/































































































































































































































































Author: Горбунова Н.П.
Tags: систематика растений ботаника водоросли пособие для вузов
ISBN: 5-06-000641-7
Year: 1991




Text
Н.П. Горбунова
АЛЬГОЛОГИЯ
Допущено Государственным комитетом СССР
по народному образованию
в качестве учебного пособия
для студентов
высших учебных заведений,
обучающихся по специальности
«Ботаника»
Москва
«Высшая школа» 1991
ББК 28.591
Г 67
УДК 582.26
Рецензенты:
кафедра ботаники Горьковского государственного университета
им. Н. И- Лобачевского (зав. каф. доц. Г. А. Юлова); докт. биол.
наук Н. П. Масюк (Институт ботаники им. Н. Г. Холодною
АН УССР)
Горбунова Н. П.
Г 67 Альгология: Учеб, пособие для вузов по спец.
«Ботаника» — М.: Высш, шк., 1991.—256 с.: ил.
ISBN 5-06-000641-7
Рассмотрены морфология, ультраструктура, онтогенез, систематика,
филогения и экология водорослей. Приведены новые данные о цитологии,
жизненном цикле и систематике, обусловленные применением электронной
микроскопии и новыми принципами культивирования водорослей. Обо-
бщены сведения по родам водорослей.
1906000000 (4309000000)—248
Г----------1---------------13—91
001(01)—91
ББК 28.591
582
Учебное издание
Горбунова Нина Павловна
Альгология
Редактор Н А. Соколова
Младшие редакторы Е. В. Бурова и Е. И. Попова
Художник В. Н. Хомчков
Художественный редактор Т. А. Коненкова
Технические редакторы Л. А. Муравьева, Г. А. Фетисова
Корректор С. К. Завьялова
ИБ№8176
Изд. № Е-568. Сдано в набор 26.07.90. Подп, в печать 22.03.91. Формат 60x88Tj<>. Бум.
офс. № 2. Гарнитура Таймс. Печать офсетая. Объем 15,68 усл. псч. л. 15,68 уел. кр.-отт.
17,97 уч.-изд. л. Тираж 5200 экз. Цена 1 р. 50 к.
Издательство «Высшая школа», 101430, Москва, ГСП-4, Неглинная у.г., л. 29/14.
Отпечатано с диапозитивов ордена Октябрьской Революции и ордена Трудового Крас^
ного Знамени МПО ,,Первая Образцовая типография” Госкомпечати СССР, 113054,
Москва, Валовая, 28 в Московской типографии N4 8 Госкомпечати СССР, 101898,
Москва, Центр, Хохловский пер., 7, Тип. зак. 1210.
ISBN 5-06-000641-7 € н. П. Горбунова, 1991
ОГЛАВЛЕНИЕ
Предисловие ........................................................... 4
Введение .............................................................. 5
Глава 1. Систематика водорослей ................................... 11
Глава 2. Морфология ............................................... 15
Глава 3. Цитология эукариотических водорослей ..................... 17
Глава 4. Размножение .............................................. 41
Глава 5. Отдел синезеленые водоросли .............................. 54
Отдел прохлорофитовые водоросли ............................ 7]
Глава 6. Отдел красные водоросли ................................... 72
Глава 7. Отдел золотистые водоросли ................................ 96
Глава 8. Отдел желтозсленые, или разножгутиковые, водоросли ....... 113
Глава 9. Отдел диатомовые водоросли ............................... 122
Глава 10. Отдел бурые водоросли .................................... 139
Глава 11. Отдел линофитовые водоросли .............................. 162
Глава 12. Отдел криптофитовые водоросли ............................ 179
Глава 13. Отдел рафидофитовые водоросли ............................ 183
Глава 14. Отдел эвгленофиговые водоросли ........................... 184
Глава 15. Отдел зеленые водоросли .................................. 189
Указатель латинских названий......................................... 253
Светлой памяти профессоров и преподавателей
Московского университета-.
Л. И. Куреанова, К. И. Мейера,
Н. И. Цешинской, Н. А. Комарницкого,
М. А. Пешкова
посвящается автором
ПРЕДИСЛОВИЕ
Как показывает название, водоросли живут преимущественно в воде (покрыва-
ющей 3/4 поверхности Земли), хотя некоторые из них встречаются в наземных
условиях: на почве и в ее толще, на коре деревьев и г. и. Распространенные по всему
земному шару, они играют колоссальную роль в жизни природы, которая в первую
очередь определяется их особенностями как фотоавготрофных организмов. В воде
они являются основными создателями органического вещества, поэтому все живо-
тное население водоемов обязано своим существованием водорослям — первому
звену пищевых цепей, Другое очень важное значение водорослей состоит в том, что
в процессе фотосинтеза они выделяют свободный кислород, необходимый для
дыхания водных организмов, как животных, так и растительных. Следовательно,
в водной среде водоросли определяют уровень биологической продуктивности водо-
емов. Наземные водоросли важны как пионеры растительности на бесплодных,
каменистых участках суши, как накопители первичною гумуса, подготавливающие
возможность поселения других растений. Повышая содержание кислорода, улучшая
структуру и накапливая в почвах связанный азот, водоросли способствуют и новы
шению их плодородия.
Повсеместное распространение водорослей в природе и массовое развитие в раз-
ных местообитаниях определяет большую роль их в практической деятельности
человека, роль неуклонно возрастающую. Положительная роль обусловлена участием
в создании плодородия почв, азотфиксацией, осуществляемой синезелепыми водоро-
слями. Существуют съедобные водоросли- источники белков, жиров, углеводов,
а также водоросли, идущие на изготовление лекарственных и биологически активных
веществ и т. д. Отрицательное значение водорослей связано с цветением воды
в водохранилищах и каналах, засорением агрегатов электростанций, фильтров водо-
проводных систем, замором рыб. Встречаются токсические водоросли.
Колоссально возросло и теоретическое значение водорослей в качестве объектов
исследования при решении физиологических, биохимических, биофизических, эколо-
гических и общебиологических проблем; они являются также объектами космических
исследований. Таким образом альгологические знания становятся необходимыми
для специалистов самых разных областей народно, о хозяйства.
Между тем наблюдается острый дефицит в отечественной учебной алыологичес-
кой литературе, особенно такой, которая отражала бы современное состояние этой
науки.
Стремительное развитие электронной микроскопии и прогресс в культивировании
водорослей в 40 — 50-х годах внесли принципиальные изменения в представления
о цитологии, онтогенезе и систематике водорослей. Эта новая информация пока
слабо отражена в тех немногочисленных изданиях по водорослям, которые у нас
имеются.
Предлагаемое учебное пособие преследует цель восполнить это, пробел, Изложе-
ние материала как в общей, так и в специальной части (при описании отдельных
родов) основано на обобщении данных последних десятилетий.
Автор
ВВЕДЕНИЕ
^Альгология1 (иначе фикология)— наука о водорослях. Крупные,
главным. образом морские, водоросли были известны человеку
с древнейших времен, и в приморских странах, особенно в Китае,
Японии, они издавна использовались в хозяйстве человека. Однако
начало альгологии как науки можно датировать только серединой
XVIII в., точнее 1753 годом, когда К. Линней в своем 24 классе
Cryptogamia выделил порядок Algae (водоросли). В линнеевском
понимании порядок Algae включал печеночные мхи, лишайники,
губки и собственно водоросли. Среди последних он различал всего
четыре рода: Chara, Fucus, Ulva и Conferva. Огромный авторитет
Линнея в какой-то мере затормозил развитие альгологии, так как
еще на протяжении более 50 последующих лет открываемые много-
численные новые виды водорослей относились ботаниками-альго-
логами все к тем же чет ырем родам Линнея. Только в начале XIX в.
появляются описания новых родов, и с 1800 по 1875 г. рядом
авторов, среди которых отец и сын Агарды (К. A. Agardh,
I. G. Agardh), немецкий исследователь Кютцинг (F. Kutzing), из-
вестный своим 20-томным атласом, содержащим до 2 тыс. таблиц
с изображениями морских и пресноводных водорослей и др.,— было
описано подавляющее большинство известных в настоящее время
родов водорослей. Имена этих авторов стоят после соответству-
ющих родовых названий. Установление большого количества но-
вых родов вызвало необходимость группирования их в таксономи-
ческие единицы более высокого ранга. Первые попытки сгруппиро-
вать роды в семейства и порядки базировались исключительно на
внешних признаках таллома; так, нитчатые водоросли (тогда их
называли членистыми) группировались вместе независимо от их
окраски. Первым, кто выдвинул окраску таллома в качестве фун-
даментального признака для установления больших таксономичес-
ких групп, был английский ученый В. Гарвей (W. Н. Harvey), уста-
новивший в 1836 г. такие крупные ряды, как Chlorospermeae — зеле-
ные водоросли, Melanospermeae — бурые . водоросли, Rhodosper-
meae — красные водоросли, позднее переименованные соответст-
венно в Chlorophyceae, Phaeophyceae, Rhodophyceae. Как известно,
этот признак пигментации не утратил своего значения и в наши дни,
1 От лат. alga - водоросль и греч. logos—наука.
'5
так что различные крупные группы водорослей распознаются в пер-
вую очередь по набору пигментов, что и отражено в их на-
именованиях. В гарвеевском понимании Chlorophyceae включали,
помимо зеленых, синезеленые и разножгутиковые, или желтозе-
леные водоросли, позднее выделенные в самостоятельные классы
Cyanophyceae и Heterocontae (Xanthophyceae). Диатомовые водо-
росли (Bacillariophyceae) еще во времена Гарвея рассматривались
как отдельная группа — класс Bacillariophyceae, сначала включа-
вший и десмидиевые водоросли, впоследствии отнесенные к зе-
леным водорослям.
В начале XIX в. альгология, как и вся ботаника, носила чисто
описательный характер. Однако в середине XIX в. наряду с чисто
описательной — классификаторской ботаникой1 начинает бурно раз-
виваться новое направление, заключающееся в изучении клеточного
строения растений и особенно их онтогенеза - направление, кото-
рому его приверженцы дали наименование «научной ботаники».
40—50-е годы прошлого века знаменуются крупнейшими открытия-
ми в области онтогенеза водорослей, важная составная часть кото-
рого — бесполое и половое размножение. Лишь в середине XIX в.
У было твердо установлено значение зооспор как нормальных элемен-
тов размножения водорослей. Впервые зооспоры наблюдались у во-
шерии (Vaucheria): в 1807 г. Трептеполем (Trentepohl) и в 1843 г.
Ф. Унгером (F. Unger), но были истолкованы не как клетки беспо-
лого размножения, а как «превращение растения в животное».
Аналогичная ситуация имела место и при истолковании феноме-
на полового воспроизведения. Еще в конце XVIII в. у спирогиры
(Spirogyra) впервые наблюдалось образование зигот, а в 1803 г.
Ж. Воше (J. Vaucher) то же самое отметил у спирогиры, а также
выявил половые органы у водоросли, названной по его имени
вошерией. Тем не менее вплоть до середины XIX в. господствовало
мнение, что половой процесс у водорослей отсутствует, причем это
выдвигалось даже как характерный для талломных растений
признак. Переворот в таком подходе наступил только в 1854 -
1855 гг. после появления замечательных работ французского
альголога Ж. Тюрэ (G. Thuret) и немецкого — Н. Прингсгейма
(N. Pringsheim). В 1854 г. Тюрэ экспериментально доказал оплодот-
ворение у рода фукус (Fucus): он выяснил, что яйцеклетка у фукуса
развивается в ооспору только в том случае, если она соединяется со
сперматозоидом. За этим последовало наблюдение Прингсгейма
в 1855 г. над процессом проникновения сперматозоида в яйцо
Vaucheria sessilis, а спустя год он же увидел слияние сперматозоида
с яйцом у зеленой водоросли эдогониума (Oedogonium). Таким
образом было неопровержимо доказано слияние различных в поло-
вом отношении — мужских и женских — клеток и возникновение
1 Эколого-флористичсские работы по о i дельным регионам продолжаются
и в настоящее время; в СССР они даже преобладают.
6
I
зигот как прямого следствия этого полового акта. Ряд авторов,
в том числе А. де-Бари (A. De Вагу), подтвердили наблюдения
Прингсгейма над половым процессом у вошерии и эдогониума.
В 1856 г. Ф. Кон (F. Cohn) обнаружил оогамию у вольвокса
(Ио/гох), а в 1867 г. Э. Борнэ и Ж. Тюрэ (Е. Bornet, G. Thuret)
описали строение половых органов и половой процесс у красных
водорослей.
Новый шаг вперед в изучении полового воспроизведения у водо-
рослей опять связан с именем Н, Прингсгейма, который в 1870 г.
описал у пандорины (Pandorina тогит) половой процесс в виде
слияния гамет одинакового размера. Это окончательно поколебало
господствующую долгое время точку зрения, что половой процесс
обязательно связан с морфологической дифференцировкой гамет на
мужские и женские. После открытия изогамии у пандорины, а спу-
стя год К. Крамером (С. Cramer, 1871) — у улотрикса (Ulothrix)
конъюгация у спирогиры и других конъюгат окончательно признана
в качестве полового процесса. Вскоре (1876—1879) у кутлериальных
(Cutleriales) был обнаружен половой процесс, связующий изогамию
и оогамию и заключающийся в копуляции макро- и микрогамет,—
гетерогамия.
В России онтогенетическое направление в изучении водорослей
успешно развивалось И. Н. Горожанкиным и его многочисленными
учениками'.
В конце XIX в. было установлено, что важнейшим этапом поло-
вого процесса является слияние ядер, т. е. кариогамия, после чего
возник интерес к наблюдениям за поведением ядер при половом
процессе. В 70—80-е годы XIX в. в альгологии начал быстро рас-
пространяться цитологический метод исследования. Первым слия-
ние ядер в зиготе спирогиры наблюдал Ф. Шмитц (Fr. Schmitz,
1879). Он также установил многоядерность клеток кладофоры
(Cladophora) и некоторых сифоновых водорослей. В России цитоло-
гический метод к изучению онтогенеза у водорослей впервые при-
менил ученик И. Н. Горожанкина Л. И. Курсанов. Поскольку поло-
вой процесс сопровождается слиянием ядер и удвоением числа
хромосом, в цикле развития водорослей с половым размножением
неизбежен процесс возникновения гаплоидных ядер из диплоидных,
и в конце прошлого века начались и до сих пор продолжаются
исследования по обнаружению момента редукционного деления
в жизненном цикле той или иной водоросли. На протяжении второй
половины прошлого века вырабатывается и принятая в настоящее
время терминология (зооспоры, гаметы, изо- и оогамия, ооспора,
спорофит, гаметофит и пр.).
Большой материал, накопленный в ходе изучения цитологии
и онтогенеза водорослей, подготовил почву для перехода альгологии
1 Содержание этих работ приводится ниже при описании соответствующих
родов.
7
в новый период ее исюрии, который начинается с самых первых лет
XX в. В области систематики он знаменуется, во-первых, разработ-
кой учения о происхождении различных групп водорослей из разных
групп окрашенных жгутиковых и, во-вторых, объяснением явления
параллелизма в развитии отдельных классов и типов водорослей.
Разработка того и другого вопроса неразрывно связана с именем
чешского альголога А. Пашера (A. Pascher). который, по существу,
создал современную систематику водорослей.
До начала XX в. все передвигающиеся с помощью жгутиков
окрашенные одноклеточные и колониальные организмы относили
к отдельному классу Flagellatae среди Protozoa или же выделяли
в отдельный тип или класс Flagellatae. Исключение составляли лишь
зеленые жгутиконосцы — вольвокальные, которые еще Л. Рабен-
хорст (L. Rabenhorst, 1863) включил в систему зеленых водорослей.
Кроме того, когда из зеленых водорослей был выделен класс жел-
тозеленые, то к нему были отнесены и некоторые пигментирован-
ные разножгутиковые подвижные формы. Между тем еще в конце
XIX в. Г. Клебсом (G. Klebs) и др. была продемонстрирована гете-
рогенность группы окрашенных жгутиконосцев, включающей не-
сколько различных линий или рядов, постоянно различающихся
числом и расположением жгутиков, природой пигментов, формой,
в которой накапливаются в клетке продукты фотосин теза, и други-
ми менее константными и потому менее важными признаками.
Ф. Ольтманнс (F. Oltmanns, 1904) ликвидирует единый класс
Flagellatae, разбивая его на четыре класса, которые он поместил
среди водорослей, подчеркивая этим филогенетические связи жгут и-
коносцев с водорослями. Полную ясность в понимание отношений
между жгутиконосцами и водорослями внес А. Пашер (A. Pascher,
1914, 1931), который разработал учение о происхождении различ-
ных групп водорослей1 от различно окрашенных жгутиконосцев.
Одно из самых веских доказательств такого происхождения, поми-
мо сходства в строении и химизме клетки,— существование подви-
жных (жгутиковых) репродуктивных стадий у неподвижных в веге-
тативном состоянии водорослей. В целом это было установлено
в ходе онтогенетических исследований водорослей еще в середине
XIX в. Так, зооспоры неподвижных зеленых водорослей подчас
неотличимы от жгутиконосцев зеленого цвета; зооспоры разнож-
гутиковых водорослей имитируют жгутиконосцев с неравными и не-
одинакового строения жгутиками; зооспоры золотистых неподвиж-
ных водорослей очень сходны с одноклеточными жгутиконосцами,
имеющими золотистую окраску; наконец, зооспоры динофитовых
водорослей, неподвижных, нитчатых или иной формы весьма напо-
минают своеобразных жгутиконосцев, у которых на теле имеются
1 Однако необходимо помнить, что современные жгутиковые сами являются
продуктом длительной эволюции и, несомненно, построены гораздо сложнее, чем тс
первичные жгутиконосцы, которые и стали предками большинства водорослей.
8
1
ложбинки или бороздки (поперечная и продольная). Факты подо-
бного рода привели к тому, что во всех современных системах
отдельные группы окрашенных жгутиковых включены в отделы
водорослей в качестве самых примитивных их представителей1.
К ранее указанным классам водорослей Пашер добавил еще ряд
классов: Chrysophyceae. Dinophyceae, Cryptophyceae,
Euglenophyceae, исходные формы которых — подвижные жгутико-
вые. различно окрашенные организмы, прежде объединяемые в од-
ну таксономическую группу. Таким образом. Пашер заложил ос-
новы современной систематики водорослей. (Об объяснении им
параллелизма в разви тии отдельных классов водорослей см. гл. 2.)
С 1945 г. начинается современный период в истории альгологии.
Для этого периода характерно, во-первых, использование элект-
ронной микроскопии для изучения ультраструктуры водорослей, а,
во-вторых, бурное и чрезвычайно продуктивное развитие методов
искусственного выращивания как пресноводных, так и морских
водорослей в условиях лаборатории. Именно этими факторами
объясняется «взрыв» информации, относящейся к ультраструктуре,
а также к вопросам размножения и циклов развития самых разных
водорослей, нередко побуждающей пересматривать давно устано-
вившиеся представления. Новая информация по ультраструктуре
водорослей в свою очередь тесно коррелирует с усовершенствовани-
ем методов электронной микроскопии. Первый «взрыв» информа-
ции такого рода наблюдался в конце 40-х и начале 50-х годов, когда
для изучения жгутиков различных водорослей был применен недав-
но разработанный метод напыления. Следующий «взрыв» инфор-
мации в конце 50-х годов связан с усовершенствованием микроско-
пической техники и внедрением ультрамикротомов для получения
ультратонких срезов, позволяющих изучать детали внутреннего
строения клетки. С середины 60-х годов введение техники замора-
живания— скалывания сделало также возможным изучение поверх-
ностей клеток, стенок мембран и органелл. В последние годы боль-
шое развитие получила сканирующая электронная микроскопия,
ставшая важным орудием исследования панцирей диатомовых во-
дорослей, изучения поверхностей клеток десмидиальных, диноф-
лагеллят и многих других водорослей. Один и тот же объект
нередко изучается комплексно, с привлечением всех вышеназванных
методов. Если первые два десятилетия исследования тонкой струк-
туры водорослей носили описательный характер, то в последнее
время данные электронной микроскопии широко используются для
расшифровки функций клетки и органелл.
Важно отметить, что полученные в ходе этих исследований
данные отражены в предлагаемом пособии. Однако небольшой
объем пособия позволяет сделать это только в отношении
' При этом окрашенные жгутиковые (Flagellatac) как особая систематическая
группа (прежде они трактовались как особый тип) отпадают.
9
сравнительно немногих, самых широко распространенных и из-
вестных родов. По этой же причине соответствующий иллю-
стративный материал, который можно найти в любом бота-
ническом курсе, в частности в «Курсе низших растений» (1981),
не приводится.
В предисловии указывалось на значение водорослей в качестве
объектов исследований при решении физиологических, биохимичес-
ких, общебиологических и других проблем. Ниже достаточно под-
робно приводятся результаты некоторых исследований, полученных
на альгологических объектах и представляющих общебиологичес-
кий интерес (работы по химической природе аттрактантов, диф-
ференцировке клеток, регенерации талломов, получению вегетатив-
ных гибридов и др.). Изложение же работ физиологических, биохи-
мических, генетических выходит за рамки настоящего учебного
пособия1.
Данные по распространению, экологии, практическому исполь-
зованию водорослей излагаются в систематической части при опи-
сании соответствующих таксонов.
’ Интересующимся можно порекомендовать сводки: «Algal Physiol, and Biochem.
(1974) и «The Genetics of Algae» (1976).
Глава 1
СИСТЕМАТИКА ВОДОРОСЛЕЙ.
КЛАССЫ, ОТДЕЛЫ ВОДОРОСЛЕЙ
К началу XX в. различали следующие классы водорослей (по
А. Пашеру):
Синезеленые водоросли — Cynophyceae
Красные водоросли — Rhodophyceae
Зеленые водоросли — Chlorophyceae
Золотистые водоросли- Chrysophyceae
Желтозеленые, или Разножгутиковые, водоросли —
Xanthophyceae, или Heterocontae.
Диатомовые водоросли--Bacillariophyceae, или Diatomeae.
Бурые водоросли — Phaeophyceae
Динофициевые водоросли — Dinophyceae
Криптофицисвые водоросли — Cryptophyceae
Эвгленофициевые водоросли — Euglenophyceae
Каждый класс характеризуется специфическим набором пигмен-
тов, запасным продуктом, откладывающимся в клетке в процессе
фотосинтеза, и если есть жгутики, то их строением. Эти постоянные
различия между большими группами водорослей побудили рас-
сматривать их как самостоятельные по своему филогенезу группы,
не связанные родством, и отказаться от понятия водоросли--Algae
как определенной таксономической единицы (класса, типа). Таким
образом, название Algae так же, как Flagellatae, утратило свое
прежнее значение определенной таксономической единицы (в лин-
невском понимании — порядок) и сохранилось лишь как биологи-
ческое понятие для обозначения низших растений, содержащих
в подавляющем большинстве хлорофилл, способных к фитоавтот-
рофному питанию и живущих преимущественно (но не всегда)
в воде.. ~
Что касается классов, то большинство современных авторов
следует системе А. Пашера. Пашеровская концепция классов, кото-
рая выдвигала требование единообразия таких признаков, как пиг-
ментация, характер запасных продуктов, строение монадных "кле-
ток, и допускала неограниченные вариации в строении таллома,
сохранилась и до настоящего времени. Принятие этой концепции не
препятствует объединению и расчленению конкретных классов си-
11
стемы Пашера в соответствии с новыми данными по биохимии
и цитологии водорослей. И в последние два десятилетия был
установлен целый ряд новых классов, например, из зеленых водо-
рослей выделен класс Prasinophyceae. из желтозеленых —класс
Eustigmatophyceae, из золотистых Haptophyceae и др. Однако
основные классы системы Патера остались прежними. Что же
касается распределения этих классов по стволам, или отделам, то
тут мнения разных авторов сильно расходятся, и современные
системы водорослей различаются главным образом по числу
и объему стволов, или отделов. Число последних варьирует от 4 до
10—12. Эти колебания определяю тся гем, какое место в системе
занимают харовые водоросли, каким образом группируются клас-
сы, характеризующиеся желто-бурой окраской, и отношением ав-
торов к сипезеленым водорослям. Согласие достигнуто относите-
льно красных водорослей, которые, по мнению большинства авто-
ров, невозможно поместить в один отдел ни с одним из других
классов водорослей. Они составляют отдел Rhodophyta. Эвглено-
фициевые водоросли также во многих системах относят к отделу
Euglenophyta. Окрашенные в зеленый цвет водоросли с крахмалом
в качестве запасного продукта в одних системах распределяются
между двумя отделами: Chlorophyta и Charophyta, как это делал
и Пашер (A. Pascher, 1931), однако многие авторы включают
харовые водоросли в качестве класса, а в некоторых системах как
порядок в отдел Chlorophyta, а в системах последних лет (В. Foil,
1971; Д. К. Зеров, 1972) большой ствол Chlorophyta объединяет не
только все окрашенные в зеленый цвет водоросли, но и высшие
растения.
Еще большее разнообразие наблюдается в системах разных
авторов относительно группирования классов водорослей, имею-
щих золотисто-бурую окраску. Еще в 1914 г. Пашер впервые об-
ратил внимание на ряд сходных признаков у золотистых, желтозе-
леиых и диатомовых водорослей. Эти сходные признаки следу-
ющие: 1) сосл ав пигментов с преобладанием ксантофиллов над
хлорофиллами; 2) запасные продукты; хризоламинарин, масло, от-
сутствие крахмала; 3) оболочки, состоящие из двух и более частей;
4) частое окремнение клеточных покровов; 5) сходство монадных
клеток; 6) образование особых эндоплазматических цист. На основе
сходства этих признаков Пашер счел эти группы родственными
и объединил их в один большой отдел Chrysophyta* что было
принято в большинстве систем зарубежных авторов. Однако
Ф. Э. Фрич (F. Е. Fritsch, 1935, 1945) воздерживается от установле-
ния отделов и каждый из приводимых им 11 классов предпочитает
рассматривать как отдельный эволюционный ряд. Того же, но
существу, придерживаются и вдэусской альгологической литературе,
где почти каждому из указанных выше классов соответствует отдел.
Наоборот, в зарубежной литературе последних лег наблюдается
тенденция к еще большему укрупнению отделов и соответственно
12
уменьшению их количества. Так, в ряде систем к классам отдела
Chrysophyta (по Пашеру) добавили еще классы Phaeophyceae,
Dinophyceae, Chyptophyceae, Raphidophyceae, создав крупный так-
сон Chromophyta? Общий признак входящих сюда классов - отсут-
ствие хлорофилла h\ имеется только хлорофилл а и в большинстве
случаев с. Это сходство пигментов коррелирует со сходством
субмикроскопического строения фотосинтезирующего аппарата,
жгутики в этих группах также обнаруживают сходное строение.
Большой таксон Chrotnophyta некоторыми альгологами принима-
ется (М. Chadefaud, 1960; В. Fott, 1971), другие эту группу отвер-
гают.
Классы Dinophyceae и Cryptophyceae Пашер объединил в отдел
Pyrrophyta, что принято в русской альгологической литературе, но
не прижилось в иностранной, где чаще всего различают два отдела:
Dinophyta и Cryptophyta.
Наконец, необходимо остановиться на положении синезеле-
ных водорослей. Как уже отмечалось, синезеленые водоросли
были выделены из зеленых и во многих системах до сих пор
фигурйруют в качестве класса Cyanophyceae, составляющего
отдел Cyanophyta, равноценный другим отделам водорослей.
Однако в последнее время синезеленые водоросли во многих
руководствах стали сближать с дробянками и противопоста-
влять царство прокариот (Procaryota) царству эукариот
(Eucaryota), куда относятся все остальные отделы водорослей5.
Эта точка зрения не нова. Сходство в организации клетки очень
давно побудило исследователей к сближению сипезеленых водоро-
слей с бактериями. Еще Ф. Кон (F. Cohn, 1854) в середине прошло-
го столетия относит бактерии к рйститёльнбм"у царству и объеди-
няет их в одну группу с синезелеными водорослями под общим
названием Schizophyta. Несколько позднее К. Нэгели (С. Nageli,
1857), основываясь на отсутствии хлорофилла у бактерий, выделя-
ет их в самостоятельную группу Schizomycetes, и большинство
последующих авторов (A. Pascher, 1931; М, Chadefaud, 1960;
Н. С. Bold, 1973 и др.) также рассматривают синезеленые водорос-
ли и бактерии как две самостоятельные группы одинакового ранга
(отдела).
Наряду с такой трактовкой синезеленых водорослей и бактерий
как самостоятельных отделов, объединяемых в группу Procaryota,
в последние годы у бактериологов наметилась тенденция придавать
еще большее значение сходству ультраструктуры у этих двух групп,
незаслуженно пренебрегая такими важными и существенными раз-
1 Для прокариот в первую очередь характерно отсутствие в клетке обосо-
бленных, одетых элементарными мембранами ядра, митохондрий, хлороплас-
тов. Клеткам эукариот присущи хорошо оформленные ядро, пластиды, мию-
хондрии, отграниченные от основной цитоплазмы системой элементарных мем-
бран.
13
линиями между этими организмами, как: 1) более высокий уровень
морфологической дифференциации таллома синезеленых водорос-
лей; 2) наличием у них пигментной системы, в принципе аналогич-
ной таковой у эукариотических водорослей и существенно отлича-
ющейся от пигментной системы фотосинтезирующих бактерий;
3) оксигенный (т. е. сопровождающийся выделением кислорода)
фотосинтез синезеленых. К этому можно добавить еще полное
отсутствие жгутиковых стадий у этих водорослей в отличие от
бактерий.
Синезеленые водоросли стали называть синезелеными бактери-
ями— цианобактериями и в некоторых системах низвели их до
ранга класса (класс синезеленые фотобактерии в отделе
Photobacteria наряду с классами красные и зеленые фотобактерии)
или даже порядка Cyanobacteriales в классе Photobacteria.
На основании принадлежности синезеленых водорослей наряду
с бактериями к прокариотам (Procaryota) многие бактериологи
сделали вывод о синонимичности терминов Procaryota и Bacteria
и выдвинули требование подчинить номенклатуру Cyanophyta пра-
вилам Международного кодекса номенклатуры бактерий
(R. I. Stanier et ai., 1978).
Против этих предложений выступили многие альгологи, в том
числе и русские, наример Н. В. Кондратьева (1981), подчеркива-
ющие вышеперечисленные существенные различия между этими
двумя группами, и то, что синезеленые водоросли до сих пор
подчинялись Международному кодексу ботанической номенклату-
ры, по которому создание нового таксона в случае водорослей
базируется на латинском описании, сопровождаемом рисунками
и фотографиями. Подчинение же Cyanophyta кодексу номенклатуры
бактерий означало бы необходимость описания новых таксонов
синезеленых водорослей (и пересмотр более 2000 уже описанных
таксонов) на основании чистых — аксеничных культур. Последние
известны сейчас для сравнительно очень немногих синезеленых
водорослей. Кроме того, использование штаммов Cyanophyta (как
того потребовал бы кодекс номенклатуры бактерий) в качестве
номенклатурных типов видов, затрудняется способностью многих
синезеленых водорослей при определенных условиях задержи-
вать свое развитие на промежуточных стадиях, а также вероят-
ностью появления мутаций под воздействием экстремальных
факторов, используемых при очистке культур. Адаптация бакте-
риологического кодекса к синезеленым водорослям сделала бы
невозможным какое-либо экологическое исследование их в при-
роде.
В дальнейшем изложении мы принимаем, что царство прокари-
от (Procaryota) имеет три ветви (отдела): I. Schizophyta.
II. Cyanophyta, III. Prochlorophyta.
Более частные вопросы классификации будут рассмотрены ниже
при описании сответствующих отделов.
14
Глава 2
МОРФОЛОГИЯ (СТУПЕНИ МОРФОЛОГИЧЕСКОЙ
ДИФФЕРЕНЦИАЦИИ ТАЛЛОМОВ ВОДОРОСЛЕЙ)
Как известно, во многих классах (и отделах) водорослей (зеле-
ные, желтозеленые, золотистые, динофитовые) простейшие пред-
ставители —это одноклеточные жгутиковые формы.
В дальнейшей эволюции этих отделов наблюдается замечатель-
ный параллелизм, который выражается в том, что в разных, само-
стоятельных по своему происхождению отделах водорослей наблю-
даются аналогичные типы или ступени морфологической дифферен-
циации таллома. Эти ступени следующие:
1. Монадная организация характеризуется активной подвижно-
стью с помощью жгутиков. Она присуща прежде всего одноклеточ-
ным жгутиконосцам, которые являются начальными звеньями эво-
люции многих отделов водорослей. Разновидность монадной ор-
ганизации -- подвижные (с помощью жгутиков) колонии и ценобии1.
У выше организованных водорослей монадной структурой облада-
ют лишь клетки, служащие для размножения - - бесполого (зооспо-
ры) и полового (гаметы).
2. Ризоподиальная (амебоидная) организация наблюдается у неко-
торых лишенных твердой оболочки форм, которые развивают ци-
топлазматические отростки—ризонодии.
3. Пальмеллоидный, или капсальный, тин организации представ-
лен неподвижными, одетыми оболочками или голыми клетками,
погруженными в общую слизь. Такие слизистые талломы имеют
различные размеры и очертания.
4. Коккоидная организация характеризуется неподвижными,
одетыми оболочками клетками, одиночными или соединенными
в колонии и ценобии.
5. Нитчатая ступень организации представлена клетками, со-
единенными в нити, простые или разветвленные. Клетки нити не-
прерывно делятся поперечными перегородками, обусловливая этим
ее нарастание в длину.
6. Гетеротрихальная или разнонитчатая организация сложнее
предыдущей и характеризуется двумя системами нитей: стелю-
щимися по субстрату и отходящими от них вертикальными ни-
тями.
7. Пластинчатая (тканевая) организация также легко выводится
из нитчатой: в результате делений клеток нити не только в попереч-
ном, но и в продольных направлениях возникают талломы в виде
паренхиматозных пластинок.
1 Ценобиями называют колонии, в которых число клеток определяется на
ранних стадиях развития и не меняется до следующей репродуктивной фазы.
15
8. Сифональная, или сифоновая, организация отличается отсутст-
вием поперечных клеточных перегородок, так что талломы часто
крупных размеров и со значительной внешней дифференцировкой
формально представляют собой одну клетку с непрерывной ваку-
олью клеточного сока и обычно большим количеством клеточных
ядер. В этом случае часто говорят о неклеточном строении.
9. Сифонокладалъная организация отличается от сифоновой тем,
что сифоновый вначале таллом рано или поздно делится на много-
ядерные участки, или сегменты.
Перечисленные ступени организации таллома наиболее полно
представлены в отделах зеленых, золотистых, желтозеленых и дино-
фитовых водорослей. В этих отделах типы организации таллома
кладутся в основу деления на порядки, последовательность которых
в каждом отделе отражает действительную картину его филогене-
тического развития.
Для эвгленофитовых водорослей, помимо монадной, пока не
установлены другие ступени дифференцировки таллома. Наоборот,
у бурых водорослей неизвестны примитивные одноклеточные пред-
ставители, здесь преобладают высшие ступени морфологической
дифференцировки таллома - - гетеротрихальная и тканевая.
Удивительное на первый взгляд явление параллелизма в эволю-
ции разных отделов водорослей объясняется довольно просто.
Многократно на многочисленных примерах из разных отделов во-
дорослей было показано, что монадная, т. е. снабженная жгути-
ками, клетка довольно легко может терять их и временно перехо-
дить в одних случаях в ризоподиальное, в других — в пальмеллоид-
ное, в третьих- —в коккоидное состояние. Обычно из этих времен-
ных состояний возврат к исходной монадной форме более или менее
легкий. Если же эти преходящие случайные состояния продлеваются
во времени, так что на них будет приходиться большая часть
жизненного цикла, а к монадному состоянию организм будет воз-
вращаться только в момент размножения, то получатся соответст-
венно ризоподиальный, пальмеллоидный и коккоидный типы тал-
ломов. Нитчатый таллом также без труда можно вывести из монад-
ного: такой переход каждый раз можно наблюдать при прорастании
зооспоры в нить, а из нити легко может возникнуть пластинчатый
таллом. Таким образом, монадная организация--исходная для
всех остальных типов организации талломов водорослей. Посколь-
ку начальные ступени эволюции во многих отделах водорослей
представлены довольно сходно устроенными монадами1 (различа-
ющимися, однако, по окраске, продуктам метаболизма и строению
жгутиков), параллельный ход эволюции в разных отделах водорос-
лей объясняется параллельным развитием из монадной остальных
типов морфологической организации таллома. Усложнение стро-
1 Некоторые отделы, однако, совсем лишены жгутиковых стадий (красные,
синезеленые водоросли), и для них невозможно установить родство со жгутиковыми.
16
ения и переход к многоклеточное™ сопровождались потерей подви-
жности в вегетативном состоянии. Исходная монадная организация
сохранялась только у репродуктивных клеток, и то не всегда. При
этом подвижная монадная стадия, все более сокращаясь, удержала
основные черты родоначальной формы. Вот почему монадные ре-
продуктивные клетки водорослей из разных отделов столь полно
воспроизводят признаки, присущие одноклеточным монадным
представителям данного отдела.
Другой причиной, обусловившей параллельное развитие разных
групп водорослей, могло быть большее однообразие водной среды
по сравнению с местообитаниями суши, что и определило более
однотипный ход эволюции у водорослей по сравнению с ходом
эволюции у организмов, покинувших водную среду.
Глава 3
Цитология эукариотических водорослей1
Протопласт как совокупность живых компонентов клетки (ядра,
цитоплазмы, пластид, митохондрий, вакуолей) лишь у сравнитель-
но немногих более примитивных водорослей ограничен снаружи
только цитоплазматической мембраной (клеточной мембраной,
нлазмалеммой). Такие клетки называют «голыми». Голыми явля-
ются, как правило, репродуктивные клетки водорослей—зооспоры
и гаметы (исключение представляют многие вольвокальные, монад-
ные клетки которых, как вегетативные, так и репродуктивные,
одеты клеточной оболочкой).
КЛЕТОЧНАЯ ОБОЛОЧКА
У большинства водорослей протопласты полностью окружены
клеточными стенками (оболочками)2. Как и у высших растений,
клеточная оболочка водорослей всегда образуется кнаружи от
нлазмалеммы, защищает протопласт и служит как бы наружным
скелетом клетки. Оболочка состоит из двух компонентов: аморф-
ного слизистого матрикса и погруженного в него фибриллярного
опорного (скелетного) компонента. В состав матрикса входят
гемицеллюлозы и пектиновые вещества, образуемые аппаратом
Гольджи внутри клетки и выделяемые через плазмалемму путем
экзоцитоза. Что касается химического состава фибриллярного
1 Описание прокариотной клетки будет дано при рассмотрении соответству-
ющих групп.
' Клеточная оболочка и клеточная стенка - синонимы, одинаково широко упот-
ребляемые в литературе.
компонента клеточных стенок, то в отличие от высших растений,
где фибриллы представлены целлюлозой, у водорослей наблю-
дается большее разнообразие полисахаридов, образующих ске-
летные фибриллы. «Создается впечатление, что природа экспе-
риментировала с разнообразными типами полисахаридов до тех
пор, пока отбор не показал, наконец, что именно цепи целлюлозы
(0-1,4-полиглюкан) могут служить наилучшим материалом для
укрепления клеточных оболочек в борьбе растений за переход
к наземному образу жизни» (А. Фрей-Висслинг К. Мюле галер.
1968). К настоящему времени у водорослей на молекулярном
и ультраструктурном уровне детально описаны три главных типа
скелетных полисахаридов: целлюлоза, маннан и ксилан.
Целлюлоза обнаружена у многих зеленых водорослей (тетрас-
пора, хлорелла, анкистродесмус, сценедесмус. недиаструм, улот-
рикс, ульва, монострома, энтероморфа, эдогониум, кладофора, хе-
томорфа, валония и др.), у желтозеленых (трибопема, вошерия),
динофитовых (пироцистис). в небольших количествах -- у красных
(гриффитзия и др.) и бурых (ламинария, пельветия, аскофиллум).
Ряд зеленых водорослей с сифоновой организацией таллома не
содержит в стенках целлюлозы. Главные составные части клеточ-
ных стенок у них—ксилан или маннан. Так, ксилан обнаружен
в клеточных стенках бриопсиса, каулерпы, галимеды, удотеи, а ман-
нан— у ацетабулярии и кодиума. Это различие в химическом со-
ставе оболочек пытались использовать для установления родст вен-
ных связей в пределах этой группы зеленых водорослей. Пред-
лагалось делить их на «ксилановую» и «маннановую» группы.
Однако позднее ксилан и маннан были обнаружены в стенках одних
и тех же водорослей на разных стадиях их жизненного цикла: споро-
фиты дербезии, бриопсиса, псевдобриопсиса содержали маннан,
а гаметофиты — ксилан и целлюлозу. В ряде случаев состав микро-
фибрилл точно не установлен.
Фибриллы — скелетные волокна — ориентированы в матриксе
по-разному. Они могут быть рассеяны по нему беспорядочно или же
они располагаются параллельно друг другу. И то и другое рас-
положение микрофибрилл может встречаться в пределах одной
систематической группы, в разных частях одного и того же таллома
и даже в разных слоях клеточной стенки одной и той же клетки.
Например, среди зеленых водорослей беспорядочное расположе-
ние фибрилл наблюдается у родов, клеточные стенки которых хара-
ктеризуются обильным аморфным матриксом (ульва, энтероморфа,
улотрихальные, хетофоральные, эдогониальпые, спирогира, педи-
аструм и др.). У сифонокладальных (кладофора, хетоморфа, вало-
ния) стенка содержит лишь небольшие количества аморфного мате-
риала. Она состоит из многочисленных ламелл, в которых фибрил-
лы ориентированы более или менее параллельно друг другу. В со-
седних ламеллах (слоях) фибриллы располагаются почти под
прямым углом. ,
18
Первый слой новой стенки, образующийся на остановившихся
зооспорах этих водорослей, состоит из беспорядочно расположен-
ных микрофибрилл.
Помимо скелетных полисахаридов, матрикс клеточных стенок
может содержать инкрустирующие вещества — кремнезем, споро-
полленин, карбонат кальция. Есть данные о присутствии в клеточ-
ных стенках протеинов (например, у хламидомонады, хлорококка).
В 1972 г. у ряда видов хлореллы, сценедесмуса, прототеки в их
оболочках впервые был обнаружен спорополленин—чрезвычайно
стойкое по отношению к ферментам вещество, характерное для
оболочек спор и пыльцевых зерен высших растений. За последние
годы число водорослей, в оболочках которых найден споропол-
ленин, заметно возросло. Он обнаружен в клеточных стенках
Phycopeltis epiphitoit, ряда видов требуксии, в стенках цист гемато-
кокка, в стенках зигот спирогиры. У некоторых водорослей (кладо-
фора, питофора, эдогониум) имеется хитин в виде топкого наруж-
ного слоя. Оболочка может быть цельной или состоять из двух
и более частей, может быть пронизана порами, нести различные
выросты. Примеры будут приведены при описании соответству-
ющих родов.
Формирование новых оболочек исследовалось с применением
различных методов электронной микроскопии (на ультратонких
срезах, путем исследования реплик — отпечатков плазмалеммы).
Объектами служили голые, одетые только цитоплазматической ме-
мбраной естественные стадии развития разных водорослей: цисты
ацетабулярии, только что оплодотворенные яйцеклетки фукуса, син-
зооспоры вошерии, зооспоры педиаструма, кладофоры, хетоморфы;
автоспоры хлореллы, ооцистиса, сценедесмуса. В других исследова-
ниях по регенерации стенки использовались искусственно получен-
ные протопласты (например, у хламидомонады). Результаты этих
исследований несколько противоречивы; по-видимому, сборка кле-
точных стенок происходит неодинаково у разных водорослей,
и в этот процесс вовлечены разные органеллы.
В ряде случаев констатируется участие в синтезе клеточных
стенок аппарата Гольджи (Acetahularia mediterranea, Vaucheria
fontinalis, Fucus vesiculosus, F. distichus, F. inflatus. Напротив, при
изучении регенерации клеточной стенки протопластами
Cldamydomonas smilhii, полученными под воздействием гаметного
автолизина, активность аппарата Гольджи на срезах протопластов,
регенерирующих клеточную стенку, не отличалась от таковой кле-
ток, обладающих стенкой.
Высокая степень упорядоченности целлюлозных микрофибрилл,
их правильная ориентация в оболочках таких водорослей, как кла-
дофора, хетоморфа, послужила основой для гипотезы «упорядо-
ченных гранул» биосинтеза целлюлозы, при котором гранулы эн-
зима, расположенные определенным образом, осуществляют все
процессы от полимеризации глюкозы до ориентации микрофибрилл
19
в ламеллах стенки. В пользу этой гипотезы свидетельствуют данные
по изучению плазмалеммы методом отпечатков в процессе био-
синтеза микрофибрилл на голых зооспорах кладофоры и хетомо-
рфы. Вскоре (через 24—48 ч) после остановки зооспор возникает
оболочка. При этом на плазмалемме этих голых клеток видны
гранулы с той же ориентацией, что и микрофибриллы в зрелых
оболочках клеток этих водорослей. Был сделан вывод, что эти
гранулы плазмалеммы участвуют в биосинтезе целлюлозных мик-
рофибрилл. Сходные данные были получены для автоспор Oocystis
apiculata — водоросли, также имеющей упорядоченную текстуру
микрофибрилл в оболочке зрелых клеток. Исследование реплик
плазмалеммы голых автоспор этой водоросли показало, что сна-
ружи плазмалемма несет спаренные ряды мелких гранул, соеди-
няющиеся в широкие полосы. На конце рядов может находиться
более крупная гранула. Между полосами гранул располагаются
отдельные гранулы разного диаметра. Ориентация полос гранул
точно соответствует ориентации микрофибрилл в оболочке.
С внутренней стороны плазмалемма несет отдельные гранулы.
Было высказано предположение, что полосы гранул являются ме-
стом синтеза целлюлозных микрофибрилл, которые вставляются
в глубокую выемку между парами рядов гранул. С другой стороны,
из анализа электронных микрофотографий кортикального слоя ци-
топлазмы и клеточной стенки Oocystis solitaria сделан вывод, что
упорядоченная организация микрофибрилл обусловлена микротру-
бочками, сосредоточенными в примембранном слое цитоплазмы.
В пользу этой гипотезы свидетельствуют опыты с ингибиторами,
действующими на микротрубочки, например с колхицином. Об-
работка клеток колхицином приводит к деполимеризации микро-
трубочек и нарушает ориентацию микрофибрилл. У водорослей
(особенно с монадной организацией), помимо цитоплазматической
мембраны и клеточных стенок, наблюдается большое разнообразие
клеточных покровов. Эти покровы можно распределить на две
группы: формирующиеся под цитоплазматической мембраной и на-
ходящиеся кнаружи от нее. К первым относятся: перипласт крип-
тофитовых, пелликула эвгленофитовых и тека динофлагеллят; ко
вторым — различного рода чешуйки, образующие покров снаружи
от цитоплазматической мембраны, особенно характерные для золо-
тистых и гаптофициевых водорослей, но встречающиеся и у зеле-
ных. Особые покровы - панцири диатомовых водорослей, непо-
лные оболочки—домики золотистых, эвгленофитовых водорослей.
ЦИТОПЛАЗМА
Цитоплазма—часть клетки, остающаяся после удаления раз-
личимых в световом микроскопе органелл (ядра, пластид, митохон-
дрий, вакуолей). Основная плазма — мелкогранулированный мат-
рикс— содержит ультраструктурные органеллы цитоплазмы: плаз-
20
малемму, эндоплазматическую сеть, аппарат Гольджи, микротру-
бочки и пр. (см. рис. 5).
Мембраны. Электронный микроскоп показывает, что цитоплаз-
ма пронизана многочисленными мембранами, имеющими на срезе
вид двойных темных линий, заключающих светлое промежуточное
пространство. Снаружи клетка ограничена такой типичной элемен-
тарной, мембраной—плазмалеммой, которая в живой клетке об-
ладает свойством полупроницаемости и выполняет барьерную фун-
кцию. Все клеточные органеллы ограничены такой элементарной
мембраной. Ядра, пластиды, митохондрии отделены от основной
плазмы оболочками, состоящими из двух (в случае пласа ид и боль-
шего числа) мембран.
Внутри клетки цитоплазма отграничена от вакуолей вакуоляр-
ной мембраной—тонопластом, имеющим ту же природу, что
и плазмалемма. Наряду с перечисленными стабильными мембрана-
ми, отграничивающими клетку от наружной среды и органеллы от
основной плазмы, существует еще чрезвычайно изменчивая внут-
ренняя система мембран—эндоплазматическая сеть. Эта система
образована ограниченными элементарной мембраной плоскими по-
лыми цистернами и трубочками, связанными в сеть. Наружная
поверхность эндоплазматической сети часто (но не всегда) покрыта
рибосомами. Соответственно различают шероховатую и гладкую
эндоплазматическую сеть, которая может встречаться в одной и той
же клетке. Эндоплазматическая сеть играет роль в передвижении
и распределении веществ внутри клетки. Рибосомы, связанные с ме-
мбранами шероховатой эндоплазматической сети, осуществляют
синтез белка.
Аппарат Гольджи. Аппарат Гольджи (см. рис. 5) состоит из
диктиосом, одной (в мелких клетках) или многих (в более крупных
клетках многоклеточных водорослей, например в ризоидах хары, их
насчитывается более 25 000). Каждая диктосома представляет собой
стопку плоских блюдцевидных цистерн, ограниченных элементар-
ной мембраной. Число цистерн в стопке, т. е. приходящихся на одну
диктиосому, варьирует от 3-7 до 30. От краев цистерн, большей
частью вздутых, отшнуровываются мелкие пузырьки — везикулы
Гольджи. Вся органелла носит полярный характер; в ней имеются
проксимальный, или формирующий, полюс (расположенный ближе
к ядру) и дистальный, или секретирующий, полюс (удаленный от
ядра), где цистерны образуют обильные пузырьки и фрагментиру-
ются. Таким образом цистерны постоянно перемещаются от форми-
рующего полюса к секретирующему. Образование новых цистерн
Гольджи, по-видимому, происходит за счет слияния пузырьков,
которые отпочковываются от эндоплазматической сети или наруж-
ной мембраны ядерной оболочки. Если основной функцией эндо-
плазматической сети является полимеризация аминокислот в специ-
фические полипептиды, то в аппарате Гольджи из простых сахаров
синтезируются олигомерные углеводы. По мере созревания цистерн
21
диктиосомы (от проксимального ее полюса к дистальному) водя-
нистая жидкость эндоплазматической сети или перинуклеарного
пространства, как правило, обогащается углеводами, что можно
выявить гистохимически. Функции аппарата Гольджи многообраз-
ны. Он участвует в образовании матрикса клеточной оболочки.
В результате слияния пузырьков Гольджи возникает клеточная
(срединная) пластинка, принимающая участие в цитокинезе многих
зеленых водорослей. Срединная пластинка представляет собой по-
лужидкий слой из гемицеллюлозы и пектиновых веществ, состав-
ляющий матрикс клеточной оболочки, в котором позднее появля-
ются фибриллы целлюлозы. Синтез фибрилл, как правило, осущест-
вляется плазмалеммой. Мембраны же пузырьков Гольджи преоб-
разуются в плазмалемму. Аппарат Гольджи поставляет также
материал матрикса для растяжения клеточной стенки. Пузырь, где
откладывается кремнезем и в котором формируется новая створка
диатомовых водорослей, также возникает за счет слияния пузырь-
ков Гольджи. То же относится и к мембране — силиколемме, окру-
жающей пузырь, в котором закладывается кремнеземная оболочка
эндоплазматических статоспор золотистых водорослей. В цистер-
нах аппарата Гольджи наблюдаются не только растворенные веще-
ства, но и оформленные чешуйки, составляющие клеточный покров
многих монадных золотистых и некоторых зеленых водорослей.
Чешуйки выделяются на поверхность клетки путем экзоцитоза-
при слиянии мембраны цистерны, содержащей готовую чешуйку,
с плазмалеммой. В этой связи заслуживают внимания данные
Р. Броуна с сотрудниками (R. М. Brown et al., 1970), относящиеся
к гаптофициевой водоросли- Pleurochrysis scherffeli.. На попереч-
ном разрезе через зрелую вегетативную клетку этой водоросли
можно видеть, что клеточный покров образован многими слоями
рыхло расположенных чешуек, погруженных в материал, по-види-
мому, пектиновой природы (рис. 1.). На тангентальном срезе клетки
чешуйки представляются в плане: их остов образован целлюлоз-
ными микрофибриллами, из которых одни располагаются концент-
рическими кругами, другие расходятся радиально из центра че-
шуек. Эти чешуйки формируются в цистернах аппарата Гольджи.
Процесс начинается в проксимальных (расположенных ближе к яд-
ру) цистернах: центральная расширенная часть таких цистерн явля-
ется местом полимеризации глюкозных мономеров в кристалличес-
кую сеть целлюлозных цепей. В дистальных (расположенных ближе
к поверхности клетки) цистернах можно видеть уже зрелые чешуй-
ки; их остов образован сетью целлюлозных микрофибрилл, одна
часть которых расположена концентрическими кругами, а другая
расходится из центра по радиусам. Мембраны самых наружных
цистерн с уже готовыми чешуйками, по-видимому, сливаются с ци-
топлазматической мембраной, принимая, таким образом, участие
в выделении готовых элементов клеточно! о покрова па поверхность
клетки, где они располагаются наподобие черепиц. Подсчитано, что
22
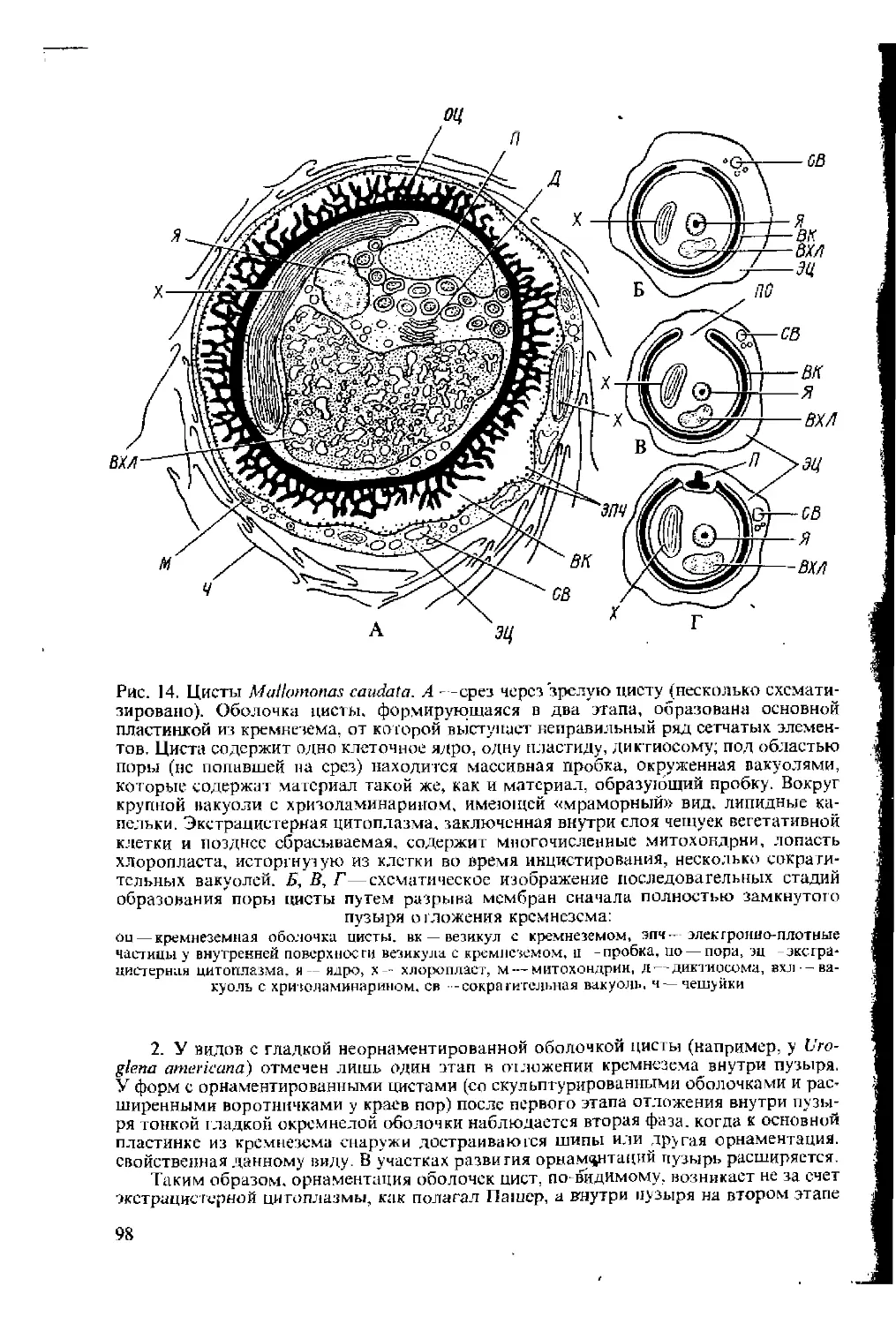
Рис. 1. Схема формирования чешуек Pleurochrysis scherffcli. Центральная расширен-
ная часть в проксимальных (расположенных ближе к ядру) цистернах аппарата
Гольджи является местом полимеризации глюкозных мономеров в кристаллическую
сеть целлюлозных цепей. В дистальных (расположенных ближе к поверхности клет-
ки) цистернах уже находятся зрелые чешуйки; мембраны самых наружных цистерн
с уже готовыми чешуйками сливаются с плазмалеммой. выделяя чешуйки на поверх-
ность клетки;
пц — проксимальные цистерны, ДЦ — дистальные цистерны с чешуйками, ч—чешуйки на поверх-
ности клетки
у Pleurochrysis scherffeli аппарат Гольджи каждые две минуты про-
дуцирует одну чешуйку. Таким образом, скорость продукции чешу-
ек достаточно велика, что, возможно, связано с большой потерей
рыхло расположенных чешуек.
Было доказано, что основной строительный полисахарид чешуек
плеврохризиса - целлюлоза. Этим было продемонстрировано, что
аппарат Гольджи участвует не только в синтезе аморфного
23
матрикса клеточной стенки, но и в синтезе целлюлозы, принимая
на себя морфогенетические функции, обычно выполняемые пла-
змалеммой. До этого исследования клеточного покрова Р. scherffel'l
общепризнанным считалось, что опорные фибриллы целлюлозы
образуются всегда вне протопласта благодаря направленной
наружу активности плазмалеммы. Возникновение чешуек, кок-
колитов в цистернах аппарата Гольджи к настоящему времени
описано у многих представителей.
В ряде случаев, например у золотистой водоросли охромонас,
было показано, что в сборку и выведение на поверхность клетки
мастигонем вовлечены пузырьки Гольджи. Сами мастигонемы, бу-
дучи белковой природы, образуются не в пузырьках Гольджи,
а в перинуклеарном пространстве, т. е. в системе синтезирующей
белок эндоплазматической сети.
Одна из функций аппарата Гольджи — регуляция содержания
воды в клетке. У рафидофитовой водоросли Vacuolaria virescens
было продемонстрировано возникновение пульсирующих вакуолей
из аппарата Гольджи. Сократительная вакуоль этой водоросли
представляет собой разбухшую цистерну Гольджи. При «сокраще-
нии» она сливается с плазмалеммой, и ее жидкое содержимое
удаляется из клетки путем экзоцитоза. Вакуоль, по существу, и не
является сократительной, так как она полностью исчезает и заменя-
ется новой цистерной. Это процесс, обратный процессу активного
поглощения воды путем пиноцитоза (эндоцитоза). Было показано,
что каждые 75 с из клетки выталкивается новая вакуоль и за 30 мин
выделяется количество воды, соответствующее полному объему
клетки. Аппарат Гольджи связан с секреторной деятельностью
клетки, при его участии образуются, по-видимому, трихоцисты,
эджектосомы, возможно, выделяется клеющая субстанция, способ-
ствующая прикреплению спор к субстрату, и т. д.
Микротрубочки. Помимо эндоплазматической сети, в цитоплаз-
ме клеток водорослей обнаруживаются трубчатые элементы — ци-
топлазматические микротрубочки. Их следует отличать от микро-
трубочек, составляющих аксонему ресничек и жгутиков. В отличие
от последних образование цитоплазматических микротрубочек по-
давляется действием колхицина, они исчезают из цитоплазмы при
низкой температуре и высоком гидростатическом давлении (все эти
воздействия не оказывают влияния на микротрубочки аксонемы
жгутиков). Цитоплазматические микротрубочки—крайне динами-
ческие клеточные структуры: они то появляются, го исчезают,
уменьшаются или увеличиваются в числе, перемещаются из одной
части клетки в другую. Стенки микротрубочек образованы глобу-
лярными макромолекулами. Хотя цитоплазматические микротру-
бочки пока не удается выделить из цитоплазмы и анализировать, на
них обычно распространяются данные, полученные в отношении
микротрубочек жгутиков, поддающихся изолированию и биохими-
ческому анализу. Принимается, что цитоплазматические микротру-
24
бочки, как и микротрубочки жгутиков, образованы белками тубули-
нами, по аминокислотному составу напоминающими актин мышц.
Цитоплазматические трубочки различаются по положению в клетке
и назначению: одни участвуют в образовании ядерного веретена
(так называемые «волокна» митотического веретена не что иное, как
волокнистые элементы, возникающие в результате агрегации мик-
ротрубочек при фиксации); другие связаны с цитокинезом — уча-
ствуют в образовании фрагмопласта и фикопласта; третьи, со-
бственно цитоплазматические микротрубочки, представляют собой
скелетную систему клетки. Последняя особенно свойственна голым
клеткам, форма которых определяется микротрубочками, образу-
ющими периферический микроскелет. Например, асимметричная
форма клетки голой золотистой водоросли охромонаса определяет-
ся присутствием цитоплазматических микротрубочек: при высоком
гидростатическом давлении или обработке колхицином (воздейст-
виях, подавляющих образование цитоплазматических микротрубо-
чек) клетки охромонаса принимают сферическую форму. Своеоб-
разная тетраэдрическая форма зооспор золотистой водоросли гид-
руруса также определяется системой микротрубочек. Микротрубоч-
ки ответственны не только за поддержание, но и за изменение
формы голых клеток, например зооспор водяной сеточки, педиаст-
рума во время формирования их дочерних ценобиев, а также в про-
цессе инверсии при образовании дочерних сфероидов вольвокаль-
ных. Микротрубочки, локализованные на переднем конце спермато-
зоидов эдогониума, возможно, наряду со жгутиковыми корнями
и фиброзным кольцом ответственны за те изменения формы кон-
чиков сперматозоидов, которые наблюдаются перед самым процес-
сом плазмогамии. В ряде случаев (гименомонас, дихотомосифон,
вольвокс, эдогониум, хламидомонада) микротрубочки наблюда-
лись по соседству с хлоропластами, текальными цистернами у ди-
нофитовых водорослей, пелликулой эвгленофитовых и пр.
ЖГУТИКИ
Строение жгутиков. Монадные клетки снабжены жгутиками.
Жгутики всех водорослей имеют единый план строения.
Жгутик можно подразделить на наружный и внутренний — эст-
ра- и интрацеллюлярный отрезки. Наружный отрезок, или свобод-
ная часть жгута, в свою очередь, состоит из трех частей: кончика,
главного стержня и переходной зоны. Внутренний- интрацеллю-
лярный— оз резок охватывает основание жгута (базальное тело)
и корни жгута.
Свободная часть жгута всегда одета жгутиковой мембраной,
которая не что иное, как приподнятая (выпяченная) цитоплаз-
матическая мембрана. Так, у бурой водоросли диктиоты един-
ственный жгутик (второй редуцирован) несколько раз свернут по
спирали вокруг молодого сперматозоида. На срезе через такой
25
сперматозоид под мембраной, одевающей клетку, можно видеть
микротрубочки жгута, характерным образом расположенные
(9 — 9 + 2), без какой-либо собственной мембраны. После освобо-
ждения сперматозоида первоначально закрученный по спирали
жгутик разматывается, приподнимая при этом цитоплазматическую
мембрану клетки, которая на этой стадии должна быть в очень
лабильном состоянии. Наоборот, при втягивании жгутиков, на-
пример при остановке синзооспор вошерии, их мембрана ста-
новится частью цитоплазматической мембраны. Кнаружи от жгу-
тиковой мембраны находится чехол — слой материала, покрыва-
ющего жгут на большем протяжении его длины.
Под мембраной жгута находится матрикс, образованный бел-
ками. Весь жгутиковый аппарат пронизан пучком белковых фиб-
рилл или микротрубочек — аксонемой, поперечный разрез которой
выглядит неодинаково на разных уровнях жгута: в его кончике,
главном стержне, переходной зоне и базальном теле (рис. 2).
На поперечном срезе главного стержня видно типичное для всех
водорослей (и вообще эукариотических растительных и животных
организмов) расположение микротрубочек или фибрилл: кольцо из
девяти пар (дублетов) микротрубочек окружает две одинарные цен-
тральные микротрубочки—структура (9 —9)+ 2. Каждый дублет
образован «А» и «В» микротрубочками, «А» трубочка каждого
дублета снабжена двумя короткими выступами—боковыми «ру-
ками», обращенными к следующему дублету, и радиальным тяжем
(спицей), соединяющим периферический дублет с центральными
микротрубочками, которые могут быть заключены в общий чехол.
Кроме того, имеются интердублетные соединения — тяжи, соединя-
ющие дублеты, и ряд дополнительных образований.
Рис. 2. Схема электронно-микроскопического строения жгутиков Chlamydomonas;
А — продольный разрез жгутиков, Б, В—поперечный разрез через кончик жгута.
Г—поперечный срез через стержень жгута, Д- переходная зона, Е - поперечный
разрез через основание жгута — базальное тело (см. текст)
26
Рис. 3. Схема строения дублетов в жгутике
Chlamydonionas reinlnirdtir. А последователь-
ное! ь растворения микротрубочек жгутика: уда-
ление мембраны жгутика оставляет нетронутой
аксонему, содержащую девять наружных дубле-
тов и две центральные трубочки (9 — 9 + 2). При
обработке аксонемы возрастающими концентра-
циями саркозила удаляется одна нз центральных
трубочек, затем растворяется оставшаяся цент-
ральная трубочка, часто вместе с наружной ча-
стью трубочки В. Б- интактные наружные дуб-
леты растворяются в такой последовательности
(сверху вниз): наружная часть трубочки В, оста-
вшаяся часть трубочки В, наружная часть трубо-
чки А, А —остаток трубочки А, за исключением
грех первичных нитей, составляющих перегород-
ку между трубочками А и В. В схема, иллюст-
рирующая гипотетическое расположение тубули-
на 1 (белые субъединицы) и тубулина 2 (черные
субъединицы) в наружном дублете
Тонкая структура микротрубочек, составляющих наружные дублеты жгутиков
и ресничек, стала объектом многочисленных исследований. Подробно были изучены
и дублеты жгутиков простейшей монадной зеленой водоросли хламидомонаса. Дуб-
леты образованы субъединицами первичными нитями, в свою очередь состоящими
из одного ряда сферических субъединиц. Каждый дублет содержит 23 первичные
нити: микротрубочка «А» образована 13 первичными нитями, из которых 3 рас-
положены медианно. создавая перегородку между просветами «А» и «В» микротру-
бочек. «В» микротрубочка включает 10 первичных нитей (рис. 3, В).
Г. Витман с соаэт. (G. Witman el al., 1972) разработали методы изоляции жгути-
ков хламидомонаса и их фракционирования на мембрану, мастигонемы (волосовид-
ные образования, прикрепленные к жгутиковой мембране), матрикс и компоненты
аксонемы. Наружные дублеты затем были разделены на «А» и «В» микротрубочки
и выделена перегородка между ними, образованная тремя первичными нитями.
Каждый и: компонентов был изучен электронно-микроскопичес-
ки и биохимически. Изолированные мастигонемы образованы эллипсоидными субъ-
единицами, соединенными конец к концу, и содержат один гликопротеин. Фракция
матрикса включает ряд белков. Изолированные аксонемы удерживают сначала
интактную (9 — 9 + 2) структуру, а дальнейшее их фракционирование осуществляется
путем нагревания или под действием детергента. Возрастающие концентрации детер-
гента саркозила растворяют микротрубочки аксонемы в следующем порядке: одна
из двух центральных микротрубочек, вторая центральная микротрубочка и наружная
27
стенка «В» микротрубочки дублета, остальные части «В» микротрубочки, наружная
стенка «А» трубочки, остаток «А» трубочки, за исключением трех первичных нитей,
составляющих «перегородку» между просветами «А» и «В» микротрубочек
(рис. 3, А, Б).
Анализ наружных дублетов показал, что они образованы двумя белками-
тубулином 1 и тубулином 2 соответственно с молекулярной массой 56 000 и 53 000
и разным аминокислотным составом.
Как в микротрубочках «А», так и в микротрубочках «В» присутствуют оба
тубулина (1 и 2), в трех же первичных нитях, образующих стенку между просветами
«А» и «В» .микротрубочек, обнаружен только тубулин 1. В остальных частях микро-
трубочек каждая первичная пить содержит только один из тубулинов и они череду-
ются парами: пары первичных нитей, образованных тубулином I, перемежаются
с парами нитей, содержащими тубулин 2 (рис. 3, В).
Кончик — это участок жгута, тупой или слегка заостренный, у ко-
торого нарушается описанное для главного стержня жгута нормаль-
ное (9 —9)+ 2 расположение микротрубочек. По мере приближения
к кончику утрачивается материал матрикса и периферические дуб-
леты один за другим постепенно теряют одну из микротрубочек,
становясь одинарными, но сохраняя при этом изначальное круговое
расположение. Затем постепенно сокращается число периферичес-
ких микротрубочек, пока не останутся только центральные микро-
трубочки, которые сохраняются почти до самого кончика жгута (см.
рис. 2).
У гамет бурой водоросли эктокарпуса оба жгутика несут длин-
ный, часто закрученный спирально терминальный придаток — ак-
ронему, в который из стержня жгута переходят две центральные
микротрубочки (см, рис, 31, А, В).
Переходная зона — между свободной частью жгута и базальным
телом — внешне легко отличается от стержня, так как здесь жгутик
перетянут: жгутиковая мембрана плотно подходит к дублетам ак-
сонемы. У проксимального — морфологически нижнего конца на-
ружной части жгута находится базальный диск, у которого окан-
чиваются две центральные микротрубочки, Периферические дуб-
леты продолжаются ниже базального диска, где к ним вскоре
добавляются дополнительные микротрубочки, превращающие дуб-
леты в триплеты базального тела (см. рис, 2).
Базальное тело, находящееся в основании каждого жгутика,
расположено в цитоплазме вблизи поверхности клетки и представ-
ляет собой короткий цилиндр, по периферии которого располагает-
ся девять триплетов микротрубочек. Триплеты соединяются друг
с другом тонкими нитями между «А» и «С» — микротрубочками
соседних триплетов (см, рис. 2). По-видимому, жгутики всех водо-
рослей имеют «корни». прикрепленные обычно к их базальным
телам, закрепляющие эти тела в протопласте. Обычно корни быва-
ют двух типов: группы исчерченных полосатых волокон и группы
микротрубочек. Ориентация корней по отношению к базальным
телам и в клетке различна. Например, у хламидомонады базальные
тела двух жгутиков связаны друг с другом широким полосатым
корнем, от которого отходят четыре четырехчленных микротруб-
28
чатых корня, идущих назад в клетку непосредственно под цитоплаз-
матической мембраной.
Такая комбинация исчерченных и трубчатых корней встречается
часто.
Совсем иной тип «корневой системы» у представителей празино-
фициевых, хризофитовых, рафидофитовых водорослей. У этих ор-
ганизмов широкая исчерченная лента—ризопласт (см. рис. 19) —
тянется непосредственно от базальных тел к ядру, где она, по-
видимому, прикрепляется к ядерной оболочке.
В некоторых группах водорослей, например зеленых, особен-
ностям жгутиковых корней в последнее время придается важное
значение в связи с выявлением родственных отношений между
видами и родами.
При едином общем плане строения жгутики разных водорослей
могут отличаться в деталях, причем эти различия весьма констант-
ны и характерны для больших групп — отделов.
Эти постоянные различия касаются как особенностей тонкого
строения жгута, так и таких признаков, как число жгутиков, место
прикрепления, длина, характер поверхности и пр. Так, у диатомо-
вых водорослей в жгутике сперматозоидов отсутствуют централь-
ные микротрубочки и вместо обычного строения (9 — 9) + 2 трубочки
расположены по формуле (9 — 9) +0, а у зеленой водоросли голенки-
нии в стержне жгутика сперматозоида имеется только одна цент-,
ральная микротрубочка (9 — 9) + 1.
Особенно вариабильной сказалась переходная зона, которая
у зеленых водорослей характеризуется так называемым звездчатым
телом (см. рис. 2). У золотистых водорослей между периферичес-
кими дублетами и центральными трубочками в этой зоне имеется
спиральное тело, а у динофитовых переходная зона благодаря
диафрагмам разделена на камеры.
Число жгутиков может варьировать от одного до многих, хотя
преобладают двужгутиковые формы; прикрепление их к клетке — к
концу (терминальные) или сбоку (латеральные).
Если у двужгутиковых клеток оба жгутика одинаковой длины,
то они изоконтные, если разной длины,— то гетерокоптные. Одина-
кового строения жгутики называют изоморфными, различающиеся
по форме— гетероморфными. В последнем случае один из жгу-
тиков (обычно более короткий) может быть гладким, а другой несет
прикрепленные к его мембране волосовидные образования — ма-
стигонемы. Мастигонемы имеют сложное и разнообразное стро-
ение и расположение. Поверхность жгутиков бывает покрыта раз-
личной формы чешуйками или шипами.
Регенерация жгутиков. Перед клеточным делением у одних мо-
надных форм (хламидомонада, охроманас) старые жгутики сбрасы-
ваются; при этом связь с базальным телом прерывается у прокси-
мального конца переходной зоны в месте превращения триплетов
в дублеты. В других случаях (например, у дюналиеллы) половина
29
старого кинетического (жгутикового) аппарата может, переходить
к дочерним клеткам, а отсутствующие половинки образуются зано-
во. Жгутики не способны делиться и всегда берут начало от базаль-
ных тел, репликация которых предшествует клеточному делению:
от дистального конца базального тела вырастает стержень жгута,
приподЕшмающий плазмалемму, которая по мере вытягивания жгу-
тика также вытягивается и превращается в его мембрану. При
репликации базальных тел формирование нового базального тела
индуцируется уже имеющимся, по соседству с которым образуются
девять новых триплетов микротрубочек. Базальные тела гомологи-
чны, а в некоторых случаях идентичны с центриолями — центрами,
от которых во время митоза у многих (но далеко не у всех)
водорослей расходятся нити веретена. Центриоли располагаются
парами, образуя диплосому с осями, ориентированными под пря-
мым углом друг к другу, в особом участке гомогенной основной
цитоплазмы — центросфере, где пег ни рибосом, ни эндоплазмати-
ческой сети, ни каких-либо иных клеточных органелл. Центриоли,
как и базальные тела, образованы девятью триплетами микротру-
бочек, расположенными по кругу. При возникновении из единичной
центриоли диплосомы (во время поздней телофазы или даже ин-
терфазы) в гомогенной центросфере перпендикулярно к оси име-
ющейся центриоли появляются девять очень коротких триплетов
микротрубочек, которые растут, пока не достигнут длины трубочек
материнской центриоли. Никаких контактов между двумя система-
ми микротрубочек обнаружить нельзя—г^ежду ними всегда со-
храняется расстояние. По-видимому, как и в случае репликации
базального гела, каждая существующая центриоль является цент-
ром дифференцировки микротрубочек. Во время митоза центриоли
расходятся в противоположные стороны, образуя полюса ядра,
к которым сходятся нити веретена. Центриоли могут располагаться
вне ядра или же при образовании в оболочке ядра полярных отвер-
стий они вместе с микротрубочками проникают внутрь ядра. Цент-
риоли встречаются не только у монадных форм и водорослей,
жизненный цикл которых включает репродуктивные подвижные
клетки, они были обнаружены и у водорослей, лишенных подви-
жных стадий (кирхнериелла, тетраадрон, сценедесмус и др.). В од-
них случаях центриоли мигрируют от ядра к поверхности клетки
и начинают функционировать в качестве базальных тел, например
у хорды, У хары в вегетативных клетках центриоли отсутствуют,
однако имеются в клетках сперматогенных нитей, где они связаны
с веретеном, а позднее принимают участие в образовании жгутиков.
Во многих других случаях базальные тела функционируют во время
митоза в качестве центриолей (так базальные тела жгутиков зоо-
спор стигеоклониума после оседания зооспор мигрируют к ядру).
Псевдоцилии. У зеленых водорослей с пальмеллоидной струк-
турой клетки многих представителей несут неподвижные отрост-
ки— псевдоцилии. Электронно-микроскопическое исследование по-
30
казало, что эти структуры — производные жгутиков, базальные
тельца и корневые системы которых почти идентичны таковым
хламидомонады; переходная зона также имеет звездчатый узор,
характерный для жгутиков зеленых водорослей. Однако свободная
часть органеллы не имеет центральных микротрубочек, и «В» тру-
бочки периферических дублетов большей частью очень коротки.
Таким образом, под мембраной псевдоцилий микротрубочки име-
ют расположение 9 —0 вместо характерного для типичных жгутиков
(9 —9)4-2.
Гаптонема, Ряд золотистых водорослей (род хризохромулина,
кокколитофоридовые), помимо двух жгутиков, обладает особой
органеллой — гаптонемой. Это расположенный между двумя жгути-
ками обычно более короткий отросток; он может находиться в скру-
ченном или выпрямленном состоянии. У одного из видов хризох-
ромулины с помощью цейтраферной киносъемки удалось опреде-
лить скорость скручивания гапотнемы (этот процесс, оказалось,
занимает менее 1/50 с) и ее вытягивания (длится от 2 до 10 с).
Существует мнение, что гаптонема может служить для временного
прикрепления клетки к субстрату. С другой стороны, у видов
с очень длинной гаптонемой она может служить приспособлением,
облегчающим парение в толще воды, как и всякий вырост клетки.
На эту мысль наводят наблюдения за поведением гаптонемы: если
клетка движется с помощью жгутиков, гаптонема свернута, если же
жгутики перестают работать, гаптонема оказывается далеко вытя-
нутой.
Как и в жгутиках, внутри гаптонемы проходят микротрубочки,
расположение которых иное. Гаптонема, как правило, содержит
лишь один круг из 5—8 простых микротрубочек. Центральные
микротрубочки полностью отсутствуют. У вхождения гангонемы
в клетку (у ее основания) добавляются еще 2—3 микротрубочки, так
что число их достигает 9. Кроме того, в отличие от жгутиков кольцо
трубочек гаптонемы окружено тремя концентрическими обвертка-
ми, из которых самая наружная служит продолжением плазмалем-
мы клетки. Очевидно, микротрубочки это те структурные элемен-
ты, которые обусловливают движение как жгутиков, так и гап-
гонемы; корней у гаптонемы нет.
ХЛОРОПЛАСТЫ
В клетках эукариотических водорослей из органелл особенно
заметны хро.матофоры (х.юропласты)' —носители окраски, кото-
рые в отличие от хлоропластов высших растений чрезвычайно
разнообразны по форме (см. рис. 5). Хроматофоры. занимающие
в клетке в большинстве случаев постенное положение (париеталь-
1 Термин «хлоропласты» сейчас широко распространен, однако в более старой
литературе обычно приводится термин хроматофор.
31
ные хроматофоры), могут быть чашевидными (большинство видов
хламидомонады), в виде кольца, опоясывающего клетку (улотрикс),
в виде полого цилиндра, продырявленного многочисленными от-
верстиями (эдогониум), одной или многих идущих по спирали лент
(спирогира), одной-двух крупных париетальных пластинок (пенна-
тофициевые диатомеи), У многих водорослей хлоропласты много-
численны и имеют вид зерен или дисков, сосредоточенных в постен-
ной цитоплазме (зеленые водоросли, имеющие сифоновую органи-
зацию, харофициевые, центрофициевые диатомовые, динофитовые,
бурые, красные водоросли). Реже хроматофор занимает в клетке
центральное положение, тогда чаще всего он состоит из массивной
центральной части, от которой к периферии клетки отходят лопасти
или гребни (зигнема и большинство десмидиальных).
Среди зеленых водорослей, обладающих сифоновой организаци-
ей, различают две большие группы. Водоросли, относящиеся к пер-
вой группе (кодиум, бриопсис, дербезия), обладают только одним
типом пластид—хлоропластами. Это гомопластидные формы. Вто-
рая группа (дихотомосифон, каулерпа, галимеда, удотеа) гетероп-
ластидная. У них, помимо хлоропластов, имеются амилопласты.
Гомопластидия, гетеропластидия, так же как морфология и положе-
ние в клетке пластид,— важные таксономические признаки.
Субмикроскопическое строение хлоропластов водорослей в ос-
новных чертах сходно. У эукариотических водорослей хлоропласты
ограничены оболочкой, под которой находится тонкозернистый
материал матрикса (стромы), заключающий уплощенные, одетые
мембраной мешочки, или пузыри,— тилакоиды, или диски, содер-
жащие хлорофилл и каротиноиды.
Кроме того, в матриксе хлоропласта находятся рассеянные хло-
ропластные рибосомы, фибриллы ДНК, липидные гранулы и осо-
бые включения — пиреноиды. Однако в деталях тонкого строения,
касающихся оболочки, расположения тилакоидов и фибрилл ДНК,
места образования и отложения зерен запасных полисахаридов
и формы пиреноидов, хлоропласты водорослей обнаруживают весь-
ма постоянные различия, что и позволяет использовать их наряду
с набором пигментов, продуктами запаса и строением жгутикового
аппарата в качестве таксономических признаков, характеризующих
большие группы — отделы водорослей.
Оболочка хлоропласта — перманентная структура, всегда прису-
тствующая даже в тех случаях, когда тилакоидная система не
дифференцируется. Например, у хлоропластных мутантов хламидо-
монаса, не способных образовывать хлорофиллы в темноте, тилако-
идная система не наблюдается, а оболочка хлоропласта всегда
присутствует. Бесцветная водоросль политома содержит пластиду
без ламелл, но с оболочкой.
" У зеленых и красных водорослей оболочка хлоропласта об-
разована только двумя параллельными мембранами, у динофито-
вых и эвгленофитовых водорослей — тремя, а у золотистых, желто-
32
зеленых, диатомовых, криптофитовых, рафидофитовых и бурых
водорослей, вокруг одетых четырехмембранной оболочкой хлороп-
ластов, имеется сложная система мембран, находящаяся в прямой
связи с мембраной ядра,— хлоропластная эндоплазматическая сеть.
Расположение тилакоидов в матриксе хлоропласта также неодина-
ково в разных отделах водорослей, при этом хлоропласты водорос-
лей со сходными пигментами характеризуются и сходным располо-
жением тилакоидов (рис. 4).
Наиболее простое расположение наблюдается у красных водорос-
лей, у которых тилакоиды лежат в матриксе поодиночке (рис. 4, А, Б).
У остальных эукариотических водорослей тилакоиды группируются,
образуя ламеллы, причем число тилакоидов, входящих в состав одной
ламеллы, постоянно в пределах больших групп, объединяющих
родственные водоросли. Есть водоросли (криптофитовые), у которых
тилакоиды соединяются по два (рис. 4, В). У золотистых, желтозеле-
ных, диатомовых, бурых, динофитовых и эвгленофитовых водорослей
тилакоиды располагаются преимущественно по три (рис. 4, Г). Обыч-
но между соседними тилакоидами внутри ламеллы имеются проме-
жутки. У зеленых водорослей наиболее вариабельное расположение ;
тилакоидов: число их в ламеллах может колебаться от 2 до 6, а иногда >
до 20, в таких случаях стопки тилакоидов так тесно прижаты друг
к другу, что пространство между соседними тилакоидами исчезает >
и тогда эти стопки называют гранами (рис. 4, Е). Помимо зеленых J
водорослей, граны имеются у эвгленофитовых. В хлоропластах \
золотистых, желтозеленых, диатомовых и бурых водорослей с трехти-
лакоидными ламеллами находятся так называемые периферические
Рис. 4. Схема расположения тилакоидов в хлоропластах водорослей. А—модель
хлоропласта красной водоросли; Б — одиночное расположение тилакоидов в хлороп-
ластах красных водорослей; В — двухтилакоидные ламеллы криптофитовых; Г, Д—
трехтилакоидные ламеллы в хлоропластах бурых и динофитовых водорослей; Е—
граны зеленых водорослей:
ох— оболочка хлоропласта, т — тилакоиды, от- опоясывающий тилакоид, г—граны
2 - 1210
33
(опоясывающие) ламеллы, расположенные параллельно оболочке
j хлоропласта и окружающие остальные ламеллы, пересекающие хло-
ропласт (рис. 4, Г, Д). Среди красных водорослей у одних опоясыва-
I ющие тилакоиды имеются, у других-- отсутствуют. У зеленых,
jl-^эвгленовых водорослей опоясывающие ламеллы в хлоропластах не
обнаружены.
Хлоропласты водорослей из разных отделов различаются и по
расположению в них генофоров. Генофоры—это электронно-про-
зрачные участки, содержащие фибриллы ДНК. Их существование
впервые было продемонстрировано в 1962 г. у хламидомонад.
В хлоропластах Ch. reinhardtii, Ch. eugametos, Ch. moewusii наблюда-
лись области, которые окрашивались, по Фёльгену. так же интен-
сивно, как и ядро. Обработка клеток дезоксирибонуклеазой снимала
положительную реакцию Фёльгена, а также восприимчивость
к окраске акридиноранжем как в хлоропласте, так и в ядре. Элект-
ронные микрофотографии вскрыли в этих участках хлоропласта,
обнаруживающих положительную реакцию Фёльгена, присутствие
чувствительных к ДНК-азе фибрилл. Позднее и у других водорос-
лей было доказано существование хлоропластной ДНК. У красных
водорослей, например у Laurencia spectabilis, электронно-прозрач-
ные участки, содержащие фибриллы ДНК, как и у хламидомонад,
рассеяны беспорядочно по матриксу хлоропласта. Такое же бес-
порядочное расположение генофоров в хлоропластах свойственно
и динофитовым водорослям. Напротив, у исследованных бурых
водорослей (хорда, фукус, алария, сфацелярия, эгрегия) кольцевид-
ный генофор располагается под опоясывакйцей ламеллой. Такое же
локализованное под опоясывающей ламеллой расположение гено-
фора описано у желтозелепых и диатомовых водорослей. У золо-
тистых водорослей существование хлоропластной ДНК доказано
ауторадиографически, у эвгленофитовых как ауторадиографичес-
ки, так и с помощью методов экстракции.
Только у зеленых водорослей крахмал откладывается в матриксе
хлоропласта между ламеллами и вокруг пиреноида, у зеленых
водорослей, лишенных пиреноида, крахмал откладывается, как пра-
вило, в строме хлоропласта. Лишь у некоторых дазикладальных (без
пиреноидов) крахмальные зерна обнаружены как в хлоропластах, так
и свободно в цитоплазме. Кроме того, выявлено, что у Caulerpa
prolifera при разрушении оболочки амилопластов крахмал переходит
в цитоплазму. У зеленых сифоновых водорослей, помимо хлоропла-
стов, имеющих амилопласты, крахмал откладывае тся в обоих типах
пластид: в амилопластах зерна крахмала крупнее, чем в хлоропла-
стах. Амилопласты отличаются от хлоропластов по у.чьтраструкту-
ре. Все тело амилопласта обычно заполнено одним (редко двумя)
крупным крахмальным зерном, тилакоиды практически отсутствуют.
У всех остальных водорослей запасные вещества (хризоламинарин,
ламинарин, крахмал динофитовых водорослей, парамилон, багрян-
ковый крахмал) откладываются вне хлоропласта — в цитоплазме.
34
Хлоропласты эукариотических водорослей содержат особые
включения — пиреноиды. Это не означает, что пиреноиды встреча-
ются у всех представителей данного отдела. Так, среди зеленых
водорослей пиреноиды отсутствуют у всех дазикладальных, у ряда
гетеропластидных бриопсидальпых (галимеда, удотеа, большинство
видов каулерпы). Среди бурых водорослей пиреноид имеется у ви-
дов с изогамным и гетерогамным половым процессом, у более
высокоорганизованных форм с оогамным половым процессом пи-
реноид, как правило, отсутствует. В ряде случаев пиреноиды при-
урочены лишь к какой-либо стадии жизненного цикла. Например,
у некоторых фукальных пиреноид встречается в хлоропластах яйце-
клеток, но отсутствует в пластидах вегетативных клеток; наоборот,
у эустигматофициевых пиреноиды имеются в пластидах вегетатив-
ных клеток, но не зооспор. Пиреноиды могут находиться как внутри
хлоропласта, так и выдаваться за его пределы (сидеть на одной или
многих ножках), но и в последнем случае они заключены в оболочку
хлоропласта. Пиреноид имеет гранулярную белковую строму, в ко-
торую у одних видов ламеллы внедряются, а у других этого не
наблюдается. Расположение тилакоидов в строме пиреноида неоди-
наково.
У зеленых водорослей матрикс пиреноида отделяется от ма-
трикса собственно хлоропласта гранулами или пластинками крах-
мала.
Подобно ядру и митохондриям, хлоропласты, содержащие свои
собственные ДНК, являются самовоспроизводящимися органелла-
ми, которые размножаются путем деления. В ряде случаев передача
потомству пластидного аппарата осуществляется только через одну
из гамет, например у зигнемы и других копьюгатофициевых. У этих
водорослей наблюдается физиологическая анизогамия, при которой
хлоропласты мужской (отдающей свое содержимое) гаметы отми-
рают. У эдогониума передача хлоропласта также происходит через
яйцеклетку, так как хлоропласт сперматозоида крайне редуцирован.
То же относится к вошерии, сперматозоиды которой лишены хло-
ропласта.
Во многих случаях наблюдаются перемещения хлоропластов
в клетке. Так, при высокой интенсивности света хлоропласт мезоте-
ниума поворачивается боковой частью к свету. В клетках мужоции
изменение ориентации хлоропластов от фронтальной к профильной
также обусловлено светом, причем имеет значение не только его
интенсивность, но и спектральный состав. Вызываемая светом миг-
рация хлоропластов наблюдалась и у Eremosphaera viridi.s: при низ-
ких и средних интенсивностях света хлоропласты располагаются по
периферии клетки, в темноте и на ярком свету они собираются
вокруг крупного центрального ядра, образуя плотную сферу,
у Dietyota dichotoma также отмечено движение хлоропластов, вы-
званное светом. На слабом свету хроматофоры располагаются
вдоль клеточных стенок, которые перпендикулярны к направлению
35
света. При ярком же освещении они перемещаются на боковые
стенки, параллельные световому лучу. Такова же реакция хромато-
форов коровых клеток Fucus vesiculosus, где расположение пластид
определяется направлением падающего света.
ГЛАЗОК —СТИГМА
Монадные клетки обычно имеют кирпично-красный глазок, или
стигму, представленный рядами осмиофильных гранул, содержа-
щих пигмент астаксантин (гематохром). У водорослей разных групп
глазки в деталях различаются; в большинстве случаев они являются
частью хлоропласта. Число рядов гранул варьирует от 1 —2 до 9 (у
колониальных вольвокальных). Если слой гранул только один, то
они располагаются непосредственно под оболочкой хлоропласта,
а если слоев много, то слои разъединяются одним вздутым тилако-
идом. В каждом слое гранулы тесно упакованы и имеют гексаго-
нальные очертания. У организмов с однослойными глазками на
тангенталыюм срезе через клетку, а также при применении техники
замораживания — скалывания можно достаточно точно подсчитать
число гранул. Например, у Chlamydomonas eugametos глазок об-
разован 800 гранулами. Глазок в женских гаметах бриопсиса (мужс-
кие гаметы лишены глазка) содержит около 3000 гранул. В много-
слойном глазке Volvox aureus на вертикальном срезе подсчитано
примерило 100 гранул, и таким образом весь глазок (9 слоев) может
содержать приблизительно 1000 гранул. У эвгленофитовых, эвстиг-
матофициевых водорослей стигма находится вне пластиды. Среди
тех водорослей, у которых гранулы стигмы заключены в оболочку
хлоропласта, у одних нет связи глазка со жгутиковым аппаратом
(зеленые водоросли), у других (золотистые, желтозелеиые, бурые)
такая связь есть, и тогда основание заднего жгута, расположенное
вблизи глазка, несет характерное вздутие. У эвгленофитовых водо-
рослей глазок располагается вне хлоропласта, но связан со жгути-
ком. Особенно сложно устроенные глазки встречаются среди дино-
фитовых.
МИТОХОНДРИИ
Митохондрии сильно варьируют по числу и форме (удлиненные,
нитевидные, разветвленные, почти округлые, неправильных очерта-
ний) не только у разных водорослей, но и в одной и той же клетке
на разных стадиях жизненного цикла (см. рис. 5). Так, у эвглены
в фазе клеточного деления митоходрии имеют вид мелких немного-
численных телец, рассеянных в цитоплазме, а во время фазы роста
они принимают форму сети, занимающей в клетке периферическое
положение. В конце фазы роста в клетках эвглены, хламидомонады
образуются гигантские митохондрии за счет слияния более мелких.
Перед делением клетки они снова распадаются на нормальные
36
мелкие митохондрии. Расположение в клетке митохондрий строго
не фиксировано: они могут находиться в ней в центре под хлороп-
ластами (центральные митохондрии), по периферии клетки у наруж-
ной поверхности хлоропластов (периферические митохондрии);
у монадных форм сравнительно крупные митохондрии часто лока-
лизованы в непосредственной близости от основания жгутиков. Как
и у других растительных и животных организмов, митохондрии
водорослей ограничены оболочкой из двух элементарных мембран.
Внутренняя мембрана образует складки-к/шс/ны, число которых,
а соответственно и расположение — более тесное или свободное —
сильно варьирует. Матрикс митохондрий содержит рибосомы и эле-
ктронно-прозрачные участки, в которых находятся фибриллы ДНК.
Размножаются митохондрии посредством деления или почкования.
Митохондрии являются источниками энергии в клетке, ее дыхатель-
ными центрами.
ВАКУОЛИ
В цитоплазме находятся отграниченные от нее элементарной
мембраной — тонопластом — и наполненные гомогенной жид-
костью полости — вакуоли. Они бывают разных типов. Взрос-
лые клетки водорослей, одетые клеточной оболочкой, как и клет-
ки высших растений, содержат центральные вакуоли с клеточным
соком, занимающие большую часть клетки и оттесняющие ци-
топлазму с ее включениями к клеточной стенке. В других слу-
чаях клетки могут содержать много более мелких вакуолей с кле-
точным соком. Вакуоли иногда пересекают цитоплазматические
тяжи. Особенно крупными вакуолями с клеточным соком отлича-
ются водоросли, имеющие сифоновую организацию таллома.
У концов клетки некоторых десмидиальных (клостериум, пениум,
плеуротениум) имеются вакуоли, которые содержат кристаллы
гипса, обнаруживающие броуновское движение. В клетках бу-
рых водорослей, особенно меристематических, репродуктивных,
вокруг ядра или пластид группируются мелкие вакуоли — физоды,
включающие фукозан (вещество, обнаруживающее свойства та-
нина).
В монадных клетках, а также в клетках ризоподиальных и паль-
меллоидных форм находятся пульсирующие или сократительные
вакуоли, отличающиеся от остальных вакуолей ритмической де-
ятельностью; через правильные интервалы времени они увеличива-
ются в объеме, воспринимая жидкость из окружающей цитоплазмы
(диастола), а затем быстро уменьшаются, освобождая содержимое
наружу (систола). Это органеллы, осуществляющие осморегуля-
цию, они свойственны в первую очередь пресноводным формам,
реже встречаются у морских. В большинстве случаев, если главная
вакуоль растянута, то представлена шаровидным пузырем, ограни-
ченным одной мембраной, и окружена многочисленными мелкими
37
дополнительными везикулами, которые, сливаясь С тонопластом
главной вакуоли, изливают в нее свое содержимое. Положение
пульсирующих вакуолей в клетке весьма стабильно. Так, у боль-
шинства вольвокальных и в подвижных репродуктивных клетках
зеленых водорослей на переднем конце клетки наблюдаются две
пульсирующие вакуоли, у хлорогониума 12—16, а у гемотококка
30—60 сократительных вакуолей рассеяны по клетке беспорядочно;
у золотистых водорослей, например у малломонаса, сократитель-
ная вакуоль находится в заднем конце клетки. У подвижных дино-
фитовых водорослей сократительные вакуоли отсутствуют, имеют-
ся особые органеллы, несущие функцию осморегуляции,— пузулы.
Они представляют собой впячивание (инвагинацию) цитоплазмати-
ческой мембраны, глубоко вдающееся от основания жгутикового
канала внутрь клетки и тесно соприкасающиеся с вакуолями с кле-
точным соком (подробнее о различных типах пузул см. гл. 11).
ЯДРО
Клетка эукариотических водорослей содержит одно или много
ядер (до нескольких тысяч — в крупных старых клетках красной
водоросли гриффитзии). Ядро различной формы (шаровидное, линзо-
видное, эллипсоидальное и пр.) располагается в постенной цитоплазме
глубже хлоропластов (улотрикс, эдогониум и др.), реже в центре
клетки; тогда оно заключено в цитоплазматическую обвертку и подве-
шено на цитоплазматических тяжахфспирогира) или находится в цито-
плазматическом мостике (пеннатофициевые диатомеи). Ядро окруже-
но ядерной оболочкой, образованной двумя мембранами, из которых
наружная связана с мембранами эндоплазматического ретикулума
клетки. Между двумя мембранами ядерной оболочки расположена
узкая перинуклеарная полость или перинуклеарное пространство,
которое в некоторых случаях (например, при формировании мастиго-
нем) может заметно расширяться. Ядерная оболочка, как правило,
пронизана многочисленными порами, размеры, строение и располо-
жение которых варьирует (беспорядочное, в прямых рядах, гексаго-
нальными группами и пр.). Ядерная оболочка заключает нуклеоплаз-
му, в которой находятся ядрышки (одно — несколько) и хроматин.
У большинства водорослей в интерфазном ядре хроматин имеет вид
равномерно рассеянного тонкогранулированного материала. Исклю-
чение составляют эвгленофитовые и динофитовые, у которых хромо-
сомы остаются спирализованными на протяжении всего ядерного
цикла и могут быть различимы и подсчитаны в интерфазном ядре.
Деление ядра. Ядра водорослей делятся, как правило, митотичес-
ки. Однако в разных группах водорослей было обнаружено большое
разнообразие в организации митотического аппарата (примеры
будут даны при описании соответствующих групп). Здесь лишь
укажем основные признаки, по которым различаются митозы в раз-
ных группах водорослей. Центриоли встречаются далеко не у всех
38
водорослей, они описаны у хлорококкальных, рафидофитовых, жел-
тозеленых, бурых.
У золотистых водорослей имеется ризопласт, у красных — так
называемые полярные кольца — короткие полые цилиндры, около
которых иногда (не всегда) могут собираться микротрубочки. У од-
них водорослей (клебсормидиум, конъюгатофициевые, харофици-
евые, золотистые, криптофитовые, диатомовые) веретено открытое,
т. е. ядерная оболочка полностью исчезает и отсутствует на протя-
жении некоторых стадий митотического цикла (с поздней профазы
до поздней анафазы — ранней телофазы). У других (эдогониальные,
эвгленофитовые, хлоромонадофитовые, желтозеленые, динофито-
вые) ядерная оболочка остается интактной на протяжении всего
митоза (закрытое веретено). У вольвокальных, хлорококкальных,
ульвовых, бурых, красных водорослей в ядерной оболочке возника-
ют полярные отверстия (полузакрытое веретено). Происхождение
микротрубочек веретена или его эквивалента различно: у динофито-
вых они полностью цитоплазматические, у эвгленофитовых, желто-
зеленых— полностью внутриядерные, у большинства водорослей
(вольвокальные, хлорококкальные, ульвовые, эдогониальные, зиг-
немальные, рафидофитовые, золотистые, криптофитовые, бурые,
красные) — отчасти цитоплазматического, отчасти ядерного проис-
хождения. Кинетохоры описаны у эндогониальных, зигнемальных,
рафидофитовых, красных, в остальных группах водорослей они
отсутствуют.
Деление клетки. Цитокипетический аппарат у водорослей также
отличается большим разнообразием. Протопласт может разделять-
ся или путем центрипетального разрастания борозды дробления,
или путем образования центрифугально разрастающейся клеточной
пластинки. И тот и другой тип разделения протопласта может
сочетаться с образованием как фрагмопласта — системы микротру-
бочек, возникающей в плоскости, перпендикулярной к плоскости
будущего цитокинеза, так и фикопласта — системы микротрубочек,
параллельных плоскости цитокинеза. Оболочка материнской клетки
либо участвует в образовании оболочек дочерних клеток, либо не
принимает в этом процессе никакого участия. В первом случае
дочерние протопласты сохраняют родительскую клеточную стенку,
с которой непосредственно связаны новые стенки (так называемый
десмосхизис), например, при вегетативном делении клеток у боль-
шинства нитчатых водорослей. Во втором случае (этот тип деления
называется элевтеросхизис), наблюдающемся, например, при форми-
ровании зооспор, гамет вольвокальных, при автоспорообразова-
нии многих водорослей их оболочки возникают независимо от
оболочки материнской клетки, которая часто подвергается авто-
лизу. В большинстве случаев цитокинез непосредственно следует
за митозом. У водорослей, имеющих сифонокладальную структу-
ру, оба эти процесса разобщены, что приводит к многоядерности
клеток.
'39
Рис. 5. Схема строения клетки Chlamydomonas по электронной микрофотографии:
кс — клеточная стенка, цм— цитоплазматическая мембрана, ох — оболочка хроматофора, т —
тилакоиды, с—стигма, п — пиреноиД, к — крахмал, м-.--митохондрии, д—диктиосомы, р —
рибосомы, эс—эндоплазматическая Ьеть, я — ядро', пв~-пульсирующие вакуоли, ж —жгутик
Особенности цитокинетического аппарата подробно изучались
у зеленых водорослей; ему придается немалое филогенетическое
значение.
На основе объемной реконструкции серий ультрагонких
срезов клетки и количественных измерений было установле-
но, что у Chlamydomonas reinhardtii около 40% объема клетки
заняты хлоропластом, около 10% — ядром, узкая область ци-
стерн Гольджи охватывает около 1%, митохондрии — 3%, ваку-
оли - 8%, липидные тельца — приблизительно 0,5%. Сходный
морфометрический анализ клетки сделан и для Chlorella vul-
garis.
На рис. 5 дана схема строения клетки эукариотической во-
доросли на примере хламидомонады по данным электронной мик-
роскопии.
40
Глава 4
РАЗМНОЖЕНИЕ
Различают вегетативное, бесполое и половое размножение.
ВЕГЕТАТИВНОЕ РАЗМНОЖЕНИЕ
К вегетативному можно отнести те процессы размножения, при
которых части таллома отделяются без каких-либо заметных изме-
нений в протопластах.
Вегетативное деление клеток десмидиальных, диатомовых, мно-
гих одноклеточных синезеленых водорослей, продольные деления
монадных представителей, распад — диссоциация колоний на две
или большее число частей (например, у синуры, диктиосфериума) —
примеры вегетативного размножения. Распространенным приме-
ром вегетативного размножения нитчатых и более высокоорганизо-
ванных форм является фрагментация—разрыв талломов на от-
дельные участки. Этот способ наиболее обычен среди зеленых водо-
рослей и достигает кульминации у таких родов, как клебсормидиум
(нити нередко распадаются на отдельные клетки) и особенно у рода
стихококкус, который встречается в виде отдельных клеток (нити
даже из нескольких клеток редки). Диссоциация нитей на корот-
кие отрезки или даже отдельные клетки легко происходит у зиг-
немальных.
Вегетативное размножение широко распространено и среди бу-
рых водорослей. Гаметофиты рода ламинария, происходящие от
одной зооспоры, могут распадаться на неограниченно большое
число особей. Среди фукальных крупные скопления поддерживают-
ся исключительно путем процесса регенерации участков таллома, то
же относится к пелагическим видам рода саргассум.
Лежащая в основе вегетативного размножения способность реге-
нерации целого таллома из отдельных частей или даже клеток
экспериментально продемонстрирована и для красных водорослей
(гриффитзия, родохортон). Этим объясняется их встречаемость
в природе там, где внешние условия не благоприятствуют прораста-
нию спор.
Более специализированные структуры, служащие для вегетатив-
ного размножения,— выводковые почки или пропагулы. Они могут
быть одноклеточными, многоклеточными, сифонового строения,
обычно содержат обильные запасные вещества и вырастают не-
посредственно в новый таллом. В качестве примера можно привести
многоклеточные выводковые почки бурой водоросли сфацелярия
с довольно сложной морфологией, различающейся у разных видов:
У одних они двухраздельпые, у других — трехраздельные. Пропа-
гулы сифонового строения, по форме поразительно похожие на
41
трехраздельные пропагулы сфацелярии, описаны для зеленой бри-
опсидальной водоросли дербезии. У другой бриопсидальной водо-
росли— кодиума — пропагулы в виде легко отделяющихся специ-
ализированных веточек. С многоклеточными пропагулами можно
сблизить клубеньки харофициевых водорослей, которые образуют-
ся у некоторых видов на нижних, погруженных частях таллома
и на ризоидах. У ряда сифоновых и сифонокладальных водорослей
(бриопсис, галицистис, валония) голые участки цитоплазмы -про-
топласты, вытекающие из таллома при его повреждении, очень
быстро (в течение нескольких минут) секретируют новые клеточные
стенки и развиваются в соответствующие талломы.
Специализированным типом вегетативного размножения можно
считать и образование структур, предназначенных для перенесения
периодов, неблагоприятных для обычного вегетативного роста.
В этих случаях в клетках утолщаются оболочки, в протопласте
накапливаются запасные вещества — жир, крахмал. Такие клетки,
называемые акинетами, встречаются у многих нитчатых форм (уло-
трикс, микроспора, кладофора).
БЕСПОЛОЕ РАЗМНОЖЕНИЕ
Бесполое размножение осуществляется посредством особых
спор. Самый обычный способ бесполого размножения у многих
водорослей — посредством зооспор, как правило, голых монадных
клеток. Из содержимого каждой клетки может сформироваться
только одна зооспора (например, у эдогониума) или чаще содер-
жимое клетки делится на две, четыре, восемь и более частей и про-
дуцируется соответствующее количество зооспор. У многих водоро-
слей клетки, в которых формируются зооспоры, специально не
дифференцированы, однако у трентеполиевых зеленых водорослей
и у всех бурых есть спорангии, образующиеся только во время
бесполого размножения и отличающиеся по форме и размерам от
обычных вегетативных клеток.
В ряде случаев выяснены (экспериментально) условия внешней
среды, стимулирующие переход к зооспорообразованию. Для мно-
гих нитчатых зеленых водорослей таким фактором служит повы-
шенное содержание в воде свободной углекислоты. Это объясняет
старые наблюдения над массовым зооспорообразованием при пере-
несении нитей из текучей воды в сгоячую, со света в темноту, т, е.
в условия, когда в воде накапливается углекислота.
В связи с зооспорообразованием уместно остановиться на воз-
никновении так называемых синзооспор. Синзооспоры — образова-
ния, соответствующие комплексу двух или многих сросшихся в раз-
личной степени зооспор,— наблюдались у разных водорослей, но
особенно часто у разножгутиковых. Нередко при выходе из мате-
ринской клетки такие зооспоры остаются соединенными по две или
по нескольку лишь посредством тонких цитоплазматических нитей
42
и позднее разъединяются. Однако часто зооспоры оказываются
связанными своими задними концами гораздо прочнее и на
большем протяжении, так что свободными остаются только их
передние концы со жгутиками или же, наконец, они образуют
сплошную цитоплазматическую массу, из которой выступают пары
жгутиков, указывая на число соединившихся зооспор. Такие стадии
очень похожи на синзооспоры вошерии, которым они полностью
соответствуют.
Синзооспоры часто встречаются у бочридиопсиса. трибонемы,
ботридиума и др., однако, как правило, эти водоросли образуют
двужгутиковые зооспоры. Эти случайные отклонения стабилизиро-
вались лишь у вошерии. Ее крупные многожгутиковые зооспоры
уже давно рассматривали как сложную зооспору, образующуюся
внутри спорангия, в котором деление спорангиального протопласта
па одноядерные двужгутиковые споры не наблюдается.
Синзооспоры имеются также у зеленых (сфероплея, кладофора,
хламидомонас и др.) и бурых водорослей (Ectocarpiis siliculosus).
У значительного числа водорослей вместо, зооспор образуются
апланоспоры—неподвижные, лишенные жгутиков споры. Примера-
ми их могут служить моно- и тетраспоры красных водорослей,
тетраспоры диктиотовых из бурых водорослей. В тех случаях, когда
апланоспоры, будучи заключенными в оболочку материнской клет-
ки, принимают все отличительные черты этой клетки (характерные
очертания, особенности оболочки), их называют автоспорами (на-
пример, у многих хлорококкальных из зеленых водорослей).
У ценобиальных (монадных и коккоидных) зеленых водорослей
при бесполом размножении образуются дочерние колонии. В ряде
групп водорослей бесполое размножение отсутствует: у конъюгато-
фициевых, харофициевых, многих зеленых водорослей сифонового
строения, у всех диатомовых, фукальных из бурых водорослей.
На примере некоторых водорослей удалось глубже проникнуть
в механизмы освобождения клеток бесполого размножения и их
последующего прикрепления к субстрату. Вот некоторые примеры.
Для многих (более 40) видов хламидомонады было показано, что
молодые клетки выделяют литический фактор — автолизин споран-
гиальной стенки, вызывающий лизис клеточной стенки спорангия,
что способствует освобождению зооспор. Фермент действует толь-
ко на спорангиальную стенку, но пе на стенки вегетативных клеток
и зооспор и обладает групповой специфичностью, т. е: влияет
только на морфологически сходные виды, а в некоторых случаях —
только на спорангии вида-продуцента.
У вольвокса клетки также образуют и выделяют в окружающую
среду ферменты, лизирующие инволюкрум материнского ценобия,
облегчая этим освобождение дочерних сфероидов. Из культураль-
ной среды ChloreUa fusca var. vacuolata был выделен фермент (от-
сутствующий в среде до начала споруляции), разрушающий кле-
точную стенку материнских клеток автоспор и не действующий
43
на клеточные стенки самих автоспор, У вошерии в. кончике синзо-
оспоры были обнаружены многочисленные везикулы, по-видимому,
дериваты диктиосом, содержащие предположительно целлюлазу
и другие энзимы, разрушающие полисахариды. Высказано пред-
положение, что они ответственны за разрушение стенки в апи-
кальной области спорангия — необходимое условие выхода синзо-
оспоры. В зооспорах зеленой водоросли Oedogonium cardiacum, бу-
рой— Chorda tomentosa, тетраспорах красной водоросли Palmaria
palmata обнаружены обильные везикулы, содержащие клеющий ма-
териал, который, возможно, способствует прикреплению спор к суб-
страту. То же, по-видимому, справедливо и для тетраспор и ка-
рпоспор Ceramium rubrum,
ПОЛОВОЕ РАЗМНОЖЕНИЕ j y
Половое размножение (воспроизведение), сущность которого за-
ключается в слиянии — копуляции двух различных в половом от-
ношении клеток-гамет с образованием зиготы, известно у большин-
ства эукариотических водорослей. Во многих случаях, особенно
у водорослей с монадной организацией, обнаружению полового
процесса способствовала разработка методов их культивирования
в лабораторных условиях. Например, при определенных условиях
культивирования в последние годы был описан половой процесс
у ряда динофлагеллят, ранее считавшихся лишенными его.
Половое размножение — сложный многоступенчатый процесс,
начинающийся с половой дифференцировки клеток—гаметогенеза,
включающий слияние гамет (происходящее в несколько этапов
и приводящее к образованию зиготы) и завершающийся мейозом.
Условия, вызывающие половой процесс. Переход к половому
размножению и дифференцировке гамет зависит не только от внут-
ренних причин, нередко определяемых возрастом водоросли, но
и от факторов внешней среды. Среди этих факторов для многих
зеленых водорослей ведущую роль играет содержание в среде азота.
Исчерпание в среде азота и наступающее азотистое голодание слу-
жит сигналом для начала полового размножения многих видов
Chlamydomonas (Ch. reinhardtii, Ch. moewusii var. rotunda,
Ch. chlamydogama, Ch. eugametos), Pandorina morum, P. unicocca,
Chlorococcum echinozygotum, Golenkinia minutissima, Scenedesmus
obliquus, Oedogonium cardiacum и др. Хорошим примером зависимо-
сти половой дифференцировки от содержания в среде азота служит
зеленая водоросль Sphaeroplea annulina. Эта азотолюбивая водо-
росль встречается у нас преимущественно ранней весной в мелких
водоемах и уже в мае при понижении содержания в воде азота
переходит к половому воспроизведению. В водоемах с более посто-
янным подтоком азотистых веществ, например на полях орошения
Москвы, сфероплея вегетирует иногда почти все лето. В лаборато-
рии также можно поддержать ее в вегетативном состоянии, культи-
44
вируя в среде со значительным содержанием азота; перенесением же
в среду, бедную солями азота, легко можно вызвать половой про-
цесс.
У перечисленных и многих других видов источники азота (нит-
раты, аммоний, мочевина, глютамин и пр.), поддерживающие рост,
препятствуют половой дифференцировке, которая наступает в усло-
виях, неблагоприятных для вегетативного роста. Рост и размноже-
ние находятся как бы в антагонистических отношениях друг с дру-
гом. В отличие от большинства зеленых водорослей у вольвоксов
половое размножение с последующим образованием покоящихся
зигот происходит в то время, когда условия среды оптимальны для
вегетативного роста. Это объясняется тем, что половой процесс
у этого рода сопряжен с продукцией специальных половых ценоби-
ев, в которых и формируются гаметы. В менее оптимальных для
вегетативного роста условиях половые сфероиды уменьшались бы
в размерах, понижалась бы их витальность. Половая дифферен-
цировка у вольвоксов наступает под действием видоспецифичных
половых гормонов — индукторов, выделяемых мужскими ценоби-
ями, появляющимися в популяции спонтанно (см. гл. 15).
Дефицит в среде азота вызывает половой процесс и у многих
динофлагеллят.
Для индуцирования полового процесса у десмидиальных водо-
рослей наиболее важными факторами оказываются свет и тем-
пература, хотя в некоторых случаях формирование конъюгацион-
ных выростов и образование зигот наблюдалось при недостатке
в среде азота. Свет и температура являются также решающими
факторами среды, оказывающими воздействие на гаметогенез мно-
гих бурых водорослей. Так, для многих ламинариальных водорос-
лей было показано, что переход их гаметофитов к гаметогенезу
контролируется синим светом достаточно высокой интенсивности
при низких (5—10° С) температурах (см. гл. 10). У организмов,
приуроченных к водам с высоким содержанием солей, например
у Dunaliella salina, понижение концентрации солей в среде вызывает
половой процесс.
Формы полового процесса. Форма полового процесса может быть
различной. У ряда лишенных клеточной оболочки монадных во-
дорослей сливаются, (копулируют) клетки, не отличимые от ве-
гетативных. Такая форма полового процесса называется хологамией
(напрймёр, у зеленой водоросли дуналиелла). У многих водорослей
гаметы, как и зооспоры, образуются внутри материнских клеток
и, выйдя из них, копулируют попарно. Если обе сливающиеся
монадцые гаметы не различаются морфологически, половой про-
цесс называют изогамным — изогамией, при копуляции монадных
гамет, рдзличающи^я. по., размерам, имеет место гетерогамия или
анизогамия. Более крупные макрогаметы трактуются как женские,
более мелкие — микрогаметы ---как мужские. При оогамном поло-
вом процессе — оогамии—более крупные женские гаметы лишены
45
жгутиков, неподвижны и называются яйцеклетками или яйцами,
более мелкие мужские гаметы со жгутиками подвижны и носят
название сперматозоидов или антерозоидов.
У красных водорослей яйцеклетки оплодотворяются мужскими
клетками, лишенными жгутиков, — спермациями.
У зеленых водорослей за единичными исключениями наблюда-
ется истинная оогамия, для которой характерно оплодотворение
яйца внутри оогония — на месте образования (in situ) в отличие от
примитивной оогамии, когда яйцеклетка покидает оогонии до опло-
дотворения, осуществляющегося в окружающей среде. Примитив-
ная оогамия, встречающаяся среди зеленых водорослей лишь в виде
исключений, является правилом для оогамных бурых водорослей.
У центрофициевых оогамных диатомовых наблюдается как истин-
ная, так и примитивная оогамия, Есда сливаются протопласты
неподвижных клеток, не отличающихся от вегетативных, говорят
о конъюгации.
Обычно предполагается, что форма полового процесса эволюци-
онировала в следующем направлении: изогамия->анизогамия->
оогамия. Эта прогрессивная эволюция началась с возникновения
различающихся по размерам гамет у изогамных предковых форм.
Образование мелких гамез повышает их нумерическую продуктив-
ность— их число, но за счет редукции количества цитоплазмы.
С другой стороны, большая масса цитоплазмы и резервных матери-
алов у более крупных гамет (возникающих в материнских клетках
в меньшем числе) необходимы для начального развития будущей
зиготы. Таким образом, раз возникший диморфизм гамет дает
основу для избирательного слияния между гаметами, различающи-
мися по размерам: гаметы минимальных размеров получают ком-
пенсацию в количестве цитоплазмы посредством слияния с более
крупными гаметами. Под эти чисто теоретические рассуждения
в последнее время была подведена экспериментальная основа.
Л. Визе с соавт. (L. Wiese, W. Wiese, D. A. Edwars, 1979) удалось
экспериментально получить фенотипическую анизогамию у трех
изогамных гетероталличных видов Chlamydomonas (Ch. reinhardtii.
Ch. chlamydogama, Ch. moewusii ). У этих видов каждый тип спарива-
ния (« + » и « —») имеет два пути гаметогенеза, которые индуциру-
ются подбором условий культивирования. Первый путь гаметогене-
за заключается в том, что протопласт вегетативной клетки в резуль-
тате внутриклеточной перестройки непосредственно превращается
в одну крупную гамету. При втором способе гаметогенеза в мате-
ринской клетке происходят два или три митоза, и образуется 4—8
гамет меньшего размера. Комбинированием совместимых гамет,
сформировавшихся разными способами, может быть получена фе-
нотипически анизогамная копуляция. Высказано предположение,
что любая мутация, которая зафиксировала бы тот или иной способ
гаметогенеза (блокировала бы один путь гаметогенеза в пользу
другого), дала бы пепосредствешю штаммы, производящие исклю-
46
чительно или микро-, или макрогаметы. Таким образом могли
возникнуть уже генетически анизогамные линии. В появившемся
в результате мутации анизогамном таксоне при дальнейшей эволю-
ции может быть утеряна подвижность у более крупных гамет.
Сначала жгутики еще сохраняются, как у Ch. suboogama, полная же
их потеря привела бы к настоящей оогамии, как у Ch. coccifera.
Ch. pseudogigantea.
Эволюция формы полового процесса происходила независимо
от эволюции таллома. Это впервые продемонстрировал русский
альголог И. Н. Горожанкин в конце прошлого века, обнаружив
у разных видов хламидомонады (стоящих на одном уровне мор-
фологической дифференцировки таллома) все основные типы поло-
вого процесса: наряду с изогамией, преобладающей у одноклеточ-
ных вольвокальных, гетерогамию у Ch. braunii и оогамию
у Ch. coccifera.
Гаметогенез. У форм с изогамным и гетерогамным половым
процессом гаметогенез в большинстве случаев не связан с форми-
рованием каких-либо обособленных морфологических структур:
изо- и гетерогаметы возникают в обычных вегетативных клетках,
как и зооспоры, но обычно в большем числе и соответственно
меньших размеров. Исключение составляют изогамные и гетеро-
гамные бурые водоросли, где гаметы образуются в специальных
органах: многогнездных или многокамерных спорангиях, в каждой
камере (клетке) которых формируется по одной гамете. Оогамия
у монадных и коккоидных форм обычно также не сопряжена с из-
менением клеток, производящих гаметы: они не отличаются от
вегетативных. Наоборот, у водорослей с нитчатой и тканевой ор-
ганизацией таллома яйцеклетки и сперматозоиды развиваются в га-
метангиях, как правило, резко отличающихся по форме от вегета-
тивных клеток и носящих специальные названия оогонии и антери-
дии (сперматогонии).
Растения, производящие гаметы, могут быть обоеполыми (од-
нодомными) и раздельнополыми (двудомными). В первом случае
гаметы разного пола возникают на одном и том же талломе,
во втором--на разных. Кроме понятий «однодомность» и «дву-
домность» существуют понятия «гомоталлизм» и «гетероталлизм».
При гомоталлизме половой процесс имеет место в пределах кло-
нальной популяции (клон — культура, полученная из одной клетки).
При гетероталлизме половой процесс возможен только при сме-
шении клеток из разных клонов противоположного типа спари-
вания или знака; в случае гетероталличных изогамных форм гаметы
разного типа спаривания обозначаются знаками « + » и « —». По-
нятия «гомоталличный» и «гетероталличный» не всегда совпадают
с понятиями «однодомный» и «двудомный». Гак. среди вольвоксов
имеются гомо галличные и гетероталличные виды, включающие
как однодомные, так и двудомные штаммы. Например, внутри
клональной популяции могут развиваться особи как однодомные.
47
с яйцами и сперматозоидами в одном сфероиде, так и двудомные,
у которых половые органы находятся в разных индивидах.
Освобождение репродуктивных клеток. Некоторые морские водо-
росли, обитающие в верхней части литоральной зоны, обнаружива-
ют периодичность в формировании и освобождении репродуктив-
ных клеток как бесполого (спор), так и полового (гамет) размноже-
ния. Так, у берегов Англии у водоросли Dictyota dichotoma форми-
рование половых органов (антеридиев и оогониев) приурочено
к летним месяцам и происходит периодически с двухнедельными
интервалами, что соответствует таковым между двумя сигизий-
ными приливами1, а освобождение гамет — яиц и сперматозоидов —
наступает через несколько приливов после наивысшего из последу-
ющих квадратурных приливов1 2 и продолжается 1—9 дней. Таким
образом, периодичность находится в строгом соответствии с лун-
ным ритмом. Периодичность сохраняется и при культивировании
талломов в лаборатории. У одного и того же вида ритм освобожде-
ния гамет зависит от географического положения местообитания.
Так, у D. dichotoma в Северной Каролине периодичность наблюда-
лась, но иная, чем у берегов Англии: гаметы освобождались не через
двухнедельные интервалы, а через месячные, а у берегов Ямайки
половые органы формировались периодически, но период созрева-
ния гамет растягивался настолько, что выход их происходил почти
непрерывно.
Наряду с полулунным ритмом в освобождении яйцеклеток
у D. dichotoma была описана и дневная периодичность. При культи-
вировании в условиях смены света (14 ч) и темноты (10 ч) вскоре
после начала светового периода наблюдался активный выход яйце-
клеток, который продолжался в течение часа, после чего освобожде-
ние яиц прекращалось до начала следующего светового периода,
когда вновь начиналось освобождение яиц. Дневной выход яиц
может быть индуцирован очень слабым кратковременным освеще-
нием. Так, 50% яиц зрелого оогония освобождается после 20-се-
кундного освещения белым светом в 1 Лк. Выход яиц зависел также
от температуры: если при 20* С максимум освобождения яиц дости-
гается через 25—30 мин после начала дневного светового периода,
то при 10 С этот максимум наступал только через 100 мин. Эффект
света данной интенсивности также зависит от температуры. Кроме
того, вскрытие полностью созревшего оогония может быть ускоре-
но или замедлено путем изменение осмотического давления среды.
Специальное исследование по выяснению влияния некоторых
внешних условий на периодичность освобождения гаме было про-
ведено в Японии над популяциями обычной в прибрежных водах
1 Два раза в месяц в новолуние и полнолуние происходят наибольшие приливы
и отливы, называемые сигизийными.
2 В фазе первой и последний четвертей Луны амплитуда приливов и отливов
наименьшая (квадратурные приливы и отливы).
48
водоросли — Monostroma nitidum, занимающей второе (после видов
порфиры) место в качестве объекта промышленного культивирова-
ния. Это культивирование осуществляется обычно двумя методами:
на хиби-сетках, расположенных горизонтально на такой глубине,
при которой талломы обнажаются во время отлива приблизитель-
но на 4 ч, и на бета-сетках с рас тениями, постоянно погруженными
в морскую воду вблизи ее поверхности. Образование гамет легко
распознается невооруженным глазом благодаря желтоватой окра-
ске плодущей части таллома. Были проведены сравнительные на-
блюдения над талломами, произрастающими поблизости друг от
друга, но в различающихся условиях: на прибрежных скалах — в
естественных местах обитания, на хиби-сетках и бега-сетках. Пери-
од плодоношения (с февраля по июнь) во всех грех случаях был
сходным, а периодичнос ть в освобождении гамет оказалась разной.
Более четко (с двухнедельными интервалами, во время каждого
квадратурного прилива) периодичность была выражена у растений,
растущих на скалах. Менее четко она обнаруживалась у талломов
на хиби-сетках, а у постоянно погруженных талломов на бета-сетках
никакой периодичности не наблюдалось. Продемонстрировано так-
же, что периодичность в образовании гамет зависит от освещения
(интенсивность света и продолжительность фотопериода) и тем-
пературы.
Периодичность наблюдалась и в спороношении Ulva pertusa,
в освобождении яйцеклеток у Sargassum muticum.
В последние годы в ряде работ прослежена дифференцировка
и механизм освобождения спермациев у красных водорослей на
электронно-микроскопическом уровне. Так, у Ptilota plumosa спер-
матогонии, в которых образуются мужские гаметы — спермации,
обладают ясно выраженной полярностью, заключающейся в том,
что клеточное ядро занимает апикальное положение, а в базальной
части молодого сперматогония находится одна или несколько ваку-
олей. В зрелых сперматогониях вакуоли замещаются крупными
фиброзными исчерченными вакуолями, занимающими приблизите-
льно половину объема клетки и возникающими, возможно, за счет
слияния пузырьков, происходящих от диктиосом. Фиброзные ваку-
оли содержа! мукополисахарид. Вокруг клеточного ядра и окружа-
ющей цитоплазмы, по-видимому, образуется новая плазмалемма,
не включающая базальных фиброзных исчерченных вакуолей. Та-
ким образом формируется спермации, который покидает оболочку
сперматогония. Одновременно с разрывом клеточной стенки спер-
матогония опорожняются базальные крупные фиброзные вакуоли,
что способствует освобождению спермация. Спермаций без клеточ-
ной оболочки одет тонкой слизистой обверткой, возможно проис-
ходящей от матрикса фиброзной вакуоли. Эта обвертка играет
важную роль во время прилипания спермация к трихогине. Сходные
процессы наблюдаются и при дифференцировке и освобождении
спермация у Polysiphonia hendryi. У ламинарии при образовании
49
сперматозоидов аппарат Гольджи выделяет большое количество
слизи, способствующей разрыву антеридия и освобождению спер-
матозоидов.
У вошерии везикулы, возможно, содержащие целлюлазы, наблю-
дались в апикальной части оогониального клювика, у места, где
сперматозоиды входят в оогоний вскоре после того, как клювик
образует пору. Такие же везикулы наблюдались вблизи кончика
антеридия, где выходят сперматозоиды.
Копуляция. Слияние — спаривание гамет, приводящее к возник-
новению зигот, происходит в несколько этапов. Первый из этих
этапов — образование групп, в которых гаметы разного пола всту-
пают в первоначальный контакт. Если в капле воды или культу-
ральной жидкости наблюдать поведение снабженных жгутиками
гамет одного пола (в случае изогамет — одного типа спаривания
или одного полового знака « + » или « —»), можно убедиться, что
гаметы равномерно рассеяны по всему полю зрения микроскопа.
Стоит к капле с такими гаметами добавить гаметы противополож-
ного пола (знака), как сейчас же образуются скопления — группы.
Это впервые наблюдалось в конце прошлого века у изогамной
бурой водоросли Ectocarpus siliculosus, где много гамет одного пола
окружают одну гамету противоположного пола. Это было проде-
монстрировано с применением витального окрашивания гамет про-
тивоположных полов в разные цвета. Сходным образом образуют-
ся группы гамет у Chlamydomonas раирега.
У других водорослей каждая группа содержит по многу гамет
обоих полов, что показано у Dunalielld salina путем смешения
окрашенных в зеленый цвет гамет одного пола (образовавшихся
в культуре с избытком азота и фосфора) с окрашенными в оран-
жевый цвет гаметами противоположного пола (такая окраска воз-
никает в культурах с недостатком в среде азота и фосфора). Среди
анизогамных водорослей, например у Ulva lobata, микроскопические
наблюдения показывают, что в одной группе имеется много гамет
обоих размеров.
Образование групп уже давно связывалось с выделением каких-
то половых веществ (аттрактантов), хемотаксически привлекающих
гаметы противоположного пола. На такой вывод наталкивали и са-
мые ранние наблюдения за половым процессом у водорослей (фу-
куса, эдогониума).
В ряде случаев присутствие привлекающих половых веществ
было продемонстрировано экспериментально, например, у зеленой
водоросли Sphaeroplea annulina. У нее наблюдается оогамный поло-
вой процесс: в одних клетках (оогониальных) образуется много
яйцеклеток, в других (антеридиальных)— колоссальное количество
двужгутиковых сперматозоидов золотистого цвета. При созревании
половых клеток в стенках оогониев и антеридиев возникают отвер-
стия. Вышедшие сперматозоиды скапливаются у отверстий, веду-
щих в оогонии, врываются в оогоний и своими движениями приво-
50
дят в движение яйцеклетки. Если среди нитей сфероплеи со зрелыми
оогониями поместить хлопчатобумажные ниточки, они абсорбиру-
ют предполагаемый аттрактант и, перенесенные в равномерную
суспензию сперматозоидов, привлекают их: сперматозоиды скап-
ливаются вокруг таких ниточек. Выделение аттрактантов гаметами
Tetraspora lubrica было продемонстрировано иным способом: при
добавлении в жидкость из-под гамет одного знака, удаленных цент-
рифугированием, гамет противоположного знака, последние обра-
зовывали группы. Такая изоагглютинация наблюдается и у других
водорослей.
Делались также попытки установить химическую природу ат-
трактантов (иначе гамонов). Однако все более ранние попытки
оказались тщетными, что в значительной мере объясняется ограни-
ченными в то время возможностями аналитической химии. Лишь
в 70-е годы XX в. для нескольких морских бурых водорослей уда-
лось установить точную химическую природу аттрактантов. Это
стало возможным только благодаря развитию методов аналитичес-
кой химии, а также в связи с возможностью культивирования
водорослей в условиях лаборатории, позволяющей накопить боль-
шую биомассу, необходимую для биохимических исследований.
У некоторых водорослей аттрактанты, по-видимому, отсутству-
ют. Например, у Volvox rousseletii глобоиды сперматозоидов, осво-
бодившиеся из мужских ценобиев. плавают в воде быстро, и в при-
сутствии женских ценобиев могут прикрепиться к ним. Однако
специфическое привлечение отсутствует, скорее, имеет место слу-
чайное столкновение. Нередко наблюдаются глобоиды сперматозо-
идов, прикрепленные к вегетативным и мужским сфероидам.
В группах осуществляется первичный контакт гамет и их по-
парное слияние, начинающееся со слияния их цитоплазм — пла-
змогамии.
Использование альгологически чистых и аксеничных культур
водорослей и развитие методов, позволяющих индуцировать га-
метогенез, сделали возможным более тщательное и детальное
исследование процессов оплодотворения у водорослей как в све-
товом, так и в электронном микроскопе, а иногда даже с при-
менением микрокипематографии (например, у Prasiola scipitata,
Aseophyllum nodosum, Halidrys siliquosa, Oedogonium cardiacum). В ря-
де случаев процесс действительного слияния половых клеток уда-
лось наблюдать при использовании штаммов, специально полу-
ченных в условиях культуры и не встречающихся в природе. Так,
слияние сперматозоида с яйцеклеткой у вольвокса (Volvox carteri
f. nagariensis) удалось наблюдать у женского мутанта, характе-
ризующегося большим числом яиц и небольшим (по сравнению
с диким штаммом) количеством соматических клеток — обилие
их затрудняло наблюдение этого процесса. У полученного в куль-
туре женского диплоидного штамма Oedogonium cardiacum более
крупные, чем у гаплоидного (обычного в природе), оогонии.
в которых, как правило, обнаруживается большее пространство
между поверхностью яйца и окружающей оогониальной стенкой,
что дает явное преимущество для наблюдения начальных ста-
дий слияния гамет в живом материале. Конкретные примеры
плазмогамии и следующего этапа полового слияния— кариогамии
будут даны при описании родов, у которых этот процесс подробно
исследовался.
В результате полового процесса образуется зигота, содержащая
одно купуляционное диплоидное ядро — продукт слияния двух гап-
лоидных ядер, происходящих из той и другой гаметы. Обычно
зигота окружается толстой оболочкой, переполняется запасными
продуктами и растворимым в жирах пигментом кирпично-красного
цвета — астаксантином. Зигота прорастает или непосредственно по-
сле образования, или после более или менее продолжительного
периода покоя.
Жизненный цикл. Соотношение диплоидной и гаплоидной фаз
в жизненном цикле разных водорослей неодинаково. В одних случа-
ях прорастание зиготы сопровождается редукционным делением
копуляционного ядра (зиготическая редукция) и развивающиеся при
этом растения оказываются гаплоидными. Так, у многих зеленых
водорослей (вольвокальные, большинство хлорококкальных, ко-
нъюга тофициевые, харофициевые) зигота — единственная диплоид-
ная стадия в цикле развития, вся вегетативная жизнь проходит у них
в гаплоидном состоянии, они являются гаплонтами.
У других водорослей, наоборот, вся вегетативная жизнь осуще-
ствляется в диплоидном состоянии, а гаплоидная фаза представлена
лишь гаметами, перед образованием которых и происходит редук-
ционное деление ядра (гаметическая редукция). Зигот а без редукци-
онного деления ядра прорастает в диплоидный таллом. Эти водо-
росли— диплонты. Таковы многие зеленые водоросли, имеющие
сифоновое строение, все диатомовые, из бурых — представители
порядка фукальные.
У третьих редукционное деление ядра предшествует образова-
нию зооспор или апланоспор, развивающихся, как правило, на
диплоидных талломах {спорическая редукция). Эти клетки беспо-
лого размножения вырастают в гаплоидные растения, размножа-
ющиеся только половым путем. После слияния гамет зигота раз-
вивается в диплоидное растение, несущее только органы бесполого
размножения. Таким образом, у этих водорослей имеет место чере-
дование форм развития (генераций): диплоидного бесполого споро-
фита и гаплоидного полового гаметофита. Оба поколения могут
быть одинаковы морфологически {изоморфная смена генераций)
или же резко различны по внешнему виду {гетероморфная смена
генераций). Изоморфная смена генераций характерна для морских
видов ульвы, энтероморфы, кладофоры, хетоморфы из зеленых
водорослей, для ряда порядков бурых и большинства красных
водорослей (несколько усложненная). Гетероморфная смена генера-
52
ций особенно распространена среди бурых водорослей, но встреча-
ется у зеленых и красных. Следует отметить, что у ряда этих
водорослей, имеющих смену генераций, помимо двух кардинальных
моментов в их жизненном цикле — копуляции (сингамии), сопрово-
ждающейся удвоением числа хромосом в копуляционном ядре,
и мейоза, приводящего к редукции числа хромосом,— могут проис-
ходить и иные процессы: спонтанное умножение числа хромосом,
партеногенетическое развитие гамет, явление гетеробластии. В ре-
зультате способ размножения (бесполое или половое) далеко не
всегда коррелирует с плоидностью таллома, т. е. спорофиты могут
быть не только диплоидными, но и гаплоидными и тетраплоид-
ными, а гаметофиты обнаруживают не только гаплоидный, но
и диплоидный наборы хромосом и т. п. Такие сложные циклы
развития были установлены, например, у эктокарпуса, ульвы. Кро-
ме того, при определенных внешних условиях из жизненного цикла
одной и той. же водоросли могут выпадать какие-либо стадии,
например, у эктокарпуса происходит репликация спорофита посред-
ством нейтральных зооспор, и в некоторых местообитаниях извест-
ны только диплоидные талломы этой водоросли.
У одного и того же вида наблюдаются различные жизненные
циклы. Так, у зеленой водоросли Bryopsis hypnoides встречается как
однофазный цикл, так и двухфазное развитие (см. гл. 15), у красных
водорослей Gigartina stellata, G. agardhtii отмечается как половой
цикл, так и апомиктическое (без слияния половых клеток) развитие
(рм. гл. 6). Даже у таких высокоорганизованных форм, как ламина-
рия, где, как правило, регулярная смена спорофита и гаметофита
носит облигатный характер, возможны отклонения в жизненном
цикле: партеногенетическое (из неоплодотворенной яйцеклетки) раз-
витие спорофитов и апоспорическое (из вегетативных клеток споро-
фита) развитие гаметофитов.
Жизненные циклы водорослей очень сложны и разнообразны.
Особенно это относится к красным водорослям, исследование
развития которых позволило обнаружить гораздо больше типов
жизненных циклов, чем эго было известно ранее. Культураль-
ные исследования позволили установить связи между водорос-
лями, долгое время считавшимися самостоятельными видами
и оказавшимися лишь стадиями в жизненном цикле одной и той
же водоросли, ,т. е. значительно умножились известные нам слу-
чаи гетероморфной смены генераций. Это относится не только
к красным водорослям, но и к зеленым — улотрихальным, бриопси-
дальным.
Наконец, у немногих водорослей, например у водоросли прази-
олы (из зеленых) — леманеи, батрахоспермума (красные), мейоз
происходит в некоторых вегетативных клетках диплоидного тал-
лома {соматическая редукция), из которых затем развиваются гап-
лоидные талломы. Подробнее жизненные циклы будут разбираться
при описании соответствующих групп.
53
ЦАРСТВО ПРОКАРИОТЫ- PROCARYOTA
Глава 5
ОТДЕЛ СИНЕЗЕЛЕНЫЕ ВОДОРОСЛИ —CYANOPHYTA
К данному отделу относят одноклеточные, колониальные и нитча-
тые водоросли, различно окрашенные в зависимости от соотношения
пигментов, представленных кроме хлорофилла а и каротиноидов еще
синими пигментами — фикоцианином и аллофикоцианином и красным
фикоэритрином. Характерно также полное отсутствие жгутиковых
стадий. Пщювояпромесс у синезеленых водорослей не наблюдался.
Цитология. При рассматривании клетки синезеленых водорослей
в оптическом микроскопе можно видеть, что окруженная оболочкой
цитоплазма, как правило, лишена вакуолей с клеточным соком,
окрашена в периферических частях (хроматоплазма) и бесцветна
в центре (центроплазма). Резкой границы между хроматоплазмой
и центроплазмой нет. В центроплазме локализирована ДНК, что
позволяет рассматривать ее как гомолог клеточного ядра, хотя
ядерная оболочка и ядрышки отсутствуют. Кроме того, в цитоплаз-
ме находятся включения запасного характера (гликоген, волютин,
цианофициновые зерна) и часто — полости, наполненные смесью
газов, по составу близкой к воздуху,— газовые вакуоли, или псев-
довакуоли. У одних видов псевдовакуошГвстречаются на протяже-
нии всей вегетативной жизни, у других — только на определенных
стадиях развития.
В электронном микроскопе клетки синезеленых водорослей име-
ют следующее строение (рис. 6). Клеточная стенка состоит из четы-
рех четко разграниченных слоев, обозначаемых Lx, L?. L3, L4.
Непосредственно кнаружи от цитоплазматической мембраны рас-
положен электронно-прозрачный слой LXy за которым следует элек-
тронно-плотный слой Ь2, состоящий из муреина1—основного ком-
понента клеточной оболочки (стенки) бактерий. У эукариотических
водорослей и грибов это вещество не обнаружено. Таким образом,
и по химизму клеточной оболочки синезеленые водоросли, нередко
называемые теперь синезелеными бактериями (цианобактериями),
более близки к бактериям, чем к эукариотическим водорослям.
Именно этот муреиновый слой L2 определяет прочность стенки.
Будучи изолирован, этот слой клеточной стенки способен сохранять
форму всей клетки. Следующие за муреиновым слоем слои—элек-
тронно-прозрачный L3 и мембраноподобный Т4:—образованы уг-
леводами и в отличие от слоя L2 гибкие, пластичные.
1 Мукополимер, в состав которого входят два аминосахара и три аминокис-
лоты. Растворяется под действием фермента лизоцима.
54
Рис. 6. Схема строения клетки синезеленых водорослей на примере нитчатой водоро-
ели Lyngbya',
цм—цитоплазматическая мембрана, L] — L4—слои клеточной стенки, ч — чехол, фч — фибрил-
лы чехла, по — пора в клеточной стенке, ц—цитоплазма, р~-рибосомы, гв — газовые везикулы,
г — гликоген, т — тилакоиды, ф—фикобилисомы, нп — нуклеоплазма, содержащая миги ДНК,
цг - цианофициновые гранулы, в--во;1ЮТИн4 с—септа
Обусловленная этими слоями пластичность клеточной стенки
допускает изменения формы клетки. На уровне слоя £3 у осциллато-
рии были обнаружены непрерывные параллельные ряды, по-видимо-
му, протеиновых микрофибрилл, которые по спирали окружают
трихом. Ориентация этих микрофибрилл совпадает с направлением
вращения трихома во время его скольжения. Было высказано пред-
положение, что волнообразные изгибы этих рядов микрофибрилл
могут обусловливать скользящее движение трихома на твердом
субстрате. Упомянутые четыре слоя наблюдаются в продольных
стенках нитчатых синезеленых водорослей. Поперечные стенки, или
септы, нитчатых синезеленых водорослей образованы только слоями
Lr и L2. У одноклеточных форм наружные слои Г,3 и /,4 образуются
на септах только после начала разъединения клеток. Как в продоль-
ных стенках, так и в септах нитчатых (гормогониофициевых) синезе-
леных водорослей имеются поры, через которые соединяются цитоп-
лазматические мембраны и протопласты соседних клеток; эти цитоп-
лазматические тяжи называются микроплазмодеемами.
55
Подсчеты показали, что между двумя вегетативными клетками
одного из видов анабенопсиса в септе может быть до 4000 микро-
плазмодесм.
У многих синезеленых водорослей клеточные стенки покрыты
слизистым слоем, который может быть толстым и плотным и об-
разует чехлы или капсулы, обычно заключающие несколько клеток,
или же слизь представлена в виде тонкого жидкого слоя. Слизь
предохраняет клетки от высыхания и, по-видимому, принимает
участие в процессе скользящего движения. Тонкая структура сли-
зи— фибриллярная (волокнистая).
Преимущественно в периферической цитоплазме—хроматопла-
зме—локализованы тилакоиды, способные иногда проникать во
все части клетки. Тилакоиды не отграничены от цитоплазмы мемб-
ранами, как у других хлорофиллоносных растений, т. е. истинные
хлоропласты, одетые оболочкой, здесь отсутствуют. Периферичес-
ки расположенные тилакоиды обычно ориентированы параллельно
продольной клеточной стенке, но иногда они расположены перпен-
дикулярно к продольной стенке. Изредка тилакоиды рассеяны по
всей клетке.
Тилакоиды синезеленых водорослей никогда не образуют групп,
как это свойственно эукариотическим водорослям, за исключением
красных, где тилакоиды также расположены одиночно. Добавочные
пигменты сипезеленых водорослей (фикоцианин, аллофикоцианин
и фикоэритрин) в форме гранул — фикобилисом—локализованы на
поверхности тилакоидов.
Центр клетки, обычно свободный от тилакоидов, занят нуклеоп-
лазмой, не отграниченной от остальной цитоплазмы ядерной обо-
лочкой' В нуклеоплазме находятся фибриллы ДНК. Для ядерного
материала синезеленых водорослей, как и для бактерий и фагов,
характерно отсутствие гистонов, чем они отличаются от остальных
эукариотических организмов, ядерная субстанция которых содер-
жит помимо ДНК, ядерный белок.
В цитоплазме, не занятой тилакоидами и нуклеоплазмой, нахо-
дятся рибосомы и запасные вещества: гликоген, волютин (метах-
роматин), цианофициновые гранулы.
Газовые вакуоли, рассеянные по всей клетке или располагающи-
еся у поперечных перегородок (септ), состоят из тесно упакованных,
наподобие сот, полых, одетых мембраной субъединиц - газовых
везикул (газовых пузырей). Эти везикулы имеют форму полых
цилиндрических трубок с коническими шапочками у концов. Они
могут быть изолированы из клетки без изменения их формы. Под-
вергнутые определенному давлению цилиндрические части газовых
везикул спадаются, при этом конические шапочки могут отламы-
ваться. Химические анализы изолированных газовых везикул пока-
зали, иго их мембраны отличаются от типичной элементарной
мембраны отсутствием липидов. Мембраны газовых везикул состо-
ят только из белков.
56
Размножение. Перед клеточным делением количество ДНК удва-
ивается, а по мере деления клетки удвоенное количество ДНК
разделяется пополам. Клетки синезеленых водорослей делятся та-
ким образом, что на боковой стенке их возникает кольцевая склад-
ка, образованная цитоплазматической мембраной и внутренними
(Lj и Т2) слоями клеточной оболочки. Разрастаясь в центростреми-
тельном направлении, ла складка смыкается наподобие ирисовой
диафрагмы микроскопа, формируя поперечную перегородку — сеп-
ту, перерезающую тилакоиды и содержимое клетки.
В середине поперечно перерезанной септы в процессе утолщения
муреинового слоя сравнительно рано становится различимым слой
более низкой плотности — срединная ламелла (рис. 7, Л). У одно-
клеточных форм и многих ностокальных, у которых образование
клеточной перегородки сопровождается появлением более или ме-
нее глубокой перетяжки между дочерними клетками, клетки разъ-
единяются путем центрипетального врастания в плоскость средин-
ной ламеллы между слоями муреина [L2) наружных (L, и Т4) слоев
клеточной оболочки (рис. 7, К). Таким образом, у многих носто-
кальных разрыв трихома при образовании гормогониев происходит
между клетками — интерцеллюларно. Совсем иначе разъединяются
Рис. 7. Деление и разъединение клеток синезеленых водорослей.
Л — деление клетки: образование септы; Б—ишерцеллюларное разъединение клеток; В—транс-
целлюларное разъединение клеток путем разрыва вдоль цепочек соединительных пор в продоль-
ной клеточной стенке «жертвенной» клетки:
цм — цитоплазматическая мембрана. L2— муреиновый слой клеточной стенки, с — септа» сл—
срединная ламелла, сп — соединительные поры
57
клетки у многих осциллаториальных и сцитонемовых. Хотя муреи-
новые слои зрелой септы разъединены срединной ламеллой, расщеп-
ления септы и расхождения клеток в этой области не происходит,
так как муреиновые слои закреплены у периферического утолщения
края диска септы. Слабым местом в муреиновом одеянии клеток
у форм с нерасщепляющейся септой, по-видимому, является набор
многочисленных мелких соединительных пор, расположенных по
окружности по обе стороны от септы — вдоль соединения ее с про-
дольными стенками. Разрыв трихомов при образовании гормогони-
ев происходит вдоль соединительных пор — поперек клеток —
трансцеллюларно и связан с гибелью клетки, «приносимой в жерт-
ву»— «жертвенной клетки». При этом ненарушенные септы между
«жертвенной клеткой» и двумя соседними с ней клетками становятся
стенками конечных клеток сформировавшихся гормогониев. Оста-
тки клеточных стенок «жертвенных клеток» могут служить доказа-
тельством того, что разрыв произошел не благодаря расщеплению
септ между живыми клетками, а трансцеллюларно. Такие остатки
оболочек отмерших клеток, по которым происходит разрыв, часто
можно видеть в оптическом микроскопе на концах трихомов осцил-
латорий (рис. 7, В), Как интерцеллюларным, так и трансцеллюлар-
ным разрывам способствуют многократные изгибы трихомов.
У одних нитчатых синезеленых водорослей все клетки нити
одинаковы — это гомоцитные. .талломы; у других нити состоят из
вегетативных клеток и гетероцист, нередко наблюдаются и споры
(акинеты) — eetnepotfumubie формы. Гетероцисты и споры — это спе-
циализированные клетки, образующиеся путём” дифференцировки
вегетативных клеток. Один из наиболее заметных признаков гетеро-
цист (даже при наблюдении в оптическом микроскопе)—их сильно
утолщенные стенки. Клеточные стенки гетероцист, как и вегетатив-
ных клеток, состоят из слоев —L4, но кнаружи от них развива-
ются еще три специальных слоя оболочки гетероцист. Непосредст-
венно кнаружи от клеточной стенки располагается пластинчатый
слой, особенно толстый у глубокой перетяжки между гетероцистой
и соседней вегетативной клеткой, где имеется поровый канал —«по-
ра», пересеченная септой, пронизанной микроплазмодесмами, по-
средством которых сообщаются гетероциста и вегетативная клетка.
Кнаружи от пластинчатого слоя находится гомогенный слой, кото-
рый без резкой границы переходит в самый наружный фибрилляр-
ный слой. Благодаря толстой обвертке, образующейся кнаружи от
клеточной оболочки, гетероцисты выдерживают механические воз-
действия, разрушающие более тонкостенные вегетативные клетки.
Другой (помимо толстых оболочек) признак зрелых гетероцист,
легко наблюдаемый в оптический микроскоп,—гомогенность кле-
точного содержимого, которая обусловлена исчезновением грану-
лярных включений, характерных для вегетативных клеток. Ни во-
лютина, ни цианофициновых зерен в полностью развитых гетероци-
стах нет. Единственные гранулярные структуры, которые можно
58
Рис. 8. Схема строения гетеро-
цисты:
цм- цитоплазматическая мембрана,
—ЪА—слои клеточной стенки,
пл—пластинчатый слой стенки гете-
роцисты, гс — гомогенный слой стен-
ки гетероцисты, фс — фибриллярный
слой стенки гетероцисты, мп- мик-
роплазмодесмы, п—пора гетероци-
сты, с —септа гетероцисты, пр —
пробка, замыкающая канал поры ге-
тероцисты, м — мебраны гетероци-
сты, яд — рассеянные нити ДНК, р
рибосомы, вк—вегетативная клетка,
гц— гетероциста
различить на электронных микрофотографиях зрелых гетероцист,—
рибосомы. Газовые вакуоли, если они были в вегетативных клетках,
при дифференцировке их в гетероцисты также исчезают.
Наконец, дифференцировка гетероцист сопровождается реорга-
низацией мембранной системы клетки: тилакоиды разрушаются
и формируются новые плотно упакованные мембраны. Преобразо-
вание мембранной системы сопряжено с изменениями в пигментном
составе: в гетероцистах обнаруживаются хлорофилл и каротино-
иды, но почти нет фикоцианина, аллофикоцианина и фикоэритрина.
Фибриллы ДНК, в вегетативных клетках обычно сконцентриро-
ванные в нуклеоплазме (центроплазме), в гетероцистах рассеяны по
всей цитоплазме (рис. 8).
В многочисленных исследованиях было показано, что образова-
ние гетероцист, как и процесс фиксации атмосферного азота, ин-
гибируется присутствием в питательной среде связанного азота,
особенно аммонийного1. Исходя из этих данных стало возможным
при выращивании на средах, содержащих связанный, особенно
ам’монийный азот, искусственно получать трихомы (нити) без
1 Помимо связанного азота на образование гетероцист оказывают влияние
и другие факторы внешней среды, в частности свет, его интенсивность и спектраль-
ный состав.
59
гетероцист у родов, в норме характеризующихся наличием гетеро-
цист, например у анабен, ностоков и др. При перенесении такого
безгетероцистного материала в среду, свободную от связанного
азота, можно проследить все стадии развития гетероцист из вегета-
тивных клеток. Дифференцировка гетероцист начинается с образова-
ния самого наружного фиброзного слоя ее оболочки кнаружи от
клеточной стенки дифференцирующейся вегатативной клетки. Обра-
зование фиброзного слоя и следующего гомогенного слоя сопровож-
дается разрушением гранулярных цитоплазматических включений
и тилакоидов. На конечной стадии развития гетероцист откладывает-
ся самый внутренний пластинчатый слой оболочки гетероцисты,
особенно толстый вокруг порового канала, и формируется развитая
мембранная система, по-видимому, путем соединения мелких пузы-
рьков, возникающих на местах предшествующих тилакоидов.
Давно известно, что у таких родов, как анабена, носток, харак-
теризующихся интеркалярными гетероцистами, в гетероцисты пре-
вращаются не любые вегетативные клетки, а лишь некоторые,
расположенные на определенном расстоянии друг от друга. Срав-
нительно недавно было высказано предположение, что гетероцисты
анабены выделяют ингибитор, подавляющий дифференцировку ве-
гетативных клеток в гетероцисты. Последние возникают из вегета-
тивных клеток, расположенных вне ингибирующих зон, окружа-
ющих ранее образовавшиеся гетероцисты.
В ряде работ эту гипотезу удалось подтвердить эксперименталь-
но. Так, Д. Тивари (D. N. Tiwari, 1977) из дикого штамма Nostoc
linckia, способного осуществлять процесс азотфиксации (а местом
нитрогеназной активности, как было доказано в многочисленных
исследованиях, являются как раз гетероцисты), выделил мутанты,
не фиксирующие атмосферный азот, по сохранившие возможность
осуществлять развитие, ведущее к дифференцировке гетероцист. На
средах, не содержащих азота, у этих ппаммов появлялись иптерка-
лярные гетероцисты. Однако в отличие от дикого исходного штам-
ма вегетативные клетки по соседству с возникшими (не функци-
онирующими здесь — не фиксирующими азот) гетероцистами также
превращались в гетероцисты, что приводило к образованию цепо-
чек гетероцист1. Напрашивается вывод, что ингибитор—это дери-
ват какого-то непосредственного продукта азотфиксации. Он мак-
симально концентрируется вокруг функционирующих гетероцист,
регулируя распределение гетероцист вдоль нити. У других родов
гетероцисты занимают терминальное положение в трихоме; быва-
ют и латеральные гетероцисты.
У очень многих гетероцитных форм, в зрелом состоянии харак-
теризующихся интеркалярными гетероцистами, их молодые про-
ростки имеют терминальные гетероцисты.
' Цепи интеркалярных гетероцист наблюдались и у аналогичного мутанта
Gloeotriehia ghosei.
60
Рис. 9. Схема строения споры (акинеты)
синезеленых водорослей:
цм--цитоплазматическая мембрана, Z.2 — i4 —
слои клеточной стенки, ос — обвертка споры,
т—тилакоиды, ф—фикобилисомы, ц — цитоп-
лазма, нп — нуклеоплазма с нитями ДНК, р —
рибосомы, г гликоген, цг — цианофициновые
гранулы, в — волютин
Относительно функции гетероцист высказывались разные пред-
положения. У гетероцитных форм распад нитей на отдельные участ-
ки (гормогонии), из которых развиваются новые талломы, обычно
происходит по гетероцистам, соответственно им приписывалась
и известная роль в вегетативном размножении. Наблюдаемые ино-
гда случаи деления содержимого гетероцист позволяли предполо-
жить их первоначальную (впоследствии утраченную) роль в качестве
клеток, предназначенных для размножения, И действительно, при
культивировании в определенных условиях описано довольно много
фактов прорастания гетероцист. Так, у Anabaena cylindrica, выращен-
ной в безазотистой среде, при перенесении ее иа среду с хлоридом
аммония 3 —10% гетероцист прорастали. У неспорулирующего му-
танта одного из видов Gloeotrichia при культивировании в среде
с хлоридом аммония прорастало 83% гетероцист. Сравнительно
недавно (1980) было даже высказано предположение, что исчезнове-
ние гетероцист, наблюдаемое на средах, в состав которых входит
связанный азот, объясняется как раз их прорастанием. Однако
в естественных условиях прорастание гетероцист — редкое явление.
Против их роли в качестве клеток бесполого размножения свиде-
тельствует не только то, что они лишены запасных продуктов,
которые могли бы поддержать начальные стадии прорастания, но
и то, что при отделении их от нити, они, как правило, быстро
разрушаются, возможно, оттого, что в отличие от спор не защищены
со всех сторон оболочкой. Согласно последним данным в гетероци-
стах происходит фиксация атмосферного азота в аэробных условиях.
Другими специализированными клетками, которые также об-
разуются путем дифференцировки вегетативных клеток, являются
споры — акинеты (рис. 9). Это обычно более крупные, чем вегета-
тивные, клетки, толстостенные, причем споровый покров окружает
акинету полностью в отличие от гетероцист, которые через поровый
61
канал сохраняют контакт с соседними вегетативными клетками.
При дифференцировке спор из вегетативных клеток заметно утол-
щается L2 (муреиновый) слой и, кроме того, кнаружи от клеточной
оболочки образуется широкая обвертка. В отличие от гетероциш
зрелые споры переполнены запасными гранулярными включени-
ями, особенно цианофициновыми зернами. Расположение и струк-
тура тилакоидов в спорах те же, что и в вегетативных клетках.
Содержание ДНК в спорах заметно возрастает, иногда в 20—30 раз
ио сравнению с содержанием ее в вегетативных клетках. Как и в слу-
чае гетероцист, вегетативные клетки, превращающиеся в споры,
утрачивают газовые вакуоли.
Спорообразование у синезеленых водорослей стимулируется
разными факторами среды, но особенно фосфором. Споры могут
выдерживать высыхание и другие неблагоприятные условия. Они
могут прорастать сразу же после образования, без какого-либо
периода покоя, при помещении в благоприятные условия. При
отсутствии таких условий споры долго сохраняют жизнеспособ-
ность. Например, споры Anabaena spiroides, A. flos-aquae,
Aphanizomenon flos-aquae, выделенные из донных осадков, возраст
которых составлял от 18 до 64 лет, хорошо прорастали на мине-
ральной среде в лабораторных условиях. При прорастании споры
образуется один проросток, освобождающийся через разрыв обо-
лочки.
Большинство одноклеточных и колониальных синезеленых водо-
рослей размножаются посредством деления клеток пополам, у не-
которых— посредством мелких клеток — «гонидий», образующихся
или внутри материнской клетки (эндоспор), или отшнуровывающих-
ся от верхушки материнской клетки (экзоспор). Подавляющее боль-
шинство нитчатых синезеленых водорослей размножается посред-
ством гормогониев — участков, па которые распадаются нити.
Синезеленые водороли подразделяют на три класса: хроокок-
кофициевые (Chroococcophyceae), хамесифонофициевые
(Chamaesiphonophyceae) и гормогониофициевые (Hormogoni-
ophyceae).
КЛАСС ХРООКОККОФИЦИЕВЫЕ —CHROOCOCCOPHYCEAE
Объединяет колониальные, реже одноклеточные формы, причем
клетки, как правило, не дифференцированы на основание и вершину.
Размножение—делением клеток пополам. Если клетки после деле-
ния не расходятся, то обычно возникают слизистые колонии, форма
которых определяется как формой самих клеток, так и способом их
деления, в одних случаях приводящим к возникновению объемных
колоний, в других—плоских, пластинчатых.
Род микроцистис (Microcystis), широко распространенный в пре-
сноводном планктоне, где, развиваясь в массе, вызывает «цветение
62
воды», характеризуется объемными сферическими или неправиль-
ной формы слизистыми колониями, образованными шаровидными
клетками. Некоторые виды токсичны.
Род мерисмоиедия (Merismopedid), обычный в пресных водах
в планктоне и среди других водорослей, представлен плоскими,
в виде табличек, колониями, образованными шаровидными клет-
ками.
КЛАСС ХАМЕСИФОНОФИЦИЕВЫЕ — CHAMAESJPHONOPHYCEAE
Сюда относятся эпифитные одноклеточные водоросли, диффере-
нцированные на вершину и основание, которым они прикрепляются
к субстрату, а также нитчатые формы, сложенные из изолированных
толстостенных клеток. Размножение- посредством эндоспор и эк-
зоспор.
КЛАСС ГОРМОГОНИОФИЦИЕВЫЕ —HORMOGONIOPHYCEAE
Этот класс объединяет нитчатые многоклеточные синезеленые
водоросли, у которых протопласты соседних клеток сообщаются
посредством плазмодесм. Такие нити носят специальное название
трихом. Размножение — посредством гормогониев, которые обыч-
но определяются как участки трихомов, способные к активному
движению и прорастанию в новые особи. У многих представителей
известны споры. Они прорастают также гормогониями, которые
в отличие от вторичных гормогониев, развивающихся из взрослых
трихомов, называются первичными. Из входящих сюда порядков
рассмотрим осциллаториальные (Oscillatoriales), ностокальные
(Nostocales) и сгигонемальные (Stigonematales).
Порядок осциллаториальные—Oscillatoriales
Порядок объединяет нитчатые гомоцитные формы. Представи-
телем может служить род осциллатория (Oscillatorid). Его виды
часто образуют синезелепые пленки, покрывающие влажную зем-
лю, подводные предметы, или плавают в виде толстых кожистых
лепешек на поверхности стоячих водоемов. Длинные нити осцил-
латории сложены из цилиндрических клеток, совершенно одинако-
вых, за исключением верхушечных, которые по форме могут неско-
лько отличаться от остальных. Рост происходит в результате попе-
речных делений клеток. Для нитей характерно своеобразное колеба-
тельное (осциллаторпое) движение, сопровождающееся вращением
нити вокруг собственной оси и ее поступательным движением.
Размножение — путем трансцеллюларного распада трихомов по
«жертвенным» клеткам на более короткие подвижные участки, ча-
сто определяемые как гормогонии. Они вырастают в новые нити.
63
Во многих случаях трансцеллюларный распад происходит по уже
мертвым клеткам — некридиям.
Близок к осциллатории род спирулнна (Spirulina), нити которой
скручены в правильную спираль. Некоторые виды спирулины, на-
пример S', maxima, содержат много протеинов (60—80% от сухой
массы) и с давних времен используются в пищу населением некото-
рых районов Африки. В последние годы в ряде стран, в том числе
в СССР, стали заниматься массовой культурой видов спирулины.
Порядок иостокальные — Nostocales
Порядок объединяет водоросли с гетероцитными неразветвлен-
ными нитями или нитями с ложным ветвлением.
Род анабена (Anahaena)— обычно одиночные или собранные
в неправильные скопления нити. Нити симметричные, одинаковой
ширины на всем протяжении, состоят из округлых или бочонкооб-
разных вегетативных клеток и промежуточных (интеркалярных)
гетероцист. Нити прямые или изогнутые. Виды анабены встречают-
ся как в бентосе (придонные), так и в планктоне. У планктонных,
видов клетки содержат газовые вакуоли. Размножение — посред-
ством подвижных нитей, часто называемых гормогониями, образо-
вание которых сопровождается ингерцеллюларными разрывами
обычно между вегетативными клетками. Таким образом, на протя-
жении активно ползающих «гормогониев» анабен (например,
A. cylindrica, A. hallensis, A. variabilis) имеются интеркалярные гете-
роцисты родительского трихома, чем они отличаются от гормого-
ниев других ролов, как правило, гомоцитных. Реже у тех же видов
происходит интерцеллюларный разрыв между вегетативными клет-
ками и гетероцистами, сначала еще живыми (с содержимым) и лишь
позднее отмирающими, возможно, в результате нарушения микро-
плазмодесм, связывающих ге тероцисты с соседними вегетативными
клетками. «Гормогонии» растут только за счет поперечных делений
клеток. Большинство видов имеет споры, обычно резко отлича-
ющиеся по форме и размерам от вегетативных клеток. Их прораста-
ние сопровождается только поперечными делениями клеток. Пер-
вые формирующиеся в проростке гетероцисты часто термальные.
Для ряда видов (A. cylindrica, A. orycae, A. variabilis и пр.) доказана
способность к фиксации атмосферного азота.
Род восток (Nostoc) характеризуется слизистыми или студенис-
тыми колониями различных размеров и форм: от микроскопически
мелких до крупных, достигающих величины сливы; от сферических
или эллипсоидальных до неправильно распростертых и нитевидных.
Слизь разной консистенции содержит массу извитых нитей (три-
хомов) или беспорядочно перепутанных, или расходящихся более
или менее радиально из центра колонии; трихомы похожи на три-
хомы анабены; наряду с вегетативными клетками содержат ин-
теркалярные гетероцисты. Размножение — посредством гормогони-
64
ев. Последние возникают в результате интерцеллюлярных разрывов
между вегетативными клетками и гетероцистами; таким образом,
гормогонии всегда гомоцитные. Отмирание гетероцист, по-види-
мому, предшествует разрыву; выпадают из трихома лишь пустые
оболочки гетероцист, в результате чего трихомы распадаются на
гормогонии. Это наблюдается у таких видов, как Nostoc puncliforme,
N. pruniforme и др. Образование гормогониев сопровождается
внутриклеточными перестройками, внешне выражающимися в том,
что в большинстве случаев изменяются размеры и форма клеток.
Очень часто у бентосных форм, в вегетативном состоянии лишен-
ных газовых вакуолей, эти вакуоли появляются в клетках гор-
могониев. Гормогонии приобретают подвижность и покидают ма-
теринскую колонию, слизь которой к этому времени расплывается,
а если колония покрыта плотным наружным слоем - перидермом,
то этот слой разрывается. Обычно гормогонии служат и для рас-
пространения вида благодаря собственному скользящему движе-
нию и вследствие лучшей плавучести, обусловленной развитием
в клетках газовых вакуолей. После некоторого периода движения
прямые гормогонии останавливаются, теряют газовые вакуоли (у
бентосных видов) и прорастают в спирально извитые нити. При
этом клетки гормогония делятся косыми или продольными пе-
регородками. Во втором случае сначала возникает двурядная нить,
а затем при разъединении определенных клеток этой нити форми-
руется уже зигзагообразная нить, свойственная ностокам. Обычно
конечные клетки молодой нити превращаются в гетероцисты, вы-
деляется обильная слизь и таким образом возникает молодая ко-
лония, в которой нити дальше растут только за счет поперечных
делений клеток. У N. puncliforme—вида, лишенного крепкого пе-
ридерма, дочерние колонии возникают за счет развития многих
гормогониев, У сферических ностоков с крепким перидермом
(N. coeruleum, N. microscopicum, N. pruniforme) дочерние колонии об-
разуются в результате прорастания только одного гормогония.
Длинные гормогонии ностоков могут непосредственно прорастать
в извитые трихомы, но в некоторых случаях гормогонии прете-
рпевают предварительную фрагментацию на короткие отрезки;
фрагментация сопровождается многократными изгибами длинных
гормогониев. Эти изгибы способствуют интерцеллюларным раз-
рывам. Фрагментация заметно повышает энергию размножения.
Это относится и к самому крупному сферическому ностоку —
AL pruniforme, развивающемуся в наших северных водоемах в ко-
лоссальных количествах. В ряде случаев у сферических ностоков
(например, N. microscopicum) может быть подавлена подвижность
гормогониев, которые не покидают слизи материнской колонии
и развиваются внутри нее с образованием эндогенных колоний,
позднее освобождающихся в результате разрушения перидерма ма-
теринской колонии. Иногда у ностоков наблюдаются ветвящиеся
гормогонии, которые прорастают, как и обычные неразветвленные
3 - 1210
65
гормогонии. Сферические ностоки нередко размножаются почко-
ванием колоний. Помимо гормогониев, у многих ностоков обра-
зуются споры, обычно мало отличающиеся по форме и размерам
от вегетативных клеток. Нередко все вегетативные клетки превра-
щаются в споры, остающиеся связанными в цепочки. При про-
растании их оболочки разрушаются, и проросток (первичный го-
рмогоний), еще не поделившийся или уже многоклеточный, осво-
бождается. Как у вторичных (развившихся из взрослых трихомов),
так и у первичных гормогониев иногда наблюдается полная потеря
подвижности, и они прорастают на месте образования внутри обо-
лочек спор. У N. linckia концы проростков, развившихся при про-
растании таких неподвижных первичных гормогониев, соединяются
с образованием как бы омоложенного нового трихома, и на месте
спороносной колонии, содержащей цепочки спор, формируется ве-
гетативная. Самый крупный N. pruniforme, а также носток с ко-
лониями в виде слизистых тяжей, в которых нити расположены
более или менее параллельными рядами (У. flagelliforme), съедобны.
Ностоки способны фиксировать атмосферный азот.
У ряда других ностокальных трихомы асимметричны. Напри-
мер, у рода калотрикс (Calofhrix) трихомы бичевидные:- на рас-
ширенном конце дифференцируется базальная гетероциста, проти-
воположный конец утончается и часто заканчивается многоклеточ-
ным волоском из отмерших клеток. Трихом одет чехлом и может
ветвиться. Ветвление ложное и обусловлено тем, что на протяжении
трихома в результате отмирания промежуточной клетки происхо-
дит разрыв. Расположенный ниже разрыва участок, прорывая чехол,
продолжает расти вбок, образуя ветвь. Конечная клетка верхнего
фрагмента может превратиться в базальную гетероцисту или же
базальный конец верхнего фрагмента может дать вторую боковую
ложную ветвь. В первом случае ложное ветвление одиночное, во
втором--двойное. При размножении С. stellaris и других видов
волоски сбрасываются, клетки трихома, начиная с его верхушки,
переполняются газовыми вакуолями, несколько меняются их форма
и размеры. Эти изменения не затрагивают лишь несколько базаль-
ных клеток. Возникает длинная гомоцитная нить — прогормогоний,
который обычно покидает чехол материнского трихома. Прогор-
могопий способен к движению, по не к непосредственному прора-
станию в новые трихомы. Прогормогонии выполняют функцию
распространения вида, которая осуществляется не столько благо-
даря собственному активному скользящему движению, сколько за
счет газовых вакуолей, переполняющих их клетки и способству-
ющих их лучшей плавучести. Такие прогормогонии закономерно
претерпевают фрагментацию на более короткие участки — гормого-
нии, также сначала подвижные и способные прорастать с образова-
нием трихома. Фрагментация активная, сопровождается многократ-
ными изгибами прогормогония и может продолжаться несколько
часов. Иногда подвижность прогормогония подавлена и распад его
66
на фрагменты происходит пассивно внутри влагалища материнс-
кого трихома, из которого выходят уже подвижные фрагменты —
гормогонии. При прорастании гомоцитных гормогониев возникают
асимметричные (за счет расширения клеток на одном конце и вытя-
гивания их на другом), гетероцитные, с терминальной гетероцистой
на расширенном конце трихомы. У некоторых видов калотрикса
известны споры.
У родов ривулярия {Rivalaria) и глеотрихия (Gloeotrichia) тал-
ломы студенистые: в слизи, расходясь из центра по радиусам,
располагаются асимметричные, как у калотрикса, нити. К центру
колонии обращены расширенные концы с базальной гетероцистой,
к периферии — волоски. Размножение ривулярии и глеогрихии —
посредством гормогониев. У всех видов глеотрихии известны спо-
ры, у ривулярии споры отсутствуют. Размножение и развитие коло-
ний подробно изучено у многих видов глеогрихии. В вегетативных
(неспороносиых) колониях из трихомов, как и у калотрикса, сначала
формируются длинные симметричные гомоцитные черные (от пере-
полняющих их клетки газовых вакуолей) прогормогонии. Они поки-
дают слизь материнской колонии, образуют плотные пучки
и всплывают (за счет газовых вакуолей) на поверхность воды.
В таких пучках прогормогопиев вскоре наступает активная, со-
провождающаяся коленообразными изгибами ломка их на более
короткие фрагменты. Фрагментация, по-видимому, трансцеллюляр-
ная, обычно происходит одновременно во всех прогормогониях,
слагающих снопик, и минут через 10—20 на его месте свободно
лежат короткие фрагмешы (подвижные и черные от газовых ваку-
олей) и вытолкнутые «жертвенные» клетки. Через некоторое время
зги клетки теряют содержимое и опускаются на дно сосуда. Распад
прогормогониев продолжается до образования гормогониев — ко-
ротких участков, при дальнейшем развитии которых возникает
только один трихом, или образуются более длинные бигормогонии,
дающие два трихома. В первом случае, характерном для Gloeotrichia
pisum, совершенно прямые и гомоцитные вначале гормогонии, сло-
женные из одинаковых черных (от газовых вакуолей) клеток, оста-
навливаются и начинают медленно изгибаться дугой, В них проис-
ходит дифференцировка клеток: па одном конце намечается буду-
щая гетероциста, на противоположном конце — едва заметное утон-
чение и удлинение клеток. По мере дальнейшего развития
асимметричного трихома газовые вакуоли из клеток исчезают. Во
втором случае, свойственном Gl. echinulata, бигормогонии сначала
образуют симметричный, утончающийся к обоим концам трихом,
впоследствии распадающийся на два асимметричных трихома
благодаря формированию двух средних гетероцист и последую-
щему разрыву между ними. Поскольку этот вид планктонный,
газовые вакуоли в клетках сохраняются. По мере дифферен-
цировки клеток трихомов выделяется слизь, в которой трихомы
располагаются более или менее по радиусам. Таким образом
67
формируются молодые колонии, как правило, за счет развития
многих гормогониев, чему способствует то, что с самого начала
прогормогонии собраны в плотные пучки. У глеотрихии длинная,
с зернистым содержимым спора образуется за счет слияния не-
скольких клеток, расположенных над базальной гетероцистой.
К концу вегетационного периода все нити в колонии образуют
споры, вегетативные клетки отмирают и в слизи остаются только
споры; у шаровидных видов (например, у Gl. pisum) споры рас-
ходятся из центра колонии по радиусам. При помещении таких
спороносных колоний в благоприятные условия споры более или
менее одновременно прорастают. Содержимое их делится с об-
разованием длинных симметричных многоклеточных нитей — про-
гормогониев, в клетках которых обычно появляются обильные
газовые вакуоли. Оболочка на верхушке споры разрывается и более
легкие, чем вода (из-за газовых вакуолей), обычно соединенные
пучками прогормогонии всплывают на ее поверхность, где и пре-
терпевают дальнейшее развитие, совершенно такое же, как это
было описано для прогормогониев, образующихся из взрослых
вегетативных колоний.
Порядок стшонсмальные—Stigonematales
Этот порядок характеризуется гетероцитными нитями с насто-
ящим ветвлением. При настоящем — истинном — ветвлении отдель-
ные клетки трихома образуют выступы, которые отделяются пере-
городкой и затем, повторно делясь поперечными перегородками,
развиваются в ветви. Примерами могут служить роды стигоиема
(Stigcmema) и мастигокладус (Mastigociadus). Нити стигонемы об-
ладают верхушечным ростом. Несколько отступая от верхушки
ветвей, клетки претерпевают продольные деления, и нить становит-
ся многорядной. Мастигокладус с единственным видом
М. laminosus имеет сложный ветвящийся гетероцитный таллом и яв-
ляется типичной термальной водорослью, распространенной по
всему свету. Образование гормогониев, в которые превращались
концы боковых ветвей, у этой термофильной водоросли наблюдает-
ся при 40—45° С, хотя талломы не теряют жизнеспособности и да-
же обнаруживают слабый рост при температурах ниже 30° С. В ста-
реющих культурах возникают споры, |расположенные цепочками.
У них нет периода покоя и в благоприятных условиях они часто
прорастают in situ, нередко имитируя при этом боковые ветви
(истинные или ложные).
Таким образом, процесс образования гормогониев у разных
представителей существенно различается. Лишь у осциллатори-
альных гормогонии отвечают вышеприведенному определению их
как участков трихомов, способных к активному движению и про-
растанию в новые особи. Разрыв гомоцитных трихомов осцил-
латории на гормогонии, отличающиеся от зрелых особей только
68
меньшей длиной, можно рассматривать как типичный пример
вегетативного размножения — распада таллома на отдельные
участки без каких-либо перестроек в содержимом клеток. У пред-
ставителей вышестоящих порядков гормогониофициевых—носто-
кальных и стигонемальных, имеющих гетероцитные трихомы, об-
разование гормогониев нельзя свести к простому разрыву трихомов
на отдельные участки. Здесь формирование гормогониев — гораздо
более сложный процесс, как правило, сопровождающийся внутри-
клеточными перестройками, о которых пока мало известно. Обычно
внешне это выражается в появлении в клетках гормогониев новых
органелл, отсутствующих в материнском трихоме,- - газовых ва-
куолей, а также в значительном изменении формы и размеров
клеток. Здесь гормогонии, почти всегда (за исключением рода
анабена) гомоцитные и симметричные, резко отличаются от ге-
тероцитных, часто асимметричных зрелых трихомов. Оба признака
гормогониев осцилляториальных—подвижность и способность
к прорастанию в новые трихомы — часто бывают разобщены в про-
гормогониях, возникающих у многих представителей ностокальных
при их размножении и часто называемых гормогониями. В связи
с этим необходимо более резко разграничивать образование гор-
могониев от случайных разрывов материнских трихомов на более
короткие участки и согласиться с несколько иным, чем было приве-
дено выше, определением самого понятия «гормогонии». Так,
Р. Ринпка и др. (R. Rippka et al., 1981) определяют гормогонии
как трихомы, отличающиеся от родительских отсутствием гете-
роцист (даже при выращивании на средах свободных от связанного
азота), а также одним или совокупностью таких признаков, как
быстрое скользящее движение, иные размеры и форма клеток, при-
сутствие газовых вакуолей в клетках (не только у планктон-
ных, но и у бентосных форм). Этому определению отвечают
структуры, служащие для размножения родов носток, калогрикс,
глеотрихия, мастигокладус и др. Размножение же осциллатори-
альных и анабен (согласно данным Р. Риппка и др.) следует рас-
сматривать как случайный разрыв (трансцеллюларный у осцил-
латории, интерцеллюларный у анабен) родительского трихома на
участки, отличающиеся от трихома только меньшей длиной. Ос-
новное различие между родами носток и анабена указанные авторы
видят именно в способах размножения: в наличии гормогониев
(наряду со случайными разрывами трихомов) у востока и от-
сутствии гормогониев у анабен. Следует, однако, добавить, что
фрагменты, на которые распадаются трихомы анабен, отличаются
от материнских подвижностью и часто присутствием газовых ва-
куолей (у бентосных видов). Что касается критериев для раз-
граничения подов анабена и носток (между неоформленными сли-
зистыми массами анабены и слабо оформленными мягкими ко-
лониями некоторых видов ностока трудно установить резкую гра-
ницу), то полезен и такой признак, как характер деления клеток
69
проростков, развивающихся из спор: у анабены происходят только
поперечные деления клеток, у разных видов ностока — косые и про-
дольные.
Подвижность прогормогониев и гормогониев сильно варьирует
у разных видов и штаммов и зависит от условий внешней среды.
Так, потребность в свете для движения гормогониев у разных
гормогониофициевых неодинакова. Здесь можно выделить по край-
ней мере три группы видов:
1. Виды и штаммы, гормогонии и прогормогонии которых
одинаково подвижны как на свету, так и в темноте; некоторые
штаммы N. punetiforme, Calothrix elenkini, С. braunii.
2. Виды, гормогонии которых для приобретения подвижности
нуждаются в свете (миграция по агару в темноте наблюдается лишь
изредка и в очень незначительной степени). Однако при добавлении
в среду глюкозы движение гормогониев в темноте не отличалось от
движения их на свету (один из штаммов TV. punetiforme).
3. Виды, гормогонии которых обнаруживаю! подвижность то-
лько на свету, в темноте движения не наблюдается даже при добав-
лении в среду углеводов. Сюда в первую очередь относится тер-
мальная водоросль Mastigocladus laminosus, для образорания гор-
могониев которой необходимы высокие температуры (38—48е' С).
Кроме того, .для приобретения подвижности и миграции по поверх-
ности агара образовавшихся гормогониев необходим свет. В темно-
те при оптимальных температурах гормогонии остаются неподвиж-
ными. Добавление же в среду различных доз глюкозы не оказывало
никакого воздействия на подвижность гормогониев. Движение на-
чиналось только после выставления культур на свет: через непро-
должительный промежуток времени наблюдалась миграция гормо-
гониев от места посева по поверхности агара. На свету глюкоза
(0,25%-ная) оказывает некоторый стимулирующий эффект на интен-
сивность движения гормогониев, которые расползаются от посева
на большую дистанцию, чем в отсутствие глюкозы.
Свет необходим также для приобретения подвижности теми
участками, на которые распадаются трихомы анабен (A. cylindrica,
A. hallensis, A. variabilis).
В ряде случаев подвижность гормогониев может быть утрачена
полностью.
При прорастании гормогониев во взрослые трихомы ход их
развития в значительной мере обратный ходу развития гормогониев
из трихомов, г. е. из гомоцитной нити при подходящих внешних
условиях в конце концов развивается нить, свойственная данному
роду.
В процессе индивидуального развития гормогониофициевых во-
дорослей они проходят ряд неодинаковых морфологических стадий
или статусов, сходных со зрелыми особями других родов. Так, все
гормогониофициевые водоросли в состоянии гормогониев отвеча-
ют диагнозу рода осцилл<1тория (симметричная гомоцитная нить).
70
При дальнейшем развитии разные роды различаются, по статусам,
и число их тем больше, чем сложнее организация таллома данного
рода. Так, у мастигокладуса со сложным гетероцитным талломом
с дифференцировкой вегетативных клеток и разными типами вет-
вления (истинного, ложного) в жизненном цикле удается установить
десять статусов.
На разных ростовых фазах или статусах гормогониофициевые
водоросли могут задерживать свое развитие, вегетировать длитель-
ное время и даже размножаться посредством гормогониев.
Это послужило причиной того, что отдельные статусы в цикле развития носто-
кальных водорослей неоднократно описывались как самостоятельные таксоны. По-
этому система гормотониофициевых водорослей нуждается в пересмотре на основе
уточнения их онтогенеза. Онтогенез удобнее всего изучать в условиях культуры.
Однако при этом не следует игнорировать наблюдения в естественных местообита-
ниях. как это делают некоторые бактериологи. Так, Р. Риппка и др, (R. Rippka,
J. В. Waterbury, R. J. Starrier, 1981), изучая в культуре синезеленыс водоросли глеот-
рихию и ривулярию, постоянно существующие в природе в виде слизистых колоний,
но не образующие их при некоторых условиях культивирования, приходят к выводу
об упразднении этих родов. Для цианобактерий с асимметричными трихомами
и базальной гетероцистой они оставляют один род калотрике.
* * *
Синезеленые водоросли — очень древние, известны из докемб-
рийских отложений. Распространены они повсеместно, растут там,
где не могут обитать никакие другие растения и первыми заселяют
вновь образующиеся поверхности Земли, например вулканические
острова. Большинство синезелепых водорослей встречается в пре-
сных водах, меньше — в море. В планктоне прудов, озер, медленно
текущих рек виды родов микроцистис, анабена, глеотрихия часто
вызывают «цветение» воды.
Синезеленые водоросли имеют большое значение в жизни чело-
века, как положительное (азотфиксации, съедобность), так и от-
рицательное (токсичность, порча воды при отмирании возбудителей
«цветения»),
ОТДЕЛ ПРОХЛОРОФИТОВЫЕ ВОДОРОСЛИ PROCHLOROPHYTA
Это новый отдел растительного царства, предложенный Р. Леви-
ным (R. A. Lewin, 1976) для одноклеточных сферических водорос-
лей, размножающихся путем деления клетки посредством экватори-
альной перетяжки, относимых к новому роду прохлорон —
Prochloron. Эти водоросли живут на поверхности или в клоаке
некоторых асцидий. Замечательная особенность их — сочетание
прокариотной организации клетки с пигментным составом, свойст-
венным зеленым водорослям: доказано присутствие у прохлорона
хлорофиллов а и 6,отсутствие фикобилиновых протеиновых пигмен-
тов, характерных для синезеленых водорослей. Фотосинтетические
ламеллы состоят большей частью из двух сближенных тилакоидов
с гладкими мембранами, лишенными фикобилисом.
71
ЦА РСТВО Э У КА РИО ТЫ—EUCAR YO ТА
Глава 6
ОТДЕЛ КРАСНЫЕ ВОДОРОСЛИ — RHODOPHYTA ?
В хроматофорах красных водорослей, помимо хлорофиллов (й
и d, но не Ь) и каротиноидов (0-каротин, зеаксантин, антераксантин,
криптоксантин, лютеин, неоксантин), содержатся еще водораст-
воримые пигменты — фикобилины: фикоэритрины красного цве-
та, фикоцианины и аллофикоциан синего цвета. От соотношения
пигментов зависит окраска таллома, варьирующая от малиново-
красной (преобладание фикоэритрина) до голубовато-стальной (при
избытке фикоцианина). Хлоропласты одеты оболочкой из двух
мембран (хлоропластной эндоплазматической сети нет) и содержат
одиночные тилакоиды, на поверхности которых локализованы фи-
кобилисомы. Генофор рассеянный. Периферические тилакоиды
у более примитивных форм со звездчатыми или сильно лопастными
хроматофорами с пиреноидами отсутствуют, а у более высокоор-
ганизованных красных водорослей с дисковидными хлоропластами
без пиреноидов имеются. Запасной продукт — полисахарид «багрян-
ковый крахмал», от иода приобретающий буро-красный цвет. Зерна
багрянкового крахмала откладываются в цитоплазме всегда вне
связи с пиреноидами и хроматофорами. Для жизненного цикла
характерно полное отсутствие жгутиковых стадий и особая форма
оогамного полового процесса. Как и бурые, красные водоросли —
почти исключительно морские.
Строение таллома красных водорослей довольно простое: изве-
стны даже одноклеточные коккоидные формы (порфиридиум),
у многих талломы гетеротрихальные и в виде разветвленных нитей,
прикрепленных к субстрату с помощью ризоидов. Однако подав-
ляющее большинство имеет псевдопаренхиматозные талломы, воз-
никающие за счет переплетения и срастания боковых ветвей либо
одной оси, неограниченно нарастающей с помощью верхушечной
клетки, либо многих таких осей; в первом случае говорят об одно-
осевом строении, во втором — о многоосевом или мультиосевом.
Пластинчатые талломы истинно паренхиматозного строения, полу-
чающиеся в результате как поперечных, так и продольных делений
клеток, встречаются среди красных водорослей редко (порфира).
Клетка красных водорослей одета оболочкой, пектиновые и ге-
мицеллюлозные компоненты которой сильно набухают и часто
сливаются в общую слизь мягкой или хрящеватой консистенции,
заключающей протопласты. Нередко в оболочках откладывается
известь. При делении клеток у большинства красных водорослей
(класс флоридеофициевые) в результате неполного смыкания цент-
72
рипеталъно растущей поперечной стенки образуются первичные
поры. Поровый канал сначала открыт, но затем замыкается проб-
кой. По этим порам можно восстановить генетическую связь между
клетками в многоклеточном талломе. Клетки одно- и многоядер-
ные, хроматофоры, как правило, париетальные, многочисленные,
в виде зерен или пластинок. У бангиофициевых встречаются звезд-
чатые хроматофоры с центральными пиреноидами.
Размножение. Вегетативное размножение — путем фрагментации
таллома; особенно часто наблюдается в местообитаниях, где фак-
торы внешней среды (температура, свет) неблагоприятны для раз-
множения спорами.
Бесполое размножение осуществляется посредством неподвиж-
ных клеток, развивающихся из содержимого спорангия в числе
одной (моноспоры) или четырех (тетраспоры).
Тетраспоры формируются на диплоидных бесполых растени-
ях—тетраспорофитах (тетраспорангиальная фаза). В тетраспоран-
гиях перед образованием тетраспор происходит мейоз. Расположе-
ние тетраспор в тетраспорангии может быть крестообразным (гели-
диум, хондрус), рядовым (кораллина, дюренея) или по углам тетра-
эдра (церамиальные).
При крестообразном расположении содержимое тетраспорангия
сначала делится в горизонтальном направлении, а затем каждая
половина, в свою очередь, делится вертикально. При рядовом или
зональном расположении тетраспор содержимое вытянутого тет-
распорангия делится поперечными перегородками на четыре клет-
ки, расположенные друг над другом. Если тетраспоры расположены
по углам тетраэдра, содержимое тетраспорангия сразу распадается
на 4 тетраспоры. Гаплоидные тетраспоры прорастают в гаплоид-
ные растения — гаметофиты (гаметангиальная фаза) того же или
иного, чем тетраспорофиты, облика. Они размножаются половым
путем: на них развиваются половые органы — гаметангии.
Половой процесс оогамный. Женский орган —карпогон—у бо-
льшинства красных водорослей (флоридеофициевые) состоит из
расширенной базальной части —брюшка, заключающего яйцеклет-
ку, и отростка—трихогины (последняя отсутствует у бангиофици-
евых). Трихогина или вообще не содержит ядра, или оно дегенери-
рует к моменту оплодотворения. Карпогон обычно развивается на
особой короткой, состоящей из 3—4, реже из 7—9 клеток карпогон-
ной ветви, которая в свою очередь сидит на поддерживающей (несу-
щей) клетке. Антеридии (сперматангии) — обычно мелкие бесцвет-
ные клетки, содержимое которых освобождается в виде мелких
голых, лишенных жгутиков мужских оплодотворяющих элемен-
тов— спермациев. Выпавшие из антеридиев спермации пассивно
переносятся токами воды и прилипают к трихогине. В месте контак-
та спермация и трихогины их стенки растворяются, и ядро спер-
мация по трихогине перемещается в брюшную часть карпогона, где
сливается с женским ядром. Эти данные классических исследований
73
конца прошлого века, сохранившие значение до наших дней, можно
дополнить некоторыми наблюдениями последних лет. Так, в 1984 г.
получены новые данные о динамике полового процесса у некоторых
красных водорослей. В условиях эксперимента у Callithamnion
cordatum удалось показать, что зрелые карпогоны (созревание их
происходит в течение четырех дней после заложения карпогонной
ветви) восприимчивы к спермациям в течение 3 ч. Сам процесс
слияния гамет, включающий известные из классических работ фазы
(прикрепление спермация к трихогине, плазмогамия, перенос ядра
спермация вниз по трихогине и кариогамия), длится 5—10 ч. После
оплодотворения базальная часть карпогона отделяется перегород-
кой от трихогины, которая отмирает, и претерпевает дальнейшее
развитие, приводящее у большинства красных водорослей к об-
разованию карпоспор. Детали этого развития имеют важное систе-
матическое значение. У одних красных водорослей содержимое
зиготы (оплодотворенного карпогона) непосредственно делится
с образованием неподвижных голых спор карпоспор, у других из
оплодотворенного карпогона (или из его дочерней клетки) выраста-
ют многоклеточные ветвящиеся нити — гонимобласты, клетки кото-
рых превращаются в карпоспорангии, производящие по одной кар-
поспоре. Наконец, у большинства красных водорослей гонимоб-
ласты развиваются не непосредственно из брюшка оплодотворен-
ного карпогона, а из особых вспомогательных—ауксилярных
клеток. Последние могут быть удалены от карпогона или же рас-
полагаются на талломе в непосредственной от него близости. Если
ауксилярные клетки удалены от карпогона, из его брюшка после
оплодотворения вырастают гифообразные, без поперечных клеточ- 1
ных перегородок соединительные или ообластемные нити. Их воз- 3
никновению предшествуют митотические деления копуляционного )
ядра и таким образом они содержат диплоидные ядра. Ообластем-
ные нити подрастают к ауксилярным клеткам, в точке контакта
оболочки растворяются, и кончик ообластемной нити и ауксиляр-
ная клетка сливаются. Диплоидное ядро ообластемной нити делит- ;
ся, одно из дочерних ядер передается ауксилярной клетке, а другое !
остается в ообластемной нити, которая может расти к следующей ?
ауксилярной клетке, и г. д. Таким образом, все большее число
ауксилярных клеток обеспечивается диплоидным ядром - потом- <
ком диплоидного копуляционного ядра одного-единственного 1
оплодотворенного карпогона. Слияние кончика ообластемной нити |
с ауксилярной клеткой не сопровождается слиянием их ядер— ;
диплоидного ообластемной нити и гаплоидного ауксилярной клет-
ки. Диплоидизированная ауксилярная клетка, отделяясь перегород- j
кой от ообластемной нити, делится на базальную клетку и цент- 1
ральную. Первая получает первоначальное гаплоидное ядро аук- j
силярной клетки, вторая—диплоидное ядро и становится центром 1
развития гонимобластов, клетки которых содержат диплоидные I
ядра и продуцируют диплоидные карпоспоры. Гонимобласты, раз- I
74 j
вивающие карпоспоры, рассматриваются как особая генерация —
карпоспорофит (карпоспорангиальная фаза).
У вышестоящих порядков красных водорослей ауксилярные
клетки формируются рядом с карпогоном до оплодотворения
карпогона или только после того, как оно произошло (у наиболее
высокоорганизованных красных водорослей). Совокупность аук-
силярной клетки (или клеток) с карпогоном называется прокарпием.
Здесь нет необходимости в образовании длинных ообластемных
нитей, связывающих карпогоп и ауксилярные клетки, ауксилярная
клетка просто сливается с брюшком оплодотворенного карпогона,
после чего из нее развиваются гонимобласты с карпоспорами.
Карпоспораигии часто располагаются тесными группами — цисто-
карпиями, которые у многих представителей одеты псевдопарен-
химатозной оболочкой, развивающейся из соседних с карпогоном
клеток.
Жизненные циклы красных водорослей отличаются большим
разнообразием, что в последнее время было продемонстрировано
на основе следующих данных, полученных разными методами. На-
блюдения в природе дают представление о встречаемости мор-
фологических фаз и репродуктивных органов, на них развивающих-
ся. В условиях лабораторной культуры можно проследить прора-
стание репродуктивных клеток и развитие из них талломов до
состояния, когда они дают репродуктивные органы, т. е. в культуре
можно строго установить последовательность морфологических
фаз (например, тетраспоры полисифонии, прорастая, дают гамето-
фиты, несущие половые органы; карпоспоры прорастают в тетра-
спорофиты, развивающие тетраспорангии). Особенно плодотворным
оказался метод лабораторных культур для установления связей
между гаметангиальной и тетраспорангиальной фазами, различа-
ющимися морфологически и долгое время рассматриваемыми как
самостоятельные таксоны. Цитологический метод позволяет уста-
новить число хромосом каждой морфологической фазы, место
в жизненном цикле сингамии (копуляции), мейоза и других цитоло-
гических деталей. Ниже приводятся жизненные циклы, обоснован-
ные совокупностью вышеперечисленных данных,
У красных водорослей наиболее распространен жизненный цикл,
состоящий в последовательности гаметангиальной, карпоспоранги-
альной и тетраспорангиальной фаз, причем первая и последняя
морфологически сходны, а карпогониальная фаза развивается на
гаметангиальной (рис. 10, А). Такая изоморфная смена гаметофитов
и тетраспорофитов (усложненная развитием особой генерации —
карпоспорофита) впервые была описана для Polysiphonia violaceae.
Поэтому такой жизненный цикл, присущий большинству красных
водорослей, обычно называют Polysiphonia-тип. Его можно встре-
тить помимо порядка церамиальных, куда относится род полисифо-
ния, и у представителей порядков криптонемиальиые, гигартиналь-
ные, родимениальные и (предположительно) гелидиальные.
75
Рис. 10. Жизненные циклы красных водорослей: А — Polysiphonia; Б—Bonnemaisonia
hamiferiT, В - Lemanea; Г—Liagora tetrasporifera; Д—Palmaria palmate (Подробнее см.
в тексте):
К—копуляция, Р—редукционное деление, ка карпоспоры, 2л — диплоидная фаза, и—iaiuio-
идная фаза
Предположение (на основе наблюдений в природе тетраспоран-
гиальных растений, морфологически неотличимых от гаметангиаль-
ных) о жизненном цикле Polysiphonia-типа у представителей выше-
стоящих порядков класса флоридеофициевых в последние годы
подтверждено строгими культуральными экспериментами.
Цикл развития Bonnemaisonia hamifera-типа (рис. 10, Б), который
можно назвать гетероморфной сменой генераций (усложненной кар-
иоспорофитом), характеризуется последовательностью гаметанги-
альной, карпоспорапгиальной и тетраспорангиальной фаз, причем
все три фазы морфологически неодинаковы. Карпогониальная фаза
развивается на гаметангиальной. В последние годы такой жизненный
цикл был описан не только для многих представителей порядка
немалиальных, но и для криптонемиальных и гигартинальных.
У Liagora tetrasporifera в жизненном цикле чередуются гаметан-
гиальная и тетраспорангиальная фазы, морфологически различа-
ющиеся, причем тетраспорангиальная фаза развивается на гаметан-
гиальной; мейоз осуществляется в тетраспорангиях, карпоспорофиз
отсутствует (рис. 10, Г).
У Palmaria palmata также отсутствует карпоспорофит и наблю-
дается чередование гаметангиальной и тетраспорангиальной фаз,
причем вторая развивается на первой (рис. 10, Д). Однако женский
гаметофит здесь крохотный, а мужской—крупный, морфологичес-
ки неотличимый от тетраспорофита.
76
Приведенные жизненные циклы характеризуются спорической
редукцией, так как мейоз происходит при образовании тетраспор.
Жизненный цикл, описанный пока только для двух родов —
леманеа и батрахоспермум (порядок немалиональные), отличается
соматической редукцией. В апикальной клетке предростка, развива-
ющегося из диплоидных карпоспор, происходит мейоз и из нее
вырастает гаплоидная гамегангиальная фаза (рис. 10, В).
Сказанным не исчерпывается все разнообразие жизненных цик-
лов красных водорослей (см. ниже).
Красные водоросли делятся на два класса: бангиофициевые
(Bangiophyceae) и флоридеофициевые (Florideophyceae)
КЛАСС БАНГИОФИЦИЕВЫЕ — BANGIOPHYCEAE
Клетки часто со звездчатым хроматофором, заключающим пи-
реноид. Поры между клетками, как правило, отсутствуют. Кар-
погон без трихогины, после оплодотворения содержимого карпого-
на непосредственно делится с образованием карпоспор. Бесполое
размножение обычно моноспорами.
Представителями могут служить роды порфира и бангия.
Род порфира (Porphyra) характеризуется листоватыми таллома-
ми истинно паренхиматозного строения, прикрепленными своими
основаниями к субстрату и достигающими 50 см в длину, редко
более (у Р. nereocystis до 2 м). Пластинка состоит из одного слоя
клеток или двухслойная. Клетки содержат по одной или по две
пластиды. Пластинки у разных видов неодинаковой формы (окру-
глые, яйцевидные, удлиненные и пр.) с цельными или зазубренными
краями, различной окраски (красноватой, буроватой, голубоватой)
и толщины. Проростки сначала представлены однорядными нитя-
ми. Последовательные продольные деления клеток этих нитей при-
водят к образованию листовидных талломов. Порфира распрост-
ранена как в южных, так и в северных морях в прибрежной лито-
ральной зоне.
Род бангия {Bangid), включающий как морские, так и пресновод-
ные виды, имеет таллом, представленный неразветвленными нитя-
ми, вначале прикрепленными к субстрату базальной клеткой; с воз-
растом от нижних клеток развиваются несептированные ризоиды.
Нити сначала однорядные, но в более старых участках клетки
претерпевают продольные радиальньге деления, так что на попереч-
ном срезе таллома видны сектора.
Зрелые талломы порфиры и бангии размножаются половым
путем. У порфиры при образовании антеридиев клетки талломов
делятся во взаимно перпендикулярных направлениях на ряд мелких
клеток, каждая из которых образует по одному спермацию. При
разбухании ин герцеллюлярной слизи спермации выдавливаются
и освобождаю гея. Карпогоны лишь незначительно отличаются от
77
вегетативных клеток. Типичная трихогина отсутствует. После опло-
дотворения зигота (оплодотворенный карпогон) непосредственно
делится на 2—32 карпоспоры, которые освобождаются так же. как
и спермации,— благодаря набуханию межклеточной слизи. Сход-
ным образом осуществляется половое размножение бангии. Так.
Е. Мань (Е. Magne, 1972) утверждает, что талломы бангии гаплоид-
ны и представляют собой гаметофиты. Они двудомны: мужские
талломы продуцируют спермации, которые копулируют с клетками
женских нитей, функционирующими как каргюгоны. В результате
возникает диплоидная зигота, которая делится на карпоспоры.
Цикл развития. Цикл развития порфиры и бангии интенсивно
изучался в последние десятилетия как в природе, так и в искусствен-
ных культурах. Для многих видов порфиры и бангии в их жиз-
ненном цикле были установлены Conchocelis-стадии: карпоспоры,
сначала голые, выделяют оболочку и прорастают униполярно
в нитчатые растеньица, образующие на поверхности и внутри изве-
стковых раковинок моллюсков розовые пятна1. Эта стадия долгое
время считалась самостоятельной водорослью Conchocelis rosea.
Нити конхоцелиса сложены из удлиненных клеток с париетальным
хроматофором. Ультраструктура клеток типична для красных во-
дорослей из класса флоридеофициевых: слоистая клеточная оболоч-
ка , одиночные тилакоиды в хлоропластах, окруженных двухмемб-
ранной хлоропластной оболочкой: периферические тилакоиды; пи-
реноид, в который проникают одиночные тилакоиды; рассеянные
между тилакоидами фибриллы ДНК; в цитоплазме—гранулы баг-
рянкового крахмала. Между соседними клетками Conchocelis-нитей
имеются закупоренные пробками поры, сходные с таковыми в клет-
ках флоридеофициевых.
В деталях Conchocelis-стадии разных видов порфиры и бангии
обнаруживают постоянные различия: морфологические (в толщине
клеточных стенок, гладких или извилистых), ультраструктурные,
касающиеся числа и расположения тилакоидов, пронизывающих
пиреноиды, пор и замыкающих их пробок и др. Имеются постоян-
ные различия в характере росла в культуре и физиологические,
в частности, каждый вид по-своему реагирует на температуру и фо-
топериод. Зрелые Conchocelis-нити образуют специализированные
веточки — конхоспорангиальные ветви, сложенные из изодиаметри-
ческих толстостенных клеток со звездчатыми осевыми х^оматофо-
рами без периферических тилакоидов. Клетки конхоспорангиаль-
ных ветвей освобождают конхоспоры, прорастающие биполярно
и образующие проростки, которые вырастают в новые макроскопи-
1 В культуре нити конхоцелиса можно выращивать и без раковин в качестве
субстрата, однако в природе они приурочены в основном к раковинам различных
моллюсков.
’ Клеточная стенка Conchocelis-стадии Bangia fuscopurpurea, Porphyra umbi'icalis.
P. leucosticta содержит целлюлозу, ранее неизвестную у бангиофициевых, хотя цел-
люлоза—обычный компонент стенок флоридеофициевых.
78
ческие листовые талломы. Для некоторых видов (Porphyra ка-
ncikaensis. Р. perforata, Р. schizophylla) было показано, что карпоспо-
ры и вегетативные клетки конхоцелис-стадии диплоидны. Ввиду дип-
лоидности конхоцелис-стадия может быть сравнима с тетраспо-
рофитом, хотя она и не образует тетраспор. Прежние утверждения,
что мейоз у порфиры происходит при прорастании зиготы, не под-
твердились, так же как и у так называемых гаплобионтных немали-
альных. Для Р. gardneri описано оплодотворение карпогона спер-
мацием и высказывалось предположение, что мейоз имеет место при
образовании конхоспор. Недавние цитологические исследования
Р. yezoensis дают возможность предположить, что мейоз осуществля-
ется при образовании проростков из конхоспор. Однако у очень
многих видов цитологические детали жизненного цикла неизвестны.
Каждая из морфологических фаз (макроскопические листоватые
растения, микроскопическая нитчатая Conchocelis-фаза) способны
к саморепродукции. Так, молодые талломы Р. yezoensis, Р. tenera
продуцируют обильные моноспоры (нейтральные споры), а у Р. ки-
niedai моноспоры образуются обильно на протяжении всего периода
жизни, даже во взрослых талломах, которые формируют спермации
и карпоспоры. Прорастая, моноспоры дают листоватые талломы
порфиры. Однако у Р. pseudolinearis размножение посредством моно-
спор отсутствует. Наоборот, некоторые виды порфиры (Р. argentinen-
sis) размножаются только бесполым путем — посредством аплано-
спор, образующихся по 8—32 в клетках по краю таллома. Эти споры
имеют звездчатый хлоропласт с центральным пиреноидом и прора-
стают биополярно, давая начало новому листоватому таллому.
На Conchocelis-нитях в качестве одиночных боковых ветвей воз-
никают моноспорангии. Они снабжены ножками (Р. miniata,
Р. brumalis, Р. perforata, Р. kanakaensis) или сидячие, как у Bangia
fuscopurpurea, располагаются поодиночке (Р. miniata) или образуют
цепочки (Р, nereocystis), Униполярно прорастающие моноспоры
вновь дают Conchocelis-стадию. Помимо моноспор, Conchocelis-
нити могут размножаться вегетативно — фрагментацией таллома.
При культивировании некоторых видов порфиры (например,
Р. yezoensis) в качестве посевного материала используются культи-
вируемые Conchocelis-нити.
Описанный выше и приводимый во всех учебных пособиях половой процесс
бангиофициевых в виде слияния спермация с карпогоном, вследствие которого
образуются карпоспоры, не удавалось наблюдать в ходе многочисленных исследова-
ний онтогенеза родов порфира и бангия как в культуре, так и в полевых условиях1.
' Однако совсем недавно (Т. Y. Chang et В. F. Zheng, 1984) описана плазмога-
мия (переход содержимого спермация в карпогон) и деление оплодотворенного
карпогона на 32 карпоспоры у Р. haitanensis — вида, культивируемого в Китае в ком-
мерческих целях. М. Куроги (М. Kurogi, 1972), подробно исследовавший японские
виды порфиры в полевых условиях, также обнаруживал у многих видов оплодот-
воряющие трубки между карпогонами и спермациями, однако цитологических дан-
ных он нс приводит.
79
Ряд авторов поставили под сомнение даже оплодотворение спермацием карпогона
и образование карпоспор как прямого следствия половою процесса1. Это повлекло
к изменению терминологии. Например, карпоспоры и сттермации были обозначены
соответственно как к- и р-споры. tx-Споры прорастают непосредственно в Conchocelis-
стадию, p-споры к прорастанию не способны. Эта новая терминология широко
применяется не только в многочисленных специальных работах, посвященных родам
порфира и бангия, но проникла и в некоторые учебные руководства. Это побуждает
несколько подробнее остановиться на новой терминологии спор бангиофицисвых и их
классификации. Так, Н. Ричардсон (N. Richardson, 1972) предложил классификацию
спор, обнаруживаемых в цикле развития бангия и порфира, с учетом в первую очередь
характера их прорастания и их дальнейшей судьбы. Он различает споры, прораста-
ющие униполярно, биполярно и не прорастающие совсем. К униполярно прораста-
ющим спорам относятся, например, споры, образующиеся путем повторного деления
материнских клеток макроскопического таллома на 16 или менее частей и которые
при определенных условиях освещения дают начало конхоцелис-фазе (а-слоры, карпо-
споры). Униполярно прорастают также моноспоры, репродуцирующие конхоцелис-
стадию. К спорам, прорастающим биполярно, относятся моноспоры, возникающие
на макроскопическом талломе, и необходимые для репродукции этой фазы, конхоспо-
ры, развивающиеся на Conchocelis-нитях и дающие начало макроскопической (листо-
ватой) фазе, а также апланоспоры, репродуцирующие у некоторых видов порфиры
листоватые талломы. Для прорастания всех этих спор требуется определенный
фотопериод. Споры, не прорастающие, образуются путем повторных делений содер-
жимого'материнской клетки па более чем 16 продуктов деления (спсрмации).
Порфира — важная промысловая водоросль, она высоко ценится
как продукт питания с высоким содержанием протеина и витаминов
(А, В, С и др.), а также применяется в медицине.
КЛАСС ФЛОРИДЕОФИЦИЕВЫЕ —FLORIDEOPHYCEAE
Клетки большей частью с париетальными хроматофорами без
пиреноидов. Между клетками имеются поры. Карпогон с трихоги-
ной. После оплодотворения развиваются гонимобласты или непо-
средственно из брюшка оплодотворенного карпогона, или из аук-
силярных клеток после слияния их с ообластемными нитями или
оплодотворенным карпогоном. Бесполое размножение большинст-
ва— с помощью тетраспор.
Дел али развития карпоспорофита (из оплодотворенного карпо-
гона или из ауксилярных клеток), а также время дифференцировки
ауксилярных клеток (до или после оплодотворения), их положение
на талломе и т. п. положены в основу деления флоридеофициевых
на порядки. » 1 2
1 То, что во многих исследованиях оплодотворение непосредст венно не наблю-
далось, еще нс свидетельствует об его отсутствии и как всякий негативный результат
нуждается в особенно тщательной проверке. Так, К. Коле (К. Cole. 1972). не наблю-
давшая оплодотворения в культурах Bangia fuscopurpwea, не исключает возможности
оплодотворения в исходном — природном материале до его изолирования в куль-
туру, т. е. ot-слоры, появившиеся в культуре, могли быть результатом полового
процесса, осуществлявшегося в природе до выделения таллома в культуру! Позже
этот же автор (К. Cole, Е. Conway, 1980) при описании способов размножения
бангиофициевых придерживается более традиционных воззрений и пишет, что из
оплодотворенных карпогопов у Bangia atropurpurea и 24 видов Porphyra развиваются
2 — 64 карпоспоры, прорастающие униполярно в конхопелис-нити.
80
ПОРЯДОК НЕМАЛИАЛЬНЫЕ —NEMALIALES
Этот порядок, включающий формы одноосевого и многоосевого
строения, характеризуется отсутствием ауксилярных клеток; гони-
мобласты развиваются непосредственно из оплодотворенного кар-
погона или из его дочерней клетки. Долгое время считалось, что
у представителей этого порядка мейоз совершается непосредствен-
но после оплодотворения при прорастании зиготы и называли их
гаплобионтными флоридеями, противопоставляя всем остальным
диплобионтным порядкам флоридей, у которых мейоз происходит
при образовании тетраспор. Однако Е. Мань (Е. Magne, 1961, 1967)
продемонстрировал цитологически отсутствие мейоза в карпогоне
вслед за оплодотворением и диплоидность гонимобластов, карпо-
спор и продуктов их прорастания для ряда представителей порядка
немалиальных (немалион, сцинайя, боннемайсопия). Он выдвинул
гипотезу существования у них спорофитной генерации или ее эк-
вивалента, где обязательно должен происходить мейоз. Исследова-
ния двух последних десятилетий подтвердили гипотезу, выдвину-
тую Е. Мань, и, кроме того, обнаружили заметное разнообразие
жизненных циклов в пределах этого порядка. Здесь можно раз-
личить по крайней мере три типа циклов развития, хорошо обосно-
ванных как с точки зрения последовательности морфологических
фаз, так и цитологических данных о числе хромосом в каждой
морфологической фазе и месте сингамии и мейоза в жизненном
цикле. Первый тип цикла развития — тип Lemanea. В этом типе две
морфологические фазы совмещены в одной структуре, где нижняя
базальная часть диплоидна, а верхняя часть гаплоидна и представ-
ляет собой гаметангиальную фазу, на которой развивается карпо-
спорангиальная фаза. Мейоз происходит соматически во время
развития этой структуры (рис. 10, В).
Как уже отмечалось, этот тип развития установлен пока только
для родов леманеа (Lemanea) и батрахоспермум (Batrachospermum).
Род леманеа (Lemanea) встречается в быстро текущих речках
с холодной водой. Таллом, имеющий вид неветвящейся щетинки
10—15 см длиной и 1 мм толщиной, темно-фиолетового или
оливково-бурого цвета с узловатыми вздутиями; прикрепляется
к субстрату при помощи подошвы из стелющихся нитей. По
оси таллома проходит одна нить из вытянутых бесцветных клеток
(одноосевое строение). От верхней части каждой клетки цент-
ральной оси отходит мутовка обычно из четырех расходящихся
по радиусам ветвей. Базальные клетки каждой такой ветви крупные,
вытянутые. От их дистального (морфологически верхнего) конца
отходят ветви второго порядка, которые, в свою очередь, мно-
гократно ветвятся. Конечные разветвления срастаются в мно-
гослойную кору. Ее наружные клетки мелкие и заполнены хро-
матофорами, внутренние — крупные и почти бесцветные. От про-
ксимальных (морфологически нижних) концов основных клеток
81
радиальных ветвей отходят коровые нити, окутывающие цен-
тральную нить. Антеридии образуются группами из поверхностных
клеток коры. Карпогонные ветви, несущие карпогоны, возникают
от внутренних клеток коры. Брюшко карпогона находится около
внутренней поверхности коры, а трихогина прорывает кору и вы-
совывается наружу. После оплодотворения из брюшной части
карпогона развиваются пучки гонимобластов, врастающие в по-
лость таллома. Карпоспорангии формируются целыми четками,
карпоспоры скапливаются внутри полости таллома и освобо-
ждаются после разрушения коры таллома. Согласно исследованиям
Е. Мань, при развитии из зиготы (оплодотворенного брюшка
карпогона) гонимобластов мейоз не осуществляется и, таким
образом, карпоспорофит и карпоспоры диплоидны. Карпоспоры
прорастают с образованием диплоидного нитчатого, состоящего
приблизительно из 20 клеток предростка, ядро в верхушечной
клетке которого редукционно делится. Из апикальной клетки
после мейоза развивается гаплоидное растение -гаметофит (га-
метангиальная фаза) леманеи; на нем образуются половые органы.
Таким образом, базальная часть таллома леманеи диплоидна.
У рода батрахоспермум (Batraehospermum) цикл развития такой
же, как у леманеи. Его представители встречаются в реках с чистой
прозрачной водой и озерах. Таллом имеет вид сильно разветвленно-
го, слизистого иа ощупь кустика оливково-зеленого или стального
цвета (избыток фикоцианина). Ось, неограниченно нарастающая за
счет деятельности верхушечной клетки, состоит из одного ряда
вытянутых бесцветных клеток. От каждой из них непосредственно
под поперечой перегородкой возникает мутовка ветвей ограничен-
ного роста, которые сложены из мелких, богатых хроматофорами
клеток. Конечные клетки ветвей могут вытягиваться в длинные
волоски. Из базальных клеток боковых ветвей (ассимиляторов)
развиваются ветви неограниченного роста. Кроме того, базальные
клетки мутовок ветвей дают начало кортикальным (коровым) ни-
тям, которые растут вдоль клеток главной оси, совершенно скрывая
их, и у некоторых видов, переплетаясь, образуют многослойную
обвертку. Из клеток коровых нитей возникают вторичные мутовки
боковых ветвей, вставляющиеся между уже имеющимися. На боко-
вых ветвях ограниченного роста (ассимиляторах) формирую гея по-
ловые органы. После оплодотворения из брюшка карпогона выра-
стают ветвящиеся гонимобласты; конечные клетки их образуют
карпоспоры, собранные в тесную группу — цистокарпий. Из карпо-
спор развиваются ползучие нити, от которых отходят вертикаль-
ные, ветвящиеся нити совсем иного внешнего вида, чем таллом бат-
рахоспермума. Эта стадия была описана под родовым названием
шантранзия — Chantransia. Она может размножаться посредством
моноспор. При благоприятных условиях из верхушечных клеток
этого Chantransia-предросгка вырастают типичные талломы батра-
хоспермума.
82
Развитие на нитях шантранзии таллома батрахоспермума зави-
сит от продолжительности и интенсивности освещения. Так, при
культивировании Batrachospernntm moniliforme при фотопериоде ме-
нее 14 ч Chantransia-стадия дает только моноспоры. При освеще-
нии ниже 750 Лк наблюдалась только Chantransia-стадия. При дли-
не дня Мчи освещении свыше 750 Лк до 4% Chantransia-нитей
образовывали гаметофиты, т. е. развивали талломы батрахоспер-
мума. При фотопериоде 15—16 ч и освещении 750 и 1500 Лк раз-
витие гаметофитов наблюдалось у 40% Chantransia-нитей. У неко-
юрых видов Batrachospermum (В. vagum, В. moniliforme) в лабора-
торных условиях онтогенез от прорастания карпоспор до развития
гаметофитов прослежен цитологически. Ввиду мелких размеров
ядер содержание ДНК в клетках гаметофитов и Chantransia-нитей,
покрашенных по Фёльгену, определялось цитофотометрическим
методом. Показано, что Chantransia-стадия диплоидна, а гамето-
фит (батрахоснермум) гаплоиден. Таким образом, мейоз Не мог
произойти во время первого деления ядра зиготы, как принималось
ранее, а иногда утверждалось и в более современных исследованиях.
Можно принять, что мейоз имеет место в переходной области
между Chantransia-стадией и гаметофитом (батрахоспермум). Со-
гласно Мань, Chantransia-стадию батрахоспермума, так же как
и базальную часть таллома леманеи, следует рассматривать как
диплоидное образование, соответствующее тетраспорофиту других
красных водорослей.
Другой тип жизненного цикла, хорошо обоснованный,— это тип
Bonnemaisonia hamifera (водоросль с одноосевым строением), состо-
ящий из последовательности гаметангиальной, карпоспорангиаль-
ной и тетраспорангиалыюй фаз, причем все три морфологически
неодинаковы и имеют карпогониальную фазу, развивающуюся на
гаметангиальной (рис. 10, Б). Такой тип, иначе гетероморфная сме-
на форм развития, наблюдался у некоторых видов морского рода
немалион (Nemaliori), отличающегося от леманеи и батрахоспер-
мума многоосевым типом таллома. В слабоветвящихся розовых
слизистых талломах немалиона центральная часть занята целым
пучком продольных нитей, состоящих из вытянутых бесцветных
клеток, от которых радиально расходятся обильно ветвящиеся,
богатые хроматофорами нити — ассимиляторы, соединенные сли-
зью мягкой консистенции.
Половые органы, как и у батрахоспермума, развиваются на
ассимиляторах. Карпоспорофит и карпоспоры у немалиона, как
у леманеи и батрахоспермума, диплоидны. У некоторых видов
Nemalion (N. vermieulare. N. multifidum) наблюдалась гетероморфная
смена генераций: макроскопического гаметофита (гаметангиальной
фазы) и микроскопического нитчатого Acrochaetium-подобного тет-
распорофита (тетраспорангиалыюй фазы).
Такой же тип развития, в подтверждение гипотезы Мань,
описан и для некоторых других представителей немалиальных,
83
обладающих, как и немалион, многоосевыми талломами, но с более
компактной структурой и нередко с заметной кальцификацией. Тако-
вы Scinaia furcellata, S. turgida, Helminthora divaricata, Helminthocladia
calvadosii, Chaetangium fastigiatum. Описанные ранее как два самосто-
ятельных вида рода Acrochaetium (с талломом из свободно разветв- ‘
ленных однорядных нитей) A. polyblasum и A. hallandicum оказались
соответственно тетраспорофитом и гаметофитом в жизненном цик-
ле одного организма. То же относится и к сильно различающимся
морфологически красным водорослям, ранее относимых к разным
родам: Falkenbergia rufolanosa и Asparagopsis armata.
Третий тип цикла развития—тип Liagora tetrasporifera1—скла-
дывается из последовательности гаметангиальной и тетраспоранги-
альной фаз, морфологически неодинаковых, причем тетраспоранги-
альная фаза развивается на гаметангиальной; мейоз происходит
в тетраспорангиях (рис. 10, Г). Такой цикл описан для
Helminthocladia agardiana, Н. senegalensis.
Наконец, недавно (1983) в культуре Acrochaetium pectinatum впер-
вые для красных водорослей наблюдалось свободное развитие зи-
готы: содержимое карпогона после оплодотворения выходило из
оболочки, прикреплялось к субстрату и прорастало в спорофит.
Иногда зигота развивалась в нити гонимобласта с терминальными
карпоспорангиями.
Порядок гелндиальные — Gelidiales1 2
Этот порядок выделил из порядка немалиальных Г. Кюлин
(Н. Kylin, 1923) на основе наличия тетраспорангиальной фазы, мор-
фологически сходной с гаметангиальной, что позволяло предполо-
жить здесь цикл развития типа Polysiphonia. Кюлин придерживался
мнения, что, как и у немалиальных, у гелидиума отсутствуют
ауксилярные клетки и гонимобласты развиваются непосредственно
из брюшка оплодотворенного карпогона. Мнения последующих
исследователей относительно существования ауксилярных клеток
у гелидиума расходятся. Одни утверждают, что поддерживающая
клетка, с которой сливается оплодотворенный карпогон, может
рассматриваться как ауксилярная, другие же, указывая на то, что
это слияние не носит облигатного характера (его может и не быть),
не признают эту клетку ауксилярной.
Талломы всех гелидиальных имеют одноосевое строение. Все
они — обитатели теплых морей. Gelidium cartilagifieum и другие виды
этого рода—важные промысловые водоросли: из них в Японии
получают кантен (агар-агар).
1 Род Liagora близок к Nemalion, имеет многоосевой таллом, местами пропитан-
ный известью.
2 Некоторые авторы, например П. Диксон (Р. S. Dixon, 1982), рассматривают
гелндиальные в качестве семейства в пределах порядка Nemaliales.
84
Порядок криптонемиальные—Cryptonemiales
В отличие от предыдущих порядков здесь представители имеют
ауксилярные клетки, которые развиваются до оплодотворения кар-
погона и обычно рассеяны по таллому на известном расстоянии от
карпогона. Связь между ауксилярными клетками и карпогоном
осуществляется с помощью более или менее длинных соединитель-
ных, спорогенных или ообластемных нитей.
Для представителей порядка криптонемиальных (Cryptonemiales)
долгое время предполагался жизненный цикл Polysiphonia-типа, при
котором гаметангиальная и тетраспорангиальная фазы морфологи-
чески идентичны. Такой жизненный цикл был давно описан для
Corallina officinalis, тогда как для остальных представителей порядка
это предположение не сопровождалось строгими доказательствами,
и лишь сравнительно недавно жизненный цикл Callophyllis firma был
прослежен в лабораторной культуре и продемонстрировано разви-
тие этой водоросли по Polysiphonia-типу. Гаметангиальные растения
несли зрелые карпоспорофиты. Освобождавшиеся карпоспоры про-
растали с образованием многослойных базальных дисков, из кото-
рых вырастали вертикальные оси и развивались листовидные пла-
стинки, характерные для этого таксона. Приблизительно через три
месяца листовидные талломы, возникшие при прорастании карпо-
спор, продуцировали многочисленные тетраспорангии с крестообраз-
ным расположением тетраспор. Из тетраспор, освобождавшихся из
тетраспорангиальных растений, в культуре развились гаметангиаль-
ные растения со зрелыми карпоспорофитами. Карпоспоры прораста-
ли в новые тетраспорангиальные растения. Гаметангиальная и тет-
распорангиальная фазы морфологически идентичны. Исследования
других представителей порядка криптонемиальных выявили иной
жизненный цикл, при котором гаметангиальная и тетраспорангиаль-
ная фазы морфологически различались (Acrosymphyton purpuriferum,
Gloiosiphonia capillaris, Pikea californica, Gloeophycus koreanum).
Среди криптонемиальных, как и среди немалиальных, встреча-
ются формы не только одноосевого (дюренеа), но и многоосевого
(кораллина) строения.
Порядок гигартипальные—Gigartinales
Порядок характеризуется тем, что ауксилярные клетки диф-
ференцируются до оплодотворения в непосредственной близости
от карпогона, так что прокарпий имеется; в качестве ауксилярной
функционирует клетка, участвующая в формировании таллома,
а не специальная — добавочная. Размножение можно рассмотреть
на примере рода анфельция (Ahnfeltiaf. Зрелые мужские и женские
гаметофиты представлены вертикальными, в различной степени
разветвленными многоосевыми талломами, вырастающими от
1 Виды этого рода используются для получения агар-агара.
85
хорошо развитых базальных корочек. На мужских талломах в плоду-
щих участках зрелые антеридии образуют один ряд вытянутых
клеток. При помещении живого материала в морскую воду под
микроскопом можно наблюдать, как вытянутые спермации с силой
отбрасываются от таллома на значительное расстояние. На женских
талломах плодущие участки находятся вблизи верхушки. Прокарпий
Ahnfeltia concinna состоит из крупной поддерживающей клетки и трех-
клеточной карпогонной ветви, верхняя клетка которой представляет
собой карпогон. Поддерживающая клетка функционирует как аукси-
лярпая. После оплодотворения карпогона и слияния его с поддержи-
вающей клеткой, одновременно являющейся ауксилярной, развива-
ются гонимобласты — карпоспорофиты, вращ ающие в ткань сердце-
вины таллома, где они могут сохраняться (по полевым наблюдени-
ям) дольше года до того, как освободятся карпоспоры.
При прорастании карпоспоры образуется корковидный таллом,
морфологически отличный от гаметангиальных талломов-гамето-
фитов в виде вертикальных разветвленных или неразвегвленных
пластинок многоосевого строения, растущих от базальных корочек.
Приблизительно через три недели после прорастания карпоспор на
корковидном талломе начинается формирование тетраспорангиев,
в которых развиваются крестообразно расположенные тетраспоры.
Тетраспоры прорастают как карпоспоры, образуя сначала корочки,
на которых, однако, возникают мпогоосевые талломы анфельции,
сходные с встречающимися в природе. Корочки, развившиеся из
тетраспор, никогда не образуют тетраспорангиев, а корочки из
карпоспор никогда не дают вертикальных талломов анфельции.
Таким образом, у Ahnfeltia concinna гетероморфная смена генера-
ций. У ряда представителей (Phyllophora pseudoeeranoides, Ph. crispa,
Ph. heredia, Stenogramma interrupta, Petroglossum pacificum, Gracilaria
verrucosa и др.) (в исследованиях, сочетающих морфологические,
культуральные и цитологические данные) был продемонстрирован
жизненный цикл Polysiphonia-типа.
У ряда видов из класса флоридсофициевых было описано аномальное сочетание
репродуктивных ортанов; среди этих сообщений многие касаются представителей
порядка гигартинальных. Например, у Cystodinium purpureum, в норме характеризу-
ющегося изоморфной сменой поколений и морфологически сходными гаметофитами
и тетраспорофитами, у некоторых растений на одном и том же талломе присут-
ствовали и тстраспорангии (по размеру и расположению нс отличающиеся от
таковых «нормальных» тстраспорофитов), и цистокарпии.
В ряде случаев у одного и того же вида обнаруживаются разные циклы развития.
Например, у Gigartina stellata наблюдается или половой цикл с Petrocellis cruenta
в качестве тстраспорофита, или же бесполый цикл, по-вт^1имому, с апогамным
развитием цистокарпиев.
Порядок родимениальные — Rhodymeniales
Порядок объединяет формы многоосевого строения. Ауксиляр-
ные клетки отчленяются от клетки, получившейся в результате
деления несущей (поддерживающей) клетки до оплодотворения кар-
86
погона. Ауксилярные клетки — добавочные, специальные клетки, не
принимающие участия в построении вегетативного таллома. Есть
прокарпии, в состав которых в одних случаях, кроме карпогона,
входит одна ауксилярная клетка, в других — две. У одних форм
тетраспорангии с тетраэдрическим расположением тетраспор,
у других — с крестообразным. Представителем служит род ломен-
тария (Lomentaria), имеющий трубчатый, полый с перетяжками или
без них таллом с боковым ветвлением.
До сравнительно недавнего времени (середина 70-х годов) к это-
му порядку относили широко распространенную по берегам Европы
водоросль родимению пальчатую (Rhodymema palmata). Таллом
многоосевого типа имеет вид листа, сидящего на суженном основа-
нии. В настоящее время эта водоросль под названием пальмария
пальчатая (Palmaria palmata) выделена в новый порядок пальмари-
альные (Palmariales) главным образом на основе строения тетраспо-
рангиев, с характерными клетками-ножками. Тетраспорангиальный
зачаток формируется из коровой клетки таллома, которая при этом
увеличивается, претерпевает митоз и неравновеликий цитокинез
с образованием крупного тетраспорангия и более мелкой клетки-
ножки. Ядро спорангия делится мейотически, вслед за чем содержи-
мое спорангия распадается на четыре крестообразно расположенные
гетраспоры. Тетраспоры содержат в цитоплазме везикулы, возмож-
но с клеющим материалом, способствующим прикреплению осво-
бодившихся тетраспор к субстрату. После освобождения тетраспор
клетка-ножка увеличивается и делится с образованием новой клет-
ки-ножки и нового тетраспорангия. Долгое время женский гамето-
фит был неизвестен, наблюдались лишь мужские индивиды, образу-
ющие мелкие сорусы антеридиев по поверхности таллома. Однако
в 1980 г. в лабораторной культуре Р. palmata, полученной из тетра-
спор этой водоросли, собранных вдоль берегов Канады, был обнару-
жен женский микроскопический гаметофит. Таким образом, мужс-
кие и женские гаметофиты Р. palmata морфологически различаются.
После оплодотворения тетраспорофит начинает развиваться на
женском гаметофите непосредственно из зиготы, из тетраспор обра-
зуются гаплоидные гаметофиты — мужские и женские (рис. 10, Д).
Порядок церамиальные—Ceramiales
Этот порядок, наиболее эволюционно продвинутый и богатый
видами, характеризуется одноосевым строением таллома, наличием
прокарпия и тем, что ауксилярные клетки дифференцируются толь-
ко после того, как произошло оплодотворение карпогона. Кар-
погонная ветвь всегда четырехклеточная, неразветвленная, несущая
клетка—перицентральная, т. е. всегда отчленяется от клетки цент-
ральной оси плодущей ветви. После оплодотворения ауксилярная
клетка отделяется от несущей (поддерживающей) карпогонную
ветвь клетки.
87
Рис. i 1. Polysiphonia. A — продольный разрез таллома: клетки центральной оси и пе-
рицентральные, образующие однослойную кору; f> — последовательные стадии
в формировании вторичных пор между перицентральными клетками разных сегмен-
тов; В— поперечный разрез ветви с многослойной корой; Г—Ж — плодущая часть
женского трихобласта; Г—молодой прокарпий; Д—образование ауксилярной клет-
ки и развитие стенки цистокарпия; Е- - слияние оплодотворенного карпогона и аук-
силярной клетки; Ж—зрелый цистокарпий; 3— начальная стадия развития мужского
88
Как было недавно показано для Callithamnion cordatum, необ-
ходимая предпосылка образования ауксилярной клетки после опло-
дотворения— контакт (не слияние) карпогона и клетки, несущей
карпогонную ветвь, от которой отчленяется ауксилярная клетка.
Без такого контакта ауксилярная клетка не образуется, что позволя-
ет предположить участие в ее формировании одного или нескольких
экзогенных гормонов. Образование ауксилярных клеток наблюда-
лось приблизительно через 40 ч, а диплоидизация их — через 72 ч
после соединения в культуре мужского и женского гаметофитов.
Брюшко оплодотворенного карпогона или непосредственно сли-
вается с рядом расположенной ауксилярной клеткой, или слияние
происходит через посредство коротких выростов.
В качестве примера лучше всего рассмотреть широко распрост-
раненный в южных и северных морях род полисифония
(Polysiphonia), который еще в начале XX в. был подробно изучен
цитологически и с точки зрения чередования морфологических фаз.
Как указывалось, жизненный цикл этой водоросли получил назва-
ние Polysiphonia-тип (см. рис. 10, А) и позднее был обнаружен
у многих представителей вышестоящих порядков флоридеофици-
евых. Талломы полисифонии представлены темно-малиновыми раз-
ветвленными кустиками, ветви которых увенчаны верхушечными
клетками. Верхушечная клетка посредством нескольких косых сте-
нок отчленяет клетки-сегменты. Эти клетки на стороне, где их
высота больше, отделяют обычно инициальную клетку боковой
ветви еще до деления сегмента на центральную и перицентральную
клетки. Инициальная клетка боковой ветви развивается или в моно-
сифоновые, состоящие из одного ряда клеток ветвящиеся трихо-
бласты, или же дает начало боковым ветвям, повторяющим стро-
ение главной оси. После этого клетка-сегмент претерпевает после-
довательные деления продольными перегородками, которые отчле-
няют периферические (перицентральные) клетки (в числе 8—12) от
центральной (рис. 11, А). Перицентральные клетки связаны с цент-
ральной посредством первичных пор, а между перицентральными
клетками соседних сегментов образуются вторичные поры. При
этом на морфологически нижнем конце перицентральной клетки
отшнуровывается маленькая клетка с одним ядром, естественно
соединенная порой с клеткой, от которой отделилась. Эта малень-
кая клеточка сливается с соответствующей перицентральной клет-
кой ниже расположенного сегмента. В результате две друг над
другом расположенные перицентральные клетки оказываются со-
единенными порой (рис. 11, Б). У некоторых видов перицентраль-
ные клетки делятся с образованием многослойной коры (рис. 11, В).
трихобласта; й—собрание спермациев; К—Л—образование тетраспорангия (К—
на продольном срезе, Л — на поперечном):
к —кариогой, к.в —карпогоннця ветвь, нк—несущая клетка, ок — клетки центральной оси, пк —
перицентральные клетки, тс—тетраспорапгий, ак — ауксилярные клетки, оц —оболочка цистока-
рпия. п — пора, я — ядро, т-— трихогиаа
89
На гаметофитах (мужском и женском) половые органы возника-
ют на трихобластах— веточках, имеющих моносифоновое стро-
ение, т. е. сложенных из одного ряда клеток и не окруженных
перицентральными клетками. Плодущая часть мужского трихо-
бласта становится полисифоновой в результате отчленения продоль-
ными перегородками перицентральных клеток. Они становятся ма-
теринскими клетками спермациев1 (рис. 11, И, К}. Спермации от-
почковываются последовательно: одновременно каждая материнс-
кая клетка спермация (например, у Polysiphonia hendryi) может нести
2—6 спермациев различного возраста. Спермации соединены с со-
ответствующими материнскими клетками посредством пор. Заново
отчленившийся спермаций содержит митохондрии; ядро располага-
ется в его базальной части — ближе к материнской клетке, дикти-
осомы и вакуоли вначале отсутствуют. По мере отпочковывания от
материнской клетки последующих спермациев содержимое ранее
отчленившегося спермация претерпевает изменения: ядро мигриру-
ет и занимает апикальное положение; в базальной части спермация
возникают цистерны шероховатого эндоплазматического ретикулу-
ма, которые сливаются с образованием двух «спермациальных»
вакуолей. Сначала эти вакуоли мелки и имеют гранулярное содер-
жимое, позднее превращающееся в фибриллярный материал (из
полисахаридов). После формирования вакуолей в спермации появ-
ляются диктиосомы, везикулы которых секретируют материал стен-
ки спермация. Образующаяся вокруг спермация слизистая стенка
содействует разрыву септальной пробки, что облегчает освобожде-
ние спермация. При этом спермациальные вакуоли (сильно увеличи-
вшиеся) выделяются путем экзоцитоза. Кроме того, по мере от-
членения от материнской клетки последующих спермациев и их
увеличения в объеме они оказывают давление на более старые
спермации сбоку и снизу, выталкивая их наружу. В результате этого
спермации энергично извергаются, а в пространство, остающееся на
месте вытолкнутого спермация, может врастать добавочный спер-
маций. Дифференцировка спермация, включающая развитие «спер-
мациальной» вакуоли (как деривата эндоплазматической сети) и ее
экзоцитоз, а также образование диктиосомами слизистой стенки
спермация, возможно, имеющей значение для прилипания его к три-
хогине, наблюдалась и у других церамиальных, например у Ptilota
plumosa.
Плодущий сегмент женского трихобласта формирует пять пери-
центральных клеток: две на нижней стороне, по одной с боков
(справа и слева) и, наконец, одну перицентральную клетку сверху —
она и является поддерживающей — несущей клеткой, на которой
развивается четырехклеточная каппогонная ветрь. Карпогон одно-
ядерный, трихогина безъядерна. После оплодотворения от несущей
' Именно спермациев, а не спсрматогониев, так как продуктом деления этих
клеток являются спермации, а не сперматангии (Р. Kurgens, 1980).
90
г ----------------------
клетки отчленяется ауксилярная клетка, диплоидное ядро оплодот-
воренного брюшка карпогона делится митотически и одно из дочер-
них ядер переходит в ауксилярную клетку при непосредственном
слиянии карпогона и ауксилярной клетки. Собственное гаплоидное
ядро ауксилярной клетки дегенерирует, а диплоидное делится и за-
тем от ауксилярной клетки наверх и кнаружи отчленяется первая
гонимобластная клетка, из которой развиваются гонимобластные
нити: их конечные клетки представляют собой карпоспорангии. Из
боковых перицентральных клеток плодущего и базального сегмен-
тов трихобласта еще до оплодотворения развиваются нити, кото-
рые образую! вокруг прокарпия и развивающихся карпоспор (ци-
стокарпия) псевдопаренхиматозную оболочку с отверстием на вер-
шине (рис. И, С—3). Через него по созревании выпадают карпо-
споры. Диплоидные карпоспоры прорастают в диплоидные тетра-
спорофиты, морфологически сходные с гаметофитами, но несущие
органы бесполого размножения — тетраспорангии (рис. 11,Д М).
Тетраспорангии возникают на ветвях полисифонового строения
таким образом, что плодущая перицентральная клетка сначала
отчленяет налево и направо две покровные клетки, а затем делится
косо на нижнюю более мелкую и верхнюю более крупную клетку,
которая и становится спорангием. После мейоза в спорангии фор-
мируются четыре тетраэдрически расположенные тетраспоры. Тет-
распоры прорастают в гаплоидные гаметофиты, несущие половые
органы.
Среди церамиальных, как и у гш аргинильных, в последние годы описаны случаи
аномального сочетания репродуктивных органов. Так, у Polysiphonia harlandii 14%
образцов, собранных в мае 1984 г. в литоральной зоне Гонконга, несли одновремен-
но и тстраспоры, и цистокарпии.
Одновременное развитие цистокарпиев и тстраспорангиев на некоторых тал-
ломах отмечено и для Р. тасгосагра. Ранее у другой церамиальной водоросли
Antithamnion tenuissimum были обнаружены некоторые мужские растения, несущие
тетраспорангии. При спаривании этих мужских растений с нормальными женскими
получены карпоспоры, развившиеся в нормальные тетраспорофиты. Последние,
в свою очередь, дали начало нормальным мужским и женским растениям и некото-
рым мужским растениям, несущим тетраспоры.
Заслуживает внимания род гриффитзия, виды которого благо-
даря простоте строения таллома и очень крупным клеткам стали
объектами интересных экспериментов. Некоторые из них приводят-
ся ниже.
Род гриффитзия (Griffithsia) отличается очень крупными, не оде-
тыми корой многоядерными клетками, легко различимыми нево-
оруженным глазом. Таллом вертикальный, разветвленный, прикре-
пленный к субстрату ризоидами. Весь таллом заключен в обильную
слизь. Короткие веточки, несущие репродуктивные органы, возника-
ют от верхушек более молодых клеток, то же положение занимают
обильно разветвленные волоски (рис. 12, Л), Спорангии развивают-
ся на специальных боковых веточках, которые образуют мутовки
у верхушек сегментов. Каждая фертильная боковая ветвь состоит из
91
Рис. 12. Griffiihsia. А-. 1 — несколько сегментов G. corallina с плодущими побегами
(пн) и волосками (в); 2 — 3—G. pacifica: 2—одна плодущая ветвь с тетраспоранги-
ями и тетраспорами (тс), J - женская плодущая ветвь из трех клеток с двумя
прокарпиями (кв — карпогонная ветвь, т — трихогина, нк — несущая клетка); 4--
кончик мужского растения с аптсридиальными сорусами (ас); 5 — одна из антериди-
альных ветвей, образующих сорус (а антеридии). Б—развитие растения G. pacifica
нз одной изолированной клетки. В — схема опыта по слиянию соматических и полу-
чению гибридных клеток G. tenuis. Интеркалярные фрагменты' мужских и женских
растений гриффитзии помещались в пустые оболочки клеток междоузлий нителлы
таким образом, чтобы основание одного фрагмента было обращено к верхушке
другого фрагмента (/). Спустя 4 — 8 ч верхний фрагмент продуцировал у своего
основания ризоид, а нижний фрагмент отчленял от своей верхушки специальную
восстанавливающую— репарационную клетку (2). Еще через 12— 16 ч ризоид от
верхнего фрагмента сливался с репарационной клеткой от нижнего фрагмента с об-
разованием одной гибридной клетки (3)
/
одной клетки, несущей несколько спорангиев, которые защищены
выростами, возникающими от той же клетки или от родительского
сегмента (рис. 12, А). Женские плодущие веточки состоят из не-
92
скольких (трех — у G. corallina, пяти—у G. setaceae) многоядерных
клеток и берут начало от верхушки крупных осевых клеток. Из
самой нижней клетки веточки позднее возникает обвертка цистокар-
пия, а из субтерминальной клетки—прокарпий. Она отчленяет три
(у G. g/obifera—лишь две) перицентральные клетки: те из них, кото-
рые отчленяются направо и налево, развиваются в двухклеточные
веточки; их базальная клетка несет карпогонную ветвь. Осуществ-
ляется непосредственное слияние ауксилярной клетки и карпогона:
специальная соединительная клетка не образуется. Ауксилярная
клетка продуцирует несколько зачатков гонимобластов, которые
последовательно развиваются в обильно разветвленные гонимоб-
ласты; большинство их клеток формирует одноядерные карпо-
споры. Наблюдается заметное увеличение пор и слияние между
клетками плодущей веточки, ауксилярной клетки с несущей и др.
Несколько одевающих цистокарпий нитей имеют только две клет-
ки. Антеридиальные сорусы состоят из плотно разветвленных вето-
чек, сформированных главным образом одноядерными клетками.
Они образуют «шапочки» на терминальных сегментах таллома или
окружают перетяжки между более молодыми клетками (рис. 12, Л).
Хотя у G. pacifica вертикальные талломы и ризоидальные нити на-
растают только за счет деления их апикальных клеток, любая изо-
лированная интеркалярная клетка таллома способна регенерировать
весь таллом. Пусковым механизмом этого процесса служит удаление
примыкающих клеток. Так, в опытах по регенерации для изолирова-
ния отдельной клетки соседние с ней клетки отрезались. Такая живая
клетка помещалась на питательную среду и при определенных внеш-
них условиях (температура, фотопериод) уже в течение первых суток
отчленяла от своей верхушки клетку будущего вертикального побега,
а у своего основания—ризоидальную клетку. Дальнейшее удлинение
побега осуществляется за счет деления апикальной клетки, когда нить
достигает длины 4—5 клеток, возникают первые боковые ветви. На
5-й день изолированная из побега клетка уже образует растение,
различимое невооруженным глазом (рис. 12, Б).
Если в нити G. pacifica отмирает интеркалярная клетка, то выше
ее расположенная клетка делится с образованием ризоидальной
клетки, а клетка, находящаяся ниже погибшей клетки, отчленяет
специализированную репарационную клетку. Последняя растет на-
встречу ризоидальной клетке и сливается с ней, восстанавливая
таким образом целостность нити.
Недавно у этого же вида удалось установить локализацию и свойст-
ва гормона, названного родоморфином, который индуцирует деление
клеток и способствует репарации (S. D. Waaland, А. В. Watson, 1980).
У другого вида — G. tenuis С. Д. Вааланд (S. D. Waaland, 1978)
получила соматическое слияние между вегетативными клетками
в разных сочетаниях: между клетками одного пола (женская —
женская, мужская—мужская) и клетками разного пола (женская —
мужская). В эксперименте использовалась следующая методика:
93
длинные клетки междоузлия нителлы (харовой водоросли) разреза-
лись лезвием бритвы на сегменты, из которых стеклянной палочкой
удалялось их цитоплазматическое содержимое. В получившиеся
цилиндры вставлялись изолированные двуклеточные интеркаляр-
ные участки растений G. tenuis. Нити в трубочках располагались
так, чтобы основание одной контактировало с верхушкой другой
нити. Спустя 4—8 ч верхняя нить продуцировала у основания ризо-
идальную клетку, а нижний фрагмент отчленял у своей верхушки
специальную добавочную клетку. Еще через 12—16 ч ризоид верх-
ней нити и добавочная клетка нижнего фрагмента сливались с обра-
зованием одной гибридной клетки (рис. 12, Б). Слившиеся клетки
содержали более 100 ядер. Эти гибридные клетки изолировались,
и из них в результате регенерации получали новые гибридные
растения, которые почти все давали репродуктивные структуры.
Характер этих структур определялся характером сливающихся кле-
ток. Гибридные клетки, возникшие при слиянии женских клеток
с женскими (вегетативных клеток женских изолятов), мужских
с мужскими (клеток мужских изолятов) при регенерации формиро-
вали нити, которые продуцировали только половые структуры —
женские в первом случае, мужские — в последнем. Если гибридные
клетки получали при слиянии женских и мужских клеток (вегетатив-
ных клеток из женских и мужских изолятов), то приблизительно
50% регенерирующих нитей давали репродуктивные структуры,
характерные для тетраспорангиальной (диплоидной) фазы водорос-
ли. При дальнейшем удлинении нити, образовавшей тетраспоранги-
альные ветви, добавляющиеся к ней сверху за счет деления апикаль-
ной клетки новые клетки начинают продуцировать половые струк-
туры. Пол последних коррелирует с полом клетки, из которой
возник апикальный конец гибридной клетки. Например, если верх-
няя нить при получении гибридной клетки была женской, а нижний
фрагмент — мужским, апикальный конец гибридной клетки будет
образован женской клеткой, а базальный—мужской. Когда нить,
полученная в результате регенерации такой клетки, начнет давать
ветви с половыми органами, они скорее всего будут женскими.
Каждая нить продуцирует только один тип половых ветвей (или
женские, или мужские).
Клетки, из которых возникали тетраспорангиальные ветви, были
вновь изолированы и при репарации давали нити, в одних случаях
образующие тетраспорангиальные ветви, в других — половые стру-
ктуры, а третьих — вообще не формирующие репродуктивных стру-
ктур. Гибридных нитей, которые продолжали бы непрерывно да-
вать тетраспорангиальные ветви, получено не было.
Тот факт, что соматические гибриды между гаплоидными клет-
ками одного и того же пола никогда не образуют репродуктивные
структуры, характерные для диплоидных растений (тетраспоранги-
альные веточки) и что последние возникают при слиянии гаплоид-
ных соматических клеток разного пола, служит указанием па то, что
94
в гибридных клетках может быть слияние ядер или же они суще-
ствуют в виде гетерокарионов. Более вероятно последнее пред-
положение, так как гибридные растения после формирования
репродуктивных структур, характерных для диплоидной фазы,
в конце концов возвращались к образованию половых структур.
Даже клетки, которые сами непосредственно продуцировали тетра-
спорангиальные веточки, будучи изолированными, впоследствии
регенерировали растения, рано или поздно дававшие половые
структуры.
Филогенез красных водорослей. Красные водоросли представля-
ют собой естественную единую, несмотря на их многообразие,
весьма древнюю группу: остатки их известны из силура и девона. Из
всех эукариотических отделов водорослей по набору пигментов,
одиночному расположению тилакоидов, отсутствию жгутиковых
стадий красные ближе всего к синезеленым, от которых они, одна-
ко, резко отличаются строением клетки и наличием полового про-
цесса.
Что касается эволюции в пределах самого отдела красных водо-
рослей, то из двух классов — бангиофициевых и флоридеофицие-
вых— первый, несомненно, более примитивный. Здесь у карпогона
еще не выработалась типичная форма и он мало отличается от
обычных вегетативных клеток. После оплодотворения содержимое
карпогона непосредственно делится на карпоспоры. Флоридеофици-
евые представляют собой более эволюционировавшую группу, у ко-
торой карпогон снабжен органом улавливания спермациев — трихо-
гиной. Из порядков класса флоридеофициевых простейшими счита-
ются немал и опальные, гелидиальные, у которых пет ауксилярных
клеток и гонимобласты с карпоспорангиями образуются непосред-
ственно из брюшной части оплодотворенного карпогона. Следу-
ющий шаг прогрессивной эволюции — криптонемиальные: они име-
ют ауксилярные клетки, способствующие увеличению продукции
карпоспор, так как образуется не один цистокарпий на месте опло-
дотворенного карпогона, а много — по числу этих клеток. При
беспорядочно разбросанных по таллому ауксилярных клетках
необходимы более или менее длинные ообластемные нити. Еще
более прогрессивны те порядки (гигартинальные, родимениальные,
церамиальные), у которых имеются прокарпии. Непосредственное
соседство карпогона и ауксилярной клетки (клеток) в прокарпии
облегчает образование цистокарпиев. Наивысшей ступени эволю-
ции достигли церамиальные, у которых есть прокарпий и ауксиляр-
ные клетки дифференцируются только после того, как произошло
оплодотворение. Этот порядок наиболее богат видами.
Красные водоросли находят довольно большое применение в хозяйстве: из них
вываривают студенистое вещество агар-агар, применяемое в кондитерской и ряде
других отраслей промышленности, а также в лабораторной микробиологической
технике для приготовления твердых питательных сред. Некоторые, например порфи-
ру, употребляют в пищу.
95
Глава 7
Рис. 13. Пространстаен-
ная реконструкция спи-
рального тела и аксонемы
в проксимальной части
стержня жгута Uroglena
americana:
д—периферические дубле-
ты, пм—центральные ми-
кротрубочки, ст—спираль-
ное тело
ОТДЕЛ ЗОЛОТИСТЫЕ ВОДОРОСЛИ — CHR YSOPHYTA |
Для водорослей этого отдела характерны хроматофоры, окра- Я
шенные в золотистый цвет благодаря тому, что, помимо хлорофил- Я
лов а и с, имеется избыток каротиноидов — 3-каротина и ксан- Я
тофиллов (фукоксантин, антераксантин, лютеин, зеаксантин, не- я
оксантин, виалоксантин). Для ультраструктуры хлоропластов Я
характерны сложная оболочка, трехтилакоидные ламеллы, ло- Я
кализованный генофор; опоясывающая ламелла у одних имеется, Я
у других отсутствует. Запасные продукты — хризоламинарин (лей- ’
Козин), откладывающийся вне хлоропласта (в цитоплазме) и жир.
Глазок, заключенный в пластиду, связан со жгутиковым аппаратом. ;
У одних форм жгутики гетероконтные и гетероморфные, у других —
изоконтные, изоморфные; у многих одножгутиковых (в оптическом -
микроскопе) форм электронная микроскопия выявила второй руди-
ментарный жгутик. Для переходной зоны жгутика характерно спи- i
ральное тело, состоящее из 4—5 витков электронно-плотного мате- ;
риала, расположенных между периферическими дублетами и цент-
ральными микротрубочками (рис. 13). Золотистые водоросли, сто-
ящие на разных ступенях морфологической дифференцировки
талломов, помимо указанных выше признаков, характеризуются
особыми эндогенными или эндоплазматическими окремнелыми ци-
стами или статоспорами, образующимися
летом, реже поздней осенью, и служащие
для переживания неблагоприятных условий
существования. Своеобразие процесса их
формирования заключается в том, что обо-
лочка развивается не вокруг протопласта |
(как у других водорослей), а закладывается |
внутри него. Протопласт пространственно 1
разрезается оболочкой на интрацистерную 1
внутреннюю и экстрацистерную наружную J
части. Экстрацистерная плазма в большей
своей части переходит в эндогенно заложив- 'Я
шуюся спору, после чего спора закупорива- Я
ется пробочкой или крышечкой, а оставщая- Я
ся экстрацистерная плазма разрушается. Во Я
многих случаях края поры вытягиваются Я
в виде трубки или расширяются в виде во- Я
ронки. В то время как оболочка цисты (ста- Я
тоспоры) обычно сильно окремпевшая, 9
окремнение пробочки значительно меньше, 1
а иногда и отсутствует. При прорастании 1
пробочки растворяются и поделившийся 1
96
или неподелившийся протопласт споры выходит большей частью
в виде монады, снабженной жгутиками, реже в виде безжгутиковой
амебы.
Впервые этот процесс эндогенного образования статоспор был
описан русским исследователем Л. С. Ценковским в 1870 г. у рода
хромулина. Позднее А. Пашер (A. Pascher, 1932) показал, что этот
своеобразный способ образования спор свойствен всем золотистым
водорослям и встречается не только у монадных и ризоподиальных
форм, но и у тех представителей, протопласты которых одеты
оболочкой (феотамнион, феодермациум, нематохризис).
В последние годы процесс образования цист (статоспор) у хризомонад, различа-
ющихся организацией таллома, был изучен на электронно-микроскопическом уров-
не. Д. Хибберд (D. J. Hibberd, 1977) исследовал образование цист у голой формы
Ochromonas tuberculaia, К. Сэндгрен (С. D. Sandgren, 1977, 1980, 1983) у двух видов
Dinobryon (£). cylindrieum, D. divergens) (их клетки заключены в целлюлозные доми-
ки), у двух колониальных форм — Uroglena volvox, U.americana и у Mallomonas
caudata — одноклеточной хризомонады, клетка которой имеет покров из окремнелых
чешуек.
Образование цист происходит в основном сходно у всех изученных до сих пор
форм, однако имеются и особенности, которые можно использовать в систематике.
Во всех случаях кремнеземная оболочка цист (статоспор), как и створки диато-
мей, образуется внутри пузыря, мембрана которого силиколемма — возникает за
счет слияния пузырьков Гольджи. Пузырь (везикул) в виде сферы, со стенкой,
образованной двумя мембранами, в пространстве между которыми и осаждается
кремнезем, закладывается внутри цитоплазмы, деля ее на интрацистерную (с глав-
ными органеллами клетки) и экстрацистерную (содержащую обычно сократитель-
ную вакуоль, иногда митохондрии, пластиду).
Когда оболочка цисты сформировалась, внутренняя мембрана пузыря (внутри
которого отложилась кремнеземная оболочка цисты) становится функционирующей
плазмалеммой цисты; растягиваясь, она перегораживает пору. Наружная мембрана
пузыря утрачивается.
Цисты всех хризомонад имеют пору, в зрелом состоянии замкнутую пробкой.
Пробка при всем разнообразии структуры у разных видов всегда образуется из
материала, освобождающегося из пузырьков Гольджи, которые сливаются с мемб-
раной цисты под порой.
Процесс созревания цист у всех изученных видов сопровождается возрастанием
электронной плотности всего содержимого цисты, накоплением липидных гранул.
Крупная вакуоль с хризоламинарином приобретает крапчатый («мраморный») вид.
Наряду с перечисленными признаками процесса инцистирования, присущими всем
хризомонадам, существуют особенности, различающиеся у разных видов. Некото-
рые из них, по мнению Сэндгрена, имеют даже филогенетическое значение. Эти
особенности касаются: 1) способа развития поры в пузыре, откладывающем кремне-
зем; 2) отложения кремнезема в пузыре в один или в два этапа в зависимости от того,
являются ли оболочки цист гладкими или орнаментированными; 3) судьбы экстраци-
стерной цитоплазмы и присутствия или отсутствия цитоплазматических микротрубо-
чек, проходящих через пору в экстрацистерную цитоплазму; 4) структуры пробки;
5) числа ядер и пластид в цисте сравнительно с таковым в вегетативных клетках.
1. Пора может возникать двумя путями: или посредством разрыва мембран
полностью замкнутого шаровидного пузыря, в котором остается нсокремнелый
участок, или же еще во время развития пузыря пора оказывается детерминированной
и ее границы очерчены кольцом электронно-плотного' материала (последний отсут-
ствует у форм, где пора появляется в результате разрыва цельного пузыря). Первый
способ образования поры можно наблюдать у Ochromonas tuberculaia (возможно,
и у Mallomonas caudata, рис. 14), второй — у Dinobryon ID. cylindrieum, D. divergens)
и Uroglena (U. volvox, U. americana, рис. 15).
4 - 1210
97
Рис. 14. Цисты Mallomonas caudata. А - -срез через зрелую цисту (несколько схемати-
зировано). Оболочка цисты, формирующаяся в два этапа, образована основной
пластинкой из кремнезема, от которой выступает неправильный ряд сетчатых элемен-
тов. Циста содержит одно клеточное ядро, одну пластиду, диктиосому; под областью
поры (нс попавшей на срез) находится массивная пробка, окруженная вакуолями,
которые содержат материал такой же, как и материал, образующий пробку. Вокруг
крупной вакуоли с хричоламинарином, имеющей «мраморный» вид, липидные ка-
пельки. Экстрацистерная цитоплазма, заключенная внутри слоя чешуек вегетативной
клетки и позднее сбрасываемая, содержит многочисленные митохондрии, лопасть
хлоропласта, исторгнутую из клетки во время икцистирования, несколько сократи-
тельных вакуолей. Б, В, Г—схематическое изображение последовательных стадий
образования поры цисты путем разрыва мембран сначала полностью замкнутого
пузыря отложения кремнезема:
оц — кремнеземная оболочка цисты, вк — везикул с кремнеземом, эпч — элекгроиио-плотные
частицы у внутренней поверхности везикула с кремнеземом, ц -пробка, по — пора, эц -экстра-
цистерная цитоплазма, я— ядро, х— хлоропласт, м — митохондрии, д — дикзиосома, вхл--ва-
хуоль с хризоламинарином. св -сократительная вакуоль, ч— чешуйки
2. У видов с гладкой неорнаментированной оболочкой цисты (например, у Uro-
glena americana) отмечен лишь один этап в отложении кремнезема внутри пузыря.
У форм с орнаментированными цистами (со скульпгурироваиными оболочками и рас-
ширенными воротничками у краев пор) после первого этапа отложения внутри пузы-
ря тонкой гладкой окремнелой оболочки наблюдается вторая фаза, когда к основной
пластинке из кремнезема снаружи достраиваются шипы или другая орнаментация,
свойственная данному виду. В участках развития орнаментаций пузырь расширяется.
Таким образом, орнаментация оболочек цист, по-йидимому, возникает не за счет
экстрацистерной цитоплазмы, как полагал Пашср, а вгнутри пузыря на втором этапе
98
Рис. 15. Цисты Dinobryon divergens. А -срез через зрелую цисту (несколько схемати-
зировано). Оболочка цисты с детерминированной норой, границы которой очерчены
кольцом элскгронно-плотного материала, содержит два ядра, два хлоропласта (ре-
пликация ядра и хлоропласта имеет место до инцистирования), диктиосому, мито-
хондрии, вакуоль с хризоламинарином, сократительную вакуоль. Эксграцистсрная
цитоплазма с включениями (сократительная вакуоль и пр.) втягивается внутрь
заложившейся оболочки цисты в этом процессе, возможно, принимают участие
микротрубочки, выступающие через пору в экстрацистерную цитоплазму. Б, В. Г—
схема последовательных стадий образования детерминированной поры с элект-
ронно-плотными краями и цитоскелстными микротрубочками, выступающими через
пору в экстрацистерную цитоплазму, которая в конце концов втягивается вместе
с сократительной вакуолью в цисту.
оц—оболочка цисты, я ядро, х—хлоропласт м — митохондрии, д диктиосома, св —сокра-
тительная вакуоль, вхл вакуоль с хризоламинарином, ж —жир, вк—везикул с кремнеземом,
кэцм — кольцо электронно-плотного материала, п -пробка, по — пора, мт—микротрубочки
окремнения. Так, у Dinobryon cylindrieum и D. divergens экстрацистерная цитоплазма,
содержащая сократительную вакуоль, втягивается внутрь оболочки цисты до заку-
порки поры пробкой, и только после этого наступает второй этап окремнения,
ответственный за образование шипов на оболочке цист у этих видов. У Mallomonas
caudata кнаружи от стенки цисты остается большое количество цитоплазмы, заклю-
ченной в слой чешуек вегетативной клетки. Эта экстрацистерная цитоплазма с мно-
гочисленными митохондриями, пластидой, сократительными вакуолями не втягива-
ется внутрь оболочки цисты до образования пробки, замыкающей пору. Она сбра-
сывается, когда циста созреет. Оболочка зрелой цисты Mallomonas состоит из
базальной пластинки кремнезема, от которой выступает неправильный ряд сетчатых
элементов, возникающих во время второго этапа окремнения. Вдоль внутренней
поверхности наружной мембраны пузыря, откладывающего кремнезем, можно ви-
деть плотные гранулы (рис. 14).
3. У изученных видов Dinobryon и Uroglena экстрацистерная цитоплазма, содер-
жащая пульсирующую вакуоль, втягивается вну1рь эндоплазматически заложившей-
99
ся оболочки цисты через пору до образования пробки и таким образом сохраняется.
В процессе втягивания экстрацистерной цитоплазмы внутрь оболочки цисты, воз-
можно, играют роль цитоскелетные микротрубочки, выступающие через пору в экс-
трацистсрную цитоплазму (рис. 15). Напротив, у Ochromonas tuberculata и Mallonionas
caudata экстрацистерная цитоплазма не втягивается внутрь цисты и таким образом
не сохраняется: при созревании цисты она сбрасывается.
4. Структура пробки, всегда образующейся в результате активности аппарата
Гольджи, у разных видов неодинакова. Пробка может быть составлена из разного
количества слоев (от одного до трех) морфологически различающихся материалов,
Каждый слой заключает несколько компонентов. Так, пробка у Uroglena
(О. americana, U. volvox) и, возможно, у Mallomonas caudata содержит только первич-
ный слой. У Ochromonas tuberculata имеется периферический вторичный слой, состоя-
щий из фиброзного материала, заключенного внутри везикул. У Dfnobryon
cylindricum, D. divergens пробка цисты также включает второй периферический слой,
но состоящий из гранулярного материала. У D. cylindrieum имеется еще и третий слой
пробки фиброзный. В химическом отношении пробка представляет собой полиса-
харид и не содержит вообще или содержит лишь небольшие количества кремнезема.
5. Цисты Ochromonas tuberculata и Mallomonas caudata заключают по одному
ядру и одной пластиде, как и вегетативные клетки. Цисты Dinobryon и Uroglena
в отличие от вегетативных клеток содержат по две пластиды и по два ядра.
Репликация этих органелл у изученных Сэндреном видов предшествует образованию
цист, которые таким образом возникают бесполым путем. Это не исключает воз-
можности появления цист у других видов в результате полового процесса. Такие
случаи на уровне оптического микроскопа неоднократно описывались.
При прорастании цисты у Ochromonas tuberculata пробка исчезает и содержимое
выходит через пору.
Отдел золотистые водоросли (Chrysophyta) включает невколько
классов, из которых ниже приводятся два: класс хризофициевые,
или гетерохризофициевые (Chrysophyceae, Heterochrysophyceae),
и сравнительно недавно установленный класс гаптофициевые, или
примнезиофициевые (Haptophyceae, Prymnesiophyceae), для золотис-
тых водорослей с гаптонемой.
КЛАСС ХРИЗОФИЦИЕВЫЕ —CHRYSOPHYCEAE
Класс объединяет водоросли с различными типами организации
таллома, которые и кладутся в основу деления его на порядки:
хризомонадальные (Chrysomonadales), объединяющие мопадные
формы, ризохризидальные (Rhizochrysidales), представленные золо-
тистыми водорослями с ризоподиальной структурой, хризокапсаль-
ные (Chrysocapsales), включающие пальмеллоидные формы, хризо-
сферальные (Chrysosphaerales) —коккоидные, феотамниальные
(Phaeothamniales)—нитчатые. Ниже приводятся только два поряд-
ка: хризомонадальные и хризокапсальные.
Порядок хризомонадальные нли хромулинальные—
Chrysomonadales, Chromulinales
Это самый обширный порядок, включающий водоросли монад-
ной структуры, одноклеточные и колониальные*
Систематика хризомонад базируется на числе и строении жгути-
ков и особенно на характере клеточных покровов. Имеются одно-
100
и двужгутиковые формы, и прежде считалось, что первые являются
исходными, более примитивными. Однако с помощью электронно*
го микроскопа обнаружено, что у кажущихся (в оптическом микро-
скопе) одножгутиковых форм имеется второй маленький боковой
жгутик. Было высказано предположение, что двужгутиковые хризо-
монады с гетероконтными и гетероморфными жгутиками могли
быть исходными, а одножгутиковые формы возникли в результате
редукции более короткого жгута. Клеточные покровы хризомонад
различны: имеются формы голые, одетые лить плазмалеммой,
у других клетки заключены в целлюлозные домики, у третьих
поверх плазмалеммы находится покров из окремнелых чешуек.
Размножение осуществляется делением клеток, у некоторых (ди-
нобрион, малломонас) описан половой процесс.
Хризомоиады— преимущественно пресноводные формы, чаще
всего приуроченные к чистым водам. Обычно они встречаются
в холодное время года, ранней весной и поздней осенью, некото-
рые— в зимнее время подо льдом. Однако, как было показано для
синуры, температура воды здесь имеет лишь косвенное значение.
Решающим же фактором является химизм воды, меняющийся на
протяжении года: в холодное время года из-за отсутствия другой
растительности в воде содержится больше железа и азота. Боль-
шинство этих водорослей живут в планктоне, имеют специальные
приспособления к планктонному образу жизни и некоторые (синура,
малломонас, динобрион) вызывают «цветение» воды, окрашивая ее
в бурый цвет.
Ниже будут рассмотрены представители трех семейств: охромо-
надовые, или хромулиновые (Ochromonadaceae), динобриевые
(Dinobryaceae) и синуровые (Synuraceae).
Семейство Охромонадовые — Ochromonadaceae. Сюда
относятся голые формы, клетки которых покрыты только цито-
плазматической мембраной с одним или двумя неодинаковыми
жгутиками (при наблюдении в оптическом микроскопе).
Род охромонас (Ochromcmas), виды которого обычны в планк-
тоне и нейстоне пресных вод, реже солоноватоводных водоемов,
представлен одиночными, окрашенными в золотистый цвет клет-
ками с двумя гетероконтными и гетероморфными жгутиками. Клет-
ка охромонас голая, снаружи одетая только цитоплазматической
мембраной. Каплевидная форма ее поддерживается цитоскелетом
из периферически расположенных микротрубочек. В центре клетки
находится клеточное ядро, окруженное ядерпой оболочкой из двух
мембран. В расширении между мембранами ядерной оболочки (в
перинуклеарном пространстве) заключены два пластинчатых хро-
матофора с типичной для всего отдела ультраструктурой (имеется
опоясывающая ламелла). В задней части клетки находится крупная
вакуоль с хризоламинарином, в цитоплазме рассеяны митохондрии,
в передней части клетки — аппарат Гольджи (рис. 16, А). От перед-
него конца клетки отходят два неодинаковых по длине и форме
101
Рис. 16. Ochromonas danica. A — клетка с двумя гетероконтными и гетероморфными
жгутами; Б—часть длинного жгута с мастигонемами — трубчатыми и фиброзными.
Трубчатые мастигонемы располагаются на жгутике в два неодинаковых ряда: с од-
- »
102
жгутика. Г. Бак (G. Bouck, 1971) проследил тонкое строение и про-
исхождение мастигонем у Ochromonas danica. Эта водоросль удобна
для изучения динамики развития мастигонем, так как ее клетки
легко теряют жгутики, а затем образуют их заново. Таким образом
можно изучить материал на разных стадиях регенерации жгутико-
вого аппарата. Оказалось, что сами мастигонемы у охромонас
сложнее, чем у фукуса и аскофиллума, и в их сборке и выведении на
поверхность клетки участвует не только перинуклеарное простран-
ство, но и элементы аппарата Гольджи. Готовые трубчатые масти-
гонемы охромонас состоят из базальной части, прикрепляющейся
к мембране жгута, микротрубчатой части и 1 —3 терминальных
нитей. Кроме этих компонентов, сходных с таковыми мастигонем
бурых водорослей (фукус, аскофиллум, эктокарпус), трубчатый
стержень мастигонемы по всей поверхности несет длинные и корот-
кие латеральные нити. Трубчатые мастигонемы располагаются на
длинном жгутике в два неодинаковых ряда: с одной стороны масти-
гонемы прикрепляются к специальным участкам жгутиковой мемб-
раны поодиночке, а на противоположной стороне жгута — пучками
по три — пять (рис. 16, Б). Помимо трубчатых мастигонем, длин-
ный жгут несет тонкие простые нитевидные фиброзные мастигоне-
мы. Короткий жгутик имеет только фиброзные мастигонемы
(рис. 16, А). На рис. 16, В представлена схема возможной последо-
вательности сборки мастигонем и их освобождения при регенерации
жгутика у охромонас. По мере удлинения жгута новые мастигоне-
мы и, возможно, мембрана, к которой они прикрепляются, добав-
ляются к основанию жгута.
Такое же образование промастигонем (в расширениях перинук-
леарного пространства) и выделение готовых мастигонем пооди-
ночке или в группах, наружу к основанию жгутика в цистернах
аппарата Гольджи наблюдалось и у Ochromonas minuta. Жгутико-
вый корень (ризопласт) у охромонаса состоит из поперечно исчер-
ченных пучков фибрилл, которые, разветвляясь, достигают поверх-
ности клеточного ядра.
Перед митозом базальные тела двух жгутиков реплицируются,
хлоропласт делится перетяжкой, во время профазы реплицируются
ной стороны они прикрепляются к специальным участкам жгутиковой мембраны по
одиночке, а на противоположной стороне жгута пучками по 3*—5. Стержень
трубчатой мастигонемы несет на конце 1 — 3 терминальные нити, а по всей поверх-
ности боковые (латеральные) нити двух родов: длинные и короткие; В—схема
стадий сборки мастигонем и их освобождения:
1 - предположительные мастигонемы образуются внутри перинуклеарного простра-
нства, 2 — предположительные мастигонемы в цистернах аппарата Гольджи, где
к ним присоединяются боковые нити, 3 --из цистерн аппарата Гольджи с помощью
пузырьков мастигонемы переносятся к поверхности клетки, 4— выделение мастиго-
нем на поверхность клетки, 5 прикрепление их к основанию жгута:
я -ядро, х — хлоропласт, ни — перинуклеарное пространство, д - диктиосома, м — митохонд-
рию вхл— вакуоль с хризоламинарином, дж — длинный жгутик, кж—короткий жгутик, гм—
1рубчатые мастигонемы, фм — фиброзные мастигонемы, тн — терминальные нити, клн—корот-
кие латеральные нити, длн—длинные латеральные нити
103
диктиосомы, появляется второй ризопласт. Вокруг ядра увеличи-
вается число микротрубочек, ядерная оболочка начинает разру-
шаться у полюсов, что позволяет микротрубочкам войти внутрь
ядра и образовать веретено. С каждым полюсом ядра ассоциирует-
ся по одному ризопласту, к которому, по-видимому, прикреплены
трубочки веретена. Одни трубочки веретена простираются от полю-
са к полюсу, другие присоединяются к хромосомам (кинетохоры
отсутствуют). Остатки ядерной оболочки разрушаются, за исключе-
нием участков, соседних с хлоропластами. Хлоропластная эндо-
плазматическая сеть остается интактной на протяжении всего мито-
за. В анафазе микротрубочки удлиняются и хромосомы разъединя-
ются на две группы. В телофазе появляются ядрышки и ядерные
оболочки. Ци гокинез происходит путем продольного раскола, ко-
торый начинается у переднего конца клетки между двумя парами
жгутиковых оснований (рис. 17). Об образовании цист у Ochromonas
сказано выше.
Род хромулина (Chromulina), как и охромонас, представлен оди-
ночными голыми клетками, но с одним (в оптическом микроскопе)
жгутиком.
Род уроглена (Uroglena). Виды этого рода (U. voivox,
U. атегкапа) обычны в пресноводном планктоне озер и прудов,
представлены шаровидными или эллипсоидальными слизистыми
колониями, в которых протопласты находятся на концах слизистых,
дихотомически разветвленных 1яжей.
Семейство динобриевые (Dinobryaceae) объединяет сво-
бодно живущие или прикрепленные, одиночные или колониальные
формы, клетки которых заключены в домик— мертвое, выделяемое
протопластом образование. Типичный представитель — род диноб-
рион (Dinobryon), виды которого широко распространены в пресно-
водном планктоне и при массовом развитии нередко вызывают
цветение воды в озерах и прудах. Голая, одетая цитоплазматичес-
Рис. 17. Схема митоза у Chrysophyceae (Ochronionaa danica).
А — профаза, Б—метафаза, В—анафаза. Г—телофаза и цитокинез:
я — ядро, р — ризопласт, мт — микротрубочки, д— диктиосомы, п — пластида, эс—эндоплаз-
матическая сеть, дя—дочерние ядра
104
кой мембраной клетка заключена в бокаловидном домике, образо-
ванном целлюлозными микрофибриллами с петлеобразным рас-
положением; базальный конус домика состоит из продольно ориен-
тированных микрофибрилл. В состав домика входит и большое
количество аминокислот. Протопласт прикреплен к одной стороне
основания домика базальной ножкой, способной к удлинению, со-
кращению или закручиванию, что позволяет клетке двигаться внут-
ри домика: в этой части клетки находятся многочисленные микро-
трубочки. Клетка овальных очертаний содержит одно ядро, рас-
положенное между хлоропластами. Два хроматофора различаются
по длине, меньший располагается сзади. В хлоропласте можно
видеть тилакоиды, жировые капли. На переднем конце более круп-
ного хроматофора имеется глазок, состоящий из рядов электронно-
плотных гранул несколько различной величины. В цитоплазме на-
ходятся эндоплазматический ретикулум, полирибосомы, митохонд-
рии, в передней трети клетки — одна крупная диктиосома из при-
близительно 12 диктиосомальпых цистерн, от которых по краям
отшнуровываются многочисленные пузырьки. В различных местах
цитоплазмы содержатся свободные липидные гранулы, в задней
части клетки — крупная вакуоль с хризоламинарином. Клетка имеет
два жгутика: длинный (с мастигонемами) и более короткий (глад-
кий). Проксимальная часть короткого гладкого жгута несет вздутие,
расположенное в непосредственной близости от стигмы. От базаль-
ного тела к ядерной оболочке тянется поперечно исчерченный ко-
рень (ризопласт), кроме того, имеются 7 микротрубчатых корней,
распространяющихся на периферию клетки. Клеточное тело сопри-
касается со стенкой домика не только базальной ножкой, но и срав-
нительно широкой экваториальной областью, в которой в молодых
и делящихся клетках наблюдаются обильные пузырьки—дериваты
аппарата Гольджи. Это заставляет предположить, что материал
домика секретируется именно этими пузырьками. При продольном
делении клетки один из дочерних протопластов или оба покидают
материнский домик и, оседая у его края, вращаясь вокруг своей оси,
формируют новый домик. Таким образом возникают древовидные
колонии различной формы. При образовании статоспоры базальная
ножка, по-видимому, отделяется от стенки домика, и клетка миг-
рирует к его отверстию, где округляется, теряет жгутики и образует
цисту или статоспору (см. с. 99). По крайней мере часть статоспор
Dinobryon cylindrieum— результат гологамного полового процесса.
Семейство Синуровые Synигасеае включает одноклеточ-
ные и колониальные формы, у которых клетки кнаружи от цитоп-
лазматической мембраны несут покров из окремнелых чешуек.
Род малломонас (Mallomonas}, обычный в пресноводном пла-
нктоне, наиболее богат видами (более 80). Клетки различной формы
покрыты окремнелыми чешуйками или чешуйками со щетинками
(рис. 18). В качестве представителя рода можно рассмотреть один
из наиболее крупных видов — Mallomonas caudata (рис. 19), для
105
Рис. 18. Чешуйки Mai- Рис. 19. Maltomonas caudata. А — продольный
кнпопая со щетинкой разрез клетки; Б—поперечный разрез двух
(схематизированно) жгутиков (/—короткого с аксонемой 9 — 9 + 0
и 2— -длинного с аксонемой 9 — 9 + 2): В - ба-
зальные тела жгутиков, соединенные тяжем
электронно-плотного материала:
я — ядро с 4 ядрышками, ж — Ж1угики, р ризоп-
ласт (исчерченный корешок), гк— кольцо грануляр-
ного материала, окружающее ризопласт на лолпути
от основания жгутиков до места его раздвоения,
д — диктиосома, х — лопасти одного-единствсшюго
хлоропласта, п — пиреноид, св—сократительная ва-
куоль, вхл вакуоль с хризоламинарином, мз --
микротрубочки, м —мйтохондрии, л липидные
капли, хв—«хвост», ч- чешуйки
которого (D. Е. Wujek, 1978) подробно описана не только уль-
траструктура содержимого клетки, чешуек и щетинок (что пра-
ктически сделано для всех видов рода), но и механизм их об-
разования, освобождения и отложения на поверхность клетки
(D. Е. Wujek, J. Kristiansen, 1978). Такого рода исследований сра-
внительно немного.
М. caudata имеет два жгутика, из которых в оптическом микро-
скопе различим только один'. Этот жгутик нормального строения
106
(аксонема 9 — 9 + 2) и несет два ряда волосовидных мастигонем.
Второй жгутик, не различимый в световом микроскопе, выступает
из клетки на короткое расстояние и скрыт покровом чешуек; он
обладает аксонемой 9 — 9 + 0 (у подрода Mallomonopsis длинный
и короткий жгутики имеют нормальную конфигурацию аксоне-
мы: 9 — 9 + 2). Два базальных тела соединены тяжем электронно-
плотного материала, Ризопласт (исчерченный корешок) соединяет
базальные тела с ядром, делясь непосредственно над ядром на две
ветви. Несколько выше разветвления поперечно исчерченный корень
окружен кольцом гранулярного материала, имеются и микротруб-
чатые корни. Одно ядро, содержащее до четырех ядрышек и рас-
положенное по соседству с основаниями жгутиков, окружено обо-
лочкой из двух мембран; наружная мембрана соединена с эндоплаз-
ма гической сетью вокруг пластиды. Вблизи ядра — аппарат Гольд-
жи. В клетке один двухлопастный хроматофор (а не два, как
считалось ранее при исследовании в оптическом микроскопе). Он
окружен двойной мембраной и хлоропластной эндоплазматической
сетью, содержит трехтилакоидные ламеллы. Имеется одна опоясы-
вающая ламелла, в одной из лопастей хлоропласта находится пире-
ноид, который не удается наблюдать в световом микроскопе. Треть
или половину заднего конца клетки занимает крупный пузырь с хри-
золаминарином. В цитоплазме содержатся еще жир, сократитель-
ные вакуоли (у заднего конца клетки), митохондрии, микротрубоч-
ки (особенно обильные в заднем выступе клетки — «хвосте»), слизе-
вые тельца, которые располагаются непосредственно под цитоплаз-
матической мембраной и состоят из темноокрашенной сердцевины,
заключенной в мембрану. Предполагают, что они ответственны за
слипание чешуек у М. caudata.
Клетка покрыта кремнеземными чешуйками, расположенными
по спирали. Каждая чешуйка отчасти окружена загнутым краем,
отсутствующим в передней части, с многочисленными мелкими
порами, почти в центре чешуйки — продолговатое отверстие. Ще-
тинка имеет вид полой трубки с расщелиной на верхушке и несколь-
кими зубцами по одному краю. Своим изогнутым основанием
щетинка укреплена в дистальной части чешуйки, где отсутствуют
поры. Место образования чешуек и щетинок — пространство между
наружной поверхностью хлоропласта и плазмалеммой. Они форми-
руются внутри плоских пузырьков — везикул, которые отпочковы-
ваются от хлоропластной эндоплазматической сети и сливаются
с везикулами, являющимися дериватами аппарата Гольджи. Концы
плоских везикул, производящих чешуйки, вздуты. Примордии (за-
чатки) и чешуек и щетинок представляют собой плоские пластины.
На более поздних стадиях плоские пузыри с пластинами скручива-
ются в разной степени: в случае щетинок закручивание сильнее. Во
время закручивания к везикулам прикрепляются трубчатые элемен-
ты, возможно принимающие участие в механизме закручивания.
Следствием этого процесса является то, что в трубчатой части
107
щетинки остается щель вдоль одной ее стороны. Клетка продуцирует
или чешуйки, или щетинки, но не обе структуры одновременно.
Мембрана везикула, содержащего зрелую чешуйку, сливается с плаз-
малеммой (процесс экзоцитоза) и чешуйка выталкивается в любом
месте клеточной поверхности под существующим слоем чешуек.
Возможно, чешуйки фиксируются в определенном положении сли-
зью, которая вырабатывается и извергается наружу тельцами, распо-
ложенными непосредственно под плазмалеммой. Каким образом
чешуйки выстраиваются в правильном порядке по поверхности
клетки, пока не ясно. Освобождение щетинок происходит также
посредством экзоцитоза. Первой выталкивается из клетки верхушка
щетинки; у более длинных щетинок верхушка должна быть вытолкну-
та из клетки еще тогда, когда ее базальная часть находится в процессе
формирования. (Об образовании цист у малломонас см. с, 98.)
У малломонас описан и половой процесс — гологамия, при этом
клетки сливаются задними концами.
Род синура (Synura) характеризуется шаровидными или эллипсо-
идальными колониями из грушевидных клеток, задними, иногда
очень длинными концами соединяющимися в центре колонии. Кна-
ружи от цитоплазматической мембраны клетки одеты спирально
расположенными, черепитчато перекрывающими друг друга окре-
мнелыми чешуйками, форма и ультраструктура которых, как
и в случае малломонас, имеют важное таксономическое значение.
Например, у 5. sphagnicola базальная пластинка на поперечном срезе
имеет одинаковую толщину, т. е. плоская. Она пронизана мелкими
перфорациями. Передний (апикальный) край снабжен утолщенной
апикальной кромкой, базальный край загнут, окружая базальную
пластинку наподобие скобы. Полый отогнутый наружу шип прикре-
плен несколько отступя от переднего края пластинки, у основания
его находится пора (рис. 20, А). Чешуйки других видов, например
S. peterscnii, сложнее. Поверх базальной мелко перфорированной
пластинки располагается полый медиальный гребень — апикальный,
заостренный или тупой, конец которого может выступать за фрон-
тальный край чешуйки, имитируя тип. В передней части медиально-
го гребня находится крупная пора. Базальный конец чешуйки подко-
вообразно загнут и нависает над телом чешуйки. Передние и задние
чешуйки, покрывающие тело клетки, несут поперечные ребра, отхо-
дящие от медиального гребня, срединные чешуйки, помимо попе-
речных, имеют еще продольные ребра (рис. 20, Б). Чешуйка не
лежит на клетке плоско, а по-видимому, прикреплена только кон-
цом, противоположным шипу. Профили чешуек тела у 5. sphagnicola
можно наблюдать в цитоплазматических везикулах, большей ча-
стью расположенных вблизи наружной поверхности хлоропласта
или между хлоропластом и везикулами с хризоламинарином.
Из углубления на переднем конце клетки отходят два гетеро-
контных и гетероморфных жгутика. Более длинный жгут — с масти-
гонемами, которые состоят из трубчатой части, несущей терминаль-
108
2
Б
Рис. 20. Схематическое изображение строения чешуйки Synura sphagnicola (Л)
и S. petersenii (Б):
1—продольный, 2 — поперечный разрез чешуйки
ные нити; короткий жгут — гладкий, с латеральным вздутием. На
мембране жгута, как длинного, с мастигонемами, так и короткого,
гладкого, располагаются чешуйки жгутиков, размеры и форма ко-
торых специфичны для каждого вида и наряду с чешуйками тела
клетки служат важным таксономическим признаком. Так,
у S. petersenii чешуйки обоих жгут иков кольцевидные,
у S', sphagnicola на обоих жгутиках чешуйки палочковидные,
у 5. uvella чешуйки (только на длинном жгутике) полукруглые,
подковообразные, у S. echinulata на длинном жгутике наблюдается
смесь булавовидных и кольцевидных чешуек, а на коротком глад-
ком жгуте — только кольцевидные чешуйки. Прикрепляются чешуй-
ки следующим образом: кольцевидные — своими краями, полукруг-
лые- выпуклой стороной, палочковидные — одним концом. В от-
личие от пропитанных кремнеземом чешуек тела чешуйки жгутиков
не минерализованы. Местом образования и сборки чешуек жгути-
ков у .S'. sphagnicola является, по-видимому, аппарат Гольджи,
у .S'. petersenii чешуйки жгутиков наблюдались в пузыре, окружен-
ном расширенной цистерной эндоплазматической сети. Клетка со-
держит два париетальных хлоропласта, каждый из которых окру-
жен слоем хлоропластной эндоплазматической сети, ламеллы трех-
тилакоидные, имеется опоясывающая ламелла, глазок отсутствует.
Пространство между передними концами хлоропластов занимает
одно клеточное ядро, в передней части клетки—аппарат Гольджи,
задний узкий конец клетки заполнен сократительными вакуолями,
между хлоропластами и поверхностью клетки находятся крупные
пузыри, по-видимо.му, с хризоламинарином.
Клетки внутри колоний делятся продольно. Молодые колонии
образуются за счет распада разросшихся старых колоний. Часто
109
одновременно во всех клетках колонии возникают цисты. Известно
и пальмеллевидное состояние. У S', petersenii описан половой про-
цесс— гологамия. Копуляция клеток осуществляется задними кон-
цами.
Порядок хризокапсальные— Chrysocapsales
Порядок включает пальмеллоидные формы. Наиболее дифферен-
цированная из пальмеллоидных форм среди водорослей - - род гидру-
рус (Hydrurus) с единственным видом Н. foetidus, довольно обычным
в быстро текущих холодных горных источниках и представленным
оливково-зелеными слизистыми сильно разветвленными тяжами до
30 см длиной, часто издающими очень неприятный запах. Отсюда
и видовое название — зловонный. Плотная слизь, образующая цилин-
дрические тяжи, заключает многочисленные грушевидные клетки
с заостренным концом, содержащие один стенкоположный хромато-
фор, пульсирующие вакуоли и хризоламинарин. Клетки плотно
расположены по периферии и более рыхло в центре слизи. На конце
каждого тяжа клетка ведет себя как апикальная, одна половинка
которой после продольного деления вновь становится апикальной,
а другая входит в вещество тяжа. Размножение осуществляется
зооспорами характерной тетраэдрической формы. Такая необычная
форма зооспор гидруруса определяется сложной системой скелетных
микротрубочек, о тходящих от жгутикового корня и простирающихся
в три передних отростка. Задний отросток также поддерживается
сетью микротрубочек. От переднего конца зооспоры отходя! два
неравной длины жгутика, из которых длинный служит для движения,
а более короткий — остаточный — лишен центральных микротрубочек
(9 — 9 — 0). Зооспоры образуются путем продольного деления клеток
более тонких ветвей, клетки более толстых тяжей несут, по-видимому,
лишь опорную функцию, не участвуя в размножении. Цисты образу-
ются в некоторых клетках, выступающих из слизистых тяжей,
КЛАСС ГАПТОФИЦИЕВЫЕ, ИЛИ ПРИМНЕЗИОФИЦИЕВЫЕ
HAPTOPHYCEAE, PRYMNESIOPHYCEAE
Этот класс выделен из золотистых водорослей в первую очередь
на основе строения монадных клеток, обладающих помимо жгути-
ков гаптонемой (см. гл. 3). Включает три порядка, из которых ниже
рассматривается один.
Порядок примиезиальные — Prymnesiales
Порядок характеризуется обычно длинной гаптонемой и двумя
изоконтными и изоморфными жгутиками. Поверхность клеток кна-
ружи от плазмалеммы покрыта органическими неминерализован-
110
ними чешуйками (семейство примнезиевые — Prymnesiaceae) или
известковыми телами—кокколитами, в совокупности образующи-
ми вокруг клетки коккосферу (семейство кокколитофоровые—.
Coccolithophoraceae).
Семейство примнезиевые — Prymnesiaceae. Род хризохро-
мулииа (Chrvsochromulina) представлен как в морях, так и в пресных
водах. Сферические или овальные клетки с двумя равной длины
гладкими жгутиками и гаптонемой кнаружи от цитоплазматической
мембраны покрыты органическими неминерализованными чешуй-
ками обычно двух типов, различающимися или только размерами,
или же формой.
Так, у Chrysochromulina birgeri два типа чешуек, покрывающих
тело, различаются только по размерам. Оба они представлены
овальными пластинками с рисунком из радиальных гребней, рас-
положенных в квадрантах. Имеются два центральных выступа в ви-
де рогов. У других видов клеточная поверхность покрыта чешуй-
ками, более или менее резко различающимися морфологически.
Например, у Ch. cyanophora внутренние чешуйки, округлые, плос-
кие, с тонкими концентрическими гребнями, перекрывают друг
друга, образуя вокруг клетки футляр, и обычно скрыты под много-
численными наружными цилиндрическими чешуйками.
У Ch. meyacylindra также есть два рода чешуек—пластинки и ци-
линдры. Цилиндры, каждый прикрепленный нижним концом
к своей собственной базальной пластинке, распределены по клетке
более или менее равномерно, почти касаясь один другого своими
боковыми сторонами. Под ними имеется много слоев плоских
чешуек с ободками.
'У Ch. chiton наблюдаются три типа чешуек с характерным рас-
положением: в группах чешуек 6 крупных без ободка располагаются
вокруг одной крупной чешуйки с ободком, а все промежутки запол-
нены самыми мелкими чешуйками.
Семейство кокколитофоридовые — Coccolithophora-
ceae. К этому семейству относятся почти исключительно морские
формы, исключение — пресноводный род гименомонас (Нутепо-
топах). Монадные клетки снабжены двумя одинаковыми жгути-
ками и обычно хорошо заметна гаптонема. Однако у ряда кок-
колитофорид гаптонема, по-видимому, редуцирована: так,
у Н. coronata явной гаптонемы не наблюдалось, у Ochrosphaera
neapolitana гаптонема представлена только в виде группы микро-
трубочек, связанных с основаниями жгутиков, и не выступает из
клетки. Внутреннее строение клетки такое же, как у других га-
птофициевых: ядро, хлоропласты, окруженные хлоропластной эн-
доплазматической сетью, содержат трехтилакоидные ламеллы, опо-
ясывающая ламелла отсутствует; пиреноид, выступающий от вну-
тренней поверхности хлоропласта и пересеченный парными тила-
коидами; аппарат Гольджи, митохондрии и пр. Характерен
клеточный покров, расположенный кнаружи от цитоплазматической
111
мембраны и состоящий из пропитанных карбонатом кальция чешу-
ек— кокколитов, в совокупности образующих вокруг клетки кокко-
сферу. У ряда форм, помимо кокколитов, имеются пеминерализо-
ванные органические чешуйки. Тонкая структура кокколитов отли-
чается большим разнообразием и представляет важный таксоно-
мический признак. Ниже ограничимся лишь некоторыми приме-
рами кокколитофорид, исследованных в последние годы с помощью
электронного микроскопа. Кокколиты, покрывающие клетки
Calyptrosphaera pirus, при изучении в сканирующем микроскопе
имеют следующее строение: куполообразная верхняя часть отчасти
погружена в более широкую, слегка воронковидную базальную
часть. На границе между этими двумя частями имеется ложбинка,
в которой почти всегда наблюдаются 7—10 перфораций; на куполо-
образной части кокколита иногда есть углубления. Кокколиты,
предположительно окружающие жгутиковый полюс клетки, отлича-
ются от остальных коническим выступом на куполообразной части
(рис. 21, Б). У Papposphaera lepida кокколиты состоят из базальной
части с краевым кольцом из пятиугольных элементов; от креста на
базальной пластинке отходит вертикальный расширяющийся на
конце придаток (рис. 21, Л, Л). У пресноводного рода гименомонас
(Hymenomonas) клетки кнаружи от плазмалеммы несут покров из
органических чешуек и кокколитов. Последние короновидные, сло-
женные из 11 —12 пятиугольных элементов, расположенных по
периферии базальной пластинки (рис. 21, В).
Изучение некоторых коккофорид в лабораторной культуре позво-
лило обнаружить в цикле их развития неподвижную нитчатую стадию.
Например, подвижная одноклеточная стадия Cricosphaeraroscoffensis
var. haptonemofera (два жгутика, гаптонема, клеточный покров образо-
ван тонкими органическими чешуйками, поверх которых расположе-
ны кокколиты) может размножаться путем деления пополам, а при
определенных условиях культивирования развивается нитчатая непо-
движная стадия, сходная с водорослью Apistonema submarinum.
Как и в случае окремнелых чешуек хризофициевых водорослей,
таких, как малломонас, синура и др., неминерализованные органи-
ческие чешуйки и кокколиты гаптофициевых водорослей в послед-
ние десятилетия изучены не только с точки зрения их ультраструк-
туры, таксономическая значимость которой не вызывает сомнения,
но и с точки зрения их генезиса. Из последних исследований в пер-
вую очередь заслуживает упоминания работа Р. Броуна с соав-
торами (1970) о синтезе чешуек Pleurochrysis scherffeli (см. гл. 3).
В 1975 г. были изучены формирование и химический состав
чешуек Chrysochromulina chiton. Было показано, что все три типа
чешуек разного размера образуются в одно и то же время внутри
аппарата Гольджи, затем транспортируются к поверхности клетки,
где и располагаются характерным для этого вида образом. То же
относится к обоим типам (плоским и цилиндрическим) чешуек
Ch. megacylindra.
112
Рис. 21. Кокколиты: А - Pupposphuera lepida; Б—Calyplrosphaera pirns'. В —
Hymenomonas roseola; Г коккосфера Papposphaera lepida
Эндогенное (в цистернах аппарата Гольджи) образование кок-
колитов и выведение их на поверхность клетки посредством процес-
са экзоцитоза продемонстрировано для Hymenomonas coronata
и других кокколитофоридовых.
Глава 8
ОТДЕЛ ЖЕЛТОЗЕЛЕНЫЕ, ИЛИ РАЗНОЖГУТИКОВЫЕ,
ВОДОРОСЛИ— XANTHOPHYTA, HETEROCONTAE
Для этих водорослей характерны следующие пигменты: хло-
рофиллы а, с, е (хлорофилл Л отсутствует), 0-каротин, £-каротин,
ксантофиллы: антераксантин, лютеин, зеаксантин, неоксантин,
виолаксантин, вошериаксантин, гетероксантин, диадиноксантин,
Ш
Рии, 22. Схема строения монадных клеток ксантофициевых
(Л) и эвстигматофициевых (F) водорослей:
гж — главный длинный жгут с мастигонемами. бж— короткий бо-
ковой жгут со вздутием у основания, от базальные тела, х •—
хлоропласт, с — стигма, м—'Митохондрии, вж— вздутие у основа-
ния жгута
диатоксантин. Клетка содержит обычно по два пластинчатых
хроматофора, которые лежат в цистерне эндоплазматической
седи, находящейся в прямой связи с оболочкой ядра, в матри-
ксе хлоропласта расположены трехтилакоидные ламеллы; почти
всегда (ксантофициевые) имеется опоясывающая ламелла (от-
сутствует у эвстигматофициевых), внутренние пиреноиды кса-
нтофициевых содержат трехтилакоидные ламеллы. Глазок (стиг-
ма) является частью пластиды (ксантофициевые) или же гра-
нулы стигмы располагаются вне пластиды (эвстигматофицие-
вые).
Запасные продукты: волютин, жир, часто хризоламинарин; крах-
мал не образуется. Более длинный жгутик несет мастигонемы,
короткий — гладкий. Клеточные оболочки часто состоят из двух
и более частей, цисты (статоспоры), как и у золотистых водорослей,
эндоплазматические, их оболочки окременелые.
На рис. 22 приведены схемы монадных клеток желтозеленых
водорослей.
В большинстве современных систем различают два класса: ксан-
тофициевые (Xanthophyceae) и эвстигматофициевые (Eustigmato-
phyceae).
114
КЛАСС КСАНТОФИЦИ ЕВ ЫЕ — XANTHOPHYCEAE
Этот класс включает водоросли, стоящие на самых разных
ступенях дифференцировки таллома: монадные, ризоподиальные.
пальмеллоидные, коккоидные, нитчатые, сифоновые. В соответствии
с типами организации таллома различают порядки, параллельные
порядкам золотистых, динофитовых, собственно зеленых водорослей.
Порядок гетерохлоридальные (Heterochloridales) объединяет монад-
ные формы, порядок ризохлоридальные (Rhizochloridales) включает
ризоподиальные, порядок гетероглеальные (Helerogloeales) — паль-
меллоидные, порядок мисхококкальные (Mischococcales)— коккоид-
ные, порядок трибонемальные (Tribonematales) — нитчатые, порядок
ботридиальные (Botrydiales)—сифоновые водоросли.
Ниже рассматриваются три последних порядка.
Порядок мисхококкальные — Mischococcales
Порядок включает многочисленные одноклеточные и колони-
альные коккоидные формы. Клетки различных очертаний одеты
клеточной стенкой, часто состоящей из двух частей. Размножение
зооспорами или апланоспорами.
Род ботридиопсис (Botrydiopsis) широко распространен в пресно-
водных водоемах (прудах, канавах и пр.), представлен одиночными
шаровидными клетками, одетыми клеточной оболочкой, под кото-
рой располагается постенная цитоплазма, содержащая многочис-
ленные дисковидные хлоропласты, а в зрелых клетках — много
клеточных ядер. Центр клетки занят крупной вакуолью с клеточным
соком, пересеченной тонкими тяжами цитоплазмы. В цитоплазме
рассеяны капли масла и глыбки хризоламинарина. Размножение
зооспорами и автоспорами, образующимися в клетке в больших
количествах (до 300). У В. arrhiza при зооспорообразовании, как
и у многих других желтозеленых водорослей (харациопсис, трибо-
нема, ботридиум и др.), наряду с типичными зооспорами нередко
наблюдаются синзооспоры (см. также гл. 4).
Порядок трибонемальные—Tribonematales
Объединяет нитчатые формы. В качестве представителя может
быть рассмотрен широко распространенный в пресных водах род
трибонема (Tribonema). Таллом представлен неразветвленной, сло-
женной из одного ряда клеток нитью.
Цилиндрические, часто слегка бочонковидные клетки, обычно
одноядерные, большей частью содержат несколько хлоропластов.
В цитоплазме -жир, хризоламинарин. Клеточная стенка всегда
состоит из двух половинок, краями находящих друг на друга в сре-
динной плоскости клеток. Каждая половинка оболочки составлена
из ряда налегающих друг на друга слоев.
115
Еще до деления ядра клетки (на стадии интерфазы) в экватори-
альной области под старой клеточной оболочкой закладывается
новый цилиндрический кусок оболочки — промежуточное кольцо.
На более поздней стадии деления материнской клетки, во время
цитокинеза, в середине этого полого цилиндра образуется попереч-
ная перегородка. Новая оболочка, имеющая в продольном оптичес-
ком сечении Н-образную форму, при разрастании раздвигает друг
от друга обе более старые половинки материнской клеточной стен-
ки, вставляясь между ними. Таким образом оболочки соседних
клеток соединяются друг с другом; соответственно каждая клетка
окружена двумя половинками соседних Н-образных фигур. При
освобождении репродуктивных клеток или под воздействием неко-
торых агентов (например, крепкой хромовой кислоты) оболочка
клеток распадается на Н-образные участки. Фрагменты нитей все-
гда оканчиваются пустыми половинками Н-образных фигур, кото-
рые в профиль выглядят как два острия. Размножение нитей, непре-
рывно нарастающих за счет поперечных делений клеток, осуществ-
ляется вегетативно — фрагментацией и бесполым путем — посред-
ством зооспор, амёбоидных клеток, апланоспор, акинет.
Порядок ботридиальные—Botrydiales
Объединяет ксантофициевые водоросли, имеющие сифоновую
организацию.
Представители рода ботридиум (Boirydium) живут на сырой зем-
ле. Таллом сифоновый, дифференцированный на надземную часть
в виде сферического грушевидного пузыря диаметром 1 —2 мм,
и погруженную в субстрат ризоидальную часть, обычно разветвлен-
ную. Эта нормальная форма таллома в зависимости от внешних
условий может заметно видоизменяться. Например, при культиви-
ровании ботридиума в погруженном в воду состоянии наблюдаются
талломы в виде разветвленных нитей. Стенка многослойная, с раз-
лично ориентированными в каждом слое целлюлозными микрофиб-
риллами. Цитоплазма сосредоточена в постенном слое и окружает
непрерывную вакуоль с клеточным соком. Во взрослом талломе
цитоплазма содержит многочисленные ядра, в надземной части —
обильные дисковидные хлоропласты с пиреноидами и прочие ор-
ганеллы. Размножение осуществляется одноядерными зооспорами
с двумя гетероконтными и гетероморфными жгутиками, возника-
ющими при погружении пузыря в воду (после дождя и т. п.).
Наблюдались и синзооспоры. Оседая затем по краю лужи, на
подсыхающей земле, зооспоры развиваются в новые растения. При
более сухой погоде вместо зооспор образуются апланоспоры. По
достижении известной внутренней зрелости ботридиум переходит
в стадию покоя. Русский исследователь В. В. Миллер, подробно
изучивший род ботридиум в культуре, описал различные способы
образования покоящихся цист. В одних случаях все содержимое
116
пузыря дает начало одной крупной цисте с толстой оболочкой.
Такие цисты В. В. Миллер назвал «макроцистами» (В. wallrothi,
В. tuberosum, В. pachidermum). В других случаях содержимое надзем-
ной части делится с образованием многих многоядерных спороцист
(В. wallrothi). Наконец, все содержимое пузыря или часть его может
переходить в ризоиды и там образовывать ризоцисты. Последние
или располагаются по многу в ряд, например, у В. granulatum, или
же содержимое переходит во вздутые концы ризоидов, в каждом из
которых образуется циста, например у В. tuberosum. У одного и то-
го же вида могут наблюдаться различные формы цист, которые при
неодинаковых внешних условиях заменяют друг друга. Все формы
цист для своего прорастания не нуждаются в периоде покоя, они
могут прорастать и непосредственно после своего возникновения.
Мелкие цисты (расположенные в ряд ризоцисты, спороцисты) или
непосредственно прорастают в новые индивиды, или образуют
зооспоры. Крупные цисты (макроцисты, образующиеся поодиночке
ризоцисты В. tuberosum) прорастают обычно зооспорами или ап-
ланоспорами. Ни у одного из исследованных видов ботридиума
В. В. Миллер не наблюдал полового процесса. Однако имеются
данные других авторов о существовании у В. granulatum полового
процесса, изо- и гетерогамного у разных рас. Эти данные нуждают-
ся в подтверждении.
У рода вошерия (Vaucheria) (известны как пресноводные, так
и морские и солоноводные виды) таллом в виде разветвленных
сифоновых нитей, образующих ватообразные скопления в воде или
обширные дерновинки темно-зеленого цвета на влажной земле.
Рост нитей апикальный. В вегетативных нитях вошерии, начиная
с верхушки, можно различить три зоны: апикальную, субапикаль-
ную и вакуолизированную. Апикальный отрезок представляет со-
бой активно растущую часть сифонового таллома: здесь находятся
многочисленные пузырьки и митохондрии; хлоропласты и клеточ-
ные ядра отсутствуют. Везикулы содержат фиброзный материал,
возможно, являющийся предшественником материала (целлюлозы)
клеточной оболочки. В субапикальной зове количество пузырьков
уменьшается, появляются хлоропласты и клеточные ядра. Хлоро-
пласты многочисленные, дисковидные, лишены пиреноидов. Выда-
ющийся в виде почки пиреноид наблюдается только в проростках
вошерии. С каждым клеточным ядром связана пара центриолей,
которые во время профазы митоза мигрируют к противоположным
полюсам удлиняющихся ядер. Оболочка ядра во время митоза
остается интактной, внутри нее образуется внутриядерное веретено,
центромеры отсутствуют; между расходящимися хромосомами
ядерная оболочка перешнуровывается и окружает телефазные до-
черние ядра. Нормальный митоз, происходящий внутри оболочки
материнского ядра, у ряда видов вошерии наблюдал еще
Л. И. Курсанов (1911). У всех исследованных видов Л. И. Курсанов
отметил интересное распределение ядерных делений на протяжении
117
Рис. 23. Схема развития, выхода, оседания и прорастания син-
зооспоры Vaucheria.
А — конец вегетативной нити; Б—Е—образование синзооспо-
ры: Б - во вздутом кончике нити постепенно исчезает вакуоль
с клеточным соком, скапливаются ядра и образуются внутрен-
ние жгутики, В— внутренние жгутики группируются вокруг
возникающих пузырьков—«везикул жгутиков», Г, Ц, везику-
лы жгутиков мигрируют к плазмалемме, с которой сливаются,
при этом жгутики достигают поверхности развивающейся син-
зооспоры, а у основания спорангия образуется септа, Е —зре-
лая синзооспора внутри зооспорангия, в периферической цитоп-
лазме, помимо ядер, находится множество везикул с фиброз-
ным материалом; Ж—освобождение синзооспоры из зооспо-
рангия; 3—синзооспора, покрытая парами гладких, очень не-
значительно различающихся по длине жгутиков; И—у осевшей
на субстрат синзооспоры перемещаются ядра в субповерхност-
ную зону и втягиваются жгутики, мембраны которых становят-
ся частью плазмалеммы; К—перемещение ядер завершено,
с ними связаны сохраняющиеся некоторое время аксонемы
жгутиков, появляется вакуоль с клеточным соком, выделяется
клеточная оболочка, при этом из периферического слоя цитоп-
лазмы исчезают везикулы с фиброзным материалом (возмож-
ным предшественником клеточной стенки); J7— прорастание
синзооспоры, возникновение выступа — нового кончика нити,
в котором образуются обильные везикулы,- центральная ваку-
оль начинает удлиняться:
кс — клеточная стенка, п — плазмалемма, я—ядро, внж — внутренние
жгутики, вж — пузырьки жгутиков, ж—жгутики, в — везикулы с фиброз-
ным материалом, вак — вакуоль с клеточным соком, аз — апикальная
зона, сз—субапикальная зона, вакз — вакуолизированная зона, м—ми-
тохондрии, х -хлоропласты, эс — эндоплазматическая сеть
нити: начавшись в одном каком-нибудь месте, митозы мало-по-
малу распространяются на соседние участки и т. д. Таким образом,
на препарате, фиксирующем известный момент этого процесса,
получается то, что всего лучше назвать волной делений, где отдель-
ные, следующие друг за другом во времени фазы деления на протя-
жении нити расположены в правильной последовательности. В апи-
кальной и субапикальной зонах еще отсутствует центральная ваку-
оль. Лишь в более старой — вакуолизированной — части нити появ-
ляется вакуоль с клеточным соком (рис. 23, А). Размножение беспо-
лое, посредством синзооспор и апланоспор.
Образование синзооспор, их выход, оседание и прорастание
были прослежены у Vaucheria fontinalis па электронно-микроскопи-
ческом уровне (рис. 23, Б—Е). При образовании синзооспоры на
конце нити, несколько вздутом, центральная вакуоль исчезает,
и здесь скапливаются все органеллы. Связанные с клеточными
ядрами пары центриолей формируют внутренние жгутики. Клеточ-
ные ядра и внутренние жгутики группируются вокруг возникающих
пузырьков, в которые вдаются жгутики. Пузырьки мигрируют к по-
верхности протопласта и сливаются с плазмалеммой. Таким об-
разом, жгутики достигают поверхности будущей синзооспоры.
Синзооспоры вошерии покрыты многочисленными гладкими,
очень незначительно различающимися по длине парами жгутиков.
Возникает септа, отделяющая спорангий с синзооспорой от вегета-
тивной нити. Выход синзооспоры, по-видимому, осуществляется
после энзиматического растворения верхушки зооспорангия. Недав-
но остановившаяся синзооспора сферическая, крупная, центральная
вакуоль отсутствует, жгутики втягиваются, сохраняя сначала типич-
ную (9 — 9 + 2) конфигурацию аксонемы; их мембраны становятся
частью плазмалеммы. ядра занимают субповерхностную зону. Кле-
точной стенки сначала нет, но в периферическом слое цитоплазмы
находятся обильные везикулы с фибриллярным материалом, кото-
рые, по-видимому, содержат предшественников целлюлозы. Их ис-
чезновение совпадает с отложением на поверхности синзооспоры
тонкой стенки. Ядра смещаются, в периферическую область втор-
гаются хлоропласты и другие органеллы. Аксонемы втянутых жгу-
тиков разрушаются, центриоли занимают свое обычное положение
рядом с клеточными ядрами. Образуется крупная центральная ваку-
оль. С одного или с обоих концов инцистированной зооспоры
возникает выпуклость, в которой скапливаются везикулы с фибрил-
лярным материалом, митохондрии, диктиосомы, хлоропласты, яд-
ра, как это наблюдается в кончике вегетативной нити. Центральная
вакуоль инцистированной зооспоры начинает удлиняться
(рис. 23,Ж—Л).
Половой процесс оогамный. Пресноводные виды вошерий боль-
шей частью однодомные, морские виды- двудомные.
Половые органы возникаю! в виде боковых выростов нити,
содержащих многочисленные ядра: более или менее шаровидной
120
формы в случае оогония и в форме цилиндрической трубки, закру-
ченной наподобие рога - при развитии антеридия. Оба типа поло-
вых выростов сначала сообщаются с вегетативной нитью, на кото-
рой заложились, а позднее отделяются от нее поперечной стенкой.
В случае оогония до образования поперечной перегородки все ядра,
кроме одного, становящегося ядром единственной яйцеклетки, миг-
рируют с очень подвижной цитоплазмой обратно в вегетативную
нить. По другим данным, лишние ядра дегенерируют после отделе-
ния оогония от нити поперечной перегородкой. В антеридии, кото-
рый отделяется поперечной перегородкой от кончика мужского
выроста, сохраняются многочисленные ядра и возникает большое
число сперматозоидов. Сперматозоид вошерии без хлоропласта
и стигмы, с крупными митохондриями, снабжен двумя гетероконт-
пыми жгутиками, причем передний жгутик с мастигонемами короче
гладкого заднего. В передней части сперматозоида имеется вырост
с микротрубчатыми скелетными элементами, напоминающий так
называемый хоботок сперматозоида фукуса. Половые органы
вскрываются, сперматозоиды проникают в оогоний и оплодотворя-
ют яйцеклетку, которая после оплодотворения выделяет толстую
оболочку. Копуляционное ядро в прорастающей после периода
покоя ооспоре редукционно делится.
Некоторые исследователи, придавая большое значение различи-
ям в строении жгутикового аппарата у сперматозоидов и синзоо-
снор вошерии, выделили этот род даже в отдел Vaucheriophyta,
поместив остальные желтозеленые водоросли в качестве класса
Heteroconlae в отдел Chrysophyta. Другие авторы объединяли роды
ботридиум и вошерия в отдел Siphonophyta, позднее понизив этот
таксон до ранга класса Xanthosiphonophyceae наряду
с Xanthophyceae в пределах отдела Chrysophyta. Сегрегация рода
вошерия от других желтозеленых водорослей на столь высоком
уровне, по-видимому, не встретила поддержки у многих исследова-
телей. Однако выделение вошерии в отдельный порядок
Vaucheriales принято многими авторами.
Класс эвстигматофициевые (Eustigmatophyceae)
Этот класс был выделен из ксантофициевых па основе строения
монадных клеток (зооспор), в первую очередь их глазка. Глазок
представляет собой крупное оранжево-красное тело у крайнего
переднего конца зооспоры, не зависимое от единственной пластиды,
и состоит из неправильной группы капель без ограничивающих
мембран и без мембраны вокруг всего комплекса. Таким образом,
в отличие от ксантофициевых стигма здесь расположена вне
пластиды. Другой отличительный признак эвстигматофициевых —
то, что утолщение жгута, перекрывающее стигму, находится у про-
ксимального конца, направленного вперед и несущего два ряда
волосков жгутика. Гладкий, направленный назад жгутик, как
121
правило, отсутствует. Кроме того, у эвстигматофициевых трех-
тилакоидные ламеллы часто образуют граноподобные стопки,
периферическая опоясывающая ламелла отсутствует, пиреноид,
обнаруженный только в вегетативных клетках (в зооспорах от-
сутствует), выступает от внутренней поверхности пластиды и не
пересечен тилакоидами.
Все эвстигматофициевые — одноклеточные коккоидные формы
(прежде относимые к порядку мисхококкальные из класса ксан-
тофициевые).
Глава 9
ОТДЕЛ ДИАТОМОВЫЕ ВОДОРОСЛИ —
BACIACILLARIOPHYTA, DIATOMЕАЕ
Сюда относятся коккоидные — одноклеточные и колониальные
водоросли с хроматофорами, в живом состоянии окрашенными
в желто-бурый цвет благодаря преобладанию каротиноидов (р-
карогин, е-каротин, ксантофиллы: неоксантин, фукоксантин, неофу-
коксантин, диадиноксантин, диатоксантин) над хлорофиллами а,
с (хлорофилл b отсутствует). Хлоропласты обычно париетальные,
в виде одной-двух крупных пластинок (многие пеннатофициевые)
или многочисленных дисков (большинство центрофициевых). Мат-
рикс хлоропласта пересекается трехтилакоидными ламеллами, име-
ется опоясывающая ламелла. Пиреноид центральный, в большинст-
ве случаев окружен специальной мембраной и пронизан различным
у разных видов числом (1—3) тилакоидов. Запасные продукты —
хризоламинарин, масло, волютин; крахмал отсутствует. Клеточное
ядро локализовано или в постенной цитоплазме, окружающей одну
центральную вакуоль с клеточным соком, или в цитоплазматичес-
ком мостике, пересекающем вакуоль. Цитоплазма содержит пери-
нуклеарный комплекс диктиосом, митохондрии. Монадные клетки
(сперматозоиды) одножгутиковые, в аксонеме отсутствуют цент-
ральные микротрубочки (9 — 9 + 0).
Клеточный покров. Диатомовые характеризуются особым кле-
точным покровом, состоящим из двух главных компонентов: крем-
неземного панциря и тонкого органического пектинового слоя, ко-
торый полностью (и снаружи и изнутри) обрамляет панцирь. Для
изучения панциря (что необходимо для флористических и система-
тических исследований) диатомовые обрабатывают таким образом,
чтобы разрушить протопласт, при этом удаляют и поверхностный
органический материал. Чтобы изолировать второй компонент кле-
точного покрова диатомовых — органический наружный слой, на
клетку воздействуют фтористоводородной кислотой, растворя-
ющей кремнезем. Топография такого органического покрова оказа-
122
лась видоспецифичпой и идентичной с топографией панциря. Пока
неясно, отражает ли рисунок органического покрова рисунок пан-
циря или же, наоборот, является матрицей для панциря. Органичес-
кий слой при формировании клеточного покрова развивается инт-
рацеллюлярно, у одних диатомей он может утрачиваться, у дру-
гих— сохраняется. Кроме того, через отверстия в панцире (шов,
ареолы, поры) может выделяться слизь, которая в зависимости от
вида формирует слизистые чехлы, трубки, ножки, подушечки, тяжи,
сосочки и пр.
Панцирь диатомовых водорослей состоит из двух половинок:
большей эпитеки и меньшей гипотеки. Каждая половинка образова-
на с/ийрркощ.которая своим загибом соединяется с поясковым коль-
цом или ободком. Свободный край пояскового кольца эпитеки
надвинут на свободный край пояскового кольца гипотеки, как кры-
шечка на коробочку. У многих диатомей между загибом створки
и поясковым кольцом возникают вставочные ободки различной
формы и в разном количестве (у рабдонемы до 28). Эти ободки
появляются последовательно, причем каждый вставочный ободок
формируется между загибом створки и последним из ранее образо-
вавшихся вставочных ободков. Таким образом самый молодой
вставочный ободок всегда граничит с загибом створки, а самый
старый — с поясковым ободком. На вставочных ободках развива-
ются септы — неполные кремнеземные перепонки, расположенные
или вдоль всей внутренней поверхности вставочного ободка, или на
одном из его полюсов (рис. 24). Септы всегда имеют одно или
несколько отверстий, перегораживая полость клетки на сообщаю-
щиеся между собой отсеки. Образование вставочных ободков обус-
ловливает возможность роста клетки вдоль ее центральной (пер-
вальварной) оси, соединяющей центры двух створок. Различные
части панциря плотно сцементированы вместе, иногда связаны
сложными закраинами. Раздвигание эпи- и гипотеки наблюдается
только при делении клетки и во время образования ауксоспор.
Створки у одних диатомовых, объединяемых в класс центрофицие-
вые (Centrophyceae), обычно округлых очертаний, например у рода
циклотелла (Cyclotella), и панцирь в целом может быть уподоблен
круглой коробочке. Реже створки треугольные, как у трицера-
циума (Triceratium), или эллиптические, как у рода биддульфия
(Biddulphia). Элементы скульптуры, представленные в первую оче-
редь порами различных типов (отверстия, альвеолы, локулы), раз-
мещаются на створках вокруг центральной точки, а у родов с окру-
глыми створками обычно располагаются радиально. У других диа-
томовых, составляющих класс пеннатофициевых (Pennatopbyceae),
створки эллиптические или продолговатые, например у родов пин-
нулярия (Pinmdaria) и навикула (Navicula), у некоторых, как у синед-
ры (Synedra). длина их во много раз превосходит ширину.
У перечисленных родов концы створок одинаковые — изополь-
ные створки. У родов гомфонема (Gomphonema}, ликмофора
123
Рис. 24. Элементы панциря диатомовых водорослей.
А, Б- элементы половинки панциря (А—вид со створки, Б—вид с пояска);
В, Г, Д—формы вставочных ободков (В—кольцевидный, Г—полукольцевидный.
Д— чешуевидный):
с—створка, зс — загиб створки, во — вставочный ободок, се—септа, по — поясковый ободок
(Liemophora) створки гетерополъные, т. е. суживаются к одному
концу. Могут быть створки и иной формы: S-образно изогнуты-
ми— у плевросигмы (Pleurosigma), гиросигмы (Gyrosigma), некото-
рых видов рода ницшия (Nitzschia), полулунными с прямым или
вогнутым брюшным и выпуклым спинным краями, как у цимбеллы
(Cymbella) и т. д. У всех пеннатофициевых скульптурные элементы
на створках располагаются в два продольных ряда по бокам от
бесструктурной срединной—медианной линии, или осевого поля.
Положение клетки диатомовых, при котором видна одна из ство-
рок, называется видом панциря со створки. Кроме того, различают
вид панциря с пояска, при котором видно, как поясковое кольцо
эпитеки надвинуто па поясковое кольцо гипотеки. У громадного
большинства диатомовых, центрофициевых и пеннатофициевых
клетка со стороны пояска имеет форму четырехугольника, но по-
ясок может быть и клиновидным (гомфонема, ликмофора), S-образ-
но изогнутым (некоторые виды ницшии) с более или менее выра-
женным изгибом посередине —у рода ахнантес (Achnanthes) и пр.
У некоторых пеннатофициевых, объединяемых в порядок фраги-
ляриальные (Fragilariales), осевое поле — бесструктурная часть
124
створки — гладкое. Этим характеризуются роды синедра, меридион
(Meridian), астерионелла (Asterionella), фрагилярия (Fragilaria), диа-
тома (Diatoma). У большинства других пеннатофициевых эта об-
ласть створки занята особым образованием — швом (рафе), име-
ющим вид двух продольных, слегка изогнутых щелей в толще
створки. Это щелевидный шов. В середине створки, где панцирь
изнутри утолщен, образуя центральный узелок, обе ветви шва погру-
жаются в две центральные поры в толще центрального узелка.
У полюсов клетки каждая ветвь шва заканчивается внутри поляр-
ных— конечных узелков. Последние, как и центральный узелок,
являются местными, но менее заметными утолщениями створки
На поперечном разрезе створок можно видеть, что шов представля-
ет собой V-образную или коленовидно изогнутую трещину, состо-
ящую как бы из двух щелей или каналов, из которых одна (внутрен-
няя щель) открывается в полость клетки, а другая — наружная —
сообщается с внешней средой. Наружная и внутренняя щели каждой
ветви шва соединяются в центральном узелке посредством вер-
тикального канала. В центральном же узелке обе ветви шва сообща-
ются с помощью горизонтального канала (рис. 25, А).
Внутри полярных узелков наружная щель шва переходил в слег-
ка скрученную (конечную) трещину (щель), а внутренняя щель
заканчивается в виде воронковидного образования.
Описанный выше щелевидный шов развивается на обеих или
только на одной створке пеннатофициевых, объединяемых в по-
рядок навикулальные (Maviculales). Каналовидный шов характерен
для диатомовых, относящихся к порядку суриреллальные
(Surirellales). Он не связан с осевым полем створки, обычно рас-
полагается в более или менее выступающем гребне или киле,
идущем продольно вдоль середины створки или лежащем экс-
центрично ближе к одному из ее краев (нищпия). Каналовидный
шов представлен обычно непрерывным трубковидным каналом
в толще створки (возможно, занятым цитоплазмой), сообщаю-
щимся с наружной средой посредством тонкой щели, гомологичной
наружной щели щелевидного шва. С внутренней полостью клетки
канал соединяется через отверстия или поры, гомологичные вну-
тренней щели щелевидного шва (рис. 25, Б). Диатомовые, имеющие
шов (щелевидный или каналовидный), способны к движению. Ме-
ханизм движения окончательно не выяснен. Имеется ряд гипотез.
По одной, наиболее распространенной гипотезе движение клетки —
следствие трения о субстрат цитоплазмы, циркулирующей в шве.
Благодаря внутриклеточному тургору цитоплазма вгоняется в ве-
ртикальный канал центрального узелка, откуда по внутренней
щели шва гонится к полярным узелкам, а з ам по воронкообразному
каналу передвигается в конечную щель полярного узелка и попадает
в наружную щель шва, по которой течет по направлению к цент-
ральному узелку. В центральном узелке по вертикальному ка-
налу, соединяющему наружную и внутреннюю щели, этот ток
125
Рис. 25. Строение швов в панцире диатомовых водорослей: А — шов шелевидный
{1 — разрез в области центрального узелка, 2 — шов в районе конечного узелка);
Б—часть створки Surirella с каналовидным швом:
нщ - наружная щель, вщ — внутренняя щель, к канал, соединяющий наружную и внутреннюю
щели, с — канал, соединяющий две ветви шва в центральном узелке, кщ конечная щель,
в — конеп внутренней шелн шва
цитоплазмы может перейти во внутреннюю щель этой же ветви
шва, ч тобы повторить пройденный путь, но может попасть и не-
посредственно в полость клетки или же через горизонтальную
борозду, соединяющую в центральном узелке обе ветви шва,
переходит во внутреннюю щель другой ветви шва и начинает
там круговой путь в противоположном направлении. Трение стру-
ящейся цитоплазмы о субстрат и развивает моторную силу, дви-
126
жущую клетку в направлении, противоположном току цитоплазмы.
Движение форм, имеющих каналовидпый шов без центрального
узелка, значительно интенсивнее благодаря непрерывному току
цитоплазмы.
Согласно другой гипотезе скольжение диатомовых обусловлено
движением жидкости в шве, представляющем собой пластинчатый
капилляр, в который жидкость втягивается под действием капил-
лярных сил.
У имеющих шов пеннатофициевых хлоропласты (обычно в виде
одной-двух крупных пластинок) располагаются на поясковой сторо-
не клетки, лишь слегка заходя на створковую. Перед делением
клетки хлоропласты перемещаются на створку. У бесшовных форм
(синедра и др.) хлоропласты располагаются со стороны створки.
Ничего сравнимого со швом не наблюдается у представителей
цен грофициевых. Они все неподвижны.
Многие диатомеи в течение всей жизни встречаются в виде
отдельных клеток, однако известны и колониальные формы, как
свободноплавающие, так и прикрепленные. Колонии берут свое
начало от одной клетки, которая в результате ряда делений дает
свойственную данному виду форму колонии. Как правило, колонии
образуются у видов, не способных к движению, т. е. совсем не
имеющих шва (фрагиляриальные из пеннатофициевых и центрофи-
циевые). Виды же с вполне развитым щелевидным или каналовид-
ным швом формируют колонии редко, и при этом клетки, входящие
в состав колонии, обычно не теряют способности к движению
(Bacillaria paradoxa, виды родов навикула, ницшия, цимбелла, живу-
щие в слизистых трубках).
Клетки прикрепляются к субстрату и соединяются в колонии
с помощью слизи, выделяемой через сравнительно крупные поры на
поверхности створки. Иногда в скреплении клеток в колонии, поми-
мо слизи, принимают участие различные выросты створок—рога,
щетинки, зубчики и пр. Все громадное разнообразие форм колоний
можно свести к трем основным типам. К первому типу относятся
студенистые колонии, в которых клетки целиком погружены в вы-
деляемую ими слизь. Колонии имеют вид бесформенных комочков
или трубок, неветвящихся или разнообразно разветвленных.
Второй тип включает нитевидные, лентовидные, цепочковидные
и звездчатые колонии, форма их зависит от способа скрепления
клеток в колонии и от формы створок. Во всех случаях клетки
соединяются только створками, соединения поясками никогда не
бывает, так как в плоскости пояска клетки делятся. Если смежные
клетки соединяются всей поверхностью створки, то в зависимости
от формы створок образуются колонии нитевидные, как у некото-
рых видов циклотеллы и у рода мелозира (Melosira), лен-
товидные— например у фрагилярии, вееровидные — у меридион.
Если же клетки смыкаются только концами створки, то возникают
или зигзагообразные цепочки, как у рода диатома (Diatoma), или
127
звездчатые колонии, характерные для рода астерионелла
{Asierionella).
У различных планктонных центрофициевых клетки соединяются
выростами створок (рогами, щетинками), слизистыми тяжами и то-
гда образуются не плотные, а рыхлые цепочки, в которых клетки
более или менее удалены друг от друга. Таковы роды хегоцерос
(Chaeloceros), скелетонема {Skeleionema. рис. 27, Л—В), талассиози-
ра (Thalassiosira. рис. 27, Г—Ж).
Третий тип колоний наблюдается у форм, прикрепленных к суб-
страту слизью, которая выделяется локально на базальном конце
клетки через поры. Слизь имеет вид тяжей неразветвленных или
древовидно ветвящихся, на концах которых сидят клетки — по од-
ной или помногу. Примерами могут служить виды синедры, гом-
фонемы, ликмофоры.
Электронно-микроскопические исследования панциря диатомо-
вых водорослей показали, что кремневые стенки панциря бывают
однослойными (ламинарные) и двухслойными (локуларные) незави-
симо от принадлежности данного вида к центрофициевым или
пеннатофициевым диатомовым. В первом случае стенка панциря
состоит из одной пластины, во втором—из двух пластин: наружной
и внутренней, пространство между которыми разделено вертикаль-
ными перегородками на камеры. Кроме того, было показано, что
створки панциря всегда пористые, и один из основных элементов
структуры панциря (помимо выростов, шипов и пр.) — именно по-
ры. По классификации X. Окуно (Н. O^uno, 1975) различают три
основных типа пор: отверстия, локулы, альвеолы. Отверстия — это
мелкие, в виде следа булавочного укола округлые или щелевидные
перфорации, возникающие в более или менее тонких однослойных
(ламинарных) стенках (например, у хетоцерос). Отверстия лишены
замыкающих ситовидных мембран (рис. 26, А). В двухслойных (ло-
куларных) стенках встречаются или округлые и полигональные
перфорации локулы, или более или менее вытянутые перфора-
ции— альвеолы. И те и другие снабжены замыкающими ситовид-
Рис. 26. Структура стенок панциря по данным электронной микроскопии. А — лами-
нарная стенка с отверстиями; Б—Д — локуларные стенки (Б. В локулы с ситовид-
ными, пронизанными ситовидными порами мембранами, расположенные снаружи
или у основания локул; Г Д -альвеолы, замкнутые ситовидными мембранами
с их наружной или внутренней стороны)
128
ними мембранами, перфорированными, в свою очередь, ситовид-
ными порами. Локулы могут иметь хорошо выраженные высокие
латеральные стенки, которые вдаются глубоко внутрь, и тогда
ситовидная мембрана находится снаружи локулы (рис. 26, Б); если
латеральные стенки высоко выступают наружу, то ситовидная мем-
брана расположена у основания локулы (рис. 26, В). В первом слу-
чае изнутри, а во втором--снаружи локул обычно наблюдается
покровная мембрана, перфорированная всегда одной крупной по-
рой— покровной порой. В том и другом чипе локул покровная
мембрана может отсутствовать. Латеральные стенки бывают с ла-
теральными порами или без них. В толстых стенках, перфорирован-
ных удлиненными альвеолами, как и в случае локул, латеральные
стенки вдаются внутрь или выступают кнаружи, соответственно
ситовидная мембрана располагается снаружи от альвеолы или у ее
основания (рис. 26, Г, Д). Покровные мембраны могут присутство-
вать или отсутствовать. Пор в латеральных стенках альвеол до сих
пор не наблюдалось.
Ситовидные мембраны, которые замыкают локулы или альве-
олы на их наружной или внутренней поверхностях, имеют характер-
ную тонкую структуру. Ситовидная мембрана перфорирована од-
ной или многими крошечными ситовидпыми порами округлых,
эллиптических, прямоугольных или полигональных очертаний. Си-
товидные поры, в свою очередь, часто замкнуты тонкой вторичной
мембраной с ультратонкими вторичными ситовидными порами.
Форма, размер и расположение ситовидных пор могут служить
в качестве надежного признака для идентификации видов.
Несколько подробнее можно рассмотреть структуру панциря
у рода пиннулярия (Pimularia). В панцире пинпулярии альвеолы
поперечно вытянуты, простираясь почти от шва до края створки.
Они представлены корытообразной (или в форме перевернутого
желобка) полостью, замкнутой снаружи только пористой ситовид-
ной мембраной. У некоторых видов пиннулярии альвеолы, лишен-
ные покровной мембраны, полностью открыты внутрь, у других
видов альвеолы изнутри загорожены тонкой покровной мембраной
с отверстием— покровной порой. Покровные поры могут быть
округлыми, эллиптическими или другой формы, они расположены
на створке в две продольные линии. Эти ряды покровных пор
различимы и в световом микроскопе. Поперечные боковые (лате-
ральные) стенки альвеол более или менее глубоко вдаются внутрь
и лишены пор. Таким образом, соседние альвеолы полностью раз-
делены этими боковыми стенками.
Электронно-микроскопическая гонкая структура пор панцирей
диатомовых водорослей строго видоспецифична. Согласно X. Оку-
но, «надежность тонкой структуры пор панциря для идентификации
видов диатомовых водорослей может быть сравнима с надежно-
стью отпечатков пальцев для идентификации человеческих индиви-
дуумов».
5 - 1210
129
Значение пор, по-видимому, заключается в том, что через них
содержимое клетки сообщается с окружающей средой.
Во многих руководствах и исследованиях можно встретиться
и с несколько иной терминологией. Различают два основных элемен-
та структуры панциря: поры и ареолы. В однослойных — ламинар-
ных стенках образуются только поры, которые могут быть откры-
тыми в виде сквозного отверстия или полузамкнутыми мембранами
разного типа. Ареолы образуются только в двухслойных—-локу-
лярных стенках и обычно затянуты снаружи или внутри тонкой
перфорированной пленкой—«велумом» или «крибрумом» (сито,
решето). Помимо пор, как уже указывалось, на створках имеются
различного рода выросты, характер и расположение которых слу-
жат важными таксономическими признаками. Так у ряда центрофи-
циевых диатомей (например, у талассиозиры, скелетонемы, стефа-
нодискуса) существуют вырост ы: центральные, краевые (маргиналь-
ные), промежуточные между ними и одиночный или добавочный
вырост, расположенный или между маргинальными выростами,
или же между центром и краем створки. Г. Хасле (G. R. Hasle, 1972)
подробно изучила их строение и назначение. Центральные выросты
оказались полыми трубками (цилиндрами), иногда расширенными
на вершине, основание их окружено ободком в виде муфты, пре-
рванной в четырех (реже трех) местах, что позволяет видеть стенки
цилиндров до самого основания. Хасле назвала эти трубки «подпер-
тыми трубками». Их функция состоит в выделении плотной слизи,
образующей студенистые тяжи, соединяющие смежные клетки в це-
почковидную колонию. Краевые и промежуточные выросты также
трубчатые, открытые наружу и внутрь клетки, через них выделяется
слизь, покрывающая всю поверхность створки. Эти выросты свой-
ственны только планктонным видам. Крупный одиночный вырост
тоже трубчатый, иногда па вершине он расширен, а внутрь клетки
открывается узкой щелью с утолщенными краями в виде двух губ.
Хасле назвала эти выросты двугубыми; функция их пока не ясна.
Примерами строения и назначения этих выростов могут служить
выросты у представителей родов талассиозира (Thalassiosira) и ске-
летонема (Skeletonema).
У большинства видов талассиозиры клетки образуют рыхлые
цепочки посредством слизистого тяжа, соединяющего центры смеж-
ных створок (тяж виден в оптическом микроскопе в виде тонкой
нити). С помощью сканирующего электронного микроскопа (СЭМ)
показано, что эта соединительная нить между соседними панцирями
образована несколькими нитями, которые выдавливаются из 7—12
(и более) «подпертых трубок», расположенных в центре створки.
Помимо центральных подпертых трубок, на створке имеются более
мелкие подпертые трубки и один двугубый отросток. У разных
видов они расположены по-разному (рис. 27, Г—Ж). Отличитель-
ный признак рода скелетонема (Skeletonema) — соединительные от-
ростки— маргинальные подпертые трубки в числе 7—9 со срав-
130 *
Рис. 27. Skeletonema (Л —В): А — общий вид колонии; Б—S. subsalsum (соединитель-
ные отростки); В—S. eostatum (соединительные отростки с продольной щелью);
Thalassiosira (Г - Ж); Г—общий вид колонии; Д — створка с маргинальными и цен-
тральными отростками; Е—часть створки: виден соединительный тяж, выходящий
через центральные подпертые трубки; Ж—часть створки с краевыми отростками
нигельно длинными наружными частями, которые смыкаются обы-
чно по одному с такими же отростками смежных клеток, образуя
рыхлые цепочки (рис. 27, А). Структура соединительных отростков
у разных видов неодинакова. Так, у S. subsalsum (рис. 27, Б) они
трубчатые, а у S. eostatum расщеплены в виде желоба почти на
протяжении всей длины (рис. 27, В).
Помимо трубковидных, створки могут нести и иные выросты,
часто также служащие для объединения клеток в колонии. Напри-
мер, у рода хетоцерос (Chaetoceros) с цепочкообразиыми колониями
клетки соединяются с помощью длинных негнущихся щетинок.
Овальные створки биддульфии тоже снабжены выростами, которые
объединяют клетки в рыхлые цепочки.
Размножение. Обычный способ размножения у диатомей — кле-
точное деление. В результате этого процесса оба образующихся
индивида получают одну створку ’Ът родительской клетки, тогда
как другая возникает заново. Две створки в делящейся клетке
слегка расходятся так, что остаются соединенными только края их
131
поясковых ободков. Ядро претерпевает митоз. Расщепление прото-
пласта начинается с периферии, идет ковнутри и всегда происходит
в плоскости, параллельной створкам. На заново образовавшейся
свободной поверхности обоих дочерних протопластов формируют-
ся новые створки, а затем около каждой из новых створок
возникает поясковый ободок и всегда ковнутри от поясковых
ободков первоначального индивида. Таким образом, поясковый
ободок каждой родительской створки перекрывает поясковый
ободок заново образовавшейся створки, и гипотека родительской
клетки становится эпитекой одного из дочерних индивидов.
Возникновение новых створок можно проследить только с помо-
щью электронного микроскопа. Электронно-микроскопические
исследования (первая работа появилась в 1964 г.) показали, что
в каждой полученной в результате деления клетке, окруженной
плазмалеммой, недостающая створка формируется внутри перифе-
рического плоского, окруженного мембраной пузыря — «пузыря
отложения кремнезема», который служит матрицей, определяя
окончательную морфологию створки. Пузырь отложения крем-
незема возникает под плазмалеммой, по-видимому, благодаря
слиянию пузырьков Гольджи. Мембрана, ограничивающая этот
пузырь, получила название силиколеммы; возможно, она ответст-
венна за отложение кремнезема, т. е. за транспорт иона SiO^2
в пузырь и его полимеризацию там до кремнезема. По достижении
новой створкой зрелости и ее освобождении силиколемма, об-
ращенная внутрь, становится плазмалеммой' и, возможно, состав-
ляет часть органического покрова, возникающего в основном за
счет слияния силиколеммы и плазмалеммы, обращенных кнаружи
клетки.
Ниже (рис. 28) приводится несколько упрощенная схема процес-
са образования новых половинок панциря (створки и первого по-
яскового кольца) в делящейся клетке Gomphonema parvulum. Срав-
нительно недавно у Fragilaria capucina было прослежено возникнове-
ние интеркалярных — вставочных поясков. Было показано, что они
также формируются внутри пузырей отложения кремнезема, воз-
никающих непосредственно под плазмалеммой, по-видимому, в ре-
зультате слияния везикул- дериватов аппарата Гольджи. Ободки
покрыты матриксом (являющимся, возможно, продуктом слияния
плазмалеммы и силиколеммы), который удерживает их вместе.
Таким образом, элементы панциря (створки, поясковые кольца,
вставочные ободки) являются независимыми единицами, которые
и формируются в сепаратных «пузырях отложения кремнезема».
Этим подтверждаются более ранние данные световой микроскопии:
поясковые и вставочные ободки не образуются до тех пор, пока не
будет завершено формирование створки.
132
' Однако в некоторых случаях плазмалемма дочерних клеток формируется,
по-видимому, заново, без участия силиколеммы.
/
Рис, 28. Схема образования створки (включая образование первого пояскового коль-
ца Gomphonema parvulum). А — деление ядра, клеточные органеллы распределяются
на две приблизительно равные части, клетка растягивается в первальварной плоско-
сти; Б- ядро поделилось; инвагинация плазмалеммы и отложение новой плазмалем-
мы из пузырьков аппарата Гольджи (получаются два дочерних протопласта, окру-
женных полностью плазмалеммой и наследующих по одной родительской створке);
В -непосредственно под плазмалеммой, одевающей свободные поверхности дочер-
них клеток, формируется «пузырь отложения кремния», пузырьки Гольджи сливают-
ся с образованием силикалеммы; Г—в пузыре произошло отложение кремнезема,
новая створка (вальва) стала электронно-плотной (у места возникновения первого
пояскового кольца начинают скапливаться пузырьки Гольджи, которые, сливаясь,
дадут пузырь, ограниченный силиколеммой, в котором отложится поясковое коль-
цо); Д — в каждой из двух дочерних клеток образовалось по поясковому кольцу,
которые выстроились под покровом новых вальв (снаружи от новых створок видны
остатки плазмалеммы и наружной силиколеммы. Внутренняя силиколе.мма стано-
вится частью плазмалеммы дочерней клетки):
я— ядро, а — аппарат Гольджи, пг — пузырьки Гольджи, л — плазмалемма, си—ситиколемма,
вк — везикул, где будет отложен кремнезем, по — поясковые кольца, э—эпитека, г— гипотека,
р -рафе
В результате деления клетки одна из дочерних клеток, полу-
чившая от материнской эпитеку, точно повторяет размеры мате-
ринской клетки. Вторая дочерняя клетка, получившая от материнс-
кой гипотеку, которая стала для нее эпитекой, несколько меньших
133
размеров: на двойную толщину пояскового ободка. Из-за окре-
мнения панциря клетки могут увеличиваться только вдоль оси,
соединяющей две створки. В результате размеры клеток в попу-
ляции диатомовых прогрессивно уменьшаются. Скорость этого
уменьшения размеров не так уж велика, некоторое растяжение
панциря у слабо окремнелых диатомей все же наблюдается и ча-
стота делений, особенно меньших из двух дочерних клеток, не-
велика. Специальное исследование показало, что уменьшение раз-
меров клеток до минимальных может растягиваться на несколько
лет: на два — пять у Stephanodiscus hantzschii, на четыре года
у Tabellaria fenestrata. Затем следует краткий период образования
растущих спор — ауксоспор, возникающих в результате полового
процесса. Зигота, одетая неокремнелой растяжимой оболочкой, пре-
вращается в ауксоспору, которая растет, пока не достигнет ма-
ксимальной для данного вида величины, после чего образуется
панцирь и начинается ряд последовательных вегетативных делений
клеток.
У пеннатофициевых (за одним исключением) половой процесс
напоминает конъюгацию, когда сливаются протопласты вегетатив-
ных клеток. В качестве примера можно рассмотреть половой про-
цесс у Gomphonema geminatum, подробно изученный русским аль-
гологом К. И. Мейером. Две клетки сближаются и окружаются
толстой слизистой обверткой. Ядро каждой из конъюгирующих
клеток претерпевает редукционное деление, а протопласт делится
с образованием двух гамет одинакового размера. Гаметы сначала
содержат по два ядра, одно из которых остается жизнеспособным,
а другое дегенерирует. После этого створки конъюгирующих клеток
расходятся, и одна гамета из каждой клетки переползает в клетку
партнера с тем, чтобы слиться там с другой гаметой. Возникшие
в результате конъюгации две зиготы начинают быстро расти и пре-
вращаются в ауксоспоры. Ауксоспоры сначала двухъядерные, но
вскоре происходит кариогамия и более зрелые ауксоспоры содержат
по одному диплоидному копуляционному ядру. Разрастаясь, они
растягивают, а затем и разрывают общую слизистую обвертку,
одевающую конъюгантов, в которой можно видеть створки конъю-
гирующих клеток, значительно меныпих размеров, чем ауксоспоры.
Достигшая максимальных размеров ауксоспора образует панцирь
со створками, несколько различающимися между собой: появля-
ющаяся первой — более толстая и выпуклая, другая створка—плос-
кая. Только после деления ауксоспоры дочерние клетки приобрета-
ют признаки, свойственные вегетативным клеткам данного вида.
У большинства пеннатофициевых половой процесс протекает таким
же образом, т. е. в каждом из конъюгантов образуется по две
гаметы и в результате возникает по две ауксоспоры. В деталях
могут быть и различия. Например, у Anomoeoneis sculpta деления
в конъюгантах неравновеликие, образуются по две гаметы, различа-
ющиеся по размерам. Затем гаметы разного размера из разных
134
конъюгантов попарно сливаются. Реже у пеннатофициевых (напри-
мер, у суриреллы, некоторых навикул) редукционное деление ядра
в конъюгирующих клетках не сопровождается цитокинезом и таким
образом образуется по одной гамете, содержащей сначала четыре
ядра, из которых три затем отмирают. В результате такого полово-
го процесса формируется только одна зигота, соответственно — ау-
ксоспора. У некоторых пеннатофициевых известны случаи апогамии
(аутогамии), когда ауксоспоры возникают из одной-единственной
родительской клетки. Здесь сливаются два ядра из четырех получив-
шихся в процессе редукционного деления в неразделившихся прото-
пластах, которые превращаются в зиготы, вырастающие в ауксо-
споры.
Таким образом, у пеннатофициевых имеет место гаметическая
редукция; перед образованием гамет вся вегетативная жизнь их
проходит в диплоидном состоянии — они являются диплонтами (то
же относится к центрофициевым).
В начале XX в. проблема изучения жизненного цикла цент-
рофициевых усложнилась благодаря обнаружению сначала у рода
биддульфия (Biddulphia), а позднее и у других родов монадных
клеток — микроспор. Ряд авторов приписывали им роль гамет
(хотя никто не наблюдал слияния их) и соответственно отрицали
половой акт при образовании ауксоспор. В 1929 г. русский ис-
следователь Б. М. Персидский, соглашаясь с тем, что ауксоспоры
центрофициевых возникают из одной клетки—материнской клетки
ауксоспор, без слияния ее с другой клеткой, отрицает бесполый
характер ауксоспор. У двух видов рода хетоцерос и у Melosira
varians Б. М. Персидский наблюдал в материнской клетке ауксоспор
редукционное деление ядра и образование четырех гаплоидных
ядер. Он утверждал, что два из четырех ядер отмирают, а два
оставшихся сливаются в молодой ауксоспоре, которая возникает
таким образом в результате апогамного полового процесса. По-
зднее то же наблюдалось у Cyclotella meneghiniana. На основе
этих данных до 1950 г. господствовало мнение, что у центро-
фициевых ауксоспоры образуются из одной клетки, в которой
происходит редукционное деление ядра, и два ядра сливаются
в созревающей ауксоспоре; этот процесс сравнивался с встреча-
ющейся у некоторых пеннатофициевых аутогамией. Однако
в 1950—1954 и последующие годы появляются работы, в которых
у различных представителей центрофициевых описывается оогам-
ный половой процесс. Первая работа Г. Штош (Н. A. Stosch, 1950)
относилась к пресноводной водоросли Melosira varians — виду,
изученному Б. М. Персидским. Штош показал, что ауксоспора
М. varians является разросшейся зиготой, получающейся в резуль-
тате слияния сперматозоида с материнской клеткой ауксоспоры.
Согласно Штош, антеридии возникают в результате двух, реже
одного, трех или четырех делений из вегетативных клеток. В ан-
теридиях происходит редукционное деление, вокруг получившихся
135
четырех гаплоидных ядер обособляется цитоплазма и образуются
одножгутиковые, лишенные хроматофоров сперматозоиды. Анте-
ридии и оогонии (материнские клетки ауксоспор) могут встречаться
на протяжении одной и той же нити. По мнению Штоша,
Б. М. Персидский ошибочно отождествил антеридии с четырьмя
ядрами как стадию тетрады, предшествующую автогамной кари-
огамии при образовании ауксоспор у мелозиры.
Обнаружение подвижных монадных сперматозоидов у мелозиры
позволило предположить, что хорошо известные микроспоры мно-
гих морских центрофициевых также являются сперматозоидами
и что и у них образованию ауксоспор предшествует оогамия
(Н. A. Stosch, 1954). Это предположение подтвердилось и в последу-
ющие годы. Штош устанавливает оогамию у целого ряда морских
центрофициевых: Lithodesmium, Bellarochea, Streptotheca, Biddulphia
rhombus, В. granulata, В. mobiliensis, Cerataurus, Aulacodiscus. Л. Гей-
тлер (L. Geitler, 1952) описал оогамию у циклотеллы.
В деталях половой процесс и ово- и сперматогенез у разных
видов неодинаков, В материнской клетке ауксоспор — оогонии по-
сле редукционного деления ядра могут образоваться два яйца (ка-
ждое сначала содержит одно ядро генеративное и одно отмира-
ющее) или же в оогонии формируется только одна яйцеклетка
(жизнеспособным сохраняется одно ядро, три остальных дегене-
рируют). Первый путь овогенеза характерен, например, для В.
mobiliensis, В. granulata, Lithodesmium. Второй тип (с образованием
только одного яйца в оогонии) наблюдается у Melosira varians,
Cyelotella tenuistriata, Streptotheca. Промежуточное положение за-
нимает В. rhombus, у которой в оогонии имеет место неравное
деление протопласта, в результате которого образуется крупная
яйцеклетка и мелкое, лишенное хлоропластов так называемое на-
правляющее тельце. В первом случае в результате полового про-
цесса формируются две ауксоспоры, во втором — только одна. Яй-
цеклетки, возникающие в оогонии в числе двух или одной, в одних
случаях (например, у биддульфии, мелозиры) оплодотворяются
внутри оогония, т. е. происходит истинная оогамия, в других (у
литодесмиума, стрептотеки, циклотеллы) — только после выхода
яйцеклеток из оогония, т. е. наблюдается примитивная оогамия
(рис. 29). В антеридиях все четыре ядра, возникающие в результате
редукционного деления исходного диплоидного ядра, остаются
жизнеспособными и вокруг них обособляются четыре спермато-
зоида. Однако дифференцировка самих антеридиальных клеток
(сперматогониев) у разных представителей происходит неодинако-
во. У Cyelotella tenuistriata вегетативные клетки непосредственно
становятся антеридиями (сперматогониями). У Melosira varians из
одной вегетативной клетки возникают 4—8 более мелких, с не-
сколько упрощенными панцирями, бедных хлорофиллом сперма-
тогониев. У Biddulphia rhombus и В. gramdata сперматогонии, об-
разующиеся из вегетативной клетки (у первого вида 4, у второго —
136
Рис. 29. Оогонии (слева), спермато! онии (справа). А—Biddulphia mobiliensis; Б—
В. granulate; В—rhombus; Г—Lithodesmium; Д—Streptotheca (объяснение в тексте)
8), более редуцированы: одеты сильно упрощенными панцирями
и до мейоза заключены в материнской клетке. У В. mobiliensis
после ряда митозов из вегетативной клетки появляются 16 ан-
теридиев, которые освобождаются после расхождения створок, об-
разующихся только после первого митоза (после остальных ми-
тозов створки не формируются). То же имеет место и у лито-
десмиума, стрептотеки и некоторых других, у которых отсутствуют
все створки, кроме сформировавшихся после первого митоза, так
что голые сперматогонии, возникающие в больших количествах,
137
заканчивают свое развитие (мейоз, формирование сперматозоидов)
свободно в окружающей среде (рис. 29). '
Штош наблюдал слияние сперматозоида с яйцом у В. mobiliensis
на живом материале, а также представил цитологические доказате-
льства слияния ядер. Он также продемонстрировал эксперименталь-
но. что яйца способны прорастать в ауксоспоры только в присутст-
вии сперматозоидов. В 70-х годах подробно описан жизненный цикл
и оогамный половой процесс у нескольких видов хетоцерос. Оога-
мия описана лишь у одного вида пеннатофициевых диатомовых —
Rhabdonema adriaticum, Однако мужские клетки здесь лишены жгу-
тика и они передвигаются посредством тонких псевдоподий.
Вегетативные клетки большинства диатомовых водорослей спо-
собны выносить высыхание, низкие температуры и прочие не-
благоприятные условия. Однако у некоторых форм, преимуще-
ственно морских центрофициевых, реже у пресноводных видов,
известны покоящиеся споры или цисты. Образованию их обычно
предшествует деление клетки, при этом дочерние клетки покры-
ваются толстостенными выпуклыми створками, смыкающимися
своими краями, поясковые кольца отсутствуют. Створка, возни-
кающая первой, покрыта шипами или выростами, створка, об-
разующаяся позже, гладкая.
* * *
Критерии, которые кладутся в основу систематического деления
диатомовых водорослей на классы, порядки, семейства и т. д., каса-
ются формы створок, деталей их структуры, отсутствия или нали-
чия вставочных ободков, шва, их формы и т. п. Как уже упомина-
лось, отдел диатомовые водоросли делится на два класса: центро-
фициевые и пеннатофициевые.
КЛАСС ЦЕНТРОФИЦИЕВЫЕ —CENTROPHYCEAE
Створки обычно округлых очертаний и радиального строения,
шов отсутствует, половой процесс — оогамия. Примерами могут
служить роды циклотелла, мелозира, талассиозира, скелетонема,
биддульфия, хетоцерос.
КЛАСС ПЕННАТОФИЦИЕВЫЕ — PENNA TOPHYCEAE
Створки вытянутые, с двусторонней симметрией. У одних шов
щелевидный, что характерно для порядка навикулальные
(Naviculales), или каналовидный (порядок суриреллальиые —
Surirellales), у других шов отсутствует — порядок фрагиляриальные
(Fragilariales). Половой процесс (за одним исключением) по типу
конъюгации. К пеннатофициевым относятся роды пиннулярия, на-
викула, цимбелла, гиросигма, гомфонема, ахнантес (порядок нави-
138
кулальные); ницшия, сурирелла (порядок суриреллальные); синедра,
фрагилярия, диатома, астерионелла, меридион (порядок фрагиляри-
альные).
Оба класса—центрофициевые и пеннатофициевыс, по-видимому, произошли от
одного общего предка, в пользу чего говорит диплоидная природа обеих групп,
сходство строения панциря, пигментации и обмена вешсств. Центрофициевые и ли-
шенные шва пеннатофициевыс могут представлять собой дивергирующие линии
эволюции от этого общего ствола, в то время как остальные пеннатофициевыс,
главным образом бентосные формы, эволюционировали в сторону выработки слож-
ного строения шва. Из двух классов центрофициевые больше сохранили черт,
связывающих их с какими-то отдаленными жгутиковыми предками, в пользу чего
свидетельствует наличие у них жгутиковых стадий (сперматозоидов).
Диатомовые встречаются во всех видах вод, а также во многих типах почв.
Мншие несущие шов виды являются донными формами, живущими в пресной
и соленой воде, другие -эпифиты — на более крупных растениях. Диатомовые,
лишенные шва, играют большую роль в планктоне, причем в море преобладают
центрофициевые, а в пресноводном планктоне фрагиляриальные. В качестве созда-
телей органического вещества в водоемах диатомовые занимают среди всех водорос-
лей первое место. Поскольку сильно окремневшие оболочки сохраняются неизмен-
ными после отмирания водорослей, то в местах массового развития диатомей
образуются обширные отложения, известные как диатомовая земля или кизельгур,
имеющие важное экономическое значение: как полировочный материал, для тепло-
вой изоляции, как связующая масса при изготовлении динамита. В меловых отложе-
ниях диатомовые представлены большим количеством видов. Совершенство стро-
ения меловых диатомей заставляет предполагать, что они уже до мелового периода
прошли длительную эволюцию.
Глава 10
ОТДЕЛ БУРЫЕ ВОДОРОСЛИ—PHAEOPHYTA
Водоросли этого отдела характеризуются хроматофорами,
окрашенными в бурый цвет благодаря тому, что кроме хлорофил-
лов (а и с; хлорофилл h отсутствует), Р-каротина имеется избыток
бурых ксантофиллов, особенно фукоксантина.
Хлоропласты, помимо оболочки, окружены сложной системой
мембран, находящейся в прямой связи с мембраной ядра—«хло-
ропластной эндоплазматической сетью». Матрикс хлоропласта пе-
ресекается параллельными трехтилакоидными ламеллами, окру-
женными одной или несколькими периферическими (опоясывающи-
ми) ламеллами. Свободный от тилакоидов пиреноид выступает
из хлоропласта в виде почки (рис. 30). Генофор кольцевидный,
под периферическими ламеллами. Запасной полисахарид — лами-
нарии— откладывается вне хлоропласта, в цитоплазме. Помимо
ламинарина, запасными продуктами служат шестиатомный спирт
Маннит и жиры, Монадные клетки (зооспоры и гаметы) имеют
глазок и жгутики. Глазок является частью пластиды и связан со
139
у
Рис. 30. Схема строения клетки бурых водорослей:
кс—клеточная стенка, м — митохондрии, эс фрагмент канала эндоплазматической
сети, д диктиосомы, в — вакуоль, я — ядро, яо — ядерная оболочка (в перинуклеар-
ном пространстве заключен хлоропласт), х — хлоропласт с трехтилакоидными ламел-
лами, п —пиреноид с обкладкой, г —кольцевидный генофор, перерезанный поперек
жгутиковым аппаратом. Жгутиков, как правило, два (у диктио-
тальных один) — гетероконтный и гетероморфный. Мембрана пе-
реднего (обычно более длинного) жгута на своей поверхности несет
сложно устроенные мастигонемы (рис. 31, В), Задний (обычно более
короткий) жгут гладкий, его основание, прилегающее к глазку,
несет вздутие (рис. 31, Г). У гамет эктокарпуса каждый жгутик
несет длинный, часто спирально закрученный терминальный при-
даток - - акронему, в которую из стержня жгута переходят две
центральные микротрубочки (рис. 31, Л, Б).
За немногими исключениями бурые водоросли — морские, осо-
бенно богато представленные в холодных водах северного и южно-
го полушария. Таллом самых простейших из ныне живущих бурых
водорослей гетеротрихальный, громадное же большинство их име-
ют крупные компактные талломы ложно- или истиннотканевого
строения.
Размножение — вегетативное, бесполое и половое. Вегетативное
размножение происходит участками таллома. У некоторых име-
ются специализированные веточки (выводковые почки), которые
легко отламываются и вырастают в новые талломы. Бесполое
размножение у большинс!ва бурых водорослей осуществляется по-
средством зооспор, возникающих в одногнездных или однокамер-
140
ных спорангиях, которые развива-
ются на диплоидных растениях
(спорофитах) и в ко торых перед об-
разованием зооспор ядра редукци-
онно делятся. У представителей
порядка диктиотальных (Dictyota-
les) в одногнездных спорангиях (те-
траспорангиях) вместо зооспор, та-
кже после редукционного деления
ядра, формируются четыре непо-
движные апланоспоры — тетраспо-
ры. Гаплоидные зооспоры и тетра-
споры прорастают в гаплоидные
половые растения — гаметофиты;
на них развиваются половые орга-
ны. У простейших бурых водорос-
лей половой процесс изогамный,
гаметы образуются в многогнезд-
ных или многокамерных споранги-
ях, состоящих из большого числа
(до нескольких сотен) мелких куби-
ческих клеток. Протопласт каждой
клетки превращается в одну гаме-
ту. У ряда бурых водорослей на-
блюдается гетерогамия: гаметы та-
кже возникают в многокамерных
спорангиях двух типов: одни состо-
ят из большего числа мелких кле-
ток, другие — из меньшего количе-
ства более крупных клеток. Соот-
ветственно в первых образуют-
ся мелкие микрогаметы, а во
вторых — крупные макрогаметы.
У наиболее высокоорганизованных
бурых водорослей половой процесс
оогамный. В оогониях и антериди-
ях, как правило, формируется по
одной гамете (яйцеклетке и спер-
матозоиду соответственно). Ис-
ключение составляют представите-
ли порядка фукальные, в антериди-
ях которых развиваются 64 спер-
Рис. 31. Схема строения монадных кле-
ток бурых водорослей. А -гамета
Ectocarpus', передний жгутик с мастиго-
нсмами (тм) и акронемой (ак), задний
жгутик гладкий с акронемой (ак); Б—
переход от стержня жгута к акронеме,
содержащий только две централь-
ные микротрубочки аксонемы жгута;
В схема строения мастигонемы
у Ascophyllum (ом- основание масти-
гонемы, тм - трубчатая часть, тн — те-
рминальная нить); Г—сперматозоид
Eucus со вздутием у основания заднего
жгута, прилегающего к стигме (с).
матозоида, а оогонии у некоторых родов содержат более одной
(2, 4, 8) яйцеклетки; например, у рода фукус в оогонии образуются
8 яйцеклеток. Антеридии диктиотальных имеют вид многокамер-
ных спорангиев, в каждой камере образуется по одному спермато-
зоиду, снабженному в отличие от монадных клеток всех остальных
141
f
i бурых водорослей только одним жгутиком. Яйцеклетка оплодот-
воряется, как правило, вне оогония (примитивная оогамия). Зигота
; без периода покоя прорастает в диплоидное растение.
i Все бурые водоросли, за исключением представителей порядка
1 фукальных, у которых отсутствует бесполое размножение и ко-
। торые являются диплонтами, обнаруживают смену генераций:
одни — изоморфную, другие — гетероморфную. Эти разные типы
жизненного цикла положены в основу деления отдела бурых
водорослей (Phaeophyta) на три класса: изогенератофициевые
(Isogeneratophyceae), характеризующиеся изоморфной сменой гене-
раций, гетерогенератофициевые (Heterogeneratophyceae), включа-
ющие бурые водоросли с гетероморфной сменой генераций, и цик-
лоспорофициевые (Cyclosporophyceae) с одним порядком фукаль-
ные, в котором смена генераций отсутствует.
КЛАСС ИЗОГЕНЕРАТОФИЦИЕВЫЕ —ISOGENERATOPHYCEAE
Порядок эктокарпальные—Ectocarpales
Представителем этого порядка может служить род эктокарпус
{Ectocarpus'). Многочисленные виды его распространены во всех
морях, особенно холодных, растут на различных подводных объект-
: ах и на других более крупных водорослях. Таллом (как спорофита,
, так и гаметофита) имеет вид желтовато-бурых кустиков, достига-
ющих в длину нескольких сантиметров. Он состоит из стелющихся
по субстрату нитей, от которых отходят вертикальные нити из
: одного ряда клеток, обильно ветвящиеся. Ветви вертикальных ни-
тей часто оканчиваются многоклеточными бесцветными волосками.
Стелющиеся по субстрату нити обнаруживают верхушечный рост.
Рост вертикальных нитей у разных видов неодинаков. У большин-
ства видов рост диффузный и лишь у некоторых видов дифферен-
i цируется ясно выраженная интеркалярная вставочная меристема
у основания волосков; еще реже наблюдается верхушечный рост.
Жизненный цикл этой водоросли оказался гораздо более слож-
ным, чем он обычно описывался на основе наблюдений в природе.
Бесполое размножение — зооспорами. Они развиваются на дип-
лоидных растениях в одногиездных спорангиях, представляющих
конечные клетки коротких боковых ветвей. Содержимое их распада-
ется на ряд голых зооспор, выходящих через разрыв оболочки
спорангия. Образованию зооспор предшествует редукционное деле-
ние ядра и гаплоидные, таким образом, зооспоры вырастают в гап-
лоидные растения, размножающиеся половым путем. Гаметы об-
разуются в многогнездных спорангиях. Хотя гаметы внешне оди-
наковы и половой процесс у эктокарпуса изогамный, по поведению
можно различить женские и мужские гаметы. Первые быстро теря-
ют подвижность, вторые могут оставаться подвижными до 24 ча-
142
сов. Они роем окружают женскую гамету и прочно прикрепляются
к ее мембране с помощью акронемы переднего жгута. Затем одна
из мужских гамет сокращает свой передний жгутик, приближаясь
к женской гамете, с которой и сливается. Остальные мужские гаме-
ты открепляются и уплывают.
Зигота без периода покоя прорастает в диплоидное бесполое
растение. Таким образом, эктокарпус обнаруживает изоморфную
смену генераций, от которой в зависимости от климатических усло-
вий могут быть отклонения. Так, в некоторых местах обитания на
спорофитах эктокарпуса, помимо одногнездных спорангиев, наблю-
даются многогнездные—нейтральные спорангии, которые в отли-
чие от многогнездных спорангиев, развивающихся на гаметофитах
и являющихся гаметангиями, образуют в своих клетках зооспоры —
нейтральные споры. Эти споры диплоидны, так как перед их возник-
новением редукционного деления ядра не происходит, они прора-
стают в диплоидные растения—спорофиты.
В 1972 г. Д. Мюллер (D. G. Muller, 1972) опубликовал резуль-
таты своих многолетних исследований размножения Ectocarpus
siliculosus в культуре. Он показал, что, помимо сингамии (копуля-
ции) и мейоза, которые связывают названные выше стадии гамето-
фита и спорофита, имеется ряд добавочных путей развития, явля-
ющихся результатом процессов гетеробластии, спонтанного умно-
жения хромосом и партеногенеза.
Как и в природе, в культуре Е. siliculosus на спорофитах, помимо
одногнездных, развиваются многокамерные — многогнездные (ней-
тральные) спорангии. При образовании в этих спорангиях нейтраль-
ных зооспор мейоза не происходит и таким образом диплоидные
спорофиты размножаются посредством нейтральных спор из мно-
гогнездных (нейтральных) спорангиев. Из зооспор, возникающих
в результате мейоза на диплоидных спорофитах, вырастают не
только гаплоидные гаметофиты, но и гаплоидные спорофиты. Это
явление называется гетеробластией. Гаплоидные спорофиты раз-
множаются посредством спор, образующихся в многогнездных
(нейтральных) спорангиях, но в потомстве постоянно находится
небольшой процент диплоидных спорофитов, по-видимому, воз-
никающих в результате спонтанного умножения числа хромосом.
Образование одногнездных спорангиев на гаплоидном спорофите
не связано с мейозом, но потомство зооспор из этих одногнездных
спорангиев снова обнаруживает гетеробластию, т. е. образуются не
только гаплоидные спорофиты, но и гаплоидные гаметофиты.
Партеногенетическое развитие гаплоидных гамет приводит к об-
разованию исключительно спорофитов, большей частью гаплоид-
ных, но иногда и диплоидных. Иногда зооспоры из многогнездных
спорангиев гаплоидных спорофитов развиваются в тетраплоидные
спорофиты. Они могут размножаться посредством многогнездных
спорангиев, а в результате мейоза в одногнездных спорангиях дают
начало диплоидным спорофитам и диплоидным гаметофитам.
143
Термин «спорофит» Мюллер относит к растениям, несущим
одногнездные спорангии независимо от числа хромЬсом (п или 2л).
Пути развития Е. siliculosus в культуре представлены на рис. 32.
В ходе исследования Е. silkulosus было замечено, что суспензия
женских гамет издает очень приятный сладкий запах. Это летучее
вещество оказывает действие на мужские гаметы, изменяя направле-
ние их движения и обусловливая их аккумуляцию в месте наивысшей
концентрации вещества вокруг женских гамет. В эксперименте мужс-
кие гаметы собирались также вокруг капелек минерального масла,
содержащего это вещество. Через суспензию, содержащую женские
гаметы, и летучее вещество, ими выделяемое, пропускали очищен-
ный воздух, который затем проходил через холодильник при - 80’ С.
При этом вещество конденсировалось. В течение двух лет было
собрано 92 мг вещества (для получения 80 мг необходим 1 кг живого
материала), которое далее подвергалось очистке, газовой хромато-
графии и другим методам, позволившим впервые у водорослей
химически идентифицировать молекулы аттрактанта. Это углеводо-
род с эмпирической формулой CiiHie, названный эктокарпеном.
Порядок сфацеляриальные—Sphacelariaies
Род сфацелярия (Sphacelarlu), как и эктокарпус, широко рас-
пространен во всех морях и имеет небольшие гетеротрихальные
талломы. Однако рост как распростертых, так и вертикальных
нитей в талломе сфацелярии строго верхушечный. Сказанное от-
носительно размножения и цикла развития эктокарпуса в большой
степени приложимо и к сфацелярии. Диплоидные бесполые расте-
ния некоторых видов сфацелярии на одном и том же индивиде
могут образовывать как одногнездные, так и многогнездные (ней-
тральные) спорангии. При морфологической изогамии сфацелярии
ее гаметы различаются по поведению: мужские гаметы дольше
остаются подвижными, и копуляция происходит только после того,
как женские гаметы остановились. Для целей вегетативного раз-
множения у сфацелярии имеются специализированные веточки —
выводковые почки, которые отламываются и прорастают в новые
талломы. Кроме того, от краев диска вырастают столоны, раз-
вивающие на концах новые диски, которые в свою очередь дают
начало вертикальным талломам.
Порядок кутлериальные— Cutleriales
Порядок включает только два хорошо известных рода: кутлерия
(Cutleria) и занардиния (Zanardinia).
Кутлерия распространена по всему побережью Европы, занарди-
ния— преимущественно в Средиземном море, а также в Черном.
Оба рода обнаруживают чередование поколений: у кутлерии смена
генераций гетероморфная, у запардинии — изоморфная.
144
145
Гаплоидные половые растения кутлерии (С. multifida) высотой
около 20 см прикреплены к субстрату с помощью ризоидов,
растущих от базальных клеток. Растения повторно дихотомически
ветвятся и лентовидные ветви заканчиваются пучками волосков,
клетки которых содержат многочисленные хроматофоры. У ос-
нования этих волосков располагается интеркалярная меристема. Ее
клетки, делясь, отчленяют сегменты в двух направлениях: отделя-
ющиеся кнаружи (дистально) обусловливают рост в длину воло-
сков, а отделяющиеся внутрь (проксимально) способствуют росту
собственно таллома. Отделяющиеся внутрь сегменты плотно со-
единяются боками и претерпевают как поперечные, так и продоль-
ные деления. В результате получается компактный паренхиматоз-
ный таллом, в более старых частях дифференцированный на
наружную мелкоклеточную кору, клетки которой богаты хромато-
форами, и внутреннюю крупноклеточную сердцевину. На поверх-
ности уплощенного таллома кутлерии возникают пучки разветв-
ленных нитей, на которых латерально образуются многогпездные
спорангии или гаметангии. У кутлерии анизогамия, гаметангии
двух типов, развивающиеся на разных индивидах (двудомность):
одни (женские) состоят из небольшого числа крупных камер,
другие (мужские) образованы гораздо большим числом мелких
клеток. В первых образуются женские макрогаметы, во вторых —
мелкие мужские микрогаметы. Женские гаметы, помимо того что
крупнее мужских, содержат больше хроматофоров и имеют более
короткий период подвижности, чем мужские. Оплодотворение
обычно происходит после того, как макрогамета становится непо-
движной.
Зигота сразу прорастает в корковидный, достигающий размеров
человеческой ладони таллом совсем иного облика, чем описанная
кутлерия. Долгое время эти талломы рассматривались как самосто-
ятельный род аглаозония (Aglaozonia). Аглаозония—стадия кут-
лерии - размножается только бесполым путем: на ее поверхности
образуются группы—сорусы одногнездных спорангиев, в которых
после мейоза формируются от 4 до 32 гаплоидных зооспор. Попла-
вав некоторое время, они вырастают в гаплоидное растение кут-
лерии с половыми органами. Таким образом, здесь чередуются
половое и бесполое поколения, внешне различающиеся. Цитологи-
ческое изучение показало, что при развитии зооспор в одногнездных
спорангиях первое деление ядра носит редукционный характер.
В 1974 г. Д. Мюллер (D. G. Muller, 1974), ободренный успешной
идентификацией аттрактантов Ectocarpus siliculosus и Fucus serratus,
обратился к аналогичным исследованиям Cutleria multifida.
Макрогаметы из женских гаметангиев С. multifida, собранных
у берегов французского Средиземноморья, спонтанно прорастали.
Один из таких проростков послужил исходным для получения
в культуре женского клона. Далее он поддерживался путем партено-
генетического развития женских гамет.
146
Среди культивируемых вегетативных фрагментов мужских рас-
тений (микрогаметы спонтанно не прорастали) в одной аберрант-
ной культуре мужской гаметофит рос в виде плотных пучков одно-
рядных разветвленных нитей, на которых развивались мужские
гаметангии с микрогаметами. Таким образом, удалось получить
культуры, продуцирующие как макро-, так и микрогаметы.
Макрогаметангии кутлерии издавали приятный запах, похожий
на запах, исходящий от женских гамет эктокарпуса. При смешении
гамет разного пола микрогаметы, привлеченные остановившимися
макрогаметами, окружали их ореолом, а затем одна из мик-
рогамет сливалась с макрогаметой. Зигота всегда развивалась
в аглаозония-стадию, остававшуюся в условиях культуры сте-
рильной.
Хемотаксическую активность извлеченных из макрогамет со-
единений испытывали, используя технику вазелиновых капель или
шариков из пористого стекла, выдержанных в парах аттрактанта.
Если шарики содержали активный материал, то вокруг них кон-
центрировались микрогаметы. Аттрактант кутлерии — мультифи-
ден — так же, как эктокарпен, оказался низкомолекулярным уг-
леводородом.
У другого рода—занардиния {Zanardinia} и гаметофит и споро-
фит внешне неотличимы: они имеют форму кожистого диска до
20 см в диаметре, по краю которого располагаются волоски. У ос-
нования волосков находится меристема, рост таллома такой же, как
у половой генерации кутлерии.
Поскольку у кутлерии обе генерации различаются морфологи-
чески, естественно, может возникнуть вопрос: почему этот род
относят к классу изогенератофициевых, объединяющему бурые во-
доросли с изоморфной сменой генераций. По мнению ряда авторов,
первоначально у кутлерии чередовались одинаковые поколения,
имеющие на первых этапах эволюции гетеротрихальное строение.
Таллом обеих генераций обнаруживал распростертую аглаозоние-
подобную систему и вертикальную кутлериеподобную часть. В про-
цессе эволюции половые индивиды утратили распростертую систе-
му, а бесполые — вертикальную. В пользу этого свидетельствует
то, что аглаозония-стадия (спорофит) в онтогенезе обнаруживает
явную гетеротрихальность: зигота, прорастая, дает вертикальную
нить, клетки которой делятся продольно с образованием так назы-
ваемой колонки. Колонка дальше не развивается, а из ее основания
вырастает крупный корковидный галлом аглаозонии. Иногда из
гаплоидных зооспор образуются индивиды с признаками обеих
генераций. Наконец, наблюдались случаи, когда растения кутлерии
возникали вегетативно на аглаозонии и таким образом один ин-
дивид совмещал обе части гетеротрихального таллома. Такие слу-
чаи, по-видимому, воссоздают картину прежнего предкового состо-
яния, когда у кутлерии оба поколения были внешне одинаковы
и имели гетеротрихальное строение.
147
Порядок диктиотальные — Dictyotales
Представители этого порядка характеризуются апикальным ро-
стом талломов и обычно дихотомическим ветвлением в одной
плоскости. Бесполое размножение посредством неподвижных ап-
ланоспор— гетраспор, половой процесс оогамный, смена генераций
изоморфная.
В качестве представителя можно рассмотреть распространенный
по берегам Европы род диктиоту (Diciyora). Таллом лентовидный,
дихотомически разветвленный, ветви обычно располагаю гея в од-
ной плоскости. Таллом берет начало от цилиндрического корневи-
ща, приклепленного к субстрату ризоидами.
Зрелый таллом диктиоты состоит только из трех слоев клеток:
средний слой из крупных, бесцветных или с немногими хроматофо-
рами клеток окружен сверху и снизу слоем мелких клеток с много-
численными хроматофорами. На спорофитах из поверхностных ко-
ровых клеток развиваются только одногнездные спорангии—тет-
распорангии, в которых образуются по четыре неподвижные тетра-
споры. Тетраспоры прорастают в гаметофиты.
Диктиота — двудомное растение. Различают мужские гаметофи-
ты, несущие антеридии в виде многокамерных спорангиев, каждая
камера которых продуцирует по одному одножгутиковому спер-
матозоиду, и женские гаметофиты, на поверхности которых груп-
пами (сорусами) развиваются оогонии. Образование оогониев
у D, dichotoma начинается у основания таллома, продвигается к его
верхушке и происходит периодически с интервалами в 14—17 дней.
(Подробнее о периодичности в формировании и выходе яиц см.
гл. 4.) Опорожнение оогониев начинается с растягивания их оболоч-
ки, которая разрывается у верхушки, после чего яйцо как бы вытал-
кивается, что сопровождается заметными изменениями его формы.
Опорожнение одного оогония занимает менее 1 мин, процесс выхо-
да яиц из всех оогониев на талломе растягивается на более длитель-
ный промежуток времени — более 60 мин с явным максимумом
через 25—30 мин после начала светового периода. Это было уста-
новлено в опытах, в которых интенсивность света достигала
1000 Лк.
Недавно (1981 г.) было показано, что яйцеклетки D. dichotoma
выделяют вещество, привлекающее сперматозоиды. Оно было иден-
тифицировано и оказалось близким к аттрактантам других бурых
водорослей. Оплодотворение начинается с прикрепления жгутика
сперматозоида к поверхности яйца. После оплодотворения ооспоры
сразу прорастают с образованием спорофитов.
КЛАСС ГЕТЕРОГЕНЕРАТОФИЦИЕВЫЕ— HETEROGENERATOPHYCEAE
Этот класс, характеризующийся гетероморфной сменой генера-
ций, включает ряд порядков, из которых одни образуют спорофиты
148
за счет плотного переплетения боковых ветвей одной или многих
осей неограниченного роста, т. е. имеют псевдопаренхиматозное
строение — одноосевое или многоосевое, другие же порядки объеди-
няют водоросли, чьи спорофиты имеют истинно паренхиматозное
строение. Из второй группы наибольший интерес представляет
порядок ламинариальные — Laminariales.
Порядок ламинариальные — Laminariales
Строение и размножение гаметофита. После открытия К. Соважо
(С. Sauvageau, 1915.) гетероморфного жизненного цикла у Saccorhiza
bulbosa развитие гаметофитов было прослежено в культуре у многих
ламинариальных водорослей. Гаметофиты различных родов и ви-
дов сходны. Они состоят из большего или меньшею числа клеток,
неразветвленные или ветвящиеся, что,по-видимому, определяется не
столько видовой принадлежностью, сколько условиями произраста-
ния. Мужские гаметофиты отличаются от женских более мелкими
клетками и более обильным ветвлением. Во многих случаях наблю-
дается редукция женского гаметофита до одной клетки. У 5. bulbosa
женская зооспора непосредственно дает начало оогонию без какой-
либо вегетативной фазы.
В последние десятилетия интенсивно изучалось влияние разно-
образных факторов внешней среды на рост и фертильность гамето-
фитов ламинариальных водорослей. Ведущими оказались свет и те-
мпература. Эти исследования представляют не только теоретичес-
кий, но и большой практический интерес, так как многие ламинари-
альные, будучи промысловыми водорослями, являются объектами
культивирования (например, Laminaria digitata, L. japonica и др.).
Оптимальное сочетание основных факторов среды — температуры
и освещения — необходимо для повышения продуктивности таких
культур.
Развитие гаметофита L. saccharina от зооспоры до зрелости при
благоприятных условиях может быть завершено в течение 8 дней.
Эмбриоспора, развивающаяся из зооспоры после ее остановки и вы-
деления клеточной оболочки, прорастает в течение 24 ч. Таким
образом формируется первая клетка гаметофита — «первичная
клетка», которая в случае женского гаметофита иногда остается его
единственной клеткой. Далее следует фаза вегетативного роста,
у женского гаметофита выражающегося в двух-, трехкратном уве-
личении диаметра первичной клетки, а у мужского—лишь в незна-
чительном увеличении «первичной клетки» и последующих ее деле-
ниях. Вегетативный рост гаметофитов L. saccharina и других лами-
нариальных имеет место при любом спектральном составе света
и при сравнительно низкой его интенсивности. Потребность гамето-
фитов в свете (для их вегетативного роста) сравнивают с потреб-
ностью глубоководных красных водорослей. Однако созревание
и переход к фертильности контролируются синим светом и требуют
149
гораздо большей интенсивности его. Так, если вегетативный рост
с образованием нитчатых гаметофитов происходит нри освещении
в 1000 Лк, то для индуцирования фертильности у'большинства
гаметофитов необходима интенсивность облучения в 2—3 раза боль-
шая. Если иррадиация низка (соответствует 300 Лк белого света),
то и на синем свету гаметофиты остаются чисто вегетативными —
стерильными. Таким образом, для перехода к гаметогенезу важны не
только определенные длины световых волн (синий свет), но и интенси-
вность света. На красном свету гаметофиты остаются стерильными,
продолжая вегетативный рост. Такое же действие синего света было
продемонстрировано и на других видах ламинариальных.
Еще одним фактором внешней среды, оказывающим решающее
воздействие на рост и фертильность гаметофитов ламинариальных,
является температура. Низкие температуры способствуют гаметоге-
незу: при 10е и 5 JC даже на красном свету могут быть получены
фертильные гаметофиты. По-видимому, необходимостью низких
температур для размножения гаметофитов объясняется приурочен-
ность ламинариальных к холодным водам.
Созревание и освобождение половых клеток у L. saccharina под-
робно прослежено К. Люнингом (К. Liming, 1981). Признаком того,
что оогоний созрел и готов высвободить яйцо, является выпячивание
несколько удлиненной клетки оогония, указывающее на место буду-
щего разрыва его оболочки. Спустя 5 с после разрыва оболочки одна
треть—половина цитоплазменного содержимого оогония выступает
из его оболочки. Через 10 с большая часть цитоплазменного содер-
жимого оставляет оогоний, и весь процесс завершается приблизите-
льно через 30 с. На этой стадии яйцо, сидящее на верхушке оогония,
имеет несколько треугольную форму, но через 3 мин после начала
освобождения яйцо округляется. Было показано, что когда яйцо
L. digit ata покидает оогоний, продуцируется вещество (СнН14О),
вызывающее освобождение из антеридиев сперматозоидов и привле-
кающее их к яйцам, которые они окружают. Массовое освобождение
сперматозоидов наблюдается через 8—12 с после того, как зрелые
мужские гаметофиты оказываются в сфере действия этого вещества.
Таким образом достигается синхронизация освобождения яиц и спер-
матозоидов, что несомненно способствует оплодотворению.
Строение и размножение спорофита. Взрослые спорофиты разных
родов ламинариальных резко различаются. Это самые крупные
водоросли, достигающие в длину 60—100 м с морфологическим
расчленением и сложным анатомическим строением. В большинстве
случаев спорофиты расчленяются на листовидную пластинку, ствол
(стебель) и ризоиды, с помощью которых все растение прикрепля-
ется к подводным камням и скалам. Развиваются они следующим
образом.
Возникшая в результате оогамного полового процесса зигота
ламинариальных сначала сферическая, затем удлиняется и делится
поперечными перегородками. В образовавшейся таким образом
150
однорядной нити клетки претерпевают не только поперечные, но
и продольные деления, причем продольные чаще происходят на
апикальном конце, чем на базальном. На этой стадии молодой
спорофит представлен плоским однослойным образованием, более
широким на переднем и более узким на базальном конце. Из
базальных клеток развиваются ризоиды, прикрепляющие растение
к субстрату. Вскоре спорофит становится многослойным благодаря
периклинальным (параллельным плоскости таллома) делениям кле-
ток. Периклинально делящиеся клетки базальной части спорофита
образуют будущий стебелек, после заложения которого базальный
конец спорофита превращается в подошву, от которой отходят
прицепки, служащие для прикрепления растущего спорофита к суб-
страту. Часть спорофита выше стебелька становится пластинкой
«листа». После дифференцировки первичных органов (подошвы,
стебелька, пластинки) рост спорофита в длину (за исключением
роста в результате вытягивания клеток) постепенно локализуется
в переходной зоне —в месте соединения стебелька и пластинки. Эта
интеркалярная меристема ответственна за рост в длину как стебель-
ка, так и листа у более взрослых спорофитов. На сравнительно
ранней стадии, особенно в стебельке, начинается дифференцировка
тканей. От центральных клеток, далее не делящихся и составля-
ющих первые сердцевинные элементы, посредством тангентального
деления отчленяется поверхностный слой клеток, претерпевающих
активные тангентальные и радиальные деления. Этот меристемати-
ческий слой клеток называется меристодермом. Клетки, отчленя-
ющиеся от клеток меристодерма внутрь, образуют вокруг сердцеви-
ны кору. По мере деятельности меристодерма коровые клетки,
граничащие с первыми сердцевинными элементами, превращаются
в сердцевинные клетки. Срезы через стебелек и пластинку более
старых спорофитов, например у L. saccharina, обнаруживают таким
образом три слоя тканей — меристодерм, кору, сердцевину, перехо-
дящие один в другой без резкой границы.
Срезы через переходную зону показывают, что сердцевина и кора стебелька
непрерывны с таковыми листовой пластипки. Меристодермальные клетки, кубичес-
кие. с уплощенной наружной стенкой, содержат большое число хлоропластов. За счет
их деления стебель растет в толщину и длину, и все время пополняются клетки
наружной коры, тогда как внутренние клетки коры сильно вытягиваются.
На вытянутых внутренних коровых клетках L. saccharina дифференцируются
сосочковидные выросты, которые образуют анастомозы (соединения) с такими же
выступами, возникающими на соответствующих уровнях на других клетках. Таким
образом устанавливаются цепочки поперечных соединений между вертикальными
рядами внутренних кортикальных клеток. Если эти нитчатые выросты не контак-
тируют друг с другом, они проходят сквозь медуллярную сердцевинную часть
стволика, которая образована этими внутренними нитями, цепочками поперечных
соединений и дериватами внутренней коры1. Из внутренних кортикальных клеток
дифференцируются и ситовидные элементы, сравнимые с таковыми у представителей
семейства лессониевых (роды нереоцистис. макроцистис). Ситовидные элементы, как
1 Такая неплотная, пронизанная нитями сердцевина свойственна многим лами-
нариальным, однако у рода саккориза (Saccorhiza) сердцевина плотного строения.
151
и клетки внутренней коры, не делятся и по мере удлинения стволик сильно вытяги-
ваются, становятся узкими, с расширениями у поперечных стенок, боковые же стенки
утолщаются. Конечные стенки кортикальных клеток и ситовидных элементов оста-
ются тонкими. На конечных стенках кортикальных клеток присутствуют поровые
поля — тонкие участки в стенке, несущие поры. Напротив, в ситовидных элементах
поры более крупные (средний диаметр пор в ситовидных элементах в два раза более
такового плазмодесм, обнаруживаемых в кортикальных клетках) и распределены
равномерно по всей поверхности конечных стенок. Поры округлые, открытые, без
каллозы. Они содержат тяж цитоплазмы, окруженный плазмалеммой, и связывают
друг с другом соседние ситовидные элементы. В каждом ситовидном элементе
L. saccharina находится крупное лопастное ядро с заметным ядрышком, диктиосомы,
шероховатый эндоплазматический ретикулум. Ситовидные элементы L. saccharina,
L. groenlandica, Alaria marginata, содержащие клеточное ядро, митохондрии и харак-
теризующиеся сравнительно мелкими норами, менее специализированы по сравне-
нию с таковыми Nereocystis luetkeana и особенно Macrocystis piryfera, у которых поры
гораздо более крупные и отсутствуют клеточные ядра. От ситовидных элементов
сосудистых растений, также энуклеированных (лишенных ядра), ситовидные элемен-
ты макроцистиса отличаются наличием многочисленных митохондрий. Если мито-
хондрии и присутствуют в ситовидных элементах высших сосудистых растений, то
в дегенерирующем состоянии. Благодаря ситовидным элементам ламинариальные
способны к транспорту ассимилятов (маннита, некоторых аминокислот) на большие
расстояния. Скорость передвижения колеблется от 10 до 60 см/ч. По-видимому,
между скоростью транспорта и диаметром ситовидных пор есть прямая зависи-
мость. Многие ламинариальные обнаруживают особые слизевые каналы или npojo-
ки и секреторные клетки как в листовой пластинке, так и в стволе.
Спорофиты ламинариальных образуют только одногнездные
(однокамерные) спорангии. Начало их возникновения знаменуется
переходом меристодерма от пополнения клеток вегетативного тал-
лома к образованию двух новых типов клеток: парафиз и материнс-
ких клеток спорангиев. У большинства изученных представителей
ламинариальных водорослей меристодермальные клетки удлиня-
ются, образуя палисадный слой, затем каждая клетка делится пери-
клинально на крупную верхнюю клетку — зачаток парафизы и не-
сколько более мелкую изодиаметрическую базальную клетку. По
мере созревания парафиз базальные клетки обнаруживают плоско-
стной рост, в результате чего парафизы у основания разъединяются.
От свободных поверхностей базальных клеток образуются высту-
пы, которые отчленяются периклинальной стенкой и становятся
материнскими клетками спорангиев. Этот ход развития несколько
отличается у рода хорда, где деления меристодермальных клеток,
продуцирующих парафизы, косые. Апикальный выступ, который
отчленяется в виде материнской клетки спорангия, уже присутствует
на меристодермальной клетке ко времени образования парафиз.
Парафизы быстро растут, принимают характерную «булавовид-
ную» форму и возвышаются над материнскими клетками споранги-
ев. Спорангии увеличиваются в объеме, становятся эллипсоидаль-
ными, на верхушке возникает характерная «шапочка». Ядро споран-
гия претерпевает мейоз, после которого следуют синхронные мито-
тические деления. Число зооспор, образующихся в спорангиях,
неодинаково у разных родов и видов, по более или менее постоянно
у данного вида. Оно колеблется от 16 до 128. Освобождение зооспор
изучено на электронно-микроскопическом уровне у Chorda
152
tomentosa. У этого" вида вокруг незрелых, т. е, заключенных еще
в спорангии, зооспор (в числе 128) находится большое количество
альгиновой кислоты, которая постоянно давит на спорангиальную
стенку и апикальную шапочку спорангия. Дополнительное давление
на спорангии, возможно, оказывают тесно расположенные и пере-
мешанные с ними парафизы. Во время особождения зооспоры,
по-видимому, секретируют энзим (или энзимы), который селектив-
но переваривает апикальную шапочку, возможно, образованную
сульфатированными полисахаридами. Начало выхода зооспор но-
сит взрывной характер — в виде моментального извержения. Пер-
вые вышедшие из спорангия споры окружены и соединены вместе
клейкой альгиновой кисло гой, которая по мере разбухания вытяги-
вает из спорангия оставшиеся споры. Когда слизь растворяется,
зооспоры становятся подвижными. После краткого периода движе-
ния зооспоры прикрепляются к субстрату, выделяют оболочку
и прорастают в растеньица гаметофитов. Гаметофиты всегда дву-
домные и определение пола, по-видимому, генотипическое: при
последующей культуре зооспоры из одного спорангия дают мужс-
кие и женские гаметофиты в одинаковом числе.
Большой порядок ламинариальные делится на ряд семейств, из
которых будут рассмотрены хордовые (Chordaceae), ламинариевые
(Laminariaceae), лессониевые (Lessoniaceae) и алариевые (Alariaceae).
Семейс тво хордовые (Chordaceae) отличается от всех дру-
гих ламинариальных водорослей простейшей формой таллома спо-
рофита: трубковидной, без расчленения на стволик и листовую
пластинку. Сюда относится только один род хорда (Chorda).
C.filum — однолетняя водоросль, широко распространенная в се-
верном полушарии. Бичевидные полые талломы, слабо опушенные
волосками, достигают длины 8 м при ширине не более 0,5 см
и прикрепляются к субстрату посредством маленького диска. Верх-
няя часть таллома плавает у поверхности воды. Меристема рас-
полагается несколько отступя от верхушки. Спорангии покрывают
большую часть таллома и развиваются снизу вверх.
Семейство ламинариевые (Laminariaceae). Спорофиты
относящихся сюда родов дифференцированы на ризоиды, стебли
(стволы) и листовые пластинки, которые у одних родов и видов
цельные, у других—рассеченные. В последних случаях на началь-
ных стадиях развития пластинка всегда цельная, как у Laminaria
saccharina, лишь позднее на ней появляются продольные трещины,
которые, однако, рассекают только листовую пластинку, не рас-
пространяясь на переходную зону и не затрагивая меристему. Груп-
пы (сорусы) зооспорангиев развиваются на листовых пластинках.
Род ламинарии (Laminaria) распространен (за исключением трех
видов) в северном полушарии, особенно в Тихом океане. Спорофи-
ты, достигающие нескольких метров в длину, многолетние. Много-
летние только стволик и органы прикрепления (ризоиды, подошва),
пластина же меняется ежегодно. Она разрушается и с началом
153
вегетационного сезона отрастает вновь. На новой пластине часто
можно видеть остатки старой. У одних видов (L.Jsdccharina) пла-
стинки остаются цельными, у других (L. digitata) с возрастом раз-
деляются на многочисленные сегменты.
Семейство лессониевые (Lessoniaceae) включает самые
крупные, достигающие 50—60 м водоросли. От предыдущего се-
мейства ламинариевых отличается тем, что процесс расщепления
пластинки в молодом спорофите распространяется на переходную
зону — разделяется и меристема. Таким образом, каждая вторичная
пластинка снабжена частью первичной меристемы и может разви-
вать свои собственные вторичные стебельки. Спорангии образуются
на поверхности пластин. Из представителей могут быть названы
роды лессония (Lessonia), макроцистис (Macrocystis), нереоцистис
(Nereocystis).
Род лессония (Lessonia), распространенный в основном в южном
полушарии, на молодой стадии развития имеет вид Laminaria
saccharina. Вскоре у основания пластины появляется срединная
щель, раскалывающая пластинку на два сегмента, снабженных ко-
роткими стебельками, которые кажутся развилками нижележащего
стволика. Таким образом, первоначальная меристема поделйлась
на две меристемы, каждая из которых способствует удлинению как
вторичных пластинок, так и нижележащих стебельков. Вторичная
пластинка расщепляется так же, и это повторяется снова и снова.
Обычно один развилок (сегмент) развивается сильнее другого и, так
как вторичные стебельки более развитых сегментов располагаются
в одну линию, получается симподиальная ось, на которой более
слабые сегменты кажутся боковыми ветвями.
Род макроцистис (Macrocystis), также распространенный глав-
ным образом в южном полушарии, имеет талломы до 60 м в длину.
Длинный тонкий ствол несет ряд рассеянных пластинок и воло-
чится косо к поверхности воды, где горизонтально плавает оста-
льная часть таллома. Плавучая часть несет плотный двойной ряд
листовых пластинок (1 —1,5 м длины), сидящих на коротких вто-
ричных стебельках. У основания каждой пластинки имеется сфери-
ческий или грушевидный воздушный пузырь. Таллом развивается
в общем так же, как у лессонии. Растение многолетнее.
Род нереоцистис (Nereocystis)—однолетнее растение, достига-
ющее 50 м. Молодые растения обычного типа (имеют вид Laminaria
saccharina), но вскоре обнаруживается очень быстрое удлинение
ствола, который остается гонким и очень гибким. У верхушки
ствола под переходной зоной возникает сферический воздушный
пузырь, а в пластинке образуется первая щель, простирающаяся
вниз до пузыря и делящая пластинку приблизительно на две равные
половины. Затем последовательные расщепления пластинок приво-
дят к тому, что у взрослых растений на поверхности пузыря (кото-
рый плавает на воде) располагаются многочисленные, сидящие на
коротких вторичных стебельках ланцетовидные пластинки.
154
Семейство алариевые (Alariaceae) характеризуется тем,
что органы размножения — одногнездные спорангии—образуются
не на вегетативных пластинках, а, как правило, на специальных
спорофиллах. Спорофиллы возникают в виде выростов или на
стволе, или по краю пластинки.
Род алария (Alaria)— многолетнее растение, распространенное
в северном полушарии. Сравнительно короткий стволик несет удли-
ненную пластинку часто с волнистыми краями и срединной жилкой.
На протяжении лета стволик у своего верхнего конца образует два
ряда толстых (в виде языков) спорофиллов, которые после созрева-
ния сорусов спорангиев (осенью и зимой) сбрасываются, оставляя
на стебле мелкие рубцы1. Пластинка рвется на сегменты перпен-
дикулярно к срединной жилке и в конце концов разрушается. В на-
чале следующего сезона пластинка возобновляется за счет деятель-
ности меристемы. Новые спорофиллы возникают над спорофил-
лами предыдущего года.
КЛАСС ЦИКЛОСПОРОФИЦИЕВЫЕ — CYCLOSPOROPHYCEAE
Класс представлен лишь одним порядком фукальные (Fucales)1 2.
Порядок фукальные — Fucales
Порядок характеризуется отсутствием смены генераций; расте-
ния диплоидны, мейоз предшествует гаметогенезу, половой про-
цесс— оогамия; бесполое размножение спорами отсутствует. Рост
талломов верхушечный. Фукальные не достигают таких больших
размеров, как ламинариальные: самые крупные талломы дурвиллеи
(Durvillea) достигают 10 м. Однако по степени внешней дифферен-
цировки истинно паренхиматозных талломов фукальные не уступа-
ют ламинариальным. Распространены они очень широко и наиболь-
шего развития достигают в северных широтах, где образуют зарос-
ли в литоральной зоне (зона прилива — отлива). Самые обычные
роды здесь — роды фукус (Fucus), пельвеция (Pelvetid), аскофиллум
(Ascophyllum), гиманталия (liimanthalia).
У рода фукус (Fucus) плоский ремневидный таллом, достига-
ющий 0,5—1,0 м, дихотомически ветвящийся, вдоль лопастей тал-
лома проходит срединная жилка, переходящая в нижней части
в черешок, который прикреплен к субстрату с помощью диска.
У F. vesiculosus и ряда других видов по бокам от срединной жилки
расположены вздутия, наполненные воздухом,— воздушные пузы-
ри, которые играют роль плавательных пузырей.
1 Однако недавно на Тихоокеанском побережье Японии было обнаружено много
талломов Alaria sp., имеющих сорусы спорангиев не только на спорофиллах, но и на
вегетативных пластинках. Сорусы можно было наблюдать осенью и зимой, сначала
на спорофиллах, а позднее на листовых пластинках.
2 Некоторые авторы разбивают этот порядок на ряд порядков.
155
У F. serratus лопасти, имеющие зазубренные края, лишены воз-
душных пузырей. * J
Род пельвеция (Pelvetia} встречается в Белом море, растет на
поверхности и в щелях скал у самой верхней границы литоральной
зоны, довольствуясь лишь брызгами воды: узкие сегменты дихото-
мически ветвящегося таллома, без воздушных пузырей и срединной
жилки, согнуты по длине в виде желобка, где и задерживается во
время отлива влага. У пельвеции, как и у фукуса, концы основных
ветвей таллома со временем становятся плодущими — превращают-
ся в рецептакулы, на которых развиваются половые органы, погру-
женные в сообщающиеся с наружной средой с помощью узкого
отверстия полости — концептакулы, или скафидии.
У рода аскофиллум (Ascophyllum), распространенного вместе
с фукусом в верхней части литорали, довольно узкий и сравнитель-
но толстый таллом, достигающий 1,5 м высоты. Он лишен средин-
ной жилки и дихотомически ветвится. Расположенные вдоль тал-
лома воздушные пузыри поддерживают таллом во время прилива
вблизи поверхности воды. В отличие от фукуса и пельвеции рецеп-
такулы развиваются на специальных боковых веточках.
Встречающийся по берегам Северной Европы род гимантаЛия
(Himanthalia) имеет таллом, дифференцированный на стерильную
чашечковидную на гибкой ножке часть, прикрепленную к субстрату
диском и достигающую нескольких сантиметров в диаметре, и воз-
никающие от ее внутренней поверхности длинные лентовидные,
дихотомически разветвленные рецептакулы. Рецептакулы возника-
ют на 2—3-й год жизни в числе 1—4 (обычно 2), достигают 2 м
в длину при ширине 1 —2 см и несут по всей своей длине (за
исключением базальной узкой и особенно гибкой части) концеп-
такулы.
У атлантических берегов Европы встречается род галидрис
(Halidrys), кустистые талломы которого с уплощенными ветвями
несут воздушные пузыри, поделенные внутри поперечными перего-
родками на камеры. У близкого рода цистозейра (Cystoseira), произ-
растающего в СССР в Черном, Японском, Охотском морях, моно-
подиально ветвящиеся талломы несут веретеновидные рецептакулы
и однокамерные воздушные пузыри.
В тропических и субтропических областях доминирует род сар-
гассум (Sargassum), отличающийся наибольшей внешней дифферен-
цировкой таллома: стволик, прикрепленный к субстрату подошвой,
моноподиально ветвится и несет уплощенные листовидные образо-
вания, шаровидные воздушные пузыри на специальных стебельках
и особые разветвленные плодущие веточки — рецептакулы.
На поперечных срезах взрослых талломов (например, фукус,
саргассум) легко различаются эпидермис, к которому изнутри при-
мыкает многослойная кора, незаметно переходящая в сердцевину.
Размножение. Размножение вегетативное и половое. Первое при-
водит к образованию плавучих масс — неприкрепленных, стериль-
156
ных, более или менее полностью утративших способность к полово-
му размножению талломов, которые распространены в некоторых
закрытых морях, например Балтийском.
Колоссальные плавучие массы стерильных талломов саргассума
наблюдаются в Саргассовом море, открытом еще Колумбом. По-
ловой процесс, как правило,— примитивная оогамия, причем поло-
вые органы — оогонии и антеридии — развиваются в специальных
углублениях таллома — концептакулах, или скафидиях. Скафидии
развиваются из одной инициальной клетки, обычно поверхностной.
Благодаря росту и делениям соседних клеток инициальная клетка
погружается в таллом, хотя всегда остается отверстие, ведущее
наружу,— будущее отверстие концептакула. Инициальная клетка
делится на наружную языковидную (язычковую) клетку и внутрен-
нюю— базальную. Язычковая клетка или дегенерирует, или остает-
ся неактивной, или же, наконец, делится, образуя короткую одно-
рядную нить-волосок с базальной меристемой. Благодаря верти-
кальным делениям базальной клетки и ее дериватов в расширя-
ющейся полости возникает выстилающий слой клеток1. У рода
фукус и некоторых других дериватами базальной клетки выстлано
лишь основание (пол) концептакула, а с боков выстилающий слой
развивается из клеток, соседних с инициальной клеткой концеп-
такула. Клетки выстилающего слоя в верхней части зрелого концеп-
такула развивают волоски, выступающие из отверстия. Из клеток
в базальной части концептакула образуются разветвленные (иногда
не разветвленные), содержащие хлоропласты нити, называемые па-
рафизами. Растения в зависимости от рода и даже вида могут быть
двудомными или однодомными. Так, одни виды саргассума дву-
домные (5. fulvellum), другие (S. kjellmanianum)— однодомные. То
же относится и к роду фукус. У однодомных форм оогонии и ан-
теридии развиваются в одном и том же концептакуле (Fucus
evanescens, виды цистозейра, саргассум) или же в разных концеп-
такулах (S. kjellmanianum). Антеридии обычно образуются на пара-
физах. Первые два деления первичного ядра антеридия носят мейо-
тический характер, затем следуют еще четыре митоза. Таким об-
разом, в зрелых антеридиях возникают 64 ядра, которые все функ-
ционируют, и образуются 64 сперматозоида. Сперматозоиды
гетероконтные и гетероморфные; в отличие от монадных клеток
остальных бурых водорослей задний жгут длиннее переднего. Роды
фукус и аскофиллум были первыми бурыми водорослями, у кото-
рых на электронно-микроскопическом уровне был подробно про-
слежен сперматогенез и исследовано тонкое строение мастигонем,
а также их происхождение, относительно которого ранее высказы-
вались лишь предположения.
1 Инициальную клетку концептакула некоторые авторы называют проспорой,
а развивающийся из нее выстилающий слой концептакула трактуют как диплоидный
гаметофит.
157
На ранних стадиях формирования сперматозоидов, когда оба
жгута еще лишены мастигнонем, в цитоплазме наблюдались обиль-
ные, окруженные мембраной пузырьки-везикулы, по-видимому, от-
членяющиеся от перинуклеарного пространства, в которых находи-
лись микротрубочки. Эти микротрубочки отличались от цитоплаз-
матических и от трубочек, составляющих аксонему жгута, мень-
шими размерами. Размеры и внешний вид их были сходны со
жгутиковыми волосками—мастигонемами. Г. Бак (G. В. Bouck,
1969) назвал их «предположительными мастигонемами». На более
поздней стадии развития сперматозоида, когда передний жгут уже
несет на своей поверхности мастигонемы, «предположительные ма-
стигонемы» в цитоплазме не обнаруживаются. Это позволило Баку
высказать предположение, что образование и сборка мастигонем
происходят внутри цитоплазмы, а не на поверхности жгута, как
полагали ранее. Везикулы, содержащие мастигонемы, мигрируют
на поверхность клетки к месту отхождения жгута, мастигонемы
выделяются на поверхность кле тки и по мере роста переднего жгута
приклеиваются к его мембране. Сами мастигонемы в готовом виде
состоят из трех частей: 1) базальной части (не трубчатой), которая
прочно приклеивается к мембране жгута; 2) трубча той части, стен-
ки которой образованы рядами глобулярных субъединиц, и 1) 1 —3
терминальных нитей из одного ряда глобулярных субъединиц (см.
рис. 31, В). Такое же строение имеют мастигонемы, покрывающие
передний жгут гамет эктокарпуса.
При наблюдении в оптическом микроскопе у зрелых антеридиев
видна двухслойная стенка. Наружный слой рвется первым, и масса
сперматозоидов освобождается, заключенная сначала во внутрен-
нюю стенку, которая быстро ослизняется.
Оогонии обычно образуются непосредственно на стенке концеп-
такула и снабжены короткой ножкой. Как и в антеридиях, деление
первичного ядра оогония редукционное, после чего следует еще
одно митотическое деление. Таким образом, всегда образуются
восемь ядер, из которых не все оказываются жизнеспособными.
Лишь у родов фукус и нотейа (Notheia) все восемь ядер остаются
жизнеспособными, и в оогонии формируются восемь яйцеклеток.
У других родов часть ядер дегенерирует и соответственно в оогонии
образуются четыре яйца (роды аскофиллум, дурвиллея и некоторые
др.), два яйца (например, у пельвеции), одно яйцо (у саргассума
и многих др.). Яйцеклетки фукальных — одни из самых крупных
репродуктивных клеток, обнаруженных среди морских водорослей;
они видимы невооруженным глазом.
По специальным подсчетам из одного растения Sargassum horneri
освобождаются 1—6 млн. яйцеклеток.
Стенка оогония (при рассматривании в оптическом микроскопе)
дифференцирована по крайней мере на три слоя: внутренний —
эндохитон, наружный — экзохитон передний — мезохитон. Обычно
оплодотворение у фукальных описывается следующим образом:
экзохитон разрывается и яйца, окруженные средним и внутренним
слоями, выдвигаются из отверстия концептакула, мезохитон лопа-
ется на верхушке и скользит назад, эндохитон растворяется и яйца
освобождаются в воду, где и оплодотворяются сперматозоидами.
Однако у Fucus evanescens, у которого яйца и сперматозоиды образу-
ются в одном и том же концептакуле, яйца оплодотворяются в са-
мом концептакуле и уже оплодотворенные освобождаются в море.
У Pelveda canaliculata при освобождении двух сформировавшихся
в оогонии яйцеклеток разрывается эндохитон, а остальные слои
набухают в воде. Антерозоиды проникают через мезохитон в оого-
нии, когда яйцеклетки находятся еще на разных стадиях развития.
У F. spiralis большая часть яйцеклеток покидала оогонии уже буду-
чи оплодотворенными — в виде ооспор, и только 5% вышедших
яйцеклеток привлекало сперматозоиды.
У Halidrys siliquosa яйца и сперматозоиды одновременно выхо-
дят из одного и того же концептакула; оплодотворение может
произойти до освобождения яйца через отверстие концептакула.
У Ascophyllum nodosum яйца появляются из отверстия концет акула,
заключенные в массу слизи, из которой они впоследствии освобож-
даются в морскую воду, где и оплодотворяются сперматозоидами.
У некоторых родов мезо- и эндохитон остаются на яйцах, образуя
слизистую ножку, посредством которой яйца прикрепляются к кон-
цептакулу. В таком состоянии происходят оплодотворение и первые
стадии развития зародыша. Таким образом, оплодотворение яйца
сперматозоидом у фукальных не обязательно происходит вне кон-
цептакула, у некоторых видов оно имеет место внутри концептакула
и даже внутри оогония.
Фукус — первая водоросль, у которой в середине прошлого сто-
летия был описан половой процесс и наблюдалось скопление спер-
матозоидов вокруг яйцеклеток. В настоящее время выяснена хи-
мическая основа этого процесса. F. serratus явился вторым после
Ectocarpus siliculosus видом бурых водорослей, у которого в 70-х
годах была установлена химическая природа аттрактанта. Испыта-
ния проводились с использованием летучести аттрактанта. Вазели-
новые капельки легко экстрагировали вещество из потока воздуха,
проходящего над ними и содержащего аттрактант. При помещении
затем таких капель в суспензию активных сперматозоидов они
воздействуют на сперматозоиды как яйцеклетки.
Привлекающее сперматозоиды вещество, выделяемое яйцеклет-
ками F. serratus и названное фукосерратеном, было идентифициро-
вано как транс-цис-1,3,5-октатриен. Было испытано действие девяти
других конъюгированных изомеров октатриена, синтезированных
в лаборатории. Количественное изучение хемотаксиса сперматозо-
идов с использованием этих синтетических окгатриенов показало,
чю мужские гаметы F. serratus наиболее чувствительны к фукосер-
рагену и транс-цис-2,4,6-окгатриену, тогда как остальные октатри-
ены менее эффективны. Аттрактант F. vesiculosus (как и ряда других
159
фукальных: F. .spiralis, F. evanescens, Pelvetia wrightii, Sargassum
horneri) также оказался фукосерратеном. Этим сходством аттрак-
тантов F. serratus и F. vesiculosus, по-видимому, объясняется извест-
ный факт, что их сперматозоиды не отличают яйцеклетки своего
вида от чужого. Тем не менее полевые исследования показывают,
что гибридизация между этими двумя видами пе наблюдается, хотя
в природе они живут рядом и могут освобождать гаметы в одно
и то же время. Барьер нескрещиваемости. очевидно, действует не на
уровне хемотаксиса сперматозоидов, а на более поздней стадии
полового процесса. Недавние эксперименты по гибридизации свиде-
тельствуют, что носителем механизма узнавания «вид-вид» может
быть мембрана яйцеклеток.
Само оплодотворение начинается с прикрепления переднего
жгутика сперматозоида к поверхности яйцеклетки. Это наблю-
далось у F. serratus, F. spiralis var. platicarpus, F. vesiculosus,
Cysioseira barhata, Pelvetia canaliculata, Ascophyllum nodosum, Halidrys
siliquosa. В двух последних случаях начальные этапы оплодот-
ворения были изучены с применением киносъемки и фазового
контраста, которые позволили, кроме того, проследить за изме-
нением формы сперматозоидов и продемонстрировать различный
характер движения жгутиков. Так, сперматозоиды Ascophyllum
nodosum, только что освободившиеся из антеридия и свободно
плавающие, имеют каплевидную форму, движение переднего жгу-
тика быстрое, колебательное, тогда как задний обнаруживает лишь
медленное покачивание. Перед слиянием с яйцом сперматозоид
удлиняется, принимает форму запятой, 'медленно ползает по по-
верхности яйца, затем прикрепляется к его поверхности посред-
ством переднего жгутика и продолжает вращаться вокруг точки
его прикрепления. У Halidrys siliquosa сперматозоиды не изменяют
формы, в остальном поведение их (движения жгутиков, начальные
этапы оплодотворения) те же, что и у Ascophyllum nodosum. Затем
эластичные мембраны жгутика сперматозоида и яйца слипаются
(сливаются), жгутик проникает в цитоплазму яйца и протопласты
половых клеток сливаются.
У всех фукальных талломы прикрепляются посредством ризоидов, которые J
образуются на протяжении всей жизни растения, формируя в конце концов подошву. ]
Однако промежуток времени от оплодотворения яйца до появления первых ризоидов
у разных видов и родов варьирует. Так, у видов фукуса развитие ризоидов очень
быстро следует за оплодотворением: у Halidrys siliquosa — через 5. а у Pelvetia
canaliculaia —через 13 17 дней после оплодотворения; зигота может даже достичь
многоклеточной стадии раньше, чем дифференцируется первый ризоид. Между тем
свободноплавающие зиготы (ооспорьГ) фукальных должны прикрепиться к субстрату |
вскоре после оплодотворение, иначе значительное движение воды на литорали унесет I
их в море, где последующие прорастание и рост невозможны. Исследования с при- |
менением сканирующего электронного микроскопа показали, что после оседания на 1
субстрат оплодотворенная яйцеклетка очень быстро покрывается слоем слизистого I
клейкого вещества, особенно обильного у видов, у которых образование ризоидов 4
задерживается. Это клейкое вещество продуцируется аппаратом Гольджи, по-види- 1
мому, еще до оплодотворения, а затем выделяется из клетки наружу. Слизь медлен- 1
160 1
но стекает —«сбегает» с поверхности зиготы на субстрат, заполняя все пространство
между зиготой и субстратом, делая как бы точный слепок контуров субстрата. Там,
где контур субстрата неровный, будет большая поверхность склеивания и более
прочное прикрепление зиготы к субстрату, чем на гладкой поверхности. Под слизью
закладывается стенка зиготы, ясно различимая уже через 24 и 48 ч после оплодот-
ворения. Слизь — временное образование, служащее для прикрепления зиготы, пока
не образуются ризоиды, после чего она сбрасывается в виде шапочки (она может
также удерживать воду вокруг зиготы, что важно для литоральных водорослей).
Кончик образующегося первичного ризоида (отрицательно фототропичного)
сразу окружается толстым слизистым эксудатом, который при контакте ризоида
с твердым субстратом распространяется по нему, образуя липкую «подошву». По
.мере удлинения ризоида его кончик ползет по субстрату, продолжая выделять
клейкий эксудат, пока не найдет в субстрате отверстия, в которое можно проникнуть.
Из соседних клеток у основания зародыша развиваются вторичные ризоиды, также
ползущие по субстрату, пока не натолкнутся на расщелину. Клейкое вещество,
продуцируемое ризоидами, и проникновение их в субстрат столь эффективны, что
невозможно вырвать молодое растение без удаления поверхностных слоев камней.
Скорость роста ризоидов зависит от внешних условий, особенно от освещения
и температуры. Обычно интенсивный свет ингибирует удлинение ризоидов.
Филогенез. Бурые водоросли — естественная группа, по своей
морфологии хорошо отличающаяся от остальных отделов водорос-
лей. Однако по ряду признаков (наличию хролофиллов а и с,
отсутствию хлорофилла Ь, расположению тилакоидов в хлоро-
пластах, по сходным продуктам запаса и строению монадных клеток
с гетероконтными и гетероморфными жгутиками) бурые водоросли
сходны с золотистыми, желтозелеными, диатомовыми и динофито-
выми водорослями. Некоторые авторы на этом основании низводят
бурые и остальные перечисленные группы водорослей до ранга
классов в одном большом отделе Chromophyta. Наличие монадных
клеток в жизненном цикле бурых водорослей позволяет выводить
их, так же как остальные перечисленные группы, из каких-то первич-
ных фотосинтезирующих жгутиконосцев с преобладанием бурых
пигментов. Эти жгутиконосцы эволюционировали несколькими пу-
тями, один из которых и привел к возникновению бурых водорос-
лей. Однако у бурых водорослей связь со жгутиковыми более
отдаленная, чем у сходных по ряду признаков с ними золотистых,
разножгутиковых, диатомовых, динофитовых, а также зеленых во-
дорослей, так как прямых переходных форм здесь пет. Среди бурых
водорослей не наблюдается примеров последовательного развития
организации таллома от монадного к нитчатому и пластинчатому,
как это легко удается проследить у зеленых, желтозеленых, золотис-
тых и динофитовых водорослей. Как уже отмечалось, среди бурых
водорослей представлены только высшие ступени морфологической
дифференцировки таллома — гетеротрихальная и пластинчатая.
Возможно, более простые формы (монадпые, коккоидные, простые
нитчатые) полностью утрачены на протяжении длительной эволю-
ции, которую проделали бурые водоросли — древняя группа, из-
вестная уже из отложений силура и девона.
Относительно родственных связей порядков в пределах самих
бурых водорослей и их системы единого взгляда нет. Согласно
6 - 1210 161
одной из схем филогенеза, построенной с учетом сходства строения
таллома на самых ранних стадиях развития, бурые водоросли
группа дифилетическая: одна линия эволюции объединяет порядки
сфацеляриальные, диктиотальные, фукальные, другая — эктокар-
пальные, кутлериальные, ламинариальные. Эти группы порядков
различаются и по способу роста талломов: первая характеризуется
верхушечным ростом, вторая — интеркалярным. Другие авторы от
общего предка бурых водорослей выводят три эволюционные ли-
нии, которые представлены как классы изогенератные, гетерогене-
ратные и циклоспоровые, различающиеся по жизненным циклам
и морфологическому строению талломов.
* * *
Крупные бурые водоросли широко применяются в хозяйстве. Из
них добиваю i альгин — клеющее вещество, которое используется
в текстильной, пищевой и ряде других отраслей промышленности.
В приморских странах выбросы морских водорослей, богатые кали-
ем и азотом, употребляют как удобрения, а также используют на
корм скоту. Некоторые, например ламинарии (морская капуста),
съедобны.
Глава 11
ОТДЕЛ ДИНОФИТОВЫЕ ВОДОРОСЛИ —DINOPHYTA
В подавляющем большинстве представителей—это монадные
формы, реже ризоподиальные, пальмеллоидные, коккоидные, нит-
чатые. Принадлежность неподвижных форм так же, как многих
бесцветных сапротрофных и паразитных форм к динофитовым во-
дорослям, легко устанавливается по совершенно уникальному стро-
ению клеточного ядра и по подвижным репродуктивным стадиям,
повторяющим характерные признаки монадных представителей от-
дела. Обычно линзовидные хлоропласты чаще всего окрашены в бу-
рый цвет благодаря преобладанию каротиноидов (p-каротин, у-
каротин, фукоксантин, диадипоксантин, диатоксантин, диноксан-
тин, перидинин и др.) над хлорофиллами а и с (хлорофилл b отсут-
ствует). Оболочка хлоропластов состоит из трех мембран, ламеллы,
как правило, трехтилакоидные, опоясывающая ламелла отсутству-
ет, генофор рассеянный. Пиреноиды различной формы: простой
внутренний пиреноид, гранулированный матрикс которого не пере-
секается тилакоидами; сложный внутренний голый пиреноид, про-
низанный более или менее параллельными ламеллами; пиреноиды,
выступающие из хлоропластов в виде почки и сидящие на одной
или многих ножках. Продукты ассимиляции — экстрапластидный
162
(откладывающийся вне хлоропластов, в цитоплазме) крахмал
и жир. Крахмал встречается преимущественно у пресноводных
форм, жир—у морских.
Ядерный аппарат. Все динофитовые водоросли имеют уникаль-
ное клеточное ядро, носящее специальное название динокарион, для
которого характерно конденсированное состояние ДНК на протя-
жении всего митотического цикла, включая интерфазу. Таким об-
разом. число хромосом может быть подсчитано на препаратах
интерфазных ядер (за исключением, по-видимому, ядра в вегетатив-
ных клетках ночесветки. где хромосомы в интерфазе ясно не раз-
личаются). Хромосомы имеют вид многочисленных палочковидных
телец, имеющих исчерченную (полосатую) структуру, кинетохоры
отсутствуют. Тесты с энзимами, такими, как ДНК-аза, в более
ранних работах показали, что хромосомы динофитовых водорослей
в отличие от хромосом эукариотических организмов состоят из
ДНК-фибрилл без какого-либо заметного количества гистона, т. е.
как у всех прокариотических организмов. Это побудило назвать
динофитовые водоросли мезокариотическими организмами, зани-
мающими по организации их ядра промежуточное положение меж-
ду прокариотическими и эукариотическими водорослями. Позднее
были представлены убедительные аргументы (в частности, размеры
ДНК-фибрилл) в пользу отнесения динофитовых к эукариотическим
организмам, а недавно были обнаружены и низкие концентрации
хромосомных белков (гистонов). Таким образом, от термина мезо-
кариотические следует отказаться. Ядра у различных представи-
телей динофитовых водорослей помимо крупных размеров отлича-
ются большим разнообразием формы: сферические, полукруглые,
эллипсоидальные, четырехугольные, треугольные, почковидные.
Они окружены ядерной оболочкой, как правило с порами, и содер-
жат ядрышко.
Митоз имеет ряд особенностей. Центриоли отсутствуют. При
делении ядро увеличивается. Его оболочка остается интактной на
протяжении всего митоза и возникают глубокие инвагинации (впя-
чивапия) ее внутрь ядра; некоторые из них пронизывают ядро
насквозь, образуя каналы или туннели. В этих цитоплазматических
каналах, и таким образом вне ядра, возникают микротрубочки —
эквивалент веретена. Сегрегация хроматид, возможно, осуществля-
ется благодаря прикреплению их к ядерной оболочке, выстилающей
туннели, которые заметно вытягиваются. В конце концов ядерная
оболочка перетягивается по экватору и разделяет ядро (рис. 33)
и ядрышко, сохраняющееся на протяжении всего митоза, на две
части.
Другие органеллы клетки — аппарат Гольджи, митохондрии — в
общем обычного строения. Аппарат Гольджи состоит из стопок
уплощенных мембран, вздутых на концах, с многочисленными терми-
нальными везикулами. Часто тела Гольджи располагаются по сосед-
ству с ядром. Митохондрии — вытянутые, иногда разветвленные
163
Рис. 33. Схема митоза у динофитовых водорослей.
А — ранняя стадия деления ядра; Б—более поздняя стадия; В — продольный разрез
через лопасть ядра: хромосомы прикреплены к ядерной оболочке, выстилающей
канал с микротрубочками; Г—образование дочерних ядер:
мт—микротрубочки, я- ядро, яо — ядерная оболочка, дя—дочерние ядра, х- -хромосомы
структуры, округлые на поперечном срезе, ограничены двойной
мембраной; кристы короткие, трубчатые. Митохондрии рассеяны по
цитоплазме. У Gonyaulax polyedra описано слияние отдельных мито-
хондрий, приводящее к образованию гигантских митохондрий.
Монадные клетки динофитовых водорослей имеют ряд специфи-
ческих особенностей, касающихся клеточного покрова, трихоцист,
строения жгутикового аппарата, осморегуляторной системы клетки,
фоторецепторного органа.
Клеточный покров. У монадных клеток динофитовых водорослей
имеется сложный клеточный покров — тека или амфиезма, пред-
ставленный цитоплазматической мембраной, под которой в один
слой располагаются компоненты теки — уплощенные пузыри (вези-
кулы, цистерны), окруженные одиночной мембраной. Число текаль-
ных цистерн в клетке варьирует от нескольких сотен (гимнодиниум)
до двух (пророцентрум). Расположение текальных везикул под плаз-
малеммой неодинаково. Например, у рода оксирис они разъедине-
ны друг от друга (рис. 34, А), а у гимнодиниума, амфидиниума
и других родов везикулы соприкасаются друг с другом у своих
краев и приобретают многоугольные очертания. Везикулы у одних
представителей, таких, как гимнодиниум, клетки которых в оптичес-
ком микроскопе выглядят как голые, не содержат текальных пла-
стинок, в них наблюдается только зернистое содержимое. У других
в цистернах находятся целлюлозные пластинки (по одной на цистер-
ну), различающиеся по толщине. Имеется обратная зависимость
между толщиной пластинок и их числом. Чем толще пластинки, тем
их меньше. У катодиниума везикулы содержат тонкие пластинки,
а у волошинскии пластинки достаточно толстые, чтобы их можно
было различить в оптическом микроскопе (рис. 34, Б, В, Г). У ро-
дов гленодиниум, перидиниум, цератиум число пластинок редуци-
руется приблизительно до двадцати, они очень толстые (рис. 35)
и могут нести шипы, различные выросты, иметь сетчатый узор.
164
Рис. 34. Схема различных типов тек динофлагеллят.
А— Oxyrrhis', Б—Amphidinium', В—Katodinltum; Г—
Woloszynskia
Рис. 35. Схема теки СегаНипт.
пл — плазма лемма, нтм — наруж-
ная текальная мембрана, втм —
внутренняя текальная мембрана,
тп—текальная пластинка, пт —
пора трихоцистная, тц -текаль-
ная цистерна
Пластинки вокруг клетки создают прочный целлюлозный панцирь.
У пророцентрума в клетке только две главные толстые пластинки
(в виде часовых стекол) формируют целлюлозный панцирь. Во всех
случаях в норме непосредственно под текой находятся микротру-
бочки.
В месте соприкосновения соседних текальных пузырей (цистерн)
их мембраны располагаются очень тесно, образуя шов. Рост пласти-
нок происходит в двух областях. Посредством отложения стро-
ительного материала у краев пластинок вдоль шва возникают ин-
теркалярные полосы, обусловливающие плоскостной рост пласти-
нок. У Gonyaulax polyedra вдоль мембран швов по обе стороны
наблюдается электронно-плотный слой, который, возможно, при-
нимает участие в плоскостном росте пластинок. В результате от-
ложения материала вдоль внутренней и наружной сторон пластинок
они утолщаются и на верхней стороне пластин развивается харак-
терная для каждого вида скульптура. Необходимый для роста
в толщину материал, возможно, доставляется в текальные пузыри
с помощью специальных шаровидных везикул и от внутренней
стороны пластин на их наружную сторону — через трихоцистные
поры.
Иногда у отдельных клеток динофлагеллят наблюдается процесс
сбрасывания панциря (экдизис), который можно искусственно
стимулировать и затем наблюдать регенерацию панциря. У G.
polyedra экдизис вызывается охлаждением клеток до 4 °C в течение
20 мин. При переносе в обычные условия культуры панцирь
регенерирует. Этот процесс занимает немного времени; уже через
165
'•у-'
4 ч на поверхности клетки можно обнаружить элементы панциря
(кольцевые поры, швы и пр.), а через 24 ч клетки, ’округлившиеся
после сбрасывания панциря, принимают характерную для них
полиэдрическую форму.
В текальных пластинках есть специальные отверстия — поры для
трихоцист. Трихоцисты располагаются по периферии клеток. Число
их варьирует от немногих (Gymnodinium simplex) до сотен (Oxyrrhis
marina).
Трихоцисты состоят из узкой области — шейки, прикрепляющей-
ся к теке в месте нахождения округлой поры с приподнимающимся
краем, и более широкой части — тела; и шейка, и тело окружены
одной мембраной. На поперечном срезе тело имеет квадратные или
ромбические очертания и состоит из протеиновых субъединиц,
упакованных в форме кубической решетки, шейка же образова-
на пучком спирально закрученных фибрилл. Выбрасываемые
при возбуждении через поры в теке трихоцисты значительно
длиннее и уже. Они представляют собой белковые нити с пери-
одической исчерченностью под прямым углом к их длинной
оси. Есть данные, что примордиальные трихоцисты возникают
в центральной части клетки в везикулах — дериватах аппарата
Гольджи.
Жгутиковый аппарат. Два жгутика различаются по форме (см.
рис. 40): один «продольный», обычного строения, содержи! ак-
сонему 9 — 9 + 2, на мембране его во многих случаях в два ряда
располагаются волоски; второй — «поперечный» — заключает вну-
три растянувшейся мембраны, помимо аксонемы, еще исчерченный
тяж; мембрана его несет односторонний ряд длинных тонких
волосков (мастигонем). Как и у других водорослей (золотистых,
бурых), мастигонемы возникают интрацеллюлярно: везикулы, со-
держащие промастигонемы, в цитоплазме можно видеть часто
вблизи ядра и в тесной связи с дикгиосомами и эндоплазма-
тическим ретикулумом.
Пузулы. Большинство монадных динофитовых водорослей об-
ладает (по-видимому, для регуляции 'осмотического давления) вме-
сто отсутствующих пульсирующих вакуолей особой органеллой —
пузулой (см. рис. 40). Пузула представляет собой инвагинацию
(впячивание) цитоплазматической мембраны в форме мешка или
трубки, глубоко вдающуюся внутрь клетки и непосредственно со-
прикасающуюся с вакуолью клеточного сока (рис. 36). Вакуоль
окружает пузулу, стенка которой становится таким образом двух-
мембранной (цитоплазматическая мембрана и тонопласт). Пузулы
изливают свое содержимое наружу через жгутиковые каналы. В слу-
чае крупной мешковидной пузулы поверхность контакта между
плазмалеммой и тонопластом увеличивается благодаря инвагина-
циям разветвлений вакуоли внутрь пузулы (рис. 36, А). То же
относится и к трубковидным пузулам. Если приходящее в кон-
такт с вакуолярной системой впячивание плазмалеммы сообща-
166
Рис. 36. Различные типы пузул динофитовых водорослей. А —простая мешковидная
пузула; Б— мешковидная пузула с инвагинациями; В — простая трубчатая пузула;
Г—собирательная пузула:
цм— цитоплазматическая мембрана, г тонопласт, в —вакуоль с клеточным соком
ется с наружной средой не непосредственно, а с помощью со-
бирающего пространства, говорят о собирательной пузуле
(рис. 36, Г). Расположение в клетке пузырей, собирательных ка-
мер, выводковых трубок у разных представителей варьирует.
Часто около мешковидной находится собирательная пузула,
изливающаяся или в мешковидную, или через свой собственный
проток.
Глазки (стигмы), расположенные вне хлоропластов у основания
жгутиков, также отличаются большим разнообразием: от простей-
шего глазка в виде массы глобул (Woloszynskia coronatd) до сложной
светочувствительной органеллы, состоящей из стекловидной, пре-
ломляющей свет линзы и пигментного тела, например у рода
эритропсис. У нематодиниума сгигма тоже состоит из прозрачного
линзовидного тела, окружающей его снизу пигментной чашечки
и расположенного на дне этой чашечки светочувствительного рети-
ноидного тела (рис. 37). Линза точно фокусирует свет на дне
пигментной чашечки. С физической точки зрения была доказа-
на возможность получения изображения объекта на ретиноидном
слое.
Размножение вегетативное, бесполое, половое, ' ,,,
167,
Рис. 37. Стигма
Nernalodbuum:
лт — линзовидмое тело, ич —
пигментная чашечка, р — ре-
тиноид
* * *
Отдел Dinophyta делится йа три класса:
десмофициевые, динофицйевые и бласто-
динофициевые. Представители класса дес-
мофициевых (Desmophyceae) — монадные,
голые или одетые панцирем, преимущест-
венно морские формы со жгутиками, от-
ходящими от переднего конца клетки.
Класс динофициевые (Dinophyceae) объ-
единяет монадные, ризоподиальные, паль-
меллоидные, коккоидные и нитчатые
формы. Монадные клетки дорзивентраль-
ные, с поперечной и продольной ложбин-
ками (бороздками), в которых располага-
ются жгутики, отходящие в месте пересече-
ния ложбинок на вентральной стороне
клетки. Класс бластодинофициевые (Blas-
todinophyceae) — искусственная группа,
объединяющая паразитные формы.
КЛАСС ДЕСМОФИЦИЕВЫЕ DESMOPHYCEAE -
Представителем может служить преимущественно морской род
пророцентрум (Prorocentrum), иногда размножающийся столь обиль-
но, что вызывает цветение прибрежных вод. Клетки более или менее
овальные, с боков несколько уплощенные, лишены ложбинок или
бороздок. Две главные пластинки тек блюдцевидные, смыкаются
вдоль всего края, за исключением их переднего конца, где в одной
из них имеется выемка. В апикальной обласги располагается ряд
(4—6) мелких толстых пластинок, довольно прочно соединенных
друг с другом так, что апикальная структура имеет тенденцию
отделяться как одно целое. От апикальных пластинок отходит
и характерный для рода апикальный шип, или зубец,— трубчатая
или сдвоенная структура. Главные текальные пластинки гладкие
или орнаментированные (сетчатые или с шипиками), перфорирова-
ны рядом трихоцистных пор. На переднем конце клетки имеются
два жгутика, выходящие через индивидуальные каналы в апикаль-
ных пластинках. Один жгут («продольный») направлен вперед, пре-
вышает по длине клетку, имеет заостренный конец и, по-видимому,
не несет волосков. Второй жгут, несмотря на то что клетка лишена
каких-либо следов поперечной борозды, свойственной монадным
клеткам динофициевых водорослей, является по структуре типич-
ным «поперечным» жгутом. Он закручен и внутри свободной Мемб-
раны, помимо аксонемы, содержит исчерченный тяж; мембрана его
несет односторонний ряд мелких волосков. Основания жгутов в от-
личие от динофлагеллят параллельны друг другу и располагаются
168
J
каждое в цитоплазматическом выступе в полости под текой. При
вхождении жгутика в цитоплазматический выступ находится толс-
тый трехслойный базальный диск, замыкающий центр аксонемы,
под которым периферические дублеты превращаются в триплеты.
Оба жгутика различаются по расположению и структуре своих
корней (какая из этих корневых структур относится к «продоль-
ному». а какая к «поперечному» жгуту — установить не удалось).
Один жгутик в месте выхода из цитоплазмы имеет «воротничок»,
и несколько микротрубочек (трубчатый корень) тянутся от основа-
ния жгутика к «воротничку». У другого жгута проксимальный конец
базального тела заключен в электронно-плотный материал, от ко-
торого отходят вперед трубчатый корень и корень с поперечной
исчерченностью (рис. 38).
В протопласте находится один желтовато-бурого цвета пари-
етальный сетчатый хлоропласт, окруженный трехмембранной обо-
лочкой и содержащий ламеллы из двух-трех тилакоидов. Пиреноид
ограничен растяжением оболочки хлоропласта, матрикс его обычно
пересечен несколькими двухтилакоидными ламеллами. Гранулы
крахмала откладываются вне хлоропласта. В задней половине клет-
ки расположено ядро («динокарион»), более или менее сферическое,
с ядрышком, оболочка ядра с многочисленными порами. Деление
ядра такое же, как у других динофлагелля г: ядро становится пер-
форированным туннелями, которые содержат многочисленные мик-
ро трубочки и ориентированы большей частью поперек ядра — под
прямым углом к плоскости деления. Тела Гольджи, митохондрии,
Рис. 38. Схема расположения оснований жгутиков и корней Prormvntrum (подробнее
см. в тексте):
ап — апикальная пластинка, бд— батальный диск, в — «воротничок», тк -трубчатый корень,
пк — полосатый—исчерченный корень
/
169
мешковидная пузула — как и у других динофитовых водорослей.
Предшественники трихоцист находятся вблизи центра клетки, а зре-
лые трихоцисты — под текой. Размножение осуществляется путем про-
дольного деления клетки, причем дочерние протопласты получают
одну половину материнского панциря, а другую выделяют заново.
КЛАСС ДИНОФИЦИЕВЫЕ — DINOPHYCEAE
В зависимости от типа организации таллома класс делится на
следующие порядки: перидиниальные, или динофлагелляты -
Peridiniales, Dinoflagellatae (включает монадные формы), динамёби-
диальные — Dinamoebidiales (ризоподиальные, или амебоидные),
глеодиниальные — Gloeodiniales (пальмеллоидные), динококкаль-
ные — Dinococcales (коккоидные) и динотрихальные — Dinotrichales
(нитчатые).
Ниже рассматриваются порядки неридиниальные и динококкаль-
ные.
Порядок перидиниальные, или динофлагелляты—
Peridiniales, Dinoflagellatae
Строение клетки. Это самый обильный представителями поря-
док динофитовых водорослей. Монадные клетки яйцевидно-шаро-
видные, более или менее сжатые дорзивентрально, так что при
рассматривании с полюсов они имеют эллиптическую или почко-
видную форму. Нередко клетки несут выросты в виде рогов' или
шипов. Большей частью это микроскопически мелкие формы, лишь
морская ночесветка видна невооруженным глазом, достигая 2 мм
в диаметре. Для тела динофлагеллят чрезвычайно характерны две
бороздки. Поперечная бороздка хорошо заметна и обычно полно-
стью опоясывает клетку, отделяя апикальную половину клетки —
эпивальву. направленную при движении вперед, от антапикальной —
задней половины — гиповальвы. Поперечная бороздка идет или по-
чти горизонтально, как у рода перидиниум (Peridinium), или чаще
несколько по спирали, иногда делая не один, а несколько оборотов,
как у рода варновия (Warnowia). Очень редко поперечная борозда
опоясывает лишь часть клетки, например у гсмидиниума
(Hemidinium). Поперечная борозда чаще проходит по середине клет-
ки (перидиниум), но может быть смещена к переднему концу, как
у рода амфидиниум (Amphidinium), реже к заднему, как у катодини-
ума {Katodinium). От положения поперечной борозды зависит высо-
та эпи- и гиповальвы. Эпивальва спереди может быть закруглена
(перидиниум) или вытянута в длинный рог, как у церациума
(Ceratium), гиповальва у церациума также несет один или два рога.
Кроме поперечной борозды есть продольная борозда, выраженная
менее резко и проходящая перпендикулярно к поперечной по вент-
170
ральной (брюшной), несколько сплющенной стороне клетки. Про-
дольная борозда очень немного заходит на эпивальву и часто
ограничивается одной гиповальвой — антапикальной стороной
клетки.
Однако у рода гоннаулакс (Gonyaulax) продольная борозда про-
стирается от переднего до заднего конца клетки. У некоторых
морских перидиней продольная борозда не прямая, а изогнутая или
спирально извитая.
Два жгутика динофлагеллят выходят на вентральной стороне
клетки через индивидуальные отверстия в текальных пластинках.
Поперечный лентовидный жгут опоясывает клетку и скрыт в попереч-
ной борозде, лишь дистальный конец его выступает наружу. Жгути-
ковая мембрана поперечного жгута растянута и, как правило, заклю-
чает узкий негибкий тяж, имеющий правильную поперечную полоса-
тость. Этот тяж находится в состоянии натяжения, что, по-видимому,
и удерживает поперечный жгут в борозде. Гораздо более длинная
аксонема (9 + 9 — 2) идет по спирали вокруг тяжа. Поперечный ленто-
видный жгут несет односторонне расположенные волоски одинако-
вой длины (Gonyaulax polyedra, Gymnodinium breve и др.) или же
волоски могут различаться по длине. Например, поперечный жгут
Gyrodinium lebouriae несет длинные и короткие волоски и еще третий
гип придатков (пока описанных только у этого вида) в виде сплош-
ных, изогнутых по направлению к кончику жгута шипов. Все три типа
отростков располагаются вдоль одного спирального ряда. На самом
кончике жгута волоски и острия отсутствуют (рис. 39).
Продольный жгут значительно длиннее продольной борозды,
высгупает из нее и направлен по движению клетки назад. Тонкие
волоски располагаются на нем в два ряда (Gyrodinium lebouriae,
Woloszynskia micra, Oxyrrhis marina). Жгутиковые волоски образу-
ются в расширенной перинуклеарной полости. Расположение жгути-
ковых корней очень варьирует.
На рис. 40 приведена схема строения монадной клетки динофи-
циевых водорослей.
Размножение динофлагеллят вегетативное, бесполое и поло-
вое. Вегетативное размножение осуществляется делением клетки
пополам, например у гимнодиниума, цератиума, перидиниума, го-
ниаулакса.
Клеточный покров динофлагеллят—тека — интенсивно изучал-
ся у взрослых, более старых клеток. Гораздо меньше сведений
о покровах во время и вскоре после деления клетки (G. Durr, 1979).
У панцирных динофлагеллят можно различить два разных типа
деления клетки. При одном типе, характерном для видов цератиум
(Ceratium hirundinella, С. tripos), гониаулакс (Gonyaulax polyedra) ста-
рый панцирь материнской клетки распределяется на две дочерние
клетки, а недостающие половинки панциря вырабатываются зано-
во. При другом типе деления клетки, характерном для рода периди-
ниум (Peridinium einctum), дочерние клетки образуют новый панцирь
171
Рис. 39. Схема поперечного жгута
G} то dintшп /еЬ о иг iае:
а —аксонсма, кв короткие волоски, лв—
длинные волоски, о - острия, с — стержень
Рис. 40. Схема строения клетки
Woloszynskia micra:
х—хлоропласт, л—диктиосома, м — мито-
хондрии, я — ядро, п — пиреноид, т трихо-
цисты, пу пузулы, пж—поперечный жгут,
прж—продольный жгут
полностью, а старый при этом погибает. Г. Дюр исследовал оба
типа: первый на примере G. polyedra, второй — у Р. cinctum.
У G. polyedra после деления ядра появляется борозда дробления.
Эта борозда закладывается почти одновременно по всей теке, сле-
дуя линии швов, мест соединения — стыка мембран текальных ци-
стерн. Мембраны шва разъединяются на две мембраны соседних
лекальных цистерн. Возникающий в этой области пузырь, одетый
мембраной (повой плазмалеммой), распространяется в плоско-
стном направлении и внедряется в соседние текальные цистерны под
находящиеся там пластинки. Этим текальные цистерны расщепля-
ются на две части: наружную с текальными пластинами, и внутрен-
нюю, содержащую лишь зернистый строительный материал для
новых пластин. Плоские пузыри, заключенные в текальные мемб-
раны, затем впячиваются по направлению к центру клетки под
несколько перекрывающими друг друга пластинами панциря и сли-
ваются. Так возникает борозда дробления, снаружи (с поверхности
клетки) еще не различимая. Одновременно внутрь клетки смещают-
ся и внутренние, нижележащие части текальных цистерн. Таким
образом, в борозде дробления под плазмалеммой уже с самого
начала находятся уплощенные пузыри — цистерны, содержащие
строительный материал для новых пластинок панцйря. Под ними
находится слой цитоплазмы с микротрубочками. Позднее борозда
прорывается наружу: мембраны шва расходятся, внутренняя новая
плазмалемма пузыря соединяется с наружной старой плазмалем-
мой и обе дочерние клетки отклоняются друг от друга (рис. 41).
172
Рис. 41. Схема образования борозды дробления Gonyaulax polyedra.
.А —в области шва — на стыке двух текальных цистерн — образуется пузырь, одетый
новой плазмалеммой. Он обнаруживает плоскостной рост и распространяется в со-
седние текальные цистерны, деля их на наружную (содержащую текальную пластин-
ку) часть и внутреннюю часть (без текальной пластинки) с хлопьевидным материа-
лом— возможным предшественником пластинки. S'-- мембраны шва расходятся.
Заключенный в текальных цистернах плоский пузырь, ограниченный новой плаз-
малеммой, впячивается внутрь клетки. В результате образуется борозда дробления,
еще не различимая снаружи, так как слегка разошедшиеся мембраны шва перекрыты
непрерывной старой плазмалеммой. В дальнейшее расхождение мембран шва,
разрыв старой плазмалеммы в области шва и соединение ее с новой плазмалеммой,
окружающей пузырь. Закладывающиеся уже в борозде дробления новые текальные
цистерны содержат строительный материал для образования новых пластинок. Под
внутренней мембраной цистерн в цитоплазме находятся микротрубочки:
ил— плазмалемма, мш — мембраны шва, нмц наружная мембрана циег.-риы. вмц внутрен-
няя мембрана цистерны, тп--текальная пластинка, в — везикул, одетый повой плазмалеммой,
мт — микротрубочки
В результате деления возникают две дочерние клетки, теки которых
состоят на одну половину из старых текальных пузырей с пластина-
ми, а на другую — из текальных пузырей, еще не содержащих
пластин. Возможно также, что обе половины теки уже с пластина-
ми, но пластины различаются по степени развития скульптуры,
однако эта разница между двумя половинками теки в молодых
дочерних клетках очень быстро исчезает, и уже вскоре после деления
почти все клетки Gonyaulax polyedra обнаруживают однородно
скульптурированный панцирь. Такая однородная структура пан-
циря молодых клеток достигается в результате обновления старой
половины панциря, унаследованной от материнской клетки.
Еще во время деления клетки в текальных цистернах и отчасти
в пластинах появляются везикулы с двойной стенкой. При увеличении
этих везикул в направлении, параллельном поверхности, как и при
образовании борозды дробления, текальные цистерны расщепляются
на наружную часть, содержащую пластинку, и внутреннюю, возмож-
но, с остатками старой пластинки. При разрыве пузыря-везикула
у расширенных краев кольцевых пор, выстланных плазмалеммой
и наружной мембраной цистерны, внутренняя мембрана пузыря
соединяется с плазмалеммой, а наружная — с наружной мембраной
текальной цистерны. Наружная часть текальной цистерны, содержа-
щая старую пластинку, пронизанную порами, изолируется и сбрасы-
вается, а во внутренней части возникает новая пластинка (рис. 42).
173
Рис. 42. Последовательные стадии сбрасывания старой, пронизанной порами текаль-
ной пластинки Gonyaulax polyedra (схема).
А— участок старой теки между тремя кольцевыми порами старой пластинки: сле-
ва -кольцевые поры (п). выстланные плазмалеммой (пл) и наружной мембраной
текальной цистерны (нмц); справа к началу деления клетки наружная мембрана
текальной цистерны и плазмалемма распространяются под текальной пластинкой
(тп), образуя под ней кольцевую перетяжку, так что поровые поля соответственно
расширяются. На срезе под пластинкой (отчасти в пластинке) можно видеть пузырь-
везикул (в), ограниченный двойной мембраной. Б— поверхностное разрастание nyj
зыря до стыка его с сжимающимися под пластинкой мембранами.?—плазмалеммой
и наружной мембраной текальной цистерны. В—пузырь разрывается. Его мемб-
раны— внутренняя и наружная — сливаются соответственно с плазмалеммой и на-
ружной мембраной цистерны, ограничивающих расширенные поровые поля под
текальной пластинкой. Таким образом, наружная часть цистерны со старой пластин-
174
Рис. 43. Возникновение плазмалеммы при де-
лении Peridinium ctnctum.
А -под внутренней мембраной текальных
цистерн появляются плоские пузыри (везику-
лы), окруженные мембраной. Л'- благодаря
слиянию пузырей возникают непрерывные
наружная и внутренняя мембраны пузыря.
Внутренняя мембрана пузыря становится но-
вой плазмалеммой дочерних клеток:
п — плазмалемма, нмц — наружная мембрана цисте-
рны. тп - текальная пластинка, вмц—внутренняя
мембрана цистерны, в—везикулы (пузыри), ныв —
наружная мембрана везикула, ни— новая плазма-
лемма
Таким образом, старые пластинки панциря сбрасываются в ре-
зультате перешнуровки текальных пузырей в плоскости, параллель-
ной поверхности. Внутренняя часть перешнурованных текальных
пузырей, возможно, содержит еще старый материал для образова-
ния новых пластинок. Эти пластинки не будут отличаться от новых
пластинок, возникающих в пузырях, выстилающих борозду дробле-
ния, т. е. в половинке клетки, образующей панцирь заново.
У Peridinium cinctum клетка делится внутри старого панциря,
текальные пластинки которого отчасти растворяются. Дочерние
клетки удерживаются некоторое время вместе внутри разрушающего-
ся панциря, заключенные в общий фиброзный слой; после его разрыва
клетки освобождаются. Еще внутри старого панциря в дочерних
клетках начинается отложение текальных пластинок. Ультратонкие
срезы через делящиеся клетки позволили проследить возникновение
новой плазмалеммы дочерних клеток (рис. 43). Под внутренней
мембраной текальных цистерн старого панциря появляются плоские
везикулы, которые сливаются с образованием наружной и внутренней
мембран. Внутренняя мембрана пузыря (везикула) становится плазма-
леммой дочерних клеток. Из содержимого пузыря, возможно, возника-
ет фиброзный слой. Наружная мембрана везикула остается с внутрен-
ней мембраной текальных цистерн, старыми текальными пластинка-
ми, наружной мембраной текальных цистерн и старой плазмалеммой.
У Woloszynskia apiculata при бесполом размножении клетки теря-
ют подвижность, округляются и превращаются в зооспорангии.
При этом под текой выделяется тонкая целлюлозная оболочка
собственно зооспорангия. Содержимое зооспорангия делится на
две, четыре, восемь клеток, которые освобождаются при растворе-
нии целлюлозной стенки собственно зооспорангия и разрыве и сбра-
сывании теки исходной материнской клетки.
кой отшнуровывается от ее нижней части без пластинки (или с остатками се — они не
изображены). Старая пластинка, пронизанная отверстиями порами, окруженная
плазмалеммой и наружной мембраной текальной цистерны, сбрасывается. В нижней
части текальной цистерны в дальнейшем закладываются поры и возникает новая
пластинка, не отличимая от таковой в цистернах, выстилающих борозду дробления:
п- - поры, пл - плазмалемма. нмц наружная мембрана цистерны, вмц— внутренняя мембрана
цистерны, тп — текальная пластинка, в- везикул
V
175
На протяжении двух последних десятилетий у многих дино-
флагеллят рядом ученых обнаружено половое размножение, ранее
известное лишь у одного из видов гленодиниума. Успех этих ис-
следований в большой степени обусловлен развитием методов
культивирования на искусственных питательных средах и накопле-
нием данных о том, что половой процесс у многих водорослей
индуцируется недостатком в среде азота. Это оказалось справед-
ливым и для многих динофлагеллят. Так, при помещении в среду,
свободную от азота, или в старых культурах, где уровень азота
низок, половой процесс наблюдался у ряда видов Peridinium
(Р. Umbatum, Р. cinctum, f. ovoplanum, Р. willei, Р. faeroense)
Woloszynskia apiculata, Glenodinium lubiensiforme, Gonyaulax moniliata,
Crypthecodinium cohnii, Protogonyaulax catenella и др. Сливаются
голые гаметы (отличающиеся от вегетативных клеток меньшими
размерами и более светлой окраской) одинакового размера, т, е.
имеет место изогамия, и зиготы у Р. cinctum, Р. willei, Р. Umbatum,
Gymnodinium pseudopalustre, Crypthecodinium cohnii возникают в пре-
делах одного клона (гомоталлизм).
Woloszynskia apiculata изогамна, но гетероталлична, у этого вида
наблюдалось образование групп. Гаметы сливаются своими бо-
ками. Напротив, у видов рода цератиум (Ceratium horridum,
С. cornutum) копуляция гамет постоянно анизогамная, и сам про-
цесс слияния иной: мужская гамета (меньших размеров) входит
в женскую (большую) через отверстие поблизости от места прикреп-
ления жгутиков.
Отличительной чертой всех до сих пор изученных в этом от-
ношении динофлагеллят (за исключением Amphidinium carteri, у ко-
торого зигота не увеличивается и становится неподвижной непо-
средственно после полового слияния) является долго продолжа-
ющийся рост планозиготы. Так, планозиготы Gymnodinium
pseudopalustre, прежде чем превратиться в покоящуюся зиготу, рас-
тут 11 дней при 15G С или 8 дней при 2Г С. У Woloszynskia apiculata
планозиготы также сильно вырастают и остаются подвижными
в течение 8—10 или 3—4 дней при температурах 15 и 21° С соответ-
ственно. Планозиготы передвигаются с помощью двух сохраня-
ющихся задних жгутиков гамет, в некоторых случаях остаются
и поперечные жгутики, по крайней мере один из них. Динофлагел-
ляты с «двойными» задними жгутиками описывались более 100 лет
тому назад и в материале из природы: по-видимому, в этих случаях
речь идет о планозиготах, которые нередко таким образом возника-
ют и в природных условиях, а не только в культурах. После
длительного движения, сопровождающегося ростом, планозиготы
(одно- или двухъядерные в зависимости от времени, когда проис-
ходит кариогамия) превращаются в неподвижные толстостенные
(одетые трехслойной оболочкой) гипнозиготы.
Г. Дюр (1979) исследовал строение оболочек гипноспор (цист)
в электронном микроскопе. Наружная оболочка (экзоспорий) похо-
176
жа на панцирь вегетативных клеток, отличаясь от него большей
толщиной пластинок и более широкими швами. Образование сред-
ней (мезоспорий) и внутренней (эндоснорий) оболочек, по-видимо-
му, сходно с образованием теки, т. е. они состоят из пластинок,
которые возникают в текальных цистернах, однако швы отсутству-
ют. Это заставляет предположить, что цистерны, в которых образу-
ются пластинки мезоспория и соответственно эндоспория, сливают-
ся с образованием непрерывных наружной и внутренней мембран:
затем между ними возникают мезоспорий и эндоспорий.
Через некоторый период покоя зиготы прорастают: так, отдель-
ные зиготы Gymnodinium pseudopalustre и Woloszynskia apiculata спо-
собны прорастать через несколько недель на свету при 15u С. Одна-
ко более обильное и синхронизированное прорастание бывает при
охлаждении зигот по крайней мере в течение 4 недель при 6 С для
Woioszynskia и при ?г С для Gymnodinium и последующем переносе
их на свет в условия более высокой температуры. При прорастании
зигот всех изученных до сих пор динофлагеллят происходит мейоз,
и таким образом динофлагелляты —гаплонты, т. е. их ядра в веге-
тативных клетках имеют гаплоидный набор хромосом.
* * *
Упомянутые выше роды динофлагеллят распределяются по мно-
гим семействам. Кроме них можно назвать еще род ночесветка
(Noctiluca) и ряд родов, объединяемых в семейство динофизовые
(Dinophysidaceae), заметно о тличающиеся от вышеописанных дино-
флагеллят.
Род ночесветка (Noctiluca) — крупная, распространенная по все-
му свету морская бесцветная голозойная динофлагеллята. Клетка
сферическая, достигающая 1 - 2 мм в диаметре, покрыта снаружи
полиэдрическими, одетыми мембраной текальными везикулами,
снабженными грихоцистными порами. Изнутри везикулы выстланы
рядами микротрубочек, тянущихся параллельно поверхности клет-
ки. Ци топлазма образует гонкий периферический слой, от которого
отходят тонкие тяжи, пересекающие крупные вакуоли с клеточным
соком. Постенная цитоплазма содержит многочисленные липидные
везикулы; некоторые из них, возможно, являются «микроисточ-
никами» биолюминесценции. На одной стороне сферической клетки
имеется глотка, в которой расположен один короткий жгутик.
Из этой части клетки образуется крупное поперечно исчерченное
подвижное щупальце, функция которого — улавливать необходи-
мые для питания организмы (диатомеи, перидинеи, мелкие жи-
вотные и их личинки) в свой слизистый эксудат и увлёкать их
в глотку, где они заключаются в пищеварительные вакуоли. В ци-
топлазме находятся ядро и крупные митохондрии, трихоцисты.
По соседству с местом прикрепления щупальца расположены пучки
исчерченных волокон, возможно, контролирующих его движение.
|1 177
’ Наряду с делением — наиболее обычным способом размножения
у ночесветки описаны: образование многих (до 5Q0) зооспор, по-
: хожих на гимнодиниум, и половой процесс в, виде слияния од-
ii ножгутиковых изогамет, возникающих внутри материнской клетки
в числе 1024. Есть данные, что перед образованием гамет про-
исходит мейоз, за которым следуют многие синхронные митозы.
Если эти данные подтвердятся, то ночесветка будет представлять
исключение среди других динофлагеллят, у которых редукция зи-
готическая.
Семейство динофизовые (Dinophysidaceae) включает
главным образом обитателей теплых тропических морей. Клетки
дорзивентральпые, сжатые с боков, снабжены, как и другие диноф-
лагелляты, бороздками, причем поперечная борозда сильно смеще-
на вперед. Таким образом, эпивальва небольшая и жгутики отходят
I в месте пересечения борозд также недалеко от переднего конца
клетки. Клетка одета текой, в везикулах которой находятся толстые
целлюлозные текальные пластинки. Текальные пузыри с пластин-
ками, соприкасаясь краями, образуют панцирь, который здесь в от-
личие от других динофлагеллят разделен на две боковые половинки,
। соединенные продольным (сагиттальным) швом. Края как продоль-
ной, гак и поперечной борозд несут крыловидные выросты. Раз-
множение— продольным делением клетки пополам в плоскости
сагиттального шва: каждая дочерняя клетка получает половинку
панциря материнской клетки, а недостающую строит заново.
Род динофнзис (Dinophysis). Края поперечной борозды направ-
; лены вперед, образуя воронки, скрывающие небольшую эпивальву.
Продольная борозда несет крылья, несколько различающиеся по
i размерам и форме. Некоторые виды встречаются и в холодных
морях (/). norvegica) и лиманах (D. ovum).
। Род орнитоцеркус (Ornithocercus) характеризуется эпивальной,
’ редуцированной до маленьких пластинок, соединенных продоль-
I ным швом. Крыловидные выросты очень сильно развиты как у про-
дольной, так и у поперечной борозды.
Перилинеи распространены в планктоне как морей, так и пресных вод. Они
занимают второе (после диатомовых) место как продуценты органического вещества
в морс.
Наряду с пользой псридинеи могут приносить и вред. Среди них имеются
паразиты, некоторые из них нападают на личинки устриц. Есть виды, вырабатыва-
ющие яд, смертельный для рыб. Кроме того, разлагаясь после своего массового
развития, они могут отравлять воду на многие километры вредными продуктами
распада, вызывая замор рыб и других водных животных.
Порядок динококкальные — Dinococcales
Объединяет коккоидные формы — неподвижные, одетые оболоч-
кой клетки с типичными для динофитовых водорослей внутренним
строением (динокарион, бурые хлоропласты и пр.). Вегетативное
деление отсутствует, размножение зооспорами, автоспорами. В ря-
178
де случаев замена зооспор автоспорами может быть прослежена
у разных видов в пределах одного и того же рода.
У рода тетрадиниум (Tetradinium) клетки имеют форму тетраэд-
ра, углы которого вытянуты в простые или двойные шипы.
При зооспорообразовании протопласт слегка сокращается и де-
лится пополам, после чего у продуктов деления формируются бо-
роздки. характерные для монадных клеток динофициевых водорос-
лей, появляются стигмы, В конце концов в растянувшейся оболочке
материнской клетки можно видеть две гимнодиниумподобные зоо-
споры. позднее освобождающиеся через разрыв оболочки.
Род цистодиниум (Cystodinium), встречающийся в болотах, харак-
теризуется веретеновидными или полулунными клетками. У одних
видов (С. bataviense, С. steinii) размножение зооспорами: содержи-
мое клетки отстает от оболочки и еще до деления принимает форму
динофлагелляты, т. е. развивает систему бороздок, появляется стиг-
ма; однако жгутики на этой стадии не образуются. Такая клетка
делится на две гимнодиниумподобные зооспоры, вырабатывающие
уже жгутики и покидающие оболочку материнской клетки. После
краткого периода движения зооспоры останавливаются, бороздки
пропадают, протопласт вытягивается и выделяется новая оболочка.
У Cystodinium lunare наблюдается редукция подвижной стадии; при
размножении протопласт сокращается, но, не вырабатывая бороз-
док и стигмы, претерпевает деление; продукты деления, также не
образуя бороздок и стигмы, несколько вытягиваются, принимают
очертания материнской клетки, одеваются собственной оболочкой
и освобождаются в виде мелких клеток цистодиниума. Таким об-
разом, здесь вместо образования зооспор происходит автоспорооб-
разование.
Глава 12
ОТДЕЛ КРИПТОФИТОВЫЕ ВОДОРОСЛИ—CRYPTOPHYTА
В подавляющем большинстве это монадные формы, реже кок-
коидные. Для хлоропластов представителей отдела характерны
хлорофиллы а и с, каротиноиды (а- и £-каротин, зеаксантин, диа-
гоксантин), а также синий фикоцианин и красный фикоэритрин.
Соответственно окраска хлоропластов варьирует от синей, сине-
зеленой, оливково-зеленой до желто-бурой, бурой и красной. Обы-
чно один или два хлоропласта, одетые оболочкой из двух мембран,
окружены слоем эндоплазматического ретикулума. Строма соде-
ржит обычно вздутые тилакоиды, расположенные по два (иногда
по три). Фикобилины (фикоцианин, фикоэритрин) в отличие от
таковых синезеленых и красных водорослей локализованы не в фи-
кобилисомах, на поверхности тилакоидов, а внутри тилакоидов.
179
В качестве запасного продукта в клетке синтезируется крахмал,
который откладывается вне пластид—между оболочкой хлоро-
пласта и слоем эндоплазматического ретикулума, а также наблю-
дается свободно в цитоплазме. Если есть пиреноиды (выступающие
из хлоропласта и сидящие на одной ножке—у Cryptomonas
cryptophila или многих — у родомонас, С. sp., Chroomonas
mesostigmatica—ножках, с матриксом, не пересеченным тилако-
идами или пересеченным двухтилакоидной ламеллой), то они окру-
жены двумя полусферическими или несколькими пластинками кра-
хмала, расположенными кнаружи от оболочки хлоропласта и пи-
реноида, но под чехлом эндоплазматической сети. У С. sp.
и С. reticulata в пиреноиде наблюдались пальцевидные цитопла-
зматические инвагинации. Стигма (наблюдаемая у немногих видов)
располагается внутри пластиды без какой-либо связи со жгути-
ковым аппаратом (см. рис. 44). Жгутиков два, базальные тела
Рис. 44. Схема строения клетки
криптофитовой водоросли:
я — ядро, х хлоропласт, с стиг-
ма, к — крахмал, св - сократитель-
ные вакуоли, б — борозда, д—дикти-
осомы, э — эектосомы, ж — жгуты
Рис^45. Схема эектосомы крип-
* тофитовой водоросли.
А—до раскручивания. Б—в
процессе раскручивания
180
их соединены волокнистым тяжем. Жгутики различаются по длине
очень незначительно (отношения их длин колеблются от 3:5 до
9:10). Более длинный несет два ряда жестких волосков, которые
имеют компактное шаровидное основание и оканчиваются тонкой
волосовидной частью; кроме того, длинный жгут может нести
дополнительный пучок из приблизительно 45 волосков (рис. 44).
Короткий жгут имеет лишь один ряд волосков, несколько более
коротких и нежных, чем волоски на длинном жгуте. У некоторых
видов короткий жгут свободен от волосков, оба жгутика закан-
чиваются акронемой — более узкой частью, в которую заходят
лишь центральные трубочки аксонемы; периферические дублеты
в ней отсутствуют.
Строение клетки. Клеточный покров монадных клеток представля-
ет собой перипласт, образованный плазмалеммой, под которой
располагается слой тонких пластинок или мембран. Они состоят из
протеинов; у рода криптомонас — гексагональные или полигональ-
ные, у рода хроомонас — четырехугольные. Вблизи перипласта обыч-
но находятся микротрубочки. От косо срезанного переднего конца
дорзивентральных монадных клеток отходит борозда, косая или
спирально завитая, не достигающая заднего конца клетки. От бороз-
ды внутрь клетки вдаются два (вентральный и дорзальный) отростка,
от одного из которых у разных форм отходят жгутики, и, кроме того,
узкий канал — глотка, хотя это образование не имеет никакого
отношения к восприятию пищи. Под перипластом, выстилающим
глотку (в случае недоразвития глотки — борозду), располагаются
сильно преломляющие свет цилиндрические структуры — эектосомы.
На периферии клетки могут также находиться эектосомы, но более
мелких размеров. Эектосомы окружены мембраной и внутри состоят
из двух главных компонентов; большая часть пространства занята
цилиндром из намотанного по спирали лентовидного мембранного
материала, который окружает узкий канал и V-образное углубление.
В V-образном углублении и несколько выступая из него находится
второй цилиндр меньших размеров, также образованный лентой
мембранного материала, свернутой в тугую спираль. Обе эти части,
по-видимому, связаны посредством волокон. При физических или
химических раздражениях происходит взрывной процесс раскручива-
ния свернутых туго лент, который можно сравнить с растягиванием
рулона бумаги. При этом разрываются мембрана эектосомы и пери-
пласт, свернутые ленты выбрасываются, превращаясь в длинные
раскрученные спирали (рис. 45). Сначала выбрасывается центральный
рулон более мелких размеров, который, покидая клетку, выдергивает
ленту из более крупного наружного цилиндра. Эектосомы образуют-
ся в везикулах аппарата Гольджи. На ранних стадиях везикулы
содержат рулон, состоящий из нескольких оборотов, и центральное
тело. По мере развития число оборотов и ширина образованного
свернутыми мембранами цилиндра увеличивается. Крупное сферичес-
кое ядро с ядрышком располагается у заднего полюса клетки.
181
До начала митоза в цитоплазме, снаружи от ядра, имеются
микротрубочки. К метафазе ядерная оболочка полностью разруша-
ется, но область, в которой происходит митоз, ограничена слоями
эндоплазматического ретикулума. В экваториальной области хро-
мосомы образуют плотную метафазную пластинку. Одни трубочки
веретена проходят сквозь пластинку, другие могут быть прикреп-
лены к ней. Центриоли не наблюдаются. В анафазе дочерние хромо-
сомы разъединяются в виде двух глыбок, расходящихся затем к хло-
ропластной эндоплазматической сети, которая, по-видимому, дает
начало ядерным оболочкам дочерних ядер (рис. 46).
В цитоплазме находятся удлиненные митохондрии, аппарат Го-
льджи, в передней части клеток — пульсирующие вакуоли, излива-
ющие свое содержимое в бороздку (см. рис. 44).
КЛАСС КРИПТОФИЦИЕВЫЕ—CRYPTOPHYCEAE
Криптофитовые водоросли включают один класс — криптофици-
евые (Cryptophyceae) с двумя порядками: криптомонадальные
(Cryptomonadales) и криптококкальные (Cryptococcales). Первый
объединяет монадные, второй — коккоидные формы.
Порядок криптомонадальные — Cryptomonadales
Порядок криптомонадальные, включающий большинство крип-
тофитовых водорослей, распространенных как в пресных водах, так
и в морях, делится на ряд семейств, различающихся по направле-
ние. 46. Схема митоза у Cryptophyceae.
А профаза; Б—метафаза; В- анафаза—те-
лофаза; Г— телофаза, цитокинез (стрелка
указывает направление борозды дробления):
бт— базальные тела, мт микротрубочки, я—яд-
ро, эс — эндоплазматическая сеть, х— хромосомы.
□ --пластиды, хэс — хлоропластная эндоплазмати-
ческая сеть, дя дочерние ядра
182
нию жгутиков (оба направлены вперед или в разные стороны)
и наличию или отсутствию глотки. Наиболее важно из них —
семейство криптомонадовые (Cryptomonadaceae), характеризующе-
еся направленными вперед жгутиками и присутствием глотки. Оно
включает как окрашенные, так и бесцветные формы. Представи-
телями первых могут служить роды: родомонас (Rhodomonas) с хро-
матофорами обычно красного цвета, хроомонас (Chroomonas) с го-
лубыми, синими или синезелеными хроматофорами и криптомонас
(Cryptomonas) с хлоропластами преимущественно бурыми или олив-
ковыми. Среди бесцветных криптомонадовых наиболее известен
часто встречающийся в загнивающей воде род хиломонас
(Chilomonas). В его клетке находятся бесцветный хлоропласт (лей-
копласт) и зерна крахмала (несмотря на отсутствие пигментов
и осмотрофное питание).
Обычный способ размножения криптомонадовых — продольное
деление клетки в подвижном или неподвижном состоянии. Половой
процесс неизвестен. У Cryptomonas ovata развиваются цисты с глад-
кой оболочкой.
Порядок криптококкальные — Cryptococcales
Представителем криптококкальных (Cryptococcales) может слу-
жить род тетрагонидиум (Tetragonidium). Бурые коккоидные клетки
имеют форму тетраэдра, одеты целлюлозной оболочкой, утолщен-
ной на углах. Размножение посредством зооспор, сходных с крии-
томонадальной водорослью, относящейся к роду криптохризис
(Cryptochrysis), который характеризуется направленными вперед
жгутиками и отсутствием глотки. После периода движения зооспо-
ры тетрагонидиума останавливаются, выделяют оболочку и приоб-
ретают тетраэдрическую форму.
Глава 13
ОТДЕЛ РАФИДОФИТОВЫЕ ВОДОРОСЛИ—RAPHIDOPHYT А
Небольшая группа жгутиконосцев. Крупные дорзивентральные
с бороздой вдоль вентральной стороны клетки одеты плазма-
леммой и содержат мелкие многочисленные дисковидные или
эллипсоидальные париетальные хлоропласты, окрашенные в свет-
ло-зеленый цвет. Цвет обусловлен хлорофиллами а и с, р-ка-
ротином, антераксантином, лютеином, диадиноксаптином, диа-
гоксантином. Запасной продукт — масло, рассеянное по клетке
в виде капель. От углубления борозды на переднем конце клетки
отходят два неравных жгутика, из которых один направлен вперед
и несет волоски, а другой — гладкий, направлен назад и лежит
183
I
1 в вентральной борозде. В центре клетки расположено крупное
клеточное ядро. В цитоплазме — митохондрии, диктйосомы, пуль-
сирующие вакуоли. Непосредственно под поверхностью клетки
находятся трихоцисты в виде палочковидных или шаровидных
образований. Размножение — продольным делением клеток в под-
вижном состоянии или в состоянии пальмеллы. Распространены
рафид офитовые водоросли в пресных водах, встречаются в море.
Род вакуолярия (Vacuolaria) с округлыми трихоцистами, содер-
жимое которых при возбуждении взбухает в слизь.
Род гониостомум (Goniostomum) характеризуется палочковидны-
ми трихоцистами, собранными в группы или рассеянными по телу.
Глава 14
ОТДЕЛ ЭВГЛЕНОФИТОВЫЕ ВОДОРОСЛИ —
EUGLENOPHYTA
Отдел представлен преимущественно монадными, реже ризопо-
диальными и капсальными формами, для которых характерны хло-
рофиллы а и b (как у зеленых водорослей) и каротиноиды: р-
каротин, антераксантин, лютеин, виолаксантин и др. Хлоропласты
веретеновидные или лентовидные, дисковидные, звездчатые, пла-
стинчатые, париетальные или расположенные в радиальных, напра-
вленных внутрь клетки рядах, одеты оболочкой из трех мембран.
Ламеллы двух-, трехтилакоидные. Опоясывающая ламелла, по од-
ним данным, отсутствует, по другим — имеется. Генофор (молеку-
лы ДНК) рассеян по матриксу хлоропластов. Пиреноид выступает
за поверхность пластиды в виде почки на ножке, так что запасной'
полисахарид парамилон, хотя и образуется в цитоплазме (вне хло-
ропласта), но у поверхности выступающего пиреноида. У видов,
лишенных пиреноида, зерна парамилона разной формы (плоские
диски, выпукло-вогнутые диски, палочки, кольца и т. п.) свободно
рассеяны в цитоплазме. Стигма экстра пластидная (т. е. расположен-
ная вне пластиды, в цитоплазме), состоит из варьирующего числа
(от 1—2 у эвтрепции и хавкинеи до 20—60 у эвглены) расположен-
ных в один слой (трахеломонас) или плотно скученных глобул,
которые поодиночке или по 2—5 окружены элементарной мембра-
ной, Стигма находится в непосредственной близости от базального
вздутия жгута, где, по-видимому, содержится фоторецептор, ответ-
ственный за распознавание клеткой интенсивности и направления
света при фототаксисе. Стигма функционирует как абсорбирующая
свет ширма, служащая для затенения фоторецептора.
Исходными формами среди эвгленофитовых, по-видимому, яв-
ляются двужгутиковые. Два жгутика (один направлен вперед, дру-
гой— вбок и назад) отходят от находящегося на переднем конце
184
г
г клетки углубления — глотки. Последняя состоит из узкой трубки
| и расширяющейся задней части — резервуара. Мембраны жгутов
! заключают, помимо аксонемы (9 — 9 + 2), парафлагеллярное тело
с кристаллической структурой, которое тянется от основания жгута
до его конца. На поверхности жгута по спирали в один ряд рас-
J полагаются волоски, У эвглены, трахеломонаса и близких родов
направленный назад жгутик редуцирован до нефункционирующего,
находящегося в глотке короткого рудимента с аксонемой, в которой
обе центральные трубочки отсутствуют, а периферический круг
образован девятью одиночными трубочками, а не дублетами.
I Клетка эвгленофитовых имеет особый покров—пелликулу, со-
стоящую из расположенных под непрерывной плазмалеммой узких,
опоясывающих клетку по спирали полос более плотного протеина.
Пелликулярные полосы (они различимы и в световом микроскопе —
спиры, стрии) своими краями находят друг на друга, соединяясь
наподобие суставов (рис. 47), У краев полос, особенно там, где
Рис. 47. Разрез через пелликулу
Euglena:
им— цитоплазматическая мембрана,
пп-—пелликулярные полосы
соседние полосы перекрывают друг друга, лежат группы радиально
и про только ориентированных микротрубочек, образующих добавоч-
ный ске icr. Кроме того, у мест перекрывания полос изнутри рас-
полагаются с шзевые тела, изливающие свое содержимое через узкий
канал наружу между пелликулярными полосами и, возможно, связан-
ные с механизмом движения. Если пелликулярные полосы гибки и по-
движны одна относительно другой, то клетка способна к изменению
формы...-метаболии (эвгленоидному движению). Например, у Euglena
gracilis клетка может вытягиваться змеевидно и сокращаться до пол-
ного округления. Эта метаболия может сопровождаться поступатель-
ным движением всей клетки с помощью жгутика. Уплотнение пел-
ликулярных полос и их взаимное «цементирование» у многих родов
и видов ведет к отсутствию метаболии. Такие ригидные клетки харак-
терны, например, для рода факус. У нескольких родов (например,
трахеломонас) имеются домики, в которых клетки помещаются свобо-
дно. Домики снабжены отверстием, через которое выступает жгутик,
а при делении клетки выползает одна из дочерних особей.
В цитоплазме находится диктиосома, состоящая из располо-
женных одна над другой стопок цистерн, число которых может
достигать 30. От краев цистерн, большей частью вздутых, отшну-
ровываются мелкие везикулы. Митохондрии обычного строения.
Во время фазы роста у эвглены наблюдались гигантские непра-
вильно разветвленные митохондрии с кристами только по пери-
ферии. По-видимому, они возникают путем слияния многих мелких
митохондрий и перед клеточным делением снова разделяются на
185
митохондрии нормальных размеров. В митохондриях, так же как
в хлоропластах эвглены, обнаружена ДНК. Вокруг главной пуль-
сирующей вакуоли, находящейся в передней части клетки, около
резервуара глотки, расположены дополнительные вакуоли, в ко-
торых, возможно, скапливается жидкость, прежде чем излиться
в главную вакуоль. Последняя затем сливается с плазмалеммой,
освобождая содержимое в резервуар.
Клеточное ядро, как и у динофитовых водорослей, имеет хромо-
сомы, сохраняющиеся в интерфазе в конденсированном состоянии.
Хромосомы, помимо ДНК, содержат нуклеогистоны. Отклоняю-
щийся от нормы митоз происходит внутри интактной ядерной
оболочки. Во время профазы в кариоплазме появляются микротру-
бочки; ядрышко (эндосома) удлиняется в направлении плоскости
деления, хромосомы располагаются в форме рыхлой ленты вокруг
него—эквивалент метафазы. После того как хромосомы собрались
у полюсов, делится эндосома. Центриоли и центромеры отсутству-
ют. Оболочки дочерних ядер возникают путем перешнуровки ста-
рой оболочки материнского ядра. Одновременно с делением ядра
удваиваются жгутики — еще до начала цитокинеза и развития резер-
вуаров будущих дочерних клеток.
На рис. 48 приведена схема строения клетки эвгленофитовых
водорослей на примере рода трахеломонас. Размножение--про-
дольным делением клетки пополам в подвижном или неподвижном
состоянии. Деление пополам у форм, живущих в домиках (напри-
мер, у трахеломонаса), происходит внутри них: один из дочерних
протопластов выходит через маленькое отверстие в домике наружу
и через некоторое время выделяет свой собственный домик. Неред-
ко клетки переходят в пальмеллевидное состояние: они округляют-
ся, выделяют слизь и, делясь, образуют неправильные слизистые
Рис. 48. Схема строения клетки
Trachetomonas:
до — домик, tie — пелликула, я — ядро,
М— МИТОХОНДРИИ, Д "ДИКТИОСОМЫ, х —
хлоропласт, пи — пиреноид, св — сокра-
тительные вакуоли, р- резервуар, дж- •
длинный жгут, кж- короткий, не высту-
пающий из резервуара жгут, вж— взду-
тие длинного жгута, ож — основание
жгута, с—стигма, п — парамилон
186
скопления. При неблагоприятных условиях формируются покоящи-
еся цисты с толстыми оболочками.
Половой процесс не доказан, хотя указания на него имеются
(Phacus pirum, Hyalophacus ocellatuP).
Среди эвгленофитовых, как и в других отделах, наряду с окра-
шенными фотосинтезирующими имеются параллельные им бес-
цветные формы. Например, бесцветный род хавкинея параллелен
роду эвглена, бесцветный род гиалофакус параллелен окрашенному
роду факус. Бесцветные формы питаются сапротрофно, т. е. осмо-
тически поглощают растворенные в среде органические вещества,
или фаготрофно (голозойно), захватывая и переваривая оформлен-
ные частицы (диатомеи и др.). У многих фаготрофных форм для
этого служит находящееся на переднем конце клетки и вдающееся
внутрь ее воронковидное углубление—фарингиальный аппарат
(сифон); у его основания располагается палочковидный орган.
Этот орган состоит из трех коротких головчато расширяющихся
палочек. Путем выворачивания фарингиального аппарата палоч-
ковидный орган выходит наружу и захватывает подвижную до-
бычу. Если добыча слишком велика, палочковидная органелла раз-
рывает ее покровы, а мягкое содержимое всасывается клеткой.
Глотка не принимает участия в захвате и переваривании оформлен-
ных частиц. При культивировании эвглены в темноте и обильном
органическом питании, а также под воздействием многих других
агентов (ультрафиолетовое облучение, антибиотики и пр.) количест-
во хлорофилла и число хлоропластов сильно редуцируется. Потеря
всех хлоропластов может дать начало бесцветному штамму. В ряде
случаев экспериментально было продемонстрировано, что продук-
ция обесцвеченных клеток базируется на замедлении скорости ре-
пликации хлоропластов по сравнению со скоростью деления клеток.
Так могли возникнуть многочисленные бесцветные эвгленофито-
вые, которые следует рассматривать как формы вторичные, произо-
шедшие от зеленых.
Распространены эвгленофитовые почти исключительно в пре-
сных водах.
КЛАСС ЭВГЛЕНОФИЦИЕВЫЕ—EUGLENOPHYCEAE
Отдел эвгленофитовые (Eugienophyta) включает один класс —
эвгленофициевые (Euglenophyceae), деление которого на порядки
различно у разных авторов. Одни делят класс на 6 порядков,
главным образом на основе строения двигательного аппарата;
другие низводят эти порядки до статуса семейств в пределах
одного порядка; наконец, в русской альгологической литературе
принято распределение тех же семейств по двум порядкам: эв-
гленальные (Euglenales), объединяющие зеленые фототрофные
и бесцветные сапротрофные формы с двумя или одним жгутиком,
и перанемальные (Peranemales), куда относят бесцветные организмы
187
с голозойным (фаготрофным) питанием, реже сапротрофные
формы. У ряда форм для захвата пищи существует*специальный
(независимый от глотки) фарингиальный аппарат с-палочковидным
органом.
Порядок эвгленальные — Euglenales
Представителями порядка могут служить роды: эвтрепция
(Eutreptia), дистигма (Distigma), эвглена (Euglena), трахеломонас
(Trachelomonas), факус (Phacus).
Роды эвтрепция (Eutreptia) и дистигма (Distigma) имеют сильно
метаболитные, чаще веретеновидные клетки с двумя жгутиками,
далеко выступающими из глотки, одинаковыми у эвтрепции и не-
сколько различающимися по длине у дистигмы. Для эвтрепции
характерны хлоропласты, дистигма — бесцветная форма: хлоро-
пласты и глазок отсутствуют. Остальные перечисленные роды из
порядка эвгленальные, по-видимому, вторично одножгутиковые.
Род эвглена (Euglena) имеет клетки веретеновидные, вытянутые
в длину, более или мепее спирально скрученные, метаболичные
в разной степени, без домиков; один длинный жгут выщупает из
глотки, второй жгут редуцирован. Самый распространенный вид —
Е. viridts— вызывает цветение воды в загрязненных мелких водо-
емах, имеет веретеновидные метаболичные клетки с лентовидными,
расходящимися по радиусам из центра клетки хроматофорами.
Е. gracilis- с длинной цилиндрической клеткой, с дисковидными
хроматофорами, содержащими пиреноиды, сильно метаболична.
У ряда видов: Е. sanguinea, Е. haematodes, Е. ruhida, обитающих
в водоемах, богатых органическими веществами, в клетках содер-
жатся многочисленные, кроваво-красные от гематохрома гранулы,
которые при сильном освещении и температуре выше 30° С равно-
мерно рассеяны по всей клетке и маскируют зеленые хлоропласты:
при этом клетка имеет красную окраску. При более низкой тем-
пературе (10е С) и при меныпей освещенности гранулы гемато-
хрома концентрируются в центре клетки, не перекрывая перифери-
чески расположенные хлоропласты, и тогда клетка имеет зеленую
окраску. При массовом развитии эти виды образуют на поверх-
ности воды налет: красный — на солнечном свету, зеленый —в тени
или после захода солнца.
Род трахеломонас (Trachelomonas, рис. 48) характеризуется нали-
чием домика. Он может быть гладким, различным образом скульп-
турированным, с отверстием простым или окаймленным кольце-
видным выростом (воротничком). При массовом развитии вызыва-
ет «цветение» воды, окрашивая ее в коричневый г£вет.
Род факус (Phacus) имеет сплюснутое, более или менее скручен-
ное штопором по продольной оси неметаболирующее тело, обычно
заканчивающееся на заднем конце бесцветным прямым или изогну-
тым отростком.
188
Порядок перанемальные — Peranemales
Род перанема (Регапета)—двужгутиковая бесцветная форма
с анимальным — фаготрофным способом питания; имеется фарин-
гиальный аппарат с палочковидной органеллой.
Глава 15
ОТДЕЛ ЗЕЛЕНЫЕ ВОДОРОСЛИ —CHLOROPHYTA1
Представители отдела (стоящие на самых разных ступенях мор-
фологической дифференцировки таллома) характеризуются травя-
нисто-зеленой окраской хлоропластов, зависящей от преобладания
хлорофиллов а и b над каротиноидами: а- и p-каротином, лютеи-
ном, неоксантином, виолаксантином, зеаксантином, антераксанти-
ном. Хлоропласты окружены оболочкой из двух мембран, хлоро-
пластная эндоплазматическая сеть отсутствует. Ламеллы состоят из
2— 6 или многих слившихся тилакоидов, образующих «граны»,
периферические ламеллы отсутствуют, генофоры рассеянные. Пире-
ноид, погруженный в строму хлоропласта, пронизан тилакоидами.
Запасной полисахарид крахмал откладывается внутри хлоропла-
ста— вокруг пиреноида и в строме. Глазок располагается внутри
хлоропласта и не связан со жгутиковым аппаратом. Жгутики в чис-
ле 2,4, реже многих, одинаковой длины (изоконтные) и одинакового
строения (изоморфные), гладкие или покрыты тонкими мастигоне-
мами, у некоторых — чешуйками. Клетка подавляющего большин-
ства зеленых водорослей поверх цитоплазматической мембраны
покрыта целлюлозной оболочкой.
Клетки зеленых водорослей одно- или многоядерные. Митоз
у них в деталях протекает несколько по-разному. У одних це-
нтриоли имеются (кирхнериелла, тетраэдрон, хлорелла, гидроди-
ктион, клебсормидиум, сценедесмус), у других (эдогониум, конъю-
гатофициевые) отсутствуют. То же касается и кинетохоров (уча-
стков хромосом, к которым прикрепляются микротрубочки ве-
ретена), Они описаны у кладофоры, эдогониума, спирогиры,
гидродикциона и не наблюдались у хламидомонады, ульвы, У эдо-
гониума, улотрикса, стигеоклониума, микроспоры, ацетабулярии
ядерная оболочка на протяжении всего митоза интактна и веретено
полностью интрануклеарное (закрытое); у других зеленых водо-
рослей— кирхнериеллы, тетраэдрона, гидродиктиона, хламидомо-
нады— в оболочке ядра возникают полярные отверстия, через
1 Некоторые авторы дают название Chlorophycophyta, так как Chlorophyta, по
их мнению, имеет более широкое значение и должно обьединять зеленые водоросли
и высшие растения.
189
которые центриоли и экстрануклеарные вначале микротрубочки
перемещаются в ядро и веретено становится ищрануклеарным
(полузакрытое веретено); наконец, у стихококкуса„клебсормидиума,
колеохете, спирогиры, клостериума, космариума, микрастериаса,
нетриума, харофициевых, как и у высших растений, ядерная обо-
лочка полностью исчезает к поздней профазе и отсутствует до
ранней телофазы, веретено открытое. Для изученных до сих пор
хлорококкальных характерно развитие еще до митоза вокруг ядра
перинуклеарной обвертки (чехла эндоплазматической сети), которая
сохраняется до телофазы.
Электронно-микроскопическими исследованиями обнаружено
значительное разнообразие в структуре ци гокинетического аппара-
та. У сравнительно немногих зеленых водорослей (стихококкус,
клебсормидиум, колеохете, спирогира, харофициевые), как и у вы-
сших растений, после анафазы митоза, когда хромосомы движутся
к полюсам и телофазные ядра, далеко раздвинутые, реконструиру-
ются, между полюсами веретена остаются натянутыми непрерыв-
ные, так называемые интерзональные волокна. В экваториальной
плоскости веретена образуются микротрубочки, расположенные па-
раллельно этим волокнам и перпендикулярно плоскости будущего
цитокинеза. Эта система микротрубочек называется фрагмоплас-
том. Сохраняющееся интерзональное веретено и фрагмопласт удер-
живаю! дочерние ядра далеко раздвинутыми в тот момент, когда
между ними образуется поперечная перегородка.
У большинства других до сих пор исследованных зеленых водо-
рослей иная цитокинетическая система: после телофазы интерзо-
нальные волокна разрушаются, далеко раздвинутые сначала дочер-
ние ядра сходятся, так что оказываются прижатыми друг к другу.
Между ними возникают микротрубочки, ориентированные под пря-
мым углом к митотическому веретену и, следовательно, параллель-
но плоскости предстоящего цитокинеза. Система таких микротру-
бочек называется фикопластом. Ее назначение — определись пра-
вильное направление растущей перегородки, чтобы она прошла
между дочерними ядрами и разделила дочерние клетки.
Фикопластная система обнаружена у вольвокальных (хламидо-
монада), тетраспоральных (тетраспора), хлорококкальных (хлорел-
ла, кирхнериелла, тетраэдрон, анкистродесмус, сценедесмус), эдого-
ниальных (эдогониум, бульбохете), некоторых представителей улот-
рихальных (улотрикс, микроспора), хетофоральных (стигеоклони-
ум). Независимо от того, какая система микротрубочек — фрагмо-
или фикопласт — развивается, сам цитокинез происходит двояко:
или посредством инвагинации плазмалеммы и центрииетального
(от периферии внутрь клетки) разрастания борозды дробления, или
же путем образования срединной (клеточной) пластинки — полу-
жидкого слоя, возникающего за счет слияния пузырьков — дерива-
тов диктиосом. Клеточная пластинка разрастается центрифугаль-
но — по направлению к периферии клетки благодаря включению
190
в нее дополнительных пузырьков Гольджи и, наконец, достигает
продольной стенки материнской клетки, сливаясь с ней. Позднее на
эту срединную пластинку откладывается клеточная стенка, завер-
шая деление клетки. Цитокинез у спирогиры занимает промежуточ-
ное положение между этими двумя типами деления. После деления
ядра новая поперечная стенка закладывается в виде кольца клеточ-
ной стенки, которое распространяется центрипетально, перерезая
вакуоль с клеточным соком. Когда септа наполовину готова, цито-
плазматические тяжи, соединяющие дочерние ядра, вздуваются в це-
нтре, соединяются с цитоплазмой, смещаемой растущей септой, и,
сливаясь, образуют между ядрами цилиндр цитоплазмы. Поперек
отверстия септы в цитоплазме собираются пузырьки, в результате
слияния которых формируется клеточная пластинка, разрастающа-
яся центрифугально. В цитоплазматическом тяже, связывающем
дочерние ядра, наблюдаются продольно ориентированные микро-
трубочки — фрагмопластоподобное образование.
Фрагмопласт в сочетании с клеточной пластинкой наблюдается
у харофициевых водорослей; системой клеточная пластинка—фико-
пласт обладают Ulothrix fimbriata, Siigeoclomum helveticum,
Fritschiella tuberosa (фикопластные микротрубочки у F. tuberosa не
описаны, но близкое расположение телофатических ядер предпола-
гает их присутствие), а также тетраспора, эдогониум, хлорелла,
сценедесмус и др. Фрагмопласт и центрипетально разрастающаяся
борозда дробления описаны у ряда видов клебсормидиума
(К. flaccidum, К. subtilissimum). Сочетание борозды дробления и фи-
копласта наблюдается у микроспоры, уроспоры. У некоторых водо-
рослей (пилиндрокапса, ульва, диктиосферия) борозда дробления
при делении клетки врастает без участия каких-либо микротрубочек,
у них не отмечалось образования ни фикопласта, ни фрагмопласта.
У водорослей с сифонокладиальной организацией таллома на-
блюдается особый тип септирования таллома, известный как сег-
регативное клеточное деление. Например, у диктиосферии много-
ядерный протопласт в первоначально сифоновом талломе (пузыре)
раскалывается на многочисленные многоядерные цитоплазматичес-
кие массы различного размера, из которых каждая секретирует
клеточную стенку, а затем увеличивается в размерах до тех пор,
пока не придет в соприкосновение с такими же соседними клетками
внутри материнской клеточной стенки.
Размножение вегетативное, бесполое, половое. Имеются всевоз-
можные циклы развития (гаплонты, диплонты, изо-гетероморфная
смена генераций).
Этот отдел занимает особое положение. Окраска зеленых водо-
рослей такая же, как у высших растений. В качестве запасного
вещества в клетках, как и у высших, обычно откладывается крах-
мал. У многих зеленых водорослей наблюдается свойственное вы-
сшим расстениям правильное чередование поколений: полового
и бесполого. И наконец, ряд этих водорослей покинули водную
191
среду и перешли к наземному существованию. Все сказанное застав-
ляет именно среди зеленых водорослей искать родоначальников
высших растений.
Представители этого отдела большей частью распространены
в пресных водах, хотя имеются и морские.
Подразделение зеленых водорослей на классы весьма варьирует.
Прежде (приблизительно до второй половины XX в.) системы зеле-
ных водорослей различались главным образом по тому месту,
которое авторы отводили харовы.м водорослям, включая их в каче-
стве класса (порядка) в отдел Chlorophyta или выделяя их в са-
мостоятельный отдел Charophyta. Во многих руководствах по аль-,
гологии, особенно монографиях, порядки класса Chlorophyceae
(Volvocales, Ulotrichales, Bryopsidales, Oedogoniales) возводились
в ранг класса (Volvocophyceae, Ulotrichophyceae, Bryopsidophyceae,
Oedogoniophyceae). Конъюгаты, как и харовые водоросли, в ряде
систем возведены в ранг отдела. При этом родовой состав этих
групп, какой бы ранг им ни приписывался, существенно не менялся.
В 60 — 80-е годы были установлены новые классы, в основу выделе-
ния которых авторы кладут разные критерии (главным образом
детали ультраструктуры). Для иллюстрации можно привести неко-
торые примеры. Так, Т. Христенсен (Th. Christensen, 1962) выделил
из класса Chlorophyceae класс Prasinophyceae на основе строения
монадных клеток, имеющих жгутики, покрытые органическими че-
шуйками и отходящие от углубления на переднем конце клетки.
В классе Chlorophyceae у монадных клеток (шаровидных или эллип-
соидальных) жгутики гладкие или покрыты мастигонемами, но без
чушуек (этот автор рассматривает харовые водоросли, у которых
жгутики сперматозоидов покрыты чешуйками, как самостоятельный
отдел). К. Стюарт и К. Мэттокс (К. D. Stewart, К. R. Mattox, 1975)
наряду с местом прикрепления жгутиков (латеральное, терминаль-
ное) придают главенствующее значение и биохимическим критериям
(наличие или отсутствие гликолатоксидазы), деталям цитокинеза
(сохраняется или не сохраняется иптерзональное веретено во время
цитокинеза). На основе этих признаков отдел Chlorophyta делится на
два класса. Класс Charophyceae с порядками Klebsormidiales,
Zygnematales, Coleochaelales, Charales объединяет представителей,
у которых интерзональное веретено во время цитокинеза сохраняет-
ся, имеется гликолатоксидаза и жгутики (если они есть) латеральные.
Класс Chlorophyceae с порядками Volvocales Chlorococcales,
Microsporales, Ulvales, Chaetophorales, Oedogoniales характеризуется
тем, что интерзональное веретено не сохраняется, жгутики монад-
ных клеток терминальные и гликолатоксидаза отсутствует,
П. Корнманн (Р. Kornmann, 1973) выделил самостоятельный
класс Codiolophyceae с порядками Ulotrichales, Monostromatales,
Codiolales, Acrosiphonales, у которых существует гетероморфная
смена генераций с одноклеточным спорофитом (Codiolum—ста-
дия). Остальные зеленые водоросли распределяются по классам
192
Chlorophyceae, Oedogoniophyceae, Bryopsidophyceae, Zygne-
matophyceae. Г. Слюиман и др. (H. J. Sluiman et al., 1980) выде-
лили сепаратный класс Ulvophyceae на основе ультраструкту-
ральных деталей жгутикового аппарата, в частности корневых
систем.
Установление этих классов повлекло значительную перегруп-
пировку ряда родов: например, объединение родов Klebsormidium,
Coleochaete, родов зигнемовых и харовых водорослей, ко торые в бо-
лее ранних системах помещались в разные классы и даже отделы!
В настоящем руководстве мы будем придерживаться более тра-
диционной системы, принятой во многих общих руководствах по
альгологии, с делением отдела Chiorophyta на классы
Chlorophyceae, Prasinophyceae, Conjugalophyceae, Charophyceae. По-
рядки, где это возможно, устанавливаются в зависимости от типов
организации таллома.
Класс собственно зеленые водоросли, или равножгутиковые,—.
Chlorophyceae, Isocontae— характеризуется бесполым размножени-
ем обычно зооспорами с двумя—четырьмя, реже многими изоконт-
ными и изоморфными жгутиками, обращенными вперед, реже апла-
носпорами. Половой процесс—гологамный, изогамный, гетерогам-
ный и оогамный. В этом классе весьма полно представлены ступени
морфологической дифференцировки таллома (все, кроме ризоподи-
альной), которые обычно кладутся в основу деления его на порядки.
Класс празииофициевые водоросли (Prasinophyceae) характери-
зуется монадными клетками и жгутиками, покрытыми органичес-
кими чешуйками. Жгутики отходят со дна углубления на переднем
конце клетки. У представителей класса конъюгат официевые —
Conjugalophyceae в жизненном цикле отсутствуют жгутиковые ста-
дии, так как бесполое размножение отсутствует, а половой про-
цесс— конъюгация.
Класс харофициевые — Charophyceae включает крупные водоро-
сли со сложной морфологической дифференцировкой нитчатого
таллома. Бесполого размножения зооспорами или апланоспорами
нет. Половой процесс оогамный. Половые органы — оогонии и ан-
теридии сложного строения.
КЛАСС СОБСТВЕННО ЗЕЛЕНЫЕ, ИЛИ РАВНОЖГУТИКОВЫЕ,
ВОДОРОСЛИ —CHLOROPHYCEAE, ISOCONTAE
Класс занимает центральное положение среди всех зеленых во-
дорослей. В соответствии со ступенями морфологической диффе-
ренцировки таллома класс делится’на порядки.
Порядок вольво кал ьные — Volvocales
К порядку относятся монадные, т. е. снабженные жгутиками,
одноклеточные, колониальные и ценобиальные зеленые водоросли,
подвижные в течение вегетативной жизни.
7 - 121Q
193
IIf 1 Типичный представитель — одноклеточная водоросль из рода Я
хламидомонада (Chlamydomonas), многочисленные Ниды которого я
обитают в лужах, канавах и других мелких пресных водоемах. При я
$?.! их массовом развитии вода нередко принимает зеленую окраску, Я
'(:: Сферическая или эллипсоидальная клетка, одетая клеточной обо- fl
I лочкой, плотно прилегающей к протопласту или (у более старых Л
I1 особей) несколько отстающей от него в задней части, на переднем Ш
, конце несет два жгутика; с их помощью она плавает в воде. Прото- fl|
: пласт содержит одно ядро, большей частью чашевидный хромато-
фор, в который погружены пиреноид и глазок, в передней части '
клетки находятся сократительные вакуоли (см. рис. 5).
।' j При подсыхании водоема хламидомонады легко переходят в так
;||'|. называемое пальмеллевидное состояние: при этом клетки теряют г
'|1 . жгутики, оболочка их ослизняется, и в таком неподвижном состоя-
I.'' нии клетки размножаются. Оболочки дочерних клеток также ослйз-
‘ । няются, и таким образом получается система вложенных друг в дру-
! | га слизистых обверток, в которых группами расположены непо-
|'1 1 движные клетки, причем клетки сохраняют свои пульсирующие
М вакуоли и глазки. Попадая в воду, клетки снова вырабатывают
jl I;, . жгутики и возвращаются к монадному состоянию. Однако легкость
И J, перехода из пальмеллевидного состояния в монадное неодинакова
У у разных видов. Есть хламидомонады (Ch. kleinii), которые боль-
|'1: шую часть жизни проводят в пальмеллевидном состоянии.
ji1 Бесполое размножение. В благоприятных условиях хламидомона-
j' да интенсивно размножается бесполым, путем. Начало ядерного
й| деления у хламидомонады знаменуются репликацией базальных
тел, теряющих связь со своими жгутиками. Жгутики дегенерируют
Ы и сбрасываются. После утраты связи со жгутиками пары базальных
||- тел, соединенные исчерченными волокнами, могут свободно пере-
мещаться в цитоплазме. Хотя отделившиеся от жгутиков базальные
Ш тела неотличимы от центриолей, они не функционируют в качестве
| таковых, не располагаются у полюсов митотического аппарата и не
принимают участия в митозе. Скорее, они имеют отношение к по-
следующему цитокинезу, располагаясь по сторонам борозды дроб- j
[ ления, причем одно базальное тело каждой пары находится по одну
if сторону борозды, а другое — по другую, у переднего конца клетки
(рис. 49). Ядрышко исчезает, ядро принимает веретеновидную ;
! > форму, хромосомы конденсируются и в метафазу располагаются ;
|| внутри срединного вздутия ядра; в нуклеоплазме видны микротру-
I; . бочки веретена, кинетохоры не наблюдаются. Ядерная оболочка I
1| сохраняется интактной, за исключением отверстий у полюсов. По у
j мере расхождения хромосом в анафазе ядро удлиняется. В телофазе
г микротрубочки веретена исчезают, ядерная оболочка расширяется,
j! образуя складки между разошедшимися наборами хромосом, раз- ,
деляя дочерние ядра. Между дочерними ядрами под прямом углом
I! к оси веретена появляются интерядерные микротрубочки. В конце f
i телофазы возникает специальный набор микротрубочек, формиру- |
194
клцих аппарат раскола. Эти микротрубочки раскола отходят от
базальных тел и тянутся вдоль обеих сторон борозды дробления
под плазмалеммой. Борозда возникает вблизи переднего конца
клетки и распространяется вокруг всей клетки. После деления мате-
ринского протопласта (оболочка материнской клетки не принимает
никакого участия в этом процессе) на два процесс повторяется еще
один или два раза, давая соответствующее число дочерних прото-
пластов. Протопласты вырабатывают жгутики, окружаются кле-
точной оболочкой и превращаются в зооспоры, которые освобож-
даются благодаря ослизнению оболочки материнской клетки.
Ослизнение (лизис) оболочки материнской клетки (спорангиальной
оболочки) происходит под действием специального литического
агента — автолизина спорангиальных стенок, выделяемого молоды-
ми клетками. Он вызывает лизис только спорангиальной стенки, но
не стенок вегетативных клеток, зооспор, гамет.
При культивировании Chlamydomonas girus, особенно в средах
с высокой концентрацией солей, наряду с нормальными двужгути-
ковыми зооспорами нередко встречаются синзооспоры. После из-
вестного периода движения они, как и зооспоры, останавливаются,
теряют жгутики и приступают к размножению. При этом их содер-
жимое делится с образованием различного числа клеток: зооспор
или же опять синзооспор.
Половой процесс у большинства видов хламидомонады изогам-
ный. Гаметы образуются внутри материнской клетки так же, как
зооспоры, но в большем количестве (32 — 64) и соответственно
меньших размеров. У многих видов хламидомонады наблюдается
гетероталлизм: половой процесс возможен только при смешении
« + » и « —» гамет и начинается с образования групп — скопления
гамет противоположных знаков. В группах гаметы соединяются
попарно благодаря хемотаксическому привлечению. Сначала они
склеиваются своими жгутиками, после чего активируется автолити-
ческая система клеток: выделяется «гаметный автолизин», вызыва-
Рис. 49. Деление протопласта
Chlamydomonas (подробнее см. в тек-
сте):
бт— базальные тела, мт — микротрубочки
Рис. 50. Соединение гамет посредством
оплодотворяющей трубки (от)
у Chlamydomonas reinhardtii
195
ющий автолиз оболочек гамет (гаметы вольвокальных одеты обо-
лочками в отличие от голых гамет большинства водорослей). Толь-
ко гаметы, освободившиеся от своих клеточных етенок, сливаются,
образуя зиготы. Некоторые вещества, ингибирующие автолиз кле-
точных стенок гамет хламидомонады, подавляют и образование
зигот. Как показали электронно- микроскопические исследования,
у некоторых видов (Ch. reinhardtii) гаметы соединяются посред-
ством оплодотворяющей трубки (вырастающей от переднего конца
между жгутиками одной из сливающихся гамет, рис. 50), у других
видов (Ch. moewusii, Ch. chlamydogama) такая трубка отсутствует
и слияние гамет начинается со слияния цитоплазматических сосоч-
ков в один цитоплазматический мостик.
Вслед за плазмогамией происходит слияние ядер — кариога-
мия— путем слияния ядерных оболочек. Связанный с наружными
мембранами ядерных оболочек эндоплазматический ретикулум обе-
их гамет внедряется в цитоплазматический мостик, где сливается,
устанавливая таким образом контакт между наружными ядерными
мембранами обоих гамегных ядер. При сокращении эндоплазмати-
ческого ретикулума ядра сближаются до соприкосновения их на-
ружных ядерных мембран, вслед за слиянием которых начинается
слияние внутренних мембран ядерных оболочек гаметных ядер.
Таким образом, возникает копуляционное ядро зиготы с диплоид-
ным набором хромосом. В последние годы с помощью элек тронно-
го микроскопа удалось проследить дальнейшее поведение пластид
после копуляции гамет. Зиготы получались в результате скрещива-
ния дикой исходной формы Ch. reinhardtii, хлоропласты которой
плотно заполнены стопками тилакоидов с желтым мутантом, име-
ющим пластиду, совершенно лишенную тилакоидов. Таким обра-
зом, благодаря четким различиям структуры пластид обеих роди-
тельских форм можно было проследить их дальнейшее поведение
после капуляции гамет.
После слияния клеток и ядер наблюдалось слияние пластид:
в основании пластид, в участке, близком к пиреноиду, возникал
тонкий мостик, а затем пластиды сливались. Сначала в слившемся
хлоропласте зиготы можно увидеть две морфологически различа-
ющиеся части: одна часть с развитыми тилакоидами от хлоропласта
дикого типа, другая часть без тилакоидов, принадлежавшая пластиде
желтого мутанта. Позднее в хлоропласте зиготы уже не удается
различить морфологически разных частей родительских пластид, так
как мембранные структуры — тилакоиды хлоропласта дикого типа
равномерно распределяются по всему хлоропласту зиготы. При
прорастании зиготы до мейоза хлоропласт делился на четыре части,
и все сформировавшиеся зооспоры имели хроматофоры дикого типа.
Уже на ранних стадиях образования зиготы выделяется первич-
ная оболочка, после кариогамии возникает вторичная оболочка,
состоящая из двух или трех слоев с характерной для вида скульп-
турой.
196
У некоторых видов хламидомонады наблюдается гетерогамный
и оогамный половой процесс. Так, у Ch. braunii русским ботаником
И- Н. Горожанкиным был описан гетерогамный половой процесс,
при котором крупные женские гаметы, образующиеся в материнс-
кой клетке, в числе четырех сливаются с мелкими мужскими гаме-
тами, возникающими в клетке в числе восьми. Оогамия впервые
была описана также И. Н. Горожанкиным у Ch. coccifera. У этого
вида одни клетки теряют жгуты и превращаются в неподвижные
яйцеклетки, в других клетках образуется большое количество мел-
ких двужгутиковых мужских гамет, которые и оплодотворяют яй-
цеклетки.
Близки к хламидомонаде род картерия (Carterictf клетка которо-
го несет не два, а четыре жгутика, и род хлорогониум (Chlorogonium),
имеющий сильно вытянутое в длину, веретеновидное, иногда иг-
ловидное тело и два жгутика. Размножение этих родов такое же, как
у хламидомонады. У некоторых видов — Ch. oogamum, С. iyengarii
описана примитивная оогамия.
У ряда одноклеточных вольвокальных в отличие от хламидомо-
нады и близких к ней родов отсутствуют клеточные оболочки. Так,
род дюналиелла (Dunaliella), встречающийся в соленых озерах с вы-
сокой концентрацией солей, по форме клетки и внутреннему стро-
ению очень похож на хламидомонаду, но отличается от нее отсут-
ствием обособленной клеточной оболочки. Размножение—про-
дольным делением клетки. Известен и половой процесс в виде
хологамии, когда копулируют клетки, внешне не отличающиеся от
вегетативных. D. salina гетероталлична, т. е. половой процесс на-
блюдается при смешении клеток из разных клонов, различающихся
в половом отношении (+) и ( —). Культивируя оба клона в разных
условиях (с избытком или, наоборот, с дефицитом азота и фосфора
в среде), удается получить неодинаково окрашенные клетки: зеле-
ные при достаточном количестве азота и фосфора и оранжевые при
их недостатке в среде. При смешении клеток из двух таких клонов
(различающихся по половому знаку и окраске) можно наблюдать
образование групп и копуляцию в них неодинаково окрашенных
клеток. Получающаяся зигота также окрашена наполовину в зеле-
ный, наполовину в оранжевый цвета.
У многих вольвокальных двужгутиковые (как и у хламидомона-
ды) клетки соединяются в подвижные ценобии: чаще всего встреча-
ются роды гоииум (Gonium), пандорииа (Pandorina), эвдорииа
(Eudorinaf вольвокс (Polvox).
Гониум образует пластинчатые колонии из клеток, расположен-
ных в один слой. У наиболее широко распространенного вида Gonium
pectorale колонии состоят из 16 клеток, которые соединяются от-
тянутыми концами своих оболочек, формируя пластинку или таб-
личку, снаружи окруженную бесцветной слизью. Клетки в таком
пластинчатом ценобии ориентированы таким образом, что все пе-
редние концы, несущие жгутики, обращены в одну сторону.
197
У остальных перечисленных родов ценобиальных вольвокаль-
ных колонии представляют собой слизистые эллийсоиды или сфе-
ры, по периферии которых в один слой располагается различное (в
зависимости от рода или вида водоросли) число клеток.
У пандорины и эвдорины ценобии микроскопически мелкие.
Повсеместно встречающийся вид Р. ntorum представляет собой
слизистый эллипсоид, по периферии которого под плотным слоем
слизи (инволюкрумом) находится 16 клеток. Клетки лежат в общем
инволюкруме очень тесно, давят Друг друга, вследствие чего имеют
граненую форму; только часть их поверхности, обращенная кнару-
жи, выпуклая. Общая полость колонии очень мала.
Эвдорина характеризуется более крупными, также слизистыми
эллиптическими ценобиями, в которых по периферии располагается
большее число клеток (32 — 64), лежащих в слизи свободно. Центр
ценобия занят жидкой слизью. Жгутики, отходящие от передних,
направленных к периферии ценобия концов клеток, проходят сквозь
одевающую всю колонию плотную слизистую обвертку через нахо-
дящиеся в ней тонкие канальцы и работой своих свободных частей
приводят колонию в движение.
Наиболее высоко организован вольвокс (рис. 51). Он представ-
ляет собой крупный, достигающий 2--3 мм в диаметре шар, оде-
тый тонким инволюкрумом, под которым по периферии шара
в один слой располагаются клетки. Число их колеблется от 500 до
60 000. Внутренняя полость шара занята жидкой слизью. Для кле-
ток вольвокса характерно, что внутренние слои оболочки с возрас-
том очень сильно ослизняются, в результате чего протопласт каж-
дой клетки значительно удален от самого наружного, не ослизнен-
ного слоя оболочки. Более плотные наружные оболочки соседних
клеток, соприкасаясь, образуют полигональный узор, иногда хоро-
шо заметный, если смотреть на ценобий с поверхности. Протопла-
сты же соседних клеток разъединены друг от друга. От протопласта,
занимающего центр клетки, в одной плоскости, параллельной пове-
рхности всей колонии, по радиусам отходят более или менее много-
численные цитоплазматические отростки (у некоторых видов от-
ростки отсутствуют), проходящие сквозь ослизненные внутренние
слои оболочки до самых наружных плотных слоев оболочки. Цито-
плазматические отростки соседних клеток соответствуют друг дру-
гу и связаны между собой плазмодесмами. У одних видов (Volvox
globator, V. rousseletii) цитоплазменные отростки толстые, грубые,
с заходящими в них лопастями хроматофора; при рассматривании
таких ценобиев с поверхности протопласты (в оптическом разрезе
конические) кажутся звездчатыми. У других видов (V. aureus) окру-
гло-яйцевидные протопласты имеют тонкие, едва различимые цито-
плазматические отростки. Наконец, такие виды, как V. tertius,
V. gigas, V. africanus, V. carteri, V. powersii, вообще лишены цитопла-
зматических отростков. На наружной поверхности клетки несут по
два жгутика, выходящих наружу через канальцы в инволюкруме.
198
Рис. 51. Volvox carteri. A — бесполый сфероид, содержащий 16 крупных вакуолизиро-
ванных гонидиев; Б— женский сфероид, содержащий около 40 яиц; В -мужской
сфероид, содержащий в равном числе соматические и антеридиальные клетки (анд-
рогонидии); в каждом антеридии формируется пакет из 64 двужгутиковых спер-
матозоидов; Г— прорастание зиготы с образованием одной нормальной зооспоры:
ск—соматические клетки, г - гонидии, о —оогонии, ai аидрогониднй, пс — пакет сперматозо-
идов
Образование дочерних ценобиев. При бесполом размножении це-
нобиальных вольвокальных образуются дочерние колонии. При
этом протопласт клеток, продуцирующих дочерние колонии, прете-
рпевает продольные деления и формирует полный комплекс клеток
будущего ценобия, после освобождения которого клетки не делятся
до тех пор, пока не наступит новая репродуктивная фаза. В даль-
нейшем дочерний ценобий, как правило, растет только за счет
увеличения размеров клеток, но не их числа. Давно замечено, что
у гониума, пандорины, Eudorina elegans каждая клетка ценобия
способна давать дочерний ценобий, а у вольвокса репродуктивными
становятся только 8 —10 клеток, остальные — чисто соматические.
Таким образом, было обращено внимание на дифференцировку
клеток в ценобиях вольвокальных. Однако ход делений, приводя-
щих к возникновению внутри материнской клетки нового дочернего
199
ценобия, и, что особенно важно для сферических ценобиев эвдори-
ны, пандорины, вольвокса, возникновение внутренней полости этих
ценобиев до 1875 т. выяснены не были.
В 1875 г. И. Н. Горожанкин подробно проследил весь ход деле-
ний при развитии дочерних колоний у Е. elegans, Pandorina tnorum,
Volvox globator и впервые показал, что возникновение полости в сфе-
рических ценобиях происходит в результате загиба краев тоническо-
го состояния. Особенно подробно было прослежено развитие дочер-
них ценобиев у Е. elegant. перед делением цитоплазма материнской
клетки отстает от оболочки, и первая продольная перегородка
возникает перпендикулярно к общей поверхности ценобия. Через
несколько минут возникает вторая продольная перегородка, перпен-
дикулярная к предыдущей и к поверхности ценобия: получается
стадия квадранта. После получасовой паузы в каждой клетке квадра-
нта появляется еще по одной продольной перегородке, стоящей под
прямым углом к направлению наибольшего роста этих клеток
и к поверхности материнского ценобия. Возникает восьмиклеточный
комплекс, в котором четыре клетки соприкасаются в центре, образуя
внутренний крест, а четыре другие лежат в промежутках креста.
Через час с небольшим наступает следующее деление в клетках
креста на периферическую и центральную. Затем возникают перего-
родки в клетках, расположенных по углам первоначального креста,
и возникает табличка из 16 расположенных в один слой клеток. Эта
стадия из-за ее сходства с G. perforate получила название «тоническо-
го состояния». Спустя 2,5 ч в 16-клеточном тоническом состоянии
происходит еще одно деление и формируется пластинка из 32 клеток.
Затем края этой пластинки заворачиваются, образуя полую сферу
с маленьким отверстием, обращенным к наружной поверхности
родительского ценобия. Такое же происхождение центральной поло-
сти сфероидов — в результате загиба краев тонического состояния —
имеют пандорина и вольвокс. Эти классические данные И. Н. Горо-
жанина вошли во все альгологические сводки и полностью сохрани-
ли свое значение до настоящего времени. И. Н. Горожанкин пола-
гал, что края тонического состояния сразу срастаются и получается
замкнутая сфера. На самом деле на стадии незамкнутой сферы,
возникающей в результате загиба краев тонического состояния,
клетки ориентированы так, что передние ядерные концы их, на
которых должны возникнуть жгутики, направлены к центру сферы.
Нормальная ориентация клеток с передними несущими жгутики
концами, обращенными кнаружи сферы, достигается путем полного
выворачивания наизнанку (инверсии) незамкнутых сфероидов, после
чего отверстие молодого сферического ценобия замыкается.
Приведенные выше данные были получеры с помощью оптичес-
кого микроскопа и на материале, взятом'из природных водоемов.
В последние десятилетия процесс развития дочерних ценобиев
и дифференцировки в них клеток на чисто соматические и репродук-
тивные, а также процесс инверсии были изучены на более высоком
200
методическом уровне в условиях чистых культур и с применением
электронной микроскопии (а на уровне оптического микроскопа на
помошь рисовальному аппарату пришла микрофотография). Осо-
бенно много подобных исследований проведено на видах вольвокса.
В 1959 г. была разработана питательная среда для культивиро-
вания вольвоксов. Получение и поддержание бактериально чистых
(аксеничных) культур дали возможность изучить дифференцировку
клеток у разных видов вольвокса. В ряде случаев соматические
клетки и репродуктивные гонидии разъединяли, действуя на цено-
бии вольвокса некоторыми ферментами. Такие изолированные го-
нидии далее претерпевали деления с образованием зародыша —
дочернего ценобия. В таких изолированных гонидиях прослеживать
и фотографировать весь ход делений, процесс инверсии гораздо
легче, чем в составе крупной сферической колонии (рис. 52, А,Е).
Развитие зародыша (дочернего ценобия) из гонидия протекает
в общем одинаково у всех видов. Однако в отношении последовате-
льности роста и деления гонидия виды вольвокса могут быть
распределены на две группы: 1) у таких видов, как Г. globator,
V. rousseletii, V. aureus, деления начинаются тогда, когда гонидий
еще очень мал, и за каждым делением следует период роста; 2) у
других видов (И. carteri, Г. africanus, V. gigas) гонидии претерпева-
ют заметное увеличение размеров до начала серии делений без
видимого роста между последующими делениями.
Разные виды различаются также по времени наступления види-
мой дифференцировки на соматические и репродуктивные клетки
в процессе формирования зародыша (дочернего ценобия). Так, у ря-
да видов репродуктивные клетки (гонидии) могут быть различимы
в молодом сфероиде до его освобождения из родительского. Осо-
бенно рано происходит дифференциация гонидиев в молодых заро-
дышах V. obversus и К. carteri f. nagariensis, у первого на стадии 16,
у второго на стадии 32 клеток.
У V. obversus во время деления 16-клеточной стадии впервые
становятся морфологически различимыми обычно 8 инициальных
клеток гонидиев благодаря неравному делению 8 клеток, окружа-
ющих фиалопору (остиум, отверстие незамкнутой сферы).
У У. carteri f. nagariensis также во время эмбриогенеза все де-
ления клеток равновелики до стадии 32 клеток. На стадии 32-
клеточного зародыша появляется первая морфологическая диффе-
ренцировка: передние 16 (при менее оптимальных условиях 10 — 15)
клеток зародыша претерпевают неравные дифференцирующие де-
ления, давая начало мелким соматическим инициальным клеткам
и крупным инициальным клеткам гонидиев. Соматические иници-
альные клетки передней и задней части молодого зародыша про-
должают синхронно делиться 11 или 12 раз (таким образом, ко-
нечное число клеток в сфероиде 2000 или 4000) и становятся все
мельче и мельче. Гонидии же (способные отчленять мелкие клетки,
но, по-видимому, претерпевающие лишь немного таких делений)
201
Рис. 52. Изолированные из сфероида гонидии Volvox carter! на разных стадиях
развития бесполого зародыша; А увеличившийся гонидий до начала делений;
А— двуклетный зародыш, заключенный в оболочку гонидия; В чстырехклетный
зародыш; Г - Е--более поздние стадии развития зародыша, видна фиалопора меж-
ду четырьмя клетками переднего конца зародыша (фп); Ж—зародыш в начале
инверсии: губы, окружающие фиалопору, отогнулись назад на поверхность зароды-
ша, все клеточные деления завершены; 3—зародыш, наполовину вывернувшийся
наизнанку; И—вывернувшийся зародыш с гонидиями, обращенными теперь внутрь
сфероида, а Ж1 утиковыми концами соматических клеток - кнаружи
оказываются сравнительно крупными и более удаленными друг
от друга, всегда сохраняя свое первоначальное положение — в пе-
редней половине молодого зародыша. К окончанию клеточных
делений соматические клетки в сферическом зародыше своими ядер-
ными (а следовательно, и жгутиковыми) концами обращены внутрь
сферы, а 16 или менее (10 —15) крупных гонидиев заметно вы-
даются на наружной поверхности сферы. Вскоре начинается процесс
202
инйерсии: в области фиалопоры четыре губы загибаются назад
над поверхностью зародыша, и в течение 45 мин зародыш вы-
ворачивается наизнанку (рис. 52, Ж— И).
Дифференцировка гонидиев в молодых сфероидах до их выхода
из родительского наблюдается также у Й rousseletii, V. aureus,
V. gigas, И. tertius. У других видов, например у V. powersii и, по-
видимому, у И. globator, гонидии дифференцируются поздно, толь-
ко после освобождения дочернего ценобия из родительского шара.
Ряд исследований с привлечением электронной микроскопии
посвящен инверсии — в высшей степени координированному про-
цессу, вовлекающему многие клетки. Напомним, что быстрые син-
хронные серии митозов и цитокинезов дают начало чашевидно
изогнутой пластинке, в которой ядра и центриоли в каждой клетке
расположены на внутренней — вогнутой стороне, а пластиды — с
выпуклой наружной стороны. Далее следует процесс инверсии. Уль-
траструктуральные исследования колонии, претерпевающей выво-
рачивание наизнанку, позволили обнаружить некоторые черты, воз-
можно, объясняющие механизм этого сложного процесса.
В клетках как вольвокса, так и эвдорины при образовании зародыша
происходят неполные цитокинезы и таким образом они остаются
соединенными посредством цитоплазматических мостиков, которые
могут функционировать как каналы для межклеточного сообщения,
обеспечивая слаженность в поведении многих клеток (рис. 53, А, Ь). Все
клетки пластинки одновременно удлиняются; также одновременно
расширяется «ядерный» конец клетки и сокращается конец, несущий
пластиду, где клетки остаются соединенными между собой цитоплазма-
тическими мостиками, действующими как шарниры. Результатом
является инверсия развивающихся ценобиев, после которой ядра
и центриоли оказываются лежащими на выпуклой наружной поверхно-
сти (рис. 53, В). Удлинение клеток и расширение их «ядерного» конца
(вообще изменение формы клетки), по-видимому, осуществляется при
участии микротрубочек, расположенных по периферии клеток. В пользу
этого свидетельствует тот факт, что обработка пластинок колхици-
ном— ядом, специфически блокирующим полимеризацию микротру-
бочек, препятствует изменению формы клетки и нарушает инверсию.
Для освобождения дочерних сфероидов вольвокса инволюкрум
(чехол) материнского ценобия должен быть отчасти или полностью
лизирован. Как у хламидомонад для освобождения зооспор, так
и у вольвоксов для освобождения дочерних сфероидов клетки выра-
батывают гидролизирующие энзимы (лизины, автолизины), обла-
дающие видовой специфичностью.
Половой процесс у ценобиальных вольвокальных, как и у одно-
клеточных, различается по форме: у гониума и пандорины наблюда-
ется изогамия, у эвдорины — гетерогамия, у вольвокса — оогамия.
У гониума в каждой клетке образуются по 16 гамет, которые
выходят в воду, где и копулируют. У пандорины вегетативные
клетки выходят из слизи колонии и функционируют как изогаметы.
203
Рис. 53. Разрез через тоническое состояние и недавно претерпевшую инверсию коло-
нию Eudorina elegant.
А — восьмиклетная стадия. Деления неполные, клетки соединяются посредством
цитоплазматических мостиков. Б—разрез через 16- или 32-клетные пластинки (тони-
ческое состояние), некоторые клетки еще соединены широкими цитоплазматичес-
кими мостиками, ядра расположены на вогнутой стороне, пластиды — на выпуклой
стороне изогнутой пластинки. Б — недавно вывернувшаяся колония
У Р. unicocca сначала образуются группы колоний (до 20 и более),
после чего гаметы начинают выходить из них, выдавливаясь из
своих клеточных оболочек через крошечное отверстие. Когда перед-
ний конец гаметы начинает выскальзывать через отверстие, прото-
пласт временно становится перетянутым. Поскольку здесь гаме-
ты—это вегетативные клетки, вышедшие из колониального мат-
рикса, то их морфология та же, что и у вегетативных клеток, только
жгутики, более не ограниченные колониальной обверткой, движутся
от точки их прикрепления к протопласту.
У Р. unicocca первоначальный мостик слияния образуется между
передними концами двух совместимых гамет. Затем происходи!
быстрое (в течение 3—4 мин) боковое слияние. Сливающиеся гаме-
ты обычно сходны по размерам, но могут быть и неодинаковыми.
Размер гамет зависит от размера клеток колоний, из которых они
выходят, колонии обоих типов спаривания продуцируют гаметы
разных размеров.
Эвдорина (Е. elegans), как правило,— форма двудомная (гетеро-
талличная), т. е. различают колонии женские и мужские. В женских
204
остановившихся колониях клетки разъединяются, сбрасывают свои
оболочки и превращаются в крупные, снабженные жгутиками гаме-
ты, которые, однако, неподвижны и остаются в слизи женской
колонии (ввиду неподвижности женских гамет половой процесс
эвдорины часто трактуют как оогамию). Каждая клетка мужской
колонии в результате продольных делений образует изогнутую
пластинку, претерпевающую инверсию. Полушаровидные пакеты из
16 — 32 двужгутиковых веретеновидных сперматозоидов освобож-
даются из слизистой обвертки родительской колонии, подплывают
к женским ценобиям и распадаются на отдельные сперматозоиды.
Сперматозоиды имеют стигму, две сократительные вакуоли, два
жгутика одинаковой длины и во время движения меняют свою
форму. У основания жгутиков сперматозоид несет тонкий цитоплаз-
матический выступ, возможно (точно не установлено) принима-
ющий участие в плазмогамии. Проникая в слизь женской колонии,
сперматозоиды оплодотворяют женские клетки.
Действительное слияние сперматозоида (антерозоида) с женской
гаметой (плазмогамию) у эвдорины не удавалось непосредственно
наблюдать И. Н. Горожанкину, который лишь косвенно признал
оплодотворение у Е. elegans на том основании, ч то при культуре во
влажной камере женские колонии, изолированные от мужских (со
сперматозоидами), более или менее скоро отмирали, а в присутст-
вии последних давали ооспоры (зиготы). Слияние сперматозоида
и женской гаметы у Е. elegans впервые увидел ученик И. Н. Горожа-
нина русский альголог К. И. Мейер (1935). Согласно К. И. Мейе-
ру, антерозоид прикрепляется к яйцеклетке носиком, прикладывает-
ся к ней боковой стороной и сливается с ней. Весь процесс длится
2 — 3 с. После К. И. Мейера еще два автора наблюдали и описали
этот процесс: Н. Покок (N. А. Рососк, 1937) и М. Айенгар
(М. Iyengar), причем данные Покок совпадают с таковыми
К. И. Мейера, а наблюдения Айенгара отличаются: по Айенгару,
у Е. elegans антерозоид входит в яйцеклетку не передним, а узким
задним концом и сливается с яйцом, медленно погружаясь в него.
Недавно X. Нозаки (Н. Nozaki, 1983) проследил плазмогамию
у Е. elegans на массовом материале в лабораторной культуре. Со-
гласно Нозаки, плазмогамия у Е. elegans начинается с вхождения
переднего конца мужской гаметы в переднюю область женской
гаметы. Сферическая четырехжгутиковая планозигота имеет два
коротких жгутика, принадлежащих мужской гамете, и два длинных
жгутика от женской гаметы. Вскоре жгутики разрушаются, сек-
ретируется гладкая клеточная стенка, и зигота, принимающая крас-
новато-бурую окраску, переходит в состояние покоя.
Как упоминалось (гл. 4), вольвоксы переходят к половому раз-
множению под действием специальных индукторов — веществ, вы-
деляемых при освобождении и дезинтеграции пакетов сперматозо-
идов и споврбных далее индуцировать образование половых сферо-
идов как в мужских, так и в женских клонах, до этого размножав-
205
шихся бесполым путем. Считается, что первые мужские сфероидь),
выделяющие индукторы, возникают в популяции’ до того размно-
жающейся бесполым путем, спонтанно. В случае V. carteri индуктор
был идентифицирован как гликопротеид с молекулярной массой
около 30000.
Различные виды и штаммы вольвокса характеризуются неодина-
ковым распределением половых органов. Так, V. carteri,
V. rousseletii, V. gigas двудомны (мужские и женские половые клетки
образуются в разных сфероидах) и гетероталличны, т. е. в пределах
одного клона половой процесс невозможен. V. aureus тоже двудом-
ный, но гомоталличный, т. е. половой процесс может происходить
внутри одного клона. У этого вида сперматозоиды оплодотворяют
гонидии, которые в присутствии мужских половых клеток функци-
онируют в качестве яиц. V. powersii тоже двудомный и гомоталлич-
ный, но у него в отличие от V. aureus имеются специальные женские
сфероиды. Примером вида однодомного (мужские и женские ре-
продуктивные клетки формируются в одном и том же сфероиде)
и гомоталличного может служить V. globator.
В отношении дифференциации пола наиболее хорошо изучен
V. carteri f. nagariensis. Под воздействием индуктора, продуциру-
емого мужскими половыми сфероидами, гонидии женского клона
образуют половые сфероиды с яйцеклетками (см. рис. 51, Б). При
развитии сексуальных сфероидов деления в гонидии до 32-клеточ-
ной стадии идентичны с таковыми при развитии бесполого зароды-
ша. Клеточные деления на 32-клеточнрй стадии равновелики, и об-
разуется 64-клеточный зародыш, в котором все клетки одинакового
размера. На этой стадии наступают дифференцирующие клеточные
деления: 32 передние клетки и несколько клеток в задней половине
зародыша претерпевают неравновеликие деления. В результа те воз-
никает зародыш с 35 45 крупными инициальными яйцеклетками.
Дальнейшее деление соматических клеток женского зародыша и ин-
версия происходят так же, как и при бесполом эмбриогенезе: об-
разуются сфероиды обычно из 2—4 тыс. клеток. Гонидии мужского'
клона реагируют на индуктор (из половых мужских сфероидов)
образованием мужских зародышей (сфероидов). До 32-клеточной
стадии деления в развивающемся мужском зародыше те же, что
и при развитии бесполого зародыша. Дальнейшие деления с об-
разованием 64, 128 и 256 клеток равновелики и при оптимальных
условиях в результате еще одного деления может образоваться 512
клеток. Это финальное деление неравновеликое во всех клетках
зародыша; в результате возникает мужской сфероид с крупными
андрогонидиями (клетками, продуцирующими сперматозоиды)
и мелкими соматическими клетками в отношении 1:1 (256 анд-
рогонидий и 256 соматических клеток). Таким образом, в отличие
от бесполых и женских мужские половые сфероиды — карликовые,
состоящие всего из 512 клеток. В менее оптимальных условиях
дифференцирующее окончательное деление может наступить на
206
более ранних (32—64- или 128-клетных) стадиях, но соотношение
мелких соматических клеток и крупных андрогонидий будет то
же—1:1 (см. рис. 51, В).
После инверсии мужского сфероида андрогонидии увеличивают-
ся и последовательно делятся, образуя изогнутые пластинки (прете-
рпевающие инверсию) из 64 или 128 двужгутиковых сперматозо-
идов (см. рис. 51, В). Мужские сфероиды ко времени своего освобо-
ждения из родительского индивида1 содержат пакеты из 64 —128
сперматозоидов. Освободившиеся мужские сфероиды эфемерны
и в течение нескольких часов разрушаются, освобождая интактные
пакеты сперматозоидов, которые плавают в среде, пока не придут
в контакт с женскими сфероидами. Растворяя отверстие в женском
сфероиде, пакеты распадаются на отдельные голые вытянутые дву-
жгутиковые сперматозоиды, которые устремляются к многочислен-
ным яйцеклеткам. Как было показано в последние годы, дезинтег-
рация пакетов сперматозоидов и проникновение сперматозоидов
сквозь слой чехла матрикса и покровы яйцеклеток обусловлены
лизинами.
У Volvox rousseletii, как и у V. carteri, индуктор продуцируется
мужскими половыми культурами. Однако действие индуктора здесь
несколько отличается от того, что наблюдалось у V. carteri
f. nagariensis. Молодые сфероиды V. rousseletii ко времени их осво-
бождения из родительского содержат репродуктивные клетки двух
размеров: несколько клеток значительно более крупных, чем сома-
тические, и клетки промежуточного размера. В отсутствие индук-
тора только крупные репродуктивные клетки развиваются и путем
последовательных делений дают бесполые зародыши (молодые сфе-
роиды). Если же в момент выхода молодого сфероида индуктор
присутствует, то как крупные, так и более мелкие репродуктивные
клетки в мужском клоне превращаются в андрогонидии и продуци-
руют пакеты сперматозоидов, а в женском клоне увеличиваются
в размерах и непосредственно трансформируются в яйцеклетки без
каких-либо делений.
Таким образом, индукционные системы V. carteri и V. rousseletii
различны, хотя в обоих случаях к действию индукторов восприим-
чивы молодые сфероиды после выхода их из родительского.
У V. carteri (и у К. obversus) сексуальные сфероиды формируются
в следующем поколении в результате изменения характера делений
гонидиев, воспринявших индуцирующие стимулы. У V. rousseletii (и
V. gigas) половые клетки возникают непосредственно в сфероиде,
подвергшемся влиянию дндуктора. Индуктор изменяет направление
развития клеток, уже имеющихся в данном сфероиде.
У однодомного вида V, capensis половые индивиды развиваются
под действием низких концентраций L-глютаминовой кислоты —
! В освобождении как мужских, так и женских сфероидов, по-видимому, участву-
ют лизины (автолизины).
207
продукта переваривания гликопротеинового матрикса родитель^
ких сфероидов при выходе из них дочерних. *' ।
Как и в случае Eudorina elegans, у вольвоксов очень долго не
удавалось наблюдать непосредственно процесс плазмогамии - ‘сли-
яние сперматозоида с яйцеклеткой. Лишь сравнительно недавно это
удалось у V. carteri f. nagariensis при использовании женского мутан-
та с гораздо большим, чем в норме, числом яйцеклеток и с мень-
шим количеством соматических клеток, естественно затрудняющих
наблюдение.
Образующаяся в результате полового процесса зигота одевается
толстой многослойной оболочкой и у всех вольвокальных перехо-
дит в состояние покоя. При прорастании ее диплоидное копуляци-
онное ядро редукционно делится, образуются четыре гаплоидных
ядра и, как правило, четыре зооспоры. Так, у многих видов хлами-
домонады зигота прорастает с образованием четырех зооспор.
У гониум (G. pectorale) при прорастании зиготы возникает малень-
кая четырехклетная ювенильная колония. Из каждой клетки такой
колонии при дальнейшем бесполом размножении формируется уже
16-клетная колония, типичная для этого вида. В других случаях не
все из четырех ядер, возникающих в результате редукционного
деления, остаются жизнеспособными, часть их дегенерирует. На-
пример, у пандорины, эвдорины при прорастании зиготы остается
только одно жизнеспособное ядро и соответственно развивается
только одна здоровая зооспора. Она выделяет слизистую обвертку,
и внутри нее делится с образованием зародышевой— ювенильной
колонии.
Формирование только одной жизнеспособной зооспоры при про-
растании зиготы описано и у некоторых видов вольвокса: например,
у V. carteri f. nagariensis прорастающая зигота продуцирует одну
двужгутиковую зооспору, содержащую одно жизнеспособное гапло-
идное ядро, получившееся в результате мейотического деления ко-
пуляционного ядра зиготы (см. рис. 51, Г). Зооспора некоторое вре-
мя плавает, а затем претерпевает деления с образованием маленько-
го зародышевого индивида, который обычно содержит 8 гонидиев.
Цитологических исследований редукционного деления ядра
в прорастающей зиготе сравнительно немного (например, у воль-
вокса). Гораздо больше косвенных доказательств такого деления
получено с помощью генетического анализа зооспор, образующих-
ся при прорас гании зиготы. Так, А. Пашеру удалось скрестить два
вида хламидомонад, различающихся по строению вегетативных
клеток и зигот. Получившиеся в результате скрещивания зиготы
имели строение, промежуточное между зиготами родительских ви-
дов. При прорастании таких гибридных зигот в каждой образова-
лось по четыре зооспоры. Каждая из них была изолирована и при
дальнейшем их бесполом размножении были получены четыре
культуры, из которых две оказались тождественны одному из роди-
тельских видов, а две другие — другому. Этот опыт показал, что
208
i
разъединение признаков гибрида наблюдается при прорастании
зиготы, согласно же цитогенетическому учению такое разъединение
происходит при редукционном делении диплоидного ядра. Еще
больше аналогичных данных получено относительно разделения
полов у раздельнополых или гетероталличных форм. Для них в ряде
случаев также было показано, что половые локусы разъединяются
в момент прорастания зиготы. Для гониума (G. pectorals) это было
продемонстрировано опытами Е. Шрейбера. Он изолировал четы-
рехклеточные колонии, развивающиеся при прорастании зиготы.
В клетках такой колонии через несколько дней формировалась
нормальная 16-клетная колония. Каждая из четырех первых вегета-
тивных колоний также изолировалась и служила для получения
клона. Таким образом, Шрейбер получил четыре клона из четырех
клеток, образовавшихся в прорастающей зиготе. Попарное смеще-
ние этих клонов позволило установить, что два из них относятся
к одному полу, два - -к другому. Ввиду изогамии гониума особи
противоположного пола обозначаются как «+» и « —». В той
и другой паре при смешении половой реакции не наступает, копуля-
ция наблюдается лишь при соединении особей разного пола. Итак,
у гониума уже при прорастании зиготы в равном числе устанавлива-
ются оба пола. Разделение полов в момент прорастания зигот —
косвенное доказательство редукционного деления; при этом одни
ядра получают потенцию одного пола ( + ), другие—потенцию
противоположного ( —) пола. При дальнейшем бесполом размноже-
нии знак пола не меняется. Определение пола в момент прорастания
зиготы, связанное с редукционным делением, называется генотипи-
ческим (или гаплогенетическим). У гомоталличных обоеполых ви-
дов при прорастании зиготы тоже происходит редукционное деле-
ние (например, у вольвокса это было доказано цитологически), но
оно не влечет за собой разделения полов. Выходящие из зиготы
зооспоры потенциально содержат в себе оба пола, лишь в их
потомстве наступает половая дифференцировка, которая не связана
с редукционным делением и скорее определяется внешними услови-
ями. Такое определение пола называется фенотипическим (или фи-
зиологическим).
Порядок хлорококкальные—Chlorococcales
Порядок объединяет коккоидные, одноклеточные, колониальные
и ценобиальные формы.
Бесполое размножение — посредством зооспор и апланоспор
(автоспор). У ценобиальных форм зооспоры и автоспоры обычно
еще внутри оболочки материнской клетки слагаются в дочерние
ценобии, которые, как и у вольвокальных, растут только за счет
увеличения размеров клетки, число же клеток остается постоянным.
Половрй процесс изо-, гетеро- и оогамия. В зависимости от
способа бесполого размножения — зооспорами или автоспорами —
209
хлорококкальные нередко делятся на две большие группы: хлорсн^И
коккальные зооспоровые и хлорококкальные автосгюровые. В каче- “Н
(стве представителя первой группы можно рассмотреть род гидроди- >
кцион, или водяную сеточку Ulydrodietyon). Это макроскопическая,
достигающая 30 см и более ценобиальная водоросль. Цилиндричес-
кие или широкоовальные клетки соединяются концами большей
частью по три, образуя сеть. У широко распространенной в водах,
богатых азотом. Н. reticidatum сеть представляет собой замкнутый
мешок. Цилиндрические с заостренными концами клетки имеют
крупную центральную вакуоль с клеточным соком.
Постенная цитоплазма содержит сетчатый хроматофор с много-
численными пиреноидами и многочисленные ядра. Митоз протекает
следующим образом: центриоли реплицируются и мигрируют к по-
люсам; в профазе формируется перинуклеарная оболочка — чехол
эндоплазматической сети, который заключает ядро, центриоли
и цитоплазматические микротрубочки, находящиеся вокруг ядра;
в ядерной оболочке на полюсах, вблизи центриолей, образуются
; полярные отверстия, через которые экстрануклеарные микротру-
бочки вторгаются в ядро; одни микротрубочки непрерывны и пере-
секают хромосомы в метафазной пластинке, другие — хромосо- Я
мальные -связывают кинетохоры с полюсами; расхождение хро- Л
j мосом в анафазе сопровождается значительным удлинением верете- И
i на внутри ядерной оболочки, которая остается интактной, за Я
j исключением полярных отверстий; в телофазе перинуклеарная обо- Я
| лочка рассеивается, ядерная оболочка восстанавливается. Л
При бесполом размножении после митозов вблизи тонопласта Я
появляются микротрубочки, ориентированные преимущественно
параллельно длинной оси клетки. Среди микротрубочек возникают
пузырьки, которые, сливаясь, дают начало вытянутым трещинам
(бороздам), изолирующим тонкий непрерывный слой цитоплаз-
I мы — вакуолярную оболочку, обычно содержащий лишь мелкие
органеллы — рибосомы, микротрубочки. Вследствие этого тоно-
пласт изолируется от тех мембран, которые участвуют в распаде
цитоплазмы при образовании зооспор. Цитоплазма, заключенная
между вакуолярной оболочкой и клеточной стенкой, претерпевает ,
радиальные расколы: между ядрами появляются микротрубочки, |
ориентированные радиально, и среди них образуются пузырьки,
удлиняющиеся и сливающиеся друг с другом и с растущими ин-
вагинациями плазмалеммы. Таким образом цитоплазма распадает-
ся на все более мелкие единицы вплоть до образования одноядер-
ных зооспор, выраба тывающих жгутики (рис. 54). Зооспоры, огра-
ниченные небольшим пространством между вакуолярной оболоч-
кой и клеточной стенкой, некоторое время движутся, а затем
останавливаются и каждая зооспора обычно соединяется с четырь-
мя другими. При этом первоначально овальная форма их меняется
на ромбическую. Вакуолярная оболочка, образованная по существу
плазмалеммой и тонопластом, ограничивающими очень тонкий
210
Рис. 54. Схема развития зооспор Hydrodictyon reliculatum.
.4— микротрубочки (мт) появляются вблизи тонопласта (т) и ориентированы в про-
дольном направлении. Б—между этими микротрубочками возникают пузырьки-
везикулы (в), которые удлиняются и сливаются, отграничивая от тонопласта узкий
слой цитоплазмы вакуолярную оболочку (во). Между клеточными ядрами появля-
ются радиально идущие микротрубочки. В цитоплазма, заключенная между ваку-
олярной оболочкой и клеточной стенкой, раскалывается радиально идущими трещи-
нами на участки, в конце концов заключающие только по одному ядру. Г зрелые
со жгутиками зооспоры, заключенные между вакуолярной оболочкой и клеточной
стенкой
слой цитоплазмы, играет существенную роль в формировании дочер-
ней сеточки, как бы прижимая зооспоры, расположенные в один слой,
к клеточной стенке. Когда зооспоры соединяются друг с другом, они
автоматически принимают конфигурацию цилиндрической полой
сети из одного слоя клеток, столь характерную для Н. reticulatum.
Если по каким-либо причинам эта вакуолярная оболочка оказывает-
ся поврежденной, то возникают неправильные, уродливые сеточки.
Соединяются зооспоры не в любом месте их поверхности, а лишь
в тех участках, где под плазмалеммой располагаются микротрубоч-
ки. Зооспоры, подвергнутые действию колхицина — яда, разруша-
ющего цитоплазматические микротрубочки, в ценобий не объединя-
ются1. Молодые сеточки растут затем только благодаря росту
слагающих их клеток, причем число ядер в них увеличивается.
Половой процесс, наступающий при недостатке в среде азота,
изогамный. Двужгутиковые изогаметы образуются так же, как зоо-
споры, но в большем числе и соответственно они мельче. Гаметы
освобождаются через отверстие в оболочке материнской клетки.
Н. reticulatum— форма однодомная (гомоталличная: даже гаметы
из одной клетки способны копулировать). Хотя у гидродикциона
1 Аналогичные опыты с колхицином показали, что образование микротрубочек
необходимо не только при аггрегации зооспор в колонию, но и для формирования
самих зооспор и последующего изменения их формы.
211
изогамия, под электронным микроскопом удалось установить раз-
ницу в строении сливающихся гамет: одна из них hSl переднем конце
между жгутиками несет электронно-плотную Молярную шапочку,
другая такой шапочки лишена. Полярная шапочка вытягивается
в виде оплодотворяющей трубки, кончик которой и сливается с ме-
мбраной, находящейся между жгутиками гаметы без апикальной
шапочки (рис. 55). Зигота одевается оболочкой, в ней накапливается
жир, окрашенный гематохромом в кирпично-красный цвет. Посте-
пенно она увеличивается в размерах и переходит в состояние покоя.
Прорастает зигота с образованием четырех крупных двужгутико-
вых зооспор, при этом ядро редукционно делится. Зооспоры, про-
плавав некоторое время, останавливаются, и каждая развивается
в многоугольную клетку — полиэдр. Полиэдр разрастается, стано-
вится многоядерным, и содержимое его распадается на двужгутико-
вые зооспоры, которые слагаются в молодую зародышевую плос-
кую сеточку, освобождающуюся через разрыв оболочки полиэдра.
Полиэдры, оболочки которых несут отростки, а содержимое
богато жировыми включениями, по-видимому, имеют значение для
распространения водоросли, так как крупные зооспоры, освобожда-
ющиеся при прорастании зигот, малоподвижны и не могут выпол-
нять эту функцию. Кроме того, полиэдры выносят высыхание,
губительное для ценобия, и, таким образом, наряду с зиготой могут
представлять покоящуюся стадию в жизненном цикле водяной се-
точки.
У хлорококкальных, размножающихся посредством автоспор,
формированию их предшествуют митозы, которые сопровождают-
ся репликацией центриолей и образованием перинуклеарного чехла
Рис. 55. Детали слияния гамет у Hydrodietyon reticulatum.
А —гамета с полярной шапочкой (пш); Д’-—гамета без полярной шапочки; В — гаме-
та с полярной шапочкой дает оплодотворяющую трубку (от); Г—соединение гаме!
посредством оплодотворяющей трубки
эндоплазматической сети. Ядерная оболочка остается интактной, за
исключением полярных отверстий, через которые в ядро перемеща-
ются микротрубочки, принимающие участие в образовании верете-
на. У ряда автоспоровых хлорококкальных (кирхнериелла, тетраэд-
ров, анкистродесмус) фикопластные микротрубочки проникают
между дочерними ядрами, тогда как другие микротрубочки ограни-
чивают протопласты, обозначая плоскости будущего распада цито-
плазмы по бороздам, по-видимому, возникающим в результате
инвагинации плазмалеммы. Однако у других родов (хлорелла, сце-
недесмус) цитокинез сопровождается образованием клеточной пла-
стинки.
Род хлорелла (Chlorella) широко распространен в пресных водах,
на сырой земле, коре деревьев и т. д. Описан ряд морских видов.
Шаровидные клетки одеты гладкой оболочкой, содержат обычно
чашевидный хроматофор и одно клеточное ядро.
Электронно-микроскопические исследования позволили обнару-
жить сложное строение клеточной стенки у некоторых видов хло-
реллы. Наружный трехслойный - триламинарный компонент стен-
ки, образованный двумя электронно-плотными зонами, заключа-
ющими электронно-прозрачную центральную зону, содержит спо-
рополленин— чрезвычайно устойчивое к действию различных
ферментов вещество, встречающееся в пыльцевых зернах и спорах
высших растений. Внутренний более толстый компонент содержит
целлюлозные микрофибриллы. У ряда видов хлореллы в стенке
имеется только микрофибриллярный слой, а трехслойный наруж-
ный компонент из спорополленина отсутствует.
Автоспорообразованию у хлореллы предшествуют митозы, со-
провождающиеся дупликацией центриолей и миграцией их к проти-
воположным полюсам ядра, после чего образуется интрануклеарное
веретено. У Ch. pyrenoidosa цитокинез начинается на стадии четырех
ядер. Между соседними близко расположенными ядрами появляют-
ся микротрубочки, и вдоль плоскости их ориентации по направле-
нию к периферии клетки центрифугально растягивается клеточная
пластинка. Последняя появляется сначала в виде мелких элект-
ронно-прозрачных капелек между двумя дочерними ядрами, кото-
рые сливаются.
Развитие упомянутых выше компонентов стенки происходит
следующим образом. Кнаружи от цитоплазматических мембран
(плазмалемм) автоспор, заключенных еще в оболочку материнской
клетки, появляется первый предшественник клеточной стенки в виде
мелких трехслойных изогнутых дисков. Эти диски увеличиваются
в числе и размерах, пока не сольются краями с образованием вокруг
автоспоры сплошного трехслойного наружного компонента, в кото-
ром откладывается спорополленин. Как только сформировался
сплошной трехслойный наружный компонент стенки автоспор, на-
чинается ^растворение внутренней микрофибриллярной части стенки
материнской клетки, и между образовавшимся трехслойным чехлом
213
Рис. 56. Схема последовательной
го (А — Г) развития разных сло-
ев стенки Chtorella при автоепд-
рообразовании:
сп- спороиоллениновый слой мате-
ринской стенки, ц - целлюлозный
слой материнской стенки, нм ннто-
плазматическая мембрана
автоспор.
спи— спорополленшювыс лиски ав-
тоспор, сна — сплошная спорополлс-
ииновая стенка автоспор, ца цел-
люлозный слой стенки автоспор
и цитоплазматической
мембраной автоспоры откладывается мик-
рофибриллярный компонент стенки автоспор. Стенка материнской
клетки, от которой остается только тонкий наружный споропол-
лениновый компонент, очень стойкий по отношению к химическим,
но не механическим воздействиям, разрывается, и автоспоры осво-
бождаются (рис. 56).
Другие близкие к хлорелле роды характеризуются иной формой
клеток и тем, что наряду с одиночно живущими клетками у них
в результате неполного расхождения автоспор нередко возникают
скопления клеток или колонии из неопределенного числа клеток.
Таковы, например, роды анкистродесмус (Ankistrodesmus) и кирх-
нериелла (Kirchneriella); первый—с веретеновидными, прямыми или
изогнутыми клетками, второй — с полулунными или дуговидными
клетками.
Род эремосфера (Eremosphaera), обычный на сфагновых болотах,
отличается очень крупными (до 150 мк) одиночными шаровидными
клетками с одним ядром и многочисленными дисковидными хрома-
тофорами с пиреноидами. Автоспоры в числе двух или многих
освобождаются через разрыв стенки. Сравнительно недавно (э
1967 г.) у Е. viridis в экспериментальных условиях наблюдался поло-
вой процесс: в одних клетках (оогониях) развивалась одна яйцеклет-
ка, в других (сперматогониях) в результате последовательных деле-
214
ний образовалось от 16 до 64 двужгутиковых сперматозоидов.
Ооспора внутри стенки оогония покрывалась скульптурированной
оболочкой.
Оогамный половой процесс, помимо эремосферы, гораздо рань-
ше1 наблюдался у родов голенкиния (Golenkinia), микрактиниум
(Micraclinium) и диктиосфериум (Dictyosphaerium).
У диктиосфериума яйцеклетка выходит из оогония до оплодот-
ворения и сливается со сперматозоидом вне оогония, т. е. имеет
место примитивная оогамия.
Из родов, характеризующихся ценобиями, сложенными из опре-
деленного числа различно расположенных клеток, можно рассмот-
реть род сценедесмус (Scenedesmus), у которого эллипсоидальные
или веретеновидные клетки соединены боками в простые или двой-
ные ряды. У самого обычного вида S', quadricauda обычно четырех-
клеточные ценобии, причем краевые клетки несут выросты стенки —
шипы.
Электронно-микроскопические исследования многих видов рода
Scenedesmus выявили весьма сложное строение клеточной стенки.
Она состоит из трех слоев: внутреннего толстого целлюлозного
слоя, очень тонкого среднего слоя, соответствующего трехслойному
компоненту стенки хлореллы и также содержащего спорополленин,
и наружного пектинового слоя. Два первых слоя окружают каждую
клетку ценобия, а пектиновый наружный слой одевает весь ценобий,
объединяя все его клетки в единое целое. Различная орнаментация
клеточных стенок видов сценедесмуса, различимая в оптическом
микроскопе и используемая как диагностический признак, обуслов-
лена сложным строением наружного пектинового слоя. Этот слой
образован гомогенным матриксом, на поверхности которого нахо-
дится сеть, сложенная из шестиугольных ячеек. Сеть расположена
на цилиндрических трубчатых подпорках; основание их прикрепля-
ется к поверхности среднего (спорополленинового) слоя. Пектино-
вый слой и сеть местами (обычно на концах клеток и вдоль гребней)
прерываются округлыми отверстиями, окаймленными подпорками.
Эти подпорки сливаются боками, а со дна отверстий отходят
короткие, тоже трубчатые подпорки, от которых, возможно, берут
начало длинные щетинки, пучком выходящие из отверстий. Щетин-
ки имеюз белковую природу и сложную микроструктуру, они спо-
собствуют лучшей плавучести ценобиев: если их удалить центрифу-
гированием, ценобии быстро оседают на дно сосуда. Видимые
в оптический микроскоп гребни и шипы также сформированы под-
порками разной длины. Самые длинные подпорки образуют шипы
(рис. 57).
Клетка сценедесмуса содержит париетальный хлоропласт с пире-
ноидом, ядро, митохондрии, диктиосому, связанную с ядерной
1 Оогамный половой процесс у хлорококковых впервые описал русский альголог
А. А. Коршиков (1937) у новых описанных им видов голенкиния и микрактиниум.
215
Рис. 57. Схема строения стенки ;
Scene desmus^quadricauda:
цм — цитоплазматическая мембрана, ц— 1
целлюлозный внутренний слой стенки» сп— j
средний спороиоллениновый слой, нсл—не- 1
ктиновый слой, пс пектиновая сеть, о— -я
отверстие в пектиновом слое, п— подпорки, .]
ыа которых покоится пектиновая сеть, ш— |
шип, образованный самыми длинными под- I
порками, кн -- короткие подпорки, полнима- I
ющиеся со дна отверстий ’
оболочкой и окруженную цилиндром эндоплазматического ретику-
лума, пару центриолей. При развитии автоспор материнская клетка
делится два или три раза с образованием соответственно 4—8
дочерних протопластов. Первое деление поперечное, второе и тре-
тье— продольные. Митозам предшествует репродукция диктиосо-
мы, пар центриолей и, по-видимому, хлоропласта; пиреноид не
делится, а исчезает между первым и вторым делениями и образует-
ся заново в каждой дочерней клетке — автоспоре. Ядерная оболочка
сохраняется во время митоза, за исключением отверстий на полю-
сах. Ядрышко разрушается. После митоза между дочерними яд-
рами появляются микротрубочки и клеточная пластинка. Эта пла-
стинка образуется за счет слияния мелких везикул, располагающих-
ся вдоль микротрубочек. Еще до завершения формирования первой |
клеточной пластинки наступает второй митоз, после которого появ-
ляется вторая клеточная пластинка и т. д. После того как сфор-
мировались все клеточные пластинки, дочерние клетки смещаются,
располагаясь рядом друг с другом и параллельно длинной оси
материнской клетки, под плазмалеммой возникают продольно ори-
ентированные микротрубочки, а в хлоропластах образуются пире- :
ноиды. Клеточные стенки развиваются одновременно вокруг всех I
дочерних клеток. Как и у хлореллы, первым откладывается средний
слой в виде трехслойных дисков, которые растут и, смыкаясь,
полностью окружают клетку. Затем начинается формирование внут-
реннего целлюлозного и наружного пектинового слоев. Автоспоры
внутри оболочки материнской клетки слагаются в молодую коло-
нию, которая позднее освобождается через разрыв материнской
оболочки и растет только за счет роста клеток, но не изменяя их
числа. В последние годы появились сообщения о существовании
у 5. obliquus изогамного полового процесса: при снижении в среде
содержания азота вегетативные клетки превращаются в гаметангии,
в которых образуется по восемь гамет. На гаметогенез влияют
также температура и освещение.
Хлорококкальные— почти исклеочитслыю пресноводные орга!!измы, особенно
богато представленные в планктоне. Отростки клеточных стенок у многих видов
сценедесмуса. характерные для Е олепкинии, микрактиниума и др., шипы, щетинки,
увеличивая трение о воду, поддерживают водоросль во взвешенном состоянии
и служат приспособлениями к планктонному образу жизни.
Хлорококкальные легко культивируются в искусственных условиях и давно
используются в качестве объектов физиологических и биохимических исследований,
особенно при изучении фотосинтеза. Массовое культивирование допускает их прак-
тическое применение в качестве источника пищи для человека и сельскохозяйстве!!-
ных животных, для получения витаминов, хлорофилла, стеринов и пр.
Порядок улотрихальные — Ulotrichales
Порядок объединяет зеленые водоросли, имеющие таллом в ви-
де неразветвленной нити, сложенной из одноядерных клеток. Реже
таллом пластинчатый или трубчатый, но в онтогенезе развива-
ющийся из нитчатого. Представителем может служить распрост-
раненный как в пресных водах, так и в морях род улотрикс (Ulothrix)
с наиболее известным видом V. zonata.
Нить состоит из одного ряда клеток, одетых толстой оболочкой,
под которой находится постеппая цитоплазма, содержащая пла-
стинчатый хроматофор, опоясывающий клетку изнутри в виде неза-
мкнутого кольца. В хроматофоре несколько пиреноидов, в постен-
ной цитоплазме одно ядро, центр клетки занят вакуолью с клеточ-
ным соком. Все клетки нити одинаковые, за исключением базаль-
ной, которая Вытягивается в короткий ризоид и служит для
прикрепления нити к субстрату. Все клетки, кроме базальной, спо-
собны делиться, обусловливая непрерывное нарастание таллома
в течение вегетативной жизни, и формировать репродуктивные
клетки. При бесполом размножении во всех зеленых клетках образу-
ется от 2 до 16(32) четырехжгутиковых зооспор. Обычно этот
процесс начинается в апикальной (конечной) клетке и продвигается
к основанию нити. Зооспоры, заключенные в слизистой обвертке,
освобождаются через боковое отверстие в клеточной стенке. После
периода движения (имеет место положительный фототаксис) зоо-
спора останавливается, сбрасывая один за другим жгутики, прикре-
пляется боком к субстрату и прорастает в нить.
При половом размножении в клетках возникают двужгутиковые
гаметы в числе 8(4)—32(64). Гаметы выходят из клетки заклю-
ченные в слизистый пузырь, в котором они двигаются, пока
не освободятся. Половой процесс изогамиый. У U. zonata наблю-
дается гетероталлизм, т. е. копулируют гаметы из разных нитей,
отличающихся в половом, отношении (« + » и « —»). В результате
копуляции сначала образуется подвижная четырехжгутиковая зи-
217
гота (планозигота) с отрицательным фототаксисом. После неко- 1
торого периода движения она прикрепляется к субстрату, окру-
гляется и втягивает жгутики. При определенных условиях зигота
прорастает в одноклеточный спорофит. Сначала появляется тру-
бковидный вырост, в который переходит ее содержимое. Этот
вырост позднее вздувается, образуя на вершине грушевидные или .
дубинкообразные спорофиты, содержимое которых при созревании I
распадается на 4—16 (большей частью 8) четырехжгутиковых зоо-
спор. Они выходят через отверстие, возникающее в результате
ослизнения части стенки спорофита, и заключены еще в слизистую
обвертку, быстро исчезающую. Зооспоры оседают на субстрат
и прорастают так же, как зооспоры, возникающие при бесполом
размножении нитей улотрикса.
Сходный цикл развития установлен для других видов улотрикса
и некоторых других родов. Дубинкообразные одноклеточные спо-
рофиты улотрикса и других родов прежде рассматривались как
самостоятельный род кодиолюм (Codiolwn). Выше уже указыва-
лось, что П. Корнман для зеленых водорослей с одноклеточным
спорофитом установил специальный класс кодиолофициевые
(Codiolophyceae).
В качестве отдельного семейства к порядку улотрихальных от-
носят ульвовые (Ulvaceae), иногда их выделяют в особый порядок
Ulvales. Представители этой группы отличаются тем, что имеют не
нитчатый, а пластинчатый таллом. Однако на ранних стадиях раз-
вития талломы их состоят из прикрепленной к субстрату нити,
очень похожей на улотрикс, но рано или поздно в такой нити,
помимо поперечных делений клеток, наступают продольные, веду-
щие к формированию двухслойной пластинки. У рода ульва (Ulva)
оба слоя клеток остаются плотно сомкнутыми. Таллом достигает
крупных размеров (кисти руки), имеет гофрированные края и при-
крепляется к субстрату суженным в короткий черешок основанием.
Клетки одноядерные, с постенным хроматофором. У близкого
к ульве рода энтероморфа (Enteromorpha) оба слоя клеток пластинки
на ранней стадии разъединяются, и талломы, также крупные, при-
нимают вид кишки или трубки с однослойной стенкой. Ульва
и энтероморфа — в основном морские водоросли, приуроченные
к прибрежной (литоральной) зоне как в северных, так и в южных
морях. Многие виды обоих родов выносят заметное опреснение
воды и часто поднимаются в устья рек, а некоторые виды эн-
тероморфы обычны и в пресноводных водоемах. Вегетативное раз-
множение осуществляется отделившимися участками таллома. Бес-
полое размножение — зооспорами, как правило, четырехжгутиковы-
ми, которые образуются путем последовательного деления прото-
пласта любой клетки на 4—8 частей. Половое размножение— у
двужгутиковыми гаметами (имеет место гетероталлизм). У разных *
видов обоих родов наблюдается изогамия или гетерогамия. У гете- а
рогамных видов талломы разного знака можно различать и нево-
218 Я
I
оружейным глазом по окраске плодущих участков — желтоватой
у мужских и темно-зеленой у женских. Оба рода детально изучены
с точки зрения их цитологии и циклов развития. У них установлена
изоморфная смена генераций. Изучение в культуре показало, что
как и у эктокарпуса, у ульвы чередование спорофита и гаметофита
не всегда коррелирует с чередованием в плоидности. Так, наряду
с диплоидными спорофитами в жизненном цикле ульвы можно
встретить гаплоидные спорофиты, возникающие в результате пар-
теногенетического развития гамет. Помимо гаплоидных гаметофи-
тов, продуцирующих гаплоидные гаметы, попадаются редкие дип-
лоидные гаметофиты, в клетках которых образуются диплоидные
гаметы и т. п. Всего было установлено 12 различных (по характеру
спороношения, числу хромосом) типов морфологически идентич-
ных растений.
Порядок хетофоралытые — Chaetophorales
Сюда относят зеленые водоросли с гетеротрихальным строени-
ем, имеющие нитчатые талломы, дифференцированные на распро-
стертую по субстрату систему нитей, от которых отходят вертика-
льно стоящие более или менее разветвленные нити. Порядок вклю-
чает ряд семейств.
Семейство хетофоровые (Chaetophoraceae) включает сле-
дующие роды.
Род стигеоклоииум (Stigeoclonium') — центральный род семейства,
у которого легко различимы обе части гетеротрихального талло-
ма— обычен в стоячей или медленно текущей воде на подводных
объектах, на их верхней освещенной стороне. От нитей, стелющихся
по субстрату, отходят вертикально стоящие ветвящиеся нити. Ветви
обычно заканчиваются многоклеточными волосками из вытянутых
бесцветных отмерших клеток. Строение клеток стигеоклониума
в общем такое же, как у улотрикса. Размножение бесполое — по-
средством четырехжгутиковых зооспор, половое — изогамия (гаме-
ты двух- или четырехжгутиковые — у разных видов).
У разных видов стигеоклониума стелющаяся по субстрату систе-
ма нитей может быть развита неодинаково: от рыхло разветвленной
нити, часто состоящей из сравнительно немногих клеток, до ком-
пактного диска, образованного срастанием многочисленных ветвей.
Развитие вертикальной системы обычно находится в обратной зави-
симости от развития распростертых нитей: у форм с хорошо разви-
той подошвой или диском вертикальная система может быть реду-
цирована до нескольких коротких веточек или. представлена только
волосками. Наоборот, при слабом развитии основания нижние
клетки хорошо развитых вертикальных нитей дают ризоиды, спо-
собствующие прикреплению растения к субстрату.
От форм, подобных стигеоклониуму, эволюция пошла в двух
направлениях: в одних случаях эволюционировала восходящая
219
часть таллома, а стелющаяся редуцировалась, в других—наблюда-
лась редукция вертикальной системы, а стелющаяся ’Тасть получала
наибольшее развитие. '
Первый путь эволюции приводит к таким формам, как род
драпарнальдия (Draparnaldia), у которого распростертая система
практически отсутствует и таллом прикрепляется с помощью ризо-
идов, отходящих от нижних клеток восходящей системы. В отличие
от большинства видов стигеоклониума у восходящей системы дра-
парнальдии резко выражена дифференцировка на длинные и корот-
кие ветви. Длинные, неограниченно нарастающие, слабо ветвящиеся
оси образованы крупными, часто слегка бочонковидными клетками
со слаборазвитым хроматофором, который представлен экватори-
альным пояском с изрезанными краями. Короткие боковые ветви
ограниченного роста, часто называемые ассимиляторами, возника-
ют пучками на длинных осях. Они обильно ветвятся и состоят из
коротких клеток с хорошо развитым постенным хроматофором,
занимающим всю поверхность клетки. Концы этих ветвей часто
превращаются в длинные многоклеточные волоски. Размножение
драпарнальдии такое же, как и у стигеоклониума. Репродуктивные
клетки (зооспоры, гаметы) образуются только в ассимиляторах,
длинные оси несу г главным образом опорную функцию.
Еще большая дифференцировка таллома наблюдается у рода
драпарнальдиопсис ( Draparnaldiopsis), у которого длинные оси состо-
ят из чередующихся коротких и длинных клеток. Ассимиляторы
отходят только от коротких клеток длинных осей. У D. indica (а также
у ряда видов стигеоклониума) отмечена изоморфная смена генераций.
Изоморфная смена генераций установлена также у рода фричиел-
ла (Fritschiella), единственный вид которого F. tuberosa найден до
сих пор только в Африке, Индии, Японии и США, где он растет на
сравнительно сухих почвах. Он имеет сложный гетеротрихальный
таллом, состоящий из подземной разветвленной системы нитей,
которая образует первичные вертикальные нити. Эти нити растут
сквозь тонкий слой почвы и развивают над ее поверхностью кустис-
тые побеги.
Другой эволюционный ряд форм характеризуется более или
менее полной редукцией вертикальной системы. Примером таких
большей частью энифитных форм могут служить роды хетонема
(Chaetonema) — редкий эпифит в слизи разных пресновидных водо-
рослей (тетраспоры, батрахоспермума) и афанохете
(Aphanochaete)— обычный пресноводный эпифит на водорослях.
У хетонемы вертикальная система представлена короткими нитями,
несущими на конце одноклеточный волосок, у афанохете, как пра-
вило,— только волосками. Помимо образования четырехжгутико-
вых зооспор, у обоих родов известен и половой процесс.
У афанохете половой процесс обычно описывается как резко
выраженная гетерогамия: в крупных оогониях образуется по одной
четырехжгутиковой макрогамете, которая выходит из оогония, не-
220
продолжительное время движется, затем теряет жгутики, останав-
ливается и оплодотворяется четырехжгутиковым сперматозоидом
(микрогаметой). Сперматозоид образуется в единственном числе
в мелких, почти бесцветных антеридиальных клетках. Однако
в 1981 г. появилось сообщение, иначе освещающее этот процесс:
четырехжгутиковым сперматозоидом оплодотворяется неподвиж-
ная, лишенная жгутиков яйцеклетка, которая выходит из оогония
наружу до оплодотворения. Если это действительно так, то у афа-
нохете имеет место примитивная оогамия, свойственная и другому
роду — хетонеме.
Половой процесс у хетонемы оогамный. Он был описан русским
ботаником К. И. Мейером (1930) у Ch. irregulare. У этого вида
можно различить мужские и женские растения. Некоторые клетки
мужских нитей превращаются в антеридии, в которых образуется по
8 сперматозоидов овальной формы с двумя жгутиками. Оогонии
образуются на женских нитях, обычно на концах коротких боковых
ветвей. Все содержимое оогония превращается в одну яйцеклетку
без жгутиков, которая выходит через отверстие в оболочке оогония.
Оплодотворяется яйцеклетка вне оогония, т. е. имеет место прими-
тивная оогамия, впервые описанная у зеленых водорослей
К. И. Мейером как раз у Ch. irregulare.
Семейство трентеполиевые — Trentepohliaceae объединя-
ет в основном гетеротрихальные зеленые водоросли, как правило,
без волосков. Зооспорангии в отличие от большинства зеленых
водорослей по форме иные, чем вегетативные клетки. Половое
размножение—путем копуляции двужгутиковых изогамет.
Род трентеполия (Trentepohlia) интересен тем, что приспособлен
к наземным условиям существования. Виды его особенно широко
распространены в тропиках и субтропиках, где растут на камнях,
скалах, стволах деревьев и как эпифиты на листьях. Некоторые
входят в состав лишайников. У нас талломы трентеполии образуют
кирпично-красные или желтые налеты на стволах деревьев. У боль-
шинства видов таллом гетеротрихальный, дифференцированный на
стелющиеся по субстрату нити и отходящие от них вертикально
стоящие нити; у других видов вертикальная система нитей развита
слабо. Более или менее шаровидные клетки толстостенные, прото-
пласт без вакуоли с клеточным соком, содержит много дисковид-
ных или лентовидных хроматофоров, зеленая окраска которых, как
правило, маскируется желтым или кирпично-красным пигментом —
гематохромом, растворенным в каплях масла. Более старые клетки
обычно многоядерные. Основной способ размножения трентепо-
лии— вегетативный: нити легко разламываются на отдельные
участки или клетки, которые переносятся ветром на новый субстрат.
Чезырехжгутиковые зооспоры образуются в специальных
крючковидных спорангиях, сидящих на клетках-ножках. Эти
спорангии легко отделяются и разносятся ветром в целом
виде. Как показал К. И. Мейер, при созревании зооспорангии
221
становятся многоядерными, и стоит им попасть на каплю
воды, как через несколько (3—5) минут содержимой их распадается
на одноядерные участки, которые вырабатывают жгутики и пре-
вращаются в зооспоры. В шаровидных сидячих гаметангиях,
которые также разносятся ветром в целом виде, образуются
(в присутствии воды) двужгутиковые гаметы. Однако копуляция
гамет наблюдается очень редко, гаметы большей частью раз-
виваются пар геногенетически, прорастая, как и зооспоры, на
влажной коре в новые нити.
Из других представителей семейства трентеполиевых можно назвать роды цефа-
леурос (Cephaleuros) и фикопельтис (Phycopeltis), встречающиеся в тропиках и суб-
тропиках на листьях различных растений, вызывая в некоторых случаях опасные
заболевания. Например, цефалеурос является возбудителем заболевания чая. Как
и у трентеполии, у этих родов зооспорангии, морфологически отличающиеся от
обычных вегетативных клеток, отделяются целиком и распространяются в целом
виде с помощью ветра или дождя, что, несомненно, служит приспособлением к на-
земному существованию. Процесс отделения зооспорангия от клетки-ножки был
подробно изучен с применением сканирующего электронного микроскопа
у С. virescens и Ph. epiphiton. Сначала формируется пора, через которую будут
выходить зооспоры. Затем происходят кольцевой разрыв боковой стенки в области
септы, разделяющей зооспорангий и клетку-ножку, и расщепление самой септы.
Септа как на зооспорангии, так и на клетке-ножке выпячивается, и это выпячивание,
так же как выступы, возникакицие в центре выпячивающихся септ, способствует
отделению спорангия от клетки-ножки.
Семейство колеохетовые (Coleochaetaceae) включает род
колеохете (Coleochaete), виды которого — пресноводные эпифиты —
поселяются главным образом на высших водных растениях. У од-
них видов типичный гетеротрихальный таллом, погруженный
в слизь, образует полусферические подушечки, у других видов про-
изошла редукция восходящих нитей, и таллом представлен только
распрос тертой по субстрату системой нитей, которая может состо-
ять из рыхло расположенных разветвленных нитей или нити могут
срастаться боковыми частями, формируя компактный диск. Одно-
ядерные клетки с крупным постенным хроматофором обычно несут
характерные, одетые воротничком щетинки, которые возникают на
клетке в виде выступа, одетого сначала двухслойной стенкой. Затем
наружный слой стенки разрывается и остается в виде воротничка
у основания щетинки, в которую вырастает выступ, одетый только
внутренней стенкой. Бесполое размножение колеохете осуществля-
ется двужгутиковыми зооспорами. Половой процесс оогамный.
Оогонии и антеридии обычно развиваются на одном талломе, хотя
есть и двудомные виды. Оогоний бутылковидный, со вздутой ба-
зальной частью, содержащей хлоропласт, которая продолжается
в длинную шейку—трихогину с бесцветной цитоплазмой. При со-
зревании кончик шейки разрушается, а содержимое базальной части
округляется, причем образуется одна яйцеклетка.
В антеридиях — маленьких бесцветных клетках—формируется
по одному бесцветному двужгутиковому сперматозоиду. После
оплодотворения ооспора одевается толстой оболочкой, а из сосед-
222
них с оогонием клеток вырастают ветвящиеся нити, которые, пере-
плетаясь, образуют вокруг ооспоры псевдопаренхиматозную одно-
слойную кору.
Ооспора зимует и весной прорастает. При этом содержимое ее
делится на 8—32 клетки, каждая из которых затем продуцирует по
одной зооспоре. Зооспора развивается в таллом. Первое деление
копуляционного ядра в прорастающей ооспоре носит редукционный
характер. Таким образом, растение колеохете гаплоидно, диплоид-
на только покоящаяся ооспора. Тем не менее ряд ультраструктур-
ных особенностей этой водоросли позволяет некоторым авторам
рассматривать ее в ряду тех зеленых водорослей, предки которых
могли дать начало высшим наземным растениям (см. ниже).
Порядок эдогониальные—Oedogoniales
Порядок включает нитчатые водоросли, одноядерные клетки
которых делятся с образованием на их стенках особых «колпачков».
По колпачкам эдогониальные водоросли легко отличить от всех
остальных водорослей. Монадные клетки (зооспоры, сперматозо-
иды) эдогониальных в отличие от монадных клеток других зеленых
водорослей многожгутиковые: жгутики располагаются венцом вок-
руг переднего конца клетки. Такие монадные клетки называют
стефанокоптными.
Стефаноконтные зооспоры, помимо эдогониальных, имеются
только у бриопсидальных водорослей (дербезия, бриопсис, псевдо-
бриопсис). Базальные тела, или основания жгутиков, удерживаются
вместе фиброзным (волокнистым) кольцом, находящимся на гра-
нице бесцветной передней (апикальной) части — «головки» зооспо-
ры (сперматозоида) и остальной окрашенной части, занятой пари-
етальным сетчатым хлоропластом — «тела». От фиброзного кольца
между базальными телами, чередуясь с ними, отходят корни жгути-
ков (рис. 58): восходящие исчерченные корни в основном
Рис. 58. Часть жгутикового аппарата
зооспоры Bulbochaete hiloensis:
* -жгут, фк фиброзное кольцо, пм-
поддерживающий материал, вк —восходя-
щий корень, нк - нисходящий корень
ВК
223
простираются в головку, а трубчатые нисходящие — в тело зооспо-
ры (сперматозоида). Основания жгутиков и корни прочно соедине-
ны с фиброзным кольцом и сохраняются вместе с ним при обработ-
ках (необходимых для электронно-микроскопических наблюдений),
при которых свободные части жгутиков утрачиваются. Эго позво-
лило достаточно точно подсчитать жгутики у монадных клеток
эдогониальных водорослей, которые различаются только по числу
жгутиков, что, несомненно, связано с размерами самих монадных
клеток. Так, в зооспорах эдогониума (наиболее крупных) насчиты-
вается около 120 жгутиков, в более мелких зооспорах бульбохете
жгутиков 37—49, а у сперматозоидов эдогониума и бульбохете —
соответственно 30 и 6—9. Жгутиковый аппарат монадных клеток
(зооспор, сперматозоидов) эдогониальных водорослей был изучен
не только в готовом, сформированном состоянии, но был просле-
жен и его генезис. В клетках, первоначально лишенных центриолей,
центриоли появляются заново. Так, при сперматогенезе у Oedo-
gonium cardiacum вблизи ядерной оболочки, снаружи, возникает
20—30 центриолей, которые располагаются в два ряда. Затем два
ряда центриолей мигрируют к поверхности клетки, где разъединя-
ются в центре и соединяются на своих концах, давая начало коль-
цевой конфигурации. Эти расположенные кольцом центриоли ста-
новятся базальными телами жгутиков. Одновременно развиваются
микротрубчатые корни, которые простираются в заднюю часть
сперматозоида и, по-видимому, функционируют, как скелетные эле-
менты. Исчерченные волокна и фиброзное кольцо образуются после
возникновения кольца из базальных телец. Во время формирования
жгутикового аппарата передняя часть цитоплазмы клетки диффере-
нцируется в Куполообразную область -головку, содержащую мно-
гочисленные пузырьки — производные аппарата Гольджи. Половой
процесс оогамный.
Представителем порядка может служить род эдогониум
(Oedogonium) — распространенная во всех частях света пресновод-
ная водоросль. Талломы имеют вид неветвящихся нитей, обычно
в зрелом состоянии свободноплавающих, а в молодом прикреплен-
ных к субстрату (подводным высшим растениям, сваям и пр.)
специальными базальными клетками. У видов, живущих в текучей
воде, нити остаются прикрепленными всю жизнь. Цилиндрические
клетки, иногда вздутые в верхней части, одеты тонкой оболочкой,
под которой располагается постенная цитоплазма, содержащая од-
но крупное, хорошо заметное даже в живом состоянии ядро и сет-
чатый цилиндрический хроматофор со многими пиреноидами.
Центр клетки занят крупной вакуолью.
Деление клетки. Некоторые клетки нити делятся. Перед началом
деления у одного конца клетки под поперечной перегородкой из
внутренних слоев оболочки образуется кольцевое утолщение — ва-
лик, вдающийся в полость клетки. Края валика сверху и снизу
образованы более плотным материалом, выступают в виде «губ»
224
и прикреплены к наружной стенке клетки. Ядро мигрирует в об-
ласть валика и делится (ядерная оболочка остается интактной на
протяжении всего митоза, имеются кинетохоры). Между дочерними
ядрами появляются пузырьки и микротрубочки, ориентированные
в плоскости будущей септы. В области валика между двумя его
выступами — «губами» — уплотненного материала происходит ко-
льцевой разрыв старой стенки клетки. Разрыв этот носит взрывной
характер — происходит судорожно, нередко приводя в движение
всю нить. Растяжение более мягкого материала валика, в резуль-
тате чего между участками старой оболочки вставляется цилинд-
рический кусок новой, тоже осуществляется быстро -в течение
полминуты. В эту часть клетки переходит одно из дочерних ядер.
Септа, сформированная пузырьками и микротрубочками, передви-
гается наверх в виде диафрагмы, пока не достигнет уровня нижней
губы растянувшегося валика. В септе пузырьки сливаются с об-
разованием новой поперечной стенки, прикрепляющейся к нижней
губе валика. Таким образом, стенка верхней из двух дочерних
клеток в основном представляет собой растянувшийся валик, лишь
сверху, выше кольцевого разрыва первичной оболочки, несколько
выступает часть материнской оболочки. Она и образует то, что
называют колечком или колпачком. Клетки с колпачком обычно
делятся повторно и обнаруживают столько колпачков, сколько
было, делений.
Бесполое размножение эдогониума осуществляется с помощью
крупных зооспор, образующихся по одной из всего содержимого
клетки.
Оболочка клетки, в которой сформировалась зооспора, разрыва-
ется около верхнего конца, обе ее половинки расходятся, и зооспора
медленно выплывает, сначала одетая тонким слизистым пузырем,
который вскоре исчезает. После некоторого периода движения зоо-
спора передним концом прикрепляется к субстрату, теряет жгутики,
выделяет оболочку и вытягивается в нить с ризоидами у основания.
Первое же деление молодого проростка обычно сопровождается
возникновением колпачка.
Половое воспроизведение эдогониума оогамное. Оогонии —
крупные, большей частью шаровидно или эллипсоидально вздутые
клетки, содержащие по одной яйцеклетке. Антеридии — короткие
дисковидные клетки с желтеющими редуцированными хроматофо-
рами. Каждый антеридий обычно развивает по два желтоватых
сперматозоида с венцом жгутиков на переднем конце. Распределе-
ние половых органов у видов эдогониума различно. Существуют
виды обоеполые, или однодомные, у которых мужские и женские
органы развиваются на одной нити, и раздельнополые, или двудом-
ные, у которых оогонии и антеридии развиваются на разных нитях.
К двудомным видам принадлежат макрандриевые с оди-
наковыми мужскими и женскими нитями, и наннандриевые (анд-
роспоровые) с половым диморфизмом: женские нити обычные,
8 - 1210
225
а мужские — карликовые (наннандрии), часто состоящие только из
одной вегетативной клетки, несущей антеридиальные. Наннандрии
вырастают из специальных спор—андроспор, оседающих на женс-
кие нити поблизости от оогония — на субоогониальной клетке или
на стенке самого оогония. Они, по-видимому, хемотаксически при-
влекаются к оогониям: для О. borisiamim было доказано, что мате-
ринская клетка оогония выделяет аттрактант, привлекающий анд-
роспоры.
Одно из самых ранних наблюдений над слиянием гамет — плаз-
могамией не только у водорослей, но и у растений вообще—от-
носится к эдогониуму (N. Pringsheim, 1855). В 1973 г. Л. Гофман
(L. R. Hoffman, 1973) проследил этот же процесс у О. cardiacum
с применением современных методов исследования, включая элект-
ронную микроскопию и микрокиносъемку, используя не только
встречающиеся в природе гаплоидные мужские и женские штаммы
этой гетероталличной водоросли, но и специально полученный
в культуре женский диплоидный штамм, о преимуществах которого
для наблюдения слияния сперматозоида с яйцом сказано выше (см.
гл. 4). Для анализа движения сперматозоидов перед и во время
плазмогамии были использованы микрофильмы, продемонстриро-
вавшие 26 полных актов слияния гамет. У О, cardiacum в оогоний
цревращается верхняя из двух дочерних клеток, получающихся в ре-
зультате деления вегетативной клетки нити. Оогониальная клетка
быстро увеличивается, достигая окончательных размеров часто
в течение 12 ч. Молодой оогоний заполнен протопластом, и стенка
его сначала не обнаруживает никаких признаков поры. Однако еще
до созревания яйца оогониальная стенка вблизи места нахождения
клеточного ядра претерпевает, по-видимому, локальную деграда-
цию, приводящую к образованию поры. Эта область заполнена
тонкой волокнистой субстанцией — «субстанцией поры», которая
интерпретируется как «переваренный» материал оболочки оогония.
Вскоре после образования поры через нее выделяются остатки
«поровой субстанции»—то, что ранними исследователями описы-
валось как гиалиновая слизь.
По мере развития поры происходит созревание яйца. Превраще-
ние оогониального протопласта в яйцо сопровождается заметным :
уменьшением крупной центральной вакуоли и соответствующей
потерей воды. Это, возможно, связано с деятельностью обильных
сократительных вакуолей, присутствующих в периферической цито-
плазме. Зрелое яйцо (сферическое, темно-зеленое, с многочислен-
ными зернами крахмала и жировыми включениями) имеет меньший
объем, чем оогоииальный протопласт, из которого оно образова-
лось; между поверхностью яйца и оогониальной стенкой, как пра-
вило, существует свободное пространство (особенно большое у дип-
лоидного женского штамма). Во время созревания оогония выделя-
ется аттрактант, привлекающий сперматозоиды к области поры
в стенке оогония.
226
Недавно освободившиеся из антеридиев наиболее активные спер-
матозоиды веретеиовидной формы, у многих задний конец остро
суживается в «хвост». По мере старения сперматозоиды увеличива-
ются в размерах, становятся более сферическими и вялыми. Они
редко осуществляют плазмогамию. Сперматозоид содержит ядро,
заметную вакуоль, митохондрии в переднем конце, небольшое ко-
личество эндоплазматического ретикулума, несколько мелких телец
Гольджи. Хлоропласт сильно редуцирован, крахмала мало. На
апикальном конце находится венец из 25—35 жгутиков.
Большинство оогониев ко времени, когда спермотозоиды
достигают поры, утрачивает «поровую субстанцию». В этих
случаях вход сперматозоида в пору—дело секунд. Сначала спер-
матозоид приближается к оогониальной поре со жгутиками,
направленными вперед. Часто во время быстрого движения
сперматозоида назад и вперед у отверстия поры наблюдается
контакт жгутиков с краем поры. Когда кончик сперматозоида
прошел через пору, жгутики большей частью волочатся своими
концами вне оогония. Если в оогониальной поре присутствует
«поровая субстанция», она мешает непосредственному прони-
кновению сперматозоида в оогоний. Тем не менее сперматозоиды
в оогонии продвигаются. Многочисленные жгутики распласты-
ваются по поверхности поровой субстанции, очерчивая таким
образом ее границы. При этом они быстро колеблются и весь
сперматозоид после нескольких легких движений делает сильный
толчок вперед в «поровую субстанцию». В присутствии спер-
матозоидов выделившаяся из поры часть «поровой субстанции»
быстро исчезает, очевидно, благодаря растворению. Оогонии,
изолированные от сперматозоидов или жидкости из-под них,
дольше сохраняют выделившуюся «поровую субстанцию». Это
наводит на мысль, что сперматозоиды могут продуцировать
диффундирующее вещество — лизирующий агент, помогающий
растворению .поровой субстанции. На основании двух микро-
киносъемок время исчезновения поровой субстанции определено
как 21 и 14 с. После исчезновения «поровой субстанции» снаружи
некоторое количество ее остается внутри оогония, где сперматозоид
прокладывает свой путь к яйцу через эту субстанцию. По данным
двух киносъемок, миграция сперматозоида от края поры до
поверхности яйца занимает 58 и 37 с. Перед плазмогамией
несущий жгутики передний конец сперматозоида вытягивается,
приобретая способность к быстрым изменениям формы, что
происходит более или менее независимо от остальной части
клетки. Эти быстрые изменения длины и диаметра переднего
конца сперматозоида, возможно, связаны с кольцевидной фи-
брозной лентой, соединяющей базальные тела находящихся здесь
жгутиков. Растягивания и сокращения этой волокнистой ленты,
по-видимому, осуществляют наблюдаемые изменения диаметра
переднего фончика сперматозоида. Кроме того, в переднем конце
сперматозоида, непосредственно под плазмалеммой, располага-
ются цитоплазматические микротрубочки, ориентированные боль-
шей частью параллельно продольной оси клетки. Сокращение
этих микротрубочек может обусловить изменение длины кончика
сперматозоида, связанное с изменением его диаметра. Наконец,
в механизме изменений переднего конца сперматозоида известную
роль, возможно, играют жгутиковые корни, ориентированные
продольно в передней части клетки. Сперматозоид соприкасается
с поверхностью яйца всегда посредством подвижного переднего
конца. Перед самой плазмогамией сперматозоид как бы подметает
поверхность яйца посредством боковых движений своего переднего
конца. Гаметное слияние начинается с сильных вибрирующих
движений клетки сперматозоида, жгутики сильно колеблются,
а затем сбрасываются. Мембраны гамет сливаются, и их про-
топласты соединяются посредством тонкого цитоплазматического
мостика, который постепенно увеличивается в размерах по мере
того, как цитоплазма сперматозоида изливается в яйцо. Среднее
время, необходимое для полного слияния, определенное на ос-
новании 25 актов плазмогамии, зарегистрированных киносъемкой,
составляет 30,5 с. В случае диплоидных оогониев слияние уско-
ряется. Во время плазмогамии с яйцом сливается вся клетка
сперматозоида целиком, за исключением жгутиков. В двуядерной
копуляционной клетке ясно различимы гаметные ядра, а также
митохондрии сперматозоида и остатки его жгутиковых аппаратов.
Непосредственно после плазмогамии и -до кариогамии кнаружи
от копуляционной клетки — продукта слияния — образуется тонкий
фиброзно-волокнистый слой.
Кариогамия следует вскоре после плазмогамии и оба события
могут происходить в течение 15 мин после смешения яиц и спер-
матозоидов. Ооспора после периода покоя прорастает с образова-
нием четырех зооспор, которые дают начало новым растениям. При
прорастании ооспоры имеет место мейоз, и таким образом вегета-
тивная фаза жизни эдогониума проходит в гаплоидном состоянии.
Другой род эдогониевых, также обычный в пресных водах,
бульбохете (Bulbochaete)— отличается тем, что нити обильно вег-
вятся и каждая клетка несет длинный бесцветный волосок со взду-
тым основанием в виде луковицы.
Порядок бриопсидальные—Bryopsidales
Сюда относятся сифоновые — многоядерные на протяжении ве-
гетативного роста водоросли. Таллом часто крупных размеров и со
значительной внешней дифференцировкой формально представляет
собой одну клетку, одетую толстой, часто слоистой оболочкой, под
которой находится постенный слой цитоплазмы, изнутри гранича-
щий с непрерывной вакуолью. В цитоплазме находятся многочис-
ленные дисковидные хроматофоры, которые наряду с хлорофил-
228
нами и каротиноидами, известными для зеленых водорослей, содер-
жат два особых ксантофилла — сифонеин и сифоноксантин, встреча-
ющихся лишь у очень немногих других зеленых водорослей. Под
хроматофорами в постенной цитоплазме, как правило, располага-
ются многочисленные ядра. Размножение вегетативное и половое.
Бесполое размножение известно не у всех представителей. Половой
процесс гетерогамный, реже изогамный. Большинство этих водоро-
слей диплонты, редукционное деление ядра имеет место перед об-
разованием гамет, у некоторых обнаружена гетероморфная смена
генераций. Подавляющее большинство их — обитатели тропических
морей, лишь немногие растут в Черном море и севернее.
Семейство бриопсидовые (Bryopsidaceae) объединяет че-
тыре рода, оказавшиеся тесно связанными между собой. Прежде их
нередко относили к разным семействам.
Род дербезия (Derbesia), встречающийся не только в тропических,
но и в морях умеренных широт, представлен стелющимися по
субстрату и укореняющимися с помощью ризоидов нитями. От этих
нитей отходят пучки вертикально стоящих обычно ветвящихся ни-
тей. Строение нитей типично сифоновое: в постенной цитоплазме,
окружающей непрерывную вакуоль с клеточным соком, находятся
многочисленные ядра и вере геновидные хлоропласты с 1 — 3 пире-
ноидами. Размножение осуществляется посредством стефаноконт-
ных зооспор, образующихся в овальных спорангиях. Спорангии
возникают в качестве боковых выростов вертикальных нитей, от
которых они отделяются сложной перегородкой. Большинство ядер
в спорангии дегенерирует, а вокруг оставшихся ядер цитоплазма
распадается с образованием 8—20 одноядерных стефаноконтных
зооспор.
Род халицистнс {Halicystis)— морская форма, растущая эпифит-
но на известковых красных водорослях (Lithothamnion, Lithop-
hyllum). Таллом — в виде шаровидного пузыря, достигающего
в диаметре 3 см, в нижней части переходящего в тонкий бесцветный
ризоид, глубоко погруженный в субстрат. Пузыревидная часть тал-
лома каждую осень отмирает и сбрасывается, а ризоид зимует
и затем развивает новый пузырь.
Размножение. Халицистис (Н. parvula) может размножаться
посредством протопластов: цитоплазматические массы, получа-
емые в результате мягкого плазмолиза пузырей, секретируют
клеточные стенки и разрастаются в новые растения. Капельки
цитоплазмы получались также путем разрыва клеточных стенок
халицистиса, выращенного в условиях длинного или короткого
дня и раздельно из пузыревидной и ризоидальной частей таллома.
Голые протопласты округлялись и продуцировали новые клеточные
стенки. Однако в отличие от бриопсиса, протопласты которого
всегда развиваются в нормальные растения, способ развития
голых протопластов халицистиса определяется условиями осве-
щения: условия длинного дня обычно способствуют развитию
229
протопласта в полное (пузыревидное с ризоидом) дли сферическое
растение, тогда как условия короткого дня — развитию в нитчатые
ризоидоподобные растения. Местоположение выделенных про-
топластов в исходных талломах халицистиса также, по-видимому,
определяет их дальнейшее развитие. Протопласты, полученные
из сферических частей таллома, развиваются почти исключительно
в пузыри без ризоидов; протопласты, выделенные из ризоидальной
части исходных талломов, в условиях длинного дня, как правило,
превращаются в полные пузыревидные растения с ризоидами.
Кроме вегетативного размножения у халицистиса известно по-
ловое— гетерогамия (анизогамия). Двужгутиковые гаметы неоди-
наковых размеров образуются на разных индивидах (гетеро- |
таллизм). Формирование гамет начинается со скопления цито- |
плазмы с ядрами и хлоропластами в ограниченных участках, й
обычно на верхушке пузыря. Мужские и женские пузыри легко |
отличить еще до выхода гамет по окраске их плодущих участков: |
желтовато-оранжевых, продуцирующих микрогаметы, и зеленых, • I,
в которых формируются макрогаметы. Плодущие участки (га-
метангии) отделяются от остальной вегетативной части пузыря
только тонким слоем цитоплазмы; содержимое распадается на
двужгутиковые гаметы, которые с силой выпрыскиваются (вы- Jj
талкиваются) через одну или несколько пор, возникающих в то- fl
лстой стенке пузыря над плодущими участками. Содержимое Я
вакуоли через эти поры не выходит, так как цитоплазматическая fl
обвертка опустевшего гаметангия (плодущего участка) прижи- j
мается к стенке и таким образом закупоривает пору. Освобождение -
(выстрел) гамет происходит так стремительно (если наблюдать
этот процесс в чашке Петри), что пузырь халицистиса при
отдаче отшвыривается в противоположном выстрелу направлении
через всю культуральную чашку. Содержимое пузыря, не ис- njfl
пользованное на образование гамет, может снова продуцировать
их через правильные интервалы в две недели.
Еще в 30-е годы было обнаружено и многократно подтверждено
в лабораторной культуре, что Halicystis parvula и Derbesia tenuissima 1
являются соответственно гаметофитом и спорофитом одного и того 1
же организма, обнаруживающего таким образом гетероморфную
смену генераций. Зигота, возникающая в результате слияния анизо- я
гамет Н. parvula, вырастает в нити D. tenuissima. Стефаноконтные Я
зооспоры D. tenuissima вырастают в пузыри Н. parvula. Как и в слу- fl
чае голых протопластов Н. parvula, развитие стефаноконтных зоо- Л
спор D. tenuissima, по-видимому, зависит от длины дня. В условиях 9
длинного дня каждая зооспора дает начало пузыревидному тал- 9
лому халицистиса с ризоидами, в условиях короткого дня — только Я
тонкому ризоидальному растению. Кроме того, при культивирова- fl|
нии во взвешенном состоянии зооспоры D. tenuissima давали ризо- Я
идальные нити лишь с местными расширениями. Для получения Я
в культуре растений Н. parvula нормальной формы, не отличимых Я
230
от растений в естественных условиях, зооспоры D. tenuissima нужда-
ются в кальциевых субстратах.
Такая же гетероморфная смена генераций была установлена
и для другого вида Halicystis — Н. ovalis, который представляет
собой гаметофит, чередующийся со спорофитом—D. marina.
Род брнопсис (Bryopsis). В. plumosa широко распространен в Че-
рном море, где растет на небольшой глубине, прикрепляясь к кам-
ням. Растение состоит из ползучего малоразветвленного корневи-
ща, прикрепленного к субстрату ризоидами, от которого вверх
отходят относительно толстые нити, в верхней части несущие пери-
сто расположенные боковые ветви. Эти ветви в свою очередь могут
перисто ветвиться один или два раза, обычно в одной плоскости.
У основания каждого «перышка» имеется заметная перетяжка
и оболочка в этом месте утолщена, но до момента образования
репродуктивных органов настоящих поперечных перегородок не
образуется, и все растение бриопсиса представляет собой одну
громадную клетку с непрерывной вакуолью. У других видов
(В. hypnoides) боковые ветви расположены радиально — вокруг
главной оси.
Размножение. Вегетативное размножение бриопсиса легко осу-
ществляется посредством отделения «перышек», которые закупори-
ваются в месте отделения и могут развивать базальные ризоиды
даже до отделения от материнского таллома. Кроме того, участки
протопласта, вытекающие из таллома при его повреждениях, окру-
гляются и в течение нескольких минут синтезируют новые оболоч-
ки. Затем они развиваются в новые растения бриопсиса. На ранних
стадиях регенерации протопласты могут сливаться. При смешении
протопластов двух разных видов: В. plumosa и В. maxima — были
получены вегетативные гибриды.
Помимо вегетативного размножения, играющего очень боль-
шую роль в распространении растений, у бриопсиса существует
и половое. Половой процесс аиизогамный (гетерогамный): мужские
и женские гаметы различаются по морфологии, окраске, размерам
и поведению. Мужские гаметы мельче женских, они светло-желтые
(хлоропласт редуцирован), без глазка, большую часть клетки зани-
мают митохондрии. Женские гаметы светло-зеленые, имеют нор-
мально развитые хлоропласты и заметный глазок. Мужские гаметы
движутся быстрее женских, которые нередко лишь вращаются вок-
руг собственной оси. Одни виды бриопсиса {В. plumosa,
В. halimeniae) двудомны: мужские и женские гаметы развиваются
в разных гаметангиях, в которые превращаются «перышки». При
этом у основания перышек возникают сложно устроенные попереч-
ные перегородки, хотя иногда они и не образуются, и гаметы могут
формироваться даже в главной оси. Другие виды (В. hypnoides)
однодомны; мужские и женские гаметы развиваются в одном и том
же гаметангии: мужские — у конца гаметангия, а женские — у его
основания. Вышедшие из гаметангиев мужские и женские гаметы
231
попарно копулируют. На процесс оплодотворения в.^тяет темпера-
тура: зиготы образовывались только при 18 и 24е.,, но не при 10 °C.
Зигота немедленно прорастает, давая, однако, не непосредственно
гаметофит (растение бриопсиса), как считали ранее, а нитчатое
малоразветвленное сифоновое растеньице, распростертое по суб-
страту,— протонему. Протонема содержит одно гигантское ядро, по
диаметру значительно (в 8—20 раз) превосходящее ядра гаметофи-
та. Через много недель или месяцев роста протонема может дальше
развиваться двумя способами: или из нее непосредственно вырастает
новая особь бриопсиса— гаметофит (однофазный цикл развития),
или же содержимое протонемы распадается на многочисленные
стефаноконтные (с венцом из многих жгутиков) зооспоры, которые
уже после выхода через разрыв протонемы дают начало талломам
бриопсиса. Это двухфазный цикл развития с гетероморфной сменой
генераций между перисто-разветвленным талломом бриопсиса (га-
метофит) и протонемой (сильно редуцированный спорофит). Оба
типа развития протонемы наблюдаются у одного и того же вида
(В. hypnoides)', при этом известную роль играет географическое про-
исхождение растений: так, в северных водах Европы (Северное,
Балтийское море) встречается только однофазный цикл, а на южных
берегах Атлантики и в Средиземном море наряду с однофазным
циклом наблюдается и двухфазное развитие (рис. 59).
Гетероморфный двухфазный цикл с мейозом, предшествующим
образованию стефаноконтных зооспор в микроскопическом споро-
фите (прот онеме), выявлен и у В. plumosa:
Наконец, еще у одного вида — В. halimeniae зигота вырастает
в разветвленную нитчатую сифоновую водоросль, известную ранее
под названием Derbesia neglecta. В спорангиях, развивающихся на
нитях D. neglecta и отделяющихся от остального таллома перего-
родками, образуются стефаноконтные зооспоры, при прорастании
которых возникают растения В. halimeniae. Таким образом, здесь
также наблюдается двухфазное развитие с чередованием В. hali-
meniae (гаметофита) и D. neglecta (спорофита).
Вопрос о месте мейоза в жизненном цикле бриопсиса остается
спорным. Так, в 70-х годах было высказано мнение, что мейоз
осуществляется не перед образованием гамет, как считалось ранее,
а, по-видимому, перед образованием стефаноконтных зооспор. Од-
нако в 1980 г. в работе, специально посвященной этому вопросу,
утверждается, что у В. plumosa и В. hypnoides мейоз имеет место при
формировании гамет, т. е. гаметофит находится в диплоидной фазе
так же, как и спорофит (микроталлом, протонема). Таким образом,
морфологические фазы жизненного цикла не связаны с ядерными.
Изучение жизненных циклов различных видов выше перечислен-
ных родов {Derbesia, Halicystis, Bryopsis) позволяет расположить их
в ряд, в котором в одном направлении редуцируется спорофит,
а в другом — гаметофит. Исходным типом, занимающим середину
ряда, можно считать почти изоморфную смену генераций между
232
Рис. 59. Схема циклов развития Bryopsis.
А однофазный цикл развития В. hypnoides; Б- -двухфазный цикл развития
В. hypnoides; В — двухфазный цикл развития с чередованием В. halimeniae (гаметофи-
та) и Derbesia neglecta (спорофита):
1 — проросток из зиготы (протонема), 2—протонема, содержимое которой распалось на зооспо-
ры, 3 — стефаноконтпая зооспора, 4— перисторазветвленный таллом, 5 — гаметангий, содер-
жащий мужские (в апикальной части) и женские (в базальной части) гаметы, 6 — мужской
гаметангий, 7—женский гаметангий, 8 —гаметы, 9— зигота, /0—D. neglecta (спорофит), 11—
спорангий
В. halimeniae и D. neglecta. Редукция спорофита наблюдается
у В. plumosa и В. hypnoides, где при двухфазном цикле обнаружива-
ется спорогенная стадия проростка, которая при однофазном раз-
витии редуцируется дальше путем утраты спорообразования, так
что сильно редуцированный спорофит представлен вегетативной
протонемой. Редукция гаметофита наблюдается у D. marina, где
имеется сравнительно малодифференцированный пузыревидный га-
метофит Н. ovalis, а у одной разновидности — D. marina var.
lamouroxii гаметофит вообще отсутствует и размножение проис-
ходит только за счет стефаноконтных зооспор. Существование не-
зависимых видов Derbesia и Bryopsis можно объяснить обособлени-
ем одной фазы первоначального двухфазного цикла путем редукции
или утраты другой фазы. Сходные с бриопсисом жизненные циклы
наблюдались и у другого близкого рода псевдобриопсис
(Pseudobryopsis), который отличается от бриопсиса тем, что гамето-
генез происходит в специализированных гаметангиях, развиваю-
щихся на перышках. Кроме того, стефаноконтные зооспоры псев-
добриопсиса каплевидные или овальные и снабжены длинным
«хвостом» в отличие от сферических с несколько выступающим
передним концом стефаноконтных зооспор бриопсиса и дербезин.
233
Семейство каулерповые (Caulerpaceae). Наиболее сложные
и морфологически дифференцированные талломы встречаются у ка-
улерпы (Сaide гр а), виды которой распространены в тропических
морях, и лишь несколько видов, в том числе и классическая
С. prolifera, живут в Средиземном море. Общий план строения у всех
видов один и тот же: горизонтально стелющееся, более или менее
разветвленное цилиндрическое корневище, которое у видов, обитаю-
щих на песке, может достигать 1 м в длину, снизу несет обильные
разветвленные ризоиды, прикрепляющие растение к субстрату, а на
своей верхней стороне—ряд вертикально сл оящих ассимилирующих
побегов. У простейших видов (С. fastigiata) вертикальный побег
представлен неправильно разветвленными нитями. У других видов
отходящая от корневища цилиндрическая вертикальная ось обычно
образует уплощенные боковые ветки — ассимиляторы. Их форма
и расположение у разных видов варьирует. Так, у С. verticillata
ассимиляторы цилиндрические и располагаются мутовками, прида-
вая растению сходство с хвощами или харой. У С. taxifolia ассимиля-
торы сидят на вертикальной оси в два ряда, так что эти талломы
напоминаю!' побеги тиса. У С. prolifera ассимиляторы плоские,
листообразные, на коротких стебельках, растут прямо из корневища.
На их поверхности обычно образуются вторичные ассимиляторы.
Как и у бриопсиса, у каулерпы полностью отсутствуют попереч-.
ные перегородки, постенная цитоплазма содержит многочисленные
клеточные ядра и хроматофоры, как правило, лишенные пирено-
идов; кроме того, имеются лейкопласты (амилопласты), в которых'
образуется крахмал. Различные части таллома растут своими вер-'
хушками, где скапливается цитоплазма. Каулерпа отличается'
обильным развитием более или менее цилиндрических скелетных,
тяжей — целлюлозных балок, пересекающих центральную полость^
во всех частях таллома. Тяжи эти связаны с оболочкой или свобод-
ны с обоих концов. Каждый тяж одет цитоплазмой. Таким образом,-
помимо увеличения механической прочности клетки, балки заметно)
увеличивают поверхность, выстланную цитоплазмой. Широко рас-?
пространен© вегетативное размножение: при отмирании более ста-’,'
рых частей корневища отдельные участки его с вертикальными
ассимиляторами становятся независимыми растениями. Кроме то-
го, известен половой процесс. Содержимое ассимиляторов скап-
ливается в отдельных плодущих участках, которые не отделяются
от остальных частей таллома перегородкой. Сконцентрированная
в этих участках цитоплазма, включающая многочисленные ядра
и хроматофоры, распадается на одноядерные участки, превраща-
ющиеся в гаметы, которые освобождаются в слизи через специаль-
ные сосочки или бугорки, возникающие в области плодущих участ-
ков ассимиляторов. У большинства видов копулирующие гаметы
несколько различаются по размерам (гетерогамия); перед образова-
нием гамет ядра редукционно делятся, зигота сразу вырастает
в диплоидное растение.
[
Семейство к о дие вне (Codiaceae). Если талломы бриопсиса
и каулерпы можно представить себе как сильно усложненные и диф-
ференцированные нити, то у представителей семейства кодиевых
наблюдается совсем иной тип строения. Здесь макроскопически
крупные талломы возникают в результате плотного переплетения
простых нитей типичного сифонового строения. Так образуются
крупные компактные псевдопаренхиматозные талломы.
Род кодиум (Codium) характеризуется таким строением. Один из
видов этого рода — С. tomenloaum — встречается в Черном море на
глубине 10 м и более. Таллом, прикрепленный к субстрату базаль-
ным диском, имеет вид цилиндрических разветвленных темно-
зеленых шнуров, достигающих длины 50 см при диаметре 8 мм.
Анатомически таллом представлен сердцевиной, состоящей из тон-
ких, идущих в продольном направлении нитей, от которых ответв-
ляются крупные дубинкообразные пузыри, плотно сгруппирован-
ные на одном уровне и образующие периферическую кору. Кор-
тикальные (коровые) пузыри и возникающие на них волоски особен-
но богаты хроматофорами и играют роль ассимиляторов.
Помимо широко распространенного вегетативного размноже-
ния, у кодиума наблюдается половой процесс. Гаметы образуются
в специальных гаметангиях, возникающих как боковые выросты
кортикальных пузырей и отделяющихся от них перегородкой.
У кодиума — гетерогамия. Как у халицистиса и бриопсиса, у ко-
дума мужские и женские гаментангии различаются еще до того, как
образуются гаметы благодаря золотисто-желтой окраске первых
и темно-зеленой окраске вторых.
Как показали недавние цитологические исследования
С. dwarkense, в молодых гаметангиях содержится всего несколько
ядер, которые делятся путем митоза. В зрелых гаметангиях ядра
апикальной области делятся мейотически, а базальной - -митоти-
чески. Часть ядер в центральной части находится в состоянии
интерфазы: по-видимому, они готовятся для вступления в мейоз.
Таким образом, у Codium, как и у Caulerpa, мейоз предшествует
гаметогенезу, и кодиум является диплонтом. Возникающая в ре-
зультате слияния гамет зигота непосредственно прорастает в новое
диплоидное растение. Партеногенетическое развитие женских гамет
(макрогамет), по-видимому, послужило поводом для описания в не-
которых популяциях кодиума бесполого размножения посредством
двужгутиковых монадных клеток.
Порядок дазикладальные — Dasycladales
Порядок объединяет сифоновые водоросли, для которых харак-
терно мутовчатое ветвление таллома, одпоядерность на протяже-
нии вегетативной фазы развития, отложение карбоната кальция
в клеточных стенках, хлоропласты, лишенные пиреноидов. Пред-
ставителем может служить род ацетабулярия (Acetabularia). У аце-
; I1
235
( . табулярии вертикальная ось, часто называемая стебельком и до-
р стирающая 3—5 (у некоторых видов до 18) см вдлину, прикрепля- ;
!' ется к субстрату с помощью лопастного ризоида. На вершине ее j
сначала образуются мутовки стерильных веточек, а в конце цикла i
развития1 возникает плодущая мутовка из гаметангиев. У средизем- i
номорской A, mediterranea гаметангии срастаются своими боками,
образуя шляпки или зонтики. У другого вида A. crenulata, встреча-
ющегося в Карибском море, зонтик состоит из долек (гаметангиев),
имеющих форму банана, радиально расходящихся от верхушки 1
стебелька. Лучи зонтика (шляпки)—гаметангии—сначала сообща- |
ются с полостью стебелька. До этой стадии ацетабулярия не только
одноклеточное, но и одноядерное растение. Одно первичное ги- 1
гантское ядро находится в ризоиде и во время вегетативного роста 1
увеличивается в диаметре —с 3 до 100 мкм, претерпевая более чем |
। тысячекратное увеличение в объеме. Ядрышко, мелкое и округлое I
t в зиготе, во время вегетативного роста также становится крупным I
и колбасообразным. Через несколько недель после формирования |
шляпки первичное ядро уменьшается в размере (от 100 мкм в диа- 1
метре до приблизительно 15 мкм), утрачивает заметное разветвлен- ]
ное ядрышко и в течение 24 ч распадается, отпочковывая большое I
количество мелких вторичных ядер, которые с цитоплазматичес- 1
кими токами перемещаются в гаметангии — лучи зонтика или ’
[ шляпки. Гаметангии отделяются от стебелька пробкой, и их много- 1
ядерное содержимое распадается на одноядерные цисты. В цистах, I
, освобождающихся из гаметангиев, позднее после ряда ядерных ?
делений формируются двужгутиковые изогаметы. Цисты вскрыва-
ются крышечкой, через которую выходят гаметы, зигота развивает- I
ся в диплоидное растение. |
До сравнительно недавнего времени оставался спорным вопрос 1
о том, когда в жизненном цикле ацетабулярии происходит мейоз: до I
или после формирования цист1 2. Были получены данные, что мейоз I
происходит после формирования цист3. Если первоначальное един- j
ственное ядро цисты диплоидно и претерпевает мейоз внутри ци-
сты, должны продуцироваться гаметы обоих типов спаривания, j
и гаметы из одной и той же цисты должны были бы копулировать. I
Между тем наблюдения показали, что гаметы из одной цисты не
копулируют, будучи, по-видимому, все одного и того же типа I
спаривания. Для подтверждения этих наблюдений сравнительно
недавно был предложен простой способ. В пробирки с морской
водой помещалось по 1, 2, 4, 12 и 30 цист. В трех отдельных
экспериментах, проведенных над A. mediterranea, из 240 пробирок,
1 В природе никл развития растягивается на несколько (2- 3) лет, в лаборато-
рии он может быть сокращен до 6 месяцев.
2 Первичное ядро ацетабулярии не окрашивается ядерными красителями, не
дает реакции Фельгена.
3 Долгое время считалось, что мейоз происходит в цистах перед образованием
изогамет.
236 '.
содержащих по одной цисте, все (кроме одной — возможна ошибка
эксперимента) не содержали проростков. В пробирках с двумя
цистами, где, возможно, присутствовали цисты противоположных
типов спаривания, 11 из 40 содержали проростки. Пробирки, содер-
жащие более двух цист, пропорционально продуцировали больше
проростков. Эти данные можно объяснить только исходя из того,
что мейоз, сопровождаемый сегрегацией — разъединением половых
локусов, происходит до обособления единственного вторичного
ядра цисты. Примененный метод количественной микроспектрофо-
тометрии также продемонстрировал в ядре цисты то же количество
ДНК, что и в ядре гамет, и половину количества ДНК, наблюда-
емого в ядре зиготы. Это подтверждает заключение, что мейоз
имеет место до образования цист, возможно, во время распада
первичного ядра.
То, что ацетабулярия на протяжении большей части жизненного
цикла — до наступления размножения представляет собой не только
одноклеточный, но и одноядерный организм (причем ядро локали-
зовано в строго определенном, легко распознаваемом месте (в
ризоиде), сделало ее идеальным объектом для изучения взаимоот-
ношений между ядром в живой клетке и окружающей его цитоплаз-
мой. Этому благоприятствовали также способность к регенерации
(место пореза быстро затягивается, и клетка начинает регенериро-
вать недостающую часть) и исключительная живучесть гигантского
ядра ацетабулярии. Его можно извлечь из клетки, отмыть в рас-
творе сахарозы и снова имплантировать в клетку. Для получения
безъядерной (энуклеированной) клетки и прослеживания ее даль-
нейшей судьбы у ацетабулярии достаточно отрезать ризоид. Ядра
(в различных количествах) могут пересаживаться из клетки одного
вида в энуклеированный ризоид другого вида и после приживления
его к стебельку можно проследить влияние ядра на характер раз-
вивающейся шляпки и т. п. Ацетабулярия, как и другие сифоновые
водоросли,— излюбленный объект изучения движения цитоплазмы.
Порядок сифоиокладальные—Siphonocladales
Сюда относятся водоросли, имеющие сифонокладальную струк-
туру. Порядок включает ряд семейств.
Семейство кладофоровые— Cladophoraceae. Представи-
телями могут служить роды кладофора (Cladophora) и ризоклониум
(Rhizoclonium).
Род кладофора {Cladophora), виды которой распространены как
в море, так и в пресных водах, имеет нитчатый, обильно разветв-
ленный таллом, состоящий из вытянутых цилиндрических сегмен-
тов. Под толстой и слоистой не ослизняющейся оболочкой находит-
ся стенкоположная цитоплазма, содержащая многочисленные кле-
точные ядра и много мелких дискретных хлоропластов, располо-
женных таким образом, что при рассматривании в оптическом
237
микроскопе производят впечатление сети. Таллом*! кладофоры,
первоначально прикрепленные к субстрату, позднее могут отры-
ваться и, разрастаясь, образовывать скопления «тины» грязно-зеле-
ного цвета и не слизистые на отцупь. Бесполое размножение осуще-
ствляется зооспорами, образующимися в большом количестве в сег-
ментах, по форме неотличимых от обычных вегетативных. Зооспо-
ры двух- или четырехжгутиковые, выходят через боковое отверстие
в оболочке. Половой процесс изогамный, гаметы двужгутиковые,
мельче зооспор. У ряда морских видов кладофоры установлена
изоморфная смена генераций.
Род ризоклониум (Rhizoclonium) отличается от кладофоры почти
полным отсутствием ветвления.
Семейство сфероплеевые (Sphaeropleaceae). Встречающа-
яся ранней весной в мелких водоемах сфероплея (Sphaeroplea) имеет
нитчатый неразветвленный таллом, с самого начала неприкреплен-
ный. Нить состоит из очень сильно вытянутых цилиндрических
сегментов, содержащих большое количество (до 70) париетальных Д
хроматофоров в виде колец с гладкими или изрезанными краями;
несколько пиреноидов располагаются в них в один ряд. Каждый
кольчатый хроматофор занимает периферию цитоплазматического
диска, тянущегося поперек сегмента. Эти диски делят центральную
полость сегмента на ряд крупных вакуолей, чередующихся с хрома-
тофорами. Каждый диск обычно содержит немного ядер — два,
иногда одно. Таким образом, весь сегмент многоядерный.
Бесполого размножения у сфероплеи нет. Помимо фрагмента-
ции нити, наблюдается половой процесс, к которому сфероплея
легко переходит при понижении в среде содержания азота. Половой
процесс оогамный. В отличие от других оогамных нитчатых зеле-
ных водорослей сегменты сфероплеи, превращающиеся в оогонии
и антеридии, не изменяют своей формы. У Sph. annulina нити обое-
полые. В оогониях рано нарушается кольчатое расположение хро-
матофоров, вся цитоплазма постепенно распадается на многочис-
ленные сферические яйцеклетки с темно-зеленым содержимым. Во 1
время образования яйцеклеток (одноядерных) ядра в оогонии не
делятся. В антеридиях кольчатое расположение хроматофоров со-
храняется до поздних стадий. Хроматофоры желтеют, ядра много-
кратно делятся, цитоплазма раскалывается на несколько тысяч а
одноядерных участков, которые превращаются в вытянутые дву- ЯВ
жгутиковые сперматозоиды желтого цвета. Сперматозоиды выхо-
дят через небольшие отверстия в боковых стенках антеридия. Такие ,^В
же отверстия возникают и в стенках оогония: через них в воду вВ
диффундируют половые вещества, хемотаксически привлекающие
сперматозоиды (см. гл. 4). После слияния со сперматозоидом каж-
дая яйцеклетка одевается гладкой бесцветной тонкой первичной
оболочкой, под которой позднее вырабатывается более толстая ^В
вторичная оболочка с неровными звездчатыми контурами; под ее Я
давлением первичная оболочка разрывается и сбрасывается. В эре- Я
238 1
лых оогониях можно наблюдать в больших количествах такие
пустые оболочки, лежащие вдоль зрелых ооспор. Содержимое оос-
пор окрашено в кирпично-красный цвет. Ооспоры в течение лета
переносят высыхание, зимуют и весной прорастают (по-видимому,
после редукционного деления ядра) с образованием, как правило,
четырех двужгутиковых зооспор, которые еще во время движения,
не достигнув субстрата, выделяют оболочку. После остановки зоо-
споры вытягиваются с образованием веретеновидного проростка,
вырастающего в двух противоположных направлениях в нить, с са-
мого начала не прикрепленную. У других видов сфероплеи половой
процесс анизогамный.
Семейство валониевые (Valoniaceae) характеризуется тем,
что у его представителей первичный многоядерный сифоновый
пузырь с возрастом становится многоклеточным образованием.
Сюда относится распространенный в тропических и субтропических
морях род валонии (Valonia). Молодые растения валонии представ-
лены микроскопическим пузырем обычного сифонового строения,
прикрепленным к субстрату несептированными ризоидами.
В любом месте от поверхности пузыря перегородкой в виде
часового стекла отчленяются маленькие линзовидные клетки, осо-
бенно мелкие у основания пузыря. Эти так называемые маргиналь-
ные (краевые) клетки у основания пузыря вырастают во вторичные
ризоиды, часто с лопастными окончаниями. У многих видов неко-
торые из более крупных маргинальных клеток на верхушке пузыря
вырастают в ветви, которые повторяют строение первичного пузы-
ря и могут дать начало пузырям третьего порядка. Бесполое раз-
множение— зооспорами, половой процесс— гетерогамия; редукци-
онное деление происходит перед образованием гамет.
Некоторые виды валонии, у которых большая часть таллома
представлена пузырем, достигающим нескольких сантиметров в диа-
метре и содержащим громадную вакуоль с клеточным соком, являют-
ся излюбленными объектами для изучения клеточной проницаемости,
аккумуляции веществ в клетке, а также биоэлектрических явлений.
КЛАСС ПРАЗИНОФИЦИЕВЫЕ — PRASINOPHYCEAE
Класс объединяет подвижные—монадные и неподвижные — ка-
псальные и коккоидные — формы, одиночные или образующие ко-
лонии. Монадные клетки на переднем конце имеют углубление, из
которого выступают жгутики в числе 2—4 (редко 8). Как клетки,
так и жгутики одеты покровом из органических чешуек.
Порядок пирамимоиадальные—Pyramimonadales
Порядок представлен монадными формами. В качестве примера
можно рассмотреть род пирамимонас — Pyramimonas, виды которо-
го встречаются как в морях, так и в пресных водах. Клетка имеет
239
обратнопирамидальную, обратногрушевидную иЛ'и овальную
форму, в поперечном сечении (по крайней мере й передней части)
квадратную. Поверх клеточной мембраны клетки покрыты несколь-
кими слоями (2—5) чешуек с характерной для каждого вида струк- ли
турой. Например, у Р, grossii три слоя чешуек: внутренние — мелкие,
в виде коробочки с ободком, средний слой из чешуек в виде
плоских с низенькими ободками перфорированных квадратов, че-
шуйки наружного слоя — сложные ажурные образования (рис. 60, Б,
НЧЖ 8ЧЖ
Рис. 60. Pyraminionas. А — схема строения клетки; Б—Р. grossii: I — 3—чешуйки
тела (I — внутренняя, 2 — средняя, 3— наружная), 4 — 5 — чешуйки жгутика, внутрен-
няя (4) и наружная (5):
нчт наружная чешуйка тела, вчт внутренняя чешуйка тела, ечт—средняя чешуйка тела,
ж жгутик, вчж — внутренняя чешуйка жгу,ика, нчж — наружная чешуйка жгутика, бт базаль-
ные тела, жк- жгутиковый корень, рч — резервуар чешуек, д—диктиосома, я—ядро, х—хло-
ропласт, и — пиреноид, с — стигма, м — митохондрии, т — трихоциста
)
240
1—5). Жгутики в числе четырех выходят из углубления на переднем
конце клетки и покрыты двумя слоями чешуек: внутренние пяти-
угольные, расположенные тесными спиральными рядами, поверх
них — наружный слой, образованный плоскими чешуйками, несущи-
ми шип, который тянется вдоль чешуйки и выступает от пластинки
к дистальному концу жгута (рис. 60, А, Б, 4, 5). Помимо чешуек
жгутики несут волосовидные отростки. Чешуйки всех типов и во-
лоски формируются в цистернах аппарата Гольджи, и в пузырьках
Гольджи транспортируются к специальному резервуару чешуек,
который открывается протоком в жгутиковое углубление. Цистер-
ны с чешуйками могут и непосредственно сливаться с плазмалем-
мой, выстилающей жгутиковое углубление, освобождая чешуйки на
поверхность клетки. Основания четырех жгутиков связаны попереч-
но-исчерченной структурой и соединены с пиреноидом посредством
ризопласта. Ядро занимает в клетке боковое эксцентричное положе-
ние. Хлоропласт разделяется продольно на 4, 8 или 12 лопастей,
которые простираются почти до его основания. В задней части
хлоропласта расположен пиреноид, пересеченный одиночными ти-
лакоидами и окруженный крахмальными зернами. Ламеллы состо-
ят из 2—5 тилакоидов, стигма—из 2—4 рядов липидных гранул.
Трихоцисты извергаются из клеток в форме длинных нитей. Раз-
множение— путем деления клетки, причем деление как хлоропла-
ста, так и цитокинез начинается с заднего конца клетки. Наблюдал-
ся и половой процесс — гологамия. Зигота прорастает четырьмя
зооспорами, сходными с клеткой Pyramimonas, но сначала меньших
размеров.
Порядок галосферальные — Halosphaerales
Порядок включает коккоидные формы. Сюда относится широко
распространенный в морском планктоне род галосфера
(Halosphaera). В жизненном цикле этой водоросли можно различить
неподвижную фазу— вегетативные шаровидные клетки, одетые
оболочкой, часто трактуемые как цисты, и подвижную фазу в виде
четырехжгутиковых монадных клеток, похожих на пирамимонас,
способных размножаться путем деления. Вегетативные клетки (ци-
сты) заметно увеличиваются в размерах, пока не достигнут зре-
лости. При этом оболочка, пектиновая внутри и, возможно, спо-
рополлениновая снаружи, растягивается по мере возрастания объ-
ема клетки и сбрасывается лишь при освобождении подвижных
монадных клеток. Клетка содержит многочисленные хлоропласты
(каждый с пиреноидом, окруженным крахмальной сферой) и в мо-
лодох£состоянии — одно клеточное ядро. Когда прекращается рост
клетки (цисты), начинается деление ядра; содержимое делится на
мелкие округлые или овальные диски, каждый со своим ядром,
располагающиеся под внутренней поверхностью оболочки. Эти дйс-
ки, называемые «розетками», число (у Н. viridis образуется до 512
241
«розеток»), размеры и внешний вид которых неодинаковы у разных
видов, при дальнейшем делении дают подвижную* фазу. Освобо-
ждение монадных клеток происходит следующим' образом: наруж-
ная оболочка клетки (цисты) разрывается щелью, через которую
выходит содержимое, окруженное внутренним слоем оболочки; этот
слой сильно растягивается, превосходя в несколько раз диаметр
первоначальной наружной оболочки. Внутри растянувшейся вну-
тренней оболочки монадные клетки продолжают делиться, так что
число их возрастает и в конце концов они освобождаются при
разрыве внутренней оболочки. После периода движения, во время
которого возможно размножение путем деления, монадные клетки
округляются, теряют жгутики и превращаются в неподвижные,
одетые оболочкой клетки (цисты).
КЛАСС КОНЪЮГАТОФИЦИЕВЫЕ—CONJUGATOPHYCEAE
Для относящихся сюда одноклеточных и нитчатых зеленых во-
дорослей характерно полное отсутствие в жизненном цикле жгути-
ковых стадий, так как бесполого размножения с помощью зооспор
у них нет, а половой процесс — конъюгация — заключается в слия-
нии амебоидных протопластов обычных вегетативных клеток. По-
мимо конъюгации, широко распространено вегетативное размноже-
ние. Для подавляющего большинства конъюгат характерен цент-
ральный осевой хроматофор; париетальный хроматофор встречает-
ся сравнительно редко. Пресноводные формы; некоторые живут на
почве и в солоноватых водах.
Класс включает четыре порядка, из которых главнейшие: мезо-
телиальные (Mesotaeniales), зигнемальные (Zygnematales) и десмиди-
альные (Desmidiales). Для представителей первых двух порядков
характерна цельная сплошная оболочка без пор, у десмидальных
клеточная оболочка состоит из двух или более частей и пронизана
сложно устроенными порами.
Порядок мезотениальные — Mesotaeniales
Одноклеточные формы. Клетки обычно цилиндрические, с за-
кругленными концами, в поперечном сечении всегда округлые.
Клетка одета сплошной гладкой оболочкой, поверх которой имеет-
ся слизистая обвертка. Одноядерные клетки содержат хромагофо-
ры, различающиеся по форме у разных родов. У рода мезотениум
(Mesotaenium) хроматофор в виде осевой пластинки с одним или
несколькими пиреноидами: в зависимости от положения клетки
хроматофор можно видеть или с его широкой стороны, или в про-
филь.
У рода спиротения (Spirotaenid) хроматофоры париетальные,
в виде спиральных лент с пиреноидами. У родов цилиидроцистнс
(Cylindrocystis) и нетриум (Netriutri) в клетке имеются по два цент-
242
ральных, в поперечном сечении звездчатых хроматофоров. состо-
ящих из массивной центральной части, содержащей пиреноид,
и расходящихся по радиусам пластинок (у нетриума сильно изрезан-
ных по краям).
Размножение вегетативное путем деления клеток в поперечной
плоскости. Половой процесс- -конъюгация1. Конъюгирующие клет-
ки, обычно погруженные в общую слизь, располагаются своими
длинными осями или параллельно друг другу, или под прямым
углом и в месте контакта образуют навстречу друг другу отростки,
которые по мере удлинения несколько раздвигают конъюгирующие
индивиды. При растворении в месте стыка отростков оболочек
возникает конъюгационный канал, внутри которого. встречаются
и сливаются протопласты конъюгирующих клеток. Зигота одевает-
ся толстой многослойной оболочкой и переходит в состояние покоя.
При прорастании зиготы копуляционное диплоидное ядро претер-
певает майоз. Как правило, все четыре гаплоидных ядра остаются
жизнеспособными и возникают четыре новых индивида, освобожда-
ющиеся через разрыв в оболочке зиготы. Однако у нетриума из
четырех гаплоидных ядер, как правило, остаются жизнеспособными
только два и соответственно при прорастании зиготы образуются
только два, реже четыре проростка.
Мезотениальные распространены в торфяных болотах, некото-
рые виды мезотениума и цилиндроцистиса встречаются на сырой
земле, образуя слизистые скопления.
Порядок зигнемальные—Zygnematales
Сюда относятся самые обычные нитчатые пресноводные водо-
росли. Все они, как правило, неветвящиеся, состоят из одного
ряда цилиндрических клеток, одетых цельной оболочкой без пор
и слизистым чехлом. Благодаря чехлу нити и их скопления на
ощупь слизистые в отличие от скоплений (тины), образованных
другими нитчатками, например кладофорой. Центр клетки занят
крупной вакуолью с клеточным соком, цитоплазма занимает по-
степенное положение. Хроматофоры тех же типов, что и у ме-
зотениальных. У рода мужоция (Mougeotia) хроматофор в виде
плоской осевой пластинки с несколькими пиреноидами. Пластинка
своей широкой стороной может быть повернута к свету, если же
освещение чрезмерно, она поворачивается на 90 и встает к свету
в профиль. Такой поворот занимает около 30 мин. Два осевых
звездчатых хроматофора, каждый с крупным центральным пире-
ноидом, наблюдаются у рода зигнема (Zygnema). Хроматофоры
в виде одной или многих лент, расположенных в постенной ци-
топлазме и опоясывающих клетку по спирали, характерны для
' У одних разновидностей N. digitus наблюдается гомоталлизм, у других -гете-
роталлизм.
243
—. 11
наиболее широко распространенного рода спирогира (Spirogyra).
Лентовидные хроматофоры спирогиры обычно имеют неровные
городчатые края и у многих видов вдоль ленты посередине про-
ходит гребень, вдающийся внутрь клетки, темная полоса. По
средней линии хроматофора расположены пиреноиды, окруженные
крахмальными зернами. Одно ядро всегда находится в центре клет-
ки: у мужоции оно прижато к одной стороне хроматофора. у зи-
гнемы заключено между двумя хроматофорами, а у спирогиры
ядро, окруженное слоем цитоплазмы, подвешено на цитоплазма-
тических тяжах, отходящих от постенной цитоплазмы, в центре
вакуоли с клеточным соком. Любая клетка нити способна к росту
и делению. За делением ядра следует образование поперечной пе-
регородки, разрезающей у спирогиры хроматофоры пополам; у му-
жоции и зигнемы деление хроматофоров предшествует делению
протопласта.
Размножение. Вегетативное размножение осуществляется благо-
даря разрыву нитей (в результате отмирания промежуточных кле-
ток) на отдельные фрагменты, даже отдельные клетки могут выра-
сти в новые нити.
Половой процесс — конъюгация. Наиболее обычна так называ-
емая лестничная конъюгация, которая происходит между клетками
двух нитей. Нити располагаются параллельно друг другу и вначале
тесно склеиваются слизью. Затем противолежащие клетки навстре-
чу друг другу образуют выросты, которые соприкасаются и сраста-
ются своими концами. Удлиняясь, выросты постепенно раздвигают
нити, так что возникает фигура в виде лестницы.
Стенки на соприкасающихся концах отростков растворяются
и возникает узкий канал, соединяющий полости конъюгирующих
клеток. У всех видов спирогиры и у многих видов зигнемы прото-
пласт одной из конъюгирующих клеток (отдающей) сокращается,
отстает от стенок и постепенно проталкивается через канал в дру-
гую (воспринимающую) клетку, где и сливается с ее содержимым,
образуя зиготу. Сокращение протопласта обусловлено, по-видимо-
му, тем, что жидкость из центральной вакуоли диффундирует в по-
являющиеся в цитоплазме многочисленные пульсирующие вакуоли,
которые, в свою очередь, выпрыскивают жидкость в пространство
между протопластом и клеточной оболочкой.
Появление сократительных вакуолей и отставание протопласта
от стенки наблюдается и в клетке воспринимающей, но значительно
позднее, чем в клетке, отдающей свое содержимое. Помимо лест-
ничной, у тех же видов можно наблюдать иной тип конъюгации —
боковой. При боковой конъюгации отростки, а затем канал воз-
никают между соседними клетками одной и той же нити.
Поскольку поведение'кднъюгирующих клеток у спирогир и мно-
гих зигнем несколько различно (клетки воспринимающие, пассив-
ные можно назвать женскими, а клетки отдающие, активные —
мужскими), половой процесс в этих случаях можно определить как
244
физиологически анизогамный, хотя морфологически он изогамен,
так как конъюгирующие клетки морфологически одинаковы. У дру-
гих видов зигнем и у всех видов мужоций слияние протопластов
и образование зиготы происходят в конъюгационном канале, как
у мезо! ениальных. Здесь наблюдается физиологическая изогамия,
которая, как правило, сопровождается и морфологической изогами-
ей. Описан лишь один вид мужоции, у которого конъюгирующие
клетки и нити различались морфологически, т. е. наблюдалась мор-
фологическая анизогамия, и у этого вида зигота возникала всегда
в конъюгационном канале.
Образовавшаяся в результате слияния протопластов конъюгиру-
ющих клеток зигота округляется, выделяет толстую трехслойпую
оболочку и переходит в состояние покоя. В молодой зиготе еще
можно различить хроматофоры обоих слившихся протопластов;
позднее, в случае физиологической анизогамии, мужские хромато-
форы разрушаются и остаются одни женские, при физиологической
изогамии сохраняются все хроматофоры. Ядра сливаются незадол-
го до прорастания зиготы. При прорастании зигот зигнемальных,
подробно изученном у многих видов зигнем русским ботаником
Л. И. Курсановым, происходит редукционное деление, причем из
четырех гаплоидных ядер остается жизнеспособным только одно
и соответственно развивается только один проросток.
Порядок десмнднальные — Desmidiales
Порядок, насчитывающий до нескольких тысяч видов, включает
одноклеточные, реже нитчатые водоросли, для которых характерны
клетки, состоящие из двух симметричных половинок — полуклеток.
У большинства родов в плоскости симметрии клетки наблюдается
более или менее заметная перетяжка, исключение составляют род
клостериум и некоторые виды рода пениум, у которых перетяжка
отсутствует. Полуклетки могут быть самой разнообразной формы:
цилиндрическими (с заостренными у клостериума, закругленными
у пениума или срезанными у плеуротениума концами), а также
уплощенными (космариум, эуаструм) или даже дисковидными (мик-
растериас). В разных положениях клетки выглядят неодинаково.
Полуклетки могут иметь более или менее глубокие надрезы, деля-
щие их на ряд симметрично расположенных лопастей, углы их
могут нести различные выросты.
Клеточная стенка десмидиальных всегда состоит из двух поло-
винок, имеющих скошенные края, плотно находящие друг на друга
в плоскости симметрии клетки. При рассматривании в оптический
микроскоп оболочка двухслойная, наружный слой часто импрег-
нирован соединениями железа, желтоватого цвета; он может нести
шипы, бородавочки и другие выступы, определенным образом рас-
положенные. В оболочке находятся сложно устроенные поры, часто
группирующиеся вокруг основания шипов, бородавок и других
• 143
выступов оболочки. Через поры выделяется слизь. Слабое движе-
ние, которое часто наблюдается у десмидиальных,-контактирующих
с субстратом, обусловлено местной секрецией слизи через специаль-
ные, особенно крупные поры, расположенные на концах клетки.
В течение часа в этих местах могут выделиться неправильные
червеобразные массы слизи, в 2 — 3 раза превышающие длину
индивида.
Одно ядро находится в цитоплазме в области перешейка, соеди-
няющего две полуклетки. В большинстве случаев хроматофоры
осевые, по одному в каждой клетке, хотя у многих видов космари-
ума и некоторых других родов в каждой полуклетке может быть по
два хроматофора. Такие осевые хлоропласты состоят из централь-
ной-- осевой части, имеющих у удлиненных форм, например у кло-
стериума, палочковидную форму. От осевой части хлоропласта
радиально расходится ряд пластинок или отростков. Пиреноиды
обычно располагаются в центральной части хлоропласта. Реже
хлоропласты постенные — париетальные; по-видимому, они про-
изошли от центральных при расширении концов радиально рас-
ходящихся отростков в париетальные лопастные пластинки и при
редукции осевой части хлоропласта. У форм с массивными осевыми
хлоропластами вакуоли с клеточным соком небольших размеров.
У клостериума и плеуротениума на концах клетки имеются тер-
минальные вакуоли, содержащие кристаллы гипса. Последние об-
наруживают броуновское движение.
Размножение вегетативное — делением клетки в плоскости сим-
метрии, половой процесс — конъюгация. При делении дочерние ин-
дивиды получают одну пол у клетку от материнской клетки, а недо-
стающую достраивают заново.
При конъюгации две клетки сближаются1 и одеваются общей
слизью. У некоторых,, например у клостериума, в плоскости сим-
метрии от каждой конъюгирующей клетки возникают отростки,
которые соединяются с образованием конъюгационного канала,
внутри которого протопласты конъюгирующих клеток сливаются.
У многих десмидиальных половинки конъюгирующих клеток рас-
ходятся в области перетяжки, протопласты освобождаются и слива-
ются (конъюгационного канала здесь не образуется). У некоторых
видов (Closterium lineatum и др.) протопласт каждой из конъюгиру-
ющих клеток предварительно делится с образованием двух гамет,
которые освобождаются и попарно сливаются. Таким образом
возникают «двойные» зиготы. Зрелые зиготы обычно одеты трех-
слойной оболочкой. При прорастании зиготы копуляционное ядро
редукционно делится, и из четырех гаплоидных ядер, как правило,
два жизнеспособны; поэтому для десмидиальных характерно об-
разование двух проростков, хотя есть и исключения (у гиалотеки
только один проросток).
1 Одни виды гомоталличны, другие — гетероталличны.
246
Примерами нитчатых десмидиальных, возникающих в резуль-
тате того, что дочерние клетки при делении не разъединяются,
а остаются плотно соединенными, могут служить роды десмидиум
(Desmidium) и гиалотека (Hyalolheca). Клетки, как и у одноклеточ-
ных десмидиальных, состоят из двух симметричных полуклеток,
в плоскости симметрии — неглубокая перетяжка. Нити обычно
скручены по спирали и одеты более или менее мощным слизистым
чехлом. У D. swartzii клетки при рассматривании сверху треуголь-
ные и соответственно нить имеет форму трехгранной призмы. По-
скольку нити скручены, выдающиеся ребра клеток у D. swartzii
выглядят темно-зеленой полосой, идущей косо вдоль нити.
У гиалотеки (Hyalotheca) нить цилиндрическая, и клетки при
рассматривании сверху округлые. Клетки гиалотеки отличаются
очень слабой перетяжкой, нить окружена мощным слизистым чех-
лом. У большинства нитчатых форм до конъюгации пить распада-
ется на отдельные клетки, однако у D. swartzii конъюгируют клетки,
соединенные в нить. При конъюгации образуется конъюгационный
канал. Десмидиевые особенно распространены в торфяных болотах.
КЛАСС ХАРОФИЦИЕВЫЕ —CHAROPHYCEAE
Класс харофициевых водорослей представлен всего одним по-
рядком Charales с тремя семействами, два из которых объединяют
только ископаемые формы, и лишь одно семейство Characeae наря-
ду с ископаемыми включает и ныне живущие шесть родов. Наибо-
лее обычны роды хара (Chara) и нителла (Nitella).
От других зеленых водорослей они отличаются сложно устроен-
ными половыми органами и расчленением своего нитчатого тал-
лома.
Нитчатые талломы харовых, всегда вертикально стоящие, до-
стигают высоты около 20 — 30 см, иногда и метра, и обнаруживают
мутовчатое расположение ветвей ограниченного роста, что придает
им сходство с хвощами. Как оси, неограниченно нарастающие
(«стебли»), так и боковые ветви ограниченного роста («листья»)
дифференцированы на узлы и междоузлия. Первые возникают из
одной клетки в результате деления ее продольной стенкой и затем
отчленения продольными перегородками ряда (6 - 8) периферичес-
ких клеток от двух центральных. От периферических клеток узла
«стебля» отделяются верхушечные клетки боковых ветвей ограни-
ченного роста — «листьев», благодаря этому расположенных всегда
мутовками. Узлы чередуются с междоузлиями, представленными
у нителлы одной вытянутой (у самых крупных видов до 25 см)
клеткой, хары клетка междоузлия снаружи одета специальной
корой.
Рост строго верхушечный: куполообразная апикальная клетка
отчленяет перегородкой, параллельной ее плоскому основанию,
сегменты. Каждый сегмент делится на двояковогнутую верхнюю
247
клетку и двояковыпуклую нижнюю клетку. Двояковогнутая клетка
дает начало узлу: она делится пополам продольной перегородкой,
после чего ряд изогнутых перегородок отделяет серию перифери-
ческих клеток от двух центральных, позднее иногда претерпева-
ющих еще несколько делений. Двояковыпуклые клетки не способны
к делению, они только растут в длину, образуя междоузлия. Боко-
вые ветви, или «листья», обнаруживают тот же способ роста, что
и главная ось, но клетки междоузлиев обычно остаются довольно
короткими, а апикальная клетка сравнительно рано прекращает
делиться, принимая вытянутую заостренную форму. От перифери-
ческих клеток узлов, расположенных у основания «листьев», воз-
никают ветви неограниченного роста; таким образом «стебель»
ветвится. Базальные или основные узлы «листьев» служат также
местом возникновения характерной коры, свойственной большинст-
ву видов хары. Из некоторых клеток каждого базального узла
возникают две нити: одна из них растет вверх, другая — вниз. Эти
коровые нити плотно прилегают к клетке междоузлия и растут с той
же скоростью, что и клетка междоузлия; таким образом, междоуз-
лие с самого начала покрыто корой. Растущая вверх нить одного
узла встречается с нитью, растущей вниз от вышерасположенного
узла. Нити коры, одевающие стеблевые междоузлия, имеют то же
строение и тот же способ роста, что и другие ветви растения. У них
также наблюдаются апикальный рост и сегментация на узлы и меж-
доузлия. Первые делятся двумя радиальными стенками на медиан-
ную и две латеральные. Медианная клетка узлов коровых нитей
делится тангентальной стенкой на более мелкую внутреннюю и бо-
лее крупную наружную клетку, которая может дать либо только
маленький выступ, либо развиться в шип или даже поделиться на
ряд клеток (эти образования сравнимы с боковыми ветвями ограни-
ченного роста). У многих видов хары «листья» также одеты корой,
нити которой, однако, не дифференцированы на узлы и междоузлия.
Прикрепляются талломы посредством многоклеточных разветв-
ленных ризоидов, возникающих из периферических клеток нижнего
узла главной оси.
Строение клетки. Клетки одеты целлюлозной оболочкой, в на-
ружных слоях которой откладывается карбонат кальция. В стен-
коположной цитоплазме находятся многочисленные мелкие диско-
видные хроматофоры, лишенные пиреноидов. В удлиненных клет-
ках междоузлий хроматофоры расположены хорошо заметными
продольными рядами в самом наружном слое цитоплазмы.
Более глубокий слой цитоплазмы, граничащий с громадной
вакуолью клеточного сока, обнаруживает интенсивное движение,
В каждой клетке существует восходящий ток цитоплазмы, с одной
стороны клетки, и нисходящий — с другой. Линия раздела заметна
по бокам клетки по бесцветной полосе, где нарушено плотное
расположение хроматофоров и где оболочка образует гребень, вда-
ющийся внутрь клетки. Все клетки, способные к делению (т. е.
248
апикальные, сегменты и клетки узлов), одноядерные, ядра делятся
митотически. Не способные к делению клетки междоузлиев во
взрослом состоянии многоядерные, содержат крупные лопастной
формы ядра, делящиеся амитотически.
Размножение. Бесполого размножения у харовых tier. Вегетатив-
ное размножение—с помощью клубеньков, возникающих из ниж-
них стеблевых узлов, половой процесс оогамный. Половые органы
имеют весьма сложное строение. Как правило, оогонии и антеридии
развиваются в непосредственной близости друг от друга, однако
имеются и двудомные виды. Оба вида половых органов возникают
на вторичных боковых ветвях ограниченного роста, образующихся
из верхних узлов «листьев». При возникновении антеридия боковая
ветвь формирует базальный узел, после чего апикальная клетка
делится с образованием антеридия. Апикальная клетка боковой
ветви, продуцирующая антеридий, отчленяет у своего основания
одну или две дисковидные клетки, принимает сферическую форму
и претерпевает деления двумя продольными стенками и одной
поперечной с образованием восьми клеток — октантов. Затем в каж-
дом октанте последовательно возникают по две параллельные стен-
ки, так что получается восемь диагональных рядов, каждый из трех
клеток. Восемь периферических клеток обнаруживают плоскостной
рост и развиваются в плоские изогнутые клетки — щитки с выпуклой
стороной, обращенной кнаружи. Их оболочки несут радиально рас-
ходящиеся складки, а содержимое в зрелом состоянии оранжево-
красного цвета. Они образуют стенку шаровидного антеридия, кото-
рая у основания замыкается одной из дисковидных клеток, отчлени-
вшейся вначале и обычно вдающейся в полость антеридия. Средние
клетки значительно удлиняются в радиальном направлении, каждая
образует клетку — рукоятку, которая таким образом отходит от
центра щитка и на внутреннем конце несет шаровидную клетку —
головку, образовавшуюся из самой внутренней клетки первоначаль-
ного диагонального ряда. Клетки головок (каждая) дают начало
шести вторичным головкам, из которых развиваются длинные нити,
обычно раздвоенные у основания. Эти сперматогенные нити запол-
няют полость антеридия в виде плотного клубка. Каждая спермато-
генная нить состоит из 100 — 200 дисковидных клеток, в которых
образуется по одному спирально изогнутому двужгутиковому1 спер-
матозоиду. Сперматозоиды освобождаются при расхождении щит-
ков антеридиальной стенки и ослизнении стенок материнских кле-
ток. Клетка базального узла антеридия на стороне, обращенной
к оси, у однодомных форм хары дает начало оогонию. Таким
образом, у хары оогоний направлен вверх, а антеридий — вниз.
У однодомных видов нителлы оогоний образуется под антеридием.
Клетка базального узла антеридия, которая развивается в оого-
ний, претерпевает два поперечных деления с образованием ряда из
' Жгутики на своей поверхности несут чешуйки.
249
трех клеток. Самая верхняя из них дает оогоний, <при этом она
сильно увеличивается. Самая нижняя дает одноклеточную ножку,
а средняя образует узел, который делится на центральную и пять
периферических клеток. Эти клетки вырастают в пять коровых
нитей, которые уже на ранних стадиях одевают оогоний и вскоре
отчленяют на своих концах одну (у хары) или две (у нителлы)
клетки коронки. При дальнейшем развитии нити, не делясь, удлиня-
ются и принимают спиральное расположение. Клетки коронки оста-
ются мелкими и вертикально стоящими. В оогонии образуется одно
яйцо. Под коронкой, клетки которой плотно соединены, нити слегка
разъединяются, так что образуется щель; через нее проникает спер-
матозоид. Верхушка оогониальной стенки ослизняется и спермато-
зоид оплодотворяет яйцо. Вокруг оплодотворенного яйца выделя-
ется целлюлозная оболочка, кнаружи находится стенка оогония и,
кроме того, коровые нити. Внутренние оболочки нитей утолщают-
ся, пробковеют, в них может откладываться кремнезем, а в поло-
стях коровых нитей еще и известь. Таким образом, вокруг ооспоры
образуется очень твердая оболочка, и ооспоры переходят в состоя-
ние покоя. При прорастании ооспоры копуляционное ядро редукци-
онно делится, после образования четырех гаплоидных ядер появля-
ются перегородки, делящие ооспору на две неравные части: верхняя
клетка получает одно из четырех гаплоидных ядер, три остальных
ядра оказываются в нижней клетке, богатой запасными веществами.
Эти три ядра впоследствии дегенерируют. Оболочка ооспоры раз-
рывается на верхушке, и верхняя клетка делится продольно. Полу-
чившиеся две клетки растут в противоположных направлениях:
образуются первый ризоид и вертикальная нить — предросток или
протонема, на которой развивается затем нормальный побег.
Сложность половых органов, стадия протонемы, или предростка, при развитии
зиготы, сложное вегетативное строение харовых значительно отличают их от других
зеленых водорослей, и ряд авторов рассматривают цх не только как отдельный
класс, но даже как самостоятельный отдел. Однако состав пигментов, присутствие
крахмала, гаплоидный характер, относительно простое строение нителлы скорее
свидетельствуют о том, что харовые водоросли представляют собой лишь очень
специализированную ветвь зеленых водорослей.
Распространены харофициевые преимущественно в пресных водах, хотя некото-
рые виды встречаются в солоноватых. Обычно они обитают в спокойных водах, где
нет сильных движений воды и где имеется песчаный или илистый грунт, в котором
укореняются ризоида.
Харовые водоросли представляют немалый теоретический интерес: на них издав-
на изучали движение цитоплазмы. Кроме того, крупные клетки междоузлий этих
водорослей, в которых цитоплазма ограничена постенным слоем, а центр клетки
занят громадной вакуолью с клеточным соком, служат объектами для исследования
биоэлектрических явлений.
* * *
Как уже отмечалось в общей характеристике зеленых водорос-
лей, они единодушно признаются предками высших наземных рас-
тений. Относительно же того, какой порядок, семейство включает
в настоящее время роды, предки которых могли быть среди воз-
250
можных прародителей высших растений, мнения несколько рас-
ходятся. Согласно некоторым авторам, исходным пунктом для раз-
вития высших растений могли быть только те зеленые водоросли,
которые лучше всего приспособлены к наземному образу жизни.
Еще Ф. Фрич (F. Е. Fritsch, 1916, 1945) высказал мысль о возник-
новении наземных растений из зеленых водорослей, обладающих
нитчатыми гетеротрихальными талломами, дифференцированными
на стелющиеся по субстрату нити и нити, приподнимающиеся
вверх, т. е. имевших примерно такую же дифференцировку, какая
наблюдается у многих современных хетофоральных. В дальнейшей
эволюции этих водорослей — предков архегониат — из стелющейся
части таллома возник гаметофит, а из восходящей — спорофит. Тех
же взглядов придерживается и Б. Фотт (В. Fott, 1971), приводящий
в качестве примеров водорослей, хорошо приспособленных к назем-
ному образу жизни, трентеполиевые и род фричиеллу из семейства
хетофоровых. К этому следует добавить, что среди хетофоровых (у
стигеоклониума драпарнальдиопсиса, фричиеллы) установлена сме-
на генераций, свойственная высшим растениям; есть данные
и о приуроченности зооспорангиев и гаметангиев к разным ин-
дивидам трентеполии. Приверженцем этой гипотезы является
и Т. Стефуреак (Т. Y. Stefureac, 1972, 1982), считающий, что высшие
растения произошли от некоторых хетофоровых водорослей, об-
ладавших гетеротрохиальным талломом, достигших высокого
уровня организации в палеозое (далее они мало изменялись), и ада-
птировавшихся к наземному образу жизни.
Другие авторы (К. D. Stewart, К. R. Mattox, 1975, 1978;
J. D. Pickett-Heaps, 1979; Н. W. Johnston, 1979; Н. Y. Sluiman, 1983;
L. Е. Graham, 1985) при поиске среди зеленых водорослей возмож-
ных предшественников наземных высших растений руководствуют-
ся иными критериями, придавая большое филогенетическое значе-
ние некоторым цитологическим признакам, полученным в ходе
электронно-микроскопических исследований зеленых водорослей за
последние десятилетия. Исходя из того, что всем высшим растени-
ям свойствен оогамный половой процесс, причем сперматозоиды
мохообразных и низших сосудистых растений имеют субапикаль-
ное— латеральное прикрепление жгутиков и многослойную струк-
туру, связанную с основанием жгутиков, и что во время цитокинеза
развивается фрагмопласт, эти авторы склонны выводить высшие
растения из тех зеленых водорослей, у которых обнаруживаются те
же черты. В этой связи особое внимание привлекает род колеохете
(порядок хетофоральные, семейство колеохетовые). Этот род харак-
теризуется оогамным половым процессом, при котором зигота,
защищенная псевдопаренхиматозным чехлом, сохраняется и прете-
рпевает свое дальнейшее развитие на родительском гаплоидном
растении; обнаружено и присутствие плацентарных передающих
клеток, напоминающих таковые семенных растений. Зооспоры
и сперматозоиды с латеральным прикреплением жгутиков имеют
251
многослойную структуру, при цитокинезе сохраняется межзональ-
ное веретено и формируется фрагмопласт. Правда, у колеохете
установлена зиготическая редукция. Согласно Л. Грэхэм
(L. Е. Graham, 1985), переход к высшим растениям мог произойти
путем смещения мейоза на все более поздние стадии развития
зиготы и тем самым удлинения диплоидной фазы. Образование
8—32 мейоспор, согласно этому автору, можно рассматривать как
преадаптацию, приведшую ко все большему смещению мейоза. Эти
взгляды поддерживают гипотезу А. Бауэра (A. Bower, 1908), соглас-
но которой родоначальниками наземных растений были зеленые
водоросли из класса равножгутиковых с зиготической редукцией.
При выходе на сушу в цикл их развития вставилась диплоидная
фаза — спорофит, благодаря чему сдвинулся и момент редукции
(мейоз), связанный с образованием спор.
Таким образом, в наиболее разработанных гипотезах происхож-
дение наземных растений связывается с разными представителями
из разных семейств, но относящихся к одному порядку хетофораль-
ные (по традиционной классификации).
Ультраструктурными признаками, которым в последнее время
придается столь важное значение, обладают и некоторые другие
зеленые водоросли. Так, род клебсормидиум (порядок улотрихаль-
ные) характеризуется зооспорами с латеральным прикреплением
жгутиков и широким пучком тесно собранных микротрубочек, с ко-
торым соединены базальные тела жгутиков, у него сохраняется
интерзональное веретено и образуется фрагмопласт при цитокинезе.
Однако оогамия отсутствует. Другая группа зеленых водорослей,
обладающая необходимым для возможных предков высших расте-
ний набором признаков,— эго харовые водоросли (класс харофици-
евые). Они имеют те же пигменты, что и высшие растения, в стенках
клеток содержатся целлюлоза и предшественники лигнина, при
делении клетки наблюдается открытое веретено и образуется фраг-
мопласт, половой процесс оогамный, у спирально закрученных
сперматозоидов- латеральное прикрепление жгутиков и много-
слойная структура и пр. Однако вся морфология харовых водорос-
лей с их крупными клетками междоузлий, согласно Г. Джонстону
(Н. W. Yohnston, 1979), исключает их существование вне водной
среды и делает маловероятным их выход на сушу.
Гипотезы происхождения наземных растений от иных, кроме
зеленых, водорослей, например взгляды А. Черча (A. Church, 1919),
согласно которому наземные растения развились из высокооргани-
зованных морских бурых водорослей, в настоящее время представ-
ляют лишь исторический интерес.
ОСНОВНЫЕ РУКОВОДСТВА И ПОСОБИЯ
Громов Б. В. Ультраструктура синезеленых водорослей. Л., 1976.
В книге приводятся современные данные о строении клетки синезеленых водорослей.
Жизнь растений. М., 1977. Т. 3.
Разделы, посвященные водорослям, охватывают как общие вопросы (цитология, морфоло-
гия, размножение и циклы развития, физиология, эколошя), так и систематику,
Кондратьева Н. В. Морфогенез и основные пути эволюции гормогониевых водо-
рослей. Киев, 1975.
В монографии содержатся исчерпывающие данные о морфологических особенностях ин-
дивидуального развития гормогониофициевых водорослей.
Куре низших растений/Под ред. М. В. Горленко. М., 1981.
Разделы, посвященные водорослям, могут служить полезным дополнением к настоящему
пособию, особенно в части иллюстративного материала.
Седова Т. В. Основы цитологии водорослей. Л., 1977.
В книге даются сведения о субмикроскопической организации клеток водорослей.
Топачевский А. В., Масюк Н. П. Пресноводные водоросли Украинской ССР. Ки-
ев, 1984.
В общей части этого руководства рассмотрены цитология, морфология, размножение,
экология, значение пресноводных водорослей. В специальной части даны таблицы для определе-
ния отделов, классов, порядков, семейств, родов и приведены характеристики этих таксонов.
Carr N. G. The biology of blue-green algae. B. A. Whitton (cd.). London — N.-Y.,
1973.
В кпите изложены различные аспекты изучения сииеЗеленых водорослей: нх биохимия,
физиология, экология, а также вопросы цитологии, такие, как тонкая структура и химический
состав оболочек, структура и расположение тилакоидов, строение газовых вакуолей и т. д.
Отдельные главы посвящены гетероцистам, фиксации азота, движению и пр.
Dodge Y. D. The fine structure of Algae cells. London — N.-Y., 1973,
Капитальная сводка по улы paci рукт уре эукариотических водорослей.
Ettl Н. Grundriss der allgemeinen Algologie. Jena, 1980.
Кпщ а посвящена в основном проблемам общей альгологии, цитологии, морфологии,
размножению (только эукариотические водоросли). Самая большая глава, в которой рассмот-
рена цитология, содержит богатейший материал по ультраструктуре водорослей. Описание же
циклов развития кратко и ни в какой мере не отражает' современное состояние Этого вопроса.
Fogg G. Е. The Blue-green algae. W. D. Stewart (ed.). London — N.-Y., 1973.
Это сборник статей по различным вопросам биохимии, физиологии, цитолот ин синезеленых
водорослей.
Foil В. Algenkunde. Jena, 1971.
Наряду с систематическим описанием всех отделов водорослей это капитальное руководство
содержит главы по экологии водорослей. В книге отражены данные 60-х годов по ультраструк-
туре и циклам развития водорослей.
Fritsch F. Е. The Structure and Reproduction of the Algae. Cambridge, 1935. T. I.;
1945. T. 2.
Наиболее ценное и полное руководство по морфологии и размножению водорослей, со-
хранившее свое значение до настоящих дней.
Manual of Phycology. G. M. Smith (ed.), Valtham. Mass.. 1951.
Разными авторами изложены морфология, размножение, классификация различных отделов
водорослей. Кроме того, отдельные главы посвящены общим вопросам: цитологии, половому
процессу, пигментам, физиологии и биохимии, экологии, методам культивирования.
Oltmanns Fr. Morphologic und Biologic der Algen. Jena, 1922—1923.
Содержит богатьщ. материал как по морфологии, так и по экологии и физиологии
водорослей.
Warienberg /t'Systematik der niederen Pflanzen. Stuttgart, 1979.
В разделах, посвященных водорослям, широко используются данные электронно-микроско-
пического изучения их. Несмотря на то что число обьектов очень ограничено, это полезное
руководство отражает современное состояние изучения низших растений.
253
УКАЗАТЕЛЬ ЛАТИНСКИХ НАЗВАНИЙ .
Aceiabularia 235
— crenuiaia 235
mediterranea 19, 235
Achnanthes 124
Acrochactntm 84
— • haHundieum 84
• — pecrinawm 84
polyblasum 84
Acrosiphonales 192
Aglaozonia 146
Ahnfeltia 85
— concinna 85
Alarm 155
— marginata J 52
Alariaceae 153, 155
Algae 5, 11
Amphidimum 165, 170
— carter/ 176
Anabaena 64
— cylindrica 61. 64, 70
— hallenxis 64, 70
— oryzae 64
— spiroides 62
- variubilis 64, 70
Ankistrodcsmus 214
Anomoeoneis scidpta 134
Antiihamnion tenuissimum 91
Aphanizomenon flax-aquae 62
Aphanochaete 220
Apistonema xubmarinum 112
A SCoph yllum 141, 155, 156
nodosum 51, 159, 160
Asparagopsis armata 84
Asierioneila 125, 128
Aulacodiseus 136
Bacillar io paradoxa 127
Bacillariophyceae 6, 11
Bacillariophyta 122
Bacteria 14
Bangia 77
• • atropurpurea 78. 80
- fuscvpurpurea 80
Bangiophyceae 77
Batrachospermum 81, 82
— monilifurmc 83
— vtrgwm 83
Bellarochea 136
Biddulphia 123, 135
— granulata 136, 137
- mobiliensis 136, 137
rhombus 136. 137
Blastodinophyceae 168
Bonnemaisonia hamifera 16. 83
Botrydiales 115. 116
Botrydlum 116
— granulatum 117
— pachidermum 117
tuberosum 117
— wallrothi 117
Bryopstdaceae 229
Bryopsidales 192, 228
Bryopsidophvceae 192, 193
Bryopsis 231,'232, 233
— halimeniae 231
— hypnoides 53, 231. 232, 233
— maxima 231
— pluntosa 231. 232, 233
Bulbochaete 228
— hiloensis 223
Callithamnion cordatum 74, 89
Callophylks firma 85
254
Calothrix 66
— braunii 70
— eienkim 70
— stellaris 66
Caiyptrasphoera pirus 112. 113
Carteria 197
Caulerpa 234. 235
fasligiata 234
- prolifera 34. 234
• taxifolia 234
eerticillata 234
Caulerpaceae 234
Ccphtdeuros 222
- virescens 222
Centrophyceae 123. 138
Ceramiales 87
Ceramium rubrum 44
Cerataurus 136
Ceratlum 165, 170
— comutum 176
hirundinella 171
— horridum 176 *
“ tripos 171
Chaetoceros 128, 131
Chaetonema 220
— irregulare 221
Chaetophorales 192, 219
Chainaesiphonophyceae 62, 63
Chanlransia 82, 83
Chara 247
Characeae 247
Charales 192
Charophyceae 192, 193. 247
Charophyta 12, 192
Chilomonas 183
Chiamvdomonas 26. 40. 44. 46., 194.
196
~ braunii 47. 197
- eocdfera 47, 197
— ehlamydogama 44, 46, 196
— eugametos 34, 36, 44
gfrus 195
kleinii 194
moewusii 34, 46, 196
“ var. rotunda 44 -
- paupera 50
— pseudogigantea 47
— rcinhardfii 27, 34, 40, 44, 46.
195, 196
— лтяйн 19
suboogama 47
Chioreila 213, 214
fusca var. vacuolata 43
pyrenoidosa 213
vulgaris 40
Chlorococcales 192, 209
Chloroeoccum echinozygotum 44
Chlorogomum 197
— iyengarii 197
bogamum 197
Chlorophyceae 5, 6, 11. 192, 193
Chlorophycophyta 189
Chlorophyta 12, 189, 192
Chlorospermcae 5
Chorda 153
fllum 153
" tomeniosa 44. 152
Chordaceae 153
Chromophyla 13, 161
Chromultna 104
Chromulinales 100
Chroococcophyceae 62
Chroomonas 183
mesostigmata 180
Chrysocapsales 100, I l!l
Chrysothromulina 111
- birgeri 111
chiion 111, 112
cyanophora 111
megaeylindra 1! 1. 112
Chrysomonadales 100
Chrvsophvceae 9, 11, 13. 100, 104
Chrysophyta 12, 13. 96, 100. 121
Chrysosphaerales 100
Cladophora 7, 237
Cladophoraceac 237
Clostenum imeatum 246
Coccolothophoraceae 111
Codiaccae 235
Codiolales 192
Codiolophyceae 192. 218
Codiolum 192, 2f 8
Codium 235
dwarkense 235
— tumentosutn 235
Coleochaetaceae 222
Colcochaetales 192
Coieochaete 222
Conchacelis 79- 80
— rosea 78
Conjiigatophyceae 193, 242
Corallina officinalis 85
Cricosphaera roscojfensis var. hap-
tonemofera 112
Crypthecodinium cohnti 176
Cryptochrysis 183
Cryptococcales 182. 183
Cryptophyceae 9. 11. 13, 182
Cryptophyta 13, 179
Cryptomonadaceae 183
Crypiomonadales 182
Cryptomonas 183
— cryptophila 180
- ovata 183
reticulata 180
Cryptoncrmaies 85
Cutleria 144
multifida 146
Cutleriales 7, 144
Cyanohacteriales 14
Cyanophyceae 6, 11, 13
Cyanophyta 13, 14, 54
Cyclosporophyceae 142. 155
Cyclotella 123
— meneghiniana 135
• -'.tenuistriata' 136
Cylindrocystis 242
Cymbeila 124
Cystodinium 179
bataciense 179
lunare 179
purpureum 86
steinii 86
Cystoseira 156
— barbata 160
Dasycladales 235
Derbesia 22% 232, 233
— marina 231
-----var lamouroxti 233
- neglecta 232. 233
Desmarestiales 242, 245
Desmidium 247
strarizii 247
Desmophyceae 168
Diuioma 125, 127
Dialumeae 11, 122
Dictyasphaenum 215
Diciyota dichotoma 35, 48, 148
Dictyolales 148
Dinamoebidiales 170
Dinobryaceae 101, 104
Dinobryon 99, 100, 104
cylindricum 97, 99
- • divergens 97, 99
Dinococcales 170, 178
ritnoflagellalae 170
Oinophyceac 9, 11, 13, 168, 170
Dlnophysidaceae 177, 178
Dinophysis 178
- norvegica 178
ovum 178
Dinophyta 13. 162, 168
Dinotricbates 170
fristigma 188
Praparnaldia 220
Draparnaldiopsix 220
— indica 220
Dunaliella 197
- salina 45, 50, 197
Durvillea 155
Ectocarpales 142
Ectocarpus 141, 142
- siliculoxus 43, 50, 143, 144, 145,
146, 159
Enteromorpha 218
Eremospftaera 214
vindis 35, 214
Eucaryota 13, 72
Eudorma 197
- elegant 199. 200, 204, 205, 208
Euglena 185. 188
gracilis 185. 188
- haematodes 188
- rublda 188
sanguinea 188
viridli 188
Euglenalcs 187, 188
Euglenophyceae 9, 1], 187
Eijglenophyta 184, 187
Eustigmatophyceae 12, 114, 121
Eutreptia 188
Falkenbergia rufblanosa 84
Flagellatae 8, 9, II
Florideopbyceae 77, 80
Fragdaria 125
- capudna 132
Fragilariales 124
Fritschiella 220
- tuberosa 191, 220
Fucales 155
Fucus 6, 141, 155
distiehus 19
— evanescens 157, 159. 160
— inflatus 19
— serratus 146, 156, ] 59, 160
- spiralis 159, 160
- - var. piafiearpus 160
- vesicutosus 19. 36, 159. 160
Gelidiales 84
Gelidium cartilagineitm 84
Gigartina 53
figardktti 53
stellata 53. 86
Gigartinales 85
Glenodinium iubiensifornte 176
Gioeodin>ale& 170
Gfoeophycus korneunum 85
^ioeotriehsu 61. 67. 68
echinuluta 67
- pisum 67
Giafosiphonia eapillaris 85
Golenkinia 215
- miniitissima 44
Gomphonema 123
geminatum 134
— parvulum 132, 133
Gonioxtumum 184
Gonium 197
— pectorale 197, 200. 208, 209
Gonyauiax 171
— moniliala 176
— polvedra 164, 165, 171, 172,
173.' 174
Gracilarta verrucosa 86
Grtffiihsia 91. 92
— coraliina 92
— gfobifera 93
— pacifica 92. 93
— setaceae 92
— tenuis 92. 93. 94
Gymnodinium 177
— breve 171
- pseudopalustrc 176, 177
— simplex 166
Gyrodiniunt lebouriae 171, 172
Gvi'osigma 124
Haltcystis 229. 132
— ovalis 231, 233
parvula 229, 230
fiatidrys 156
— siliquoxa 51. 159, 160
Halosphaera 241
— viridis 241
Halosphaeraies 240
Haptophyccae 12, 100, 110
Nelminthocladia 84
— agaroides 84
— calvadosii 84
— senegaiensis 84
Helminthnra divaricata 84
Hcmidinium 170
Heterochloridalcs 115
Heterachrysophyecae 100
Heterocoittae 6, II. 113. 121
Hcterogeneratophyccae 148
Heterogloeales 115
Himanthalia 155, 156
Hormogoniophyceae 62, 63
Hyalop/racus otellofus 187
Hyulotheca 24?
Hydrodictyvn 210
— reticulatum 210. 211, 212
Hydrurus 110
— foetidus 110
ffymenomanas 111
— coronata 111, 113
roseola 113
Isocontae 193
Isogeneratophyceae 142
Katodinium 165. 170
Kirchner ielitt 214
Klebsormidiales 192
Klebsormidium 191
— flaccidum 191
— xubtdirsimum 191
Laminaria 153
— digitata 149, 150. 154
— grnenlandiea 152
— japtmica 149
— saceharina 149. 150, 151, 152,
153, 154
Laminariaceae 153
Laminanalcs 149
Laurencia spectabilL 34
Lemanea 76, 81
Lessonia 154
Lessoniaceae 153, 154
Liagora tetrasporifera 76, 84
Licnwphora 124
Liihodesmium ) 36. 137
Lithophyllum 229
Lifhmhamninn 229
Lomentaria 86
Lyngbya 55
Macrocysds 154
— piryfera 152
Mallomonus 99, 105. 106
caudata 97. 98, 99, 100, 105.
106. 107
Mallomonopsix 107
Mastigocladus 68
lamin wax 68, 70
Melanospenneae 5
Melatoit 127
- variant 135, 136
Meridian 125
Merismopedia 63
Mesotaeniales 242
Mesotaenium 242
Micraetinium 215
Microcystis 62
Microsporales 192
Mischococeales 115
Monostrama nilidum 49
Monosiromaiates 102
Mougeotia 243
Navicula 123
Naviculales 125
Nemaliales 81. 84
Nemalion 83, 84
— muiufldum 83
— vermiculare 83
Nemalvdinium 168
Nereocysfix 154
— luetkeana 152
Neirium 242
— digitus 243
Nitella 247
Nltzschia 124
Noctiluca 177
Nostoc 64
—• coeruleum 65
— flagellifornic 66
— ffnckia 60, 66
— microscopicum 65
— pruniforme 65. 66
— punetiforme 65, 70
Nostocales 64
Nothe.ia 158
Ochromonadaceae 101
Ochromonas 101
— danica 102. 103, 104
— minuta 103
— tuberculata 97, 100
Ochrosphaera neapaiitana 111
Oedogoniales 192, 223
Oedogoniophyceae 192, 193
Oedogonium 6. 224
— fneisianum 226
— cardiacum 44, 51, 224, 226
Ooeystls 20
— apiculata 20
- so/itoria 20
Ornithacercus 178
Oscillatoria 63
Oscillaioriales 63
Oxyrrhis 165
- marina 166, 171
Pafrnaria palrwta 44, 76, 87
Palmariuics 87
Pandorina 197
— 7, 44. 198. 200
255
— unit Осс а 44. 204
Papposphaera lepida 112, 113
Pelvetia 155. 156
— canaliculate 159, 160
— nrightii 160
Pennalophyceae 123, 138
Peranema 189
Perancmaies 187, 189
Peridimalcs 170
Peridin ium 170
— cinctum 171, 172. 175
---f. oroplanum 176
— faeroense 176
— limbarum 176
— willei 176
Petroeellix eruenta 86
Petrogloxsum pacifteum 86
Phacus 188
— pirum 187
Phaeophyceae 5, II, 13
Phaeophyia 139. 142
Phaeot ham males 100
Photobacteria 14
Phyco pelt is 222
epiph it on 19, 222
Phyllophora 86
erf spa 86
— heredia 86
- pseudoceranoides 86
Pikea caiiforntca 85
Pinnularia 123, 129
Pleurochrysix seherffeli 22, 23,24, 112
Pleura sigma 124
Polysiphonia 76. 84. 85, 86, 88, 89
— harlandii 91
— hendryi 49, 90
— maemcarpa 91
— violctceae 76
Porphyra 77, 80
— brumafis 79
— gardneri 79
— haitanensis 79
— kanakaensis 79
— kuniedai 79
— leucwticta 78
— miniata 79
— nereocystis 71, 79
— perforata 79
— schizophylla 79
— tenera 79
— umbiliealis 78
— yezoensis 79
Prasinophyceae 192, 193, 239
Prochltiron 71
Pnrhlorophyta 14, 71
Ptilota phmiosa 49
Prasiola scipilata 51
Procaryota 13, 14, 54
Prochlorophyta 14
Prorocentrum 168, 169
Protogonyaulax eatenella 176
Prymnesiaceae 111
Prymnesiales 110
Prymnesiophyceae 100, 110
Pseudohryopsis 233
Pyramimonadates 239
Pyramimunas 239, 241
gross»’ 240
Pyrrophyta 13
Raphidophyceae 13
Raphidophyta 183
Rhodanema adriaticum 138
Rhizochloridales 115
Rhizochrysidales 100
Rhizoclonium 1 38, 237
Rhodophyceae 5, 11
Rhodophyta 12, 72
Rhodomonas 183
Rhodospermeae 5
Rhodymenia palmata 87
Rhodymeniales 86
Rivularia Ы
Saechorhiza bulbosa 149, 151
Sargassum 156
— fu helium 157
horneri 158, 160
kjellmanianum 157
muiicum 49
Scenedesmus 215
obliquus 44, 217
— quadricauda 215, 216
Schizophyta 13, 14
SchizomyceteS 13
Seinaia 84
furcellata 84
turgida 84
Siphonocladus 237
Si phonophyla 121
Skeletunema 128, 130, 131
coxtatum 13 I
— subxalsutn 131
Sphaeelaria 144
SphaceJarialcs 144
Sphaernplea 238 r
— annulina 44, 50. 238
Sphaeropleaceae 238
Spirugyrtt 6, 244
Spirtnaenia 242
Spirulina 64
Stephanodiscus hantzsckii 134
Stenogramnta interrupta 86
Stigeoc/onium 219
— helreticum 191 '
Sligonema 68
Stigonematales 68
Streptotheca 136, 137
Surirella 126
Surirellales 125
Synedra 123
Syniira 108
— echinulaia 109
— petersenii 108. 109
sphagnicola 108. 109
- uvella 109
Synuraceae 101, 105
Tubeliaria fenestra la 134
Tetradinium 179
Tetragonidium 183
Tkalassioxira 128. 130 131
Trachemmonas 186, 188
Trgntepohlia 221
Trentepohliaceae 221
Tribonema 115
Tribonemalales 115
Triceratium 123
Ulotrichales 192, 217
Ulotrichophyceae 192
Ulothrix 7, 217
— fimbriaia 191
— zonata 217
Utva 218
— lobata 50
— pertusa 50
Ulvaceae 218
Ulvates 192. 218
Uroglena 99. 100, 104
— americana 96. 97. 98. 100, 104
— roll-ox 97, 100, 104
Vacuolaria 184
virescens 24
Vakmia 239
Valoniaceae 239
Vaucheria 6. 117, 119
— fominalis 19. 120
sessi/is 6
Vaucherialcs 121
Vaucheriophvta 121
Volvocales 192. 193
Volvocophyceae 192
Voltax 7. 197
— africanus 198. 201
— aureus 36. 198- 201. 203, 206
— capensis 207
— carteri 198. 199. 200, 201, 202,
206, 207
f. nagariensis 51. 201, 206, 207
gigas 198. 199. 203. 206
globator 198. 200. 201. 203
— abverstts 201, 207
— powersii 198, 203, 206, 207
— rousseletii 51, 198, 201, 203.
206, 207
— teriius 198, 203
Wdrnowia 170
Woloszynskia 165, 177
— apiculura 175, 176, 177
coronata 167
micro 171. 172
Xanthophyceae 6, It, 114, 121
Xanthophyta 113. 121
Xanthosiphonophyceae 121
Zanardinia 144, 147
Zygnema 243
Zygncmatales 192, 242, 243
Zygnema toph yceae 193