамнегативних бак-
терш е лшопо.дюахаридами — складними бгопол!мерами. Здатшеть спричинювати
гарячку мае лгпщна частина лшополгеахаридгв (лшощ А). Вивчення трогенних
властивостей останнього показало, що вже в юлькост! 0,0001 мкг на 1 кг маси тша
вш може зумовити гарячку у кроля. Людина, а з тварин собака та кгнь приблизно
однаково чутлив. до пгрогенног дй лшощу А.
Пгсля того як було помгчено. що пщ впливом пгрогеюв у тварин г людини
зменшуеться тяжкгеть перебггу багатьох хвороб, у тому числ! 1 злоякгених новоут-
ворень, сифшгсу тошо. гх почали застосовувати в клгнщг (шрогенал — отримано з
Рхеи&нпопаз аепррпоза, пгрексаль — гз 8а1топе11а аЬопизедш). Пщвищення темпе-
ратури т!ла у хворих виникае через 40—90 хв шсля парентерального введения
вказаних препаратов ! тривае 6—9 год.
Останшм часом шрогени почали добувати 1 синтетичним способом. При цьо-
му було шдтверджено, що 1х бюлопчна актившсть зумовлена лшщним складни-
ком (лшощом А).
Первинш шрогени можуть утворюватися в самому оргашзм! незалежно вщ
шфекцшних збудниюв (гарячка шсля перелому исток, пщ час шфаркту мюкарда,
гомологичного кризу тощо). Ц1 речовини утворюються внаслщок ушкодження або
руйнування власних тканин 1 впливають на оргашзм як первинш екзогенш, тобто
бактерхальш шрогени.
ПАТОГЕНЕЗ
Патогенез гарячки починаеться з того, що пщ впливом первинних шрогешв утво-
рюються вторинн!. Цей процес вщбуваеться насамперед у макрофагах (фпссова-
них 1 в!льних), а також у нейтрофыьних гранулоцитах 1 деяких шших юптинах.
Синтез вторинних шрогешв закодований у геном! юптин-продуценпв. Якщо
блокувати процес транскрипци РНК на ДНК-матриц! актиномщином В або пу-
ромщином — процес трансляцн б!лка на РНК, то синтез шроген!в не вщбуваеть-
ся. Бюсинтез шрогешв вщбуваеться шсля того, як первинш (бактероальш) шроге-
ни подпоть на клггини-продуценти 1 тим самим активують метаболочш процеси в
них. Ця д!я реал!зуеться через рецептори на мембранах юптин або в раз! проник-
нення токсину всередину макрофапв шляхом фагоцитозу чи шноцитозу. Зазначе-
ний процес, очевидно, не е суто специф!чним, осюльки синтез вторинних шро-
гешв може бути шдукований також впливом нешфекцшних шрогешв, тобто ре-
човин, яю проникають в оргашзм 31 стерильним материалом або утворюються
внаслщок ушкодження оргашзму поза шфекцшним процесом.
Доведено, що властиысть шдвищувати температуру пла та спричинювати
шип (м!сцев! та системш) ефекти мають БАР, вщом! пщ загальною назвою ци-
токлни. Серед них чи не найбшьше значения мае 1Л-1 — гормонопод!бний б!лок,
який вид!ляеться юптинами макрофагально-моноцитного ряду, ендотелюцитами
та деякими спещальними юптинами (кератиноцитами, дендритними юптинами)
шсля !х активацп. До вторинних шроген!в, кр!м того, належить 1Л-6 (продукуеть-
ся моноцитами, ф!бробластами, ендотелюцитами), ФНП-а (кахексии, мюцем ут-
ворення якого е моноцити, тканинш макрофаги та базофыьш гранулоцити, л!м-
фоцити), катюнн! быки (продукт специфтчних гранул нейтрофыьних грануло-
цитов), штерферони, колошестимулюючий фактор (КСФ) тощо. Шрогенна
здатшсть вторинних шрогешв неоднакова: найбыьша вона в 1Л-1, приблизно
вдвочо менша у ФНП-а, ще менша — в 1Л-6. Характер взаемовщношення м!ж
наведеними мед!аторами гарячки досить складний. Доказом цього можуть бути
стамулюючий вплив ФНП-а (пщ час пухлин, бакгер!альних о паразитичних ш-
фекцш), у-штерферону (у раз! ворусних шфекцой) на утворення 1Л-1 моноцитами
та ендотелюцитами, вплив 1Л-1 о ФНП-а на утворення 1Л-6.
До клотин-мошеней 1Л-1 та шших цитокшов належать нейрони, а також лом-
фоцити, ештелюцити печшки (гепатоцити), фобробласти, синовщити, хондроци-
та, мооцити та ш. Побутуе думка, що на мембранах цих юптин е вщповщш рецеп-
Схема 11. Патогенез гарячки
тори, Д1я на як! ЦИТОК1Н1В або Тх посереди глав зумовлюе пщвишення температури
тьда (схема 11). Зокрема. коли вдеться про ппоталамус, де розмпцений центр тер-
морегуляцп. то безпосередньо в контакт гз нейронами нього центру 1Л-1, 1Л-6,
ФНП-а не вступають, осюльки не проникають крТзь гематоенцефалгчний бар’ер.
Однак пш Тх впливом на ргвш мозкових артерюл Т капыярТв утворюються ПГЕ. 1
ПГЕ2, як! проходять кр!зь бар’ер 1 безпосередньо впливають на центр терморегу-
ляцц. Як наслщок зм1нюеться установочна точка центру, шо встановлюе темпера-
туру тгла на вищий рТвень, на якому вона 1 лишаеться доти, доки тривае синтез
наведених медТаторТв гарячки.
Гарячка — патолопчний процес, до якого залучена не лише система терморе-
гуляцп. а й пшп системи, насамперед гмунна. Це й зрозумыо. якщо врахувати. шо
гарячка в еволюцн виникла ж вщповщь на гнфекцно. Привергае увагу також
зв’язок мхж гарячкою х западениям. Можна сказати, що гарячка, хмуштет х запа-
лення — це своерщна трхада, яка визначае вщповщь на шфекцшний вплив х тка-
нинне ушкодження шшох природи. Зв’язок мхж ними трьома реакщями такий
пений, що вони хстотно доповнюють одна одну. Цей зв’язок найбихыи чггко про-
стежуеться у виникненю “гострофазно! вцщовцп”, медхаторами яко! е 1Л-1, 1Л-6,
ФНП-а 1 простагландини. П виникнення та розвиток пов’язують хз синтезом у
печппц та макрофагах “гострофазних” бхлкхв (СРП, компонента системи компле-
менту, транспортах быки, у тому чиелх й церулоплазмш, быки системи коагуляци
та антикоагулянтах быки, шпбхтори протешаз тощо).
Клхнхчними х патофхзюлогхчними проявами реакцп “гострофазно! вщповщх” е
збыыпення ШОЕ, агрегацшних властивостей клггин кров! та попршання реоло-
гхчних властивостей останньох в цхлому, порушення в систем! гемостазу, посилен-
ня антиоксидантно! та антимикробно! здатносп. 1ншими важливими ознаками
“гострофазно! вщповщх” е лейкоцитоз унаслщок стимуляцн гранулоцитопоезу та
моноцитопоезу в червоному юстковому мозку пщ дхею 1Л-1, 1Л-6 та КСФ, поси-
лення адгезп вказаних видхв лейкоципв, !х мхграцп до мхеця ушкодження, акти-
вацхя фагоцитозу. При шфекцхйнш гарячщ посилюеться також лтмфопоез, вщбу-
ваеться активацхя клхтинно! та гуморально! ланок хмунхтету.
Нарешп, до проявхв “гострофазно! вщповщх” пщ час гарячки належить також
посилення (пщ дхею Ш-1) продукцп колагенази хондроцитами та руйнування
хряща, активацхя протешаз х ушкодження мхоцитхв посмуговано! м’язово! ткани-
ни, деградацхя протеоглгканхв ! демхнералхзацхя кхстково! тканини, б!ль у суглобах,
кхстках ! м’язах. 1з загальних клхнхчних проявхв гарячки слщ вщзначити також
втрату апетиту та порушення травления в юпиках (зумовлеш дхею ФНП-а), ви-
никнення сонливост! та маячних симптомхв унаслщок дп опхощних пептидхв,
синтез яких у рхзних дшянках мозку пщ впливом 1Л-1 також посилюеться.
РОЛЬ ЦЕНТР1В ТЕРМОРЕГУЛЯЦИ У РОЗВИТКУ ГАРЯЧКИ
Головним центром терморегуляци е гшоталамхчна дшянка, хоча ппнх вщдши цен-
трально! нервово! системи, починаючи хз сегментних центрхв спинного мозку !
закхнчуючи корою великого мозку, також беруть участь у хпдтриманнх стану го-
мойотерми. Установлено, що термочутливх нейрони (холодов! й тепловх) роз-
мццеш переважно в надзоровому ядрх передньо! дхлянки (поля) гшоталамуса,
куди надходить хнформацхя вщ периферичних (поверхневих ! глибоких) терморе-
цепторхв. Ця зона безпосередньо чутлива до температурних коливань. Доказом
цього е результати дослщжень з термодами — тонкими трубочками, як! вводили
у певний центр мозку х пропускали через них теплу або холодну воду. Пщ час
проходження тепло! води спостерхгали перебудову терморегуляци, спрямовану на
виведення тепла, унаслщок чого температура тиха знижувалася. У раз! охолоджен-
ня мозкового центру холодною водою температура тша, навпаки, пщвищувалася.
Роль задньо! дхлянки гшоталамуса проявляеться штегращею температурно!
шформацп та формуванням ефекторних стимулхв, що керують фхзичною х ххмхч-
ною терморегуляцхею. Руйнування и або всього гшоталамуса робить тварин пой-
кыотермними. Шсля руйнування передньо! дхлянки гшоталамуса через деякий
час вцщовлювалася здатшсть до виникнення гарячки.
Мал. 30. Учаси певних вцццлн) центрально! нервово! системи в терморе! утящ! (пе-
рер!зи позначено римськими цифрами):
1 — смугасте тыо; 2 — таламус; 3 — М1ст; 4 — довгастий мозок; 5 — слияний мозок
Олерацп, виконант на центральны нервовш систем!, довели значения й ш-
ших и вщдьтгв. Шсля псрертзування головного мозку вище гшоталамуса тварина
зберпала здатшсть до гарячки. Перер!зування, за якого гшоталамус вщокремлю-
вався вщ стовбура головного мозку, позбавляло тварину щет здатность Нарешп,
у разг перер1зування спинного мозку в груднш частиш здатшсть дс гарячки вщ-
новлювалася шсля виходу тварини з! стану сигнального шоку (мал. 30).
Перертз I (талам1чна клика) — терморегулящя збер)гаеться; перертз II (деие-
ребрована юшка) — терморегуляцн немае; перер1з III на р)вн! СУ11 (сшнальна юш-
ка) — терморегуляцтя вщсутня; перертз IV на р!вш Т, — фтзична терморегуляция
зникае, х1М1чна — залишаеться.
Центр терморегуляцн таким чином забезпечуе температурний гомеостаз, ур1в-
новажуючи процеси теплопродукш! та тепловщдачт. Це е можливим завдяки тому,
що центр терморегуляцн працюе як юбернетичний пристрЫ у точно заданому
режим!, 1 коливання температури (добов!) допускаються лише у вузьких межах вщ
установочно! точки.
Отже, оргашзм теплокровних е бюлопчним термостатом, температура якого
залежить вщ того, на яку точку виставлено терморегулятор, тобто вщповщний
центр головного мозку. Ця усгановочна точка може бути змшена в двох випадках:
у раз! надзвичайного впливу (перегртвання, гшотермтя, замерзания, ппокс!я),
коли цей мехашзм повшстю або частково виводиться з ладу, або пщ д!ею шро-
гешв, коли установочний мехашзм не руйнуеться, а змшюеться так, що устано-
вочна точка перемнцуеться на вищий р!вень.
Результата тонких електрофтзюлопчних дослщжень дають змогу уявити цей ме-
хашзм так: у ппоталамтчному центр! терморегуляцн е нейрони зрьох титв — чутлив!
до тепла, чутлив! до холоду ! '‘глух!” до коливань температури. Припускають, що
головну роль вщтграють останн). Вони генерують сигналя стандартного характеру,
як! с сигналом поргвняння для гермочутлиьих нейрошв. За будь-яко! змши темпе-
ратури т!ла вона повертаеться до нормального ршня завдяки “глухим” нейронам.
6 й !шш пояснения формування установочного механизму температури, згщ-
но з яким установочна точка визначаеться функшею тепло- т холодочутливих
нейротнв. Доведено, шо вони е двох видтв — з лшшною ! нелшшною функшею.
Дй
Термочутлив! нейрони з лппйною функщею — це нейрони, у яких кглькгсть де-
поляризацгй прямо пропорцшна коливанням температури тгла. Термочутлив!
нейрони з нелпййною функцгею на змшу температури т!ла вщповщають непро-
порщйно, наприклад, зменшенням приросту к!лькост! деполяризаций у раз! пщви-
щення температури т!ла. Установочна точка температурного гомеостазу фор-
муеться нейронами з нелпййною функщею г саме на них впливають медгатори
гарячки. Термочутлив! нейрони з нелпййною функщею встановлюють нову, вищу
установочну точку температурного гомеостазу. Нормальна температура сприй-
маеться як знижена. Тод! перекриваються шляхи вщдачг тепла, посилюються ме-
хашзми теплопродукцп, унаслщок чого температура тгла пщвищуеться ! на цьому
ргвнг лишаеться протягом деякого часу (на перюд гарячки).
Кргм шрогенгв у формуванш гарячково! реакцп певну роль вщгграють також
пши речовини, насамперед гормони. В ос!б зг зниженою функщею щитоподгбног
залози або гшофгза супутн! гнфекщйнг хвороби супроводжуються меншою га-
рячкою.
Глюкокортикощи (гщрокортизон) галъмують розвиток гарячково! реакцп,
очевидно, унаслщок того, що вони е гнтгбгторами експресп штерлейкгнових генгв,
пригшчують метаболгчнг процеси в клгтинах — продуцентах вторинних шрогенгв,
блокують синтез простагландин !в ! зменшують чутливгсть клгтин г тканин до вто-
ринних шрогенгв.
СТАДП ГАРЯЧКИ. СП1ВВ1ДНОШЕННЯ
М1Ж ТЕПЛОПРОДУКЦИЮ IТЕПЛОВ1ДДАЧЕЮ
У гарячковому процесг розргзняють три стадп: пщвищення температури тша (81. гп-
сгепгепй); збереження температури тгла на пщвшценому ргвнг (81. Газйёгг); знижен-
ня температури тгла до вихщного ргвня (81. бесгетепй).
Стада» пщвищення температури тгла. Пщвищення температури тша в цгй стадп
вщображуе перебудову терморегуляцп: змгну теплопродукцп г тепловщцачг. При
цьому теплопродукцгя перевищуе тепловщдачу. Головне значения мае обмеження
тепловщдачг, що не тшьки ефективнгше в розумгннг шввдкостг розггргвання тгла,
а й економнгше для органгзму, оскгльки не потребуе додаткових енергозатрат.
Тепловгддача зменшуеться внаслщок звуження периферичних судин г змен-
шення притоку кровг до шкгри, одночасного гальмування потовидглення г змен-
шення невидимого випаровування води. У тварин спостерггаеться скорочення
м’язгв волосяних цибулин г скуйовдження шерстг, що збглыпуе теплогзоляцпо.
Еквгвалентом цгег реакцп в людини е “гусяча шкгра”.
Важливе значения для пщвищення температури тгла пщ час гарячки мае г
збщьшення теплопродукцп за рахунок скоротливого та нескоротливого термоге-
незу. Провщну роль у зростанш теплопродукцп шляхом скоротливого термогене-
зу вщгграють пщвищення терморегуляторного м’язового тонусу г м’язове тремтш-
ня (приргст виробленого тепла може становити 35—45 %). М’язове тремтгння
пов’язане зг спазмом периферичних судин. Через зменшення припливу кровг тем-
пература шкгри знижуеться гнколи на кглька градусгв. Терморецептори збуджу-
ються, виникае вщчуття холоду — озноб. У вщповщь центр терморегуляцп посилае
гмпульси до рухових нейрошв — виникае тремтгння. Одночасно збглыпуеться не-
СТАДЙ ГАРЯЧКИ. СШВВЩНОШЕННЯ М1Ж ТЕПЛОПРОДУКЦ1СЮ I ТЕПЛОВЩДАЧЕЮ
скоротливий термогенез (так званий загальний калоригенний ефект) у внутрешнлх
органах, таких як печшка, легеш, мозок тощо. Це е наслщком стресових нейро-
ендокринних мехашзмхв, зокрема гормошв 1 мед!атор!в симпатико-адреналово!
системи, гормошв щитоподхбног залози, глюкагону тощо, коли активуються фер-
менти клети иного дихання, збшьшуеться споживання кисню та утворення тепла.
У розбалансувашп теплового гомеостазу певну роль можуть вщегравати деяю
бактергальш токсини, здатш роз’еднувати окиснення 1 фосфорилювання 1 тим
самим збшьшувати утворення тепла. Цей додатковий термогенез може прискори-
ти пщвищення температури тша у першш стадп гарячки.
Стадая у гримання температури тша на шдвищеному рхвш. Шсля того як темпе-
ратура тша пщвищилася, вона утримуеться на цьому ргвш протягом деякого часу
(дш, години). Осюльки при цьому дещо збшьшуеться тепловщдача, то подальше
пщвищення температури тша не вщбуваеться. Тепловщдача здшснюеться завдяки
розширенню периферичних судин; блща дос1 шюра стае пперемшованою, гаря-
чою на дотик. Виникае вщчуття жару.
Утримання температури тша на пщвшценому р!вш пояснюеться тим, що пщ
впливом вторинних шрогешв зменюеться установочна точка центру терморегуля-
цп, за яко! тепловщдача 1 теплопродукцёя стають збалансованими. На цьому ргвн!
вщновлюеться мехашзм пщгримання сталост! температури ёз характерними коли-
ваннями вранц! ё ввечерё, амплётуда яких значно бёльша, нёж у норм! (мал. 31).
За ступеней пщвищення температури тша в друпй стадп гарячки розрёзняють
таю Г! види: субфебрильну — пщвищення температури т!ла до 38 °С; пом1рну — 38—
39 °С; високу — 39—41 °С; гтергйретичну — понад 41 °С.
Дне
хвороби
Стадп
гарячки
123456789 10
О
О
«г
41
40
39
а 38
<и
I
37
36
35
Пульс
Дихання
Д!урез
М'язове
тремпння
Мал. 31. Змши температури тёла, пульсу, ди-
хання та дёурезу в резнях стадиях гарячки
Мал. 32. Критичне (А) е лггичне (Б) знижен-
ня температури пла в Ш стаде! гарячки
I
П
ш
Новий ргвень температури т!ла, п коливання протягом доби визначаються
певними факторами, серед яких вирпиальне значения мае характер ! кыыпсть
шрогешв, чутлив1сть до них центргв терморегуляцн. Кр!м того, мають значения
потужшсть системи вщведення тепла, точшсть 1 надшшсть функцюнально! 1 тро-
ф1чно1 шнервацй, утворення речовин-роз’еднувачхв 1, наренгп, наявшсть в орга-
шзм! запасу енергетичного матерхалу, насамперед жиру. У виснажених людей пе-
реб1г шфекцшних хвороб може вщбуватися без гарячки. У дхтей вона розвиваеть-
ся швидко, в ос1б лггнього вхку — повыьно 1 е помхрною.
Стадхя зниження температури ттла. Шсля причинения ди шрогенгв центр тер-
морегуляцй набувае попереднього стану, установочна точка температури зни-
жуеться до нормального рхвня. Тепло, що накопичилося в оргашзм!, виводиться
за рахунок розширення шкхрних судин, збгльшення потовидхлення х втрати рщи-
ни через шкхру, слизов! оболонки й органи дихання. Зниження температури тыа
може бути поступовим, лйпичним (протягом кхлькох дхб), або швидким, критич-
ным (мал. 32). В останньому випадку може вщбутися дуже рхзке розширення су-
дин, у раз! поеднання якого з штоксикащею виникае небезпечний для життя
колапс.
ТИПИ ТЕМПЕРАТУРНИХ КРИВИХ
Температурна крива пщ час гарячки складаеться з трьох частин — пщвищення,
утримання ! зниження. Однак у кожно! з них, як х у криво! в щлому, можуть бути
сво! особливост!, як! дають лхкаревх шформацпо про стан хворого х мають дифе-
решцально-дхагностичне значения.
На характер температурно! криво! можуть впливати особливост! збудника,
наприклад циюичшсть його розвитку в кров!. Щодо цього показовою е темпера-
турна крива у хворих на малярпо (ГеЬпз пйег-
пнйепз — перемгжна гарячка). При цш хво-
роб! напади пщвищення температури т!ла
можуть вщбуватися щоденно, через день на
третей (триденна маляр!я), через два дш на
четвертий залежно вщ типу збудника. Пщ
час нападу гарячки температура тхла р!зко
пщвищуеться й утримуеться на високому
рхвш вщ 30—60 хв до 2—3 год, а пот!м зни-
жуеться до вихщного рхвня ! навггь нижче вщ
нъого (мал. 33).
Залежшсть температурно! криво! вщ збуд-
ника добре простежуеться також на приклад!
гарячки в раз! поворотного епщемхчного тифу
(ГеЬпз гесиггепз — поворотна гарячка). Сшро-
хета поворотного тифу фагоцитуеться макро-
фагами 1 розмножуеться в них. У м!ру нако-
пичення в юптинах сшрохети проникають
через бар’ер мононуклеарних фагоципв 1
надходять у кров. Це спричинюе черговий
Мал. 33. Динамхка температурно! кри-
во'! у хворих на малярпо
------------Температура---------------Пульс
Мал. 34. Динамша температурно! ( пульсовоТ криво! у хворих на поворотний гиф
Мал. 35 Крив! температури ттла. пульсу та дихання в раз! постншо! гарячки
(крупозна пневмошя)
напад гарячки тривалктю 6—8 дшв. шсля чого температура тша критично зни-
жуеться 1 настае перюд ашрекси, який також тривае 6—8 дшв. Напади можуть
повторюватися (мал. 34).
Гарячка щд час крупозно! пневмонп мае характер постшно! (ГеЬпь сопйлиа —
постшна гарячка). Спочатку' температурна крива р!зко пщвищуеться, потгм н!би
сягае свого максимуму й утримуеться 7—9 дшв, коливаючись у межах одного гра-
дуса, а попм р1зко знижуеться (мал. 35). При сепсис! (ГеЬпз Ьеспса — ^иснажлива
гарячка) добов! коливання температури пла сягають 2—3 °С.
Гарячков! бюритми залежать не лише вщ збудника, дози 1 циклу утворення
шрогешв, а й вщ оргашзму хворого, його пщившуально! реактивносп.
Слщ зазначити, що остан нш часом унаслщок широкого застосування анти-
бютиюв температури! крив! втратили свою типовкть.
Г1ПЕРТЕРМ1Я ТАЙ В1ДМ1НН1СТЬ В1Д ГАРЯЧКИ
Вщ гарячки слщ вщр!зняти перегршання, або гшертермпо. Обидва процеси посд-
нуються лише кшцевим результатом — пщвишенням температури пла. а мехашз-
ми !х прямо протилежш. Перегр!вання не пов’язане з д!ею шрогенно! речовини.
Температура тгла пщвищуеться внаслщок або зовшшнього впливу, який обмежуе
гепловщдачу, або первинного порушення роботи центру терморегуляцп. Перше,
тобто перегр!вання оргашзму внаслщок затримання тепла в ньому, спостерйаегь-
ся на пшприемствах э високою температурою виробничого середовиша або в ре-
понах !з жарким климатом. Перс1р!ванню в цих випадках сприяе зростання теп-
лопродукцй у зв’язку !з роботою м’яз!в.
Компенсащя пщ час перегр!вання спрямована на усунення утруднень щодо
видшення тепла ! збереження теплового гомеостазу. Оск!льки за температури
навколишнього середовища близько 33 °С выдача тепла шляхом випромшюван-
ня та конвекцп практично припиняеться, цей процес здшснюеться тыьки через
випаровування поту ! вологи пщ час дихання. Однак якшо волопсть повггря
висока, зазначений шлях також перекриваеться г вс! компенсаторш мехашзми
виявляються неефективними. Температура тша пщвищуеться, проте це не га-
рячка.
Температура т!ла може шдвишуватися 1 без впливу факторов навколишнього
середовища. Це так звана ендогенна гтертерм/'я, або гарячкопошбний стан.
Розр!зняють так! ендогенш гшертермп:
1) неврогенш: а) центрогенш (у раз! прямого ушкодження р!зних вщдвнв го-
ловного мозку, насамперед гйюталамуса); б) психогенн! (у раз! неврозы, псих!ч-
них розлад!в, значного емоцшно-розумового напруження);
2) рефлексогенн! (у раз! сечо- та жовчнокакГяно! хвороби, подразнення оче-
рсвини, катетеризапй сеч!вника);
3) ендокринш (у раз! гипертиреозу,, унаслщок дц стерощних гормошв — спо-
холанолону. прогестерону);
4) зумовлеш ентеральним 1 парентеральним уведенням деяких фармаколопч-
них препараты: кофсгну, ефедрину, метиленового синього, гйгеросмолярних роз-
чишв, адреналшу, фенампту. агрошну. стрихшну, сальварсану, сульфазину.
Описано так звану злояклсну гтертерм1ю (пщвищення температури тша сягае
44,4 °С). Вона виникае у вшповшь на введения засобгв для наркозу (галотан,
еф!р, метоксифлуоран), м'язових релаксанпв (суксэметонш). Спостерйаеться 1з
частотою 1 випадок на 1500 осУб молодого вгку, персдаеться як автосомно-домУ-
нантна ознака, зумовлена порушенням внутрУшньоклУ тинного метаболизму юшв
кальцпо в скелетних 1 серцевому м’язах. Смерть настае внаслщок ацидозу та зу-
пинки серця
ЗМ1НИ В ОРГАНАХ I СИСТЕМАХ У РА31 ГАРЯЧКИ
ОкрУм розладтв терморегуляци пщ час гарячки спостерггаються також Унпп пору-
шення, насамперед обмшу речовин, дУяльностУ серпево-судиннох 1 дихально! сис-
тем, секреторних У екскреторних функпш. Виникае комплекс симптомтв, у якому
слщ розрУзняти, як1 симптоми залежать вщ шрогеюв, а яю вщ само) хвороби
(пневмошя, шфаркг, гепатит).
Найвираженппими е змши в систем! кровообзгу. За правилом ЛУбермейстера,
пщвищення температури тша на 1 °С супроводжуеться прискоренням пульсу на
8—10 за 1 хь. Осюльки локальне зУгрУвання водхя ритму серця зумовлюе приско-
рення скорочень серця, то тахгкардпо на тл! гарячки пояснюють так само. КрУм
того, мае значения пУд вишен ня тонусу симпатичних нерыв. Збшьшуються удар-
ний 1 хвилинний об'ем серця. У першш стадзУ гарячки може шдвищуватися ар-
герУальний тиск. Вщбуваються звуження судин шгари У розширення судин внут-
рУшнзх органзв. У третгй стадГУ в разз критичного зниження температури тша може
виникнути колапс унаслщок рУзкого зниження тонусу артерш.
Тах У кардзя пщ час гарячки виникае не завжди. При деяких УнфекцУйних хво-
робах температура тша пщвищуеться У водночас спостерУгаеться брадикардУя.
Прикладом е черевний, а також поворотний тиф — хвороби, перебУг яких вщбу-
ваеться зУ значною Унтоксикашею, коли серце реагуе не стшьки на високу тем-
пературу, скшьки на дУю токсичних речовин екзогенного й ендогенного поход-
ження.
Зовшшне дихання в першш стадп гарячки дешо сповшьнюеться. ПУсля досяг-
нення максимально! температури тша дихання прискорюеться, УнодУ у 2—3 рази.
Осюльки при цьому глибина дихання зменшуеться, то легенева вентиляцУя Ус-
тотних змУн не зазнае. Прискорсння дихання (тахУпное) е наслшком пщвищення
температури головного мозку.
Система травления пщ час гарячки зазнае значних змш — втрачаеться апетит,
зменшуеться секрецУя слинних залоз (язик сухий, обкладений), знижуються юль-
юсть У кислотшсть шлункоьогэ соку, порушуються пристУнкове травления, пе-
ристальтика кишок. Однак ступУнь них змш неоднаковий У здебшыного залежить
вщ характеру хвороби. Наприклад, при гриш вУн менший, нУж при черевному
тиф1.
ПорУвнюючи змУни в органУзмУ, спричиненУ дУею високоочищених бактерУаль-
них шрогенУв, з! змшами, яю вщбуваються в разУ природного розвитку шфекцш-
но‘1 хвороби, П.М. Веселкш дУйшов висновку, що змУни у травнш систем! вини-
кають не стУльки через гарячку, скшьки внаслщок голодування, Унтоксикаци та
нетермогенних впливУ в бакл ерУальних токсинУв.
-----------------------------------------------------------------Гг —
287
Гарячка супроводжуеться змшами ендокринно! системи: активуеться гшоф1-
зарно-ндцнирникова система, спостерхгаються ознаки стресу. При шфекцшнш
гарячщ збшыиуеться викид гормошв ищтоподхбно! залози, що зумовлюе пщви-
щення основного обмшу.
У центральны нервовш систем! виникають змши процесхв збудження х галъму-
вання. На електроехщефалограмх з’являеться повшьний альфа-ритм, характерний
для гальмування кори великого мозку. Пхсля введения шрогешв у людини можуть
спостерхгатися безсоння, вщчуття втоми, головний б!ль. При шфекцшних хворо-
бах ступить цих проявхв вищий. Можливх галюцинацц, потьмарення свщомостх,
запаморочення. Осюльки щ симптоми з’являються х в раз! пом!рного пщвищення
температури тша, то, очевидно, вони пов’язаш не стшьки з гцдвищенням темпе-
ратури пла, скшьки з штоксикащею.
Основний обмш пщ час гарячки пщвищений, хоча прямого зв’язку м!ж акти-
вацию обмшу та гцдвищенням температури пла немае. Дихальний коефпцент у
першш стадп гарячки наближаеться до единиц!, що евщчить про збшъшення
окиснення вуглеводхв, а згодом — жир!в, особливо коли резерви вуглевод!в уже
вичерпан!. У таких випадках окиснення жирхв вщбуваеться не до кшцевих про-
дукпв 1 в кров! накопичуються кетонов! пла. 1з сечею видшяеться ацетон. Якщо
хворому з гарячкою вводити достатню кшыйсть вуглевод!в у легкозасвоюванш
форм!, то ц! порушення можна призупинити.
При деяких шфекцшних хворобах порушуеться бхлковий обмш. Азотистий ба-
ланс стае негативним, збшьшуеться видшення хз сечею продукт! в азотистого об-
мхну, зокрема й сечовини, що евщчить про збшыиення розпаду бхлюв. Однак це
спостерхгаеться не завжди. Так, при гриш обмш, як правило, не порушуеться.
Водночас для деяких хнфекщйних хвороб, що мають безгарячковий перебтг, ха-
рактерне рхзке пщвищення катаболхзму бшкхв. Це евщчить про те, що причина
порушень не в гарячщ, а в ступеш штоксикацц, розвитку запальних х дистрофхч-
них змш у тканинах, нарепгп в голодуванш внаслщок втрати апетиту та порушен-
ня всмоктування в юпиках.
Характерними для гарячки е змши водно-мшерального обмшу. У першхй стади
спостерхгаеться збшъшення дхурезу внаслщок пщвищення артерхального тиску 1
притоку кров! до внутрхшнхх органтв, зокрема до нирок. У друпй стадп гарячки
внаслщок пщвищеного синтезу альдостерону 1 вазопресину у тканинах затриму-
ються натрш х вода. Дхурез зменшуеться. У трепй стади збшыиуеться виведення
хлоридхв, зокрема х натрпо хлориду, вщбуваеться зневоднення тканин, збшьшуеть-
ся дхурез х потовидшення.
ЗНАЧЕНИЯГАРЯЧКИ
Пщ час гарячки, як х пщ час шших типових патолопчних процесхв, шкщливе ! ко-
рисне нерозривно пов’язаш м!ж собою. Гшершретична температура тша перешкод-
жае розмноженню багатьох збудникхв — кокхв, сшрохет, вхрусхв. Рхзко галъмуеться
репродукцхя вхрусу полхомхелхту за температури тша понад 40 °С. За такох темпера-
тури багато мткрооргашзмхв збертгають здатшсть розмножуватися, однак втрачають
стшюсть до лхкарських засобхв. Так, чутливхсть мжобактерш туберкульозу до стреп-
томщину при температур! тша 42 °С майже в 100 разхв вища, н!ж при 37 °С.
Гарячка позитивно впливае на низку функцгй оргашзму: пгдвшцуе актившсть
фагоцитозу, стимулюе вироблення антиты, гнтерферону. Штучне пригнгчення га-
рячки призводить до зменшення в кровг юлькосп нейтрофыьних гранулоцитов,
зниження функцгй макрофапв. Вивчення мехашзмгв впливу гарячки на гмунну
систему показало, що пщвищення активност! мехашзмгв !мунгтету пов’язане не
стыьки з щдвищенням температури тыа, скыьки з безпосередшм впливом вто-
ринних шрогешв (цитокйпв) на 1 муноцити та гепатоцити.
Привертас увагу зв'язок мгж гарячкою 1 запаленням. Макрофаги не тгльки пог-
линають г знешкоджують бактерц, а й синтезують шроген (ТЛ-1). Якщо врахувати,
що макрофаги разом з лгмфоцитами беруть участь у виробленн! антиты (юптин на
взаем од гя), то зв’язок мгж гарячкою, гмунпетом г запаленням стае очевидним.
У позитивному вплив! гарячки на перебгг основного захворювання важливого
значения набувае стрес (активацгя гшоталамо-гшофгзарно-наднирниковог систе-
ми), який гцдвшцуе неспецифгчну резистентшсть оргашзму.
Захисно-пристосувальш реакцп оргашзму, яю мобппзуються пщ час гарячки.
що супроводжуе гнфекцшну хворобу, можуть значною мгрою “знецшюватися”
внаслщок розвитку гнгоксикацп та ушкодження життево важливих оргашв Тод!
пщвищення температури тша може мати негативн! наслщки, оскыьки гарячка
пов’язана з додатковим навантаженням на органи, насамперед на серпе 1 судини.
Розробляючи тактику лгкувальних заходов пщ час гарячки, лгкар повинен ви-
ргшити, що в клип41 пй картин! хвороби “е наслщком ушкодження г що наслщком
протидп органгзму цьому ушкодженню” (ГП. Павлов).
Лжувальне застосування гарячки спочатку грунтувалося на випадкових споете -
реженнях. Одеський лгкар А.С. Розенблюм (1876) помгтив, що стан хворих на
сифшс полшшувався, коли вони захворювали на поворотний тиф. Згодом зара-
жения поворотним тифом, а частице — маляргею почали застосовувати для л!ку-
вання хворих з шзшми стадгями сифыгсу (Вагнер — Яурегг, 1917). Пгзшше для
цього застосовували високоочищеш препарати шрогешв т!рогенал). Тепер у
клгшщ з шею метою використовують ГЛ-1 та пип! цитокши. 1х псревага над гйро
генами бактергального походження полягае в тому, що введеш ззовнг, вони не
спричинюють поб1чну дгю. а справляють стимулюючий ефект на гмунну систему.
Пгрогерашю призначають не пльки при сифшс!, а й при шших шфекцшних
хворобах, зокрема при к!стково- суглобовому губеркульозг, коли .пкувальне засто-
сування гарячки поеднують з лгкуванням специфгчними антибак тергальними за-
собами. П!ротерап!я ефективна також у раз! гшертензивно! хвороби ниркового
генезу, оскыьки розширення судин оргашв черевно* порожни ни, яке виникае
при цьому, посилюс кровот!к у нирках г знижуе загальний артер!альний тиск.
Оскыьки активацгя гмунпету залежить вщ дц цитокгнгв на гмунну систему, а
пщвищення температури пла при гарячш безпосередньо пов’язане гз синтезом пщ
впливом питокппв ПГЕ та гх впливом на нейрони герморегуляторного центру, щ два
прояви гарячки можна роздшити. Якщо застосувати нестерощнг протизалальн! засо-
би, як! блокують фермент циклооксигеназуг тим самим синтез ПГЕ, то температура
т!ла знизиться, а позитавний ефект спгмулягш гмунпету залишиться. Це добре вщо-
мо вс!м. хто при застудг споживас малину, калину-, червсне вино, як! мгсгять значку
юльюсть ацетилсалщилово! кислоти. Якшо ж застосовувата для зниження темпера-
тури тгла глюкокортикощи, якг одночасно блокують синтез 1 дгю як простагл андин !в.
так 1 цитокппв то позитивний ефект гарячки буде лгквщоваво разом гз гарячкою.
Розд1Л XIV
ПУХЛИНА
Пухлина — це типовий патолог/чний процес, який характеризуешься неконтрольова-
ним I потенцшно безмежним размножениям клгтин унас/йдок мутацшного та / або
етгеномного порушення активност! гешв, як1 регулюють клйпинний поди, що приз-
водить до розростання тканини, не пов’язаного з п загальною структурою та функ-
ц1ями.
Особливост! росту пухлинних КЛ1ТИН:
— пухлина виникае в оргашзм! внаслщок перетворення нормально! юптини
на пухлинну, яка втрачае здатшсть реагувати на гальмхвш впливи кейлошв ! кон-
тактно гальмування юптинного под!лу та набувае здатност! до самойщтримуючо!
стимуляцн подщу;
— пухлинн! юштини втрачають лхмхт Хейфлгка х мають потеншйно безмежний
рхст, що закхнчуеться лише зх смертю оргашзму;
— здатнють пухлинних юптин безмежно розмножуватися успадковуеться як
домхнантна ознака соматично! спадковост! х виявляеться не лише в оргашзм!, а й
у культур! пухлинно! тканини, а також пщ час трансплантацн пухлини.
Класифшацхя. За характером росту пухлини бувають доброяюсними х злоя-
кхсними. Доброяюсю пухлини мають експансивний рхст, за якого прилегла здо-
рова тканина з ростом пухлини розсуваеться. Злояюсн! пухлини мають хнфшьтра-
тивний р!ст — пухлинн! юптини проростають мхж нормальними клхтинами х крхзь
базальш мембрани та судинну стхнку. Потрапляючи в лхмфу або кров, вони пере-
носиться в ппш органи х можуть утворювати нов! вогнища пухлинного росту —
метастази.
За гхстолопчною будовою клхтин, з яких вони походять, розрхзняють пухлини
з ештелпо, сполучно! тканини, паренххматозних клхтин певних оргашв. Злояюсн!
пухлини з ештелхальних клхтин отримали назву раку (вщ лат. сапсег), злоякхсш
пухлини зх сполучно! тканини — саркоми (вщ лат. загсота). У назвах пухлин ви-
користовують також суфхкс “ома”, що означае “пухлина”, наприклад лейомхома
матки — доброяюсна пухлина матки, яка розвиваеться з непосмуговано! м’язово!
тканини матки.
ЕКСПЕРИМЕНТАЛЬНЕ В1ДТВОРЕННЯ ПУХЛИНИ
Незважаючи на те що пухлина як хвороба вщома давно, п довго не вдавалося
вщтворити експериментально. Ось чому вщтворення в експеримент! цього пато-
лопчного процесу стало на початку XX ст. значним науковим досягненням. Екс-
периментальш моделх пухлини дають змогу вивчати етхологпо х патогенез пухлин-
ного процесу, розробляти новх методи його профхлакгики та лхкування.
Методами експериментального моделювання е Ёндукщя, експлантацЁя 1 транс-
плантацЁя пухлини.
1ндукц1я пухлини хшгчними речовинами. У 1775 р. хЁрург лондонського гостта-
лю П. Потт описав професшну злояюсну хворобу — рак шюри калитки у са-
жотрусЁв. Однак, незважаючи на очевидний зв’язок раку сажотрус!в ёз забруднен-
ням шюри сажею ё смолою, спроби вщтворити таку пухлину в експеримент!
тривалий час були невдалими. У 1915 р. японсью вченЁ 1ипкава та ЯмагЁва впер-
ше змогли спричинити розвиток пухлини у тварин. Протягом 6 мёс. вони змащу-
вали шкЁру кролЁв кам’яновупльною смолою, ё лише шсля цього у тварин розви-
нувся рак шк!ри. ШзнЁше було видыено канцерогенн! речовини у чистому виг-
ляд!, встановлено канцерогешпсть речовин, що належали до р!зних клас!в хёмёч-
них сполук.
1ндукц1я пухлини вирусами. У 1908 р. Еллерман 1 Банг уперше змоделювали
лейкоз курей за допомогою безклпинного фшьтрату з лейкозних лейкоцитов.
Його отримують, фыьтруючи екстракт подрЁбнено! пухлинно! тканини кр!зь фар-
форов! фшьтри, яю не пропускають клггини, а у фшьтрат проходить тшьки в!руси,
що мають молекулярну будову ё пор!вняно менш! розм!ри. У 1910 р. Раус за допо-
могою безклпинного фшьтрату, добутого Ёз саркоми в курки, спричинив розвиток
саркоми у здорових курей. Так уперше було одержано докази вЁрусно! епологц
лейкозу! пухлин.
Однак у наступи! десятилгття викликати безюйтинним фшьтратом рак у до-
рослих ссавщв не вдавалося. Виняток становила пашлома Шоупа. Тшьки в 1950 р.
Л. Гросс шсля багатьох невдалих спроб спровокувати лейкоз у дорослих мишей
уперше вв!в безюптинний фшьтрат з лейкозних юптин кров! новонародженим
мишам 1 спричинив у них лейкоз. Отже, було добуто прям! докази в!русно! етЁо-
лоп! пухлини у ссавщв, а 40 рок!в невдалих спроб теля вщкритгя Рауса поясню-
ються ошршетю оргашзму дорослих ссавщв до в!рус!в. Але Шоуп виявив у диких
крол!в бородавчастЁ розростання на шюр! (пашломи), яю вдалося перещепити
дорослим тваринам за допомогою безклпинного фшьтрату.
€ лёнё! мишей високораков! (з високою захворювашетю на рак молочно! за-
лози) й низькораковг Однак якщо у самки високораково! л!нп забрати новона-
роджених мишенят вщ першо! годёвл! 1 вщдати !х на вигодовування самщ низько-
раково! лшп, то частота захворюваностЁ на рак у перших р!зко знизиться. I, нав-
паки, у раз! вигодовування самкою високораково! лшп мишенят вщ самок низь-
кораково! лёнй частота виникнення пухлин у других значно пщвищуеться. Бптнер
у 1936 р. довёв, що в молощ високоракових мишей е фактор молока, який зумо-
влюе у потомства рак молочно! залози. Шсля вщкритгя Л. Гросса було встанов-
лено, що фактор молока Битнера — це пухлинотв!рний в!рус. Стало зрозумшим,
що новонароджен! мишенята заражаються цим в!русом з молоком матер!.
1ндукц1я пухлини ф1зичними факторами. Пухлину вдаеться вщтворити за допо-
могою юшзуючого випромшювання, у тому числ1 й рентгенЁвського, радюактив-
них !зотошв, а також ультрафюлетового випромшювання.
Експлантащя пухлини — вирощування пухлини в культур! тканини поза ор-
ган!змом. Цей метод усшшно застосовував О.Д. Тимофеевський. Культура ткани-
ни, взята безпосередньо з пухлини тварини або людини, називаеться первинною.
Кр!м того, у лаборатор!ях е велика юльюсть штамЁв пухлинних клётин, яю постёй-
но пасеруються 1 властивостЁ яких добре вивчено, що дае змогу проводити дослё-
ди на однаковому матер!ал!. Культура тканин дае можливють шдукувати пухлину
поза оргашзмом хшгчними онкогенами та онкогенними в!русами. Цей метод
особливо шнний тим, що можна вивчати шдукцпо пухлин 1 пухлинотвхршгх В1РУС1В
на тканинах оргашзму людини. Пасероваш або шдуковаш в культур! тканини
пухлинн! клетини шсля тдсаджування здоровш тварин! ростуть в Г! оргашзм! й
утворюють злояюсну пухлину.
Трансплантация пухлини. Уперше М.0 Новинський у 1876 р. усшшно транс-
плантував пухлину дорослого собаки цуценятам. Фактично цим досл!дом було
покладено початок експериментально! онкологи. Метод трансплантаци широко
використовуеться й ниш. € штами пасерованих пухлин з добре вивченими влас-
тивостями: асцитна карцинома Ерл!ха у мишей, куряча саркома Рауса, саркома
Тенсена у щур!в, карцинома Брауна — Шрса у крол!в 1 т. п. Алогенна трансплан-
тац!я пухлини (тобто пересаджування пухлини нешбредним тваринам того самого
виду) проходить успппно, тод! як така сама трансплантащя нормальних тканин
без !мунодепресп не вдаеться. Причинами вдалих пересаджувань алогенних пух-
лин е антигенне спрощення пухлин у м!ру !х малптпзацц, маскування антиген!в у
пухлинах, а також !х гмунодепресивна д!я Уведення невелико! юлькост! пухлин-
них клггин (400 000) зумовлюе пригшчення !мунно! системи 1 ргст пухлини (зга-
даемо, що в 1 мл кров! мгститься 5 млн еритроципв). Ильки !н’екц!я ще меншо!
юлькост! пухлинних юптин може призвести до (мушзащ! 1 подалыпого вщторг-
нення трансплантовано! пухлини.
ЕТЮЛОПЯ
Причинами розвитку пухлини е р!зш фактори, здатн! зумовлювати перетворення
нормально! юптини на пухлинну. Вони називаються онкогенними, або бластомо-
генними. Причини пухлин, р!зш за своею природою 1 способом впливу на ор-
гашзм, але здатш порушувати регуляцпо юптинного подйту, — х!м!чш, фхзичн! та
бюлопчш.
Онкогент фактори мають таю власти вост!:
— справляють мутагенну дно — здатшсть прямо чи опосередковано впливати
на геном юптини, що призводить до мутацш. Таку властивгсть мають хгмгчш ре-
човини (вуглеводш, н!трозам!ни тощо), ф!зичш (юшзуюче випромпповання) та
бюлопчш (в!руси) фактори. Вгруси можуть спричинювати пухлини також епгге-
номним шляхом. М!сцем взаемодн х!м!чних онкогешв з нуклешовими кислотами
е, очевидно, гуашн;
— здатн! проникати крхзь зовшшн! та внутршпп бар’ери. Так, у раз! потрап-
ляння на шюру розвиток пухлини викликатимуть лише т! потенцшш х!м!чш он-
когени, яю проникають крхзь зрогов!лий еп!дерм!с. Осюльки бюлопчш мембрани
складаються з лшопротеццв, крхзь них проникають насамперед лшщорозчинш
речовини, до яких належать 1 онкогенн! вуглеводш;
— дозована д!я онкогенних факторгв, яка забезпечуе незначне ушкодження
юптини, що дае !й змогу вижити. У зв’язку !з цим для досягнення онкогенного
ефекту мають значения доза 1 ступгнь токсичносп онкогенного фактора. Незнач-
не пщвищення дози призводить до збшыпення юлькост! пухлин, випадюв захво-
рювань тварин 1 зменшення термппв розвитку пухлин. Подальше збшыпення дози
супроводжуеться переважанням токсичного ефекту 1 загибеллю тварин, перш нгж
утвориться пухлина. Зменшення дози онкогену дало змогу встановити, що:
• субпорогових онкогенних доз немае (у дослщах виявляеться канцероген-
шсть дуже низьких доз), однак при цьому збгльшуеться тривалгсть виникнення
нових пухлин);
• дгя онкогешв необоротна;
• онкогенам властивг ефекти сумащг та кумуляцп. Така сама залежнгсть спос-
терггаеться у випадку дп юшзуючого випромшювання: висок! дози зумовлюють
променевий опгк г загибель тканини г лише вщносно невисокг дози створюють
умови для виникнення пухлин;
— дгя вгрусгв з абортивним перебггом, а не в раз! гостро! шфекцп (загибель
клгтин) найчастгше спричинюе онкогенез. Вгрогщнгсть розвитку онкогенезу
пщвищуеться в м!ру тривалост! впливу онкогенного фактора;
— в органах г тканинах з ргзними особливостями проникност! й метаболгзму
можуть створюватися умови, сприятлив! для прояву канцерогенност! одних фак-
торгв г несприятливг для прояву шших. Цим можна пояснити гснування органо-
тропних онкогешв;
— онкогени пригшчують тканинне дихання та гмуннг реакцп;
— утворення пухлини пщ дгею кглькох онкогенних факторгв посилюеться
(синканцерогенез). 1нодг фактори, якг сам! не е онкогенними, можуть посилюва-
ти дгю онкогешв. Такс явище отримало назву коканцерогенезу, а фактори, якг його
спричинюють, — коканцерогешв.
Було вщкрито таю групи причин пухлин, здатних впливати на гени мутацгй-
ним або епггеномним шляхом:
— хгмгчш онкогеннг фактори: полгциклгчнг ароматичн! вуглеводш (ПАВ),
амгноазосполуки, штрозамгни, дгоксини, оксиди азоту;
— онкогеннг фактори бголопчного походження: продукти грибгв г рослин,
гормони ссавцгв, зокрема статев! гормони, онкогеннг вгруси;
— фгзичнг онкогеннг фактори: гонгзуюче випромшювання, г навгть механгчнг
фактори при введеннг в тканини неживих тш (пластмасовг плгвки г грудочки).
Хгмгчнг онкогеннг фактори. Через 15 рокгв пгсля дослщгв Ямаггви та 1шгкави в
1930 р. Кук, Хюгтт г Хайджер гз 2 т кам’яновуггльног смоли добуди 50 г хгмгчно
чистого 3,4-бензшрену, який виявився активним онкогеном. Ще рангше, в 1929 р.,
Кук синтезував дуже активний онкоген 1-,2-,5-,6-дибензантрацен (ДБА). Вщтодг
почалося вивчення хгмгчно чистих канцерогенних речовин. Було встановлено
канцерогеншсть багатьох сполук, якг належать до ргзних класгв.
Полщиклгчш ароматичн! вуглеводш. До них належать 3-,4-бензпгрен, ДБА г
9-,10-диметил-1,2-бензантрацен (ДМБА). Шляхом обробки жовчних кислот було
одержано активний онкоген — метилхолантрен. Синтез цього онкогену вперше
навгв на думку про можливу канцерогеншсть деяких бголопчних продуктгв, якг
виробляються самим органгзмом, що згодом пщтвердилося.
Онкогеннг ПАВ виявляють здебгльшого мгсцеву канцерогенну дгю: введен!
пщ шкгру вони спричинюють саркому, нанесен! на шкгру — рак. У разг введения,
яке забезпечуе поширення онкогену в органгзмг, ПАВ спричинюють утворення
пухлин у тих органах, де накопичуються: при видшеннг з молоком утворюються
пухлини молочних залоз, гз сечею — пухлини нирок г ниркових мисок, при видг-
леннг сальними залозами шкгри — пухлини шкгри.
Деяк! ПАВ е дуже сильными онкогенами: 0,2—0,5 мг ДМБА теля пщшк!рно-
го введения мишам спричинювали розвиток пухлин майже в ус!х тварин. Актив-
нппим онкогеном е 20-метилхолантрен.
Пюльман, користуючись методами квантово! органично! Х1ми, визначав густи-
ну електронно! хмари в молекулярных структурах ПАВ. З’ясувалося, що коли яке-
небудь похщне антрацену або фенантрену мае канцерогенну актившсть, то быя
певного радикала в одному й тому самому м!сщ основно! цикл!чно! структури
спостерйаеться пщвшцена густина електронно! хмари, яка сягае 1,2 е. Цю зону в
молекул! вуглеводню було названо К-районом (вщ шм. КгеЬз — рак). Попм тео-
ретично було обчислено, у яких похщних антраценового ряду електронна густина
у К-райош мае значения, що вщповщае канцерогенност!. Експериментально цей
розрахунок пщтвердився переважно для антраценового та фенантренового ряд!в.
Онкогены! ПАВ дуже поширеш в середовипц кнування людини, оскйтьки
вони часто е продуктами неповного згорання. ПАВ утворюються за температури
400—600 °С (температура горшня тютюну в сигарет!), мктяться в дим! 1 смол!
тютюну, пересмаженш олп, выхлопных газах, копчених продуктах, а також у на-
фт!, б!тум1, асфальт!. У щур!в, як! перебували пщ час експерименту на асфальто-
вашй мапстралц частице розвивалися пухлини легень пор!вняно з тими, як! були
на польовгй дороз!.
Пщ час тривалих спостережень за людьми, як! курять, було встановлено, що
частота раку легень 1 верхшх дихальних шлях!в у них пропорцшна юлькост! вику-
рених сигарет з латентным перюдом, що становить 10 роюв. Вона в к!лька десят-
ые раз!в вища в тих, хто курить, пор!вняно з тими, хто не курить.
Канцерогенн! амшоазосполуки й амши мають значку органотрошпеть. Диме-
тиламшоазобензол (ДАБ) в експеримент! спричинюе рак печшки у 80 % выпалив
незалежно вщ того, яким шляхом в!н потратив в оргашзм. Аналопчну дно справ-
ляе ортоамшоазотолуол. р-Нафтиламш у людини 1 тварин спричинюе рак сечово-
го м!хура. Органотропшеть онкогенних речовин пояснюють утворенням в ураже-
ному орган! активних речовин з менш акгивних попередникгв. Канцерогеннкть
р-нафтиламшу проявляеться дкю його метаболит — 2-амшонафтолу-1 1 2-на-
фтилокс!ам!ну.
Нпрозамши. Особливктю цих речовин також е органотропшеть, яка може
зм!нюватися через вщносно незначн! перебудови в молекул!. Так, й!етилн!троз-
ам!н спричинюе переважно рак печшки 1 стравоходу, метилнпрозосечовина —
пухлини головного мозку, триметилнпрозосечовина — пухлини головного мозку
та периферично! нервово! системи.
Нпрозамши утворюються в шлунку людини !з неканцерогенних попередни-
к!в (нпритш та амш!в) за наявносп хлоридно! кислоти. Иприты, наприклад на-
трпо иприт, та амши (амшокислоти, амщошрин), потрапляючи в оргашзм !з
!жею, утворюють нпрозамши, спричинюючи у 80—100 % пщдослщних тварин
розвиток пухлин.
Дюксини — продукт окиснення деяких пол!мер!в 1 пластичных мае, хгхпчно
стопа 1 высоко канцерогены!; утворюються зокрема пщ час спалювання пластмас.
Оксиди азоту — висококанцерогешп.
Онкогенш фактори бюлоггчного походження. 1х виробляе оргашзм, вони вхо-
дять до складу !ж!! мктяться серед речовин, як! використовують у медицин! та на
виробництв!. У цю трупу входять також в!руси.
Гриб Ахрег^Шиз /1сгиит синтезуе афлатоксин — речовину з рхзко вираженими
канцерогенними властивостями. У дощове лето весь урожай земляних гор!х!в
(арахзсу) бувае заражений грибом, який продукуе афлатоксин. Гриб паразитуе
також на кукурудз!, рис!, яйцях, порошковому молощ. Ще бглыпе поширений
А$рег%111и8 п1(1и1ап8, який виробляе канцерогенний стеригматоцистин.
Канцерогенну дпо гонадотропних гормошв гшофхза було виявлено при про-
веденш доелщв 1з пересадки яечника. Щ гормони спричинюють у яечнику про-
Л1ферацпо клхтин фолпсула. Естрогени, що продукуються цими юптинами за при-
нципом зворотного зв’язку, гальмують вироблення фолггрошну. Шсля пересад-
ження яечника в селезшку з одночасним видаленням другого яечника перший
постшно зазнае впливу !нтенсивно! стимуляцн фолггрошном, який спричинюе в
ньому у 80 % випадюв розвиток пухлини. Це евщчить про те, що власш гормони,
якщо вони утворилися в бьтышй юлькост! 1 бЬгьше, Н1Ж звичайно, атакують ор-
ган-м1шень, можуть спричинити утворення в ньому пухлини.
Пухлинотв!рш В1руси. У класифжацп онкогенних в!рус!в враховано таю озна-
ки: тип нуклешово! кислота, яка входить до складу в!русу (РНК або ДНК); м!сце
1 спос1б розмноження в!русу в юптин!; форму:
• РНК-вм!сн! в!руси сгпралепод!бно! форми, що розмножуються в цитоплаз-
ме в!руси лейкозу мишей 1 курей, саркоми Рауса, в!рус молока Бггтнера тощо;
• РНК-вм1сш в!руси ползедрально! форми. В!руси обох цих труп, як! мтстять
РНК, отримали назву онкорнав!рус!в (онкогенних в!рус!в, що мютять РНК), або
ретров]рус!в (через передавания шформацп у зворотному напрямку — в!д РНК до
ДНК);
• ДНК-вмюш в!руси пол!едрально! форми; розмножуються в ядрах юптин. До
них належать в!руси пашломи крол!в, полюми, бородавки людини, вакуолхзую-
чий в!рус мавп — 8У40. Властивост! цих в!рус!в настшьки типов! для вс!е! групи,
що !х об’еднали п!ц загальною назвою Рароха, що походить в!д початкових лггер
назв пухлин 1 функцюнальних змш (рарШота, роНота, уасиоНгайоп);
• велик! ДНК-вмюш вхруси; розмножуються в цитоплазм!, утворюючи харак-
терн! юптинш включения. До ще! групи належить в!рус ф!броми Шоупа, в!руси
Яба । контапозного молюска. Ус! вони дуже под!бш до в!руса в!спи 1 спричиню-
ють здебптыпого дОброяюсн! пухлини.
Великий штерес становить в!рус полюми, детально вивчений Стюартом 1
Едд!. Цей в!рус мютить одну молекулу ДНК, що мае вигляд подвшного юльця або
подвшно! лппйно! молекули. Вш може зумовлювати близько 27 тишв пухлин у
р!зних тканинах у семи видгв ссавц!в (мишей, щур!в, крол!в, хом’яюв та !н.). При
введенш в!русу новонародженим тваринам захворювашеть сягае 100 %. 3 вгком
чутливгсть до в!русу знижуеться: якщо в!рус уводять мишам шсля 14 дшв, пухли-
на не розвиваеться. 3 кров! шфпсованих тварин можна видшити в!русну ДНК,
здатну до шдукцп пухлин.
Серед пухлин, яю утворюються в людини 1 спричинеш, очевидно, в!русом, ведо-
ма л!мфома Берютга, що уражуе гпднижньощелепн! л!мфатичн! вузли в детей. Вона
поширена в низиннш мтсцевосп Африки, де переносниками шфекцп е мосюти.
Установлено зв’язок вхрусгв з виникненням 1 розвитком деяких шших поши-
рених пухлин людини: в!русу герпесу й раку шийки матки; в!русу гепатиту В 1
гепатоцелюлярно! карциноми (рак печшки, який походить з гепатоцитгв); аде-
нов!рус!в 1 пухлин ештелпо верхних дихальних шляхгв, з якими, як з’ясувалося,
пов’язаний також в!рус Епстайна — Барр (назофарингеальн! пухлини), щентиф!-
кований ранние як збудник В-юптинно! лшфоми Берютга. Видшено ретровхрус
(онко-РНКовий С-типу) з лейкозних юптин людей, хворих на Т-лшфому людини, —
форму Т-л!мфоцитного лейкозу. В!рус названо НТЬУ (вщ англ. кипит Т-1утркота
уп-из). Т-клпинний лейкоз — шфекцшна хвороба людей, заражения якою вщбу-
ваеться пщ час переливания кровь Установлено четкий зв’язок мгж онкогенними
ДНК-вмгсними вирусами папшом 1 пухлинами статевих орган1в людини.
Ф1зичн1 онкогент фактори. Онкогенну мутацшну дпо мають таю ф!зичн! фак-
тори, як юшзуюче та ультрафюлетове випромшювання, можливо, теплова енер-
пя, ультразвук. Кр!м того, фгзичн! фактори можуть вщггравати роль син- або ко-
канцерогеюв.
Онкогенну дпо ультрафюлетового випромшювання спостерггали в дослщ! на
тваринах. Щодня п’ятигодинне перебування лабораторних щур!в на яскравому
сонщ через 10 м!с. призвело до розвитку у багатьох тварин пухлини шюри. Часто
пухлина виникае пщ впливом рентген!вського випромшювання 1 шсля введения
в оргашзм радюнуклцпв.
У людини спостерггають професшш пухлини! хвороби, спричиненг впливом
юшзуючого випромшювання: рак у рентгенолопв, рак легень у шахтар!в, як! пра-
цюють у копальнях з радиоактивною рудою.
Трапчними були наслщки атомних вибухш у Хгросгмг й Нагасаю. Серед 18 тис.
жителю Хгросгми, яю перебували поблизу вщ епщентру вибуху 1 лишилися живи-
ми, !стотно збглыпилася захворювашсть на лейкоз.
З’ясовано, що д!я одночасно юшзуючого випромшювання та хгмгчних онко-
гешв у низьких дозах зумовлюе надзвичайно сильну шдукцпо пухлин, непропор-
цшну дозам цих факторгв, яю спричинюють розвиток пухлини лише в незначшй
юлькосп випадюв, коли дпоть окремо.
Доведено також канцерогенну роль тривалого мехашчного впливу на ткани-
ну. У 1948 р. Б. Оппенгеймер, Е. Оппенгеймер ! Стоун виявили, що у щур!в,
яким, щоб створити ниркову ппертензгю, обгорнули нирки целофаном, розвину-
лася саркома. В експеримент! з гмплантащею пластмасових пластинок установле-
но, що бщя пластинок розм!рами 0,5x0,5 см 1 б!льше шдукуються злояюсш спо-
лучнотканинш пухлини, тод! як уведення порошку з ще! пластмаси не спричиню-
вало утворення пухлин. Можливо, металев! або пластмасов! пластинки
перешкоджають завершению пролгферативно! стадп запалення, що призводить до
надм!рного нагромадження шдукторш розмноження, яю викликають утворення
пухлини.
ПАТОГЕНЕЗ
У патогенез! пухлинного росту розр!зняють три етапи: трансформацпо нормально!'
клггини в пухлинну (!нщ!ац!я), промошю (“пщбурювання”) г прогреспо пухлини.
Трансформация полягае в набутп первюно нормальною клпиною здатносп
необмежено 1 неконтрольовано розмножуватись 1 передавати цю здатшеть дочгр-
шм клпинам у спадок. Трансформация може вщбуватися двома шляхами — мута-
цшним та еппеномним. Обидва шляхи включають мехашзми порушення системи
регуляцп юптинного подшу: системи регуляторних гешв, яю в норм! збуджують
(стимулюють) або гальмують юптинний подш та !х диференщащю, здатшсть до
апоптотично! загибел! мутованих юптин ! юльюсть мттотичних подоив (лгмег Хей-
фл!ка), яю можуть вщбутися з юптиною без термшально! диференщацй обох
дочтрнтх клггин. Якщо юптина буде здатна до апоптозу, то будь-яка мутащя спри-
чинить п загибель. Якщо юптина не буде здатна до апоптозу г набуде здатностг до
неконтролъованого подтлу, але не скасуе лтмтт Хейфлтка, то тз тако! клггини не
зможе вирости пухлина, осюльки подш швидко припиниться.
Узагалт трансформация (тшщацтя) — процес багатокроковий з поступовою
змшою ртзних гешв, яю беруть участь у регуляцц юптинного подшу, однак пер-
шою и ланкою с тмморталтзацтя, тобто набуття юптиною здатностг до безмежного
розмножування.
Втрата пухлинами лйшту клтинного подшу Хейфлиса. У 1961 р. Хейфлгк г
Мурхед вщкрили процес обмеження подшу нормальних фгбробластгв людини;
шсля того як клгтини ембрюна дшяться 50 разгв, вони втрачають здатшсть до
подальшого розмноження. Автори також виявили залежнгсть мгж вгком донора
фгбробластгв г кшьюстю подвоень цих клттин: що старгший донор, то менше разгв
можуть подшитися його клгтини в культур! тканини фгбробластгв.
Оллсопп зт спгвавторами з’ясували, що лтмтт юптинного подшу пов’язаний зт
зменшенням довжини теломертв, яю е юнцевими дшянками ланцюпв ДНК хро-
мосом; що коротки теломери хромосом клгтини, то менше разгв вона здатна подт-
литися, тому що за кожним подшом клгтини вщ теломери вщр!заеться певна юль-
юсть нуклеотидних послщовностей. Розмноження клггини припиняеться, коли
вщ теломера майже нгчого не залишаеться.
Поргвняно з нормальними диференщйованими клгтинами, в ембрюнальних г
ракових юптинах за кожним подшом клттини теломера не скорочуються, а добу-
довуються, г тхня довжина не зменшуеться. Грейдер г Блекберн виявили в таких
юптинах фермент теломеразу, яка е термшальною трансферазою з властивостями
зворотно! транскриптази. Теломераза на матриц! РНК добудовуе в теломерах
втрачеш дшянки ДНК г таким чином скасовуе лтмтт Хейфлтка, а юптини пухлин
стають безсмертними (гмморталтзованими). У нормальних ембрюнальних клтти-
нах шсля синтезу теломера розраховано! довжини ген теломерази блокуеться, а в
юптинах пухлин залишаеться активним. Таким чином, у цих пухлинах здатшсть
до безмежного розмноження досягаеться втратою репресора гена теломерази.
У частиш пухлин теломерази немае, а вщновлення довжини теломертв дося-
гаеться альтернативними мехашзмами — АКТ (вщ англ. аИетаНуе тесИататх /ог
1еп%1кепт§ о/ 1е1отег$ — альтернативш мехашзми подовження теломер). Цими ме-
хашзмами е рекомбтнацгя та ретротранспозишя, за рахунок яких частина довгих
теломертв приеднуеться до вкорочених ! вщновлюе здатшсть клгтини розмножу-
ватися.
Другою обов’язковою ланкою трансформацп нормально! клттини в пухлинну
е втрата здатностг до апоптозу, який у нормальних юптинах включаеться в разг
змши !х геному.
Апоптоз — запрограмована смерть юптини. Ця програма може бути вв!мкнена
геном р53, який активуеться в разт накопичення в ядрт нерепарованих ушкодже-
них ДНК, зокрема з розривами н ланцюггв. При апоптоз! активуються ферменти,
якт руйнують бюлопчш макромолекули, дезштегрують субюптинш органели. Ак-
тиващя апоптозу залежить також вщ спгввщношення продукттв гешв Вах, яю пщ-
тримують апоптоз, 1 проти-
апоптичних вс]-2. Крш того,
апоптоз може бути активо-
ваний ФНП, який вироб-
ляеться лейкоцитами та ф1б-
робластами. У пухлинах ус!
щ мехашзми апоптозу не
дпоть, осюльки при онкоге-
нез! втрачаеться ген р53, шо
унеможли влюс акт и ва шю
апоптозу цим супресором.
Порушення збуджуваль-
но! системи регуляторних
гешв. Збуджувальна система
Фактоэ эозмножечня
Цитоплазматичний передавая
Ядерш регуляторш битки
Ген - ппщатор фази 5
Внутртшня частина рецептора
фактора розмноження
Зовжшня частила репеггтора
фактора розмноження
Схема 12. Система збудження пятинного подглу
регуляторних гешв, яка ак-
тивуе розмноження юптини, мае зовнппньо- 1 внутрпцньоюнтинну частини. У нор-
М1 активашя розмноження юптини починаеться з дп позаклггииного фактора росту
на зовшшньоюптинну час!ину рецептора в мембраш юптини. Дай збудження пе-
редаеться на внутршшьоюйтинну частину рецептора. вщ яко! активуються цито-
плазматичш рецептори збудження розмноження. Щ рецептори активують в ядр! кй-
тини 1ени, шо стимулюють синтез ДНК, який зумовлюе мггоз юпгини (схема 12).
Мутацшний механизм порушення збуджувально! системи геп!в полягае в тому,
шо один 13 них унаслщок мутацп змшюе свою структуру так, що на ньому почи-
нае синтезуватися вщповщний фактор — шщгатор подшу в незворотно актившй
форм!, який одразу активуе поды юптини, тощ як у норм! вш синтезуеться в не-
активней форм! 1 його нормальна активащя вщбуваеться тыьки пед д!ею нормаль-
них стимулов юйтинного подыу. У нормально! юноши ця активащя завжди е
зворотною ! може бути зупинена вщповщно до ф1зюлопчних потреб. Зокрема
нормальна юптина перетворюеться на пухлинну внаслщок мутацп гена зовнппньо!
або внутрплньо! частини рецептора фактора розмноження, що призведе до син-
тезу цих рецепторе в в актившй форм! без дп стимулятора розмноження (фактора
росту) на рецептор. Оскыькп в пронес] розмноження тако! клпини цей ген вщ-
творюеться вже з мутацшними змшами, то дочтрня юйгина також одразу починае
процес редупйкацп ДНК 1 м1тоз. Як наслщок, в оргашзм! з’являеться клон (ро-
дина) пухлинних клггин, у яких здатшсть нерегульовано розмножуватися пере-
даеться як фактор соматично! спадковосп.
Ус! онкогени пухлин, шдукованих онкогенними факторами, а також В1русш
онкогени виявилися власними генами оргашзму — регуляторами росту юптин,
тыьки змшеними до стану постшно! активность Тому нормалый гени, що ре-
гулюють юптинний поды, з погляду потенцшного перетворення !х на онкогени
отримали назву протоонкогешв. Онкогени — пе мутацгйно або ешгеномно акти-
рован! протоонкогени. а синтезоваш на них б!лки — онкобыки
Етгено.мний механизм порушення збуджувальна! системи гешв полягае в тому,
що в юптиш з являеться фактор, який не належить до геному ще! юптини ! не
спричинюе мутацп, однак зумовлюе еййке порушення нормально! регуляцн гено-
му, шо призводить до неконтрольованого синтезу онкобыюв та росту (розмно-
ження) юптин.
Ешгеномний вплив, який передаегься з поколшня в поколения, може сфор-
муватися щц д!ею вгрусу, що шфткуе первюно норматы ту юптину 1 потрапляе в
кожну новоутворену юптину щл час мггозу. Доведено, що пухлинотв!рн! втруси
мають у своему геном! онкогени, як! е регуляторними генами юптин. В!руси ко-
лись узяли цт гени у юптин, у яких вони паразиту вали, але зумовили в них таку
мутацпо. шо призвела до вироблення факторов кл! тинного подшу саме в постшно
активнш форм!. Це своею чертою спричинито трансформашю нормальних юптин
у пухлинш. Наприклад, деяк! пухлинотвгрш в!руси раку молочно! залози як онко-
ген використовуютъ внутринньоклттинну частину рецептора фактора розмножен-
ня, що е ферментом протешкшазою. Цей захоплений вирусом ген внутр!шньоюп-
тинно! частини рецептора змшений втрусом так, що закодована на ньому про-
тешкшаза синтезуеться постшно в активнш форм! т стимулюе розмноження
юптини без взаемодп рецептора юптини з фактором розмноження, поки в!рус
перебувае в кяттиш.
З’ясувалося, що онкогеню в!руси можуть мати в такш постшно активнш
форм! будь-який ген збуджувально! системи розмноження — 1 цитоплазматичний
передавач, < ядерн! активатори. Загалом сгимуляц!я розмноження пухлинних юп-
тин вщбуваеться на таких етапах регулящ! топтанного под!лу: посилення ди фак-
тор!в розмноження, збшьшення мембранних рецепторов факторгв росту, збшь-
шення передач! сигналу, шо активуе юптинний под!л, посилення процесу транс-
крипцп ген!в.
З’ясувалося, що як онкобшки взруси продукують ус! зазначен! нижче види
молекул, як! в нормг стимулюють мехашзми юптинного подшу.
1. Фактори росту. Онкобиок ворусу саркоми мавп виявився аналогом нор-
мального ФРТ, який стимулюе розмноження ф!бробласт!в. 81з е геном, який ко-
дуе Р-ланцюг ФРТ. На вщмшу вщ нормальною ФРТ, який активуеться, з’сднуточись
1з рецептором на мембран! юптини, продукт цього в!русного онкогену вироб-
ляеться вже в активному стан! ! поеднання з рецептором юптини для активацп не
потребуе. Мутованими генами факторгв росту е також онкогени 1п1-2, ЮР-1.
2 Рецептори юптинних мембран до фактор!» росту. Аналогами рецепторов юп-
тинних мембран е продукта вгрусних онкогенов зге вгрусу саркоми Рауса. егЬВ
в!рус!в АЕУ, як! спричинюють у птахш саркому 1 рак. З’ясовано, що егЬВ е анало-
гом юптинного рецептора для ЕФР, його внутршшьоюптинною частиною, а за
дгею — тирозиновою протешкшазою (приеднуе до тирозинових залишюв про-
теши фосфата! групи, вщ чого функцоональна актившсть фосфорильованих бш-
юв розко змшюеться) Поровняно з нормальним рецептором продукт ворусного
онкогену виробляеться в активному сташ й !мотуе для клотини дйо ЕФР. Протеш-
коназами е також рецептори фактоотв росту — онкобшки Гшз. тег, тгк, гет
3. Передавач! сигнал!в з рецепторгв. Прикладами втрусних онкогешв, як! коду-
ють аналоги передавали сигнал!в у юптинах, е газ т РКС. Онкогени газ — аналоги
гени О-битов М1ТОТИЧНОЮ циклу. Вони кодують б!лок (21 кДа), який при-
кр!плюеться до внутр!шньо! поверхш цитоплазматично! мембрани 1 бере участь у
проведенн! сигналу, шо стимулюе мттоз. Цей бшок у поеднаннт з гуанозинтри-
фосфатом (ГТФ) стае активним, але пщ час поедкання з гуанозиндифосфатом
(ГДФ) шактивуеться. Унаслщок мутацп протоонкоген втратив ферментну актив-
шсть ГТФази т став онкогеном гаг, який не здатен розшеплювати молекулу ГТФ
! утворювати ГДФ, що постшно стимулюе мггоз юптини.
4. Цитоплазматичш т ядерш быки. Цитоплазматичними передавачами сигналтв
е онкобыки онкогешв га/, тоз, ртз, яю мають властивостт цитоплазматичних ю-
наз. Вони е похщними протоонкогешв нормальних цитоплазматичних передава-
ч!в — серин-треоншових юназ МАР (вщ англ. тИореп асПуа1е(1 рго1е1п ктазез —
активоваш митогеном протеппонази), як! в активному стан!, зокрема кшази ЕКК1
! ЕКК2 (ех!гасе1и1аг зщпа! — ге§и1а1еб ргоСет ктазез 1,2 — кшази 1 т 2, що тх ре-
гулюе позаюптинний сигнал), !з цитоплазми перемпцуються в ядро, де фосфори-
люють ядерш быки — активатори гешв юптинного подшу.
У ядрах клггин виявлено продукта онкогешв у-тус, у-туЬ втруств, як! спричи-
нюють мтелощний лейкоз у птах!в, у-/оз втрусу остеосаркоми у мишей. Продукта
д!яльност! багатьох онкогешв ДНК-умтсних втруств виявляються в ядрах юптин,
яю трансформуються: продукт Е1А — регулятор транскрипцп в ядр! та цитоплаз-
м!; великий биток Т-в!русу 8У-40 у ядр! шщпое синтез ДНК, впливае на транс-
крипцгю, шактивуе супресор р53.
За бюлопчною актившстю продукттв синтезу онкогени втруств 1 вщповщш !м
юптанш прогоонкогени можуть бути подшеш на такт групи:
• тирозинов! протешюнази: уез, /§г, /рз, гоз, /тз, егЬВ, зег, аЬЦ
• серин-треошновт протешкшази (фосфорилюють быки за радикалами амт-
нокислот серину т треоншу): тоз, тП, га/;
• родина газ онкогешв, яю походять в!д дуже поширених регулятортв розмно-
ження т е навтть факторами росту дртжджтв; активують адентлатциклази та гу-
аншатциклази;
• шдуктори подтлу, як! дгють через ядернт б!лки (тус, туЪ, /оз, ]ип/, у/ип / у-/оз
утворюють гомо- т гетеродимери й активують синтез РНК на певних генах ядра;
• аналоги фактортв росту: з1з (аналог ФРТ);
• промотори гешв. Онкоген тус може виконувати роль промотора, тобто мо-
бильного гена, який активуе РНК-полтмеразу в дшянц! гена, до якого в!н приед-
нуеться. З’ясувалося, коли онкоген тус мтститься в геном! втрусу раку молочно!
залози, у мишей, яю отримали такий втрус, у пер! од лакгацй штенсивно утво-
рюеться пухлина молочно! залози. Однак коли онкоген тус приеднуеться до гентв,
яю контролюють вироблення тмуноглобулппв, то у мишей розвиваеться лтмфо-
лейкоз.
Як формуютъся вгрусш онкогени та пухлинотв1рн(стъ е(рус1в?
1. Змши структури протоонкогешв, яю призводять до !х неврегульованост!,
вщбуваються пщ час неповного захоплення цих протоонкогешв втрусами, коли
здшснюеться вщрив протоонкогену вщ його власних регуляторних дшянок або
регуляторних дшянок шших юптинних гешв. Вщмтншстю втрусних онкогешв вщ
протоонкогентв е те, що протоонкогени в геном! клттини представлен! двома ге-
нами — штроном т екзоном, а у втрусних геномах виявляються тшьки екзони
протоонкогешв, тобто вщбуваеться втрата значно! регуляторно! частили юптин-
них гентв.
З’ясувалося також, що втрусна зворотна транскриптаза “припускаеться” по-
милок у зчитуванш геному. Точкова мутацтя в онкоген! втрусу, що зумовив розви-
ток доброяюсно! пухлини, перетворюе втрус на шдуктор злояюсних пухлин. Тол-
ков! мутацп в онкогенах втруств тстотно впливають на ртзю ланки онкогенезу.
2. Особливим механизмом пщвищення онкогенност! е супершфекцтя втрусом
клттин, заражених ранппе слабким штамом онкогенного втрусу. Мтж втрусами може
вщбуватися обмш генетачною шформащею, 1 дефектний в!рус може огримати ген,
якого не вистачае, або стимулятор своеТ активность Супергнфекцзя може зумовити
розмноження слабкого онкогенного в!русу або активащю функшй його гешв.
Порушення галышвшн системи репресорних гешв. Кожна клиппа мае автоном-
ну гальмгвну систему гешв, завдяки як!й вона в культур! тканини поза оргашзмом
розмножуеться нормально. Онкогенез у раз! порушення системи репресорних гешв,
як! в норм! гальмують клгтинний пол!л, полягае в тому, що ген ще! системи втра-
чае здатшсть виконувати свою репресорну функщю ! перестав галъмувати клпин-
ний под!л.
Системи гальмзвних гешв функцтонують здебьтьшого в контрольних пхнктах
на меж! фаз мистичного циклу О, / 3, 8 / С_, Сг2/ М. Особ.тиве значения мають
галъм!вн! гени КЬ та р53, як! функшонують на меж! фаз С^/ 8.
Продукта д! ял вноси гальмтвних ген!в входять до складу мембранних. цито-
плазматичних 1 ядерних бьтктв. Окремо генуе мехашзм обмеження (л!мпу) клпин-
ного подшу Хейфл!ка, який реалтзуеться шляхом зменшення довжини теломергв.
Супресори — мембраны быки. У мембрану кштин вбудований продукт гальмгв-
ного гена ОСС. Його втрата призводить зокрема до розвитку раку товсто! кишки.
Цей ген моститься в довгому плеч! хромосоми 18. У норм! вш зумовлюе утворення
мгжклпинних зв'язктв. Кр!м того, в мембрану вбудован! рецептори для ФНП 1
рецептори СО95-К. Унаслщок дп на сво! рецептори ФНП 1 СО95 вмикають ме-
хашзм апоптозу.
Супресори — иртоплазматичш быки. Продукт гальмгвного гена ЫЕ-1 е цито-
плазматичним бьтком у поеднанн! з ГТФ, який у норм! акгивуеться ГТФазою 1
блокуе цитоплазматичну передачу сигналу активацп розмноження клпин.
Групи цитоплазма!ичних б!лк!в Вах 1 Вс1-2 у певних спзввщношеннях можуть
блокувати розмноження 1 зумовлювати апоптоз.
Супресори — ядерш быки. Це продукта ген!в КЬ 1 р53, як! регулюють експреспо
гешв, беруть участь у вибор! шляху пролзферащ! або диференцповання клпини, а
головне — контролюють бтохтмзчний мехашзм, який регулюе шпцащю синтезу
ДНК Ген р 53 може вмикати апоптоз — залрограмован) загибелъ клггини. Ген КЬ
моститься в людини у хромосом! 13 1 зчеплений !з геном естерази О. У раз! вграти
цього гена клпинами ставки ока розвиваеться пухлина — ретинобластома.
З'ясувалося, шо в!н мае велике значения в регуляцн клпинного подьту та запобт-
ганн! розвитку пухлин. Продуктом синтезу гена КЬ е биток з молекулярною масою
105 кДа — р105-В.Ь, який угворюе комплекс з шпцаторами транскрипц!! Е2Р 1
Г)Р1, що гальмуе мпотичний цикл, осюльки блокуе транскрипцпо гешв 8-фази.
Супресорна д!я протешу КЬ залежить вщ сгупеня його фосфорилювання, яке
здшенююгь циклшзалежн! кшази сёк-2 1 сёк-4 у поеднанн! з пиклшами Ц 1 Е.
Ген р53 кодуе фосфопротеш, який побудований !з 375 амшокислот 1 функшо-
нуе в ядр! клпини. Як 1 проте!н КЬ, р53 репресуе гени — зшшатори редуплткацп
ДНК, блокуючи фазу 8 мпотичного циклу. Особливкть репресорно! дп гена р53
полягае в тому, шо за нормального процесу розмноження вш не чинить свою д!ю,
але швидко блокуе процес розмноження в раз! появи ушкоджених молекул ДНК,
у тому числ! в раз! загрози розвитку неконтрольованого росту'. Зугшнка шппаторзв
транскрипцп ДНК 1 початку фази 8 дае змогу клггин! репарувати ушкоджен! гени.
Встановлено, що юлыасть р53 значно збьтьшуеться в клзтинах пщ д!ею юшзую-
чого випром!нювання та шших мутагенних факторгв.
Онкогенез тип час порушення гальмгвно! системи гешв вщбуваеться, як г при
порушеннях збуджувально! системи гешв, двома шляхами — мутацшним г епгге-
номним.
Мутацшний мехашзм порушення галъмгвшп системи гешв. Мутагенш фактори
можуть спричинити мутацпо всгх гальмгвних гешв, що кодують: мембрани!, ци-
топлазматичш, ядерш быки. При цьому гальмгвнг гени можуть повшстю втрати-
тись або синтезуватимуться неактивш репресори. Унаслщок тако! мутацп здоров!
клггини перетворюються на пухлинш.
Доведено, що, на вщмгну вщ гешв збуджувально! системи, як! е здебшьшого
домшантними, гальмгвн! гени клггинного под!лу успадковуються за рецесивним
типом. Быышсть спонтанних або !ндукованих канцерогенами пухлин виникае в
раз! порушення генгв збуджувально! системи, спадков! види пухлин — гальмгвних
гешв. При цьому для мутацгйного порушення клггинного подшу достатньо мута-
цп або одного збуджувального гена, або обох алельних гальм!вних генгв.
У бшыпост! випадкгв мутоваш гени в клгтинах успадковуються соматично.
Але мутоваш гальмгвнг гени виявляються в статевих клпинах 1 зумовлюють успад-
кування пухлин. Так успадковуеться недостатшсть р53, яка призводить до розвит-
ку в таких родинах остеогенно! саркоми, раку молочних залоз, глгоми, а також
синдрому Лг-Фраумеш, при якому в нащадк!в уже в молодому виц виникають
злоякгсш пухлини молочних залоз, головного мозку, кгрково! речовини наднир-
кових залоз. Спадковими можуть бути пухлини, що з’являються внаслщок мутацп
шших ген!в — супресор!в клггинного подшу, а саме: ретинобластома, остеогенна
саркома в раз! мутацп гена КЬ, полшоз товсто! кишки при спадковгй недостат-
ностг супресора АРС, меланома — супресора МТС-1, який у норм! приеднуеться
до циклгнзалежног кгнази 4 1 запобггае ц активацп циклгном П1, що блокуе пере-
ход фази мггозу СП у фазу 8.
Ешгеномний мехашзм порушення галъм^вшй системи ген1в. Функцгя генгв — су-
пресоргв клгтинного подшу може порушуватися ешгеномно онкогенами в!рус!в.
Б!лок КЬ втрачае актившсть у раз! поеднання з онкогеном Е7 вгрусу папшоми
людини, що й призводить до онкогенезу. 1нш! онкогени ДНК-вгрусгв, зокрема й
онкоген Е1А аденовгрусгв, великий Т-антиген вгрусу 8У40, також гндукують пух-
лини, гнактивуючи бшок КЬ.
Епггеномним шляхом постгйну гнактивацгю гена р53 зумовлюють вгруси, он-
когени яких кодують саме такт бшки, що руйнують або блокують р53. Так, р53
блокуеться продуктом онкогену тЗт-2 вгрусу, який у такий спосгб спричинюе
розвиток пухлин.
Ешгеномний мехашзм установлений для бшыпост! генгв — супресоргв клг-
тинного подшу.
Сучасн! данг про мехашзми онкогенезу свщчать, шо цг репресорш мехашзми
виявилися набагато складнгшими, але центральним об’ектом усгх систем регуля-
цп розмноження клгтин е гени — гнщгатори транскрипцп ДНК, якг вмикають
перехгд фаз мггозу О, / 8, причому безпосереднгми генами-репресорами, здатни-
ми заблокувати гени — ппцгатори транскрипцп, е гени КЬ г р53.
Мутацшний г епггеномний мехашзми онкогенезу можуть бути пов’язанг мгж
собою. У клгтинг е спецгальнг регуляторн! гени, якг репресують геном пухлино-
твгрного вгрусу. Тому мутацгя може вгдбутися з репресорним геном клгтини, унас-
лщок чого активуеться онкоген пухлинотвгрного вгрусу, який вийшов з-щд конт-
ролю, 1 вщбуваеться ешгеномна трансформашя клггини. Таким чином, хгмгчнг та
фгзичн! фактори можуть не спричинювати трансформацгю безпосередньо, а спри-
нта активацп вирусного онкогенезу.
Дослщи Гердона, пов'язанг з пересадженням ядер диференщйованих юптин у
цитоплазму зиготи, було використано для вивчення регуляцгг подшу в ядрах пух-
линних юптин. Якщо 13 зашпднених яйцеюптин жаби видаляють ядро г замгсть
пього гадсаджутоть ядро зг спешалгзованог юптини з подвгйним набором хромо-
сом, наприклад, ядро юптини кишкового ештелпо, то розвиваеться пуголовок з
усгма органами, шо диференшюються. Отже, у ядрах спещалгзованмх юптин ор-
гашзму зберггаеться весь набгр гешв, який був у ядрг зиготи Коли в цитоплазму
зиготи пщсадили ядро раковог клгтини з нирково! пухлини Л юкке жаби, також
розвинувся пуголовок, а не пухлинна тканина. Шсля пересадження ядра юптини
такого пуголовка в цитоплазму юптини Люкке утворилася нормальна ниркова
тканина.
Отже, здорова цитоплазма зиготи зумовила повнс ещновлсння регуляци в
пересадженому ядр! пухлинног юптини 1 закргллення його як успадковано! влас-
ти вост!. Осюльки пухлина Люкке жаби мае вгрусне походження, то йдеться про
вгдновлення функцп гешв, якими репресуеться втрусний геном.
Трансформоваш юптини починають синтезувата нов! фак гори росту; яю
впливають на пухлинш юптини автокринно г паракринно. Фактори росту, гндук-
цгя яких починаеться юптинами п!д час перетворення гх на пухлинш, належать до
двох груп:
— 1рупа А — фактори росту, шо дпоть на сам! юптин и-продуценти г шдтри-
мують !х розмноження: глгкопротегн р52, !нсул!нопод!бн! фактори росту — ШФР-1
та ШФ Р-11, аналог ФРТ онкоген р28*, у-газ та шли онкогени 1 протоонкогени. У
юптинах, як! зазнали онкогенезу, встановлюеться автокринна секрещя цих фак-
торгв росту, тобто клгтини залишають гх у собц г цим шдтримуеться безперервне
розмноження. Невелика частина екскретованих факторгв росту впливае на сусгдгп
клгтини цгег само! тканини;
— група Б — фактори росту, призначен! для юптин гншого типу, насамперед
для юптин строми ! судин. За допомогою цих факторгв росту пухлинна тканина
спричинюе вростання гнших юптин у пухлинний вузол. Для ф!бробласт!в вироб-
ляеться згаданий ФРТ або його вгрусний аналог р28Ь1\ а також особливий фактор
росту, який стамулюе синтез колагену ф1бробластами — С55Р (вщ англ. соИа^еп
фп1Ие$18-8йти1аНп8 §гом>1И /асюг). Для судин пухлини! клгтини виробляють стиму-
лятор росту анпогенгн, який проявляв актившсть у надзвичайно низьких дозах, а
також шсу.тгноподгбш та гнш! фактори росту. Секрецгя факторгв росту для клггин
гншого типу отри мала назву паракринно'1.
У пухлинних юптинах ргзко збгльшуються синтез 1 екепреегя рецепторгв. на-
самперед для фактор!в росту, наприклад у-егЬВ для ЕФР.
У пухли нах культурах пухлинних юптин спостерггаеться зниження або вгд-
сутнгсть контактного гальмування. Коли в культур! тканини нормальш клггини
двох сусгднгх дглянок, розмножуючись у напрямку росту, торкаються одна одног,
ргст тканини ! подш юптин у дгй дьлянцг приштняються. Пухлинш юптини в та-
ких випадках продовжують роста, утворюючи багатошаров! дглянки.
Промоцгя (активацгя) е другим етапом у мехашзмг онкогенезу. Трансформо-
ванг клгтини можуть залишатись у тканин! тривалий час у неактивнгй форм!.
ЗоГ
Додатковий вплив канцерогенного фактора, який сам не зумовлюе трансформа-
ццо, але стимулюе юптини до розмноження, призводить до того, що пухлинш
юптини, яю перебувають у латентному сташ, починають дшитись, утворюючи
пухлинний вузол.
Бшышсть онкогешв е повними, тобто такими, що спричинюють 1 трансфор-
мацпо, 1 активацйо. Однак в експеримент! онкогенез можна перетворити на дво-
етапний процес, коли трансформацпо 1 промоцпо можна вивчати окремо. У до-
слщ! Беренблюма — Моттрама мишам наносили на шюру 25 мкг метилхолантре-
ну, що було недостатшм для вщтворення пухлини протягом життя тварини. Потгм
змащували ту саму дшянку шюри кротоновою ол!ею, що сама по соб! школи пух-
лини не зумовлюе, однак в умовах цього дослщу активувала подш юптин, транс-
формованих онкогеном. У тварин почали утворюватися пухлини.
Тснування латентних (“що сплять”) трансформованих юптин можна виявити
1 в дослщ! Фппера. У вену ввели 50 пухлинних юптин карциноми Уокера. Ц!е!
дози недостатньо для шдукцл пухлини, 1 протягом багатьох мтсяцгв теля ш’екцп
у щур!в пухлини не розвивалися. Однак якщо цим щурам юлька раз!в зробити
розтин черевно! порожнини 1 доторкнутися до печшки, то в печшщ у них розви-
неться карцинома Уокера.
Не вс! речовини, яю спричинюють запалення, е активаторами онкогенезу.
Так, розведен! розчини шриту та кантаридину, яю спричинюють лише слабке
подразнення шюри, справляють активну антиканцерогенну д!ю в умовах дослщу
Беренблюма — Моттрама.
Можливим молекулярним мехашзмом промоцп е ввгмкнення трансмембран-
но! сигнально! системи, яке закшчуеться акгиващею и внутршшьоюптинно! части-
ни — протешкшази С. Сигнал вщ рецепторгв ще! системи, розмйцених на поверх-
ш юптинно! мембрани, активуе обмш фосфатидилинозитолу в мембраш, генеращю
дгацилглщеролу, який стимулюе протешкшазу С. Промотори пухлин — еф!ри фор-
болу — прямо стимулюють протешкшазу С. Можливо, протешюназа С передае дпо
також шших промоторов, у тому числ! й факгор!в росту, продуктов онкогешв. Од-
ночасно в пухлинах актившсть сигнально! системи другого типу — адешлатциклаз-
но! — знижуеться. Вище було описано мехашзм промоцп, який полягае у прямому
вбудовувашп промоторного гена в геном юптини (онкогену тус).
Прогресгя — третш етап мехашзму онкогенезу. Це стшю яюсш змши влас-
тивостей пухлини в бхк малишзацп в процес! п росту. Доброяюсш пухлини мо-
жуть перетворюватися на злояюсш, а злояюсн! — на ше б!льш злояюсн! пщ впли-
вом кшькох факторгв:
• постшних спонтанних мутащй пухлинних клггин у раз! зниження в них ак-
тивност! репаративних ферменпв. Унаслщок мутацш з’являються агресившпп
юптини пухлини;
• змш гено- та фенотипу клгтин, що призводять до прогреей та можуть бути
пов’язаш з продовженням ди на геном пухлинних юптин канцерогенного факто-
ра, або супершфекцп пухлинотв!рними та непухлинотв!рними втрусами, полепле-
но! в пухлинних клгтинах;
• до первинного онкогенезу, як правило, залучаеться не одна юптина, а юлька,
що спричинюе формування в пухлин! юлькох сублшш юптин. У пухлиш, що росте
в мшливих умовах (харчування, кровопостачання, шнервашя), постшно вщбуваеть-
ся вщб!р найбшьш життездатних юптин. Певш юптини при цьому переважають.
Стшкктъ пухлин до хьмюпрепарагтв. При лгкуванн! цитостатичними або кан-
церол1тичними хтмгчними сполуками у багатьох хворих з’являються юптини, стш-
К1 до них сполук, яю зумовлюють нове розростання пухлини. Ц1 юптини утворю-
ються шляхом мутацп або ешгеномно з попереднгх юптин пухлини. Яю саме ме-
хашзми стшкосп до хтмтопрепарапв виникають у мутованих юйтинах пухлин?
Один 13 таких мехашзм тв — пе зв'язування чужорщних речовин (ксенобютиюв) 1
викидання !х гз юптини, що здшснюе Р-гл!копротеш. Вш е ! в нормальних кл!ти-
нах. але пщ час прогресп пухлин з’являються юптини, у яких юльюсть Р-пп-
копроте'шу значно зб!льшена. Кртм того, у прогресп кожно! пухлини можуть фор-
муватися сво! особлив! мехашзми стшкост! до конкретних х!м!опрепарат!в.
ОСОБЛИВОСТ1 ПУХЛИННОТТКАНИНИ
У процес! онкогенезу та прогресп юптини втрачають здатшсть до диференцпован-
ня, повертаючись шби до ембрюнального стану. Це явище огримало назву анап-
лази. Ознаки анаплазп е в бюх!м!чних пропесах пухлинних юптин (бюхтмтчна
анаплаз!я), у ф!зико-х!м!чному !х еташ (ф1зико-х!м!чна анаплазгя), у будов! та
функцп (морфологтчна ! функцюнальна анаплазтя). Вщбуваеться також метаплазия —
перетворення на нов! юптин ш форми.
Бюхтмтчн! особливост! пухлинно! тканини. Основою бтох!м!чних особливостей
пухлиннот тканини е змши генетично! регуляии кл!тини. Унаслщок репресп одних
гешв припиняеться синтез пов’язаних !з ними ферменпв, структурних б!лк!в
тошо, дерепрестя шших ген!в призводить до того, що в юптин! з’являються нов!
типи бтлюв, тзоферменпв. Як правило, репресуеться вироблення ферменпв 1 бтл-
юв, яю дають змогу юптин! виконуъати спец!ал!зовану функщю, й активуються
шляхом дерепресп п ферменти, яю забезпечують клгтинний поды.
Онкогени! фактори потенщйно здатш зумовлювати не лише розлад регуляцн
юптинного под!лу, а й порушення функцш шших структурних 1 регуляторних
гешв. Тому в пухлиннш юптин! спостерйаеться раптова дерепрестя синтезу речо-
вин, наприклад гормошв, як! в норм! не утворюються в ц!й тканин!. Так, у пер-
виннш карцином! легень може бути виявлений синтез кортикотрошну або гона-
дотрошну, у пухлинах нирок — тироксину. 3 пет само’! причини в пухлинах може
втрачатися вироблення якого-небудь ферменту або здшснюватись утворення но-
вого. Характерно, однак. що р1зш пухлини наближаються одна до одно! за комп-
лексом ферменпв, яю входять до гх складу, причому цей комплекс гим менший.
чим бьтыпою е дедиференщашя кл!тин.
Найважлившгою бюхгмтчною особливгстю пухлиннот клпини е активащя
синтезу нуклешових кислот. У пухлинних юптинах поргвняно з нормальними зм!-
нюеться набтр ДНК-полгмераз. Серед трьох вищв ДНК-полгмераз у пухлинах
зменшуеться ктльюсть ДНК-пол!мерази 3, яка використовуе як матрицю нативну
ДНК, ! збьтьшуеться юльюсть ДНК-пол!мерази 2, здатно! будувати ДНК не лише
за нативною, а й за денатурованою матрицею.
У пухлинних юптинах яюсно та юльюсно змшюеться синтез бтлюв. До бтлгав,
синтез яких у пухлинних юптинах р!зко збшыпуеться, належать протеши м! готич-
ного апарату, у тому числ! великомолекулярнии бшок веретена. У норм! вмгст
бтлюв у мистичному апарат! становить близько 11 % тх юлькост! в юптин!, а в
пухлинт !х юльюсть збйгьшуеться до 30 %.
Змшюеться обмш ботов. Знижуеться здатшсть пухлинних клттин до переамг-
нування та дезамтнування амшокислот, школи не утворюються деяю ферменти,
яю беруть участь в обмш! амшокислот. У бтлыпост! пухлин зростае захоплення
амшокислот гз кровт т синтез ботов. Катаболтзм бтлка знижуеться настигьки, що
навггь в оргашзм!, який голодуе, бглок пухлини не бере участт в загальному об-
мш1. Кргм того, радюлопчним методом було доведено, якщо тканини оргашзму,
який голодуе, втрачають амшокислоти, пухлина “привласнюе” гх собг Усе це
дало змогу схарактеризувати пухлину як “пастку для азоту”. Так само пухлини
захоплюють г “привласнюють” вггамши та мгкроелементи.
Особливе мтсце в бтохгмп пухлин посщае обмш вуглеводтв 1 вироблення енер-
гп. У пухлинах нерщко значно збшьшуеться швидюсть глтколтзу. 1нтенсивний
глгколгз не е спепифгчною особливгстю пухлин. Однак коли поргвняти актившсть
фермента глтколтзу в пухлин! т первиннш тканиш, то вдаеться виявити збгльшен-
ня активност! основних фермента гл!кол!зу: гексоюнази, фосфофруктоюнази та
шруваткшази. У пухлинах вщбуваеться аеробний глгколгз, тобто розпад вуглеводтв
до шрувату 1 перетворення його на молочну кислоту за наявностг кисню — нега-
тивный ефект Пастера. Водночас у бшьшост! нормальних тканин спостерггаеться
гальмування перетворення шрувату на молочну кислоту за наявностг кисню та
зниження гнтенсивностг глтколтзу. Варбург сформулював гшотезу про те, що при-
чиною злояюсно! трансформацп клттин е фактори, яю перешкоджають одержан-
ию клгтинами енергп вщ окиснення, унаслщок чого вони переходить на глгколгз.
Ниш посилення глтколтзу слщ розглядати не як причину виникнення пухлин, а
як наслщок онкогенезу, складно! перебудови синтезу та регуляцп функцп фер-
менттв.
Пухлина гнтенсивно захоплюе глюкозу з кровт. Навпъ у разг пщвищення
вмтсту глюкози в кровт до 16,7 ммоль/л (300 мг%) кров, що вщтгкае вщ пухлини,
не мтстить глюкозу (В.С. Шапот). Цю здатшсть пухлини пов’язують з! зниженням
активностт регульовано! глюкоюнази 1 ртзкою активащею гексоюнази, що менш
чутлива до гормонально! регуляцп.
Енергтя, яку одержують пухлинш клттини вщ глтколтзу, достатня для забезпе-
чення синтезу нуклегнових кислот г юптинного подшу.
У пухлинах також змшюеться окиснення (тканинне дихання). Загалом е тен-
денцгя до зниження дихання пропорцшно ступеню дедиференцгювання клттин.
При цьому спостерггаеться ефект Кребтрт: пригнтчення окиснення в раз! наванта-
ження глюкозою, що може бути наслщком “боротьби” глтколттично! системи
ферменттв пухлини з и окисними ферментами за неоргантчний фосфат, шш! суб-
страта та кофермента.
Фгзико-хгмгчш особливост! пухлинног тканини. Змша фтзико-хгмтчних власта-
востей пухлинних клттин е здебглыпого наслщком бгохтмтчно! перебудови пухлин-
но! тканини. Тнтенсивний глгколгз призводить до накопичення молочно! кислота.
У разг навантаження вуглеводами рН у пухлиннш тканин! може знизитися до 6,4.
У пухлин! пщвищений вмгст води, а школи г деяких електролгта, зокрема солей
калпо. Вмгст кальцпо г магнпо знижений, сшввщношення К+/ Са2+ пщвищене. За
рахунок гщратацп та пщвищення вмтсту тошв водню, а також деяких електролгта
електропровщюсть пухлинног тканини збшьшена. При цьому зменшена в’язюсть
колоццв. Спостерггаеться збшьшення негативного заряду клттин пухлини, вели-
чина якого наближаеться до величини заряду лтмфоцита. Припускають, що через
подгбшсть зарядгв лгмфоцити меншою лирою здатн! контролювати пухлинну тка-
нину, юж нормальну. тому не атакують пухлини, мутанта. Збщьшення негатив-
ного заряду пухлинних кл!тин вщбуваеться за рахунок збит мнения юлькосп елек-
тронегагивних радикатгв нейрамшовог кислота, у зовшшнш мембран! юптин.
Ступшь ф1зико-х1м1чно1 анаплазн вщповщае ступеню дедиференцповання та
швидкосп росту.
Антигени! особлипостг пухлини. За свогм антигеннггм складом пухлинна ткани-
на вщргзнясться вщ нормально! тканини, з яко! вона походить. Так, у пухлинах
можуть бути виявлен! антигени, власти в! ембргональним тканинам (пуклинно-
ембрюнальнг антигени). Г.1. Абелев показав це на приклад! гепатом и, у якш в!н
виявив бгток а-фетспротеш. За наявшстю цього б!лка в кров! можна дгагностува-
та пухлину печшки до появи клгнгчних гг ознак. У пухлинах вгрусного походжен-
ня з'являються шдукован! вгрусами антигени, специфгчнг для цього вгрусу й од-
наков! в ргзних пуки инах г в ргзних шдивщгв.
Випадков! антигени виникають в шдукованих пухлинах унаслщок мутацш.
Якщо пщ впливом одного канцерогенезу в ор]ан!зм1 виникае кыька первинних
пухлин, то вони можуть вироблята ргзш випадков! антигени так само, як г ргзш
комплекси ферменпв.
Поява в оргашзм! пухлинних юптин не обов’язково спричинюе розвиток пух-
линного пронесу. Клони пухлинних клгтин потрапляють пщ контроль гмуноком-
петентног тканини, г внаслщок гмунних реакцш клон з якими-небудь антигенни-
ми вщмшностями усуваеться.
Ргст пухлинно! тканини спосгерггаеться тод!, коли пухлинн! юптини здатн!
уникати гмунологгчного контролю.
Мехашзми. яю дають змогу уникати гмунолопчного контролю:
• прогресгя пухлини супроводжуеться втратою частини антигенгв, якг е в нор
мальнгй клгтинг, — гак званим антигенним спрощенням, що може сприяти вижи-
ванню пухлинних клгтин г досягати такого ступени, коли повюстю втрачаються
тканиноспецифгчнг й гндивщуальш антигени Лишаються лише видоспецифгчнг
антигени, до яких у кожного оргашзму певного виду генуе толерантшеть,
• поява в пухлиннш тканин! фетальних антигешв не зумовлюе гмунну реак-
цию внаслщок того, що до цих антигенгв е гмунолопчна толерантнгсть;
• маскування антигенгв пухлин. Так, юптини хоргонештелгоми мають ней-
тральну полгеахаридну капсулу;
• у деяких пухлинах виявлено антагенш детермшанти, якг запобгжно ста-
мулюють Т-супресорну функдгю, шо призводить до гальмування гмунну реакцпо
проти пухлини:
• канцероген нг фак гори можуть спричинювати гмунодепреегю;
• перевантаження гмунно! системи та пригнгчення гмунно! вщповщг пухлин-
ною тканиною пгсля досягнення певно! маси.
Однак, незважаючи на вс! мехашзми уникнення пухлиною гмунологгчного
нагляду, трансформоваш юптини руйнуються в органгзмг г клшгчний прояв пух-
лини вгцбуваеться значно рдше, нгж трансформапгя клгтин.
Мехашзми гмунного захисту органгзму вщ пухлини. Для боротьби з пухлиною в
органгзмг е мехашзми. якг подгляють на адалтивгп та неадаптивш.
Аоаптивнг мехашзми — це класичнг гмуннг реакцп. шо здшснюються Т- г В-лгм-
фоцитами проти пухлинних клгтин, якщо на цих клгтинах е пухлинш антигени
Псява нових антигешв пухлини може бути зумовлена коъкома факторами;
— мутащею пщ впливом мутагенгв;
— !ндукшею пухлини в!русами (вгрусш антигени);
— дерепресгею гешв, шо призводить до появи в пухлинних клпинах незви-
чайного для вихщно! тканини ферменту, гормону або зародкового антшену;
— змшою антигешв ГКГС унаслщок мутацп або порушення регуляцп гешв.
В гмуннгй вщповщ! можуть брати участь лзмфоцити як обмежував! антигена-
ми ГКГС, гак 1 необмежуваш, причому наслщки залежать вщ шлькостг та якост!
експресованих антиген!в ГКГС. Якщо в процес! онкогенезу в пухлинних клпинах
змшюються антигени пстосумгсностг I класу (НЬА-А, НЬА-В, НГА-С), таю юп-
тини будуть розшзнаш г знишеш Т-кглерами без лопередньо! !мунно! реакцш Те
саме вщбуватиметься у раз! змши антигенш ггстосумкност! II ктасу (НЬА-О). Але
в цьому випадку антигени, що не належать до ГКГС, не будуть розшзнан! на пух-
линних юптинах, оскшьки Т-хелпери мають антигенне обмеження г розшзнають
антигени тыьки на клпинах, у яких однаков! з Т-хелперами антигени II класу
(НЬА-О). Якщо антигени II класу на пухлинних клпинах та !муноцитах однаков!
г не мае додаткових антигенних ознак, блокуеться можливють 1муннО1 атаки г вми-
кання мехашзмгв адаптивного гмунгтету Антигени пстосумгсностг III класу (ком-
поненти комплементу) потргбн! для активацц цитолгзу клттин.
До неадаптивних (що здшснюються без участт антитш) .мехашзмгв захисту
проги пухлин належать:
• НК;
• ФНП лтмфонипв, який руйнуе юптини та судини пухлин дгя якого поси-
люеться у-1Ф:
• лгмфощний токсин (ЛТ) лгмфоцивв;
• цитотоксичний фактор натуратьних юлергв (ЦФНК);
• Л13ОСОМШ ферменти лейкоципв. Кргм того, неадаптивш мехагпзми залучають-
ся до адаптивних через Рс-фрагменти тмуноглобулппв ! активац!ю комплементу.
Функпюнальга особливост! пухлинно! тканини. Функциональна анаплазгя прояв-
ляеться втратою функцш, якг юйгини здатш були виконувати до трансформацп.
Наприклад, у гепатом! припиняеться синтез жовчних шгменпв, значно дедифе-
ренщйован! пухлини, як! швидко ростуть, втрачають первинш специфтчн! функ-
цп. Часгково диференщйован! пухлини, як! зберегли здатшсть здшснювати деяк!
специфтчш для первинно! тканини процеси, втрачають контроль над ними. Так,
у пухлинт мозково! речовини надниркових залоз (феохромоцитома) спостершаеть-
ся пеконтрольований синтез адреналтну. у пухлинах статевих органгв може част-
ково збереггися чутливгсть до гормонально! регуляцп. Порял з дедиферевдпюван-
ням г зниженням ефективностг контролю в пухлинних клпинах може вщбуватися
незвичайний для первинно! тканини процес, наприклад синтез глгкозамшогл!-
канш або гормошв. зокрема глюкокортикоццв, пухлиною легень.
Злоякгсшсть пухлини. Здатшсть пухлинних юптин до безмежного неконтрольо-
вапого розмноження ще не означае неминучзсгь загибел! оргашзму в раз! росту
пухлини, осюльки хгрурпчне видалення пухлинного вузла забезпечуе вовне вилг-
кування. Однак цьому може перешкоджати злояктсшсть пухлини, для яко! харак-
терними е шфйгътративний (Ывазивний) ргст / здатшсть метастазувати. 1нфшь-
гративний ргст г утворення метастазгв пов’язаш з порушенням у пухлинвш тка-
нинг мгжюптинних взаемозв’язюв.
Для злоякхсних пухлин характерным е також значнппий, шж у доброякхсних,
тканинний атитзм, який проявляеться порушенням нормального спхввщношен-
ня тканинних структур. Тыьки у злоякхсних пухлин з’являеться клтинний ати-
п1зм, який проявляеться клхтинним ! ядерним полиморфизмом зх збхльшенням
ядерно-цитоплазматичного спхввщношення та посиленою базофйпею ядра х ци-
топлазми, що виникае внаслщок значного збыыпення юлькост! ДНК! РНК. Кр!м
того, злоякхсш пухлини здатш спричинювати загальне глибоке виснаження оргашз-
му — кахекаю.
Метастазування мае так! етагги: вщрив пухлинно! клггини вщ сусщнхх клггин;
рух у тканини, розплавляння при цьому компонента сполучно! тканини х стшки
судини; перенесения з кров’ю або лхмфою скупчення пухлинних клхтин; прикрш-
лення до стшки судини в новому мхсцх; шдукц!я росту сполучно! тканини 1 судин
у новоутворену пухлинну тканину. Серед мехашзмхв х факторхв, яхс! зумовлюють
метастазування на рхзних його етапах, можна видшити основш:
— припинення мхжклхтинних контакта, змша рецепторхв мембрани х набуття
рухомосп значною мхрою пов’язаш зх змшами бхлкхв цитоскелета, зокрема з !х
фосфорилюванням протешкшазами, якими е значка кшькхсть продуктхв жит-
тедхяльностх онкогешв х фактори росту. Вщбуваеться також змша регуляцн ген!в,
як! кодують быки цитоскелета х рецептори мембран;
— у клхтинах, як! трансформуються, вщбуваеться синтез активатора про-
фхбринолхзину — ферменту, що штенсивно руйнуе компонента основно! речови-
ни сполучно! тканини х судиннох стшки, а також активуе ферменти шших бюло-
пчно активних систем, зокрема профхбринолхзин. У пухлинних клхтинах утворю-
ються колагенази, якх руйнують колаген рхзних типхв, у тому числх IV типу, з
якого складаеться базальна мембрана судин. Пухлинн! клхтини, що не мають про-
фхбринолхзину, виробляють фактор, який залучае моноцита, ферменти яких роз-
рщжують матрикс х сприяють метастазуванню пухлинних клхтин. Аналопчно пух-
линн! клхтини залучають базофыьш гранулоцита, ферменти яких, зокрема сери-
нова протеаза та металпротешаза, також сприяють розщепленню матриксу, а
гепарин посилюе дпо анпогеншу х вростання судин у пухлинну тканину;
— хснують катепсини як вбудоваш в мембрану пухлинних клхтин, так 1 у выь-
ному стан! в мхжклггиншй рщиш пухлинно! тканини;
— пухлинн! клхтини мають набхр факторхв, якх активують у сполучнотканин-
них клхтинах синтез колагену, глгкопротещхв та шших компонента основно! ре-
човини х розмноження цих клхтин, вростання в пухлинний вузол;
— пухлинн! клхтини видшяють анпогенш та шгш фактори росту судин, що
забезпечуе кровопостачання пухлинно! тканини;
— у мембранах пухлинних клхтин, на вщмшу вщ нормальных, радикалы нейр-
амхново! кислота, глхкопротещхв, а-В-глюкошранозиду ! КГ-ацетил-В-галактоз-
амхну залишаються вщкритими. Быок конканавалш А, а також лектини завдяки
наявност! вщкритих радикалхв аглютинують пухлинн! клхтини. Якщо пухлинн!
клхтини обробита розщепленим конканавалшом А, який блокуе вщкритх радика-
ли мембран, у тому числ! аглютинацпо, то вони деякий час ростуть так само, як х
нормальш. Усе це дае пщетави припустита, що порушення мембран пухлинних
клхтин х поява в них вщкритих радикалхв перешкоджають утворенню щыьних
контакта мхж пухлин ни ми клхтанами х сприяють шфшьтратавному росту й утво-
ренню метастазгв. Однхею з причин розкритгя в пухлинах радикалхв х порушення
функцп мембран е збольшення вмосту соалтрансферази, що переносить радикали
глокопротещов.
У разо змшгування в культур! тканини невелико! юлькосп пухлинних юптин
з нормальними перш! доляться ! ростуть, як нормально. Очевидно, пухлинно кло-
тики при цьому втрачають здатность посилати сигналы про гальмування шшим
клотинам, але здатн! певною морою сприймати гальмовш сигнали, яю посилають
нормально клггини. У пухлинному вузло створюються умови для переважання пух-
линних клотин та ох онвазивного росту.
ВПЛИВ ПУХЛИНИ НА ОРГАН13М
Залежно вод локалозацп пухлини та п метастазов можуть виникати розно патолопч-
но процеси в органозмо. Так, пухлини органов травления супроводжуються тяжки-
ми порушеннями травления о живлення, рак шлунка — пригноченням секреторно!
функцп цього органа. Унаслщок недостатнього надходження о засвоення ожо при
цьому розвиваеться голодування органозму.
Кром прямого ураження органов пухлиною та п метастазами е й шшо шляхи
впливу злояюсних пухлин на органозм, наприклад токсичними речовинами, що
зумовлюють порушення обмшу речовин, яке нерщко призводить до глибокого
виснаження органозму — раковог кахексп. Значке схуднення хворих зо злоякосни-
ми пухлинами зокрема пояснюеться пригночуючою доею на центр голоду кахек-
сину (ФНП-а), який виробляеться пухлинними клотинами.
В органозмо в разо пухлинного процесу знижена активность каталази — фер-
менту, що каталозуе реакцою розпаду пероксиду водню. Вивчаючи причини цього
явшца, Накахара о Фукуока видшили з пухлини людини фракцою, яка шсля вве-
дения и мишам зумовлювала зниження ровня каталази в печонцо. Ця речовина
отримала назву токсогормону. Згодом було видшено високоактивний кристалоч-
ний полшептид з молекулярною масою 4000 Да. Очищений препарат токсогормо-
ну зумовлюе у хворих з пухлинами зниження вмосту каталази в печонцо о нирках,
а також ровня залоза в крово, на який токсогормон впливае у 200—500 разов силь-
нппе, нож на активность каталази, розвиток анемо! через приппчення еритронито-
поезу, гопертрофпо надниркових залоз та онволюшю загруднинно! залози, зболь-
шення селезшки о печшки.
1з пухлини в органозм надходять недоокиснено продукта обмшу. У нейтрало-
зованому сташ вони виводяться через нирки. У норм! в сечо споввщношення кшь-
косто вуглецю й азоту (С / М) становить 0,7, у раз! пухлинного процесу — 0,9 о
больше, що евщчить про збольшене видолення оз сечею недоокиснених продуктов
(дизоксидативна карбонургя).
Частина ферментов пухлини переходить у прилегло тканини внаслщок пщви-
щеноо проникност! клотингшх мембран, а також некрозу пухлинно! тканини. При
цьому в крово або онопих родинах органозму виявляються синтезовано пухлиною
ферменти, а також шшо болки, у тому число емброонально. Так, ровень лужноо фос-
фатази в крово у хворих з остеогенною саркомою зростае у 20—40 разов; пщвищен-
ня й активносто спостерогаеться також у процес! росту гепатоми. Збольшення вмос-
ту у крово кисло! фосфатази спостерогаеться при пухлин! передмохуровоо залози, а
пщвищення активносто глюкозофосфатозомерази — у хворих на рак молочно! за-
31Г
лози. При рхзних пухлинах збшылуеться вм!ст у кров) альдолази, деяких !зоформ
лактатдег'щрогенази, знижуеться актившсть холшестерази та рибонуктеази. Сл!д
зазначити, що змгни юлькост! ферментов кров! не завжди специф!чш для того або
шшого виду пухлин
Особлив! зм1ни б оргашзм! хворих 13 пухлинами спостерйаються тод!, коли
вщбуваеться безконтрольний синтез гормошв або шших БАР. У раз! синдрому
Золлшгера — Еллйона в пухлин! тдшлунково! залози спостерйаеться штенсив-
ний синтез гастрину (гормону7 слизово! оболонки шлунка, активного стимулятора
секрецй шлункового соку). Синтез гастрину, не властивий пщшлунковш залоз!, е
наслщком аномально! функпп вщповщного гена, який функцюнуе безконгроль-
но, поза зв'язком з природними мехашзмами регуляпп синтезу шлункового гаст-
рину. Безперервна стимуляция шлунково! секрецй пухлинним гормоном призво-
дить до розвитку виразки шлунка. Аналопчно у феохромоцитом) секретуеться
адреналш, шо спричинюе розвиток артергально! гшертензп.
РОЛЬ ОРГАШЗМУ В ПУХЛИННОМУ ПРОЦЕС1
Оргашзм справляс дно на пухли нний пронес на вс!х етапах. На етапах онкогенезу
д!ють:
— антиоксиданта! та шш! ферменти, як! знешкоджують канцерогенн! факто-
ри, що п!дтверджено виведенням шбредних лш!й тварин (одержаних унаслщок
близькоспорщненого схрсщування). спйких до дц певних онкогешв;
— ферменти репарацп ДНК;
— спещальш гени — репресори клгтинного подглу;
— гени — репресори вгрусного геному 1 синтезу РНК, зокрема штерферони;
— мутован! юптини перевщяються захисними системами оргашзму, зокрема
!мунною системою, 1 в раз! появи в цих клпинах пухлинних антиген 1в вщбуваеть-
ся гх знишення. За кожно; ново1 мутацп клпини весь мехашзм боротьби вми-
каеться знову.
Отже, у б!льшост! випадкгв онкогенезу оргашзм л!кв!дуе мутаппо в клгтин!
або саму мутовану клпину, 1 пухлина не розвиваеться. Пухлина росте в тому ви-
падку, коли вег захисш системи е недостатшми. Однак кр!м захисних механ!зм!в
в оргашзм! можуть спостерйатися процеси 1 стани, як! сприяють виникненню
пухлин.
Вщкрито спадков! порушення регуляторних гешв клпини, зокрема спадкову
недостатшсть гешв — репресор!в юптинного подглу р53 га КЬ, яка зумовлюе роз-
виток ргзних пухлин у родинах.
Вщомг передраков! стани — хвороби, при яких значно зростае частота розвит-
ку пухлин, наприклад рак шийки матки у хворих, у яких дгагностують ерозгю ц
слизово! оболонки.
Видов!, статев!, тканинш та !нш! особливост! оргашзму визнанаюзь варианта
метаболгзму 1 ди х!м!чних онкогешв, а також вщм!нност! !мунних реакшй проти
пухли нотвгрних в!русгв 1 мутантних клоыв пухлинних клгтин. Як наслщок, деяю
вида органгзмш нечутлив! до впливу пухли нот вгрного вирусу, в шших цей в! рус
спричинюе розвиток пухлини. У чоловгкш частице спостерйаеться рак шлунка, у
жшок — рак статевих оргашв.
Значений вплив на пухлинний процес чинить гормональна регуляция. Гормони
можуть ёндукувати розвиток пухлини або полеппувати п перебёг (див. вище). Пух-
лини, що ростуть, нерёдко проявляють особливу чутливёсть до гормонально! реак-
цц. Так, рёст рёзних пухлин гальмуеться пщ впливом ёнсулёну, дефшиту соматот-
ропёну, посилюеться при ппофункцп щитоподёбно! залози, а також унаслщок дц
деяких статевих гормонёв. Варёабельнёсть змш у пухлинних клётинах зумовлюе
рёзнё !х реакцп на гёпер- або гёпосекрецёю гормонёв. Тодё як при гёперсекрецп ён-
сулёну розвиток багатьох пухлин пригнёчуеться, в експериментё спостерёгали ён-
сулёнозалежну карциному молочно! залози, яка була ёндукована ДМБА. Без ён-
сулшу ця пухлина рости не може.
Тканина злоякёсних пухлин у бёльшостё випадкёв не ённервуеться. Нервовё
закёнчення розмёщуються в стромё — нормальней сполучнёй тканинё. Однак ё тут
ённервацёя недостатки.
Проте, оскёльки дёяльнёсть нервово! системи пов’язана з ендокринною регу-
ляцёею, порушення п може спричинити змёни гормонально! регуляцё!, що зумо-
влюе розвиток пухлин. Так, при нервово-емоцёйному стресё вёдбуваеться значке ё
три вале збёлыпення в кровё рёвня глюкокортикоёдёв, якё зумовлюють масову апоп-
тотичну загибель лёмфоцитёв, що викликае пригнёчення клётинного ёмунётету ё тим
самим порушуе ёмунний контроль над мутованими клётинами.
Кёлькёсть мётозёв у тканинё перебувае щд контролем репресорних гормошв —
кейлонёв, якё утворюються в клётинах, що дёляться. Що бёльше в цёй тканинё
клётин, якё дёляться, то вищою е концентрацёя кейлонёв ё сильнёше гальмуеться
мётотична актившсть решти клётин. Припускають, що в такий спосёб в органёзмё
пёдтримуеться постёйна кёлькёсть клётин. У пухлиншй тканинё цей механёзм регу-
ляцп подёлу клётин порушуеться.
Однак, як зазначалось више, органёзм мае засоби захисту в1д канцерогенних
факторгв. Це насамперед функцёя оргашв ё систем, якё захоплюють, знезаражують
ё виводять бёологёчнё та хёмёчнё канцерогени, захищають клётини ё макромолекули
вёд дп активних кисневих рад икал ёв, що утворюються при радёацёйному ураженнё,
а також ёмунна система, яка знешкоджуе вже мутоваш ё трансформоваш клётини.
Крём того, е особливё мехашзми захисту: система репаративних ферментёв, якё
усувають порушення генёв ё вёдновлюють 1х нормальну структуру шсля мутацп
(ендонуклеази); клётинш ёнпбётори синтезу власних ё вёрусних нуклегнових кис-
лот (а-1Ф ё Р-1Ф); гени, якё репресують вёрусний геном та ён. Зрештою, слёд виз-
нати, що стан антибластомно! резистентностё оргашзму мае бёльше значения для
утворення пухлин, нёж наявшеть канцерогешв.
ПРИНЦИПИ Л1КУВАННЯ ПУХЛИНИ
С.А. Розенберг з колегами застосовували для стимуляцё! автогенних лёмфоцитёв
лёмфокён — трансгенний 1Л-2, синтезований мёкроорганёзмами, яким вбудовува-
ли його людський ген. 3 видалено! пухлини хворого висёвали лёмфоцити, якё
емёгрували в тканину пухлини, ё розмножували гх пщ впливом 1Л-2. Крём того,
Ш-2 додавали до лёмфоцитёв кровё цих самих хворих з ёноперабельними формами
рёзних пухлин ёз метастазами. Пёд дёею Ш-2 лёмфоцити розмножувалися, активу-
валася !х функцёя. Потём кожному хворому вводили його власнё активованё лём-
фоцити, видшеой з охньо! пухлини о крово (1,8—18.4*10:0 клггин) разом з 1Л-2.
Поеднане введения активованих автогенних лшфоцитов 1 лшфокшу зумовило
регреспо пухлин, у тому числ! метастазов, в 11 оз 25 хворих. В одапе! хворо!' на
меланому досягнуто повного вилокування, у тому числ! зникнення метастазов у
костках.
Автори выкрили при цьому нову субпопуляцпо лимфоцитов, названу ними
лшфоктнактибованими талерами (ЛАК), вплив на яко за допомогою 1Л-2 пщвищу-
вав цитотоксичну активность шодо пухлин, стойких до природиих колеров у зви-
чайних умовах. Ефеосгивность такого локування пщтверджено в розних кражах на
сотнях онопе рабел ьн их хворих. Довезено, що метод оз використанням 1Л-2 особ-
ливо доевош при меланомо та раку нирок.
Поряд з ПТ-2 проводять дослодження омунологочних методов та ох комбшацш
для локування пухлин. Доведено, шо а-1Ф, який посилюе експресою антигенов
ГКГС I класу та мае цитотоксичну дпо, здатен забезпечувати оюдовжену ремоегю
волосато-клотиннооо лейкозу о гальмуе рост пппих пухлин. у-1Ф зболыпуе пред-
ставництво на мембранах клотин антигенов ГКГС I о II класов, активуе макрофаги
1 Т-колери. Цей у-1Ф виявився спроможним гальмувати розвиток перитонеатьних
пухлин яечников у раз! його мосцевого введения у черевну порожнину. ФНП-а
активуе макрофаги о ломфоцити, спричинюе злипання клггин пухлин о гальмуе
рост перитонеальних злоякосних пухлин.
1нший метод було застосовано групою Т. Таквороан у хворих на лейкоз. У них
хворих брагой костковий мозок, вид клял и моноклоналъно антитола до лейкозних
клотин (добирали хворих, чи! лепкозно клотини експресували антиген В1), очища-
ли за допомогою антитол костковий мозок вод лейкозних клотин, опромонювалоо
хворих випалювальною дозою оонозуючого випромшювання о вводили кожному
його вл асний костковий мозок. Результат: у 34 !з 45 хворих с постер! галася рсмосоя
без подтримуючоо терапп, яка тривала близько 52 мос., у середньому — 11 мос.
Кром омунологочних методов, за допомогою яких улерше досягнуто повного
вилокування окремих хворих оз пухлинами з лоширеними метастазами, ведуться
поогоуки й онших методов патогенетичного локування, спрямованих на блокаду дп
онкогенов, вщновлення регуляцп розмноження трансформованих клотин о запобо-
гання утворенню метастазов.
Роздал XV
ППОКС1Я
ПпоксЫ, або кисневе голодування, — типовий патологичный процес, який развиваешь-
ся внаслгдок недостатнього забезпечення оргашзму киснем або порушення його вико-
ристання клйпинами, через що знижуеться енергоутворення в тканинах.
Гепоксёя — найпоширенгший патолопчний процес. Народження, смерть, бёль-
ппсть хвороб охоплюють фактор гёпоксп. Нестача кисню ё зниження утворення
АТФ зумовлюють порушення метаболизму й енергозалежних процесёв в оргашзм!.
Класифёкапёя. За локал ёзашею розр!зняють ппоксёю м1сцеву та загальну, за
перебегом — гостру ё хрошчну, за причинами ё мехашзмом розвитку — гтоксичну
(екзогенну), дихальну (респёраторну), циркуляторну, гем1чну (кров’яну), тканинну
(пстотоксичну) та змйиану.
ЕТЮЛОПЯ
Численн! причини ппоксп об’еднують за одшею загальною властигястю — пору-
шенням транспорту кисню в оргашзм! або утилёзацп його тканинами. Саме за
причинами ппоксп видыено п види.
Причиною ппоксичноё ппоксп е зниження парщального тиску кисню у вдиху-
ваному повётрё (прська або висотна хвороба, аварп киснезабезпечення на вироб-
ництвё). В експеримент! цей вид ппоксп моделюють за допомогою дихальних
сумёшей, бгдних на кисень, або барокамер.
Причинами дихалыюё ппоксп е тага, що призводять до порушення газообмену
в легенях ё недостатностё дихання (див. роздёл XXVII “Патолопчна фёзёологёя зов-
нппнього дихання”).
Гемёчиа ппоксёя розвиваеться внаслщок зменшення кисневоё емност! кровь 1ё
своею чертою подёляють на анем1чну й гтокст внаслгдок шактивацП гемоглобшу.
Анемпо як причину ппоксп описано в роздёлё “Патолопчна ф!з!олог!я системи
кров!”. Щодо гемоглобену, то в патолопчних умовах можливе утворення таких
його сполук, яга не можуть виконувати дихальну функцею. Так, песля отруення
оксидом вуглецю (чадним газом СО), спорёднешсть до якого в гемоглобену у 300 ра-
з!в вища, нёж до кисню, утворюеться карбоксигемоглобин, який не здатен перено-
сите кисень, а також ушкоджуються залезовмесш дихальнё ферменти. У разё от-
руення нётратами ё нётритами, похёдними анелёну, деякими лёкарськими засоба-
ми (з окисною актившстю) утворюеться метгемоглобгн, у якому тривалентне
залтзо не приеднуе кисень.
Причинами циркуляторног гшоксн е таю, що призводять до мтсцевих (гшемтя,
тромбоз, емболтя, стаз) т загальних (гостра та хрошчна недостатшсть серця або
судин, шок, колапс) порушень кровообпу .(див. роздш IX “Патолопчна фгзюлопя
периферичного кровообтгу”, роздш XXVI “Патолопчна фгзюлопя системного
кровообпу” та роздш ХХХИ1 “Патолопчна фгзюлопя екстремальних сташв. Шок,
Колапс. Кома”). У раз! порушень у судинах великого кола кровообпу утруднюеть-
ся доставка кисню у тканини, а якщо порушення гемодинамгки виникають у
систем! малого кола кровообпу, зазнае негативного впливу оксигенац!я артерталь-
но! кров!. Причиною циркуляторно! ггпоксп може бути не тшьки абсолютна, а й
вщносна недостатшсть кровообпу, коли потреба тканин у кисш перевищуе нор-
мальну його доставку з кров’ю.
Тканинна (синонгми: пстотоксична, клггинна, цитотоксична) ппоксш е наслщ-
ком дп факторгв, яю безпосередньо ушкоджують у клттин! систему утилтзаци кис-
ню й енергоутворення. У цьому випадку ппокстя виникае на тл! достатнього забез-
печення тканин киснем. Так, теля отруення щашдами гнактивуеться система
цитохромтв; отруення деякими наркотичними засобами та алкоголем пригшчуе
депдрогенази циклу Кребса. Антибютик олтгомщин ушкоджуе фермент АТФ-син-
тетазу, юни важких металтв — функщональш групи дихальних ферменттв. Зни-
ження синтезу дихальних ферменттв спостерггаеться при дефщит! вттамштв, якт
входять до складу окисно-вщновних ферментних систем. Важливими з цього пог-
ляду е рибофлавш т шкотинова кислота: перший е простетичною групою флавгно-
вих ферменттв, друга входить до складу кодепдрогеназ. 1ошзуюче випромшювання
призводить до утворення вшьних радикалтв, якт ушкоджують мттохондрп т тканин-
не дихання. Так зван! роз’еднувачт окиснення г фосфорилювання (токсини мгкро-
оргашзмгв, адреналтн, тироксин у високих дозах, 2-4-диштрофенол) сприяють
витгканню протонтв, зниженню ефективносп бюлопчного окиснення, активацп
ПОЛ, розегюванню енергп у вигляд! вшьного тепла т дефщиту АТФ.
Змтшана ппокстя спостерггаеться найчастгше, осюльки зазначею вище види
кисневого голодування в гзольовашй форм! трапляються рщко, часттше вони на-
бувають вигляду ртзних комбшацш. Наприклад, пщ час пневмонп розвиваеться г
дихальна, т циркуляторна недостатшсть. Хрошчна гшокстя будь-якого генезу уск-
ладнюеться ураженням дихальних ферменттв (тканинним компонентом).
Гшокстя навантаження виникае тодт, коли пперфункцтя оргашв г велика пот-
реба в кисш (наприклад пщ час надмтрного навантаження у спортсмешв) не за-
безпечеш киснем навтть на тлт пщвищеного його постачання. Тодт з’являються
метаболтчн! порушення, характерш для справжньо! киснево! недостатносп. Такий
стан е пусковим механтзмом розвитку втоми.
ПАТОГЕНЕЗ
Гшокстя мае двт стадп розвитку — компенсации декомпенсаци. Спочатку завдяки ком-
пенсаторно-пристосувальним реакщям гцдтримуються нормальне тканинне дихан-
ня та енергоутворення. У разг виснаження пристосувальних мехашзмгв розвиваеться
стадгя декомпенсаци, або власне кисневе голодування.
КОМПЕНСАТОРНО-ПРИСТОСУВАЛЬН! РЕАКЦП
Оскыьки гшоксхя е одним !з найпоширенгших патолопчних процесгв, в еволюци
сформувалося багато пристосувальних реакцш, яю охоплюють майже вег системи
оргашзму — системи транспорту й утшпзацп кисню. Але вег вони розвиваються
не одночасно. Щодо стратеги пристосування, то компенсаторш реакцп подыяють
на негайш (аваршш, але короткочасш та неекономш, оскыьки самг потребують
кисню) 1 тривалг (шертнг, але економш). Кргм того, видыяють мехашзми “бо-
ротьби за кисень” 1 мехашзми пристосування до умов його дефщиту.
Негайш компенсаторно-пристосувальн! реакцп
Ц1 реакцп розвиваються головним чином у динамгчних системах транспорту кисню.
Ггпервентилягця в раз! ппоксп вщбуваеться внаслщок рефлекторного збуд-
ження дихального центру гмпульсами з хеморецепторгв довгастого мозку та су-
динного русла, переважно синокаротидно! та аортально! зон, яга реагують на
змшу х!м1чного складу кров!! насамперед на накопичення СО2 та юн!в водню.
У випадку гшоксично! ппоксп патогенез гшервентиляцп дещо шший — збуд-
ження каротидних та аортальних хеморецептор!в вщбуваеться у вщповщь на зни-
ження в кров! напруження кисню (рО2). Гшервентиляцгя е, безперечно, позитив-
ною реакщею оргашзму на гшокспо, але в умовах гшоксично! ппоксп вона мае й
негативш наслщки, оскыьки ускладнюеться пщвищеним виведенням вуглекисло-
ти (СО2) г зниженням п вм!сту в кров! (ппокапшею). Якщо врахувати вплив СО2
на мозковий г вгнцевий кровообгг, регулящю тонусу дихального г судинорухового
центргв, пщтримання кислотно-основного стану (КОС), дисошащю оксигемог-
лобину, то стане зрозумыим, що в патогенез! екзогенно! ппоксп ппокапнгя мае не
менше значения, н!ж дефщит кисню.
Г!первентиляц!я супроводжуеться мобыгзащею резервних альвеол г полш-
шенням дифузп газгв кр!зь альвеолярно-капыярш мембрани.
Посилення кровообггу також спрямоване на адекватну доставку кисню у ткани-
ни г реалгзуеться через пщвищення артер!ального тиску, аваршну пперфункцпо
серця, збыьшення швидкостг току кров!, розкриття нефункцюнуючих капыярних
судин. Не меню важливою характеристикою кровообпу в умовах ппоксп е пере-
розподы кров! в бгк переважно! доставки й життево важливим органам 1 пщтри-
мання оптимального кровотоку в легенях, серцг, головному мозку за рахунок змен-
шення кровопостачання шюри, селезшки, м’язш, кишок. Змши кровообггу ре-
гулюються рефлекторними 1 гормональними мехашзмами. Мають значения також
тканини! продукт ендотелгального походження (оксид азоту, простациклгн), як!
справляють судинорозширювалъну дпо.
Реакцгя регуляторных (нервово! та ендокринно!) систем на гшокспо прояв-
ляеться збудженням дихального та судинного нервових центр!в, вивыьненням
катехоламгнгв, що забезпечуе гшервентиляцгю, а також аваршну пперфункцпо
серця г системи кровообггу.
Збглыиення кглькостг еритроцитгв г гемоглобину в кровг збыыпуе кисневу емшсть
кровг та забезпечуе швидке пристосування до гшоксп завдяки викиду кровг з депо.
Описанг вище негайш пристосувальш змши розвиваються у найбыып реак-
тивних системах, однак не можуть забезпечити стшке г тривале пристосування до
Мал. 36. Змши криво! дисощаци оксигемоглобшу у процес! пристосування оргашзму
до ппоксп
ппоксп, осюльки сам! потребують додаткових енергетичних витрат, супроводжу-
ються збгльшенням штенсивносп функцюнування структур (1ФС) 1 посиленням
розпаду субстрата.
Кргм них е негайш пристосувальш реакцп, яю не потребують додаткового
кисню.
Змша криво! дисощаци оксигемоглобшу полягае в посиленш здатносп молеку-
ли гемоглобшу приеднувати кисень у легенях 1 вщдавати його тканинам. Це
пов’язано з конформащйними змшами молекули гемоглобшу. Юлька варианта
цього пристосування наведено на мал. 36.
Змпцення криво! дисощацп в дглянщ верхньо! шфлексп лгворуч свщчить
про здатшсть гемоглобгну щдвищувати спорщнешсть до кисню в судинах мало-
го кола кровообпу в раз! його зниженого парщального тиску у вдихуваному
повггр! й альвеолах. Змгщення криво! дисощаци в д!лянц! нижньо! шфлексп
праворуч е ознакою здатносп гемоглобшу знижувати споргднешсть до кисню г
полеплено! його в!ддач! в раз! зниженого напруження кисню в тканинах. У та-
кий спосгб тканини можуть отримувати достатню юльюсть кисню при р!зних
формах ппоксп.
Система утимзаци кисню й енергоутворення також мае негайне пристосуван-
ня. Мпохондрп — досить мобйтьна система утворення АТФ. 1хня висока реактив-
шсть забезпечуеться алостеричною активащею ферменпв, за рахунок чого спор!д-
нешсть цитохромоксидази (юнцевого ферменту дихального ланцюга) до кисню
може збериатися в раз! меншого вмгсту кисню в тканин!. Процес енергоутворен-
ня в цикл! Кребса перебудовуеться на штенсившший, яким е окиснення буршти-
ново! кислоти (сукцинату), кшетичш переваги якого зумовлюють збыьшення
швидкосп фосфорилювання АДФ (цей шлях транспорту електрон!в стшкипий до
ппоксп, н!ж НАД-залежний, г через нього штермедгати циклу Кребса — сукцинат
! а-кетоглутарат — одержують вих!д у термшальне окиснення). Катехоламши та
оксид азоту активують цей процес.
У м!тохондр1ях головного мозку посилюеться поеднання фосфорилювання з
окисненням.
У таких структурах, як мембрани, теж гснують швидк! пристосувальнт змши.
Пщ час гшоксп дифузтя газтв (О2та СО2) кртзь мембрану може посилюватися за-
вдяки конформацшнш перебудовт мембранних быкових молекул т змшам власти-
востей лшщного шару.
Тривал) компенсаторно-пристосувальш реакцп
Тривалг компенсаторно-пристосувальн! реакци розвиваються в системах, як! несуть
найбтльше навантаження пщ час гшоксп, а також у системах, яга бшыпе за шип
потерпають вщ дефщиту кисню. Вони полягають у розвитку гтертрофп та гтер-
плази, 1 таким чином аваргйна гшерфункцтя одержуе пластичне та енергетичне
забезпечення (мал. 37). На додаток до цього ще низка тривалих пристосувальних
змш розвиваеться на метаболгчному ргвш.
Пперплаз1я юсткового мозку / посилення еритроцитопоезу е одним гз найефек-
тивнптгих пристосувань (показником е збшьшення юлькостт ретикулоциттв у пе-
рифериями кровт). Роль стимулятортв еритроцитопоезу виконують продукта роз-
паду еритроциттв, еритропоетини нирок, а також продукт глтколтзу 2,3-дифос-
фоглтцерат (2,3-ДФГ), вмгст якого в кровт шдвищуеться (ретикулоцита також
мтстять бтлыпе 2,3-ДФГ). З’являеться фетальний гемоглобш, який мае бтлыпу
спорщнешсть до кисню.
Ппертрофгя мтохондрш (активацтя тх бюгенезу) розвиваеться в доповнення
до тих змш у мттохондртях, яга описаю вище, як негайнт компенсаторш реакцп.
Ппертрофтя мгтохондрш пщкршлюе потужнтсть системи енергоутворення. Почат-
ковою ланкою в патогенез! цих явшц е ослабления окисного ресинтезу АТФ г
зростання юлькостт продукттв розпаду макроерпв (сптввщношення (АДФ)+(Фн)/
(АТФ), що позначаеться як потенцтал фосфорилювання, збтльшуеться). Це е сти-
мулом для генетичного апарату клгтини, активацтя якого зумовлюе активацпо
синтетичних процеств у систем! мгтохондрш г зростання юлькостт дихальних фер-
менттв (дихальних ланцюпв) (схема 13).
Ппертроф1я дихальних м’язгв / мюкарда забезпечуеться посиленням кровопос-
тачання цих органтв за рахунок
утворення нових кашляртв пщ
дтею фактортв росту (тромбоци-
тарного та ендотелтального).
Збтльшуеться маса легеневих аль-
веол. У м’язах пщвищуеться вмгст
мтоглобшу, який е додатковою
кисневою емнтстю г мае антиок-
сиданту актившсть.
Аналопчний процес ппертро-
фп спостерггаеться в системах ре-
гуляци — вщбуваеться ппертро-
фтя юрково! речовини наднирко-
вих залоз, гшоталамуса г навтть
нейронтв дихального центру.
Метабол1чн1 пристосувальш
реакци. Активацтя безкисневого
Кагплярш
судини
Еритроцити,
втддача О2
МИохондрп
Мюгпобш
Норма
Адаптацгя
0000-0 ООООР
ООО 000
ооооо ООООО ООООО
оооо оооо ооЪо
Мал. 37. Тканинш мехашзми адаптацп до гшоксп
Схема 13. Мехашзми адаптацп до ппоксп (за Ф.З. Меерсом, 1978)
утворення енергп шляхом глгколозу вщбуваеться при ппоксп завдяки активацп
його ферменпв продуктами розпаду АТФ.
Спостерйаеться активац1я гексозомонофосфатного шунта як шляху утворен-
ня енергп у вигляд) сполук, що вщновлюють НАДФ до НАДФН (донаторов Н+).
Кром того, утворюються пентози, потробно для синтетичних о репаративних про-
цесов (боосинтезу нуклешових кислот, нуклеотидов, стерощов, жирних кислот), а
також ппертрофп органов.
Вщбуваеться посилення глюконеогенезу (утворення глюкози з невуглеводних
попередников) унаслщок активацп корково’о речовини надниркових залоз о синтезу
глюкокортикощов.
Збхльшуеться вироблення ферментов антиоксидантного захисту (каталази, пер-
оксидази, супероксиддисмутази), простагландинов (як! мають вазоактивно власти-
восто), а також цитохрому Р450.
Пщвищуеться експресоя МО8, що призводить до додаткового утворення МО,
який е не лише вазоактивною речовиною, а й регулятором багатьох функцш (кло-
тинноо активносто, кооперацп клггин крово, гемостазу тощо).
Важливу роль у здойсненно пристосування до гшоксп, у розвитку мехашзмов
як негайноо, так о тривалоо адаптацп вщйрають киснечутливий проте'шовий ком-
плекс оз функщею транскрипцойноУ активносто — ппоксое-шдуцибельний фактор
(ПФ, Нурохоа-шбюсоЫе ГасХог, ШР), а також металотранскрипцойний фактор
(МТФ, теОа! Ггапзспрйоп ГасОог, МТР). ПФ е основним транскрипцойним регуля-
тором генов, яко вщповщають за реакцпо ссавцов на дефщит кисню (див. схему
13). В1н забезпечуе швидку й адекватну вщповщь на гшоксичний вплив шляхом
залучения генов, яко регулюють мотохондроальний енергетичний метаболозм, вазо-
моторной контроль, процеси еритропоезу, ангюгенезу, гшертрофою о гшерплазпо
клотин о навоть процес апоптозу. Пщ конролем ПФ перебувае синтез катехола-
монов, ЫО8, еритропоетину, ендотелону-1, анпотензину II, факторов неспецифоч-
ного та специфочного захисту вщ онфекцп тощо.
Отже, якщо пщ час тривалоо ппоксп гшерфункщя систем транспорту кисню
лишаеться недостатньою, змши на ткани иному о метаболочному ровнях довершу-
ють пристосувалыгий процес.
3 усох видов гшоксп гшоксична гшоксоя як така спостерйаеться у людей в
умовах високопр’я та в експерименто. У клйшц переважають шшо види. Природ-
но, що спгввщношення мгж описаними пристосувальними реакщями буде ргзним.
Так, пщ час дихальног або серцевог недостатносп обмежуються можливост! при-
стосування в систем! зовнгшнього дихання або кровооб!гу. При тканиннгй гшок-
сп неефективними е пристосувальн! мехашзми в систем! транспорту кисню.
Адаптащя. Наведен! пристосувальн! метаболгчнг г тканинн! змши здатн! забез-
печити не тгльки виживання, а й можливгсть активно! фгзично! г розумово! пращ
в умовах тривало! пом!рно! ппоксп. А це вже адаптация — економне, стгйке, три-
вале пристосування. Для аналгзу цих явищ вивчають умови життя мешканщв ви-
сокопрних райошв, ггрських тварин, тих тварин, що шрнають, а також дослщжу-
ють експериментальних тварин, як! пристосувалися до ппоксп протягом кглькох
поколшь. Активацгя ппоталамо-ггпофгзарног системи г кгрковог речовини над-
ниркових залоз вщгграють важливу роль.
Варто зазначити ще деяк! важливг позитивы змши, як! вщбуваються в процес!
адаптацп до ппоксп: пщвищення гмунгтету, стшкосп до шфекцп, радгацп, шфар-
кту мгркарда, стресу, тобто пщвищення загально! неспецифгчна! резистентностг
органгзму. Саме тому пом!рну гшокспо, що сприяе пщвищенню стшкосп тканин
до ппоксп, використовують тодг, коли необхщно стимулювати резистентшсть ор-
гангзму (спортивнг тренування, лгкування хрошчних хвороб тощо).
ПАТОЛОПЧН1 ПОРУШЕННЯ ПРИ ППОКСГГ
У раз! виснаження або недостатносп пристосувальних мехашзмгв патогенез п-
поксп вступае в другу стадно — стадгю декомпенсацп, г розвиваються патолопчн!
порушення, характерш для гшоксп.
Порушення на молекулярному (метаболгчному) ргвгп. Провщною ланкою гх па-
тогенезу е зниження енергоутворення. Синтез АТФ гальмуеться внаслщок знижен-
ня напруження кисню в кров!, ушкодження мембран г ферментних систем мгто-
хондргй. У разг роз ’еднання окиснення та фосфорилювання потгк електронгв вже не
генеруе синтез АТФ, ушкоджуються шляхи транспорту АТФ з мгтохондргй до
мгсць п використання. Причому це часто передуе ушкодженню систем утворення
АТФ, що пояснюе можливгсть розвитку гшоксп на тлг досить високого ргвня
аденшнуклеотидгв. Здатшеть тканин до споживання АТФ також зменшуеться
внаслщок ослабления активностг АТФаз. Отже, дефгцит кисню призводить до
енергетичного голодування тканин, яке е основою бгльшостг порушень у разг п-
поксп.
Як наслщок енергодефщиту порушуються метаболгчнг енергозалежш проце-
си: ослаблюеться синтез бглкгв, гормонгв, РНК, ДНК, фосфолшщгв, а також
нейромедгаторгв. Через порушення функцп енергозалежних мембранних гон-
них каналгв г насосгв змгнюеться баланс електролгтгв, зменшуеться кглькгсть внут-
ргшньоклгтинного калгю, порушуеться кальщевий гомеостаз. Накопичення г ци-
тозол! вшьних гонгв кальцпо та порушення гх виведення з клгтин (через дефгцит
АТФ, зниження трансмембранного потенцгалу) е одшею з провщних ланок ггпок-
сичного ушкодження (кальцгеве ушкодження).
Порушення окисно-вщновних реакцш призводить до накопичення продуктов
неповного окиснення, багато з яких е токсичними. Промгжнг продукти обмгну
лшщгв (ацетон, ацетоопдова г р-гщроксимасляна кислоти) е мембранотоксични-
ми. Збтлыпуеться вмгст малонового дтальдегщу (продукту розпаду пероксидтв
жирних кислот). У печшщ 1 м'язах зменшуеться юльюсть глткогену, а глюкоза не
окиснюеться до кшця. Лактат, шруват та шли недоокиснеш продукта можуть
змшювати КОС у бгк метаболгчного ацидозу, який е додатковим фактором ушкод-
ження клттин шляхом шпбхвання ферменттв тканинного дихання, посилення гщ-
ролпичного ефекгу фосфолшаз. активацп ПОЛ, ушкодження мембран (аиидо-
тичне ушкодження). Осюльки внаслщок активацц глтколтзу зменшуеться надхо-
дження в цикл Кребса шрувату 1 вйтьних жирних кислот (^-окиснення останнтх
гальмуеться), то залучаються бтлки як джерело субстрату для апетил-КоА. Поси-
лення катаболтзму б!лктв т порушення сечоутворення призводять до накопичення
в кров) амошю. нейтралтзашя якого вщбуваеться шляхом зв’язування з а-кетоглу-
таровою кислотою, шо вилучае а-кетоглутарат з циклу Кребса т також порушуе
енергоутворення.
Особливо слщ вщзначити активащю ПОЛ при гшоксп. Причиною утворення
активних форм кисню й активацп’ втльнорадикального окиснення в умовах гшок-
сп е гальмування транспорту електронтв по дихальному ланцюгу. Якщо виника-
ють проблеми з дихальними ферментами, самс активацтя радикалоутворення роз-
вантажуе дихальний ланцюг вщ електронтв. Поява продуктов ПОЛ — важливий
мехашзм гшоксичного ушкодження. Окиснюються мем брани) лтпщи, гемоглобш
перетворюеться на метгемоглобтн. Нейтралгзацтя ироду к пв ПОЛ реалтзуеться за-
собами природного антиоксидантного захисту, який пщ час гшоксп, враховуючи
ослабления активностт глутаттонредукгази, теж порушуеться. Беручи до уваги те,
шо на початкових етапах гшоксп проникшсть мембран для кисню шдвищуеться,
може виникнути ситуацтя “вщносно) ппероксп”, коли утшйзашя кисню менша
вщ його надходження. Тобто до гшоксичного ушкодження приеднуеться перок-
сидне ушкодження.
Порушення на клттинному р1внт проявляються ушкодженням енергозалежних
клттинних функщй. Осюльки споживачем АТФ е мембрани, то тх ушкодження —
суттева ланка патогенезу гшоксичних порушень. Мембранозалежн) ферментнт бтл-
ки та рецептори шсля тимчасового посилення активностт в мтру подальшого роз-
витку гшоксп шгтбуютъея або тнактивуюгься (внаелшок активацп ПОЛ, ацидозу,
гщролтзу мембранних лштдтв та фосфол)пщ)в етщогенними, у тому числт лтзосом-
ними, лшазами та фосфолшазами). Енергодефщит справляе ушкоджувальну дно
на функщю 1 структуру мембран через порушення тонних насоств, дефосфорилю-
вання мембранних сполук, активащю фосфолшаз т протешаз. Змшюеться текуч) ст ь
як важлива фтзико-хтмтчна властивтсть мембран. Перерозподтт натрпо призводить
до набухания клттин (пщ час елекгронно-мткроскошчного дослщження виявля-
ються розриви та фрагментация мембран, аномальн) канали в лшщному бтшарт).
Пор1внюючи метаболии) та мембрани) мехашзми гшоксичного ушкодження клт-
тин, учен) дтйшли висновку, шо ушкодження мембран первтент.
1з клттинних органел пщ час гшоксп найбтльшот уваги заслуговують мнпохон-
дри. Надмтрне накопичення тонтв Са в них призводить до роз’еднання процеств
дихання т фосфорилювання. Мткроскоптя виявляе ушкодження мгтохондрш у
виглядт набухания, вакуолтзацп т деградацтт (руйнування та часткова редукщя
крист). Щодо ядра, то структури) порушення в ньому проявляютья гшерхромато-
зом т руйнуванням. У раз) ушкодження мембран лЬзосом вщбуваеться вихщ актив-
них протеолттичних ферменттв у цитозоль, руйнгвча дтя яких посилюеться на тлт
ацидозу. Гальмуеться поды юптин, послаблюються регенеративнз процеси. Дефз-
цит енергп та надлишок Са спричинюють апоптоз з некроз юптин.
Порушення в органах I фЫологгчних системах. В умовах гшоксп ушкодження
окремих оргашв ! систем залежить вш зхньо! чутливост! до ппоксп, яка зумовлена:
1) знтенсивнзстю обмшу речовин, тобто потребою тканин у кисш; 2) потужнзстю
глзколзтично! системи, тобто здатшстю виробляти енерпю без участз кисню; 3) за-
пасами енергп у вигляд! макроерпчних сполук; 4) потенцшною можливзстю гене-
тичного апарату забезпечувати гзпертрофзю та пластичне закршлення гшерфункцп.
3 огляду на це найбыьшого впливу зазнае нервова система. Осюльки процеси
внутрзшнього гальмування е бзлып енергозалежними, то першими розвиваються
порушення психзчноз (вищо! нервово!) дзяльностз та емоцзйнз розлади (ейфор!я).
При тривалзй гшоксп порушуються рефлекторна дзялыпсть, регуляция дихання з
кровообзгу. Потьмарення свщомостз та судоми е загрозливими симптомами тяж-
кого перебзгу кисневого голодування.
Порушення збудливост!, провздностз та скоротливостз мюкарда клшзчно про-
являються аритмзею, причому скоротлив! елементи стзйкзшз, н!ж провщна систе-
ма серця. Кальцзевим з пероксидним механизмам належить провщна роль у цих
порушеннях. Недостатшсть серця та ослабления тонусу судин зумовлюють за-
гальш порушення кровообзгу, що значно утруднюе перебзг ппоксп, якою б не
була першопричина.
Порушення легенево'1 вентиляцП при гшоксп супроводжуеться змшою ритму
дихання, що набувае характеру перзодичного дихання Чейна — Стокса. При
хрошчнзй ппоксп внаслщок застшних явищ у легенях стовшуеться альвеолярно-
кашлярна мембрана, попршуеться дифуззя газзв.
При всзх видах хронзчноз ппоксп зменшуеться секреторна актившсть шлунка,
кишок з пщшлунковоз залози. Спостерзгаеться пригнзчення рухово! функцц трав-
ного каналу. У печ1нц1 порушуються система мзкросомального окиснення, деток-
сикацзя ендогенних метаболзтзв з лзкарських засобзв, гальмуються синтетичнз про-
цеси.
Початкова ползурзя змшюеться порушенням функцп нирок з сечоутворення.
За тяжкого перебзгу ппоксп знижуеться температура тша внаслщок ослаблен-
ия обмгну речовин з порушення терморегуляци.
У кзрковзй речовиш надниркових залоз первична активацзя !х функцш змз-
нюеться виснаженням.
Пригнзчуеться 1мунолог1чна реактившсть, порушуеться система гемостазу.
Глибший аналзз описаних вище змш дае змогу зробити висновок, що при
ппоксп однз й тз самз явища, з одного боку, е патолопчними, з зншого — можуть
бути оцзнеш як пристосувальнз. Так, гальмування нервово! системи е охоронним
(пщвизцуеться вмзст ГАМК), оскзльки ослаблюе п чутливзсть до подальшого роз-
витку кисневого голодування. Зниження температури тша та обмшу речовин може
бути оцзнене так само. Навзть перзодичне дихання Чейна — Стокса (перзодична
затримка вуглекислого газу) е механззмом пристосування, оскзльки таким чином
пщтримуеться тонус дихального центру. Певний захисний ефект дають навзть
помзрний ацидоз з активацзя ПОЛ: перший пригшчуе актившсть фосфолшаз, по-
силюе в мембранах актившсть Са2+-АТФази, активуе окиснення сукцинату, а
продукта ПОЛ спочатку пздвищують проникшсть мембран, полегшують роботу
мембразгних бзлкзв). Ушкодження з захист пщ час гшоксзз тзсно пов’язаш. Так,
322 —
саме зниження рО2 у кровг (гшоксемгя) е початковою ланкою компенсаторного
пристосування через подразнення хеморецепторгв г мобйпзацгю зовнгшнього ди-
хання та кровообпу, Саме дефщит АТФ сприяе процесам, якг активують бгогенез
мгтохондргй,
Чутливгсть до гшоксп. В еволюцп чугливгсть до гшоксп посилюеться 1 водно-
час формуються складншп пристосувальн] мехашзми. Новонароджеш (люди 1
тварини) поргвняно з дорослими особинами витривалшп щодо гшоксп завдяки
наявностг фетального гемоглобшу та гзоформ ферментгв.
Тндивщуальна чутливгсть до гшоксп коливаеться в дуже широких межах. Мар-
керами високог чутливосп до гшоксп е гшерглгкемгя, гшерлшщемгя, ацидемгя,
зменшення в кровг ргвня гнсулгну, збщьшення — тироксину, гнтенсивнппий енер-
гообмш, гшерфункцгя щитопод1бног залози, симпатикотонгя, зниження фонду
ключового ферменту антиокисного захисту еритроципв (супероксиддисмутази),
високий ргвень пероксидного метабол гзму, астенгчний тип конституцп.
Маркерами резистентносп до гшоксп е ггпоглгкемгя, гшолшщемгя, пгдвищен-
ня вмгсту шсулгну, знижений ргвень тироксину та соматотропгну, менш штенсив-
ний енергообмш, ваготонгя, а також гшерпггментовангсть шкгри (пггменти е ан-
тиоксидантами), ггперстешчний тип конституцп.
ПРИНЦИПИ Л1КУВАННЯ ТА ПРОФИЛАКТИКИ ППОКСП
Лгкарсыа засоби, як! компенсують гшоксичне ушкодження, отримали назву ан-
тигтоксанпйв.
Заушена тератя полягае в додаваннг кисню у випадках, коли його утилгзацгя
не порушена (гнгаляцгйна, пероральна). При деяких хворобах застосовують ки-
сень пщ пщвищеним диском (гшербарична оксигенацгя), що створюе запаси кис-
ню, фгзично розчиненого в кровг г тканинах. Це бувае корисним у разг отруення
СО г барбгтуратами, при вроджених вадах серця. Проте, коли утилгзацгя кисню
порушена, штучна оксигенацгя може бути небезпечною та призводити до утво-
рення акгивних форм кисню, стамуляцп ПОЛ (“кисневий парадокс”). Кргм того,
за таких умов порушуеться кальщевий гомеостаз, пщвищуеться проникшсть плаз-
матичних мембран для цього гона г транспорту його з позаклгтинно'г рщини в
клгтину г дал1 в мггохондрп (“кальщевий парадокс”). Це звужуе показания до ок-
сигенотерапп, яка може обтяжити гшоксичне ушкодження, що набувае необорот-
ного характеру саме в той час, коли постачання кисню штучно посилене.
Ефекгивним у боротьбг з гшоксгею е застосування антиоксидант!в, тобто спо-
лук, якг мгстять лабшьнг атоми водню, що несуть неспарений елекгрон. При взае-
модп з активними кисневими радикалами та лшщними пероксидами вони гх ней-
тралгзують. До природних антиоксиданпв належать вгтамгни С, Е, А, Р г лшоева
кислота, дуже важливим антиоксидантом е глутапон, що мае в своему склад!
выьну сулъфгщрильну групу, та пов’язанг з ним мгкроелемент селен г амшокис-
лота цистегн. Розробляють штучш антиоксиданта. Пропонують стимулятори глг-
колгзу (пананггн), а також блокатори кальцгевих каналгв.
Кргм того, для усунення ппоксп корисним е активацгя системи мгкросомаль-
ного окиснення в печгнщ арггнгном. Глюкокортикощи стабглгзують мембрани лг-
зосом, активують деякг фермента дихального ланцюга. Застосування простаглан-
дишв иод! дае захисний ефект — оптим!зуе м!кроциркуляцпо, запобйае аритмп,
гальмуе набряк.
Варто зазначити, що антигшоксанти не справляють вузькоспрямовану дпо.
Вони водночас е адаптогенами 1 навпаки, рхзн! адаптогени пщвшцують стшюсть
до гшоксп.
Проте на практищ застосування антигшоксант!в супроводжуеться непередба-
ченими труднощами, зумовленими дуже широким коливанням ч угли вост! до п-
поксп. Залежно в!д !ндив!дуалъно'! чутливост! медикаментозний ефект може бути
рхзним чи нав!ть прямо протилежним (антиоксидант стае прооксидантом). Ефект
лткарських засоб!в залежить вщ виду 1 стадп гшоксп (саме в стадп декомпенсаци
ефекту може не бути). Кр1м того, унаслщок порушення микросомального окис-
нення в печ!нц! зм!нюеться фармакодинамика лгкарських засобгв. Тому в юпшч-
нш практищ основне значения мае л!кування хвороби, що спричинила ппокспо,
а не застосування так званих антигшоксаипв.
Чутлив!сть оргашзму до дефщиту кисню можна зменшити шляхом моделю-
вання деяких стан!в, як! супроводжуються гальмуванням центрально! нервово!
системи та зниженням обмшу речовин (наркоз, гшотерм!я).
Тренування в умовах барокамери або високопр’я сприяе пщвищенню ст!й-
кост1 до гшоксп та ширшому розвитку пристосувальних реакцш (активацп анти-
оксидантних систем, вивыьненню ендотел!ального МО). Заслуга в розробленш
методу ступенево! аюпматизацп до високопрного климату належить М.М. Сиро-
тиншу.
Хоч як це не парадоксально, гшокспо застосовують як нефармаколопчний
терапевтичний 1 профшактичний зас!б: акл!матизащя в горах, барокамери, так
зване зворотне дихання (дихання пов!трям в обмеженому простор!). Тренування
в умовах помтрно! гшоксп пщвищуе ст!йк!сть оргашзму не лише до не!, а й до
багатьох шших несприятливих факторхв — ф!зичного навантаження, змш темпе-
ратури навколишнього середовища, шфекцп, отруення, юшзуючого випромшю-
вання тощо. Дихалью методики, спрямоваш на зниження вентиляцн 1 затримку
дихання, використовують при лгкуванш деяких хрошчних захворювань, у патоге-
нез! яких е фактор гшоксп (броюпальна астма). Пом!рна г!покс!я захищае ткани-
ни вщ променевого ураження, осюльки радючутливють юптин залежить вщ вм!с-
ту кисню в тканинах.
ПОРУШЕННЯ ЕНЕРГЕТИЧНОГО ОБМ1НУ
Порушення регуляцн обмшу речовин. Обмш речовин, або метаболизм, в ор-
гашзм! визначаеться спадковими факторами 1 регулюеться ендокринною та веге-
тативною нервовою системами. Вщповщно до цього 1 порушення обмшу речовин
можуть мати спадковий характер або виникати внаслщок порушення функцп ре-
гулюючих систем.
Провщна роль у здшснешп внутршшьоюптинно! саморегуляци належить ге-
нетичнш шформацп. Бшышсть спадкових дефекпв обману речовин зумовлена
мутащею генгв, як! кодують синтез ферменпв (спадков! ферментопатп).
Для координацп метаболхчних реакцш у клхтиш потрхбним е поспйне надхо-
дження шформацп, що забезпечують мед!атори нервово! системи 1 гормони. Для
сприйняття шформацп клпини мають специфхчн! рецептори. Порушення рецеп-
торного механизму також може бути одшею з причин розвитку патолопчних про-
цес!в, наприклад у раз! цукрового 1 нецукрового дхабету.
Окргм внутршшьокгптинних мехашзмхв саморегуляци оргашзм мае й склад-
шпп нервово-ендокринш мехашзми саморегуляци.
Гормональна регуляцхя на клтгинному р!вш може вщбуватися також за допо-
могою генетичного апарату шляхом хндукци утворення ферменпв (наприклад ш-
сул!н шдукуе синтез ферменпв ппколхзу) або змши активносп наявних ферменпв
(адреналш активуе фосфорилазу, шсулш — гексокшазу).
Нервова система виконуе троф!чну функщю, тобто контролюе тканинний
обмш за допомогою мед!атор!в, а також аксоплазматично! течи. У раз! порушення
ще! функцн розвиваеться нейродистроф!чний процес.
Порушення обмшу речовин на вищому р!вн! бюлопчно! оргашзаци — рхвш
оргашв 1 оргашзму — значною мхрою залежить вщ стану нейроендокринно! регу-
ляцп. Так, емоцшне збудження супроводжуеться змшою гарково! регуляцн тепло-
продукци, вуглеводного обмшу та ш.
Багато порушень обмшу речовин, терморегуляци, статевого 1 фхзичного роз-
витку зумовлеш ураженням промхжного мозку. Особливо великою е роль ппота-
ламуса, який за допомогою рил!зинг-факгор!в через гшофхз або парагшофхзш
проводников! шляхи дае на метаболхзм. Порушення автономно! (вегетативно!) не-
рвово! системи також зумовлюють змши обмшу речовин. 3 ураженням симпатич-
них вузл!в, спинного 1 промхжного мозку пов’язане виникнення хвороби Барра-
кера — Слмонса.
Отже, м!ж внутрииньоклггинними мехашзмами саморегуляци, пов’язаними з
генетичною шформащею, 1 нервово-гормональною регулящею обмшу речовин е
пений зв’язок, 1 порушення з будь-якого боку спричинюе розвиток патологи.
Порушення обмшу енергп е основою бЁлыпосп функщональних 1 органЁчних
порушень органЁв ё тканин. Воно може вин и кати на всгх етапах енергетичних пе-
ретворень унаслщок вщсутностЁ або дефЁциту енергетичного субстрату, змёни
кёлькостё або активностЁ ферментгв, у зв’язку з генетичними дефектами, впливом
ЁнпбЁторЁв ферменпв ендо- й екзотенного походження, недостатшм надходжен-
ням в органЁзм незамЁнних амшокислот, жирних кислот, впамЁнЁв, мЁкроелемен-
тёв та Ёнших речовин. якё беруть участь у метабол Ёчних процесах, або через ушкод-
ження регуляторних систем.
Нормальний перебЁг обмЁну енергп на молекулярному р1вн1 зумовлюе дина-
мЁчну взаемодЁю процесЁв катаболЁзму й анаболвму.
КатаболЁзм може вЁдбуватися поза клётиною за допомогою травних ферментЁв
1 внутрЁшньоклЁтинно за участю лЁзосомальних гщролаз. ВнутрЁшньокиЁтанного
розпаду зазнають макромолекули, якё мають конформапЁйнЁ порушення, набуп
внаслЁдок розладу синтезу або Ёнших ушкоджень, зокрема й пероксидного окис-
нення. Продукта Ёх розпаду використовуються клётиною для синтезу Ёнших ком-
понентЁв. Генетична недостатнЁсть лЁзосомальних ферментЁв зумовлюе виникнен-
ня хвороб накопичення (мукополЁсахаридоз, сфЁнголЁпщоз, глЁкогеноз).
Окремим прикладом позаклЁгинного розпаду макромолекул е протеолЁз. що
забезпечуе пщвищення функцЁонально'Ё активностЁ ферментЁв, гормонЁв, якё спо-
чатку синтезуються як попередники з быыиою молекулярною масою, нёж основ-
на функциональна молекула (наприклад проЁнсулЁн-ЁнсулЁн). Ферментативиий
процес такого типу отримав назву обмеженого протеолау. Характерним його при-
кладом е функшонування каскадних систем (система комплементу, системи зсё-
дання кровЁ, фЁбринолЁзу, кЁнЁнова система).
Найефективншшм з енергетичного погляду е окиснення продуктов обмЁну в
циклё трикарбонових кислот (Кребса), менш ефектавним — глгколёз.
У разЁ порушення катаболЁчних процссгв передусЁм зазнае впливу синтез АТФ,
а також надходження субстратЁв, позрЁбних для бЁосинтезу (анаболЁзму). Своею
чертою порушення анаболЁчних процесЁв призводить до розладу вщтворення фун-
кцЁонально важливих сполук (ферментЁв, гормонЁв), потрЁбних для катаболЁзму.
НайвираженЁшЁ змёни анаболЁзму спостерЁгаються в разЁ порушення системи бю-
логЁчного окиснення або механЁзмЁв зв’язку дихання й окисного фосфорилюван-
ня. Приблизно на двё третини зменшуеться вироблення енергЁЁ в раз1 блокування
циклу трикарбонових кислот (пригнЁчення ферменту цитратсинтетази, дефЁцит
пантотеново] кислота, гёпоксёя).
Тяжкё роздали енергетичного обмЁну виникають у перебЁгу пукрового дЁабету,
коли значно зменшуеться вироблення макроерпчних сполук у зв’язку з порушен-
ням дихального ланпюга, зумовленого обмеженням потужностЁ циклу7 грикарбо-
нових кислот (дефЁцит щавлево-оптово! кислота), а також ослаблениям глЁколЁзу.
Порушення глёколётичних процесЁв негативно позначаеться на можливостё ор-
ганизму адаптувагися до ппоксп.
Взаемозв’язок дихання Ё фосфорилювання в клЁтинах е регульованим проие-
сом, який залежить вщ стану мЁтохондрЁй. У патолопчних умовах у разЁ порушен-
ня функцЁй мЁтохондрЁй ш органели триватий час перебувають у набухлому станЁ.
Це призводить до зменшення окисного фосфорилювання та стимулягш глЁко-
лЁзу.
За деяких умов, особливо пов’язаних з потребою пщгримання постойнох тем-
ператури тща, наприклад в умовах холоду, оргашзм термшово мобыхзуе тепло. Це
вщбуваеться шляхом роз’еднання окисного фосфорилювання х посилення винно-
го окиснення (до факторов роз’еднання належать адреналш, паратирин, прогесте-
рон, соматотропш, вазопресин, диштрофенол, урамщидин тощо). При роз’еднанн!
енергетичний обмш клггини спрямовуеться в бхк посиленого утворення тепла за
рахунок тимчасового зниження специфично! функцп клгтини х пластичних про-
цесхв.
Неабиякий штерес становлять даш про вплив на енергетичний обмш бак-
терхальнох штоксикацп. Дослщження довели, що дифтерхйному токсину, живим х
вбитим культурам золотистого стафйюкока властива роз’еднувальна дхя.
Калоригенний ефект тироксину, який рашше пов’язували з роз’еднанням
окисного фосфорилювання внаслщок набухания мггохондрхй, ниш пояснюють
збгльшенням маси мхтоховдрш та активацхею окисних ферменти, а також голо-
вним чином посиленням роботи №+, К+-АТФази.
Окисне фосфорилювання значно порушуеться при авггамшозц особливо гру-
пи В, осюльки вггамши цхе! групи входять до складу коферментов циклу трикар-
бонових кислот ! залучен! до перенесения електронхв у дихальному ланцюгу.
При гшовтгамшоз! В1 (хвороба берх-берх) порушуеться цикл Кребса ! тим са-
мим зменшуеться кыьюсть субстратного матер!алу для дихального ланцюга. Су-
доми 1 психози, як! спостерйаються у хворих, е клхшчними симптомами пору-
шення бюлопчного окиснення в головному мозку. Порушення в дихальному лан-
цюгу, пов’язаш з вщсутшстю нхкотинамщних х флавшових дегщрогеназ, характер-
ш для гшовхтамшозхв РР (пелагра) та В2 (арибофлавшоз).
Про стан енергетичного обмшу кожного шдивщуума можна дхзнатися, вимх-
рявши його основный обмш (ОО) — лабораторный показнык, що в1дображае наймен-
шу киыасть енергп, яку споживае I вивиьнюс у виглядг тепла людина для шдтри-
мання базовых вкцеральных процессе, що забезпечують и життед^яльтсть. ОО
вимхрюють у стандартних умовах:
• через 14 год теля останнього споживання !жх;
• зранку шсля пробудження в положенш пащента лежачи;
• пщ ковдрою в умовах температурного комфорту (18—21 °С);
• у стан! повного ф!зичного ! психолопчного спокою.
За цих умов бхльша частина енергп витрачаеться на роботу юнних насосгв, що
забезпечуе функцюнування насамперед нервово! та серцево-судинно! систем. Для
дорослого чолов!ка хз середньою масою тола ОО становить приблизно 1785 ккал
на добу, а для жшки — 1679 ккал на добу, що евщчить про бхльшу ефектившеть
енергетичних процесхв жшочого оргашзму. ОО також залежить вщ питомох по-
верхнх х температури т!ла, вхку, стану гормонально! та нервово! систем х взагал!
стану здоров’я, у тому чиелх наявностх шфекцшних захворювань.
Що бхльша хстота, то менше вщношення поверхнх п тхла до маси х меншим п
ОО, якщо розраховувати його на 1 кг маси т!ла. При розрахунку ОО на 1 м2 по-
верхш пла рхзниця виявляеться не дуже значною. Пщвищення температури тша
на 1 °С за правилом Р.М. Рассела збшьшуе ОО на 13 %.
Змши ОО, як правило, е наслщком захворювання. Особливо чхтко це вияв-
ляеться в умовах патологп, що порушуе нейрогормональну регуляцпо обмхну. Так,
при прогресуючому паралхчх, старечому недоумств! ОО дещо знижуеться. Вира-
женшп порушення спостерйають у раз1 ураження всгетативних центров гшотала-
муса (пгготаламгчний синдром Пейджа., пухлини, крововилив у мозок).
Найбглыпий вплив на ОО справляють гормони. Пщвищують ОО за опи сани-
ми више механизмами тироксин, трийодтиронш, кортикотропш, глюкокортико!-
ди, соматотропш, прогестерон, тестостерон, катехолам!нм. Знижують ОО гнсулш
за рахунок збёльшення частки енергп, шо запасаеться у вигляд! АТФ, та сома-
тостатин ! ошощи, як! гальмують обмяли процеси. В1ков! змгни ОО значною
м!рою залежать вщ змш гормонально! секрет!.
Збптьшення ОО на 20 % 1 б!льше е важливою д!агностичною ознакою тирео-
токсикозу, а його зменшення свщчигь про ггпофункцпо щитоподабно! залози.
Статев! стеро’щш гормони 1 глюкокортико’щи теж активують вгльне окиснен-
ня, а разом з альдостероном — натрйв! канали 1 тим самим опосередковано по-
силюють роботу Ка+-, К+-АТФази, що сприяе витрат! енергп’. У раз1 ппофункш!
гшофгза, надниркових 1 статевих залоз (ппофвектомы. негормональш пухлини.
крововиливи, хвороба Ащпсона, кастращя, нецорозвинення, кл!макс) штенсив-
нйть енергетичних процеств знижуеться, що нерщко супроводжуеться ожирш-
ням.
Пщвищення основного обмшу спостерйаеться в раз! посилення роботи серця
та органтв дихання. У початковгй стали серне во! недостагност! пщвишення основ-
ного обм,ну становить 30—50 %. У патогенез! дього явиша мае значения ппокс!я,
що зумовлюе компенсаторне посилення функшй орган !в дихання ! кровообпу.
Молочна кислота, яка утворюеться при цьому, частково окиснюсться з додатко-
вими затратами кисню. Гшеркапшя теж збуджуе дихання 1 посилюе роботу' серця
з! збгпьшенням ОО. Пщвишення ОО шд час гарячки пояснюють як роз'еднанням
окиснення 1 фосфорилювання пщ впливом адреналину, так 1 посиленням обм!н-
них процес!в ! функцш серцево-судинно! системи та системи зовшшнього дихан-
ня внаслщок пщвищення температури пла.
Збудники шфекцш 1 !хн! токсини прямо або за допомогою цитокпйв роз’ед-
нують окиснення та фосфорилювання 1 пщвищують ОО. Цей самий вплив пи-
токпйв спостерйаеться при багатьох захворюваннях, якг супроводжуються запа-
ленням, гшокс!ею, гарячкою. пухлинами.
ГОЛОДУВАННЯ
Голодування — це стан, що виникае в тому рал, коли оргашзм зовам не одержуе
харчових речовин або одержуе недостатню Тх киькгсть, або не засвоюе Тх унаслТ-
док хвороби.
Голодування як социальна проблема, а також як патолопчний процес, що
супроводжуе деяк! хвороби, особливо травно! системи, здавна привертае увагу
досл1дник1в. Значний внесок у вивчення голодування зробили В.О. Манасеш
(1869), В.В. Пашутш (1902) та його учш. 3!брано багато факпв, як! свщчать про
те, що при голодуванш насамперед активуються пристосувальн! мехашзми, вщбу-
ваються своерщна ферментативна адаптац!я оргашзму до вщсутност! поживних
речовин 1 перехщ до ендогенного живлення. Разом з тим доведено, що порушен-
ня задоволення потреб оргашзму в !ж! призводить до хвороб харчово! недостат-
ност!, як! проявляються змшами ферментних констелящй тканин 1 розладом об-
мшних процеав.
Нин! голодування розглядають як стан тривалого стресу, пов’язаного з адап-
тивною активащею бюсинтезу гормонгв надниркових залоз, як! справляють пря-
му (активуючу) 1 непряму (зберггальну) дпо на житгево важлив! ферментш систе-
ми оргашзму.
Голодування за походженням може бути ф!зюлопчним 1 патолопчним. Ф!з!о-
лопчне голодування перюдично повторюеться в деяких вид!в тварин у зв’язку з
особливими умовами !х кнування або розвитку. Прикладом такого голодування е
зимова сплячка деяких ссавц!в (байбакгв, ховрахгв), риб, плазун!в. Найпоширеш-
шим е патолопчне голодування. Розр!зняють голодування повне, неповне (юльюс-
не недощання) 1 часткове (якгсне). Повне голодування можливе без обмеження води
1 з обмеженням або зовс!м без води (абсолютно голодування). Неповне голодування
розвиваеться в тому раз!, коли в оргашзм надходять ус! поживш речовини, але в
недостатшй за енергетичного цшшстю юлькосп. Часткове голодування спостер!-
гаеться при недостатньому надходженш з !жею одного або кшькох харчових ком-
поненпв (бшки, жири, макро- ! мгкроелементи, вггамши, вуглеводи) за нормаль-
но! енергетично! цшносп !ж!. Ц! дв! форми голодування часто комбшуються.
ПОВНЕ ГОЛОДУВАННЯ
Причини розвитку повного (як ! шших вид!в) голодування можуть бути зовюшш-
ми 1 внутршгшми. Зовнйиш причини — вщсутшсть 1ж1. Голод виникае пщ час воен,
е постшним супутником людей низького социального статусу (безробггних, без-
хатчешов тощо). Щор!чна смертшсть вщ голодування у свш перевищуе 50 млн
ос!б. Найбшьшо! шкоди голод ! недощання завдають дням.
Внутрйин! причини — вади розвитку в д!тей, хвороби оргашв травно! системи,
шфекцшш процеси, анорексгя (патолопчна вщсутшсть апетиту).
Умови. У розвитку голодування, його тривалост! !, отже, тривалост! життя
оргашзму велике значения мають зовншпп 1 внутрппш умови. Тривалгсть життя
при голодуванш зменшують т! зовнйшй умови, яю збптьшують втрату тепла, шдви-
щуючи енергетичш затрата оргашзму на пщтримання температури пла (низька
температура навколишнього середовища, висок! волопсть 1 швидюсть руху
пов1тря, активш рухи). 1з внутр1шн1х умов на тривал!сть життя год час голодування
впливають стать, вне, загальний стан оргашзму, кшьюсть 1 яюсть жирових 1 бшко-
вих резервгв, а також !нтенсившсть обм!ну речовин.
Тривал1сть. Величина ОО, як вщомо, певною м!рою залежить вщ питомо! по-
верхш, тобто вщ сп!вв!дношення м1ж поверхнею тша 1 його масою. Що б!льша
питома поверхня, то значнпш витрати енергп !, отже, напруженшп обмгнш про-
цеси у тканинах для забезпечення гомойотермп. Цей закон поверхш пояснюе,
чому тривалють життя при голодуванш у др!бних тварин менша, шж у великих.
Так, др!бш птахи при голодуванш живуть лише 1—2 дш, а кондор — до 40 дшв,
мини — 2—4 дш, щури — 6—9 дшв, собаки — 40—60 дн1в, кош — до 80 дшв.
Граничним термшом повного голодування для людини вважаеться 65—70 дшв.
Невелик! розм!ри тша 1 менш досконала регуляцхя обм!ну й теплообмшу по-
яснюють швидку загибель новонароджених пщ час голодування. Так, триденне
цуценя може голодувати лише 3 дш 1 загинути вщ втрата 17—20 % маси тша, а
дорослий собака — 40—60 дшв до втрати близько 50 % маси тша. Впсове знижен-
ня р1вня ОО визначае бшьшу тривалють голодування в людей лпнього 1 старечо-
го в!ку 1 дорослих тварин. Кр1м усгх цих факторгв тривалють голодування визна-
чаеться також шдивщуальними особливостями, пов’язаними !з характером нерво-
во-гуморально! регуляцп та реактившстю оргашзму шдивщуума.
Перюди. За клйачними проявами повне голодування можна подшита на чота-
ри перюди: байдужост!; збудження, яке наростае з посиленням вщчуття голоду;
пригшчення (найдовший); парал!ч!в 1 загибел! тварини.
Краще уявлення про особливост! р!зних перюд!в голодування дае патоф1з1о-
лог1чна характеристика, що враховуе стан обмпту речовин 1 енергп. На основ! ц!е!
характеристики голодування можна подшита на три перюди: неекономного вит-
рачання енергп; максимального пристосування; тканинного розпаду, штоксикацп
та загибел! (термшальний перюд).
Тривалють кожного перюду р!зна залежно вщ виду тварин. У людини перший
перюд тривае 2—4 дш, другий фактачно визначае термш голодування 1 може три-
вата 40—50 дшв, третш — 3—5 дшв.
Основш прояви. Одним !з раннгх проявхв голодування е в1дчупгтя голоду у
зв’язку з! збудженням травного центру. Це зумовлено зниженням р!вня глюкози й
амшокислот у кров!, вщсутшстю нервових !мпульс!в вщ шлунка, зниженням сек-
рецц шсулшу та дуоденального холецистоюшну, який стимулюе центр ситост!.
Мае значения активащя секрецп нейропептиду У, мотилшу, як! стимулюють трав-
ний центр. Надал! вщчуття голоду пщтримуеться зниженням продукцп адипоци-
тами кахексину та лептину (див. роздш XIX “Порушення жирового обмшу”).
Вщчуття голоду у випадку повного голодування може зникнути через кшька
дн!в шсля його початку, причому пригшчення травного центру надал! може бути
настшьки глибоким, що його збудження потребуе спещальних заходгв. У раз! не-
Жиров! депо 97 %
Селез1нка 60 %
Печ1нка 53,7 %
Яечка 40 %
М'язи 30,7 %
Кров 26 %
Нирки 25,9 %
Шюра 20,6 %
Кишки 18 %
Деген! 17,7 %
Шдшлункова залоза 17 %
К1стки 13,9 %
Нервова тканина 3,9 %
Серне 3,6 %
0 20 40 60 80 100%
Мал. 38. Стутнь зниження маси окремих оргашв 1 тка-
нин у раз! повного голодування
повного голодування збудження травного центру весь час пщтримуеться 1 вщчут-
тя голоду перюдично вщновлюеться.
Голодування супроводжуеться зниженням маси пила. У перший перюд маса
тша знижуеться значно за рахунок неекономного витрачання енергп 1 видшення
екскременпв. У другий перюд темп зниження маси тша зменшуеться 1 становить
щодня 0,5—1 %. У третш перюд зниження маси тша знову посилюеться.
Маса рхзних оргашв зменшуеться неоднаково (мал. 38). Найштенсившше
зменшуеться маса жирово! тканини (97 %), а найменше — серця (3,6 %) та нерво-
во! тканини (3,9 %). Ще Л.О. Тарасевич (1898) звернув увагу на значну стшюсть
головного мозку до голодування. ГПзшше ГП. Разенков (1949) 1 Л.С. Галеева
(1952) довели, що в молодих тварин, незважаючи на зниження маси вс1х оргашв,
головний мозок продовжував рости. Дуже незнание зниження маси мозку 1 серця
при голодуваню свщчить про те, шо в оргашзм!, який голодуе, продовжуе здшс-
нюватися складна регулящя пром1жного обмшу речовин 1 процес1в М1Ж органами,
що забезпечуе насамперед пластичним 1 енергетичним матерхалом житгево важ-
лив1 органи, яю постшно працюють.
ГП. Разенков (1949), вивчаючи стан шлункового соку в собак, яю голодува-
ли, звернув увагу на те, що на 6—8-й день голодування в шлунковому соку збшь-
шуеться юльюсть азотистих речовин — альбумш!в 1 глобушшв. Було з’ясовано,
що бшки шсля розщеплення всмоктуються в кров 1 використовуються на побудо-
ву життево важливих оргашв. Цей процес е наслщком уведення в дно пристосу-
вальних мехашзм!в, яю забезпечують повторне використання бшюв для синте-
тичних процесхв.
П1д час голодування спостерггаються й шш! процеси, яю характеризують
адаптацйо оргашзму до незвичайних умов юнування 1 свщчать про перехщ до ен-
догенного живлення. Так, у ранш термши голодування активуються гл1кол1тичш
та лшолгтичш ферменти, пот1м посилюеться актившсть фермента переам^нуван-
ня. До юнця голодування шдвишуеться актившсть лхзосомних гщролаз, що свщ-
чить про порушення цшост! лшопротещно! мембрани цих органел.
Зм1на ферментно! активности в органах щд час голодування може бути
пов'язана зё змёною ёзоферментних спектрёв. Таю даш е про глюкозо-6-фосфат-
дегщрогеназу (Г-6-ФДГ), фосфоглюконатдегёдрогеназу (ФГД), яю посщаютъ клю-
чове мгеце в пентозофосфатному цикл!. П1Д час голодування знижуеться як за-
гальна акгившсть цих ферментгв. так ! активность !хшх окремих гзоферментёв. При-
чини зниження активност! гзоферментёв, очевидно, пов'язан! 31 змшою
мехашзм ёв генетично! регуляцп 1х синтезу
Про змши тд час голодування в генетичному апарапи юптини, який контрол юс
процеси белкового синтезу, евщчить порушення первинно! структури ферменпв.
Особливост! бтлкового синтезу пёд час голодування: частково можна поясните
змшами, яких зазнае в цих умовах транспорта РНК (тРНК), акцепторяа здат-
нёсть яко’! вёдносно таких аминокислот, як тирозин, лейцин, метёошн, ппцин,
знижуеться.
Обмш речовин. Перший перёод голодування характеризуеться плсиленим вит-
рачанням вуглевод1в, у зв’язку ёз чим дихальний коефгцёент шдвигцуеться, наближа-
ючись до 1,0. Рёвень глюкози в кровё знижуеться (нижче 3 ммоль/л), шо зумовлюе
зменшення секрецп ёнсулёну. посилення активност! а-ендокриноцит!в острёвцёв
щдшлунково! залози ё видшення глюкагону. Стимул юеться глюкокортикоид на
функшя юрково! речовини надниркових залоз, що викликае посилення катаболизму
бйиав ё глюконеогенезу. Запаси гл!когену в печёнщ швидко зменшуються, однак не
зникають повшетю за рахунок посилення процесёв глюконеогенезу. Унаслёдок при-
ппчення секрецп шеулёну в печёнцё ослаблюелься ефективнёсть циклу Кребса,
знижуеться р!венъ окисного фосфорилювання, шо позначаеться на енергетичному
обм!ш клётин. Гальмування глюкокортикоидами швидкостё гексою назло! реакцп
знижуе засвоення глюкози юптинами печшки.
На початку першого перёоду голодування ОО може бути дещо пёдвищеним.
При переход! до економного витрачання енергп вён знижуеться на 10 -20 % 1
залишаеться на цьому рёвнё до юнця другого перюду, дешо збёльшуючись лише
в третш. Зниження ОО пёд час голодування вёдображае значку перебулову обмён-
них процесёв. спрямовану на економне витрачання енергетичних ресурсёв. Певну
роль при цьому втдтграе пригшчення функцп щигоподтбно! залози.
Показово, що паралельно зниженмю маси тша ! печшки знижуеться вмтст
мётохондрёйного бёлка. Однак пояснюегься пе не лише зменшенням кёлькостё
мгтохондргй. Пёд час голодування системи окиснення й акумулювання енергп’ мо-
жуть перебудовуватися на економнёше витрачання енергетичних ресурсов ёз пёдви-
щенням сфективностё гх використання.
Видёлення азоту ёз сечею зменшуеться вже на 2—3-й день голодування (з
12—14 г до 10 г за добу). Потгм на 5—6-й день, коли запаси вуглеводёв вичерпу-
ються ё вёдбувасться перехёд обмёну на жири, спостерггаеться короткочасне пёдви-
щення видшення азоту ёз сечею, теля чого воно знижуеться до 7—4 г, вёдобража-
ючи економне витрачання бшкёв. Зменшуються штенсившсть дезамёнування г
переам!нування амшокислот у печёнш, бюсинтез амёнокислот з а-кетокислот та
амонёю. Однак усё цё процеси не можуть збалансувати розпад б!лк!в — розви-
ваеться негативний азотистий баланс.
У другий, найдовший, перёод голодування дихальний коефщгент знижуеться
до 0,7, шо вёдображае переважне окиснення жирев Майже 84 % енергёё оргашзм
одержуе за рахунок окиснення жир!в, 3 % — глюкози ё 13 % — бёлюв.
ЭЕ
Увесь запас глжогену забезпечуе енергетичнх потреби оргашзму протягом не
бшьше 6 год голодування. Потхм спостерхгаеться активацы обмшу в жировш тка-
нин!. Унаслщок низького рхвня хнсулшу знижуеться доставка глюкози в лшоцити х
виникае дефщит глщерину для синтезу триглщерхщхв. Тим часом переважання дп
глюкагону х катехоламшхв посилюе лхполхз. Вшьш жирнх кислоти надходять у
кров (л!пем!я) та хншх органи.
У тканинах, у яких транспорт глюкози крхзь клхтинш мембрани залежить вщ
шсулхну (мхокард, скелетах м’язи, жирова тканина), змешпуеться вмхст глюкози.
У печшцх та м’язах пщвищуеться рхвень вхльних жирних кислот 1 стимулюеться
хх транспорт крхзь мхтохондрхйш мембрани до дщянок окиснення. Лшогенез х
синтез жирних кислот у печшцх загальмований, однак через дефщит бшкхв х не-
достатке утворення лхпопротехнхв триглщерхщи затримуються в печшцх х розви-
ваеться и жирова шфыътращя.
У разх вичерпання запасхв глгкогену починаеться продукцхя кетонових тш
унаслщок значного збшьшення утворення ацетил-КоА в процесх р-окиснення жир-
них кислот. За умови штенсивного лпхолхзу весь ацетил-КоА не може зразу бути
використаним у циклх трикарбонових кислот х частина його ще на синтез у печхн-
цх кетонових тш. Цьому зокрема сприяе те, що в умовах голодування в гепатоци-
тах збхльшуеться спхввщношенххя НАДН / НАД+. Протягом 12—24 год голодуван-
ня штенсифхкащя процесхв р-окиснення виражаеться в наростанн! концентраци
кетонових ты у кров! вщ 0,2 до 0,3 ммоль/л. Цх змгни забезпечують збшьшення
потреби периферичних тканин у додатковому джерелх енерпТ. При голодуваннх
протягом 48—72 год рхвень кетонових тш сягае 3 ммоль/л.
Катаболхзм ендогенних енергетичних запасхв спрямований на забезпечення
енерпею функшй житгево важливих органхв, особливо головного мозку, який
споживае 1600—1800 кДж за добу, що забезпечуеться розпадом 100—150 г глюко-
зи. Пщ час голодування утилгзащя глюкози мозком може знизитися до 1/4 норми
х 40 % потреби в субстратах забезпечуеться за рахунок метаболгпв жирних кислот.
У людей з надмхрною масою тша на 3-у тижнх голодування ацетоацетат х р-гщрок-
сибутират забезпечують бшьше половини енергетичних витрат мозку.
Оскшьки посилення кетогенезу може призвести до кетоацидозу, вмикаються
регуляторш мехашзми, якх гальмують продукцхю кетонових тш. Пщвищення
рхвня кетонових тш у кровх за принципом зворотного зв’язку справляе антил!-
пол!тичний вплив на жирову тканину.
У нирках хнтенсивно вщбуваеться глюконеогенез. Щодня утворюеться майже
80 г глюкози, причому половина — за рахунок амхнокислот (бшковий катаболхзм)
х глщерину (катаболхзм жирхв). Амонш, який вивхльнюеться пщ час дезамхну-
вання глютамхну в процесх ниркового глюконеогенезу, витрачаеться на зв’язування
надлишку кетонових тш, якх потрапляють у первинну сечу у виглядх натрхевих
солей. Утворення амоншних солей дещо зменшуе негазовий ацидоз, оскшьки
супроводжуеться регенеращею гщрокарбонатного буфера (див. роздш XXI “По-
рушення кислотно-основного стану”).
ОО в цей перход, як уже зазначалося, знижений, азотистий баланс негатив-
ний. Тим часом зберхгаеться можливхсть синтезу життево важливих бшкових
структур за рахунок розпаду шших бшкхв. Вщбуваеться перебудова обмшних
процесхв, спрямована на ращональне використання резервних речовин, обмежен-
ня потреби тих оргашв. якх мають менше значения для збереження життя.
Третш, термшальний, перюд голодування мае характер ргзкого посилення
розпаду б1лк1в життево важливих органов, як! витрачаються як енергетичний ма-
тергал Дихальний коефппент становить 0,8. Збшыпуеться виведення гз сечею азо-
ту, сгрки, калпо, фосфору; сшввщношення азоту, кал!ю г фосфору в сеч! таке
саме, як 1 в цитоплазм! м’язових волокон. Це евщчить про розпад не лише бшкгв,
що легко мобшзуються, а й стабшьних бпткДв м’язово! тканини. Виникають де-
структивн! змши в м!тохондр!ях. У зв’язку 31 зменшенням кшькост! бшкгв плазми
кров! та зниженням п онкотичного тиску вщбуваеться вихщ води !з судин у тка-
нини з розвитком голодных набряклв. Порушення трофгки тканин 1 зниження
загально! резистентност! супроводжуються шод! наявшетю пролежшв 1 дшянки
некрозу на шкгр!г слизовш оболонщ, кератитом.
Безпосередньою причиною порушення обм!ну речовин пщ час голодування
е розлад ферментных систем, що пов’язано з! складшстю вщновлення бшкгв-фер-
ментгв, як! руйнуються в процес! голодування. Причому швидюсть розпаду 1 вщ-
новлення р!зних ферментних систем неоднакова 1 змша ’гх функцш вщбуваеться
неравномерно. Наприклад, актившсть катепсинов майже не змшюеться, а окисних
ферменттв порушуеться вже в перший перюд. Виникае дискоординац!я у процесах
обмену, якг суворо узгоджен! в норм!. Можливим е накопичення промгжних про-
дуктов токсично! дп.
Стан оргашв г систем. Теплопродукщя тцдгримуеться протягом усього термшу
голодування на мппмальному ргвш й ослаблюеться наприкшщ третьего периоду.
Тепловщдача дещо зменшуеться. Температура т!ла змшюеться, залишаючись на
нижшй меж! норми, 1 лише наприкшщ знижуеться до 30—28 °С, що було встанов-
лено в дослщ! В.В. Пашутша.
1нш! функцп оргашзму протягом першого г другого перюдгв голодування та-
кож зберггаються в межах, близьких до фгзголопчних. 3 боку нервово! системи у
перший перюд спостерггаеться збудження, особливо травного центру. Згодом
розвиваеться його пригшчення, рефлекси ослаблюються, деяк! умовш рефлек-
си зникають. Пщ час голодування легше розвиваеться шок. Розумова дгяльшеть
зберйаеться, однак школи можливий розвиток психозу.
На початку голодування шдвищуеться функция щитопод!бно! залози, гшоф!-
за, збшыиуеться секреция кортикотрошну ! тиротропшу, що своею чертою сти-
мулюе наднирковг залози. У другий перюд голодування функц!я бшьшост! ен-
докринних залоз приппчуеться. Основним фактором у цьому процес! е зниження
нейросекрецн в ядрах ппоталамуса. У 5 раз!в знижуеться секрепгя соматолгбери-
ну, що на 40—50 % зменшуе р!вень у кров! соматотропшу. Так! сам! змши вини-
кають з боку тиротропшу, фол!тропшу й лютропшу гшофгза.
У систем! кровообпу 1 дихальшй систем! особлив! порушення не спостергга-
ються. Функцш травно! системи пригшчуеться. Хоча за оптималъних умов основ-
ш житгев! функцп голодуючих не вщхиляються вщ норми, недостатшсть гх регу-
ляцп, яка прогресивно наростае, обмежуе адаптацшн! можливосп оргашзму, що
вдаеться виявити за допомогою ртзних навантажень. Патоморфолопчш змши в
тканинах 1 органах оыб, як! померли вщ голодування, незначш г зазвичай зво-
дяться до атрофп (переважно в паренхгматозних органах).
Вщгодовування навгть на початку останнього перюду голодування повшстю
вщновлюе функцп оргашзму. Це евщчить про те, що повне голодування не при-
зводить до необоротних змш. Процес вщновлення вщбуваеться дуже швидко.
~334
Втрата 40—50 % маси тгла за мгсянь вщновлюеться за 2 тиж. Виникае апетит,
прискорюються окиси! пронеси, стимулюеться пронес асимыяцп, всгановлюеть-
ся позитивний азотистий баланс. Однак, враховуючи стан травног системи у
випадку голодування (слабка перистальтика1 низька секреторна актившсть), вщ-
годовування сл!д проводити обережно.
У раз! повторного голодування пристосувальн! мехашзми, спрямован! на еко-
номне витрачання енергп, реалгзуються швидше, ! перший перюд скорочуеться
до мшгмуму. Однак бататоразове голодування поступово виснажуе вщновн! мож-
ливост! оргашзму, 1 настае момент, коли в!дгодовування неефективне 1 тварина
гине наыть за наявност! жирових вщкладень пщ шк!рою г в чепщ.
Перебп повного голодування без води такий самий, як г щд час голодування з
водою, однак тяжчий г не такий триватий (3—6 дшв). Якшо вода не вводиться
ззовн!, оргашзм черпае ц 13 тканин — не оксидацгйна вода. Найбыьша кглькгсть
води вивыьнюеться гз жирових вщкладень: 100 г п!д час окиснення вщдають
112 г води, а бглки й вуглеводи — приблизно вдвгчг менше. При цьому утво-
рюеться багато продуктов обмшу, для виведення яких потребно ще бшьше води —
створюеться порочне коло, шо прискорюе загибель тварини.
НЕПОВНЕ ГОЛОДУВАННЯ
Неповне голодування (недощання) споетерпаеться часпше, юж повне. Нимало
патолопчних стангв, особливо пов’язаних гз порушенням функщй органгв трав-
ления, супроводжуеться голодуванням того чи шшого ступеня. Сощальн! катаст-
рофу так! як вшна, а також безробгггя призводягь до недощання.
Неповне голодування виникае тод!, коли оргашзм хронгчно недоотримуе з
гжею належну для енергетичних затрат к!льк!сть енергп. Наприклад, замгсть
10 467—10 454 кДж (2500—3500 ккал) лише 8374—6280 кДж (2000—1500 ккал) 1
менше. Оскгльки таке голодування тривале, розвиваються пристосувальн] ме-
хашзми. Так, ОО знижуеться сутгевгше, нгж пщ час повного голодування (на 30—
35 % замють 10—20 %). Оргашзм надзвичайно економно витрачае енергетичш
ресурси. Повшьно знижуеться маса тгла, що школи маскуеться затриманням
води. Тим часом у тканинах розвиваються дегенеративнг процеси, виникають ви-
раженпш змши, нгж пщ час повного голодування, оскгльки воно тривае довше.
Тварини гинуть теля втрати близько 40 % маси тша. Унаслщок зменшення
вмгсту бык!в у кров! знижуеться онкотичний тиск, шо може зумовити виникнен-
ня набрякгв.
Дихальний коефшгент зменшуеться незначно. Нерщко виникае анемгя. 3 боку
системи кровообггу спостсрпаються брадикардгя, зниження артергального тиску.
Ослаблюеться дихання. Пригнгчуеться статевий шетинкт.
ЧАСТКОВЕ ГОЛОДУВАННЯ
У разг дефициту в гжг вуглеводгв основю порушення пов’язанг з посиленням кето-
генезу в печшцг, куди транс портуються жири внаслщок зменшення вмгсту глгко-
тену. Вуглеводи можуть утворюватися в процесг глюконеогенезу. Проте для ней-
тралгзацп та виведення азотистих шлакгв необхщю здоров! печшка г нирки. Недо-
статне надходження жирш в енергетичному аспект! можна вщкоригувати за рахунок
вутлеводтв 1 б'лкдв. Однак для забезпечення пластичних процеств потребно вводи-
те хоча б мшшальну кьтьктсть жир! в (5—6 г на добу), як! мтстять незамгнш жирш
кислоты: арахщонову, лшолеву та лтноленову, Варто також враховувати, шо !з
жирами надходять жиророзчиюп впамши, т тому жирове голодування не-
розривно пов’язане з вттамтнним.
Б!лкове голодування настае в тих випадках, коли юльюсть бтлюв, яю надхо-
дять тз !'жею, не забезпечуе в оргашзм! азотисту ртвновагу, необхтдний ртвень
пластичних процес!в. Потреба в бътках не стала т залежить вщ фтзюлопчного ста-
ну оргашзму, а також в!д якост! бтлюв. яю надходять тз тжею.
Якшо в склад! тжт немае навтть одтпе! з незамтнних аминокислот (арпнтну,
петидину, лейцину, тзолейцину, лтзину, триптофану, треошну, фентлалашну.
метюшну. валтну), азотисгий батане стае негативным.
Тривале недошання з переважним дефшитом у рашош бтлюв призводить до
бтлково-енергетичн от недостатностт.
1снуе зв’язок мтж споживанням енергп та азотистим балансом. Зниження
споживання енерп! збтльшуе витрати азоту т затри муе фтзичний розвиток.
Клпйчш та патофтзюлопчю прояви бтлково-енергетично! недостатностт в до-
рослих т дттей в блокадному Леншград! отримали назву алиментарно! дистрофа.
Хвороба починал ася в разт зниження енергетично! цшност! тжт на 50 %. Нес гача
повноцгнного бтлка, холод, фтзичне т нервове перенаируження — основш еттоло-
пчш фактори алтментарнот дистрофп.
У трошчних! наптвтрошчних крашах Африки, Азп, Центрально! та Птвденнот
Америки цей патолопчний процес огримав назву алиментарного маразму I ква-
иаоркору. В умовах цих кратн хронтчна бтлково-енергетмчна недостатшсть пог-
либлюсться п!д впливом несприятливих природних фактортв (сильна шеолящя,
високт температура 1 волопсть повттря). важко! фтзично! працт, шфекцшних хво-
роб особливо тих, шо уражують органи травления.
У розвитку ал!ментарного маразму видтляють тривалий перюд “збалансовано-
го голодування”, коли гомеостаз пщтримуеться за рахунок зменшення енергетич-
них затрат т зниження основного обмтну на 15—20 % т навтть 30 %. Однак унаслт-
док того, шо затрата енергп п!д час виконання фтзично! робота не можуть ком-
пенсуватися енергетичною цтншетю Тж!, витрачаються власн! запаси енергп
оргашзму. Знижуеться ршень глюкози, холестерину та нейтрального жиру в кровт.
Розвиваються анемтя, брадикардтя, плотен зтя, порушуеться секреторна т моторна
дтяльшеть травного каналу. Поступово розвиваеться дистрофтя оргашв 1 гканин.
Характеры ими е ртзке зменшення маси тша. атрофтя м’язтв т вщсутшсть пщ пиар-
ного жиру. У кров! — р!зко збьтылений ртвень глюкокортикощтв т глюкагону, як!
стимулюють глюконеогенез. 3 боку нервово! системи при алтментарному маразм!
можуть спостертгатись я винта парктнеошзму. зниження пам’ят!, змгни психтчного
стану, особистость Ослаблюеться слйюсть оргашзму до в!русних т грибкових тн-
фекцтй.
Алтментарний маразм у дттей розвиваеться швидше, шж у дорослих, т нерщко
е безпосереднъою причиною смерть Дтти вщетають у фтзичному та псих!чному
розвитку, у них вщзначаються детцгментацгя волосся, шкгри, м’язове виснаження.
Обтяжуе розвиток захворювання гшовттамшоз А, Е), В.
Найбшьш згубно впливае ця форма голодування на розвиток нервово! систе-
ми в дггей в1ком вщ 6 мю. до 3 рокгв, коли штенсивно ростуть нервов! клггини,
вщбуваеться мхелгшзащя ! глюгенез, установлюються нервов! зв’язки.
Быкова недостатшсть у дней у раз! достатньо! енергетично! цгнност! !ж! при-
зводить до розвитку кваипоркору, який супроводжуеться порушенням процесу
зроговгння та штенсивним злущуванням еп!дерм!су, що дало назву шй патологи —
“червоний хлопчик”. Саме слово “кваппоркор” перекладаеться з мови захщно-
африканського народу га як “хвороба первктка шсля народження молодшо! ди-
тини”. Це означае, що шсля того як перша дитина залишаеться без грудного
молока, вона позбавляеться джерела бык!в, що 1 спричинюе хворобу. У д!тей не-
рщко розвиваеться жирова гнф!льтрац!я печшки, можливим е виникнення циро-
зу. Характерним е розвиток набряк!в 1 асциту внаслщок значно! ггпоальбумшемп
та гшеральдостерошзму. Пщшлункова залоза зазнае впливу палшозу й ф!брозу,
унаслщок чого зменшуеться утворення травних ферменпв, школи розвиваеться
цукровий д!абет. Дистроф!чш змши поширюються на цирки, серце. Уникнути
цих змш можна лише завдяки ращональному харчуванню.
Про вггамшне голодування вдеться в роздш! XXIV “Патоф!зюлопя обмшу
вггамппв”, а про недостатшсть макро- 1 мгкроелеменпв — у роздшах XXII “Пору-
шення водно-електролггного обмшу” г XXIII “Патоф!з!олопя обмшу мгкроеле-
менпв”.
Р03Д1Л XVIII
ПОРУШЕННЯ ВУГЛЕВОДНОГО ОБМШУ
Вуглеводний обмш — сукуонктъ процессе споживання, перетворення, новс-утворення
и утимэацН вуглеводк. основным призначенням якого е тдтримання постшно! кон-
центрат! глюкози в кров! в межах 3,9—6,1 ммоль/л.
Це необхщно для нормального функцюнування нейрошв головного мозку,
яко за нормал ьних умов як енергетнчний субстрат можуть використовувати титьки
глюкозу, яку вони не здатн, аю депонувати, аш утворювати в процес! глюконео-
генезу.
За умови нормального харчування вуглеводи становлять 3/4 району 1 забез-
печують 50 % енергетичних потреб оргашзму. Серед вуглевод!в, якг споживають-
ся, частка моно-о дисахаридов не мае перевищувати 25 %, решту становлять пол!-
сахариди. Надлишок моно- 1 дисахаридов спричинюе ожир!ння, розвиток атеро-
склерозу та зумовлюе ман офсета щю цукрово го д!абету.
Вуглеводи не е незамшними речовинами, але при глюконеогенез!, який акти-
вуеться в раз! зниження р!вня глюкози в кров!, руйнуються болки й утворюеться
багато токсичних речовин. для знешкодження о виведення яких потробно снерпя о
нормальне функцюнування печшки та нирок.
Полосахариди крахмаль т глйсоген розщеплюються ферментом амолазою до
глюкози та мальтози в ротовой порожнин! о тонкой кишю. Дисахариди сахароза,
лактоза, мальтоза розщеплюються в тонкш кишю. Сахарозу розщеплюе сахараза
на глюкозуо фруктозу, лактозу (молочний цукор) — лактаза на глюкозу г галакто-
зу, мальтозу — мальтаза на дв! глюкози.
Порушення катаболозму вуглеводав може виникати внаслшок розладу, пере-
травлювання ! всмоктування вуглевод!в у кишках. Порушення ферментативного
розщеплення полосахаридов у кишках спостерогаеться поровняно рщко, оскольки
амгтаза виообляетъся слинними. кишковими та подшлунковою залозами. У раз!
ахолоо доя ам!лази слини тривае о в шлунку Процес завершуеться за допомогою
олпосахаридаз мембранного травления.
Синдром порушеного всмоктування вуглеводов (мальабсорбцгя) може бути спри-
ч мнений дефщитом специф!чних дисахаридаз еппелпо кишкових ворсинок або
недостатносто систем транспорту моносахаридов. В обох випадках вуглеводи на-
копичуються в просвото кишок, шдвищуючи осмолярность кишкового соку й тим
самим додагково залучаючи в г.росвот кишок воду. Хвор! скаржаться на вщчуття
болю, здуття живота, пронос, спостерогаеться затриомка росту о розвитку.
Найболыпе проблем виникае через порушення метаболозму лактози (5 % у
коров’ячому молош) г фруктози.
118
Порушення перетравлювання та всмоктування лактози в багатьох дорослих
людей пов’язане з припиненням з вёком експресп гена, що кодуе лактазу у 10 %
дорослих бёлих европейцёв 1 пёвшчноамериканщв, 70 % арабёв, 80 % китайцёв, 95 %
афроамериканцев.
Спадковий (вроджений) дефщит лактази, або низька ц актившсть, призводить
до неспроможностё кишкового соку дитини розщеплювати молочний цукор, вмёст
якого в жёночому молощ становить 6—7 %>. Таке явище називають непереносим1с-
тю лактози, що в раз! споживання грудного молока спричинюе мальабсорбцёю,
яка проявляеться диспепсёею.
Неспроможшсть бёохёмёчних систем оргашзму перетворювати галактозу на
глюкозу внаслщок спадкового дефекту синтезу галактозо- 1-фосфат-уридилтранс-
ферази проявляеться галактоземоею. Споживання дитиною грудного молока при
цёй патологп зумовлюе накопичення галактози в кров! та внутрёшнёх органах, що
проявляеться помутшнням кришталика, збшьшенням печенки та затримкою ро-
зумового розвитку.
Непереносимость фруктози, яка пов’язана з автосомно-рецесивною спадко-
вою недостатшстю фруктозо-1-фосфатальдолази, призводить до накопичення в
тканинах фруктозо-1-фосфату, що блокуе ферменти, необхёднё для розщеплення
глёкогену. Така патолопя проявляеться фруктозем!ею, фруктозур!ею, тяжкою п-
поглёкемёею ёз загальною слабёстю, тремтённям, пётливёстю, потьмаренням свёдо-
мостё в разё споживання фруктових сокёв або фруктёв.
При недостатност! фруктоконази виникають доброякёснё фруктоземёя та фрук-
тозурёя без виражених клёнёчних проявёв.
Молекули глюкози та ёнших моносахаридёв не здатш вёльно проходити крёзь
лёпщний бар’ер мембран. Для ёх транспорту необхёднё спецёальнё бёлки-перенос-
ники, якё мёстять 492—524 амшокислоти та мають форму зигзага, що 12 разёв
перетинае мембрану.
Клётини тонко! кишки ё нефронёв на апгкальнёй мембранё мёстять натрёйза-
лежний котранспортер моносахаридёв (симпорт). Цей переносник транспортуе
глюкозу, фруктозу або галактозу за мехашзмом вторинно активного транспорту,
тобто проти градёента концентрацёё разом з ёонами натрёю, якё переносяться за
градёентом концентрацп, що утворюеться внаслёдок роботи Ыа+-, К+-АТФази на
базальнгй мембранё цих клётин. Натрёй, що перемёщуеться завдяки енергп АТФ,
тягне за собою через клётинну мембрану моносахариди, навёть у разё !х дуже низь-
коё концентрацёё в травному каналё або первиннёй сечё.
1ншё переносники глюкози, фруктози ё, мабуть, галактози позначаються як
ГЛЮТ (глюкозш транспортери) ё забезпечують полегшену дифузёю моносахаридёв
через клётинну мембрану за градёентом концентрацёё:
• ГЛЮТ-1 мёститься у великёй кёлькостё в ендотелёё судин мозку ё в меншёй
— в ёнших органах ё тканинах;
• ГЛЮТ-2 виявляеться в клётинах оргашв, якё здатш не тёльки всмоктувати, а
й виводити глюкозу в кров (ентероцити тонко! кишки, гепатоцити та нефроцити).
За умови ацидозу нирки видёляють бёльше глюкози, нёж печёнка. У В-клётинах
острёвцёв Лангерганса цей переносник е сенсором глюкози;
• ГЛЮТ-3 мае найбёлыпу спорёдненёсть до глюкози ё мёститься в нейронах
головного мозку;
• ГЛЮТ-4 — шсулгнозалежний. За звичайних умов цей переносник мгститься
в мгкровезикулах цитоплазми м’язових клггин ! адипоципв. Титьки в раз! дп ш-
сулгну на шсулшов! рецептори ГЛЮТ-4 експресуеться на клпини ш мембран!. У
раз! глюкозного навантаження 80 % усге! глюкози входить у м’язов! клпини;
• ГЛЮТ-5 транспортуе фруктозу в травному тракт! та сперматозоидах.
Порушення транспорту вуглевод!в може бути пов’язане зг спадковою недо-
статшстю переносникгв або порушенням роботи Ма+-, К+-АТФази, що найчасп-
ше е наслщком зниження мпохондргального синтезу АТФ.
Для пщтримання постшно! концентрацп глюкози в кров! и надлишок, що
виникае теля прийому !ж!, вщкладаеться у вигляд! глгкогену в печищ! та м’язах
(глгкогенез) г у вигляд! лшщгв — в адипоцитах (лшогенез). При зменшенш кон-
центрацп глюкози в кров! глгкоген печшки розпадаеться до глюкози (глткоге-
нолгз), яка виходить у кров. Кргм того, у печгнцз г нирках глюкоза утворюеться тз
лактату, птрувату та глщерину, постачальником якого е лшолгз, а також з амгно-
кислот, кр!м лейцину, у процес! глюконеогенезу.
Для порушення анаболтзму вуглеводш е характерними розлад синтезу та депо-
нування глгкогену в печшцг, наприклад у раз! тяжкого ушкодження ештелюципв
печшки (гепатоципв) при гепатитах, отруешп чотирихлористим вуглецем, хлоро-
формом, пщ час гшоксп.
Спадково знижена актившсть будь-якого з ферменпв розпаду глгкогену спри-
чинюе розвиток гл1когеноз1в, як! належать до хвороб накопичення. Найпоширенг-
шим ппкогенозом е хвороба Прке (гткогеноз I типу), що виникае внаслщок дефг-
циту глюкозо-6-фосфатази. У дней хвороба проявляеться надмгрним вщкладен-
ням глгкогену в печшщ та нирках, ггпоглгкемгею. Спостерггаеться затримка
фгзичного та психгчного розвитку. При хвороба Герса (глгкогеноз IV типу) вияв-
ляеться низька актившсть фосфорилази ! вщкладення глгкогену в печшц! та лей-
коцитах. Для глгкогенозу III типу (хвороба Форбса) характерною е поява глгкогену
з короткими численними зовнгшнгми плками. Цю хворобу пов’язують зг зниже-
ною активнгстю ам!ло-1,6-глюкозидази. У титей, у яких вщмгчають цю форму
глгкогенозу, спостерггають набряки, гепатомегалгю, м’язову ггпотошю, кровото-
чивгеть. Глгкоген депонуеться в печшц!, м’язах, еригроцитах, лейкоцитах.
Значно змшюеться вуглеводний обмш у разг ггповгтамшозгв, особливо групи
В, оскгльки ц! вгтамгни е коферментами багатьох ферменпв (див. роздщ XXIV
“Патофгзгологгя обмшу вгтамгнгв”).
Депонування, вивйльнення, утшйзащя / новоутворення глюкози регулюютъея 1н-
сулшом I контршеулярними гормонами.
Гормони, що регулюють вуглеводний обмш
1.1 нсулЫ, який знижуе концентрацгю в кров! глюкози г посилюе и утилгзацгю
клгтинами за рахунок:
• активацп ГЛЮТ-4, що посилюе захоплення глюкози м’язовою г жировою
тканинами;
• активацп в усгх тканинах, кргм мозку, гексокшази та глюкокшази, як! пере-
творюють глюкозу, що потрапила в клгтину, на глюкозо-6-фосфат;
• стимуляцн ферменпв глгкогенезу в печгнцг та м’язах;
• стимуляцн ферменпв литогенезу в адипоцитах;
• стимуляцн ферменпв циклу трикарбонових кислот (ЦТК), пентозо-фос-
фатного шунта (ПФШ), глтколззу, що прискорюе утилгзанпо глюкози;
• блокування ферменпв глюконеогенезу, ппкогенолгзу 1 лшол!зу.
Кр!м того шсулш:
• активуе №+-, К+-АТФазу, що сприяе переходу юнгв калпо в юптини;
• зб!льшуе синтез бшктв;
• прискорюе клггинний р!ст.
II. Контршсулярнг гормони пщвищують концентрацию глюкози в кров!, але
вони е функцюнальними агошстами шсулшу, осюльки зб!льшують дифуз!ю глю-
кози в кл1тини в умовах дефщиту шсулшу за рахунок пщвищення градиента його
концентрацп.
1. Глюкагон:
— активуе розщеплення глгкогену в печшц!;
— активуе глюконеогенез;
— активуе лшолтз;
— активуе утворення кетонових пл !з жирних кислот у печищ;
— блокуе ЦТК, ПФШ, глгкол!з.
2. Катехоламши:
— стимулюють гл!когенол!з;
— стимулюють ЛШОЛ13.
3. Глюкокортикощи:
— блокують гексо- 1 глюкокшазу;
— активують глюконеогенез, переважно за рахунок розпаду бшюв м’язтв 1
катабол!зму амшокислот;
— дуже слабко активують лшол!з.
4. Гормони щитопод1бно! залози:
— активують ГЛ1КОЛ13, ЦТК, ПФШ;
— стимулюють усмоктування глюкози в кишках;
— активують л!пол!з;
— стимулюють глпсогенолтз.
5. Соматотрошн:
— !нг!буе глюко- 1 гексокшазу;
— стимулюе л!пол!з;
— активуе глюконеогенез;
— активуе протешази, що прискорюе розщеплення шсулшу та знижуе його
концентрацпо;
— стимулюе синтез 1 секрецпо глюкагону.
Порушення нервово-гормоналыюТ регуляцп е найчаспшою причиною патологи
вутлеводного обмшу. Об’ектами регуляцл е вщкладення вуглевод!в у печищ! 1
м’язах у вигляд! глгкогену 1 перетворення тх на жири та депонування як джерела
енергп в лшоцитах, а також глгкогенолтз т глюконеогенез. Ц! процеси псно
пов’язан! м!ж собою. Порушення зазначено! координацп проявляються гшер- або
ппогл!кем!ею I глюкозур!ею.
Наслщки порушення нервово'1 регуляци вуглеводного обм1ну вперше вивчив К. Бер-
нард (1855). Вш установив, що укол у дтлянцт дна IV шлуночка зумовлюе розви-
ток гшерглткемц. Такий ефект може спричинити подразнення строго горба, гшо-
таламуса, сочевицеподтбного ядра та смугастого пла базальних ядер великого
мозку. Кеннон спостертгав. як унаслщок психтчного перенапруження, емощйних
ГздГ
1нсулш
Проникшсть КЛ1ТИННИХ мембран
Швидюсть гексокшазно! реакци ,
-> Синтез ферменттв глтколтзу
Синтез ферменттв глюконеогенезу'
Утворення глюкозо-6-фосфату
Посилення
Ослабления
Антагонтсти
тнсултну
Мал. 39. Вплив шсулшу та його антагошстш на метаболтзм глюкози
в клттиш .
впливтв пщвищувався ртвень глюкози в кровт. Пперппкемтя виникае також год
час болю, нападу ешлепсп та тн.
Вщомий шший шлях впливу центрально! нервово! системи на вуглеводний
обмш: на остртвщ годшлунково! залози через парасимпатичнт волокна.
Порушення гормонально! регуляцп вуглеводного обмшу може виникати внаслщок
не тшьки розладу ц центральних механизмов, а й ураження ендокринних залоз або
розладу периферичних механтзмтв дп гормошв.
Провщним фактором у порушенш гормонально! регуляцп обмшу вуглеводтв е
змша сптввщношення мтж актившстю шсулшу та контршсулярних гормошв.
На мал. 39 показано вплив шсулшу та його антагошспв на процеси, як! зу-
мовлюють утшпзащю глюкози в тканинах т визначають певною мтрою показник
глткемп.
Дефщит шсулшу спричинюе гтперглткемпо. Гт мехашзм пояснюеться знижен-
ням проникносп клттинних мембран для глюкози, сповшьненням швидкост! гек-
сокшазно! реакцп й утворення глюкозо-6-фосфату, а отже, ! подальшого мета-
болтзму глюкози, посиленням процеств глюконеогенезу.
Гшерглткемтя спостертгаеться також у раз! надмтрного вмтсту глюкагону, ад-
реналшу, тиреощних гормонтв, глюкокортикощтв, соматотропшу та кортикотропт-
ну в кровт.
Патология вуглеводного обм1ну проявляеться гшерглткемтею, глюкозуртею або
гшоглткемтею.
Гшерглткемтя — пщвищення ртвня глюкози в кровт понад 6,1 ммоль/л. Залеж-
но вщ еттологп можна видшити таки вида гшерглткемп:
— алиментарна — короткочасне пщвищення ртвня глюкози при вживанш по-
над 75 г моно- т дисахаридтв;
— при абсолютнш чи в1дноснш недостатност/ шсулшу,
— ендокринно — при надмтрнш продукцп (синдром т хвороба 1ценка — Кунин-
га, гтпертиреоз, глюкагонома, феохромоцитома, акромегалтя) чи надлишковому
введенш якогось !з контршсулярних гормонтв;
— стресорна — сумарна д!я катехоламтнтв т глюкокортикощтв, що вившъню-
ються при дп стресорних фактортв;
— при патологи печшки шсля споживання !жт.
Глюкозуртя — поява глюкози в сечт. Розртзняють:
— дгабетичну глюкозурйо, що е наслщком збшъшення в кровт ртвня глюкози
при цукровому дтабетт понад нирковий пород тобто понад концентрацию, до яко!
нирки здатнх П0ВН1СТЮ реабсорбувати профхльтровану глюкозу (для неушкодже-
них нирок поргг становить 11 ммоль/л, але при дхабетичнш нефропатп вш зни-
жуеться до 7—8 ммоль/л);
— ниркова глюкозур1я виникае на тлх нормоглхкемп при порушены мехашзмхв
реабсорбцн глюкози в нирках унаслщок спадково! недостатностх або блокади
флоридзином натршглюкозного переносника, блокади №+-, К+-АТФази уабаи-
ном чи порушення утворення в нефроцитах АТФ. Вхдоме рщкхсне автосомно-ре-
цесивне захворювання, зумовлене муташею гена (хромосома 6) Ыа-залежного
флоридзинчутливого переносника моносахарид хв, яке супроводжуеться значним
зниженням ниркового порога реабсорбцп глюкози та глюкозурхею.
Ппоглхкемхя — зниження рхвня глюкози кров! нижче 3,8 ммоль/л. Основн!
причини ппоглхкемп:
— голодування',
— значне ф1зичне навантаження',
— ниркова глюкозур1я (нирковий дхабет);
— гтергнсулмешя (доброяюсна пухлина хз В-клпин або передозування шсулшу);
— недостатшсть продукцп контршсулярних гормошв (мхкседема. хвороба Ад-
д!сона тощо);
— патолог1я печшки — у стан! натще нездатысть утворювати глюкозу в печш-
цх шляхом глхкогенолхзу та глюконеогенезу;
— глшогенози — порушення синтезу або розщеплення глхкогену.
Значне зниження рхвня глюкози в кров! (менше 2,5 ммоль/л) шсля введения
шсулшу натще без глюкози спричинюе розвиток гшоглшемхчно! коми. Гшогл!-
кемхчнш комх передують загальна слабхсть, дратхвливхсть, вщчуття голоду у зв’язку
зх збудженням вентролатеральних ядер гшоталамуса, посилене потовидхлення,
таххкардхя (пперпродукцхя адреналхну), тремор; можливх судоми. Усе це е наслщ-
ком порушення енергетичного забезпечення нейронхв.
1. а-Ендокриноцити острхвцхв тдшлунково’Г залози
2. Юркова речовина надниркових залоз
3. Мозкова речовина надниркових залоз
4. Аденоппофхз
Мал. 40. Нормальна регуляхця вуглеводного обмхну та його ймовхрш
порушення в разх змши сшввцщошення мхж гормонами-антагошстами
Мал. 41. Крив!, шо в!дображають результата
навантаження глюкозою, у здорово! людини
(А), у разг знижено’! толерантност! до глюко-
зи (Б), у хворого на цукровий даабет (В)
Кома — це патолопчне гальмування
функцп центрально!’ нервово’! системи, для
якого характерш потьмарення свщомостг,
вщсутнгсть рефлексов ! розлад регуляцп
життево важливих функщй оргашзму.
У патогенез! гтоглисешчно! коми основ-
ного значения набувае енергодефгцит юп-
тин ГОЛОВНОГО МОЗКу, ДЛЯ ДгЯЛЬНОСП яких
глюкоза е основним джерелом енергп.
Спгввщношення мгж гормонами, яю
беруть участь у регуляцп р!вня глюкози в
кров!, наведено на мал. 40.
За станом регуляцц вуглеводного об-
мшу, здатшстю органгзму засвоювати пев-
ну кглькгсть вуглеводгв роблять висновок
про толерантшсть до вуглеводгв, яку виз-
начають за допомогою навантаження глю-
козою. Швидюсть зниження ргвня глюко-
зи багато в чому залежить вщ функцгй
остргвцгв пщшлунково! залози. На мал. 41
наведено крив! в норм! г в раз! знижено!
толерантност! до вуглеводш, яю можуть
бути при цукровому дгабет!, акромегалп, тиреотоксикоз!, синдром! 1ценка —
Кушшга.
У цих випадках ргвень глюкози може перевшцувати 11 ммоль/л. Пщвищена
толерантшсть до вуглеводгв спостериаеться при пперфункцп В-клгтин остргвцгв
Лангерганса, мгкседем!, гшофгзектомп. Глгкемгчна крива у цих випадках характе-
ризуеться незначним пщвищенням 1 ргзким зниженням протягом друго! години
спостереженшг.
Толерантшсть до вуглеводгв визначае та максимальна кгльюсть глюкози, яку
може засвог'ти оргашзм без виниюгення глюкозурп, тобто появи глюкози в сечг:
у здорово! людини — 160—180 г глюкози, спожитог натще. Толерантшсть до глю-
кози знижуеться при цукровому та цирковому дгабет!.
ЦУКРОВИЙ Д1АБЕТ
Цукровий д1абет — стан хрошчноТ гтерглйсеми, зумовлений абсолютною чи в1днос-
ною недостатшстю шсулшу.
Проявами цукрового дгабету е порушення обмгну речовин, особливо вугле-
водного, кетоацидоз, прогресуюче ураження периферичних нервгв, капгляргв ни-
рок, сгтювки, а також атеросклероз (ВООЗ, 1980).
Цукровий дгабет дгагностують у 4—5 % населения розвинутих краш, а серед
осгб вгком понад 65 рокгв — у бглыпе нгж 20 % випадкгв. У ргзних репонах поши-
ренгсть хвороби неоднакова: вщ нуля у жителгв високопрних районгв Папуа-Но-
во! Гвгне! до 85 % серед гндганцгв пгма (США). На юнацький дгабет найбгльше
хворгють скандинавщ, а найменше — корейщ.
За даними ВООЗ, у 2010 р. юльккть хворих на цурковий дгабет становила
240 млн; кожнг 15 роив захворювашсть на цю недугу збыьшуеться вдвгчг.
Основними проявами цукрового даабету е ггперглгкемгя, яка сягае школи по-
над 25 ммоль/л, глюкозургя з умктом глюкози в сеч! до 555—666 ммоль на добу
(100—120 г на добу), пол!ур!я (до 10—12 д сеч! на добу), полщипс!я ! полгфапя.
Класифгкацк. Комитетом експерпв ВООЗ у 1999 р. запропоновано еполопчну
класиф’кащю цукрового диабету, наведену нижче з деякими модифхкашями, шо
вщповгдаюгь Мгжнародшй статистичнгй класифгкан!! хвороб 10-го перегляду
(1998):
ЕПервинний щабет (95 %) — переважно генетично зумовлене первинне пору-
шення мехашзмгв шсулшово! регуляцп вуглеводного обмшу, що подгляеться на:
1. Цукровий дгабетп типу 1 (10—20 %) — генетична дисфункщя або, часпше,
генетично зумовлена автоёмунна деструкщя В-клггин остр)вц!в Лангерганса, шо
виникае шсля провокуючо! дп зовнппнгх факторгв або спонтанно з розвитком
абсолютно! шсулшово! недостатносп переважно в юнацькому вщ! (синошми: ди-
тячий, юнацький, дгабет худих, шсулшозалежний; 13ЦД).
2. Цукровий 01абет типу 2 (80 %) — дшбет дорослих гз надмгрною масою пла,
спричинений первинною шсулшорезистентшстю або зниженою чутливгстю В-
кл!тин до глюкози, що зумовлюе вщносну шсулшову недостатшсть, тобто гшер-
глгкемпо на тлг гшершсулшемп (синонгми: д!абет старих, дгабет товстих, шсулшо-
незалсжний; 1НЦД)
Р1зновиди цукрового д!абету 2 типу:
а) 31 збшьшенням маси пла;
б) з нормальною масою пла,
в) МООУ (ташгпу опкес сИаЬеГе^ оГ Гйе уоип§, 1НЦД старих у молодих).
3. 1нш1 специфгчш типи цукрового дшбету, що включають р!зн! моногенн! де-
фекти В-кл!тин чи шсулшу, а також хвороби екзокринно! частини пщшлунково!
залози.
4. Гестицшний цукровий дшбет (дгабет вагйпних) — зниження толераптност!
до глюкози, яке вперше проявляеться шц час ваптносп.
II. Вторинний дгабет — ппергл!кем!чш синдроми, що виникають у раз! вто-
ринного порушення мехашзмш вуглеводного обм!ну при хворобах, якг первин-
но не ушкоджують систему шсулшово! регуляпн 1 не мають генетично! зумовле-
ност!:
• ендокринопатп, що супроводжуються надм!рною секретею контршсуляр-
них гормошв (хвороба г синдром 1ценка — Кушшга, феохромоцитома, глюкаго-
нома, акромегал!я, гггантизм, тиреотоксикоз);
• порушення КОС (див. роздал XXI “Порушення кислотно-основного стану”);
• ускладнення медикаментозно! тераш! (глюкокорти кощи, ооальш контра-
цептиви, антидепресанти, сечопнш засоби, пропранолол, фенопазини тощо);
• системш автошунш пронеси;
• генетичн! синдроми (Лу!-Ьар, Вернера, Да^тта, Шерешевського, Клайнфел-
тера тощо).
Експериментальний цукровий дгабет. Основнг дан! про еполопю г патогенез
цукрового дгабету стали вщомими завдяки дослщженням на тваринах. Першу його
експериментальну модель виконали Мершг г М!нковський (1889) шляхом вида-
лення у собак умей або бглыио! частини (9/10) тдшлунково! залози.
Дия те! форми експериментального дхабету характеры вег ознаки, що хх спос-
терхгають у людини, однак перебхг и був тяжчим, завжди ускладнювався високою
кетонемхею, жировою шфшьтрашею печхнки, розвитком дхабетично! коми. Унас-
лщок видалення всхех пщшлунково! залози оргашзм страждав вщ дефициту не
тхльки шеулхну, а й травник ферменпв.
Великого поширеннянабула модель алоксанового диабету, який виникае теля
введения тваринам алоксану Ця речовина ушкоджуе переважно В-клпини ост-
рхвцхв пщшлунково! залози, унаслщок чого знижуеться секрецхя шсулшу. 1ншою
ххмхчною речовиною. шо спричинюе цукровий дхабет, е дитизон, який зв’язуе
цинк, що бере участь у депонуваннх та секрецй' шеулхну. Бщып специфшно уш-
коджуе острхвш пхдшлунково! залози антибютик стрептозотоцин. Цукровий дха-
бег у тварин можна вщтворити за допомогою антипйл до 1нсул1ну. Такий дхабет
виникае у раз! як активно!', так х пасивно! хмунхзацй.
Експеримент альний цукровий дхабет розвиваеться також теля введения контр-
(нсулярних гормошв. Так, теля тривалого застосування гормон!в аденоппофхза
(соматотропшу, кортикотрошну), як зазначалося више. може розвинутися гтофг
зарний дшбет. Унаслщок уведення глюкокортикощхв можна досягти розвитку
стероидного иукрового дгабету.
Спонтанний цукровий дхабет спостерхгають у китайських хом’ячкхв, мишей
деяких лшгй (“колючих”, новозеландських, МОЕ), об / оЬ та ш.) У МОЭ-мишей,
наприклад, виявляють генетично зумовлеш порушення хмунно! системи у вигляд!
надлишку Т-ефектор1в ! дефициту супресорно! функцн Т-льмфоцитхв.
Усх форми експериментального пукрового дхабету чггко заевщчують. шо ос-
новою його розвитку е абсолютний або вщносний дефщит шеулхну.
Абсолютна (панкреатична) шеулшова недостатшсть пов’язана з генетичними
або набугими порушеннями бюсинтезу х видщення шсулшу, вщносна (позапанк-
реатична) виникае за нормального утворення шсулшу пщ впливом факторхв, якх
пригшчують його дно або прискорююхь його катаболхзм.
Цукровий дЗабет типу 1
Етюлопя. 13ЦЦ виникае в людей вхком до 30 рокх'в, частпне восени чи взимку, у
бхльшостх випадкхв теля перенесено! в! рус но! шфекцп.
Реовхруси III типу, цитомегаловхруси, вхруси Коксакх В3! В4, кору, вродженох
краснухи, вхспи, епхдемхчного паротиту, ентеровхруси та ххмхчнх дхабетогени (нхт-
розамши, штрозосечовина. вакор, алоксан, сечова кислота, стрегпозотоцин, пен-
гамщин. бичачий сироватковий альбумш) здатнх спричинити ушкодження В-клх-
тин острхвцхв Лангерханса з подальшим розвитком автохмуннохо процесу.
Проте така провокашя автохмуннохо цитолхзу можпива тыьки в оргашзм! лю-
дей з певними генетичними особливостями ретуляци хмунно! вщповщх. Таке яви-
ще мае назву “адитивно-полхгенне успадкування з пороговим ефектом I провоку-
ючим впливом факторхв на вколи шньохо середовища”. Характерно, що до остан-
нього найбьтып схильнх дни раннього вхку.
Вважають, що найбшыпий внесок у генетичну схильшсть до цукрового дхабе-
ту тапу 1 здхйснюе комплекс генхв ГКГС у короткому плеч! хромосоми 6 мхж
локусами ОК х В, Наявшсть гена збщыпуе ймовхршеть захворювання на цук-
ровий дгабет типу 1 у 5 разгв, ВК(— у 7 разгв, а у гетерозигот ЭК, / ПК4вона вища
в 14,3 раза.
Наявшсть певних антигешв ГКГС ще не означае, що з!гхворювання розви-
неться обов’язково. ате збыьшуе ризик його виникнення Наприклад, локус Эр
при зчепленнг з локусами ОК. зумовлюе розвиток пукрового дгабету типу 1, якщо
в 57-у положены! р-ланцюга быка ОО с серин, валш або алашн. Наявнгсгь же
аспарапново! кислота зменшуе ризик виникнення цукрового дгабету. Таким чи-
ном, 13ЦД належить до антигенасоцгйованих хвороб.
Значения генетичних факторгв в етголота цукрового дгабету шдтверджуетъся
наявшстю роди иного дгабету, коли захворювання реестру еться в кглькох члешв
родини, шод! в 3—4 поколшнях: высокою конкордантнгстю в однояйцевих близ-
нюктв (у 4 рази вище, нгж у двояйцевих). Очевидно, генуе низка мутантних гешв,
якг визначають розвиток цукрового дгабету. Успадкування вщбуваеться полтген-
ним, школи автосомно-рецесивним або автосомно-домгнантним шляхом.
Патогенез. Взаемозв’язок генетичних та тмунних мехашзмгв пояснюеться роз-
мпценням генгв системи ГКГС у локус!, який контролю»: гмунну вщповщь ор-
гангзму, на хромосом! 6. Очевидно, наявшсть певних антигенгв евщчить про особ-
ливост! тмунного статусу органгзму.
Вгруси та х!м!чш дтабетогени, проникаючи в В-клпини остргвцгв Лангерганса
(р-ендокриноцити), можуть безпосередньо токсично впливати на них або шщпо-
вати автогмунну реакцпо на быки цих клгтин у носив певних антигешв ГКГС
Незалежно вщ початкового мехашзму деструкнтя В-клзтин спричинюе абсолют ну
шеулшову недостатшсть (схема 14).
Генетичнг особливост! гмунно! системи у вигляд! пщвишено! хелперно! актив-
ное тг та пригшчено! супресорно! функцп визначають мехашзм розвитку автогмун-
ного ушкодження В-клгтин.
Якщо в оргашзм проникае вгрус з антигеном, лодгбним до бшкгв р- ендокри-
ноцитгв, наприклад быок з молекулярною масою 64 кДа (глютаматдекарбоксила-
за). то починаеться автогмунний процес за участю клгтанних г гуморальних ме-
хангзмгв гмунгтету. Макрофаги поглинають антиген г презенгують його на свогй
поверхш разом з быками ГКГС II класу хелперним лгмфоцитам. Кргм того, вони
секретують 1Л-1, який стимулюе лгмфоцити (Т-хелпери, Т-к!лери, НК, В-лгмфо-
цита). Лгмфокши, особливо ФНП-а та у-(Ф. беруть безпосередню участь у де-
струкпи р-ендокриноцит!в.
Мають значения також активацгя системи комплементу та утворення проста-
гландишв. Останшм часом значну роль у руйнуваннг В-клпин придыяюзь МО,
який утворюеться за допомогою МО-синтетаз з Ё-арпнгну в моноцитах-макрофа-
гах г р-ендокриноцитах, що зумовлюе гх деструкцгю та розвиток цукрового дгабе-
ту типу 1.
Розргзняють так! опади патогенезу 131Щ'.
1) стад! я генетично! схильностг, яка може три вата вш кглькох мгсяшв до де-
сяткгв рокгв,
2) провокуючог дп — може тривати 1—2 дат;
3) первинних автогмунних уражень В-клггин — тривае вщ 2—3 мгс. до 3—
4 рокгв. Вщбуваеться руйнування В-клгтин головним чином за мехашзмом ппер-
чутливостт сповыьненого типу, але недостатшсть гнеулгну ще не вгдчуваеться (мае
загинути понад 90 % В-кл!тин);
Схема 14. Етюлопя 1 патогенез цукрового даабету типу 1
4) стадщ латентного д!абету. Клинчних симптомгв немае, але при наванта-
женш глюкозою виявляеться штолерантшсть до не!. За тривалютю вщповщае 3-й
стадп;
5) стадия явного дгабету, що проявляеться вираженою клцйчною картиною цук-
рового ддабету: ппергл1кем1ею, глюкозур!ею, пол!ур1ею, полщишлею, кетонемгею.
Цей перюд мае гострий початок. Згодом з’являються ознаки мГкроанпопатп.
Пщгрунтям патогенезу цукрового дхабету типу 1 е абсолютна шсулгнова недо-
статшсть 1 порушення паракринно! взаемодп М1ж юптинами остргвшв Лангерганса.
Остргвгц Лангерганса за нормальних умов на 70 % складаються з В-клггин, що
секретують шсулш, на 20 % — з А-клпин, яю секретують глюкагон 1 на 10 % — гз
О-юптин, яю продукують соматостатин. При цьому глюкагон стимулюе утворен-
ня та секрешю шсулшу г соматостатину. Соматостатин гальмуе утворення г сек-
рещю глюкагону та шсулшу. Тнсулгн своею чертою паракринно гальмуе утворен-
ня та секрешю глюкагону. Тобто у ф!зюлопчних умовах функцюнальна актив-
ность А-юггтин бггормонально гальмуеться шсулшом 1 соматостатином.
При нукровому дгабет! типу 1 унаслщок автогмунно! загибел! В-юптин частка
А-юптин в остргвцях збглыпуеться до 70—75 %, а частка О-юптин — до 20—25 %.
Б|гормональне гальмування А-юитин втрачаеться, що спричинюе надлишковс
вироблення глюкагону. Спгввщношення мгж. глюкагоном та шсулшом збшывуеть-
ся, що й визначае подалыш порушення обмшу речовин.
Гормоналънг мехашзми порушення обмшу речовин при цукровому д1абет! типу 1:
• у гепатоцитах, м юнитах 1 лшоцитах дефщит шсулшу пригнгчуе гексокшазну
(глюкокшазну) реакцию, що зменшуе утворення глюкозо-6-фосфату, без чого
значно менше глюкози потрапляе в щ юптини;
• зазначене вшце, а також те, шо без шсулшу не активуються ферменти гл!-
колгзу, никну Кребса г пентозного шунта, призводить до дефщиту АТФ;
• у печшш без шсулшу блокуеться глгкогенез та активуеться шд впливом
глюкагону 1 катехолам1шв глткогенолгз;
• у гепатоцитах г нефроцитах пщ впливом глюкагону, глюкокорти копив г со-
матотропшу активуеться глюконеогенез, що збшьшуе ргвень глюкози в кров! та
прискорюе розпад бглкгв;
• без шсулшу порушусться транслокащя ГЛЮТ-4 в мембрану мю- 1 лшо-
цитлв, що перешкоджае поглинанню надлишку глюкози м’язами 1 жировою тка-
ниною;
• без шсулшу блокуеться литогенез, а шд впливом контршсулярних гормон! в
активуеться лшолтз у жировш тканиш та кетогенез у печшщ.
Усе вищенаведене спричинюе гтергл1кем1ю, глюкозурйо, кетонемпо, кето-
нурт.
Ппергткем1я е наслщком збыыпеного утворення глюкози (глгкогенолтз, глю-
конеогенез), зменшеного п поглинання тканинами через порушений транспорт
вуглевод1в до юптини (зменшена транслокашя в юптинну мембрану' й активацгя
ГЛЮТ-4), а також зниженого утворення глюкозо-6-фосфату (Г-6-Ф), а потгм !
використання цього метаболиту обмгну глюкози на всгх шляхах перетворення його
в юптин!. Пщвищення активност! фосфорилази ! глюкозо-6-фосфатази в печшц!
спричинюе посилене утворення глюкози г зниженс — глгкогену.
При цукровому дгабет! типу 1 порушуеться перетворення вуглеводгв на жири,
пригшчуються ГЛ1КОЛ13 ] мегабол 1чн1 процеси в цикл! Кребса
Акгивашя глюконеогенезу пояснюеться вщносним переважанням глюкокор-
тикощгв, як! шдукують синтез потр!бних для цього ферментгв.
Пперглткемгя, що е наслщком зазначених порушень, зумовлюе глюкозур!ю га
неферментативне глгктрування бглкгв шо е важливим мехашзмом патогенезу д!а-
бетичних ангто- та нейропат! й.
Порушення водно-мшерального обмшу при цукровому д!абетг е наслщком гшер-
глгкемп та глюкозурп. Пперглгкемгя, шо перевищуе нирковий поргг, спричинюе
глюкозурйо. Глюкоза як осмотично активна речовпна сприяе розвитку осмотич-
ного дгурезу та полгурЦ, яка зумовлюе зневоднення, що своею чертою активуе
ренш-альдостеронову систему ! затримку натргю. Гшергл!кем1я 1 гшернатр1ем!я
пр изводить до гшеросмп, яка посилюе спрагу та спричинюе полгдипст.
Для порушення жирового обмшу в раз! дефщиту шсулшу характерно зменшен-
ня утворення жиру з ву1левод:в ! ресинтезу триглшерид!в !з жирних кислот у
жировш тканини Посилення пшол!зу ; вих!д жирних кислот 13 жирово! тканини
пов’язаш з вщносним збы мнениям лшолггичного ефекту глюкагону, катехола-
мине, соматотропшу, тиреощних гормошв. Посилюеться також окиснення жир-
них кислот у тканинах. Кр!м пперлшацидеми для иукрового д!абету характерною
е дислшопротегдемтя у зв’язку з! зменшенням унаслщок дефщиту битка р!вня Л1-
попроте!д1в високо! щыьност! (ЛПВЩ) 1 вщносним збгтьшенням у крою вмкту
лшопротерцв низькоТ щыьност! (ЛПНЩ) ! лшопротещдв дуже низько! щыьност!
(ЛПДНШ).
Жирн! кислота у збыьшенш кшькост! надходять у печшку, яка збериас здат-
шсть до синтезу триглщерщцв при зменшенш утворення лшопротешв, що ! ство-
рюе передумови для ожир!ння печшки.
При цукровому д!абет! вщбуваються посилене утворення в печшщ та накопи-
чення в оргашзм! кетонових т!л (апетоапетат, р-г!дроксибутират. ацетон). Це по-
яснюеться тим, що ацетил-КоА, який у велиюй юлькост! утворюеться в процес!
розщеплення жирних кислот, не може повшстю перетворитися на цитрат! “згор!-
та” в цикл! трикарбонових кислот, осюльки метабошчна потужшсть останнього
обмежена ! введения в нього активно! оцтово! кислота порушене. Крш того, в!л-
буваеться сповыьиення ресинтезу жирних кислот з ацетил-КоА в печшщ, жи-
ровгй та шших тканинах унаслщок дефщиту7 НАДФН2, який виникае внаслщок
зменшення при недостатност! шсулшу швидкост! пентозофосфатного циклу. При
цьому ослаблена також актившсть ферменпв, як! синтезують жирн! кислота через
малоны-Ко А. Тому не використаний у синтез) жирних кислот ! не залучений до
циклу Кребса ацетил-КоА е джерелом посиленого кетогенезу. € дан! про участь у
цьому процес! глюкагону, який стимулюе фермент карштин-ацетилтрансферазу,
що прискорюе окиснення жирних кислот до утворення кетонових ты.
Порушення КОС у хворих на цукровий д!абег пов’язане з розвитком негазово-
го метабол!чного ацидозу через накоггичення кетонових ты. Частим 1 небезпеч-
ним гострим ускладненням иукрового щабегу типу 1 е кетоацидотична кома. При
цукровому д!абет! не тыьки пщсилюеться вироблення кетонових ты, але й пору-
шуеться !х утилтзащя. На тл! не дуже високо! гшерглгкем!! (18—50 ммоль/л) рН
знижуеться до 7,2—6,8. Вщчуваеться запах ацетону в сеч! та видихуваному повпр!.
Спостерпаються гшеросм!я, гшотенз!я, ол!гур!я. дихання Куссмауля. Сопор пере-
ходить у глибоку кому
Коли цукровий дхабет поеднуеться !з хворобами, як! спричинюють гшокспо,
може виникнути лакгат-апидемтчна кома (р!венъ молочно! кислота понад 2 ммоль/л).
Крим пригшчення свщомост! при цгй ком! спостерггаються ознаки серпево-судин-
но! недостатност! (тах!кард!я, артер!альна гшотенз!я); можливий розвиток шоко-
вого стану.
Лгкування !з застосуванням !нсул!ну супроводжуеться ризиком набухания
клггин мозку, осюльки може виникнути ГШ00СМ1Я позаюптинно! рыини в раз!
переходу глюкози в клггини гад впливом шсулшу. Це дуже часто е основною при-
чиною смерт! при д!абетичних кето- 1 ла ктат-аци дозах.
Порушення битового обмшу при цукровому д!абеп проявляеться не т!льки
пригншенням анаболгчних процеств, а й посиленням катаболизму бшкгв з вико-
ристанням дезамшованих амшокислот для утворення вуглевод!в (глюконеогенез).
Сповыьнення синтезу б!лка та прискорення глюконеогенезу зумовлюють нега-
тивний азотистий баланс. У зв’язку з порушенням белкового обмгну пригшчують-
ся пластичн! процеси, загоювання ран. утворення антитш. Пригшчення лшогене-
зу, активац1я лшолгзу 1 використання бглюв м’язгв у процес! глюконеогенезу
спричинюють зниження маси т!ла. Порушення синтезу б!люв на тлг мобйпзапгй-
но! гшерлшемп може спричинити жирову шфйгьтращю печшки.
Порушення гормонально!регуляцп при цукровому' д!абет! може позначитися на
псребпу ваптносп, призвести до патологп плаценти, мимовьзьних абортгв. У
зв’язку з порушенням проникносп плаценти для гормошв, ппоксгею виникають
ембрюпатн, шо спричинюють тяжк! вади розвитку плода.
Значно зменшують тривалють життя хворих на 13ЦД мйсроангюпати, яю уш-
коджують судини мгкроциркулягорного русла. Частине це судини нирок1 ставки
очей. М!кроанг!опат!я проявляеться склерозом, обл!терац!ею та !ншими уважен-
иями кровоносних судин. Провщними факторами в генез! цих ускладнень еме-
таболгчн! порушення. Так, гшерппкемгя супроводжуеться пшвищенням концент-
рат"! глгко- й мукопротеццв. яю накопичуються у сполучшй тканин!, зумовлюючи
утворення палшу й ураження судинно! стшки.
Тривала гшерглгкемгя спричинюе глпарування б!дюв плазми г стшки судин,
що порушуе гх функцп. Це стосуеться 1 колагену базально! мембрани судин. Кр!м
того, при цьому активуеться сорбгголовий шлях перетворення глюкози з утворен-
ням осмотично активних сорбгголу 1 фрукгози, шо спричинюе набухания та уш-
кодження юптин. Активуються макрофаги, як! секретують цитокши. Можливе
утворення 1мунних комплексов
Основними про я вами дгабетично! м1кроанпопата е нефропания з розвитком
хрошчно! нирково! недостатност! як основна причина передчасно! смерт! хворих
на 13ЦД, ретинопагтя, що часто зумовлюе розвиток слшоти.
3 м!кроангюпат!ями псно пов’язаш дгабетичш нейропати, яю проявляються
уражсннями рухових, чутливих 1 вегетативних нерв!в. головним чином у зв’язку з
ушкодженням швашвських юптин 1 дем!ел!шзап!ею нервових проводники. Ме-
хашзми нейропатш зумовлеш м!кроанпопат!ею хаьа пегуошш та утворенням в
умовах д!абетично! гшерглпсемп токсичного для шваншських юптин сорбполу.
Цукровий д!абет типу 2
Ет1Олое!я । патогенез. 1НЦД — це велика грука лахворювань р1зних за етюлогкю /
патогенезом, як! характеризуються гтергл1кем1ею на тл! гтерйзеулшеми, що, як
правило, не шддаеться корекцц шеулшотерагаею. В основ! ШЦД лежить первин-
на резистентшсть до шсулшу. тобто знижена чутливкть печшки та м’яз!в до ш-
сул!ну, та/або знижена чутливкть В- ! А-юптин остр!вщв Лангерганса до глюкози.
1нсул!но- та глюкозорезистентшсть може бути генетичною або набутою, рецеп-
торною, пострецепторною 1 дорецепторною.
Незважаючи на вщеутшеть генетичних маркертв у вигляд! антигешв ГКГС,
цукровий д!абет типу 2 мае значно б!лыпу спадкову зумовлешеть, юж 13ЦД (схе-
ма 15). Конкордантшсть у монозиготних близнююв сягае 100 %, тод! як у раз!
Схема 15. Етюлопя 1 патогенез цукрового дгабету типу 2
цукрового диабету типу 1 — лише близько 50 %. Антитгл до В-ендокргшоцитгв та
(нсулшу немае. Спадкова схильшсть до цукрового дгабету типу 2 рсалгзуеться при
перещанш, алгмеНтарному ожиршнг.
Хвороба виявляеться шсля 40 рокгв г характеризуеться вщносним дефгпитом
шсулгну та мппмальними метаболгчними порушеннями.
Основш причини шсулгно- та глюкозорезистентностг:
1) рецепторнг: недостатшсть шсулшових рецепторгв або гх нечутливгсть до
шсулгну;
2) пострецепторш: порушення передач] сигналу вгд шсулшових рецепторгв до
внутргшньоктгтинних ферментних систем;
3) дорецепторнг:
а) панкреатичнг:
• МО ОУ (2-й пгдтип) пов’язаний з автосомно-домгнантним успадкуванням
дефекту глюкокшази у В-клгтинах, що зумовлюе зниження гх чутливостг до глю-
кози;
• зниження чутливосп В-клггин до глюкози може бути пов’язане г з вгдсутнгс-
тю або дефектом ГЛЮТ-2 на гх мембранг;
ИГ
• накопичення амЁлЁну й амЁлоТду в острЁвцях знижуе чутливЁсть В-клётин до
стимулюючих впливёв глюкози Ё, можливо, А-клётин — до ц гальмЁвних впливёв;
• вщсутшсть О-клётин усувае гальмування А- Ё В-клггин;
• нечутливЁсть А-клЁтин до гальмЁвних впливёв шсулшу чи глюкози;
• порушення структури прошсулшу.та / або порушення його протеолЁзу зё
зменшенням кёлькостё активного шсулшу;
• генетичнЁ аномалп структури шсулшу, що зменшують його зв’язування з
рецептором на клЁтинах-мЁшенях;
б) позапанкреатичнЁ:
• вироблення антитЁл до шсулшу;
• активащя протешаз 1 прискорений протеолЁз ЁнсулЁну;
• дефЁцит хрому, селену та ванадЁю (див. роздЁл XXIII “ПатофЁзЁологЁя обмЁну
мЁкроелементгв”).
Рецепторна резистентшсть до ЁнсулЁну найчастЁше пов’язана зё спадковим
зменшенням кёлькостё рецепторЁв або спорЁдненостЁ Ёх до ЁнсулЁну. Ця форма зок-
рема спостерЁгаеться при ЁнсулЁнорезистентному цукровому дЁабетЁ, поеднаному з
пЁгментно-судинною дистрофЁею шкЁри. Рецепторна резистентшсть виникае та-
кож у разЁ утворення антитЁл, якё блокують зв’язування ЁнсулЁну з рецептором, при
генетичному порушеннЁ амЁнокислотного складу ЁнсулЁну або його рецептора.
Велика, але досё маловивчена роль у виникненнЁ ЁнсулЁнорезистентностЁ на-
ложить генетично зумовленим порушенням передач! стимулюючих сигналЁв вёд
зовнёшньоё частини ЁнсулЁнових рецепторЁв до внутрЁшньоклЁтинних ефекторних
систем. Так, надлишкова експресЁя гена Кас! призводить до блокади ЁнсулЁновоЁ
активацЁЁ ё транслокацп на поверхню цитоплазматичноЁ мембрани ГЛЮТ-4, що
порушуе транспорт глюкози в м’язовё та жировЁ клётини. Надлишковий синтез
а2-8Н-глЁкопроте‘Ёну або глЁкопротешу РС-1 Ёнпбуе в скелетних м’язах тирозин-
кЁназну актившсть ЁнсулЁнових рецепторЁв ё вёдповёдно блокуе ефекти ЁнсулЁну.
Припускають можливёсть зменшення кёлькостё ГЛЮТ-2 в печЁнкових клЁтинах,
що обмежуе потрапляння в них глюкози.
Факторами, що провокують виявлення генетичних порушень вуглеводного
обмену Ё розвиток 1НЦД, е перещання й ожирЁння. Проте остаточно не з’ясованим
залишаеться питания про те, що е первинним — ожирЁння чи ЁнсулЁнорезистент-
нёсть. 3 одного боку, передумовою ожирЁння мае бути пперглЁкемЁя, гЁперЁнсулЁн-
емЁя та достатня чутливЁсть адипоцитЁв до ЁнсулЁну. ГЁперЁнсулЁнемЁя е наслЁдком
ЁнсулЁнорезистентностЁ, яка при цукровому дЁабетЁ типу 2 зменшуе вплив ЁнсулЁну
на глЁкогенез у печЁнцЁ та м’язах ё майже не впливае на стимуляцЁю ЁнсулЁном
лЁпогенезу. Тому за умови ЁнсулЁнорезистентностЁ гепатоцитЁв ё скелетних м’язёв ё
вЁдсутностЁ ЁнсулЁнорезистентностЁ жирово'1 тканини виникае ожирЁння, яке спри-
чинюе ЁнсулЁнорезистентшсть адипоцитЁв, оскёльки збЁлыпуе вЁдстань мёж рецеп-
торами до ЁнсулЁну на адипоцитах, що пригшчуе гх чутливЁсть до гормону Ё е за-
хистом вёд подалыпого ожирЁння. При первиншй ЁнсулЁнорезистентностЁ адипо-
цитёв 1НЦД не супроводжуеться ожирЁнням. 3 Ёншого боку, адипоцита при
ожирЁннЁ виробляють велику кёлькёсть ФНП-а, який блокуе тирозинкЁназну ак-
тивнёсть ЁнсулЁнових рецепторЁв м’язових ё жирових клётин Ё мае цитотоксичну
Д1Ю на В-КЛЁТИНИ острЁвцЁв пЁдшлунковоЁ залози.
У значшй частиш випадкЁв цукрового дЁабету типу 2 первинною е не ЁнсулЁ-
норезистентнЁсть, а знижена чутливЁсть В-клётин до глюкози (див. вище). При
виникненш глюкозорезистентност! В-кл1тини починають секретувати шсулш
тшьки тод!, коли ртвень глюкози пщвшцуеться значно вище нормального. Якщо
при вьому (наприклад у раз! накопичення в остртвцях амшшу) зменшуеться галь-
мтвний вплив глюкози та / або шсулшу на синтез глюкагону, ртвень глюкози
посттйно залишаеться високим, що стимулюе секреппо шсулшу. Гшертнсулшемтя
г гшсрглткемтя спричинюють ожиртння й шсулшорезистентшсть. Це саме вщбу-
ваеться 1 в раз] зменшення кшькостг або активностт Э-кгитин.
Порушення функцп панкреатичних ендокриноципв та тнсулшорезистент-
н.сть, яка посилюе навантаження на шсулярний апарат, призводять до того, що
вщносна шсулшова недостатшсть, яка е характерною для цукрового дтабету типу
2, згодом переходить в абсолютну (див. схему 15).
Для ШЦД харакгерним гострим ускладненням е ггаерглткемтчяа (пперосмоляр-
на) кома, що розвиваеться в людей лпнього вгку, як не отримують необхщш лт-
кування 1 догляд. Кома виникае на тл! значно! гшерппкеми (55—200 ммоль/л) !
ппернатртем!!. Осмолярнтсгь позакл!тинно! рщини збшыпуеться до 500 мосмоль/л,
що е наслщком гшерглткемл, ипернатртемп та згушення кров! за неможливостт
адекватно! компенсащ! втрат води при цтабетичюй полтуртт. Гшеросмолярюсть
позаклтгинно! рщини зумовлюе втрату води кл(тинами, що порушуе Гх дтяльшсть
1 в першу черту нормальне функцюнування нейрошв головного мозку.
Мггогенний вплив ппершсулшемй на фтбробласти, макрофаги та
гладком’язов! клттини стшки судин, гшерхолестеринем1я, гшерлшщемтя за раху-
нок ЛПНЩ т ЛПДНЩ при зниженнт ртвня ЛПВШ та глтктруванн! апопротетшв
лшопроте'цив, шо збшьшуе !х атерогеннкть, а також активан1я агрегацп тромбо-
циттв спричинюють прискорений розвиток атеросклерозу, який уражуе судини
великого д таметр а — макроангюпати.
1шем1чна хвороба серця, порушення мозкового кровообпу та облттеруючий
атеросклероз аргертй нижнтх кшщвок, який зумовлюе !х штемтю та некроз, слос-
герпаються за будь-якого типу цукрового .набегу. але особливо тяжких форм т
фатальних наслщкпв вони набувають при цукровому дтабет! типу 2. Дуже часто
хвор] на 1НЦД вмирають вщ шфаркту мюкарда або шсульту.
Основним принципом пагогенетично! терага! цукрового дтабету типу 1 е зам!сна
гераптя шляхом парентерального введения шсулшу. Здшснено спроби трансплан-
тацц р-ендокриноцитгв або пщшлунково! залози. Однак вони дають тимчасовий
ефект, як ! використання штучно! пщшлунково! залози. При цукровому дтабетт
типу 2 ефективними е пероральнт глюкозознижувальнт препарата (сульфоншсе-
човина, б(гуанщи), якт збыьшують секрецпо шсулпгу, потеннпоють дно власного
шсул!ну, знижують актившсть протешаз, звтльняють шсулш вщ зв’язк’:в з битками
плазми.
Нов! даш про тмунн! мехашзми ушкодження р-ендокриноципв за участю
МО зумовили спроби лткувати цукровий дтабет вщповщно тмунодепресантами, а
також нткогинамщом, амшогуанщином, якт гнпбують експрешю МОЗ й 5творен-
ия МО.
Розд1Л XIX
ПОРУШЕННЯ ЖИРОВОГО ОБМ1НУ
Порушення жирового (лгпщного) обмгну, зокрема обмшу триглщеридш г виших
жирних кислот, може виникати внаслщок розладу всмоктування, видшення г
транспорту жиргв, надлишкового накопичення жир)в у тканинах, що не належать
до жирово! (жирова шфгльтрацгя 1 жирова дистрофгя), порушення промгжного
жирового обмшу, розладу обмшу жир) в у жировгй тканин! (наддишкове або недо-
статке гх утворення й вщкладення).
РОЗЛАД УСМОКТУВАННЯ I ВИДШЕННЯ ЖИР1В
Одюею з основних умов, що забезпечують нормальне всмоктування жиргв, е гх
емульгування, розщеплення на глщерин г жирш кислота й утворення сполук гз
жовчними кислотами (холешатгв). Емульгування жиргв вщбуваеться за певного
спгввщношення жовчних, жирних кислот г моноглщеридгв. Два останы компо-
ненти утворюються внаслщок розщеплення жиргв лшгвальною, шлунковою г
панкреатичною лгпазами. Отже, дефщит лшази, що виникае в першу черту в раз)
хвороб пщшлунковог залози (панкреатит, гострий некроз, склероз), чи недостат-
шсть п активацп при патологи печшки, жовчного м)хура (обтурацгйна жовтяниця,
бшгарний цироз печгнки) супроводжуються порушенням усмоктування жиргв. У
цьому випадку вмгст жиргв у калг ргзко збшьшуеться, спостерггаеться стеаторея.
Стеаторея може виникати г пщ час лгкування антибготиками (неомщин, хлортет-
рациклш), якг притнгчують лшолгз. У разг надлишку в !жг кальцпо г магнгю пору-
шуеться всмоктування жирних кислот — утворюються нерозчиннг у водг солг
жирних кислот (мило), якг виводяться через кишки.
Усмоктування жиргв гальмуеться у випадку ураження епгтелгю тонко! кишки
шфекцшними ) токсичними факторами, авгтамшозу А г В через порушене утво-
рення ферментгв, якг беруть участь у ресинтезг триглщеридгв. На резорбцп жиргв
позначаеться г дефщит холшу.
За нормальних умов в органгзмг зазвичай засвоюеться майже 95 % жиргв зг
спожитих. Приблизно 5 % жиргв виводиться в основному через кишки г меншою
игрою — сальними г потовими залозами. У разг споживання велико! юлькостг
жирно! гжг, а також при розчавленнг кгсткового мозку, ушкодженнг великих дшя-
нок жирово! тканини, лгпощному нефрозг спостерггаеться лгпургя.
Наслщком порушення всмоктування жиргв е якгсне голодування.
ПОРУШЕННЯ ТРАНСПОРТУ IДЕПОНУВАННЯ ЖИР1В
Ресинтезоваш в кишковш стшщ жири надходять у л!мфатичну систему, попм
плечо-головш вени 1 циркулюють у кров! у вигляд! х!лом!крон!в, що мктять
близько 1 % бшюв 1 99 % л!пщ!в. Перший орган, у якому частина хыомпсрошв
затримуеться, — леген!. 1м властива лшопектична функц!я, що регулюе надхо-
дження жирш в артергальну кров. Якщо дихалъна функщя легень обмежена (пнев-
моторакс, емф!зема), жири затримуються в них. Збшьшення дихально! поверхн!
легень, прискорення кровотоку (наприклад у сшваюв) у раз! недостатнього роз-
витку мезенхгмних елеменпв легень спричинюють посилене надходження лшщГв
в артер!альну кров 1 вщкладення !х у жировш тканин!. Частина х!лом!крон!в
розщеплюеться лшопротещною лшазою (фактор просвгглення), що синтезуеться
в ендотелп кап!ляр!в жирово! тканини, легень, серця, м’яз!в, печшки, у молочнш
залоз! при лактацп та в меншш юлькост! в шших органах 1 надходить у кров пщ
впливом жиргв 1 гепарину. Неестерифшован! жирн! кислоти (НЕЖК), що утворю-
ються при цьому, адсорбуються на альбумин та р-лшопротещах 1 транспортують-
ся в органи 1 тканини. Частина НЕЖК у печшц! ресинтезуеться в триглщериди,
а частина використовуеться органами 1 тканинами, переважно серцем 1 частково
скелетними м’язами як джерело енергп. Жири транспортуються кров’ю не т!льки
1з кишок до органы 1 тканин, а й вщ жирово! тканини до печшки та шших ор-
ганов.
Таким чином, кров мае х!лом!крони, що надходять 1з кишок 1 мктять тригл!-
цериди, НЕЖК — !з жирово! тканини, легень та шших органы з високою актив-
Н1СТЮ лшопротещно! лшази та лшопротещи (ЛП) р!зно! щыьност!, що надходять
з печшки. Це ЛПДНЩ, що транспортують ендогенш триглщериди до оргашв !
тканин, ЛПНЩ 1 лшопротещи помхрно! щыьност! (ЛППЩ), як! переносить хо-
лестерин до периферичних тканин, ЛПВЩ, як! транспортують надлишок холе-
стерину 13 судин 1 тканин до печшки.
Одним 13 показники порушення жирового обмшу е пперлшемы (понад 3,5—
8 г/л). Вона може бути ал!ментарною (харчовою), що супроводжуеться хыомжро-
немкю, транспортною, або мобшзацшною (у раз! перемпцення жирних кислот !з
жирово! тканини в печ!нку), 1 ретенцшною (унаслщок затримки розщеплення
лшопроте'ццв у кров! в раз! нестач! секрецп або втрати лшопротещно! лшази, на-
приклад при постгеморапчнш анемп або нефротичному синдром!).
Частине гшерлшемП е тимчасовими. Бшьше значения мають тривал! пперлшоп-
ротещеми, пов’язаш з! спадковим збшьшенням р!вня ЛПДНЩ 1 ЛПНЩ, що мають
атерогенш властивосп. У 1970 р. ВООЗ ухвалила класифгкацпо гшерлшопротещемп
за п’ятьма класами залежно вщ типу лшопротещы, причин 1 мехашзмы виникнен-
ня, !мов!рност! розвитку атеросклерозу (див. “Атеросклероз”, роздш XXVI).
Кр!м гшерлшопротещемп спостеркаються гшо- та дислшопротещемп. Прикла-
дом може бути абетсшпопротегдешя, або акантоцитоз (синдром Бассена — Корн-
цвейга), що виникае як наслщок автосомно-домшантного порушення утворення
апопроте'шу В та ЛПДНЩ, ЛППЩ, ЛПНЩ 1 супроводжуеться невролопчними
порушеннями, мальабсорбщею й акантоцитозом (див. “Гемолпичш анемп”, розды
XXV). Аналъфапротегнешя — автосомно-рецесивна хвороба, пов’язана з вщсутшстю
синтезу апопротешу А та ЛПВЩ (тенжерська хвороба). При щй хвороб! порушуеть-
ся транспорт холестерину вщ тканин до печшки, виникае гепатоспленомегал!я.
ПпобетампонротеЮемш успадковуеться за автосомно-домшантним типом,
супроводжуеться зменшенням ртвня ЛПНЩ 1 гальмуванням розвитку атероскле-
розу
Жирова шфпьтрашя 1 дистрофгя. Жири, то надходять у тканини, окиснюють-
ся або депонуються- Якщо накопичення вщбуваеться поза кштинами жирово)
тканини, це евщчить про жпрову шфгльтращю. Поеднання шфгльтрацп з лору-
шенням структури цигоплазми цих кштин визначасгься як жирова дистрофгя.
Причиною жирово! шфшьтрацн нерщко е зниження активност! г!дрол!тичних
або окисних ферментгв (у разг отруення фосфором, миш’яком, хлороформом,
вгрусно! шфекцп, авгтамгнозу). Найчастгше спостерггаеться жирова шфтътрацы
печшки До не: може призвести будь-яке порушення, що дезштегруе обмш г син-
тез лгтпдгв у печшцг: 1) посилений лшогенез; 2) знижене окиснення жирних кис-
лот; 3) пщвищений лшолгз жировог тканини; 4) сповгльнене виведення ЛПДНЩ.
Продукцгя ЛПДНЩ у печпщт потребуе поеднання процесгв лшщного 1 бшкового
синтезу. Порушення синтезу бшка в разг навпъ мгнтмального синтезу триглще-
ридгв зумовлюе акумуляшю жиргв у печшцг, оскгльки порушуеться утворення гх
водорозчинних транспортних форм. Неповноцгнне харчування, дефгцит амино-
кислот । АТФ порушують синтез апопротегнгв г знижують утворення лшопро-
теццв. До такого самого наслщку призводить посилений лшолгз у жировгй тканин!
пщ час голодування або цукрового дгабету, коли порушуеться введения лшщних г
бшкових попередникгв в обмш лшопротещгв.
У патогенез! жировог шфшьтрацп великого значения набувае порушене утво-
рення фосфол гпгдгв. Достатнгй вмгст гх у печгнц! забезпечуе тонке диспергування
жиру г можливгсть видалення його з печшки. Фосфолшщи входять до складу р-
лшопротецпв г полегшують вихщ гх гз клгтин печгнки. Частина жирних кислот
бере участь в утворенш фосфолггццгв г в гх склад! виводиться з печгнки. Кргм того,
у молекул! фосфолшшгв жиров! кислота краше окиснюються. Необхщними ком-
понентами основного фосфолшщу печгнки — лепитину — е холш ! метгонгн, який
вщдае сво'г метильн! групи для його утворення. Тому нестача в !жг холшу, метгонг-
ну та шших лгпотропних речовин (гнозит, нуклегнов! кислота) спричинюе розви-
ток алтотропно! жирово! шфшътрацп печшки. До такого самого наслщку призво-
дить дефгцит ендогенного лшотропного фактора — лшокагну. який утворюеться
в епгтелп дргбних проток пгдшлунково! залози. Лшокаш активуе утворення фос-
фолшцйв у печшцг, окиснення в нш жирних кислот запобггае ожиршню печш-
ки. Дефгцит цього фактора вщгграе важливу роль у патогенез! ожиргння печгнки
у хворих на цукровий дгабет.
Порушення синтезу холшу можливе в раз! дефщиту шанокобаламшу. фолге-
во! та пантотеновог кислот.
ПОРУШЕННЯ ПРОМ1ЖНОГО ЖИРОВОГО ОБМ1НУ
Одним !з найважлившгих порушень промгжного обмгну жиргв е посилення кето-
генезу. Кетонов! тша, якг утворюються в пропес! [3-окиснення жирних кислот,
посглають одне з центральних мгспъ у систем! забезпечення органгзму енерпею,
конкуруючи щодо цього з глюкозою. У раз! унеможливлення використання глю-
кози як джерела енергп в органгзмг посилюються лгполгз г кетогенез, розвиваеться
кетоацидоз. Такий кетоацидоз можна спостертгати й у фтзюлопчних умовах (ф!-
зична праця, емоцгйне напряжения, шзн! термши ваптностг), але тодт вш не е
тривалим, ртвень кетонових ттл у кровт не перевищуе 0,1 мкмоль/л, осюльки вщ-
буваеться швидка утилтзацтя тх як енергетично! сировини (ф1зюлог1чний кетоаци-
доз). У раз! патолог1чного кетоацидозу утворення кетонових пл перевищуе утилт-
зацпо. Зазвичай це вщбуваеться внаслщок посилення лшолтзу в жировш тканин!,
коли печшка не використовуе вс! жирш кислоти для синтезу триглщеридтв 1 час-
тина !х залучаеться до процесу р-окиснення та кетогенезу. Таким е походження
кетоацидозу пщ час голодування, цукрового д!абету. За значного накопичення
кетонових т!л у кров! (понад 0,1 мкмоль/л, а шодт — до 20 мкмоль/л) виникае
загроза житгю внаслщок метабол!чного ацидозу.
ПОРУШЕННЯ ОБМ1НУ ЖИРШ У ЖИРОВ1Й ТКАНИН!
Жирова тканина характеризуемся високим р!внем метаболизму, штенсивним кро-
вопостачанням 1 е свого роду енергетичним акумулятором, що саморегенеруе.
Накопичення в шй енергп у вигляд! нейтральних жиртв вщбуваеться шсля кожно-
го споживання тжт, а мобтлтзацтя енергп — у будь-який час пщ впливом тмпульств,
що вившьнююм жирш кислоти.
Роль нервово! системи в регуляцп жирового обмшу пщмерджують дан! про
те, що тривале емощйне напруження зумовлюе мобшзацпо жиру тз жирових депо
1 зниження маси т!ла. Те саме спостерггаеться т гцд час подразнення симпатичних
нервгв. Десимпатизащя перешкоджае виходу жир!в !з депо. Подразнення пара-
симпатичних нервгв супроводжуеться вщкладенням жиру.
Якщо протягом тривалого часу накопичення енергп перевищуе п витрати,
розвиваеться ожиргння.
Жировим клттинам властив! вег три шляхи метаболтзму — глгколгз, пентозний
цикл г цикл Кребса, у яких здшснюються синтез жирних кислот, лшогенез г лг-
полтз. На мал. 42 показано роль нервово! системи г гормонтв у депонуваню жир!в
т лшолтзт.
Лшолтз запускаемся адреналшом т глюкагоном за допомогою цАМФ, що ак-
тивуе триглгцеридну лшазу. 1з гормональних факторгв, яю справляють жиромо-
бпйзуючий ефект, слщ також вщзначити соматотропш, тиротропш, трийодти-
ронш т тироксин. Вщомо, що в перюд посиленого росту, а також у раз! пперти-
реозу настае значне зниження маси тша. Стимулюе лшолтз також видшений з
аденогшофтза р-лшотропш.
Глюкокортикощи сприяюм посиленню мобйпзацп жиру гз жирово! тканини т
гальмуюм литогенез. Але цей ефект може перекриватися гншими: здатшетю спри-
чинювати гшерглткемпо та стимулювати секрецпо шсулшу, який активуе лшогенез
в адипоцитах та накопичення глткогену в печшцт, що зумовлюе гальмування мо-
бйпзацп жиртв т вщкладення тх у жировш тканинт; спроможшстю у високих дозах
затримувати жиромобштзуючу т стимулюючу окиснення жиртв дно соматотропшу.
Цим можна пояснити накопичення жиру в жирових депо у раз! гшеркортицизму
(синдром Кушшга та хвороба Тценка — Кушшга). Кортикотропш, стимулюючи
секрецпо глюкокортикощтв, може впливати на жировий обмш у тому самому на-
прямку, але, кртм того, справляе ще й екстраадреналову жиромобштзуючу дно.
Мал. 42. Нейрогуморальна регулящя жирового обмшу: фактори, що спри-
яють депонуванню (а), 1 фактори, що зумовлюють мобшзащю жир!в (в)
Литогенез (анаболтчш процеси в лшоцитах) регулюеться шсулшом. 1нсулш
стимулюе синтез нейтральних жиртв з глюкози 1 жирових кислот, гальмуе лйтолтз,
знижуе ртвень глюкози в кров!, що посилюе апетит. На мал. 43 показано роль
тнсултну й адреналшу в метаболтзм! жиру в лшощгп ! сшввщношенн! процеств
литогенезу та лшолтзу.
Вплив гормошв на метаболтзм жиру в лшоцитах здшснюеться через специфтч-
нт рецептори. Дефтцит рецепторхв для шсулшу або зниження тх чутливост! може
бути причиною резистентност! тканин до нього та шдукувати гшерглткемпо г п-
першсулипзм, як! будуть супроводжуватися зниженням маси ттла.
Синтез г розпад триглщеридтв регулюються також ртвнем глюкози в кровь
Якщо створюеться надлишок глюкози, збйтыттуеться секрецтя шсулшу г частина
НЕЖК як друге джерело енергп вилучаеться з обпу 1 вщкладаеться в жирових
депо; при дефщит! глюкози з депо мобшзуються жири. Цей процес саморегуляци
е однтею з ланок у складнш систем! регуляцн жирового обмшу, що здшснюеться
нервовою та ендокринною системами.
Мал. 43. Вплив шсулшу й адреналГну на лГгогенез 1 лшолГз у лшо-
цит1 в норм) (а) та при ожиршш ((Г) (за Я. Татонь, 1981):
ВЖК — в1льн1 жирн! кислоти; Жк — жирш кислота, Тг — триглще-
риди: Глю — глюкоза; Гл1 — глщерин; X — лшогенез, У — лшолГз
Ожиршня — надмГрне вщкладення жиру в жировгй тканиш. Серед дорослого
населения економгчно розвинених крапг вщ 30 до 60 % осгб мають масу пла, що
на 20 % г бгльше перевищуе норму. Ожиршня частгше розвиваеться в жзнок, пе-
реважно теля 50 рокгв.
Класиф|кацш. Залежно вщ того який фактор е провщним у виникненю ожи-
ршня, розргзняють ожиршня переинне (алгментарно-конституцюнальне) 1 вторин-
не, ибо симптоматичне (церебральне й ендокринне). За сучасними уявленнями
первинне ожиршня пов'язане зг спадковим порушенням синтезу адипоцитами леп-
тину або зг зниженням чутливосп до нього ппоталамуса.
За пагогенетичною класифгкашею ожиршня, що Грунтуеться на критергях
розмгргв г кшькосп лшоцит1в, видшяють два види ожиргння: ппертрофгчне й п-
перпластичне.
Ппертрофгчне ожиршня залежить вщ юлькосп жиру в кожному лшоцип, шо
взаемопов язано з пщвищеною коннентрашею шсулшу, пперлшемгею та ппер-
глгкемгею. Збщьшення маси пла при такому ожиршш обмежене кгльюстю лшо-
цитгв та !х максимачьним об'емом.
Пперпластичне ожиршня пов’язане зг збгльшенням кшькосп лшоципв, що
залежить вщ генетичних факторов або впливгв, якг регулюють морфогенез жирово!
тканини в ембргональний перюд г ранньому дитинсгвг, наприклад пперглгкемгя у
ваптно!' або перегодовування дитини в перпп мгсяцг життя. Новоутворенггя ади-
попитгв можливе також г в препубертатний та преклгмактеричний перюди. Гшер-
пластичне ожиршня виникае рангше, може супроводжуватися дуже значним нал-
лишком маси тша та е бглып резистентним до щетотератш.
За клшгчними ознаками видгляють: андроТдний (яблучнии) тип ожиршня, коли
жир вщкладаеться на верхнш половинг гулуба та животг. а також у чепцг та брижг
"зйГ
(спостерхгаеться частице в чоловххов) та гшсйдний (грушопод!бний) — у нижшй
половин] ту.туба, стегнах (у жшок). Андрощний тип ожиршня частгше усклад-
яюеться атеросклерозом ! цукровим дебетом типу 2.
Експерименталъне ожиршня. € кхлька лшхй мишей, у яких ожиршня вияв-
тяеться як генетичне порушення, що зазвичай успадковуеться за автосомно-реце-
сивним типом. У мишей зх спадковим ожирхшхям спостерхгаються гшерглхкемхя,
гшердшемхя, посилення лшогенезу, гшершсулшемхя; виявлено пщвищення чут-
ливост! до гшерглхкемхчнох дп соматотрошну х глюкагону. Ферментативш змши в
ештелюцитах печшки (гепатоцитах) свщчать про збхльшене використання фосфа-
тоглщеролу для лшогенезу.
Списано л! нею жовтих мишей, у яких поеднан! двх властивосп: жовте забар-
влення шерсп х схильшсть до ожиршня. Для мишей характерними е шдвищене
сложивання вуглеводхв. резистентшсть до шсулшу за нормально? глхкемп. У ми-
шей друго’х лшп ожиршня । гшерглхкемхя, яю розвиваються на другому мхсяц)
життя, поеднуються з пперхолестеринемхею, тпдвищенням чутливост! до дп сома-
тотропшу та глюкагону. У цих гварин виявляютъся ппертроф)я острхвщв п!д-
шлунково] залози та ппершсулшемхя.
Порушення балансу мхж надходження м ! витрачанням енергп може бути
спричинене впливом на центри гшоталамуса, з якими пов’язане вщчуття голоду
та ситост!. Електролхтичне руйнування вентро медхальн их ядер — центру ситостх,
який пригшчуе дхяльнхсть травного центру у щурхв, юшок х мавп, спричинюе п-
перфапю з подальшим розвитком ппоталамхчного ожиршня. Такий саме ефект
зумовлюе аурот’юхлюкоза. яка вибхрково уражуе вентромедхальн! ядра. Експери-
ментальне гшоталамхчне ожиршня е аналогом гшоталамхчного ожиршня в людей,
яке може розвиватися внаслщок перенесеного енцефал!ту, меншпту, травми го-
ловного мозку.
] ормональне ожиршня в експеримент! розвиваеться теля видалення щито-
подхбно! залози Ожиршня шсля кастрацп пов’язане з гшертрофхею кхрково! речо-
вини надниркових залоз, що розвиваеться через потребу в компенсаци 1 ормональ-
них порушень, 1 гхпершеулшхзмом. При цьому активуються процеси глюконеоге-
незу х псретворення вуглеводш на жири. Можливо, таке саме походження мае
ожиршня в жшок у перюд кд!максу.
Екзогенне ожиршня моделюеться обмеженням рухово! активност! тварин !
перегодовуванням. особливо вуглеводним.
Етюлопя. Ожиршня е наслщком порушень енергетичного обмхну. У виник-
ненш його вщхграють роль внутрхшш та зовнхшн! фактори, якх змшюють поведш-
ку людини шодо харчування. Надларне споживання 1ж1 е основною умовою виник-
нення ожиршня, а причиною — порушення регуляцн харчовоТ поведшки. Фактори, що
регулюють алхментарну поведшку людини, визначаються и генетично-конститу-
цхональними особливостями. Мають значения також харчування матер! в лерход
ваптностх, дитини — у рахшьомудитинствх; типи безумовних рефлекс в, пов’язаних
!з харчування м; родиннх, нащональн! традицп; рхвень материального забезпечен-
ня; рухова актившсть.
Надмхрне споживашгя хжх, зумовлене посиленням апетиту, може бути наслщ-
ком тдвишено! збудливост! травного центру, розмхщеного в передньобхчних яд-
рах задньох гшотатамхчно! дезянки.
Змши, на якё реагуе травний центр, можуть бути причиною тривалого харче-
вого збудження ё внаслщок цього — алЁментарного ожиршня. Так, будь-який
стан, коли стёйко знижуеться рЁвень глюкози в кровЁ, наприклад посилення фун-
кци острЁвцЁв пщшлунково! залози, супроводжуеться вщчутгям голоду, що зумо-
влюе можливёсть пере'щання.
У функцп травного центру мае значения також ЁмпульсацЁя, яка надходить з
рецепторЁв травного каналу. Розтягнення шлунка до певного ступеня гальмуе ак-
тившсть травного центру. У раз! зниження чутливост! нервових закЁнчень у стёнцё
шлунка гальмування центру вщбуваеться лише теля надм!рного його розтягнен-
ня, що також створюе передумови для пере’щання й ожир!ння.
Порушення балансу енергп можливе 1 теля переходу вщ фЁзично! пращ до
способу життя, який не потребуе значного ф!зичного напруження, якщо поперед-
нш ступЁнь збудливост! травного центру зберЁгся. За нормально! функцп травного
центру ожиршня може бути пов’язане з порушенням мобЁлЁзащ! жирЁв Ёз жирових
депо — джерела енергп.
Як зазначалося вище, регуляция процесу мобйпзацй та вщкладення жирЁв
здшснюеться нервовою та ендокринною системами. Так, зниження тонусу сим-
патично! нервово! системи може спричинити затримання мобйпзацп та виходу
жирЁв Ёз жирово! тканини. У кёшок теля однобЁчного перер!зування черевного
нерва в раз! голодування кёлькёсть жиру в навколонирковш клётковинё виявилася
бЁльшою на денервованому боць
При хвороб! Барракера — С!монса — прогресивнш лшодистрофп, що пов’язана
з ураженням центров промгжного, спинного мозку ё вузлЁв симпатичного стовбу-
ра, спостерггаеться зникнення жиру в жировш тканин! в дёлянцё голови ! трудно!
кл!тки з одночасним вщкладанням його в нижнш половин! т!ла.
Порушення мобЬпзуючого впливу гормошв спостерЁгаються при патологп п-
поф!за, щитоподЁбно!’, надниркових Ё статевих залоз.
Однак спЁввщношення мёж генетичними та зовнёшнёми факторами за р!зних
вид!в ожиршня неоднакове. Первинне ожиршня мае в основ! полЁгенне успадку-
вання ! спричинюеться надмЁрним сложиванням !жё та гЁподинамЁею. Успадкува-
тися можуть структура ! функцп систем, як! регулюють алЁментарну поведшку,
особливост! метаболЁзму лшоципв Ё мёоцитёв. Про це евщчать результата спосте-
режень за розвитком ожиршня в юлькох поколениях одше! родини. Виявлено
високу конкордантшеть за щею ознакою в однояйцевих близнюкЁв. Вщкрито “ген
ожирЁння” та його продукт — лептин — пептидний гормон адипоцитЁв (ФрЁдман
та спЁвавт., 1994), який видЁляеться за достатньо! кёлькостё в них жиру, зв’язуеться
з клЁтинами вентромедЁальних ядер ппоталамуса Ё зумовлюе вЁдчуття ситостё. За
допомогою глюкагоноподЁбного пептиду I вён зменшуе секрецЁю нейропептиду У
у вентролатеральних ядрах ппоталамуса, що пригнЁчуе апетат.
У дЁтей у родинах з ожирЁнням лептинову недостатшсть виявляють у 70—80 %
випадкЁв: у 20 % спостерЁгають абсолютну лептинову недостатшсть, у 80 % — п-
перлептинемЁчну форму з вёдносною лептиновою недостатнЁстю. Це збЁгаеться з
даними експериментЁв. Зокрема Ёснують двё форми генетичного ожирЁння в ми-
шей: лептинопенЁчна, пов’язана з дефектом гена лептину, та гЁперлептанемЁчна,
за яко! рЁвень лептину нормальний або пЁдвищений, але на нейронах ядер гЁпота-
ламуса недостатня кёлькёсть рецепторЁв до лептину.
В адипоцитах виявлено також цитокш кахексии (ФНП-а). Це протишсулшо-
вий фактор, який гальмуе литогенез, пригнгчуе апетит, активуе катабол1зм, бере
участь у мехашзм! шсулшорезистентносп.
Лептин, кахексии, а також гормони травно! системи (грелш та пт.), що ре-
гулюють апетит, е складниками нейрогуморально! регуляцп маси пла, яка отри-
мала назву “лптостат”.
Лтостат — це саморегуляц!я маси тша шляхом взаемодп гшоталамуса з ади-
поцитами та гормонами ентерально! ендокринно! системи. Виходячи з цього по-
ложения, основним мехашзмом первинного ожиршня е генетично зумовлене по-
рушення взаемодп мтж центральними та периферичними ланками лшостату,
унаслщок чого установочна точка маси ттла перемпцуеться на вищий ртвень, !
нормальна маса тша сприймаеться як знижена, що активуе алтментарну поведпт-
ку людини. Нашмовтрнппе, що генетичний дефект лшостату пов’язаний з абсо-
лютною чи вщносною лептиновою недостатшстю. При цьому людина втрачае
вщчуття ситосп, активуеться секрецтя нейропептиду У в травному центр! (вент-
ролатералып ядра гшоталамуса), що призводить до надм!рного споживання !ж!
(схема 16).
Вторинне ожиршня теж пов’язане з чинниками, шо порушують баланс енер-
гп, але воно виникае за первинно нормальних взаемозв’язктв гшоталамуса й ади-
ПОЦИТ1В.
Схема 16. Мехашзм розвитку первинного ожиршня
Основш вида вторинного ожиршня — це ендокринне та церебральне. Ендо-
кринне ожиршня розвиваеться при деяких ендокринних захворюваннях унаслщок
зниження обмшу речовин (ппофункщя щитопод!бно! 1 статевих залоз), надм!рно-
го перетворення вуглеводтв на жири (гшершсулпмзм), порушення жиромобппзу-
ючого впливу гормошв (гшофункц1я ппоф1за, надниркових залоз).
Церебральне ожиршня спостерггаеться при травмах, пухлинах мозку, ениефал!-
тах, меншптах.
Ожиршня можливе також шсля отруення колощним золотом (пщ час лгку-
вання ревматоидного артриту), миш’яком, у раз! тривалого вживания р-адренобло-
каторгв, сульфаншамщних препаратгв. Колощне золото впливае на центри си-
тост!; мехашзм дц шших речовин не з’ясовано.
Основним у патогенез! ожиршня е первинне або вторинне порушення лшо-
стату. Ожиршня несприятливо впливае на життедгяльшсть оргашзму. У молодому
виц, коли адаптащйю можливосп виражеш краше, негативна д!я ожиршня про-
являеться меншою м!рою. 3 вжом кшыастъ ускладнень, пов’язаних з ожиршням,
зростае. Смертшсть серед хворих з ожиршням у вщ! 20—24 роив на 3 %, а в
40—45 роив — на 50 % вища, н!ж в ос!б !з нормальною масою тша.
При ожиршю порушуеться обмш речовин, особливо в жировш тканиш, де пщ-
вищуеться швидкгсть синтезу триглщерщцв, порушуеться здатшсть до мобшзащ!
жирових резерв1в, спостершаються гшерлшем1я, пщвищення р!вня вщьних жир-
них кислот, г!перхолестеринем!я.
У людини з ожиршням можлива жирова шф!льтрац!я печшки.
Порушення вуглеводного обмшу проявляються обмеженням обману глюкози,
пщвищенням р!вня глткогену в печшпд. У м’язовш тканиш порушуеться утилиза-
ция глюкози, незважаючи на гшершсулппзм. Зниження дихального коефпцента
до 0,70—0,74 евщчить про те, що як джерело енергп використовуються переважно
жирш кислоти.
Порушення бокового обмшу визначаеться зниженням р!вня загального бшка за
рахунок альбум1шв, пщвищенням тромбогенних властивостей кров!, загальноТ ко-
агулящйно! активност! та ртвня фтбриногену, припиченням фтбринолтзу. Усе це
зумовлюе виникнення тромбозу.
У зв’язку з вщкладенням велико! кшькост! жиру 1 пщвищенням навантаження
на бшышеть життево важливих органш ожиршня спричинюе функщональш зм!-
ни в них, а також порушення метаболтзму.
Вщкладення жиру в мюкардт викликае значке зниження скоротливо! функцп
серця. Ожиршня часто супроводжуеться атеросклерозом, пщвищенням артерталь-
ного тиску, зсщання кров!, тромбозом. При цьому погтршуеться легенева венти-
ляц!я, зменшуеться життева емшеть легень, виникае схильшсть до застою кров! й
розвитку в дихальних шляхах хрошчного запалення. Задатка виникае навпъ пщ
час незначного ф!зичного навантаження. Розвиваеться циркуляторна й дихальна
ГШОКСТЯ.
Поеднання ожиршня тз цукровим д!абетом спостерггаеться в раз! шеулшоре-
зистентносп, пов’язано! зт збшьшенням продукцй адипоцитами кахексину, який
зменшуе чутливтсть шеулшових рецептсдав печшки, м’яз1В та шших органтв. Ком-
пенсаторна гшертрофгя 1 гшерплазтя В-клггин остртвшв пщшлунково! залози, за-
безпечуючи пщвишену секрецтю шсулшу (гшершеулштзм) для подолання резис-
тентностт, з часом змшюетъся !х виснаженням. Припускають. що в таких випад-
ках ожиртння е фактором ризику виникнення иукрового дтабету типу 2.
Еттотропна терашя хворих з ожиршням шляхом корекш! ендокринних т не-
вролопчних порушень ефектившша в разт вторинного ожиршня
Пато генетичну терашю, що полягае в норматтзацп енергетичного балансу,
застосовують за ртзних видтв ожиршня. Це зокрема пригнтчення апетиту (вико-
ристання аноректичних засобгв, я кт гальмують травтшй центр), збшьшення фтзич-
ного навантаження. зниження енергегично! щнностт тжт за достатнього п об’ему.
При абсолютнш лептиновш недостатностт ефективним е введения хворим лепти-
ну. Останнтм часом вдаються до хтрургтчних вгручань для зменшення об'ему
шлунка. Використовують також лткувальне голодування
Схуднення (виснаження) — це зниження жирового запасу оргашзму Крайнтй
ступтнъ виснаження, що супроводжуеться втратою 50 % маси ттла. називають ка-
хекаею- Екзогенна кахекстя виникае в разт голодування, ендогенна спостертгаеть-
ся при нейрогеннтй аноре ксп, пперпродукцп кахексину й шших речовин, що
пригнтчують центр голоду (травний центр) т стимулюють катаболтзм. Так, ракова
кахекстя, ранове та опткове виснаження пов’язаш з впливом кахексии)', цукровий
дтабет типу 1 — з втдсутнтстю анаболтчно! дц шсулшу та катаболтчним впливом
глюкокортикотдтв.
Виснаження спостерйаеться також при глюкагономт, соматостатином! як на-
слщок збтлынення лшолтзу в адипоцитах, а в раз! гшокортицизму — за умови
вщсутностт ди глюкокортикотдтв.
Л1П1ДОЗ
Лгтдоз — загальна назва спадкових або набутих хвороб, що належать до хвороб на-
копичення, характеризуютъся порушенням лпидного обмшу / зумовлеш дефектами
специфичных л!зосомних г'юролаз.
Хвороба Вольмана (хвороба накопичення ефтртв холестерину) — рщктсна авто-
сомно-рецесивна спадкова хвороба, яка в перпп тижнт супроводжуеться б.пован-
ням, дтареею зт стеатореею, гепатоспленомегаттею 1 двобтчним кальцинозом над-
ниркових залоз. Дтти здебтльшого помирахль у вщ! 6 мтс. Генетичний дефект —
втдсутнтсть кислот лшази лтзосом, шо призводить до накопичення ефтртв холесте-
рину т триглщеридтв у лтзосомах печшки, селезшки, надниркових залоз, гемопо-
етичнот тканини 1 тонко! кишки.
Хвороба Шюллера — Христгана харакгеризуеться втдкладенням холестерину та
його ефтртв у клттинах гранулящйнот тканини, шо розростаеться в ктстках т бтль-
шостт вттутртшшх органтв. Харакгерними е деструктивш змши в ктстках.
За деяких патолопчних стантв, якт успадковуються за автосомно-рецссивним
типом, спостерпаеться надмтрне вщкладання у тканинах фосфолпцдтв. Так, при
хвороб! Гоше у зв’язку зт спадковою вщсутшстю глюкоцереброзидази цереброзиди
(трупа глткосфшголтпщтв) вщкладаються в макрофагах селезшки, печшки, лтмфа-
тичних вузл1В т ктсткового мозку. Ключовими симптомами хвороби е спленомега-
лтя, збшьшення печшки, а також змши в ктстках (остеопороз).
У разт хвороби ГПманна — П1ка фосфатид ефшгомтелш вщкладаеться в моно-
нуклеарних фагоцитах ртзних оргашв т центральнш нервовш системт. Генетичний
дефект — дефтцит сф1нгом!елгнази. Характеризуеться значним збшьшенням пе-
чшки та селезшки, затримкою психхчного розвитку’ дитини, розвитком слшоти 1
глухоти. Д1ти здебшьшого помирають у вщ) до 2 роюв. Успадковуеться за авто-
сомно-рецесивним типом.
Амавротична 1дюпия — загальна назва спадкових хвороб 1з групи хвороб нако-
пичення, що виникають унаслщок вщкладання танглюзидгв у клпинах централь-
но! нервово! системи. Харакгерним е зниження штелекту, зору внаслщок атрофп
зорових нерв1в Успадковуеться за автосомно-рецесивним типом.
Порушення холестеринового обмшу описано в роздш XXVI “Патолопчна
ф1з1олопя системного кровообггу”.
Розд1л XX
ПОРУШЕННЯ Б1ЛКОВОГО ОБМЕНУ
Оскгльки белки посЁдаютъ головне мЁсце в процесах життедЁяльносп оргашзму
(“життя — це спосЁб Ёснування бЁлкових тел”), порушення Ёх обмгну е елементом
патогенезу всех патолопчних пропесш. Щоб отримати загалъне уявлення про по-
рушення белкового обмену, слёд розглянути поняття азотисто! ргвноваги. У здоро-
во! доросло'1 людини кёлькёсть азотистих речовин. якё вона одержуе з ежею. дорЁв-
нюе тёй кёлькостё. яка виводиться з оргашзму. В оргашзм;, що росте, пёд час ва-
гётностё, надмЁрно! секрецп або гпеля введения гормонЁв анаболечно) дп, а також
у разг посиленого харчування теля виснажливих хвороб азоту виводиться менше,
нёж надходить, тобто анаболЁчнЁ процеси переважають над катаболЁчними. Це
позитивный азотистый баланс. Негативный азотистый баланс спостерЁгаеться в
раз; втрати 6ёлкёв або збЁлыпеного витрачання ёх органЁзмом пёд час голодування.
у разЁ втрати бЁлкЁв через нирки (протеТнурЁя), шкЁру (ошки). кишки (пронос),
при тиреотоксикозЁ. гарячцЁ тощо.
Порушення белкового обмЁну можливе на всЁх етапах, починаючи зё всмокту-
вання ё закЁнчуючи виведенням з оргашзму кЁнпевих продуктЁв.
ПОРУШЕННЯ ПЕРЕТРАВЛЮВАННЯ I СИНТЕЗУ Б1ЛК1В
Оскёльки в органЁзмЁ практично немае депо бЁлкЁв, а джерелом амЁнокислот для ёх
синтезу е переважно компоненти ёжё, то в разЁ порушення перетравлювання ё
всмоктування оёлхёв розвиваеться алиментарна быкова недостатшсть. Вона спос-
терЁгаеться год час голодування, незбалансованого за амёнокисл одним складом
харчування, при запальних Ё дистрофЁчних процесах у кишках, лЁдшлунковёй за-
лоз] (хронЁчний панкреатит).
Перетрантювання йлкёв порушуеться в разЁ зниження секрецп пептидаз (пеп-
син, гастриксин, хёмозин) клЁтииами шлунка абс при зниженнЁ кислотностё шлун-
кового соку (збЁльшення рН до 5,0—6,0). На наступному етаго — кишковому —
починають прапювати проте'Ёнази пЁдшлунковоЁ залози (трипсин, хЁмолрипсин,
еластаза, карбоксипептидаза А та В). Усё зазначеш ферменти секретуються у виг-
ляд] неактивних ироферментЁв ё в разЁ недостатностЁ кишковего ферменту енте-
рокЁнази (описано спадковий дефект цього еязиму) або зменшення рН Ёх пере-
творення на активнЁ проте’Ёнази порушуеться Це призводить до пригнЁчення по-
рожнинного перетравлювання бЁлкЁв ёз розвитком креаторе'1 (див. роздш XXVIII
“Патолопчна Фёзёологёя травления”).
Пёсля розщеплення бЁлкЁв на ди- Ё трипептиди й амЁнокислоти мае вЁдбутися
‘ёх усмоктування. ОстаннЁй процес забезпечуеться пермеазами — групоспепифЁч-
ними котранспортерами (переносниками), що здшснюють перенесения речовин
разом 13 натрхем за мехашзмом вторинно активного транспорту. Усього е 5 груп
пермеазних систем, яю транспортують близью за х!м!чною будовою амшокисло-
ти. Наприклад, видшяють пермеази, що забезпечують перенесения нейтральних
моноамшо-монокарбонових (аланш, валш), двоосновних (арпнш, л!зин), дикар-
бонових (аспарагш, глутамш) амшокислот. Описано селективш та трупов! пору-
шення всмоктування амшокислот у кишках унаслщок спадкових дефекта перме-
азних систем, як! призводять до дефщиту в оргашзм! певних амшокислот за ра-
хунок порушення як кишкового транспорту, так 1 реабсорбцн амшокислот у
нирках (див. роздш XXX “Патолопчна фгзюлопя нирок”). Тому щ захворювання,
як правило, супроводжуються пщвищенням виведення амшокислот !з сечею, тоб-
то амтоацидургею (л!зинур!я, цистинур!я, цистиноз тощо).
Достатня юлыасть амшокислот забезпечуе синтез бщюв за умови збережено-
го генетичного коду 1 нормального функцюнування систем транскрипцп ! транс-
ляцп. Порушення продукцп бшка може бути набутим 1 спадковим. Воно прояв-
ляеться змшою юлькост! синтезованих молекул або утворенням аномальних за
своею структурою бшюв. Збшъшення або зменшення синтезу бшка пов’язане пе-
реважно 31 змшою регуляторних вплив!в певних гормошв, нервово! системи. Кр!м
того, до порушення синтезу бшка може призвести неврегульована швидюсть син-
тезу мРНК унаслщок розладу функцюнування гена (наприклад у пухлинних юп-
тинах), а також дефекти в структур! рибосом, що виникають зокрема пщ впливом
стрептомщину.
Синтез бшюв 31 змшеною структурою зазвичай е наслщком помилок у геном!.
Це проявляеться порушенням амшокислотного складу бшкових молекул (напри-
клад молекула гемоглобшу при серпопод!бно-кл!тинн!й анеми), вкороченням мо-
лекул (коли транскришця ДНК вщбуваеться т!льки до дефекту в шй), а також
синтезом аномально довгих бшюв при динам!чних мутац!ях або якщо мутащя
сталася в “стоп-кодон!” гена. Прикладом цього можуть бути аномальш бшки у
випадку хоре! Гентшгтона, при спадковому амшощоз! (мутац!я в “стоп-кодон!”
гена аполшопротешу) та !нших патолопях. Продукщя бшюв з! змшеною структу-
рою можлива також унаслщок порушення трансляцп або посттрансляцшно! мо-
дифжацГ! молекул. 31 збшьшенням частота виникнення помилок транскрипцп 1
трансляцп протягом життя пов’язують стар!ння оргашзму.
ПОРУШЕННЯ РУЙНУВАННЯ Б1ЛК1В
Руйнування бшюв е не менш важливим завданням для юптин оргашзму, шж !х
синтез. Видшяють два основних шляхи деградац!! б!люв — л!зосомний 1 протеа-
сомний. Перший забезпечуеться гщролггичними ферментами л!зосом (протеша-
зами) 1 мае значения в розщепленш бшюв з тривалим термшом життя, що повшь-
но оновлюються, та бшюв, яю потрапили до юптини через фагоцитоз. Другий
шлях деградацп бшюв реал!зуеться за участю мультикатал!тичного макромолеку-
лярного комплексу пщ назвою “протеасома” ! забезпечуе ел!мшаццо бшюв з ко-
ротким перюдом життя.
Порушення л1зосомного протеолгзу виникають у раз! спадкових дефекта гщ-
ролггичних фермента або пщ впливом факторгв, потр!бних для !х спрямування в
лхзосоми, та внаслщок дп рхзних ххмхчних речовин (хлорохтн, амонпо хлорид
тощо). Наслщком пригшчення лхзосомного протеолхзу е накопичення в клхтин!
быкових агрегапв ! порушення спегщфхчних функцш. Надмхрна активацхя лхзо-
сомного протеолхзу також може спостерхгатися при деяких патолопчних проце-
сах. Наприклад, пщ час голодування для забезпечення клхтини потрхбххими для
синтетичних процесхв аминокислотами в лхзосоми потрапляють не тыьки окремх
быки, а й цы! органели — вщбуваеться автофагхя.
Порушення протеасомного протеошзу. Протеасома, яка порхвняно з лхзосома-
ми е мобыьнппою структурою та може виконувати свою функцпо як у цитоплаз-
м!, так х в ядрх, мае унхверсальне значения х бере участь у регуляцн багатьох важ-
ливих внутр!шньоклхтинних процесхв (поды клхтини, диференцхащя, апоптоз,
оновлення репепторхв, юнних каналхв, ферменпв тощо). Тому порушення актив-
ной! протеасоми чи факторхв, як! забезпечують пщготовку або убгквхтинхзацпо
(послщовне приеднання низькомолекулярного быка убхквхтину до протешу, що
мае гщролхзуватися) субстрапв для протеасомного протеолхзу, мае тяжкх наслщки
для клхтини. Рхзке пригнхчення активностх протеасоми пщ час оксидативного
стресу, дефххцпу АТФ призводить до запуску апоптозу за рахунок накопичення
быкхв з проапоптичними властивостями (наприклад р53). Велике значения мае
порушення протеасомного протеол1зу в патогенез! нейродегенеративних хвороб.
Доведено, що при хвороб! Альцгеймера, амыощозх, хоре'! Генпнгтона, латераль-
ному амютрофхчному склероз!, прюнових шфекцхях у клхтин! накопичуються
убхквхтишзован! протеши, якх не гщролхзувалися протеасомою. Крхм того, пору-
шення протеасомного протеолхзу негативно впливае на утворення пептщцв, що
презентуються в склад! быкхв ГКГС I класу. Доведено зокрема, що генетичнх де-
фекти в катал ггичн их субодиницях протеасоми пов’язанх з розвитком низки авто-
хмунних захворювань (цукровий дхабет, ревматощний артрит та ш.).
ПОРУШЕННЯ ОБМ1НУ АМ1НОКИСЛОТ
Порушення трансамшування й окисного дезамшування. Процеси трансамшування !
дезамхнування мають унхверсальне значения для всхх живих оргашзмхв: трансамх-
нування сприяе синтезу амхнокислот, дезамшування — ’хх руйнуванню.
Суть реакцц трансамшування полягае у зворотному перенесенн! амшогрупи
вщ амшокислоти на а-кетокислоту без промхжного утворення выьного йону амо-
нпо. Реакщя каталхзуеться специфхчними ферментами амшотрансферазами (тран-
самшазами), кофакторами яких е фосфорильованх форми шридоксину (пхридок-
сальфосфат х шридоксамшфосфат).
Порушення реакцш трансамшування можуть виникати з кыькох причин х
передусхм унаслщок дефщиту шридоксину (ваптнхсть, пригнхчення сульфаныа-
мщними препаратами кишково! мгкрофлори, гальмування синтезу тридоксаль-
фосфату пщ час лхкування фтивазидом). Зниження активностх амшотрансфераз
вщбуваеться також у раз! обмеження синтезу быкхв (голодуванххя, тяжкх хвороби
печшки). Якщо в деяких органах виникае некроз (йхфаркт мхокарда або легень,
панкреатит, гепатит та ш.), то внаслщок руйнування клхтин тканини! амшотран-
сферази надходять у кров, х пщвищення ххньо'х активностх в кров! за тако! патоло-
ги е одним хз дхагностичних тестхв. У змш! швидкостх трансамшування важливу
роль вщгграють порушення спгввщношення мгж субстратами реакцп, а також
вплив гормонгв, особливо глюкокортикощгв г гормонгв щитоподгбног залози, якг
стимулюють цей процес.
Пригнгчення окисного дезамшування, яке спричинюе руйнування невико-
ристаних амгнокислот, зумовлюе бглыпу концентрацгю амшокислот у кровг — г1-
перамшоацидемйо. Наслщками цього е посилена екскрецгя амгнокислот нирками
(амиюацидурм) г змгна спгввщношення окремих амгнокислот у кровг, що створюе
несприятливг умови для синтезу бглкових молекул. Дезамшування порушуеться в
разг дефгциту компонентгв, якг прямо або опосередковано беруть участь у цгй ре-
акцп (пгридоксин, рибофлавш, шкотинова кислота), а також пщ час гшоксп, го-
лодування (бглкова недостатшсть).
Порушення декарбоксилювання. Декарбоксилювання, яке е важливим, хоча й
не унгверсальним напрямом бглкового обмгну, вщбуваеться з утворенням вугле-
кислого газу г бгогенних амгнгв. Декарбоксилювання зазнають лише деякг амшо-
кислоти: петидин — з утворенням ггстамгну, тирозин — тирамгну, у-глутамгнова
кислота — у-амгномасляног кислоти, 5-ггдрокситриптофан — серотоншу, похщнг
тирозину (3,4-дгоксифенглалангн) г цистину {(Ь-цистегнова кислота — вщповщно
3,4-дгоксифенглетиламшу (дофамш)} г таурину. Бгогеннг амгни, як вщомо, мають
специфгчну бголопчну актившсть, г збгльшення гх кглькостг може спричинити
певнг патолопчнг змши в органгзмг. Зростання кглькостг бгогенних амгнгв може
бути наслщком не тгльки посиленого декарбоксилювання вгдповгдних амшокис-
лот, а й пригнгчення окиснення амгнгв г порушення зв’язування гх быками. Нап-
риклад, при гшоксичних станах, гшемп г деструкцц тканин (травма, опромшення
тощо) ослаблюються окиенг процеси, що сприяе посиленню декарбоксилювання.
Поява великог кглькостг бгогенних амгнгв (особливо ггстамгну та серотонгну) у
тканинах може зумовити значке порушення мгецевого кровообггу, пщвищення
проникностг стшки судин г ушкодження нервового апарату.
Спадков! порушення обмшу деяких амшокислот. Проходження амгнокислот че-
рез певнг метаболгчнг шляхи детермгнуеться кглькгетю й активнгстю вгдповгдних
ферментгв. Спадковг порушення синтезу ферментгв призводять до того, що вщ-
повщна амшокислота не включаеться в метаболгзм, а накопичуеться в бголопч-
них середовищах органгзму: кровг, сечг, калг, потг, спинномозковгй рщинг. Клппч-
на картина в таких випадках визначаеться, по-перше, появою надто великог кгль-
костг речовини, що мала метаболгзуватися за участю заблокованого ферменту,
по-друге, дефщитом речовини, яка мала утворитися.
Генетично зумовлених порушень обмгну амшокислот вщомо досить багато,
усг вони успадковуються рецесивно. Деякг з них наведено в табл. 13.
Порушення обмшу фешлалангну. Фенглаланш у норм! незворотно окиснюеться
в тирозин. Якщо в печшцг порушуеться синтез потргбного для цього ферменту
фенглаланшгщроксилази (схема 17), то окиснення фешлалангну вщбуваеться
шляхом утворення фенглшровиноградног та феншмолочног кислот — розвиваеть-
ся фен1лкетонур1я. Однак цей шлях мае малу “пропускну” здатнгсть, тому велика
кглькгсть фешлалангну накопичуеться у кровг, тканинах г спинномозковгй рщинг,
що в пергш ж мгсяцг життя спричинюе тяжке ураження центрально! нервовог сис-
теми г невилгковне недоумство. Через недостатшсть синтезу тирозину зменшуеть-
ся утворення мелашну, що зумовлюе поевгтлгння шкгри г волосся. Кргм того,
Таблиця 13. Спадков! порушення обм!ну амшокислот унаслщок дефщиту або низькоТ
активност! ферменпв
Амшокислота ................. . - . . . .. .. < - Фермент - - .. .. КЛ1Н1ЧН1 ознаки
Феншалажн Феншалашштдроксилаза Феншкетонуртя, феншпрови- ноградна ол1 гофрен (я
Тирозин Оксидаза гомогентизиново! кислоти Тирозиназа Алкаптонурш Альбгшзм Тирозиноз
АрГШТН Ксантиноксидаза Аргшшсу книпаза Ксантину р1я Арп ншеу кт1,инатур1я
унаслцюк збглыиеного утворення феншпровиноградно! кислоти гальмуеться ак-
тившсть ферменту (дофамшгщроксилази), нотрхбного для синтезу катехолампив
(адреналшу, норадренал1ну). Тяжк1сть спадково! хвороби визначаеться комплек-
сом ус!х цих порушень. Хвор1 помирають у дитинств!, якщо не проводиться спе-
щальне лткування, що полягае в постшному, але обережному (контроль за амлно-
кислотним складом кров!) обмеженш надходження фсюлаланшу з гжею Ранню
диагностику захворювання потребно проводити одразу теля народження дитини.
Для цього застосовуютъ рлзш бюх1М1чш тест-системи.
Порушення обмшу тирозину. Обмш тирозину вщбуваеться юлькома шляхами.
У раз! недостагнього перетворення тирозину на гомогентизинову кислоту (див.
схему 17), що може бути пов’язано з дефектом рлзних ферменпв. тирозин нако-
пичуеться в кров! та виводиться и сечею. Це порушення називаеться тирозинозом
1 супроводжуеться печшковою 1 кирковою недостатшстю та рапньою смертю ди-
Феылалатн
Тирозин
Фен1'1кетонур(я
3,4-Дипдрооксифешлаланш (ДОФА)
Альбйазм
Мелаши
Фен1Г|П1ровино-
градна кислота
1 щроксифешлтро-
виноградна кислота
Фенгшактат
2, 5-Дипдроокситровиноградна
кислота
Тирозиноз
Фешлоцтова
кислота
Гомотентизинова
кислота
Л зкапгпонурт
С0. + Н.0
Схема 17. Блокада шляхтв обмшу фешлаланшу г тирозину
тини або .лише затримкою психомоторного розвитку. Коли порушення обмшу
тирозину вщбуваеться в момент окиснення гомогентизиново! кислота (див. схему 17),
розвиваеться алк.аптонур1я. Фермент, який окиснюе гомогентизинову кислоту
(гомогентизиноксидаза), утворюеться в печшщ. У норм! вш настыьки швидко
розривае п пдрохшоновс к!льце, що кислота “не встигае” пограпити в кров, а
якщо 1 потрапила, то швидке вид!ляеться пирками. У раз! сдадкового дефекту
цього ферменту гомогентизинова кислота у велиюй юлькост! виявляеться в кров!
! сеч!. Сеча хворих на алкапгонур!ю на пов!тр! або шсля додавав ня л ул в стае
чорною. Це пояснюеться окисненням гомогентизиново!’ кислота киснем пов!тря
й утворенням у нш алкаптону. Гомогентизинова кислота з кров! проникае в тка-
нини — хрящову. сухожилии, зв’язки, внутрпшпй шар стшки аорта, унаслщок
чо го утворюються теми! плями в д!лянщ вух, носа, ш!к, на склерах. Алкаптон
робить хрящ! та сухожилки крихкими. що шод! призводить до тяжких змш у суг-
лобах.
Тирозин е вихцшим продуктом для утворення шгменту меланшу, який м!с-
титься в шюр! га волосе!. Якщо перетворення тирозину на мелашн зменшене
через спадковий дефщит тирозинази (див. схему 17), виникае алъбтзм, який суп-
роводжуеться пщвищенням чутливост! шюри до сонячного свила 1 порушенням
зору.
Нареши, тирозин е попередником тироксину. У раз! недостатнього синтезу
ферменту, який катал!зуе взаемод!ю тирозину з в!льним йодом, порушуеться ут-
ворення гормошв щитопод!бно1 залози.
Порушення обмшу триптофану. Основний шлях метабол!зму триптофану, як I
шкотиново! кислота (див. розд!л XXIV “Латоф.’зюлопя обм)ну в!тамш!в”), приз-
водить до синтезу НАД 1 НАДФ, яю вйцграють важливу роль у житгед!яльност!
оргашзму, будучи коферментами багатьох реакщй обм!ну, а значний дефщит цих
речовин спричинюе розвиток пелагри. Порушення обмшу триптофану може суп-
роводжуватися також змшою кшькост! серотоншу, який утворюеться з нього.
ПОРУШЕННЯ К1НЦЕВИХ ЕТАП1В
Б1ЛК050Г0 0БМ1НУ
На кшцевих етапах бокового обмшу можуть порушуватися процеси утворення
азотистах продукпв (сечовини, амошю. сечово’1 кислота) 1 виведення !х з ор-
гашзму. Основним показником розладу утворення 1 видщення сечовини та шших
азотистах продуктов обмшу е змша р!вня 1 складу залишкового (небшкового)
азоту в кров! (норма — 14,3—21,4 ммоль/л, або 20—30 мг%). Залишковий азот на
50 % складаеться з азоту сечовини, близько 25 % його припадае на амшокислоги.
решта — на шил азотис г! продукта. Несечовинна частина його отри мала назву
резидуального азоту. Збшьшення залишкового азоту в кров! — гтеразотелпя —
може бути наслщком порушення синтезу в печшц! сечовини з амошю (продук-
цшна пперазотемтя) 1 розладу видшьно!’ функцц нирок (ретенщйна гшеразо-
темЗя).
Порушення синтезу сечовини спостерйаютъ при деяких патолопчних станах
(дисгрофтчн! змши в печшщ, гшокс!я), а також можуть бути зумовлеш спадковим
дефектом. СпадковЁ порушення синтезу сечовини виявляють у раз) недостатнього
утворення аргЁнЁнсукцинатлЁази (аргМнсукцинатургя), карбамоЁлфосфатсинтетази
й орштинкарбамоЁлтрансферази (а.мон1ем1я), а також арпнЁнсукцинатсинтетази
(цитрумнургя).
НайчастЁшим наслЁдком порушення синтезу сечовини е накопичення амонт в
крови Амонёй спричинюе токсичне ураження насамперед центрально! нервовоЁ
системи. Токсична дёя його пов’язана зё зв’язуванням його а-кетоглутаровою кис-
лотою з утворенням ЕлугамшовоЁ кислоти, яка приеднуе ще один юн амонЁю ё
перетворюеться на глутамЁн. Вилучення внаслЁдок цього а-кетоглутаровоЁ кисло-
ти з циклу Кребса призводить до порушення синтезу АТФ ёз розвитком енергоде-
фЁциту нервових клётин.
ПОРУШЕННЯ Б1ЛКОВОГО СКЛАДУ КРОВ1
ЗмЁнами кёлькёсного та якёсного спЁввЁдношення бЁлкгв кровЁ супроводжуються
майже всё патолопчнЁ стали, а також уродженЁ аномалЁЁ синтезу бЁлкЁв. Порушен-
ня вмЁсту бЁлкЁв плазми кровЁ може проявлятиея змёною загально! кёлькостё бЁлкЁв
(гЁпопроте’ЁнемЁя, гЁперпроте’ЁнемЁя) або спЁввЁдношення мёж окремими бЁлковмми
фракциями (диспротеЁнемЁя) на тлё нормального загального вмЁсту бЁлюв.
ГгпопротеТнемЁя виникае здебЁльшого за рахунок зменшення кёлькостё аль-
бумЁнЁв, е набутою (у разЁ голодування, хвороб печЁнки, порушеного всмоктуван-
ня бЁлкЁв) ё спадковою. ГЁпопроте'ЁнемЁю може спричинити також вихёд бьткЁв ёз
кровоносного русла (крововтрата, плазмовтрата, ексудацЁя, транссудацЁя) ё втрата
бЁлкЁв Ёз сечею (протешурЁя).
ГЁперпротешемЁя бувае переважно вёдносною (згущення кровЁ). Абсолютна гЁ-
перпроте’ЁнемЁя зазвичай пов’язана зё збЁльшенням рЁвня глобулгнЁв, як правило,
у-глобу.тЁнЁв унаслЁдок посилення синтезу ЁмуноглобулЁнЁв.
ДиспротеЁнемЁя мае як набутий, так Ё спадковий характер. Умовно розрЁзня-
ють дисглобулЁнемЁю, дисгаммаглобулЁнемЁю ё дисЁмуноглобулЁнемЁю. За остан-
ньоё бгтковий склад кровЁ е лише вЁдображенням загально! перебудови ЁмунноЁ
системи, у першу чергу и гуморальноЁ ланки. Прикладами диспротеЁнемЁЁ. яку
виявляють найчастЁше, може бути збЁльшення вмгсту у-глобулЁнЁв, зменшення
рЁвнЁв а- Ё р-лЁпопроте’ЁдЁв унаслщок порушення функцЁй печЁнки, змЁна кёлькостё
та структури фЁбриногену.
Змёни у-глобулЁнЁв можуть бути КЁЛЬКЁСНИМИ та ЯКЁСНИМИ. ЯкЁСНО ЗМЁНСНЁ у-
глобулЁни мають назву парапротеЁнЁв. Вони належать до ЁмуноглобулЁнЁв Ё е зазви-
чай продуктами патолопчних клонёв плазмоцитЁв, що вироб.впоть антитьта або
окремЁ Ёх танпюги (важкЁ, легкЁ). ЗбЁльшення Ёх кёлькостё в кровЁ — моноклоналъна
г1пергамиаглобул1нем1я — спостерЁгаеться здебЁльшого в разЁ пролЁферацй вёдповщ-
них клонёв, зумовленоЁ пере важно пухлинною природою патологЁчного процесу
(мЁсломна хвороба, макроглобулЁнемЁя Вальденстрема). РЁзновилом парапротегнЁв
е також крЁоглобулЁни — патолопчнЁ протеЁни з властивостями ЁмуноглобулЁнЁв,
якё преципЁтують пёд час охолодження.
ПОРУШЕННЯ МЕТАБОЛИЗМУ НУКЛЁ1НОВИХ ОСНОВ
Порушення обмшу пуринових основ. Кшцевим продуктом обмшу пуриневих основ
(адешну та гуаншу), яка набежать до структуры нуклетнових кислот, е сечова кис-
лота. Порушення н синтезу 1 видтлення призводить до гтерурикеми, що е досить
поширеним патолопчним синдромом (у ртзних популящях частота виникнення
гшерурикемп становить вщ 2 до 18 %).
Найяскравштим прикладом порушень синтезу т виведення сечовот кислоти та
найтяжчим вартантом первинно'1 гшерурикемп е подагра (вщ грец. уражена нога,
капкан).
Подагра була вщома ще в лавнину й описана Гтппокрагом. Вивчення п поча-
лося в 1860 р , коли А. Гаррод, який сам страждав на подагру, зробив класичне
описания хвороби т встановив, що в устх хворих на подагру в суглобах 1 першрти-
кулярних тканинах втдкладаються урати — солт сечовот кислоти. Подагра — полт-
генна хвороба, розвиток якот залежить вщ способу життя людини (споживання
велико! юлькостт м’яса, алкоголю, гшодинамтя) та дп додаткових фактортв навко-
лишнього середовища (геохтмтчнт особливостт територн та ш.).
Генетична схильнтсть до подагри визначаеться кшькома генами, серед яких
найбтльше значения мають патолопчш алелг гптоксантингуашнфосфорибозил-
трансферази (ГФРТ) т фосфорибозилптрофосфатсинтетази (ФРПС). ГФРТ за
нормальних умов регулюе вмгст фосфорибозилшрофосфату, перетворюючи його
на гуанозинмонофосфат та шозинмонофосфат, т таким чином зменшуе утворення
сечовот кислоти з ппоксантину. У хворих на подагру часттше спостерпають вартан-
ти гена ГФРТ (ниш описано близько 60 алельних варганттв цього гена), шо кодуе
фермент зт зменшеною активнтстю, а це призводить до утворення надлишковот
юлькостт сечовот кислоти. Алельний полтморфтзм гена ФРПС (вщомо 8 вартан-
ттв), навпаки, спричинюе збыыпення активностт цього ферменту, унаслщок чого
утворюеться надлишкова юльктсть фосфорибозилшрофосфату, який згодом пере-
творюеться на сечову кислоту. Важливо, шо обидва гени розташоваш в Х-хромо-
сомт. Цим здебглыпого пояснюеться пщвищена схильнтсть чоловтктв до подагри
(близько 95 % хворих).
Вторинна гшерурикемгя, шо спричинюе розвиток подагричного синдрому,
пов’язана переважно з порушенням видшення сечовот кислоти цирками. Хрошч-
на ниркова недостатшсть, полтктстоз нирок, нефропаття ваптних досить часто
призводять до затримки в органтзмт сечовот кислоти. Зниження секрепп сечовот
кислоти в нирках спостерггаеться в разт ацидозу будъ-якот еттологп, ппотиреозу,
свинцевого отруення тощо.
Патогенез ураження оргашв 1 систем при подагрт пов’язаний з вгдкладанням
ураттв, ушкодженням тканин кристалами солей сечовот кислоти та розвитком
запального процесу. Найуразливштою в цьому аспект! е хряшова тканина, фтзико-
хтмтчнт особливостт (температура, рН тошо) якот сприяють переходу ураттв з роз-
чинного стану в нерозчинний. Вщкладання солей спричинюе гостре подагричне
запалення, яке супроводжуеться болем, гарячкою 1 заюнчуеться утворенням по-
дагричних вузлтв, деформащею суглобтв. Не меншу небезпеку становить т вщкла-
дання ураттв у нирках з розвитком сечокам’яно! хвороби (уролтттазу) та подагрич-
но'т нефропатГ!
Порушення обмшу шримтдинових основ. Катабол!зм шримгци нових основ (ци-
тозину, тимшу та урацилу) призводить до утворення р-амшокислот. Описано
юлька вар!ант!в порушень шримщинового обмшу. Наиб!лыпе клпичне значения
мае оротацидургя — спадкова автосомно-ревесивна хвороба, пов'язана з дефщи-
том двох ферменпв: оротатфосфорибозилтрансферази та оротидин-5'’-фосфатде-
карбоксилази. За нормальних умов щ ферменти забезпечують утворення уридину,
а при оротаццщдш уридин не утворюеться 1 накопи чуеться його поперед ник —
оротова кислота. Ця речовина е малорозчинною 1 легко кристашзуеться !з форму-
ванням конкрементов у сечових шляхах. Однак найнебезпечншп патолопчн! зм!-
ни в раз! оротацидурй вщбуваються в систем! кров!. Порушення подглу еритро-
бл аст!в призводить до гяжко! металобластно!' анемп, рефрактерно! до лгкування
щанокобаламшом (вотам он В12). Також розвиваеться гмунодефщит, пов’язаний !з
недостатшм утворенням Т-л!мфонит!в.
Розд1Л XXI
ПОРУШЕННЯ КИСЛОТНО-ОСНОВНОГО СТАНУ
Кислотно-основний (кислотио-лужний) стан — тдтримання концентрации /ошв
водню (протонов, Н+) у бюлогёчних р/динах на дуже низькому р1вн1 та в дуже вузъких
межах коливань.
Нормальна концентрацхя !ошв водню (Н+) у позаклпшппй рщин! становить
40 некв/л, що вщповщае рН 7,4. Така конпентрашя Н+ у 100 тис. раз!в менша, шж
концентращя К+, 1 в 1 млн разгв менша, юж концентрация Нормальн! коли-
вання рН вщ 7,35 до 7,45 вщповщають змгнам концентрацп Нт т!льки на 10 некв/л,
а змши рН у межах вщ 7,0 до 7,8, як! е сумгсними гз життям, — на 86 некв/л. Це
пов’язано з тим, шо гони водню мають дуже високу бгологгчну актившсть. Пору-
шення КОС, тобто зрушення рН за меж! норми, зумовлюе зм!ни активносп фер-
менпв, проникносп мембран, чутливосп рецепторгв, активноеп БАР, секрецп
гормошв, стану колощгв, нервово-м'язовог збудливосп, порушення синтезу АТФ
тошо.
Тим часом у процес! метаболгзму жир!в, б!лк!в ! вуглевощв щодоби утво-
рюеться 15 000—20 000 ммоль вуглекислого газу. Частково розчиняючись у вод!,
вш утворюе вуглекислоту. Кр!м того, у реакциях пром!жного обману за нормаль-
них умов утворюеться приблизно 50—100 мскв нелетких кислот, !з яких 1/3 ста-
новить сульфатна (с!рчана) кислота, що синтезуеться пщ час мегабол!зму метюш-
ну га цистешу, 1 /3 — молочна кислота, ацетоацетат 1 0-гщроксибутират, що утво-
рюютъея в раз! неповного окиснення вуглеводгв 1 жир!в. Ще третина нелетких
кислот утворюеться пщ час обм!ну фосфолшщгв, фосфопротещгв 1 солей з ор-
гангчними капонами, як! метабол!зуються. Кзльюсть кислот, як! утворюються,
залежить вщ вмгсту бьзка в гж! та р!зко зменшуеться в раз! дотримання молочно-
рослинно! д!ети. Значне збщьшення вироблення органгчних кислот вщбуваеться
при гшоксп, голодуваннг та цукровому дгабеп.
Мехашзми пщтримання сталосп* КОС. У норм! рН мгжклггинно! рщини та
кров! становить 7.4, цитоплазми — 7.0—7,2, активованих л!зосом — 5,0—5,5. При
цьому рН дистильовано! води за температури 37 °С сягае 6,8. Отже, рН позаюп-
тинно! рщини мае бглып лужну реакшю. шж води, яка е ушверсальним бюлопч-
ним розчинником. Наявшсть град!ента концентрацп !ошв водню М1Ж р!зними
компартментами (водними секторами) оргашзму та гх суворе пщтримання в дуже
вузьких межах потргбш, з одного боку, у зв’язку з тим, що ф!з!олог!чн! реакцп
можуть вщбуватися тгльки за певних значень рН, з шшого, вони забезпечуються
вщповшними ф!зико-х!м!чними (буфери та гонообмш)! фгзюлопчними (леген! та
нирки) механизмами.
Головний ф1зико-хш1чний мехашзм пщтримання сталосп рН — буферт сис-
теми, яю е сумшшпо слабко! кислота або слабко! основи з !хшми солями. Шсля
додавання до буферного розчину сильно'! кислота быышсть юнгв водню зв’язуються
31 сполученою основою, утворюють нейтральну с!ль 1 слабку кислоту, яка не ди-
соцпое. Це зменшуе юльюсть вшьних юшв водню та запобггае зрушенню рН, як,
наприклад, у таюй реакцп:
НС1 + ИаНСО3 = Н2СО3 + ИаС1.
Якщо прийняти всю буферну емшсть кров! за 100 %, то пдрокарбонатний
(бжарбонатний) буфер становись 53 %, 13 яких пдрокарбонат плазми — 35 %, а
еритроцити — 18 %; 35 % становись емшсть гемоглобшового буфера, 7 % — бо-
кового буфера плазми, 3 % — оргашчних фосфата еритроципв, 2 % — неоргашч-
них фосфата плазми та еритроципв.
Отже, найважлив!шим буфером кров! е пдрокарбонатний буфер. У м1жюп-
тиншй рщиш. об’ем яко! втрич! перевищуе об’ем кров!, буферна емшсть майже
виключно визначаеться пдрокарбонатним буфером. Кргм того, унгкальною влас-
тив1стю цього буфера е його вщкритий характер. Це видно з рхвняння Гендерсона —
Гассельбалха:
рН = рК + 1§
[(ошзована форма]
[неюшзована форма] ’
яке для системи НСО3 — Н2СО3 набувае такого вигляду:
рН = 6,1 +1§
[НСО3 ](метабол 1чний компонент; контролюеться нирками)
[Н2СО3](респ!раторний компонент; контролюеться легенями)
Осюльки Н2СО3 утворюеться пщ час пдратаци розчиненого СО2, а концентращя
останнього дор!внюе рСО2, помноженому на коефпцент розчинносп (0,03 ммоль/л
на 1 мм рт. ст.), то ртвняння набуде такого вигляду:
рН = 6,1 +1§
[нсо;]
[0,03 рСО2]’
Напруження вуглекислого газу в артер!альн!й кров! людини перебувае в р!вно-
ваз! з парщалъним його тиском у альвеолярному повггр! — приблизно 40 мм рт. ст.
за температури 37 °С. Таким чином, юльюсть вуглекислого газу, розчиненого в
плазм! кров!, а отже, 1 концентращя Н2СО3 дор!внюють:
[СО,]розч. = 0,03 рСО2 = 0,03 • 40 = 1,2 ммоль/л.
За рахунок того, що леген!, виводячи вуглекислий газ (ресшраторний компо-
нент буфера), щдтримують юльюсть розчиненого вуглекислого газу, а отже, 1
Н2СО, на поспйному р!вш, можна легко розрахувати концентраццо гщрокарбона-
ту в позаюптиннш рщиш, потр!бну дтя пщтримання нормального рН.
Так як вщомо, що 7,4 = 6,1 + 1§
(КаНСО3)
(н2со3)
(№НСО3)
(Н2СО3)
, то вщповщно 1§
= 1,3.
Осюльки 1,3 = 1§ 20, то за нормального рН кров! сшввщношення
(МаНСО,)
(Н,со,)
Звщси, знаючи вмгст у кров! Н2СО3, ргвняння можна записати у такому вигляд!:
_ , , , ,24 ммоль/л №НСО,
7 4 = 61 + 12---------------.
1,2 ммолъ/л Н2СО3
Отже, ефекти внють роботи пдрокарбонатного буфера щодо пщтримання рН
на нормальному р!вш залежить вщ збереження сшввщношення [гщрокарбонат] /
[СО2розч ] у межах, близьких до 20,0. Резервш ж можливосп цього буфера за нор-
мального рСО2 будуть найбшьшими при концентрацп гщрокарбонату в плазм!
близько 24 мекв/л. Стал!сть ресшраторного компонента цього буфера залежить
вщ роботи легень 1 дихального центру, метаболхчного — визначаеться д!яльшстю
нирок. Вщкритий характер 1 швидка регенерац!я пдрокарбонатного буфера забез-
печують те, що переважна бшышсть кислот 1 основ, як! з’являються в оргашзм!,
нейтралтзуються саме цим буфером.
ЕТЮЛОПЯ ТА ФОРМИ ПОРУШЕНЬ КОС
Осюльки буферна емшсть позакштинно! рщини, а отже, ! п рН визначаються
головним чином станом гщрокарбонатного буфера, очевидно, що змгна сшввщ-
ношення ресшраторного 1 метабол!чного компонент цього буфера зумовить по-
рушення КОС оргашзму у форм! ацидозу чи алкалозу.
Ацидоз — патологгчний стан, для якого характерном е зниження рН або тен-
денция до зниження рН унаслгдок зменшення ствв1дношення м1ж метабол1чним г
рестраторним компонентами г1дрокарбонатного буфера.
Алкалоз — патолог1чний стан, що характеризуешься тдвищенням рН або тен-
денщею до тдвищення рН унаслгдок збиыиення сп1вв1дношення М1Ж метаболгчним I
рестраторним компонентами ггдрокарбонатного буфера.
Якщо внаслщок порушення виведення легенями вуглекислого газу збшьшить-
ся рСО понад 40 мм рт. ст., це спричинить розвиток газового ацидозу. Якщо ж
унаслщок гшервентиляцц рСО2 зменшиться, виникне газовий алкалоз. Вщповщно
в раз! первинного зменшення концентрацп гщрокарбонату в плазм! кров! нижче
24 мекв/л виникне негазовий ацидоз, а в раз! первинного збшъшення вмгсту гщро-
карбонату — негазовий алкалоз. Якщо при цьому рН завдяки робот! мехашзм!в
компенсац!! буде залишатися в межах норми, ацидоз 1 алкалоз будуть називатися
компенсованими. Якщо внаслщок недостатносп мехашзм!в компенсац!! рН вийде
за меж! норми, ацидоз 1 алкалоз будуть декомпенсованими.
Взаемозв’язок КОС 1з мшеральним обмшом. Концентращя Н2СО3 у норм! зале-
жить, як вщомо, вщ рСО2 в альвеолярному пов!тр! та вщ газообмшу в легенях.
Виникають запитання: вщ чого залежить концентрац!я гщрокарбонату, чому вона
саме така 1 чому може змшюватися? Для того щоб вщповкти на ц! запитання,
потр!бно згадати, що зпдно !з законом електронейтральносп, граф!чним вира-
женням якого е юнограма Гембла (мал. 44), сума катюн!в у бюлопчних рщинах
завжди доргвнюе сум! анюнгв, а вс! катюни й ашони подшяються на: 1) фпссоваш
(вм!ст !х у позаюптиннш рщиш оргашзму вщповщае !х концентрацн у водах пер-
в!сного океану, де зародилося життя на нашш планет!, 1 може змшюватися тшьки
внаслщок !х введения в оргашзм або виведення; до них належать №+, К+, Са2+,
М§2+, СГ, НРО42 тощо); 2) нашвфгксоваш (утворюються 1 метаболтзуються в про-
ПгтГ
Мал. 44. Граф!чне зображення залежност! концентрацп НСО'3 вщ сшввщношення концентрацп
шших 1ошв ! катюшв (юнограма Гембла) у бюлопчних рщинах:
а — норма; б — ппонатр!ем1чний ацидоз (вшцльний); в — лактат- або кетоацидоз (метабол)чний);
г — пперхлорем)чний ацидоз (екзогенний); Г — ппохлоремзчний алкалоз (видшьний); д — ппер-
натр1ем1чний алкалоз (екзогенний)
цес1 обмшу вуглеводш, жир!в ! б1люв з невисокою швидюстю — >Ш4+, лактат,
шруват, кетонов! тша, бшки); 3) нефгксоваш (утворюються 1 зникають у процес!
метабол!зму практично миттево — гщрокарбонат та юн водню).
Якщо вмгст у бюлопчних рщинах гонгв водню цщком визначаеться станом
буферних систем, головним чином пдрокарбонатного буфера, то концентрацгя
гщрокарбонату залежить вщ сшввщношення у бюлопчних рщинах фгксованих г
напгвфгксованих кат!он!в та ангошв. Саме швидка зм!на концентрацп гщрокарбо-
нату забезпечуе електронейтральшсть бюлопчних рщин за умови змши кглькостг
шших ангонгв або катюшв. Надлишок катюшв, насамперед натр по, збшьшуе вмгст
нефгксованого ангона — гщрокарбонату, а гх дефщит — знижуе його концентра-
цгю. Виходячи з цього, можна вважати, що негазовий ацидоз виникае внаслгдок
зменшення вмюту г1дрокарбонату, спричиненого зменшенням спгввгдношення м1ж
фгксованими катюнами та анюнами, а негазовий алкалоз — через збшьшення р1вня
г1дрокарбонату, спричиненого збиьшенням ствв1дношення м1ж ф1ксованими катиона-
ми й анюнами.
Етюлоггя та класифйсащя негазових зрушень КОС. Залежно вщ причини, що
зумовлюе порушення сшввщношення м!ж катюнами та анюнами позаклггинно!
рщини, негазовий ацидоз подшяють на: 1) метабол!чний; 2) видшьний; 3) екзо-
генний.
Метабол1чний ацидоз виникае при надапрному утворенш або порушенш мета-
болизму оргашчних кислот, наприклад, лактату пщ час гшоксп та при цукровому
ддабет!, кетонових тш при голодуванн! та цукровому д!абетц шровиноградно! та
а-кетоглутаровоТ кислот при дефщит! вггамшу В, тощо.
Негазовий видшьний ацидоз може бути нирковим 1 виникае або через порушен-
ня виведення ашошв нелетких кислот у раз! значного зменшення клубочково!
фшьтрацц, або ж порушення функцш ниркових канальцев. Канальцевый ацидоз
подшяеться на к!лька тишв: I — гтокал1ем1чний дистальний канальцевый ацидоз
виникае в раз! порушення реабсорбцп юшв калпо в обмш на юни водню, що
секретуються вставними юптинами А-типу зб!рних ниркових трубочок ! прояв-
ляеться неспроможшстю знижувати рН сеч! менше 5,3 пор!вняно з нормою, за
якою рН може знижуватися до 4,5; II — проксимальный канальцевый ацидоз спос-
терггаеться в раз! порушення реабсорбцп профшьтрованого натрпо гщрокарбона-
ту; III — ацидоз I типу в дней; IV — гтеркал1ем1чний дистальний канальцевый
ацидоз, зумовлений порушенням секрецй К+ 1 Н+ в обмш на Ма+ у головних клх-
тинах збхрних ниркових трубочок при гшоальдостерошзм! або резистентност! до
мшералокортико'щ!в. Канальцевий ацидоз може виникати також пщ час пору-
шення синтезу та секрецй юн!в амонпо в проксимальних канальцях, що характер-
но для хрошчно! недостатност! нирок.
Кргм ниркового до видшьного належить також ацидоз, який виникае в раз!
пщвищеного виведення з кишок гщрокарбонату внаслщок д!аре!. Велик! юлькост!
№НСО3 та КНСО3 утворюються в кишках при метабол!зм! натр!евих 1 кал!евих
солей оргашчних кислот.
Негазовий екзогенний ацидоз виникае теля введения в оргашзм кислот (нап-
риклад хлоридног або оцтово!) або солей сильних кислот з катюнами, що мета-
болхзуються, зокрема МН4С1, який шсля проходження кр!зь печ!нку втрачае юн
амонпо пщ час бюсинтезу сечовини, унаслщок чого в кров потрапляе юн хлору й
утворюеться НС1. Надходження в оргашзм етиленгл!колю зумовлюе утворення
щавлево! кислоти, метанол перетворюеться на мурашину кислоту, етанол — на
оцтову.
Негазовий алкалоз подшяють на: 1) видиьний, який виникае внаслщок пщви-
щення в нирках реабсорбцп юшв натрпо без супутнгх юн!в хлору при гшеральдо-
стерошзм! та в раз! тривалого введения мшерало- або глюкокортикоццв або пору-
шення реабсорбцп юшв хлору теля застосування хлорних дхуретиюв чи пщ час
блювання, коли оргашзм втрачае СГ; 2) екзогенний, який виникае теля введения
в оргашзм основ або солей з оргашчними анюнами, що метабол!зуються, напри-
клад натрпо сукцинату або натрпо лактату, пщ час метабол!зму яких на завер-
шальному еташ утворюеться МаНСО3.
МЕХАН13МИ КОМПЕНСАЦ» ПОРУШЕНЬ КОС
Мехашзми тимчасовог компенсац» порушень КОС. Негазов! ацидоз 1 алкалоз мо-
жуть тимчасово компенсуватися за рахунок змши роботи легень (табл. 14). Так, у
випадку негазового ацидозу внаслщок стимуляцп юнами водню дихального цен-
тру виникае гшервентилящя, що призводить до зниження рСО2, а в раз! негазо-
вого алкалозу спостерггаеться деяка затримка дихання, що сприяе незначному
пщвишенню рСО2. При зниженн! концентрацп гщрокарбонату на 1 мекв/л рСО2
зменшуеться на 1,2 мм рт. ст. Це дае змогу до певно'1 мгри зберггати нормальне
сшввщношення метаболгчного 1 газового компонента пдрокарбонатного буфера,
а отже, 1 пщтримувати рН у межах норми, однак зменшуе резервш можливосп
буферное системи, осюльки юльюсть \таНСО3 при цьому не вщновлюеться. Та-
кий стан вщповщае компенсовангй стадп негазового ацидозу. Повну нормализа-
цию КОС позаюптинно! рщини й усього оргашзму в раз! негазових ацидозу й
алкалозу здатш забезпечити пльки цирки.
Таблица 14. Основн! параметри КОС у раз! його порушення,
при компенсаторних реакц!ях! в!дновленн!
Вид порушення РН [Н’1 Основн! порушення Тимчасова компенсация Остаточна корекц!я
Негазовий ацидоз 1 Т ЦНСО;] 1рСО2 Т[НСО3]
Негазовий алкалоз Т Т[НСО3] ТрСО2 ЯНСО;]
Газовий ацидоз 1 т )рСО2 Т[НСО3] 1рсо2
Газовий алкалоз Т 1 1рСО2 1[НСО3] ТрСО2
У випадку газового ацидозу нирки залучаються до компенсаторно! реакцп,
посилюючи реабсорбцию натргю гщрокарбонату. Однак цей мехашзм менш ефек-
тивний 1 б1лыц латентний, шж гшервентиляц!я. Так, у раз! гострого ресшратор-
ного ацидозу концентрация гщрокарбонату плазми збглыпуеться пльки на 1 мекв/л
на кожних 10 мм рт. ст. пщвищення рСО2, у раз! хрошчного — приблизно на
3,5 мекв/л. Повну й остаточку нормалгзашю КОС при газовому ацидоз! може
забезпечити лише нормалхзащя системи зовшшнього дихання.
Швидке часткове зменшення порушення нормального сшввщношення ф!к-
сованих капошв 1 анюшв, а отже, ! концентрацп гщрокарбонапв кров! досягаеть-
ся за рахунок ще одного ф!зико-х!м!чного мехашзму компенсац!!' — юнообмшу. У
раз! позаюптинного ацидозу з клхтин виходять гони калпо, а з исток — юни каль-
цпо, магнпо та натрпо. При цьому в юптини входить юни хлору. Кожна змша рН
на 0,1 супроводжуеться протилежною змшою концентрацц калпо в плазм! при-
близно на 0,6 мекв/л. Винятком е лише окрем! рхзновиди ниркового канальцево-
го ацидозу та д!арея, за яких оргашзм втрачае разом з натр!ем велику юльюсть
калпо. Пщ час позаюптинного алкалозу вщбуваються прямо протилежш змши.
Виникае враження, н!би при ацидоз! зайв! йони водню заходить у юптини в обмш
на юни калпо. Насправд!, цей процес значно складнппий, 1 точний мехашзм його
й дос! залишаеться невстановленим, хоча, ймов!рно, вш пов’язаний з! змшою
активност! №+-, К+-АТФази, Иа+/ Н+ ! С1_/ НСО3" проти переносниюв ! селек-
тивних кал!евих канал!в.
Значения нирок в остаточшй корекци негазових порушень КОС визначаеться
тим, що вони забезпечують виведення нелетких кислот, регенерують метаболгч-
ний компонент пдрокарбонатного буфера, який був витрачений на нейтралгзащю
сильних нелетких кислот, реабсорбують увесь гщрокарбонат, який профшьтру-
вався в первинну сечу. Проте дуже часто пщ виведенням кислот по милко во ро-
зумпоть екскрешю !он!в водню. Однак виведення катюшв без анюшв, по-перше,
неможливе, а по-друге, екскрещя Н+ не мала б жодного сенсу, адже !х кшьюсть в
оргашзм! залежить тгльки вщ стану буфергв. До того ж кислоти, яю вводиться в
оргашзм або утворюються в процес! ъгетаболгзму, потрапляють у нирки переваж-
но в забуференому стан! у вигляд! нагргевих солей. Тому немае можливост! здш-
снювати екскрешю кислот як таких. Не можна виводити кислоти з оргашзму г у
вигляд! натр!евих солей, осюльки це може призвести до втрати води гз подальшим
розвитком гшоволемц.
Виходячи з того, шо основною патогенетичною ланкою нересшраторних по-
рушень КОС оргашзму е порушення спгввщношення в позаклгтиншй рщинг фгк-
сованих г напгвфгксованих катюнгв й ашонгв, треба визнати, що роль нирок у
компенсаторных реакциях оргашзму при негазових ацидоз! та алкалоз! полягае у в!д-
новленн! нормального стввгдношення м!ж ф'иссованими катюнами й ашонами плазми
кровг. У цьому процес! роль нирок е визначальною, оскыьки телки вони здатн! забез-
печити !зо!он!ю кровз.
У раз! ацидозу нирки виконують функцию повернення в кров дещо бьтылог
ктлькостг натрпо. н!ж анюнгв нелетких кислот, при алкалоз! — протилежну д!ю.
Цю функцгю нирки здгйснююгь за допомогою мехашзм!в селективно! реабсорбцй
та секрецп катюшв г ашонгв. Селективна реабсорбнгя натрпо забезпечуеться обмг-
ном його на гони амошю, якг утворюються в проксимальних канальиях нефрона,
на !они водню, що секретуються в проксимальних канальцях, товстгй висхщнгй
частин 1 петлг нефрона (Генле) г у вставних клгтинах А-типу зб!рних трубочок, а
також на йони кал1ю, яю секретуються в головних клгтинах кгркових збгрних тру-
бочок. При алкалоз! вставш кл!тини В-типу кгркових збгрних трубочок секретують
у сечу гщрокарбонат в обмш на хлор . реабсорбують у кров йони водню та хлору
У цьому обмгнг беруть участь селективнг юннг канали та переносники (схема 18).
Натрш, бгльша частина якого реабсорбуеться в проксимальному в!дд!л! неф-
рона, повертаеться в кров у вигляд! натрпо гщрокарбонату. Останнгй та юни вод-
ню, шо секретуються в проевгт канальцгв в обмш на натр!г!, утворюються в про-
цес! реакцп, що катал!зуегься нирковою карбоангщразою. Секрецгя гошв водню в
проксимальному' вщд!л! та петлг нефрона забезпечуе реабсорбшю КаНСО3, який
профшьтрувався з кровг в первинну' сечу. Частина гошв волню. взаемолгючи з
№2НРО4, який профшьтрувався, перетворюе його гга №Н,РО4, що дае змогу ре-
абсорбувати юн натрпо у вигляд! МаНСО,.
Збереження основноТ юлькосп натрпо та регенерацгя тим самим гщрокарбо-
натггого буфера, витраченого на нейтралгзацгю нелетких кислот при ацидоз!, за-
безпечуетъея нирковим амонюгенезом. Ранние було прийнято вважати, шо в про-
цес! ниркового амошогенезу з глутамшу г частково з !нших амшокислот утво-
рюеться амгак (>Ш3), який у проевгп ниркових канал ьщв з’еднуеться з юном
ьодню й утворюе амонгй (МН4Т), що е засобом виведення з органгзму гонгв водню.
Однак тепер вщомо, шо рН системи ам!ак — амонгй доргвнюе 9,2. Пе означае, що
при рН 7,4 понад 99 % амгаку перебуваютъ у протонованому сташ Осюльки внуг-
182
Проксимальный Первинна сеча
каналець ИаНСО,
нефрона Ка.НРО'
№80.‘
4
№+-лактат (ацетат)
№С1
нсо;+н+
V
н2со3
Карбоанпдраза
V
| н2о + со2
Глутамш
№+
Глутамш
№+
лактат
№+
Глюкоза
Збгрна трубочка
ЫН*( К+) (ацетат)
КС1(8О2’)
ХаН,РО,
№ЛС1
Юнцева сеча
К4
К+
Схема 18. Мехашзми, що забезпечують ниркову регулящю КОС:
1 — первинно активн! переносчики та протипереносники, що використовують АТФ;
2 — вторинно активы! ко- та протипереносники, що використовують концентращйний
град1ент одюй речовини для транспорту шшоц 3 — селективны юнн1 капали
ршшьоюптинний рН — 7,0—7,2, то всередиш юптин ниркових канальщв вм!ст
амонпо в 10 тис. разгв б!льший, нгж амгаку.
Отже, амошогенез не може бути засобом забуферування та виведення 13 се-
чею юшв водню. Нирковий амошогенез е засобом, що дае змогу виводити з оргашзму
надлишок анюшв нелетких кислот у вигляд! амоншних солей без втрати юн!в на-
трйо. Амошогенез ! карбоанггдразна реакщя забезпечують таким чином регенерацйо
г!дрокарбонатного буфера за рахунок в!дновлення нормального сп!вв!дношення м!ж
катюнами й ашонами плазми кров!.
Частина оргашчних ашошв реабсорбуеться ештелгем ниркових канальщв г
метаболгзуеться в них до глюкози, що потрапляе в кров. Решта оргашчних ашошв
1 надлишок усгх неоргашчних ашошв виводяться з оргашзму як амонгйнг та калгев!
сол1, а також у меншгй юлькостт у вигляд! так званих титрованих кислот, що €
здебглыпого дигщрофосфатами.
Регуляцгя ниркових ацидо-, амошо- та глюконеогенезу, а також активност!
селективних !онних переносникгв 1 канал!в здгйснюеться як м!сцево, п!д впливом
метабол (я них факторгв (пперкалгемгя, рН, рСО2 та !н.), так г дистантно — за до-
помогою нейрогуморальних фактор!в, серед яких зараз в!дом! катехоламши, до-
памш, альдостерон, анпотензин II, глюкокортикощи, шсулш, вазопресин, парат-
гормон, мехашзми впливу яких на нирков! процеси в раз! порушення КОС вив-
чаються.
Вщновлення завдяки робот! нирок нормального сшввщношення фгксованих
катюнгв 1 ашошв плазми кров! автоматично нормал!зуе концентрацгю гщрокарбо-
нату, а отже, ! КОС оргашзму.
ПАТОГЕНЕЗ ПОРУШЕНЬ КОС
Надмгрна концентращя вуглекислого газу (пперкапшя) при газовому ацидоз! так
само, як 1 юни водню та калпо, справляе прямий судинорозширювальний ефект у
бгльшосп тканин, особливо в головному мозку, унаслщок чого пщвищуеться внут-
ртшньочерепний тиск. Кр!м того, пперкапшя стимулюе вазомоторний центр у дов-
гастому мозку. Вплив активацп нейрошв вазомоторного центру, опосередкований
симпатичним судинозвужувальним вщдшом нервово! системи, зумовлюе у перифе-
ричних тканинах спазм артерюл. Цей централ ьний судинозвужувальний ефект вуг-
лекислого газу потужшший, н!ж прямий периферичний судинорозширювальний
вплив вуглекислого газу, що спричинюе пщвищення при гшеркапнп системного
артер!ального тиску, рефлекторний розвиток брадикардп через барорецептори ар-
тергй та може зумовити виникнення лгвошлуночково! серцево! недостатносп.
Гшокапн!я при газовому алкалоз! спричинюе звуження судин мозку, що про-
являеться головокружениям, запамороченням 1 парестезгями. При цьому гальму-
вання вазомоторного центру знижуе тонус периферичних судин г, вщповщно,
артергальний тиск.
При декомпенсаци газових 1 негазових порушень КОС ! зсув! рН за меж! нор-
ми виникае полюрганна недостатшсть, зумовлена змшою активност! ферментгв,
проникносп мембран, порушенням синтезу АТФ, змшою нервовом’язово! збуд-
ливосп, гуморально! регуляцп тощо. Порушення функцп мозку спричинюе сплу-
тангсть свщомосп, запаморочення, ступор, кому та смерть.
Патогенетична роль механгзмгв компенсаци порушень КОС. Численн! порушення
функцш оргашзму виникають ще на етат компенсованого ацидозу й алкалозу, ос-
юльки мехашзми компенсаци, що включаються при порушеннях КОС, можуть сам!
вццгравати патогенетичну роль. Так, у раз! негазового ацидозу виникають гтер-
калгемгя, гтеркалыремгя та ггпокапнгя, негазового алкалозу — ггпокалгемгя та гтокаль-
цгемгя. Гшеркал!ем!я може призвести до зупинки серця, гшокал!ем!я — до м’язово!
слабкосп, артер!ально! ппотензп, аритмп. Гшокальщем1я здатна спричинити судоми
1 тетанпо. Гшокапшя супроводжуеться шгем!ею та ппокаею мозку. запамороченням,
сонливютю 1 може спричинити потьмарення свщомосп. Виведення кальцпо 13 юсток
у раз! тривалого ацидозу сприяе розвитку остеопорозу 1 нефролтазу. Видалення пщ
час алкалозу з оргашзму надлишку ИаНСО3 може викликати гшоволем!ю.
Напруження регуляторних систем оргашзму, яке супроводжуе порушення
КОС, зумовлюе !х дисбаланс, що спричинюе розвиток р!зних патолопчних змш.
Так, негазовий ацидоз сприяе викиду катехолам1шв 1 активацп симпатично! час-
тини автономно! (вегетативно!) нервово! системи з одночасним пщвищенням то-
нусу парасимпатично! частини 1 зниженням чутливост! а- 1 р-адренорецептор!в.
Усе це спричинюе вегетативно-судинну дистонпо, бронхоспазм, пщвищення сек-
рет! слизу, блювання, пронос. При негазовому алкалоз! пщвищуеться збудливють
р-адренорецептор!в серия, судин, бронхов, кишок, знижуеться тонус блукаючого
нерва, що супроводжуеться тахпсард!ею, зниженням артер!алъного тиску, уповшь-
ненням перистальтики кишок.
Патологгчний вплив порушень КОС на обмш вуглеводгв. Помхрний компенсова-
ний ацидоз значно шдвищуе зв’язування шсулшу з рецепторами 1 знижуе спорщ-
нешсть тканин до глюкагону (Н.В. Крышталь, 1989). Значний декомпенсований
ацидоз спричинюе шсулшорезистентюсть. Як хрошчний негазовий ацидоз, так 1
хрошчний негазовий алкалоз зумовлюють пперппкемпо натще та знижують толе-
рантшсть до глюкози. Першопричиною д!абетогенно! дп хрошчного ацидозу е
виснажлива стимулящя р-ендокриноцитгв (р-клггин) острхвцгв пщшлунково! за-
лози гшергл!кем!ею, яка е наслщком посилення неоглюкогенезу, зумовленого п-
перпродукщею глюкокортикоццв. Алкалоз спричинюе первинну пригшчувальну
дпо на р-ендокриноцити й активуе а-ендокриноцити (а-клггини) остр!вц!в пщ-
шлунково! залози (Н.В. Крышталь, Э.Г. Гареева, 1993).
Патологгчний вплив ацидозу на бглковий обмш. Нирковий синтез юшв амонпо,
який забезпечуе виведення з оргашзму переважно! юлькост! анюн!в нелегких
кислот у вигляд! амошйних солей, як субстрат використовуе глутамш, що утво-
рюеться в раз! переамшування р!зних амшокислот. Основним джерелом амшо-
кислот для киркового амошогенезу е АТФ-залежний уб!кв!тин-протеасомальний
протеол!з, який при ацидоз! стимулюеться глюкокортикощами. Активац!я проте-
олизу при хрошчному ацидоз! стимулюе катаболизм бшюв м’язгв г юсток, що спри-
чинюе втрату м’язово! маси й остеопороз юсток на тл! негативного азотистого
балансу (М.В. Кришталь, 2003).
Принципа корекци порушень КОС полягають у л!куванш хвороб, яю стали при-
чиною порушення рСО2 або [НСО~3]. При негазовому ацидоз! чи алкалоз! необ-
хщно виявити причину порушення нормального сшввщношення фгксованих
катюшв й анюшв та усунути п. Якщо це не вдаеться зробити швидко, то доц!ль-
ними будуть нормал!зац!я буферних систем оргашзму, усунення супутшх розлад!в
водно-мшерального обмшу та симптоматичне лгкування.
Р03Д1Л XXII
ПОРУШЕННЯ ВОДНО-ЕЛЕКТРОЛ1ТНОГО ОБМШУ
Водно-електролТтний обмш (ВЕО) — пТдтримання постшно! киькосггй води в ор-
ганизм! з певним перерозподиом н мТж «одними секторами (компартментами) !з
суворим шдтриманням Тх Тонного складу й осмолярностТ.
Нормальна житгедгяльнгсть клггин, якг е основною морфофункцюнальною
единицею людського органгзму, може здгйснюватися лише за умови оточення гх
водним розчином постгйного складу, який К. Бернар назвав внутрппнгм середо-
вищем органгзму. Вода, що е унгверсальним бгологгчним розчинником, мгститься
також г всерединг клгтин, що забезпечуе обмгн речовин г метаболгзм. Причому,
якщо склад внутргшньоклгтинних розчингв дещо вгдргзняеться в ргзних органах г
органгзмах, то йонний склад позаклгтинног рщини завжди однаковий г подгбний
до океанськог води (табл. 15). Це пов’язано з тим, що першг клгтини виникли г
мшгарди рокгв розвивались у первгеному океаш, неоргангчний склад якого бага-
токлгтиннг органгзми навчилися вщтворювати штучно в мгжклгтиннгй рщинг, щоб
забезпечити гомеостаз г нормальну життедгяльнгсть сво'гх клгтин.
Таблице 15. Середый склад основних южв у бюлопчних рщинах людини
— .-Г* 1- - ~ Рщина Иа*, мекв/л К+, мекв/л СГ, мекв/л НСО3, мекв/л
Внутр1шньокл1тинна 12—38 100—150 4—50 12
Плазма 142 4,5 104 24
Мгжклгтинна 140 4,4 117 27
ШлуНКОВИЙ С1К 60 15 130 0
Вмгст ТОНКО! кишки 130 10 70 80
ВМГСТ товстоГ кишки 30 80 20 25
Дгарейна рщина 60 45 40 50
Пгт 30 5 35 0
Вмгст води в оргашзмг та П перерозподгл мгж водними секторами. Загальна кгль-
кгсть води в новонароджених (табл.Чб) сягае 75—80 %>, а в дгтей вгком 5 рокгв г у
дорослих чоловгкгв — 60 %. У жшок 1 чоловгкгв з надмгрною масою тша вмгст води
в органгзмг становить 50—55 %, а в осгб лгтнього вгку — 45—55 %. Уся вода роз-
подшяеться мгж двома основними рщинними компартментами (секторами, про-
сторами): внутршгньоклгтинним г позаклгтинним. У дорослог людини 2/3 води
шститься всередин! юптин, у дитини грудного вгку — близько половини, а в но-
вонародженого — ще менше. При цьому внутршшьоклггинний об’ем найбшьш
стадий, а змша загально! юлькосп води вщбуваеться за рахунок позаюптинного
сектора. Позаюптинна рщина подшяеться на внутршшьосудинну, штерстищаль-
ну 1 трансцелюлярну, до яко! належать сщгнномозкова, плевральна, перикард!аль-
на, внутр!шньоочеревинна, синовхальна рщини, а також вода, що мгститься в
травному канал!. Найбшьш сталим з ус!х позаклггинних комнартменттв е об’ем
внутрппньосудинно! рщини, вщ якого залежить нормальна робота серцево-судин-
но! системи, а отже, й кнування оргашзму. Вш майже не вщр!зняеться в дней 1
дорослих. Якщо в дорослих плазма кров! становить близько 5 % маси тша, а сама
плазма на 90 % складаеться з води та на 7 % з бшка, то внутрпиньосудинна вода
становить близько 4,5 % маси тша. Менш стабшьною е штерстишальна, або мгж-
юптинна, рщина, до яко! належить 1 лгмфа. У новонароджених вона становить
близько 1/2 вше! води, а в дорослих — лише 1/4. Частка трансцелюлярно! води за
нормальних умов становить 1—3 % маси тша, але може значно збшыпуватися,
наприклад, при асцит!.
Таблица 16. Середжй загальний вмгст води в оргажзм!
та деяких його компартментах вщносно маси т!ла (%)
В1к, стать, юлыастьжиру Загальна вода Внутршньо- клпгинна вода Позаклпинна вода
плазми 1нтерстиц1К>
Новонароджен! 75—80 34 5,5 34—39
Доросл! 60 40 4,5 14
Худорляв! чолов1ки 65 41 5 17
Чоловпси та ж!нки з надонрною масою пла або похилого в!ку 50 36 4 9
Осмотично активы! речовини та осмотичний тиск у норм! та при патолоп!. Вода
утримуеться в рщинних компартментах завдяки розчиненим у н!й речовинам, як!
подшяються на електролгги та неелектролпи. Елекгрол!ти — це речовини, що
дисоцпоють у розчин! на заряджен! катюни й ан!они, а неелектрол!ти — це речо-
вини, яю не дисоцпоють, наприклад глюкоза, сечовина, креатинш, бшрубш.
Концентрац1я розчинених речовин зумовлюе осмотичний тиск розчишв, тому щ
речовини називаються осмотично активными. Концентращя осмотично активних
речовин, або осмотичний тиск, тобто сила з якою притягуеться й утримуеться
вода, визначаеться не розмтрами, а кшьюстю юшв, молекул, колощних частинок.
Осмотичний тиск виражаеться осмоляршстю. тобто кшьюстю мйпмолей розчине-
них речовин в 1 л розчинника (одиниця вим!ру — мосмоль/л), або осмоляльшс-
тю, тобто кшьюстю мйпмолей розчинених речовин в 1 кг розчинника (одиниця
вим1ру — мосмоль/кг). Хоча завдяки наявност! в плазм! бшка п осмоляльшсть
трохи вшца за осмоляршсть, р!зниця м!ж ними в бюлопчних рщинах несутгева.
Кожний водний компартмент вщокремлений вщ пшгого перегородками, як!
е проникними для води завдяки тому, шо спнки капшяр!в легко пропускають
М1Ж ендотелюцитами воду 1 сол!, а лшщш мембрани бшьшост! юптин, у тому
чишп ендотелюципв, мають бшков! водн! канали — аквапорини. Тому осмотич-
ний тиск однаковий в усгх водних секторах г в норм! становить 290± 10 мосмоль/
л, що вщповщае осмолярноста первгсного океану. За нормалъних умов збщьшен-
ня або зменшення осмолярност! не перевищуе 1 %. Пщвищення осмолярност!
плазми понад 300 мосмоль/л називають гшеросмоляршстю, а зменшення нижче
280 мосмоль/л — гшоосмолярнгстю. Збщьшення осмолярност! понад 340 мо-
смоль/л або зменшення нижче 260 мосмоль/л спричинюе розвиток коми.
Ефективш осмотично активн! речовини як головна руппйна сила перемгщення
води мгж водними компартментами. Ус! осмотично активн! речовини подыяються
також на ефективш та неефективш. Ефективними речовинами е так!, що не мо-
жуть выьно перетинати межу м!ж двома водними компартментами, а тому утри-
мують воду в одному з них. На меж! мгж кров’ю та гнтерстицгем ефективними
осмотично активними речовинами е лише быки плазми, оскыьки солг, глюкоза
та сечовина выьно проходять кр!зь стшку капшяра. Тиск, що створюеться быка-
ми плазми (альбумшами, а,-глобул!нами), називаеться онкотичним. Його частка
в загальному осмотичному тиску кров! незначна, усього 2 мосмоль/л, але саме
його змгна спричинюе перемгщення води мгж судинним руслом 1 тканинами.
Кр!зь плазматичну мембрану клггин выьно проникають алкоголь та гншг жи-
ророзчиннг речовини. За наявноста на мембран! клгтин аквапорину-1 через клгги-
ни проходять вода г фгзголопчна кглькгсть сечовини, а в раз! наявноста достатньог
кыькоста ГЛЮТ — нормальна кыыасть глюкози, а сол! та надлишок глюкози —
ш. Тому на меж! м!ж позаклгтинною та внутргшньоклгтинною рщиною ефектив-
ними осмотичними речовинами е !они солей г надлишок глюкози. Оскыьки, як
видно з табл. 16, осмолярнгсть плазми г м!жкл!тинно! рщини на 90 % залежить вщ
натр!ю 1 супутшх йому анюн!в, то й об’ем позаклгтинног рщини залежить у першу
черту вщ кыькоста в шй натрпо.
Збыьшення осмолярноста позаклгтинног рщини при гшернатргемгг або ггпер-
глгкемгг спричинюе пасивний вихщ води з клгтин за осмотичним градгентом, що
зумовлюе зморщування клгтин, а зменшення осмолярноста позаклгтинног рщини —
набухания клгтин. Це особливо небезпечно для функщонування клгтин мозку !
може спричинити розвиток коми. Тому для забезпечення оптимальних умов гсну-
вання клгтин оргашзм у першу черту пщтримуе сталгсть осмолярноста, а не об’ему
позаклгтинног рщини.
Мехашзми пщтримання нормально! осмолярноста позаклгтинног рщини. Збыъ-
шення осмолярност1 позаклтиюип р1дини вщ нормальних коливань лише на 2 %
подразнюе осморецептори гшоталамуса, що спричинюе акгивацгю секрецп анти-
дгуретичного гормону (АДГ) нейронами супраоптичного та паравентрикулярного
ядер гшоталамуса через задаю частку ггпофгза. Пщ впливом АДГ у люменальну
мембрану нефроцитгв збгрних трубочок вбудовуеться трансмембранний бшок ак-
вапорин-2, що збгльшуе проникшсть цього вгддглу нефрона для води г сечовини
й активуе гх реабсорбцгю. При цьому АДГ, стимулюючи синтез у нирках проста-
гландишв, зумовлюе г натргйуретичний ефект, що прискорюе лгквщацгю ггпер-
осмг'г. Пперосмоляршсть також через, централып осморецептори викликае вщчут-
тя спраги, яке змушуе людину вживати воду. Кргм того, збыьшення концентрацп
Ка+ г СГ у канальцевгй рщинг в зонг таси1а Зепза спричинюе гальмування продук-
цп ренгну, що зменшуе синтез ангготензину II г альдостерону, а це своею чертою
зменшуе реабсорбцгю та збгльшуе екскрецпо №+ нирковими канальцями.
Зменшення осмолярностг позаклтинног ргдини, навпаки, зумовлюе блокування
секрецй АДГ, зникнення вщчутгя спраги й активацпо ренш-ангютензин-альдо-
стероново! системи (РААС). Усе це спричинюе прискорене виведення з оргашзму
води, затримку натрпо 1, як результат, пщвищення осмол ярносто позаклотинно!
рщини до норми. „
Мехашзми пщтримання сгалосп об’ему 1 тиску кровь 1ншими стимулами для
збшьшення спраги ! секрецй АДГ с зменшення ОЦК або зниження артергального
тиску. Хнформацпо про змши об’ему 1 тиску кров! сприймають барорецептори,
розташован! в стонках аорти та сонних артерш, у передсердях, шлуночках, в!нце-
вих судинах о легеневих венах. Ц1 рецептори являють собою механорецептори, що
реагують на розтягнення стшок судин або серця. Для мехашзмов регуляцн об’ему
рщини, омоворно, найважлившшми е серпево-легенев! барорецептори, що реагу-
ють на змшу центрального венозного тиску й об’ему кров!, а тому називаються
волюморецепторами. Зниження об’ему кров! або артер!алъного тиску не менш
н!ж на 10 % знижуе штенсивиость !мпульс!в, яю надходять вщ волюморевепторов
у гшоталамус, шо збшьшуе спрагу 1 секрецпо АДГ, а вщ барорецепторов аорти та
сонних артерш — у судиноруховий центр довгастого мозку, що посилюе актив-
шсть симпатико-адреналово! системи. Активацоя симпатико-адреналово! системи
через 0-адренорецешори спричинюе тахокардно, через а-адренорепептори збшь-
шус тонус судин, через ^-адренорецептори гранулярних клотин юкстагломеру-
лярного апарату нирок стимулюе секрецпо реши у о, впливаючи переважно через
с^-адренорецептори на №+ / Н^-антипорт (протипереносник), безпосередньо
сгимулюе реабсорбцпо натрою в проксимальних канальцях нефрона. Кром того,
гранулярно юптини, шо синтезуютъ о секретують реши, само дпоть як барорецеп-
гори о збольшують секрещю в разо зниження артероалыюго тиску. -Активащя РААС
збшьопуе реабсорбцпо 1Ма+, шо пшвишуе осмоляршсть крово, а це своею чертою
ше бшьше стимулюе секрещю АДГ о, вщповщно, збшьшуе реабсорбцпо води.
Кром того, активована симпатико-адреналова система о АДГ, звужуючи артерюли
ниркових клубочков, зменшують клубочкову фольтрацпо о. вщповщно, екскрещю
натрою та води. Анпотензин II до того ж е погужним стимулятором нейронов то-
пот аламуса, як! вщповщають за вщчутгя спраги.
При збглыиенш об’ему позаклйпинноТ ргдини, особливо ОЦК, або при пЮвищенш
артергального тиску больше нож на 10 % реакцп органозму будуть, навпаки, спря-
мовано на виведення оз сечею натрою та води. Кром гальмування РААС о зменшен-
ня секрецн АДГ розтягнення передсердь у разо збшьшення об’ему кров! спричи-
нюе секрещю кардюмюцитами передсердного (атроальното) нагршуретичного
пептиду (передсердною наоршуретичного фактора, атроопептину), який знижуе
реабсорбшю натрою канальцями нефрона, гальмус синтез реншу, альдостерону й
АДГ та збшьопуе клубочкову фольтрапою. Кром того, натршуретичну дно мають ПГЕ,
о ПП2 та МО, як! утворюються в нирках при зболыпенн! артергального тиску.
Шляхом згегпавлення механгзмгв пгдтримання осмолярностг з механизмами за-
безпечення сталостг об’ему встановили, шо, по-перше, для оргашзму прооритет-
ошм е пщтримання осмолярносто, а, по-друге, в обох випадках задояно однаково
доурети чно. антидоуретичш, натройуретичш й антинатр!йуретичн! мехашзми. Вщ-
моншеть полягае в тому, що при забезпеченн! сталост! об’ему’ чи тиску кров! ак-
тивацоя або блокада реабсорбцп води збогаеться з активацию чи пригшченням
реабсорбцп натрпо, тод! як при порушеннях осмолярносп все вщбуваеться навпа-
ки. Так, при пщвищенш осмолярност! позаклпинно! рщини збыьшення реабсор-
бцп води супроводжуеться зменшенням реабсорбцп натрпо, а при зниженн! —
зменшення споживання 1 прискорене виведення води 13 сечею вщбуваеться на тл!
посилено! реабсорбцп натрпо.
Р13НОВИДИ, ПРИЧИНИ ТА НАСЛ1ДКИ ПОРУШЕНЬ
ВОДНО-ЕЛЕКТРОЛ1ТНОГО ОБМ1НУ
Порушення балансу м!ж споживанням 1 виведенням води спричинюе зневоднення
(дегидратация, ексикоз, гтоггдратащя, гтоггдргя) або накопичення води в оргашзли
(гтерггдратащя, г1перг1др1я). Залежно вщ змш осмолярност! позаюптинно! рщини
ппо- та гшергщратац1ю подыяють на 1зоосмолярну, гтоосмолярну та гтеросмоляр-
ну (мал. 45).
Депдратащя розвиваеться в тих випадках, коли видыення води перевищуе
надходження и в оргашзм (негативний водний баланс). Причиною може бути
порушення надходження води в оргашзм (водне голодування, розлад ковтання,
атрез!я стравоходу, коматозний стан та пт.) або пщвищена п втрата (пронос, блю-
вання, крововтрата, полтуртя, гшервентиляцгя, збыьшене потовидшення, втрата
рщини з ексудатом ! транссудатом — отк тощо), а також поеднання цих пору-
шень.
Зневоднення зумовлюе тяжк! наслщки, пов’язаш з! зменшенням ОЦК (ппо-
волемтя) ! пщвищенням п в’язкостт Порушення кровооб!гу спричинюе гшокспо
тканин, яка впливае передуам на центральну нервову систему. Це може проявля-
тися потьмаренням свщомост!, Об’ем
галюцинащями, розвитком ко-
матозного стану. Порушуються
також функцп нервових центр!в,
ритм дихання, шдвищуеться тем-
пература т!ла. Значне зниження
артер!ального тиску може супро-
воджуватися порушенням ф!ль-
трацп в клубочках нефрошв, олх-
гур!ею, пперазотем!ею 1 негазо-
вим ацидозом.
Добре вщомо, що при нор-
мальшй глпсемп осмоляршсть
позаклпинно! рщини досить точ-
но можна розрахувати за форму-
лою: Р (мосмоль/л) = 2 • {[ТЧа+] +
[К+]} (мекв/л). Отже, зрозумыо,
що осмоляршсть плазми та м!ж-
клггинно! рщини залежить пере-
важно вщ вмтсту натрпо. Оскыь-
ки вм!ст травного каналу (див.
табл. 15), сеча при нецукровому
Норма
1зоосмолярна депдратащя
Нестача Ха+ та води
Гшеросмолярна депдратащя
Нестача переважно води
Гшоосмолярна депдратащя
Нестача переважно №+
1зоосмолярна пперпдратащя
Надлишок №+ та води
позаклт-
тинний
внутр^шньо-
КЛ1ТИННИЙ
Гшеросмолярна пперпдратащя
Надлишок переважно Иа+
Гшоосмолярна пперпдратащя
Надлишок переважно води
Мал. 45. Сшввщношення позаклггинно! (злтва) та внут-
рцпньоклггинно! (справа) рщини в норм! 1 при патоло-
пчних процесах
390
_______
(порушення секрецп або рецепцп АДГ) г цукровому дгабет! та, особливо, шт 1 пара
мктять солей нагр!ю менше, Н1ж плазма, блювання. пронос, лперсашвашя. гшер-
вентиляцгя, надмгрне потовидыення та полгургя спричинюють гтеросмолярну де-
г!дратац1ю Гшеросмолярнкть (гптернатртемгя) особливо виражеш в щтей ран-
нього В1 ку. Це зумовлено високим вмтстом у кров! дггей раннього в!ку анпотснзи-
ну II 1 альдостерону, зменшеною швидкктю клубочковот фгльтрацп, низькою
конпенгращйною здатнктю нирок, вщмосно великою площею поверхн! тша, ви-
сокою частотою дихання 1 недосконалктю регуляторних механгзм!в, як! забезпе-
чують екскрецпо надлишку натрпо. Унаслщок цього в дней перших рокгв життя
гшеросмолярне зневоднення у випадку' гострих шлунково-кишкових захворювань,
якг супроводжуються блюванням г / або дгареею, та в разг збглыденого потовидт-
лення та гшервентиляцп, що спостерггаються при гарячш та в спеку, настае часть
ше, Н1ж у дорослих, ! е тяжким ускладненням, яке може призвссти до смерть
Пщвищення осмотичного тиску позаклгтинно']’ рщини спричинюе вихщ води
з нервових клгтин за осмотичним градкнтом та гх зморщування, що зумовлюе вщ-
чутгя спраги, дралвливкть. сонливкть г судом и. Якшо доросла людина при ньому
п'е воду, гшеросмоляршсть змшюетъся на нормоосмоляршсть, а при надлишковому
споживанш безсольових напо1в — на гтоосмоляршстъ, яка викликае перемгщення
води всередину клгтин. Дгтям грудного вгку рщину слщ давати часто, але дргбни-
ми порцгями, шоб запобпти як ггперосмолярнгй дегщратацп, так г ппоосмолярнгй
пперпдратацй. Дорослим у спеку слщ пити пщсолену воду.
Ппоосмолярна дег1драташя виникае також унаслщок втрати назргю гз сечею
при недостатносп кгрковог речовини надниркових залоз (хвороба Двдгсона), при
штенсивному застосуваинг дгуретикгв г в полгуричну фазу гостро! нирковог недо-
статностг. Осюльки при ппоосмолярнгй тшогщратацп основн! втрати рщини вщ-
буваються гз позаклгтииного сектора, то швидко розвиваються ознаки дегщратацп
зг зменшенням ОЦК: зниження тургору шкгри, сухгсть слизових оболонок, тахг-
кардгя, ортостатична ппотензгя, слабка пульсацгя яремних вен та олггургя. Зни-
ження осмотичного тиску в позаюптиннгй рщин! зумовлюе перехщ води в кл!ти-
ни та гх набухання з розвитком симптомгв пщвищеиого внутрппньочерепного тис-
ку (блювання, головний б!ль, потьмарення свщомостг, браликардгя, апапя).
Перемгщення води у внутр!шньоклгтинний сектор посилюе пповолемпо, згушен-
ня кровг га порушення кровообггу.
Тзоосмолярна дег!дратащя, класичними прикладами яког е гостра крововтрата
та значний опгк шюри, за яких оргашзм ргвномгрно вграчае сол! та воду, супро-
воджуеться зменшенням об'ему позаклгтинно!’ ргдини без змши кглькостг води у
внутр!шньоклгтинному сектор!.
Лповолемя (зменшення ОЦК), яка супроводжуе значну дегщратащю будь-яко!
осмолярностг, може спричинити розвиток колапсу 1 шоку з гострою нирковою
недостатнктю.
Гшергщраташя бувае загальяою та з переважним накоггиченням рщини в ок-
ремих компартментах. Так, накопичення води у мгжклгтинному сектор! нази-
ваеться набряком (оейета), внутршшьосудинному сектор! — пперволемкю, внут-
ргшньоклпи иному компартменп — набуханиям клгтин, а в серозних порожнинах —
водянкою (ИудгорЦ. водянку внутргшньоочеревинно! порожнини називають
асцигом (азсИея), плеврально! порожнини — ИуагогИогах. шлунолкгв мозку —
1тус1госерИа1их, перикардхально! порожними — ИуйгорепсагсНит. Накопичення неза-
пально! рщини мае назву “транссудат”, а запально! — “ексудат”.
1зоосмолярна г1перг1дратац1я, яка можлива при надмгрному введенш !зотошч-
ного розчину №С1 або шших сольових кровозамшниюв, харакгеризуеться збшь-
шенням юлькост! води в позаюптинному сектор! без суттево!' змши об’ему внут-
ршшьоюптинного сектора.
Пперосмолярна гтерг1дратац1я в дорослих спостерйаеться при вторинному п-
перальдостерошзм! (серцева або печшкова недостатшсть, нефротичний синдром)
та дуже рщко при вживанш сольових розчишв, а в дней можлива в перюд ново-
народженосп та у вщ! до 1 року при надмгрному вживанн! натрпо. При цьому
вода накопичуеться переважно в позаюптинному сектор! з розвитком генералтзо-
ваних набряюв.
Ппоосмолярна гтерггдратащя (водне отруення) може виникати або в дней ран-
нього впсу у зв’язку з недорозвинешстю нирок 1 систем !х регуляцн, або в дорос-
лих 1 датей р!зного в!ку при неадекватно збшьшенш секрецй АДГ або при надмгр-
ному парентеральному введенш розчишв, як! не мктять електролгти (наприклад
розчин глюкози, який шсля утшпзацн глюкози перетворюеться на дистильовану
воду). При цьому вода потрапляе в юптини, що спричинюе !х набухания.
В експеримент! на тваринах гтоосмолярну гтерггдратацт можна спричинити
повторним уведенням води в шлунок. Одноразове водне навантаження у здоро-
вих тварин зазвичай не зумовлюе тяжк! наслщки. Згщно з результатами дослщ-
жень при надмтрному введенш води в кров, навпь в об’ем!, який дор!внюе мае!
кров!, масова частка води в кров! мало змшюеться (М.Н. Зайко). Це зумовлене
затримкою води в печшц!, м’язах, селезшц!, шюр!, а також посиленим виведен-
ням ц з оргашзму. Однак у раз! порушення регуляцн водного обмшу вже незнан-
ие водне навантаження може призвести до гшергщратацп. Так, експериментальне
водне отруення можна спричинити водним навантаженням з одночасним уведен-
ням вазопресину, альдостерону або видаленням надниркових залоз. Та обставина,
що адреналектомоваш тварини, яю зазвичай гинуть вщ втрати солей натрпо !
зневоднення, погано переносить водне навантаження, пояснюеться зниженням
артергального тиску, а отже, г клубочково! ф!льтрацй шсля видалення наднирко-
вих залоз.
Пперволелйя при значнш гшергщратацп будь-яко! форми може спричинити
гостру серцеву недостатшсть 1 набряк легень.
Набухания кмтин можливе 1 без змши осмолярност! позаклпинног рщини. Це
пов’язано з тим, що плазматичш мембрани не е абсолютно непроникними для
юшв. Вони можуть проходити кр!зь клггини! мембрани через селективш юнш ка-
нали. 1они калпо за градиентом концентрацп постшно “виходять” з клпин, а юни
натрпо — “заходять” у юптини. Це зумовлюе збшьшення юлькост! Иа+ в клпинах
лише за умови недостатност! №+, К+-АТФази, яка виникае при енергодефщит!
клпин, наприклад, в умовах гшоксп. Осюльки кожний юн натрпо зв’язуе 14 моле-
кул води, а юн кал!ю — лише 10, накопичення натрпо в клпинах спричинюе гх
набухания, яке збшьшуе проникшсть цитолеми через механо- й АТФ-залежш ка-
пали та прискорюе вхщ натрпо ! кальцпо всередину клпин 1 вихщ кал!ю з клпин,
що своею чертою порушуе бюелектричш процеси в нервових г м’язових юптинах.
Набряки, згщно з теоргею Е. Старлшга (1896), виникають або при збшыпеню
фшьтрацп води гз капшяр!в в штерстищальний сектор пщ впливом пщвшценого
Мал. 4о. Мехашзм розвитку набряку в раз) змши гщ-
роста!ичною (А) й онкотичного (Б) тиску (за Стар-
. питом):
1 — артер1альна дьтянка катлярно! судини (рщина
входить у судини); 2 — зона ршноваги: 3 — зенозна
дьтянка кагплярноЁ судини (рщина налходить и ткани-
ни в кров); пунктир!й лшК: а — гпдвищення пдроста-
тичною тиску; б — зниження онкотичного тиску кров1
при патологи; суцщыи лнш — нормально величина
вщповщних ТИСК1В
гщростатичного тиску кров! на венозному кёнщ каппяра, або при зменшенн: ос-
мотичного висмоктування штерстишальноТ рщини бьтками плазми на тлё зни-
ження онкотичного тиску кров) га / або пщвишення онкотичного тиску тканин
(мал 46). Найчаспше причиною пщвищення гщростатичного тиску кров] в умо-
вах патологи е вади клапанов серця, як! зумовлюють застигну правошлуночкову
серцеву недостатшсть ё застиг кров) у венах. Венозний тиск пгдвищуегъся також
вщ здавлювання га закуп орюван ня (тромбоз) вен, порушення гх клапанного апа-
рагу, пщ час тривалого стояния.
Пщвишення гщростатичного тиску кров! спостерггаеться також при збгль-
шенш об’ему кров!. ГЁперволемЁя своею чертою може бути наслщком затримки
натрпо та води при нирковгй недостатностЁ або в разг посилення гх реабсорбцп.
Осганне зокрема спостерггають при склероз) судин нирок, лшошлуночковЁй сер-
цевш недостатносп та заспингй серпевгй недостатносп, коли зменшуеться при-
ток артергальноТ кровг до юкстагломерулярного апарату нирок знижуеться ар-
тергальний тиск без зменшення ОПК. Усе це ЁмЁтуе крововтрату г спричинюе, за
описаними вище мехашзмами, стимуляшю РААС, розвиток вторинного гшераль-
достеронгзму та збшъшення секрецп АДГ, що посилюе ниркову реабсорбшю на-
трию г води з розвитком пперволемп. Так виникае “помилка регулящТ". яка поси-
люе набряк. Г. Селье назвав такт явища хворобами адаптацгг.
Локальне збшъшення гщростатичного тиску кровЁ в систем) ворЁтно'Ё вени при
цирозг печшки та заспйнгй серцевгй недостатносп спричинюе розвиток асциту.
Зменшення онкотичного тиску кровг виникае або внаслщок порушення син-
тезу бшкЁв плазми при голодуванш та печшковгй недостатносп. або в раз! гх втра-
ти ёз сечею при нефротичному синдром), або внаслщок виходу бшкгв ёз кров) в
ЁнтерстицЁв при збшьшенш проникностЁ капЁлярног стёнки. Остан не виникае в
разЁ впливу на судинну стЁнку кёнёнёв, амЁвгв, ХО ё ейкозанощЁв (простагландини
та лейкотрЁени) при алерпчних ё запальних процесах, тнфекцп, ггпоксп, ацидоз) Ё
токсикоз). БЁлки плазми кровЁ, якё виходять у мЁжклЁтинну рЁдину, можуть повер-
нутся в кров лише через лЁмфатичну систему. Оскшьки лЁмфатичнЁ капЁляри при
набряку перетискаються, бьчки кров! затримуються в Ёнтерсппш, що зумовлюе
гпдвищення онкотичного тиску тканин. При запатеннЁ, алергп та ппоксп виникае
мЁспевий ) / або системний ацидоз, який активуе протеолЁтичш ферменти, шо
руйнують макромолекули в тканинах Ё тим самим збглъшують кёлькёсть осмотич-
но активних речовин. Це своею чертою зумовлюе пЁдвишення онкотичного тиску
тканин
Зниження онкотичного тиску кров’ та пщвищення онкотичного тиску тка-
нин спричинюе вихщ води Ёз судин у мёжклётинний сектор, нас лёд ком чого е
зменшення ОЦК. Це сприймаеться регуляторними системами як ознака кро-
вовтрати та спричинюе посилення реабсорбцп натрпо ! води нирками за вище-
описаними механизмами. Таким чином, у патогенез! системних (генерайзованих)
набряюв будь-яко! етюлоп! мае м!сце надапрна затримка нирками натр!ю 1 води.
Осюльки в дйей раннього вгку спостерйаеться тенденц!я до затримки натрпо при
нездатносп виводити надлишок води 1 натр!ю, про що йшлося вище, немовлята
в цьому впц схильш до розвитку системних набряюв, яю прийнято називати пас-
тозшстю.
Перелечен! фактори беруть участь у мехашзм! розвитку всгх вщцв набряку,
однак значения !х у цьому процес! залежить в!д походження набряку.
Залежно вщ причини ! мехашзму виникнення розргзняють системы! набряки
внаслщок ураження серця (недостатност! кровообпу), нирок, печгнки, при голо-
дуванн! (кахектичний) 1 переважно локальш (запальний, токсичний, нейроген-
ний, алерпчний, л!мфогенний та ш.).
Набряк унаслгдок серцево! недостатност! (заспйний набряк) виникае внасль
док здебшыпого венозного застою 1 пщвищення венозного тиску, що супровод-
жуеться посиленням фщьтрацп плазми кров! в каголярах. Гшокс!я, що розви-
ваеться, зумовлюе порушення троф!ки 1 посилення проникносп стшки судин.
Велике значения при цьому мае також затримка натрпо ! води через активащю
РААС (схема 19).
Набряк унаайдок ураження нирок. У патогенез! набряку при гострш нирковш
недостатност! первинного значения набувае зменшення швидкост! клубочково!
фщьтрацп, що зумовлюе затримку води в оргашзм!. Пщвищуеться також реабсор-
бц!я натр!ю в канальцях нефрошв, у чому, очевидно, певна роль належить вто-
ринному гшеральдостерошзму, осюльки антагошст альдостерону — сшронолак-
тон (синтетичний стеро'щ) — справляе дгуретичний ! натршуретичний ефекти. У
раз! нефротичного синдрому основным мехашзмом набряку е зниження онкотич-
ного тиску кров! внаслщок протешурп, що спричинюе виникнення гшопроте!-
немп.
У розвитку набряку внаслгдок ураження печ!нки також найважлив!шу роль вцц-
грае гтопротешем!я, зумовлена порушенням синтезу б!лк!в у печшц!. Певне зна-
чения мае пщвищення продукцп або порушення шактивацп альдостерону. У роз-
витку асциту при цироз! печшки вирппальну роль вщйрають утруднення крово-
обйу в печшщ ! пщвищення гщростатичного тиску кров! в систем! ворйно!
вени.
Причинами кахектичного (голодного) набряку е алиментарна дистрофия (голо-
дування), гшотрофы в дйей, злояюсн! пухлини та !нш! виснажлив! хвороби. Най-
важливппим фактором його патогенезу е зниження онкотичного тиску кров!
внаслщок гшопротешемп, зумовлено! порушенням синтезу б!лк!в 1 пщвищенням
проникносп стшки капыяр!в, пов’язаним 13 порушенням троф!ки судин.
У патогенез! запального, алерг!чного 1 токсичного набряк!в, яю е переважно
локальными (мйцевими) основну роль вщйрае зменшення онкотичного тиску
кров! 1 збшьшення онкотичного тиску тканин унаслщок пщвищення проникносп
стшки кап!ляр!в 1 активацп протеолйичних ферменпв в ушкоджених тканинах.
Певне значения в патогенез! цих набряюв мае пщвищення гщростатичного тиску
кров! внаслщок розвитку артер!ально! та венозно! гшеремп, яю виникають у вог-
нигщ ураження пщ впливом вазоактивних медиаторы (БАР 1 метаболии).
Катлярнг. ппертензся
Зменшення хвилипно го об'ему серця
Недостатшсть серця
Венозний застш
Г1ПОКС1Я тканин
Подразнення
волюморецел тор1в
Зменшення ниркового
кровотоку
Збътыпенпя вироблення
ретпну
С гимуляшя секрецн
альдостерону,
вторивнид альдостеронизм
Пщвищення реабсорбцн №
в цирках 1 затримка води
Гшернатр1тм1я
Подразнення осморецепюрш
Посилення вироблення вазоиресину
Збщьшення реабсорбцн води
> Накопичення води в тканинах
(порожнинах)
V
Ацидоз
Набряк, водянка
Схема 19. Патогенез набряк} внаслщок ураження серця
Нейрогенний набряк розвиваеться внаслщок порушення нервово! регуляцп
водного об и! ну. трофгки тканини ! судин (анпотрофоневроз). Це набряк кпщгвок
у раз! гемшлегп 1 сирингомгелГ!, набряк обличчя при невралти тршчастого нерва.
У походженш нейрогенного набряку важливу роль вщгграе пщвищення про-
никност! стшки судин 1 порушення обмшу в уражених тканинах.
У розвитку набряку разного походження слщ розргзняти дв! стадп. У першгй
стадп надлишкова рщина, яка налходить у мгжтканинний простгр, накопичуеться
переважно в геле под! бних структурах (колагенов! волокна й основна речовина
сполучно! тканини), збшьшуючи масу немобтльног, фтксовано!, м!жтканинно! р!-
дини. Коли маса фшсовано! рщини збшыиуеться приблизно на 30 %. а тиск сягае
атмосферного, починаеться друга стащя, для яко! характерно накопичення вшь-
нот мтжюптинно! рщини (мал. 47). Ця рщина
здатна перемтщуватися в мьжклпинному про-
стор! пщ дтею сили тяжтння 1 дае симптом ямки
шсля надавлювання на набряклу тканину.
Наслщки набряку залежать вщ його ступеня
та локалтзацп. Знание накопичення рщини спри-
чинюе здавлювання тканин, порушення тх тро-
ф1ки ! функцш (див. мал. 47).
Особливо небезпечним е набряк мозку 1 ле-
гень. Скупчення рщини в серозних порожнинах
тша порушуе функцпо прилеглих органтв (утруд-
нення дихання внаслщок водянки плеврально!
порожнини).
ОБМ1Н ЕЛЕКТРОЛ1Т1В
ТА ЙОГО ПОРУШЕННЯ
Дост ми розглядали електролтти виключно як ос-
мотично активы речовини, юльюсть яких визна-
чае бшыпу частину осмолярност!, а отже, й об’ем
рщинних компартмент!в. Ця функцтя електро-
лгпв е дуже важливою, але не единою. Зокрема
вщ град!ента концентрат! катюшв мтж цитоп-
лазмою та штерстищем залежать збудливтсть т
Тиск М1ЖКЛ1ТИНН01 рщини, кПа
Мал. 47. Залежшсть об’ему выьно!
(1) । фжсовашй (2) мжклгпшноТ рь
дини вщ п тиску, що характеризуе дв!
стадп розвитку набряку (за А.С. Сиу-
Топ, 1975)
провщшсть у нервовш ! м’язовш тканинах, а вщ сптввщношення м!ж фгксовани-
ми каттонами й анюнами — КОС оргашзму. Тому здатшсть щдтримувати постш-
нкть концентрацп електролтв у поза клттин ному сектор! (тзоюшя) е одшею з
найважлив!ших функцш оргашзму.
Обмш натрпо та його порушення. Натрш е головним катюном позаюптинно!
рщини, який забезпечуе понад 90 % Г! каттоннот осмолярност!. У середньому ор-
гашзм людини мктить близько 60 мекв №+ на 1 кг маси тша, що становить
близько 0,15 % вщ загально! маси тша: 50 % натрпо мктяться в позаклттинному
сектор!, 43 % — у юстках ! лише 7 % — внутртшньоюитинно. При цьому 70 %
натрпо здатш в!льно обмшюватися. У норм! концентрацтя в плазм! кров! ста-
новить 135—145 мекв/л, тобто и коливання не перевищують 10 %>. Водночас ртз-
ниця у внутртшньоклттинних концентрациях натрпо в ртзних тканинах перевищуе
50 %. Потреба оргашзму в натри, який дуже добре всмокгуеться в кишках, стано-
вить 1—1,5 мекв/кг на добу. Проте доросла людина традицтйно вживае 2,5—
3,1 мекв/кг або 4—5 г натрпо на добу. Потужна система регуляцп натртевого об-
мтну, про яку йшлося вище, дае змогу дорослим збертгати баланс натрпо при
значних змтнах у його споживаннт. Виведення наддишку натрпо, а за необхщностт
т його затримка в норм! здшснюютьсд переважно нирками. Тонка регуляцтя цтет
дтяльност! нирок забезпечуеться альдостероном, який стимулюе реабсорбцпо №+
в збтрних трубочках. Вщ альдостерону залежить реабсорбцтя лише 2 % натрпо, що
профшьтрувався, але це становить близько 30 г №С1 на добу. У разт необхщностт
нирки здатнт реабсорбувати та повертати в кров 99,9 % натрпо, що профшьтрував-
ся. Кргм того, альдостерон стимулюе всмоктування натргю в товстгй кгипщ. Тому
дефшигу натрпо в доросло! людини не спостерггаеться навгть у раз! вживання 0,5 г
Т9а+ на добу
Вище згадувалося про те, що нирки дней раннього вгку не здатн! виводити
надлишок нагргю. Це може спричинити розвиток набряюв. Але навгть за помгр-
ного споживання натргю в дгтей спостерггаеться позигивний натргевий баланс,
оскгльки збыьгдення маси тгла на 1 кг потребуе затримки 3 г натрию.
Патологична затримка натрпо в органгзмг виникае при значному зменшеннг
швидкостг клубочковог фглырацп або при надмгрнгй реабсорбцп натргю пгд вггли-
вом анпотензину И та альдостерону при вторинному гшеральдостерогггзмг, який
мае мгеце при серцевгй ! ггечгнковгй недостатностг та при нефротичному синдро-
ме. Вторггнним такий гшеральдостеронгзм називаеться тому, що йому гтередуе
збыьшення продукцгг анпотензину II, який стимулюе реабсорбцгю №+ в прокси-
мальних канальцях Реабсорбцгя 1 мекв К’а+спричинюе затрим ку 6 мл Н2О у по-
заклгтинному секторг, що при значному накопи чеши натргю в органгзмг призво-
дить до розвитку набрягсу.
Первинний гшеральдостеронгзм, який пов'язаний гз альдостеронпродукую
чою пухлиною кгрковог речовини надниркових залоз, не спричинюе значку за-
тримгсу натргю г води. Це пов'язано з тим, що за умови стабыьног роботи серця г
нормального онкотичного тиску кровг затримка Ма+ г Н2О збгльшуе ОЦК г ар-
тергальний тиск, що пригнгчуе актившсть симпатико-адреналово! та ренгн-антго-
тензиново! систем г пшвищуе секрешю ПНП, просгагландингв та КО. Зниження
активностг симпатичних нервгв г зменшення кониенграцн анпотензину II зумо-
влюс пригнгчення проксимально! реабсорбцг! Ь!а'Г, що компенсуе посилену аль-
достеронзалежну реабсорбцгю натргю у збгрних трубочках. Кргм того, ПНП, нир-
ковг ПГЕ2 г 12та ендотелгальний релаксуючий фактор (МО) пригнгчують реабсор-
бцгю натргю г води переважно у збгрних трубочках.
Дефщит натрпо виникае внаелгдок або порушен гля його нирково! реабсорбцг!
при недостатностг кгрково! речовини надниркових залоз г введеннг дгуретикгв, або
його вграти при дгарег та гюговидгленш. Наследии натргевого дефщиту залежать
вгд характеру змш при цьому осмолярностг позаклгтинног ргдини, про що йшлося
вище.
Обмш калгю та його порушення. Калгй е основним катгоном внутргшньоклг-
тинног ргдини. Його вмгст в органгзмг людини — близько 54 мекв/кг, шо стано-
вить 0,2 % усге! маси тгла; 98 % калгю мгстяться всерединг клгтин г лише 2 % — у
позаклгтиннгй ргдинг; 95 % усьото калгю здатнг выьно обмтнюватися. Концентра-
цгя калгю всерединг ргзних клгтин значно вгдргзняеться В еритроцитах вона най-
менша (100—110 мекв/л), у м’язах — найбыьша (140—160 мекв/л). У плазм! кровг
в дорослих концентрацгя К+становить у норм! 3,8—5,1 мекв/л. Протягом лершо-
го тижня житгя спостерггаеться пперкатгемгя, тобто пщвищення ргвня калгю
плазми вгд 5,7 до 8 мекв/л. Це пов'язано з тим, що кгльюсть К+у позаклгтиннгй
ргдинг визначаеться не стгльки його вмгстом в органгзмг, скгльки КОС. Ацидоз,
який мае мгспе в цьому вгцг, як правило, супроводжуеться пперкалгемгею, про шо
йде мова у вгдповгдному роздглг, а алкалоз — ггпокалгемгею, тобто зменшенням
концентрацп К+ у плазмг кровг нижче 3,5 мегсв/л. Кргм того, оскгльки висока
внутргшньоклггинна конценграшя калгю створюеться лише завдяки активностг
№+-, К+-АТФази, навгть незнание п пригнгчення при гшоксп або ушкодженнг
клхтин здатне суггево збшыпити концентрацию К+ у позаюптинному сектор! без
змши загально! юлькост! цього канону в орган 1зм1. Навпаки, при первинних змх-
нах юлькост! калпо в позаклхпгннгй рщиш вш активно закачуеться в клпини №+-,
К+-АТФазою або пасивно виходить !з них через селективш калхев! канали. Ма+-,
К+-АТФаза у м’язах 1 деяких шших клхтинах активуеться шсулпхом х адреналшом,
що дае змогу запобхгти гшеркалхемп шсля щи та пщ час фхзичного навантаження.
Головним регулятором балансу калпо в оргашзм!, як х для шших ф!ксованих
йонхв, е нирки. Вони здатш реабсорбувати близько 90 % калпо, що профшьтру-
вався. Обов’язков! втрати калпо !з сечею х калом становлять 1,2 мекв/кг на добу.
Тому добова потреба оргашзму доросло! людини в калл становии, 1,2—1,5 мекв/кг
на добу. У д!тей, як! ростуть, потреба в калн становить 3—7 мекв/кг на добу. Голов-
ними джерелами калпо для оргашзму е м’ясо, фрукти й овочк
Ппокал1см1я може виникати при: недостатньому надходженн! калпо з !жею;
первинному ! вторинному гшеральдостерошзм!; надмхрному виробленш (хвороба
1ценка — Кушшга) чи введенш глюкокортикоида; втрап калпо пщ час блювання,
дхарех, лхкування д!уретиками та нирковому канальцевому ацидоз! I типу; пере-
мхщенш калпо всередину клггин алкалоз! та введенш шсулшу. Осюльки мембран-
ний потешцал спокою нервових ! м’язових клхтин прямо залежить вщ спхввщно-
шення мхж внутр хшньо клхтин ною та позаклхтинною концентрацхею калхю, зни-
ження концентрацп К+ у позаклхтиннш рщиш зумовлюе гшерполяризацпо
клхтинно! мембрани та зниження п збудливосп. Тому ппокалхемхя проявляеться
швидкою втомлювашстю, парестезхями, м’язовою слабкхстю, гшорефлексхею,
спазмом м’язхв нижнгх кшцхвок, порушенням перистальтики хз закрепами х па-
ралхтичною непрохщшстю кишок, пщвищенням чутливост! серцевого м’яза до
дипталхсу. На ЕКГ характерними е подовження хнтервалу ()— Т, сплощення зубця
Т х поява зубця !/; можливими е атрховентрикулярна блокада, таххаритмхя. Зни-
ження концентрацп К+ у плазм! нижче 1,5 мекв/л спричинюе паралхч м’язхв х
зупинку серця.
Пперкал1€м1я виникае при нирковш недостатност!, гшоальдостерошзм! та п-
покортицизмх, ацидозх, шсулшовхй недостатност! та надмхрному введенш калхю.
Збшьшення концехгграцп калпо в позаклхтиннш рщиш знижуе мембранний по-
тенцхал спокою, що збшьшуе збудливхсть нервових х м’язових клхтин. При ще
бшьшому пщвищенш рхвня калпо в позаклхтиннш рщиш потенцхал спокою може
досягги рхвня порогового потенщалу, що спричинить рхзке зниження збудливосп,
загальну слабхсть х паралхч. Пперкалхемхя проявляеться дратхвливхстю, дхареею,
кишковими спазмами, патолопчним впливом на серцеву дхяльшсть. Оскшьки
пщвищення рхвня К+ порушуе провщнхсть х серцевий ритм, на ЕКГ зубець Т стае
високим х загостреним, штервал Р—<2 збшыпуеться, комплекс ()К5 розширюеть-
ся, можуть розвинутися блокада пазухо-передсердного вузла та брадиаритмхя.
При збшыпеннх рхвня калпо в плазм! понад 7,5 мекв/л можуть виникнути парок-
сизмальна таххкардхя, фхбриляцхя шлуночкхв х асистолхя.
Обмш магнпо та його порушення. Магнпо в оргашзм! людини мхститься менше
0,036 % маси пла, але вш е другим за значениям внутрхшньоклгпшним капоном
1 четвертом за абсолютним вмхстом в оргашзм!. Близько 60 % магнпо мхститься в
кхстках, третина з якого здатна до обмхну. Лише 1 % магнпо мхститься в позаклх-
тинному сектор!, де його концентращя становить 0,75—1,25 ммоль/л. Близько 30 %
магнпо плазми зв’язаш з битками та хншими речовинами. Решта магнпо юнхзова-
на. Саме ця його частина е бюлопчно активною. У м’язах концентращя магшю
становить близько 10 ммоль/л, а в еритроцитах — близько 2,5 ммоль/л. Як 1 у
випадку з калыцем, при гшоальбумшемп загальна юльюсть магнпо буде знижена,
але водночас р)вень юшзованого магнпо може 1 не змшюватися. Зниження рН
збшьшуе юльюсть юшзованого магшю.
Людина отримуе М§2+ переважно !з зеленими овочами, горками. бобовими,
арахтсом, питрусовими, шоколадом, чаем, морепродуктами. У норм! в тонкш
кишщ абсорбуеться близько 35 % М§2+ з того, шо надходить з !жею. Жирш кис-
лоти, фттин 1 алкоголь зменшують абсорбцпо Добова потреба у магнп до-
росло! людини — 0,4 ммоль на 1 кг маси тша, у ваптних ! датей — у 1,5—2 рази
бшьше. Нирки здатш реабсорбувати близько 95 % М&2* переважно в товстому
висхщному колпп петл1 Генле. Алкоголь, мшералокортикоУди та внутр! шньовенне
введения кальцпо 1 глюкози зменшують реабсорбшю М§-+. Абсопбц’я М§2+ в киш-
ках 1 його реабсорбщя в нирках, як т кальппо, активуються паратгормоном 1 вгта-
мшом В. Тнсулш стимулюе переход М&2+ !з позакл!тинно! рщини в юптини.
Внутрппньоюптинний М§2т е кофактором 1 активатором багатьох ферменпв
Позаюптинний М§2+ бере участь у нейром’язовому збудженш, д!ючи на цей про-
цес подтбно Са2+ । протилежно К+.
Ппомаен1ем1я виникае при змсншенш надходження М§1+ в оргашзм, знижен-
ш всмоктування в кишках 1 збшьшенн! втрати його тэ сечею. Дефшит магнпо
спостерггаеться при бшково-кал орш ному голодуванш, ентеритах, ураженнях нир-
кових канальщв, уведенн! д!уретик!в, пперальдостерошзмь У дорослих основною
причиною ппомагшемп е алкоголтзм. Симптоми ппомагшемп проявляються при
знижешп концентрацп магшю нижче 0,5 ммоль/л. Зменшення кттькосп М§2+ в
плазм! кров! зумовлюе збшьшення нервово! та м'язово! збултивосп, шо прояв-
ляеться тремором, судомами шт, атакшею, головокружениям, безсонням, де-
пресёею, шод! галюцинащями та психозом, анорекс!ею, нудотою, блюванням,
проносами, парестез1ями. Часто розвиваються артертальна гшертензтя, тахжардтя
та екстрасистолп.
Ппермагшешя найчастйпе бувае наслщком гшервггампюзу О. Вона також спос-
терпаеться при нирковш 1 наднирковш недосгатносп та при застосувашп препа-
ратов, як! мютять М§2+. Симптоми виникають при збшьшенн! р!вня магнпо в плаз-
М1 понад 2 ммоль/л. Спостерггаеться гальмування проведения нервового тмпульсу,
що проявляеться зниженням сухожилкових рефлексов, м’язовою слабюстю, гшо-
гошею, нудотою, блюванням, почервоншням шюри з вщчуттям жару, штливютю,
СОНЛИВ1СТЮ. При збшьшенн! концентрацп М§2+ в плазм! понад 5 ммоль/л розвива-
ються пригшчення дихального центру, порушення провшност! мюкарда 1 кома, а
при концентрацп понад 7,5 ммоль/л — зупинка серия.
Обмш хлору та його порушення. Хлор — головний ашон поза юптинно! рщини
1 шлункового сою'. Його загальна юлькёсть в оргашзм! становить близько 0,12 %
маси т!ла. У внутршгньоюптинному сектор! мютиться всього 12,4 % хлору. При-
чому його концентрация в р!зних тканинах неоднакова. У скелетних м’язах вона
становить 4 мекв/л, у серцевому м'яз! — 7 мекв/л, у печ!нц! — 12 мекв/л 1 в срит-
роцитах — 50 мекв/л. Ргвень хлору в плазм! кров! мало залежить вш в!ку 1 стано-
вить 99—105 мекв/л. 1они хлору вщпрають важливу роль у гццтриманн! осмоляр-
носп та КОС позаюптинно! рщини. Зменшення спгввщношення мгж Ма+ та СГ
зумовлюе розвиток негазового ацидозу, а збшьшення — негазового алкалозу.
Можливгсть змши сшввщношення м!ж СГ та Иа+ зумовлена тим, що абсорбцхя х
секрецхя хлору в травному тракт! та його реабсорбщя в нирках вщносно незалеж-
ш вщ транспорту натрпо. Так, у ворсинках кишок частина СГ всмоктуеться через
мхжклггинн! щхлини за електроххмхчним ! концентрацшним градхентами, створе-
ними при активнш реабсорбцп натр!ю. 1нша частина хлору абсорбуеться в киш-
ках ! секретуеться у шлунку трансцелюлярно за допомогою СГ / НСО3_-антипор-
ту. Реабсорбщя хлору в канальцях нефрона також частково натршзалежна (Яа+-,
К+-, 2СГ- та Ка+-, СГ-котранспорт, парацелюлярний транспорт), а частково не-
залежна вщ транспорту натрпо (СГ / НСО3_-антипорт у вставних клгтинах В-типу,
К+-, СГ-симпорт у проксимальних канальцях х петл! Генле та селективш хлорнх
канали).
Ппохлорелйя виникае найчастгше при блюванш та посиленому потовидшенш.
Кргм того, причиною гшохлоремп може бути застосування хлорних д!уретик!в х
муковхсшздоз. Зниження р!вня СГ у плазм! бшьше н!ж на 20 мекв/л спричинюе
розвиток тяжкого алкалозу.
Пперхлорелйя виникае при нирковш недостатност! або за вщносно б!льшо!
втрати натрпо 1 води, шж хлору, що спостершаеться при канальцевому ацидоз!,
недостатност! мшералокортико'щхв та при дхарех. Головним наслщком гшерхлор-
емп е розвиток ацидозу.
Обмш кальщю та його порушення. Кальцш — найпоширешший канон оргашз-
му людини, вмгст якого становить близько 2 % маси тша; 99 % усього кальщю
м!стяться в метках х зубах переважно у форм! кристалхв гщроксиапатиту. Не мен-
ше 1 % юсткового кальщю здатн! до обмшу, а за 5—6 роюв увесь кальцш юсток
оновлюеться. Близько 1 % кальцпо мхститься в позаклгтинному сектор!. Його
концентрация в плазм! кров! дорослих у норм! — 2,2—2,5 ммоль/л. Внутрпиньо-
клтгинна концентращя кальщю в 10 000—100 000 разгв менша. У плазм! кальцш
перебувае в трьох формах: 40 % його зв’язано з бшками, 13 % поеднано з цитра-
тами, фосфатами, карбонатами х 47 %— у йошзованому стан!. Фхзхолопчну актив-
шсть мае лише хонхзований кальцш, концентрац!я якого в дорослих у норм! ста-
новить 1,05—1,15 ммоль/л, або 2,1—2,3 мекв/л. Кшьюсть юнхзованого кальщю
збшьшуеться при ацидоз! та зменшуеться при алкалоз!. При цьому ргвень загаль-
ного кальцпо може сутгево не змшюватися. Це пов’язано з тим, що збщьшення
рН переводить додаткову кшьюсть бшюв у форму полханхошв, а зменшення рН —
навпаки. Крхм того, ацидоз безпосередньо активуе резорбцгю юсток остеокласта-
ми. Усе це дае змогу при ацидоз! вившьняти частину Са2+ як вщ зв’язку з бшками
плазми, так х з юсток. Вмхст загального кальщю в новонароджених становить
2—2,4 ммоль/л, у дггей вгком до 1 року х старших — 2,5—2,8 ммоль/л. Пперкаль-
цхемхя у дней раннього вгку, хмовгрно, пов’язана зх схильнхстю хх оргашзму до
ацидозу та пщвищеною чутливхстю до вхтамгну О.
Кальцш надходить в оргашзм з хжею, перевалено з молоком г сиром. У норм!
в кишках всмоктуеться лише 1/4 частина кальцпо. Цей процес активний х регу-
льований, а отже, може значно змшюватися залежно вщ вмхету кальщю в !ж!. У
нирках може реабсорбуватися до 99 % профшьтрованого кальщю; 85 % Са2+ реаб-
сорбуеться в проксимальному вщдшх та в товстгй висхщнш частин! петл! Генле
пасивно х парацелюлярно. Решта Са2+ реабсорбуеться в дистальному вщдшх неф-
рона активно х трансцелюлярно за допомогою Са2+-АТФази та Са2+ / Ма+-проти-
переносника. Ргвень хошзованого кальцпо в плазм! пщтримуеться завдяки взае-
модй паратгормону, який виробляегься прищ1ттопод!бними залозами, активова-
ного вггамшу В (1,25-(ОН)2О3, калыгигрюл) та гормону парафол1кулярних
С-клтгин шитоподгбно!’ залози — кальцитошну. Секретя паратгормону безпосе-
редньо стимулюеться зменшенням коштенграцй Са2+ в позаклггиннш рщиш. Па-
ратгормон стимулюе активацпо вгтамшу В 1 разом з ним посилюе резорбцпо ис-
ток остеокластами, усмоктування Са2+ в кишках 1 реабсорбцпо його в нирках, а
також пригшчуе ниркову реабсорбцпо фосфату. Кальцитонш зменшуе резорбцпо
юсток, але його вплив на гомеостаз Са2+ обмежений поргвняно з паратгормоном
1 в!там!ном В.
Юльюсть кальцпо, шо екскретуеться гз сечею за добу, у дорослих доргвнюс
юлькост! кальцпо, який абсорбувався в кишках, тобто його баланс е нульовим.
Навпаки, у ваптних 1 дней баланс кальцпо е позигивним, осюльки Са2+ погр!б-
ний для побудови скелета дитини. Потреба в кальцй у дорослих становить 0,8—
1,5 г на добу, або 0,3—0,5 ммоль/кг на добу. В останньому триместр! ваптност!
потреба в кальцп збгльшуеться вдвгчь У немовлят потреба в кальцп становить
у середньому 2,5 ммоль/кг на добу, у дней в!ко.м вщ 1 до 5 рок!в — приблизно
2 ммоль/кг на добу, старших — 1,5 ммоль/кг на добу.
У процес! перенесения ! вщкладення кальцпо важливу роль вщпрають
кальцгйзв’язува'гын бглки (СаЗвБ). Нин! описано понад 70 СаЗвБ: кальмодулш,
парвальбумтн, СаЗвБ кишок, б!лок 810и мозку, протромбш, амелогенш ! енамелш
емал! зуб!в, остеокальцин тощо. Порушення активност! СаЗвБ вццграють важливу
роль у розладах транспорту кальцпо кр!зь мембрани, скоротливо! функцп серце-
вого, скелетних ! непосмугованих м’яз!в, зсщання кровг, мшерашзацп юсток ! в
мехашзм! деструкцй твердих тканин зуб!в при кар!ес!.
Роль кальцпо в оргашзм! не обмежуеться побудовою скелета. Вщ концентра-
цй позаклггинного Са 2+ залежать генерашя потенщалу дй та зсщання кров!, а !з
внутргшньоюптинним кальщем пов’язаш поеднання збудливосп та скорочення
м’яз!в, активация ферменпв, регуляшя секрецй гормонгв, передача гормонального
! нервового сигналов, мехашзми апоптозу 1 некробюзу.
ГтокальщемЫ властива вс!м новонародженим у зв’язку з ргзким припинен-
ням його надходження вщ матер!. Зниження р!вня кальцпо виражешше в недоно-
шених, а також у новонароджених, яю перенесли сильний стрес п!д час полопв,
спричинений асф!кс!ею або травмою. У таких дгтей ппокальц!ем!я пов’язана, по-
перше, з абсолютною 1 вщносною парат иреошною недосгатшстю в цьому вшк
по-друге — з гшерпродукшею глюкокортикощгв, як! зменшують реабсорбцию
Са2+; по-трете — з пперфосфатем!ею. яка е наслщком интенсивного дефосфори-
лювання АТФ 1 креатинфосфату. У новонароджених можуть спосгерпатися лише
летарпя 1 погане смоктання та ознаки пщвищено!' збудливосп (дргбнорозмашис-
тий тремор шдборщдя 1 пальщв, прискорене дихання з апное). Легкий перебй
гшокалыцсмй в цьому вш! пов’язаний з гим, шо недостатшсть загального каль-
цию компенсуеться збгшьшенням частки юшзованого кальпйо, шо е наслщком
ацидозу. Пгсля п’ято! доби життя в дней, яю отримують сум!ш! на основ]
коров’ячого молока, може розвинутися теташя, повязана з гшерфосфатемгсю,
шо стимулюе вщкладення в юстках кальцпо фосфату. Гшерфосфатемгя в них д!-
гей спричинена тим, що !х нирки нездатш екскретувати надлишок фосфате, яко-
го багато в коров’ячому молоцг. У старших дней г дорослих гшокальщемы спо-
стершасться при абсолютшй чи вщносшй недостатност! паратгормону га / або
активно! форми вггамшу О, нирковш 1 печшковш недостатност!, ентерип, панк-
реатит!, переливанш цитратно'1 кров!, уведенш петльових д!уретик!в 1 глюкокор-
тикоццв, алкалоз!, пщвищеному р!вн! фосфору та зниженн! концентрацн магнпо
в плазм! кров!. Гшокальщемгя супроводжуеться щдвищенням нейром’язово! збуд-
ливосп, яка проявляеться гшеррефлекс!ею, судомами, теташею, спазмофйпею
(дитяча тетан!я в дггей вгком вш 2 м!с. до 2,5 року), бронхо- 1 ларингоспазмами.
Характерними е периоральш та периферичн! парестезп. Можуть виникати серцев!
аритмп, пшороспазм з блюванням, бшь у живот!. Недостатке всмоктування Са2+з
кишок при пповггамшоз! О може спричинити виникнення остеомалящ! — пщвище-
ного виходу кальцпо з юсткового депо. Хрошчна ппокальщемгя зумовлюе остео-
пороз 1 перелом юсток.
Пперкалыцелйя виникае при гшерпаратиреоз!, ппервггамшоз! В, злояюсних
новоутвореннях, тривалш нерухомосп або невагомосп та гострому ацидоз!. П-
перкальц!ем!я знижуе нейром’язову збудливють, що проявляеться нудотою, блю-
ванням, анорекс!ею, закрепами, гшотошею м’яз!в, аритм!ею, швидкою втомлю-
вашстю, депрес!ею 1, як наслщок, комою. Пперкальщемгя та гшеркальц!ур!я мо-
жуть спричинити виникнення нефропатп, що проявляеться зниженням швидкосп
клубочково! фшьтрацп, натршурезом, зниженням чутливост! ниркових каналыцв
до АДГ 1 пол!ур!ею, дегщратащею 1 полщипыею. У раз! тривало! дп патогенних
чинниюв пщвищена актившсть остеокласте призводить до остеопорозу з перело-
мами юсток I за вщсутносп гйюфосфатемп — до кальциф!кацц судин, нирок,
ропвки й утворення камешв у нирках.
Обмш фосфору та його порушення. Фосфор в оргашзм! людини становить
близько 1 % маси тша 1 перебувае в оргашчних 1 неоргашчних сполуках переваж-
но у вигляд! залишюв ортофосфатно! кислоти (Н3РО4); 85 % ус!х фосфапв м!с-
тяться у юстках 1 зубах, де 2/3 !х представлен! важкорозчинними фосфатами каль-
щю, а 1/3 — розчинними з’еднаннями, яю становлять депо фосфапв кров!. Усьо-
го 1 % фосфат!в моститься в позаюптиннш рщин!; 67 % фосфапв плазми
представлено фосфол шщами. Концентращя неоргашчних фосфапв плазми в до-
рослих становить 0,9—1,5 ммоль/л, або 1,7—2,6 мекв/л, з яких 10—15 % зв’язаш
з бшками. У д!тей першого року життя р!вень неоргашчних фосфат!в плазми ся-
гае 1,3—2,3 ммоль/л, а з 2 до 14 рокгв — близько 1,6 ммоль/л. При нормальному
рН 80 % усгх неоргашчних фосфапв плазми представлено НРО42’ 1 20 % Н2РО4~.
Внутршшьоюптинна концентращя фосфапв становить 65—100 ммоль/л, з яких
близько 98,5 % входять до складу оргашчних сполук.
Фосфата потрапляють в оргашзм !з тваринними 1 рослинними продуктами.
За нормальних умов у кишках усмоктуеться 70 % фосфате, тод! як при голоду-
ванн! може абсорбуватася до 90 %. У нирках реабсорбуеться 70—90 % неоргашч-
них фосфапв, переважно у проксимальному вщдш нефрона за мехашзмом
котранспорту з натрием. Потреба дорослих у фосфор! становить 10—15 мг/кг на
добу, або 0,3—0,5 ммоль/кг на добу. У ваптних мш!мальна потреба у фосфатах
становить 0,6 ммоль/кг на добу, а в немовлят — 0,8 ммоль/кг на добу.
Усмоктування фосфапв у кишках стимулюеться 1,25-(ОН)2В3 1 меншою
м!рою паратгормоном. Ниркова реабсорбц!я фосфапв активуеться вггамшом В 1
гальмуеться паратгормоном. За нормальних умов зменшення надходження фос-
фапв з !жею 1 зниження концентрацп неоргашчних фосфат!в у плазм! збшыпують
утворення 1,25-(ОН)2В3, що посилюе абсорбщю з кишок 1 вихщ !з юсток фос-
фатгв 1 кальщю. Збшьшення концентрацп Са2+ своею чертою зменшуе секрешю
паратгормону, шо щдвищуе реабсорбцпо г знижуе екскрещю фосфатаз гз сечею.
Навпаки, при дгетг з пщвишеним вмгстом фосфатов утворення 1,25-(ОН)2О, змен-
шуеться, що зумовлюе зниження мобшгзащ! з кгсток г всмоктування в кишках
фосфапв г кальщю. Розвиток гшокальщемп при цьому збшьшуе секрецпо парат-
гормону, який посилюе екскрешю фосфалв гз сечею.
Пщтримання нормально! концентрацп неоргашчних фосфалв у плазм! кров!
перш за все необхщне тому, що вони постгйно переходят ь у середи ну клттин, де
беруть упасть у процесах окисного фосфорилювання, тобто переходять тз неор-
ганичного пулу в органтчний. Фосфата вхоцять до сктаду фосфолшщв, фосфо-
протегшв, нуклеотидов, нуклегнових кислот, беруть участь у глгкогенезг, глгкоге-
нол13г, глгколгзг, неоглюкогенезг гошо. Найважлив1шою е роль фосфапв у проце-
сах зберггакня й обмшу енергп, потргбно! для здшснення руху, активного
транспорту та бгосинтезу. Вшьна енерггя, джерелом яко! е окиснення харчових
субстрат!в, не може безлоссредньо використовуватася организмом. Обов’язковим
етапом використання енергп е утворення макроерпчних фосфатних сполук —
безпосередюх донатортв вшьно! енергп, найважлившшм тз яких е АТФ. Перене-
сения фосфорильно! групи вщ АТФ до шших сполук щд впливом кшаз е засобом
обмшу енергп, а вщщеплення ортофосфату пщ впливом фосфатаз призводить до
вивЬьиення енергп.
Ппофосфагпем'ы виникае при голодуванш, мальабсорбцп, недостатносп втта-
мгну О. гшерпаратиреозг, вживавнг анташцвв, хрошчному алкоголгзмг, недостат-
носп ниркових канальщв, зокрема при синдром) Фанкош, ппофосфатемгчному
рахгп, проксимальному канальцевому ацидозг, цистинозг (див. роздш XXX “Па-
то логочна фгзюлопя нирок). Зменшення неоргашчного фосфату в плазм г також
може бути наслщком поглинання фосфату клгтанами при введенш шсулшу з
глюкозою або при гостро му ресшра горному алкалозг. Гшофосфатем1я спричинюе
зниження синтезу АТФ, що гюрушуе функцпо оргашв з штенсивним енергетич-
ним обмшом. Зменшення синтезу в еритроцитах 2,3-дифосфоглщерату знижуе
надходження кисню в тканини, що зумовлюе гшокспо Симптомами гостро! гшо-
фосфатемп е запаморочення, ошмтння га парестезп кшчиктв палыив, судоми.
м’язова слабкгсть, стенокардгя, бшь у м’язах, гемолгтична анемгч. ]мунодспрссив-
ний стан. Хрошчна гшофосфатемгя спричинюе попршення пам’ятт, летаргтю, ос-
теомаляцию, б!ль у ктстках, артралпю, гематоми, затримку росту.
Пперфосфателия спостерггаеться при штоксикацп вггамшом □, зокрема в
раз; передсзування ультрафзолетового опромшення, а також при гшопарагиреозг,
гострш кишковтй непрохщностг, нирковш недостатносп, переломах исток, ново-
утвореннях у кгстковгй тканиш. Гшерфосфатемтя призводить до вщкладання фос-
фату кальцпо в м’яких тканинах, суглобах 1 артергях, шо може спричинити розви-
ток нирково! недостатносп, аритмп, помутнпшя ропвки. Значка гшерфосфатемгя
в тяжких випадках може супроводжуватася гшокальцгемгею, яка проявляеться
анорексзею, нудотою, блюванням, гшеррефлексгею, гахгкардгею, геташею, а в дг-
тей раннього вгку — спазмофплею.
1з порушенням кальшй-фосфорного обману пов'язане виникнення дистро-
фтчних уражень системи звапнших тканин. Провщне мгсце серед них посщае ос-
теопороз. який ВООЗ оцшюе як трегю за значениям проблему' (уражено 11—12 %
населения розвинених крагн) шсля серцево-судивних 1 онколопчних хвороб Ос-
теопороз — це втрата маси юсток 1 розвиток порозност! (д!рчастост!) трабекуляр-
них структур 13 порушенням нормального ремоделювання орган!много та мине-
рального матриксу юстково! тканини.
Швидкому розвитку остеопорозу сприяе нездоровий способ життя сучасно.
людини з характерними !й гшодинамтею та нерашональним харчуванням, яю е
етюлопчними факторами. Роль ал!ментарного фактора полягае в недостатньому
або незбалансованому надходжешп в оргашзм кальшю та фосфору (нормал ьне
сшввщношення цих елемештв у рашон) рхзних в!кових груп може коливатися вщ
1 : 1,5 до 1,5 : 1), регулярному вживанн! переварено! води, злакових, сол! та цукру,
а також продуктов, бщних на таю втгамши, як аскорбшова кислота (впамш С),
ретинол (в1там1н А), групи О 1 К (фьлохшони). Етюлопчну роль вщтграють також.
стресорш впливи 1, особливо, вжовим фактор Розвитку остеопорозу сприяють
також ал1‘ментарний дефщит цинку, мар ганию, кобальту 1 надлишок алюмппю та
залгза.
У патогенез! остеопорозу велике значения мають порушення гомеостазу каль-
цпо 1 дисбаланс м!ж резорбщею 1 формуванням юстково! тканини. Зменшення
ф!зичного навантаження та р!вня кальцпо в кров! (недостатке вживания та пору-
шене всмоктування в кишках) збшьшуе процукцпо паратгормону (б)лково-пеп-
тидний гормон пришит оподтбних залоз). який сприяе мобшзац!! кальшю з! ске-
лета. Быково- зернова д!ета сприяе розвитку апидозу, що також посилюе мобш!-
зацпо кальшю. Гшовггамшоз С зменшуе синтез колагену, який становить основу
оргашчного матриксу юстки. Дефицит ретинолу зменшуе синтез глткозамшопп-
кан!в, як! вщпрають важливу роль у м!нерал1занп юсток, а дефщит ф!лох1нон!в
зменшуе утворення основного кальщйзв’язувального быка юстково! тканини —
остеокальцину. Стреси супроводжуються посиленням продукцп глюкокорти-
кощ!в, а останнт, як вщомо, спричинюють деградацию быкового матриксу юстко-
во! тканини та втрату солей катьшю. Дефщит в!там!ну О, порушуе всмоктування
Са2+ в кишках.
Патогенез вжового остеопорозу пов’язаний !з гшогонадизмом 1, особливо, з!
зниженням продукпп естрогешв. як! чинять значний остеопротекторний ефект.
Близько 23 % жшок шсля 50 роюв хворпоть на остеопороз, а в ж!нок в!ком понад
70 роюв ця хвороба призводить до переломов шийки стет на. хребшв 1 променево!
юстки.
У профшактищ остеопорозу значну роль вщпрають ф!зичне навантаження,
обмеження споживання солодопцв, сол! й бтлого хл!ба з проектного борошна,
перехщ на лакто-ово-вегетар!анський ращон з! споживанням молока, сиру, яень
1 соевих продуктов, багатих на калъщй, быок 1 фггоестрогени, яю сприяють ф!к-
сацГ! кальшю в юстках.
Розд!л XXIII
ПАТОЛОПЧНА Ф13ЮЛ0ПЯ
ОБМШУ М1КРОЕЛЕМЕНТЕБ
3 усгх хгмгчних елементгв, якг кнують у природ], в оргашзм! людини вже виявилв
81. Оргашзм людини на 99 % складаеться з вутленю, кисню. водню, азоту, каль-
цпо, фосфору, калпо, натргю, магнпо, Орки, хлору. Це так зван! макроелементи.
Ус! х1м1чн1 елементи, концентрация яких в оргашзм! перебувае в межах 10~3—10~12 %,
называются лакроелементами ! подгляються на есенщальш 1 токсичш. До есен-
щальних, тобто абсолютно необх!дних оргашзму, належать 15 — залгзо, м!дь,
цинк, кобальт, хром, молгбден, нгкель, ванадш, марганець, селен, йод, фтор,
кремюй, миш’як, лгтгй. Ус! щ мгкроелементи, утворюючи комплекси з фермента-
ми, гормонами, вггамгнами, транспортними 1 структурними бглками, справляюсь
значили вплив на вс! аспекта житгедгяльностг оргашзму. Загальнов1домими ток-
сичними мпсроелементами е ртуть, свинець, кадмгй, беритш, алюмгшй, миш’як,
фтор, хром. Под!л мгкроелементгв на есенщальш ! токсичш значною м!рою е
умовним. Так, з одного боку, бйтышсть, а можливо г вег, токсичш мгкроелементи
е есенщальними. 3 шшого боку, передозування життево необхшних дйкроелсмен-
ив спричинюе розвиток гнтоксикапп.
Мгкроелементи потрапляють в оргашзм людини переважно з !жею, але мо-
жуть надходити 1 через повттропровгдш шляхи та шкгру. Бгльша частина гх усмок-
туеться в проксимальному вгддглг кишок. Причому абсорбцгя катгошв-металгв е
специфгчною г селективною та потребуе наявностг спещалъних бглкгв-переносни-
к!в г ферментгв. Виводяться вони головним чином зг злущеним ештелкм травно-
го тракту, жовчю, епгдермком г волоссям. Ангони-неметали всмоктуються вщнос-
но легко г виводяться переважно гз сечею.
Кглькгсний склад мгкрослемент!в в органгзмг людини значною игрою визна-
чаеться гхнгм вмгстом у навколишньому середовишг, у першу черту, у рослинах,
гваринах г водг. Проте вибгрковгеть абсорбцп бгльшостг мгкроелементгв, здатнгсть
до гх депонування та селектившсть виведення демонструють здатшеть зрелого
здорового органгзму пгдтримувати гомеостаз мгкроелеменпв за рахунок авторегу-
ляцгг. Це, а також те, що в оргашзмах генуе виб!ркова концентрация мгкроелемен-
пв в окремих органах, тканинах г клгтинних органелах, а ёх дефгцит або надлишок
зумовлюе тяжкг патолопчнг наелгдки, евгдчить про значку участь мгкроелеменпв
у систем! регуляцн функцгй оргашзму.
В ембргональний пергод плгд отримуе мгкроелементи вгд матер!г здатен нако-
пичувати гх. Концентрангя деяких мгкроелементгв у деяких органах г тканинах
доношеного новонародженого перевищуе таку в органгзмг доросло! людини. У
немовлят баланс мгкроелеменпв залежить вщ гхнього вмгсту в грудному молош
або молочних сумгшах, а в дорослих — у продуктах харчування Значний дефщит
мгкроелеменпв виникае при хронтчних дгаре!, мальабсорбпп, патологп печшки,
ахолп, панкреатит], дуодент. Абсорбшя деяких мткроелеменпв порушуеться при
надлишку в !жг шших макро- 1 м!кроелемент!в. Порушення обмшу мшроелемен-
пв може бути наслщком спадкових хвороб 3 гншого боку, дефщит або наллишок
деяких мткроелеменпв в органтзмт матер! с правд я? тератогенну дно на потомство,
що спричинюе ембрю- та фетопатп з появою вроджених вад
На прикладах деяких найважливипих для розвитку г функцюнування людсь-
кого оргашзму есенщальних мткроелеменпв розглянемо загальн законом грносп
фтзюлоги та патофгзтологи цих складових гомеостазу.
ОБМ1Н ЗАЛ13А ТА ЙОГО ПОРУШЕННЯ
В оргашзм! доросло! людини мгститься 3—5 г залгза, що становить 0.006% маси
пла. За норма,!ьних умов близько 70 % залгза входить до складу гемоглобтну ерит-
роципв, близько 10 % — мюглобтну, близько 20 % — депонуеться у вигляд! фери-
тину га гемосидерину т менше 1% входить до складу транссрерину, лактоферину 1
понад 70 ртзних ферменпв. Майже все заттзо перебувае всередин! клпин. У плаз-
м1 кровт залгза мгститься менше 0,01 % вщ його вмтсту в оргашзм!. Там воно лише
транспортуеться у вигляд! трансферину г феритину.
Залтзо е життево необхщним для оргашзму металом у зв’язку з його вмгстом
у великтй юлькостт залтзовмгених ферменттв т неферменгних металопротешж
Прозе не зв’язане з битками юшзоване зал!зо е дуже токсичним, тому що у форм!
Ре2+ воно каталгзуе утворення гщроксильних радикалтв у реакци Фентона. Ре2+ +
Н,О, = Ре3т + ОН” + ОН. 3 цте! причини оргашзм змушений дуже точно регулю-
вати концентрашю втльного юшзованого залгза в цитозол! клттин. Мехашзми аб-
сорбци залпа через етигелш кишок, його транспорту в сироватщ кров!, доставки
до клттин, депонування, мобйпзацп 1 реутилгзаци е дуже складними г водночас
ефективними.
Джерела г мехашзми надходження залгза в оргашзм та шляхи його виведення.
Органтзм людини не здатний швидко й ефективно виводити залтзо. Ус1 дли до
пубертатного втку т чоловтки втрачають не бГттьше 1 мг залгза на добу переважно
з! злутценими кл!тинами слизово! оболонки кишок 1 шюри. Д!вчата г жшки додат-
ково пщ час менструацп втрачають гз кров’ю 15—40 мг залгза. Гомеостаз цього
мгкроелемента забезпечуеться регуляшею його всмоктування у верхнш частинг
тонко! кишки, переважно у дванадцятипалш кишщ.
1снують два мехашзми всмоктування залгза. Легко г прямо, за допомогою
спецтального переносника — гемтранспортера (ЬешсГгапзройег) абсорбуегься за-
кисне (Ре2+) фери-залгзо у склад! гему г окисне (Ре3+) фери-зал!зо у склад! гемшу,
як! мктяться лише в гваринних продуктах (печшка, м’ясо, нирки). Ц! продукти
не входягь до рацюну немовлят ! вегетартанцгв. Негемшове окисне залтзо, що
моститься в молоц!, овочах, фруктах г мтнеральних речовинах, перед усмоктуван-
ням мае вивтльнитися з оргашчних комплексов. вшновитися за допомогою фери-
редуктази та вттамтну С до Ре2+. Цей пронес зна’шо полегшуеться при пщвишенш
розчинностт залгза пщ впливом хлоридно! кислоти шлункового соку. Механтзм
усмоктування негемового Ре2+, який нин! уже достатньо вивчено, пов’язаний тз
роботою апикального мембранного переносника залгза, що мае назву транспорте-
ра двоваиентних метал) в 1 (йтуакгншегакгапяройег 1, ОМТ 1) або транспортера
двовалентних каттонгв 1 (Дп аТептсапопггапзропег 1, ОСТ1), або макрофагального
быка, пов’язаного з природною резне гентшстю 2 (пашга1гез)81апсе-а8&ос)агес1гпас-
горйа^ергогет 2,14КАМ Р 2). Цей мембранний глткопротеш, який мзетить 12 гранс-
мембранних доментв, виявляють у посмутовашй облямтвщ ентероципв ворсинок
дванадцятипало! кишки, ендосомах некишкових юптин т фагосомах макрофапв,
як) срагоцитують стар) еритроцити. ОМТ 1 за допомогою активного ко-транспор-
ту з протонами переносить як Ре2+, так 1 2п2+, Мпн, Си2+, СД2+, Со2\ 14г2+ ) РЬ2+ з
проевпу кишок у цитоплазму ентероципв або з ендосом 1 фагосом у нитозоль
шших клпин. В ентероцитах гемове 1 гемшове залтзо вивыьнюються гем-
оксигеназою 1 разом з Ре2+. транспортованим ОМТ 1, за допомогою битка з умов-
ною назвою мобыферин транслортуеться до базальнот мембрани або у везикули.
Останнт дант евщчать, шо мобыферин, 1мов)рно, е Ре-АТФазою На базальшй
мембран) або у везикулах за допомогою гефестину (йерйаезбп) — битка, под)бного
до сироваткового быка церулоплазмшу, що мае власти востт фероксидази, Ре2+
окиснюеться до Ре3т. Це необхтдно тому, що подальший експорт зал)за в кров або
його депонування в ентероцитах у вигляд) феритину можливе лише в окиенш
(Ре3+) ферт-форм). Транспорт Ре3+ через базальну мембрану ентероципв, а в по-
дальшому 1 з гепатопиттв та макрофапв, здшснюеться спещальним переносни-
ком, який отримав назву феропортин 1 (ГеггоротТт 1), або быок — транспортер
металу 1 (тетайгапзропегргогепг 1, МТР 1). Ртвень МТР в ентероцитах, гепатоци-
тах ) макрофагах регулюеться спещальним быком — гепсидином (йерсгсйп), який
синтезуеться в печшц). Ц)каво. що утворення гепсидину значно збыынуеться при
запаленн). Дал) на зовншгнтй поверхн) базальнот мембрани Ре3+ зв’язуеться з
трансферином плазми кров), молекула якого здатна переносили 2 атоми Ре3+, )
транспортуеться з кров’ю. Частина Ре3+ тимчасово депонуегься в ентероцитах у
вигляд) феритину. При дефшит) в оргашзм) залгза збыьшуються синтез 1 актив-
шсть ОМТ 1 । МТР 1, а синтез феритину знижуеться. При надлишку залтза все
вщбуваеться навпаки: усмоктування залгза зменшуеться, а чистка феритинового
залтза в ентероцитах збыьшуеться, 1 воно поступово втрачаеться оргашзмом при
злущуванн) ентероципв ворсинок дванадцятипалог кишки. 3 усього залгза, яке
надходить з тжею, усмоктуеться не быьше 10 %. 1з залтза, яке мгститься у м’яст,
абсорбуеться 13—22 %, )з залтза грудного молока — 49 %, з коров’ячого молока —
нс быьше 10 %, а тз залгза рослинних продукты — лише 1—3 %. Особливо погано
засвоюеться залтзо зернових продукты, у яких воно зв’язане з фпиновою кисло-
тою Знижують абсорбцию Ре2+ фосфата, оксалата та клггковина. Хлоридна кис-
лота шлунка, аскорбшова та )нш) оргашчш кислота сприяють вгдновленню Ре3+
до Ре2+, полптшуючи тим самим усмоктування залгза. Сприяють абсорбцп залтза
також аминокислота з тваринних быкгв, просп вуглеводи й алкоголь. Осюльки
ОМТ 1 здатен переносита ргзш двовалентн) метали, надлишок будь-якого з них
може перешкоджати абсорбцй шших. Водночас вщсутшсть мцц зумовлюе пору-
шення синтезу полтмщного быка гефестину й унеможливлюе всмоктування за-
лтза.
Залежшеть потреби в харчовому залтзг вгд вшу г стан. Потреба доросло) людини
в залтз), яка становить 20—25 мг на добу, задовольняеться в основному завдяки
реутилтзаш) залгза, що вивыьняеться при мегаболтзм) гемоглобшу. Шодобоьа по-
треба в харчовому залгзг в дорослих чолов)к)в — 10 мг. що забезпечуе всмоктуван-
ня 1 мг, або 4—5 % потреби. У дзтей з жшок потреба в харчовому залззз значно
вища. Воно мае забезпечити близько 30 % добовоз потреби в залззз дзтей раннъого
взку. У перюд ваптносп г лактац!! жзнки потребують 30—60 мг харчового залзза на
добу.
Мехашзми реутилззащ! залзза. Шсля фзззолопчного або патолопчного гемолззу
еритроципв гем, який потрапляе в цитоплазму макрофапв, гепатоципв, нефро-
цитзв 1 деяких шших юптин, пщ дзею ферменту гемоксигенази розщеплюеться на
взльне двовалентне залззо Ре2+, СО з бызвердин. Далз Ре2+ за допомогою нещодавно
вщкритого быка — переносника Ре-АТФази переноситься в ендоплазматичний
ретикулум, де за допомогою гефестину окиснюеться до Ре3+, зв’язуеться з апо-
трансферином з через апарат Гольджз виводиться в кров у виглядз трансферину.
Мехашзми депонування залзза. Надлишок залзза головним чином у макрофагах
юсткового мозку, селезшки, печшки та в сидеробластах юсткового мозку депо-
нуеться у вигляд! феритину з гемосидерину. Кожна молекула апоферитину мае
порожнину, у яюй може перебувати близько 4300 атомзв Ре3+ у форм! кристалу
ферипдрату (5Ре2О3 9Н2О). У норм! ступзнь насиченосп феритину залззом стано-
вить близько 2000 атомзв на молекулу. Феритин розчиняеться у вод! й легко вщдае
залззо. У раз! перенасиченосп оргашзму зал!зом молекули феритину агрегуються,
частково протеолззуються з перетворюються на нерозчинний гемосидерин.
Мехашзми обмшу залзза в некишкових клгтинах. Зал!зо зз макрофапв, як з хар-
чове, транспортуеться до кровотворних та шших клиин трансферином. У некиш-
ковз юптини зал!зо потрапляе шляхом рецептор-опосередкованого ендоцитозу.
Специфзчш трансферинов! рецептори на зовшшньому боц! плазматичноз мембра-
ни зв’язують трансферин, ! комплекс трансфериновий рецептор — трансферин
штерналззуеться (захоплюеться клзтиною). У цитоззлазмз цей комплекс потрапляе
вендосоми, уякихрН становить 5,5—6,0. У кислому середовищз залззо вщ’еднуеться
вщ трансферину, з комплекс апотрансферин — рецептор повертаеться назад до
плазматичноз мембрани, де апотрансферин звзльняеться вщ рецептора. Взльний
Ре3+ вщновлюеться оксидредуктазою до Ре2+ з за допомогою ОМТ 1 транспор-
туеться в цитоззлазму, де включаеться в гемоглобш, мюглобш або ферменти.
Мехашзми регуляцп обмшу залзза. Усмоктування, мобыззацзя та депонування
залзза регулюються концентрацзею вшьного залзза в цитоплазм! через быки, чут-
ливз до залзза (згоззгекропззуергоГезпБ, 1КРз), як! вшзззвають на елементи, що ре-
гулюють залззо (!гопге§и1а!огуе1етеЩ8, 1КБз), у нетранслюючих дыянках кшькох
мРНК, яю кодують протеши, що беруть участь у метаболззмз залзза. Наприклад,
взаемодзя 1КР8 з 1КБ8 при зменшеннз пулу взльного цитоплазматичного залзза
зумовлюе збыьшення абсорбцн Ре2+ ентероззитами завдяки пщвищенню синтезу
ОМТ 1 та активацпо захоплення трансферину некишковими юптинами шляхом
посттранскрипщйноз стабыззацп мРНК трансферинового рецептора. Водночас
запаси феритинового залзза змензззуються внаслщок блокування трансляцп фери-
тинового мРНК.
Роль залзза в оргашзм!. Залззо бере участь у транспорт! та депонуваннз кисню,
окисному фосфорилювашц, окисно-вщновних реакцзях тощо. Окисне Ре3+ справ-
ляе антиоксидантну, а закисне Ре2+, яке каталззуе реакцпо Фентона, — проокси-
дантну дно.
Залгюдефщит у чоловзюв виззикае дуже рщко: унаслщок хрошчних крововтрат
при гемофзлп, виразцз шлунка, а також при ахолп, дуодент та в разз суворого
вегетарианства. Навпаки, за останюми даними ВООЗ, 46 % дней вгком 5—14 рок!в
г 48 % ваптних в усьому свттт мають знижений запас залгза; у 50 % жтнок виявля-
ють запас залгза, який можна вважати ранньою стадгею залгзодефщиту. У дгтей до
пубертатного перюду залгзодефшит е наслщком недостатнього надходження залт-
за з !жею в раз! неповнощнного харчувдння або д!ареТ. Можливо, що випадки
вроджених залгзодефщиту й анемш е наслщком генетичного порушення синтезу
одного з бшкгв — переносниюв залгза. У статевозрших дтвчат г жшок залгзодефт-
цит виникае через надмгрну втрату залгза при менораггях, метрораггях, пологах та
пщ час лактацп. Спочатку зменшуються запаси гемосидерину г феритину, а потгм
знижуеться концентрацтя залгза в плазм!, що супроводжуеться низьким насичен-
ням трансферину залтзом г пщвищенням залгзозв’язуючог здатностг сироватки
кровг. Пгзнгше виникають зниження ртвня гемоглобгну в одиницг об’ему кровг,
гшохромгя г мгкроцитоз еритроципв. Дефщит залгза, спричинюючи зменшення
юлькостт гемоглобгну г мгоглобгну, призводить до вторинно! тканинно! гшоксп, а
зумовлюючи зниження ргвня залгзовмгсних мгтохондргальних цитохромгв, ци-
тохромоксидаз г ферменттв ЦТК, — до первинног тканинног гшоксп. Гшокстя
проявляеться швидкою втомлювангстю, м’язовою слабкгстю з нгчним нетриман-
ням сечг у щвчаток. Дефщит залгза порушуе утворення колагену, що зумовлюе
ураження шюри, нггггв г слизових оболонок. Характерними е тргщини в кутах
рота, червоног облямгвки губ, атрофгя ниткоподгбних сосочюв язика, що робить
поверхню язика гладкою г болючою. Спостерггаеться стоматит, ларингофаринго-
трахегт з пперемгею задньог стгнки глотки, захриплгстю голосу, дисфапею з вщ-
чуттям застрягання гжг (синдром Пламмера — Угнсона). Часто виникае атрофгч-
ний рингт, гнодг — озена. У багатьох випадках спостерггаеться атрофгчний гастрит
з ахглгею, а це значно посилюе залгзодефгцит. Нестача залгза порушуе дгяльшсть
центрально! та периферично! нервово! системи, що проявляеться порушеннями
та змшою смакових вщчуттгв г парестезиями.
Надлишок зал1за, який частппе спостерггаеться в чоловгкгв, може бути як спад-
ковим автосомно-рецесивним захворюванням, яке мае назву спадковий (первин-
ний) гемохроматоз, так г наслщком залгзорефрактерних г гемолгтичних анемгй або
хронгчного алкоголгзму (вторинний гемохроматоз).
Спадковий гемохроматоз, як правило, пов’язаний з мутацтею гена НРЕ, який
належить до родини генгв ГКГС I класу. Нормальний бглок НРЕ приеднуеться до
частини трансферинових рецепторгв, що зменшуе зв’язування насиченого залгзом
трансферину зг свогми клгтинними рецепторами. Також е данг про те, що бшок
НРЕ негативно модулюе ендоцитоз комплексу трансферин — рецептор. Модифг-
кований бглок НРЕ, що утворюеться при мутацп гена НРЕ, на це не здатен. Тому
при спадковому гемохроматоз! клгтини починають захоплювати бглыпе залгза г
накопичувати його, збшьшуючи юльюсть апотрансферину. Це супроводжуеться
щодобовою абсорбцгею в кишках 3—4 мг залгза замгсть 1 мг, що зумовлюе акуму-
ляцгю 500—1000 мг додаткового залгза в оргашзм! щороку. Збшьшення абсорбцп
залгза спостерггаеться на тлг нормального його надходження з гжею г непоруше-
ного еритропоезу, що спричинюе прогресивне збшьшення запасгв залгза в ор-
гангзмг з вщкладанням його в печшцт, пщшлункозгй залозг, серцг та ендокринних
органах. Оскшьки Ре2+ активуе процеси пероксидного окиснення, первинний г
вторинний гемохроматоз спричинюе розвиток цирозу печшки, цукрового дтабету
з потемншням шюри (бронзовий дтабет), ураження мюкарда, шфантилтзм г гшо-
гонадизм. НРЕ-асоцшований гемохроматоз — спадкове автосомно-рецесивне за-
хворювання, що виявляеться в европейськйй популяцй йз частотою 1 : 400. Спос-
терйгаються й шшй форми спадкового гемохроматозу. Наприклад, ювеныьний
гемохроматоз пов’язують з мутащею гена НРЕ-2. Бувае й автосомно-домйнантний
гемохроматоз.
Перевантаження клггин Ре2+ з их ушкоджен ням спостерйаеться також при
тканиннш гшоксп внаслщок пригнйчення роботи Ре-АТФази. Гостре отруення
зализом виникае в раз! перорального вживання понад 900—1500 мг Ре2+ у вигляд!
таблеток й призводить до смерт! внаслщок некротичного гастроентериту та шоку.
ОБМ1Н М1Д1 ТА ЙОГО ПОРУШЕННЯ
Вмйст 1 розподйл мщ! в оргашзм!. В оргашзм! доросло! людини мйститься 0,1—0,25 г
мйдй, що становить близько 0,0002 % маси т!ла. Практично вся мйдь перебувае внут-
рйппньоклйтинно в склад! ферменпв, переважно оксидредуктаз. Лише 1—3 % мйдй
мйстяться в плазм! кров!; п концентращя в чоловйкйв становить 10—20 мкмоль/л, у
ж!нок — 13—25 мкмоль/л; 95 % мщ! плазми входять до складу церулоплазм!ну, а
решта зв’язана з альбуминами. У новонароджених концентрация мщ! в плазм! —
лише 3—10 мкмоль/л. Водночас у тканинах новонароджених коищентрацйя мщ! в
галька разйв бйльша, н!ж у дорослих, й знижуеться до их рхвня лише у вщ! 6—12 мйс.
Мйсце ! мехашзми всмоктування мйдй. Мщь усмоктуеться в шлунку й прокси-
мальному вйддйлй тонко! кишки переважно за допомогою ПМТ 1 та, можливо,
транспортера мщ! 1 (соррегйгапзроПег 1, СП 1), надал! зв’язуеться в цитоплазм!
гастро- й ентероципв з бйлком металотйонешом. Одна молекула апопротеину цього
внутрйшньоклйтинного бйлка здатна зв’язати 7—10 атом!в мйдй, цинка, кадмйю, зо-
лота, ср!бла або ртуть Тому щ метали конкурують мйж собою при абсорбцц в
травному тракт!. Знижують усмоктування мщ! також фйтинова кислота й високий
р!вень бшка в !жй. 1з харчово! мщ! зазвичай усмоктуеться близько 30 %, а у вагйт-
них — близько 50 %.
Мехашзми транспорту, депонування та виведення з оргашзму мйдй. На базально-
му полюс! ентероципв мщь вившьнюеться вщ металотйонешу 1 тимчасово
зв’язуеться з альбумшами та деякими аминокислотами й транспортуеться в печ!нку
та ппш органи й тканини. У плода функцию транспорту мщ! замйсть альбумйнйв
виконуе а-фетоглобул!н.
Ключову роль у метаболйзмй мщ! вйдйграе печйнка, у якйй мйдь спочатку
зв’язуеться металотйонешом, шо виконуе функцп детоксикации та внутрйшньоклй-
тинного транспорту м!дй. Синтез металотйонешу регулюеться вмйстом мйдй та цин-
ку в клйтинах на рйвнй транскрипцй! мРНК. Пидвиицення вмйсту Си2+ або 2п2+ може
спричинити амплйфйкацйю генйв, яга кодують синтез металотюнешу, що зумовлюе
рйзке пидвиицення синтезу цього бйлка. Далй в лизосомах мйдь звйльняеться вйд ме-
талотюнешу й потрапляе в апарат Гольджй, де за допомогою Си-АТФази або ви-
водиться в жовчнй протоки, або транспортуеться до ензимйв, що синтезуються,
або включаеться в склад церулоплазмину, який синтезуеться в гепатоцитах. Цей
мультифункцйональний а2-глйкопротеш мйстить 6 атомйв мйдй й транспортуе их йз
печйнки в органи й тканини, де вони включаються в склад внутрйшньоклйтинних
ферменпв, переважно оксидредуктаз.
Доставку мгдг до Си-залежних ферменпв. таких як цитохромоксидаза. су-
пероксиддис мутаза, амшоксидаза, безпосередньо здшснюють маленьк! цитоплаз-
чатичн! протегни, якг називають шаперонами м1д1.
В ентероцитах, гепатоцитах г деяких пил их клгтинах плода г новонароджсних
концентращя металотгонепгу значно бьтыпа, а синтез церулоплазмгну менший, нгж у
дорослих, що г пояснюе плокупремгю та накопичення в тканинах мгдг в цей перюд.
Надлишковий церулоплазмш вграчае сгалову кислоту, зв язуеться зг спещаль-
ними рецепторами на поверхш гепатоиитш, шляхом ендоцитозу потрапляе в ге-
патоцита г частково разом гз зайвим комплексом мщь — металотюнегн захоп-
тюеться лгзосомами, де вщбуваеться гх протеолгз. Асгалоцерулоплазмш цитогглаз-
ми гепатоципв разом гз вмгстом лгзосом вивыьнюються через апарат Гольдж! в
жовчнг протоки, кон’югуеться з бпирубшом 1 високомолекулярними б!л ками та
виводиться з оргашзму гз калом. Усього гз жовчю виводиться 80 % мгдг, ще 16 % —
у складг випорожнень зг злущеним еппелгем шлунка г кишок. У норм! гз сечею
виводиться лише близько 4 % мгдг. Щодоби в оргашзм г доросло! людини вклю-
чаеться в склад церулоплазмгну । виводиться гз жовчю близько 0,5 мг мгдг.
Оснсвш джерела надходження мин в оргашзм г добова потреба дорослих г дгтей
у харчовш мцц. Оскгльки оргашзм людини не здатен до реутилгзацп м!дг, потреба
в цьому мгкроелеменп за повод ьняеться лише його надходженням з гжею. Основ-
ними джерелами мгдг для доросло! людини е печгнка I м’ясо тварин. риба, бобов!,
горгхи. а для немовлят — грудне молоко. Добова потреба в харчовгй м!д! для до-
росло! людини становить 2—5 мг, для немовляти — 0.6—0,7 мг.
Роль мгдг в оргашзм! людини. Значения мгдг для органгзму визначаеться тим,
що вона е необхгдним складовим компонентом дуже важливих оксидредуктаз.
Так. головний мгдьумгсний метало протегн коовг церулоплазмш окр!м виконання
транспорт™! функцп е оксидазою катехоламгнгв, аскорбшовог кислота та гнших
сполук. Особливо важливим е те, що церулоплазмш у плазм!, а подгбний йому
гефестин у цитоплазмг окиснюють Ре2+у Ре3"1. Це, по-перше, дае змогу апотранс-
фсрину мобглгзувати залгзо гз клгтин г перенести його в гем, а по-друге, блокуе
реакцию Фентона г зменшуе ПОЛ. Антиоксидантну та протизапальну д!ю чинить
г мщь- та щшкумгсна супероксиддисмутаза, шо активуе реакцгю: 20/ + 2Н+=
Н2О. + О,. Цитохромоксидаза — термшальний фермент дихального ланцюга, не-
обхгдний для синтезу АТФ. Тирозиназа катапгзуе утворення мсланшу а дофамш-
р-пдроксилаза — катехоламгнгв. Лгзилоксидаза необхгдна для утворення попереч-
ник зшивок колагенових г еластичних волокон. Ггсгамгназа та шил амгнооксидази
шактивують бюлопчно акта в ш амгни.
Недостатшсть лад/ можлива в д!тей при повному парентеральному або не-
збалансованому за мгкроелементами штучному молочному вигодовуваннг При
цьому дефщит трансферину й АТФ спричинюе виникнення мгкронитарно! залг-
зодефгцитног анемп. Нестача мгдг супроводжуеться порушенням синтезу норадре
налгну г дофамгну в головному мозку. зниженням синтезу мгелшу й актавашею
ПОЛ, що в ноеднанн! зумовлюе порушення дгяльносп центрально! нервово! сис-
теми. В останнгй час з’явилися данг про те, шо хвороби Атьцгеймера 1 Паркгнсо-
на пов’язанг з оксидатавними процесами, якг зокрема стимулюються недостатньою
активнгстю мгдьумгсних ферменпв супероксиддисмутази та церулоплазмшу. Зни-
ження активностг тирозинази може спричинити депггментзцпо шкгои г волосе я.
Дефектний синтез колагену й еластину може стати причиною деформапп та лам-
костт ктсток, порушення зроговшня шктри, вазопатгй т розшаровуючо! аневризми
аорти, що найхарактерншге для синдрому Марфана, який становить автосомно-
домтнантну недостатшсть лтзилоксидази.
Дефщит мщ! супроводжуеться пперхолестеринемгею, яку пояснюють знижен-
ням активност! лецитинхолестеринацилтрансферази, лшопротешлшази та деяких
шших ферменпв. Гшерхолестеринемтя та вазопаття при дефщит! мщт може стати
причиною прискорбного розвитку атеросклерозу та ппемтчно! хвороби серця.
Хвороба Менкеса, яка успадковуеться рецесивно г зчеплено з Х-хромосомою
та виявляеться тз частотою 1 випадок на 35 тис. новонароджених, характеризуеть-
ся дефщитом мщт в печшщ, кров!, судинах, мозку та порушенням функцш мщь-
залежних ферменттв. Водночас ентероцити, нирки, м’язи та фтбробласти шк!ри
мтстять у 3—10 раз!в бшыпе мщт, шж звичайно. Характерними е дешгментащя та
кучеряв!сть волосся. Порушення стабшьност! колагену та еластину спричинюе
розвиток аневризм, остеопорозу, емфтземи. Спостерггаеться порушення розвитку
мозку, виникае нейродегенерацтя тз високою летальшстю в ранньому дитинств!.
Як нещодавно встановлено, патогенез ще! хвороби пов’язаний гз мутащею
гена АТР 7 Ав Х-хромосомц який кодуе синтез Си-АТФази, або бшка Менкеса в
багатьох органах, кргм печшки. При цьому виведення мцц з оргашзму в склад!
жовчт зберггаеться, а абсорбцтя через ентероцити т транспорту у печшку порушу-
ються, що ! спричинюе негативний баланс м!д! в оргашзм!.
Надм1рне потрапляння м1д1 в оргашзм можливе при споживанш кислих про-
дукттв, що збертгалися в мщному посуд!, при отруенш пестицидами, фунгщидами
та при контакт! з мщними рудами. Але в педтатрп причиною збшьшення концент-
рацп мцц в органах 1 тканинах найчастгше е хвороба Вшьсона — Коновалова. Це
захворювання мае автосомно-рецесивний тип успадкування т виявляеться тз час-
тотою 1 : 30 000. Гетерозиготне носгйство патолопчного гена ще! хвороби спос-
тертгаеться гз частотою вщ 1 : 500 до 1 : 200.
Пщгрунтям патогенезухвороби Вшьсона — Коновалова е мутация гена АТР 7
В у хромосом! 13, який кодуе синтез ще одше! Си-АТФази або бшка Вшьсона. У
норм! цей ген значно експресуеться в печшщ, нирках г плацент! та слабко — в
мозку, серщ, м’язах 1 шдшлунковш залоз!. Мутащя цього гена перешкоджае ви-
веденню мцц тз жовчю та утворенню церулоплазмшу з апоцерулоплазмтну, що
зумовлюе накопичення мцц в печищ! та зменшення вмтсту церулоплазмтну в
кров!. Хоча екскрецтя мцц !з сечею при цьому збшьшуеться в 30 разгв, вщбуваеть-
ся затримка мщ! в органтзмт за нормального п всмоктування. Накопичення ютп-
зовано! мцц в гепатоцитах спричинюе генеращю втльних радикалтв, якт руйнують
клггинш мембрани, ДНК, фермента! та структурнт бшки. Розвиваються стеатоз г
цироз печтнки. Пертодично при руйнуванн! гепатоциттв значна кшьктсть тошзова-
но! мцц потрапляе в кров, виникають напади гемолттично! анемп та вщбуваеться
вщкладання мщ! в нирках, мозку та шших органах з !х ураженням. Характерним
для ще! хвороби е вщкладання по периферт! роггвки кшьця Кайзера — Флейшера
золотисто-коричнюватого або зеленкуватого кольору.
Клгшчн! ознаки хвороби Вшьсона-— Коновалова починають з’являтися в 4-
ртчному вщ!, а середшй втк установления ддагнозу становить 15,5 року. Раншми
ознаками захворювання е збшьшення печшки й селезшки, жовтяниця й анорек-
с!я. Без лгкування неминуче розвиваеться цироз печшки. У вщ! 6 рокгв т старше
виникають нейром’язовт порушення: тремор кшщвок, дизартртя (скандована
мова) 1 дистошя (порушення м’язового тонусу) Одночасно вщбуваеться ушко-
дження проксимальних ниркових канальщв. що проявляеться втратою хз сечею
глюкози, амшокислот, фосфатхв х розвитком циркового канальцевого ацидозу.
ГИзнхше превалюють нейропсиххчш ускладнення: змши особи с госи та поведшки,
зниження здатносп до навчання.
Для Л1кування застосовують хелатори м!д), наприклад Э-пенщиламж, а та-
кож вилучають 1з рацюну м’ясо, горххи, боби, шоколад.
ОБМ1Н ЦИНКУ ТА ЙОГО ПОРУШЕННЯ
Вмхст 1 розподш цинку в оргашзм!. В оргашзм! здорово! доросло) людини мхститься
1,5—2,5 г цинку, що становить близько 0,003 % маси тша. Вш наявний в ус!х ор-
ганах 1 тканинах, переважно у склад! металоферменпв. Близько 63 % усього цин-
ку мхститься в скелетних м’язах. Значка юльюсть 7п2+мхститься в печ’шш, мозку,
щитопод1бн!й залоз!, гхпофхзх, молочних залозах х грудному молод! Проте най-
5!лыпа його концентращя спостерйаеться в передмххуровш залоз!, сперматозоидах
та р-юптинах острхвшв Лангерганса шдшлунково’! залози. У кров1 мютиться лише
1 % цинку. У плазм!, де 7п2+на 70% зв’язаний з альбумшами, його концентращя
становить 46—61 мкмоль/л, а в еритроцитах — 153—214 мкмоль/л.
Мехашзми всмоктування, транспорту, депонування та виведення цинку. Перед аб
сорбшею в тонкш кишщ 7п2+ мае звшьнитися в!д оргашчних носив, а дал) частина
цього пдрофшьного металу проникае кр!зь алгкальну мембрану ентероципв за допо-
могою ОМТ 1, про який шшося вище. Кр!м того, транспорт 7п2+ в ентероцитах, як
х в шших клхтинах, забезпечуеться двома родинами селективних транспортерхв цин-
ку: родиною бшкхв хмпоргерхв цинку (хтсхтрогХегрхоГешз, 7п1Рз), яю хмпортують
його у цитозоль клхтин, 1 родиною транспортерхв цинку (ДпсГгапзрохТегз, 7пТз), якх
видаляють його хз клхтин або хзолюють у внутршшьоклхтинних везикулах.
Так, 2п1Р I сприяе захопленню клхтинами цинку х залхза, 2п1Р 4 селективно
абсорбуе в кишках 2п24, а 2п1Р 3 вившьняе його хз внугршшьохсптинпих компарт-
ментхв у цитозоль. Своею чертою 7пТ 1, шо розташований на базолатеральшй мем-
бран! ентероципв, нефроцит!в дистальних канальшв нирок ! на плазматичних мем-
бранах кл)тин шших органхв, вившьняе цинк хз клхтин у кров. 2пТ 2 1 7пТ 3 пере-
м)шають цинк хз цитозолю клхтин у специфхчнх везикули. 7пТ 4 забезпечуе секрещю
2п2+ в молоко 2пТ 5 транспортуе цинк у секреторн! грану ли р-клхтин остр)вц)в Лан-
герганса. 7пТ 6 може транспортувати 2п24 як у везикули, так х в апарат Гольджь
Потрядивши в цитозоль ентероципв та шших клггин, цинк або депонуеться
у везикулах у зв'язаному з металопонешом станх, або використовуеться для син-
тезу металоферменпв, або виводиться за допомогою 7пТ 1 у кров, де зв'язуеться
з альбумшами та, частково, з петидином ) цистешом. Цей процес може регулю-
ватися кошхентращею цинку в цитозоль Збшьшенххя рхвня 7п2+ пшвшцуе екс-
пресхю гешв, як! кодують синтез металотюнешу х 7пТ 1, через фактор транскрип-
цц, шо регулюеться металрегульованим фактором транискрипцп 1 (хпе1а1-ге5и1ато
гутгапзспрнопГасгог 1, МТЁ 1), локашзованим у хромосом! 1.
Така складна система обмшу пинку не ильки дае змогу пщтримувати концен-
трашю пього метату в щхтозолх на поспйному ргвш, а й здатна регулювати )нтен-
сивнхсть його всмоктування в кишках ! реабсорбцп в нирках. Так, з хжх, б)днох на
цинк, засвоюеться близько 85 % 7п2+, а зг звичайног гжг — лише 10—30 %. Виво-
диться цинк з органгзму переважно гз калом, у який потрапляють металофермен-
ти пишглунково! залози та ентероцилгв, а лакож злушен! ентеропити.
Джерела надходження циню в оргашзм та погреба в ньому залежно вщ вшу та
стать У немовлят потреба в пинку задовольняеться його надходженням гз грудним
молоком. У грудному молоцг цинк зв’язаний з альбумшом г лактоферином, тодг як
у коров’ячому — з казегном, який перетравлюетъся значно прше. Ним г пояснюеть-
ся бгльша поступи гсть цинку грудного молока Цинк мгститься як у тваринних, лак г
рослинних продуктах. Проте засвоенню його з останнгх перешкоджае фггинова кис-
лота . клгтковина. Усмоктуванню цинку перешкоджэюль також мщь, кальцш, кад-
мш, фосфати. 1Л-1, алкоголь : дефшит впамшу А. Твариннг битки 1 таю амгнокисло-
ти, як Л13ИН, цистеш, глгцин г глутамат, навпаки, сприяють усмоктуванню /и2’’.
Щодобова потреба в харчовому цинку в дорослих чоловгкгв становить 15 мг,
у жгнок — 12 мг, у ваггтних —15 мг, у перюд лактацп — 19 мг. У немовлят потре-
ба в 2п2+ становить 5 мг на добу, а у сгаргггих дгтей — 10 мг на добу.
Значения цинку' для оргашзму визначаеться тим, шо вш входить до складу
близько 200 ферментгв, яю належать по всгх вщомих класгв. Цей мгкроелемент
здатний зв’язуватись одночасно гз 4 г бгльше лггандами та мае велику споргд-
ненгсть до електронгв, шо забезпечуе стабглънгстъ сполук, до яких вгн входить.
Тому в металоферментах цинк може виконувати не тгльки каталгтичну. а й струк-
турну функцгю. До найважливгших цинкумгсних ферментгв належать алкогольде-
гщрогеназа, супероксиддисмутаза. амшопептидаза, карбоксипептидаза, анпотен-
зиназа, лужна фосфатаза, карбоанггдраза, колагеназа тощо. Цинку належить важ-
лива роль у синтез! ДНК, РНК 1 бглка. Вш необхгдний для регуляпн клгтинного
циклу, пролгферацп 1 диференцповання клгтин. Цинк входить до складу всгх вгдо-
мих лранскритщгйних факторгв, необхщних для функцюнування ДНК- 1 РНК-
полгмераз. Цинкумгснг протегни беруть участь у генетичнгй експресй ргзних фак-
торгв росту, модуляцгг протегнкгназ.
Цинк входить до складу гормону загруднинно! залози тимулгну г е необхгдним
для дозргвання лгмфоцитгв. Вгн бере участь в упаковуваннг шсулгну, гонадо-
грошнгв та шших гормонгв, входить до складу глюкокортикогдних рецепторгв
Цинк вщновлюе ретинол у сгткгвцг ока. Вш е необхгдним складником бглка Гус-
тину, гцо виробляеться в привушних слинних залозах. Цей бглок потргбний дня
росту смакових сосочкгв. а отже, г для нормально! смаково! чутливосп.
На вгдмгну вщ зал!за, мгдг, марганпю. хрому, ваналгю. берилгю, миш’яку, 2п2+
не змгнюе свою валентнгсть ла безпосередньо не бере участь в окисно-вгдновних
реакщях, а отже, не стимулюе утворення акгивних кисневих радикалгв. Це, а та-
кож те, що вш входить до складу тгитозольног супероксиддисмутази, робшь цинк
мгнеральним антиоксидантом.
Причини та насмдки дефициту цинку. Недостатшсть цинку значно поширена
в усьому свт, особливо в реггонах, де в рацюнг населенггя переважають злаков!
протеши. Стан цинкового дефщиту супроводжуе захворювагшя травного тракту г
печшки, а також може бути наслщком; спадкових захворювань.
Значний дефгцит цинку у ваптно! може спричинити гщроцефалгю, зменшен-
ня об'ему мозку та загально! кглькостг нервових клгтин, мгкро- г анофгапьмгю.
розколину пгднебгння, вади серця у плода 1 новонародженого. У немовлят дефг-
цит цинку може бути наслщком або нестачг цього мгкроелсмента в органгзмг ма-
4141
герЁ, або недостатшстю переносника 7.пТ 4 в молочних залозах, або спричинений
переведениям дитини на штучне вигодовування. У ранньому дитинствё га шзнЁшс
дефшит цинку найчастЁше е наслщком або недостатнього споживання тваринних
бшкгв, або ентериту, або хронЁчного гемолизу еритроципв. У дорослих основною
причиною нестачг цинку може стати алкоголизм. Цей дефЁцит зумовлюе затримку
росту та недостатке розвинення гонад у хлопчикёв, розумове недорозвинення,
зниження апетиту, вштадЁння волосся, шчну слшоту, зниження ё зникнекня сма-
кового сприйнягтя, пригшчення клётинного ниуштету, порушення сну. Раншми
ознаками цинкового дефшиту можуть бути прищавЁсть ё поява б!лих плям на
нитях, ёх ламкЁсть. У щдлтткгв ё дорослих, якё з дитинства страждали на дефщит
цинку, крЁм вищезазначеного спостерггаються карликовЁсть, гшогонадизм без
вторинних статевих ознак, безшпднЁсть, гшохромна мЁкроцитарна анемЁя. пору-
шення загоення ран. Патогенетичною складовою бЁлыпоЁ' частини пих наслЁдкгв
дефЁциту пинку е вЁдсутнЁсть функцЁЁ цинкумЁсних ферментЁв, необхЁдиих для
процесу пролЁферацп та диферснцЁювання клётин. Нестача лужно’1 фосфатази при
цинковому дефЁцитЁ може порушувати остеогенез. АгевзЁя (втрата смаку) спричи-
нюеться нестачею цинку мёсного бЁлка густину, а ЁмунодефЁцит е наслщком пору-
шення синтезу тимулЁну разом з пригнЁченням синтезу клётин.
ДефЁцит цинку може бути ё наслЁдком ентеропатичного акродерматиту —
рЁдкЁсного автосомно-рецесивного захворювання, яке проявляеться везикуляр-
ним ё псорЁ азоформним дерматитом на лёктях, у пах вин них складках ё навколо
природних отворЁв, а також алопешею (випадЁнням волосся) ё дЁареею. За вЁдсут-
ностё лЁкування препаратами цинку це захворювання спричинюе дуже значний
дефЁцит цинку Ё смерть. ОстаннЁми роками було визначено патогенез ентеропа-
тичного акродерматиту. З’ясувалося, що його причиною е мутацЁя гена 8ЬС 39 А 4
в 15 ё, можливо, 8 хромосомах, який у нормЁ кодуе 2п1Р 4, Цей бьток, що мае 8
трансмембранних доменЁв. мёститься у великЁй кёлькостё в апЁкальних мембранах
ентеропитЁв Ё нефроциттв дистальних канальщв ё забезпечуе кишкову абсорбшю
та ниркову реабсорбцЁю пинку. Нестача 7п1Р 4 ё зумовлюе фатальний дефЁцит
цинку.
Надлишок цинку при поступовому його надходженнЁ мало токсичний, оскёль-
ки цинк не кумулюеться ё швидко виводиться. УзагалЁ вмёст цинку в органах ё
тканинах завдяки досконалЁй системЁ регулящ’1 мало змшюеться в раз! надлишко-
вого чи помЁрно недостатнього надходження цинку в оргашзм. Змёни вЁдбувають-
ся лише в кровЁ. Проте в раз! рЁзкого надходження велико) кёлькостё цинку спос-
терЁгаються сонливёсть, головокружЁння, блювання. галюцинапп, анемЁя. У разЁ
вживания 45 г Хп8О4 зареестровано отруення, що закЁнчилося смертто.
ОБМ1Н МАРГАНЦЮ ТА ЙОГО ПОРУШЕННЯ
Вмёст I розподЁл марганцю в оргашзмЁ. В оргашзмЁ здоровоТ .людини мёститься близь-
ко 20 мг марганцю, концентращя якого найбЁльша в кЁстках, печЁнпЁ, мозку, гона-
дах. нирках та ендокринних залозах. Навпаки, вмёст марганцю в скелетних м’язах
ё плазм! кровт дуже низький. ПечЁнка плода не накопичуе цей мЁкроелемент.
Споживання, усмоктування, транспорт 1 виведення марганцю. Основним джере-
лом марганцю для людини е рослиннё продукт, передусЁм чай, з яких Мп2+
вивыънюеться ! в невеликш юлькосп всмоктуеться у проксимальному вщдцп тон-
ко! кишки. У грудному молош марганцю менше, н!ж у коров’ячому. але всмок-
туеться В1Н значно краще саме !з грудного молока. Кальцш I фосфати утруднюють
абсорбцйо марганцю, а алкоголь ц пщвишуе.
Переносниками Мп2т через ашкальну мембрану ентероципв е ПМТ 1 та спе-
циф!чш транспортери, яю одночасно здатш переносити кадмш. Марганець транс-
портуеться в кров! спешальним (3,-глобулшом. Вш ефективно захоплюеться пе-
чшкою. Невелика кыьюстъ Мп2+ окиснюеться до Мп3+1 транспортуеться транс-
ферином в ппш органи ! тканини, де б!льша його частина включаеться в
мгтохондрп.
Лобова потреба в марганщ для доросло! людигш становить 2—5 мг, у немо-
влят — 0,3—0,6 мг. Быьша частина марганцю екскретуеться !з жовчю ! соком
пщшлунково! залози.
Значения марганцю для оргашзму визначаеться тим, що вш входить до актив-
них центр!в низки ферменпв. а також здатен активувати багато шших ферменпв
Це робить марганець необхщним для полгмеризацп ДНК. транскрипцн, синтезу
гангл1озид1в г глутамшу. Лшотропна й антиатеро генна ды марганцю забезпечуеть-
ся тим, що в!Н активуе ферменти, необхщн! для синтезу фосфолшцпв I холестери-
ну. Його участь у синтез! глжозамшоглгкашв забезпечуе остео-1 хондрогенез. Цей
м!кроелемент необхщний для глюконеогенезу, обмшу катехолам!н!в й акгивац!! X
фактора зсщання кров!. Дуже важливу роль в оргашзм! в ил грае м!тохондр!альна
суперокспдцисмугаза, що мгстать марганець. Цей фермент захищае м!тохондрп
вщ руйшвно! дп супероксидного ан!он-радикала та запобггае апоптозу клпин у
норм! 1, особливо, при опромшенш, гшероксп та синдромг гшемн-реперфузи.
Недостатшсть марганцю, що виникае при порушенш його всмоктування,
зумовлюе затримку росгу, аномал!! розвитку юсток 1 хряпцв, гптогонадизм, зни-
ження маси пла, гшохолестеринемпо, норушуе зсщання кровь Зниження актив-
ност! глутамшсинтетази спричинюе штоксикащю центрально! нервово! системи
амон!ем 1 пригшчуе актившсть глутаматерпчних нейронш, а недостатшсть м!то-
хондр!ально! супероксиддисмутази зумовлюе ушкодження мггохондртальних мем-
бран, шо е причиною зниження синтезу АТФ. Усе це порушуе нормальну дгяль-
шсть мозку г проявляеться ешлсптичними судомами. Недостатня актившсть су-
пероксиддисмутази в мпохондртях р-юптин осгр!вц!в Лангерганса зумовлюе !х
руйнування супероксидом та розвиток иукрового дгабету. При дефщит! марганцю
у внутршшьоутробний перюд можуть порушуватися структура 1 функцгя вестибу-
лярного апарату.
Надлишок марганцю виникае при щдвищенш його концентраци у вод! та про-
дуктах харчування. Марганець може потрапляти в оргашзм також при вдиханн!
марганпевого пилу п!д час робот» на шахп. У пщвищешй концентраци в оргашзм!
цей м!кроелемент справляе прооксидантну дно 1 здатен ушкоджувати мембрани
мпохондрш та шших органел. Головною мшгенню для токсично! ди марганцю е
ЦНС 1, особливо, базальн! гангли. У першу чергу ушкоджуються астроцита та по-
рушуються синтез г депонування в нейронах дофамшу, шо супроводжуеться не-
вролопчними порушеннями, подтбними до хвороби Паркшсона. Можлив! також
психгчш розлади, порушення функцн гонад, рахгтопошбш змши скелета, анем!я,
анорекстя та дизартрш. Менш тяжке хрошчне отруення марганцем проявляеться
зменшенням у дней здатносп до концентраци уваги та зниженням пам’яп.
ОБМШ ХРОМУ ТА ЙОГО РОЛЬ В ОРГАЖЗМ1
Джерела надходження хрому га його обмш в оргашзм]. В оргашзм! доросло! людини
мгститься приблизно 0,6 мг хрому. Концентращя цього мокроелемента в органах 1
клпинах значно перевищуе його р!вень у сироватш кров!. У тканинах новонарод-
жених концентращя хрому в середньому в 2,5 рази болыпа. шж у дорослих. Усмок-
туеться вш переважно в порожнш кишщ, транспортуеться в кров! трансферином
видтляеться в основному нирками. Джерелом харчового хрому е м’ясо, печшка,
яйця, пивш дргждж!, фрукти й овоч!. 3 харчового хрому в норм! всмоктуеться не
б!льше 1 %. Добова потреба в цьому мокроелеменп вщ народження 1 до 6 м!с.
становить 10—40 мкг, в!д швроку до 3 рокгв — 20—80 мкг, в!д 4 до 6 рок!в — 30—
120 мкг, а в д!тей вгком понад 7 рок!в 1 дорослих — 50—200 мкг.
Форми кнування хрому та особливостт Тх впливу на органтзм. У природ! хром
звичайно генуе у двох станах: Сг64! Сг34. Тривалентний хром е х!м!чно стаб!льним
1 нетоксичним дня оргашзму. Хоч в!н 1 не входить до складу ферменпв 1 кофак-
торов, його все-таки вважають есеншатьним нутроенгом, осюльки в!н бере участь
у метаболозмо вуглевод!в, лшод!в 1 б!лк!в, посилюючи дно шеулону.
Шестивалентний хром оснуе лише у вигляд! окислив о оксо-ашонов, як! утво
рюються при обробш та зварюванш металтв, виготовленн! нержавпочо! стад! та
сплавов, а також фарб ! латав тощо. Останньм часом забруднення грунтових вод г
повотря сполуками, шо мктять Сг64 , прогресивно збттыпуеться. У бюлопчних
рщинах Сг64легко вщновлюеться через пром!жш форми Сг44! Сг54 до х!м!чно стой-
кого Сг34. При цьому утворюеться велика юльюсть дуже токсичних активних кис-
невих радикалов, яю спричинюють оксидативний стрес з пероксидним окиснен-
ням лопйцв, ДНК о болков, шо активуе апоптоз клотин 1 генш та хромосомно
мутацп. У травному траки болыпа частина Сг6’швидко вщновлюеться о потрапляе
в кров переважно у вигляд! Сг34. Водночас з аерозолями Сг61 проникав в легеш, де
спричинюе розвиток фоб розу о раку.
Встановлено, шо дефщит хрому знижуе толерантность до глюкози, зб!ль-
шуе потребу в шсулш!. шдвишуе ртвень у кров! холестерину о прискорюе роз-
виток атеросклерозу. Вщомо, що у хворих на цукровий доабет типу 2 Сг34 зни-
жуе р!вень глокемп га зменшуе конпентрашю в кров! холестерину та шсулшу.
Проте у здорових людей вон таку дою не чинить. Водночас у хворих на цукро-
вий д!абет типу 2 всмоктування хрому оз кишок збглыпуеться в 3 рази, ртвень
його в сироватш кров! пщвищуеться, а концентращя в клпинах зменшуеться.
Це евщчить про можливость спадкового порушення при цукровому доабет! типу
2 транспорту Сг34 у клпини, де вон необхщний для забезпечення нормально!
чутливосто до шеулону. Дефщит хрому в Ъю посилюе шсулшорезистенгшсть,
що прискорюе розвиток цукрового доабету типу 2 й атеросклерозу. Враховуючи
те, шо у близько 50 % населения о в першу черту в дней спостерогаеться суб-
нормально споживання тривалентного хрому, можна вважати, що поморю до-
бавки Сг34 з нетоксичними логандамп здатю справляги профшактичну дпо
щодо виникнення цукрового дтабету типу 2, шсулшорезистентного ожиршня й
атеросослерозу.
ОБМ1Н ВАНАДПО ТА ЙОГО РОЛЬ В 0РГАН13М1
Джерела надходження та шляхи обмшу ванадию. Вм!ст ванад!ю в оргашзм! не пере-
вищуе 100 мкг. У кишках усмоктуеться близько 1 % харчового ванадпо, переваж-
но у склад! ашонгв. Транспортуеться цей мгкроелемент у кровг трансферином 1
виводиться в основному гз сечею. Накопичуеться ванадгй у клетках, жировш тка-
ниш, печгнцг, нирках, селезгнщ, яечках. В еритроцитах його бгльше, нгж у сиро-
ватцг кровг. Харчовими джерелами ванадпо е рослинш жири, гречка, петрушка,
овес, рис, морква, капуста, крш, редиска. Добова потреба у ванадй для доросло!
людини становить 12—30 мкг.
Ванадгй дуже поширений у навколишньому середовигш та широко викорис-
товуеться у виготовленш стал! та сплавгв, скла, гуми, фарб, лакгв г фотоматер!ал!в.
Промисловг викиди, вулкани, згоряння нафтопродуктгв г випаровування з повер-
хш океану пщтримують його значку концентрации в повггрг. Побутове емгття е
основним джерелом накопичення ванадпо у грунтових водах.
Значения ванадпо для оргашзму та наслщки його дефщиту г надлишку. За свогми
хгмгчними 1 бюлопчними властивостями ванадш мае багато епшьного гз хромом.
3 одного боку, цг важкг метали е необхщними компонентами жиггедтяльносп
оргашзму, з шшого — у високих концентрацгях вони можуть гндукувати гостру чи
хрошчну штоксикацгю, що ушкоджуе структуру клгтин г спричинюе патолопчш
наслщки. Токсичшсть цих мгкроелементгв значною м!рою визначаеться ступенем
гх окиснення, тобто валентшетю. Найчаст!ше ванадш входить до складу ванадатгв
гз п’ятивалентним або чотиривалентним ванадгем.
При штрацелюлярному вщновлешп У5+ до У4+ утворюються активн! киснев!
радикал и, яю спричинюють запалення г пухлини через ушкодження ДНК, пост-
транскрипцшну модифгкацпо бшюв або лйггдну пероксидацгю. У раз! надлишко-
вого надходження в оргашзм ванадпо виникають гематолопчнг, бюхгмгчнг, не-
фролопчн!, гмунолопчш та репродукщйш порушення, затримуеться розвиток
оргашзму та виникають змши поведгнки у вигляд! машй 1 депресп. Водночас ос-
таншми роками ванадш використовують у лгкуваннг цукрового дгабету, атеро-
склерозу г пухлин.
У 1899 р., ще до вщкритгя шсулгну, було виявлено, що ешь ванадгю полпппуе
перебтг цукрового д!абету в людини. Проте лише останнгми роками вдалося
з’ясувати мехашзм цього феномену. Ванадгй, так само як ! хром, пщвищуе чут-
ливгеть м’яз!в, адипоциттв г, можливо, гепатоцитгв до шсулшу. Молекулярною
основою цього е стимуляцгя шеулгном тирозинкгнази, яка, фосфорилюючи тиро-
зин у бглку, необхщному для транслокагщ в мембрану ГЛЮТ-4, переводить його
в активний стан. Своею чертою ванадгй г хром пригшчують актившсть тирозино-
во! фосфатази, яка вщокремлюе фосфорильну групу вщ тирозину. Це подовжуе
перебування бшка, який транслокуе ГЛЮТ-4 у мембрану, в активному сташ, а
отже, ! активну дгю ГЛЮТ-4. Таким чином, ванадгй 1 хром зменшують шеулшо-
резистентшсть при цукровому дгабет! типу 2, збшыпуючи час 1 силу ди шсулшу.
Пптвищення чутливосп тканин до шсулшу лежить ! в основ! антиатерогенно! дй
ванадгю г хрому. Для профшактики та лткування цукрового дгабету типу 2, ожи-
ршня й атеросклерозу найперспективнгшими е низькг дози ванадпо сульфату в
комбшагщ з антиоксидантами.
Ванадш дае також парадоксальней протипухлинний ефект, який пояснюеться
хндукщею апоптозу пухлинних юптин х / або активащею гешв — супресоргв юп-
тинного подглу. Причиною тако! дп може бути блокада тирозин-фосфатази та / або
активащя тирозин-фосфорилази. Крхм того, ванадш може шдукувати затримку
клхтинного циклу та / або справляти цитотоксичний ефект на пухлинш клггини,
ушкоджуючи ДНК, ядерш транскрипщйш фактори х лшщш мембрани акгивними
кисневими радикалами. Ванадш, можливо, також зменшуе метастатичний потен-
цхал пухлинних клхтин через модуляцпо юптинних адгезивних молекул.
Проте даю про здатшсть ванадпо блокувати №+-, К+-АТФазу на сучасному
еташ розвитку науки не шдтвердилися.
ПАТОЛОПЧНА Ф13ЮЛ0Г1Я ОБМ1НУ МОЛ I БД ЕНУ
Кхлъюсть молхбдену, що мхститься в оргашзм! доросло! людини, за рхзними даними,
коливаеться вщ 20 до 70 мг. Його концентращя найбшьша в печшцх та нирках. У
кровх анхоннх комплекси молхбдену зв’язаш з быками. Вш добре всмоктуеться в тон-
кхй кишщ як з неоргаючних сполук, розчинених у водх, так х з продукгхв харчування.
Багато молхбдену в зелених овочах, зернових, бобових, у нирках х печшщ тварин.
Пригшчують кишкову абсорбцхю молхбдену мщь1 сульфата. Добова потреба доросло!
людини в цьому мхкроелеменп становить 75—250 мкг, а в немовлят — 15—30 мкг.
В оргашзм! людини молхбден входить до активних центрхв трьох важливих
ферменпв: альдегщоксидази, ксантиноксидази х сульфхтоксидази. Першх два фер-
менти е флавопротещами, якх крхм молхбдену мхстять залхзо х схрку. Альдепдокси-
даза окисшое альдегщи до карбонових кислот. Ксантаноксидаза виконуе важливу
роль у кшцевому етат обмшу пурингв, окиснюючи гхпоксантин ! ксантин вщ-
повхдно до ксантину 1 сечово! кислота. Окиснення здшснюеться завдяки тому,
що Мо6+ вщновлюеться в цих реакцхях до Мо5+ х далх до Мо4+. При цьому погли-
наеться кисень х утворюються супероксидний анхон х пероксид водню.
При генетичному дефект! ксантиноксидази виникае ксантинурхя з утворен-
ням ксантинових камешв у сечових шляхах. У репонах з геоххмхчним надлишком
молхбдену збхльшуеться синтез ксантиноксидази, що спричинюе розвиток подаг-
ри, яка харакгеризуеться вщкладанням солей сечово! кислоти в сухожилках х суг-
лобах. Спостерхгаються також затримка росту кхсток у дхтей, функщональш змши
печшки, ппотонхя, анемхя та лейкопенхя. Аерозолх молхбдену можуть спричинити
ураження бронххв х пневмокошоз.
Сульфхтоксидаза перетворюе сульфхт на сульфат переважно в мхтохондрхях
печшки. Генетичний дефект сульфхтоксидази харакгеризуеться аномалхями роз-
витку мозку та розумовою вщсталхстю. Причина цього, можливо, полягае в недо-
статньому синтез! сульфолшццв х быкхв.
ПАТОЛОПЧНА Ф131ОЛОПЯ ОБМ1НУ СЕЛЕНУ
Вмхст I розподхл селену в оргашзм!. Концентрапхя селену в оргашзм! людини значно
вщрхзняеться за географхчними регюнами, що визначаеться його вмхстом у Грунт! та
рослинах. Концентращя селену в плазм! кровх, де вш мхститься у вигляд! як орга-
шчних, так 1 неоргашчних сполук, у рхзних регхонах становить вщ 50 до 820 мкг/л.
У нирках, як! е основним органом виведення селену, його концентрац!я коли-
ваеться вщ 640 до 6170 мкг/кг. В шших органах 1 тканинах селен перебувае у
склад! б!люв, 1 його концентращя стабыъшша. Так, у м’язах вона становить ПО—
430 мкг/кг, у легенях ! серщ — 570—800 мкг/кг, а в печшц! — 370—720 мкг/кг.
Велика юльюсть селену в юптинах !мунно! системи, сперматозоидах 1 мозку, де вш
зберйаеться навггь за умови селенодефщиту. У новонароджених 1 дггей перших
м!сяц!в життя р!вень селену досить низький 1 з в!ком поступово збыыпусться.
Джерела 1 форми надходження селену в оргашзм людини. Хоча селен може пот-
рапляти в оргашзм через шюру 1 леген!, переважна його юльюсть надходить !з
!'жею. Вш може засвоюватись у форм! неоргаючних сполук, таких як натрпо се-
деют №28еО3 ! натрпо селенат №28еО4. Проте оргашчн! форми селену е б!лып
бюпридатними 1 менш токсичними. Тваринн! продукта 1 грудне молоко м!стять
селен переважно у вигляд! селеноцистешу. Велика його юлыасть моститься в пив-
них др!жджах, злакових, хл!б! та бобових, де вш перебувае переважно у вигляд!
селенометююну, який е основною формою надходження селену в оргашзм до-
росло! людини 1 дггей старшого вгку.
Мехашзми всмоктування селену. Розчинш форми селену всмоктуються в тон-
кий кишш на 80—100 %. Селенометюнш 1 селеноцистеш абсорбуються за допомо-
гою натршзалежно! амшокислотно! транспортно! системи. Неоргашчш сполуки
селену в кишках частково вщновлюються !з селенатов до селентв, а останн! до
натрпо селенщу №28е ! абсорбуються вторинно актавним транспортом.
Шляхи бютрансформацп та виведення селену. Осюльки вищ! тварини не здат-
ш синтезувати 1 вщр!зняти один вщ одного селенометюнш 1 метюшн, частина
селенометююну, який усмоктався в кишках, неспецифгчно вбудовуеться в р!зю
быки на м!сця, призначеш для метююну. Така замша метюшну на селенометюнш
сутгево не позначаеться на функцп быюв, але може слугувати засобом депону-
вання селену в оргашзм!.
Бюлопчна функщя селену здшснюеться лише тими селенопротешами, яю
м!стять у свою первиншй структур! селеноцистеш. Ця 21-а ампюкислота мгстить-
ся в активних центрах селеноензим!в. Включения селеноцистешу здшснюеться на
основ! генетично! експресп за допомогою уникально! селеноцистош!л-тРНК, яка
розюзнаеться специф!чними УГА-кодонами в мРНК. Таким чином, селен е еди-
ним м!кроелементом, який перебувае пщ прямим генетичним контролем.
Хоча быыпа частина селенометююну шляхом транссулъфатування перетво-
рюеться на селеноцистеш, вш безпосередньо не включаеться в бщки, а перетво-
рюеться в печшщ на серин 1 водню селеюд Н28е за участю ферменту р-л!ази. Се-
лении своею чертою за участю глутатюново! системи перетворюються на Н28е в
еритроцитах 1 плазм! через селенодиглутатюн г глутатюнселеноперсульфщ. Водню
селеюд е ключовим метаболитом, який або утворюе активну форму селенофосфату
для включения в селенопротеши у вигляд! селеноцистешу, або метилюеться за учас-
тю селенаденозилметюншу до метилселенолу СН38еН, диметилселенщу (СН3)28е 1
триметилселеноюуму (СН3)38е+. Диметилселенщ може виводитася з повггрям, що
видихаеться легенями, а триметилселеноюум — !з сечею. Незначна частина селену
виводиться через кишки, але за нормальних умов гомеостаз цього мжроелемента
регулюеться в основному екскрещею його похщних !з сечею.
Селенопротеши 1 роль селену в норм! та при патолог!!. На сьогодю щентифгко-
вано понад 30 селенопротеппв, як! м!стять у свош первинюй структур! селено-
цистегн. Близько половини з них очищено, а для 9 селенопротешгв секвеновано
га клоновано ДНК, що дало змогу вивчити г схарактеризувати гх функцпо.
Першими було вивчено ферменти родини глутатгонпероксидаз, серед яких
розргзняють гастрогнтестинальну, цитозольну, позаклгтинну та зв’язану з плазма-
тичними мембранами.
Усг селеновмгсш глутатюнпероксидази за участю вщновленого глутатгону ка-
талгзують реакцгю вщновлення г детоксикацп пероксиду водню та оргашчних пе-
роксидгв г гщропероксщцв до нетоксичних сполук:
Н2О2 + 2Г-8Н-+ 2Н2О + Г-8-8-Г;
КД, + 2Г-8Н -> 2КОН + Г-8-8-Г;
КООН + 2Г-8Н -> КОН + Н2О + Г-8-8-Г.
Водночас субстратом каталази г глутатгонпероксидази, що не мгстить у свой
структур! селен, е лише пероксид водню.
Гастрогнтестинальна глутатюнпероксидаза утворюеться виключно в травному
тракт! та функщонуе як бар’ер, що запобггае всмоктуванню гщропероксщцв з гжг.
Причому низьког, але стабильно! концентрацп цього ферменту, яка спосгерггаеть-
ся при дефщит! селену, достатньо для запобггання абсорбцц гщропероксщцв г
ушкодження ними ештелгю.
Цитозольна глутатюнпероксидаза захищае внутргшньоклгтинш бглки печш-
ки, серця, мозку, легень г клгтин кров! вщ ушкодження пероксидом водню, лгпо-
пероксидами г лшогщропероксидами, вщновлюючи гх до нетоксичних сполук.
Позаклгтинна глутатюнпероксидаза е секреторним ферментом, який вщнов-
люе оргашчш пероксиди та гщропероксиди в материнськгй децидуальнгй оболон-
щ, ембрюнальшй ентодерм!, жовтковому мгшку, мюкардг плода, делянках скос-
тенгння, ембрюнальшй жировгй тканин!, епггелп та шкгрг. Дефщит селену в жшок
спричинюе безплщшсть г спонтаннг аборти, що евщчить про захисну роль селе-
нопротегшв в ембрюнальному розвитку. У дгтей г дорослих позаклгтинна глу-
татюнпероксидаза виявляеться у значшй кшькостг в проксимальних ниркових
канальцях г в меншш концентрацп — в мюкард! передсердь, кишках, шкгрг та жи-
ровш тканиш.
Зв’язана з плазматичними мембранами глутатюнпероксидаза захищае гх вщ
пероксидного окиснення мембранних лгпщгв.
Тгоредоксинредуктаза являе собою, як г глутатюнпероксидаза, селеновмгс-
ний антиоксидантний фермент. Вона вщновлюе за допомогою НАДФН тгоредок-
син та гншг сполуки. Видгляють три гзоформи тгоредоксинредукгази: цитозольну,
мгтохондргальну г позаклгтинну, розташовану переважно в яечках. Цей фермент
захищае клггини вщ оксидативних ушкоджень, гнггбуе апоптоз, впливаючи на сиг-
нальну кгназу 1 апоптозу, регулюе клгтинш процеси, захищаючи вщ ушкодження
ргзнг транскрипцгйнг фактори г вщновлюючи нуклеотиди до дезоксинуклеотидгв.
У цглому селенумгсш тгоредоксинредукгази сприяють виживанню клгтин.
Три типи селеновмгсних йодтиронгндейодиназ каталгзують перетворення тет-
райодтирошну (тироксину) в актившший трийодтирошн.
Селенопротегн Р, молекула якого мгстить 10 атомгв селену, переносить селен з
печгнки в гншг органи, у першу черту в мозок 1 яечка. У перюд ваптностг вгн транс-
портуе селен до плода. Приблизно 60 % селену плазми мгститься в цьому бглку.
Вщсутшсть селенопротегну Р у тварин спричинюе накопичення селену в печшцг та
зниження його концентраци в плазмо й шших органах. Кром транспортноТ функцп
вш виконуе ще й антиоксиданту. Вважають, що недостатшсть цього селенопроте!-
ну може бути патогенетичною основою одного оз субтигпв шизофренп.
Селенопротеш XV е необходним для нормального метабол!зму м’язов.
Селенопротеш капсули сперматозоидов з молекулярною масою 17 000 кДа бере
участь у формуванно джгутиков та стабшозуе ох цшсть, що, омоворно, зумовлюе
властивють селену подтримувати високу фертильность (плодшсть) чоловоков.
Кром того, за допомогою неводомих досо селенопротешов чи низькомолеку-
лярних неоргашчних форм селену стимулюеться мембранний транспорт глюкози
завдяки збшыпенню юлькост! ГЛЮТ-1 у плазматичних мембранах адипоцитов о
фобробластов, нав!ть у раз! !х шсулшорезистентносто. Селен також справляе ш-
сулшоподобну дою, пригночуючи лшол!з в адипоцитах о глюконеогенез у печшц! та
стимулюючи глоколоз, пентозофосфатний шунт о синтез жирних кислот. Мехашзм
шсулшоподобного впливу селену не до конця з’ясовано, проте встановлено, що
селен активуе ключов! бшки, як! входять до каскаду шсулшових сигналов.
Селен регулюе активний стан омунно! системи, а також здатен захищати ор-
ганозм вод запальних факторов, зокрема при бронхоалыпй астмо. Вш за фозоологоч-
них ровопв пригшчуе активность транскрипцшного ядерного фактора КР-кВ, який
контролюе експреспо гешв, що кодують синтез запальних цитокшов. Це призво-
дить, наприклад, до того, що селен значно зменшуе синтез печонкою СРП у вщ-
повщь на дою ФНП-а.
Антиканцерогенна доя селену. Завдяки численним епщемоолопчним спостере-
женням о дослщженням на тваринах о культурах пухлинних клггин було встанов-
лено антиканцерогенно ефекти селену. Статистичний аналоз зв’язку мож вмостом
селену в грунто та !ж!, його добовим споживанням о концентрацоею в крово, з од-
ного боку, о частотою вошикнення раку, з шшого, переконливо довов негативну
кореляцою мож ними. Зокрема в Черновецькш областо Б.П. Сучковим ще у 1981 р.
було встановлено, що значно пщвищена захворюваность на злоякосно пухлини
сшьського населения передпрсько! лосостепово! зони областо пов’язана зо зниже-
ним вмостом селену в грунт! мосцевих полов о пасовищ.
Позшше у Великой Британоо та шших крашах було виявлено, що додаткове
споживання сполук селену людьми у виглядо харчових добавок сприяе значному
зниженню загально! захворюваносто на рак. Причому антиканцерогенний ефект се-
лену був набагато потужношим, нож такий у ретинолу, р-каротину або вотамшу Е.
Нещодавно встановлено, що антибластомна доя селену не пов’язана оз син-
тезом селенопротешов та !х антиоксидантною доею. Серед усох х!мочних сполук
селену найбшьшу канцеропревентивну дно мае монометильована його форма —
метилселенол СН38еН, який утворюеться при метилюванно водню селенщу. Щка-
во, що таю рослини, як астрагали, особливо &епи&, о добре водомий нам
часник кром селенометооншу синтезують у 100—1000 разов больше, нож шшо рос-
лини, селенметилселеноцистешу, який за допомогою р-лоазноо реакцо! перетво-
рюеться на метилселенол.
Метилселенол у мокромолярних дозах ондукуе апоптоз трансформованих кло-
тин незалежно вод ушкодження ДНК о без участо бшка р53. Ця доя селену, можли-
во, пов’язана з тим, що його монометильована форма здатна онпбувати протеш-
коназу юптинного циклу сбк-2 о протеонкшазу С за механизмом модифокацо! цистео-
нових залишктв з утворенням похщних селену (селеногрисульфшу або селенш-
сульфщу) або шляхом катал]заш! утворення дисульфиду.
Причини г наслгдки селенодефщиту. Основною причиною дефщиту селену в
людини 1 тварин е недостатке його надходження з тжею в репонах тз низьким
вм1стом секеншв г селенаттв у труни. СЧльськогосподарська дтялыйсть людини
спричинюе поступове зниження концентраци селену як у Грунт) т рослинах, так I
в кров) людини. У дней селенодефщит може бути також спричинений штучним
або парентеральним вигодовуванням або ж виникае внаслщок мальабссрбцп.
Дефщит селену зумовлюе кардюмюпатпо г значно збгтыпуе ризик смерп вщ
шфаркту мюкарда. Ендем1чна форма мюкардюпатп вшома в деяких провшщях Ки-
таю ше з 1935 р. як хвороба Кешана. Вона супроводжуеться фокальними некрозами
мюкарда, аризмоями 1 розвитком серцево! недостатносп. Застосування натрпо се-
ленпу запобйае розвитку цтет хвороби, яка е найтиповтшою для дггей г ваптних. При
хвороб! Кешана також виявляють дегенеративн! змши м’язгв нижнтх ктншвок, що
нагадуе алтментарну м’язеву дистроф!ю (бшом’язов) хворобу) телят, лошат, поросят,
ягнят I молодняка свшських птахтв, яка теж виникае при недостатносп селену. Де-
генерация серцевого та скелетних м’яз!в зумовлена вшьнорадикальним ушкоджен-
ням мембран т бшкгв за умови недостатнього утворення селеновмтсних антиокси-
дантних ферменпв, а також, можливо, пов’язана з нестачею селенопротешу УУ.
Дефщит селену змешпуе кшьюсть г функцюнальну актившсть Т-лтмфоцитгв
та, як згадувалося вище, у 2—3 рази збшьшуе ймовгрнтсть захворювання на рак,
спричинюе внутршшьоутробну загибель плода, а в чолов!к!в спричинюе азоос-
перм!ю або знижуе рухомгсть сперматозоиде унаслщок вщриву хвоста (джгугика)
вщ головки,
Селенодефшит у перинатальний перюд спричинюе виникнення муковтеци-
дозу в д!тей молодшого в!ку. Це захворювання характеризуеться юстозно-фтброз-
ним переродженням вивщних проток пщшлунково! залози.
При нестач! селену спостертгаються також гепатози та некрози печшки, мож-
лив1 емоцшш та психгчн! розлади.
Токсична дйг селену проявляеться в репонах тз високим його вм!стом у Груни
га вод! год!, коли добове споживання цього мгкроелемента перевищуе рекомендо-
вану дозу в понад 5 разгв. При хронтчному отруенн! селеном його концентрацтя в
сеч! може збшьшуватися в 100 раз!в поршняно з такою в людей з оптимальним
споживанням пього мжроелемента т в 400 разгв пор!вняно з вмтстом селену в сеч!
при хвероб! Кешана, що евщчить про значну алапташйну спроможюсть нирок у
пщтриманн! гомеостазу зазначеною мгкроелемента. Для селенового токсикозу
найхарактерншшми симптомами е ураження нггпв ! випадшня волосся. Також
спостерггають порушення формування в дней емал! та исток. Можливт хроючнг
дерматит, артрит, анемгя, дегенерашя печшки, гастроентерит, депресивш стани.
Хоча мехашзми цитотоксично! дц селенпу та шших неоргашчних сполук се-
лену залишаються нез'ясованими, при пускают ь, що вони Груитуються на здат-
ност! цих сполук катанзувати окиснення тюл!в ! одночасно продукувати суперок-
сидний ашон-радикал. Це зумовлюе загибель кл!тин шляхом апоптозу або некро-
зу. Кргм того, Н,8е здатен зв’язуватися з металами з утворенням нерозчинних
комплекса. Ця властивють селену може бути корисною при детоксикацп отруень
важкими металами. Проте гоксичш дози селену здатн! вивьзьнюваги метали з
металопротенпв, що спричинюе порушення .хн!\ функщй.
ПАТОЛОПЧНА Ф13ЮЛ0ПЯ ОБМ1НУ ЙОДУ
Вмгст । розподол йоду в оргашзм!. У тыг доросло! людини за нормальних умов мгс-
титься близько 20—30 мг йоду, з яких 30 % — у щитоподгбшй залоз!. Пор!вняно
з м’язами та деякими шшими органами концентращя цього мгкроелемента пщви-
щена також у печпщг, нирках, слинних, надниркових залозах, ппофгзг, яечниках,
макрофагах г нейтрофыьних лейкоцитах. У кров! здорово! людини мгститься 50—
120 мкг/л йоду, причому 40—80 мкг/л представлено його оргашчними сполуками,
зв’язаними з быками плазми, а 85—90 % йоду плазми кров! входить до складу
тетрайодтирошну (Т4, тироксин) г лише 4—5 % — трийодтиронгну (Т3). Дуже не-
значна частина йоду мгститься в кров! у вигляд! выьних Т4гТ3, концентращя яких
становить 20 нг/л г 6 нг/л вщповщно. Вмгст неоргашчних йодидгв у плазм! кров!
коливаеться в межах 0,8—6 мкг/л.
Обмш йоду в природ! та джерела надходження його в оргашзм. Неоргашчш спо-
луки йоду добре розчиняються у водг, тому з дощами ! таненням сшгу поступово
перемпцуються в моря г океани, де концентрацгя цього мгкроелемента с тановить
50—60 мкг/л. Завдяки випаровуванню з поверхш светового океану йод знову пот-
рапляе на суходш, причому основна його частина припадав на приморськ! репо-
ни. Тому вмгст йоду в питшй водг, рослинних г тваринних продуктах визначаеться
вщдалешстю вщ моря. Найбыьш бщними на йод е прсью мгсцевосп.
Основними джерелами цього мгкроелемента для людей е морська риба, мо-
люски, морськг водоросп та йодована сыь. Немовлята отримують йод гз грудним
або коров’ячим молоком. Вмгст йоду в грудному молощ становить 40—80 мкг/л,
а в коров’ячому, де вш мгститься лише у вигляд! йодидов, — 70—140 мкг/л.
Добова потреба в Йодо залежно вщ вгку та стат!. Довгий час вважали, що добо-
ва потреба дорослих г дгтей вгком понад 11 рокгв у йодг становить 100—150 мкг.
Дням вщ народження до 6 мгс. рекомендували вживати 40 мкг на добу, вщ 6 мгс.
до 1 року — 50 мкг, вщ 1 до 3 рокгв — 70 мкг, вщ 4 до 6 рокгв — 90 мкг. Ваптним
радили вживати 175 мкг йоду на добу, а в пергод лактацп — 200 мкг на добу. Зараз
таю дози вважають мгнгмальними, а оптимально споживання йоду, на думку аме-
риканських фахгвцгв, мае становити 200—600 мкг на добу. Причому найбыьшим
споживання цього мгкроелемента мае бути в пергод статевого дозргвання, пщ час
ваптносп г лактацп.
Усмоктування та виведення йоду. 3 гжею йод потрапляе переважно у виглядг
йодидгв г добре всмоктуеться в кишках у йоннгй формг шляхом полеплено! ди-
фузц з використанням переносникгв та юнних каналгв. Йодованг амшокислоти
абсорбуються за допомогою натргйзалежного вторинно-активного транспорту.
1ншг сполуки йоду перед усмоктуванням вщновлюються до йодидгв.
Виведення йоду з органгзму можливе з молоком, потом г калом, але основна
його юльюсть виводиться гз сечею.
Фгзюлопчна г патолопчна роль йоду головним чином зумовлена його наявшс-
тю в структур! тирео'щних гормонгв, необхщних для нормального росту, розвитку
нервовог та юстково! тканини, синтезу бшюв, енергетичного обмгну та репродук-
тивно! функцп. Кргм того, йод, як г шип галощи, здатен утворювати в мгкро- 1
макрофагах йодиди й оксийодиди, якг мають бактерицидну дгю.
Дефщит йоду, спричинений недостатнгм надходженням його з гжею г водою,
може зумовити порушення гормонально! функцгг щитоподгбно! залози, яка для
ИГ
нормального синтезу х секрецй гормошв мае щодоби захохшювати хз кров! близь-
ко 60 мкг йоду. Ппойодоз спочатку компенсуеться гшертрофхею 1 гшерплазхею
тирео'1'дного ештелхю та загальним збшьшенням маси щигоподхбно! залози, що до
певно! мхри забезпечуе нормальний рхвень синтезу Т41 Т3. Проте подальше змен-
шення надходження йоду спричинюе гшофункцпо щигоподхбно! залози.
Таке захворювання називають ендемхчним зобом. Воно мае масовий характер
в Альпах, на Алта!, в Андах, Пмалаях, на Кавказ!, у Карпатах, Кордильерах х на
Тянь-Шань В Украпп кр!м Карпат ендемгчний зоб спостерггають на Полюс!.
Усього в свхтх, за даними ВООЗ, на це захворювання страждають 200—400 млн
осхб. Хвороба харакгеризуеться хрошчною втомлювашстю, гальмуванням обмш-
них 1, у першу черту, окисних процесхв, зниженням температури т!ла х основного
обмшу. Тривалий дефщит йоду в дитячому вщ! спричинюе розвиток кретшпзму.
При гшотиреозх в дорослих розвиваеться мхкседема (слизовий набряк). У раз!
недостатност! йоду в оргашзм! ваптно! в новонародженого можуть виявлятися
ознаки вроджено! мхкседеми.
Кр!м того, хснують спадков! та набут! дефекти здатносп щитоподгбно! залози
захоплювати 1 концентрувати йод, а також синтезувати або секретувати тирео'щн!
гормони (детально патофхзюлопю синтезу, обмхну х впливу на оргашзм гормонхв
щитоподхбно! залози розглянуто в роздшх XXXI “Патолопчна фхзюлопя ендок-
ринно! системи”).
Надлишок йоду в орган1злй може бути наслщком ятрогенно! або професшно!
штоксикаци йодидами. Проникнення йоду через шкхру ! слизов! оболонки може
спричинити алерпчний дерматит, який отримав назву “йодизм”. Парентеральне
введения препарапв йоду також може порушувати функцпо щитоподгбно! залози.
При гшертиреозах, якх характеризуються значшхм збшьшенням продукцп гор-
монхв щитоподхбно! залози, концентрацхя йоду в кровх може перевищувати
1000 мкг/л. Проте це не е наслщком пщвищеного надходження йоду в оргашзм, а
спричинено надмхрним захопленням його щитоподхбною залозою та збшьшенням
синтезу х секрецй тиреощних гормонхв у кров, де вони зв’язуються з бшками плаз-
ми, що унеможливлюе хх фхльтрацхю в клубочках нефрона та екскрещю хз сечею.
ПАТОЛОПЧНА Ф13ЮЛ0ПЯ ОБМ1НУ ФТОРУ
Розподш фтору в оргашзм!. Майже весь фтор мхститься в твердих тканинах у склад!
гщроксиапатиту, який мае формулу Са10(РО4)6(ОН)2. Хон фтору мае властивхсть
замхщати юн гщроксилу як у молекул! гщроксиапатиту, так х в активних центрах
метало-, а хнколи х неметалоферменпв 1 неферментних бхлкхв. Спхввщношення
м!ж ОН- 1 Р'у твердих тканинах у середньому становить 40 : 1, але може значно
змхнюватися залежно вщ бюдоступностх фтору. У зубах його середня концентра-
ц!я становить 560 мг/кг, а в ктстках — 490 мг/кг. Серед твердих тканин зуба най-
бшьше фтору мхститься в цемент!, менше — у дентинх й емалх. Губчастх кхстки
мхстять фтору бшьше, нхж трубчастх. Причому в новонароджених вмхст фтору в
ктстках мхшмальний т поступово збшьшуеться до старечого в!ку, коли його нако-
пичення припиняеться. Кхсткова тканина певною мхрою регулюе концентраций
фтору в позаклхтиннш рхдинт, швидко зв’язуючи його при надлишку ! вщдаючи в
кров при дефщит!.
У м’яких тканинах кшьюсть фтору дуже мала, але стабшьна. При кальцифг-
кацг! стгнок судин та шших оргашв у них зростае г вмгст цього мгкроелемента.
У кровг людини концентрацгя фтору коливаеться вщ 0,03 до 0,15 мг/л. У плаз-
мг вш мгститься в гоннш г оргашчнш формах. Як г в юстках, у плазм! кровг вмгст
фтору збшьшуеться з вгком.
Шляхи надходження, усмоктування та виведення фтору. 1он фтору всмоктуеть-
ся в шлунку Г тонкш кишцг шляхом просто! дифузп, хоча, можливо, кнують 1
мехашзми полеплено! дифузп. У норм! засвоюеться близько 80 % фтору, що над-
ходить у травний тракт переважно у вигляд! розчинних неоргашчних сполук з
водою та !жею. Немовлята отримують фтор гз материнським молоком. Кр!м того,
цей м!кроелемент може швидко проникати в оргашзм через леген! та шкгру. Го-
меостаз фтору, як 1 всгх шших ангонгв, регулюеться переважно нирками, як! ви-
даляють близько 90 % його надлишку. Решта виводиться гз калом г лише незнач-
на частина — з потом.
Джерела та добова потреба у фтор! залежно вщ вгку. Фтор досить поширений
у природ! та широко використовуеться в дгяльностг людини. Найбглыпе його там,
де були або кнують вулкани чи залягають родовища фосфапв. Значнг юлькосп
фтору потрапляють у землю гз фосфатними добривами, але найнебезпечшшими
для навколишнього середовища е викиди в повпря сполук фтору при виробниц-
тв! алюмгшю. Вщ народження 1 до 0,5 року добова потреба д!тей у фтор! становить
0,1—0,5 мг. Дорослг незалежно вщ стат! повинш споживати цей мгкроелемент у
юлькосп 1,5—4 мг на добу.
Важливо ВЩМ1ТИТИ, що поргвняно з шшими мгкроелементами особливктю
фтору е вщносно мала р!зниця м!ж фгзголопчною 1 токсичною дозами.
Роль фтору в оргашзм! дгтей полягае в тому, що вш сприяе первиннш мше-
ралгзацп юсток г зуб!в, особливо емалг, формуючи центри кристалгзацп гщрокси-
апатиту. Велике значения мае здатшеть цього мгкроелемента запобггати розвитку
карксу зубгв. Мехашзм такого ефекту фтору дос! достатньо не вивчено. Саме по
собг фторування гщроксиапатиту не збшыцуе мщнкть твердих тканин зуба. Про-
те наявшсть фтору, можливо, зумовлюе шактиващю металоферменпв бактерш,
яю спричинюють демшералгзащю емал! з розвитком карксу.
Дефщит фтору, що виникае при зменшенш концентрацп цього мгкроелемен-
та в питнш вод! нижче 0,5 мг/л, проявляеться в дней г дорослих збгльшенням
захворюваносп на каргес. Вроджений ппофтороз, а також нестача фтору в немо-
влят 1 дгтей раннього вгку характеризуються також затримкою росту г зашзненням
прор!зування зубгв. Застосування профглактичного фторування води та фторумк-
них зубних паст у репонах гз низысим вмктом фтору в питнш вод! значно змен-
шуе ураження карксом.
Надлишкове споживання фтору з раннього дитинства, яке спостерггаеться в
раз! гцдвищено! його концентрацп в питнш вод! (понад 1,5 мг/л), зокрема в де-
яких репонах пгвденного сходу Украши, зумовлюе виникнення в дней ендемгч-
ного флюорозу. Це захворювання характеризуеться появою на емал! зубгв плям
молочно-матового або жовтуватого кольору. Зуби стають крихкими г швидко руй-
нуються без запального процесу. При цьому спостеркаються ламюсть юсток,
м’язова слабюсть, зменшення маси тша й анемгя. Вважають, що вег щ прояви е
наслщком гнггбування ферментгв при зв’язуванш фтору з гонами залгза, мгдг, цин-
ку, марганцю та шших металгв або гз залишками деяких амгнокислот в акгивних
центрах цих ензим!в. Фтор може також шактивувати ферменти, яю не м!стять
метали, але активуються ними, що найхарактернппе для магнпо. Зокрема шакти-
вац!я фосфатаз в остеобластах 1 одонтобластах спричинюе порушення структури
та мщносй меток 1 зуб!в на тл! високо! резистентност! до кархесу.
Водночас фтор, активуючи О-б!лки, стимулюе так! ферменти, як адешлат-
циклаза та фосфолшаза С, що через вторинн! месенджери — цАМФ, шозитолтри-
фосфат, дхацилглщерол 1 кальцш — зумовлюе численш порушення юптинних
функцш. Зокрема змшюеться мембранний транспорт юшв, посилюеться скоро-
чення юптин непосмугованих м’яз!в, порушуеться бар’ерна функция ендотелпо.
Кр!м того, фтор здатний, незалежно вщ О-протешового мехашзму, активувати
кальщев! канали в саркоплазматичному ретикулум! та !нпбувати серин- або тре-
аншов! фосфатази.
Для хрошчно! фтористо! штоксикацц характерш таю ознаки: астешчш стани
з нейроциркуляторною дистошею, артер!альна ппертенз!я, атеросклероз судин,
токсичн! гастрит, кол!т, гепатит, нефропатп, неврит, ради кул и, ознаки гшофунк-
цц ендокринних залоз. Бшя шдприемств, що виробляють суперфосфата й алю-
м!шй, найчаепше спостерйають захворювання оргашв дихання; носов! кровотеч!,
ришт ! бронхгг з астматичним компонентом.
Гостре отруення фтором не проявляеться специф!чними ознаками 1 характе-
ризуемся нудотою, блюванням, проносами, шлунковими колпсами. Значне гос-
тре отруення, як правило, спричинюеться натр!ю фторидом, який використову-
ють у побут! для боротьби з гризунами, комахами 1 грибами. При цьому внаслщок
утворення нерозчинного СаР2 можуть виникати гшокальщем!я !з розвитком су-
дом, пригшчення дихально! та серцево! д!яльност!.
Р03Д1Л XXIV
ПАТОЛОПЧНА Ф131ОЛОПЯ ОБМШУ В1ТАМ1Н1В
КЛАСИФ1КАЦ1Я ТА ЗАГАЛЬНА ХАРАКТЕРИСТИКА
Б1ТАМ1Н1В ЛЮДИНИ
За фхзико-млячними властивостями впамши под!ляють на водо- та жиророзчинш
(табл. 17).
Водорозчинш впамши, за винятком вгтамппв В121 В9, не здатн! накопичуватись
у тканинах ! тому, щоб уникнуги гшовпамшозу, !х потр!бно постшно споживати.
Надлишок водорозчинних вгтамппв легко виводиться з оргашзму, а тому !х пере-
дозування неможливе.
Жиророзчинш впамши, навпаки, здатш накопичуватись у тканинах, що до
певно! м1ри запоб!гае !х дефщиту. Надмгрне споживання вгтамппв А, О, Е може
спричинити розвиток гшервпамшозу.
За функщональними властивостями бшышсть водорозчинних вгтамппв (за
винятком впамппв С 1 Р) 1 жиророзчинний впамш К належать до ензимовта-
мййв, осюльки !хш активш форми вщгграють роль коферментов. Впамши А, О2 1
О. за своею функшею нагадують стерощш гормони, а похщш вггамшу Р е потуж-
ними БАР, що дае пщстави зарахувати гх до гормоновйпамййв. Впамши С, Р, Е, А
беруть участь у регуляцн редокс-стану юптин 1 у вщновлешй форм! виконують
роль антиоксидантов, тобто сполук, яю мають лабшьш атоми водню та неспаре-
ний елекгрон. Взаемодпочи з активними кисневими радикалами та лшоперокси-
дами, антиоксиданти нейтралгзують !х. При взаемоди з окисненими формами
шших антиоксидантов вони вщновлюють 1 реактивують !х. Тому, виходячи 1з су-
часних позицш, !х можна зарахувати до втамшв-антиоксиданпйв або редокс-в1-
тамиив.
За лгкувально-профшактичною д!ею впамши можна класифгкувати як анти-
анелпчш (В6, В9, В12, С), антигемораг1чн1 (С, Р, К), антиг1поксичн1 (В6 С, Н, Р, Р),
антисклеротичш (В3, В6, Р, Р), антитоксичн! (В3, В5, В6, В12, С), анти1нфекцшн1
(С, В, Р, А, Р), антидерматитш (А, В2, В3, В5, В6, Н), антиневритичних (В,, В12),
антирахйпичш (О2, В3), антигемералотчний / антиксерофталъм1чний (А).
Бшышсть впамгшв представлена не одшею, а юлькома сполуками — впаме-
рами, що мають под!бш бюлопчш властивосто Наприклад, термш “шридоксин”
(впамш В6) об’еднуе три близью за д!ею та взаемно перетворюваш в оргашзм!
людини сполуки: шридоксол, шридоксаль 1 тридоксамш. У табл. 17 впамери
видшено курсивом.
Таблица 17. Класиф|кац>я В1таги1жв та Тхн! властивост!
Буквене позна- чення Х!м1чна назва В1ТЭМ1Н1В таТхв1тамер1в Активн! форми I ?;д'" = - с-.. — ) Ф|зюлопчн1 властивост! Г ..А* Ж >
Водорозчиннт в 1 т а м1н н
в. Ттамш Ттамтндифосфаг Спамшшрофоефат, кокарбоксилаза) Як кофермент забезпечуе Ключовт реакцп аероб- ного обману вуглеводтв, втдпрас важливу роль у метаболтзм! нервово! системи
Вг Рибофл автн Ф. тавтнадентнди- нуклеотид. флавтн- мононуклеогид Як кофермента анаеробних т аеробних деггдроге- наз та оксидаз берут ь участь в окисненнт вугле- водтв амшокислот, жиртв т синтез) сгеротдтв
в. Пантотенова кислота Коензим А(КоА- 8Н), дефосфо-КоА, 4-фосфопантететн Забезпечують метаболтзм вутлеводв, амшокислот 1 жиртв, синтез жирних кислот т сгеротдтв; беруть участь у процесах детоксикацп тощо
В5(РР) Нтацин, нгко- тинова кис- лота, Н1КО- тинамгд Н1котинам1даденш- динуклеотид, шко- тинамхдаденшди- нуклеотидфосфат НАД г коферментом анаеробних дегюрогеназ, якт беруть участь в окисненнт глюкози, жирних кис- лот, глтцерину, амшокислот. переносить протони га електрони в дихальному ланцюгу мтгохондртй, бере участь у синтез! та репарацн ДНК. НАДФН — донор водню в реакщях синтезу жирних кислот, холестерину, стеротдних гормонтв та при втднов- ленн) тлутатюну, а також е компонентом моноок- сигеназних реакцш мткросомального окиснення ксенобтогиктв та ендогенних токсиюв
вь ПфИДОКСИН. тридоксол, п(ридоксаль, тридоксамш Птридоксаль- фосфат, шридоксамтн- фосфат Як кофермента беруть учаегь у трансамшуванш, декарбо ксилюваню, тзомеризацй амшокислот, бюсинтез! гему, гормонтв щитоподтбнот залози, пета мшу, серотонтну, ГАМК та угвореннт нжотин- амтду з триптофану
В,(Вс) Фолат (фолт- сва кислот а, фолацин, пте- ротлтутамт- нова кислота) Тетрапдрофолтева кислота (ТГФК) Як кофермент ТГФК забезпечуе мтжмолекулярне перенесения одновуглецевих фрагменттв, що ви- користовугться в реакцтях обмшу амшокислот, бюсинтезт фтзтолопчно актавних сполук, нуклео- тидтв 1 нуклетнових кислот
В12 Кобаламтн Метилкоба та мш, дезоксиаденозил- кобаламтн Метвлкобаламтн € коферментом гомоцистелн- метилтрансферази, яка каталтзуг перенесения метильнот трупи з №-метил-ТГФК на гомоцистеш з утворенням меттонтну 1 ТГФК, Це одночасно зменшуе сироватковий ртвень гомощтстетну, який токсично впливае на серце. утьорюе меттонш — донор метильних грун у метилюванн! нуклеотидтв нуклетнових кислот т поповнюе пул ТГ ФК, яка далт у форм! №-, Н’0-метентл-ТГФК т 5Г11-формтл- ТГФК стае донором вщпов1дних одновутлецевих радикашв при синтез! пуринових нуклеотидтв, а у формт №-, тЖ-метилен-1 ГФК бере участь у пере- гворенн! дезоксиурт’дин 5 -монофосфату в дезок- ситимт дин-5 -монофосфат, необхтдшгй для синтезу ДНК. Дезоксиаденозилкобаламш е коферментом метизмат-оны-КоА-мутази, що каталтзуе перетво- рення метилмалонш-КоА на сукцинтп-КоА
Продовження табл. 17
Буквене позна- чення Х1м1чна назва В1там<н!в та IX вггамер!в ' Активн) форми ф|зюлопчн1 властивосп
с Аскорбшова кислота Аскорбшова 1 де- гщроаскорбшова кислоти Бере участь у реакции пдроксилювання при бю- синтезг колагену, серотоншу, катехоламгнгв, сте- рогдав. У дихальному ланцюгу мгтохондргй е доно- ром електрошв для цитохрому С. Бере участь в антиоксидантному захистг як донор протонгв 1 електрошв для нейтралгзаци акгивних кисневих радикалов у воднгй фазг. Регенеруе глутатюн г токо- ферол, яю своею чертою беруть участь у вщнов- ленш дегщроаскорбшово!' кислоти. Сприяе вщнов- ленню Ре3+ до Ре2+, що необхццю для всмоктування в кишках або для використання юптинами залгза
н Б1ОТИН Карбоксибютин Як кофермент ацетил-КоА-карбоксилази бере участь у бюсинтез! жирних кислот. У склад! про- тон1л-КоА-карбоксилази сприяе окисненню жир- них кислот 13 нефгзголопчною непарною кглькгс- тю атомгв вуглецю. Як кофермент труваткарбо- ксилази сприяе перетворенню шрувату на оксалоацетат, необхгдний як для функцюнування циклу Кребса, так г для глюконеогенезу
р Рутин, бюфла- воногди та полгфеноли БюфлавоноИди та пол1феноли: квер- цетин, рутин, гес- перидини, катехг- ни, лирецетин, ге- нистрин, бетагн, г!дрокситирозол, ферулова г голова кислоти тощо Антиоксиданты, дгя яких пояснюеться здатшстю перехоплювати акгивш кисневг радикали, зв’язувати метали з перемшною валентнгстю, вщ- новлювати сульфпдрильш сполуки, вгтамгн С г глутатюн. Вщома здатшеть рутину змщнювати судинну стшку та зменшувати гт проникшсть (тер- мш “вггамш Р” походить вщ слова регтеаЬИИу — проникшсть) пояснюеться реакгивацгею аскорбг- новог кислоти та зменшенням активностг ггалу- ротдази. Седативна, знеболювальна г ппотензивна дгя бюфлавоноУдгв зумовлена гх здатшстю зв’язуватися з бензодгазепшовими ре- цепторами мозку. Бюфлавонощи, зокрема кверце- тин, блокують лтоксигеназу, що запобггае про- дукт! прозапальних г проппемгчних лейкотр!ешв
Жиророзчинн! виакпни
А, а2 Ретинол Дег1дрорети- нол Ретинол, ретиналь, ретиноева кислота, дегщроретинол Беруть участь у забезпечешп шчного зору, росту г диференщаци, мають антиоксидантну, антиканце- рогенну та гмуностимулюючу дно
О, Ч Ергокалъци- ферол Холекальци- ферол Кальцифедюл 25(ОН)-О3, кальцитрюл 1, 25(ОН)2-О3, секальциферол 24, 25(ОН),-О3 1ндукують бюсинтез остеобластами мшорних кальцшзв’язуючих бшюв юсткового матриксу (ос- теокальцину, остеонекгину тощо); пщвищують усмоктування в кишках г реабсорбцию в нирках кальщю г фосфапв; стимулюють резорбццо хря- ща й остеоща, не спричинюючи резорбщю юсток; стимулюють остеогенез г посилюють мшералгза- цгю юсток; пригнгчують продукцпо паратгормону
Закгнчення табл. 17
Букве** позна- чення Юм!чнаназва в|там!н1В таТх вггамер!в • Активн» форми Ф131ОЛОПЧН1 ВЛаСТИВОС!!
Е А-, р- у-, 8- токофероли А- р-. у-, 8- токотрзеноли А-, р-, у-, 8-токофе- роли А-, р-? у-, 8-токо- Тр1€НОЛИ Антиоксиданти з антирадикальними та мембра- ностабъизуючимя властивостями
ч. В Незалпнт омега-6 полшенаси- чен1 жирш кислоти (а>-6 ПНЖК): лша- лева, арах1до- нова Незамшт ш-3 ПНЖК. а-лЫоленова, ейкозапен- таенова, до- козагексаено- ва ЕйкозаноЁди — похёднё арах : до но- во: кислота: (ПГ) 2-1 серп’, тромбоксани 2-1 сери, лейкотрЁсни 4-1 серЁТ ПГ 3-1 серп, тромбоксани З-Т серЁ1, лейкотр1ени 5-1 серп Похзднё ш-6 ПНЖК: Ш Е,а, тромбоксан Аг лей- котрЁен В4 е бронхо-1 вазоконстрикторами та ли- нять переважчо прозапальну дёю, стимулюючи хемотаксис нейтрофиЁв 1 фагоцитоз, адгезЁю й агрегащю тромбоципв, посилюючи больовий ефект гЁстамЁну Ё кёншёв, разом з 111Е2 збЁльшу- ють проникшсть стёнки судин. Вазодилататорна Ё антиагрегантна дёя 1111, та 111Е вёдносно незнания. Похё.дн! ш-З ПНЖК: тромбоксан А3 Ё лейкотрЁен В5 не виявляють виражеш прозапальш властивостЁ, а ПГ13 е потужним антиагрегантом та вазодилата- тором. Загалом похёднё ю-3 ПНЖК мають гЁпокоа- гуляцЁйиу, пполЁшдемЁчну, антиЁшемЁчну та Ёму- номодулюючу ДЁЮ
К К, К. НафтохЁнони Фиохтони Менахшони (6. 7, 9) МенахЁнон-4 Як кофермент у-глутаматкарбоксилази бере участь в утворенш Са2+-зв’язуючоТ у-карбоксиглу- тамЁновоТ кислоти, необхЁдноЁ' для функцЁопуваи- ня таких кальцЁйзв’язуючих оёлкёв, як фактори коагуляпй' кровЁ II, VII, IX, X, антитромботичпЁ б1лки С Ё 8, остеокальцин та ш.
ВЁтамЁни не постачають енерпю дгя оргашзму ё не мають великого пластич-
ного значения. Обов'язковими ознаками випамййв е б1окатамтична аборегулятор-
на роль / нездатнгсть органззму забезпечити себе цими сполукими за рахунок бюсин-
тезу. Речовини, яю не мають усЁх основних ознак вЁтамЁшв, але яким притаманш
тё чи ёншё гх власгивостЁ, називають вп амшоподЁбними речовинами Зокрема орото-
ва ё лЁпоева кислоти, а гакож карштин синтезуються в органЁзмЁ у досгатнЁй кёль-
костё. Тнозит Ё холён входять до складу фосфолЁпЁдЁв Ё виконують в органЁзмЁ плас-
тичну функшю Пангамова кислота не е незамЁнною ё не виконуе роль кофермен-
ту. ПараамЁнобензойна кислота е вггамЁном тёльки для мЁкроорганЁзмЁв. 3 ёншого
боку, аскорбЁнова кислота вважаеться вЁтамЁном тёльки для людини, морсько’1
свинки, деяких приматЁв 1 кажашв, а в бЁлыпостт тварин вона синтезуеться в до-
статшй кёлькостё КобаламЁн ё рибсфлавЁн не е вЁтамЁнами у жуйних тварин, а
людина повинна Ёх споживати, осюльки в неЁ вЁтамгни В12Ё В2 не всмоктуються Ёз
товстот кишки, де мЁкрофлорою синтезуеться велика юльюсть цих вЁтамЁнЁв.
Деяю вЁтамЁни можуть синтезуватися в органЁзмЁ зё своёх попередниюв — про-
вЁтамЁнЁв. ПровЁтамЁни А — жовн шгменти рослин (а-, р- та у-каротини) при пл-
розды ххгу. патолопчна фтзюлопя обмшу вггамппв
ролУзУ за учасгю ферменту р-каротинази стшки гонко) кишки га печшки узворю-
ють цв1 молекули вггамшу Аг ПровУтамшом холекальциферолу (вУтамш Ц) е 7-
депдрохолестерин, який перетворюеться в ыкУрУ на вУтамш пУд дУсю ультрафУоле-
тового випромшювання.
ДЖЕРЕЛА НАДХОДЖЕННЯ В1ТАМ1Н1В
ТА IX МЕТАБ0Л13М В 0РГАН13М1
ОсновнУ харчов) джерела постачання вУтамУнУв в оргашзм людини наведено в габл 18.
Продукти розташоваш в порядку зменшення концентрацп в них вУтамУнУв. АналУз
даних табл. 18 доводить, шо б Ул вплоть вггамшУв мУстигься в продуктах як рослин-
ного, так У аварийного походження. Винятком е лише вУтамш В|2, який мктиться
виключно у продуктах аварийного походження, та вУтамш Р, джерелом якого е
лише продукти рослинного походження. Таким чином, цУлком зрозумшо, шо
нормальне забезпечення оргашзму вУгамУнами можливе лише за умови ртзно-
манггного та збалансованого харчування.
СлУд пам’ятати, що вУтамши В3, В,, С, К руйнуються пУд час термУчноУ оброб-
ки. вУтамУни В9 У С значно втрачаються при консервуваннт, вУтамУни В2, В6, Вд, В12,
С, А, Е, Р, К швидко руйнуються пУд дтею ультрафюлетового випромшювання.
вУтамУни Вр В2, С 1 К не стшкт до лугУв.
Таблица 18. Основн! харчов) джерела надходження в!там1жв
Буквене позна- чення Х!м!чна назва В1ТЭМ1Н1В та Тх вггамер1в — - . .......—. Продукти, ЩО МкСТЯТЬ В1ТЭМ1Н у найбй1ьш1й кшькосп
В, ТУамш Житнш хл1б, соя, дрЬкджц квасоля, горох, вгвсяна та гречана крупи, печшка, нирки
в2 РибофлавУн Печзнка. нирки, дрУжджУ, яйця, сир, м’ясо
В. Пантотенова кислота ПечУнка, дрУжджУ, нирки, яйця, сир, соя. квасоля, горох, гпечка, овес
В,(РР) Шанин Печшка, дрУжджУ, м’ясо, нирки. соя, квасоля. горох, рисова крупа, хлтб, картопля
В„ ГПридоксин Квасоля, соя, печшка, нирки, дрЬкджц банани, м’ясо, риба, яйця, кар- топля, СВ1Ж1 ОВОЧ1
В9(Ве) Фолат (фолзе- ва кислота) ДрУжджУ. печшка. нирки. серпе, сир. зелен! листянУ овочц зелений го- рошек, баклажани, фрукти
В12 КобаламУн Печшка, нирки, серпе, м’ясо, сир, молоко
с Аскорбшова кислота Шипшина. волоський горУх, перець солодкий червоний, смородина чорна, цитрусов), капуста бУлокачапна, яблука, печшка, томати, зеле- ний горошок, картопля. нирки
н БУотие Нирки, печшка, серце, соя, жовток курячого яйця, риба, соя, дрУжджУ, шоколад, арахУс, вУвсява крупа, значно менше — у молоцУ, хлУбУ га овочах
Зактчення табл. 18
БуквеНе позна- чення Х1мнна назва в!там1Н1в таТхвпамер1в ... Продукти, що мютять впамТн у найб1льш1й юлькост!
р Рутин, бюфла- еоноТди та гюйфеноли Черноплодна горобииа, смородина черна, шипшина, щавель, часник, цитрусовт, вишня, брусниця, ат рус, яблука, чайне листа, шоколад, кава какао, винограда соки та вина, карюпля
А Провт- тамтн А Ретинол р-Каротин Жир глибоководних морсыжх риб, печшка, вершкове масло, сметала, сир, яйця Морква, гомати, облшиха, горобина, шиишина, перець солодкий чер- воний, гарбуз, печшка
Ц'Т ц Ергокальцифе- рол Холеканъцифе- рол Дртждж!, гриби, рослинш олп Жир 1 печшка глибоководних морських риб, вершкове масло, жовток яець
Е у А-, Р- у-, д- токофероли та токотрге- ноли Соева, кукурудзяна га соняшникова олп, соя, квасоля, злаков!, вершко- ве масло, м’ясо, морська риба. овоч!
Р о-бПНЖК: лиюлева, арахгдонова а>-3 ПНЖК: а-дшоленова, ейкозапента- енова, докоза- гексаенова Рослинш олп Глибоководна морська риба та и свтжий жир, лляна ол!я
К, Ц Ф1ЛОХ1НОНИ Менахшони Шпинат, капуста, зелен) томати, салат, I оробина, кропива Печшка, м’ясо, яйця, продукти життгдтяльностт мткрооргантзмтв
Бшышсть в1там1Н1В усмоктуеться в проксимальных вцтдтлах тонко! кишки, а
вттамш В12 — у клубовш кинтцт. Усмоктування встх водорозчинних вттамппв вщбу-
ваеться за допомогою спещальних бшктв-переносниюв. Абсорбщя вггамшу В]2 т
фолтевот кислоти е Ма+-незалежним пронесом, проте вст шип водорозчиннт втта-
мши всмоктуються за допомогою переносниктв, якт е ^+-котранспортерами.
Жиророзчинн! вттамши всмоктуються разом тз жирами у склад! мщел. Абсорбитя
жиророзчинних вттамппв обмежуеться нестачею панкрсатичних ензимтв т жовч-
них кислот. Пронеси, що вшбуваються пщ час усмоктування жиртв. недостатньо
сформован! в новонароджених, тому в немовлят абсорбуеться лише 10—15 % жи-
ртв т, вщповтдно, жиророзчинних вттамтшв вщ устх спожитих.
Мткрофлора кишок людини в значнтй юлькостт синтезуе фолацин, вттамши
К, Вг В„ В3, В6, В]2, РР, проте всмоктування цих вттамштв дуже обмежене або
зовстм немождивс- переносники водорозчинних вттамштв розташован! в тонктй
китшп, а бтльша частина мткрофлори, що синтезуе вттамтни, — у товстш. Усмок-
тування жиророз чинного вттамшу К з товстот кишки також не може бути ефек-
тивним, осктльки воно залежить втд абсорбцп жиртв. якт майже повшстю всмок-
туються в тонктй кишш. Нездатнтсть органтзму засвотти вттамши, як! виробляють-
ся кшпковою мткрофлорою, пояснюе той факт, шо концентрацтя цих вттамштв у
калт бувае бтльшою. юж у тжт.
Шсля всмоктування водорозчиню впамши транспортуються кров'ю у выьно-
му стан! або в комплекс! з быками (впамши Н. В,2, С) у клпини печ!нки та шших
органгв 1 тканин. Серед ус!х водорозчинних впамшгв лише В , 1 значно менше
фолква. кислота здатш депонуватися в печшц! та нирках, де вони перетворюють-
ся на сво! коферментн! форми, як! поступово !з током кров! розносяться по вс!х
тканинах оргашзму. Виводятъся водорозчинн! впамши або !хю метабол !ти пере-
важно 1з сечею.
Жиророзчинш впамши в ентероцитах включаються у склад х!лом!крон!в 1 в
такому вигляд] потрапляють у кров. Дал! вони зв'язуютъся з быками 1 потрапля-
ють у печшку. Провпамши А — каротини — в ентероцитах кишок перетворюють
ся на ретинол 1 теж включаються у склад х!лом!крон!в. Впамш А депонусться в
печшц! у вигляд! жирнокислотних еф!р!в, а поим розноситься до оргашв 1 тканин
у поеднанн! з ретинолзв’язуючим быком. Впамш К депонуеться в печ!нш, се-
лезшц! та серц!, звщки транспортуеться разом з альбумшами. Впамши Е ! Р у
печшщ включаються у склад ЛПНЩ чи ЛПДНЩ 1 транспортуються до орган!в 1
тканин, де включаються в склад фосфол! пццв юптинних мембран, у яких вони
депонуються перед виконанням сво!х фхзюлопчних функцш.
Найскладшшим е метабол!зм впамшу I). За умови нормально! шеоляцц
близько 80 % впамшу Е\ утворюеться в макрофагах 1 адипонитах дерми 13 7-де-
гщрохолестерину пщ впливом ультрафюлетового випромшювання. Зв!дси ендо-
генний впамш разом 1з харчовими впамшами П31 О2 транспортуеться в печшку
в поеднанн! з П-зв’язуючим а1 -глобулшом, де гщроксилюються за 25-м атомом
вуглецю з утворенням кальпифедюлу 25(ОН)-П3, який дал! з О-зв’язуючим бы-
ком транспортуеться в нирки, у яких утворюеться найактившший кальцитрюл
1,25(ОН),-О3. Таке ниркове гщроксилювання стимулюеться гтокатьц!ем!ею че-
рез паратгормон або безпосередньо гшофосфатемюю. Проте за нормально! кон-
центрат! в кров! Са2+ 1 фосфату нирки утворюють матоактивний секальциферол
24Г25(ОН)2-П3. Кальцитрюл може утворюватися також у плацент!. Основне депо
впам!ну розташоване в адипоцитах.
Впамши Е) 1 А шактивуються в печшш, а продукти шактивацп виводяться тз
жовчю у вигляд! г.тюкурошд!в. Продукти метаболизму вгтамппв К т Е виводяться з
оргашзму тз сечею.
ОСНОВН1 ПРИЧИНИ ВИНИКНЕННЯ В1ТАМ1ННО1 НЕДОСТАТНОСТ1
Оргашзм людини не здатен запасати впам!ни на тривалии час. Тому вони мають
надходити регулярно, у повному набор! та в юлькост!, що выповыае ф!зюлопч-
ним потребам. Потреба у впамшах може змшюватися пщ впливом р!зних фак-
гор!в Значний вплив на потребу у впамшах чинять в!к ! стать людини. характер
та !нтенсившсть пращ, особливосп д!еги. Необхщшсть у впамшах значно зб!ль-
шуеться в жшок пщ час вагпност! та в перюд лактацп, у дпей — у перюд штен-
сивного росту. Потреба у впамшах пщвишуеться при первохолодженнг та перегр!-
ванш оргашзму, интенсивному фгзичному навактаженш та нервово-психтчному
перенапруженн!, гшоксп й штоксикацп, ггпотиреоз! та недостатност! к!рково! ре-
човини надниркових залоз. У похитому 1 старечому вщ! пывищена потреба у в!-
тамшах зумовлена попршенням усмоктування й утишзапп впамш!в, а в новона-
роджених ё в ранньому дитячому виц — недорозвиненням механгзмгв травления ё
всмоктування. частями диспепсиями 1 дисбактертозом.
Недостатке надходження в оргашзм вётамёнёв з ёжею спричинюе патолопчна
стани, якё мають назву игтов1там1нози”х “авйпамтози ”. Значний дефицит впамгну
в органёзмё з вираженою клёшчною картиною його недостатносп отримав назву
"авётамшоз” (рахгг, цинга, пелагра, берё-берё, пернпцозна анемёя. гемералопёя тощо).
Стан помтрного дефщиту впамгну коли спостерёгаються лише деякг, незначно
виражеш специфгчш симптоми юпнёчноё каргини авётамшозу, а в основному пе-
реважають загальш неснецифгчнг прояви недостатносп вётамёнм (затримка росту 1
розвитку-. швидка фёзична ё розумова втомлюванёсть, порушення концентрат! ува-
ги, драттвливтсть, зниження пам'яп та здатносп до навчання, головокружения та
головний бьть, апатёя, втрата апетиту тошо). називають пповёгамёнозом.
Зменшення надходження вётамёну в оргашзм мае назву екзогенного гтовтамшо-
зу. Кр)м того, розргзняють ендогенш гтовтамшози, основними причинами яких е:
• порушення всмоктування вётамёнёв у тонкш кишцё при захворюваннях трав-
ного тракту, гелатобгпарног системи ё пёдшлунковоё залози;
• порушення всмоктування й утилёзацц вётамшш пёд впливом хёмёчних речо-
вин, у тому числе фармаколопчних препараттв. Наприклад, неомёцин порушуе
всмоктування вётамёну А, авщин — глёкопротеён сирого яечного быка — мёцно
зв’язуе бютин ё перешкоджае його абсорбцц, ацетилсалщилоьа кислота пригшчуе
утилёзащю фолату, нёкотин ё закис азоту ёнактавують вётамён В12;
• нездатшеть бёохёмёчних систем органгзму включати вётамёни в обмённё про-
неси, зокрема внаслщок наявноси ёх структурних конкурентёв — антивташшв
Часто це похёдш вётамёнгв, але такт, що не здатн 1 виконувати ёх функцп у фермен-
тативнихреакцёях. Наприклад, антимёкобакгерёальне препарата — пдразиди ёзонё-
котиновоё кислоти — включаються замтсть вётамшу ₽Р у структуру НАД ё 11АДФ.
при цьому кофермента втрачають здатнёсть брата участь у метаболёзмё тих клётин,
у якё проникають цё препарата, зокрема в мёкобактергю туберкульозу; транквёлё-
затори трё оксазинового ряду приппчують утилёзащю рибофлавёну, порутпуючи
синтез його коферментноё форми; протираковё препарата (амтноптерин 1 ме-
тотрексат) витёсняють фолёеву кислоту з фолатзалежних ферментёв, блокуючи
синтез нуклегнових кислот ё подёл клётин; сульфанёламщи включаються замёсть
параамшобензойноё кислоти в структуру фолгевоё кислота, що робить ёё фгзёоло-
пчно неактивною; антикоагулянта (кумарини) е антивётамшами нафтохёнонёв
• порушення транспорту вётамёнёв кров’ю при зменшеннё або вёдсутносп син-
тезу бёлкгв-транспортерш печёнкою, а також унаслёдок ёх втрати при нефротично-
му синдромё,
• порушення депонування вётамгнёв при патолопё печптки ё нирок;
• конкурентна уталёзацёя вётамгнёв кишковими паразитами ё патогенною
мёкрофлорою;
• пёдвишена потреба у вётамгнах (ваптшеть, лактацтя, фёзичне ё нервово-
психёчне перевантаження, несприятливё клёматичнё га виробнич] умови тощо);
• вродженё порушення обмёну вётамёнёв (порушення синтезу або структури
бёлкёв-переносникёв, бёлкёв-транспортергв, ферментёв, необхёдних для щворення
коферментних форм вётамёнёв, або апоферментёв. посилення катаболёзму впа-
мёнёв або порушення ёх реабсорбцн в нирках тощо);
• прмгшчення сапрофётноё кишковоё мшрофлори, шо продукуе деякг вгтамёни.
Класичн! авхтамшози розвиваються луже рщко. в основному в умовах трива-
лого голодування, коли вхгамшна недостатшсть супроводжус алхментарну дисгро-
фйо. 1ншими причинами авггамхххозхв можуть бути масивне потрапляння в ор-
гашзм антивггамхшв, а також деяю спадков! ферментопатп та тяжкх захворювання
травно! системи, що супроводжуютъся синдромом малъабсорбцп. Поширешшими
е пловггамшози, особливо серед ваптних 1 дггей. Дуже часто спостерггаеться не-
достатшсть одразу кхлькох вхтамхнхв. Так! сгани мають назву “потгтовипалпнози”.
В умовах сучасно! Украши полшповхтамшози залишаються поширеним яви-
щем серед дорослих ! дней. Це зумовлено низкою обставин. По-перше, у рашош
мшького населения збхлыххилася частка консервованих 1 рафшованих продукта
харчування та белого хлхба, що мають зменшену вхтамхнну хцншсть. По друге,
значна кшькхсть вхтампйв втрачасться при термхчнш обробщ Ью, а також унаслх-
док ди кисню та свила при тривалому зберхганш. По-трете. рхзке зниження енер-
говитрат сучасною людиною призвело до значного зменшення загального об’ему
!ж!, шо споживаеться. Це своею чертою знизило введения в оргашзм вхгамхшв !
мхкроелемента. що особливо посилюеться в раз! дотримання популярних у наш
час вегетархансько! дхети, “лхкувального голодування” та шших екзотичних ! мод-
них дхет, що характеризуються незбалансовашстю та неповноцшшстю. I нарешп,
найважлившгою причиною вхтамхнного дефщиту для значно! частини населения
нашо! кра'хни залишаеться низький р!вень життя, який унеможливлюе придбання
необхщно! кшькосп свхжххх овочхв, фрукта, м’яса та риби.
Походження ! розвиток вггамшнох недостатност! в дней мае деяк! особли-
вост!. У ново народ жен их х дхтей раннього в!ку вггамшну недостатшсть споете рхга-
ють часттше. Вона може бути наслщком недостатнього надходження впамшхв до
плода в перюд внутршшьоутробного розвитку та недостатнього вмхсту деяких вх-
тамипв у молохи матер! в раз! Г! пера тонального харчування. Недостатня забезпе-
чешсть вхтамшами ваптних х жшок, якх годуютъ груддю, завдае шкоди здоров’ю
матер! та дитини, е одшею з причин недоношування, вроджених вад, порушення
фхзичного х розумового розвитку дней. В’тамшна недостатшсть новонароджених
х дхтей раннього вхку посилюеться в раз! застосування при штучному вигодовуван-
ш неадапгованих для дитячого харчування сумшхсй хз коров’ячого молока, нера-
цюнального харчувашхя дней раннього вхку. спадкових ! набузих хвороб, при
яких порушуються надходження в оргашзм дитини в!там1Н1в. !х депонування та
метаболхзм. Частими причинами вхтамшно! недостатносп в дхтей бувають дисбак-
терхоз х пщвищехха потреба у вхтамшах при гшоксп новонароджених та шфекцхях,
дхатезах, алерпчних захворюваннях ! залхзодефхххитних станах дхтехх раннього вхку.
Особливо велика схильнхсгь до розвитку пповхтамшозхв у першх мхсяц! життя в
недоношених дхтей унаслщок меншого депонування в органхзмх вггамшхв А, Ц, Е,
В9. В.,, з одного боку, х бхлыпох погреби у вхтамшах — з шшого. що визначаеться
вищою захворювашстю та штенсивним лхкуванням.
Недостатшсть антикевритного в1там!ну В
Гшовхтамшоз памшу (вхтамш В]) прояв.хяеться головним чином ураженням перифе-
рично! та центрально! нервово! системи. Розвиваеться полшеврит хз порушенням пе-
риферийно! чутливосп, що супроводжуеться вщчуттям повзання комах х виникнен-
ням невралгп. Характерними е втрата пам'ятх вхдносно нещодавних подш, головний
бшь, драпвливкль. зниження апетиту, нудота. закрепи. Кр1м того, розвиваеться слаб-
юсть мюкарда. яка проявляеться тахгкардгею навпь при незнаниях навантаженнях.
Патогенна дтя нестачг впамшу В, пояснюеться передуетм порушенням окис-
ного декарбоксилювання шрувату 1 а-кетоглутарату з частковим блокуванням об-
ману вуглеводш, що супроводжуеться енергодефшитом г наксличенням тнрови-
ноградноТ та а-кетоглутаровоё кислот гз розвитком метабол гчного ацидозу. Пору-
шення метаболтзму вуглеводш активуе катабо.тгзм быкгв т жиргв, то викликае
зменшення маси тша г м’я зову атрофйо, а в дней — затримку росту. Особлива
чутлиБтсть нервово! тканини до нестачт ттам1ну пояснюеться тим, шо основним
енергетичним джерелом для нервових клттин е глюкоза. Харчування переважно
гжею, збагаченою вуглеводами, 1 вживания на цьому тл! морепродуктов, якт мтс-
тять фермент памшазу, що руйнуе ттамтн, посилюе недостатшсть впамшу В,.
Авггамшоз В, описано як хворобу бер1-бер1, що в переклад) з шдшсько') мови
означае “вовця”. Це пов язано зт змшою ходи хворих. 1нша назва — кайданова
хвороба — виникла тому, що на нет часто хворый в’язнт в швденних крашах, яких
годували майже виключно очищеним рисом, шо не мгстить ттамш. Видыяють дво
форми хвороби: суху (нервово-парайтичну) 1з розвитком парал1Ч1в г атрофию
м’язтв та набрякову (серцеву) залежно вщ переважання ураження нервово! або
серцево-судинно! системи.
До вроджених порушень обману па мшу належать:
• синдром Вершке — Корсакова, щдгрунтям якого е зменшення спорщненосп
ферменту транскетолази до ттамшптрофосфату. При цьому синдром! вщбуваються
крововиливи в мозок гз розвитком офтальмоплегп. шетагму, атаксп. апати, дез-
ортентацй, шо характеризуе енцефалопатйо Вершке. У раз! прогресування хвороби
приеднуються ретроградна амнезтя, неспроможшсть засвоювати нову шформа-
цпо, батакучгсть, що щентиф!куеться як психоз Корсакова Набута форма цього
синдрому часто спостерйаеться у хворих на хрошчний алкоголхзм;
• тдгостра некротизуюча енцефалопапйя виникае при порушены утворення в
мозку ттамгатрифосфату. Хвороба проявляеться втратою апетиту, блюванням, ут-
рудненням смоктання. Немовлята вграчають здатшсть утримдвати голтвку, у них
розвиваються численш невролопчш роздали. Без лткування хвороба заюнчуеться
летально протягом перших роюв життя.
Недостатшсть рибофлавшу
Патогенна д!я недостатност) впамшу В, (рибофлавшу) пов’язана з порушенням
процесгв бголопчного окиснення, що спричинюе енергодефщит, який уже на
раншх етапах проявляеться загальною слабгстю, пщвищеною втомлювашетю г
схилъшстю до застуди их захворювань
Арибофлавшоз клппчно проявляеться ушкодженням високоаеробних м>то-
тичних тканин, до яких належи гь епггелгальна, кровотворна та лгмфощна. Спо-
стерггаеться хейлоз (трпцини губгв г кут!в рота), ангулярний стоматит 1 глосит
(язик яскраво-червоного кольору), себорейний дерматит, особливо носогубного
трикутника. Кон’юнктива очей втрачае блиск унаслщок закупорки слъозового
каналу ештелгем, шо злущився. Ропвка проростае судинами ) згодом мулле.
Може виникнути катаракта. У дгтей часто спостерн аються нормохромна гшореге-
нераторна анем!я г лейкопенгя.
Недостатшсть антипелагричного впамшу РР
В основ! патогенезу недостатност! вгтамшу РР лежать зменшення енергетичного
забезпечення клггин, порушення синтезу та репарацп ДНК, пригнгчення синтезу
жирних кислот, холестерину I стеро'щгв, а також зниження антиоксидантних мож-
ливостей клгтин. Усе це порушуе прол!феративну та функцюнальну актившсть
мгтотичних тканин г спричинюе дегенеращю провщних шляхгв, загибель нейронгв
г порушення функщй периферично! та центрально! нервово! системи.
Клппчна картина нгациново! недостатност! характеризуеться симптомокомп-
лексом “трьох Д”: дерматит, дгарея, деменщя, як! в комплекс! формують клгнгчну
картину пелагри (в!д !тал. ре Не а§га — шорстка шк!ра). Це захворювання було по-
ширене в XVI ст. в Ггали, Тспангг, Румунп, Захщнгй Грузи, де в рацгонг населения
на таг нестач! тваринного б!лка знание мгеце посщала кукурудза, у якгй вггамш РР
мгститься у важко засвоювангй для людини форм!. Сама назва вгтамгну РР озна-
чав протипелагричний (реПа^га ргеуепйуе).
Дерматит виникае на вщкритих дшянках шкгри пщ дгею сонячних промен!в.
Шкгра симетрично червонге, виникають гшеркератоз, злушування, трпцини, гг-
перпггментащя та вторинна гнфекщя. Уражуються слизов! оболонки (стоматит,
пнггвгт, глосит, уретрит, валют, проктит).
Д!арея спричинюеться атрофгею ештелгю шлунка та кишок г супроводжуеться
нудотою г втратою апетиту.
Деменщя починаеться з головокружения, головного болю, апатп, неврастенгч-
ного синдрому та закшчуеться розумовою в!дстал!стю 1 розвитком психозгв гз дез-
оргентащею та галюцинацгями. Ураження периферично! нервово! системи харак-
теризуеться сильним нервово-м’язовим болем г парестез!ями у вигляд! печшня.
До шших проявгв хвороби належать анемгя ! мгокардгодистрофгя.
Недостатшсть антидерматитного в1там1ну В.,
Основною ланкою патогенезу недостатност! шридоксину (вггамш В6) е блокуван-
ня перетворення амшокислот шляхом трансамшування, декарбоксилювання й
гзомеризацп, що порушуе синтез бшюв, гормонгв шитоподгбног залози, бгогенних
амгнгв г вгтамшу РР гз триптофану, спричинюе розвиток пперамшоацидемп, амг-
ноацидури г негативний азотистий баланс. Кргм того, без вгтамшу В6 неможлива
актившсть деяких синтетаз, що перешкоджае бгосинтезу гему г сфшголшщгв, по-
рушуе перетворення гомоцистегну на цистегн.
Клгнгчна картина недостатностг шридоксину часто спостерггаеться в дитячому
вгцг при одномаштному харчуваннг штучними сумгшами г стерилгзованим моло-
ком, у ваптних при токсикоз!, а також у дорослих при лгкуванш антимгкобак-
тергальними препаратами г деякими антибготиками. Характерною для ггповгтамг-
нозу В6 е гтохромна, мйсроцитарна, сидеробластна (сидероахрестична або замзореф-
рактерна) анем1я, що виникае внаслщок порушення синтезу гемоглобгну в
еритропитах в разг одночасного пщвищення вмгсту залгза в сироватцг кровг та мак-
рофагах, розвитку гемохроматозу з ураженням Ре2+ серця, печгнки, шдшлунковог
залози та шших паренхгмних органгв (див. попереднш роздал). Кргм того, вщмг-
чаеться розвиток сухого себорейного пелагроподгбного дерматиту, стоматиту г гло-
ситу, що частково пояснюеться зменшенням ендогенного синтезу вгтамшу РР.
Часто виникають судоми, зумовлеш недостапнм утворенням у центральны не-
рвовш систем! гальм!вного мещатора ГАМК. Порушення синтезу бшюв ! сф!н-
голппщв спричинюе в дггей затримку росту ! розвиток лейкопенп.
Дефщит тридоксалю зумовлюе виникнення гтергомоцистешемп, яку остан-
нтм часом розглядають нартвнт з атерогенними лшопротещами як фактор ризику
розвитку серцево-судинних захворюванъ. Гомоцистеш — сульфоамшокислота з
високою прооксидантною актившстю, яка зумовлюе функщональш розлади ен-
дотелтальних юптин, пролтферащю гладком'язових юптин артер!альних судин,
посилення агрегацп тромбоцилв. Гшергомоцистешемтя спричинюе прискорений
розвиток атеросклерозу, значно збшьшуе ризик цереброваскулярных захворювань
та шфаркту мюкарда.
До вроджених порушень обмшу шр ид оксину належать:
• гомоцистинуры, шо зумовлена недостатшсгю цистапошнсинтази — вттамгн-
В6-залежного ферменту, який катал!зуе конденсацпо гомоцистешу 1 серину з ут-
воренням цистатюшну та цистешу. У кров1! тканинах хворих накопичуеться го-
моцистеш, який екскретуеться цирками у вигляд! окиснено! форми — гомоцис-
тину. Захворювання проявляеться в долей г шдлпктв симтомокомплексом, який
включае: ектоппо кришталика; генералгзований остеопороз г деформацпо скелета
у вигляд! “павучих” кшшвок (довп та тоню), викривлення хребта, плоскостопое
тт, неправильного прикусу; своерщну (негнучку) ходу; затримку фхзичного роз-
витку. Характерними е блакигний кол гр очей I свттле волосся внаслщок порушен-
ня шгментапп. Часто спостерггаеться затримка розумового розвитку. Усе це,
гмовгрно, е наслщком нестач! цистешу. Основною причиною смерт: хворих е
тромбоз артергй або вен, спричинений гшергомоцистешсмгею,
• цистат1оншур1я, яка зумовлена генною мутащею апоферменту циста гюна-
зи, що спричинюе екскрешю гз сечею цистатгонгну. Хвороба проявляеться за-
три мкою розумового розвитку;
• спадкова ксантуренургя (синдром Кнаппа), яка пов’язана з недостатнгстго вт-
тамш-В6-залежного ферменту — кгнуреншази, що спричинюе порушення пере-
творення триптофану на вгтамш РРг накопичення в кровг г тканинах ксантурено-
вот кислоти г кгнуреншу. Хвороба проявляеться у 0,5— 1 % дней такими симпто-
мами, як гетер]я, невмотивоваю вчинки, фобп, сомнамбулгзм, судоми. Часто
виникають алерпчш стани:
• тридоксинзалежний судомний синдром, який виникае на тлт нормально! за-
безпеченосп оргашзму ьгтамшом В6, що пов’язано зг зменшенням спорщненостг
генетично дефектно! глутаматцекарбоксилази до глутамату.
Недостатшсть фолъевоТ кислоти
Природна фолгева кислота (вгтамш В9) погано всмоктуеться г дуже швидко руй-
нуеться при термгчнш обробщ, консервуванш г тривалому збергганнг продует! в хар-
чування Здатшсть до депонування обмежуеться в дорослих 5—20 мг, чого вистачае
на 1—3 мгс. Оргашзм плода здатен активно вилучати фолацин !з материнськс! кровт,
тому його концентрацтя в кровх плода в 2—8 разгв витца. нтж у ваггтног. Проте в
недоношених дггей запаси фол к во! кислота дуже обмежею. У кровг фолацин пере-
носиться спещалышм транспортером у вшлядг №-метилтетрагщрофол!ево! кисло-
ти (№-метил-ТГФК), яка в юйтанах деметалюеться за участю вггамшу В]2 до ко-
ферментно! форми фолхево! кислоти — тетрагщрофолату, який один здатен депону-
ватися в цитоплазм!. Без впамшу В)2 фолхева кислота втрачаеться оргашзмом.
В основ! патогенезу недостатност! фолхевох кислоти лежить порушення М1Ж-
молекулярного перенесения одновуглецевих фрагменпв, що призводить до бло-
кади реакцн перетворення гомоцистешу на метюнш хз виникненням гшергомо-
цистешемп, а також до зниження синтезу нуклеотидгв ! нуклешових кислот, що
викликае порушення подыу клпин.
Основними КЛ1Н1ЧНИМИ проявами недостатност! впамшу В9 е гтерхромна, ме-
галобластична, макроцитарна анем1я; лейко- I тромбоцитопенгя з появою в кров!
гшерсегментованих багатоядерних лейкоципв; запально-атроф!чн! процеси у сли-
зовш оболонц! травного каналу (глосит, стоматит, езофапт, ахшчний гастрит,
ентерит). Спостерйаються загальна слабость, доарея, стеатоз печшки, раннш роз-
виток атеросклерозу, дратовливость, попршення пам’ятх, ворожость.
До вроджених порушень обмшу шридоксину належать:
• мегалобластична анем1я внасгидок вродженого порушення всмоктування фо-
лапйв /з порушенням психочного розвитку;
• мегалобластична анемгя внасл1док дефекту фолатредуктази, що порушуе пе-
ретворення фолхевох кислоти на №-формхл-ТГФК. Захворювання прогресуе
повыьно без невролопчноо симптоматики;
• мегалобластична анем1я внаслгдок недостатност! формгмшотрансферази, при
яюй порушуеться утворення з петидину 5-формхмшо-ТГФК. Захворювання про-
являеться протягом першого року житгя дитини та харакгеризуеться затримкою
розумового 1 фхзичного розвитку.
Недостатшсть антианем1чного впамшу В12
Впамш В12 (кобаламш) не утворюеться ан! рослинами, ан! тваринами. Лише мхк-
роорганхзми синтезують кобаламш, у тому чиелх 1 в товстш кишщ людини. Проте
людина не може скористатися ендогенним впамшом В12, осюльки його всмокту-
вання можливе лише в клубовш кишщ. Единим джерелом цього впамшу для
людини е тканини жуйних тварин, тварин-копрофапв х хижакзв. Печшка людини
теж накопичуе кобаламш у киькост!, яка забезпечуе оргашзм на 3—4 роки. Про-
те люди, що дотримуються суворо! вегетар!ансько! дхети, обов’язково страждають
на гшовпамшоз В12. Впамш стшкий до термообробки, але розкладаеться пщ д!ею
свила. Пар!етальн! юптини шлункових залоз виробляють глхкомукопротеш (внут-
рнннш антианемхчний фактор Касла), який зв’язуе х зберхгае впамш в!д руйну-
вання панкреатичними ферментами, а утворений комплекс абсорбуеться через
слизову оболонку клубово! кишки за допомогою спещального рецептора-пере-
носника. Порушення вироблення цього фактора, його структурний дефект, руй-
нування антиплами або дефект кишкового рецептора-переносника спричинюють
розвиток ендогенно! недостатност! кобаламшу. Утворення внутршшього фактора
Касла порушуеться при автохмунному ураженн! пар!етальних юйтин, атрофхчному
гастрит!, у тому числ! при недостатност! Дюлхево! кислоти, при пухлинах х внаелх-
док резекщй шлунка. Можлив! й шли причини виникнення ендогенно! недостат-
ност! кобаламшу. Наприклад, гельмшти можуть конкурентно споживати впамш,
ентерити порушують його всмоктування. Зменшення утворення транскобаламхну
II порушуе транспорту впамхну, а пригшчення синтезу транскобаламшу I переш-
коджае його збергганню г депону ванню. При захворюваннях печшки 1 нирок по-
рушуеться як депонування вгтамшу, гак г перетворення його на коферментн! фор-
ми Ваптшсть 1 лактацгя значно збгльшують потребу в кобаламгнг.
В основ! патогенезу кобаламшовог недостатност! лежить порушення двох ре-
аки !й. У лерппй реакцп метиткобаламгн необхгдний для метилювання гомоцистег-
ну в метгонш. При цьому донатором метильног групи е №-мезил-ГГФК. УнаслЁ-
док реакцп зменшуеться вы Ёст токсичного гомоцистешу г збЁльшуеться кёлъкёсть
тетрагЁдрофолату, необхЁдного для синтезу нуклешових кислот. Таким чином, не-
достатнЁсть вЁтамЁну В,2, як Ё вЁтамгнгв В6 ё Во. зумовлюе стергомоцистешемио. що
спричинюе ушкодження серпе во-судинног системи. Зменшення метилкобаламшу
також приппчус, як ё нестача фолЁевог кислоти, подёл клётин. Останне порушуе
структуру Ё функцЁю М1ТОТИЧНИХ тканин Ёз розвитком мегалобластично! анеми, лей-
копенИ, тромбоцит опеки, атрофи етте.йю травного тракту. Якщо недостатнЁсть
кобаламЁну спричинена порушенням утворення або структури внутргшнього ан-
тианемЁчного фактора Касла, то тодё анемЁя мае назву першцюзно? (злоякЁсна,
хвороба АддЁсона — БЁрмера).
У другой реакцгг дезоксиаде нозиткобаламЁв бере участь в утворенш сукшгнЁл-
КоА з метилмалонЁл-КоА, шо продукуеться при карбоксилюванн! прошонгл-КоА.
Блокування цЁег реакцЁЁ в разЁ недостатностЁ вЁтамЁну В]2 зумовлюе накопичення
метилмалоновоЁ та прогпоново! кислот, що спричинюе утворення жирних кислот
ёз непарним числом вуглеводних а том! в. Включения таких жирних кислот у лёпёди
нейронгв г клётин Шванна спричинюе жирову дистрофгю нервових клётин ё де-
мгелгнЁзацгю нервових волокон. Ураження нервово! системи з розвитком фунику-
лярного мгелозу (дегенерацгя г склероз заднгх ! бгчних стовпёв спинного мозку) й
ушкодженням черепних ё периферичних нервгв проявляеться полЁневритом, па-
рестезгями, а пот!м Ё парапЁчами з порушенням функцш органгв малого таза Така
неврологЁчна симптоматика не спостерЁгаеться при недостатностЁ фолЁевог кисло-
ти, що дае змогу встановити правильний дгагноз.
До вроджених порушень обмЁну кобаламЁну належать:
• мегалобластичш анеми при спадкових дефектах утворення або структури
внутриинього антианемгчного фактора Касла',
• мегалобластично анем1я при порушены всмоктування вйпамшу В12, спричине-
на вродженим дефектом механЁзму вившьнення вЁтамЁну в кров;
• хвороба 1мерслунда — Гресберка, шо зргдка спостерЁгаеться в кра'гнах Скан-
динавського п! вострова, яка пов'язана зё спадковим автосомно-реиесивним де-
фектом кишкового рецептора до комплексу вЁтамЁн — фактор Касла та прояь-
ляеться мегалобластичною анемЁею ё протегнурЁею;
• мегалобластичш анеми, зумовлеш вроОженим дефектом утворення транскоба-
ламтв. За вЁдсутностЁ в кровЁ транскобаламгну II з перших гижнёв життя в дитини
розвиваеться тяжка анемгя;
• вроджена метилмалонатацидем1я, що може бути наелгдком спадкового де-
фекту утворення як дезоксиаденозилкобаламЁну, так г апоферменту. Хвороба про-
являеться в перший рЁк життя дитини нестримним блюванням, кетоацидозом,
нейтропенгею, тромбоцитопенгею ё затримкою психомоторного розвитку. Мега-
лобласти в кровг не виявляються.
Недостатшсть антицинготного впам!ну С
В!тамш С синтезуеться !з глюкози веша рослинами 1 переважною бьлышетю тва-
рин. Проте в органгзм! людини вш не утворюеться, а його надлишок повшстю
виводиться з оргашзму за 4 год. Тому людина повинна постшно споживати ас-
корбшову кислоту, яка мютиться лише у св!жих або пщкислених продуктах.
В основ! патогенезу недостатносп вггамшу С лежить передуйм пригшчення
реакцш гщроксилювання, що спричинюе порушення синтезу колагену, серотош-
ну, катехоламгшв, стеро’щ!в, карштину. Уже незначний дефщит аскорбшово! кис-
лоти зумовлюе вщчуття втоми, зниження апетиту, ослабления 1муштету до застуд-
них захворювань, зменшення спйкосп до стресу. Вщсутшсть нормального пдрок-
силювання проколагену перешкоджае стабшзацп сшралепод!бно! конфггурацп
колагенових молекул. Такий неповноцшний колаген погано секретуеться ф1броб-
ластами, швидко розчиняеться 1 не мае достатньо! мехашчно! мщносп. У першу
черту уражуеться найнасичешший пдроксипролшовими залишками колаген кро-
воносних судин. Саме тому цинга (скорбут), що розвиваеться при нестач! вггамшу
С, характеризуемся вазопапями, як! проявляються петех!ями, екхгмозами, син-
цями, кровоточивхстю ясен. Внутршгньочерепш та внутршшьоперикардгальш
крововиливи в разг тяжкого перебйу цинги можуть спричинити смерть. Ослаб-
ления прикрепления оюстя до исток ! фгксацц зуб!в у ком!рках призводить до
пщоюсних крововилив1в, субпериостальних перелом1в за типом “зелено! гы оч-
ки”, набряюв 1 болю в суглобах, розхитування 1 випадшня зуб1в. Цинга в дитячо-
му оргашзм! затримуе утворення кзетки на м1сщ хряща, який при цьому
розростаеться 1 мшерайзуеться. Виникае деформащя трудно! кйтки з чоткопод!б-
ними болючими кшцями ребер 1 коритопод!бним западаниям груднини, а також
варусне викривлення трубчастих меток нижшх кшц!вок. Недостатшсть карштину
спричинюе м’язову слабйсть у хворих на цингу.
Недостатшсть аскорбшово! кислоти викликае розвиток зайзодефщитно! ане-
мй через ослабления всмоктування негемшового зайза 1 зниження споживання
його клпинами юсткового мозку при синтез! гемоглоб!ну.
Аскорбшова кислота е важливим компонентом у систем! антиоксидантного
захисту оргашзму, проте за умови надлишку вгльних Ее2+ або Си2+, нестач! вггам!-
ну Е 1, особливо, за наявносп микросомального цитохрому Р450 вццграе роль про-
оксиданта, сприяючи утворенню активних кисневих радикал!в. Це, з одного боку,
забезпечуе участь впамшу С у знешкодженш токсишв, антибютийв та шших
ксенобютийв, а з шшого — активуе реакцп ПОЛ.
Недостатшсть бютину
Недостатшсть бютину (в!тамш Н) спостерхгаеться в людей, так само як ! нестача
пантотеново! кислоти, дуже рщко, але може виникати при парентеральному хар-
чуванш, мальабсорбцп або за надопрного споживання сирих курячих яець. В ос-
нов! патогенезу гшовпамшозу Н лежить пригшчення активност! трьох ферменпв,
для яких карбоксибютин е коферментом. Зниження активност! шруваткарбокси-
лази спричинюе блокування циклу Кребса ! глюконеогенезу, а пригшчення аце-
тил-КоА-карбоксилази 1 прошошл-КоА-карбоксилази — порушення бюсинтезу
нормальних жирних кислот ! ослабления руйнування неф!зюлопчних жирних
кислот з непарним числом атом!в вуглецю.
Недостатшсть впамшу Н проявляеться лускатим себорейним дерматитом но-
согубного трикутника 1 волосисто! частини голови (звщси назва впамшу: з н!м.
Иаи1 — нлара), алопещею, гшерестез!ею, кон’юнктивпом, м!алпею, атакс!ею. У
дней виникае затримка росту.
До вроджених порушень обмшу бютину належать:
• десквамативна еритродерлия Лейнера, пов’язана з недостатшстю панкреа-
тично! бютишнази, яка звшьняе впамш вщ бшка перед усмоктуванням у порож-
шй КИПЩ1. Хвороба проявляеться в новонароджених проносом ! характерною се-
бореею;
• вроджений дефект протоны-КоА-карбоксилази, який призводить до пщви-
щення в тканинах 1 кров! концентраци прошоново! кислоти, що спричинюе в
перш! тижн! життя дитини кетоацидоз, який супроводжуеться блюванням, зне-
водненням оргашзму, артер!альною гшотенз!ею, м’язовою слабкктю. Вшбуваеть-
ся утворення аномальних жирних кислот з непарним числом вуглеводних атомгв.
Накопичуючись у фосфолшщах мозку, так! жирн! кислоти спричинюють тяжкх
невролопчш порушення. У бхльшостх випалктв хвороба закшчуеться летально;
• вроджений дефект активност! ^-метилкротоноы-КоА-карбоксилази зумо-
влюе порушення розпаду лейцину в стадп карбоксилювання р-метилкротоно'ш-
КоА, що спричинюе розвиток метабол!чного ацидозу, який супроводжуеться
нестримним блюванням. Через екскрецпо аномальних продуктов перетворення
р-метилкротоно!л-КоА сеча набувае своерщного запаху котячох сечь Хвороба,
хмовхрно, пов’язана зх зменшенням спорщненосп апоферменту до карбоксибюти-
ну, оскшьки висок! дози бютину полшшують стан хворого.
Недостатжсть рутину
Патогенез недостатност! впамшу Р (рутин), який являе собою рослинн! бюфла-
вонощи х пол!феноли, значною м!рою зумовлений зменшенням реактивацп ас-
корбшовох кислоти, що проявляеться пщвищенням проникност! та ламкост! ка-
п!ляр!в.
Недостатн!сть антиксерофтальм!чного в1там1ну А
Ретинол (впамш А) надходить в оргашзм хз продуктами тваринного походження,
а каротини — рослинного. Причому в норм! засвоюеться не бшьше 1/6 провпа-
мшу. Впамш А всмоктуеться значно краше, але при стеаторе! порушуеться за-
своення 1 ретинолу, 1 каротишв. Купфер!вськ! кл!тини печшки депонують у до-
рослих значну кмыасть еф!р!в ретинолу, яких вистачае на 2—3 роки. Новонарод-
жеш не мають такого запасу впамшу, що зумовлюе швидкий розвиток
авпамшозу в раз! недостатнього його надходження. Недостатшсть цинку порушуе
вившьнення ретинолу з печшки.
Оскшьки альдегщна форма ретинолу — 11-цис-ретиналь — бере участь в ут-
воренш фоточутливого зорового шгменту паличок ставки ока родопсину, при
гшовпамшоз! А спостерйаеться дегенеращя зовшпшхх сегмент!в паличок хз ви-
никненням порушення сутшкового (гемералопия, “куряча слтота”) х шчного (н!к-
талотя) зору. У раз! виражешшо! недостатност! ретинолу порушуеться 1 фуякшя
колбочок.
Из
Дпочи як стероТдний гормон, ретиноева кислота бере участь у рост! га дифе-
ренппованш ештелУальних клгтин, запобпае метаплазп залозистого ештслУю в
зроговыий. стимулюе синтез глУкопроте'шУв, якУ становлять основу муцин 1 в, що
захищають слизовУ оболонки. Тому нестача вУтамУну А спричинюе кератишзашю
епУтелУю слизових оболонок У залоз. Зроговшня епУтелУю проток сльозових залоз
зумовлюе сухУсть очного яблука (ксерофтальмия), що спричинюе запалення У
розм'якшення ропвки (кератомаляция) 13 подальшим утворенням “бУльма”. Мета-
плазУя ештелУю ргзних слизових оболонок 1 залоз знижуе Ух бар’ерну функцпо, що
одночасно 31 зниженням Умуштету спричинюе розвиток ларинпту, трахелу, брон-
хиту, пневмонп, гастриту, ентериту, шеллу, циститу, юст ргзних залоз. Зниження
антиоксидантних потужностей оргашзму при нестачг карагиноццв зумовлюе ви-
никнення катаракт.
У дгтей недостатшсть вгтамшу А проявляеться затримкою росту- (звщси назва —
“впамш росту”), що зокрема пояснюють порушенням синтезу хондроУтинсуль-
фапв.
До вроджених порушень обмшу вггамУну А належать.
• гтеркаротинелпя, що виникае за вУдсутност! кишкового ферменту р-каро-
тин-15-15 -дюксигенази, який каталУзуе перетворення каротишв на ретинол. Хво-
роба проявляеться гемералошею та помутнУнням ропвки;
• спадковий фомкулярний кератоз Дар’е проявляеться затримкою розумового
розвитку У психозами на тл) уражень шк!ри та слизових оболонок. Типовими е
поздовжня покреслешсть У зазубрешстъ ютив. Тривале вживания вУтамУну А у
високих дозах дае позитивний ефект.
Недостатжсть антирахггичного в!там!ну □
Гщроксильован! форми вУтамУну О — це стеро'щш гормони, рецептори до яких е
в клгтинах багатьох оргашв У тканин. Основними мишенями для цих гормоновпа-
мппв е травний тракт, нирки 1 юстки, а основною функщею — пщтримання ра-
зом з паратгормоном У кальцитоншом кальцш-фосфорн от о гомеостазу.
Екзогенна недостатнють впамшу В виникае за вщсутностУ впливу на шшру
ультрафиолетового випромшювання та в раз! зменшення в рацУонУ юлькосп каль-
циферолУв. Ендогенна недостатшсть вггамУну може бути спричинена порушенням
перегравлення У всмоктування лУпщУв, печшкового синтезу холестерину, змен-
шенням гщроксилювання вУтамУну О при печУнковш У / або нирковУй недосгат-
ностУ, зниженням синтезу або втратою Уз сечею вУтамш-О-зв’язуючого от-глобулУ-
ну, пригшченням реабсорбцп в ниркових канальцях У / або всмоктування в киш-
ках фосфатУв, спадковим дефектом клУтинних рецепторгв до кальцитрУолу.
Дефшит вУтамУну Е» проявляеться значною ппофосфатемтею У помУрною ппо-
калыпемУею внаслУдок недостатностУ кальцитрУолу, який стимулюе всмоктування
Са2+ 1 фосфату в кишках, та Ух реабсобцпо в ниркових канальцях, ГшокальцУемУя
стимулюе секрецпо паратгормону. Вторинний гшерпаратиреоз посилюеться тим,
що в норм! кальцитрУол пригшчуе продукцпо паратирину ВисокУ концентрат"]
гормону прищитоподУбних залоз активують остеокласти, що призводить до осте-
олгзу, розчинення гщроксиапатиту 1 вивьтьнення кальцпо Уз юсток. КрУм того,
паратгормон стимулюе абсорбцУю кальцУю в кишках, збУльшуе його реабсорбпУю
в нирках, але посилюе екскреппо фосфату7 Уз сечею. НевУдповщшсть достатньо
високой концентрацп кальцпо й гшофосфатемп при дефицит! вггамшу С и вторин-
ному гиперпаратиреоз! порушуе утворення фосфатив кальцию. Мшералйзацйя юс-
ток порушуеться ще й тому, що в норм! вгтамш В сприяе окисненню шрувату в
цитрат, який е необхщним для утворення нерозчинних солей кальцпо. Кр!м того,
зменшення утворення моногщроксильованои форми вггамшу В — кальцифедюлу —
пригшчуе биосинтез остеобластами мшорних битв юсткового матриксу (остео-
кальцину, остеонектину, остеопонтину, тромбоспондину тощо), необхщних для
фиксаций кальцпо на колагенових б!лках юсток. Таким чином, недостатшсть пдро-
ксильованих форм витамину В на тли вторинного гиперпаратиреозу зумовлюе пере-
важання остеолизу та деминерализации над остеогенезом и мйнералйзациею юсток, що
становить патогенетичну основу остеомаляци (деминерализация юсток без их струк-
турной перебудови) та остеопорозу (деминерализация юсток из перебудовою их струк-
тури). Гшовйтамшоз В проявляеться остеомаляцйею в дитей старшого ппсыьного
вику та в дорослих.
У рази недостатности витамину В у период интенсивного росту организму до завер-
шения енхондрального скостеншня метаепйфизарних хрящйв виникае рахйп. Кл!-
нйчнй прояви захворювання найвираженйшй в немовлят та в дйтей раннього вику.
Перши ознаки екзогеннои недостатности витамину проявляються на 2—3-у мйсяци
життя дитини. Спочатку з’являються неспецифични симптоми: порушення сну,
плаксивисть, пидвищена питливисть, зниження апетиту тощо. У кров! шдвищуеться
ривеииь дужной фосфатази.
Осюльки кальцитриол сприяе диференцпованню моноцитив й макрофагив в ос-
теокласта, необхидни для резорбцп хряща и надлишкового остеоиду, при дефицит!
витамину В утворення кйсток на мйсцй хряща вщбуваеться без адекватной резорбций
хряща й остеойду та без достатнього их звапнення. У рази надлишковои гиперплазий
хряща й остеоидной речовини утворюються безболйсни стовщення на кистках (“брас-
лета” у дилянци епифйзив трубчастах юсток, “чотки” на ребрах). Пласта остеоидной
тканини нашаровуються периостально, утворюючи стовщення в дшянках точок
скостеншня лобових и тйм’яних юсток (“лоб Сократа”, “квадратна голова”). Недо-
статки минерализация и прискорена декальцификация спричинюють пом’якшення та
механйчну деформацию юсток (кранютабес, “груди шевця”, “курячи груди”, запа-
дания перешсся, недорозвинення нижньой щелепи, високе пщнебшня, кифоз або
сколиоз хребта, викри влення нижних кинцивок, плоский рахитичний таз). Затриму-
ються закриття тйм’ячка и проризування зубив, спостерйгаеться схилыисть до роз-
витку кариесу. Гипотония м’язив передовой черевной стшки розвиваеться на тли гипо-
тоний кишок, що зумовлюе значне збильшення живота. Деформация грудной клйтки
та гипотония м’язив порушують легеневу вентиляцию, що поприцуе функцию серця.
Тяжкий рахит може супроводжуватися спазмофшею (дитячою тетаниею), в ос-
нов! патогенезу якой лежить системна гипокальциемия, що пщвищуе нервово-м’язову
збудливйсть. Хвороба проявляеться в дйтей виком вид 2 мис. до 2 роюв нападами
тошчних и клоничних судом, яки можуть супроводжуватися ларингоспазмом.
До вроджених порушень обмину витамину В належать:
• сшейний г'пюфосфатемгчний в1там1н-О-резистентний рах1т. Захворювання
успадковуеться доминантно, зчеплено з Х-хромосомою. В основ! патогенезу — по-
рушення структури бшюв — переносниюв фосфату в ниркових канальпях, у де-
яких хворих — у тонкий кишци. При нормальному ривни калъцитриолу и кальцию в
крови фосфор втрачаеться из сечею и, можливо, з калом. Виникае гипофосфатемия,
що зумовлюе появу симптомгв рахггу без м’язово! гшотонп вже на 1—2-у рощ
життя. Затримка росту ! стовщення меток спричинюють розвиток псевдоатлетич-
но! будови пла;
• спадковий псевдодефщитний випамш-В-залежний рах1т I типу. В основ! за-
хворювання — автосомно-рецесивний дефект нирково! сц-пдроксилази, необхщ-
но! для пдроксилювання в нефроцитах кальцифедюлу в кальцитрюл. На першому
роц! житгя з’являються симптоми “класичного” рахггу на тл! гшокальщемп, не-
значно зниженого або нормального рхвня фосфату, високо! активност! лужно!
фосфатази, генера/пзовано! гшерамгноацидурп та вторинного гшерпаратиреощиз-
му. Для лгкування застосовують кальцитрюл;
• спадковий псевдодефщитний втам1н-Д-залежний рахт II типу, пов’язаний
!з дефектом юптинних рецептор!в кальцитрюлу. Клппчна картина рахггу при ць-
ому захворюванш поеднуеться з алопещею (вогншцеве облисшня) та м’язовою
слабюстю. Позитивний ефект дають висок! дози впамшу Е);
• спадковий автосомно-рецесивний синдром Де Тон1 —Дебре — Фанкош, пщгрун-
тям якого е порушення канальцево! реабсорбцп фосфату, пдрокарбонату, амшо-
кислот 1 глюкози. Ппофосфатем!я при нормокальщеми спричинюе розвиток раха-
ту на тл! пол!урп, ппотонп та полщипсп;
• спадкова автосомно-рецесивна гтофосфатазгя, пов’язана з р!зким знижен-
ням активност! лужно! фосфатази, необхщно! для утворення фосфату кальцпо,
побудови та мшералхзацн юсток. Для цього захворювання вже на першому рощ
життя характерн! виражеш рах!тичш юстков! деформацн, остеопороз, “куляста”
форма черепа, вщетавання в рост!, гшотон!я м’яз!в на тл! пперкальщеми, нормо-
або гшерфосфатемп.
Недостатжсть вгтамжу Е
Гшовпамшоз Е найчастппе виникае в недоношених 1 дней з гшотроф!ею, у яких
спостерйають зменшене депо токоферол!в ! погане !х усмоктування. Екзогенна не-
достатшсть впамшу Е розвиваеться в малююв при штучному вигодовуванш, особ-
ливо в раз! надлишку полшенасичених жирних кислот 1 дефщиту б!лка в ращош.
Ендогенну недостатшсть токоферол!в зумовлюють ус! захворювання печшки, пщ-
шлунково! залози! тонко! кишки, яю попршують перетравлювання 1 всмоктування
ЛШЩ1В. Розвиток пповпамшозу Е можливий 1 в раз! порушення його транспорту.
Пщгрунтям патогенезу пповпамшозу Е е зниження антиоксидантного захис-
ту юптинних мембран, активашя ПОЛ, зменшення вщновлення впамшу А й уб!-
хшону, що спричинюе ушкодження клпин, у яких висока актившсть окисних
процесхв 1 як! утворюють велику юльюсть активних кисневих радикалхв. Пере-
дус!м до таких юптин належать еритроцити, нейрони та мюцити, а також сперма-
тогенний еппелш, клпини ембрюна, гепатоцита 1 нефроцита.
Клхшчно гшовпамшоз Е проявляеться в новонароджених гемол!тичною ане-
мхею, а в д!тей — мюдистрофхею з креатанур!ею х вщкладанням ефшголшщхв у
м’язах, демхелипзацхею аксошв ЦНС, що супроводжуеться атаксхею, порушенням
пропрюцептивно! чутливост!, гшоестезхею, дизартр!ею, ппорефлексхею. У раз!
значно! недостатност! впамшу можливий розвиток некрозу печшки та розм’якшення
дхлянок мозку. В експериментальних тварин виникае безплщдя, яке лхкують хз
застосуванням токоферолу (в!д грец. 1окоз — потомство, рйего — несу).
До вроджених порушень обмшу впамшу Е належать:
• акантоцитоз (еритроцит мае форму листя аканта; шпороподгбна клпина) —
спадкова гемолгтична анемгя. шо супроводжуеться м’язовою слабюсгю, атаксгею
1 пггментною ретинопапею. В основ! патогенезу цього захворювання лежить спал-
кове порушення синтезу одного гз структурних бщкгв хгломгкрошв г лшопротещв,
якг транспортують вггамш Е Вщсутшсть хшомгкрошв порушуе всмоктування вг-
тамшу. а нестача ЛПДНЩ — транспорт токоферолгв до тканин;
• спадковий дефект токоферолзв ’язуючого бглка, який в печшш зв’язуе вггамш
Е та включае його у склад лшопротсшв. Хвороба проявляеться атаксге ю та
м’язовою дистрофгею.
Недостатшсть вЕтамЕну Р
Основою етюлогп та патогенезу недостатностг вгтамшу Е е обмеження надход-
ження 1 порушення спгввщношення в органгзмг, з одного" боку, переважно проза-
пальник, проагрегангних, бронхо- та вазоконстрикторних похщних ш-6-ПНЖК,
з пшгого — переважно протизапальних, антиагрегантних, вазодилататорних, гшо-
лшщемгчних похщних го-З-ПНЖК.
При тотальнгй недостатностг незамшних полшенасичених жирних кислот,
яка може бути наслщком недоношеностг, голодування або порушення всмокту-
вання жиргв, слоегерцамився затримка росту, алопешя. дерматит гз розвитком
фолгкулярного гтеркератозу (надлишкове зроговшня епщермгсу навколо волося-
них фолгкулш), гшерхолестеринемгя г стеатоз печшки.
Частные виникае недостатшсть ш-З-ПНЖК, оскгльки гх природними джере-
лами е жир глибоководних морських риб I лляна олгя (в гнших рослинних олгях
переважають ю-6-ПНЖК). Зменшення утворення при цьому гшокоагулянтних.
гшолгпщемгчних г антигшемгчних ейкозано'глгв зумовлюе передчасний розвиток
атеросклерозу та збгльшуе ризик виникнення тромбозгв. тромбоемболш, шфаркту
мюкарда й гшемгчного шеульту Встановлено зворотну залежшеть мгж смертшстю
вщ ускладнень гшемтчног хвороби серця та вживаниям и-З-ПНЖК.
Проте в разг вщносно!" недостатностг в рашош ю-б-ПНЖК, шо спостерггаеть-
ся при персважаннг в продуктах ю-З-ПНЖК (нормальне спгввщношення мгж иг-6-
ПНЖК 1 ш-З-ПНЖК — 4 : 1), можуть з’являтися кровоточивгеть 1 крововиливи.
Зокрема це спостерггаеться в аборигешв Крайньог Пгвночг та японшв, як! харчу-
ються переважно морепродуктами. Цгкавим г красномовним е той факт, що в
Ялонп геморалчний шеульт рееструють у 4—5 разгв частгше. нгж у США г Фгн-
ляндп, хоча племгчна хвороба серця менш поширена.
Усе це ще раз переконливо евщчить про те, що харчовий рацюн людини мае
бути ргзномангтним 1 збалансованим.
Недостатшсть антигеморапчного вЕтамЕну К
В1тамш ЬС,, який виробляетъея мгкрофлорою кишок 1 мгститься в продуктах тва-
ринного походження, на вщмгну вщ вгтамшу Кг який утворюеться в рослинах,
для всмоктування в кишках не потребуе специального переносника г може абсор-
буватися не тгльки в порожюй, а й у клубовгй кипшг га проксимальней частиш
товсто! кишки. Це, а також те, шо вгтамгн К мгститься в багатьох продуктах, сприяе
низкий поширеностт гшовггамшозу К у дорослих. Недостатшсть вггамшу в цьому
вщ! може виникати лише при вживанш антивнамштв, яки ми е антикоагулянта
тривалому Л1'куванш ангибютиками 1 сульфашламщними препаратами та при сте-
аторет. Проте в новонароджених недостатшсть вггамшу К спостерггаеться часто,
що зумовлено йоге незначним запасом у печшщ, особливо в недоношених дней
Останне супроводжуеться низьким вмтстом втгамгн-К-залежних фактор! в коагу-
ляцц в кров! 1 визначаеться дуже низькою концентращею впамгну в грудному та
коров’ячому молощ, несформовангстю мгкрофлори кишок 1 частими порушення-
ми функцп травного тракту.
В основ! патогенезу гшовггамшозу К, який проявляеться щдвищеною крово-
ТОЧИВ1СТЮ 1 порушенням остеосинтезу, лежить пригшчення активност! ферменту
у-карбоксиглутаматкарбоксилази, коферментом якого е вгтамш К. Унаслщок цьо-
го блокуеться утворення у-карбоксиглутамшовоТ кислоти, необхшно! для фхксаци
Са2+ кальшйзв'язуючими бшками кровт (II. VII, IX. X фактори зсщання кровт.
антикоагулянтш протеши С 1 8) 1 юсток (у-карбоксиглутамгиовий протеш, остео-
кальцин, остеонектин тощо). Вгтамш К, кр!м того, активуе ртст гладком’язових
клттин 1 в!тамш-К-залежний скоротливий биток хвоста сперматозоцпв.
Г1ПЕРВ1ТАМ1НОЗИ
У наш час в економтчно розвинутих крашах тшовпамшози спостерпаються лише
серед малозабезпечних прошаркш населения та у хворих на мальабсорбцпо й ал-
коголтзм. Проте актуальною стала проблема передозування деяких внамппв.
Мегадози водорозчинних вггамгшв школи не спричинюють виникнення за-
хворювань г пом1тних неспецифгчних побгчних проявш. Винятком е лише впамт-
ни РР, В6 т С. Так. у раз! введения понад 50 мг нгкотиново! кислоти вщбуваеться
неспецифтчна дегрануляцтя лаброципв, що супроводжуеться свербежей, кропи-
в’янкою, гшеремгею шюри, жаром ! гшотошею. П!стя вживания понад 2000 мг
шрпдоксину виникають ошмшня шюри, особливо, навколо рота, вщчуття поко-
лювання. короткотривала втрата втбращйног чутливость При гемолттичних анемтях
т гемохроматоз!, що супроводжуються поя вою в оргашзм! надлишку вгльного за
л!за, висою дози впамшу С посилюють ПОЛ. У хворих 31 спадковим дефщитом
Г-6-ФДГ мегадози аскорбшово! кислоти спричинюють розвиток кризового ге-
молтзу еритроципв.
Здатшсть жиророзчинних вттамштв накопичуватись у тканинах оргашзму зу-
мовлюе можливтсть розвитку тх гшервпамшозтв, Ц! захворювання, спричинен!
тривалим передозуванням, найхаракгернпш для вггам!н!в А г Э.
Хрошчне отруення вггамшом А спостерггаеться при рег/лярному вживанш
ретинолу у високих дозах. Гостр! отруення 31 смертельними наслщками виникали
при споживанш печшки акули, бшого ведмедя та !нших морських тварин. Го-
стрий г'тервнпалйноз А у немовлят проявляеться гщроцефалгею з випинанням
пм’ячка, гарячкою. втратою апетиту. блюванням. Хрошчний гтервйпамйюз А у
дней 1 дорослих супроводжуеться головним болем, нудотою. втратою апетиту,
зниженням маси тиа, безсонням, драттвливтстю, гепатомсгалгсю, гптеркератозом,
западениям ропвки, сухтстю 1 ламюстю шгпв 1 волосся, перюсталытим гшеросто-
зом, деструкшею хрящовоУ та юстково! тканини, що спричинюе бшь у юстках т
сутлобах. Водночас надмтрне вживания каротишв проявляеться лише пожовтш-
ням долонь, шдошов 1 слизових оболонок без ознак штоксикацп.
Ппервйпалпноз О виникае в Д1тей при “штенсивному” лткуванш або надмтршй
профшактищ рахату 1 може проявлятися вже на 3—4-у мзсящ житгя. Патогенез
цього захворювання пов’язаний 1з посиленням усмоктування кальцпо ( фосфапв
у кишках 1 збшьшенням !х реабсорбцп в нирках. Пе спричинюе виникнення п-
перкальшемн, гшерфосфатеми. вторинного ппопаратиреозу та зниження актив-
ностг лужно! фосфатази. Ранньми ознаками гшерЕгтамшозу Б е млявгсть, сон-
ливгсть 13 перюдами збудження г драНвливостт, головний бгть, нудота, втрата апе-
титу, зменшення маси тша. Налли шки кальшю I фосфапв утворюють у тканинах
нерозчинш сош, що зумовлюе вогнищеву кальцифжашю нирок, судин, печшки,
легень 1 серця з порушенням !х структури та функшй. Виникають пошуртя, спра-
га 1 полцшпстя, шдвищуеться артерзальний тиск 1 розвиваються аритмй, ниркова
1 серцева недостатшсть. У дней припиняеться рют скелета в довжину, стовщують-
ся ешфтзи та д1афтзи юсток. Через лередчасне заростання тьм'ячка розвиваеться
мгкроцефалтя з пщвищенням внутршшьочерепного тиску.
Частина третя
ПАТОЛОПЧНА Ф13ЮЛ0ПЯ
ОРГАШВ I СИСТЕМ
Розд1л XXV
ПАТОЛОПЧНА Ф13ЮЛ0ПЯ СИСТЕМИ КРОВ1
ЗАГАЛЬНА ХАРАКТЕРИСТИКА ПАТОЛОПЧНИХ ЗМ1Н
У СИСТЕМ! КРОВ!
Патолопчнх зм1ни можуть виникати в будь-якш складовш системи кров!: у кро-
вотворних органах, кров], що циркулюе або депонована в судинах, в органах !
тканинах, де кров руйнуеться, а також у скупи еннях лхмфощво! тканини, шо мхс-
титься в некровотворних органах. Ц] складом псно взасмопов’язан], унаслщок
чого патолопчний пронес, як правило, не бувае суворохзольованим, 1 на нього
реагус система кров! в щлому, хоча прояви реакцп з боку окремих и компонент)в
бувають рвними.
Основними ознаками порушень у систем] кров! е змши:
• загального об’ему кров!;
• кшькосп, структури х функцш клхтин кровх (еритроципв, лейкоиитхв, тром-
бопитхв) унаслщок патологи кровотворення або кроворуйнування (див. схему
нормального кровотворення, кольор. вкл., мал. 1);
• гемостазу;
• бюххмхчних показникхв х фхзико-ххмхчних властивостей кровь
Пх змши виникають пщ впливом дц патогенних фактор)в на саму систему
кров], у разх порушення и нейрогуморально! регуляцн, а також унаслщок уражен-
ня шших систем х органхв.
Висока частота розвитку вгоринних змш у систем! кров! зумовлена п функ-
шональними особливостями. Так, збшьшення продукцц еритроципв, якх забезпе-
чують дихальну функшю кровх як переносники кисню та вуглекислого газу, е
компенсаторною реакщею системи кров! на гшоксйо. Пщ час шфекцшно! хворо-
би посилюеться лейкопоез хз податыпим збшьшенням у кровх кшькост! лейко-
ципв, яю беруть участь у захисних, у тому7 числх й змунних реакнхях органхзму.
Пщ час ушкодження судин передусгм активуеться зсгаальна система кров!, баланс
мгж якою г протизсщальною та фгбринолпичною системами забезпечуе гемостаз
Патолопя системи кров! може проявлятись у вигляд] як самостгйних хвороб
(наприклад, хвороба Аддгсона — Бгрмера, або пернщюзна анемгя, лгмфобластний
лейкоз, гемофглгя), так 1 гематологгчних синдромгв, як! супроводжують захворю-
вапня !нших оргашв г систем (наприклад, еритроцитоз при деяких уроджених
вадах серця, нейтрофшьний лейкоцитоз при пневмонн).
Серед патолог!! системи кров!часто спостерйаються так! хвороби, як пухлини
(гемобластози), авто!мунна патолопя, спадков! порушення, депреегя кроветво-
рения (гшопластичн! елани). Це зумовлено тим, що гемопоетичшй тканин!, яка
входить у систему кров!, властива висока м!тотичяа актившсть, !з чим пов’язана
п! дви щена чутливгеть п до ушкоджувальних впливгв р!зних факторгв: мутагенгв
(вгруси, хгм!чнг речовини, юнгзуюче випромшювання), лжарських засоб!в, як! ли-
нять вплив на обм!н речовин або пригшчують клгтинний подгл, дефщиту ллас-
тичних матер!ал!в (бглки, залгзо), вгтамгнгв (цганокобаламш, фолгева кислота)
ПОРУШЕННЯ ЗАГАЛЬНОГО ОБ’ЕМУ КРОВ!
Класиф!кап!я. Зпдно з Броуд! га Раунтрг (1853), порушення об’ему кров! про-
являються гтоволемйло або ггперволемйю — зменшенням або збгльшенням об'ему
кров! поршня но з нормою (нормоволемию), яка становить 6—8 % маси тага або
близько 1/14 маси тша доросло! людини (у новонародженого — 1/7). Своею чер-
тою гто- ! гшерволемйо подшяють на нормоцитемгчну (просту), полгцитемгчну й
олйоиитедйчну залежно вш того, чи зберггаеться при цьому нормальне сшввщно-
шення плазми г кч1тин (48—64 % об’ему кров! становить плазма, 36—52 % — юп-
тини кров!), чи змгнюеться в бгк переважання клгтин (полщитемгчна форма) або
плазми кров! (ол!гоцитем!чна форма). Кр!м того, до порушень об’ему кров! нале-
жать змши об’емного стввщношення м!ж клпинами кров! та плазмою в разг
нормального загального об’ему кров! — олгго- г полщитем/чна нормоволелйя
Показником об’емного спгввщношення е гематокритне число, яке визначае
вмгст клгтин кроВ1 (переважно еритроципв) у заг альному об’емг кров! (норма для
ж!нок — 0,36—0,46 л/л за С1, або об’емна частка 36—46 %; для чоловгкгв — 0,4—
0,52 л/л).
Етгологгя. Пповолелая нормоцитемчна (зменшення об’ему кров! без змши
гематокритного числа) виникае одразу теля гостро! крововтрати ! зберггаеться
доти, доки рщина не перейде з тканин у кров.
Пповолелйя ол1гоиитем1чна (зменшення об’ему крош з переважним зменшен-
ням у нгй клгтин — еритроцитгв) спостерггаеться в раз! гостро! крововтрати в тих
випадках, коли надходження кровг г тканинно! рщини у кровоносне русло не
компенсуе об’ем 1, особливо, склад кровг.
Пповолемгя пол1цитем1чна (зменшення об’ему кровг внаелгдок зменшення
об’ему плазми за вгдносного збщьшення вмгсту еритроцитгв) розвиваеться в разг
зневоднення органгзму (пронос, блювання, посилене потовидшення, гшервенти-
лящя), втрати плазми (плазморапя в раз! великих ошкгв). При шоку кров депо-
нуетъея в розширених судинах черевно! порожнини, що зумовлюе зменшення
ОЦК. вихщ рщини в тканини внаелгдок пщвищення проникност! ст!нки судин
зумовлюе згущення кровг та виникнення полгцитемгчно! гшоволемп.
Пперволемгя нормоцитемгчна (збшьшення об’ему кров! в раз! збереження нор-
мального сп!ввщношення м!ж еритроцитами 1 плазмою кров!) виникае одразу
шсля переливания велико! юлькосп кров!. Проте невдовз! рщина виходить з кро-
воносного русла в тканини, а еритроцити залишаються, що спричинюе згущення
кровь Нормоцитем!чна пперволемгя пщ час гостро! гшоксп та надмгрного фгзич-
ного навантаження зумовлена надходженням у загальний кровотгк кров! з депо.
Пперволемгя олггоцитемгчна (збшьшення об’ему кров! за рахунок плазми) роз-
виваеться в раз! введения кровозамшникгв, плазми, затримки води в оргашзм!
внаслщок гшерпродукцп вазопресину (АДГ), при гшеральдостеронгзм!, хворобах
нирок, а також у процес! !х л!кування, коли швидко зменшуються набряки.
Пперволемгя полщитемгчна (збшьшення об’ему кров! за рахунок збшьшення
кшькост! еритроципв) спостерггаеться в раз! зниження атмосферного тиску, а
також при ргзних хворобах, пов’язаних гз кисневим голодуванням (вади серця,
хронгчнг хвороби легень), г розглядаеться ж компенсаторне явище. При мгело-
пролгферативнгй хвороб! — справжнш полщитемИ, або хвороб! Вакеза, полщи-
темгчна пперволемгя е наслщком пухлинного розростання клттин еритроцитарно-
го ряду юсткового мозку.
Нормоволем'гя олггоцитемгчна виникае при анеми внаслщок втрати кров!
(об’ем кров! нормал!зувався за рахунок тканинно! рщини, а юльюсть еритроципв
ще не вщновлена), гемолгзу еритроципв, порушення гемопоезу.
Нормоволемгя полщитемгчна спостерггаеться пщ час переливания невелико!
юлькостг еритроцитно! маси, при еритроцитоз! в раз! хронгчно! гшоксп.
Патогенез. Пповолемгя супроводжуеться порушенням основних функцш кров!:
транспортно! та пов’язаних гз нею дихально!, трофгчно!, екскреторног, регуляторно!
(гуморальна регуляцгя, терморегулящя), а також захисно! функцш. При цьому змен-
шення ОЦК спричинюе органо- г мгкроциркуляторнг порушення, мгкротромбоз (у
раз! полщитемгчно! форми), гшоксгю (особливо при олггоцитемгчнгй форм!).
Пперволелйя зумовлюе пщвищення навантаження на серце збшыпеним
об’емом кров!. Одночасне зростання гематокритного числа (полщитемгчна ппер-
волем!я) спричинюе збшьшення в’язкостг кров! та посилення тромбоутворення,
що попршуе порушення кровообпу в органах.
У патогенез! порушень у випадку олггоцитемгчно! нормоволеми основну роль
вщгграють ослабления дихально! функцп кров! та розвиток гшоксп.
Крововтрата
Крововтрата — патологгчний процес, що виникае внасл1док кровотечг (гемораги) /
характеризуешься складним комплексом порушень / компенсаторних реакцш оргашз-
му у вгдповгдь на зменшення ОЦК та гшоксгю, зумовлену ослаблениям дихально!
функцй крови
Етгологгя. До етюлопчних факторгв, яю зумовлюють кровотечу, належать:
• порушення шлостг судин пщ час поранения (травми) або ураження патоло-
пчним процесом (пухлина, туберкульоз);
• ушкодження стшки судин (гостра променева хвороба);
• зниження зсщання кров! (гемофшгя, тромбоцитопенгя).
П атогенез. Видшяють три стадп крововтрати: початкову, компенсаторну 1
термшальну.
Початкова стадия, що виникае в перни годный шсля крововтрати, коли ком-
пенсаторы! реакцп оргашзму ще не можуть забезпечити гомеостаз, характеризуеть-
ся зменшенням ОЦК (нормонитемзчна гшоьолемзя), систол!чного об’ему серця,
зниженням артерзального тиску, ппоксзею, переважно циркуляторного типу.
У разг значно! крововтрати (25—40 % ОЦК) може розвинутися гемора&чний
(постгемораггчнии) шок, у патогенез! якого основним ланцюгом е гемодинамзчн!
порушення внаслщок швидкого зниження ОЦК (первинний гшоволемгчний шок),
що супроводжуеться розладами мзкроциркуляцп, недостатшм кровопостачанням
органзв з тканин, гшоксзею, диезрофзчними змшами в тканинах. Зменшення ОЦК е
також наслщком депонування кров1 в раз! патолопчного розширення судин кров’яних
депо. Остан не виникае пщ впливом вазоактивных речовин, як! вивзльняються з!
зруйнованих тканин пщ час крововтрат, що супроводжують травму. Крзм того,
пщвищення проникност! судинних стшок пщ дзею БАР. а також кислих метаболыгв
зумовлюе вихщ рщко! частини кров; в тканини, що ще сутгевтше зменшуе ОЦК.
Компенсаторна стадия зумовлена комплексом захисно-пристосувалышх реак-
цш. спрямованих на вшновлення об’ему та складу кров!, нормалгзаигю гемодина-
мзки з кисневого забезпечення оргашзму. Тривае ця стадзя вЩ кглькох годин до
галькох тижшв залежно вщ об’ему втрачено! кровз та реакгивност! оргашзму.
Терм1нов1 (негайш) мехашзми компенсац '1 тага:
• спазм кровоносних судин, зумовлений активащею симпатико-адреналово!
системи з д!ею катехоламш!в. шо призводить до пщвишення опору току кровз в
судинах внутрпшпх органзв (за винятком головного мозку з серця) та шгари, вихо-
ду кров! з депо в кровоносне русло, унаслщок чого пщвищуеться артерзальний
тиск, певною м!розо вщновлюеться ОЦК з кровопостачання житгево важливих
орган зв;
• рефлекторне прискорення та посилення скорочень серця;
• затримка води в оргашзм! внаслщок стимуляцз! секрецй вазопресину й аль-
достерону (у зв’язку з активащею ренш-ангзотензиново! системи пирок);
• надходження мзжтканинно! рщини в судини;
• рефлекторне прискорення та поглиблення дихання, що сприяе усуненню
дефщиту кисню в оргашзм!;
• збшьшення здатност! гемоглобшу вщдавати кисень тканинам (посилення
дисощаш! оксигемоглобзну пщ час ацидозу);
• пщвищення зездання кров! з тромбоутворенням, що еззиняе кровотечу.
Нетермтоы (травам) мехашзми компенсаци проявляються в пззшшз термшп у
вигляд! посиленого кровотворення та вщновлення бзлковоз о складу кров Протя-
гом 5-1 доби збзльшуеться гальгасть ретикулоцитзв у кровз, шо пов'язано з пщви-
щенням гемопоетично! активност! кзеткового мозку внаслщок збзльшено) иродук-
цп еритропоетину1 з внутрзшнього фактора Касла Бзлковий склад кров! нормалз-
'Еритропоетин — основний регулятор еритропоезу. Це глюкопроте'щ, який виробляеться в пост-
натальний перюд здебьтьшого в знтерстицзйних юптинах нирок. у незнаний юлькост! — в печзнцз,
макрофагах. аденогшофзз!, у плода 1 гал час патологи нирок — у печшщ та селезшщ. Стимуляторами
продукцв ЕП е зменшення вмзету кисню в нирковш тканизп, пщвищення ргвня молочно'; кислоти й
оксиду вуглепю. андрогени. ЕП шдукуе пролгферашю та еритроГдне дифереишювання еритропоетин-
чутливих юптин юсткового мозку, гальмуе Зхнзй апоптоз шляхом взаемоди з рецепторами мембран 1
зумовлюе посилення експреси гензв 13 подальшим збшьшенням синтезу ДНК з РНК, залЗзотранспор-
тних систем з гемоглобшу. У разз пщвишення ршня ЕП у плазм! прискорюються лро.пферапгя та
цозрзвання еритроцитзв. що сприяе збшьшенню юлькост! ретикулоцитзв у юегковому мозку г кров.
зуеться починаючи з 5—10-го дня шсля крововтрата завдяки збшьшенню синтезу
бшкгв у печшщ.
У компенсаторних реакциях оргашзму в раз! крововтрата беруть участь нерво-
ва система (рефлекси з рецепторних зон аорта, сонно! пазухи, збудження симпа-
тичного вщдшу автономно! вегетативно! нервово! системи), ендокринна система
(вазопресин, катехоламши, альдостерон), ренш-анпотензинова система, як! за-
безпечують часткову або повну нормалтзащю об’ему кров!, тонусу судин та шших
показниюв гемодинамгки.
ТермЫальна стад1я може настата в раз! недостатнього виявлення компенса-
торних реакцш унаслщок змши реактивност! оргашзму, а також при великгй 1
швидкш крововтрат! (залежно в!д виду ушкоджено! судини, кшькост! втрачено!
кров!) на тл! дп несприятливих екзогенних г ендогенних фактор!в (охолодження,
велика травма, серцево-судинш хвороби), за вщсутност! л!кувальних заход!в За
цим патолопчш змши в оргашзм! наростають аж до настання смерть Летальний
наслщок настае в раз! швидко! втрата 33 % ОЦК 1 повшьно! та тривало! втрата
50 % ОЦК.
Патолог1чн1 зм1ни внаслщок втрати кров! в термшальнш стади подгбш до та-
ких у початковш стади. Вони проявляються:
• зменшенням ОЦК гз розвитком постгеморапчно! анеми (ггповолемгя ол!го-
цитем!чна);
• розладом гемодинамики (зменшення венозного надходження кров! до сер-
ця, серцевого викиду, об’ему вгнцевого кровотоку, артер!ального тиску), арит-
м!ею, порушенням мшроциркулящ! в тканинах;
• недостатшстю зовшшнього дихання;
• розвитком циркуляторно!, гемгчно! т тканинно! гшоксп;
• порушенням тканинного метабол!зму 1 КОС (негазовий ацидоз);
• розладом нейрогуморально! регуляцп найважлившшх функцш оргашзму;
• зниженням зсщання кров! та розвитком ДВЗ-синдрому (особливо при знач-
них травмах).
ПАТОЛОПЧШ ЗМ1НИ ЕРИТР0ЦИТ1В
Патолопчш змши еритроцитгв можуть виникати в будь-яких кгптинах еритрону.
Еритрон — це сукупшсть нерухомих г циркулюючих клггин еритроцитарного ряду,
як! розташован! штра- та екстравазально г перебувають в ус!х стад!ях розвитку:
утворення (пролтферуючий, дозргваючий, резервний пули в кровотворнш тка-
нин!), функцюнування (циркулюючий пул у кров!) та !х загибел! (у макрофагоци-
тарних органах кроворуйнування).
Кл ас и фхкацхя. Патолопчш змши еритроцитгв подшяють на киыасш (у раз!
змши !х кшькост! в кров! поргвняно з нормою1) та як1сн1. Останн! можуть прояв-
лятися збшьшенням у кров! ргвня незрглих або навггь патолопчних, невластивих
нормальному еритропоезу еритроцитгв; змгною структури (форми, розмгру), хгмгч-
ного складу, метаболтзму, осмотично! та мехашчно! резистентностт, функцп ерит-
роцитгв, що супроводжуеться порушенням транспорту кисню.
‘У норм! юлыасть еритроципв у чоловцов становить 4—5,5 Т/л (Т за Мокнародною системою
одиниць — тера — 1012/л), або 4,0—5,5 • 1012/л, у ж!нок —3,7—4,7 Т/л (3,7—4,7 • 1012/л); юльюсть гемо-
глоб1ну в чолоыюв — 8,4—10,5 ммоль/л (135—170 г/л), у жшок — 7,4—9,9 ммоль/л (120—160 г/л).
Кыыасш змши еритроципв — це збщьшення (еритроцитоз) ! зменшення
(еритропешя, анемгя) вмгсту еритроципв в единиц! об’ему кров! вище або нижче
норми.
Яюсш змши еритроципв проявляються:
• збгльшенням або зменшенням у кров! юлькосп регенеративных форм ерит-
роцитгв (клйпини фгзюлоггчног регенерацп), яю в норм! м!стяться в кров! (ретикуло-
цита', полгхромагпофиьнг еритроцити1), а також появою в н!й незр!лих клгтин
еритроцитарного ряду, як! в норм! мктяться лише в юстковому мозку, — ацидо-
ф!льних, пол!хроматоф!льних нормобластгв (кольор. вкл., мал. II);
• клгтинами патологгчног регенераци еритроцитгв (у раз! мегалобластного
еритропоезу) у юстковому мозку та кров!, до яких належать базофшьнц пол!хро-
матофыьш, ацидофшьн! мегалобласты 1 мегалоциты (див. кольор. вкл., мал. II);
• дегенеративными змшами еритроцитгв (дегенеративнг формы), до яких нале-
жить ангзоцитоз, пойкглоцитоз, змгни забарвлення еритроцитгв, патологгчнг вклю-
чения (див. кольор. вкл., мал. II).
Ангзоцитоз — зм!на розм!р!в еритроципв пор>вняно з нормальними кл!ти-
нами (середнш д!аметр нормоцита становить близько 7,2 мкм), коли з’являються
мгкроцити (д!аметр менше н!ж 6,5 мкм), макроциты (д!аметр понад 8 мкм) 1 мега-
лоциты (дгаметр быьше н!ж 9 мкм).
Пойкглоцитоз — змша форми еритроципв, коли в мазку кров! виявляють-
ся еритроцити грушоподтбн!, видовжен!, серпопощбш (дрепаноциты), овальн!
(овалоцити), сферично! форми (сфероциты), шистоцити (фрагментованг), стома-
тоцити (ротоподгбнг) та 1н.
Змгни забарвлення еритроцитгв залежно в!д вмкту в них гемоглобину:
ггперхромгя — переважання в мазку кров! 1нтенсивно забарвлених ггперхром-
них еритроципв;
гтохромгя — переважний вм!ст блщо забарвлених еритроципв та анулоцитгв
(у вигляд! юльця забарвлена лише периферична частина еритроцита, де розмпце-
ний гемоглоб!н, а в центр! е незабарвлене просветления);
ангзохромгя — значна вщмшнкть у забарвленн! еритроципв.
Патологгчнг включения в еритроцитах:
тгльця Жоллг — утвори розмграми 1—2 мкм, що е залишком ядра, здебшьшо-
го мегалобласта;
кгльця Кебота — залишки ядерно! оболонки, яю мають форму юльця, вклмки;
базофгльна зернистгсть — гранули в еритроцитах (прециштати рибосом) синь-
ого кольору, що виявляються шсля забарвлення мазка за Романовським 1 евщчать
про токсичне ушкодження юсткового мозку;
тйгъця Паппенгейма — гранули тривалентного зал!за темно-блакитного кольо-
ру в сидеробластах 1 сидероцитах;
тгльця Гейнца — Ерлгха — син! прециштати гемоглобшу (виявляються в раз!
суправшального фарбування), юльюсть яких перевищуе чотири при спадковш
‘У норм! вмгст ретикулоципв — незрглих попередниив еритроципв — становить 0,2—2 % загаль-
но! юлькост! еритроцитгв; виявляються шсля суправггального фарбування мазка кров! брилганткре-
зилблау у вигляд! без’ядерних клггин синьо-зеленого кольору гз синьою зерниспспо (комплекс рибо-
сом з мРНК) 1 низьким вмгстом гемоглобшу.
ШолЬсроматофгиьш еритроцити — еквгваленти ретикулоципв; виявляються у вигляд! без’ядерних
клгтин бузкового кольору (сприймають киелг та основш барвники) пгсля фарбування мазка за Рома-
новським.
ферменто- 1 гемоглобшопатн з порушенням антиоксидантних властивостей ерит-
роципв, та ш.
Ет1олог1я. К1лыасн1 змши еритроципйв можуть бути зумовлеш:
• посиленням або ослаблениям еритропоезу в истцовому мозку;
• посиленим руйнуванням еритроципв (еритрод!ерез);
• зменшенням юлькосп еритроципв у раз! порушення цшосп судин (кро-
вовтрата);
• перерозподшом еритроципв у кровоносному русл!.
Як1сш змши еритроцит!в виникають унаслщок:
• порушення дозргвання еритроципв у истцовому мозку або пщвищення
проникносп юстковомозкового бар’ера, теля чого збшыпуеться надходження в
кров незрших клпин еритроцитарного ряду з низьким вмютом гемоглобшу (реге-
неративных форм еритроципйв)',
• зм1ни типу кровотворення в истцовому мозку з еритробластного на мегало-
бластний, цоли в ньому 1 цров! з’являються клтини патологично! регенерацй ерит-
роцитарного паростка;
• набутих 1 спадцових порушень обмшу речовин, складу ! структури еритро-
ципв, у тому числ! синтезу гемоглобшу (зменшення утворення або синтез ано-
мального гемоглобшу), що зумовлюе появу в кров! дегенеративных форм еритро-
ципв.
Еритроцитоз
Еритроцитоз — збшьшення в кров! юлькосп еритроципв з 6 Т/л (6,0 • 10’2/л) та
бшьше, гемоглоб!ну понад 10,85 ммоль/л (175 г/л), гематокритного числа — бшь-
ше 0,52.
Класиф!кац!я. За еттологгею видшяють еритроцитоз набутий 1 спадковий.
За механизмом виникнення еритроцитоз подшяють на:
• абсолютний (справжшй), зумовлений посиленням еритропоезу в кютко-
вому мозку I збшьшенням маси циркулюючих еритроципйв (у норм! — 28—36 мл/кг);
• вгдносний (несправжшй) — перерозпод!льний !гемоконцен-
трацшний.
У випадку вщносного еритроцитозу збшьшення юлькосп еритроципв в еди-
ниц! об’ему кров! е наслщком зменшення об’ему плазми або виходом кров! з депо
1 тому не супроводжуеться збшьшенням маси циркулюючих еритроципв. Абсо-
лютний еритроцитоз своею чертою подшяють на первинний (у вигляд! само-
стшних хвороб) 1 вторинний (симптоматичний), а останнш — на ф!з!о-
лог1чний (компенсаторний) 1 патолопчний.
Ет1олог1я. Причиною виникнення первинного абсолютного еритроцитозу
може бути спадкове, генетично детермшоване посилення еритропоезу. Такий
еритроцитоз може бути спричинений генетично зумовленим дефицитом в еритро-
цитах 2,3-дифосфоглщерату (регулятор оксигенацп та дезоксигенацц гемоглоб!-
ну). При цьому пщвищуеться спорщнеюсть гемоглобину до кисню 1 зменшуеться
передача його тканинам. Розвиваеться тканинна гптокоя, стимулюеться продук-
ция еритропоетину, пщ впливом якого посилюеться еритропоез.
Вщомий родинний еритроцитоз !з доброяюсним перебиом 1 домшантним або
рецесивним успадкуванням.
Причинами вторинного абсолютного (набутого) еритроцитозу, що виникае в
разг посилення еритропоезу при шдвищенш продукцп еритропоетину переважно
в нирках. е:
• гшоксична, ресшраторна, циркуляторна, гемгчна гшокстя при висотнш хво-
роб), хрошчних хворобах органов дихання та серцево-судинно! системи, мет- г
карбокси гемоглобшемд;
• локальна гшокстя нирок унаслщок !х штемп (гщронефроз, полткгстоз нирок,
стеноз ниркових артерш);
• гшерпродукшя еритропоетину деякими пухлинами (гшернефрома, рак пе-
чшки, фгбромгома матки, гемангиобластома мозочка);
• збудження симпатичного вщдыу нервово! системи, гшерфункщя деяких ен-
докринних залоз1 (шитоподюно!, надниркових та гн.).
Кргм того, можлива пухлинна трансформашя еритрощних ктттин з гюсилен-
ням !х пролгферацп, незалежно! вщ еритропоетину (при хвороб! Вакеза).
Хвороба Вакеза (справжня полщитемтя) належить до м1елопрол1феративних
хвороб з одночасним збьтьшенням ктлькоетг еритроштпв. гранулоциттв 1 тромбо-
цитгв, що пов'язано з пухлинним ураженням клттини — попередника мгелопоезу,
але з переважною диференшащею в еригрогдному напрямку.
Ыдносний еритроцитоз виникае пщ дтею причинних фактортв, якт зумовлю-
ють зневоднення органгзму (пщвищене потовидшення пщ час гшертермп, трлвале
блювання в разт кишково! непрохщностг, пронос) або внааидок перерозподигу кров!,
що спричинюе полщитемтчну ппово.темгю (шок), а також у раз! виходу кровт з
депо при гостргй гшоксп, коли спостерггаеться полщитемгчна гшерволемтя.
Патогенез. У випадку абсолютного еритроцитозу пщвищення утворення
еритропоетину зумовлюе посилення еритропоезу з подальшим збгльшенням у
кров) юлькостт циркулюючих еритроцитгв, вмтсту гемоглобгну, гематокритного
числа. При цьому спостерггаеться збгльшення ОЦК (полщитемгчна гшерволемгя),
ц в’язкостг, сповгльнення швидкостг кровотоку, порушення функцт! серцево-су-
динно! системи. Аргертальний тиск пщвищуеться. спостерггаються повнокргв’я
внутршштх органгв, пперемтя шюри г слизових оболонок, посилюеться тромбоут-
ворення, гнодг виникае ДВЗ-синдром, порушуеться мгкроциркуляшя.
Змши в кровг в разг набутого абсолютного еритроцитозу в умовах гшоксп
часто мають компенсаторний характер, сприяють полшшенню постачання тка-
нин киснем. 3 припиненням втиву етюлопчного фактора ктлъкгсть еритроципв
г гемоглобгну нормалгзуеться. Однак еритроцитоз, який виникае внаслщок пух-
линно! пролтферацй клггин еритропитарного ряду, не мае компенсаторного зна-
чения.
АнемЕя
Анемгя (вщ грец ярягд/я г?) — гематологгчний синдром або самостгйна хвороба, для
яко г характерно зменшення вмгсту гемоглобгну в единиц! об’ему кровг нижче вгд
норми, часто зг зменшенням кглькостг еритроцитгв та гх якгсними змшами.
’Катехоламши, андрогени, кортикотропш, тиреошн! гармони, глюко кортикощи. що продукують
ендокринш залози, носилююгь упвизацно кисню, що сприяе розвитку ппоксн та утворенню ЕП
у нирках. Кр1м того, ц! гармони щдвищують чутпивють еритропоетинчутливих клпин йсткового моз-
ку до ЕП 1 таким чином активують вплив на еритропоез.
Класифшацпо анемп наведено в табл. 19.
Таблица 19. Класиф1кац1я анемГГ
Локазник, покладений в основу класиф1каци Вид анемп
Етюлопя Спадкова Набута (гмунна, радгацшна, алгментарна, медикаментозна, ш- фекцшна та гн.)
Патогенез Анемгя внаслщок крововтрати (постгеморапчна) Анемгя внаслщок пщвищеного кроворуйнування (гемолгтична) Анемгя внаслщок порушення еритропоезу
Тип кроветворения Анемгя з еритробластним типом еритропоезу (еритробластна) Анемгя з мегалобластним типом еритропоезу (мегалобластна)
Здатшеть юсткового мозку до регенерацп Регенераторна Гшеррегенераторна Гшо-, арегенераторна
Колгрний показник (КП) Нормохромна (КП — 0,85—1,15) Гшохромна (КП < 0,8) Гшерхромна (КП > 1,15)
Дгаметр еритроципв Нормоцитарна (середнш д!аметр еритроципв — 7,2 мкм) Мгкроцитарна (середнш д!аметр < 6,5 мкм) Макроцитарна (середнш д!аметр > 8 мкм)
Клшгчиий переби Гостра Хронгчна
Кр!м того, видгляють анемгю справжню, зумовлену порушенням утворення
або збёлыпеною втратою та руйнуванням еритроципв, та несправжню, гемодилю-
щйну, при якш зменшення юлькост! еритроцитгв 1 гемоглобшу пов’язане 31 збгль-
шенням ОЦК (ггперволемгя олггоцитемгчна) 1 суггутшм зменшенням концентрацп
еритроципв та вщповщно гемоглобшу г гематокритного числа. Це може спостер!-
гатися при серцевш недостатносп, р!зкому зменшенш набрякгв унаслщок штен-
сивного лпсування вщповщно! патолоп!.
Ет1олог1Я анемп розглядаеться у вщповщних пщроздглах.
Патогенез анемп окргм власне патолопчних змш еритрону та порушення
функцш органгв г систем охоплюе також захисно-компенсаторш реакцп оргашзму.
До патологичных змш насамперед належать порушення еритропоезу, еритродгерезу
(руйнування еритроципв), кглькгсного та якгсного складу еритроципв 1 гемоглоби-
ну, якг зумовлюють розвиток ппоксичного синдрому (гемгчного типу), пов’язаного
31 зменшенням основно! функцп еритроципв — транспорту кисню.
Компенсаторы реакцп при анемп спрямоваш на усунення гшоксп. Вони про-
являються посиленням еритропоезу (лейкоеритрощне вщношення в юстковому
мозку змгнюеться з 3 : 1 до 1 : 11 навпъ 1 : 3, у кров! збглыцуеться кщькгсть клгтин
фгзголопчно! регенерацп), змгщенням криво! дисощацп оксигемоглобгну, змгною
функцгй оргашв кровообггу г дихання, тканинного метаболгзму.
Постгеморапчна анемгя
ПостгеморогЫна анемгя — анемгя, що розвиваеться внаелгдок гострсг або хрошчног
втрати крови
КласифЁкацЁя. За клёнёчним перебЁгом розрЁзняють гостру Ё хрошчну пост-
геморапчну анемЁю.
Етёологёя. Гоетра постгемораггчна анемгя виникае пёсля швидкоё масивно"
втрати кровЁ внаслЁдок поранения судин збо руйнування стёнки судин патологёч-
ним процесом.
Хронгчна постгемораггчна анемгя розвиваеться внаслЁдок повторних крововтрат,
зумовлених ураженням кровоносних судин при певних хворобах (виразкова хво-
роба шлунка. гемсрой тошо) або порушенням тромбоцитарно-судинного ё коагу-
ляцёйного гемостазу (геморагЁчний синдром).
Патогенез. Гоа пра постгемораггчна анемгя в першЁ годи ни пёсля втрати кровЁ
за достатнём проявом компенсаторних реакшй (спазм судин, вихёд кровЁ з депо)
харакгеризуеться нормальною кшькёстю еритроцитЁв ё гемоглобЁну в одиницё
об’ему кровЁ на тлё нормоцитемЁчно) гЁповолемп та зменшення абсолютно! кёль-
кост) еритроцитЁв. У разг дуже сильно) крововтрати вже в 1—2-у добу може розви-
нутися нормохромна анемЁя. Якёсних змён еритроцитЁв у мазку кровЁ немае.
Через 2—3 днё пёсля спинення кровотечЁ кётькёсть еритроцитЁв ё гемоглобЁну
в одиницё об’ему кров1 дещо зменшуеться за рахунок надходження тканинно) рЁ-
дини в судини (гемодил юцЁйна анем1я); КП залишаеться нормальним.
На 4- 5-й день пёсля крововтрати посилюеться прол1ферадЁя клётин еригро-
иитарного паростка кёсткового мозку пёд впливом еритропоетину, продукта яко-
го збЁльшуеться внаслЁдок гёпоксёё. У кров] збЁльшуеться кёлькёсть полЁхромато-
фшьних еритроцитЁв, ретикулоцит!в, з’являються поодинокё нормобласти (реге-
нераторна анем1я). КП знижуеться (гЁпохромна анемия), оскёльки
прискорена регенерацЁя випереджае дозрЁвання клетин, якё не встигають втратити
ознаки незрЁлостЁ (ядро, зернистЁсть) ё насититися гемоглобЁном.
Хронгчна постгемораггчна анемгя супроводжуеться зменшенням запасЁв за лё за в
органЁзмЁ в разЁ повторних крововтрат ё, отже, виникненням залЁзодефЁцитно)
анемн з низьким вмёстом гемоглобЁну, зниженим КП I появою в мазку кровЁ гё-
похромних еритроцитЁв, мЁкропитЁв. У разЁ приппчення кровотворення така анемЁя
е ппорегенераторною (рЁзке зменшення регенерат ивних форм еритроцитЁв).
Гемолгтичнэ анемгя
Гемолгтична анемгя — анемгя, що виникае внаелгдок пгдвищеного еритродгерезу, коли
руйнування еритроципйв (гемолгз) переважае над гхнгм утворенням.
КласифЁкашя. За епйологгею видёляють гемол'гтичну анемйо набуту г спад-
кову
Набуту гемолгтичну анемгю залежно вёд етюлогЁчних факторЁв, шо зумовили
гемолЁз еритроцитЁв, подЁляють на.
• пюксичну, за яко) гемолЁз еритроцитЁв зумовлений дЁею ЁнфекцЁйних фак-
торЁв, хёмёчних речовин рЁзного походження;
• гмунну (ёзо-, гетеро-, автоЁмунну), коли гемолЁз виникае пщ впливом комп-
лексу антиген — антиеритропитарне антитъто,
• механгчну — у разг мехашчного ушкодження еритроципв;
• набуту мембранопатгю, пов’язану гз соматичною мутацгею, при якш у юст-
ковому мозку утворюеться клон еритроципв гз дефектом структури мембрани.
Спадкову гемолйпичну анемт, залежно вгд характеру генетичного дефекту
структури мембрани, синтезу ферментгв еритроцитгв та гемоглобшу, що зумовлюе
посилення гемолгзу, подшяють на:
• мембранопатй',
• ферментопатгг,
• гемоглобшопати, яю гснують у двох ргзновидах:
— анемп, зумовлеш генетичним дефектом первинно! структури ланцюггв глобшу;
— анемп, зумовлеш порушенням синтезу а- або $-ланцюггв глобшу.
Залежно вщ того, чим спричинений гемолгз, розргзняють таю гемолггичш
анемп:
• ендоеритроцитарну, пов’язану з внутргшшми аномалгями еритроципв (до
не! належить спадкова гемолгтична анемгя);
• екзоеритроцитарну, що виникае пщ дгею екзогенних факторгв, розташова-
них поза еритроцитом (набута гемолгтична анемгя, див. вище).
За мгсцем, де вгдбуваеться гемолгз, гемолгтичну анемгю подшяють на:
• гемолгтичну анемт з внутргшнъосудинним гемолизом (токсична, гмунна, ме-
ханична гемолгтична анемгя);
• гемолгтичну анемт з внутргшньоклгтинним гемолгзом, перебгг якого вщбу-
ваеться в макрофагах шляхом фагоцитозу (частппе це спадкова гемолгтична анемгя
з аномалгею еритроцитгв, так звана ендоеритроцитарна гемолгтична анемгя).
За перебггом гемолгтична анемгя може бути гострою та хрошчного (переважно
спадкова гемолгтична анемгя).
Етгологгя набутог гемолгтично! анемп. Токсична набута гемолгтична анемгя
може розвинутися пщ дгею:
• гемолгтичних отрут (сполуки миш’яку, свинцю, штробензол, феншгщразин,
алкоголь, жовчш кислоти, токсичш продукти азотистого обмгну, змпна, бджоли-
на, грибна отрути);
• збудниюв шфекцшних г паразитичних хвороб (гемолгтичний стрептокок,
анаеробш мгкрооргашзми, маляршний плазмодш, лейшмангя, бартонела).
1мунна (гетеро-, 1зо-, автогмунна) гемолгтична анемгя виникае при:
• переливагшг видо-, групо- г резус-несумгсног кровг;
• резус-несумгсностг матерг та плода;
• утвореннг автоантитгл до власних еритроцитгв у випадку змгни гх антиген-
них властивостей (внаелгдок дгг лгкарських засобгв, вгрусгв, радгацп);
• виникненш мутантного клона лгмфоцитгв, яю утворюють антитгла до нор-
мальних еритроцитгв (при лейкозг, системному червоному вовчаку тощо).
Мехашчна гемолгтична анемгя спостерггаеться в разг:
• мехашчного ушкодження еритроцитгв протезами кровоноеггих судин г кла-
пангв серця;
• тривалого маргпу або бггу по твердому Грунту (маршева гемоглобшургя)’,
• егшеномегалп, коли еритроцити проходять кргзь звужеш синусогднг капгля -
ри збгльшено! селезгнки.
Гемолгтична анемгя може виникати пщ дгею шших фгзичних факторгв, напри-
клад, юшзуючого випромгнювання, ультразвуку.
Набута мембранопатгя спричинюеться мутащею юптини — попередника мге-
лопоезу пщ впливом втруств, лжарських засобтв, гнших мутагенних фактортв з
утворенням патолопчного клона еритроципв, гранулоцитов г тромбоцитов.
При пароксизмалънгй нгчниг гемоглобгнури (хвороба Марклафави — Мгкелг) унас-
лщок мутацп в аномальних еритроцитах порушуеться структура мембрани, при-
пиняеться синтез глгканшозитолфосфатиду, через що з мембранами не може
зв’язатися сироватковий шпбггор комплементу (бглок ПАЕ). Це спричинюе ге-
молтз таких еритроципв у шчш години, коли зниження рН 1 пперкапшя викли-
кають активащю комплементу.
Патогенез. Механизм гемолгзу при набутш гемолгтичнгй анеми полягае в
ушкодженш структури мембрани еритроцитгв. Одш гемолттичш фактори справля-
ють пряму ушкоджувальну дпо на мембрани еритроцитгв (механгчний гемолиз при
маршевой гемоглобшури), шпп (арсин — гщрид миш’яку, ютрити) як силып окис-
ники зумовлюють спочатку метаболочш, а потом структурно змши в мембранг та
стромт еритроцитгв (окисний гемолгз). Багато гемолгтичних отрут бюлопчного по-
ходження ферментативно активнг, руйнують лецитин мембран (лецитиназна ак-
тивнгсть стрепто-, стафглолгзингв, отрути комах г змш). У разг омунноо гемолгтичног
анеми 1§О г 1§М приеднують до мембрани еритроцитгв комплемент, який при
цьому активуеться г зумовлюе лгзис мембран (гмунний комплементзалежний ге-
МОЛгз).
У випадку холодовог омунноо гемолгтичног анеми, яка спостерггаеться при
деяких шфекцгях, зумовлених вгрусом Епстайна — Барр, Мусор 1азта рпеитопгае,
а також у пгзнгх стадтях сифглгсу, у разг хронгчного лгмфолейкозу, гемолтз еритро-
цитгв вщбуваеться за температури кровг 4—18 °С (часттше в судинах кгнцгвок). Це
пов’язано зг здатшстю 1§М (холодов! антитгла) активувати комплемент за зниже-
ног температури.
Пщ дтею гемолгтичних факторгв у мембранг еритроцита утворюються пори,
кргзь якг з клгтини виходять гони калпо, фосфата, а гони натрпо надходять у клг-
тину. Унаслщок тонного дисбалансу вода проникае в еритроцит, який набухае,
набувае сферичнот форми. Його здатшсть змшювати конфпурацпо знижуеться.
Коли об’ем еритроцитгв сягае критичного (146 % початкового), а розмгр пор гх-
ньо’т мембрани перевищуе 6 нм, настае гемолгз гз виходом гемоглобгну в плазму.
Гемолтз еритроцитгв при набупй гемолгтичнш анемн вщбуваеться переважно
в кровоносному русл! (внутрпиньосудинний гемолгз). Проте в разг резус-конфлгкту
(гемолгтична хвороба новонароджених) антарезусш аглютинтни, що утворилися в
органтзмт жшки з резус-негативною кров'ю, зумовлюють гемолтз резус-позитив-
них еритроцитгв плода або новонародженого не тшьки всередиш судин, а й у
печшщ та селезтнщ (внутргшнъоклтинний гемолгз).
У разг спадково! гемолгтичног анеми гемолгз зумовлений зменшенням осмотич-
нот та мехашчнот резистентностт еритроципв гз генетично детермшованими пору-
шеннями структури мембрани, метаболтзму, синтезу гемоглобгну.
Так, при спадковш мембранопати (мгкросфероцитарна гемолйпична анемгя, або
хвороба Мгнковського — Шоффара, частгше з автосомно-домгнантним типом ус-
падкування) генетичний дефщит у мембранг еритроцитгв бтлктв спектрину й анкг-
рину спричинюе змши цитоскелета мембрани еритроцитгв, якт втрачають здат-
шсть пщтримувата двояковвггнуту форму. Одночасно гццвищуеться проникшсть
мембрани, у юптинах накопичуеться надлишок юшв натрпо та води. Еритроцита
перетворюються на сфероцити з ргзко зниженою осмотичною резистентшстю та
здатшстю деформуватися пщ час проходження кр!зь синусощш кашляри селезш-
ки. Вщрив частини оболонки в таких еритроципв призводить до утворення
мйсросфероципйв з! зменшеною тривалктю життя (8—14 дшв замгсть 90—120 дшв
у норм!) у зв’язку з передчасним захопленням !х макрофагами селезшки й печш-
ки (внутр!шньокл!тинний гемолхз).
У випадку спадкова! ферментопатн, наприклад у раз! дефщиту Г-6-ФДГ (не-
повне домшантне успадкування, зчеплене з Х-хромосомою), гострий гемол!з ви-
никае при споживашп бобових (У!с!а Гауа) або шсля вживания л!карських засобш
13 високою окисною здатшстю (протималяршш препарата, фтивазид тощо). Ге-
мол13 зумовлений ушкодженням юптинних мембран пероксидами, осюльки в
еритроцитах з дефщитом Г-6-ФДГ знижений вм!ст вщновленого глутатюну (ан-
тиоксиданту). Кр!м того, пщ д!ею пероксид !в виникае окисна денатурация гемог-
лобшу з утворенням пйлецъ Гайнца — Ер.йха.
За спадково! гемоглобйюпати внутрпнньоюптинний гемол!з пов’язаний !з син-
тезом аномального або невластивого певному в!ку гемоглобшу1.
У випадку серпопод!бно-клтинно! анеми утворюеться НЬ8 (у р-ланцюгу глоб!-
ну глутамшова кислота замшена на валш), який у вщновленому еташ випадае в
кристалл 1 спричинюе серпопод!бну деформацпо еритроципв (дрепаноциты)-, п-
покс!я сприяе посиленшо гемолгзу таких еритроципв. Серпоподгбно-юптинна
анем!я успадковуеться за неповним дом!нуванням. Тому гомозигота за НЬ8, в
еритроцитах яких мктаться пльки НЬ8, хворпоть на тяжку форму анемп; гетеро-
зиготи (!хш еритроцити м!стять 22—45 % НЬ8 1 55—78 % НЬА,) — носп серпо-
подобно! аномалп еритроципйв, гемолгз яких вщбуваеться в умовах гшоксп (пщ час
тяжко! пневмонп, польоту в негерметизованих лиаках, у стан! наркозу) 1 тод! вже
виникае серпопод!бно-кл!тинна анем!я.
При а-таласеми (генетичний дефект синтезу а-ланцюпв глобшу) вщбуваеть-
ся гемол!з еритроципв з аномальним гемоглобшом — Ваг!-НЬ (у4) у новонародже-
них 1 НЬН (Р4) у дорослих. У раз! $-таласеми (неповне домшантне успадкування),
коли порушений синтез р-ланцюпв ! не утворюеться НЬА] (а2Р2), гемолгзуються
еритроцити, яю мктять шдвищену юльюсть НЬР (а2у2) або НЬА/а^). У гетеро-
зигот ступшь гемолизу незначний, виникае помгрна мпсроцитарна анемгя (мала
$-таласем!я), тод! як у гомозигот дуже значний гемол!з спричинюе розвиток
тяжко! анемп (велика ^-таласемоя, або анем!я Кул!).
Насл!дком масивного гемолизу еритроципйв е:
• гемолггична анем!я з порушенням дихально! функцп кров! та розвитком
гемгчно! ппоксп;
• гемоглобшемы — циркулящя в кров! гемоглобшу, що вивыьнився внасл!-
док розпаду еритроципв 1 з’еднуеться з гаптоглобшом у великомолекулярний
комплекс, що не проходить кр!зь нирковий фшьтр;
• гемоглобшур!я — видщення !з сечею гемоглобину, не зв’язаного з гапто-
глобшом, що спостерйаеться в раз! його низького вихщного р!вня або тод!, коли
вмкт выьного гемоглобшу в плазм! кр<5в! перевишуе 2482 нмоль/л (0,04 г/л);
’ У норм! еритроцити плода мктять переважно фетальний гемоглобш НЬР (а2у2) I невелику к!ль-
юсть гемоглобшу дорослих НЬА1 (а2р2), синтез яких починаеться теля 8-го тижня ембрюналъного
перюду; еритроцити новонародженого — 70—90 % НЬР 1 10—30 % НЬА1. На кшець першого року
життя 1 в доросло! людини еритроцити мктять 96—98 % НЬА1, 2—3 % НЬА2 (а252) та 1—2 % НЬР.
• гемолгтична жовтяниця як наслщок пщвишеного утворення з гемоглобшу
жовчних шгменпв;
• холелгтгаз унаслщок утворення пггментних каменгв;
• гемосидероз селезшки, нирок, печшки, юсткового мозку через реактивне
розростання сполучно! тканини в цих органах, де гемоглобш поглинаеться клгтина-
ми системи мононуклеарних фагоципв г розщеплюеться в них до гемосидерину;
• спленомегалгя — збыьшення розмгргв селезгнки пщ час хрошчного внутргш-
ньоклгтинного гемолизу;
• тромбоутворення — унаслщок внутргшньосудинного розпаду еритроципв гз
подальшим порушенням кровопостачання тканин г розвитком трофгчних виразок
кшцгвок, дистрофгчних змш у селезшцг, печшш, нирках;
• ДВЗ-синдром у разг надходження в судинне русло велико! юлькосп еритро-
цитарного тромбопластину;
• пщвищення температури пла пщ час масивного гемолгзу, що пов’язано з
посиленим утворенням 1Л-1 акгивованими макрофагами.
Картина кровь Набута гемолгтична анемгя за типом кровотворення е ерит-
робластною, за ступенем регенерацп юсткового мозку — регенераторною, за КП —
нормо- або ппохромною, рщше — псевдопперхромною (унаслщок абсорбцп ге-
моглобину на еритроцитах). Ступшь зменшення юлькосп еритроцитгв г гемоглобг-
ну залежить вщ штенсивносп гемолгзу. У мазку кровг виявляються клггини
фгзгологгчно! регенерацп. особливо характерним е високий ретикул оцитоз,
та дегенеративно змшенг еритроцити (пойкглоцитоз, розгрванг, фрагментованг
еритроцити — шистоцити, ангзоцитоз). Велика кглькгсть еритробластгв г
нормобласпв можлива при гемолгтичнш хворобг новонароджених.
При спадковш гемолгтичнш анеми спостерггаеться посилена регенерацгя ерит-
роцитарного паростка, часто з неефективним еритропоезом, коли в юстковому
мозку руйнуються ядерш форми еритроципв. У мазку кровг одночасно з регене-
ративними формами (високий ретикулоцитоз, полгхроматофглгя, по-
одинокг нормобласти)виявляютьдегенеративнозмшенгклгтини(мгкросфе-
роцити при хворобг Мшковського — Шоффара, дрепаноциты — серпо-
подгбнг еритроцити — при НЬЗ-гемоглобгнопатп, мгшенеподгбнг ерит-
роцити, а також еритроцити з базофгльною зернистгстю — при
таласемп). У зв’язку з частими гемолгтичними кризами можуть виникнути ггпо-
та арегенераторна анемп.
Анемгя внаслщок порушення еритропоезу
Класифгкацгя. За етюлоггею анемгя внаслщок порушення еритропоезу може
бути набутого та спадковою.
За механгзмом порушення еритропоезу розргзняють такг види цгег анемп:
• дисрегуляторну, зумовлену порушенням регуляцп еритропоезу внаслщок
зниження синтезу еритропоетину або пщвищення синтезу його гнггбгторгв (при
хронгчних хворобах нирок, гшофункцц ггпофгза, щитоподгбно! залози);
• дефщитну внаслщок дефщиту речовин, потргбних для утворення еритроци-
тгв (залгзо-, В12-, фолгево-, бглководефщитна анемгя);
• ферментопатт, зумовлену порушенням активностг ферментгв, якг беруть
участь в еритропоезг (наприклад, у синтезг порфгрину г гема);
• г1по(а)пластичну в разг ураження еритроцитарного паростка юсткового моз-
ку на тлг ргзкого пригшчення юстковомозкового кроветворения;
• метапластичну (мгелофтизичну) шсля зампцення або виттснення еритроци-
тарного паростка шшою тканиною;
• дисеритропоетичну, коли порушений подш ядер еритрокарюцитгв юстково-
го мозку та збтльшуеться внутрштньоюстковомозкове руйнування клттин еритро-
цитарного паростка.
За типом еритропоезу цт анеми можуть бути еритробластними або мегалоблас-
тними, за клийчним перебегом, як правило, хронгчними.
Залгзодефщитна анем1я (ЗДА) — анемгя, зумовлена нестачею залгза (див. роздгл
XXIII “Патологична фгзюлоггя обмшу мйероелеменпйв”) в оргашзмг внаелгдок пору-
шення балансу м1ж його надходженням, використанням I втратою.
Це найпоширенштий вид анеми (80 % захворюваностг на анемтю; у ртзних
кратнах страждае на ЗДА вщ 15 до 50 % населения, особливо жшки та дгти).
Етгологгя. Причинами розвитку ЗДА е:
• повторит кровотечт, якт спричинюють втрату залгза разом з еритроцитами
(переважно це матков! кровотечт, рщше — шлунково-кишковт, нирковт, легеневт,
кровотечт при геморапчному синдром!); це найчасттша причина ЗДА;
• недостатке надходження залтза з тжею (тз м’ясними продуктами);
• пщвищент витрати залгза в перюд росту, дозртвання оргашзму, пщ час вапт-
ностт, лактаци;
• зниження всмоктування залгза при хворобах травного каналу (ппоацидний
гастрит, хрошчний ентерит) або шсля резекци його вщцшгв;
• порушення транспорту залгза (гшотрансферинемтя через ураження печшки,
спадкова атрансферинемтя);
• недостатки утилгзацтя залгза з резервтв (у разт шфекцп, штоксикацп, глистя-
но! швазн) та його депонування (при гепатит!, цироз! печгнки).
Патогенез. Основною ланкою в патогенез! ЗДА е дефщит зал1за екзогенного
та ендогенного походження, що зумовлюе порушення утворення гема гемоглобг-
ну, до складу якого входить залтзо. Це своею чертою спричинюе зменшення юль-
костт гемоглобгну та зниження киснетранспортно! функцп кровт з наступним роз-
витком гемгчно! гшоксп.
В оргашзм! вщбуваються компенсаторы реакци:
• збшьшення мобштзацп залгза з депо у вигляд! феритину та гемосидерину;
• посилення абсорбцп залгза в травному канал!;
• гшерплазтя еритроцитарного паростка;
• збшьшення штенсивност! глтколтзу та вмтсту 2,3-дифосфоглгцерату в ерит-
роцитах, що полегшуе дисощацпо оксигемоглобшу т вщцачу кисню тканинам.
Коли компенсаторш реакцп, спрямоваш на лщвщащю дефщиту залгза, вияв-
ляються недостатшми, в органтзм! з розвитком ЗДА наростають патологгчш змши.
До них належать:
• зменшення резервтв залгза та цоступове гх вичерпування, що проявляеться
зникненням гемосидерину в макрофагоцитах печшки та селезшки, зменшенням
феритину в печшщ та м’язах, зниженн! до 2—5 % у юстковому мозку кшькост!
сидеробласттв (еритрокарюцитгв, якт мтстять зашзо у вигляд! гранул феритину !
юльюсть яких у норм! становить 20—40 %);
• зменшення в сировапц кров! концентраци залоза (ггпосидеремгя, що стано-
вить 1,8—5,4 мкмоль/л зам!сть 12,5—30,4 мкмоль/л у норм!), зниження р!вня фе-
ритину (нижче вш 12 мкг/л за норми 12—200 мкг/л) та коефщоента насичення
трансферину залозом (менше юж 20 % за норми 30—50 %), що призводить до
зменшення транспорту залоза в костковий мозок;
• порушення надходження залоза в еритроцити о зниження синтезу гема в
гемоглобин! та деяких залозовмосних о залозозалежних ферменпв (каталази, глу-
тапонпероксидази) в еритроцитах, що шдвищуе !'хню чутливость до гемолозуючоо
ди оксидантов;
• неефективний еритропоез унаслщок посилення гемолозу еритрокарооципв у
костковому мозку й еритроцитов у крово, що зумовлюе зменшення тривалосто жит-
тя еритроципв (до 20—30 дшв у разо тяжких форм ЗДА);
• морфолопчн! змони в еритроцитах косткового мозку: ггпохромгя, пов’язана з
недостатньою гемоглобонозащею еритроцитов, переважанням базофгльних нормо-
бластов над ацидофыьними в костковому мозку, мгкроцитоз о деструкцгя ядерних
клгтин у вигляд! карюрексису, вакуолозацп цитоплазми еритробл астов о нормо-
бласпв тощо;
• розвиток у разо дефщиту залоза в органозмо гемгчно'1 (унаслщок зменшення
кисневоо емносто крово) о тканинно'г (через зменшення колькосто мооглобону й ак-
тивносто залозовмосних ферментов тканоотного дихання) гтоксГг,
• атрофочн! та дастрофочш змши в тканинах о органах як наслщок гшоксп,
особливо в травному канало (глосит, гшговот, кароес зубов, ушкодження слизово!
оболонки стравоходу, порушення ковтання, атрофочний гастрит з ахолоею) о серп!
(дистрофоя моокарда).
Картина крово. ЗДА — анемоя з еритробластним типом кровотворення, п-
похромна (КП становить 0,6 о менше, до 0,4). Колькость гемоглобону знижена
большою морою, н!ж колькость еритроцитов, яка може бути в межах норми в почат-
ковш стадп розвитку ЗДА. Для мазка кров! характерно гшохромоя, “тпп” еритро-
ципв, наявшсть анулоцитов, микроцитоз, пойколоцитоз. Колькость ретикулоцитов
залежить вод регенераторноо здатносп еритроцитарного паростка, але частппе це
гшорегенераторна анемоя.
Мегалобластна анемгя — анемгя, пов’язана з порушенням синтезу нуклегнових
кислот г замгною еритробластного типу кровотворення на мегалобластний.
Класифокацоя. За епиологгею мегалобластна анемгя може бути набутого о спад-
кового.
Залежно вщ того, який патогенетичний мехашзм лежить в основ! розвитку
анемп, видоляють три основно вида мегалобластноо анемп:
• В 12-дефщитну,
• фолгеводефщитну,
• анемйо внаслгдок поругиення синтезу ДНК у разо дефщиту або гальмування
активносто ферментов синтезу пуринових о шримщинових нуклеотидов.
Етоологоя. Загальнг причини виникнення набутих В12- о фолгеводефщитног ане-
мш (унаслщок тосного зв’язку метаболозму щанокобаламшу — вотамону В12 та фо-
лоево'о кислоти) описано в роздал XXIV “Патолопчна фозоологоя обмшу вотамшов”.
П атогенез. Через дефщит коферменту цганокобаламгну мепиглкобаламгну в раз!
В12-дефщитноо анемп не вщбуваеться перетворення фолоево'о кислоти на п кофер-
ментну форму ТГФК, без яко! неможливий синтез тимщин монофосфату, що вхо-
дить до складу ДНК. Те саме спостерггаеться г при дефщит! фол!ево! кислоти.
Порушення синтезу ДНК при В12- I фолгеводефщитнш анеми спричинюе:
• порушення подглу клггин, насамперед клгтин кроветворно! тканини г трав-
ного каналу, як! активно розмножуються;
• затримання дозр!вання еритрокарюцитгв у юстковому мозку;
• замшу еритробластного типу кроветворения на мегалобластний;
• зростання неефективност! еритропоезу та скорочення тривалост! життя
еритроципв, !х пщвищений гемол!з;
• розвиток анемп, за яко! клгтани патолопчно! (мегалобластно!) регенерацп з
ознаками дегенерацп з’являються не ильки в юстковому мозку, а й у кров!;
• порушення лейко- г тромбоцитопоезу;
• розвиток запальних г атроф!чних процесгв {гласит Гунтера з лакованим язи-
ком через вщеутшеть сосочюв, стоматит, езофагт, ахийчний гастрит, ентерит)
унаслщок атипових м!тоз!в г появи пгантських юптин ештелпо травного каналу,
що утруднюе первинне порушення секрецп та всмоктування внутргшнього факто-
ра 1, отже, посилюе дефгцит вггамппв (порочне коло).
Унасл1док дефщиту дезоксиаденозилкобалам1ну — !ншого коферменту щаноко-
баламшу, який бере участь в утворенш бурштиново! кислоти з метилмалоново!, в
оргашзм! вщбуваеться:
• накопичення метилмалоново! кислоти, яка е токсичною для нервових клгтин;
• порушення утворення мгелгну й ушкодження аксонгв через синтез у нерво-
вих волокнах жирових кислот зг змшеною структурою;
• дегенеративнг процеси у заднгх г бгчшгх стовпах спинного мозку (фунгку-
лярний мгелоз), ураження черепних г периферичних нервгв гз розвитком ргз-
номангтно! невролопчно! симптоматики.
Отже, для В12-дефщитно! анемп е характерною клиична тр1ада з ураженням
трьох систем — системи кровг, травно! г нервовог систем; у разг фолгеводефгцит-
но! анемп невролопчних порушень немае.
Картина кровг. В12- г фолгеводефгцитна анемп визначаються мегалобласт-
ним типом кроветворения, гшерхромгею, макроцитозом. Вмгст еритроцитгв 1 ге-
моглобшу в кровг може ргзко знижуватися, проте КП (ступгнь насиченостг гемо-
глобгном кожного окремого еритроцита) вищий 1,15 (1,4—1,8) за рахунок наяв-
носп у кров! великих за розмграми мегалобластгв г мегалоципв, насичених гемог-
лобшом.
У мазку кров! з’являються клггини патолопчно! регенерацп юсткового мозку
(див. кольор. вкл., мал. II) — мегалоцита (штенсивно забарвлегй клгтани дгамет-
ром 10—12 мкм 1 бгльше, без центрального проевгтлення, дещо овально! форми) г
поодиною мегалобласта (клгтани дгаметром 12—15 мкм з базофгльною, полгхро-
матофгльною або ацидофильною цитоплазмою г нгжносгтчастим, зазвичай ексцен-
трично розмпценим, ядром). Вважають, що мегалобласта г мегалоцита при В12- та
фолгеводефгцитнгй анемгях не гдентичнг ембргональним клгтанам еритроцитарно-
го ряду г ильки зовнг подгбн! до них. У'кровг виявляеться багато ознак дегенера-
тавних змш еритроцитгв: пойюлопитоз, ангзоцитоз !з макроцитозом, гшерхромгя,
мегалоцита з включениями у виглядг тглець Жоллг, гшгець Кебота, еритроцити з
базофгльною зерниспстю. Зменгггуеться кгльюсть юггтин фгзголопчно! регенерацп
(ретикулоципв, полгхроматофгльних клгтин) унаслщок переважання мегалобласт-
ного еритропоезу над еритробластним. Спостерггаеться лейко- 1 тромбоцитопешя
з атиповими, великими за розмграми юптинами кров! (наприклад пгантськг пол!-
сегментоядерш нейтрофшьш гранулоцита з 8—10 сегментами 1 дааметром 20—
30 мкм).
Анемгя, зумовлена порушенням активностг ферментгв, як1 беруть участь в
еритропоезг (сидеробластна анемгя). До ще! групи можна зарахувати багато вид!в
анеми, у тому числ! й тг спадковг мембрано-, ферменто- ! гемоглобшопатп, як!
входять у групу гемолггично! анеми', осюльки при цьому також порушуеться
еритропоез унаслщок генетичного дефекту синтезу тих чи шших ферменпв. Про-
те коли наслщком порушення еритропоезу е значно посилений гемолгз, який стае
основним патогенетичним механизмом у розвитку цих видгв анеми, тод! це дае
змогу зарахувати гх у групу як гемолггично! анеми, так г анеми, пов’язано! з пору-
шенням еритропоезу.
Зменшення активност! ферменпв, яю беруть участь в еритропоез!, зумовлюе
виникнення анемш, основою розвитку яких е порушення синтезу порфгрингв г гема.
Ц! анеми мають ргзнг назви, що вщображають гхнг характернг особливост!: сидеро-
ахрестична {анемгя внаслщок невикористання залгза для синтезу гема), сидеро-
бластна (анемгя, пщ час яко! збшьшення юлькосп залгза в кров! супроводжуеться
вщкладанням його у вигляд! ферифосфатних комплекс!в у мгтохондргях 1 цито-
плазм! бластних еритро'щних юптин юсткового мозку з утворенням сидероблас-
та), зал1зорефрактерна анемгя (стгйка до лгкування препаратами залгза).
Класиф1кац1я. За етюлоггею сидеробластна анем!я може бути спадковою г
набутою.
Ешолопя. Сидеробластна анем!я може виникнути в раз! порушення в будь-
яюй ланцг бгосинтезу гемоглобгну, що катал!зуеться тими чи гншими фермента-
ми, починаючи вщ взаемодн глщину з бурштиновою кислотою, через послщовне
утворення Д-дельта зам!сть 3-амгнолевулгново! кислоти (АЛК), порфобшшогену,
уро-, копро-, протопорфириногену, протопорфирину, гема г до формування мо-
лекули гемоглобгну. Причини тако! анеми видшяють:
• дефщит шридоксину (вгтамгну В6), з якого в оргашзм! утворюеться шри-
доксальфосфат (кофермент синтетази АЛК) г який спостерггаеться в раз! нестач!
шридоксину в гжг (пщ час штучного вигодовування немовлят), а також при лгку-
ванн! засобами, яю пщвищують споживання шридоксину (наприклад, деякими
протитуберкулъозними препаратами);
• побутове та виробниче отруення свинцем, що спричинюе порушення син-
тезу порфирингв (свинець блокуе сульфгщрильш групи у ферментах, яю беруть
участь в утворенш гема, 6-депдрази АЛК, декарбоксилази уропорф!риногену,
гемсинтетази);
• хромосомш мутацп в юстковому мозку, внаслщок яких виникае клон ано-
мальних юптин зг зниженою актившстю синтетази АЛК (рефрактерна анемгя з
юльцеподгбними сидеробластами — ргзновид мгелодиспластичного синдрому,
див. с. 440);
• генетачний дефект синтезу АЛК або протопорфгрину (успадкування реце-
сивне, зчеплене з Х-хромосомою).
Пато генез. Зменшення актавносп фермента, що беруть участь в утворенш
порфгрингв г гема, зумовлюе ослабления утилгзацп залгза й порушення синтезу
гемоглобгну, що призводить до:
• ппохромноТ анемп з низьким вмгстом гемоглобшу в еритроцитах за одно-
часного пщвищення вмгсту залгза в сироватш кров! (до 54—98,5 мкмоль/л);
• появи в юстковому мозку ознак подразнення еритроцитарного паростка,
збщьшення юлькосп базофшьних нормобластгв г сидеробластгв та одночасного
зменшення кглькостг гемоглобппзованих форм;
• неефективносп еритропоезу, що зумовлюе гемолгз г скорочення тривалостг
життя еритроцитгв;
• розростання сполучно! тканини в разг вщкладання залгза у внутргшнгх орга-
нах (гемосидероз печгнки, серця, пщшлунковог залози та шших органгв).
Картина кровг. Для сидеробластног анемп характерним е зменшення кгль-
костг гемоглобшу до 2,48 ммоль/л (40 г/л) та колгрного показника до 0,6—0,4
(ппохромна анемгя). У мазку кровг переважають ппохромнг еритроцити, сидеро-
цити, еритроцити з базофгльною зернистгстго, ангзоцитоз, гнодг спостерггаються
мгшенеподгбнг еритроцити; кглькгсть ретикулоципв нормальна або знижена, пщ
час свинцевого отруення — збгльшена.
Анемгя внаслщок ураження еритроцитарного паростка юсткового мозку
Апластична анемгя (АА) — анемгя, при якш уражуеться еритроцитарний паросток
кгсткового мозку на тл1 глибокого пригнгчення кроветворения, ргзко знижуеться про-
букцгя еритроцитгв, а також гранулоцитгв г тромбоцитгв (панцитопенгя).
АА, як г агранулоцитоз та амегакаргоцитна тромбоцитопенгя, належить до
синдрому недостатностг кгсткового мозку.
Класифгкацгя. За етюлоггею розргзняють спадкову г набуту АА.
Набуту АА своею чергою подгляють на токсичну, медикаментозну, радга-
цшну, гнфекцгйну, а також гдгопатичну в разг невщомог етгологг!.
За патогенезом розргзняють дисрегуляторну, мгелотоксичну, гмунну на-
буту АА.
Етгологгя. Серед екзогеннгзх етюлоггчних факторгв, що зумовлюють розви-
ток АА, видгляють такт:
• хгмгчнг: лгкарськг засоби (сульфангламщнг, цитостатачнг, антимгкобак-
тергальш та протисудомнг препарата, гндометацин, левомщетин), бензол г його
похщнг, шсектациди, пестициди;
• гонгзуюче випромшювання;
• шфекцгйнг: вгруси гмунодефщиту людини, гепатиту тапу Е, грипу, шфек-
цгйного мононуклеозу, парвовгрус В19, мгкобактергг туберкульозу тошо.
До ендогенних етюлоггчних факторгв належать:
• генетичнг дефекта еритропоезу;
• автоантитгла до клгтин еритропоетичног тканини;
• гшофункцгя ендокринних залоз (ггпофгза, загруднинног, щитоподгбно! за-
лоз, яечникгв);
• порушення синтезу еритропоетцну пщ час хвороби нирок г збщьшення
вмгсту шггбгторгв еритропоетину.
Серед спадкових АА частгше трапляеться анемгя Фанконг, яка належить до
синдрому хромосомно! нестабгльностг з генетичним дефектом у систем! репарацп
ДНК (не синтезуеться у-ендонуклеаза). Це призводить до накопичення мутацш,
Тб8
___<_
наслщком чого е апластичш змши в кгстковому мозку з панцитопешею (автосом-
но-рецесивне успадкування).
У патогенез! АА можна видтлити основнг мехашзми, яю зумовлюють пригш-
чення гемопоезу, у тому числг еритропоезу (можлива вибтркова депрестя титьки
еритропоетичнот тканини при так званщ парцгальнтй червоноклгтиншй
аплазп):
• зменшення юлькосп клггин — попередниюв еритроцитарного ряду шд пря-
мою ушкоджувальною дкю еполопчних фактори на стовбуровт клггини кгстково-
го мозку або !х мутацп, унаслщок чого ртзко зменшуеться мгтотична актившсть
кровотворних клгтин, а тнодт в них порушуеться синтез гемоглобшу (наприклад
пщвищення вмгсту фетального гемоглобшу при анемп Фанконт);
• тмунне ушкодження антиплами г Т-лтмфоцитами еритропоетинчутливих
клгтин або клгтин — попередниюв мтелопоезу в кгстковому мозку, а також еритро-
поетину г його рецепторгв;
• неповнощншсть стромального мткрооточення еритропоетичних клгтин, уна-
слщок чого порушуеться гх пролтферативна функцгя г здатшсть до диференщацп
(це доведено в експеримент! на мишах зт спадковою АА, у яких вщновлювався
подгл трансплантованих клгтин ктсткового мозку титьки шсля введения тканини
селезшки як кровотворнот строми).
Усг цг мехашзми пщ час апластичнот анемп призводять до:
• порушення утворення клгтин еритроцитарного, а також гранулоцитарного 1
тромбоцитарного ряду (якщо одночасно уражуеться загальна для трьох рядтв клг-
тина-попередник) 1 ргзкого збщнення ктсткового мозку на клгтинш елементи;
кроветворна тканина зампцуеться жировою;
• появи при тмуннш форм! апластичнот анемп в юстковому мозку активова-
них цитотоксичних Т-лгмфоципв СЭ8+, яю синтезують гемопоезблокуючг ци-
токши (1Ф, ФНП), що тндукують апоптоз кровотворних клгтин;
• неефективносп еритропоезу зт зменшенням тривалосп життя еритроципв;
• поеднання тнтрамедулярного руйнування еритрокарюциттв з посиленням
гемолгзу еритроципв у кровг, селезшш та печшщ;
• ускладнення АА кровоточивгстю внаслщок тромбоцитопенп (див. с. 428), а
також шфекцшними процесами, яю розвиваються на тлг гранулоцитопенп;
• збглыиення в кров! вмгсту залгза, щанокобаламшу, еритропоетину внаслт-
док нездатносп кровотворних клгтин засвоювати потргбш для тх пролгферацп
речовини.
Картина кровг. АА здебтльшого нормохромна, норме- або макроцитарна з
панцитопешею — ртзким зменшенням юлькосп еритроципв (до 0,6 Т/л), гемо-
глобшу (до 1,86—1,24 ммоль/л, або 30—20 г/л), лейкоципв, особливо грануло-
ципв (до 0,2 Г/л) г тромбоципв (менше 20 Г/л). Кзльюсть ретикулоципв у мазку
кровг ртзко зменшена, до повно'т тх вщсутносп в термшальний перюд (арегенера-
торна анемгя).
Метапластична (мгелофтизична) анемгя — анемгя, за яког порушення еритро-
поезу зумовлене випгисненням або зам 'гигенням еритропоетичног тканини пухлинни-
ми метастазами (рак молочнот залози, легень, шлунка, передмтхуровог залози
тощо), лейкозними клгтинами, волокнистою сполучною (мгелофтброз) г жировою
тканинами.
Назва “мгелофтизична анемгя” пов’язана з тим. шо пщ час вторинно! ме-
тапластично! анеми витгснясться не тшьки еритропитарний, а й ус! шш! паростки
кроветворения, виникае спустошсння юсткового мозку — панлйелофпшз (сухоти
юсткового мозку).
ПАТОЛОПЧШ ЗМ1НИ ЛЕЙКОЦИТ1В
Класифгкагия. Патолопчнг змши лейкоцитгв проявляюгъся у вигляд!:
• порушення лейкопоезу;
• киыасних / яклсних злин лейкоцитгв кров!.
За походженням ш змши можуть бути первинними внаслщок ураження клггин
лейкоцитарного ряду в кровогворнгй тканин! га кровоносному русл! пщ д!ею р!з-
них патогенних факторгв. Вториннг змши лейкоциттв виникають як вщповщна
реакшя на патолопчш процеси в органах г тканинах шших систем оргашзму.
Ет1олог1ю розглянуто в окремих пщроздыах пього роздшу.
Патогенез. Основною ланкою в патогенез! порушень у разг патологи лейко-
цитгв с змша реактивност! оргашзму, у тому числ! !мунолопчно! та алерпчно'!. Це
пов’язано з функцюнальними особливостями лейкоцитов: !х участю в реакщях
КЛ1ТИННОГ та гуморально! ланки гмунггету, пропесах фагоцитозу, синтез!, депону-
ванш й шактивацп БАР (штерлейюни, г!стам!н, серотонш гошо). Патолопчнг
змши лейкоципв можуть супроводжуватися зниженням захисту оргашзму вщ мгк-
роорган!зм!в (бактерш. в1рус!в, гриб!в. найпростгших), гельмшпв г таким чином
негативно впливати на переб!г хвороб.
Порушення лейкопоезу
Класиф1кап!я. Видшяють гаю порушення лейкопоезу:
• посилення або пригшчення утворення лейкоцитгв у гемопоетичнш тка-
ниш;
• порушення дозр!вання лейкоцитгв у кровотворних органах;
• продукцгя патолопчно змшених лейкоцитгв.
Певш види порушень лейкопоезу часто поеднутоться.
Посилення лейкопоезу своею чертою може бути реактивного та пухлинного
походження. За ступеней поширення розргзняють парщалъне порушення лейкопоезу,
коли спостерггаеться ураження певних паросткгв (мгелощного, лгмфоцитарного)
лейкопоетично! тканини, г тоталъне.
Етголошя. Порушення лейкопоезу виникае п!д вптивом низки екзогенних
факторгв'. бюлопчних (бактери, в!руси, найпроепгш), фгзичних (юшзуюче, ультра-
фголетове випромшювання). хгмгчних, алгментарних. До ендогенних факторгв пору-
шення лейкопоезу належать генетичш дефекта утворення та диференщювання лей-
коцитов, д!я токсичних метаболтв (при уремп), антатгл. метастаз!в пухлин тощо
Патогенез. Посилення лейкопоезу реактивного характеру може вини кати
внаслщок пщвищення вироблення гуморальних стимуляторов лейкопоезу (ко-
лонгестимулююч! фактори’, 1Л-1, 1Л-3 та ш.) або зменшення продуют гх гнггб!-
1 КСФ — шггокш. який стимулюе прол1ферашю га дозр1вания клггин м!елопоезу. За х1М1чиою
природою — це глгкопроте'гд (з ргзною молекулярною масою. що утворюеться моноцитами, макрофа-
гами, Т-лимфоцитами, ендотелюцитами, фибробластами.
торге (кейлони, ПГЕ, г ПГЕ2, лактоферин, гзоферитин тощо). При цьому в юст-
ковому мозку виявляеться пролгферащя клгтин — попередникгв млело- та лгмфо-
поезу з прискоренням гх диференщювання в зрш лейкоцити. Збгльшуеться
надходження лейкоцитгв у кров, тобто виникае лейкоцитоз.
Вгд етголопчного фактора залежить, якг клгтини лейкоцитарного ряду зазна-
ють пперплазп. Так, бактергальнг ендотоксини (стрептококовий, стафглококо-
вий), деякг продукти розпаду тканин (наприклад пщ час гемолгзу, ппеми) зумо-
влюють переважне збгльшення секрецп ростового фактора, що стимулюе пролг-
ферацгю та диференщювання попередникгв нейтрофгльних гранулоцитгв. Це
своею чертою викликае пщвищення ргвня нейтрофгльних гранулоцитгв у кровг
(нейтрофгльний лейкоцитоз). Посилення еозинофглопоезу г прискорення виходу
еозинофгльних гранулоцитгв гз кгсткового мозку у кров, що спостерггаеться при
алерпчних хворобах, пов’язане зг збгльшенням синтезу КСФ у лгмфоцитах пгсля
стимуляцгг гх антигеном, а також гз пгдвищенням проникностг кгстковомозкового
бар’ера пщ дгею ггстамгну та гнших БАР, якг вивгльнюються пщ час реакцп анти-
ген — антитшо.
Посилення лейкопоезу пухлинноо природи вщбуваеться внаелгдок дп онкогенних
факторгв, якг спричинюють трансформацпо нормальних гемопоетичних клгтин у
лейкознг, що мають шдвищену пролгфератавну здатшеть.
Пригшчення лейкопоезу може зумовлюватися порушенням регуляцп утворення
лейкоцитгв (у разг зменшення продукцп КСФ або збгльшення продукцп гнггбг-
торгв лейкопоезу), дефгцитом пластичних факторгв, потргбних для лейкопоезу
(при бшковому голодуваннг, нестачг цганокобаламгну г фолгевог кислоти). Лейко-
поез знижуеться внаелгдок спадкового або набутого ураження клгтин — попере-
дникгв грануло- та агранулоцитгв, а також клгтин строми, що визначають у норм!
диференцповання стовбурових клгтин у напрямку мгело- та лгмфоцитопоезу, або
при генералгзованому ураженнг лейкопоетичног тканини. Таке зменшення лейко-
поезу спостерггаеться у випадку спадковог нейтропенп, пщ впливом гонгзуючого
випромшювання, цитостатичних препаратгв, при метастазах пухлин г лейкозних
гнфгльтратах, що витгеняють нормалью продуцента лейкоцитгв, а також пгдвище-
ному руйнуваннг клгтин лейкоцитарного ряду в кровотворних органах автоанти-
тглами, токсичними речовинами, вгрусами.
Пригнгчення лейкопоезу, як г посилення його, охоплюе або вег види лейко-
цитгв, або переважно один гз них.
Порушення дозргвання лейкоцитов зумовлюеться блокадою диференщювання
на тому чи гншому ргвнг розвитку клгтин. Цей процес регулюеться генетично г
забезпечуеться певними метаболгчними реакцгями. До його змгни призводять му-
тацй (при лейкозг, спадковому дефектг дозргвання лейкоцитгв), дгя екзогенних та
ендогенних факторгв (збудники гнгйних г вгрусних шфекцгй, лгкарськг засоби,
гнтоксикацгя). Порушення дозргвання лейкоцитгв часто трапляеться внаелгдок
пухлинног пперплазп гемопоетичног тканини як у разг збгльшення продукцп лей-
козних лейкоцитгв, так г пригнгчення лейкопоезу. Кргм того, прискорений вихщ
незрглих лейкоцитгв гз кгсткового мозку в кров пов’язують зг змгною проникностг
кгстковомозкового бар’ера гад впливом КСФ, глюкокортикощгв, 1Л-1,1Л-8, ФНП,
бактергальних ендотоксингв.
Поява незрглих клгтин у кровг спостерггаеться гад час лейкемощних реакцш г
ядерного зеуву нейтрофгльних гранулоцитгв улгво.
Продукция ооатологочооо змонеошх лейкоцитов у юстковому мозку може виникну-
ти внаслщок пухлинно! трансформацп при лейкоз!, генетично зумовлених пору-
шеннях структури й обм!ну речовин у лейкоцитах. Наприклад, у раз! домшантно
успадковано! пельгер1вськоо аномала лейкоцитов утворюються нейтрофшып грану-
лоцита, яю 1 шсля дозрхвання, надгйшовши у кров, зберйають кругле або палич-
коядерне, або двосегментне ядро (кольор. вкл., мал. III).
Спадковий дефщит м!елопероксидази, глюкозо-6-фосфатдегщрогенази приз-
водить до зниження фагоцитарно! активност! лейкоципв (синдром Чед!ака — XI-
гас!). Може спостерйатися неефектившсть лейкопоезу з! скороченням тривалосп
життя лейкоципв. Патолопчш клони лгмфоципв можуть продукувати антипла до
тканин власного оргашзму, що зумовлюе розвиток автогмунних хвороб.
Кшьюсн! зм1ни лейкоципв у кров!
Класиф!кац1я. Змши юлькосп лейкоципв у кров! полягають у збшыпенш (лей-
коцитоз) або зменшенн! (лейкопеноя) пор!вняно з нормою (4—9 Г/л, 9,0«109/л; за
М!жнародною системою ф!зичних одиниць (С1) Г — пга = 109).
Лейкоцитоз
Лейкоцитоз — зболъоиення загалъноо киькост! лейкоцитов у крово понад 9 Г/л
(9,0 • 1(Г/л).
Класиф!кац1я. За механизмом виникнення розр!зняють лейкоцитоз абсолют-
ний 1 водносний.
Абсолютний лейкоцитоз (справжнш) — пщвищення абсолютно! кшькосп лей-
коципв у кров! внаслщок посиленого лейкопоезу реактивного чи пухлинного ха-
рактеру або збшыпеного надходження лейкоципв !з юстковомозкового депо в
кровоносш судини. За патогенезом абсолютний лейкоцитоз може бути ре-
активним 1 пухлинного походження (неопластичним).
В1дносний лейкоцитоз (несправжнш) — збшьшення юлькосп лейкоципв у
кров! внаслщок переходу лейкоципв !з приспнкового пулу в циркулюючий (л е -
рерозподолъний лейкоцитоз) або в раз! зменшення об’ему плазми та згущен-
ня кров! (гемоконцентрац1йний лейкоцитоз).
У зв’язку з там що зростання загально! юлькосп лейкоципв зазвичай поед-
нуеться з переважним збшьшенням абсолютно! юлькосп окремих видгв лейко-
ципв, лейкоцитоз подшяють на нейтрофЫьний, еозиноф1льний, базофтьний,
лгмфоцитарний 1 моноцитарний. Часто спостерйаеться поедания окремих форм
лейкоцитозу, наприклад, нейтрофшьно-еозинофшьний, л!мфоцитарно-моноци-
тарний лейкоцитоз.
Ет1олопя. Нейтрофыьний лейкоцитоз (нейтрофиъоз) зумовлюють шфекцш-
ш агента (стрептококи, стафшококи, гриби), продукти розпаду тканин (у випадку
шфаркту мюкарда, гострого гемол!зу, злояюсних пухлин), токсичш метаболии
(при уремп, печшковш ком!), фззичш (холод, тепло) 1 психзчш (страх, лють) фак-
тори; високий нейтрофшьоз е характерним для хрошчного м!елолейкозу.
Еозинофйоьний лейкоцитоз (еозинофшя) спричинюють !мунна патолопя (брон-
х!альна астма, автогмунш хвороби), паразитичш гнфекцп (гельмгнти, амеби, лям-
бли). В!н також спостерйаеться пщ час хвороби Леффлера (еозинофшьш шфшь-
трати в легенях, серцг, шших органах), пухлин (хронгчний мгелолейкоз), у перюд
видужання вщ бактергальних шфекцш (скарлатина).
Базофгльный лейкоцитоз (базофшя) спостерггаеться в разг анафшаксп, авто-
гмунног патологп, колагенозгв, хрошчного мгелолейкозу, вгрусних шфекцш,
хрошчного колгту, ппотиреозу.
Лгмфоцитарний лейкоцитоз (тмфоцитоз) спричинюють деяю вгруси (шфек-
цшного мононуклеозу, гепатиту, кору), бактергалью (збудники кашлюку, тубер-
кульозу, сифглгсу) та паразитичнг шфекцп, гмунопатолопя (автогмунш ендокри-
нопатп, сироваткова хвороба), гриби; надвисокий лгмфоцитоз спостерггаеться щд
час хрошчного лгмфолейкозу та лгмфом.
Моноцитарный лейкоцитоз (моноцитоз) розвиваеться пщ дгею на оргашзм
вгрусгв (кору, краснухи, шфекцгйного мононуклеозу), збудниюв туберкульозу, си-
фглгсу, лепри, протозойних шфекцш (малярп, лейпгмашозу), у випадку дифузних
хвороб сполучно! тканини, автогмунног патологп та гемобластозу.
Патогенез. Видгляють таю механгзми виникнення лейкоцитозу:
• пщвищення продукцп лейкоцитгв у кровотворних органах реактивного ха-
рактеру (пщ час шфекцп та некрозу тканин) або при пухлиннш гшерплазп лейко-
поетичног тканини, коли збгльшуеться мгтотичний, дозргваючий г резервний пул
лейкоцитгв у юстковому мозку;
• прискорення виходу лейкоцитгв гз кгсткового мозку в кров,
що може бути наслщком пщвищення проникносп кгстковомозкового бар’ера пщ
впливом медгаторгв запалення, глюкокортикощгв, а також посилення протеолгзу
оболонки, що оточуе остргвець гранулоцитопоезу, при септичних станах;
• перерозподгл лейкоцитгв за рахунок мобглгзацп !х гз пристшкового
(марггнального) пулу в циркулюючий (пщ час стресу, коли в кровг пщвищуеться
вмгст адреналшу та глюкокортикощгв, у разг сильних емоцш, болю, переохолод-
ження або перегргвання, дп ендотоксингв мгкрооргангзмгв), унаслщок пере-
розподщу кровг (у зв’язку з шоком, колапсом);
• зменшення об’ему плазми та згущення кровг.
Лейкоцитоз часто поеднуеться з порушенням дозргвання клггин лейкоцитар-
ного ряду г продукцгею патолопчно змшених лейкоцитгв.
У випадку лейкоцитозу, що виник унаслщок реактивно!' гшерплазп лейко-
поетичног тканини, як правило, пщвищуеться функщональна актившсть лейко-
цитгв, що зумовлюе посилення захисних реакцгй оргашзму. Так, гз розвитком ней-
трофглъного лейкоцитозу та моноцитозу одночасно посилюеться фагоцитарна ак-
тивнгсть лейкоцитгв. Еозинофглышй лейкоцитоз завдяки антипстамшнгй функцп
еозинофщьних гранулоцитгв вщгграе компенсаторну роль пщ час алерпчних реак-
цш. Водночас лейкоцитоз при лейкозг може супроводжуватися зменшенням захис-
них властивостей клгтин лейкоцитарного ряду, що зумовлюе розвиток гмунолопч-
ног гшореактивностг, за яко'г оргашзм зазнае впливу вторинних та автошфекцш.
Картина кровь Пщ час лейкоцитозу збшьшення загальног кглькостг лейко-
цитгв супроводжуеться змшою лейкоцитарног формули (вщсотковий вмгст окремих
форм лейкоцитгв, що визначаеться пщрахунком 100—200 клгтин у пофарбованому
мазку кровг; табл. 20). Для встановлення абсолютного характеру окремих видгв
лейкоцитозу розраховують абсолютный вмгст ргзных форм грануло- та агрануло-
цитгв в 1 л (на шдставг знания загальног кглькостг лейкоцитгв в 1 л кровг та лей-
коцитарног формули), осюльки сама лейкоцитарна формула дае уявлення лише
про вщносний (вщсотковий) вмгст лейкоцитгв та вщображае спгввщношення мгж
ргзними гхнгми формами.
Пщ час лейкоцитозу, особливо нейтрофшьного, у кров! нерщко з’являються
незрш клгтини (ядерний зсув нейтрофшъних гранулоцитгв улгво). Велика кгль-
кгсть дегенеративно змгнених лейкоцита на тлг лейкоцитозу характерна для сеп-
сису, гншних процесгв, шфекцшних хвороб, розпаду злоякгсног пухлини.
Таблиця 20. Лейкоцитарна формула й абсолютний вм!ст лейкоцитгв у норм!
Лейкоциты Кшыасть клггин
% в 1 л в 1 мкл
Базофшьт гранулоцита 0—1 0—0,0910’ 0—90
Еозинофгльш гранулоцита Нейтрофшьш гранулоцита: метамгелоцити (кип) паличкоядерш сегментоядерш 2—5 0—1 51—67 0,08—0,4510’ 0—0,0910’ 0,04—0,54 10’ 2,04—6,03 10’ 80—450 0—90 40—540 2040—6030
Моноцита 4—8 0,16—0,7210’ 160—720
Лгмфоцити 20—40 0,8—3,610’ 800—3600
Лейкопен/я
Лейкопения — зменшення загально! киъкоспй лейкоцитгв у кров! нижче 4 Г/л (4,0*1(Г/л).
Класифгкацгя. Лейкопенгя, як г лейкоцитоз, за механизмом виникнення може
бути абсолютною, або справжнъою (унаслщок ослабления утворення лейкоцита,
затримки 1х виходу з юсткового мозку, пщвищеного руйнування або видшення з
оргашзму), г вгдносною, або несправжньою (перерозподыъною, або гемодилюцшною).
За переважним зменшенням абсолютно! кйгькост! окремих форм лейкоцита
розргзняють нейтро-, еозино-, л!мфо-, моноцитопент.
Ет1олог1я. У виникненнг нейтропени може вщггравати роль шфекцгя (вгруси
грипу, кору, шфекцгйного мононуклеозу, гепатита, В1Л, збудники черевного г
висипного тифу), юнгзуюче випромшювання, лгкарськг засоби (сульфаншамщнг,
антитиреощш, цитостатичш препарати, барбггурати тощо), бензол, дефщит цга-
нокобаламгну та фолгевог кислоти, бшкове голодування, анафглактичний шок,
пперспленгзм, гемодгалгз, а також генетичний дефект пролгферацп та диферен-
щювання нейтрофгльних гранулоцита (спадкова нейтропенгя).
Еозинопенгя й анеозинофиия (вщсутнгсть еозинофгльних гранулоцита у кров!)
спостерггаються в разг пщвищеног продукцп глюкокортикощгв (стрес, хвороба чи
синдром 1ценка — Кушгнга), уведення кортикотрошну та кортизону, гострих ш-
фекцгйних хвороб, агранулоцитозу, лгмфобластного лейкозу.
Л1мфоцитопен1ю може спричинити сепсис, гмунодефщит спадкового чи набу-
того характеру (у разг променево)' хвороби, В1Л-шфекцн, мглгарного туберкульозу,
бглкового голодування, лгкування гмунодепресантами), хронгчний стрес г ггпер-
кортицизм.
Моноцитопешя спостерйаеться за всхх станхв, яю супроводжуються припп-
ченням мхелощного паростка юстковомозкового кровотворення (наприклад про-
менева хвороба, тяжю септичн! стани, агранулоцитоз).
Патогенез. Основою розвитку лейкопени е таю мехашзми:
• пригнхчення лейкопоезу;
• порушення виходу зрхлих лейкоцитхв хз кхсткового мозку в
кров;
• руйнування лейкоцитхв у кровотворних органах х кровх;
• перерозподхл лейкоцитхв у судшшому руслх;
• пхдвищене видхлення лейкоцитхв з оргашзму.
Мехашзми, що зумовлюють пригнгчення лейкопоезу, розглянуто вище. Сповиъ-
нення виходу гранулоцитгв гз Мсткового мозку у кров спостерйаеться пщ час спад-
кового синдрому “ледачих ” лейкоцитгв (р!зке зниження рухово! активност! лейко-
ципв унаслщок генетичного дефекту бшкхв мткровщростюв клхппшо! мем-
брани).
Руйнування лейкоцитгв у кровг вщбуваеться пщ впливом тих самих факторов,
яю спричинюють лхзис юптин лейкоцитарного ряду в кровотворних органах (пе-
реважно хмунн! мехашзми цитолхзу), а також у зв’язку зх змшою ф!зико-х!м!чних
1 метаболхчних властивостей 1 проникносп мембран самих лейкоцит!в унаслщок
неефективностг лейкопоезу, що й зумовлюе посилення л!зису цих клхтин у кровх х
макрофагах селезшки. Одним 1з мехашзмхв збшьшення руйнування СО4+-лхмфо-
ципв при ВГЛ-хнфекцп, що призводить до лхмфопенн, е активацхя апоптозу. Ос-
таннш мехашзм лежить частково в основ! лхмфопешх та еозинопени в раз! збшь-
шення в оргашзм! вмхсту глюкокортикощхв (тривале лххсування цими гормонами,
пперкортицизм, хрошчний стрес).
Перерозподгльний мехашзм лейкопени частице полягае в тому, що змшюеться
сшввщношення мхж циркулюючим х приспнковим пулом лейкоципв. Останне
спостерхгаеться в раз! переважання тонусу парасимпатично! частини автономно!
(вегетативно!) нервово! системи над симпатичною, запальних процесхв, перегрх-
вання, деяких бакгерхальних хнфекцхй. Так, при сальмонельоз! ендотоксини мхк-
роорганхзмхв сприяють активацп' синтезу адгезивних бшкхв та хх включению в
мембрани лейкоципв (нейтрофхльних гранулоципв) та ендотелхоцитхв. Це спри-
чинюе пщвищену маргшацпо гранулоципв та хх прилипания до ендотелхоципв,
що зумовлюе розвиток перерозподхльно! нейтропенн. Стимуляторами маргшаци
лейкоципв можуть бути компонент системи комплементу, 1Л-1, лейкотрхени,
ФНП та ш. Зрщка лейкопенхя може зумовлюватися тдвигценим видиенням лейко-
цитгв з оргашзму (при гншному ендометрип, холецистоанпохолхп). Значна заги-
бель гранулоципв у великих вогнищах гшйного запаленння в раз! абсцесхв ! флег-
мони також може викликати лейкопенпо.
Основном наслгдком лейкопени е ослабления реактивносп оргашзму, зумовле-
не зниженням фагоцитарно! активностх нейтрофшьних гранулоципв та участю
лхмфоципв у гуморальних х клхтинних Гмунних реакцхях за рахунок не тшьки
зменшення хх загально! кшькосп, а й можливого поеднання лейкопени з продук-
цхею функщонально неповноцшних лейкоципв. У таких хворих вщзначаеться
схильшсть до шфекцшних х пухлинних хвороб, особливо за спадково! нейтропе-
нп, а також дефщиту Т- х В-лшфоципв. Яскравими прикладами тяжко! ареактив-
ноете оргашзму е синдром набутого ёмунодефёциту вёрусного (СН1Д) ё рал1ашино-
го походження, агранулоцитоз ё алёментарно-токсична алейкёя.
Агранулоцитоз — ргзке зменшення кмькоспй (до 0,75 Г/л I менше) нейтрофиь-
них гранулоципйв у кров! на пш лейкопени (1 Г/л I менше).
Класифёкацёя. За ет!олог1ею агранулоцитоз поделяють на набутий ё спадко-
вий, за мехашзмом виникнення — на мгелотоксичний (ураження юсткового мозку)
та гмунний (руйнування клггин гранулоцитарного ряду антилейкоцитарними анти-
телами).
Етёологёя. В етёологп набутого агранулоцитозу основну роль вёдёграють тё
самё фактори, що спричинюють нейтропешю: деяю лёкарськё засоби (особливо
амёдошрин, сульфанёламёднё, цитостатичш препарати, амёназин), бензол, споро-
фузарин, дефщит бшюв, цёанокобаламёну, фолёевоё кислоти, ёонёзуюче випромё-
нювання, вёруснё ёнфекцп, колагенози, антинейтрофёльнё антитела, ёзоёмунний
конфлёкт матерё ё плода за антигенами нейтрофёльних гранулоцитёв, метастази
пухлин, лейкознё клони в юстковому мозку.
Причиною спадкового агранулоцитозу е генетичний дефект продукцп та доз-
рёвання гранулоцитёв у юстковому мозку або ёх виходу з нього у кров.
Патогенез. Основою набутого м!елотоксичного агранулоцитозу можуть бути
таю мехашзми:
• пряма ушкоджувальна дёя етёологёчних факторёв (ёонёзуточе випромёнюван-
ня, вёруси) на клётини — попередники гранулоцитопоезу або на ёх мёкрооточення
в юстковому мозку;
• детальна мутацёя в клётинах гранулоцитопоезу лед впливом радёацп, вёрусёв,
токсичних похёдних бензолу;
• тяжкё порушення метаболёзму леёгкопоетичних клётин (синтезу ДНК) пед
дёею цитостатичних препаратёв, дефёциту вётамёнёв (В12та фолёевоё кислоти), по-
силення вёльнорадикального окиснення, у тому числе за рахунок пригшчення
систем антиоксидантного захисту.
Мехашзмами виникнення набутого 1мунного агранулоцитозу е:
• гаптенасоцёйоване утворення антител до нейтрофельних гранулоцитёв кровё,
а також до клётин — попередникёв гранулоцитопоезу в юстковому мозку (при
цьому найчастёше роль гаптенёв виконують лёкарськё засоби — амёцопёрин, аналь-
гён, ацетилсалёцилова кислота та ён.);
• асоцёйоване з автоантитёлами ослабления гранулоцитопоезу пёд час вёрус-
них ёнфекцёй, гёперпролёферацёё великих гранулярних лёмфоцитёв — природних
кёлерёв (ВГЛ-лейкоз), дифузних хвороб сполучноё тканини (ревматоедний артрит,
системний червоний вовчак);
• утворення антилейкоцитарних антител до нормальних антигешв неёггрофёлёв
у разё мутацёй в ёмуннёй системё, коли виникае заборонений клон лёмфоцитёв.
Механёзм розвитку спадкового агранулоцитозу пов’язаний з особливёстю гене-
тичного дефекту гранулоцитопоезу.
Наприклад, у хворих на синдром'Костманна (автосомно-рецесивне успадку-
вання) порушенё дозрёвання ё диференцповання нейтрофельних гранулоцитёв у
стадёё промёелоцит — мёелоцит. Мутацёя зумовлюе втрату чутливостё промёело-
цитёв до деё КСФ. У разё циклгчного агранулоцитозу (автосомно-домёнантне успад-
кування) кёлькёсть нейтрофёльних гранулоцитёв коливаеться вед нормальное вели-
176
чини до повного Тх зникнення в периферичнгй кров! з тривалгстю нейтропени
протягом 3—10 днгв, що повторюеться кожы 21—28 дшв. В основ! ще! хвороби
лежать гшерпродукгця Т-лгмфоцитами шгобггоров подшу клиин — попередникгв
гранулоцитопоезу г зниження чутливост! цих клгтин до КСФ. Накопичення фак-
торов росту вщновлюе нормальну колькгсхь нейтрофгльних гранулоцитгв.
Клгычы прояви агранулоцитозу, як г нейтропени, пов’язано з р!зким змен-
шенням фагоцитарно! функцп нейтрофгльних гранулоцитгв. Тяжка форма аграну-
лоцитозу характерна для алейки.
Алешин — ураження кгсткового мозку з ргзким пригнгченням / навгть повною
вгдсутнгстго мгелопоезу г лгмфопоезу.
Алгментарна токсична алейкгя розвиваеться внаслщок використання в рацооы
зерна, що перезимувало на пол! о заражене плоснявими грибами, яко утворюють
токсичнг речовини (наприклад спорофузарин). При цьому спостерогаеться пан-
цитопенгя — розке зменшення кглькосто лейкоцитов (алейкоя), еритроцитов
(анемгя) г тромбоцитов (тромбоцитопенгя). Усунення захисноо функцп лейкоцитгв
зумовлюе виникнення некротично-запальних уражень мигдаликов (ангша), шко-
ри, слизових оболонок рота, носа, глотки, кишок, бронхов. У разг значного зни-
ження гмунгтету (пщ дгею ерготоксину — алкалощу маткових рожков — гриба, що
паразитуе на зерно жита, шших злаков), може розвинутися некротичне ураження
м’яких тканин обличчя ! рота — нома, або водяний рак.
Яюсж зм1ни лейкоцитов у кров!
Класифгкацоя. Якгсш змши в кровг проявляються як:
• дегенеративно змгн лейкоцитгв;
• порушення сшввщношення зрглих г незрглих форм лейкоцитгв у крово як у
бок збгльшення кглькостг незрглих (дозргваючих клгтин V класу), так о переважан-
ня зрших форм лейкоцитгв (VI клас) на тлг вщсутносто гх попередникгв;
• цитохгмгчнг, цитогенетичш та антигеннг змши лейкоцитгв (особливо харак-
теры для лейкозов);
• функционально порушення лейкоцитов (передусгм фагоцитарно! активностг
нейтрофгльних гранулоцитгв та учасп лгмфоцитгв в гмунних реакцгях).
Дегенеративнг змши лейкоцитгв. Ознаками дегенеративних змгн лейкоцитгв е:
• ангзоцитоз (розы розмгри клгтин);
• токсогенна зернисттсть (числены азурофгльнг гранули) у нейтрофгльних
гранулоцитах, тгльця Князькова — Деле (велико базофольно забарвлеш грудочки
цитоплазми), зерна Амато (овально утвори блщо-блакитного кольору, що мостять
червоно та червоно-фоолетово зерна); вакуолг в цитоплазмг та ядре,
• зникнення звичайног зернистостг в гранулоцитах',
• пгкноз або набухання ядра, його ггпер- або гтосегментацгя, кольцеподобна
форма ядра, вщмшност! в ступено зролосто ядра о цитоплазми, каргорексис;
• цитолгз (див. кольор. вкл., мал. III).
Дегенеративних змгн часто зазнають нейтрофольш гранулоцита о моноцита.
Причиною ох появи е продукцгя лейкоцитов оз порушеним обмгном речовин (при
лейкоз!, спадковш ферментопатп), що й зумовлюе структуры аномали, а також
’ 47Г
ушкопження лейкоципв у кровотворних органах 1 кров! шд впливом ргзних пато-
генних факторов (бактерш. вгрусгв. антитгл).
Наявнтсть дегенеративних змш лейкоштпв (токсогенно! зернистости вакуолз-
зацп тощо) може бути одним тз дгагностичних показниюв тяжко! (нтоксикацв
оргашзму (при алкоголизм!, гнгйно-сешичних хворобах, крупознгй пневмонп,
скарлатин!, гангренозному апендицип. гострш дистрофг! печшки та ш.). Ггпер-
сегменгашя нейтрофгльних гранулоципв характерна для мегалобластно! анемп.
Юльцепощбна форма ядра спостерйаеться у випадку тяжкого алкоголгзму.
Порушення стввгдношення зрыих г незрглих форм лейкоцит!в у крови Поява не-
зр!лих форм лейкоципв у кров! зумовлена порушенням дозргвання !х у крово-
творнш тканин! та пгдвишенням проникносп юстковомозкового бар’ера. Цг зм!-
ни зазвичай вщбуваються на тл! посилено! продукты лейкоципв унаслщок як
реактивно!, так г пухлинно! гшерплазп лейкопоетично! тканини.
Переважання в кров! зрглих, сегментоядерних, клггин гранулоцитарною ряду
(насамперед нейтрофгльних гранулоцитгв) за вщсутност! !х попередниюв (палич-
коядерних гранулоцитгв г мегамгелоцитгв) 1 наявносп дегенеративних змш у клг-
тинах пов’язане з пригшченням юстковомозкового кровотворення.
Класифгкацгя. До порушень сшввщношення зрглих г незрглих форм лейко-
ципв належать:
• зсув лейкоцитарной формулы (або ядерныы зсув нейтрофыьних гранулоцитгв);
• лейкемогдна реакц1я.
Ядерний зсув нейтрофгльних гранулоципв. На пщставг розмпцення нейтроф!ль-
них гранулоципв у лейкоцитарной формул! за Арнетом г Шиллингом (табл. 21; у
л!вш частит формули розташован! незрглт несегментованг клгтини — паличко-
ядернг нейтрофыьнг гранулоцити, метамгелоцити, мгелоцити, або юнг, у правы
частит — зрыг сегментоядернг нейтрофыьнг гранулоцити. ядра яких мають два г
быьше сегменпв) видыяють:
• ядерний зсув неытрофыъних гранулоцитгв улгво — у разг збшьшення юлькосп
молодих, незрглих форм нейтрофгльних гранулоципв;
• ядерний зсув управа — у разг переважання зрыих нейтрофгльних гранулоципв
гз п’ятьма-шгстьма сегментами (пперсегментацгя ядер) на тлг вщсутност! поперед-
шх молодих форм нейтрофгльних гранулоципв
Виокремлюють юлька видгв ядерного зсуву влгво:
• регенеративный зсув — показиик реактивно! активацп грануловитопое-
зу в раз! гострого запалення; на тлг пом!рного лейкоцитозу (до 20 Г/л) — пщви-
шений вм!ст паличкоядерних гранулоципв г метамгелоципв; спостерйаеться год
час гншних шфекцш, гемолгзу еритроципв;
• глперрегенератывниы зсув (лейкемгчний) вштворюе надмгрну гпгер-
плазпо лейкопоетично! тканини з порушенням дозргвання клгтин г значним омо-
лодженням складу кровг; значно зросгае юльюсть паличкоядерних нейтрофгльних
гранулопипв г метамгелоцитгв, з’являються мгелоцити, поодинок! промгелоцити,
загальна юльюсть лейкоципв спочатку може бути збшьшена понад 20 Г/л, поим
знижуеться до норми та нижче: таким зсув характерний для гострого запалення,
лейкемощних реакдгй мгслощного типу, хрошчнего мгелолейкозу;
• регенеративно-дегенеративный зсув спостерйаеться в разг поруше-
ного дозргвання лейкоципв г пперпродукш! в юстковому мозку патолопчно змг-
Таблиця 21. Ядерне зрушення нейтрофшьних гранулоципв
Лейкоцита Загальна юльюсть лейкоципв, (Г/л) —— ———.—,—. ————— - Нейтрофшьж гранулоцита, % Дегене- ративн) зм)ни нейт- роф)л)в
Пром1е- лоцити М!ело- цита Метамгело- цита (юж) Паличко- ядерн! Сегменто- ядерж
Норма 4—9 . — — 0—1 1—6 51—67 —
Ядерне зрушення вл!во
Регенеративна > 9 до 20 . - — — Т т ё —
Пперрегенера- тивне >20 4—9 <4 + + тт и ё —
Дегенеративне <4 — — — тт ёё +
Регенеративно- дегенеративне > 9 до 20 — — т т ё +
Ядерне зрушення вправо <4 — — ё ё т +
нених лейкоципв; проявляеться помёрним лейкоцитозом, енодё нормальною кёль-
кёстю лейкоципв, у мазку кров! — збёлъшенням кёлькостё паличкоядерних грану-
лоцитёв, метамёелоцитёв з ознаками дегенерацп, що е можливим у разё тривалого
перебегу ёнфекцёё (сепсису), коли ресурси мёелопоезу виснажуються;
• дегенеративный зсув евщчить про пригшчення 1 глибокий розлад лей-
копоезу, коли збёлыпуеться юльюсть паличкоядерних нейтрофыьних грануло-
цитёв !з дегенератпивними зм1нами на тл! лейкопени, зменшення кёлькостё сегменто-
ядерних форм ё вёдсутшсть метамёелоцитёв; такий зсув може виникати при сеп-
сис!, тяжкёй ёнтоксикацц оргашзму, коли порушуеться як поды, так ё дозревания
лейкоцитгв.
Ядерний зсув управо (дегенеративний зсув) з переважанням зрёлих сегментоядер-
них нейтрофыьних гранулоцитёв з великою кёлькёстю сегментёв ё за зменшення
кёлькостё чи за вёдсутностё ёхнёх попередникёв — паличкоядерних нейтрофёльних
гранулоцитёв ё метамёелоцитёв; спостерёгаеться на тлё лейкопенёё ё е показником
значного пригшчення лейкопоезу (променева хвороба, мегалобластна, апластич-
на анемёя, агранулоцитоз, алейкёя). У гёперсегментованих нейтрофёльних грану-
лоцитёв часто виявляються рёзш дегенеративнё змёни цитоплазми.
Киькосну оцшку ядерного зеуву лейкоцитарное формули здёйснюють шляхом
обчислення ёндексу ядерного зеуву за формулою:
Паличкоядернё нейтрофельнё гранулоцита + метамёелоциТи + меелоцити _
Сегментоядернё нейтрофыьнё гранулоцита
= 0,05 — 0,10 (норма).
У разё ядерного зеуву влево ёндекс буде бёльший нёж 0,10, у разё зеуву вправо —
менший нёж 0,05.
Лейкемоодна реакцоя — це змши в кров! та костковому мозку у вигляд! зболъшення
киькосто незролих форм лейкоцитов, подобно до лейкозу. В!др!зняетъся вод нього'.
• етоологоеоо (причина часто вёдома);
• патогенезом (реактивна пперплазёя лейкопоетичноё тканини ё вёдсутнёсть нео-
пластичноё грансформаци в гу пухлину крове, з якою е морфологёчна подебнесть);
• клш1чним перебггом (часто мають гимчасовий оборотный характер)
Класифёкацёя. Затежно вщ того, пролёферашя яких паросткёв кроветво-
рения спричинила ведповёднё змши картин и крове, розрёзняють лейкемоиМ. реакци
м1ело1дного та лимфоцитарного тише
До лейкемоёдних реакцш м1ело!дного типу належать псевдобластична, пролйсло-
цитарна, нейтрофйшна, еозинофшъна (‘‘велика еозинофиия”), базофиьна, моноцитар-
на реакци.
Серед лейкемоедних реакцш л1мфоцитарного типу видгляють лейкемогдн!реак-
ци з переважанням атипових лимфоцит! або лшфоципйв типовое структури.
Етёологёя. Причинами виникнення лейкемоедних реакцш лаелоедного типу е
в основному тё, що викликають нейтрофётьний, базофшьний. еозеенофёльний лей-
коцитоз.
Псевдобластична I пром1елоцитарна лейкемощна реакцея м1ело'1дного типу
з’являеться в стали завершения ёмунноё о агранулоцитозу, коли вщновлюсться
мёелопоез. У кровё мёстягься клётини, морфотогёчно подёбнё ло бластёв промее-
лоцитёв. Такт саме змёни спостерёе ають ё в кёстковому мозку. Це три вас вёд кшькох
годин до доби. Така клёнёчна картина ёнодё бувае на перелому мёсяцё життя в детей
ёз хворобою Дауна, але потём склад кровё нормалёзуеться.
Моноцитарш лейкемоТднг реакци чаете ше перебёеають як моноцитарно-макро-
фаговё реакцёё. Вони виникають у випадку тяжкое дисемёнованоё форми туберку-
льозу, при брупельозё, сифёлёсё. токсоплазмозе, лейшманёозё та ёнших бактерёаль-
них, мекозних ё протозойных ёнфекцёях. При цьому высокий моноееитоз у кровё
погднуеться з граиульоматозом, коли в рёзних органах е тканинах формуються
гранульоми (горбики), що склэдаються з макрофапв га ёхнёх похщних — еш-
гелёоёдоподёбних клётин, яке перетворюються на гёгантськё багатоядернё клётини.
Одиёею з поширених причин виникнення лейкемоедних реакцш лимфоцитарно-
го типу е вёруси. Так, вёрус Епстайна — Барр ё цитомегаловёрус спричинюють
ёнфекцейний мононуклеоз, ентеровёрус родили Коксакё —гострий шфекцёйний
лёмфоцитоз (хвороба Смёта). Вёруси кору ё краснухи гакож можуть зумовити ви-
никнення лёмфоцитарноё лееТкемощноё реакцёё.
Причиною лейкемоёдноё реакцп цього тешу е також порушення ёмунноё сис-
теми при автоёмуепшх хворобах (ендокринопатёё, сеестемний червоний вовчак та
ён.), реакпёё “трансплантат проти хазяёна”, резус-несумёсностё матере га плода, що
зумовлюе гемолётичну хворобу новочароджених.
Патогенез. Основний мехашзм виникнешея лейкемоедних реакцёй резного
типу — це пёдвищена пролёферашя вёдповщного паростка гемопоезу кёстковоео
мозку полёклоналъного характеру, що вщрёзняе лейкемоёднё реакцёё вёд ггухлинних
хвороб — лейкозёв, для яких на першому етагп розвитку характерна моноклональ-
на пролёфераиея неопластичних клётин. Бластнё клётини пед час лейкемоёдних
реакцей не мають антигенних маркерёв ё хромосомних аномален, а також не зазна-
ють цитохемёчних змён, властивих лейкозам. При деяких видах лейкемощних ре-
акции мёелоедного типу поява незрелих форм нейтрофёльних гранулоецетёв у крове
може бути пов’язана зё ееевидким ё значним виходом ёх ёз кёсткового мозку.
Картина кровё. Для лейкемоёдних реакцш мёелоедного ё лёмфоцитарного
типе в характерним е рёзке збёлыпення загальноё кёлькостё лееёкоцитёв, частеше до
I
П
III
Стовбурова кроветворна юптина
Юйтина - попередниця лхмфопоезу
Колон
Юптина - поперед-
ниця Т-Л1МфоЦИТ!В
Юптина - поперед-
ниця В-лшфоцит1в
хсутворювальн! единиц!
Юптина - попередниця мзелопоезу
IV
В-лхмфобласт
Плазм облает
Монобласт
Базоф1льний
бласт
М1слобласт
культур1
Еритробласт
К Л 1 т и н
Мегакарюбласт
Т-л1мфобласт
Т-прол1мфоцит
VI
Т-л1мфоцит
В-прол1мфоцит
В-Л1мфоцит
Проплазмоцит
Плазмоцит
Промоноцит
Про
1 € Л О
Еозинофгльний
бласт
Пронормоцит
।
Моноцит
м
Ц
И Т И
и
Нормобласт базофыьний
Нормобласт пол^хроматофйтьний
Мет
ц и т и
а
М 1 € Л О
Нормобласт ацидоф1льний
*Г'
Пал ичкоядерн1 гранулоцити
•к. ’
Сегменто ядерн 1 гранулоцити
базофыьний нейтроф1льний еозинофыьний
Промегакарюцит
№
И--------Мегакариоцит .
ПолГхромагофгп Ретикулоцит |
Еритроцит
Тромбоцит
Мал. I. Схема кроветворения (за 0.1. Воробйовим, 1981):
I — клас стовбурових клггин; II — клас частково детермшованих полшотентних кштин; III — клас уншотентних клггин (кшти-
ни—попереднищ Т-, В-л1мфоцит1в, колошеутворювалып одиниц! — моноцитш, гранулоцити, еритроцита!, мегакариоцита!);
IV — клас прол!феруючих кл!тин (бласти); V — клас клгтин, як! дозргвають; VI — клас зрших кштин
Юптини ф131олопчноТ регенерацп
Дегенеративш змши еритроци пв
Клпини патолопчно! регенерацп
Змша розм1ру (ашзоцитоз)
Ретикулоцита
(суправпальне забарвлення)
Пол4хроматоф1льш еритроцити
(забарвлення за Романовським)
Змша форми (пойюлоцитоз)
Змша забарвлення
13 14 15
Мал. II. Змши еритроципв при анемп:
1 — еритроцит; 2 — ретикулоцит; 3 — пол1хроматофш; 4 — еритробласт; 5 — базофшьний нормо-
бласт; 6 — пол1хроматофшьний нормобласт; 7 — ацидофшьний нормобласт; 8 — нормоцит (серед-
Н1й д1аметр [б] — 7,2 мкм); 9 — мшроцит (4 — менше 6,5 мкм); 10 — макроцит (б — 8 мкм 1 бшь-
ше); 11 —- мпсросфероцит; 12 — серпоподобний еритроцит (дрепаноцит); 13 — нормохромний еритро-
цит; 14 — анулоцит; 15 — гшохромний еритроцит; 16 — пперхромний еритроцит; 17 — базофшьна
зерниспсть; 18 — кшьце Кебота; 19 — пльце Жоллц 20 — мегалоцит; 21 — пол1хроматофшьний
мегалобласт; 22 — ацидофшьний мегалобласт
Мал. III. Дегенерат ивнт змжи лейкоципв.
7 — токсична зерниспсть; 2 — вакуолизация
ядра 1 цитоплазми; 3 — пльця Князькова-
Деле; 4 — аномалзя Иельгера; 5 — ппер-
сепиентацхя ядра; 6 — антзоцитоз
Мал. ГК Картина кровз в разз гострого лейкозу:
1 — бласгш КИ1ТИНИ
Мал. V. Картина кров! в раз! хрошчного кпело-
лейкозу:
1 — мгепобласт; 2 — пром1слоцит; 3 — нейтрофшь-
ний шелоцит; 4 — нейтроф1льний метамхелоцит; 5 —
паличкоядерний нейтроф1льний гранулоцит; 6 — сег-
ментоядерний нейтроф1льний гранулоцит; 7 — еози-
нофшьний М1елоцит; 8—еозинофшьний гранулоцит;
9 — базофшьний гранулоцит; 10 — Л1мфоцит
Мал. VI. Картина кров! в раз! хрошчного Л1мфо-
лейкозу:
1 — лгмфоцит; 2 — великий Л1мфоцит; 3 — л1мфо-
бласт; 4 — Т1ш Гумпрехта
50 Г/л ё бёльше, з абсолютным нейтрофёчьозом. еозинофёдёею, базофшею. моно-
цеггозом ё лимфоцитозом вёдповёдно до типу лейкемощноё реакш’Г.
У випадку М1€ло1дно1 лейкемоёдноё реакци спостерегають гёперрегенератавний
зсув лейкоцитарное формулы влево (до мёелоцитёв 1 промёелоцитёв) або дегенера-
та вний ядерний зсув (до 40 % паличкоядерних нейтрофельних гранулоцитёв при
токсикоёьфекцёях, спричинених Е. соИ). До* найпоширешших дегенеративных
змён лейкоцитёв належать вакуолёзацёя ядер ё цеггоплазми, токсична зернистёсть у
нейтрофёльних еранулоцитах.
При лёмфоцитарнёй лейкемо'ёднёй реакцп загальна кёлькёсть лейкоцитёв ёнодё
сягае 200 Г/л (ёнфекцейний лёмфоцитоз), а вмёст лёмфоцитёв у лейкоцитаретёй
формуле — 90 %. Окрем морфолотёчно типовых лёмфоцитёв за цим типом лейке-
моедноё реакпёё можуть з’являтися, наприклад при ёнфекцейному мононуклеозё.
моноцитоподёбне лёмфоцити — вёроцити. Вони е носёями геному ДНК-вмёсного
вёрусу Епстайна — Барр ё здебельшого належать до Т-лёмфоцитёв. Лёмфопитарну
лейкемощну реакцёю при краснусё характеризуюсь подёбнё до плазмобластёв кле-
тини з базофельною цитоплазмою ё круглим ядром, у якому мёститься близько
чотирьох нуклеол.
Гемобластоз. Лейкоз
Гемобластоз — клоналина пухлинна хвороба, яка виникае. з кровотворних клгтин.
Класифёкацёя. В основу класифёкацёё пухлинних хвороб кровотвор-
ноё тканини (ВООЗ, 2008) покладено морфолопчнё, питохёмёчнё, ёмунологёчнё,
пито-, молекулярно-генетичнё особливостё гемобластозе в. Браковано також кло-
налънесть е ревень диференпёювання неопластичних клетин. Выделено такё основнё
види гемобластозу.
• лейкози;
• лёмфоми.
Лейкоз
Лейкоз — пухлина, що виникае з кровотворних клётин кёсткового мозку.
Класифёкацёя. Розрёзняють гострий ё хронёчний лейкоз не тётьки за тривалёс-
1ю хвороби (сучаснё методи лёкування дають змогу продовжувати життя хворих на
гострий лейкоз ёнодё до кётькох рокев), а й за там. наскёльки лейкозне клётини
здатнё диференцеюватися до зрелых, шо визначае морфологёчний склад пухлини.
Гострий лейкоз — це злоякёсна пухлина. яка складаеться з бластных е<лётин I,
И, П1, IV класёв гемопоезу. що втратили здатнёсть до дозрёвання (кольор вкл.,
мал. IV).
Хронёчний лейкоз — пухлина, що складаеться переважно з дозрёваючих ё зрё-
лих клётин V та VI класёв, оскётьки основна маса лейкозних клётин диферен-
цеюеться до зрёлих форм.
На лёдсгавё морфологёчних, питохемёчних та ёмунологёчних показникзв розрёз-
няють таки вещи гострих ё хронёчних лейкозёв:
Гострё лейкози:
• мёелобластнё гострё лейкозп;
• недиференшйований гострий лейкоз;
• мхслобластнихй гострий лейкоз хз рхзним ступенем дозрхвання бластхв х рхз-
ними цитогенетичними аномалиями;
• промхелоцитарний гострий лейкоз;
• мхеломонобластний 1 монобластний гострий лехгкоз;
• мегакарюбластнихх гострий лейкоз;
• еритролейкоз (гострий еритромхелоз).
Лхмфобластш:
• В-клхтинного походження лейкоз (з попередниюв В-клхтин; до щех групи
належи ть лхмфома Берютта);
• Т-клпинного походження лейкоз (хз попередниюв Т-клхтан).
Хрошчш лейкози
Хронхчнх мхслопролхфератавнх лейкози (захворювання)1:
• хронхчний мхелолейкоз (кольор. вкл., мал. V);
• хронхчний нейтрофхльний лейкоз;
• хронхчний еозшофхльнийлейкоз;
• еритремхя;
• есенщальна тромбопитемЫ
Хрошчнх лхмфопролхферативнх лейкози (захворювання):
• хронхчний лхмфолейкоз (В - х Т-клхтиннх лейкози, що складаються
з1 зрхлих В- х Т-лхмфоцитхв; кольор. вкч., мал. VI);
• волосатоклпинний лейкоз;
• мхеломна хвороба;
• макроглобулшемхя Вальденстрема,
• хвороба важких ланцюпв.
Мхеломну хворобу, макроглобулхнемпо Вальденстрема та хворобу важких лан-
цюпв об’еднують у трупу парапроте1нем1чних гемобластозов — пухлинних захво-
рювань зрхлих В-клхтин, яю продукують значну юльюсть моноклональних хму-
ноглобулпйв (патолопчних) або фрагмента хх молекул.
1снуе трупа захворювань, яю об’еднують назвою "мгелодиспластичний синд-
ром” (МДС). Це клоналый новоутворення з первинним ураженням полшотентно!
стовбуровох кроветворно! клхтини юсткового мозку, втратою здатносп дозрхвати
до зрхлих юптин, аномальним ростом (диспластичнх змгни) мхелохдних клхтан
одночасним пригнхченням грануло-, еритро- та хромбоцитопоезу. Р1 знобил а ми
МДС е рефрактерна анемхя, яка не коригуеться звичайними лгкувальнимп засо-
бами, хз кьтьцеподхбними сидеробластами х без них, рефрактерна анемхя з над-
лишком бластхв у юстковому мозку. При МДС шдвишеним с ризик розвитку го-
строго лейкозу
Крхм того, розрхзняють форми лейкозхв за к1лък1стю лейкоцигтв у кров! (Даме-
шек, 1958): лейкопен1чну (кхлькхсть лейкоцитхв менша нхж 4 Г/л), алейкем1ч-
ну з нормальним вмхстом лейкоцитхв у межах 4—9 Г/л; сублейкем1чну (умхст
лейкоцитхв бътьнхе 9 Г/л — до 50 Г/л); лейкем1чну з рхвнем лейкоххипв понад
50 Г/л (може бута 200 -500 Г/л х бйгыпе).
'Мхелопролхферативн! хвороби — це хвороби. основою яких е пухлинна трансформация стовбу-
рових клхтин — попередниюв мхелопоезу зх збереженням ххнъоГ здатносп диференшювэтися до зрхлих
клхтин (про.пферац1я може бути переьажно як одного, так 1 всхх трьох паросткш кровотворення — гра-
нулоцитного, еритроцитного. тромбоцитного).
Лгмфома
Лгмфома— загальна назва злояюсних пухлин, що утворюються з лгмфоцгних клг-
тин, яю мгстяться переважно поза юстковим мозком (у лгмфатичних вузлах, се-
лезшцг, скупченнг лгмфощног тканини в ргзних органах, наприклад, у кишках,
приносових пазухах та ш.) г мають здебгльшого локальний ргст. Ураження юстко-
вого мозку та кровотворних органгв можливе при лейкемгзацп лгмфоми. Зггдно гз
сучасними уявленнями, окремг види лгмфоми г лгмфолейкози е однгею хворобою
з ргзними гематологгчними проявами, що залежать вгд стадп.
Класифгкащя злояюсних лгмфом
1. Лгмфогранульоматоз (хвороба або лгмфома Ходжкгна).
2. Неходжюнсью злояюсн! лгмфоми.
За морфюлоггею розргзняють таю неходжюнсью злояюснг лгмфоми: лгмфому
(лейкоз) В- г Т-клйпинну, яка своею чертою дглиться на лгмфому з клгтин — попе-
редникгв В- г Т-клгтин або зг зрглих В- г Т-лгмфоцитгв. € юлька морфолопчних
варгантгв неходжюнських злояюсних лгмфом.
Лгмфогранульоматоз (хвороба Ходжкгна) — злояюсна пухлинна хвороба лгм-
фатично'г системи з утворенням у лгмфатичних вузлах полгморфноклгтинних гра-
нульом, у яких мгстяться атиповг багатоядернг юптини Березовського — Рща —
Штернберга, невстановленог природи (припускають гх як макрофагально-моно-
цитарне, так г лгмфоцитарне походження).
Етгологгя. Причини виникнення пухлинних хвороб кровг розглядають
здебгльшого на прикладг лейкозгв г лгмфом. В етюлогп лейкозгв г лгмфом установ-
лено роль онкогенних вгрусгв, юнгзуюного випромгнювання, хгмгчних онкогенних речо-
вин, генетичних факторгв.
Онкогеннг вгруси зумовлюють:
• Т-клгтинний лгмфолейкоз (лгмфому) людини (ретровгрус типу С — НТЬУ-
I), вгрус г антитгла до якого виявляються не тгльки у хворих, а й у здорових осгб,
яю контактували з ними (можлива передача вгрусу Т-клгтинного лейкозу пгд час
переливания кровг, статевого контакту, трансплацентарно, подгбно до вгрусу
СН1Д - НТЬУ-Ш);
• злояюсну лгмфому Берютга (ДНК-вмгсний герпесвгрус Епстайна — Барр);
• лейкоз птахгв, мишей, котгв, великог рогатог худоби, мавп та шших тварин
(С-тип РНК-умгсних вгрусгв), лгмфопролгферативну хворобу Марека у курей
(ДНК-умгсний герпесвгрус) г лгмфому щургв, мишей (аденовгруси); вгруси переда-
ються через кал, сечу, видшення з носа г глотки та вгд матерг нащадкам (напри-
клад при вгсцеральному лгмфоматозг курей);
• лейкоз в експериментальних тварин у разг введения здоровий тваринам без-
клгтинних фгльтратгв лейкозних клгтин хворих тварин г людей.
Непрямим показником етголопчног ролг РНК-умгсних онкогенних вгрусгв у
виникненнг лейкозгв людини е наявшсть у лейкозних клгтинах зворотног транс-
криптази (РНК-залежног ДНК-полгмерази). Ген, що кодуе цей фермент, мгстить-
ся в геномг ретровгрусу г забезпечуе синтез ДНК-копп РНК-вмгсного вгрусу. Кргм
того, у структур! нормально! ДНК людини виявлено понад 60 протоонкогенгв,
гомолопчних вгрусним онкогенам, яю акгивуються пгд час хромосомних г генних
мутацш г зумовлюють розвиток лейкозу, причому самг вгруси можуть гндукувати
цг мутацп.
1оюзуюче випромшювання е причиною радтапшного лейкозу, доказом чого слугуе:
• тдвишена захворювашсть на гострий т хронтчний мтелолейкоз жителтв XI-
ростми т Нагасакт, яю пережили атомний вибух у 1945 р. (так зван! хтбакуст),
особливо тих. яю пере бу вали в радтуст 2 км вщ епщентра;
• зростання поширеност! лейкозу у хворих. яю отримали променеву терашю
з приводу злояюсних хвороб ртзнот локалтзацц, лтмфогранульоматозу, тнших видтв
гемобластозу, а також анктлозуючого спондилоартриту, у дттей, опромшених т
итего. а також у радюлопв 1 рентгенолопв,
• розвиток лейкозу в лабораторних тварин (мишей, щуртв) за умови однора-
зово! дп високих доз т гривало! дц низьких доз юшзуючого випромшювання.
Хгмгчш мутагени можуть спричинити гострий лейкоз т лтмфому:
• п!д час профестйного контактування людей з бензолом, фарбами, мастила-
ми, пестицидами тощо;
• у зв’язку з лткуванням лейкозтв деякими лткарськими засобамя шо мають
мутагеннудтю (цитостатичн! препарата, тмуноделресанти, бутадюн, левомицетин),
може виникати вторинний гострий лейкоз, частице мтелобластний, а також шсля
лжування гемобластозу алюлуючими препаратами (мтелосан. мелфалан, цикло-
фосфамщ);
• лейкоз в експериментальних тварин п1сля введения бензолу т його похщних,
бензантрацену (екзогенних мутагешв), похщних триптофану, тирозину, шдолу (ен-
догенних мутагешв), а також бензольних екстракпв тз тканин хворих на лейкоз.
Генетичш фактори також вциграють еттолопчну роль у розвитку лейкозу !
лтмфоми, про шо евщчать:
• родинний лейкоз (хрошчний лтмфолейкоз тз домшантним 1 рецесивним ус-
падкуванням);
• висока захворювашсть на лейкоз у деяких етнтчних трупах населения;
• конкордантшеть за формою, юпшчною т гематологтчною картиною лейкозу
в 1/3—1/4 однояйцевих близнюктв:
• пщвищена частота розвитку лейкозу у хворих тз хромосомними аномалтями
(при синдромах Клайифелтера, Тернера; у дттей 13 хворобою Дауна частота во-
строго мтелобластного лейкозу в 20 раз!в вища, тттж серед остб тз нормальною
ктльктстю хромосом 21), спадковими дефектами тмуннот системи (синдроми Втс-
котта — Олдртча, Л у!-Бар);
• хромосомш мутацп (транслокашя. делешя, тнверстя) у 80—90 % хворих на
гострий т хрошчний мтелолейкоз та у 50 % хворих на хрошчний лтмфолейкоз;
• так, для хрошчного мтелолейкозу характерною е аномальна ф!ладельф!йсь-
ка (РЬ7) хромосома (вщкрита в 1960 р. Ноуелом т Хангерфордом у ФтладельфтТ),
що утворилася внаслщок транслокацп сегмента хромосоми 9 на хромосому 22
(спостертгаеться у 90—97 % хворих);
• Рй-хромосому виявляють у 2 % виттадктв гос грота мтелолейкозу, а також
при гострому лтмфолейкоз! (прогноз у таких хворих полршуеться);
• при лтмфомт Беркттта часто вщбуваеться транслокашя сегмента хромосоми 8
на хромосому 14, !мов!рио, пщ дтею втрусу Епстайна — Барр;
• експериментальнт дант про високу (60—90 %) захворювашсть на лейкоз в
тнбредних мишей генетично чистих лппй.
Патогенез. Пщ впливом етюлопчних фактортв (онкогенних втруств, юнтзу-
ючого випромшювання. хтмтчних мутагешв) вщбуваеться пухлинна трансформация
кровотворних клипин, переважно стовбурових, напхвстовбурсвхгх, клхтин — поперед-
ниюв лхмфо- та м!елопоезу, наслщком чого е виникнення пухлини.
Для лейкозу характерно первинне утворення неопластичного клона в кхстко-
вому мозку, для лхмфоми — у лгмфатичшхх вузлах х лхмфощшй тканин! рйзних
органов.
Найпоширенхпхими е мехашзми пухлинное трансформацп кровотворних юптин:
• вбудовування вирусного онкогену ДНК-вм!сних вхрусхв або ДНК-копхх рет-
ровхрусхв у геном юптини з подалылим синтезом онкоб!лк!в;
• перетворення протоонкоген!в на клхтинш онкогени та хх щдвищена екс-
пресхя або дерепрес!я внаслщок точково! мутацп, транслокацп, утворення химер-
них генгв, амплхфхкашх тошо;
• репресхя антионкогешв, як! кодують антионкопротеши — шдуктори апоп-
тозу, адгезивш протеши (потрхбш для вмикання контактного гальмування пролх-
фераци клхтин), за участю мутащиного та епхгеномного мехашзмхв онкогенезу.
Конкретний мехашзм пухлинно! трансформацп кровотворних клхтин розхля-
нсмо на приклад! злоякхснох лхмфоми (лейкозу) Беркхтта, де можна видыяти так!
етапи:
• вбудовування ДНК-герпесвхрусу Епстайна — Барр у геном В-лхмфоципв
призводить до гпхерекспресп гена Ьс)-2 (блокатор апоптозу) та хмморталхзацп
(безсмертя) шфхкованого клону В-клхтин; однак ще вщсутня автономшсть росту,
властива злоякхсним пухлинам;
• хрошчне шфхкування малярхйним плазмодхем спричинюе ослабления клх-
тинного хмунхтету х перехпкоджае знищенню патолопчного клону В-лхмфоцитхв;
• порушення регулядп ххролхферацп (посилення пролхферацп) В-лхмфоцитхв
полегшуе виникнення в них додатково'х мутацп;
• транслокащя дшянки хромосоми 8 з протоонкогеном с-тус на хромосому 14
б!ля гена, що кодуе синтез важких ланцюпв антитш (при цьому с-глус пперекс-
пресуеться), завершуе пухлинну трансформашю В-клхтин. Клон В-лхмфоцитхв на-
бувае здатност! до автономного росту; виникае длоякхсна лхмфома Беркхтта
Пщ час хранённого мёелолейкозу основою пухлиннох трансформапп е трансло-
кацхя частки хромосоми 9 (мхстить онкоген с-аЫ, що кодуе одну з протешхДназ —
регуляторов стану рецепторхв до КСФ) у ддлянку розриву хромосоми 22 (Рй-хро-
мосома), де мхститься онкоген Ьсг. Поеднання двох онкогенгв на хромосом! 22
зумовлюе утворення химерного гена Ьсг-аЫ ! подалыпу пперекспресхю химерного
онкобшка — аномально! тирозиновох протешинази р210 з надмхрним подшом
клхтин.
Розвиток гострого лёмфолейкозу та деяких лёмфом спричинюе делещя корот-
кого плеча хромосоми 9 або 12, шо зменшуе або повшстю усувае вплив антионко-
гешв.
Механхзм пухлиннох трансформацп п!д час Т-кмтинного лёмфолейкозу мае
сво! особливостх. Вш зумовлений вхрусом НЕТУ-), який не мае вхрусних онко-
генхв — аналопв клхтшших протоонкох енхв, на вщмхну вщ хвших РНК-вмхсних
вхрусхв. Цей вхрус не спричинюе мутанпо хпляхом включения його ДНК-копп в
геном юптини, але вш мхстить ген (а/, що посилюе транскрипцхю. Спочатку в
уражених вхрусом Т-лхмфоцитах експресуеться велика кьтькхсть 1Л-2 1 його рецеп-
торхв, що призводить до прогрссуючо! пролхферапхх лхмфопитхв. Наступи!, додат-
ков! мутацп в клонх Т-лхмфощнхв змхнюють характер хх росту; вш стае автоном-
ним, злоякгсним. Процес лейкозогенезу в такому випадку7 тривалшшй, шж у разг
первгсно-муташйного механизму.
У патогенез! лейкозу можна видыитидвгстадп — мсноклоновута по.тгкло-
яову (0.1. Воробйов, 1985).
Для моноклоновог стадИ лейкозу (вщносно доброякгсног) характерними
е так! особливост!:
• активно прол!феруюч! клгтини утвореного лейкозного клону вщ значаються
однаковим гено- I фенотипом, мають одш й т! самг маркери — антигенш, хромосом-
на бгохгмгчш. За хронгчног форми лейкозу вони зовш подгбнг до нормальних юп-
тин кров! Пщ час гострого лейкозу (кр!м окремих, так званих недиференцшова-
них форм) можлива цитох!м!чна та (мунолопчна !дентиф!капгя бластних юптин;
• найхарактернгшою цитохгмгчною особливгстю лейкозних бласт>в у раз! го-
строго промгелоцитарного лейкозу е позитивна реакшя на мгелопероксидазу га
лппди гз Суданом чорним: гострого мон облает його лейкозу — на кислу фосфатазу,
неспецифгчну естеразу (а-нафгилацетатестеразу); гострого лгмфобластного лей-
козу — РА8-реакцпо (Ш1К-реакшю) на глгкоген,
• вщомг гмунологгчнг маркери стовбурових юптин за деяких форм гострого мге-
ло- 1 л!мфобластного лейкозу (антигени СОЗЗ, СЭ34), бластних юптин у раз! го-
сгрото мгел о лейкозу (СОЗ, СЕ) 14, СВЗЗ), В-лгмфоггитгв — В-лгмфолейкозу
(СЭЮ, СП 19, СО20), Т-л!мфоцит1в — Т-л!мфолейкозу (СПЗ, СЭ5, СБ7);
• вгдеутшеть або знижена здатшеть лейкозних клгтин до дозргвання зумовлюе
накопичення тд час лейкозу пухлинних клтин, якг активно про.йферуютъ (нормаль-
н! юптини кров! шсля 4—6 подпив починають диференцгюватися до зрыих юп-
тин, уже позбавлених мистично! активност!). Порушення в лейкозних клгтинах
апоптозу (наприклад, усунення гена р53) сприяе збыьшенню в лейкозному клон!
довгоживучих клтин, для розмноження яких не погргбн! ростов! стимулятори,
тому що вони автономно регулюють евгй подгл. Це зумовлюе гой факт, що клон
пухлинних юптин, який мае переваги над нормальними кровотворними кинина-
ми, заповнюе юстковий мозок, поступово тх витгеняючи. Однак на першому ета-
ш розвитку лейкозу (який може бути дуже коротким пщ час гостро! його форми)
одночасно з пухлинними юптинами утворюються 1 нормальна клпини кров!;
• лейкозю клгтини рано метаетазують гз кгсткового мозку в кров г поширю-
ються по веш систем! кровг (принцип “вгдчуття дому'’ — йопппё-ейест). Вони
потрапляють у лгмфатичнг вузли, селезгнку, печгнку, як! можуть збгльшуватися в
розмграх унаслщок пперплазп в них лейкозних клгтин (лгмфаденопатгя, гепаго-
спленомегалгя). За межами системи кров! лейкозних гнфыьтратгв немае;
• вгдсутнгстъ значних змгн з боку шших систем органгзму, кр!м наявност! харак-
терних для певно! форми лейкозу гематологгчних порушень.
Полгклонова стадгя розвитку лейкозу (злояюсшша, термтнальна) змгнюе
моноклонову стад по внаслщок пухлинног прогреси, власти во! будь-якгй пухли нь
Для цге! стад!! характерним е:
• виникнення спонтанно! та шдуковано! мутацп внаслщок нестабгльност! ге-
номгв пухлинних клгтин шд дгею первинного або нових мутагенних фак гор! в. Це
зумовлюе утворення замгсть одного клону лейкозних юптин кглькох нових клонгв.
Пухлина стае гено- та фенотипово неодноргдною;
• лейкозш клгтини стають злоякгешшими, морфололчно г цитохгмгчно неди-
ференшйованими. повнгстю втрачають функцп (синтез гмуноглобулгнгв, фермен-
Т1в, фагоцитоз), що були певною игрою 1м власти в! в моноклон овш стадп. У
кровотворних органах г кров! збглыпуеться кьтыасть бластних клгтин гз дегенера-
тивними змшами ядра г нитоплазми (бластный криз);
• здшснення добору, внаслщок якого знищуються юптини тих клошв, на яю
Д1Яли фактори гмунного захисту, гормони, цитостатичн! препарата, гошзуюче
випромшювання (у пронес! променево! терапп). Домтнування клонгв пухлинних
клггин, найбгльш резистентних до вплив!в цих факторгв, зумовлюе неефектив-
шстъ лйсування лейкозу в полклоновш стадп;
• витг'снення гнтенсивно пролгферуючими лейкозними юптинами в юстково-
му мозку нормальних кровотворних клгтин, що пригшчуе нормальне кроветво-
рения га гмунну систему г призводить до розвитку анемичного, гемораг1чного й 1му-
нодеф'щитного синдрому;
• поширення лейкозних клгтин за меж! системи кров! з утворенням лейкозних
1нф1лътрагтв у резных органах.
Кл1н1чн1 прояви лейкозу
Гематологгчний синдром
• Загальна юльюсть лейкоцитгв у кров! залежить вгд виду, стадгг лейкозу (мо-
ноклонова чи полгклонова) та особливостей перебйу хвороби. Тому можуть бути
лейкемгчна, сублейкемгчна, алейкемгчна та лейкопеючна форми лейкозу.
• Гострий лейкоз супроводжуеться поя вою в кров! лейкозних бластних юптин
(вгд 10 до 90 %), якг диференшюють за допомогою цитохгмгчних га гмунних ме-
тодгв доелгдження. Спостерггаеться лейкемгчний провал — вгдсутнгсть пере-
хгдних форм мгж бластними г зрглими юптинами. Це пов’язано з ргзким зменшен-
ням здатностг пухлинних клгтин до диференщювання.
• Хрошчний миелолейкоз характеризуеться збгльшенням у лейкоцитарной фор-
мулг кглькостг нейтрофгльних гранулоцитгв (метамгелопитгв, паличкоядерних,
сегменгоядерних) з ядерним зеувом лейкоцитарной формули влгво до мгелоцитгв,
промгелоцитгв та поодиноких мгелобтастгв. Може збгльшуватися юльюсть еози-
нофьтьних базофгльних гранулоцитгв (еозинофгльний г базофгльний лейкоци-
тоз). Спостерпаються дегенеративно змши клгтин. У термональнгй стадо'! настае
бластный криз, за яким у кров! ргзко зростае вмгст бластних юптин — моелоб-
ластгв г недиференцгйованих бластов.
• Хрошчний л1мфолейкоз (частгше трапляеться В-л!мфонитарний варгант лей-
козу) характеризуемся лгмфопитозом. за якого 80—98 % лгмфоиитгв становлять
малг зргл! юптини; спостерггаються поодиною проломфоцити ! лгмфобласти, тгнг
Гумпрехта (лгмфоцити в станг лгзису). Ьластний криз за цгег форми лейкозу вини-
кае вгдносно пгзно.
• При вах видах лейкозу порушуеться нормальний гемопоез. тому що лейкоз-
нг клгтини не тгльки замощують кровотворну паренхгму юсткового мозку, а й
гальмують диференщювання нормальних стовбурових клгтин. Наслгдком пригш-
чення гемопоезу под час гострого лейкозу е паниитопенгя — зменшення юль-
кост! вегх нормальних клгтин кровг.
• Анем/я при лейкозе за механгзмом свого виникнення е переважно ггпоплас-
тичною । метапластичною, частково — гемогнтичною. Автогмуиний гемолгз ерит-
роцитгв спостерггаеться при хрошчному лгмфолейкозг. Кргм того, у зв’язку з ге-
мораггчним синдромом може розвинутися постгеморапчна анемгя.
• Гемораг!чний синдром у випадку лейкозу зумовлений не ильки тромбоци-
топен!ею,ай функциональною неповношншстю лейкозних тромбоципв (тром-
боцитопатгя) та ураженням стшок кровоносних судин лейкозними хнфхльтра-
тами (гемораг!чна вазопапия). Нерщко сильна кровотеча е причиною летального
наслщку.
• ДВЗ-синдром при лейкоз! е наслщком значних порушень у систем! гемоста-
зу. Лейкозш юптини здатш до секрецп прокоагулянтхв. Так, у раз! гострого про-
м!елоцитарного лейкозу пухлинн! клхтини продукують тромбопластин, який зу-
мовлюе внутрхшньосудинне зсхдання кровх, а також протешази, шо шп-
бують фактори VII, XII, XIII, 1 активатори плазмшогену. Усе це спричинюе
спочатку пщвищене тромбоутворення, а пот!м кровотечу.
Синдром 1мунолог!чно1 недостатност! та зниження неспециф!чно1 протимис-
робно! резистентност!
• Депресхя нормального грануло-, моно- ! лгмфоцитопоезу при лейкоз!, а та-
кож функщональна неповноцшшсть лейкозних лхмфоципв (при лхмфолейкозх) х
гранулоципв (при мхелолейкозх) зумовлюють порушення фагоцитозу, пригшчен-
ня гуморальних ! юптинних реакцш хмунхтету, що сприяе приеднанню вториннох
бактерхальнох та мхкознох хнфекцп, активацп автошфекцп.
• Втрата лейкозними лимфоцитами функцп хмунолопчного нагляду спричи-
нюе утворення заборонених клошв, здатних синтезувати антипла до власних тка-
нин; розвиваються автохмуннх процеси.
Синдром позагастковомозкових проявив лейкозу
• Механхзмами розвитку ххъого синдрому е ураження лейкозними шфхльтра-
тами рхзних органхв, а також секрецхя пухлинними клхтинами токсичних речовин,
якх в значнш юлькосп вившьняються пщ час хх розпаду.
• Загальними проявами синдрому е гпдвищена стомлюванхсть, загальна сла-
бхсть, зниження маси тша, гарячка. Клппчнх прояви залежать вщ локалхзацп лей-
козних хнфшьтрапв у полхклоновхй стадп лейкозу. До них належать: нейролейкоз у
раз! шфнхьтрацп оболонок головного мозку, рхзних дхлянок головного та спинно-
го мозку, ядер х стовбурхв черепних нервгв, шо зумовлюе пщвищення внутрхшнь-
очерепного тиску, парез, паралхч та шшу невролопчну симптоматику; ураження
шк!ри та слизових оболонок (пнпвхт, стоматит тощо); ураження суглоб!в ! к!сток,
шших оргашв (легень, серця, нирок, яечка) з вщповщною симптоматикою та оз-
наками хх функцхонально! недостатност!.
ПОРУШЕННЯ ГЕМОСТАЗУ
Система гемостазу (вщ грец. аща — кров усерединх судин та отаот? — застш, сто-
яния, зупинення) — комплекс мехашзмхв, спрямованих на спинення крововтрати
шсля ушкодження судин шляхом утворення згусткхв кровх та пщтримання рщкого
стану кровх у кровоносних судинах. До системи гемостазу належать судинно-тром-
боцитарний гемостаз, який забезпечуеться судинною стшкою, тромбоцитами та
хншимхх клхтинами кровх, та коагуляцшний гемостаз, який забезпечуеться плазмо-
вими х тканишшми факторами зсщання кровх х калхкрехн-кппновою системою.
Система гемостазу функщхонуе в баланс! з протизс!далъною та ф!бринол!тичною
системами, якх належать до системи антигемостазу. Такий баланс забезпечуе рщ-
кий стан крово в судинах, а також оборотный о локальный характер процесу тром-
боутворення. Порушення в будь-якш ланцо щео системи спричинюе патолопю
гемостазу.
Класифокацоя порушень гемостазу.
— за етгологгею — набутг о спадковё, -
— за патогенезом:
• порушення тромбоцитарных механгзмгв гемостазу (тромбоцитопеноя, тромбо-
цитопатоя, тромбоцитоз);
• порушення судинного компонента гемостазу (вазопатоя);
• порушення коагуляцшних I антикоагуляцшних механ1зм1в гемостазу (коагуло-
патия з порушенням I, II, III фаз зсщання крово);
• комбгнован! — за участю ргзних мехашзмов гемостазу (ДВЗ-синдром, хвороба
Вшлебранда);
— за спрямовашстю змш:
• гтокоагуляцгя (зниження зсщання крово);
• ггперкоагуляцгя (пщвищення зсщання крово);
• комбгнованг порушення (тромбогеморапчно порушення при ДВЗ-сштдромо,
геморапчному васкулото).
Зниження зсщання кров! (ппокоагуляц1я)
Основным проявом ппокоагуляцш е геморапчний синдром.
Гемораггчний синдром (застарший термон — “геморапчний доатез”) — це пато-
логгчний стан, який характеризуеться пгдвищеною кровоточивгсгпю, повторними,
часто тривалими кровотечами, крововиливами, що виникають як довгльно, так г в
разг незначно! травми.
Гэморапчн) синдроми внаслщок порушення
тромбоцитарного механизму гемостазу
Тромбоцитопешя — зменшення вмгсту тромбоцитгв у кровг до 100 Г/л I менше (нор-
ма — 150—400 Г/л, або 150,0—400,0-109/л). Проте спонтанно кровотечо, небезпеч-
но для життя, виникають лише тодо, коли кглькгсть тромбоцитов становить 20 Г/л
1 менше.
Класифокацоя. За етгологгею видгляють набуту о спадкову тромбоцитопенйо,
за механизмом виникнення — абсолюгпну о вгдносну (перерозподйгыгу та гемодилю-
цшну).
Етоологоя. Причини виникнення абсолютно! тромбоцитопенП:
• фактори, що уражують мегакарюцитарний паросток кгсткового мозку та
пригшчують тромбоцитопоез — оошзуюче випромшювання, воруси омунодефщиту
людини, гепатиту С; хомочш речовини, особливо цитостатичш препарата, тоазидно
доуретики; алкоголь; лейкозно шфшьтрати; дефщит щанокобаламшу та фолосво!'
кислоти; дисфункщя гемопоетичних стовбурових клгтин (при мгелодиспластич-
ному синдром!);
• фактори, що спричинюють руйнування тромбоцитгв унаслгдок гмунних реак-
цгй, пов’язаних з утворенням антитш до тромбоцитов, у яких антигенна структура
змшена пщ впливом в1рус1в, лгкарських засобтв, а також до незмтнених антигенш
тромбоципв унаслщок зняття 1мунолопчноТ голерантносп (авто1мунна або ццопа-
тична тромбоцитопеючна пурпура, або хвороба Верльгофа); при несумкносп
громбоцитарних антигешв матер! 1 плода;
• мехашчне руйнування тромбоцитов при спленомегалп. штучних судинах «
клапанах серця;
• посилене споживання тромбоцитов при ДВЗ-синдром1, множив ному утво-
ренш тромбоцитарних тромбов у судинах нирок при гемолличному урем1чному
синдром! в дней;
• спадковий дефект утворення тромбоцитов (у тому числ! в раз! дефщиту тром-
боцитопоетину, при комбшованому змунодефщип Вгскотта — Олдрича).
Причиною виникнення перерозподольноо форми водносноо тромбоцитопени може
бути спленомегалш, коли в селезгнщ депонуегься приблизно 90 % тромбоципв
(норма — до 30 %).
Причиною гемодилюишно! форми водносноо тромбоцитопени е пперпдратащя
р1зного генезу.
П атогенез. Видшяють так1 механозми виникнення абсолютно! тромбоцито-
пеног.
• зменшеооня продукии тромбоцитов;
• посилене руйнування',
• подвшцене споживання тромбоцитов пщ час тромбоутворення.
Мехашзмами розвитку водносноо тромбоцитопени можуть бути перерозподгл
тромбоцитов 1 гемодилюцоя.
Порушення гемостазу о кровоточивость при тромбоиитопени зумовленг
• порушенням формування тромбоцитарного тромбу внаслщок зменшення юль-
кост1 тромбоцитов 1, отже. гх учасп в судинно-тромбоцитарному гемостаз);
• водсутшстю спазму судин, порушенням адгези й агрегацп тромбоцитов о змен-
шенням зеодання крово, якз зумовлеш порушенням вившьменмя з тромбоципв ад-
реналину, АДФ 1 ссротонгну, фактора Вылебранда, тромбоксану А?, ТФ зсщання
кров! (мембранного фосфолшопротешу. або ТФ-3, ф!бриногену, антигепариново-
го фактора ТФ-4) тощо,
• порушенням ретракци згустка внаслщок зменшення вмкту скоротливого
бшка тромбонипв тромбастен1ну;
• зменшенням ангютроф'очно! функци тромбоцитов, шо зумовлюе порушення
Ц1ЛОСТ1 стшки М1кросудин, гх ламккть. унаслщок чого еритроцита та ипш с клац-
ни ки кров) переходить у навколосудинний проспр (доапедезна геморагоя з утворен-
ням петехш).
Кл1Я1чн1 прояви тромбопитопенп:
• крововиливи у вигляд! петехш (точков!) та пурпур (петехи, що злилися) на
шкдл та слизових оболонках:
• носов), шлунков! кровотеч), М1крогематур1я (як правило, у раз) зменшення
кшькосп тромбоципв нижче 50 Г/л);
• гию- або гшерплазш юсткового мозку з вщповщними змшами мегакарюци-
тарного паростка:
• позитавн! ендотелзальш проби (поява петехш теля стандартного наванта-
ження);
• збольшений час кровотечо (за методом Айво норма — 3—8,5 хв, за Дьюком —
3—5 хв); зазвичай нормальш показники коагулограми.
Тромбоцитопатия — як1сна неповноцшшсть (бсохшсчна, морфологгчна) I дис-
функция тромбоципнв.
Класиф|кац1я. За егтологгею тромбоцитопатия може бути спадковою о набу-
тою.
Спадкову тромбоцитопатою подоляють залежно в1д порушення основ-
них функцш тромбоципйв на таю види:
• дисадгезивну, наприклад, при макроцитарнгйтромбоцитодистрофп (хво-
роба Бернара — Сулье) генетичний дефект утворення глокопротешу (ГП) 1Ь —
мембранного рецептора до фактора Вшлебранда — порушуе початковий етап ад-
гези тромбоципв до колагену;
• дисагрегацойну, наприклад, при хвороб! (тромбастеноя) Гланцманна —
Негело спадковий дефект мембранного рецептора ГП ПЬ — Ша зумовлюе немож-
ливость поеднання фобриногену з мембраною тромбоципв, що спричинюе пору-
шення ох агрегацп',
• деф!цитну та дисдегрануляцойну в разо дефщиту гранул о порушення
ох секрецп; це спостерогають, наприклад, при синдром! с!рих тромбоципв, коли
в!дсутн! а-гранули о порушене вившьнення фактора Вшлебранда, тромбоспонди-
ну, антигепаринового фактора ТФ-4, факторов росту, а також у випадку синдрому
(омунодефщиту) Вюкотта — Олдр!ча, при якому дефщит а-гранул поеднуеться з
порушенням вивольнення АДФ, серотоншу з а-гранул (щшьних т!лень) о змен-
шенням адгезп тромбоцитов до колагену, ох агрегацп, ослаблениям коагуляцп
кров! та ретракцц згустка.
Етоологоя. Найпоширеношо причини виникнення набуто! тромбоцито-
пати:
• д!я деяких локарських засобов, особливо ацетилсалщиловоо кислоти (необо-
ротно шпбуе циклооксигеназу тромбоцитов, що зумовлюе припинення синтезу
простагландинов, наслщком чого е порушення секрецц гранул о агрегацп тромбо-
цитов);
• доя токсичних речовин екзогенного (етанол) й ендогенного походження
(при уремп, цирозо печонки);
• оонозуюче випромшювання;
• моелопролоферативно та моелодисотластично хвороби;
• дефщит цоанокобаламону, фолоевоо та аскорбшово! кислот, залоза,
• автоомунно хвороби;
• вплив продуктов деградацп фобрину (при екстракорпоральному кровообпу,
ДВЗ-синдромо).
Причиною виникнення спадкова! тромбоцитопати е генетично дефекти струк-
тури мембрани, гранул о боохомочного складу тромбоооитов.
Патогенез. € три основы мехашзми розвитку тромбоцитопати:
• продукцоя патолопчно змонеооих тромбоооитов у костковому мозку;
• блокада тромбоцитарних рецеготоров токсичними метаболотами, макрогло-
булшами тощо;
• часткова деструкцоя тромбоооитов у систем! крово, а також у разо екстракор-
порального кровообпу.
Мехашзм порушення судинно-тромбоцитарного гемостазу ! кровоточивост! при
тромбопитопатп визначаеться характером функционально! недостатност! тромбо-
цита ! с таким самим, як 1 при тромбоцитопенП.
Окремо слщ розглянути функцюнальну недостатшсть тромбоцита пщ час та-
ко! спадково! тромбопитопатп, як анг!огемоф!л!я, або хвороба Виллебранда, що фак-
тично належить до комбшовано! патологи з порушенням судинно-тромбоцитарного
! коагуляцшного гемостазу. Геморапчний синдром спричинений мутащею гена
фактора Вщлебранда, що мгститься в хромосом! 12 1 кодуе синтез цього фактора
в ендотелюцитах 1 мегакариоцитах. Фактор Вщлебранда в норм! накопичуеться в
тромбоцитах 1 вивщьнюетъся у раз! !х дегрануляци. За його вщсутносп не вщбу-
ваються адгезия тромбоцита до колагену судинно! спнки, !х агрегация 1 не фор-
муеться тромбоцитарний тромб. Кр!м того, хвороба Вщлебранда одночасно е 1
коагулопапею з дефщитом фактора VIII, 1 порушенням першо! фази зсщання
кров!. Це пов’язано з тим, що фактор Вщлебранда утворюе комплекс !з фактором
VIII, бере участь у його стабйпзацп 1 транспорт!, а також захищае вщ руйну-
вання.
Набута хвороба Вгллебранда частице зумовлена утворенням автоантитш до
фактора Вщлебранда (при системному червоному вовчаку) або зменшенням його
вмгсту внаслщок абсорбцп фактора Вщлебранда патолопчними бшками (при м!е-
ломшй хвороб!).
Геморапчн! синдроми внаслщок порушення
судинного компонента гемостазу
Геморог!чна вазопапйя — ураження судин у вигляд! змши !х структура, х!м!чного
складу та функцюналъшл неповноцшност!, що зумовлюе порушення судинно-тромбо-
цитарного гемостазу г кровоточивост!.
Класифгкацгя. Геморапчна вазопапя за етюлоггею може бути спадковою 1
набутою, а остання — !нфекц!йною, гмунною, токсичною, рад!ацшною, медикаментоз-
ною, в!там!н-С-дефщитною, зумовленою пухлинним ураженням судинно! спнки.
Етгологгя. Причини виникнення набуто! вазопатп:
• !нфекщйн! фактори (юр, скарлатина, в!спа, геморапчна гарячка);
• токсичш речовини (при уремп, недостатност! печ!нки; отрути комах, зм!й);
• юшзуюче випром!нювання;
• гмунш реакцп;
• ппо- й авггамгноз С;
• пухлинне ураження судинно! спнки лейкозними шфшьтратами, шд час ге-
манпом тощо.
Спадков! вазопатп спричинеш генетичними дефектами синтезу б!лк!в, що
становлять структуру судинно! спнки.
Патогенез.Видшяють таю мехашзми виникнення вазопап!:
• !мунниш, наприклад, при хвороб! Шенляйна — Геноха (пурпура анафшактич-
на, геморапчний васкулит) !мунш комплекси ушкоджують стпгку мгкросудин шю-
ри, суглоб!в, травного каналу, нирок;
• запальний у раз! ушкодження ендотелпо судин вирусами, бактер!ями та пой-
ми токсинами;
• метапласт ичний у випадку геманпоми та метастазування пухлин у судинну
станку;
• диспластичний у раз! порушення структурио трофоки судинно! станки; напри-
клад, при алтментарному дефицита аскорбшово! кислоти порушуеться синтез ко-
лагену; судинна станка ушкоджуеться в раз! посилення пероксидного окиснення
мембранних фосфолопщов при променевой хвороб!; при синдром! Ослера — Рандю
(спадкова геморагочна телеанпектазоя, хвороба Рандю — Ослера — Вебера) у разт
мутацп гена глокопротеону ендоппну судинна станка фактично мае тшьки ендо-
телоальний шар, шо зумовлюе розширення кшцевих воддшов судин (телеанпекта-
зи) з наступним !х розривомо кровотечею; при спадковому синдром '! Еяерса — Дан-
лоса розрив судин зумовлюе порушення перетворення прокола гену на колаген
унаслщок дефщиту проколагеново! амшопротеази;
• дистрофЫний, наприклад у раз! ослабления анпотрофочно! функцп тромбо-
цитов при тромбоцитопени о тромбоцитопати.
Клоночно прояви:
• телеангоектазц (розширення кшцевих вщдолов др!бних судин у раз! стоншен-
ня ох станок);
• петехо! ! пурпура, що виникають унаслщок порушення структур судинно!
стшки;
• кровотеч! р!зно! локалозацо!;
• зазвичай нормально показники коагулограмм.
Геморапчн) синдроми внаслщок порушення коагуляцшннх
I антикоагуляц!йних механ/эм/в гемостазу
Коагулопатия — геморагзчний синдром, спричинений порушенням коагуляцшних з ан-
тикоагуляищних мехашзмзв гемостазу.
Класифокаоия. За етюлог1ею розрозняють коагулопатйо спадкову та набуту.
За патогенезом залежно вщ порушення I, II о III фаз зсщання крово видшяють
протромбшазопатпо, або тромбопластинопатио, тромбшопатт о ф1бринопатйо.
Ет1олог1я. Причини коагулопати з порушенням першо'1 фази задания кров) —
утворення протромбоназного комплексу (активно! протромбо-
нази);
• патолоооя печонки (зниження продукт'! факторов VII, IX, X);
• гшовггамшоз К (фактори VII, IX, X синтезуються в печонцо за участю фшо-
хшонов — вотамппв групи К);
• дефщит кальщю (фактор IV);
• блокада факторов VIII о IX антитшами тд час гемобластозу о дифузних хво-
роб сполучно! тканини, у хворих на гемофолою Л;
• тромбощггопеноя о тромбопитопатая (дефщит ТФ-3);
• надлишок екзо- й ендогенних антикоагулянтов (антитромбопластини);
• генетично дефекта синтезу факторов VIII, IX, XI при гемофшо! (вщповщно
гемофоло! А, В о С), факгораХП, плазмового калокрешогеозу (фактора Флетчера),
фактора VII (ппопроконвертинемоя).
Причини коагулопати з порушенням другоТ физи задания кровз — утворення
тромбту (фактора II):
• хвороби печшки 1 дефщит фшохшошв (у раз! мехашчно!' жовтянищ, енте-
риту, дисбактерюзу, шсля резекцп тонко! кишки), що зумовлюе недостатнш син-
тез у печшщ вхтамш-К-залежного фактора II (протромбшу);
• омунн! шпбггори факторов II, V, VII (наприклад, пщ час локування стрепто-
мщину сульфатом);
• протешуроя, яка спричинюе виведення факторов зсщання кров! нирками;
• онактиващя тромб!ну компонентами антизсщальноо системи — антитромбо-
нами, гепарином (у раз! анафшактичного шоку, передозування гепарину);
• спадковий дефект синтезу фактора II пщ час спадково! гопопротромбшемп,
фактора V — при парагемофшп.
Причини коагулопати з порушенням третып фазы задания кров! — у творен -
ня ф!брину (фактора I):
• патолопчш процеси в легенях, печшц!, яю супроводжуються зменшенням
синтезу фобриногену;
• травми (операцоя) легень, матки, пщшлунково! залози, р!зн! види шоку, що
зумовлюють посилення ф!бринол!зу у зв’язку з утворенням фоброшолозину1 (плаз-
мону);
• передозування ф!бринол!гичних о деф!бринуючих препаратов;
• спадково порушення синтезу фобриногену, ф!бринстаб!л!зуючого фактора
(фактора XIII).
Патогенез. До основних механгзмгв виникнення коагулопати належать:
• набуте о спадкове зменшення, а також якосне порушення синтезу фаосгоров
зсщання кров!, тромбоципв о компонентов калокреш-кшшовоо системи;
• пригшчення факторов зсщання кров! омунними шпботорами — антит!лами;
• надморие споживання цих факторов у раз! посиленого тромбоутворення;
• збшьшення синтезу або активацоя ендогенних антикоагулянтов;
• активацоя фобринолотичноо системи;
• передозування антикоагулянтов, фобринолотичних о дефобринуючих препа-
ратов.
Провгдними ланками в патогенез! гемораггчного синдрому под час коагуло-
патоо, незалежно вщ того, яка фаза зсщання крово порушена, е хронгчна кро-
вовтрата та п наслщки (анемоя, гшоксоя), а також морфолопчн! й функцоонально
змши в мосцо офововиливов (у суглобах, внутршшох органах, шкоро тощо).
Клоночно прояви:
• тривала кровотеча пщ час травми, хорургочних втручань (видалення зубов);
• крововиливи (часто у виглядо гематом — великих скупчень крово, що ви-
йшла оз судини) у пщшкорну жирову клотковину, м’язи, суглоби та шли органи о
тканини, найнебезпечнпш — у головний мозок;
• гематуроя;
• подовжений час кровотечо;
• змши коагулограми залежно вщ вощу коагулопати.
'Посилення фобринолозу зумовлене надходженням у кров активаторов профобринолозину (плазми-
ногену) — тканинних (продукуються в ендотелоальних клотинах, моноцитах, мегакарюцитах), бак-
тероальних компонентов (стрепто- та стафолоконаз) о компонентов калгкре'он-кононовоо системи о систе-
ми комплементу. Кром того, надморна активность фобринолозину може бути спричинена ослаблениям
доо його шгоботоров (а,-антиплазмшу, с^-макроглобулону, 0,-антитрипсину, аоотитромбшу III).
Пщвищення зсщання кров! (пперкоагуляц)я)
Гнтеркоагуляпхя, основою яко! е порушення судинно-тромбоцитарного ! коагуля-
шйного гемостазу, проявляеться локальным тромбозом, тромбофыгчним синдромом,
дисемпюваним внутрйиньосудинним задацрям кровг (ДВ3-синдром).
Локальный тромбоз — це утворення артериальных, венозных, внутр'ниньосериевих
тромбгв, а також тромбгв у мйсроциркуляторному рус.и (визначення тромбозу,
класифгкашю, мехашзми розвитку' див. у роздш ГХ “Патолопчна фхзюлопя пери-
феричного кровообну”).
Тромбофшчний синдром (тромботичний синдром, гематогенна тромбоф!л!я.
клгшчна назва — тромбоембо.шчна хвороба) — це тдвшцена схильшсть до тромбо-
утворення, зумовлена порушенном мех ан 1зм1в гемостазу й антигемостазу.
Класиф»ка.п!я. Залежно в!д етюлогн тромбофшчний синдром може бути спад-
ковим (первинним) 1 набутим (вторинним, симптоматичним).
Ет1Олог1Я.До найпоширешших причин виникнення спадкового тромбо-
фшчного синдрому належать:
• генетичний дефект проакцелерину (фактора V) зумовлений тим, що внасл!-
цок так звано! лейденсько! мутацп хромосоми 1 фактор V стае нечутливим до
шактивацп ангигромботичним комплексом тромбомодулш — протеш С, що зни-
жуе здатшеть судинно! стшки обмежуваги утворення фибрину (5 % населения
€вропи з такою аномалию, яка мае автосомно-домшантне успадкування, страж-
дають на флеботромбоз нижнгх кшщвок);
• тшергомоцистешемтя, що виникае при порушен ш метаболизму гомоцистешу,
зумовленого спадковим дефектом цистатюнсинтетази, гомоцисте!нметитгрансферази
та !нших ферменпв, спричинюе ушкодження судинно! стшки. Це зумовлюе зни-
ження п ангикоагулянтних втастивостей, що викликае пщвищення агрегацп тром-
боцитов, активацпо утворення тканинного тромбопластину, наслщком чого е ар-
тер!альний 1 венозний тромбоз, посилення атерогенезу пщ впливом тромбоцитар-
них циток!н!в, своею чертою атеросклероз також зумовлюе тромбоз артергй;
• генетичн! аномали синтезу антитромбшу ПТ, антикоагулянтних протешгв С
‘ 8, шо спричинюють венозний тромбоз;
• спадков! гшерлшопротещемц, особливо типу I (гшерхьтомгкронемгя) 1 типу
V (пщвишений вм!ст у кров! хптомгкрошв 1 ЛПДНЩ), як! зумовлюють розвиток
атеросклерозу 1 пщвишену схильшсть до тромбоутворення.
Етюлопчн! фактори набутого тромбофшчного синдрому.
• гшергомоцистешемзя, спричинена недостатшстю в!тамгн!в В6, В,! Вг (див.
розд!л XXIV “Патолопчна фгзюлопя обмшу вггампйв”);
• атеросклероз;
• цукровий диабет;
• сепсис (порушення тромборезистентност! судинно! стшки й активашя ге-
мостазу гпд дгею бактерТальних токсишв);
• пгеляоперащйний тромбоз як наслщок травми, стресу й плотинами, що
сприяють активацп зсщання кровх;
• нефрогичний синдром (втрата антитромбшу 111 за значно! протегнурп);
• гшерестрогешзм (лгкарськг засоби, контрацептиви, гшергонадизм) ослаб-
люе синтез антикоагулянт (в ! зб!льшуе синтез прокоагулянпв;
• тромбоцитоз ( збшьшення кшькост! тромбоципв до 600 Г/л 1 бшьше). особ-
ливо щд час гемобластозу (есешцатьна тромбопитем!я, гострий мегакарюбласт-
ний лейкоз, справжня полшитем!я), теля спленектомп;
• авготмунна громбофьйя, коли автоантитша до фосфолшццв мембрани фтк-
суютъся на тромбоцитах 1 ендотелюцигах т активують гемостаз (при колагенозах,
В1Л-шфекци, лгкуванш хлорпромазином);
• злояюсн! пухлини, яю продукують прокоагулянта (фактор Ш), протешази —
активатори фактора X (аденокарцинома молочно! залози, легень та ш.).
Патогенез. Основними мехашзмами, яю можугь зумовити розвиток тром-
бофштчного синдрому, е:
• зниження ангитромботичних властивостей стшки судин;
• збшьшення вмюту в кров! тромбоципв (тромбоцитоз);
• пщвищення функцюнально! активносп зещально! системи кров! за рахунок
збшьшеного надходження в кров прокоагулянт, в та активатор,в зсщання кров!;
• зменшення антикоагуляшйно! активносп кров! й актавносп ф!бринолггич-
но! системи.
Клтатчн! прояви:
• флеботромбоз, тромбофлебт, варикозне розширення вен !з подалыиим розвит-
ком набряДв I трофзчних порушень у тканинах та органах;
• тромбоз 1 тромбоембо.йя артер!альних 1 венозних судин ртзних оргашв (ле-
гень, серця, головного мозку, кишок та тн.) з розвитком некрозу (шфаркту) 1
функшонально! недостатносп цих орган!в,
• зменшена тривалтсть кровотеч!;
• порушення коагулограми — збшьшення вмтсту прокоагулянпв ! зменшення
компоненпв системи антигемостазу.
ДВЗ-синдром — це патологгя гемостазу, що характеризуеться фазовыми змша-
ми гтер- та гиюкоагуляци: починаеться з генерализованы активацп задания кровг,
що зумовлюе утворення множинних мйеротромб'гв у судинах мпсроциркуляторного
русла; виникнення тромбоцитопени I коагулопати споживання, активащя системи
антигемостазу спричинюють подалыиий розвиток гтокоагуляци та гемораги.
Класиф|кац!я, За перебйом ДВЗ-синдром подшяють на гострий ! хрошчний,
за поширенням — на генераллзований т локальний (рщко).
Еттологтя. Причини виникнення гострого генералгзованого ДВЗ-синдрому.
• генералтзоваш шфекцп та септичш стани;
• уст види шоку т термшалънт стани;
• травм и (також хтрурпчн!);
• акушерська патолог!я (передчасне в!дшарування плаценти, ручне вщокрем-
лення плацента тощо);
* гострий внутршптьосудинний гемолиз;
• гостр! запальш процеси з некрозом (гострий панкреанекроз, перитошт).
Хрошчний ДВЗ-синдром спричинюють злояюсш пухлини, у тому числ! гемо-
бластоз, тяжи хвороби й ураження печшки (цироз, некроз), затримка в матщ
мертвого плода тощо.
Локальний ДВЗ-синдром виникае при хворобах, за яких на певних дшянках су-
динного русла теля тромбоутворення розвиваються тромбоцитопентя т коагуло-
папя споживання з геморапею (аневризма аорги, геманпома, патологтя нирок).
патогенез. Пров!дною ланкою в патогенез! ДВЗ-синдрому е порушення ба-
лансу м!ж кал!кре1н-к!н!новою, зс!далъною, протизс!далъною ! ф!бринол!тичною сис-
темами кров! та фазов! змши гемостазу.
Перша фаза ДВЗ-синдрому — фаза гтеркоагуляци — харакгеризуеться посиле-
ним тромбоутворенням у судинах мткроциркуляторного русла. Це пов’язано з
активащею системи зсщання кровг починаючи з п першо! фази, перебгг яко! може
вщбуватися здебшыпого за зовн!шн!м (при травмах) ! внутрппшм мехашзмами
зсщання (наприклад пщ час внутр, шньосудинного гемошзу).
Активащя зс!дання кров! може виникати'.
• пщ час надходження в судинне русло велико!’ юлькост! прокоагулянпв (на-
самперед тканинного тромбопластину) та !хшх активатор!в (у раз! травми, некро-
зу печшки, гемолизу);
• у раз! первинного порушення ендотелпо судинно! стшки (бактер!альними
токсинами, вирусом герпесу, ошками), коли колаген субендотелгального шару зу-
мовлюе активацпо адгезп тромбоципв;
• унаслщок початкового ушкодження тромбоципв (безпосередня д!я вгрусгв,
бактерш, !мунних комплекс!в або за рахунок стимуляцн макрофапв). Таким чи-
ном порушуеться така важлива функция системи гемостазу, як збереження нор-
мального агрегатного стану кров!.
Фгбринов! мжротромби зумовлюють блокаду мпсроциркуляцп та розвиток
тяжких дистрофгчних 1 функщональних порушень в органах ! тканинах (недостат-
шсть нирок, надниркових залоз, легень, мозкового кровообпу та ш.), нерщко не
СУМ 1С них 13 життям.
У друпй фаз! ДВЗ-синдрому — фаз! гтокоагуляци — виникае геморапчний
синдром 1з сильною кровотечею, яку складно спинити. Основою ц!е! фази е втра-
та здатносп кров! до зсщання й агрегацп тромбоципв, юльюсть яких, як 1 фак-
тор!в зсщання кров!, р!зко зменшуеться завдяки !х використанню для утворення
тромб 1 в у попереднш фаз! (коагулопагтя споживання). Кр!м того, пщ впливом
тканинних фактор!в активуються протизсщальна 1 ф!бринол!тична системи кров!,
що посилюе кровотечу.
Трети фаза ДВЗ-синдрому може бути в!дновною (фаза нормал!заци гемостазу) 1
терм!нальною. В останньому випадку повне виснаження систем гемостазу, анти-
гемостазу 1 ф!бринол!зу зумовлюе тяжку крововтрату, постгеморапчну анемпо,
некроз тканин, недостатшсть функцп оргашв 1 систем (нирок, серця, мозку, ле-
гень та !н.), у яких виникли розлади кровообпу в раз! тромбозу або крововтрати.
Природно, що так! змши мають необоротний характер.
Р03Д1Л XXVI
ПАТОЛОПЧНА Ф13ЮЛ0ПЯ
СИСТЕМНОГО КРОВООБ1ГУ
За нормальних умов координована робота серця та судин забезпечуе кров’ю орга-
ни о тканини вщповщно до !хнох потреб. У стан! повного спокою сумарна потреба
в крово становить у доросло! людини близько 3 л/хв • м2. У разо значного фозично-
го навантаження вона може зрости у 3—4 рази, а у спортсменов ще больше.
Велик! функцоональш можливост! системи кровообогу й адекватне присто-
сування Г! до потреб органозму забезпечуються тим, шо серце о судини мають
тонку о водночас стойку регуляцою. Ця регулящя, як внутрппньосерцева, так о
загальна нервово-гуморальна, забезпечуе не лише координовану роботу розних
воддолов серця, його зв’язок оз судинами, а й з шшими системами — дихання та
крово. Тому на пщвищення вимоги до кровообпу реагуе не лише серце (зболь-
шенням ХОС) або судини (змоною тонусу, перерозподолом штенсивност! ре-
понарного кровообогу), а й система дихання (збольшенням легенево! вентиля-
цп, утилозацп кисню тканинами) о система кровотворення (активацоею еритро-
поезу).
Патолопю кровообогу, яка може виникнути внаслщок ушкодження серця чи
судин або порушення !х регуляцп, також супроводжуе розвиток перелочених вище
пристосувань. Завдяки цьому надовго можуть компенсуватися порушення в той
або оншой ланцо кровообпу, проте, якшо ушкодження надто значне, а компенса-
торно можливост! органозму зменшено або вогчерпан!, то розвиваеться недостат-
шсть кровообогу.
Недостатшсть кровообггу — це порушення гемодинамики, яке проявляеться в
тому, що органи / тканини не забезпечуються потргбною киькгстю циркулюючо'1
кровг, а це своею чергою спричинюе недостатне забезпечення Тх киснем, поживними
речовинами, порушення видалення кгнцевих продуктгв обмгну.
Недостатшсть кровообпу може виникнути внаслщок попршення роботи сер-
ця (серцева недостатшсть) або змони функцой судин (недостатшсть судин). Часто
спостерогаеться комбшована серцево-суцинна недостатшсть. Як правило, будь-
яка !зольована форма недостатносто надало стае змошаною.
Кожна оз цих форм недостатност! може перебогати гостро або хроночно о мати
розний ступшь прояву, набуваючи вигляду компенсовано! (приховано!), субком-
пенсованоо або декомпенсовано! (явно!) недостатносто.
РОЗЛАДИ КРОВ00Б1ГУ, ЗУМОВЛЕН1 ПОРУШЕННЯМ
ФУНКЦП СЕРЦЯ
Робота серця харакгеризуеться особо ивостями, пов'язаними з його функщону-
ванням, метаболтзмом, кровопостачанням та шнервацтею. яю зумовлюють яюсн!
вщмшносп патолопчних процеств, що розвиваються в ньому.
До них належать безперервшсть функшонування серия за рахунок спецталтзо-
ваного апарату автоматизму та високий аеробний метаболизм. Навить за макси-
мального напруження глткошзу вш не може забезпечити больше шж 10—20 %
енергетичнот потреби серця, що зумовлюе високу чутливтсть серцевого м’яза до
нестач! кисню.
Серце добре постачаеться кров’ю. Особ.гивостями судин серця е високий то-
нус, який у разт збтльшеного навантаження дае !м змогу розширюватись у 5—6
раз1в, наявшсть анастомоз!в мтж артер1ями четвертого-п'ятого порядку, а також
мтж артергями 1 кашлярами та невелика юльюсть анастомозтв мтж вгнцевими ар-
тертями. Тому, коли виникае о клюзы мапстральноУ артери, внутртшньосерцевт
анастомози нездатш забезпечити нормальну цирку ляцтю кровт, осюльки ними
надходить не быьше 1/4 ц вихщно! юлькосп.
Осюльки мтокард навтть у стант спокою бере з кровт, що надходить, 3/4 кис-
ню (скелетний м’яз у стаю спокою. наприклад, потребуе лише 20—30 % кисню),
единим способом забезпечити птдвищену потребу серця в кисш е збтлыиення
втнцевого гоку кровт. Це робить серце, якжоден тнший орган, залежним вщ стану
судин, механтзмтв регуляцн втнцевого току кровт т здатносп втнцевих артерш адек-
ватно реагувати на змтни навантаження.
Серце дуже чутливе до порушень обмтну електролтттв, вщ якого залежать ав-
томатизм, провщюсть, взаемозв'язок збудження та скорочення, а також стан де-
яких ферментних систем.
Серпе тннервуеться симпатичною т парасимпатичною частицами автономно!
(вегетативно!) нервово! системи, а за насичентстю адренерпчними нервовими во-
локнами, а також за вмтстом норадреналпзу не мае соб! ртвних серед шших ор-
таюв. Втдомо, що медтатор симпатично! нерьовот системи пщвищуе напруження,
яке розвивае м’язове волокно серця, посилюе обмтн речовин, споживання кисню
т жирних кислот, обмтн юнтв кальцпо та калпо. Таке переважання симпатично!
щнервацп створюе т теред умови для пщвтпценот уразливостт мтокарда.
Встановлено, що м'язов! клттини серця дорослого оргашзму не дтляться, не
здатш до регенерацп. Замещения функцп клггин, яю загинули. т пристосування до
тривалого пщвищеного навантаження вщбуваеться пльки за рахунок збшьшення
внутрт'шньоклттинних структур неушкоджених клггин — тх гшертрофп. Певна ре-
генерацтя може втдбутися за рахунок стовбурових юптин мтокарда.
Незалежно вш причини виникнення патолопчного процесу в серш наслщки
його типов! — це порушення кровообтгу ! кровопостачання оргашв т тканин.
Недостатшсть серця
Недостатшсть серця розвиваеться в разт невщповщностт навантаження на серце,
його здатносп виконувати роботу, яка визначаеться ктльюстю кровт. що надхо-
дить до серця, опором вигнанню кровт в аорт! та легеневому стовбурт. Отже, не-
достатнгсть серця виникае тощ, коли серце не може за наявного опору перекача-
ти в артерп всю кров, що надшита венами.
Розргзняють три патофгзгологгчних варганти серцево! недостатност!.
1. Серцева недостатшсть в/д перевантаження, що розвиваеться при: хворобах,
якг спричинюють збыьшення опору серцевому викиду або притоку кров! до пев-
ного вщдглу серця, наприклад у раз! вади серця; гшертензп в судинах великого
або малого кола кровообггу, артерговенозних ф!стулах або в раз! надмгрного фг-
зичного навантаження.
2. Серцева недостатшсть через ушкодження мюкарда, спричинене гнфекщею,
гнтоксикацгею, гшоксгею, авгтамшозом, порушенням вгнцевого кровообггу, фг-
зичним та психгчним перенапруженням, деякими спадковими дефектами обмгну.
У таких випадках недостатшсть розвиваеться навгть за нормального чи зменше-
ного навантаження на серце.
3. Змниана форма серцево!' недостатност!, що виникае в разг поеднання уш-
кодження мюкарда та його перевантаження, наприклад пгд час ревматизму, коли
спостерггаеться комбгнацгя запального ушкодження мюкарда та порушення кла-
панного апарату серця. Такий варгант недостатност! серця виникае г в тих випад-
ках, коли внаслщок дистрофгчних змгн або загибелг частини м’язових волокон
серця на решту припадае шдвищене навантаження.
Серцева недостатшсть вщ перевантаження, мехашзми компенсацп. Пщвищене
навантаження на серце може бути наслщком збыьшення кглькостг кровг, що при-
тгкае, або посилення опору кровг, що вгдтгкае. Перше спостерггаеться пщ час фг-
зично! роботи або вад серця, що супроводжуються недостатнгстю клапанного апа-
рату. За таких вад пщ час дгастоли в порожнину серця потрапляе не лише та кров,
яка надходить нормальними шляхами, а й та, яка внаслщок неповного замикання
клапашв викинута з порожнини пщ час систоли. Те саме спостерггаеться й у раз!
вроджених дефекпв перегородок серця. Причиною пщвищеного навантаження на
серце е також збгльшення опору вщтоку кровг, що виникае внаелгдок звуження
(стенозу) вихщного отвору з порожнини серця (наприклад отвору легеневого
стовбура або аорти, передсердно-шлуночкового отвору), артергально! гшертензп,
генералгзованого атеросклерозу, пневмосклерозу.
В експеримент! ргзш види порушення дгяльностг серця вивчають шляхом
створення штучно! вади клапашв або звуження великих вщвщних судин — аорти
та легеневого стовбура.
Серце здатне швидко пристосовуватися до пщвищеного навантаження г, вико-
нуючи пщвищену роботу, компенсувати можливг розлади кровообггу. При цьому,
залежно вщ виду навантаження, спрацьовуе той чи гнший мехашзм компенсацп.
У разг перевантаження об’емом кровг спрацьовуе гетерометричний мехашзм
компенсацп (Франка — Старлгнга). Пщ час дгастоли пщвищуеться кровонаповнен-
ня порожнин (або однге! порожнини) серця, шо зумовлюе збыьшене розтягнення
м’язових волокон. Наслщком такого розтягнення е посилене скорочення серця
пщ час систоли. Цей мехашзм зумовлений властивостями клгтин мюкарда. У в!-
домих межах навантаження генуе лгншна залежшеть мгж кыькгетю кровг, що над-
ходить, г силою скорочення серця (мал. 48, а).
Проте, якщо ступгнь розтягнення м’язового волокна перевищуе допустим!
меж!, сила скорочення зменшуеться. Зменшення напруження вщбуваеться в разг
розтягнення сегмента мюкарда понад 25 % його вихщног довжини, що вщповщае
Мм. 48. Залежшсгь хвтинного об’ему серця (Р) ащ його роботи (И)
при 1'етерометрнчному (а) 1 гомеоме триному (б) мехашзмах компенса-
цп (частота скорочень серця постшна):
1, 2 — нижнгй 1 верхней ртвш об’ему кровь шо припливае. та опору п
вщтоку, поза якими компенсашя не вщбуваеться
збшъшенню об’ему порожнини левого шлуночка приблизно на 100 %. За допус-
тимих псревантажень лилию розмтри серця: збшыпуються не бшьше н!ж на 15—
20 %. Розширення порожнин серця при цьому супроводжуеться збшьшенням
ударного об’ему । називаеться: тоногенною дилатащею.
За пщвищеного опору вщтоку кров! спрацьовуе гомеометричний мехашзм ком-
пенсаци. У цьому випадку довжина м’язового волокна серця збгльшуеться не так
ргзко, але пщвищуються тиск 1 напруження. шо виникли внаслщок скорочення
м’яза в кппц д!астоли. Сила скорочень серця збшьшуеться не одразу, а поступово
з кожним наступним скороченням серця, доки не сягне ргвня, потр!бного для збе-
реження сталосп ХОС. У вщомих межах навантаження потужнють, шо розвиваеть-
ся пщ час скорочення серця, лгшйно пов’язана з величиною опору вщтоку кров>
У раз! виходу за щ меж! сила скорочення серця зменшуеться (мал. 48, б).
Енергетично обидва мехашзми компенсац!] пщвищеного навантаження не-
ргвношнн! Так, у раз! однакового збшьшення зовшшньо!' роботи серця, розрахо-
ваного за добутком ХОС на середнш систол!чний тиск в аорт!, споживання кис-
ню серцем змшиться неоднаково. залежно вщ того, чим зумовлене посилення
роботи — збшьшенням надходження кров! до серця або збшьшенням аортального
опору. .Якщо робота подвошася за рахунок збшьшення вдв!чг ХОС, то споживан-
ня кисню зростае лише на 1/4, якщо ж робота подвошася за рахунок збшьшення
вдвгч! опору вщтоку кров!, то споживання кисню мюкардом збгльшуеться на
200 %. Це пояснюеться тим, що за гомеометричним мехаюзмом компенсацп для
подолання пщвищеного опору вщтоку кров! потр!бне значне пщвищення сис-
толгчного тиску, якого можна досягти шляхом збшьшення величини г швидкост!
напруження м’язового волокна. А саме фаза !зометричного напруження е най-
б!льш енергоемною ! визначае витрагу АТФ 1 спожгшання кисню мюкардом.
Отже, гетерометричний мехашзм компенсацп е економгчнппим, юж гомео-
метричний, чим, можливо, г пояснюеться сприятливппий перебт тих патолопя-
Мал. 49. Змша тривалоеп серцевого циклу,
систоли та дгастоли за р1зно) частота серцевих
скорочень у людини (штрихом позначено
систолу передсердь)
динамично! характеристики серця: шд
них процесхв, що супроводжуються роз-
витком механизму Франка — Старл!нга,
наприклад, недостатност! клапашв по-
р1вняно 31 стенозом отвору.
Компенсаторним мехашзмом, що
забезпечуе сталхсть ХОС, також може
бути прискорення скорочень серця —
тахйсардгя. Вона може виникати як за
рахунок прямого впливу пщвищеного
тиску кров! в порожниш правого перед-
сердя на вод!я ритму — пазухо-перед-
сердний вузол, так 1 за рахунок нервових
1 гуморальних екстракард!альних вплив!в.
3 енергетичного погляду це найменш
вигщний мехашзм компенсацп, ос киш-
ки вш супроводжуеться, по-перше, вит-
рачанням велико! юлькосп кисню, по-
друге, значним скороченням д!астоли —
перюду вщновлення ! вщпочинку
мюкарда 1, по-трете, попршенням гемо-
: д!астоли шлуночки не встигають запов-
нюватися кров’ю, систола стае меню повноцшною, осюльки за цим неможлива
моб!л!защя гетерометричного механизму компенсацп. 3 мал. 49 видно, що 31 ско-
роченням серцевого циклу (верхня крива) тривалгсть систоли зменшуеться мен-
ше, н1ж тривалгсть дгастоли. Момент початку скорочення передсердь (штрихова
л1Н1я) наближаеться до кпщя систоли шлуночюв, доки при частот! 170 за 1 хв не
зб1житься з ним (“закупорювання передсердь”). На ЕКГ при цьому зубець Р на-
шаровуеться на зубець Т.
Описан! мехашзми компенсацп шд час перевантаження серця можна проде-
монструвати 1 на !зольованому, позбавленому регуляторних зв’язюв з оргашзмом
серц!. Вони зумовлеш властивостями серцевого м’яза, його провщно! системи !
певною м!рою функщею внутр!шньосерцево! нервово! системи. Остання пред-
ставлена нейронами, розмщеними в серц! до р!вня передсердно-шлуночково! пе-
регородки, яю утворюють рефлекторш дуги в межах серця. Вважають, що функ-
щя внутршшьосерцево! нервово! системи полягае в пристосуванш д!яльност! сер-
ця до навантаження 1 координацп роботи передсердь 1 шлуночюв серця, л!во! 1
право! його половин.
На внутрппньосерцев! мехашзми регуляцп впливають позасерцев! (нервов! й
гуморальш) мехашзми. Серед них особлива роль належить симпатичшй частин!
автономно! нервово! системи, 1! мед!аторам норадреналшу й адреналшу. Перший
видыяеться нервовими заюнченнями, другий — юптинами мозково! речовини
надниркових залоз. Щ мед!атори (катехолам!ни) взаемодпоть з рецепторами на
поверхш серцевого мюцита. Рецептори симпатично! частини нервово! системи
подыяють на два класи — а- 1 р-рецептори, кожен з яких подыяеться на шдкласи
ар а2, рр Р2. У серц! ссавцш мгстяться переважно р,-рецептори, а в непосмуго-
вашй м’язовш тканин! судин — о^- 1 Р2-рецептори. Внутрппньоклпинш ефекти
стимуляцп рецепторгв зумовлеш пщвищеним вмгстом цАМФ, збыьшенням ак-
тивностт цАМФ-залежно! протешюнази, змшою потоков юшв Са+ т зв’язування
Са24 клттинними структурами. У раз! збудження симпатичного вщдшу нервово!
сисгеми значно збшьшуються сила ! швидюсть скорочень серця, зменшуеться
об’ем залишково! кров! в порожнинах серця за рахунок повинного вигнання кров!
пщ час систоли (при звичайному навантаженн! приблизно половина кров! в шлу-
ночку залишаеться в юнц! систоли), збтлыпуеться частота серцевих скорочень У
раз! пщвищення тонусу симпатичних нершв г видпення велико! юлькост! катехо-
ламш!в ефектившше компенсуеться перевантаження за рахунок внутрштньосер-
цевих регуляторних мехашзмгв.
Порушення симпатично! пшервацп серця, зокрема т шсля введения деяких фар-
маколопчних засоб!в або застосування експериментально! си млате ктомп, утруд-
нюе мобшзашю компенсагорних механгзмгв, що знижуе функшональю можли-
вост! серця.
Якшо навантаження на серце надм!рне. компе нсаторн! мехашзми не снрав-
ляються з ним. розвиваеться гостри серцева неоостатшсть. При цьому в серцево-
му м’яз! виникають змши у вигляд! внутрштньоклпинного накопичення юшв на-
трпо 1 кальшю, порушення синтезу макроерпчних сполук, закиснення внутрйпньо-
КЛ1ТИННОГО середовища з подалыпим порушенням процеств скорочення !
розслаблення серцевого м’язового волокна. Це зумовлюе зниження сили та швид-
кост! скорочення серцевого м’яза, збшьшення залишкового систол!чного об’ему г
д!астол!чного тиску, розширення порожнин серця. Гостра сернева недостатшсть
супроводжуеться тяжкими порушеннями: нщвищенням венозного тиску, змен-
шенням ХОС, г!покс!ею тканин. У серцевому м’яз! кр!м обмшних можуть вини-
кати 1 структури! змши, через яю навпь у раз! зменшення подальшого каванта-
ження функщю серця нормалгзуваги не вдаеться.
Гостра недостатшсть серця розвиваеться гакож пщ час фтбриляцт! шлуночюв,
ттароксизмально! тахткардн, шфаркту мтокарда, мюкардиту, тромбозу клапанного
отвору, емболи легенево! артери, тампонада сердя. При цьому спостерйаеться
недостатке наловнення кров'ю артертально! системи, що зумовлюе пнемпо голо-
вного мозку з тяжкими порушеннями його функцп як! нагадують клтшчну кар-
тину шоку ! нерщко супроводжуються потьмаренням свщомосп та судомами.
У раз! гривалого перевантаження серця, наприклад пщ час вади клапан!в, п-
пертензивнот хвороби, розвиваються гривалт мехашзми компенсаци у вигляд!
специфгчних обмшних т структурних змш у мтокард!, шо зумовлюе збшьшення
маси серця, посилюе його функшю.
Гшертрофтя мтокарда. Тривале значне навантаження на серцевий м’яз супро-
воджуеться збшьшенням навантаження на одиницю м’язово! маси! потребуе бтль-
шо! тнтенсивностт функшонування п структур. У вщповщь на гаке навантаження
активуеться генетичний апарат м язових т сполучнотканинних юптин. Гак. у тпд-
доелшно! тварини через юлька годин птсля звуження аорти в клттинах серця вияв-
ляються ознаки посилення функцц ядра, збшьшення синтезу РНК т юлькост! ри-
босом. Пщ юнець першо! доби посилюеться синтез бтлюв, шо спричинюе швидке
збшьшення об’ему м’язового волокна, його ггпертрофтю !, як правило, супровод-
жуеться гшертрофтсю того вщдшу серця, на який припадае збшьшене навантажен-
ня. При цьому збшьшуеться об’ем кожного серцевого м’язового волокна, загальна
ж юльюсть волокон залишаеться незмшною. За рахунок гшертрофп мтокарда на-
вантаження на одиницю м’язово! маси знижуеться до нормального ртвня.
У разт зменшення навантаження, наприклад шсля лтквщаци стенозу, вщнов-
лення клапантв маса мюкарда зменшуеться до норми. Це вказуе на те, що штен-
сившсть синтезу бтлктв у кл (тинах мюкарда значною мтрою регулюеться ртвнем
навантаження. Кртм того, цей процес контролюеться мехашзмами нервово-гумо-
ральнот регуляцп.
Гшертрофтя мюкарда — явшце пристосувальне, спрямоване на виконання
пщвищенот роботи без тстотного збтлыиення навантаження на одиницю м’язовот
маси серця. Це досить досконале пристосування. Так, гшертрофтя мюкарда у
спортсменов дае змогу серию виконувати надзвичайно велику роботу. При цьому
змшюеться т нервова регуляцтя серця, що значно розширюе дтапазон його адап-
таци та можливосп витримувати значно навантаження. У разо патолопчних про-
цесс гшертрофтя серця довго компенсуе порушення, що виникають. Так, напри-
клад, пщ час розтинтв було виявлено, що майже 4 % людей мають вади клапанов,
яко супроводжуються ппертрофоею серця, о лише в 0,5—1 % особ захворювання
проявлялося клштчно.
З’ясуванню мехашзмгв розвитку гшертрофп серця, особливостей обмшу та
функцш гшертрофованого мюкарда, причин виникнення декомпенсаци серця
сприяло вивчення процесу в експеримент!. Велика заслуга у створенно розних ек-
спериментальних моделей хвороб серця, зокрема й експериментальних вад серця,
належить О.Б. Фохту.
Пщ час експериментального втдтворення розриву клапана або звуження аор-
ти гостро збольшуеться навантаження о змшюеться гемодинамтка. Це може спос-
терггатися т в людини, наприклад пщ час травматичного ушкодження клапанного
апарату, гострого перевантаження серця, гшертензивного кризу. Експерименталь-
на модель гострого перевантаження серця дае змогу з’ясувати послщовшсть змш,
як! вщбуваються, визначити тх причинно-наслщковий зв’язок.
Гшертрофоване серце вщрозняеться вщ нормального низкою обмшних, фун-
кцюнальних о структурних ознак, яко е вщображенням, з одного боку, можливосто
протягом тривалого часу долати пщвищене навантаження, з шитого — наявносп
передумов для виникнення патолопчних змш.
Збтльшення маси серця вщбуваеться за рахунок стовщення кожного
м’язового волокна, що супроводжуеться змшою сшввщношення внутршшьокл!-
тинних структур. Об’ем клттини при цьому збшыиуеться пропорцтйно кубу лтнтй-
них розмтртв, а поверхня — пропорщйно тхньому квадрату, що зумовлюе змен-
шення клттинно! поверхнт на одиницю маси клттини. Вщомо, що через
поверхню клттини вщбуваеться п обмш тз позаклттинною рщиною — поглинання
кисню, поживних речовин, виведення продукпв метаболтзму, обмш води й елек-
тролтв. Зазначент змши створюють умови для попршення постачання м’язового
волокна, особливо його центральних втддтлтв.
Клттинна мембрана вщтграе важливу роль у проведенн! збудження 1 в поед-
нанн! процеств збудження та скорочення, що здшснюеться через тубулярну сис-
тему т саркоплазматичну сттку. Оскщьки ртст цих утвортв пщ час гшертрофп
м’язового волокна також вщстае, створюються передумови для порушення
скорочення 1 розслаблення серцевих мтоциттв: унаслщок сповтльиен-
ня виходу юшв кальщю в саркоплазму попршуеться скорочення, а внаслщок ут-
руднення зворотного транспорту юшв кальцпо — розслаблення. 1нодт можуть ви-
никати локалъш контракгури окремих серцевих мюципв.
Мал. 50. Сшввщношення мЬк м’язовими во-
локнами, кашлярними судинами 1 нервами
серця в немовляти {а, г), здорово! доросло!
людини (б, П маса серця 310 г) I доросло!
людини з ппертрофованим мюкардом (в, д;
маса серця — 540 г)
У раз! ппертрофц збгльшення
об’ему клётини перевищуе збыьшен-
ня об’ему ядра. Здатшеть ядра висо-
кодиференцшовано! клётини до подё-
лу рёзко обмежуеться. Оскыьки роль
ядра полягае в забезпеченш белково-
го синтезу, а отже, ё процесёв вёднов-
лення внутрёшньоклётинних структур,
то вёдносне зменшення ядра може
спричинити порушення синтезу
бёлкёв ё погёршення пластич-
ного забезпечення клётини.
О - М’язове • - Кашлярна ▼ - Нервове
волокно судина
волокно
У процесг розвитку гёпертрофп маса мётохондрёй спочатку збёлыпуеться швид-
ше, нёж маса скоротливих бёлкёв, створюючи умови для достатнього енергетично-
го забезпечення ё повноё компенсацп функцп серця. Проте згодом, у мёру усклад-
нення процесу, збёльшення маси мётохондрёй починае вёдставати вёд росту маси
цитоплазми. Мётохондрёё починають зазнавати граничного навантаження, у них
розвиваються деструктивнё змёни, знижуеться ефективнёсть гхнъоё’ роботи, пору-
шуеться окисне фосфорилювання. Це спричинюе погёршення енергетично-
го забезпечення гёпертрофованоё клётини.
Збёльшення маси м’язових волокон серця часто не супроводжуеться адекват-
ним збёльшенням капёлярноё сётки, особливо у випадках швидкого розвитку його
гёпертрофп. Великё вёнцевё артерп також не здатнё до потрёбного пристосувально-
го росту. Тому пёд час навантаження погёршуеться судинне забезпечення
гёпертрофованого мёокарда (мал. 50).
У ппертрофованому серцё порушена структура вставних дискёв ё 2-лёнёй,
унаслёдок чого змёнюеться електрична активнёсть мёокарда, погёр-
шуеться координованёсть скорочення серця в цёлому.
До процесу гёпертрофп мёокарда залучаеться нервовий апарат серця. Спос-
терёгаеться посилене функцёонування внутрёшньо- ё позасерцевих нервових еле-
ментёв. Проте ёх рёст вёдстае вёд росту маси скоротливого апарату мёокарда. Вёдбу-
ваеться виснаження нервових клётин; порушуються трофёчнё впливи, зменшуеть-
ся вмёст норадреналёну в мёокардё, що спричинюе погёршення скоротливих
властивостей, утруднення мобёлёзацп його резервёв. Отже, порушуеться ё ре-
гуляторне забезпечення серця.
Гёпертрофоване серце за рахунок збыьшення маси скоротливого й енергоза-
безпечувального апаратёв здатне тривалий час виконувати значно бёлыпу роботу,
нёж здорове серце, зберёгаючи при цьому нормальний метаболёзм. Проте здат-
шеть пристосовуватися до змёни навантаження, дёапазон адаптацёйних можливос-
тей у гёпертрофованого серця обмежеш.
Зменшений функцгональний резерв. Незбалансовашсть внутршшьо-
клттинних г тканинних структур зумовлюе бшыпу уразливтсть гшертрофованого
серця в разг впливу несприятливих факторгв.
Тривале штенсивне навантаження на серцеве м’язове волокно зумовлюе вис-
наження його функцц. Причиною може бути розлад скоротливо! функцп м’язового
волокна внаслщок зменшеного утворення енергп мгтохондргями г порушення п
використання скоротливим апаратом. За ртзних форм серцево! недостатносп один
13 патолопчних варганив може переважати. Зокрема в раз! тривало! гшерфункцп
серця провщним е порушення використання енергп. Одночасно зт зменшенням
скоротливо! функцц утруднюеться розслаблення м’язового волокна, виникають
локальн! м’язов! контракгури, що надал! призводить до дистрофц 1 загибел! сер-
цевих МЮЦИПВ.
Збшыпене навантаження нер!вном!рно розподшяеться мтж р!зними трупами
м’язових волокон. Т! волокна, що штенсившше функцюнують, швидше виснажу-
ються, гинуть 1 замтщуються сполучною тканиною, а решта перебирае на себе
навантаження, яке зростае. Замццення ж серцевих мюципв сполучнотканинними
елементами спричинюе стиснення сусщнгх клттин, змши механгчних властивос-
тей серця, подальше попршання дифузп, поглиблення порушень обмшу. Вважа-
ють, що нормальна робота серця унеможливлюеться, якщо зам!на сполучною
тканиною становить 20—30 % його маси.
Дистрофтчнт змши серцевого м’яза супроводжуються розширенням порож-
ний серця, зменшенням сиди його скорочень — виникае мюгенна дилатацгя
серця, за яко! збшыпуеться об’ем кров!, що залишаеться пщ час систоли в по-
рожнинах серця, переповнюються вени. Пщвищений тиск кров1 в порожнин!
правого передсердя г в отворах порожнистих вен безпосередньо (через вплив на
пазухо-передсердний вузол) г рефлекторно (рефлекс Бейнбрщжа) зумовлюе тахт-
кардпо, яка ускладнюе порушення обмшу в мюкард!. Тому розширення порожнин
серця I тахйсардгя е загрозливими симптомами розвитку декомпенсаци.
Оцшюючи бюлогтчне значения гшертрофп мюкарда, слщ звернути увагу на
внутршшю суперечлив!сть цього явища. 3 одного боку, це досить досконалий
пристосувальний мехашзм, який забезпечуе протягом тривалого часу виконання
серцем пщвищено! роботи в нормальних 1 патолопчних умовах, з пппого — особ-
ливосп структури 1 функцп гшертрофованого серця е передумовою для розвитку
патологп. Переважання першого чи другого в кожному конкретному випадку виз-
начае особливостт перебтгу патолопчного процесу.
За динамткою змш обмшу, структури г функцп мюкарда у фаз! компенсатор-
но! пперфункцп серця видшяють три основнт стадп (Ф.З. Меерсон).
1. Аваршна стад1я розвиваеться безпосередньо теля пщвищення навантажен-
ня; характеризуеться поеднанням патолопчних змш у мюкард! (зникнення глтко-
гену, зниження ртвня креатинфосфату, зменшення вмтсту внутрштньоклттинного
калпо 1 пщвищення вмтсту натрпо, мобшзацтя глтколтзу, накопичення лактату) з
мобйпзашею резервтв мюкарда та органтзму в цтлому. У ц!й стад!! — тпдвищеш на-
вантаження на одиницю маси мюкарда. штенсившсть функцюнування структури
(1ФС), вщбуваеться швидке, протягом тижшв, збшьшення маси серця за рахунок
посиленого синтезу бшкгв т стовщення м’язових волокон.
2. Стадия завершения гтертрофи та в1дносно стшко! гтерфункци. У ц!й стадп
маса мюкарда збшьшена на 100—120 % 1 бтлыпе не зростае, 1ФС нормалтзуеться.
Патолопчш зм1ни в обмш! та структур! мтокарда не виявляються, споживання
кисню, утворення енергй, вмтст макроерпчних сполук не вщртзняються вщ нор-
ми. Нормалтзуеться гемодинамика. Гшертрофоване серце вже пристосувалося до
нових умов навантаження 1 протягом тривалого часу компенсуе його.
3. Стпадгя поступового виснаження I прогресуючого кардиосклерозу; характери-
зуеться вираженими обмшними т структурними змшами в енерготвтрних т скоро-
тливих елементах мтокарда. Частина м’язових волокон гине т замтщуеться сполуч-
ною тканиною, 1ФС зростае. Порушуеться регуляторний апарат серця. Прогресу-
юче виснаження компенсаторних мехашзмтв спричинюе розвиток хрошчно'т сер-
цевот недостатност!, надал! призводить до недостатност! кровооб!гу.
Хрошчна, або застшна, недостаппсть серця розвиваеться поступово, переваж-
но внаслщок метаболтчних порушень у мюкард! за тривало!’ гшерфункцп серця
або рхзних вид!в ураження мтокарда. Через недостатшсть викиду кров! !з серця
зменшуеться кровонаповнення оргашв на шляхах вщтоку. Водночас унаслщок
нездатност! серця перекачали всю кров, що надходить до нъого, розвиваеться за-
стш на шляхах п притоку, тобто у венах. Осюльки об’ем венозного русла при-
близив в 10 раз!в перевишуе об’ем артер!ального, у венах накопичуеться значка
юльюсть кров!.
У раз! порушення роботи переважно одного шлуночка серця недостатшсть
кровообпу набувае деяких специф!чних ознак ! виникае вщповщно недостат-
шсть за лтвошлуночковим або правошлуночковим типом. У пер-
шому випадку засттй кров! спостерггаеться у венах малого кола кровообпу, що
може спричинити набряк легень, у другому — у венах великого кола кровообпу,
за якого збшьшуеться печшка, з’являються набряки на ногах, асцит.
Порушення скоротливо'т функцц мтокарда не одразу спричинюе розвиток не-
достатност! кровообпу. Як пристосувальний мехашзм спочатку рефлекторно зни-
жуеться периферичний ошр в артертолах великого кола кровообтгу, що полегшуе
приплив кровт до бшыпост! оргашв. Рефлекторно звужуються артертоли в малому
колт кровообтгу, унаслщок чого зменшуеться приплив кров! до лтвого передсердя
т водночас знижуеться тиск у систем! легеневих кашляртв. Останне е мехашзмом
захисту легеневих кашляртв вщ переповнення кров’ю т запобтгае розвитковт на-
бряку легень.
Характерною е певна послщовнтсть розладу функцп ртзних вщд!лтв серця.
Так, декомпенсацтя функцц сильного л!вого шлуночка швидко зумовлюе пору-
шення функцц лтвого передсердя, застш кров! в малому колт кровообтгу, звужен-
ня легеневих артерюл. Поттм дещо слабший правий шлуночок змушений долати
пщвищений ошр у малому кол!, що призводить до його декомпенсацп та розвит-
ку недостатностт за правошлуночковим типом.
Гемодинамтчнт показники шд час хронтчнот серцевот недостатностт змшю-
ються таким чином: зменшуеться ХОС (з 5—5,6 до 3—4 л/хв); у 2—4 рази зни-
жуеться швидюсть току кровт; артертальний тиск змшюеться несутгево, венозний —
пщвищуеться; каптляри ! посткапйтярн! вени розширюються, кровоттк у них
сповшьнюеться, тиск пщвищуеться (схема 20).
Виникають патолопчш змши в тнших органах т системах. Сповшьнення току
кров! в судинах великого кола кровообтгу т порушення кровообпу в легенях зумо-
влюють збтльшення юлькост! вщновленого гемоглобшу в кров!. Унаслщок цього
шюра т слизова оболонка набувають характерного синюшного кольору (щаноз).
Схема 20. Патогенез освовних порушень у раз! серцево! недостатност!
Тканинам не вистачае кисню. Г! покоя супроводжуеться накопиченням недо-
окиснених продукпв обмшу I СО2 — розвиваеться ацидоз. Ацидоз 1 ппокс!я
зумовлюють порушення регуляцГ! дихання, виникае задишка. Шоб компенсувати
гшокспо, стимулюеться сритронитопоез, збшьшуеться загальний ОЦК 1 вцшос-
ний вмкт юптин кров!, що спричинюе пщвищення в’язкосп кров! га попршуе
гемоди нам !чш ц властивосп.
Унаслщок пщвищення тиску у венозних дшянках кап!ляр!в та ацидозу в тка-
нинах розвиваеться набряк, який своею чертою посилюе гшокспо, осюльки при
цьому’ збшьшуеться дифузгйний шлях вщ капйтяра до клггини. Розвиток застшно-
го набряку зумовлюе загалып порушення обмшу води й електролтв (затримка в
оргашзм! натргю 1 води) унаслщок порушення д!яльност! РААС, а також видшен-
ня натр!йуретичного гормону (пептиду) передсердями. Це е ще одним доказом
внутрштньо! суперечливосп механ!зм!в компенсацп пщ час патолопчного проце-
су. Мехашзми, яю еволюшйно виникли для забезпечення достатнього вмгсту в
оргашзм! солей ! рщини в разг загрози зневоднення або втрати кров!, при серцевш
недостатносп завдають оргашзму шкоди. У хворих гз серпевою недостатшстю
ж
надлишок вживано! сол! не вищляеться нирками, як це вщбуваеться в здорово!
людини, а затримусться в оргашзм! разом з еквтвалентною юльюстю води. Пору-
шення живлен ня тканин за тривало! недостатност! кровообтгу спричинюе тяжкий
1 необоротний розлад вн уз ршшьоюпти иного метабол !зму, шо супроводжуеться
порушенням синтезу бпгюв, у тому числг дихальних ферменпв, розвитком гшоксп
пстотоксичного типу. Ц1 явища характерш для термшальног фази недостатност!
кровообпу. У поеднанн! з! значним порушенням функцй травного каналу прогре-
суюча недостатшсть кровообпу спричинюе тяжке виснаження оргашзму — серце-
ву кахексйо
Серцева недостатшсть у раз! ушкодження мюкарда. Як уже зазначалося, !ншим
патоф]з!олог!чним мехашзмом виникнення серцево! недостатност! е ушкодження
серцевого м’яза. Воно може бути запального або дистроф!чного характеру, на-
слщком генетичних дефекпв, шфекцй штоксикацй, !мунопатолог!чних процесш,
хвороб, шо зумовлюють гшоксйо мюкарда або призводять до порушення боково-
го, лшщного, минерального та впамшного обм!ну.
При цьому може порушуватись утворення макроерпчних фосфат!в у серцевих
мюцитах або використання Тхньо! енергп. Процеси першого роду виникають у
раз! недостагнього надходження кисню в серцевий мюпит, зменшення вмюту
його в кров! або !шемй, а також порушення надходження субстрат!в окиснення,
функшонування м!тохондр!й, системи креатинюназа — креатинфосфат; другого
роду — унаслщок ушкодження бпгюв мюфгбрил, саркопдазматично! с!тки 1 розла-
ду обм!ну основних юн!в кальшю, калпо. нагр!ю (мал. 51).
Одним 13 мехашзмгв ушкодження серцевого мюпита може бути ушкодження
його мембранних структур унаслщок пероксидного окиснення лйгщгв. що входять
до ёх складу, вшьними радикалами ! гщропероксидами. Пщвищення ж ртвня в!ль-
норадикалыгого окиснення своею чертою може виникнути в раз! порушення
окисною метабол!зму в серцевому мюцип або внаслщок недостатност! антиокси-
дантних систем. Насамперед порушуються функцй спепиф!чних мембранних на-
сос!в (Ма+-, К*-, Са2ч-АТФаза), поступово збшьшуеться проникшсть мембрани,
пот!м вщбуваеться ушкодження фосфолшццв у шй 1 виникають дефекти. Пору-
шення мембрани спричинюе зм!ни потоки юшв натрпо, калпо, хлору 1 води, що
зумовлюе набухания клггини, а також значне надходження юн!в кальцпо з роз-
витком токсичних ефекпв цього канона. Можуть збьтыпуватися юльюсть виявле-
них а- та р-адренорецепторш 1 вившьнення катехолампцв !з нервових заюнчень,
що поглиблюе первинне ушкодження.
У випадках виражених порушень обмшу можлива загибелъ серцевих мю-
цит!в.
Некоронарогенш ушкодження серця. € юлька експериментальних моделей не-
крозу серцевого м’яза, виникнення якого не пов’язане з патолопею судин серия.
Ц! модел! певною м!рою вщображають ситуацпо, шо спостерйаеться за природ-
них умов.
Ппоксичний некроз мюкарда може бути вщтворений за допомогою ргзних вид!в
гшокей — нетоксично!, гем!чно/ На тл! загально! недостатност! кисню в орган!з-
М1, шо зумовлюе пщвищення навантаження на систему кровообйу. розвиваеться
некротичне ушкодження м’язових волокон серця. Розвитков! некрозу сприяе
фгксашя тварини в незручшй поз!, наприклад розгягування у станку, або додат-
кове навантаження — бй у тредбан!.
Мал. 51. Порушення обмшу речовин у серцевому м’язъ
Види порушень: I — дефщит кисню; надлишок молочно!' кислоти; зм!на юлъкосп позакштичних
юнш. !х спшвщношення; порушення сгпввщ ношения жирна кислота / глюкоза; Н — порушен-
ня синтезу окисних фермента (гтовггам)нози Вг 1Г та 1н., отруення наркотич ни ми препарата-
ми); порушення процесш окисного фосфорилювання: порушення перенесения макроерпчних
фосфата; Ш — порушення структури скоротливих бшюв; порушення саркоплазматично! с1тки;
порушення обмшу Са2*, ЬГа+, К*
Електролтно-стероТдна кард1опат1я з некрозом. За спостереженнями Г. Селье,
у раз! введения щурам значно) ктлькост! солей натрпо в поеднанш з деякими ашо-
нами (сульфатами, фосфатами) у серщ з’являються вогнища ушкодження дегене-
ративно-некротичного типу, як! часто супроводжуються палшозом судин шших
оргашв. Ц] ушкодження поширюються або виникають шсля застосування меншоТ
ктлькосп солей, якщо одночасно вводиться деяк! стеротдш ! гормони наднирко-
вих залоз. На такому тл1 легше розвиваються ушкодження серця, зумовлен! иппи-
ми причинами, а також спостсрпаеться тяжчий 1х перебтг. Наприклад, навтть не-
висок! дози норадреналшу, похщних кальциферолу, гшокФя, м’язове напруження
або, навпаки, значне обмеження рухомост) спричинюють поширення некрозу
мюкарда. Сол! калтю 1 магнпо при цьому справляють захисну дно.
1мунн1 ушкодження серця можлив! шсля введения в оргашзм експерименталь-
но'! тварини гетерогенно! сироватки, шо мктить антитша до бтлктв серця гварини
цього виду (кардюпитотоксини). Доведено також, що в оргашзм! можуть утворю-
ватись антипла та сенсибннзован! л!мфоцити, спрямоваш проти тканин власного
серця. Цьому сприяе проникнення в кров денатурованих компонент!в некрозизо-
ваних серцевих мюципв. В експеримент! аналопчний процес можна спричинити
шляхом уведення твариш суспензг мюкарда з! стимулятором тмунно! реакцп
(ад’ювант Фрейнда). Ушкодження серця може зумовлюватась г циркулюючими
гмунними комплексами антиген — антитшо — комплемент.
Коронарогенш ушкодження серця. 1шем1чна хвороба серця, шфаркт мюкарда. Як
зазначалося ранние, особливост! функцюнування, метаболизму та кровопостачан-
ня серця роблять його надзвичайно вразливим у разг порушення вщповщностг
потреби мюкарда в киснг ргвню притоку кров! через вгнцевг артери.
Хвороби г патологгчнг стани, що супроводжуються порушенням кровообггу в
мюкард,, до якого призводять ушкодження вшцевих артергй переважно атеро-
склеротичного характеру, об’еднано в особливу нозологгчну одиницю, що отри-
мала назву “йием1чна хвороба серця ” (1ХС). 1ХС може проявлятися переважно
функцгональними розладами г больовим синдромом (стенокардия) або зумовлюва-
ти некротачнг змгни мюкарда. Останш можуть мати велико- г дргбновогнищевий
характер, гострий або хронгчний перебгг. При цьому виникае комплекс мета-
болгчних змгн, спрямованих на пгдтримку життедгяльностг мюкарда. Цг змгни
можна роздшити на два етапи — короткочасну захисну реакцпо г тривалу фазу
виживання. На першому еташ вичерпуються макроерггчнг фосфата, спочатку КФ,
потгм АТФ, пщвищуеться ргвень неоргангчного фосфату; посилюеться анаероб-
ний глгколгз, зменшуються рН г скоротливгсть серцевих мгоцитгв, що сприяе гх
виживанню. Наслгдками цього е активацгя генетично детермпюваних захисних
механгзмгв, так звана метаболгчна адаптацгя мюкарда (прекондицгювання), що
залежить вщ змгни активностг АТФ-залежних калгевих каналгв, збереження об’ему
мгтохондргй, запобггання кальцгевому перевантаженнго. Прекондицгювання
пщвищуе адаптацгю мюкарда до повторно! гшемп теля короткочасного поперед-
нього обмеження вгнцевого кровообггу.
Проте навгть короткочасна гшемгя мюкарда супроводжуеться порушенням
його локально! скоротливостг, що зберггаеться деякий час пгсля повного вщнов-
лення кровотоку вшцевими артергями. Ця постгшемгчна дисфункцгя мюкарда от-
римала назву приголомшеностг (зШппгпё). Вона спостерггаеться шсля короткочас-
ног (5—15 хв) гшемп в експеримент!, а також при стенокардй, у разг вщновлення
кровотоку при хгрурпчнгй реваскуляризацй серця або успгшног тромболгтичног
терапгг вшцевих артергй, а також пгсля значного фгзичного навантаження. В ос-
нов! патогенезу приголомшеностг мюкарда лежать надлишок гонгв кальшю в ци-
тозолг й ушкоджувальна дгя вгльних радикалгв.
Пгсля значного тривалого зменшення коронарно! перфузп виникае феномен
“засинання” (ггбернацп) мюкарда, пщ час якого спостерггаеться вщсутнгсть ско-
ротливог функцг! певних дглянок мюкарда, якг, однак, у разг вщновлення крово-
постачання здатн! вщновити свою скоротливу активнгсть — “прокинутися”. Вва-
жають, що пбернацгя мюкарда виникае як захисна реакцгя на недостатшсть його
кровопостачання. Вона допомагае йому вижити в несприятливих умовах, захища-
ючи тим самим вщ розвитку некротичних ушкодженъ. 1снування дглянок ггберну-
ючого мюкарда мае велике прогностичне значения пщ час вщновног терапгг.
Наслщком закупорювання вшцевих артергй е зниження постачання кисню
серцю, яке, якщо воно досить значне г тривале, спричинюе ушкодження мемб-
ран, некроз г втрату клгтин мюкарда. Проте в разг швидкого вщновлення поста-
чання кисню та нормалгзацп коронарного кровообггу в гшемгзованому мюкард!
виникають патологгчнг змгни, що називаються реперфузшним ушкодженням г якг
додатково збгльшують зону шфаркту мюкарда. При цьому, якщо сама по собг
1шем1я спричинюе загибель серцевих мюципв за рахунок некрозу, то релерфузгя
призводить до смерт! клгтин шляхом апоптозу. Вважають. що швидке вщновлен-
ня постачання кисню мюкарду супроводжуеться “вибухом” вироблення вглъних
радикалов, ушкодженням ДНК г м)тохондрш гз вивгльненням з останнгх в цито-
плазму цитохрому С й шших апоптозшдгтсуючих речовин, як! активують каспаз-
ний каскад 1з подальшою апоптотичною загибелтю серцевих мюципв. Серед
форм 1ХС за особливостями патогенезу та клгнгчним значениям слщ видглити
гнфаркт мюкарда.
1нфаркт мюкарда — вогншцева !шемгя г некроз м’яза серця, що виникають
унаслщок припинення притоку кровг через одну з плок вшцевих артерш або в
разг надходження п в кглькостг. недостатнш для забезпечення енергетичних пот-
реб мюкарда. Здебглыпого ушкодження стшки вшцевих артерш спричинюе атеро-
склероз.
Останюм часом порушення кровопостачання серця стало настшьки пошире-
ним 1 мае таку високу питому вагу серед шших видгв патологи в людини, що
варто говорите про своерщну ещдемпо 1ХС, яка охопила промислово розвинуп
крагни, де смертность вгд хвороб оргашв кровообпу пос фае перше мюце. На ш-
фаркт мюкарда хворпоть переважно люди похилого 1 старечого вгку, проте ч!тко
простежуеться тенденцгя до збьтьшення показниюв по шире ноет! шфаркту мю-
карда г захворюваносп на нього серед особ молодшого в1ку.
Чинники, шо створюють передумови для виникнення шфаркту мюкарда, на-
зивають факторами ризику. Ними е: спадкова зумовлешеть; ппертензивна
хвороба, цукровий диабет, подагра; фактори навколишнього середовища — мало-
рухливий, емощйно напружений спосгб життя, надморие харчування зг споживан
ням велико'] кглькостг жир!в; куршня. У биоьшосп випадкгв гнфаркт мюкарда роз-
виваеться внаслщок кальцифокащТ та виразкових змш атеросклеротичног бляшки
з подальшим закупорюванням судини тромбом. Закупорювання одше! з плок
вшиево'о артерп здебыыпого не супроводжуеться мобйпзацгею колатеральних су-
дин, осюльки атеросклерозом певною мгрого ушкоджеш й шип судини серця.
Стенозуючий склероз судин створюе жорсткий лгмгт надходження поживних
речовин до серцевого м’яза, коли навпъ дещо больший ступгнь звуження судини
або пщвищення потреби м’яза в кисш може зумовити некроз. Слгдом за цим у
вогнищ] гшемц виникають порушення мокроциркуляцц у вигляд! паралотичного
розширення капглярних судин. стазу, набряку. Вони ускладнюютъ циркуляторн!
порушення.
Можлив! так! патогенетичн) вароанти розвитку шфаркту мюкарда (мал:
52): 1) закупорювання судини, яке зумовлюе абсолютне зменшення величини
вшцевого току крово нижче критичного ргвня (звичайнс понад 3/4 первосного);
2) стенозування, яке не проявляеться у стан! спокою, а июля незначного навэн-
таження (фгзичного або псих!чного) спричинюе гшемгю серцевого м’яза; 3) знан-
ие ф1зичне навантаження або емоцгйне напруження, яке г без атеросклеротичних
ушкоджень може зумовити невгдповщшеть _мгж потребою мюкарда в кисш та
можтивгетю надходження кров!.
В остан ньому випадку велику роль вгдгграе посилення секрецп катехол аминь
1 гормошв юрково! речовини надниркових залоз. Кргм того, е дан! про те, що
судини, навпъ незначною игрою склерозоваш, можуть вщповщати спазмом тодт,
Причини Мехашзми Синдроми Юмшчш ознаки Ускдаднення
Бшь
Резорбци
продукте
розпаду
Ектотпчв!
вогнища
збудження
Порушення
лровздносп
Збшьшення
вмюту жиро-
вих кислот
V кров'
Звуження,
закупорюван-
ня вншевих
артерш
Пщвищення
витрача <ня
кисню
Пшвшцення тонусу симпатично!
частини автономно!
нервово!' системи
Викид катехоламппв
Звуження
периферичних
судин
Ослабления
насосноТ
функцц
серця
Авто!му 1 пзащя ->
2
х
Лейкоцитоз
Гарячка
Пшвишенпя
ШОЕ
Поява ферменпв
мюкарда у кров1
Б1ль тз характер-
ною зррадзашею
Нелосзапнсть
серця
х
Екстрасистолзя,
пароксизмальна
тах1кард1я
Блокада
провшно!
системи серця
Лйи. 52. Причинно-наслшков! зв’язки мехаквмш порушень при шфаркп мюкарда
Поспнфаркт-
синдром
Карлюгенний
шок
Ф1брИЛЯЦ1Я
шлуночюв
РОЗЛАДИ ХРОВООБ1ГУ, ЗУМОВЛЕНГ ПОРУШЕННЯМ ФУНКЦП СЕРЦЯ
коли нормальш судини розширюються, наприклад шд впливом катехоламгнгв. Це
може бути наслщком порушення секрецп (або дй) так званого ендотелгального
фактора розслаблення, яким виявився оксид азоту, шо синтезуеться в клгтинах
ендотелгю судин (аналоги оксиду азоту, наприклад нггрити, застосовують у тера-
певтичнгй пракгицг як судинорозширювальш засоби).
€ калька скспериментальних моделей шфаркту мюкарда: перев’язування од-
шег з пл о к вгнцевих артергй у гострому або хронгчному експеримент!; закупорю-
вання артери за допомогою катетера або введения емболгзуючих часгинок (ртутг,
агару); перфузгя вгнцево! артери через катетер кров’ю, позбавленою кисню, або
кров’ю, яка мгстить антимюкардгальвд антитгла.
Пгсля порушення кровообггу вже протягом перших хвилин виникають змгни
на електрокардгограм! у вигляд! змгщення сегмента 57, змши комплексу (?Л5 1
зубця Т.
Морфолог гчно найбьтьш рано можна вгдзначити порушення структури мгто-
хондргй, потгм вгдбуваеться набухания або пгкноз ядер, зникас поперечна посму-
говангсть м’язових волокон. Серцевг мгоцити вграчають глгкоген г калгй, у них
збшьшуеться кглькгсть лгзосом.
1нфаркт розвиваеться в дглянщ, кровопостачання яко! здгйснювала ушкодже-
на судина. Основними наслгдками шфаркту е локатьний коагуляцгйний некроз,
лгзис серпевих мгоцитгв. набряк мюкарда. Розргзняють кглька зон у вогнищг ш-
фаркту. У центральнгй, переважно субендокардгальнгй, зонг переважають необо-
ротнг ушкодження (псрерозтягненг мгофгбрили, грудкопод!бний ядерний хрома-
тин, мггохондрп з аморфними ушшьненнями матриксу, дефекта плазматичног
мембрани). У промгжнгй зонг виявляють некротизован! м’язовг клгтини з ознака-
ми кальцгевого навантаження (перескорочення мгофгбрил, контракгури, вгдкла-
дання кадьпгю фосфату в мгтохондргях), аморфнг ущгльнення матриксу, грудко-
подгбний хроматин, краплг жиру. У зовнгшнш зон! шфаркту в м’язових клгтинах
переважае накопичення крапель жиру, некротичних змш немае. Спгввщношення
розмгргв цих зон мае велике значения для прогнозування перебггу хвороби та ви-
бору тактики .пкування. Клгтини. що загинули, швидко огочуються нейтрофгль-
иими гранулоцитами, як! гпзншге замшюються макрофагами, лгмфоцитами г плаз-
моцитами. Згодом серцевг мгоцити розсмоктуються г замгщуються фгбробластами,
утворюеться сполучнотканинний рубець.
Вогнище некрозу в мюкард! порушуе роботу серця в шлому, що проявляеться
розладом ритму г зниженням насосног функцп серця. Стуш’нь г характер пору-
шень залежать вгд локалгзацп та поширення шфаркту.
Пгд впливом гшемп серцевг мгоцити можуть набувати здатностг до автоматиз-
му, г тодг з’являеться ектопгчне вогнище збудження, що спричинюе виникнення
екстрасистолгг. Зниження провгдностг в уражених дглянках серця, а гнодг бло-
када разом гз МНОЖИНЯ1СТЮ ектошчних вогнищ створюють умови для повторно!
циркулящг збудження г виникнення пароксизмальног гахгкардгг, а також
такого тяжкого ускладнення, як фгбрцляцгя шлуночкгв — основног причини
ранньо! смертг внаслщок шфаркту мюкарда.
1нфаркт мюкарда може супроводжуватися гострою або хронгчною серцевою
нсдостатнгстю, причому попршення гемодин амгки тим значнгше, чим бгльша
зона шфаркту. При цьому пщвищуеться тиск кровг на шляхах пршгливу и до сер-
ця г зменшуеться ХОС. Одним гз найтяжчих ускладнень шфаркту мюкарда е кар -
дгогенний шок, за якого зменшуеться серцевий викид на тлг значного пщви-
щення загального периферичного опору судин унаслщок збшьшення активносп
симпатико-адреналово! та ренш-анпотензиновог систем. Приеднання розладгв
мгкроциркуляцп в тканинах зумовлюе гщокспо, ацидоз, порушення функщй го-
ловного мозку та шших оргашв, смерть.
Для шфаркту мюкарда характерними е больовий 1 резорбцгйно-некротичний
синдроми. Б г ль мае типову локалгзацгю (л!воруч у верхнш частит тулуба 1 за
грудниною), а також тяжке емоцгйне забарвлення. Це пояснюеться гррадгацгею
збудження в спинному мозку з в!сцеральних нейрошв на вщповщнг проекцгйнг
зони соматичних чутливих нейрошв. Проте можливий г безбольовий перебгг ш-
фаркту мюкарда.
Гострий гнфаркт мюкарда в людини часто супроводжуеться пщвищенням
функцп симпатико-адреналово! системи г викидом у кров високих доз катехола-
мгнгв. Це своею чертою призводить до пщвищення функцп серця, ргвня в кровг
вшьних жирних кислот, що зумовлюе зменшення транспорту глюкози в серцевг
мгоцити й штенсивностг глгколгзу в них, збшьшення витрат кисню, посилення
метаболгчних порушень г, як наслщок, ускладнення перебггу шфаркту. У таких
випадках захист серця вщ впливу катехоламшгв (наприклад шляхом застосування
р-адреноблокаторгв) дае позитивний результат.
Резорбшя з некротизованих дшянок мюкарда у кров вмгсту ушкодже-
них клгтин зумовлюе появу в кровг внутргшньоклгтинних ферментгв (креатин-
кшази, аспартатамшотрансферази, серцевих гзоферментгв лактатдегщрогенази), а
також мгоглобгну, що може бути використано з дгагностичною метою. Резорбцгя
клгтинних бшкгв супроводжуеться лейкоцитозом, гарячкою, збшьшенням ШОЕ.
Надходження в кров внутргшньоклгтинних мгокардгальних бшкгв може супро-
воджуватись автогмушзацгею з утворенням протисерцевих антитш г сенсибглгзова-
них до серцевих антигешв лгмфоцитгв, еозинофшгею та ггпергаммаглобулшемгею.
Природно, що така гмунна реакщя поглиблюе ураження мюкарда, спричинюе
появу вторинних вогнищ некрозу. Кргм того, з утворенням автоантитш пов’язують
розвиток постгнфарктного синдрому (синдрому Дресел ера), який характе-
ризуеться запаленням серозно! оболонки серця, легень, суглобгв. При цьому лгку-
вання антибютиками неефективне, позитивний результат спостерггаеться в разг
введения кортикостеро'щгв.
Нейрогенш ушкодження серця. Дистрофгчнг змши г некроз мюкарда можна
спричинити шляхом гострого або хрошчного подразнення шийно-грудного вузла
симпатичного стовбура, блукаючого нерва, ппоталамуса, стовбура або шших вщ-
дшгв головного мозку, уведення у кров адреналшу або норадреналшу у високих
дозах. Основою нейрогенних ушкоджень е невщповщнгсть мгж функцюнальними
можливостями, метаболгзмом г кровопостачанням серця. Пщ час подразнення
симпатичних нервгв серця споживання кисню мюкардом пщвищуеться бшьшою
мгрою, юж вшцевий кровотгк, унаслщок чого розвиваеться ппокегя мюкарда. У
разг склерозування вгнцевих артерш ргзниця мгж ргвнями кровотоку й обмгну
значшша, що може мати загрозливг наслщки. Подразнення блукаючого нерва
зумовлюе протилежнг змши у епгввщношеннг мгж ргвнем обмшу та величиною
вгнцевого кровотоку, полшшуючи умови кровопостачання серця. У спортсмена
тонус серцевих плок блукаючого нерва пщвищений, а в людини, яка веде мало-
рухливий спос1б життя (“детреноване серце” за В. Раабом), переважають симпа-
тичнг впливи. Це, можливо, зумовлюе щдвищену уразливгсть серця сучасноТ лю-
дини, яка веде малорухливе, емоцшно насичене життя, на вщмшу вщ життя п
далеких предав, пов’язаного зг значними фгзичними навантаженнями.
Порушення серцевого ритму
Робота серця як единого насосного пристрою залежить вщ узгодженост! робота
м’язових волокон кожного його вщдглу, послщовностг, ритму г частота скорочень
цих вщдглгв. Ш вимоги, як вщомо, забезпечуються основними властивостями сер-
ця: автоматизмом, збудливгстю, провщнгстю г скоротливгстю. У нормальних умо-
вах автоматизм забезпечуеться водгем ритму — пазухо-передсердним вузлом, про-
вщнгсть — провщною системою серця, що складаеться з провщних пучкгв перед-
сердь, передсердно-шлуночкового пучка, передсердно-шлуночкового вузла г
провщних серцевих мюцитгв (волокон Пуркшье), з яких збудження передаеться
на клггини скоротливого мтокарда. Попри те, що здатшсть до автоматизму влас-
тива й шшим вщдглам провщно! системи серця, частота гмпульсгв, яга генерують-
ся, зменшуеться в напрямку вщ передсердь до шлуночюв (закон градгента серия)
1 за нормальних умов здатшсть нижчерозташованих вщдглгв серця до виявлення
автоматизму пригшчуеться утворами, розташованими вище.
Порушення автоматизму, провщностг г здатносп серця до засвоення ритму збуд-
ження зумовлюють порушення частота, ритму, узгодженосп або послщовностг
скорочень серця — аритмйо.
Порушення серцевого ритму виникають у раз! запального, гшемгчного або ток-
сичного ураження мюкарда, змши сшввщношення мгж внутргшньо- та поза-
клгтинним вмгстом юшв калпо, натр(ю, кальщю г магшю, у зв’язку з гормональ-
ними дисфункщями, а також можуть бути наслщком порушення взаемоди симпа-
тично! та парасимпатично!' шнервацп серця. Пщ впливом зазначених етюлоггчних
факторгв можуть змшитись актившсть нормального водгя ритму, рефрактерний
перюд ргзних збудливих структур або порушитися проведения збудження мгж ргз-
ними ланками провщно! системи г мгж провщною системою г скоротливим мю-
кард ом, виникнути ектогпчш вогнища збудження. Усг щ змши, кожна окремо або
в поеднанш, зумовлюють аритмпо. У ц виникненнг, кргм того, значну роль може
вщггравата наявшсть структур гз ргзною швидгастю проведения збудження (у виг-
ляд! певно! структурно! аномали або вогнищевого патолопчного процесу), що
створюе умови для безперервно! циркуляцп хвил! збудження.
Порушення автоматизму. Здатшсть до автоматичного утворення !мпульс!в, як
вщомо, залежить вщ юптин провщно! системи серця (р-клггини), у яких вщбу-
ваеться спонтанна повгльна деполяризац!я клгтанног мембрани в перюд дгастоли.
Як наслщок, теля досягнення певного критичного ргвня виникае потенцгал дп.
Частота генераци гмпульсгв залежить вщ максимального дгастолгчного потенщалу
цих клгтин, ргвня того критичного потенщалу на мембраш, шсля якого виникае
потенцгал ди, г швидкосп повгльног дгастолгчног деполяризащ! (мал. 53).
Змша ргвня максимального дгастошчного потенщалу, критичного потенщалу
або швидкосп дгастолгчног' деполяризацп в той чи шший бгк спричинюе змши
частота генерацц гмпульсгв або появу шших джерел гмпульсацп, якщо цг змгни
Мал. 53. Спонтанш змши мембранного потенщалу клггини пазухо-
передсердного вузла
виникають в шших, здатних до збудження дглянках серця г зумовлюють появу там
потенщалгв дп. У разг зменшення ргвня максимального дгастолгчного потенщалу
клгтин пазухо-передсердного вузла, наближення до нього порогового критичного
потенщалу або збгльшення швидкостг повгльног дгасто.нчног деполя ризацп гм-
пульси генеруються частгше, розвиваеться тах!кард1я. Це спостерггаеться пгд
впливом пщвищеног температури тгла, симпатичного медгатора. розтягнення дг-
лянки пазухо-передсердного вузла. I навпаки, зменшення швидкостг повгльног
дгастолгчноТ деполяризанп, гшерполяризацгя в дгастолг та вщдалення критичного
порогового потенщалу, як це спостерггаеться пгд час подразнення блукаючого
нерва, супроводжуюгься сповгльненням генерацп гмпульегв. а отже, г скорочень
серця — брадикардгею. Коливання тонусу блукаючого нерва пщ час дихання мо-
жуть спричинити дихальну аритмт (прискорене серцебиття пгд час вдиху, сповгдь-
нене — пгд час видиху). Дихальна аритмгя в норм] бувае в дгтей, але зрщка може
спостерггатись г в дорослих.
За патолопчних умов може проявитися власнии автоматизм нижчерозташо-
ваних вгадглгв провщног системи серця (потенцшних водив ритму). Такт умови
можуть виникнути пщ час зниження автоматизму пазухо-передсердного вузла або
в разг пщвищення здатностг до генерацп гмпульегв в шших дглянках мюкарда У
цих випадках частота гмпульегв, якг генеруються нормальним водгем ритму, вияв-
ляегься недостатньою для пригнгчення автоматизму шших вгддглзв, що зумовлюе
появу додаткових гмпульегв з ектошчних вогнищ збудження.
Гншим механгзмом появи ектошчних вогниш збудження може бути виник-
нення ргзницг потенщалгв мгж розгашованими поряд мгоцитами внаслщок, на-
приклад, неодночасного закгнчення реполяризацн в них, що може спричинити
збудження у волокнах, якг вже вийшли з фази рефрактерностг. Це явище спо-
стерггаеться шд час локально^ гшемп мюкарда 1 в разг отруення серцевими глгко-
зидами.
В усгх згаданих випадках виникае позачергове скорочення серця або тгльки
шлуночкгв — екстрасистола.
Мал. 54 Елекгрокардюграма лад час екстрасис-
толи иередсердноЕ (а), лшоадлуночково! (б). пра-
вошлуночьово) (в):
1 — пазухо-передсердний вузол; 2 — передсердно-
шл}ъочковий вузол: 3 — передсердно-шлуночко
вий пучок; Е — екстрасистола; С — компенсатор-
на пауза; 1—111 — вщведенпя
Залежно вщ локал!запН вогнища, з
якого виходить позачерговий шпульс,
розр!зняюзь кыька видтв екстрасистоли:
синусну (або нормотопну), псредсердну.
переде ердно-шлуночкову ) шлуночкову
Оскшьки хвиля збудження, що виникла
в незвичному шещ, поширюсться В ЗМ1-
неному напрямку, змшюеться структура
електричного поля серця, що вщобра-
жаеться на електрокардюграмь Кожний
вид екстрасистоли мае свою електро-
кард1ограф1чну картину, яка дае змогу
визначши м1сце ектошчного вогнища
збудження. Калька еле ктрокардюграм, харакгерних для р!зних видхв екстрасисто-
ли наведено на мал. 54.
Синусова екстрасистола виникае внаслщок передчасного збудження
частини юптин пазухо-передсердного вузла. Електрокард!ограф!чно вона не
вщр'хзняеться вщ нормального скорочення за винятком зменшення щастол1чного
шгервалу Т — Р Унаслщок укорочения д!астоли 1 зменшеного наповнення шлу-
ночюв пульсова хвиля при екстрасистоли зменшена.
Передсердна екстрасистола спостерггаеться за наявносп вогнища ек-
топ1чного збудження в р!зних дшянках передсердь. Характеризуемся викривлен-
ням форми зубця Р (знижений, двофазний, негативний) при збереженому комп-
лекс! 0&8Т! дешо подовженому д!астол!чному штервал! шсля екстрасистоли. Це
зумовлено тим, що збудження, яке передаеться ретроградно, передчасно розряд-
жае нормальний синусний 1мпулъс, що збпаеться з! збудженням шлуночкгв. На-
ступний персдсердний !мпулъс, що виникае через нормальний интервал, за часом
дещо вщетае вш моменту закшчення збудження шлуночшв — неповна компенса-
торна пауза.
Передсердно-шлуночкова екстрасистола спостерхгаеться враз! ви-
конання додагкового шпульсу в передсердно-шлуночковому вузлк Хвиля збуд-
ження, шо виходить !з верхньоТ га середньо! частин вузла. поширюеться у двох
напрямках: у шлуночках — у нормальному, у передсердях — у ретроградному.
При цьому негативний зубепь Р може збпатися з комплексом ()К8 Дшстолхчний
штервал шсля екстрасистоли дещо подовжении. Екстрасистола може супрово-
джуватись одночасним скороченням передсердь 1 шлуночюв. У раз) передсердно-
шлуночково! екстрасистоли, що виходить !з нижньо! частини вузла, виникае така
сама компенсаторна пауза, як 1 при шлуночковш екстрасистол!
Для шлуночково! екстрасистоли характерною е наявшетъ повно! ком-
пенсаторно! паузи шсля позачергового скорочення. Вона виникае внаслщок того,
що збудження, яке охоплюе шлуночки, не передаеться через перед сердно-шлу-
ночковий вузол на передсердя г черговий нормальний гмпульс збудження, що йде
з пазухо-передсердного вузла, не поширюеться на шлуночки, якг перебувають у
фаз! рефрактерностг. Наступне скорочення шлуночюв виникае ильки теля чер-
вового нормального гмпульсу. Тому тривалгсть компенсаторно! паузи разом з ин-
тервалом, що передуе гй, доргвнюе тривалостг двох нормальних дгастолгчних пауз.
Проте якщо скорочення серця настшьки рщкг, що на момент надходження черво-
вого нормального гмпульсу шлуночки встигають вийти зг стану рефрактерност!,
то компенсаторно! паузи не бувае. Позачергове скорочення потрапляе в штервал
мгж двома нормальними г в цьому випадку мае назву вставно! екстрасисто-
л и. Оскыьки хвиля збудження гид час шлуночково! екстрасистоли поширюеться
в шлуночках як у нормальному, так г в ретроградному напрямках, то це супрово-
джуеться значним викривленням форми комплексу ОК8.
Позачергов! скорочення можуть виникати поодинцг або групами. У разг ви-
никнення групи екстрасистол, що швидко повторюються г повнгстю пригигчують
фгзгологгчний ритм, розвиваеться пароксизмальна тах1кард1я. Нормальний ритм
серця раптово перериваеться нападом скорочень частотою вщ 140 до 250 за 1 хв.
Тривалгсть нападу може бути р!зною — вщ кшькох секунд до кглькох хвилин, те-
ля чого вш так само раптово припиняеться г налагоджуеться нормальний ритм.
Здебшьшого спостерггаеться передсердна форма пароксизмальног таю кард!!.
А оскыьки тривалгсть потенщалгв дп г рефрактерних пергодгв збглыпуеться за хо-
дом провщног системи, то дистально розташоваю дглянки ц не завжди здатнг
вщтворити частоту гмпульсацц, що йде з проксимальних вщдглгв. Тому быьша
частина гмпульегв шд час передсердно! тахгкардгг не може проводитися передсер-
дно-шлуночковим вузлом. Оскгльки тривалгсть рефрактерних пергодгв г потенцга-
ли дп у волокнах право! нгжки передсердно-шлуночкового пучка бглыш, нгж у
лгвш, частгше порушуеться за високог частота гмпульегв проведения збудження до
правого шлуночка.
Порушення провщностг. Аритмгя, зумовлена порушенням проведения гмпуль-
су, називаеться блокадою. Причиною блокада може бути ушкодження провщних
шляхгв, що зумовлюе подовження рефрактерного перюду, попршення гнших
функщональних характеристик г супроводжуеться сповшьненням або повним
припиненням проведения гмпульсу. Порушення провщностг можуть виникати
мгж пазухо-передсердним вузлом г передсердям, усерединг передсердь, мгж перед-
сердями г шлуночками г в однгй гз нгжок передсердно-шлуночкового пучка. У разг
внутргшньопередсердно! та внутргшньошлуночковог блокади час-
тота скорочень серця не змгнюеться, а порушення проявляеться змгною форми
елекгрокардгограми. Змгною ритму г частоти скорочень серця може супроводжу-
ватися передсердно-шлуночкова блокада.
Передсердно-шлуночкова, або поперечна, блокада серця може
бути повною г неповною. Видгляють три ступенг неповно! блокади серця (мал. 55).
Неповна передсердно-шлуночкова блокада I ступеня характеризуеться збыь-
шенням часу проведения гмпульсу вщ передсердь до шлуночкгв, що супрово-
джуеться подовженням гнтервалу Р—0 (0,2—0,5 с). Для блокади II ступеня (пе-
ргоди Самойлова — Венкебаха) характерним е прогресуюче збгльшення гнтервалу
Р— 2 до тих пгр, поки одне зг збуджень, зазвичай восьме—десяте, не проводиться.
Пгсля випадгння скорочення шлуночка штервал Р-0 вщновлюеться, поступово
Мая. 55. Електрокард’ограма п!д час передсердно-шлуночково'!
блокади:
а — I ступшь (половження штервалу Р—<2У. б — II стушнь (по-
С1упове подовження гнтервалу Р— Оу низш наступних скорочень
з виляниям одного чергового скорочення — перюди Самойлова —
Венкебаха); в — III ступшь (проводиться кожне друге збудження);
г — повна поперечна блокада (передсердя 1 шлуночки збуджують-
ся та скорочуються кожний у своему ритм:)
подовжуючись кожним скорочснням серця. Вважають, що цей феномен пов'язаний
13 наростаючим утрудненням проведения !мпульс!в через вузол. Пщ час блокади
III ступени (Мобтгца тип блокади) спостерйаеться випадйтня кожного другого-
третього скорочення або, навпаки, проводиться тыьки кожне друге, трете або
четверте збудження передсердь.
У раз! повно! передсердно-шлуночково!' блокади передсердя 1 шлуночки ско-
рочуються незалежно одне вы одного, кожний у своему ритм!, передсердя 1з час-
тотою приблизно 70 за 1 хв. шлуночки — приблизно 35 за I хв (!д!овентрику-
лярний ритм).
Особливе значения мае момент переходу неповно! блокади в повну, коли до
шлуночюв не надходять тмпульси вы передсердь. Повыьна Д1астол!чна деполяри-
зашя в потенцшних вод!ях ритму виникае тыьки через деякий час теля припи-
нення надходження !мпульс!в вщ пазухо-передсердного вузла. Цей перюд мае
назву преавтоматично! паузи, пщ час яко спостерйаеться асистолы шлуночюв.
При цьому внаслщок припинення притоку кров) до головного мозку виникають
непритомюсть, судоми (синдром Морганы — Адамса — Стокса), можлива смерть.
Проте зазвичай у раз! поновлення скорочень шлуночюв зазначен! симптоми ми-
нають. Напади можуть повторюватися багаторазово.
У раз! порушення провщвосп в одни !з н!жок передсердно-шлуночкового
пучка частота серцевих скорочень не змппоеться, але скорочення вщповщного
шлуночка зашзнюеться внаслщок того, що хвиля збудження доходить до нього
кружним шляхом.
Порушення засвоення ритму. Аритм!я може полягати в тому, що порушуеться
вщтворення частота збудження (трансформация ритму, подщ частота) або потен-
3®
щали ди га скорочення, як) йдугь один за одним, виявляються неоднаковими
(альтернащя).
Трансформа ц1я ритму може спостертгатися в раз? порушення проведения збу-
дження через ртзн! д!лянки провщно! системи серия або шд час переходу збуджен-
ня 13 серцевих провщних мющтв на м’язов) волокна. Вона ч)тко проявляеться в
разт порушення функционального стану серця внаслщок пттоксикацн, ппоксп або
ппемп в поеднанн) з тахткард)ею. При цьому частота збуджень мюкарда може не
вщповщати частот) скорочень. Наприклад, за кожним другим потеншатом ди не
настае скорочення. Пояснюеться це тим, що скоротливий апарат юптини, система
поеднання збудження г скорочення мають тривалшгий перюд вщновлення, нгж
збудлива мембрана серцевого мюциту. Тому це явише виникае в раз! таких уражень
мюкарда, коли функдюнальн) властивост) мембрани ще збережею, а скоротливий
апарат уже порушений, 1 розглядаеться як несприятлива прогностична ознака.
Альтернашя харакгеризуеться вщмшшстю за амплитудою ) тривал)стю збуд-
жень 1 скорочень, шо йдугь одне за одним. Можлива альтернащя тёльки збуджень
або пльки скорочень, або одночасно тих 1 тих. Це здебыьшого пов’язане з тим,
шо пщ час ураження мюкарда у вщповщь на один тмпульс збуджуються ) скоро-
чуються вс) волокна, а у вщповщь на наступний — лише частина Ьс. Тому потен-
шали дц й амгпптуда скорочень неоднаков). Проте можлив) альтернуючт скоро-
чення кожного м’язового волокна.
Порушення засвоення ритму свшчать про глибокий розлад обмшу г часто
спостерпаються в термшальних станах.
Аритмтя унаслщок одночасного порушення автоматизму 1 провщкост). За наяв-
ностз численних ектовдчних вогнищ збудження ) тако! змши проведения импуль-
су, при яюй порушуеться швидюсть проведения його через ртзн) дптянки мюкар-
да або тмпульс поширюеться титьки в одному напрямку, створюються умови для
тривало! циркуляцп хвил) збудження в певному вщдйп серця, виникають розлади
ритму: тремзтння г мерехтшня.
За нормальних умов хвиля збудження, виникиувши в одному мтсц), поши-
рюеться в обидва боки серцево! камери. Досягнувши протилежно! стшки, вона
згасае, зустртвшись з )ншою хвилею. яка залишила за собою зону' рефрактерност)
(мал. 56, а). Якщо ж унаслщок виникнення тимчасово) блокади або затримки
надходження 1мпульс)в деякими волокнами мюкарда збудження проводиться до
мтсця, яке вже вийшло зт стану рефрактерност), то створюються умови для грива -
лот ииркуляш! (мпульсу (мал. 56. б).
У низтл випадктв частота скорочень передсердь сягае 250—400 за 1 хв. Такий
стан визначають як тремгтння передсердь, яке може тривати талька мгсятцв ) роюв
При пьому внаслщок нездатност) шлуночюв вщтворювати високий ритм передсердь
розвиваеться вщносна блокада серця; шлуночки вщповщають скороченням на кож-
не друге, трете або четверге скорочення передсердь, оскшьки решта хвиль збуджен-
ня потралляе у фазу рефрактерност) Скорочення шлуночюв може виникати ранппе
вщ достатнього наповнення !х кров'ю. що зумонтюе тяжк) порушення кровообтгу.
Якщо юльюсть збуджень передсердь сягае 400—600 за 1 хв, (деться про мерех-
пйння, або фьбриляцно, передсердь При цьому скорочуються лише окремт м’язов)
волокна, а все передсердя перебувае в стан! неповного скорочення, його участь у
перекачуванш кров) припиняеться. Ылышсть )мпульс)в, шо хаотично надходять
до передсердно-шлуночкового вузла через окрем) м’язов) волокна передсердя, не-
" _52[
Мал. 56. Рз’х хви.п збудження по мюкарду в норм! (я) ! при ме-
рехтливш аритмн (б) (за Б. Фолковим)
здатш зумовлюваги його збудження, осктльки застають вузол у стан! рефрактер-
ност! або не досягають пороговою ртвня. Тому передсердно-шлуночковий вузол
збуджуеться нерегулярно ! скорочення шлуночктв мають випадковий характер. Як
правило, ктльктсть скорочень шлуночктв за 1 хв перевищуе нормальну. Нерщко
скорочення шлуночктв вшбуваються до наповнення тх кров’ю ! не супроводжу-
ються пульсовою хвилею. Тому частота пульсу виявляеться меншою. н!ж частота
скорочень серия — дефщит пульсу Такий патолопчний стан серця мае назву
мерехтливог аритми. В1н виникае здебшьшого в раз! стенозу л!вого передсердно-
шлуночкового от вору, тиреотоксикозу, кардюсклерозу.
За певних патогенних впливтв (проходження електричного струму кр!зь сер-
це, наркоз хлороформом або циклопропаном, закупорення вшцевих артерш чи
тнш! випадки резко! гтпоксп мюкарда, травма серця, д!я токсичних доз препарат] в
наперстянки ] кальщю) виникае ф1бриляц1я шлу нониле. При цьому через хаотичне
скорочення окремих м’язових волокон пропульсивно! сили скорочень практично
немае, кровообп припиняеться, швидко виникае стан неттритомност! т настае
смерть. До фтбриляцп призводить зменшення концентрацп внутрштньоклттинно-
го кал1ю. шо зумовлюе зниження мембранного потенщалу серцевих мюципв 1
полетшуе виникнення в них деполяризацп та збудження, а також зм!на вмтсту
нервоьих медтатортв, особливо катехол амий в.
Найефектившший застб усунення фтбрилящ! шлуночктв — електрична дефтб-
риляц!я (дтя на серце короткочасним електричним розрядом), за яко! вщбуваеться
одночасна деполяризация вс!х волокон мюкарда т припиняються асинхроннт збуд-
ження м’язових волокон. Дтя запобйання розвитку фтбриляцп вдаються до ко-
рекцц сольового складу кровт.
Недостатшсть кропообку б раз! порушення
надходження крэв! до серця
Цей вид недостатносп розвиваеться в тих випадках, коли до серця надходить
мало кров! венами або коли серце не може прийняти всю кров, що надходить.
Перше спостерггаеться в раз! пповолемп (крововтрата) або рхзкого розширення
судин (колапс), друге — теля накопичення рщини в порожнин! перикарда, шо
утруднюе розширення порожнин серця пщ час дтастоли.
Накопичення рщини в порожнин! перикарда може бути швидким ! повхльним.
Швидке накопичення виникае внаслщок крововиливу теля поранения або розри-
ву серця або в раз! перикардиту, шо швидко розвиваеться. Унаслщок недостатньоТ
розтяжност! перикарда в порожни нх пщвищуеться тиск, що перешкоджае дхас-
холхчному розширенню серця. Виникае гостра тампонада серця. В експеримент!
цей процес моделюеться введениям рщини в порожнину перикарда (О.Б. Фохт),
що дае змогу докладно вивчити патолопчш та компенсаторш мехашзми, яю при
цьому виникають. Передуши зменшуеться кровонаповнеиня порожнин серця,
ударний об’ем серця та артерхальний тиск. Мхж цими показниками х внутр!шньо-
перикард1альним диском хенуе чхтка зворотна залежи есть: шо вищий внутр! шньо-
перикардхальний тиск, то нижчий артерхальний. Венозний тиск при цьому пщви-
шуеться.
Вмикання компенсаторних механхзмхв при перикардит! вщбуваеться рефлек-
торно за участю сигналхв, яю надходять хз трьох рецепторних зон: 1) !з зони от-
ворхв порожнистих 1 легеневих вен — унаслщок щд вишен ня тиску на шляхах
притоку7 кров); 2) з аорти х сонних пазух (синокаротидш зони) — унаслщок зни-
ження тиску на шляхах вщтоку кров! та подалъшохо зменшення депресорного
ефекту; 3) !з перикарда — пщ впливом пщвищення внутрхшньоперикардхального
тиску. Пхсля перерхзання блукаючих х депресорних нервхв, а також у раз! пригнх-
чення рецепторних зон за допомогою новокашу пристосувальн! мехашзми не
функщонують х порушення кровообпу супроводжуеться значними ускладнення-
ми. У раз! тампонади серця мобшзацхя погужних механхзмхв компенсацп для
посилення скорочень серия (гомсо- та гетеро метричш мехашзми, гнотропний
ефект кате ходамхшв) малоефективна або неможлива. Тому працюе тшьки порхв-
няно малопотужншг х енергетично неекономний мехашзм компенсацп х пщтри-
мання артерхального тиску — прискорення скорочень серця. до якого пот!м при-
еднуеться звуження периферичних судин. Цим х пояснюеться тяжкий клшхчний
перебхг гостро! тампонади серця.
У раз! повхльшшого накопичення рщини в порожшхн! перикарда робота при-
стосувалх.них механхзмхв виявляеться ефектившшою, пщвишення внутрхшньопе-
рикардхального тиску протягом певного часу може компенсуватися. Повхльне на-
копичення рщини, яке спостерггаеться пщ час хрошчного ексудативното пери-
кардиту х гщроперикарду, супроводжуеться поступовим розтягненням перикарда
1 збхлыпенням об’ему його порожнини. Унаслщок цього внутрхшньоперикардхалъ-
ний тиск змхнюеться порхвняно мало, а порушення кровообхху не виникае протя-
гом тривалого часу.
РОЗЛАДИ КРОВООБ1ГУ, ПОВ’ЯЗАН!
3 ПОРУШЕННЯМ ФУНКЦП СУДИН
Кровообхг здхйснюеться завдяки тхенхй взаемодц серця х кровоносних судин. Ос-
ноьне завдання останнхх полягае в тому, шоб регулювати об’ем периферичного
русла х його вхцповхднхеть об’ему кров!, а також сталхсть ! адекватшеть крово-
постачання орган!в х тканин. Усу це стае можливим завдяки специфхчиим функ-
цхональним особливостям судин (еластичнхсть, скоротливхсть. тонус, проникнхеть
стшки). Попри те шо ш особливост! притаманш практично всем вхддхлам судин-
Мал. 57. Диференгцащя системи кровообйу за функциональною ознакою I змша тиску вздовж судин-
иого русла (за Б. Фолковим, 1967)
ного русла, можна видщити такт, де та або 1нша особямвкть переважае. Зпдно 13
цим судини подшяють на компенсашшп, резистивш, судини обману та емнкн)
(мал. 57).
Компенсац1йн1 судини — це аорта, артерп еластичного й еластично-м’язового
типу. 1хня функцтя по.>1ягае передуем в тому, щоб перетворювати лоштовхоподхб-
Н1 викиди кров) 13 серця в р1вном)рний ток кровт Еластичш та колагенов) структу-
ри цих судин визначаютъ напруження 1хн)х стшок, потртбне для протидй значив!
розтягувальшй ди кровт Важливо при иьому те, шо гпдгримання постшного фун-
кционального напруження за рахунок зазначених структур не потребуй енергп.
Резистивш судини, або судини опору, — артерюли 1 венули, розмпцеш в пре - 1
пост кап ктярних дщянках суди иного русла Основний ошр току кров) припадав на
артерхальну дщянку судинного русла (66 %), решта — на кашлярну (27 %) : веноз-
ну (7 %). Ошр току кровт в резмстивних судинах здшснюеться завдяки !х струк-
турним особливостям (досить товста стшка порхвняно з величиною просвэту), а
також здатност] м’язових структур стшки перебувати в стан) постшного тонусу й
активно змпповати величину простату у вщповщь на додатков) нейрогуморалып
впливи. Це забезпечуе вщповщшсть просв)ту резистивних судин об’ему кров! в
них, а також контроль надходження кров) в кашляри оргашв 1 тканин. Не можна
повшстю виключити можлив1сть активноТ регуляцп просвету 1 в кашлярах. Цю
Л24.
роль виконують першндотелхалью кл!гини, що за сво!м розм!щенням у стшщ та
функц1ями нагадукпь клггини непосмуговано! м’язово! тканини, проте менш спе-
шалхзован]
Згщно 1з сучасними уявленнями, тонус судин складасться з двох основних
компонент — базального 1 вазомоторного. Базальний компонент судинного то-
нусу визначаеться структурними особливостями (пильною судинною “сумкою”,
у твореною колагеновими волокнами) та мюгекним фактором — т!ею частиною
скорочення судинно! ствпси, яке виникае у вщповщь на розтягнення и кров'ю 1
зумовлене змшами обмшу, зокрема й обмшу катююв, у м’язовш тканин!.
Вазомо горний компонент тонусу судин залежить вщ судинозвужувально!
симпатично! шнерваци та ди на гладком’язов! клхтини судинно! стшки гормонш,
БАР 1 мюцевих метабол !чних факторш.
Судини обмшу — катляри 1 вснули. На дшянках цих судин здшснюеться
двоб!чний обмш М1Ж кров’ю ! тканинами водою, газами, елекгролггами, пожив-
ними речовинами й метаболггами.
Смшсн! судини (переважно др!бн! вени) депонують кров для розподшу и та
повернення до серця. Основна маса кров! (60—80 %) зосереджена саме в цих су-
динах, Викид кров! з емнгсних судин здшснюеться завдяки як насивно-еластич-
н!й вщдач! й активним скороченням непомугованих м'яз!в ст!нки вен, так ! ско-
роченню поперечно посмугованих скелетних м’яз!в, при якому кров витискаеться
з др!бних 1 середн!х вен у бшыш в напрямку серця.
Наведена характеристика судин дае змогу послщовно розглянути !х па голо-
г!ю. Так, атеросклероз, що харакгеризуеться дистроф1чно-шфйътративно-прол1фе-
ративними змшами в артерсях еластичного й еластично-м ’язового тите (тобто су-
динах компенсацшного типу), можна розглядаги як хворобу з переважним пору-
шенням еластичних властивостей судин, хоча певне значения можуть мати !
порушення !хнього тонусу та проникност!.
Вщповщно артер!альну ппер-! гшотензпо слщ зарахувати пере важно до пато-
ло1н резистивних судин, а порушення проникносп стшки судин — до харам ер-
них ознак патологи судин обм!пу.
Патолопчн! зм!ни а судинах компенсац|йного типу.
Атеросклероз
Патолопчн! змши стшки артерш !з вщкладанням у них лшщ!в патологи описува-
ли ще з XVIII ст., а сам термш “атеросклероз” (вщ грец. адцр — каша 1 аккорде —
твердий) запропонував Ф. Мартан у 1904 р. Це поняття визначало наявшеть двох
тип!в змш: скупчення лшщних речовин у вигляд! кашкопод1бних мае у внутрпи-
нш оболонщ артерш 1 власне склерозу — сполучноткани иного уппль нення спнки
артерш.
Атеросклероз е р!зновидом ширшого поняття — артериосклерозу, який гакож
включае артерюсклероз (медшкальциноз) Мьонкеберга ! артерюлосклероз, яю уш-
коджують м’язову оболонку середшх 1 др!бних аргерш та не супроводжуються
вщкладанням лппдгв. Артерюлосклероз чаепше е наслщком хрошчно! ппертензи
та цукрового д!абету типу 1 1 спричинюе мгкроанпопати нирок, Фгктвки ока
тощо
Осюльки саме атеросклероз г його ускладнення е найбыьшим ризиком зрос-
тання захворюваностг та смертносп в економгчно розвинутих кра'шах, вивчення
його етюлогп та патогенезу г досг залишаеться однгею з найважлившгих проблем
медицини.
За сучасними уявленнями, атеросклероз — хрошчне захворювання гнтими великих
г середнгх, еластичних та м’язово-еластичних артерш, що виникае внаелгдок адгезп
тромбоцитгв / моноцитгв, мгграци, пролгфераци та фагоцитарно-синтетичног актива-
ци моноцитгв-макрофаггв г гладком’язових клгтин у вгдповгдь на ушкодження ендотелгю
екзогенними та ендогенними факторами, у тому числг якгено змгненими або геглькгено
надлишковими лгпопротегдами, з подалыиим утворенням атером г фгброатером.
Атероми та фгброатероми всерединг мгстять переважно холестерин г його ефг-
ри та мають вигляд бляшки, яка може перешкоджати кровотоку г е джерелом
вазоконстрикторних ейкозанощгв. Унаслщок цього, а також дисфункцн ендо-
телгоцитгв та зниження еластичних властивостей, ушкодженг атеросклерозом ар-
тери мають схильшсть до спазмування г не можуть адекватно змпповати евгй
проевп залежно вщ потреби органа або тканини в кровопостачанш.
Спочатку функцюнальна неповноцшшетъ склеротично змгнених судин, а
отже, оргашв г тканин проявляеться лише в раз! пщвищення вимог до них, тобто
збыьшення навантаження. Подальше прогресування атеросклеротичного процесу
може призвести до зниження функщональних можливостей г в стан! спокою.
Гострий атеросклеротичний процес, як правило, супроводжуеться звуженням г
навпъ повним закритгям проевпу артерш. У раз! повыьного склерозування артерш
в органах гз порушеним кровопостачанням вщбуваються атрофгчш змши з посту-
повим замнценням функцгонально активно! паренхгми сполучною тканиною.
Склеротично змшенг судини (найчастгша локалгзашя — аорта, артерп серця,
головного мозку, нижнгх кшцгвок) виргзняються пщвищеного щыьнгстю та лам-
кгетю. Швидке звуження або повне перекриття проевпу артерп у випадку розриву
або вщриву атеросклеротичног бляшки, що зумовлюе тромбоз, тромбоемболпо
або крововилив у бляшку, спричинюе розвиток гшемп г некрозу дыянки органа,
тобто гнфаркт або гшемгчний шеульт.
1нфаркт мюкарда — найчастппе та найнебезпечшше ускладнення атероскле-
розу вшцевих артерш.
Бляшки, кргм того, ушкоджують г ослаблюють медпо, що може спричинити
розвиток аневризми та п розрив.
Експериментальнг моделг атеросклерозу. У 1912 р. М.М. Аничков г С.С. Хала-
тов запропонували спосгб моделювання атеросклерозу в кролгв шляхом уведення
гм холестерину (через зонд або у виглядг добавок до звичайного корму). Чгткг
атеросклеротичнг змши розвивалися за кыька мгсяцгв пгсля щоденного введения
0,5—1 г холестерину на 1 кг маси тгла. Як правило, вони супроводжувалися щдви-
щенням р!вня холестерину в сироватц! кровг (у 3—5 разгв поргвняно з початковим
ргвнем), що стало пщетавою для припущення про провщну патогенетичну роль
гшерхолестеринемп в розвитку атеросклерозу. Ця модель може бути легко выдво-
рена не лише в кролгв, а й у курей, голубгв, мавп, свиней.
У собак, щургв та шших м’ясощних тварин, резистентних до дп холестерину,
атеросклероз вщтворюють лише шляхом комбшованого застосування холестерину
г метилтгоурацилу, який пригнгчуе функцгю щитоподгбног залози. Таке поеднання
двох фактор!в (екзогенного й ендогенного) зумовлюе тривалу та виражену ппер-
холестеринемгю (понад 26 ммоль/л — 1000 мг%). Додавання до !'ж! вершкового
масла 1 солей жовчних кислот також сприяе розвитку атеросклерозу.
Пгзшше С.С. Халатов, його учш та !нш! дослщники довели, шо ендогенна
гшерхолестеринемгя, яка виникае при тоивалому стрес! або введенш стеро’щних
гормон!в без екзогенного вживания холестерину, здатна спричинити атероскле-
роз у будь-яких тварин.
Я. Ватанабе (1980) на кролях г В Кумар (1997) на мишах виявили, що гене-
тичш дефекти обмгиу лптопротеццв, що супроводжуються значною гшерхолесте-
ринем!ею, спричинюють виражений атеросклероз та 1ХС в експериментальних
тварин.
X. Баумгартнер (1964) викликав розвиток атеросклерозу шляхом деендотел!-
заш! судин на тл! нормального ргвня холестерину в кров!.
Дуже важливим для розумшня етюлогп та патогенезу атеросклерозу е те, що
ознаки пього захворювання, як! стабптьно виникають в штактних тварин як за
моделлю М.М. Аничкова — С.С. Халатова, так г за моделлю X. Баумгартнера,
слабко проявляються або зовсгм не вщтворюються при порушенн! функцГ! тром-
боципв. Це зокрема довели в доел щах з використанням антитромбоцитарно! си-
роватки Т. Шимамото (1974)! X. Мур (1976).
Етюлопя. Наведен! експериментальн! дан!, а також спостереження за спон-
танним атеросклерозом у людини ! його епщемюлопею евщчать, що цей патоло-
пчний процес розвиваеться внаслщок комбшованог ди низки факторов (навко-
лишнього середовища, генетичних, харчових). У кожному окремому випадку один
!з них е основним.
Нин! вщомо понад 200 фактор!ь ризику атерогенезу. Частина з них (спад-
ковфть, стать, в!к) належать до конституцюнальних або ендогенних, яю дуже
складно або неможливо контролювати. Вони виявляють свою дно вщ моменту
народження (стать, спадковгстъ) або на певному етап! постнатального онтогенезу
(в!к). 1нш1 фактори належать до набутих або екзогенних. 3 !х даю оргашзм люди-
ни стикаеться в р!зн! висов! перюди 1 потенцшно може !х уникати.
Роль спадкового фактора у виникненш атеросклерозу пщтверджують статис-
тичн! дан! про високу частоту виникнення 1ХС у деяких родинах, а також в одно-
яйцевих близнююв. (деться про спадков! форми пперлшопротещемй, генетично
зумовлену неповнощншеть лшопротещлшази, ферменпв 0-окиснення жирних
кислот, генетичн! аномал!’! юптинних рецепторгв для аполшопроте'Щв. Кр!м того,
доведено значения в розвитку артер!ально! ппертензй та атеросклерозу полгмор-
фгзму гешв, яю кодують анпогензиноген, анпотензинперетворювальний фер-
мент, рецептори до ангютензину II, дефекти генгв Ь1О8. Нареши, у розвитку
атеросклерозу важливу роль можуть вщггравати дефекти ген!в, як! регулюють кл!-
тинний цикл за участю тромбоцитарного фактора росту, фактора росту ф!броб-
ласив, ендотелтнгв г специф!чних рецепторов до них. Сюда ж варто зарахувати 1
генетичну схилыисть до шсулшорезистентност! та цукрового д!абету.
Стать. До настання менопаузи жшки за вщсутносп цукрового дтабету. спад-
ково! гшерхолестеринеми чи тяжко! артершльноз гшертензп хвор!ють на атероск-
лероз та гнфаркт мюкарда не так часто, як чоловжи (у середньому в 3—5 разтв
менше). ГПсля 70 роюв захворювашсть на атеросклероз серед чоловгюв 1 ж!нок
приблизно однакова Зазначен! вщмшносп пов’язаш, з одного боку, з нижчим
вихщним р!внем холестерину 1 вшстом його здебшьшого у фракцп неатерогенних
а-лшопроте'щдв сироватки кров! жшок, з шшого — з антисклеротичною д!ею ж!-
ночих статевих гормонов. Зниження функцп статевих залоз у зв’язку з висом або з
будь-яко! шшо! причини (видалення чи опромшення яечникзв) зумовлюе пщви-
щення в сироватщ кров! р!вня холестерину 1 ргзке прогресування атеросклерозу.
Припускають, що захисна д!я естрогешв зводиться до регуляцп не тглъки
вмгсту холестерину в сироватщ кров!, а й шших вид!в обмшу в артер!альнш стш-
щ, зокрема й окисного. Така антисклеротична д!я естрогешв виявляеться пере-
важно щодо вшцевих артерш.
В1к. 3 огляду на ргзке збщьшення частоти 1 тяжкост! атеросклеротичного ура-
ження судин у зв’язку з В1ком, особливо шсля 30—40 рокгв, деяк! дослщники д!-
йшли висновку, що атеросклероз — функция в!ку 1 е винятково бюлопчною про-
блемою (1.В. Давидовський, 1966). Цим пояснюеться песим!стичне ставлення до
практичного вирппення проблеми. Бшышсть досл!дник!в, проте, дотримуються
думки, що В1ков! й атеросклеротичю змши судин, особливо в шзшх стадшх свого
розвитку, — це р!зш форми атеросклерозу, але вгков! змши судин зумовлюють
його прогресування.
Вплив в!ку на розвиток атеросклерозу проявляеться у вигляд! мюцевих струк-
турних, ф!зико-х!м!чних 1 бюх!м!чних змш артер!ально! стшки та загальних пору-
шень обмшу речовин (гшерл!пем!я, гшерлшопротещем!я, гшерхолестеринем!я) 1
його регуляцп.
Пперхолестеринелия, гтер- та дисл1попроте1дем1я е одними з найважлившшх
факторов ризику розвитку атеросклерозу. ЕкспериментальшдослщженняМ.М. Анич-
кова та С.С. Халатова дали змогу припусти™ важливу етюлопчну роль у виникненш
спонтанного атеросклерозу надм!рного харчування, зокрема 1 споживання !ж!,
багато! на жири. Досвщ краш !з високим р!внем економхчного розвитку доводить:
що бптыпе задовольняеться потреба людини в енергп за рахунок тваринних жир!в
’ продукт!в, як! м!стять холестерин 1 насичен! жирн! кислота, то вищий вмгст хо-
лестерину в кров! та бшьша захворювашсть на атеросклероз. Навпаки, у крашах,
де жири тваринного походження становлять незначну частку енергетично! щн-
ност! добового рацюну (приблизно 10 %), а споживання омега-3-ненасичених
жирних кислот, харчових волокон ! антиоксиданпв збшьшене, захворювашсть на
атеросклероз низька (Аз!я, Африка, Латинська Америка, Швдень Свропи). Вщ-
повщно до розроблено! в США програми, що грунтуеться на цих фактах, змен-
шення вживания тваринних жир!в, пропаганда здорового способу життя, заняття
спортом, контроль артер!алъного таску та р!вня глюкози кров!, боротьба з шфек-
щйними хворобами дали змогу значно знизита смертнють вщ шфаркту мюкарда
в друпй половин! XX ст.
Проте останшм часом значения екзогенного холестерину в атерогенез! пере-
глядаеться. У багатьох людей, що споживають дуже багато тваринного холестери-
ну, зокрема в африканських маса!в, не помечено розвитку атеросклерозу та його
ускладнень. В Украпп у 1999 р. споживання холестерину було вдв!ч! меншим, шж
у мешканцгв Шмеччини, а серцево-судинна захворювашсть, пов’язана з атеро-
склерозом, — удв1ч! бшьша. Водночас у Францп, де не спостерйалося зменшення
споживання продукпв тваринного походження, частота серпево-судинних захво-
рювань останшм часом знизилася на 30 %. Усе це евщчить про те, що атероскле-
528~ "
роз не стшьки “дхетична”, скшьки “соцхально-стресова” хвороба (Ж. Медведев,
2008).
Дослши, якх провхв у США на здорових волонтерах професор Ф. Керн (1991),
показали, що тривале надлишкове споживання холестерину не супроводжуеться
пщвищенням його рхвня в кров!. Це пов’язано з тим, що бшышсть зайвого холес-
терину в людини взагалх не всмоктуеться в кишках, а бшыпа частина того, що
всмокгався, у норм! перетворюеться в печшщ на жовчш кислоти ! виводиться в
кишки. Крхм того, холестерин, який усмоктався, знижуе синтез ендогенного хо-
лестерину в печшц!.
3 шшого боку, повна вщсутшсть у харчовому ращош холестерину не гарантуе
захисту вщ гшерхолестеринемп та розвитку атеросклерозу, оскшьки бшьша частхх-л
на холестерину кров! синтезуеться в печшц! з вуглеводхв. Зокрема, патологоан*
томхчнх спостереження Ф. Благе на матерхал! померлих вщ голоду 1 стресу в’язнхь"
нацистського концтабору Дахау показали значне ураження судин атеросклерозом.
Генетичю порушення обмшу лшццв, стрес ! гормоналып порушення вщпрають
суттевхшу роль у розвитку гшерхолестеринемп, нхж споживання жирхв. Так, на
Пхвночх Европи, в США, Канад!, Австралп, Новш Зеландц висока захворюванхсть
на атеросклероз корелюе зх значною поширешстю спадкових ггпер-хдислшопротещ-
емш, патолопчних ген!в апопротещхв, зокрема апопротещу (а), та хх рецептор!в.
Водночас у монголощгв так! патолопчн! гени майже не виявляють, так само як !
гшерхолестеринемш з атеросклерозом.
У кров! холестерин е складником х!лом!крон!в (др!бнодисперсних частинок,
нерозчинних у плазм!) х лшопротеццв — надмолекулярних гетерогенних комплек-
сов триглгцеридгв, ефхрхв холестерину (ядро), фосфолхпщхв, холестерину х спе-
цифхчних бшкхв (аполшопротещи: апо-А, апо-В, апо-С, апо-Е), що утворюють
поверхневий шар.
Лшопротещи вщрхзняються за розмхрами, сшввщношенням ядра й оболонки,
якхсним складом ! атерогеннхстю. Видшено чотири основш фракцп лхпопротещхв
плазми кров! залежно вщ щшьностх й електрофоретично! рухомост!. Визначено
високий вмхст б!лка та низький вм!ст лпгщхв у фракцп ЛПВЩ (а-лшопротещи)!,
навпаки, низький вмхст бшка 1 високий вм!ст лшщхв у фракцхях хшомгкронхв,
ЛПДНЩ (пре-р-лшопротещи) х ЛПНЩ (р-лшопротещи).
Лшопротещи плазми кров! доставляють синтезован! й одержан! з !жею холес-
терин 1 триглщериди до м!сць хх використання та депонування. ЛПВЩ справля-
ють антиатерогенну д!ю шляхом зворотного транспорту холестерину з клхтин, у
тому числх стшки судин, до печшки з подалыпим виведенням з оргашзму у фор-
м! жовчних кислот. 1ншх фракцп лшопротещхв (особливо ЛПНЩ) е атерогенними
х зумовлюють надлишкове накопичення холестерину в стшщ судин.
У табл. 22 наведено класифгкацпо х характеристику первинно! (генетично зу-
мовленох) х вториннох (набуто!) гшерлшопротещемп з тим чи тим ступенем проя-
ву атерогеннох дп. Як видно з табл. 22, у розвитку атероматозних змш судин ос-
новну роль вщхграють ЛПНЩ 1 ЛПДНЩ, пхдвищена концентрацхя хх у кровх,
надлишкове надходження до внутрхшньох оболонки судин.
Надмхрний транспорт ЛПНЩ ! ЛПДНЩ до судинно! стшки може ушкодити
ендотелш.
За концешцею американських дослщникгв Голдстайна! Брауна (1985), ЛПНЩ
х ЛПДНЩ надходять до клхтини шляхом взаемоди зх специфхчними рецепторами
530
Таблица 22. Класиф1кац1я первинноТта вторинно! пперлшопротеТдемГГ
(ВООЗ, 1970)
Тип пперлто- протеТдемГГ Пошире- жсть Мехашзм пперл1пщемй
вмгст у плазм! кров» Етюлопя |МОВ1РЖСТЬ розвитку атеро- склерозу ’-Г-.ь
холестерину ТрИГЛ!- церид!в
I (гшерхшо- кпкронем1я) Спостерг- гаеться рщко Нормальний або дещо пщвшцений Р13КО шдвище- ний Уроджене збшьшення синтезу шпбпора лшопротещлшази, спадковий дефГцит лшопротещлшази (рецесивний тип ус- падкування); дефщит апопротещу С-П; надмГрне споживання харчових жирГв; системний червоний вовчак Недостатшсть лшопротещ- лшази плазми кровт Дещо пщвищс- на
ПА (гшер- беталшо- протещемГя) Спостерь гаегься часто Пщвищений Нормаль- ний Генетична аномалия рецепторгв для ЛПНЩ (передаеться за домшантним ти- пом); споживання ГжГ з високим вмГстом холестерину; цукровий дГабег; гшоти- реоз; нефротичний синдром; синдром Кушшга; мехашчна жовтяниця Знижений катаболвм ЛПНЩ; збщьшена секрецгя ЛПДНЩ та ЛПНЩ Значно пщвище- на
ПБ (гшербе- та-1 пребе- талшопро- теТдем1я) Те саме Те саме Дещо пщвище- ний
III (дисбе- талшопро- те'ГдемГя) СпостерГ- гаеться рщко Те саме Пщвище- ний Генетична аномалГя апопротещГв; спожи- вання 1Ж1 високо! енергетично'Г цшносп 13 високим вм1стом холестерину; ожирш- ня; ппотиреоз; кетоацидоз 1муноглобул1ни утворюють комплекси з ЛПДНЩ, пору- шуючи розщеплення Гх; зни- жена актившсть лшопро- тещлшази; збшьшена сек- реция ЛПДНЩ Те саме
IV (гшер- пребеталь попроте'Г- демгя) Спостерь гаеться часто Дещо шдви- щений Те саме Комбшована сГмейна гшерлппдемГя, С1- мейна ппертриглщеридемГя (успадко- вуеться домшантно); споживання ГжГ з високим вмхстом тваринних жирГв; алко- голГзм; ожирГння; гепатит; цукровий дГа- бет; уремГя; подагра; голодування Те саме + зменшена актив- шсть лецитинхолестерин- ацилтрансферази Пщвище- на
V (пперпре- беталшо- протещемГя 1 пперхыо- М1кронем1я) Досить пошире- НИЙ Дещо пщви- щений або нормальний Значно пщвище- ний СТмейна ппертриглГцеридемГя; дефщит апопротещу С-П; надмГрне вживания жирГв Г простих цукрГв; алкопмпзм; ппо- тиреоз; прийом Д1уретичних засоб^в 1 0- адреноблокаторхв тощо Зменшена швидюсть ката- болизму ЛПДНЩ; надлишок хшомГкронГв 1 ЛПДНЩ Дещо шдвище- на
РОЗД1Л XXVI. ПАТОЛОПЧНА Ф13ЮЛОПЯ СИСТЕМНОГО КРОВООБПУ
(апо-В, апо-Е-рецехтгори-глткопротещи), шсля чого вщбуваються !х захоплення
(ендоцитоз) 1 злитгя з лхзосомами. При цьому ЛПНЩ розщеплюються на проте!-
ни й ефгри холестерину. Своею чертою протеши розщеплюються на выъш амшо-
кислоти, яю виходять з юптини, а ефхри холестерину зазнають гщролгзу з утворен-
ням вгльного холестерину, який надходигь з лгзосом у цитоплазму з подалыпим
використанням з тхею чи нею метою (утворення мембран, синтез стерощних гор-
монхв тощо). Важливо, що цей вхльний холестерин пригшчуе синтез холестерину
з ендогенних джерел, у раз! надлишку створюе “запаси” у форм! ефхргв холестери-
ну 1 жирних кислот, та найголовшше — за механизмом зворотного зв’язку пригш-
чуе синтез нових рецепторхв для атерогенних лшопротещгв г подальше !х надход-
ження до юптини. Поряд з регульованим рецепторопосередкованим механгзмом
транспорту лшопротеццв, що забезпечуе внутрхшш потреби юптин у холестерин!,
описано мгжендотелхальний транспорт, а також так званий нерегульований ендо-
цитоз, який е трансцелюлярним (у тому числ! трансендотелхальним) везикуляр-
ним транспортом ЛПНЩ х ЛПДНЩ з подалыпим екзоцитозом у внутргшню обо-
лонку артергй ендотелхем, макрофагами, гладком’язовими юптинами).
Отже, у мехашзм! початкових змш при атеросклероз!, що характеризуються
надмхрним накопиченням лшщгв у внутршшш оболонщ артерш, можуть вщггра-
вати роль кхлька факторгв.
I. Генетична аномалгя рецепторопосередкованого ендоцитозу ЛПНЩ (вщ-
сутшсть рецепторхв (менше 2 % норми), зменшення хх юлькосп (2—30 % норми),
наявшсть яко! властива родиншй гшерхолестеринемп (ппербеталшопротещемхя
типу ПА) у гомо- х гетерозигот.
2. Перевантаження рецепторопосередкованого ендоцитозу п!д час гшерхолес-
теринем!!.
У перших двох випадках ргзко посилюеться нерегульований ендоцитоз части-
нок лшопротеццв макрофагами, юптинами ендотелпо х гладком’язовими юптина-
ми стшки судин унаслщок значно! гшерхолестеринемп.
3. Сповхльнення видалення атерогенних лшопротещхв зх стшки судин через
лхмфатичну систему у зв’язку з гшерплазхею, артерхальною гшертензхею, запаль-
ними змшами.
Сутгевим доповненням до наведеного е р!зш якхсш перетворення (модифхка-
цп) лшопротещхв у кров! х судиннш стгнщ. Глгюроваш, окиснеш, ацетильоваш
лшопротещи х комплекси лшопротещхв !з 1§6, ендо- й екзотоксинами, глгкозам!-
ноглгканами, фхбронектином, колагеном ! еластином активнппе, нхж нормальн!
лшопротещхв, захоплюються макрофагами I мюцитами за допомогою апо-рецеп-
тор-незалежного мехашзму ендоцитозу за участю “смхттевих” рецепторгв.
Артер1альна гтертензы е важливим фактором ризику атерогенезу, особливо
при тиску понад 160/95 мм рт. ст. За такого тиску захворювашсть на шфаркт м!о-
карда у 5 разгв вища, нгж за нормального артерхального тиску. В експеримент! на
кролях, до гж! яких додавали холестерин, атеросклеротичш змхни розвивалися
швидше 1 досягали бхлыпого ступеня на тлх артерхально! гшертензп. Це можна
пояснити ушкодженням ендотелпо, стимулюванням пперплазп гладком’язових
юптин судинно! стшки та посиленням шфыьтрацп лшопротещхв в штиму ар-
терш.
Цукровий д1абет е одним !з найважливхших факторхв ризику розвитку атеро-
склерозу, оскгльки 75—85 % хворих на цукровий дхабет страждають на атероскле-
рознил ххи. патолопчна фтзюлопя системного кровообтгу
роз । помирають вщ нього, а у 4/5 хворих на атеросклероз встановлено зниження
толерантност! до глюкози. При шших р!вних умовах ризик виникнення шфаркту
мюкарда вдвхчх виший у хворих на цукровий дхабет, а шдукована атеросклерозом
гангрена нижшх кшшвок виникае в 100—150 разгв частице.
За будь-якого типу иукрового дхабету виникають пперхолестеринемхя х гшер-
глхкемхя, якх ушкоджують ендотелгй. Крхм того, пперг.и!кем!я зумовлюе модифика-
цию лшопротещгв завдяки !х глхюруванню. шо сприяе 1х захопленню клттинами
судинно! ст1нки шляхом нерегульованого ендоцитозу. Проте найбпьше значения
для розвиткут атеросклерозу х його ускладнень мае цукровий дхабет типу 2. Це
зумовлено тим. що гшершсулшемхя, яка притаманна цьому типу захворювання,
активуе пролхферащю юптин судинно! стшки, оскшьки шсулпх е фактором росту,
1 сприяе синтезу ахерогенних лшопроте!д!в.
Стрес. € спосгереження, що свщчагь про вишу хахворювашсть на атероскле-
роз у людей ‘"стресових професш”, тобто таких, яю потребують тривалого х силь-
ного нервового напруження (лхкарх, вчителх, пшоти, працхвники управлтнського
апарату7 таги.).
У цшому захворювашеть на атеросклероз вища серед мхського населения. Це
можна пояснили тим, що в умовах великого мтста людина частхше зазнае нейро-
генних стресових вплив!в. Експерименти пщтверджують можливу роль нервово-
психхчного стресу у виникненн! атеросклерозу. Це не дивно, осюльки стрес суп-
роводжуеться артериальною ппертензхею, пперхолестеринемхею ! гшерглхкемхею.
1нтоксикаи1я. зумовлена рхзними хтмтчними речовинами (фториди, СО, Н.8,
бензол, свинець, кадмш, сполуки ртулг, гомоцистеш, шкотин, гшервпамшоз Э,
гпхерурхкемхя), також е фактором, який зумовлюе виникнення атеросклерозу.
Бшышсть штоксикащй спричинюють типов! дистроф!чн! та шфьтьтративно-про-
лтферативн! змши в артерхалыпй стшцх.
Найбшьше ушкодження судинно! стшки зумовлюють палшня цигарок (бшьше
однхе! пачки в день) х гтергомоцистешелия.
Згщно з результатами юпшчних х епщемюлопчних спостережень останнхх
роюв, пщвищення р!вня гомоцистешу в кровт дае виражений атерогенний ефект.
Ця амшокислста мае сильну прооксидантну актившсть. яка спричинюе ушкод-
ження енд отел дальних клхтин, агрегашю тромбоцит! в х посилену пролхферашю
гладком’язових юптин артерхально! стшки. Своею чертою ппергомопистешемхя е
наслщком нестачх в оргашзм! вкамппв В (пхридоксину), В, (фолхево! кислоти) х
В (кобаламшу) (див. роздш XXIV “Патофтзюлопя обмшу вхтамхшв”).
1нфекц1я. Бактериальна, паразитарна та втрусна шфекшя за рахунок фактора
штоксикацц може спричинити ушкодження ендотелхалытих юптин х неклпинних
утвор!в стшки великих артерхальних судин. подалыпий розвиток запального про-
цесу 1, як наслщок, атеросклерозу. У деяких популящйних дослщженнях установ-
лено. що в раз! хрошчно! бактерхально! шфекцп (ресгпраторно!, гастроштести-
нальнох, урогенхтально! та ш.) ризик прогресування атеросклерозу великих артерхй
збшыпуегься в 4 рази. Останшм часом особливого значения в розвитку атероск-
лерозу та 1ХС надають латентним персистуючим шфекшям, зумовленим питоме-
гал о вхрусом, вхрусом герпесу, гепатиту А, СМаткИа рпеитотае, НеНсоЬааег ру1оп.
Жив! хламщп виявлено безпосередньо в атероматозних бляшках (ендотелюцитах,
непосмуговашхх мюцитах ! макрофагах). 1мунопозитивними щодо хлам!л!йно1 ш-
фекц!х е 75 % осгб хз вираженим атеросклерозом, щодо хелхкобактернох та цитоме-
галовхрусно! шфекцш — приблизно 20 % хворих на атеросклероз. Особливе зна-
чения в ушкодженш ендотелпо. активацп адгез!!, мхграцп. пролхферацп й еластолх-
зу в артерхальнш стшщ мають бактерхальн! екзо- й ендотоксини (М.М. Цветкова.
В П. Захарова, М.В. Кришталь, 2011).
1нш1 фактори. У деяких випадках атеросклероз виникае на тлх попередшх
гормональних порушень (ппотиреоз, зниження функцц статевих залоз) або хво-
роб обмшу речовин (подагра, ожир!ння, ксантоматоз). € дан! про те, що розвитку
атеросклерозу сприяють гпходинамхя, надмхрне споживання солодопцв, нестача в
рацюш ванадию, селену, хрому, споживання м’якох води, юнхзуюче випромшю-
вання тощо
Патогенез. Наявш хеорй патогенезу атеросклерозу можна звести до двох, при-
нципово вщмшних за вхдповщдю на запитання: що первинне, а що вторинне при
атеросклероз!, шшими словами, що е причиною, а що наслщком — лшохдоз внут-
рхшньох оболонки артерш чи дегенеративно-пролхферативн! п змши. Це запитан-
ня вперше поставив Р Вхрхов (1856;. Вш першим х вшповхв на нього, зазначив-
ши, шо “за вс!х умов провес, (мовхрно. починаеться з того, що основна речовина
сполучно! тканини, з яко! переважно складаеться внутр! шшй шар артерш, стае
крихкою”. Учень Р. Вхрхова Ю. Конгейм (1877) назвав атеросклероз хрохпчпим
запаленням, що завершуеться жировою дистрофхею, склерозом 1 кальнинозом.
Вщтод! й бере початок уявлення нхмецько! школи патолопв та ц послщовни-
кхв в хнших кра!'нах, зпдно з яким пщ час атеросклерозу спочатку розвиваються
дистрофин! змхни внутр!пхньох оболонки артерхй, а вщкладення лнпдхв ! солей
кальцпо — явище вторинне. Перевага зазначено! концепцп в тому, шо вона здат-
на пояснити розвиток спонтанного й експериментатьного атеросклерозу як у тих
випадках, коли е значнх порушення холестеринового обмшу, так х в тих, коли
таких порушень немае, а таких випадкхв близько 50%. Основну роль автори кон-
цепцп вщводять стану артерхально! спнки. гобто субстрату, який безпосередньо
залучаеться до патолопчного процесу. “Атеросклероз е не тшьки х навпь не стхль-
ки вщображенням загальних обмшних порушень (лабораторно вони можуть на-
вив не визначатися), скхльки похщним власних сгруктурних, фхзичних х ххмхчних
перетворень субстрату артерхально! стшки... Первинний фактор, що зумовлюе
атеросклероз, лежить саме в артерхальнхй стшщ, у п структур! х в 'Гх ензимшй сис-
тем!” (I В Давидовський, 1966).
На сучасному етат вчення про атеросклероз збагатилося додатковими експе-
риментальними фактами 1 клхшчними спостереженнями на користь зазначено!
концепцп. З'ясовано причини та мехашзми ушкодження ендотелхальних клттин
спнки артерш як пускового мехашзму подальше! вщповщх оргашзму на ендо-
телхальне ушкодження (Р. Росс та спхвавт.., 1976—1999). Важливо вхдзначити, що
ушкоджуючу дио на ендотелюцита чинять не пльки екзо-, а й ендогенн! фактори
з навсденого више перелхку. Особливо сильно ушкодження ххмхчш фактори спри-
чхшюють у дхлянках аргерш з турбулентним кровотоком х високим артерхатьним
тиском.
Далх в мхеххях ушкодження вщбуваеться експресхя на поверхш ендотелхоципв
молекул клхтиннох адгезп, що спричинюе прилипания до судинно’! спнки тром-
боципв х моноцитхв, а поим х Т-лхмфоцит!в (мал. 58). Вщбуваеться мхграшя в
пхтиму моноципв 1 перетворення ’хх на макрофаги. Мед!атори запалення, що иро-
ду куються активованими тромбоцитами х лейкоцитами, шдукують хм.грацхю в ш-
Мал. 58. Патогенез атеросклерозу:
а — ушкодження ендотелто при артерОальнш гшертензп,
цукровому дОабето, куршнц гшерхолестеринемп, гшергомо-
цистешемп, шфекцп, 1нтоксикацп; б — емггращя । прош-
ферашя клггин: 1 — адгез!я тромбоцитов та моноцитов;
2 — емпращя моноцитов в штиму; 3 — емпрашя поперед-
ник1в гладком’язових клиин 13 кров: в штиму; 4 — мнращя
гладком’язових клиин 13 меди в 1нтиму; 5 — прол!ферац1я
гладком’язових клггин; в — утворення тпнистих юптин
унаслщок захоплення лшопротеццв макрофагами та мюш-
тимоцитами; г — формування атеросклеротииноТ бляшки:
1 — лшщне ядро, шо мютить позаюптинний холестерин 1
юптинний детрит; 2 — ф1брозна кришка
тиму гладком’язових юптин 13 медп та Тх попе-
редниюв 1з кров!, стимулюють Тх пролТферацпо
1 перетворення на макрофагоподТбш мюшти-
моцити, що здатш разом 13 макрофагами до не-
регульованого захоплення атерогенних лшо-
протецпв за допомогою “смггтевих” рецепторгв
1 перетворення на пшисл клгтини. Нагадаемо,
що кр1м медТаторТв запалення пролгферащю
гладком’язових юптин посилюють, зокрема,
шсулш, анпотензин II й ендотелш.
Захоплення лшопротещхв мюштимоцитами
1 макрофагами вщбуваеться 1 за нормального Тх
р1вня в кров!, але значно посилюеться при п-
перлшопротещемп. Фагоцитозу лшщгв сприяе 1
те, що активоваш макрофаги 1 нейтрофши
видгляють значну кшьюсть акгивних кисневих
радикалТв, яю окисно модифгкують лшопротеТ-
ди. Окиснеш ЛПНЩ 1 ЛПДНЩ е цитотоксичними щодо ендотелюцилв I мю-
цит1в, що спричинюе Тх дисфункции з! зменшення синтезу МО та додатковим
синтезом хемоюнТв 1 факгорш росту. Окиснеш ЛП активно поглинаються макро-
фагами 1 мюштимоцитами шсля зв’язування 31 “смптевими” рецепторами. Може
також приеднуватись гмунний мехашзм додаткового ушкодження юптинних 1 по-
заюптинних структур стшки артерп. Джерелом антигенгв е мйфооргашзми, про-
дукти !х житгед1яльносп, антигени з ушкоджених власних структур, а також мо-
дифгковаш лшопротещи.
Цитокши — медТатори запалення — активують не тшьки миращю 1 пролгфе-
рацпо мюципв, а 1 !х синтетичну актившсть. Мюштимоцити синтезують велику
кшьюсть таких компонентов екстрацелюлярного матриксу, як глгкозамшоглткани,
соалоглгкопротещи й еластичн! та колагенов! волокна.
На завершальному етагп апоптоз 1 некроз переповнених лшщами шнистих
юптин сприяе екстрацелюлярному вщкладанню волокон 1 лшщ1в. Формуеться
ф1броатерома з лшщним ядром 1 ф1брозною кришкою. Частка холестерину та
його ефхргв у лшщному ядр1 поступово збшьшуеться. Пщ впливом циклооксиге-
нази 1 лшоксигенази макрофапв лшщне ядро фтброатероми стае джерелом ейко-
занощ1в, що активують спазм судин 1 утворення тромбов. Шсля чого настае стадгя
Ендотелоалше ушкодження
Високий ровенъ ЛПНЩ у плазм!
Прилипания громбоципв
Вившьнення ТФР
Тнфътьтрашя ЛПНЩ у плазм!
Окиснен! ЛПНЩ
Макрофаги
П1НИСТ1 клггини
у -—- 1нш1
Клггинна пролофераоця фактори
। росту
Задавненс ушкодження
Жирова смута
I I
Про1ресуюча атеросклеротичпа бляшка
I |
Ускладнене ураження Фоброзна бляшка
Схема 21. Патогенез атеросклерозу: зв’язок мЬк гшотезами лшдноТ !н-
фыьграцп та л1дповцц на ушкодження (адаптовано за О. 81ешЪег§, 1988)
ускладнень (тромбоутворення. кальцифокашя, емболоя. шфаркт, шсульт тощо;
схема 21).
Доведено, шо при атеросклероз! кром мосцевих проявив запалення слостерна-
ютьсяо сист емы! прояви запального процесу. Найважливппими з них е: эболынен-
ня в кров! юлькост! нейтрофгльних гранулоципв, моноципв о пщвищення ’охньоо
функционально!’ активностг; збельшення в плазм! кров! вм!сту лрозапальних ци-
ток!н1в, молекул клпинног адгезп та б!люв гостроо фази запалення; серопозитив-
ность до хламщойноо та хелгкобакгерно!’ шфекцп; збшьшення вмосту та змши про-
ф!лю лшопроте’щов, охн!х модифжованих форм у плазм!, пщвищення !х атероген-
ност! за умови шфекцц, ендотоксемп о екзотоксемп; збшьшення вмосту в крово
вольнорадикальних продуктов о зменшення Г! антиоксидантного лотенцоалу, зболь-
шення титру антитш до модифжованих лшопроте’щов та кшькосп Т-цитотоксич-
них клпин
Мож мосцевими та системными проявами запалення пщ час атеросклерозу
оснуе взаемозв’язок. Кожну атеросклеротичну бляшку можна розглядати як вог-
нище запалення, спричинене мосцевим ушкодженням ендотел!ю. Порушення
бар’срноо функцп запального процесу зумовлюе надходження в кров медоаторов
запалення, а також продуктов з автоантигенними властивостями з подальшим
впливом на судинну спнку системних факторов 1 розширенням зони впливу. Сис-
темный вплив медоаторов запалення збшьшуе адгезивность ендотелпо до тромбо-
цитов о лейкоцитов, шо формуе ново вогнища ушкодження ендотелпо о розвиток
новш атеросклеротичних бляшок. Додаткове уооокодження ендотел!ю може в!дбу-
ватися о пщ впливом омунних механозмов. Кром того, антипла, киснев! радикали,
ппергл!кемоя, екзо- й ендотоксини, бшки гостроо фази зумовлюють модифжацою
л!попротещ!в, що прискорюе ох ендоцитоз макрофагами о мюштимоцитами су-
динноо стшки. Так чи шакше протизапалъна терапоя при атеросклероз!, апробо-
вана останнгм часом, довела свою певну ефектившсть.
На противагу цим поглядам з часу дослщжень М.М. Аничкова та С.С. Хала-
това штенсиво розвивалася концепцоя про головну роль у розвитку атеросклерозу
загальних метабол!чних порушень в оргашзм), яю супроводжуються пперхолесте-
ринемгею, гшер- 1 дислшопротещем!ею. 3 цих позищй, атеросклероз — наслщок
первинно! дифузно! шфшьтрацц лшццв, зокрема й холестерину, у внутрппню
оболонку артерш. Подалыш змши в судиннш стшщ (ендотелпо 1 юптинних еле-
ментгв субендотел!ального шару, мукощний набряк, дистрофочн! змши волокнис-
тих структур, продуктивно змши) розвиваються через наивность у ной лшццв, тоб-
то е вторинними.
Спочатку провщну роль у гцдвищенш ровня лшццв, особливо холестерину, у
крово належала алгметарному фактору (надлишковому харчуванню), що дало на-
зву 1 водповщной теорп виникнення атеросклерозу — алшентарна. Проте дуже швид-
ко п довелося доповнити, осюльки стало очевидним, шо не в усох випадках атеро-
склерозу можна встановити ггричинний зв’язок з алтментарною пперхолестерин-
емоею. Зпдно з шфыътрацшно-комбшацшною теоргею С.С. Халатова (1946), у роз-
витку атеросклерозу кр!м аломентарного фактора мають значения ендогенно пору-
шення лшщного обмшу та його регуляцп, механочний вплив на стшку судин, змши
артероального тиску, головним чином його пщвищення, а також дистрофочно змши
в самой артероальнш стшщ. У цой комбшацп причин о мехашзмов атерогенезу одно
(ал!ментарна та / або ендогенна гптерхолестеринемоя) вщограють роль пищального
фактора, ппш або забезпечують збшьшене надходження холестерину в стшку суди-
ни, або зменшують його екскрецпо з не! через ломфатично судини.
Особлива роль на початковому еташ гшерлшопротещемй належить ушко-
дженню ендотелпо, на поверхно якого за таких умов фоксуються моноцити о тром-
боцита. Моноцита в подальшому могрують у субендотелоальний шар 1 перетворю-
ються на шнисп клггини; тромбоцита видшяють ФРТ, який стимулюе пролофе-
рацпо фобробластов 1 гладком’язових клотин.
Могращю макрофапв кров! у внутропшю оболонку судин забезпечуе моноци-
тарний хемотаксичний фактор, що утворюеться пщ впливом лшопротещов, та
1Л-1, який видшяеться оз макрофапв.
На завершальному етат процесу формуються фоброзно бляшки як пролофера-
тивна вщповщь гладком’язових юптин, фобробластов о макрофапв на ушкоджен-
ня, стимульована факторами росту тромбоципв.
Наведен! вище концепцп патогенезу атеросклерозу на сучасному етап! вщроз-
няються лише там, що за 1нф1льтрац1йно-комб1нацшною теоргею атеросклероз суп-
роводжуеться пперлшопротещемоею та г!перхолестеринем!ею, а теория в1дпов1д1
на ушкодження (Р. Росс, А. Гломсет, А. Готто, Р. Джексон, 1976) пояснюе появу
атеросклерозу ! без пперхолестеринемп та первинним вважае вплив на судинну
стшку будь-яких чинниюв, здатних ушкодити ендотелш о спричинити запальний
процес, а не тшьки атерогенних о модифокованих лшопротещов.
Про те що ушкодження ендотелпо о розвиток запалення е первинним, пере-
конливо евщчить той факт, що модель М.М. Аничкова о С.С. Халатова, як о модель
X. Баумгартнера, не вщтворюеться при хвороб! фон Вшлебранда чи при блокад!
функцп тромбоципв, яю е джерелом прозапальних медоаторов о в першу черту фак-
торов росту для гладком’язових юптин судинно! спнки (див. мал. 58). Таким чи-
ном, концепц!я первинного значения мосцевих змш в артероальнш стшц!, яку було
висловлено понад 150 роков тому, успшшо повертаеться.
Патолспчж зм1ни в судинах резистивного типу
Резистивн! судини бер>ть участь у пщтриманш на нормальному ртвш артериального
тиску. Отже, патолог1я резистивних судин проявляеться передус!м значними в!дхи-
леннямп вщ норми р!вня артериального тиску. Вщновщш патолопчш стани нази-
вають артергальною гшер- 1 ппотензгею. У першому випадку ргвш систолхчного
(САТ) 1 Д1астол1чного (ДАТ) артериального тиску становлять вщповщно 140 мм рт. ст.
190 мм рт. ст. 1 вище, у другому — вони менпй за нижет границ] (1001 60 мм рт. ст.)
допустимих у норм! коливань.
Артер)альна ппертенз)я
Артер'юльна гтертенля — це стиле тдвищення системного артериального тиску.
Системний артертальний тиск (сАТ, або Р) — це середнш тиск в артериях,
завдяки якому кров розноситься до оргашв 1 тканин протягом усього серцевого
циклу: сАТ = ДАТ + 1/3 (САТ — ДАТ). Вт е однаковим в аорт! та шших великих
артер1ях тому, що вс1 вони мають великий дтамегр 1 незначний ошр.
У бйтьшосп випадюв першопричина артер!ально! гшертензп залишаеться нев!
домою, 1 таку гшертензто називають первинного. Первинно артергальна гтертенмя
мае назву есешцально! (Е. Ргапк, 1911), або щюпатично!, гшертензп, що вказуе на
нез’ясовашсть ц походження. На частку первинно! гшертензп припадае 95 % ус)х
випадюв захворювання. Решту (5 %) становить вторинна артер1альна гтертензгя. 1з
них 2/3 випадюв пов’язаш з хворобами паренхтми нирок або ьх судин.
Вторинне пщвищення артериального тиску е симптомом (симптоматична п-
пертенз1я), наслщком шшого захворювання (гострий гломерулонефрит, тело-
нефрит, хрошчна ниркова недостатшсть, полшстоз нирок, стеноз ниркових ар-
терш, нирковий васкул1т, реетнпродукуюча пухлина, звуження аорти, гшер- або
ппотиреоз, аденома ппоф1за, пухлини мозково! або к1рково1 речовини наднирко-
вих залоз, пухлини головного мозку, пщвищення внутрппньочерепного тиску,
вживания глюкокортикоццв. естрогешв, стерощних протизапальних 1 конграцеп-
тивних засоб1в тощо).
Вщповщно до рекомендацш ВООЗ (1999) втщшяють кыька ступешв (р^вшв)
пщвищення артертального тиску (табл. 23).
Таблице 23. Класиф)кац<я артер!альнсго тиску
залежно вщ його р!вня I ступеня артеркально!ппер'гензГГ
У'-'.'и5- • -Я--’ '.V’-; И- Категов1Я САТ (мм рт ст.) ДАТ (мм р т п.;
Оптимальний <120 <80
Нормальний < 130 <85
Високий нормальний 130—139 85—89
Гшертензш: I ступшь (м’яка) I] ступшь (пом1рна) Ш ступшь (тяжка) 140—159 160—179 >180 90—99 100—109 >110
1зольована систсгачна >140. < 90
Експерименталыи модем. В експеримент! на тваринах сталого пщвишення ар-
тер!ального тиску можна досягги, послщовно впливаючи на р!зш ланки системи
нейрогуморально! регуляцн тонусу судин.
Значне 1 тривале пщвищення артер!ального тиску розвиваеться в раз! одно- та
двоб!чно! ппемп головного мозку, яка настае теля перев’язування артерш, що
живлять мозок (хребетних, сонних або !хн!х плок). Це центрально-ниелнчна гтер-
тенз1я, зумовлена зниженням кровотоку в мозковш тканин! ! накопиченням унас-
лщок цього вуглекислого газу в стовбуровш частит головного мозку, що зумо-
влюе потужну стимуляцпо симпатичних центр!в пшервацп серця 1 судин.
Аналопчного ефекту можна досягти шляхом уведення в мозочково-мозкову
(велику) цистерну мозку тварин каол!ну, частники якого блокують зворотний ток
спинномозково! рщини через периневральн! та периваскулярш лтмфатичш шля-
хи. 3 одного боку, виникае пщвищення внутрппньочерепного тиску, з шшого —
спостерйаеться деякий ступшь ппемп.
У високооргашзованих тварин (собаки, мавпи) вдалося спричинити ппертен-
зпо шляхом зггкнення процессе гальмування 1 збудження (вплив диференщюваль-
ного подразника одразу за позитивним без звичайного м!ж ними штервалу,
зггкнення харчового й захисного рефлекыв). Гшертензы, що розвиваеться, е на-
слщком неврозу. Виникнення артер!ально! гшертензп можливе в раз! психоемо-
цшного стресу, в умовах зоосощального конфликту.
Модел! друго! групи вщтворюють артергальну гшертензпо шляхом ушкоджен-
ня депресорних систем.
Гейманс у 1931—1937 рр. ! М.М. Горев у 1939 р. одержали рефлексогенну г1-
пертензт (“гшертошю розгальмування”) шсля двобгчного перер!зання в крол!в 1
собак депресорних нервгв 1 пазухових плок язико-глоткового нерва. Мехашзм Г!
виникнення пов’язаний з! зниженням гальм!вно! (“стримувально!”) гмпульсацн з
рефлексогенних зон дыянки дуги аорти та сонно! пазухи. Руйнування ядра солй
тарного тракту шляхом електролгтично! дп дае такий самий результат. До депре-
сорних фактор!в належать ПГА, ПГЕ(, ПГЕ2, ПП2. Природно, що пригючення !х
синтезу (шдометацином) супроводжуеться пщвшценням артер!ального тиску.
Розроблено також нирков! модел! вщтворення артергально! гшертензп. У
1934 р. Гольдблатт вщтворив хрошчну артер!альну гшертензпо шляхом часткового
звуження просвету обох ниркових артерш (реноваскулярна гтертенз1я). Ця модель
мае так! особливост!: по-перше, вона можлива лише за часткового звуження про-
евпу ниркових артерш; по-друге, ппертензгю можна вщтворити лише в раз! обме-
ження надходження кров! до обох нирок. Одноб!чне порушення ниркового кро-
вообйу спричинюе, як правило, нестале пщвищення артергального тиску, проте,
якщо при цьому видалити другу (нормальну) нирку, розвиваеться стшке пщви-
щення артериального тиску.
Нарепгп, тривалу гшертензпо можна зумовити видаленням обох нирок (ре-
нопривна г1пертенз1я), перевшши тварин на гемод!ал!з або перитонеальний дгалгз
для запобйання уремп.
Пресорну дно на судини справляють гормон мозково! речовини надниркових
залоз адреналш 1 гормон гшоталамуса вазопресин, що виробляеться нейронами суп-
раоптичного ! паравентрикулярного ядер. Якщо щ гормони вводили в оргашзм про-
тягом тривалого часу, а головне регулярно, у шддослщних тварин розвиваеться ппер-
тензы. Г! виникнення пов’язують переважно з прямим впливом адреналшу та ва-
зопресину на м'язовг слементи стшки артертальних судин. Кргм того, псвнс значения
при цьому мае стимуляцтя адреналшом решн-анпотензиновог системи (РАС).
В експеримент! на тваринах доведено також роль гормошв ктрково! речовини
надниркових залоз у розвитку артергально’! ппертензп (Селье, 1943). Особливе
значения при цьому мають мгнералокортикоцти — дезоксикортикостерон г альдо-
стерон. Постшне Ёх уведення у помгрних дозах чутливим тваринам (щурам, соба-
кам, кролям) з одночасним призначенням !м замтсть питно! води розницу натрпо
хториду зумовлюе значний гшертензивний ефект. У раз! вилучення натрпо хло-
риду з води 1гжт артертальний тиск у вщповщь на введения дезоксикортикостеро-
ну чи альдостерону не пщвищуеться. Вважають, що безпосередньою причиною
гшертензй е збьтьшення вмтсту натрпо в стшщ судин, який пщвищуе шло чут-
ливгеть до вазопресорних вплив! в. Уведення натрпо хлориду не тыьки зумовлюе
розвиток м1нералокортико1дно1 ггпертпензН, а й здатне спричинити Г! без будь-яких
додаткових вплив,в (солъова г1пертенз1я). Важливо зазначити, що в 2/3 тварин
(шуртв) птсля вщмши сольово! дтети гшертензтя залишалася
1снуе пряма залежшетъ мгж ргвнем артериального тиску г добовою дозою на-
трпо хлориду, гривалтстю його вживания, веком тварин (молод! тварини быьш
схильш до розвитку сольоьо! ппертензп) г спадковою схильшстю. Виведено лшп
щуртв, схильних або резистентних до розвитку сольово! ппертензп. Цю схильшсть
у тварин зумовлюють генетичиг дефекти функцп нирок або синтезу гормонтв, що
спричинюе затримку натрпо в оргашзмЕ 1нпп варганти вщтвореноЁ генетично зу-
мовлено! артергально! ппертензп Грунтуються на схильносп певних лтшй тварин
(щуртв) до частих тнеульттв (до 80 %), артерюлшщозу судин головного мозку, гг-
перчутливосп а-адренергтчних структур м’язових елеменпв спнки судин до кате-
холамштв, наявностг спадкових дефекты синтезу простагландишв.
Е лО.Ю1Т я. Специфтчну причину первинно! артергально! ппертензп щентиф!-
кувати не вдалося г, мабуть, п не генуе, тому що первинна артертальна ппертензтя
ш за етюлопею, ш за патогенезом не е единою хворобою. У кожному7 конкретно-
му випадку и розвиток визначаеться спадковими або набутими факторами, а
часттше !х комбгнащею з ргзними вартантами порушення пресорних або депресор-
них мехашзмгв. быыпою чи меншою участю в цьому нервового. ниркового г гор-
мональних факторгв.
С падкое! фактори. Ниш вважають, що близько 40% випадкгв первинно! ар-
тер!ально! ппертензп спричинеш спадковими факторами. Спадкова схильшсть до
ппертензп полтгенна, тобто може були пов’язана з дефектами або полгморфгзмом
ргзних гешв. Ыльша частина цих гентв пов’язана з нирковою екскрешею натрпо,
а серед них найбыьша кыькгеть мае вщношення до РААС. Сюди належать дефек-
та гентв, що кодують рецептори до анпотензину II, синтез ренгну, анпотензино-
гену й анпотензинперетворювалъного ферменту. Кргм спадково! патологп систе-
ми РААС вивчено кыька гешв, якт впливають на нирковий транспорт сол! г води
та клгтинний мембраннии транспорт натрпо г кальщю. До них зокрема належать
дефекта! тени, що кодують Ха+- / ЕГ-протипереносник. Ма+-, К+-, 2СР-когранс-
портер, Яа+-, К+ АТФазу, атртальний натргйуретичний фактор, адреномедулш
тошо. У випадку синдрому Лщдла мутащя гентв, як! кодують р-субодиницю або
у-субодиниггю натргевого каналу7 в алЁкальнтй мембран! нефрощгпв збтрних тру-
бочок спричинюе !х посилену актившсть, що зумовлюе затримку натрпо в ор-
гангзмт та розвиток артергально! ппертензп. 1нпп тени, мутащя або полгморфтзм
яких причеты! до розвитку артергально! гшертензп шляхом модуляцп судинного
тонусу, мають вщношення до синтази ендотел!ального 1ЧО, кгнази рецептора О-
протегну, адренерпчних рецепторгв, спадково! симпатикотони. Важливим спадко-
вим фактором, причетним до розвитку атеросклерозу г гшертензп, ниш вважають
гнсулшорезистентшсть з асоцгйованою ендотел!альною дисфункцгею. Гг пер гл г-
кемгя на тлг ппершсулшемг! ушкоджуе ендотелш судин зг зменшенням синтезу г
секрецп МО г простациклшу, порушуе структуру г функцгю нирок, стимулюе тка-
нинний синтез анпотензину II, сприяе стовщенню судинно! стшки г зменшенню
диаметра артерюл.
В1к. Артер!альна гшертензгя — це “хвороба осенг життя людини, яка позбавляе
п можливост! дожиты до зими” (О.О. Богомолець). Отже, в!к людини мае велике
значения в походженн! хвороби. Зокрема в ос!б лгтнього в!ку (понад 60 роюв)
частота виникнення артергально! гшертензп становить 60 %. Проте г в молодому
вщг первинна ппертензгя спостерйаеться досить часто.
Стать. До 40 рокгв, починаючи з п!дл!ткового вгку, чоловгки хворгють частг-
ше, нгж ж!нки, а теля 50 роюв — щ вщмшност! згладжуються г шсля 70 роюв
бглышеть хворих на артер!альну гшертензш становлять жгнки.
Стрес. Не мае сумнгву, що певне значения у виникненнг артергально! гптер-
тензп мае перенапруження нервових процесгв пщ впливом надм!рних емоцгй,
стресових впливгв, що спричинюе гшерактивацпо симпатичного вцщглу вегета-
тивно! нервово! системи та гшоталамо-гшофтрно-наднирниково! системи. Про
це евщчить “ещцемгя” розвитку гшертензп в людей, що пережили Ленгнградську
блокаду, П высока поширеюсть у людей “стресових” професгй. Особливе значен-
ия при цьому мають негативн! емоцп, пригшчений настргй, вщчуття тривоги, на
як! не може вщбутися реакцгя в руховому актг, коли вся сила гх патогенно! дг!
припадае на систему кровообггу. На цш пщетавг Г.Ф. Ланг назвав гшертензгю
“хворобою невщреагованих емоцгй”.
Споживання сол1. Проведен! серед населения деяких краги (Япошя, Китай), а
також негритянського населения Багамських остров! в, деяких районгв Закарпат-
ськог областг Украши епщемголопчн! дослщження показали наявшсть тгеного
зв’язку мгж ргвнем артергального тиску г юльюстю спожитог солг. Вважають, що
тривале споживання понад 6 г солг на день зумовлюе розвиток первинно! арте-
ргально! гшертензп в людей, як! мають до не! спадкову схильшсть. Успгшне екс-
периментальне моделювання солъово! гшертензп пщтверджуе значения надлиш-
кового споживання солг. 3 наведеними спостереженнями добре узгоджуються
клшгчш даш про позитивный терапевтичний ефект низькосольовог дгети в па-
цгентгв гз деякими формами первинно! гшертензп. Тим часом е даш, яю евщчать,
що зменшене надходження з гжею калпо, кальцпо та магшю при пщвищеному чи
нормальному споживанш натргю також е фактором ризику первинно! артергаль-
но! гшертензп.
Нарештг, слщ зазначити генування певно! залежност! мгж пщвищенням ар-
тергального тиску г надмгрною масою угла, зменшенням фгзично! активностг, зло-
вживанням алкогольними напоями, кургнням тощо.
Отже, встановлено юлька етюлоггчних факторгв артергально! гшертензп. У
кожному випадку хвороби важливо з’ясувати, який гз них е причиною, а який
вщгграе роль умови п виникнення, особливо за наявностг спадково! схильностг.
Патогенез. Зпдно з ос-
новним ровнянням гемодина-
мики р—ДР/К, яке випливае
хз закону Ома (1=Е/К. для
електричною струму), об’ем -
на швидюсть системного кро-
вотоку, або юльюсть кров»,
шо викидае серце за хвилину
(Ц, ХОС), прямо пропорщй-
на р1знищ тиску (АР) мгж
аоотою та порожнистими ве-
нами й обернено пропоршй
на пдродинамзчному опору в
суди нах (К) або загальному
периферийному опору (ЗПО).
Осюльки тиск у порож-
нистих венах становить
Мал. 59. Тиск I загальний периферитний ошр у великому
ВОЛ) кровообпу
О мм рт. ст., можна вважати,
що ДР = САТ, а САТ = ХОС • ЗПО або Р = Р В. ХОС (Р) своею чертою зале-
жить вщ ударного об’ему серця (УОС) 1 частота серцевих скорочень (ЧСС) за 1 хв
Р = УОС ЧСС, тобто вш робот и серця. Величина УОС, або систол^чногс об’ему
серця (у норм! — 70 мл) визначаеться ОЦК (у норм)— 4—5 л), швидюстю веноз-
ного повернення кров! до серця та силою його скорочення.
Осюльки ХОС мае бути однаковим у кожному вщдйп судинно) системи, зни-
ження тиску кров! вщ левого шлуночка до правого передсердя залежить виключно
вщ загального периферичного опору. Осюльки в артерюлах створюеться найбшь-
ший ошр току кров) поршняно з 1ншими вщдшами судинно! системи, то ! К зале-
жить переважно вш !х структури 1 функцп (мал. 59).
Згщно з ршнянням Пуазейля К = в-туЯ/я-г4, пдродинам!чний ошр току рщини
прямо пропорщйний ц в’язкосп (г]) та довжин! судини (1) й обернено пропорши-
ний четвертому ступеню внутршшього рашуса судини (1/г4). Осюльки в’язюсть
кровт та довжина судин швидко не змшюються, зрозумшо. шо змша опору артерюл
переважно визначаеться змшою !х радиуса (К ® 1/г4) за рахунок скорочення або роз-
слаблення м’язово! стшки. Так, зменшення дгаметра артерюли лише вдв1ч! призведе
до пщвищення опору в шй у 16 раз!в, а збшьшення диаметра в 4 рази, що е щлком
можливим, зменшить ошр 1 шдвищить мюцевий об’емний кровотж у 256 разш.
Таким чином, артерхальний тиск визначаеться в основному двома факторами;
величиною ХОС (ф), що залежить вщ частота 1 сили серцевих скорочень, < опо-
ром, якого ток кров! зазнае в судинах (К), шо переважно залежить вы д!аметра
артерюл.
Залежно вщ того, збшьшення якого з двох параметров (Ц або В.) визначае
пщвищення артериального тиску, розр!зняють таю гемодинамгчн! типи артершлъ-
ног гтертензи: 1)г!перк1нетичний— збшьшений О, не змшений або трохи
знижеяий К: 2) еук!нетичний — спостерогаеться збшьшення як Ц, так 1 К;
3) г1пок1нетичний — величина О не змшена або дешо зменшена, К — р!зко
збшыпена
Вторинна артерхальна гшертензгя може мати рхзш гемодинам1чн1 типи. Нап-
риклад, гшертиреоз завдяки посиленню роботи серця пщ впливом тиреощних
гормон1в 1, вщповщно, збшьшенню 9 вщповщае пперкшетичному типу. Гшоти-
реоз, навпаки, супроводжуеться вщкладанням мукополкахаридхв у стшщ артерюл,
!х звуженням, збшьшенням К, ослаблениям серцево! дгяльносп та зменшенням
0, що вщповщае гшокшетичному типу.
Важливо вщзначити, що на початковому етапг первинно! артергально! гтер-
тензи в молодому та середньому виц основною ц ознакою е пщвищення арте-
риального тиску, яке зумовлюеться не пщвищенням тонусу судин, а посиленою
скоротливою дхяльшстю мюкарда, що вщповщае пперкшетичному типу артергаль-
но! гшертензн (схема 22). У разг подалыпого розвитку захворювання гшеркшетич-
Схема 22. Патогенез первинноТ артергально! гшертензн
ний тип переходить в еуюнетичний 1 ггпокшетичний. На запитання, чи стосуеть-
ся це I так звано! гзольовано! систолгчног артергально! гшертензп, яка виникае в
ос!б лгтнього вгку (теля 65 рокгв) на тлг знижено! еластичноет! судин, сьогоднг
ствердно вгдловгеги неможливо.
Спадкова. стресорна чи центратьно-ппемгчна гтерактивац!я симпатично! не-
рвово! системи (схема 23) прямо та за допомогою стимуляцп секрецп мозковою
речовиною надниркових залоз норадреналгну г адреналину через (^-адренорецеп-
тори зумовлюе збгльшення частота га сиди серцевих скорочень, а через а-адрено-
рецептори спричинюе спазм непосмугованих м’язгв артерш, що щдвищуе пери-
феричний ошр у деяких органах. Завдяки одночасному судинорозширювальному
впливу адреналину на Р2-адренорецептори судин скелетних м’язгв, як! становлять
бшьшу часгину судин тша, активацгя симпатико-адреналово! системи призводить
до збгльшення ХОС за практично незмгнено! величини ЗПО. Кргм того, симпати-
ко-адреналова система через а-адренорецептори стимулюе вазоконсгрикцпо вен,
що збшьшуе приплив венозно! кровг до серця. Завдяки впливу на р,-адренорепеп-
тори гранулярних клгтин юкстагломерулярного апарату7 нирок пперактивацгя
симпатичного вгддглу нервовог системи стимулюе секрецгю ренгну, що активуе
РААС г, вцщовгдно, ниркову реабсорбцгю натргю г води. Через активации (^-адре-
норецепторов симпатико-адреналова система стимулюе К1а+-, Н+-протиперенос-
ники на апгкальнш мембран) нефропитгв, що безпосередньо збгльшуе реабсорб
цгю натргю в проксимальних канальцях нефрона. Усе це спричинюе гшерволемгю
г збгльшуе приплив кров! до серця. Проте найсильнцпе ОЦК збгльшуеться при
спадковш чи набутгй стимуляцп РААС.
Найзначвгшу роль у кардговаскулярнгй патологи вщгграе ангготензин II (схе-
ма 24). Вгн може бути як пиркулюючим гормоном — компонентом РаАС, так г
мгецевим медгагором, що продукуеться баг атьма тканинами, у тому числг серцем
! судинами, у вщповщь на !х ушкодження. Ангготензин II через АТ(-рецептори
збгльшуе вазомоторний тонус завдяки прямому стимулюванню скорочення не-
посмугованих м’язгв судин, а також через пригнгчення вившьнення ендотелгем
Схема 23. Симпатична серцево-судинна регуляцГя (за В. Ра1ш, 1993)
Анпотенз иноген
А нпотензин (
Анпотензин П
Стимуляция
фактор!в
росту
Порушення
ендотетальноТ
функцп
(дефицит КО)
▼
Гшертроф1я
судинно! стшки
Вазоконстрикция
Активащя
пресорних
факторТв
(ендотелш-1)
Посилення
симпатично!
активноси
Посилення
розпаду
брадикнпну
Стимулящя
секрецй
альдостерону
Пщвищення АТ
Затримка №
1 рщини
Схема 24. Мехашзм пресорноТ дп анпотензину II
МО 1 простациклшу та стимулящю секрецй ендотелталъними клггинами потужно-
го вазоконстриктора — ендотелшу. Анпотензин II безпосередньо стимулюе реаб-
сорбщю натр1ю в проксимальних канальцях 1 через активащю секрецй альдосте-
рону клубочковою зоною юрково! речовини надниркових залоз збшьшуе реабсор-
бщю натрпо в збтрних трубочках.
Збгльшення надходження кровг до правого передсердя призводить до пщвищення
ЧСС 1 без попередньо! активацп симпатично! нервово! системи. Частина цього
ефекту зумовлена прямим розтягненням 1 подразненням атипових клгтин пазухо-
передсердного вузла. Бшьша ж роль у прискоренш серцевих скорочень при цьому
належить розвитку рефлексу Бейнбриджа: передсердш рецептори розтягнення по-
силають гмпульси по аферентних волокнах блукаючого нерва в довгастий мозок,
а еферентш сигнали передаються симпатичними нервами до серця, що збшьшуе
частоту I силу серцевих скорочень. Кргм того, збшьшення наповнення л!вого
шлуночка та його розтягнення за законом Франка — Старлшга (гетерометричний
мехашзм компенсаци) додатково збшьшуе силу серцевих скорочень 1 УОС. Усе це
пщвищуе ХОС 1 артергальний тиск за гшеркшетичним типом.
У здорово! людини, у яко! немае порушень нейроендокринно! регуляцп та
функцп нирок, безперервно здшснюеться контроль над системним артергальним
диском. Так, раггтове пщвищення тонусу симпатичного вщдшу нервово! системи
й артериального тиску при психтчному чи ф!зичному навантаженш спричинюе
подразнення синокаротидних 1 аортальних барорецепторгв, гмпульси вщ яких по
аферентних волокнах язико-глоткового 1 блукаючого нерв!в надходять до довгас-
того мозку, де зумовлюють гальмування тошчно! активностх симпатичних суди-
нозвужувальних нерв!в ! збуджують парасимпатичш центри блукаючих нервхв.
Унаслщок цього вщбуваеться розширення артерюл 1 вен у периферичних вщдшах
великого кола кровообпу 1 зменшення частота 1 сили серцевих скорочень, що
знижуе артергальний тиск. Якщо ж чинник пщвищення артериального таску про-
довжуе дгята понад 1—2 доби, барорецептори звикають до пщвшценого тиску,
перебудовуються 1 починають сприймата високий тиск як нормальний.
Тривала регуляцгя артериального тиску неможлива без нормального функцю-
нування нирок г!х адекватно! нейрогормонально! регуляцп, осюльки сталгсть тис-
ку залежить в!д тпдтримання постойного об’ему кров!, який своею чертою визна-
чаеться здатшстю нирок забезпечити нормальний ргвенъ натрпо ! води в оргашзм!
за допомогою описаних вшце мехашзмгв (див, роздш XXII “Порушення водно-
електролгтного обмшу”). Якщо ж через наявшсть спадкових або набутих порушень
захисш рефлекторш мехашзми не здатш швидко вщновити нормальний ргвень
артериального тиску, то тод! артергальна гшертензгя набувае хрошчного перебггу.
На другому етат патогенезу первинно! гтертензи хрошчне пщвищення ар-
тергального тиску призводить до вазоконстрикцп, ушкодження судинно! стшки з
п подальшим перманентним стовшенням (ремоделюванням), звуженням просвпу
артерюл 1 збшьшенням периферичного опору (див. схему 22). Це вщповщае закону
Лапласа: Т=Р г, який стверджуе, шо напруження судинно! стшки (Т) прямо про-
порцгйне тиску (Р) в судин! та п радгусу (г). Для кровоносних судин це означае, що
якщо пщвищуеться артергальний тиск, то для збереження цшост! судинно! стшки
1 забезпечення нормального обмшу речовин, що прямо залежить вщ сталост! Т,
необхщно зменшити г. Ц! теоретичш мгркуваггня шдтверджуються законом Остро-
умова — Бейлгса, який було сформульовано на пщставг клпичних спостережень.
Зазначений закон стверджуе, що пщвищення артергального тиску призводить до
генералгзованого спазму артерюл г збшьшення ЗПО. Це забезпечуе сталгсть крово-
току в периферичних органах ! тканинах г запобйае розриву дргбних судин.
Звуження артерюл вщбуваеться поетапно завдяки наступним механизмам. У
раз! збшьшення серцевого викиду г ргзкого пщвищення артергального тиску, що
супроводжуеться прискоренням кровотоку, зпдно з мюгенною теор!ею авторегу-
ляцп кровотоку вщбуваеться розтягнення гладком’язових клгтин стшки артерш г
артерюл, що за законом Франка — Старлгнга спричинюе гх подальше спазмуван-
ня, зменшення радгуса судин, збглъшенггя ЗПО г зменшення швидкостг кровотоку
до нормального ргвня. Мехашзм носиленого скороченггя пгсля поперелнього роз-
тягнення як непосмуговагшх м’язгв, так г кардгомющгпв, за сучасними уявлення-
ми, пов’язаний зг швидким наростанням кальцгевого току, направленого гз позак-
лгтинно! рщини в саркоплазму, унаслщок активацп механозалежних кальщевих
каналгв сарколеми.
Пролонговане г значне пщвищення артергального тиску ушкоджуе стшки су-
дин, що стимулюе мгсцеве утворення г секрецпо антютензину II й ендотелшу-1,
яю е потужними вазоконстрикторами. Як стало вщомо останнгм часом, вони е
факторами росту, що стимулюють мгграцгю, ггпертрофпо г гшергшазпо
гладком’язових клгтин г фгбробластгв судинно! стшки, синтез ними колагену,
еластину, протеоглгкашв, що призводить до потовгцення меди та штими. Анпо-
тензин II також зумовлюе виникнення гшертрофп ! фгброзу мюкарда пгсля
гшемгчного його ушкодження або у вщповщь на постгйне пщвищення артергаль-
ного тиску. Пролгферативне потовгцення артерюл значно посилюеться за наяв-
ностг гнсулгнорезистентностг, оскгльки ггперглгкемгя додатково ушкоджуе ендо-
телгй, а надлишок гнсулгну е фактором росту. Кргм того, ггпертензивне та гшерг-
лгкемгчне угггкодження ендотелпо спричинюе адгезгю до стгнгси судин тромбоципв
г моноцитгв з розвитком запального процесу, який супроводжуеться набряком г
пролгферащею фгбробластгв штими судин. Пролгферативне потовгцення меди та
штими залишае артерюли перманентно звуженими, незалежно вгд наявностг або
вщсутноси стимултв. якг зумовлюють вазоконстрикппо. ВЦ цього моменту ЗПС
залишаеться постшно пщвишеним, а артертальна ппертензтя набувас ознак еук!-
нетичного, а шсля гшертрофп мюкарда, кардюсклерозу та виснаження серця т
ппокшетичного типу.
Наслтдки артерталыю! гшертензп. Рання стадтя гшертензп, як правило, не мае
клштчних проявив. окр]'м тпдвишеного артериального тиску, т тому називаеться
латентною. Вона е небезпечною. оскйгьки хвор! не звертаються по медичну допо-
могу ! хвороба прогресуе. Близько 5 % хворих тз первинною або вторинною арте-
рхальною ппертензтею схильш до швидкого г значного гцдвищення артериального
тиску (САТ > 200 мм рт. ст., ДАТ — > 120 мм рт. ст.). Ця патолопя називаеться
злояк!сною гтертенз1ею 1 може спричинити смерть без локування за 1—2 роки.
У бйтьшосп хворих КЛГН1ЧН1 прояви гшертензп пов’язаш з ремоделюванням
стшки артерюл 1 пщвищенням ризику розвитку атеросклерозу. Перманентне зву-
ження просвету артерюл зумовлюе штемтчне ураження серия, нирок, центрагьно!
нервово! системи, с!тк1вки ока тощо.
Орган, що страждае найбелыие, — серце. Кртм абсолютно! та вщносно! коро-
нарно! недостатносп, яка проявляеться стенокардтею та шфарктом мюкарда, ар-
терхальна гшертензтя супроводжуеться розвитком гшертрофп мюкарда, оскйгьки
лтвий шлуночок поспйно перевантажуеться збйтьшеним объемом кров! та / або
збьтьшеним опором току кровг. Гшертрофтя мюкарда поступово зумовлюе розви-
ток кардюсклерозу 1 хрошчно! серцево! недостатносп. При значному та ртзкому
пьцвищеню артертального тиску може виникнути гостра лгвошлуночкова серцева
недостатшсть, що супроводжуеться набряком легень.
Ппертошчне порушення кровооб!гу в нирках не пйлъки призводить до хрошчно!
нирково! недостатност! внася!док зменшення к!лькост! функщонуючих нефрошв, а й
спричинюе значне прогресування артерсалъно! гтертензп.
Вщомо, що нирки можуть сприяти як пщвшценню, так т зниженню артериаль-
ного тиску. Перше залежить вщ функцп юкстагломерулярних (епттелющних, мюепт-
телтальних) клпин. шо у ын ,тяд1 манжетки оточують приносну клубочкову артерюлу
юкстагломерулярного (приктубочковою) апарату нирок. У юкстагломерулярних кл1-
тинах утворюеться ренш — протео.штичний фермент, субстратом для якого е а2-гло-
булш плазми кров! — ангтогензиноген. Збыьшення секрецп реншу супроводжуеться
ферментативним перетворенням анпогензиногену кров! на анпотензин I. Цей пол!-
пептид (декапептид) п!д впливом перетворювального ферменту кровх перетворюегь-
ся, иереважно в легенях. на анпотензин II, який кртм багатьох шших ефекттв, опи-
саних више, стимулюе синтез альдостерону. Оскйгьки секретя реншу регулюеться
зокрема ступенем розтягнення приносних артерюл клубочктв 1 партцальним тиском
кисню, зрозумыо, що ппергошчне звуження ниркових аргерш г артерюл призводить
до гшемтчно! стимуляцп вироблення реншу, анпотензину II й альдостерону.
Вивчено хтмтчну структуру анпотензину II (вш е октапептидом), здтйснено
його штучний синтез. Перюд твжиття анпотензину II, видйгеного тз сироватки
кров! коня, свин! або бика, становить у середньому 200 с. Таке швидке зникнення
анпотензину II тз системи кровообпу пояснюеться його руйнуванням у кров! !
тканинах (переважно в нирках) пщ впливом ферменту ангютензинази. Здорова
нирка (у раз! порушення кровообпу в друпй нирщ) утворюе велику кйтьктсть ан-
гютензинази, тим самим запобшаючи розвитку спйко! гшертензп. У раз! пору-
шення гемодинамики в обох нирках, шо спостертгастъся при аргертальнш гшер-
тензп, Тх здатшсть виробляти анпотензиназу р!зко знижуеться, що зумовлюе ста-
де пщвищення артерхальнохо тиску. Отже, у раз! порушення кровообпу в нирках
артергальна гшертензхя частково зумовлена активащею РААС, частково — змен-
шенням утворення анпотензинази.
Проте пщвищена секрещя рехгхну хшемгзованими нирками вияьляеться лише
в перш! дш шсля експериментального звуження ниркових артерш, тод! як гшер-
тензхя утримуеться на високому рхвн! протягом тривалого часу. Цей факт, а також
стала гшертензхя п!сля видалення обох нирок дали змогу припустили, що хрошч-
на гшертензхя може бути зумовлена порушенням якоТсь депресорно! функцп ни-
рок (ренопривна гшертензхя) та / або дхею якихось позаниркових речовин (Гролд-
мен). Це припущення повшстю шдтвердилося. Ниш загальновизнано, шо нор-
мальш нирки здатн! контролювати дхю як ниркових, так 1 позаниркових пресорних
речовин (анпотензин II, вазопресин, катехоламши), тобто мають антипресорну
(протиппертеизивну) функшю. Вважають, що ця функция здшснюеться за допо-
могою синтезу нирками простагландин !в г к!н]н!в.
Особливий интерес становлять простагл андини — похгдш полхненасичених жир-
них кислот. Основним мхсххем гх утворення в нирщ е хнтерстицшш зхрчаст! клггини
мозково! речовини, що мають мезенхгмну природу. Контроль за секрецхею проста-
гландиюв здшснюеться за допомогою ангиотензину II, катехоламшгв г вазопресиггу.
ГИдвищена хх юльюсть у плазм! кров! стимулюе утворення простагландишв.
ПГЕ, ! ПГ1, справляють як виражений мкцевий, так г помгрний системний
вазодилатацшний ефект. Кр!м розширюватьно! дп на артерюли простагландин и
блокують реабсорбцпо натр!ю в ниркових канальцях х спричинюють посилене
видыення його гз сечею, що також запобхгае розвитков! гшертензп. Аналопчна
дгя властива нирковгй калгкреш-кгшновгй систем!.
Отже, пригнхчення антипресорно! функцп нирок при ппемгчному гх ушко-
дженш, як г системне зменшення тцд да ею анпотензину II продукт! ендотелюци-
тами оксиду азоту та простациюпну, зумовлюють переважання вазопресорних
мехашзмгв над депресорними г виникнення гшертензп.
Розвиток хрошчно! нирково! недостатностт (синдром лервинно-зморщено!
чирки) унеможливлюе виведення з оргашзму надлишку натрпо г води, що спри-
чинюе гшерволемхю, яка е основною передумовою стабильного пщвищення ар-
териального тиску. Таким чином, наведен! даш переконливо евщчать про важливу
патогенетичну роль ниркового фактора у виникненш, розвитку 1 прогресуванш
артерхальнох гшертензп.
Ппертензивне звуження мозкових судин зумовлюе !шем!ю мозку, активащю сим-
патичного в!дд!лу нервово! системи та посилення артергальна! гтертензи за механиз-
мом порочного кола. Дуже небезпечними для життя та здоров’я людини е також
таю усктаднення артерхальнох гшертензп, як гшертензивна енцефалопатхя та ш-
сульт, крововиливи в ехтювку ока, набряк зорового нерва та пт.
Легенева гшертензхя, шо харакгеризуеться пщвшценням сАТ у легеневих супи-
нах понад 25 мм рт. ст., е причиною розвитку легеггевого серия (сот рШтопа1е)!
недостатност! кровообхгу за правошлуночковим типом.
Швидюсть кровообпу через легенх так само, як ! через велике коло кровооб!-
гу, доргвнюе ф При цьому сАТ у малому кол! кровообпу становить приблизно
15 мм рт. ст., а у великому — близько 100 мм рт. ст. Ця рхзниця зумовлена тим.
шо стгнка легеневих судин мктить поргвняно менше м’язових кл!тин, дгаметр
легеневих артерюл та !х здатшсть розтягуватися бглыш, а ошр значно менший.
Тому за нормально! структури 1 функцп легень збглыпення щд час фгзичного на-
вантаження ХОС у 4—6 разгв знижуе ошр легеневих судин за рахунок !х розши-
рення 1 суттево не збглыпуе тиск у легеневш артерп.
Етюлопя. За походженням розргзняють первинну г вторинну легеневу гшер-
тензпо. Виникнення вторинно! легенево! гтертензи пов’язують гз хворобами, що
зменшують резерви судинного русла, первинно ушкоджуючи бронхюли й альвео-
ли разом з альвеолярними капглярами (емфгзема легень, хрошчний бронхгг, брон-
хгальна астма, хрошчна пневмошя, фгброз легень, ателектаз 1 пухлини легень,
бронхоектази) або судини легень (вузликовий пергартерпт, тромбоз дргбних кро-
воносних судин, тромбоемболгя легенево! артерп), а також гз хворобами, що збшь-
шутоть тиск у лгвому передсерд! (стеноз лгвого передсердно-шлуночкового отвору,
дефект мгжпередсердно! та мгжшлуночково! перегородок, ураження аортальних
клапашв, кардюмюпатгя, обструкщя лгвого передсердя, констриктивний пери-
кардит тощо).
Первинна легенева г!пертенз1я, яку спостерйають значно рщше, розвиваеться
без попереднього ушкодження структури легень г розвитку лгвошлуночково! сер-
цево! недостатность Основною причиною п виникнення е зниження паршально-
го тиску кисню (рО2) в альвеолярному повпрь Природно, що в умовах високопр’я
вплив цього фактора виявляеться сильшшим.
Патогенез. За первинним мгсцем збшьшення опору току кров! та кров’яного
тиску легеневу ппертензпо подгляють на прекапшярну 1 посткапшярну.
Прекапыярна легенева гтертензгя, до яко! належить г первинна легенева ппер-
тензгя, характеризуемся первинним збшьшенням тиску в легеневих артер!ях. 5
основ! патогенезу первинно! легенево! гтертензи лежить наявшсть у гладком’язових
клпинах спнки легеневих артерюл ушкальних киснечутливих кал!евих канал!в.
Непосмуговаш м’язи судин усгх органгв г тканин, кргм легень, мгстять АТФ-
залежнг калгевг канали, якг за умови гшоксп та, вщповщно, зниження синтезу
АТФ розкриваються. Це зумовлюе вихщ К+ гз саркоплазми, пперполяризацпо сар-
колеми г розслаблення гладком’язових клгтин. Дгаметр артерюл збгльшуеться,
кровопк посилюеться, що збшьшуе доставку кисню г синтез АТФ. Гладком’язовг
клгтини легеневих артерш г артерюл реагують на зниження рО2 в альвеолярному
повггрг закриттям О2-чутливих К+-каналгв, шо спричинюе деполяризацию сарко-
леми, збшьшення надходження Са2+ в саркоплазму г спазм непосмутовано!
м’язово! тканини легеневих судин (артерюл), зумовлюючи стшке пщвищення !х-
нього тонусу. Цей мехашзм необхщний для збереження нормального стввщно-
шення мгж вентиляцгею окремих альвеол ! перфузгею судин, що !х омивають. При
тотальшй екзогеншй чи вентиляцгйшй ггпоксп зазначений захисний механгзм
стае патогенним г зумовлюе застшну правошлуночкову серцеву недостатшсть г
зменшення надходження кров! в л!ву половину серця.
Вазопресорний ефект гшоксп посилюеться в разг розвитку ацидозу та щд час
фгзичного навантаження. Кргм того, вважають, що гони водню, ендотелгни, вазо-
активш амгни (пстамш, серотонгн), як! утворюються при гшоксп в легеневих су-
динах, а також надходять до них гз периферичних органгв г тканин, вгдгграють роль
посередникгв у пресорному вплив! гшоксп на легеневг судини (А.1. Хомазюк, 1978).
Алкалоз ослаблюе вазопресорний ефект гшоксп. Судинорозширювальний
вплив на легеневг артерп справляють ПП2, МО, аденозин, АМФ.
Посткатлярна легенева гтертенз1я найчастппе е наслщком лхвохххлуночковох
серцевох недостатност!, що спричинюе пщвищення тиску в лхвому передсерд!,
легеневих венах х кашлярах х лише вторично призводить до збшьшення тиску в
легеневих артер!ях. За нормальних умов тиск кров! в л!вому передсерд! становить
2—8 мм рт. ст., а в легеневих капшярах — 7 мм рт. ст. Коли при гострш лхвошлу-
ночковш недостатност! тиск у л!вому передсерд! пщвищуеться понад 30 мм рт. ст.,
виникае набряк легень.
Артер!альна гшотензйх. на вщмшу вщ гшертензп, е сталим зниженням ар-
териального тиску (САТ < 100 мм рт. ст., ДАТ < 60 мм рт. ст.), зумовленим або
ослаблениям насосно! функцп серця, або переважно зниженням тонусу резистив-
них судин. Спостерйаеться частгше в ос!б астешчнох конституцп, проявляеться
зниженням фхзичного розвитку, загальною адинам!ею, швидкою стомлювашстю,
таххкардхею, задишкою, запамороченням, головним болем, непритомшстю.
Видшяють артер!альну гшотензпо фхзюлопчну (не супроводжуеться хвороб-
ливими симптомами) х патолопчну (хз харакгерним симптомокомплексом). Ос-
тання бувае гострою 1 хрошчною. Гостра патологгчна артергальна гтотенз1я ви-
никае при колапс!, шоку (див. розд!л XXXIII “Патолопчна фхзюлопя екстремаль-
них станхв. Шок. Колапс. Кома”).
Хротчну патолог1чну артер1алъну гтотензйо подшяють на вторинну (симпто-
матичну) 1 первинну.
Враховуючи, що рхвень артерхального тиску визначаеться роботою серця, кшь-
кхстю циркулюючо! кров!! тонусом резистивних судин, розрхзняють три гемодина-
м!чн! форми артерхальнох гшотензп, патогенетичну основу яких становхггь у пер-
шому випадку недостатшсть скоротливо! функцн серця, у другому — зменшення
кхлькостх циркулюючо! кровх, у третьему — зниження тонусу резистивних судин.
Симптоматична хронхчна артерхальна гхпотензхя (вторинна) е на-
слщком деяхеих соматичних гострих 1 хрошчних хвороб серця (вада, мхокардит,
шфаркт мюкарда), головного мозку (струс), легень (крупозна пневмонхя), печшки
(гепатит, мехашчна жовтяниця), кровх (анемхя), ендокринних залоз (ппофункщя
надниркових залоз, гшофхза), а також екзогенних х ендогенних штоксикацгй.
Що стосуеться походження нейроциркуляторно! (первиннох) ар-
терхально! гхпотензп, то вважають, що провщним етхолопчним х патогене-
тичним фактором е перенапруження основних процесхв кори великого мозку
(збудження 1 гальмування). Проте, на вщмхну вщ первиннох гшертензп, переважа-
ють процеси гальмування з поширенням на пщкхрковх вегетативн! утвори, зокре-
ма судиноруховий центр.
Ослабления внаслщок цього ефективних судинозвужувальних впливхв на тлх
властивого астешчному типу конституцп переважання холшерпчних процесхв над
адренерпчними е безпосередньою причиною зниження тонусу резистивних су-
дин, периферичного опору та артерхального тиску. Певну роль у розвитку первин-
нох артерхальнох ппотензп вщхграють набут! ! спадковх порушення продукцп та
рецепцп фхзюлогхчно активних речовин вазоконстрикторнох дп (катехоламхшв,
вазопресину, анпотензину II, ендотелшхв, ПГР2а, тромбоксану А, тощо), з одного
боку, та пперпродукцхя оксиду азоту х простациклхну — з шшого. Наявшсть збшь-
шенох кхлькостх оксиду азоту в ендотелп, крхм того, посилюе ефекти ендотелшза-
лежних вазодилататорхв (ацетилхолш, брадикхнш тощо).
РОЗД1Л XXVII
ПАТОЛОПЧНА Ф13ЮЛ0ПЯ
ЗОВН1ШНБОГО ДИХАННЯ
Зовнгшне дихання — сукупнгсть процессе, якг вгдбуваються в легенях г забезпечують
нормальний вм1ст у кров! кисню та вуглекислого газу.
У легенях перебггають три основш процеси: вентилящя альвеол, дифуз!я мо-
лекулярного кисню та вуглекислого газу кр!зь альвеолокапшярну мембрану 1 пер-
фузия — проходження вщповщно! юлькосп кров! через легенев! кашляри. Завдяки
чпкому взаемозв’язку цих процес!в забезпечуеться нормальний газовий склад ар-
тер!ально! кров!. Хвороби, як! розвиваються в дихальшй систем!, можуть зумови-
ти порушення цих процес!в, зниження ефективност! дихання 1 розвиток його
недостатност!.
Недостатшсть дихання — патологгчний процес, що розвиваеться внаелгдок по-
рушення зовнгшнього дихання I при якому не забезпечуеться пгдтримання адекват-
ного потребам органгзму газового складу артергально! кровг в станг спокою або пгд
час фгзичного навантаження.
За основними мехашзмами розвитку недостатносп дихання можна видщити
так! ц форми: 1) вентиляцшну, що виникае внаслщок порушення вентиляцп аль-
веол; 2) дифузшну, яка розвиваеться внаслщок порушення дифузп газ!в кр!зь аль-
веол окашлярну мембрану; 3) перфузшну, що виникае внаслщок порушення кро-
вопостачання альвеол; 4) змппану.
ПОРУШЕННЯ ВЕНТИЛЯЦП АЛЬВЕОЛ
Альвеолярна вентилящя полягае в регулярному оновленш альвеолярних газ!в вщ-
повщно до потреб оргашзму поглинати кисень 1 виводити вуглекислий газ. Що-
хвилини до альвеолярного простору в людини в стан! спокою надходять 4,5—5 л
повпря, завдяки чому й оновлюеться газовий склад альвеол. У цьому процес! бе-
руть участь центральна нервова система (дихальний центр головного мозку 1 мо-
тонейрони спинного мозку), периферичш нерви (рухов! та чутлив! — соматичш 1
вегетативш), верхи! дихальн! шляхи, трудна клггка, дихальш м’язи та легень
Вщомо, що д!яльшсть дихального Центру забезпечуе певний ритм дихання —
його частоту 1 глибину. Дихальними шляхами здшснюеться рух пов!тря до альве-
ол ! в зворотному напрямку. Трудна клика мае здатшеть активно змшювати свш
об’ем, що дае змогу змшювати свш об’ем 1 легеням, як! характеризуються розтяж-
нтстю певного ступеня. Уст щ процеси потртбш для нормально! вентиляцп альве-
ол; у разт !х порушення розвиваеться вентилящйна недостатшсть дихання.
Залежно в!д мехашзмгв, що е основою розвитку вентиля щй но! недостатносп
дихання, видшяють так! Г! види: дисрегуляторну, обструкгивну, рестриктивну.
Дисрегуляторна вентилящйна недостатшсть дихання
ГПдТрунтям дисрегуляторно! вентиляцшно! недостатносп дихання е порушення
нервово! регуляцп функцш дихальних м’яз!в трудно! клики та дтафрагми, яка за-
безпечуе тх пертодичне скорочення. Це може бути пов’язано з такими мехашзма-
ми, як порушення функцц дихального центру ! порушення передач! еферентних
впливтв дихального центру на дихальнт м’язи.
Порушення функци дихального центру. Дихання е автоматичним процесом, т
ця автоматичнтсть забезпечуеться ритмтчною тмпульсацтею нейрон!в, як! утворю-
ють так званий генераторний комплекс регулювання дихання т мктяться в дов-
гастому мозку. До цього комплексу належать дорсальна 1 вентральна групи ней-
рошв довгастого мозку, а також група синаптично з’еднаних пейсмекерних клг-
тин, як!у творюють пре-бутзшгеровий комплекс, що мктиться мгж подвшним т
бтчним ядрами сттчастого утвору.
У мосту головного мозку мктяться групи нейрошв, як! утворюють апнейстич-
ний т пневмотаксичний центри. Апнейстичний центр розташований у нижнш
частин! мосту 1 забезпечуе достатню тривалкть вдиху, пневмотаксичний розмтще-
ний у верхшй частин! мосту т бере участь у припиненнт вдиху.
За нормальних умов у стан! спокою людина дихае без будь-яких видимих
зусиль, здебшьшого не помтчаючи цього процесу. Такий стан називають дихаль-
ним комфортом, а дихання, що спостертгаеться при цьому, — евпное.
У раз! впливу на дихальний центр ушкоджувальних фактортв або рефлекторно
може змшюватися ритм дихання, його глибина й частота, виникае задишка. Ц!
змши можуть бути проявом компенсаторних реакцтй оргашзму, спрямованих на
пщтримання сталосп газового складу кров!, або виникають при розладах регуля-
цп дихання ! спричинюють порушення альвеолярно! вентиляцй 1, як наслщок,
недостатшсть дихання.
Змши частоти т глибини дихання зумовлюють або зменшення вентиляцп аль-
веол (гшовентиляцтя), або 1"! надм!рне збшьшення (гшервентиляцтя).
Пповентилящя е причиною розвитку в органтзм! гшоксп, гшеркапнп 1 газово-
го ацидозу.
Ппервентилящя, що виникае не як пристосувальна реакцтя при фтзичному
навантаженнт т не супроводжуеться вщповщним збшьшенням ХОС, призводить до
зменшення парщального тиску вуглекислого газу (рСО2) у кров! (ппокапнп) т
розвитку газового алкалозу. Це своею чертою може спричинити зменшення кро-
вопостачання головного мозку т серця через пщвищення тонусу тхнтх судин, змши
електролттного складу кров! (гшокальщемпо, ппокалкмгю), попршення дисоцта-
цй оксигемоглобшу, зменшення утилтзацн тканинами кисню та шшт змши.
Порушення функцш дихального центру можуть виникати пщ дкю ушкоджу-
вальних фактортв, як! безпосередньо впливають на метаболтзм, структурнт та фун-
кшональш властивостт його нейрошв. Такими факторами можуть бути гшокстя,
ппоглткемтя, штоксикацтя (наприклад, пщ дтею наркотичних засобтв, шкщливих
продуктов метаболгзму в разг недостатност! печгнки г нирок тощо), запалення го-
ловного мозку, здавлення стовбура головного мозку (при набряку, пухлинах, ге-
матом!), травма мозку (наприклад у новонароджених п(д час полопв), порушення
мозкового кровообггу.
Функцгя дихального центру г, вщповщно, характер дихання можуть також
змшюватися рефлекторно внаелгдок дп патолопчних факторгв на велику трупу
рецепторгв, розташованих у судинному русл!, дихальних шляхах, паренхгмг ле-
гень, довгастому мозку.
У разг порушення нервовог регуляцп можуть виникати такт типи дихання.
Брадипное — рщке дихання. Виникнення рщкого дихання спричинюють змгни
характеру нервово! гмпульсацп, яка йде вщ ргзних рецепторгв до дихального цент-
ру, або первиннг порушення дгяльностг самих дихальних нейронгв. При цьому
ступшь гшовентиляцгг визначатиметься як частотою, так г глибиною дихання.
Рефлекторне зменшення частота дихання спостерггаеться при пщвищеннг ар-
тергального тиску (рефлекс гз барорецепторгв дуги аорти г сонно! пазухи), у разг
ггперокегг (унаслщок пригнгчення “ггпоксичного драйва” — пергодичного збуд-
ження хеморецепторгв, чутливих до зниження парцгального тиску молекулярного
кисню в артергальнгй кровг).
Глибоке рщке дихання може розвинутися у зв’язку з щдвищенням опору ру-
ховг повгтря у верхнгх дихальних шляхах — стенотичне дихання. У цьому випадку
вдих г вид их здгйснюються повгльншге, нгж звичайно. У виникненнг такого типу
дихання певну роль вщгграють гмпульси, що надходять до дихального центру вгд
мгжребрових м’язгв, якг працюють гз пщвищеним навантаженням. Кргм того, мае
значения запгзнювання в цьому випадку гальмгвного рефлексу Гергнга — Брейе-
ра. Вилученням аферентног ланки цього рефлексу пояснюеться дуже рщке, але
глибоке дихання, що спостерггаеться в експеримент! у тварин шсля перергзання
обох блукаючих нервгв (вагусне дихання). Глибоке рщке дихання може бути
пов’язане також зг зменшенням активност! пневмотаксичного центру.
Тахшное (полшное) — чаете поверхневе дихання. За таким типом дихання
вентилюеться переважно мертвий простгр легень г зменшуеться надходження
повгтря до альвеол (гшовентиляцгя).
У розвитку тахшное можлива участь пневмотаксичного центру, пщвищення
активностг якого спричинюе скорочення вдиху г збгльшення частоти дихання.
У деяких тварин (наприклад у собак) чаете поверхневе дихання е компенса-
торною реакцгею на дно високог температури повгтря (одним гз механгзмгв термо-
регуляцп). У людини тахшное може бута пов’язане з функцгональними змгнами в
централыпй нервовгй систем! при гарячцг, гстерп, деяких оргашчних ураженнях
головного мозку.
Чаете поверхневе дихання може виникати в разг подразнення в дихальних
шляхах чутливих закгнчень вагусних мгелгнгзованих волокон гритантних рецеп-
торгв або рецепторгв, що швидко адаптуються. Вони стимулюються пщ час вди-
хання таких шкщливих чинникгв, як пил, деякг гази, холодне повгтря тощо.
Тнгтгу трупу гритантних рецепторгв становлять нервовг закгнчення вагусних
немгелгшзованих С-волокон, що мгстяться в паренхгмг легень, повгтропровщних
шляхах г кровоносних судинах, Вони можуть активуватись як екзогенними под-
разнювальними хгмгчними речовинами та фгзичними факторами, так г ендоген-
ними (наприклад медгаторами запалення). Активацгя С-волокон також призво-
152
лить до розвитку частого поверхневою дихання, що обмежуе поширення шкщли-
вих факторов по дихальних шляхах. Тахшное може спостерёгатися при емболи
-еликих плок легенево! артерй, шо пов'язано !з впливом на нервов! закшчення
блукаючого нерва серотоншу, який впвётьнюеться !з тромбоципв.
У паренхём! легень бшя легеневих капёлярёв мтстяться особлив! закшчення С-
волокон — 3-рецептори, що стимулюються розвитком штерстицшного набряку.
1х активащя (наприклад, при запаленн! легень, лгвошлуночковш недостатност!
серця) теж здатна спричинювати тахшное.
У раз! ателектазу в легенях збуджуються механорецепгори, розташоваш в
бронхёолах. яю реагуюгь на спадання легень. 1х подразнення активуе вдих. Але
пщ час вдиху неуражен! альвеоли розтягуються швидше, н!ж звичайно, що зумо-
влюе передчасне виникнення ёмпульсаци з боку рецептор!в розтягнення, яка
гальмуе вдих.
До розвитку тахшное може призвести бёть певно! локапзацй (у грудшй клёгщ,
черевшй стшп!, плевр!). Б!ль обмежуе глибину дихання 1 зумовлюе збёльшення
його частота (щадне дихання).
Гшерпное — глибоке чаете дихання. За ф!з1олог!чних умов пперпное виникае
як реакщя дихально! системи, спрямована на збёльшення вентиляцп легень 1 при-
ведения ц у вщповщшеть до потреб обмшу речовин, що посилюеться. наприклад,
шд час м’язово! робота. Водночас полёпшуеться оксигенашя кров! 1 пщтримуеть-
ся КОС в оргашзм! шляхом виведення надлишково! илькосп СО2.
Пперпное розвиваеться внашцдок посиленого подразнення хеморедепторгв
сонно! пазухи ! дуги аорти, як! реагують на збёльшення напруження СО3ё зменшен-
ня О, у кров!! на пщвищення в ши концентраци !онгв водню. За умови патологи
це може спостер!гатася при всех видах ппокси, недостатност! серця, анеми, ацидоз!
тощо Найвищий стушнь рефлекторного збудження дихатьного центру проявляеть-
ся у вигляд! дихання Куссмауля, яке здебьлмного може спостерёгатися у хворих у
стан! дёабетично! коми. Це шумне прискорене дихання, коли шсля глибокого вди-
ху вщбуваеться посилений видих за активною учаслю ексшраторних м’язёв.
1нший мехашзм рефлекторного розвитку пперпное пов’язаний з! збудженням
барорецепторёв сонно! пазухи 1 дуги аорти в раз! зниження артер!ального тиску
(наприклад пщ час крововтрати).
Гшерпное може розвиватися гакож при емоишному збудженш, подразненш
екстерорсиептор!в шк!ри — больових, темпсратурних (наприклад пщ час занурен-
ня в гарячу чи холодну воду). Це пов’язано зё збудженням сётчастого утвору, у
якому м!стяться нейрони дихального центру. М’язова робота також рефлекторно
активуе нейрони с! гчастого утвору.
Гикавка. Цим термином позначають виникнення клошчних судом дзафрагми,
пщ час яких пов!тря втягуеться в легень Вона розвиваеться в раз! подразнення
аферентних закёнчень у дёафрагмц органах черевно! порожнини. Особливо тривка
гикавка спостерёгаеться саме шсля операцш на цих органах, що може сутте во
порушувати ритм дихання ё призводити до зменшення алъвеолярно! вентиляцп.
Термён “апное” дословно перекладаеться як вщсутшсть дихання, але зазвичай
цим словом позначають тимчасову зупинку дихання. Апное може спричинити
порушення газообм!ну в оргашзм!, тяжкёсть якого залежить вш частота виникнен-
чя 1 тривалосп апное, шо своею чертою визначаеться причинами, ёз яких воно
оозвинулося.
Тимчасова зупинка пихания може бути пов’язана 31 зменшенням рефлектор-
но! або безпосередньо! х!м!чно! стимуляш! дихального центру. Наприклад, апное
виникае теля пасивно! ппервентиляцп у тварин або людини пщ наркозом унас-
лщок зменшення в артер!альнш кров! напруження вуглекислого газу 1 припи-
няеться одразу теля нормалтзацй його вмкту В експеримент! у тварин можна
снос тер! гати рефлекгорну гимчасову зупинку дихання в раз! швидкого пщвищен-
ня артер!ального тиску (наприклад гасля введения у кров адреналшу), шо пов’язано
з! стимулящею барорецепторш дуги аорти 1 сонно! пазухи.
В умовах патологп апное найчастние виникае пщ час порушень функцш ди-
хального центру, зокрема в раз! розвитку перюди чного дихання.
Кашель ! чхания належать до рефлекторних акт!в, як! виникають у вщповщь
на подразнення певних рецепторних зон, переважно верхюх дихальних шлях!в, 1
супровопжуються змшою ритму ! глибини дихання.
Кашель найчастпне виникае внаслщок подразнення закшчень язико-глотко-
вого 1 блукаючого нерв!в у слизовш оболонц! глотки, горташ, трахе! (найчутлив!-
ша д!лянка — роздвоення (б!фуркац!я) трахе!) та бронх!в. Кр!м того, його може
спричинити подразнення чутливих закшчень плсври. Кашель складаеться з ко-
роткого вдиху, шсля якого негайно змикаеться голосова щшина ! водночас вщбу-
ваеться ексшраторне посилення роботи дихальних м'яз!в. Унаслщок цього р!зко
пщвищуеться тиск у дихальних шляхах, легеневих альвеолах ! плевральной порож-
нин!. Голосова щшина пот!м раптово розкриваеться 1 пов!тря з великою силою 1
швидкктю виходить 13 дихальних шляхтв, виносячи !з собою частники з поверхн!
слизово! оболонки.
Чхання виникае у вщповщь на подразнення закшчень тршчастого нерва, шо
мктяться в слизовш оболонц! носа (особливо середньо! носово! раковини 1 пере-
городки). На вщмшу вщ кашлю, пщ час чхання форсований видих, який виникае
шсля вщкривання голосово! щшини, вщбуваеться не через рот, а через шс.
I кашель, 1 чхання е захисними реакц!ями, спрямованими на очищения ди-
хальних шляхш вщ слизу, мокротшшя, р!зних х!м!чних речовин ! мехашчних час-
тинок. Виникаючи етзодично, вони не впливають на газообмш у легенях. Проте
част! напади кашлю зумовлюють тривале пщвищення внутр!шньогрудного тиску,
полршують вентиляццо альвеол 1 кровообтг особливо в судинах малого кола.
Перюдичне дихання — дихання, яке характеризуемся чергуванням перюдтв
дихання з перюдами його вщсутносп (апное), 1снують два типи периодичного ди-
хання — дихання Чейна — Стокса ! дихання Бюта.
Дихання Чейна — Стокса характеризуемся поступовим наростанням ампл!ту-
ди дихання, а пот!м зменшенням п до апное, теля чого знову настае цикл ди-
хальних рух!в, яю закшчуюмся також энное (мал. 60).
Циюпчш зм1ни дихання в людини можуть супроводжуватися потьмаренням
свщомост! в перюд апное 1 нормал!зан!ею и з! збшьшенням вентиляцн. Артер!аль-
ний тиск при цьому також коливасться, як правило, пщвищуючись у фаз! поси-
Мал. 60. Дихання Чеиаа — Стокса в кивки теля крстортряти
(за Я. Бритваном, 1966)
лення дихання г знижуючись у фаз! його ослабления. Вважають, шо в бшьшослг
випадков дихання Чейна — Стокса е ознакою гшоксп головного мозку. Його мож-
на спостерпати у здорових людей на великгй висотг (особливо гид час сну), у не-
доношсних дзтей, що, можливо, пов'язано з недорозвиненгстю нервових центргв.
Дихання Чейна — Стокса у тварин можна вгдтворити шляхом зменшення
забезпечення головного мозку киснем (шгаляцгя газових сумйпей гз низьким
вмгстсм кисню, порушення кровопостачання головного мозку) або перергзання
стовбура головного мозку на ргзних ргвнях.
Патогенез дихання Чейна — Стокса недостатнъо з’ясовано. Деяю дослщ-
ники пояснюкль його мехашзм таким чином. Клгтини кори великого мозку г
пщкгркових утворгв унаслщок гшоксп пригнгчуються — дихання зупиняегься,
пригшчуетъся дгяльнгсть судинорухового центру, Однак хеморецептори при цьо-
му ще здатн! реагувати на змгни вмгсту газгв у кровг Значне посилення гмпульса-
цц з хеморецепторгв разом з одночасною прямою дгею на центри високог концен-
трат!' СО, г стимулами з барорецепторгв унаслщок зниження артергального тиску
виявляеться достатнгм, щоб збудити дихальний центр — дихання поновл юеться.
Поновлення дихання сприяе оксигенанп кровг, що зменшуе гшоксгю головного
мозку г полшшуе функцгю нейронгв судинорухового центру. Диханггя стае глиб-
шим, пщвищуеться артергальний тиск, полгпшуеться наповнення серця, але вгд-
новлена вентиляцгя зумовлюе пгцвишення напруження кисню та зниження: на-
пруження вуглекислого газу в артергальнш кровг. Це своею чертою спричинюе
ослабления рефлекторно!' та хгмгчног стимуляцп дихального центру, дгяльнгсть
якого починае згасати, — настае апное
Варто зазначити, що дослщи з вгдтворення пергодичного дихання у тварин
шляхом перетяну стовбура головного мозку на ргзних ргвнях дають змогу деяким
дослщникам стверджувати, шо дихання Чейна — Стокса виникае внаслщок змгни
ргвноваги мгж гальмгвними г стимулюючими процесами в цьому вглдглг мозку.
Дихання Бюша вщргзшгеться вгд дихання Чейна — Стокса тим, що дихальнг
рухи, якг характер изуються постгйною амплпудою, так само раптово припиня-
ються, як г починаються (мат 61).
Здебшьшого дихання Бюта спостерггаеться гпд час меншггту, егщефачгту та
гнших хвороб, якг супроводжуються ушкодженням центрально!' нервовог системи.
Апнейстичне дихання характеризуеться тривалим судомним зусиллям вдихну-
ти, що зрщка перериваеться видихом.
В експеримент! апнейстичне дихання спостерггаеться шсля перергзання в
тварини обох блукаючих нервгв г стовбура головного мозку мгж пневмотаксичним
та апнейсгичним центрами. Вважають, що апнейстичний центр мае здатшеть
збуджувати гнешраторш нейрони, якг пергодично гальмуються гмпульсами з блу-
каючого нерва ; пневмотаксичного центру. Перергзання зазначених структур зу-
мовлюе постшну шепграторну актившсть апнейстичного центру.
Мал. 61 Дихання Бюта в кппкп шсля перергзання головного мозку на ргвгн моста
(за Я. Бритваном, 1966)
Гасшнг-дихання (вщ англ, &азр— ловити повггря, задихатися) — це поодиною
“зпхання”, що спадають за частотою х глибиною х спостерхгаються пщ час агонп,
наприклад у термшальнгй стадп асфхксй. Таке дихання називають також термг-
нальним, або агональним. Зазвичай “зхтхання” виникають теля тимчасовох зу-
ггинки дихання (претермшально! паузи).
Порушення передавания еферентних впливгв дихального центру на дихальш м’язи
можуть виникати з низки причин. Так, вони можуть бути пов’язаш з ушкоджен-
ням нервових провщних шляххв, що з’еднують нейрони дихального центру з мо-
тонейронами спинного мозку, яю шнервують дихальш м’язи. Якщо в експери-
мент] перетнути провщнх шляхи, яю з’еднують нейрони пре-бутзшгерового ком-
плексу довгастого мозку г мотонейрони, що шнервують дхафрагму, дихання
припиняеться. Ураження цих провщних шляххв призводить до втрати дихального
автоматизму, х залишаеться лише довшьний контроль дихання (синдром проклят-
тя ундши).
Вентиляцшна недостатшсть дихання може виникати в разх порушення функ-
цц мотонейрошв спинного мозку, що шнервують дихальш м’язи. Збудження або
гальмування цих нейрошв може порушуватися внаслщок розвитку в спинному
мозку запальних 1 дистрофхчних процесхв (бхчний амютрофхчний склероз, полю-
мхелп, сирингомхелхя), а також штоксикацп (стрихншом, правцевим токсином).
Порушення вентиляцн альвеол можуть виникати унаслщок ушкодження не-
рвгв, яю шнервують дихальш м’язи (запалення, авпамхноз, травма), порушення
передавания на м’язи нервового хмпульсу (при мхастенп, ботулхзмг, уведенш мю-
релаксанта). До значного розладу дихання може призвести ураження п. /гетсиз х
паралхч дхафрагми, унаслщок чого виникають й парадоксальш рухи: вгору — пщ
час вдиху, униз — пщ час видиху.
Обструктивна вентиляцшна недостатшсть дихання
Обструктивна недостатшсть дихання виникае внаслщок звуження повггропровщ-
них шляххв (вщ лат. оЬзГгисйоп — перешкода) та пщвищення опору в них руху
повггря. При цьому перешкоди руху повггря можуть утворюватись як у г верхнхх,
так х в нижн1х дихальних шляхах.
Причинами звуження проевпу верхнхх дихальних шляххв або навпь повного
закупорювання гх можуть бути: 1) сторонш предмети, що потрапляють у проевхт
верхшх дихальних шляххв; 2) стовщення стшок дихальних шляххв (запальний на-
бряк слизово! оболонки носа, горташ, трахе!, розвиток пухлини); 3) спазм м’язхв
горташ (ларингоспазм); 4) здавлення стшок дихальних шляхгв ззовш (пухлина
тканин, яю оточують дихальш шляхи, заглотковий абсцес, збшьшення прилеглих
оргашв, наприклад щитоподхбно! залози тощо). В уехх цих випадках виникае так
звана фхкеована обструкция, за яко! просвет дихальних шляхгв майже не змшюеть-
ся пщ час вдиху або видиху г спостерхгаеться стенотичне дихання.
Нижш дихальш шляхи (дхаметром менше 2 мм) у здорово! людини створюють
незначний ошр диханню. Проте саме йони е мхецем виникнення перешкоди руху
повггря при розвитку бшьшостх хвороб легень. Це зумовлено тим, що ошр потоку
повхтря зростае пропорцхйно четвертому ступеню зменшення проевпу бронха,
через який проходить повхтря. Якщо дгаметр проевпу бронха 1 мм з будь-яких
причин зменшився до 0,5 мм (тобто у 2 рази), ошр потоку повхтря зросте в 16 разхв.
Вщповщно до звуження бронхи збшыиуеться г швидюсть руху в них повттря, а це
також супроводжуеться пщвищенням опору повггряному потоку (пропорцшно
квадрату збшьшення швидкосп руху повттря).
Причини обструкцп нижнтх дихальних шляхтв: 1) надходження в просвгт дртб-
них бронхтв 1 бронхюл рщини — води, блювотних мае, гною, транссудату; 2) стов-
щення слизово! оболонки нижнтх дихальних шляхтв, що спосгертгаеться зазвичай
пщ час запальних процеств (бронхтт) ! у виникненш якого вщгграють роль таю
фактори, як пперемтя слизово! оболонки, п набряк, гшертрофтя ! гшерплазтя юп-
тин слизових залоз; 3) накопичення в бронхах мокротиння, чому сприяе гшерсек-
рецтя слизу, попршення його реолопчних властивостей, порушення мукоцилиар-
ного транспорту, розлад кашльового мехашзму; 4) спазм непосмугованих м’язтв
бронхюл; 5) пщвищена розтяжшеть легень унаслщок втрати ними еластичность
Накопичення в бронхах мокротиння найчасттше спостерггаеться у випадку
тнфекцшно-запальних процеств дихальних шляхтв. Кргм того, це може розвивати-
ся при деяких видах спадково! патологп, пов’язаних тз порушенням структури !
функцп втйчастого егптелтю. Наприклад, надлишкове накопичення мокротиння
вщбуваеться при синдром! Картагенера (спадково-родинна аномалтя), за якого
втрачаеться рухомтсть вшок через недостатшсть у них бшка динешу, що мае
АТФазну актившсть.
У раз! тншо! спадково! патологи — муковтецидозу — порушуються процеси
видтлення води секреторними юптинами вс!х екзокринних залоз оргашзму, у тому
числ1 й ештелтальними юптинами бронх!в через вщеутшеть у мембран! цих клттин
особливого быка — трансмембранного регулятора муковтецидозу, який е компо-
нентом каналу для тонтв хлору. Унаслщок цього вони затримуються в юптинах, а
разом з ними затримуються тони натрпо т вода, що зумовлюе дегщратацпо встх
секреттв, зокрема г слизу, унеможливлюе транспорт слизу втйчастим ештелтем.
Спазм непосмугованих м’язтв бронхюл — один тз найважливштих мехашзмгв
обструкцп у випадку бронхтально! астми г запальних процеств у дихальних шляхах.
У розвитку спазму можуть брати участь гуморальш т нервов! фактори.
Серед гуморальних факторгв, здатних спричинювати скорочення непосмуго-
ваних м’язтв дихальних шляхтв, е велика трупа речовин — медгаторгв запалення й
алергп: пстамтн, серотонш, лейкотртени, юнгни, простагландини, тромбоксани.
Кртм того, у разг розвитку запалення ушкоджуеться ештелтй дихальних шляхтв 1
може знижуватися вироблення ештелгального фактора розслаблення — речовини,
що сприяе розелабленню непосмугованих м’язтв бронхтв т бронхюл.
Щодо нервових мехашзмгв потргбно зауважити, що величина проевпу ди-
хальних шляхтв регулюеться блукаючим ! симпатичними нервами. У склад! блука-
ючого нерва видшяють ртзш типи нейрошв. Холгнерпчш мотонейрони вившьню-
ють ацетилхолтн, який стимулюе скорочення непосмугованих м’язтв дихальних
шляхтв. Другий тип нейрошв вившьнюе речовину Р, яка також спричинюе спазм
непосмугованих м’язтв дихальних шляхтв.
Ще один тип нервових клгтин вившьнюе вазоактивний штестинальний пеп-
тид (ВШ), який розелабляе непосмуговант м’язи. Факторами розслаблення м’язтв
дихальних шляхтв е також норадреналш г адреналш, що виробляються симпатич-
ними нервами г наднирковими залогами.
Роль нервових мехашзмгв у розвитку бронхоспазму може полягати або в над-
мтршй активностт холптерпчних чи пептидерпчних нейрошв блукаючого нерва,
що вивглъняють речовину Р (вщомо, що подразнення гритантних рецепторгв блу-
каючого нерва в легенях може спричинювати спазм бронхгол), або в недостатнш
активностг ВШ-ерпчних нейронгв блукаючого нерва чи симпатико-адреналово!
системи.
Зниження еластичних властивостей легень спостерггаеться при емф1зем1 —
хворобг легенево! паренхгми, яка супроводжуеться руйнуванням тонко! сгтки ле-
геневих капгляргв г альвеолярних перегородок, а також звуженням просвгту термг-
нальних бронхгол. Важливим фактором у розвитку емфгземи е ферментативний
розпад волокнистих структур легенево! тканини внаслщок збгльшення активностг
протеолгтичних ферментгв (еластази, колагенази, трипсину), що синтезуються пе-
реважно нейтрофгльними гранулоцитами й альвеолярними макрофагами. Цг клг-
тини в легенях можуть стимулюватися мгкробною мгкрофлорою, тютюновим ди-
мом, частками пилу (пилу вугглля, азбесту, цементу тощо). Про роль посиленого
протеолгзу розвитку емфгземи легень евщчить гг виникнення у пщдоелгдних тва-
рин, яким тривалий час уводили гнтратрахеально протеолгтичнг ферменти. У роз-
витку емфгземи можуть вгдгтравати роль г спадковг фактори: дефгцит у кровг а,-
антитрипсину, дефекта в легенях структури колагену, еластину, протешглгкангв,
атонгя м’язових клгтин, якг оточують реепграторну бронхголу.
У механгзмг виникнення обструктивних явищ при емфгземг вщгграе роль той
факт, що проевгт бронхгол, якг мають м’якг г тонкг стшки, пгдтримуеться транс-
пул ьмональним тиском. Бшьша еластичнгсть легенг потребуе створення бглыпого
розрщження у плевральнгй порожнинг пгд час вдиху (що й пщвищуе транспуль-
мональний тиск), щоб подолати еластичну тягу легень г розтягнути гх. Якщо ж
легенг втрачають еластичнгсть, то вони розтягуються значно легше, тобто потре-
бують значно меншого транспульмонального тиску. Це зумовлюе зменшення
сили, що мае дгяти на стгнки бронхгол зсередини г розправляти гх: стшки бронхгол
спадаються, проевгт гх зменггтуеться. Кргм того, у нормг алъвеоли, сполученг з
бронхголами, сприяють розправленню останнгх унаслщок радгально! тракцп. Пгд
час емфгземи внаслщок втрати альвеол зменшуеться радгальна тракцгя.
Зменшення просвгту нижнгх дихальних шляхгв значно пщвищуе опгр руху
повгтря г заважае ргвномгрному розподшу його в альвеолах. Особливо порушуеть-
ся акт видиху. Пояснюеться це тим, шо при емфгземг пгд час вдиху транспульмо-
нальний тиск, хоч г менший за нормальний, проте е достатнгм для того, щоб
розправити стгнки бронхгол. Пщ час видиху (який при емфгземг стае активним,
оскгльки еластична тяга легень слабшае, а опгр руху повгтря зростае) тиск у плев-
ральнгй порожнинг пщвищуеться г вщповщно в мгру видихання збглыпуеться сила,
що дге на бронхголи ззовнг. Стгнки бронхгол поступово спадаються, г настае такий
момент, коли подальше видихання стае неможливим. У такий спосгб стгнки брон-
хгол вщгграють роль клапана, який пщ час видиху закриваеться г повгтря опи-
няеться нгби у пастцг. Унаслщок цього альвеоли залишаються постшно роздути-
ми, у них збшьшуеться кглькгсть залишкового повгтря. Аналопчно передчасне
ексшраторне закриття дихальних шляхгв може виникати г за шших обструктивних
процесгв: бронхгально! астми, бронхгту тощо. Подгбнг обструктивнг порушення
вентиляцгг можуть спостершатися 1 при бронхоектатичнгй хворобг, за яко! внаелг-
док хронгчно! шфекцп та запального процесу розвиваеться деструкцгя м’язового
й еластичного компонентгв стшок бронхгв, а також за наявностг вродженог були
легень.
Рестриктивна вентиляцшна недостатшсть дихання
Цей вид недостатност! дихання пов’язаний хз порушенням вентиляцп альвеол
унаслщок обмеження розтягнення легень (в!д лат. геМпсИо — обмеження). Вщц-
ляють двх форми рекстриктивно! вентилящйнох недостатност! дихання: позалеге-
неву 1 легеневу.
Позалегенева рестриктивна вентиляцшна недостатшсть дихання розвиваеться
внаслщок таких причин: 1) порушення структури х функцц дихальних м’яз!в;
2) порушення (утруднення) рухомост! трудно!' клхтки ! дхафрагми; 3) пщвищення
тиску в плевральнш порожний.
Порушення структури I функцн дихальних м ’язгв можуть спостерхгатася в раз! !х
запалення (мюзит), при хворобах м’язово!' системи, наприклад при м’язовш дистро-
фц Дюшенна. В усхх цих випадках розвиваеться м’язова недостатшсть, коли змен-
шуеться сила 1 швидюсть скорочення м’язхв, що обмежуе рух трудно! клхтки х
дхафрагми х, вщповщно, розправлення легень. Навхть незначй додатковх наванта-
ження на дихальш м’язи (наприклад год час фхзично! робота) можуть призвести до
!х швидкох втоми, що ще бшьше пришвидшуе розвиток недостатност! дихання. Щоб
оцхнита функфональнх можливостх дихальних м’язхв ! ступшь ххньо'х втоми, визна-
чають тиск у дихальних- шляхах пщ час вдиху х споюйного дихання (Тсп), тиск при
максимальному шсйраторному зусиллх (Тмакс) та ххне спхввщношення (Т^/Т^). У
норм! це спхввщношення становить приблизно 0,05. Ступшь недостатност! м’язхв,
ххньо! втоми харакгеризуеться збшьшенням ще! величини понад 0,4.
Порушення рухомост! грудно'1 клйпки / дгафрагми можуть спостерхгатась у раз!
вроджених або набутих деформайй ребер х хребетного стовпа (юфосколюз), за-
пальних х дистрофхчних процесхв у мхжхребцевих х реброво-хребцевих суглобах,
скостенхння ребрових хрящхв, травматичних ушкоджень трудно! клхтки. Утруд-
нюеться рухомхсть трудно! клхтки х в раз! зрощення вхсцерального ! пархетального
листкхв плеври.
Рухомхсть дхафрагми може обмежуватися при вроджених вадах розвитку, де-
яких патолопчних процесах у черевйй порожний — накопиченй в нхй рщини
(асцит), надмхрнох кшькостх жиру в жирових депо, при метеоризм!, за наявност!
великих пухлин, спленомегалп, гепатомегалп.
У випадку всхх цих процесхв порушуються повноцшй екскурси трудно! клхтки
та / або дхафрагми х, вщповщно, розправлення легень, що призводить до гшовен-
тиляцп альвеол.
Шдвищення тиску в плевральнш порожним може спостерхгатася в разх пору-
шення цшостх трудно! клхтки х плеврально! порожнини. Цшхсть гшеврально! по-
рожними забезпечуе певний транспульмональний тиск (рхзниця мхж тиском
повхтря всередий альвеол х тиском у плевральнш порожний), який пщтримуе
легеш в розправленому станх. Пщ час вдиху, коли об’ем трудно! клхтки збшь-
шуеться, транспульмональний тиск зростае дота, поки не подолае еластичну тягу
легень, унаслщок чого альвеоли розширюються. Якщо цшхсть плеврально! порож-
нини порушуеться х в не! потрапляе атмосферне повхтря, транспульмональний
тиск знижуеться, а легеня спадаеться. Скупчення повхтря в плевральнш порож-
ний та пщвищення тиску в нхй мае назву пневмотораксу.
Повхтря може потрапита в порожнину плеври внаслщок проникаючого пора-
нения трудно! клхтки, розхрвання емфхзематозних альвеол на поверхй легей, роз-
паду легенево! тканини (туберкульоз, пухлина, абсцес). У таких випадках порож-
нина плеври може сполучатися з легенею та 1ншими повгтропровщними органа-
ми — стравоходом, шлунком, кишкою. 1нод1 повттря вводять у плевральну по-
рожнину з лгкувальною метою.
Якшо повгтря, яке потрапило в плевратьну порожнину. не сполучаеться з
атмосферним пов!грям, пневмоторакс називають зэкритим, якщо сполу-
чаеться — в'дкритим. Нареши, якщо особливоси входного отвору допускають
потрапляння повпря в порожнину плеври щд час вдиху, аде перешкоджають його
виходу пщ час видиху, пневмоторакс називають клапанним, або вентиль-
ним,якийе особливо небезпечним, бо зумовлюе прогресуюче накопичення
повттря в плевральшй порожнинт, значке пщвищення в шй тиску, зсув леген!, що
спалась, у бтк середостшня т змтщення розташованих там оргашв ! судин.
Спатання легень спостерггаеться також у разт накопичення в плевральнш
порожнинт транссудату або ексудату (гщроторакс, шоторакс), кров! (гемоторакс).
У раз! спадания (колапсу) легень посилюеться вдих, зросгають екскурсп грудноТ
клики для створення потргбного транспульмонального тиску, щоб розтягнути ле-
геш (хоча при вщкритому 1 особливо вентильному пневмоторакс) цього повшстю
досягти не вдаеться). Тим самим збшьшуегься навантаження на дихальш м’язи.
що може призвести до гхвьо! втоми.
Легенева рестриктивна вентилящйна недостатшсть дихання розвиваеться внаслт-
док зменшення розтяжност! легень. Розтяжшеть легень можуть порушувати за-
пальш та застшн! процеси в легенях. При цьому переповнеш кров’ю легеневг
кашляри, а також набрякла штерстишйна тканина здавлюють альвеол и 1 заважа-
югь гм розправитися повною мтрою. Кргм того, за цих умов знижуеться розтяж-
шеть самих каш л яр! в та гнтерстицшног тканини.
Пневмосклероз (фгброз легень) також призводить до зменшення розтяжност!
легень. Цим прочесом завершу ються ртзш хвороби легень. До пневмосклерозу
призводять хрошчш запальш хвороби (шфекщй н!, алерпчш), токсичнт ураження.
гривале запилення легень (пневмоконюз), емфгзема. дифузнз хвороби сполучно!
тканини, застгй кровг в легенях унаслщок недостатност! кровообпу, вплив юшзу-
ючого випромшювання, тютюнокуршня. Для пневмосклерозу характерним е над-
лишкове розростання волокнисто! сполучно! тканини на м!сщ ушкоджених па-
ренхгматозних елеменпв, капглярних судин 1 еластичних волокон.
Порушення сурфактантной системи легень. У норм! внутршгня поверхня альвеол
вкрита як ПЛ1ВК0Ю сурфактантами — поверхнево-активними речовинами, основ-
ну масу яких становлять лшопротещи. До сурфактантно! системи легень належать
юптини альвеолярного ештелпо II типу, яю. беруть участь у синтез! фосфолшцпв
1 бшково! частини сурфактантов, депонувашп готового матер!алу, а також макро-
фаги, за допомогою яких вщпрацьований сурфактант видатяеться з поверхн! аль-
веол. За рахунок високог поверхнево! активносп сурфактант!в поверхневий натяг
у легенях близький до нуля, шо запобпае транссудащ! рщини в проевгг альвеол гз
капгляргв 1 захишае алъвеоли вщ спадания. Здатшсть легень розтягуватися на двг
третини забезпечуеться саме сурфактантами.
Дефщит сурфактанпв зумовлюе пщвищення повер.хневого натягу альвеол,
унаслщок чого шдвищуеться ошр легень розтягненню. Вш може виникнути в разг
порушення гх синтезу, прискореноге видалення з поверхш альвеол або руйнуван-
ня. Наприклад, у новонароджених (частгше недоношених) у випадку так звано!
хвороби палшових мембран утворення сурфакгантгв р!зко зменшуеться. Водночас
вщбуваються часткове руйнування альвеол ! накопичення в ’гхньому просвт без-
структурно! речовини (палшових мембран), у якш спостерпаються залишки кл!-
тин ештелно, кров!. Ця хвороба спричинюе асфгксзю 1 смерть дитини.
Вм!сз сурфактант!в може змеишуватися внаслщок гшоксп, ацидозу, запален-
ня легень 1 порушення кровообпу в них, асшрацп блювотних мае та шших рщин,
шд час штучною кровообпу7. промивання легень. Сурфактант легко руйнуеться
пщ впливом високих концентрашй кисню, тому застос^’вання чистого кисню в
раз! недостатност! дихання може шод! зумовити п посилення.
Ателектаз легекь — патолопчний процес, за якого припиняеться вентиляц!я
альвеол ! вони спадаються внаслщок роземоктування в них повпря До ателектазу
здебшьшого призводить порушення прохщносп бронха, що супроводжуеться спа-
даниям легенево! тканини дисталышие м!сця закупорювання (обтурацгйний
ателектаз), а також здавлювання легенево! тканини ззовш ексудатом, пухли-
ною тощо (компресшний ателектаз). У розвитку ателектазу важливе зна-
чения може мати зменшення вмзету сурфактантов.
Слщ зазначити, що при обструкцГ!, яка виникае, наприклад, унаслщок за-
пальних проиесгв у дихальних шляхах (обструктивний бронхи) 1 яка спричинюе
обтурацпо бронх! в, розвитков! множинних ателектазгв перешкоджае колатеральна
вентилящя через мгжальвеолярю анастомози (пори Кона). Тх роль у забезпеченн!
газообмшу незначна, але вони мають важливе значения у пщгриманш стабшь-
носп просвету альвеол.
Пухлини о мсти легень також призводять до зменшення розтяжност! легень !
виникнення легенево! рестриктивно! вентиля цгйно! недостатност! дихання.
Змши показниюв зовжшнього дихання
п1д час обструктивних»рестриктивних процес!в
Обструктивн! ! рестриктивн! процеси в легенях спричинюють пповентилящю
альвеол.
Одним !з найважлившшх показниюв, що характеризуй вентилящйну здат-
шеть легень, е об'емна швидкзеть повггряного потоку, яка визначаеться пщ час
форсованого видиху, що здшснюеться шсля максимального вдиху, — так звана
форсована життева емнгсть легень (ФЖСЛ) 1 витрачений на це час. Пщ час виз-
начення об’емно! швидкост! повггряного потоку важливо враховувати кшьюсть
повгтря, що видихасться за першу секунду, — об’ем форсованого видиху за 1 с
(ОФВ,). Здоров! люди за першу секунд)’ видихають щонайменше 70 % ФЖСЛ.
На мал. 62 представлено сшрограми здорово! людини г хворого з обструктив-
ними 1 рестриктивними провесами в легенях. Якшо паренхгма легень нормальна,
але звужеш лов!тропровщн! шляхи, то ФЖСЛ може збершатися нормальною,
проте об’емна швидкзеть повггряного потоку зменшуеться, особливо за 1 с. При
рестриктивних процесах легеш стають жорсткпними г погано розправляються, але
функшя повгтропровщних шляхов зазвичай не порушуеться, тому швидкзеть
ловотряного потоку не змгнюеться. I хоча ФЖСЛ га ОФВ, зменшуються, !х сшв-
вщношення залишаеться нормальним.
Мал. 62. Сшрограми здорово! людини (а), хворо! на пневмосклероз (б),
броняальну астму (в) 1 емфгзему легень (г):
3€Л — загальна емшсть легень; Ж€Л — житгева емшсть легень; РОВД — ре-
зервний об’ем вдиху; РОвил — резервний об’ем видиху; ЗО — залишковий
об’ем
Обструктивю та рестриктивн! процеси супроводжуються пщвищеною робо-
тою дихальних м’яз!в, збшьшенням енергетичних витрат на дихання, так звано!
киснево! щни дихання, тобто збшьшенням вщсотка молекулярного кисню, який
використовуеться дихальними м’язами п!д час утворення АТФ.
Тривале перевантаження дихальних м’язгв може призвести до !х утоми 1 по-
глиблення недостатност! дихання.
ПОРУШЕННЯ ДИФУЗП ГА31М У ЛЕГЕНЯХ
Рух газУв кргзь альЕсолокашлярну мембран}- вщбуваеться шляхом дифузп У зале-
жить, за законом Фиса, вщ дифузшноУ здатностУ мембрани У рУзницУ парциального
тиску газгв з обох бою в мембрани. Дифузшна здатшсть мембрани складаеться з
юлькох компонентов, що охоллюють розчиннкть газу в тканинах (насамперед у
лбищах 1 вод!), площу поверхш мембрани, й товтцину, молекулярну масу газу 1
коефппент дифузп,
Через те що дифузУя газУв у легенях вщбуваеться з альвеол у кров, що рухаеть-
ся, У кисень у кровУ з’еднуеться з гемоглобином, дифузшна здатшсть легень зале-
жить також вщ об’ему кагплярноУ кров! У швидкостт реакцп кисню з гемоглобином
Виходячи з цього, утруднення дифузп газгв у легенях, й зменшення можна
пов’язати з такими факторами: 1) зменшення площУ альвеолокапглярноУ мембра-
ни; 2) стовщення альвеолокапУлярног мембрани; 3) зменшення трансмембранно-
го градюнта парцУального тиску газу в систем! альвеола — кров, шо спостерУгаеть-
ся при всУх патолопчних процесах, якУ характеризуються зменшенням альвеоляр-
ное вентиляцп та / або зменшенням об’ему капУлярног кров! в легенях; 4) змен-
шення швидкост! реакцп кисню з гемоглобшом 1 киснево’У емностУ кров!, шо
вщбуваеться при анемп, шактивашУ гемоглобгну; 5) зменшення об’ему капУлярног
кровУ в легенях (при емфУземУ легень, недостатносп серця, запаленнУ легень тощо);
6) зменшення експозицп контакту' еритроципв з альвеол я рним ловпрям.
Дифузшна новерхня може змепшуватися внаслщок резекцп частини леген!
або видалення шло'У легеш, загибелт частини альвеол пщ час хрошчних запальних
процеств. туберкульозу. припинення вентиляцп певноУ дУлянки леген! (ателектаз)
тощо.
Патолопчш процеси в легенях можуть супроводжуватися стовшенням стгнок
альвеол I капыярУв, збгльшенням кшькосп сполучноУ тканини мгж ними, рщини
в альвеолах У Унтерстицй легень. Це призводить до утруднення дифузп газгв у ле-
генях — виникае алъвеолокатлярна блокада (мал. 63).
Альвеолокагплярну блокаду можуть спричинювати таю дифузш ураження ле-
гень, як саркощоз, пневмоконУоз рУзноУ еполопУ, пневмосклероз, склеродерм!#,
емфУзема, пневмонУя.
Важливу роль у розвитку альвеолокатплярноУ
блокади вщУграе набряк легень. ВУн виникае тодУ,
коли рщина фшьтруеться через легеневе мжро-
циркуляторне русло швидше, нУж видаляеться
через лУмфагичнУ судини. Спочатку рщина нако-
пичуеться в Унтерстицшному простору мУж: ендо-
телУем капшярУв У альвеолярним ештелУем (Ун-
терстишйний набряк). Бътъша частина ще'У
Мал. 63. Причини порушення аифузп:
1 — нормальне сшввщношенвд мЬк кагплярними судинами
га альвеолами; 2 — потовщення спнки альвеол: 3 — потоь-
щення спнки кашлярник судин: 4 — внщршньоальвеоляр-
ний набряк; 5 — штерстишйний набряк; 6 — розширення
кашлярних судин
рщини перемпцуеться в 1нтерстиц1йний простщ пщ плеврою 1 б!ля бронх!в, що
перешкоджае виходу рщини в альвеоли. Але якщо резервш можливосп лёмфатич-
но! системи легень та ппш захисш фактори вичерпутоться, рщина потрапляе |
повгтряний проспр альвеол (альвеолярний набряк), шо спричинюе порушення
гаюобмшу.
Причинами набряку легень можуть бути гемодинамёчш фактори, яю зумо-
влюють пщвишення тиску в кашлярах до 3,3—4 кПа (25—30 мм рт. ст). Це спос-
терпаеться у випадку лёвошлуночково! недостатност! серця, деяких уроджених
або набутих вад серця, наприклад при стеноз! лшого передсердно-шлуночкового
отвору.
Серед !нших причин набряку легень слщ зазначити ушкодження ендотелёю
легеневих мёкросудин, шо сприяе щдвишенню !хньо! проникносп для быкёв. Цей
мехашзм е основою рес гираторного дистрсс-синдрому дорослих. Пщвишення про-
никност! капёлярёв легень може спостериатися пщ час вдихання токсичних речо-
вин. у тому числ! 1 високих концентрашй кисню; при системному запально-реак-
тивному синдром!, коли в раз1 розвитку запального процесу, сепсису в кров! нако-
пичуютъся БаР (медтатори запалення); при бактер!альшй з в!русшй пневмонп:
у раз! швидкого зниження атмосферного тиску внаслщок перерозтягневня легень
ё гостро! ппоксп; емболи легеневих судин, зокрема жирово! емболи, що може ви-
никати теля перелому трубчастих юсток. Виникнення респираторного дистрес-
синдрому може загрожуъати також реципёентам легеневого трансплантата.
За нормальних умов у станё спокою час проходження червоних юптин через
катпляр становить приблизно 0.75 с. Чи буде його достатньо для нормально! ок-
сигенат! кров!, залежить вщ факторгь, яю визначають дифузшну здатшсть мем-
брани.
У здорово! людини р!вновага альвеолярного ! катлярного рО2 настае не п!з-
н!ше, нёж еритроцит подолае третину довжини капётяра. У раз! патолог!! легень т
помёрного порушення !х дифузшно! здатносп ця рёвновага настае шзшше, але ще
в межах капьляра. Пщ час тяжко! патологи часу .для досягнення рёвноваги може
забракнути, 1 рО2 в юнпё капьляра може залишатися нижчим, юж в альвеолах.
Варто зазначити, що патологёчш процеси в легенях, як! утруднюють дифузёю,
спричинюють порушення, передусём дифуз!ю кисню, осюльки вуглекислий газ
дифундуе у 20—25 разёв легше Залежно вщ того як змёнюеться газовий склад ар-
терщчьно! кровё видщяють два види недостатност! дихання — гёпоксемтчну ё п-
перкапшчну.
Пперкапшчна недостатшсть дихання харакгеризуеться збыьшенням напру-
ження вуглекислого газу в артерёальнёй кровё понад 6 кПа (45 мм рт. ст.) 1 вини-
кае при зменшеюп альвеолярно! вентиляцё!, що призводить до накопичення СО
в альвеолярному газ! !, вщповщно, в кров!.
Ппоксемгчну недостатшсть дихання харакгеризуе напруження кисню в ар-
тертальнш кровё, величина якого менша нёж 7,33 кПа (55 мм рт. ст). Причомх
хворий повинен у цей час дихати киснево-повётряною сумпшпю, яка мтстить
понад 60 % кисню. ГЦдгрунтям гшоксемёчно! недостатност! е ушкодження па-
ренхгми легень ё порушення через це дифузй кисню крхзь альвеолокапётярну
мембрану.
ЗМЕНШЕННЯ ЗАГАЛЬНОГО ТОКУ КРОВ1 В ЛЕГЕНЯХ
У норм! величина загального кровотоку в легенях вщповщае ХОС г становить 4,5—
5 л/хв. Зменшення перфузи легень можуть зумовити таю патологгчнг стани: 1) пору-
шення скоротливо! здатностг правого шлуночка (гнфаркт мюкарда, мюкардит, кар-
диосклероз, ексудативний перикардит тощо); 2) порушення скоротливо! здатносп
Л1вого шлуночка, яке спричинюе розвиток застгйних явищ у легенях; 3) деякг врод-
жеш та набуп вади серця (стеноз або атрезгя легеневого стовбура, стеноз правого
передсердно-шлуночкового отвору, атрезгя правого передсердно-шлуночкового кла-
пана); 4) судин на недостатшсть (шок, колапс); 5) ембол!я легенево! артерп.
В усгх перелгчених випадках тгк кров! в легенях уповгльнюеться, зменшуеться
зрошення кров’ю легеневих альвеол. Водночас вентилящя альвеол переважае над
перфузгею, що зумовлюе, як правило, достатке насичення киснем кров!, яка вщ-
тгкае вщ легень, г зменшення в нй вмгсту вуглекислого газу (гшокапнгя). Але
осюльки ХОС у цих умовах зменшуеться (до 2—4 л/хв)г сповшьнюеться рух кров!
в судинах великого кола кровообггу, тканини вщчувають кисневе голодування, а
в кров! визначаються недостатшсть кисню г надлишок вуглекислого газу.
ПОРУШЕННЯ ЗАГАЛЬНОГО I РЕПОНАРНОГО ВЕНТИЛЯЦ1ЙНО-
ПЕРФУ31ЙНОГО СП1ВВ1ДНОШЕННЯ У ЛЕГЕНЯХ
Для нормального газообмшу в легенях дуже важливим е правильне спгввщношен-
ня вентиляцн г перфузи. У сташ спокою в здорово! людини ефективна альвеоляр-
на вентиляция (АВ) становить 4,0—5,0, ХОС — близько 5 л, а сшввщношення
АВ / ХОС — 0,8—1,0. Саме таке спгввщношення забезпечуе нормальний газовий
склад кров!, що вщтгкае вщ альвеол. У тих випадках, коли вентиляцгя переважае
над током кров! (АВ / ХО> 1,0), з кров! видаляеться бшьша, шж зазвичай, кгль-
кгсть вуглекислого газуг розвиваеться г1покапн1я. Якщо вентиляцгя вщстае вщ току
кров! (АВ / ХОС < 0,8), в альвеолярному повгтрг пщвищуеться парщальний тиск
вуглекислого газу г знижуеться рО2, що спричинюе гтоксемио та г1перкапн1ю.
Для того щоб газообмгн у легенях вщбувався нормально, оптимальне спгввщ-
ношення мгж вентилящею альвеол г гх кровопостачанням мае шдтримуватися в
усгх альвеолах. Це означае, що повгтря, яке вдихаеться, г вщповщно ток кровг
мають ргвномгрно розподглятися по всгх легеневих альвеолах. Однак повнгстю ця
умова не виконуеться навгть у норм!; у здорово! людини спостерггаються вщхи-
лення в розподглг як вентиляцн, так г перфузи альвеол у ргзних дглянках легень:
показники вентиляцн та перфузг! в нижшх вщдглах легень вшцг, нгж у верхнгх,
кргм того, у верхнгх вщдглах вентиляцгя перевищуе перфузгю, а в нижшх — пер-
фузгя переважае над вентилящею.
Пщ час хвороб легень до фгзголопчног нергвномгрностг вентиляцп та перфузи
може приеднатися й патолопчна. Наприклад, у разг пневмоюг, емфгземи, ателек-
тазу або пневмосклерозу внаслщок залучения до патолопчного процесу крово-
носних судин ток кров! в кашлярах альвеол на одних дглянках легенг ргзко зни-
жуеться, а на гнших — посилюеться. Змгни еластичностг легень або порушення
проходження повгтря дихальними шляхами, що спостерггаються при цьому, у ргз-
них дглянках легень можуть бути неоднаково виявленими, що спричинить нергв-
Мал. 64. Емовгргп репонарю вентиляцшно-перфузшш взаемо-
зв’язки в легенях та 1х вплив на оксигенащю кровк
УЛ— альвеолярна вентилящя; <2 — юлыасть кров!, яка протпсае
через легенев! судини за 1 хв (за С. ХУезг, 1988)
ном!рну вентиляцгю альвеол. Кргм того, значш порушення газообмшу можуть
виникати 1 внаслщок того, що в добре вентильованих альвеолах ток кров! може
бути слабким, а альвеоли, яю штенсивно постачаються кров’ю, можуть погано
вентилюватися. Загальна альвеолярна вентиляцгя г загальний кровотгк у легенях
при цьому 1стотно не змшюються. На мал. 64 показано, як значно може змшюва-
тися вептиляцшно-перфузшне сшввщношення в подгбних випадках. Видно та-
кож, що високе вентиляцшно-перфузшне спгввщношення лише дещо збгльшуе
оксигенащю кров!, тод! як низьке зумовлюе значке и зменшення, що викликае
зниження концентрацп кисню в кров), яка вщпкае вщ певно! дшянки легеш.
Варто зауважити, що в легенях гснують мехашзми, здатш пщтримувати по-
тргбне сп1ввщношення мгж вентилящею альвеол та !х перфузгею в раз! його вщхи-
лення вщ нормально! величини. Так, пщ час обструкцп окремих бронхов або брон-
хгол попршуеться вентилящя певно! частки альвеол г вмгст кисню в альвеолярно-
му газ! зменшуеться. Вщомо, що непосмугованг м’язи легеневих артерш мгстять
О2-чутлив! К+-канали. Зменшення надходження до них кисню призводить до за-
криття К+-каналгв ! деполяризацп мембрани м’язових юптин, що зумовлюе над-
ходження до юптин Са2+ та !х скорочення. Тому недостатшсть кисню спричинюе
спазм непосмугованих м’язгв судин малого кола кровообпу (гшоксична вазокон-
стрикщя), унаслщок чого приплив кров! до гшовентильованих дглянок легенг
зменшуеться. Кргм того, накопичення вуглекислого газу призводить до збщьшен-
ня концентрацп Н+, що також зумовлюе вазоконстрикцгю в легенях (на вщмгну вщ
шших органгв, у яких цей процес сприяе вазодилатащг). 3 шшого боку, якщо кро-
вопостачання окремих дглянок легенг зменшуеться, це зумовлюе зниження пар-
щального тиску вуглекислого газу в альвеолах, що спричинюе спазм непосмугова-
них м’язгв бронхгв г бронхгол г, як наслщок, зменшення вентиляцп альвеол, що
погано перфузуються. Усе це сприяе пщтриманню нормального вентилящйно-
перфузгйного спгввщношення в окремих дглянках легень.
ВПЛИВ НЕДОСТАТНОСП ДИХАННЯ НА ОРГАНОМ
Недостатшсть дихання е одшею з причин розвитку гшоксп та ацидозу. Проте при
багатьох розладах системи зовнппнього дихання, коли порушуються процеси вен-
тиляцп, дифузп та перфузп, у стан! спокою недостатшсть дихання може не вини-
кати, осюльки при цьому пщтримуеться нормальний газовий склад кровь Це
пов’язано, по-перше, з наявшстю в легенях тако! кшькост! функцюнальних еле-
ментгв (альвеол ! кашляргв), яка в багато разгв перевищуе потребу оргашзму, що
перебувае в стан! спокою, у газообмгш. По-друге, досконала система регуляцп
зовшшнього дихання реагуе на найменш! коливання напруження кисню та вугле-
кислого газу в кров!! в раз! потреби збшьшуе роботу дихальних м’яз!в, змшюючи
глибину ! частоту дихання. Вщповщно до дихання змшюеться г кровообгг. Усе це
сприяе пщтриманню сталостг газового складу кров! в стан! спокою в раз! патоло-
ги органов дихання. Проте за цих умов компенсашя досягаеться за рахунок мо-
бпизацп резервгв дихально! системи, тому ознаки недостатносп дихання
обов’язково виявляться пщ час фгзичного навантаження, осюльки резервш мож-
ливостг зовшшнього дихання в хворо! людини будуть вичерпаю ранппе, нгж у
здорово!.
Розлади процесгв вентиляцп, дифузп або перфузп зумовлюють посилення ро-
боти дихальних м’язгв у стан! спокою, зростання навантаження на систему кро-
вообггу, посилення обм!ну речовин. Потреба оргашзму в кисн! постгйно зростае.
Зрештою настае такий момент, коли г в сташ спокою пщтримання нормального
газового складу кров! стае неможливим. Тод! провщними ланками в розвитку
подалышгх порушень в оргашзм! стають ггпоксгя, пперкапн!я г газовий ацидоз.
Якщо недостатшсть дихання виникае гостро чи пщгостро 1 сягае такого сту-
пеня, що в кров перестае надходити кисень, а з кров! не виводиться вуглекислий
газ, розвиваеться асфгкс1я (вщ грец. асщю&а — вщсутшсть пульсу, ядуха). Здебшь-
шого асфгксгя виникае в раз! здавлення дихальних шлях!в (задушения), закупо-
рювання гхнього просвпу (сторонш предмета, запальний набряк), наявностг рщи-
ни в дихальних шляхах г альвеолах (утопления, набряк легень, блювотш маси),
двобгчного пневмотораксу. Кргм того, до розвитку асфгксп може призвеста р!зке
пригшчення дихального центру, порушення проведения нервових гмпульсгв до
дихальних м’язгв, значке обмеження рухомостг трудно! клики.
У перебггу асфгксп видшяють три перюди, як! можна спостерпати у тварини
в експеримент!, наприклад шсля перетиснення трахе!.
Перший пергод характеризуемся швидко наростаючим збшьшенням гли-
бини г частоти дихання з переважанням фази вдиху над фазою видиху. Спостерг-
гаються загальне збудження, пщвищення тонусу симпатично! частини автономно!
(вегетативно!) нервово! системи (розширення згнипь, тахгкардгя, пщвищення ар-
тергального тиску), можливг судоми.
Удругий пергод частота дихання поступово зменшуеться за збереження
максимально! амплгтуди дихальних рухгв, посилюеться фаза видиху. Переважае
тонус парасимпатично! частини автономно! нервово! системи (згницг звужуються,
знижуеться артергальний тиск, розвиваеться брадикардгя).
Утретгй пергод асфгксп спостерггаються поступове зменшення амплпуди
дихання, його частоти г нарештг зупинка дихання. Артергальний тиск значно зни-
жуеться. Пгсля короткочасно! зупинки дихання зазвичай з’являються юлька судо-
мних дихальних руххв (гаспшг-дихання), шсля чого дихання припинясться оста-
точно.
Явища, як! спостерггаються пщ час асф!кси, пов’язаш спочатку з накопичен-
ням в оргашзм! вуглекислого газу. Дпочи рефлекторно 1 безпосередньо на дихаль-
ний центр, вутлекислий газ збуджуе його. Максимально можливих величин дося-
гають глибина 1 частота дихання. Кр1м того, дихання рефлекторно стимулюеться
зниженням у кров! напруження молекулярного кисню. 31 збшьшенням у кров!
вмюту вуглекислого газу пщвищуеться 1 артер!альний тиск. Експерименти !з вди-
ханням газових сум!шей, як! мктять 10—20 % вуглекислого газу, показали, що це
пщвищення пов’язане, по-перше, з рефлекторним впливом хеморецепторгв на
судиноруховий центр, по-друге — з посиленим викидом адреналшу в кров, по-
трете — 31 збшьшенням ХОС унаслщок пщвищення тонусу вен 1 збшьшення над-
ходження кров! шд час посиленого дихання.
У раз! подальшого збщыпення концентраци вуглекислого газу в кров! почи-
нае проявлятися його наркотична д!я, рН кров! знижуеться до 6,8—6,5. Поси-
люеться гшоксем1я !, вщповщно, гшоксгя головного мозку. Це спричинюе при-
гшчення дихання, зниження артер!ального тиску. Як наслщок, настае парал1ч
дихання та зупинка серця.
Задшпка, або диспное. При недостатност! дихання та деяких шших патолопч-
них процесах у людини може виникнути вщчутгя нестач! повгтря 1 пов’язана з
ним потреба посилити дихання. Це явище мае назву задишки, або диспное. Вщ-
чуваючи нестачу повгтря, людина не лише мимовыьно, а й свщомо збшьшуе ак-
тившсть дихальних рух!в, намагаючись усунути це вщчутгя, наявшсть якого 1 е
найсуттевнпою вщм!ншстю задишки вщ шших вид!в порушення дихання (гшер-
пное, тахшное тощо). Тому в людини в непритомному стан! задишки не бувае.
У здорово! людини задишка може виникнути пщ час виконання важко! ф!-
зично! роботи, якщо вона потребуе значних зусиль на меж! ф!зичних можли-
востей.
У патолопчних умовах задишку можуть спричинити: недостатня оксигенац!я
кров! в легенях (зниження паршалъного тиску молекулярного кисню в повггр!,
що вдихаеться, порушення вентиляцп альвеол, дифузп газ!в 1 кровообпу в леге-
нях); порушення транспорту газ!в кров’ю (анемгя, шакгиващя гемоглобшу, недо-
статшсть кровооб!гу); утруднен ня рухгв трудно! клики та д!афрагми; ацидоз;
пщвищення обмшу речовин; функцюнальш й оргашчш ураження центрально!
нервово! системи (силып емоцшш впливи, !стер!я, енцефалгг, порушення мозко-
вого кровообпу тощо).
В експеримент! вщчутгя нестач! повггря у здорово! людини виникае в раз!
тривалого довыьного затримання дихання. 1мов!рно, це вщчутгя 1 пов’язане з ним
нездоланне бажання зробити вдих зумовлеш сильним збудженням шсшраторних
нейрошв, яю рефлекторно стимулюються зниженим вы ютом молекулярного кис-
ню в кров!, а також рефлекторним 1 прямим впливом високо! концентраци вугле-
кислого газу. Зробивши один-два вдихи, людина вщчувае полегшення, оскшьки
напруження газ!в у кров! нормалгзуеться 1 дихальний центр знову збуджуеться
звичайними за силою подразниками.
Задишку в людини можна спричинити г за допомогою вдихання сумшп газ!в
з високою концентращею вуглекислого газу. У цьому випадку вщчутгя нестач!
повгтря також пов’язане з штенсивним збудженням дихального центру, але поси-
лення дихальних рухгв не дае полегшей ня, осюльки вплив збуджувальних сти-
мулов на дихальш нейрони е постгйним г не зменшуеться наприкгнщ вдиху.
Очевидно, розвиток задишки пщ час патолопчних процесг в також пов’язаний
оз надмгрною стимуляцгею гнспграторних нейронов. Збудження вш них поширюеть-
ся не тшьки на периферпо. до дихальних м’язов. а й у вищерозташоваш вгддгда
центрально-! нервово! системи, де, ймоворно, г формуеться специфгчне вгдчутгя.
У розвитку задишки беруть участь численш фактори. Вона виникае, якщо
линники, ям збуджують вдих. переважають над тими, яю гальмують його, або
якщо чутливость до них дихального центру подвишена. Серед найважлившгих гз
цих факторов видгляють:
1) збудження рецепторов, шо стимулюють центр вдиху, яко активуються в разо
зменшення об’ему легеневих альвеол (один гз рефлексов Геронта — Брейера —
рефлекс на спаданья легень). За деяких видов патологи може виникнути постоина
мпульсацоя з цих рецепторов. Наприклад, у разо застшних явищ у легенях (серце-
ва недостатшсть, пневмоноя) переповнеш кров’ю судини, оцо оточують альвеоли.
здавлюють гх, емность альвеол зменшуеться, шо спричинюе збудження рецепторов
спадания. Осюльки розтяжность гшеремойованих дглянок легенг значно знижена,
активность рецепторов, шо збуджують вдих, не знижуеться навоть щд час сильного
вдиху. В усох цих випадках гнший рефлекс Геронта — Брайера — гнсшраторно-
гальмгвний — виявляеться менш ефективним, оскгльки легенево альвеоли не рсз-
тягуються достатньою игрою;
2) збудження рецепторов гнтерстищйно! тканини легень О-рецепторов). Пато-
лопчно процеси, пов’язано гз застгйними явищами в легенях о розвитком гнтерсти-
цгйного набряку (пневмоноя, недостатшсть серия), можуть спричинювати тривале
збудження 3-рецепторов о посилення стимуляцц дихальних нейронов;
3) збудження гритантних рецепторгв, розташованих в епгтелг! о субештелгально-
му шар! всгх повгтропровщних шляхгв У разг подразнення гритантних рецепторов
трахе! виникае кашель, а якшо подразнюються рецептори бронхов, посилюеться
шспораторна активность, що може спричинити задатку. Значне збудження гритан-
гних рецепторов може спосгерггатися в раз) бронхгту, пневмонп. бронхоально! аст-
ми, пневмотораксу, ателектазу, застою кровг в малому коло кровообпу;
4) рефлекси з барорецепторов аорти о сонно! пазухи. Цо рефлекси залучаються
до патогенезу задишки у зв’язку з втратою кровг. шоком, колапсом. Артергалъний
тиск 70 мм рт. ст. (9,1 кПа) 1 нижче спричинюе розке зменшення потоку гмпуль-
егв, яю в нормг справляють гальмгвну дгю на гнспграторну активность;
5) рефлекси з хеморецеггторгв аорти г сонно! пазухи. Зниження в кровг напру-
ження кисню, пщвищення напруження вуглекислого газу або ж збгльшення кон-
центрат! гонгв водню посилюють збудження рецепторгв, розташованих в аорталь-
ному г сонному югубочках, що пщвищуе актившсть центру вдггху. Цей механгзм
вщгграе важливу роль у розвитку задишки при ацидозг, недостатностг дихання,
анемп, недостатностг кровообггу тошо;
6) стимуляцгя нейрошв довгастого мозку7. якг вибгрково чутливг до вуглекис-
лого газу г силе не збудження яких у разг гшеркапнн спричинюе гнтенсивну сти-
муляцпо дихального центру;
7) рефлекси з дихальних м’язгв, Вщчуття недостатностг дихання може виник-
нути внаелгдок надмгрного навантаження на мгжребровг м’язи г сильного збуд-
ження м’язових рецепторгв розтягу, гмпульсацгя з яких надходить у вищг вшдгли
центрально! нервово! системи. Цей мехашзм д!е пщ час виконання важко! ф!зич-
но! роботи, яка потребуе значного напруження шсшраторних м’язгв для забезпе-
чення вщповщно! вентиляцп, а також у раз! зменшення здатност! легень до роз-
тягування, звуження верхшх дихальних шлях!в. В експеримент! така задишка ви-
никае в дослщжуваних пщ час дихання через маску або в одяз!, який ш!льно
прилягае й утруднюе рухи трудно! клггки;
8) стимуляция дихального центру продуктами власного метаболтзму. 1деться
про накопичення вуглекислого газу, кислих продуктов обмшу та зниження тиску
кисню безпосередньо в нервових центрах унаслщок порушення мозкового крово-
обпу (спазм або тромбоз судин головного мозку, набряк мозку, колапс).
Дихання пщ час задишки, як правило, е частим ! глибоким. Посилюеться як
вдих, так ! видах, який мае активний характер г здшснюеться за участю ексшра-
торних м’язтв. Проте в деяких випадках може посилюватися переважно або вдих,
або видах, т тод! говорить про шстраторну (посилений вдих) або екстраторну
(посилений видих) задшику. Тнсшраторна задишка спостерггаеться, наприклад, у
першш стадп асфгксп, пщ час збудження центрально! нервово! системи (стрес,
тстертя), ф!зичного навантаження у хворих тз недостатшстю кровообпу, при пневмо-
торакс!. Ексшраторна задишка трапляеться переважно у хворих на бронх!альну
астму, емф!зему, кода пщ час видиху збшыпуеться ошр потоку повггря в нижнгх
дихальних шляхах.
За наявносп перешкоди продолжению повггря у верхних дихальних шляхах
спостерггаеться стенотичне дихання.
ПОРУШЕННЯ НЕРЕСП1РАТОРНИХ ФУНКЦ1Й ЛЕГЕНЬ
У легенях здшснюеться не лише газообмш мтж кров’ю та пов!трям, !м властив! 1
шш! — нересшраторш функцп, як! можуть порушуватися пщ час р!зних патоло-
пчних процеств (запалення, набряк, емф!зема, склероз тощо).
Захисна функщя. Поверхня легень, що контатуе з навколишшм середовищем,
яке в сучасних умовах стае все агресившшим, мае найбйтыпу в оргашзм! сумарну
площу (50—100 м2). У легенях затримуються мехашчш частники (приблизно 90 %
частинок дтаметром понад 2 мкм), шкщлив! токсичш продукти, що надходять з
повттрям пщ час вдиху. Осш частники видаляються зт стшок бронх!в шляхом
висхщного просування слизу (мукоцилтарний транспорт). Слиз просуваеться вго-
ру 31 швидюстю близько 20 мм/хв завдяки ритм!чним рухам численних тонких
вгйок ештелпо. Частники, що потрапляють до альвеол, здебтлыного поглинаються
макрофагами, як! в подальшому “полишають” леген! з кров’ю та лтмфою.
Фтльтрацшна функщя. Леген! здатш очищати кров вщ деяких мехашчних до-
мшгок — крапель жиру, др!бних тромбтв, бактерш тощо. Усе це затримуеться в
легенях, зазнае деструкцп та р!зних перетворень.
Екскреторна функщя легень виявляеться у виведенш деяких летких метаболгпв
(наприклад ацетону, ам!аку тощо) або екзогенних речовин (алкоголю, бензолу),
що мае значения в раз! штоксикацц.
Важливою е також усмокгувальна функщя легень. Багато жиро- ! водорозчин-
них речовин, переважно летких, аерозол!в здатш з великою швидюстю всмокту-
ватися легенями. На цш властивост! грунтуеться шгаляцшний шлях уведення де-
яких лгкарських засоб!в.
Метаболхчна функхця. Леген! вщгграють важливу роль в обмхнх бшкхв, лпхщхв 1
вуглеводхв. Вони багап на лхполхтичш та протеолхтичнх ферменти х за штенсивхпс-
тю обмшу лшщхв подхбнх до печшки. Легеш регулюють надходження лпхщхв в
артерхальну кров, осюльки в них затримуеться х залучаеться до метаболизму час-
тина хшомхкрошв, яю надходять хз кишок через лхмфатичш судини. У легенях
синтезуються жиров! кислоти I фосфолхпщи, зокрема й дипальмгтиллецитин, що
входить до складу сурфактанпв. Важливу роль вщпрае також синтез бхлюв, ос-
юльки структурною основою легень е колаген х еластин. Порушення синтезу,
посилений розпад або гшерпродукцхя цих бшюв можуть бути причинами розвитку
емфхземи та пневмосклерозу. Велике значения мае також обмш вуглеводхв, особ-
ливо утворення глхкозамшоглхканхв, що входять до складу бронххального слизу.
У легенях вщбуваеться обмш багатьох речовин, яю впливають на судини.
Адже саме через легеш проходить уся кров х саме в них зосереджена значна час-
тина ВС1Х судинних ендотелхальних клпин оргашзму. Багато вазоактивних речо-
вин повшстю або частково втрачае актившсть пщ час проходження судинами
легень, наприклад брадикшш може шакти вуватися на 80 %>. Легеш е головним
органом, який звшьняе кров вщ серотоншу шляхом його захоплення х депонуван-
ня. При цьому серотонш частково надходить у тромбоцита х в раз! розвитку ана-
фшаксп вившьнюеться. Тут е також ферменти, що шактивують ПГЕр ПГЕ2 та
ПГЕ2а, частково (до 30 %) захоплюються норадреналш х пстамш. Альвеолярний
еттелш мае особливу трупу рецептор!в, яю активуються протешазами, наприклад
тромбшом або трипсином. Пщ д!ею протешаз частково перетравлюються лгганди,
з’еднанх хз цими рецепторами, що призводить до хх активацп. У раз! активацп цих
рецептор!в вившьнюеться ПГЕ2, який пщвищуе резистентшсть ештелхальних юп-
тин до патогенних факторхв.
У легеневих судинах анпотензин I пщ впливом АПФ змшюеться на анпотен-
зин II. Леген! вщхграють важливу роль у пщтриманш фхбринолхтично! та протиз-
сщальнох активност! кров!. В штерстицшнш тканиш легень е велика юльюсть
базофщьних гранулоцитхв (тканинних базофшв), що мхстять гепарин. Леген! бе-
руть участь у детоксикацц деяких лхкарських засобхв (амшазину, сульфаншамццв
тощо).
Вщома також роль легень у терморегуляци. За низько! температури повхтря,
що вдихаеться, у легенях активуються процеси бхолопчного окиснення (зокрема
й р-окиснення лшщхв) х збхльшуеться теплопродуктця. Водночас вщбуваеться зни-
ження капшярного току кровх в малому колх кровообхгу та змехгшенххя тепловщ-
дачх через легень
Нарештх варто зазначити, що легеш е резервуаром кровх завдяки здатностх су-
дин малого кола кровообпу змшювата свою емшсть навггь за умови незначно!
змши тиску в них. Леген! вщхграють певну роль у пщтриманш водного балансу,
оскшьки з повхтрям, яке видихаеться, видаляеться з оргашзму х вода.
Порушення всхх цих важливих для оргашзму функцш легень може сприяти
розвитку найрхзноманхтнппих патолопчних процесхв.
Р03Д1Л XXVIII
ПАТОЛОПЧНА Ф13ЮЛ0ПЯ ТРАВЛЕНИЯ
Травления — це сукупшсть процессе, якг забезпечують мехашчне подргбнення та
хгмгчне (головном чином ферментативне — г1дрол1тичне) розщеплення харчових ре-
човин на компонента, позбавлеш видово! специфгчноспй / придатш до всмоктування
та учаспй в облша речовин (О.М. Уголев).
Основне у вищих оргашзмгв позаклпинне травления за локалтзашею травно-
го процесу под!ляють на порожнинне (дистантне), мембранне (контактне) та про-
м!жне м!ж ними травления в шар! слизових накладень (премембранне). За похо-
дженням ферменпв його подигяють на власне, автолтгичне 1 симбюнтне, питомий
внесок яких гстотно змшюеться при порушенш продукцй ферменпв травними
залогами (власне травления), змшах мгкрофлори травного каналу (симбюнтне),
термгчнш обробш !ж! (зниження автолпичного травления внаслщок шактивацп
лгзосомальних ферменпв продукпв рослинного 1 тваринного походження). М1Ж
названими процесами та р!зними вщдитами системи травления юнуе взаемозв’язок
1 етапюсть у робоп, що забезпечуе висок! компенсаторн! можливосп ще! системи
в умовах патологи (Ю.С. Лондон). Однак коли щ компенсаторн! можливосп ви-
черпуються, виникае недостатшсть травления — стан, за якого травна система не
забезпечуе адекватш потребам оргашзму перетравлювання, усмоктування та засвоен-
ня поживних речовин (нутргенпв). Основним наслщком недостатносп травления е
розвиток голодування (головним чином неповного) р!зного ступеня.
ПОРУШЕННЯ АПЕТИТУ
Вщповщшсть мгж енергетичними г пластичними потребами ! надходженням по-
живних речовин до оргашзму пщтримуеться вщчуттям голоду та спраги г може
порушуватись у випадку розладу апетиту.
Вщчуття голоду залежить вщ наповнення шлунка г складу кров! (ппоглгкемгя).
Його стимулюють збудження латерального ядра ппоталамуса, гшоталамгчний
нейропептид У, ендорфши та кортикостеро'щи. Гальмують апетит наповнення
(розтягнення) шлунка, “сита” кров, вентромед!альне ядро ппоталамуса, холецисто-
кгшн, а-меланоцитостимулювальний гормон (а-МСГ), лептин (гормон жирово!
тканини), кахексии та амши.
Зниження апетиту — анорексгя — спостерггаеться при хворобах оргашв трав-
ления, гарячщ, шфекцшних хворобах та интоксикациях, злояк!сних пухлинах, авг-
тамгнозах, зневодненнг та хрошчшй втомг. Пщвищення апетиту — гтерорексы,
або булгмгя, — виникае при ураженнях центрально! нервово! (психгчш хвороби,
енцефалгт, пухлини) та ендокринно! (цукровий дгабет, гшергнсулгшзм, ггперти-
реоз) систем. В експеримент! анорексгю вщтворюють руйнуванням латеральних,
а булгмгю — вентромедгальних ядер гшоталамуса.
ПОРУШЕННЯ ТРАВЛЕННЯ В ПОРОЖНИН1 РОТА
Основш розлади травления в порожнинг рота: 1) порушення подргбнення !жг (жу-
вання); 2) розлади слиновидшення; 3) порушення ковтання.
Порушення жування виникае внаслщок ушкодження або втрати зубгв, розла-
ду функцг! жувальних м’язгв чи ураження скронево-нижньощелепних суглобгв г
запалення слизово! оболонки ротово! порожнини. Головними причинами ушкод-
ження г втрати зубгв е каргес 1 пародонтит.
Мгкрофлора, що розвиваеться при каргесг зубгв, пародонтит! або тонзилгп,
може зумовлювати сенсибглгзацгю органгзму, спричинювати ендокардит, гломеру-
лонефрит г навгть оральний сепсис. Джерелом захисних факторгв ротово! порож-
нини е слина та лейкоцита, що емггрують. Захисну функцгю виконують гмуно-
глобулгни та ферменти слини, якг справляють окисну, антабактергальну та про-
тавгруснудгю (пероксидаза, лгзоцим, РНКаза, ДНКаза). Цг речовини взаемодгють
з цитокгнами г захисними факторами лейкоцитгв (мгелопероксидаза, лгзоцим, лак-
тоферин, катепсин О, катюши бглки).
У ротову порожнину щохвилини мггрують десятки тасяч лейкоцитгв, пере-
важно нейтрофглгв. Головним мгсцем гх емгграци е ясенна борозна. Втрата зубгв
зменшуе емгграцгю, а запальнг процеси (ггнггвгт, пародонтит) — гстотно шдвищу-
ють. Розроблено кглькгснг методи визначення мгграцц лейкоцитгв до ротово! по-
рожнини (М.О. Ясиновський; 0.1. Сукманський та ш.), якг застосовують у клгшч-
нгй практицг.
Кар!ес зубгв
Каргес зубгв (англ. — с1еп1а1 сапен) — патолопчний процес, що руйнуе тверд! тка-
нини зубгв (емаль г дентин) з утворенням порожнини. Вщбуваеться як деструкцгя
мгнерально! фази (оксиапатату), так г розпад оргашчних речовин твердих тканин
(протеолгз). Каргес може ускладнюватася пульпгтом г пергодонтитом. Поши-
ренгсть каргесу в деяких реггонах земно! кулг рангше сягала 100 %. Однак ефек-
тавна профилактика (фторист! зубнг паста, фторування води)! змгни харчування
сприяли гстотному зниженню за останнг 15—20 рокгв поширеност! каргесу в де-
яких европейських крагнах.
Основн! етюлопчш фактори каргесу — алгментарний г мгкробний. Головна
причина його виникнення г поширення — неприродне харчування сучасног лю-
дини, споживання гжг, оброблено! мехашчно г термгчно, г (особливо) чаете спо-
живання солодощгв. Важливу етголопчну роль вщгграють мгкрооргангзми зубного
нальоту, особливо 81гер1ососсин тШапн, 81г. нап&йз г АсИпотусен уюсонин Роль мгк-
робного фактора шдтверджують епщемгологгчнг доелгдження та дослщи на без-
мгкробних тваринах (гнотобгонтах), у яких каргес змоделювати не вдаеться.
У патогенез! кар!есу важливе значения мають процеси, що вщбуваються на
поверхш емаль Слина справляе на емаль зуб!в захисний вплив (очисний, буфер-
ний, антибактерхалышй, ремшералхзуючий). Порушення слиновидхлення сприяе
утворенню на поверхн! емал! зубного нальоту (бляшки), який складаеться пере-
важно з адгезивних полхглхкашв, що синтезуються з вуглеводхв гж1. Мхкрооргашз-
ми нальоту метаболхзують вуглеводи (особливо цукор) в оргашчш кислоти, пере-
важно молочну, як! демхнералхзують емаль. Розчиненню мшеральних солей часто
передуе руйнування мхкрооргашзмами оргашчних елеменпв емал! (пластинки,
оболонки призм). Утворюваш продукти гщролхзу быкхв можуть виконувати роль
хелаторгв (комплексоутворювачхв), яю забирають калъцш з кристалгв гщроксиа-
патиту емалх та дентину. Певну роль у патогенез! кархесу вщгграе розлад ендоген-
них процесхв на шляху пульпа — тверд! тканини зуба, зокрема порушення цирку-
ляцп зубно! лхмфи.
Руйнування зуба кархозним процесом сповхльнюе захисна реакцхя — утворен-
ня одонтобластами з боку пульпи вторинного, замхнного дентину. Так, повтори!
травми рхзцхв у гризунгв рефлекторно зумовлюють гшертрофхю слинних залоз,
прискорення росту зубхв (рхзц! в гризушв ростуть постшно), новоутворення де-
нтину й емал! та пщвищення стшкост! хх до кархесу (0.1. Сукманський). Можна
припустите, що за подхбним мехашзмом вщбуваеться захисне новоутворення вто-
ринного дентину у випадку руйнування твердих тканин зуба кархозним процесом.
Певну роль у патогенез! кархесу зубхв вщхграе також спадкова схильшсть х зни-
ження хмунно! та неспецифхчно! резистентност! оргашзму.
В експеримент! на щурах х хом’яках кархес моделюють шляхом утримання на
дхетах хз високим вмхстом сахарози (50 % х быьше).
Пародонтит
Пародонтит — запально-дистрофхчний процес, що уражуе пародонт, тобто ткани-
ни, як! оточують х фхксують коршь зуба (перходонт, альвеолярна кхстка й окхстя,
ясна). Проявом пародонтиту е резорбцхя зубних альвеол, гноетеча з ясенних кар-
машв, розхитування х випадшня зубхв.
Пародонтит спостерхгаютъ у 30—50 % осхб вгком понад 30 рокхв. Останшм ча-
сом усе частппе пародонтит виявляють в осхб молодого вхку 1 навггь пщлггкхв.
В етюлоте пародонтиту важливу роль вщхграють:
1) емоцшне перенапруження й ниш надзвичайш (стресовх) впливи;
2) зниження загального фхзичного (гшодинамхя) 1 жувального навантаження;
3) мгкрофлора зубного нальоту ! ясенних карманхв;
4) неповноцхнне харчування, а надто недостатшсть антиоксидантних вхта-
мшхв — аскорбшово! кислоти, бхофлавонощгв х токоферолу (вхтамшхв С, Р х Е).
У патогенез! пародонтиту важливу роль вщхграють нервово-дистрофхчний фак-
тор, розлади ендокринно! регуляцн, порушення кровопостачання тканин паро-
донта, гмунний чинник та порушення функцц слинних залоз. В умовах неадекват-
ного трофхчного забезпечення тканини пародонта ушкоджуються ферментами
слини (калхкреш, РНКаза тощо), а також цитокгнами (1Л-1, ФНП та ш.), як! спри-
чинюють запалення х сприяють розсмоктуванню кхстки. Виникнення запалення
зумовлюе активацхя пероксидного окиснення ! посилення синтезу ейкозанощхв
(простагландишв, лейкотрхешв). Мгкрофлора х розлади слиновидыення спричи-
174
нюють утворення м’якого зубного нальоту, а дал! — зубного каменя, який пору-
шуе кровопостачання тканин пародонта. Важливого значения надають тмунному
ушкодженню тканин, ди бактергально! та лейкоцитарно! колагенази, а також ен-
докринним розладам (гшогонадизм, гшогнсулшгзм, пперпаратиреоз. ппотиреоз,
зниження шкреторно! функпи слинних залоз).
В експерименп пародонтит вцп ворюють шляхом нейрогенних, ендокринних,
алгментарних та тмунних впливгв.
Порушення функцн слинних залоз
Слина бере участь у травлен нт — змочуе й ослизнюе !жу. розчиняе г ферментуе
деяю сполуки, разом тз жуванням девтталтзуе гжу. Кр1м того, вона виконуе захис-
ну 1 троф1чну функцп, необхщна для вщчуття смаку, а завдяки змащувальним
властивостям сприяе жуванню. ковтанню та мовленню.
Слина (ротова р!дина) мтстить близько 50 ртзних ферменпв. Найбшьшою е
концентрацтя а-ам!лази. пероксидази та РНКази. Помггну юльюсть лшази проду-
куе шд’язикова заноза. Вмгст шших ферменпв (а-глгкозидаза, лтзоцим, калгкреш,
ДНКаза, кисла та лужна фосфатази, ренш тощо) е незначним, але вони вццгра-
ють важливу роль у девггалгзадп !жт, захисних реаки тях, регуляцп крозообйу й
очищенш слизово! оболонки рота г зуб1в. Загальний вмгст мтнеральних речовин т
вмтст Ма+1 СГу един! нижчий, а К+г НСО3_ — вищий, юж у кровь Цс пояснюеть-
ся тим, шо продукта слини е двостадшною. У кшцевих вццплах (ацинусах) 1
вставних протоках утворюеться первинна слина, близька за структурою до уль-
трафьтьграту плазми кров!. У внутртшньочасточкових (посмуговаттих) протоках
первинна слина модифткуеться шляхом реабсорбцп №+ т СГ та секрецп К* 1
НСО,-. Секрешю води й електролтттв стимулюють парасимпатичш (М-холшергтч-
ш), а секрецпо оргашчних речовин — симпатичш (ферменттв — переважно (3-ад-
ренерпчш, а гормонтв — а-атренерпчш) впливи.
Пщвищення слиновидтлення — г1персал1ваи,1я — виникае внаслщок запалення
слизово! оболонки ротово! порожнини (стоматит, гшпвгг), при отруеннях фосфор-
органтчними сполуками (пестициди, шсектициди), мухоморами г ртутпо, при
сказт, ящур! та невролопчних порушеннях (хвороба Паркшсона). Збшьшення
швидкосп секрецп супроводжуеться пщвищенням загалъно! концентрацп мтне-
ральних речовин (закон Гейденгайна), концентрацп №+ т СГ, а також зниженням
концентрацп К+т НСО3“у слинт.
Падмтрна секрещя слини може порушувати травления в шлунку,
Зниження секрецп слини — глпосалгващя — може супроводжуватися ксеросто-
м(ею (сухтсть слизово! оболонки ротово! порожнини). Спостерггаеться при запален-
нях слинних залоз (вгрусний паротит, субмаксилтт), муковтсцидозг, гарячщ г зневод-
ненш, прийомт деяких лгкарських засобгв (атроглн. транквьпзатори, антипстамгнш,
антигшертензивш та протиблювотн! засоби). Тяжка ксеростомия при автотмунному
сталоадент (синдром Шегрена) та опромгненнг слинних залоз супроводжуеться
зниженням експресп в них аквапоринтв (АКП-5 г АКП-1). В експеримент! на твари-
нах установлено можливтсть вщновлення слиновидтлення шляхом уведення в оттро-
-тшент залози гена АКП-1 за допомогою аденовтрусного вектора (ВаитеТа!. 2009).
Ксеростомгя утруднюе жування г ковтання, спричинюе запалил процеси в
слизовш оболонш рота, а також розвиток каргесу зубш.
1з слинних залоз видщено гнкрет — паротин, який знижуе ргвень кальцпо е
кров!г сприяе росту та звапненню юсток г зубгв (У. Ио, 0.1. Сукманський). Кргм
паротину слиннг залози продукують шсулш; нейротрофгчш фактори (фактор рос-
ту нервгв г нейролейкгн); фактор росту епщермгсу (урогастрон), що активуе роз-
виток тканин епгтелгального походження ! гальмуе шлункову секрецгю; еритропо-
етин, колонгестимулювальний 1 тимотропний фактори, що впливають на систему
кровг; калгкреш 1 реши, яю регулюють тонус судин г мгкроциркуляцпо. Паротин
та гншг шкрети видгляються не ильки в кров, а й у слину. Тому порушення сли-
новидьчення позначаеться на гнкрецп слинних залоз. 31 зниженням синтезу паро-
тину пов’язують розвиток хондродистрофп плода, деформуючого артриту г спон-
дилпу, пародонтиту 1 хвороби Кашина — Бека. Дефщиту фактора росту нервгв
надають значения в розвитку родинног дизавтономп, хвороби Альцгеймера (пре-
сешльна деменцтя) г хоре! Гентйптона, а його ггперпродукцп — у виникненш
нейрофгброматозу (хвороби Реклгнгхаузена). Зниження продукцп фактора росту
епщермгсу зумовлюе розвиток виразки дванадцятипало! кишки. До проявгв п-
персгалоаденгзму належить симетричне незапальне припухання слинних залоз
при цукровому дгабет!, гшогонадизм! та шших ендокринних розладах. Таку гшер-
трофгю слинних залоз розщнюють як компенсаторну.
Порушення ковтання
Розлади ковтання — дисфагия — порушують надходження гжг ! води з ротово! порож-
нини до шлунка. Вони можуть бути пов’язанг з розладом функцп тргйчастого.
пщ’язикового, блукаючого г язико-глоткового нервгв, порушенням роботи ков-
тальних м’язгв, спастичними скороченнями м’язгв глотки при сказ!, правцг, гс-
терп. Утруднення ковтання виникають при вроджених г набутих дефектах твердо-
го г м’якого пщнебгння, а також у зв’язку з ураженням дужок пщнебшня г мигда-
ликгв (анггна, абсцес). Остаточним (мимовгльним) етапом ковтання е просування
харчових мае через стравохщ завдяки перистальтичним скороченням його м’язгв
Цей процес може порушуватися в раз! спазму або парал!чу м’язово! оболонки
стравоходу, його звуження (ошк, запалення, дивертикул), а також при ахалазп —
обмеженнг чи втратг здатност! до розелаблення. Зниження тонусу нижнього стра-
вохщного сфгнктера спричинюе рефлюкс шлункового вмгсту в стравохщ 1 виник-
нення печп (дулауй).
ПОРУШЕННЯ ТРАВЛЕНИЯ В Ш Л УН КУ
Основними функцгя ми шлунка е секреторна г рухова (моторна), яю забезпечують
хгмгчне та фгзичне оброблення !жг, депонування хгмусу г транспорт його до кишок.
Шлунок виконуе також гнкреторну, усмоктувальну та екскреторну функцп.
Порушення секреторно! функцн шлунка
Порушення шлунково! секрецп виникае внаслщок оргашчних уражень шлунко-
вих залоз (змгна юлькосп, гшертрофгя або атрофгя клгтин), а також при змгн! гх
функцгонально! активностг. Так, середня кглькгсть паргетальних екзокринощгпв
шлункових залоз становить у здорово! людини 0,82 109. Вона збглыпена у хворих
гз виразкою дванадцятипало! кишки х зменшена при виразщ шлунка, раку шлун-
ка 1, особливо, при пернщхознш анемп.
Шлункову секрецпо стимулюють: нейрогенно — блукаючий нерв (ацетил-
холш), гормонально — гастрин ! пстамш. Вироблення гастрину О-клпинами сли-
зово! оболонки шлунка активуеться блукаючим нервом, а також шляхом х!м!чно!
та мехашчно! стимуляцн воротарно! печери (апггит ру1опсит) !жею та продуктами
п перетравлення. Стимулюючу дно справляють мал! пептиди, феншаланш, трип-
тофан, а також кальцш !жх. Р!вень гастрину в сироватц! кров! пщвищуеться при
синдром! Золлшгера — Еллхсона, а також при першцюзшй анемп. Секрецпо пс-
тамшу ЕСЬ-клггинами шлунка активуе гастрин. Пстамш активуе пархетальнх кл!-
тини через цАМФ.
Стимулюють шлункову секрецпо також пощбний до гастрину холецистокшш,
гастринолхберин, соматотропш, кортикотропш, шсулш, глюкокортикощи, парат-
гормон 1 тироксин.
Гальмують секрецпо шлунка соматостатин 1 гастрошпбуючий пептид (глюко-
зозалежний шсулшотропний пептид). Гастрошпбуючий пептид пригшчуе також
моторику шлунка, активуе вившьнення шсулшу, кишкову секрещю. Видшяеться
гастрошпбуючий пептид унаслщок пщвищення ргвня глюкози в кров! та надход-
ження триглщеридхв у тонку кишку. Гальмують шлункову секрецпо також фактор
росту епщермхсу (урогастрон), секретин х под!бн! до нього В1П х глюкагон, каль-
цитонш, а також зниження рН у воротарнш печерь
За нормальних умов шлунок секретуе близько 2 л шлункового соку на добу.
Основн! мшеральш компоненти соку — Н+ та СГ — продукуються пар!етальними
клхтинами. Концентрацхя хлоридхв у соку перевищуе таку в плазм! кров!. Вона, як
1 концентрация Н+, прямо залежить вщ швидкосп секрецц. Тому збщыпення юль-
кост! соку супроводжуеться пщвищенням його кислотность Концентращя натрпо
1стотно нижча, нхж у кров!, х зменшуеться при збшьшенш швидкосп секрецц.
Ферментний спектр шлункового соку представлений головним чином протешаза-
ми, такими як пепсин (оптимальний рН 1,5—2,0), гастриксин (рН 3,0—3,5) 1 хи-
мозин (рН 7,0). Останнш е головним протеолхтичним ферментом у немовлят. Сгк
мхстить також незначну юльюсть шших ферменпв (лшаза, л!зоцим, РНКаза
тощо), як! не вщхграють хстотну роль у травленш.
Порушення видщення шлункового соку може проявлятися як ппер-, так х
гшосекрещею.
Пперсекрецгя шлункового соку харакгеризуеться не ильки збщьшенням кщь-
костх секрету, а й пщвищенням утворення хлориднох кислоти (гшерхлоргщрхя).
Збщыпуеться як базальна, так х максимальна секрецхя шлунка, тобто секрецхя,
стимулъована внутршшьом’язовим уведенням пентагастрину (6 мкг/кг) або пста-
мхну дигщрохлориду (24 мкг/кг; табл. 24).
Пперсекрещя спостерхгаеться при пептиднш виразщ дванадцятипало! киш-
ки, антральному гастрит!, хплороспазм! та пхлоростеноз!, охолодженш, вживаннх
алкоголю, гарячо! хжх та Деяких лхкарських препаратхв (салщилати, бутадхон, ш-
сулш, кортикостерощи.тощо). Вона супроводжуеться пщвищенням перетравлю-
вальнох здатносп соку, що може зумовлювати ушкодження слизово! оболонки 1
виникнення ерозш чи навтть виразок.
Ппосекрещя шлункового соку спостерхгаеться при гострому х хронхчному гас-
трит!, пухлинах шлунка, пернхщознш анемп, гарячщ х зневодненнх оргашзму.
Таблиця 24. Кшыасна характеристика нормальной патолопчно!секрецп шлунка
——— ... - ->• -т-с-л'. т Секрефя —— — - —... : —— ——, . . ... ... .у-г- Показники секрецп
базально! максимально?
Кшьюсть секрету, мл/год НС1 Кшьюсть секрету, мл/год НС1
ммоль/год ммоль/л ммоль/год ммоль/л
Нормальна 60±25 3±2,5 45±15 200±55 20±4 100±15
Гшерсекрецгя (виразка дванад- цятипало! кишки) 100 6 60 250 27,5 ПО
Гшосекрецгя (рак шлунка) 40 0,5 12,5 80 4 50
Вона зазвичай поеднуеться зг зниженням кислотност! шлункового соку (гшохлор-
ПДрГЯ).
Анацидний стан визначаеться тодг, коли шлунковий сгк не мгстить в!льну хло-
ридну кислоту (ахлоргщргя). Коли залози шлунка не видгляють хлоридну кислоту
г ферменти, евщчить про ахшю (вщеутшеть шлункового соку).
Припускають, що порушення секрецп шлунка можуть бути зумовлеш змша-
ми експресп АКП, особливо АКП-3 г АКП-4, якг локалгзуються на базолатераль-
нш мембран! секреторних клгтин.
Гшосекрецгя супроводжуеться зниженням перетравлювально! здатностг шлун-
кового соку. Унаслщок ослабления бактерицидно! дй хлоридно! кислоти можуть
розвиватися процеси бродшня г гниття.
Розлад руховоТ функцп шлунка
Розлад рухово! функцп шлунка проявляеться змгною його перистальтики (ппер-г
ппокгнез) г м’язового тонусу (ппер- г гшотошя; мал. 65), що супроводжуеться
сповыьненням або прискоренням евакуацй харчових мае зг шлунка.
Посилення перистальтики — гтерктез — зазвичай зумовлене збгльшенням
кглькостг гмпульегв, якг надходять до м’язово! оболонки шлунка волокнами блу-
каючого нерва. Подразнення симпатичних нервгв, а також катехоламгни ослаблю-
ють силу г ритм скорочень непосмугованих м’язгв шлунка. Згщно гз сучасними
уявленнями, деякг еферентш волокна ослаблюють моторну актившсть шлунка
також шляхом вившьнення ИО та В1П.
У мехашзм! порушення рухово! функцп шлунка важливу роль вщгграють роз-
лади гуморально! регуляцп. З’ясовано, що моторику шлунка г кишок активуе гор-
мон мотилш — пептид гз 22 амгнокислот. Вгн виробляеться у тонкш кишцг (пере-
важно в дванадцятипалш кишцг). Продукцгя мотилшу посилюеться в разг олуж-
нення вмгсту дванад пяти пало! кишки г гальмуеться при низьких значениях рН.
Ргвень мотилгну пщвищуеться також при подразненнях блукаючого нерва. Вш
збшьшуеться пщ час проносу. Рухову актившсть шлунка пщвищують також серо-
тошн, пстамш, холецистокшш, субстанцгя Р та деяк! ошати.
Посилення перистальтики
шлунка — гтеркшез — може
спостерггатися при виразках
шлунка । дванадцятипало! киш-
ки, а також у раз! нирково!
або печшково! кольки 1 суп-
роводжуватися спастичними
скороченнями м’язових во-
локон 1 появою болю в шлун-
ку. Посилюють перистальтику7
груба Ужа, алкоголь, а також
гривога, гшв.
Посилення перистальтики
шлунка зазвичай поеднуетъся
з пщвищенням КИСЛОТНОСТ!
Мал. 65 Форма шлунка залежно вш його м’язового тонусу
(за Д.1. Сгепановим, 1963):
а — гшертошчпа; б — нормотошчна; в — ппспогпчна; г —
атошчна
шлункового соку. Кисл1 харчов! маси, потрапляючи в дванадпятипалу кишку,
спричинюють тривале закриття воротаря 1 сповЁтьнення евакуацй шлункового
вм1сту (ентерогастральний рефлекс).
Гальмують рухову актившсть шлунка гастрошпбуючий пептид, В1П, сома-
тостатин, секретин, глюкагон, кальцитонш, пептид УТ, нейротензин ! ПГЕ2.
Ослабления перистальтики шлунка — гтокгнез — спостерйаеться при гаст-
рит!, а також пщ час спланхно- 1 гастроптозу, схуднення та ослабления загально-
го тонусу оргашзму. Сповпгьнюють перистальтику шлунка жирна тжа, неприемш
смаков! в!дчуття, страх, печаль В експеримент! гшокшез шлунка вщтворюють у
собак за допомогою ваготомп або видалення воротарно! частини з накладанням
зуоденогастрального або еюногастрального анастомозу (Ю.С. Лондон). Особливо
слщ вщзначити розслаблення нижнього сгравохщного сф!нктера (шлунково-стра-
вохщного з’еднання), яке може виникати в раз! зменшення вироблення гастрину,
пщвищення продукцл секретину ! ошо'щних пептидов. Таке розслаблення зумо-
г.тюе рефлюкс шлункового вмкту в стравохщ ! виникнення печи, яка вщчуваеться
в надчеревшй д!лянщ ! за грудниною.
Тяжким розладом моторики, шо зумовлюе порушення евакуащ! харчових мае
31 шлунка, е пиюроспазм. В!н спостерйаеться при пперхлоргщрп, деяких формах
гастриту 1 гплородуодешту, полшозу шлунка ! супроводжуеться компенсатормим
посиленням перистальтики та ппертрофкю м’язово! оболонки шлунка. До гплоро-
спазму може приеднатися г!пертроф!чний шлоростеноз (у д!тей вхком до 4 м!с.).
Порушення рухово! функш! шлунка може проявлятися блюванням 1 вщрижкою.
Блювання (уотНиз) — складний рефлекторний акт, наслщком якого е вивер-
ження вм!сту шлунка (1 кишок) назови! через рот Блюванню зазвичай передуе
нудота (паизеа), салгващя, прискорене дихання 1 серцебиття. Блювання часто мае
захисне значения — зв!льнення шлунка вщ недоброяюсно! 1ж!, токсичних рсчо-
вин. Однак триватс невтримне блювання зумовлюе втрату оргашзмом води та
.он!в хлору, унаслщок чого розвиваеться зневоднення й алкалоз, а в хрошчних
випадках — виснаження оргашзму.
Вгдрижка (егисгасю) зумовлюеться виходом з! шлунка повггря або газ!в (СО2,
арководень, метан тощо), що утворилися внаслщок бродильних або гнильних
процес!в.
Пептична виразка
Пептична виразка (англ. — рерНси1сег) шлунка ! дванадцятипало! кишки е хрошч-
ним захворюванням, що характеризуеться виникненням виразкових дефекпв у
слизовш оболонц! шлунка чи дванадцятипало! кишки, тобто там, де може дгяти
шлунковий спс, що мгстить хлоридну кислоту 1 пепсин. Поширенгсть хвороби в
деяких крашах сягае 10—15 % дорослого населения. Прийнято розргзняти пер-
винну виразку 1 вторинн! (симптоматичн!) виразки, хоча такий подш е дещо
умовним. Вторинн! виразки можуть бути наслщком тривалого вживания деяких
препаратов (НПЗЗ, глюкокортикощи, резертн тощо), ендокринних розладгв (пух-
лина з О-клгтин, що продукуе гастрин, — синдром Золлгнгера — Елл!сона; гипер-
паратиреоз, гшеркортицизм, гшершсулгнгзм), тяжких огшав, обструктивно! леге-
нево! недостатносп та гн.
В етюлоп! пептично! виразки видшяють роль таких факторгв:
• аламентарний — нерегулярне харчування, погане пережовування !ж!, спожи-
вання надмгрно гарячог, гостро!, солоно! !ж!, уживання алкоголю тощо;
• психосощальний — хрошчна нервово-психгчна травма, тривал! невирппу-
ваш конфликта, емоцшне г психгчне перенапруження, стресов! стани;
• шфекцгйний — заражения грамнегативною бактергею НеПсоЬас1ег ру1оп\
певну еполопчну роль надають також в!русу герпесу (Негрез з1тр1ех у1гиз), цито-
мегалов!русу, Н. /геПтаппи та грибам роду СагиМсг,
• медикаментозний — тривале вживания салщилапв та шших НПЗЗ, глюко-
кортикоццв, б1фосфонат!в та шших ульцерогенних препаратов;
• куршня;
• генетичний — полотенца схильшсть до розвитку виразок, особливо дуоде-
нальних, зокрема в ос!б гз 1(0) групою кров!, а також автосомно-домшантна ггпер-
пепсиногенемоя о спадкове пщвищення продукцй юнгв водню паргетальними кл!-
тинами.
Еполопчна роль алгментарного фактора пщтверджуеться численними клшгч-
ними спостереженнями. Ульцерогенну роль грубого неадекватного корму було
доведено в експеримент! Л. Ашоффом.
Роль психосощального фактора, нервово-емоцшного перенапруження о стре-
сових впливгв уперше було встановлено в роки Друго! свггово! войни пщ час гн-
тенсивних бомбардувань Лондона шмецькою авгащею. Вона пщтверджуеться
одержаниям виразок в експеримент! за допомогою стресових впливгв (знерухом-
лення, больова травма тощо). Уявлення про провщну роль психосощального чин-
ника лягло в основу кортиковгсцерально! теори етголоп! та патогенезу виразково!
хвороби (К.М. Биков, 1.Т. Курцин, 1949).
Вщкритгя у 1983 р. спгралеподгбно! бактерп, вщомо! сьогодн! як НеНсоЬас1ег
ру1оп, гстотно змгнило уявлення про еполопю ! патогенез пептично! виразки. Не-
дарма за вщкритгя Н. ру1оп та п рол! у розвитку гастриту 1 пептично! виразки
австралшським ученим Б. Маршаллу 1 Р. Уоррену у 2005 р. було присуджено Но-
белгвську премию.
Ця бактергя синтезуе велику юлькгсть уреази, яка розщеплюе сечовину на
амгак 1 вуглекислий газ. Хмарка амгаку захищае бактерио вщ хлоридно! кислоти
шлункового соку. Бактергя також виробляе протешази, що руйнують глисопроте!-
ди слизу, дезгнтегрують захисний слизовий бар’ер шлунка та спричинюють розви-
ток хрошчного запалення 1 виразок. Кргм того, лшополюахарид оболонки Н. ру1оп
взаемодте з ламппном базально!’ мембрани ештелЁю шлунка, шо призводить до
оуйнування його зв’язку з Ёнтегрином 1 порушення цёлост! ештелпо. Ушкодження
ештелно спричинюе мЁсцеву гмунну реакцш, секрецпо прозапальних питокппв.
мобйпзацш нейтрофЁлЁв Ё розвиток запалення. При ньому виникають ушкоджен-
ня ендотелш мЁкросудин, розлдти мЁкроциркуляцц 1 трофЁки слизово! оболонки,
що зумовлюе утворення виразок. У випадку виразки шлунка запалення мае ди-
фузний характер 1, як правило, спричинюе виникнення атрофп ! зменшення к!ль-
костё парЁетальних клётин, а отже, зниження продукцп хлоридноТ кислоти. При
цьому три вал а гелЁкобактерна 1нфекц1я сприяе також розвитку раку шлунка
Заселению Н. ру1оп у дванадцятипалш кишцц розвитку запалення слизово!
оболонки зазвичай передуе “шлункова метаплазия” егптелпо кишки, яка розви-
ваеться пщ впливом високо! кислотност! шлункового вм!сту, що надходить у не!.
Пщвищення кислотност! зумовлюе розвиток антрального гастриту, галъмування
продукт! соматостатину (який гальмуе секрешю хлоридно! кислоти) 1 пщвищен-
ня продукцп юн1в водню парЁетальними юптинами.
У патогенез! пептично! виразки вирппальну роль вщгграе порушення ршноваги
и!ж ушкоджувальними (агресивними) та захисними факторами. До факторЁв аг-
респ належать кислий шлунковий сёк (пептичний фактор), мехашчне, терм!чне,
х!м!чне та бактерЁалъне ушкодження слизово! оболонки шлунка !жею, л!карськими
засобами тощо. Захисну роль вшйрають сл изовий бар’ер, який перешкоджае зво
ротнЁй дифузй юн!в водню та гхнгй ушкоджувальшй дп, адекватне кровопостачан-
ня Ё високий регенеративний потентат слизово! оболонки. Таку саму роль вщЁгра-
ють нейгралЁзацзя кислого секрету гщрокарбонатами, якг разом з! слизом виробля-
ються юптинами поверхневого ештелно, ростов! й трофгчш фактори, що сприяють
регенерацп, простагландини та ёншё гормони (див. нижче). Гщрокарбонати вироб-
ляються ё парЁетальними клЁтинами, як! секретують СГё Н+з ашкально! поверхш
у шлунок, а НСО3 ! К+ — з базально! поверхш в пщслизовий прошарок.
Роль пептичного фактора гпдтверджуеться пщвищенням кислотност! при ви-
разках дванадцятипало! кишки, добре вщомим правилом Шварца “без кислоги
немае виразки”, численними екепериментами, а також усшшним: застосуванням
у л!куванн! виразок, особливо дванадцятипато! кишки, операщй (ваготомЁя, вида-
лення воротарного вшдгпу шлунка), лЁкарських засобш (блокатори протоново! пом-
пи, Н2-антагошсти, антихол шерпчн! пре-
парата), яю знижують кислота ют ь шлун-
кового соку. В той же час виразка шлунка
частгше локал!зуеться в тих вгддгпах (мала
кривизна, прешлорична д!лянка), що м!с-
тять пор!вняно мало парЁетальних клггин,
якё продукують хлоридну кислоту (мал 66,
67) 1 одночасно гщрокарбонат. Таким чи-
ном, щ вёддёли шлунка менше захищен! вщ
дп кислого шлункового соку. Як правило,
виразка дванадпятипало! кишки виникае
на тлё гЁперацидного гастриту, а пептична
виразка шлунка — гшоацидного гастриту
або нормально! секрецп. у — ЮО; 2 — 75; 3 - 50; 4-0—1
Мал. 66. Ильгасть парктальних екзокри-
ноиит1в (%) у слизовш обо.тонщ шлунка
Мал. 67. Частота локалЬ
зацп виразки
Важливу захисну роль вццграе слиз, що вкривае сли-
зову оболонку шлунка. В експеримент! на тваринах ут-
ворення виразок можна спричинити, вводячи у шлунок
салщилати чи жовчш кислоти, як! порушують слизовий
бар’ер. У фглогенез! в людини сформувався слабкий сли-
зовий бар’ер шлунка, властивий траво'щним ссавцям.
Проте за високим кислотовцщленням (шдукованим м'яс-
ною !жею) людина близька до м’ясощних.
Вирппальну роль у збереженн! високо! резистент-
ност! слизово! оболонки шлунка вщгграе адекватне не-
рвово-троф!чне забезпечення, яке зумовлюе швидке (за
4—6 дн1в) оновлення поверхневих екзокриноципв шлун-
ка. Важливе значения нервово-троф!чного фактора п!д-
тверджене кл!шчними (М.Н. Бурденко 1 Н.Б. Могильни-
цький) та експериментальними (О.Д. Сперанський) до-
сл!дженнями розвитку виразок шлунка 1 кишок у раз!
ушкодження гшоталамуса, у якому мхстяться вищ! цент-
ри автономно! (вегетативно!) нервово! системи.
Згщно з кортиковюцеральною теор!ею, пусковим фактором у розвитку пеп-
тично! виразки е порушення дхяльносп вищих нервових центров унаслщок трива-
лих несприятливих екстеро- та штероцептивних впливхв, шо своею чертою спри-
чинюе порушення взаемозв’язюв м!ж корою великого мозку ! пцпарковими вуз-
лами. Як наслщок, вщбуваеться дезштеграц!я автономно! нервово! системи,
пщвищуеться секрец!я шлункового соку, настае тривалий спазм судин 1 м’язово!
оболонки шлунка та розвиваються дистроф!чн! процеси в слизовш оболони!.
Важливого значения в патогенез! пептично! виразки надають гуморальним
порушенням: збйгыиенню концентраци пстамшу в кров! та слизовш оболонщ
шлунка, а також зниженню активност! пстамшази. За концепщею Б.П. Бабкша,
надм!рне вившьнення пстамшу пщ час шлунково! секрецй збитыпуе продукщю
хлоридно! кислоти пар!етальними клхтинами, пщвищуе проникшсть кап!ляр!в 1
порушуе мгкроциркулящю, що спричинюе виникнення ерозш та виразок слизо-
во! оболонки. Правильшсть ще! концепцй пщтверджуеться експериментально, а
також успшшим застосуванням блокатор!в Н2-рецептор!в у лгкуванш виразки.
Доведено, що тривале застосування з лгкувальною метою глюкокортикоццв
або кортикотрошну спричинюе загострення виразок або виникнення нових у
шлунку або дванадцятипалш кишщ. У дослщах на собаках показано, що щ гормо-
ни пщвищують секрецпо ! кислотшсть шлункового соку та знижують вм!ст слизу,
який вщхграе захисну роль. Вони також гальмують синтез б!лк!в 1 регенерацпо
кл!тин. Добре вщомо про збшьшення продукцй цих гормошв теля стресових
ВПЛИВ1В, що вццграють роль еполопчного фактора в розвитку пептично! виразки.
З’ясовано, що ПГЕ, та ПГЕ2, синтез яких гальмують глюкокортикоши, при-
гшчують шлункову секрецпо 1 запобпають розвитку експериментальних виразок
шлунка у щур!в. Д1я простагландишв грунтуеться на приппченш активност! аде-
н!латциклази й утворення цАМФ у слизовш оболонщ шлунка. Гальмують ульце-
рогенез також естрогени, секретин 1 епщермальний фактор росту. Протективний
ефект ЕФР пов’язаний 13 трофхчною (що стимулюе пролгферащю) д!ею, гальму-
ванням секрецй хлоридно! кислоти ! стимулящею синтезу глжозамшопикаюв
слизу. У хворих з активним виразковим процесом концентращя ЕФР в един! зни-
жена.
Отже, у патогенез] пептично! виразки важливу роль вщпрають нейрогенш (а
надто трофгчш) та гуморальш порушення Прояв цих порушень у вигляд! виразки
шлунка або дванадцятипало! кишки зумовлений мзсцевими несприятливими
гтливами та спадково-конститушональними особямвостями.
ПОРУШЕННЯ КИШКОВОГО ТРАВЛЕНИЯ
Кишки виконують сскреторну. рухову, усмоктувальну, шкреторну та екскреторну
функцц. У кишках здшснюеться порожнинне (дистантне) ! мембранне (пристпз-
кове) травления. Останшм часом видыяють також етап, пром!жний м!ж порож-
ни нним 1 мембранним травлениям — пдрол!з харчових речовин у шар] слизових
щкладень. Порожнинне травления вшбуваеться в просвт кишок 1 полягае в
инуванш надмоле кулярних систем 1 великих молекул, мембранне — на мемб-
ран! стовпчастих еп!тел!оцит!в кишкових ворсинок. Тут здшснюються завершаль-
ш етапи гщрол!зу харчових речовин 1 перехщ до всмоктування. Порушення по-
рожнинного травления залежить передус!м вщ розладу видыення панкреатичного
соку та жовч!. У патологи мембранного травления вир!шальну роль вы (грае пору-
шення продукц!! ферменпв стовпчастимн ештелюцитами.
Вщповщно до сучасних уявлень кишков! крипти (залози Л(беркюна) беруть
участь у видыенш рщкого компонента кишкового соку 1 не секретують ферменти.
Ферменти виробляються стовпчастими ештелюцитами кишкових ворсинок. Вшц-
тяючись на поверхню клггин м!ж мпсроворсинками, ферменти беруть участь у
мембранному травленш (О.М. Уголев). Проникнення !х у кишковий спс вшбу-
ваеться в основному внаслщок вщторгнення 1 розпаду стовпчастих клгтин (за нор-
мальних умов цикл !х оновлення становить 3 дн!). Отже, слизову оболонку кишок
у щлому можна розглядати як единий залозистий апарат.
Найважлившгим вщдшом тонко! кишки е дванадцятипала кишка, у яку над-
ходить секрет дуоденал ьних залоз. жовч, спс пщшлунково! залози.
У дослщах Ю.С. Лондона з повним або частковим видаленням дванадцягипало!
кишки у тварин уперше встановлено и житгево важливе значения для оргашзму.
З’ясовано, що екстирпащя дванадцятипало! кишки в кот!в г собак спричинюе гли-
бок! порушення обмшу речовин, функцш нервово! та ендокринно! систем, зумов-
леш переважно випаданням шкреторно! функцй кишки (О.М. Уголев). У дванадця-
типалш кишц! виробляються секретин, холецистоюнш, мотилш, гасгрошпбуючий
нтестинальний пептид 1 В1П та ппш шкрети, що регулююгь д!яльн!сть системи
травления.
ПОРУШЕННЯ ТРАВЛЕНИЯ, ПОВ’ЯЗАНЕ 3 РОЗЛАДОМ
ВИД1ЛЕННЯ ЖОВЧ11 ПАНКРЕАТИЧНОГО СОКУ
Вщсутшсть жовч! (ахол/я) або знижене утворення п (гтахамя) з недостатшм над-
ходженням у дванадцятипалу кишку е наслщком порушення функцш утворення
та видыення жовч! 1 супроводжуеться розладом перетравлюванвя та всмоктуван-
ня жирш, сповгльненням перистальтики кишок г посиленням у них пропеспз
гниття 1 бродгння (див. роздш XXIX "Паголопчна фгзюлопя печшки”).
Тяжк! порушення травления спричинюються змшою панкреатично! секрецп,
осюльки гццшлункова залоза виробляе вег основю ферменти травления.
За добу пшшлункова залоза секретуе близько 1,5 л соку, що мае лужну реак-
цш (рН 7,5—8.8). Г! забезпечуе високий вмгст гщрокарбонату (до 150 ммоль/л),
концентращя якого прямо иропорцшна швилкостг секрецп. Концентрация натрпо
сягае 140 ммоль/л, а вмкт хлоридов е поргвняно низьким. Основну масу бглк!в
панкреатичного соку (понад 70 %) становлять протеолгтичн! ферменти: трипсин,
хгмотрипсин, еластаза, карбоксипептидаза (А 1 В) та калгкреш. Усг цт ферменти, а
також фосфолшаза А виробляються в неактивному стан! (у вигляд! зимогенгв).
Решта ферменпв — лшаза. а-ам!лаза. РНКаза га ДНКаза — секретуеться в акгив-
нш форм). У забезпеченш секрецп певну роль вщпрають АКП-1, АКП-5 г АКП-8.
Так, блокатор аквапоришв хлорид ртуп майже повшстю гатьмуе секрецпо рцшни
в мтжчасточкових протоках залози, де експресуеться АКП-1
Порушення секрецп панкреатичного соку спостерггаеться в раз! закупорювання
або здавлення протоки пщшлунково! залози. при муковгсцидозг (кгстозиий фгброз
пщшлунково! залози), гострому та хроючному панкреатит! чи дуоденит, порушенн!
нервово-гуморальних мехашзмгв регуляцп панкреатично! секрецп. Секрегорним
нервом пщшлунковог залози е блукаючий нерв, гуморальна регуляпгя здшснюеться
за допомогою секретину, який активуе видшення води г гщрокарбонатгв, холецис-
токгшну (панкреозимгну), що стимулюе продукцию ферменттв, панкреатичного
пол! пептиду г соматостатину, як! гальмуготь п. Посилюють панкреатичну секрецпо
також гастрин, гастринолгберин, гнсулгн, В1П г речовина Р, а глюкагон, кальпи-
тонш, гастрошпбуючий штестиназьний та ошощн! пептили гальмують п.
У разг дефшиту панкреатичного соку значна частина жиртв не перетравлюсть-
ся 1 виводиться з калом (стеаторея) Порушення перетравл ювання бшюв виникае
внаслщок недостатнього вироблення пептидаз пщшлунковою залозою, а також у
разг порушення 1х активацп. Так. трипсиноген активуеться ентерокгназою кишко-
вого соку й автокататгтично, решта протеолгтичних ферменпв г фосфолшаза А
активуються трипсином. У разг зниження панкреатично! секрецп порушуеться
гщролгз нуклегнових кислот !жг 1 меншою мгрою — розшеплення крохмалю
Панкреатит
Запалення пщшлунково! залози нерщко мае гострий перебег г може супроводжу-
ватися розвитком панкреатичного шоку, небезпечного для життя
В етголоп! панкреатиту гстотного значения надають: зловживанню алкоголем
г перещанню. що його супроводжуе; надмгрному споживанню жирно! !жт; жовч-
ним каменям г полшам протоки пгдшлунково! залози; мехаючному ушкодженню
пщшлунковог залози 1 м’яза — замикача печшково-пщшлунковог ампули пщ час
травм г хгрурпчних втручань; шфекшйному фактору (вирусам паротиту, гепатиту,
Коксакт, бактергальнш шфекцп), штоксикацп, у тому чиелг деякими лгкарськими
засобами (гмуяодепресанти, пазиди, кортикостерощи тощо).
У патогенез! панкреатиту важливу роль вщгграють: посилення пщ впливом
етюлопчних факторгв секрецп панкреатичного соку; порушення вщтоку секрету;
пщвищення тиску в протоцг пщшлунково! залози; потрапляння в не! жовч! та
Е-полопчш фактори
-> Затримання евакуацп секрету
Ушкодження юптин
Вившьнення велико!'
юлькосп трипсину
Рефлюкс жовчт
або дуоденального вмхсту
Проеластази
Активашя
Профосфолшази А
Активац1я
т
Еластаза
Кал1кре!ногену
Фосфолшаза А
Калткреш, брадиюшн
Лхзолецитин
Лшази
Ушкодження судин,
крововилив
Набряк, бхль,
Пстамш у
/ Некроз
паренх1ми органа
Некроз
жирово'! тканини
втрата плазми, запалення
Схема 25. Патогенез гострого панкреатиту (за К.П. Веремеенком, 1977)
дуоденального х!мусу (шо мхстить ентероюназу); порушення мгкроциркуляцп,
трофхки 1 бар’ерних властивостей екзокринних панкреоципв. Основною ланкою
патогенезу панкреатиту е передчасна активацхя ферменпв (трипсину, калхкрешу,
еластази, фосфолшази А) безпосередньо в протоках х клпинах залози, яка вщбу-
ваеться пщ дхею ентерокхнази, жовчх або автокаталхтично, при ушкодженш панк-
реоцитхв (схема 25).
Наслщком цього е автолхз тканини залози, некроз окремих ц дхлянок, утво-
рення токсичних речовин (лхзолецитину) х БАР, у тому числх кппнхв, якх справля-
ють сильну ппотензивну дцо. Вихщ пептидаз та шших панкреатичних ферментхв
у кров зумовлюе тяжкх розлади гемодинамхки, дихання, шших життево важливих
функцш (панкреатичний шок). Велике значения в патогенез! цих порушень мае
змша рхвноваги мхж протеолхтичними ферментами та хх шгхбпорами. Останн! ви-
робляються самою пщшлунковою залозою та шшими органами (слинш залози,
легеш) х з успехом застосовуються для лпсування панкреатиту.
Певну роль у патогенез! панкреатиту, особливо хронхчного, вщхграють пору-
шення кровообхгу в пщшлунковхй залозх (при атеросклероз!, гшертензп), а також
хмунн! (автоалерпйн!) фактори, про що свщчить виявлення протипанкреатичних
антиты у кров! деяких осхб, хворих на панкреатит.
Порушення мембранного травления
Мембранне (пристхнкове) травления здшснюеться ферментами, фхксованими на
поверхш посмугованох облямгвки, утворено'х мхкроворсинками стовпчастих еш-
телхоципв кишкових ворсинок. Харакгеризуеться поеднанням процесхв фермен-
тацп та всмоктування харчових речовин, високою шввдкхстю гщролхзу х стериль-
шстю, зумовленою невеликими розмхрами пор мхж мхкроворсинками (10—20 нм),
у як! не можуть проникнута мгкрооргашзми. Ферменти мембранного травления
(ологосахаридази, ологопептидази, фосфатази тощо) синтезуються всередин!
стовпчастих ештелооцитов о транслокуються на поверхню клггинно! оболонки, а
також частково адсорбуються з хомусу (панкреатична амшаза, лшаза тощо). Трав-
ления в шар! слизових вщкладень (глгкокалокс) характеризуеться частковою фок-
сацоею й ороентованостю ферменпв та субстратов о за ефектившстю займае про-
можну позицпо мож порожнинним 1 мембранним травлениям.
Розлади мембранного травления о травления в шар! слизових вщкладень
спричинюють так! фактори; порушення структури ворсинок та ультраструктури
поверхш стовпчастих ештелюципв; змша ферментативного шару кишково! по-
верхно та сорбцшних властивостей клггинно! оболонки, розлади перистальтики,
за яких порушуеться перенесения субстратов оз порожнини кишки на п поверхню.
Так, зменшення поверхно травления за рахунок атрофп та скорочення юлькосп
ворсинок або мокроворсинок виявлено при холер!, спру, ыеоеюнт, шсля онтен-
сивно! антибоотикотерапп (неомщин), гастроеюностоми о резекцн шлунка. При-
кладом порушення ферментативного шару кишково! поверхно може бути непере-
носимость молока у випадку дефщиту лактази ф-галакгозидази, КФ 3.2.1.22) або
непереносимость сахарози при дефщито сахарази (р-фруктозидази).
Для виявлення порушень з боку ферментов мембранного травления в клошщ
використовують методику аспорацойно! бюпсп слизово! оболонки тонко! кишки з
подалыпим пстохомочним (пстоензимолопчним) дослщженням.
Порушення синтезу ферментов стовпчастими епотелюцитами може позначи-
тися також на порожнинному травленю. Особливо це стосуеться ентерокшази,
що активуе панкреатичний трипсиноген.
Порушення всмоктувально'Гта вид1льноГ функщй кишок
Усмоктування харчових речовин, пдролозованих, як правило, до стадп мономеров,
здшснюеться головним чином у тонкой кишцо. У процесо мембранного травления
гщролоз харчових речовин о перенесения ох крозь мембрану клотин псно пов’язано.
Тому порушення всмоктування спричинюють усо фактори, яю зумовлюють розла-
ди мембранного травления.
У кишках виявлено юлька форм аквапоришв (водних каналов): АКП-1, АКП-3,
АКП-4, АКП-6, АКП-8, АКП-9 о АКП-10. Вони беруть участь у всмоктуванно о
секрецп води, а також у забезпеченно бар’ерних властивостей ештелою кишок. Зок-
рема порушення всмоктування води в кишках виявлено в трансгенних мишей з
нокаутом генов АКП-3 о АКП-4. Змши активносто та локалозацп аквапоринов
спостерогають при деяких формах онфекцойного проносу, а у хворих оз синдромом
подразнено! кишки з переважанням доаре! виявлено зниження експресп АКП-8 в
ештелп товсто! кишки.
Синдром малъабсорбци (порушеного або неадекватного всмоктування пожив-
них речовин у кишках) характеризуеться доареею, зменшенням маси тша, бшко-
вою недостатносто та ознаками гшовотамшозов. Вш може бути первинним (спад-
ковим) або вторинним (набутим).
Спадковий синдром мальабсорбцн е одним оз найчастнпих автосомно-рецесив-
них порушень (1 : 3000) о здеболыпого мае характер селективного дефщиту фермен-
тов або порушення мембранного транспорту нутроенпв. Унаслщок цього порушуеть-
ся всмоктування одное! або юлькох близьких за структурою харчових речовин. До
ша групи належать: непереносимгсть моносахаридов — глюкози, фруктози, галак-
:эзи (вщсутшсть переносниюв): дисахарид!в — лактози, сахарози, озомальтози (де-
фщит дисахаридаз); целоакш, або глутенова хвороба (дефщит пептидаз); порушення
тгоринно-активного транспорту амшокислот (цистинуроя, триптофанматьабсорб-
шя, мепоншмальабсорбщя) 1 вгтамппв (шанокобаламшу, фолювоТ кислоти).
Набутий синдром мальабсорбцп спостерйаеться шсля гастрекгомп, при хво-
робах кишок (ентероколп, хвороба Крона, або репонарний ентерит, тощо), тд-
шлунковоо затози (панкреатит, муковисцидоз), печшки, а також теля тривало!
роменевоТ та медикаментозно) терагш
Розлад усмоктування в тонкш кишщ може спостерпатись у раз1 ослабления
"эрожнинного травления, яке готуе харчов! речовини до за вершат ьного гщролтзу
ча мембран! клпин, а також пщ час порушення рухово! функцц кишок 1 рухомосп
: орсинок, при розладах крово- та л!мфообпу. Розлади кровообпу порушують ви-
: едення речовин. як! всмокгуються, !хш концентранйпп градтенти та енергетичне
забезпечення активного транспорту. Ослабления активного транспорту поживних
речовин зумовлене також впливом токсинов, як! блокують актившсть ферменпв.
в раз! порушення водно-електролпно! р!вноваги. Особлива роль належить ак-
тивному транспорту натр!ю за допомогою енергп АТФ у вторинно-активному
транспорт! глюкози, амшокислот га шших сполук.
Ба)ато з наведених механ!зм!в беруть участь у порушенн! всмоктування в раз!
запалення тонко! кишки (ентерит), кишково! непрохщносп, гшов!там!ноз!в тошо.
Шцвищення проникносп судин кишково! стшки внаслщок н запалення та гшер-
еш! може супроводжуватися всмоктуванням речовин антигенно! природи ! сен-
сибпизащею оргашзму
3! всмоктувальною функшею кишок псно пов’язана !х екскреторна (видшьна)
функшя. Через кишки виводяться кшцев! продукта обмшу гемоглобшу та холесте-
рину, сол! металгв, молочна кислота, пурини, деяк! гормони, феноли, салшилати,
д.тьфанйтамщи, барвинки тощо. У раз! недостатност! нирок компенсаторно поси-
юеться видщення продуктов азотистого обм!ну (сечовина, сечова кислота тощо).
Розлад рухово! функцн кишок
Розлад рухово! функцп кишок може проявлятася посиленням або ослаблениям
перистальтичних 1 локальних (сегментуючих ! маятникопод!бних) рух!в. Перис -
тальтичн! скорочення забезпечують просування (транзит) вм!сту по кишках. При
цьому транзит по говспй кишц! (у середньому 36 год) займае бшьше 90 % часу
перебування !ж! в кишках.
Пщвищення рухово! функцн кишок виникае в раз! запальних процеств (енте-
рит, кол!т), розлад 1в нервово! та гуморально! регуляцн, пщ впливом мехашчних
або х!м!чних подразнень недостатньо перетравленою !жею, унаслщок дп бак-
тертальних гоксишв. Скорочення м’язово! оболонки кишок посилюе блукаючий
нерв, а симпатичн! (черевн!) нерви гальмують и. Серотонш, речовина Р, гастрин,
мотилш активують перистальтику кишок, а ВШ, КО, нейропептид У, глюкагон 1
секретин — гальмують.
Прикладом порушення нервово! та гуморально! регуляцн рухово! функш! ки-
шок е синдром подразнено! кишки, який спостерйаеться майже у 30 % населения !
харакгеризуеться болем, метеоризмом 1 проносом, шо часто змшюеться запором.
У патогенез! цього синдрому провщну роль вщгграе пщвищення психтчно! та
нервово-вегетативно! лабшьност!, а також негативш емоцй. В!н може бути наслщ-
ком перенесено! рашше дизентерп або виникати пщ впливом алтментарних чи фт-
зичних (перевантаження, охолодження) фактортв. Певне значения в його розвитку
може мати алергшний компонент. У слизовш оболонщ порожньо! кишки, отри-
мангй за допомогою асшращйно! бюпсп, виявляють пщвищений вмгст мотилгну.
Посилення перистальтики зазвичай спричинюе прискорене пересування хар-
чових мае у кишках, попршуе перетравлювання ! всмоктування, призводить до
розвитку проносу (сНаггИоеа). Сприяючи звшьненню оргашзму вщ токсичних ре-
човин (у раз! харчових отруень) або надлишку неперетравлено! гжг, дтарея може
вщггравати захисну роль. Проте тривалий пронос, особливо в дитячому вщ!, спри-
чинюе зневоднення оргашзму г втрату електролтв (№+, К+), розвиток гшово-
лемн, а в тяжких випадках — кардиоваскулярного колапсу.
За патогенезом д!арею подшяють на осмотичну г секреторну. Осмотична д!арея
виникае в раз! потрапляння у травний тракт речовин, не здатних перетравлюватися
та / або всмокгуватися. Окртм того, порушення перетравлювання нормальних компо-
нента !ж] також призводить до накопичення в кишках осмотично акгивних речовин,
перемгщення в них води г до появи проносу. Голодування усувае осмотичну щарею.
Секреторна д!арея е наслщком активацп секрецп СГ у криптах т гальмування
абсорбцп №+ у ворсинках у раз! накопичення в цитоплазм! ентероципв цАМФ
або цГМФ, що може бути наслщком ди бактер!алъних ентеротоксишв (холерного
тощо), гормошв системи травления (В1П тощо) та деяких рослинних г морських
продукта, лгкарських препарата тощо.
Непрохщжсть кишок
Непрох1дн1сть кишок (Иеиз, вщ грец. еИео — повертаю, зачиняю) — синдром, що
характеризуеться припиненням транзиту кишкового вмгсту, зневодненням, што-
ксикащею, розладами кровооб!гу та мгкроциркуляцп. За етюлопею п подшяють
на вроджену г набуту. Непрохщшсть можуть спричинювати вроджеш аномалп,
гельмштоз, вона може розвиватися як шеляоперацгйне ускладнення, а також у
раз! недощання та споживання недоброякгено! !ж!. За патогенезом непрохщшсть
подшяють на мехамчну г динашчну. Мехашчна може бути обтурацшною — унаслГ
док закупорки кишки каловими масами, клубком гельмшта чи сторонюми тша-
ми, компресшною у раз! здавлювання кишки ззовн! гематомою, кгетою яечника,
пухлиною ! странгуляцшною у раз! перетяжки спайками, завороту, ущемления в
грижових воротах. Динам!чну непрохщшсть подшяють на спастичну1 паралбпичну
(унаслщок спазму або парагачу м’язово! оболонки кишок).
Початков! ланки патогенезу непрохщност! багато в чому визначаються причи-
нами, що зумовили п. Так, паралтичну непрох1дн1стъ (тсляоперацгйна або пщ час
перитониту) часто спричинюе активащя а- та р-адренерпчних рецепторгв, як!
гальмують скорочення м’язово! оболонки кишок. Спастична непрохгдшетъ при
карциноид (серотошнпродукуюча пухлина з аргентафшоцита тонко! кишки)
може бути пов’язана з пщвищеного актившетю м’язово! оболонки кишок пщ
впливом надлишку серотошну.
Подальший розвиток порушень мае багато стильного пщ час непрохщност!
ргзно! етюлогп.
Слщ ВИД1ЛИТИ три основш групи порушень: 1) зневоднення, порушення гщро-
електролггного балансу та КОС; 2) штоксикацгя та посилена продукция БАР;
3) посилена больова !мпульсап!я, розлад нервово! та нейрогуморально! регуляцп.
Вирпиальну роль ввдграе розлад обмшу води й електромпйв, зумовлений пору-
шенням секрецп (зазвичай пщвищенням) 1 зворотнсго всмоктування сок!в трав-
ления. Виникають блювання, зневоднення (за добу може втрачатися до 5—7 л
травних секрепв), втрата юн!в натр!ю, катпо т хлорщцв. Розвиваються гшовс-
тем!я, гшотензхя ! гемоконпентращя, унаслщок чого порушуеться кровообгг 1 ви-
никае клппчна картина, яка нагадуе шок. Втрата юшв калпо спричинюе атонйо
кишок.
Нелрохщшсть кишок призводить також до порушення КОС. Виведення пере-
лажно натрию гщрокарбонату7 (панкреатичний 1 кишковий соки) призводить при
низыой непрохщносп до розвитку негазового ацидозу. Якщо переважае виведен-
ня хлоридно! кислоти при висоюй непрохщносп. виникае негазовий алкалоз.
Значну роль у патогенез! непрохщносп кишок вщпрае 1нтоксик,ац1я, шо ви-
никае внаслщок порушення перетравдювання гжг, процеств бродгння ! гниття,
утворення токсичних речовин 1 всмоктування !х у кров. Велике значения при цьому
чае утворення пщвищено! кшькост! БАР, особливо кшш!в, що зумовлене перед-
часною активашею панкреатичних ферменпв (потрапляння вмосту кишок !з пе-
реповнено! кишки в протоку пщшлунково! залози).
У розвитку описаних вище змш важливе м!сце посщають розлади нейрогумо-
ралъно! регуляцп, що виникають рефлекторно пщ впливом 1мпульс!в 13 репептор!в
уражено! кишки (розтяг кишки, быв тощо). Вони особливо значш в раз! странгу-
ляшйно! непрохщносп (заворот, грижа), що супроводжуеться перетисканням
бриж! та порушенням кровопостачання уражено! д!лянки кишки.
Кишкова авт оштоксикафя
У кишках людини, особливо в товспй 1 нижнш частин! клубово! кишки, р!зно-
маштна мткрофлора представлена в основному облйатно анаеробними безспоро-
впми паличками Васгегоюдез 1 В1]й1оЬас{епит. Факультативно анаеробн! кишкова
паличка, молочнокисл! бактерп, стрептококи тощо становлять близько 10 %.
Нормальна мпсрофлора кишок в!д!грае певну зависну роль, гальмуючи розвиток
патогенних м!крооргашзм!в ! сприяючи виробленню природного !муштету, що
довели дослши на гнотобюнтах (безмшробних тваринах).
Однак вмгст кишок може справа яти токсичну дпо, зумовлену наявшстю про-
дукт гниття (!ндол, скатол, протешогенн! ам!ни тошо). У норм! ш продукти утво-
рюються в невеликих кьтькостях 1 не чинять помгтну токсичну дпо на оргашзм
завдяки бар’ершй функцп кишково! стшки 1 печшки. Посилення процеств гниття
внаслщок запалення товсто! кишки (кол!т), закрепу, непрох!дност! кишок 1 дис-
бактерюзу (паголопчна змша складу г розподдлу мгкрофлори в кишках) супровод-
жуеться порушенням бар'ерних функшй та штоке и кашею оргашзму.
1.1. Мечников уперше запропонував використовувати мжробний ангагошзм
для боротьби з кишковою авто!нтоксикац!сю. 1де! 1.1. Мечникова набули розвит-
ку з погляду лгквщащ! кишкового дисбактерюзу, шо розвиваеться за експеримен-
галъних умов, у раз! ослабления оргашзму, а шодг пщ впливом антибактергальних
препаратов.
ПОРУШЕННЯ СИНТЕЗУ ГОРМОН1В СИСТЕМИ ТРАВЛЕНИЯ
Нин! ведомо понад 20 речовин, яю е доведенный або передбачуваними гормонами
системи травления. Разом з регуляшею процесев травления ! всмоктування гормони
травно! системи впливають на кроьообгг. обмш речовин, апетит, дгяльнкть нервово!
та ендокринно! систем. Тому гх с.шд зараховувати до групи регуляторних пептцдёв.
Для гормонёв системи травления характернг певнё особливост], яю визначають
!х роль у патологи органгв травления. До таких особливостей належить подгбнгсть
хёмёчно! структури 1 бюлопчно! дп деяких гормонгв. За такою подгбнгстю нине
видглено ролини гастрину (ргзнг форми його г холецистоюнгну), секретину (секре-
тин, глюкагон, гас троен пбуючий шгестинальний пептид г ВШ, кальцитонш, каль-
цитонгн-ген-пов'язанг пептиди I г II, паратиреощний гормон, кортиколёберин
тощо). панкреатичного полшептиду (включае пептид УУ г нейропептид У), мо-
тилгну (включае грелён та гн.), тахёюншу (включае також речовину Р, гастринолг-
берин, нейромедини га багато пептидгв молюскгв г амфгбш). Таким чином, подгб-
нг порушення травления та ёнших функцгй органгзму можуть виникати внаелгдок
надмтрног або знижено! продукц!! ргзних гормонгв системи сравнения (табл. 25).
Гормони виробляються клгтинами дифузно! нейроендокринно! системи (тер-
мит уведено в 1977 р. Пгрсом), що походять з нервовою гребшця г локалгзован!
в слизовёй оболонщ травного каналу г травних залоз. Це зумовлюе !х пений
зв’язок гз нервовою системою. Так, нейропептиди (речовина Р, нейропептид У,
ВШ, гас трин олгберин тощо) виробляються в органах травления 1 беруть участь у
регуляцп секреторное функпп та кровообггу цих органгв. У центральней нервовш
систем!, як г в травнгй, функшонують речовина Р, ВШ, гастрин, холецистокшён,
нейротензин та тн. Деякг трави! гормони одночасно е нейротрансмётерами ё мо-
жуть досягати органе в-мёшеней як з кров’ю. так г нервовим шляхом. Одержано
також дан! про можливу роль порушення синтезу гормонгв травления при пато-
логи нервово! системи, у зв’язку гз чим говорить про “нейрокишкову вгеь’’.
1снуе пений зв’язок м!ж гормонами гравног та ендокринно! систем г функ-
щею плот аламо-ппофгзарно! системи. Соматостатин, який гальмуе вироблення
соматотропшу, пригнгчуе також продукцпо деяких гормонгв травления. Зв’язок гз
пгдшлунковою залозою проявляеться четким впливом гастрогнпбуючого гнтести-
нального пептиду 1 секретину на продукцгю шсулгну га глюкагону, шо зумовлюе
важливу роль гормонгв травления в патогенезе ожирения та схуднення. Стимуля-
цгя гастрином, холеиисгоюнгном г глюкагоном вироблення кальцитоншу визна-
чае участь ёх у патогенез! порушень кальцеевого обмшу.
Важчивою особлив!стю гормонгв травное системи е надходження ёх у кров е
зовннпн! секрети. Це зумовлюе можливгсть впливу на клгтини слизово! оболонки
з боку просвгту органгв травления г пений зв’язок мёж порушеннями внутрппньо!
та зовигшньог секрецп цих органгв.
За бага тьма ознаками перелечен! гормони подгбне до гнкретгв слинних залоз,
як! також варто зарахувати до гормонгв сисгеми травления.
З’ясовано, що порушення продукт! гормонгв травно! системи зумовлюе тяжел
розлади травления, метаболгзму та дтяльностг ргзних органгв 1 систем органгзму. Най-
бёлъше вивчено патологечнё процеси, пов’язанг з виникненням пухлин, джерелом яких
е гормонпродукуюч! клётини органгв травления: гастринома (синдром Золдёнгера —
Еллёсона). вшома (синдром Вернера — Морргсоиа). соматостатинома (табл. 25).
Таблица 25. Характеристика тормоша системи травления
Назва Основна локализация Молёку- лярн! форми* Д|Я Р1вень секрецй, роль у патологи
Гастрин Антральний вцццл шлун- ка 17, 34 Стимулящя секрецй кислоти в шлунку 1 ферменпв шд- шлунковоТ залози, скорочен- ня нижнього стравох.дного сфшктера, прол1ферацп сли- зово! оболонки, вившьнення кальцитоншу Знижений при ахлор- пдрй, ваготомп; шдви- щений при перющоз- н.и анемп, синдром! Золлшгера — Еллкона (гастринома— виразки шлунка та дванадцяти- пало! кишки)
Холецистокппн гпанкреозимш) Тонка кишка 8, 33, 39, 58 Стимулящя скорочення жовч- ного мтхура 1 секрецц ферменпв шдшлунковою за- лозою, троф1чна д1я на шд- шлункову залозу, пригшчен- ня апетиту Пщвищений у раз! не- достатност! пщшлун- ково!' залози, знижений при цел!акй
Секретин Тонка кишка 27 Стимуляция секрецп' пдро- карбонату шдшлунковою за- лозою, гальмування секрецй кислоти в шлунку, розслаб- лення нижнього стравохт,дно- го сфшктера Знижений при цсл1акп
ВШ Тонка кишка 28 Зниження тонусу судин, ак- тивация панкреатично!’ та кишково!’ секрецй Опосередковуе синд- ром Вернера — Морри- сона (водна Д!арея, п- покал!СМ1я, ахлор- пдртя)
Гастрошпбую- чий (глюкозо- залежний ш- сулшот ропиий) пептид Тонка кишка 42 Стимулящя секрецй’ шсулшу, гальмування секрецй кисло- ти 1 моторики шлунка, вики- ду глюкози пешнкою, усмок- тування жиру Знижений у раз! цел1акй, припускають участь у патогенез! ожиршня
Ентероглюка- гон Тонка 1 товс- та кишка 69 Трофхчна Д1я на слизову обо- лонку кишок Шдвищений у раз! ви- далення або ушкоджен- ня слизово!оболонки
Мотилш Тонка кишка 22 Стимулящя моторики шлун- ка 1 кишок Шдвищений при д1аре1
Нейротензин Тонка кишка 13 Стимуляцгя секрецй" в тонкш кишщ, пригшчення секрецй кислоти в шлунку га шсулшу ГКдвищеннй у раз! син- дрому мальабсорбцй
Гастринол1бе- рин Шлунок, тон- ка кишка, шдшлункова залоза 27 Стимуляция секрецй’ гастри- ну, моторики шлунка 1 панк- реатично! секрецй, вивйгь- нення ХЦК, мотилшу, нейро- тензину, шсулшу, речовини Р, соматостатину Не виз качено
Зактчення табл. 25
Назва Основна локал1зац1я Молеку- ЛЯРН1 форми* Р1вень секрецн, роль у патологп
Речовина Р (нейрокшш А) Кишки, гшо- таламус, ешф!з 11 Стимуляция секрецп пхд- шлунково! 1 слинних залоз, розширення судин, мед1ац1я болю Не визначено
Панкреатичний полшептид ГПдшлункова залоза 36 Гальмування секрецп фер- менте шдшлунковою зало- зою 1 скорочення жовчного мххура, моторики шлунка й кишок Знижений у разг недо- статносп шдшлунково! залози, пгдвищений при д!аре!
Соматостатин Гшотала- мус, шд- шлункова залоза, шлу- нок 14, 28 Гальмування шлунково! та панкреатично! секрецп, ви- роблення соматотрошну, гас- трину, гастрошпбуючого ш- тестинального пептиду, ш- сул1ну, глюкагону тощо Опосередковуе синд- ром соматостатиноми (диспепсия + цукровий д!абет + холелтаз)
Ендогенн! оша- ти (ендорфши) Головний мо- зок, кишки, шлунок 5, 16, 17,31 Гальмування секрецп шлун- ка, активащя моторики ки- шок, розслаблення нижнього стравохлдного сф!нктера, сти- мулящя апетиту Знижений у раз! ожи- ршня, тдвищений при голодуванш; припуска- ють участь у патогенез! синдрому подразнено! кишки
* Юльюсть амшокислотних залишктв у молекул!.
Роздал XXIX
ПАТОЛОПЧНА Ф1310Л0ПЯ ПЕЧ1НКИ
ЗАГАЛЬНА ХАРАКТЕРИСТИКА
ПАТОЛОПЧНИХ ПРОЦЕСС У ПЕЧ1НЦ1
Печшка виконуе в оргашзм! багато функцш, серед яких найважлившшми е участь
в обмин речовин, гравленш, жовчоутворенш та жовчовидйтенш (екскрегорна
функптя), гемодинамщх, кровотворенн]. Значка також захисна (бар’ерна) роль
пього органа. Ус! ш функцп, спрямоваш на збереження гомеостазу, порушуються
в разт виникнення патологи печшки, що проявляеться як самостшними хворобами
(наприклад втрусний гепатит), так 1 печшковими синдромами (жовтянипя, холестаз,
холемгя, портальна ппертензтя, печшкова кома тощо). Останш спостертгаються
не титьки при хворобах само! печшки, а й супроводжують хвороби шших оргашв
1 систем (наприклад жовтяниця у випадку гемолттично! хвороби новонародже-
них). У зв’язку 13 цим розртзняють первинш I вторинш ураження печшки.
До патолог1чних процессе, що найчастйие е основою хвороб печшки, належать:
• запалення (гепатит);
• порушення кровообпу в судинах печшки (тромбоз, ембол!я вортто! вени,
печшкових вен, печшково! артерп);
• набут] та спадков] порушення обмшу речовин в ештелюцитах печшки (ге-
патоцитах) з розвитком дистрофп та некрозу (гепатоз);
• дифузне розростання сполучно! тканини (ф!броз) на тлт дистрофп, некрозу
еттелюцитпв ] перебудови сгруктури органа (пироз);
• пухлини.
Еттологтя. Етюлопчними факторами, що спричинюють ураження печшки, е
такт:
• шфекцшнп — втруси (втрусних гепатиттв А, В, С, О, Е, С, шфекцшного
мононуклеозу), збудники туберкульозу, сифйпсу, найпростил! (лямбли. амсби),
гриби (акпшомщета), гельмшти (ехшокок, аскариди);
• токсичн! — речовини екзогенного (адкоголь; лткарськ! засоби — ПАСК-на-
трш, сульфанитамтди, питостатичн! препарата, салщилати, бюмщин, тетрациклш;
промислоь] отрути — чотирихлористий вуглець, хлороформ, миш як, фосфорор-
гашчн! речовини, шеектофунгщиди; рослинш отрути — афлатоксин, мускарин,
алкалощи гелютропа) та ендогенного (продукти розпаду тканин щд час ошку, не-
крозу; токсичш речовини в разт уремп, токсикозу ваптних) похооження',
• фтзичш — юшзуюче випромшювання (променевий гепатит), мехашчна
травма;
• ашментарш — бщкове, вггалянне голодування, жирна !жа;
• алерпчш реакцп — на введения вакцин, сироваток, харчових 1 лпсарських
алергешв;
♦ порушення кровообггу в печшщ хпсцевого (!шем!я, венозна пперемгя, тром-
боз, емболхя) та загального (при недостатносп кровообпу) характеру;
• ендокринн! та обмшю порушення в оргашзм! (цукровий д!абет, голодуван-
ня, ожиршня);
• пухлини — первиню (гепатоцелюлярний рак) ! метастатичш (унаслщок раку
шлунка, легень, молочно! залози, лейкозш шфшьтрати);
• генетичш дефекта обмгну (спадкова ферментопатгя), вроджеш вади анато-
мгчного положения 1 структури печшки.
У патогенез! уражень печшки р!зно! етюлогп варто вщзначити деякг загаль-
ш закономгрностг:
• наявшсть двох основних механгзмгв виникнення патологи печшки:
— пряме ушкодження печгнки (вирусами, хгмгчними речовинами, отрутами,
унаслщок порушення кровотоку), що проявляеться дистрофгчними эхи-
нами в шй, аж до некрозу;
— автогмунне ушкодження внаслщок появи автоантитенгв (патолопчно змг-
нених компонента ештелюципв печшки, що частгше утворюються в раз!
прямого ушкодження печшки) г розвитку автогмунних реакцш гумораль-
ного г клгтинного типу, як! поглиблюють ураження печгнки шляхом {мун-
ного цитолгзу за участю Т-кглергв г цитотоксичних антитш;
• певний взаемозв’язок патолопчних процесгв у печшцг: як запальш (гепа-
тит), так г обмшно-дистрофгчн! (гепатоз) ураження печшки можуть закшчитися
розвитком цирозу;
• розвиток недостатносп печшки унаслщок часткового або повного пору-
шення функцгй печшки;
• висока частота поеднання патологи печгнки з патолопею органгв травлен-
ия, селезшки, що зумовлено анатомгчними г функщональними зв’язками мгж
цими органами; так, утворювана в печшщ жовч бере участь у перетравлюванш та
всмокгуваннг компонента !ж! в кишках, забезпечуе рухову функшю кишок; у
печгнцг вщбуваеться промгжний обмш продукта гщролгзу гжг, багато з яких депо-
нуються в нт; унаслщок того що печшка г травний канал е единою гепатоенте-
ральною системою, ураження однгег з частин цгег системи зумовлюе залучения до
патолопчного процесу й шшо! (гепатоентеральний синдрому, спшьшсть крово- й
лгмфообггу, гннерваци та деяких функцгй (кроветворения г кроворуйнування,
бар’ерна функцгя, депонування кровг) зв’язуе печгнку гз селезгнкою ! зумовлюе
чаете епгльне ураження цих органгв (гепатол1еналъний синдром)-,
• наявшсть одночасно зг структурними та функщональними порушеннями
печгнки компенсаторних реакцш у нш, спрямованих на обмеження ! повну лгквг-
дацгю патолопчних змш в орган!; до таких компенсаторних реакцш належать
посилення метаболгчних процесгв (енергетичних, дезгнтоксикацгйних тощо) у пе-
чшщ, фагоцитозу, збгльшення екскреци токсичних речовин, перерозподгп кров!,
змша судинного тонусу, розвиток анастомозгв, висока здатшеть печгнки до реге-
нераци, що проявляеться як пщ час резекци, так г дифузного ураження г! ткани-
ни; при цьому вщбуваеться пролгферацгя ештелюципв з повним або частковим
вщновленням структури органа;
• участь печхнки в адаптивних реакциях оргашзму в раз! крововтрати, висо-
тнох хвороби, дй токсичних речовин екзо- та ендогенного походження, хвороб
шших оргашв 1 систем.;
• ускладнення перебггу патолопчних процесхв в шших органах п!д час пато-
лап! печшки (ендокринопатй внаслщок. порушення метабол!зму гормонхв у пе-
чшцх, посилення недостатност! серця в раз! портально! гшертензп) та зниження
гфективностх дй лгкарських засобхв у разх порушення детоксикацшно! функцн
печшки.
НЕДОСТАТШСТЬ ПЕЧ1НКИ
Недостатшсть печшки — патологичный стан, за якого функциональна дгяльнгсть
печшки не забезпечуе п'гдтримання гомеостазу, що призводить до порушення жипг-
тсдгяльностг оргашзму.
Класифхкацхя. На пгдставг механгзмгв виникнення (патогенезу) розрхзхтяють
так! види недостатност! печхнки:
• печшково-клгтинну (гепатоцелюлярну) при дистрофхчних х некротичних ура-
женнях ештелюципв печшки;
• екскрегпорну або холестатичну1 (унаслщок порушення жовчоутворювально! х
жовчовидхльно! функцш печшки);
• печгнково-судинну (у раз! порушення кровообну в печшцх внаслщок тромбо-
зу або емболи ворйно! вени, печхнково! артерй, печшкових вен або загальних
циркуляторних порушень, наприклад при недостатност! хдювообхгу, шоку);
• комбгновану (унаслщок поеднахшя кгтькох мехашзмхв у процес! розвитку
функцхонально! недостатност! печшки, наприклад холестаз, шо може супрово-
джуватися печхнково клхтиниою та судинною недостатшстю печхнки).
Печшкова недостатшсть може бути:
• абсолютною, основою розвитку яко! е зменшешхя функцюнальних можли-
юстей органа;
• вгдносною, що виникае внаслщок невщповщностх функцхонально! активностх
печхнки пщвищеним потребам оргашзму в й функцй (наприклад пщ час посиле-
ного гемолхзу еритроцитхв, коли, незважаючи на значне збшьшення жовчоутво-
рювально! функцй’ печгнки, виникае гемолпична жовтяншгя).
За ступенем порушень функцш недостатшсть печшки бувае:
• тотальною (порушен! вех функцй);
• парцгалъною (порушен! окрем! функцй).
За клйачним перебггом печгнкова недостатшсть подхляеться на:
• блискавичну (розвиваеться за кхлька годин);
• гостру (протягом кзлькох д!б х тижнхв);
• хрошчну (упродовж кхлькох мхсяцхв ! рокхв).
Етхслопя.Етхолопчш фактори, якх спричинюють недостатшсть печгнки,
можна подшити на печшковг, що зумовлюють первинне ураження печхнки, та по-
запечгнковг, пов’язаш !з загальнопатолопчними процесами (гшоксхя, ггповггамшо-
’Холестаз — порушення вщтоку жовчх з накопиченням п компонент Хв у печшщ та кровх внаслщок
зниження екскреторно'! функцп егптелхецитхв печшки або механхчно! перешкоди вщтоку жовчх у жовч-
них пробочках X протоках печшки (внутршньопечтковий холестаз), а також через механхчне порушен-
ня вхдтоку жовчх в позапечшкових жовчних протоках (позапечшковий холестаз).
Мал. 68. Накладення судинних ф!стул (за К.М. Биковим):
а — судини до операцп; б — пряма ф1стула (Екка); в — зворотна ф1стула (Екка — Павлова)
зи, голодування) або патолопею окремих органов 1 систем (недостатшсть серия та
нирок, ендокринопатп, гемобластози).
Експериментальн! модель Печшково-клйпинна недостатшстьпечшкимо-
делюеться:
• Х1рурпчним видаленням печшки (повним або частковим);
• токсичним ураженням печшки гепатотропними отрутами.
Повне видалення печшки здшснюють у юлька етап!в:
I — накладають зворотну фхстулу Екка — Павлова (анастомоз м!ж ворггною 1
нижньою порожнистою венами 1 перев’язування останньо! над анастомозом (мал. 68),
щоб печшка переповнилася кров’ю в!д усге! нижньо! половини т!ла, а це спричи-
нюе розвиток колатералей, як! вщводять кров у верхню порожнисту вену через
ут. 1когас1сае 1п1етае);
I I — через 4—6 тиж. перев’язують вор!тну вену вище анастомозу;
I II — видаляють печшку.
Через 3—8 год теля видалення печшки в собаки вш гине вщ ппогл!кем!чно!
коми. Уведення глюкози зб!лыпуе тривал!сть життя тварин до 40 год; смерть на-
стае при симптомах недостатносп кровооб!гу 1 дихання. На пщетав! виявлених
при цьому змш бюх1М1чного складу кров! доведено участь печшки в обмш! речо-
вин, зещанн! кров! та детоксикацшних процесах.
Часткове видалення печшки дае змогу вивчити особливост! регенерацп цього
органа. Видалення 65 % маси печшки зумовлюе п функцюнальну недостатшсть,
але протягом 4—8 тиж. вона повшстю вщновлюе свою масу за рахунок гшерплазп
не! частини ештелюцитгв, яю збереглися.
Токсичного ушкодження печшки досягають уведенням гепатотропних отрут
(чотирихлористий вуглець, хлороформ, фосфор, тршптротолуол), як! спричиню-
ють некроз I тслянекротичний (токсичний) цироз печшки.
Холестатичну форму недостатноспй печшки моделюють перев’язуванням жовч-
них проток. Так, пщпечшкова (мехашчна) жовтяниця, що супроводжуеться поз-
апечшковимхолестазом, вщтворюеться шляхом перев’язування сшльно! жовч-
но! протоки. Для моделювання внутрпиньопечшковогохолестазу тваринам уво-
дять а-нафтил!зот1ошонат та шип речовини, як! утруднюють в!дт!к жовч! з
печхнки. В обох випадках розвиваеться холестатичне ушкодження ештел!оцит!в
печшки, а згодом — бшарний цироз.
Судинну форму недостатностг печгнки моделюють у раз! порушення в нш
кровообпу шляхом накладання фгстули Екка та Екка — Павлова (див. мал. 68),
перев’язування або часткового звуження печшково! артерп, вор!тно! та печшково!
вен.
Накладання прямо! фгстули Екка полягае в створешп анастомозу мЬк ворпною
. нижньою порожнистою венами 1 перев’язуванн! ворхтно! вени над анастомозом.
Як наслщок, припиняеться надходження кров! 13 судин травного каналу в печшку,
ток кров! в нш зменшуеться вдвхч!, порушуеться метаболхчна функшя (особливо
бшковий обмш; “м’ясне отруення”, за 1.П. Павловим).
Перев’язування печшково! артерп може спричинити смерть собак, осюльки
внаслщок ппемп печшка шфшуеться анаеробною мгкрофлорою кишок 1з розвит-
ком масивного некрозу. Уведення антибютиюв запобтгае виникненню некрозу.
Через 10 дшв шсля перев’язування артерхальш колатерал! стають настхльки роз-
винутими, що кровообхг у печшщ вщновлюеться, надходження кисню стае до-
статнхм для того, щоб запобпти розвитку шфекцп.
Моделювання портально! гшертензп (синдрому, що супроводжуе судинну фор-
му недостатност! печшки) здшснюють шляхом часткового звуження воргтно! вени,
печшкових вен або нижнъо! порожнисто! вени вище впадання в не! печшкових вен.
Наслщками цих судинних порушень е пщвищення тиску в систем! ворхтно! вени,
розвиток портокавальних анастомозхв, асциту, ппоксичного ушкодження еш-
телхоципв печшки х портального цирозу.
В умовах хрошчного експерименту для вивчення функщонально! недостат-
ност! печшки (метаболхчно!, захисно! тощо) застосовують метод анггостомп
(Ю.С. Лондон).
До стшок печшково! артерп, ворхтно! та печшково! вен пщшивають металев!
канюл!, вхльний кшець яких виводять назови! крхзь передаю стшку живота. У
хрошчному дослщ! через щ канюл! здшснюють узяття кров!, що надходить до
печшки 1 вщходить вщ не!.
Для вивчення функщй печшки в експеримент! застосовують також метод
перфузи гзольовано! печгнки, проводить клшхчш дослщження (пункцшна бхопсхя
печхнки, радхонуклщне сканування, УЗД, бюххмхчш та хмунолопчш дослщження
тощо).
Порушення метабол1чноТ функцп печшки
Порушення вуглеводного обмшу пщ час ураження печшки полягае в гальмуванш
таких процесхв, як:
• утворення з глюкози глюкозо-6-фосфату х його подальша утилхзацхя р!зни-
ми шляхами (перетворення глюкози на глхкоген, окиснення в цикл! трикарбоно-
вих кислот ! пентозофосфатному цикл!);
• бюсинтез 1 розщеплення гликогену;
• глюконеогенез;
• перетворення на глюкозу галактози та хшпих моносахарвдхв (фрукгози, ма-
нози);
• утворення глюкуроново! кислоти.
За ет10.юг1€Ю порушення вуглеводно! функцп печшки можуть бути набути-
ми 1 спадковими
Основними механизмами розвитку наведенихпорушень. якз е загальними
1 для шших порушень метабол!чних процесов у печшщ, с такт
• ушкодження паголопчним процесом (запалення, гепатоз, цироз. пухлина,
!шем!я) ештелюципв печшки, у яких здшснюються метаболзчш перетворення
вутлеводгв;
• дефпшт субстрапв (у раз! голодування. порушення перегравлювання 1
всмоктування вуглевод!в у кишках);
• деф1цит енергп (пФ час ппоксп, пповпамшозу, особливо дефщиту- тридок-
сину, рибофлавину);
• знижештя активност! ферменпв, як1 катанзують р!зш ланки вуглеводного
обм1ну, унаслщок зменшення (хнього синтезу (у разт спадково!’ ферментопати,
бпткового голодування. гшоксп. ушкодження мпохондрш при гепатитах, цироз));
• розлади нейрогуморально! регуляцн вуглеводного обмшу (при цукровому
д!абеп, патологи ппофзза, надниркових залоз та ш.).
Основа) КЛ1Н1ЧН1 прояви порушень вуглеводного обмшу в печшц!:
• змши вм!сту глюкози в кров! (г!пер- та г!погл!кем!я);
• зменшення запас1в глгкогену в печшц! — головному депо глюкози в оргашзм!;
так, при цукровому доабетт. для якою характерна г!пергл!кем!я, глгкогениий ре-
зерв може зменшуватися в 50 раз!в; р!зке його виснаження, але на тл! гшоглисем!!,
також спостерйаеться пщ час повного голодування, при гепатит!, цироз!, печ!н-
ковш ком!, спадковому агл!когеноз! (пов’язаний тз дефщитом глжогенсинтетази),
який може супроводжуватися розвитком коматозного стану;
• хвороби накопичення гмкогену в печшц!; у випадку спадкових гл!когеноз!в деф!-
цит глюкозо-6-фосфагази (хвороба Прке) 1 печшково! глокогенфосфорилази
(хвороба Герса) зумовлюе р!зке зростання запасхв глгкогену в печшц! внасл!-
док його нездатност! розщеплюватися з вив!льненням молекул глюкози; це приз-
водить до гепатогенно! гтогпйсеми, яка спричинюе енергетичне голодування голо-
вного мозку з подалыпим розвитком розумово! вщсталосп в дхтей;
• спадков! ферментопатп з порушенням метаболхзму фруктози (фруктозем!я) та
галактози (галактоземия) (див. розды XVIII “Порушення вуглеводного обмшу”);
• метабол!чний ацидоз у раз! посилення анаеробного ппкол!зу в печшц! з на-
копиченням метаболгпв — лактату, шрувату, кислот !з ЦТК тощо (при голоцуван-
ш, цукровому д!абег!. гепатитах, синдром! портально! гшертензп).
Порушення жирового (лшыного) обмшу при хворобах печшки можуть перебь
гати у вигляд!:
• зниження гщролтзу 1 всмоктування лппгцв !ж! в кишках унаслщок недостат-
яього надходження в них жовчних кислот пщ час патологи жовчоутворення !
жовчовидыення (при печшковш 1 пщпечшковш жовтяниш (див. ‘‘Ахол!чний син-
дром”);
• порушення синтезу I розщеплення жирових кислот, григлшеридгв (ней-
трал ьних жир!в), фосфолшщш, глгколшщгв, лшопроте’щов, холестерину (при гепа-
титах, цироз!);
• збшьшення синтезу та порушення утшпзацп кетонових т!л (див. розды ХЕХ
“Порушення жирового обмшу”).
Основы КЛ1Н1ЧН1 прояви розладу жирового обмшу в печшщ:
• змгна лгтдного складу’ кровг у вигляд!:
— транспортноЧ гтерлтеми, що спостерггаеться в раз! зменшення запаств глг-
когену в печшщ при цукровому дгабетт, голодуванш;
— ггперлтопротегдеми зт збшьшенням вмтсту в кров! ртвнгв ЛПНЩ, ЛПДНЩ
унаслщок посилення тхнього синтезу в ештелюцитах печшки тз жирових кислот,
як! надходять у печшку тз кишок у разг надлишкового гх вмтсту в гжт (такий вид
пперлшопротещемп зумовлюе розвиток атеросклерозу);
— ггперхолестеринеми внаслщок порушення виведення холестерину тз жовчю
у випадку холемтчного синдрому (при механтчнш та печшковш жовтянищ);
— гтохолестеринеми при патолопчних процесах у печтнцт (гепатит, цироз),
коли знижуеться бтосинтез холестерину з ацетил-КоА (цей процес здебшыпого
вщбуваеться в печшцт), що впливае на процеси утворення тз холестерину жовчних
кислот, холекальциферолу (вттамгну О3), кортикостеро'щгв, статевих гормонтв !
спричинюе вщповщнт порушення;
• ггперкетонемгя ! кетонургя внаслщок пщвищеного синтезу в печшц! кетоно-
вих тщ з ацетил-КоА, а також зниження тх утилгзаци (кетонемтчна кома — при
цукровому дтабетт, кетоз — при голодуванш, печшковш комг);
• жировий гепатоз (дистрофтя печшки жирова, ожиршня печшки), для якого
характерно накопичення жиру в ештелюцитах г мтжклггиншй речовин! печшки,
дифузне або вогнищеве п ожиршня;
• стеаторея — збшьшення виведення жиргв тз калом у разг порушення тх ус-
моктування в кишках пщ час ахолтчного синдрому.
Порушення бшкового обмшу в раз! патологп печшки проявляеться розладом:
• синтезу бшкгв (у тому числ! бшкгв плазми кровг, зсщальнот, протизсщально!
та фтбринолттично! систем кровг);
• дезамгнування, трансамшування та декарбоксилювання амшокислот;
• утворення сечовини з продукттв азотистого (головним чином бшкового) ка-
таболгзму.
Основы! КЛ1Н1ЧН1 прояви порушення бшкового обмшу в печшщ:
• гтопротегнемгя внаслщок зменшення синтезу альбумтшв та а- ! р-глобулштв
(у норм! в ештелюцитах печшки синтезуеться весь альбумш плазми кровт, 75—
90 % — а-глобулгнтв 150% — 0-глобул!н!в);
• диспротегнемгя — наслщок порушення спгввщношення мгж окремими фрак-
щями проте'ппв сироватки кровт;
• печгнковий набряк, розвиток якого зумовлений зниженням онкотичного тис-
ку кровт в раз! гшоальбумшемп;
• гемораггчний синдром при зменшенш синтезу бшкових факторгв зсщання
кровг (ф. I, II, V, VII, IX, X, XI, XII, XIII);
• пгдвищення ргвня вгльнглх амгнокислот у кровг (амгноацидемгя) та сечг (амгно-
ацидургя), змши яктсного складу амшокислот сироватки кров! в разг дифузних г,
особливо, некротичних уражень печшки, коли в шй порушуються окисне дезам!-
нування! трансамтнування амшокислот;
• ггперазотемгя (збшьшення в кровт концентрацп залишкового азоту) за раху-
нок амшокислот г гтерамонгеми (збшьшення ртвня амонпо в кровг) унаслщок по-
рушення синтезу сечовини г бшкгв (продукцшна азотемгя) — показник тяжко! не-
достатностг печшки, як правило, у раз! ушкодження 80 % (г бшьше) п паренхтми;
• тдвищення вмгсту в кровг деяких ферментгв (у-глутамглтранспептидази, амг-
нотрансфераз та ш.) унаслщок руйнування епгтелгоцитгв печгнки при гепатит!,
цирозг, пухлин!.
Порушення обмшу вгтамшгв (див. роздгл XXIV “Патофгзголопя обмгну вгта-
мшгв”) при патологи печшки проявляеться:
• зменшенням усмоктування жиророзчинних вгтамшгв (А, В, Е, К) унаслщок
розладу жовчовидгльног функцй печшки (вщсутнгсть або недостатке надходження
жовчних кислот у кишки через механгчну та паренхгматозну жовтяницю);
• порушенням синтезу вгтамшгв г утворенням гхнгх бюлопчно акгивних форм
(ретинолу з каротину, шридоксальфосфату — активно! форми впамшу В6, коен-
зиму А з пантотеновог кислоти та гн.);
• порушенням депонування вгтамшгв (ретинолу, щанокобаламшу, фолгевог та
шкотиновог кислот тощо) та виведенням гх з оргашзму.
Основн! КЛ1Н1ЧН1 прояви порушень метаболгзму вгтамшгв у печшщ:
• ггповгтамгнози з вщповщною симптоматикою в раз! дефициту жиророзчин-
них вгтамшгв (наприклад геморапчний синдром при гшовгтамгнозг К);
• порушення бгологгчного окиснення й утворення енерги пщ час дефициту вгта-
мшгв групи В, бглышсть яких виконуе коферментну функцпо;
• посилення пероксидного окиснення лгтдгв з подальшим ушкодженням парен-
хгми печшки при дефщит! антиоксиданту вгтамшу Е;
• порушення еритроцитопоезу в раз! дефщиту В121 фолгевог кислоти (дефщит-
н! анеми).
Порушення обмшу гормошв г БАР у раз! патологи печшки можуть бути наслщком:
• порушення в печшщ синтезу гормошв (наприклад тирозину, що утворюеть-
ся в ештелгоцитах печшки з фенглаланшу 1 е попередником тироксину, трийод-
тирошну, катехоламгнгв) г транспортних б!лк!в гормонгв (наприклад транскорти-
ну, що зв’язуе 90 % глюкокортикощгв);
• порушення шактиваци гормонгв шляхом, наприклад, кон’югаци стерощних
гормошв з глюкуроновою г сульфатною (сгрчаною) кислотами, ферментативного
окиснення катехоламгнгв пщ впливом амшоксидаз, розщеплення гнсулшу шсулг-
назою та гн.;
• зниження шактиваци БАР (наприклад окисного дезамшування серотошну,
ггстамгну).
Основнг КЛ1Н1ЧН1 прояви:
• ендокринно патологгя',
• посилення алергп в разг зниження дезамгнування БАР у патолопчно змгнених
епгтелгоцитах печгнки.
Порушення обмгну мгкроелементгв (див. роздгл XXIII “Патофгзголопя обмшу
мгкроелементгв”) при хворобах печгнки пов’язанг зг змгною:
• депонування в печшцг залгза (у форм! феритину), мщг, цинку, кобальту,
мол!бдену, марганцю тощо;
• синтезу транспортних бглкгв мгкроелементгв (трансферину, церулоплазмгну);
• екскрецг! мгкроелементгв гз жовчю.
Клгнгчними проявами цих порушень е:
• гемохроматоз, гемосидероз у разг накопичення в печгнцг залгза (з подалыпим
розвитком цирозу г гепатомегалп);
• залгзо-, В!2- I фолгеводефщитнг анеми внаслщок тяжких уражень печгнки.
Порушення захисноТ функцн печ!нки
Порушення захисно! функцп печшки проявляеться зниженням:
• фагоцитно! та цитотоксично! активност! ззрчастих макрофапв (макрофаго-
цитзв, яю етановлять 85 % загально! юлькост! макрофапв организму);
• цитотоксично! активност! великих грануловмюних л!мфоцит!в (рН сеПз, або
ямков! юптини печенки, як! здшснюють противтрусну та протипухлинну !мунну
вщповщь);
• детоксикацшно! (знешкоджувааьноТ) антитоксично! функцп печшки;
• юиренсно! функцп — виб!ркового захоплення еттел юнитам и печшки окре-
мих речовин !з кров! без !х х!мтчного перетвсрення 1 подальшого виведення !з
жовчю;
• екскреторно! функцц — виведення токсичних речовин 1з жовчю.
Функциональна недостатшсть з!рчастих макрофапв 1 рП-ювтин при хворобах
печшки (цироз, гепатит, злояюсна пухлина) може розвиватися внаслщок:
• змш структури печшки;
• ппоксп в раз! порушення кровотоку ! дп токсичних речовин — продуктов
розпаду клггин 1 метабол!т!в. шо утворюються в самгй печшц!.
Основ ними КЛ1Н1ЧНИМИ проявами недостатност! фагоцитно! функцц з!р-
частих макрофапв ! цитотоксично! активност! лимфоцитов (рН се!1з) печшки е:
• зниження сгтйкост1 организму до [нфекцшних факторов',
• септицелия (до шфекцшно-токсичного шоку);
• метастатичнеураження печшки;
• збыыиення частота розвитку алергп та посилення й прояв!в, зумовлене по-
рушенням захоплення з кров! ! руйнуванням макрофагами печшки речовин з
антигенними властивостями та гмунних комплекс!в.
Ослабления детоксикащйно! функцн печшки — бюгрансформацп ксенобютигав
та ендогенних токсишв1 у менш токсичн! для оргашзму речовини — пов’язане з
порушенням таких и метабол!чних функцш:
• синтезу сечовини, у процес! якого в!дбуваеться знешкодження амонпо;
• реакшй микросомального окиснення (бютрансформашя полщикшчних аро-
матичних вуглеводшв !з канцерогенними властивостями, пестицид!в. амщошри-
ну, ефедрину тощо) та вщновлення (нейтрал1зап!я нггро- та азосполук);
• реакшй ацетилювання (знешкодження сулъфаншамшних препаратов) г пд-
рошзу (розщеплення алкалойпв, серцевих гл!козид!в);
• реакц!й метилювання (похщних пурин!в);
• реакщй кон’югацй з утворенням нарних сполук !з глюкуроновою кислотою,
гл 1 цином, цистешом, таурином (вщбуваеться детоксикац!я непрямого бгпрубшу,
скатолу, фенолу, шдолу, серотоншу тощо);
• перстворення водонерозчинних речовин у розчиюп сполуки. яю можуть
виводитися з оргашзму !з жовчю 1 сечею та шшими шляхами екскрецц.
Ксевобютикм (вш грец. хепоз — чужий, Ыт — життя) — речовини разного походження, яю ток-
сично впливають на оргашзм в цЬтому або окрем! його тканини. До них належать фармацевт ичш та
косметичш тасоби, промислов) отрута, пестициди, радюнуклщи, харчов) добавки (барвинки, консер-
ванта), засоби побутово! х!мн. Еидогенш токсини — токсичш продукти метаболизму, яю утворюються
в оргашзм! (наприклад жовчн! ппменти), а також речовини, що утворюються виаслшок гнитгя б!лк!в
у товстш книги) (феноли, шдол) пш впливом мткрооргашзмгв.
Ослабления детоксикацгйно! функцц печшки спричинюеться:
• ушкодженням ештелюципв печшки — основного мгсця знешкодження ксе-
нобютиюв та ендогенних токсишв;
• зменшенням активносп ферменпв, шо каталгзують реакцп детоксикацп;
• дефщитом енергп.
Основними КЛ1Н1ЧНИМИ проявами ослабления детоксикацшно! функцп
печшки може бути:
• печткова енцефалопапйя, для яко! характерними е порушення психгки, св!-
домосп, рухов! розлади (тремпння, атаке! я, рипдшсть м’язгв) 1 яка за тяжкого
перебггу може перейти в печгнкову кому,
• пгдвищення чутливоспй организму до ргзних лгкарських засобгв — хгшну, морф!-
ну, барбгтурапв, препарапв наперстянки тощо, яю через недостатшсть гхньо! де-
токсикацп в печшщ накопичуються в оргашзм!, спричинюючи медикаментозну
штоксикацпо;
• утворення ще токсичшших речовин у процес! метаболгчних перетворень ток-
сичних сполук в ештелюцитах печшки (метабол!™ деяких лгкарських засобгв,
наприклад гзошазиду).
Печ?нкова кома — патологгчний стан, зумовлений токсичном уроженкам цент-
рально? нервово? системи з глибокими розладами ?? функщй (потьмарення св?до-
моспн, вгдсутшсть рефлексов, розвиток судом), порушенням кровообггу, дихання, що
виникае внаслгдок тяжко? недостатност? печшки.
Класиф!кац!я. За причинами та мехашзмами розвитку видшяють три вида
печшково? коми:
♦ печ?нково-кл?тинну, або паренхгматозну (унаслщок масивного некрозу еш-
телюципв печшки, наприклад, при вгрусних гепатитах, токсичнгй дистрофп пе-
чшки, цироз!, гострих порушеннях печшкового кровотоку);
• портокавальну, або “шунтову ” (у раз! надходження портокавальними анас-
томозами в загальний кровопк речовин, як! в норм! знешкоджуються в печшщ),
що спостерггаеться при синдром! портально! гшертензн;
• змйиану (у раз! некрозу паренхгми печшки 1 наявносп портокавальних анас-
томозгв у хворих на цироз).
В експеримент! печшкову кому моделюють за допомогою методики видалення
1 токсичного ураження печшки, прямо! фгстули Екка (“м’ясне отруення” собак). ।
Патогенез. Основними патогенетичними ланками печшково! коми е:
• збшьшення вм?сту в кров? нейротоксичних речовин, яю утворюються у сами
печшщ пщ час руйнування п паренхгми або надходять гз кишок у печшку г не
знешкоджуються в н!й через порушення детоксикацгйно! функцц ештелюципв
печшки; у раз! наявносп шунпв м!ж воргтною ! порожнистими венами ц! речови-
ни надходять !з кишок у загальний кровопк, минаючи печшку (при синдром!
портально! ппертензп).
До нейротоксичних речовин належать:
• юни амонйо (?Ш4+); при патолргг! печшки порушуеться ц сечовиноутворю-
вальна функщя: амошй, який утворюеться в оргашзм! в результат! метабол!зму
бглюв, не бере участь в орштиновому цикл! та не перетворюеться на сечовину,
через що надмгрно накопичуеться в кров! {г?перамон?ем?я — понад 40 мкмоль/л).
У мгтохондршх клгтин головного мозку амошй з’еднуеться з а-кетоглутаровою
I
кислотою (утворюеться глутамгнова кислота, а потгм глутамш). Це спричинюе
тефшит а-кетоглутарово! кислоти та порушення ЦТК, що зумовлюе зниження
активностг аеробного окиснення глюкози та синтезу АТФ г, отже, зниження енер-
гетичного забезпечення нейронгв. Це спричинюе деполяризацию мембран нерво-
вих клгтин 1 порушення синаптичног передач! з подалыпим розвитком невроло-
ггчно! симптоматики;
• бьгкое! метабол)ти (феноли, гндол, скатол, амши, похгднг меткий ну), що
утворюються в кишках г не гнактпвуються при патологи печгнки;
• низъкомолекулярн) жирн) кислоти (масляна, капронова, валерганова), якг,
взаемодгючи з лппдами мембран нейронгв, гальмують передання збудження;
• пох)дн) тровиноградно'1 та молочно) кислот (ацетогн, бутиленглгколь);
• посилення синтезу несправжнйс нейромедгаторгв (октопамгну, р фенглетил-
амгну, тирамшу); несправжнг кейромедгатори утворюються з ароматичних амшо-
кислот (фешлалангну, триптофану, тирозину), якг у великш юлькосп потрапля-
ють у головний мозок через недостатшсть гхнъого метаболгзму шд час патологи
печгнки; вони гальмують ферменти, що в нормальних умовах забезпечують пере-
творення тирозину на ДОФА (дгоксифешлалангн), а потгм на дофамгн та норад-
реналш; зменшення юлькосп фгзголопчних нейромедгагоргв (норадреналш, до-
фамгн) зумовлюе тяжке порушення функцгй ЦНС, яке посилюеться також унас-
лгдок надмгрного синтезу серотошну з триптофану (серотонгн е нейромед гатором,
що впливае на психоемоцгйш реакнп);
• гтогл)кем1я < унаслщок ослабления глюконеогенезу в патолог гчно змгнених
епгтелгоцитах печгнки), що посилюе дефгцит енергп в клгтинах головного мозку;
• гтокс'ш циркуляторного г тканинного типу;
• порушення КОС I водно-електрол/тного обмшу., у разг порушення метаболгч-
ног функцп печгнки нергдко спостерггаеться негазовий ацидоз через накопичення
пгро вин оград ног, молочног, трикарбонових, жирних кислот, кетонових тш; за цим
пгдвищуеться проникшсть мембран епгтелгоцитгв г збшьшуеться надходження
нейротоксичних речовин у кров, а потгм у клгтини мозку; своею чертою ушко-
дження мембран нервових клгтин сприяе посиленому проникненню в них води,
що спричинюе гх набухания; подразнення дихального центру кислими речовина-
ми та гонами амонгю може зумовити ппервентиляцпо з поддавшим розвитком
газового алкалозу г негативним впливом гшокапш! на мозковий кровотгк, що по-
силюе ппоксло мозку г нервов! порушення при печгнковш ком!; вторинний гтер-
алъдостерошзм, який виникае в разг ослабления метаболгзму альдостерону в пато-
лопчно змгненгй печпщ!, спричинюе порушення водно-минерального обмгну
(збгльшення реабсорбцп №+ в канальцях нефрона) г КОС (негазовий алкалоз).
Порушення жовчоутворювальноТ
та жовчовид1льно[' функцш печгнки
Печгнковг клгтини секретують жовч, до склада’ яког входять вода, жовчн! кислоти
(як жовчн! солг), жовчш пггменти. холестерин, фосфолшгди, жиров! кислоти, му-
цин та гншг речовини.
Печшка бере участь в утворенн!, мегаболгзмт й екскрецп жовчних пггменпв
(схема 26).
Система
мононуклеарних
фагоцита
Кров
Гемолгз еритроцита
I
НЬ
I
Бшрубш (непрямий)
Б1п грубш-ал ьбумг новий
комплекс
Печшка
Жовчнг шляхи
Тонка кишка
Товста кишка
Кал
Нирки
Сеча
Уробшш Стеркобппн
Ентерогепатична
циркулящя
Нижш
прямокишковг вени
Схема 26. Утворення г видыення жовчних шгменттв
У зтрчастих макрофагах печшки, макрофагах юсткового мозку, селезгнки з
гемоглобгну зруйнованих еритроцитгв утворюеться вердоглобш, який шсля вгд-
щеплення атома залгза г глобгну перетворюеться на бшвердин, а той, вцщовлюю-
чись, на бшрубгн. У кровг бшрубш з’еднуеться переважно з альбумшом, утворюю-
чи комплекс, що називаеться некон’югованим непрямим бглгрубгном
(непрямий, осктльки забарвлення з дтазореакгивом Ерлгха дае тшьки шсля осад-
ження бтлка). Вш нерозчинний у водт, у нормт становить 75 % загального бшрубт-
ну кровг (1,7—20,5 мкмоль/л у разг визначення за методом €ндраппка), нетоксич-
ний, не проникае в головний мозок. Вгльний бглтрубтн, не включений у
быгрубш-альбумшовий комплекс, е .тшщорозчгшною речовгшою, яка легко про-
шкае кргзь гемагоенцефалгчний бар'ер 1, взаемодгючи з фосфолптщами мембран
нейронгв. надходить у клггини ЦНС ! може уражувати Ёх. Проте в норм! концент-
ащя вгльного бш!рубшу в кров! наспльки низька, а здатшеть алъбумшу зв’язувати
його така висока, шо вш не справляе токсичну дгю.
Б1ля судинного полюса сштелгоцитгв печшки вщбуваегься захоплення !з кров!
непрямого бппрубшу, вш якого в цитоплазматичнш мембран! вщокремлюеться
альбумш. У перенесенн! бглгрубшу кргзь мембрану клгтини, а потгм з цитоплазми
в мембрану ендоплазматично! егтки ештелюципв печгнки1 беруть участь бглки У
' пгандин) та 2, як! е глутатюнтрансферазами. У мембран! ендоплазматичног сет-
ки вщбуваеться кон’югацгя бглгрубгну з уридиндифосфоглюкуроновою кислотою
год дгею ферменту УДФ-глюкуронозилгрансферази й утворюеться моноглюку-
ровдд бшгрубгну. Бгля бинарного полюса еппелгоцита моноглюкурошд бглгрубшу
переходить кргзь гелатоканалгкулярну мембрану в жовч проти градгента концент-
рацш який утворюеться жовчними кислотами (енергозалежний процес). У жовч-
них канальцях гз двох молекул моноглюкуронщу бглгрубшу утворюеться диглюку-
ронщ бглгрубшу. Таким чином:, у жовч! утворюеться кон’югований з ури-
диндифосфоглюкуроновою кислотою б!л!руб!н (в основному диглю-
куронщ бшгрубшу), який ще мае назву пря мого, оскыьки вгн дае пряму реакцгю
з дгазореактивом Ерлгха. Прямий бшрубш (0,86—5,1 кмоль/л) становить близько
25 % загального бглгрубшу сироватки кров!.
У позапечшкових проточках, жовчному мгхург Ё головним чином у тонкгй
кишц! шд дгею ферменпв кишково! мпсрофлори в!д диглюкурогцду бглгрубшу
ащщеплюетъея глюкуронова кислота (декон’югацгя), быгрубш проникае в кров, а
диглюкуронщ б!л!руб!ну, шо залишився, вщновлюеться до мезобшногену (уробш-
ногену), частина якого всмоктуеться кргзь кишкову стшку в кров г з воргтно! вени
надходить у печЁнку (ентерогепатична циркуляция), де руйнуеться до шрольних
сполук. Оскыьки в норм! уробшгноген не потрапляе в загальний кровотгк, його
немае г в сечь Быьша частина уробышогену в товстгй кишцг вщновлюеться до
стеркобшногену, що виводиться гз калом у вигляд! окисненог форми — стеркобш-
ну (пггмент жовтото кольору). Незначна кглькгсть стеркобышогену проникае в
кров у нижньому вёддыё товстог кишки Ё кргзь нижш прямокишков! вени надхо-
дить у систему нижньо! порожни сто! вени, пгсля чого виводиться Ёз сечею. У
норм! в сеч! мгстяться слщи стеркобшну (1—4 мг на добу). У клшгчшй практищ
цей стеркобыш прийнято називати уробиином як продукт обмгну бшгрубгну, що
виводиться гз сечею.
Порушення жовчоутворювальноТ функцп печшки проявляеться збыьшенням
або зменшенням секрецп жовчг епгтелгоцитами органа, реабсорбщею п компопен-
тгв, шо впливае на кглькгсть г склад жовчё.
Етгологгя. Зменшення жовчоутворення Ё змша складу жовчг можуть спричи-
нити.
• ураження печгнки г жовчних шляхгв у виглядг гх дистрофй та деструкцн (ггри
гепатитг, гепатозг, холецистит!), що зумовлюе роздал утворення компоненпв жов-
‘Еюхгмгчним еквгвалентом мембран ендоплазматичног егтки ештелюципв печшки с м.кросомна
фракцгя. отримана штатом дифере ншалъного центрифугування Це стало основою класифгкаци видгв
печшково! жовтяниш.
I
41 (бйпрубшу, жовчних кислот), секреторно! функцй ештелюципв печ!нки та по-
рушення реабсорбцп компонента жовч!;
• екзогенш й ендогенш фактори, що порушують енергетичний обмш в ор-
гашзм! (ппоксгя, перегргвання, ппотерм!я, отруення щанщами) 1 впливають на
синтез компонент! в жовч!, !х секрешю ештелюцитами та реабсорбцпо складники
жовч! (енергозалежш процеси);
• змши нейрогуморально! регуляцн в раз! посилення тонусу симпатично! час-
тини автономно! нервово! системи, зниження секрецц секретину;
• патолопчн! процеси в тонкш кишщ, доя антибютикгв, що пригшчують фер-
ментативну актившсть кишково! мйрофлори, зменшують ентерогепатичну цир-
куляцпо компонента жовч! 1, як наслщок, концентрацпо !х у жовч!;
• бщкове голодування;
• л!карськ! засоби (естрогени у високих дозах, антим!кобактер!альш, суль-
фаныамщш препарата тощо), що гальмують пдроксилювання жовчних кислот !
!х секрецпо.
Збмьшення секрецй жовч! спостерйаеться в раз!:
• посилення тонусу блукаючого нерва, пщвищення !нкрецп секретину;
• впливу алгментарних фактор!в (жири, яечний жовток);
• застосування деяких л!карських рослин 1 препарата (настш кукурудзяних
приймочок, сорбй, алохол тощо).
Патогенез. До мехашзми, як! зумовлюють яюсш та юльюсш порушення
жовчоутворення, належать:
• змша секреторно! активност! ештелюципв печшки;
• порушення реабсорбцп компонента жовч! у жовчних шляхах 1 кишках та
ентерогепатично! циркуляцп;
• зм1ни транс- та штерцелюлярно! фщьтрацп деяких речовин !з кров! в жовч-
н! капыяри печ!нки, пов’язаш з коливанням осмотичного град!ента 1 станом про-
никносп мембран.
Вщ ентерогепатично! циркуляцп залежить юльюсть реабсорбованих у кишках
жовчних кислот, що визначае штенсившсть !х бюсинтезу в печшц!! впливае на
холатохолестериновий шдекс жовч! — важливий показник п лпогенних властиво-
стей.
Порушення жовчовид!лення
Е т1олог1я. Причинами порушення надходження жовч! в дванадцятипалу
кишку можуть бута:
• мехашчна перешкода вщтоку жовч! — здавлення жовчних шлях!в !ззовю
(наприклад пухлиною головки пщшлунково! залози, запаленою тканиною, руб-
цем) або закупорювання !х (каменем, гельмштами, густою жовчю);
• порушення шнервацп жовчних шлях!в — гшер- або гшокшетачна дисю-
нез!я (наприклад зменшення жовчовидолення внаслщок скорочення м’язових еле-
мента шийки жовчного м!хура);
• розлади гуморально! регуляцн жовчовидыення (наприклад посилення жо-
вчовидщення внаслщок пперпродукц!! секретину, холецистокшшу, мотилшу);
• холецистектом!я (теля видалення жовчного м!хура жовч постшно вид!-
ляеться безпосередньо у дванадцятипалу кишку).
Патогенез порушення жовчовидолення розглянуто нижче (див. “Пгсляпе-
чшкова жовтяниця”).
ПатофЫологгчн! синдроми, зумовлеш порушенням жовчоутворення I жовчовид!-
сння.
Порушення жовчоутворення г жовчовидиення проявляют вся такими синдро-
мами, як жовтяниця, холелня, ахомя, дисхо ия
Жовтяниця (1с1егия) — синдром, що виникае в раз! збтьшення вм!сту в кров!
Ямрубшу (г!перб1г!руб!нем!я) ! для якого характерным е жовте забарв/ення итри,
с шзових оболонок, особливо склер, унасл!док в!дкладення в них жовчних шгментёв.
Класифгкапгя. Залежно в!д первинно!локал1зацН патологичного процесу г ме-
ан!зму виникнення видгляють такт види жовтянищ:
• передпечгнкову, спричинену тдвишеною продукцгею бшрубшу здебшыпого
через посилення розпаду еритроципв (гемолтична жовтяниия). 1 рщше — пору-
шення плазмового транспорту бшрубшу та його надлишкове утворення додатко-
ими (шунтовими) шляхами,
• печшкову (паренх!матозну), зумовлену порушенням захоплення, кон’югацп
та екскрецп бшрубгну ештелюцитами печшки внаслщок ушкодження гх пщ час
чзних патолопчних процесгв (гепатоцелюлярна жовтяниця), внутршшьопечшко-
эго холестазу (холестатична жовтяниця), а також спадкових г набутих дефекпв
структури мембран ештелюципв г ферменпв, яю беруть участь у метаболгзмг г
ранспорп бшрубшу в юптинах печшки (ензимопатична жовтяниця);
• тсляпечшкову (мехашчну), що виникае при утрудненш вщтоку жовч! позапе-
чшковими жовчними протоками.
Передпечшкова жовтяниия
Класифгкацг'я. За механгзмом виникнення розргзняють так! види передпе-
:шковог жовтянищ, як.
• гемолтична жовтяниця, яка розвиваеться внаслщок пщвищеного розпаду
(гемо.пзу) еритроцитгв;
• исунтова жовтяниця (шунтова гтербшрубшем1я) — унаслщок надлишкового
утворення так званого шунтового (некон’югованого) бшрубшу з гемоглобгну ано-
гальних еритроципв через неефекгивний еритропоез або з гема таких гемопро-
тегшв, як мюглобш, цитохроми, каталаза, пероксидаза;
• транспортна жовтяниця, зумовлена порушенням плазмового транспорту
бшрубшу в разг недостатнього утворення бшрубш-альбумшового комплексу че-
рез ргзке зменшення вмгсту альбумгчу в кровг або через розрив зв’язку мгж бглгрубг-
ном та альбумшом.
Етгологгя. Причинами виникнення гемолгтично! жовтянищ е тг сам! факто-
ри, що зумовлюють гемолгз еритроцитгв г розвиток гемолгтичног анемгТ.
Причиною розвитку’ шунтово! передпечшково! жовтянищ може бути:
• неефективний еритропоез у випадку В12-дефщитноТ анемп, який супрово
ркуеться посиленим гемолгзом мегалобластгв юсткового мозку, а також мегало-
цитгв у кровг й селезгнщ, що зумовлюе пщвищений синтез некон’югованого
бшрубшу;
• значнг травми, гематоми, шфаркти в разг порушення кровотоку в ргзних
органах (наприклад легенях, серщ), коли бглгрубш утворюеться з гема мюгло-
бшу.
Одшею з поширених причин виникнення транспортна! передпечшково! жое-
тянищ е дефгнит альбумшу при бглковому голодуванш, у недоношених дней.
Патогенез (розглядаеться на приклад! гемолйпичног жовтянищ). Унаслщок
посиленого внутрёшньосудинного та позасудинного гемолёзу еритроципв у зёр-
частих макрофагах печшки, макрофагах селезенки, кзеткового мозку утворюеться
така велика кёлькесть некон’югованого (непрямого) бёлёрубёну, що ештелёоцити
печшки не здатш повшетю вилучити його з кров! е зв’язати з уридиндифосфоглю-
куроновою кислотою (ыдносна недостатшсть печшки). Кр!м того, гемолетичш от-
рути часто е й гепатотоксичними речовинами, а в уражених ештелёоцитах печшки
метаболизм ё транспорт бйпрубшу порушен!. У кров! збельшусться вмёст непрямого
бшрубшу, який не виводиться ёз сечею, оскшьки зв’язаний з альбумгном плазми.
Може педвищитися р!вень кон’югованого (прямого) бйпрубшу в раз! його зворот-
ноё дифузёё в кров шсля того, як ештелёоцити печшки виявляються вже не здат-
ними екскретувати прямий бёлёрубён у жовч.
За дуже високо! непрямо! гёпербёлёрубёнемп (260—550 мкмоль/л), коли не
весь бйпрубш залучаеться до бйпрубгн-альбумгнового комплексу, розвиваеться
так звана ядерна жовтяниця (забарвлення жовчними шгментами — вшьним
бйпрубшом ядер головного мозку) з ураженням ЦНС ! неврологёчною симптома-
тикою (енцефалопагтя). Це особливо характерне для гемолётично! хвороби (анемп)
новонароджених у раз! Кй-антигенноё несумёсностё еритроципв матер! 1 плода.
Токсична дёя вёльного бйпрубшу на нервову систему може виявитись ё в раз! не-
значного збшьшення ргвня бйпрубшу в кров! на тл! ппоальбумёнемёё, пщвищення
проникносп мембран нервових клгтин (наприклад при гшоксп, ацидоз!).
У випадку гемолётично! жовтянице в печшц!, жовчних шляхах ё кишках син-
тезуеться надлишкова келькёсть глюкурошдёв бйпрубшу, уробйгшогену, стеркобйп-
ногену, у кров! пщвищуеться ргвень стерко- й уробёлёногенёв. Спостерегаеться г1-
перхол1я (збйгьшене утворення жовч! та екскрецп её в кишки), яка спричинюе над-
мёрне виведення стеркобшгну й уробйпну ёз калом г сечею, що зумовлюе гх темне
забарвлення. Але при цьому немае холемгчного синдрому (жовчнё кислоти у кров не
надходять) ё розладу кишкового травления як у разг ахолёчного синдрому.
До гемолгтичног жовтяницё може приеднатися печёнкова (якщо одночасно з
гемолёзом вёдбуваеться ураження ештелёоцитгв печёнки) або механёчна жовтяниця
внаслёдок закупорювання жовчних шляхёв жовчними тромбами г каменями з
бёлёрубёну, холестерину й кальцёю.
Печшкова жовтяниця
Класифгкацгя.За вёдношенням до мембрани ендоплазматично! сётки печён-
кових епгтелгоцитёв, бёохгмгчним еквёвалентом яко! е мгкросомна фракцёя, виделя-
ють кйгька патогенетичних вар1ант1в печёнковог жовтяницё:
• жовтяниця внаслщок порушення захоплення бёлгрубёну ештелёоцитами печён-
ки (премёкросомна абсорбцёйна жовтяниця);
• жовтяниця внаслёдок порушення кон ’югацН бёлгрубёну з уридиндифосфоглю-
куроновою кислотою в мембране ендоплазматично! сётки егптелёоцитев печёнки
(мгкросомна кон’югацгйна жовтяниця);
• жовтяниця через розлад екскрецп бёлгрубёну з епгтелгоцитёв печёнки у жовчнг
шляхи (пгслямёкросомна екск-рецёйна жовтяниця);
• жовтяниця, зумовлена комбшованим порушенням захоплення, кон’югацёё
та екскрецгг бёлгрубёну епётелёоцитами печёнки.
Етёолопя. Причиною виникнення печёнковог жовтяницё е передусгм вплив
факторёв, що зумовлюють ушкодження епётелёоиитёв — ёнфекцгя, токсичш, у тому
числе лгкарсью, речовини, внутрЁшньопечЁнковий холестаз, а також спадковий
дефект мехашзмгв захоплення, кон’югацп г виведення бшрубшу з ештелюципв
печшки. Деяю причини зазначено щд час розгляду патогенезу окремих видев пе-
чшково! жовтяниш.
Патогенез. Жовтяниця внаелгдок порушення захоплення бшрубшу епгтелюци-
тами печшки. У п виникненш мають значения таю мехашзми:
• зменшення вмгсту в епгтелгоцитах печгнки транспортних У- та 2-бглюв, якг
забезпечують перенесения бглгрубшу кргзь цитоплазматичну мембрану з кровг до
клётини (причиною може бути бглкове голодування, а також недостатнгй синтез у
новонароджених у першг днг життя бглка У як головного переносника бглгрубшу);
• конкурентне гальмування захоплення бглгрубшу (рентгенконтрастними ре-
човинами, деякими лгкарськими, наприклад антигельмгнтними, засобами);
• генетично детермшоване порушення структури мембрани судинного полю-
са ештелюцитгв печшки (це призводить до змши проникностг мембрани), розщеп-
лення в нш бглгрубгн-альбумшового комплексу г перенесения бглгрубшу в еш-
телгоцит; при спадковому синдром! Жгльбера — Мейленграхта, який спричинюеться
ргзними генетичними дефектами, спостерггаеться також г порушення здатностт
ештелюцитгв до захоплення некон’югованого бглгрубшу — так звана абсорбцшна
жовтяниця).
При цьому виде жовтяниш некон’югований бйпрубш не потрапляе в ештелЁо-
цити печшки, не вщбуваеться його кон’югацЁя, наслщком чого е г!пербгл!руб!нем!я
за рахунок збгльшення непрямого бшрубшу в кровг, а також зменшення вмгсту стер-
кобшну в сеч! та кал! через порушення утворення глюкуронщгв бглгрубшу в еш-
телюцитах г, отже, ёхнёх похщних у жовчних проточках Ё кишках (зменшуеться
штенсившсть забарвлення калу).
Жовтяниця внаелгдок порушення кон’югацп бшрубшу з УДФ у мембран! ендоп-
лазматично! егтки ештелюцитгв печгнки. Основним механгзмом виникнення ще!
жовтяниш е зниження активностг УДФ-глюкуронглтрансферази, яка каталгзуе
кон’югацгю бйпрубгну.
Причинами розвитку цього виду печшково! жовтяницг може бути:
• зашзнення експресп генгв, що кодують УДФ-глюкуронглтрансферазу в не-
доношених дгтей;
• пригнгчення естрогенами активностг УДФ-глюкурошлтрансферази внаелг-
док п конкуренцп з бшрубшом за зв’язок з ферментом, що спричинюе жовтяни-
цю в немовлят, яю отримують материнське молоко з високим вмгстом естро-
гешв;
• застосування деяких лгкарських засобгв (вЁкасолу), яю зумовлюють конку-
рентне гальмування активност! ферменту;
• спадковий дефгцит УДФ-глюкурошлтрансферази (синдром Кргглера — Найя-
ра, синдром Жгльбера — Мейленграхта з генетичним дефектом синтезу цього фер-
менту — кон ’югацшна жовтяниця).
ВщсутнЁсть або зниження активностЁ УДФ-глюкурошлтрансферази зумовлюе
порушення утворення прямого бшрубгну в епгтелгоцитах печгнки г пгдвищення в кровг
концентрацп некон’югованого (непрямого) бшрубгну. Одночасно в жовчг зменшуеть-
ся юльюсть диглюкуронЁдгв бЁлЁрубгну, що призводить до зниження виведення
стеркобшну з калом г сечею.
Високий ртвень непрямого бшрубтну, наприклад при синдром! Крюгера —
Найяра, спричинюе тяжку енцефалопатпо через розвиток ядерно! жовтянищ
внаслщок надмтрно! кшькост! втльного бшрубшу, його здатност! проникати через
гематоенцефалгчний бар’ер у головний мозок ! вщкладатися в стрш речовин!,
особливо у швкулях великого мозку ! стовбур! головного мозку.
Жовтяниця внаслгдок розладу екскрецп бшрубшу з ештелюцитгв печшки у жовч-
нг шляхи. Основний мехашзм розвитку ц!е! жовтянищ полягае в порушенн! транс-
порту глюкурошдгв бшрубшу з ештелюципв у жовч.
Причиною цього виду жовтянищ може бути як порушення проникносп ци-
топлазматично! мембрани в зон! бшарного полюса ештелюципв печшки (в раз!
цитолтзу ештелюципв, розриву жовчних проточив, згущення жовч! та обтурацп
внутр!шньопеч!нкових проток у випадку внутрпиньопечшкового холестазу), так г
генетичн! дефекти ферменпв транспорту глюкуронццв бшрубтну з ештелюципв
печтнки у жовчнг каптляри.
1зольоване порушення виведення прямого бшрубтну спостерггаеться при
спадкових синдромах Дубгна — Джонсона I Ротора (екскрецшна жовтяниця).
При цих синдромах розвиваються внутршшьопечтнковий холестаз тз потрап-
лянням у кров надмтрно! кшькост! кон’югованого (прямого) бшрубшу, бйпрубш-
ургя 1 ппохолтчний синдром, за якого знижуеться виведення стеркобшну з калом
1 сечею. Для синдрому Дубша — Джонсона, кргм того, характерне вщкладення в
печгнц! лшохромного шгменту.
Жовтяниця внаслгдок комбшованого порушення захоплення, кон’югаци та екс-
крецп бшрубшу ештелюцитами печшки. Основы мехашзми виникнення ще! жов-
тянищ — ушкодження клгтин печшки {гепатоцелюлярна жовтяниця) г внутргшньо-
печшковий холестаз {холестатична жовтяниця).
Причинами розвитку комбшованого (за патогенезом) виду печшково! жовтя-
ницг, яка спостерггаеться найчаспше, можуть бути гепатити втрусного, парази-
тичного (токсоплазмоз), токсичного (у тому числт медикаментозного й алкоголь-
ного) походження, гепатози, цирози печшки (наприклад первинний бшарний
цироз), шфгльтрацгя печшки при лейкоз! та гн.
3 ушкодженням печшкових клгтин пов’язане виникнення сполучення мгж
жовчними шляхами, кровоносними г лгмфатичними судинами, що спричинюе
потрапляння жовч! у кров (холемгчний синдром)! частково — у жовчн! шляхи. При
цьому жовч частково або повшстю не надходить у кишки {г'гпохолгчний або ахолгч-
ний синдром). Набряк у дшянцг перипортального простору також може спричини-
ти зворотне всмоктування жовчг тз жовчних проточкгв у кров. Набухл! клгтини
здавлюють жовчнг протоки, створюючи механгчну перешкоду вщтоку жовчг; роз-
виваеться внутргшньопечшковий холестаз. Метаболтзм г функцц печшкових клттин
порушуються.
При гепатоцелюлярнгй т холестатичнш печтнковш жовтянищ спостерггаються
такт змши вмгсту жовчних тгментгв г жовчних кислот у кровг, сеч!I каш:
• ггпербшрубшемгя', у кровг шдвищуеться ргвень некон’югованого (непрямого)
бшрубшу внаслщок зниження його захоплення г кон’югацп в ештелюцитах печш-
ки в разг ушкодження мембрани ендоплазматично! сттки 1 зниження активностт
УДФ-глюкуронштрансферази; збгльшуеться вмгст кон’югованого (прямого) бшрубг-
ну, який пщ час холестазу ! руйнування ештелюципв потрапляе в склад! жовчг у
кров замтсть його звичайного транспорту в жовчн! кашляри;
1Й ~
• уробшногенемиг, зумовлена нездатшстю ушкоджених ештелюципв печшки
метаболгзувати уробшноген (мезобшшоген), який надходить у печшку завдяки
ентерогепатичнш циркуляцп, що зумовлюе збшьшення його ргвня в кров!;
• холалем1я; поява жовчних кислот у кров! зумовлюе загальну симптоматику,
характерну для холемгчного синдрому)',
• бшрубшур1я', наслщок надходження кон’югованого бшрубшу через мембра-
ни нирок у сечу, що разом з уробшном надае сеч! насиченого жовтого кольору;
• уробйшур1я; е наслщком уробшногенемп;
• зменшення кллъкоспи (сл1ди) стеркобшну в кал! (знебарвлений кал) I сече,
чов’язано з! зменшенням або припиненням надходження жовч! в кишки (гто-
хол1я, ахолгя), що спричинюе зниження утворення похщних глюкуронццв бглгрубг-
ну та появу симптомгв гтохол1чного (ахол1чного) синдрому.
Ушкодження печшкових юптин запально-дистроф!чного характеру в раз! ге-
патоцелюлярно! та холестатичног печшково! жовтянищ супроводжуеться розвит-
ком недостатност! печшки з порушенням ус!х п функшй. При цьому в кров!
з’являються ферменти — показники руйнування ештелюципв печшки (АлАТ,
АсАТ), нерщко порушуеться зсщання кров! (гемораг1чний синдром), можуть вини-
кати печшковг (гтопротешем1чн1) набряки, енцефалопагтя, печшкова кома.
Шсляпечшкова (механична, обтурацшна) жовтяниця
Ет!олог1я ще! жовтянищ пов’язана з порушенням жовчовидглення.
Патогенез. Механгчна перешкода вщтоку жовч! спричинюе застш жовч!
позапечшковий холестаз) г пщвищення Г! тиску понад 2,7 кПа (270 мм вод. ст.),
розширення г розрив жовчних капшяргв, надходження жовч! у кров прямо або
через лгмфатичш шляхи; виникае холем1чний синдром. Ненадходження жовч! в
гшгки через мехашчну перешкоду в жовчних шляхах зумовлюе виникнення
ахол1чного синдрому, найвираженшюго в раз! повно! обтурацн жовчних шляхгв.
Холемочний синдром — це комплекс порушень в орган1зм1, що виникае внаслодок
чадходження жовчних кислот та шших компонентов жовчо у кров.
Вш спостерйаеться не ильки в раз! мехашчно!, а й гепатоцелюлярног та хо-
юстатично! печшково! жовтянищ.
Холемоя — поява жовч! в кров!, що зумовлюе:
• гёпербшрубшемио — збшьшення в кров! вмгсту кон’югованого (прямого)
бшрубгну з подальшим забарвленням шкгри г слизових оболонок у жовтий кол!р
унаслщок вщкладення в них бйпрубшу;
• бшрубшурт — появу кон’югованого бшрубшу в сечг, яка набувае темно-
жовтого забарвлення (“кол!р пива”);
• холалемио (збшьшення вмгсту жовчних кислот у кров!) та холалурио (появу
жовчних кислот у сеч!);
• гтерхолестеринемйо, що зумовлюе утворення на шкгрг обличчя холестери-
нових бляшок (ксантелазм) навколо очей г ксантом на кистях г стопах.
Холалемгя зумовлюе такт патолопчн! змши та симптоми, що визначаються
шею жовчних кислот:
• з боку серцево-судинноо системи: брадикардио, зниження артергального тис-
ку, що пояснюеться дгею жовчних кислот на рецептори г центр блукаючого нерва,
пазухо-передсердний вузол г кровоносю судини; геморагп внаслщок порушення
шлосп судивших стшок;
• з боку центрально! нервово! системи'. загальну астешзацгю, пщвищену стом-
лговангсть, емоцгйнг порушення — дратгвливгсть, яка змгнюеться депресгею, сон-
ливгсть удень г безсоння вночг, головний бить у разг токсично! дп жовчних кислот
на головний мозок;
• з боку шкгри'. свербгж, зумовлений подразненням чутливих нервових закгн-
чень шкгри жовчними кислотами;
• у кровг. гемолгз еритроципв, лейко-, тромбоцитолгз унаслщок детергентно!
дц жовчних кислот на мембрани клгтин;
• у тканинах, ушкоджених жовчними кислотами, — запалення (перитонгг, го-
стрий панкреатит), некроз печгнки.
Лхолгчний (ггпохолгчний) синдром — зумовлений вгдсутнгстю або зниженням
надходження жовч! в кишки внасл!док обтураци жовчних шляхгв або порушення ек-
скреторно! функцп ештелюцитгв печгнки (при механгчшй г печгнковгй жовтяницг).
Для нього характерш такт прояви:
• розлад кишкового травления; через вщсутшсть у кишках жовчних кислот не
може проявитись актившсть панкреатичног лгпази, не емульгуються жири, не
утворюються розчинш комплекси жовчних кислот гз жирними, через що 60—70 %
жиргв не перетравлюються, не всмоктуються г виводяться з органгзму разом з ка-
лом — стеаторея;
• порушення всмоктування жиророзчинних в!там!н!в (А, О, Е, К); призводить
до розвитку вщповщних пповгтамгнозгв, а також геморапчного синдрому при гг-
повгтамгнозг К унаслщок недостатнього синтезу протромбгну та шших факторгв
зсщання кровг;
• порушення рухово! функцп кишок; ослабления тонусу г перистальтики кишок
у разг вщсутностг жовчних кислот спричинюе розвиток закрепу, який нерщко
змгнюеться проносом унаслщок посилення гнильних 1 бродильних процесгв у
кишках, зниження бакгерицидних властивостей жовчг;
• знебарвлення калу, осюльки в кишках не утворюеться стеркобипн;
• вгдсутнгсть стеркобшну в сеч!.
Дисхолгя — синдром, за якого жовч набувае лгтогенних властивостей, що зумо-
влюс утворення жовчних каменгв (холелгтгаз) у жовчному мгхург г жовчних прото-
ках, сприяерозвитку жовчнокам’яно! хвороби.
Етгологгя. Причини дисхолн ргзш:
• запалью процеси, дискгнезгя жовчного мгхура г жовчних проток;
• хвороби шлунка й кишок;
• споживання гжг, багато! на холестерин;
• порушення обмгну речовин (особливо обмгну холестерину, бшгрубгну), у
тому числг спадкового походження, при ожиршш г цукровому дгабетг та пов’язанг
з лгтнгм вжом (пщвищення вмгсту холестерину в жовчг).
Патогенез. Основними мехашзмами набутгя жовчю лгтогенних властивостей
е змгни спгввщношення (концентрацп) компонентгв жовчг та п застш у жовчному
мгхург, жовчних шляхах унаслщок порушення рухово! функцп жовчного мгхура.
Найчастгше (70—90 % випадкгв) унаслщок збгльшення насиченостг жовчг хо-
лестерином г випадшня його в осад у виглядг кристалгв утворюються холестеринов!
камею. Це може бути наслщком зменшення концентрацп в жовчг жовчних кислот
г лецитину, якг в нормг забезпечують розчинний стан холестерину. Зменшення
щтохолестеринового ё лецигинхолестеринового ёндексёв (стввёд ношен ня жовч-
-пх кисло! ё лецитину до холестерину жовчг) може зумовлюватися:
• зменшенням гепатоентерального кругообёгу жовчних кислот у раз) патологи
. шток ё змши ёхньоё мёкрофлори;
• пригшченням синтезу жовчних кислот у печёнш внаслщок зниження актив-
: стё 7а-гёдроксилази;
• прискоренням усмоктування жовчних кислот слизовою оболонкою щд час
-щалення жовчного м1хура;
• зменшенням вмгсту лецитину ё збёльшенням синтезу холестерину.
Тнфекцёя. заспи жовч! також сприяють процесу утворення каменев, осктльки
троводжуються змшою фёзико-хёмёчних властивостей жовч!: зеувом рН у кис-
ти бёк, зниженням розчинностё солей, вин ад ён ням ёх в осад, коагуляцию бшкгв
клётин, шо розпадаюгься.
Крём холестеринових утворюються тгментн! (при гемолёзё еритроцитгв), вапня-
та змгшан! камет (наприклад. холестериново-пёгментно-вапняковё). Наявнёсть
.дменгв зумовлюе порушення жовчовидёлення ё розвиток механёчноё жовтянищ.
"орушення гемсщинамёчноТ функцп печёнки
Для порушення гемодинамёчноё функцц печёнки характерним е:
• зменшення або збйгыиення ОЦК в разё змёни кёлькостё депонованоё в печёнцё
ров: (у нормё близько 800 мл); при шоку, патологёё серцсво-судинноё системи
бётьшення депонування кровг в печёнцё зумовлюе гтоволемйо', вихёд кровг ёз су-
тн печёнки в загальний кровотёк е компенсаторною реакцёсю при крововтратё,
исотнёй хворобё;
• змёни судинного тонусу генератёзованого та локального характеру в системё
юрётноё вени, що проявляеться артерзальною гтотенз!ею та синдромом портально!
•пертензи.
Артерёальна ппотензёя може розвиватися внаслгдок:
• зменшення синтезу в печёнцё ангёотензиногену — субстрату для утворення
шпотензину лёд впливом ренёну;
• гёпотензивноё дц жовчних кислот (при холемёчному синдромё, характерному
для печёнковоё (паренхёматозноё) та механёчноё жовтянищ).
Синдром портально! гтертензи характеризуешься тдвищенням тиску в сис-
тем! вортгноТ вени.
Залежно вёд локалёзашё перешкоди вщтоку кровё видшяють такё види пор-
•палъно! гтертензи'.
• надпечткову (при тромбозё. емболёё. здавленнё нижньоё порожнистоё ё печгн-
.ових вен, правошлуночковш недостатностё серця, перикардии);
• внутрйиньопечшкову (при цирозё, хронёчному алкогольному гепатитё, мета-
газах пухлин у печёнку, коли порушений кровотёк у самёй печёнцё);
• тдпечшкову (у разё тромбозу, здавлення ворётноё вени);
• змйиану (унаслёдок порушення кровотоку як у самёй печёнцё, так ё в позапе-
чёнкових вёшшгах ворётноё або печгнкових вен).
Провёдними ланками в патогенез! портально) ггпертензёё е:
• застш кров! та збыыиення тиску в систем! вортно! вени понад 20 мм рт. ст.
300 мм вод. ст.) за норми 5—10 мм рт. ст. (70—140 мм вод. ст.);
• розвиток колатерального кровообзгу, який опечатку мае компенсаторно зна-
чения для забезпечення вщтоку кров! з печшки через анастомози ворйно! вени з
порожнистими венами, печшковою артерхею х через новоутвореш колатерал! з
печшковими венами (клппчним проявом портокавальних анастомозхв е “голова
медузи” — розширення вен на переднш стшц! живота навколо пупка);
• пгдвищена здатшсть внутрзшнъопечшкових судин до збыъшення в них тиску в
раз! будь-яких структурних змш у печшц! внаслщок вздсутностг клапангв у пор-
тальнш венознш системе,
• варикозне розширення вен стравоходу, кард!ального в!дд!лу шлунка з можли-
вим !х розривом х розвитком тяжких кровотеч',
• недостатшсть кровопостачання паренх1ми печшки, тому що близько 50 с.
кров! пропкае через портокавалън! анастомози, не потрапляючи в паренххму пе- !
чшки, що призводить до порушення вс!х функцш печшки (печшкова недостат-
шсть з можливим розвитком бактерхемп, енцефалопатп та печшково! коми);
• асцит (накопичення рщини у внутрппньоочеревиннш порожнин!);
• мехашзм виникнення асциту:
— збшьшення фшьтрацп плазми в раз! пщвищеного гщростатичного тиску в (
систем! ворхтно! вени, а також зниженого онкотичного тиску внаслщок гптоаль-
бумшемп (порушена бхлоксинтетична функцхя печшки);
— активацхя РААС (секрецхя ренхну пщвищуеться в умовах зниженого цирко-
вого кровотоку у випадку значного депонування кровх в судинах черевно! порож-
нини), що зумовлюе посилення реабсорбцп натрхю в ниркових канальцях х нако-
пичення води в органхзмх, чому також сприяе розвиток вторинного гшеральдосте-
ронхзму як наслщок зменшення шактивацп гормону при порушенш метаболхчно!
функцп ештелюципв печшки;
— збшьшення лхмфоутворення в печшц! пщ час застою в шй кровх х транссу- ]
дащх рщини з лхмфатичних судин у черевну порожнину;
• гепатолгенальний синдром зх спленомегалхею внаслщок застою кров! та по-
далъшого фхброзу в селезшщ, а також 1з пщвищеним руйнуванням еритроцитхв
розвитком анемп.
Порушення кровотворноТ функцн печшки
Порушення еритропоезу в патолопчно змгненш печшцх (гепатит, цироз) пов’язане з! I
зменшенням депонування в нш таких важливих для кровотворення факторхв, я:
щанокобаламш, фолхева кислота, залхзо, що зумовлюе розвиток дефхпцтно! анемп. .
Зниження задания кровз при патолопчних змшах ештелюципв печхнки вини-
кае через зменшення синтезу фактора II (протромбшу), факторхв V, VII, IX, X, XI,
XII, XIII1 фактора I (фхбриногену). Крхм того, при печшковхй х мехашчшй жовтя-
ниц! виникае пповггамхноз К — наслщок ппо- й ахолп, порушення всмоктування
жирхв х жиророзчинних вхтамшхв. Унаслщок дефхциту цього вхтамшу в органхзмх
знижуеться утворення протромбшу та шших факторхв зсщання кровх. Усе це по-
яснюе частоту розвитку геморапчного синдрому при хворобах печшки.
Пздвищеннзо зс1дання кровз, що може спостерхгатись у фазу гшеркоагуляцп ДВЗ-
синдрому, сприяе вцдшення з ушкоджених ештелхоцитхв печхнки у кров прокоагу-
лянтхв, а також зниження синтезу в раз! патологи пе'йнки компонент! в протизсь
дально! та фхбринолхтичнох систем кровх (антиплазмшу, антитромбшу, гепарину).
Р03Д1Л XXX
ПАТОЛОПЧНА Ф13ЮЛ0ПЯ НИРОК
Нирки вщгграють важливу роль у пщтриманш гомеостазу, для якого в норм! е
арактерною сталгсть об’ему рщин (гзоволемгя), гх осмотично! концентрацп
!зоосмгя), генного складу (гзогонгя), концентрацп гошв водню (гзогщргя). Тому тг
ти гншг порушення дгяльностг нирок можуть спричинити виникнення змш зазна-
тених показникгв. Вихщ !х за меж! норми евщчить про порушення основних го-
неостатичних констант, як! неможливо компенсувати, тобто про недостатшсть
-ирок. Водночас змши внутргшнього середовища органгзму, наприклад втрата або
накопичення електролгтгв, води, гонгв водню, можуть гстотно утруднювати або
"орушувати функцп нирок (диселектролгтна нефропатгя — гшокалгемгчна, гшо-
-атргемгчна, у тяжкгй форм! — з ознаками недостатностг нирок).
Не менш важливою е роль нирок щодо виведення з оргашзму продуктгв азо-
тистого обмгну, стороншх речовин. Тому порушення екскрецп речовин е одним з
сновних проявгв недостатностг нирок у разг безпосереднього гх ушкодження, а
-акож унаслщок дп позаниркових факторгв.
Для нирок характерними е штенсивне кровопостачання, високий ргвень
енергетичного обмгну, що визначае пщвищену чутливгеть гх до порушень
кровообггу, дц ргзних токсичних речовин г навгть лгкарських засобгв гз розвитком
патолопчних синдромгв, зокрема гостро! та хронгчно'г недостатностг нирок.
Нирки е важливим не тшьки екскреторним, а й гнкреторним органом, що
сере участь у регуляцп тонусу судин (РАС, простагландини) та еритропоезу,
еритропоетин, гнпбгтор еритропоезу), зсщання кровг (урокшаза). 3 цим пов’язана
висока частота розвитку г тяжкгеть гшертензивного й анемгчного синдромгв на тлг
патологи нирок.
Быковий склад нирковог тканини, особливо клубочкгв, визначаеться ан-
тигенною спорщненгстю до бглкгв сполучног тканини та деяких мгкрооргангз-
пв, зокрема стрептококгв. Це зумовлюе взаемозв’язок деяких хвороб нирок (го-
.трий г хронгчний дифузний гломерулонефрит) з дифузними ураженнями сполуч-
чог тканини стрептококовог природи, а також вщгграе важливу роль у патогенез!
гострого г хрошчного гломерулонефриту за рахунок гмунного й автогмунного
механгзм!в.
Нарештг, оскгльки сеча може бути живильним середовищем для мгкроор-
гашзмгв, а опгрнгсть окремих частин нирок (мозковог речовини) до патогенних
мгкрооргангзмгв невисока, то виникнення деяких хвороб нирок часто пов’язане
з гематогенною або висхщною шфекщею (гострий г хронгчний пгелонефрит).
ПОРУШЕННЯ ОСНОВНИХ НИРКОВИХ ФУНКЦ1Й
ТА ПРОЦЕС1В
Патолопя нирок супроводжуеться порушенням основних ниркових функщй. Ос-
танн! подшяються на функцц, пов’язаш !з сечоутвореиням, 1 таю, що не мають до
нього прямого вщношення.
Функцп, пов’язаш в сечоутвореиням:
1) шдтримання нормально! осмолярносп бюлопчних рщин оргашзму (осмо-
ре гулю юча функщя);
2) пкггримання сталост! ОЦК (волюморегулююча функщя);
3) шдтримання норма!ьного юнного складу кров! (юнорегулююча функция);
4) регуляшя КОС позаюнтинно! ршини;
5) виведення шлаюв !з оргашзму (екскреторна функщя);
6) тдтримання нормального артертального тиску кров!,
Функцп, що не пов’язаш тз сечоутвореиням:
1) утворення еритропоетину та регуляшя еритропоезу;
2) утворення урокшази 1 регуляшя зсщання кров!;
3) метаболтзм витамину О (див. роздал XXIV "Патолопчна фтзюлопя обману
впамгшв”);
4) участь в обмпй вуглевод!в, бонов, жир!в, мйкроелсменттв, вттамппв. гор-
мошв.
Функцц, пов’язан! тз сечоутвореиням, грунутуються на трьох основних нир-
кових процесах: клубочковш фгльтраци, каналъцевш реабсорбиИ та каналъцевш сек-
реци. Порушення будь-якого з них унеможливлюе здшснення одночасно майже
вс!х зазначених ниркових функцш. Своею чертою нормальний перебтг ниркових
процеств т функцш забезпечуеться 1нкрец1ею — виробленням I видтленням пирка-
ми гормонтв 1 БАР.
Порушення процесу клубочковоТфЕльтрацн
Клубочкова фтльтращя, тобто перенесения рщини в!д гломерулярних каптляртв у прс-
спр Боумена за нормальних умов здшснюеться в малъптпевих ттльцях (клубочках,
юркових нефрошв (2 млн) через фильтрапшний бар’ер, що складаеться з феиестро-
ваного ендотелпо. базальной мембрани 1 подошгпв капсули Шумлянського — Бо-
умена. У норм! фыьгруегься близько 20 % плазми, що надходить до клубочка.
Клубочкова фыьтраптя характеризуемся двома параметрами: об’емом филь-
трату, який утворюеться за одиницю часу, що позначаеться як швиднлстъ клубоч-
ковоТ ф1льтрацН (ШКФ), та проникшстю клубочкового фильтра.
ШКФ визначають за кл!ренсом (очищения, с1еагапсе, С) або ендогенного
креатишну, або екзогенного птултну. яю фильтруються. але не реабсорбуються 1 ни
секретуються в нефронах. ШКФ розраховують за формулою:
Ц У Ц-У
ШКФ = або Ссг = —*—, або С =—,
р 7 ,п р
г ст * * т
де Ц— конпентрац!я в сеч! або креатишну. або 1нул!ну; Р — концентращя в плаз-
М1 кров! або креатишну, або шултну; V — дтурез.
За нормальних умов у здорово!' людини ШКФ становить близько 180 л на
добу, або 125 мл/.хв. Це означае, шо вся плазма фцтьтруеться в нирках близько
*ч) разгв на добу, що забезпечуе виведення токсичних продуктов обмшу речовин.
В1д чого залежить ШКФ, видно гз формули:
ШКФ = РГ- КГ,
де Р}'— ефективний фшьтрашйний тиск; К[ — коефппент ультрафшьтрапп.
РГ = Рг.кр. — (Рг.кап. +- Ронк.кр.),
де Рг.кр. — гшродинамхчний тиск кровт в капшярах клубочкгв (у норм! —
'тизько 59 мм рт. ст.), Рг.кап. — пдростатичний тиск первинно! сеч! в капсул!
Шумлянськохо — Боумена (у норм! — близько 15 мм рт. ст.), Ронк.кр. — онкотач-
-ий тиск кровх (у норм! — близько 27 мм рт. ст.).
Таким чином, за нормальних умов РГ® 17 мм рт. ст.
Своею чертою:
КГ= 8 • т|,
ае 5 — плота фФьтращйно! поверхн! клубочкових капхлярхв, яка залежить вщ
ллькост! кхркових нефронхв 1 кшькосп функшонуточих капьтярЩ у кожному клу-
гочку 1 за нормальних умов дорхвнюе 1,6 м2 в обох нирках; г] — проникшсть клу-
бочкового фитьтра для певних речовин.
Зниження ШКФ, а отже, ! зменшення утворення первинно! сеч! з порушен-
ням очищения кров! в!д шлакхв спостерйаеться при зменшенн! або ефективного
фьтьтрацгйного тиску (РГ), або плохш фшырацшно! поверхн! (8).
Зниження РГ спостерхгаеться при зменшенн! Рг.кр або при збшыпенн! Рг.кап.
чи Ронк.кр.
Зниження Рг.кр. своею чертою може бути наслщком як зменшення систем-
ного артерхального тиску, так х спазмування аферентно! артерюли клубочка п!д
пливом анпотензину 1! чи аденозину або тромбозу, або перетискання нирково!
артерц.
Пщвишення Рг.кап. виникае при обструкш! сечових шляххв, а збшьшення
Ронк.кр. — при згущенш кров! чи мхеломнхй хворобе
Зменшення 8 виникае внаслщок загибелх нефронхв, видалення нирки або при
спазмуванн! зхрчастих мезанпальних клхтин, наприклад пхд впливом анпотензину
И, що призводить до зменшення функщонуючих капшяр!в у клубочках.
Порушення клубочково! фыьтрашх знижуе екскреторну функтпю нирок, зумо-
д.тюе затрпмку в кров! продуктов азотистого обмхну 1 пщвищення концентрат! в
хдховх залишкового азоту переважно за рахунок сечовини I сечовох кислоти (ретеп-
ц1йна гтеразотелйя), що спричинюе уремхю (сечокрхв’я) розвиток уремхчно! коми.
Проникшсть клубочкового ф1льтра залежить як вщ його структури, так х вщ не-
гативного елекгричного заряду’ полхашонтв (глткозамхноглхкангв ; схалоглткопро-
теппв) його базальнох мембрани. Тому через клубочкову мембрану проходять неза-
рядженх молекули дхаметром до 4—8 нм, а негативно заряджеш — лише до 2,5 нм.
Пщвищення проникност! клубочкового фыьтра прояв.хяеться клубочковою про-
тешурхею 1 нирковою гематурхею. У норм! через клубочковий фшьтр в!льно про-
ходить лише низькомолекулярнх бшки з молекулярною масою близько 7000 Да,
якх реабсорбуються в проксимальних ниркових канальпях У кгнцеву сечу потрап-
хяе не бшьше 100 мг бшка на добу. Патолопчна клубочкова протешурхя супровоц-
жуеться втратою бшьше нгж 10 г бшка на добу. При зменшеннх негативного елек-
тричного заряду базатьно! мембрани виникае селективна клубочкова протеТнургя з
появою в- сечх бшка з низькою молекулярною масою до 40 000 Да х дхаметром до
4 нм. Такх змхни з боку клубочково! мембрани можуть спостерхгатася в разх фун-
"бгГ
кщональних порушень нирково! гемодинам!ки, спричинених посиленням про-
дукцп адреналину та норадреналшу; у положены! стоячи в дгтей (ортостатична
проте!нургя), при важкгй робот! (маршова протешургя), у раз! втрати р!дини в
немовлят (депдратацшна протегнургя), пщ час охолодження. Проте!нур!я може
виникати шсля споживання гжг з великим вм!стом битка, особливо в д!тей (ал!-
ментарна протегнургя). У быьшост! таких випадк!в функциональна протешургя не-
значна (зазвичай до 1 г/л) г минае шсля усунення п причини.
П!сля ушкодження ниркового фыьтра (у раз! гострого 1 хрошчного уражен-
ня клубочково! мембрани при гломерулонефрит! та шших захворюваннях ни-
рок) виникае неселективна клубочкова протегнургя, що характеризуеться значною
втратою быка (до 120 г на добу) з молекулярною масою понад 70 000 Да т розвит-
ком нефротичного синдрому.
Промтжне положения посщае протешургя, зумовлена недостатнгстю кро-
вообггу, шфекцгйними хворобами, деякими токсичними станами, тиреоток-
сикозом, мехашчною 1 паренхиматозною жовтяницею, ентероколггом, непрохщ-
н!стю кишок, ошками.
У тих випадках, коли быки не видшяються нирками, але домппуються до сеч!
пщ час запального процесу в сечових шляхах, виникае позаниркова, або несправж-
ня, протешургя (быюв зазвичай не быьше 1 г/л).
Ушкодження клубочково! мембрани, зокрема стшки гемокапыяртв, може
супроводжуватися гематургего — виходом еритроципв у проевгт канальцгв г по-
явою у св!ж1й сеч! вилужених еритроцитгв (ниркова клубочкова гематургя), яю
часто набувають вигляду тшей унаслщок часткового гемолгзу. Така гематурия е
одним гз провщних симптомгв вогнищевого нефриту, гострого г хрошчного гло-
мерулонефриту.
Ниркову гематурпо потр!бно диференщювати вщ позанирковог, зумовленог
травмою, пухлинами або запаленням сечових шляхгв. Вщмшними ознаками по-
занирково! гематурц е наявшсть у сечовому осад! велико! юлькосп свгжих неви-
лужених еритроципв 1 быково-еритроцитарна дисоцгацгя (вщсутшсть значно!
юлькосп бглкгв у сечг).
Порушення канальцевих процес!в
Канальцева реабсорбщя. За нормальних умов у канальцях нефрона реабсорбуеться
100 % профыътрованих глюкози г амшокислот, 99,5 % натр!ю, 99 % води г близь-
ко 50 % калпо. Причому в проксимальних канальцях реабсорбуються вся глюкоза,
амшокислоти I быки та 65 % натр!ю г води. Натргй тут реабсорбуеться пщ впли-
вом анпотензину II, катехоламгнгв тощо за мехашзмом первинно активного
транспорту за допомогою №+-, К+-АТФази, а амшокислоти, глюкоза, ангони,
впамши, лактат, мгкроелементи реабсорбуються через люменальну мембрану не-
фроципв за допомогою ко-переносникгв разом з гонами натрпо за мехашзмом
вторинно активного транспорту, пов’язаного з! створенням за допомогою №+-,
К+-АТФази натргевого град!ента. При цьому натр!й перемгщуеться за сво!м кон-
центрацгйним гращентом, а супутнг речовини — проти градгента. Вода г сечовина
реабсорбуються за мехашзмом полеплено! дифузп. Слщ нагадати, що лпгщна
мембрана клиин не пропускав воду г водорозчинн! речовини. Мембрани стають
проникними для води тгльки за наявностг водних каналгв, роль яких виконують
. ки аквапорини, як! також транспортують сечовину, вутлекислий газ, глщерин.
рвинно 1 вторинно активний транспорт дас змогу реабсорбувати речовини про-
га конце нтраишн ого градгента, а дифузгя води здгйснюеться пльки в напрямку
гсмотичного градгента, створеного головним чином первинно активним транс-
:этом натрпо.
В ашкальну мембрану нефроггитгв проксимальних канальцгв, як г в бгльшгсть
"тин органгзму, вбудована велика кгльюсть быка — переносника води — аква-
адину-1, а в базальну мембрану — аквапорину-3. Завдяки цьому сеча тут гзоос-
:.тярна кровг.
У низхгднш частинг пет л г Генле реабсорбуеться 10 % води, шо профгльтрува-
ися, без реабсорбцп №+, а у висхщнгй частин), де немае аквапорингв, — 25 %
'-.а без Н,О. Це зумовлюе когщентрування сечг.
У збгрних трубочках пщ впливом альдостерону, шо вгдкривае натргевг канали
нюменальнгй мембранг нефроцитгв, натргй пограпляе в цитозоль нефроцитгв за
. еангзмом полегшейО1 дифузп. Це зумовлюе активацгю №4-, КД-АТФази на ба-
_1ьнгй мембранг. Унаслщок цього через базолатеральну мембрану збгрних трубо-
к за механгзмом первинно активного транспорту реабсорбуеться лише 4—5 %
зтргю, але це дае змогу точно пщтримувати натргевий баланс.
Вода г сечовина в них канальцях, у яких немае аквапорину-1, реабсорбуеться
з механгзмом полеплено! дифузп лише пгд впливом вазопресину (антидгуретич-
ни гормон), який активуе аквапорин-2. Цей быок за вщсутносп вазопресину
.титься в цитозол! у вигляд! пухиргггв 1 вбудовуеться в мембрану нефроцитгв
нше пщ впливом вазопресину. Тому без антидгурегичного гормону стгнки збгр-
их трубочок не пропускають воду г сечовину. Завдяки цьому сеча концентруеть-
я або розводиться, що пщтримуе нормальна осмолярнгсть г об ем позаклгтинног
дини.
1ншим прикладом полеплено! дифузп е перемгщення глюкози через базальну
ембрану нефроггитгв за допомогою быка-переносника ГЛЮТ-2. Так само, за
эпомогою спецгальних переносникгв, через базатьну мембрану транспортуюгься
игнокислоти га ггшпг волорозчиннг сполуки.
Бглки реабсорбуються через люменальну мембрану проксимальних нефро-
_итгв за допомогою енергозалежного ендоцитозу. Ендогхитознг пухирцг зливаютъ-
. я з лгзосомами, вщбуваеться розщепленггя бглкгв до амгнокислот, яю транспор-
туються через базолатеральну мембрану за допомогою кглькох переносникгв г
.•траггляють у кров.
Канальцева секретя також здгйснюеться за механгзмами (див. схему 18 у XXI
щздглг) первинно-активного транспорту (Н+-АТФаза, К+-, Н+-АТФаза), вторин-
но активного транспорту (К+/ С1_-котранспортер г С1“ / НСО.-- та №+/ ЫН/-
лротипереносники) 1 полегшено! дифузп (калгевг та хлорнг канали). У прокси-
мальних каналыхях секретуються переважно катгони (К1, Н+, МН/) в обмгн на
Иа+, а в дистальних каналъггях г збгрних трубочках — переважно ангони.
За етюлопею порушення канатьцево! реабсорбцп та секрецп подгляються на
спадковг г набутг.
Спадкова недостатшсть реабсорбцг! може бути пов’язана з вщсутнгстто певних
переносникгв або селективних гонних каналгв. Частгше спадковг тубулопатг! су-
проводжуються парцгальним порушенням реабсорбцг! або секрецп речовин. При-
чалами таких порушень е спадковг фосфатургя, амгноацидурц, глюксзургя тошо.
Слмейний гтофосфателйчний втамш-В-резистентний рахйп (див. с. 445)
проявляеться значним пщвищенням добовоё екскрецёё фосфатов ё, як наслёдок,
зниженням ревня ёх у плазм! кров!, нормальним або дещо зниженим р!внем каль-
щю в крове, пщвищеного кальщуреею, демёнералёзацёею юсток, рёзними формами
рахёту (у детей) та остеомаляцёею (у дорослих), резистентними до лёкування ерго-
кальциферолом у високих дозах.
Для цього типу рахёту характерним е первинне порушення здатностё нирок
реабсорбувати фосфати в проксимальней частин! канальцев нефронев або педви-
щення чутливосте канальцев до паратгормону (псевдогшерпаратиреоз).
За спадково) остеодистрофи (псевдогёпопаратиреоз) спостерёгаються так!
саме змёни в крове, як ё за справжнъого гёпопаратиреозу, а саме гёпокальцёемёя,
геперфосфатемёя. Проте застосування паратгормону не усувае це змёни ё не пёдви-
щуе екскрецёео фосфатёв ёз сечею внаслёдок резистентностё канальцев нефронев
до дп гормонёв. Вважають, що така резистентнёсть зумовлена вёдсутнёстю або де-
фектом вёдповёдних рецепторёв до паратгормону в канальпях нефронёв.
Основним механезмом виникнення спадково'1 амшоацидури е спадково зумо-
влений дефёцит натрей-амёнокислотних переносникёв, що беруть участь у вторин-
но активному транспортё амёнокислот. Описано спадковё тубулярнё синдроми,
для яких характерними е цистинурёя, глёцинурёя тощо. Бёльшёсть ёз них супро-
воджуеться симптомами сечокам’яноё хвороби та её ускладнень.
Комбёнованё тубулопатП. Анатомёчна подёбнёсть структур ё певна бёохёмёчна
спёльнёсть, особливо енергозалежнесть механезмёв, що забезпечують реабсорбцею
глюкози, фосфатёв, амёнокислот, спричинюють комбёнованё порушення канальце-
вих процесёв ё появу вёдповёдних тубулярних синдромев. Найвёдомеше з них —
глюкофосфатний деабет, глюкозурёя з амёноацидуреею, комбёнованё порушен-
ня реабсорбцеё амёнокислот ё гёдрокарбонатёв, фосфатёв е деяких аменокислот.
Найскладнешим за проявом е найтяжчим за клёнёчним перебегом е синдром Фан-
кони Для цього генетично зумовленого патологёчного синдрому характерне одно-
часне порушення реабсорбцеё глюкози, фосфатёв, гёдрокарбонатёв, аменокислот, а
також розвиток канальцевого ацидозу (унаслёдок втрати гёдрокарбонатёв) е гёпо-
калёемёя. У деяких випадках спостерёгаються порушення концентрацёйноё здат-
носте нирок е зневоднення оргашзму внаслёдок осмотичного дёурезу. Як правило,
цей стан супроводжуеться гёпофосфатемёчним рахётом з нормокальцеемёею
резистентним до лёкування ергокальциферолом.
Набутё порушення реабсорбцее можуть бути пов’язанё з недостатнёстю секрецёё
гормонёв (альдостерону, вазопресину) або з вёдсутнёстю рецепторёв до цих гор-
монёв (наприклад ниркова форма нецукрового деабету за вёдсутностё або при бло-
кадё рецепторёв до аеггидёуретичного гормону). Для такого стану характерне видё-
лення велико! кёлькостё сечё низькоё вёдносноё густите (сёеаЬеёез ёпяёрёсёиз — неееук-
ровий дёабет, або нецукрове сечовиснаження). Можлива також блокада аквапо-
ринёв-2, що забезпечують трансцелюлярешй транспорт води через нефроцити
збёрних трубочок за наявностё аеггидёуретичного гормону. Наприклад, так деють
ртутне дёуретики, що блокують 8Н-групи аквапоринёв.
Надлишок гормонёв, навпаки, може патолопчно посилювати парцёальну ре-
абсорбцёю натрёю (альдостерон, анпотензин II при вторинному гшеральдосте-
ронезмё) або води (гшерпродукцёя антидеуретичного гормону при синдроме Пар-
хона) та знижувати реабсорбцею фосфатёв (паратирин).
1шемгчнг або токсичн! ураження каналъщв нефрошв порушують синтез у не-
фроцитах АТФ з блокадою роботи Ха"-, К1 -АТФази, шо зумовлюе розлад первин-
-о активно! реабсорбцп натрпо. Це своею чертою блокуе роботу вторин но актив-
ого транспорту, що здшснюеться натршзалежними переносниками глюкози. амг-
аокислот, ангонгв, мгкроелеменпв тощо Тобто виникае тотальне порушення
-лнальцево! реабсорбцп. Клшгчну картину, под!бну до тако! при синдром! Фан-
той!, спостерп'али в раз) отруення солями важких металгв (ртуть, свинець, уран).
Амшоапидургя може розвиватися також як наслщок посиленого катаболгзму
люв, порушення промгжного обмшу амшокислот за умов гшоксп, голодування,
ефшиту нгкотиново! кислоти, вгтамппв групи В, шд час ошковог хвороби, шфарк-
ту мтокарда, тяжких уражень печшки. Виникнення п пов;язане з гшерамшоацид-
ешею 1 перевантаженням транспортного механгзму канальи1в нефрошв унаслщок
збшьшеного надходження амшокислот до ультрафильтрату.
Екстраренальна амгноацидургя в деяких випадках е наслщком спадкових
вороб обмшу речовин г вторичного ураження канальщв через збшьшення вмгсту
в кров! амшокислот (феншкетонургя, лейпиноз, гшерпролшемгя. пперглшинемгя.
цистиноз, оксалоз тощо).
Набуте порушення реабсорбцп може бути 1 парцгальним. як, наприклад. при
етруеню глгкозидом флоридзином, що мютиться в смол! фруктових дерев, вини-
кае глюкозур!я внаслщок селективно! блокади натр!й-глюкозного переносника.
Порушення функцй проксимальных канальщв, що розташован! в кгрковш речо-
зинг нирок, проявляеться зниженням реабсорбцп глюкози, амшокислот, фосфа-
ту, пдрокарбонату, бшка ! появою гх у сеч!. Виникае селективна канальцева про
тешур1я. шо харакгеризуеться появою в сеч! невелико! юлькосп б!лка з низькою
поле куля рною масою (вш 7 000 до 40 000 Да), який може фшьтруватися ! в норм!
майже весь реабсорбуеться.
У раз! ушкодження та руйнування клпин канальшв у сеч! з являеться велико-
молекулярний быок разом гз зернистими, палшовими й епгтелгальними цилпщра-
ми, що е злшками просв!ту канальщв. Це неселективна канальцева протегнурш.
При ушкодженш та порушены функцн прямих канальщв петлг Генле / зб1рних
трубочок, шо розташован! в мозковш речовин! та сосочках нирок, порушуеться
гормонально-залежна факультативна рсабсорбцгя натр!ю г вещи. Унаслщок цього
нирки втрачаюгь здатшсть концентрувати ! розводити сечу. Виникае йостенурш,
за яко! вщносна густина сеч!, яка в норм! коливаеться вы 1,002 до 1,035, станс-
вить 1,010, як г у первиннш сеч!,! не змшюеться залежно в!д режиму споживання
.подиною води. 1зольоване порушення реабсорбцп води при недостатност! вироб-
лення або рецепцп вазопресину (антидгуретичного гормону) або ушкодження ме-
хангзму концентрування сечг за допомогою протиплинно-поворотно-мн ожильно!
системи, яка залежить вщ ргзниц! проникност! для натрпо, води г в першу черту
сечовини стшок ргзних дшянок прямих каналыггв, — гтостенурш. Вона характе-
ризуеться зниженням вгдггосног густини сеч! нижче 1.010.
Порушення секреш’1. Спадковими або набутими можуть бути також порушен-
чя секрецц, що спричинюють канальцев! форми негазового ацидозу:
• ушкодження проксимальных канальшв порушуе в них процеси канальцевого
ацидо- й амонгогенезу, що знижуе секрещю йонгв амонпо ! водню в обмш на
йони натрпо, якг реабсорбуються. Це зумовлюе виникнення проксимального ка-
нальцевого аппдозу (див. роздгл XXI “Порушення кислогно-основного стану”);
• при ушкодженш збгрних трубочок порушуеться секрешя юнш водню в обмш
на юни калпо або натрпо (див. роздш XXI “Порушення кислотно-основного ста-
ну”), що спричинюе розвиток дистального канальцевого ацидозу.
Порушення секреци сечовог кислоти супроводжуеться пщвищенням концен-
трацц в кров! сечово! кислоти та ц солей (г!перурикем!я), розвитком нирково'1
форми подагри. Такий стан часто е спадково зумовленим 1 передаеться за дом1-
нантним типом.
Порушення секреци стороншх речовин: лжарських засобгв (антибютики), барв-
ник1в (фенолрот), йодовмкних рентгеноконтрастних препарата тощо, що ушко-
джують нирков! каналъщ, особливо проксимальш — спостерхгаеться у раз! ура-
ження нирок 13 тяжким тубулоштерстицшним синдромом. Затримання в кров!
цих речовин, зокрема пешцилшу ! продукта його перетворення, може супрово-
джуватися гх токсичною дхею.
ПАТОГЕНЕЗ НИРКОВИХ СИНДРОМ1В
Через обмежешсть ниркових пропета, що забезпечують численш функцп нирок,
клппчна картина ниркових захворювань, як! спричинюються р!зними етюлопч-
ними факторами, мае багато спыьного 1 проявляеться чотирма великими ниркови-
ми синдромами, а саме:
• нефротичним синдромом;
• построю нирковою недостатшстю;
• хрошчною нирковою недостатшстю;
• тубулоштерстицшним синдромом.
Ц1 синдроми можуть супроводжувати один одного або переходити один в
шший, змшюючи картину перебпу захворювання. Своею чертою вони характери-
зуються виникненням на певних етапах шших ниркових синдромов, до яких на-
лежать:
• сечовий синдром;
• набряковий синдром;
• ппертошчний синдром;
• анем!чний синдром;
• урем!чний синдром.
Сечовий синдром — це змши кшыасного та яюсного складу сеч!, що виника-
ють унаслщок ушкодження клубочк!в 1 канальщв: ол!гур!я, анур!я, пол!ур!я, про-
тешур!я, гшо!зостенур!я, глюкозурхя, амшоацидур!я, фосфатур!я, еритроцитур!я,
лейкоцитурхя, цилшдрур!я.
Слщ зазначити, що за винятком анурп, еритоцитурп та клубочково! протеш-
ури, ус! ппш змши в сеч! пов’язаш з ушкодженням ниркових канальц!в. Визна-
чення сечового синдрому належить до найпоширешших лабораторних дослщжень
1 е важливою д!агностичною ознакою ушкодження нирок. Потр!бно також зазна-
чити, що змши величин и д!урезу можуть бути не пов’язаш 1з функщональним
станом нирок ! залежать вщ водного балансу оргашзму, який завжди потр!бно
враховувати.
Набряковий синдром. Патолопя нирок часто супроводжуеться розвитком на-
бряюв, як! спочатку проявляються характерними “мшгками” пщ очима, а згодом
.причинюють накопичення рщини в плевр!, перикард! тошо Збыьшення загаль-
но! юлькосп рщини в оргашзм!, особливо в позаюптинному сектор!, розвиваеться
як наслщок зросгания реабсорбцн натр!ю (первинно) та води (вторинно). Слщ
зазначити, що при патологи' нирок гол овну роль у пщвищенш реабсорбцп натрпо
ыдгграе пперсекрещя альдостерону внаслщок активацп РААС. Своею чертою при-
чини цього можуть бути як позанирковими, так 1 нирковими. Пперсекрещя атьдо-
стерону може бути наслщком первинного або вторинного гшеральдостерошзму.
Первинвий гшеральдостерошзм виникае при доброякгсних пухлинах клубоч-
ково! зони надниркових залоз, як! синтсзують надлишок альдостерону. У цьому
ипадку набряки не виникають. Це пов’язано з тим, що збыьшення реабсорбцн
натрпо в дистальному вщдцп нефрона 1 в зб!рних трубочках при цьому компен-
суеться зменшенням реабсорбпп натрпо в проксимальному вщды! нефрона. Це
зумовлено тим, що при первиннш пперпродукцн альдостерону приппчуеться
синтез реншу й анпотензину II.
Набряки з’являються тыьки при вторинному тшеральдостерошзм!, за якого
здночасно збыьшуеться продукшя решну, анпотензину III альдостерону, а отже,
посилюеться реабсорбщя натрпо як у проксимальних канальцях. так 1 в збгрних
трубочках. Це спостерггаеться зокрема в раз! серце во-судин но! недостатност!.
коли зменшення УОС та ХОС спричинюе зниження артер!атьного тиску й акти-
ващю РаАС. При цьому зростае реабсорбцгя натр!ю, за яким реабсорбуеться вода,
шо г зумовлюе появу набряк!в унаслщок збыьшення об’ему позаклгтинно! рщини.
Другою причиною вторинного пперальдостеронгзму е зменшення онкотичного
тиску кров! та об’ему внутр!шньосудинно! рщини при ппопротешемп, що вини-
кае при голодуванш, патологи печшки г, нарешп, при нефротичному синдром!
(див. патогенез нефротичного синдрому у роздгл! XXX).
Синдром артергальноТ ппертензн розвиваеться внаслщок або активацп РАС !
утворення потужного вазоконстриктора — анпотензину II, який здатен впливати
на спстемний кровообгг, або зменшення продукцп в мозковш речовин! нирок
вазодилататорних простагландин! в при ушкодженн! и та сосочктв нирок при
хрошчному шелонефрит!. Акгиващя РАС вщбуваеться при порушенн! ниркового
кровотоку, здебшьшого в раз! гострого або хрошчного ураження клубочюв нир-
кових т!лець (вогнищевий ! дифузний гломерулонефрит, дгабетичяий гломеруло-
склероз), звуження або закриття проевпу ниркових артерш або гхнгх плок через
аномал!!' розвитку, атеросклероз, тромбоз, емболпо, здавлення рубцем або пухли-
ною, при гострш ! хронгчнш недостатност! нирок.
Мехашзми виникнення нирково! гшертензп викладено в роздш XXVI “Пато-
лопчна фгзюлопя системного кровообггу”.
Анем!чний синдром. При хворобах нирок, яю супроводжуються порушенням
функцп клубочюв г канальцы (гострий г хронгчний гломерулонефрит, дгабетична
нефропапя, амыо’щоз, шелонефрит тошо), часто спостерйаеться анемы. Як пра-
вило, вона е нормоцитарною, нормохромною, гшорегенераторною.
Патогенетично виникнення таких вид!в анемп пов’язують здебгльшого з! зни-
женням продукцп еритропоетину на тл! збыыпено. секрецп шпбтгора еритропое-
зу. Наслщками зменшення або випадання еритропоетично! функцп нирок е галь-
мування синтезу ДНК в еритропоетинчутливих клгтинах кгсткового мозку, пору-
шення !х пролгферацп та диференщювання, приличен ня дозргвання нормопипв
! виходу ретикулоциты з юсткового мозку в кров.
Додаткову роль у розвитку нирковс! анемп вгдгграють пригнгчення функшй
ктсткового мозку азотумгсними речовинами, гематурия, наявшсть геморапчного
диатезу, а також зумовленг цими факторами г зниженою реабсорбшею дефшит за-
лгза, в грата трансферину 13 сечею за значно! протешурп, дефщит щанокобаламгну.
Уремгчний синдром е наслщком значного зниження ШКФ г накопичення у
кровг токсичних продукпв обмшу речовин. Уремгя (сечокргв'я) зумовлена здебыь-
шого накоттиченням у кровг сечовини, сечово! кислоти, креатиншу, меншою
игрою — амшокислот, а також токсичних продуктгв (шдикану. фенолгв, гндолу.
скатолу, молекул еередньо! маси — продуктгв ггдролгзу быка), шо утворюються
внаслщок гнитгя в кишках.
Рзвень азотемп може бути ргзним: вщ 35,7 ммоль/л (дещо псревищуе верхню
межу норми залишкового азоту) до 142.8—357 ммоль/л. Визначальним фактором
е ступшь зниження ШКФ
Унаслщок порушення екскреторно! функш! нирок при зменшенш ШКФ за-
тримуеться також виведення фосфатгв, сульфалв та оргашчних кислот, пщви-
щуеться концентрация !х у кровг.
При ретенцшнш азотемп та уремп виникае штоксикашя оргашзму, описана
нижче (див. патогенез хронгчно! нирково! недостатностг). Смерть настае вщ уре лич-
но! коми, що розвиваеться внаслщок отруення мозку азотистими шлаками.
Нефротичний синдром
Нефротичний синдром еупроводжуе ргзш за етголопею захворювання нирок, яю
проявляюгься ушкодженням клубочкового фгльгра г масивною неселективною
клубочковою проте!нур1ею, гшопротегнемгею, гшоонюею, набряками, диелротеш-
емгею г гшерлшщемгею. Характерним прикладом такого захворювання е гломеру-
лонефрит, за якого вщбуваеться гмунокомплексне або цнтотоксичне автогмунне
ураження клубочюв нефрона.
Етголопя. За походженням нефротичний синдром подшяють на первинний
г вторин ний.
Первинний нефротичний синдром не пов’язаний тз жодною попередньою хво-
робою нирок. Здебыгьшого причиною його е генетично зумовлений дефект обмг-
ну речовин (лшощний нефроз) або трансплацентарне перенесения специфтчних
лротинирковггх антитш вщ матер! до плода (вроджений родинний нефроз) тощо.
Вторинний нефротичний синдром зумовлений деякими хворобами нирок (гло-
мерулонефрит) або шших органгв (нефропатгя вагпних, цукровий дгабет, амыо-
щоз, системний червоний вовчак, сироваткова хвороба, стафшококовий сепсис
тощо). Спостерйаеться також теля отруення солями важких металгв, у разг знач-
них опгкгв, променевого ураження, вщторгнення ниркового трансплантата, засто-
сування деяких лгкарських засобгв (сульфаншамщш препарати, пеншилгн, корти-
костерощн1 гормони), порушення кровопостачання нирок.
Патогенез. У бывшосп випадктв розвиток нефротичного синдрому зумовлений
гмунними механгзмами, здебтл ьшого ггперчутливтсгю сповшьненого типу. При цьому
антигени можуть бути як екзогенного (бакгергальнг, втрусш, паразитичю, лгкарськг.
харчовг, сполуки важких металгв тощо), так г ендогенного (ДНК, денатуроваш нуклео-
вротещи, быки пухлинного походження, тиреоглобулш) походження. Антипла, шо
утворюються у вщповщь на надходження антигентв. належать переважно до 1§М.
Ураження клубочки ниркових тыець пов’язують Уз вУдкладенням на поверхш
або в сами базалънУй мембран! кагплярУв амУлощу, глУко- У лшопротещтв, фтбри-
ногену з актива шею гуморальних т клттинних мехашзмгв запальнот реакци. Як
наслщок, втрачаеться структурна цшснУсть базальноУ мембрани. змшюються и
склад У фУзико-хУмУчнУ властивостУ, рУзко гпдвищуеться проникшсть клубочковото
фУльтра для бУлю в плазми кровт.
У розвитку нефротичного синдрому беруть участь Т-лУмфоцити, якУ продуку-
ють фактор, який активуе хемотаксис нейтрофшв У Ух ушкоджуючу дпо, шо спри-
чинюе пщвищення проникностУ каптляртв клубочктв. Одним Уз механУзмУв може
бути втрата негативного заряду базалъно'У мембрани кашляртв. шо сприяе полег-
шешй фгльтрапи дрУбнодисперсного бУлка через кашлярнт мембрани клубочктв до
ультрафУльтрату, що своею чертою зумовлюе значку селективну клубочкову протеш-
уртю (понад 3,5 г на добу). Основним ланцюгом патогенезу нефротичного синдро-
му е масивна неселективна клубочкова проте'Унуртя та спричинена нею ппоать-
бумшемУя (менше нУж 35 г/л). Зменшення концентрацУУ альбумине у плазмУ кровт
призводить до зниження ртвня онкотичного тиску, шо е причиною переходу час-
тини ртдини. яка перебувае в судинному руслУ, у позасудинний проспр.
Зниження внаслУдок цього ОЦК зумовлюе зменшення юлькостт кровт, що
надходить до серця, У, як наслУдок, зниження УОС Це е проявом серцево'У недо-
статностт, для лУквщаци яко’У необхУдно збУльшити об'ем кровт за рахулток затрим-
ки нирками натртю та води. Така адаптивна реакцтя нирок вщбуваеться за раху-
нок активашт РААС унаслщок пщвищення активностт симпатичного вУддтлу не-
рвово‘1 системи та зменшення ниркового кровообтгу. ОкрУм того, зменшення
притоку венозноУ кровт до серия супроводжуеться зниженням розтятування пе-
редсердь, особливо правого, що спричинюе гальмування секреци цими вщдУлами
передсердного натрУйуретичного пептиду, унаслщок чого його пригнУчувальна дтя
на секрецтю альдостерону також знижуеться. Усе це за механУзмом порочного
кола призводить до затримки в органУзмУ натрпо У води та посилення гУпоонки У
набрякУв.
ОкрУм змш водно-електролУтного обмУну при нефротичному синдромУ вини-
кають мегаболтчнУ порушення. Основним Уз них е гУперлУпУдемУя — пщвищення в
кровт ртвня винного й етерифткованого холестерину, УнодУ триглУцеридтв, що
пов’язують також Уз ппоальбумУнемУею У зниженням у кровт вмУсту ферменттв,
нсобхщних для розщеплення лшоттротещУв та мегаболУзму лшщУв. При нефротич-
ному синдромУ порушуеться У гемостаз Уз проявами пперкоагуляшт.
Гостра ниркова недостатшсть
Гостра ниркова недостатнУсть (ГНН) — патолопчний синдром рУзнот етУоло-
ги, який характеризуеться значним У швидким зменшенням ШКФ, що порушуе
здатшсть нирок пщтримувати нормальний об’ем У склад рщин органУзму.
ЕтУологтя. Фактори, що зумовлюють розвиток ГНН, можна об'еднати в три
групи: преренал ьш, ренальнУ, постренальнУ.
Пререналый фактори пов’язаш зУ зниженням системного артертальною тиску
внаслщок:
• зменшення ОЦК теля крововтрата, оптку. невтримного блювання, профуз-
ного проносу, рУзкого зменшення об’ему внутртшньосудиннот та лозаклУтиннот
рщини в раз1 застосування д!уретик!в, значного потовидшення чи гтервенти-
ляцп;
• ргзкого розширення щд д!ею вазодилататоров об’ему циркуляторного русле
при судинних формах шоку (септичний. анафыактичний), колапс!;
• порушення насосно! функцп серия при гострш (шфаркт мюкарда. емболк
легенево!' артерп) та хрошчшй недостатносп серця.
Реналыа фактори:
• мгсцевх порушення кровооб)гу в нирках (тромбоз, емболк нирковсЛ артери,
тромбоз ниркових вен, ппемк. зумовлена преренальними факторами, синдром
дисемшованого внутршшьосудинного зсщання кров!);
• гостро запал ьн! хвороби нирок (гострий штерстишйний нефрит, гострии
гломерулонефрит, васкулп);
• нефпотоксичш впливи (антибютики, сол! важких металгв. органично роз-
ни нники. рентгенконтрастш речовини, грибна 1 змпна отрути, анаеробна шфек-
щя, ендогенна 1нтоксикашя в раз! токсикозу ваптних, д!абетично! коми, сепсису,
перитошту, недостатност! печшки);
• ушкоджувальна ды шгментш (гемоглобшу — в раз! масивното внутр!шньо-
судинного гемолозу, мюглобшу — масивного травматичного г нетравматичного
рабдомюлгзу).
Постреналът фактори:
• обструкшя сечо водш (камен!, пухлина, згустки кров!, некротичш маси —
зсередини; пухлина, збгльшен! лгмфатично вузли, спайки — зовн!);
• затримка видшення сеч! на ровш виходу !з сечового м!хура (аденома, пухли-
на передмохурово! затози, камею).
Патогенез ГНН головним чином пов’язаний зг спазмуванням приносних
артерюл клубочков пщ д!ею анпотензину II, що е наслщком активацн РАС через
зниження аргергального тиску й активацп у зв’язку !з ним симпатико-адреналово!
системи. Другою причиною активацп утворення анпотензину II е порушення ре-
абсорбцп натр!ю нефропитами щшьно'о паями (таси!а <1епза) при ураженнях ка-
нал ьшв, що сприймаеться секреторними клотинами юкстагломерулярного апарату
як недостатшсть натр!ю в оргашзм!. Спазмування аферентних артерюл зумовлюе
зменшення ШКФ, виникнення шгемп к!рково! речовини нирок, п гшоксп та не-
крозу. Додатковими факторами зменшення ШКФ при ГНН е зменшення плош!
функшонуючих кашляр!в клубочков при спазмуваню мезанпальних клотин пщ
впливом анпотензину II, мокротромбоз ниркових судин г пщвищення тиску в
канальцях нефрона.
У клипчному перебггу ГНН видшяють 4 стадп:
• почат кову, або латентну, що тривае вщ кшькох годин до кшькох д!б.
• ол!гурй (зменшення д!урезу нижче 500 мл на добу), анурп (зменшення дгу-
резу нижче 100 мл на добу);
• полгурп (збыьшення дгурезу понад 1500 мл на добу), що частшге евщчить не
про вщновлення фшьтрацп, а про порушення канальцево! реабсорбцп;
• завершальну (смерть або одужання).
Значш розлади гомеостазу спостерогають у стадп ол!го- та анурп. Унаслщоя
розкого зменшення (або вщсутност!) д!урезу розвиваеться пперазотемы, пору-
шуеться обмш води, електролтпв 1 КОС. виникають систе.мш набряки. Основш
клинчи! прояви ще! стадп: набряк головного мозку, штерстищйний набряк ле-
126~
~ень, тяжкх порушення дхяльносп оргашв кровообпу (зниження насосно! функ-
Ш1 мюкарда, порушення ритму серця на зразок екстрасистолп, брадикардп, бло-
зди, артерхальна гйютензхя з подалыпим переходом у гшертензпо), дихання Кус-
мауля, тяжкх розлади функцш нервово! системи (головний бшь, блювання,
.гефлексхя, порушення свщомосп, судоми, кома), прогресуюча анем!я тощо.
Бшышсть хворих помирають у II стадп ГНН вщ пперволемп та р!зкого пщви-
хення артерхального тиску, що спричинюе гостру серцеву недостатшсть хз набря-
еом легень або мозку.
Якщо полхурхя, що розвиваеться внаслщок ушкодження проксимальних ка-
вальщв 1 порушення реабсорбцп, зменшуе гшерволемпо й артерзальний тиск, то
юдина може уникнути швидко! смерт!, але за збереження низько! ШКФ помре
грез кшька тижшв вщ уремп, яка е наслщком ппемхчного ураження кхркових
зефрошв 1 посилення зменшення ШКФ.
Патогенетична терапхя при ГНН полягае в термшовому вщновленн! ШКФ за
рахунок медикаментозно! блокади РАС х усунення причини И активацп.
Хрожчна ниркова недостатжсть
Хрожчна ниркова недостатшсть (ХНН) — полхетюлопчний синдром, що характе-
-1зуеться повшьно прогресуючим, тривало непомхтним зниженням ШКФ унаслх-
_ок зменшення кшькостх (маси) дпочих нефрошв.
Етхологхя. У розвитку ХНН вщпрають роль хрошчш прогресуюч! хвороби:
• пререналый — тривале пщвищення артерхального тиску або стеноз нирково!
артери;
• реналый — хрошчш захворювання нирок запально! (хронхчний гломеруло-
нефрит, хронхчний шелонефрит), алерпчно! (ревматощш васкулгги, системний
ервоний вовчак) х метаболхчно! (дхабетична нефропатхя, амшощоз, подагра)
~рироди, полпастоз та хн.;
• постренальш — тривала обструкцхя сечових шляххв.
За клшчним переб1гом видшяють так! стадп ХНН:
• латентну, що може тривати багато рокхв без будь-яких клхшчних проявхв;
• початкову, або стадно полхурп, що розвиваеться шсля загибелх понад 50 %
-ефрошв х харакгеризуеться полхурхею внаслщок порушення реабсорбцп натрпо
та води;
• олпо- й анурп, що виникае при загибелх понад 70 % нефронхв х зменшення
ШКФ нижче 20 % вщ норми;
• уремп, що розвиваеться при зменшенш ШКФ нижче 10 %.
Патогенез ХНН зумовлений прогресуючим зменшенням кшькостх функ-
шонуючих нефронхв. Початков! ознаки ХНН спостерпаються в разх зменшення
илькосп нефронхв до 50—30 % вщ вихщнох. Клшхчно виражена картина розви-
ваеться тодх, коли зменшення становить 70—90 %, а величина клубочково! фшь-
трацп — менше 20 %. Подальше зменшення клубочково! фщьтрацп (нижче 10 %)
зумовлюе виникнення уреми (термпхальна стадхя недостатност! нирок).
При ХНН дхурез змхнюеться не суттево, а в початковш стад!! впх може бути
дещо збшыпеним (полхурхя), що зумовлено обмеженням реабсорбцп води з дис-
тально! частини канальшв нефронхв х збхрних трубочок х зниженням концент-
ращйнох здатносп нирок (гшо-, хзостенурхя).
Порушення осмотичного й об’емного гомеостазу при ХНН не вщтграють
вирштальну роль, як при ГНН. Ц! ознаки, а також азотемтя виникають лише в
термшальшй стадп ХНН, коли значно зменшуеться фшыращя та виникае олгг-
ур!я. У раз! пол!ури можливт також гшоволемтя, внутр!шньо- та позакл!тиннз
дег!дратац!я, гшонатр!ем!я, бшьш стал! ппокашемтя, гшокальщемтя, гшермагшемтя
Набряки в шй стади вщсутш. В олтгоануричшй стади ХНН спостерпаються п-
перволем!я, пперпдратащя поза- 1 внутрппньоклттинного простору, тобто
кл!шчна картина водного отруення (набряк головного мозку, легень), гшона-
тртемтя, гшеркалгемтя, гшокальщемтя, з якою пов’язують розвиток остеодистро-
фи та остеомаляци. 1стотного значения набувають також порушення КОС на
зразок ренального (азотемтчного) ацидозу.
Урем1я (сечокрш’я) — це термшальна стад!я ХНН, що характеризуеться зни-
женням маси дпочих нефронтв 1 ШКФ нижче 10 %. Основш прояви уреми зумо-
влен! передуши азотем!ею внаслщок зниження екскреци кшцевих продукттв азо-
тистого обмшу. Стугинь азотемп вщцзеркалюе ступшь зменшення кшькост! не-
фровдв. Виявлено понад 200 токсичних речовин, накопичення яких у кров! пр;
ХНН визначае штоксикацпо оргашзму та пов’язаш з нею анорекспо, диспепспо
(блювання, пронос), зменшення маси тша, загальну слабгсть, головний бшь
апатпо, порушення смаку, слуху, нестерпний свербгж шк!ри, полшеврит, роздал
дихання, прогресуючу анемпо, уремгчний перикардит, мюкардит, плеврит, арт-
рит, судоми, кому. Основним показником, оборотно пропорцшним зменшенню
кшькост! нефрон!в, е збшьшення ргвня креатиншу в плазм! кров!.
Наростаюча штоксикацгя ЦНС супроводжуеться розвитком уремгчно! ком
(потьмарення св!домоет!, вщеутшеть активних рух!в 1 рефлекств).
У розвитку цих порушень важливу роль вщтграють розлади метаболтзму: п-
перкал!ем!я, пперфосфатемгя, гшермагшемтя, ппо- чи гшернатртемгя, метаболтч-
ний ацидоз, гшергщратацгя, що виникають унаслщок порушення встх функцш
нирок. Важливу роль вщтграють гормональн! порушення у зв’язку з ренальною
затримкою в оргашзм! гормону росту, глюкагону, шсулшу, пролактину тощо. Не-
достатшстъ утворення в нирках еритропоетину е причиною анеми, що одночасно
з порушеннями функцп серцево-судинно! системи призводить до гшоксп. У нир-
ках менше утворюеться кальцитрюлу, що зумовлюе виникнення гшокальщемп.
Патогенетичною основою прогресування ХНН е поступове т незворотне
зменшення кшькост! функцюнуючих нефронтв унаслщок !х загибел!. На сьогодн!
вщомо два основних мехашзми прогресуючо! загибел! нефронтв.
Перший — тмунолопчний — як наслщок авто!мунного ушкодження нефрон!в.
коли внаслгдок первинного етюлопчно зумовленого ураження в нирках з’являються
эмитент за структурою бшки-автоантигени, що надал! стають об’ектом гмунного
ушкодження, передустм клубочктв.
Другий — за мехашзмом пперфшьтраци дйочих нефрошв, яка е мехашзмом
компенсац!! при зменшенш кшькост! нефронтв, але сама по собт внаслщок функ-
цюнального перевантаження канальцевого вщдшу (зростаюча реабсорбигя натр!к?
б!лка тощо) призводить до виснажелня канальщв та !х ушкодження з подальшою
загибеллю нефрошв або розвитком тубулоштерстицшного синдрому, що супро-
воджуеться дистрофтчно-атрофтчними проявами в ниркових канальцях з утворен-
ням сполучно! тканини в шерстили нирок, особливо в мозковш речовин!.
~убуло1нтерстиц1йний синдром
.'убулоштерстицшний синдром е окремою формою ураження нирок, для якоТ ха-
_ктерна атрофЁя ештелио канальшв нефрошв у поеднанн] з вираженим склеро-
юм с громи га порушенням основних канальиевих функцш, шо виникае у вгд-
зёдь на р!зш токсичш, метаболтчш, фгзичш (юшзуюче випромЁнювання) та Ён-
екцгйш впливи
Для тубулоЁнтерстицЁйного синдрому харакгерним е значне порушення в дёючих
ефронах найважливЁших канальцевих функцЁй, яю регулюють водно-електролп-
.-.ий обмЁн, що зддйснюеться переважно дистальними канальиями 1 збЁрними тру-
ояками. До них функшй належать осмотичне концентрування Ё розведення сечЁ.
При губулоЁнтерстицЁйному синдром] можлив! також порушення прокси-
чальних канальцЁв нефрона, якё проявляються ренальною глюкозургею, реналь-
ною фосфатургею, нирковим канальцевим ацидозом, синдромом Фанконг тошо.
Детальний опис етЁсшогц га патогенезу тубулоштерстишиного синдрому вихо-
тить за рамки пропонованого пЁдручника ё розглядатиметься в курсЁ нефрологи.
ЕТЮЛОПЯ I ПАТОГЕНЕЗ ОСНОВНИХ ХВОРОБ НИРОК
1вороби нирок рЁзног природи спосгерЁгаються в 1,5—2 % населенна, становлять
близько 5,5—6 % загально! захворюваност! та харакгеризуються тяжким перебггом
ё високою летальшстю.
Причому осганнгм часом спостерйаеться зростання поширеностг хвороб ни-
рок, а також гх фЁнальног стадп — ХНН.
Умовно основн! хвороби нирок можна роздЁлити на две групи. По-перше, це
шорой и, при яких патологгчний процес первинно перебЁгае безпосередньо в нир-
ках: гломерулонефрит, шелонефрит, токсичнё нефропатЁЁ, спадкова патологЁя ни-
рок. До другог групи належать ураження пирок при патологи Ёнших органЁв Ё
зистем: цукровий дЁабет, гшертошчна хвороба, амглогдоз. Як наслщок на початку
Ехворювашгя патогенез цих нозологёчних форм мае суттевг вгдмённостё.
Проте хронгчний перебгг усгх хвороб супроводжуеться поступовим перева-
жанням загальних механгзмЁв патогенезу, шо в клгшчшй практиш призводить до
розвитку ХНН.
Гломерулонефрит
Гломерулонефрит — дифузна хвороба нирок запально? природи з переважним
смунним ураженням клубочк1в ниркових милець.
Рёзнё змёни в клубочках зумовленЁ поеднанням ексудат явного та пролЁферативно-
го процесЁв, якё розвиваються ЁнтракапЁлярно. У разг тяжкого перебггу хвороби з
розвитком некротичних змён вщбуваеться повна деструкция клубочкЁв. До патоло-
пчного процесу залучаеться вся маса клубочкЁв обох нирок. Розргзняють гострий
. хрошчний гломерулонефрит.
Гострий (дифузиий) гломерулонефрит. Експерименталыи модели У 1901 р. В.К. Лён-
деман у лабораторн 1.1. Мечникова спостерЁгав основш прояви нефриту в кроля шсля
внутрЁшньовенного введения йому нефротоксично] сироватки гвЁнейськог свинки,
попереднъо гмуюзовано! суспензгею нирки кроля. Застосовуючи аналоггчку схему,
японський учений Мазнет в 1933 р. вгдтворив клгнгчну картину нефриту в кроля шля-
хом уведення йому сироватки кровг качок. гмунгзованих тканиною нирки кроля. 1н-
ший варгант моделг нефриту ?Ла8и§1 здгйснив за схемою щур — кргль — щур.
У 1909 р. росгйський хгрург П.О. Герцен, намагаючись з’ясувати мехашзм ви-
никнення “окопного” нефриту (нефрит военного часу), одержав його експери-
мснтальну модель у кроля шляхом охолодження (заморожування хлоретилом)
нирки. При цьому в кров! шддослгдних тварин було виявлено специфгчш проти-
нирковг антитгла. Особливютю щег моделг виявилося ушкодження не пльки охо-
лодженог, а й штакгно! нирки. Ця модель уперше навела на думку про можливгсть
автогмунног природи зазначено! хвороби.
1тал!йськими вченими. подружжям Ковелл (1945) було вщтворено гломеруло-
нефрит у кролгв шляхом уведення !м у черевну порожнину кптинно! суспензп нир-
ковог тканини та культури стрептококгв. 1з достатньою надгйнгстю гострий гломеру-
лонефрит вгдгворюеться шляхом уведення тваринам чужорщного бглка. сироватки.
ЗгаЫау (1962) вщтворив гломерулонефрит в овець гмушзащею !х базальними
мембранами клубочкгв нефрошв нирки людини з повним ад'юваггтом Фрейнда.
Неутапп 1 спгвавгори (1965) викликали експерименгальний нефрит у щур1в
шляхом активно! гмушзацп суспензгею гомологгчно! або автол опчно'г нирки з
повним ад'ювантом Фрейнда. У новозеландських мишей лгнп К7В/ВЬ визначено
спонтанне виникнення серолопчних 1 морфолопчних ознак автогмунного гломе-
рулонефриту та гнших авю!мунних хвороб: системного червоного вовчака. авто-
гмунно! гемолпичног анемп. На базальшй мембран! уражених клубочюв виявлено
автолопчн! у-глобулгни, що е автоантитгтами до ниркових автоанти генгв. Розви-
ток хвороби рееструеться з 3—5-мгсячного вгку.
Етгологгя. Гострий гломерулонефрит виникае на тлг (або пгсля) будь-яко!
гнфекцп, переважно стрептококово; Вважають, шо гемолггичний стрептокок гру-
пи А (типи 4, 12) е специфгчним “нсфритогенним" штамом. Певну роль вцпгра-
ють 1 шшг гнфекцп (вгруснг, паразитичнг). Доведено етгологгчну роль у виникнен-
нг гломерулонефриту охолодження. опгку, дифузних уражень сполучног тканини
(системний червоний вовчак, ревматоцгний артрит, вузликовий пергартерпт), по-
переднгх вакпинацгй або застосування гетерол опчн их сироваток.
Патогенез. Вгдсутнгсть нерщко гнфекцгйного фактора, наприклад стрепто-
кокк, у нирках пгд час розвитку в них дифузного запального процесу, наявнгстъ
латентного перюду (1—3 тпж.) у перебггу постшфекцгйного гломерулонефриту,
виникнення його на тл! хвороб автогмунног природи, сироватковог хвороби, пгсля
вакпинацп, пщ час експериментального моделювання за допомогою гмунолопч-
ного втручання, результата серологгчних та гмунофлюоресцентних дослцтжень да-
ють змогу припустите алерггчну й автоалерпчну природу цгег хвороби. Вгдповщно
до дього видгляють два основних мехашзми ураження клубочюв:
• ураження батально!' мембрани клубочкгв антитглами до и антигенгв — нефро-
токсичный гломерулонефрит (виргзняеться швидко прогресуючим перебшом)
(мал. 69, а, б). Носгем антигенних властивостей базально! мембрани е глюкопротещ:
• розвиток запального процесу в клубочках унаслшок фгксацп на базатьнш
мембранг та штрамембранно гмунних комплексгв — 1мунокомплексний гломеруло-
нефрит (мал. 69, в).
Мал. Ь9 Взаемоди антиты (АТ) з антигенами (АГ) базально!
мембрани клубочшв (БМК):
а, 6 — нефрогоксичиий гломерулонефрит; в — (мунокомплекс-
ний глумерулонефрит
Як антиген при ньому мехашзм! виступае екзогенна (шфекцшного або неш-
фекцшного походження) або ендогенна (тканинний бшок, ДНК) субстанщя. Ан-
типла СёО, 1§М), шо утворюються безпосередньо в сироватщ кров! взаемошють
з антигенами, поим у вигляд) змунних комплекс!в (антиген — антипло — комп-
емент) надходять у клубочки, вщкладаючись на гхнш базальшй мембран!. Уш-
коджувальна д1я (мунних комплексов, як 1 нефротоксичних антитш, реа’пзуеться
шляхом ондукцп !мунного запалення.
У бщьшост! випадюв (не менше 80 %} гломерулонефрит е !мунокомплекс-
ним, у тому числ! 1 гломерулонефрит, шо розвиваеться внаслщок стрептококово!
|нфекпл, у раз! системного червоного вовчака, сироватково! хвороби тощо.
Клппчн) та патофозюлопчн! прояви гострого гломерулонефриту вщображають
зм!ни ниркових, в основному клубочкових, ! позаниркових функцш. Характерни-
м ознаками класичного перебпу хвороби е стргмкий початок, ол±гур!я, протегн-
урзя, азотемзя, артерзальна гзпертенззя, набряк (унаслщок затримання натр!ю, п-
попроте'шеми, пперволемп, пщвишення проникност! стшки кашлярзв), гематурзя.
порушення з боку ЦНС.
Хрошчний (дифузний) гломерулонефрит — тривала прогресуюча хвороба, для
яког характерном е дифузне двобгчне ураження нирок запальной' природи, неоднор1дне
за походженням, клтчними проявами I патогенезом.
Етзологзя. Хрошчний гломерулонефрит може виникнути як наслщок го-
строго гломерулонефриту, проте здебзльшого розвиваеться первинно. Розрззня-
ють таю форми хрошчного гломерулонефриту.
• шфекцшну (постстрептококовий, у раз! затяжного бактергального ендокар-
диту, малярп, сиф!л!су, туберкульозу та шших шфекпш);
• нешфекцшну (сироватковий, поствакцинальний, медикаментозной, травма-
тичний, у разг отруення ргзними отрутами, охолодження, тромбозу ниркових вен,
дифузних хвороб сполучно! тканини — ревматоидного артриту, системного черво-
ного вовчака, геморапчного васкулгту тощо);
• окрему (постеклампсичний, променевий, спадковий тощо).
Патогенез. Загальновизнаною е гмунологгчна концепция розвитку хронгч-
ного гломерулонефриту. Окргм двох основних механгзмгв, гз якими пов’язують
розвиток як гострого, так г хронгчного гломерулонефриту, певне значения мае
гшерчутливгсть сповгльненого типу.
За клгнгчними ознаками функцгонально-компенсованог фази хвороби виргз-
няють чотири форми хронгчного гломерулонефриту:
• латентну (65 % усгх хворих гз хронгчним гломерулонефритом), що прояв-
ляеться гзольованим сечовим синдромом — помгрною протегнургею, гематургею.
У частини хворих (20—25 %) спостерггаються набряк г транзиторна гшертензгя;
• гтертензивну (32 % хворих), що проявляеться стгйким пщвищенням ар-
териального тиску. В 1/3 хворих спостерггаються набряк, у 2/3 — гематургя, у по-
ловили — цилгндрургя г лейкоцитургя, в усгх хворих — протегнургя;
• нефротичну (2—4 % хворих), яку виргзняють набряковий синдром (у 2/3
хворих), значна протегнургя г цилгндрургя (в усгх хворих), характерш змгни в кровг
(ппопротегнемгя, гшерлшгдемгя);
• змииану, або нефротично-гтертензивну (2,4 % хворих), для яког характерш
набряк г гшертензгя (в усгх хворих), змгни в сечг, як г в разг нефротичног форми,
проте, на вщмгну вщ нег, немае характерних змгн у кровг.
Маса дгючих нефрошв у бглъшостг хворих збережена або дещо зменшена за
рахунок гмунопатолопчних механгзмгв.
Подальший розвиток хвороби вщбуваеться за рахунок як гмунопатолопчних,
так г негмунних механгзмгв патогенезу. До перших слщ зарахувати розвиток авто-
гмунних механгзмгв ушкодження нирок, серед яких найбыьшу роль вщгграють
мехашзми клгтинного гмунгтету з появою сенсибглгзованих до антигенгв нирки
Т-лгмфоцитгв.
До негмунних механгзмгв ураження нирок належать протегнургя при ушко-
дженнг клубочкгв гз подальшим накопиченням профгльтрованого бглка в прокси-
мальних канальцях, ушкодження яких при цьому пов’язане з активащею перок-
сидного окиснення лшщгв г лгзосомальних ферментгв.
Активацгя внутргшньонирковог РАС, що спостерйаеться при гломеруло-
нефрит!, пщукуе розвиток сполучно! тканини в паренхгмг нирок гз поширенням
патолопчного процесу на гншг структури нирки, особливо гнтерстицгй. Це пов’язано
з тим, що анпотензин II е фактором росту, який активуе пролгферацгю клгтин.
Одним гз провщних механгзмгв прогресування ушкодження нирок е пперфгль-
трацгя дгючих нефрошв. Збгльшення фгльтрацп в дпочих нефронах дае змогу пщ-
тримувати нормальний ргвень загальнонирковог ШКФ в умовах скорочення кгль-
костг функцгонуючих нефрошв унаслщок гх загибелг. Але водночас пперфгльтра-
цгю розглядають як основний механгзм’негмунного ушкодження дгючих нефронгв
та гх загибелг, що зумовлюе постшне зменшення функцгонуючих нефронгв.
Клгшчнг прояви гломерулонефриту виявляються у виглядг всгх ниркових син-
дромгв: сечового, набрякового, нефротичного, гшертонгчного, анемгчного, пору-
шень зсщання кровг, канальцевого ацидозу г, нарештг, нирковог недостатност!.
У фаз! декомпенсацп через значнг склеротична змши г зниження внаелгдок
дього маси дпочих нефрошв хронгчний гломерулонефрит проявляеться синдро-
ом хрошчно! нирково'Г недостатност!.
ГИелонефрит
Шелонефрит — шфекцшно-запальна хвороба, за якоё до патологичного процесу залу-
чаешься слизова оболонка сечових шляхёв / паренхёма нирок (одночасно або последов-
ав) з переважним ураженням ттерстицёйноё тканини нирок.
Ет1олог1я 1 патогенез. Тнфекцгя потрапляе в нирки гематогенним шля-
ам або поширюеться у висх!дному напрямку сечовими шляхами. Збудниками
дебгльшого е кишкова паличка, коки.
Виникненню хвороби, переходу гострого шелонефриту в хрошчний сприяють
'1зш фактори, що спричинюють застш сеч! (звуження, закупорювання сечоводгв,
аденома передм!хурово! залози), порушення троф!ки сечових шляхгв, хвороби та
гтани (цукровий дгабет, атеросклероз, ожиршня, хронгчна гнтоксикацгя тощо),
якг знижують реакгившсть органгзму.
Починаеться пгелонефрит гостро г найчастгше (за винятком випадкгв повного
дужання) через латентну, малосимптомну фазу переходить у хрошчну форму,
жа закшчуеться зморгггуванням г недостатнгстю нирок.
Процес ушкодження починаеться з епгтелгю мгхурово-мисково! системи з по-
дальгпим переходом на епгтелгй канальцгв, спочатку тих, якг розташованг в мозко-
вш речовинг нирок, згодом — на штерстицгй ще! дглянки. Надалг уражуеться г
кгркова речовина. Зрештою частина уражених нефрошв гине.
Основш порушення нирок пов’язанг гз двома видами ушкоджень: зменшен-
ням кглькостг дпочих нефрошв та ураженням переважно прямих канальцгв, якг
розташованг в сосочковгй г мозковгй речовинг ггирок г забезпечують переважно
осморегулюючу функцгю. Згодом у патогенез! та клшгчшй картин! основним стае
меншення кглькост! функцюнуючих нефрошв, тобто формуеться ХНН.
Клшчними проявами шелонефриту е ознаки тяжкого шфекцгйного про-
цесу: пгтоксикащя (особливо в гострий пергод); артергальна гшертензгя (у 25 %
випадкгв у початковш стадп г в 75 % — у стадп розвитку); помгрний набряковий
синдром та анемгя; сечовий синдром (полгургя, у шзгггй стадп — олггургя, по-
лакгургя — чаете сечовипускання, ггпостенургя, у завершальшй стадп — гзостен-
ургя, лейкоцитургя, гематургя, помгрна (5—10 г/л) протегнургя, ггилшдрургя).
Порушення клубочково-канальцево! ргвноваги проявляеться переважанням
канальцевих дисфункцгй над ютубочковими, особливо в початкових стадгях хво-
роби (своергдна функцюнальна дисоцгацгя). Про це евщчать зниження здатностг
нирок до концентруванггя сечг внаелгдок порушення процесу реабсорбцп води у
збгргшх трубочках, раннгй г тяжкий канальцевий ацидоз, пов’язаний гз порушен-
ням секрецп гонгв водню в них г ацидо- та амошогенезу в проксимальних каналь-
пях, втрата солей, в основ! яко! — ргзке зниження канальцево! реабсорбцг! гонгв
натрпо г кальцпо. Як наелгдок, можуть розвиватися небезпечнг для життя пору-
шення водно-мгнерального обмшу г КОС.
Прогресування шелонефриту супроводжуеться наростанням отшеаних пору-
шень, зменшенням маси дпочих нефронгв г переходом тубулоштерстицшног недо-
статностг в ХНН.
РОЗД1Л XXXI
ПАТОЛОПЧНА Ф13ЮЛ0ПЯ
ЕНДОКРИННО! СИСТЕМИ
ГОРМОНИ ТА1Х роль В 0РГАН13М1
За допомогою гормонтв ендокриниа система разом тз нервовою та 1мунною сис-
темами забезпечуе в оргашзм! регулярно фтзтологтчних т бюхтмтчних процеств
Гормони справляють генералтзований тривалий вплив на органтзм. Вони беруть
участь в ембртогенезт та подалыпому розвитку оргашзму, збереженн! гомеостазу,
пропесах розмноження, пристосування (адаптацц) до умов тснування. а також
впливають на обмгн речовин, поведшку та шип психофтзтолопчю функцп.
Окртм гормонтв (тироксин, кортизол, шсулш тощо), якт виробляють щито-
подтбна залоза, наднирковт залози, остртвцт птдшлунковот залози та шшт залози
внутоштньо1 секреци, нимало гормонтв продукуеться тканинами, окремими кгпти-
нами або тхнтми скупченнями незалозисто! будови (схема 27).
Зокрема в органах травления утворюються травит гормони, такт як гастрин,
секретин, холецистокшш та тн. Нирки е мтецем синтезу еритропоетину, серце —
иередсердного натршуретичного пептиду, жирова тканина — естрогешв т нещо-
Клггини
Схема 27. Джерела утворенкя гормогав
давно вщкритих гормошв лептину, адипонектину тощо. Важливим джерелом гор-
мошв е клггини дифузно! нейроендокриино! системи АР1Ю (див. роздш XXVIII
[ “Патолопчна фззюлопя травления”). Така р!зномаштшсть структурно-функцю-
нально! оргашзацп ендокринно! системи надзвичайно утруднюе д!агностику та
лгкування ендокринних хвороб.
Класичш гормони вщповщають таким критериям: 1) синтезуються спещалззо-
ваними ендокринними органами або юптинами; 2) видшяються в циркулюючу бю-
[ лопчну ршину — кров, лхмфу, спинномозкову рщину (внутршшя секретя); 3) дпоть
дистантно, тобто на вщсташ в!д м!сця утворення; 4) мають надзвичайно високу
фгзюлопчну актившсть, яка виявляеться в концентращях 10-11—10-9 моль/л;
5) здшснюють вибзрковий ! специфзчний вплив на тканини.
Проте варто зауважити, що ш критерп дещо умовш. Досить послатися на
естрогени, яю частково утворюються жировою тканиною та нейронами головного
мозку 1 здатш дзяти в мгсц! утворення на сусщш юптини (паракринна д!я) або
навпъ на и сам! юптини, де виробляються, наприклад, на нейрони чи гранульоз-
| ш юптини фолжулгв яечниюв (автокринна д!я). Деяю !з тканинних гормошв,
зокрема простагландини, дпоть не тшьки локально, а й циркулюють у кров!,
впливаючи на вщдалено розташоваш органи ! тканини. 1х вщр!зняе широкий
спектр прояв!в ф!з!олопчно! активност!. Таю гормонопод!бш речовини отримали
назву г1стогормон1в, або гормоноТдгв. Нин! налзчують майже 100 гормонтв 1 гормо-
но'цпв (табл. 26, 27). 1хня найголовнппа функц!я — передача шформацшного сиг-
налу в систем! гуморально! регуляцп.
Таблиця 26. Гормони, що синтезуються завозами внутрданьоТ секреци
Залоза(и) Гормони
Ппоф1з Аденогшоф1з Кортикотропш, лютропш, фолпрошн, пролактин, тиротрошн, сома- тотропин, р-лшотрошн, Р-ендорфш
Пром1жна частина аденоппофхза Нейрогшоф!з Еп1ф1з Наднирков1 залози Юркова речовина Меланотропш, Р-ендорфш Вазопресин, окситоцин’ Мелатонш, ешф!зш пептида Кортизол, кортикостерон, альдостерон, дегццдазоандростерон, анд- ростендюн, прогестерон
Мозкова речовина Статев! залози Яечко Адреналш, енкефалши Тестостерон, андростендюн, естрадаол, шпбш, активш, фактор ре- гресн парамезонефричних проток (мюлерових канал1в)
Яечник Естрадюл, естрон, прогестерон, 17-пдроксипрогестерон, андростен- дюн, тестостерон, шпбш, акгивш, релаксин, вазопресин
Щитоподтбна залоза Прищитопод1бш залози Щдшлункова залоза Загруднинна залоза Т4, Т3, кальцитонш Паратгормон 1нсул1н, глюкагон, амппн, соматостатин, ВШ Тимопоетин
‘Гормони, що утворюються в ппоталамус1 та накопичуються в нейроппоф131 до видыення !х у
кров’яне русло.
Таблица 27. Гормонита гормонощи, що синтезуються поза залозистими органами
Анатаючний утвХр Гормони, гормоноТди
Гипоталамус та шгш Д1ЛЯНКИ головного мозку Система гравлення Печ1нка Серпе Нирки Передм1хурова залоза Кров, псриферичгп ткашши Жирова тканина Макрофаги, лхмфоци- ти, макро гл хя Тромбоцита Фхбробласти, нейрони га хнш! клпини Плацента Рхзнх орган! 1 Ршйзинг-гормони. кортикол1берин, лкмпберин, тиролхберин, соматош- берин, соматостатин; вазохгресин, окситоцин, ошохди, галанхя, нейро- пептид V та шгш нейропептиди, естрогени, катехолестрогени, 5а- дипдротестостерон, дофамш Гастрин, секретин, холецистокшш, апелш, ентероглюкагон, ВГП, грели, галашн, нейропептид V, речовина Р та 1н. 1нсулхнопод)бн1 фактори росту, анпотензиноген, 25-п цроксихолекать- циферол Передсердний натрйТуретичний пептид Еритропоетин, 1,2,5- диг) дроксихо. хе кальциферол 5а-Дигидротестостерон Анхтогензини Лептин, резистин, адипенекгин, естрон. естрадход Цитохани (щтерлейкши), кортпкотрошн. Р-енлорфгн Фактор росту Р тромбоципв 1нсул1нопод1бн1 фактори росту, трансформуючип фактор росту а, еп)- дермальний фактор росту, фактори росту нервхв, ф)бробласт)в Хорютчний гонадотропхн, хорюхпчний соматомамотрогин (плацентар- ний лактоген) Простагландина
Пщ дхею гормон!в у клхтинах-мхшенях розвиваеться низка функцхональних х
морфолопчних змхн, що стосуються процеств пролхферацп, апоптозу, атрофхх, п-
пертрофп, секрецц тощо. Змшюються електричний потенцхал юптинних мемб-
ран, Гхня проникшсть для хонхв, глюкози, амшокислот, жирових кислот та шших
речовин, актившсть гешв. Синтезуються структури 61лки, нуютехновх кислоти,
ферменти та шпп речовини.
Бьтышсть цих ефекпв зумовлена акгивуючим або гальмхвним впливом гор-
мошв на процеси транскрипцп в ядерному геном! юптини за класичною трхадою
гормон — ген — фермент. Але чимало з них чинять позагеномний вплив Так,
збхльшення швидкосп проникнення глюкози й амшокислот крхзь плазмолему в
м’язов! та жиров! юптини тд дхею шеулхну вщбуваеться завдяки прям1й стимуля-
цц синтезу быкхв-транснорт ерхв на полхеомах. До негеномних ефекпв належить х
майже митгеве збудження нейронхв унаслщок дм стерощних хормонхв, а також
посилення внутрхшньох та зовшшньох секрецй за механизмом екзопитозу, який
шщпоеться пщвищенням внутрхшньоюптинноУ концентраци юшв кальщю.
ЕТЮЛОПЯ I ПАТОГЕНЕЗ ЕНДОКРИННИХ РОЗЛАДIВ
Ендокринна патологхя виникае як наслщок оргашчних ушкоджень або функцю-
нальних порушень. якх стосуються синтезу та секрецц гормонхв, Тх метаболхзму,
мехашзмхв дп на клхтини х тканини. За етхолопею резрхзняхоть набут! та спадков!
ендокринхд хвороби (ендокринопатп). Основними еполопчними чинниками ен-
докринних хвороб е бактерхалвш га вхрусш шщекцп. вроджеш хромосомш ано-
< ,ш, мутацп генгв, психоемошйш травми, пгтоксикацп, алгментарн: фактори,
пнгзуюче випромгиювання, мгсцевг порушення кровопостачання (крововилив,
гбоз), ураження пухли ин и-г або запатьним процесом.
Неабияку роль у виникненш ендокринних хвороб вщгграе спадковгсть. Але
частшге вдеться не про пряме чи рецесивне !х успадкування, а про схильшсть
ендокринно! патологи, яка може реалгзуватися за певних умов, зокрема при
яровому дгабетг, синдром! полгкгстозних яечникгв. Прикладами вроджених ен-
сринних порушень, пов’язаних гз хромосомними або гешгими аномалгями, е
огонадизм у чоловгкгв, хворих на синдром Клайнфелтера, г несправжнгй гер-
фродитизм у жгнок з адрен огенгтальним синдромом,
Основгп патогенетичгп форми ендокринних розладгв. Залежно вгд патогенезу ен-
сринопатп, вона може бути занозистою, периферичною або дисрегуляторною. У
разг первинного ушкодження залози внутр!шньо! секрецп розвиваеться первинна
- юкриняа патолопя, а у випадках, коли органгчнг ураження або функшоналып
глади спочатку виникають за и межами (наприклад у ппоталамгчнгй дглянцг
юзку), — вгоринна. Найчастппе первинна ендокринна патология розвиваеться як
эгмунний процес (наприклад автогмунний тиреощит), пухлина (аденома гшо-
га, кортикостсрома), некроз або крововилив. вродженг вади бюсинтезу гор-
•нгв (вроджена пперплазгя кгрковог речовини надниркових залоз).
У виникненш вторинно'1 ендокринно! патологи переважне значения мають роз-
_ди нейрогормонально! регуляпп секрецп гормонгв (наприклад псевдоспгввщно-
_ення продукт! лютротну г фолггропгну аденоппофгюм при синдром! лолгкге-
озних яечникгв). До групи вторинних гормональних порушень належать ! пери-
еричш, тобто тг, якг е наслщком розладгв рецепцп, метаболгзму гормонгв або
г’язування гх гз битками плазми кровт.
Основою патогенезу биты пост! ендокринних хвороб е недостатня (гтофунк-
?) або надмзрна (гтерфункцгя) актившсть ендокринних залоз г клгтин. Ш понят-
"т стосуються не тгльки залоз внутргшньо! секрецп, а й окремих гормонгв. (деться
.ро те, що будь-яка залоза е джерелом двох або бгльшо! кглькостг гормонгв. В
жому лише ппофгзг мгститься не менше десяти ргзних гормошв бшково! приро-
и. 3 к!рково! речовини надниркових залоз ввдглено близько п’ятдесяти стерощ-
их сполук, багато з яких мають гормональну актившсть. У випадку тотального
раження залози внаслщок крововиливу або запального процесу водночас припи-
няеться або ослаблюеться продукпгя всгх гормонгв. Наприклад. некроз передньо!
г детки гшоф!за (аденогшофгза) призводить до генерализовано! аденогтофнзарно! не -
•. статности Проте вщомо нимало ендокринних розладгв, для яких харакгерним
е гзолъоване порушення секрецп того або шшого гормону, що визначають як пар-
Аалъну (часткову) ендокринну гтер- або гтофункшю. Це видно на прикладг хворо-
би Хценка — Куштнга, зумовленог надмгрною секрецгею кортикотропгну (адрено-
кортикотропного гормону), та синдрому Кальмана, тобто вродженого вторинного
ппогонадизму, коли гзольована недостатшсть продукцг! лютротну аденогшофг-
зом е причиною слабкого розвитку статевих органгв г вторинних статевих ознак.
Поняття ендокринно! гшо- або гшерфункцй не вичерпуе всього ргзномангт-
тя ендокринопатш, перебгг деяких з них вщбуваеться за типом дисфункци. Вона
зумовлена ргзноспрямованими змшами синтезу гормошв г близьких до них фгзго-
логгчно акгивних сполук в одному 1 тому самому ендокринному ортанг або ж ут-
воренням г надходженням у кров аномальних за молекулярною будовою г бголо-
пчними властивостями гормональних речовин. Так, патогенез уродженого адре-
ногенггального синдрому, шо зумовлюе лояву жшочого несправжнього гермафро-
дитизму, пов’язаний гз блокадою певних ферменпв биосинтезу стероццв у юрков!?
речовин! надниркових залез. Це зумовлюе р!зке зменшення утворення кортизол
(гщрокортизону) з подалыпим збшьшенням продутой! надниркових андрогеюв.
Аденопп<?ф!з може виробляти структурно змшен! бшков] гормони, шо спричиню*
повну або часткову втрату ними специфично! бюлопчно! активносп.
Подобно до бьтьшост! шших хвороб, ендокринн! хвороби перебпають у го-
стрш або хронЫнш форм!.
За морфолопчною ознакою — юльюстю уражених патолопею ендокринни
залоз — ендокринопатн бувають моногландулярними та, рщше, полггландулярними.
Патогенез енлокринних порушень вивчають на експериментальних моделях
яю сгворюють у лабораторних тварин шляхом видалення, руйнування залоз, уве-
дення гормональних 1 фармаколопчних речовин, що вибтрково впливають на
синтез, транспорт, секрецпо гормонтв або !х взаемодпо з кл!тинами-м!шенями.
Осташйм часом значного поширення набувають так зван) альтернативн! методи
дослщження т гиго (“у скляному посуд!”) на культивованих ендокринних юпти-
нах людини ! тварин, комп’ютерне моделювання га ш., що дае змоту зберегти
життя багатьом тваринам. Нимало даних про патогенез ендокринних розлад!в от-
римано п!д час проведения дослщжень на хворих, чому сприялэ поява високочут-
ливих гормондльних методик.
Роль ендокринних взаемозв’язкгв у патогенез! ендокринопатш. Ус! ланки ендо-
кринно! системи функцюнують у йсному взаемозв’язку. Пряма стимулювальнз
д!я гшофтзних тропних гормошв на пщлегл! ендокринн! залози (тиротропшу — на
щитопод!бну залозу, кортикотропшу — на юркову речовину надниркових залоз.
гонадотроп!н1в — на статев! залози) вщображае так зван! вертикалью регуляторю
зв’язки в ендокринн! й систем!. О крш них !снуе ендокринна регуляшя за гори зон-
тал ьним типом. Зокрема естрогени яечниюв стимулюють утворення тиреощних
гормон!в у щитоподзбшй залоз! (дистантна д!я), а шсулш, що утворюеться в р-
ендокриноцитах остр!вц!в пщшлунково! залози, пригшчуе секрешю глюкагону з
розташованих ноблизу а-ендокриноцийв (паракри нна регуляц!я). Тому розлад
функцп одше! ендокринно! залози неминуче зумовлюе ланцюгову реакцпо гормо-
нальних змш. Так виникають поеднаш (корелятивш) ендокринш розлади: недостат-
н!сть статевих залоз при цукровому л!абей, функщональне перенапруження, а
пойм виснаження р-ендокриноцийв панкреатичних остр!вц!в у випадку ппер-
функци юрково! або мозково! речовини надниркових залоз. В експеримент! вида-
леиня щитоноддбно! залози спричинюе пригшчення активноей статевих залоз !
юрково! речовини надниркових залоз. Шсля кастрацп розвиваеться ппер'грофп.
юрково! речовини надниркових залоз. До цього типу ендокринопайй належить
також поеднання пперандрогенй з ппершсулшем!ею та шсулшорезистентшстю у
хворих на метабо.йчний синдром, або синдром Штейна — Левенталя, для якого
характерними е пол!юстоз яечниюв, аменорея, ожиршня, безплщшеть.
Реакшя ендокринно! залози на первинне ушкодження шшо! ланки ендокрин-
но! системи нерщко е компенсаторною 1 спрямованою на збереження гомеостазу.
Гормони подщяють на синерггсти, як! впливають на ф!з!олопчю та б!ох!м!чн!
процеси в однаковому напрямку, й антагон1сти, що д!ють протилежно. Так, п-
погл!кем!чний ефект шсулшу можна нейтрал!зувати введениям кортизолу або ад-
эеналпху. Якщо в собак з видаленою пщшлунковою залозою видалити також над-
жирковх залози, прояви иукрового дхабету будуть значно слабшими. Деяк! гормо-
ч, характеризуються односпрямовашгм впливом на обмш речовин, наприклад
щреналхн х глюкагон, якх зумовлюють пперглхкемйо.
Такий подпт гормонхв значною мхрою умовний. Хоча шсулш х кортизол по-
еному впливають на рхвень глюкози в кров!, хх сумхсна дхя спрямована на полш-
лення постачання тканин вуглеводами: кортизол сприяе пхдвищенню концентра-
глюкози в кров!, шсулш полегшуе проникнення п всередину юптини. Глюко-
ортико'щи х соматотропгн (гормон росту) дпоть як антагошсти вщносно
ежового обмшу: перший сприяе розщепленню бппов, тодх як другий мае ана-
ххчну властивхсть. Проте за впливом на енергетичний обмш хх можна характе-
гнзувати як синерпсти, тому що глюкокортикоида спричинюють гхперглжемпо, а
матотропш мае лшолхтичну дхю, тобто загальний ефект цих гормонхв полягае в
восиленн! окиснення вуглеводхв х жирхв.
Спхввщношення мхж гормонами — антагошстами х синерпстами — посщае
сновне мхсце в патогенез! деяких ендокринних розладхв. Так, порушення толе-
рантностх до глюкози ! ггперглхкемхя можуть розвинутися внаслщок посиленого
видхлення антагонхстгв шсулшу: глюкагону, соматотрошну або кортикостерощёв.
Роль автохмунних процесхв у патогенез! ендокринних порушень. Вщхилення в
хнстемх хмунного гомеостазу при ендокринних хворобах проявляються утворен-
- чм антитхл до антигешв тканин ендокриннох залози (наприклад у щитоподхбшй
залоз! — до тироглобулшу, рецепторхв тиротрошну), шших тканин (м’язово! та
хировох — до рецепторхв шсулшу) або гормонхв, як! циркулюють у кровх. Змшю-
ються також вмхст х функцюнальний стан Т- х В-лхмфоцитхв, Ц1К. Прикладом
автохмунного захворювання е автогмунний тиреощит, до якого призводить уш-
одження щитоподхбно! залози органоспецифхчними антиплами. Пхсля травми
течка шодх розвиваеться автохмунний орхгг, що пов’язують хз порушенням про-
никностх гематотестикулярного бар’ера.
Якщо алерпчна реакцхя активуе секреторш клхтини залози, наслщком авто-
гресй е посилення п функцп. Стимулювальний вплив антитш на продукцпо гор-
'онхв в експеримент! доведено пщвищенням функцхонально! активностх статевих
алоз пхсля введения тваринам невеликих доз вщповщних цитотоксичних сирова-
гок у невисоких дозах, якх мхстять антипла до антиген!в яечка або яечника.
Автоантитхла до гормонхв здатнх нейтралхзувати ххню бхолопчну актившсть. У
। ровх хворих нерщко виявляють антипла до шсулшу, соматотропшу, тиротроппху.
Основы! мехашзми ендокринних хвороб. Регуляцхя секрецй гормонхв спрямова-
на на оптимально забезпечення фхзхолопчно! потреби оргашзму в гормонах. Вона
дшснюеться гуморальними та нервовими шляхами. Найскладнхше зоргашзована
.ндокринна функцхя у тих випадках, коли секрецхя гормонхв регулюеться гйтота-
ламо-гшофхзарною системою (схема 28).
Залежно вщ рхвня ураження ендокриннох системи, розрхзняють так! мехашзми
. ндокришхих хвороб: 1) порушення центрально! нервово! та нейроендокринно! регу-
яцп ендокринних функцш; 2) порушення бюсинтезу х секрецй гормонхв; 3) пору-
лення транспорту, метаболхзму та реалхзацй бюлопчно! дй гормонхв.
Порушення центрально! регуляцн ендокринних функцш реалхзуеться двома шля-
хами: нервово-провщниковим х нейрогуморальним (нейроендокринним). Цх сам!
шляхи задхяш в нормальней регуляцй ендокринних функцш.
Метаболии Метаболии
Виведення з оргашзму
Нирки, печхнка, залоза зовшшньо"! секрецп
Схема 28. Ф1зюлопчна оргашзашя ендокринно! функцп
Нервова регулящя може бути прямою 1 непрямою. Прямим нервовим шляхом
регулюеться секрещя гшоталам!чних рил!зинг-гормон!в, катехолам!н!в мозково!
речовини надниркових залоз, етф1зного мелатошну. Пщ непрямою регулящею
розумпоть нервовий вплив на кровопостачання ендокринно! залози та синтез 1
секрецпо тропних гормонов.
Порушення нервово! регуляцп нерщко призводить до розвитку ендокрино-
патп. Так, психзчна травма може стати причиною розвитку ендокринно! безплщ-
ност!, цукрового дгабету, тиреотоксикозу. 1снування психогенних ендокринопатш
пщтверджуе важливе значения ЦНС у патогенез! порушень дгяльност! ендокрин-
них залоз.
Руйнування або стимуляция певних дшянок л!мб!чно! системи, сггчастого ут-
вору (речовини) стовбура мозку, промгжного мозку порушуе секрецио гормонгв.
Розлади нейроендокринно'1 регуляцп зумовлеш первинним ураженням гшотала-
муса або пов’язаних !з ним вищерозташованих вщдшв головного мозку. Саме в
ппоталамус! зосереджеш головш центри регуляцп секрецп гормонгв гшофгза г,
отже, низки пщпорядкованих йому ендокринних залоз. Тому в раз! ураження
центральних механгзмгв регуляцГ! можуть виникати плюригландулярш (множинш)
хвороби ендокринно! системи.
Нейроендокринна регуляцгя е таким типом регуляцп, який здгйснюе гнтегра-
цпо нервових г гормональних сигнал!® г гцдтримуе гормональну ргвновагу за при-
нципом зворотних зв’язкгв. Анатомгчним субстратом нейроендокринно! регуляцп
е гшоталамо-гшофгзарний комплекс та деяк! позаппоталамгчнг утвори: морський
коник (гшокамп), мигдалеподгбш ядра та гн. Здебшьшого вдеться про саморегу-
ляцпо дгяльностг залежних вщ гшофгза залоз шляхом негативного зворотного
Мал. 70. Регулящя гормонально! р!вноваги в норм! (а) ! при порушеннях у
систем! зворотного зв’язку (б — у раз! зниження, в — пщвищення збудли-
восп гшоталамуса):
1 — гшоталамус; 2 — ппоф!з; 3 — периферична залоза; Г — гормон пери-
ферично! залози; РГ — рил!зинг-гормон; ТГ — тропний гормон гшоф!за
» ’язку. Це означае, з одного боку, що пщвищення вмгсту гормону “пщлеглог”
залози в циркулюючш в оргашзм! кров! гальмуе секрецпо вщповщного тропного
ормону гшофгза, г в такий спосгб нормалгзуеться функцгя периферично! залози.
3 гншого боку, ослабления секрецп гормону зумовлюе автоматичне посилення
тпофгзарно! стимуляцп, унаслщок чого знову-таки вщновлюеться гормональна
^гвновага (мал. 70).
Один гз численних експериментальних доказгв гснування негативного зворот-
аого зв’язку полягае в тому, що видалення в щура чи собаки одшег наднирково!
залози спричинюе внаслщок пщвищення секрецп кортикотрошну ппертрофгю
тег залози, що залишилася. Тривале введения тестостерону у високих дозах зумов-
тое атрофгю яечок, а кортизол в аналопчних дослщах спричинюе атрофпо кгрко-
воТ речовини надниркових залоз. В обох випадках це пов’язано з пригнгченням
’родукцг! вгдповгдних тропних гормонгв гшофгза.
В окремих випадках гормон периферично! залози здатний стимулювати
секрецпо вщповщного гормону гшофгза (позитивный зворотний зв’язок). Напри-
<тад, обов’язковою передумовою овуляцп (виходу яйцеклгтини з фолгкула) в яеч-
ику е багаторазове пщвищення ргвня гонадотропних гормонгв у кровг, яке вщбу-
\аеться завдяки стимулювальному впливу естрогенних гормонгв яечника на сек-
рецпо гонадотропних гормонгв.
У нейросекреторних клгтинах гшоталамуса утворюються рилгзинг-гормони
яейрогормони пептидно! будови), якг стимулюють (лгберини) або гальмують (ста-
тини) вивпгьнення гормошв аденогшоф!за та промжно! частей гшоф!за у кров I
До л!берин!в належать кортикол!берин, тиролаберин, соматолгберин, гонадолгбе- ।
рин, пролактол1берин, мел анол 1берин, до статингв — соматостатин, меланоста I
тин. Вони видйгяються з термшалей аксошв нейросекреторних клгтин у кров .
через шжку ппофгза потрапляють до його передньо! та промгжно! часток.
Гормони вазопресин 1 окситоцин, як! секретуються 13 задньо! частей гшофаъ I
(нейрогшофхза), утворюються у велико юптинних ядрах передньо! гшоталам!чнсг I
дыянки. Звщси вони по аксонах нейросекреторних юптин надходять у нейроппс- I
ф1з, де 1 депонуються. Переносниками вазопресину й окситоцину е спещальв
быки — нейроф1зини. Секрец1я рюпзинг-гормошв регулюеться 1 модулюетьс- I
численними нейромед гаторами, нейропептидами, шшими регуляторами. До них I
належать норадреналш, серотошн, дофамш, юспептин, опющи, КО, галашн 3
нейропептид V тощо. Наприклад, доведено, що до регуляцп секрецп гонадол!бе- 1
рину причетш щонайменше 25 фхзюлопчно активних речовин мозку.
Кр!м пщтримки гормонально! ргвноваги бюлопчна дощлъшсть нейроендок-
ринно! регуляторно! системи полягае в тому, що вона здшснюе каскадне поси- 1
лення нервових сигналов 1 генерал!защю !х шляхом надсилання гормональны’ '
сигналов до вс!х тканин оргашзму.
Ураження нейроендокринних структур спричинюють розлади гормонально' |
р1вноваги. Наприклад, ураження надзорового (супраоптичного) та пришлуночко- I
вого (паравентрикулярного) ядер ппоталамуса супроводжуеться порушенням ]
синтезу ! транспорту вазопресину й окситоцину. Розлади секрецп ппофгзарних |
гормошв можуть виникати внаслщок порушення регуляторних функцш ендоген-
них ошоццв, катехоламшгв та шших нейромед!атор!в
Порушення нормального функцюнування мехашзму зворотного зв’язку ме?
гшоталамо-гшофхзарною системою та пщконтрольними ш залогами е найваж-
ливппою патогенетичною ланкою деяких ендокринних розлад1в. Так, для патоге-
незу хвороби Тценка — Кушшга е характерною змша чутливосп гшоталам!чни
нейрошв до гальм1вно! дп кортикостероида, що своею чертою спричинюе ппер-
секрецпо кортикотропшу 1 вторинну гшерплазпо юрково! речовини надниркових
залоз.
Порушення швидкоспи утворення (синтезу) / видыення (секреци) гормошв е од-
ним гз головних мехашзмгв виникнення 1 розвитку ендокринно! патологп. Синтез (
! секрещя гормошв сутгево вщр!зняються для похщних амшокислот, бглково-пеп-
тидних, стерощних та шших гормонгв. Так, синтез гормошв щитопод!бно! залози
вщбуваеться у склад! тиреоглобулшу, що мгститься в колощ! у порожнин! фол!-
кул!в, шляхом приеднання йоду до тирозину та конденсацй йодованих залипла?
ще! амшокислоти. Попм ештелш фол!кул!в поглинае шляхом пшоцитозу крапл!
колощу. Процес гормоноутворення завершуеться протеолггичним розщепленням
тиреоглобулшу, що зумовлюе вивыьнення тироксину та трийодтироншу 1 вщц-
лення цих гормошв у кров.
Быково-пептидш гормони утворюються за загальновщомою схемою балково-
го синтезу внаслщок активацп вщповщних генгв. Як правило, спочатку син-
тезуеться велика молекула прогормону. Послщовне ферментне розщеплення !
змша просторово! конф!гураци ц!е! молекули вщбуваються в цистернах ендоплаз-
матично! с!тки (ретикулуму), що зумовлюе утворення спочатку прогормону, г
пот1м власне гормону.
Початковим субстратом для синтезу стеро'щних гормошв е холестерин, що
хадходить до гормоноутворювалъних юптин хз кров! або синтезуеться в них з оц-
говох кислоти. Вщ молекули холестерину вщокремлюеться бхчний ланцюг, а зали-
лгок молекули зазнае численних ххмхчних модифхкацш за участю ферменпв. За
такою схемою утворюються кортикостерощи, естрогени, андрогени, прогестини.
Стерощш гормони не затримуються в залозах х надходять у кров шляхом зви-
чайно’х дифузп. Быково-пептидш гормони накопичуються в юптинах, де вони
. интезуються, у вигляд! гранул х секретуються шляхом екзоцитозу пщ впливом
. гимулювальних нервових або гуморальних факторхв.
Вщом! певш ритми секрецц гормошв залежно вщ часу доби, освгглення, пори
року, коливань фхзхолопчно! акта вноси оргашв х систем. Так, максимальна сек-
тещя мелатошну спостерйаеться вночх за умов мппмального освгглення (фотопе-
чодичшсть). Найвища концентрацхя в кров! тестостерону, глюкокортикощхв, кор-
тикотрошну визначаеться в ранковх години, гормону росту — у першх шчш годи-
ли 1 т. п. Секрещя жшочих статевих гормошв змппоеться не ильки протягом
доби, а й щомхсяця вщповщно до стадш менструального циклу.
Мхсцевх патолопчш процеси в ендокринних залозах змппоють ххню функцю-
:альну актившсть. Гшофункцхя розвиваеться внаслщок зменшення маси паренххми
алози (атрофхя, некроз), недостатностт ферментних систем 1 кофакторхв бюсинтезу
юрмонхв (фолхева кислота, мхкроелементи тощо), блокади мехашзмхв депонування
та секрецц. Навпаки, джерелом надлишковох продукцц гормошв (гшерфункцц) е
«хльшення маси занозистого еттелпо (гшерплазхя, гшертрофхя, доброякзсш та зло-
якхснх пухлини), активащя ферменпв бюсинтезу гормошв тощо. Особливхстю гор-
монально-активних пухлин залоз, що за нормальних умов контролюються гшофх-
зом, е 1хня автономшсть, тобто незалежний характер секрецц гормонхв. Тому змен-
шення секрецц вщповщного тропного гормону аденогшофхзом за механизмом зво-
ротного зв’язку не зменшуе секрецпо гормошв у пухлинних юптинах периферично!
ендокринно! залози.
Ендокринна дисфункцхя нерщко е наслщком спадкового дефекту ферменпв
бюсинтезу гормошв (адреногештальний синдром).
Порушення швидкосп утворення та видщення гормошв можуть виникати
внаслщок змгни чутливост! залоз до нервових 1 гуморальних регуляторхв. Так,
зменшення юлькосп рецепторхв лютрошну на мембранах штерстицхйних юптин
яечок, що спричинюеться тривалою стимулящею статевих залоз препаратом цьо-
го гонадотрошну, зумовлюе недостатню вщповщь на ендогенний гонадотропш !
зниження утворення тестостерону. Надлишок прогестерону або андрогешв у кров!
пригшчуе чутливхсть гонадотропоцитхв ппофхза до рилхзинг-фактора лютрошну.
Причиною змшенох чутливост! залози може стати порушення трофхчно! шнерва-
цц. Наприклад, пхсля перер!зання симпатичних нервхв змхнюеться реакцхя юрко-
во! речовини надниркових залоз на стимуляцию кортикотропшом.
Розрхзняють абсолютну та вгдносну (приховану) недостатшсть ендокринних
залоз. Остання виявляеться в разх пщвищено! потреби оргашзму в гормонах, тодх
як за умови фхзхолопчного спокою показники утворення та секрецц гормонхв
мають нормалью величини. Невщповщшсть продукцц кортикостерощхв потребам
оргашзму нерщко вщзначають, наприклад, при бронххальшй астм! та шших алер-
пчних хворобах.
Неповноцгншсть гормональних резервгв визначають за допомогою спещаль-
них функщональних проб. Зокрема навантаження глюкозою з подалыпим визна-
ченням вмгсту шсулшу в кров! дае змогу виявити в частини хворих на цукровий
дгабет виснаження резервних можливостей клгтин, яга виробляють шсулш.
Порушення транспорту, метаболгзму та бюлоггчног ди гормошв. Цг порушення
формують позазалозистий мехашзм ендокринних розладгв, якг виникають на тлг
нормально! секрецп гормонгв.
Транспорт гормонгв в органгзмг здшснюеться бголопчними ргдинами (кров’ю
лгмфою, спинномозковою ргдиною). У кровг гормони перебувають у вгльному
стан! або утворюють комплекси з бглками. Саме вгльнг молекули проявляють гор-
мональну актившсть, що е важливим складником гормонально! регуляцп. Особ-
ливо це стосуеться стерогдних г тиреогдних гормонгв. Наприклад, вгльна фракцгя
тиреогдних гормонгв становить лише 1 % загального вмгсту гх у кровг, андрогенгв —
1-3 %.
Транспорта! бглки утворюються в печшцг, звгдки надходять у кров. Вони вибгр-
ково зв’язують певнг гормони. Так, транскортин утворюе комплекси з глюкокор-
тикощами та прогестинами, тестостеронестрадюлзв’язувальний глобулш взае-
модге з естрогенами та андрогенами, тироксинзв’язувальний глобулш — з гормо-
нами щитоподгбно! залози тощо. Саме цг гормони регулюють штенсившсть
синтезу в печшцг вгдповцших зв’язувальних бглкгв.
Зв ’язувалънг бглки виконують не лише транспорту функщю. Вони утворюють
депо, з якого гормони гггвидко вивгльнюються в разг термшово! потреби. Суттева
роль бглкгв полягае г в тому, що вони утримують гормони вгд передчасно! мета-
болгчно! деградацц та подовжують термш гхнього перебування в кров!. Кргм того,
вони зв’язують надмгрну кглькгсть гормонгв г в такий спосгб запобггають гормо-
нальному “отруенню”. Цей захисний мехашзм спрацьовуе зокрема пгд час ваггт-
ност!, коли значно зростае секрецгя кортикостерощгв, естрогешв г прогестингв.
Вщом! гормональш порушення, зумовлеш змгною концентрацп зв’язувальних
бглкгв плазми кровг. Симптоми ггпертиреозу можуть розвинутися внаелгдок змен-
шення концентрацп тироксинзв’язувального глобулгну. У жшок з ознаками вг-
рилгзацг! (ргст волосся за чоловгчим типом, низький голос, недорозвинен! вто-
риннг статевг ознаки) нерщко виявляють зменшення тестостеронестрадголзв’язу-
вально! здатностг плазми кровг.
Метаболгзм ! виведення гормонгв з органгзму е важливими механгзмами збе-
реження гормонально! ргвноваги. Зазвичай швидкгеть цих процесгв змгнюеться
вщповщно до гнтенсивност! секрецп гормошв. Час перебування гормонгв у судин-
ному русл! коливаеться в!д хвилин (наприклад для ггпоталамгчних рилгзинг-гор-
монгв) до кглькох дгб г навгть тижнгв (для тиреощних гормонгв).
Метаболгчнг перетворення гормонгв вщбуваються в печшцг, нирках, кишках
та шших органах. Бглково-пептидш гормони пгддаються розщепленню протеша-
зами, тиреогднг — дейодгшазами. Стерогднг гормони шактивуються шляхом при-
еднання ггдроксильних груп та ш.
Метаболгзм гормонгв супроводжуеться втратою гхньо! активностг та утворен-
ням водорозчинних сполук, як! легко виводяться гз сечею, жовчю, единою. Проте
внаелгдок метаболгзму гормонгв можуть утворюватися нов! речовини, якг шод!
навгть перевигцують свого попередника за силою гормонально! дц.
ма 29^ Патогенез резистентност! до шсулшу в раз!
ожиршня (за Каком ( Ротом, 1975)
У раз! порушення процеств
метабол 1зму гормонш (цироз, ге-
патит, низька або висока актив-
Н1сть ферменпв метаболизму) та-
кож виникають гормональн! роз-
дали.
Реалгзащя бюлоггчно! ди гор-
монш починаеться з !х взаемодп
13 вйщов] дними к/итинними ре-
цепторами — быковими молеку-
лами, як мають значну х!м!чну
спорщнен!сть до певних гор-
монгв 1 винятково або переваж-
40 взаемодпоть саме з ними. Ф!з!олопчна реакщя клггини на гормон значною
рою залежить в!д кшькост! гормональних рецепторш.
За мюцем розташування розргзнякль мембрани! рецептори, вбудован! в юп-
дгу оболонку, ! внутр! шньоюптинн!, шо мктяться в цитоплазм! та ядрь 1з мем-
шними рецепторами взаемодпоть катехоламши (адреналш, дофамш), бшково-
гттидн! гормони (шсулш, ппоф!зарн! гормони). 1з мембранних рецептор!в юрмо-
-1 ,ьш сигналя надходять до внутр!шньокл!тшших бюх!м!чних систем 1 генш.
Принципово (ншими е мехашзми ди стерощних ! тиреощних гормонш. Завдя-
малим розм1рам молекул вони легко проникають усередину юптини 1 там ут-
. зюють комплекси з вшповщними цитоплазм ат ич ними ! ядерними рецепторами,
рецептори функшонують як регулятори активносп ген!в, а роль гормошв по-
; де у забезпеченш пшьово! доставки рецепторних молекул до вщповщних ген!в.
щодавно виявлено наявшсть мембранних рецепторш стерощних гормонш, сиг-
али з яких також надходять до ядра юптини-мппен!. Але деяк! змши, що Тх спри-
глнюють стероццп та тиреощн! гормони у клггин!, не потребують учасп ядерного
.ному. Зокрема вони можуть безпосередньо впливати на актившсть ферменпв.
Порушення гормонально! рецепцп в юптинах зумовлюе низку патолопчних
лхилень. Наприклад, за вроджено! вщсугносп клттинних рецепторов андрогенш у
ювшв розвиваеться синдром тестикулярно) фемшТзацп. Вш характеризуеться
твою жТночих вторинних ознак, попри те що яечка продукують достатню к!ль-
сть тестостерону. Прсутизм (оволосшня в жшок за чолов!чим типом) може бути
5» (явлений шдвишеною чутливктю волосяних фолжулш до ендогенних андро-
вшв. В!дом1 випадки жшочого ппогонадизму. зумовленого нечутливктю яечни-
№ до гонадогропних гормонТв (синдром резистентних яечниюв).
Хнеулшов! рецептори вщповщальн! за розвиток нечутливосп тканин до ш-
ину при ожиршш. що спричинюе виникнення цукрового д!абету. На схем! 29
сазано мехашзм цього явища. Пщвищення вм!сту шсулшу в кров!, що виникае
щповщь на гшерглткемпо, зумовлену надмтрним уживанням вуглеводш, знижуе
ьюсть рецепторш шсулшу. Як наелшок, зменшуеться чутлив!сть тканин до ш-
"лну, що сприяе пщвищенню ргвня глюкози ь кровг з подальшим зменшенням
ькост! рецепторгв шсулшу.
Отже, характер основних проявив ендокринно! патологп (ппо- та пперфункцп)
; жить вщ направленост! функцюнальних змш (ушкодження) на вс!х ргенчх регуля-
— вщ утворення т секрецп гормошв до реа/пзацп !хньо! бюлопчно! дат (табл. 28).
Таблица 28. Патогенез ендокринних порушень
- —1 Рхвень ушкодження ендокринних функшй Характер змш
при ппофункцн при пперфункшх
Центральна регуляция 1нкреторно1 функцп: • стимуляция Ослаблена Посилена
• гальмування Посилене Ослаблене
Утворення 1 секрецхя гормону Зменшен! Збхльшенх
Зв’язування гормону з бшками кровх Пхдвищене Знижене
Тканинна рецешця, чутливхсть органхв-мхшеней Зниженх Пхдвищенх
Метаболхчна хнактивацхя гормону Збхльшена Зменшена
Компенсаторно-пристосувалън1 мехашзми. У патогенез! ендокринних, як х ш-
ших, хвороб розрхзняють, з одного боку, мехашзми ушкодження, тобто виник-
нення 1 розвитку гормональних порушень, а з шшого — мехашзми компенсаци та
вщновлення. До останшх нал ежить, наприклад, компенсаторна гхпертрофхя яеч-
ника або яечка теля видалення одше! з пари залоз. Гшертрофхя 1 гшерплазй.
секреторних клетин кхрковох речовини надниркових залоз здатш повшетю ком-
пенсувати и недостатшсть при ххрурпчному вилученнх бхльшо! частини залози. Ц1
компенсаторш реакцп регулюються негативним зворотним зв’язком гшофхза и
периферичними ендокринними органами.
Один тз ефекгивних шляххв пристосування оргашзму до патолопчних роз-
ладхв секрецц гормонхв полягае в регуляцн зв’язування гормонхв быками кров!.
Так, пщвищене зв’язування глюкокортикощхв хз транскортином плазми, що спо-
стерхгаеться в раз! наднирниково! недостатност!, захищае щ гормони вщ мета-
болхчно! деградацп х тим самим сприяе щдтриманню нормально! концентраци п
у кровх. Усунення несприятливого впливу гшо- та гшерсекрецп гормонхв дося-
гаеться також змшою активностх ферментхв печшки та шших органхв, у яких к
гормони руйнуються й шактивуються.
ПОРУШЕННЯ ФУНКЦ1Й
ППОТАЛАМО-ППОФ13АРНОТ СИСТЕМИ
Порушення функцш ппоф!за
Панппопхтухтаризм, або повну недостатшсть ппофхза, вщтворюють в експеримен-
тх видаленням усхех залози (гшофхзектомхя) або передньо! и частки. Найдокладш-
ше наслщки гшофхзектомн вивчено в собак х щурхв. У молодих тварин пхсля ви-
далення ппофхза розвиваеться складний симптомокомплекс, зумовлений дезор-
гашзащею обмшу речовин ! регуляцн ендокринних функцш. Найпомхтшшим
зовшшшм проявом цього стану е рхзке затримання росту х загального розвитку
тварин. Спинення росту пов’язане головним чином !з порушенням соматотроп-
но! х тиреотропно! функцш ппофхза. Бхлышсть шших порушень зумовлена недо-
статшм утворенням гормонхв у периферичних залозах: щитоподхбшй, наднирко-
вих х статевих, якх зазнають атрофй внаслщок розладу стимулювально! функцй
тропних гормонхв гшофхза.
У гшофгзектомованих тварин спостерггаються млявгсть г малорухливгсть, зни-
ц ення основного обмшу, схильшсть до гшоглгкемп, зниження температури тгла
ппотермгя), м’язового тонусу г резистентносп до р!зних несприятливих впливгв.
У людини повна недостатшсть функшй гшофгза бу вас вродженою г набугою.
йчаспше причиною хвороби с пухлина, некроз гшофгза шсля полопв, травма
снови черепа, запалення, тромбоз, вгрусна гнфекпгя. Ураження залози в ембруо-
-тьний пергод або перед статевим дозргвання м призводить до карликовость ста-
рого недорозвинення, ослабления функцгй щитоподгбног залози, ендокринно-
бмгнних порушень, зниження загально! реактивности
У разг руйнування понад 95 % маси залози в дорослих розвиваеться г1поф1зар-
а кахекая. або хвороба Сгммондса, для яко! характерними с загальне виснажен-
я, атрофгя щитоподгбног, надниркових г статевих залоз, м’язово! тканини, внут-
лнгх органгв. руйнування кгстковог тканини, випадшня зубгв г волосся, розлад
нкцгй автономно: (вегетативно!) нервово! системи, гшоглгкемгя Ылышсть по-
тень пов’язана з припинснням синтезу соматотропгну. тиротропзну г корти-
отропгну.
Пвофупкшя аденогшофгза. Патогенез г симшоми гшофункцп аденогшофгза
жтежать вгд того, секрецгя якого саме гормону або гормонгв е порушеною
Для частково! недостатностг аденолпофгза с характерним зменшення продук-
та! якогось одного гормону. Наприклад, ранне випадання або пригнгчення сома-
“ тропно! функцп спричинюе розвиток гтоф1зно1 карликовости Ослабления син-
езу бглкгв зумовлюе атрофгю м’язово! г сполучног тканин, що зови! проявляеться
дальним постаршням, зморшкамп, млявгстю шкгри. Унаслщок випадання лг-
агптично! дп соматотролшу може збгльшуватися вгдносиа маса жирових вщкла-
ень. Через те що статевий розьиток пов’язаний гз соматичним, статев! органи
залишаються в гнфантильному сташ (ппогонадизм).
Гонадотропна недостатшсть зумовлюе вторинний ппогонадизм, фгзичне не-
розвинення, затримку статевого розвитку — гиюфпний тфантшйзм. У дгвчаток
.постерггаються вгдсутнгсть менструашй. безшпдшсть, у хлопчикгв — затримка
лущения яечок у калитку (крипгорхгзм) та гх гшоплазгя.
Гшерфункщя аденолпофгза. Ыльшгсть патолопчних змгн, що спостерггаються
гюдини при гшерфункцп аденолпофгза, можна вщтворити в експеримент! вве-
-чням тваринам екстракпв гшофгза або окремих гормошв.
Лперпродукцгя соматотропгну в людини проявляеться як гтоф1зний г1гантизм
бо акромегалия, залежно вщ того, у якому вщг виникла патолог гя. Гшофгзний п-
.гятизм розвиваеться за надлишково! секрецп соматотропгну в молодому вщг, до
1криття етфгзних зон росту. Аналопчнг гормональнг порушення в старшому
шг, шсля закриття ешфгзних швгв г припинення росту, е причиною акромегалп.
Джерелом пгдвищено! секрецп соматотропгну у випадку акромегалп е еозино-
иыга аденома гшофгза. Окремг частини тгла при цьому непропоршйно збглыпу-
ються, риси обличив також збгльшенг. Одночасно спостерггають збгльшення
нутргшнгх органгв — спланхномегалгю (збгльшення печшки, селезшки, серця
ошо). Змгни зовггшпгього вигляду хворого зумовлеш посиленням пергостального
осту ! розростанням м’яких тканин.
У разг акромегалп концентращя соматотропгну в кровг може перевишувати
- трмальнг показники у 100 разгв г бгльше. Патолопчна д!я надлишку соматогрот-
су на оргашзм визначаеться здатшстю гормону пщвишувати проникшсть клгтин-
' 4.47
них оболонок для амшокислот, прискорювати включения хх у синтезований бхлок
гальмувати розпад бшкхв. Унаслщок посиленого лпхолхзу х гальмування утворення
жиру з вуглеводхв збшыиуються мобинзацхя жиру !з депо, вмхст неестерифхкованих
жирних кислот у кровх, окиснення хх у печппц й утворення кетонових тш. Через
вплив соматотропшу на рхзнх ланки регуляцн вуглеводного обмхну при акромегал'
часто спостерхгаються гшерглхкемхя, зменшення чутливост! тканин до шеулхну.
Патолопчна дхя соматотрошну на сполучну, кхсткову ! хрящову тканини
випадку акромегалн зумовлена зокрема здатшстю гормону стимулювати утворен-
ня оксипролшу (найважливхшого компонента колагену) х хондротгинсульфату. Ш
та низка шших ефекпв соматотрошну опосередковуються особливим б 1л ковш
фактором — соматомедином (шеулшоподхбним фактором росту), який утво-
рюеться в печхнцх пщ дхею соматотрошну.
1нша поширена форма пперфункцн аденогшофхза — хвороба 1ценка — Куийн-
га. Для не! характерне збшьшення секрецй кортикотропшу, зумовлене доброякхс-
ною кортикотропшпродукуючою базофхльною пухлиною гшофхза х, як наслщок,
надмхрною секрещею глюкокортикощхв наднирковими залозами.
Пперпролактинемгя, причиною яко! здебшьшого е доброякхена пухлина з лак-
тотропоцитхв аденогшофхза, спричинюе в жшок синдром галакторех-аменоре
Його наслщками е стимулящя утворення ! видшення молозива, а також порушен-
ня дозрхвання фолгкулхв у яечниках х приппчення овуляцп. У чоловхкхв пщвищенз
секрецхя пролактину може бути причиною приппчення сперматогенезу х безплщ-
носп. Через те що головним природним чинником, який гальмуе секрешю про-
лактину, е дофамш, препарата його агошспв з успххом використовують для лхку-
вання гхперпролактанемхх.
Ефекти пщвищення секрецй кортикотропшу, тиротропшу х гонадотрошшв
зводяться переважно до пперфункцн надниркових, щитоподхбно! х статевих зале,
(див. вщповщш роздши).
Порушення функцш нейроппофхза. Збшьшення секрецй вазопресину, яком)
притаманна антидхуретична дхя, сприяе накопиченню води в оргашзм!, вщхграе
важливу роль у патогенез! рефлекторно! анури, тобто затримки утворення сечх
(наприклад при больовому шоку), х набряк1в, зокрема х при цироз! печхнки, коля
порушуеться шактиващя гормону. Наслщками надмхрно! секрецц вазопресину <
також пщвищення артерхального тиску, гшонатрхемхя та зменшена осмоляршсть
сироватки кровх.
Дефхцит вазопресину виникае внаслщок ураження дшянок нейроендокрин-
но! системи: надзорового (супраоптичного) х пришлуночкових (паравентрикуляр-
них) ядер гшоталамуса або гшоталамо-ппофхзарного нервово-провщниковоп
шляху. Унаслщок цього розвиваеться нецукровий д1абет, або нецукрове сечовис-
наження.
У деяких хворих нецукровий дхабет виникае на тлх нормального або навгг
пщвищеного вмхету вазопресину в кровх. У таких випадках патогенез хворобя
пов’язаний !з пщвищеною шактиващею гормону або, що бшып гмовхрно, хз недо-
статньою чутливхстю до нього дистальних частан канальщв нефронхв, у яких вщ-
буваеться зворотне всмоктування води.
I абсолютний, х вщносний дефицита вазопресину порушують реабсорбщк
води в канальцях нефронхв, що спричинюе збшьшення дхурезу (полхурпо), спрап
зниження артергального тиску. Хвор! видшяють за добу 3—8 л сеч! низько! вщ-
осно! густини. Нередко дхурез становить 10—12 л, а в окремих випадках — 40—
43 л.
Деякт патолопчш змши, шо спостерггають шсля видалення нейрогшофтза, по-
чзаш, гмовгрно, з дефщитом окситоцину. Вважають, шо змша секреци оксито-
лшу вршграе роль у патогенез! порушення пологово! дгяльносп г лактаци, диск!-
ези жовчних шлях! в.
-ейроендокринж хвороби
>дш й тт сам! патолопчш пронеси можуть розвиватись як при ураженш гшотала-
уса, так ! в разг первинного порушення функцш ппофхза. I при нецукровому
абеп, 1 в разг порушення статевого розвитку чи росту нерщко виявляють ознаки
ши нейросекреци, розладу функщй гшоталамо-плофгзарно! системи. Навпъ у
их випадках, коли первинне патолопчне ураження з’являеться в гшофгзг, до про-
есу завжди залучаеться ппоталамус. Так, аденома гшофгза, яка е джерелом
двищенох продукт!' соматотротну! причиною пгантизму або акромегали, може
чроводжуватися нецукровим дгабетом. Багато розладтв обмшу речовин, гермо-
егуляцп, статевого г фгзичного розвитку, що ускладнюють перебтг основно! ен-
окринно! хвороби, зумовлеш ураженням промгжного мозку. 1нод! порушення
ейроендокринно! регуляци виникають як наслшок ураження позаг!поталамтчних
.трукгур — мигдалепод1бного пла, морського коника (гшокампа) та ш.
Ендокринно-обмшш порушення гшоталамо-гптофгзарного походження вщ-
-осять до групи нейроендокринних хвороб. Типовим прикладом е адипозогеш-
лальна дистрофия, яку супроводжують ураження ппоталамуса, ослабления гона-
отропног активносп гшофгза т порушення г!поталам!чно! регуляцп енергетично-
₽о обмшу. Це зумовлюе затримку статевого розвитку. недорозвинення статевих
оргашв, крипторх1зм, припинення менструашй (аменорея). Розвиваеться ожирш-
я з харакгерним вщкладенням жиру в дшянщ плечей, живота, газа г стегон.
ктновний обмш 1 температура тша знижеш, у деяких випадках спостерггають
фигурно, спрагу г поспйне вщчуття голоду В експериментах на щурах 1 мишах
йнування дутоподгбного (аркуатного) ядра або вентромедтальних ядер гшотала-
са призводить до посилення апетиту. гшер.тшщемп, ггперглгкемв, шсулшоре-
гстентностт й ожиршня.
Порушення нейроендокринно! регуляцп секрецп гонадотропних гормонтв е
ровщним мехашзмом синдрому Штейна — Левенталя, для якого характерш
олгктстоз яечниюв, безплщшсть, порушення менструального циклу, часто ожи-
1ння, ознаки гшерандрогешзацп оргашзму. Узагалг 90 % випадктв аменоре! в
жшок репродуктивного вгку повязан! з нейроендокринними порушеннями
знаслшок оргашчних або функшоналъних ура жен ь пром!жного мозку, сильних
сихоемоцшних впливгв. Вщомо, шо в жшок тривалий 1 значний стрес або вжи-
п. ння наркотичних засоб!в спричинюе безплщшсть нейроендокринного похо-
_ження через гальмування ендогенними або екзогенними ошатами нейроендок-
ннних мехашзмгв овуляцп.
Експериментально доведено можтивкть формування нейроендокринно! па-
тологи внаслгдок неадекватного впливу на мозок ембрюна та плода нейромедга-
оргв, ендогенних ошатгв, статевих гормошв, гшококортикощгв. тироксину та гн-
ших гормошв, а також тривалого стресу пщ час ваптносп. Отже, нейроендокрин-
Н1 порушення можуть бути наслщком патолопчних впливгв на головний мозок 5
ранньому онтогенез! — пщ час ваптносп або в ранньому дитин спи. Клппчн
спостереження також свщчать про пщвищений ризик народження хлопчикгв,
яких у майбутньому виявляеться гомосексуальна поведшка, якщо гхш матер! пщ
час ваптносп переживали сильний стрес. Деяк! форми ендокринно! безплщност
в жгнок, зумовлеш порушенням статево! циюпчносп, пов’язують гз уживанняк
1ХШМИ матерями пщ час ваптносп гормональних та ипих лжарських засобгв. й
експеримент! зазначену патолопю вщтворюють одноразовим уведенням тестосте-
рону новонародженим самкам щур! в, мишей або ваптним самкам крол!в, гвшей-
ських свинок. До пубертатного вгку тварини за сво!м зростом 1 розвитком шчим
не вщргзняються вщ штактних тварин того самого виплоду. Проте тод!, як у кон-
трольна щур1в статевозрыого вгку спостерйаються регулярний 4—5-денний ест-
ральний цикл, у андрогешзованих тварин виявляються поспйна пчка, полжгсто
яечникгв, безплщшсть. Рання кастращя новонароджених самцгв щур!в зумовлкх
прояви гомосексуально! поведшки в зршому вщг, незважаючи на замкну терапж
тестостероном. Такий самий патолопчний вплив на плоди чоловгчог етап маг
стрес ваптно! самки. До того ж у них порушуеться реактившстъ ппоталамо — п-
пофгзарно — наднирниково! системи на стресовг подразнення.
Перелгчен! порушення зумовлеш первинним впливом андрогешв, кортико-
стеро'щ!в 1 нейромедгаторгв на структури! утвори гшоталамуса, вщповщальш за
нейроендокринну регуляшю статево! повед!нки й адаптивних ендокринних реак-
цш у самщв 1 овуляцп у самок. Виявлення цих явищ дало змогу розробити при-
нципи превентивно! нейроендокринологп.
ПОРУШЕННЯ ФУНКЦ1Й ШШИХ ЕНДОКРИННИХ ОРГАН1В
Порушення функц|й щитопод!бно'Г залози
Гшофункшя щитоподабно!’ залози. Ппотиреоз вщтворюють у тварин повню--
або частковим хгрурпчним видаленням залози, уведенням тиреостатичних препа-
ратов, якг перешкоджають синтезу г видшенню тиреощних гормошв.
Стан вираженого ппотиреозу, що його спричинюе тотальна тиреощекгомгя
позначають як тиреопривну кахекейо. Наслщки тиреощектоми тим тяжчг, чи
рашше зроблено операцгю. У молодих собак, щур!в, крол!в та шших тварин спос-
терйають значну затримку росту (мал. 71), статеве недорозвинення, розлад усгх
вид1в обмшу речовин, трофгчш порушення.
Унаслщок зниження ргв-
ня окисних процесгв основ-
ний обмш зменшуеться на
25—40 %, знижуеться темпе-
ратура тша, спостерггаеться
схильшсть до гшоглгкемп. Че-
рез зменшення розпаду холес-
терину й утилгзаци його в тка-
нинах у 2—3 рази пщвищуеть-
Мал. 71. Затримка росту щура (праворуч) шсля тиреощек-
ТОМ11 в молодому ВЩ1
концентрация холестерину в кров!. На ньому тлх посилюеться лхя атерогенних
кторхв, Результати радхологхчних дослщжень свщчать про гальмування вклю-
шя мхчених амшокислот у бттки, тобто приппчення бшкового синтезу. Спос-
-ер.гають затримку води в тканинах (набряки).
Тиреощектомованх тварини малорухливх. Розлади функцш виших вцщхлхв
. 4 С проявляються недостатшстю утворення умовних рефлекс хв, переважанням
роцесхв гальмування.
Етюлопчними факторами розвитку гпхотиреозу в людей е: вроджеш дефекти
|Осинтезу тиреощних гормошв; вроджена ппогхлазхя або аплазш щитоподхбно!
_ юзи; автохмунн! га хнфекцшш процеси в залоз!; видалення велико! юлькост!
мренххми залози гид час ххрурпчного втручання; ушкодження залози тиреоста-
шими засобами, радхоакгивнххм йодом унаслщок перевищення допустимих лх-
зальних доз або техногенно! катастрофи (наприклад п!д час авар!! на Чорно-
тьськхй АЕС), а також п!д впливом шкорпорованих радюнуклщхв або локаль-
го зовншшього опромшення. Але найчастхше причиною гшотиреозу е
достатке надходження в оргашзм йоду з !жею та питною водою (див. розды
-XIII “Патолопчна фхзюлопя обмшу м!кроелемент!в”).
У разх тяжко! недостатност! щитоподхбно! залози (якщо захворювання виник-
ч) в ранньому дитинств! або е вродженим) розвиваеться кретишзм, у дорослих
юней — микседема (ппотиреоз, який супроводжуеться слизовим набряком шкх-
"н|. Порушення психхчно! дхяльностх. трофхки, водно-мшерального, бшкового й
пхдного обмшу, росту х статевого розвитку. терморегуляци та хнших функцхй
анхзму, шо е характерными для гшотиреозу. при кретишзм! сягаюгь крайнього
гупеня. Навпь незначний дефхцит йоду в органхзм! матер! пщ час вагхтностх зда-
ен спричинити неповноцшне формування ! дозрхвання головного мозку плода,
-ху шо тиреоцгнх гормони вщхграють ключову роль у процесах формування не-
вовох системи. Через це вже в дитинств! виявляють зниження розумового потен-
шалу, здатностх до навчашхя та шш! вади.
Для мхкседеми типовими е зниження обмхну речовин, ожирхння, малорухливхстъ,
-шження температури тхла. Унаслщок посиленох гщратацв шкхри х пхдшкхрнох жи-
во! клхтковини та надлишкового накопичення в них гщрофыьних слизових речо-
лхн обличчя хворого стае одутлим, з бщною мхмхкою, стовщеними носом х губами,
^остерхгають ламкхсть нхгпв, випадхння волосся та !шш грофхчш порушення. Ста-
;ва функцхя поступово згасае. знижуеться штелекг, попршуеться пам'ять, що суп-
роводжуеться апаттею, сонлив!стю, а в шзн!й перюд хвороби — недоумством.
Збхльшення об’ему щитоподхбно! залози внаслщок дефщиту йоду вщоме пщ
язвою ендем'тний зоб, Ця хвороба поширена в Альпах, Карпатах, Пмалаях, Андах
шших прських районах земно! кулх, де грунт х вода мхстять мало йоду. Дефхцит
оду зумовлюе зниження синтезу тироксину х грийодтироншу. унаслщок чого в
’ пофхзх посилюеться вироблення тиротрошну. Це своею чертою зумовлюе ппср-
газхю щитоподхбно! залози, маса яко! хнодх сягае кхлькох кыограм’в.
Етюлопчну роль дефхциту йоду ь розвитку ендемхчного зобу пцхтверджуе та-
ий експеримент. Якщо собаку протягом перших хпвтора року життя напувати
водою, яка не мхстить йод, маса щитоподхбно! залози сягне 100 г за норми 1 г.
-айбхльш переконливим доказом е успхшна профхлактика йодом ендемхчного зобу
зсередках його поширення. Як свщчить досвщ багатьох ендокринологхв, дода-
шхя солей йоду до ххитно! води або харчово! сол! запобхгае розвитков. хвороби.
Гшерфункщя ШИТ0П0Д10Н01 залози. Пщвишення продукта! тиреощних гормона
ослабления мщност! зв'язку тироксину з тироксинзв'язувальним глобулшом, по-
рушення метабол 13му гормонш шитопод)бноГ залози або пщвишення чутливост
тканин-мшгеней до !хньо! ди зумовлюють розвиток гтертиреозу. Найпоширень
шим його прояво.м е дифузний токсичный зоб (базедова хвороба, хвороба Грейвса).
Типовим симптомокомплексом для дифузного токсичного зоба е збшьшенш
щитоподдбно! залози, екзофтальм, пщвишення основного обмшу, посилення теп-
лопродукции, тахЁкарлЁя та (нив ознаки порушення серцево! щяльносп (“тиреоток-
сичне серце”), гремтшня палыцв рук, пщвищення психёчно! збудливосп. Ц! тг
багато шших патолопчних проявив зумовлен) токсичною д!ею тироксину 1 трийод-
тирон!ну, яю виробляються в надлишков,й КШЬКОСТ].
Розвитков) хвороби сприяють шфекшя, переохолодження, а також фгзюло-
пчн! коливання функщондтьно! активност! залози, пов язан) з менструальшг
циклом. У жшок ппертиреоз спостерйають частйпе, н!ж у чоловгюв. Це поясню-
ють прямою стимулювальною д)ею естроген!в на фолжулярний етателш щито-
подгбно! залози. Провокуючим фактором розвитку ппертиреозу в людини може
бути псиючна травма.
Уявлення про гшерсекретаю тиротропшу як провщну патогенетичну лашс
дифузного токсичного зоба ниш переглядають. Основного значения надають по-
рушенню !мунних пропесзв, насамперед. появ! в кров! таких хворих антитш до •
рецепторов тиротропшу на мембранах фол!кулярних ендокриноципв, а також ан-
титш до тиреоглобулшу, мЁкросомних та шших антигешв шитопопдбно! затози.
Доведено також патогенну роль простагландин!в у патогенез) ппертиреозу та йог,,
ускладнень. Екзофтальм зумовлен ий екзофтальм)чним фактором, який мае ппо-
ф)зне походження 1 под!бний за низкою ознак до тиротропшу. Ц! фактори про-
довжують д)яти 1 шсля хгрурпчного видалення залози 1 можуть спричинити пост-
тиреотоксичну енцефалоофтальмопатЁю.
Патогенез ппертиреозу вивчати не тшьки на людях, а й на лабораторних тва-
ринах, яким уволили препарата тиреощних гормошв або додавали в ’гжу висушену
щитопод)бну залозу При цьому в собак, щур)в, мишей, крол)в спостерйалися
втрата маси тша, збшьшення ЧСС 1 дихальних рухзв. основного обмшу. темпера-
тури тша. трофЁчш розлади, пронос, блювання. Посилювалися чутливйть до п-
поксп, збудливЁсть усьх ланок рефлектопно! дуги, збсльшувалася рухова актив-
шсть.
Провщним мехашзмом токсично! дп надтишку тироксину 1 трийодтирошну е
збшьшення проникносп мггохондр!ально! мембрани. Порушення функционально) ]
шгеграцп мггохондргй зумовлюе роз’еднання окисного фосфорилювання, унаслЁ-
док чого зменшуеться акумуляшя енерп! у фосфатних зв’язках аденозинтрифос-
форно! кислоти та шших макроерпчних сполук. В!льна енерпя окиснення при
цьому видшяеться у вигляд! тепла.
Вщ’емний азотистий баланс пщ час ппертиреозу евщчить про стимуляцию
розпаду бшюв. Спостерхгаеться гшерглЁкемЁя внаслщок посиленого розпаду гл)ко-1
гену в печшщ та м’язовш тканин!. Утил!зац!я глюкози тканинами прискорена.
актившеть гексоюнази пщвищена.
Надлишок тиреощних гормошв гальмуе перехщ вуглеводгв у жири. прискорюе
розпад холестерину та його утшпзацпо в тканинах, штенсифжуе окиснення жир!В
у печшщ, а також пщвишуе чутлив Ёсть жирово! тканини до лпюлпично! дп адре-
яалгну. Усе це призводить до збщьшення енергетичних витрат оргашзму. Наслщ-
ком зазначених змш е посилена мобшзацгя жиру з депо, що пояснюе зменшення
маси тгла хворих на тиреотоксикоз, ггпохолестеринемгю та пперкетонемгю.
Тиреощш гормони порушують метаболгзм серцевого м’яза. Спостерггають
дистрофгчш змгни в мгокардг, порушення атрговентрикулярног провщностг, пере-
ъантаження лгвого шлуночка. Порушуються пероксидне окиснення лгпщгв мемб-
ран серцевих мгоцитгв, енергетичне г пластичне забезпечення дгяльностг серця.
Тиреотоксичне серце” неадекватно реагус на холгнерпчш й адренерпчш впливи.
Порушення секрецп кальцитонгну. Деякг наслщки тиреощектомп, очевидно,
зумовлеш вщсутшстю секрецп бглкового гормону щитоподгбно'г залози — кальци-
гонгну. Утворення кальцитонгну порушуеться також у разг ггпофункцп гцитоподгб-
ног залози, спричиненог тиреостатичними засобами, г ппертиреозу ендогенногс
та екзогенного походження.
Надлишкова секрещя кальцитонгну шодг спричинена пухлинами, що походять
з розташованих мгж фол скулами С-клгтин щитоподгбно'г залози, у яких синтезуеть-
ся цей гормон. Можливо, зг збщьшенням п пов’язане походження несправжнъого
гипопаратиреозу — хвороби, при якгй, незважаючи на нормальне функцюнування
прищитоподгбних залоз, спостерггаються ппокальцгемгя та гнил розлади фосфор-
но-кальцгевого обмгну.
Порушення функщй прищитоподгбних залоз
Ппофункцы прищитоподгбних залоз. Вщсутшсть функцп прищитоподгбних залоз,
тобто тяжкий гипопаратиреоз, зумовлюе розвиток паратиреопривно! тетани. В ек-
спериментг и вщтворюють видаленням залоз у собак г кппок. Через одну-двг доби
пгсля операцп тварини стають млявими, вщмовляються вщ гжг, у них спостергга-
ються спрага, зниження температури тгла, задишка. Унаслщок зниження концен-
трацц кальцпо в кровг змгнюеться спгввщношення одновалентних (Иа+, К’) г дво-
валентних (Са2+, М§2+) гошв. Наслщком цього е ргзке пщвищення нервово-м’язовог
збудливостг. З’являеться м’язова ригщнгсть, порушуеться хода. При цьому спос-
терггаються множиннг фгбрилярш скорочення м’язгв усього тгла, до яких потгм
приеднуються напади судом. Останнг переходят у тонгчнг судоми, розвиваеться
опгстотонус (ргзке вигинання тулуба гз закинутою головою). Судомнг скорочення
можуть поширюватись г на внутргшнг органи (пглороспазм, ларингоспазм). Пщ
час одного з таких нападгв тварина гине, як правило, унаслщок спазму дихальних
м’язгв.
На тлг гшокальцгемп у кровг збгльшуеться вмгст неоргангчного фосфору. Роз-
лади мгнерального обмгну зумовлеш гальмуванням резорбцп кгстковог тканини,
усмоктуванням кальцгю в кишках г збшьшенням реабсорбцп фосфата у каналь-
цях нефрошв.
У патогенез! паратиреопривног тетангг певного значения надають порушенню
дезштоксикацгйног функцп печгнки. Годування м’ясом собак, яким видалено
прищитоподгбнг залози, посилюе тетанпо через недостатке знешкодження про-
дукта азотистого обмгну, зокрема й ослабления здатностг печгнки перетворювати
амонгй на сечовину.
За наявностг додаткових прищитоподгбних залоз (у кролгв, шургв) або збере-
ження пщ час операцп часточки прищитоподгбног залози в тварини розвиваеться
хрошчний гшопаратиреоз, клшочна картина якого вщома як паратиреопривна ка-
хекая. Для не! характерними е схуднення, вщмова вщ Ько (анорексоя), пщвищення
нервово-м’язово! збудливосто, пронос та розн! трофочш розлади.
Гшопаратиреоз у людини розвиваеться найчастппе внаслщок випадкового
ушкодження або видалення прищитоподобних залоз пщ час оперативного втру-
чання на щитоподобной залозо. Вщносна гшофункцоя залоз спостерйаеться в раз:
пггенсивного росту, пщ час ваптносто, у перюд лактацп та за шших станов, для
яких характерна пщвищена потреба оргашзму в солях кальцпо.
Патогенез I клшочна картина гшопаратиреозу в людини подобно до тих, що
спостерогають в експеримент!. Пщвищення нервово-м’язово! збудливост! вста-
новлюють за появою м’язових скорочень пщ час подразнення рухових нерв!в
гальвашчним струмом певно! сили, здавлювання руки вище локтя або легкого
постукування по шюро в мосщ виходу лицевого нерва спереду вщ зовншшього
слухового ходу.
Гшерфункцоя прищитоподабних залоз. При гиперпаратиреоз! внаслщок шдвище-
но! секрецц паратгормону посилюються утворення й актившсть остеокластов, що
здшснюють розсмоктування (резорбцпо) юстки, о гальмуеться утворення остео-
бластов, яю беруть участь у новоутворенш юстково! тканини. Разом з тим пщви-
шуеться всмоктування кальцпо в кишках, зменшуеться зворотне всмоктування
фосфатов у канальцях нефрошв, пщвищуеться вмост розчинних солей кальщю в
юстковш тканин! та нерозчинного фосфату кальцпо в розних органах, у тому чис-
ло й нирках (див. роздол XXII “Порушення водно-електролотного обмшу”).
Гшерпаратиреоз в експериментальних тварин вщтворюють уведенням екстра-
кту прищитоподобних залоз або очищеного паратгормону. Пщ впливом високих
доз гормону ровень кальцпо в крово сягае 5 ммоль/л, тобто вдвочо поровняно з нор-
мою; концентрацоя неоргашчного фосфору зменшуеться; посилюеться виведення
фосфору оз сечею. Хоча паратгормон о посилюе дещо канальцеву реабсорбцпо
основ кальцою, виведення ’ох оз сечею пщвиоцуеться за рахунок значно! гпоеркаль-
щеми. Виникають зневоднення оргашзму, блювання, гарячка, ГНН, унаслщок
чого тварина гине.
Експериментальний хрошчний гшерпаратиреоз вщрозняеться вщ гостро! ш-
токсикацп паратгормоном. При цьому спостерогають прогресуюче розрщження
юстково! тканини (остеопороз), вщкладення солей кальцою в нирках, легенях,
серцо та шших внутрошнох органах аж до повноо !х кальцинацц. Стшки судин ста-
ють твердими о ламкими, тиск крово пщвищуеться. Тварини гинуть, як правило,
вщ ураження нирок.
Походження гшерпаратиреозу в людей пов’язують з аденомою або гшер-
плазоею прищитоподобних залоз. Для генератзовано! фгброзно! остеодистрофи, що
розвиваеться при цьому, характерш боль у м’язах, юстках о суглобах, розм’якшення
юсток, розка деформацоя скелета. Мшерально компонента вимиваються з костко-
воо тканини о вщкладаються у м’язах та внутрошнох органах (це явище образно
називають перемощениям скелета в м’яко тканини). Розвиваються нефрокальци-
ноз, звуження проевпу канальщв нефронов о закупорювання ’ох каменями (не-
фролтаз), а як наслщок — тяжка недостатшсть нирок. Унаслщок вщкладень со-
лей кальцою в стшках мапстральних судин порушуються гемодинамока о крово-
постачання тканин.
_________________________________________________________________________
| 654
Порушення функщй надниркових залоз
Ппофункцгя кгрково! речовини надниркових залоз. Видалення обох надниркових
залоз (адреяатектомгя) спричинюе в оргашзм! патолопчн! змгни, не сумгснг з
житгям. При цьому вирипатьне значения мае вщсутшсть функцп кгрково. речо-
вини, тодг як недостатшсть мозково! речовини компенсуеться виробленням кате-
|.олам!н1В хромафшними клгтинами симпатично! частини автономно/ нервовог
системи.
Гостра недостатшсть кщковоТ речовини надниркових залоз розвиваеться в лю-
дини, а також у собак, кшюк г гвгнейських свинок, тобто в тих тварин, як! не
мають додаткову юркову речовину надниркових залоз.
За один-два дш пгсля адреналектоми спостерггають м’язову слабк/сть, ано-
рексгю, блювання, пронос. Тварини апатичш, слабко реагують на зовнгшнг под-
разники. Температура т!ла знижуеться. Адреналектомоваш тварини досить чут-
:ивг до гнфекци, штоксикацн, переохолодження тощо. Симптоми гостро" недо-
статност! надниркових залоз прогресують гз часом.
Пгсля адреналектоми виснажуеться депо глгкогену в печгшц та м’язах. Унас-
лщок зменшення активностг гдюкозо-6-фосфатази знижуеться швидкзеть глгкоге-
нолгзу в клгтинах печгнки. Водночас уповгльнюеться утворення глюкози з амгно-
кислот (глюконеогенез), що призводить до гшоглгкемп.
Для порушення обмгну бглкгв г нуклешових кислот е характерними перева-
кання процесгв дисимгляци, гидвищення в кровг ргвня азотумюних сполук — про-
дуктгв розпалу бглкгв.
У шзшй стали гостро! недостатност! надниркових залоз знижуеться артергаль-
ний тиск. Гшотензгя зумовлена зменшенням ОЦК, брадикардгею й ослаблениям
судинозвужувальног ди катехоламгнгв.
У генезг гостро! недостатностг надниркових залоз найважпивгшу роль вщггра-
ють порушення водно-мгнерального обмгну. У фгзюлопчних умовах альдостерон
забезпечуе реабсорбцгю гонгв нагргю в дистальних вщдглах канадъшв нефронгв, а
: дюкокортикощи пщвищують фгльтрашю рщини в клубочках нирок. Пгсля вида-
зення надниркових залоз унаслщок втрати гонгв натргю гз сечею знижуеться його
вмгст у плазм! кровг Спочатку спостерггаеться полгургя, а потгм олггургя й анургя.
Одночасно розвиваеться “водне отруення”, тобто посилена гщратацгя клгтин. Це
пояснюегься тим, що внаелгдок порушення роботзг натргевого насоса збшьшуеть-
ся внутргшньоклгтинна концентращя гонгв натргю г пщвищуеться осмотичний
тиск. Ргзке пщвищення концентрацп гонгв качгю у плазмг кровг е причиною по-
пущения бгоелектричних процесгв, сили г ритму скорочень серця (аж до мерехт-
-ивог аритмп), ослабления скоротливо! здатностг м'язгв.
У термгнальнгй стад!! гостро! недостатностг надниркових залоз повнгстго при-
диняеться сечовилускання, сповгльнюеться пульс, дихання. Тварина впадае в ко-
чатозний стан г гине. Тривалгсть життя адренатектомованих собак г козгв коли-
.аеться вгд 2—3 до 9—11 дгб.
Вщсутшсть мозково! речовини надниркових затоз г негативнг наелгдки опера-
дгйно! травми утруднюють патофгзголопчний анатгз змгн, зумовлених вщсутшстю
функци кгрково! речовини, в алренатектомованих тварин. Тнший метод експери-
гентального моделювання гшофункцгг кгрково! речовини надниркових залоз Грун-
туеться на застосувангп хгмгчних речовин (гнггбгторгв), якг шсля введеггня в орга-
I ЙГ
Н1зм виб!рково гальмують секрецпо кортикостероцЦв, Найефектившшим е введен-
ия собакам хлодитану.
Гостра недостатшсть юрково! речовини надниркових залоз у людини може
виникати внаслщок крововиливу в них.
Хрошчна недостатшсть кгрково! речовини надниркових залоз (гтоадренокорти-
цизм) у людини вщома щд назвою хвороби Алдтсона. або бронзово! хвороби. Вона
здебшыпого е наслщком губеркульозного ураження надниркових залоз, а також
атрофп юрковт речовини внаслщок перенесених тяжких шфекцшних хвороб або
тривалого лгкування коргикостерощними препаратами
Для хрошчного гптоадренокортипизму характерш зниження маси пла. швид-
ка фгзична 1 псих!чна стомлювашсть, знижений апетит. дисфункщя травного ка-
налу. артер!альна гшогензхя, прогресуточа гшерпггменташя шюри. Мехашзм п-
пертгменташ! пов’язаний 13 посиленням меланопитстимулюючо! активносп п-
поф!за, яка супроводжуе збшьшення секрецп кортикотропшу, що виникае в рал
ппоадренокортицизму. Ргзн! патогенш впливи (травма, шфекшя, крововтрата
навпъ екстракщя зуба) у патента 13 хворобою Ададсона можуть спричинити гос-
тру недостатшсть юрково! речовини надниркових залоз.
Гшерфункщя 1 дисфункщя юрково! речовини надниркових залоз. Провщнимн
патогенетичними ланками, шо визначають клппчну картину пперфункцп 1 дис-
функцй юрково! речовини надниркових залоз, е надмзрна секретя корпполу (хво-
роба 1 синдром Шенка — Кушшга), альдостерону (пперальдостерошзм), андрогеш:
(.ядрен огеютальний синдром, андростерома) або естрогешв (кортикоестрома).
Еполопя хвороби та синдрому 1ценка—Кушшга р!зна, проте клппчш симптоми
1 патогенез порушень под!бш. Хвороба е наслщком надм!рно! секрецп корт»-
котротну через пухлину аденогшофзза або порушення гшоталамгчно! регуляцп.
синдром — наслщком первинного ураження юрково! речовини надниркових залоз
пухлиною. шо походить !з юркових ендокринопипв. Хвороба супроводжуеться
збшьшенням у кров! р!вшв кортикотропшу ! меланотропшу з потемншням шюри
а синдром — зменшенням 1х концентрат!.
Пщ впливом наданрно! юлькосп глюкокортикоцпв виникають тяжю ендокрин-
но-обм!нн! розлади. Розвиваеться ожиршня з переважним вщклалеиням жиру на
ту луб), обличия стае м!сяцепод!бним, пперемшованим, на шюр! живота з'явля-
ються синьо-багрян! дистроф!чш смути розтягнення. Негативний азотистий баланс
евщчить про переважання катабол!чних процеств. Пщвищення розпаду бшюв при-
зводить до дистроф!чних змш у м’язах, юстках 1 суглобах. Остеопороз може дося-
гати такого ступеня, за якого виникають спонтанш компреейш переломи хребщв.
П1д д!ею глюкокортикощ!в у печшш пщвищуеться вм!ст гликогену, розви-
ваеться стшка готерппкемтя (стерогдний д!абет). Останн я е наслщком штенсявно-
го глюконеогенезу 1 пшвишено! активносп глюкозо-6-фосфатази в печшщ.
Унаслщок збшьшення об’ему кров! та сенсибпизапп судинно! стшки до да
катехолам!шв пщвищуеться артер!альним тиск. У л!мфо’!дн!й тканиш вщбуваеться
посилений розпад нуклеопроте!д!в Надлишок глюкокортикошв зумовлюе л!м-
фоцито- та еозинопенпо. пригшчуе продуюпю антитш 1 реакцп юптинного !мун!-
тету. Нершко спостерггають множиню крововиливи та виразков! ураження слизо-
во! оболонки шлунка 1 кишок.
Аденома, яка походить 1з юптин клубочково! зони юрково! речовини наднир-
кових залоз, може стати джерелом над мерного утворення альдостерону, шо спри-
Ефективний розтягнення
э^ем кровх
Рецептори Анпотензин П
/Надниркова
залоза (бхо-
синтез альдо-
" стеР0НУ)
ха ^—^Альдостерон
Обм^н^а+,К+
№+
Мал. 72. Взаемодхя кирок з наднирковими за-
логами в норм! (в), при первинному (б) X вто-
ринному (в) пперальдостерошзм! (за Дж. Кон-
ном, 1975)
чинюе первинний гтеральдостерон1зм,
або хворобу Конна. Секрецхя глюко-
кортикоида при цьому в норм!.
П!д впливом надлишку альдосте-
рону в оргашзм! затримуються юни
натрпо 1 вода. Збхлыпення концент-
раци юшв натрхю в клхтинах. зокрема
1 судинно! стшки, пхдвшцуе чут-
ливхсть !х до катехолампда, що зумо-
влюе розвиток артерхальнох гшертен-
зи. Втрата значно! юлькост! юнхв
калхю ! хлору е причиною виникнен-
ня мхастенИ та парезов, нападхв судом
скелетних м’язхв, порушення скорот-
ливо! функцц мюкарда. Канальщ не-
фронхв зазнають дистрофхчних змш х
втрачають здатнхсть реагувати на вазо-
-ресин. Так виникае полхурхя, що частково пояснюе вхдсутнхсть набряюв при пер-
инному пперальдостеронхзм! (див. роздхл XXX “Патолопчна фхзюлопя нирок”).
Патолопчн! стани (недостатшсть кровообхгу, асцит при цироз! печхнки, нефро-
ичний синдром тощо), яю спричинюють звуження ниркових артерш або змен-
лення ниркового кровотоку, нерхдко викликають вторинний гтеральдостерошзм.
альдостерон якнайпсшше пов’язаний !з ниркою: його продуюця залежить в!д ста-
ну кровотоку в нирщ, а функщя нирки (реабсорбцхя юшв натрпо) своею чертою
алежить в!д вмхсту альдостерону. Отже, розвиток вторинного гшеральдостерошзму
«гоже вхдбуватися в таюй послхдовносп: ипемхя — реши — анпотензин II — альдо-
стерон — звивист! канальцх нефрохда (мал. 72).
Адреногешталъний синдром у дхтей х дорослих е клхшчним проявом уроджено!
пперплази юрково! речовини надниркових залоз. Цей синдром пов’язують хз ге-
-етично зумовленою недостатшстю синтезу кортизолу (пдрокортизону). При цьому
озгальмовуеться секрецхя кортикотропшу, який стимулюе вироблення андро-
ешв наднирковими залозами. Останш справляють вхрилхзуючу дно (поява й по-
-илення ознак чоловхчо! стат! в жшок) у внутрппдаоутробний перюд. Наслщки
гормональних порушень рхзш — вщ легко! маскулшхзацп до грубих анатомхчних
шомалш фхзичного х статевого розвитку (схема 30).
У хлопчикхв така патолопя зумовлюе передчасний розвиток вторинних стате-
зих ознак, дхвчатка часто народжуються з ознаками псевдогермафродитизму.
Один !з вархантхв адреногештального синдрому в дхтей визначаеться вира-
женппим ушкодженням ферменпв синтезу стерохдних гормошв 1 супроводжуеть-
з тяжкими порушеннями водно-мшерального обмшу {синдром втрати аш). Без
замкно! тераш! кортикостерощами дхти помирають у ранньому вщх.
г
Якшо синтез гормошв блокований на за-
вершальшй стадн, утворюеться надлишок дез-
оксикортикостерону — стероТду з властивостя-
ми мшералокортикощгв. Унаслщок цього роз-
виваеться тяжка артергальна гшертензгя.
Гигерандрогенгзацпо та ггперестрогешза-
цгю оргашзму дгтей г дорослих може спричи-
нити пухлинне переродження сгтчасто! (внут-
ршгньо!) зони кгрково! речовини надниркових
залоз. Залежно вщ характеру гормонально!
секрецп та вщповщностг п стат! хворого спос-
терггаються вгрилгзацгя (у жшок), фемппзацгя
(у чоловгкгв) або передчасний статевий розви-
ток в осгб обох статей.
Порушення функцш мозково! речовини над-
ниркових залоз. Надмгрна секрецгя катехола-
мгшв спостерггаеться при феохромоцитом! —
доброякгснш пухлинг, що походить гз мозково!
речовини надниркових залоз. Вона супровод-
жуеться артергальною гшертензгею, нападами,
пщ час яких спостерггаеться тахгкардгя, пщви-
Жшочий псевдогермафродитизм
Вгрильний синдром
Прсутизм
Аменорея
Безплщшсть
Схема 30. Розвиток пперандрогенного
стану в жшок з адреногештальним синд-
ромом
щуеться артергальний тиск, з’являються ргзкий бщь у надчеревнш дшяшц, рясне
потовидглення. Напади пояснюють масивним викидом у кров адреналгну й нор-
адреналгну пщ впливом психгчного або фгзичного навантаження та гнших фак-
торгв, що гх провокують.
Порушення функцш статевих залоз
Гшофункщя статевих залоз. Недостатшсть ендокринно! функци статевих залоз (а-
погонадизм) може бути зумовлена видаленням гх хгрурпчним шляхом (кастрацгя ,
руйнуванням запальним процесом, токсинами, гонгзуючим випромшювання^
розладами ппоталамо-ппофгзарно! регуляцп, нечутливгстю залоз до гонадо-
тропних гормошв. Окре му трупу становлять уродженг порушення статевого роз-
витку, зумовлеш хромосомними аномалгями, мутацгею генгв або впливом пато-
генних факторгв (гормональний дисбаланс, стрес, вгрусна гнфекцгя тощо) на ней-
роендокринну систему плода. Неопущення яечок у калитку (крипторхгзм), гшо-
плазгя або вщсутшсть яечок г яечникгв, псевдогермафродитизм — усг цг вродженг
аномалгг розвитку супроводжуються ппогонадизмом. Основнг симптоми гшого-
надизму майже однаковг в разг первинного ураження статевих залоз г вториннс
ггпофункцгг, зумовлено! гонадотропною недостатнгстю.
Гормональну недостатшсть статевих залоз вщтворюють в експериментальних
тварин шляхом кастрацп, уведення лгкарських препаратгв, якг пригшчують утво-
рення гормонгв у статевих залозах або блокують гхнг клгтиннг рецептори.
Кастрацгя тварин, якг ростуть, спричинюе змгну зовнгшнього вигляду, видов-
ження г стоншення кгсток, зменшення маси м’язово! тканини, надмгрне вщкла-
дання жиру. Останне було помгчено ще в стародавш часи, кастрацпо г досг вико-
ристовують у продуктивному тваринництвг для збгльшення маси тварин. У самщв
158
-г.остерггаються недорозвинення вторинних статевих ознак, статевих орган!в (ста-
"евого члена, сш’яних м!хурц!в 1 передм!хурово! залози), безпл)дшсть, у самок —
тедорозвинення матки та и придатков, згодом — вщсутшсть циюпчних змш у
“атевш систем!, безплщшсть. Ц1 порушення зумовлен) випаданням ди статевих
.’терощхв.
Кастращя, виконана шсля статевого дозргвання, позначаеться переважно на
' <пш речовин ! функшях статевих оргашв, спричинюючи !х атроф!ю, зникнення
зтевих шстинкпв, припинення циюпчних процеств у матш, молочних залозах,
аволющю вторинних статевих ознак.
Змши, зумовлен! кастращею або гшогонадизмом у людини до настання ста-
тье! зршосп, дуже под!бш до тих, що спостерггаються в експеримент!. Вщповщ-
йий юпшчний синдром у чолов1к!в отримав назву евнухоидизму. Вш може бути
рмлщком не лише вщсутност! або зменшення продукцп статевих гормошв у яеч-
IX, а й наслщком вроджено! нечутливосп тканин до тестостерону {синдром тес-
.кулярно'1 фемтзаци) або дефекту ферменту, який вщповщае за перетворення
ещостерону на акгивний метаболит — дигщротестостерон.
Вщсутшсть функци яечок у зр!лому вш! внаслщок травми або хзрурпчного
тручання, так само як пригшчення !хньо! ендокринно! функци за допомогою фар-
«аколопчних засоб!в, що !х застосовуютъ для лгкування злояюсних пухлин перед-
«курово! залози, спричинюе синдром кастраци. У чоловиов поступово розвиваеть-
ся 1мпотенц!я, порушуеться обмш речовин, що призводить до збшьшення маси
-тирово! тканини, гшерхолестеринемп, остеопорозу. Так! сам! розлади виникають 1
дорослих жшок шсля овархектоми, унаслщок рдщащйного або х!м!чного уражен-
ня яечниюв. I в жшок,! в чоловиов спостерггаеться комплекс судинних! психоемо-
.гних розлащв, пов’язаних з! зменшенням вщповщно естрогенно! та андрогенно!
- лсиченосп оргашзму. Це призводить до надм!рного збудження 1 дезоргашзаци п-
"оталамхчних нервових 1 нейроендокринних центр!в мед!аторами симпатично! не-
рвово! системи, що супроводжуеться нападами рясного потовид!лення, пщвищен-
чям температури тша, артергального тиску, загального слабктю, тахгкарщею, болем
суглобах 1 юстках, головним болем, безсонням тощо.
У зв’язку з тим що тестостерон е вкрай потр!бним для нормального спермато-
-енезу в шм’яних каналъцях, гормональна недостатшсть яечок спричинюе пору-
чения цього процесу аж до його повно! зупинки, тобто призводить до безплщ-
остг
Зменшення синтезу прогестерону в жшок унаслщок гормонально! недостат-
-ост! жовтого тша (як правило, гшоталам!чного генезу) може бути причиною
торушень менструального циклу, раннього невиношування ваптносп, осюльки
прогестерон потр!бен для и нормального перебйу. Застосування в цих випадках
-.репарапв прогестерону дае змогу зберепи ваптшсть.
Зниження гормонально! активност! статевих залоз хз вхком е основною озна-
ою ктмаксу в жшок 1 векового ппогонадизму в чоловиов. Кл!мактеричний син-
ром шодх набувае патолопчного перебйу 1 потребуе лгкування. Патогенез 1 його
сновн! симптоми дуже подхбш до таких шсля овархекгоми. Через те що пщГрун-
ам юймактеричних проявгв та ускладнень у жшок старшого вйсу (остеопороз,
_ртер!альна гшертензгя, нейродегенеративн! процеси, психотичш розлади, пору-
шення сечовипускання) е естрогенний дефщит, призначення гормональних пре-
парата 1з замгоною метою виявилося дуже ефекгивним.
Гшерфункщя статевих залоз. Посилення гонадогропно!
функцй гшофтза або гормонально-активна пухлина яечок 1
яечниктв у ранньому виц зумовлюють нередчасне статеве
дозревания. Пщ дтею надлишково! кшькосп андрогентв
хлопчики перетворюються на "маленьких геркулес!в” гз
короткими юншвками т сильными м’язами (мал. 73).
У раз] збшьшено! секрецц гонадотрогпшв у стм’яних
канальцях яечка выявляюсь зргл) сперматозоши. Зовнштн!
сгатев! органи, а також яечка гшертрофованг шо дае змогу
вшр1знити цю патологпо вщ адреноген стального синдрому.
Гшерандрогентзащя спричинюе ранню появу чоловтчих
вторинних статевих ознак: низький голос, оволооння лоб-
ка, обличчя, пахвових ямок тошо. У дтвчаток пщ впливом
надлишку есгрогенхв дуже рано з’являегься менструашя та
с постер! гаються передчасний рют молочних затоз. ово-
лосшня лобка, вщкладання жиру в д!лянш таза < стегон.
Пщвищена секреция гормон!в яечниюв у дорослих ж!-
нок зумовлюе порушення менструального циклу, масто-
патно га пшп розлади. Так, наданрне надходження в кров
естрогентв унаслщок персистенци фолгкула (стан, за якого
фолткул не досягае повного дозр!вання 1 не настае овуля-
ц!я) часто спричинюе матков! крово гечс Пшвишена сек-
рецтя прогестерону супроводжуеться аменореею, збшьшен-
ням об’ему матки ! молочних залоз. Якшо яечники стають
Мал. 73 Передчаснест»
теве дозртваштя в хлоп-
чика ежом 2,5 року
джерелом надлишку андрогенних гормошв, розвиваеться гшерандрогенний стат
проявами якого е оволосшня за чоловшим типом, пперхолестеринемтя, ожир!-1-
ня, шсулшорезистентшсть, артертазьна гшергензш, порушення менструально
циклу, безплщшсть тощо.
Порушення ендокринно) функцн загруднинно? залози
Гормон загруднинно! залози (тимуса) — тимопоетин — вщтграе роль паракринно
го регулятора дозр!вання Т-л1мфоципв. Водночас в!н надходить у загальний к]
вообп т здшснюе дистанщйний вплив на шш! органи та кл!тини тмунно! систему
Патогенез тмунних розладтв, як! спостерйають пхсля тимекэомп, унаслшок де
структивного процесу в залоз! або ц гптерплазп, значною мтрою пов’язаний
порушеннями ендокринно! функцй загруднинно! залози.
Порушення функций епгф!за
Для гормону мелатоншу характерний широкий дгапазон ф!з!олопчно! активное г
тому гшофункцтя або гтерфункшя ет'фгза (шнеально! залози) мае р!зноманп-
прояви. Х!рурпчне видалення затози в експерименталъних тварин або введенн
мелатон!ну змгнюютъ затежн! в!д фотоперюдичност! хроноритми щяльност! ег
докринних залоз (щитопод!бнот, надниркових, статевих). нирок та шших органи
стан пероксидного окиснення лшцвв 1 бъдктв. Надмтрна секрецхя мелатоншу мохь
бути причиною затримки статевого дозртвання (через антигонадотропну даю г<г
иону), зниження емоцгйного тонусу, аж до розвитку депресивних стангв. В осгб,
що передали на постшне мгеце проживания з ГПвдня на Пгвшч, часто спостерг-
'ають апапю, сонливють, швидку втомлювангсть, що, гмовурно, пояснюеться п-
персекрешею мелатонину внаслщок скорочення тривалосп свгтлого часу доби.
Причинами гшошнеалгзму бувають оргашчне ураження залози, судинна па-
тололя, деструктивна пухлина. 3 гшофункщею ешфг'за пов’язують деяк1 випадки
передчасного статевого дозргвання, прискореного постаргння, що може поясню-
’атись ослаблениям антиоксидантног дй гормону.
СТРЕС IЗАГАЛЬНИЙ АДАПТАЦ1ЙНИЙ СИНДРОМ
, Одним гз досягнень сучасног медицини е розкриття важливо! рол! ендокринних
залоз, зокрема системи гшофгз — кгркова речовина надниркових залоз, в аданта-
цп органгзму до дй' патогенних факторгв. Широко вщомою е теоргя канадського
вченого Г. Селье про стрес
Термитом “стрес” (вш англ, зггга — напруження) позначаюгь неспецифгчну
реакшю оргашзму, шо виникае год впливом будь-яких сильних подразник)в (стре-
соргв) г супроводжуеться перебудовою захисних систем оргашзму. Г Селье звер-
нув увагу на те, що, незважаючи на розмагття стресоргв (травма, пгфекшя, пере-
лхолодження, огруення, наркоз, м’язове навантаження, сильнг емоий тощо), усг
вони зумовлюють однотип нг змгни в загрудниннш г надниркових залозах. лгмфа-
шчних вузлах, склад) кров1 й обмин речовин. У дослгдах на щурах вгн спостер|гав
тппову тр)аду: гшертрофгю кгрковог речовини надниркових залоз, гнволюцйо
тимгко-лгмфатичного апарату та геморапчш виразки на ели зов) й оболонц) шлун-
ка г д ванад пятипал о)' кишки.
Стрес проявляеться як загальний адапташйний синдром, який мае три лосл)-
довн1 стадп: тривоги, резистентност) та виснаження. Стадгя тривоги означав не-
.-айну мобпггзаггйо захисних сил органгзму. Для не)' притаманнг фаза шоку г фаза
протишоку. У фаз) шоку спостер)гаються гшотошя м’язгв г артергальна плотензш,
ппотермгя, ГШОГЛГКСМ1Я, згущення кровз. еозинопешя. пщвищення проникност)
тонки капйгяргв. (нволюшя лгмфо'щног тканини, негативний азотистий батане,
виразков) ураження шлунка евгдчать про переважання пронес 1в катаболгзму. Ха-
рактерними ознаками фази протишоку е змши у зворотному напрямку (шдви-
щення артериального тиску, м’язового тонусу, вмгсту' глюкози в кровг), що зумо-
елюють розвиток наступног стали — резистентность Основна патогенетична лан-
ка фази протишоку — стгйке посилення секрецп кортикотрошну 1 глюкокортикощгв
(кортизолу). У стадп резистентностг кгркова речовина надниркових залоз ггпер-
трофуеться । секретуе велику кглькгсть гормонгв, активутоться анаболзчнг пронеси,
посилюеться глюконеогенез
Барто зауважити. шо здатшеть кгрковог речовини надниркових залоз реагува-
ти посиленням функцгональног активностг на патогенний фактор уперше вщкрив
О.О. Ьогомолепь на початку XX ст. Проте на той час ньому явишу ше не було
чадано значения узагальнювально) закономгрностг.
У раз) тривалог дй' ушкоджувального фактора адапташя порушуеться. Ргзке
зменшення функщональних резервов г атрофгя кгрковог речовини надниркових
залез, зниження артергального тиску, розпад бьткових речовин характеризуюгь
перехЩ стадй резистентносгг в стадно виснаження.
I
Стресорний агент
Схема 31. Основш шляхи вйшовщ! на вплив стресорного фактора (за Г. Селье);
КРГ — кортикотропний рилгзинг-гормон, або кортикол!берин
Наслщки стресу залежать вщ сили 1 тривалост! дп стресора та потенцшних
можливостей захисних сил оргашзму.
Бюлопчне значения адаптацшного синдрому полягае не лише в тому, що в
друпй, найтривалппш, стадп пщвищуеться резистентшсть оргашзму до фактора,
який спричинив стан стресу, а й у тому, що не дуже сильний ! тривалий стрес
здатний створити або пщвигцити неспециф!чну резистентшсть оргашзму до ш-
ших факторгв. Це проявляеться пщвищенням виживаносп внаслщок ди леталь-
них фактор!в або ж зменшенням запалення, запобгганням гшерерпчним реак-
щям, ураженню серця, нирок та шших оргашв пщ д!ею патогенних факторхв.
Наприклад, кровопусканиям (стресор) можна ослабити запалення, зумовлене
внутр! шньошкфним уведенням формалшу. М’язове напруження або травма захи-
щае тварину вщ некрозу в мюкард!, спричиненого внутр!шиъовенною ш’екщею
протеолггичних ферменттв. Кортикотрошн 1 кортикостерощи вшграють провщну
роль у формуванш неспепифгчно! резистентност!, тому !х названо адаптативними
гормонами (схема 31).
Систематичний вплив на оргашзм слабких ё помёрних подразни к! в (напри-
клад, холодний душ. фёзичш влрави) п.тгрим1 с готовнёсть ендокринно! системи
до адаптивних реакцёй. На вщмшу вщ цього, стресовий стан, який виникае пщ
шею факторгв надмёрпоё сили, визначаеться як дистпрес. Тодё стрес замёсть захис-
ного стае руйшвним для организму.
Недостатшсть адаптацп або п вёдхилення в протилежний бёк е, за Г. Селье,
причиною розвитку хвороб адаптици. Експериментатьне пёдтвердження цього по-
ложения полягае у виникненш артерёальноё ппертензёт, нефрокальцинозу, гёалё-
нозу оргашв, посиленнё запальних реакщй шсля введения у високих дозах дезок-
сикортикостерону, який мае прозапал ьш власти вост:. У ведения тваринам гчюко-
кортикоццв (протизапальних гормонш) гальмуе запалення, але водночас пригшчуе
1унш реакцп, зумовлюе виразкове ураження шлунка ё дванадцятипало! кишки,
сгворюе умови для виникнення некрозу мёокарда. Недостатшсть секреиё! глюко-
кортикощёв сприяе алергёчним розладам, знижуе стёйкёсть до хвороботворних фак-
торёв.
Хворобами адаптацё!, за Г. Селье, е ревматизм, бронхёальна астма, деяю хво-
роби нирок, серця ё судин, шкёри та ён. У !х виникненш великого значения нала-
гать кондицёоналъним (зумовлюючим) факторам. Цими факторами можуть бути
переохолодження, перегревания, фвична перевтома, обтяжена спадковёсть, над-
т ишкове споживання солё. Одночас не введения кортикостеронов ё натрпо хлори-
ду створюе тло, на якому рёзнё подразники зумовлюють некротичнё змши в сер-
цевому м’язё.
Г. Селье сформулював свою теоргю на основё вивчення ендокринних механёз-
чёв пристосувальних реакцёй оргашзму. Проте ще ранёше працями У. Кеннона,
Л.А. Орбелё та ёхшх учнёв було створено вчення про адаптацёйно-трофёчну роль
симпатично! частини автономно! нервово! системи в захисно-комлеиса горних
реакпёях. Деякё ознаки стресу (поява виразок) спостерё] аються в гёпофёзектомова-
них тварин. Отже, неможливо зволити численнё прояви стресу до однёе! лише
гормонально! перебудови. Стрес е складною реакцёею, у розвитку яко! беруть
участь нервова система й ендокриннё залози. Разом з тим вэрто зауважити, що
теорёя Г. Селье справила ё продовжуе справляти великий вплив на розвиток ме-
дицини. Вона дала теоретичне обгрунтування кортикостерощно! терапёё. На ос-
новё ше'ё теорп розроблено новий пёдхёд до проблеми старёння. пояснено неспе-
цифёчну терапёю (кровопускания, автогемотерашя, голковколювання).
Основнё положения цёе! теорё! успёшно розробляються ниш. Зокрема дослёд-
ження ультраструктури клётин, персбёгу молекулярно-бёологёчних ё бёохёмёчних
процеств у них дали змогу дёйти висновку про ёснування так званого клётинного
стресу. [деться про сукупнёсть стереотилних змён у клётинё пёд дёею стресових
факторёв. До них належать порушення ПОЛ, структури та функцёё мётохондрёй,
властивостей клётинних мембран, посилене утворення лейкотрёенёв тощо. Виник-
ло поняття про оксидантний стрес, у якому провёдну роль вёцёграе порушення
вёльнорадикальних процесёв. Вважаюгь, шо супсроксидш радикали, якё дуже
швидко утворюються в надмёрнёй кёлькостё пщ дёею стресових фактор<в. е пуско-
вими факторами в збудженнё симпатичного вёддёлу нервовоё системи й активаш!
системи гёпоталамус — гёпофёз — наднирковё залози.
РОЗД1Л XXXII
ПАТОЛОПЧНА Ф13ЮЛ0ПЯ НЕРВОВО! СИСТЕМИ
ЕТЮЛОПЯ I ПАТОГЕНЕЗ
ПОРУШЕНЬ ФУНКЦ1Й НЕРВОВО! СИСТЕМИ
Порушення функцзй нервово/ системи можуть виникати внаслщок впливу на ор-
ганизм рззних екзогенних 1 ендогенних факторзв, що спричинюють порушення
метаболизму, структури з функци нервових юптин.
Етюлопя. Насамперед варто зазначити, що нервова система з передусзм п
центральнз взддыи дуже чутливз до гтокси. Головний мозок споживае близькс
20 % усього кисню, який потрапляе. в оргашзм. У разз рантового припинення
надходження кисню до мозку (вдихання безкисневих газових сумзшей, порушен-
ня мозкового кровообегу) людина через 6—7 с може знепритомшти, а через 15 с
припинясться нормальна бюелектрична актившсть мозку. Повне вщновлення
функцш мозку можливе тод!, коли кровообзг припинився не бзльше нзж на 5—
6 хв. Якшо зшемзя мозку тривае довше, память та штелект необоротно порушу-
ються. Варто зауважити, що р!зш взддзли ЦНС неоднаково чутливз до кисневогс
голодування. Фзлогенетично стар! структури е стшкими до гшоксзз. Наприклад.
нейрони стовбура головного мозку. як! входять до складу дихального та судино-
рухового центр!в, можуть переносити анокспо тривалзстю до 30 хв.
Патогенно впливати на нервову систему можуть також фгзичш фактори: зонь
зуюче випромшювання, електричний струм, шум. взбрацзя, електромагштне поле,
механзчна травма, висока та низька температури. Пщ час пщвищення температу-
ри т!ла (перегрзвання, гарячка) посилюються обмш речовин, процеси збудження
в нервовзй систем! з подалыпим розвитком гальмування з виснаженням енерге-
тичних ресурс!в у нервових клзтинах. У раз! охолодження (загального або м!сце-
вого) сповзльнюеться швидкзеть обмшних процеезв у нейронах, знижуеться зх
здатнзеть генерувати потеншал дц та проводите його через нервов! вщростки.
Функцп нервово! системи порушуютъея п!д впливом рззних токсичных речо-
вин природною або штучного походження. Взздзляють велику трупу нейро гропних
огрут, як! можуть вибзрково порушувати бюенергетичш процеси в нервових кл!-
тинах, утворення, транспорт, видшення з метаболззм нейро медзаторзв, впливати
на проникшсть зонних канал!в у нейронах, футзкцпо синапсзв.
Розлалом дзяльност! нервовоз системи, насамперед и центральних взддзлзв
можуть супроводжуватися порушення обмшу речовин в органззмз.
Головний мозок дуже чутливий до гйиалйсемп. Практично весь кисень, який
. дожпваеться головним мозком. витрачаеться на окиснення глюкози. Ртзке зни-
-ення ртвня глюкози в кров! зумовлюе порушення лтячьност! нейрошв. бюелект-
ччно! активносп мозку ] може призвести до знепритомнення. Тривала тшог.ч!-
ем!я спричинюе необоротю змши в корт великого мозку. У разт значно! гшопй-
емп порушуються функцц, яю регулюються стовбуровими мехашзмами.
Розлади дтяльност! нервово! системи спостертт аються внаслщок змши власту
кров/ електролтйв та юн/в водню.
Голодування. зокрема вгтамшне, нерщко зумовлюе порушення нервово! дтяль-
остт. Насамперед це стосуеться вттамппв групи В.
До тяжких порушень функцш нервово! системи, особливо ц вищих вщдшв,
оже призвести патолог/я ендокринних залоз.
Важливу роль еттолопчного фактора в патологи нервово! системи вщтграють
ирекцшю збудники. Деякт з них. видтляючи токсичнт речовини, здатнт порушувати
:етаболтзм ! функшт нервових клттин, передачу нервових тмпульств на ефекторнт
клттини (наприклад, правцевий токсин, ботулотоксин та ш.). Серед шфекцшних
эудниктв потртбтю втщшити такт групи втруств, якт можуть безпосередньо ушкод-
увати нервов! клггини, — втруси сказу, полтомтелтту.
Визначним науковим досягненням у галузт молекулярно! бтологы та медици-
и стало вщкритгя шфекцтйних фактортв нового типу, якт отримали назву пртонтв,
зумовлен! ними хвороби — пртонових. Пртони — це певна груда бтлктв, як! ут-
орюються нервовими клпинами т потртбш для тхньот нормально! життедтяль-
ностт. Унаслщок ще не з’ясованих причин оргашзм може почати продукувати
1Гутантш, конформацшно змтнеш пртони. Важливим е те, що щ патогеннт моле-
.ули прюнтв здатш до самореплткацц без участт нуклетнових кислот. Можна уяви-
ги, що вони вщтграють роль своерщно! матриц!, на я кт и формуються нов! моле-
кули патогенних пртошв Це зумовлюе дистрофтчш змши т загибель у головному
чозку частини нейрошв з утворенням пустот, як! на зртзах нагадують губку, т тому
такт хвороби отримали назву губчастих (спонгюформних) енцефалопатш. Подтбн!
процеси можуть вщбуватись ! в головному мозку велико! рогато! худоби. У цьому
випадку патогеннт пртони тварин можуть набувати значения шфекцтйних фак-
тортв. якт, ттотраштяючи в органтзм людини, створюють умови для перетворення в
ц головному мозку нормальних прюнтв на патологтчш.
Безперечним е вплив фактора вису на будову 1 функшт нервово! системи. Ме-
хашзм пього впливу вивчено недостатньо, однак його наслщком е зменшення
кшькост! клттин в устх вузлах периферично! нервово! системи ! головного мозку,
у тому числ! й кори. Побутуе думка, згщно з якою в людини, починаючи при-
близно з 30 роктв, регулярно вщмирае певна кйтьктсть нейротттв, причому в мтру
старшня темп загибел! нервових клттин зростае. Однак вважають також, що з вт-
ком нейрони не гинуть, а лише атротруються, шо спричинюе зменшення маси го-
ловного мозку. При ньому в ртзних дшянках мозку маса нейрон!в зменшуеться з
неоднаковою швидктстю, т починаеться цей процес у ртзний час.
З’ясовано, що в певних дшянках головного т спинного мозку мтстяться ней-
рогенш стовбуровт клттини, здатнт мтгрувати т, диференцтюючись у зр!лт клттини,
ьщновлювати ктльктсний склад нейрон!в у мтецях !х ушкодження.
У забезпеченш нормально! життептялъност! нервових клттин велике значения
мають глтальн! клттини, кшыасть яких у 10—50 разтв перевищуе кшьктсть ней-
I Г665
ронхв. Зокрема астроцити беруть участь у формуванш гематоехщефалхчного
бар’ера, видхляють нейротрофхчш фактори, забезпечують сталкть потребно! кон-
центращТ юнхв (здатш акумулювати юни калпо) х деяких нейротрансмхтерхв (глу-
тамату, у-амшомасляно! кислоти). Впалый клхтини мктять глхкоген, що розкла-
даеться до глюкози, яка потрапляе в нейрони. Тому порушення функцй нервово!
системи може зумовлюватися також структурними х функщоналъними змшами
гл1алышх кл1тин. Наприклад, у раз! генетичного дефекту, який призводить до при-
гшчення утворення нейротрофхчного фактора, що продукуеться глхальними клх-
тинами, у тварин порушуеться розвиток ентерично! нервово! системи.
Функцн нервово! системи можуть порушуватися рефлекторно тд впливом
сильних або незвичайних подразните на зовнйпнх х внутргшш рецептори.
Серед причин порушення функцш нервово! системи важливе мкце посщають
соц1алън1 фактори. Для людини характерною е друга сигнальна система. За допо-
могою образхв, символов, понять у и уявх вибудовуеться модель навколишнього
свпу. Друга сигнальна система тхсно пов’язана хз сощальним життям людини ! е
наслщком стосункхв ххщивща !з сощальним середовищем. Вплив цього середови-
ща, який здшснюеться переважно через словесну сигналхзацпо, на психхку люди-
ни, п штелект, емоцшний стан надзвичайно великий. Людина постшно оцшюе
свое становище в суспыьств!, ступшь свое! вол! в ньому, можливост! задоволення
свохх потреб, ставлення до не! шших людей тощо. Усе це, а також штенсивна
трудова д!яльн!сть вимагають вщ людини емоцшно! та розумово! напруженостх.
Конфлхктш ситуацп, яю виникають часто або кнують тривалий час ! пов’язаш як
з особистктю шдивща, так х з характером його социального оточення та устроем
сустпльства в щлому, з умовами пращ, побуту, можуть призводити до надм!рного
збудження його емощйних центрхв х порушення психхчно! дхяльност!, розвитку
невротичних станхв, психхчних хвороб х пов’язаних хз ними рхзних психосоматич-
них розладхв. Особливо великого значения набувають слова, нав!ювання (у тому
числ! й самонавповання), як! ктотно впливають на перебк фхзюлопчних процесхв
у нервовш систем! х можуть справляти як патогенний, так х сприятливий, лхку-
вальний вплив.
Патология нервово! системи може бути спадковою. Це проявляеться незавер-
шенктю розвитку окремих структур нервово! системи х порушеннями метаболхз-
му в рхзних трупах нейронхв. 1нколи ураження нервово! системи при спадкових
хворобах мають вторинний характер. Наприклад, при феныкетонурп первинним
е порушення обмшу феншаланшу, а нервова система уражуеться внаслщок шток-
сикаци продуктами порушеного обмшу ще! амшокислоти.
Розлад функцш нервово! системи можливий унаслщок розвитку типовых пато-
лог1чних процессе — запалення, пухлини, мхсцевих порушень кровообпу, набряку.
Пухлина подразнюе той чи той нервовий центр, спричинюючи його надмхрне
збудження. 3 ростом пухлини атрофуються нервов! клхтини х волокна, а це приз-
водить до вщсутност! ххньо'х функцн. Крхм того, збшьшення маси пухлини супро-
воджуеться пщвищенням внутрхшньочерепного тиску, зменшенням кровонапов-
нення головного мозку та його хшемкю.
Причиною порушень функцш нервово! системи досить часто е запалення.
Запалення периферичних нервових волокон зумовлюе порушення чутливост!, ру-
хомостх або функшонування внутрхшнхх органхв. Запальний процес у ЦНС, най-
частйпе в оболонах головного мозку, спричинюе порушення продукцц та вщтоку
спинномозково! рщини, пщвищення внутр!шньочерепного тиску, розлад мозко-
^ого кровообпу, може охоплювати 1 речовину головного мозку (енцефалгг). У
-атогенез! енцефалпу ктотну роль вщпрае автоалерпчна реакцш в раз! порушен-
ня гематоенцефал!чного бар’ера, осюльки оргашзм не мае гмунолопчно! толеран-
пюсп до власно! нервово! тканини.
У головному мозку може розвиватися набухания нервових юптин, пов’язане
токсичним впливом на нейрони, розвитком тяжко! гшоксп. За цим порушуеться
’етаболззм у нервових юптинах, зменшуеться утворення АТФ, що зумовлюе при-
гшчення д!яльност! №+-, К+-насоса 1 збшьшення юлькосп внутр!шньоюптинних
Ма+, а з ними 1 води.
Патогенез розладав функцш нервово! систем насамперед пов’язаний !з ме-
ашзмами порушення функцш окремих нейрошв.
Тнтегративна й аналитична здатносп нервово! системи багато в чому визнача-
ються множинними контактами нервових клиин одна з одною. Вщомо, що в де-
яких вщцшах головного мозку юльюсть синаптичних контакпв на одному ней-
ош може сягати 2*105. Зменшення юлькосп контакт!в мгж нейронами за деяких
атолопчних процес!в також е одним гз суттевих мехашзмгв порушення функщй
-ервово! системи.
Важливою ланкою в патогенез! багатьох розладав дгяльносп нервово! системи
може бути порушення синтезу, видшення 1 розпаду мед!атор!в. Кр!м того, е чис-
енш дан! щодо того, що на дгяльнгсть нервово! системи, особливо п виших вщ-
длгв, дуже впливають речовини пептидно! природи (нейропептиди), як! виробля-
ються як нервовими, так г шшими юптинами г можуть виконувати медгатори! та
немедгаторш функцп. У мозку людини г тварин виявлено десятки олйопептидав,
ведения яких у шлуночки мозку або безпосередньо в нервов! центри може спри-
чинювати ргзш емоцгйнг стани г реакцп поведанки, впливати на вироблення умовних
рефлексов, здатшеть до запам’ятовування, навчання тощо. Очевидно, мехашзм по-
рушення функцш нервово! системи може зумовлюватися недостатшм або надмгр-
ним утворенням нейропептидав, змшою чутливосп до них нервових юптин.
Варто зазначити, що в патогенез! функщональних порушень центральних вщ-
_тпв нервово! системи можуть мати значения нейрони, яю з’являються в нерво-
зих центрах, працюють певною м!рою автономно г е джерелом надмгрного збуд-
кення. Таю нейрони називають генераторами патолопчно посиленого збудження
(Г.М. Крижановський). У норм! актившсть нейрон!в або нервових центр!в конт-
ролюеться й обмежуеться вщповщними мехашзмами гальмування. У раз! ушкод-
кення цих механ!зм!в г виникае генератор патолопчно посиленого збудження,
ткий залежно вщ його локалгзацгг може бути причиною чутливих, рухових г веге-
гативних розладгв, а також порушень психгчног дгяльностг.
ПОРУШЕННЯ ФУНКЦ1Й НЕРВОВО! СИСТЕМИ
Порушення функцгй нервових шптин । пров!дник1в
Найважливгшими функщями нервово! юптини е генерування потенщалу дц, про-
ведения збудження через нервов! волокна й передача його на шшу юптину (нер-
ову, м’язову, залозисту). Функцгя нейрона забезпечуеться обмшними процеса-
ш, яю вщбуваються в ньому. Одним гз призначень метаболгзму в нейрон! е ство-
> | 667 {
д
§
а
о
а
й
I
а
л
§
о
—1----1----I---I------1---к-----1----1----------I------>
0,01 0,03 0,1 0,3 1 3 10 30 100 300 Т, мс
Мал. 74. Крива “сила — тривалтсть” у р1зш перюди госля перер1зу-
вання рухового нерва:
I— одразу теля перервування; II — через 16 тиж.: III — через 20 тиж.,
IV— через 24 тиж.; V —у норм! (за ВИсЫе)
рения асиметричного розподйту юнтв на поверхш й усередин! клттини, що визначае
потенщал спокою т потеншал дц. У ход! обмшних процессе Ка+-, К+-насос набу-
вае енергп, потребно! для активного подолання електрох!м!чного градюнта на
мембран!.
Отже, вст речовини ! процеси, яю порушують мегабол!зм т зменшують вироб-
лення енергп в нервовш клттин! (гшоксемтя, отруення щанщами, дишгрофено-
лом, азидами тощо), здатш пригшчувати збудливтсть нейрон!в.
Функцтя нейрона порушуеться 1 в раз! змши вмтсту одно- т двовалентних юшв
у середовищ!, що його оточуе. Зокрема нервова кл!тина цшком втрачае здатшсть
до збудження, якщо помютити и в середовише, позбавлене юнш натрпо. Значний
вплив на величину мембранного погеншалу нейрона мають також юни кал!ю !
кальшю. Мембранний потентат, який визначаеться ступенем проникност! для
№+, К+т СГ та тх концентрашею, може нщгримуватися лише тодт, коли мембра-
на стабштзована кальцтем. Як правило, пщвищення ртвня кальщю в середовитш.
де перебувають нервов! клттини, призводить до тхньот пперполяризацтт, а його
часткове або повне видалення — до деполяризацп.
Функцтя нервових волокон — провщниктв збудження — може порушуватися
в раз! дистрофтчних змш мтелтновот оболонки (наприклад унаслщок дефициту тта-
м!ну або цтанокобаламшу), здавлювання або охолодження нерва, запатення, гт-
поксп, щд впливом деяких отрут т токсинтв м1кроорган!зм!в.
Як вщомо. збуцлив!сть нервово! тканини визначаеться кривою “сила — трива-
лтсгь”, яка вцюбражае залежшсть порогово! сили подразнювального струму вщ три-
ватост! його впливу. У раз! ушкодження нервово! клттини або дегенерацн нерва кри-
ва “сила — тривалтсть” значно змшюеться, зокрема збшьшуеться хронакстя (мал. 74).
Унаслщок дп ртзних патогенних фактортв на нерв може розвинутись особли-
вий стан, який М.€. Ввеленський назвав парабюзом. Залежно вш ступеня ушкод-
ження нервових волокон розртзняють кыька фаз парабюзу. Птд час вивчення
проявтв парабюзу в руховому нерв! на нервово-м’язовому препарат! визначено,
ло в разг незначного ушкодження нерва настае момент, коли на сильне г слабке
? дразнения м’яз вщповщае однаковими за силою теташчними скороченнями.
Це зргвняльна фаза. 3 поглибленням альтерацп нерва виникае парадоксальна
аза. коли на сильне подразнення нерва м’яз вщповщае слабкими скороченнями,
тодг як на помгрне подразнення м’яз реагуе енерпйшше. Нарепт, в останнгй фазг
щрабгозу — фазг гальмування — жодш подразнення нерва не можуть спричинити
.корочення м’яза.
Якщо внаслщок ушкодження нерва втрачаеться його зв’язок з тглом нейрона,
ерв зазнае дегенераци. Основним мехашзмом, який спричинюе дегенерацйо не-
вового волокна, е припинення аксоплазматичного току г транспорту речовин
ксоплазмою. Процес дегенераци, детально описаний Уоллером, полягае в тому,
шо вже через добу шсля травми нерва мгелш починае вщходити вщ перехватав
вузлгв нервового волокна (перехватав Ранв’е). Потам мгелш збираеться у велик!
крагой, якг поступово розсмоктуються. Нейрофгбрили зазнають фрагментацц. Вщ
черва залишаються вузьк! трубочки, утворен! нейролемоцитами. Через кглъка дшв
!сля початку дегенерацп нерв втрачае збудливгсть. У ргзних трупах волокон втра-
га збудливостг настае в ргзш термши, що, очевидно, залежить вщ запасу речовин
• аксон!. У нервових закгнченнях дегенеруючого нерва змши настають тим швид-
ше, чим ближче до закшчення перергзаний нерв. Невдовз! шсля перер!зування
нейролемоцити починають виявляти фагоцитну актившсть щодо вшьних нерво-
вих закшчень — гхш вщростки проникають у синаптичну щшину, поступово вщо-
кремлюючи нервов! закшчення вщ постсинаптичног мембрани г фагоцитуючи гх.
Пгсля травми нерва починаються змши г в проксимальному вщцглг нейрона
червинне подразнення), ступшь яких залежить вш виду та штенсивностг ушко-
дження, його локалгзацп (вгддаленгсть вщ тша нейрона), типу г вгку нейрона.
У раз! поранения периферичного нерва змши в проксимальному вщдш! нейрона,
як правило, мгшмальш, г надал! нерв регенеруе. У центральны! нервовш систем!
нервове волокно, навпаки, дегенеруе ретроградно на значн!й довжинг, г нерщко
нейрон гине.
Порушення функций синапов
Синапси — це спецгалгзоваш контакт, через як! здшснюеться передача збуджу-
вальних або гальмгвних впливгв (збуджувальш та гальмгвш синапси) вщ нейрона
до нейрона або гншог клгтини (наприклад м’язово'!). У ссавцгв цей процес у си-
напсах вщбуваеться хгмгчним шляхом — за допомогою медгаторгв. На мал. 75
схематично показано основнг структури! компоненти холшерпчного синапсу г
процеси, якг вщбуваються в ньому.
Порушення синтезу медгатора. Синтез медгатора може порушуватися внаслщок
зниження активностг ферментгв, якг беруть участь у його утворенш. Наприклад,
синтез одного з медгаторгв гальмування — у-амшомасляног кислоти — може при-
гнгчуватися дгею семикарбазиду, який блокуе фермент, що каталгзуе перетворен-
ня глутамшовог кислоти на ГАМК. Синтез ГАМК порушуеться г в разг дефщиту
в гжг шридоксину, який е кофактором цього ферменту. У таких випадках у ЦНС
порушуються процеси гальмування.
Процес утворення медгаторгв пов’язаний гз затратами енергп, яку постачають
мгтохондргг, що мгстяться у великгй кглькостг в нейронг г нервових закгнченнях.
Пресинаптичний Пресинатична Постсинаптична
елеменч мембрана мембрана
Мал. 75. Холшерпчний синапс:
АХ (ацетилхолш) синтезуеться н цитоплазм! кл1тин, попм концентруеться в нервовому
зак!нченн1 та збсрцаеться в пресинаптичних мзхурцях. Пцз вшивом потенщалу дп мзх-
урш поблизу пресинаптично'1 мембрани розкриваються 1 медаатор виходигь у синап-
тичну щиину. Июля взаемоди з холзнорепептором ацетилхолш руйнуеться естеразою.
Холш реабсорбуеться нервовим закшченням 1 бере участь ) синтез! ацетилхолз ну (поз-
начено р1вш, на яких можливе порушення цих процеств;
Отже, порушення цього процесу може виникати внаслщок блокади метаболтчних
процеств у мпохондртях т зниження вмтсту макроерпв у нейрон! (пщ час гшоксп,
ТОКСИЧНИХ ВПЛиВТВ тошо).
Порушення транспорту меддатора. Мед!атор може синтезуватись як у т!ш ней-
рона, так 1 безпосередньо в нервовому закшченн!. Мед!атор, шо утворюеться при
цьому, транспортуеться по аксону в пресинаптичну частину. У мехашзм! транс-
порту' значну роль вЩграють цитоплазматичн! мткротрубочки, побудоваш з особ-
ливого битка тубулшу, який за сво!ми властивостями близькии до скоротливого
бшка актину. За участю мткротрубочок у нервове закшчення надходять медтатори,
ферменти, що беруть участь в обмш! мед!атор!в, та ш. Микротрубочки легко роз-
падаються п!д дтею знеболювальних засобгв. пщвишено! температури, протеоль
тичних ферменпв. речовин типу колхщину, що зумовлюе зменшення кшькост!
мед!атор!в у пресинаптичних елементах. Наприклад, гемохолш блокуе транспорт
ацетилхолтну в нервов! закшчення, тим самим порушуючи передачу нервових
впливтв у холшерпчних синапсах
Порутпения депонування медтатори в нервових закшченнях. Медтатори зберпа-
ються в пресинаптичних мтхурцях, у яких мтститься комбшащя молекул медта то-
ра, АТФ т специфтчних бйпав. Мтхурц! утворюються в цитопдазм! нейрона, а
попм транспортуються по аксону до синапса. Нин! вщомо, шо в окремих нерво-
вих волокнах можуть синтезуватись т накопичуватись у м!хурцях нервових закш-
чень не один, а два мсшатори, наприклад, у деяких волокнах блукаючого нерва
утворюються ацетилхолш т ВШ.
Порушення видалення медаатора в синаптичиу щёлину. Процес виходу медёатора
синаптичну щетину може порушуватися пёд впливом деяких лёкарських засобёв
токсишв. Зокрема правцевий токсин перешкоджае виходу медёатора тальму ван-
я тлёцину, ботулёшчний токсин блокуе викид апетилхолёну. Механёзм бпокуван-
я цими токсинами видалення нейротрансмётера пов’язаний з ёнактивацёею бшюв
выбрани синаптичних мтхурцёв ё бёлкёв пресинапгично! мембрани, що забезпе-
ють процеси злиття обое мембран ё подал ьший екзоцитоз меддатора. Так, прав-
евий токсин дёе на бшок мембрани мёхурцёв синаптобревён, а деяк! види бо-
лёнёчного токсину, шо прилиняють видёлення ацетилхолёну в мгсщ нервово-
[’язового сполучення, можуть впливати як на синаптобревён, так ё на бшок
-есинаптично'1 мембрани синтаксин.
У секрецп медёатора вёдёграе роль скоротливий бшок тубулён, який входить до
кладу преси нал тично!’ мембрани. Блокада цього бшка колхёцином пригшчуе
идёлення ацетилхолшу.
Секрецёя мед!атор!в нерьовими закёнченнями може також порушуватись
наслёдок шактивацп кальщевих канал!в у пресинаптичшй мембранё.
Порушення взаемодп медаатора з рецептором. € багато речовин, якё впливають
-а зв’язок медёаторёв зё специфёчними рецепторними бёлками, локатёзованими на
остсинаптичнёй мембранё. Це здебёлыпого речовини, для яких властивий конку-
ентний тип дц’, тобто вони легко зв’язуються з рецепторами. До таких речовин,
априклад, належитьтубокурарин, який блокуе Н-хол!норецептори, стрихнён, що
:-токуе рецептори, чугливё до глёцину, тощо. Цё речовини блокують вплив медёа
эра на ефекторну клттину.
Тривале зв’язування рецепторёв ёз нейротрансмётером або ёншим лёгандом
юже призвести до зменшення чутливостё мембрани ефекторно! клётини до дц
_ього лёганда — розвиваеться десенсибёлёзацёя. Один ёз ц механёзмёв полягае в
эму, шо при цьому посилюеться ендоцитоз релепторёв 1 пригнёчуеться процес !х
/творения. Це зумовлюе зменшення кёлькостё рецептортв на поверхш клётинно!
мембрани.
До рецепторних бёлкёв постсинаптично! мембрани можуть утворюватися ан-
тптёла.
Можливё також генетичнё дефекти, унаслёдок яких у структур! рецептора мо-
..уть замёнюватись одна або кшька амёнокислот, що впливае на спорёднешсгь
рецептора до вёдповёдного лёганда.
Порушення видалення медаатора ёз синаптично) шпини. Для того шоб синапс
функцёонував нормально, медёатор шсля взаемодп з рецептором мае бути видале-
ний ёз синаптично! щёлини. 6 три мехашзми видалення: 1) руйнування медёатора
ферментами, локалёзованими на постсинаптичнёй мембранё; 2) зворотне захоп-
тення медёатора нервовим закёнченням: 3) дифузёя медёатора ёз синаптично! щё-
’ини в позаклётинну рёдину. Ацетилхолён, наприклад, руйнуеться в синаптичшй
щшинё холёнеегеразою. Порушення цього процесу може зумовлюватись ёнактива-
цёею холёнестерази, зокрема фосфорорганёчними сполуками. При цьому ацетил-
холён на тривалий час зв’язуеться з великою кёлькёстю холёнорецепторге, що
спричинюе порушення функцц багатьох оргашв.
Доведено, що вмёст у нервових закшченнях амтнокислотних нейромед!атортв.
"аких як норадреналён, дофамш, глёцин, ГАМК, серотонён, залежить вёд !х зво-
ротного поглинання ёз синаптично! щёлини до цитоплазми пресинаптичного ней-
рона. Холшерпчнг нервов! закшчення захоплюють також продукт гщролгзу аце-
тилхолшу холш для ресинтезу медгатора. Таке зворотне поглинання нейротран-
смгтергв вщбуваеться за участю спещальних б!лк!в-транспортер!в. Деяк! речовини
можуть порушувати процес депонування мед!атор!в, блокуючи щ транспорта! мо-
лекули. Наприклад, резерпш здатний блокувати два моноамшов! м!хурцев! транс-
портери, що може зумовити зменшення вм!сту в нервових закшченнях норадре-
налшу, дофамшу, серотоншу.
Тим часом шггбування зворотного захоплення нейротрансмгтера подовжуг
його вплив на постсинаптичну мембрану, що посилюе його дпо на ефекторщ
клгтину аж до и ушкодження (у деяких випадках). Вважають, що в раз! ггпоксй
головного мозку частина нервових клгтин ушкоджуеться тому, що пригнгчуеться
зворотне поглинання деяких медгаторгв, зокрема г глутамату, який у раз! йог:
накопичення справляв нейротоксичну дпо.
Порушення чутливосп
Усг види чутливост! вщ шкгри, м’яз!в, суглоб!в г сухожилкгв (соместезгя) переда-
ються в ЦНС трьома нейронами. Перший нейрон мгститься у спинномозкових
вузлах, другий — у заднгх рогах спинного мозку (больова г температурна чут-
ливгсть) або в тонкому г клиноподгбному ядрах довгастого мозку (глибока г так-
тильна чутливгсть), трети — у таламус!, вщ якого аксони спрямоваш до чутливих
зон кори великого мозку.
Патологгчнг процеси г пов’язанг з ними порушення чутливост! можуть локал!-
зуватись у будь-якгй д!лянц! сенсорного шляху. У раз! ушкодження периферичних
нервгв (перергзування, запалення, авгтамшоз) у вгдповщнгй зон! порушуються вс
види чутливост!. Втрату чутливост! називають анестез1ею, зниження — гтестезгею
пщвищення — г1перестез1ею. Залежно вщ характеру втраченог чутливост! розргз-
няють анестезпо тактильну (власне анестезпо), больову (аналгезгю), термгчнх
(термоанестезпо), а також втрату глибоког, або пропргоцептивног, чутливост!.
Якщо патолопчний процес локалгзуеться в спинному або головному мозку
порушення чутливост! залежить вщ того, якг саме висхщнг шляхи ураженг.
€ двг доцентровг системи чутливостг. Одна з них отримала назву лемшсково!
мгстить нервовг волокна великого дгаметра, якг проводять гмпульси вщ пропргоре-
цепторгв м’язгв, сухожилкгв, суглобгв г частково вщ шкгрних рецепторгв дотику I
тиску (тактильних рецепторгв). Волокна цгег системи входять у слияний мозок !
спрямовуються у складг заднгх стовшв у довгастий мозок. Вщ ядер довгастого моз-
ку починаеться медгальна петля (лемнгсковий шлях), яка переходить на протилеж-
ний бгк г закшчуеться в задньобгчних вентральних ядрах таламуса, нейрони яких
передають отриману гнформацпо в соматосенсорну зону кори великого мозку.
Друга висхщна система — це спгноталамгчний (переднш г бгчний) шлях, який
несе больову, температурну г частково тактильну чутливгсть. Його волокна спря-
мовуються догори в склад! переднгх г бгчних канатикгв спинного мозку г закшчу-
ються в клгтинах ядер таламуса (антеролатеральна система).
Характерш змши чутливостг спостерпають пгсля перергзування право! або
лгвог половини спинного мозку (синдром Броун — Секара): на боцг перергзування,
нижче вщ нього зникае глибока чутливгсть, тод! як температурна г больова чут-
ливгсть зникають на протилежному боцг, оскгльки провщн! шляхи, якг належать
^672
до антеролатерально! системи, перехрещуються в спинному мозку. Тактильна
чутливйть частково порушена з обох боюв.
Порушення лемшсково! системи можливе в раз! ушкодження периферичних
,ерв1в (товстих м!елшових волокон), а також унаслщок р!зних патолопчних про-
цесгв у спинному мозку (порушення кровообпу, травма, запалення). 1зольоване
шкодження задшх канатиюв спинного мозку спостерйаеться рщко, однак нар!в-
Н1 з шшими провщними шляхами вони можуть ушкоджуватися пухлиною або
внаслщок травм.
Порушення провщносп у волокнах медгально! петл! спричинюе ртзш розлади
чутливост!, характер яких залежить вщ ступени ушкодження системи. При цьому
може втрачатися здатшсть визначати швидюсть 1 напрямок руху кйпцвок. Значно
лорушуеться вщчутгя роздшьного сприйняття дотику одночасно у двох М1сцях, а
акож здатшсть вщчувати вгбращю й оцшювати масу предмета, який пщшмають.
Дослщжуваний не може на дотик визначити форму предмет! в та щентиф!кувати
.итери й числа, якщо !х написати на шюр!. Вш вщчувае лише мехашчний дотик 1
:е може точно визначити його м!сце 1 силу. Вщчутгя болю 1 температурна чут-
;ив!сть при цьому зберйаються.
Ушкодження зацентрально! звивини пм’яно! частки великого мозку. У мавп ви-
алення зацентрально! звивини спричинюе розлади чутливост! на протилежному
чип пла. Певною м!рою характер цих розладш можна визначити на основ! знань
"ро функцп лемшсково! системи, пов’язавши таку операцпо з лемшсковою де-
ерващею на протилежному бощ, на якому, однак, зберйаються елементи анте-
"юлатерально! системи. Розлад полягае, очевидно, у втрап м’язово-суглобово! чут-
ивосп. Тварина часто перестае рухатися, залишаючись у незручюй поз! протягом
ривалого часу. Тактильна, больова! температурна чутлив!сть на цьому бощ збер!-
аеться, хоча п поргг може пщвищуватися.
У людини !зольоване ушкодження зацентрально! звивини спостерйаеться
туже рщко. Наприклад, х!рурги шод! видаляють частину ще! звивини для лгкуван-
-я ешлепс!! юркового походження. У такому раз! спостерйають уже описан! роз-
ади: втрату вщчуття положения кшщвок у простор!, здатносп на дотик визнача-
и форму предмепв, !хш розм!ри, масу, характер поверхш (гладка, шорстка), дис-
кримшащйно! чутливосп.
Ушкодження пм’яно! частки швкул! великого мозку. Вщомо, що за участю
пм’яно! частки здшснюються штегративш функцн вищого порядку. Вважають,
що штегращя чутливо! нервово! д!яльносп у пм’янш частщ сприяе свщомому
прийняттю зовншшього свггу 1 здатносп ор!ентуватися в ньому. Зокрема нор-
мально сприйняття людиною форми свого т!ла та положения в простор! залежить
шд штеграцц в щй дглянщ птформацц, що надходить кшькома сенсорними кана-
лами: соматосенсорним, зоровим, вестибулярним, слуховим. Ушкодження пм’яно!
-стки спричинюе комплекс порушень, що мае назву аморфосинтезу. У чистому
зигляд! аморфосинтез у людини (але не в шульги) спостерйаеться шсля видален-
1Я кори право! швкул! великого мозку. Людина при цьому втрачае уявлення про
тросторове розташування частин тша на протилежному бощ. Вона не може одяг-
нутися або впорядкувати одяг на цьому бощ, не може поголитися або зачесати
волосся з цього боку. Вона навпь може заперечувати наявшсть ще! половили пла
не помтчати патолопчш процеси на шй, наприклад супутшй парал!ч. Але така
людина здатна вшзнаваги частини тЁла 1 називати ёх. Якщо ж ушкодженою е
тш'яна частка з лЁвого боку (тобто в домЁнуючЁй швкул!), то до аморфосинтез}
додаегься агнозгя — нездатшсть вшзнавати частини тша, предмета, Ёхш зображен-
ня ё розташуваняя в простор!.
Б!ль
Поняття болю охоплюс. по-перше, своерщне вшчуття, по-друге — реакцЁю на не
В1дчуття, яка проявляешься певним емоцшним забарвленням, рефлекторними змн
нами функщй внутр! шшх орган ёв, руховими безумовними рефлексами 1 вольови-
ми зусиллями, спрямованими на звыьнеиня вщ больового фактора. Ця реакщя за
своём характером близька до вшчуття страждання, якого зазнае людина в раз
загрози й житпо, ё надзвичайно гндившуальна, осюльки залежить вш фактор,в.
серед яких основне значения мають м!сце ! стутнь ураження тканин, вихованш
людини, емощйний стан у момент нанесения больового подразнення.
Спостереження евщчать, що щд дЁею ушкоджувального фактора людина мож
вщчувати два види болю. Якщо, наприклад, гарячим сЁрником доторкнутися де
шюри, то спочатку7 виникае вшчуття, подЁбне до уколу, — ‘"перший” бшь. Цей бшь
ч!тко локализуется 1 швидко згасае. Через деякий час виникае дифузний пекучи.
“другий” бшь, який може бута досить тривалим. Такий подвЁйний характер болк-
спостериасться в раз! ушкодження шюри , слизовоё оболонки деяких оргашв
Великого значения у симптоматищ рЁзних хвороб набувае вгецералъний биь
тобто бшь, який виникае у внутршш!х органах. Цей б!ль важко гцддаеться л окал,-
заци, мае розлитий характер, супроводжуеться тяжкими переживаниями, пригш-
чешетю, змшою дёяльностё автономно! нервово! системи. ВЁсцеральний бпп
под!бний до “другого” болю.
Дослщження, проведен! переважно на людях пщ час оперативник втручань.
показали, що не вс, анатом!чн! утвори можуть бута джерелом больових вщчутпв.
Органи черевноЁ порожнини не чутлив! до звичайних хЁрургЁчних вплив!в (розрЁ-
зування, зшивання). болючими е лише брижа , пристшкова (партетальна) очере- ]
вина. Однак ус! внутрЁшн! органи з непосмугованою м’язового тканиною болюче
реагують на розтягнення. спазм або судомне скорочення.
Дуже чутлив! до болю артерп. Звуження артерш або раптове Ёх розширенн’
спричинюе гострий бшь.
Тканина легень 1 нутрошева (висцеральна) плевра не чутлив! до больовоп
подразнення, дуже чутлива до цього парЁетальна плевра
Результата операшй на людях ё тваринах показали, що серпевий м’яз не чут-
ливий до механЁчноЁ травм и (укол. розр!з). Якшо ж у тварини потягти одну
в!нцевих артерш, виникае больова реакщя. Чутливим до болю е перикард.
Кшьюсть больових рецепторЁв у вЁсцеральних структурах менша, нёж у сома-
тичних, 1 це е однкю Ёз причин, чому вюцеральний б!ль погано локалЁзований
Больобё Ёмпульси вщ внутрЁшшх органЁв дося гаюгь ЦНС симпатичними або пара-
симпатичними провщними шляхами, а в ЦНС проходить тими самими шляхами
шо й Ёмпульси вш соматичних структур.
Складним е питания про мехашзми розвитку больового вшчуття. Доведено^
що !снують спецЁальш больов! рецептори, аферентнЁ шляхи, як! передають болъс-
ье подразнення, ё спецёальш структури в головному мозку, якё переробляють бо-
ьову ёнформацёю.
На пёдставё результатов дослёджень установлено, що рецептори шкгри ё види-
;о! слизово! оболонки, якё реагують на больов! стимули, належать до чутливих
эолокон антеролатерально! системи двох типёв — тонких мёелёнових А5-волокон
зё швидкёстю проведения збудження 10—30 м/с ё безмёелёнових С-волокон зё
швидкёстю проведения 0,5—2 м/с. Актившсть в А5-волокнах спричинюе вёдчуття
□строго колючого болю, тодё як збудження повёльнопровёдашх С-волокон зумов-
дое вёдчуття печёння.
Доведено, шо синагпичним трансмёгером, який видёляеться первинними афе-
рентними волокнами, що забезпечують вщчуття швидкого болю. е глутамат, а
трансмётером, який передае повёльний бёль, — речовина Р
Важливим е питанья про мехашзм активацп больових рецепторёв. Факти
гвёдчать про те, що значна деформацёя вёльних нервових закёнчень (наприклад лёд
час здавлення або розтягнення тканини) та щцвищення температури шкёри (по-
пал 45 °С) або зниження п (нижче 15 °С) е адекватними стимулами для рецептор]в
долю, впливають на проникшсть ёхньо! мембрани для ёошв ё зумовлюють виник-
нення потенщалу дп. Больовё рецептори реагують також на електричну енергёю 1
хёмёчш подразники.
€ певш припущення, що вёльнё нервовё закёнчення, якё належать до Ао- або
С-волокон, мёстять одну або калька специфёчних речовин, шо видёляються пёд
пливом ушкоджувальних фактор!в, взаемодёють з рецепторами зовнёшньо! по-
ьерхнё мембрани нервових закёнчень ё спричинюють ёх збудження. Одним ёз таких
ёмёчних факгорёв може бути АТФ, який, вшгиваючи на вёдповёдш рецептори не-
чвових закёнчень, вёдкривае хемочутливё ёоннё канали Вёдомо, що АТФ зумовлюе
шикнення болю в разё внутрёшньошкёрного введения.
На мембранё нервових закшчень виявлено капсащиновё рецептори, з якими
взаемодёе капсаё'цин — речовина, шо мёститься в червоному пркому перцё ё зумо-
..тюе пекучий бёль. Цё рецептори з’еднанё з неселективними ёонними каналами,
якё забезпечують проникнення натрёю ё катьцёю всередину ноцицепторёв ё спри-
-инюють ёх деполяризашю. € припущення, шо пёд час подразнення больових за-
шчень ними видёляеться ще неёдентифёкований лёганд капсаё’цинових рецепторёв,
кий ёх активуе. Крём того, капсащиновё рецептори активуються ёонами водню.
Активаторами больових рецепторёв можуть бути пстамён, серотонён, бради-
пнён, соматостатин, субсганнёя Р, простагландин и ё деякё ёншё БАР. Не всё цё
'ечовини виявлено в нервових закшченнях, однак багато з них утворюються в
канинах пёд час ушкодження клётин ё розвитку запалення, ё з ё'х накопиченням
чов’язують виникнення болю.
Припускаютъ також, що ендогеннё БАР у невеликих (пёдпорогових) кёлькос-
’ях знижують порёг реакцёё’ больових рецепторёв на адекватнё стимули (механёчнё,
-ермёчнё), шо е фёзёологёчним пёдгрунтям для стану' пёдвишено’ё больово! чутли-
востё (гёпералгезщ пперпатн), який супроводжуе деякё патолопчнё пропеси.
В органёзмё виявлено лриродний полёпептид, який мёстить 17 амёнокислот ё
гае назву нопицептину. Ноцицептин та його рецептори е в багатьох дёлянках
головного мозку, зокрема й гёпоталамусё, стовбурё головного мозку, а також у
зднёх рогах спинного мозку. Пёс.тя внутрёшньомозкового введения ношшептину
експериментальних тварин спостерёгаеться гёлералгезёя.
I "б7Г
Мал. 76. Функщя воротного механизму в спинному мозку (за Ме1хаск I УУаП):
80 — нейрон лраглистоТ речовини; Т — релейний нейрон задн1Х роив спинного
мозку; М — товст! м1ел1нов! волокна лемшсково! системи; А — тонк! волокна
антеро. 1атерально! системи
ЧУаЛ । Ме!хаск запропонували концепцию, вщповшно до яко! у виникнеш
больового вшчуття важливу роль в!д!грае функцюнування у спинному мозку та
званого ворггного механизму.
Одним 1з основних положень ще! концепцп е те, шо передача нервових 1м-
пульсш вш аферентних волокон до нейрошв спинного мозку, як! несуть сигнали ь
головний мозок, регулюеться системою нейрошв драглисто! речовини (мал 76).
Припускають, що бшь виникае шсля дп високо! частота розрядш на нейрон
Т. На тш цих нейрошв заюнчуються як товст! мйелшов! волокна (М), шо належат
до лемшсково! системи, так 1 тонк! волокна антеролатерально! системи (А). Кр!
того, колатерал! обох волокон утворюють синаптичн! зв’язки з нейронами дра-
глисто! речовини. Вшростки нейронш 80 своею чертою утворюють аксо-аксонт
синапси на закшченнях волокон М 1 V, здатш гальмувати передачу 1мпульс1в ь
волокон обох вид1в на нейрони Т. Сам! нейрони 86 збуджуються импульсами, яй
надходять волокнами лемшсково! системи, : гальмуються в раз! активацп тонки
волокон (на мал. 76 збудливий вплив позначено знаком а гальмйвний — зна-
ком
Отже, нейрони 86 можуть вццгравати роль вор!т, що вщкривають або закри-
вають тлях !мпульсам, якими збуджуються нейрони Т. Вор!тний мехашзм обме-
жуе передачу нервових !мпульс1в до нейрони Т у раз! збудження аферентних вс-
локон лемшсково! системи (“закривае ворота’’) !, навпаки, полсгшуе проходжен-
ня нервових !мпульс!в до нейрошв Т у випадках, коли зростае аферентний поп
через тонк! волокна (“вщкривае ворота”).
Коли збудження нейрон!в Т перевищуе критичний р!вень, 1мпульсац!я !х
спричинюе збудження системи дп. До ще! системи належать нервов! структур!'
як! забезпечують форми поведшки щд час дп больового подразника, рухов!, веге-
тативн! й ендокринш реакцп, 1 т! структури, де формуються вшчуття, характерш
для болю.
Функцию сигнального воротного мехашзму контролюють р!зн! вцщши голо-
дного мозку, чи! впливи передаються нейронам спинного мозку через волокна
низхщних шляхтв (докладшше див. антиноцицептивш системи мозку). Система
антрального контролю болю активуеться !мпульсами, яю надходять товстими
волокнами лемшсково! системи.
Теоргя вхщних ворй дае змогу пояснити природу фантомного болю 1 каузал-
гп. Фантомный б1ль спостерйаеться в людей шсля ампутацп юнц!вок. Протягом
три вал ого часу хворий може вщчувати ампутовану юнщвку ! сильний, йюд! не-
стерпний бшь у нш. Пщ час ампутацп зазвичай перер!зують велию нервов! стов-
эури з безл!ччю товстих нервових волокон, переривають канали для надходження
мпульсацп з перифери. Нейрони спинного мозку стають менш керованими 1 мо-
жуть зумовлювати спалахи на р!зш несподйаш стимули.
Каузалггя — нестерпний бшь, який спостерйаеться в раз! ушкодження сома-
тичного нерва. Будь-який, навпъ найменший вплив на хвору юнщвку спричинюе
-1зке посилення болю. Каузалпя виникае частице внаслщок неповного перертзу-
ання нерва, коли ушкоджуеться бшыпа частина товстих м!елшових волокон. За
дим збшыиуеться потгк !мпульс!в до нейрошв задай ропв спинного мозку — “во-
рота вщкриваються”.
Отже, у раз! 1 фантомного болю, 1 каузалгй у спинному мозку або вище з’яв-
дяеться генератор патолопчно посиленого збудження, утворення якого зумовлене
позгальмуванням групи нейрошв унаслщок порушення зовнппнього апарату кон-
-ролю, локал!зованого в ушкодженш структур!.
Запропонована теор!я дае пщстави пояснити ! той давно вщомий у л!куваль-
шй практищ факт, що бшь помгтно згасае в раз! застосування вщволйальних
процедур: з!гр!вання, розтирання, масажу, накладання прчичниюв тощо. Ус! ц!
«ходи посилюють гмпульсацпо в товстих мгелшових волокнах, що зменшуе збуд-
кеиня нейрошв антеролатерально! системи.
У раз! розвитку в деяких внутрццнй органах патолопчних процесй може ви-
чикати в1дбитий (рррадшований) б1лъ, який здебшьшого поширюеться до структу-
ри, яка розвиваеться з того самого ембрюнального сегмента (дерматома), що й
.рган, який е джерелом болю. Наприклад, хвороба серця супроводжуеться болем
дшянц! л!во! лопатки ! зон! шнервацй лйсгьового нерва л!во! руки; у раз! розтяг-
чення жовчного мйура б!ль локал!зуеться м!ж лопатками; проходження каменя
терез сечоводи супроводжуеться болем, який з дшянки попереку перемйцуеться в
пахвинну дшянку. 1ррадшований бшь пояснюеться тим, що ушкодження внут-
’1шшх оргашв зумовлюе збудження, яке аферентними волокнами автономних
вегетативних) нерв!в досягае тих самих нейрошв задай рог!в спинного мозку, на
ких заюнчуються аферентш волокна вщ шюри. Посилена аферентна !мпульсац!я
вщ внутрппнй оргашв знижуе порй збудження так, що подразнення вщповщно!
дшянки шюри сприймаеться як бшь.
Експериментальн! та клйпчш дослщження евщчать про те, що у формуванш
юльового вщчуття ! реакци оргашзму на б!ль беруть участь багато вптдйпв ЦНС.
Таламус забезпечуе анал!з якост! больового вщчуття (штенсившсть, локалгза-
!ю тощо).
Больова шформацы активуе нейрогенш 1 нейрогуморальш структури гшотала-
муса. Це супроводжуеться розвитком вегетативних, ендокринних ! емощйних ре-
акцш, спрямованих на перебудову вей систем оргашзму в умовах дп больових
стимулов. Больове подразнення, яке йде вщ зовнхшнього покриву, а також вщ де-
яких оргашв пщ час травми, супроводжуеться загальним збудженням х активащею
симпатично! нервово! системи — посиленням дихання, пщвищенням артерхально-
го тиску, таххкарщею, гшерглхкемхею тощо. Активуеться гхпофхзно-наднирникова
система, спостерхгаються вс! компоненти стресу. Надмхрний больовий вплив може
призвести до розвитку шоку. Бшь, який ще з внутршпйх оргашв х за свош харак-
тером подхбний до “другого” болю, найчастхше супроводжуеться загальним при-
гшченням х вагусними ефектами: зниженням артерхального тиску, гхпоглхкемхех
тощо.
Лхмбхчна система вщхграе важливу роль у створеннх емоцшного забарвлення
поведшки людини у вщповщь на больову стимуляцпо.
Мозочок, шрамщна й екстрашрамщна системи здшснюють програмування
рухових компонент реакцш поведшки пщ час появи больового вщчутгя.
Щодо кори великого мозку, то сприйняття больових подразнень зберхгаеться
в експериментальних тварин навхть за ц вщсутност!. Вважають, що за участю корн
вщбуваються уточнения й штерпретацхя деяких характеристик больового вщчут-
тя, а також реалхзуються свщомх компоненти поведшки пщ час болю.
Через спинний мозок реалхзуються руховх та симпатичнх рефлекси. Там же
вщбуваеться первинне оброблення больових сигналхв.
Рхзнх функцп з переробки больовох шформацп виконуе схтчастий утвхр (ретику-
лярна формацхя). До цих функцш належать: пщготовка х передача больовох шфор-
мацн у вихщ соматичш й автономш вщдши головного мозку (таламус, гптоталамус,
лхмбхчну систему, кору); полегшення захисних сегментних рефлексхв спинного
мозку 1 стовбура головного мозку; залучения до рефлекторно! вщповщх на больо!
стимули автономно! нервово! системи, дихального х судинорухового центр!в.
Антиноцицептивш (аналгетичш) системи мозку. На пщставх експериментальних
дослщжень доведено, що в нервовхй систем! е не лише больовх центри, збудження
яких зумовлюе формування больового вщчутгя, а й структури, активацхя яки
може змшити больову реакцпо у тварин аж до й повного зникнення. Показано
наприклад, що електрична стимулящя або ххмхчне подразнення деяких зон цент-
рально! схро! речовини, покриву мосту, мигдалеподхбного тша, ппокампа, яде;
мозочка, схтчастих ядер середнього мозку зумовлюе чхтку аналгезхю. Вщомо та-
кож, що емоцй людини впливають на характер реакци-вщповщх на больовий сти
мул: страх посилюе реакцпо на бшь, знижуе порхг больово! чутливост!, агресив-
нхсть х лють, навпаки, рхзко зменшують реакщю. Цх та шшх спостереження сфор-
мували уявлення про те, що в оргашзм! е антиноцицептивш системи, якх можуть
пригшчуваги сприйняття болю. € докази того, що таких систем у мозку чотирх
нейронна ошатна, гормональна ошатна, нейронна неошатна, гормональна не-
опхатна.
Нейронна огйатна система локалхзована в середньому, довгастому ! спинном
мозку. Доведено, що центральна схра речовина, ядра шва х схтчастий утвхр мхстят
тша 1 закшчення енкефалшерпчних нейрошв. Частина цих нейрошв посилае с во
аксони до нейрошв спинного мозку. У заднхх рогах спинного мозку також вияв-
лено енкефалшерпчш нейрони, якх розподшяють сво! закхнчення на нервови
провщниках чутливост!. Енкефалш, що видшяеться, гальмуе больову передач
через синапси до нейрошв спинного мозку. В експеримент! показано, що ця сис-
тема активуеться п!д час больово! стимуляцн тварини.
Функцгя гормонально')' огиатно) системи полягае в проведенн! аферентног гм-
"ульсацп 31 спинного мозку до гшоталамуса 1 плоф!за з видгленням кортикол!бе-
ину, кортикогропгну г р-лшотрошну, з якого утворюеться сильней аналгезуючий
олшептид р-ендорфш. Останнгй, потрапивши в кровоносне русло, гальмуе ак-
ившсть нейронгв больовоТ чутливосп в спинному мозку г таламус! 1 збуджуе
ейрони центрально! сгрог речовини, яю гальмують бьть.
Нейронна неотатна система охоплюе ссротоншерпчнг. адренерпчнг й дофа-
днерпчш нейрони, яю утворюють ядра в сговбур! головного мозку. Стимуляцгя
Шважливгших моноамшерпчних структур стовбура (ядер шва, блакитного мгс-
пя, чорно! речовини, центрально! сгрог речовини) зумовлюе значну аналгезгю. Усг
_ у твори мають прямий вихгл на нейрони больовог чутливостг спинного мозку. г
шд впливом серогонгну та норадренал1ну, шо видгляюгься ними, вщбуваеться
тачне пригнгчення рефлектор] тих больових реакщй.
Гормоналъну неотатну систему пов'язують переважно з функщею ппоталаму-
с а г гшофгза та гхшм гормоном вазопресином. Вщомо, що у щургв гз генетично
орушеним синтезом вазопресину пщвишена чутливгсть до больових стимулгв.
ведения вазопресину в кров або шлуночки мозку спричинюе у тварин стан гли-
’око! । тривалог аналгезп. Кргм того, вазопрссинерпчн! нейрони гшоталамуса по-
илаютъ сво! аксони до ргзних структур головного г спинного мозку, у тому числ!
до нейронгв драглието! речовини, г можуть впливати на функцию сигнального
ьоргтного механгзму та шших аналгетичних систем. € даш про те, шо кргм ва-
эпресину антиноцицептивну д!ю маюгь також гнил гормони ппоталамо-ппоф!-
зарног системи, зокрема соматостатин.
Ус! аналгетичн! системи взаемодгють одна з одною, забезпечують можливгсть
керувати больовими реакщями й ослаблювати гх негативна наслщки. У раз! пору-
дення функцгй цих систем можуть виникати ргзш больов! синдроми. Тому одним
з ефективних напрям!в подолання болю е розроблення способгв активацп анти-
ноцицеттгивних систем (акупунктура, навгювання, застосування лвкарських за-
.об!в тощо).
Значения болю для оргашзму. Ешь так часто виникае у повсякденному житп
юдини, шо ввгйшов у и свщомгсть як неминучий супутник генування. Однак варто
чам’ятати про те, шо це явище не фгзголопчне, а патолопчне. Ыль спричинюють
р!знг фактори, единою загальною властивгстю яких е здатнгсть ушкоджувата ткани-
.ди органгзму. Як 1 будь-який патолог гчний процес, бить суперечливий за сво!м
.петом г мае, з одного боку, захисно-пристосувальне, а з шшого — патолопчне
значения. Залежно в!д характеру болю. причини, часу г мгеггя його виникнення
южуть переважати або захиенг, або власне патолопчн! елемента. Захисн! власти во-
ст! болю вщгграють значну роль у житп людини г тварин: вгн е сигналом небезпеки,
нформуе про розвиток патолопчного процесу. Однак бщь надалг сам стае компо-
нентом патолопчного процесу, шод! дуже загрозливим.
Порушення рухово! функцп нервовоТ системи
За участю нервово! системи вщбуваються здгйснення г регуляцгя соматично! рухо-
во! активност! — як мимовшьно! (рефлекторно!), так г довгльног.
Як вщомо, скорочення скелетних м’язгв. його сила, тривалгсть. швидкгеть
залежать вщ ргвня збудження а-мотонейронгв спинного мозку. До кожного мого-
нейрона надходять численно гмпульси вщ чутливих нервов, нейрошв. розтаююва-
них у розних сегментах спинного мозку, стовбуро головного мозку, мозочку, ба-
зальних ядрах, кор! великого мозку. Ид нервово структури регулюють активност
мотонейронов як безпосередньо, так о влливаючи на штернейрони спинного моз-
ку (зокрема на гальмовно клгтини Реншоу) ! у-мотонейрон и, шо активують скоро-
чення внутрошньоверетенних (онтрафузальних) м’язових волокон, Цо впливи ре-
гулюють поставу о координують дгяльшсть р1зних груп м’язов под час виконанн
щлеспрямованого руховою акту.
Рухов! розлади виникають як унаслщок ушкодження зазначених вщдолгв ЦНС
так о внаслщок порушення проведения омпульсов руховими нервами, передач! ом-
пульсов вщ нерва до м’яза.
Порушення руховоо активност! супроводжуються розними проявами ппоко-
незп та гшерконезй.
Формами гопоконезп е параич (втрата рухов) о парез (ослабления рухов). Па-
рал оч м’язгв одноео полови ни тола мае назву гемшлегп, обох верхнгх або нижн
кони шок — параплегп, усох кшповок — тетраплеги. Залежно вод патогенезу парато-
ну тонус уражених м’язов може бути або втрачений — млявий паралоч, або пщви-
щений — спастичний паралоч. Кром того, розрозняють паралоч периферичний
центральний (унаслщок ураження вщповщно периферичних або центральних ру-
хових нейронов), а також рефлекторный парая1ч, зумовлений тим, шо в раз! ушко-
дження пугливого нерва гмпульси, яко йдуть вщ нього, можуть пригючувати мо-
тонейрони вщповщного м’яза.
Рухово розлади внаслщок патологи нервово-м’язових синапсов о рухових нервов
Нервово-м’язове сполучення е холонерпчним синапсом. У ньому можуть виника-
ти вс! паголопчн! пронеси, розглянуто ранппе.
Одним оз найвщомшоих прикладов порушення нервово-м’язовоо передач! в умо-
вах патологп е лйастешя. Якшо хворого на моасгенпо попросите калька раз!в пос-
пйть стиснута руку в кулак, йому вдасться це зробити тольки перший раз. 3 кожною
наступною спробою сила скорочень швидко зменшуеться. Така м'язова слабоасть
швидка стомлюваность стосуеться багатьох скелетних м’язов хворого, у тому чисо
момочних, окорухових. ковтальних та он. Зпдно з результатами елекгромоограф!чно-
го дослщження, у таких хворих порушуеться нервово-м’язова передача.
Хвороба виникае внаслщок утворення антитол до нокотинових холонорецеп- ।
торов. Цо антитша зв’язуються з рецепторами поетсинаптичноУ мембрани мионев-
рального синапсу, посля чого рецептори усуваються з поверхно мембрани шляхом
ендоцитозу.
М'язова слабкость спостерйаеться також у разо синдрому Ламберта — 1тона.
за якого утворюються антитиа до одного з калъщевих каналов нервових закончена
концевоо пластинки. Це зумовлюе зменшення надходження Са2+ до нервового за-я
к!нчення о видолення останном меншо'У колькосто ацеталхолону.
У разо ушкодження рухових нервов розвиваеться паралоч м’язов, яко оннерву-
ються цими нервами (периферичного типу), зникають усо рефлекси, м’язи атоноч-
но (млявий паралоч) о з часом атрофуються. В експеримент! такий тип рухови
розладов зазвичай можна спостерогати шсля перерозування переднох коронцк
сшонномозкових нервов або периферичноо о нерва.
Рухово розлади внаслщок порушення функцш спинного мозку. В експеримент
порушення функцш спинного мозку можна вщтворити його перерозуванням. У
Ж! ' I
.ребетних це спричинюе глибоке прхггшчення рефлекторно! активное!!, пов'язано!
нервовими пентрами, розмпценими нижче вщ м!сця перерхзування, — стналъ-
•ий шок. При пьому розвиваеться гшерполяризащя спхналыгих мотонейрошв.
Тривалхсть 1 тяжкхсть пього ставу в рхзних тварин рхзнх, однак вони тим бхлыш,
тим вищий розвиток тварини. У жаб шок гривае кмька хвилин. у собак, котж —
глька годин, у мавп — кптъка дшв, у людини — приблизно 2 гиж.
Клхнхчна картина шоку залежить вщ ргвня перерхзування. Перерхзування, зробле-
не нижче вщ довгастого мозку, спричинюе повне припинення дихання х рхзке зни-
кення тиску кровх. осюльки в такому разх життево важлив! центри повшетю в!ло-
кремлюються вщ ефекторних оргашв. Перерхзування спинного мозку на рхвш п’ятого
щийного сегмента не порушуе дихання Це пояснюеться тим. шо й дихальний центр,
ядра, як! хннервують дихальш м’язи, залишаюхься вище вхд мхеця перерхзування,
1днак не втрачають зв’язок хз ним, пщтримуючи його через дхафрах моих нерви.
Спхнальний шок не е простим наслщком травми, остдльки пхсля вщновлення
ефлекторххих функшй повторне перерхзування, виконане нижче порхвняно з по-
переднхм, не спричинюе шок. € рхзнх припушення стосовно патогенезу сигналь-
ного шоку. Одш дослщники вважають. шо шок виникае внаслщок випадання
буджувального впливу з боку вищих нервових центрхв на актившсть нейрошв
спинного мозку. На думку хнших, перерхзування усувае пригнхчувальну дно ви-
щих рухових центрхв на спхнальне гальмування.
Через деякий час пхсля зникнення ознак сигнального шоку рефлехегорш реак-
дц вцхновлюютъея х навпь посилюються. Рхвень порога рефлекс! в вщемикування,
розтягуванмя неухильно знижуеться. У разх подразнення шкхри в експеримен-
альних тварин унаслщок хррадхацп збудження в еххинному мозку рефлекси вгра-
ають нормальну обмежешеть х локалхзашю. Спостерхгаеться хррадхавхя хмпульсхв
.о автономиях цеххтрхв, що може спричинити опорожнения сечового мххура х пря-
юх кишки, коливання кров’яного тиску (мае- рефлекс) тощо.
Вщновлення рефлексхв х гпхеррефлексхя можуть бути пов’язап! з розвитком
надмхриох денервапхйнох чутливостх до нервових медхаторхв, шо хх видхляють ст-
нальш нейрони. Можливим е також утворенххя додаткових колатералей вщ наяв-
них нервових клхтин спинного мозку з формуванням збуддивих закхнчеиь на хи-
ер- х мотонейронах.
Руховх розлади в разх порушення функцш стовбурово! частини головной» мозку.
Для вивчешхя рухових розладхв. зумовлених порушеннями функцш рхзних струк-
тур головного мозку, якх здхйснююгь вищий руховии контроль, найчастхше пере-
эхзають мозок на рхзних його рхвнях.
Пхсля перерхзування мозку м!ж нижшми ! верхнхми горбками покриву серед-
нього мозку спостерхгаеться рхзке пщвищення тонусу м’язхв-розгиначхв — деце-
ебрацшна риг1дн1стъ. Шоб зхгнути кхнцхвку в сутлобх, потрхбно докласти значних
усиль. У певнхй стали згинання ошр раптово слабне — це реакшя подовження.
Якщо пхсля реакцп подовження трохи розхгнути кхнцхвку, ошр згинанню вщнов-
доеться — реакшя вкорочення. Розвиток деперебрапшно. ригхдностх пов’язаний
з рхзким посиленням хмгхульсацп мотонейронами. Посилення тонусу м’язхв мае
•ефлекторне походження: пхсля перерхзування заднхх канатиюв спинного мозку
тонус м’язхв вщповщно) хехнцхвки ослаблюеться.
Патогенез децеребрацхйнох рипдностх складний. Вщомо, що х тонхчнх, х
разовх рефлекси регулхоюгься схтчастим утвором, у якому е дв! рхзш за своею
функщею зони. Одна з них, бгльша, охоплюе дглянку вщ гшоталамуса до довгас-
того мозку. Подразнення нейронгв ще! зони посилюе скорочення скелетнил
м’язгв, що пояснюеться активащею у-мотонейронгв. Друга зона займае лише пе-
редньомедгальну частину довгастого мозку. Збудження нейронгв ще! зони спри-
чинюе гальмування спинномозкових рефлексгв г зниження м’язового тонусу
Функцгя нейрошв друго! зони пщтримуеться гмпульсацгею вщ мозочка, а такой
вщ кори великого мозку через екстрашрамщн! шляхи. Актившсть першо! зонт
забезпечуеться аферентною гмпульсацгею вщ чутливих нейрошв спинного ! вес-
тибулярних ядер довгастого мозку. Цг ядра вщгграють важливу роль у пщтриман-
ш м’язового тонусу, 1 в разг !х руйнування у пщдослщно! тварини децеребрацшна
ригщшсть м’язгв на вщповщному бощ ргзко слабне.
Шсля перетину стовбура головного мозку м!ж нижнгми г верхшми горбками,
покриву середнього мозку двг з трьох дглянок, яю керують шпбувальним центров
спчастого утвору (ретикулярно! формацп), залишаються вище вщ мгсця перетин
1 гхнгй вплив припиняеться. Вплив дглянки, яку активують, зберггаеться !, осюль-
ки актившсть гальмгвних нейрошв спчастого утвору знижена, баланс мгж актив) -
вальними г гальмгвними гмпульсами, шо впливають на у-мотонейронгв, змгщуетъ-
ся в б!к !х стимулювання. Наслщками цього е посилення гмпульсацп з нервово-
м’язових веретен, активацгя а-мотонейрошв 1 пщвищення тонусу м’язгв.
Руховг розлади внаслщок порушення функцш мозочка. Мозочок е високооргаш-
зованим центром, який чинить регулювальний вплив на функцпо м’язгв. До ньо-
го надходить потгк гмпульегв вщ рецепторгв м’язгв, суглобгв, сухожилкгв г шкгри
а також вгд органгв зору, слуху й ргвноваги. Вщ ядер мозочка нервовг волоки
пролягають до гшоталамуса, червоного ядра середнього мозку, вестибулярни.
ядер г спчастого утвору стовбура головного мозку. Цими шляхам и мозочок впли-
вае на рухов! центри, починаючи з кори великого мозку г закгнчуючи спинномоз-
ковими мотонейронами. Мозочок визначае цглгсшсть рухового акту, коригуе ру-
ховг реакцп органгзму, забезпечуючи !х точнгсть, що особливо яскраво прояв-
ляеться пщ час мимовгльних рухгв. Основна його функцгя — узгодження фазових
г тошчних компонентгв рухового акту.
Пгсля ураженггя мозочка в людини або видалення його в експериментальнш
тварин виникае низка типових рухових порушень. У пергггг дш шсля видалення
мозочка ргзко пщвищуеться тонус м’язгв, особливо розгинальних. Однак поп
вгн, як правило, ргзко знижуеться, г розвиваеться атон1я. Через тривалий ча
атонгя може змгнитися ппертошею. Отже, у цьому випадку йдеться про порушен-
ня м’язового тонусу, що, очевидно, зумовлено вщсутшстю регулювального впли-
ву мозочка, зокрема його передньо! частки, на у-мотонейрони спинного мозку.
У тварин, позбавлених мозочка, м’язи не здатн! до теташчного скорочення.
Це проявляеться постгйним тремтгнням г похитуванням тулуба та кгнцгвок твари-
ни (астазгя). Мехашзм цього порушення полягае в тому, що за вщсутносп мозоч-
ка не вщбуваеться гальмування пропрюцептивних рефлексгв г кожне м’язове ско-
рочення збуджуе пропрюрецептори, що викликае новий рефлекс.
У таких тварин порушуеться г координацгя рухгв (атакс1я). Порушення в лю-
дини здатностг виконувати тоню й точШ рухи позначаеться на функцпо мовленнт
воно стае монотонним, уривчастим — скандованим. Рухи втрачають плавюсп
(асинергия), стають незграбними, дуже мшними, розмашистими, що евщчить про
розлад взаемозв’язку сиди, швидкосп з напрямком руху (дисметр1я). Розвиток
атакой та дисметрп зумовлений порушенням регулювального впливу мозочка на
гктивнгсть нейрошв кори великого мозку. За цим змшюеться характер гмпульсгв,
яю кора посилае кгрково-спинномозковими шляхами, унаслщок чого юрковий
ехашзм мимовгльних рух!в не може досягти потртбного обсягу.
Одним 1з характерних симптомгв порушення функцп мозочка е сповшьнення
мимовшьних рухгв спочатку 1 ргзке посилення гх пщ кшець. У раз! намагання ви-
конати певний довшьний рух виникае тремор кппцвок. Втрачаеться здатшсть
раптово припиняти виконуваний рух, що лежить в основ! адгадохокгнезп, — не-
ожливост! швидко змгнювати напрямок руху кшцгвки на протилежний.
У раз! видалення клаптиково-вузликово! частки мозочка в мавп порушуеться
лвновага. У положенн! лежачи в тварини не виявляють жодних порушень. Однак
сидгти вона може, лише притулившись до стгни, а стояти зовс!м неспроможна
абаз1я). Спшальш рефлекси, рефлекси положения тша та мимовшью рухи при
цьому не порушуються.
Нареши, харакгерним симптомом, який спостерггаеться шсля видалення мо-
зочка, е астешя (легка стомлювашсть).
Рухов! розлади внаслщок порушення функцш шрамщних (юрково-спинномозко-
вих) та екстрашрамщних шлях!в. Як вщомо, шрамщним шляхом !мпульси надхо-
дять вщ великих шрамщних клггин кори великого мозку до мотонейрошв спин-
ного мозку. В експеримент! для того, щоб звшьнити мотонейрони вщ вплив!в
шрамщних клгтин, виконують одно- або двоб!чне перергзання шрамщних шлях!в.
Легше робити таке гзольоване перерезания стовбура головного мозку на р!вш тра-
пец!епод!бних тш. При цьому, по-перше, у тварини втрачаються або значно по-
рушуються поставн! ! стрибков! рефлекси; по-друге, порушуються деяю фазов!
рухи (дряпання, удари лапою тощо). Одноб!чне перер!зування шрамщного шляху
у мавп показало, що тварина дуже рщко 1 начебто неохоче користуеться кгнщв-
кою, яка втратила зв’язок з шрамщного системою. Уражена юнщвка починае
рухатися лише шсля сильного збудження г виконуе прост!, стереотипн! рухи
(ходгння, лазания тошо). Порушуються тоню рухи пальцгв, тварина не в змоз!
взяти предмет. Знижуеться тонус м’яз!в в уражених кшцгвках. Порушення фазо-
вих рухгв одночасно з гшотошею м’яз!в евщчить про зниження збудливостг мото-
нейрон!в спинного мозку. Шсля двоб!чного перергзання шрамщних шлях!в ми-
мов!льн! рухи можуть здгйснюватися тшьки екстрашрамщною системою. При
цьому спостерггаеться ппотошя м’язгв як кшцгвок, так г тулуба: змшюеться по-
става, випинаеться живгг. Через юлька тижнгв рухов! реакцп в мавпи вщновлю-
ються, однак ус! рухи вона виконуе дуже неохоче.
Екстрашрамщш шляхи заюнчуються на базальних ядрах кори великого моз-
ку, червоному ядр! середнього мозку, клпинах сгтчастого утвору 1, очевидно, на
шших шдюркових структурах. Вщ них 1мпульси числении ми нервовими шляхами
передаються мотонейронам довгастого г спинного мозку. Вщсутшсть симптомгв
гальмування пгсля перергзання шрамщних шлях!в дае пщетави припустили, що
вс! гальмгвнг впливи кори великого мозку на мотонейрони спинного мозку здгй-
снюються через екстрашрамщну систему. Ця дгя поширюеться як на фазов!, так 1
на тонгчш рефлекси.
До базальних ядер належать так! структури, як хвостате ядро, души ина, блща
куля, ггпоталамгчне ядро (тшо Льюгса) г чорна речовина. Хвостате ядро г лушпина
утворюють смугасте тшо, а лушпина г блща куля — сочевицепод!бне ядро. У ба-
зальних ядрах дтють три бюхЁмтчно вщмгнш системи: дофамгнерпчна — мгж чс”
ною речовиною 1 смугастим тйгом: внутршгньосмугаста холшерпчна систем*
ГАМК-ерпчна система, шо сполучае смугасте тою з блщою кулею г чорною реъ
виною.
Одшею з функцш бл1до‘1 кулЁ е гальмування ядер екстратпрамшмо'1 системи
розташованих нижче, зокрема 1 червоного ядра середнього мозку. У раз] ушк.
дження блщог кул1 значно пщвищуеться тонус скелетних м’яз]в, що пояснюеп .
звшьненням червоного ядра вш гальмЁвних впливёв блшот куль Оскшьки через бл
ду кулю проходить рефлекторш дуги, як! зумовлюють допомёжнё рухи, що супр
воджують руховий акт, то в разЁ и ушкодження розвиваеться г1пок1нез1я — рух.
стають скутими, незграбними, одномаштними, зникае актившсть мёмёчних м’язи
Смугасте тёло посилае еферентш Ёмпульси переважно до блщо'Ё кулЁ, регулк
ючи 1 частково гальмуючи и функцй. Цим, можливо, пояснюеться те, що в раз
його ушкодження виникають явища, протилежш тим, якё споетерйаються пщ ча
ураження блЁдог кулЁ. Виникають рЁзш вили гтерктези, зокрема й посилення дс-
пом1жних рухгв пщ час складного рухового акту. КрЁм того, можуть розвинутис
атетоз ё хорея. Для атетозу характерними е повёльнё червоггоцЁбш рухи, переваж-
но верхнЁх юнщвок, передусЁм пальцЁв. При цьому в скороченш одночасно берут
участь м’язи-агошсти ё м’язи-антагонЁсти. Хорея визначаеться швидкнми розма-
шистими неритмЁчними рухами юн цёвок, голови й тулуба.
Таю рухов] розлади спостерЁгаються зокрема пщ час хорет Гентингтона, як
успадковуеться за автосомно-домЁнантним типом Ё симптоми яко) починають про-
являтися переважно у вёцё вш 30 до 50 роюв. За шет хвороби ушкоджуютьс
ГАМК-ергЁчш та холшерпчш нейрони хвостатого ядра । лушпини сочевипеподЁб-
ного ядра.
Чорна речовина бере участь у регуляцй пластичного тонусу Ё мае значения
для виконання дрЁбних рухтв пальшв рук, яю потребують великот точностт ё тон-
кот регуляцн тонусу. У разЁ ушкодження чорно'т речовини м’язовий тонус пщвн-
щуеться
Дегенеращя дофам Ёнерпчних нейронЁв нтгросгртарнот системи с причин гс
дрижальний паралхч (хвороба Парюнсона), який розвиваеться переважно в люде
лггнього втку, коли виникае стЁйке зменшення юлькост] дофамЁну 1 дофамЁнови
рецептор!в у базальних ядрах. При цьому утворюеться дисбаланс мёж процесам!
збудження та тнпбування у базальних ядрах, шо виникае внаслщок втрати дофа-
мЁнергЁчного тнпбування лушпини.
Для паркшсошзму характерними е не тёльки гЁпоюнетичш, а й гшеркшетичн
прояви рухових порушень. Гшоюнетичними проявами е аюнезЁя (утруднен ня по-
чатку руху й ослабления спонтанних рухЁв) ё брадикшезтя (сповшьнення рухЁв) на
тлё пщвищення м’язового тонусу (ригЁднЁсть м’язёв). Пперкшезтя супроводжуеть-
ся тремором ЮНЦ1ВОКё тулуба, що спостерйаеться в стан! спокою Ё зникае шд час
рухЁв, зумовлених регулярними скороченнями м’язЁв-антагошстЁв.
Осюльки пщ час парюнсошзму збшыпуеться актившсть блщот култ (унаелшо?
зменшення галъмЁвно! дн з боку лушпини), то руйнування шляхёв. яю проводят*
тмпульси вщ блщог кулЁ, або патщотомЁя, усувають пщвищення тонусу м’язтв 1
тремор.
РуховЁ розлади внаелшок порушення функцш кори великого мозку. В експери-
ментЁ Ёзольованс ушкодження чутливо-рухово! дшянки кори, а гакож повна де-
а
Мал. 77. Елекгрична актившсть крко-
всго нейрона в епътептичному вогнипп
(за Шмщтсм):
а — посилення спайковоГ активностг з
повиъниме милями; б — збшыпення
частота розвиткл спайкГв гид час напа-
ду; в — збГлъшення юлькосп повшьних
хви.ть, г — збереження хвилъ у клошч-
шй фазг
кортикацгя тварин призводять
до двох основних наслщкгв: по-
рушення дифере нцшован их ру-
хгв г пщвишення тонусу м’язгв.
Пщвищення тонусу м’язгв у де-
кортикованих тварин зумовле-
не екстирпащею певного кгр-
кового поля, з якого через сг-
тчастий утвгр вщбуваеться
гальмування у-мотонейронгв.
Дуже важливою е проблема
вщновлення рухових функшй у
тварин гз видаленими дглянка-
ги рухових зон кори. Пгсля вндалення веге! кори великого мозку собака або кгт
дуже швидко вгдновлюе здатнгсть прямо стояти, бггати, хоча такт порушення, як
-щсутнгсть стрибкового г поставного рефлексгв, залишаються ггазавжди. Двобгчне
’идалення дглянок рухових зон кори в мавп робить гх неспроможними гцднгмати-
;я, стояти г навгть гсти, вони безпорадно лежать на боцг.
У декортикованих тварин втрачаються набутг завдяки доевгду умовно-рефлек-
орнг рухов! реакпгг.
3 порушенням функцгй кори великого мозку та шших структур головного
мозку пов’язаний ще один тип рухових розладгв — судами, якг спостерггаються щд
ас епглеиегг. У тонгчнш фазг епглептичного нападу ноги хворого ргзко розггнутг,
а руки зггнутг. Ригцгнгсть при цьому частково нагадуе децеребрацгйну. Потгм на-
-тае клонгчна фаза, яка проявляеться чергуванням мимовгльних, переривчастих
. корочень м’язгв кгнцгвок з гхнгм розелабленням. Пгдгрунтям епглептичного на-
аду е надмгрна синхронгзашя розрядгв у нейронах кори великого мозку. Елект-
гоеннефалограма. зроблена пгд час судомного нападу, складаеться з гпкових
розрядгв великог амплгтуди, якг ритмгчно вдуть один за одним г дуже поширенг в
корг (мал. 77). Така патолопчна синхронгзацгя залучае до посиленог активностг
безлгч нейронгв, унаслщок чого вони припиняють виконувати звичайнг для них
шференцгйованг функцп.
Причиною судомних нападгв можуть бути пухлина абс рубцевг змгни, як! ло-
алгзуються в руховгй або чутливгй дшянцг кори. У деяких випадках у патолопч-
нгй синхронгзацгг розрядгв бере участь таламус. Вщомо, шо неспецифгчнг ядра
таламуса в норм! синхронгзують розряди клгтин кори великого мозку, що й зумов-
гюе характерний ритм елеклроенцефалограми. Очевидно, пщвищена актившсть
цих ядер, пов’язана з виникненням у них генераторгв патолопчно посиленоп
збудження, може супроводжуватися розрядами в корь
В експеримент! судомн! розряди можна спричинити л!карськими засобамв
як! дпоть безпосередньо на поверхню кори великого мозку. Наприклад, стрихнш
зумовлюе серп розряд!в велико! амплпуди, як! евщчать про те, що в !х генерапь
синхронно бере участь багато клттин. Судомну актившсть можна викликати та-
кож, подразнюючи кору сильним електричним струмом.
НЕРВОВАТРОФ1КАIДИСТРОФ1ЧНИЙ ПРОЦЕС
Мал. 78. Дослщ Ф. Мажанд!:
троф!чна виразка на ропвц! ока кроля через 4
тиж. шсля перер!зання тршчастого нерва. У
центр! ропвки — кратерошмибна виразка, на пе-
риферГГ — вросл! вщ л!мба кровоносш судини
Нервова троф!ка — вплив нерв!в на тканину, що зумовлюе змши обм!ну речовин
у нш вщповщно до потреб у певний момент. Це означае, що троф!чна д!я нервг
тгено пов’язана з шшими шпми функ-
цхями (чутливою, руховою, секретор-
ною) 1 разом з ними забезпечуе оггги-
мальну функцпо кожного органа.
Перш! докази того, що нерви маютт
трофгчну функщю, отримав ще в 1824 р.
французький учений Ф. Мажанд!. В ек
спериментах 13 перер!занням тр!йчасто-
го нерва у крол!в вш виявив утворенш1
виразок у зон! чутливо! денервацп (оке
мал. 78).
Згодом модель нейрогенно! виразкь
багаторазово вщтворювалась ! не лише
зон! розм!щення плок тршчастого не-
рва. Троф!чю розлади виникають у будь-
якому орган!, якщо порушуеться йоп
шнервашя втручанням на нервах (афе-
рентних, еферентних, автономних) або нервових центрах. Медична практика евщ-
чить також про те, що ушкодження нервгв (травма, запалення) загрожуе виник-
ненням виразки або шшими розладами (набряк, ероз!я, некроз) у вщповщнь
зон!.
БЮХ1М1ЧН1, структура । функцюнальн! зм!ни
в денервованих тканинах
3 огляду на доевщ патогенн! впливи на периферичний нерв завжди зумовлюють
змгну обмшу речовин (вуглевод!в, лшщ!в, быюв, нуклегнових кислот тощо)
вщповщному орган!. Ц! змши мають не лише юльюсний, а й яюсний характер
Наприклад, мюзин у денервованому м’яз! втрачае АТФазш властивост!, а пико-
ген за своею структурою стае простёшим, елементарнйпим. Спостерггаеться пе-
ребудова ферментативних процесгв, наприклад, !зоферментний спектр лактатде-
пдрогенази (ЛДГ) змшюеться в б!к переважання ЛДГ41 ЛДГ3, тобто тих молеку-
лярних форм ферменпв, як! адаптоваш до анаеробних умов. Знижуетьс;
актившсть ферменту сукцинатдегщрогенази. Загальна тенденщя метабол!зму по-
ягае в тому, що вш набувае ембргонального характеру, тобто в ньому почина-
эть переважати глгколгтичнг процеси над окисними. Ослаблюеться потужнгсть
шклу Кребса, зменшуеться вихщ макроерпв, знижуеться енергетичний потен-
_гал.
У тканинах у разг порушення гннервацп виникають гстотш морфолопчш змг-
ш. Якщо йдеться про ропвку, шкгру або слизову оболонку, то в них послщовно
розвиваються вег стадп запалення. Усунення шфекцп, травм 1вного фактора, виси-
,ання не запобггае процесовг, хоча й сповглыпое його розвиток. Як наслщок, ут-
ворюеться виразка, яка не мае тенденцп до загоювання. У детальних дослщжен-
чях встановлено змши органонов, зокрема зменшення кглькостг мгтохондргй,
".освгтлшня гхнього матриксу. Очевидно, гз цим пов’язане порушення окисного
фосфорилювання г Са2+-акумулюючог здатностг мгтохондргй, а одночасно й енер-
гетичних можливостей клпини. У денервованих тканинах знижуеться мистична
актившсть.
Щодо функщональних розладгв, наслщки денервацп залежать вщ виду ткани-
ш. Наприклад, скелетний м’яз пгсля денервацп втрачае свою головну функшю —
щатшсть скорочуватися. Серцевий м’яз скорочуеться навгть у разг перергзання
всгх екстракардгальних нервгв. Слинна залоза секретуватиме слину, однак харак-
тер слини не залежатиме вщ виду гжг.
Денервована тканина реагуе на багато гуморальних факторгв шакше, нгж нор-
мальна. 1деться насамперед про медгатори нервовог системи. В. Кеннон (1937)
становие, що скелетнг м’язи, позбавлеш в одному випадку симпатичних, а в гн-
шому — холшерпчних нервгв, реагують вщповщно на адреналш г ацетилхолш
сильнгше, нгж у нормг. Так було вщкрито закон денервацп — тдвищено! чутливост!
денервованих структур. Зокрема це пов’язано з тим, що холгнорецептори, якг в
нормг зосереджеш лише в дглянцг нервово-м’язових синапсгв, пгсля денервацп
з’являються на всгй поверхнг мембрани мгоцита. Незвичайнгсть вщповщг денерво-
ваних структур полягае не лише в п посиленш, а й у перекручуваннг, коли, на-
приклад, замгсть розслаблення м’язгв судин вщбуваеться гх скорочення, шо може
сутгево позначитися на сташ судин, кровообггу тканин тощо.
Важливим е питания, чи гснують спецгальнг трофгчш нерви.
Свого часу Ф. Мажандг зробив припущення, що кргм чутливих, рухових г
секреторних нервгв е ще особливг трофгчш, якг регулюють живлення тканини.
Пгзнгше Т.П. Павлов (1883) в експериментг на тваринах серед нервгв, якг
вдуть до серця, виокремив таку гглочку, яка, не впливаючи на кровообгг, шдви-
щувала силу його скорочень. Цей нерв вш назвав посилюючим г визнав його суто
трофгчним. Повну г гармонгйну гннервацгю органа, на думку 1.П. Павлова, забез-
печують три види нервгв: функцгональш, судиноруховг (що регулюють надход-
ження поживних речовин) г трофгчш (що визначають остаточну утилгзацйо цих
речовин).
Таког самог думки дотримувався г Л.А. Орбелг, який разом з О.Г. Ггнецинсь-
ким у 1924 р. довгв, що гзольований (без кровообггу) м’яз жаби, виснажений три-
валим подразненням рухового нерва, знову починае скорочуватися, якщо стиму-
лювати симпатичний нерв. Трофгчна функцгя симпатичного нерва — це вплив на
метаболгзм, шдготовка органа до дп й адаптацгя його до майбутньог роботи, яка
здшснюеться завдяки руховому нерву.
Викладене вище, однак, не дае пщстав стверджувати, шо симпатичш нерв
не справляють шххгу, крхм трофшно!. дно на тканину або шо руховий нерв н
впливае на обмш речовин. О.Д Сперанський (1935) вважав. що вс‘ нерви впли
вакпъ на метаболизм, нетрофхчних нервхв немае, “нерв тхльки через те 1 функд :
нальннй, що вш грофхчний”.
Мехашзми троф!чного впливу нерв!в
Нервов! ’мггульси, приводили в дно орган (наприклад м’яз), одночасно змшюки
обмш речовин у клпин! за схемою: мещатор - активащя вторинних посередникхв —
активацхя генетичного апарату, ферменпв. Обмш речовин у клхтинах зм1нюетьс
також гоп впливом судинорухових нерв1в, яю розширюютъ або звужують судинн
таким чином зм1нюють приплив поживних речовин. Крш них двох (функшонатх-
ного (импульсного) та судинного) вплив! в нервово! системи на обмш речовин
непвовох клпини е третш — нехмпульсний. або власне трсф’чний. Вш забезпе
чуеться рухом аксоплазми як вщ нейрона до ефекторно'] клетини (ортоградно), таг
х в зворотному напрямку (ретроградно). За допомогою ортоградного аксотоку ш-
нервоваш клхтини отримують гроф!чш речовини, шо продукуються нейронами,
шляхом ретроградного аксотоку клпини-мштеш (м’язов], ештелхаоьн!) пос гач
ють так! речовини нейронам. Щ речовини отримали назву нейро хрофхчн их фак-
гор!в. або нейро трофшхв.
Нин! з рхзних нервових стр} ктур. клпин-сателп!в (глхальш клхтини, лемоци-
ти), а також !з тканин-мппеней 1 деяких оргашв видхлсно низку нейротрофшп
розшифровано ххню структуру та вивчено б!олог!чну дпо. Це фактор росту нерви
! спорщнен! з ним пептида, таю як мозковий нейротрофгший фактор, нейротро-
фши-3, -4, -5, -6.
Мозковий нсйротрофхчний фактор утворюеться безпосередньо в нейронах
транспортуеться до нервових зашнчень 1, видшяючись звщти, пщтримуе нормаль-
ний стан постсинаптичного нейрона.
1нпп нейротрофши зв’язуються з рецепторами нервових закжчень, потрапля-
ють у нейроплазму ! ретроградно перемпцуюгься до т!ла нейрона, де активукл
синтез речовин, потрхбний для життедхялвноси нервово! клхтини.
До ще! родини нейротрофшхв певною мхрою належать фактор росту епщерм!-
су, трансформуюч! фактори росту (а х р), шсулшоподхбш фактори росту 1 та II.
До нейротроф!чних факторхв належать нейролейкш, цилхарний I глхальнхг.
нейротрофхчш фактори, тромбоцитарний фактор росту, а також кислий та основ-
ний фактори росту ф!бробласт!в. Нейротрофхчш властивосп виявлено у речовин
Р, опхо'щних пептидхв, атриального натршуретричного пептиду. Крхм того, ней-
ротрофхчну тхю справляють глхколппда — ганглхозида, а також деяю гормони —
тироксин, тестостерон, кортикотрошн, хнсулш.
Найбшьше вивченим нейротрофшом е фактор росгу нервхв. Вш мхститься в
рхзних тканинах тварин х людини, але найбйыпу кьтькхсть йохо виявлено в слин-
них залозах самцхв мишей. Сприяе ембрюнальному розвитку та виживанню сим-
патичних х деяких сенсорних нейрошв, а також холшерпчних нейрошв централь-
но! нервово! системи, вщповщальних за пам’ять. Якщо отримати антипла до фак-
тора росгу нервхв 1 ввести хх новонародженим тваринам, то можна викликатв
майже повну деструкшю симпатичних вузлхв (гмуносимпатектомш)
Головними об'ектами да фактора росту епщермЬсу е ппальш юптини (астро-
иити), лемоцити, юптини ЦНС. яю своею чертою продукують таю нейротроф!чш
фактори, як гл1альний, цшпарний 1 фактор росту- нервов та ш.
Цилтарний нейротроф1чний фактор створюе умови для виживання моторних,
еенсорних 1 симпатичных нейрошв. Нейролейюн впливае як на руховй так 1 на
г-тлив! нейрони 1 продукуеться слинними залозами, скелетними м’язами га сти-
[ульованими Т-л1мфоцитами.
Експерименталый дослщження довели, шо дефщит нейрогроф] н!в або Тхшх
рецепторш може зумовлювати розвиток нейродегенеративних хвороб. Наприклад,
.лефщит мозкового нейрогрофтчного фактора у мишей спричинюе вграту перифе-
’ичних чутливих нейрошв 1 дегенеративна змши в нейронах вестибулярних нер-
я1в. У тварин з1 спадковим порушенням утворення нейротрофшу-З спостернаеть-
.я втрата механорецептор1в шюри.
У патогенез! нейрогенно! дистрофп визначальну роль вЩ^грае порушення син-
тезу й аксонального транспорту нейротроф1чних фактор!в. Однак, анал!зуючи
процес, слщ керуватися тим, що троф1чна функцтя здшснюеться за принципом
рефлексу 1 потр1бно оцшювати значения кожнот його ланки, роль розвитку у цьо-
у мехашзм! дистрофичного процесу.
Чутливий нерв, очевидно, вщ! грае в цьому особливу роль, осюльки, по-пер-
ше, перериваеться передача шформацп в нервовий центр !з зони денервацп, по-
руге — ушкоджений чутливий нерв е джерелом патолопчноТ шформацп, у тому
Схема 32. Патогенез троф4чних порушень у раз! ушкодження периферичних (чутливих, еферент
них, змпнаних) нерв1Е
числй й больовой, по-трете — йз нього виходять центрифугальнй (вйдцентровй) впли-
ви на тканину. Доведено, зокрема, що через чутливй нерви з аксоплазми в ткани
ну надходить речовина Р, яка впливае на метабол!зм й мйкроциркуляцйю.
Про значения нервових центров у розвитку дистрофйй свйдчать дослйди О.Д. Спе-
ранського з вибйрковим ушкодженням центров гшоталамуса. Наслщком цього е
утворення трофйчних виразок у рйзних органах на периферйй.
Роль еферентних нервйв у дистрофйй полягае в тому, що припиняеться а&
перекручуеться йхня функшя (рухова, секреторна). Припиняються ймпульсна ак-
тивнйсть, синтез мед!атор1в (адреналину, серотонйну, ацетилхолину тощо), змйню-
ються синтез й аксональний транспорт нейротрофйнйв. До процесу залучаетьс
геном, порушуеться синтез ферменпв, зменшуеться вихщ макроерпв, обмш наб}
вае примйтивнйшого характеру. Зазнають змш транспорта! функцп мембран юп-
тин. Орган йз порушеною йннервацйею може стати джерелом автоантигенйв. Про-
цес ускладнюеться тим, що до суто нейротрофйчних змйн додаються порушеннч
крово- й лймфообйгу (мйкроциркуляцйя) з розвитком гшоксп.
Отже, нейрогенна дистрофйя — це складний багатофакторний процес, який
починаеться з того, що нервова система перестае керувати обмйном речовин
тканинах, й, як наслйдок, виникають складнй порушення метаболизму, структури 1
функцй'й (схема 32).
ПОРУШЕННЯ ФУНКЦ1Й
АВТОНОМНО! НЕРВОВО! СИСТЕМИ
До автономной (вегетативной) нервовой системи належать Гй симпатична й парасим-
патична частини. Симпатичнй нерви починаються у вузлах, розмйщених уздовх
хребтового стовпа. До клйтин вузлйв пйдходять волокна вйд нейрошв, розмйщених
у грудних й поперекових сегментах спинного мозку. Центри парасимпатичной час-
тини автономной нервовой системи мйстяться у стовбурй головного мозку й крижо-
вих сегментах спинного мозку. Нерви, що вйдходять вйд них, спрямованг до внут-
рйшнйх органйв й утворюють синапси на вузлах, якй мйстяться бйля або всередин.
цих органйв.
Бйлышсть органйв йннервуеться й симпатичними, й парасимпатичними нерва-
ми, якй протилежно впливають на них.
У центрах автономной нервовой системи пйдтримуеться постййний рйвень ак-
тивностй, тому внутрйшнй органи безперервно отримують вйд них гальмйвнй абс
збуджувальнй ймпульси. Якщо орган з будь-якой причини позбавляеться йннерва-
цйй, наприклад симпатичной, усй функцйональнй змши в ньому визначаються пере-
важним впливом парасимпатичних нервйв. У разй парасимпатичной денервацйй
спостерйгаеться протилежне.
В експериментй для порушення автономной йннервацйй того чи того органа пе-
рерйзують вйдповйднй симпатичнй й парасимпатичнй нерви або видаляють вузли. КрЬ
того, знизити актившсть будь-якой частини автономной нервовой системи або на
деякий час повнйстю припинити н можна за допомогою фармакологйчних засобйв —
антихолйнергйчних (холйнолйтичних), антиадренергйчних (симпатолйтичних).
€ також метод “ймунологйчной екстирпацйй” симпатичной частини автономно;
нервовой системи. Якщо ймунну сироватку, яку отримують шляхом ймунйзацйй тва-
рин фактором росту нервгв, що мёститься в слинних залозах мишей, увести ново-
народженим тваринам, вузли симпатичного стовбура припиняють розвиватись г
легенерують. У цих тварин зникають усг перифериям прояви активностт симпа-
। тично! частини автономно! нервово! системи, вони мляв! й апатичш, в умовах,
шо потребують напруження оргашзму (перегргвання, охолодження, крововтрата),
виявляють меншу витривалтсть. У них порушуеться система терморегуляцн, г для
шдтримання температури тша на нормальному ргвш потр!бно щдвищити темпе-
ратуру навколишнього середовища. Система кровообтгу при цьому втрачае влас-
тивгсть пристосовуватися до змш у потреб! оргашзму в кисш у вщповщь на пщви-
щення фгзичного навантаження. У таких тварин знижуеться резистентнгсть до
ппоксг! та шших сташв, що в умовах стресу може призвести до загибел!.
Дуги автономних рефлекс! в замикаються у спинному, довгастому г середньо-
му мозку. Ушкодження цих вщдшгв центрально! нервово! системи може зумовити
порушення функцш внутршппх органгв. Так, у раз! сигнального шоку кргм рухо-
вих розладгв спостерггаеться ргзке зниження артергального тиску, порушення тер-
морегуляцп, потовидглення, рефлекторних актгв дефекацй та сечовипускання.
Пгсля ушкодження спинного мозку на ргвнг останнього шийного г двох верх-
нгх грудних сегментгв спостерггаеться звуження зшиць (мгоз), повгковог щглини г
западания очного яблука (енофтальм).
Пщ час патолопчних процеств у довгастому мозку ушкоджуються нервов!
центри, якг збуджують сльозовидшення, секрецпо слинних, пщшлунковог залоз г
залоз шлунка, зумовлюють скорочення жовчного мгхура, шлунка г тонко! кишки.
Ушкоджуються також дихальний г судиноруховий центри.
Автономна нервова система пщпорядкована вищим центрам, розмгщеним у
сгтчастому утворг, ггпоталамусг, таламус! та корт великого мозку. Вони штегрують
взаемозв’язок ртзних частин само! автономно! нервово! системи, а також зв’язки
автономно!, соматично! й ендокринно! систем.
1з 48 ядер г центртв, якг мгстяться в сгтчастому утворг стовбура головного моз-.
ку, бгльгшсть бере участь у регуляцп кровообпу, дихання, травления, екскрецг! та
лших функцш. Наргвнг гз соматичними елементами спчастого утвору вони за-
безпечують автономний компонент для вс!х видгв соматично! дгяльностг органгз-
му. Прояви порушення функцш спчастого утвору ргзномаштш г можуть стосува-
тися розладу дгяльностг серця, тонусу судин, дихання, функцш травного каналу
тшцо.
У разг подразнення ппоталамуса спостерггаються ефекти, подгбнг до виявле-
них пщ час стимуляцн парасимпатичних г симпатичних нервгв. 3 урахуванням
дього в ньому видшяють две зони. Подразнення однге! з них — динамогенног, яка
хоплюе задню, бгчну г частину промгжно! дшянки ппоталамуса, — спричинюе
тахгкардгю, гпдвищення артергального тиску, мщргаз, екзофтальм, пглоерекцгю,
трипинення перистальтики кишок, ггперглгкемпо та гншг ефекти симпатично!
нервово! системи. Подразнення друго! зони — трофогенног, до яко! належать пе-
редзоровт ядра г передня гшоталамгчна дшянка, — зумовлюе протилежш реакцп,
щрактерш для збудження парасимпатичних нервгв.
На функци ппоталамуса значно впливають розмпценг вище вгддши ЦНС.
Пгсля гх видалення автономн! реакци зберггаються, однак втрачаються стушнь
1хньо! активност! г тонка диферешцащя контролю.
Структури лхмбхчнох системи зумовлюють ефекти автономно! нервово! систе-
ми, яю виявляються в органах дихання, травления, кровообпу, систем! регуляцн
величини зшиць, терморегуляци. Таю ефекти спричинюють бшьше збуджуваль-
них, Н1ж гальмгвних впливхв на щ структури.
Мозочок також бере участь у контрол! функцш автономно! нервово! системи.
Подразнення мозочка спричинюе здебшыпого ефекти симпатично! нервово! сис-
теми — пщвищення артерхального тиску, розширення згниць, вщновлення скоро-
тливо! здатносп стомлених м’язхв. Пхсля видалення мозочка порушуеться регуля-
цхя кровообггу, функцш травного каналу.
Кора великого мозку хстотно впливае на регулящю автономиях функцш. То-
пографхчна подхбнхсть автономиях х соматичних центргв на рхвнх як чутливо!, так
х рухово! зон кори зумовлюе одночасну штеграцпо в нш автономних х соматичних
функцш. У разх електричного подразнення моторно! х премоторнох дхлянок кори
спостерхгаються змши в регуляцн дихання, кровообхгу, потовидшення, дхяльносп
сальних залоз, рухово! функцп травного каналу, сечового мгхура.
До автономно! нервово! системи належить також ентерична, або метасимпа-
тична, нервова система, утворена нервовими клхтинами, що мхстяться в стхнц!
травного каналу х формують мхентеричне (мхж шарами непосмугованих м’язхв) х
пщслизове (мхж коловим шаром х слизовою оболонкою) сплетения. У людини ця
система налхчуе понад сто мшьйонхв чутливих, рухливих х промхжних нейрошв.
На дхяльшсть ентерично! системи впливають симпатичш х парасимпатичн! нерви
травного каналу, але вона може дхяти автономно х без цих впливхв, здшснюючи
рефлекторну регуляцйо переважно моторно! та секреторно! функцш травного ка-
налу. Великою е юльюсть нейротрансмхтерхв, що утворюються х видшяються ней-
ронами ентерично! системи, серед яких ацетилхолш, адреналш, серотонш, АТФ,
у-амшомасляна кислота, бшьше десяти полшептидхв, а також КО ! СО.
У разх порушення функцш ентерично! системи змшюються структура х фун-
кц!я окремих дшянок травного каналу. Прикладом таких порушень е хвороба
Пршпрунга, яка виникае внаслщок того, що пщ час ембрюнального розвитку не
вщбуваеться мпрацхя нервових клхтин у певш дхлянки товстох кишки й утворю-
ються аганглюнарш зони, де немае нейронхв ентерично! системи або !хня юль-
юсть значно менша. За вщсутност! нервових впливхв в аганглюнарних зонах по-
рушуеться розвиток м’язово!! слизово! оболонок товстох кишки. Цх дхлянки киш-
ки розширеш, атонхчш, в них немае перистальтики.
ЕКСПЕРИМЕНТАЛЬНЕ ВИВЧЕННЯ
ЕМОЦ1ЙНИХ РЕАКЦ1Й 11Х ПОРУШЕНЬ
Факта, яю мають принципове значения для розумшня рол! емоцш в оргашзацп
поведшки вищих тварин, отримано експерименталышм шляхом. Насамперед
хдеться про метод подразнення електричним струмом через вживлен! в рхзнх вщ-
д!ли головного мозку електроди. 1нформативною виявилася також методика са-
моподразнення, суть яко! полягае в тому, що електроди, вживлен! в мозок, вми-
каються самою пщдослщною твариною. Доведено, що в мозку е центри, подраз-
нення яких зумовлюе задоволення, осюльки тварина намагаеться багато разхв
подразнювати мозок, шодх до виснаження. Але е й таю центри, яю тварина уни-
Мал. 79. Струкгурш утвори л!мб!чноТ системи:
1 — передне ядро таламуса; 2 — д1агокальна
смута нюхового нерва; 3 — б1чна нюхова смута;
4 — сосочкове того ппоталамуса; 5 — лрпсеред-
Н1й пучок кпшевого мозку: 6 — присередня ню-
хова смута; 7 — нюхова цибулина; 8 — прозора
перегородка: 9 — медулярна смута, 10 — нюхо-
вий горбок, И — Лобова частка; 12 — мозолисте
тою; 13 — склегання; 14 — стовбур головного
мозку, 15 — термшальна пластинка: 16 —
морський коник (ппокамп); 17— мпгдалепод1б-
не тсто
кае подразнювати, центри “незадо во
лення”, або точки “покарання”. Ц1
дан! свщчать про наявшсть псного
зв’язку емошйних реакцш з шяльшс-
ю певних структур головного мозку, насамперед гшоталамуса, льмбгшо! системи
кори великого мозку (зокрема лобово! частки). Схему основних структур лшб1ч-
зо! системи 1 зв'язюв мок. ними подано на мал. 79.
Експериментальне ушкодження або подразнення ргзних утворгв л!мб!чно!
.истеми, гшоталамуса або ново!' кори призводить до порушення таких найваж-
лив1ших реакцш иоведшки, як статева, зравна, захисна, 1 пов’язаних 13 ними
емоцшних сташв.
Значш емоцшн! порушення сиостерггають у раз1 ушкодження гшоталамуса,
що зумовлюе у самщв 1 самок втрату статево! активност!, порушення травно! ре-
кци в поведшц!. Коти шсля ушкодження передньо! дыянки гшоталамуса пере-
ьорюються на диких тварин, схильних до реакцш розлюченост!! страху. У раз!
шкодження обох б!чних дыянок гшоталамуса емоцшн! реакци слабшають. тва-
-,ини стають млявими.
У людини з пухлинами гшоталамуса видыяють дв! групи емощйних розладгв.
Перша трупа — розлади, пов’язан! з ушкодженням заднього ядра, шо проявля-
ються мляв1стю, зниженням гнгщативи, втратою !нтересу до всього, шо оточуе;
друга трупа — розлади, як! спричинен! ушкодженням передньо! дыянки гшотала-
муса 1 супроводжуються збудженням, ейфоргею, сексуальною розгальмовашетю.
йзн! емоцшн! розлади виникають унаелцюк ушкодження л!мб1чно! системи.
У раз! двоб!чного видалення скроневих часток, у тому числ! морського коника та
игдалепод!бного тыа, у поведши! тварин виявляюзь харакгерн! змиш (синдром
Клюве ра — Бюсг). Зокрема у мавп спостерггаються виражена тшерфаггя, ггперсек-
суальшсть 1 суттев! змши в поведений пов’язан! з втратою реакцш агреси г страху.
Руйнування мигдалеподгбного тыа призводить до змш внутр!шньогрупових в!д-
носин у мавп. Сами!, як! ранние панували в гурт!, переходить у щдпорядковане
становище. С також численн! даш про перетвоиення диких тварин на ручних
унаслщок руйнування мигдалепод!бного тыа Елекгричне подразнення цього ут-
вору 1 гшокампа, навпаки, спричинюють лють у тварин.
Руйнування центральных 1 присередшх ядер мигдалеподгбного т!ла супровод-
жуеться розладом регуляцп травних поведшкових реакцш — тварина повшетю
перестае споживати корм. Подразнення р!зних вгддшгв гшокампа призводить як
до шдвищення. так 1 до зниження травних реакшй.
У щур1в ушкодження дшянки, що вщповщае пояснш звивиш, зумовлюе по-
рушення поведшки, пов’язано! з материнством, — самки перестають доглядат
новонароджених 1 не оберп'ають !х вщ небезпеки.
У раз! патологи л!мб!чно! системи порушуються й шли функци нервово'1 сис-
теми, У норм! емоцшн! мехашзми беруть участь у концентраци уваги, сприяют
тому, що мислення зосереджуеться на розв’язанш завдань доти, доки не буде до-
сягнуто задоволення потреб. Ураження поясно! звивини та порушення и зв’язк
з корою лобово! частки великого мозку спричинюють у тварин порушення увагт
Кр1м того, утруднюються процеси навчання, що евщчить про розлад мехашзм!:
пам’ять Груб! порушення пам’ят! на поточи! поди в людей виникають шел*'
двобхчного видалення гшокампа 1 мигдалепод!бного тша.
Важливу роль у формувашп емоцшних реакцш вщгграе кора лобових часто!
великого мозку. Про це евщчать результати спостережень над людьми, яким я
певними показаниями перервзали бшу речовину мозку в дшянц! лобових часто!
(лейкотомгя). У таких людей почутгя втрачають св!ж!сть, силу й гостроту. Ц1 люд?
стають безтурботними, ус!м задоволеними, и! до чого не прагнуть, не вщчувають
мук сумлшня, не мають вщчуття страху, печал!, не можуть виявляти сшвчутге
симпатпо або антипатпо, вщчувати ненависть, втрачають творч! зд!бност!. Зника
ють вищ! емоцп, 1 вс!ею поведшкою хворих керуе принцип “задоволення — неза-
доволення”.
Нервов! та соматичш порушення пщ час емощйного стресу. Неврози. Емоп
завжди супроводжуються автономними й ендокринними реакщями. Це пояс-
нюеться тим, що будь-яке емощйне збудження пов’яЗане насамперед з гшотала-
мусом. Як вщомо, ппоталамус — це, по-перше, вищий автономний центр, яки
оргашзовуе вс! автономш компоненти емоцш (змшу артергального тиску, частот
скорочень серця, функцш оргашв травления тощо), по-друге — самоспйно 1 че-
рез гшоф!з керуе багатьма ендокринними залогами, що й зумовлюе виникнення
ендокринного компонента емошй (викид кортикотропшу, соматотрогану, стате-
вих гормонов, кортикостерощ!в тощо). Сутшсть цих реакций полягае в тому, щоб
пщготувати оргашзм до майбутньо! м’язово! роботи, пов’язано! з добуванняк
корму, втечею вщ ворога тощо.
У норм! вс! емоцшн! реакцп мають певний ступшь прояву, адекватний жит-
тевш ситуаци, у якш перебувае оргашзм. Процеси збудження в емоцгйних цент-
рах визначаються певною силою 1 тривалютю. Вони контролюються 1 своечасш
припйчуються вщповщними гальм!вними структурами. Якщо з якихось причин
настае надапрне збудження емоцшних центртв, що отримало назву емоцшног-
стресу, може виникнути стшке порушення функцш ЦНС, яке клппчно прояв-
ляеться неврозом.
Експериментальш прийоми вщтворення емощйного стресу 1 неврозу буле
розроблен! в лаборатори 1.П. Павлова. 1х сутшсть полягае в тому, щоб створити
важк! умови для двяльност! головного мозку, за яких вщбуваеться перенапру-
ження процесхв гальмування 1 збудження в нервових центрах. Перенапруження
процесу збудження досягають шляхом застосування потужних подразниюв (евгг-
лових, звукових, больових тощо), одночасного застосування багатьох сильних
подразнивав, тривалого впливу цих подразнивав, застосування незвичайних по-
дразнивав.
Розке посилення процесов збудження спостерогають о шд час моделювання
неврозу вщчаю, наприклад, коли тварина в процесх експерименту позбавлена
можливосп уникнути повторних ударов електричного струму.
Перенапруження гальмхвного процесу можна досягги шляхом вироблення
дуже тонких х складних диференцповань, застосування велико! юлькосп дифе-
ренцповань в одному стереотиш. Сюди варто додати х модель неврозу очхкування,
коли тварина пщцаеться больовим впливам (наприклад удари струму) через рхзш
штервали часу.
Класичним прийомом вщтворення неврозу у тварин е часта х раптова змша
' збудження емощйних центров, одним хз вароантов якого е раптове подання елек-
тричного струму в годхвницю в момент шдкрполення травного рефлексу. Якщо
। багаторазово повторювати таку ситуацпо, виснажуються гальмхвш процеси, що
вимикають роботу травного центру в момент дп больового подразника. Це зумо-
влюе стойке некероване збудження захисного центру х порушення функцш
цнс.
У тварин можна спровокувати невроз, якщо помостити ох в умови одночасно-
го впливу трьох факторов, яю часто трапляються о в житто людини: 1) великий
обсяг шформацп, яка пщлягае обробщ х засвоенню (у тому число прийняття рх-
шень); 2) постшний дефщит часу, водведеного на таку роботу мозку; 3) високий
рхвень мотивацп в досягненно позитивного результату.
Дуже часто причиною розвитку емоцшного стресу е конфлоктна ситуацоя, за
| яко! людина або тварина под впливом рхзних обмежувальних факторов не може
задовольнити сво! головш бюлопчш та сощальш потреби. Наприклад, значний
емоцшний стрес можна вщтворити, стимулюючи емоцюгеннх центри в умовах
хммоболозацп, обмежешхя часу на задоволення потреб ! т. п. У конфлхктнш ситуа-
цц, особливо тривалш або такой, що часто повторюеться, наростае емощйне на-
пруження, яке внаслщок недостатност! процесхв гальмування може перейти в
стшке збудження певних нервових центров.
Болышсть патогенних факторхв, яю впливають на нервову систему, ушкоджуе
насамперед функцц тих п структур, яю вщповщають за процеси гальмування.
Тому голодування, хнтоксикацхя, шфекцхя, гшоксхя, хошзуюче випромшювання,
травма головного мозку, ендокринн! розлади сприяють розвитково неврозу.
Невроз проявляеться рхзними соматичними розладами. Функшонування не-
керованого вогнища в ЦНС через автономну нервову х гшоталамо-ппофхзну сис-
теми спричинюе порушення дхялъностх внутрхшнхх оргашв х залоз внутрхшньо!
секрецц, виникнення стойко! артерхальнох’ гшертензп, 1ХС, виразкового ураження
травного каналу, иукрового дхабету, тиреотоксикозу, порушення менструального
циклу тощо.
Емоцшний стрес може призвести до порушень функцой ЦНС, характер яких
залежить вщ типу нервово! системи, способу вщтворення емоцшного стресу, фун-
кцш ендокринних органов та онших факторхв. Дослщження показали, що перебог
таких порушень в експерименталышх тварин вщбуваеться з переважанням або
процесов збудження, або процесов гальмування. У першому вшхадку характерними
е рухове збудження, агресивна поведгнка, втрата раноше виробленого диферен-
цповання х здатностх виробляти нове. Цо розлади спостерогаються здебхлыпого у
тварин хз силышм типом нервово! системи.
У тварин 31 слабким типом нервово! системи дуже швидко виснажуються
процеси збудження 1 розвиваеться гальмування. Тварини стають млявими, мало-
рухливими. У них згасають умовш рефлекси, а в деяких випадках зникають без-
умовш — тварини не приймають корм ! т. п. У людини щд час неврозу пору-
шуеться емоцшна сфера. Спостерггаються емоцшна нестшюсть, швидка змша
настрою, шод! тривал! перюди пригшченого стану. Можуть виникати 1 незвичай-
н1, неадекваты емоцшн! стани фобп (страху) 1 т. п. Часто порушуються пам’ять.
сон, здатшеть до концентрацп уваги. Можуть спостерйатися р!зн! рухов! розлади
(за!куват!сть, гшеркшез тощо).
Р03Д1Л XXXIII
ПАТОЛОПЧНА Ф1310Л0Г1Я
ЕКСТРЕМАЛЬНИХ СТАН IВ
Екстремалый стани — це дуже тяжкё, фактично критичнё стани, оскиыси безпо-
середньо загрожують життю оргашзму, став.зять його на межу життя / смеряй.
Екстремальш стани розвиваються внаслщок впливу на оргашзм надзвичай
них за силою або тривал!стю ди фактор!в. як! виходять за мехе фшогенетично
передбачсних можливостей оргашзму, у раз! несприятливого леребпу хвороби або
тяжких морфолопчних ! функцюнальних порушень. До таких сташв належать
шок, колапс, кома. Хвор), як! перебувають у такому еташ, потребують негайного
медичного втручання.
Критичн! стани можуть спричинити розвиток термшальних сташв (див. розди I
"Загальне вчення про хворобу”), шо е кшиевими стад1ями життя, перехщними
\ож життям ! бюлопчною смертю.
ШОК
Шок (вщ англ. зИоск — удар, струе або франц. скос — поштовх, удар) — екстре-
малышй стан, який виникае внаслёдок дёТ на органёзм патогенних факторёв надзви-
чайное сили ! для якого характерними е порушення гемодинамёки з критичном змен-
шенням капёлярного кровообёгу (тканинноё перфузёз) та прогресуючим порушенням
усёх систем життезабезпечення оргашзму.
Основн! прояви шоку включають порушення мйероциркуляци та периферий-
ного кровообпу (блща або мармурова. холодна, волога шк!ра), центрально!' гемо-
динамгки (зниження артер!ального тиску), змши з боку ЦНС, псюйчного статусу
загальмовашеть. прострашя), порушення функцш шших оргашв (нирок, печш-
ки, легень, серия тощо) !з законом 1рним розвитком ! прогресуванням недостат-
ност! багатьох оргашв, якщо не буде надано негайну медичну допомогу.
Етюлопя. Шок можуть спричинювати 6удь-як! патогенш фактори, здатш по-
рушувати гомеостаз. Вони можуть бути екзогенними й ендогенними, але надзви-
чайно силъними. Д1я таких фактор!в 1 змши, що виникають у вщповщь на них в
орган1зм!, е потенцшно смертельними. Ц! фактори за силою або тривалгетю ди
перевишують певну межу, яку можна назвати '‘порогом шоку”. Так, пщ час кро-
вотеч! — це втрата понад 25 % ОЦК, у раз! оппйв — ураження бшьше н!ж 15 %
поверхн! тша (якщо понад 20 % — шок розвиваеться неодмшно). Проте, оишю-
ючи д!ю шокогенних фактор!в, обов’язково потр!бно враховувати попереднш стан
оргашзму, який може ютотно вплинути на щ показники, а також наявшсть
впливгв, здатних посилювати дпо патогенних фактор!в.
Залежно вщ причини, що зумовлюе шок, описано близько 100 р!зних йоге
варганив. Найпоширенгшими е такт види шоку: первинний гтоволелйчний (у том
числ1 гемораггчний), травматичний, кардюгенний, септичний, анафиактичнии
ошковий (комбустшний) (схема 33).
Патогенез. Шокогенний фактор спричинюе в оргашзм! змши, що вихо-
дять за межг пристосувальних г компенсаторних можливостей його органов г сис-
тем, унаслщок чого виникае загроза житгю оргашзму. Шок — це “героина бо-
ротьба 31 смертю”, що здшснюеться шляхом максимального напруження всЬ.
компенсаторних мехашзмгв, гх р!зко! системно! акгиваци. За звичайного ргвня
патолопчних впливгв на оргашзм компенсаторн! реакцп нормалгзують вщхилен-
ня, що виникли; системи реагування “заспокоюються”, гх активацгя припиняеть-
ся. В умовах дц факторов, що зумовлюють шок, вщхилення так! значш, що ком-
пенсаторн! реакцп не в змоз! нормалгзувати параметри гомеостазу. Активащя
адаптащйних систем продовжуеться г посилюеться, стае надм!рною. Порушуеться
збалансовашсть реакцш, вони стають несинхронними, а на певному еташ сам!
спричинюють ушкодження та попршують стан оргашзму. Утворюються численю
порочн! кола, процес набувае тенденцп до самопщтримки та стае спонтанно не-
оборотним (мал. 80). У подальшому вщбуваються прогресуюче звуження дгапазо-
ну пристосувальних реакщй, спрощення та руйнування функцюнальних систем,
як! забезпечують компенсаторн! реакцп. Наслщком цього е перехщ до “екстре-
мального регулювання” — поступове вщключення ЦНС вщ аферентних вплив!ь
якг в норм! здгйснюють складну регуляцпо. Зберйаеться лише мгнгмум аферента-
цц, потр!бний для забезпечення дихання, кровооб!гу та небагатьох шших життево
важливих функцш. На певному етап! може вщбутися перехщ регуляцп життедгяль-
носп на гранично спрощений метабол!чний р!вень.
Схема 33. Види шоку залежно вщ причин 1 мехашзм!в !х виникнення
Мал. 80. Реакщя оргашзму яа дпо патыештх факторов звичайноТ сили 1 таких, якг
спричинюють розвиток шоку (адаптовано за Ю. Шутеу та ш., 1981):
1 — Д1я патогенного фактора; А — коливання параметров гомеостазу без дц пагоген-
них факторов за умови здоров’я; В — реакщя оргашзму на дгю патогенного фактора
звичайног сили (змши в органозм) не виходять за максимально допустимо межг при-
стосувальнил можливостей органозму; завдяки компенсаторним реакпоям параметри
гомеостазу вгдновлююгься); С — реаюия органозму надою шокогенного фактора (змо-
ни, що виникають, перевишують межо пристосувальних можливостей органозму о мо-
жуть призвесои до його загибелз): О — патогенний фактор, який зумовлюе ураження,
шо спричинюе загибель органгзму без розвитку шоку; Е — межа пристосувалыгих
можливостей органозму
Для розвитку бгльшост! видов шоку потробен певний штервал часу шсля дц
агресивного фактора, оскшьки якщо оргашзм гине одразу, стан шоку не встигае
розвинутися. Для розгортання компенсаторних реакцш шд час шоку необхщними
також е початкова анагом1чна й функцюнальна цйпсть нервово! та ендокринно! сис-
тем, тому зазвичай черепно-мозковг травми та первинний коматозний стан не
супроводжуються клшгчною картиною шоку.
На початку дп шокогенного фактора ушкодження ше локалг зо ване, збер!-
гаеться специфшшсть вщповщ! на етголопчний фактор. Проте з появою систем-
чих реакцш ця специфика втрачаеться, шок розвиваеться певним шляхом, сшль-
ним для ргзних його видгв. До нього лише цодаються особливост!, властивг цим
зкремим видам. Такими загалышми ланками патогенезу шоку е (за Ю Шутеу та
ш., 1981, гз доповненнями):
1) дефщит ефективно циркулюючого об’ему кров! (ЕПОК), що поеднуеться з!
зниженням серцевого викиду 1 пщвишенням загального периферичного опору
-удин;
2) над.шрне вид1лення катехоламине, яке стимулюеться невщкорегованими п-
повочем!ею, ппотензгею, гшокс!ею, ацидозом тошо;
3) генерал 1зоване видшення й активацгя велико! юлькосп БАР:
4) порушення мгкроциркуляци, шо е провщною пагогенетичною ланкою за шо-
кового стану;
5) зниження артериального тиску (однак тяжюсть стану пщ час шоку залежить
не вщ р!вня тиску, а здебьтьшого вщ ступеня порушення перфузи тканин);
6> гтокс1я, наслщком яко! е недостатке енергоутворення й ушкодження клг-
тин в умовах гх пщвишеного навантаження;
7) прогресуючий ацидоз',
8) розвиток дисфункцп та недостатност! багатьох оргашв (пол1органно! недо-
статност!).
У розвитку шоку схематично можна видыити так! основы етапи:
1) нейроендокринний етап, що складаеться зе
• сприйняття шформацп про ушкодження;
• центральних штеграцшних циюпв;
• нейрогормональних еферентних впливгв;
2) гемодиналтний етап, який охоплюе:
• зм1ни системно! гемодинам!ки;
• порушення мгкроциркуляцп;
• штерстицшно-лгмфатичш порушення;
3) клтинний етап, що подщяеться на стани:
• метабол хчного напруження;
• метаболхчного виснаження;
• необоротних ушкоджень юптинних структур.
Ц1 етапи зумовлюють один одного 1 можуть вщбуватись одночасно. У розвит-
ку кожного етапу розр!зняють фази.
• функцюнальних змш;
• структурних оборотних порушень;
• необоротних змш.
Нейроендокришп реакцп. У розвитку шокового стану завжди вщбуваються зли
ни функцш нервово! системи, як! мають певну послщовшсть 1 циюпчшсть. До не
рвово! системи надходить шформац!я про вщхилення, що виникли внаслщок д'
шокогенного фактора. Запускаються реакцп, спрямован! на порятунок житгя ор
гашзму, однак вони е надзвичайно штенсивними, стають несинхронними, розба-
лансованими. Спочатку розвиваеться збудження кори великого мозку внаслщо
дп масивно! аферентно! !мпульсацп, що надходить у ЦНС з периферп (еректилъъ-
стад!я). Кора спричинюе збудження пщюркових структур, а вони своею чертою —
кори; утворюються позитивш зворотш зв’язки. Збудження надм!рно посилюеть-
ся. Цьому сприяють також висхщн! активуюч! впливи сггчастого утвору. Одночас-
но 1стотно знижуеться синтез ГАМК, змшюеться вм!ст опющних пептид!?
(ошаттв). Наданрне тривале збудження може спричинити виснаження ЦНС 1 по-
яву необоротних структурних ушкоджень, як! посилюються також унаслщок гу
морального впливу на мозок. Под!бним чином у високих концентращях дпот
ацетилхолш, адреналш, вазопресин, кортикотрошн, пстамш, серотонш, анало-
пчно впливае зниження рН, зменшення вмюту кисню. Якщо нейрони кори бу
дуть здатними розвинути активне захисне гальмування, тод! кора буде захишеног
1, можливо, п функцп вщновляться в раз! сприятливого виходу оргашзму з! стан
шоку. На тл! гальмування в кор! збер!гаетъся домшуюче вогнище, до якого над-
ходять стимули вщ шокогенного ураження, що пщтримуеться. У цьому перена-
пруженому вогнищ! виникають явища парабюзу. Якщо стан оргашзму не ног
мал!зуеться, то метабол!чш запаси кори великого мозку виснажуються, порушен-
ня прогресують, виникае фаза зовшшнього пасивного гальмування з подалышг.-1
структурними ушкодженнями нейрошв 1 можливим розвитком смерп головного
мозку. Фаза гальмування отримала назву тортдно! стадп 1 проявляеться змша’
ПСИХ1ЧНОГО статусу — загальмовашстю, простращею.
Початкове збудження охоплюе також елементи лгмбгчног системи, у якгй вщ-
буваеться штеграцгя гуморально! вгдповщг на шокогенний вплив. Проте, якщо в
корг розвиваеться захисне гальмування, то шдкгрковг центри залишаються в збуд-
женому сташ, г лгмбгчна система забезпечуе ргзке пщвищення тонусу симпатико-
адреналово! системи (вм!ст катехоламгнгв може бути пщвищений у 30—300 разгв),
яке передаеться на гшоталамо-ппофгзарно-наднирникову систему з видшенням
вщповщних гормошв. За всгх видгв шоку визначаеться пщвищений вм!ст у кров!
бглыпостг гормошв: кортикотрошну, глюкокортикощгв, тиротропшу, тиреощних
гормошв, соматотропшу, вазопресину, альдостерону, катехоламгшв, а також ан-
потензину II, ендогенних опгатгв.
Реакщя ендокринно! системи пщ час шоку вибухова, концентрацп гормошв
зростають стргмко г сягають надзвичайно високих ргвнгв. Найшвидше збыьшуеть-
ся вмгст катехоламгшв, вазопресину, кортикотропшу г кортизолу. Тим часом спо-
стерпаються порушення ритмгв видглення гормошв, коливання гормонально! ре-
акцп, змши концентрацп гормошв. У цшому реакцп ендокринно! системи пщ час
шоку спрямоваш на збереження життя оргашзму: забезпечення енергогенезу, пщ-
тримання гемодинамгки, об’ему кровг, артергального тиску, гемостазу, елекгролгт-
ного балансу. Проте ендокринна вщповщь е надзвичайно сильною, тому вона
зумовлюе виснаження оргашв-ефекторгв г стае руйшвною.
Гемодинамгчн! змши (схема 34). Провщною ланкою в патогенез! шоку е пору-
шення гемодинамики, насамперед зниження ЕЦОК. Такий розлад може бути спри-
чинений:
• втратою организмом рщини (кров!, плазми, води), що характерне для пер-
винного гшоволемгчного, а також геморапчного, травматичного, опгкового шоку;
• перемгщенням рщини гз судин в ппш вщдши оргашзму, наприклад накопи-
чення води в серозних порожнинах, штерстицшному простор! (набряки), внут-
ргшньокишково (так! шоки називають перерозподиышми, або дистрибутивними,
що характерно, наприклад, для септичного, анафшактичного шоку);
• розвитком серцево! недостатносп, що зумовлюе зменшення серцевого ви-
киду (кардгогенний шок).
У раз! зменшення ЕЦОК 1 зниження артер!ального тиску через баро-, волю-
мо-, осморецептори долучаються мехашзми корекцн цих параметргв. Активують-
ся симпатико-адреналова, РААС, гшоталамо-гшофгзно-наднирникова системи,
посилюеться викид вазопресину. У судини надходить кров гз депо, штерстицгйна
рщина; вода затримуеться нирками. Розвиваеться генералгзований спазм перифе-
ричних судин. Це забезпечуе певне пгдтримання тиску в центральних судинах за
рахунок обмеження надходження кров! в мгкроциркуляторне русло паренхгматоз-
них органов, тобто вщбуваеться центрамзащя кровооб1гу. Саме тому ргвень ар-
тергального тиску при шоку не вщображае стан кровопостачання оргашв г стушнь
тяжкосп стану хворого. Якшо тиск не нормалгзуеться в процесг подальшого роз-
витку шокового стану, то активацгя вазоконстрикторних систем не тшьки продов-
жуеться, а й посилюеться за рахунок ргзко збшьшеного викиду катехоламгнгв.
Вазоконстрикцгя стае надмгрною. Вона генералгзуеться, але нергвномгрно за ш-
тенсившстю г тривалгстю в ргзних органах. Це пов’язано з особливостями регуля-
цп окремих дглянок судинного русла, такими як: наявшсть ргзного типу г кгль-
костг адренорецепторгв, ргзна реактившсть судинно! стшки, особливост! мета-
болгчно! регуляцп. Тому в умовах дефщиту кровопостачання деяю органи стають
Схема 34. Змши гемодинам1ки за рвиих видш шоку
ура длившими 1 швидше ушкоджуються, "приноситься в жертву” (органи травно-
го каналу, нирки, печшка) для годтримання мозкового г вшцевого кровообпу.
Критичний тиск “закриття” руху кров! в кишках, нирках становить 10,1 кПа
(75 мм рт. ст.), у серц! та легенях кровообгг порушуеться за зниження тиску нижче
4,7 кПа (35 мм рт. ст.), у мозку — нижче 4 кПа (30 мм рт. ст.), а за тиску нижче
2,7 кПа (20 мм рт. ст.) не перфузуеться жодна тканина
Паралельно розвиваються порушення мжроциркуляцН (схема 35). Тут також
можна видътиги кьтька етапгв. Спочатку п!д д!ею вазоконстрикторних речовин
(катехоламппв через а-адренорепепгори, вазопресину, анпотензину II, ендо-
тел!н!в, тромбоксангв тошо) розвиваеться спазм судин ммроциркуляторного русла —
артерюл, метартерюл, прекапитяоних ефшктерш. Спазмуються також венули.
Розкриваються артерюловенулярш шунти (найбыьше в легенях, м’язах). кров про-
ходить, оминаючи капьтярн! судини. Змшюються реолопчн! власти вост! кров!,
розвиваеться сладж-синдром. Спостерггаеться 1 центральна веноконстрикщя, яка
зумовлюе шдвишення центрального венозного тиску, збитыпення венозного по-
вернення кров! до серця, шо може маги компенсагорне значения. Тривалий спазм
судин 1 порушення перфузи оргашв спричинюють розвиток гтокси тканин, пору-
шення метаболгзму клгтин, ацидоз. Ацидоз зншае спазм прекатлярних сфшктер!в
Схема 35. Порушення мхкроциркуляцн при рхзних видах шоку
/ закривае сфшктери артериоловенулярных шунтгйв. У мщроциркуляторне русло над-
ходить велика киыасть кров!, проте носткатлярно-венулярн1 сфтктери менш чут-
лив! цо анидозу 1 залишаютъся спазмованими Унаслщок цього в систем! мткроцир-
кулящ! накопичуеться велика юльшсть застшно! кисло! кров!. Г! юльюсть може в
3—4 рази перевишувати ту, яка мктиться там за фтзюлот хчних умов. Таке явите
отримало назву рооИпр.
Водночас пщвищуеться проникшсть судин, р!дина виходить у тканини, що
збшьшуе дефщит ОЦК. Набряк, шо розвиваеться, у свою черту утруднюе поста-
чання кисню. Вщбуваеться згущення кровг, порушуються ц реолопчн! властиво-
стг, унаслщок чого рух кровг сповгльнюеться. Створюються умови для розвитку
ДВЗ-синдрому. зменшуеться тромборезистентшсть судинно! стшки, виникае дис-
баланс зсщально! та протизсщально! систем кровт, вщбуваеться активащя тромбо-
цита. Унаслщок цього ще бптыпе порушуеться кровооби, фактично закупорюеть-
ся мгкроциркуляторне русло, що зумовлюе подальше посилення гшоксп, уражен-
ня оргашв, прогресування шокового стану. Артергалыи судини втрачають
здатшсть тдтримувати свШ тонус, перестають реагувати на вазоконстрикторш
впливи; розширюються г посткапглярш вгдцши судинного русла. Застш кровт вщ-
буваеться здебгльшого в легенях, кишках, нирках, печшщ, ппарт, що г зумовлюе
зрештою ушкодження цих органтв г розвиток Тхньо! недостатносп.
Отже, на ргвш мгкроциркуляторного русла простежуються числешп порочит
кола, якт значно посилюють розлади кровообггу.
Одночасно вщбуваються змши в ммфообггу. Коли розвиваеться блокада мткро-
циркуляторного русла, лгмфатична система посилюе свою дренажну функцпо за
рахунок збшьшення пор у лтмфокапглярних судинах, венулолгмфатичного шунту-
вання. Це суттево збшьшуе лгмфовщпк вщ тканин, г таким чином значка частина
штерстицгйно! рщини, що накопичуеться внаслщок порушення мгкроциркуляцп,
повертаеться в системний кровообгг. Це е доцгльним компенсаторним мехашз-
мом у разг зменшення венозного повернення кров! до серця. На шзнгх етапах
шоку лгмфовщпк ослаблюеться, шо спричинюе штенсивний розвиток набрякгв.
особливо в легенях, печшщ, нирках.
Гемодинамтчнг розлади значною мтрою пов’язаш з порушенням функци серия
(схема 36). Ушкодження серця може бути причиною розвитку шоку (кардюген-
ний шок) або виникае в процес! розвитку шокового стану г поглиблюе розлади
гемодинамгки. В умовах шоку ушкодження серця спричинюються порушеннями
вгнцевого кровообггу, гшоксгею, ацидозом, надлишком вгльних жирних кислот
ендотоксинами мгкрооргантзмтв, реперфузгею, катехоламшами, д!ею цитокпив.
Сироватка хворого в шоковому сташ мае кардюдепресорну дпо, мтстить речо-
вини, що пригшчують дгяльшсть серця. Найбшыпе значения серед них мае ФНП-а.
Його кардгодепресорний ефект може зумовлюватися здатшстю запуска™ апоптоз
клгтин через вщповщн! рецептори, впливом на метаболтзм сфшголшщгв, що спри-
чинюе збшьшення сфгнгозину, здатного прискорювати апоптоз (рани! ефекти), а
також шдукцгею МО8 г утворенням велико! кшькост! МО (пгзш ефекти). Збшь-
шення р!вня МО8 спричинюють також Ш-1, лшополтсахариди. Може мати зна-
чения утворення пероксиштриту за наявностг кисневих радикалтв г МО. ФНП-а
також ушкоджуе мгтохондрп. Кргм ФНП-а кар дю де пресор ш ефекти спричиню-
ють ФАТ, Ш-1, Ш-6, лейкотргени, пептиди, що утворюються в ппемгзовашй пщ-
шлунковгй залозг. Кардюдепресорш фактори можуть порушувати внутргшньоклг-
тинний калыцевий обмш, впливати на поеднання збудження г скорочення; мож-
лива гх безпосередня дгя на скоротливу актившсть. До цього слщ додати, що
лейкотргени справляють дуже сильну вазоконстрикторну дпо на вгнцевг артерп,
спричинюють аритмц, знижують венозне повернення кровг до серця, а СЗа ком-
понент комплементу шдукуе тахгкардпо, поггршуе скоротливу функцпо мюкарда
г також спричинюе вгнцеву вазоконстрикцпо.
Метаболгчш змши та клггиннг ушкодження. Розлади кровообггу пщ час шоку
обов’язково спричинюють порушення метаболтзму клттин, !х структури та функ-
Схема 36. Фактори ушкодження серця при розних видах шоку
цц, що мае загальну назву “шокова клйпина”. На першому еташ для клпини ха-
рактерним е стан пперметаболозму, що розвиваеться внаслщок нервових та ен-
докринних впливов. Швидкость обмшу зростае в 2 рази 1 больше. Органи о тканини
потребують значно большого надходження субстратов о кисню. Вщбуваеться роз-
пад онкогену, посилюеться глюконеогенез. Формуеться онсулшорезистентность. У
м’язових та шших тканинах здшснюеться розпад бшков оз використанням амшо-
кислот як субстратов для глюконеогенезу. Це зумовлюе розвиток м’язово! слаб-
косто, у тому числ! дихальних м’язов. Створюеться негативний азотистий баланс.
Амоной, який утворюеться пщ час розпаду болков, недостатньо знешкоджуеться в
печонцо, що перебувае в умовах шокового стану. Вш своею чертою справляе ток-
сичну дпо на клотини. Порушення мокроциркуляпоо на тло збольшено! потреби в
кисно спричинюють розкий дисбаланс мож потребою о надходженням кисню о по-
живних речовин, накопичення продуктов обмшу. Окром того, деяко цитокони, зок-
рема ФНП-а, ендотоксини мокроорганозмов (лшополосахариди) значною морою
ушкоджують дихально ланцюги, порушуючи окисш процеси, тим самим значно
посилюючи гшоксичне ушкодження тканин.
Тнтегральним показником ступеня порушення енергетичного обмшу тканин
в умовах обмеження ох кровопостачання та гшоксп може бути поступове зболь-
шення вмосту молочно! кислоти до 8 ммоль/л (у нормо < 2,2 ммоль/л), що е не-
сприятливою прогностичною ознакою. Розвиваються виснаження о розлад кло-
тинного обмшу, яко зумовлюють функцоональш змши о структурно ушкодження
тканин, виникнення недостатносто органов (легень, нирок, печшки, органов трав-
ного каналу), що о спричинюе смерть хворого. Варто зауважити, що причинами
клотинно! загибело е не лише метаболочно порушення внаслщок гшоксп, а й уш-
кодження пщ дхею активних кисневих радикалхв, протеаз, лхзосомних факторхв,
цитокппв, токсишв мхкрооргашзмхв та хн.
Роль цитокпйв 1 БАР. Принщшово важливими у виникненш 1 прогресуваннх
патолопчних змш пщ час шоку е видшення й активацхя велико! кшькостх ци-
токшхв та шших БАР. Вони взаемодпоть один з одним, утворюючи цитокшову
сггку, 1 з клхтинами (евдотелхю, моноцитами, макрофагами, нейтрофшьними гра-
нулоцитами, тромбоцитами та ш.). Особливхсть ще! взаемоди полягае в тому, шо
цитокши стимулюють видшення один одного (ФНП-а, ФАТ, штерлейкши тоща
х навхть власне утворення. Формуються самогенеруючх з позитивним зворотним
зв’язком цикли, що спричинюе рхзке збшьшення кшькостх цих речовин.
Тим часом е й шпбхторш впливи, якх обмежують рхвень активацп х цитоток-
сичну дно БАР. У разх вщповщх оргашзму на патогенш дп звичайно! штенсивносп
пщтримуеться баланс мхж цитотоксичними та шпбхторними мехашзмами, конт-
ролюються мхсцевх та загалып прояви запального процесу, що запобхгае ушкод-
женню клхтин ендотелхю та ш. Пщ час розвитку шокового стану вщбуваютьсг
форсування подш, надмхрна продукцхя медхаторхв, яка здшснюеться на тлх кри-
тичного зниження вмхсту шпбхторхв, позитивш зворотш зв’язки стають нерегу-
льованими, реакци — генералхзованими, системними. Кшыасть БАР може збшь-
шуватися в сотнх разхв, х тодх вони хз “захисникхв” перетворюються на “агресорхв”
За рхзних видхв шоку хх активацхя може починатися з рхзних ланок х в рхзний час
але потхм, як правило, вщбуваеться системна активацхя БАР, розвиваеться ССЗВ‘
(див. розды XI “Запалення”, розды XIII “Гарячка”). У разх подальшого розвитку
шокового стану гшоксхя, накопичення продуктхв обмхну, змхни хмунно! системи
токсини мхкрооргашзмхв посилюють цей “медхаторний вибух”.
Найважливхшу роль на початкових етапах “медхаторного вибуху” вщхграють
ФНП-а, ФАТ, 1Л-1, потхм залучаються ппш цитокхни та БАР. Через це ФНП-а
ФАТ, 1Л-1 належать до “раннхх” цитокшхв, 1Л-6, 1Л-8, 1Л-9, 1Л-11 та шшх БАР —
до “пхзнхх”.
ФНП-а визнано центральним медхатором шоку, особливо септичного. Утво-
рюеться ФНП-а головним чином макрофагами пхсля хх стимуляцн, наприклад.
компонентами комплементу СЗа, С5а, ФАТ, при ппемп та реперфузп. Дуже силь-
ними стимуляторами е лпхополхсахариди грамнегативних мхкрооргашзмхв. ФНП-а
мае широкий спектр бюлопчних ефекпв:
• е пхдуктором апоптозу, зв’язуючись зх специфхчними рецепторами на ци-
топлазматичних мембранах х мембранах ендоплазматичного ретикулуму;
• спричинюе депресивний вплив на мхокард;
• пригшчуе внутрхшньоклггинний обмш кальцпо;
• посилюе утворення активних кисневих радикалхв, стимулюючи ксантин-
оксидазу;
• прямо активуе нейтрофшьш гранулоцити, стимулюе видшення ними протеаз.
• впливае на ендотелхальш клхтини: зумовлюе скспресхю адгезивних моле-
кул, стимулюе синтез х викид ендотелхальними клхтинами ФАТ, 1Л-1, 1Л-6, 1Л-8:
стимулюе прокоагулянтш функцп ендотелхю. Може спричхшити ушкодження ци-
тоскелета ендотелхальних клггин х збшьшити проникшсть судин;
'Основними проявами ССЗВ е: температура пла > 38 °С або < 36 °С, частота серцевих скорочень
> 90 за 1 хв, частота дихання > 20 за 1 хв, юльюсть лейкоципв > 12,0-10’/л або< 4,0-10’/л , юльюсг»
молодих форм > 10 %.
• активуе систему комплементу;
• спричинюе розвиток дисбалансу прокоагулянтног та фгбринолгтичног сис-
тем (ослаблюе фгбринолпичну систему й активуе систему зсщання кровг).
ФНП-а може дгяти мгсцево г виходити в циркуляцию. Вш дге як синерпст з
1Л-1, ФАТ. У такому раз! гх вплив р!зко посилюеться навпъ у мгкроюлькостях, як!
самостгйно виражених ефекпв не дають.
У раз! введения ФНП-а тваринам спостерггаються генерализован! ефекти:
системна артер!альна гшотензгя, легенева ппертензгя, метаболгчний ацидоз, п-
перглгкемгя, ггперкалгемгя, лейкопен!я, петехгальнг геморагй в легенях г травному
канал!, гострий тубулярний некроз, дифузна легенева шфшьтрацгя, лейкоцитарна
шфшьтращя.
Важливу роль у цитоюнових взаемод!ях при шоку вщгграе ФАТ. В!н син-
тезуеться г секретуеться р!зними типами юптин (ендотелгем, макрофагами, тка-
нинними базофшами, юптинами кров!) у вщповщь на вплив мед!атор!в! цитокгнгв,
особливо ФНП-а. Ендотелгальнг юптини видгляють ФАТ у вщповщь на тромбш,
пстамш, лшополгсахариди. Це видшення швидке г короткочасне. А внаслщок дц
1Л-1 видшення ФАТ здшснюеться за рахунок стимуляцп синтезу б!лкаг е тривалг-
шим. ФАТ д!е на нейтрофши, моноцити, макрофаги, ендотелгальнг клгтини, тром-
боцита через вщповщш рецептори. Вш спричинюе тага ефекти:
• е сильним стимулятором адгезп та агрегацп тромбоцит! в, сприяе тромбо-
угворенню;
• збшыпуе проникшсть судин, осюльки зумовлюе входження кальцпо в ендо-
телгальн! клпини, гх скорочення, можливе ушкодження клгтин;
• гмовгрно, опосередковуе дгю лшополгсахаридгв на серце; гастроштестиналь-
ш ушкодження;
• спричинюе ураження легень: збшыпуе проникшсть судин, сприяючи на-
эряку, пщвищуе чутливгсть до пстам!ну, збшыпуе утворення Тх 1 простаглан-
лгшгв;
• е сильним хемотаксичним фактором для лейкоцитгв, стимулюе вившьнен-
ня протеаз, супероксиду. Зумовлюе продукцпо супероксиду в нейтрофшах, стиму-
тьованих ФНП-а;
• справляе виражену дгю на макрофаги: навгть у мгкрокшькостях запускае або
збгльшуе утворення 1Л-1, ФНП-а, ейкозанощгв (Тх).
Таким чином, ФАТ опосередковуе частику сво'гх бюлопчних ефекпв через
макрофаги. Своею чертою макрофаги, видшяючи ФАТ, модулюють свою функ-
по. В експеримент! у тварин уведення ФАТ вщтворюе стан шоку. У собак пгсля
цього спостерггаються зниження артергального тиску, зменшення коронарного
кровотоку, зниження скоротливостг мюкарда, змши в судинах (системних, леге-
невих), гемоконцентрацгя, розвиваються метаболгчний ацидоз, ренальна дисфун-
кщя, лейкопенгя, тромбоцитопешя.
Хоча ФНП-а вважають центральним медгатором, шшг цитокши, тага як 1Л-1,
Ы-6, 1Л-8, метаболии арахщоновог кислоти, також вщгграють важливу роль у
розвитку ушкодження органгв при шокових станах. При ргзних типах шоку спо-
стерггаеться збыьшення утворення зазначених штерлейкгнгв.
Хнтерлейкши утворюються головним чином макрофагами шсля гх стимуляцп.
Тншим важливим джерелом циркулюючих цитокгнгв е ендотелгальнг клгтини.
Вони здатнг видшята штерлейгани пгсля стимуляцп ФАТ, ФНП-а. 1х видшення
може бути зумовлено також гшемгею, брадикгншом, компонентами комплемент
(С5а спричинюе продукцпо 1Л-1р, 1Л-6), ендотоксином. Високий р!вень 1Л-6
1000 разгв вищий, юж звичайний) спостерггали у хворих гз септичним шоком
Бюлопчш ефекти 1Л-6 подабш до ефектгв 1Л-1. Вш справляе також виражен;
кардюдепресорну дпо. Велике значения мае утворення Ш-8 (ЯАР-1 — нейтро-
фшактивуючий пептид). Його основна бголопчна функщя — сильна хемотаксич
на Д1я на нейтрофши г Т-л!мфоцити. Високий ргвень у кров! ФНП-а та гнтерлей-
кшгв корелюе з тяжким перебит» м шоку ! смертшстю.
Порушенню мпфоциркуляци й ушкодженню тканин значною м!рою сприя
утворення ейкозаноццв. В умовах шоку генуе багато шлях!в запуску гх синтез)
фосфолшаза А2 активуеться в раз! збшьшення кальцпо в кгптиш, що е можливим
унаслщок ппоксн, при порушенн! клггинних мембран мгкробними токсинами
комплементом. Синтез метаболтв арахщоново! кислоти запускаеться пстамшом
серотоншом, кшгнами, анпотензином II, СЗа, С5а. Лейкотргени, що утворюють-
ся у великгй кшькост!, спричинюють вазоконстрикцпо, особливо посткапшярни'
венул, пщвищення проникносп судин, набряк. Вони зумовлюють хемотаксис
хемокшез г активацпо лейкоципв, макрофапв, тканинних базофшв, гх адгезпо.
дгапедез, дегранулящю. Лейкотргени (особливо В4) посилюють продукцпо актив
них кисневих радикал!в нейтрофшьними гранулоцитами, макрофагами.
Надзвичайно сильне ушкодження спричинюють активн! выьш радикали кис-
ню (див. роздш XI “Запалення”), найстабшьшшим 1 найтоксичнгшим гз яких с
супероксидний анюн. Вони утворюються в дуже великих кшькостях у разг иле’.,
та реперфузп, при “кисневому вибуху” у фагоцитах. Одночасно в раз! розвитк
шоку знижуеться ргвень антиоксиданпв. Активн! киснев! радикали спричинюют.
деполяризацгю й ушкодження мембран, збшьшення проникносп, розвиток на
бряку, дисбаланс кальцпо. Порушують утворення АТФ, ушкоджують ДНК. Впли-
вають на вироблення ендотелгем ЫО г сприяють його руйнуванню, тим сами},
дпочи на судинний тонус. Киснев! радикали можуть пщукувати утворення ФНП-1
стимулюють вироблення ендотелгем ФАТ, сприяють адгезп лейкоцит!в до ендо-
телпо судинно! спнки.
В умовах шоку активуються вс! плазмов! протеолпичн! системи. Велике зна-
чения мае активац!я системи комплементу. При шоку вона можлива як альтерна-
тивним, так ! класичним шляхом, може запускатися шшими протеазами, м!кро-
оргашзмами або гхнгми токсинами при септичному шоку, комплексами антиген —
антитшо в раз! ушкодження тканин, гшоперфузп та пиеми. Утворюютьсв
бюлопчно активн! анафшотоксини СЗа, С5а 1 термшальний комплекс С5Ь-
Анафшотоксини, особливо С5а, значною м!рою впливають на лейкоцити: справ-
ляють сильну хемотаксичну дпо, сприяють адгези лейкоципв до ендотелпо, л
агрегацп. Вони зумовлюють активацпо лейкоципв гз вившьненням лгзосомальни
ферменпв, кисневих радикал!в, еластази, сприяють експресц рецептор!в для ком-
плементу СК-1 та СК-3. Така активация лейкоцитгв значно ушкоджуе судини п
прилепи тканини. Анафшотоксини спричинюють дегранулящю тканинних базо-
фшв 1 посилюють дпо пстамшу, сприяють утворенню метаболтв арахщоновс
кислоти. Кргм того, вони прямо впливають на судини: пщвищують проникшсть:
С9 стимулюе видшення ендотелгем 1Л-8, сильного хемоатракганта для нейтро
фшгв, моноципв, експреспо адгезивних молекул (Р- г Е-селектингв), секреци
фактора Вшлебранда. Анафшотоксини спричинюють скорочення непосмугова-
них м’язхв бронхового дерева х травного каналу. СЗа хндукуе коронарну вазокон-
стрикцию, таххкардпо, ослабления функцц серця.
Таким чином, БАР дпоть на рхзнх клпини: макрофаги, ендотелш, нейтрофшь-
н! гранулоцити та ппш клпини кровь Для розвитку шокового стану найважливх-
шою е дхя цих речовин на ендотелш судин х лейкоцити. Крхм того що ендо-
телхальн! клхтини сам! е джерелом видшення низки цитокшхв (1Л-1, 1Л-6, 1Л-8,
ФАТ), вони виступають мпненню для дп цих самих речовин. Вщбуваються акти-
вацхя контрактильних елемента ендотел!альних клпин, порушення цитоскелета,
ушкодження ендотелпо, що спричинюе рхзке пщвищення проникностх судин.
Водночас стимулюеться експресхя молекул адгезп, якх забезпечують фхксацпо лей-
коцитхв на судиннш стппц. Накопиченню нейтрофшьних гранулоцита сприяе
також велика кшькхсть речовин хз позитивною хемотаксичною дхею — компонен-
ти комплементу СЗа х особливо С5а, 1Л-8, ФАТ, лейкотрхени. Лейкоцити вщхгра-
ють надзвичайно важливу роль в ушкоджених судин х тканин при шоку. Активо-
ванх цитокшами нейтрофшьш гранулоцити видшяють лхзосомнх ферменти, велику
кхлькхсть протеолпичних фермехпхв, серед яких провщну роль вщхграе. еластаза.
Одночасно посилюеться актившсть лейкоцитхв щодо генерацн та вившьнення ак-
тивних кисневих радикалхв. Вщбуваються масивне ураження ендотелхю, рхзке
пщвищення проникностх судин, що сприяе розвитку зазначених порушень мпсро-
циркуляцп. Ш сам! речовини ушкоджують не лише судини, а й клпини паренхх-
матозних органхв, посилюють ураження, спричинеш гшоксхею, сприяючи розвит-
ку хх недостатност!. Ушкодження, особливо судин, спричинюють також компо-
ненти комплементу, ФНП-а, ФАТ тощо. Цитокхни мають значения х для
розвитку ДВЗ-синдрому в разх шокового стану. За ’хх впливу зменшуеться тромбо-
резистентшсть судинно! стшки, стимулюються прокоагулянтах функцп евдотелхю,
що сприяе тромбоутворенню. ФАТ, ФНП-а активують тромбоцита, спричиню-
ють ’хх адгезхю, агрегацхю. Розвиваеться також дисбаланс мхж актившстю системи
зсщання кровх, з одного боку, та протизсщально! х фхбринолпично! систем, з ш-
шого.
Недостатшсть оргашв х систем. Описанх впливи (ппоксхя, ацидоз, вплив ак-
тивних кисневих радикалхв, протешаз, цитокшхв, БАР) зумовлюють масивш уш-
кодження клхтин. Розвиваються дисфункцхя х недостатшсть одного, двох ! бшьше
органхв х систем. Такий стан отримав назву синдрому полюрганно'1 недостатност!
(ПОН), або синдрому мультиорганно! дисфункци (МОД; гпиШр1е ог^ап бузйхпсбоп
зупбготе, МОО8). Стушнь функцхонально! недостатност! органхв залежить вщ
тривалостх та тяжкост! шоку. Пщ час розвитку шоку в людини насамперед ушкод-
жуються леген!, потхм розвиваються енцефалопатхя, ниркова та печшкова недо-
статшсть, ушкодження травного каналу. Можливе переважання проявхв недостат-
ност! того або шшого органа. Унаслщок дисфункци печшки, нирок, кишок вини-
кають нов! патогеннх фактори: шфекцхя з травного каналу, висок! концентраци
токсичних продукта нормального ! патолопчного обмхну. Показник смертностх
таких хворих дуже високий: у разх недостатност! за однхею системою — 25—40 %,
за двома — 55—60 %, за трьома — понад 80 % (75—98 %), а якщо розвиваеться
дисфункцхя чотирьох х бшьше систем, смертшсть наближаеться до 100 %.
Одним хз органхв, як! найпершими уражуються в умовах шоку в людини, е
легеш. Ушкодження може розвинутися за кшька годин або дхб вщ початку розвит-
ку шоку як гостра легенева недостатшсть, яка отримала назву гострого рестра-
торного дистрес-синдрому дорослих (ГРДСД; АсиСе гезрйаФгу сНыгевз зупйготе,
АКВ8); також застосовують терлин ‘‘шоков! пегеш”. Рання стащя ГРДСД, яка
вир!зняеться меншим ступснем ггпоксемп, мае назву синдрому гострого легеневого
ушкодження (СГЛУ). Провзднилги факторами розвитку легенево! недостатност! е
р1зке пщвищення проникност! альвеолокаппгярно! мембрани, ушкодження ендо-
тел!ю судин, легенево! парен хгми. ше зумовлюе вихщ рщини за меж! судинно!
стгнки 1 розвиток набряку легень.
Ргзке пщвищення проникност! стшки судин спричинюють БАР. як! у велик!й
юлькосп потрапляють у леген! з кров] або утворюються м!сцево з р!зних кл!тин:
легеневих макрофапв, нейтрофшышх гранулоципв, клггин ендотелпо судин, еш-
телпо нижшх дихальних шляхгв. Ц! речовини недостатньо там гнактивуються, ос-
кальки в умовах шоку дуже рано порушуються нереспграторш функцп легень
Велике значения л(ае активац!я комплементу, кшшовох системи.
У легенях секвеструеться значна кшьюсть лейкоципв, спостерггаеться лейко-
ишпарна шфиьтращя. Скупченню лейкоципв сприяе велика юльюсть хемоатрак-
танпв у легенях — компонента комплементу, лейкотр!ен!в, ФАТ, 1Л-8 (видь
ляеться з легеневих макрофапв г альвеолой ипв II типу). Лейкоцита додатково
активуються ФНП-а, ФАТ, лшопол!сахаридами. 3 них видыяються протеази, ак-
тивн! киснев! ради кал и, як! ушкоджують спинку судин. Вщбуваються також вихщ
лейкоципв за меж! судинно! стшки ! ушкодження легенево! тканини. Руйнуються
колатен, еластан, фибронектин. Багатий на б!лки ! фгбрин ексудат виходить в ш-
терстицгйний проспр г альвеоли, вщбуваеться позасудинне вщкладання фибрину,
шо надал! може спричинити розвиток фгброзу.
Ушкодження посилюються внаслщок порушення кровообггу, мгкротромбгв.
що утворюються внаслщок розвитку ДВЗ-синдрому. До пього призводить пору-
шення гемостазу в легенях — збщьшення прокоагулянтно! га зниження ф!бри-
нолгтично! акгивносп органа. Збшьшуеться продукция ! зменшуеться руйнування
ендотелшу в легенях, шо сприяе розвитку бронхоконстрикци. Знижуеться розтяж-
шсть легень. Зменшення утворення сурфактанту зумовлюе спадання альвеол 1
розвиток множинних ателектазов Вщбуваеться шунтування — скидання кров!
справа нал!во, шо викликае подальше попршення газообмшно! функцГ! легень
(венгилящйно-перфузшного сп!вв!дношення). Ушкодженню може сприяти та-
кож реперфузгя, що виникае на тл! .пкування. Усе це спричинюе тяжку прогре-
суючу гтоксемою, яку складно нормалгзувати навгть гз використанням гшероксич-
них газових сумгшей. Збшьшуються енергетичш витрати на дихання, дихалью
м’язи починають споживата близько 15 % ХОК. Найважливгшими показниками,
що св!дчать про розвиток легенево! недостатностг, е парщальний тиск кисню в
артер1альнш кров! < 71 мм рт. ст., зниження ресгпраторного гндексу РаО. / РЮ
< 200 мм рт. ст., при СГЛУ — < 300 мм рт. ст. На рентгенограм! визначають
двоб!чн! шфьтьтрати в легенях, таек заклинювання кашляр!в легенево! артерп
(ТЗКЛА) < 18 мм рт. ст.
У раз! розвитку ГРДСД стан хворих значно попршуеться. Смертшсть за не-
сприятдивого перебггу може сягати 90 %
Значну роль у розвитку критичних сташв вциграс ушкодження кишок. Слизо-
ва оболонка кишок постшно оновлюеться, мае високу метабол!чну актившсть,
тому дуже чутлива до гшоксп. Унаслщок порушення мгкроциркуляпп та дп шших
факторгв гинуть клттини кишок, порушуеться цшсть слизово’]' оболонки, розвива-
ються ерозивш змши. Спостерггаеться кровоточа, мгкрооргашзми т токсини з ки-
шок потрапляють у мезентер1альш лтмфатичш судини, ворттну систему та загаль-
ний кровооб1г. Виникае ендогенна токсем!я. шо може зумовлювати розвиток
нирково! та печпгково! недостатносп в шзнш перюд шоку. Перебтг шоку усклад-
нюеться розвитком септичного стану.
Прояви ураження печшки розвиваються зазвичай за кшька дтб вщ початку ос-
новного захворювання Це можуть бути енцефалопаття, жовтяниця, коагулопапя
та ДВЗ-синдром. Кргм того, при печшковш недостатносп порушуеться клтренс
циркулюючих ЦИТОК1И1В, шо сприяе тривалш пщтримш тх високого ргвня в кровг.
Великого значения набувае порушення дезштоксикацтйно! функци, особливо на
тл1 надходження значно! кглькосп токсичних речовин 1з кишок 1 метаболтв. Пщ
час шоку порушуеться синтез бйгкгв у печгнцг. Особливо проявляеться дефщит
синтезу короткоживучих бгткгв. таких як фак гори зсщання кровг, що призводить
до виснаження системи коагулянт! та переходу ДВЗ-синдрому в стадно гшокоагу-
ляцп. На метаболтзм ештелюципв печшки суттево вплггвають ФНП-а, 1Л-1, 1Л-6.
Зменшення ОЦК, зниження артергального тиску. граничний стушнь спазму
приносних артерюл спричинюють зменшення ШКФ г розвиток ГНН. Пщ час
тяжкого шоку перфузтя нирок ослаблюеться 1 нерщко припиняегься. Розвивають-
ся одно- та ануртя, у кровт збгльшуеться вмгст креатишну, сечовини, наростае
азотемия. Тшемы. шо тривае понад 1,5 год, зумовлюе ушкодження нирково! тка-
нини, розвиваеться гломерулярна, а поттм губулярна недостатшсть, пов’язана з
некрозом канальцевою ештелпо. У такому разт ниркова недостатшсть може зберт-
гатися шсля виведення хворого тз шокового стану.
Про наявнгсть полторганно! дисфункцн та недостатносп евщчать певш клшг-
ко-лабораторнт показники: при печ!нков!й недостатност! — ргвень бшрубтну кровт
понад 34 мкмоль/л, спостерггаеться збшьшення вмтсту АсАТ, нужно! фосфатази в
2 рази 1 бшьше вщ верхньо! межт норми; при нирковш недостатност! — ргвень кре-
атишну кров! перевищуе 176 мкмоль/л, дтурез знижуеться нижче 30 мл/год; у раз!
дисфункци е систем! гемостазу — збшьшення продукттв деградаш! фтбрину /фиб-
риногену та О-димеру, протромбшовий шдекс < 70 %, ртвень тромбоципв
< 150.0* 109/л, фтбриногену < 2 г/л; при дисфункш! центрально! нервово! ситеми —
менше 15 баш в за шкалою Глазго.
Особливост) розвитку Р13НИХ ВИД1В шоку
Гшоволемтчний шок. Первинний гшоволемтчний шок розвиваеться внаслщок
втрати рщини та зменшення ОЦК. Це може бути в разг:
— втрати кровг пш час кровотеч як зовшшюх, так г внутрштшх (такий вид
шоку мае назву гемораггчногоу,
— втрати плазми пщ час огокгв, ураження тканин тошо;
— втрати рщини пщ час профузних проноств, невтримного блювання, унас-
лщок пол гур п при цукровому або нецукровому дгабеп.
Гшоволемтчний шок починае розвиватися, коли об’ем внутрштньосудиннот
рщини зменшуеться на 15—20 % (1 л на 70 кг маси тша). У людей молодого в1ку
класичю прояви гшоволемгчного шоку з’являються в разг втрати 30 % ОЦК. Якщо
втрата становить 20—40 % ОЦК (1—2 л на 70 кг маси тша), розвиваеться шок
Г ‘ рп8
середнього ступени тяжкости, понад 40 % ОЦК (быьше юж 2 л на 70 кг маси тша) —
тяжкий шок. Розвиток шокового стану залежить не лише вщ того, наскйльки
зменшився ОЦК, а й вщ швидкосп втрати рщини. Саме йнтенсивнйсть, швидюсть
й тривалисть кровотечй перетворюють кровотечу на геморапчний шок.
У вщповщь на зменшення ОЦК виникае стандартний набйр компенсаторних
реакщй. Вщбуваеться перемищення рщини йз позасудинного простору в судини,
тому втрата ОЦК супроводжуеться дефицитом позаклитинно! рщини, ривноцйнним
дефщиту плазми. Спостерйгаються затримка води нирками, вихйд крови з депо.
Розвиваються спазм судин мйкроциркуляторного русла, централизация кровообйгу.
Зменшення венозного повернення кров! до серця зменшуе серцевий викид, рано
виникае недостатшсть центрально! гемодинамики. До головних гемодинамйчних
показникив, що характеризують гйповолемйчний шок, належать: низький ТЗКЛА,
низький серцевий викид, високий загальний периферичний судинний ошр. На-
дал! розвиток шокового стану продовжуеться за загальними закономйрностями.
Тривала централизация кровообйгу спричинюе ушкодження органйв й розвиток
МО ОБ. Пйд час лйкування гйповолемйчного шоку потрйбно швидко вйдновити де-
фицит ОЦК та усунути вазоконстрикщю.
Кардйогенний шок. Кардйогенним називають шок, причиною розвитку якого е
гостра недостатшсть серця з рйзким зменшенням серцевого викиду, що можуть
спричинювати:
• зниження скоротливо! здатноспй серця пйд час инфаркту мйокарда, тяжкого
мйокардиту, кардйомйопатй!, ускладнень тромболйтичнои терапй! з розвитком репер-
фузййного синдрому;
• тяжк! порушення ритму серця',
• зменшення венозного повернення кров1 до серця',
• порушення внутриинъосерцево'1 гемодинамиси, що спостерйгаються в разй тяж-
ких вад й розривйв клапанйв, сосочкових м’язйв, мйжшлуночковои перегородки, ку-
леподйбного тромбу передсердя, пухлин серця1;
• тампонада серця, масивна тромбоембол1я легенево! артери або напружений
пневмоторакс. Такий вид шоку називають обструктивным. Вйн розвиваеться внаслй-
док порушення наповнення серця або вигнання кровй из серця. У разй тампонады
серця механйчна перешкода розширенню його камер пйд час дйастоли порушуе их
наповнення, рйзко знижуеться також венозне повернення крови до серця.
Тромбоембол1я легеневих артерш зумовлюе обмеження припливу кровй до лйвих
вйддйлйв серця, що е наслидком комбйнацй! механйчного фактора пид час закупорю-
вання великим тромбоемболом й спазму легеневих судин у разй емболйи численни-
ми невеликими тромбоемболами. За напруженого пневмотораксу зростання тиску
в плевралыпй порожнинй спричинюе змищення середостйння та перегин порож-
нистих вен на ривнй правого передсердя, що блокуе венозне повернення кровй до
серця.
Найпоширенйшою причиною розвитку кардйогенного шоку е шфаркт мюкар-
да, який у 5—15 % хворих ускладнюеться шоком. Розрйзняють окремй клйнйчнй
варйанти кардйогенного шоку при инфарктах — рефлекторний, аритмйчний,
справжнйй кардйогенний. У розвитку рефлекторного кардюгенного шоку великого
значения набувають реакции на рйзкий бйль, рефлекторнй впливи (рефлекс Бецоль-
'Найчастнце це миксома передсердя, яка виявляеться здебыьшого в ливому передсерд! з можли-
вим закупорюванням передсердно-шлуночкового (митрального) отвору.
да — Яриша) з вогнища некрозу на серце 1 на хону с судин хз депонуванххям кров1
в мхкроииркуляторному русл). Унаслщок патолопчних рефлекторних впливш,
особливо при шфарктх мюкарда задньох стшки, можуть розвиватися брадикардия,
значно знижуватися артерхальний тиск.
Аритшчний кардюгенний шок пов'язаний хз приеднанням тяжких порушень
ритму серця, що значно знижують серневий викид. Найчастппе це пароксизмаль-
на шлуночкова таххкардхя хз дуже високою частотою скорочення шлуночюв. трх-
попння передсердь або значна брадикардхя, наприклад, пщ час повно! атрювент-
рикулярно! блокади.
Справжнш кардюгенним шоком називають шок, що розвиваеться внаслщок
рхзкого зменшення скоротливо)' здатност! мюкарда. Це частице виникае в раз!
шфаркпв. як! перевишують 40—50 % маси лхвого шлуночка, трансмуральних,
передньобхчних ! повторних на тлх попередньо зменшено!' скоротливосп мюкар-
ца, артергально!' ппертснзп, цукрового дхабету. в оехб вхком понад 60 роюв.
Початковою ланкою патогенезу кардюгенного шоку е ргзке зменшення серцевого
викиду, зниження артергалъного тиску (систолхчний артергальнил тиск < 90 мм рт. ст.,
середнш артерхальний тиск < 60 мм рт ст. (7,9 кПа) або зниження бьльше нхж на
30 мм рт. ст.). При ньому збхлыцусться тиск наповнення шлуночюв серия !, вщ-
повщно, ТЗКЛА становить > 20 мм рт. ст., серцевий шдекс < 1,8—2 л/хв • м2.
Долучаюхься компенсаторн! реакцп. спрямован! на нормалхзашю артерхального
тиску: активашя симпатико-адреналово!' системи, РААС тошо. Р1зко збшьшуеться
периферичний судинний отр, шо створюе додаткове навантаження на серце х
водночас попршус перфузхю тканин. Катехоламши безпосередньо впливають на
серце — виявляеться ’ххня шо- 1 хронотропна дхя, що збшьшуе потребу серця в
киснх, а одночасне зниження тиску в аорт! перешкоджае надходженню лотрхбно'1
юлькосп кров! в коронарнх судини. Це посилюе недостатнхсть кровозабезпечення
мюкарда. До попршення метаболгзму серця призводить х таххкардхя. В хшемхзова-
ному мюкардх активуеться утворення метаболхпв арахщоново! кислоти, особливо
лейкотрхенхв, продукт пероксидного окиснення, видхляються лейкопитарш фак-
тори. Усе це додатково ушкоджус серце. У такий споФб виникають порочнх кола
Ураження серххд ! тяжкхсгь стану хворого нароста ють. Приеднання пооушень ле-
геневого кровообггу, розвитку набряку легень спричинюе тяжку артерхальну п-
поксем!ю. Надатх шоковий стан розвиваеться за загальними закономхрностями.
Смертнхсть у раз! кардюгенного шоку становить 50—80 %, а за деяких його видхв
сягае 100 %.
Септичний шок ускладнюе перебхг рхзних шфекцшних хвороб, спричинених
переважно грамвегативними бактерхями, Проте почаспшали випадки септичних
станхв х в разх грампозитивнох х грибково! шфекцн.
Розвиток шокового стану пщ час грамнегативного сепсису головним чином
пов’язаний хз дгею ендотоксину, який вившьнюеться в раз! подшу або руйнування
м1кроорганхзм!Б. у тому числ! х при застосуванш антибактерхально! терапх!'. Ендо-
токсин е лхлогюлхсахаридом, який мае здатшеть самостхйно або в комплекс! з
лхпополхеахаридзв’язувальним бшком кровх1 (ЕВР) зв’язуватися хз репепторним
'Лпгополгсахаридзв’язувальний биток плазми крова — бгток гостро! фази запалення, що пооду-
куегься гепатоцитами й ентероцитами 1 мае високу спорщненХсть (афшнХсть) до лХпополХсахаршпв.
забезпечуе зв’язування його 1з СЭМ
комплексом, шо складаеться з СБИ, МД2 1 ТЕВ.-4-рецептораии1 на моноци-
тах / макрофагах та шших клпинах — ендотелпо, тромбоцитах. Окргм того, деяк!
бактер!альн! молекули розшзнаюзься питоплазматичними рецепторами МОБ-1 та
МОО-2. Надал! запускаеться внутрпшгьоклггинний каскад з активашею фактора
транскрипцп МРкВ, наслщком чого е синтез ФНП-а. Тндукуеться також видшен-
ня шших ЦИТОК1Н1В, прозапатьних БАР. стимулюеться утворення молекул адгезц,
шдуковано] ИО5 тощо. Експрес!я ТЕК I, отже, реакцш оргашзму на ендотоксин
значно посилюеться одним 1з цитокштв — фактором, що шпбуе мгграшю макро-
фапв (Ф1ММ), який у великш юлькосп виявляеться у хворих !з септичним шо-
ком. В)н вившьнюеться ендотелгем та шшими клггинами пщ дтею мгкрооргашзмФ
1 прозапальних цитокипв. Лшополгсахарид активуе також плазмов! протеолтичш
системи.
На початку розвитку шфекцшного процесу БАР утворюються у вогнигш ш-
фекцшного запалення. У раз) надмхрно! вщповщ!, недостатносп мюцевих захис-
них мехашзмгв 1 нестабшьносп бар'ера можлив, тх надходження в кров, некон-
грольоване поширення медтатортв т генералнацтя процесу тз розвитком ССЗВ.
При цьому бактертемтя може бути короткочасною або взагал! вщсутньою. Цт ре-
човини справляють системний вплив насамперед на мткротшркуляторне русло, а
також пот ужну пряму ушколжувальну дно на тканини. Тому гемодинамтчнт змши
при септичному шоку починаються з порушень мткроциркулящ! з подальшим
приеднанням змш центрально! гемодинам тки.
Септичний шок е найбшьш “клтинним "типом шоку, при якому ураження
тканин виникають дуже рано т стушнь 1х набагато вищий, нгж можна очткува-
ти внаслшок лише гемодинамтчних змш. Ендотоксин (лшопол!сахарид) зумо-
влюе швидку шактиващю цитохрому а, а3 (нитохромоксидази). ФНП-а також
дуже ушкоджуе дихальнт ланцюги, що порушуе мпохондртальне окисне фос-
форилювання, незалежно втд вмтсту оксигемоглобшу або штенсивносп крово-
току в органах. Через дисфункцию на клттинному р!вн! поглинання кисню з
кровт попршуеться, що проявляеться зменшенням артертовенознот розниц! за
киснем.
Найважливтшими серед цитоктшв у разт септичного шоку вважають ФНП-а,
ФАТ. Можливо, шо саме ФНП-а вщтграе провщну роль у тих випадках шоку, якт
закшчуються смертю, осктльки разом тз лшополт сахар илом вони справляють дуже
сильну дпо, значно посилюють ефекти один одного, навпъ у низьких дозах. Саме
тому пщ час розвитку септичного шоку спостерггаеться дуже ранне ушкодження
ендотелтю судин тз ртзким щдвишенням проникностт, виходом бтлка т велико!
кшькост! рщтши в штерстищйний просттр т зменшенням ЕОЦК. Тому такий шок
отримав назву дистрибутивного, або перерозподыьного. Ушкодження судин т тка-
нин спричинюють також актавован! лейкоцита. Ще одшею особливтстю септич-
ного шоку е ранне т стшке розширення судин мткроциркуляторного русла, що
'Родина ТЬК опосередковуе реакцп ьроджечого тмуштету. що е “першою лтнтею” захисту вщ
шфекшйних агентов. Вони реагують на илька консервативних структур мткрооргантзмтв (патогенасо-
циюваш молекулярш образи), для яких харакгерним е те, шо вони власгивт лише тнфекщйним фак-
торам 1 не утворюються юптинами оргашзму, тобто тх поява сигналтзуе про виникнення шфекцп.
Кртм того, вони е найбшьш загальними, характерними для низки груп мткрооргантзмтв, наприклад
лшополтсахариди характерш для вс о грамнегативних шфекцш, лшотейхоева кислота — д ля встх грам •
позитивнпх. Активашя ТЁК через факт ор транскрипцп ТхР-кВ стимулюс синтез бататьох цитоктшв.
Ж
разом хз секвестрах, хею 1 виходом рщини в тканини зумовлюе значне зниження
артериального тиску, яке не пщдаеться корекцп
Мехашзмхв рхзко! вазодилатацй е кшька. Так, лшополхсахариди, цитокхни (особ-
ливо ФНП-а), ендотелш-1 стимулюють утворення макрофагами, ендотелхальними
1 непосмугованими м’язовими юптинами 1МО8. яка продукуе дуже велику кхлькхсть
МОБ, унаслщок чого знижуеться тонус 1 резистивних судин, ! венул. У процес! екс-
периментального модел юванпя септичного шоку спостерхтають дв1 фази зниження
тиску у вщповщь на дно евдотоксину — фазу негайного зниження, пов’язану з ак-
тиващею конститутивно! МОЗ, 1 пхзнхххху фазу, спричинену утворенням 1МО8. Крш
вазодилататорно! дц МО, реагуючи з великою кшькхстю вшьних кисневих ради-
калов, утворюе високотоксичнии пероксинприт (ОМ'ОО), який ушкоджуе клхтинш
мембрани, ДНК ендотелпо 1 юптин прилеглих тканин. Ослаблению судиниого то-
нусу сприяе також вшкритгя АТФ-залежних калхевих канашв1, вихш унаслщок цьо-
го К4 з юптин. Спостерхгаютъ зменшення вмхсту вазопресину (виснаження його
запасов у ппофхз! внаслщок попереднъого надмхрного викиду) Вщбуваються шакти-
ващя катехолампйв сунероксидними радикалами, що утворюються у велиюй кшь-
кост1, зменшення реакцп судин на катехолам!ни. Як наслщок, ослаблюеться скоро-
тлива здатшсть непосмугованих м’язов судин, знижуеться тонус х розвиваеться реф-
рактерна вазодилатафя. Судини втрачають чутливхсть до ди судинозвужувальних
впливхв. Порушення мпсроциркуляш! набувають неоднорщного характеру — поед-
нуються зони вазодилатаци та вазокочетрикци. Характерним е також вщкриття ар-
тер! олове нулярних шунтхв.
Септичний шок у разх трам позитивно! шфекцп пов’язаний хз прямою дхею як
токсинхв, так х БАР, що утворюються. Токсини грампозитивних мжроорганхзмхв
(лхпотейхоева кислота, пептидоглгкани, флагелш тощо) також зв’язуються з в!д-
повщними ТЬК (ТЬК-2, ТЪК-5, ТЬК-6, ТЬК-9), що зумовлюе видшення ни-
токхшв. Токсини, що мають властивостх суперанта гешв (токсин синдрому ток-
сичного шоку, стафшококовий ентеротоксин, стрептококовий шрогенний екзо-
гоксин), спричинюють неспецифхчну активацпо велико! кшькостх лхмфоципв
також хз видшенням БАР
На початкових етапах розвитку септичного шоку пщ дхею катехоламппв збхль-
шуеться ЧСС та УОС. Проте надалх вщбуваеться ушкодження мюкарда кардю-
депресорними факторами (ФНП-а, ФАТ, лейкотрхени тощо), дхя яких значно
посилюеться лхпополхеахаридами. Приеднуеться серцева недостатшсть, що знач-
но поглиблюе порушення гемоди намхки.
Оскшьки пщ час септичного шоку спостерхгаеться значне ураження тканин,
рано розвиваеться недостатшсть рхзних оргашв, насамперед легень 1 нирок. Особ-
ливхстю розвитку ГРДСД в умовах септичного шоку е те, що до його патогенезу
долучасться дхя лшополхеахаридхв, якх стимулюють видшення та посилюють ефек-
ти цитоюшв х лейкоцитхв. Це спричинюе швидке й хнтенсивне ушкодження ен-
дотелпо, набряк легень х розвиток гостро! легеневох недостатност!.
Нирки уххгкоджуються ендотоксином 1 внаслщок порушення хх кровопоста
чання. Розвхттку ппеми нирки додатково сприяе стимуляпхя ендотоксином видх-
лення ренхну з подалыпим утворенням анпотензину II х спазмом ниркових судин.
Розвиваеться гострий канальцевий некроз.
‘Канали для; проходження юшв калхю; вщтр.'шньоклггинна АТФ 1х блокуе, а в раз! зменшення
рХвня АТФ канали вщкриваклься.
Для септичного шоку характерним е раннгй розвиток ДВЗ-синдрому. Ушко-
джуеться також ЦНС аж до розвитку коматозного стану.
Головнг гемодинамгчш характеристики септичного шоку такт: низький ТЗКЛА,
високий серцевий викид, низький загальний периферичний опгр судин.
Септичний шок е одним гз найтяжчих вид! в шоку. Смертшсть унаслщок сеп-
тичного шоку дотепер залишаеться високою — 40—60 %, а в раз! шоку внаслщок
абдоминального сепсису може сягати 100 %. Септичний шок — найчастпиа при-
чина смертг у вщдгленнях загально! реашмаци.
Анафьгактичний шок. Цей вид шоку, як г септичний, також належить до су-
динних форм шоку. До його розвитку може призвести алерггчна реакцгя ан-
афглактичного типу в раз! Г! генералгзацп. При цьому вщбуваеться поширення
БАР, як! видшяються з тканинних базофглгв, а саме: ггстамгну, гепарину, лей-
котргешв, ФАТ, протеолгтичних ферментгв, факторгв хемотаксису, а також шших
БАР, цитокгнгв. Значно знижуеться судинний тонус, розширюються судини
мгкроциркуляторного русла, пщвищуеться гх проникшсть. Кров накопичуеться в
мгкроциркуляторному русл!, рщина виходить за меж! судин, зменшуються ЕЦОК,
венозне повернення кров! до серця. Робота серця попршуеться також унаслщок
порушення вшцевого кровообггу, розвитку тяжких аритмгй. Так, лейкотргени
(С4, 04) г пстамш спричинюють коронароспазм. Пстамш (через Н,-рецептори)
пригнгчуе роботу пазухо-передсердного вузла, зумовлюе (через Н2-рецептори)
гншг види аритмгй аж до розвитку фгбриляцгг шлуночкгв. Унаслщок зменшення
ЕЦОК г порушення роботи серця знижуеться артерхальний тиск, порушуеться
перфузгя тканин. Дгя ггстамгну, лейкотргешв на непосмугованг м’язи бронхгалъно-
го дерева спричинюе спазм бронхгол г розвиток обструктивног недостатностг ди-
хання. Це значно посилюе ппоксгю, зумовлену розладами гемодинамгки. Окргм
типового перебггу анафглакгичного шоку можливг деякг шшг його клгшчнг варган-
ти. Так, може спостерггатися гемодинамгчний ргзновид, при якому на перший план
виступають порушення гемодинамгки з ураженням серця, аритмгями аж до асис-
толп, розвитком гостро! серцево! недостатностг. Наявшсть у людини хрошчних
захворювань дихально! системи може сприяти розвитку асфгктичного варганта
анафглакгичного шоку, у клгнгчшй картинг якого домгнуе гостра недостатшсть
зовнгшнього дихання, зумовлена набряком дихальних шляхгв, бронхоспазмом,
набряком легень. Особливгстю анафглакгичного шоку е можливгсть його стргмко-
го, блискавичного розвитку, коли смерть хворого може настати впродовж кглькох
хвилин. Тому медичну допомогу потргбно надати негайно за появи перших ознак
шокового стану. Це мають бути швидке масивне введения рщини, катехоламгнгв,
антипстамгнних препаратгв та гншг протишоковг заходи, спрямованг на вщнов-
лення роботи дихально! 1 серцево-судинног систем.
Опгковий шок розвиваеться як наслщок великих термгчних уражень шкгри та
пщлеглих тканин. Перпп реакци органгзму на ошк пов’язанг з дуже сильним бо-
льовим синдромом г психоемоцгйним стресом, що е пусковим механгзмом для
ргзко! активацп симпатико-адреналово! системи зг спазмом судин, тахгкардгею,
збгльшення м УОС г ХОС, можливим пщвищенням артергального тиску. Надалг
розвиваеться стандартна нейроендокринна вщповщь. Одночасно на великих пло-
щах, ушкоджених пщ час ошку тканин, починаеться запалення з видгленням усгх
його медгаторгв. Ргзко зростае проникшсть судин, бшок г рщка частини кровг ви-
ходить 13 судинного русла в мскклггинний простхр (у раз! огпкгв з ураженням
бшьше Н1Ж 30 % поверхш тша — 4 мл/кгтод); рщина втрачаеться також через
обпалену поверхню назови!. Це спричинюе значне зменшення ОЦК, шок стае
г1поволем1чним. Ппопроте'шемхя, що виникае внаслщок втрати б!лк!в, посилюе
розвиток набряку в неуражених от ком тканинах (особливо в раз! огню в з уражен-
ням понад 30 % поверхн! тша). Це своею чертою поглиблюе гшоволем!ю. Змен-
шуеться серцевий викид, значно пщвищуеться затальний периферичний ошр су-
дин, знижуеться центральний венозний тиск, таким чином посилюються гемоди-
намхчн! розлади. Мед!атори потрапляють у затальний кровообп, вщбуваються
генерализована активацхя БАР I розвиток ССЗВ. Унаслщок руйнування тканин,
розпаду бшгав утворюеться велика кшьюсть токсишв, яю також надходять у сис-
темний кровооб!г 1 спричинюють додаткове ушкодження тканин. Подальший пе-
ребп шоку вщбуваеться за затальними закономхрностями. Можливе приеднання
шфекцп з розвитком сепсису, шо значно попршуе стан хворого.
Травматичний шок виникае як наслщок тяжких мехашчних ушкоджень — пе-
реломов юсток, розчавлювання масиву тканин, травмування внутршппх оргашв,
значних поранень. Шоковий стан може розвиватися одразу шсля травми або че-
рез галька годин теля не!. Спричинюють розвиток шоку, як правило, сильна
больова реакшя, рхзке подразнення 1 нав!ть ушкодження екстеро-, штеро- та про-
пр!орецептор!в 1 порушення функцй ЦНС.
У розвитку травматичного шоку чтгко розр!зняють стадно збудження (ерек-
тильну) ! гальмування (торпщну). Яскравий опис травматичного шоку належить
М.1. Пирогову.1 Еректилъна стад1я зазвичай короткотривала (5—10 хв), спричине-
на ргзким збудженням ЦНС з ознаками рухового, мовленневого збудження та
больових реакщй на дотик. Вщбуваеться значна активация ендокринно! системи з
викидом у кров велико! юлькост! катехоламшгв, кортикотропшу 1 гормошв Юрко-
во! речовини надниркових залоз, вазопресину. Посилюеться функцш дихально! та
серцево-судинно! систем: пщвищуеться артер!альний тиск, збЁльшуеться ЧСС,
частота дихання. Потём настае тортдна стадия — стад!я гальмування ЦНС, яке
поширюеться на вщдши гшоталамуса, стовбура головного мозку, спинного мозку.
Для не! характерш адинамия, загальна загальмовашеть, хоча хворий 1 притомний,
проте вкрай мляво реагуе на зовшшш подразники; знижуеться артер!альний тиск,
спостерЁгаються ознаки порушення перфузп тканин, зменшуеться дхурез. Унасл!-
док кровотеч!, що супроводжуе травму, приеднуються прояви пповолем!чного
шоку. У будь-якому випадку розвиваються характерш для всёх видёв шоку пору-
шення гемодинамики.
3 ушкоджених 1 прилеглих тканин, з юптин кров! вившьнюеться значна юль-
юсть медЁаторхв запалення 1 розвиваеться ССЗВ. Кр!м того, у кров надходить ве-
лика юльюсть токсичних продукпв, яга утворюються внаслщок розпаду тканин,
'“3 вщрваною рукою або ногою лежить такий закляклий на перев’язочному пункт! нерухомо; вш
не кричить, не волае, не скаржиться, шчого не вимагае; тою його холодне, обличчя блще, як у трупа;
погляд нерухомий та спрямований удалину, пульс — як нитка, ледь вщчутний год пальцем 1 з частими
промгжками. На запитання закляклий або зовам не вщповщае, або лише про себе, ледь чутним голо-
сом (пошепки); дихання також ледь помгтне. Рана та шюра майже зовом нечутливг; але якщо великий
нерв, що звисае з рани, буде чим-небудь подразнений, то хворий одним ледь поманим рухом м’язхв
виявляе ознаки вщчуття. 1нколи такий стан минае через юлька годин вщ застосування збуджувальних
засобхв, гнколи ж вш тривае без змш аж до само! смерть Закляклий не те щоб зовом не уевщомлюе
свого страждання, вш начебто весь у нього занурився, затих 1 закляк у ньому”.
13 продукта порушен ого обмшу. Значка штоксикац1я посилюе ушкодження вщ-
дачених вщ мзспя гравми оргашв. Для травматичного шоку характерною е значнг
(мунодепресзя, на тл1 яко! можтиве виникнення шфекцшних ускладнень 1з не-
сприятливим перебхгом. Уст ц! змши. як 1 в раз! шших вид1в шоку, зумовлюють
виникнення ПОН.
Р1зновидом травматичного шоку е шок, що розвиваеться внаслщок комп
ресшно! травми, — синдрому тривалого здавлювання (у раз! закрито! травми) абс
розчавтвания (вгткрито! травми), сгаяк-синдрому. Вш розвиваеться теля велико!
та тривало! (понад 2—4 год 1 бгдьше) комлресп м’яких тканин 13 перетисненняч
великих судин, коли людина потрапляе пщ завати в раз» катастроф, обвал!в будг-
вель, землетрусёв, аварий. Наичасйше зазнають здавлювання кшщвки. Аналопч-
ний стан виникае шсля зняття джгута, накладеното на тривалий термш (гуршкет-
ний шок).
У патогенез! сгазй-синдрому головними факторами е порушення кровообггу з
значним ступеней 1шемн в здавлених тканинах, ушкодження нервових стовбурш.
розвиток больово! реакци, мехашчне ушкодження масиву м’язово! тканини з
вивьтьненням велико! кгтькоей токсичних речовин. П1сля звгльнення тканин вщ
здавлення за калька годин розвиваеться ! наростае набряк на мгсщ ушкодження ;
в дистально розташованш цшянтц тканин, що зумовлюе зменшення ОЦК, пору-
шення реолойчних властивостей кровт. 1з травмованих тканин у загальний крово-
об!г надходить дуже багато токсичних речовин — продукти розпаду тканин, що
були накопичеш в ушкоджених дыянках, креатинш, молочна кислота, продукти
порушеного обмшу. Вившьнюються калш. фосфор, розвиваеться гшеркагпемтя
Особливктю сгакй-синдрому е надходження в кров велико! кыькоей мюглобшу з)
зруйновано! м’язово! тканини. що е доцатковим фактором ушкодження нирок
зумовлюе розвиток ГНН (мюренальний синдром), йзко активуються цитокшп.
БАР. Шоковий стан розвиваеться за загальними законом!рностями.
Загальш принципи протишоково! терапп. Прогноз для хворого в шоковому'
стан; значною м!рою визначаеться своечасним ужиттям реашмашйних заходов.
Головне завдання Л1кування — стабйпзувати гемодинам1ку та вщновити перфузию
оргашв для шдтримки адекватного системного 1 репонарного транспорту киенк’
У раз! розвитку' шокових станш доцщьними е таю загальш заходи:
• припинення або ослабления да! шокогенного фактора (наприклад спинення
кровотечО;
• знеболювання за наявносп сильних больових вщчутпв — при травмах, ошках;
• забезпечення прохщноей дихальних шляДОв 1 роботи системи зовшшнього
дихання — штучна вентиляция легень, застосування вщповщних газових сумипей:
• вщновлення перфузп оргашв 1 тканин, для чого потр]бш нормалгзашя ОЦК
(шфузшна терашя — уведення рщин), вщновлення I шдтримка гемодинамгки —
роботи серця, нормалгзашя судинного тонусу;
• нормал1защя системи гемостазу (у зв'язку з розвитком або загрозою розвит-
ку ДВЗ-синдрому);
• корекшя ацидозу, гшоксп, електролггного балансу, гшотермн;
• летоксикашйнг заходи, можливо. гз застосуванням екстракорпорально! де-
гоксикацп (плазмаферез, гемосорбчгя, л1мфосорбшя, гемодгадаз, ульграгемофгть-
тращя), уведення антидотних засоб1в;
• боротьба з шфекцгею (септичний шок, отковг ураження, вщкритг травми,
а також у разг приеднання сепсису за шших видав шоку).
Розробляються методи боротьби з надаирною кглькгстю цитокгнгв та шших
БАР — застосування шггбгторгв протеаз, моноклональних антиты, наприклад до
ФНП-а, блокаторгв деяких рецепторгв, у тому числ! й ТЬК, у випадку септичного
шоку, ендотелшових рецептор!в, уведення розчинних рецепторгв, наприклад СП-
14, антиты до молекул адгезп та гн. Деяк! з ефект!в ФНП-а блокуються шпбгто-
рами циклоксигенази, глюкокортикощами.
КОЛАПС
Колапс (вщ лат. соИарзиз — раптово падати) — форма гостро! судинно! або серце-
во-судинно! недостатност!, що проявляеться швидким /' значним зниженням арте-
риального тиску нижче певного критичного р!вня (САТ до 10,7—7,9 кПа, або 80—
60 мм рт. ст., ДАТ — до 5,3 кПа, або 40 мм рт. ст. г нижче).
Переб!г колапсу може бути короткочасним, але з тяжкими порушеннями ге-
модинамгки практично в ус!х органах г тканинах, що призводить до порушення
гхнгх функцш. Насамперед зазнае впливу кровопостачання головного мозку з
раптовим нетривалим потьмаренням свщомостг. Деякг форми колапсу можуть
прогресувати з подалыпим розвитком шоку г бути складовою системних пору-
шень гемодинамгки пщ час шоку.
Залежно вщ причини, що зумовила розвиток колапсу, розргзняють такт най-
поширенгшг його види: постгемораг!чний, дег!дратац!йний, токсичний, токсико-
!нфекц!йний, кардюгенний (гнодг аритшчний), ортостатичний, панкреатичний,
рад!ацшний та гн. За основними патофгзгологгчними механгзмами розвитку колап-
су, тобто за гемодинамгчними варгантами його походження, розргзняють гто-
волем1чний, кардюгенний та ангюгенний (анпоппотонгчний, або вазодилатацгйний)
колапс.
Колапс, що виникае внаслщок зменшення ОЦК, отримав назву гтоволемгч-
ного.
Етгологгя г патогенез. Причиною, розвитку колапсу е дисоцгацгя (пору-
шення спгввщношення) мгж ОЦК г емшетю судинного русла, яка розвиваеться
так швидко, що компенсаторы мехашзми не встигають нормалгзувати зниження
артергального тиску.
Причиною виникнення г!поволем!чного колапсу (схема 37) можуть бути гпвид-
ка штенсивна кровотеча, у тому числг г внутргшня (наприклад, у разг позаматковог
ваптностг, перфорацц виразки шлунка або дванадцятипалог кишки, розриву анев-
ризми), втрата плазми, рщин гз травного каналу, штенсивне потовидыення в разг
критичного пщвищення температури тыа пщ час гарячки. Можливою е також
втрата внутргшньосудинног рщини у випадку и виходу в позасудинний простгр
унаслщок ргзкого поширеного пщвищення проникностг судинно! стгнки. Це
спричинюеться впливом токсичних речовин, у тому числг токсишв мгкрооргангз-
мгв, БАР у разг гх масивного видыення та надходження в системний кровообп. У
таких випадках мехашзми компенсацп (надходження в судини гз мгжклгтинного
простору води, лгмфи, бглкгв, затримка води цирками та судинозвужувальний
вплив) або не встигають спрацьовувати, або е недостатнгми.
Схема 37. Мехашзм в|1никкеш1я гшоволем)чыого колапсу
Кард'югенний колапс (схема 38) розвиваеться як лрояв гостро) недостатност)
серця, пов’язано! з рззким ослаблениям систол)1! но) функцп левого шлуночка та
зменшенням серцевого викиду в раз) )нфаркту, аритмш, таких як раптова значна
брадикардгя в раз) повно) передсердно-шлуночково) блокади, пароксизму мерехт-
ливо) аритми, ф)бриляпп шлуночк)в тощо. Р1зке зменшення ХОК 1 зниження
артерхального тиску виникають також в умовах зменшення кровонаповнення сер-
ця в раз) обмеженого венозного повернення кров) до серия, порушень у судинах
малого кола кровообггу, зокрема емболи, або наявност) перешкод для викиду
кров) )з серця.
Ангюгтотошчний, або вазодилаташйний, колапс (схема 39) розвиваеться внасл)-
лок раптового збшьшення емност) судиннего русла внаслщок первинно) вазо-
констрикторно) недостатност) з) значним зменшенням тонусу венозного в)дцщу
судинного русла або )э системною гшогонгею артерюл, що зумовлюе швидке на-
копичення велико) юлькост) кров) в емюсних судинах. За таких умов знижуеться
централъний венозний тиск, зменшуються повернення венозно) кров) до серця
серцевий викид, знижуеться артер)альний тиск. Системна вазодилатащя виникае
в раз) порушення нейрогенно) або гуморально) регуляцн тонусу судин )з перева-
жанням судинорозширювальних впливгв. Нейрогенну вазодилаташю спричиню-
ють змпги робота нейтральних механ)зм)в регуляш) гемодинамики, зокрема й ро-
бота судинорухового центру з ослаблениям актпвност) пре сорного в)ддьту або
збудженням депресорного, нацм)рною активащею а,-адренорецепторгв у стовбу-
рових дьтянках мозку, у тому числ) теля застосування )х агон)ст)в. У таких випад-
ках зменшуеться вазоконстрикторний вплив симпатичного вщдшу нервово) сис-
геми на судини з генерал)зованим )х розширенням ) зниженням артериального
тиску. Змгни стану судинорухового центру можуть спричинювати оргаючнг ура-
ження вщповщних дшянок мозку, гнтоксикащя, отруення або надмгрна (мпульса-
ц)я з депресорних рефлексоген них зон, у тому числ) у раз) синдрому пперчутли-
вост) барорецепторгв сонно) пазухи.
Схема 38. Мехашзм виникнення кардюгенного колапсу
Системне розширення судин виникае також унаслщок появи велико! кшь-
кост! гуморальних вазодилататор!в (пстамшу, кппнтв, простагландишв тощо) у
раз! генерал!зованих алерпчних реакцш, ССЗВ, гострого панкреатиту 1 т. п. Кр!м
того, розширення судин може зумовлюватися зм!ною !х реактивносп, зменшен-
ням реакци на катехолам!ни або шпп судинозвужувальш впливи. Гшо- або ареак-
тившсть судин виникае в раз! зменшення кшькост! рецептор!в, зокрема а-адрено-
рецепторгв, !хньо! чутливосп (десенситизац!я), або неможливост! непосмугованих
м’яз!в судин адекватно вщповщати на стимуляцпо — у раз! порушень процес!в
збудження, блокування скоротливо! активносп. У таких випадках розвиваеться
рефрактерна вазодилатац!я, судини не реагують на вазоконстрикторн! впливи, у
тому числ! й на судинозвужувальну тераппо. Ослабления тонусу судин виникае
пщ д!ею токсингв м!крооргашзм!в, у раз! неправильного застосування лжарських
засобхв (гангл!облокатор!в, а-адреноблокатор!в, симпатолпикгв, блокаторгв каль-
щевих каналов та шших судинорозширювальних засоб!в). Вазодилатацио можуть
спричинити або посилити порушення функцш ендокринно! системи — гостра або
хрошчна недостатшсть надниркових залоз, гшофункщя щитопод!бно! залози.
Якщо в патогенез! колапсу переважае зниження тонусу вен, то це може спричи-
нити розвиток ортостатичного колапсу, який виникае часпше у вертикальному по-
ложенш тша, особливо теля р!зкого переходу з горизонтального положения у верти-
кально, або пщ час тривалого напруженого стояния з обмеженням рухово! активносп
(на парадах, почесних вартах, п!д час масових вистушв). Шсля переведения хворого
в горизонтально положения артергальний тиск стабшзуеться. Якщо колапс виникае
внаслщок гшоволемн, серцево! недостатносп або зниження тонусу артерюл, то ар-
тер!альний тиск може знизитися 1 в горизонтальному положенш пла. Основним на-
Пору шення нейрогенно!
регуляцп судинного тонусу
Порушення гуморально!
регуляцп судинного тонусу
Порушення
реактивносп судин
Подразнення
деггресорних
рефлексоген-
них зон
Отруення,
штокси-
кацп
Оргашчш
ураження
мозку
Системи!
алерпчн!
реакцп
ссзв
Гострий
панкреатит
Зменшення
кшькостз або
блокада
ц-адрено-
рецепторзв
Порушення
скоротливо!
здатност!
1.1ЯДКИХ
м’язтв судин
Вазодилат атори:
пстамш, юшки,
простат ландини,
ФНП, ФАТ та ш.
Зменшення
пресорних
НПЛИВ1В
У ражеиня га / або порушення
функцп судинорухового
центру
Збудження
депоесорчого
в1ддшу
Зниження артершльного тиску
Генералвоване розширення судин
венозного В1дд1лу га / або артерюл
Анг 1огшотон1чний (вазодилатацшний)
колапс
±
Схема 39. Мехашзми розвитку анпоппотошчного колапсу
слщком гостро! недостатносп судин е порушення кровопостачаяня всгх органгв 1
систем 13 розвитком циркупяторно! гшоксп та з подалыпим ураженням мгтохондргй 1
приеднанням тканинно! ппоксп (див. роздгл XV “Гптокая”). Сгушнь тяжкост) ппок-
сп та ураження оргашв залежагиме вщ р)вня зниження артергального тиску г трива-
лоси колапсу. Колапс може стати причиною розвитку ГНН з олпо- та анур!ею.
Загалып принципи лгёування. Хворим у стаю колапсу допомогу потр:бно нада-
вали негайно. Бш швидкосп и ефективносп надання медичнот допомоги залежить
подальший стан хворого (вщ повного вщновлення гемодинамгки до можливого
розвитку шоку або навпь смерп хворого).
За можливосп потр!бно л!квщувати дпо патогенного фактора, що зумовив
розвиток колапсу: спинити кровотечу, усунути втрату рщини, змшити положения
тша на горизонтально, застосувати антидоти в раз! дп токсичних факторов або
вжити дезштоксикацшних заходив. Варто також вщновити ОЦК у випадку гшово-
лешчного колапсу, нормал!зувати судинний тонус 13 застосуванням судинозвужу-
вальних препарапв, ужити заход!в щодо нормал!зацп та шдтримки роботи серця,
лпсвщацц гшоксп.
КОМА
Кома (вщ грец. кедра — глибокий сон) — екстремальний стан, який розвиваеться
внаслёдок ушкодження головного мозку I для якого характерними е глибоке пригшчен-
ня свёдомостё, порушення функцш систем життезабезпечення та аналёзаторёв.
Основним проявом коматозного стану е значний ступшь приппчення свщо-
мост! — п повна втрата. Порушення дихання, кровообпу, температурного гомео-
стазу, рефлекторно-тошчю розлади, зшичн! порушення, окорухов! розлади е на-
слщками мозково! недостатност!, що розвиваеться пщ час коми.
Свщошсть — це такий стан, за якого людина усвщомлюе, що вщбуваеться з
нею та з оточенням, здатна адекватно реагувати на зовшшн! стимули. Свщомють
визначаеться двома компонентами — станом пилънування, тобто ступеней (р!внем)
активацп свщомосп, та зм1стом свёдомостё, тобто тзнавальними функциями голов-
ного мозку, його псих1чною д!яльн!стю. Стан пилънування (неспання) залежить вщ
висхщних активуючих впливгв спчастого утвору на вищ! вщдщи кори великого
мозку. Ц1 впливи генералхзоваш та проявляються десинхрошзащею к!рково! елек-
трично! активност!, властивою пробудженню. Зм!ст свщомост! пов’язаний !з функ-
щонуванням швкуль великого мозку, певних функцюнальних зон, як! взаемодпоть
одна з одною та з глибинними активуючими системами. Ушкодження цих вщд!л!в
ЦНС (стовбура головного мозку 1 кори великих гпвкуль) проявлятиметься пору-
шеннями вщповщних компонент свщомост! (пилънування та / або психдчно! д!яль-
ност!) 1 розвитком коматозного стану, а також супроводжуватиметься розладом
автономних (вегетативних) функцш, залежно вщ того, як! вщдщи мозку ушкод-
жен!. Для того щоб розвинулася кома, ушкодження кори великого мозку мають
бути дифузними, ДВ061ЧНИМИ.
Тривалий несвщомий стан — ознака вкрай тяжкого ураження мозку, що е
загрозою для житгя хворого ! потребуе негайного лжарського втручання, оскшьки
змши функцш головного мозку обороти! лише в певних межах його структурних
1 метабол!чних порушень.
Розвитку коми можуть передувати прекоматознё стани, за яких спостерша-
ються р!зш ступен! приппчення свщомост! — потьмарення, оглушения (легке,
глибоке), сопор. Для оглушення характерними е стан торпщносп, або притуплен-
ия ПСИХ1ЧНИХ процес!в, пщвищена сонлив!сть. Сопор (вщ лат. зорог — зацшеншня,
млявгсть) — це стан глибокого сну або аналопчного йому за поведшковими реак-
Ц1ями стану ареактивносп !з втратою дов!льно! та збереженням рефлекторно!
дтяльносп, координовано! рухово! реакцп на больов! подразники.
Кома — це таке глибоке порушення свщомост!, за якого неможливо вивести
хворого з непритомного стану жодними зовншпйми стимулами, навпь больови-
ми, його неможливо “пробудити”. Кома може бути ргзного ступеня тяжкост! — I,
II, III, вщповщно, пом!рна, глибока, позамежова, або термшальна. 1нод! видщя-
ють ще легкий ступшь коми. Позамежова кома — еквгвалент смерт! мозку. Смерть
мозку — стан, пщ час якого вс! функци, у тому числ! й юрков!, пщюрков!, стов-
буров!, необоротно втрачеш. Ушкодження мозку в таких випадках необоротне з
обов’язковим ураженням стовбурових структур, що унеможливлюе самостгйне
дихання, яке пщтримують лише штучно. Кровообгг може зберйатися.
Коматозний стан може закшчуватись або смертю, або розвитком вегетатив-
них сташв, при яких наявний ритм сон — пильнування, проте повшстю вщсутш
ознаки шзнавально! д!яльносп, псюочних функщй. Можлив! також р!зш ступен!
вщновлення псюпчно! д!яльност! та рухово! активност!, що зумовлюе або повне
повернення хворого до вихщного стану, або р!зш ступен! швалщносп.
Етюлопя. Причини, що зумовлюють кому, можуть мати генерал!зований вплив,
викликають дифузн!, множинш др!бновогнищев! або локальш ушкодження.
Розр!зняють екзогенш й ендогенш етюлопчш фактори. Ендогенн! фактори,
що спричинюють ушкодження головного мозку, можуть бути штрацеребральни-
ми й екстрацеребральними як наслщок позамозково! патологп. Залежно вщ ло-
калхзацп патолопчного процесу в мозку !х подшяють на супратентор!альш, суб-
тентор!алъю, дифузн! (метабол!чш) або множинш др!бновогнищев1
Екзогенш фактори:
• черепно-мозкова травма;
• екзогенш отруення (алкоголь, наркотачш засоби, седативш препарата, на-
приклад барбпурата та !шш снодшш засоби, транквйпзатори, бромщи, отати, пси-
хотропна речовини, протасудомш препарата, метиловий спирт, еталенппколь, хло-
рид амонпо, паральдепд, важю метали, чадний газ, метгемоглобшоутворювач!);
екзогенна (ппоксична) ппокс!я;
• дефщит впамгюв — памшу, шридоксину, щанокобаламшу, фол!ево! кислота;
• шфекшя;
• фтзичш фактори: температури! фактори — тепловий удар, гшотерм!я; елек-
трична травма головного мозку; юшзуюче випромшювання.
Ендогенш фактори
А. Екстрацеребралъш фактори:
• дефщит кисню, субстратов або метабол!чних кофактор!в;
• ендогенн! !нтоксикаци (у раз! недостатносп печшки, нирок — урем!я
тощо);
• патолопя ендокринно! системи: г!поф!за, щитопод!бно! залози, наднирко-
вих залоз (гостра 1 хрошчна недостатшсть, хвороба 1ценка — Кушшга, феохромо-
цитома), пщшлунково! та прищитопод!бних залоз;
• шип хвороби — пухлини, патолопя легень, пщшлунково! залози (екзокрин-
на панкреатична енцефалопат!я);
• патолопя водно-мшерального обмшу (ппо-, ппергщратац!я, ппо-, гшер-
осмоляршсть);
• порушення КОС (ацидоз, алкалоз);
• порушення терморегуляцп.
Б. Тнтрацеребральш фактори:
• порушення мозкового кровообпу;
• пухлини;
• шфекцп, запальш процеси — менгнггти, мешнгоенцефалгт;
• егплепсгя;
• первиню ураження нейрошв або глп.
Патогенез. Етгологгчш фактори, що спричинюють розвиток коми, мають за-
гальнг мехашзми ураження головного мозку. Це можуть бути механгчю ушко-
дження, порушення функцш нервових клггин г мгжнейронних взаемозв’язкгв,
пов’язан! з формуванням потенщалгв дц нейрон! в, утворенням г вид!ленням ней-
ромедгаторгв або гх дгею на рецептори.
Механгчне ушкодження тканин головного мозку виникае внаслщок травми,
пщвищення внутргшньочерепного тиску, здавлювання через появу додаткових
об’емгв у порожнинг черепа, якг утворюються в разг крововиливу, пухлин, абс-
цесгв, набряку. За локалгзацгею первинного патолопчного процесу в мозку щодо
намету мозочка (вище вщ нього в передаю черепнш ямцг або нижче — у заднгй
черепнш ямцг) розргзняють кому супратенторгальну та субтенторгалъну.
Оскгльки внутрппньочерепний об’ем постшний, додатковг об’еми в порож-
нинг черепа можуть розвиватися лише за рахунок гснуючих структур мозку. До-
датковий об’ем спричинюе здавлення розташованих поряд г вщдалених дглянок
головного мозку. Якщо ш патологгчнг утворення розташованг супратенторгально,
то вони зумовлюють змгщення (дислокацп) частини мозково'г тканини, видавлю-
вання п в отвори, зокрема г в тенторгальну виргзку. Частина мозку, що вкли-
нюеться, змггцуе, здавлюе й ушкоджуе розташованг нижче дглянки стовбура голов-
ного мозку, спчастого утвору. Це зумовлюе порушення його функцгй, зокрема й
активуючих впливгв на кору, розлади пильнування г свщомостг, розвиток коми з
появою невролопчних симптомгв вщповщно до ушкоджених дглянок мозку. Уш-
кодження мозку посилюються судинними розладами, що супроводжують такг па-
толопчнг процеси, та порушеннями шляхгв вщтоку спинномозковог рщини. По-
ширеною причиною тако’г коми е крововиливи внаслщок розриву судин, анев-
ризм. Майже чверть усгх крововиливгв супроводжуеться розвитком коматозного
стану. Навколо крововиливу розвиваеться запалення, набряк, ще бгльше пщви-
щуеться внутргшньочерепний тиск.
Стовбур головного мозку може ушкоджуватися також у разг субтенторгальног
локалгзацн патолопчних процесгв. Виргзняють компресШнг ушкодження, при яких
зона патологи розташована зовю стовбура мозку г здавлюе його, та деструктивы,
якщо стовбур головного мозку ушкоджуеться безпосередньо патолопчними про-
цесами, що локалгзуються в його межах. Такими е мгсцевг порушення кровообггу
(гшемгя, крововиливи), абсцеси, пухлини, травми, демгелгшзуючг процеси в стов-
бург мозку. Проте якщо патолопчний процес розвиваеться поволг, наприклад при
пухлинах, що повгльно пролгферують, то кома може не виникнути завдяки адап-
тацгйним можливостям спчастого утвору. Кргм потьмарення свщомостг ураження
стовбура головного мозку супроводжуються ргзними порушеннями вщповщно до
ушкодження розташованих там центргв.
Одним гз найважливппих видгв коми е кома внаелгдок черепно-мозкових травм.
Ушкодження мозку при закритш черепно-мозковш травмг пов’язанг з раптовим
стргмким пщвищенням тиску в порожнинг черепа, розвитком гшемп та набряку
мозку, деполяризацгею нейронгв унаслщок раптового викиду ацетилхолшу. Пгсля
травми черепа стан потерпглого може попршуватися через прогресуючг судиннг
порушення г набряк мозку, що спричинюе подальше пщвищення внутршгньо-
черепного тиску о розвиток дислокацш мозково! тканини. Спостерогають поширеш
ураження катлярних судин. Таю змши найбшьш характерно для спчастого утвору,
стовбура головного мозку. Шсля травми можуть виникати о мають велике додатко-
ве значения в розвитку коматозного стану внутршшьочерепно крововиливи.
Велика група етоолопчних факторов спричинюе порушення метаболизму нерво-
во! тканини, що зумовлюють приппчення и функцой. Серед метаболочних ушко-
джень мозку найбшыпе значения мае недостатке утворення АТФ, що порушуе
синтез нейромедоаторгв, попршуе роботу енергозалежних оонних насосов, ство-
рення потенцоалгв на мембранах нервових клотин. Порушуються генерацоя о пере-
дача нервових гмпульсов, знижуеться збудливость, розвиваеться гальмування ней-
ронов, що й зумовлюе порушення функщонування нервовоо системи з розвитком
прекоматозних станов о коми. Недостатке енергоутворення може бути наслщком
гтоксп мозку, дефщиту субстратов для утворенооя енергп, зокрема о глюкози, фер-
ментных порушень утворення АТФ.
Нервова система дуже чутлива до гтокси не лише через недостатке утворення
енергп в умовах дефщиту кисню. У разо дефщиту кисню в головному мозку нако-
пичуеться велика колькость збудливих амшокислот (багато з них видшяеться з
нервових закшчень пщ час деполяризацп о не поглинаеться назад через дефщит
енергп), зокрема й глутамату, яю з’еднуються з МВА-рецепторами (М-метил-
Б-аспартат-рецептори). При цьому вщкриваються Са2+-, Иа+-канали, у результат!
чого в нервово клггини надходить багато натрпо о разом з ним води, а також каль-
цш. Надходження кальцою пов’язане з вщкритгям потенщалзалежних кальцоевих
каналов (пщ час деполяризацп мембран через потрапляння натрою в клотини) о
ММБА-потенцоалнезалежних каналов. Накопичення кальцою зумовлене також
зволъненням його оз внутрошньоклотинних структур о недостатком поверненням у
внутрппньоклотинн! депо та позаклотинне середовище (через низьке енергозабез-
печення). Розвиваеться гшеркальщеве ушкодження нейронов, апоптоз клотин
(див. роздол “Патофозоолопя клотини”). Кром того, в умовах ппоксп накопичуеть-
ся лактат, розвиваеться як внутрппньоклотинний, так о позаклотинний ацидоз, що
посилюе ушкодження нервовоо тканини.
Нестача вменю в нервовш тканин! виникае внаслщок розних видов гшоксп (п-
поксично'о, дихальноо, гемочно'о, циркуляторноо, пстотоксичноо). У разо загально!'
гшоксп мозковий кровоток може компенсаторно навоть зростати (удвоч!), щоб за-
безпечити доставку кисню до мозку. Енпефалопатоя розвиваеться внаслщок зни-
ження напруження кисню в артероальнш крово до 2,7—4 кПа (20—30 мм рт. ст.),
коли навоть збщьшений кровообог не забезпечуе доставку до мозку потробно! коль-
косто кисню. Таке суггеве обмеження забезпечення головного мозку киснем спо-
стерогаеться при тяжкой серцевш недостатност! з розким зменшенням серцевого
викиду (шфаркт, кардюгенний шок, аритмп, асистолоя), у разо значного о тривалого
зниження артероального тиску. При анемп киснетранспортна функцоя крово повин-
на зменшитися вдвочо, щоб спричинити ппоксою мозку. Патологоя мозку частице
розвиваеться в разо поеднання розних видов гшоксп, особливо за обмежених ком-
пенсаторних можливостей з боку мозкового кровотоку. Наприклад, шфаркт мю-
карда на тло анемп та мозкового атеросклерозу часто зумовлюе розвиток енцефа-
лопатп навоть за поморного впливу кожно! оз зазначених причин.
Гшоксично ушкодження головного мозку, що призводять до розвитку сопору
о коми, можуть виникати також у результат! первинних порушень мозкового крово-
обггу. Це або дифузш порушення у великих зонах кори обох пгвкуль великого
мозку, або локалып ушкодження з локалгзашею в зонах, яю вщповщають за стан
пильнування.
Дифузш порушення кровопостачання мозку спричинюють мозкову гшокспо
в тих випадках, коли значно збшьшуеться опгр судин головного мозку. Це гшемгч-
ш стани, зумовлеш спазмом судин або !х системним склерозуванням. Важливим
з цього погляду е ураження судин мозку при гшертошчшй хвороб!. Порушення
мозкового кровообггу можуть зумовлювати також тромбоз у раз! розвитку ДВЗ-
синдрому, емболгя (жирова емболгя, емболгя з порожнин серця при ендокардитг).
Спостерггаеться множинне ураження мозкових судин при системному червоному
вовчаку. Локальнг ушкодження — це здебгльшого гшемгчний або геморапчний
гнсульт. 1шемгчнг ушкодження нейронгв можуть виникати в разг зниження мозко-
вого кровообггу нижче нормального ргвня (55 мл / 100 гхв1). Доведено, що коли
внаелгдок значного зменшення енергоутворення мозковий кровотгк становить
20 мл / 100 г-хв, втрачаеться елекгрична актившсть нейронгв, проте гх ушкоджен-
ня ще може бути оборотним (верхнш гшемгчний поргг). У разг зниження кровотоку
до 10 мл / 100 гхв ушкодження нейронгв стають необоротними — вони гинуть
(нижнш гшемгчний поргг).
Чутливгсть ргзних вщдглгв ЦНС до ппоксп неоднакова. Першими гинуть ней-
рони кори великого мозку, особливо морського коника, г кори мозочка (грушо-
подгбнг нейрони — клгтини Пуркгнье). Тому пщ час хрошчног гшоксп, якщо
розвиваеться патолопя мозку, вона починаеться з проявгв порушення змгсту евг-
домостг: втрати правильно! оцшки навколишнього середовища, дезоргентацц,
сплутаностг свщомостг, розвитку сонливосп з можливими сопором г комою. До-
лучаються порушення дихання та гншг розлади.
Метаболгчна кома може розвиватися також вгд нестачг головного субстрату
для енергетичного метаболгзму мозку — глюкози. Ггпоглгкемгчна кома виникае в
разг зниження ргвня глюкози в кров! до 2,5 ммоль/л г нижче (див. роздгл XVIII
“Порушення вутлеводного обмгну”). У разг ггпоглгкемп уражуються насамперед
швкулг великого мозку.
Дгя багатьох токсичних речовин, здатних у високих дозах спричинювати роз-
виток коматозних стангв, також пов’язана з порушенням енергетичного обмгну в
мозковш тканин!. Так, цгангди блокують транспорт електрошв у дихальних лан-
цюгах мгтохондргй; опгати пригнгчують дихальний центр, зумовлюючи розвиток
ппоксп; амгак, барбгтурати за високог концентрацп блокують мгтохондргальнг
ферментш комплекси, пригнгчують центри в гшоталамусг, стовбур! головного
мозку, довгастому мозку, можуть спинити дихання.
Великого значения в патогенез! ком, що розвиваються в разг ргзних екстраце-
ребральних патолопчних процесгв, набувають порушення водно-мгнералъного обмг-
ну. Найтяжчг наслщки спостерггають, якщо змша осмолярностг вщбуваеться швид-
ко, оскгльки не можуть бути повною мгрою реалгзованг компенсаторш мехашзми
мозку щодо пщтримки нормального мшерального складу та пдратащ! клгтин. Гг-
поосмолярнгсть зумовлюе накопичення води в головному мозку, розвиток набря-
ку мозку, перехщ води в нервовг клгтини, гх гшергщратацгю. Це збгльшуе об’ем
мозку, вщбуваеться його здавлення. Збгльшення об’ему внутргшньоклгтинног води
'У сташ спокою мозковий кровотгк становить 55 мл / 100 гхв, тобто 15—20 % серцевого викиду
у споког, причому в сгрш речовин! вш у 3—4 рази вищий, н!ж у бшй.
спричинюе структури! т функщональш порушення нервових юптин. Змша балан-
су внутргшнъо- та позакл!тинного натрпо 1 калпо зумовлюе порушення електрич-
них потенщал1в, збудливост! мембран. Розвиваеться гшоосмолярна енцефалопапя
аж до сопору 1 коми. Гшоосмолярнгсть виникае, як правило, на тлт ппонатртемп,
що може розвиватися в раз! порушення функцш нирок, надлишку вазопресину,
великого 1 ртзкого водного навантаження (див. розд!л XXII “Порушення водно-
електролггного обману”). У раз! коми р!вень натрпо в плазм! кров! може знижува-
тися до 95—110 ммоль/л (у норм! — 135—147 ммоль/л).
Г!перосмолярн!сть (пщвищення осмотичного тиску) також е небезпечним для
клттин головного мозку, осюльки зумовлюе перехщ води з клггин у позаюптин-
ний проспр, зневоднення юптин з порушенням тх функцюнування (див. роздал XXII
“Порушення водно-електролггного обмшу”).
Пщвищення осмотичного тиску, що може спричинити розвиток коматозного
стану, найчастппе виникае за ппернатртеми або значно! пперглткемп. Збшьшення
вмтсту натрпо понад 160 ммоль/л (осмоляршсть понад 340 мосмолъ/л) зумовлюе
зниження пильнування, сонливгсть, а подальше збшьшення до 180 ммоль/л (при
осмолярност! 360—380 мосмоль/л) — розвиток сопору та коми. Такий вмгст на-
тртю в кров! можливий, як правило, при патологп ппоталамуса, порушенн! вщ-
чуття спраги 1 вмтсту вазопресину, тнтенсивно! втрати води з неможливгстю и
адекватного поповнення.
Гшеросмолярнгсть може розвиватися також за рахунок шших осмотично ак-
тивних речовин, зокрема глюкози, що зумовлюе розвиток коми при цукровому
д!абет! типу 2, коли ртвень глюкози може сягати 40—60 ммоль/л т вище, а осмо-
лярнгсть перевищуе 350 мосмоль/л.
У патогенез! енцефалопатш, яю призводять до розвитку коматозного стану,
мають значения порушення КОС, розвиток ацидозу або алкалозу. Проте сопор т
кома виникають найчастппе лише за умови тяжкого ресшраторного ацидозу на
тлт дихально! недостатносп, осюльки вуглекислий газ легко проникае кр!зь гема-
тоенцефалтчний бар’ер ! вщбуваеться змша рН мозку, що спричинюе порушення
мозкового кровотоку, ферментативно! активносп, енергетичного обмшу. Серед
метаболтчних ацидозгв, яю вщтграють певну роль у розвитку коматозного стану,
найбшьше значения мае ацидоз унаслщок накопичення кетонових пл при цукро-
вому дтабет! чи голодуванш або внаслщок пщвищення ртвня молочно! кислоти в
умовах гшоксп. Решта вартанпв порушення КОС рщко сам! по собт зумовлюють
розвиток коми.
У патогенез! коматозного стану, який розвиваеться за ртзних патолопчних про-
цеств, може спостертгатися поеднання юлькох патогенетичних мехашзмтв, яю ушко-
джують мозок. Так, мехашзмами розвитку коми, що ускладнюе цукровий дтабет, е
значна депдратащя, гемоконцентрацтя, що виникають унаслщок полтурп. Розви-
ваеться ппокстя — унаслщок зменшення ОЦК т порушення кровообггу, зниження
артертального тиску, змши властивостей гемоглобшу. Спостерггаеться ацидоз за
рахунок накопичення кетонових пл головним чином при цукровому дтабет! типу 1
(кетонелична кома) або молочно! кислоти переважно при цукровому щабеп типу 2
{лактацидемгчна кома). У разт лакгацидемтчно! коми ртвень лактату у венознш кровт
може пщвитцуватися понад 5 ммоль/л (у норм! 0,55—2,2 ммоль/л), розвиваеться
декомпенсований ацидоз. Дефщит шсулшу та внутр!шньоклттинний ацидоз пору-
шують роботу ферменттв та утилтзацгю глюкози, що посилюе дефщит АТФ, спри-
чинений ппоксгею. Виникають змши минерального обмшу: в раз! дефщиту шсулшу
клгтини втрачають калгй, збшьшуеться його вмгст у кров!, що стимулюе видшення
альдостерону, який сприяе виведенню калпо нирками. Поки е д!урез, генуе г небез-
пека втрати калпо у зростаючих кшькостях. У деяких випадках цукрового д!абету
типу 2 виявляеться дуже високий ргвень глюкози в кров!, 1 тод! до патогенезу коми
додаються значит ггперосмолярнг ушкодження головного мозку.
Дгабетична кетоацидотична кома супроводжуеться порушеннями дихання
(розвиваеться дихання Куссмауля), судинним колапсом, зниженням артергально-
го тиску, ослаблениям м’язового тонусу, олгго- або анургею, низьким р!внем рН
кров! (зазвичай у межах 6,8—7,3), електрокардгографгчними ознаками спочатку
гшеркалгемй, а надал! гшокалгемгг.
Пперосмолярна кома частгше розвиваеться при цукровому дгабет! типу 2, в осгб
лгтнього вгку, пщ впливом факторгв, якг зумовлюють видшення контршеулярних
гормонгв (стрес, травми, шфекцп) або депдратацгю (кровотечг, втрата води через
травний канал, отки). Зневодненню сприяе також полгургя (осмотичний дгурез).
Якщо неможливо забезпечити адекватне поповнення рщини, розвиваеться ргзке
зневоднення, пщвищуеться осмотичний тиск (понад 360 мосмоль/л). Ргвень глю-
кози в кровг пщвищуеться до 50 ммоль/л г вище г може сягати 200 ммоль/л. При-
еднуються гшернатргемгя 1 гшерхлоремгя внаслщок надмгрного видшення альдо-
стерону у вщповщь на дегщратацгю та ггповолемгю. Знижуеться артергальний
тиск. Порушення гемодинамгки сприяють тромбоутворенню. Розлади кровообггу
можуть призвести до гшоксп та лактатацидозу, що поглиблюе ураження мозку.
Кома розвиваеться дуже повгльно, протягом кшькох дгб. Смертнгсть може сягати
60 %.
Розвиток гтерлактацидем1чно'1 коми у хворих на цукровий дгабет 2 типу часто
зумовлюють анемгя, недостатшсть серця, хвороби легень, печгнки, нирок, над-
мгрне фгзичне навантаження, шфекцгйш хвороби. Початок коми зазвичай го-
стрий, упродовж кглькох годин. Супроводжуеться болем у м’язах, серцг, живот!,
диспепсгею. Розвиваеться дихання Куссмауля, швидко прогресуе недостатшсть
кровообггу, знижуеться артергальний тиск. Дгагноз пщтверджуе високий ргвень
молочно! кислоти в кровг. Смертнгсть унаслщок ггперлактацидемгчно! коми може
сягати 80 %.
У патогенез! печшково! енцефалопатгг з розвитком прекоматозних г коматоз-
них стангв також поеднуються кшька механгзмгв ураження ЦНС. Насамперед це
токсична дгя амгаку, кглькгсть якого значно збшьшуеться внаслщок порушення
функцп печгнки (див. роздгл XXIX “Патолопчна фгзголопя печгнки”). Амонгй
вгльно проходить кргзь цитоплазматичш та мгтохондргальнг мембрани нервових
клгтин, блокуе роботу ЦТК (зв’язуеться з а-кетоглутаратом, вилучаючи його гз
циклу) г таким чином сприяе зниженню енергозабезпечення головного мозку.
Кргм того, е припущення, що глутамш, який утворюеться пщ час реакцп амгаку з
глутаматом, може справляли токсичну дпо на нервов! клгтини. Багато шших ток-
сичних речовин, якг надход ять з травного каналу, справляють пряму токсичну дгю
на мембрани нервових клгтин, змшюють роботу гонних насосгв, що порушуе
електричну актившсть головного мозку. Мае значения порушення медгаторного
обмгну за рахунок накопичення несправжшх нейромедгаторгв, наприклад октопа-
мшу. Гальмування може розвиватися також унаслщок накопичення великог кгль-
костг ГАМК. Окргм дп нейротоксичних речовин у патогенез! печшково! коми
। 729 ’
певну роль вщтграють гшоглткемтя, порушення КОС, водно-електролггного обма-
ну (затримка натрпо та води, втрата калпо, осюльки в разг недостатност! печшки
збшыиуеться вмтст альдостерону, який за нормальних умов руйнуеться в пе-
ЧТНЦТ).
У патогенез! урем1чно1 коми, що може розвинутися внаслщок хрошчно! недо-
статностт нирок, також домшуючим етюлопчним фактором е вплив на мозок
велико! кшькост! токсичних речовин, здебшьшого кшцевих продуктов азотистого
обмшу та багатьох !нших, що не виводяться з оргашзму в раз! патологп нирок.
Збшьшенню !х кшькост! сприяе ушкодження травного каналу. Мехашзми !х дц на
нервову систему аналопчш таким при недостатносп печшки. Значним е вплив
гшоксп, що виникае внаслщок ураження мюкарда, недостатносп зовшшнього
дихання з можливим розвитком набряку легень, анеми (и ступшь пропорцшний
азотемп) та ушкодженням дихальних ферменпв мттохондртй. Ушкоджують не-
рвов! клттини порушення водного, мшерального обмшу — калпо, натрпо, каль-
щю, а також магнпо (збшьшення вмтсту магшю понад 2 ммоль/л сприяе значному
пригшченню нервово! системи), ацидоз.
Загальш принципи лткування. Потр!бно забезпечити достатке постачання кис-
ню, вщновлення 1 тдтримання функцш серцево-судинно! системи, нормалтзацпо
водно-електрол!тного обм!ну, КОС. Лгкувальн! заходи полягають у лгквщацп па-
толопчних процеств у головному мозку: зниженш внутршшьочерепного тиску,
знятп набряку. метабол!чному пщтриманю мозку. Заходи специфтчно! терапп ма-
ють бути спрямоваш на лткування хвороб т екзогенних штоксикацш, яю е причи-
нами розвитку коматозного стану, на запобпання виникненню ДВЗ-синдрому,
шфекцшних ускладнень, трофтчних уражень.
ВИБУХОВА ШАХТНА ТРАВМА
Людина може опинитися в зон! вибуху на виробництв! (вибухова шахтна травма),
а також у побуп, транспорт! та пщ час бойових дш. Проте ВШТ мае низку особ-
ливостей. Щороку на шахтах Украши гинуть 200—250 прниюв; 1 млн тонн видо-
бутого вуплля коштуе життя чотирьох прниюв.
Еполопя. У момент пилогазових вибух!в у пщземних виробках вупльних шахт
виникають таю фактори ушкодження: вибухова ударна хвиля (тиск досягае 3922 кПа),
полум’я високо! температури (до 2650 °С), висока концентращя оксиду вуглецю,
метану, продукттв неповного згорання цитотоксичних полщиюпчних ненасиче-
них вуглеводнтв т низький вм!ст кисню. Ц! патогеннт фактори взаемообтяжлив!,
хоча причиною загибел! потерпших може бути будь-який тз них.
Умови. Еколопчне середовище, виробнич! фактори вупльних шахт мають по-
тенцшно небезпечш властивост!, яю особливо проявляються пщ час вибухтв у
шахтах горючих газгв г вупльного пилу. Основними виробничими факторами, що
формують несприятливе преморбщне тло ртзних форм шахтно! травми, у тому
числт т вибуховот, е висока температура повттря, пщвищений барометричний тиск,
шум, втбрацтя, пщвищена волопсть ! запорошешсть повттря, пщвищений вмтст у
повттр! токсичних речовин (радон, метан, оксид вуглецю, оксиди азоту) т зниже-
ний вмгст кисню, природна радюактившсть вугшля т порщ, важка напружена
праця (частково у вимушеному положенш), нервово-емоцшне напруження тощо.
Зё збёльшенням кёлькостё ушкодж^ъальних факторхв змёнюеться леребхг шахт но!
травми, пояснениям чому е виражена поломка мехашзмёв адаптацп. що призво-
дить до стрёмкого розвитку критичних СТЗН1В.
Пёд кёнець дй комплексного шахтного преморбшного навантаження оргашзм
вёдчувае дефщит маси тёла (унаслёдок значного потовидёлення при ёнтенсивному
фхзичному навантаженнё), погёршуеться системна ё церебральна гемодинамёка,
вщбуваеться репонарний перерозподйт кров! в мезенгерылыпй дыянцё ё ссквест-
рацёя кровё в н!й, пёдвишуються температура пла ё проникшсть гематоенцефалм-
ного бар'ера, ёмовёрно. унаслщок гшервентиляшйного синдрому.
Таким чином, робота в нагрхваючому мхкрокчёматё глибоких вугёльних шахт
на тлё дп шших перерахованих факторёв супроводжуеться напруженням механёз-
мёв адаптацп ё певними втратами маси пла. Останш е тим несприятливим пре-
морбщним тлом, яке спричинюе збёльшення тяжкостё ушкоджень при ВШТ.
Патогенез. Преморбшне тло ВШТ обтяжуе розвиток й основних патогенетич-
них мехашзмёв: мехашчного (контузёя) ё тсрмёчного (опёк), що зумовлюе пору-
шення гемодинамёки на трьох рёвнях (системному, органному, мёкроциркуляцй) 1
зрештою спричинюе виникнення циркуляторно! гёпоксп. Токсичний компонент
зумовлюе гёпоксичну (метаном), гемёчну (карбоксигемоглобёном) ё тканинну (чад-
ний газ блокуе Ре2+, а цёанёди Ре34 у цитохромах тканинного дихання) ппоксёю.
Таким чином, гёпоксёя стае змёшаною за своём генезом, обтяжуючи перебёг пато-
логёчного стану.
Складнёсть патофхзёологёчних процесхв, якими харакгеризуеться патогенез
ВШТ, полягае в тому, що ураження виникають у бёльшостё оргашв. Проте най-
важливёшими патогенетичними ланками е ушкодження легеш>, нервово! системи
ё кровообёгу. Патофхзёологёчнё змёни в органёзмё одразу пёсля да ударно! хвилё
(якщо не настае мигтева смерть) характеризуються потьмаренням свёдомосп, ко-
роткочасним апное, частим поверхневим диханням, збёльшенням частота пульсу
ё зниженням артерхального тиску. Знижуеться вмёст кисню як в артерхальнёй, так
ё у венознёй кровё. Порушення гематоенцефалёчного бар’ера зумовлюе формуван-
ня набряку-пабухапня головного мозку. У потерпёлих при пилогазових вибухах у
шахтах спостерхгають контурнё опёки II—III ступени площх тёла, не захишеноё
одягом. Зазвичай це обличчя та кистё, а у випадках. коли хёрники прапюють ого-
леними по пояс,— голова, шия, груди, спина, руки. Часто приеднуеться термогн-
галяцёйна травма. Опёковё поверхнё ёмпрегнуються вугёльним пилом, особливо на
боцё, звсрненому до епёцентру вибуху (вторинна дёя ударнох хвилё). Роговий шар
епёдермёсу на ошкових ранах часто майже вщсутнёй, а мёсцями звисае у вигляд!
шматкёв рёзно! величи ни. Обпалюються вн, брови, волосяний покрив. Швидко
набрякае обличчя. Набряк обпечених вёй нерхдко перешкоджае розкриттю пов.-
кових щетин. Згёдно з результатами клёнёко-гёстологёчних дос/.ёджень, при пило-
газових вибухах виникають переважно поверхневх дермальнё опхки, рёдше глибокё,
тодё як у потерпёлих, що вижили. глибокё опёки майже нхколи не спостерёгають.
Рёзна глибина ураження хххкёри пов’язана з рёзною тривалёстю впливу полум’я та
рёзною його температурою. Незважаючи на високу температуру полум’я вибуху.
короткочасна експозицёя його зумовлюе виникнення лише поверхневих опёкхв на
обпечених дёлянках тёла (за типом митгевого опёку). Гёстохёмёчно миттевий опёк
харакгеризуеться наявнёстю чётко виражено! демаркащйно! лёнп мёж ушкодженою
ё здоровою шкёрою. У глибоких шарах шкёри лёнёя демаркапй пролятае у криптах
1 волосяних фолпсулах. Для миттевого ошку типобими е еритема 1 набряк тканин
безпосередньо лщ ушкодженими вщ тепла тканинами 1 до перифери ввд них. При
звичайних ошках кровоносн! судини шири ф!ксуються тепловим впливом, 1 су-
динш реакцп вщсугш. При мпневому оп!ку загоення вщбуваеться шляхом сек-
вестрацн поверхневих шар!в шири, п!д яким настае реештелгзашя. Обидва фено-
мени — демаркашя 1 загоення ищ секвестром — вщбуваються в умовах. коли
тепло передаеться кргзь тканину протягом дуже короткого промЬкку часу. Мит-
тевтсгь гмпульсу не дае змоги теплу поширюватися по перифсри ! всередину,
створюючи крайову межу ураження. Тому уражеш тканини не вщновлюються,
але глибш! шари залишаються неушкодженими. здатними як до судинно’1 реакци,
так 1 до звичайного процесу репараци. При звичайних оптках зберпаеться посту-
повий перехщ вщ нормальних клиин до ушкоджених. Загоення при миттевому
ошку вщбуваеться завдяки тому, що ештелш в!льно виростае 13 крипт, во лося них
фолжул'.в г неушкоджених кратв шири. Однак при глибших теплових ураженнях
шири з руйнуванням ештелно в криптах 1 волосяних фолткулах еппелтальних ос-
тршцгв росту буле недостагньо. 1 загоення вщбувати меться повально, шляхом гра-
нулювання. Подтбш рани, шо гранулюються. нерщко зумовлюють утворення ке-
лощних рубшв Е експерименгальш докази того, шо регенерация шири можлива
ильки в тому випадку, якщо буде збепежено ретикулярний шар У раз! видалення
ретикулярного шару шири разом з основою волосяних фол псу шв рана загоювати-
меться шляхом утворення рубля. У вупльних шахтах у потергплих у зон! пожеж!
займання одягу може спричинити глибой ураження шири. Тяжйсть стану хво-
рих у гострий перюд ошково! травми пов’язана не з ушкодженням шйрного по-
криву, а 13 системними змшами, яй терм!чний фактор спричинюе в оргашзм!.
Виражешсть змш залежить вщ сили впливу температурного факгора, м!сця 1 пло-
щ! ураження, реактивност! организму, в!ку хворого, часу, який минув вщ моменту
отримання ошку до початку надання медично! допомоги.
Терм 1чний фактор зумовлюе еферентну !мпульсац!ю з обпалено! поверхш,
порушення центрального рсгулювання та функцюнальну недостатшсть шйрного
покриву. Ослабления та дезоргашзашя таких функлтй шйри, як бар’ерна, дихаль-
на, видшьна, спричинюють посилення змш гемоди нам! ки та мжропиркуляцп,
лавинопод!бне наростання патолопчних змш з боку житгево важливих органгв 1
систем, що зумовлюе генералззашю патологи.
Подразнення нервових зайнчень у ран! е причиною раншх ! виражених змш
функщонально! активност! ЦНС у вигляд! порушень функпюнальних взаемовщ-
ношень центрального апарату регуляцп з периферичними ланками нервово! сис-
теми. Аналопчним чином змшюеться 1 функцюнальний стан пшйркових ' пери-
феричних вщдцпв нервово! системи. Функшональна перебудова апарату нейроген-
но! стимуляцп мае виражений компенсаторно-пристосувальний характер, спрямо-
ваний на запобшання виснаженню центральних нервових структур.
Предметний покажчик
Абаз1я 683
Абеталшопротещемтя 356
Автоантигени 690
Автоштоксикацгя кишкова 589
Автоматизм серця 516
— порушення 516
Автофапя 369
Агноз1я 674
Агранулоцитоз
— гмунний 476
— класифпсащя 476
— м!елотоксичний 476
- ЦИКЛ1ЧНИЙ 476
Агрегащя тромбоцит!в 205
Адаптация 49, 319, 661
Адгезившсть 78
Адгезия 205
- молекул 237
- оборотна 239
Аденог1поф1з
— гшерфункщя 647
— ппофункщя 647
АФадохокшези 683
Ад’ювант Фрейнда 511, 630
Азотемш 617
Азотистий баланс
— негативний 351, 367
— позитивний 367
Акантоцитоз 356
Аквапорини 387, 388, 586, 619
Акромегал1я 647
Алейюя
— ал1ментарна токсична 477
Алергени 254
— класифпсащя 255
Алерпйш реакцп 254
— анаф!лактичного типу 257
— гальм!вш 260, 261
— класифгкацгя 257
— мехашзми формування 267
— негайного типу 257
— несправжн! 271
— перехресш 272
— сповшьненого типу 260
— справжш 271
— стадо 256
---б1ох1м1чна 261
---1мунолопчна 256
---патоф1зюлог1чна 266
---патох1м!чна 261
---стимулююч! 260
Алерпя 254
— етюлопя 254
— запобшання 272
Алкалоз
— газовий 378
— декомпенсований 378
— компенсований 378
- негазовий 378
---видшьний 380
---екзогенний 380
Алкаптонургя 372
Алоантигени 261
Альбппзм 372
Альтерац1я 222, 224
- вторинна 224
— первинна 222, 224
Амавротична щюпя 366
Амппн 353
Амшощоз 85, 353, 369
— старечий 131
Ам1ноацидем1я 599
Амшоацидур1я 368, 370, 599
— ренальна 620
Амошогенез
- нирковий 382
— посилення 384
Аморфосинтез 673
Анальгез1я 672
Анальфапроте!нем1я 356
Анаплаз1я
— антигенна 307
— бюх1м1чна 305
— клзтинна 309
— тканинна 309
— ф1зико-х1м1чна 306
— функщональна 308
Анафйтатичш реакцп 257, 268
Анафзлаксзя 258
— загальна 258, 716
— мзсцева 258
Ангзогемофшя 492
Анпопатзя дзабетична 138
Анпоспазм 198
Анпотензин II 198, 534, 539, 543—547
Анпотрофоневроз 395
Аневплоздзя 92
Анемзя 455, 457, 623
— апластична 46ч, 468
— аретенераторна 458, 469
— гемодилюцзйна 458, 459
— гемолпмчна 459
--- госзра 460
---екзоеригроцитна 460
---ендоеритронитна 460
---змунна 460
---мехашчна 460
---мзкросфероцитна 461
---набута 4б0
---спадкова 460
---токсична 460
---хронзчна 460
— ппохромна 459, 465
— дисеритролоетична 464
— дисрегуляторна 463, 468
— еритробластна 458
— залззодефщитна 409, 464
— залззорефрактерна 467
— Кулз 462
— мегалобластна 465
---В1:!-деф1цитна 441, 466
---фолзеводефзцитна 440, 466
— метапластична 469
— мзелофтизична 469
— пернзцзозна 441
— постгеморапчна 459
— регенератор на 459
— серпоподзбно клзтанна 462
— сидероахрестична 467
— сидеробластна 467
— справжня 558
— Фзнкон) 468
Анестеззя 672
Анззохромзя 455
Анззоцитоз 455, 463
Аномал1я лейкоципв пельгерзвська 472
Анорексзя 330, 572, 628, 654
Антигени 166. 169, 254
— демаскування 269
— ксеногенн! 261
- маркернз клзтин СВ 167
---неповнз 254
---повн! 254
Антаппоксанти 323
Антиоксиданта 136, 323
Антаоксидаптнз системи 64, 101
Антиретикулярна цитотоксична сироватка
(АИС) 139
Азгурзя 626
— рсфлекторна 648
Аззное 553
Апоптоз клзтин 91, 96, 297. 486
— порушення в лейкозних клпинах 486
АРЬЮ-система 198. 590. 635
Арахзлоновз похзднз (ейкозанозди) 231, 263
Арззтмзя 516—522, 708
— дихальна 517
— мерехтлива 521, 655
Артерзальна ппертенззя 537
Асинергзя 682
Астаззя 682
Астензя 683
Асфзксзя 567
Асцит 614
Атаксзя 682
Ателектаз легень
— компресзйний 561
— обтурацзйний 561
Атеросклероз 525
— експериментальнз модел! 526
— георп патогенезу 533
— факзори ризику 527
Атетоз 684
Атонзя 682
Атрзальний натрзйуретичний петзтид 194. 389,
539, 636
Ахалаззя 576
Ахзлзя 578
Ахлорпдрзя 578
Ахолзя 583, 612
Ацетилхолш 188, 192
Ацидоз 378
— газовий 378
— декомпенсований 378
— компенсований 378
— лактацидемзчний 380
— негазовий 378, 380
---видзльний 380
---екзоз-енний 380
---каназьпевззй 380
---метаболзчний 380
— ренальнии азотемзчний 380
Базофзлзя 473
Баротравма 55
Берз-бер! 437
Ылзрубш 604
— взльниИ 604
— зв’язаний з альбумгнами непрямий 604
— кон’югований з глюкуроновою кислотою
прямий 605
— некон’югований непрямий 604
Бйпрубшуртя 611
Б1лки
— гостро! фази 233, 280, 713
— катюши 230, 278, 573
— теплового шоку 93
— шляхи руйнування 94
Б1лъ 46, 515 674
— вщбитий 677
— втсцеральний 674
— значения для оргашзму 679
— тррадтйований 677
— фантомний 677
Бюлопчний вгк людини 133
Бюлопчш бар’ери
- внутрипш 164
— гематоенцефалтчш 164
- пстогематичн! 164
— зовнппш 163
Бюлопчно активн! речовини (БАР) 192, 198,
203, 205, 226-234, 249, 261-265, 267, 706
- мехашзми дезактивацп 244,265
Бластний криз 487
Бластопат1я 120, 122
Блокада
- альвеолокап!лярна 563
— серця
----внутрппньопередсердна 519
----внутрппньошлуночкова 519
----передсердно-шлуночкова
----неповна 519
----повна 519
Брадикардтя 517
Брадипное 552
Бронхтальна астма 258, 557
Бултмтя 573
Вазопаття геморапчна
— етюлопя 492
— класифткацтя 492
— патогенез 492
Вентиляцшно-перфузшне сптввщношення в
легенях
— порушення 565
Вентиляция альвеолярна 565
Видужання 35
— мехашзми 36
— неповне 36
— повне 36
В1дрижка 579
Втльнорадикальне окиснення 133, 224—226
В1рил!зац1я 657
Втроцити 481
Втрулентшсть 78
Втруси пухлинотвгрю 295, 299
— роль в онкогенез! 299, 300, 483
Вттамши 428
— джерела надходження 432
— класифткацтя та властивост! 429
— недостатшсть 434
---бютину (вгтамш Н) 442
---вттамшу С 442
---вггамшу Б 444
---вттамшу Е (токофероли) 446
---вггамшу Р 447
---вггамшу К 447
---кобаламшу (вгтамш В12) 440
---шридоксину (вгтамш В6) 438
---ретинолу (вгтамш А) 443
---рибофлавгну (вгтамш В2) 437
---рутину (вгтамгн Р) 443
---тгамшу (вгтамш В^ 436
---фолгевог кислоти (вгтамш В9) 439
Внутргшньосудинне зсщання кровг
- дисемшоване 204, 496
Внутргшньоутробного розвитку порушення
119
— класифткацтя 120
— причини 120
Вуглеводтв порушення
— анаболтзму 340
— катаболгзму 338
— нервово-гормонально! регуляцп 341
Галактоземгя 339, 598
Гаметопаття 120
Гаптени 166, 254
Гарячка 276
— види 284
— виснажлива 285
— експерименталъне вивчення 276
— значения 288
— лткувальне застосування 289
— медтатори 277—280
— перемгжна 284
— поворотна 284
— посттйна 285
— стадп 282
Гастрин 577, 591
Гейденгайна закон 575
Гематокритне число 451
Гематуртя
— ниркова клубочкова 618
— позаниркова 618
Гемералогпя 443
Гемобластоз
— класифткацтя 481
Гемоглобшешя 462
Гемоглобшопаты 104
— спадкова 462
Гемоглобшур1я 462
— маршева 460
— пароксизмальна шчна 461
Гемолхз
— внутршпгьокштинний 461
— внутрйпньосудинний 461
— 1мунний комплементзалежний 461
— мехашчний 461
— окисний 461
Гемостаз 204, 488
— коагуляцшний 205, 488
— порушення 488
— судинно-тромбоцитний 204, 489
Генетичне обстеження
— значения 110
Гепатоз жировий 599
Гер1атр1я 124
Гершга — Брайера рефлекс 569
Гермафродитизм несправжшй 658
Геронтолопя 124
Гетероалерпя 271
Гетероантигени 261
Гетерометр1я 31
Гетеротоп1я 31
Гетерохрония 31
Пбернащя мюкарда 511
Пгантизм ппоф1зний 647, 649
ГшеразотемЗя
— продукцшна 372, 599
— ретеншйна 372, 617
Гшералгез1я 675
Гшеральдостеронгзм
— вторинний 394, 397, 603, 623, 657
— первинний 657
Гшерамшоацвдемзя 370
Пперамошемзя 599, 602
Гшерб1л1руб1нем1я 607, 610
— шунтова 607
Ппервентилящя 316, 381, 551, 603
Ппервггамшози 448
Гшерволемзя
— нормоцитем1чна 452
— ол1гоцитем1чна 452
— полшитем1чена 452
Ппергаммаглобул1нем1я моноклональна 373
Г шерпдратащя
— пперосмолярна 392
— гшоосмолярна 392
— гзоосмолярна 923
Гшерпя 159
Пперппкемгя 342, 349, 354, 532, 598, 648, 652,
656
Гшергомоцисте!нем1я 441, 532
Гшерем1я
— артер!альна 190
---нейрогенна нейропаралпичного типу 192
---нейрогенна нейротошчнсго типу 192
---патолопчна 191
---фгзюлопчна 191
— венозна 196
— реактивна 191
— робоча 191
Гшерерпя 159
Пперестез1я 672
Гшершсулгшзм 364
Гшеркалкм1я 385, 398
Гшеркетонем1я 333, 350, 358, 380, 599
Гшеркшез 578
Гшеркшез1я 684
Пперкоагуляшя 495
Гшеркортицизм 358, 656
Гшерлшемхя 356
- ал!ментарна 356
— ретеншйна 356
— транспорта 356, 599
Гшерлшопроте1дем1я 356, 528, 599
— класифшащя 356, 530
Гшермаппемзя 399
Гшерокая 56
Гшерорекс1я 573
Гшеросмолярн1сть 388, 729
Пперпаратиреоз 654
Гшерпное 553, 568
Гшерпролактинем1я 648
Пперпротешемзя 373
Гшерсалгвашя 575
Ппершалоадешзм 576
Гшертенз1я
— артер!альна 537, 656
---вторинна 537
---експериментальш модел! 538
---есенщальна 537
---ццопатична 537
---мшералокортикощна 539
--- реноваскулярна 538
---ренопривна 538
---рефлексогенна 538
--- сольова 539
---типи 537
---центрально-нпемхчна 538
— легенева 547
— портальна 597, 613
---внутршгнъопечшкова 613
---змппана 613
---надпечшкова 613
---щдпечшкова 613
Гшертермзя 53, 285
— ендогенна 286
— злояккна 287
Гшертрофхя мюкарда 503, 546
Гшерурикемхя
— вторинна 374
— первинна 374
Гшерхолестеринешя 528, 530, 534, 536, 659
Гшестезгя 672
Гшерхол!я 608
Пперхлорпдрхя 577
Гшерхромгя 455
Гшерчутливгсть сповшьненого типу 260
Гшоадренокортицизм хрошчний 656
Гшеральдостерошзм 657
Гшобархя 55
Гшобеталшопроте!'дем1я 357
Гшовентиляцхя 551
— альвеол 561
Гшоволемхя 451
— нормоцитешчна 451
— ол1гоцитем1чна 451
— пол!цитем!чна 451
Гшогаммаглобулптемхя
— Брутона 177
— транзиторна 176
— фхзюлопчна 176
Гшог1дратац1я 391
Гшогл!кем!я 343, 598, 603, 650, 655
- гепатогенна 598
Гшогонадизм 658
— вторинний 647
Гптодинашя 533
Гшокал!ем1я 398
Ппокальщемхя 401
Ппокапшя 316, 565
Гшоюнезхя 684
Ппокоагулящя 489
Гшоксемхя 565
Гшокс1я 314, 603
— гешчна 314, 465
— ппоксична 55, 314
— дихальна 314
— класифйсашя 314
— мюкарда 509
— патолопчш порушення 320
— тканинна 315, 409, 465
— циркулярна 315
Гшоосмолярюсть 388
Гшопаратиреоз 653
— несправжшй 653
— хрошчний 654
Гшоплазхя загруднинно! залози 176
Гшопротешемхя 373, 394, 599
Г1посал1вац1я 575
Гшосидеремхя 465
Гшостенурхя 621
Гшотензхя артерхальна 549, 613
— вторинна 549
— патолопчна гостра 549
— патолопчна хрошчна 549
— первинна 549
— симптоматична хрошчна 549
Гшотермхя 52
Гшотиреоз 650
Ппотрансферинемхя 464
Гшохлорпдрхя 578
Гшохолестеринемхя 599
Гшохолхя 611
Гшохромхя 455, 459
Гл1когеноз(и) 340
— спадков! 340, 598
— типу I 340
— типу VI 340
Глхкозамшоглпсани 137, 140, 141
Глюкозурхя 342
— дхабетична 342
— ниркова 343
— спадкова 343
Глхкопротещи
— структури! 142
Гломерулонефрит 629
— гострий 629
--експериментальш модел! 629
--хмунокомплексний 630
--нефротоксичний 630
--патогенез 630
— хрошчний 631
--фази 632, 633
--форми 632
Глосит Гунтера 466
Гнотобюнти 577, 589
Голодування 329, 380, 594, 665
— абсоютне 329
— бьткове 336
— жирове 336
— неповне 335
— патолопчне 329
— повне 329
--основш прояви 330
— перюди 330
— причини 329
— умови 330
— повне без води 335
- ф!з!олопчне 329
— часткове 335
Гомеостаз 29, 386
Гомойотермхя 162, 276, 330
Гормони 634
— адаптацию 661
— антагошсти 638
— регуляция секреци 640
— синерпсти 638
— синтез 1 секрецтя 642
Гормоноцщ 590, 635, 636
Гранулоцити 227
— базофтльнт 140, 227, 229
--дегрануляшя 227
Гранульоматоз 480
Дегенерацтя нерва 669
Депдратацтя 390
— гшеросмолярна 391
— гшоосмолярна 391
— гзоосмолярна 391
Декарбоксилювання порушення 370
Делецтя 92, 100
Десатуращя 55, 57
Десенсибшзацгя 181
— специфгчна за методом Безредки 182
Детермппзм 40
“Детреноване серце” 516
Дефензини 230
Дефтбрилящя електрична 522
Дефщит пульсу 522
Дизерпя 159
Дилатац1я 501
— серця мюгенна 506
— тоногенна 501
Дисбактерюз 589
Дискшезгя жовчних шляхтв
— гшерктнетична 606
— ппокшетична 606
Дискразтя 45, 115
Дисметр1я 682
Диспное 568
Диспротешемгя 373, 599
Дистрес 663
Дистрофия
— адипозогештальна 649
— алтментарна 336
- жирова 357
— нейрогенна
--патогенез 689
Дисфапя 576
Дисхолгя 612
Дифузтя 190
— газгв у легенях порушення 563
Дифузна нейроендокринна система (АРГЮ)
198, 590, 635
Дихання
— апнейстичне 555
— Бита 555
— вагусне 552
— гасшнг 556
— зовнпнне 550
змши показниктв 561
— Куссмауля 350, 553, 729
— перюдичне 554
— стенотичне 552, 556
— Чейна — Стокса 554
— щадне 553
Дтабет
— алоксановий 346
— вторинний 345
— класифжащя 345
- МОЭУ 345, 352
— нецукровий 390, 620, 648
---ппофтзна форма 648
---нефрогенний 620, 648
— стерогдний 346, 656
- цукровий 344
---гестацгйний 345
---експериментальний 345
---типу I 345, 346
---типу II 345, 351
---ускладнення 350, 354
Дгатез 118
— астешчний 119
— ескудативно-катаральний 118
— лгмфатико-ппопластичний 119
— нервово-артритичний 119
Дтурез осмотичний 349
Д1я атмосферного тиску
— зниженого 55
— пгдвищеного 56
— йошзуючого випромшювання 57
— бюлопчних факторгв 77
— шфекцтйних факторгв 77
— факторгв космгчного польоту 68
— хтмтчних фактортв 69
— температури
---високот 53
---низькот 52
Домшанта 48
Дослщ(и)
— Клода Бернара 192
— Фппера 304
— Ф. Мажандт 686
— Ю. Конгейма 234
— Ю.С. Лондона 583
Ейкозано'щи 231, 263, 708
Екзофтальм 652, 691
Експресившстъ гена 102
Екстрасистола
' — вставна 519
— передсердна 518
— передсердно-шлуночкова 518
— синусна 518
— шлуночкова 518
Екстре мальш стани 51, 697
Екструзгя 236
Ексудащя 222, 235
Емболгя 206
— вор'.тног вени 211
— газова 55, 209
— екзогенного походження 207
— ендогенного походження 208
— жирова 208
— навколоптгдними водами 208
— парадоксальна 207
— ретроградна 207
— судин великого кола кровооб!гу 210
— судин малого кола кровообггу 209
— ткан инна 208
Ембргопатгя 120, 122
Емггращя лейкоципв 222, 236
Емф1зема 558
Ендотелши 198, 545
Ендоцитоз 531
Ензимопапг 103
Енкефалш 678
Енофтальм 691
Ентерогепатична циркуляция 605
Енцефалгг 667
Енпефалопатгя 85. 547. 608, 610, 611, 726
— гшоосмолярна 728
— губчаста 6б5
— печгнкова 602, 729
Еозинофшя 472
Еритрон 454
Еритропешя 455
Еригропоетин 456, 457, 615
Еритроцити
- змши 454
---дегенеративн! 455
---забарагення 455
---кгльюснг 455
---ягаснг 455
— форми дегенеративн! 456
---регенеративнг 456
— серпоподгбн! 455, 462
Еритроцигоз 455, 456
— етгологгя 456
— класифгкашя 456
Етгологгя 40
— психосоматичний напрям 42
Ефект кисневий 59
Ефект Пастера негативний 306
Евнухогдизм 659
Жиосна шфгльтрапгя 333, 357
— печ1нки 357, Зб4
---алгпотропна 357
Жовтяниця 607
— абсорбцшна 607
----премгкфосомна 608
— гемолпична 607
— гепатоцелюлярна 607. 610
— екекрепгина пгслямгкфосомна 608, 610
— ензимопатична 607
— кон’гогапгйна 609
— механгчна 607
— передпечгнкова 607
— печгнкова 607. 608
— пгеляпешнкова 6й?
— транспорта 607
— холестатична 607, 610
— шунтова 607
— ядерна 608, 610
Загальна нозолопя 29
Задишка 210, 568
— ексшраторна 570
— гнепгра горна 570
Закон градгента серця 516
Закон денервацп 687
Запалення 219
— експериментальне вщтворення 220
— загальнг прояви запалення 219
— значения для оргашзму 219
— класифгкашя
----альтеративне 250
----геморапчне 251
----ггперерпчне 250
----гните 251
----гостре 251.
----патогенез 221
---- ексудагивне 251
— змлпане 251
----катарачьне 251
----нормерпчне 250
----продуктивна 251
----пролгферапгвне 251
---- серозне 251
----фгбринозне 251
----хрошчне 251
— клггини запалення 226
— медгатори 229
----впливу на
----гшоталамус 233
----печгнку 233
----гуморальш 229, 232
----клггинш 230
— мгецев! ознаки запалення 219
— стадп 222
Здоров’я
— визначення 29
Зневоднення 54, 390
Зоб
— дифузний токсичний 260, 652
---ендешчний 425, 651
[дюпатичний прсутизм 261, 645
1зоволем1я 615
1зопдр!я 615
1зоюн1я 615
1зоосм1я 615
1зостенур!я 621
1ммортал!зац!я клггини 297
1мунггет 165, 166
Хмунна агрес1я матер! до плода 121
Тмунна система 165
— пперфункщя 172
— ппофункщя 173
---експериментальне вщтворення 173
— дисфункщя 173
— значения 165, 166
— порушення 172
---органогенезу 179
— регуляц1я 180
- функцн 169
1мунн1 реакцп проти трансплантата 261
1муноглобулщи 168
— будова 168
— особливост! 168
1мунодепресивш стани 179
1мунодефщити 173
Хмунокомплексш реакцп 259
1нваз1йн1сть 78
1ндекс жовч!
— лецитинохолестериновий 613
— холатохолестериновий 613
Тндекс фагоцитний 140
1ндекс ядерного зсуву 479
1нтегрини 237
Хнтерлейкш-1 167, 226, 228, 231, 233, 236, 238,
278-280
Хнтерлейкши 81, 167, 231, 240, , 244, 278, 707
1нтоксикац1я 72, 532
— гостра 72
— легка 73
— перюди 72
— шдгостра 72
— середньо! тяжкост! 73
— тяжка 73
— хронхчна 72
1нфантил1зм 647
Хнфаркт 201
— мтокарда 511, 512, 515
---експериментальн! модел! 509. 514
---насл!дки 514
---патогенетичн! вар!анти розвитку 512
---фактори ризику 512
Хнфекцшний процес 77
— системн! прояви 86
Хнфекпдя (!) 79
— бактер!альна 82
— в!русна 83
— грибна 82
— паразитична 80
— пов!льна 85
— прюнов! 85
— протозойна 80
— рикетсшна 82
— хламщшна 82
Тритантн! рецептори 552, 569
1шем!чний порп-
— верхней 727
— нижнлй 727
1шем!я 197, 594
— ангюспатична 198
— зм1ни в тканинах 200
— компресшна 198
— обтурацГйна 198
— повна 199
Хошзуюче випром!нювання 54, 484, 664
— етюлопя 57
— патогенез 58
1онограма Гембла 379
Хонообмш 376, 381
Калгкреш 232, 263, 585
Кальцитошн 653
Каналопата 89
Капсащинов! рецептори 675
Кар!ес зубхв 573
Карюрексис 465, 477
Карликовость гшоф!зна 647
Катехоламши 502, 515, 523, 543, 547, 701
Каузалпя 677
Кахексии 330, 363
Кахекс!я
— гшоф!зна 647
— екзогенна 365
— ендогенна 365
— паратиреопривна 654
- ракова 309
— серцева 509
— тиреопривна 650
Квашюркор 336, 337
Кейлони 129, 247
Кератомаляция 444
Кетоацидоз
— патолопчний 350, 358, 380
— фгзюлопчний 358
Кетоз д!абетичний 350
Кетонур!я 349
Кислотно-основного стану порушення 376
— мехашзми компенсац!! 381
— патогенез 384
Кинни 188, 194, 231. 232, 244, 263, 585, 721
Юлмакс 659
Кттпенс фагоцитний 140
Клгтини
— Березовськото — Р!да — Штернберга 483
— дендритш 226, 249
— дифузно! нейроендокринно! системи
(АРШ) 198, 590, 635
— загибель
----ентерфазна 61
----мт готична 61
----репродуктивна 61
---- типи 95
— запалення 226
— к!лери 167
— мттотичн! 129
---стар!ння 129
— необоротно постмттотичнт 129
---старшня 130
— оборот но постмгготичн! 129
---старгння 130
— патолопчно! регенерацп еритроштпв 455,
466
— порушення функщй 88
— розмноження у культур! г29
— фгзюлопчно! регенерапп еритроцитгв 455
Коагулолапя 104, 493
— еполоня 493, 494
— класифткаптя 493
— патогенез 494
- споживання 496
Когорта
- фази 124
Коканнерогенез 293
Колаген
— антигенш властивосп 147
- бтосинтез 143
---егапи 144
----порушення 145
— глткозилювання 144
— молекулярна тетерогеншеть 146
— експериментальне моделювання хвороб 147
— роздал 147
— типп 145, 146
Колапс 522, 719
— анпогшотошчний 720
— вазодилатащйний 720
- види 719
— гшоволемтчний 719
— кардтоваскулярний 588
- кардюгеннпй 720
— ортостатичний 721
Колектини 249
Кома 350, 354, 388, 723
— пперкетонемтчна 350
— гшерлактацидемтчна 729
— гшеросмолярна 388, 729
— ппотлткемтчна 343, 727
— дтабет ич на 350, 354, 729
— кетоаттидотична 350, 729
— кетонемгчна 350, 728
— лактацидемтчна 728
— патогенез 725
— печшкова 602
--змштанэ 602
--паренхтматозна 602
--печшково-клпинна 602
--портокавалъна 602
--шунтова 602
— субтентортальна 725
— супратентортальна 725
— уремтчна 730
— фактори
--екзогенш 724
--екстраиеребролью 724
--ендогенн! 724
--штраиеребральш 724
Компартменти 386
Компенсаторна гшерфункцтя серия
- стадп 506
Компенсац! я
- мехашзми 49
— принципи 50
Комплемент 182, 233
Кона пори 561
Копдищоналгзм 41
Конституцтоналтзм 42, 114
Констигуцюнальн! типи
— аноплексичний 115
- астешчний 116
— атлетичний 116
— г!перстен!чний 116
— ппостеючний 116
— дихальний 115
— класифткашя 115
— лшоматозний 117
— мозковий 115
— м’язовий 115
— нормостентчний 116
— пастозпий 117
— пткшчний 116
— розумовий 117
— силъний повтлъний уртвноважений 117
— силъний рухтивий невртвноважений 117
— силъний рухливии уртвноважений 117
— слабкий 117
- травний 115
— фтброзний 117
— фтизичний 115
— художшй 117
Конститущя 114
— роль у патологи 114
Конформацгя 85
Концепц1я
— адитивно-полйенного успадкування з по-
роговим ефектом 107
— нервтзму 46
Кортико-вюцерально! динамтки порушення
48
Кортикотропш 635
Кразис 45
СгасЬ-синдром
— патогенез 718
Креаторея 367
Кретишзм 651
Крипторхизм 647, 649, 658
Критичш перюди 119, 120
Крововтрата 51, 452, 522
- мехашзми компенсацп 453
— стади 452—454
Кровоносне русло мткроциркуляторне 212
Ксенобютики 70, 261, 601
— бютрансформац!я 76, 601
Ксеростомтя 575
Ксерофтальмтя 444
Лаброцити 227, 229
Латиризм 147
Латирогени 147
Легенева гшертенз1я 547
Лейкемхчний провал 487
Лейкемощна реакщя 479—481
Лейкоз 481
— гострий 481
— стади патогенезу 486
— хрошчний 481
— форми 481, 482
Лейкопен1я 474
— етюлопя 474
— класифжащя 474
— патогенез 475
Лейкопоез
— порушення
---парщальне 470
---тотальне 470
— посилення
---пухлинно! природи 471
---реактивного характеру 470
— пригшчення 471
Лейкотртени 231, 263, 431, 557
Лейкоцитарно! формули змши 473
Лейкоцити
— змши юлыасш 472
— змши як1сш 477
---дегенеративн! 477
Лейкоцитоз 472
— абсолютний 472
— вщносний 472
— етюлопя 472
— класиф1кац!я 472
— патогенез 473
Лептин 330, 360, 362, 572, 635, 636
Лецитин 357
Л1бератори пстамшу 271, 273
Л1М1Т Хейфлгка 290, 297
— втрата 297
Лгмфогранульоматоз 483
ЛГмфокши 364
— групи 265
Лхмфолейкоз хрошчний 482, 487
Лтмфома 483
— Беркпта 296, 483
— Ходжкша 483
Лтмфоцитоз 473
Л1мфоцитопен1я 474
Лшем1я 333
Лппдоз 365
Литогенез 359
Лшощ А 277
Лшокаш 357
Лйюпроте'нш
— фракци 356, 529
Лшостат 363
Лшофусцин 126, 131, 134
Лшур1я 355
Макроанпопати 354
Макрофаги 139, 226, 228, 229, 265, 601, 707
Макроелементи 405
— калгй 397
— кальцш 400
— магнш 398
— натрш 396
— фосфор 402
— хлор 399
Маразм алментарний 336
Мастоцити 227, 229
Махизм 41
Медицина
— модел!
— генетична 42
— шволюцшна 43
— метабол1чна 43
— молекулярна 46
— онтогенетична 42
— психосоматична 42
Мед1атори
— пперчутливосп сповиьненого типу 264, 265
- запалення 229—233, 237-242, 244-250
---шоку (септичного) 706—709
‘Мещаюрний вибух” 706
Мезобппноген 605
Мембранопапя 104
— набута 460. 461
— спадкова 461
Метаболии аномальш
— лшщш 59
— хшонсв! 59
Метаплазгя 305
Метастази 290, 308
Мгастенгя 680
М1елолейкоз хрошчний 485, 487
Мгкроашюпатп 351
Мткровезикулярний транспорт 190
Мгкроелементи 405
— ванадш 418
— залгзо 406
— йод 424
— марганець 415
— мгдь 410
— молгбден 419
— селен 419
— фтор 425
— хром 417
— цинк 413
М1кросфероцити 462
Мткроциркуляшя 212
— порушення типов! 212, 702
---внутрштчьосудиню 213
---позасудинш 216
---проникносп спнки судин обм!ну 189,
214, 235
Микроцитоз 455, 465
Мгкседема 651
Мпохондрп 90, 317
Модель неврозу оч!кування 695
Модель нейрогенно'1 виразки 686
Молекули клпинно! адгезп 237
— апресини 238
— быки суперродини гмуноглобулшгв 237, 238
— тнтегрини 237, 238
— селектини 237, 238
Монокаузалтзм 40
Моноюни 140
Моношггоз 473
Моноцигопешя 475
Мотилш 578, 583, 587, 588
Муковгсцидоз 557
Мукопол (сахарид оз 141
Мукоцилгарний транспорт 570
Мутагени
— бюлопчш 99
— фгзичн! 99
— Х1М1ЧН1 99
Мутащяф 91, 99. 100
— бюхтмгчш наслщки 103
— гаметична 100
— герм (нативна 100
— генш 100
— шдукована 100
— корисна 100
— методи виявлення НО
---бюммгчний 111
---близи юковий 111
---генеалопчний 111
---генний ПО
---1мунолог1чний 111
---популяшйно статистичний 111
---нитолопчний 111
— соматичнг 100
— спонтанна 100
— хромосомш 100
— шкщлива 100
“М’ясне отруення” собак 597
Набряк 391, 392
— алерпйний 394
— альвеолярний 564
— запальний 394
— штерстищйний 563
- кахектичний 394
— Кв1нке 271
— легень 563, 627, 710
— мозку 603, 627
---нейрогенний 395
— пешнковий 599
---токсичний 394
---унаслщок ураження
---нирок 394
---печшки 394
---серця 394
— фактори патогенезу 392
Невагомгсть 69
Невроз 694
Недосконалий остеогенез 146
Недостатшсть
— абсолютна шеулшова 345, 348
— алшентарна быкова 367
— быково-енергетична 336
— дихання 489, 510
---вентиляшйна 550
---дисрегуля горна 551
--- обструктивна 556
---рестриктивна 559
---вплив на оргашзм 567
---гшеркалшчна 564
---гптоксемтчна 564
---форми 550
— експресп антшетпв ГКГС класу II 176
— ендокринних залоз 643
— шсултнова 345
— кагилярно-троф!чна 214
— юрково! речовини надниркових залоз гос-
тра 655
---хрошчна 656
— кровообггу 498, 522, 523
— лтмфатичнот системи 217
— лтмфообтгу 217
---динамтчна 218
---механтчна 218
---резорбцтйна 218
— нирок 615, 616, 618, 622
---гостра 625
--- стади 626
---фактори 626
---хрошчна 627
---стали 627
— печшки
---абсолютна 595
---блискавична 595
---в!дносна 595
---гепатоцелюлярна 595
--- гостра 595
---екскреторна 595
---експериментальш модел! 596
---комбшована 595
---парцтальна 595
— течтатоъо-юпта&та 5%
---печшково-судиняа 595
---'’отальна 595
---холестатична 595
---хрошчна 595
— серия 499
---вар!анти 500
--- гостра 503
---застойна 507
---мехашзми компенсацн 500—507
---хрошчна 507
— Т-лтмфоииттв первинна 176
— травления 572
— фуюоцональна зтрчастих макрофапв т рй-
клттин 601
Нейролейкоз 488
Нейропати дтабетпчш 351
Нейропептид V 330, 363, 572, 590, 636, 642
Нейротрансмттери 590, 666
Нейротрофши 688
Нейтропения спадкова 474
Нейгрофтльоз 472
Некроз 95, 334, 509, 510, 686
— аденог!поф1за 637
— канальцевий 718
— мюкарда гшоксичний 509
— пшистих клини 535
Непрохтднтсть кишок
— динамична
---парал1тична 588
---спастична 588
— мехашчна
---компрестйна 588
---обтуращйна 588
---с грангуля цтйна 588
Нткгалоптя 443
Норме вол ем!я
— олтгоцитемтчна 451
— полтттитемтчна 451
Ноцицептин 675
Нугртенти 572
Обмшу типов! порушення
— амшокислот 369
— бшкового 367
— вттамтшв 428
— водно-електрол1тного 386
— вуглеводного 338
— енергетичного 325
— жирового 355
— мткроелсменттв 405
— нуклетнових основ 374
— основного 327
— пром1жно]'о жирового 357
— тирозину 371
- ТП
Одиниця багатоклттинна основна (ОБО) 152
Ожиршня 360, 361
— алтментарно-конституцтональне 360
— андрощне 360
— вгоринне 360, 363
— пношне 361
— ггперпластичне 360
— ппертрофтчне 360
— гормональне 360
— експериментальне 361
— ендокринне 360. 364
— первинне 360, 362
— церебральне 360, 362
Олтгуртя 626
Онкогенез
— мехашзми 296—305
Онкогенш фактори 292—296
— бюлопчного походження 294—296
— ендогенш 295
— органотропн! 293, 294
— фтзичш 296
— хтмтчн! 293, 294
Онтогенез 161
Опасистт клттини 227, 229
Огак 54
Огастотонус 653
Опромшення
— внуршгне 58, 65
— зовнштне 58
— 1нкорпороване 65
Опотацидурш 375
Остеодистрофтя
— генералвована ф1брозна 654
— спадкова 620
Остеомалящя 155, 445
Остеопетроз 154
Остеопороз 154, 403, 404, 445
— локальний ’54
— патогенез 404, 444—446
— постме нопаузний генералтзований 154, 404
Пангшоштутгаризм 646
Панкреатит 584
Панмтелофтиз 470
Панцитопетпя 477
Параалерпя 27]
Парабюз 48, 668
— фази 669
Парал1ч 680
— Фелтона 272
Парез 680
Парюнсошзм 411, 684
Пародонтит ювешльний 574
Паротин 576
Патогенез 44
Патогеншсть 78
Патолопчна фтзюлопя 15
— визначення 15
- зв’язок з шшими медичними науками 15,
16
— метод 17
Патолопчний пронес 34
Патолопчна реакшя 34
Патолопчний стан 34
Патолопя
— ендокринна
---вторинна 637
---множинна 640
---первинна 637
- загальна 15
— К1стково7 тканини 152
— молекулярна 46
— напрям функцюнальний 46
— спадкова нервово! системи 666
— теорн
---гуморальна 45
---солидарна 45
---целюлярна 45
Патофтзюлопчний експеримент 17
— етапи 19. 20
- методики 18, 19
---введения в оргашзм ртзних речовин 18
---вилучення 18
---зольованих оргашв 1 тканинних культур
19
---моделювання генетично зумовлено! пато-
лога 19
—- подразнення 18
---портвняльно! патолога 19
Пелагра 438
Пенен гратшсть гена 102
Пентраксини 233
Пептид на виразка 580
Перевантаження 68
Перев’язування
— печшково! артери 597
- спйишоТ жовчно! протоки 596
Перекисне окиснення л1п1Д1в (ПОЛ) 59, 226,
263
- активашя 225, 321
- арахщоново') кислоти 263
Пермеази 367, 368
Персистенщя фолпсула 660
Печшки видалення
— повне 596
— часткове 596
Печ1я 579
Птелонефрит 633
П порос пазм 579
П1ротерашя 289
Пневмосклероз 500, 560
Пневмоторакс 559
— вентилъний 560
— вщкритий 560
— закритий 560
— клапанний 560
Подагра 374
Пойилоцитоз 455
Пол1дипс1я 350
Полтмеразна лани югова реакщя 19
Полшлоцця 92
Пол1ур1я 349, 390, 626, 648, 729
Полщикл1чю ароматичы вуглеводи! (ПАВ)
293
Пори мпохонпр1альгп 224
Пород не коло 46
Прекоматозш стали 723
Приводомшешсть 511
Прюни 85. 665
Прогери 133
Протрес1я 304
Пролтферашя 121. 222, 244, 534, 545
Промене ве ураження 57, 58
— корекдтя 67
Промошя 303
Простагландины 192, 205, 231, 263, 431, 547,
571
Протеасома 94, 369
Прогепгурщ 617, 618, 621
— ал'ментарна 618
— депдраташина 618
— маршова 018
— несправжня 618
— ортостатична 618
— поза ниркова 618
— тубулярна 621
— функцюнальна 618
Протеоглпсани 141
Протеошзу порушення
— гнзосомного 95, 368
— протеасомного 94, 369
Протоонкогени 298
Процесинг 167
Пухлина 290
— антигенш особливосп 307
— вплив на оргашзм 310
— експериментальне вщтворення 290
— експланташя 291
— злоякгсшсть 308
— шдукшя 291
— класиф1кац1я 290
— особливосп росту 290
— спйюсть до юмюпрепарапв 305
— транс плантацгя 292
--алогенна 292
Радюлгз води 58
Радюнуклщи 65
Радюпротектори 67
Радютоксини 59
Рак 290
Рах1т 445, 620
Реагши 258
Реактившсть 157
- бюлопчна 157
- видова 157
— 1рупова 158
— еволющя 161
— гмунолопчна 158
— шдивщуальна 158
— клгтинна 160
— мехашзми 162
— на молекулярному ргвш 160
— на р1вш органа 161
— неспепнфггна 158
— патолопчна 158
— первинна 157
— системи оргашв 1 оргашзму в щлому 161
— специфична 158
- фгзюпопчна 158
Реакцп нейроендокринш 700
— стадп
--- еректильна 700
---торпщна 700
Реакшя Артюса 259
Реакшя •‘трансплантат проти хазяпга” 261
Реан1мац;я 38
Резистентшсть 159
— активна 160
— вторинна 159
— неспециф1чна 160
— пасивна 159
— первинна 159
— специфгчна 160
Ремоделювання йстково! тканини 152
Ретикулоцити 455
Ретракщя згустка 206
Рефрактерна вазодилаташя 715
Рецидив 37
Речовини шрогеню 277
Рибосоми 93
Рипдшсть децеребрацшна
- патогенез 681
Ритму серця порушення 516
— альтернащя 521
— засвоення 520
— провщносп 519
— трансформащя 521
Самойлова — Венкебаха перюди 519
Саркома 290
Сатурашя 56
Свщомють 723
Секретин 577, 591
Селектини 237
Сенсибкчгзащя
- активна 256
- пасивна 257
Септицем:я 601
Синапси 669
Синдром(и)
— адаптащйний загальний
---стадц 661
— адреногештальний 657
— ахол!чний 610
— Бассена — Корнцвейга 356
— Броун-Секара 672
- Брутона 177
— Вернера 133
— Вернера — Моррюона 590. 591
— Вернгке — Корсакова 437
- Вюкотга — Олдр1ча 178. 484, 490, 491
- втрати сол! 657
- галактореТ — аменорег 648
— гематолопчний 62, 487
— геморапчний 62, 488, 493, 599
— гепатаентеральний 594
— гепатолтенальний 594. 614
— Гетчшсона — Гыфорда 133
— Г1ПОХОЛ1ЧНИЙ 6 .0
— Гудпасчера 259
— Гурлера 142
— Дауна 109. 484
- ДВЗ 204, 463, 489, 491, 495, 696, 710, 711
---еполопя 496
---класиф1кац1Я 496
---патогенез 497
---фази 497
— декомпресп 55
---вибухово'1 56
— днстрес гострий ресшраторний дорослих
(ГРДСД) 709
— Д1 Джордж! 176
— Дресслера 515
— Дубша — Джонсона 610
— гострого легеневого серця 210
— госгрого легекевсго ушкодження 710
— Елерса - Данлоса 145, 146. 493
— ендокринно! недостатносп 63
— Жыьбера — Мейленграхта 609
— Золлшгера — Еллкона 577, 580. 590. 591
— зниження )мунолопчно! реактивносп 63
— 1мунолопчно! недостатносп 488
— 1ценкэ — Кушшга 345, 656
— Картагенера 557
— Кастрацн 659
— кл!мактеричний 659
— Клайнфелтера 108, 484, 637
— Клюве ра — Бюс1 693
— Кнаппа 439
— Костманна 476
— Крйлера — Найяра 609
— Кушшга 358
— Ламберта — Иона 680
- Лцщла 539
- Лу!-Бар 178, 484
— мальабсорбцп 339, 586
— Маркю 142
— Марфана 146. 412
— Морганы — Адамса — Стокса 520
— мульти орган ко! дисфункцн (МОД) 709
— набутого !мунодефшиту (СН1Д) 181
— недостатносп нирок хротпчно! 627
— нервово-церебрапьний 63
— нирков!
---анем1чний 623
---артергально! ппертензп 623
---набряковий 622
---нефротичний 624
---сечовий 622
---урем!чний 624
---тубулоштерстицшним 629
— Ослера — Рандю 493
— шридоксинзалежний судомн ий 439
— подразнено! кишки 587
— позаюстковомозкових проявш лейкозу 488
— полюрганно! недостатносп 709
— портально! ппертензп 597, 602, 613
— постшфарктний 515
— резистентних яечниюв 645
— резорбцшно-некротичний 515
— реперфуз1йний 202
— Ротора 610
— системно! запально! вцщовип 219, 717
— спонтанно! хромосомно! несгабьтьносп 104
— Тернера 108, 48ч
— тестикулярно! фемш’ваш! 645, 659
— тривалого розчавлювання 51, 718
— тромбогеморапчний 204
— тромбофшчний
---етюлопя 495
---класиф1кап1я 495
---патогенез 49ь
— губулошгерстицшний 629
— Фан кон) 620
— холем)чний 610, 611
— Чед!ака — Х1гас! 183, 472
— Шерешевськот о — Тернера 108
— шлунково-кишковий 63
— Штейна — Левенталя 638, 649
Син канцерогенез 293
Система(и)
— антеролагеральна 672
— анта гемостазу 495
— антаноципептавш 678
— гемостазу 488
- ентерична нервова 666. 692
- комплементу
---активашя 262, 708
---порушення 182
— лемшскова 672
---порушення 673
— л!мб1чна 678. 692
---ушкодження 693
— метасимпатична 692
— плазмов) протеол1тачн! 708, 714
— симпатико-адреналова 389, 543, 701
- ферменпв реларацп 101
Скреп! 85, 93
Сладж-феномен
— необоротний 213
— оборотиий 213
Смерть мозку 724
Соматомедин 648
Соместез1я 672
Сопор 723
Сонячний удар 54
Спадкова схильшсть 109
Спадковгстъ 98
— епггеномна 92
Сшнальний воргтний мехашзм 676, 679
Спланхномегалгя 647
Сполучна тканина як фгзюлоггчна система
137
Стадп компенсаторно! лперфункцп серця
— аваргйна 506
— завершения гшертрофп та вщносно стшко!
пперфункцп 506
— поступового виснаження г прогре суюч ого
кардюсклерозу 507
Сгаз 202, 234
— венозний 202
— гшемгчний 202
— справжшй 202
Старгвня 123, 154, 663
— гетеротопнгстъ 126
— гетерохроннгсть 126
— змгни
— клгтин 129
---молекул 131
---систем г оргашв 126
— мехашзми розвитку 134
— причини 133
— шляхи впливу 135
Статеве дозргвання передчасне 660
Стеаторея 355, 584, 599, 612
Стенокардия 511
Стеркобглгн 605
Сгеркобглгноген 605
Стрес 48, 180, 532. 540, б61
— емоцгйний 694
Судини
— емнгснг 525
— компенсашйнг 524
---патологгчнг змгни 537
— обмгну 525
— О1 гору 524
— резистивш 524
---патолопчнг змгни 537
Судоми епыептичнг
— фази 685
Супресорна функцгя лгмфоцитгв 168
Счрфактанти 571
— дефгцит 560
Сурфактанта э система легень
— порушення 560
Схуднення 358. 365
Таласемгя 462
Тампонада серця 503
— гостра 523
Тахгкардгя 502, 506, 517, 652, 715
— пароксизмальна 519
Тахшное 552, 568
Тварини
— гомойотермш 276, 280
— зрглонароджеш 161
— незрглонароджеш 161
— пойкглотермнг 161, 280
Телеанггекгазгг 493
Теоргя (!)
- атеросклерозу
---апгментарна 536
---гнфшьтрацггЧно-комбгнацгйна С С. Хала-
това 536
- вщповгдг на ушкодження 536
-запалення
---бголопчна 252
---бюхгмгчна 253
---еволюцгйна 221
---судинна Ю. Конгейма 252
---фгзико-хгмгчна 252
— гмовгрнгена 64
— Селье 661
— мгшею 59,64
— Старлшга 189
Тепловий удар 54
Термшальнг стани 37
— агонгя 37
— преагогпя 37
— смерть
---бголопчна 37
---клгнгчна 37
Термогенез
— нескоротливий 283
— скоротливип 282
Тетангя паратиреолривна 653
Типов! патологгчнг процеси 16, 34, 186
Тиреотоксикоз 522, 652
Тирозиноз 371
Ткашгню базофгли 227, 229
Токсигеннгсть 78
Токсогормон 310
Толерантшсть
— високодозова 171
— до вуглеводгв 344
— гмунолопчка 171, 269
- низьколозова 171
Травления
— мембрани е 583
< — порожнин не 583
— порушення 513
- у порожнинг рога 573
---у дыунку 576
---кишковог о 583
---мембранного 585
Травма
— електрична 67
— мехашчна 51
- променева 57
— тсрмгчна 51
Трансамшування порушення 369
Транслокащя 92
Транспортери
— глюкози 339, 340. 349, 352, 388, 418, 422,
619
— зашза 406, 407
— М1Д1 410
— цинку 413
Транспорт
— активний 189
---первинно активний 618, 619
---вторично активний 618, 619
— пасивний 189
---проста дифуз1Я 189
---полегшена дифузы 618, 619
Транспульмональний тиск 558, 559
Трансферини 407. 464
Трансформащя 296
- мехашзми 296—303
— пухлина кровотворних юптин 484
---мехашзм 485
Трисомтя
— автосом 109
— за Х-хромосомою 108
Тр1ада Верхова 203
Тромбоемболш 208
Тромбоз 203, 495, 593
— локал ьний 495
— наслщки 206
Тромбоутворення 463
— юптинна фаза 204
- плазматична фаза 205
Тромбоцитопати 102, 491
Тромбоцитопешя 489
- класифткащя 489
— патогенез 490
Трофогени 48
Туберкулшова реакщя Манту 260
Тубулопати комбгноваш 620
Тучн! юптини 227, 229
Убгкв-тиюзащя 369
Умирания 37
Уремия 624, 628
Уробшн 605
Уробйтшоген 605
Уроб1л1ногенем1я 611
Уроб)Л1нур1я 611
Успадкування
— зчеплене 31 статевою хромосомою 106
— типи
---автосомно-домшантний 106
---домшантний 106
---пол ненний 107
---рецесивний 106
Ушкодження
— мембран 321
— серця
---1мунн1 510
---коронарогенш 511
---нейрогент 515
---некоронарогенш 509
Фаю.изосома 242
Фаюсома 242
Фагоцитоз 183. 221, 241
— порушення 183
— стадп 241
Факгор(и)
— алерпзуюч! 254
— атерогенезу 527
- Вшлебранда 431, 492, 708
- Касла
---внутртштй 440, 441,453
— ендотелшзалежний вазорслаксацп (МО) 194
— етюлопчш 51
- канцерогенш 180
— кондищональш 663
— мозковий нейротрофгчний 688
— молока Б1ттнера 291
— мутагеюн 180
— некрозу пухлин 152, 231, 233, 238, 310, 353,
706
— онкогенш 292
— пептичний 581
— перенесения 265
— постренальн! 626
— преренальш 625
— просвгглення 356
— реналью 626
- ризику 109
- росту 246-248, 299, 303
---егндермальний 246, 248, 583, 688
---шсулшоподтбш 246, 688
---нервов 246. 688
---тромбоцитний 246, 248, 688
— ф1зичш 664
- флогогешп 221, 250
— Хагемана 262
---активащя 232, 235
- цилтарний нейротрофгчний 689
- цитостагичю 180
Фемтшзащя 659
Фешлкегонургч 370
Фенокопп 98, ПО
Феномен
— агрегацп еритроципв 203
— пбериаци мюкарда 511
— крайового стояния лейкоципв 236
— Санарелл! 272
— Швартцмана 272
Феохромоцитома 658
Ферментопатц 103, 460, 462
- спадков! 460, 462, 598
Фегопапя 120. 122
— неспеиифзчна 122
— специф!чна 122
Ф1бриля1йя
— передсерд ь 521
— шлуночкгв 522
Фибробласт и 139, 229
Ф1броплаз1я 247
ФЬюгенез 161
Ф1стула
— Екка — Павлова 596
— Екка пряма 596, 597
— зворотна 596
Фоби 696
Фруктозсмтя 339, 598
Фруктозуртя 339
Фунгкулярний М1ЕЛОЗ 441
Хвороба(и) 30, 33, 35, 40. 44
— автоатерпчш 269
- автозмунн! 142, 148, 259. 269, 369, 459. 469,
491, 639
— апапташ! 663
— Ащпсона — Ырмера 441
— Ашцсона (бронзова) 656
— Альдера 184
— Атьпгеймера 369, 411, 576
— антигенасоцшован! 184
— Барракера — Симонса 362
— берьбер! 437
— Бернара — Сулье 491
— Вакеза 457
— Вальденстрема 482
— вари козна 196
— Верльгофа 259
— визначення 30
— Вылебранда 492, 536
— В1льсона — Коновалова 412
- Вюкотга — Олдртчэ 178. 490. 491
— Вольмана 365
— вроджен! 98
— гемолггичча новонароджених 461, 480, 608
— генетично детерм1нован1 98
— Герса 340, 539
— Герстманна — Штрауслера 85
— палгнових мембран 561
— гшерпчн! 159
— гшерерпчн! 159
- Прке 340, 598
— Прпгпрунга 692
— Гланцманна — Негел! 491
— гострого легеневого серця 210
— Гоше 365
— Грейвса 652
— Дауна 109. 484
— дифузн! сполучно! тканини 148
— ендокринн]
----мехашзми 639
— Ыерслунда — Гресберка 441
— 1мучодеф1цитн1 173
— 1ценка — Куппнга 345, 358, 398. 642, 648, 656
— Кальмана 637
— Кашина — Бека 576
— кератоз Дар’е 444
— комбшован! гмунодефшитш 177
— Конна 657
- Крейтцфельдта — Якоба 85, 93
- Крона 586
— кригерП клаеифжацй 33
— Леффлера 472
— л1мфопрол1феративш 482
— Марека 483
— Маркзафави — М1кел1 461
— Менкеса 412
— м!елопрол1феративь1 482
— Мшковського — Шоффара 461
— молекулярно-геню 102
- накопичення 95, 103, 140, 326, 340, 365
— нейроендокринш 649
- ЕИманна — П1ка 365
— ошкова 54
— основн! перюди
----виражених проявш 35
----завершения 35
----кгнець 35
----латентний 35
---- продромальний 35
— Паркшсона 411. 684
— Педжета 155
— прюнов! 85, Зб9, 665
- променева 65
----гостра, форми 65
----кишкова 66
----юстковомозкова 65
----перюди 65
----церебральна 66
----хрошчна 66
— Рандю — Ослера — Вебера 493
- Реклшгхаузена 106, 576
— серця гшемзчна 511
— сироваткова 260
— Сгммопдса 647
— Слая 142
— спадковг 98, 102
--вггамш залежш 104
--експериментальне моделювання 113
— тромбоемболгчна 495
— умови виникнення 44
— фшософське узагальнення 31
— Форбса 340
— Ходжкгна 483
— хромосоми! 108
— Шенляйна — Геноха 492
— Шюллера — Христгана 365
Хемоатрактанти 239—241
Хемотаксис лейкоцитгв 239, 241
Хтломгкрони 213, 356, 529
Холалемгя 611
Холал ургя 611
Холелгтгаз 612
Холемгя 611
Холестаз
- внутршшьопечшковий 595, 596, 607, 610
— позапечшковий 595, 596, 611
Хорея 684
— Гентшгтона 102, 576, 684
Хромосомна аберашя 100
Хромосомна аноматгя 100, 108
Целгаюя 587
Централгзацгя кровообггу 701
Циклгни 92
Цилгндрургя 621
Цинга 442
Цироз
— бшарний 597
--первинний 610
- птслянекротичний печгнки 596
- портальний 597
— токсичний 596
Цитокгни 231, 233, 240, 706-709, 714, 715
— пгзнг 706
— грозапальчг 231, 233, 240, 714
— ранит 706
Цитолгз 89, 225, 226, 259
— мехашзми 224—226, 259, 262, 263
— реакцгя 259
Цитотоксичн! реакци’ 259
Шсшоз 507
Шлункового соку
— гшерсекрецтя 577
— гшосекрецгя 577
Шок 697
— анафглактичний 258, 716
--мехашзми 257
— види 698
— геморапчний 453, 711
— пповолемгчний 391, 711
- дистрибутивний 701
— етапи розвитку 699
— етгологгя 697
- кардюгений 515, 712
--аритмтчний 713
--патогенез 713
--рефлекторний 712
--снравжнгй 713
— обструктивний 712
— опгковий 716
— нанкреатичний 585
— нерерозподьтьний 701
— септичний 713
— слшатьний 681
— травматичний 51, 717
--стадп 717
Шоковг органи 709
Ядерний зсув нейтрофгльних гранулоцитгв
478
— кгльктсна оцтчка 479
— влгво 478
— вправо 479
Навчальне видання
Биць Юрш Викторович
Бутенко Геннадш Михайлович
Гоженко Анатолш Тванович та ш.
ПАТОФ13ЮЛОГ1Я
Пщручник
За редакщею
професора М.Н. Зайка,
професора Ю.В. Биця,
професора М.В. Кришталя
Подписано до друку 25.11 2013.
Формат 70x100 1/16. Патр офсет.
Гарн. Тппез. Друк офсет.
Ум.-друк. арк. 61,1.
Зам. № 13-552.
ВСВ “Медицина”
01030, м. Ки!в, вул. Стрыецька, 28
Свщоцтво про внесения до Державного реестру видавщв,
вигопвникш 1 розповсюджувашв книжково! продукцц
ДК № 3595 вщ 05.10.2009
Тел.: (044) 581-15-67, 234-58-11
Е-таП: те<1@8ос1е1у.1аеу.иа
Вщдруковано на ПАТ“Б1лоцергавська книжкова фабрика”,
09117, м. Б1ла Церква, вул. Леся Курбаса, 4.
Свщоцтво сер1я ДК № 4063 вщ 11.05.2011р.
Впроваджена система управлпшя яюстю
згщно з мгжнародним стандартом ПП4 ЕМ 180 9001:2000
ПАТ0Ф13ЮЛ0ПЯ
Шдручник подготовлений авторським колективом сшвроботниюв
вищих медичних навчальних закладов IV ровня акредитацп 1 академоч-
них науково-дослодних заклада Украши, яю е визнаними фаховцями
з розних питань патофозоолопГ, що дае змогу вважати цей базовий под-
ручник нащональним.
Поровняно з останшми виданнями суттево перероблеш таю роздши,
як “Патолопчна фозюлопя омунно! системи”, “Алерпя”, “Запалення”,
“Пухлина”, “Порушення вуглеводного обмшу”, “Порушення кислотно-
основного стану”, “Порушення водно-електролИного обмшу”, "Розла-
ди кровообпу, пов’язаш з порушенням функцп судин” та ш. Заново на-
писано роздолоо “Патофозоолопя обмшу мо'крослементов”, “Патофозооло-
пя обмшу вотамонов”, “Вибухова шахтна травма”.
Матероал подручника викладено водповодно до ново! навчальноТ
програми з патолопчно! фозюлогн, складено! на осново кредитно-мо-
дульно! системи оргашзащ! навчального процесу.
Для студентов вищих медичних навчальних закладов IV ровня акре-
дитацп. Подручник може бути корисиим для викладачов о локаров усох
спещальностей.