/













































































































































































Author: Рундина Л. А..
Tags: систематика растений ботаника водоросли гидробиология
ISBN: 5-02-026092-4
Year: 1998




Text
Л.А. РУНДИНА
ЗИГНЕМОВЫЕ
ВОДОРОСЛИ
РОССИИ
РОССИЙСКАЯ АКАДЕМИЯ НАУК
БОТАНИЧЕСКИЙ ИНСТИТУТ им. В.Л. КОМАРОВА
RUSSIAN ACADEMY OF SCIENCES
KOMAROV BOTANICAL INSTITUTE
L.A. RUNDINA
THE ZYGNEMATALES
OF RUSSIA
(Chlorophyta: Zygnematophyceae)
Й
SANKT-PETERBURG
"N A U К A"
1998
Л.А. РУНДИНА
ЗИГНЕМОВЫЕ
ВОДОРОСЛИ
РОССИИ
(Chlorophyta: Zygnematophyceae,
Zygnematales)
в
САНКТ-ПЕТЕРБУРГ
“НАУК А"
1998
УДК 582.262
ББК 28.591
Р 86
Рунднна Л. А. Зигнемовые водоросли России (Chlorophyta: Zygnematophyceae, Zygne-
matales). — СПб.: Наука, 1998. — 351 с.
ISBN 5-02-026092-4
Первая в России обобщающая сводка по систематике, таксономии, синонимике, мор-
фологии, экологии и географии нитчатых зеленых водорослей — зигнемовых — одного из
4 порядков в классе конъюгат. Сообщаются общие сведения, преимущественно по морфо-
логии, с таксономической оценкой признаков в вегетативной и фертильной стадиях. В
Систематической части даны таблицы для определения и диагнозы таксонов всех рангов —
от отдела до внутривидовых форм. Приведены соответствующие сведения, с критическими
примечаниями к иим, о 360 видовых и внутривидовых формах, потенциальных и обнару-
женных в России, с привлечением данных о их распространении в республиках бывшего
СССР и странах Европы. Для редких видов указываются все находки в мире. Особо вы-
делены роды редкие, недостаточно изученные и сомнительные; сведены воедино сомни-
тельные и ошибочные указания видов в родах Zygnema, Zygogonium, Mougeotia, Spirogyra.
Для широкого круга ботаников-систематиков, гидробиологов, экологов, преподавателей и
студентов биологических факультетов. Библиогр. 265 иазв. Ил. 142.
Rundina L. A. The Zygneipatales of Russia (Chlorophyta: Zygnematophyceae). — St. Pe-
tersburg: Nauka, 1998. — 346 p.
ISBN 5-02-026092-4
This monograph of filamentous green algae of the order Zygnematales for the first time
for Russia presents the data on their taxonomy, synonyms, morphology, ecology, and geogra-
phy. It opens with a discussion of the morphology of Zygnematales, in particular the taxonomic
value of characters in both vegetative and reproductive stages. Systematic part presents the
tables for indentification of all the taxa and their diagnoses from division to all the intraspecific
forms. The monograph summarizes all the data on the morphology of 360 species and intra-
specific forms found and potentially occurring in Russia, and presents the data on geographic
distribution of these taxa in the former USSR and Europe. All findings of rare species are
indicated on a global scale. Author pays special attention to the poorly studied and questionable
genera and summarizes all doubtful and erroneous occurrences of taxa from the genera Zyg-
nemai Zygogonium, Mougeotia, and Spirogyra. The book is addressed to a wide audience of
systematic botanists, hydrobiologists, ecologists, teachers and students of biology. Bibliogr.
265 ref. Ill. 142.
Ответственный редактор К. Л. Виноградова
Рецензенты: И. В. Макарова, И. С. Трифонова
Издание осуществлено при поддержке
Российского фонда фундаментальных исследований
по проекту № 97-04-62040
Р
И
ТП-98-П-№ 270
ISBN 5-02-026092-4
© Л. А. Рунднна, 1998
© А. Т. Пожванов, оформление, 1998
© Российская академия наук, 1998
ПРЕДИСЛОВИЕ
Зигнемовые — нитчатые зеленые водоросли, макроско-
пические, широко распространенные в континентальных,
преимущественно в пресных и чистых водоемах и на на-
земных субстратах. Их насчитывается свыше 700 видов, и
они встречаются на всех широтах всех континентов, вклю-
чая Антарктиду. В вегетативном состоянии эти нитчатки
изумрудно-зелеными шелковыми космами устилают дно рек
и прибрежья озер и водохранилищ, в фертильной стадии
они довольно крупными скоплениями буровато-желтого цве-
та плавают на поверхности воды. Зигнемовые являются су-
щественным компонентом водной растительности, в массе раз-
виваясь в теплые сезоны и даже вызывая цветение воды.
Нередко их используют в качестве очистителей на фильтра-
ционных установках и как экологические индикаторы и стра-
тиграфические маркеры при гидрологических и геологических
изысканиях. Некоторые из зигнемовых (Spirogyra, Mougeotia,
Zygnema) давно служат классическими объектами для изу-
чения различных процессов в растительной клетке. Всесто-
ронние знания о зигнемовых необходимы также для пост-
роения гипотетических схем филогении и выяснения во-
просов эволюции и происхождения растений.
Зигнемовые водоросли в ранге порядка входят в обшир-
ную (около 6000 вадов) группу конъюгат (класс Zygnema-
tophyceae), четко очерченную по вегетативным признакам
и выделяющуюся среди зеленых водорослей характерным
типом полового процесса — конъюгацией. Между тем на-
учное определение зигнемовых крайне затруднено. Главная
трудность состоит в том, что точная идентификация зигне-
мовых возможна лишь при наличии зрелых зигот, в отличие
от представителей других порядков конъюгат — мезотени-
5
евых, гонатозиговых и десмидиевых, определяемых в основ-
ном по вегетативным признакам. Процесс же конъюгации
протекает, как правило, кратковременно, требует специаль-
ных условий, поэтому найти зрелые зиготы в природе бы-
вает нелегко. Исследователю в поисках зигоспор следует
учитывать как температурные и световые условия, так и
особенности гидрологического и гидрохимического режимов
водоемов, влияющих на половой процесс. Сложность раз-
работки систематики зигнемовых водорослей усугубляется
еще и тем, что российских определителей и региональных
флористических сводок по этой группе водорослей нет, за
исключением работы Воронихина (1954) по флоре конти-
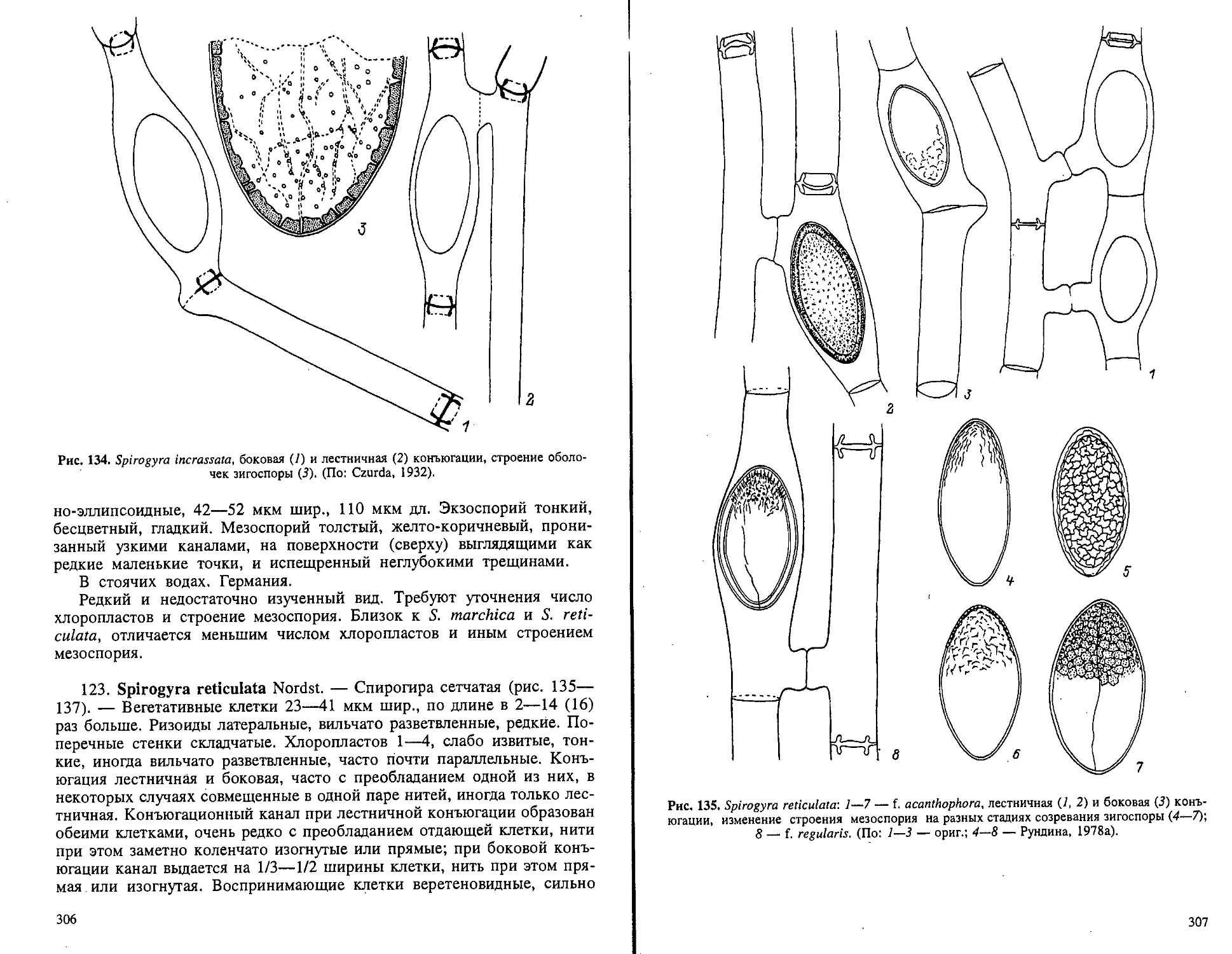
нентальных водоемов европейского Севера.
Представляемая монография — первая обобщающая сводка
по систематике, таксономии, синонимике, морфологии, эко-
логии и географии зигнемовых водорослей, потенциальных
и обнаруженных на территории России. Она написана на
основе сравнительного критического изучения обширного
материала — оригинальных данных и литературных источ-
ников. В Общей части монографии кроме общей характе-
ристики зигнемовых подробно рассматриваются основные
морфологические признаки, необходимые исследователю для
определения, и этим признакам дается авторская таксономи-
ческая оценка. В связи с переоценкой значимости ряда морфо-
логических признаков вид у зигнемовых водорослей автором
монографии понимается в широком объеме, и этим объясня-
ются введение в систематику зигнемовых таксономической
категории формы и критическое переосмысление некоторых
положений в их классификационной системе. В Системати-
ческой части монографии приведены сведения о 360 видовых
и внутривидовых формах, потенциальных (найденных в рес-
публиках бывшего СССР и странах Европы) и обнаруженных
в России. Для видов редких или представляющих особый так-
сономический интерес указываются все находки в мире. Опи-
сания видов и внутривидовых форм даются с исчерпывающей
полнотой, по литературным источникам и оригинальному ма-
териалу; при этом описания большинства таксонов сопро-
вождаются примечаниями, облегчающими понимание таксо-
нов различного ранга и их определение. Кроме того, в при-
мечаниях даются ссылки на близкородственные таксоны, не
встреченные до сих пор в России, республиках бывшего
СССР или Европе, но имеющие вероятность потенциального
обнаружения.
Территория России столь обширна, а водоемы ее столь
разнообразны, что до сих пор нельзя считать хорошо изу-
6
ченной флору зигнемовых водорослей. Флористические ис-
следования сводятся обычно к упоминанию отдельных пред-
ставителей, в лучшем случае — к инвентаризации видового
состава без критического изучения их систематики. Так как
в большинстве случаев видовое название не сопровождается
описанием и рисунком, то, естественно, достоверность оп-
ределения подвергается подчас оправданному сомнению.
Разрозненность и труднодоступность сведений о местона-
хождениях зигнемовых и отсутствие обобщающих сводок
по этой теме в отечественной литературе привели к досад-
ным пробелам и в монографиях зарубежных авторов, в ко-
торых данные по России чаще всего отсутствуют. Наиболее
значимыми флористическими работами, содержащими не
только списки обнаруженных видов, но и описания редких
и новых таксонов и критические замечания к видам, явля-
ются исследования Скворцова (1917, 1927а, 19276, 1946),
Скворцова и Ноды (Skvortzov, Noda, 1973) — по Сибири
и Дальнему Востоку, Воронихина (1926, 1954) — по Кав-
казу и европейскому Северу России, Полянского (1951,
1955, 1959) — по северо-западу и центру европейской
части России, Рундиной (1993) — по Северной Осетии. Из
сопредельных с Россией стран (республик бывшего СССР)
флора зигнемовых наиболее полно изучена в Латвии и Эс-
тонии (Skuja, 1924, 1926, 1928, 1929, 1932а), на Украине
(Рундина, 1988) и в Казахстане (Красноперова, 1966, 1970,
1971а, 19716; Воронихин, Красноперова, 1970; Рундина,
1977а, 19776, 1978а, 19786, 1981).
В представляемой работе использованы многочисленные
личные сборы, проведенные автором во многих областях и
краях России и республиках бывшего СССР, сборы геобо-
таников и альгологов, а также коллекции, хранящиеся в
Отделе альгологии Ботанического института РАН и альго-
теке Института ботаники АН Украины. Основой для учета
литературных данных о видовом разнообразии зигнемовых
водорослей России и сопредельных стран послужили 2
сводных Указателя по водорослям к отечественным библи-
ографиям (Голлербах, Красавина, 1971; Водоросли, 1983).
При обобщении литературных сведений и критической ре-
визии приведенных в Указателях таксонов 'большинство из
них либо ушло в синонимы, либо было таксономически
преобразовано; многие были выделены в особые разделы
«Сомнительные и ошибочные указания видов» (в родах Zyg-
пета, Zygogoniutn, Mougeotia, Spirogyra). Так как из 16
родов зигнемовых водорослей, предложенных в монографи-
ях зарубежных авторов, в России обнаружены представите-
7
ли только 8 родов, то для полного представления о системе
зигнемовых водорослей в каждом из трех семейств (Zygne-
mataceae, Mougeotiaceae, Spirogyraceae) особо рассматрива-
ются роды редкие (представители которых не найдены в
России и в Европе), недостаточно изученные (как правило,
синонимы в предлагаемой монографии) и сомнительные (ис-
ключаемые, по мнению автора, из пор, Zygnematales'). Об-
ширные таблицы для определения видовых и внутривидовых
форм даются с привлечением всего комплекса морфологи-
ческих признаков зигнемовых водорослей в вегетативной и
фертильной стадиях.
Работа выполнена в Отделе альгологии Ботанического
института РАН. Большинство рисунков подготовлено к пе-
чати художницей И. Г. Гай. Всем коллегам и друзьям, ока-
завшим мне помощь в работе, приношу искреннюю благо-
дарность. Выражаю глубокую признательность Кире Леони-
довне Виноградовой за советы, поддержку и редактирование
монографии. Не могу не вспомнить с благодарностью моих
учителей: Максимилиана Максимилиановича Голлербаха,
первым прочитавшего рукопись и сделавшего ценные заме-
чания, и Владимира Ивановича Полянского, мною не виден-
ного, но своими работами всегда вдохновлявшего. Сердечно
благодарю моих голландских друзей: Дольфа ван Рехтерна
(Dolph van Rechteren) — за бескорыстную материальную по-
мощь, Мадлен и Мартина Инген Хуз (Madeleine, Maarten J.
Ingen Housz) — за предоставление принтера и компьютера.
I. ОБЩАЯ ЧАСТЬ
1. ОБЩАЯ ХАРАКТЕРИСТИКА
ЗИГНЕМОВЫХ ВОДОРОСЛЕЙ
Основные признаки зигнемовых водорослей. Нитчатые зеленые во-
доросли порядка Zygnematales относятся к классу конъюгат (.Zygnema-
tophyceae), отличающемуся от других зеленых водорослей специфи-
ческим половым процессом — конъюгацией. Многоклеточные нити
довольно длинные (до 15 см), обычно неразветвленные или с корот-
кими ответвлениями из 1—10 клеток, свободноплавающие или при-
крепленные ризоидами к субстрату, изумрудно-зеленого или нежно-
оливкового цвета, обычно слизистые на ощупь (исключение — род
Sirogonium). Они широко распространены в континентальных, преиму-
щественно пресных, редко в слабосоленых или соленых водоемах всех
широт и предпочитают чистую, хорошо прогреваемую воду. Предста-
вители родов Zygogonium и Sirocladium, некоторые виды Zygnema и
Spirogyra образуют хорошо заметные глазом дерновинки и налеты на
увлажненной почве, техногенном грунте, на мхах, на орошаемых водой
скалах и даже на стволах деревьев. Конъюгация с последующим об-
разованием зигоспор протекает в теплое время года, преимущественно
в весенне-летний период; сроки и продолжительность ее зависят от
климатических, гидрологических, температурных и световых режимов.
Клетки, составляющие нить, цилиндрические, до 200 мкм шириной,
одноядерные, довольно плотно соединенные друг с другом. Распадение
нитей на отдельные фрагменты и клетки, некоторое время самостоя-
тельно существующие, происходит лишь при резко изменившихся в
неблагоприятную сторону условиях среды. Поперечные оболочки
(септы) вегетативных клеток во всех родах гладкие, только в роде
Spirogyra они складчатые или гладкие, редко (секция Colligata) с до-
полнительной кольцевой накладкой. Продольные оболочки вегетатив-
ных клеток двух- или многослойные. Наружный слой пектиновый, сли-
зистый и аморфный, отсутствующий только у рода Sirogonium-, внут-
ренние слои целлюлозные, фибриллярные, без пор и орнаментаций.
Зеленая окраска нитей определяется основными пигментами — хло-
рофиллом а и хлорофиллом Ь, сосредоточенными в хлоропластах. В
них же находятся крупные и мелкие пиреноиды, от 1 до многих,
иногда пиреноид отсутствует (род Mougeotiopsis). Основной продукт
ассимиляции — крахмал, в виде глыбок откладывающийся вокруг цент-
9
рального тела пиреноида. Различают 3 основных типа хлоропласта —
по форме и положению его в клетке: осевой (центральный) звездчатый,
осевой пластинчатый, пристенный спирально-лентовидный. Очень
редко встречается пристенный пластинчатый хлоропласт (род Sirocla-
dium). Количество хлоропластов в клетке бывает различным: звездча-
тых — обычно 2, пластинчатых — 1—2, спирально-лентовидных —
от 1 до 16.
Для зигнемовых водорослей присуще вегетативное, бесполое и поло-
вое размножение. Вегетативное размножение происходит поперечным
делением клеток надвое, чем достигается рост нити в длину, и фраг-
ментацией нити; бесполое — акинетами, апланоспорами и партено-
спорами (азигоспорами в широком смысле); половое — характерной
для всего класса Zygnematophyceae конъюгацией. Специфика полового
процесса у зигнемовых — наличие лестничной и боковой конъюгаций,
иногда совмещенных в одной паре нитей. При конъюгации специали-
зированных подвижных гамет нет, и имеет место слияние протопластов
двух клеток, выполняющих функцию гамет. При лестничной конъюга-
ции клетки одной нити функционируют как мужские гаметы, а дру-
гой — как женские, морфологически оставаясь неразличимыми. Ти-
пичной формой конъюгации является изогамная, при которой форми-
рование зиготы в конъюгационном канале происходит в результате
слияния двух совершенно одинаковых по морфологии и скорости дви-
жения друг к другу изогамет. Но у целого ряда зигнемовых наблюдается
также и функциональная анизогамия (гетерогамия), при которой мор-
фологически одинаковые гаметы различны по «поведению», т. е. по
активности движения. В этом случае формирование зиготы в женском
гаметангии (воспринимающей клетке) происходит в результате слияния
активно движущейся мужской гаметы с неподвижной женской гаметой.
Такой процесс, скорее, напоминает оогамию как разновидность ани-
зогамии. У родов Spirogyra, Sirogonium и Sirocladium таковая является
наследственным родовым признаком, а у родов Mougeotia и Zygnema
характер полового процесса служит стабильным видовым признаком.
Главными особенностями конъюгации и зиготообразования, характер-
ными для некоторых родов у зигнемовых водорослей, являются: запол-
нение гаметангиев коллоидным веществом, окружение зигоспоры ос-
татком цитоплазмы в виде мантии, предварительное деление вегетатив-
ной клетки перед конъюгацией на собственно гаметангиальную и
стерильную, отсутствие или слабая выраженность конъюгационного ка-
нала, появление спорангия — пространства для зигоспоры, отделенного
перегородками от гаметангия.
Происхождение и родственные связи. Широко распространено
мнение о том, что отсутствие у конъюгат дифференцированных гамет
монадной структуры и характерный для них половой процесс (конъ-
югация) представляют собой вторичное явление. Обычно это связывают
с обмелением и высыханием водоемов и со сменой водного место-
обитания на наземное и аэрофильное. Согласно этой точке зрения, конъю-
гаты происходят от предков монадной структуры, но их эволюция
пошла в направлении утраты подвижных стадий размножения, замены
этих стадий интенсивным делением вегетативных клеток, частым об-
ю
разованием акинет и апланоспор и появлением половых протопластов,
лишенных органов передвижения. В качестве довода в пользу проис-
хождения конъюгат от жгутиковых предков приводятся случаи нахож-
дения в конъюгирующих клетках Spirogyra пульсирующих вакуолей
(Lloyd, 1924; Матвиенко, 1941) и появление на полюсах сливающихся
протопластов вытянутых носиков или папилл, наподобие таковых у
Chlamydomonas eugametos. По последним данным (Hoek et al., 1995),
предполагаемый монадный предок конъюгат имел два боковых жгутика,
a Zygnematophyceae на особом ответвлении филогенетического древа
были предшественниками высших растений (Sphenopsida и Magnoliop-
sida) и ряда зеленых водорослей (Klebsormidiophyceae: Klebsormidiales,
Coleochaetales).
Противоположных взглядов придерживаются Штрём (Strom, 1944)
и Топачевский (1952, 1953, 1954), полагающие, что подвижных пред-
ков у конъюгат вообще не было и они возникли на том этапе эволю-
ции, когда жгутиковая форма еще не выработалась, так что отсутствие
ее здесь первично. Штрём идет настолько далеко, что считает конъ-
югаты самостоятельной труппой, равноценной «настоящим» зеленым
водорослям (Chlorophyceae, равнозначным современному отделу Chlo-
rophyta) и произошедшей независимо от них. На современном этапе
фикологи не сомневаются в принадлежности конъюгат к зеленым во-
дорослям; вопрос, очевидно, сводится к тому, обособились ли конъ-
югаты от главного ствола зеленых водорослей после того, как у пос-
ледних выработалась монадная структура, или ранее того. Что касается
непосредственно зигнемовых водорослей, то их широкое географичес-
кое распространение, приуроченность к различным местообитаниям,
большая внутривидовая изменчивость и морфологическая пластичность
видовых и родовых признаков характеризуют эту группу как находя-
щуюся в разгаре эволюции.
По представлениям отечественных фикологов, в группу конъюгат в
ранге самостоятельных порядков входят Mesotaeniales, Gonatozygales,
Zygnematales и Desmidiales. Выяснение родственных связей этих по-
рядков, заключающееся в сравнении эволюционного развития их веге-
тативных и генеративных признаков, дает основание построить следу-
ющую гипотетическую схему их филогении (Yamagishi, 1963а, с ав-
торскими коррективами):
1) пор. Mesotaeniales (род Mesotaenium род Cylindrocystis —ь род
Netrium) пор. Desmidiales-,
2) пор. Mesotaeniales -ь пор. Gonatozygales —> пор. Zygnematales.
В каждой из этих ветвей эволюция талломной организации идет от
одноклеточной формы (мезотениевые и большинство десмидиевых)
через форму нестойких нитей, легко распадающихся на отдельные
клетки, существующие независимо (гонатозиговые и у десмидиевых
роды Desmidium, Gymnozyga, Hyalotheca), к форме настоящего много-
клеточного нитчатого таллома (зигнемовые). Вместе с прогрессивным
усложнением вегетативных признаков у конъюгат намечается и эволю-
ция их генеративной стадии развития. Десмидиевые водоросли (за ис-
ключением родов Closterium, Penium, Hyalotheca, Gymnozyga и Desmi-
dium) по межклеточному, свободному положению зиготы резко обо-
11
соблены от мезотениевых, гонатозиговых и зигнемовых, у которых зи-
гота формируется, как правило, внутриклеточно, замкнуто — в конъ-
югационном канале или в гаметангии.
Мезотениевые водоросли являются наиболее примитивными в груп-
пе конъюгат как по чрезвычайной простоте обычно одиночных клеток,
так и по строению однослойных, лишенных скульптуры и пор, не
сегментированных оболочек. Гонатозиговые водоросли занимают в сис-
теме конъюгат спорное положение, совмещая одновременно признаки
мезотениевых, зигнемовых и десмидиевых водорослей. Они очень близ-
ки к зигнемовым водорослям по типу хлоропластов и нитчатому стро-
ению таллома, но хорошо отличаются от них более хрупкими, легко
распадающимися на отдельные клетки нитями и сегментированными,
с порами и орнаментацией оболочками вегетативных клеток. Кроме
того, в конъюгации у гонатозиговых водорослей принимают участие
только две, свободные и вполне изолированные клетки, в то время
как у зигнемовых конъюгация происходит всегда между клетками в
целых, не распадающихся нитях. Что касается десмидиевых водоро-
слей, то можно с достоверностью предположить, что они произошли
не от нитчатых зигнемовых, дегенерировав в одноклеточную форму
клеток, а от Mesotaenium-поцобных. одноклеточных предков водоро-
слей, параллельно с эволюционным развитием Zygnematales.
Предполагается, что три семейства внутри порядка Zygnematales:
Zygnemataceae, Mougeotiaceae и Spirogyraceae, различающиеся формой
и положением в клетке хлоропласта (осевой звездчатый, осевой плас-
тинчатый, пристенный спирально-лентовидный), произошли от общего
предка (Mesotaenium) и развивались параллельно:
1) пор. Mesotaeniales (род Mesotaenium —> род Cylindrocystis) —> ? пор.
Gonatozygales —> сем. Zygnemataceae (род Zygnema —> ? род Neozygne-
та —> род Zygnemopsis —> ? род Zygogonium —> ?);
2) пор. Mesotaeniales (род Mesotaenium) —> пор. Gonatozygales (род
Gonatozygon) —> сем. Mougeotiaceae (род Mougeotiopsis —> ? род Мои-
geotiella —> род Debarya —> род Mougeotia —> род Temnogametum —> род
Sirocladium)',
3) пор. Mesotaeniales (род Mesotaenium) —> пор. Gonatozygales (род
Genicularia) —> сем. Spirogyraceae (род Spirogyra —> ? —> ? род Тетпо-
gyra род Sirogonium).
Конъюгаты хорошо сохранились в геологических напластованиях
разных эпох в форме микрофоссилий (аналогов современных зигоспор)
благодаря спорополленину в их оболочках. Проведенное сравнение
микрофоссилий с морфотипами зигоспор современных конъюгат поз-
волило установить между ними сходство, в основном на родовом уров-
не, а в некоторых случаях и на видовом. В кайнозойских отложениях
(средний олигоцен—ранний миоцен) были определены разнообразные
зигнемовые, относящиеся к родам Spirogyra, Mougeotia, Zygnema, а
также десмидиевые, сближаемые с современными родами Cosmarium,
Closterium и Penium. Микрофоссилии форм-рода Tetrapidites были оп-
ределены как зигоспоры Mougeotia gracillima, М. punctata, М. produc-
ta, а микрофоссилии форм-рода Rundinella — как зигоспоры видов из
группы Mougeotia laetevirens. В мезозое (ранний мел) зигоспоры из
12
формального рода Schizocystia подушкообразной формой сходны с зи-
госпорами Mougeotia или Mesotaenium. Морфотип зигоспор форм-рода
Lecaniella напоминает зигоспоры Zygnema или Zygnemopsis, а гладкие
эллипсоидные микрофоссилии Ovoidites близки зигоспорам с гладкой
оболочкой Spirogyra. В палеозое (пермь) микрофоссилии таких родов,
как Haplocystia, Mehlisphaeridium и Peltacystia, поразительно схожи с
зигоспорами современных родов Spirotaenia, Cylindrocystis и Debarya.
Карбонские отложения в различных угленосных бассейнах не только
России, но и других регионов мира содержат большое количество мик-
рофоссилий форм-рода Tetraporina, необычайно разнообразных по
форме. С большой долей вероятности они могут рассматриваться как
прямые предшественники современных видов рода Mougeotia. По мере
продвижения вглубь геологических эпох наблюдается явное уменьшение
морфологического разнообразия микрофоссилий — аналогов современ-
ных Zygnematophyceae.
Распространение. Зигнемовые являются одними из наиболее мно-
гочисленных и широко распространенных в пресных водоемах. За не-
которым исключением, их можно считать космополитами, так как они
встречаются на всех континентах, включая Антарктиду. Зигнемовые
водоросли — по преимуществу обитатели пресных вод, но среди них
есть представители с наземным и аэрофильным местообитаниями.
Массовое развитие зигнемовых чаще можно наблюдать в стоячих или
слабопроточных водах, сравнительно чистых, хорошо прогреваемых и
освещенных: в мелких прудах, озерцах, канавах, лужах, ручьях, при-
брежье рек и рисовых полях. В больших озерах зигнемовые развива-
ются в литоральной зоне. В реках и водоемах с заметным течением
часто обнаруживаются прикрепленные формы, такие как Spirogyra flu-
viatilis. Морских видов среди зигнемовых нет, но можно назвать не-
скольких представителей (Spirogyra subsalsa, S. salina, S. cateniformis,
S. tenuissima, S. decimina f. juergensii), которые выдерживают широкую
амплитуду колебаний солености воды — от опресненной до 8.5—
9.7 %о. В прибрежье морей они попадают со стоками речных вод, а
потом разносятся течением на довольно значительные расстояния. К
примеру, в Аральском море стерильные нити Mougeotia найдены во
всех районах моря, за исключением центрального, на глубине до 26 м,
при солености 8—11 %о и температуре 10.4—24.2 °C. Зигнемовые
достигают большой биомассы во всей акватории опресненной и с не-
большими глубинами северной части Каспийского моря, куда влива-
ются воды Волги, Урала, Эмбы.
Рисовые поля в течение вегетационного периода представляют
собой водоемы, исключительно благоприятные для зигнемовых водо-
рослей. Здесь хорошо прослеживается четкая закономерность в их раз-
витии, и к тому же конъюгация у зигнемовых на рисовых полях про-
исходит чаще и обильнее, чем в других местообитаниях.
В минеральных источниках (радоновые, серные, некоторые сазовые
водоемы) зигнемовые никогда не развиваются в массе, и для каждого
из специфических водоемов характерен определенный, хотя и немно-
гочисленный набор видов. Так, в водах с примесью радона хорошо
вегетируют Mougeotia genuflexa f. genuflexa, Spirogyra tenuissima f. re-
13
nuissima, S. hassallii. В сильно минерализованных водоемах (с концен-
трацией минеральных веществ свыше 2000 мг/л) хорошо развиваются
Spirogyra decimina f. communis и f. longata, S. gracilis, S. heeriana,
S. inflata, S. cateniformis, S. varians f. varians.
Сильное течение в реках, особенно в горных районах, разруши-
тельная сила воды в прибойной полосе больших озер в общем пре-
пятствуют росту и вегетации зигнемовых водорослей, подавляющий
процент которых предпочитает стоячие мелкие водоемы. Однако в ро-
дниковых топях и по руслу большинства, преимущественно мелких
рек все же могут интенсивно вегетировать некоторые виды зигнемовых,
в основном Spirogyra, имея вид нежно-зеленых шелковых косм, пере-
плетающихся между собой и устилающих дно на многие метры.
Большинство зигнемовых — олиготрофы, однако у представителей
рода Spirogyra отмечен сравнительно широкий экологический спектр.
Замечено также, что виды Mougeotia предпочитают щелочную воду, а
Zygogonium ericetorum распространен в кислых водах, главным образом
в верховых болотах, где образует при высыхании так называемую ме-
теорную бумагу. Зигнемовые водоросли часто развиваются на увлаж-
ненной почве, выбросах техногенного грунта, на пустошах, где в виде
налетов покрывают большие пространства. Их можно встретить и на
скальных плитах с проточной водой, на известковых и песчаниковых
откосах и вблизи термальных источников. В горных районах зафик-
сированы своеобразные рекорды высоты, достигнутые зигнемовыми:
Spirogyra varians найдена на высоте 3400 м над ур. м. в Заилийском
Алатау (Казахстан), a S. borgeana — свыше 5600 м над ур. м. в Ти-
бете (Китай).
Благоприятные для развития различных видов Zygnematales пределы
концентрации водородных ионов различны. Так, например, представи-
тели рода Zygogonium растут при pH 3.5—5.5, Spirogyra varians и S. mi-
rabilis — при pH 6.0—7.4, a S. crassa и S. maxima — при pH 7.8—8.5.
Большинство зигнемовых нуждается для своего развития в интенсивном
освещении, встречаясь лишь в летние месяцы в хорошо освещенных по-
верхностных слоях водоемов. Температурный оптимум для большинства
видов лежит между 14 и 22 °C. Некоторые виды выдерживают без ясно
выраженных повреждений более или менее продолжительное повышение
температуры до +35 °C или понижение до - 8 °C. Некоторые из зигнем
вмерзают в лед, долгое время оставаясь живыми.
Из зигнемовых водорослей три рода: Spirogyra, Mougeotia, Zygnema
имеют наибольшее количество видов, подавляющий процент которых
относится к повсеместно распространенным. Другие роды характери-
зуются сравнительно малым количеством видов и ограниченным рас-
пространением: виды Temnogametum найдены только в тропических и
субтропических областях, а виды Sirocladium описаны только для почв
Индии и Кубы. У родов Zygogonium и Sirogonium соответственно 22
и 15 видов имеют узко очерченные ареалы с определенной экологи-
ческой приуроченностью, и только Zygogonium ericetorum и Sirogonium
sticticum широко распространены на всех континентах с большим диа-
пазоном экологических условий. Несомненно, что с ростом исследо-
ваний в природе и дальнейшим изучением особенностей развития зиг-
14
немовых водорослей границы ареалов многих видов и даже родов будут
расширяться и уточняться.
Практическое значение. Клетка зигнемовых водорослей с хорошо
заметным крупным ядром, ядрышком и интенсивным движением про-
топлазмы давно считается классическим объектом для цитологов и био-
физиков. Зигнемовых, как представителей зеленых водорослей, легко
демонстрировать студентам и школьникам: живой материал весной и
летом есть везде, а зимой они хорошо растут в склянках с природной
водой. Теоретический интерес зигнемовые несомненно вызывают у эво-
люционистов и морфологов — в построении филогенетического древа,
в изучении приспособительных морфологических признаков, возника-
ющих у зигнемовых при различных местообитаниях и протекании
столь необычного для других водорослей полового процесса, при раз-
работке гипотезы о возникновении высших растений от нитчатых зе-
леных водорослей. В геологии аналоги зигоспор современных конъюгат
успешно используются для целей стратиграфии и распознавания воз-
раста пород. Палинологи, без знания современных зигнемовых водо-
рослей, определяли прежде микрофоссилии как пыльцу магнолиевых
или пыльцу голосемянных без воздушных мешков. Экологи широко
используют зигнемовые водоросли как прекрасные показатели чистоты
воды, в некоторых случаях решая чрезвычайно важные проблемы ан-
тропогенного загрязнения. Заслуживает внимания вопрос о значении
зигнемовых в создании первичной биопродукции и определении сап-
робности воды. В массе развиваясь, часто вызывая цветение воды в
мелководных водоемах и прибрежных участках больших озер и опрес-
ненных участках морей, они способствуют образованию высокопродук-
тивных илов. По данным ряда авторов, зигнемовые водоросли приоб-
ретают особое значение в питании бентосоядных рыб: маринки и са-
зана (в устье Волги и на северном побережье Каспийского моря),
воблы, частично усача, белоглазки и шемаи (в прибрежных районах
и заливах Аральского моря). Зигнемовые водоросли являются харак-
терным кормом для молоди сазана и усача на рисовых полях. Так,
частота встречаемости нитчатых зеленых водорослей, среди которых
преобладали зигнемовые, в желудках сазана колеблется от 34.5 до
54.55 %.
Прямое использование зигнемовых водорослей человеком невелико
и определяется местными обычаями и привычками: в Китае из них
пекут лепешки, в Чехии их добавляют в муку для десертного печенья,
а в России, в Новгородской области, их в высушенном виде подме-
шивают зимой в корм скоту — овцам и свиньям, на севере (Архан-
гельская область) ими, тоже в высушенном виде, прослаивают бревна
в избах, подметив с давних пор их хорошие антисептические свойства.
Методы сбора и изучения. Стадии вегетативного развития у зиг-
немовых водорослей наблюдаются в течение всего года с двумя нерав-
номерными пиками — наивысшим в весенне-летний сезон и мень-
шим — теплой осенью. Доказано, что многие нити Zygnema и Spiro-
gyra благополучно перезимовывают, вмерзая в лед или оседая на дно
водоемов. Процессы конъюгации и зиготообразования сравнительно
кратковременны и непостоянны и зависят, во-первых, от температур-
15
ных и световых условий сезона, а во-вторых -г- от особенностей гид-
рологического и гидрохимического режимов каждого водоема. Поэтому
сбор материала обычно приурочен к наиболее теплым месяцам с уче-
том климатических особенностей области и района. Материал соби-
рается вручную, обычно в прибрежной полосе водоемов, где зигнемо-
вые развиваются обильно и хорошо видны невооруженным глазом. Же-
лательно брать несколько образцов из разных местообитаний одного
и того же водоема — со дна и с поверхности, среди высших растений,
из свободноплавающих скоплений (так называемой тины) и прикреп-
ленные дерновинки. Следует по возможности брать нити разного воз-
раста, захватывая у прикрепленных форм ризоидную часть. Главным
условием успеха должен быть сбор материала в вегетативном и гене-
ративном состояниях. Поэтому следует быть особенно внимательным,
выискивая конъюгирующие нити: как правило, они хорошо отличаются
от изумрудно-зеленых вегетативных нитей своим желтовато-бурым
блеклым оттенком.
Сбор материала в каждом, отдельно взятом биотопе имеет свои
особенности. К примеру, пробы для определения видового состава в
родниках собираются не непосредственно в источниках, где конъюга-
ция невозможна из-за слишком низкой температуры воды (5—8 °C),
а в ручьях, топях и заводях от родников, где вода хорошо прогревается
(до 25—28 °C) и глубина незначительная.
По руслу небольших, преимущественно мелких рек прикрепленные
формы зигнемовых интенсивно вегетируют, но в большинстве остаются
стерильными (исключение составляет Spirogyra fluviatilis). Преимущест-
венно конъюгируют только нити, всплывшие на поверхность воды, где
они, спирально переплетенные, постепенно приобретают вид рыхлого,
с пузырьками воздуха внутри, желтовато-бурого скопления. Конъюгация
протекает в прогреваемых, более мелких, чем по руслу, местах, при
температуре воды 18—30 °C, pH 6.9—8.2.
Иногда местообитания зигнемовых водорослей бывают необычными,
развитие водорослей бурным и кратковременным, и поэтому от иссле-
дователя требуются особое внимание и осторожность для распознава-
ния объекта исследования. Например, типовая форма Zygnema insigne
была собрана мною в водах временного водопада 10—12 м высоты,
образовавшегося от ливней в горах Кызыл-Рай (Прибалхашье, Казах-
стан). Водоросль развивалась в течение 2—3 дней, росла на скалах
при очень сильном течении и нити ее были темно-зелеными, почти
черными, они устилали дно потока от основания водопада и были
переплетены в плотные пучки, окруженные толстой стекловидной
слизью, напоминая своим внешним видом синезеленые водоросли (Mic-
rocoleus). Вегетируя обильно под сильным напором воды, конъюгиро-
вать они начали позднее, спустя неделю после начала вегетации, в
ручье от водопада, на глубине 0.2—0.5 м.
Собранный живой материал желательно просматривать под сильной
лупой или полевым микроскопом на месте сбора. Просмотр и опре-
деленная фиксация материала ведутся в зависимости от цели сбора.
Свежесобранные водоросли, необходимые для занятий в школах и
вузах, должны быть в хорошем состоянии: хлоропласты интенсивной
16
окраски (яркого изумрудно-зеленого цвета у спирогиры, оливково-зе-
леного — у зигнемы и мужоции) и свободного (не плотного) распо-
ложения в довольно крупного размера клетке. Для первоначального
знакомства с клеткой зигнемовых желательно выбрать экземпляры спи-
рогиры, ширина вегетативных клеток которых была бы от 40 до
70 мкм, с наличием 3—4 неплотно расположенных хлоропластов, что
позволяло бы отчетливо видеть крупное ядро в центре клетки.
Удачно собранным считается материал, в котором представлены
последовательные стадии конъюгации и зрелые зиготы. Для цитологи-
ческих целей фиксировать его рекомендуется в хромово-уксусной смеси
с последующим промыванием в дистиллированной воде и осторожным
проведением через каждые 5—10 минут через шкалу спиртов, начиная
с 20°-ного до 75—80°-ного спирта, в котором.можно хранить материал.
Следует избегать непосредственного перенесения материала из крепкого
спирта в воду, лучше провести его через спирты крепостью 50, 40,
30° и тогда уже поместить в воду. Фиксация формалином для цито-
логических целей считается грубой.
Фиксированные формалином водоросли достаточно хорошо сохра-
няются для определения их систематической принадлежности, которое
основывается исключительно на морфологических признаках. Фикси-
рование обычно производится 4 %-ным формалином в небольших по
объему склянках, желательно темного стекла. Если формалин более
крепкой концентрации, то он каплями добавляется в склянку с водой
и материалом до слабого запаха. Для нейтрализации хорошо добавить
щепотку соды. При наличии в пробе не только стадий конъюгации,
но и зрелых зигот рекомендуется наряду с фиксированием материала
иметь гербарный образец — для последующего выращивания водоро-
слей в культуре непосредственно из зигоспор, которые в высушенном
состоянии сохраняют свою «всхожесть» до 20—30 лет. Для гербари-
зации на плотном листе белой бумаги в воде равномерно распределя-
ются скопления нитей, затем этот лист осторожно, в горизонтальном
положении, поднимают из воды, он прокладывается фильтровальной
бумагой и просушивается сменой бумаг в затененном (не на солнце)
месте. Одновременно со сбором материала обычно измеряют темпера-
туру воды и pH (по колориметрической шкале или лакмусовой бума-
гой).
Пробы зигнемовых водорослей, наземных или аэрофилов, растущих
на сырой почве, горных скалах, песчаниковом или техногенном грунте,
пустошах, тоже в виде хорошо заметных глазом дернинок и налетов,
наряду с фиксацией формалином собираются лопаточкой или ножом,
предварительно простерилизованными спиртом и прокаленными непос-
редственно перед сбором материала, и помещаются либо как гербарный
материал в почвенные конверты, либо в чашки Петри для исследования
в культуре.
В лабораторных условиях собранный материал, весь или по частям,
предварительно просматривается под бинокулярной лупой для визуаль-
ной оценки видового обилия, стадий конъюгации и зрелости зигот.
При наличии фертильной стадии и зигоспор материал исследуется под
микроскопом при постоянном применении водной и масляной иммер-
2 Рундина Л. А.
17
сий. Для просветления скульптуры на оболочках зигоспор применяются
кислоты различной концентрации: соляная, уксусная, молочная и азот-
ная, а также щелочь (едкий натр). Для изучения строения оболочек
на сканирующем электронном микроскопе применяется особая мето-
дика для растворения тех или иных слоев у зигоспоры, с применением
ферментов пектиназы и целлюлазы.
Каждая проба состоит, как правило, из серии образцов; средние
величины различных параметров вычисляются по выборке, состоящей
из 10—50, иногда 100 экземпляров.
Методика культивирования зигнемовых на питательных растворах
довольно сложна, так как у разных видов неодинакова потребность в
биодобавках, минералах, солях, микроэлементах, различна реакция на
температуру, освещенность, pH. Неплохие результаты дает культивирова-
ние водорослей в водной вытяжке из почвы, через которую рекомен-
дуется пропускать некоторое количество углекислого газа (Голлербах,
Полянский, 1951). В подходящих условиях температуры и освещения
некоторые экземпляры свежесобранной спирогиры могут продержаться
неделю-две в более или менее удовлетворительном состоянии. В этом
случае рекомендуется осторожно поместить (не набивая!) несколько
пучков нитей в просторную и незакупоренную склянку (колбу, лабо-
раторный стакан) с водой из природного водоема и поставить ее на
северное окно, время от времени, по мере испарения, доливая воду
по стенке сосуда.
2. МОРФОЛОГИЯ ЗИГНЕМОВЫХ ВОДОРОСЛЕЙ
Строение нитей и вегетативных клеток. Нити зигнемовых водоро-
слей, как правило, неразветвленные, состоящие из одного ряда цилинд-
рических клеток, иногда с короткими ответвлениями (веточками) из
1—10 клеток у наземных форм. Нити свободноплавающие, часто об-
разующие ватообразные скопления на поверхности воды; иногда они
прикреплены ризоидами к субстрату, и тогда имеется ясное различие
между основанием и вершиной. Рост нитей в длину осуществляется
поперечным делением клеток. Нити макроскопические, от нескольких
миллиметров до 8—15 см длиной, окруженные общей слизью. Пере-
плетенные между собою и прикрепленные к субстрату, они хорошо
заметны глазом — в виде дерновинок или пучков, при наземном мес-
тообитании — в виде налетов и обрастаний. У нитей, прикрепленных
к различного рода субстратам, наземным или водным, развиваются
ризоиды — своеобразные выросты клетки, чаще базальной. Для ризо-
идов характерны утолщение оболочки и отсутствие четко оформлен-
ного хлоропласта. Ризоиды имеют крайне разнообразный вид — от
слегка изогнутых неразветвленных выростов до значительно развет-
вленных, розетковидных и лопастевидных (рис. 1). Ризоиды характер-
ны для значительного числа видов, их образование и форма зависят
от экологических условий среды и местообитания водоросли.
18
Рис. 1. Ризоиды у некоторых представителей зигнемовых водорослей: 1—4 — Spirogyra flu-
viatilis; 5 — S. tenuissima f. cylindrical 6 — S. occidentalism 7 — Zygnema melanosporum. (По:
I-—4 — Rieth, 1976; 5 — Рундина, 1988; 6 — Gauthier-Lievre, 1965; 7 — Skuja, 1964).
19
Оболочка вегетативных клеток довольно тонкая, не сегментирован-
ная, без пор и орнаментаций, гладкая, двух- или многослойная. Внешний
слой студенистый и аморфный, состоящий из пектиновых веществ. Ис-
ключение составляет род Sirogonium, у представителей которого отсутст-
вует пектиновый слой оболочки, что обусловливает некоторую жесткость
его нитей. Внутренние слои оболочки целлюлозные, фибриллярные. По
последним ультраструктурным исследованиям, сборка целлюлозных мик-
рофибрилл происходит в терминальном комплексе, находящемся в плаз-
малемме и имеющем вид розетки из 6 цранул в форме кольца и цент-
ральной, более крупной глобулы. Такие четкие картины розеток и от-
печатков микрофибрилл, связи их с концами фибрилл целлюлозы, как
у зигнемовых, были найдены также у десмидиевых, харовых и хлоро-
кокковых водорослей. Внешний студенистый покров нитей достигает
иногда значительной толщины и придает зигнемовым водорослям харак-
терную для них шелковистость. Толщина этого покрова, как правило,
зависит от условий местообитания, в некоторых случаях она видоспеци-
фична. Обычно с увеличением концентрации водородных ионов (pH)
толщина покрова уменьшается. В случае длительной приостановки про-
цессов размножения клеток делением и роста клеток, вызванной подсу-
шиванием субстрата или повышением кислотности среды, клеточные
оболочки утолщаются иногда весьма значительно. При повторных пре-
кращениях и возобновлениях указанных процессов внешний слой обо-
лочки отделяется от внутреннего и из-за роста клеток в длину разрыва-
ется на 2 или 3 части, в результате чего на концах клетки образуется
шапочка, а по длине нити происходят довольно значительные разрывы
наружного слоя (рис. 2, 5, 6). Прилегающие друг к другу шапочки двух
соседних клеток обычно образуют при этом одно целое, имея на про-
дольном сечении Н-образную форму. Такие образования наблюдаются у
зигнемовых водорослей при наземном местообитании (Zygogonium) и у
некоторых представителей Spirogyra и Zygnema. Установлено также, что
слизь, окружающая нити Zygnema в фертильной стадии, обладает гор-
мональными свойствами, повышающими половую активность гамет.
Поперечные перегородки клеток, иначе называемые септами, у
большинства зигнемовых водорослей гладкие (рис. 3, 1, 2). На границе
двух примыкающих друг к другу клеток они выглядят в обычном со-
стоянии как две параллельные плоскости с тонкой пектиновой про-
слойкой между ними. Когда тургорное давление в клетках уменьшается,
поперечные перегородки прогибаются, как бы втягиваются внутрь кле-
ток. Виды Spirogyra имеют три формы поперечных перегородок: глад-
кие, гладкие с дополнительной кольцевой накладкой и складчатые
(рис. 3). Складчатые перегородки образуются путем врастания септ в
форме загнутого языка внутрь клеток. Предполагается, что такие врас-
тания септы возникают при увеличении поверхности поперечной пе-
регородки в результате роста вегетативных клеток. При повышении
тургорного давления в клетках, имеющих складчатые септы, а это про-
исходит обычно при половом процессе, врастания в перегородках рас-
правляются. Таким образом, при конъюгации в нитях могут встречаться
как складчатые, так и гладкие септы, причем их правильное чередо-
вание или частота встречаемости одной из них зависит от стадии конъ-
20
Рис. 2. Строение нитей и клеток некоторых представителей зигнемовых водорослей: 1 —
Zygnema pectinatum-, 2—4 — Spirogyra borysthenica- 5, 6 — Mougeotia genuflexa. (По: 1, 5,
6 — De Bary, 1858; 2—4 — Казаковский, Смирнов, 19136).
югации и степени участия в ней той или иной клетки (рис. 2, 2—4).
У некоторых видов Spirogyra собственно складчатые септы модифици-
руются и приобретают разнообразную форму. Только у двух видов
Spirogyra (секция Colligata) имеется дополнительная накладка в виде
довольно широкого кольца, обхватывающего соприкасающиеся друг с
другом клетки с гладкой поперечной перегородкой. Диаметр кольца
почти равен ширине клетки, и он не увеличивается с ростом клетки.
По окружности кольца на одинаковом расстоянии от его ободов за-
метно выделяется более темная полоса. При определенном навыке и
легком усилии кольцо может быть снято. Предполагается, что кольце-
вая накладка препятствует распаду нити на отдельные клетки.
Протопласт зигнемовых водорослей по своей общей организации
и набору органелл не отличается от других представителей зеленых
21
водорослей. Цитоплазма обычно располагается постенно, одним слоем,
а в клетках большого диаметра образует несколько слоев. Даже при
небольшом увеличении микроскопа заметно ее активное движение. От
прилегающего к стенке слоя цитоплазма дает тяжи через центральную
полость клетки, заполненную клеточным соком. У спирогиры на таких
тяжах подвешено ядро в центральной части клетки, у зигнемы тяжи
тянутся к двум центрам — звездчатым хлоропластам, а у мужоции
через центральную полость клетки проходит цитоплазматическая плас-
тинка, в которую погружен пластинчатый хлоропласт. Клеточный сок
в вакуоли обычно бесцветный, но у некоторых видов Zygnema и всех
представителей с наземным и аэрофильным местообитаниями он бывает
окрашен в красный, фиолетовый или розовый цвета разной интенсив-
ности. Окраска обусловливается действием дополнительного пигмента
фикопорфирина, легко растворимого в воде, а ее интенсивность —
степенью светового и теплового воздействий.
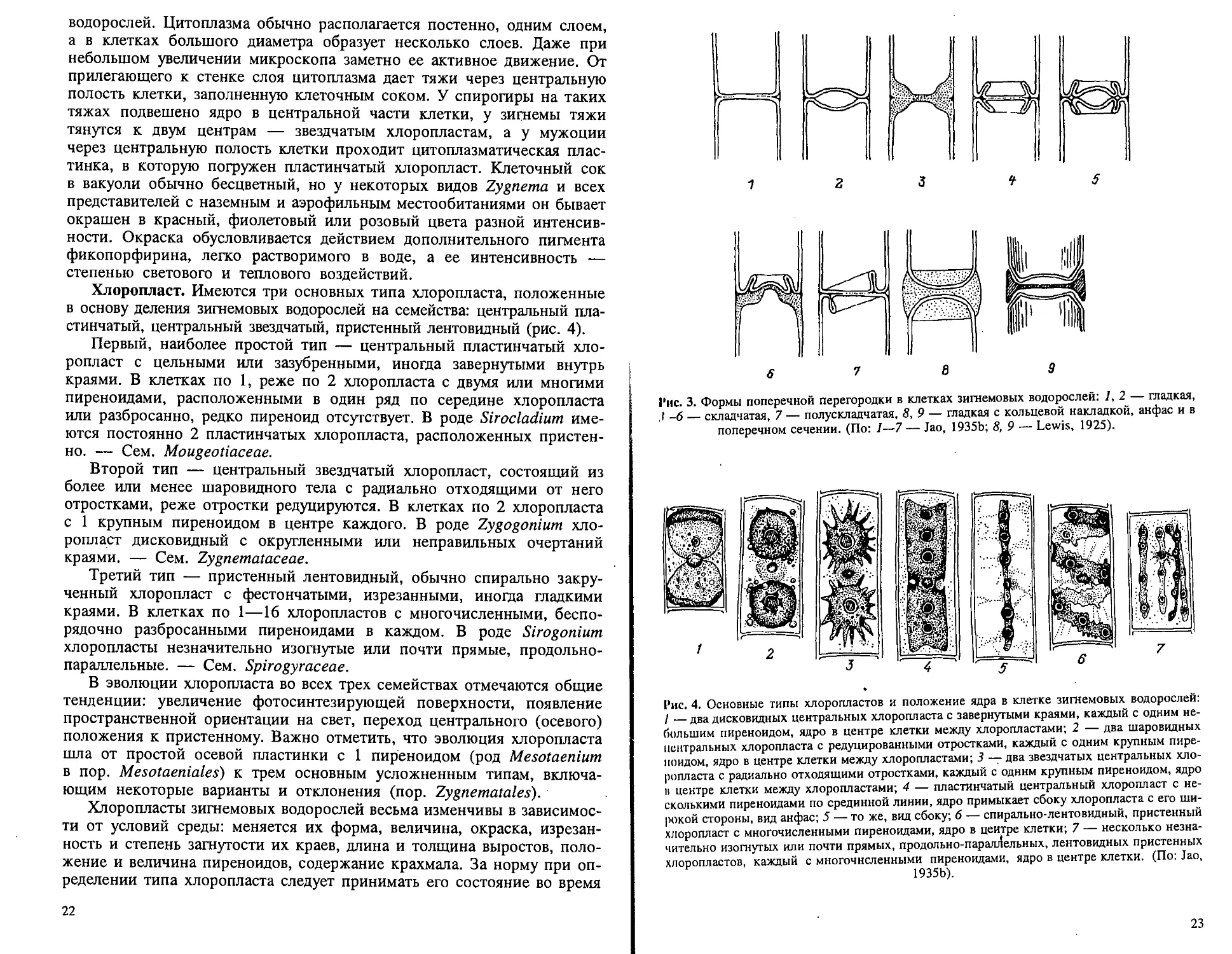
Хлоропласт. Имеются три основных типа хлоропласта, положенные
в основу деления зигнемовых водорослей на семейства: центральный пла-
стинчатый, центральный звездчатый, пристенный лентовидный (рис. 4).
Первый, наиболее простой тип — центральный пластинчатый хло-
ропласт с цельными или зазубренными, иногда завернутыми внутрь
краями. В клетках по 1, реже по 2 хлоропласта с двумя или многими
пиреноидами, расположенными в один ряд по середине хлоропласта
или разбросанно, редко пиреноид отсутствует. В роде Sirocladium име-
ются постоянно 2 пластинчатых хлоропласта, расположенных пристен-
но. — Сем. Mougeotiaceae.
Второй тип — центральный звездчатый хлоропласт, состоящий из
более или менее шаровидного тела с радиально отходящими от него
отростками, реже отростки редуцируются. В клетках по 2 хлоропласта
с 1 крупным пиреноидом в центре каждого. В роде Zygogonium хло-
ропласт дисковидный с округленными или неправильных очертаний
краями. — Сем. Zygnemataceae.
Третий тип — пристенный лентовидный, обычно спирально закру-
ченный хлоропласт с фестончатыми, изрезанными, иногда гладкими
краями. В клетках по 1—16 хлоропластов с многочисленными, беспо-
рядочно разбросанными пиреноидами в каждом. В роде Sirogonium
хлоропласты незначительно изогнутые или почти прямые, продольно-
параллельные. — Сем. Spirogyraceae.
В эволюции хлоропласта во всех трех семействах отмечаются общие
тенденции: увеличение фотосинтезирующей поверхности, появление
пространственной ориентации на свет, переход центрального (осевого)
положения к пристенному. Важно отметить, что эволюция хлоропласта
шла от простой осевой пластинки с 1 пиреноидом (род Mesotaenium
в пор. Mesotaeniales) к трем основным усложненным типам, включа-
ющим некоторые варианты и отклонения (пор. Zygnematales).
Хлоропласты зигнемовых водорослей весьма изменчивы в зависимос-
ти от условий среды: меняется их форма, величина, окраска, изрезан-
ность и степень загнутости их краев, длина и толщина выростов, поло-
жение и величина пиреноидов, содержание крахмала. За норму при оп-
ределении типа хлоропласта следует принимать его состояние во время
22
Рис. 3. Формы поперечной перегородки в клетках зигнемовых водорослей: 1,2 — гладкая,
1-6 — складчатая, 7 — полускладчатая, 8, 9 — гладкая с кольцевой накладкой, анфас и в
поперечном сечении. (По: I—7 — Jao, 1935b; 8, 9 — Lewis, 1925).
Рис. 4. Основные типы хлоропластов и положение ядра в клетке зигнемовых водорослей:
I — два дисковидных центральных хлоропласта с завернутыми краями, каждый с одним не-
большим пиреноидом, ядро в центре клетки между хлоропластами; 2 — два шаровидных
центральных хлоропласта с редуцированными отростками, каждый с одним крупным пире-
ноидом, ядро в центре клетки между хлоропластами; 3 — два звездчатых центральных хло-
ропласта с радиально отходящими отростками, каждый с одним крупным пиреноидом, ядро
в центре клетки между хлоропластами; 4 — пластинчатый центральный хлоропласт с не-
сколькими пиреноидами по срединной линии, ядро примыкает сбоку хлоропласта с его ши-
рокой стороны, вид анфас; 5 — то же, вад сбоку; 6 — спирально-лентовидный, пристенный
хлоропласт с многочисленными пиреноидами, ядро в центре клетки; 7 — несколько незна-
чительно изогнутых или почти прямых, продольно-параллельных, лентовидных пристенных
хлоропластов, каждый с многочисленными пиреноидами, ядро в центре клетки. (По: Jao,
1935b).
23
интенсивного размножения клеток делением. Подвижность хлоропласта,
имеющая место у представителей всех родов, особенно наглядна у му-
жоции. При ярком освещении ее Хлоропласт поворачивается на 90° во-
круг своей оси и тогда выглядит не привычной для глаза широкой плас-
тиной, а узкой полоской (рис. 4, 5). Кроме того, доказано наличие спи-
ральной структуры в постенном слое цитоплазмы клеток у мужоции,
которая при определенных условиях (продолжительность освещения и
его высокая интенсивность) приводит к спиральному закручиванию и са-
мого хлоропласта, придающему ему сходство с хлоропластом спирогиры.
Сходство усиливается, если хлоропласт в клетке мужоции делится вдоль,
полностью или частично, в результате чего внутри клетки оказываются
две спирально извитые ленты.
Как и у всех зеленых водорослей, хлоропласты зигнемовых содер-
жат в качестве основных пигментов хлорофилл а и хлорофилл b и
дополнительные пигменты: 6-каротин, волютин, лютеин, виолаксантин
и неоксантин.
Пиреноиды, как особые клеточные образования, непосредственно
связаны с хлоропластом и часто достигают у зигнемовых водорослей
очень крупных размеров. Они представляют собой шаровидные или
почти шаровидные бесцветные тельца, вокруг которых в процессе фо-
тосинтеза откладывается крахмал в форме мелких зернистых глыбок.
При усиленном фотосинтезе крахмал может откладываться и в самой
толще хлоропласта. Размеры пиреноидов, их количество и расположе-
ние по хлоропласту, хотя и зависят от экологических условий, но все
же являются систематическим признаком: у представителей сем. Zyg-
nemataceae в клетках всегда 2 очень крупных пиреноида, погруженных
по одному внутрь каждого хлоропласта, в семействах Spirogyraceae и
Mougeotiaceae пиреноиды, как правило, многочисленные, располагаю-
щиеся по средней линии хлоропласта или беспорядочно. У единствен-
ного представителя рода Mougeotiopsis пиреноида нет, а у наземных
представителей рода Zygogonium пиреноид может временно исчезать.
Запасные вещества, кроме крахмала, в клетках зигнемовых водо-
рослей представлены еще каплями масла и так называемыми кариода-
ми — каплями неустановленного характера, возможно являющимися
запасным веществом или рудиментарными пиреноидами. Из других
включений заметны в клетках блестящие капли дубильных веществ
(танины) и кристаллы щавелевокислого кальция, иногда откладываю-
щиеся на наружной оболочке клетки. В клеточной вакуоли находятся
в растворе сахар и другие вещества, которые рассматриваются как
питательные и как обусловливающие тургор клетки.
Ядро занимает различное положение в клетке в пределах каждого
из семейств пор. Zygnematales. У представителей рода Spirogyra, типо-
вого в сем. Spirogyraceae, клеточное ядро очень крупное (от 5—8 мкм
у 5. hyalina до 40 мкм у S. crassa), с ясно заметным ядрышком (до
16 мкм у S. crassa). Оно расположено в центральной вакуоли, окружено
слоем цитоплазмы и подвешено на цитоплазматических тяжах, отходя-
щих от постенного слоя цитоплазмы. У одних видов ядро имеет шаро-
видную форму, у других — линзовидную и тогда ориентировано поперек
оси клетки. Такое разнообразие формы ядра сочетается с некоторыми
24
морфологическими и физиологическими признаками, в первую очередь
с шириной и длиной клетки. Ядро у представителей рода Zygnema, ти-
пового в сем. Zygnemataceae, лежит между двумя звездчатыми хлороплас-
тами в протоплазматическом мостике; оно значительно меньше, чем у
спирогир (от 4—8 мкм у Z. cruciatum до 12 мкм у Z. insigne), и его
иногда трудно заметить из-за многочисленных радиальных выростов хло-
ропластов. Ядро у представителей рода Mougeotia, типового в сем. Mou-
geotiaceae, сравнительно небольшое, оно прилегает к центру хлоро-
пласта с его широкой стороны. У Mougeotia часто встречаются ядра с
2—3 ядрышками, и их число может доходить до 8.
Кариотипы лучше изучены у родов Spirogyra и Zygnema, значитель-
но слабее — у Mougeotia. У Spirogyra преобладают виды с четным
набором хромосом, причем не всегда кратное увеличение числа хро-
мосом можно рассматривать как полиплоидную серию. Самое низкое
их число п = 2 (£ teodorescoi, S. weberi, S. cylindrical, а самое высо-
кое n = 92 ± 2 (S. nitida). У Sirogonium pseudofloridanum отмечено наи-
высшее число хромосом для зигнемовых водорослей: п = 100 ±2. У
Zygnema самое низкое число хромосом равно 12—14 (Z. stellinum), а
самое высокое — 82, и между ними почти непрерывный ряд после-
довательных чисел. Хромосомы зигнемы значительно мельче и более
слабо дифференцированы, чем у спирогиры. В роде Mougeotia самое
низкое хромосомное число равно 12, а самое высокое — 94. Распре-
деление хромосом по видам проследить не удалось, поскольку изучено
всего два вида (М. nummuloides и М. viridis). Кариотипы зигнемовых
водорослей состоят как из мелких хромосом с размерами, не превы-
шающими 1 мкм, так и из комплекса мелких и крупных хромосом с
размерами до 3 мкм (Spirogyra submargaritata). Последний кариотип
встречается значительно чаще, особенно у Spirogyra и Sirogonium. Ка-
риотипы, состоящие исключительно из крупных, порядка 6.7—8.1 мкм,
хромосом, встречаются крайне редко (Spirogyra teodorescoi). Хромосо-
мы могут быть или мелкими, палочковидными, с хорошо выраженными,
параллельно лежащими хроматидами, что подтверждает наличие либо
полицентрических, либо диффузных центромер, или точкообразными,
в которых природа центромер пока остается неясной. Попытка выяв-
ления связи между кариотипическими и морфологическими признака-
ми, что было бы важно для идентификации видов в случае отсутствия
зигот, оказалась пока безуспешной. Экземпляры, относящиеся к раз-
личным видам, могут иметь одинаковое число хромосом и сильно от-
личаться морфологически. Не существует связи между шириной нити
и числом хромосом, числом хромосом и числом хлоропластов.
Митоз (деление вегетативной клетки) у зигнемовых водорослей про-
текает при благоприятных условиях в течение 1—1.5 ч в вечернее
время и начинается прежде всего с деления ядра (рис. 5). По ключевым
признакам — по поведению ядерной оболочки и отсутствию центрио-
лей, связанному с выпадением в жизненном цикле подвижной стадии,
митоз у зигнемовых водорослей охарактеризован как закрытый ацен-
трический (Седова, 1996).
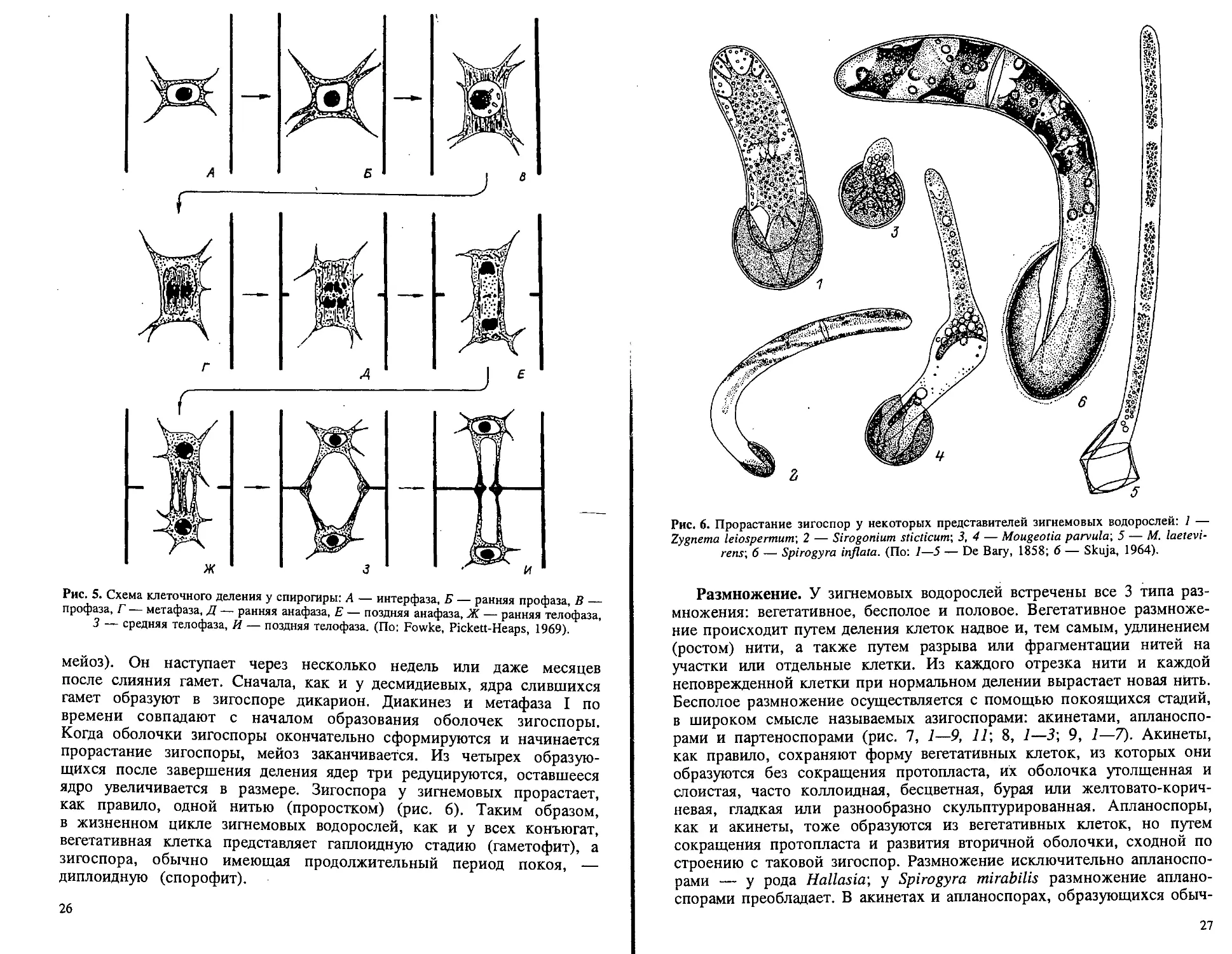
Мейоз, или редукционное деление, у зигнемовых, как и у всех
конъюгат, происходит при прорастании зигоспоры (зиготический
25
Рис. 5. Схема клеточного деления у спирогиры: А — интерфаза, Б — ранняя профаза, В —
профаза, Г — метафаза, Д — ранняя анафаза, Е — поздняя анафаза, Ж — ранняя телофаза,
3 — средняя телофаза, Й — поздняя телофаза. (По: Fowke, Pickett-Heaps, 1969).
мейоз). Он наступает через несколько недель или даже месяцев
после слияния гамет. Сначала, как и у десмидиевых, ядра слившихся
гамет образуют в зигоспоре дикарион. Диакинез и метафаза I по
времени совпадают с началом образования оболочек зигоспоры.
Когда оболочки зигоспоры окончательно сформируются и начинается
прорастание зигоспоры, мейоз заканчивается. Из четырех образую-
щихся после завершения деления ядер три редуцируются, оставшееся
ядро увеличивается в размере. Зигоспора у зигнемовых прорастает,
как правило, одной нитью (проростком) (рис. 6). Таким образом,
в жизненном цикле зигнемовых водорослей, как и у всех конъюгат,
вегетативная клетка представляет гаплоидную стадию (гаметофит), а
зигоспора, обычно имеющая продолжительный период покоя, —
диплоидную (спорофит).
26
Рис. 6. Прорастание зигоспор у некоторых представителей зигнемовых водорослей: 1 —
Zygnema leiospermum\ 2 — Sirogonium sticticum', 3, 4 — Mougeotia parvula', 5 — M. laetevi-
rens\ 6 — Spirogyra inflata. (По: I—5 — De Вагу, 1858; 6 — Skuja, 1964).
Размножение. У зигнемовых водорослей встречены все 3 типа раз-
множения: вегетативное, бесполое и половое. Вегетативное размноже-
ние происходит путем деления клеток надвое и, тем самым, удлинением
(ростом) нити, а также путем разрыва или фрагментации нитей на
участки или отдельные клетки. Из каждого отрезка нити и каждой
неповрежденной клетки при нормальном делении вырастает новая нить.
Бесполое размножение осуществляется с помощью покоящихся стадий,
в широком смысле называемых азигоспорами: акинетами, апланоспо-
рами и партеноспорами (рис. 7, 1—9, 1Г, 8, 1—3; 9, 1—7). Акинеты,
как правило, сохраняют форму вегетативных клеток, из которых они
образуются без сокращения протопласта, их оболочка утолщенная и
слоистая, часто коллоидная, бесцветная, бурая или желтовато-корич-
невая, гладкая или разнообразно скульптурированная. Апланоспоры,
как и акинеты, тоже образуются из вегетативных клеток, но путем
сокращения протопласта и развития вторичной оболочки, сходной по
строению с таковой зигоспор. Размножение исключительно апланоспо-
рами — у рода Hallasia-, у Spirogyra mirabilis размножение аплано-
спорами преобладает. В акинетах и апланоспорах, образующихся обыч-
27
Рис. 7. Образование апланоспор и партеноспор у Spirogyra mirabilis (схема): 1,2 — гипер-
трофированные апланоспоры в одногодичной культуре; 3 — нормальная апланоспора во
вздувшемся апланоспорангии; 4 — то же, с конъюгационным выростом; 5 — поделившаяся
апланоспора; 6—9 — образование партеноспор при лестничной конъюгации; 10 — образо-
вание нормальной зигоспоры при лестничной (терминальной) конъюгации; 11 — образова-
ние двух партеноспор При лестничной (терминальной) конъюгации. (По: Rieth, 1972b).
но при неблагоприятных условиях, накапливаются в больших количест-
вах крахмал и масло. Партеноспоры — это неслившиеся гаметы, воз-
никновение их связано с определенными нарушениями в процессе
конъюгации отдельных клеток; по форме, цвету и строению их обо-
лочки подобны зигоспорам, но по размерам партеноспоры вдвое мень-
ше зигоспор.
Половое размножение у всех зигаемовых — так называемая конъ-
югация, т. е. слияние двух недифференцированных протопластов, вы-
полняющих функцию гамет. Различают два основных типа конъюгации:
лестничную и боковую (рис. 10).
При лестничной конъюгации нити располагаются напротив друг
друга, обычно попарно, реже до 5 одновременно, спирально закручи-
ваясь по всей длине пучка, иногда кольцеобразно скручиваясь на кон-
цах. В начале конъюгации у взаимно противоположных клеток в двух
сближенных нитях оболочка начинает образовывать выпячивания, на-
правленные навстречу друг другу. Эти выпячивания, разрастаясь, смы-
каются, их вершинные оболочки растворяются, и в результате получа-
28
Рис. 8. Образование апланоспор и зигоспор у Mougeotia abnormis (схема): 1—3 — различ-
ные формы и положения апланоспор в разной степени изогнутых клетках; 4 — образование
зигоспоры при лестничной конъюгации; 5,6 — образование зигоспор при терминальной
конъюгации без образования конъюгационного канала. (По: Rieth, 1972а).
ется сквозной конъюгационный канал, соединяющий обе конъюгирую-
щие клетки (рис. 10, 1—5; 11; 12, 1, 2). Так как этот’процесс про-
текает во многих клетках сразу, то получается подобие лестницы, пря-
мой или причудливо изогнутой в зависимости от степени изогнутости
конъюгирующих клеток. Только в роде Mougeotia встречена псевдо-
конъюгация — многократное деление и рост клеток конъюгационного
канала, функционирующих в этом случае как обычные вегетативные
клетки.
Боковая конъюгация протекает в пределах одной и той же нити,
между соседними клетками, с образованием конъюгационного канала
наподобие изогнутого рукава, в разной степени выраженного, или без
образования конъюгационного канала — при частичном или полном
растворении поперечной перегородки (рис. 9, 6, 8; 10, б—9). Как пра-
вило, боковая конъюгация встречается реже лестничной, у некоторых
видов может быть преобладающей или отсутствует вовсе. При наличии
обоих типов конъюгации боковая конъюгация может иногда протекать
между клетками той же нити, что и лестничная. Терминальная конъ-
югация, встречающаяся редко, обычно рассматривается как разновид-
ность боковой или лестничной конъюгации (рис. 7, 10, 1Г, 8, 5, 6;
10, 7, 9).
Одновременно с образованием конъюгационного канала внутри
обеих конъюгирующих клеток происходят значительные изменения
(рис. И, 12). Протопласты клеток, выполняющие при конъюгации
функцию гамет, сжимаются, отстают от оболочки клетки, хлоропласты
теряют при этом свой обычный вид и отчетливые контуры. Слияние
29
Рис. 9. Апланоспоры, акинеты, партеноспоры и вегетативные клетки без конъюгационной
пары: 1 — апланоспоры в различной стадии формирования (Spirogyra mirabilis)-, 2—5 —
акинеты (2 — S. weberi f.farlowii, 3 — S. decimina f. juergensii, 4 — Zygnema frigidum, 5 —
Zygnema sp.); 6 — партеноспоры прр боковой конъюгации (Spirogyra majuscula)-, 7 — две
партеноспоры, зигоспора и вздувшаяся вегетативная клетка без конъюгационной пары при
лестничной конъюгации (S. polymorpha)', 8 — вздувшиеся вегетативные клетки без конъюгаци-
онной пары и зигоспора при боковой конъюгации (S. varians f. woodsii). (По: I — De Вагу,
1858; 2—5 — Transeau, 1951; 6 — Руццина, 1988; 7 — Jao, 1935b; 8 — Рундина, 1978a).
гамет с последующим формированием зиготы происходит или в конъ-
югационном канале (изогамная конъюгация) (рис. 10, 3—5, 8, 9; 11,
4—6), или в воспринимающей клетке — женском гаметангии (оогамная
конъюгация как разновидность анизогамной) (рис. 7, 70; 9, 7, 8\ 10,
7, 2, 6, 7; 12, 5). Иногда, как правило редко, конъюгационный канал
при лестничной конъюгации или поперечная межклеточная перегородка
при боковой конъюгации растворяются, и тогда зигота лежит свободно
между конъюгирующими клетками, обычно окруженная толстым слоем
слизи (рис. 10, 5, 9). Такая частная форма конъюгации именуется вне-
клеточной конъюгацией, противополагаемой нормальной внутриклеточ-
ной (рис. 10, 7—4, 6—S).
Воспринимающие клетки, в которых формируется зигота, остаются
цилиндрическими или вздуваются, шаровидно или веретеновидно, со
всех сторон или односторонне (рис. 13). Одностороннее вздутие может
зо
Рис. 10. Типы и формы конъюгаций у зигнемовых водорослей (схема): 1,2 — лестничная,
анизогамная внутриклеточная (Spirogyra); 3 — лестничная, изогамная внутриклеточная
(Zygnema); 4 — лестничная, изогамная внутриклеточная (бигаметангиальная) (Mougeotia);
5 — лестничная, изогамная внеклеточная (Mougeotia); 6 — боковая, анизогамная внутри-
клеточная (Spirogyra); 7 — боковая (терминальная), анизогамная внутриклеточная (Mougeo-
tia); 8 — боковая, изогамная внутриклеточная (Zygnema); 9 — боковая (терминальная), изо-
гамная внеклеточная (Mougeotia). (По: Rieth, 1961).
31
Рис. 11. Различные стадии конъюгации у зигнемы, зигогониума и мужоции: 1—3 — Zygne-
ma leiospermunr, 4 — Zygogonium ericetorunr, 5, 6 — Mougeotia tenuissima. (По: De Bary,
1858).
происходить как co стороны, противоположной конъюгационному ка-
налу, так и со стороны канала, иногда сильно маскируя его. Клетки,
оставшиеся без пары в конъюгирующих нитях, обычно не изменяют
своей формы, но у некоторых видов могут сильно и со всех сторон
шаровидно вздуваться, до 3—4 раз превышая ширину вегетативных
клеток (рис. 9, 7, 8). Отдающие клетки обычно остаются неизменен-
ными по форме — цилиндрическими.
Конъюгационный канал обычно формируется обеими клетками рав-
ноценно, реже с преобладанием одной из них, редко — исключительно
отдающей клеткой. Он может быть широким или узким, чаще хорошо
выраженным, реже совсем невыраженным, в некоторых случаях он
растворяется или вовсе отсутствует (рис. 10). В месте соприкосновения
двух конъюгирующих клеток при отсутствии конъюгационного канала
или его слабой выраженности появляется значительное количество
слизи — так называемая слизистая муфта (рис. 12, 3—5).
Особенности конъюгации. У большинства родов зигнемовых водо-
рослей формирование гаметы (сжавшегося протопласта) происходит без
предварительного деления вегетативной (гаметангиальной при конъюга-
32
ции) клетки. Но в родах Temnogyra, Sirogonium (сем. Spirogyraceae)
и Temnogametum (сем. Mougeotiaceae) вегетативная клетка (прогаме-
тангиальная) перед конъюгацией делится на две неравные по длине и
разные по функции клетки, и собственно гаметангиальной становится
лишь меньшая по длине, которая присоединяет к своему протопласту
весь протопласт сестринской клетки, остающейся таким образом сте-
рильной (рис. 12, 3—5). В результате такого неравномерного деления
клеток у представителей вышеназванных родов в конъюгирующих
Рис. 12. Различные стадии конъюгации у спирогиры и сирогониума: 1,2 — начальные ста-
дии конъюгации у Spirogyra decimina f. longata; 3—5 — Sirogonium sticticum: 3 — сопри-
косновение конъюгирующих клеток, 4 — предварительное деление прогаметангнальной
клетки, 5 — образование зиготы. (По: De Вагу, 1858).
3 Рундина Л. А.
33
Рис. 13. Формы воспринимающих клеток у зигнемовых водорослей: 1 — иевздутая (цилин-
дрическая), 2 — веретеновидная (вздутая в средней части), 3 — равномерно вздутая со всех
сторон, 4 — вздутая со стороны, противоположной конъюгацноиному каналу, 5 — цилинд-
рически вздутая со всех сторон, 6 — вздутая со стороны конъюгационного канала. (По: Jao,
1935b).
нитях отмечается довольно четкое чередование воспринимающих (с
зиготами) и стерильных клеток.
К особенностям конъюгации у зигнемовых водорослей надо также
отнести отсутствие или невыраженность конъюгационного канала у
родов Sirogonium (рис. 12, 3—5) и Sirocladium (соответственно семей-
ства Spirogyraceae и Mougeotiaceae), заполнение гаметангиев (при фор-
мировании в них зиготы) светлым и слоистым коллоидным веществом
у представителей родов Debarya (сем. Mougeotiaceae) (рис. 15, 6) и
Zygnemopsis (сем. Zygnemataceae), формирование остаточной после
конъюгации цитоплазмы, в виде тонкой мантии окутывающей зигоспору
и простирающейся в гаметангиях (роды Mougeotiella и Neozygnema в
семействах Mougeotiaceae и Zygnemataceae), образование спорангия у
представителей рода Zygogonium (сем. Zygnemataceae) и у некоторых
видов в роде Mougeotia (сем. Mougeotiaceae).
Зигоспора. Зигота с оформившимися слоями оболочки, приобрет-
шая определенную форму и цвет, заполнившаяся запасными питатель-
ными веществами, в частности большим количеством масла, называется
зигоспорой. Зигоспора у зигнемовых прорастает обычно одним нитча-
тым отростком после длительного периода покоя, редко — сразу после
конъюгации (рис. 6). Исключение составляет род Hallasia, у единст-
венного вида которого апланоспора может иметь как 1, так и 2—3
проростка.
34
Формирование зигоспоры в процессе конъюгации происходит либо
в конъюгационном канале, с полным или частичным его заполнением,
либо в конъюгационном канале и обоих гаметангиях, с частичным или
полным, вплоть до внешних стенок, их заполнением, либо только в
воспринимающей клетке (женском гаметангии) (рис. 10). У каждого
из родов имеется своя специфика в этом отношении: к примеру, у
родов Spirogyra и Sirogonium зигоспора формируется исключительно в
воспринимающей клетке.
Форма зигоспор очень разнообразна (рис. 14, 15): эллипсоидная
(двух-, редко трехосная), сбоку эллиптическая, сверху (в поперечном
сечении) круглая, редко эллиптическая; шаровидная; линзовидная,
анфас круглая, сбоку эллиптическая; подушковидная — особой формы
линзовидная, анфас округлая, с оттянутыми в разной степени рогами,
на концах закругленными или втянутыми внутрь, и выпуклыми или
вогнутыми сторонами, сбоку эллиптическая; короткоцилиндрическая,
редко усеченно-конусовидная (в проекции трапециевидная), с фрон-
тальной стороны (анфас) обычно с вогнутыми (в разной степени), реже
со слегка выпуклыми или прямыми боковыми сторонами, сверху и
снизу с округлыми крышечками (створками). Усложненные формы зи-
госпор (кубические, полиэдрические, в форме волчка) встречаются
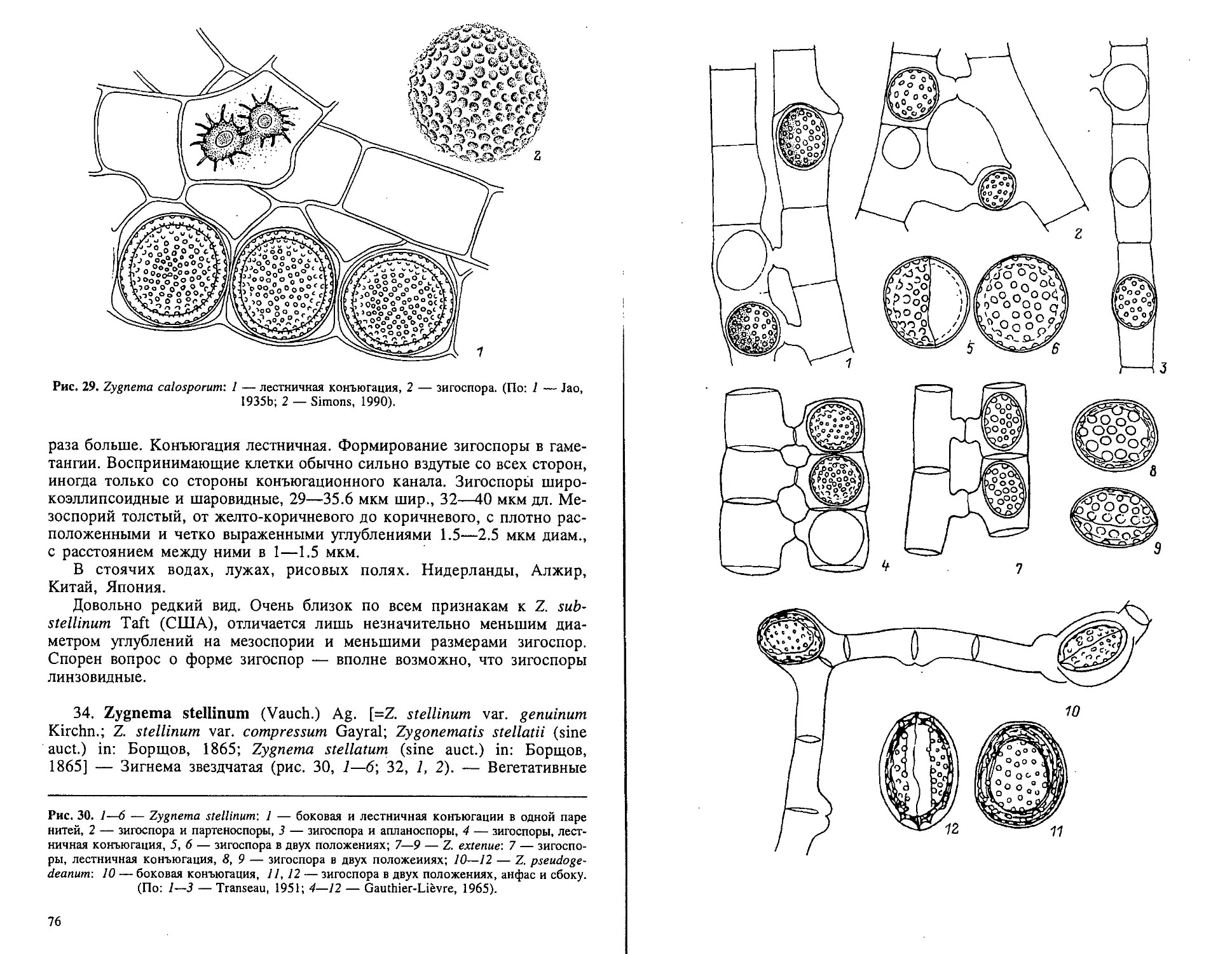
Рис. 14. Основные формы зигоспор у зигнемовых водорослей (цифровое обозначение — вид
зигоспоры анфас, обычно с широкой стороны, буквенное — вид зигоспоры в разных положе-
ниях): 1 — шаровидная; 2 — линзовидная; 3 — линзовидная с килем; 4 — короткая двухос-
но-эллипсоидная; 5 — короткая трехосно-эллипсоидная; 6 — удлиненная двухосно-эллип-
соидная с заостренными вершинами; 7 — короткая двухосная цилиндрнческн-эллипсоидная
с широко закругленными вершинами; 8 — короткая трехосная цилиидрически-эллипсоид-
ная с широко закругленными вершинами; 9 — удлиненная двухосная цилиидрически-эллип-
соидная с закругленными вершинами; 10 — четырехугольная (подушковидная) с закруглен-
ными углами и прямыми сторонами; 11 — четырехугольная (подушковидная) с вдавленными
углами и вогнутыми сторонами. (По: Jao, 1935b).
35
Рис. 15, Основные формы зигоспор на примере некоторых зигнемовых водорослей: 1 —
шаровидная (Zygnema)-, 2 — линзовндная, сбоку и анфас (Spirogyra heeriana)-, 3 — линзо-
видная с килем, сбоку (Zygogonium)-, 4 — двухосно-эллипсоидная (Spirogyra elliptica)-, 5 —
четырехугольная (подушковидная), анфас (Mougeotia); 6, 7 — сложиолинзовидная — в
форме трехплоскостного волчка, анфас и сбоку (Debarya glyptosperma). (По: 1, 3—5 — Jao,
. 1935b; 2 — Gauthier-Lievre, 1965; 6,7 — Bourrelly, 1966).
крайне редко и, как правило, бывают специфичны для одного рода
или секции в роде.
Оболочка зигоспор состоит из трех основных слоев, различных по
своему строению, толщине, цвету, скульптуре (орнаментированности),
составу и функции.
Экзоспорий (наружная оболочка) — обычно тонкий, прозрачный,
очень редко слабоокрашенный, однослойный, редко двухслойный, глад-
кий, очень редко скульптурированный слой. Экзоспорий состоит из
пектина и целлюлозы, растворим в смеси целлюлазы с пектиназой и
в хитиназе.
36
Мезоспорий (средняя оболочка) — толстый, обычно желтый или
коричневый со всеми оттенками, реже голубой, синий или синевато-
зеленый, одно- или двухслойный, реже трехслойный, очень редко че-
тырехслойный, глад,кий или разнообразно скульптурированный слой
(рис. 16, 17). В мезоспории содержатся хитин, спорополленин, хро-
Рис. 16. Строение мезоспория (с поверхности и в поперечном разрезе): 1 — гладкий; 2 —
точечный (пунктированный); 3 — гранулированный (зернистый, бугорчатый, бородавча-
тый); 4 — ямчатый; 5 — окаймленно-ямчатый; 6 — складчато-ямчатый с килем (сбоку —
складчатый с килем, анфас — ямчатый); 7 — двухслойный: наружный слой морщинистый
(волнисто-складчатый), внутренний слой гладкий; 8 — извилисто-бороздчатый; 9 — сетча-
тый; 10 — шиповидно-сетчатый; 11 —- зубчато-ячеистый (зубчато-сетчатый); 12 — игольча-
тый (тонкошиповидиый). (По: Jao, 1935b).
37
5
Рис. 17. Строение скульптурированного мезоспория (в поперечном разрезе и с поверхно-
сти): 1,3,7 — ямчатый; 2 — точечный (пунктированный); 4 — гранулированный (зернис-
тый, бугорчатый, бородавчатый); 5, 6, 8 — сетчато-ячеистый; 9 — зубчатый (шиповидный);
10 — сосочковидный; 11, 12 — извилисто-бороздчатый (морщинистый). (По: Czurda, 1931).
мовая и плавиковая кислоты; он устойчив к ферментам и ацетолизу.
Благодаря содержанию спорополленина в мезоспории зигоспоры зиг-
немовых хорошо сохранились в геологических слоях разных эпох, на-
чиная с палеозоя.
Эндоспорий (внутренняя оболочка) — тонкий, прозрачный,
обычно гладкий, трудноразличимый слой. Эндоспорий формируется
последним из оболочек, он же служит первой оболочкой проростка
зигоспоры.
Линия разрыва зигоспоры, проходящая через мезо- и экзоспорий,
обычно хорошо заметна на вполне зрелых зиготах и, естественно,
перед их прорастанием (рис. 6). Разрыв оболочки зигоспоры виден
в виде прямой* косой или слегка извилистой линии, проходящей
вдоль продольной, реже поперечной оси зигоспоры (в зависимости
от ее формы); иногда линия разрыва как бы опоясывает зигоспору
и тогда принимает форму почтового конверта. У рода Temnogametum
линия разрыва весьма усложненная и специфичная — в форме си-
нусоиды или фигурной скобки. Как правило, линия разрыва одна,
но в некоторых случаях (виды группы Laetevirens в роде Mougeotia)
линий разрыва две — по краям обеих крышечек, сверху и снизу
зигоспоры (рис. 6, 5).
38
3. ТАКСОНОМИЧЕСКАЯ ОЦЕНКА
ОСНОВНЫХ МОРФОЛОГИЧЕСКИХ ПРИЗНАКОВ
Из вегетативных признаков наличие коротких ответвлений (веточек)
при наземном местообитании водоросли является одним из хороших
родовых признаков (роды Zygogonium и Sirocladium).
Ширина клетки (средние значения) является достаточно стабиль-
ным, видовым и внутривидовым признаком. Выявление коррелятивных
связей между этим и другими признаками приобретает подчас решаю-
щее значение. Ширина вегетативных клеток всегда измеряется по по-
перечной перегородке (септе).
Отношение ширины к длине вегетативной клетки — признак до-
статочно стабильный по сравнению с признаком длины клетки, зави-
сящей от возраста и физиологического состояния клетки и факторов
среды. Величина отношения как некая видовая константа применяется
обычно при определенной математической обработке материала, но
имеет маловажное значение для видовой характеристики.
Форма поперечной перегородки клеток оценивается как видовой
признак только у рода Spirogyra. Особое внимание следует проявлять
при просмотре фертильного материала у видов, имеющих постоянную
складчатость перегородки в вегетативном состоянии и перемежающу-
юся, подчас нерегулярно, с гладкой при конъюгации. Так, у круга
форм Spirogyra reticulata складчатые поперечные перегородки в фер-
тильных нитях встречаются намного реже, чем гладкие, и тем не менее
за основную форму перегородок здесь принимается складчатая, прису-
щая вегетативным клеткам. Внимательно следует просматривать также
вегетативные клетки с гладкими септами — только у двух видов из
секции Colligata имеются на поперечных перегородках кольцевые на-
кладки.
Форма хлоропласта и положение его в клетке — важнейший ро-
довой признак. Количество хлоропластов в клетке у родов Spirogyra
и Sirogonium — один из важных и довольно стабильных видовых при-
знаков. Для разграничения видов в этих родах важны пределы коле-
баний в их количестве: постоянно 1, 1—2, 2—10, 10—12, 12—16.
Однако при совпадении комплекса прочих видовых признаков разница
в количестве хлоропластов (в известных пределах) не может считаться
решающим признаком для установления самостоятельной видовой еди-
ницы. Число витков хлоропластов и плотность их расположения в
клетке очень зависят от условий среды и возраста клетки, так что они
не могут, как правило, претендовать на таксономическую значимость.
Для подсчета числа хлоропластов применима формула п +1, где п —
число видимых, хотя и мнимых, пересечений хлоропластов по длине
одного витка.
Пиреноиды, их форма, количество и расположение не являются для
зигнемовых значимым признаком, за единственным исключением — у
рода Mougeotiopsis пиреноид отсутствует.
Положение ядра в клетке является только вспомогательным родовым
признаком.
39
Морфологические признаки, связанные с половым размножением,
у зигнемовых водорослей имеют решающее значение в их таксономии,
однако не все признаки этого цикла однозначны. Мнение о возмож-
ности определения видовой принадлежности только по вегетативным
признакам, в частности по ширине вегетативных клеток и числу хло-
ропластов, является ошибочным и устаревшим. Более того, без фер-
тильной стадии невозможно различить не только виды, но даже и
некоторые роды. Стабильными родовыми признаками, проявляющимися
только в процессе конъюгации и формирования зигоспоры, являются:
предварительное деление вегетативной (прогаметангиальной) клетки на
собственно гаметангиальную и стерильную, невыраженность или отсут-
ствие конъюгационного канала, заполнение гаметангиев коллоидным
веществом, формирование из остатка цитоплазмы мантии, окутываю-
щей зигоспору и простирающейся в гаметангиях, образование споран-
гия.
Наличие того или иного типа конъюгации (лестничной или боко-
вой) ранее считалось родовым, позднее — видовым признаком; в на-
стоящее время тип конъюгации признается одним из существенных
внутривидовых признаков. Причины возникновения того или иного
типа конъюгации или преобладания одного из них неизвестны. Следует
иметь в виду, что разные типы конъюгаций у одного и того же вида
могут быть встречены как в одном месте сбора, так и в разных. Кроме
того, лестничная и боковая конъюгации у одного вида могут встре-
чаться одновременно как в одной паре конъюгирующих нитей, так и
в раздельных нитях. Преобладание одного из типов конъюгации также
служит существенным таксономическим признаком.
Формирование зигоспоры только в конъюгационном канале, в конъ-
югационном канале с частичным или полным внедрением в гаметангии
или только в женском гаметангии — один из важнейших видовых
таксономических признаков (в родах Mougeotia, Debarya, Zygnema,
Zygnemopsis, Temnogametum) и родовой признак у Spirogyra, Sirogonium
и Sirocladium. Есть все основания считать этот признак наследственно
константным.
Особенности формирования конъюгационного канала при лестнич-
ной конъюгации обязательно учитываются при определении видов и
внутривидовых форм во всех родах, кроме Sirogonium и Sirocladium,
где канал невыраженный или вовсе отсутствует. Канал формируется,
как правило, обеими конъюгирующими клетками в равной мере, реже
с преобладанием или исключительно одной из них. Учитывая значи-
тельную пластичность формирования конъюгационного канала, связан-
ную во многих случаях со вздутием воспринимающих клеток с его
стороны, наиболее правильно давать его характеристику при окончании
конъюгации, когда все процессы формирования завершены.
Форма и степень вздутия воспринимающих клеток учитываются как
важные видовые и внутривидовые признаки. Особое внимание должно
быть здесь обращено на пластичность вздутия, коррелятивно связанную
с изменчивостью других признаков (с формированием конъюгационного
канала, размерами и формой зигоспор). Как правило, воспринимающие
клетки бывают значительно шире вегетативных, у некоторых видов
40
они приобретают при вздутии шаровидную, веретеновидную или ци-
линдрически-эллипсоидную форму.
Клетки без конъюгационной пары у большинства видов невздутые.
Однако степень вздутия, если таковая имеет место, является стабиль-
ным видовым признаком, но в пределах круга форм одного вида может
значительно варьировать.
Форма зигоспор считается одним из важнейших таксономических
признаков. У большинства видов она постоянна и только у немногих
видов и форм подвержена значительной изменчивости. Учитывая ва-
риации формы зигоспор, изучаемому образцу придают либо ранг са-
мостоятельной видовой единицы, либо ранг внутривидового таксона.
Более того, для определенных родов характерен определенный набор
форм зигоспор. Например, у видов рода Spirogyra никогда не бывает
подушковидных или короткоцилиндрических зигоспор, типичных для
видов Mougeotia. Для точного определения формы зигоспор необходи-
мо просмотреть достаточно обильный материал и постараться увидеть
их с разных сторон. Обычно это удается сделать легким постукиванием
препаровальной иглой по покровному стеклу.
Строению оболочек зигоспор: количеству слоев, цвету, деталям
скульптуры или текстуры в настоящее время придается большое зна-
чение для определения видовых и внутривидовых таксонов. Особен-
ности строения необходимо прослеживать на всех стадиях формиро-
вания зигоспор и его окончательную характеристику давать к моменту
их полного созревания. Изучение деталей строения оболочек позволяет
в большей мере уточнять и таксономическое положение, и границы
вида.
Наличие апланоспор, как таксономический признак, не имеет зна-
чения, если апланоспоры существуют наравне с зигоспорами, или имеет
решающее значение как видовой (Spirogyra mirabilis) и даже родовой
признак (род Hallasia'), если размножение осуществляется ими преиму-
щественно или исключительно.
4. СИСТЕМА ЗИГНЕМОВЫХ ВОДОРОСЛЕЙ
К группе конъюгат (Zygnematophyceae, Conjugatophyceae) причис-
ляют мезотениевые, гонатозиговые, зигнемовые и десмидиевые водоро-
сли, объединяя их в один или два класса и даже считая эту группу
подотделом Chlorophyta (Голлербах, Полянский, 1951; Косинская,
1952, 1960; Полянский, 1959; Воденичаров, 1972; Паламарь-Мордвин-
цева, 1982а, 19826, 1984, 1986). Объединение этих водорослей в са-
мостоятельную группу (любого таксономического ранга) основывается
на своеобразии их полового процесса — конъюгации, а подразделение
конъюгат на классы и порядки зависит от точки зрения авторов клас-
сификаций на ранговую таксономическую значимость некоторых мор-
фологических признаков, таких как форма клеток и строение их обо-
лочек, строение нитчатых талломов, форма хлоропластов, особенности
41
процесса конъюгации. Предметом спора в основном служит положение
в системе конъюгат порядка Gonatozygales, немногочисленные предста-
вители которого имеют близкое морфологическое сходство как с зиг-
немовыми, так и с некоторыми из десмидиевых и мезотениевых водо-
рослей.
По классификации, принятой мною, конъюгаты объединены в один
класс — Zygnematophyceae в ранге самостоятельных порядков: Meso-
taeniales, Gonatozygales, Desmidiales и Zygnematales.
При разработке системы зигнемовых водорослей с XIX века были
заложены разные принципы их подразделения. По первой классифи-
кации (De Вагу, 1858), 5 родов зигнемовых водорослей (сем. Zygne-
maceae) разделялись на два подсемейства (Mesocarpeae и Zygnemeae)
на основе разницы в процессе формирования зигот. С тех пор эта
классификация была неоднократно модифицирована многими фиколо-
гами (Wittrock, 1871, 1878; West, West, 1898; West G. S., 1904; Borge,
1913; Printz, 1927; West, Fritsch, 1927). В последней, слегка измененной
системе Фрича (Fritsch, 1935) уже 8 изученных родов зигнемовых во-
дорослей были размещены в двух семействах (Mougeotiaceae и Zygne-
maceae) с учетом разницы в строении фертильных клеток и положении
в них зигот — с наличием или отсутствием зигоспорангиальных пе-
регородок. Классификация Фрича была поддержана позднее рядом ис-
следователей (Okada, 1953; Berger, 1954; Papenfuss, 1955; Hirose, 1959).
Представители другого направления клали в основу классификации
зигнемовых водорослей, рассматриваемых как одно семейство, форму
и положение хлоропласта. Названия выделенным группам давались по
типовому роду, форма хлоропласта которого характеризовала соответ-
ствующее подсемейство. Так, по подробно разработанной классифика-
ции Веста (West G. S., 1916) сем. Zygnemaceae подразделялось на три
подсемейства (Mesocarpeae, Zygnemeae, Spirogyreae) с входящими в
них 10 родами. Того же принципа в основном придерживался и Чурда
(Czurda, 1932), признававший, однако, только 3 рода: Mougeotia, Zyg-
nema и Spirogyra. Виды этих родов объединялись им в несколько групп
(секций) нетаксономического ранга с учетом комплекса вегетативных
и генеративных признаков.
Первым, кто обосновал подразделение зигнемовых водорослей на
три семейства (Zygnemaceae, Mougeotiaceae и Spirogyraceae) по форме
хлоропласта, был Палла (Palla, 1894). Блэкмен и Тэнсли (Blackman,
Tansley, 1902) поддержали эту классификацию. Я считаю себя привер-
женцем именно этой классификации, но при выделении трех семейств
зигнемовых водорослей расширяю их характеристики, подключая все
морфологические признаки в вегетативной и генеративной стадиях.
Количество родов и видов в пор. Zygnematales в различных опре-
делителях, сводных работах и монографиях сильно варьирует, и по
мере все возрастающих флористических исследований намечается оп-
ределенная тенденция к их увеличению. В сводке Чурды (Czurda, 1932)
содержится 227 видов зигнемовых водорослей, объединенных в три
рода. Известные к тому времени роды Debarya, Mougeotiopsis, Sirogo-
nium, Temnogyra, Temnogametum, Pleurodiscus, Zygogonium, Hallasia
этим автором были сведены в синонимы Mougeotia, Spirogyra и Zyg-
42
пета. В первой мировой сводке Кригера (Kolkwitz, Krieger, 1941—
1944) приводятся уже 452 вида, распределенных в 8 родах. Кроме
основных трех родов (Mougeotia, Zygnema, Spirogyra) здесь приняты в
качестве самостоятельных роды Debarya, Mougeotiopsis, Sirocladium,
Temnogametum, Zygnemopsis на основании морфологических признаков
в вегетативном состоянии и особенностей конъюгации. Монографии
Кадлубовской (Kadhibowska, 1972, 1984), построенные по таксономи-
ческим принципам Кригера, обобщают данные о 688 видах зигнемовых
водорослей. В крупных сводках Трансо (Transeau, 1951) и Рандхавы
(Randhawa, 1959b) числятся соответственно 534 и 580 видов (в 13
родах), причем сохранены в качестве самостоятельных роды Sirogoni-
um, Pleurodiscus, Hallasia и Zygogonium, бывшие по сводке Кригера
синонимами Spirogyra и Zygnema, и признан род Entransia, В ряде
сводных работ Ямагиси (Yamagishi, 1963а, 1965b, 1966а) вновь возрож-
ден род Temnogyra и выделены некоторые представители родов Mou-
geotia и Zygnema в новые роды Mougeotiella и Neozygnema на осно-
вании некоторых усложнений в строении гаметангиальных клеток и
особенностей конъюгации. Наконец, Ахмад и Гольдштейн (Ahmad, Gold-
stein, 1971, 1972) при использовании только метода культур выделили
и описали новый род Lloydina (= Lloydiella), по ряду характерных
признаков в вегетативном состоянии имеющий большое сходство с
родами Mougeotia и Mougeotiopsis.
Таким образом, в настоящее время известно около 700 видов зиг-
немовых водорослей. Они объединены в 16 родов разной степени изу-
ченности и объема. Сравнительная и комплексная оценка родовых призна-
ков заставила меня произвести следующие перестановки. Род Pleuro-
discus и род Hallasia признаны мною синонимами родов соответственно
Zygogonium и Zygnemopsis. Род Temnogyra впредь до выяснения неко-
торых спорных морфологических особенностей в стадии конъюгации
идентифицируется мною с родом Spirogyra, род Mougeotiella — с
родом Mougeotia, род Neozygnema — с родом Zygnema. Род Lloydina
считается недостаточно описанным из-за неизученное™ процесса раз-
множения, и сама принадлежность его к зигнемовым водорослям ос-
тается спорной. Род Entransia, хотя и помещен мною в разряд сомни-
тельных родов зигнемовых водорослей, по своей морфологии необы-
чайно схож с улотриксовыми водорослями и, вероятнее всего, должен
быть исключен из пор. Zygnematales.
Определение (характеристика) рода, вида и внутривидовых таксо-
номических категорий (разновидность, форма) у зигнемовых водоро-
слей, имеющее в вопросах систематики, номенклатуры и классифика-
ции важное значение, в работах исследователей выражено неопреде-
ленно и оставляет широкий простор для субъективизма.
Как и у других водорослей, основная систематическая единица у зиг-
немовых — вид понимается исследователями то более узко, то более
широко. При сугубо практическом применении в систематике сторон-
ники узкого понимания вида понимают эту таксономическую категорию
как совокупность особей, которую отличает от других хотя бы один, но
ясно выраженный признак. При такой формулировке все существующие
внутривидовые таксоны должны быть возведены в ранг вида. Такая точка
43
зрения на объем вида у зигнемовых водорослей наиболее ярко выражена
в работах Чурды (Czurda, 1931, 1932), а затем поддержана многими ис-
следователями (Krieger in: Kolkwitz, Krieger, 1941—1944; Transeau,
1951; Воронихин, 1954; Randhawa, 1959b; Gauthier-Lifevre, 1965; Kadiu-
bowska, 1972, 1984). При более широком понимании (Рундина, 1991)
вид воспринимается как естественная целостность, характеризующаяся,
с одной стороны, определенным морфологическим, экологическим и
географическим постоянством, обусловленным наследственностью и
поддерживаемым стабилизирующими адаптациями, а с другой — состо-
янием поступательной эволюции, которое представляет собой постепен-
ный процесс дивергентного расхождения признаков во внутривидовых
группах. Таким образом, эти два состояния вида определяют и морфо-
логическое единство вида, и полиморфизм его.
В своей работе я руководствуюсь принципами внутривидовой сис-
тематики и широкого понимания вида, разработку которых излагали
в своих исследованиях А. А. Еленкин (1936) и В. И. Полянский
(1954, 1956, 1966). Основываясь на их общих положениях, считаю,
что при современном уровне знаний внутривидовое многообразие зиг-
немовых водорослей может быть достаточно полно передано посред-
ством таксономической категории формы. При этом видов становится
меньше, чем описано в настоящее время в литературе, и вид как таксон
становится намного крупнее и полиморфнее, а вся система зигнемовых
водорослей становится зримее. От понятий «подвид» и «разновидность»
я отказалась в работе, полностью согласившись с мнением Полянского
и Еленкина, что эти термины применительно к низшим растениям не
определены и не отграничены от понятия формы.
Разнообразие и дифференциация родовых, видовых и внутривидо-
вых признаков, широкий полиморфизм видов, до сих пор «становле-
ние», если так можно выразиться, некоторых родов — все это говорит
о быстроте и масштабе происходящих у зигнемовых водорослей эво-
люционных процессов. Но это и значительно усложняет систематику
и связанную с ней таксономию зигнемовых. Не исключена возмож-
ность, что некоторые из родов являются здесь экологическими моди-
фикациями (экоморфами, экородами) иных описанных родов, а неко-
торые из известных до сих пор видов — лишь отдельными морфоло-
гически различными стадиями развития.
В полном объеме система зигнемовых водорослей имеет следующий
вид:
Отдел Chlorophyta
Класс Zygnematophyceae (Conjugatophyceae)
Порядок Zygnematales
Семейство Zygnemataceae Palla
Род Zygnema Agardh, 1817
Род Zygogonium Kiitzing, 1843
Род Hallasia Rosenvinge, 1924
Род Zygnemopsis (Skuja) Transeau, 1934
Род Neozygnema Yamagishi, 1963
Семейство Spirogyraceae Palla
Род Spirogyra Link, 1820
44
Род Sirogonium Kiitzing, 1843
Род Temnogyra Lewis, 1925
Семейство Mougeotiaceae Palla
Род Mougeotia Agardh, 1824
Род Debarya Wittrock, 1872
Род Mougeotiopsis Palla, 1894
Род Temnogametum W. et G. S. West, 1897
Род Sirocladium Randhawa, 4941
Род Entransia Hughes, 1948
Род Mougeotiella Yamagishi, 1963
Род Lloydina Ahmad et Goldstein, 1972
II. СИСТЕМАТИЧЕСКАЯ ЧАСТЬ
Отдел CHLOROPHYTA —
ЗЕЛЕНЫЕ ВОДОРОСЛИ
Одноклеточные, многоклеточные, колониальные и неклеточные
формы с хлоропластами чисто зеленого цвета, с пиреноидами или реже
без них, с одним или реже с несколькими ядрами, неподвижные или
иногда активно движущиеся в вегетативном состоянии. Клеточная обо-
лочка преимущественно целлюлозная и пектиновая, в некоторых слу-
чаях с включением спорополленина. Продукты ассимиляции крахмал
и масло. Размножение делением вегетативных клеток, бесполым путем
посредством особых спор (подвижных и неподвижных) и половым
путем разного типа, у некоторых представителей с правильной сменой
половой и бесполой форм развития.
Класс ZYGNEMATOPHYCEAE
(CONJUGATOPHYCEAE) —
КОНЪЮГАТЫ, СЦЕПЛЯНКИ
Водоросли одноклеточные, колониальные и многоклеточные (нитча-
тые), простые или образующие короткие ответвления из одной-немногих
клеток, неподвижные или слабо передвигающиеся посредством выделе-
ния слизи, свободноплавающие или прикрепленные к субстрату ризои-
дами, часто наземные, реже аэрофильные. Клеточная оболочка цельная
или сегментированная, состоящая из двух и более сегментов, однослой-
ная или дифференцированная на внешний и внутренний слои, гладкая
или покрытая мелкими зернышками или шйпиками, без пор или про-
низанная порами. Хлоропласты осевые или пристенные, по одному или
по нескольку в клетке. Ядро 1. Подвижные жгутиковые стадии в про-
цессе размножения полностью отсутствуют. Вегетативное размножение
делением клеток и фрагментацией талломов у нитчатых форм. Половое
размножение — конъюгация (изо-, гетеро- и оогамия), в процессе кото-
рой происходит слияние протопластов двух клеток, одиночных или на-
ходящихся в одной или в разных нитях. Зрелая зигота с оформившимися
46
гладкими или скульптурированными слоями оболочки прорастает после
периода покоя 1, 2 или 4 проростками.
Класс включает 4 порядка: Mesotaeniales, Gonatozygales, Desmidiales
и Zygnematales, из которых в данной работе рассматривается только
последний.
I. Клеточная оболочка обычно цельная (не сегментированная), однослойная
или дифференцированная на внешний и внутренний слои, без пор или реже
с порами. Место клеточного деления не фиксировано. Клетки одиночные
или соединены в колонии и нити; клетки по форме преимущественно ци-
линдрические, без перетяжки посередине.
1. Клеточная оболочка гладкая, лишенная пор.
А. Клетки одиночные или соединены в колонии, окруженные общей, хоро-
шо заметной, иногда концентрически-слоистой слизью; очень редко со-
единены в короткие 2—16-клеточные нити (род Ancylonema). Клетки по
форме эллипсоидные, цилиндрические или веретеновидные............
............................................. пор. Mesotaeniales
Б. Клетки соединены в длинные неразветвленные нити, редко (при наземном
местообитании) с короткими ответвлениями из одной-немногих клеток.
Клетки по форме всегда цилиндрические............................
............................................. пор. Zygnematales
2. Клеточная оболочка покрыта мелкими зернышками или шипиками, реже
гладкая, с порами. Клетки соединены в хрупкие, легко распадающиеся
нити; после распада нити клетки продолжают жить самостоятельно. Клет-
ки по форме цилиндрические или цилиндрически-веретеновидные . .
..................................................пор. Gonatozygales
II. Клеточная оболочка всегда сегментированная, состоящая из двух или более
сегментов, постоянно дифференцированная на внешний и внутренний слои
и обычно пронизанная порами. Клеточное деление происходит, как правило,
по строго определенному типу (исключение — некоторые виды рода Реш-
ит). Клетки в большинстве одиночные или, реже, соединены в колонии в
форме рыхлых скоплений, цепочек или нитей. Клетки по форме очень
разнообразные, с перетяжкой посередине, реже они лишены перетяжек и
тогда серповидные ...................................... пор. Desmidiales
Порядок ZYGNEMATALES — ЗИГНЕМОВЫЕ
Нити макроскопические, длинные, из одного ряда клеток, неветвя-
щиеся или реже образующие короткие ответвления из одной-немногих
клеток, свободноплавающие или прикрепленные к субстрату ризоида-
ми, редко образующие дерновинки или налеты при обитании на сырой
земле, камнях и скалах, орошаемых водой, на влажной коре деревьев.
Ризоиды разнообразной формы, чаще от базальной клетки; их обра-
зование, форма и количество зависят от местообитания водоросли.
Клетки цилиндрические, без перетяжек посередине, часто покрытые
слизью. Клеточная оболочка цельная (не сегментированная), однослой-
ная или дифференцированная на внешний пектиновый и внутренний
целлюлозный слои, гладкая, без пор. Место клеточного деления не
фиксировано. Хлоропласты осевые и пристенные, звездчатые, диско-
47
видные, пластинчатые, лентовидные, спирально изогнутые или парал-
лельные продольной оси клетки, по 1, 2 и более (до 16) в каждой
клетке. Ядро 1, обычно в центре клетки, в протоплазматическом мос-
тике между хлоропластами или подвешено на протоплазматических
тяжах, прикрепленных к продольным стенкам клетки, или оно при-
мыкает сбоку к хлоропласту. Пиреноиды многочисленные, расположен-
ные по всему хлоропласту беспорядочно или в один ряд, иногда по
одному в каждом хлоропласте, очень редко хлоропласт без пиреноида.
Размножение вегетативное, бесполое и половое. Конъюгация лестнич-
ная и боковая, с исключительным или преимущественным развитием
одной из них, нередко совмещенные в одной паре нитей. Формиро-
вание зигоспоры в конъюгационном канале, очень редко с полным его
растворением, в конъюгационном канале с разной степенью внедрения
в гаметангии или в одном из гаметангиев (в воспринимающей клетке).
В порядке 3 семейства, в основном различающиеся формой хло-
ропласта и положением его в клетке.
I. Хлоропласт осевой, звездчатый, с радиальными выростами, реже дисковидный,
без выростов. Ядро в центре клетки........сем. Zygnemataceae (с. 48)
И. Хлоропласт осевой, реже пристенный, широкопластинчатый. Ядро примы-
кает сбоку к хлоропласту, реже в центре клетки ................
.................................... сем. Mougeotiaceae (с. 106)
III. Хлоропласт пристенный, спирально извитой, слегка извитой или почти па-
раллельный продольной оси клетки. Ядро в центре клетки ........
......................................сем. Spirogyraceae (с. 161)
Зигнемовые водоросли очень трудно определять в вегетативном со-
стоянии даже до рода, так как у ряда родов, в особенности если они
принадлежат к одному семейству, морфологические признаки в этой
стадии сходны. Их точная таксономическая идентификация возможна
лишь при наличии процесса конъюгации и зрелых зигот, которые при
сборе материала в природе встречаются далеко не часто.
Семейство ZYGNEMATACEAE Palla — ЗИГНЕМОВЫЕ
Нити неразветвленные, очень редко с короткими ответвлениями,
свободноплавающие или прикрепленные к субстрату ризоидами, у на-
земных и аэрофильных форм стелющиеся по субстрату. Поперечные
стенки клеток гладкие. Хлоропласты осевые (центральные), по 2 в
клетке, очень редко по 1 или по 3—6, звездчатые, многолучевые (с
радиальными выростами) или дисковидные, обычно без радиальных вы-
ростов. Пиреноид 1, центрально расположенный в каждом хлоропласте.
Ядро в центре клетки, в протоплазматическом мостике между хлоро-
пластами. Размножение преимущественно зигоспорами, реже аплано-
спорами или акинетами. Формирование зигоспоры преимущественно в
полости одного из гаметангиев, реже в конъюгационном канале, иногда
с частичным внедрением в полости гаметангиев. Конъюгационный
канал отчетливый, очень редко расплывающийся при формировании
зигоспоры. Дифференциация вегетативной прогаметангиальной клетки
48
на собственно гаметангиальную и стерильную клетки отсутствует. Га-
метангии при формировании зигоспоры иногда заполнены слоистым
коллоидным веществом. Зигоспоры очень редко окружены остатком
цитоплазмы в виде мантии.
I. Хлоропласты звездчатые, с радиальными выростами.
1. Гаметангии заполнены слоистым коллоидным веществом.
А. Размножение зигоспорами. Апланоспоры редкие................
........................................род Zygnemopsis (с. 92)
Б. Размножение только апланоспорами. Зигоспоры не найдены.....
...............................................род Hallasia*
2. Гаметангии без коллоидного вещества.
А. Зигоспоры окружены остатком цитоплазмы в виде мантии ......
............................................род Neozygnema*
Б. Зигоспоры без мантии .................. род Zygnema (с. 49)
II. Хлоропласты дисковидные, обычно без радиальных выростов........
...........................................род Zygogonium (с. 101)
Краткие сведения о родах, отмеченных звездочкой, помещены в
конце обзора всего семейства.
Род ZYGNEMA Ag. — ЗИГНЕМА
^Zygogonium Kutz., pr. p.; Zygnemopsis (Skuja) Trans., pr. p.; Neo-
zygnema Yamagishi
Нити преимущественно неразветвленные, свободноплавающие или
прикрепленные к субстрату ризоидами, редко коротко разветвленные, с
вертикальными и горизонтальными, стелющимися по наземному субстрату
ответвлениями, с ризоидами от базальной клетки или от клеток по длине
нити, соприкасающейся с субстратом. Вегетативные клетки до 60 мкм
шир., обычно короткие или по длине до 9 раз больше ширины. Попе-
речные стенки клеток гладкие. Продольные стенки клеток тонкие, со-
стоящие из пектинового и целлюлозного слоев, значительно ослизненные.
Хлоропласты по 2 в клетке, очень редко по 1 или по 3—4, звездчатые,
многолучевые, каждый с 1 центрально расположенным пиреноидом. Конъ-
югация лестничная, редко боковая. Конъюгирующие клетки прямые или
изогнутые. Формирование зигоспоры преимущественно в одном из гаме-
тангиев, реже в конъюгационном канале, иногда с частичным внедрением
в гаметангии. Зигоспоры линзовидные, шаровидные или эллипсоидные,
вытянутые вдоль или поперек конъюгационного канала, или коротко- или
удлиненно-цилиндрические с закругленными концами, иногда неправиль-
ной формы. Экзоспорий тонкий, очень редко толстый, бесцветный, глад-
кий, редко скульптурированный. Мезоспорий толстый, желто-коричневый
или коричневый, редко голубой или синий, однослойный или реже двух-
слойный, гладкий или разнообразно скульптурированный. Эндоспорий
тонкий, бесцветный, гладкий, обычно трудноразличимый. Бесполое разм-
ножение акинетами и апланоспорами. Акинеты цилиндрические, с утол-
щенной, гладкой или разнообразно скульптурированной оболочкой. Ап-
ланоспоры эллипсоидные и шаровидные, редко цилиндрические или
4 Рундииа Л. А.
49
цилиндрически-яйцевидные, с гладким или разнообразно скульптури-
рованным мезоспорием. Линия разрыва у зигоспор 1, продольная или
поперечная, у акинет 1, реже их 2, продольная или поперечная, косая
или в форме почтового конверта.
В роде свыше 130 видов, широко представленных на всех континен-
тах. В данной работе рассматриваются 55 видов, потенциальных и об-
наруженных на территории России. Представители рода встречены в
мелководье стоячих и проточных водоемов, прудах, озерах, реках, ри-
совых полях, болотах, реже в горных текучих водах, водопадах, скаль-
ных выбоинах, редко на влажной почве, на орошаемых водой скалах,
на влажном мху. Развиваются в массе в теплое время года, предпочитая
чистую, незагрязненную воду.
I. Зигоспоры имеются.
1. Формирование зигоспоры только в конъюгационном канале или с незначи-
тельным внедрением в гаметангии.
А. Формирование зигоспоры только в конъюгационном канале.
а. Мезоспорий голубой или синий.
а. Мезоспорий гладкий.
* Вегетативные клетки 16—20 мкм шир................
....................................1. Z. rhynchonema
** Вегетативные клетки 20—27 мкм шир. . . . 2. Z. czurdae
р. Мезоспорий скульптурированный.
* На мезоспорий имеется киль................3. Z. carinatum
* * На мезоспорий киль отсутствует.
+ Конъюгация только лестничная, вегетативные клетки 17—
21 мкм шир.............4. Z. synadelphum (частично)
++ Конъюгация только боковая, вегетативные клетки 19—
23 мкм шир.....................5. Z. pseudogedeanum
+++ Конъюгация боковая и лестничная, вегетативные клетки
23—27 мкм шир................6. Z. cyanosphaeroidica
б. Мезоспорий желто-коричневый, коричневый или оливковый.
а. Мезоспорий гладкий..............................7. Z. ralfsii
р. Мезоспорий скульптурированный.
* Мезоспорий однослойный, ямчатый, вегетативные клетки
15—39 (43) мкм шир.
+ На мезоспорий имеется киль.
х Вегетативные клетки 15—18 мкм шир............
...........................8. Z. margalefianum
хх Вегетативные клетки 20—22 мкм щир...........
...........................9. Z. circumcarinatum
++ На мезоспорий киль отсутствует.
х Мезоспорий желто-коричневый.
О Мезоспорий мелкоямчатый (углубления 1.5—
2 мкм в диам.).
Д Вегетативные клетки 16—20 мкм шир. .
.......................10. Z. decussatum
ДА Вегетативные клетки 22—27 мкм шир. .
...................11. Z. conspicuum
ОО Мезоспорий крупноямчатый (углубления 2—
4 мкм в диам.).
Д Вегетативные клетки 22—27 мкм шир. .
...................12. Z. lawtonianum
50
ДА Вегетативные клетки 26—28 (30) мкм шир.
......................13. Z. globosum
ДДД Вегетативные клетки 30—39 (43) мкм
шир. ... 14. Z. pectinatum (частично)
хх Мезоспорий оливковый, вегетативные клетки 20—
27 мкм шир.........................15. Z. skujae
** Мезоспорий двухслойный, ямчато-бороздчатый, вегетативные
клетки 27—34 мкм шир....................16. Z. laetevirens
Б. Формирование зигоспоры в конъюгационном канале с незначительным
внедрением в гаметангии.
а. Мезоспорий голубой или синий.
а. Мезоспорий гладкий, вегетативные клетки 18—24 мкм шир. . . .
............................................17. Z. cyanosporum
Р- Мезоспорий ямчатый.
* Вегетативные клетки 17—22 мкм шир., конъюгирующие клет-
ки сильно изогнутые, редко почти прямые................
............................4. Z. synadelphum (частично)
* * Вегетативные клетки (20) 23—26 мкм шир., конъюгирующие
клетки прямые или слегка изогнутые . . 18. Z. coeruleum
б. Мезоспорий желтый или коричневый.
а. Мезоспорий однослойный, ямчатый.
* Вегетативные клетки (14) 16—22 мкм шир..............
.......................... 19. Z. spontaneum (частично)
* * Вегетативные клетки 30—39 (43) мкм шир.............
...........................14. Z. pectinatum (частично)
р. Мезоспорий двухслойный, наружный слой гладкий, внутренний
слой бородавчатый, иногда морщинисто-бородавчатый, вегетатив-
ные клетки 21—35.6 мкм шир..................20. Z. pawhuskae
2. Формирование зигоспоры в одном из гаметангиев.
А. Мезоспорий желтый или коричневый, иногда с красноватым оттенком,
а. Мезоспорий гладкий.
а. Вегетативные клетки 20—24 мкм шир. . . . 21. Z. ieiospermum
р. Вегетативные клетки 26—38 мкм шир.............22. Z. insigne
б. Мезоспорий скульптурированный.
а . Мезоспорий однослойно скульптурированный.
* Мезоспорий точечный.
+ Вегетативные клетки 9—12 мкм шир..................
..........................23. Z. stagnate (частично)
++ Вегетативные клетки 14—20 (22) мкм шир.
х Конъюгация только лестничная.......................
................24. Z. cyiindrospermum (частично)
хх Конъюгация лестничная и боковая............
...............................25. Z. subtile
+++ Вегетативные клетки 20—26 мкм шир................
.....................................26. Z. argillarii
* * Мезоспорий ямчатый.
+ Мезоспорий очень мелкоямчатый, почти точечный.
х Вегетативные клетки 9—12 мкм шир............
....................23. Z. stagnate (частично)
хх Вегетативные клетки 15—18 (22) мкм шир. . .
............24. Z. cyiindrospermum (частично)
ххх Вегетативные клетки 20—24 мкм шир...........
....................27. Z. allorgei (частично)
++ Мезоспорий с углублениями 1.3—2 мкм в диам.
51
х Зигоспоры линзовидные (сжато-шаровидные): в
разных положениях круглые и широкоэллиптичес-
кие.
О Конъюгация лестничная и боковая, вегетатив-
ные клетки .24—30 мкм шир.................
.............................28. Z. vaginatum
ОО Конъюгация только лестничная.
Д Вегетативные клетки 22—25 мкм шир. .
.........29. Z. luteosporum (частично)
ДД Вегетативные клетки 27—30 мкм шир. .
...................... 30. Z. biforme
ААЛ Вегетативные клетки 30—36 (39) мкм шир.
........................ 31. Z. cruciatum
хх Зигоспоры короткоцилиндрические и широкоэл-
липсоидные .................32. Z. cylindrosporum
ххх Зигоспоры широкоэллипсоидные и шаровидные .
.................33. Z. calosporum (частично)
+++ Мезоспорий с углублениями более 2 мкм в диам.
х Мезоспорий с углублениями 2—4 мкм в диам.
О Конъюгация лестничная и боковая, вегетатив-
ные клетки (25) 27—38 мкм шир.............
...............................34. Z. stellinum
ОО Конъюгация только лестничная.
Д Вегетативные клетки (14) 16—24 мкм шир.
§ Углубления на мезоспорий 2—3 мкм в
диам....................35. Z. tenue
§§ Углубления на мезоспорий 3—4 мкм в
диам. ............................
... 19. Z. spontaneum (частично)
ДД Вегетативные клетки (20) 23—28 мкм шир.
§ Воспринимающие клетки слегка
вздутые...........36. Z. vaucheri
§§ Воспринимающие клетки сильно взду-
тые . 33. Z. calosporum (частично)
§§§ Воспринимающие клетки не вздутые .
... 29. Z. luteosporum (частично)
хх Мезоспорий с углублениями 4—9 (10) мкм в
диам.
О Вегетативные клетки 19—26 мкм шир.
Д Углубления на мезоспорий 4.5—6.5 мкм в
диам......................37. Z. extenue
ДД Углубления на мезоспорий 7—9 (10) мкм в
диам....................38. Z. hausmannii
ОО Вегетативные клетки 25—33 (35) мкм шир.
Д Воспринимающие клетки сильно вздутые
...................... 39. Z. germanicum
ДД Воспринимающие клетки слабо вздутые или
совсем не вздутые . . 40. Z. fanicum
Р- Мезоспорий двухслойно скульптурированный.
* Мезоспорий морщинистый и мелкоямчатый, вегетативные
клетки 20—23 мкм шир..............41. Z. verrucopunctatum
* * Мезоспорий бородавчатый и крупноямчатый, вегетативные
клетки 31—33.5 (38) мкм шир.............42. Z. bohemicum
Б. Мезоспорий иного цвета, преимущественно синий.
52
а. Мезоспорий гладкий, однослойный.
а. Вегетативные клетки 14—17 мкм шир. . . 43. Z. atrocoeruleum
Р. Вегетативные клетки 20—27 мкм шир.......................
..................................44. Z. chalybeospermum
у. Вегетативные клетки 30—32 мкм шир...........45. Z. cyaneum
б. Мезоспорий скульптурированный.
а. Мезоспорий однослойный, ямчатый.
* На мезоспорий имеется киль, вегетативные клетки 23—
26 мкм шир.........................46. Z. excompressum
* * На(мезоспорий киль отсутствует.
+ Вегетативные клетки 12—14 мкм шир..............
................................47. Z. mucigenum
++ Вегетативные клетки 20—30 мкм шир.
х Воспринимающие клетки сильно вздутые . . .
..................... 48. Z. peliosporum
хх Воспринимающие клетки не вздутые или незна-
чительно вздутые..........49. Z. melanosporum
Р. Мезоспорий двухслойный, ямчато-шиповатый ...............
......................................50. Z. carinthiacum
II. Зигоспоры обычно отсутствуют; размножение преимущественно акинетами,
реже апланоспорами.
1. Вегетативные клетки 14—35 мкм шир.
А. Мезоспорий ямчатый.
а. Мезоспорий с округлыми углублениями, 2—3 мкм в диам.
а. Вегетативные клетки (14) 16—22 мкм шир..................
............................. 19. Z. spontaneum (частично)
р. Вегетативные клетки 25—33 мкм шир.........51. Z. cylindricum
б. Мезоспорий с удлиненными, прямыми или слегка изогнутыми углуб-
лениями, 0.5 мкм шир., 1—2 мкм дл.................52. Z. borzae
Б. Мезоспорий мелкобугорчатый и ямчатый, вегетативные клетки 28—32
(3 5) мкм шир...............................53. Z. subcylindricum
2. Вегетативные клетки 40—54 мкм шир.
А. Вегетативные клетки 40—42 мкм шир., мезоспорий мелкобугорчатый и
крупноямчатый ..................................... 54. Z. irregulare
Б. Вегетативные клетки 44—54 мкм шир., мезоспорий ямчатый или бугор-
чатый ...............................................55. Z. sterile
1. Zygnema rhynchonema Hansg. — Зигнема клювонитчатая (рис. 18,
/). — Вегетативные клетки 16—20 мкм шир. Конъюгация боковая.
Формирование зигоспоры в конъюгационном канале. Зигоспоры шаро-
видные или эллипсоидные, 23—33 мкм шир., 30—35 мкм дл. Мезо-
спорий голубой, гладкий.
В небольших стоячих водоемах, лужах, болотах. Польша, Чехия,
Марокко, Алжир, Австралия.
Недостаточно описанный вид. Первоописание, повторяющееся от мо-
нографии к монографии, неполное и в некоторых отношениях проти-
воречивое. Совмещение двух объемных форм зигоспоры (шаровидной и
эллипсоидной) вряд ли возможно: либо зигоспоры эллипсоидные — в
поперечном сечении (вид с вершин) круглые, либо линзовидные — в
поперечном сечении (вид сбоку) эллиптические. Во всяком случае, в
описаниях даны измерения только эллипсоидной зигоспоры. Таковая
изображена и на единственном рисунке вида. Судя по этому рисунку,
53
Рис. 18. 1 — Zygnema rhynchonema, начало боковой конъюгации, незрелая зигота; 2—4 —
Z. synadelphum: 2, 3 — конъюгирующие клетки с зигоспорами, общин вид, 4 — зигоспора;
5 — Z. ralfsii, зигоспора в конъюгационном канале; 6 — Z. carinatum, зигоспора. (По: 1 —
Hansgirg, 1888; 2 — Skuja, 1926; 3,4 — ориг.; 5 — West G. S., 1904; 6 — Taft in: Transeau
et al., 1934).
зигоспора находится в гаметангии, а не в конъюгационном канале, как
указано у автора вида. Близок к Z. gedeanum Czurda (Индонезия: о-в
Ява), также имеющему только боковую конъюгацию и синий мезоспо-
рий. Отличается от него гладким мезоспорием.
2. Zygnema czurdae Randh. — Зигнема Чурды (рис. 19, 1—6). — Ве-
гетативные клетки 20—27 мкм шир., по дайне в 1.5—4 раза больше.
Конъюгация преимущественно боковая, реже лестничная. Конъюгирую-
щие клетки сильно изогнутые, особенно при боковой конъюгации на ее
поздних стадиях. Конъюгационный канал очень широкий, при боковой
конъюгации полость канала сливается с полостями гаметангиев, перего-
родки которых полностью растворяются. Формирование зигоспоры при
54
обоих типах конъюгации в конъюгационном канале, очень редко при
лестничной конъюгации с незначительным внедрением в один из гаме-
тангиев. Зигоспоры шаровидные, 30—40 мкм диам., реже и на ранних
стадиях созревания широкоэллипсоидные и тогда вытянутые вдоль конъ-
югационного канала, иногда отделяющиеся перегородкой от гаметангиев.
Экзоспорий тонкий, бесцветный или светло-синий, гладкий. Мезоспо-
рий светло-синий, иногда голубой с блеском, гладкий. Эндоспорий глад-
кий или слегка выемчатый.
В стоячих и текучих водах, прудах, реках, болотах, иногда в водах
весенних паводков, свободноплавающая. Дания, Сьерра-Леоне, Индия.
Рис. 19. 1—6 — Zygnema czurdae'. I, 3, 5 — лестничная конъюгация, 2, 4, 6 — боковая конъ-
югация; 7 — Z. carinatum’, 8 — Z ralfsii. (По: 1—3 — Kadhibowska in: Kadhibowska, Christen-
sen, 1979; 4—6 — Randhawa, 1936; 7 —Simons, 1990; 8 — Gauthier-Lievre, 1965).
55
Редкий вид, отличающийся образованием в некоторых образцах (и
только при боковой конъюгации) спорангия, что является как бы пе-
реходом к роду Zygogonium. Спорен вопрос о форме зигоспор. Вполне
возможно, что зигоспоры линзовидные — круглые и эллиптические в
разных положениях. От близкой Z. cyanosphaeroidica отличается глад-
ким мезоспорием.
3. Zygnema carinatum Taft — Зигнема килевая (рис. 18, 6; 19,
7). — Вегетативные клетки 14.3—18 мкм шир., по длине в 2 раза
больше. Конъюгация лестничная. Конъюгирующие клетки прямые.
Формирование зигоспоры в конъюгационном канале с полным его за-
полнением. Зигоспоры сжато-шаровидные (широкоэллипсоидные?),
23—32.1 мкм шир., 28.5—33 мкм дл., вытянутые поперек конъюгаци-
онного канала и окруженные слоем пектинового вещества 3—6 мкм
толщ. Экзоспорий бесцветный, гладкий. Мезоспорий голубой, ямчатый,
с четко выраженным выпуклым килем.
В стоячих (?) водах. Нидерланды, США.
Редкий вид. От видов, имеющих на мезоспорий киль, отличается
голубым мезоспорием.
4. Zygnema synadelphum Skuja — Зигнема братственная (рис. 18,
2—4). — Вегетативные клетки 17—22 мкм шир., по длине в 2—7 раз
больше. Конъюгация лестничная. Конъюгирующие клетки сильно изо-
гнутые, редко почти прямые. Формирование зигоспоры в широком конъ-
югационном канале, реже с незначительным внедрением в гаметангии.
Зигоспоры линзовидные, сбоку широкоэллиптические, (23) 27—36 мкм
шир., 29—44 (55) мкм дл., вытянутые вдоль конъюгационного канала.
Экзоспорий около 2 мкм толщ., прозрачный, с округлыми, довольно гру-
быми углублениями. Мезоспорий толстый, около 3.8 мкм толщ., голу-
бой, с округлыми углублениями 1—2 мкм в диам. Эндоспорий тонкий,
гладкий, различимый лишь на зрелых зиготах. Формирование партено-
спор в конъюгационном канале или в обоих гаметангиях.
В стоячих, реже слабопроточных, иногда заболоченных водах, в
озерах, на рисовых полях. Латвия (оз. Каниерис, Усмас), Сев. (запо-
ведник Боровое) и Южн. Казахстан.
Редкий вид. Близок к Z. coeruleum, отличается более узкими веге-
тативными и довольно сильно изогнутыми конъюгирующими клетками.
Требует уточнения строение оболочек зигоспоры: по некоторым дан-
ным, экзоспорий гладкий, а мезоспорий трехслойный, причем внешний
слой неравномерно точечный.
5. Zygnema pseudogedeanum Gauthier-Lievre — Зигнема ложноге-
денская (рис. 30, 10—12). — Вегетативные клетки 19—23 мкм шир.
Конъюгация боковая. Конъюгационный канал очень широкий, сильно
пузыревидно вздутый, конъюгирующие клетки при этом слегка или
почти под прямым углом изогнутые. Формирование зигоспоры в конъ-
югационном канале. Зигоспоры линзовидные, анфас круглые, 32—
38 мкм диам., сбоку широкоэллиптические, (25) 28—35 мкм шир.,
35—48 мкм дл. Экзоспорий тонкий, гладкий, бесцветный. Мезоспорий
56
толстый, голубой или голубовато-черный, с округлыми ямчатыми уг-
лублениями, отсутствующими вдоль линии разрыва.
В стоячих водоемах, на мелководье. Испания, Марокко.
Редкий вид. Противоречивы описания формы зигоспор: от почти ша-
ровидных до трехосно-эллипсоидных (Gauthier-Lifevre, 1965 : 53, pl. 15,
В) и эллипсоидно-яйцевидных (Aboal, 1991 : 368, fig. 1, 1, 2, fig. 2, 9).
Сопоставление размеров и изображений зигоспор в работах этих иссле-
дователей дает четкое представление о линзовидной форме зигоспор, в
разных положениях круглых и эллиптических. Наличие лестничной
конъюгации требует обстоятельного подтверждения, последним из цити-
руемых авторов она только упомянута. Непонятно отсутствие ямчатости
вдоль линии разрыва — можно предположить, что эта линия проходит
по гладкой поверхности довольно широкого кольца, опоясывающего зи-
госпору. Очень близок по морфологическим признакам, кроме ширины
вегетативных клеток, к Z. gedeanum Czurda (Индонезия: о-в Ява, вершина
вулкана Геде).
6. Zygnema cyanosphaeroidica О. et W. Bock [=Zygogonium cyano-
sphaeroidicum (O. et W. Bock) Kadi.] — Зигнема синешаровидная
(рис. 20, 1—3). — Вегетативные клетки 23—27 мкм шир., с толстыми
оболочками. Конъюгация боковая, редко лестничная. Конъюгирующие
клетки прямые или слегка изогнутые. Формирование зигоспоры в конъ-
югационном канале, при боковой конъюгации с образованием споран-
гиальных стенок. Зигоспоры утолщенно-линзовидные, сбоку широко-
Рис. 20. 1—3 — Zygnema cyanosphaeroidica-. I — боковая конъюгация, 2 — лестничная
конъюгация, зигоспора анфас, 3 — зигоспора сбоку; 4, 5 — Z. circumcarinalum-. 4 — лест-
ничная конъюгация, общий вид, 5 — строение мезоспория (фрагмент). (По: 1—3 — Воск,
Воск, 1956; 4,5 — Czurda, 1930).
57
эллиптические, 22—23 мкм шир., 31—39 мкм да. Мезоспорий темный,
черновато-синий с округлыми глубокими углублениями. Линия разрыва
зигоспоры поперечная.
На аллювии при периодическом обводнении, при pH 6.0. Германия.
Редкий вид. Характерное строение мезоспория четко проявляется толь-
ко на последних стадиях созревания зигоспоры. Образование спорангия,
послужившего поводом для некоторых исследователей отнести эту водо-
росль к роду Zygogonium, наблюдалось только при боковой конъюгации.
От сходной по многим признакам Z. czurdae отличается скульптуриро-
ванным мезоспорием, от Z randhawae Krieger (Индия) — большей ши-
риной вегетативных клеток. Крайне близок к Z. pseudogedeanum.
7. Zygnema ralfsii (Hass.) De Вагу — Зигнема Ральфса (рис. 18,
5; 19, 8). — Вегетативные клетки 14—22 мкм шир., по длине в 2—6
раз больше. Конъюгация лестничная. Конъюгирующие клетки прямые.
Формирование зигоспоры в конъюгационном канале. Зигоспоры эллип-
соидные, вытянутые поперек конъюгационного канала, 15—25 мкм
шир., 24—35 мкм да. Экзоспорий тонкий, бесцветный, гладкий. Ме-
зоспорий толстый, коричневый, гладкий.
В стоячих и текучих водах, озерах, реках, на рисовых полях, в
мелководной и опресненной зоне морей. Россия: Причерноморье, сев.
часть Каспийского моря, Республика Саха (р. Джанки, р. Тимирдях,
долина р. Яны); Эстония (Пярнуская бухта), Узбекистан (Ферганская
долина, долина р. Зеравшан).
Широко распространенный, однако еще недостаточно изученный
вид. Требует уточнения форма зигоспор: по некоторым данным, они
шаровидные. Отличается от Z. micropunctatum Trans. (США) гладким
мезоспорием.
8. Zygnema margalefianum Aboal — Зигнема маргалефская. — Ве-
гетативные клетки 15—18 мкм шир. Конъюгация лестничная. Конъ-
югирующие клетки прямые. Формирование зигоспоры в конъюгацион-
ном канале. Зигоспоры сжато-шаровидные, 33 мкм шир., 35 мкм да.,
вытянутые поперек конъюгационного канала. Мезоспорий коричневый,
ямчатый, с углублениями 1—2 мкм в диам., с килем.
В щелочной воде мелких прудов. Испания.
Редкий вид. От самого близкого вида — Z. circumcarinatum отли-
чается четко оформленным конъюгационным каналом, меньшей шири-
ной вегетативных клеток и мелкой ямчатостью мезоспория. У другого
вида — Z. excompressum, на ямчатом мезоспорий которого также есть
киль, зигоспоры формируются в гаметангии, а цвет мезоспория —
синий. Рисунок вида отсутствует.
9. Zygnema circumcarinatum Czurda [=Zygnemopsis circumcarinata
(Czurda) Krieger] — Зигнема окружнокилевая (рис. 20, 4, 5). — Вегетатив-
ные клетки 20—22 мкм шир., окруженные толстым слоем слизи 14—
16 мкм толщ. Конъюгация лестничная. Конъюгирующие клетки прямые.
Формирование зигоспоры между клетками, в конъюгационном канале, рас-
творяющемся к моменту образования зигоспоры. Зигоспоры шаровидные
58
или сжато-шаровидные, 24 мкм шир., 29 мкм дл., окруженные к момен-
ту созревания плотным слоем коллоидного вещества. Экзоспорий тон-
кий, гладкий, бесцветный. Мезоспорий толстый, коричневый или желто-
коричневый, ямчатый, с углублениями 2—2.5 мкм в диам. и расстоянием
1—2 мкм между ними, с килем, отчетливо выраженным у сжато-шаро-
видных зигоспор по их экватору. Линия разрыва по килю, отчетливая.
В стоячих водах, в лабораторной культуре. Чехия, США.
Редкий вид. Требует уточнения наличие коллоидного вещества в
гаметангиях. От близких видов, у которых есть киль на мезоспорий,
отличается отсутствием четко оформленного конъюгационного канала.
10. Zygnema decussatum (Vauch.) Ag„ поп (Trans.) Czurda — Зиг-
нема перекрестная (рис. 21, 7). — Вегетативные клетки 16—20 мкм
шир., по длине в 1.6—6.3 раза больше. Конъюгация лестничная, реже
боковая. Конъюгирующие клетки прямые или слегка изогнутые. Фор-
мирование зигоспоры в конъюгационном канале. Зигоспоры линзовид-
ные, сбоку широкоэллиптические, 24—33 мкм шир., 26—36 мкм дл.,
вытянутые вдоль конъюгационного канала. Мезоспорий коричневый, с
мелкими углублениями 1.5—2 мкм в диам.
В стоячих водах, придорожных канавах, ямах, лужах, сфагновых
болотах, реже в горных текучих водах, водопадах. Украина: горный
Крым (водопад Учан-Су).
Широко распространенный вид на всех континентах. Очень близок
к Z. conspicuum, незначительно отличается только более узкими веге-
тативными клетками. От другого близкого вида — Z. ovoidanum Trans.
(США) отличается широкими вегетативными клетками и более круп-
ными углублениями на мезоспорий. Возможно, что на территории Рос-
сии имеет широкое распространение.
11. Zygnema conspicuum (Hass.) Trans. [=Z. pectinatum (Vauch.) Ag.
var. conspicuum (Hass.) Kirchn., non sensu( Скворцов, 19276] — Зиг-
нема заметная (рис. 21, 2). — Вегетативные клетки 22—27 мкм шир.,
по длине в 2—4 раза больше. Конъюгация лестничная. Конъюгирующие
клетки прямые. Формирование зигоспоры в конъюгационном канале.
Зигоспоры линзовидные, сбоку широкоэллиптические, 24—32 мкм
шир., 26—33 мкм дл., вытянутые вдоль конъюгационного канала. Ме-
зоспорий коричневый, с небольшими округлыми углублениями 1.5—
2 мкм в диам., расстоянием в 1.5—2 мкм между ними.
В стоячей и проточной воде, на рисовых полях, в водопадах горных
районов. Россия: центр европ. части, Зап. Сибирь (Алтай) (?); При-
балтика; Украина: горный Крым (водопад Учан-Су); Узбекистан.
Широко распространенный вид, особенно в Европе. Хорошо отли-
чается от Z. pectinatum меньшей шириной вегетативных клеток и не-
значительным диаметром углублений на мезоспорий. Очень близок к
Z. decussatum, незначительно отличаясь чуть более широкими вегета-
тивными клетками.
12. Zygnema lawtonianum Taft — Зигнема лотонская (рис. 21,
3). — Вегетативные клетки 22—27 мкм шир., по длине равные ши-
59
Рис. 21. I — Zygnema decussatum, 2 — Z. conspicuum', 3 — Z. lawtonianum\ 4, 5 — Z. globo-
sum: 4 — зигоспоры, лестничная конъюгация, общий вид, 5 — строение зигоспоры. (По: I,
2 — Transeau, 1951; 3 — Taft in: Transeau et al., 1934; 4, 5 — Czurda, 1932).
рине или до 5 раз больше. Конъюгация лестничная. Формирование
зигоспоры в конъюгационном канале. Зигоспоры линзовидные, сбоку
широкоэллиптические, 33—40 мкм шир., 33—46 мкм дл., вытянутые
вдоль конъюгационного канала и отделенные со стороны гаметангиев
тонкими гиалиновыми прослойками. Экзоспорий тонкий, бесцветный,
гладкий. Мезоспорий толстый, желто-коричневый или коричневый, с
ямчатыми углублениями 3—4 мкм в диам., с расстоянием в 3—4 мкм
между ними.
В стоячих водах, глинистых ямах, озерах. Дания, США, Индия,
Мьянма.
60
Редкий вид. По первоописанию экзоспорий зигоспоры срастается
со стенками конъюгационного канала, что создает видимость образо-
вания спорангия. По этому признаку и по большему расстоянию между
ямчатыми углублениями на мезоспории хорошо отличается от близкой
Z. globosum.
13. Zygnema globosum Czurda — Зигнема шаровидная (рис. 21, 4,
5). — Вегетативные клетки 26—28 (30) мкм шир., по длине в 2.5—3.7
раза больше. Конъюгация лестничная. Конъюгирующие клетки прямые.
Формирование зигоспоры в конъюгационном канале. Зигоспоры лин-
зовидные, сбоку широкоэллиптические, 30—50 мкм шир., 42—65 мкм
дл., вытянутые вдоль конъюгационного канала. Экзоспорий тонкий,
бесцветный, гладкий. Мезоспорий толстый, коричневый, с углублени-
ями 3—4 мкм в диам., с расстоянием в 1—2 мкм между ними.
В стоячих, иногда загрязненных водах. Чехия, Центр. Индия.
Редкий вид. Близок к Z. lawtonianum, Z. adpectinatum Trans. (США,
Сьерра-Леоне), Z. sinense Jao (Китай). Отличается от них или размерами
углублений на мезоспории, или иным расстоянием между углублениями.
14. Zygnema pectinatum (Vauch.) Ag. [=Z. pectinatum f. terrestre
(Rabenh.) Hansg.; Zygogonium pectinatum (Vauch.) Kutz.] — Зигнема
гребешковая (рис. 22, 1, 2). — Вегетативные клетки 30—39 (43) мкм
шир., по длине меньше ширины или до 4 раз больше. Оболочка клеток
ослизненно-студенистая, часто значительно утолщенная. Конъюгация
лестничная, редко боковая. Конъюгирующие клетки прямые. Форми-
рование зигоспоры при лестничной конъюгации в конъюгационном ка-
нале, иногда с незначительным внедрением в гаметангии, при боковой
конъюгации — в широком конъюгационном канале. Зигоспоры утол-
щенно-линзовидные, сбоку широкоэллиптические, 35—60 мкм шир.,
40—70 мкм дл. Экзоспорий тонкий, бесцветный, гладкий. Мезоспорий
толстый, коричневый, с довольно крупными округлыми углублениями
2—3 мкм в диам. Партеноспоры шаровидные. Акинеты цилиндричес-
кие, 30—38 мкм шир., 30—60 мкм дл.
В стоячей воде мелких' водоемов, в прудах, канавках, лужах, бо-
лотцах, на рисовых полях, мелководьях озер и водохранилищ, реже в
реках и ручьях, редко на сильно увлажненной почве у родников. Рос-
сия: центр европ. части, Цимлянское водохранилище, Республика Саха
(долины рек Вилюй и Оленёк); Прибалтика, Зап. Украина, Кавказ,
Узбекистан (Ферганская долина, окрестности Самарканда, Ташкента,
Бухары, Пенджикента, долина р. Зеравшан).
Широко распространенный и часто встречающийся вид. В наземных
условиях обитания вегетативные клетки значительно укорачиваются, а
их оболочка утолщается. При этом наблюдаются интенсивное накоп-
ление в клетках масла и образование покоящихся клеток — акинет.
15. Zygnema skujae Czurda (=Z. laetevirens Klebs sensu Skuja,
1929) — Зигнема Скуй. — Вегетативные клетки 20—27 мкм шир,, по
длине в 1.5—4 (5) раза больше. Конъюгация лестничная. Конъюгиру-
ющие клетки слегка изогнутые. Формирование зигоспоры в конъюга-
61
Рис. 22. 1, 2 — Zygnema pectinatum'. 1 — лестничная конъюгация, 2 — лестничная и боко-
вая конъюгации в одной паре нитей, акинета; 3, 4 — Z laetevirens, зигоспора анфас и сбоку.
(По: I — ориг.; 2 — Transeau, 1951; 3,4 — Bock, Bock, 1956).
62
ционном канале. Зигоспоры линзовидные, анфас круглые, 40—55 мкм
диам., сбоку широкоэллиптические, вытянутые вдоль конъюгационного
канала. Мезоспорий толстый, зеленовато-коричневый (оливковый), гру-
боямчатый, с тесным расположением углублений.
В стоячих, часто заболоченных мелких водоемах, лужах, канавах.
Эстония (о-в Сааремаа).
Редкий вид. Отличается от Z. globosum цветом мезоспория и боль-
шей амплитудой ширины вегетативных клеток. Оригинальное изобра-
жение отсутствует.
16. Zygnema laetevirens Klebs, non sensu Skuja, 1929 [-Zygogonium
laetevirens (Klebs) Migula] — Зигнема ярко-зеленеющая (рис. 22, 3, 4). —
Вегетативные клетки 27—34 мкм шир., по длине в 1.5—3 раза больше.
Конъюгация лестничная. Формирование зигоспоры в конъюгационном
канале. Зигоспоры линзовидные, анфас круглые, 40—55 мкм диам.,
сбоку яйцевидно-эллиптические, 34—45 мкм шир., 51—68 мкм дл., вы-
тянутые вдоль конъюгационного канала. Экзоспорий тонкий, бесцвет-
ный, гладкий. Мезоспорий толстый, 6 мкм толщ., каштаново-коричне-
вый, двухслойный: наружный слой с углублениями 2—4 мкм в диам., с
расстоянием в 3—5 мкм между ними; внутренний слой мелкобородавча-
тый. Эндоспорий тонкий, бесцветный, гладкий. Партеноспоры сходные
по строению с зигоспорами, но меньше их по размерам.
В стоячих мелких и заболоченных водоемах, в лужах, среди нит-
чатых водорослей. Латвия.
Вид широко распространенный на всех континентах. Некоторые
авторы относят его к роду Zygogonium. Высказано предположение, что
орнаментация внутреннего слоя мезоспория возникает из гранул, сфор-
мированных между двумя слоями. Отличается от Z. adpectinatum Trans.
(США, Сьерра-Леоне) двойной скульптурой мезоспория и от Z. агео-
latum Taft et Trans. (США) небольшими углублениями на мезоспорий.
17. Zygnema cyanosporum Cleve — Зигнема синеспоровая (рис. 23,
1—3). — Вегетативные клетки 18—24 мкм шир., по длине в 2—9 раз
больше. Конъюгация лестничная, реже боковая. Конъюгирующие клет-
ки при лестничной конъюгации прямые, при боковой конъюгации
иногда довольно сильно изогнутые. Формирование зигоспоры в конъ-
югационном канале с частичным внедрением в гаметангии. Зигоспоры
линзовидные: анфас круглые, 26—40 мкм в диам., сбоку широкоэл-
липтические, 30—34.5 мкм шир., 37—44 мкм дл. Экзоспорий бесцвет-
ный, гладкий. Мезоспорий толстый, синий или голубоватый, гладкий.
В проточной и стоячей воде: прудах, озерах, родниках. Россия:
север европ. части (Карелия); Туркмения (центр. Копетдаг).
Вполне вероятно, что в пределах России распространен шире, чем
указано в литературе. Следует отметить, что на первом рисунке этого
вида хорошо заметен спорангий, не оговоренный в описании. Отлича-
ется от близких видов: Z. coeruleum, Z. synadelphum и Z. cyanosphae-
roidica гладким мезоспорием, от других видов с синей окраской ме-
зоспория — формированием зигоспоры в конъюгационном канале с
частичным внедрением в гаметангии. Близок также к Z. majus Czurda
63
Рис. 23. 1—3 — Zygnema cyanosporum: 1,2 — лестничная конъюгация, 3 — боковая конъ-
югация; 4—11 — Z. coeruleum'. 4 — различные стадии формирования зигоспоры, 5—8 —
различные положения зигоспоры, 9, 10 — различные формы зигоспор, 11 — зигоспора
анфас. (По: 1 — Cleve, 1868; 2,3 — Rattan, 1971; 4—8 — Рундина, 1977а; 9—11 — Czur-
da, 1932).
64
(Южн. Африка) и Z. indicum Misra (Индия), отличается от них узкими
вегетативными клетками.
18. Zygnema coeruleum Czurda — Зигнема голубая (рис. 23, 4—
77). — Вегетативные клетки (20) 23—26 мкм шир., по дайне в 1.5—4
раза больше. Конъюгация лестничная. Конъюгирующие клетки прямые
или слегка изогнутые, длина их в 4—7 раз больше ширины. Форми-
рование зигоспоры в конъюгационном канале с частичным внедрением
в гаметангии. Зигоспоры линзовидные, анфас круглые, (26) 32—38
(43.5) мкм диам., сбоку эллиптические, иногда яйцевидно-эллиптичес-
кие, 26—38 мкм шир., 32—43.5 (46.4) мкм да., вытянутые вдоль конъ-
югационного канала. Экзоспорий тонкий, бесцветный, ямчатый. Ме-
зоспорий толстый, голубой, с округлыми углублениями 1.5—3 мкм в
диам., с расстоянием в 3—5.5 мкм между ними. Эндоспорий тонкий,
трудноразличимый.
В мелких заболоченных водоемах, старицах рек, на рисовых полях.
Южн. Казахстан.
Редкий вид. Скульптурированность экзоспория, впервые отмеченная
в образцах из Казахстана, достаточно четко выражена, но видна лишь
при определенном положении зигоспоры и при достаточном разделении
слоев ее оболочки. По этому признаку близок к Z. synadelphum, не-
значительно отличаясь шириной вегетативных и степенью изогнутости
конъюгирующих клеток. Близок, а возможно, и идентичен Z. gorakh-
porense Singh (Индия).
19. Zygnema spontaneum Nordst. — Зигнема спонтанная (рис. 24,
7—4). — Вегетативные клетки (14) 16—22 мкм шир., по длине равные
ширине или до 5.5 раза больше. Размножение преимущественно апла-
носпорами, реже акинетами или зигоспорами. Конъюгация лестничная,
очень редкая. Конъюгирующие клетки прямые или изогнутые. Форми-
рование зигоспоры в гаметангии, иногда с частичным или полным
заполнением конъюгационного канала. Воспринимающие клетки невзду-
тые или слегка вздутые, преимущественно со стороны конъюгационного
канала. Зигоспоры шаровидные и широкоэллипсоидные, 19—36 (42) мкм
шир., 21—42 мкм да. Экзоспорий тонкий, бесцветный, гладкий. Ме-
зоспорий зигоспор толстый, от желтого до желто-коричневого, с ок-
руглыми углублениями 3—4 мкм в диам., с расстоянием в 3—5 мкм
между ними. Апланоспоры шаровидные и широкоэллипсоидные, 16—
22 мкм шир., 18—32 мкм да., очень частые, в отдельных нитях или
совместно с зигоспорами. Клетки с апланоспорами слегка вздутые.
Мезоспорий апланоспор от желтого до желтовато-коричневого, с мел-
кими углублениями 2 мкм в диам., с расстоянием в У—5 мкм между
ними. Акинеты цилиндрические, с закругленными вершинами, очень
редкие.
В стоячих водах мелких водоемов, на рисовых полях. Дания, Ан-
гола, Трансвааль, Индия, Китай, Япония, Мьянма, о-в Ява, Австралия,
Гавайские о-ва.
Широко распространенный вид. Отличается от близких видов —
Z. tenue и Z. extenue невздутыми воспринимающими клетками, час-
5 Рундина Л. А.
65
Рис. 24. 1—4 — Zygnema spontaneum: 1 — разные стадии формирования апланоспор, 2 —
зрелые апланоспоры, 3 — зигоспоры, лестничная конъюгация, 4 — зигоспора; 5—8 —
Z. stagnate’. 5, 6 — разные формы вздутия воспринимающих клеток прн лестничной конъюга-
ции, 7, 8 — зигоспора в разных положениях; 9 — Z. subtile, зигоспоры анфас и сбоку, лест-
ничная конъюгация. (По: /, 2 — Transeau, 1951; 3, 4 — Yamagishi, 1965с; 5,6 — West G. S.,
1904; 7—9— Gauthier-Lievre, 1965).
тым и обильным образованием апланоспор, от Z. hausmannii —
небольшими углублениями на мезоспории. Требует уточнения фор-
ма зигоспор и апланоспор. Вполне возможно, что они линзовид-
ные.
66
20. Zygnema pawhuskae Taft — Зигнема похаская (рис. 25, 1). —
Вегетативные клетки 21—35.6 мкм шир., по длине в 2—3 раза больше.
Конъюгация лестничная. Конъюгирующие клетки прямые или слегка
изогнутые. Формирование зигоспоры в широком конъюгационном кана-
ле с незначительным внедрением в полости гаметангиев. Зигоспоры лин-
зовидные (?), в двух положениях круглые и широкоэллиптические, 34—
48 мкм шир., 40—65 мкм дл., окруженные слоем пектинового вещества
3—6 мкм толщ. Мезоспорий сургучно-коричневый, двухслойный: на-
ружный слой гладкий, с отчетливой линией разрыва, внутренний слой
мелко и плотно бородавчатый, иногда морщинисто-бородавчатый.
В стоячих (?) водах. Нидерланды, США.
Редкий вид. Близок по ширине вегетативных клеток и двухслой-
ное™ мезоспория к Z. verrucosum Jao (Китай), отличается нескульп-
турированным наружным слоем мезоспория. Требует уточнения форма
зигоспор.
21. Zygnema leiospermum De Вагу — Зигнема гладкоспоровая
(рис. 25, 2). — Вегетативные клетки 20—24 мкм шир., по длине рав-
ные ширине или до 2 раз больше. Конъюгация лестничная. Формиро-
вание зигоспоры в одном из гаметангиев. Воспринимающие клетки
вздутые, преимущественно со стороны конъюгационного канала. Зи-
госпоры шаровидные, 23—35 мкм в диам., или почти шаровидные,
23—32 мкм шир., 23—35 мкм дл. Экзоспорий тонкий, бесцветный,
гладкий. Мезоспорий толстый, коричневый, гладкий. Партеноспоры ша-
ровидные.
Рис. 25. 1 — Zygnema pawhuskae', 2 — Z leiospermum', 3,4 — Z cylindrospermum'. 3 — цилин-
дрически-эллипсоидные зигоспоры, 4 — короткая эллипсоидная зигоспора. (По: 1 — Taft in:
Transeau et al., 1934; 2 — De Bary, 1858; 3 — West, West, 1905; 4 — Yamagishi, 1965c).
67
В стоячих водах, заболоченных мелких водоемах, канавах, плавнях,
оросительных каналах, на рисовых полях. Россия: центр европ. части,
оз. Селигер, Зап. Сибирь (Тюменская обл., пойма р. Оби), Республика
Саха; Эстония (о-в Хийумаа), Латвия, Вост. Украина, Центр. Казах-
стан, Южн. Узбекистан, Туркмения (канал от р. Мургаб).
Широко распространенный вид. Отличается от близкой Z. insigne
узкими вегетативными клетками и отсутствием боковой конъюгации.
На одном из рисунков автора Z. leiospermum мезоспорий изображен,
в отличие от описания, скульптурированным: бугорчатым или ямчатым.
22. Zygnema insigne (Hass.) Kutz. — Зигнема приметная (рис. 26,
1—6). — Вегетативные клетки 26—38 мкм шир., по длине в 1.5—3
раза больше, иногда окруженные прозрачным, слабо перешнурованным
футляром слизи 14—17.4 мкм шир. Ризоиды короткие, неразветвлен-
ные, иногда лопастные, латеральные и базальные. Конъюгация лест-
Рис. 26. 1—6 — Zygnema insigne: 1—5 — f. insigne: 1 — инцнстирование вегетативных кле-
ток, 2, 3 — зигоспоры в разных положениях, лестничная конъюгация, 4, 5 — боковая и лест-
ничная конъюгации; 6 — f. kasachstanicum. (По: 1—3 — Рундина, 1977а; 4, 5 — De Вагу,
1858; 6 — Рундина, 19776).
68
иичная и боковая. Формирование зигоспоры в одном из гаметангиев.
Воспринимающие клетки невздутые или слегка вздутые со стороны
конъюгационного канала. Зигоспоры шаровидные, 26—49 мкм в диам.,
или широкоэллипсоидные, 26—35 мкм шир., 27—40.6 мкм дл. Экзо-
спорий тонкий, бесцветный, гладкий. Мезоспорий толстый, желто-ко-
ричневый или коричневый, гладкий. Партеноспоры (апланоспоры ?)
яйцевидные или цилиндрически-яйцевидные, 28—33 мкм шир.
Широко распространенный и часто встречающийся вид. Отличается
от близкой Z. leiospermum широкими клетками и наличием боковой
конъюгации, от Z. chalybeospermum и Z. суапеит — окраской мезо-
спория.
F. insigne (рис. 26, 1—5). — Вегетативные клетки 26—32 мкм шир.
Конъюгация лестничная и боковая. Зигоспоры шаровидные, 26—38 мкм
в диам., или широкоэллипсоидные/26—35 мкм шир., 27—40.5 мкм дл.
В стоячих, реже текучих водах, в мелководьях водохранилищ и
ручьях, на рисовых полях, а также на влажных скалах, дождевых по-
токах в высокогорных районах. Россия: Республика Саха; Украина
(Прикарпатье, Причерноморье), Грузия (долина р. Белая Арагви), Сев.,
Вост, (горы Бегазы, Кызыл-Рай) и Южн. Казахстан, Южн. Узбекистан
(долина р. Зеравшан, Фархадское водохранилище).
F. kasachstanicum Rund. (рис. 26, 6). — Вегетативные клетки 35—
38 мкм шир. Конъюгация лестничная. Зигоспоры шаровидные, 43.5—
49 мкм в диам.
В чистых стоячих водах. Южн. Казахстан.
23. Zygnema stagnate (Hass.) Kutz. — Зигнема прудовая (рис. 24,
5—8). — Вегетативные клетки 9—12 мкм шир., по длине в 1.7—5.5
раза больше. Конъюгация лестничная. Формирование зигоспоры в
одном из гаметангиев. Воспринимающие клетки незначительно вздутые,
преимущественно со стороны конъюгационного канала. Зигоспоры лин-
зовидные, в разных положениях круглые и широкоэллиптические, 14—
19 мкм шир., 14—25 мкм дл. Мезоспорий коричневый, точечный или
очень мелкоямчатый.
В стоячих небольших мелких водоемах. Англия, Венгрия, США,
Марокко.
Широко распространенный, но мало изученный вид. Разными ис-
следователями указывалась разная форма зигоспор (шаровидная и эл-
липсоидная), что связано, вероятно, с различным положением линзо-
видной зигоспоры в полости гаметангия. Неясен также вопрос о стро-
ении мезоспория: мелкоямчатый мезоспорий выглядит точечным при
взгляде на зигоспору сверху.
24. Zygnema cyiindrospermum (W. et G. S. West) Krieger — Зиг-
нема цилиндроспоровая (рис. 25, 3, 4). — Вегетативные клетки 15—18
(22) мкм шир., по длине в 1.3—2.3 раза больше. Конъюгация лест-
ничная. Формирование зигоспоры в одном из гаметангиев. Восприни-
мающие клетки невздутые. Зигоспоры цилиндрически-эллипсоидные
69
или широкоэллипсоидные, 15—19 мкм шир., 23—54 мкм дл. Мезо-
спорий толстый, коричневый, мелкоямчатый (точечный ?). Линия раз-
рыва косая, поперечная.
В стоячих и временных водоемах, дождевых лужах. Великобрита-
ния: Шетландские о-ва; Южн. Африка; Индия; Япония.
Широко распространенный, но мало изученный вид. Впервые был
описан в ранге разновидности Z. stellinum, мезоспорий которой гру-
боямчатый. Требует уточнения вопрос о скульптуре мезоспория, хотя
принципиального различия между точечным и мелкоямчатым мезоспо-
рием нет: при микроскопировании без масляной иммерсии мелкие
ямки сверху выглядят точками. Отличается от близкой Z. cylindrospo-
гит узкими вегетативными клетками и относительно длинными зиго-
спорами.
25. Zygnema subtile Kutz. [=Z. stellinum (Vauch.) Ag. var. subtile
(Kutz.) Kirchn.; Z. tenue Kutz, sensu Krieger in: Kolkwitz, Krieger,
1941 —1944, pr. p.] — Зигнема изящная (рис. 24, 9). — Вегетативные
клетки 14—20 мкм шир., по длине в 1.5—6 раз больше. Конъюгация
лестничная и очень редко боковая. Конъюгирующие клетки прямые.
Формирование зигоспоры в одном из гаметангиев. Воспринимающие
клетки невздутые или незначительно вздутые, преимущественно со сто-
роны конъюгационного канала. Зигоспоры от почти шаровидных до
широкоэллипсоидных, (14) 20—29 мкм шир., 22—30 мкм дл. Мезо-
спорий коричневый, точечный.
В стоячих, преимущественно заболоченных водоемах, канавах,
лужах, прибрежье озер, редко в проточной воде. Россия: европ. часть.
Недостаточно изученный вид. Вполне возможно, что форму зиго-
спор, определяемую как шаровидная и широкоэллипсоидная, следует
считать линзовидной, видимой в двух проекционных положениях —
анфас округлой и сбоку широкоэллиптической. То же самое можно
отнести и к близкому виду Z. tenue, с которым, по-видимому, его
часто смешивали. Некоторые авторы вовсе отрицают видовую значи-
мость Z. subtile и вносят его в синонимы Z. tenue. Близок также к
Z. stellinum, в качестве разновидности которого и был впервые описан.
Отличается от него меньшей шириной вегетативных клеток и точечной
скульптурой мезоспория.
26. Zygnema argillarii Kadi. — Зигнема глинистая (рис. 27, 7). —
Вегетативные клетки 20—26 мкм шир. Конъюгация лестничная. Конъ-
югационный канал от обеих клеток. Формирование зигоспоры в одном
из гаметангиев. Конъюгирующие клетки прямые. Воспринимающие
клетки незначительно вздутые со всех сторон. Зигоспоры линзовидные,
в разных положениях круглые и широкоэллиптические, 24—28.8 мкм
шир., 33.6—48 мкм дл. Экзоспорий тонкий, бесцветный, гладкий. Ме-
зоспорий красно-бурый или желто-коричневый, точечный.
В глинистой яме. Дания.
Редкий и довольно спорный по описанию вид. Автором вида кон-
статирована только лестничная конъюгация, однако при упоминании
особо мелких по размеру зигот дается ссылка на боковую конъюгацию.
70
Рис. 27. 1 — Zygnema argillarii-, 2—4 — Z. luteosporum: 2 — лестничная конъюгация, 3,
4 — зигоспора в двух положениях; 5, 6 — Z. cruciatum (6 — акинета и зигоспоры в двух
положениях); 7—10 — Z. cylindrosporum. (По: 1 — Kadhibowska in; Kadhibowska, Christen-
sen, 1979; 2—4, 7, 8 — Gauthier-Lievre, 1965; 5 — Скворцов, 19276; 6-—Transeau, 1951;
9, 10 — Czurda, 1932).
71
Зигоспоры охарактеризованы как сферические (шаровидные), в то
время как указания длины и ширины зигоспоры противоречат этому
утверждению. Большую таксономическую значимость, по мнению ав-
тора вида, имеет особая, тонкая оболочка, окружающая зигоспору и
остающаяся в гаметангии после прорастания зигоспоры или выпадения
ее из клетки. Логичнее думать, что в данном случае речь идет об
экзоспорий: как известно, эта наружная оболочка легко сбрасывается
зигоспорой, и наличие ее в клетке после освобождения зигоспоры не
может подтверждать таксономическое родство этого вида с представи-
телями рода Zygogonium.
27. Zygnema allorgei Gauthier-Lievre — Зигнема Аллоржа (рис. 28,
1—3). — Вегетативные клетки 20—24 мкм шир., по длине в 2—2.5 раза
больше. Конъюгация лестничная. Формирование зигоспоры в одном из
гаметангиев. Конъюгационный канал от обеих клеток. Воспринимающие
клетки вздутые со стороны конъюгационного канала. Зигоспоры линзо-
видные (сжато-шаровидные), в разных положениях круглые, 30—32 мкм
в диам., и эллиптические, 28 мкм шир., 32—40 мкм дл. Экзоспорий тон-
кий, бесцветный, гладкий. Мезоспорий толстый, коричневый или желто-
коричневый, очень тонко и мелко ямчатый, почти точечный. Линия раз-
рыва четкая.
В стоячих водах, лужах. Польша (?), Алжир.
Редкий вид. По первоописанию форма зигоспор сомнительна: от ша-
ровидной до трехосно-эллипсоидной, не подтвержденная соответствую-
щими рисунками. По ширине вегетативных клеток, форме зигоспор, од-
носторонне вздутым воспринимающим клеткам этот вид сближается с
Z. perforatum Gauthier-Lievre (Марокко) и Z. reticulosporum Gauthier-
Lievre (Алжир). Хорошо отличается от них цветом и мелкой ямчатостью
мезоспория. Местонахождение в Польше сомнительно. Найденный там
вид и определенный как Z. allorgei по ширине вегетативных клеток
(20—27 мкм), размерам зигоспор (26—34 х 34—45 х 26 мкм) и, главное,
довольно крупным углублениям на мезоспории (2 мкм в диам’., с рассто-
янием в 2—3 мкм между ними), расположенным правильными рядами,
ближе к Z. vaginatum, Z. luteosporum либо к Z. biforme.
28. Zygnema vaginatum Klebs (=Z. emersum Skv.) — Зигнема вла-
галищная (рис. 28, 6—10). — Вегетативные клетки 24—30 мкм шир.,
по длине в 1.5—3.5 раза больше. Конъюгация лестничная, редко бо-
ковая. Конъюгирующие клетки при лестничной конъюгации прямые,
при боковой конъюгации прямые или слегка изогнутые, конъюгацион-
ный канал при этом выдается на 1/3 ширины вегетативной клетки.
Формирование зигоспоры в одном из гаметангиев. Воспринимающие
клетки невздутые или только слегка вздутые со стороны конъюгаци-
онного канала. Зигоспоры утолщенно-линзовидные: в разных положе-
ниях круглые, 25—32 (35) мкм в диам., и широкоэллиптические, 25—
36 мкм шир., 28—45 (46.4) мкм дл. Экзоспорий тонкий, прозрачный,
гладкий. Мезоспорий толстый, желто-коричневый или коричневый, с
небольшими, утолщенными по краям углублениями 1.3—2 мкм в диам.,
с расстоянием в 2.5—5 мкм между ними. Эндоспорий тонкий, гладкий.
72
Рис. 28. 1—3 — Zygnema allorgei: 1 — зигоспоры в разных положениях, лестничная конъ-
югация, 2, 3 — зигоспора в двух положениях; 4, 5 — Z. biforme, разные формы зигоспор;
6—10 — Z. vaginatum: 6 — лестничная конъюгация, 7, 8 — боковая конъюгация, 9, 10 —
зигоспора в двух положениях. (По: 1—3 — Gauthier-Lievre, 1965; 4—6 — Jao, 1947; 7—10 —
Рундина, 1977а).
73
В стоячих и слабопроточных водоемах, в мелководье водохранилищ,
природниковых лужах, на рисовых полях. Россия: северо-запад и центр
европ. части, Предкавказье (Сев’.Осетия); Украина (Киевское водохра-
нилище, р. Днепр в окрестностях Канева), Центр. Казахстан, Южн.
Узбекистан (Ферганская долина, Голодная степь).
Широко распространенный и довольно редко встречающийся вид.
Мезоспорий отчетливо ямчатый, но утолщенность краев углублений со-
здает впечатление легкой бугорчатости при взгляде на зигоспору сбоку.
От двух близких видов — Z. luteosporum и Z. cruciatum отличается на-
личием наряду с лестничной также и боковой конъюгации и размерами
вегетативных клеток — более широких, чем у первого вида, и более
узких, чем у второго. От Z. stellinum отличается невздутыми восприни-
мающими клетками при лестничной конъюгации. Наличие боковой
конъюгации сближает этот вид с Z. fanicum, отличающимся вздутыми
воспринимающими клетками при лестничной конъюгации, более выра-
женным конъюгационным каналом при боковой конъюгации и крупны-
ми (4—7.5 мкм в диам.) ямчатыми углублениями на мезоспорий. Очень
близок к Z. biforme, отличается существованием обоих типов конъюга-
ции и четко выраженными утолщениями по краям ямок на мезоспорий.
29. Zygnema luteosporum Czurda — Зигнема желтоспоровая (рис. 27,
2—4). — Вегетативные клетки 22—25 мкм шир., по длине в 1.7—4
раза больше. Конъюгация лестничная. Формирование зигоспоры в
одном из гаметангаев. Воспринимающие клетки невздутые или редко
незначительно вздутые, преимущественно со стороны конъюгационного
канала. Зигоспоры линзовидные, сбоку широкоэллиптические, 26—
31 мкм шир., 31—40'мкм дл. Экзоспорий тонкий, бесцветный, глад-
кий. Мезоспорий толстый, золотисто-желтый, с небольшими, грубыми,
широко разбросанными, округлыми углублениями. Линия разрыва от-
четливая с боковой стороны зигоспоры.
В небольших стоячих, иногда заболоченных водоемах, в старицах
рек, литорали озер. Чехия, Швейцария, Алжир.
Редкий вид. От Z. vaginatum отличается узкими вегетативными
клетками и отсутствием боковой конъюгации, от Z. leiospermum —
скульптурированным мезоспорием. По мнению многих исследователей,
у Z. luteosporum зигоспоры эллипсоидные.
30. Zygnema biforme Jao — Зигнема двоякая (рис. 28, 4, 5). —
Вегетативные клетки 27—30 мкм шир., по длине равны, немного пре-
вышают или до 2.5 (3.7) раза больше ширины. Конъюгация лестничная.
Формирование зигоспоры в одном из гаметангиев. Конъюгационный
канал от обеих клеток. Воспринимающие клетки невздутые или слегка
вздутые со стороны конъюгационного канала. Зигоспоры линзовидные
(сжато-шаровидные), в разных положениях от круглых и эллиптических
до цилиндрически-эллиптических, 27—37 мкм шир., 27—48 (58) мкм
дл., 25—30 мкм толщ. Экзоспорий тонкий, бесцветный, гладкий. Ме-
зоспорий толстый, 5 мкм толщ., слоистый, желтый или буроватый, с
округлыми углублениями 1.5—2 мкм в диам. и расстоянием в 2.5—4
(5) мкм между ними.
74
В стоячих водах, прудах. Польша, Китай.
Редкий вид. Наиболее близок к Z. vaginatum по ширине вегетатив-
ных клеток и особенностям ямчатого мезоспория. При нахождении у
Z. biforme боковой конъюгации, которая крайне редко наблюдается у
Z. vaginatum, следует перевести Z. biforme в синонимы последнего.
31. Zygnema cruciatum (Vauch.) Ag. (=Z. inconspicuum Czurda;
Z. inconspicuum Czurda emend. Skv.; Z. mucosum Skv.) — Зигнема
крестовидная (рис. 27, 5, 6). — Вегетативные клетки 30—36 (39) мкм
шир., по длине равные ширине или до 2 (4) раз больше. Конъюгация
лестничная. Формирование зигоспоры в одном из гаметангиев. Воспри-
нимающие клетки невздутые или слабо вздутые со стороны конъюга-
ционного канала. Зигоспоры утолщенно-линзовидные, в разных поло-
жениях круглые и широкоэллиптические, 29—40 мкм шир., 30—
60 мкм дл. Экзоспорий тонкий, бесцветный, гладкий. Мезоспорий
толстый, коричневый или бурый, с округлыми углублениями 1.5—
2 мкм в диам., с расстоянием в 3—5 мкм между ними. Партеноспоры
цилиндрически-яйцевидные, 30—35 мкм шир., 30—60 мкм дл.
В стоячей воде, лужах, прудах, канавах, болотах, реже в водоемах
со слабопроточной водой, зарастающих участках рек и озер, на рисо-
вых полях. Повсеместно.
Широко распространенный и часто встречающийся вид. Отличается
от близких видов — Z. stellinum и Z. vaginatum большей шириной
вегетативных клеток, от первого вида к тому же — невздутыми вос-
принимающими клетками. Близок также к Z. neocruciatum Trans.
(США), в отличие от него имеет более узкие вегетативные клетки,
незначительно меньшие по диаметру и более удаленные друг от друга
углубления на мезоспорий.
32. Zygnema cylindrosporum Czurda — Зигнема цилиндроспоровая
(рис. 27, 7—10). — Вегетативные клетки 30—36 мкм шир., по длине
равные ширине или до 3 раз больше. Конъюгация лестничная. Фор-
мирование зигоспоры в гаметангии. Воспринимающие клетки преиму-
щественно невздутые. Зигоспоры короткоцилиндрические с закруглен-
ными концами и широкоэллипсоидные, почти шаровидные, 30—
43 мкм шир., 42—65 мкм дл. Экзоспорий тонкий, бесцветный,
гладкий. Мезоспорий толстый, коричневато-желтый, с округлыми уг-
лублениями 1.5—2 мкм в диам., с расстоянием в 3—5 мкм между
ними. Линия разрыва поперечная, отчетливая.
В небольших стоячих водоемах, лужах. Югославия, Сев. и Южн.
Африка, Индия, Япония.
Отличается от Z. cyiindrospermum широкими вегетативными клет-
ками и относительно короткими зигоспорами. По всем морфологичес-
ким признакам, кроме формы зигоспор, сходен с Z. cruciatum. С боль-
шой вероятностью можно допустить, что зигоспоры у Z. cylindrosporum
и Z. cyiindrospermum линзовидные.
33. Zygnema calosporum Jao — Зигнема красивоспоровая (рис. 29). —
Вегетативные клетки (20) 23—26 (28.5) мкм шир., по длине в 1.5—2.5
75
Рис. 29. Zygnema calosporum: 1 — лестничная конъюгация, 2 — зигоспора. (По: 1 — Jao,
1935b; 2 — Simons, 1990).
раза больше. Конъюгация лестничная. Формирование зигоспоры в гаме-
тангии. Воспринимающие клетки обычно сильно вздутые со всех сторон,
иногда только со стороны конъюгационного канала. Зигоспоры широ-
коэллипсоидные и шаровидные, 29—35.6 мкм шир., 32—40 мкм дл. Ме-
зоспорий толстый, от желто-коричневого до коричневого, с плотно рас-
положенными и четко выраженными углублениями 1.5—2.5 мкм диам.,
с расстоянием между ними в 1—1.5 мкм.
В стоячих водах, лужах, рисовых полях. Нидерланды, Алжир,
Китай, Япония.
Довольно редкий вид. Очень близок по всем признакам к Z. sub-
stellinum Taft (США), отличается лишь незначительно меньшим диа-
метром углублений на мезоспорий и меньшими размерами зигоспор.
Спорен вопрос о форме зигоспор — вполне возможно, что зигоспоры
линзовидные.
34. Zygnema stellinum (Vauch.) Ag. [=Z. stellinum var. genuinum
Kirchn.; Z. stellinum var. compressum Gayral; Zygonematis stellatii (sine
auct.) in: Борщов, 1865; Zygnema stellatum (sine auct.) in: Борщов,
1865] — Зигнема звездчатая (рис. 30, 1—6; 32, 1, 2). — Вегетативные
Рис. 30. 1—6 — Zygnema stellinum: 1 — боковая и лестничная конъюгации в одной паре
нитей, 2 — зигоспора и партеноспоры, 3 — зигоспора и апланоспоры, 4 — зигоспоры, лест-
ничная конъюгация, 5, 6 — зигоспора в двух положениях; 7—9 — Z. extenue: 7 — зигоспо-
ры, лестничная конъюгация, 8, 9 — зигоспора в двух положениях; 10—12 — Z. pseudoge-
deanum: 10 — боковая конъюгация, И, 12 — зигоспора в двух положениях, анфас и сбоку.
(По: 1—3 — Transeau, 1951; 4—12 — Gauthier-Lievre, 1965).
76
72
77
клетки (25) 27—38 мкм шир., по длине равны ширине или до 4 раз
больше. Конъюгация лестничная, реже боковая, иногда совмещенные
в одной паре нитей. Конъюгирующие клетки прямые. Формирование
зигоспоры в одном из гаметангиев. Воспринимающие клетки невздутые
или незначительно вздутые по ширине зигоспоры, преимущественно со
стороны конъюгационного канала. Зигоспоры линзовидные, в разных
положениях круглые и широкоэллиптические, 28—42 мкм шир., 29—
48 (57) мкм дл. Экзоспорий тонкий, бесцветный, гладкий. Мезоспорий
толстый, желто-коричневый или коричневый, с окаймленными, округ-
лыми, крупными углублениями 3—4 мкм в диам., с расстоянием в
3—4 мкм между ними. Партеноспоры (акинеты ?, апланоспоры ?) ци-
линдрические, очень редко шаровидные, заполняющие всю клетку или
только ее среднюю часть.
В мелких, стоячих, реже слабопроточных водоемах, болотах, у бе-
регов водохранилищ, на рисовых полях. Есть указания на нахождение
в высокогорных районах и в солоноватой воде. Повсеместно.
Один из самых распространенных видов. Наиболее близким видом
является Z. vaginatum, но у него воспринимающие клетки постоянно
невздутые. Отличается от Z. fanicum сравнительно небольшими углуб-
лениями на мезоспории.
35. Zygnema tenue Kutz. [=Z. stellinum (Vauch.) Ag. var. tenue
(Kutz.) Kirchn.; Z. tenue sensu Krieger in: Kolkwitz, Krieger, 1941—
1944, pr. p.] — Зигнема тонкая (рис. 31, Г, 32, 3, 4). — Вегетативные
клетки 18—24 мкм шир., по длине до 4 раз больше. Конъюгация
лестничная. Конъюгирующие клетки прямые. Формирование зигоспоры
в одном из гаметангиев. Воспринимающие клетки невздутые, иногда
значительно вздутые посередине, преимущественно со стороны конъ-
югационного канала. Зигоспоры от почти шаровидных до широкоэл-
липсоидных, 25—30 мкм шир., 25—40 мкм дл. Мезоспорий толстый,
желто-коричневый, с углублениями 2—3 мкм в диам., с расстоянием
в 3—4 мкм между ними.
В мелких стоячих, преимущественно заболоченных водоемах, кана-
вах, лужах, иногда в прибрежье озер. Эстония (о-ва Сааремаа и Хий-
умаа), Латвия, Таджикистан (Памир: оз. Зоркуль).
Вероятно, распространен шире, чем указано в литературе. Очень
близок к Z. subtile по ширине вегетативных клеток, по форме и раз-
мерам зигоспор, благодаря чему Кригер (Krieger in: Kolkwitz, Krieger,
1941—1944 : 248) объединил оба вида. Возможно, идентичен ему
Z. tenue f. compressum Gauthier-Lievre (Африка), имеющий отчетливо
сжато-эллипсоидную форму зигоспор.
36. Zygnema vaucheri Ag. [=Z. stellinum (Vauch.) Ag. var. vaucheri
(Ag.) Kirchn.] — Зигнема Воше (рис. 31, 2, 3). — Вегетативные клет-
ки (22) 24—28 мкм шир., по длине в 2.5—7 раз больше. Конъюгация
лестничная. Конъюгирующие клетки прямые. Формирование зигоспоры
в одном из гаметангиев. Воспринимающие клетки слегка вздутые, пре-
имущественно со стороны конъюгационного канала. Зигоспоры линзо-
видные, сбоку широкоэллиптические, 24—36 мкм шир., 26—45 мкм
78
Рис. 31. 7 — Zygnema tenue, зигоспоры в разных положениях и акинета; 2, 3 — Z. vaucheri,
зигоспоры в разных положениях, лестничная конъюгация; 4—6 — Z. hausmannii: 4, 5 —
разная степень вздутия воспринимающих клеток, 6 — зигоспора; 7, 8 — Z. germanicum,
формирование зигоспор, лестничная конъюгация. (По: 1 — Gauthier-Lievre, 1965; 2 — Tiffa-
ny, 1937; 3 — Transeau, 1951; 4—8 — Czurda, 1932).
79
Рис. 32. 1, 2 — Zygnema stellinum: 1 — зигоспоры, лестничная конъюгация, 2 — строение
мезоспория (фрагмент); 3,4 — Z. tenue: 3 — зигоспоры, лестничная конъюгация, 4 — стро-
ение мезоспория (фрагмент); 5 — Z. extenue, вегетативная клетка, начальная стадия боко-
вой конъюгации, зигоспоры, лестничная конъюгация. (По; Jao, 1935b).
дл. Мезоспорий толстый, коричневый, с округлыми углублениями 2—
3 мкм в диам.
В стоячих мелких водоемах, канавах, лужах, болотах, реже текучих
водах. Россия: северо-запад европ. части, Зап. Сибирь; Латвия (о-в
Моритцхольм в оз. Усмас), Вост. Украина (р. Псёл) (?).
Широко распространенный, но недостаточно изученный вид. Отли-
чается от Z. stellinum более узкими, от Z. tenue — более широкими
вегетативными клетками, от Z. calosporum и Z. substellinum Taft (США),
имеющих одинаковую с Z. vaucheri ширину вегетативных клеток, —
слабым вздутием воспринимающих клеток и незначительными разме-
рами зигоспор. Местонахождение на Украине спорно.
37. Zygnema extenue Jao — Зигнема неузкая (рис. 30, 7—9; 32,
5). — Вегетативные клетки 19—25 мкм шир., по длине в 1.5—2.5 раза
больше. Конъюгация лестничная, редко боковая, иногда совмещенные в
одной нити. Формирование зигоспоры в одном из гаметангиев. Воспри-
80
нимающие клетки невздутые или незначительно вздутые со всех сторон.
Зигоспоры линзовидные, в разных положениях круглые и эллиптичес-
кие, 23—32 мкм шир., 29—39 (42) мкм дл. Мезоспорий толстый, желто-
коричневый или коричневый, с округлыми грубыми углублениями 4.5—
6.5 мкм в диам., с расстоянием в 2—3 мкм между ними.
В мелких и сильно кальцинированных прудах, лужах, на рисовых
полях. Дания, Алжир, Израиль, Китай.
Редкий вид. Отличается от близкого вида Z. tenue крупными углуб-
лениями на мезоспорий.
38. Zygnema hausmannii (De Notar.) Czurda — Зигнема Хаусманна
(рис. 31, 4—6). — Вегетативные клетки 20—26 мкм шир., по длине
в 1.5—3.5 раза больше. Конъюгация лестничная. Конъюгирующие клет-
ки часто слегка изогнутые. Формирование зигоспоры в одном из га-
метангиев. Воспринимающие клетки невздутые или значительно взду-
тые со стороны конъюгационного канала. Зигоспоры от шаровидных
до широкоэллипсоидных, 28—35 мкм шир., 32—34 мкм дл. Экзоспо-
рий тонкий, бесцветный, гладкий. Мезоспорий толстый, от желто-ко-
ричневого до коричневого, с очень грубыми углублениями 7—10 мкм
в диам. Линия разрыва поперечная (экваториальная).
В стоячих водах, в зоне рек. Австрия, Испания, Италия, Австралия.
Редкий вид. Отличается от Z. spontaneum более крупными углубле-
ниями на мезоспорий и отсутствием партеноспор (акинет ?), от Z. ex-
tenue — более грубыми, редко расположенными углублениями, от
Z. vaginatum — узкими вегетативньгми клетками, от Z. luteosporum —
часто выраженной шаровидностью зигоспор и грубой ямчатостью ме-
зоспория. Требует выяснения вопрос о форме зигоспор. Вполне веро-
ятно, что они линзовидные.
39. Zygnema germanicum Czurda — Зигнема немецкая (рис. 31, 7,
8). — Вегетативные клетки 26—28 мкм шир., по длине равные ши-
рине или до 1.5 раза больше. Конъюгация лестничная. Формирование
зигоспоры в одном из гаметангиев. Воспринимающие клетки сильно
вздутые со всех сторон. Зигоспоры шаровидные, иногда сжато-шаро-
видные, 36—38 мкм шир., 36—45 мкм дл. Экзоспорий тонкий, бес-
цветный, гладкий. Мезоспорий толстый, коричнево-черный, с крупны-
ми ямчатыми углублениями 6—7 мкм в диам.
В стоячих водоемах, старицах рек. Германия, Чехия.
Редкий вид. Спорен вопрос о форме зигоспор. Судя по размерам,
форма зигоспор, вероятно, широколинзовидная (круглая и широкоэл-
липтическая в двух положениях). Отличается от Z. calosporum и
Z. substellinum более широкими клетками и коричнево-черной окрас-
кой мезоспория, от самого близкого вида — Z. fanicum — отсутствием
боковой конъюгации, сильно вздутыми воспринимающими клетками и
цветом мезоспория.
40. Zygnema fanicum Li (=? Z. pulcherrimum Skv.) — Зигнема фан-
ская (рис. 33). — Вегетативные клетки 25—33 (35) мкм шир., по
длине равные ширине или до 1.5—2.5 раза больше. Конъюгация лест-
6 Рундина Л. А.
81
Рис. 33. Zygnema fanicum: 1,2 — зигоспоры в разных положениях, лестничная конъюга-
ция, 3 — боковая конъюгация, 4 — строение мезоспория (поперечный разрез). (По: 1,
3 — Li in: Transeau et al., 1934; 2, 4 — Jao, 1935b).
ничная, редко боковая. Формирование зигоспоры в одном из гаметан-
гиев. Воспринимающие клетки слабо вздутые или совсем не вздутые.
Зигоспоры шаровидные и широкоэллипсоидные, 30—38 мкм шир.,
34—46 мкм дл. Мезоспорий толстый, желтый, с крупными ямчатыми,
утолщенными по краям углублениями 4—7.5 мкм в диам., с расстоя-
нием в 2—3 мкм между ними. Апланоспоры эллипсоидные, редкие.
В хорошо прогреваемых мелких водоемах с чистой водой и в пойме
рек. Россия: центр европ. части (окрестности Ярославля) (?).
Редкий вид, найденный до сих пор лишь в Китае. Местонахождение
в России сомнительно. Вполне возможно, что, так же как и у Z. germa-
nium, зигоспоры по форме линзовидные, в разных положениях круглые
и широкоэллиптические. Вид имеет несомненное сходство с Z. vagina-
turn и Z. germanicum, от первого отличается более крупными углубления-
ми на мезоспорий, от второго — наличием боковой конъюгации, почти
невздутыми воспринимающими клетками и желтым цветом мезоспория.
41. Zygnema verrucopunctatum Prasad et God ward — Зигнема бо-
родавчато-точечная (рис. 34, 1). — Вегетативные клетки 20—23 мкм
шир. Конъюгация лестничная. Формирование зигоспоры в одном из
82
гаметангиев. Воспринимающие клетки слегка вздутые со всех сторон
или только со стороны конъюгационного канала. Зигоспоры широко-
эллипсоидные, 28—32 мкм шир., 40—44 мкм дл. Экзоспорий толстый,
бесцветный, гладкий. Мезоспорий желто-коричневый, двухслойный:
тонкоморщинистый и с углублениями 1 мкм в диам., с расстоянием в
3—5 мкм между ними.
В озере. Англия.
Вид редкий, неясный и противоречивый по своим морфологическим
характеристикам, особенно при сравнении описания с рисунком и даже
Рис. 34. 1 — Zygnema verrucopunctatum; 2, 3 — Z. chalybeospermum; 2 — лестничная и бо-
ковая конъюгации в одной паре нитей, 3 — лестничная конъюгация; 4, 5 — Z. суапеит,
разные формы зигоспор, лестничная и боковая конъюгации; 6—8 — Z. excompressum; 6 —
лестничная конъюгация, 7, 8 — зигоспора в двух положениях. (По: 1 — Prasad, Godward,
1966; 2 — Kniep, 1928; 3 — Gauthier-Lievre, 1965; 4, 5, 8 — Czurda, 1932 (8 — как Z. com-
pressum); 6,7 — Yamagishi, 1965c).
83
с видовым названием. Конъюгация и образование зигоспор наблюдались
только в лабораторной культуре.
42. Zygnema bohemicum Czurda — Зигнема богемская (рис. 35). —
Вегетативные клетки 31—33.5 мкм шир. (с неослизненными оболоч-
ками), реже 37—38 мкм шир. (с ослизненными оболочками), по длине
в 1.2—3 раза больше. Конъюгация лестничная. Формирование зиго-
споры в одном из гаметангиев. Воспринимающие клетки невздутые или
незначительно вздутые со стороны конъюгационного канала. Зигоспоры
шаровидные, бобовидные, правильно или неправильно эллипсоидные,
цилиндрические с закругленными вершинами, 31—52 (58) мкм шир.,
39—84 (95.5) мкм дл., вытянутые, как правило, вдоль продольной оси
клетки. Экзоспорий тонкий, бесцветный, гладкий. Мезоспорий тол-
стый, желто-бурый или коричневый, двухслойный: внешний слой с
довольно грубыми, продолговатыми, округлыми или слегка изогнутыми
бородавочками, более или менее одинаковыми по размерам; внутрен-
ний слой с правильно округлыми, иногда плотно сомкнутыми и тогда
слегка ограненными углублениями 3.8—6.3 мкм в диам., что придает
Рис. 35. Zygnema bohemicum: 1 — лестничная конъюгация, 2—8 — различные формы зи-
госпор, 9 — строение мезоспория (фрагмент). (По: 1—7 — ориг.; 8 — Yamagishi, 1965с;
9 — Czurda, 1932).
84
этому слою вид ячеистых сот. Эндоспорий тонкий, гладкий, очень
трудно различимый.
В стоячих, реже слабопроточных водах, старицах и затонах рек,
мелководьях водохранилищ и опресненных заливах морей. Россия:
центр европ. части (Рыбинское водохранилище); Вост. Казахстан
(р. Арт-Жарык), Туркмения (Аральское море, южные заливы).
Редкий вид. От ряда видов, сходных по ширине вегетативных кле-
ток, хорошо отличается формированием зигоспоры в гаметангии и ха-
рактерной скульптурой мезоспория.
43. Zygnema atrocoeruleum W. et G. S. West — Зигнема темно-
голубая. — Вегетативные клетки 14—17 мкм шир., по длине в 2.5—5
раз больше. Конъюгация лестничная. Формирование зигоспоры в одном
из гаметангиев. Воспринимающие клетки более или менее сильно взду-
тые. Зигоспоры почти шаровидные или широкоэллипсоидные, 23—
26 мкм шир., 23—29 мкм дл. Мезоспорий толстый, темно-синий, глад-
кий.
В стоячих водах. Южн. Англия.
Редкий вид. Отличается от Z. chalybeospermum узкими вегетатив-
ными клетками и постоянно вздутыми воспринимающими клетками.
Оригинальное изображение отсутствует.
44. Zygnema chalybeospermum Hansg. — Зигнема сероспоровая
(рис. 34, 2, 3). — Вегетативные клетки 20—27 мкм шир., по длине
равные ширине или до 3 раз больше. Конъюгация лестничная, редко
боковая. Формирование зигоспоры в одном из гаметангиев. Восприни-
мающие клетки невздутые или слегка вздутые со стороны конъюгаци-
онного канала. Зигоспоры широкоэллипсоидные, (22) 27—33 мкм
шир., (22) 30—38 мкм дл. Экзоспорий тонкий, гиалиновый, гладкий.
Мезоспорий толстый, темно-синий со стальным оттенком, гладкий.
В стоячих водах мелких, иногда высыхающих водоемов, лужах, бо-
лотах, вади, дренажных канавах, на литорали озер. Зап., Центр, и
Южн. Европа, США, Канада, Алжир, Тунис, Египет, Сев. Индия.
Широко распространенный вид. Близок к Z. atrocoeruleum и Z. те-
lanosporum. Отличается от первого более широкими вегетативными
клетками, от второго — гладким мезоспорием.
45. Zygnema cyaneum Czurda — Зигнема синяя (рис. 34, 4, 5). —
Вегетативные клетки 30—32 мкм шир., по длине в 1.4—2 раза больше.
Конъюгация лестничная и боковая. Конъюгирующие клетки прямые.
Формирование зигоспоры в одном из гаметангиев. Воспринимающие
клетки невздутые. Зигоспоры широкоэллипсоидные или закругленно-
цилиндрические, 30—34 мкм шир., 38—40 мкм дл. Экзоспорий тон-
кий, бесцветный, гладкий. Мезоспорий толстый, синий, гладкий.
В стоячих водах. Чехия, Индия.
Вероятно, имеет более широкое распространение, чем указано в
литературе. Отличается от Z. sphaericum Misra (Индия) наличием бо-
ковой конъюгации и формой зигоспор, от Z. chalybeospermum — круп-
ными вегетативными клетками, от Z. insigne — цветом зигоспор.
85
46. Zygnema excompressum Trans. (=Z. compressum Czurda) — Зиг-
нема несжатая (рис. 34, 6—8). — Вегетативные клетки 23—26 мкм
шир., по длине в 1.2—3.5 (5) раза больше. Конъюгация лестничная.
Формирование зигоспоры в одном из гаметангиев. Воспринимающие
клетки невздутые. Зигоспоры шаровидные или широкоэллипсоидные,
26—28 мкм шир., 28—36 мкм дл. Экзоспорий тонкий, бесцветный,
гладкий. Мезоспорий толстый, синий, с килем и ямчатыми углублени-
ями 2.5—3 мкм в диам., с расстоянием в 2.5—3 мкм между ними.
В стоячих водах. Чехия, Япония, США.
Редкий вид с широким ареалом. По ширине вегетативных клеток,
цвету и структуре мезоспория близок к Z. peliosporum, Z. melanospo-
rum, Z. carinthiacum. Отличается от них наличием киля на мезоспорий.
47. Zygnema mucigenum Randh. — Зигнема слизеродная (рис. 36,
1—3). — Вегетативные клетки 12—14 мкм шир., по длине в 3.5—8
раз больше. Конъюгация боковая, реже лестничная. Конъюгирующие
клетки прямые. Конъюгационный канал при лестничной конъюгации
от обеих клеток, удлиненный, при боковой конъюгации равный 1/3
ширины вегетативной клетки. Формирование зигоспоры в одном из
гаметангиев. Воспринимающие клетки невздутые или слабо вздутые во-
круг зигоспоры. Зигоспоры сжато-шаровидные или эллипсоидные, 20—
22 мкм шир., 30—36 мкм дл. Мезоспорий толстый, зеленовато-голу-
бой, с рельефными углублениями 1—1.5 мкм в диам., с расстоянием
в 3—4 мкм между ними, с 5—6 углублениями в ряд.
Свободноплавающая, погруженная в общую светло-голубоватую
слизь. В мелких стоячих водоемах, в рыбоводных прудах. Южн. Узбе-
кистан.
Редкий вид. Отличается от других видов с голубым структуриро-
ванным мезоспорием узкими вегетативными клетками, наличием боко-
вой конъюгации и небольшим диаметром углублений на мезоспорий.
48. Zygnema peliosporum Wittr. — Зигнема темноспоровая (рис. 37,
1—6). — Вегетативные клетки 20—30 мкм шир., по длине равные
ширине или до 5 раз больше. Оболочка клеток слизистая, часто утол-
щенная до 5—8 мкм. Конъюгация лестничная, редко боковая. Конъ-
югационный канал при лестничной конъюгации преимущественно от
отдающей клетки. Формирование зигоспоры в одном из гаметангиев.
Воспринимающие клетки сильно вздутые со стороны конъюгационного
канала, 72—90 мкм шир. Зигоспоры линзовидные, в разных положе-
ниях круглые и широкоэллиптические, (25) 30—39 (42) мкм шир., (27)
32—48 (56) мкм дл. Экзоспорий тонкий, бесцветный, гладкий. Мезос-
порий толстый, голубой или темно-синий, с грубыми углублениями
1—2 мкм в диам., с расстоянием в 2—3 мкм между ними. Партено-
споры цилиндрически-овальные, лежащие в воспринимающих клетках
или в конъюгационном канале.
В стоячих водах преимущественно мелких водоемов, родниках, при-
родниковых лужах. Россия: север и северо-запад европ. части (Карелия,
окрестности Санкт-Петербурга), Сев. Осетия; Эстония (о-ва Сааремаа
и Хийумаа), Грузия (окрестности Тбилиси, Военно-Грузинская дорога).
86
Рис. 36. 1—3 — Zygnema mucigenumt 1,2 — зигоспоры в разных положениях, лестничная
и боковая конъюгации, 3 — ранние стадии формирования апланоспор и зрелая апланоспо-
ра; 4—8 — Z. carinthiacum: 4, 6, 8 — зигоспоры анфас, 5 — зигоспора сбоку. (По; 1—3 —
Randhawa, 1938; 4, 5, 8 — Gauthier-Lievre, 1965; 6,7 — Рундина, 1977а).
Широко распространенный, но редко встречающийся вид. Конъ-
югационные каналы в некоторых случаях разрастаются в длинные, на-
поминающие ризоиды образования. По ширине вегетативных клеток и
строению мезоспория близок к Z. synadelphum, отличается положением
зигоспоры в полости гаметангия. По многим признакам неотличим от
Z. carinthiacum, однако у последнего вида мезоспорий двухслойный,
ямчато-шиповатый. По большинству признаков сходен также с Z. ve-
87
7 i
Рис. 37. 1—6 — Zygnema peliosporum: 1,2 — зигоспоры, лестничная конъюгация, 3—6 —
зигоспоры в двух положениях, анфас и сбоку; 7, 8 — Z. melanosporum: 7 — зигоспоры в
разных положениях, лестничная конъюгация, 8 — зигоспора. (По: 1—6 — Gauthier-Lievre,
1965; 7, 8 — Skuja, 1964).
nusturn Jao et Hu (Китай), но для последнего вида характерны более
крупные и тесно расположенные углубления на мезоспорий.
49. Zygnema melanosporum Lagerh. —; Зигнема черноспоровая
(рис. 37, 7, 8). — Вегетативные клетки (20) 22—27 мкм шир., по
длине до 5 раз больше. Клеточные оболочки довольно тонкие (около
0.6 мкм толщ.), без наружной студенистой обвертки. Ризоиды в виде
выростов или разветвленных лопастей от базальной клетки. Конъюга-
ция лестничная и боковая. Формирование зигоспоры при лестничной
конъюгации в одном из гаметангиев. Воспринимающие клетки невзду-
тые или незначительно вздутые. Зигоспоры от эллипсоидных до ок-
88
ругло-цилиндрических, 19—26 (30) мкм шир., 24—36 (40) мкм дл.
Экзоспорий тонкий, бесцветный, гладкий, плотно прилегающий к ме-
зоспорию. Мезоспорий относительно толстый, более или менее отчет-
ливо слоистый, в начальной стадии созревания зигоспоры оливково-
зеленый с желтовато-коричневым оттенком, позднее от синего до
черно-синего, реже черновато-зеленый, с маленькими круглыми ямоч-
ками 1—2 мкм в диам., с расстоянием в 2—3 мкм между ними. Эн-
доспорий тонкий, бесцветный, гладкий.
В стоячих водах, прудах и прибрежьях рек, на камнях. Швеция,
Алжир, Индия, США.
Вероятно, широко распространенный вид. Характеристики строения
и цвета мезоспория разноречивы. По первоначальному описанию, ме-
зоспорий гладкий, а по рисункам ямчатый. Большинство исследовате-
лей считают, что мезоспорий мелкоточечный. Наличием скульптуриро-
ванного мезоспория хорошо отличается от Z. chalybeospermum, но, по
мнению некоторых исследователей (Kadhibowska, 1984 : 202), является
его синонимом.
50. Zygnema carinthiacum Beck — Зигнема каринтийская (рис. 36,
4—8). — Вегетативные клетки (22) 25—32 мкм шир., по длине равные
ширине или до 3 раз больше. Конъюгация лестничная. Конъюгацион-
ный канал тонкий, легко разрывающийся. Формирование зигоспоры в
одном из гаметангиев. Воспринимающие клетки слабо, иногда значи-
тельно вздутые, преимущественно со стороны конъюгационного канала.
Зигоспоры от сжато-шаровидных, 28.5—44 мкм в диам., до яйцевид-
ных и широкоэллипсоидных, 27—35 мкм шир., 32—49 (58) мкм дл.
Экзоспорий тонкий, бесцветный, гладкий. Мезоспорий толстый, синий,
при полном созревании зигоспоры сине-зеленый, двухслойный: внеш-
ний слой с округлыми углублениями 3—4 (6) мкм в диам., с рассто-
янием в 3—5 мкм между ними, внутренний слой мелко-, иногда круп-
но- и часто тупошиповатый (бугорчатый), различимый только на зре-
лых зиготах.
В стоячих, реже слабопроточных водах, ямах, мелководьях, нередко
среди харовых водорослей. Россия: центр европ. части (Рыбинское
водохранилище); Азербайджан, Южн. Казахстан.
Довольно редко встречающийся, хотя и распространенный на всех
континентах вид. В образцах из Азербайджана и Казахстана обращает
на себя внимание отчетливая двухслойность мезоспория. При обработке
зигоспор концентрированной азотной кислотой оболочки расслаивают-
ся и особенно четко вырисовывается частая и мелкая шиповатость
(точечность при рассматривании сверху) внутреннего слоя мезоспория,
не отмеченная до сих пор ни одним из исследователей. При длительном
выдерживании зигоспор в азотной и молочной кислотах этот слой
быстрее прочих растворяется, поэтому вполне возможно, что он ранее
просто просматривался. По ширине вегетативных клеток и строению
мезоспория этот вид очень близок, а возможно, и идентичен Z. pelio-
sporum. По первому признаку сходен и с Z. excompressum, и с Z. ог-
natum (Li) Trans. (США, Китай), но от первого вида отличается от-
сутствием киля на мезоспорий, а от второго — небольшими по раз-
89
мерам и удаленными друг от друга углублениями. От Z. catenation
Trans. (США) его отличают меньшая ширина вегетативных клеток и
более крупные углубления на мезоспории.
51. Zygnema cylindricum Trans. — Зигнема цилиндрическая (рис. 38,
1). — Вегетативные клетки 25—33 мкм шир., по длине до 2 раз больше.
Конъюгация неизвестна. Размножение акинетами. Акинеты цилиндри-
ческие или слегка овально-цилиндрические, толстостенные, 25—33 мкм
шир., 24—58 мкм дл. Мезоспорий очень толстый, 5 мкм толщ., корич-
невый, с округлыми углублениями 2—3 мкм в диам., с расстоянием в
3—8 мкм между ними. Линия разрыва 1, реже их 2, косая, продольная
или поперечная, иногда в виде конверта.
В мелких лужах. Швейцария, Алжир, США, Чили.
По-видимому, широко распространенный вид. Представляет боль-
шой интерес разнообразие формы и положения линии разрыва. Спе-
цифической является линия разрыва в форме почтового конверта. Вид
близок к Z. subcylindricum и, вероятно, нередко смешивался с ним.
Рис. 38. 1 — Zygnema cylindricum', 2 — Z. borzae; 3 — Z. irregulare; 4, 5 — Z. subcylindri-
cum', 6 — Z. sterile. (По: 1—4 — Krieger in: Kolkwitz, Krieger, 1941—1944; 5 — Czurda,
1932 (как Z. cylindricum); 6 — Transeau, 1951).
90
52. Zygnema borzae Krieger — Зигнема Борз (рис. 38, 2). — Ве-
гетативные клетки 24—26 мкм шир. Конъюгация неизвестна. Размно-
жение акинетами. Акинеты цилиндрические, с толстой оболочкой,
24—27 мкм шир., 18—54 мкм дл. Экзоспорий тонкий. Мезоспорий
толстый, синий, с небольшими, плотно расположенными, чуть удли-
ненными, прямыми или слегка изогнутыми углублениями, 0.5 мкм
шир., 1—2 мкм дл.
На камнях, в слабопроточной воде, при pH 5.4—5.9, на высоте
2000 м над ур. м. Румыния (Трансильванские Альпы).
Редкий вид. Характерным строением мезоспория отличается от всех
видов рода, размножающихся бесполым путем. Самые близкие виды:
Z. frigidum Taft (США) и Z. hypnosporum Rich (Южн. Африка), име-
ющие, как и Z. borzae, синий цвет мезоспория у акинет. От первого
вида отличается чуть большей, от второго —- значительно меньшей
шириной вегетативных клеток.
53. Zygnema subcylindricum Krieger (=Z. cylindricum Trans, sen-
su Czurda, 1932) — Зигнема цилиндровидная (рис. 38, 4, 5). —
Вегетативные клетки 28—32 (35) мкм шир., по длине в 1.5—2.5
раза больше, иногда окруженные толстым, до 10 мкм толщ., сли-
зистым чехлом. Конъюгация неизвестна. Размножение акинетами.
Акинеты цилиндрические, 29—33 мкм шир., (50) 55—70 мкм дл.
Экзоспорий тонкий, бесцветный, гладкий. Мезоспорий очень толс-
тый, от желтого до темно-коричневого, с очень мелкими бугорка-
ми-бородавочками и неотчетливыми, равномерно распределенными и
неглубокими ямками. Линий разрыва 1—2, косых, поперечных или
в виде почтового конверта.
В стоячих водоемах, прудах, в лабораторной культуре. Дания, Гер-
мания, Испания, Чехия, Китай: Центр. Тибет (?).
Редкий вид, требующий более детального изучения. Спецификой
этого вида является линия разрыва, формой напоминающая почтовый
конверт. По этому признаку близок к Z. cylindricum, отличается от
него усложненной скульптурой мезоспория, от Z. irregulare — равно-
мерно распределенными углублениями на мезоспории.
54. Zygnema irregulare Krieger (=Z. subcylindricum Krieger sensu
Aboal et Llimona, 1984) — Зигнема неправильная (рис. 38, 3). — Ве-
гетативные клетки 40—42 мкм шир., по длине в 1.6—2 раза больше.
Конъюгация неизвестна. Размножение акинетами. Акинеты цилиндри-
ческие или цилиндрически-яйцевидные, толстостенные, заполняющие
всю клетку, 41—43 мкм шир., 70—80 мкм дл. Экзоспорий тонкий,
бесцветный, гладкий. Мезоспорий очень толстый, 5—6 мкм толщ., ко-
ричневый, с маленькими бугорками 0.5—1 мкм шир. и неравномерно
распределенными округлыми углублениями 3—6 мкм в диам., с рас-
стоянием в 4—26 мкм между ними. Линия разрыва 1, косая, попереч-
ная, отчетливая.
В стоячей воде, в канавах, ямах. Германия, Испания, Чехия (?).
Редкий вид. Отличается от Z. sterile более сложной скульптурой
мезоспория.
91
55. Zygnema sterile Trans. — Зигнема стерильная (рис. 38, 6). —
Вегетативные клетки 44—54 мкм шир., по длине до 1.6 раза больше,
с толстой оболочкой и плотным слизистым футляром 6—15 мкм толщ.
Конъюгация неизвестна. Размножение акинетами и апланоспорами.
Акинеты слоисто-толстостенные, цилиндрические, с широко закруглен-
ными вершинами, заполняющие всю клетку, 44—54 мкм шир., 22—
69 мкм дл. Мезоспорий коричневый, ямчатый (?) или бугорчатый (?).
В мелких водоемах, часто в прудах и небольших озерах с высоким
содержанием гниющей органики; легко культивируется. Испания (?),
Греция, Малая Азия, США.
По-видимому, довольно широко распространенный вид. Требуют
уточнения строение мезоспория и форма линии разрыва — по неко-
торым данным (Aboal, 1991 : 368, fig. 3, 14), она косая, в виде поч-
тового конверта. У двух близких видов, апланоспоры которых тоже
имеют ямчатый мезоспорий, — Z. schwabei Krieger (Чили) и Z. kaby-
licum Gauthier-Lievre (Алжир), ширина вегетативных клеток небольшая
(соответственно 17—20 и 20—22 мкм).
Род ZYGNEMOPSIS (Skuja) Trans. — ЗИГНЕМОПСИС
=Zygnema Ag., pr. p.; Debarya Wittr., pr. p.; Debarya sect. Zygne-
mopsis Skuja; Hallasia Rosenv.
Нити неразветвленные, свободноплавающие. Вегетативные клетки до
50 мкм шир., по длине в 2—5 (10) раз больше ширины. Поперечные
стенки клеток гладкие. Продольные стенки целлюлозные, с наружным
пектиновым слоем. Хлоропластов по 2, очень редко по 3—6 (7) в клетке,
звездчатые, многолучевые или сжато-эллипсоидные с редкими и корот-
кими лучами, каждый с центрально расположенным пиреноидом. Ядро
в протоплазматическом мостике, в центре клетки между хлоропластами.
Размножение преимущественно зигоспорами, редко только апланоспо-
рами. Конъюгация лестничная. Конъюгирующие клетки прямые или изо-
гнутые, легко отчленяющиеся. Формирование зигоспоры в конъюгаци-
онном канале с частичным или полным внедрением в полости гаметан-
гиев. Свободные полости гаметангиев при созревании зиготы заполнены
более или менее слоистым, голубовато-белым или желтоватым коллоид-
ным веществом. Зигоспоры квадратно- или прямоугольно-подушковид-
ные, трех- или четырехрогие, редко шаровидные или эллипсоидные,
иногда неправильной формы. Экзоспорий тонкий, гладкий, обычно бес-
цветный, редко голубой, сине-зеленый или коричневый, нередко отде-
ленный от мезоспория слоем, заполненным студенистым веществом. Ме-
зоспорий толстый, желтый, коричневый или синий, однослойный или
двухслойный, гладкий или разнообразно скульптурированный. Эндоспо-
рий тонкий, бесцветный, гладкий, очень редко скульптурированный,
трудноразличимый. Бесполое размножение апланоспорами, акинетами и
партеноспорами. Апланоспоры и партеноспоры разнообразной формы,
преимущественно эллипсоидные или угловато-шаровидные, расположен-
ные во вздувшихся клетках, полости которых заполнены светлым студе-
нистым веществом, редкие. Акинеты цилиндрические, очень редкие.
92
Представители рода распространены преимущественно в субтропи-
ческих и тропических областях. Они предпочитают мелкие, хорошо
прогреваемые водоемы со стоячей или слабопроточной водой: лужи,
канавы, болота, отстойники. Как и для всех зигнемовых, идеальным
местом для вегетации и конъюгации являются рисовые поля.
Скуя (Skuja, 1929 : 44), подразделивший род Debarya на две сек-
ции, в одну из них — Zygnemopsis объединил виды со зведчатым хло-
ропластом и коллоидным веществом в гаметангиях. Впоследствии Тран-
со (Transeau, 1934b) придал представителям этой секции ранг рода,
признающийся в настоящее время всеми исследователями. Род насчи-
тывает 40 видов, в вегетативном состоянии не отличимых от видов
Zygnema. В данной работе представлено 9 видов, потенциальных и
найденных на территории России.
I. Зигоспоры имеются.
1. Мезоспорий от желтого до желто-коричневого.
А. Мезоспорий точечный.
а. Эндоспорий гладкий, вегетативные клетки 8—12 мкм шир. . .
.......................................... 1. Z. desmidioides
б. Эндоспорий бородавчато-ямчатый, вегетативные клетки 17—25 мкм
шир................................................2. Z. spiralis
Б. Мезоспорий ямчатый или ямчато-бородавчатый.
а. Мезоспорий с килем.............................3. Z. quadrata
б. Мезоспорий без киля.
а. Вегетативные клетки 9—14 мкм шир., мезоспорий двухслойный
...........................................4. Z. americana
Р. Вегетативные клетки 16—20 мкм шир., мезоспорий однослой-
ный .............................,............5. Z. decussate
В. Мезоспорий ячеисто-сетчатый ........................6. Z. globosa
2. Мезоспорий сине-зеленый или синий.
А. Мезоспорий мелкоямчатый и волнисто-складчатый, экзоспорий бес-
цветный .............................................7. Z. foersteri
Б. Мезоспорий волнистый и мелкоямчатый, иногда крупноточечный, эк-
зоспорий сине-зеленый................................8. Z. lamellata
II. Зигоспоры отсутствуют, размножение апланоспорами . . . 9. Z. reticulata
1. Zygnemopsis desmidioides (W. et G. S. West) Trans. (=Deba-
rya desmidioides W. et G. S. West; D. desmidioides var. orientale
Carter; D. cruciata Price) — Зигнемопсис десмидиевидный (рис. 39,
1, 2). — Вегетативные клетки 8—12 мкм шир., по длине в 1.5—7
раз больше, закругленные на вершинах и слегка перешнурованные
у поперечных перегородок. Хлоропластов 2, звездчатые, сильно ре-
дуцированные. Размножение преимущественно зигоспорами. Конъюги-
рующие клетки изогнутые, легко отчленяющиеся от нити при фор-
мировании зигоспоры. Формирование зигоспоры в широком конъ-
югационном канале с полным внедрением в гаметангии. Зигоспоры
подушковидные, прямоугольные, с выпукло-вогнутыми сторонами и
слегка выступающими, срезанно закругленными углами или совсем
без углов, сбоку широкоэллиптические с оттянутыми концами, 18—
32 мкм шир., 14—24 мкм дл., 12—14 мкм толщ. Экзоспорий тон-
кий, прозрачный, гладкий. Мезоспорий толстый, желто- или золо-
93
тисто-коричневый, мелко- и тонкоточечный. Эндоспорий прозрач-
ный, гладкий.
В болотах, лужах, канавах. Эстония (о-в Сааремаа).
Довольно широко распространенный вид. Самый близкий вид —
Z. columbiana Trans. (Канада, Япония) имеет по сравнению с Z. des-
midioides более плотную и тонкую точечность на мезоспорий и не-
большое отслаивание экзоспория в виде едва заметных рогов. У сход-
ного по ширине вегетативных клеток Z. americana рога более выра-
жены, а двухслойный мезоспорий отличается более грубой скульптурой.
Явление конъюгации между свободноживущими одиночными клетками,
замеченное первоначально у Z. desmidioides, наблюдается и у других
видов Zygnemopsis.
2. Zygnemopsis spiralis (Fritsch) Trans. [=Debarya spiralis (Fritsch)
Trans.; Zygnema spirale Fritsch] — Зигнемопсис спиральный (рис. 39,
3, 4). — Вегетативные клетки 17—25 мкм шир., по длине равные
ширине или до 4 раз больше. Хлоропластов 2, звездчатые. Размножение
преимущественно зигоспорами. Конъюгирующие клетки прямые или
слегка коленчато изогнутые; гаметангии иногда удлиненные до 130 мкм,
легко отчленяющиеся от нити при полном созревании зиготы. Форми-
рование зигоспоры в широком конъюгационном канале и большей
части обоих гаметангиев. Зигоспоры широкоэллипсоидные, четырех-
угольные (подушковидные), с выпуклыми сторонами и широко закруг-
ленными углами, с боковой стороны широкоэллиптические, 27—36
мкм шир., 47—56 мкм дл., 25—27 мкм толщ., окруженные хорошо
заметным чехлом (отслоения экзо- и мезоспория ?), образующим по
углам зигоспор выступы в виде рогов, вдающихся в полости гаметан-
гиев. Экзоспорий тонкий, прозрачный. Мезоспорий от светло-желтого
Рис. 39. 1, 2 — Zygnemopsis desmidioides, зигоспора в двух положениях, анфас и сбоку; 3,
4 — Z. spiralis (3 — зигоспоры анфас, лестничная конъюгация). (По: Skuja, 1929 (1, 2 —
как Debarya desmidioides, 3, 4 — как D. spiralis)).
94
до коричневого, плотно- и мелкоточечный. Эндоспорий тонкий, бес-
цветный, бородавчато-ямчатый.
В болотах, лесных лужах. Россия: центр европ. части; Эстония
(о-ва Сааремаа и Хийумаа).
Отличается от Z. decussata иным строением оболочек у зигоспор,
от Z. americana, Z. columbiana Trans. (Канада, Япония), Z. sinensis
Trans. (Китай) — широкими вегетативными клетками. Строение обо-
лочек у зигоспор требует уточнения. Противоречивы описание и изо-
бражение скульптуры зигоспоры.
3. Zygnemopsis quadrata Jao — Зигнемопсис квадратный (рис. 40,
1, 2). —- Вегетативные клетки 13—18 мкм шир., по длине в 1.5—5
раз больше. Хлоропластов 2, широколопастные. Размножение преиму-
щественно зигоспорами. Конъюгирующие клетки незначительно изо-
гнутые, более длинные, чем вегетативные. Формирование зигоспоры в
широком конъюгационном канале и в обоих гаметангиях вплоть до
Рис. 40. 1, 2— Zygnemopsis quadrata: 1 — зигоспоры анфас, 2 — зигоспоры сбоку;
3—5 — Z. foersteri, строение мезоспория. (По: 1,2 — Jao, 1935b; 3—5 — Muller, 1964).
95
наружных стенок. Зигоспоры подушковидные, квадратные с вогнутыми
сторонами и закругленными углами или круглые, сбоку эллиптические,
(26) 29—48 мкм шир. (диам.), 35—60 мкм дл., 29—38 мкм толщ.,
иногда с отслоениями экзоспория в виде рогов, внедряющихся в поло-
сти гаметангиев, заполненных стекловидным желтоватым слоистым ве-
ществом. Экзоспорий тонкий, бесцветный, гладкий. Мезоспорий толс-
тый, золотисто-желтый, однослойный, с килем, хорошо заметным сбоку
зигоспоры, и с ямками около 3 мкм в диам., с расстоянием в 1.5—
2.5 мкм между ними.
В стоячих и слабопроточных водах, преимущественно на рисовых
полях. Южн. Казахстан (Алма-Атинская обл.), Ср. Азия.
Редкий вид. Хорошо отличается от всех видов Zygnemopsis нали-
чием киля на мезоспории, четко заметного при рассматривании зиго-
споры сбоку. Самые близкие виды: Z. stephensiae Trans. (Зап. и Южн.
Африка) и Z. sinensis Trans. (Китай). От первого отличается удлинен-
ными зигоспорами с закругленными вершинами (вид сбоку), от второ-
го — большими размерами клеток, от обоих — наличием желтого
слоистого коллоидного вещества, заполняющего пространство между
экзоспорием и мезоспорием.
4. Zygnemopsis americana Trans. [=Debarya americana Trans.; Zyg-
nema americanum (Trans.) Czurda] — Зигнемопсис американский (рис. 41,
1—7). — Вегетативные клетки 9—12 (14) мкм шир., по длине в 2—4
(12) раза больше. Хлоропластов 2, сжато-овальные, звездчатые, со сла-
боразвитыми радиальными отростками. Размножение преимущественно зи-
госпорами. Конъюгирующие клетки длиннее вегетативных в 1.5—2
раза, иногда сильно изогнутые. Формирование зигоспоры в широком
конъюгационном канале со значительным, часто полным внедрением
в полости обоих гаметангиев. Зигоспоры подушковидные, округло-
квадратные или прямоугольные, редко округло-треугольные, широко-
овальные или круглые, сбоку широкоэллиптические, (20) 35—43.5 мкм
шир., (30) 32—43.5 мкм дл., 27—38 мкм толщ., с вогнутыми сторонами
и закругленными или незначительно втянутыми углами, иногда с от-
слоениями экзоспория в виде рогов, незначительно внедряющихся в
полости гаметангиев, заполненные стекловидным слоистым веществом.
Экзоспорий тонкий, бесцветный, гладкий, при отслоениях до 5.8 мкм
толщ. Мезоспорий толстый, желтовато- или буро-коричневый, двух-
слойный: внешний слой мелко- и частобородавчатый (гранулирован-
ный), внутренний слой ямчатый, с углублениями меньше 2 мкм в диам.
Партеноспоры широкоэллипсоидные с вдавленными вершинами при от-
слоениях экзоспория, 15—20 мкм шир., 20—30 мкм дл.
В стоячих водах, в лужах, на рисовых полях. Южн. Казахстан
(Алма-Атинская, Кзыл-Ординская области), Южн. Узбекистан.
В первоначальном диагнозе этого вида, описанного как Debarya
americana (Transeau, 1915 : 18; 1925 : 197, tab. 1, fig. 6, 7, 16, 17),
мезоспорий охарактеризован как бородавчатый. В последующем опи-
сании (Transeau, 1951 : 53, tab. 8, fig. 18—21) он значится как ямча-
тый, но замаскированный слоем желто-коричневых гранул, располо-
женных между экзоспорием и мезоспорием. Судя по этим описаниям,
96
подтвержденным позднее Готье-Льевр (Gauthier-Lievre, 1965 : 79,
tab. 22, fig. В, а—с), мезоспорий Z. americana следует считать двух-
слойным. От видов, близких по вегетативным признакам (Z. quadrata,
Z. sinensis Trans., Z. columbiana Trans., Z. desmidioides), хорошо от-
личается двухслойным мезоспорием и отсутствием киля на нем. По
комплексу морфологических признаков наиболее близок к Z. decussa-
ta, но имеет менее широкие вегетативные клетки.
5. Zygnemopsis decussata Trans. [=Debarya decussata Trans.; Zygne-
ma decussatum (Trans.) Czurda; Z. pectinatum (Vauch.) Ag. var. decus-
Рис. 41. 1—7 — Zygnemopsis americana: 1—3, 5, 6 — различные формы зигоспор анфас, 4,
7 — то же, сбоку; 8—10 — Z. decussata: 8 — зигоспоры анфас и сбоку, 9 — зигоспора и
две партеноспоры, 10 — акинеты. (По: 1—4— Рундина, 1977а; 5—8— Gauthier-Lievre,
1965; 9, 10 — Transeau, 1925 (как Debarya decussata)).
7 Рундина Л. А.
97
satum (Vauch.) Kirchn.; Zygnemopsis pseudodecussata (Czurda) Krie-
ger] — Зигнемопсис крестовидный (рис. 41, 8—10). — Вегетативные
клетки 16—20 мкм шир., по длине равные ширине или до 4 раз
больше. Хлоропластов 2, звездчатые. Размножение преимущественно
зигоспорами. Конъюгирующие клетки слабо изогнутые, почти прямые.
Формирование зигоспоры в широком конъюгационном канале со зна-
чительным внедрением в полости обоих гаметангиев. Зигоспоры подуш-
ковидные, широкоэллипсоидные, яйцевидные, почти ланцетовидные
или асимметрично эллипсоидные, сбоку эллиптические, 22—34 мкм
шир., 26—50 мкм дл., с вогнутыми сторонами и широко закруглен-
ными или незначительно втянутыми углами, с отслоениями экзоспория
в виде рогов, значительно внедряющихся в полости гаметангиев. Эк-
зоспорий тонкий, бесцветный, гладкий. Мезоспорий толстый, желто-
коричневый, грубоямчатый. Апланоспоры яйцевидные или эллипсоид-
ные, односторонне вздутые, 17—25 мкм шир., 20—40 мкм дл. Пар-
теноспоры угловато-шаровидные, 15—20 мкм шир., 20—30 мкм дл.
Акинеты цилиндрические, неравнобокие, 18—20 мкм шир., 20—
36 мкм дл.
В небольших стоячих водоемах, лужах. Россия: Карелия (р. Умба).
Широко распространенный вид на всех континентах. Вероятно,
имеет более широкое распространение и в России, чем представлено
в литературе. Отличается от Z. americana и Z. sinensis Trans. (Китай)
широкими вегетативными клетками, от Z. quadrata, помимо этого, от-
сутствием киля на мезоспорий.
6. Zygnemopsis globosa Randh. (=Z. lamellata Randh. var. globosa
Randh.) — Зигнемопсис шаровидный (рис. 42, 3, 4). — Вегетативные
клетки 11—14 мкм шир., по длине в 6—6.5 раза больше. Хлоропластов
2, округло-звездчатые. Размножение преимущественно зигоспорами.
Конъюгирующие клетки значительно изогнутые. Формирование зиго-
споры в широком конъюгационном канале с полным внедрением в
> полости обоих гаметангиев, иногда с выходом за их пределы. Зиго-
споры от широкоэллипсоидных до шаровидных, 36—50 мкм диам.,
сбоку широкоэллиптические, часто со значительными отслоениями эк-
зоспория в виде небольших закругленных рогов. Экзоспорий темно-
желто-коричневый. Мезоспорий толстый, коричневато-зеленый или
темно-коричневый, неправильно ячеисто-сетчатый. Партеноспоры (ап-
ланоспоры ?) 30—32 мкм шир., 40—44 мкм дл., редкие.
В стоячих водах, на рисовых полях преимущественно в чеках. Южн.
Казахстан (Алма-Атинская обл.), Ср. Азия.
Редкий вид. На основании большого сходства с Z. lamellata Ранд-
хава (Randhawa, 1938 : 131, fig. 15) низвел Z. globosa на уровень
разновидности первого вида. Однако, учитывая разницу в ширине ве-
гетативных клеток и цвете мезоспория, эти два вида следует сохранить
пока самостоятельными. Отличается от Z. splendens Randh. (Сев.
Индия, Китай) формой зигоспор, немного большей шириной вегета-
тивных клеток и более крупными ячеями на мезоспорий, а от Z. iyen-
gari Randh. (Сев. Индия) — отсутствием игольчатых выростов на ме-
зоспории.
98
Рис. 42. 1,2 — Zygnemopsis reticulata, апланоспоры; 3, 4 — Z. globosa: 3 — две зигоспоры
анфас, 4 — зигоспора сбоку; 5—9 — Z. lamellata: 5, 7, 8 — зигоспоры аифас, 6 — зигоспо-
ра сбоку, 9 — разные стадии формирования апланоспоры. (По: 1,2 — Hallas, 1895 (как
Zygnema reticulatum): 3,4 — Randhawa, 1938 (как Zygnemopsis lamellata var. globosa): 5,
6 — Transeau, 1951; 7—9 — Randhawa, 1937).
7. Zygnemopsis foersteri Muller — Зигнемопсис Ферстера (рис. 40,
3—5). — Вегетативные клетки 17—19 мкм шир., по длине в 2—3 раза
больше. Размножение преимущественно зигоспорами. Конъюгирующие
клетки прямые или незначительно изогнутые. Формирование зигоспоры
в широком конъюгационном канале с полным внедрением в полости
гаметангиев. Зигоспоры от широкоэллипсоидных до шаровидных и
удлиненно-подушковидных с закругленными углами, иногда с выступа-
99
ющими рогами, 33—39 мкм шир., (33) 39—54 мкм дл. Экзоспорий
2—3 мкм толщ., двухслойный или многослойный, бесцветный. Мезо-
спорий 3—4 мкм толщ., слоистый, сначала коричневый, потом серо-
голубой, мелкоямчатый (ямки 2 мкм диам.) и волнисто-складчатый.
Партеноспоры по размерам вполовину меньше зигоспор.
В проточной воде, мелководьях, среди водных растений и камней,
в виде ватообразных скоплений. Германия.
Редкий и недостаточно изученный вид, по всей вероятности иден-
тичный Z. lamellata.
8. Zygnemopsis lamellata Randh.' — Зигнемопсис пластинчатый
(рис. 42, 5—9). — Вегетативные клетки 15—18 мкм шир., по длине
в 2—4 раза больше. Хлоропластов 2, звездчато-округлые, со слабораз-
витыми радиальными выростами. Размножение преимущественно зиго-
спорами. Конъюгирующие клетки до 98 мкм дл., изогнутые, иногда
значительно. Формирование зигоспоры в широком конъюгационном ка-
нале со значительным, иногда полным внедрением в полости обоих
гаметангиев. Зигоспоры подушковидные, четырехугольные, со слегка
выпуклыми сторонами и закругленными углами, сбоку эллиптические,
36—48 мкм шир., 36—52 мкм дл., или шаровидные, 44—52 мкм диам.
Экзоспорий довольно толстый, сине-зеленый. Мезоспорий двухслой-
ный: наружный слой синий или темно-сине-зеленый, волнистый при
полной зрелости зиготы, внутренний слой желто-коричневый, мелко-
ямчатый (сетчато-ямчатый), с ямками 1 мкм в диам., с расстоянием
3—4 мкм между ними, иногда также с крупными и редкими точками.
Апланоспоры узкоэллипсоидные, почти ланцетовидные, 24—36 мкм
шир., 40—74 мкм дл.
В небольших водоемах, прудах, промышленных отстойниках. Южн.
Узбекистан.
Редкий вид. Отличается от близких видов Z. globosa и Z. splendens
Randh. (Сев. Индия, Китай) окраской мезоспория.
9. Zygnemopsis reticulata (Hallas) Trans. [=Debarya reticulata (Hal-
las) Trans.; Hallasia reticulata Rosenv.; Zygnema reticulatum Hallas] —
Зигнемопсис сетчатый (рис. 42, 1, 2). — Вегетативные клетки 18—
20 мкм шир., по длине в 2—5 раз больше. Хлоропластов обычно 2,
к моменту спорообразования до 7, звездчатые, с длинными и тонкими
лучами. Конъюгация и зиготообразование неизвестны. Размножение
только апланоспорами. Формирование апланоспор в полости веретено-
видно расширенных и вытянутых до 240 мкм дл. клеток, заполненных
коллоидным веществом. Апланоспоры широкоэллипсоидные, иногда с
двумя оттянутыми бесцветными вершинами, 35 мкм шир., 60 мкм дл.
Экзоспорий прозрачный, гладкий, иногда значительно отходящий от
мезоспория. Мезоспорий желтый, сомкнуто-ямчатый (плотносетчатый).
В мелких водоемах. Дания.
Редкий и недостаточно изученный вид. Некоторыми исследователя-
ми по наличию нескольких (1—3) проростков из одной апланоспоры
отнесен к роду Hallasia. По ширине вегетативных клеток и размно-
жению только апланоспорами похож на ряд видов из Сев. Африки
100
(Z. ouarseniense, Z. montana, Z. mediterranea, Z. barcense, Z. akfaduen-
sis, Z. atrocoerulea), описанных Готье-Льевр (Gauthier-Lievre, 1965). По
последнему признаку сходен также с Z. transeauiana Randh. (Индия,
Япония), отличается от него формой апланоспор, отсутствием киля на
мезоспорий и более широкими вегетативными клетками.
Род ZYGOGONIUM Kutz. — ЗИГОГОНИУМ
=Conferva MU11., pr. p.; Zygnema Ag., pr. p.
Нити свободноплавающие, неразветвленные или преимущественно с
довольно короткими ответвлениями из одной или немногих клеток: го-
ризонтальными, стелющимися по наземному субстрату, и вертикальными
(аэрофильными). Ризоиды разнообразной формы, часто по всей длине
клеток от горизонтальных нитей, иногда только от базальной клетки,
многочисленные. Вегетативные клетки цилиндрические, реже слегка
вздутые, короткие, обычно не более 33 мкм шир., по длине вдвое мень-
ше ширины или чаще удлиненные, до 2—10 раз больше ширины. По-
перечные стенки клеток гладкие. Продольные стенки тонкие, двухслой-
ные, иногда на концах клеток явственно Н-образные, нередко неослиз-
ненные, при наземном местообитании часто толстые и многослойные,
иногда окрашенные в желтый или бурый цвета. Клеточный сок обычно
бесцветный, при наземном местообитании пурпурный или фиолетовый.
Хлоропластов по 2 в клетке, осевые, небольшие по размеру, дисковид-
ные, с широкой стороны округлые или глыбковидные, с неправильно
очерченными, часто загнутыми краями, обычно без выростов, очень
редко с небольшим количеством коротких выростов, сбоку удлиненные,
в виде валика, с заостренными концами, расположенные под углом друг
к другу, каждый с центрально расположенным пиреноидом. Ядро в цент-
ре клетки, между хлоропластами, линзовидное, очень мелкое. Размно-
жение преимущественно апланоспорами и акинетами, редко зигоспора-
ми. Конъюгация лестничная, реже боковая, редкая. Конъюгирующие
клетки прямые или в разной степени изогнутые. Обычно лишь часть про-
топласта гаметангиев (ядро и большая часть хлоропластов) образует га-
мету, после формирования зигоспоры отмечается наличие цитоплазма-
тического остатка в гаметангиях. Формирование зигоспоры в широком
конъюгационном канале, иногда с частичным внедрением в полости
обоих или одного из гаметангиев. Зигоспоры широкоэллипсоидные,
сжато-шаровидные, шаровидные, линзовидные, отделяющиеся от гаме-
тангиев особыми перегородками, образующими спорангий. Экзоспорий
тонкий, гладкий, иногда слоистый, бесцветный. Мезоспорий толстый,
слоистый, от желтого до коричневого, очень редко синий, гладкий или
разнообразно скульптурированный. Эндоспорий трудноразличимый. Линия
разрыва неразличимая или отчетливая, поперечная или продольная. Апла-
носпоры и акинеты по строению сходные с зигоспорами, частые.
В роде 23 вида, из которых только один, Z. ericetorum, широко
распространен во многих странах всех континентов. Именно он, един-
ственный, встречен на территории России. Для большинства видов ха-
рактерно наземное местообитание: на влажной земле, на открытых
101
грунтах, на техногенных выбросах, на болотах, на торфе, на латерит-
ной глине, на влажном мху, с предпочтением кислой среды (pH 3.4—
6). Есть указания на нахождение на высоте до 2500 м над ур. м.
Встречены также аэрофилы: на орошаемых скалах и на стволах де-
ревьев. Конъюгация и зигоспоры очень редки.
Виды этого рода в вегетативном состоянии крайне похожи на виды
Zygnema. Отличаются в основном ответвлениями при наземном место-
обитании, формой хлоропластов, часто с редуцированными радиальны-
ми отростками, образованием спорангия и наличием остатка цитоплаз-
мы в гаметангиях после формирования зигоспоры.
Zygogonium ericetorum Kutz. [=Z. ericetorum f. aquaticum Kirchn.;
Z. ericetorum f. terrestre Kirchn.; Zygnema ericetorum (Kiitz.) Hansg.;
Z. ericetorum var. terrestre (Kirchn.) Hansg.; Conferva ericetorum Kiitz.) —
Зигогониум пустошный (рис. 43, 44). — Нити неразветвленные или
чаще разветвленные на горизонтальные, стелющиеся по наземному суб-
страту, и на вертикальные ответвления, состоящие из одной или многих
клеток. Вегетативные клетки цилиндрические или слегка вздутые, 12—
26 (33) мкм шир., по длине меньше ширины вдвое или до 4 раз больше.
Рис. 43. Zygogonium ericetorum, ризоиды и ответвления при наземном местообитании
водоросли. (По: Randhawa, 1959а).
102
Рис. 44. Zygogonium ericetorum: 1—3 — различное положение хлоропластов в клетке,
4, 5 — различные стадии образования зигот и партеноспор, 6 — прорастание аплано-
спор. (По: 1—3 — Skuja, 1932b; 4—6 — Tiffany, Britton, 1952).
Оболочка клеток часто толстая и слоистая, иногда окрашенная в желтый
или бурый цвета. Клеточный сок часто пурпурного или фиолетового
цвета. Размножение зигоспорами, апланоспорами и акинетами. Конъюга-
ция лестничная, редко боковая. Зигоспоры широкоэллипсоидные, 15—
26 мкм шир., 19—36 (40) мкм дл., редкие. Мезоспорий толстый, от жел-
того до коричневого, гладкий. Апланоспоры шаровидные или широко-
эллипсоидные, 15—20 мкм шир., 15—40 мкм дл., с гладкой оболочкой,
•частые. Акинеты с сильно утолщенными оболочками, частые. Партено-
споры шаровидные, 23—26 мкм диам., редкие.
Преимущественно на кислых сырых почвах, на латеритной почве,
при pH 3.4—4.2 (6), в виде налетов или дерновинок бурого, фиоле-
тового или черного цветов. В торфяных лужах верховых, преимущест-
103
венно сфагновых болот сильно разрастается, образуя так называемую
«метеорную бумагу». Покрывает большие участки выбросов кислого
техногенного грунта угольных шахт. Встречен на открытом грунте на
высоте 1440 м над ур. м. (Индия). Задерживает в значительном коли-
честве влагу, вместе с мхами играет роль первопоселенца в колониза-
ции почв. Устойчив к высыханию, изменениям освещенности и тем-
пературы, высокой кислотности и минимальному содержанию мине-
ральных веществ. Россия: Арктика (острова Новой и Северной Земли),
Карелия, северо-запад, центр и северо-восток европ. части, Предкав-
казье, Урал; Прибалтика, Сев.-Зап. Украина, Сев. Казахстан (Кокче-
тавская обл., заповедник Боровое).
Наземное местообитание обусловливает появление характерных
морфологических признаков: утолщение оболочек вегетативных клеток
и зигоспор, подвижность и редуцированность хлоропластов, окраску
клеточного сока, появление разветвлений. Частое и многочисленное
образование акинет следует рассматривать как реакцию на подсыхание.
У наземных форм многочисленные мелкие капли масла могут сильно
маскировать хлоропласты.
СОМНИТЕЛЬНЫЕ И ОШИБОЧНЫЕ УКАЗАНИЯ
ВИДОВ РОДОВ ZYGNEMA И ZYGOGONIUM
Zygnema affine Kutz.: Алексенко, 1888 : 214. — Неполно описан-
ный вид (Kiitzing, 1849 : 445). Из описания Алексенко следует, что
по ширине вегетативных клеток этот вид легче всего укладывается в
диагнозы Z. stellinum (Vauch.) Ag. и Z. fanicum Li. Из видов, имеющих
сильное вздутие воспринимающей клетки, шаровидные зигоспоры и
мелкие углубления на мезоспорий, которые могут быть приняты за
пунктированность (точечность), более всего близок к Z. calosporum
Jao. Non Z. affine Hass.
Z. altaicum Skv. [=Z. pectinatum (Vauch.) Ag. var. conspicuum (Hass.)
Kirchn. sensu Скворцов, 19276 :. 252, fig. 2]: Скворцов, 1946 : 2, Tab. 13,
Fig. 8, с авторством «Skvortzov»; Skvortzov, Noda, 1973 : 38, fig. 1, c
авторством «Skvortzov et Noda». — Недостаточно изученный вид. Со-
мнительно разнообразие форм зигоспор: описание сделано на различ-
ных стадиях их формирования. Нет данных о строении мезоспория.
Определен неточно тип конъюгации: по описанию боковая, по рисун-
ку — лестничная. Наличие в гаметангиях коллоидного вещества (судя
по рисунку) — несомненный признак рода Zygnemopsis.
Z. anomalum (Hass.) Cooke (=Zygogonium anomalum Ralfs; Zygnema
pectinatum var. anomalum): Рябинин, 1889 : 337, с авторством «Ralfs»;
Алексенко, 1893 : 29, с авторством «Hassall». — Неполно описанный
вид (Cooke, 1882—1884 : 81, pl. 31, fig. 1).
Z. pectinatum Ag. f. conspicuum (nom. nudum, sine auct.): Lakowitz,
1929 : 85. — Узаконено: Z. pectinatum (Vauch.) Ag. var. conspicuum
(Hass.) Kirchn.
Z. stellinum (Vauch.) Ag.: Алексенко, 1888 : 214; 1894 : 98. —
Сомнительное определение, неполное описание из-за отсутствия дан-
104
ных о зигоспорах. По ширине вегетативных клеток более отвечает
диагнозам Z. tenue Kutz, и Z. vaucheri Ag.
Z. stellinum var. genuinum Kirchn.: Алексенко, 1888 : 214; Сатина,
1912 : 137. — Сомнительное определение, неполное описание из-за
отсутствия данных о конъюгации и зигоспорах.
Z. stellinum var. subtile Rabenh.: Шелль, 1883 : 49; Алексенко,
1888 : 214; Сатина, 1912 : 137. — Узаконено: Z. stellinum f. subtile
(Kutz.) Kirchn.
Z. stellinum f. subtile Rabenh.: Лобик, 1915 : 30. — Узаконено:
Z. stellinum f. subtile (Kutz.) Kirchn.
Z. vaginatum Klebs: Ширшов, 1937 : 176. — Неполное описание
из-за отсутствия данных о конъюгации, форме и строении зигоспор.
По ширине вегетативных клеток более соответствует Z. cruciatum
(Vauch.) Ag.
Z. variabile Skv.: Скворцов, 1946 : 2, Tab. 14, Fig. 1—5; Skvortzov,
Noda, 1973 : 38, fig. 2, a, b. — Вероятно, вид сборный, по ширине
вегетативных клеток включающий в себя виды Z. conspicuum (Hass.)
Trans, и Z. pectinatum (Vauch.) Ag.
Zygogonium crassum Kutz.: Змеев, 1872 : 39. — Неполно описан-
ный вид (Kiitzing, 1845 : 225; 1855 : Tab. 15, Fig. 13). Узаконено:
Z. crassum (Kutz.) De Toni.
*
РЕДКИЕ, НЕДОСТАТОЧНО ИЗУЧЕННЫЕ
И СОМНИТЕЛЬНЫЕ РОДЫ СЕМ. ZYGNEMATACEAE
1. Род Hallasia Rosenvinge (1924) преобразован из Zygnema reticu-
latum Hallas по характерной особенности размножения только апла-
носпорами. Трансо (Transeau, 1925) перенес этот вид сначала в род
Debarya, а позднее (Transeau, 1934b) — в род Zygnemopsis по наличию
коллоидного вещества в гаметангиях. В отличие от большинства видов
Zygnema и Zygnemopsis род Hallasia имеет как 1, так и 2—3 проростка
апланоспоры. Однако такой особенностью обладают и некоторые пред-
ставители Zygnemopsis. На основании большого сходства Hallasia с
Zygnema в вегетативном и с Zygnemopsis в генеративном состояниях
этот род был идентифицирован то как Zygnema (Czurda, 1932), то как
Zygnemopsis (Transeau, 1934b; Krieger in: Kolkwitz, Krieger, 1941—
1944; Yamagishi, 1963a). Позднее Трансо (Transeau, 1951), Рандхава
(Randhawa, 1959b), Готье-Льевр (Gauthier-Lievre, 1965) и Кадлубовская
(Kadhibowska, 1972, 1984) восстановили род Hallasia. Род монотипный,
с единственным видом Н. reticulata Rosenv. Мною отнесен к синони-
мам рода Zygnemopsis.
2. Род Neozygnema Yamagishi (1963а) выделен из рода Zygnema по
оригинальному описанию и рисунку Zygnema laevisporum Jao (1935а).
Отличается от прочих видов Zygnema тем, что зигоспора у этой во-
доросли формируется только из части содержимого гаметангиев, а ос-
таток цитоплазмы окружает зигоспору наподобие мантии. По предполо-
жению автора рода, эта водоросль в эволюционном отношении более
105
дифференцирована по сравнению с обычными видами Zygnema. Мною
включен в синонимы рода Zygnema.
3. Род Entransia Hughes (1948) — самый спорный и малоизучен-
ный род с 2 видами. Entransia fimbriata Hughes встречена в стоячих
водоемах (Канада, Польша), Е. dichloroplastes Prescott — на скале из
песчаника в окрестностях Панамского канала (Центр. Америка). Оба
вида описаны в вегетативном состоянии, половой процесс неизвестен.
Принадлежность к зигнемовым водорослям очень сомнительна, во-пер-
вых, из-за отсутствия генеративной стадии, во-вторых — из-за наличия
пристенного пластинчатого хлоропласта (несвойственного, как правило,
зигнемовым водорослям) с боковыми лопастями или фестончатыми вы-
ростами (так называемого незамкнутого поясковидного или кольцевид-
ного хлоропласта), в длинных стареющих клетках с многочисленными
удлиненными, пальцевидными отростками и разделенного узким пере-
шейком на 2 части. Такая форма раздвоенного хлоропласта с разрос-
шимися выростами отдаленно напоминает, по мнению автора рода En-
transia, таковую у рода Zygnema. Если принять точку зрения моно-
графов (Transeau, 1951 : 121; Randhawa, 1959b: 273), признающих
принадлежность этого рода к зигнемовым водорослям, то он в этом
случае должен занимать условное промежуточное положение между
семействами Zygnemataceae и Mougeotiaceae. По моему мнению, этот
род идентичен, скорее всего, роду Uronema Lagerh. (Ulotrichaceae) йо
поясковидному хлоропласту, особенно хорошо выраженному в молодых
коротких клетках, по ядру, примыкающему к хлоропласту, по заост-
ренной верхушечной клетке, по удлиненной и отчетливо суженной к
основанию базальной клетке, по наличию дермоида и шаровидных ап-
ланоспор. Предположение о сходстве с улотриксовыми водорослями
впервые было высказано автором рода Entransia (Hughes, 1948 : 427),
однако точно не определившим положение рода в системе зеленых
водорослей. Исходя из вышесказанного, род Entransia следует исклю-
чить из зигнемовых водорослей.
Семейство MOUGEOTIACEAE Palla — МУЖОЦИЕВЫЕ
Нити преимущественно неразветвленные, иногда с короткими от-
ветвлениями из одной-немногих клеток, свободноплавающие, прикреп-
ленные ризоидами к субстрату или стелющиеся по субстрату в виде
дерновинок или налетов при наземном местообитании. Поперечные
стенки клеток гладкие. Продольные стенки целлюлозные, с наружным
пектиновым слоем, значительно ослизненные. Хлоропласт обычно 1
(редко 2), осевой, пластинчатый, изменяющий положение при разной
интенсивности освещения, реже хлоропластов всегда 2, пристенных
или осевых. Пиреноидов от одного до многих, расположенных беспо-
рядочно или в один ряд посередине хлоропласта, иногда пиреноиды
отсутствуют. Ядро обычно примыкает сбоку к центральной части хло-
ропласта или лежит в протоплазматическом мостике между хлороплас-
тами. Размножение зигоспорами, апланоспорами и акинетами, с пре-
106
имущественным, иногда исключительным развитием одной из этих
форм. Формирование зигоспоры преимущественно в конъюгационном
канале, иногда с внедрением в полости обоих гаметангиев, вплоть до
их внешних стенок, изредка в одном из гаметангиев с частичным внед-
рением в конъюгационный канал. Конъюгационный канал в большин-
стве случаев отчетливый, иногда расплывающийся при формировании
зигоспоры, реже неотчетливый или вовсе отсутствует. Дифференциация
вегетативной прогаметангиальной клетки на собственно гаметангиаль-
ную и стерильную клетки отсутствует, редко — имеется. Гаметангии
в некоторых случаях заполнены бесцветным слоистым коллоидным ве-
ществом. Зигоспоры очень редко окружены остатком цитоплазмы в
виде мантии.
I. Хлоропласт 1 (редко 2), осевой. Ядро примыкает сбоку в центре хлоро-
пласта. Конъюгационный канал отчетливый.
1. Пиреноиды имеются.
А. Дифференциация прогаметангиальных клеток имеется ...........
.........................................род Temnogametum*
Б. Дифференциация прогаметангиальных клеток отсутствует.
а. Гаметангии заполнены бесцветным слоистым коллоидным веществом
'......................................... род Debarya (с. 157)
б. Гаметангии без коллоидного вещества.
а. Зигоспора окружена остатком цитоплазмы в виде мантии . . .
..........................................род Mougeotiella*
Р. Мантия вокруг зигоспоры отсутствует . . род Mougeotia (с. 107)'
2. Пиреноиды отсутствуют....................род Mougeotiopsis (с. 159)
II. Хлоропластов всегда 2, пристенные или осевые. Ядро в протоплазматическом
мостике между хлоропластами. Конъюгационный канал неотчетливый или
вовсе отсутствует.
1. Хлоропласты пристенные. Дифференциация прогаметангиальных клеток
имеется..............................................род Sirocladium*
2. Хлоропласты осевые. Дифференциация прогаметангиальных клеток отсут-
ствует .............................................. род Lloydina*
Краткие сведения о родах, отмеченных звездочкой, помещены в
конце обзора всего семейства.
Род MOUGEOTIA Ag. — МУЖОЦИЯ
=Serpentinaria Gray; Mesocarpus Hass.; Staurospermum Kutz.; Crate-
rospermum A. Br.; Pleurocarpus A. Br.; Debarya Wittr., pr. p.; Gonato-
nema Wittr.; Zygnema Ag., pr. p., Mougeotiella Yamagishi
Нити неразветвленные, свободноплавающие или прикрепленные ри-
зоидами к субстрату. Вегетативные клетки до 63 мкм шир., удли-
ненные, по длине до 20 раз больше ширины. Поперечные стенки кле-
ток гладкие. Продольные стенки клеток тонкие, целлюлозные, снаружи
покрытые тонким пектиновым слоем, ослизненные. Хлоропласт 1
(редко 2) в клетке, осевой, широкопластинчатый, изменяющий поло-
жение при разной интенсивности освещения, с двумя или многими
пиреноидами, расположенными в срединный ряд или разбросанно.
107
Ядро прилегает к хлоропласту в центре клетки или к перешейку хло-
ропласта, если он разделен на две пластины. Конъюгация лестничная,
реже боковая. Конъюгирующие клетки прямые или в разной степени
изогнутые. У некоторых видов наблюдается явление ложной конъюга-
ции, при которой клетки двух нитей коленчато изгибаются навстречу
друг другу и соприкасаются местом изгиба, но без образования
конъюгационного канала и без слияния содержимого клеток. Форми-
рование зигоспоры преимущественно в конъюгационном канале, иногда
с внедрением в полости обоих гаметангиев, вплоть до их внешних
стенок, изредка в одном из гаметангиев с частичным внедрением в
конъюгационный канал. Зигоспоры шаровидные и эллипсоидные, вытя-
нутые вдоль или поперек конъюгационного канала, треугольно-округ-
лые (сбоку круглые), угловато-округлые, квадратно-округлые, трапецие-
видно-округлые, короткоцилиндрические (катушковидные), с вогнутыми
или слегка выпуклыми продольными сторонами, и квадратно-подушко-
видные, с рогами или без рогов, с вогнутыми, выпуклыми или прямыми
сторонами. В некоторых случаях зигоспоры окружены слоем слизи
разной толщины и плотности, гиалиновым стекловидным слоистым ве-
ществом в виде оболочки или прослоек со стороны гаметангиев или
мантией — остатком цитоплазмы в виде тонкой сероватой мелко-
точечной и складчатой пленки, простирающейся и в полости гаметан-
гиев. Экзоспорий тонкий, бесцветный, гладкий, очень редко скульпту-
рированный. Мезоспорий толстый, желтый, коричневый или синий,
редко бесцветный, гладкий или разнообразно скульптурированный, од-
нослойный, иногда двухслойный. Эндоспорий тонкий, бесцветный,
очень редко слабоокрашенный, гладкий, редко скульптурированный,
трудноразличимый. Линия разрыва преимущественно одиночная, продоль-
ная или поперечная у эллипсоидных зигоспор, двойная у короткоцилин-
дрических зигоспор — по окружности крышечек (створок) на обоих кон-
цах цилиндра. Бесполое размножение апланоспорами, акинетами и парте-
носпорами. Апланоспоры и партеноспоры разнообразной формы, с
гладким или скульптурированным мезоспорием, трудно отличимые друг
от друга, редкие. Акинеты цилиндрические, гладкие или скульптуриро-
ванные, очень редкие.
В роде около 130 видов, распространенных на всех континентах.
В данной работе рассматриваются 56 видов, потенциальных и найден-
ных на территории России. Представители рода встречены в стоячих,
заболоченных и проточных, преимущественно чистых водах: озерах,
прудах, реках, ямах, лужах, болотах, ручьях и родниках, водохрани-
лищах, рисовых полях, на заливных участках речных долин, на скаль-
ных выбоинах, на водных растениях и среди влажного мха. Зафикси-
рованы местонахождения на высоте свыше 2000 м над ур. м., в сла-
босолоноватой воде и в минеральных источниках. Развиваются в массе
и конъюгируют в теплое время года, предпочитая хорошо прогревае-
мые, мелководные участки водоемов. В древнейших слоях зигоспоры
найдены начиная с карбона (палеозой).
При определении даже родовой принадлежности материала следует
соблюдать особую осторожность: в некоторых случаях при спиральном
закручивании хлоропласта виды мужоции становятся очень похожими на
108
виды спирогиры. При этом следует учитывать также разную степень
освещенности, при которой хлоропласт мужоции меняет свое положе-
ние.
I. Зигоспоры имеются.
1. Формирование зигоспоры в одном из гаметангиев, с частичным внедре-
нием в конъюгационный канал...........................1. М. transeaui
2. Формирование зигоспоры только в конъюгационном канале или лишь с час-
тичным, хотя иногда и значительным внедрением в гаметангии.
А. Мезоспорий гладкий.
а. Формирование зигоспоры только в конъюгационном канале.
а. Конъюгация боковая, редко лестничная .... 2. М. genuflexa
Р. Конъюгация только лестничная.
* Зигоспоры короткоцилиндрические (катушковидные).
+ Вегетативные клетки (22) 28—42 мкм шир.............
............................. 3. М. laetevirens
++ Вегетативные клетки (43) 48—53 мкм шир.............
...................................4. М. acadiana
* * Зигоспоры шаровидные, редко эллипсоидные.
+ Мезоспорий синий.......................5. М. maltae
++ Мезоспорий желто-коричневый.
х Вегетативные клетки 5—13 мкм шир.
О Зигоспоры с узкой стекловидной прослойкой
со стороны гаметангиев; . 6. М. heterogama
ОО Зигоспоры без стекловидной прослойки’ . .
.............................7. М. parvula
хх Вегетативные клетки (10) 12—35 мкм шир.
О Вегетативные клетки (10) 12—18 мкм шир.,
формирование зигоспоры только в средней час-
ти конъюгационного канала...................
. ...........................8. М. recurva
ОО Вегетативные клетки (17.5) 20—35 мкм шир.
Д Вегетативные клетки (17.5) 20—29 (34) мкм
шир., зигоспоры 27—40 (50) мкм диам. .
.............9. М. scalaris (частично)
ДД Вегетативные клетки 30—35 мкм шир.,
зигоспоры 55—70 мкм в диам...............
........10. М. macrospora (частично)
* ** Зигоспоры эллипсоидные, редко шаровидные.
+ Зигоспоры вытянуты поперек конъюгационного канала
..................................... 11. М. gelatinosa
++ Зигоспоры вытянуты вдоль конъюгационного канала.
X Вегетативные клетки 5—11.5 мкм шир............
..............................12. М. ellipsoidea
хх Вегетативные клетки 17.5—35 мкм шир.
О Зигоспоры 25—38 (46.5) мкм шир., 27—
43.5 (55) мкм дл. . . ..................
................9. М. scalaris (частично)
ОО Зигоспоры 40—48 мкм шир., 43—50 мкм
дл.............................13. М. hirnii
ООО Зигоспоры 74—84 мкм шир., 84 мкм дл.
......................10. М. macrospora (частично)
ххх Вегетативные клетки 37—40 мкм шир.............
............................14. М. ovalispora
109
б. Формирование зигоспоры в конъюгационном канале с частичным,
иногда значительным внедрением в гаметангии.
а. В гаметангиях и вокруг зигоспоры имеется мантия.
* Вегетативные клетки 14—18 (21) мкм шир................
.............................. 15. М. drouetii (частично)
* * Вегетативные клетки 17—24 (28) мкм шир...............
.......................... 16. М. sphaerocarpa (частично)
(3. В гаметангиях и вокруг зигоспоры мантия отсутствует.
* Зигоспоры шаровидные или эллипсоидные, вегетативные
клетки (8) 10—17 мкм шир.
+ Гиалиновая прослойка вокруг зигоспоры в виде двух ка-
липтр.............................17. М. bicalyptrata
++ Гиалиновая прослойка сплошная.......................
.........................18. М. calcarea (частично)
* * Зигоспоры короткоцилиндрические (катушковидные), вегета-
тивные клетки 22—27 мкм шир...............................
...........................................19. М. varians (частично)
Б. Мезоспорий скульптурированный.
а. Мезоспорий синий................................20. М. суапеа
б. Мезоспорий желтый или коричневый.
а. Формирование зигоспоры только в конъюгационном канале.
* Мезоспорий с выростами.
+ Мезоспорий бородавчатый . . . 21. М. microverrucosa
++ Мезоспорий бугорчатый и радиально исчерченный . . .
............................... 22. М. obuchoviae
+++ Мезоспорий зернистый (морщинистый?).................
.................................23. М. gotlandica
* * Мезоспорий ямчатый.
+ Мезоспорий мелкоямчатый.
х Зигоспоры вытянуты поперек конъюгационного
канала.............................24. М. depressa
хх Зигоспоры вытянуты вдоль конъюгационного канала.
О Вегетативные клетки (5) 8—16 мкм шир. . .
..........................25. М. nummuloides
ОО Вегетативные клетки 25—33 мкм шир. . .
........................... 26. М. robusta
++ Мезоспорий крупноямчатый.
х Вегетативные клетки 11—17 мкм шир.............
.................................27. М. ovalis
хх Вегетативные клетки 15—21 мкм шир.............
............................28. М. megaspora
ххх Вегетативные клетки 20—29 мкм шир.............
... г............................29. М. laevis
Р- Формирование зигоспоры в конъюгационном канале с частич-
ным, иногда значительным внедрением в гаметангии.
* Зигоспоры широкоэллипсоидные, мезоспорий крупнозернис-
тый ....................................30. М. sumatrana
* * Зигоспоры шаровидные, мезоспорий ямчатый.
+ Вегетативные клетки 11—13 мкм шир...................
.........................31. М. pseudocalospora
++ Вегетативные клетки 19—21 мкм шир...................
......................32. М. talyschensis (частично)
3. Формирование зигоспоры не только в конъюгационном канале, но всегда и
в обоих гаметангиях с частичным или полным их заполнением.
110
А. Мезоспорий и эндоспорий гладкие.
а. В гаметангиях и вокруг зигоспор имеется мантия.
а. Вегетативные клетки 14—18 (21) мкм шир.....................
...................................15. М. drouetii (частично)
р. Вегетативные клетки 17—24 (28) мкм шир.....................
............................. 16. М. sphaerocarpa (частично)
б. В гаметангиях и вокруг зигоспор мантия отсутствует.
а. Зигоспоры шаровидные или широкоэллипсоидные.
* Вегетативные клетки 7—9 мкм шир. ... 33. М. cherokeana
* * Вегетативные клетки (8) 10—17 мкм шир...............
...............................18. М. calcarea (частично)
Р. Зигоспоры короткоцилиндрические (катушковидные) .....
.......................................19. М. varians (частично)
у. Зигоспоры квадратно-подушковидные.
* Зигоспоры четырехрогие.
+ Зигоспоры с прямыми сторонами . . 34. М. elegantula
++ Зигоспоры с выпуклыми сторонами.
х Рога у зигоспор на концах плохо выраженные,
прямо срезанные . . 35.М. americana (частично)
хх Рога у зигоспор на концах хорошо выраженные,
мягко закругленные .............36. М. corniculata
+++ Зигоспоры с вогнутыми сторонами.
х Вегетативные клетки 3.5—5 мкм шир.
О Зигоспоры окружены студенистым веществом
35. М. americana (частично)
ОО Зигоспоры не окружены студенистым вещест-
вом ...................37. М. delicata
хх Вегетативные клетки 5—9 мкм шир.
О Рога у зигоспор на концах вдавленные . .
............................38. М. viridis
ОО Рога у зигоспор на концах закругленные . .
............................39. М. virescens
ххх Вегетативные клетки (И) 14—21 мКм шир.
................................40. М. capucina
* * Зигоспоры без четко выраженных рогов.................
..........................................41. М. paludosa
Б. Мезоспорий или эндоспорий скульптурированные.
а. Эндоспорий ямчатый (точечный ?), мезоспорий гладкий.............
.....................................................42. М. regeiii
б. Эндоспорий гладкий, мезоспорий скульптурированный.
а. Мезоспорий складчатый (морщинистый)..........43. М. fossa
р. Мезоспорий бородавчатый или точечный.
* Мезоспорий бородавчатый..............44. М. gracillima
* * Мезоспорий точечный.
+ Зигоспоры почти шаровидные............45. М. aspera
++ Зигоспоры квадратно-подушковидные.
х Вегетативные клетки 4—5.5 (8) мкм шир. . .
........................46. М. boodlei (частично)
хх Вегетативные клетки 7—9 мкм шир............
................................47. М. costata
у. Мезоспорий ямчатый или зернистый.
* Зигоспоры шаровидные или широкоэллипсоидные.
+ В гаметангиях и вокруг зигоспор имеется мантия ....
.........................................48. М. tunicata
111
++ В гаметангиях и вокруг зигоспор мантия отсутствует . .
.......................32. М. talyschensis (частично)
* * Зигоспоры квадратно-подушковидные.
+ Зигоспоры четырехрогие.
х Зигоспоры со слегка выпуклыми сторонами . .
...........................49. М. thylespora
хх Зигоспоры с вогнутыми сторонами.
О Рога у зигоспор на концах закругленные .
..................50. М. austriaca
ОО Рога у зигоспор на концах прямо срезанные
или вдавленные................51. М. punctata
++ Зигоспоры без четко выраженных рогов...............
...............................52. М. quadrangulata
И. Зигоспоры отсутствуют, размножение апланоспорами или акинетами.
1. Известны только апланоспоры.
А. Апланоспоры эллипсоидные или шаровидные,
а. Мезоспорий гладкий................................53. М. ventricosa
б. Мезоспорий скульптурированный.
а. Мезоспорий точечный .................46. М. boodlei (частично)
р. Мезоспорий ямчатый......................54. М. faveolatospora
Б. Апланоспоры изогнуто-цилиндрические..............55. М. notabilis
2. Известны только акинеты............................ 56. М. turicensis
1. Mougeotia transeaui Collins [=M. tenuis (Cleve) Wittr., non sensu
Kutz.; M. notabilis Hass, sensu Transeau, 1951, pr. p.; M. abnormis
Kissel.] — Мужоция Трансо (рис. 45). — Вегетативные клетки 9—
14.5! мкм шир., по длине в 4—16 раз больше. Хлоропласт с 4—8 (12)
пиреноидами. Конъюгация лестничная. Конъюгирующие клетки прямые
или слегка, иногда сильно, коленчато изогнутые. Формирование зиго-
споры в одном из гаметангиев с частичным внедрением в конъюгацион-
Рис. 45. Mougeotia transeaui, зигоспоры сбоку (7) и анфас (2—5) в нитях разной степени
изогнутости. (По: 1—4 — Рундина, 1977а; 5 — Gauthier-Lifevre, 1965).
112
ный канал. Зигоспоры треугольно-округлые, сбоку круглые, 20—30 мкм
шир., 26—36 мкм дл. Мезоспорий толстый, желто-коричневый, гладкий.
Апланоспоры яйцевидные или широкоэллипсоидные, 12—20 мкм шир.,
20—32 мкм дл., формирующиеся в середине прямых или слегка изогну-
тых клеток.
В стоячих и слабопроточных водах, на рисовых полях и опреснен-
ных морских заливах. Ср.’ Азия, Южн. Казахстан, Узбекистан (заливы
Аральского моря).
Редкий вид с широким ареалом. Хорошо узнается по характерной
форме зигоспор. Близок к М. floridana Trans. (США, Индия, Африка),
М. poinciana Trans. (США) и М. djalonensis Gauthier-Lievre (Африка:
Гвинея), треугольно-округлые зигоспоры которых также формируются
в гаметангии, но вегетативные клетки более широкие у первых двух
видов (соответственно 14—20 и 21—25 мкм) и более узкие у послед-
него вида (6—10 мкм). К тому же у М. djalonensis имеется зубчатая
оторочка вокруг зигоспоры.
2. Mougeotia genuflexa (Dillw.) Ag. — Мужоция коленчато изогну-
тая (рис. 46, 47). — Вегетативные клетки 15—40 мкм шир., по длине
в 2—9 (12) раз больше, часто коленчато изогнутые. Ризоиды от базаль-
ной клетки и боковые, чаще одиночные, слабо развитые. Хлоропласт с
(2) 4—8 пиреноидами, расположенными беспорядочно. Конъюгация ред-
кая, преимущественно боковая, реже лестничная с частым образованием
псевдоконъюгационных каналов. Формирование зигоспоры при лестнич-
ной конъюгации в конъюгационном канале, с полным или частичным
его заполнением. Конъюгирующие клетки прямые или очень незначи-
тельно изогнутые. Конъюгационный канал при боковой конъюгации ши-
роко закругленный, равный по ширине вегетативной клетке или больше
ее; нить при этом остается прямой или изгибается. Зигоспоры угловато-ок-
руглые, квадратно-округлые, трапециевидно-округлые (чаще при боковой
конъюгации), эллипсоидные или шаровидные, 24—40 мкм шир., 30—
44 мкм дл. Мезоспорий светло- или желто-коричневый, гладкий.
В стоячих и текучих водах. По-видимому, распространен шире, чем
указано в литературе.
Вид недостаточно изученный из-за редкой встречаемости зигоспор
и, вероятно, сборный из-за часто ошибочного определения образцов
только в вегетативном состоянии по коленчато изогнутым клеткам,
характерным для псевдоконъюгации, которые, как выяснилось, свойст-
венны многим видам мужоции.
F. genuflexa (=Zygnema genuflexion Ag.; Pleurocarpus mirabilis A. Br.;
Mesocarpus genuflexus Ag.; M. pleurocarpus De Вагу) (рис. 46). — Веге-
тативные клетки (24) 29—40 мкм шир., по длине в 2—5.5 (12) раза
больше. Хлоропласт с 4—8 пиреноидами. Зигоспоры 29—40 мкм шир.,
36—44 мкм дл.
В стоячих и текучих водах, осоковых болотах, в заболоченных и
мелких участках озер, прудов, рек, водохранилищ, среди высших вод-
ных растений, в обрастаниях на камнях. Есть указание на нахождение
в горных речках и минеральных (радоновых) источниках. Россия:
8 Рундина Л. А.
113
Рис. 46. Mougeotia genuflexa f. genuflexa'. 1 — ложная конъюгация, 2 — ранняя стадия лестнич-
ной конъюгации, 3,4 — лестничная конъюгация, 5,6 — боковая конъюгация. (По: 1,2 —
ориг.; 3, 4 — Krieger in: Kolkwitz, Krieger, 1941—1944; 5, 6 — Fott, 1971).
европ. часть, Зап. Сибирь, Прибайкалье; Зап., Сев. и Вост. Украина,
Крым (р. Загмату в окрестностях Ялты), Казахстан, Ср. Азия.
F. reinschii (Trans.) Rund. [=М. genuflexa var. gracilis (Kiitz.) Reinsch;
M. gracilis (Reinsch) Czurda, non sensu Kiitz.; M. reinschii Trans.]
(рис. 47). — Вегетативные клетки 15—24 (29) мкм шир., по длине в
4—9 раз больше. Хлоропласт с (2) 4—6 (8) пиреноидами. Зигоспоры
24—30 мкм шир., 26—32 мкм дл.
В мелких, стоячих, заросших высшими водными растениями водое-
мах, нередко в загрязненных, с запахом гнили водах, часто вместе с ти-
114
Рис. 47. Mougeotia genuflexa f. reinschii: 1 — ранняя стадия лестничной конъюгации, 2 —
терминальная конъюгация, 3 — рнзоид, 4 — боковая конъюгация, 5 — «раздвоение» веге-
тативной клетки. (По; J—3, 5 — Рундииа, 1977а; 4 — Transeau, 1951).
повой формой. Россия: север европ. части, Карелия, Новгородская обл. (оз.
Валдай); Прибалтика, Украина (Киевское водохранилище), Казахстан.
От типовой формы отличается меньшей шириной вегетативных кле-
ток и зигоспор.
3. Mougeotia iaetevirens (А. Вг.) Wittr. (=Craterospermum laetevirens
А. Br.; Mougeotia laetevirens var. varians Wittr. f. minor Kissel.) — My-
жоция ярко-зеленоватая (рис. 48). — Вегетативные клетки (22) 28—
42 мкм шир., по длине в 3—12 раз больше. Хлоропласты с многочис-
115
Рис. 48. 1—8 — Mougeotia laetevirens, различные положения зигоспоры, лестничная конъ-
югация. (По: I — Волкова, Данилова, 1970; 2 — Transeau, 1926; 3, 4 — Skuja, 1928; 5—
8 — Gauthier-Lievre, 1965).
ленными пиреноидами. Конъюгация лестничная. Конъюгирующие клет-
ки значительно коленчато изогнутые. Формирование зигоспоры в широ-
ком конъюгационном канале. Зигоспоры короткоцилиндрические (ка-
тушковидные), с вогнутыми, редко почти прямыми боковыми сторонами,
сверху и снизу круглые: по краю вогнутые, в центре выпуклые, 38—
60 мкм шир., (29) 36—60 (75) мкм дл. Экзоспорий очень тонкий, гиа-
линовый, бесцветный, гладкий. Мезоспорий толстый, от золотисто-жел-
того до желто-коричневого, гладкий. Эндоспорий довольно толстый,
гладкий. Партеноспоры редкие, неправильно эллипсоидные или косояй-
цевидные. Линий разрыва 2, по окружности крышечек (створок).
В стоячих, заболоченных, реже текучих водах: ямах, лужах, прудах,
болотах, водохранилищах, на рисовых полях, в планктоне рек, иногда
в солоноватых водах. Зигоспоры найдены в отложениях из нижнего и
верхнего плейстоцена, а также из голоцена. Повсюду.
116
Широко распространенный, часто встречающийся вид. Близкие
виды — М. acadiana и М. varians, но первый имеет более крупные
размеры, а у второго иное положение зигоспоры. В вегетативном со-
стоянии и начальной стадии конъюгации неотличим от М. genuflexa.
Сходная по размерам вегетативных клеток и зигоспор, форме зиго-
споры и формированию ее в конъюгационном канале М. rimulosa
Rund. (Мьянма) отличается скульптурированным мезоспорием.
4. Mougeotia acadiana Trans. [=М. laetevirens (A. Br.) Wittr. f. (sine
nomen) Skuja; non M. acadiana f. (sine nomen) Skuja] — Мужоция
акадианская. — Вегетативные клетки (43) 48—53 мкм шир., по длине
в 4—10 раз больше. Хлоропласт с многочисленными пиреноидами,
расположенными беспорядочно. Конъюгация лестничная. Конъюгирую-
щие клетки сильно коленчато изогнутые. Формирование зигоспоры в
широком конъюгационном канале. Зигоспоры короткоцилиндрические
(катушковидные), с вогнутыми боковыми сторонами, сверху и снизу
круглые: по краю слегка вогнутые, в центре выпуклые, 51—70 (73) мкм
шир., 51—78 мкм дл. Экзоспорий очень тонкий, бесцветный, гладкий.
Мезоспорий толстый, золотисто-желтый, гладкий. Эндоспорий доволь-
но тонкий, гладкий. Линий разрыва 2, по окружности крышечек (ство-
рок).
В стоячей воде, у побережья озера. Латвия (оз. Покротас), США.
Редкий вид, по размерам наиболее крупный в роде. Близок к М. lae-
tevirens, от которого отличается большими размерами вегетативных кле-
ток и зигоспор. Яо (Jao, 1988) считает М. acadiana синонимом М. lae-
tevirens. Именно как форма М. laetevirens эта водоросль была описана
Скуей (Skuja, 1932а : 63) с примечанием, что речь идет, вероятно, об
особой бивалентной расе М. laetevirens. Там же, в примечании, отмечена
шаровидная (сферическая) полость зигоспоры, размером 48—61 мкм.
Изображение вида отсутствует. Следует заметить, что рисунки в моно-
графиях Рандхавы (Randhawa, 1959b, fig. 57, а—d) и Кадлубовской
(Kadhibowska, 1972, rys. 214—217; 1984, Fig. 86) соответствуют только
форме (без названия) М. acadiana, описанной Скуей (Skuja, 1949 : 97, Taf.
21, Fig. 1—4) из Мьянмы. У этой формы мезоспорий не гладкий, как у
М. acadiana, а с тонкими и очень короткими трещинками, как бы тонко
и плотно испещренный, процарапанный. По этому признаку форма возве-
дена в ранг нового вида — М. rimulosa Rund. (Рундина, 1995).
5. Mougeotia maltae Skuja — Мужоция Мальты (рис. 49, 1—4). —
Вегетативные клетки 17—22 мкм шир., по длине в 3—6 (10) раз боль-
ше. Хлоропласт с 4—8 пиреноидами, расположенными в один ряд.
Конъюгирующие клетки прямые или незначительно изогнутые. Форми-
рование зигоспоры в конъюгационном канале с полным его заполне-
нием. Зигоспоры шаровидные, реже широкоэллипсоидные, (30) 32—35
(40) мкм диам., с прослойкой гиалинового вещества со стороны гаме-
тангиев, окруженные толстым слоем слизи 4—8 мкм толщ. Экзоспорий
бесцветный, гладкий. Мезоспорий темно-синий, гладкий.
В мелководных песчаных отмелях озер, бухт, в ручейках, среди
высших водных растений. Эстония (о-в Сааремаа), Латвия (оз. Усмас).
117
Рис. 49. 1—4 — Mougeotia maltae: 1 — вегетативная нить, 2,3 — лестничная конъюга-
ция, 4 — прорастающая зигоспора; 5 — М. macrospora. (По; 1—4 — Skuja, 1926; 5 —
Wolle, 1887 (как Mesocarpus macrosporus')').
Редкий вид. При прорастании зигоспоры в культуре чисто синий
цвет мезоспория изменялся на серо-зеленый. Наличие широкого слоя
слизи вокруг зигоспоры, который, однако, может растворяться и ис-
чезать, сближает этот вид с М, gelatinosa и М. victoriensis G. S. West
(Австралия). По цвету мезоспория и ширине вегетативных клеток бли-
зок к М. суапеа, отличается гладким мезоспорием.
6. Mougeotia heterogama Geitler — Мужоция гетерогамная (рис. 50,
5—7). — Вегетативные клетки 8—13 мкм шир., по длине в 6—25 раз
118
Рис. 50. I—3 — Mougeotia calcarea: 1 — общий вид конъюгирующих клеток, 2, 3 — зиго-
спора анфас и сбоку; 4 — М. bicalyptrata: 5—7 — М. heterogama. (По: 1 — Wittrock, 1872;
2,3 — Gauthier-Lievre, 1965; 4 — Transeau, 1926 (как М. calcarea var. bicalyptrata): 5—7 —
Geitler, 1958).
больше. Хлоропласт с многочисленными пиреноидами, расположенными
в один ряд. Конъюгация лестничная. Конъюгирующие клетки прямые
или слабо изогнутые. Формирование зигоспоры в конъюгационном ка-
нале с полным его заполнением. Зигоспоры шаровидные, реже широко-
эллипсоидные, 23—29 мкм шир., 24—36 мкм дл., с узкой прослойкой
гиалинового вещества со стороны гаметангиев. Экзоспорий тонкий, бес-
цветный, гладкий. Мезоспорий толстый, коричневый, гладкий. Аплано-
споры 12—25 мкм шир., 18—25 мкм дл.
В стоячей воде. Австрия.
Редкий вид. Автор этого вида выделяет в конъюгирующей паре
«женскую» нить, 12—13 мкм шир., и «мужскую» нить, 8—9 мкм шир.
Вполне возможно, что этот вид неправомочный, если допустить здесь
конъюгацию нитей, принадлежащих двум разным видам.
119
Рис. 51. 7—3— Mougeotia parvula f. parvula: 1,2 — зигоспоры, лестничная конъюгация,
3 — апланоспора; 4 — М. recurva. (По: 1,2 — Рунднна, 1977а; 3,4 — Transeau, 1926).
7. Mougeotia parvula Hass. — Мужоция крошечная (рис. 51, 1—3;
52, 1, 2). — Вегетативные клетки 5—13 мкм шир., по длине в 3—22
раза больше. Хлоропласт с 2—8 (9) пиреноидами, расположенными в
один ряд. Конъюгация лестничная. Конъюгирующие клетки прямые
или очень слабо и плавно изогнутые. Формирование зигоспоры в конъ-
югационном канале с полным или почти полным его заполнением.
Зигоспоры шаровидные, 7—32 (36) мкм диам., редко широкоэллипсо-
идные, почти шаровидные, иногда окруженные нетолстым слоем слизи.
Мезоспорий толстый, от желтого до темно-коричневого, гладкий. Ап-
ланоспоры широкоэллипсоидные, 16—20 мкм шир., 20—24 мкм дл.,
редкие, лежащие в слегка изогнутых клетках.
В стоячих и текучих водах. Один из самых распространенных видов.
120
Описаны формы вида, однако амплитуды колебаний ширины веге-
тативных клеток у них часто перекрывают друг друга. От М. ellipsoidea
отличается формой зигоспор, от М. recurva — узкими клетками и
положением зигоспоры в конъюгационном канале, от М. victoriensis
G. S. West (Австралия) — отсутствием очень широкой слизистой об-
вертки вокруг зигоспоры.
F. parvula (=Mesocarpus parvulus Hass.; M. parvulus var. genuinus
Kirchn.) (рис. 51, 1—3). — Вегетативные клетки (6) 8—12 (13) мкм
шир., по длине в 3—10 раз больше. Зигоспоры шаровидные, редко почти
шаровидные, (13) 16—32 (36) мкм диам.
В стоячих, заболоченных, реже текучих водах: родниках, ручьях,
на рисовых полях, часто на высших водных растениях. Хорошо раз-
вивается при pH 4.5—7.5. Зафиксировано местонахождение на высоте
2360 м над ур. м. Повсеместно.
Широко распространенная и часто встречающаяся форма.
F. angusta (Hass.) Rund. [-Mesocarpus parvulus Hass. var. angustus
(Hass.) Kirchn.; M. angustus Hass.; Sphaerocarpus angustus Hass.; Mou-
geotia angusta Hass.; M. parvula var. angusta (Hass.) De Toni] (рис. 52,
1, 2). — Вегетативные клетки 5—7 (9) мкм шир., по длине в 6—22
раза больше. Зигоспоры шаровидные, 7—9 мкм диам.
В прибрежьях рек. Россия: Прибайкалье (р. Большие Коты); Зап.
Казахстан (Актюбинская обл., р. Караганда).
Редкая форма. Описанная как самостоятельный вид (Hassall, 1843 :
187) Mougeotia angusta принимается и до сих пор почти всеми моног-
рафами в этом таксономическом ранге. Впервые Кирхнер (Kirchner,
1878 : 128) представил эту водоросль как разновидность Mesocarpus par-
vulus. В дальнейшем Де Тони (De Toni, 1889 : 714) узаконил ее как
разновидность Mougeotia parvula. Яо (Jao, 1935b : 575, pl. 2, fig. 31, 32),
не выделяя особо Mougeotia angusta в ранге формы М. parvula, описал
2 «типа» последнего вида, которые в точности, по описаниям и рисун-
кам, соответствуют М. parvula f. parvula и М. parvula f. angusta. От ти-
повой формы отличается более узкими и длинными вегетативными клет-
ками и малым диаметром зигоспор. Близка к М. ellipsoidea f. tenuissima,
отличается только формой зигоспор. Легко просматривается из-за не-
больших размеров. Вероятно, имеет более широкое распространение,
чем указано в литературе.
8. Mougeotia recurva (Hass.) De Toni (-Mesocarpus recurvus Hass.) —
Мужоция согнутая (рис. 51, 4). — Вегетативные клетки (10) 12—18 мкм
шир., по длине в 4—10 раз больше. Хлоропласт с 4—8 пиреноидами,
расположенными в один ряд. Конъюгация лестничная. Конъюгирующие
клетки слегка изогнутые. Формирование зигоспоры в расширенной сред-
ней части конъюгационного канала. Зигоспоры шаровидные, 22—
33 мкм в диам. Мезоспорий коричневый, гладкий. Апланоспоры шаро-
видные, 24—30 мкм в диам., в изгибах коленчато изогнутых клеток, или
яйцевидно-цилиндрические, 14—18 мкм шир., 28—34 мкм дл., в пря-
мых клетках.
121
Рис. 52. 1,2 — Mougeotia parvula f. angusta-, 3—6 — M, scalaris; 7 — M. gelatinosa. (По:
1, 6 — Jao, 1935b; 2, 4, 5 — ориг.; 3 — Gauthier-Lievre, 1965; 7 — Wittrock in; Wittrock,
Nordstedt, 1889).
В стоячих водах, болотистых понижениях, на заливных участках
речных долин, в дождевых лужах. Латвия, Юго-Зап. Украина (долина
Дуная в окрестностях Измаила).
Довольно редко встречающийся вид, распространенный на всех кон-
тинентах. От М. scalaris отличается узкими вегетативными клетками,
от М. parvula — широкими вегетативными клетками и положением
зигоспоры в конъюгационном канале.
9. Mougeotia scalaris Hass. (=Mesocarpus intricatus Hass.; M. sca-
laris Hass.) — Мужоция лестничная (рис. 52, 3—6). — Вегетативные
клетки (17.5) 20—29 (34) мкм шир., по длине в 2—12 раз больше.
Хлоропласт с (2) 4—8 (10) пиреноидами, расположенными в один ряд
122
или беспорядочно. Конъюгация лестничная. Конъюгирующие клетки
прямые, иногда слабо изогнутые. Формирование зигоспоры в конъюга-
ционном канале, часто с полным его заполнением. Зигоспоры шаро-
видные, 27—40 (50) мкм диам., или широкоэллипсоидные, 25—38
(46.5) мкм шир., 27—43.5 (55) мкм дл., вытянутые преимущественно
вдоль конъюгационного канала, иногда с прослойкой стекловидного
пектинового вещества со стороны гаметангиев. Мезоспорий довольно
толстый, желтовато-коричневый или желто-бурый, гладкий.
В стоячих и заболоченных водах, реже в текучих и слабопроточных,
на рисовых полях. Повсеместно.
Один из самых широко распространенных видов. Обращает на себя
внимание указываемая в литературе большая амплитуда в размерах ши-
рины вегетативных клеток и диаметре зигоспор. Возможно, вид сбор-
ный. Mougeotia ovalispora, иногда рассматриваемая как форма М. sca-
laris, отличается от последнего значительно большей шириной вегета-
тивных клеток и формой зигоспор.
10. Mougeotia macrospora (Wolle) De Toni (=Mesocarpus macrosporus
Wolle) — Мужоция крупноспоровая (рис. 49, 5). — Вегетативные клетки
30—35 йкм шир., по длине в 6—10 раз больше. Конъюгация лестничная.
Конъюгирующие клетки прямые или слегка коленчато изогнутые. Форми-
рование зигоспоры в конъюгационном канале с полным его заполнением.
Зигоспоры шаровидные, 55—70 мкм в диам., или широкоэллипсоидные
(яйцевидные), 74—84 мкм шир., 84 мкм дл., слегка вытянутые вдоль конъ-
югационного канала, иногда окруженные толстым слоем стекловидного пек-
тинового вещества. Мезоспорий желто-коричневый, гладкий.
Естественные наскальные выбоины. Россия: Сев. Осетия (Цейское
ущелье).
Очень редкий и неполно описанный вид, ранее найденный только
в Сев. Америке (США). Близок к М. hirnii и М. scalaris, отличается
от последнего вида большей шириной вегетативных клеток и крупными
зигоспорами. Возможно, является только его формой. Меньшими раз-
мерами вегетативных клеток отличается от М. subcrassa G. S. West
(Австралия) и от М. crassa (Wolle) De Toni (США). У последнего
вида близкие по размеру зигоспоры. Полное сравнение затруднено из-за
редких находок видов и их неполного, вследствие этого, описания.
11. Mougeotia gelatinosa Wittr. — Мужоция студенистая (рис. 52,
7). — Вегетативные клетки 12—18 мкм шир., по длине в 6—12 раз
больше. Конъюгация лестничная. Конъюгирующие клетки прямые. Фор-
мирование зигоспоры в середине удлиненного конъюгационного канала,
который иногда растворяется. Зигоспоры широкоэллипсоидные, вытя-
нутые поперек конъюгационного канала, 28—39 мкм шир., 38—47 мкм
дл., окруженные толстым слоем бесцветной пектиновой слизи 7—
10 мкм толщ. Мезоспорий коричневый или бурый, гладкий.
В мелких болотистых водоемах со стоячей водой, ямах, канавах,
иногда в ручьях и реках. Россия: север европ. части (арктический пояс,
Карелия — р. Умба), Республика Коми (окрестности Ухты); Эстония
(о-ва Сааремаа и Хийумаа).
123
Вид широко распространенный по всей Европе. От М. суапеа отли-
чается гладким и иначе окрашенным мезоспорием. По сравнению с
М. pawhuskae Taft (США, Пакистан), зигоспоры которого также окру-
жены толстым студенистым чехлом, имеет узкие вегетативные клетки и
гладкий мезоспорий.
12. Mougeotia ellipsoidea (W. et G. S. West) Czurda — Мужоция
эллипсоидная (рис. 53, 1—3). — Вегетативные клетки 5—11.5 мкм
шир. Конъюгация лестничная. Конъюгирующие клетки прямые или не-
значительно изогнутые. Формирование зигоспоры в конъюгационном
канале с полным или почти полным его заполнением. Зигоспоры ок-
ругло-эллипсоидные, вытянутые вдоль конъюгационного канала, 11—
26 мкм шир., 12—28 мкм дл. Экзоспорий тонкий, бесцветный, глад-
кий. Мезоспорий толстый, коричневый, гладкий.
Преимущественно в стоячих водах,
Редкий вид. Отличается от М. parvula формой зигоспор.
F. ellipsoidea (=М. parvula Hass. var. ellipsoidea W. et G. S. West)
(рис. 53, 1, 2). — Вегетативные клетки 9.5—11.5 мкм шир. Зигоспоры
16—26 мкм шир., 22—28 мкм дл.
В лужах на заболоченных лугах. Эстония (о-в Сааремаа). ‘
F. tenuissima (De Вагу) Rund. [=М. tenuissima (De Вагу) Czurda]
(рис. 53, 3). — Вегетативные клетки 5—6 мкм шир. Зигоспоры 11—
15 мкм шир., 12—14 мкм дл.
Рис. 53. 1, 2 — Mougeotia ellipsoidea f. ellipsoidea; 3 — M. ellipsoidea f. tenuissima; 4 —
M. himii; 5 — M. ovalispora. (По: I, 2 — West, West, 1907 (как M. parvula var. ellipsoidea);
3 — De Bary, 1858 (как M. tenuissima); 4 — Hirn, 1895 (как M. scalaris var. macrospora);
5 — Krieger in: Kolkwitz, Krieger, 1941—1944).
124
В стоячих водах, случайно в планктоне. Россия: Ср. Поволжье (при-
брежье Волги).
Очень редкая форма. Отличается от типовой меньшими размерами
вегетативных клеток и зигоспор; от М. parvula f. angusta, имеющей
ту же ширину клеток, — формой зигоспор.
13. Mougeotia hirnii Trans. (=Л/. scalaris Hass. var. macrospora
Hirn) — Мужоция Хирна (рис. 53, 4). — Вегетативные клетки 25—
28 мкм шир., по длине в 2—5 раз больше. Хлоропласт с 4—8 пире-
ноидами, расположенными в один ряд. Конъюгация лестничная. Конъ-
югирующие клетки прямые или почти прямые. Формирование зиго-
споры в конъюгационном канале с полным его заполнением. Зигоспоры
широкоэллипсоидные, 40—48 мкм шир., 43—50 мкм дл., вытянутые
вдоль конъюгационного канала, окруженные слоем стекловидного пек-
тинового вещества. Мезоспорий очень толстый, желто-коричневый,
гладкий.
В стоячих водах. Финляндия, США, Китай.
Спорный вид. Первоначально бьш описан Хирном (Hirn, 1895 : 5,
Fig. 1) как разновидность М. scalaris, с которым имеет несомненную
близость, в первую очередь по ширине вегетативных клеток. По опи-
санию мезоспорий гладкий, в то время как на рисунке Хирна он
выглядит то мелкоточечным (игольчатым в поперечном разрезе зиго-
споры), то морщинистым. Близок также к М. macrospora, отличается
меньшими по размеру зигоспорами.
14. Mougeotia ovalispora Krieger — Мужоция овальноспоровая
(рис. 53, 5). — Вегетативные клетки 37—40 мкм шир. Хлоропласт с
8—10 пиреноидами, расположенными беспорядочно. Конъюгация лест-
ничная. Конъюгирующие клетки преимущественно прямые. Формиро-
вание зигоспоры в конъюгационном канале, с полным или почти пол-
ным его заполнением. Зигоспоры эллипсоидные, 39—41 мкм шир.,
51—59 мкм дл., вытянутые вдоль конъюгационного канала, окружен-
ные тонкой прослойкой стекловидного пектинового вещества. Экзо-
спорий тонкий, бесцветный, гладкий. Мезоспорий толстый, желто-ко-
ричневый, гладкий.
В пруду. Германия.
Редкий вид. Отличается от М. scalaris широкими вегетативными
клетками и наличием только эллипсоидных зигоспор.
15. Mougeotia drouetii Trans. [=Mougeotiella drouetii (Trans.) Yama-
gishi] — Мужоция Друэта (рис. 54, 1, 2). — Вегетативные клетки 14—
18 (21) мкм шир., по длине в 5—10 раз больше. Хлоропласт с 4—8
пиреноидами, расположенными в один ряд. Конъюгация лестничная.
Конъюгирующие клетки почти прямые или плавно коленчато изогну-
тые, часто окруженные плотной слизью. Формирование зигоспоры в
широком конъюгационном канале со значительным, вплоть до полного,
внедрением в полости гаметангиев. Зигоспоры шаровидные, (32) 35—39
(41) мкм в диам., или широкоэллипсоидные (линзовидные ?), (32) 35—
38 (42) мкм шир., 28—39 (45) мкм дл. Экзоспорий тонкий, гладкий,
125
Рис. 54.1, 2 — Mougeotia drouetii: 1 — конъюгирующие клетки, общий вид, 2 — зигоспо-
ра, окруженная мантией; 3—6 — М. sphaerocarpa: 3,4 — зигоспоры анфас, 5 — зигоспоры
в конъюгирующих клетках, партеноспоры, 6 — апланоспора. (По: 1, 2 — Красноперова, 1970;
3 — ориг.; 4 — Transeau, 1944 (как М. africana); 5,6 — Transeau, 1926).
126
трудноразличимый. Мезоспорий толстый, до 2.8 мкм толщ., желто-ко-
ричневый или коричневый, гладкий, редко слабоволнистый. Эндоспо-
рий тонкий, гладкий, трудноразличимый. Вокруг зигоспоры и в гаме-
тангиях имеется мантия — тонкая, сероватая или слегка коричневатая,
мелкоскладчатая и тонкогранулированная.
В стоячих, хорошо прогреваемых, мелких водоемах, на рисовых
полях. Южн. Казахстан.
Редкий вид. Возможно, идентичен М. sphaerocarpa. Форма зигоспор
требует уточнения.
16. Mougeotia sphaerocarpa Wolle [=Debarya africana G. S. West;
D. immersa (W. West) Trans.; Mougeotia africana (G. S. West) Trans.;
Mougeotiella africana (G. S. West) Yamagishi;. M. sphaerocarpa (Wolle)
Yamagishi] — Мужоция круглоплодная (рис. 54, 3—6). — Вегетатив-
ные клетки 17—24 (28) мкм шир., по длине в 3—11 раз больше.
Хлоропласт с 4—16 пиреноидами, расположенными более или менее
равномерно в один ряд. Конъюгация лестничная. Конъюгирующие клет-
ки коленчато изогнутые. Формирование зигоспоры в широком конъ-
югационном канале с частичным, часто значительным внедрением в
полости гаметангиев. Зигоспоры шаровидные, 31—45 (55) мкм в диам.,
или'широкояйцевидные, 30—44 мкм шир., 35—60 мкм дл., вытянутые
вдоль конъюгационного канала. Экзоспорий тонкий, бесцветный, глад-
кий. Мезоспорий толстый, от желтого или желто-коричневого до
черно-бурого, гладкий, иногда слабоволнистый. Вокруг зигоспоры и в
гаметангиях имеется мантия — тонкая, сероватая, мелкоскладчатая и
тонкогранулированная. Апланоспоры (партеноспоры ?) эллипсоидные
или неправильно-эллипсоидные с косо срезанными вершинами, 20—
30 мкм шир., 22—50 мкм дл., редкие.
В стоячих, хорошо прогреваемых, мелких водоемах, на рисовых
полях. Южн. Казахстан, Южн. Узбекистан.
Широко распространенный вид, приуроченный к тропическим и
субтропическим областям. Отличается от М. drouetii незначительно
большей шириной вегетативных клеток. Возможно, оба вида иден-
тичны.
17. Mougeotia bicalyptrata Wittr. [=М. calcarea (Cleve) Wittr. var.
bicalyptrata Wittr.] — Мужоция двукалиптровая (рис. 50, 4). — Веге-
тативные клетки 10—12 мкм шир. Конъюгация лестничная. Конъюги-
рующие клетки слабо изогнутые. Формирование зигоспоры в конъюга-
ционном канале, изредка с частичным внедрением в один или оба
гаметангия. Зигоспоры шаровидные или Широкоэллипсоидные, 25—
28 мкм .шир., 33—38 мкм дл., вытянутые вдоль конъюгационного ка-
нала и имеющие со стороны гаметангиев гиалиновые прослойки в виде
двух колпачков (калиптр). Экзоспорий коричневый (?), гладкий. Ме-
зоспорий коричневый, гладкий.
В стоячих водах. Сев. и Южн. Европа, Марокко, США.
Спорный и недостаточно изученный вид, часто расцениваемый как
синоним М. calcarea или принимаемый за его форму.
127
18. Mougeotia calcarea (Cleve) Wittr. — Мужоция известняковая
(рис. 50, 1—3). — Вегетативные клетки (8) 10—17 мкм шир., по
длине в 5—14 (20) раз больше. Хлоропласт с 4—8 пиреноидами. Конъ-
югация лестничная. Конъюгирующие клетки сильно коленчато изогну-
тые или прямые. Формирование зигоспоры в конъюгационном канале,
иногда со значительным, вплоть до цолного, внедрением в полости
гаметангиев. Зигоспоры шаровидные, ,21—30 мкм диам., или широко-
эллипсоидные, 21—30 мкм шир., 30—50 мкм дл., редко неправильной
формы (от угловатых до кубических), окруженные толстым слоистым
гиалиновым веществом. Мезоспорий бесцветный или бледно-желтый,
гладкий. Апланоспоры преимущественно шаровидные, 17—21 мкм в
диам., или трапециевидно-округлые, 15—20 мкм шир., 20—28 мкм дл.
В слабопроточных, стоячих и заболоченных водоемах. Преимущест-
венно кальцефил. Россия: европ. часть, Вост. Сибирь; Ср. Азия, Ка-
захстан.
Широко распространенный вид на всем северном полушарии. Ха-
рактеризуется большим разнообразием формы зигоспор. В некоторых
случаях размножается исключительно апланоспорами. Близкий вид
М. cherokeana отличается незначительно более узкими вегетативными
клетками (7—9 мкм шир.) и всегда шаровидными зигоспорами, глубоко
лежащими в гаметангиях. Возможно, вид сборный, объединяющий
М. bicalyptrata и М. sphaerospora (Borge) Czurda (Бразилия, Китай:
Центр. Тибет).
19. Mougeotia varians (Wittr.) Czurda [=Л7. laetevirens (A. Br.)
Wittr. var. varians Wittr.] — Мужоция изменяющаяся (рис. 55, 1—
3). — Вегетативные клетки 22—27 мкм шир. Хлоропласт с несколь-
кими пиреноидами, расположенными беспорядочно. Конъюгация лест-
ничная. Конъюгирующие клетки сильно коленчато изогнутые. Форми-
рование зигоспоры в коротком и широком конъюгационном канале с
частичным или полным внедрением в гаметангии. Зигоспоры коротко-
цилиндрические (катушковидные) с вогнутыми боковыми сторонами,
сверху и снизу — круглые, с выпуклыми крышечками (створками),
36—60 мкм шир., 40—70 (78) мкм дл. Мезоспорий толстый, желтый
или желто-коричневый, гладкий. Линий разрыва 2, по окружности кры-
шечек (створок).
В стоячих, реже слабопроточных водах: болотах, лужах, прудах,
озерах, на рисовых полях. Россия: северо-запад европ. части (окрест-
ности Выборга), Дальн. Восток (Приморский край); Казахстан (оз.
Зайсан).
Вероятно, более широко распространенный вид, чем указано в ли-
тературе. Первоначально был описан в качестве разновидности М. lae-
tevirens, от которого его отличает меньшая ширина вегетативных кле-
ток и глубокое внедрение зигоспоры в полости гаметангиев. Сходен с
М. pseudovarians Gauthier-Lievre (Судан), отличается большими разме-
рами вегетативных клеток и зигоспор.
20. Mougeotia cyanea Trans. — Мужоция синяя (рис. 56, 7, 2). —
Вегетативные клетки (14) 16—20 мкм шир., по длине в 8—10 раз боль-
128
Рис. 55. 1—3 — Mougeotia varians: 1 — две пары конъюгирующих клеток, общин вид, 2,
3 — зигоспора в двух положениях; 4—8 — М. nummuloides: 4—6, 8 — зигоспоры, разная
степень изогнутости клеток, 7 — апланоспоры. (По: 1 — Wittrock in: Wittrock, Nordstedt,
1886 (как M. laetevirens var. varians)', 2—6 — Gauthier-Lievre, 1965; 7, 8 — Transeau, 1951).
ше. Хлоропласт с 4—10 пиреноидами, расположенными в один ряд.
Конъюгация лестничная. Конъюгирующие клетки прямые или очень не-
значительно изогнутые. Формирование зигоспоры в конъюгационном ка-
нале с его частичным заполнением. Зигоспоры широкоэллипсоидные,
(28) 30—40 мкм шир., (36) 38—48 мкм дл., вытянутые поперек конъю-
гационного канала и окруженные прозрачным слоем пектиновой слизи
4—8 мкм толщ. Мезоспорий синий, мелкоточечный. Апланоспоры
(партеноспоры ?) от угловато-эллипсоидных до неправильно шаровид-
ных, расположенные сбоку от гаметангиев, иногда в конъюгационном
канале, 30—32 мкм в диам., окруженные бесцветным слоем слизи.
, В стоячих водах: лужах, прудах, озерах. Южн. Казахстан (окрест-
ности Алма-Аты) (?).
9 Рундина Л. А.
129
Рис. 56. I, 2 — Mougeotia cyanea\ 3 — M. gotlandica-, 4 — M. depressa-, 5 — M. ovalis-,
6 — M. megaspora. (По: 7, 2, 5 — Transeau, 1926; 3, 6 — Wittrock, 1872; 4 — Kiitzing,
1855).
Редкий вид, найденный лишь в Швейцарии и США. Местонахож-
дение в Казахстане сомнительно. Вегетативные клетки у этого вида
шире, чем у М. depressa, и уже, чем у М. seminoleana Tiffany (США),
отличается от обоих видов также цветом мезоспория. Сходный по
ширине вегетативных клеток и типу формирования зигоспоры вид
М. pectosa Trans. (США) имеет гладкий и бесцветный мезоспорий. По
ширине вегетативных клеток и цвету мезоспория близок к М. maltae,
130
отличается скульптурированным мезоспорием. Апланоспоры, описан-
ные и изображенные автором вида, Кадлубовской (Kadhibowska, 1972,
1984) трактуются как партеноспоры.
21. Mougeotia microverrucosa Krieger — Мужоция мелкобородавча-
тая (рис. 57, 1, 2, 5, 6). — Вегетативные клетки 26—29 (32) мкм шир.,
по длине в 4.5—5.2 раза больше, с кристаллами звездчатой формы на
поверхности. Хлоропласт широкопластинчатый с 5—9 пиреноидами,
расположенными в один ряд. Конъюгация лестничная. Конъюгирующие
клетки прямые, короче вегетативных, иногда с остатком цитоплазмы.
Формирование зигоспоры в конъюгационном канале с полным его за-
полнением. Зигоспоры широкоэллипсоидные, иногда эллипсоидно-ло-
пастные с оттянутыми вершинами, 32—36 мкм шир., 43.5—49 мкм дл., ок-
руженные коллоидным, светлым, слегка слоистым веществом. Экзоспорий
тонкий, бесцветный, гладкий. Мезоспорий толстый, темно-коричневый, с
неравномерно расположенными, разной величины тупыми бородавочками,
не превышающими 1 мкм в диам. Линия разрыва поперечная.
В стоячих и проточных водах. Украина (Зап. Полесье: оз. Турское;
Карпаты: р. Путала).
Редкий вид, ранее найденный только в Германии. Легко узнаваем
по характерной бородавчатости мезоспория. Близок по строению ме-
зоспория к М. verrucosa Wolle (США), но вегетативные клетки по-
следнего вида уже (13—14 мкм) и зигоспора занимает конъюгационный
канал и некоторую часть гаметангиев. По форме зигоспор сходен с
М. areolata Trans. (США), отличается несколько более широкими ве-
гетативными клетками, гладким экзоспорием и иной скульптурой ме-
зоспория. Близок также к М. lamellosa Jao (Китай).
22. Mougeotia obuchoviae Rund. — Мужоция Обуховой (рис. 57,
3, 4). — Вегетативные клетки 29—32 мкм шир., по длине в 3—6.5
раза больше. Хлоропласт широкопластинчатый с (4) 6—8 пиреноида-
ми. Конъюгация лестничная. Конъюгирующие клетки прямые или слег-
ка изогнутые. Формирование зигоспоры в конъюгационном канале. Зи-
госпоры короткоцилиндрические (шайбовидные), сверху и снизу круг-
лые, 38—43.5 мкм диам., или широкоэллипсоидные, (32) 35—40.5
(46.5) мкм шир., 40.6—43.5 (55) мкм дл., слегка вытянутые вдоль
конъюгационного канала. Экзоспорий тонкий, бесцветный, гладкий.
Мезоспорий толстый, желто-коричневый, со светлыми бугорками
1.3 мкм в диам., расположенными на расстоянии 2.7—5.6 мкм друг
от друга, и исчерченный длинными и короткими ребрами, по 9—11
в 7 мкм, из них 3—4 длинных. Линия разрыва зигоспоры поперечная.
В стоячей, слегка заболоченной воде. Южн. Казахстан.
Скульптурированность мезоспория выявляется только при больших
увеличениях микроскопа, желательно с водной или масляной иммер-
сией. Следует заметить, что на крышечках (створках) зигоспоры видна
только радиальная исчерченность. По этому признаку вид близок к
М. hirnii, хотя у последнего существуют противоречия в описании и
изображении. По ряду морфологических признаков близок также к
М. scalaris, хорошо отличаясь скульптурированным мезоспорием.
131
Рис. 57. 7, 2, 5, 6 — Mougeotia microverrucosa'. 1 — зигоспоры с оттянутыми вершинами,
5 — триплоидная зигоспора; 3, 4 — М. obuchoviae: 3 — конъюгирующие клетки с зиго-
спорами, общий вид, 4 — зигоспора анфас. (По: 1,5 — Рундина, 1988; 2 — Krieger in:
Kolkwitz, Krieger, 1941—1944; 3, 4 — Рундина, 19776; 6 — ориг.).
132
23. Mougeotia gotlandica (Cleve) Wittr. (=Mesocarpus gotlandicus
Cleve) — Мужоция готландская (рис. 56, 3; 58, 4). — Вегетативные
клетки 17—24 мкм шир., по длине в 3—8 раз больше. Хлоропласт с
многочисленными пиреноидами, расположенными беспорядочно. Конъ-
югация лестничная. Конъюгирующие клетки прямые или слегка изо-
гнутые. Формирование зигоспоры в конъюгационном канале, с полным
его заполнением. Зигоспоры шаровидные, 30—34 мкм диам. Мезоспо-
рий желто-коричневый, зернистый (морщинистый ?).
Мелководные прибрежья озер, вместе с другими зигнемовыми. Эс-
тония (о-в Сааремаа).
Редкий вид, кроме Европы (Швеция, Австрия) найденный также в
Индии и Китае. Вопрос о скульптуре оболочек зигоспоры неясен. По
первоописанию Клеве (Cleve, 1868 : 31, Taf. 9, Fig. 8—10), экзоспо-
рий желто-коричневый и морщинистый, а мезоспорий гладкий. В то
же время на рисунке зигоспоры у того же автора видна крупная то-
чечность — скорее зернистость, чем морщинистость.
24. Mougeotia depressa (Hass.) Wittr. — Мужоция сжатая (рис. 56,
4). — Вегетативные клетки 7—12 мкм шир. Конъюгация лестничная
и боковая. Конъюгирующие клетки прямые или плавно и незначитель-
но изогнутые. Формирование зигоспоры при лестничной конъюгации
в конъюгационном канале с частичным его заполнением. Зигоспоры
удлиненно-эллипсоидные, вытянутые поперек конъюгационного канала,
12—14 мкм шир., 28—32 мкм дл. Мезоспорий толстый, коричневый,
мелкоямчатый.
В стоячих водах. Сев., Зап. и Центр. Европа, Африка (?), Сев.
Индия, Сев. Америка.
Широко распространенный вид, встреченный почти на всех конти-
нентах. Отличается от М. gelatinosa узкими вегетативными клетками,
от М. суапеа — коричневым цветом мезоспория, от М. ovalis и
М. operculata Trans. (США) — сравнительно мелкими углублениями
на нем.
25. Mougeotia nummuloides (Hass.) De Toni (=Mesocarpus nummu-
loides Hass.) — Мужоция монетообразная (рис. 55, 4—8). — Вегета-
тивные клетки (5) 8—16 мкм шир., по длине в 4—10 раз больше.
Хлоропласт с 2—6 пиреноидами, расположенными в один ряд. Конъ-
югация лестничная. Конъюгирующие клетки слегка, иногда значительно
изогнутые. Формирование зигоспоры в конъюгационном канале, с пол-
ным, иногда частичным его заполнением. Зигоспоры шаровидные,; (17)
22—32 (37) мкм диам., или широкоэллипсоидные, вытянутые вдоль
конъюгационного канала, 20—24 мкм шир., 20—26 мкм дл., с тонкой
прослойкой гиалинового вещества со стороны гаметангиев. Экзоспорий
тонкий, бесцветный, гладкий. Мезоспорий коричневый, ямчатый. Ап-
ланоспоры широкоэллипсоидные или шаровидные, формирующиеся в
коленчатых, изогнутых под острым углом клетках.
В мелководьях озер, в реках, прудах, заболоченных и родниковых
лужах, оросительных каналах, на орошаемых скалах и в торфяных
водоемах при pH 5.5—5:7. Россия: север и центр европ. части; При-
133
Рис. 58. 7—3 — Mougeotia robusta'. 1,3 — лестничная конъюгация, 2 — зигоспора; 4 —
М. gotlandica. (По: 1 — Yamagishi, 1965а; 2, 3 — Рундина, 1977а; 4 — Cleve, 1868 (как
Mesocarpus gotlandicus)).
134
балтика, Зап. Украина (р. Прут в окрестностях Снятына, Карпаты:
р. Путина), Ср. Азия.
Широко распространенный, часто встречающийся вид, эврибионт.
26. Mougeotia robusta (De Вагу) Wittr. [=Mesocarpus robustus De Ва-
гу; Mougeotia pulchella (Wittr.) De Toni, non sensu Hirn, 1895] — Му-
жоция мощная (рис. 58, 1—3). — Вегетативные клетки 25—33 мкм
шир., по длине в 5—10 раз больше. Хлоропласт 1, иногда их 2, с (5)
10—20 пиреноидами, расположенными беспорядочно. Ризоиды вильчато
разветвленные, от базальной клетки, редкие. Конъюгация лестничная.
Конъюгирующие клетки прямые. Формирование зигоспоры в конъюга-
ционном канале с полным его заполнением. Зигоспоры от широкоэл-
липсоидных до почти шаровидных, иногда сложной конфигурации, с
широко закругленными вершинами, 28—43.5 мкм шир., (36) 40—50
(69.5) мкм дл., вытянутые вдоль конъюгационного канала. Экзоспорий
тонкий, бесцветный, гладкий. Мезоспорий толстый, желто-коричневый,
ямчатый, с небольшими по размеру углублениями 1—1.6 мкм в диам.
В стоячих, текучих и слабопроточных водах, родниках, ручьях, на
рисовых полях. Россия: центр европ. части (окрестности Москвы);
Эстония (о-в Сааремаа), Узбекистан, Таджикистан (хребет Моголтау),
Южн. Казахстан.
Вид с широким ареалом. У некоторых экземпляров диаметр ямча-
тых углублений на мезоспорий достигает 1.8 мкм, с удаленностью уг-
лублений друг от друга 1.8—3.6 мкм, что сближает его с М. laevis.
Возможно, что М. robusta является синонимом или формой М. laevis.
Что касается формы зигоспор, то кроме типичных, широкоэллипсоид-
ных или почти шаровидных, в редких случаях зигоспоры могут быть
симметрично или неправильно лопастными — характерными для
М. oblongata Trans. (США), у которого, однако, более узкие вегета-
тивные клетки (19—22 мкм шир.). От М. ornata Jao (Китай) отлича-
ется также более широкими вегетативными клетками, эллипсоидными
зигоспорами и менее плотной и крупной пунктированностью (ямча-
тостью ?) мезоспория.
27. Mougeotia ovalis (Hass.) Nordst. [=Mesocarpus depressus Hass,
var. ovalis (Hass.) Rabenh.] — Мужоция овальная (рис. 56, 5). — Ве-
гетативные клетки 11—17 мкм шир. Конъюгация лестничная. Конъ-
югирующие клетки прямые или незначительно и плавно изогнутые.
Формирование зигоспоры в конъюгационном канале с почти полным
его заполнением. Зигоспоры широкоэллипсоидные, почти шаровидные,
вытянутые поперек конъюгационного канала, 24—38 мкм шир., 29—
40 мкм дл. Мезоспорий крупноямчатый.
В стоячих водах. Англия, Германия, Швейцария, Польша (?).
Довольно широко распространенный в Европе вид. Требует уточ-
нения форма зигоспор. Очень близок к М. depressa, формою которого
и был первоначально описан. Отличается от него более широкими
вегетативными клетками, менее вытянутыми широкоэллипсоидными зи-
госпорами и крупными грубыми углублениями на мезоспорий. Сходен
также с М. pawhuskae Taft (США, Пакистан) и М. operculata Trans.
135
(США), отличается от них чуть меньшей шириной вегетативных кле-
ток, а от первого, сверх того, отсутствием слизистой обвертки вокруг
зигоспоры.
28. Mougeotia megaspora Wittr. — Мужоция крупноспоро-
вая (рис. 56, 6). — Вегетативные клетки 15—21 мкм шир., по длине
в 10—22 раза больше. Конъюгация лестничная. Конъюгирующие клет-
ки прямые. Формирование зигоспоры в конъюгационном канале с пол-
ным его заполнением. Зигоспоры преимущественно шаровидные, 40—
50 мкм в диам., или широкоэллипсоидные, 41—44 мкм шир., 45—
48 мкм дл. Мезоспорий темно-коричневый, ямчатый, с беспорядочно
расположенными крупными углублениями.
В стоячих водах. Швеция, Япония, США.
Крупные размеры вегетативных клеток и зигоспор отличают этот
вид от близкого М. nummuloides. Возможно, что М. megaspora является
лишь формой последнего вида.
29. Mougeotia laevis (Kutz.) Arch. [-Debarya laevis (Kutz.) W. et G. S.
West; Mougeotia pulchella (Wittr.) De Toni sensu Him, 1895] — Мужоция
гладкая (рис. 59, 1—3). — Вегетативные клетки 20—32 (36 ?) мкм шир.,
по длине равные ширине или до 6 (8) раз больше. Хлоропласт с 2—6
пиреноидами, расположенными в один ряд. Конъюгация лестничная.
Конъюгирующие клетки прямые или реже очень слабо изогнутые. Фор-
мирование зигоспоры в конъюгационном канале с полным его заполне-
нием. Зигоспоры широкоэллипсоидные, часто с оттянутыми вершинами,
иногда почти шаровидные, 20—39 (40) мкм шир., 32—50 мкм дл., вы-
тянутые вдоль конъюгационного канала, нередко окруженные тонким
слоем стекловидного пектинового вещества. Экзоспорий бесцветный,
гладкий. Мезоспорий от темно-оливкового до бурого или желто-корич-
невого, с крупными ямчатыми углублениями, 3 мкм в диам.
В мелких и заболоченных стоячих водоемах, реже в текучих водах,
реках. Россия: север европ. части (реки Свирь, Мандрога); Латвия (?),
Зап. Украина (Прикарпатье), Казахстан.
Широко распространенный вид. В первоначальных описаниях вида
мезоспорий значился гладким, но позднее — ямчатым. Очень близок к
М. robusta, имеющему, однако, более мелкие углубления на мезоспорий
и более широкие вегетативные клетки. Возможна их идентичность.
30. Mougeotia sumatrana Schmidle [=Mougeotiella sumatrana
(Schmidle) Yamagishi] — Мужоция суматранская (рис. 59, 4). — Ве-
гетативные клетки 20—24 мкм шир., по длине в 4—10 раз больше.
Хлоропласт с несколькими беспорядочно расположенными пиреноида-
ми. Конъюгация лестничная. Конъюгирующие клетки прямые или слабо
коленчато изогнутые, короче и толще вегетативных клеток. Формиро-
вание зигоспоры в широком конъюгационном канале, с частичным,
иногда значительным внедрением ее в полость одного или обоих га-
метангиев. Зигоспоры широкоэллипсоидные, вытянутые вдоль конъюга-
ционного канала, 40—42 мкм шир., 52 мкм дл. Мезоспорий коричне-
вый, крупнозернистый (бородавчатый ?). . '
136
Рис. 59. 1—3 — Mougeotia laevis-, 4 — M. sumatrana. (По: 1, 2 — Gauthier-Lievre, 1965;
3 — Рундина, 1988; 4 — Schmidle, 1895).
В стоячих водах, временных мелких водоемах. Сев. Казахстан.
Редкий вид, ранее найденный на о-ве Суматра. Требуют уточнения
сведения о наличии мантии вокруг зигоспоры и о строении оболочек
у зигоспоры. Зернистостью оболочек отличается от М. talyschensis,
имеющего ту же ширину вегетативных клеток и сходный тип форми-
рования зигоспоры.
31. Mougeotia pseudocalospora Czurda [=Debarya calospora (Palla)
West] — Мужоция ложнокрасивоспоровая (рис. 60, 7). — Вегетатив-
ные клетки 11—13 мкм шир. Хлоропласт с многочисленными пире-
ноидами, расположенными беспорядочно. Конъюгация лестничная.
Конъюгирующие клетки прямые. Формирование зигоспоры преимущест-
венно в конъюгационном канале с частичным внедрением в один или
137
Рис. 60. 1 — Mougeotia pseudocalospora-, 2 — M. cherokeana\ 3 — M. americana. (По: 1 —
West, West, 1898 (как Debarya calospora); 2 — Taft in: Transeau et al., 1934; 3 — Transeau,
1926).
оба гаметангия. Зигоспоры шаровидные, 20 мкм в диам. Мезоспорий
толстый, коричневый, крупноямчатый.
В стоячих водах. Англия.
Редкий вид. Очень близок к М. nummuloides, возможно являясь его
синонимом. Положение зигоспор может значительно варьировать.
32. Mougeotia talyschensis (Woronich.) Czurda [-Debarya talyschensis
Woronich.; Mougeotia sumatrana (Schaarsch.) Schmidle var. rotundospora
Skv.; Mougeotiella talyschensis (Woronich.) Yamagishi] — Мужоция та-
лышская (рис. 61, 1—3). — Вегетативные клетки 19—21 мкм шир., по
длине в 4—5 раз больше. Конъюгация лестничная. Конъюгирующие
клетки почти прямые или плавно и незначительно изогнутые. Формиро-
вание зигоспоры в широком конъюгационном канале со значительным
внедрением, вплоть до внешних клеток, в оба гаметангия. Зигоспоры ша-
ровидные, 42—50 мкм диам. Мезоспорий желто-коричневый, со срав-
нительно небольшими углублениями, расположенными беспорядочно.
В мелкой глинистой канавке. Азербайджан (Ленкорань: долина
р. Талыш).
138
Редкий и недостаточно изученный вид, другое местонахождение ко-
торого — Сев. Маньчжурия. Требуются уточнения о наличии или от-
сутствии мантии, форме и строении зигоспор. По ширине вегетативных
клеток наиболее близкие виды — М. sphaerocarpa и М. sumatrana,
имеющие соответственно гладкий и зернистый мезоспории.
33. Mougeotia cherokeana Taft — Мужоция черокская (рис. 60,
2). — Вегетативные клетки 7—9 мкм шир., по длине в 10—17 раз
больше. Хлоропласт с 2—6 пиреноидами, расположенными в один ряд.
Конъюгация лестничная. Конъюгирующие клетки сильно коленчато
изогнутые. Формирование зигоспоры в широком конъюгационном ка-
нале и обоих гаметангиях, вплоть до их внешних стенок. Зигоспоры
шаровидные или реже почти шаровидные, 21—25 мкм диам., окружен-
ные слоем пектинового студенистого вещества, 4—6 мкм толщ. Ме-
зоспорий желтый или желто-коричневый, гладкий.
В стоячих водах, в высыхающих канавах на торфяниках. Польша,
США.
Рис. 61. 1—3 — Mougeotia talyschensis-. 1 — конъюгирующие клетки с зигоспорами, общий
вид, 2 — строение зигоспоры с поверхности, 3 — то же в поперечном разрезе; 4—6 —
М. tunicata. (По: 1—3 — Скворцов, 1927а (как М. sumatrana var. rotundospora)', 4, 5 — Ya-
magishi, 1965b (как Mougeotiella tunicata)', 6 — Bock, Bock, 1956).
139
Редкий вид. От М. calcarea отличается узкими вегетативными клет-
ками и шаровидными зигоспорами, от М. caimanii Trans. (Гаити) —
широкими вегетативными клетками и иным положением зигоспор.
34. Mougeotia elegantula Wittr. (=М. elegans Wittr.) — Мужоция
изящненькая (рис. 62, 1, 2). — Вегетативные клетки 3.5—5 мкм шир.,
по длине в 15—26 раз больше. Хлоропласт с 2—8 пиреноидами, рас-
положенными в один ряд. Конъюгация лестничная. Конъюгирующие
клетки сильно и плавно изогнутые. Формирование зигоспоры в широ-
ком конъюгационном канале и обоих гаметангиях, вплоть до их внеш-
них стенок. Зигоспоры правильно квадратно-подушковидные, коротко-
четырехрогие, с прямыми сторонами и закругленными углами, 18—
24 мкм шир., (18) 20—24 мкм дл., сбоку правильно эллиптические,
(6) 12—14 мкм шир. Мезоспорий тонкий, гладкий. Апланоспоры (?)
эллипсоидные, 6—9 мкм шир., 20—24 мкм дл.
В стоячих, заболоченных, реже текучих водах: канавах, прудах,
озерах, водохранилищах, в прибрежье рек. Россия: северо-запад и
центр европ. части, Зап. Сибирь (Тюмень); Латвия, Украина (Причер-
номорье, Днепровское водохранилище), Ср. Азия (Голодная степь) (?),
Сев. (р. Нура) и Южн. Казахстан (окрестности Алма-Аты) (?).
Широко распространенный вид. Выделяется сильно удлиненными
вегетативными клетками при малой их ширине. Отличается от ряда
видов, имеющих квадратно-подушковидные зигоспоры, прямыми сто-
ронами зигоспор и отсутствием пектинового вещества вокруг них.
Встречается редко и поэтому еще слабо изучен: неизвестен цвет ме-
зоспория и недостоверно наличие апланоспор, похожих на зигоспоры
в их боковом положении. Местонахождения в Казахстане и Ср. Азии
сомнительны: в литературе по этим территориям этот вид ошибочно
назван М. elongatum Wittr.
35. Mougeotia americana Trans. — Мужоция американская (рис. 60,
3). — Вегетативные клетки 4—5 мкм шир., по длине в 8—30 раз боль-
ше. Хлоропласт с 4—10 пиреноидами, расположенными в один ряд.
Конъюгация лестничная. Конъюгирующие клетки коленчато изогнутые.
Формирование зигоспоры в конъюгационном канале и обоих гаметанги-
ях, с полным, вплоть до внешних стенок, или частичным их заполнени-
ем. Зигоспоры неправильно квадратно-подушковидные, с вогнутыми или
выпуклыми сторонами и четырьмя плохо выраженными, иногда прямо
срезанными углами, 13—24 мкм шир., 18—32 мкм дл., окруженные пек-
тиновым студенистым веществом. Экзоспорий толстый, бесцветный. Ме-
зоспорий бесцветный, гладкий. Апланоспоры неравнобокие, эллипсоид-
ные, с прямо срезанными вершинами, окруженные пектиновым вещест-
вом, 10—14 мкм шир., 20—26 мкм дл., лежащие в середине коленчато
изогнутых, сильно удлиненных клеток.
В стоячих водах: рыбоводных прудах, хаузах, водохранилищах. Ср.
Азия (?), Южн. и Вост. Казахстан (?).
Редкий вид, ранее известный лишь из Сев. Америки (США). Очень
близок к М. elegantula, широко распространенному виду, имеющему
сходную ширину вегетативных клеток и всегда правильно квадратно-
140
Рис. 62. 1,2 — Mougeotia elegantula: 1 — конъюгирующие клетки с зигоспорами анфас,
2 — зигоспора сбоку; 3 — М. corniculata: 4 — М. delicata. (По: 1,2 — Wittrock, 1872;
3 — Hansgirg, 1886; 4 — Beck-Mannagetta, 1926).
подушковидные зигоспоры, не окруженные пектиновым веществом.
Возможно, эти виды идентичны. Указания на местонахождение в Ср.
Азии и Казахстане сомнительны.
36. Mougeotia corniculata Hansg. — Мужоция рожковидная (рис. 62,
3). — Вегетативные клетки (3) 5—7 мкм шир., по длине в 10—26
раз больше. Хлоропласт с 2 пиреноидами, расположенными в один
ряд. Конъюгация лестничная. Конъюгирующие клетки очень сильно
изогнутые. Формирование зигоспоры в широком конъюгационном ка-
нале с полным внедрением в оба гаметангия, вплоть до их внешних
141
стенок. Зигоспоры округло-подушковидные с сильно выпуклыми сто-
ронами и четырьмя хорошо выраженными, небольшими, мягко закруг-
ленными рогами, 18—26 мкм шир., 22—27 мкм дл., окруженные очень
небольшим количеством студенистого вещества, втянутого внутрь;
сбоку широкоэллиптические, с оттянутыми концами (рогами). Мезо-
спорий толстый, желто-коричневый, гладкий.
В мелких водоемах, водохранилищах)' иногда в солоноватых топях.
Узбекистан (Чардаринское водохранилище, канал от Ирмак-Узеньского
водохранилища).
Редкий вид. Описанный давно, он все еще имеет сравнительно мало
местонахождений в Европе (Дания, Германия, Франция, Австрия,
Чехия) и Сев. Африке. Самый близкий вид — М. faurelii Gauthier-Li-
evre (Алжир) имеет чуть более широкие вегетативные клетки (7—
9 мкм) и мелкоямчатый экзоспорий.
37. Mougeotia delicata Beck — Мужоция нежная (рис. 62, 4). —
Вегетативные клетки 3.5—5 мкм шир. Конъюгация лестничная. Конъ-
югирующие клетки сильно коленчато изогнутые. Формирование зиго-
споры в широком конъюгационном канале и обоих гаметангиях, вплоть
до их внешних стенок. Зигоспоры квадратно-подушковидные, с сильно
вогнутыми сторонами с четырьмя рогами, вдавленными на концах и
довольно сильно внедряющимися в гаметангии, 27.6 мкм шир. Мезо-
спорий гладкий.
В стоячих водах, на торфяниках. Австрия.
Редкий и недостаточно изученный вид. Близок к М. viridis, отли-
чается узкими вегетативными клетками и сильно вогнутыми сторонами
у зигоспор.
38. Mougeotia viridis (Kutz.) Wittr. (=Staurospermum viride Kutz.) —
Мужоция зеленая (рис. 63, 7). — Вегетативные клетки 5—10 мкм
шир., по длине в 4—20 раз больше. Хлоропласт с (2) 4—8 пиренои-
дами, расположенными в один ряд. Конъюгация лестничная. Конъюги-
рующие клетки значительно изогнутые. Формирование зигоспоры в ши-
роком конъюгационном канале и обоих гаметангиях, вплоть до их
внешних стенок. Зигоспоры квадратно-подушковидные с незначительно
вогнутыми сторонами и короткими рогами, вдавленными на концах,
16—33 мкм шир., 19—33 мкм дл.; сбоку — асимметрично эллипти-
ческие с чуть оттянутыми концами, 15—20 мкм шир. Мезоспорий бес-
цветный, гладкий. Апланоспоры (?) веретеновидные, асимметрично эл-
липсоидные с оттянутыми концами, 14—16 мкм шир., 30—36 мкм дл.
В стоячих и слабопроточных водах, в канавах, сфагновых болотах,
на лугах, рисовых полях. Россия: европ. часть, от севера до дельты
Волги.
Часто встречающийся вид. От наиболее близкого вида — М. сари-
etna отличается небольшой шириной вегетативных клеток, от М. ele-
gantula — вогнутыми сторонами зигоспор.
39. Mougeotia virescens (Hass.) Borge — Мужоция зеленоватая
(рис. 63, 2). — Вегетативные клетки 8—10 мкм шир., по длине в
142
Рис. 63. 1 — Mougeotia viridis, конъюгирующие клетки с зигоспорами в двух положениях
(анфас и сбоку); 2 — М. virescens, конъюгирующие клетки с зигоспорами анфас; 3 —
М. quadrangulata, конъюгирующие клетки с зигоспорами в двух положениях (анфас и сбоку).
(По: Jao, 1935b).
143
5—27 раз больше. Конъюгация лестничная. Конъюгирующие клетки
довольно сильно изогнутые. Формирование зигоспоры в широком конъ-
югационном канале и обоих гаметангиях, вплоть до их внешних стенок.
Зигоспоры квадратно-подушковидные со слабо вогнутыми сторонами и
очень короткими рогами, закругленными на концах, 25—36 мкм шир.,
25—36 мкм дл. Мезоспорий бесцветны^, гладкий.
В стоячих водах, болотах, озерах, на рисовых полях, возможно с
небольшим загрязнением. Узбекистан (окрестности Бухары).
Широко распространенный вид на всех континентах. По-видимому,
встречается чаще, чем указано в отечественной литературе. От М. vi-
ridis, близкого вида по ширине вегетативных клеток, отличается за-
кругленными рогами у зигоспор. Возможно, что оба эти вида иден-
тичны.
40. Mougeotia capucina (Bory) Ag. (=Staurocarpus capucinus Hass.) —
Мужоция капюшоновая (рис. 64). — Вегетативные клетки (11) 14—
21 мкм шир., по длине в 5—14 (17) раз больше. Хлоропласт 1 (2),
узкопластинчатый, с 4—10 пиреноидами, расположенными в один ряд.
Конъюгация лестничная, очень редко боковая (?). Конъюгирующие
клетки значительно коленчато изогнутые, их длина до 23 раз больше
ширины. Формирование зигоспоры при лестничной конъюгации в ши-
роком конъюгационном канале и обоих гаметангиях, вплоть до их
внешних стенок. Зигоспоры неправильно квадратно-подушковидные,
преимущественно с вогнутыми сторонами и четырьмя рогами, закруг-
ленными или прямо срезанными на концах, 45—70 (80) мкм шир.,
55—100 (117) мкм дл., окруженные слоистым пектиновым веществом,
иногда глубоко заходящим в гаметангии. Мезоспорий коричневый или
красно-фиолетовый, иногда золотисто-буроватый, редко бесцветный,
гладкий, до 5 мкм толщ. Апланоспоры эллипсоидные, неравнобокие,
окруженные пектиновым слоистым веществом, заметно выступающим
на концах, 20—36 (51) мкм шир., 45—70 (80) мкм дл., с пектиновыми
рогами до 127 мкм дл.
В стоячих, медленно текущих и проточных водах, в торфяниках,
болотах, на увлажненных скалах из песчаника, в опресненных заливах
морей. Россия: Карелия (р. Умба); Сев. Казахстан (заповедник Боро-
вое), Узбекистан (Каракалпакия, Аральское море — опресненные за-
ливы).
Широко распространенный вид. При определенных экологических
условиях (например, при наземном местообитании) клеточный сок и
оболочка зигоспоры окрашиваются в красновато-фиолетовый цвет, что
придает соответствующую окраску дерновинкам. Близкий вид М. ame-
ricana имеет более узкие вегетативные клетки.
41. Mougeotia paludosa G. S. West [-M. quadrata Randh., non sen-
su (Hass.) Wittr.; M. quadra-undulata Kissel.] — Мужоция болотная
(рис. 65). — Вегетативные клетки 9—14 мкм шир., по длине в 6—7
раз больше. Хлоропласт с (3) 4—8 пиреноидами, расположенными в
один ряд. Ризоиды от базальной клетки, неразветвленные, редкие.
Конъюгация лестничная. Конъюгирующие клетки сильно и плавно изо-
144
Рис. 64. Mougeotia capucina: 1 — вегетативная клетка, 2, 4, 5, 7 — зигоспоры анфас, 3 —
апланоспора, 6 — зигоспора сбоку. (По: 1—3 — Transeau, 1926; 4 — Топачевский, 1952;
5 — Hassall, 1843 (как Staurocarpus capucinus)-, 6,7 — Воронихин в: Воронихин, Красно-
перова, 1970).
10 Рундина Л. А.
145
Рис. 65. Mougeotia paludosa;. 1 — зигоспоры сбоку, 2, 3 — зигоспоры анфас.
(По: Рундина, 1977а).
146
гнутые, их длина в 15—20 раз больше ширины. Формирование зигос-
поры в широком конъюгационном канале и обоих гаметангиях, вплоть
до их внешних стенок. Зигоспоры разнообразной формы, от подуш-
ковидных до многоугольных, с широко закругленными углами, без
рогов, а также от широкоэллипсоидных до шаровидных, (22) 26—
38 мкм шир., (27) 32—49 мкм дл., окруженные толстым слоем студе-
нистого пектинового вещества, незначительно внедряющегося в гаме-
тангии, по углам преимущественно вогнутого или слегка волнистого.
Экзоспорий тонкий, бесцветный, гладкий. Мезоспорий толстый, желто-
коричневый, гладкий. Эндоспорий тонкий, четко различимый на зре-
лых зиготах.
В стоячей воде, на рисовых полях. Южн. Казахстан, Узбекистан.
Широко распространенный вид почти на всех континентах (Центр.
Азия, Сев. Индия, Южн. Америка), в Европе встречен в Англии, Ис-
пании и Польше. Отличается большим разнообразием формы у зиго-
спор, благодаря чему среди близких видов узнается без труда.
42. Mougeotia regelii Skuja — Мужоция Регеля (рис. 66, 1, 2). —
Вегетативные клетки 9—10 мкм шир., по длине в 5—16 раз больше.
Хлоропласт с 2—4 пиреноидами, расположенными в один ряд. Конъ-
югация лестничная. Конъюгирующие клетки слабо изогнутые. Форми-
рование зигоспоры в широком конъюгационном канале и обоих гаме-
тангиях, вплоть до их внешних стенок. Зигоспоры неправильно подуш-
Рис. 66. 1, 2 — Mougeotia regelii; 1 — конъюгирующие клетки с зигоспорами анфас, 2 —
то же с зигоспорой сбоку; 3—5 — М. thylespora; 3,4 — конъюгирующие клетки с зигоспо-
рами анфас, 5 — зигоспора сбоку. (По: 1, 2 — Skuja, 1937; 3—5 — Skuja, 1929).
147
ковидные, мягко очерченные, без рогов, 22—25 мкм шир. (выс.), 29—
30 мкм дл., 25 мкм толщ., сбоку округлые. Экзоспорий тонкий, бес-
цветный, гладкий. Мезоспорий тонкий, желто-коричневый, гладкий.
Эндоспорий довольно толстый, бесцветный или светло-желтый, густо-
и мелкоямчатый (точечный ?).
В родниковой луже, на высоте 1100 м над ур. м. Греция.
Редкий и недостаточно изученный вйд. Скульптурированность эн-
доспория — довольно редкое явление у зигнемовых водорослей вооб-
ще, у рода Mougeotia в частности. По этому признаку данный вид
достаточно резко отличается от других видов, близких по вегетативным
признакам: М. thylespora, М. austriaca, М. paludosa, М. calcarea.
43. Mougeotia fossa О. et W. Bock — Мужоция канавная (рис. 67,
2, 3). — Вегетативные клетки 5—6 мкм шир. Конъюгация лестничная.
Конъюгирующие клетки довольно сильно изогнутые. Формирование зи-
госпоры в широком конъюгационном канале и обоих гаметангиях,
вплоть до их внешних стенок. Зигоспоры квадратно-подушковидные
со слабо вогнутыми или очень редко прямыми или слегка выпуклыми
сторонами и четырьмя короткими рогами, прямо срезанными или слег-
Рис. 67. 1 — Mougeotia costata, зигоспора; 2, 3 — М. fossa-. 2 — зигоспора анфас, 3 — зи-
госпора сбоку; 4, 5 — М. gracillima: 4 — зигоспора анфас, 5 — зигоспора сбоку; 6,7 —
М. austriaca-. 6 — конъюгирующие клетки, общий вид, 7 — зигоспора анфас. (По: 1—3 —
Воск, Воск, 1956; 4, 5 — Transeau, 1926; 6,7 — Yamagishi, 1965а).
148
ка закругленными на концах, 22—27 мкм шир., 25—30 мкм дл., 16—
20 мкм толщ., сбоку асимметрично эллиптические с сильно вдавлен-
ными углами. Экзоспорий тонкий, бесцветный, гладкий. Мезоспорий
бесцветный, коротко и неправильно складчатый (морщинистый).
В канавах, рвах, болотах, преимущественно при pH 6.0. Германия.
Очень редкий вид. Морщинистость мезоспория отличает его от
близкого вида М. punctata, имеющего ямчатый мезоспорий.
44. Mougeotia gracillima (Hass.) Wittr. — Мужоция тончайшая
(рис. 67, 4, 5). — Вегетативные клетки 5—7 мкм шир., по длине в
8—20 раз больше. Хлоропласт с 9 пиреноидами. Конъюгация лестнич-
ная. Конъюгирующие клетки довольно сильно и плавно изогнутые.
Формирование зигоспоры в конъюгационном канале и обоих гаметан-
гиях, вплоть до их внешних стенок. Зигоспоры квадратно-подушковид-
ные с сильно вогнутыми сторонами и четырьмя заметными рогами, на
концах вдавленными или прямо срезанными, 18—25.5 мкм шир., 20—
28 мкм дл., сбоку правильно эллиптические со слабо вдавленными
концами. Мезоспорий бледно-серый или желтоватый, мелкобородавча-
тый. Апланоспоры (?) правильно эллипсоидные со слабо вдавленными
или прямо срезанными концами.
В стоячих, преимущественно заболоченных водах, болотах, поймен-
ных озерах, прудах, лиманах, низовьях рек. Зигоспоры найдены в от-
ложениях из голоцена. Россия: повсеместно в европ. части; Ср. Азия,
Приаралье, Казахстан.
Широко распространенный, редко встречающийся вид. От наиболее
сходной М. punctata отличается мелкобородавчатой скульптурой мезо-
спория. Наличие апланоспор спорно, так как по описанию они сходны
с зигоспорами, рассматриваемыми сбоку.
45. Mougeotia aspera Woronich. — Мужоция шероховатая. — Ве-
гетативные клетки 13—16.5 мкм шир., по длине в 5—8 раз больше.
Конъюгация лестничная. Формирование зигоспоры в конъюгационном
канале и обоих гаметангиях. Зигоспоры почти шаровидные, 36—
46 мкм диам., реже широкоэллипсоидные или яйцевидные, 46 мкм
шир., 66 мкм дл. Мезоспорий светло-коричневый, точечный.
На скалах, орошаемых родниковой водой, в реках. Грузия (окрест-
ности Тбилиси), Узбекистан.
Недостаточно описанный вид, некоторыми исследователями призна-
ваемый сомнительным. Нет оригинального рисунка, требуют уточнения
форма зигоспор и скульптура мезоспория.
46. Mougeotia boodlei (W. et G. S. West) Collins [=M. boodlei (W. et
G. S. West) Krieger; M. rectangularis Krieger] — Мужоция Будля (рис. 68,
1—3). — Вегетативные клетки 4—5.5 (8) мкм шир., по длине в 4—40
раз больше. Хлоропласт с 4—6 пиреноидами, расположенными в один
ряд. Размножение апланоспорами и зигоспорами. Конъюгация лестнич-
ная, очень редкая. Конъюгирующие клетки почти прямые. Формирова-
ние зигоспоры в широком конъюгационном канале и обоих гаметангиях,
вплоть до их внешних стенок. Зигоспоры правильно квадратно-подуш-
149
Рис. 68. 1—3 — Mougeotia boodlei'. 1 — зигоспора анфас и две апланоспоры, 2, 3 — апла-
носпоры; 4—7 — М. ventricosa, апланоспоры. (По: 1 — Transeau, 1926; 2, 3, 5—7— West G. S.,
1904; 4 — Wittrock, 1878 (как Gonatonema ventricosum')').
ковидные, преимущественно с прямыми сторонами, без рогов, 15—
18 мкм шир., 15—23 мкм дл. Мезоспорий от желтого до коричневого,
мелко- и частоточечный. Формирование апланоспор внутри клеток в
прямых или слегка изогнутых нитях. Апланоспоры правильно эллипсо-
идные или реже односторонне вздутые, с усеченными или закругленны-
ми вершинами, 12—17 мкм шир., 17—25 мкм дл., со скульптурой обо-
лочек, как у зигоспор, очень частые.
В стоячих водах, прудах. Англия, Япония, США.
Один из редких видов, у которых апланоспоры встречаются значи-
тельно чаще, чем зигоспоры. В некоторых случаях размножение ис-
ключительно апланоспорами. Отличается от двух близких видов —
М. miamiana Trans. (США) и М. prona Trans. (Сев. Америка) формой
и строением зигоспор.
47. Mougeotia costata О. et W. Bock — Мужоция ребристая (рис. 67,
7). — Вегетативные клетки 7—9 мкм шир. Конъюгация лестничная.
150
Конъюгирующие клетки сильно изогнутые. Формирование зигоспоры в
широком конъюгационном канале и обоих гаметангиях, вплоть до внеш-
них стенок. Зигоспоры квадратно-подушковидные с вогнутыми сторона-
ми и прямо срезанными и короткими рогами, с втянутыми внутрь углами,
30—34 мкм шир., 33—36 мкм дл. Экзоспорий тонкий, бесцветный,
гладкий. Мезоспорий бесцветный, плотно и равномерно точечный (пун-
ктированный), неправильно сетчато-ребристый.
На торфяниках, преимущественно при pH 6.0. Германия.
Редкий вид. Тонкую пунктированность мезоспория можно рассмот-
реть только с помощью иммерсионного объектива.
48. Mougeotia tunicata О. et W. Bock \=Mougeotiella tunicata (O. et
W. Bock) Yamagishi] — Мужоция покровная (рис. 61, 4—6). — Вегета-
тивные клетки 17—25 мкм шир., по длине в 4—8 раз больше. Хлоро-
пласт с 6—9 пиреноидами. Конъюгация лестничная. Конъюгирующие
клетки слегка коленчато изогнутые или почти прямые. Формирование
зигоспоры в очень широком конъюгационном канале и обоих гаметан-
гиях, вплоть до их внешних стенок. Зигоспоры шаровидные, 30—34 мкм
в диам., или широкоэллипсоидные, 36—44 мкм шир., 40—45 мкм дл.
Экзоспорий тонкий, прозрачный, гладкий. Мезоспорий толстый, коричне-
вый, густозернистый. Эндоспорий тонкий, гладкий. Вокруг зигоспоры име-
ется мантия, мелко- и тонкоскладчатая, бесцветная или светло-коричнева-
тая, гладкая или слегка точечная, глубоко заходящая в гаметангии.
В стоячей воде временных водоемов, лужах. Германия, Япония.
Редкий вид. От М. sphaerocarpa и М. drouetii, также имеющих ман-
тию вокруг зигоспоры и сходную ширину вегетативных клеток, отли-
чается скульптурированным мезоспорием.
49. Mougeotia thylespora Skuja — Мужоция мешочкоспоровая
(рис. 66, 3—5). — Вегетативные клетки 5—8 мкм шир., по длине в
5—10 раз больше. Хлоропласт с 4—8 пиреноидами, расположенными
в один ряд. Конъюгация лестничная. Конъюгирующие клетки незначи-
тельно изогнутые. Формирование зигоспоры в широком конъюгацион-
ном канале и обоих гаметангиях, вплоть до их внешних стенок. Зи-
госпоры правильно квадратно-подушковидные со слегка выпуклыми
сторонами и слабо выступающими, закругленными рогами, 24—32 мкм
шир., 20—29 мкм дл., 16—22 мкм толщ., сбоку эллиптические с за-
кругленными и слегка оттянутыми рогами. Экзоспорий бесцветный,
гладкий, с гиалиновыми наслоениями поверх рогов. Мезоспорий до-
вольно толстый, желтовато-коричневый, с ямчатыми углублениями раз-
ного размера, преимущественно мелкими. Партеноспоры (апланоспо-
ры ?) эллипсоидные, 18—25 мкм шир., 29—40 мкм дл.
В стоячих, часто болотистых водах: лужах, канавах, луговых по-
нижениях. Россия: Зап. Сибирь; Эстония (о-ва Сааремаа и Хийумаа).
Редкий вид, найденный также в США. От М. quadrangulata отли-
чается относительно узкими вегетативными клетками, закругленными
зигоспорами, гиалиновыми наслоениями экзоспория, окрашенным и
мелкоямчатым мезоспорием. Другие близкие виды: М. punctata и
М. producta W. et G. S. West (Мьянма, Ассам).
151
50. Mougeotia austriaca Czurda — Мужоция австрийская (рис. 67, 6,
7). — Вегетативные клетки 7—9 мкм шир., по длине в 5—20 раз боль-
ше. Хлоропласт с 2—3 пиреноидами. Конъюгация лестничная. Конъюги-
рующие клетки довольно сильно изогнутые. Формирование зигоспоры в
широком конъюгационном канале и обоих гаметангиях. Зигоспоры не-
правильно квадратно-подушковидные с сильно или умеренно вогнутыми
сторонами, четырьмя умеренно или слабо выраженными рогами с ши-
роко закругленными концами, 20—25 мкм шир., 27—30 мкм дл., сбоку
широкоэллиптические, иногда с чуть оттянутыми рогами. Экзоспорий
тонкий, гладкий, бесцветный. Мезоспорий толстый, от желтого до жел-
товато-коричневого, с небольшими углублениями, 1 мкм диам.
В стоячих водах. Австрия, Япония, США.
Редкий вид. Близок к М. punctata по ширине вегетативных клеток,
отличается более крупными углублениями на мезоспорий и мягко за-
кругленными рогами.
51. Mougeotia punctata Wittr. (=М. tumidula Trans.) — Мужоция
точечная (рис. 69). — Вегетативные клетки 6—10 (12.5) мкм шир.,
по длине в 5—20 раз больше. Хлоропласт с 4—8 пиреноидами, рас-
Рис. 69. Mougeotia punctata'. 1 — конъюгирующие клетки с зигоспорами, общий вид, 2,3 —
зигоспора анфас, 4 — зигоспора сбоку. (По: 1,2 — Yamagishi, 1965а; 3, 4 — Gauthier-
Lievre, 1965).
152
положенными в один ряд. Конъюгация лестничная. Конъюгирующие
клетки довольно сильно изогнутые. Формирование зигоспоры в широ-
ком конъюгационном канале и обоих гаметангиях, вплоть до их внеш-
них стенок. Зигоспоры квадратно-подушковидные с незначительно во-
гнутыми сторонами и четырьмя небольшими рогами, на концах прямо
срезанными или вдавленными, 18—30 мкм шир., 20—38 мкм дл., 18—
20 мкм толщ., сбоку эллиптические с прямо срезанными или чуть
вдавленными рогами. Мезоспорий (экзоспорий ?) бесцветный, мелко-
ямчатый.
В мелких, стоячих и слабопроточных водах, сфагновых болотах,
лужах, водохранилищах, на затопленных лугах, рисовых полях, при
высокой температуре воды (24.5—29.5 °C) и pH 5.2—8.0. Зигоспоры
также найдены в отложениях четвертичного периода. Россия: авандель-
та и култучная зона Волги; Узбекистан (Чардаринское и Шертское
водохранилища).
Возможно, распространен шире, чем указано в литературе. От
М. austriaca отличается прямо срезанными или чуть вдавленными на
концах рогами и более мелкими углублениями на мезоспорий. По
форме зигоспор и размерам почти не отличается от М. fossa, мезо-
спорий которого не ямчатый, а морщинистый. Близок, а возможно, и
идентичен М. quadrangulata.
52. Mougeotia quadrangulata Hass. [=Staurospermum quadratum
(Hass.) Kutz.; Mougeotia quadrata (Hass.) Wittr., non sensu Randh.; M. qua-
drangulata f. (sine nomen) Popova] — Мужоция четырехугольная (рис. 63,
3; 70). — Вегетативные клетки 7—13 мкм шир., по длине в 5—13 раз
больше. Хлоропласт с 6—16 пиреноидами, расположенными в один ряд.
Конъюгация лестничная. Конъюгирующие клетки изогнутые. Формиро-
вание зигоспоры в широком конъюгационном канале и обоих гаметан-
гиях, вплоть до их внешних стенок или с незначительным выходом за
их пределы. Зигоспоры квадратно-подушковидные со слегка выпуклыми,
вогнутыми или прямыми сторонами, обычно без рогов или с четырьмя
слабо выраженными рогами, на концах прямо срезанными или широко
закругленными, 20—40 мкм шир., 22—45 мкм дл., 18—22 мкм толщ.,
сбоку округлые или эллиптические с прямо срезанными вершинами. Ме-
зоспорий обычно бесцветный или бледно-желтый, ямчатый. Апланоспо-
ры (?) неправильно эллипсоидные, ямчатые, 20—21 мкм шир., 36—
44 мкм дл„ в слегка изогнутых клетках.
В стоячих, мелких, иногда заболоченных, реже текучих водах, бо-
лотах, озерах, лужах, на рисовых полях. Россия: европ. часть, Зап. и
Вост. Сибирь; Зап. (Карпаты, Прикарпатье, Буковина) и Вост. Укра-
ина, Казахстан, Приаралье.
Редкий вид с широким ареалом, встречающийся на всех континен-
тах. Отличается от М. punctata почти полным отсутствием рогов.
53. Mougeotia ventricosa (Wittr.) Collins (=Gonatonema ventricosum
Wittr.) — Мужоция вздутая (рис. 68, 4—7). — Вегетативные клетки
5—9 мкм шир., по длине в 11—23 раза больше. Хлоропласт с 4—5
пиреноидами, расположенными в один ряд. Зигоспоры неизвестны. Раз-
153
Рис. 70. Mougeotia quadrangulata: 1—3 — конъюгирующие клетки с зигоспорой анфас,
4, 5 — конъюгирующие клетки с зигоспорой сбоку. (По: Gauthier-Lievre, 1965).
множение апланоспорами. Формирование апланоспор в сильно колен-
чато изогнутых клетках. Апланоспоры неправильно эллипсоидные, од-
носторонне вздутые, с оттянутыми концами, лимонообразные, реже
почти шаровидные, 12—24 мкм шир., 16—34 мкм дл. Мезоспорий
толстый, желто-коричневый, гладкий.
В стоячих водах, лужах, ямах, на рисовых полях. Эстония (о-в
Сааремаа), Латвия, Узбекистан.
Вид часто встречающийся, с широким ареалом. Возможно, иденти-
чен одному из близких видов: М. tibestica Gauthier-Lievre (Африка),
М. tenerrima G. S. West (Сев. и Южн. Америка), М. rava Trans. (Сев.
Америка), имеющих сходную ширину вегетативных клеток и размно-
жающихся также только апланоспорами, мезоспорий которых тоже
гладкий.
54. Mougeotia faveolatospora Kiriakov — Мужоция мелкоячеисто-
споровая (рис. 71, 1—4). — Вегетативные клетки 8—10 мкм шир., по
длине в 8—25.5 раза больше. Хлоропласт с 3—10 пиреноидами, рас-
положенными беспорядочно. Зигоспоры неизвестны. Размножение ап-
ланоспорами. Формирование апланоспор в особых вздутиях вегетатив-
ных клеток, реже внутри самих клеток. Апланоспоры шаровидные,
17—22 мкм диам. Экзоспорий тонкий, бесцветный, гладкий. Мезоспо-
рий желто-коричневый, с небольшими ямками неправильной формы.
В стоячей воде. Болгария.
154
Рис. 71. 1—4 — Mougeotia faveolatospora, клетки с апланоспорами; 5 — М. notabilis,
то же. (По: 1—4 — Кнряков в: Воденичаров, Киряков, 1970; 5 — Hassall, 1852).
Очень редкий вид. От близких видов — М. ventricosa и М. boodlei
отличается шаровидностью апланоспор. В клетках после образования
апланоспор сохраняется бесцветный цитоплазматический остаток.
55. Mougeotia notabilis Hass. — Мужоция заметная (рис. 71, 5). —
Вегетативные клетки (9) 12—14 мкм шир. Зигоспоры неизвестны. Раз-
множение апланоспорами. Формирование апланоспор в коленчатых,
изогнутых под острым углом клетках. Апланоспоры цилиндрические,
изогнутые, расположенные в середине клеток в местах их изгибов.
В стоячих и текучих водах. Россия: Вост. Сибирь (Республика Саха:
реки Вилюй, Холомоллюх); Узбекистан (Каракалпакия).
Недостаточно изученный вид. Нет сведений о размерах апланоспор
и строении их оболочки.
56. Mougeotia turicensis Nipkow — Мужоция цюрихская. — Вегета-
тивные клетки 20—28 мкм шир. Зигоспоры и апланоспоры неизвестны.
Размножение акинетами. Формирование акинет в вегетативных клетках,
прямых или коленчато изогнутых, имеющих частые псевдоконъюгаци-
онные выросты. Акинеты цилиндрические, короче вегетативных клеток
в 3—4 раза, с бесцветной плотной оболочкой, лишенной видимой линии
разрыва, и темно-серо-зеленым, бугристым содержимым.
155
В стоячих водах небольших водоемов. Швейцария.
Редкий, недостаточно изученный вид. Отсутствует оригинальный ри-
сунок вида. По наличию псевдоконъюгационных выростов и сходной
ширине вегетативных клеток близок к М. genuflexa. Вполне возможно,
что только определенные экологические условия могут вызвать образо-
вание акинет, исключительное присутствие которых в жизненном цикле
водоросли не должно, по-видимому, рассматриваться как видовой при-
знак.
По сообщению Кадлубовской и Лангангена (Kadhibowska, Langan-
gen, 1997), поступившему ко мне уже после сдачи монографии в из-
дательство, в Норвегии найдены Mougeotia olivacea Ling и М. myso-
rensis Iyengar, новые для Европы. Эти виды следует принять как по-
тенциальные для флоры России.
СОМНИТЕЛЬНЫЕ И ОШИБОЧНЫЕ УКАЗАНИЯ
ВИДОВ РОДА MOUGEOTIA
Mougeotia caricetorum Skv.: Скворцов, 1946 : 2, Tab. 13, Fig. 9—
11. — Неполно описанный вид из-за отсутствия зигоспор. Сходен с
М. genuflexa (Dillw.) Ag.
М. condensata (sine auct., nom. nud. et nom. invalid.): Демин, 1955 :
189.
M. decussata Kiitz.: Змеев, 1872: 55. — Неполно описанный вид
из-за отсутствия зигоспор (Kiitzing, 1845 : 222; 1855 : Taf. 3, Fig. 3).
М. elongatum Wittr.: Эргашев, 1971 : 79; Музафаров, Эргашев,
1972 : 11. — Ошибочное название. Non М. elegantula Wittr.
М. gracilis Kiitz.: Волкова, Данилова, 1970 : 558. — Неполно опи-
санный вид из-за отсутствия зигоспор (Kiitzing, 1845 : 221). По ширине
и коленчатой изогнутости вегетативных клеток сходен с М. genuflexa
(Dillw.) Ag. f. reinschii (Trans.) Rund.
M. minutissima Lemm.: Levander, 1908 : 130; Schneider, 1908 : 47;
Skuja, 1928 : 118. — Неполно описанный вид из-за отсутствия зиго-
спор (Lemmermann, 1898 : 4).
М. notabilis Hass.: Абибуллаева, 1968 : 81. — Сомнительное оп-
ределение из-за отсутствия зигоспор.
М. porticalis (sine auct., nom. nud. et nom. invalid.): Демин, 1955 :
189.
M. protecta (sine auct., nom. nud. et nom. invalid.): Демин, 1955 :
189.
M. reniformis Swir. (nom. nud.): Свиренко, 1927 : 462.
M. renispora Swir.: Свиренко, 1927 : 459, 471, fig. 6. — Неполно
описанный вид из-за незрелости зигот. Сходен с М. scalaris Hass.
М. staurosperma Skv.: Скворцов, 1946 : 2, Tab. 13, Fig. 12. —
Неполно описанный вид из-за отсутствия данных о размере и строении
зигоспор. Сходен с М. americana Trans.
М. weberi (sine auct., nom. nud. et invalid.): Демин, 1955 : 189.
156
Род DEBARYA Wittr. — ДЕБАРИЯ
=Debarya sect. Mougeotiella Skuja; Mougeotia Ag., pr. p.
Нити неразветвленные, свободноплавающие. Вегетативные клетки
до 16 мкм шир., по длине до 20 раз больше. Поперечные стенки
гладкие. Продольные стенки целлюлозные, с наружным пектиновым
слоем. Хлоропласт 1, очень редко их 2, осевой, пластинчатый или
лентовидный, с 2—20 пиреноидами, расположенными в ряд или раз-
бросанно. Ядро прилегает к хлоропласту в центре клетки. Конъюгация
лестничная. Конъюгирующие клетки преимущественно коленчато изо-
гнутые, иногда значительно, реже почти прямые. Конъюгационный
канал отчетливый, иногда очень длинный, сначала узкий, при созре-
вании зиготы расширяющийся. Формирование зигоспоры в конъюгаци-
онном канале с частичным или полным внедрением в полость гаме-
тангиев. При созревании зиготы свободные полости гаметангиев запол-
няются слоистым голубовато-белым коллоидным веществом. Зигоспоры
сжато-эллипсоидные, линзовидные, реже шаровидные или квадратно-
подушковидные, часто сложной конфигурации, по форме напоминаю-
щей волчок. Экзоспорий разной толщины, бесцветный, гладкий. Ме-
зоспорий желтый, коричневый или желтовато-коричневый, редко голу-
бой, разнообразно скульптурированный, редко гладкий. Эндоспорий
трудно различимый, гладкий, очень редко ямчатый. Апланоспоры, аки-
неты и партеноспоры редкие.
Представители рода встречены преимущественно в стоячих или сла-
бопроточных водоемах: озерах, прудах, в устьях рек, мелких водоемах
оросительной системы и в болотах.
Ранние предшественники современного рода Debarya были описаны
как формальный род Peltacystia Balme et Segroves из пермских отло-
жений. Сходные по строению зигоспоры в отложениях мелового и
третичного периодов были обозначены формальным родом Lecaniella
. Cookson et Eisenack. Зигоспоры Debarya glyptosperma найдены и в
голоцене: в песках и болотистых почвах.
Род включает 10 видов, очень редко встречающихся в Азии,
Сев. Америке, Новой Зеландии и Австралии. В России, как и во
всех странах Европы, обнаружен только один, самый распростра-
ненный вид.
По ряду морфологических признаков, в первую очередь по наличию
одного пластинчатого хлоропласта, в вегетативном состоянии род De-
barya неотличим от рода Mougeotia. Однако в генеративной стадии
его виды очень сходны также с видами рода Zygnemopsis — у тех и
других полости гаметангиев при созревании зиготы заполняются кол-
лоидным веществом. Поэтому для правильного определения материала
необходимо учитывать и сравнивать морфологические признаки в
обеих стадиях — вегетативной и генеративной.
Debarya glyptosperma (De Вагу) Wittr. (=Mougeotia glyptosperma
De Bary) — Дебария бородавчатосемянная (рис. 72). — Вегетативные
клетки 9—16 мкм шир., по длине в (4) 6—12 (20) раз больше. Хло-
157
Рис. 72. Debarya glyptosperma: 1,2 — зигоспоры аифас, лестничная конъюгация, общий
вид, 3 — зигоспора анфас, 4, 5 — зигоспора сбоку. (По; 1,3,4 — Bourrelly, 1966,- 2, 5 —
Transeau, 1951).
ропласт 1, узкопластинчатый, с 8—12 пиреноидами, расположенными
в один ряд. Конъюгирующие клетки обычно сильно коленчато изогну-
тые, их длина значительно больше длины вегетативных клеток. Конъ-
югационный канал отчетливый, иногда очень длинный, до 80 мкм.
Формирование зигоспоры в конъюгационном канале и обоих гаметан-
гиях, иногда вплоть до их внешних стенок. Зигоспоры уплощенно-эл-
липсоидные (линзовидные), трехосные, с короткой осью, перпендику-
лярной к плоскости конъюгации, — сложной конфигурации, по форме
напоминающей волчок, (16) 30—50 мкм шир., (30) 42—72 мкм дл.
Экзоспорий бесцветный, гладкий. Мезоспорий желтый, коричневый
или желтовато-коричневый, сложной конфигурации, причем плоскости
между тремя поперечными килями и двумя вершинными бугорками
испещрены довольно тонкими радиальными ребрышками.
В озерах, болотах, устьях рек, водоемах оросительной системы.
Россия: Карелия; Латвия, Вост. Украина, Узбекистан.
Вероятно, более широко распространенный вид, чем указано в
литературе. Узнается без труда по характерной конфигурации зи-
госпоры.
158
Род MOUGEOTIOPSIS Palla — МУЖОЦИОПСИС
=Debarya Wittr. sensu W. et G. S. West, pr. p.; Mesogerron Brand;
Mougeotia Ag., pr. p.
Нити неразветвленные, свободноплавающие или в виде налетов и дер-
новинок при наземном местообитании. Вегетативные клетки до 18 мкм
шир., короткие, по длине равные ширине, меньше ее или до 2 (4) раз
больше. Поперечные стенки клеток гладкие. Продольные стенки клеток
целлюлозные, с наружным пектиновым слоем. Хлоропласт 1, осевой,
пластинчато-вогнутый, крупный, с утолщенными, обычно загнутыми кра-
ями. Пиреноид отсутствует. Ядро прилегает к хлоропласту в центре клет-
ки. Конъюгация лестничная. Конъюгирующие клетки прямые, длиною
значительно больше вегетативных. Конъюгационный канал отчетливый.
Формирование зигоспоры в конъюгационном канале с частичным, реже
полным внедрением в один или чаще оба гаметангия. Зигоспоры широ-
коэллипсоидные и удлиненно-цилиндрические с закругленными верши-
нами, вытянутые вдоль конъюгационного канала. Экзоспорий тонкий,
бесцветный, гладкий. Мезоспорий толстый, светло-серо-коричневый,
светло-желто-коричневый или коричневый, крупноямчатый.
Род включает только 1 вид, встреченный в Европе и Сев. Америке
(США).
4
Рис. 73. Mougeotiopsis calospora: 1 — вегетативные клетки, 2 — вегетативная клетка в
поперечном разрезе, 3, 4 — зигоспоры анфас, лестничная конъюгация. (По: Skuja, 1929).
159
Только по одному признаку — отсутствию пиреноида этот род от-
личается от рода Mougeotia.
Mougeotiopsis calospora Palla [-Mesogerron fluitans Brand; Mou-
geotia calospora (Palla) Czurda] — Мужоциопсис красивоспоровый
(рис. 73). — Вегетативные клетки 10—16 (18) мкм шир. Зигоспоры
эллипсоидные с широко закругленными вершинами и удлиненно-ци-
линдрические с тупо закругленными вершинами, 16—23 мкм шир.,
(21) 25—38 мкм дл. Мезоспорий с углублениями 1.5—2.5 мкм в диам.
В стоячих, преимущественно заболоченных водах: канавах, лужах,
осоковых и моховых болотах, на влажной земле. Эстония (о-в Сааре-
маа), Латвия, Вост. Украина, Сев. Казахстан (Кокчетавская обл., за-
поведник Боровое).
Редкий вид. Вполне возможно, что наличие или отсутствие пире-
ноида — признак непостоянный и зависит от условий обитания водо-
росли. При наземном местообитании в клетках накапливаются запасные
вещества — крахмал и масло в виде небольших вкраплений.
РЕДКИЕ, НЕДОСТАТОЧНО ИЗУЧЕННЫЕ И СОМНИТЕЛЬНЫЕ РОДЫ
СЕМ. MOUGEOTIACEAE
1. Род Temnogametum W. et G. S. West (1897) в вегетативном
состоянии неотличим от Mougeotia. В начальных стадиях конъюгации
деление прогаметангиальных клеток протекает как у рода Temnogyra.
Конъюгационный канал невыраженный или короткий и очень широ-
кий; формирование зигоспоры в обоих гаметангиях, вплоть до их пол-
ного заполнения. Зигоспоры разнообразной формы, окруженные сло-
истым пектиновым коллоидным веществом. Мезоспорий гладкий или
разнообразно скульптурированный, с уникальной для зигнемовых во-
дорослей сигмовидной или в виде фигурной скобки линией разрыва.
Кроме Чурды (Czurda, 1932), относившего Temnogametum в синонимы
Mougeotia, большинство авторов считают его самостоятельным родом.
Насчитывает 15 видов, распространенных в тропиках и субтропиках
Индии, Китая, Пакистана, Южн. Америки и Африки. В Европе не
обнаружен.
2. Род Mougeotiella (non Debarya sect. Mougeotiella Skuja, 1929)
выделен Ямагиси (Yamagishi, 1963a) из рода Mougeotia. Представители
этого рода отличаются от представителей Mougeotia только в репро-
дуктивной стадии четко оформленным остатком цитоплазмы в виде
мелкоточечной, тонкой, сморщенной оболочки (мантии), обволакива-
ющей зигоспору и простирающейся в гаметангии. Несмотря на теоре-
тическую обоснованность самостоятельного существования рода Mou-
geotiella, мною он пока признан синонимом Mougeotia, так как вопрос
о постоянном оформлении мантии у некоторых видов до сих пор
является довольно спорным и требует дальнейших проверок. В роде
9 видов, типовым является М. sphaerocarpa (Wolle) Yamagishi.
3. Род Sirocladium Randh. (Randhawa, 1941) c 5 видами: 5. vanda-
lurense Randh., S. maharashtrense Randh., S. kumaoense Randh., S. cu-
160
bense Rieth, S. cuttackense Sanna, зафиксированными на влажных поч-
вах Индии и Кубы. Хлоропластов всегда 2, пристенных, широкоплас-
тинчатых, с многочисленными разбросанными пиреноидами в каждом
из них; ядро 1, в центре клетки между хлоропластами; конъюгацион-
ный канал невыраженный; конъюгация протекает как у рода Sirogoni-
um. В Европе не обнаружен.
4. Род Lloydina (=Lloydiella) Ahmad et Goldstein (1972) с единст-
венным видом L. kankensis, изучен в культуральных условиях на ма-
териале из почвенного образца, собранного на рисовом поле в Индии.
Описан только по вегетативным признакам. Принадлежность к пор.
Zygnematales мною подвергается сомнению. В Европе не обнаружен.
Семейство SPIROGYRACEAE Palla — СПИРОГИРОВЫЕ
Нити неразветвленные, свободноплавающие, реже прикрепленные
ризоидами к субстрату. Поперечные стенки клеток гладкие, складчатые,
у немногих видов с гладкой септой — наружной кольцевой накладкой.
Продольные стенки клеток разной толщины, в период интенсивного
роста тонкие, состоят из внутренних целлюлозных и наружного пек-
тинового слоев или только из целлюлозных слоев, значительно ослиз-
ненные или неослизненные. Хлоропласты пристенные, от 1 до 16 в
каждой клетке, лентовидные, преимущественно спирально извитые или
почти параллельные продольной оси клетки. Пиреноиды многочислен-
ные, беспорядочно расположенные. Ядро крупное, с ясно заметным
ядрышком, шаровидное, эллипсоидное или линзовидное, окруженное
слоем цитоплазмы и подвешенное в центре клетки на плазматических
тяжах, соединяющихся с постенным слоем протоплазмы. Конъюгация
лестничная и боковая, встречающиеся вместе или порознь, в разных
нитях или в одной и той же конъюгирующей нити. Конъюгирующие
нити обычно спирально извитые, иногда концы их кольцеобразно за-
кручены; конъюгирующие клетки прямые или в разной степени изо-
гнуты. Размножение преимущественно зигоспорами, гораздо реже ап-
ланоспорами и акинетами, иногда зигоспорами и апланоспорами од-
новременно, очень редко — с преимущественным или исключительным
развитием апланоспор. Формирование зигоспоры в полости одного из
гаметангиев. Конъюгационный канал отчетливый, реже неотчетли-
вый — очень короткий и широкий, окруженный слоем плотной слизи
наподобие муфты, или отсутствует. В некоторых случаях формирование
конъюгирующей (гаметангиальной) клетки происходит в процессе де-
ления вегетативной (прогаметангиальной) клетки на две неравные клет-
ки — длинную, собственно гаметангиальную, и короткую, стерильную.
Иногда наблюдающееся двойное деление прогаметангиальной клетки в
мужской нити приводит к образованию трех клеток — одной гаме-
тангиальной и двух стерильных. В этом случае гамета образуется из
всего содержимого гаметангиальной и большей части стерильной (сте-
рильных) клеток. Гаметангии при формировании зигоспоры не запол-
нены коллоидным веществом. Зигоспоры не окружены остатком ци-
топлазмы в виде мантии.
11 Рундина Л. А.
161
I. Конъюгационный канал отчетливый. Хлоропласты обычно спирально изви-
тые.
1. Дифференциация прогаметангиальных клеток имеется .............
................................................род Temnogyra*
2. Дифференциация прогаметангиальных клеток отсутствует..........
.............................................род Spirogyra (с. 162)
II. Конъюгационный канал неотчетливый и^и отсутствует. Хлоропласты слабо
извитые или почти параллельные продольной оси клетки ..............
...........................................род Sirogonium (с. 323)
Краткие сведения о роде Temnogyra, помеченном звездочкой, по-
мещены в конце обзора всего семейства.
Род SPIROGYRA Link — СПИРОГИРА
=Conferva Miill., pr. p.; Mougeotia Ag., pr. p.; Zygnema Ag., pr. p.;
Rhynchonema Kiitz.; Degagnya Conard; Temnogyra Lewis
Нити неразветвленные, свободноплавающие или прикрепленные ри-
зоидами к субстрату. Вегетативные клетки до 200 мкм шир., по длине
меньше ширины, равные ей или до 25 раз больше. Поперечные стенки
клеток гладкие или складчатые, у немногих видов с дополнительной на-
ружной кольцевой накладкой; у видов с типично складчатыми попереч-
ными перегородками в фертильной стадии складчатые и гладкие пере-
городки чередуются. Продольные стенки клеток состоят из внутренних
целлюлозных и наружного пектинового слоев, значительно ослизненные.
Хлоропласты преимущественно спирально извитые или очень редко
почти параллельные продольной оси клетки, от 1 до 16 в клетке. Конъю-
гация лестничная и боковая, иногда совмещенные в одной паре нитей.
Конъюгирующие клетки прямые или в разной степени изогнутые. Раз-
множение преимущественно зигоспорами, гораздо реже апланоспорами
и акинетами. Конъюгационный канал отчетливый, при лестничной конъ-
югации обычно образован обеими клетками, реже одной, преимущест-
венно или исключительно отдающей клеткой. Воспринимающие клетки
невздутые или вздутые со всех сторон или односторонне. Отдающие
клетки, как правило, невздутые. Клетки без конъюгационной пары не-
вздутые или иногда шаровидно или бочонковидно вздутые. Зигоспоры
эллипсоидные (двухосно- и трехосно-эллипсоидные), эллипсоидно-ци-
линдрические, шаровидные, линзовидные. Экзоспорий обычно тонкий,
бесцветный, гладкий, очень редко скульптурированный, однослойный,
редко двухслойный. Мезоспорий преимущественно толстый, от желтого
до коричневого со всеми оттенками, очень редко синий, гладкий или
разнообразно скульптурированный, однослойный и двухслойный, очень
редко трехслойный. Эндоспорий тонкий, бесцветный, гладкий, преиму-
щественно однослойный, не всегда четко различимый. Партеноспоры,
обычные для многих видов, по форме, окраске и строению сходны с зи-
госпорами. Апланоспоры эллипсоидные, шаровидные, веретеновидные и
цилиндрические с закругленными концами; встречены только у 5 видов.
Акинеты цилиндрические, заполняющие всю клетку, с утолщенной,
часто слоистой оболочкой, редкие.
162
Характерный, широко распространенный и обширный род среди
зигнемовых водорослей. На всех континентах найдено свыше 350 пред-
ставителей этого рода. В данной работе рассматриваются 135 видов,
потенциальных и найденных на территории России. Представители
рода встречены в стоячих, заболоченных и проточных, преимущест-
венно чистых водах: озерах, прудах, водохранилищах, реках, болотах,
ямах, лужах, рисовых полях, заливаемых участках речных долин, ручь-
ях и родниках, на скальных выбоинах, среди водных растений. За-
фиксированы местонахождения на высоте свыше 5000 м над ур. м., в
соленой (Spirogyra salina, S. subsalsa, S. subsalso-punctulata) и в сла-
босоленой воде (S. decimina f. longata, S. decimina f. olivascens), в
минеральных источниках (S. velata f. velata, S. cateniformis, S. decimi-
na f. longata). Спирогира развивается в массе, иногда вызывая цветение
воды, и конъюгирует в теплое время года, предпочитая хорошо про-
греваемые, мелководные участки водоемов. В древнейших слоях най-
дены микрофоссилии, аналогичные зигоспорам Spirogyra, начиная с
раннего мела (мезозой).
По строению поперечных перегородок род Spirogyra условно раз-
делен на 3 секции, не имеющие таксономического значения. Они ис-
пользуются мною лишь для облегчения в пользовании ключом при
определении многочисленных видов.
I. Поперечные стенки клеток гладкие...........секция Conjugate (с. 163)
II. Поперечные стенки клеток складчатые ........секция Salmacis (с. 176)
III. Поперечные стенки клеток гладкие с дополнительной кольцевой накладкой
.............................................секция Colligata(c. 179)
Секция I. Conjugata
Поперечные стенки клеток гладкие.
I. Размножение преимущественно апланоспорами, редко зигоспорами ....
................................................1. S. mirabilis (частично)
II. Размножение преимущественно зигоспорами, редко апланоспорами.
1. Зигоспоры шаровидные или эллипсоидные.
А. Зигоспоры шаровидные.
а. Вегетативные клетки 43—45 мкм шир. . ... 2. S. sphaerospora
б. Вегетативные клетки 65—70 мкм шир.............3. S. wittrockii
Б. Зигоспоры правильно-эллипсоидные (двухосно-эллипсоидные), в попе-
речном сечении (вид сверху) круглые.
а. Экзоспорий и мезоспорий гладкие.
а. Хлоропласт постоянно 1.
* Клетки без конъюгационной пары обычно невздутые.
+ Воспринимающие клетки невздутые.
х Конъюгация лестничная.
О Зигоспоры эллипсойдногцилиндрические.
Д Вегетативные клетки (26) 29—35
(38) мкм шир.....................
. . . 4. S. decimina f. circumscissa
ДД Вегетативные клетки 40—44 мкм
шир..............................
. 5. S. porticalis f. cylindrosperma
163
ОО Зигоспоры эллипсоидные.
Д Вегетативные клетки (17) 20—38 (42)
мкм шир...........................
.... 4. S. decimina (f. decimina,
f. communis, f. juergensii) (частично)
ДД Вегетативные клетки 53—56 мкм шир.
. . . 6. S. hungarica (частично)
АЛЛ Вегетативные клетки 65—76 мкм
шир........................7. S. gallica
хх Конъюгация лестничная и боковая.
О Конъюгация преимущественно боковая, ве-
гетативные клетки 20—55 (62) мкм шир.
Д Вегетативные клетки 20—24 мкм
шир...............................
4. S. decimina f. elongata (частично)
ДД Вегетативные клетки (24) 26—38 (42)
мкм шир.............................
4. S. decimina f. longata (частично)
АЛЛ Вегетативные клетки 44—55 (62) мкм
шир. . 8. S. condensata (частично)
ОО Конъюгация преимущественно лестничная,
вегетативные клетки (34) 38—50 (55) мкм
шир........................................
5. S. porticalis f. porticalis (частично)
++ Воспринимающие клетки вздутые.
х Воспринимающие клетки вздутые со стороны,
противоположной конъюгационному каналу . .
..................................9. S. borgeana
хх Воспринимающие клетки вздутые со стороны
конъюгационного канала.
О Конъюгация лестничная.
Д Конъюгационный канал образован
равномерно обеими клетками . . .
4. S. decimina f. juergensii (частично)
ДД Конъюгационный канал образован
преимущественно отдающей клеткой.
§ Вегетативные клетки 16—26
мкм шир......................
. . . 10. S. gracilis f. gracilis
§§ Вегетативные клетки 53—56
мкм шир......................
. . 6. S. hungarica (частично)
ОО Конъюгация лестничная и боковая.
Д Конъюгация преимущественно бо-
ковая.
§ Вегетативные клетки 10—16
мкм шир............ ll.S. fla-
vescens f. flavescens (частично)
§§ Вегетативные клетки 17—22 мкм
шир............................
. . . 12. S. chenii (частично)
§§§ Вегетативные клетки 19—21
(24) мкм шир. . . 10. S. gra-
cilis f. gibberosa (частично)
164
ДД Конъюгация преимущественно лест-
ничная.
§ Конъюгационный канал образо-
ван равномерно обеими клет-
ками.
! Вегетативные клетки 10—
16 мкм шир...............
...........11. S. flaves-
cens f. flavescens (частично)
!! Вегетативные клетки 25—
33 мкм шир. . 13. S. sub-
salsa f. gracilis (частично)
!!! Вегетативные клетки (34)
38—50 (55) мкм шир. .
...........5. S. portica-
lis f. porticalis (частично)
§§ Конъюгационный канал образо-
ван преимущественно или ис-
ключительно отдающей клеткой.
! Вегетативные клетки 10—
16 мкм шир................
.... ll.S. flavescens
f. flavescens (частично)
!! Вегетативные клетки 19—
21 (24) мкм шир............
..........10. S. gracilis
f. gibberosa (частично)
!!! Вегетативные клетки 25—
33 мкм шир. . 13. S. sub-
salsa f. gracilis (частично)
ххх Воспринимающие клетки вздутые со всех сторон.
О Конъюгация лестничная.
Д Конъюгационный канал образован
равномерно обеими клетками.
§ Воспринимающие клетки сла-
бо вздутые...................
. . . . 4. S. decimina (f. deci-
mina, f. communis) (частично)
§§ Воспринимающие клетки силь-
но вздутые.
! Размножение преимущест-
венно апланоспорами . .
. 1. S. mirabilis (частично)
!! Размножение преимущест-
венно зигоспорами.
— Зигоспоры разнообраз-
ной формы . 14. S. ро-
lymorpha (частично)
= Зигоспоры только эл-
липсоидные.
/ Хлоропласт постоян-
но 1, вегетативные
клетки 25—32.5 мкм
шир............13. S.
subsalsa f. uralensis
165
И Хлоропласт 1, редко
их 2, вегетативные
клетки 22—35 (38)
мкм шир.
# Вегетативные кле-
тки 22—30 мкм
\ шир............
14. S. polymor-
pha (частично)
## Вегетативные клет-
ки 30—35 (38) мкм
шир................
... 4. S. deci-
mina f. olivascens
ДД Конъюгационный канал образован
преимущественно отдающей клеткой.
§ Вегетативные клетки 8—10 мкм
шир..........................
. . .11. S. flavescens f. parva
§§ Вегетативные клетки 22—28
мкм шир. . . . 13. S. sub-
salsa f. subsalsa (частично)
ОО Конъюгация боковая и лестничная.
Д Конъюгация преимущественно бо-
ковая.
§ Конъюгационный канал обра-
зован равномерно обеими клет-
ками при лестничной конъю-
гации.
! Вегетативные клетки 10—
30 мкм шир.
- Вегетативные клетки
10—16 мкм шир. . . .
11. S. flavescens
f. flavescens (частично)
= Вегетативные клетки
20—24 (25) мкм шир.
.... 15. S. parvula
= Вегетативные клетки
25—30 мкм шир. . . .
. . 16. S. cateniformis
!! Вегетативные клетки 44—
52 (62) мкм шир...........
8. S. condensate (частично)
§§ Конъюгационный канал обра-
зован отдающей клеткой при
лестничной конъюгации.
! Вегетативные клетки 10—
16 мкм шир................
... 11. S. flavescens
f. flavescens (частично)
!! Вегетативные клетки 17—
22 мкм шир................
. 12. S. chenii (частично)
166
ДД Конъюгация преимущественно лест-
ничная.
§ Вегетативные клетки 10—
16 мкм шир.......11. S. fla-
vescens f. flavescens (частично)
§§ Вегетативные клетки 25—
33 мкм шир.................13..
S. subsalsa f. gracilis (частично)
** Клетки без конъюгационной пары вздутые.
+ Конъюгация боковая.
х Вегетативные клетки 40—45 мкм шир..........
.......................17. S. pseudosahnii
хх Вегетативные клетки 46—72 мкм шир..........
........................................ 18. S. sahnii
++ Конъюгация лестничная.
х Воспринимающие клетки невздутые или очень
слабо вздутые
О Вегетативные клетки 14.5—17.4 мкм шир.
................19. S. bullata f. minor
ОО Вегетативные клетки 19—23 мкм шир.
........ 19. S. bullata f. bullata
ООО Вегетативные клетки 26—27.5 мкм шир.
........19. S. bullata f. major
хх Воспринимающие клетки вздутые со стороны
конъюгационного канала ... 20. S. varians (f.
varians, f. bicalyptrata, f. circumlineata) (частично)
xxx Воспринимающие клетки вздутые со всех сторон.
О Вегетативные клетки 22—30 мкм шир.
........14. S. polymorpha (частично)
ОО Вегетативные клетки 35—39 мкм шир.
...................21. S. henanensis
+++ Конъюгация боковая и лестничная.
х Воспринимающие клетки невздутые............
4. S. decimina (f. longata, f. elongata) (частично)
хх Воспринимающие клетки вздутые со стороны
конъюгационного канала.........................
20. S. varians (f. minor, f. woodsii) (частично)
xxx Воспринимающие клетки вздутые со всех сто-
рон ..............................22. S. pratensis
Р. Хлоропластов (1) 2—8 (10).
* Хлоропласт 1, изредка их 2—3.
+ Вегетативные клетки 22—30 мкм шир.
х Конъюгационный канал образован преимущест-
венно или исключительно отдающей клеткой . .
............... 13. S. subsalsa f. subsalsa (частично)
хх Конъюгационный канал образован равномерно
обеими клетками............................
............... 14. S. polymorpha (частично)
++ Вегетативные клетки (35) 38—50 (52) мкм шир. . . .
..............5. S. porticalis f. porticalis (частично)
** Хлоропластов (1) 2—10.
+ Воспринимающие клетки обычно невздутые или очень
слабо вздутые;
х Конъюгация лестничная.
167
О Хлоропластов (1) 2—4.
Д Вегетативные клетки (24) 30—42 мкм
шир.
§ Вегетативные клетки 24—30
мкм шир.......................
... 23. S. rivularis f. minor
§§ Вегетативные клетки (31) 36—
42 мкм шир....................
. . 23. S. rivularis f. rivularis
ДД Вегетативные клетки (42) 45—54 мкм
шир..................................
... 24. S. neglecta f. columbiana
АЛЛ Вегетативные клетки 82—85 мкм
шир...................................
25. S. nitida f. paradoxa (частично)
ОО Хлоропластов (2) 3—8 (10).
Д Вегетативные клетки 50—115 мкм
шир.
§ Вегетативные клетки 50—90 мкм
шир., хлоропластов (2) 3—5 (7).
! Вегетативные клетки 50—
68 (75) мкм шир............
. 24. S. neglecta f. neglecta
!! Вегетативные клетки (68)
75—90 мкм шир..............
25. S. nitida f. nitida
§§ Вегетативные клетки (85) 90—
115 мкм шир., хлоропластов (3)
4—8 (10) . . 26. S. setiformis
ДД Вегетативные клетки 115—150 (155)
мкм шир.
§ Вегетативные клетки 115—128
мкм шир., хлоропластов 4—6
.................27. S. elliptica
§§ Вегетативные клетки (120) 125—
150 (155) мкм шир., хлоропла-
стов 3—8......................
................................28. S. ellipsospora
хх Конъюгация лестничная и боковая.
О Вегетативные клетки 31—46 мкм шир.,
хлоропластов 2—4.
Д Вегетативные клетки 31—42 мкм
шир. 29. S. irregularis
ДД Вегетативные клетки 40—46 мкм
шир. . 24. S. neglecta f. fuellebornii
ОО Вегетативные клетки (45) 55—60 (67.5)
мкм шир., хлоропластов (2) 3—4 (5) .
................. 30. S. hyalina (частично)
++ Воспринимающие клетки значительно вздутые.
х Воспринимающие клетки вздутые со стороны
конъюгационного канала.........................
............. 25. S. nitida f. paradoxa (частично)
хх Воспринимающие клетки обычно вздутые со
всех сторон.
168
О Вегетативные клетки (20) 22—33 мкм
шир., хлоропластов (1) 2—3.
Д Вегетативные клетки (20) 22—26 (30)
мкм шир..........................
.....................................31. S. elongata f. elongata
ДД Вегетативные клетки 26—33 мкм
шир. . . 31. S. elongata f. rupestris
ОО Вегетативные клетки 40—52 (56) мкм шир.,
хлоропластов 2—4 (6) . . . . 32. S. dubia
ООО Вегетативные клетки (45) 55—60 (67.5)
мкм шир., хлоропластов (2) 3—4 (5) . .
.................................... 30. S. hyalina (частично)
у. Хлоропластов 12—15.
* Вегетативные клетки 100—120 мкм шир. . . 33. S. margaritata
* * Вегетативные клетки 150—189 мкм шир................
.................................... 34. S. polytaeniata
б. Экзоспорий или мезоспорий скульптурированные.
а. Экзоспорий скульптурированный, мезоспорий гладкий.
* Конъюгация лестничная..............35. S. pseudovarians
* * Конъюгация боковая ............36. S. velata f. lateralis
* ** Конъюгация лестничная и боковая...................
........................... 36. S. velata (f. velata, f. ellipsoidea)
p. Экзоспорий гладкий, мезоспорий скульптурированный.
* Хлоропластов 1—4.
+ Хлоропластов не более 2.
х Хлоропласт постоянно 1.
О Мезоспорий однослойный.
Д Скульптурные элементы мезоспория в
виде бородавок, бугорков, игл, точек.
§ Мезоспорий бородавчатый
или бугорчатый.
! Вегетативные клетки 35—
37 мкм шир..............
.... 37. S. tuberculata
!! Вегетативные клетки 42.5—
45 мкм шир................
. . .38. S. pseudowoodsii
§§ Мезоспорий мелкоточечный или
тонкоигольчатый (точечный
сверху).
I Клетки без конъюгацион-
ной пары невздутые.
- Конъюгация лестнич-
ная.
/ Вегетативные клет-
ки 20—25 (29) мкм
шир. ... 39. S.
subsalso-punctulata
И Вегетативные клетки
27—30 (39) мкм шир.
.... 40. S. pun-
ctiformis (частично)
= Конъюгация лестнич-
ная и боковая.
169
I Конъюгация преиму-
щественно лестничная,
вегетативные клетки
14.5—40 мкм шир.
# Вегетативные кле-
тки 14.5 (17.4) мкм
шир................
. 41. S. tenuispina
## Вегетативные кле-
тки 30—40 мкм
шир.................
. . . 42. S. salina
И Конъюгация преиму-
щественно боковая, ве-
гетативные клетки 24—
33 мкм шир.............
. . 43. S. lagerheimii f.
lagerheimii (частично)
!! Клетки без конъюгационной
пары вздутые.
- Воспринимающие клетки
невздутые.
/ Конъюгация только лест-
ничная ................
... 43. S. lagerheimii
f. obovata (частично)
// Конъюгация боковая и
лестничная ............
. . 43. S. lagerheimii f.
lagerheimii (частично)
= Воспринимающие клетки
вздутые со стороны конъ-
югационного канала . . .
. . . 44. S. subpapulata
ДД Скульптурные элементы мезоспория в ви-
де ямок, складок (утолщений).
§ Мезоспорий ямчатый.
! Конъюгация боковая, очень
редко лестничная ............
.................45. S. danica
!! Конъюгация лестничная.
- Клетки без конъюгацион-
ной пары невздутые.
/ Воспринимающие клет-
ки невздутые или не-
значительно вздутые.
# Вегетативные кле-
тки 22—24 мкм
шир...............
. 46. S. luteospora
## Вегетативные кле-
тки (38) 41 (45) мкм
шир.................
47. S. acumbentis
170
### Вегетативные кле-
тки (45) 50—57
(62) мкм шир. . .
48. S. rugulosa
И Воспринимающие клет-
ки сильно вздутые . .
49. S. punctata f. crassior
= Клетки без конъюгацион-
ной пары вздутые.
/ Вегетативные клетки
17 мкм шир., мезоспо-
рий мелкоямчатый . .
........50. S. sibirica
И Вегетативные клетки
30—40 мкм шир., ме-
зоспорий грубоямчатый
. . . 51. S. scrobiculata
§§ Мезоспорий складчатый или сетча-
то-складчатый.
! Конъюгация только лестнич-
ная.
- Клетки без конъюгацион-
ной пары невздутые.
/ Складки (утолщения)
на мезоспорий тонкие
и короткие ............
.... 52. S. esthonica
И Складки (утолщения)
на мезоспорий более
грубые и удлиненные.
# Вегетативные клетки
(27) 30—41 мкм шир.
® Сеть складок
рыхлая . . .
53. S. daeda-
lea f. daedalea
@© Сеть складок
плотная . . .
53. S. daeda-
lea f. africana
## Вегетативные клет-
ки 50—63 мкм шир.
. 54. S. labyrinthica
= Клетки без конъюгацион-
ной пары более или менее
шаровидно вздутые . . .
...............55. S. nodifera
!! Конъюгация лестничная и бо-
ковая.
- Клетки без конъюгацион-
ной пары невздутые.
/ Конъюгация преиму-
щественно лестничная,
вегетативные клетки
29—44 мкм шир. . . .
171
...................... 53. S.
daedalea f. daedaleoides
// Конъюгация преимуще-
ственно боковая, вегета-
тивные клетки 34—38
мкм шир. ... 53. S.
\ daedalea f. oudhensis
— Клетки без конъюгационной
пары вздутые ..........
. 56. S. papulata (частично)
ОО Мезоспорий двухслойный.
Д Скульптурированный слой мезоспория
сетчатый и угловато-ямчатый, вегета-
тивные клетки 20—35 (38) мкм шир.
§ Воспринимающие клетки невзду-
тые, вегетативные клетки 20—35
мкм шир..........................
...........57. S. pseudodaedaleoides
§§ Воспринимающие клетки вздутые
со стороны конъюгационного кана-
ла, вегетативные клетки 28—32
(38) мкм шир.......................
.................56. S. papulata (частично)
ДД Скульптурированный слой мезоспория
мелкоточечный, вегетативные клетки
32—40 мкм шир...........................
.... 58. S. aphanosculpta (частично)
хх Хлоропласт 1, изредка их 2.
О Мезоспорий однослойный.
Д Мезоспорий мелкоточечный (пунктиро-
ванный) ...............................
. 43. S. lagerheimii f. obovata (частично)
ДД Мезоспорий ямчатый.
§ Конъюгация лестничная..........
.... 49. S. punctata f. punctata
§§ Конъюгация преимущественно бо-
ковая, реже лестничная.
! Вегетативные клетки 18—22
мкм шир. Мезоспорий крупно-
ямчатый .....................
... 59. S. collinsii f. collinsii
!! Вегетативные клетки 25—29
мкм шир. Мезоспорий мелко-
ямчатый .....................
. . . . 59. S. collinsii f. ampla
AAA Мезоспорий сетчато-морщинистый с ши-
повидными выростами в узлах сети (гру-
бозернистый сверху)......................
60. S. pseudonodifera
ОО Мезоспорий двухслойный.......
....................................61. S. franconica
xxx Хлоропластов постоянно 1—2 или только 2.
О Мезоспорий мелкоточечный....................
...............40. S. punctiformis (частично)
172
ОО Мезоспорий сетчатый . 62. S. quezelii
++ Хлоропластов постоянно 1—3.
х Мезоспорий мелкозернистый . . 63. S. asiatica
хх Мезоспорий ямчатый.........................
............64. S. occidentalis f. occidentalis
+++ Хлоропластов 4..................................
......................64. S. occidentalis f. striata (частично)
** Хлоропластов 2—8.
+ Мезоспорий однослойный.
х Мезоспорий мелкозернистый..................
.................. 65. S. endogranulata (частично)
хх Мезоспорий ямчатый или точечно-ямчатый, мелко-
точечный, очень редко продольно исчерченный.
О Вегетативные клетки (36) 40—54 мкм
шир. ...................................
64. S. occidentalis f. striata (частично)
ОО Вегетативные клетки (65) 70—95 мкм шир.
Д Хлоропластов 4.
§ Конъюгация лестничная . .
66. S. punctulata f. macrospora
§§ Конъюгация лестничная и бо-
ковая ..................66. S.
punctulata f. cylindrospora
ДД Хлоропластов (3) 4—7............
... 66. S. punctulata f. punctulata
ООО Вегетативные клетки 110—123 мкм
шир. . . 67. S. trachycarpa (частично)
ххх Мезоспорий морщинистый или морщинисто-сет-
чатый, часто с бородавчатыми или шиповидными
выростами.
О Мезоспорий морщинистый.
Д Воспринимающие клетки невзду-
тые ....................... 68. S.
S. paraquayensis f. paraquayensis
ДД Воспринимающие клетки сильно
вздутые со всех сторон.
§ Вегетативные клетки 26—35
мкм шир..............69. S. flu-
viatilis f. fluviatilis (частично)
§§ Вегетативные клетки 30—50
мкм шир.......................
. . 69. S. fluviatilis f. ramosa
ОО Мезоспорий морщинисто-сетчатый.
Д Мезоспорий с бородавчатыми выро-
стами.
§ Вегетативные клетки (34) 40—52
(59) мкм шир. ... 68. S. pa-
raquayensis f. rhizobrachialis
§§ Вегетативные клетки 100—126
мкм шир. . . 70. S. verrucosa
ДД Мезоспорий с шиповидными выро-
стами.
§ Вегетативные клетки (32)
35—45 (52) мкм шир., воспри-
173
нимающие клетки сильно
вздутые со всех сторон . . .
. . . 69. S. fluviatilis f. grossii
§§ Вегетативные клетки 40.5—55
(58) мкм шир., воспринимаю-
щие клетки слабо вздутые, пре-
\ имущественно со стороны конъ-
югационного канала ....
..................... 71. S.
brunnea f. pulchrifigurata
АЛЛ Мезоспорий без шиповидных выро-
стов.
§ Вегетативные клетки 26—50
(52) мкм шир..................
. . 69. S. fluviatilis (частично)
§§ Вегетативные клетки более 50
мкм шир.
! Вегетативные клетки 56—71
(84) мкм шир..........
71. S. brunnea f. brunnea
!! Вегетативные клетки (80)
85—117 мкм шир. . . .
........72. S. reinhardii
!!! Вегетативные клетки 110—
123 мкм шир............67.
S. trachycarpa (частично)
++ Мезоспорий двухслойный.
х Внешний слой мезоспория гладкий, внутренний
мелкозернистый ..............................
.................. 65. S. endogranulata (частично)
хх Внешний слой мезоспория продольно- и корот-
коморщинистый, внутренний слой разнообразно
скульптурированный.
О Внутренний слой мезоспория густоборо-
давчатый ............73. S. mienningensis
ОО Внутренний слой мезоспория морщинис-
тый, морщинисто-сетчатый или точечно-
сетчатый.
Д Внутренний слой мезоспория ко-
роткоморщинистый или морщинис-
то-сетчатый ... 74. S. corrugata
ДД Внутренний слой мезоспория мел-
коточечный, иногда тонко- и непра-
вильно-сетчатый ...................
...........75. S. quadrilaminata
ххх Внешний слой мезоспория точечный, внутрен-
ний морщинисто-сетчатый, иногда с шиловидны-
ми выростами.
О Хлоропластов 2—3 (4), вегетативные
клетки 25—32 (35) мкм шир.
Д Воспринимающие клетки вздутые .
. . 76. S. dictyospora f. dictyospora
ДД Воспринимающие клетки невздутые
.............................76. S.
dictyospora f. subcylindrospora
174
ОО Хлоропластов (2) 3—6, вегетативные кле-
тки 26—35 мкм шир............................
69. S. fluviatilis f. fluviatilis (частично)
у. Экзоспорий и мезоспорий скульптурированные.
* Мезоспорий однослойный, волнистый ... 77. S. cavata
* * Мезоспорий двухслойный, внешний слой мелкоточечный или
мелкоямчатый .................58. S. aphanosculpta (частично)
В. Зигоспоры сжато-эллипсоидные (трехосно-эллипсоидные), в поперечном
сечении (вид сверху) эллиптические.
а. Мезоспорий гладкий.......................... 78. S. crassoidea
б. Мезоспорий скульптурированный.
а. Мезоспорий точечно-бородавчатый.............79. S. subcrassa
Р. Мезоспорий ямчатый............................ 80. S. crassa
у. Мезоспорий бугорчатый.............81. S. krubergii (частично)
2. Зигоспоры линзовидные (сжато-шаровидные, чечевицеобразные), в двух по-
ложениях (анфас и сбоку) круглые и эллиптические.
А. Мезоспорий гладкий.
а. Хлоропластов 2—4.
а. Вегетативные клетки 22—26 (29) мкм шир. ... 82. S. sinensis
р. Вегетативные клетки 30—50 мкм шир.
* Вегетативные клетки 30—40 мкм шир....................
................................83. S. pellucida f. minor
* * Вегетативные клетки 45—50 мкм шир...................
..............................83. S. pellucida f. pellucida
б. Хлоропластов (3) 5—9(10).
а. Вегетативные клетки 50—110 мкм шир.
* Вегетативные клетки 50—80 (100) мкм шир., воспринима-
ющие клетки вздутые со стороны, противоположной конъ-
югационному каналу.............84. S. majuscula (частично)
* * Вегетативные клетки 70—79 (90) мкм шир., воспринимаю-
щие клетки невздутые .... 85. S. submaxima f. lamellata
* ** Вегетативные клетки (70) 96—ПО мкм шир., воспринима-
ющие клетки невздутые или незначительно вздутые со сто-
роны конъюгационного канала ..............................
.......................85. S. submaxima f. submaxima
Р. Вегетативные клетки 145—153 мкм шир............86. S. glabra
Б. Мезоспорий скульптурированный.
а. Скульптура мезоспория однотипная (состоящая из одинаковых эле-
ментов).
а. Мезоспорий радиально исчерченный . 84. S. majuscula (частично)
р. Мезоспорий ямчатый.
* Хлоропластов 5—6 (8), вегетативные клетки (57) 65—80
(90) мкм шир..............................87. S. bellis
* * Хлоропластов 8—9, вегетативные клетки 116—132 мкм
шир........................................88. S. jassiensis
у. Мезоспорий бугорчатый или сетчатый.
* Мезоспорий бугорчатый.
+ Хлоропластов 4—7 . . 81. S. krubergii (частично)
++ Хлоропластов 7—10 ... 89. S. heeriana f. heeriana
* * Мезоспорий сетчатый.
. + Вегетативные клетки 46.4—57.6 мкм шир............
..................................90. S. czurdiana
++ Вегетативные клетки более 60 мкм шир.
х Вегетативные клетки 77—117 (130) мкм шир. .
....................91. S. maxima f. tenuior
175
хх Вегетативные клетки (115) 125—155 (170) мкм
шир................... 91, S. maxima f. maxima
ххх Вегетативные клетки (150) 170—200 мкм шир.
..................91. S. maxima f. livida
б. Скульптура мезоспория многотипная (состоящая из разных элементов),
а. Скульптура, мезоспория в основе сетчатая.
* Мезоспорий плотносетчатый и короткоребристый (морщи-
нистый) ..............'...........92. S. pseudomaxima
** Мезоспорий сетчато-бородавчатый.........93. S. oblata
р. Скульптура мезоспория другого типа.
* Мезоспорий бугорчатый и мелкозернистый...............
..............................89. S. heeriana f. ucrainica
* * Мезоспорий мелкоточечный и с короткими извилистыми
складками (морщинами), не сомкнутыми в сеть...............
..............................91. S. maxima f. cupientis
* ** Мезоспорий короткоскладчатый (морщинистый) и бородав-
чатый.
+ Хлоропластов 5—8 . . 91. S. maxima f. woronichinii
++ Хлоропластов 9—10..................94. S. lenticularis
3. Зигоспоры смешанные по форме (нескольких типов в пределах одного вида).
А. Вегетативные клетки (51) 58—78 (87) мкм шир..............95. S. ternata
Б. Вегетативные клетки 100—115 мкм шир. . . . 96. S. diversizygotica
Секция II. Salmacis
Поперечные стенки клеток складчатые.
I. Оболочка зигоспор гладкая.
1. Хлоропластов 1—2.
А. Хлоропласт постоянно 1.
а. Клетки без конъюгационной пары невздутые.........97. S. arta
б. Клетки без конъюгационной пары вздутые . . 98. S. tumida (частично)
Б. Хлоропласт 1, редко их 2.
а. Воспринимающие клетки невздутые или слабо вздутые со всех сторон.
а. Конъюгация лестничная или преимущественно лестничная.
* Вегетативные клетки 16—20 мкм шир...................
.............................98. S. tumida (частично)
* * Вегетативные клетки (19) 23—32 (36) мкм шир.........
.................................99. S. weberi f. weberi
* ** Вегетативные клетки 40—46.5 (50) мкм шир...........
..............................99. S. weberi f. tjibodensis
р. Конъюгация боковая или преимущественно боковая ...........
...........................................100. S. aequalis
б. Воспринимающие клетки вздутые со стороны конъюгационного канала.
а. Вегетативные клетки 15—20 мкм шир. . 101. S. pseudospreeiana
р. Вегетативные клетки 24—29 (32) мкм шир. . . 102. S. hopeiensis
в. Воспринимающие клетки сильно вздутые со всех сторон.
а. Конъюгационный канал при лестничной конъюгации образован
обеими клетками равномерно.
* Конъюгация лестничная.......99. S. weberi f. farlowii
* * Конъюгация лестничная и боковая.
+ Вегетативные клетки 8—24 мкм шир.
х Вегетативные клетки (8) 10—14.5 (16) мкм шир.
. . . 103. S. tenuissima f. tenuissima (частично)
хх Вегетативные клетки (14) 15—24 мкм шир.
176
О Воспринимающие клетки веретеновидно
вздутые ... 104. S. inflata (частично)
ОО Воспринимающие клетки цилиндрически
вздутые.................................
.... 105. S. groenlandica (частично)
++ Вегетативные клетки 24—35 мкм шир.
х Воспринимающие клетки преимущественно ве-
ретеновидно вздутые..........................
..........................99. S. weberi f. grevilleana (частично)
хх Воспринимающие клетки преимущественно ци-
линдрически вздутые .... 106. S. quadrata
р. Конъюгационный канал при лестничной конъюгации образован
обеими клетками, с преобладанием отдающей клетки.
* Вегетативные клетки 18—24 мкм шир...................
....................... 105. S. groenlandica (частично)
* * Вегетативные клетки 28—39 мкм шир. . . 107. S. chuniae
у. Конъюгационный канал при лестничной конъюгации образован
отдающей клеткой.
* Воспринимающие клетки преимущественно цилиндрически
вздутые....................103. S. tenuissima f. cylindrica
* * Воспринимающие клетки преимущественно веретеновидно
вздутые.
+ Вегетативные клетки (8) 10—14.5 (16) мкм шир. . .
............ 103. S. tenuissima f. tenuissima (частично)
++ Вегетативные клетки (14) 15—21 (23.5) мкм шир. . .
............................. 104. S. inflata (частично)
+++ Вегетативные клетки 25—30 (35) мкм шир..........
..............99. S. weberi f. grevilleana (частично)
2. Хлоропластов (1) 2—4.
А. Воспринимающие клетки цилиндрически вздутые ...................
...................................108. S. insignis f. rectangularis
Б. Воспринимающие клетки веретеновидно вздутые.
а. Конъюгация лестничная.................108. S. insignis f. wangii
б. Конъюгация лестничная и боковая.
а. Вегетативные клетки (26) 30—35 мкм шир. . . 109. S. hassallii
Р. Вегетативные клетки (30) 36—45 (48) мкм шир..............
................................ 108. S. insignis f. insignis
II. Оболочка зигоспор скульптурированная.
1. Экзоспорий скульптурированный, мезоспорий гладкий.
А. Воспринимающие клетки невздутые или слабо вздутые.
а. Концы стерильных клеток шаровидно вздутые...................
.................................... ПО. S. calospora f. venusta
б. Концы стерильных клеток невздутые...........................
............... 110. S. calospora (f. calospora, f. major, f. minor)
Б. Воспринимающие клетки сильно вздутые ...........111. S. areolata
2. Экзоспорий гладкий, мезоспорий скульптурированный.
А. Хлоропласт 1, редко их 2.
а. Конъюгация боковая.........................112. S. adjerensis
б. Конъюгация лестничная.
а. Вегетативные клетки 19—21 мкм шир.
* Мезоспорий точечный.................113. S. aisiophila
* * Мезоспорий сетчатый................114. S. czubinskii
Р. Вегетативные клетки 22—32 мкм шир.
* Мезоспорий мелкоточечный............115. S. australica
12 Рундина Л. А.
177
** Мезоспорий от мелкоточечного до сетчато-точечного . . .
................................................. 116. S. goetzii
в. Конъюгация боковая и лестничная.
а. Конъюгационный канал при лестничной конъюгации образован
обеими клетками равномерно.
* Вегетативные клетки 10—17 (20) мкм шир.
+ Мезоспорий мелкоямчатый..........117. S. rugosa
++ Мезоспорий сетчатый.............................
.................... 118. S. kuusamoSnsis (частично)
х Мезоспорий тонкосетчатый......................
.............118. S. kuusamoensis f. kuusamoensis
хх Мезоспорий грубосетчатый...................
...........118. S. kuusamoSnsis f. skujaensis
* * Вегетативные клетки 17—24 мкм шир. . 119. S. fritschiana
р. Конъюгационный канал при лестничной конъюгации образован
обеими клетками с преобладанием отдающей клетки.
* Вегетативные клетки 11—17 (20) мкм шир..............
......................118. S. kuusamoSnsis (частично)
* * Вегетативные клетки 18—26 мкм шир.
+ Мезоспорий мелкоточечный . . 120. S. gaterslebensis
++ Мезоспорий сетчато-шиповатый ...................
...........................121. S. dentireticulata
* ** Вегетативные клетки (25) 26—40 (52) мкм шир.
+ Мезоспорий мелкоточечный и морщинистый . . . .
.....................122. S. incrassata (частично)
++ Мезоспорий морщинисто-сетчатый и игольчатый . . .
.... 123. S. reticulata f. acanthophora (частично)
+++ Мезоспорий волнисто-морщинистый.................
.................124. S. herbipolensis (частично)
у. Конъюгационный канал при лестничной конъюгации образован
отдающей клеткой.
* Вегетативные клетки 11—17 (20) мкм шир..............
......................118. S. kuusamoensis (частично)
* * Вегетативные клетки (31) 34—40 (52) мкм шир........
......................124. S. herbipolensis (частично)
Б. Хлоропластов (1) 2—4.
а. Конъюгация боковая.
а. Воспринимающие клетки невздутые или слабо вздутые, вегетатив-
ные клетки 32—41 мкм шир...................... 125. S. costulata
р. Воспринимающие клетки значительно вздутые, вегетативные
клетки 24—33 мкм шир.
* Мезоспорий сетчатый................... 126. S. miranda
* * Мезоспорий ребристый................. 127. S. venosa
б. Конъюгация лестничная.
а. Мезоспорий с сосочковидными или шиповидными выростами.
* Мезоспорий с сосочковидными выростами...............
.................... 128. S. borysthenica f. borysthenica
* * Мезоспорий с шиповидными выростами ................
.............................128. S. borysthenica f. echinospora
р. Мезоспорий морщинисто-сетчатый.
* Хлоропластов (1)2 . . . 123. S. reticulata f. nawaschinii
* * Хлоропластов (2) 3.......123. S. reticulata f. czurdae
у. Мезоспорий морщинисто-сетчатый с шиповидными выростами.
* Хлоропластов (1) 2 .... 123. S. reticulata f. regularis
178
* * Хлоропластов 2—3...................129, S. spinescens
в. Конъюгация боковая и лестничная.
а. Мезоспорий однослойный.
* Мезоспорий мелкоточечный и морщинистый................
............................122. S. incrassata (частично)
* * Мезоспорий бородавчатый............130. S. marchica
* ** Мезоспорий сетчатый.
+ Мезоспорий ребристо-сетчатый . ..................
.............................131. S. costata (частично)
++ Мезоспорий морщинисто-сетчатый....................
...............123. S. reticulata f. reticulata (частично)
+++ Мезоспорий морщинисто-сетчатый с игольчатыми выро-
стами . . 123. S. reticulata f. acanthophora (частич-
но)
Р. Мезоспорий двухслойный.
* Внутренний слой мезоспория гладкий ...................
........................124. S. herbipolensis (частично)
* * Внутренний слой мезоспория скульптурированный.
+ Внутренний слой мезоспория сетчато утолщенный.
х Игольчатые выросты в узлах утолщений имеются .
.... 123. S. reticulata f. reticulata (частично)
хх Игольчатые выросты в узлах утолщений отсутст-
вуют ...................131. S. costata (частично)
++ Внутренний слой мезоспория зернистый и слабо мор-
щинистый ..........................132. S. granulata
+++ Внутренний слой мезоспория плотнобородавчатый
...........................133. S. pseudogranulata
Секция III. Colligata
Поперечные стенки клеток гладкие с дополнительной кольцевой на-
кладкой.
I. Вегетативные клетки 29—40 мкм шир......................134. S. colligata
II. Вегетативные клетки 43—52 мкм шир.....................135. S. silesiaca
1. Spirogyra mirabilis (Hass.) Kiitz., non sensu Petit [=5. communis
(Hass.) Wittr. f. mirabilis (Kiitz.) Kirchn.; S. polymorpha Kirchn., pr. p.;
5. polymorpha sensu Казановский, Смирнов, 1914; S. mirabilis f. minor
V. Poljansk.] — Спирогира удивительная (рис. 74). — Вегетативные клетки
(18) 20—29 (34) мкм шир., по длине в 2—10 раз больше. Поперечные стен-
ки гладкие. Хлоропласт 1, очень редко 2. Размножение преимущественно
апланоспорами, очень редко зигоспорами. Апланоспоры эллипсоидные, ша-
ровидные, веретеновидные или цилиндрические, с закругленными вершина-
ми, (18) 23—39 (45) мкм шир., 29—88 (130) мкм дл. Клетки при созревании
апланоспор довольно часто сильно вздутые до 37.7—47 мкм шир., иногда
невздугые. Конъюгация лестничная. Конъюгационный канал образован обе-
ими клетками, нечетко выраженный. Воспринимающие клетки довольно
сильно вздутые вокруг зигоспор, 29—48 (53) мкм шир. Зигоспоры эллип-
соидные, 18—41 мкм шир., (23) 29—85 мкм дл. Экзоспорий апланоспор и
зигоспор тонкий, бесцветный, гладкий. Мезоспорий толстый, от желтого до
бурого, гладкий. Эндоспорий тонкий, отчетливый. Линия разрыва на зрелых
апланоспорах и зиготах отчетливая.
179
Рис. 74. Spirogyra mirabilis: 1—4 — различные формы апланоспор на разных стадиях фор-
мирования, 5 — апланоспоры и зигоспоры, 6 — начальная стадия лестничной конъюгации,
7 — зигоспоры, 8 — партеноспоры. (По: Rieth, 1972b).
приуроченности к водоемам, богатым питательными веществами, при
pH 6.4—7.8. Россия: северо-запад и центр европ. части, Вост. Сибирь
(Республика Саха); Украина, Киргизия.
Широко распространенный вид, морфологически очень сходный с
5. subsalsa и S. polymorpha, возможно идентичный им. Близкими ви-
дами по ширине вегетативных клеток и размножению апланоспорами
являются S. maravillosa Trans. (Бразилия), имеющий в отличие от S. mi-
rabilis 2—3 хлоропласта, и S. oltmannsii Huber-Pestalozzi (Южн. Аф-
рика), зигоспоры которого, вероятно, скульптурированы.
180
2. Spirogyra sphaerospora Hirn [=S. pellucida (Hass.) G. S. West f.
sphaerospora (Him) V. Poljansk.] — Спирогира круглоспоровая (рис. 75,
У). — Вегетативные клетки 43—45 мкм шир., по длине в 4—7.5 раза
больше. Поперечные стенки гладкие. Хлоропласт 1. Конъюгация лест-
ничная. Конъюгационный канал образован обеими клетками. Воспри-
нимающие клетки очень сильно вздутые вокруг зигоспор. Зигоспоры
шаровидные, 85—88 мкм диам. Экзоспорий тонкий, бесцветный, глад-
кий. Мезоспорий толстый, коричневый, гладкий.
В стоячих водах. Финляндия.
Редкий вид. Очень близок к S. pellucida, но имеет шаровидные, а
не широколинзовидные, как у последнего вида, зигоспоры и постоянно
1 хлоропласт. От другого близкого вида — S. discoidea Trans. (Южн.
Африка) отличается шаровидными зигоспорами и немного большей
шириной вегетативных клеток.
3. Spirogyra wittrockii Alexenko — Спирогира Виттрока (рис. 75, 2). —
Вегетативные клетки 65—70 мкм шир., по длине в 3—4 (6) раз больше.
Поперечные стенки гладкие. Хлоропласт 1. Конъюгация лестничная.
Конъюгационный канал образован обеими клетками. Воспринимающие
клетки со всех сторон умеренно вздутые, иногда невздутые. Зигоспоры
шаровидные, 52—58 мкм диам. Мезоспорий гладкий (?).
В стоячих водах, в торфяном болоте. Украина (долина Днепра в
окрестностях Переяслав-Хмельницкого).
Редкий, недостаточно изученный вид, в описаниях которого отсут-
ствуют какие-либо указания на строение оболочек зигоспор. По ав-
торскому рисунку можно предположить, что мезоспорий гладкий. Об-
наруживает наибольшее сходство с S. sphaerospora, который имеет,
однако, значительно более узкие вегетативные клетки, более крупные
зигоспоры и более сильно вздутые воспринимающие клетки. Наряду с
S. gallica — видом, очень сходным по всем признакам, кроме формы
зигоспор, это самая крупная среди спирогир с одним хлоропластом.
Возможно, они идентичны.
4. Spirogyra decimina (Miill.) Kiitz. — Спирогира десятерная
(рис. 75, 3; 76; 77,1—4). — Вегетативные клетки (17) 20—38 (42) мкм шир.,
по длине в 1.5—12 раз больше. Ризоиды латеральные, разветвленные,
многочисленные, преимущественно от клеток, лишенных конъюгацион-
ной пары. Поперечные стенки гладкие. Хлоропласт 1 (2). Конъюгация
лестничная и боковая. Конъюгационный канал при лестничной конъюга-
ции образован обеими клетками, иногда с преобладанием отдающей
клетки; при боковой конъюгации канал выдается на 1/4—1/2 ширины
клетки, нить при этом прямая или слегка, изредка значительно изогнутая.
Воспринимающие клетки невздутые или слабо вздутые со стороны конъю-
гационного канала или со всех сторон, до 43.5 мкм шир. Клетки без конъ-
югационной пары невздутые, иногда сильно вздутые до 55—78 (95) мкм-
шир. Зигоспоры эллипсоидные, нередко с более или менее заостренными
вершинами, реже цилиндрически-эллипсоидные, почти шаровидные или
неправильной формы, (16.5) 18—44 мкм шир., (26) 35—83 (113) мкм дл.
Экзоспорий тонкий, бесцветный, гладкий. Мезоспорий толстый, от желтого
181
Рис. 75. 1 — Spirogyra sphaerospora, 2 — S. wittrockii, 3 — S. decimina f. decimina.
(По: 1 — Hirn, 1895; 2 — Алексенко, 1893; 3 — ориг.).
до желто-коричневого, гладкий. Партеноспоры от эллипсоидных до
почти шаровидных, 29—38 мкм шир., 32—64 мкм дл., редкие.
Широко распространенный, эврибионтный, полиморфный вид. На
основании изучения материала из разных районов, а также литератур-
ных данных диагноз этого вида пришлось несколько расширить по
сравнению с первоописанием. В. И. Полянский (1966) расширил гра-
ницы этого вида, включив ряд других видов в ранге форм. Возможно,
что в дальнейшем, при нахождении у ряда форм боковой конъюгации,
вид может рассматриваться как монотипный.
F. decimina [=Zygnema deciminum (Miill.) Ag.; Spirogyra porticalis
(Miill.) Cleve var. decimina (Hass.) Cooke (?); S. lutetiana Petit; S. sin-
gularis Nordst.] (рис. 75, 3). — Вегетативные клетки (26) 30—38
(42) мкм шир., по длине в 1.5—8 (11) раз больше. Конъюгация лест-
ничная. Воспринимающие клетки невздутые или лишь слабо вздутые.
Клетки без конъюгационной пары невздутые. Зигоспоры эллипсоидные,
реже почти шаровидные, цилицдрически-эллипсоидные или неправиль-
ной формы, 25—43.5 мкм шир., 35—82 (104) мкм дл. Партеноспоры
32—35 мкм шир., 35—41 мкм дл.
В стоячих и текучих водах, на рисовых полях, нередко в болотах, в
пойменных водоемах, водохранилищах, часто, иногда в массе. Повсюду.
Широко распространенная, эврибионтная форма. Отличается от
близкой S. porticalis меньшей шириной вегетативных клеток, отсутст-
вием боковой конъюгации и вздутия клеток без конъюгационной пары
182
Рис. 76. 1—3 — Spirogyra decimina f. circumscissa (3 — строение зигоспоры, поперечный
разрез); 4, 5 — S. decimina f. communis; 6 — S. decimina f. olivascens; 7—9 — S. decimina f.
juergensii (9 — партеноспоры). (По: 1 — Воронихин в: Воронихин, Красноперова, 1970; 2,
3 — Czurda, 1930 (как S. circumscissa); 4, 8, 9 — ориг.; 5, 7 — Jao, 1935b (5 — как S. com-
munis, 7 — как 5. juergensii); 6—Krieger in: Kolkwitz, Krieger, 1941—1944 (как S. olivascens)).
(последнее присуще в некоторых случаях и S. porticalis). От S. neglecta
f. fuellebornii отличается меньшим числом хлоропластов и меньшей
шириной вегетативных клеток. По данным СЭМ, у S. singularis, рас-
сматриваемой здесь как синоним типовой формы S. decimina, мезо-
спорий двухслойный.
F. circumscissa (Czurda) V. Poljansk. (=S. circumscissa Czurda)
(рис. 76, 1—3). — Вегетативные клетки (26) 29—35 (38) мкм шир.,
по длине в 2—8 раз больше. Конъюгация лестничная. Воспринимаю-
183
Рис. 77. 1, 2 — Spirogyra decimina f. Juergensii, лестничная конъюгация; 3, 4 — S. decimina
f. longata: 3 — боковая конъюгация, 4 — партеноспоры; 5, 6 — S. porticalis f. porticalis,
лестничная конъюгация. (По: 1 — Jao, 1935b (как S. juergensii)-, 2—4 — ориг.; 5 — Gauthier-
Lievre, 1965; 6 — Красноперова, 1968).
щие клетки невздутые. Клетки без конъюгационной пары невздутые.
Зигоспоры более или менее цилиндрически-эллипсоидные, 29—35
(41) мкм шир., (38) 40—83 (113) мкм дл.
В стоячих, реже текучих водах, в озерах, по руслу рек и затопля-
емым местам. Вместе с типовой формой, довольно часто. Россия:
европ. часть (оз. Валдайское); Сев. (заповедник Боровое, р. Джана-су)
и Южн. Казахстан (Заилийский Алатау, р. Узун-Каргалы).
184
Довольно редкая форма. Отличается от других форм вида почти исклю-
чительно формой зигоспор. По-видимому, распространена шире, чем ука-
зано в литературе. Возможно, идентична типовой форме или f. juergensii.
F. communis (Hass.) V. Poljansk. [=S. communis (Hass.) Kiitz.; S. ju-
ergensii Kiitz. sensu Skuja, 1924, non auct. pl.; S. paludosa Czurda]
(рис. 76, 4, 5). — Вегетативные клетки (17) 20—26 (28) мкм шир.,
по длине в 2—6 (12) раз больше. Конъюгация лестничная. Восприни-
мающие клетки невздутые, реже — очень незначительно вздутые. Клет-
ки без конъюгационной пары невздутые. Зигоспоры эллипсоидные,
(16.5) 18—26 (39) мкм шир., 35—69 (79) мкм дл.
В стоячих и текучих водах, на рисовых полях, преимущественно в
водах, богатых органическими веществами. Вместе с типовой формой.
Повсюду.
Широко распространенная форма, отличающаяся от типовой лишь
меньшей шириной вегетативных клеток.
F. elongata Rabenh. — Вегетативные клетки 20—24 мкм шир., по
длине в 4—12 раз больше. Длина воспринимающих клеток до 2 раз
превосходит их ширину. В остальном — как f. longata.
В стоячих и текучих водах. Швейцария, США.
Неполно описанная и недостаточно изученная форма. Отсутствует
изображение. Отличается от f. longata меньшей шириной вегетативных
клеток. Не исключено, что этим именем автор формы ошибочно обо-
значил какую-то другую спирогиру, может быть S. decimina f. communis.
F. juergensii (Kiitz.) V. Poljansk. [=S. juergensii Kiitz., non sensu
Skuja, 1924; S. quinina (Ag.) Kiitz. f. juergensii (Kiitz.) Kirchn.] (рис. 76,
7—9; 77, 1, 2). — Вегетативные клетки 24—32 (36) мкм шир., по
длине в 2—8 раз больше. Конъюгация лестничная. Воспринимающие
клетки невздутые, реже слабо вздутые, преимущественно со стороны
конъюгационного канала. Клетки без конъюгационной пары невздутые.
Зигоспоры от эллипсоидных до эллипсоидно-цилиндрических, иногда
почти шаровидные, 22—35 (38) мкм шир., (35) 40—81 мкм дл. Парте-
носпоры (32) 35—38 мкм шир., (35) 43.5—46 мкм дл.
В стоячих и текучих водах, нередко в лужах и канавах, иногда на
рисовых полях и в солоноватых водах, часто при незначительном за-
грязнении, иногда вместе с типовой формой, в массе. Повсюду.
Широко распространенная форма. Отличается от типовой меньшей
шириной вегетативных клеток. По этому признаку является промежу-
точной между f. decimina и f. communis.
F. olivascens (Rabenh.) V. Poljansk. (=S. olivascens Rabenh.; S. subsa-
lina Cedercr.] (рис. 76, 6). — Вегетативные клетки 30—35 (38) мкм
шир., по длине в 3—10 раз больше. Конъюгация лестничная. Воспри-
нимающие клетки вздутые со всех сторон до 43.5 мкм шир. Клетки без
конъюгационной пары невздутые. Зигоспоры широкоэллипсоидные,
30—44 мкм шир., (40) 48—75 (101) мкм дл. Партеноспоры 29—38 мкм
шир., 32—64 мкм дл.
185
В стоячих и текучих водах, в родниках, нередко в слегка солоно-
ватых водах. Россия: северо-запад европ. части (Онежское озеро); Сев.
(заповедник Боровое) и Южн. Казахстан (р. Талас), Узбекистан (ок-
рестности Бухары).
Редкая и еще недостаточно изученная форма. От типовой отлича-
ется постоянным и обычно более сильным вздутием воспринимающих
клеток, от S. cateniformis — большей шириной вегетативных клеток,
формой зигоспор и отсутствием боковой конъюгации.
F. longata (Vauch.) V. Poljansk. [=S. longata (Vauch.) Kiitz.; S. longa-
ta var. reversa (Hass.) Kirchn. (?); 5. indica (Rao) Krieger] (рис. 77, 3,
4). — Вегетативные клетки (24) 26—38 мкм шир., по длине в 2—5 (10)
раз больше. Ризоиды латеральные, разветвленные, многочисленные, пре-
имущественно от клеток без конъюгационной пары. Конъюгация боковая,
редко лестничная, иногда совмещенные в одной паре нитей. Конъюгаци-
онный канал при боковой конъюгации выдается на 1/4—1/2 ширины клет-
ки, нить при этом прямая или слегка, изредка значительно изогнутая. Вос-
принимающие клетки невздутые. Клетки без конъюгационной пары обычно
невздутые, иногда вздутые до 55—78 (97) мкм шир. Зигоспоры эллипсоид-
ные, нередко с более или менее заостренными вершинами, (18) 25—40
(43.5) мкм шир., (26) 41—79 (100) мкм дл.
В стоячих и текучих водах, нередко в болотах, встречена в мине-
ральных и серных источниках, на рисовых полях. Может выдерживать
небольшую соленость (0.3—0.48 %о), есть данные о приуроченности
к водоемам, богатым органическими веществами. Повсюду.
Широко распространенная, эврибионтная форма. Отличается от ти-
повой формы наличием наряду с лестничной боковой конъюгации, при-
чем боковая конъюгация может не только преобладать над лестничной,
но в отдельных случаях быть даже единственным типом конъюгации.
Близка также к f. juergensii, имеющей несколько более узкие вегетатив-
ные клетки и только лестничную конъюгацию. При вздутии клеток без
конъюгационной пары может быть смешана с S. varians f. woodsii. Из
других видов довольно близка также к S. microspora Jao (Индия, Китай)
и S. porticalis, но у первого вида 2 хлоропласта и конъюгация только
боковая, а у второго вида вегетативные клетки шире и хлоропластов
может быть больше одного.
5. Spirogyra porticalis (Mull.) Cleve — Спирогира аркадная (рис. 77,
5, 6). — Вегетативные клетки (34) 38—50 (55) мкм шир., по длине в
1.7—6 (8) раз больше. Поперечные стенки гладкие. Хлоропласт 1, редко
их 2 (3). Конъюгация лестничная, очень редко боковая. Конъюгационный
канал образован обеими клетками. Воспринимающие клетки невздутые или
слегка вздутые со стороны конъюгационного канала. Клетки без конъюга-
ционной пары невздутые, изредка более или менее вздутые. Зигоспоры ши-
рокоэллипсоидные, реже цилиндрически-эллипсоидные, 35—61 мкм шир.,
(38) 40—140 мкм дл. Экзоспорий тонкий, бесцветный, гладкий. Мезоспо-
рий толстый, желтый или желтовато-коричневый, гладкий. Партеноспоры
эллипсоидные, почти вдвое меньше зигоспор, редкие.
Весьма изменчивый, вероятно монотипный вид. Особенно сильно
186
варьируют размеры клеток, размеры и форма зигот. Чурда (Czurda, 1932)
вовсе отбрасывает этот вид, так как он, по его мнению, перекрывается
другими видами. Кригер (Krieger in: Kolkwitz, Krieger, 1941—1944) со-
храняет 5. porticalis, но ограничивает его размерами вегетативных кле-
ток в 43—52 мкм шир. Такое сужение объема этого вида является, од-
нако, произвольным, так как амплитуда колебания ширины клеток у
S. porticalis в действительности нередко выходит из указанных Кригером
пределов в сторону меньших величин. На этом основании невозможно
отграничить S. porticalis от 5. lacustris.
F. porticalis [=Zygnema quininum (Miill.) Ag. (?); Spirogyra quinina
Kiitz. (?); S. fuscoatra Rabenh.; S. lacustris Czurda] (рис. 77, 5, 6). — Ве-
гетативные клетки (34) 38—50 (55) мкм шир. Хлоропласт 1, изредка по
2 (3). Конъюгация лестничная, изредка боковая: Воспринимающие клет-
ки невздутые или слегка вздутые со стороны конъюгационного канала.
Зигоспоры преимущественно широкоэллипсоидные, изредка эллипсоид-
но-цилиндрические, 35—61 мкм шир., (38) 40—90 (115) мкм дл.
В стоячих и текучих водах, в водоемах с чистой или со слабоза-
грязненной водой (0-мезосапроб), на рисовых полях, в опресненных зали-
вах и в прибрежье морей, при солености до 5.74 %о. Россия: европ. часть,
Башкирия, Южн. Урал (окрестности Уральска), приустье Волги, зап. и сев.
побережья Каспийского моря, Сев. Осетия (окрестности Владикавказа),
Дальн. Восток (Камчатка, Приморье); Украина, Южн. Казахстан.
Широко распространенная и часто встречающаяся форма. Близка
к S. decimina f. decimina, от которой отличается главным образом
большей шириной вегетативных клеток. От S. gallica ее отграничивает,
наоборот, меньшая ширина вегетативных клеток. По наиболее часто
встречающейся ширине вегетативных клеток приближается к S. neg-
lecta f. fuellebornii, но у S. porticalis f. porticalis обычно 1 хлоропласт
и только в редких случаях их несколько, а у S. neglecta f. fuellebornii,
как правило, несколько хлоропластов и только изредка 1.
F. cylindrosperma (W. et G. S. West) V. Poljansk. [=5. cylindrosper-
ma (W. et G. S. West) Krieger], — Вегетативные клетки 40—44 мкм
шир. Хлоропласт 1. Конъюгация только лестничная. Воспринимающие
клетки невздутые. Зигоспоры эллипсоидно-цилиндрические, 39—43 мкм
шир., 92—140 мкм дл.
В стоячих водах. Англия.
Редкая форма. Аналогична S. decimina f. circumscissa, которая также
имеет эллипсоидно-цилиндрические зигоспоры, но характеризуется
меньшими размерами вегетативных клеток и зигоспор. От типовой
формы отличается меньшим количеством хлоропластов, только лестнич-
ной конъюгацией и постоянно невздутыми воспринимающими клетками.
6. Spirogyra hungarica Langer — Спирогира венгерская (рис. 78,
1). — Вегетативные клетки 53—56 мкм шир., по длине в 3—8 раз боль-
ше. Поперечные стенки гладкие. Хлоропласт 1. Конъюгация лестничная.
Конъюгационный канал образован преимущественно отдающей клеткой.
Воспринимающие клетки невздутые или слабо вздутые со стороны конъ-
187
Рис. 78. 1 — Spirogyra hungarica: 2 — S. gallica; 3,4 — S. condensata, лестничная (5) и бо-
ковая (4) конъюгации; 5—7 — 5. borgeana: 5 — партеноспоры и зигоспора, 6 — партено-
споры, 7 — зигоспоры. (По: 1 — Langer, 1932; 2 — Gauthier-Lievre, 1965; 3 — Prescott, 1962;
4 — Randhawa, 1959b; 5—7 — ориг.).
югационного канала. Зигоспоры удлиненно-эллипсоидные, с заостренны-
ми вершинами, 45—53 мкм шир., 120—148 мкм дл. Экзоспорий тонкий,
бесцветный, гладкий. Мезоспорий желто-коричневый, гладкий.
В стоячих водах. Венгрия.
Очень редкий вид. По ширине вегетативных клеток занимает про-
межуточное положение между S. porticalis и S. gallica. Отличается от
них формированием конъюгационного канала только отдающей клет-
кой и размерами зигоспор. От S. condensata отличается наличием толь-
ко лестничной конъюгации.
188
7. Spirogyra gallica Petit — Спирогира галльская (рис. 78, 2). —
Вегетативные клетки 65—75 мкм шир., по длине в 2—7 раз больше.
Поперечные стенки гладкие. Хлоропласт 1. Конъюгация лестничная.
Конъюгационный канал образован обеими клетками. Воспринимающие
клетки невздутые. Клетки без конъюгационной пары невздутые. Зиго-
споры от эллипсоидных до яйцевидно-эллипсоидных, (45) 60—66 мкм
шир., (65) 90—ПО мкм дл. Экзоспорий тонкий, бесцветный, гладкий.
Мезоспорий толстый, желтый или светло-желтый, гладкий.
В стоячих, редко в текучих водах. Россия: центр европ. части;
Центр. Украина.
Редкий вид, найденный лишь в Европе (Франция, Бельгия) и Сев. Аф-
рике (Тунис). Самый крупный вид из группы спирогир с 1 хлоропластом.
Наиболее близки к нему S. porticalis и S. hungarica, от которых он отгра-
ничен своими размерами, а от последнего вида также особенностями фор-
мирования конъюгационного канала. От S. neglecta отличается меньшим
числом хлоропластов. Сходен, а возможно, идентичен S. wittrockii.
8. Spirogyra condensata (Vauch.) Kutz. — Спирогира сжатая
(рис. 78, 3, 4). — Вегетативные клетки 44—55 (62) мкм шир., по
длине меньше ширины или до 4.5 раза больше. Поперечные стенки
гладкие. Хлоропласт 1. Конъюгация боковая, редко лестничная. Конъ-
югационный канал при боковой конъюгации часто нечетко отграничен
или выдается до 1/2 ширины клетки, нить при этом обычно прямая
или незначительно изогнутая; при лестничной конъюгации канал об-
разован обеими клетками. Воспринимающие клетки невздутые или
очень слабо вздутые, изредка вздутые со всех сторон до 78 мкм шир.
Клетки без конъюгационной пары невздутые или очень слабо вздутые.
Зигоспоры эллипсоидные, 27—52 (58) мкм шир., 50—87 (93) мкм дл.
Экзоспорий тонкий, бесцветный, гладкий. Мезоспорий толстый, желто-
коричневый, гладкий. Партеноспоры редкие.
В текучих и стоячих водах, на рисовых полях и природниковых
лужах; предпочитает воды, богатые азотом. Россия: центр европ. части,
Кавказ (Сев. Осетия), Зап. и Вост. Сибирь (Республика Саха); Эстония,
Зап., Центр, и Вост. Украина, Присивашье, Крым, Узбекистан, Вост.
Казахстан (хребет Тарбагатай).
Широко распространенный вид. Близок к S. decimina f. olivascens,
отличается большей шириной вегетативных клеток, наличием боковой
конъюгации и невздутыми или слабо вздутыми воспринимающими клет-
ками. Отсутствие вздутия отграничивает этот вид от S. sahnii, имею-
щего к тому же часто более широкие вегетативные клетки, иной цвет
мезоспория и характеризующегося только боковой конъюгацией. Бли-
зок и к S. porticalis, но у него вегетативные клетки более узкие и
длинные, боковая конъюгация наблюдается очень редко, и он обитает,
по-видимому, в менее загрязненных водах.
9. Spirogyra borgeana Trans. (=S. calcarea Trans.) — Спирогира
Борге (рис. 78, 5—7). — Вегетативные клетки (28) 30—43 мкм шир.,
по длине в 1.3—5 раз больше. Поперечные стенки гладкие. Хлоропласт
1. Конъюгация лестничная, редко боковая. Конъюгационный канал при
лестничной конъюгации образован обеими клетками. Воспринимающие
189
Рис. 79. 1 — Spirogyra gracilis f. gracilis, отчленившиеся воспринимающие клетки с зиго-
спорами, лестничная конъюгация; 2, 3 — S. gracilis f. gibberosa, отчленившиеся восприни-
мающие клетки с зигоспорами при лестничной (2) и боковой (5) конъюгациях. (По: 1 —
ориг.; 2, 3 — Jao, 1935b (как S. gibberosa)).
клетки вздутые, иногда сильно, со стороны, противоположной конъ-
югационному каналу. Клетки без конъюгационной пары невздугые. Зи-
госпоры короткоэллипсоидные, с притупленными вершинами, (25) 32—
43 (46) мкм шир., (40) 45—71 (92.5) мкм дл. Экзоспорий тонкий,
бесцветный, гладкий. Мезоспорий толстый, от желтого до желто-ко-
ричневого, гладкий. Партеноспоры преимущественно эллипсоидные,
26—35 (46) мкм шир., 35—46.4 (52) мкм дл., формирующиеся как в
мужских, так и в женских гаметангиях.
В стоячих и текучих водах, на рисовых полях, преимущественно в пред-
горных и горных районах (до 5000 м над ур. м.). Конъюгирует при pH
7.4—7.62 и температуре воды И—18 °C. Россия: Урал, Предкавказье (Сев.
Осетия, окрестности Владикавказа); Грузия (р. Иори), Сев., Центр, (пред-
горья Бегазы, Дегелен), Вост, и Южн. (предгорья Джунгарского Алатау, За-
илийский Алатау) Казахстан, Туркмения (предгорья Копетдага).
Широко распространенный и часто встречающийся вид. Вздутие
воспринимающих клеток со стороны, противоположной конъюгацион-
ному каналу, — главное отличие этого вида от S. decimina.
190
7J
Рис. 80. 1 — Spirogyra gracilis f. gracilis, лестничная конъюгация; 2 — S. gracilis f. gibbe-
rosa, боковая конъюгация; 3,4 — S. flavescens f. flavescens, боковая и лестничная конъюга-
ции; 5 — S. flavescens f. parva; 6, 7 — S. subsalsa f. subsalsa; 8 — S. subsalsa f. gracilis; 9,
10 — S. subsalsa f. uralensis; 11—13 — S. polymorpha. (По; 1, 2, 11—13 — Yamagishi,
1966b; 3, 4 — Полянский, 1955 (как S. liana}; 5 — Gauthier-Lievre, 1965 (как S. parva}; 6,
7 — Czurda, 1932; 8 — Borge, 1907; 9, 10 — Полянский, 1951).
191
10. Spirogyra gracilis (Hass.) Kutz. — Спирогира изящная (рис. 79;
80, 1, 2). — Вегетативные клетки 16—26 мкм шир., по длине в 2—9
раз больше. Ризоиды латеральные, редкие, иногда от клеток без конъ-
югационной пары. Поперечные стенки гладкие. Хлоропласт 1. Конъ-
югация лестничная и боковая. Конъюгационный канал при лестничной
конъюгации образован преимущественно отдающей клеткой, иногда
только ею, часто неотчетливо отграничен от воспринимающей клетки;
при боковой конъюгации канал выдается не более чем на 1/3 ширины
клетки, нить при этом прямая или слабо изогнутая. Воспринимающие
клетки вздутые, почти всегда со стороны конъюгационного канала,
часто отчленяющиеся друг от друга после конъюгации. Клетки без
конъюгационной пары невздутые. Зигоспоры эллипсоидные, 22—
34 мкм шир., 38—75 мкм дл. Экзоспорий тонкий, бесцветный, глад-
кий. Мезоспорий толстый, желтовато-коричневый, гладкий. Партено-
споры от эллипсоидных до шаровидных, более мелкие, чем зигоспоры.
Широко распространенный вид. Следует быть осторожным при ус-
тановлении формы конъюгационного канала, границы которого у вос-
принимающей клетки часто маскируются сильным вздутием гаметангия.
F. gracilis [-Spirogyra gracilis f. genuina (Hass.) Kirchn.; 5. gracillima
Skv.] (рис. 79, Г, 80, 1). — Вегетативные клетки 16—26 мкм шир.
Конъюгация только лестничная. Зигоспоры 22—34 мкм шир., 38—
75 мкм дл.
В стоячих и текучих водах, на рисовых полях; предпочитает воды,
богатые минеральными веществами. Повсюду.
Широко распространенная и часто встречающаяся форма. Есть данные,
что в культуре типовая форма обоепола. При нахождении в природном ма-
териале боковой конъюгации может стереться разница между типовой фор-
мой и f. gibberosa. Сходна с У decimina f. communis, отличается отчетли-
вым, часто сильным, односторонним вздутием воспринимающих клеток и
особенностями формирования конъюгационного канала.
F. gibberosa (Jao) V. Poljansk. (=S. gibberosa Jao) (рис. 79, 2, 3; 80,
2). — Вегетативные клетки 19—21 (24) мкм шир. Конъюгация лестнич-
ная и боковая. Зигоспоры 22—29 (32) мкм шир., 38—55 (60) мкм дл.
На рисовых полях, в лужах. Азербайджан.
Довольно редкая форма с широким ареалом, найдена также в
Дании, Индии, Китае и Японии. От типовой формы отличается нали-
чием наряду с лестничной боковой конъюгации. Близкий ей вид —
У chenii имеет вздутые со всех сторон воспринимающие клетки. У
сходного по морфологическим признакам У parvula конъюгационный
канал формируется обоими гаметангиями.
11. Spirogyra flavescens (Hass.) Kutz. — Спирогира желтоватая
(рис. 80, 3—5). — Вегетативные клетки 8—16 мкм шир., по длине в
2.5—14 раз больше. Поперечные стенки клеток гладкие. Хлоропласт 1.
Конъюгация лестничная и боковая, часто с преобладанием одной из них.
Конъюгационный канал при лестничной конъюгации образован обеими
клетками, часто преимущественно отдающей клеткой, иногда только ею;
192
при боковой конъюгации канал выдается слабо или совсем не выражен,
нить при этом прямая или заметно изогнутая. Воспринимающие клетки
довольно сильно вздутые со всех сторон, часто сильнее со стороны конъ-
югационного канала. Клетки без конъюгационной пары невздутые. Зи-
госпоры эллипсоидные и удлиненно-эллипсоидные, 10—29 мкм шир.,
20—58.5 мкм дл. Экзоспорий тонкий, обычно бесцветный, очень редко
коричневатый, гладкий. Мезоспорий толстый, желтовато-коричневый,
очень редко бледно-буроватый с оливковым оттенком, гладкий.
F. flavescens [=5. gracilis (Hass.) Kutz. var. flavescens (Hass.) Ra-
benh.; 5. liana Trans.; Temnogyra liana (Trans.) Yamagishi] (рис. 80, 3,
4). — Вегетативные клетки 10—16 мкм шир. Конъюгация лестничная
и боковая. Зигоспоры эллипсоидные или удлиненно-эллипсоидные,
18—29 мкм шир., 25—50 (58.5) мкм дл.
В стоячих мелких водоемах, канавах, ямах, дорожных выбоинах,
на рисовых полях, болотах, редко в текучих водах. Украина, Сев.,
Вост, и Южн. Казахстан, Узбекистан.
Необходимо отметить, что в конъюгации участвуют не все клетки
нити, наблюдается достаточно четкое чередование фертильных и сте-
рильных клеток. От самых близких видов — X chenii и X pratensis
отличается преимущественно меньшей шириной вегетативных клеток.
F. parva (Hass.) V. Poljansk. [=S. parva (Hass.) Kutz.] (рис. 80, 5). —
Вегетативные клетки 8—10 мкм шир. Конъюгация только лестничная.
Зигоспоры эллипсоидные, 10—20 мкм шир., 20—40 мкм дл.
В стоячих и текучих водах. Швеция, Англия, Чехия, Эфиопия, Судан.
Самая тонкая из спирогир. Отличается от типовой формы несколько
меньшей шириной вегетативных клеток и зигоспор и отсутствием бо-
ковой конъюгации.
12. Spirogyra chenii Jao — Спирогира Чена (рис. 81, 1). — Веге-
тативные клетки 17—22 мкм шир., по длине в 2—6 раз больше. По-
перечные стенки гладкие. Хлоропласт 1 с 1—5 оборотами спирали.
Конъюгация боковая, иногда лестничная. Конъюгационный канал при
боковой конъюгации выдается слабо, нить при этом прямая или лишь
слабо изогнутая; при лестничной конъюгации канал образован отдаю-
щей клеткой. Воспринимающие клетки сильно вздутые со всех сторон
до 42 мкм шир., часто более сильно вздутые со стороны конъюгаци-
онного канала, изредка совсем не вздутые со стороны, противополож-
ной конъюгационному каналу. Клетки без конъюгационной пары иног-
да вздутые. Зигоспоры эллипсоидные, 25—32 мкм шир., 43—61
(77) мкм дл. Мезоспорий толстый, желто-коричневый, гладкий.
В стоячей воде временных водоемов, на рисовых полях. Германия,
Китай, Япония, Африка (Алжир; Ливия, Чад: нагорье Тибести).
Редкий вид. Очень близок к X pratensis и 5. flavescens. Отличается
от первого вида несколько большей шириной вегетативных клеток и
особенностями формирования конъюгационного канала при лестничной
конъюгации, а от второго вида — в основном большей шириной ве-
гетативных клеток. Возможно, что по этому признаку не может быть
отделен от X flavescens в качестве самостоятельной видовой единицы.
13 Рундина Л. А.
193
Рис. 81. 1 — Spirogyra chenii, боковая и лестничная конъюгации в одной паре нитей; 2,
3 — S. polymorpha: 2 — вздувшаяся вегетативная клетка без конъюгационной пары, зиго-
спора и две партеноспоры во вздувшейся фертильной клетке, 3 — зигоспоры в разных поло-
жениях, лестничная конъюгация. (По: Jao, 1935b).
Близок также к S. gracilis f. gibberosa, отличаясь преимущественно
вздутыми со всех сторон воспринимающими клетками.
13. Spirogyra subsalsa Kiitz. — Спирогира солончаковая (рис. 80,
6—10). — Вегетативные клетки 22—33 мкм шир., по длине до 9 раз
больше. Поперечные стенки гладкие. Хлоропласт 1 (3). Конъюгация лест-
ничная, редко боковая. Конъюгационный канал при лестничной конъ-
югации образован обеими клетками равномерно или с преобладанием
отдающей клетки, редко только ею; при боковой конъюгации канал
слабо выражен, нить при этом прямая. Воспринимающие клетки сильно
вздутые со всех сторон, редко преимущественно со стороны конъюгаци-
онного канала, до 40—45 мкм шир. Клетки без конъюгационной пары
невздутые. Зигоспоры эллипсоидные, 17—42 мкм шир., 30—77.5 мкм
дл. Экзоспорий тонкий, бесцветный, гладкий. Мезоспорий толстый, от
желтого до коричневого, гладкий.
Установлены формы этого вида, но возможно, что он монотипен.
Степень вздутия воспринимающих клеток сильно зависит от стадий
конъюгации. На поздних стадиях конъюгации часть конъюгационного
канала от воспринимающей клетки может быть полностью деформи-
рована сильным вздутием соответствующего гаметангия.
194
F. subsalsa (рис. 80, 6, 7). — Вегетативные клетки 22—28 мкм
шир. Хлоропласт I (3). Конъюгация только лестничная. Конъюгаци-
онный канал образован преимущественно или исключительно отдаю-
щей клеткой. Зигоспоры 17—32.5 мкм шир., 30—56 мкм дл. Мезо-
спорий желто-коричневый.
В стоячих и текучих, соленых и пресных водах. Россия: Южн.
Урал; Зап. Украина (окрестности Тернополя) (?).
От близкой У decimina f. olivascens отличается меньшей шириной
вегетативных клеток и строением конъюгационного канала, от У gra-
cilis — большей шириной вегетативных клеток и характером вздутия
воспринимающих клеток. Последний признак, а также отсутствие бо-
ковой конъюгации и отсутствие вздутия у клеток без конъюгационной
пары отграничивают эту типовую форму от S. varians f. minor.
F. gracilis (Borge) V. Poljansk. [=У varians (Hass.) Kiitz. f. gracilis
Borge; 5. suecica Trans.] (рис. 80, S). — Вегетативные клетки 25—
33 мкм шир. Хлоропласт 1. Конъюгация лестничная, редко боковая.
Конъюгационный канал образован равномерно обеими клетками или
преимущественно, иногда исключительно отдающей клеткой. Зигоспо-
ры 26—39 мкм шир., 36—60 мкм дл. Мезоспорий желтый.
В стоячих и текучих водах. Швеция, Франция, Индия, США.
От типовой формы отличается большей шириной вегетативных кле-
ток и зигоспор и в некоторых случаях особенностями формирования
конъюгационного канала.
F. uralensis V. Poljansk. (рис. 80, 9, 10). — Вегетативные клетки
25—32.5 мкм шир. Хлоропласт 1. Конъюгация только лестничная.
Конъюгационный канал образован равномерно обеими клетками. Зиго-
споры 35—42 мкм шир., 52.5—77.5 мкм дл. Мезоспорий коричневый.
В сильно соленом степном озере. Россия: Южн. Урал (оз. Терен-
куль).
Редкая форма. По размерам вегетативных клеток и зигоспор сбли-
жается с f. gracilis, но в отличие от нее имеет только лестничную
конъюгацию, при этом конъюгационный канал образован обеими клет-
ками равномерно.
14. Spirogyra polymorpha Kirchn. [=S. polymorpha f. (sine nomen)
Petkoff; S. subsalsa Kiitz. sensu Krieger in: Kolkwitz, Krieger, 1941—
1944, pr. p.] — Спирогира полиморфная (рис. 80, 11—13\ 81, 2, 3). —
Вегетативные клетки 22—30 мкм шир., по длине в 2—12 раз больше.
Поперечные стенки гладкие. Хлоропласт 1 (2). Конъюгация лестничная.
Конъюгационный канал образован обеими клетками равномерно. Вос-
принимающие клетки со всех сторон более или менее вздутые до
26—43 мкм шир., иногда сильно вздутые до 65 мкм шир. Клетки без
конъюгационной пары иногда вздутые до 64 мкм шир. Зигоспоры пре-
имущественно широкоэллипсоидные, реже яйцевидные, шаровидные,
почти цилиндрические или неправильной формы, 22—36 мкм шир.,
25—56 мкм дл. Экзоспорий тонкий, бесцветный, гладкий. Мезоспорий
толстый, желтый, гладкий. Партеноспоры редкие.
195
Рис. 82. 1 — Spirogyra parvula, боковая конъюгация; 2 — S. cateniformis, боковая конъюга-
ция; 3 — S. bullata f. bullata, вздувшаяся клетка без конъюгационной пары, зигоспора, лест-
ничная конъюгация; 4 — S. bullata f. major, 5 — S. pseudosahnii; 6 — S. varians f. woodsii,
боковая конъюгация. (По: 1 — Воронихин в: Воронихин, Красноперова, 1970; 2 — Рунди-
на, 1988; 3— Рундина, 1978а; 4 — Полянский, 1959; 5 — Kadhibowska in: Kadhibowska,
Christensen, 1979; 6 — Рундина, 19786).
В стоячих и текучих водах, на рисовых полях. Латвия, Зап. Украина.
Довольно редкий вид, встреченный также в Польше, Болгарии и
Китае. Морфологически этот вид очень близок S. subsalsa и 5. mira-
bilis, возможно даже, что все они являются формами одного вида.
15. Spirogyra parvula (Trans.) Czurda [=5. arcta (Ag.) Kiitz. var.
cateniformis (Hass.) Kirchn. sensu Gutwinski, 1895a, 1895b] — Спиро-
гира крошечная (рис. 82, 7). — Вегетативные клетки 20—24 (25) мкм
шир., по длине в 2—9 раз больше. Поперечные стенки гладкие. Хло-
196
ропласт 1. Конъюгация боковая, редко лестничная. Конъюгационный
канал при лестничной конъюгации образован обеими клетками, при
боковой конъюгации выражен неотчетливо, выдается слабо, не превы-
шая 1/3 ширины клетки, нить при этом прямая или незначительно
коленчато изогнутая. Воспринимающие клетки со всех сторон равно-
мерно вздутые, до 38 (43.5) мкм шир., иногда отчленяющиеся друг от
друга после конъюгации. Клетки без конъюгационной пары невздутые.
Зигоспоры эллипсоидные, 20—30 (36.6) мкм шир., 38—67 (73) мкм
дл. Экзоспорий тонкий, бесцветный, гладкий. Мезоспорий толстый, от
золотисто-желтого до коричневого, гладкий. Партеноспоры эллипсоид-
ные, 23—25.6 мкм шир., 62—65.5 мкм дл.
В стоячих и текучих водах, на речных порогах, в родниковых ручь-
ях, на рисовых полях. Россия: Карелия, Предкавказье (Сев. Осетия),
Вост. Сибирь (Сев. Хакассия); Зап. Украина (окрестности Тернополя),
Сев. Казахстан (заповедник Боровое), Узбекистан.
Широко распространенный и часто встречающийся вид. Связан пе-
реходами с S. cateniformis и У pratensis по ширине вегетативных кле-
ток и размерам зигоспор.
16. Spirogyra cateniformis (Hass.) Kiitz. [=5. arcta (Ag.) Kiitz. var.
cateniformis (Hass.) Kirchn., non sensu Gutwinski, 1895a, 1895b; У affinis
(Hass.) Petit] — Спирогира цепочковидная (рис. 82, 2; 83, 1). — Веге-
Рис. 83. 7 — Spirogyra cateniformis, лестничная и боковая конъюгации в одной паре нитей;
2—4 — S. sahnii (2 — партеноспоры и зигоспора, боковая конъюгация). (По: 1 — ориг.;
2 — Randhawa, 1938; 3, 4 — Simons, 1990).
197
тативные клетки 25—30 мкм шир., по длине в 1.5—4.5 раза больше.
Поперечные стенки гладкие. Хлоропласт 1. Конъюгация преимуществен-
но боковая, редко лестничная. Конъюгационный канал при лестничной
конъюгации образован обеими клетками; при боковой конъюгации канал
выдается обычно слабо, не более чем на 1/3 ширины клетки или выра-
жен неотчетливо, нить при этом прямая или слабо изогнутая. Воспри-
нимающие клетки равномерно веретеновидно вздутые до 38 мкм шир.
Клетки без конъюгационной пары невздутые, Зигоспоры эллипсоидные,
26—29 мкм шир., 43—86 мкм дл. Экзоспорий тонкий, бесцветный,
гладкий. Мезоспорий толстый, от желтого до желтовато-коричневого,
гладкий. Эндоспорий тонкий, гладкий, трудноразличимый.
В стоячих и текучих водах, на рисовых полях, иногда в минераль-
ных источниках. Повсюду.
Широко распространенный и часто встречающийся вид. Отличается
от ряда близких видов: 5. decimina f. longata — вздутыми восприни-
мающими клетками, S. subsalsa — наличием боковой конъюгации и
местообитанием, 5. pratensis — главным образом большей шириной
вегетативных клеток и невздутыми или лишь слабо вздутыми клетками,
лишенными конъюгационной пары, 5. parvula — незначительно боль-
шей шириной вегетативных клеток.
17. Spirogyra pseudosahnii Kadi. — Спирогира псевдосана (рис. 82,
5). — Вегетативные клетки 40—45 мкм шир. Поперечные стенки глад-
кие. Хлоропласт 1. Конъюгация боковая. Конъюгационный канал обра-
зован обеими клетками, широко закругленный, по ширине прибли-
зительно равный 1/2 ширины вегетативной клетки. Воспринимающие
клетки вздутые со стороны конъюгационного канала. Клетки без
конъюгационной пары вздутые со всех сторон. Зигоспоры эллипсоид-
ные, 28—30 мкм шир., 35—45 мкм дл. Экзоспорий тонкий, бесцвет-
ный, гладкий. Мезоспорий желто-коричневый или красновато-бурый,
гладкий.
В мелких стоячих водоемах. Дания.
Редкий вид. Очень близок к 5. sahnii. Отличается незначительно
меньшей шириной вегетативных клеток и цветом (?) мезоспория.
18. Spirogyra sahnii Randh. — Спирогира Сана (рис. 83, 2—4). —
Вегетативные клетки 46.3—72 мкм шир., по длине равны или немного
больше ширины. Поперечные стенки гладкие. Хлоропласт 1. Конъюга-
ция боковая. Конъюгационный канал выдается на 1/4—1/2 ширины клет-
ки, нить при этом прямая. Воспринимающие клетки односторонне или
со всех сторон вздутые. Клетки без конъюгационной пары вздутые до
82 мкм шир. Зигоспоры эллипсоидные, 22—39.2 мкм шир., 36—68 мкм
дл. Экзоспорий тонкий, бесцветный, гладкий. Мезоспорий толстый,
желто-коричневый, гладкий. Линия разрыва продольная, отчетливая.
Партеноспоры эллипсоидные, 22—24 мкм шир., 22—36 мкм дл.
В стоячей и проточной воде, в ручьях. Нидерланды, Индия.
Редкий вид, требующий дополнительного изучения. В первоописа-
нии указан сине-зеленый цвет мезоспория, не встреченный прежде ни
у одного из видов спирогир. На материале из Нидерландов цвет у
198
Рис. 84. Spirogyra bullata, зигоспоры и вздувшиеся клетки без конъюгационной пары:
1 — f. bullata, 2 — f. minor. (По; 1 — Jao, 1935b; 2 — Рундина, 19776).
мезоспория был желто-коричневый. Можно предположить, что сине-
зеленый цвет мезоспория в образцах из Индии был обусловлен незре-,
лостью зигот. Кроме того, на оригинальном рисунке автора вида видна
мелкая точечность на мезоспорий, не оговоренная в первоописании.
Вид имеет несомненное сходство с 5. pseudosahnii.
199
19. Spirogyra bullata Jao — Спирогира пузырчатая (рис. 82, 3, 4;
84). — Вегетативные клетки 14.5—27.5 мкм шир., по длине в 2—9 раз
больше. Поперечные стенки гладкие. Хлоропласт 1. Конъюгация лест-
ничная. Конъюгационный канал образован обеими клетками. Восприни-
мающие клетки невздутые или очень слабо вздутые. Клетки без конъю-
гационной пары очень сильно вздутые,^13.5—64 мкм шир., шаровидные
или почти шаровидные, редко яйцевидные, расположенные по 1—25 под-
ряд. Зигоспоры эллипсоидные, 22—32 (35) мкм шир., 31—49 (61) мкм дл.
Экзоспорий тонкий, бесцветный, гладкий. Мезоспорий довольно тол-
стый, от желтого до желто-коричневого, гладкий.
Наиболее типично для этого вида сильное вздутие клеток, лишенных
конъюгационной пары, приобретающих часто шаровидную форму. Этот
признак хорошо отличает его от S. decimina f. communis. От S. pratensis,
имеющего также сильно вздутые клетки без конъюгационной пары, от-
личается формой воспринимающих клеток, размерами вегетативных кле-
ток и зигоспор, отсутствием боковой конъюгации, а также расположе-
нием вздутых неконъюгирующих клеток — иногда по многу в ряд.
F. bullata (рис. 82, 3; 84, 1). — Вегетативные клетки 19—23 мкм
шир., по длине в 2—4 раза больше. Зигоспоры 22—32 (35) мкм шир.,
31—43.5 (51) мкм дл.
В стоячих водах, прудах, на рисовых полях. Южн. Казахстан.
Типовая форма, встречающаяся вместе с двумя другими формами
вида, по ширине вегетативных клеток является промежуточной между
ними. Ширина клеток f. major и f. minor носит относительно транс-
грессивный характер.
F. major V. Poljansk. (=5. poljanskii Kadi.) (рис. 82, 4). — Веге-
тативные клетки 26—27.5 мкм шир. Зигоспоры 23.7—25 мкм шир.,
45—47.5 мкм дл.
На рисовых полях. Южн. Казахстан.
От типовой формы отличается большей шириной вегетативных кле-
ток.
F. minor Rund. (=S. subpratensis Woronich. sensu Kadhibowska, 1984, pr.
p.) (рис. 84, 2). — Вегетативные клетки 14.5—17.4 мкм шир., по длине в
4.5—9 раз больше. Зигоспоры 23—29 мкм шир., (35) 41—46.4 (61) мкм дл.
В рыбохозяйственном пруду, вместе с типовой формой, часто. Южн.
Казахстан.
От типовой формы отличается меньшей шириной вегетативных кле-
ток.
20. Spirogyra varians (Hass.) Kiitz. — Спирогира изменчивая (рис. 82,
6; 85; 86). — Вегетативные клетки 20—48 (51) мкм шир., по длине рав-
ные ширине или чаще в 2—6 раз больше. Поперечные стенки гладкие.
Хлоропласт 1, очень редко их 2. Конъюгация лестничная и боковая.
Конъюгационный канал при лестничной конъюгации образован обеими
клетками, иногда с преобладанием отдающей клетки или только от нее;
при боковой конъюгации канал часто ушковидный, реже тупо закруг-
200
Рис. 85. Spirogyra varians f. varians'. 1—4 — лестничная конъюгация (3, 4 — зигоспоры в
двух положениях), 5 — боковая конъюгация. (По: 7—4 — ориг.; 5 — Рундина, 1988).
ленный, часто неясно отграниченный, выдается на 1/2—2/3 ширины
клетки, нить при этом прямая или более или менее изогнутая. Воспри-
нимающие клетки вздутые со стороны конъюгационного канала, часто
сильно — до 41—55 мкм шир., с противоположной стороны невздутые
или реже незначительно вздутые. Клетки без конъюгационной пары
сильно вздутые, до 65 мкм шир., бочонковидные, шаровидные или не-
правильно эллипсоидные. Зигоспоры широкоэллипсоидные, очень редко
удлиненно-эллипсоидные, иногда цилиндрически-эллипсоидные, 21—
50 мкм шир., 35—125 мкм дл., изредка с темно- или черно-коричневы-
ми колпачковидными образованиями (калиптрами) на вершинах, иногда
и в других местах. Экзоспорий тонкий, бесцветный, гладкий. Мезоспо-
рий толстый, от желтого до желто-коричневого, гладкий. Эндоспорий
тонкий, гладкий, четко различимый на зрелых зиготах. Партеноспоры
26—35 мкм шир., 32—72.5 мкм дл.
Широко распространенный вид, отличающийся большим полимор-
физмом. Легко узнается по характерному вздутию клеток, лишенных
конъюгационной пары, и обычно одностороннему вздутию восприни-
мающих клеток со стороны конъюгационного канала. Ряд морфологи-
ческих отклонений описан в качестве внутривидовых форм. Изучение
зигоспор 5. varians в сканирующем микроскопе показало, что мезо-
спорий у него двухслойный: наружный слой тонкий и гладкий, внут-
ренний слой толстый, шероховатый, с маленькими разнокалиберными
бугорками на поверхности, не видимыми в световом микроскопе.
201
Рис. 86. Spirogyra varians'. 1—5 — f. bicalyptrata'. 1 — лестничная конъюгация, 2—5 — раз-
ные стадии формирования колпачков (калиптр) на вершинах зигоспор; 6,7 — f. circumline-
ata\ 8—11 — f. minor, лестничная (8—70) и боковая (11) конъюгации; 12—16 — f. woodsii:
12 — начальная стадия лестничной конъюгации, 13—16 — боковая конъюгация. (По: 1—
5 — Czurda, 1932; 6, 12, 13 — Gauthier-Lifevre, 1965 (как S. circumlineata)', 7—9, И, 14—
16 — Рундина, 1988; 10 — Воронихин в: Воронихин, Красноперова, 1970).
F. varians (рис. 85)..— Вегетативные клетки (26) 28—35 (40) мкм
шир. Конъюгация лестничная. Зигоспоры 24—44 (48) мкм шир., 35—
72.5 (100) мкм дл. Партеноспоры 26—29 мкм шир., 32—38 мкм дл.
В стоячих и текучих водах, часто в мелких водоемах, низовых
болотах, на рисовых полях, изредка на влажных скалах. Повсюду.
Широко распространенная и часто встречающаяся форма. По наблю-
дениям в культуре размножению благоприятствует pH 6.0—7.0, при pH
9.0 размножение еще возможно, при pH 5.4 клетки сразу отмирают. Тем-
пературный оптимум развития 18—20 °C. Часто и довольно значительно
202
варьирует форма зигоспор. Обнаруживает значительное сходство с S. de-
cimina f. decimina, отличается вздутием клеток без конъюгационной
пары, а также обычно односторонним вздутием воспринимающих кле-
ток. Эти отличительные признаки особенно сближают ее с формами
5. decimina (f. juergensii и f. longata), у которых эти вздутия наблюда-
ются тоже, хотя это происходит изредка и в слабой степени.
F. bicalyptrata (Czurda) V. Poljansk. (=5. bicalyptrata Czurda) (рис. 86,
1—5). — Вегетативные клетки 36—39 мкм шир. Хлоропласт 1, редко
их 2. Конъюгация лестничная. Зигоспоры 31—34 мкм шир., 55—70 мкм
дл., с темно- или черно-коричневыми утолщениями (калиптрами) на вер-
шинах, иногда и в других местах.
В стоячих и текучих водах, болотах. Португалия, Дания, Австрия,
Чехия, Венгрия, Греция.
Довольно редкая и спорная форма. Единственным значимым отличи-
ем от типовой формы, кроме изредка встречающихся двух хлоропластов,
служат своеобразные темные образования на оболочках зигоспор, одна-
ко сходная картина наблюдалась и у других видов (У nodifera, S. pseu-
donodifera, S. porticalis и т. д.). Учитывая это, можно предположить, что
f. bicalyptrata вообще не имеет самостоятельной таксономической зна-
чимости, тем более что у типовой формы очень редко, но тоже встре-
чаются подобные темные утолщения на зигоспорах.
F. circumlineata (Trans.) V. Poljansk. (=5. circumlineata Trans.) (рис. 86,
6, 7). — Вегетативные клетки (38) 40—48 (51) мкм шир. Конъюгация
лестничная. Зигоспоры (30) 35—50 мкм шир., (50) 58—125 мкм дл.
В стоячей, иногда заболоченной, реже в текучей воде, ручьях, ста-
рицах рек, нередко в горах (до 2000 м над ур. м.). Россия: Южн. Урал;
Сев. (заповедник Боровое) и Южн. (Заилийский Алатау) Казахстан.
Редкая форма, близкая f. woodsii, отличается от нее лишь отсут-
ствием боковой конъюгации. Обнаруживает также заметное сходство
с 5. porticalis, отличается постоянным наличием одного хлоропласта,
отсутствием боковой конъюгации, регулярным вздутием воспринимаю-
щих клеток и большим постоянством формы зигоспор. Фертильные
нити часто разъединяются после образования зигоспор.
F. minor Teod. (=5. teodorescoi Trans.; S. fragilis Jao; S. commutata
Skv.) (рис. 86, 8—11). — Вегетативные клетки 20—31 мкм шир. Конъ-
югация лестничная, редко боковая. Зигоспоры 21—38 мкм шир., 36—
67 мкм дл.
В стоячих, реже текучих водах, лужах, канавах, торфяных озерках,
на рисовых полях, в обрастаниях камней, в родниковых лужах. Россия:
северо-запад европ. части, Сев. Осетия (Тарское ущелье); Сев., Зап.,
Центр, и Южн. Казахстан.
По всем признакам, за исключением меньшей ширины вегетативных
клеток и редкой боковой конъюгации, подобна типовой форме. От
наиболее близкого вида — S. gracilis отличается большей шириной
вегетативных клеток и вздутием клеток, лишенных конъюгационной
пары. Частое разъединение соседних воспринимающих клеток после
203
конъюгации, по-видимому, не может быть признано таксономическим
признаком, как полагают многие авторы, так как то же самое явление
иногда наблюдается и у других спирогир. Данные сканирующего мик-
роскопа показывают, что мезоспорий 5. teodorescoi (синоним f. minor)
двухслойный, причем толстый внутренний слой имеет сетчатую струк-
туру-
F. woodsii (Hass.) V. Poljansk. [=5. woodsii (Hass.) Czurda; 5. vari-
ans (Hass.) Kutz, sensu Transeau, 1951; Волкова, 1970; Rhynchonema
woodsii Kiitz.] (рис. 82, 6; 86, 72—76). — Вегетативные клетки (32)
35—40 (47) мкм шир. Конъюгация боковая и лестничная. Зигоспоры
31—40.5 мкм шир., 46.5—90 мкм дл. Партеноспоры 29—35 мкм шир.,
37.5—72.5 мкм дл.
В стоячих и текучих водах, реках, озерах, водохранилищах, преиму-
щественно в виде обрастаний на камнях. Зап. и Центр. (Смелянское во-
дохранилище) Украина, Зап., Южн. Казахстан, Таджикистан (Памир: оз.
Яшиль-куль).
Весьма вероятно, что f. woodsii распространена достаточно широко.
Отличается от типовой формы наличием боковой конъюгации и не-
сколько большей шириной вегетативных клеток, причем различие в
размерах носит трансгрессивный характер, говорящий о возможной
идентичности обеих форм.
21. Spirogyra henanensis (Bi) Jao, non (Bi) Kadi. (=5. shanxiensis
Ling et Zheng var. henanensis Bi) — Спирогира хэнаньская (рис. 87,
7). — Вегетативные клетки 35—39 мкм шир., по длине в 1.5 раза
больше. Поперечные стенки гладкие. Хлоропласт 1 с 1.5—4 оборотами
спирали. Конъюгация лестничная. Конъюгационный канал преимущес-
твенно или исключительно от отдающей клетки. Воспринимающие
клетки со всех сторон сильно и равномерно вздутые до 55—72 мкм
шир. Клетки без конъюгационной пары со всех сторон сильно и рав-
номерно вздутые до 50—64 мкм шир. Зигоспоры эллипсоидные, 28—
40 (46) мкм шир., 45—64 мкм дл. Мезоспорий от желтоватого до
желто-коричневого, гладкий.
В стоячих водах, прудах. Испания, Китай.
Редкий вид. Близок к 5. polymorpha и У decimina f. olivascens,
отличается от первого вида большей шириной вегетативных клеток,
от второго — вздутием клеток без конъюгационной пары. От У shan-
xiensis Ling et Zheng (Китай) отличается незначительно большей ши-
риной вегетативных клеток и всесторонним вздутием воспринимающих
клеток, от У chenii — отсутствием боковой конъюгации.
22. Spirogyra pratensis Trans. [=5. pratensis f. fennica (Cedercr.) V.
Poljansk.; 5. pratensis var. minor Woronich.; 5. pratensis f. minor (Wo-
ronich.) V. Poljansk.; 5. subpratensis Woronich.] — Спирогира луговая
(рис. 88, 3—7). — Вегетативные клетки (13.7) 15.5—21 (23) мкм шир.,
по длине в (3) 4—14 (16) раз больше. Поперечные стенки гладкие.
Хлоропласт 1, редко их 2. Конъюгация лестничная и боковая, изредка
совмещенные в одной паре нитей. Конъюгационный канал при лест-
204
Рис. 87. 1 — Spirogyra henanensis-, 2—6 — S. paraquayensis: 2, 3 — f. paraquayensis, лест-
ничная конъюгация (2) и строение мезоспория (3), 4—6 — f. rhizobrachialis, конъюгирую-
щие клетки с зигоспорами (4, 5) и строение мезоспория (6); 7 — S. aria. (По: 1 — Jao,
1988; 2,3 — Borge, 1903; 4, 7 — Jao, 1935b (4 — как S. rhizobrachialis'): 5,6 — Jao, 1936
(как 5. rhizobrachialis)).
ничной конъюгации образован обеими клетками; при боковой конъ-
югации канал округленный, выдается не более чем на 1/2 ширины
клетки, нить при этом прямая или незначительно изогнутая. Воспри-
нимающие клетки вздутые со всех сторон до 35—40.5 мкм шир. Клет-
ки без конъюгационной пары часто шаровидно вздутые до 90 мкм шир.
Зигоспоры эллипсоидные, иногда цилиндрически-эллипсоидные, 19—
36 мкм шир., (34) 43—70 (100) мкм дл. Экзоспорий тонкий, бесцвет-
ный, гладкий. Мезоспорий толстый, желтый, гладкий. Эндоспорий тон-
кий, гладкий, заметный только на зрелых зиготах. Партеноспоры по
форме и размерам подобны зигоспорам, редкие.
205
В стоячих и текучих водах, в рыбоводных прудах, озерах, на ри-
совых полях, в дождевых потоках со скал. Россия: северо-запад европ.
части; Зап. Украина, Центр, (предгорья Бегазы) и Южн. Казахстан,
Узбекистан (Ферганская долина, Голодная степь).
Редкий вид. От близких S. cateniformis и S. decimina f. communis
отличается более узкими вегетативными ^летками и сильным, хотя и
Рис. 88. 1,2 — Spirogyra rivularis, разная степень вздутия воспринимающих клеток; 3—7 —
5. pratensis: 3 — вздутие клетки без конъюгационной пары, 4 — лестничная конъюгация,
5 — лестничная и боковая конъюгации в одной паре нитей, 6,7 — боковая конъюгация;
8—15 — S. neglecta: 8—11 — f. neglecta, 12, 13 — f. Columbiana, 14, 15 — f. fuellebornii.
(По: 1—3 — Gauthier-Lievre, 1965; 4—7 — Рундина, 19786; 8—11 — Рундина, 1988; 12,
13 — Gauthier-Lievre, 1965 (как S. columbianay 14 — Prescott, 1962; 15 — Schmidle, 1903
(как S. fuellebomif)).
206
нерегулярным, вздутием клеток, лишенных конъюгационной пары.
Очень близок У chenii, от которого отличается в основном лишь осо-
бенностями формирования конъюгационного канала.
23. Spirogyra rivularis (Hass.) Rabenh. — Спирогира ручейная
(рис. 88, 7, 2). — Вегетативные клетки (24) 30—42 мкм шир., по
длине в 2.5—И раз больше. Ризоиды базальные, сильно вытянутые,
узкие, неразветвленные. Поперечные стенки гладкие. Хлоропластов 2—
3. Конъюгация лестничная. Конъюгационный канал образован обеими
клетками. Воспринимающие клетки невздутые или очень слабо вздутые,
до 50 мкм шир. Клетки без конъюгационной пары невздутые. Зиго-
споры эллипсоидные, реже цилиндрически-эллипсоидные, 35—44 мкм
шир., 60—100 мкм дл. Экзоспорий тонкий, бесцветный, гладкий. Ме-
зоспорий толстый, бесцветный, гладкий.
F. rivularis (рис. 88, 1, 2). — Вегетативные клетки (31) 36—42 мкм
шир. Хлоропластов 2—3.
В текучих водах, по берегам рек и в ручьях, реже в стоячих водах,
часто прикрепленно к субстрату. Россия: центр европ. части, дельта
Волги, сев. часть Каспийского моря, Зап. Сибирь (Томск, окрестности
Колывани); Латвия, Белоруссия (окрестности Минска), Зап. Украина
(окрестности Снятына, р. Прут).
Довольно широко распространенная, но не часто встречающаяся и
недостаточно изученная форма. Многие авторы причисляют ее к си-
нонимам S. fluviatilis, от которого она отличается гладким мезоспори-
ем, меньшим числом хлоропластов, невздутыми или слабо вздутыми
воспринимающими клетками и особенностями формирования конъюга-
ционного канала.
F. minor (Hansg.) V. Poljansk. (=S. rivularis var. minor Hansg.;
S. fluviatilis Hilse sensu Krieger in: Kolkwitz, Krieger, 1941—1944, pr.
p.). — Вегетативные клетки 24—30 мкм шир. Хлоропластов 3.
В текучей воде (?). Чехия.
Слабо изученная и спорного таксономического положения форма.
Изображение отсутствует.
24. Spirogyra neglecta (Hass.) Kiitz. — Спирогира забытая (рис. 88,
8—15). — Вегетативные клетки 40—68 (75) мкм шир., по длине в
2—6 раз больше. Ризоиды латеральные, разветвленные, редкие. Попе-
речные стенки гладкие. Хлоропластов (1) 2—5 (7). Конъюгация лест-
ничная, редко боковая. Конъюгационный канал при лестничной конъ-
югации образован обеими клетками. Воспринимающие клетки невзду-
тые или слабо вздутые. Клетки без конъюгационной пары невздутые.
Зигоспоры эллипсоидные или эллипсоидно-цилиндрические, 32—
69 мкм шир., 50—160 мкм дл. Экзоспорий тонкий, бесцветный, глад-
кий. Мезоспорий толстый, желто-коричневый, гладкий. Партеноспоры
широкоэллипсоидные, редкие.
207
Вид широко распространенный и полиморфный, с сильной измен-
чивостью ширины вегетативных клеток, размеров и отчасти формы
зигоспор, степени и характера вздутия воспринимающих клеток. Есть
данные об образовании акинет при подщелачивании среды, насыщен-
ной питательными веществами.
F. neglecta [=5. neglecta f. varians (Skigh) V. Poljansk.; 5. szechwa-
nensis Jao var. varians Singh; S. emilianensis Bonhomme; S. margalefii
Aboal et Llimona] (рис. 88, 8—11). — Вегетативные клетки 50—68
(75) мкм шир. Хлоропластов (2) 3—5 (7). Конъюгация лестничная.
Зигоспоры (40) 46—70 мкм шир., (55) 65—160 мкм дл. Партеноспоры
сходные по форме и размерам с зигоспорами.
В стоячих и текучих водах, родниках, часто в мелких, хорошо
прогреваемых водоемах, нередко с сильным азотистым загрязнением,
на рисовых полях, большими скоплениями. Повсюду.
Очень близка к 5. gallica, но имеет большее число хлоропластов.
От близкого S. nitida отличается более узкими клетками, меньшим
числом хлоропластов и отчасти формой зигоспор. Обнаруживает сход-
ство с 5. ternata и 5. hyalina, отличается от первого вида невздутыми
или лишь слабо вздутыми воспринимающими клетками и формой зи-
госпор, а от второго — наличием только лестничной конъюгации и
отсутствием сильно вздутых воспринимающих клеток, хотя этот пос-
ледний признак у 5. hyalina варьирует.
F. columbiana (Czurda) Rund. [=5. columbiana Czurda; 5. neglecta
f. fuellebornii (Schmidle) V. Poljansk., pr. p., sensu Полянский, 1966]
(рис. 88, 12, 13). — Вегетативные клетки (42) 45—54 мкм шир. Хло-
ропластов (1) 2—3. Конъюгация лестничная. Зигоспоры 42—60 мкм
шир., 65—115 мкм дл.
В текучих и стоячих водах, на рисовых полях. Россия: центр европ.
части; Южн. Казахстан.
Распространенная на всех континентах; по-видимому, и в России
имеет более широкое распространение, чем известно по литературе.
Отличается от типовой формы более узкими вегетативными клетками.
Очень сходна по этому признаку с f. fuellebornii, но имеет только
лестничную конъюгацию и иногда меньшее число хлоропластов. Близка
также и к S. hyalina.
F. fuellebornii (Schmidle) V. Poljansk. [=S. fuellebornii Schmidle;
S. decimina (Mull.) Kiitz. sensu Алексенко, 1888; 5. irregularis Nag.
sensu Gutwinski, 1895a, 1895b] (рис. 88, 14, 15). — Вегетативные клет-
ки 40—46 мкм шир. Ризоиды латеральные, разветвленные, редкие.
Хлоропластов 2—4. Конъюгация лестничная, редко боковая. Зигоспоры
32—50 мкм шир., 50—80 мкм дл. Партеноспоры редкие.
В текучих и стоячих водах, на заболоченных участках, в дождевых
лужах, на рисовых полях. Россия: европ. часть — почти повсюду,
Предкавказье (Сев. Осетия, Тарское ущелье); Вост. Украина, Зап. и
Южн. Казахстан.
208
Отличается от всех других форм вида наличием боковой конъюга-
ции наряду с преобладающей лестничной. Возможно, идентична f. co-
lumbiana. Близка также к У hyalina, но имеет более узкие вегетативные
клетки.
25. Spirogyra nitida (Dillw.) Link — Спирогира блестящая (рис. 89,
1, 2, 5—7). — Вегетативные клетки (68) 75—90 мкм шир., по длине
в 1.5—3 (5) раз больше. Поперечные стенки гладкие. Хлоропластов
(2) 3—5, часто слабо извитых. Конъюгация лестничная. Конъюгацион-
ный канал образован обеими клетками, иногда с преобладанием отда-
ющей клетки. Воспринимающие клетки невздутые или едва заметно
вздутые, преимущественно со стороны конъюгационного канала. Клет-
ки без конъюгационной пары невздутые. Зигоспоры эллипсоидные,
иногда с более или менее заостренными вершинами, редко цилиндри-
чески-эллипсоидные или почти шаровидные, (50) 60—90 мкм шир.,
73—174 (210) мкм дл. Экзоспорий тонкий, бесцветный, гладкий. Ме-
зоспорий толстый, коричневато-желтый до коричневого, гладкий.
F. nitida [-Zygnema nitidum Hass.; Spirogyra nitida var. densa Ra-
benh.; 5. turfosa Gay; 5. szechwanensis Jao; S. nitida auct. non Link:
Artari, 1885; Казановский, Смирнов, 1914] (рис. 89, 1, 2, 5, 6). —
Вегетативные клетки (68) 75—90 мкм шир. Воспринимающие клетки
невздутые или только едва заметно вздутые. Зигоспоры эллипсоидные,
нередко с более или менее заостренными вершинами, редко цилинд-
рически-эллипсоидные, (50) 60—90 мкм шир., 73—174 (210) мкм дл.
В стоячих, реже медленно текучих водах, на рисовых полях, не-
редко в болотах, лужах. Отмечена приуроченность к бедным известью
водам и к развитию в весеннее время. Олигосапроб. Россия: европ.
часть — повсюду, Зап. Сибирь — местами; Украина — повсюду, Ка-
захстан (устье Сыр-Дарьи).
Возможно, эта типовая форма S. nitida вместе с S. neglecta f. neg-
lecta и У. setiformis являются формами одного полиморфного вида. По
всей вероятности, они иногда смешивались, и наиболее узкие из спи-
рогир, обозначенные именем S. nitida, в действительности относятся к
5. neglecta, а наиболее широкие — к S. setiformis.
F. paradoxa (Rao) V. Poljansk. (=5. paradoxa Rao) (рис. 89, 7). —
Вегетативные: клетки 82—85 мкм шир. Хлоропластов 3—4. Восприни-
мающие клетки вздутые со стороны конъюгационного канала или не-
вздутые. Зигоспоры от широкоэллипсоидных до почти шаровидных,
73—82 мкм шир.
В дождевой луже. Индия.
Редкая форма. Отличается от типовой формы вздутием, хотя и не-
постоянным, воспринимающих клеток со стороны конъюгационного ка-
нала и отчасти формой зигоспор. Учитывая, что некоторая тенденция
к вздутию клеток со стороны конъюгационного канала иногда свойст-
венна и типовой форме, можно предположить, следуя Кригеру (Krieger
in: Kolkwitz, Krieger, 1941—1944 : 353), что f. paradoxa — синоним
последней.
14 Рундина Л. А.
209
Рис. 89. 1, 2, 5—7 — Spirogyra nitida: 1, 2, 5, 6 — f. nitida, 7 — f. paradoxa, зигоспора в
разных положениях; 3, 4 — S. setiformis: 8, 9 — S. elliptica. (По: 1,2 — Gauthier-Lievre,
1965; 3, 4 — Рувдина, 19786; 5, 6, 8 — Jao, 1935b; 7 — Rao, 1937 (как S. paradoxa)', 9 —
Полянский, 1951).
26. Spirogyra setiformis (Roth) Kutz. [=5. jugalis (Dillw.) Kutz.;
5. wollnyi De Toni; 5. yunnanensis Li] — Спирогира щетинковидная
(рис. 89, 3, 4). — Вегетативные клетки (85) 90—115 мкм шир., по
длине меньше ширины, равны ей или до 3 (6) раз больше. Поперечные
стенки гладкие. Хлоропластов (3) 4—8 (10), часто слабо извитые.
Конъюгация лестничная. Конъюгационный канал образован обеими
клетками равномерно, иногда с преобладанием отдающей клетки. Вос-
принимающие клетки невздутые, редко очень слабо вздутые. Клетки
без конъюгационной пары невздутые. Зигоспоры эллипсоидные, с более
или менее заостренными вершинами, (66) 77—98 (112) мкм шир., (80)
87—160 (180) мкм дл. Экзоспорий тонкий, бесцветный. Мезоспорий
толстый, от желтого до коричневого, гладкий.
В стоячих и текучих водах, нередко заметно загрязненных, на ри-
совых полях. Отмечена приуроченность вида к богатым известью водам
со щелочной реакцией. Россия: европ. часть, Зап. Сибирь (окрестности
Томска), Вост. Сибирь (окрестности Иркутска, окрестности оз. Бай-
210
кал), Дальн. Восток (Приморский край); Украина — всюду, Южн. Ка-
захстан, Южн. Узбекистан.
Широко распространенный вид. Очень близок к S. nitida, и не
исключено, что эти виды иногда смешивались.
27. Spirogyra elliptica Jao, non Skv. (=S. hatillensis Trans.) — Спи-
рогира эллиптическая (рис. 89, 8, 9). — Вегетативные клетки 115—
128 мкм шир., по длине равны ширине или до 5 раз больше. Попе-
речные стенки гладкие. Хлоропластов 4—6, слабо извитые. Конъюга-
ция лестничная. Конъюгационный канал образован обеими клетками
равномерно, нередко с преобладанием отдающей клетки. Воспринима-
ющие клетки невздутые. Клетки без конъюгационной пары невздутые.
Зигоспоры эллипсоидные, часто с более или менее заостренными вер-
шинами, 82—106 мкм шир., 128.2—202 мкм дл. Экзоспорий тонкий,
бесцветный, гладкий. Мезоспорий толстый, от желтого до коричневого,
гладкий.
В стоячих водах, на рисовых полях. Украина (Полтава).
Редкий вид, приуроченный в основном к субтропической зоне. Бли-
зок к 8. setiformis, характеризующемуся меньшей шириной вегетатив-
ных клеток. Возможно, является синонимом 8. ellipsospora.
28. Spirogyra ellipsospora Trans. (=8. splendida G. S. West) — Спи-
рогира эллипсоспоровая (рис. 90, У). — Вегетативные клетки (120)
125—150 (155) мкм шир., по длине равные ширине или до 4 раз больше.
На поверхности клеток нередко имеются сдвоенные игольчатые крис-
таллы оксалата кальция. Поперечные стенки гладкие. Хлоропластов 3—
8. Конъюгация лестничная. Конъюгационный канал образован обеими
клетками. Воспринимающие клетки невздутые. Зигоспоры эллипсоид-
ные, с более или менее заостренными вершинами, реже цилиндричес-
ки-эллипсоидные, 95—140 мкм шир., (150) 160—255 мкм дл. Мезоспо-
рий толстый, от коричневато-желтого до коричневого, гладкий.
В стоячих и текучих водах, на рисовых полях. Вост. Казахстан
(оз. Зайсан), Узбекистан (отводной канал Дегрезского водохранилища,
оросительные каналы).
Редко встречающийся вид с широким ареалом. Известен также из
Нидерландов, Германии, Китая, Индии, США и Колумбии. Сходен с
8. crassa и особенно с 8. crassoidea, отличается двухосно-эллипсоид-
ными зигоспорами. Близок к 8. elliptica, отличаясь от него главным
образом большей шириной вегетативных клеток. Возможно, что оба
являются в действительности лишь формами одного вида.
29. Spirogyra irregularis Nag., non sensu Gutwinski, 1895a, 1895b
[=8. decimina (Miill.) Kiitz. var. oblonga Wang] — Спирогира неправиль-
ная (рис. 91, 1—4). — Вегетативные клетки 31—42 мкм шир., по длине
в 1.5—7 раз больше. Поперечные стенки гладкие. Хлоропластов 2—4,
слабо извитые. Конъюгация лестничная, редко боковая. Конъюгацион-
ный канал при лестничной конъюгации образован обеими клетками; при
боковой конъюгации канал выдается примерно на 1/2 ширины клетки,
нить при этом прямая. Воспринимающие клетки невздутые. Клетки без
211
Рис. 90. 1 — Spirogyra ellipsospora', 2 — S. elongata f. rupestris. (По: I — Transeau, 1914;
2 — Рундина, 1978a).
конъюгационной пары невздутые. Зигоспоры эллипсоидные или эллип-
соидно-цилиндрические, с широко закругленными вершинами, 32—40
(46) мкм шир., 34—90 (105) мкм дл. Экзоспорий тонкий, бесцветный,
гладкий. Мезоспорий толстый, от желтого до желтовато-коричневого,
гладкий. Партеноспоры сходные по форме и строению с зигоспорами.
В стоячих и текучих водах, олигосапроб. Россия: центр европ.
части, Зап. Сибирь (окрестности Томска); Зап. Украина (окрестности
Тернополя), горный Крым (водопад Учан-су), Сев. Казахстан.
Близок к S. decimina, от которого отличается большим числом хло-
ропластов. Сходен по многим признакам с S. neglecta f. fuetlebornii,
но имеет более узкие вегетативные клетки. Возможно, идентичен S. ri-
vularis.
30. Spirogyra hyalina Cleve (=5. pseudoneglecta Czurda, non Gaut-
hier-Lievre) — Спирогира гиалиновая (рис. 91, 5—9). — Вегетативные
клетки (45) 55—60 (67.5) мкм шир., по длине в (1) 2—6 раз больше.
Поперечные стенки .гладкие. Хлоропластов (2) 3—4 (5), слабо извитые
или редко почти параллельные. Конъюгация лестничная и боковая.
212
Конъюгационный канал при лестничной конъюгации образован обеими
клетками, при боковой конъюгации обычно резко выдается, приблизи-
тельно на 1/2—3/4 ширины клетки, нить при этом прямая или незна-
чительно изогнутая. Воспринимающие клетки незначительно вздутые
со всех сторон или невздутые. Клетки без конъюгационной пары не-
вздутые. Зигоспоры эллипсоидные, 45—60 мкм шир., (60) 75—160 мкм
дл. Экзоспорий тонкий, бесцветный, гладкий. Мезоспорий толстый,
красновато- или каштаново-коричневый, гладкий. Партеноспоры эллип-
соидные, редкие.
В стоячих и текучих водах, в том числе в промышленных стоках.
Россия: северо-запад и центр европ. части; Украина (окрестности
Киева).
Широко распространенный вид на всех континентах. Близок к
5. ternata и S. neglecta f. fuellebornii, отличается от первого вида на-
личием наряду с лестничной боковой конъюгации и формой зигоспор,
Рис. 91. 1—4 —• Spirogyra irregularis1. 1,4 — лестничная конъюгация, 2 — вегетативная
клетка, 3 — боковая конъюгация; 5—9 — 5. hyalina, разная степень вздутия воспринимаю-
щих клеток при лестничной (5, 7, 8) и боковой (б, 9) конъюгациях; 10, 11 — 5. dubia1, 12 —
5. margaritata1, 13—15 — S. pseudovarians: 13, 14 — лестничная конъюгация, 15 — строение
оболочек зигоспоры (поперечный разрез). (По: 1 — Jao, 1935b; 2, 3 — Wang, 1933 (как 5.
decimina var. oblonga); 4 — Yamagishi, 1966b; 5, 6, 13—15 — Czurda, 1932; 7—9 — Cleve,
1868; 10 — Рундина, 1988; 11 — Gauthier-Lievre, 1965; 72 — Transeau, 1951).
213
а от второго — часто более сильным вздутием воспринимающих кле-
ток и более широкими вегетативными клетками.
31. Spirogyra elongata (Berk.) Kiitz., non Wood — Спирогира удли-
ненная (рис. 90, 2). — Вегетативные клетки (20) 22—33 мкм шир., по
длине в 1.5—14 раз больше. Ризоиды латеральные, частые. Поперечные
стенки гладкие. Хлоропластов (1) 2—3. Конъюгация лестничная. Конъ-
югационный канал образован обеими клетками, с преобладанием отда-
ющей клетки, редко только от нее, более или менее четко отграничен-
ный. Воспринимающие клетки со всех сторон сильно вздутые, до 49—
52 мкм шир. Клетки без конъюгационной пары невздутые. Зигоспоры
эллипсоидные, иногда почти цилиндрические, с более или менее широко
закругленными, редко заостренными вершинами, 28—52 мкм шир., 48—
120 мкм дл. Экзоспорий тонкий, бесцветный, гладкий. Мезоспорий тол-
стый, от желтого до коричневого, гладкий.
Редкий вид.
F. elongata [=5. macrospora (Rao) Krieger], — Вегетативные клетки
(20) 22—26 (30) мкм шир. Хлоропластов (1) 2—3. Конъюгационный
канал образован обеими клетками равномерно. Зигоспоры эллипсоидные,
нередко с заостренными вершинами, 28—37 мкм шир., 48—120 мкм дл.
В мелких, стоячих и текучих водах, свободноплавающая или в при-
крепленном состоянии. Ископаемые находки — в карбоне. Россия:
центр европ. части, горная Башкирия, Предкавказье (Ставропольский
край).
Довольно редкая форма, распространенная также в Англии, Авст-
ралии и Сев. Америке. Сходна по ширине вегетативных клеток с
S. subsalsa, отличается обычно большим числом хлоропластов, более
крупными зигоспорами и образованием конъюгационного канала от
обеих клеток. Довольно близка к S. rhizoides Randh. (Сев. Индия), но
последний вид. имеет иной цвет мезоспория (голубоватый или бледно-
желтый — по разным сведениям), очень толстые и темные клеточные
стенки, иную форму зигоспор.
F. rupestris (Schmidle) V. Poljansk. [=5. rupestris Schmidle; S. rho-
dopea Petkoff (?)] (рис. 90, 2). — Вегетативные клетки 26—33 мкм
шир. Хлоропластов 2—3. Конъюгационный канал образован обеими
клетками, с преобладанием отдающей клетки, редко только от нее.
Зигоспоры эллипсоидные или почти цилиндрические, с широко закруг-
ленными вершинами, 36—52 мкм шир., 48—101.5 мкм дл.
В стоячих водах, прудах, на рисовых полях, влажных скалах, под-
водных известковых камнях, свободноплавающая или в прикрепленном
состоянии. Южн. Казахстан.
Редкая форма, известная также из Болгарии и Индии. Отличается
от наиболее близких видов — S. fluviatilis и S. dictyospora гладким
мезоспорием, от 5. decimina f. olivascens — большим числом хлоро-
пластов, меньшей шириной вегетативных клеток, большим вздутием
воспринимающих клеток и частой встречаемостью ризоидов, от S. ri-
vularis f. minor — сильно вздутыми воспринимающими клетками.
214
32. Spirogyra dubia Kiitz. [=S. dubia var. genuina Kirchn.; S. turfosa
Gay var. manschurica Skv.; S. adnata (Vauch.) Kiitz. var. obscura Petkoff;
S. ternata Ripart sensu Gutwinski, 1895a; S. ternata Ripart f. gagrensis
Lobik (?)] — Спирогира сомнительная (рис. 91, 10, 11). — Вегетатив-
ные клетки 40—52 (56) мкм шир., по длине до 5 (8) раз больше.
Ризоиды латеральные и базальные, редкие. Поперечные стенки гладкие.
Хлоропластов 2—4 (6). Конъюгация лестничная. Конъюгационный
канал образован обеими клетками, со стороны воспринимающей клетки
он отграничен иногда менее резко. Воспринимающие клетки вздутые
со всех сторон, до 61—81 мкм шир., редко почти невздутые или взду-
тые преимущественно со стороны, противоположной конъюгационному
каналу. Клетки без конъюгационной пары невздутые. Зигоспоры эл-
липсоидные, изредка с более или менее заостренными вершинами,
иногда почти шаровидные, 40—67 мкм шир., 57—118 мкм дл. Экзо-
спорий тонкий, бесцветный, гладкий. Мезоспорий толстый, от желто-
коричневого до коричневого, гладкий. Партеноспоры эллипсоидные,
35—46 мкм шир., 46—52 мкм дл.
В стоячих, изредка в текучих водах, старицах рек, на рисовых
полях. Россия: европ. часть — повсюду, Сев. Осетия; Зап. и Вост.
Украина, Присивашье, степной и горный Крым, Зап. и Вост. Закав-
казье, Грузия, Азербайджан, Узбекистан, Казахстан.
Вид с широким ареалом. Степень вздутия воспринимающих клеток
варьирует даже в пределах одной нити, но в большинстве случаев
вздутие выражено вполне отчетливо, что служит главным отличием от
S. neglecta f. fuellebornii. От S. elongata f. rupestris хорошо отличается
большей шириной вегетативных клеток и образованием конъюгацион-
ного канала от обеих клеток.
33. Spirogyra margaritata Wollny [=5. setiformis (Roth) Kiitz. f.
margaritata (Wollny) V. Poljansk.] — Спирогира жемчужная (рис. 91,
12). — Вегетативные клетки 100—120 мкм шир., по длине в 1.5—
4 раза больше. Поперечные стенки гладкие. Хлоропластов 13—15,
очень слабо извитые, почти параллельные. Конъюгация лестничная.
Конъюгационный канал образован обеими клетками. Воспринимаю-
щие клетки невздутые или незначительно вздутые до 165 мкм шир.,
преимущественно со стороны, противоположной конъюгационному
каналу. Клетки без конъюгационной пары невздутые. Зигоспоры эл-
липсоидные, 92—125 мкм шир., 140—200 мкм дл. Мезоспорий ко-
ричневый, гладкий.
В стоячих водах. Германия, Индия.
Очень редкий вид. Значительное количество хлоропластов отличает
его от S. setiformis.
34. Spirogyra polytaeniata Strasburger — Спирогира многоленточ-
ная. — Вегетативные клетки 150—189 мкм шир., по длине равные
ширине или до 1.5 раза больше. Поперечные стенки гладкие. Хлоро-
пластов 12—14, с половинным оборотом по спирали. Конъюгация лест-
ничная. Конъюгационный канал образован обеими клетками. Воспри-
нимающие клетки невздутые. Клетки без конъюгационной пары не-
215
вздутые. Зигоспоры эллипсоидные, 120 мкм шир., 158 мкм дл. Мезо-
спорий коричневый, гладкий.
В стоячей воде. Швейцария.
Вид редкий, требующий дополнительного изучения. Изображение
отсутствует. От близкого вида 5. margaritata, имеющего сходное ко-
личество хлоропластов, отличается значительной шириной вегетатив-
ных клеток. По тому же признаку отличается от S. desikacharyensis
Rattan (Индия) и от 5. subpolytaeniata Jao (Китай), имеющих 10—12
хлоропластов и ширину вегетативных клеток 92—105 мкм — у пер-
вого вида и ПО—125 мкм — у второго.
35. Spirogyra pseudovarians Czurda — Спирогира ложноизменчи-
вая (рис. 91, 13—15). — Вегетативные клетки 36—39 мкм шир., по
длине до 2 раз больше. Поперечные стенки гладкие. Хлоропласт 1,
редко их 2. Конъюгация лестничная. Конъюгационный канал образован
обеими клетками с преобладанием отдающей клетки. Воспринимающие
клетки преимущественно вздутые со стороны конъюгационного канала.
Клетки без конъюгационной пары невздутые. Зигоспоры эллипсоидные,
33—37 мкм шир., 47—57 мкм дл. Экзоспорий толстый, бесцветный,
однослойный, снаружи с округлыми ямчатыми углублениями, иногда
неотчетливыми, вследствие чего кажется двухслойным. Мезоспорий
тонкий, темно-коричневый, гладкий.
В стоячих водах, природниковых лужах. Дания, Австрия, Чехия.
Редкий вид и недостаточно изученный, особенно в отношении стро-
ения оболочек зигоспор. Близок к 5. varians, от которого хорошо
отличается скульптурированностью экзоспория. Этим же признаком и
отчетливым вздутием воспринимающих клеток хорошо отличается и
от S. decimina, сходного по ширине вегетативных клеток и количеству
хлоропластов.
36. Spirogyra velata Nordst., auct. non Nordst.: Рябинин, 1889 —
Спирогира укутанная (рис. 92). — Вегетативные клетки 28—50 мкм
шир., по длине в (1.5) 2—5 (7) раз больше. Поперечные стенки глад-
кие. Хлоропласт 1, очень редко их 2. Конъюгация лестничная и бо-
ковая. Конъюгационный канал при лестничной конъюгации образован
обеими клетками; при боковой конъюгации канал широко закруглен-
ный, выдается на 1/4—1/3 ширины клетки, нить при этом почти прямая
или значительно изогнутая. Воспринимающие клетки невздутые или
слабо, иногда значительно вздутые со стороны конъюгационного кана-
ла. Клетки без конъюгационной пары обычно невздутые или иногда
бочонковидные или почти шаровидные. Зигоспоры широкоэллипсоид-
ные, иногда с более или менее заостренными вершинами, реже широ-
кояйцевидные, почти шаровидные или цилиндрически-эллипсоидные,
33—48 (59) мкм шир., 50—105 (150) мкм дл. Экзоспорий бесцветный,
стекловидный, двухслойный: внешний слой тонкий, гладкий, внутрен-
ний слой очень толстый, грубоямчатый (бугристый). Мезоспорий тон-
кий, от желто-коричневого до коричневого, гладкий, иногда слоистый.
Эндоспорий тонкий, трудноразличимый. Партеноспоры редкие.
Довольно редкий вид, морфологически малоизменчивый.
216
F. velata. — Вегетативные клетки 28—41 мкм шир. Конъюгация
лестничная, редко боковая. Воспринимающие клетки невздутые или
слабо, иногда значительно вздутые со стороны конъюгационного кана-
ла. Клетки без конъюгационной пары невздутые. Зигоспоры широко-
эллипсоидные, 33—48 (59) мкм шир., 50—100 (150) мкм дл. Мезо-
спорий иногда слоистый.
В мелких стоячих водоемах, реже в текучих водах, в одном случае
в минеральном углекислом источнике при температуре воды 34—42 °C.
Россия: Зап. Сибирь (Южн. Алтай, Рахмановские ключи, долина
р. Арасан); Украина (окрестности Киева и долина р. Ворсклы).
По строению оболочек зигоспоры близка к 5. occidentalis и S. scro-
biculata, по ширине вегетативных клеток является переходным звеном
между ними.
F. ellipsoidea (Wang) V. Poljansk. (=5. velata var. ellipsoidea Wang;
5. subvelata Krieger). — Вегетативные клетки 34—50 мкм шир. Конъ-
Рис. 92. Spirogyra velata-. 1—4 — модификация формы зигоспор, лестничная конъюгация,
5—7 — боковая конъюгация (5 — зигоспора, поперечный разрез, 6 — вздутие клеток без
конъюгационной пары). (По: Рундина, 1988).
217
югация боковая, реже лестничная. Воспринимающие клетки невздутые.
Клетки без конъюгационной пары часто бочонковидно вздутые. Зиго-
споры эллипсоидные, с более или менее заостренными вершинами,
34—48 мкм шир., 50—105 мкм дл. Мезоспорий не слоистый.
В стоячих водах, лужах. Германия, Китай.
Очень редкая форма. По ширине вегетативных клеток более, чем
типовая форма, приближается к S. occiaentalis, но у последнего вида
клетки шире, число хлоропластов больше, оболочка зигоспор постро-
ена иначе, вершины зигоспор не заострены и отсутствует боковая конъ-
югация.
F. lateralis Gauthier-Lievre. — Вегетативные клетки 30—37 мкм
шир. Конъюгация боковая. Воспринимающие клетки вздутые. Зигоспо-
ры эллипсоидные, 38—42 мкм шир., 48—62 мкм дл.
В стоячих водах. Африка (Сев. Сахара).
Очень редкая форма, по всей вероятности идентичная типовой.
37. Spirogyra tuberculata Lagerh. — Спирогира бугорчатая. — Ве-
гетативные клетки 35—37 мкм шир., по длине в 2—4.5 раза больше.
Поперечные стенки гладкие. Хлоропласт 1. Конъюгация лестничная.
Конъюгационный канал образован обеими клетками. Воспринимающие
клетки вздутые со всех сторон. Зигоспоры эллипсоидные, 30—38 мкм
шир., 50—67 мкм дл. Экзоспорий тонкий, бесцветный, гладкий. Ме-
зоспорий толстый, коричневый, с бугорчатыми (бородавчатыми) воз-
вышениями разного размера.
В стоячих водах. Швеция.
Редкий вид, многими исследователями считающийся недостаточно
изученным. Оригинальное изображение отсутствует.
38. Spirogyra pseudowoodsii V. Poljansk. — Спирогира псевдовудса
(рис. 93, 1—4). — Вегетативные клетки 42.5—45 мкм шир., по длине
меньше ширины до 3 раз, реже несколько больше — до 1.5 раза.
Поперечные стенки гладкие. Хлоропласт 1, с 1/2—2 оборотами спи-
рали. Конъюгация боковая, реже лестничная. Конъюгационный канал
при лестничной конъюгации образован обеими клетками, при боковой
конъюгации нечетко отграниченный, особенно со стороны восприни-
мающей клетки, выдается приблизительно на 1/2 или несколько более
ширины клетки, нить при этом прямая или слабо изогнутая. Воспри-
нимающие клетки более или менее вздутые, особенно со стороны конъ-
югационного канала. Клетки без конъюгационной пары сильно вздутые,
иногда почти шаровидные. Зигоспоры эллипсоидные, 32.5—40 мкм
шир., 45—67.5 мкм дл. Экзоспорий тонкий, бесцветный, гладкий. Ме-
зоспорий толстый, желто-коричневатый, бугристый.
В стоячей воде. Россия: юг европ. части (окрестности Ростова-на-
Дону).
Редкий вид. Степень бугристости мезоспория сильно варьирует,
часто даже у зигоспор в одной нити; иногда бугристость отсутствует.
Главное отличие от S. varians f. woodsii — скульптурированность ме-
зоспория. В свете новых данных о бугристости мезоспория у S. varians,
218
Рис. 93. 1—4 — Spirogyra pseudowoodsii: 1 — лестничная конъюгация, 2 — боковая и
лестничная конъюгации в одной нити, зигоспоры в разных положениях, 3, 4 — строение зи-
госпоры (поперечный разрез); 5—7 — 5. tenuispina: 5 — зигоспора с фрагментом строения
мезоспория (с поверхности и в оптическом разрезе), 6 — лестничная конъюгация, 7 — бо-
ковая конъюгация; 8, 9 — S. subsalso-punctulata'. 8 — лестничная конъюгация, 9 — зиго-
спора, строение мезоспория (поперечный разрез). (По:/—4— Полянский, 1959; 5—7—
Рундина, 19776; 8, 9 — Красноперова, 1966).
219
хорошо заметной в сканирующем микроскопе, возможен пересмотр
видовой значимости S. pseudowoodsii.
39. Spirogyra subsalso-punctulata Kadi. (=5. subsalsa Kiitz. f. pun-
ctulata Krasnop.) — Спирогира солонцевато-точечная (рис. 93, 8, 9). —
Вегетативные клетки 20—25 (29) мкм шир., по длине в 4.5—6 раз
больше. Поперечные стенки гладкие. Хлоропласт 1. Конъюгация лест-
ничная. Конъюгационный канал образован обеими клетками, часто с
преобладанием отдающей клетки, редко только от нее. Воспринимаю-
щие клетки веретеновидно вздутые со всех сторон, до 46—55 мкм
шир. Клетки без конъюгационной пары невздутые. Зигоспоры широ-
коэллипсоидные, 38—43.5 (47) мкм шир., 55—61 (71) мкм дл. Экзо-
спорий тонкий, бесцветный, гладкий. Мезоспорий толстый, 2.8—
3.5 мкм толщ., желто-коричневый, тонко- и остроигольчатый (тонко-
радиальный в поперечном сечении, мелкоточечный сверху).
В стоячей воде пресных и соленых озер (сульфатно-хлоридный тип
засоления), на поверхности рапы после дождя. Сев. Казахстан (запо-
ведник Кургальджино).
Редкий вид. Экологически и морфологически, за исключением
скульптурированного мезоспория, обнаруживает значительное сходство
с кругом форм S. subsalsa, формой которого и был первоначально
описан. Близок по строению мезоспория к S. tenuispina, но имеет
более широкие клетки и только лестничную конъюгацию. Есть пред-
положение об идентичности S. hoehnei Borge (Бразилия).
40. Spirogyra punctiformis Trans. (=S. punctata Cleve sensu Kadlu-
bowska, 1984, pr. p.) — Спирогира точковидная (рис. 94, 1, 2). —
Вегетативные клетки 27—30 (39) мкм шир., по длине в 5—13 раз
больше. Ризоиды латеральные, вильчато разветвленные. Поперечные
стенки гладкие. Хлоропластов 1—2. Конъюгация лестничная. Конъюга-
ционный канал образован отдающей клеткой. Воспринимающие клетки
веретеновидно или цилиндрически-веретеновидно вздутые, 45—51 мкм
шир., расположенные попарно или одиночно. Клетки без конъюгаци-
онной пары невздутые. Зигоспоры широкоэллипсоидные, 40—48 мкм
шир., 59—110 мкм дл. Экзоспорий толстый, бесцветный, гладкий. Ме-
зоспорий желтый, мелкоточечный.
В стоячих и текучих водах, озерах, на рисовых полях, в коллек-
торно-дренажной сети. Россия: Вост. Сибирь (Республика Саха: оз.
Ирипчан); Узбекистан (Ферганская долина).
Довольно редкий и малоизученный вид. Встречен также в США.
От близких видов — S. punctata, S. aequinoctialis G. S. West (Вост.
Африка), 5. schmidtii W. et G. S. West (Норвегия; Сиам; Мьянма;
Китай; США; Африка: Гвинея, Чад) отличается толстым экзоспорием,
шириной вегетативных клеток и количеством хлоропластов. По мнению
некоторых исследователей, не является самостоятельной видовой еди-
ницей.
41. Spirogyra tenuispina Rund. — Спирогира тонкоигольчатая (рис. 93,
5—Т). — Вегетативные клетки 14.5 (17.4) мкм шир., по длине в 3—7
220
Рис. 94. 1,2 — Spirogyra punctiformis'. 1 — лестничная конъюгация, 2 — строение мезо-
спория (фрагмент); 3—5 — S. salina-. 3 — зигоспора и партеноспоры в иевздутых гаметан-
гиях, 4 — зигоспоры во вздувшихся гаметангиях, 5 — строение мезоспория, с поверхности
и в поперечном сечении. (По: 1 — Transeau, 1914; 2 — Hoek van den, 1959; 5—5 — Aleem,
1952).
раз больше. Поперечные стенки гладкие. Хлоропласт 1. Конъюгация
лестничная, редко боковая. Конъюгационный канал при лестничной
конъюгации образован обеими клетками, с преобладанием отдающей
клетки, редко только ею, конъюгирующие нити при этом прямые или
реже слегка коленчато изогнутые; при боковой конъюгации канал вы-
дается на 1/3 ширины клетки, нить при этом прямая или незначительно
изогнутая. Воспринимающие клетки веретеновидно вздутые до 26—
32 мкм шир. Клетки без конъюгационной пары невздутые. Зигоспоры
221
эллипсоидные, с более или менее заостренными вершинами, 20—23
(29) мкм шир., (40.6) 43.5—52 мкм дл. Экзоспорий тонкий, бесцветный,
гладкий. Мезоспорий толще экзоспория в 3—3.5 раза, желтоватый,
тонко- и частоигольчатый.
В рыбоводном пруду. Южн. Казахстан.
Редкий вид. По вегетативным признакам сходен с S. flavescens, от-
личается скульптурированным мезоспорйем. От 5. tuberculata, имею-
щего сходную скульптуру мезоспория, отличается меньшей шириной
вегетативных клеток. Очень близок к S, bourrellyana Gauthier-Lievre
(Африка), но имеет гладкие, а не складчатые, как у последнего вида,
поперечные стенки. Тонкоигольчатый мезоспорий у 5. tenuispina при
взгляде на поверхность зигоспоры выглядит мелкоточечным, сбоку (в
поперечном сечении) — тонкорадиальным. Мелкоточечность мезоспо-
рия характерна и для сходного по всем остальным признакам S. рег-
forans Trans. (США). Вполне возможно, что эти виды идентичны.
42. Spirogyra salina Aleem — Спирогира солоновато-водная (рис. 94,
3—5). — Вегетативные клетки 30—40 мкм шир., по длине в 5—10 (12)
раз больше. Поперечные стенки гладкие. Хлоропласт 1. Конъюгация
лестничная, редко боковая. Конъюгационный канал образован обеими
клетками, иногда с преобладанием отдающей клетки. Воспринимающие
клетки невздутые или вздутые до ширины зигоспор. Клетки без конъ-
югационной пары невздутые. Зигоспоры от коротко- и широкоэллипсо-
идных до почти шаровидных или цилиндрически-эллипсоидных, 35—
62 мкм шир., 60—85 мкм дл. Экзоспорий тонкий, бесцветный, гладкий.
Мезоспорий толстый, коричнево-красный, точечный. Партеноспоры эл-
липсоидные, 30 мкм шир., 45—80 мкм дл.
В прудах, при солености 5—17.5 %о и pH 6.6—8.2. Легко разви-
вается и конъюгирует в культуре. Конъюгирует в массе при солености
воды 3—13 %о. Франция.
Редкий вид, своеобразный по экологии. Близок к S. esthonica по
вегетативным признакам, отличается строением мезоспория.
43. Spirogyra lagerheimii Wittr. — Спирогира Лагерхейма (рис. 95,
1, 2). — Вегетативные клетки 24—33 мкм шир., по длине в 1.25—6
раз больше. Ризоиды латеральные, вильчато разветвленные. Поперечные
стенки гладкие. Хлоропласт 1, очень редко их 2. Конъюгация боковая
и лестничная, иногда совмещенные в одной паре нитей. Конъюгаци-
онный канал при боковой конъюгации выдается более или менее резко,
до 1/2 ширины клетки, иногда — слабо, редко ушковидный, нить при
этом прямая или слабо изогнутая; при лестничной конъюгации канал
образован обеими клетками. Воспринимающие клетки невздутые. Клет-
ки без конъюгационной пары иногда вздутые. Зигоспоры широко-
эллипсоидные, часто удлиненно-эллипсоидные, с заостренными вер-
шинами, 25—38 мкм шир., 30—100 мкм дл. Экзоспорий тонкий,
бесцветный, гладкий. Мезоспорий толстый, желтовато-коричневый,
мелкоточечный. Эндоспорий и линия разрыва отчетливые на зрелых
зиготах. Партеноспоры короткоэллипсоидные, 29—32 мкм шир., 38—
42 мкм дл.
222
Малоизученный вид. По наблюдениям в сканирующем микроскопе
мезоспорий двухслойный, причем внутренний слой густо- и плоскозер-
нистый.
F. lagerheimii (рис. 95, 1). — Вегетативные клетки 24—33 мкм
шир. Хлоропласт 1. Конъюгация боковая, реже лестничная, иногда со-
вмещенные в одной паре нитей. Зигоспоры удлиненно-эллипсоидные,
обычно с более или менее заостренными вершинами, 25—38 мкм шир.,
30—100 мкм дл.
В стоячих и текучих водах, водоемах ирригационной системы, на
рисовых полях, нередко в низовых болотах и ручьях со значительным
содержанием железа и несколько повышенным содержанием азота. Рос-
Рис. 95. 7, 2 — Spirogyra lagerheimii-. 1 — f. lagerheimii, боковая конъюгация; 2 — f. obo-
vata, лестничная конъюгация; 3 — S. subpapulata, лестничная конъюгация между тремя ни-
тями, зигоспоры и вздутые вегетативные клетки без конъюгационной пары. (По: 1 — Воро-
нихин в: Воронихин, Красноперова, 1970; 2, 3 — Jao, 1935b (2 — как S. obovata)).
223
сия: европ. часть, Зап. Сибирь (окрестности Челябинска и Томска),
Вост. Сибирь (Республика Саха); Эстония (о-ва Сааремаа и Хийумаа),
Латвия, Украина (окрестности Киева), Сев. Казахстан (заповедник Бо-
ровое), Киргизия (окрестности оз. Иссык-Куль), Узбекистан (?).
Широко распространенная форма. Обнаруживает значительное сход-
ство с S. decimina f. longata и S. varians. Близка к S. robusta (Nyg.)
Czurda (Южн. Африка). Указания на Местонахождения в пределах Уз-
бекистана сомнительны.
F. obovata (Jao) V. Poljansk. (=S. obovata Jao; S'. incerta Skv.) (рис. 95,
2). — Вегетативные клетки 25—29 мкм шир. Хлоропласт 1 (2). Конъ-
югация только лестничная. Зигоспоры широкоэллипсоидные, 25—
35 мкм шир., 40—73 мкм дл.
В ямах с гравием, на рисовых полях. Дания, Китай.
Редкая форма. Отличается от типовой формы отсутствием боковой
конъюгации и незаостренными зигоспорами, от S. luteospora — широ-
кими вегетативными клетками и строением мезоспория, от S. hoehnei
Borge (Бразилия) — невздутыми воспринимающими клетками и неза-
остренными зигоспорами.
44. Spirogyra subpapulata Jao — Спирогира пупырышковатая (рис. 95,
3). — Вегетативные клетки 22—31.3 мкм шир., по длине в 1.5—7.5 раза
больше. Поперечные стенки гладкие. Хлоропласт 1. Конъюгация лест-
ничная. Конъюгационный канал образован обеими клетками. Восприни-
мающие клетки вздутые со стороны конъюгационного канала. Клетки
без конъюгационной пары сильно вздутые со всех сторон. Зигоспоры эл-
липсоидные, с более или менее заостренными вершинами, 21—30
(34) мкм шир., (32) 40—61 мкм дл. Экзоспорий тонкий, бесцветный,
гладкий. Мезоспорий толстый, желтый, тонко- и густоточечный (пунк-
тированный). Партеноспоры эллипсоидные или почти шаровидные, 22—
26 мкм шир., 32—48 мкм дл.
В стоячих водах, на рисовых полях. Нидерланды, Германия, Алжир,
Марокко, Центр. Сахара, Китай, Япония.
Довольно широко распространенный вид. От близких видов —
S. papulata и S. micropunctata Trans. (США, Индия, Япония) отлича-
ется главным образом меньшей шириной вегетативных клеток и иным
строением мезоспория. Возможно, идентичен S. micropunctata.
45. Spirogyra danica Kadi. — Спирогира датская (рйс. 96, J, 2). —
Вегетативные клетки 24—32.1 мкм шир. Поперечные перегородки глад-
кие. Хлоропласт 1. Конъюгация боковая, очень редко лестничная.
Конъюгационный канал при лестничной конъюгации образован отдаю-
щей клеткой. Воспринимающие клетки сильно вздутые со стороны
конъюгационного канала. Клетки без конъюгационной пары шаровидно
вздутые. Зигоспоры эллипсоидные, с более или менее заостренными
вершинами, 25.6—32.1 мкм шир., 42.7—67.6 мкм дл. Экзоспорий тон-
кий, бесцветный, гладкий. Мезоспорий красно-бурый, с мелкими, ок-
руглыми, ямчатыми углублениями и нередко бурыми утолщениями (ка-
липтрами) на вершинах зигоспоры, часто и в других ее частях.
224
Рис. 96. 1,2 — Spirogyra danica, лестничная и боковая конъюгации, зигоспоры и вздутые
вегетативные клетки без конъюгационной пары; 3—6 — S. sibirica'. 3 — лестничная конъ-
югация, 4 — ряд вздутых вегетативных клеток без конъюгационной пары, 5,6 — зигоспо-
ры. (По: 1,2 — Kadhibowska in: Kadhibowska, Christensen, 1979; 3—6 — Скворцов, 19276).
15 Рундина Л. А.
225
В стоячих водах. Дания, Нидерланды.
Редкий вид. Близок к S. cateniformis по ширине вегетативных кле-
ток, отличается скульптурированным мезоспорием и наличием калиптр
на вершинах зигоспор. По наличию таких образований сходен с 5. va-
rians f. bicalyptrata, но имеет в отличие от последней формы меньшую
ширину вегетативных клеток и ямчаты^ мезоспорий.
46. Spirogyra luteospora Czurda — Спирогира желтоватоспоровая
(рис. 97, 7). — Вегетативные клетки 22—24 мкм шир., по длине в 2—4
раза больше. Поперечные стенки гладкие. Хлоропласт 1. Конъюгация
лестничная. Конъюгационный канал образован обеими клетками. Вос-
принимающие клетки невздутые или очень незначительно вздутые. Зи-
госпоры от широкоэллипсоидных до цилиндрически-эллипсоидных, 25—
26 мкм шир., 38—46 мкм дл. Экзоспорий тонкий, бесцветный, гладкий.
Мезоспорий толстый, желто-коричневый, с крупными, округлыми, ямча-
тыми углублениями, около 2 мкм диам.
В стоячих водах, старице реки. Чехия.
Редкий вид. На авторском рисунке на мезоспорий изображена кроме
ямчатости еще и точечность (пунктированность), не оговоренная особо
в первоописании. Отличается от S. czurdae Misra (Сев. Индия) несколь-
ко более узкими клетками и эллипсоидными зигоспорами, от 5. lager-
heimii f. obovata — меньшей шириной вегетативных клеток и строе-
нием мезоспория, от S. subpapulata — невздутыми воспринимающими
клетками.
47. Spirogyra acumbentis Vodenid. — Спирогира заостренная (рис. 97,
2—4). — Вегетативные клетки (38) 41 (45) мкм шир., по длине в 2—5
раз больше. Поперечные стенки гладкие. Хлоропласт 1. Конъюгация
лестничная. Конъюгационный канал образован обеими клетками. Вос-
принимающие клетки невздутые. Клетки без конъюгационной пары не-
вздутые. Зигоспоры широкоэллипсоидные или реже цилиндрически-эл-
липсоидные, (38) 42 (48) мкм шир., (45) 64 (88) мкм дл. Экзоспорий
тонкий, 0.5—0.7 мкм толщ., бесцветный, гладкий. Мезоспорий толс-
тый, 2—2.2 мкм толщ., оранжево-желтый, с равномерно расположен-
ными, правильной формы углублениями, 1.5—2.2 мкм диам., 0.6—
0.8 мкм глубины. Эндоспорий толстый, 1.1—1.5 мкм толщ., оранже-
во-желтый, гладкий.
В озере, свободноплавающая. Болгария.
Редкий вид. От наиболее близкого вида — S. luteospora отличается
большими размерами вегетативных клеток и зигоспор. По строению
мезоспория сходен с кругом форм S. lagerheimii и 5. czurdae Misra
(Сев. Индия), отличается от них ббльшими размерами вегетативных
клеток и зигоспор, а также грубыми и крупными углублениями на
мезоспорий. Цвет эндоспория вызывает сомнение.
48. Spirogyra rugulosa Iwan. (=5. vagans Teod.) — Спирогира мел-
коморщинистая (рис. 97, 5, 6). — Вегетативные клетки (45) 50—57
(62) мкм шир., по длине в 2—7 раз больше. Поперечные стенки глад-
кие. Хлоропласт 1. Конъюгация лестничная. Конъюгационный канал
226
Рис. 97. 1 — Spirogyra luteospora', 2—4 — S. acumbentis: 2, 3 — зигоспоры разной формы
с фрагментами строения мезоспория, 4 — строение зигоспоры с поверхности и в попереч-
ном разрезе; 5, 6 —. S. rugulosa. (По: 1 — Czurda, 1932; 2—4 — Водеиичаров в: Водеиичаров,
Киряков, 1970; 5 — Teodoresco, 1907; 6 — Иванов, 1901).
227
образован отдающей клеткой. Воспринимающие клетки невздутые или
слегка вздутые со стороны конъюгационного канала. Зигоспоры эллип-
соидные, с притупленными вершинами, 45—55 мкм шир., (55) 72—
127 мкм дл. Экзоспорий тонкий, бесцветный, гладкий. Мезоспорий
толстый, от желто-коричневого до коричневого, с мелкими, неправиль-
ной формы, тесно сомкнутыми ямчатыми углублениями, создающими
впечатление мелкой морщинистости. 41
В гипновых болотах, прудах и медленно текучих водах, на рисовых
полях. Россия: центр европ. части; Узбекистан.
Редкий вид, встреченный также в Румынии, Испании и США. Не-
достаточно изученный вид, особенно в отношении строения мезоспо-
рия, которое трактуется по-разному. По первоописанию, мезоспорий
имеет «ясную морщинистую (как у сухой сливы) или ямчатую струк-
туру», по Теодореско (Teodoresco, 1907), мезоспорий нежно точечно-
морщинистый. Если судить по авторскому рисунку, то мезоспорий
можно охарактеризовать как сетчато утолщенный, причем ямчатые уг-
лубления соответствуют участкам между сетчато расположенными утол-
щениями. От 5. punctata хорошо отличается значительно большей ши-
риной вегетативных клеток, невздутыми или лишь односторонне взду-
тыми воспринимающими клетками, участием в конъюгации всех или
почти всех клеток нити и строением мезоспория; от 5. kaffirita Trans.
(Южн. Африка) — более широкими клетками и строением мезоспория;
от S. novae-angliae Trans. (США) и S. Ьгиппеа — незначительным чис-
лом хлоропластов и иным строением мезоспория.
49. Spirogyra punctata Cleve, non Petit — Спирогира точечная
(рис. 98). — Вегетативные клетки (20) 24—40 мкм шир., по длине в
4—12 раз больше. Поперечные стенки гладкие. Хлоропласт 1 (2).
Конъюгация лестничная при участии немногих клеток нити. Конъюга-
ционный канал образован исключительно или преимущественно отда-
ющей клеткой. Воспринимающие клетки укороченные, равномерно
сильно вздутые со всех сторон. Клетки без конъюгационной пары не-
вздутые. Зигоспоры эллипсоидные, 28—66 мкм шир., 44—122 мкм дл.
Экзоспорий тонкий, бесцветный, гладкий. Мезоспорий от золотисто-
желтого до желто-коричневого, с крупными ямчатыми углублениями.
Вид достаточно интересный в эволюционно-морфологическом от-
ношении. Перед конъюгацией вегетативная (прогаметангиальная) клетка
делится на две неравные по длине клетки, причем собственно гаме-
тангиальной становится более короткая клетка.
F. punctata [-Temnogyra punctata (Cleve) Yamagishi] (рис. 98, 1,
2). — Вегетативные клетки (20) 24—32 (35) мкм шир. Хлоропласт 1
(2). Воспринимающие клетки умеренно вздутые, 40—65 мкм шир.,
73—165 мкм дл. Зигоспоры 28—53 мкм шир., 44—122 мкм дл. Ме-
зоспорий от золотисто-желтого до желто-коричневого, с округлыми
или неправильной формы ямчатыми углублениями.
В стоячих, иногда заболоченных водах, на рисовых полях, часто
на бревнах и мхах. Россия: север и северо-запад европ. части; Латвия
(окрестности Риги), Сев. Казахстан (заповедник Боровое), Узбекистан.
228
Широко распространенная форма, особенно в странах Европы. Не-
которые особенности сближают ее с S. collinsii f. ampla. Главное раз-
личие заключается в том, что у f. punctata отсутствует боковая конъ-
югация, свойственная S. collinsii f. ampla.
F. crassior Nordst. (=S. punctata var. major Hirn; S. punctata f. major
Hirn; S. suomiana Trans.) (рис. 98, 3—7). — Вегетативные клетки (30)
33—40 мкм шир. Хлоропласт 1. Воспринимающие клетки очень силь-
но вздутые. Зигоспоры 40—66 мкм шир., 60—120 мкм дл. Мезоспорий
золотисто-желтый, с крупными ямчатыми углублениями.
Рис, 98. Spirogyra punctata, лестничная конъюгация и зигоспоры: 1,2 — f. punctata', 3—7 —
f. crassior: 3, 4 — лестничная конъюгация, 5—7 — зигоспоры, вид с поверхности (5) и в
поперечном разрезе (6). (По: 1,2 — Gauthier-Lievre, 1965; 3 — Him, 1895 (как S. punc-
tata var. major}: 4—7 — Скворцов, 1927a (как S. punctata var. major}}.
229
В стоячих водах, в болоте, вместе с типовой формой. Россия: Ка-
релия (р. Умба).
Форма с широким ареалом. В Европе встречена в Англии и Фин-
ляндии. От типовой формы отличается большей шириной вегетативных
клеток, большим вздутием воспринимающих клеток и, судя по рисункам,
несколько более крупными углублениям^ на мезоспории. От близкого
по строению мезоспория 5. scrobiculata хорошо отличается характером
вздутия воспринимающих клеток, особенностями формирования конъ-
югационного канала, невздутыми клетками без конъюгационной пары и
более крупными зигоспорами.
50. Spirogyra sibirica Skv. — Спирогира сибирская (рис. 96, 3—
6). — Вегетативные клетки 17 мкм шир., по длине в 2—3 раза больше.
Поперечные стенки гладкие. Хлоропласт 1. Конъюгация лестничная.
Конъюгационный канал образован отдающей клеткой. Воспринимаю-
щие клетки вздутые со стороны конъюгационного канала. Клетки без
конъюгационной пары часто сильно вздутые, шаровидные или почти
шаровидные. Зигоспоры эллипсоидные, 20 мкм шир., 32—40 мкм дл.
Экзоспорий тонкий, бесцветный, гладкий. Мезоспорий толстый, золо-
тисто-желтый, густо покрытый мелкими ямчатыми углублениями.
В стоячей и текучей воде, озерах и реках, на сыртах, среди об-
растаний, в бентосе. Россия: Зап. Сибирь (Алтай, окрестности оз. Зай-
сан); Киргизия (Тянь-Шань, Терскей-Алатау).
Очень редкий вид. В первоописании мезоспорий значится густоям-
чатым, впоследствии автор вида охарактеризовал его крупноточечным.
Сильно вздутыми клетками, лишенными конъюгационной пары, напо-
минает S', bullata, однако хорошо отличается от него скульптурирован-
ным мезоспорием. Обнаруживает сходство также с S. subpapulata и
S. scrobiculata.
51. Spirogyra scrobiculata (Stockm.) Czurda — Спирогира ямчатая
(рис. 99; 100, 1, 2). — Вегетативные клетки 30—40 мкм шир., по длине
в 3—9 раз больше. Поперечные стенки гладкие. Хлоропласт 1. Конъ-
югация лестничная. Конъюгационный канал образован обеими клетками,
к концу конъюгации нечетко отграниченный со стороны воспринимаю-
щей клетки. Воспринимающие клетки сильно вздутые со стороны конъ-
югационного канала. Клетки без конъюгационной пары бочонковидно
вздутые, до 55—67 мкм шир. Зигоспоры широкоэллипсоидные, с при-
тупленными вершинами, 32—49 (52) мкм шир., 42—68 мкм дл. Экзо-
спорий тонкий, бесцветный, гладкий. Мезоспорий довольно толстый,
желтовато-коричневый, с плотно расположенными грубыми ямками,
5.29—7.06 мкм диам., окруженными выпуклыми кольцевыми складками.
В мелкой стоячей воде, старицах, болотах. Имеются сведения о
хорошем развитии в водах, обогащенных известью. Вегетирует в свет-
лые летние месяцы, может расти и конъюгировать теплой зимой. Легко
выращивается и конъюгирует в культуре, на почвенной вытяжке. Зи-
госпоры встречены в отложениях позднего ледникового периода и го-
лоцена, в песчаных почвах. Россия: Предкавказье (Сев. Осетия, окрест-
ности Владикавказа).
230
Рис. 99. Spirogyra scrobiculata. (По: Степняк в: Рундина, Степняк, 1979).
Вид достаточно широко распространенный в Европе (Дания, Гер-
мания, Чехия, Австрия, Италия). Близок к S. varians, с которым, ве-
роятно, часто смешивался, однако хорошо отличается от него скуль-
птурированным мезоспорием. Иное строение оболочек зигоспор и
форма клеток, лишенных конъюгационной пары, отграничивают его
от S. pseudovarians.
52. Spirogyra esthonica (Skuja) Czurda [=S. punctata Cleve var. es-
thonica Skuja; S. punctata f. esthonica (Skuja) V. Poljansk.; Temnogyra
esthonica (Skuja) Yamagishi] — Спирогира эстонская (рис. 100, 3, 4). —
Вегетативные клетки 27—32 (35) мкм шир., по длине в 3—12 раз
больше. Поперечные стенки гладкие. Хлоропласт 1. Конъюгация лест-
ничная. Конъюгационный канал образован обеими клетками, с преоб-
231
Рис. 100. 1,2 — Spirogyra scrobiculaia, лестничная конъюгация и зигоспора; 3, 4 — S. es-
thonica: 3 — лестничная конъюгация, 4 — строение мезоспория (вид с поверхности зиго-
споры); 5, 6 — S. daedalea f. daedaleoides: 5 — общий вид конъюгирующих клеток с зиго-
спорами (боковая конъюгация), 6 — две пары конъюгирующих клеток и скульптурирован-
ные зигоспоры; 7—9 — S. daedalea f. oudhensis, боковая и лестничная конъюгации и
зигоспора. (По: 1, 2 — Gauthier-Lievre, 1965; 3 — Skuja, 1928; 4—6 — Skuja, 1929 (5, 6 —
как S. daedalea); 7—9 — Randhawa, 1938 (как S. oudhensis)).
232
ладанием отдающей клетки. Воспринимающие клетки вздутые со всех
сторон, 46—61 мкм шир., 81—125 мкм дл. Клетки без конъюгацион-
ной пары невздутые. Зигоспоры эллипсоидные, 35.6—60 мкм шир.,
64—121.1 мкм дл. Мезоспорий желтый, со слабыми, коротковолнис-
тыми утолщениями, местами сливающимися в тонкую, иногда довольно
грубую сеть с ямчатыми выемками.
В стоячих водах, лужах, канавах, болотах, иногда прикреплен ко
мхам. Эстония (о-ва Сааремаа и Хийумаа).
Редкий вид. Отмечено, что в конъюгации, как и у S. punctata,
участвуют не все клетки нити. Отличается от последнего вида иным
строением мезоспория. Очень близок к кругу форм S. collinsii и S. dae-
dalea, с которыми встречается вместе. От S. lushanensis Li (Китай)
его отличает ббльшая ширина вегетативных клеток и зигоспор и мень-
шее вздутие воспринимающих клеток.
53. Spirogyra daedalea Lagerh. — Спирогира узорчатая (рис. 100,
5—9; 101, 1—3). — Вегетативные клетки 29—44 мкм шир., по длине
в 2—10 раз больше. Поперечные стенки гладкие. Хлоропласт 1. Конъ-
югация лестничная и боковая. Конъюгационный канал при лестничной
конъюгации образован обеими клетками, хорошо отграниченный; при
боковой конъюгации канал выдается не более чем на 1/2 ширины
клетки, нить при этом прямая или лишь незначительно изогнутая. Вос-
принимающие клетки обычно невздутые или очень слабо вздутые со
всех сторон, легко отчленяющиеся друг от друга. Клетки без конъюга-
ционной пары невздутые. Зигоспоры эллипсоидные, иногда со слегка
заостренными вершинами, 30—46 (58) мкм шир., (43.5) 46—98 мкм
дл. Экзоспорий тонкий, бесцветный, гладкий. Мезоспорий толстый, от
желто-коричневого до коричневого, с неправильно изогнутыми, развет-
вленными утолщениями, часто образующими широко или тесно сомк-
нутую сеть. Эндоспорий отчетливый, по толщине равный экзоспорию.
Линия разрыва отчетливая. Партеноспоры только в воспринимающих
клетках или одновременно в обоих гаметангиях, редкие.
Редкий вид. При просмотре многих образцов выяснилось, что сте-
пень сомкнутости сетчатых утолщений на мезоспорий, характеризую-
щая формы этого вида, — признак весьма относительный и сильно
зависящий от зрелости зиготы. К тому же разница в ширине вегета-
тивных клеток и размерах зигоспор у внутривидовых форм незначи-
тельна, а отсутствие или наличие боковой конъюгации — признак не-
постоянный. Все это вызывает сомнение в таксономической значимости
установленных форм вида.
F. daedalea (S. daedalea auct. non Lagerh.: Skuja, 1929) (рис. 101,
1, 2). — Вегетативные клетки 30—38 мкм шир. Конъюгация лестнич-
ная. Зигоспоры эллипсоидные, с более или менее широко закруглен-
ными вершинами, 30—44 мкм шир., 43.5—98 мкм дл. Мезоспорий с
неправильно изогнутыми, разветвленными утолщениями, образующими
широкую сеть.
В стоячих, преимущественно заболоченных водоемах. Россия: юг
европ. части (окрестности Ростова-на-Дону); Зап. Украина.
233
Рис. 101.1—3 — Spirogyra daedalea: 1,2 — f. daedalea, лестничная конъюгация и строение
мезоспория (фрагмент), 3 — f. africana: 4 — S. nodifera; 5 — S. pseudodaedaleoides. (По: 1,
2 — Полянский, 1955; 3 — Gauthier-Lievre, 1965 (как S. africana): 4 — Bock, Bock, 1956;
5 — Kadhibowska in: Kadhibowska, Christensen, 1979).
Довольно редкая форма с широким ареалом (Финляндия, Германия,
Югославия, Сев. Индия, США). При применении иммерсии становится
очевидным, что мезоспорий у зрелых зигот шиповато-игольчатый.
Шипы при взгляде сверху выглядят точками, которые при тесном и
неравномерном расположении, сливаясь, образуют неправильно изогну-
тые утолщения — линии в форме сетки.
F. africana (G. S. West) V. Poljansk. [=5. porticalis (Vauch.) Cleve
var. africana G. S. West; S. africana (G. S. West) Gauthier-Lievre, non
(Fritsch) Czurda; S. westii Trans.] (рис. 101, 3). — Вегетативные клетки
234
(33) 36—41 мкм шир. Конъюгация лестничная. Зигоспоры эллипсоид-
ные, с более или менее широко закругленными вершинами, 35—38
(42) мкм шир., 56—95 мкм дл. Мезоспорий очень плотно сетчатый.
В стоячих, часто заболоченных водоемах, постоянных и временных
лужах, топях. Алжир, Танганьика, Мадагаскар, Центр. Индия.
Редкая форма. Крайне близка к типовой, от которой отличается не-
сколько более широкими вегетативными клетками и плотносетчатым ме-
зоспорием. По последнему признаку она резко отличается не только от
типовой формы, у которой сетка на мезоспорий иногда вообще отсут-
ствует, но и от f. daedaleoides. От последней отличается также отсутст-
вием боковой конъюгации и широко закругленными вершинами зиго-
спор.
F. daedaleoides (Czurda) V. Poljansk. (=S. daedaleoides Czurda;
S. daedalea Lagerh. sensu Skuja, 1929, non auct. pl.) (рис. 100, 5, 6). —
Вегетативные клетки 29—44 мкм шир. Конъюгация лестничная и бо-
ковая. Зигоспоры эллипсоидные, с более или менее заостренными вер-
шинами, 30—46 мкм шир., 35.6—90 мкм дл. Мезоспорий сетчатый.
В стоячих, преимущественно заболоченных и текучих водах, ручьях,
канавах, приозерных лужах. Эстония (о-ва Сааремаа и Хийумаа).
Редкая форма. Отличается от типовой формы наличием боковой
конъюгации и зигоспорами с заостренными вершинами.
F. oudhensis (Randh.) V. Poljansk. (=S. oudhensis Randh.) (рис. 100,
7—9). — Вегетативные клетки 34—38 мкм шир. Конъюгация лестнич-
ная и боковая. Зигоспоры эллипсоидные, с более или менее закруг-
ленными вершинами, 30—40 мкм шир., 55—86 мкм дл. Мезоспорий
широкосетчатый, явственно отдаленный от экзоспория.
В рыбоводном пруду, свободноплавающая. Узбекистан.
Редкая форма, найденная прежде в Сев. Индии. Очень близка к
типовой форме, отличается от нее наличием боковой конъюгации. Эта
особенность сильно сближает ее с f. daedaleoides, однако f. oudhensis
отличается от нее широкосетчатым мезоспорием.
54. Spirogyra iabyrinthica Trans. (=S. rugulosa Iwan. sensu Krieger
in: Kolkwitz, Krieger, 1941—1944, pr. p.) — Спирогира извилистая. —
Вегетативные клетки 50—63 мкм шир., по длине в 3—7 раз больше.
Поперечные стенки гладкие. Хлоропласт 1. Конъюгация лестничная.
Конъюгационный канал образован обеими клетками. Воспринимающие
клетки невздутые или очень незначительно вздутые со всех сторон.
Зигоспоры эллипсоидные, с заостренными вершинами, 43—58 мкм
шир., 80—118 мкм дл. Экзоспорий тонкий, бесцветный, гладкий. Ме-
зоспорий толстый, от желто-коричневого до коричневого, покрытый
вытянутыми, неправильно изогнутыми утолщениями.
В стоячих водах. Германия.
Редкий вид. От S. daedalea, в качестве разновидности которого был
первоначально описан, отличается в основном значительно более круп-
ными размерами. Близок также к S. rugulosa, иногда рассматривается
как его синоним. Изображение отсутствует.
235
55. Spirogyra nodifera О. et W. Bock — Спирогира узлоносная
(рис. 101, 4). — Вегетативные клетки 33—42.5 мкм шир. Поперечные
стенки гладкие. Хлоропласт 1. Конъюгация лестничная. Конъюгацион-
ный канал образован обеими клетками, редко с преобладанием отда-
ющей клетки. Воспринимающие клетки вздутые со стороны конъюга-
ционного канала, редко — со всех сторон, изредка — совсем не взду-
тые. Клетки без конъюгационной пары более или менее шаровидно
вздутые. Зигоспоры эллипсоидные или цилиндрически-эллипсоидные,
27—42.7 мкм шир., 37.1—84 (112) мкм дл. Экзоспорий тонкий, бес-
цветный, гладкий. Мезоспорий толстый, желто-коричневый, неправиль-
но сетевидно-складчатый, с темно-коричневыми грубыми утолщениями
(калиптрами) на вершинах, реже на других местах зрелой зиготы.
Линия разрыва отчетливая.
В канавах, луговых озерках, рыбных прудах, родниках, при pH
6.0—7.5. Нидерланды, Германия, Польша.
Редкий и спорный вид, требующий дополнительного изучения. Схо-
ден с S. varians f. bicalyptrata, отличается скульптурированным мезо-
спорием. Возможно, идентичен 5. pseudonodifera. По данным СЭМ,
мезоспорий у 5. nodifera двухслойный: наружный слой тонкий и гиа-
линовый, внутренний — толстый, окрашенный и скульптурированный.
56. Spirogyra papulata Jao — Спирогира пупырышковая (рис. 102,
1—3). — Вегетативные клетки 28—32 (38) мкм шир., по длине в 2—6
раз больше. Поперечные стенки гладкие. Хлоропласт 1. Конъюгация
лестничная и боковая. Конъюгационный канал при лестничной конъ-
югации образован обеими клетками; при боковой конъюгации канал
выдается на 1/3—1/2 ширины клетки, нить при этом прямая. Фертиль-
ные клетки обычно укороченные. Воспринимающие клетки вздутые со
стороны конъюгационного канала, до 55 мкм шир. Клетки без конъ-
югационной пары вздутые до 64 мкм шир. Зигоспоры эллипсоидные,
22—38 мкм шир., 35—63 мкм дл. Экзоспорий тонкий, бесцветный,
гладкий. Мезоспорий толстый, желтый или золотисто-желтый, непра-
вильно сетчатый или сетчато (угловато)-ямчатый.
В небольших лужах на песке, на рисовых полях, в прудах с мощ-
ным развитием матов на поверхности воды, на увлажненных скалах,
в горном потоке. Польша, Марокко, Китай, Папуа—Новая Гвинея.
От самого близкого вида — S. subpapulata отличается большей ши-
риной вегетативных клеток, наличием боковой конъюгации и сетчатой
скульптурой мезоспория. От S. pseudodaedaleoides, близкого по ширине
вегетативных клеток и скульптуре мезоспория, отличается вздутыми
воспринимающими клетками и золотистым цветом мезоспория. Шири-
ною вегетативных клеток и вздутыми клетками без конъюгационной
пары сходен с S. scrobiculata, отличается от него сетчатым мезоспо-
рием.
57. Spirogyra pseudodaedaleoides Kadk — Спирогира ложноузор-
чатая (рис. 101, 5). — Вегетативные клетки 20—35 мкм шир. Ризоиды
латеральные, разнообразно разветвленные, от клеток без конъюгацион-
ной пары. Поперечные стенки гладкие. Хлоропласт 1. Конъюгация
236
Рис. 102. 1—3 — Spirogyra papulata- 1 '— лестничная конъюгация, 2 — боковая конъюга-
ция, 3 — строение оболочек зигоспоры (фрагмент); 4—6 — S. aphanosculpta: 4 — лестнич-
ная конъюгация, 5 — модификации формы зигоспор, 6 — строение оболочек зигоспоры
(фрагмент); 7 — S. collinsii f. ampla, лестничная н боковая конъюгации в одной паре нитей.
(По; I—3 — Jao, 1935b; 4—б — Skuja, 1937; 7 — Jao, 1935а (как S. collinsii var. ampla)).
237
боковая, очень редко лестничная. Конъюгационный канал при лестнич-
ной конъюгации образован обеими клетками, при боковой конъюгации
канал выдается приблизительно на 2/3 ширины клетки. Воспринимаю-
щие клетки невздутые. Зигоспоры удлиненно-эллипсоидные, 25—
35.2 мкм шир., 57.5—73.6 мкм дл. Экзоспорий тонкий, бесцветный,
гладкий. Мезоспорий красновато-бурый,х сетчатый и угловато-ямчатый,
нередко с бурыми пятнами (калиптрами) на вершинах или в других
местах зигоспоры.
В пруду. Дания.
Редкий вид. От близкой 5. daedalea f. daedaleoides в первую очередь
отличается более узкими вегетативными клетками. Наличие на зигос-
порах бурых пятен (калиптр) никакого таксономического значения, по-
видимому, не имеет.
58. Spirogyra aphanosculpta Skuja — Спирогира неясно скульпту-
рированная (рис. 102, 4—б). — Вегетативные клетки 32—40 мкм
шир., по длине в 2.5—5 раз больше. Поперечные стенки гладкие. Хло-
ропласт 1. Конъюгация лестничная. Конъюгационный канал образован
обеими клетками, иногда с преобладанием отдающей клетки. Воспри-
нимающие клетки невздутые или вздутые со стороны конъюгационного
канала. Клетки без конъюгационной пары иногда вздутые. Зигоспоры
коротко- или удлиненно-эллипсоидные, иногда цилиндрически-эллипсо-
идные, изредка с заострёнными вершинами, 31—46 мкм шир., 48—
81 мкм дл. Экзоспорий тонкий, бесцветный, гладкий или на внутренней
стороне ямчатый. Мезоспорий двухслойный: внешний слой довольно
толстый, желтовато-коричневый, мелкоточечный (или мелко и непра-
вильно ямчатый), внутренний слой тонкий, бесцветный, гладкий, утон-
чающийся к вершинам зигоспор.
В родниковых болотах, в зацементированных бассейнах. Греция,
Южн. Африка, Индия, США.
Довольно редкий вид. Своеобразное сложное строение оболочек зи-
госпор служит главным отличием S. aphanosculpta от других видов. Наи-
более близок S. daedalea, у которого, однако, воспринимающие клетки
если и вздуваются, то со всех сторон. Из видов, сходных по нескольким
морфологическим признакам, кроме строения оболочек зигоспор, обна-
руживает некоторую близость к 5. decimina f. longata и S. varians.
59. Spirogyra collinsii (Lewis) Printz (=Temnogyra collinsii Lewis) —
Спирогира Коллинза (рис. 102, 7; 103, 1—4). — Вегетативные клетки
18—29 мкм шир., по длине в 5—16 раз больше. Поперечные стенки
гладкие. Хлоропласт 1 (2). Конъюгация боковая, реже лестничная.
Конъюгационный канал при лестничной конъюгации образован обеими
клетками или с преобладанием отдающей клетки; при боковой конъ-
югации канал выдается до 1/2 ширины клетки, нить при этом сильно
изогнутая. Воспринимающие клетки сильно вздутые до 39 мкм шир.
Клетки без конъюгационной пары невздутые. Зигоспоры эллипсоидные,
26—40 мкм шир., (48) 52—77 (НО) мкм дл. Экзоспорий тонкий, бес-
цветный, гладкий. Мезоспорий толстый, желто-коричневый, с округлы-
ми ямчатыми углублениями.
238
Рис. 103. 1—4 — Spirogyra collinsii f. collinsii'. 1,2 — лестничная конъюгация, 3 — боковая
конъюгация, 4 — зигоспора; 5,6 — S. quezelii. (По: 1,4 — Prescott, 1962; 2, 3 — Transeau,
1951; 5, 6 — Gauthier-Lievre, 1965).
Редкий вид. Характерная особенность его заключается в том, что
на начальных стадиях конъюгации (как боковой, так и лестничной)
клетка, участвующая в этом процессе (так называемая прогаметанги-
альная клетка), делится на две неравные по длине клетки, причем
почти весь протопласт из длинной клетки переходит в короткую. Ко-
роткие клетки дают начало собственно гаметангиальным клеткам (от-
дающим и воспринимающим), тогда как более длинные клетки в поло-
вом процессе участия не принимают. Эта особенность характерна
также для двух близких видов: S. punctata и 5. corrugata.
239
F. collinsii [=S. collinsii var. (sine nomen) Skuja] (рис. 103, 1—4). —
Вегетативные клетки 18—22 мкм шир. Зигоспоры 26—37 мкм шир.,
52—62 (НО) мкм дл. Мезоспорий с крупными ямчатыми углублениями.
В стоячих водах, в лужах по краям верховых и топяных болот.
Эстония (о-ва Сааремаа и Хийумаа).
Близка к 5. punctata. От сходной S. luteospora отличается наличием
боковой конъюгации, а также сильно4, вздутыми воспринимающими
клетками.
F. ampla (Jao) V. Poljansk. (=5. collinsii var. ampla Jao; 5. ampla
Krieger) (рис. 102, 7). — Вегетативные клетки 25—29 мкм шир. Зи-
госпоры (28) 35—40 мкм шир., (48) 55—77 мкм дл. Мезоспорий с
мелкими ямчатыми углублениями.
В стоячих водах. Болгария (?), США.
По ширине вегетативных клеток обнаруживает большое сходство
с типовой формой S. punctata, однако для S. collinsii f. ampla осо-
бенно характерна боковая конъюгация, неизвестная для 5. punctata.
Кроме того, воспринимающие клетки у нее вздуты сильнее, а зи-
госпоры мельче.
60. Spirogyra pseudonodifera О. et W. Bock — Спирогира ложно-
узлоносная (рис. 104, 1—3). — Вегетативные клетки (23.5) 26—39 мкм
шир., по длине меньше ширины или до 6 (8) раз больше. Поперечные
стенки гладкие. Хлоропласт 1, редко их 2 (в проростках зигоспоры).
Конъюгация лестничная, редко боковая. Конъюгационный канал при
лестничной конъюгации образован обеими клетками с преобладанием
отдающей клетки, часто только ею; при боковой конъюгации канал
четко выраженный, выдается на половину ширины клетки, нить при
этом более или менее изогнутая. Воспринимающие клетки невздутые
или значительно вздутые со стороны конъюгационного канала, до
28.5—67.5 мкм. Клетки без конъюгационной пары иногда шаровидно
вздутые. Зигоспоры эллипсоидные, 23.5—41.5 мкм шир., 31—79 мкм
дл. Экзоспорий тонкий, бесцветный, гладкий. Мезоспорий толстый,
коричневый, плотно и правильно сетчато-морщинистый, с шиповидны-
ми выростами в узлах сети, грубозернистый (сверху), иногда с темно-
коричневыми уплотнениями на вершинах и в других местах зигоспоры.
Эндоспорий тонкий, от бесцветного до светло-коричневого. Линия раз-
рыва отчетливая. Партеноспоры шаровидные и эллипсоидные, 23—
25 мкм шир., 36 мкм дл.
В стоячих водах, воронках, сфагновых болотах, в культуре. Герма-
ния.
Редкий и спорный вид, так как имеются разногласия в описаниях.
Обнаруживает близкое сходство с S. nodifera и S. varians f. bicalypt-
rata.
61. Spirogyra franconica O. et W. Bock — Спирогира франконская
(рис. 104, 4, 5). — Вегетативные клетки 50—58 мкм шир. Поперечные
стенки гладкие. Хлоропласт 1, редко их 2. Конъюгация лестничная.
Конъюгационный канал образован отдающей клеткой. Воспринимаю-
240
щие клетки сильно вздутые со стороны конъюгационного канала, редко
невздутые. Зигоспоры эллипсоидные, преимущественно с заостренными
вершинами, 50—68 мкм шир., 80—144 мкм дл. Экзоспорий тонкий,
бесцветный, гладкий, иногда тонко исчерченный (артефакт ?). Мезо-
спорий двухслойный: внешний слой тонкий, от бесцветного до светло-
желто-коричневого, плотно сетчато-морщинистый, часто с многочис-
ленными, более или менее крупными бородавочками, иногда шиповид-
но заостренными; внутренний слой толстый, золотисто-коричневый, от
гладкого до неотчетливо плоскоямчатого. Эндоспорий тонкий, гладкий.
Линии разрыва отчетливые, иногда неправильно разветвленные.
В лесных лужах, при pH 7.7. Германия.
Редкий вид. По строению внешнего слоя мезоспория обнаруживает
большое сходство с 5. pseudonodifera, отличается значительно более
широкими вегетативными клетками. Наличие тонких штрихов на эк-
зоспории может рассматриваться как артефакт и поэтому в качестве
Рис. 104. 1—3 — Spirogyra pseudonodifera: 1 — лестничная конъюгация, 2 — боковая конъ-
югация, 3 — зигоспора; 4, 5 — S. franconica: 4 — лестничная конъюгация, 5 — зигоспора;
6, 7 — S. endogranulata: 6 — лестничная конъюгация, 7 — зигоспора. (По: Воск, Воск,
1956).
16 Рундина Л. А.
241
таксономического признака должно, по мнению авторов вида, прини-
маться с осторожностью.
62. Spirogyra quezelii Gauthier-Lievre — Спирогира Кезеля (рис. 103,
5, 6). — Вегетативные клетки 30—32 мкм шир. Поперечные стенки
гладкие. Хлоропластов 2. Конъюгация лестничная. Конъюгационный
канал образован обеими клетками с преобладанием отдающей клетки
или исключительно ею. Воспринимающие клетки веретеновидно вздутые
до 40—60 мкм шир. Зигоспоры широкоэллипсоидные, от почти шаро-
видных до удлиненных, цилиндрически-эллипсоидных, 38—52 мкм
шир., 60—ПО мкм дл. Экзоспорий тонкий, бесцветный, гладкий. Ме-
зоспорий толстый, желто-коричневый, крупносетчатый.
В прудах, небольших временных водоемах. Дания, Центр. Африка
(Ливия, Чад: нагорье Тибести).
Очень редкий вид. От S. corrugata — вида, сходного по многим
признакам, отличается меньшей шириной вегетативных клеток и не-
значительным числом хлоропластов.
63. Spirogyra asiatica Czurda — Спирогира азиатская (рис. 105,
1—4). — Вегетативные клетки 40—60 мкм шир., по длине в 2—3 раза
больше. Поперечные стенки гладкие. Хлоропластов 1—3. Конъюгация
боковая. Конъюгационный канал широкий, выдается приблизительно
на 1/2 ширины клетки, нить при этом прямая или слабо изогнутая.
Воспринимающие клетки невздутые или незначительно вздутые по ши-
рине зигоспоры. Зигоспоры эллипсоидные, яйцевидные или неправиль-
но цилиндрические, иногда в средней части неотчетливо суживающиеся
или слегка конические, с закругленными вершинами, 57.2—70 мкм
шир., 70—127.4 мкм дл. Экзоспорий тонкий, бесцветный, гладкий. Ме-
зоспорий толстый, темно- и черно-коричневый, мелкозернистый.
В реках и ручьях, в горных районах, в Центр. Тибете встречен на
высоте свыше 5000 м над ур. м. Киргизия (хребет Терскей-Алатау).
Очень редкий вид. От самого близкого вида — S. condensata от-
личается зернистостью мезоспория, размерами и вариацией форм зи-
госпор и несколько большей длиной вегетативных клеток.
64. Spirogyra occidentalis (Trans.) Czurda — Спирогира западная
(рис. 105, 5, 6). — Вегетативные клетки (36) 40—54 мкм шир., по длине
в 3—7 раз больше. Ризоиды истонченно удлиненные, разветвленные,
редкие. Поперечные стенки гладкие. Хлоропластов 1—3 (4). Конъюга-
ция лестничная. Конъюгационный канал образован обеими клетками.
Воспринимающие клетки невздутые, слабо вздутые со всех сторон до
66 мкм шир. или незначительно вздутые со стороны конъюгационного
канала. Зигоспоры широкоэллипсоидные или цилиндрически-эллипсоид-
ные, 36—61 мкм шир., (50) 57—105 мкм дл. Экзоспорий тонкий или
толстый, бесцветный, гладкий. Мезоспорий толстый, желто-коричневый,
с округлыми ямчатыми углублениями, иногда продольно исчерченный.
По данным некоторых исследователей (Transeau, 1951 : 162; Gau-
thier-Lievre, 1965 : 139), экзоспорий толстый и двухслойный, с ямча-
тым внутренним слоем, а мезоспорий гладкий.
242
Рис. 105. 1—4 — Spirogyra asiatica: 1,2,4 — боковая конъюгация, 3 — строение зигоспо-
ры (фрагмент); 5, 6 — S. occidentalis f. occidentalis-, 7 — S. punctulata f. punctulata. (По; 1,
2 — Полянский, 1951; 3,4 — Czurda, 1932; 5 — Gauthier-Lievre, 1965; 6 — Transeau, 1951;
7 — Красноперова, 19716).
F. occidentalis [=5. velata Nordst. sensu Рябинин, 1889 (?)] (рис, 105,
5, 6). — Хлоропластов 1—3. Экзоспорий толстый. Мезоспорий только
с округлыми ямчатыми углублениями.
В стоячих, реже текучих водах, реках. Вост. Украина (окрестности
Харькова) (?).
Редко встречающаяся форма с довольно широким ареалом (Герма-
ния, Тунис, Алжир, США, Канада). По ширине вегетативных клеток
и строению мезоспория очень близка к S. velata, в качестве формы
243
которого была первоначально описана (Transeau, 1914 : 294). Место-
нахождение на Украине спорно.
F. striata (Bourr. et Mang.) V. Poljansk. (=S. occidentalis var. striata
Bourr. et Mang.; non S. striata Klein). — Хлоропластов 4. Экзоспорий
тонкий. Мезоспорий продольно исчерченный, с округлыми ямчатыми
углублениями.
В лужах, периодически наполняемых морской и пресной водой.
Гваделупа.
Очень редкая и спорная форма. От типовой отличается в основном
исчерченностью, выявленной только на зрелых зиготах. Вопрос о тол-
щине оболочек зигоспоры спорен.
65. Spirogyra endogranulata О. et W. Bock — Спирогира внутрен-
незернистая (рис. 104, б, 7). — Вегетативные клетки 56—99 мкм шир.
Поперечные стенки гладкие. Хлоропластов (2) 3—5. Конъюгация лест-
ничная. Конъюгационный канал образован обеими клетками равно-
мерно, очень редко с преобладанием отдающей клетки. Воспринима-
ющие клетки невздутые, очень редко слабо вздутые со стороны конъ-
югационного канала. Зигоспоры от широкоэллипсоидных, с более или
менее заостренными вершинами, до удлиненных цилиндрически-эллип-
соидных, 69—95 мкм шир., 72—190 мкм дл. Экзоспорий тонкий, бес-
цветный, гладкий, иногда отчетливо отдаленный от мезоспория. Ме-
зоспорий толстый, желтовато-зеленый, двухслойный: внешний слой
гладкий, внутренний слой мелкозернистый. Линий разрыва от одной
до нескольких, исчерчивающих зигоспору продольно или в разных на-
правлениях.
В стоячих водах, озерах, прудах, лужах, природниковых водоемах,
при pH 5.4. Германия.
Редкий, спорный и недостаточно изученный вид, особенно в отно-
шении строения мезоспория. Мелкая зернистость, почти точечность
(пунктированность) мезоспория видна только при иммерсии. По веге-
тативным признакам обнаруживает большое сходство с S. neglecta,
S. nitida, S. setiformis. Возможно, вид сборный. Spirogyra endogranulata
f. (sine nomen), описанная теми же авторами и найденная там же, по
ширине вегетативных клеток (122—136 мкм) и постоянному вздутию
воспринимающих клеток со стороны конъюгационного канала так
сильно отличается от типовой формы, что следует, вероятно, ее вы-
делить в самостоятельный вид.
66. Spirogyra punctulata Jao — Спирогира мелкоточечная (рис. 105,
7; 106, 7). — Вегетативные клетки (65) 70—95 мкм шир., по длине
в 1.5—4.5 раза больше. На поверхности клеток изредка встречаются
кристаллы оксалата кальция, игольчатые или крестообразные. Попе-
речные стенки гладкие. Хлоропластов (3) 4—7. Конъюгация лестнич-
ная, редко боковая. Конъюгационный канал при лестничной конъюга-
ции образован обеими клетками, нередко с преобладанием отдающей
клетки. Воспринимающие клетки невздутые. Клетки без конъюгацион-
ной пары невздутые. Зигоспоры эллипсоидные, с более или менее за-
244
остренными вершинами, 64—95 мкм шир., 83—179 мкм дл., или ко-
ротко эллипсоидно-цилиндрические, с закругленными вершинами, 70—
77 мкм шир., 78—98 мкм дл. Экзоспорий тонкий, бесцветный, глад-
кий. Мезоспорий толстый, от лимонно-желтого до желтого, однослой-
ный, с небольшими, довольно грубыми углублениями, точечно-ямчатый
или мелкоточечный. Эндоспорий тонкий, гладкий, различимый только
на зрелых зиготах. Партеноспоры эллипсоидные, по размерам значи-
тельно мельче зигоспор.
F. punctulata (рис. 105, 7; 106, /). — Вегетативные клетки (65)
70—84 (86) мкм шир. Хлоропластов (3) 4—7. Конъюгация лестничная
и боковая (?). Зигоспоры эллипсоидные, с более или менее заострен-
ными вершинами, 64—74 (81) мкм шир., 83—179 мкм дл. Мезоспорий
с очень небольшими, довольно грубыми углублениями.
Рис. 106. 1 — Spirogyra punctulata f. punctulata; 2,3 — S. verrucosa: 2 — лестничная конъ-
югация, 3 — строение мезоспория (фрагмент). (По: 1,3 — Gauthier-Lievre, 1965; 2 — Rao,
1938 (как S. malmeana var. verrucosa)).
245
В стоячих водах, небольших родниковых топях. Зап. Казахстан
(предгорья Мугоджар).
Форма с широким ареалом (Дания, Алжир, Индия, Китай). Верши-
ны более заострены у удлиненных зигоспор, длина которых зависит
от длины женских гаметангиев, более коротких, чем вегетативные клет-
ки. На рисунке вида, данном его автором, мезоспорий выглядит мел-
коточечным, и большинство исследователей характеризуют мезоспорий
как тонко пунктированный (точечный). Близка по многим вегетатив-
ным признакам к S. malmeana Him (Южн. Америка, Африка), у ко-
торого в отличие от f. punctulata морщинистый мезоспорий, и к S. ni-
tida, у которого, однако, гладкий мезоспорий.
F. cylindrospora (W. et G. S. West) Rund. (=S. cylindrospora W. et
G. S. West). — Вегетативные клетки 70—77 мкм шир. Хлоропластов 4.
Конъюгация лестничная и боковая. Зигоспоры коротко эллипсоидно-ци-
линдрические, с закругленными вершинами, 70—77 мкм шир., 78—
98 мкм дл. Мезоспорий с грубыми углублениями (точечно-ямчатый).
В стоячих водах, мочажинах, лужах. Россия: центр европ. части.
Редкая форма, известная до сих пор только из Зап. Африки (Ангола).
Наиболее характерным является короткоцилиндрическая форма эллип-
соидных зигоспор, служащая главным отличием от типовой формы. От
близкой по постоянному числу хлоропластов f. macrospora отличается,
кроме упомянутой формы зигоспор, наличием боковой конъюгации и
меньшей шириной вегетативных клеток. Изображение отсутствует.
F. macrospora (Jao) Rund. (=S. punctulata var. macrospora Jao). —
Вегетативные клетки 75—95 мкм шир. Хлоропластов 4. Конъюгация
лестничная. Зигоспоры эллипсоидные, обычно с более или менее за-
остренными вершинами, 75—95 мкм шир., 100—165 мкм дл. Мезо-
спорий мелкоточечный (тонко пунктированный).
В стоячих водах, лужах. Китай.
Редкая форма. Отличается от типовой формы большей шириной
вегетативных клеток и зигоспор. Постоянное наличие 4 хлоропластов
сближает эту форму c'f. cylindrospora. Изображение отсутствует.
67. Spirogyra trachycarpa Skuja — Спирогира шероховатоплодная
(рис. 107, 1, 2). — Вегетативные клетки 110—123 мкм шир., по длине
равные ширине или до 4 раз больше. Поперечные стенки гладкие.
Хлоропластов 4—6 (8). Конъюгация лестничная. Конъюгационный
канал образован обеими клетками. Воспринимающие клетки невздутые
или очень слабо вздутые со стороны конъюгационного канала. Зиго-
споры эллипсоидные, 87—108 мкм шир:, 117—172 (204) мкм дл. Эк-
зоспорий довольно толстый, бесцветный, гладкий. Мезоспорий толс-
тый, неправильно ямчатый.
В стоячих водах, в озере, на илистом грунте, отдельными нитями
среди вошерии и кладофоры. Латвия.
Очень редкий и недостаточно изученный вид. В первоописании
отсутствует указание на цвет мезоспория и имеется противоречие
между описанием и изображением скульптуры мезоспория: в описании
246
Рис. 107. 1,2 — Spirogyra trachycarpa: I — лестничная конъюгация, 2 — строение зиго-
споры (фрагмент); 3—8 — S. fluviatilis-. 3, 4 — f. fluviatilis, лестничная конъюгация (3) и
строение зигоспоры (4); 5—8 — f. grossii, лестничная конъюгация, общий вид и ризоиды
(б), строение оболочек зигоспоры (фрагменты) (7, 8). (По: 1,2 — Skuja, 1932а; 3 — Skuja,
1928; 4 — Gauthier-Lievre, 1965; 5—8 — Rieth, 1976 (как S. fluviatilis f.)).
247
он ямчатый, на рисунке — морщинистый, с намеками на образование
плотно сомкнутой сети. Близок к S. setiformis и S. ellipsospora, отли-
чается от них скульптурированностью мезоспория и шириной вегета-
тивных клеток. Близок также к двум редким видам из Сев. Индии —
S. kundaensis Singh и S. ghosei Singh. Возможно, является синонимом
S. reinhardii, если считать мезоспорий морщинисто-сетчатым, судя по
авторскому рисунку.
68. Spirogyra paraquayensis Borge — Спирогира парагвайская (рис. 87,
2—6). — Вегетативные клетки (34) 40—-52 (59) мкм шир., по длине в
2—5 раз больше. Ризоиды латеральные и базальные, часто многочислен-
ные, лопастные. Поперечные стенки гладкие. Хлоропластов 3—4 (5), с
незначительным количеством оборотов или почти прямые. Конъюга-
ция лестничная. Конъюгационный канал образован обеими клетками.
Воспринимающие клетки невздутые или слегка вздутые, укороченные.
Клетки без конъюгационной пары невздутые. Зигоспоры широкоэллип-
соидные, 34—65 мкм шир., 58—105 (125) мкм дл. Экзоспорий тонкий,
бесцветный, гладкий. Мезоспорий толстый, желто-коричневый или
темно-коричневый, морщинистый или морщинисто-сетчатый.
Редкий, возможно монотипный, вид с 2 формами, отличающимися
друг от друга степенью сомкнутости морщин на мезоспории, зависящей,
по-видимому, от степени зрелости зигот.
F. paraquayensis (=5. elliptica Skv., non Jao) (рис. 87, 2, 3). —
Вегетативные клетки 41—45 (55) мкм шир. Хлоропластов 3—4. Зи-
госпоры 34—45 (55) мкм шир., 58—70 (125) мкм дл. Мезоспорий
морщинистый, с довольно короткими и извилистыми утолщениями
(морщинами), не сомкнутыми в сеть.
В стоячей (?) воде. Китай (Сев. Маньчжурия), Южн. Америка (Гва-
делупа, Парагвай).
Очень редкая форма. Отличается от близкой по ширине вегетатив-
ных клеток 5. fluviatilis f. fluviatilis образованием конъюгационного
канала обеими клетками без преобладания отдающей клетки, отсутст-
вием сильного вздутия воспринимающих клеток. По рисунку Борге
(Borge, 1903 : 280), тонкие, короткие и слабо разветвленные утолще-
ния иногда смыкаются, создавая впечатление слабо сомкнутой сети.
F. rhizobrachialis (Jao) V. Poljansk. (=5. rhizobrachialis Jao; S. sub-
cylindrica Jao var. major Bi; S. subcylindrica f. major Bi) (рис. 87,
4—6). — Вегетативные клетки (34) 40—52 (59) мкм шир. Хлороплас-
тов 3—5. Зигоспоры 38—65 мкм шир., 58—105 (111) мкм дл. Мезо-
спорий морщинисто-сетчатый, с короткими утолщениями (морщинами),
сомкнутыми в сеть.
В мелких стоячих водоемах, прудах, на рисовых полях. Испания,
Африка (Кот-д’Ивуар, Судан, Чад, Конго), Китай, Индия, США.
Редкая форма с широким ареалом. По первоописанию (Jao, 1935b :
599, tab. 7, fig. 81—83; 1936 : 57, fig. 18—21), мезоспорий плотно
морщинисто-сетчатый, по рисунку — мезоспорий с маленькими, изо-
гнутыми, плотно расположенными, бородавчатыми возвышениями, при
248
сомкнутости которых создается впечатление сетчатости мезоспория,
особенно при малом увеличении микроскопа. По этому признаку сбли-
жается с 5. verrucosa, отличается от него значительно меньшей ши-
риной вегетативных клеток и однотипностью скульптуры мезоспория.
69. Spirogyra fluviatilis Hilse — Спирогира речная (рис. 107, 3—8',
108). — Вегетативные клетки 26—50 (52) мкм шир., по длине меньше
ширины или до 12 раз больше. Ризоиды базальные и латеральные,
нитевидные и лопастные, часто разветвленные, многочисленные. Попе-
речные стенки гладкие. Хлоропластов (2) 3—6. Конъюгация лестнич-
ная. Конъюгационный канал образован обеими клетками равномерно,
часто с преобладанием отдающей клетки, иногда только ею. Воспри-
нимающие клетки сильно вздутые со всех сторон, до 70—93 мкм шир.
Клетки без конъюгационной пары невздутые. Зигоспоры широкоэллип-
соидные, редко удлиненно-эллипсоидные, 37—65 (75.5) мкм шир.,
41—140 мкм дл. Экзоспорий тонкий, гладкий, бесцветный. Мезоспо-
рий толстый, темно-коричневый, морщинистый или морщинисто-точеч-
ный; у зрелых зигот мезоспорий отчетливо многослойный, скульпту-
рированный слой морщинистый или морщинисто-сетчатый, с густо рас-
положенными мелкими шипами (зубчиками), при взгляде сверху —
точечный. Эндоспорий тонкий, бесцветный, гладкий. Линия разрыва
четкая.
Широко распространенный, но еще недостаточно изученный вид
из-за редкой встречаемости зигоспор. Не изучено до конца строение
мезоспория на всех стадиях созревания зиготы. Вполне возможно, что
мезоспорий четырехслойный, причем скульптурированы только два
нижних слоя. От стадии зрелости зиготы зависит высота шипов (зуб-
чиков) и формирование сетчатости на мезоспории. Данные сканирую-
щего микроскопа доказывают тонкосетчатую скульптуру двухслойного
мезоспория, причем на внутренней (изнаночной) стороне внутреннего
скульптурированного слоя выявлена губчатая (пористая, ноздреватая)
структура.
F. fluviatilis (=5. fluviatilis f. tibestica Gauthier-Lievre) (рис. 107, 3,
4; 108, 1—3). — Вегетативные клетки 26—35 мкм шир. Хлоропластов
(2) 3—6. Конъюгационный канал образован обеими клетками, с пре-
обладанием отдающей клетки, иногда только ею. Зигоспоры 37—
55 мкм шир., 65—122 мкм дл. Мезоспорий обычно однослойный, мор-
щинистый, с вытянутыми, короткими, неправильно волнисто изогнуты-
ми утолщениями, не соединяющимися в сеть; у вполне зрелых зигот
мезоспорий двухслойный; внешний слой тонкий, прозрачный, часто-
и мелкоточечный (шиповатый), трудно различимый (даже при иммер-
сии), внутренний слой морщинистый (складчатый).
В реках, озерах, стоячих и текучих водах, на рисовых полях, при-
крепленно к субстрату. Характерна для чистых и сравнительно холод-
ных вод. Россия: европ. часть — почти повсюду, Сибирь, Чукотский
авт. округ; Латвия, Украина, Узбекистан, Туркмения, Казахстан.
Широко распространенная форма. Вследствие многочисленных оп-
ределений этой водоросли в стерильном состоянии только по наличию
249
Рис. 108. Spirogyra fluviatilis-. 1—3 — f. fluviatilis, лестничная конъюгация (/, 2) и строение
мезоспория (фрагмент) (3); 4 — f. grossii, строение зигоспоры; 5,6 — f. ramosa, лестничная
конъюгация (5) и ризоиды (6). (По: 1—4 — Рундина, 1988; 5, 6 — Свиренко, 1941 (как
S. ramosa)).
ризоидов, не все имеющиеся в литературе указания на ее нахождения
можно считать достоверными. По строению мезоспория наиболее близ-
ка к S. dictyospora, иногда тоже образующему ризоиды, и к S: fossa
Jao (США), имеющему незначительно меньшую ширину вегетативных
клеток.
F. grossii (Schmidle) V. Poljansk. [=5. grossii Schmidle; 5. adnata
(Vauch.) Kutz., pr. p. (?); S. fluviatilis Hilse, pr, p.; S. fluviatilis f. (sine
nomen) Rieth; Spirogyra sp. sensu Хмелевский, 1891] (рис. 107, 5—S;
108, 4). — Вегетативные клетки (32) 35—45 (52) мкм шир. Хлоро-
пластов (2) 3—5. Конъюгационный канал образован преимущественно
или исключительно отдающей клеткой. Зигоспоры широкоэллипсоид-
ные, 40—65 (75.5) мкм шир., 61—140 мкм дл. Мезоспорий у зрелых
зигот отчетливо двухслойный, морщинисто-сетчатый и шиповидный,
сверху точечный.
В стоячих и текучих водах, реках, ручьях, озерах, на рисовых
полях, часто прикрепленно к субстрату, иногда в свободноплавающем
250
состоянии, нередко вместе с типовой формой. Россия: север, северо-
запад и центр европ. части, Сибирь (Республика Саха); Прибалтика,
Украина, Крым (окрестности Симферополя), Узбекистан, Киргизия.
Возможно, распространена шире, чем указано в литературе. Как и
типовая форма, сравнительно редко образует зиготы и часто размно-
жается только вегетативно. Близка к S. dubia, отличаясь скульптури-
рованным мезоспорием, несколько иным строением конъюгационного
канала и более сильным вздутием воспринимающих клеток. От S. af-
ricana (Fritsch) Czurda (Африка, Южн. Америка) отличается узкими
вегетативными клетками, от 5. paraquayensis — вздутыми воспринима-
ющими клетками.
F. ramosa (Swir.) V. Poljansk. (=5. ramosa Swir.) (рис. 108, 5,
6). — Вегетативные клетки 30—50 мкм шир. Хлоропластов 2—4.
Конъюгационный канал образован обеими клетками равномерно. Зи-
госпоры 41—52 мкм шир., до 120 мкм дл. Мезоспорий морщинистый.
В стоячих и текучих водах, преимущественно в береговой зоне с
замедленным течением. Эпифит, часто развивающийся в больших ко-
личествах на макрофитах, где образует кожистые густые наросты ин-
тенсивного черно-зеленого цвета. Украина (Причерноморье).
Редкая и спорная форма с узким ареалом. Возможно, является си-
нонимом f. grossii. Близка также к типовой форме, но является ис-
ключительно эпифитом, нити ее более короткие (5—20 мм дл.), в
нижней части могут разветвляться. Интересно отметить, что конъюга-
ция у этой водоросли протекает не в свободноплавающем, как у боль-
шинства спирогир, а в прикрепленном состоянии. Близка также к S. ri-
vularis.
70. Spirogyra verrucosa (Rao) Krieger (= S. malmeana Hirn var. ver-
rucosa Rao) — Спирогира бородавчатая (рис. 106, 2, 3). — Вегета-
тивные клетки 100—126 мкм шир., по длине равные ширине или до
1.5 раза больше. Поперечные стенки гладкие. Хлоропластов 4—8.
Конъюгация лестничная. Конъюгационный канал от обеих клеток. Вос-
принимающие клетки невздутые. Зигоспоры эллипсоидные, 75—
103.3 мкм шир., 115—165 мкм дл. Экзоспорий тонкий, бесцветный,
гладкий. Мезоспорий толстый, от желтого до коричневого, с малень-
кими бородавчатыми возвышениями и выпуклыми короткоизвилистыми
складками (ребрами), сомкнутыми в довольно крупноячеистую сеть.
В стоячих водах, лужах. Нидерланды, Зап. Африка (Кот-д’Ивуар),
Индия.
Редкий вид с широким ареалом. Первоначально был описан в ка-
честве разновидности S. malmeana Hirn (Бразилия, Африка), от кото-
рого отличается бородавчатостью мезоспория. Близок к S. verruculosa
Jao (Китай, Япония), отличается сетчатым мезоспорием. Так как скульп-
тура мезоспория многотипная (бородавчатая и ребристо-сетчатая),
можно предполагать, что мезоспорий двухслойный.
71. Spirogyra brunnea Czurda — Спирогира буроватая (рис. 109,
1—4). — Вегетативные клетки 40.5—71 (84) мкм шир., по длине в
251
1.5—7 раз больше. Ризоиды латеральные, преимущественно в виде ро-
зетки. Поперечные стенки гладкие. Хлоропластов (2) 3—5. Конъюгация
лестничная. Конъюгационный канал образован обеими клетками. Вос-
принимающие клетки слабо вздутые со всех сторон или преимущест-
венно со стороны конъюгационного канала, реже невздутые. Клетки
без конъюгационной пары невздутые. Зигоспоры эллипсоидные, (45)
48—66 мкм шир., (60) 70—113 (130.5) мкм дл. Экзоспорий тонкий,
бесцветный, гладкий. Мезоспорий толстый, от желто-коричневого до
коричневого, неправильно морщинисто-тонкосетчатый, иногда с шипо-
видными выростами.
Вид редкий, с ограниченным, преимущественно субтропическим
местообитанием.
F. brunnea (рис. 109, 7). — Вегетативные клетки 56—71 (84) мкм
шир. Хлоропластов 3—5. Воспринимающие клетки слабо вздутые со
всех сторон или реже невздутые. Зигоспоры 54—66 мкм шир., 70—
94 мкм дл. Мезоспорий неправильно тонкосетчатый.
В текучих, реже стоячих водах. Южн. Африка, Индия.
Редкая форма, приуроченная к тропическим и субтропическим об-
ластям. От f. pulchrifigurata отличается незначительно большей шири-
ной вегетативных клеток, от S. africana (Fritsch) Czurda (Южн. Аме-
рика, Африка) — сетчато-морщинистым мезоспорием, от S. propria
Trans. (Сев. Америка) — отсутствием боковой конъюгации, от S. mal-
теапа Hirn (Бразилия, Африка) — меньшей шириной вегетативных и
вздутием воспринимающих клеток.
F. pulchrifigurata (Jao) V. Poljansk. (=S. pulchrifigurata Jao; S. brun-
nea Czurda sensu Полянский в: Обухова, 1961) (рис. 109, 2—4). — Ве-
гетативные клетки 40.5—55 (58) мкм шир. Хлоропластов (2) 3—4. Вос-
принимающие клетки слабо вздутые до 83 мкм шир., преимущественно
со стороны конъюгационного канала или со всех сторон. Зигоспоры (45)
48—64 мкм шир., (60) 70—113 (130.5) мкм дл. Мезоспорий морщинис-
то-сетчатый, иногда с шиповидными выростами.
В текучих и стоячих водах, реках, на рисовых полях. Узбекистан
(окрестности Самарканда), Южн. Казахстан.
Редкая форма, найденная также в Китае. Очень близка к типовой
форме, однако отличается более узкими вегетативными клетками и
более крупными зигоспорами. Кроме того, у этой формы воспринима-
ющие клетки в большинстве случаев сильнее вздуты со стороны конъ-
югационного канала. Скульптура мезоспория по мере созревания зигот
заметно меняется: сначала мезоспорий продольно-морщинистый, затем
морщинисто-сетчатый, наконец в узлах сетчатых утолщений возникают
сначала бугорчатые, а потом и тонкоигольчатые образования. Этот при-
знак характерен также и для S. reticulata f. acanthophora, от которой
эта форма отличается гладкими поперечными стенками и значительно
большей шириной вегетативных клеток.
72. Spirogyra reinhardii Chm. — Спирогира Рейнхарда (рис. 109,
5—9). — Вегетативные клетки (80) 85—117 мкм шир., по длине рав-
252
Рис. 109. 1—4 — Spirogyra brunnea; 1 — f. brunnea, 2—4 — f. pulchrifigurata; 5—9 —
S. reinhardii; 5 — лестничная конъюгация между тремя нитями, общий вид, 9 — строение
мезоспория (фрагмент). (По: 1, 2, 4, 5, 7, 8 — Gauthier-Lievre, 1965 (2, 4 — как 5. pulchri-
figurata); 3 — ориг.; 6, 9 — Borge, 1913).
253
ные ширине или до 3 раз больше. Поперечные стенки гладкие. Хло-
ропластов. (3) 4—6, с небольшим количеством оборотов. Конъюгация
лестничная. Конъюгационный канал образован обеими клетками, иног-
да нерезко отграниченный. Воспринимающие клетки слабо вздутые со
стороны конъюгационного канала. Клетки без конъюгационной пары
невздутые. Зигоспоры эллипсоидные, 65—100 мкм шир., (97.5) 105—
175 мкм дл. Экзоспорий тонкий, бесцветный, гладкий. Мезоспорий
толстый, коричневый, с утолщениями, образующими неправильную ред-
кую сеть.
В стоячих, слабопроточных и текучих водах. Россия: центр европ.
части (окрестности Москвы); Вост. Украина.
Редкий и недостаточно изученный вид. Отличается от близкого
S. ghosei Singh (Сев. Индия) строением оболочки зигоспор, вздутыми
воспринимающими клетками и меньшим числом хлоропластов. Воз-
можно, что идентичен S. trachycarpa, если у последнего вида подтвер-
дится морщинисто-сетчатое строение мезоспория.
73. Spirogyra mienningensis Li — Спирогира миеннингская (рис. 110,
1—3). — Вегетативные клетки 42—49 мкм шир., по длине в 2—5 раз
больше. Ризоиды базальные, в виде плотной розетки, редкие. Попереч-
ные стенки гладкие. Хлоропластов 2—3. Конъюгация лестничная. Конъ-
югационный канал образован обеими клетками, иногда с преобладанием
отдающей клетки. Воспринимающие клетки невздутые или слегка взду-
тые, в редких случаях до 84 мкм шир. Клетки без конъюгационной пары
сильно вздутые со всех сторон, до 75.4—93 мкм шир. Зигоспоры эллип-
соидные, (49) 52—59 мкм шир., 70—88 мкм дл. Экзоспорий тонкий,
бесцветный, гладкий. Мезоспорий толстый, двухслойный: внешний слой
тонкий, бесцветный или желтоватый, широко- и короткоморщинистый,
внутренний слой толстый, коричневый, густобородавчатый. Эндоспорий
толстый, гладкий, хорошо заметный на зрелых зиготах.
В стоячей и текучей воде, в реках родникового происхождения, в
свободноплавающем или прикрепленном состояниях. Южн. Казахстан.
Редкий вид, приуроченный к субтропическим областям, найден пре-
жде в Китае. Строение экзоспория спорно. Указание на его двухслой-
ность, выдвинутое при первоописании, требует уточнения. Во всяком
случае, как показали исследования образцов из Казахстана, линия раз-
рыва зрелой зиготы проходит через два слоя мезоспория, оставляя
нетронутым однослойный экзоспорий.
74. Spirogyra corrugata Trans. [=Temnogyra corrugata (Trans.) Yama-
gishi] — Спирогира сморщенная (рис. ПО, 4—8). — Вегетативные клет-
ки (29) 32—46.5 (49) мкм шир., по длине в (3) 6—12 (17) раз больше.
Поперечные стенки гладкие. Хлоропластов (1) 2—4 (5). Конъюгация
лестничная, редко боковая. Конъюгационный канал при лестничной
конъюгации образован отдающей клеткой; при боковой конъюгации
канал четко отграниченный, выдается приблизительно на 1/3 ширины
клетки, нить при этом слегка изогнутая. Воспринимающие клетки слегка
вздутые по ширине зигоспоры, до 40—60 мкм. Клетки без конъюгаци-
онной пары невздутые. Зигоспоры эллипсоидные, 40.5—60 мкм шир.,
254
Рис. 110. 1—3 — Spirogyra mienningensis: 1,2 — лестничная конъюгация, 3 — ризоид; 4—
8 — S. corrugata: 4 — лестничная конъюгация, 5 — боковая конъюгация, 6 — зигоспора,
7, 8 — строение зигоспоры (фрагменты). (По: 1—3 — Рундина, 1978а; 4, 8 — Jao, 1935b;
5—7 — Красноперова, 1968).
80—122 мкм дл. Экзоспорий тонкий, бесцветный, гладкий. Мезоспорий
двухслойный: внешний слой тонкий, бесцветный или желтоватый, с вол-
нистыми продольными складками-морщинами, внутренний слой желто-
вато-коричневый, испещренный довольно короткими ребрами-утолще-
ниями, смыкающимися по мере созревания зиготы в неправильную сетку,
тонкую или более грубую, полностью или частично сомкнутую. Эндо-
спорий тонкий, различимый лишь на зрелых зиготах.
255
В стоячих, реже текучих водах, на рисовых полях, может вегети-
ровать при небольшой солености (0.12—5.7 %о) и на глубине 1.6—
2.5 м. Россия: устье Волги с заносом на западное и в меньшей мере
северное побережье Каспийского моря.
Довольно редкий вид с широким ареалом. Известен также из Сев.
Америки, Африки и Китая. Особенность вида состоит в том, что конъ-
югируют лишь немногие клетки, распЬложенные одиночно, парами или
редко рядами. Близкие в этом отношении виды — S. punctata и S. col-
linsii хорошо отличаются меньшей шириной вегетативных клеток и
ямчатым мезоспорием. От совпадающего по размерам и близкого по
ряду признаков S. reticulata хорошо отличается гладкими поперечными
стенками и особенностями формирования конъюгационного канала. По
своеобразному строению двухслойного мезоспория наиболее близок к
S. quadrilaminata и S. shantungensis Li (Китай, Уганда).
75. Spirogyra quadrilaminata Jao — Спирогира четырехслойная
(рис. Ill, 1, 2). — Вегетативные клетки 37—58 мкм шир., по длине
в 2—6.5 раза больше. Поперечные стенки гладкие. Хлоропластов 3—4,
с небольшим количеством оборотов. Конъюгация лестничная. Конъ-
югационный канал образован обеими клетками. Воспринимающие клет-
ки невздутые, очень редко незначительно вздутые. Зигоспоры эллип-
соидные, иногда цилиндрически-эллипсоидные, (40) 42—55 мкм шир.,
Рис. 111. I, 2 — Spirogyra quadrilaminata-. 1 — лестничная конъюгация, 2 — строение ме-
зоспория (поперечный разрез); 3—5 — S. cavata: 3 — лестничная конъюгация, 4 — боковая
конъюгация, 5 — строение зигоспоры (модель). (По: 7,2 — Jao, 1935b; 3—5 — Воденича-
ров в: Воденичаров, Киряков, 1970).
256
64—102 мкм дл. Экзоспорий тонкий, бесцветный, гладкий. Мезоспо-
рий двухслойный: внешний слой темно-коричневый, более или менее
густо покрытый продольными волнистыми морщинами (ребрышками),
внутренний слой коричневый, тонкоточечный, иногда неправильно тон-
косетчатый. Эндоспорий тонкий, гладкий.
Преимущественно в стоячих водоемах, редко в слабопроточной
воде, на рисовых полях и в культуре. Германия, Сев., Центр, и Южн.
Америка, Китай, Индия.
Довольно редкий вид с широким ареалом. От наиболее близкого
по размерам клеток S. subreticulata Fritsch (Африка, США, Китай)
отличается главным образом сложным строением мезоспория.
76. Spirogyra dictyospora Jao — Спирогира сетчатоспоровая
(рис. 112). — Вегетативные клетки 25—32 (35) мкм шир., по длине в
2—9 (12) раз больше. Ризоиды преимущественно латеральные, редко ба-
зальные, разветвленные и неразветвленные, иногда лопастные, с широ-
кой ослизненной оболочкой. Поперечные стенки гладкие. Хлоропластов
2—3 (4). Конъюгация лестничная. Конъюгационный канал образован
обеими клетками равномерно, иногда — преимущественно, редко — ис-
ключительно отдающей клеткой. Воспринимающие клетки часто сильно
вздутые со всех сторон или невздутые, или незначительно вздутые по
ширине зигоспоры. Клетки без конъюгационной пары невздутые. Зиго-
споры эллипсоидные, редко цилиндрически-эллипсоидные, с закруглен-
ными вершинами, 32—55 мкм шир., 57—ПО (116) мкм дл. Экзоспорий
тонкий, бесцветный, гладкий, иногда слоистый на вершинах зигоспор.
Мезоспорий толстый, желтовато-коричневый, двухслойный: внешний
слой тонкий, иногда неясно слоистый, нежно-желтоватый, мелко- и
плотноточечный; внутренний слой толще внешнего, желтовато-коричне-
вый, покрытый извилистыми морщинистыми утолщениями, в некоторых
случаях образующими более или менее четкую неправильную сетку.
Редкий вид, приуроченный к тропическим и субтропическим об-
ластям. Требует уточнения вопрос о скульптуре мезоспория. Точечность
мезоспория выявляется только при его рассмотрении у совершенно
зрелых зигот, и по мере созревания зигот в узлах морщинисто-сетчатых
утолщений могут возникать шиповидные выросты, которые при взгляде
сверху выглядят точками.
F. dictyospora (=S. rhizopus Jao) (рис. 112, 1—3). — Вегетативные
клетки 25—32 (35) мкм шир. Воспринимающие клетки вздутые, пре-
имущественно незначительно, со всех сторон — бочонковидно, редко
цилиндрически, до 55 мкм шир. Зигоспоры эллипсоидные, реже ци-
линдрически-эллипсоидные, 35—55 мкм шир., 53.4—124.6 мкм дл.
В текучих и стоячих водах, реках, ручьях, озерах, рыбоводных
прудах, на рисовых полях, в свободноплавающем или прикрепленном
состояниях. Узбекистан (оз. Муюнь-куль в дельте Амударьи), Южн.
Казахстан.
Редкая форма, найденная также в Нидерландах, Китае и Индии.
Наиболее близка к S. fluviatilis, отличающемуся степенью вздутия вос-
принимающих клеток и несколько иным строением мезоспория.
17 Рундина Л. А.
257
Рис. 112. Spirogyra dictyospora: 1—3 — f. dictyospora-, 4—7 — f. subcylindrospora: 4 — лест-
ничная конъюгация между тремя нитями, 5 — зигоспора, 6, 7 — строение мезоспория
(фрагменты). (По: 1,5,6 — Рундина, 19786; 2 — Jao, 1935b; 3 — Полянский, 1951; 4, 7 —
Jao, 1935b (как S. subcylindrospora)).
258
F. subcylindrospora (Jao) V. Poljansk. (=S. subcylindrospora Jao)
(рис. 112, 4—7). — Вегетативные клетки 29—32 мкм шир. Восприни-
мающие клетки невздутые или незначительно вздутые по ширине зиго-
споры. Зигоспоры эллипсоидные, 32—49 мкм шир., 57—108 мкм дл.
В текучих и стоячих водах, реках, на рисовых полях. Узбекистан
(окрестности Самарканда), Южн. Казахстан.
Редкая форма, прежде найденная в Сев. Африке, Китае, США.
Отличается от типовой формы почти исключительно невздутыми или
слабо вздутыми воспринимающими клетками.
77. Spirogyra cavata Vodenic. — Спирогира выемчатая (рис. 111,
3—-5). — Вегетативные клетки 32—39 мкм шир., по длине в 1.5—3.5
раза больше. Поперечные стенки гладкие. Хлоропласт 1. Конъюгация
лестничная, редко боковая. Конъюгационный канал при лестничной
конъюгации образован обеими клетками; при боковой конъюгации канал
четкий, выдается на 1/3—1/2 ширины клетки. Воспринимающие клетки
невздутые или изредка очень слабо вздутые. Клетки без конъюгационной
пары невздутые. Зигоспоры эллипсоидные, (30) 35 (37) мкм шир., (42)
64 (80) мкм дл. Экзоспорий толстый, 0.6—1 мкм толщ., бесцветный, уп-
лотненный снаружи и с неправильными глубокими ямками. Мезоспорий
толстый, 1—1.2 мкм толщ., бледно-желтоватый, с волнистой внутренней
поверхностью (морщинистый ?).
В луже. Болгария.
Редкий вид. Очень близок, а возможно и идентичен, S. pseudova-
rians. По строению экзоспория сходен с S. velata. Строение мезоспо-
рия требует уточнения.
78. Spirogyra crassoidea Trans. [=5. crassa Kutz. f. crassoidea (Trans.)
V. Poljansk.; S. crassa sensu Казановский, Смирнов, 1914] — Спирогира
толстовидная (рис. ИЗ, 2, 3). — Вегетативные клетки (114) 140—
150 мкм шир., по длине равные ширине или до 4 раз больше. На по-
верхности клеток имеются многочисленные кристаллы оксалата кальция.
Поперечные стенки гладкие. Хлоропластов 3—8 (10), с 0.5—3 оборота-
ми. Конъюгация лестничная. Конъюгационный канал образован обеими
клетками. Воспринимающие клетки невздутые. Клетки без конъюгаци-
онной пары невздутые. Зигоспоры сжато-эллипсоидные (трехосно-эллипсо-
идные), в поперечном сечении эллиптические, (80) 95—150 мкм шир.,
(115) 120—255 мкм дл., 90—ПО мкм толщ. Экзоспорий тонкий; бесцвет-
ный, гладкий. Мезоспорий толстый, коричневый, гладкий.
В стоячих, редко текучих водах. Украина (окрестности Киева), Сев.
Казахстан (заповедник Боровое).
Редкий вид, найденный до сих пор лишь в США. Очень близок к
S. crassa, возможно смешивался с ним, однако четко отличим от него
по отсутствию скульптуры на мезоспорий. Сходен также с S. ellipso-
spora, в качестве разновидности которого и был первоначально описан,
отличается формой зигоспор.
79. Spirogyra subcrassa Woronich. — Спирогира толстоватая (рис. 113,
7). — Вегетативные клетки 114—131.7 (140) мкм шир., по длине мень-
259
Рис. 113. 1 — Spirogyra subcrassa; 2, 3 — S. crassoidea: 2 — лестничная конъюгация, 3 —
зигоспора в двух положениях; 4—8 — S. crassa: 4 — зигоспоры в разных положениях, лест-
ничная конъюгация, 5, 6 — зигоспоры в двух положениях, лестничная конъюгация, 7 — зи-
госпора в двух положениях, справа — строение мезоспория, 8 — зигоспора в трех положе-
ниях. (По: 1 — Воронихин, 1954; 2 — ориг.; 3 — Transeau, 1951; 4, 8 — Рундина, 1988;
5,7 — Yamagishi, 1966b; 6 — Wang, 1933).
260
ше ширины, равные ей или в 1.5—2 раза больше. Поперечные стенки
гладкие. Хлоропластов (5) 7—10 (11), преимущественно с половинным
оборотом, иногда почти прямые. Конъюгация лестничная. Конъюгаци-
онный канал образован обеими клетками, с преобладанием воспринима-
ющей клетки. Воспринимающие клетки невздутые. Клетки без конъюга-
ционной пары невздутые. Зигоспоры сжато-эллипсоидные (трехосно-эл-
липсоидные), в поперечном сечении эллиптические, 108—123 мкм шир.,
142—181 мкм дл., 95—100 мкм толщ. Экзоспорий бесцветный, гладкий,
иногда хорошо заметный. Мезоспорий желтовато-буроватый, неправиль-
но и мелко точечно-бородавчатый, шагреневый.
В стоячих и слабопроточных водоемах, водохранилищах, иногда
большими скоплениями. Россия: северо-запад европ. части (окрестнос-
ти Санкт-Петербурга); Украина (Киевское водохранилище).
Редкий вид. Вместе с близким видом S. crassoidea входит в услов-
ную группу полиморфного вида S. crassa, возможно являясь либо фор-
мой последнего, либо стадией его развития.
80. Spirogyra crassa Kiitz., auct. non Kiitz.: Казановский, Смирнов,
1914; Randhawa, 1959b; Эргашев, 1974, 1976 (=S. crassa f. genuina
Kirchn.) — Спирогира толстая (рис. 113, 4—8). — Вегетативные клет-
ки 120—160 (165) мкм шир., по длине равные ширине или до 2 раз
больше. Поперечные стенки гладкие. Хлоропластов (4) 6—12, с 0.5—1
оборотом спирали или почти прямые. Конъюгация лестничная. Конъ-
югационный канал образован обеими клетками. Воспринимающие клет-
ки невздутые. Клетки без конъюгационной пары невздутые. Зигоспоры
сжато-эллипсоидные (трехосно-эллипсоидные), в поперечном сечении
эллиптические, 120—150 мкм шир., 135—190 мкм дл., 80—120 мкм
толщ. Экзоспорий тонкий, бесцветный, гладкий. Мезоспорий толстый,
коричневый, с крупными и неправильной формы, довольно плоскими
углублениями.
В стоячих и текучих водах, реках, озерах, водохранилищах, пре-
имущественно в крупных водоемах. Чувствителен к железу, предпочи-
тает щелочные воды с повышенным содержанием азота, мезосапроб.
Способен развиваться при загрязнении, часто встречается в массовых
скоплениях. Россия: европ. часть — повсюду, Сев. Кавказ, Зап. Сибирь
(окрестности Томска, Обь-Енисейский канал); Украина — повсюду,
Узбекистан.
Широко распространенный и полиморфный вид. По редко встре-
чающейся трехосно-эллипсоидной форме зигоспор наиболее близкими
видами являются S. crassoidea и S. subcrassa. Именно по этому при-
знаку четко отличается от других близких видов, сходных с ним по
многим вегетативным признакам: S. maxima, S. heeriana, S. jassiensis,
S. krubergii. На рисунках первых исследователей 5. crassa (Kiitzing,
1843; Petit, 1880) мезоспорий выглядит не ямчатым, а, скорее, боро-
давчатым или неправильно и мелкобугорчатым.
81. Spirogyra krubergii V. Poljansk. — Спирогира Круберга (рис. 114,
1—3). — Вегетативные клетки 120—135 мкм шир., по длине меньше
ширины или до 3 раз больше. Поперечные стенки гладкие. Хлороплас-
261
Рис. 114. 1—3 — Spirogyra krubergii: 1 — трехосно-эллипсоидные зигоспоры в разных по-
ложениях, лестничная конъюгация, 2а — трехосно-эллипсоидная (почти линзовидная) зи-
госпора в двух положениях, 26 — линзовидная зигоспора анфас, 3 — строение зигоспоры
(фрагменты); 4, 5 — S. sinensis: 4 — зигоспоры в разных положениях, лестничная конъюга-
ция, 5 — лестничная и «попытка» боковой конъюгации; 6, 7 — S. pellucida f. pellucida: 6 —
вегетативная клетка, 7 — лестничная конъюгация; 8—13 — S. majuscula: 8, 9 — различная
степень вздутия воспринимающих клеток, лестничная конъюгация, 10—12 — зигоспоры в
разных положениях, боковая конъюгация, 13 — зигоспора в трех положениях (схема). (По:
1—3 — Полянский, 1951; 4, 5 — Красноперова, 19716; 6, 7 — West G. S., 1899; 8, 10 —
Gauthier-Lievre, 1965; 9, 11—13 — ориг.).
тов 4—7, с 0.5—1.25 оборота спирали. Конъюгация лестничная. Конъ-
югационный канал образован обеими клетками. Воспринимающие клетки
невздутые. Клетки без конъюгационной пары невздутые. Зигоспоры сжато-
эллипсоидные (трехосно-эллипсоидные), в поперечном сечении эллипти-
ческие, 100—120 мкм шир., 120—142.5 мкм дл., 80—95 мкм толщ.,
реже линзовидные. Экзоспорий толстый, 5.9—7.8 мкм толщ., бесцветный,
гладкий, неявственно двухслойный, причем наружный слой всегда тоньше
внутреннего. Мезоспорий тонкий, бесцветный или бледно-коричневый, с
более или менее правильными бугорчатыми выпуклостями, сравнительно
пологими или крутыми, заостряющимися или закругленными.
В пруду. Россия: северо-запад европ. части (Санкт-Петербург).
Редкий вид. Строение оболочек зигоспор сходно с S. borysthenica.
От S. crassa, имеющего такие же по форме зигоспоры, отличается
строением оболочек зигоспор, более узкими вегетативными клетками
и меньшим количеством хлоропластов. От S. maxima, сходного по ве-
гетативным признакам, отличается формой зигоспор и строением их
оболочек.
82. Spirogyra sinensis Li — Спирогира китайская (рис. 114, 4, 5). —
Вегетативные' клетки 22—26 (29) мкм шир., по длине в 5—13 раз боль-
ше. Ризоиды латеральные, обычно вильчато разветвленные, многочис-
ленные. Поперечные стенки гладкие. Хлоропластов 2—4. Конъюгация
лестничная. Конъюгационный канал образован обеими клетками. Вос-
принимающие клетки веретеновидные, сильно вздутые со всех сторон
до 49—55 мкм. Клетки без конъюгационной пары невздутые. Зигоспоры
толстолинзовидные, 32—38 (43.5) мкм толщ., (38) 43.5—49 мкм диам.
Экзоспорий тонкий, бесцветный, гладкий. Мезоспорий толстый, от
желто-коричневого до коричневого, гладкий. Эндоспорий различим
только на зрелых зиготах.
В стоячих водах, в рыбохозяйственных прудах, на рисовых полях.
Южн. Казахстан.
Редкий вид, найденный также в Китае. От спирогир с линзовид-
ными зигоспорами хорошо отличается значительно меньшими разме-
рами. Наиболее близок к S. pellucida f. minor.
83. Spirogyra pellucida (Hass.) G. S. West — Спирогира прозрач-
ная (рис. 114, 6, 7). — Вегетативные клетки 30—50 мкм шир., по
длине в 3.5—9 (12) раз больше. Ризоиды латеральные, розетковидные,
немногочисленные. Поперечные стенки гладкие. Хлоропластов 2—4, с
незначительным числом оборотов спирали или почти прямые. Конъ-
югация лестничная. Конъюгационный канал образован обеими клетка-
ми. Воспринимающие клетки очень сильно веретеновидно вздутые во-
круг зигоспор, до 75.4—84 мкм. Клетки без конъюгационной пары
невздутые. Зигоспоры утолщенно-линзовидные, в двух положениях
круглые и эллиптические, 40—58 мкм толщ., 50—72.5 мкм диам., или
почти шаровидные, 77—86 мкм диам. Экзоспорий тонкий, бесцветный,
гладкий. Мезоспорий толстый, желто-коричневый, гладкий.
Редкий и недостаточно изученный вид, особенно в отношении стро-
ения оболочек зигоспор.
263
F. pellucida (рис. 114, 6, 7). — Вегетативные клетки 45—50 мкм
шир. Хлоропластов 3—4, почти прямые. Зигоспоры утолщенно-линзо-
видные, почти шаровидные, 77—86 мкм диам.
В стоячих водах, на рисовых полях, в оросителях. Россия: центр
европ. части; Молдавия (Бессарабия) (?), Узбекистан.
Довольно редкая форма. Обнаруживает большое сходство с S. sphae-
rospora, но имеет большее число хлоропластов и другую форму зиго-
спор. Отсутствуют убедительные сведения о строении оболочек зиго-
спор.
F. minor (Tiffany) V. Poljansk. [=5. pellucida Kutz.; 5. pellucida var.
minor Tiffany; S. frankliniana Tiffany; S. rotundispora Skv. (?); S. ugan-
dense Gauthier-Lievre], — Вегетативные клетки 30—40 мкм шир. Хло-
ропластов 2—4. Воспринимающие клетки вздутые до 75.4—84 мкм
шир. Зигоспоры утолщенно-линзовидные, 40—58 мкм толщ., 50—
72.5 мкм диам.
В стоячих водах, озерах, в том числе пойменных. Россия: центр
европ. части (Заволжье).
Довольно редкая форма с широким ареалом (Англия, Нидерланды,
Финляндия, Уганда, Сев. Китай, США). Отличается от типовой формы
в основном меньшими размерами клеток и меньшим числом хлоро-
пластов. Наиболее близка к 5. sinensis, но ширина вегетативных клеток
последнего вида значительно меньше.
84. Spirogyra majuscula Kutz. [=5. majuscula var. brachymeres Stit-
zenberger; S. majuscula var. genuina Kirchn.; S. orthospira Nag.; S. bellis
(Hass.) Cleve sensu Казаковский, Смирнов, 1914, non auct. pl.] — Спи-
рогира крупноватая (рис. 114, 8—13). — Вегетативные клетки 50—80
(100) мкм шир., по длине в 1.2—10 раз больше. Ризоиды латеральные,
нередкие. Поперечные стенки гладкие. Хлоропластов (3) 5—8 (10),
почти прямые. Конъюгация боковая и лестничная, с преобладанием
одной из них, иногда совмещенные в одной паре нитей. Конъюгацион-
ный канал при лестничной конъюгации образован обеими клетками; при
боковой конъюгации канал широкий, иногда ушковидный, выдается на
1/2 ширины клетки, нить при этом прямая. Воспринимающие клетки
невздутые или иногда значительно вздутые со стороны, противополож-
ной конъюгационному каналу. Клетки без конъюгационной пары не-
вздутые. Зигоспоры линзовидные, в двух положениях круглые и эллип-
тические, 35—60 (82) мкм толщ., 50—85 (98) мкм диам. Экзоспорий
довольно толстый, бесцветный, гиалиново-пектиновый, гладкий, одно-
слойный. Мезоспорий толстый, от желтого до коричневого, трехслой-
ный: внешний слой тонкий, бесцветный, гиалиновый, гладкий; внутрен-
ний слой состоит из двух окрашенных подслоев, плотно расположенных
друг к другу, первый подслой гладкий, второй подслой радиально исчер-
ченный. Эндоспорий бесцветный, гладкий, двухслойный: первый слой
очень тонкий, целлюлозный, второй слой толще первого, пектиновый.
Партеноспоры 46.6—51 мкм толщ., 54—64.8 мкм диам., довольно частые.
В стоячих и текучих, нередко загрязненных водах со значительным
содержанием железа, в водоемах оросительной системы, минеральных
264
водах, опресненных водах морей. Россия: европ. часть — повсюду,
Каспийское море (Кировский залив), Зап. Сибирь (окрестности Том-
ска, окрестности Обь-Енисейского канала), Вост. Сибирь (окрестности
Иркутска, Туркинские минеральные воды близ оз. Байкал); Украина —
повсюду, Сев. (заповедник Боровое) и Южн. Казахстан, Узбекистан.
Широко распространенный и часто встречающийся вид. Типичным
односторонним вздутием воспринимающих клеток и иным строением
мезоспория отличается от S. bellis. От S. manoromae Randh. (Сев.
Индия), имеющего сходную форму воспринимающих клеток, отличает-
ся отсутствием сетчатой структуры мезоспория. Шестислойная оболоч-
ка зигоспоры S. majuscula, хорошо изученная в сканирующем микрос-
копе, уникальна для зигнемовых водорослей. В световом микроскопе
исчерченность второго подслоя мезоспория подчас трудно заметить, в
СЭМ эта исчерченность представляется губчатой.
85. Spirogyra submaxima Trans. — Спирогира большая (рис. 115,
1, 2). — Вегетативные клетки 70—ПО мкм шир., по длине в 1.5—6
раз больше. Поперечные стенки гладкие. Хлоропластов 5—9, почти
прямые или незначительно извитые. Конъюгация лестничная. Конъюга-
ционный канал образован обеими клетками. Воспринимающие клетки
невздутые или незначительно вздутые со стороны конъюгационного ка-
нала. Клетки без конъюгационной пары невздутые. Зигоспоры линзо-
видные, в двух положениях круглые и эллиптические, 50—75 мкм
толщ., 67—100 мкм диам. Экзоспорий тонкий, бесцветный, гладкий.
Мезоспорий толстый, желто-коричневый, гладкий, иногда слоистый.
Вид малоизменчивый, возможно монотипный. Представляется так-
сономически спорным признак слоистости мезоспория.
F. submaxima (S. submargaritata Godward) (рис. 115, 7). — Веге-
тативные клетки (70) 96—ПО мкм шир. Хлоропластов (5) 8—9. Вос-
принимающие клетки невздутые или незначительно вздутые со стороны
конъюгационного канала. Зигоспоры 50—75 мкм толщ., 70—100 мкм
диам. Мезоспорий не слоистый.
В стоячей воде; Украина: Причерноморье (бассейн Днепра).
Форма найдена также в Индии, Африке (Марокко и Уганда), США,
Китае. Нити обычно окружены толстым слизистым покровом (влага-
лищем), около 17 мкм толщ. От S. bellis отличается гладким мезос-
порием и большей шириной вегетативных клеток, от S. majuscula и
S. randhawae (Randh.) Krieger (Сев. Индия) — в основном невздутыми
воспринимающими клетками.
F. lamellata (Rao) V. Poljansk. [=5. submaxima var. lamellata Rao;
S. lamellata (Rao) Krieger] (рис. 115, 2). — Вегетативные клетки 70—79
(90) мкм шир. Хлоропластов 6—8. Воспринимающие клетки невздутые.
Зигоспоры 50—60 мкм толщ., 67—83 мкм диам. Мезоспорий слоистый.
Лужи в каменоломнях. Индия.
Редкая форма. Отсутствие слизистого покрова вокруг нитей и сло-
истость мезоспория — основные, хотя и незначительные отличия от
типовой формы.
265
86. Spirogyra glabra Czurda — Спирогира голая (рис. 115, 3). —
Вегетативные клетки 145—153 мкм шир., по длине меньше ширины
или в 1.5 раза больше. Поперечные стенки гладкие. Хлоропластов 7.
Конъюгация лестничная. Конъюгационный канал образован обеими
клетками. Воспринимающие клетки невздутые. Клетки без конъюгаци-
онной пары невздутые. Зигоспоры линзовидные, в двух положениях
круглые и эллиптические, 120 мкм тоАц., 170 мкм диам. Экзоспорий
толстый, около 10 мкм толщ., бесцветный, гладкий. Мезоспорий тол-
стый, около 20 мкм толщ., желто-коричневый, гладкий.
В слабопроточных водоемах, старицах, каналах, дренажных ямах.
Россия: Предкавказье (Сев. Осетия) (?); Ср. Азия (Каракумы) (?).
Редкий и слабоизученный вид, найденный до сих пор только в
Австрии. От близкого вида — S. maxima, именем которого мог оши-
бочно обозначаться, отличается гладким мезоспорием; от S. submaxima,
имеющего также гладкий мезоспорий, — значительно более крупными
размерами.
87. Spirogyra bellis (Hass.) Cleve, auct. non Cleve: Казановский,
Смирнов, 19136, 1914 (=S. subaequa Kiitz., auct. non Kiitz.: Алексенко,
1888, 1894) — Спирогира красивая (рис. 115, 4—6). — Вегетативные
клетки (57) 65—80 (88) мкм шир., по длине изредка меньше ширины,
обычно в 1.5—5 раз больше. Ризоиды латеральные и базальные, часто
разветвленные, многочисленные. Поперечные стенки гладкие. Хлоро-
пластов 5—6 (8), почти прямые или слабо извитые. Конъюгация лест-
ничная. Конъюгационный канал образован обеими клетками, с небольшим
преобладанием отдающей клетки. Воспринимающие клетки невздутые
или незначительно вздутые со всех сторон. Клетки без конъюгационной
пары невздутые. Зигоспоры линзовидные, в двух положениях круглые
и эллиптические, 37.7—75 мкм толщ., 57.2—105 мкм диам. Экзоспо-
рий толстый, бесцветный, гладкий, двухслойный, целлюлозно-пектино-
вый. Мезоспорий толстый, коричневый, двухслойный, ямчатый. Эндос-
порий гладкий, однослойный, пектиновый.
В стоячих и текучих водах, нередко в болотах, родниках, на ри-
совых полях, на сильно увлажненной почве. Повсюду, кроме Дальн.
Востока.
Широко распространенный вид. По ширине, вегетативных клеток
близок к S. majuscula, от которого отличается вздутием воспринима-
ющих клеток со всех сторон, ямчатым двухслойным мезоспорием, тол-
стым двухслойным экзоспорием и однослойным эндоспорием. По на-
Рис. 115. 1, 2 — Spirogyra submaxima, зигоспоры в разных положениях, лестничная конъ-
югация: 1 — f. submaxima, 2 — f. lamellata-, 3 — S. glabra, зигоспоры в двух положениях,
лестничная конъюгация; 4—6 — S. bellis: 4 — вегетативная клетка и ризоид, 5 — зигоспора
в двух положениях, лестничная конъюгация, 6 — строение мезоспория (фрагмент); 7—9 —
S. Jassiensis, зигоспоры разной формы и в различных положениях, боковая конъюгация;
10—12 — S. czurdiana: 10 — вегетативные клетки и ризоиды, 11 — зигоспоры в двух поло-
жениях, лестничная конъюгация, 12 — строение мезоспория (фрагмент). (По: 1 — Tran-
seau, 1914 (как S. submaxima): 2 — Rao, 1938 (как S. lamellata)-, 3, 6 — Czurda, 1932; 4,
5 — Рундина, 1988; 7—9 — Teodoresco, 1907; 10—12 — Kadhibowska in: Kadhibowska,
Christensen, 1979).
266
9
блюдениям в сканирующем микроскопе, экзоспорий имеет два подслоя:
первый — тонкий, гиалиново-целлюлозный, второй подслой толще
первого, блестящий, пектиновый; мезоспорий тоже состоит из двух
подслоев: первый — тоньше второго, бесцветный и явственно ямчатый
(пористый, причем стенки пор приподняты над поверхностью подслоя
наподобие вулканчиков), второй подслой толстый, окрашенный, глад-
кий, с отчетливой линией разрыва.
88. Spirogyra jassiensis (Teod.) Czurda (=S. crassa Kiitz. var. jassi-
ensis Teod.) — Спирогира яссинская (рис. 115, 7—9). — Вегетативные
клетки 116—132 мкм шир., по длине до 2 раз больше. Поперечные
стенки гладкие. Хлоропластов 8—9, слегка извитые или прямые. Конъ-
югация боковая. Конъюгационный канал нечетко и слабо выраженный,
выдается на 1/5—1/6 ширины клетки, нить при этом прямая или очень
слабо изогнутая. Зигоспоры линзовидные (в укороченных воспринима-
ющих клетках), в двух положениях круглые и эллиптические, 72—
97 мкм толщ., 102—126 мкм диам., или цилиндрически-эллипсоидные
(в неукороченных воспринимающих клетках), 83—88 мкм шир., 140—
154 мкм дл. Экзоспорий тонкий, бесцветный, гладкий. Мезоспорий
толстый, коричневый, с округлыми или неправильной формы плоскими
углублениями.
В стоячих водах, преимущественно в мелких и небольших водоемах,
большими скоплениями на поверхности воды. Россия: центр европ.
части (окрестности Москвы).
Очень редкий и малоизученный вид, впервые найденный в Румынии.
Требует уточнения скульптура мезоспория, не оговоренная особо в
первоописании. Не исключено, что мезоспорий гладкий. Противоречи-
вы описания формы зигоспор: то линзовидные, то трехосно-эллипсо-
идные, то цилиндрические с закругленными вершинами. Вышеприве-
денный диагноз составлен в основном по первоописанию. Отличается
от S. bellis широкими вегетативными клетками и наличием боковой
конъюгации. Строение мезоспория у обоих видов сходное.
89. Spirogyra heeriana Nag. — Спирогира Хеера (рис. 116). —
Вегетативные клетки 120—150 мкм шир., по длине меньше ширины
или до 2 раз больше. Поперечные стенки гладкие. Хлоропластов 7—16,
иногда больше, с небольшим количеством оборотов или почти прямые,
в некоторых случаях многократно разветвленные, с анастомозами.
Конъюгация лестничная. Конъюгационный канал образован обеими
клетками, иногда с преобладанием воспринимающей клетки, очень
редко — исключительно ею. Воспринимающие клетки невздутые.
Клетки без конъюгационной пары невздутые. Зигоспоры линзовидные,
в двух положениях круглые и эллиптические, 80—130 (145) мкм толщ.,
107—195 мкм диам. Экзоспорий тонкий, бесцветный, гладкий, по кон-
туру неровный из-за особенностей строения мезоспория. Мезоспорий
толстый, до 40 мкм толщ., желто-коричневый или светло-бурый, од-
нослойный или двухслойный,, с беспорядочно расположенными бугор-
чатыми возвышениями разных размеров и форм. Эндоспорий тонкий,
бесцветный, гладкий. Линия разрыва четкая, продольная.
268
5
Рис. 116. Spirogyra heeriana: 1—4 — f. heeriana: 1,2 — зигоспоры в разных положениях,
лестничная конъюгация, 3,4 — строение мезоспория (фрагменты); 5—7 — f. ucrainica: 5 —
вегетативная клетка, 6 — зигоспоры в двух положениях, лестничная конъюгация, 7 — стро-
ение мезоспория (фрагмент, микротомный срез). (По; 1,3 — Полянский, 1955; 2, 4 —
Czurda, 1932; 5—7— Казановский, Смирнов, 1914 (как 5. ucrainica)).
Редкий, недостаточно изученный вид. Особенности формирования
конъюгационного канала находятся в прямой зависимости от стадии
конъюгации и образования зигот.
F. heeriana (рис. 116, 1—4). — Вегетативные клетки 120—150 мкм
шир. Хлоропластов 7—10, неразветвленные. Зигоспоры 80—130 (145) мкм
толщ., 120—195 мкм диам. Мезоспорий однослойный, с беспорядочно
расположенными, более или менее округлыми или удлиненными бугор-
чатыми возвышениями разных размеров.
В стоячих водах. Характерна для известковых вод. Россия: При-
черноморье (окрестности Ростова-на-Дону); Сев. Казахстан.
Известна также из Франции, Австрии, Чехии (?), Алжира и Ма-
рокко. Своеобразное строение бугорчатого мезоспория (сверху воспри-
нимается крупно- и груботочечным) служит главным отличием от близ-
ких видов: 5. maxima, S. glabra, S. jatobae Trans. (Алжир, Бразилия)
и 5. lenticularis.
269
F. ucrainica (Smirn.) V. Poljansk. (=5. ucrainica Smirn.) (рис. 116,
5—7). — Вегетативные клетки 127—143 мкм шир. Хлоропластов 12—
16, иногда больше, многократно разветвленных и анастомозирующих.
Зигоспоры 88—91 мкм толщ., 107—140 мкм диам. Мезоспорий двух-
слойный: наружный слой тонкий, с призматически-бугорчатыми выро-
стами и мелкими зернами у их основания, внутренний слой толстый,
гладкий.
В стоячих водах пойменных водоемов. Россия: европ. часть (По-
волжье); Украина (окрестности Киева).
Редкая форма. Отличия от типовой формы сводятся к большему
числу хлоропластов, их сильному ветвлению и анастомозированию,
двухслойности мезоспория и несколько иной конфигурации возвыше-
ний на нем.
90. Spirogyra czurdiana Kadt. — Спирогира Чурды (рис. 115, 10—
12). — Вегетативные клетки 46.4—57.6 мкм шир. Ризоиды латеральные,
клубочковидные или в виде неразветвленных выростов. Поперечные
стенки гладкие. Хлоропластов 5—7. Конъюгация лестничная. Конъюга-
ционный канал образован обеими клетками. Воспринимающие клетки
вздутые со стороны, противоположной конъюгационному каналу. Зигос-
поры линзовидные, в двух положениях круглые и эллиптические, 43.2—
68.8 мкм толщ., 52.8—72 мкм диам. Экзоспорий тонкий, бесцветный,
гладкий. Мезоспорий толстый, красновато-бурый, сетчатый, в попереч-
ном сечении тонко исчерченный.
В глинистой яме. Дания. .
Редкий вид. Отличается от близкой S. majuscula строением мезо-
спория.
91. Spirogyra maxima (Hass.) Wittr. — Спирогира наибольшая (рис. 117,
118). — Вегетативные клетки 70—200 мкм шир., по длине меньше ши-
рины, равны ей или до 4 раз больше. Ризоиды латеральные, простые
или разветвленные. На поверхности клеток имеются кристаллы оксалата
кальция, квадратные и крестообразные, нередкие. Поперечные стенки
гладкие. Хлоропластов (3) 5—8 (12), неизвитые, почти прямые или с
небольшим количеством, до 2, оборотов спирали. Конъюгация лестнич-
ная и боковая, с преобладанием одной из них. Конъюгационный канал
при лестничной конъюгации образован обеими клетками; при боковой
конъюгации выдается незначительно, до 1/3 ширины клетки, округлый,
нить при этом прямая. Воспринимающие клетки невздутые или очень
слабо вздутые. Клетки без конъюгационной пары невздутые. Зигоспоры
линзовидные, в двух положениях круглые и широкоэллиптические, (33)
56—122 (140) мкм толщ., (64) 70—190 мкм диам. Экзоспорий тонкий,
изредка довольно толстый, гладкий, однослойный, иногда двухслойный:
внешний слой тонкий, бесцветный, гладкий; внутренний слой до 5.8 мкм
толщ., от бледно-желтого до желтого, гладкий. Мезоспорий толстый, до
14.5 мкм толщ., от желто-коричневого до темно-коричневого, почти
черного, с извилистыми, короткими или длинными, выпуклыми утолще-
ниями (морщинами), сомкнутыми или несомкнутыми в тонкую или гру-
бую сеть, иногда складчато-бородавчатый или точечно-извилистый, двух-
270
Рис. 117. Spirogyra maxima: 1,2 — f. maxima: 1 — зигоспоры в двух положениях, лестнич-
ная конъюгация, 2 — строение мезоспория (фрагмент); 3, 4 — f. livida: 3 — зигоспоры в
двух положениях, лестничная конъюгация, 4 — зигоспора в двух положениях, сверху —
строение мезоспория (фрагмент); 5—7 — f. tenuior: 5, 6 — зигоспоры в разных положени-
ях, лестничная конъюгация, 7 — зигоспора анфас. (По: 1 — Petit, 1880 (как S. orbicularis):
2 — Borge, 1913 (как S. maxima); 3—7 — Gauthier-Lievre, 1965 (3, 4 — как S. megaspora,
5—7 — как S. moebii)).
271
Рис. 118. Spirogyra maxima: 1—3 — f. cupientis: 1 — ризоиды (справа) и ризоидовидные
конъюгационные выросты, боковая конъюгация, 2 — зигоспоры в разных положениях, лес-
тничная конъюгация, 3 — зигоспора анфас, слева — строение мезоспория; 4 — f. tenuior,
зигоспоры в двух положениях и вегетативная клетка без конъюгационной пары; 5 — f. wo-
ronichinii, зигоспора анфас, боковая конъюгация. (По: 1—3 — Водеиичаров, Киряков, 1986
(как S. cupientis): 4, 5 — Рундина, 19786).
272
слойный. Эндоспорий тонкий, светло-желтый, гладкий. Партеноспоры
эллипсоидные, 49—58 мкм шир., 70—87 мкм дл., редкие.
Вид полиморфный. Между его формами существует ряд переходов
по признакам ширины вегетативных клеток и скульптуры мезоспория.
Можно предположить, что по мере изучения скульптуры мезоспория
на различных стадиях созревания зигоспор у различных форм границы
этих форм будут постепенно размываться и из полиморфного вида, как
представляется сейчас, он превратится в монотипный или количество
его внутривидовых форм уменьшится.
F. maxima [=5. orbicularis (Hass.) Kiitz., auct. non (Hass.) Kiitz.: Ar-
tari, 1885; Иванов, 1898; S. maxima sensu Троицкая, 1930, auct. non
(Hass.) Wittr.: Скворцов, 1917; Воронихин, 1920, 1926, 1927; S. troitzka-
jae V. Poljansk.] (рис. 117, 1, 2). — Вегетативные клетки (115) 125—155
(170) мкм шир. Хлоропластов (3) 5—8 (12). Конъюгация лестничная,
очень редко боковая. Зигоспоры (60) 70—122 (140) мкм толщ., 100—150
(170) мкм диам. Мезоспорий сетчато утолщенный.
В текучих и стоячих водах, водохранилищах, на рисовых полях, в
родниках, оросительных каналах, часто при значительном азотистом
загрязнении, иногда в теплых ключах при 52 °C. Повсюду.
Широко распространенная форма. Отличается довольно значитель-
ной изменчивостью ряда признаков, таких как размеры вегетативных
клеток и зигоспор, число хлоропластов, степень сомкнутости сети на
мезоспорий. От S. crassa отличается формой зигоспор и строением
мезоспория, от S. glabra — скульптурированный мезоспорием.
F. cupientis (Vodenic. et Kiriakov) Rund. (=S. cupientis Vodenic. et
Kiriakov) (рис. 118, 1—3). — Вегетативные клетки 70—94 мкм шир.
Хлоропластов 6—8. Конъюгация лестничная. Зигоспоры 33—60 мкм
толщ., 64—86 мкм диам. Мезоспорий толстый, мелкоточечный и с
короткими извилистыми складками (морщинами), не сомкнутыми в
сеть, возможно двухслойный.
В небольшом болоте. Болгария.
Редкая форма. По совокупности признаков f. cupientis, естественно,
входит в круг форм S. maxima, являясь как бы промежуточной формой
между f. tenuior и f. woronichinii. С f. tenuior она сближается по ширине
вегетативных клеток, с f. woronichinii — по склонности к боковой конъ-
югации и двухслойности мезоспория. Однако скульптура мезоспория у
f. cupientis более тонкая, чем у f. woronichinii, и этим сходна с f. tenuior.
Возможно, идентична последней. Следует подчеркнуть склонность f. cu-
pientis к боковой конъюгации, не отмеченную в авторском описании, но
четко зафиксированную на рисунке (Водеиичаров, Киряков, 1986). При
этом конъюгационный канал образуется путем слияния ризоидовидных
выростов, иногда лопастных, от соседних по нити клеток. Канал длин-
ный, и его образование в данном случае, скорее, случайность, чем зако-
номерность.
F. livida (Vail.) V. Poljansk. (=S. maxima var. livida Vail.; S. me-
gaspora Trans.) (рис. 117, 3, 4). — Вегетативные клетки (150)
18 Рундина Л. А.
273
170—200 мкм шир. Хлоропластов 5—8. Конъюгация лестничная.
Зигоспоры 90—120 мкм толщ., 135—190 мкм диам. Мезоспорий
тонкосетчатый.
В стоячих и текучих водах. Литва.
Редкая форма, известная также из Швеции, Нидерландов, Марокко
и Южн. Америки (Уругвай). Отличается от типовой формы главным
образом большей шириной вегетативный клеток и зигоспор. При изу-
чении зигоспор 5. maxima и 5. megaspora в сканирующем микроскопе
подтверждено их большое сходство в строении мезоспория.
F. tenuior (Magnus et Wille) V. Poljansk. [=5. orbicularis (Hass.)
Kiitz. f. tenuior Magnus et Wille; 5. orbicularis sensu Artari, 1885; Ива-
нов, 1899; S. moebii Trans.] (рис. 117, 5—7; 118, 4). — Вегетативные
клетки 77—117 (130) мкм шир. Хлоропластов 4—8. Конъюгация лес-
тничная. Зигоспоры (50) 55—75 (90) мкм толщ., 70—103 (135) мкм
диам. Мезоспорий тонкосетчатый.
В стоячих и текучих водах, возможно в слабосоленых водах, в
реках, свободноплавающая или в прикрепленном состоянии. Россия:
северо-запад и центр европ. части; Южн. Казахстан.
Крайне редкая в России форма, однако довольно широко распрос-
траненная за ее пределами (Германия, Африка, Индия, Сев. и Южн.
Америка, Австралия). Отличается от типовой формы, с которой часто
вегетирует совместно, только меньшими размерами вегетативных кле-
ток и зигоспор. Наличие скульптуры на мезоспорий является главным
отличием от S. submaxima.
F. woronichinii V. Poljansk. (=S. maxima sensu Скворцов, 1917; Во-
ронихин, 1920, 1926, 1927; S. lodziensis Kadi.) (рис. 118, 5). — Веге-
тативные клетки 125—174 мкм шир. Хлоропластов 5—8. Конъюгация
боковая, очень редко лестничная, иногда совмещенные в одной паре
нитей. Зигоспоры 75—ПО мкм толщ., 100—155 (163) мкм диам. Ме-
зоспорий с короткими, выпукло-извилистыми складками (морщинами),
обычно не сомкнутыми в сеть, и густо расположенными, разного раз-
мера бородавочками.
В стоячих водах, морце, озере, пруду, канаве, иногда при довольно
сильном загрязнении. Россия: северо-запад европ. части (Санкт-Петер-
бург); Азербайджан (Ленкорань), Сев. Казахстан (оз. Кара-Камыш).
Редкая форма, известная также из Польши. Преобладание боковой
конъюгации над лестничной и морщинисто-бородавчатый (бугорчатый
и крупно-, груботочечный при взгляде на зигоспору сверху) мезоспо-
рий — основные отличия этой формы от типовой.
92. Spirogyra pseudomaxima Kadi. — Спирогира ложнонаибольшая
(рис. 119, 1, 2). — Вегетативные клетки 128—144 мкм шир. Попе-
речные стенки гладкие. Хлоропластов 5—7. Конъюгация лестничная.
Конъюгационный канал образован обеими клетками. Воспринимающие
клетки невздутые. Зигоспоры линзовидные, в двух положениях круглые
и эллиптические, 61—112 мкм толщ., 90—126 мкм диам. Экзоспорий
тонкий, бесцветный, гладкий. Мезоспорий каштановый, двухслойный:
274
Рис. 119. 1,2 — Spirogyra pseudomaxima: 1 — зигоспоры в разных положениях, лестничная
конъюгация, 2 — строение мезоспория; 3,4 — S. lenticularis: 3 — зигоспоры в разных по-
ложениях, лестничная конъюгация, 4 — строение мезоспория (вид с поверхности зигоспо-
ры). (По: 1,2 — Kadhibowska in: Kadhibowska, Christensen, 1979; 3, 4 — Transeau, 1938).
внешний слой тонкий, с разветвленными короткими ребрами (морщи-
нами); внутренний слой толстый, рельефный, неправильно плотносет-
чатый.
В глинистой яме. Дания.
Редкий, недостаточно изученный вид. Следует учесть, что орнамен-
тация внутреннего слоя мезоспория отчетливо видна только после уда-
ления внешнего слоя. Незначительно отличается от S. maxima деталями
в строении мезоспория.
275
93. Spirogyra oblata Jao — Спирогира облатковидная (рис. 120, 1,
2). — Вегетативные клетки (93) 96—119 мкм шир., по длине до 2.5
раза больше, изредка меньше ширины или равны ей. Поперечные стен-
ки гладкие. Хлоропластов 6—13, почти прямые. Конъюгация лестнич-
ная. Конъюгационный канал образован обеими клетками. Восприни-
мающие клетки невздутые. Зигоспоры лицзовидные, в двух положениях
круглые и эллиптические, 64—106 мкм толщ., 92—160.5 мкм диам.
Экзоспорий толстый, по толщине равный мезоспорию или уже его,
2—2.5 мкм толщ., бесцветный, гладкий, слоистый. Мезоспорий 8—
10 мкм толщ., на ранних стадиях созревания зиготы желтый, непра-
вильно сетчатый, в зрелом состоянии зиготы золотисто-коричневый,
бородавчатый.
В озерах. Россия: европ. часть (оз. Валдайское).
Очень редкий вид, встреченный также в Китае и Японии. Измене-
ние скульптуры мезоспория по мере созревания зиготы обусловлено,
возможно, частичным разрушением сетчатых утолщений, причем не-
редко встречаются промежуточные стадии. По ширине вегетативных
клеток близок к S. maxima f. tenuior, по скульптуре мезоспория — к
S. maxima f. woronichinii.
94. Spirogyra lenticularis Trans. — Спирогира чечевицеобразная
(рис. 119, 3, 4). — Вегетативные клетки 136—184.8 мкм шир., по
длине меньше ширины, равны ей или до 2 раз больше. Поперечные
стенки гладкие. Хлоропластов 9—10, прямые или с 1 оборотом спи-
рали. Конъюгация лестничная. Конъюгационный канал от обеих кле-
ток. Воспринимающие клетки укороченные, невздутые или слегка взду-
тые со стороны конъюгационного канала. Зигоспоры линзовидные
(сжато-шаровидные), в двух положениях круглые и эллиптические,
90—100 мкм шир., 103.3—156.4 мкм дл. Экзоспорий тонкий, бесцвет-
ный, гладкий. Мезоспорий толстый, коричневый, трехслойный: верхний
подслой с короткими извилистыми складками (морщинами), не сомк-
нутыми в сеть, средний подслой гладкий, нижний подслой мелкобо-
родавчатый. Эндоспорий гладкий.
В стоячих (?) водах. Нидерланды, Южн. Африка.
Редкий, хорошо изученный в отношении строения мезоспория вид.
Имеет несомненное сходство с кругом форм S. maxima, особенно с f.
woronichinii. Основные отличия: наличие только лестничной конъюга-
ции, большее число хлоропластов и трехслойность мезоспория. Близок
по многим признакам к S. oblata.
95. Spirogyra ternata Ripart, auct. non Ripart: Gutwinski, 1895a
[=5. neglecta (Hass.) Kiitz. var. ternata (Ripart) W. et G. S. West;
5. unocula Chm. (?)] — Спирогира тройчатая (рис. 120, 3—6). — Ве-
гетативные клетки (51) 58—78 (87) мкм шир., по длине в 1.2—2 (3)
раза больше. На поверхности клеток имеются редкие кристаллы окса-
лата кальция. Поперечные стенки гладкие. Хлоропластов (2) 3—5.
Конъюгация лестничная. Конъюгационный канал образован обеими
клетками, иногда с преобладанием отдающей клетки, нечетко отгра-
ниченный со стороны воспринимающей клетки. Воспринимающие
276
Рис. 120. 1,2 — Spirogyra oblata: 1 — зигоспоры в разных положениях, лестничная конъ-
югация, 2 — строение оболочек зигоспоры; 3—6 — S. ternata: 3—5 — разные формы зигос-
пор, лестничная конъюгация, 6 — форма зигоспор: а — трехосно-эллипсоидная, анфас и
сбоку, б — линзовидная, анфас и сбоку, в — эллипсоидная (двухосно-эллипсоидная), анфас
и сверху; 7 — 5. diversizygotica, форма зигоспор: а, б —- трехосно-эллипсоидная, анфас и
сбоку, в — линзовидная, анфас и сбоку, г —- эллипсоидная (двухосно-эллипсоидная), анфас
и сверху. (По: 1, 2 — Jao, 1936; 3—7 — Полянский, 1951 (7 — как 5. setiformis f. diversi-
zygotica)).
277
клетки более или менее, иногда значительно вздутые со всех сторон,
редко со стороны конъюгационного канала. Клетки без конъюгаци-
онной пары невздутые. Зигоспоры трех форм: двухосно-эллипсоидные
(в поперечном сечении круглые), (50) 55—75 (87.5) мкм шир., 65—
112.5 (127.5) мкм дл., широколинзовидные (в двух положениях круглые
и эллиптические), 47.5—84 мкм толщ^ 65—92.5 (95) мкм диам., и
трехосно-эллипсоидные (в поперечном сечении эллиптические), 72.5—
80 мкм шир., 60—100 мкм дл., 50—67 (75.4) мкм толщ. Экзоспорий
тонкий, бесцветный, гладкий. Мезоспорий толстый, желтовато-корич-
невый, гладкий.
В стоячих и текучих водах, в канавах, бассейнах с непроточной
водой, на рисовых полях, в эстуариях крупных рек, изредка при не-
большой солености (0.12—5.74 %о), иногда большими скоплениями на
поверхности воды. Россия: Карелия, северо-запад и центр европ. части,
приустье Волги, зап. побережье Каспийского моря, Сев. Осетия; Лат-
вия, Зап. и Вост. Украина, Причерноморье, Молдавия, Сев. и Южн.
Казахстан.
Вид широко распространенный (найден почти во всех странах Ев-
ропы), однако не часто встречающийся. Вид труден для определения
из-за сложности распознавания одновременно существующих трех
форм зигоспор. Отличается от S. neglecta значительным вздутием вос-
принимающих клеток и усложненной формой зигоспор, от S. dubia —
большей шириной вегетативных клеток, иногда меньшим числом
хлоропластов и разнообразием форм зигоспор. По последнему признаку
близок к S. diversizygotica.
96. Spirogyra diversizygotica (V. Poljansk.) Rund. [=5. setiformis
(Roth) Kiitz. f. diversizygotica V. Poljansk.] — Спирогира разнозиготная
(рис. 120, 7; 121, 1, 2). — Вегетативные клетки 100—115 мкм шир., по
длине меньше ширины или до 3 раз больше. Поперечные стенки гладкие.
Хлоропластов 3—7, с небольшим количеством оборотов спирали. Конъ-
югация лестничная. Конъюгационный канал образован обеими клетками.
Воспринимающие клетки невздутые. Клетки без конъюгационной пары
невздутые. Зигоспоры трех форм: двухосно-эллипсоидные (в поперечном
сечении круглые), 70—102.5 мкм шир., 1'02.5—175 мкм дл., линзовид-
ные (в двух положениях круглые и эллиптические), 70—85 мкм толщ.,
100—125 мкм диам., и трехосно-эллипсоидные (в поперечном сечении
эллиптические), 81.3—105 мкм шир., 100—135 мкм дл., 65—75 мкм
толщ, (не вполне зрелые). Экзоспорий тонкий, гладкий. Мезоспорий
толще экзоспория, гладкий. Партеноспоры парные, располагающиеся по
одной в отдающей и воспринимающей клетках.
В стоячих водах. Украина: Причерноморье (бассейн Днепра).
Редкий вид, встреченный только однажды. Имеет несомненное сход-
ство, особенно по вегетативным признакам, с S. setiformis, резко от-
личается от него разнообразием форм зигоспор.
97. Spirogyra arta Jao (=S. subarta Jao; S. frigida Gay sensu Kad-
lubowska, 1972, pr. p., 1984, pr. p.; Jao, 1988; S. frigida f. arta Jao) —
Спирогира узкая (рис. 87, 7). — Вегетативные клетки 16—18 мкм
278
Рис. 121. 1, 2 — Spirogyra diversizygotica-. 1 — зигоспоры в разных положениях, лестничная
конъюгация, 2 — зигоспора и две партеноспоры, лестничная конъюгация; 3—5 — S. pseu-
dospreeiana, лестничная конъюгация. (По: 1, 2 — Полянский, 1951 (как S. setiformis f. di-
versizygotica)-, 3 — Полянский, 1955; 4 — Yamagishi, 1966b; 5 — ориг.).
279
шир., по длине в 6—22 раза больше. Поперечные стенки складчатые.
Хлоропласт 1, с 3—8 оборотами спирали. Конъюгация лестничная.
Конъюгационный канал образован обеими клетками. Воспринимающие
клетки со всех сторон вокруг зигоспоры слегка вздутые, веретеновидно
или цилиндрически-веретеновидно. Клетки без конъюгационной пары
невздутые. Зигоспоры удлиненно-эллипсоидные, реже цилиндрически-
эллипсоидные, 20—29 (32) мкм шир., '§0—103 мкм дл. Экзоспорий
тонкий, хорошо заметный на вершинах зигоспоры при отслаивании
мезоспория, гладкий. Мезоспорий неярко-желтоватый или желтый,
двухслойный, гладкий.
Во временных каменистых и небольших водоемах со стоячей водой,
отдельными нитями среди скопления эдогониума, в речных долинах
горных районов, на рисовых полях. Россия: Вост. Сибирь (Алтайский
край, долина р. Катунь).
Крайне редкий вид. Найден также в Китае и Дании (?). Хотя
конъюгационный канал отчетливо формируется обеими клетками рав-
номерно, но к моменту полного созревания зиготы и вздувания вос-
принимающей клетки в некоторых случаях создается впечатление, что
он сформирован только отдающей клеткой. Отсутствие вздутия клеток,
лишенных конъюгационной пары, наличие очень узких, удлиненно-эл-
липсоидных зигоспор и большая длина вегетативных клеток служат
отличием от весьма близкой 5. tumida. От S. tandae Randh. (Индия)
также отличается удлиненностью зигоспор, большей длиной вегетатив-
ных клеток и зигоспор и, главное, нескульптурированным мезоспорием.
От S. weberi отличается в основном меньшей, а от S. tenuissima —
большей шириной вегетативных клеток и, сверх того, отсутствием бо-
ковой конъюгации. У описанной из Китая S. arta f. maxima Н. et
W. Chu вегетативные клетки незначительно шире (21—-23 мкм шир.,
107—285 мкм дл.), а зигоспоры чуть больше (28—35 мкм шир., 50—
93 мкм дл.), что не является, на мой взгляд, убедительным основанием
для выделения особой формы.
98. Spirogyra tumida Jao (=5. frigida Gay sensu Krieger in: Kolk-
witz, Krieger, 1941—1944, pr. p.; Kadhibowska, 1972, pr. p., 1984, pr.
p.; non sensu Gay, 1884a) — Спирогира вздутая (рис. 122, 4). — Ве-
гетативные клетки 16—20 мкм шир., по длине в 4.5—И раз больше.
Поперечные стенки складчатые. Хлоропласт 1, с 3—9 оборотами спи-
рали. Конъюгация лестничная. Конъюгационный канал образован обе-
ими клетками, иногда с преобладанием отдающей клетки. Восприни-
мающие клетки слегка веретеновидно вздутые со всех сторон вокруг
зигоспоры. Клетки без конъюгационной пары цилиндрически, редко
веретеновидно вздутые до 45 мкм. Зигоспоры эллипсоидные, 25—
35 мкм шир., 38—86 мкм дл. Экзоспорий тонкий, прозрачный, глад-
кий. Мезоспорий толстый, желтый, гладкий. Эндоспорий тонкий, глад-
кий.
В текучих и стоячих водах, реках, прудах, на рисовых полях. Ка-
захстан.
Редкий вид с узким ареалом (Дания ?, Индия, Китай, Япония). От
близких видов S. inflata и S. tenuissima отличается вздутием клеток без
280
Рис. 122. I—3 — Spirogyra hopeiensis; 4 — S. tumida. (По: 1 — Jao, 1935b; 2, 3 — ориг.;
4 — Рундина, 1978a).
конъюгационной пары, иной шириной вегетативных клеток и отсутст-
вием боковой конъюгации. Главные отличия от S. arta: вздутие клеток,
лишенных конъюгационной пары, укороченная форма зигоспор и более
короткие вегетативные клетки. От S. tandae Randh. (Индия) отличается
гладким мезоспорием и вздутием клеток, не участвующих в конъюгации.
99. Spirogyra weberi Kiitz. — Спирогира Вебера (рис. 123,
124). — Вегетативные клетки (19) 23—35 (39) мкм шир., по длине в
281
Рис. 123. Spirogyra weberi f. weberi'. 1—4 — разная степень вздутия воспринимающих
клеток, лестничная конъюгация, 5 — боковая конъюгация. (Ориг.).
(2) 3—16 раз больше. Поперечные стенки складчатые. Хлоропласт 1
(2). Конъюгация лестничная, реже боковая, иногда совмещенные в
одной паре нитей. Конъюгационный канал при лестничной конъюгации
образован обеими клетками или только отдающей клеткой; при боко-
вой конъюгации канал выдается на 1/2 ширины клетки, изредка от-
граничен неясно, нить при этом слабо или довольно значительно изо-
гнутая. Воспринимающие клетки при лестничной конъюгации невзду-
тые или незначительно вздутые, или более или менее сильно вздутые
до 74 мкм шир., иногда значительно, бочонковидные и веретеновидные,
реже цилиндрически вздутые до 58 мкм шир., при боковой конъюгации
вздутые значительно. Клетки без конъюгационной пары невздутые или
слегка вздутые. Зигоспоры эллипсоидные, реже цилиндрически-эллип-
соидные, с более или менее закругленными вершинами, 21—49
282
(54) мкм шир., (26) 40—105 (122.5) мкм дл. Экзоспорий тонкий, бес-
цветный, гладкий. Мезоспорий толстый, от желтого до коричневого,
двухслойный, гладкий. Партеноспоры эллипсоидные или почти шаро-
видные, 25—35 мкм шир., 32—67 мкм дл.
Широко распространенный, достаточно характерный, полиморфный
вид. Тонкий первый подслой мезоспория при обработке кислотами
легко отделяется от второго подслоя и под сканирующим микроскопом
выглядит рыхлым.
F. weberi [=5. weberi var. elongata (Spree) Rabenh.; S. weberi var.
genuina (Kiitz.) Kirchn.; S. laxa Kiitz.; S. semiornata Jao; 5. tolosana
Comere (?)] (рис. 123). — Вегетативные клетки (19) 23—32 (36) мкм
шир. Конъюгация лестничная, реже или очень редко боковая, иногда
совмещенные в одной паре нитей. Конъюгационный канал при лестнич-
ной конъюгации образован обеими клетками равномерно. Восприни-
мающие клетки при лестничной конъюгации невздутые или незначи-
Рис. 124. 1—3 — Spirogyra weberi f. farlowii, разная степень и форма вздутия воспринима-
ющих клеток, лестничная конъюгация; 4—6 — S. weberi f. grevilleana, разная степень взду-
тия воспринимающих клеток, лестничная (4, 6) и боковая (5) конъюгации. (По: 1—3 — Рун-
дина, 1978а; 4—6 — ориг.).
283
тельно вздутые по ширине зигоспоры. Зигоспоры 21—40.5 (43.5) мкм
шир., (26) 40—93.5 (120) мкм дл.
В стоячих и текучих водах, часто в родниках, заводях и старицах
рек, в болотах, соленых озерах и на рисовых полях. Развивается в
кислых, нейтральных и щелочных водах и при малом содержании
азота. Конъюгация происходит при температуре воды 10—30 °C и pH
6—7, довольно часто. Повсюду.
Широко распространенная и часто встречающаяся форма. Ампли-
туда изменчивости ширины воспринимающих клеток довольно значи-
тельная.
F. farlowii (Trans.) V. Poljansk. (=S. farlowii Trans.; S. caroliniana
Dillard) (рис. 124, 1—3). — Вегетативные клетки 23—35 (39) мкм
шир. Конъюгация лестничная. Конъюгационный канал образован обе-
ими клетками равномерно, реже только отдающей клеткой, широкий
и короткий, слабо выраженный. Воспринимающие клетки со всех сто-
рон сильно вздутые до 64—74 мкм шир., преимущественно бочонко-
видные или веретеновидные, редко цилиндрически вздутые до (35)
40.5—58 мкм шир. Зигоспоры 29—49 (54) мкм шир., (40.5) 48—105
(119) мкм дл.
В текучих и стоячих водах, реках, лужах, прудах, родниках, на
рисовых полях. Россия: Чукотка (окрестности г. Анадырь); Украина и
Крым, Зап. и Южн. Казахстан.
Довольно редкая и таксономически спорная форма. В Европе най-
дена в Нидерландах, Германии и Дании, известна также из Азии (Сев.
Индия, Китай, о-в Ява), Сев. и Южн. Америки. Отличается от типовой
формы сильным вздутием воспринимающих клеток, которое, однако,
наблюдается крайне редко, а вообще степень вздутия здесь сильно
варьирует. Крайне близка, а возможно, и идентична f. grevilleana.
F. grevilleana (Hass.) V. Poljansk. [=5. grevilleana (Hass.) Kiitz.;
5. weberi var. grevilleana (Kiitz.) Kirchn.; S. inaequalis Kiitz., non sensu
Nag.j (рис. 124, 4—6). — Вегетативные клетки 25—30 (35) мкм шир.
Конъюгация лестничная и боковая, с преобладанием одной из них,
иногда совмещенные в одной паре нитей. Конъюгационный канал при
лестничной конъюгации образован преимущественно отдающей клет-
кой, иногда обеими клетками равномерно. Воспринимающие клетки
более или менее вздутые со всех сторон, часто сильно — до 52 мкм
шир., преимущественно веретеновидные или реже цилиндрически-ве-
ретеновидные. Зигоспоры (26) 30—42 (51) мкм шир., (35) 50—85
(122.5) мкм дл.
В стоячих и текучих водах, лужах, ручейках, лесных болотцах,
часто в пересыхающих водоемах и осушительных канавах, на рисовых
полях, в теплых ручьях. Повсюду.
От типовой формы отличается частым наличием боковой конъюга-
ции и более сильным вздутием воспринимающих клеток. Оба типа
конъюгации наблюдались в клональных культурах. Ширина вегетатив-
ных клеток, особенности формирования конъюгационного канала и
степень вздутия воспринимающих клеток довольно разнообразны.
284
F. tjibodensis (Faber) V. Poljansk. (=8. tjibodensis Faber). — Веге-
тативные клетки 40—46.5 (50) мкм шир. Конъюгация лестничная. Вос-
принимающие клетки невздутые. Зигоспоры 32—48.5 мкм шир., 37—
92 мкм дл.
В стоячих водах (?). Индия, о-в Ява.
Недостаточно описанная форма, отсутствует изображение. От ти-
повой формы отличается большей шириной вегетативных клеток и
зигоспор.
100. Spirogyra aequalis Harv. — Спирогира ровная. — Вегетатив-
ные клетки 45 мкм шир., по длине в 2—7 раз больше. Поперечные
стенки складчатые. Хлоропласт 1, редко их 2. Конъюгация преиму-
щественно боковая, реже лестничная, иногда совмещенные в одной
паре нитей. Воспринимающие клетки невздутые. Зигоспоры эллипсо-
идные, 40—45 мкм шир., (40) 75—105 мкм дл. Мезоспорий от желто-
буроватого до бурого, гладкий.
В стоячих водах, лесных лужах. Польша, Сев. Америка.
Редкий и недостаточно изученный вид, игнорируемый многими мо-
нографами. Авторский диагноз следует считать неполным и неточным:
отсутствуют сведения о мезоспорий, форма зигоспор ошибочно описана
как «шаровидная до эллипсоидной». Судя по отношению ширины к
длине у зигоспор, речь идет, вероятнее всего, об эллипсоидных зиго-
спорах, круглых по очертаниям, если смотреть на них с вершины.
Кроме того, на одном из авторских рисунков (Harvey, 1892, pl. 126,
fig. 19—21) изображена, по-видимому, апланоспора, а не зигоспора.
Это предположение основывается на ее небольших по сравнению с
зигоспорой размерах и отсутствии конъюгационного канала. Из-за со-
мнительности и невыразительности авторского рисунка он в данной
работе не воспроизводится. Авторский диагноз дополнен описанием
водоросли, найденной Хмелевским (1891 : 156, табл. 4, рис. 1—7) в
Польше и не идентифицированной им. Обнаруживает большое сходство
с S. insignis f. insignis.
101. Spirogyra pseudospreeiana Jao — Спирогира ложношпреевская
(рис. 121, 3—5). — Вегетативные клетки 15—20 мкм шир., по длине в
4.5—14 раз больше. Поперечные стенки складчатые. Хлоропласт 1.
Конъюгация лестничная. Конъюгационный канал образован исключи-
тельно или преимущественно отдающей клеткой. Воспринимающие
клетки сильно вздутые посередине только со стороны конъюгационного
канала, до 40 мкм шир., иногда отчленяющиеся друг от друга. Клетки
без конъюгационной пары невздутые. Зигоспоры эллипсоидные, (22)
25—35 мкм шир., 45—78 мкм дл. Экзоспорий тонкий, бесцветный,
гладкий. Мезоспорий толстый, желтовато-коричневый, гладкий.
В текучих и стоячих водах, прудах, на рисовых полях. Южн. Ка-
захстан.
Редкий вид, найденный прежде в Китае и Дании. От близких
видов — S. hopeiensis и S. croasdaleae Blum (США) отличается не-
значительно меньшей шириной вегетативных клеток. По всей вероят-
ности, все три вида являются только формами одной видовой единицы.
285
102. Spirogyra hopeiensis Jao — Спирогира хэбэйская (рис. 122, 1—
3). — Вегетативные клетки 24—29 (32) мкм шир., по длине в 4—10
(16) раз больше. Ризоиды подошвообразные, латеральные, неразветвлен-
ные, немногочисленные. Поперечные стенки складчатые. Хлоропласт 1
(2). Конъюгация лестничная. Конъюгационный канал образован исклю-
чительно или преимущественно отдающей клеткой. Воспринимающие
клетки вздутые только со стороны конъюгационного канала, преимущес-
твенно посередине, до 55 мкм шир., иногда отчленяющиеся друг от
друга. Клетки без конъюгационной пары невздутые. Зигоспоры эллип-
соидные, 28—48 мкм шир., 54—96 мкм дл. Экзоспорий тонкий, бес-
цветный, гладкий. Мезоспорий толстый, желто-коричневый, гладкий.
В текучих и стоячих водах, реках, на рисовых полях. Конъюгация
довольно частая, с конца мая по июль, при температуре воды 13—
20 °C. Сев., Зап. и Южн. Казахстан.
Редкий вид, найденный также в Нидерландах и Китае. Односто-
роннее вздутие воспринимающих клеток хорошо отличает этот вид от
круга форм 5. weberi, в первую очередь от f. farlowii. От схожего с
ним в этом отношении S. pseudospreeiana может быть отграничен
более крупными размерами.
103. Spirogyra tenuissima (Hass.) Kiitz. — Спирогира тончайшая
(рис. 125, 1—5). — Вегетативные клетки (8) 10—16 (17.4) мкм шир.,
по длине в 4—20 раз больше. Ризоиды латеральные, в виде слабо раз-
ветвленной розетки, немногочисленные. Поперечные стенки складчатые.
Хлоропласт 1. Конъюгация лестничная и боковая. Конъюгационный
канал при лестничной конъюгации образован обеими клетками или толь-
ко отдающей клеткой; при боковой конъюгации канал отчетливо отгра-
ничен и выдается на 1/3—1/2 ширины клетки или равен ей, нить при
этом часто сильно изогнутая, иногда прямая. Воспринимающие клетки
сильно вздутые со всех сторон, веретено видные, реже цилиндрические.
Клетки без конъюгационной пары невздутые, изредка цилиндрически
вздутые. Зигоспоры эллипсоидные, 19—36 мкм шир., 33—95 мкм дл.
Экзоспорий тонкий, бесцветный, гладкий. Мезоспорий толстый, от жел-
того до коричневого, гладкий. Партеноспоры округлые, редкие.
Широко распространенный и часто встречающийся вид. Вероятно,
монотипен, так как особенности, свойственные раздельно типовой
форме и f. cylindrica, могут встречаться у одного и того же предста-
вителя.
F. tenuissima (=S. naegelii Kiitz.; S. pseudonaegelii Skv.; S. pseudote-
nuissima O. et W. Bock; S. angulata Nipkow) (рис. 125, 1, 2). — Вегета-
тивные клетки (8) 10—14.5 (16) мкм шир. Конъюгация лестничная,
редко боковая. Конъюгационный канал при лестничной конъюгации об-
разован обеими клетками равномерно, реже только отдающей клеткой.
Нить при боковой конъюгации в разной степени, часто сильно изогнутая,
иногда прямая. Воспринимающие клетки сильно веретеновидно вздутые.
Зигоспоры (21.5) 24—36 мкм шир., 40—80 (95) мкм дл.
В стоячих и текучих водах, нередко в ручьях и болотцах с железис-
той водой, часто отдельными нитями среди других водорослей. Повсюду.
286
Рис. 125. 1—5 — Spirogyra tenuissima: 1,2 — f. tenuissima, 3—5 — f. cylindrica, боковая (3,
4) и лестничная (5) конъюгации; б, 7 — S. inflata, лестничная и боковая конъюгации; 8—
10 — S. groenlandica: 8, 10 — боковая и лестничная конъюгации, 9 — апланоспора. (По: 1,
2 — Nipkow, 1962 (как 8. angulata): 3,4 — Gauthier-Lievre, 1965 (как S. cylindrica): 5 —
Рундина, 1988; б, 7 — Czurda, 1932 (как 8. pascheriana): 8—10 — Transeau, 1951).
287
Широко распространенная форма. Несмотря на способность как к
лестничной, так и к боковой конъюгации, в некоторых случаях конъ-
югирует только одним способом, что следует учитывать при определе-
нии. Наиболее близка к S', inflata, отличается меньшей шириной ве-
гетативных клеток и сравнительно более сильным вздутием восприни-
мающих клеток.
F. cylindrica (Czurda) V. Poljansk. (=S. cylindrica Czurda; S. aust-
riaca Czurda) (рис. 125, 3—5). — Вегетативные клетки (9) 10.4—16
(17.4) мкм шир. Конъюгация лестничная и боковая. Конъюгационный
канал при лестничной конъюгации образован отдающей клеткой. Нить
при боковой конъюгации в большинстве случаев сильно изогнутая. Вос-
принимающие клетки сильно цилиндрически, изредка веретеновидно
вздутые. Зигоспоры (19) 22—32 мкм шир., (33) 51—71 (75.4) мкм дл.
В текучих и стоячих водах, реках, старицах, на рисовых полях, в
опресненных участках морей при солености до 4 %о, на глубине до
5.5 м, изредка в массе. Конъюгация в апреле—августе, при температуре
воды 5.8—24 °C. Россия: приустье Волги, сев. побережье Каспийского
моря; Зап. Казахстан (р. Малая Хобда, старица и устье р. Урал).
Широко распространенная форма в Европе, Африке и Азии. От-
личается от типовой, с которой часто вместе развивается, почти ис-
ключительно формой вздутия воспринимающих клеток. Этот признак
у f. cylindrica непостоянен и можно наблюдать все переходы от вере-
теновидных до цилиндрически вздутых воспринимающих клеток, а под-
час трудно разграничить одну форму вздутия от другой.
104. Spirogyra inflata (Vauch.) Kiitz. [=S. tenuissima (Hass.) Kiitz.
f. inflata (Vauch.) Cleve; S. spreeiana Rabenh.; S. pascheriana Czurda] —
Спирогира вздутая (рис. 125, 6, 7). — Вегетативные клетки (14) 15—
21 (23.5) мкм шир., по длине в (2) 3—15 (20) раз больше. Поперечные
стенки складчатые. Хлоропласт 1, очень редко их 2. Конъюгация бо-
ковая и лестничная. Конъюгационный канал при боковой конъюгации
выдается на 1/2—3/4 ширины клетки, нить при этом слегка изогнутая,
реже изгиб или более значителен, или отсутствует; при лестничной
конъюгации канал образован преимущественно или исключительно от-
дающей клеткой, редко обеими клетками. Воспринимающие клетки
сильно веретеновидно, реже цилиндрически вздутые до 60 мкм шир.
Клетки без конъюгационной пары невздутые. Зигоспоры эллипсоидные,
21—50 мкм шир., (35) 40—105 мкм дл. Экзоспорий тонкий, бесцвет-
ный, гладкий. Мезоспорий желто-коричневый или коричневато-желтый,
гладкий. Эндоспорий тонкий, гладкий, почти всегда заметный. Парте-
носпоры эллипсоидные, 32 мкм шир., 52 мкм дл., реже шаровидные,
17 мкм диам., очень редкие.
В стоячих и текучих водах, на рисовых полях, иногда в минераль-
ных и богатых биогенными веществами водоемах. Повсюду.
Широко распространенный и часто встречающийся вид. При опре-
делении вида следует помнить, что хотя для него свойственны оба
типа конъюгации, но в большинстве случаев преобладает боковая. Оба
типа конъюгации встречаются в образцах как вместе, так и порознь.
288
Форма вздутия воспринимающих клеток — признак очень пластичный,
а следовательно, и ненадежный при определении. Типичное цилинд-
рическое вздутие клеток встречалось довольно часто вместе с типичным
веретеновидным в одной и той же нити, а наличие множества пере-
ходных форм между ними говорит о размывании формы вздутия вос-
принимающих клеток как таксономического признака. Отличается от
S. weberi f. grevilleana главным образом меньшей шириной вегетатив-
ных клеток, от 5. tenuissima — большей шириной вегетативных клеток
и менее вздутыми воспринимающими клетками, от S', groenlandica —
несколько меньшей шириной и длиной вегетативных клеток, длиной
и светлой окраской зигоспор.
105. Spirogyra groenlandica Rosenv., non sensu Hirn, 1895 — Спи-
рогира гренландская (рис. 125, 8—10). — Вегетативные клетки 18—
24 мкм шир., по длине в (15) 18—28 (33) раз больше. Поперечные
стенки складчатые. Хлоропласт 1, чаще слабо извитой или с 3—8 (?)
оборотами спирали. Конъюгация боковая и лестничная. Конъюгацион-
ный канал при боковой конъюгации выдается на 1/2—1 ширины клет-
ки, нить при этом прямая или очень незначительно изогнутая; при
лестничной конъюгации канал образован обеими клетками, иногда с
преобладанием отдающей клетки, нити при этом изгибаются в разные
стороны. Воспринимающие клетки сильно цилиндрически вздутые во-
круг зигоспоры до 51 мкм шир. Зигоспоры эллипсоидные или цилин-
дрически-эллипсоидные, 34—48 мкм шир., 100—130 мкм дл. Мезо-
спорий каштаново-коричневый, гладкий. Апланоспоры подобны зиго-
спорам, 34—44 мкм шир., 60—90 мкм дл.
В проточных и стоячих водах. Гренландия, США.
Редкий вид. От S. tenuissima f. cylindrica отличается большей ши-
риной вегетативных клеток. Очень близок к S. inflata, возможно, яв-
ляется его синонимом. Цилиндрическим вздутием воспринимающих
клеток сходен с 5. quadrata, отличается несколько меньшей шириной
вегетативных клеток.
106. Spirogyra quadrata (Hass.) Petit [=Rhynchonema quadratum
(Hass.) Kiitz.; Spirogyra quadrata f. genuina Kirchn.] — Спирогира квад-
ратная (рис. 126, 1—3). — Вегетативные клетки 24—30 (33) мкм шир.,
по длине в 2—12 раз больше. Поперечные стенки складчатые. Хло-
ропласт 1, редко их 2. Конъюгация боковая и лестничная. Конъюга-
ционный канал при боковой конъюгации выдается приблизительно на
1/2 ширины клетки, нить при этом прямая или незначительно изогну-
тая; при лестничной конъюгации канал образован обеими клетками.
Воспринимающие клетки преимущественно цилиндрически, реже вере-
теновидно сильно вздутые вокруг зигоспоры до 60 мкм шир. Зигоспоры
эллипсоидные или цилиндрически-эллипсоидные, 31—48 мкм шир.,
42—ПО мкм дл. Экзоспорий тонкий, бесцветный, гладкий. Мезоспо-
рий толстый, от желтого до коричневого, гладкий. Партеноспоры не-
редкие.
В стоячих, реже слабопроточных водах, иногда в водоемах торфя-
ных болот и в родниках, прикрепляется к камням. Россия: север (Со-
19 Рундина Л. А.
289
Рис, 126. 1—3 — Spirogyra quadrata, боковая (7, 2) и лестничная (3) конъюгации; 4 —
S. chuniae, лестничная конъюгация; 5,6 — S. hassallii, боковая и лестничная конъюгации.
(По: I — ориг.; 2, 3 — Petit, 1874; 4 — Jao, 1935b; 5, 6 — Gauthier-Lievre, 1965).
290
ловецкие о-ва) и центр европ. части (окрестности Москвы), Зап. Си-
бирь (окрестности Обь-Енисейского канала); Эстония (о-в Сааремаа),
Латвия, Литва, Украина и Крым.
Вид, широко распространенный почти на всех континентах, спор-
ный. Характер вздутия воспринимающих клеток весьма изменчив, и в
одном образце можно встретить все переходы, от сильно выраженного
веретеновидного до цилиндрического. Очень близок к кругу форм
S. weberi, особенно к f. grevilleana. Возможно, идентичен ей.
107. Spirogyra chuniae Jao — Спирогира Чун (рис. 126, 4). — Ве-
гетативные клетки 28—39 мкм шир., по длине в 1.5—4.5 раза больше.
Поперечные стенки складчатые. Хлоропласт 1, иногда их 2, с незна-
чительным количеством оборотов спирали. Конъюгация лестничная и
боковая. Конъюгационный канал при лестничной конъюгации образо-
ван обеими клетками с преобладанием отдающей, очень короткий. Вос-
принимающие клетки сильно вздутые до 80 мкм шир. Зигоспоры эл-
липсоидные, с заостренными концами, 35—43 мкм шир., (70) 73—
119 мкм дл. Мезоспорий желтый, гладкий.
В лесных лужах, на рисовых полях. Германия, Китай.
Редкий вид. Имеет несомненное сходство по многим признакам с
формами S. weberi. От S. weberi f. grevilleana отличается заострен-
ностью вершин у зигоспор и несколько ббльшими размерами клеток.
По ширине вегетативных клеток сходен с S. weberi f. farlowii, но у
последней формы менее сильно вздуты воспринимающие клетки и нет
боковой конъюгации. Исходя из этого, можно предположить, что в
дальнейшем 5. chuniae потеряет видовую значимость и в качестве
формы войдет в круг форм S. weberi.
108. Spirogyra insignis (Hass.) Kiitz. — Спирогира заметная
(рис. 127). — Вегетативные клетки 30—45 (48) мкм шир., по длине
в 3.5—12 (14) раз больше. Поперечные стенки складчатые. Хлоро-
пластов (1) 2—4, довольно тонких, слабо извитых. Конъюгация лест-
ничная и боковая с преобладанием одной из них, иногда только лест-
ничная. Конъюгационный канал при лестничной конъюгации образован
обеими клетками или только отдающей клеткой, иногда укороченный;
при боковой конъюгации канал выдается на 1/3—1/2 ширины клетки,
ушковидный или совсем не выражен, нить при этом прямая или слегка
изогнутая; концы фертильных нитей часто закручиваются. Восприни-
мающие клетки при лестничной конъюгации веретеновидно или ци-
линдрически-веретеновидно вздутые со всех сторон до 72 мкм шир.
или почти исключительно со стороны конъюгационного канала; при
боковой конъюгации клетки невздутые или веретеновидно и цилинд-
рически-веретеновидно слегка вздутые со всех сторон. Клетки без
конъюгационной пары невздутые. Зигоспоры эллипсоидные или цилин-
дрически-эллипсоидные, с более или менее широко закругленными вер-
шинами, 28—65 мкм шир., 42—140 (200) мкм дл. Экзоспорий тонкий,
бесцветный, гладкий. Мезоспорий толстый, от желто-бронзового до
коричневого, гладкий.
Широко распространенный вид.
291
Рис. 127. Spirogyra insignis; 1,2 — f. insignis, боковая конъюгация (7), лестничная и боко-
вая конъюгации в одной паре нитей (2); 3,4 — f. rectangularis, боковая (3) и лестничная
(4) конъюгации; 5 — f. wangii, лестничная конъюгация. (По: 1 — ориг.; 2 — Смирнов в:
Казаковский, Смирнов, 1914 (как S. insignis var. longispora); 3, 4 — Transeau, 1951 (как
S. rectangularis); 5 — Li, 1933 (как 5. wangii)).
F. insignis [=5. insignis var. longispora Smirn.; 5. fallax (Hansg.)
Wille, pr. p.; S. hartigii (Kiitz.) De Toni] (рис. 127, 1, 2). — Ве-
гетативные клетки (30) 36—45 (48) мкм шир. Хлоропластов (1)
2—3 (4). Конъюгация лестничная и боковая, с преобладанием одной
из них, иногда совмещенные в одной паре нитей. Конъюгационный
канал при лестничной конъюгации образован обеими клетками,
часто укороченный. Воспринимающие клетки при лестничной конъ-
югации более или менее веретеновидно вздутые со всех сторон,
при боковой конъюгации невздутые или слегка вздутые со всех
сторон. Зигоспоры эллипсоидные, 28—55 (60) мкм шир., 42—140
(200) мкм дл.
В мелких стоячих и текучих водах, на рисовых полях; по-видимому,
предпочитает воды, богатые железом. Россия: европ. часть — повсюду,
Зап. Сибирь (окрестности Томска), Вост. Сибирь (Республика Саха),
292
Дальн. Восток (Камчатка); Украина, Зап. Казахстан (окрестности
Уральска), Узбекистан.
Широко распространенная форма. Встречена также во всех странах
Европы, Китае и Сев. Америке. Близка к 5. reticulata, хорошо отли-
чается гладким мезоспорием. Из группы видов, имеющих гладкий ме-
зоспорий, обнаруживает значительное сходство с S. hassallii, отличаясь
лишь незначительно большей шириной вегетативных клеток.
F. rectangularis (Trans.) V. Poljansk. (=5. rectangularis Trans.) (рис. 127,
3, 4). — Вегетативные клетки 35—40 мкм шир. Хлоропластов 2—4.
Конъюгация боковая и лестничная. Конъюгационный канал при лест-
ничной конъюгации образован отдающей клеткой, более или менее
длинный. Воспринимающие клетки цилиндрически вздутые до 48—
70 мкм шир., при лестничной конъюгации — значительно и неотчет-
ливо, почти исключительно со стороны конъюгационного канала; при
боковой конъюгации — слабо и отчетливо, со всех сторон. Зигоспоры
эллипсоидные или цилиндрически-эллипсоидные, с широко закруглен-
ными концами, 45—65 мкм шир., 75—120 мкм дл.
В стоячих водах. Австрия, США.
Редкая форма. От типовой формы хорошо отличается цилиндри-
чески вздутыми воспринимающими клетками и в некоторых случаях
наличием цилиндрически-эллипсоидных зигоспор.
F. wangii (Li) V. Poljansk. [=5. wangii Li; 5. fallax (Hansg.) Wille,
pr. p. (?)] (рис. 127, 5). — Вегетативные клетки 30—32 мкм шир.
Хлоропластов 2—3. Конъюгация лестничная. Конъюгационный канал
образован обеими клетками. Воспринимающие клетки сильно верете-
новидно вздутые со всех сторон до 72 мкм шир. Зигоспоры эллипсо-
идные, 60—64 мкм шир., 108—124 мкм дл.
В стоячих водах, в озерных протоках. Россия: север европ. части
(бассейн р. Северная Двина).
Очень редкая форма, встреченная за пределами России только в
Китае. Отличается от типовой формы несколько меньшей шириной
вегетативных клеток (не выходящей, однако, из пределов, указываемых
для f. insignis), большим вздутием воспринимающих клеток, большей
шириной зигоспор, а также отсутствием боковой конъюгации. Близка
к S. weberi f. farlowii, отличается большим числом хлоропластов и
большими размерами зигоспор.
109. Spirogyra hassallii (Jenner) Petit — Спирогира Хассалля (рис. 126,
5, 6). — Вегетативные клетки (26) 30—35 мкм шир., по длине в 4—13
раз больше. Поперечные стенки складчатые. Хлоропластов 2—4, слабо
извитые. Конъюгация боковая, редко лестничная. Конъюгационный
канал при лестничной конъюгации образован обеими клетками, иногда
с преобладанием отдающей клетки; при боковой конъюгации канал
выдается приблизительно на 1/2 ширины клетки, нить при этом прямая
или слабо изогнутая. Воспринимающие клетки обычно более или менее
веретеновидно вздутые со всех сторон до 48—50 мкм шир. Клетки
без конъюгационной пары невздутые. Зигоспоры эллипсоидные, иногда
293
цилиндрически-эллипсоидные, (30) 35—54 мкм шир., 52—169 (223) мкм
дл. Экзоспорий тонкий, бесцветный, гладкий. Мезоспорий толстый, от
желтого до желто-коричневого, двухслойный, гладкий. Партеноспоры
38—40 мкм шир., 47—72 мкм дл., редкие.
В стоячих, реже текучих водах, заболоченных канавах и ямах, пру-
дах, минеральных источниках, в том числе радоновых, на рисовых
полях. Россия: европ. часть — почти пЬвсюду, Вост. Сибирь (Респуб-
лика Саха, оз. Байкал), Дальн. Восток (Амурская обл.); Украина (ок-
рестности Киева), Узбекистан, Вост. Казахстан.
Широко распространенный вид, встреченный также в Европе, Сев.
Африке, Азии и Сев. Америке. Близок к S. insignis f. insignis, имеет
несколько меньшую ширину вегетативных клеток и преимущественно
боковую конъюгацию. Главнейшие отличия от 5. weberi f. grevilleana
заключаются в большей ширине вегетативных клеток, большем числе
хлоропластов и менее сильном вздутии воспринимающих клеток.
110. Spirogyra calospora Cleve — Спирогира красивоспоровая
(рис. 128, 1—5). — Вегетативные клетки 20—56 мкм шир., по длине
в 4—14 раз больше. Поперечные стенки складчатые. Хлоропласт 1
(2—3). Конъюгация лестничная. Конъюгационный канал образован обе-
ими клетками. Воспринимающие клетки невздутые или слабо вздутые.
Клетки без конъюгационной пары невздутые или более или менее,
иногда сильно, бочонковидно вздутые. Зигоспоры от эллипсоидных до
цилиндрически-эллипсоидных, преимущественно с закругленными вер-
шинами, 26—60 мкм шир., 52—130 мкм дл. Экзоспорий толстый, бес-
цветный, двухслойный: внешний слой тонкий, гладкий, внутренний
слой толстый, с округлыми ямчатыми углублениями. Мезоспорий тон-
кий, от желтого до коричневого, двухслойный, гладкий.
F. calospora (=S. protecta Wood; S. punctata Petit, non sensu Cleve;
S. cleveana Trans.) (рис. 128, 7). — Вегетативные клетки (26) 29—
42 мкм шир. Зигоспоры 30—51 мкм шир., 52—125 мкм дл.
В стоячих и текучих водах, водохранилищах, озерах, прудах,
скважинных водоемах, на рисовых полях, оросительных каналах,
часто в прибрежных лужах и старицах, предпочтительно с неболь-
шим загрязнением, иногда при солености до 5.7 %о (при выносе в
море). Россия: север (Кольский п-ов, Печенга), центр европ. части
(окрестности Москвы), устье Волги, зап. побережье Каспийского
моря, Вост. Сибирь (сев. Байкал, Республика Саха), Дальн. Восток
(Чукотка: р. Колыма); Эстония (о-в Сааремаа), Латвия, Узбекистан,
Туркмения (Зап. Узбой).
По данным сканирующего микроскопа, мезоспорий двухслойный:
внешний слой тонкий, бесцветный, гиалиновый, внутренний слой
толще первого, окрашенный и плотный. Очень близка к S. areolata,
отличающемуся сильно вздутыми воспринимающими клетками.
F. major Cleve (=S. denticulata Trans.) (рис. 128, 2, 3). — Вегета-
тивные клетки 42—56 мкм шир. Зигоспоры 45—60 мкм шир., 60.5—
138.8 мкм дл.
294
Рис. 128. 1—5 — Spirogyra calospora-. 1 — f. calospora-, 2, 3 — f. major, лестничная конъ-
югация (2), зигоспора (3); 4,5 — f. venusta, зигоспора, лестничная конъюгация (4), строение
мезоспория (5); 6 — S. areolata. (По: I — ориг.; 2, 3 — Cleve, 1868 (как 5. calospora); 4, 5 —
Jao, 1935b (как S. venusta); 6 — Lagerheim, 1883).
295
В стоячих и текучих водах. Швеция, Нидерланды, Германия, США.
Редкая форма, отличающаяся от типовой более крупными размера-
ми вегетативных клеток и зигоспор.
F. minor Borge [=5. calospora sensu Krieger in: Kolkwitz, Krieger,
1941—1944, pr. p.; S. protecta Wood f. '(sine nomen) Skuja; S. latviensis
(Skuja) Czurda; S. petitiana Trans.]. — Вегетативные клетки 20—30 мкм
шир. Зигоспоры 26—45 мкм шир., 65—115 мкм дл. Воспринимающие
клетки вздутые до 30—40 мкм шир.
В болотах и прудах. Латвия.
Редкая форма, встреченная также в США. Отличается от типовой
формы более сильным вздутием воспринимающих клеток. По этому
признаку сходна с S. areolata, но у последнего вида вздутие сильнее,
а вегетативные клетки шире. Довольно близка к S. microgranulata Jao
(Китай), отличающемуся более узкими и длинными вегетативными
клетками и иным строением оболочек зигоспор. Оригинальное изобра-
жение формы отсутствует.
F. venusta (Jao) V. Poljansk. (=5. venusta Jao; S’, calospora sensu
Krieger in: Kolkwitz, Krieger, 1941—1944, pr. p.) (рис. 128, 4, 5). —
Вегетативные клетки 25—27 мкм шир. Зигоспоры 26—32 мкм шир.,
67—84 мкм дл. Концы клеток, лишенных конъюгационной пары, ша-
ровидно сильно вздутые (артефакт ?).
В стоячих водах. Китай.
Редкая форма. По ширине вегетативных клеток особенно близка к
S. calospora f. minor. Отличается от всего круга форм S. calospora
своеобразным сильным вздутием на концах стерильных клеток в фер-
тильных нитях.
111. Spirogyra areolata Lagerh. (=S'. calospora Cleve sensu Krieger
in: Kolkwitz, Krieger, 1941—1944, pr. p.) — Спирогира ареолированная
(рис. 128, 6). — Вегетативные клетки 30—36 мкм шир., по длине в
3—10 раз больше. Поперечные стенки складчатые. Хлоропластов 1—2.
Конъюгация лестничная. Конъюгационный канал образован обеими
клетками. Воспринимающие клетки обычно сильно вздутые со всех
сторон до 40—49 мкм шир. Зигоспоры эллипсоидные, иногда почти
шаровидные, реже цилиндрически-эллипсоидные, 35—47 мкм шир.,
54—97 (126) мкм дл. Экзоспорий толстый, бесцветный, двухслойный:
наружный слой тонкий, гладкий, внутренний слой толстый, с округ-
лыми ямчатыми углублениями. Мезоспорий тонкий, коричневый, глад-
кий.
В стоячих и проточных водоемах, приозерных лужах и ручьях.
Эстония (о-в Сааремаа).
Довольно редкий вид, известный ранее из Швеции и Германии. От
сходного по строению экзоспория S. calospora отличается сильно взду-
тыми воспринимающими клетками, хотя степень вздутия их у исход-
ного вида может варьировать в зависимости от стадии конъюгации и
зрелости зиготы.
296
112. Spirogyra adjerensis Gauthier-Lievre — Спирогира аджерская
(рис. 129, 1, 2). — Вегетативные клетки 22—25 мкм шир. Поперечные
стенки складчатые. Хлоропласт 1. Конъюгация боковая, нить при этом
значительно изогнутая. Воспринимающие клетки незначительно вере-
теновидно вздутые вокруг зигоспоры до 40—42 мкм. Зигоспоры эл-
липсоидные, 38—42 мкм шир., 88—100 мкм дл. Экзоспорий тонкий,
бесцветный, гладкий. Мезоспорий толстый, коричневый, с утолщения-
ми, образующими сетку с глубокими ячеями.
В стоячих водах, глинистых ямах. Дания, Центр. Африка (Алжир:
плато Тассили).
Редкий вид. Близок к S. granulata Jao (Китай), отличается мень-
шими размерами и другой орнаментацией зигоспоры. По последнему
признаку имеет расхождение и с другим близким видом — S. libyca
Gauthier-Lievre (Африка).
113. Spirogyra alsiophila Skuja ex Rund. [=S. inflata (Vauch.) Kiitz.
var. alsiophila Skuja; S. alsiophila Skuja nom. provis] — Спирогира
холодолюбивая (рис. 129, 3—5). — Вегетативные клетки 19—21 мкм
шир., по длине до 15 раз больше. Поперечные стенки складчатые.
Хлоропласт 1. Конъюгация лестничная. Конъюгационный канал обра-
зован обеими клетками, с преобладанием отдающей клетки. Воспри-
нимающие клетки значительно вздутые со всех сторон, 28—40 мкм
шир. Клетки без конъюгационной пары невздутые. Зигоспоры от эл-
липсоидных до яйцевидных, со слепка закругленными или заостренны-
ми концами, 26—36 мкм шир., 60—92 мкм дл. Экзоспорий тонкий,
бесцветный, гладкий. Мезоспорий толстый, светлый, серовато-корич-
невый, с оттенком сепии, мелкоточечный (неправильно мелкокрапча-
тый или мелкопористый), при небольшом увеличении микроскопа ка-
жущийся гладким. Эндоспорий тонкий, бесцветный, гладкий.
В холодном потоке с ледников, в роднике между дернинками мха
и нитями зигнемовых. Швеция.
Редкий вид с необычным для зигнемовых местообитанием. Особое
внимание следует уделить строению мезоспория, так как точечность его,
вернее, пористость, выявляется только при сильном увеличении микро-
скопа. Очень близок к S. inflata, в качестве разновидности которого и
был впервые описан. Отличается скульптурированным мезоспорием. По
этому признаку сближается с 5. kuusamoensis и S. fritschiana. Возможно,
является синонимом последнего вида. Значительное сходство обнаружи-
вает с 5. discreta Trans. (США, Китай), но в отличие от него имеет не-
сколько иные структуру и цвет мезоспория. Сходен по всем признакам,
кроме скульптуры мезоспория, с S. czubinskii.
114. Spirogyra czubinskii Kadi, ex Rund. — Спирогира Чубиньско-
го. — Вегетативные клетки 19—21 мкм шир. Поперечные стенки
складчатые. Хлоропласт 1. Конъюгация лестничная. Конъюгационный
канал образован обеими клетками, часто с преобладанием отдающей
клетки. Воспринимающие клетки значительно вздутые вокруг зигоспо-
ры. Клетки без конъюгационной пары невздутые. Зигоспоры эллипсо-
идные, 31—34 мкм шир., 74—91 мкм дл. Экзоспорий тонкий, бес-
297
Рис. 129. 1,2 — Spirogyra adjerensis', 3—5 — S. alsiophila'. 3,4 — изогнутые и прямые вос-
принимающие клетки, лестничная конъюгация, 5 — строение мезоспория. (По: 1, 2 —
Gauthier-Lievre, 1965; 5—5 — Skuja, 1964).
цветный, гладкий. Мезоспорий сетчатый (с утолщениями, сомкнутыми
в сеть).
В небольшом пруду. Польша.
Редкий вид. В- первом описании этого вида (Kadhibowska, 1972 :
413, rys. 1594) мезоспорий охарактеризован как правильно сетчатый —
в отличие от неправильно сетчатого во втором описании (Kadlubowska,
1984 : 466, Fig. 725). Детали строения мезоспория, весьма существен-
ные для разъяснения этого вопроса, не изображены на авторском ри-
сунке. На этом основании рисунок этого вида в данной работе не
298
воспроизведен. Цвет мезоспория неизвестен. Представляет интерес
лестничная конъюгация между клетками одной изогнутой нити. К со-
жалению, этот примечательный факт, сообщенный в первом описании,
тоже не подтвержден изображением. Близкими видами являются S. ип-
dulisepta Randh. и S. tandae Randh., оба из Сев. Индии. Они отлича-
ются от S. czubinskii чуть меньшей шириной вегетативных клеток и
зигоспор, а также веретеновидной формой вздутия воспринимающих
клеток. По многим признакам, в том числе ширине вегетативных кле-
ток, сходен с S. dentireticulata, однако у последнего вида мезоспорий
зубчато-сетчатый. Только иным строением мезоспория отличается от
S’, alsiophila, сходного по всем другим признакам.
115. Spirogyra australica Czurda (=S. protecta Wood sensu Mobius,
1892; S. weberi Kiitz. var. punctata Langer) — Спирогира австралий-
ская. — Вегетативные клетки 24—32 мкм шир. Поперечные стенки
складчатые. Хлоропласт 1. Конъюгация лестничная. Воспринимающие
клетки слегка вздутые. Зигоспоры удлиненно-эллипсоидные, 27—36 мкм
шир., 54—74 мкм дл. Экзоспорий тонкий, бесцветный, гладкий. Мезо-
спорий толстый, коричневый, с поверхности мелкоточечный, в попереч-
ном сечении радиально исчерченный,
В стоячих водах временных водоемов. Германия, Австралия.
Редкий и недостаточно изученный вид. Если сопоставить структуру
мезоспория с поверхности зигоспоры и в поперечном сечении, то
можно выразить предположение об истинном тонкоигольчатом его
строении. Близок к S. goetzii и S. fritschiana; отличается от первого
вида иным строением мезоспория и незначительно большей шириной
вегетативных клеток, а от второго — незначительным вздутием вос-
принимающих клеток и отсутствием боковой конъюгации. Оригиналь-
ное изображение отсутствует.
116. Spirogyra goetzii Schmidle — Спирогира Гётца (рис. 130, 7). —
Вегетативные клетки 22—25 мкм шир., по длине в 5—14 раз больше.
Поперечные стенки складчатые. Хлоропласт 1. Конъюгация лестничная.
Конъюгационный канал образован обеими клетками равномерно. Вос-
принимающие клетки невздутые или незначительно вздутые вокруг зи-
госпоры до 32 мкм. Зигоспоры эллипсоидные, 28—31 мкм шир., 42—
62 мкм дл. Экзоспорий толстый, бесцветный, гладкий. Мезоспорий от
коричневого до красно-коричневого, от мелкоточечного до сетчато-то-
чечного.
В озерах, прудах, ручьях. Дания, Вост. Африка, Индия (?), США.
Слабо изученный вид, особенно в отношении строения мезоспория.
По многим признакам, в том числе и по точечному мезоспорию, очень
сходен, а возможно, и идентичен S', fritschiana, но имеет только лест-
ничную конъюгацию и более нежную точечность мезоспория. Близок
также к S', australica.
117. Spirogyra rugosa (Trans.) Czurda [=S. tenuissima (Hass.) Kiitz.
f. punctata Skuja; S. tenuissima f. rugosa (Trans.) V. Poljansk.] — Спи-
рогира морщинистая (рис. 130, 2—4). — Вегетативные клетки 10—
299
Рис. 130. 1 — Spirogyra goetzii', 2—4 — S. rugosa: 2, 3 — лестничная конъюгация, 4 — зи-
госпора; 5 — S. fritschiana, боковая конъюгация. (По: 1 — Schmidle, 1901; 2—4 — Gauthi-
er-Lievre, 1965; 5 — Fritsch in: Fritsch, Rich, 1930 (как S. protecta var. inflata)).
13 мкм шир., по длине в 5—20 раз больше. Поперечные стенки склад-
чатые. Хлоропласт 1. Конъюгация лестничная и боковая. Конъюгаци-
онный канал при лестничной конъюгации образован обеими клетками.
Воспринимающие клетки веретеновидно вздутые до 30 мкм. Клетки
без конъюгационной пары невздутые. Зигоспоры эллипсоидные, 22—
32 мкм шир., (38) 55—70 мкм дл. Экзоспорий тонкий, гладкий. Ме-
зоспорий толстый, желто-коричневый, с небольшими ямчатыми углуб-
лениями (точечный ?).
Преимущественно в стоячих водах, ямах, канавах. Эстония (о-в
Хийумаа).
Редкий вид, встреченный также в Африке и Сев. Америке (США).
По ширине вегетативных клеток очень близок к 5. tenuissima, отли-
чается скульптурированным мезоспорием. По этому признаку сходен с
S. alsiophila, отличаясь от него меньшей шириной вегетативных клеток
и наличием обоих типов конъюгации.
118. Spirogyra kuusan^nsis Hirn — Спирогира кусамоская
(рис. 131). — Вегетативные клетки 11—17 (20) мкм шир., по длине
300
в 3—9 раз больше. Поперечные стенки складчатые. Хлоропласт 1.
Конъюгация боковая и лестничная. Конъюгационный канал при боко-
вой конъюгации резко выступающий, широко закругленный, выдается
на 1/2 ширины клетки, нить при этом сильно изогнутая, очень редко
почти прямая; при лестничной конъюгации канал образован преиму-
щественно или исключительно отдающей клеткой, редко обеими клет-
ками. Воспринимающие клетки сильно веретеновидно вздутые до 32—
38 (43.5) мкм. Клетки без конъюгационной пары невздутые. Зигоспоры
эллипсоидные, (23) 25—33 (37) мкм шир., (45) 49—75 мкм дл. Эк-
зоспорий тонкий, бесцветный, гладкий, возможно двухслойный. Мезо-
спорий толстый, до 2.8 мкм шир., от светло-желтоватого до желто-
коричневого, двухслойный, отчетливо сетчатый.
F. kuusan^nsis (рис. 131, 1). — Вегетативные клетки 13—17 мкм
шир. Мезоспорий тонкосетчатый.
В стоячих водах, канавах, озерцах, болотах. Россия: север европ.
части (Кольский п-ов — Печенга, поморская Карелия).
Форма редкая, найденная прежде только в Финляндии и Германии.
При малом увеличении микроскопа скульптура мезоспория выглядит
нежно-точечной, как ее описывает автор вида, но уже при увеличении
в 300—400 раз оболочка обнаруживает ясную, хотя и нежную, сетча-
тость. По ширине вегетативных клеток занимает промежуточное поло-
жение между 5. inflata и S. tenuissima, но клетки ее никогда не бывают
столь длинными, а мезоспорий гладким.
F. skujaensis V. Poljansk. [=5. inflata (Vauch.) Kiitz. var. foveolata
Trans, sensu Skuja, 1928, non sensu Transeau, 1914; S. foveolata Czurda]
(рис. 131, 2—5). — Вегетативные клетки 11—16 .(20) мкм шир. Ме-
зоспорий грубосетчатый.
В стоячих, мелких, пересыхающих, иногда заболоченных водоемах,
прудах, канавах, ямах, лужах, на рисовых полях, редко в проточной
воде. Россия: Дальн. Восток (Уссурийский край, окрестности Влади-
востока); Эстония (о-в Сааремаа), Латвия (окрестности Риги), Украина
(Карпаты, окрестности Киева).
По-видимому, распространена шире, чем описано в литературе. До-
стоверные находки в Европе (Финляндия) и в Азии (Китай). Отлича-
ется от типовой формы более грубой сетчатостью на мезоспорий.
Меньшее значение имеют другие различия: более длинные клетки,
сильный изгиб нити при боковой конъюгации. У двух близких по
вегетативным признакам видов — S. discreta Trans. (США, Китай) и
S. amplectans Skuja (Китай) иное строение оболочек зигоспоры. Близка
также к S. tandae Randh. (Индия), но у последнего вида отмечена
только лестничная конъюгация.
119. Spirogyra fritschiana Czurda (=S. protecta Wood var. inflata
Fritsch et Rich) — Спирогира Фрича (рис. 130, 5). — Вегетативные
клетки 17—24 мкм шир., по длине в 3.5—15 раз больше. Поперечные
стенки складчатые. Хлоропласт 1. Конъюгация боковая и лестничная.
Конъюгационный канал при боковой конъюгации незначительно выда-
301
Рис. 131. Spirogyra kuusamoensis: 1 — f. kuusamoensis, боковая конъюгация; 2—5 — f.
skujaensis, лестничная (2) и боковая (3—5) конъюгации. (По: 1 — Krieger in: Kolkwitz, Krieger,
1941—1944; 2 — Jao, 1935b (как S. foveolata): 3 — Skuja, 1928 (как S. inflata var. foveola-
ta): 4, 5 — Р)чгдина, 1981).
ется, нить при этом прямая или слабо изогнутая; при лестничной конъ-
югации канал образован обеими клетками. Воспринимающие клетки
значительно вздутые со всех сторон. Зигоспоры удлиненно-эллипсоид-
ные, (22) 35—40 мкм шир., 48—71 мкм дл. Мезоспорий коричневый,
груботочечный.
В лужах и реках. Нидерланды, Германия, Израиль, Южн. Африка
(ЮАР).
Очень редкий вид. Спорен вопрос о строении мезоспория: грубая
точечность может обусловливаться либо его бугорчатостью, либо ям-
чатостью.
302
120. Spirogyra gaterslebensis Rieth — Спирогира гатерслебенская
(рис. 132, 1, 2). — Вегетативные клетки 18—21 мкм шир., по длине
в 4—10 раз больше. Поперечные стенки складчатые. Хлоропласт 1,
редко их 2. Конъюгация боковая, реже лестничная, иногда совмещен-
ные в одной паре нитей. Конъюгационный канал при боковой конъ-
югации выдается на 1/2 ширины клетки, нить при этом часто довольно
сильно изогнутая; при лестничной конъюгации канал образован обеими
клетками с преобладанием отдающей клетки. Воспринимающие клетки
при лестничной конъюгации сильно веретеновидно вздутые, 41.5—52
(70) мкм шир., при боковой конъюгации иногда невздутые. Клетки без
конъюгационной пары невздутые. Зигоспоры эллипсоидные, иногда при
боковой конъюгации удлиненные, цилиндрически- или веретеновидно-
эллипсоидные, (26) 34—45 (52) мкм шир., 41.5—78 (96) мкм дл. Эк-
зоспорий тонкий, бесцветный, гладкий. Мезоспорий желто-коричневый,
мелкоточечный, на вершинах зигоспоры иногда немного морщинистый.
Линия разрыва отчетливая.
В мелкой канаве со слабопроточной водой. Германия.
Редкий вид. Конъюгация и образование зигоспор наблюдались толь-
ко в культуре. При определении следует учитывать, что часть конъ-
югационного канала от воспринимающей клетки может быть замаски-
рована сильным вздутием соответствующего гаметангия. Близок по мно-
гим признакам к S. fritschiana, отличается прежде всего более мелкой
точечностью мезоспория.
121. Spirogyra dentireticulata Jao — Спирогира зубчато-сетчатая
(рис. 133). — Вегетативные клетки 18—26 мкм шир., по длине в
4.5—14 раз больше. Поперечные стенки складчатые. Хлоропласт 1.
Конъюгация лестничная, редко боковая. Конъюгационный канал при
лестничной конъюгации от обеих клеток, с преобладанием отдающей
клетки; при боковой конъюгации канал выдается на 1/5—1/4 ши-
рины клетки, нить при этом слегка изогнутая. Воспринимающие
клетки сильно вздутые со всех сторон до 49 мкм при лестничной
конъюгации и до 43.5—46.5 мкм — при боковой конъюгации. Зиго-
споры от широкоэллипсоидных до удлиненно-эллипсоидных, с оттяну-
тыми вершинами, 29—43 мкм шир., 42—96 мкм дл. Экзоспорий тон-
кий, бесцветный, гладкий, иногда слабослоистый на вершинах зиго-
споры. Мезоспорий толстый, от желтого до желто-коричневого, грубо-
или тонкосетчатый, с мелкими или значительно удлиненными шипами
или зубчиками в местах разветвлений сетчатых утолщений. Эндоспорий
тонкий, гладкий.
В текучей и стоячей воде, в реках, на рисовых полях. Зап. и Южн.
Казахстан (р. Темир и устье р. Курты).
Редкий вид, найденный также в Китае и США. По структуре ме-
зоспория приближается к S. reticulata f. acanthophora, хорошо отли-
чается меньшей шириной вегетативных клеток и числом хлоропластов.
От близкого вида 5. lambertiana Trans. (Индия, США) отличается глав-
ным образом структурой мезоспория. Этот признак, а также форма
воспринимающих клеток и зигоспор отграничивают этот вид и от
S. laxistrata Jao (Китай).
303
Рис. 132. 1,2 — Spirogyra gaterslebensis'. 1 — лестничная н боковая конъюгации в одной
паре нитей, 2 — строение оболочек зигоспоры; 3, 4 — S. herbipolensis. (По: 1,2 — Rieth,
1973; 3, 4 — Bock, Bock, 1956).
304
122. Spirogyra incrassata Czurda — Спирогира утолщенная
(рис. 134). — Вегетативные клетки 26—30 мкм шир., по длине в 7—9
раз больше. Поперечные стенки складчатые. Хлоропластов 1—2. Конъ-
югация боковая и лестничная. Конъюгационный канал при боковой
конъюгации выдается примерно на 1/2 ширины клетки, нить при этом
сильно изогнутая; при лестничной конъюгации канал образован обеими
клетками с преобладанием отдающей клетки. Воспринимающие клетки
сильно вздутые со всех сторон, веретеновидные. Зигоспоры удлинен-
Рис. 133. Spirogyra dentireticulata: 1,2 — лестничная конъюгация, 3 — боковая конъюга-
ция, 4—6 — строение мезоспория, вид с поверхности (4, 6) и в поперечном разрезе (5). (По:
1, 6 — Jao, 1935b; 2—5 — Красноперова, 19716).
20 Рундина Л. А.
305
Рис. 134. Spirogyra incrassata, боковая (/) и лестничная (2) конъюгации, строение оболо-
чек зигоспоры (3). (По: Czurda, 1932).
но-эллипсоидные, 42—52 мкм шир., 110 мкм дл. Экзоспорий тонкий,
бесцветный, гладкий. Мезоспорий толстый, желто-коричневый, прони-
занный узкими каналами, на поверхности (сверху) выглядящими как
редкие маленькие точки, и испещренный неглубокими трещинами.
В стоячих водах. Германия.
Редкий и недостаточно изученный вид. Требуют уточнения число
хлоропластов и строение мезоспория. Близок к 5. marchica и 5. reti-
culata, отличается меньшим числом хлоропластов и иным строением
мезоспория.
123. Spirogyra reticulata Nordst. — Спирогира сетчатая (рис. 135—
137). — Вегетативные клетки 23—41 мкм шир., по длине в 2—14 (16)
раз больше. Ризоиды латеральные, вильчато разветвленные, редкие. По-
перечные стенки складчатые. Хлоропластов 1—4, слабо извитые, тон-
кие, иногда вильчато разветвленные, часто почти параллельные. Конъ-
югация лестничная и боковая, часто с преобладанием одной из них, в
некоторых случаях совмещенные в одной паре нитей, иногда только лес-
тничная. Конъюгационный канал при лестничной конъюгации образован
обеими клетками, очень редко с преобладанием отдающей клетки, нити
при этом заметно коленчато изогнутые или прямые; при боковой конъ-
югации канал выдается на 1/3—1/2 ширины клетки, нить при этом пря-
мая или изогнутая. Воспринимающие клетки веретеновидные, сильно
306
Рис, 135, Spirogyra reticulata: 1—7 — f. acanthophora, лестничная {1, 2) и боковая (3) конъ-
югации, изменение строения мезоспория на разных стадиях созревания зигоспоры (4—7);
8 — f. regularis. (По: 1—3 — ориг.; 4—8 — Рундина, 1978а).
307
Рис. 136. Spirogyra reticulata'. 1,2 — f. acanthophora, лестничная конъюгация и зигоспоры
(7), строение мезоспория (2); 3—5 — f. regularis, лестничная конъюгация (3), зигоспоры (4,
5); 6, 7 — f. czurdae, лестничная конъюгация (6), строение мезоспория (7). (По: 1, 2 —
Skuja, 1928 (как S. willei var. acanthophora); 3—5 — Gauthier-Lievre, 1965 (как S. regularis);
6,7 — Czurda, 1930 (как S. willei)).
вздутые co всех сторон до 43.5—70 (75) мкм шир., редко невздутые или
вздутые только по ширине зигоспоры. Клетки без конъюгационной пары
невздутые, иногда удлиненные. Зигоспоры от широкоэллипсоидных до
удлиненно-эллипсоидных, иногда эллипсоидно-цилиндрических с более
или менее заостренными вершинами, 30—81 мкм шир., 45—146 мкм дл.
Экзоспорий тонкий, бесцветный, гладкий. Мезоспорий толстый, от зо-
лотисто-желтого до желто-коричневого, иногда с розоватым или крас-
новатым оттенком, однослойный, морщинистый, с утолщениями, сомк-
нутыми в сеть полностью или только местами, или двухслойный, мор-
щинисто-сетчатый, с рядами более или менее острых игольчатых
(шиповидных) выростов. Эндоспорий тонкий, гладкий, различимый
только на зрелых зиготах. Партеноспоры по форме и строению подобны
зигоспорам, 35—38 мкм шир., 58—65 мкм дл., редкие.
Вид, по всей вероятности, монотипный, так как различия между
установленными формами сводятся по существу к особенностям струк-
туры мезоспория, которая сильно варьирует в зависимости от степени
308
зрелости зигот. По данным СЭМ, в центре бугорчатых утолщений на
сетчатых переплетениях мезоспория имеются поры. При определении
формы поперечной септы следует помнить, что складчатые перегородки
при изменении внутриклеточного давления во время конъюгации вы-
прямляются, становясь гладкими. Необходимо просматривать большое
количество фертильного материала, где можно найти клетки с нерав-
номерно чередующимися гладкими и складчатыми перегородками.
F. reticulata [=S. fallax (Hansg.) Wille, pr. p.; S. willei Skuja, 1928,
non sensu Czurda, 1930]. — Вегетативные клетки (28) 30—37 мкм
шир. Хлоропластов (1) 2—3 (4). Конъюгация лестничная, реже боко-
вая. Воспринимающие клетки сильно вздутые со всех сторон. Зиго-
споры эллипсоидные, 38—81 мкм шир., 55—146 мкм дл. Мезоспорий
желтый, с утолщениями в виде неправильной сетки, иногда двухслой-
ный: наружный слой тонкий, морщинистый, внутренний слой сетчато
утолщенный, иногда с игольчатыми выростами в узлах утолщений.
В стоячих водах, в мелководье водохранилищ. Россия: северо-запад
европ. части (Республика Коми); Прибалтика, Белоруссия, Украина
(окрестности Киева, Киевское водохранилище).
Широко распространенная форма, в том числе в Финляндии, Гер-
мании, Чехии, Румынии, Венгрии, Индии, Китае, Бразилии. По-види-
мому, и типовой форме иногда свойственны игольчатые выросты на
сетчатых утолщениях мезоспория, столь характерные для f. acantho-
phora. Поэтому разграничение их представляет подчас большую труд-
ность. Впервые на развитие шиповидных выростов в узлах сетчато
переплетенных утолщений обратил внимание Скуя (Skuja, 1928) на
Рис. 137. Spirogyra reticulata f. nawaschinii'. 1 — лестничная конъюгация и прорастающая
зигоспора, 2 — зигоспора (поперечный разрез) в воспринимающей клетке, 3 — строение ме-
зоспория. (По: Казановскнй в: Казаковский, Смирнов, 1914 (как S. nawaschinii)).
309
образцах S. willei Skuja, относимого мною в синонимы типовой формы
5. reticulata.
F. acanthophora (Skuja) V. Poljansk. [=S. willei Skuja var. acan-
thophora Skuja; S. acanthophora (Skuja) Czurda] (рис. 135, 1—7; 136,
1, 2). — Вегетативные клетки (25) Я?—38 мкм шир. Хлоропластов
1—4. Конъюгация лестничная и боковая, с преобладанием одной из
них, иногда совмещенные в одной паре нитей. Воспринимающие клет-
ки сильно вздутые до 43.5—70 (75) мкм, веретеновидные, редко не-
вздутые или вздутые по ширине зигоспоры. Зигоспоры от широкоэл-
липсоидных до удлиненно-эллипсоидных, с более или менее заострен-
ными вершинами, иногда эллипсоидно-цилиндрические, (35) 40—60 мкм
шир., (72.5) 80—144 (165) мкм дл. Мезоспорий от золотисто-желтого
до желто-коричневого, иногда с красноватым оттенком, двухслойный,
морщинисто-сетчатый, с рядами более или менее острых игольчатых
выростов.
В стоячих, часто заболоченных, мелких водоемах, в прибрежье озер,
прудов. Россия: северо-запад европ. части, Зап. Сибирь; Латвия, Ук-
раина, Сев. и Центр. Казахстан.
Широко распространенная, но нечасто встречающаяся форма. В
Европе найдена в Дании, Нидерландах, Германии и Польше. Наиболее
существенное отличие от типовой формы — постоянное наличие в
узлах сетчатых утолщений мезоспория игольчатых (шиповидных или
сосочковидных) выростов, частота и заостренность которых зависят от
степени зрелости зиготы. Однако эта характерная особенность иногда
наблюдается и у других форм этого вида.
F. czurdae V. Poljansk. (=S. willei Skuja sensu Czurda, 1930, non
sensu Skuja, 1928; S. willei Czurda) (рис. 136, 6, 7). — Вегетативные
клетки 32—36 мкм шир. Хлоропластов (2) 3. Конъюгация лестничная.
Воспринимающие клетки сильно вздутые. Зигоспоры удлиненно-эллип-
соидные, 54—60 мкм шир., 92—120 мкм дл. Мезоспорий коричневый,
морщинисто-сетчатый, с извилистыми утолщениями, местами смыкаю-
щимися в сетку.
В стоячих водах, в культуре. Германия.
Редкая, недостаточно изученная форма. Отличается от типовой и
других форм S. reticulata неполной сомкнутостью в сеть утолщений
на мезоспории. По наблюдениям в клональной культуре, полностью
лишена способности к боковой конъюгации.
F. nawaschinii (Kasan.) V. Poljansk. [=S. nawaschinii Kasan.; 5. fal-
lax (Hansg.) Wille var. (sine nomen) Smirn.] (рис. 137). — Вегетативные
клетки 27—41 мкм шир. Хлоропластов (1) 2. Конъюгация лестничная.
Воспринимающие клетки преимущественно слабо вздутые со всех сто-
рон до 40—45 (49) мкм. Зигоспоры эллипсоидные, 30—49 мкм шир.,
45—100 мкм дл. Мезоспорий желтовато-коричневый с розовым оттен-
ком, морщинисто-сетчатый, сетчато утолщенный.
В стоячих водоемах, на рисовых полях. Украина (окрестности
Киева), Узбекистан (Голодная степь) (?).
310
Редкая, спорная, недостаточно изученная форма. Некоторые авторы
считают ее синонимом типовой формы. В первоописании ее омонима
(.$. nawaschinii) значится, что воспринимающие клетки мало или со-
всем не вздуты, что противоречит авторским рисункам. Изображенные
там же разнообразные формы зигоспор следует рассматривать, скорее
всего, как модификации обычной эллипсоидной формы у зигоспор,
находящихся в разных положениях (вид анфас и сверху). Несомненно
также, что кроме сетчатого уплотнения мезоспорий имеет грубые ши-
повидные выросты, отчетливо видные на авторском рисунке. По этому
важному таксономическому признаку, не оговоренному в первоописа-
нии, эта форма тесно сближается с f. regularis.
F. regularis (Cedercr.) V. Poljansk. [=5. reticulata var. regularis Ce-
dercr.; S. regularis (Cedercr.) Krieger] (рис. 135, 8; 136, 3—5). — Веге-
тативные клетки 23—32 (35) мкм шир. Хлоропластов (1) 2. Конъюгация
лестничная. Воспринимающие клетки сильно вздутые со всех сторон,
веретеновидные. Зигоспоры от эллипсоидных до широкоэллипсоидных,
32—48 мкм шир., 55—94 мкм дл. Мезоспорий морщинисто-сетчатый, с
рядами туповатых шипов в узлах сети.
В стоячих и текучих водах, небольших реках. Азербайджан, Зап.
Казахстан.
Редкая форма, найденная также в Финляндии, Германии, Алжире
и Марокко. Шиповатость мезоспория отмечается нами впервые, ранее
мезоспорий характеризовался только как правильно сетчато утолщен-
ный (складчатый). От сходного в этом отношении 5. dentireticulata
отличается немного более широкими вегетативными клетками и нали-
чием почти постоянно двух хлоропластов. Близка, возможно, идентична
f. nawaschinii.
124. Spirogyra herbipolensis О. et W. Bock — Спирогира плотно-
вершинная (рис. 132, 3, 4). — Вегетативные клетки (31) 34—40
(52) мкм шир. Поперечные стенки складчатые. Хлоропластов 1—2.
Конъюгация боковая, очень редко лестничная. Конъюгационный канал
при боковой конъюгации выдается приблизительно на 1/2 ширины
клетки, нить при этом незначительно изогнутая; при лестничной конъ-
югации канал образован преимущественно отдающей клеткой. Воспри-
нимающие клетки вздутые со всех сторон, веретеновидные, редко не-
вздутые. Зигоспоры от эллипсоидных до цилиндрически-эллипсоидных,
с широко закругленными вершинами, 42—60 мкм шир., 90—170 мкм
дл. Экзоспорий тонкий, отчетливый. Мезоспорий желтовато-коричне-
вый, двухслойный: внешний слой светлый или темно-коричневый, не-
правильно волнисто-морщинистый, у зрелых зигот утолщенный на вер-
шинах, внутренний слой толстый, коричневый, гладкий. Линия разрыва
отчетливая.
В стоячих водах, при pH 6.0. Германия.
Редкий вид, требующий дополнительного изучения.
125. Spirogyra costulata Kadi. — Спирогира мелкоребристая (рис. 138,
1). — Вегетативные клетки 32—41 мкм шир. Поперечные стенки склад-
311
Рис. 138. I — Spirogyra costulata, боковая конъюгация; 2, 3 — S. miranda: 2 — боковая
конъюгация, 3 — строение оболочек зигоспоры; 4, 5 — S. venosa: 4 — боковая конъюга-
ция, 5 — зигоспора; 6—8 — S. costata: 6 — вегетативная клетка и две зигоспоры, боковая
конъюгация, 7 — зигоспора, 8 — строение мезоспория. (По: Kadhibowska in: Kadhibowska,
Christensen, 1979).
чатые. Хлоропластов 2—3. Конъюгация боковая, нить при этом прямая.
Воспринимающие клетки невздутые или слабо вздутые. Зигоспоры уд-
линенно-эллипсоидные, 40—51 мкм шир., 80—124 мкм дл. Экзоспорий
бесцветный, гладкий. Мезоспорий красновато-бурый, рельефный, непра-
вильно сетчатый.
В пруду. Дания.
Редкий вид. Близок к S. venosa, но имеет в отличие от последнего
вида более толстые вегетативные клетки и невздутые или очень слабо
вздутые воспринимающие клетки.
126. Spirogyra miranda Kadi. — Спирогира поразительная (рис. 138,
2, 3). — Вегетативные клетки 24—33 мкм шир. Поперечные стенки
складчатые. Хлоропластов 2—3. Конъюгация боковая. Конъюгационный
канал выдается на 1/2 ширины клетки, широко закругленный, нить при
этом почти прямая или незначительно изогнутая. Воспринимающие клетки
вздутые со всех сторон. Зигоспоры эллипсоидные, 43—54 мкм шир.,
312
83—ПО мкм дл. Экзоспорий бесцветный, гладкий, двухслойный. Мезо-
спорий красновато-бурый, неправильно рельефно-сетчатый, однослойный.
В стоячих водах. Дания.
Недостаточно изученный и спорный вид, требующий уточнения в
отношении строения оболочек у зигоспор. Существует противоречие
между его описанием и рисунком: изображена орнаментация экзоспо-
рия, в то время как он описывается гладким и к тому же двухслойным.
127. Spirogyra venosa Kadi. — Спирогира жилковатая (рис. 138,
4, 5). — Вегетативные клетки 25—32 мкм шир. Поперечные стенки
складчатые. Хлоропластов 2. Конъюгация боковая, нить при этом
почти прямая. Воспринимающие клетки значительно вздутые. Зиго-
споры эллипсоидные, 38—51 мкм шир., 76—114 мкм дл. Экзоспорий
бесцветный, гладкий. Мезоспорий желтый, с неправильно извилистыми
и разветвленными ребрами (утолщениями).
В глиняном карьере. Дания.
Редкий вид. Отличается от близкого S. adjerensis шириной вегета-
тивных клеток и иной скульптурой мезоспория. Близок или, возможно,
идентичен S. costulata.
128. Spirogyra borysthenica Kasan. et Smirn. — Спирогира днепров-
ская (рис. 139). — Вегетативные клетки 30—40 мкм шир., по длине в
4.5—12 раз больше. Поперечные стенки складчатые. Хлоропластов 2—4,
почти прямые или с незначительным количеством оборотов спирали.
Конъюгация лестничная. Конъюгационный канал образован обеими
клетками. Воспринимающие клетки вздутые со всех сторон. Клетки без
конъюгационной пары невздутые. Зигоспоры эллипсоидные, реже вере-
теновидные, 40—62 мкм шир., 73.7—163.7 мкм дл. Экзоспорий тол-
стый, бесцветный, гладкий, двухслойный, наружный слой тоньше внут-
реннего. Мезоспорий толстый, в 2—4 раза толще экзоспория, от желтого
до желто-коричневого, покрытый крупными, широкими в основании со-
сочковидными или удлиненными шиповидными выростами, 3—4 мкм
дл., внедряющимися во внутренний слой экзоспория. Эндоспорий тон-
кий, гладкий, трудноразличимый.
Редкий вид, найденный также в Нидерландах и США. По строению
мезоспория хорошо отграничен от близкого S. reticulata. Представляется
спорным вопрос о формах у этих двух видов, поскольку длина и заост-
ренность выростов на мезоспорий зависят от степени зрелости зигот.
F. borysthenica (=5. borysthenica var. typica Kasan. et Smim.) (рис. 139,
1—5). — Мезоспорий с крупными, широкими в основании, сосочковидны-
ми выростами; поверхность мезоспория между выростами гладкая.
В стоячих и текучих водах, осоковых болотах, на рисовых полях,
маленькими хлопьями среди других видов спирогиры. Россия: северо-
запад европ. части; Украина (берег Днепра в окрестностях Киева),
Узбекистан (Голодная степь).
F. echinospora (Kasan. et Smirn.) V. Poljansk. (=5. borysthenica var.
echinospora Kasan. et Smirn.) (рис. 139, 6—9). — Мезоспорий с сильно
313
Рис. 139. Spirogyra borysthenica: 1—5 — f. borysthenica, 6—9 — f. echinospora (/ — лест-
ничная конъюгация, 2, 6 — строение мезоспория, вид с поверхности, 3, 8 — строение обо-
лочек зигоспоры, поперечный разрез, 4, 7 — отдельные выросты мезоспория, 5, 9 — стро-
ение оболочек зигоспоры, схема). (По: Казановский, Смирнов, 19136 (/—5 — как 5. bo-
rysthenica var. typica', 6—9 — как S. borysthenica var. echinospora)).
вытянутыми шиповидными выростами; поверхность мезоспория между
выростами неровная.
В стоячих водах, большими скоплениями. Украина (остров на Днеп-
ре в окрестностях Киева).
129. Spirogyra spinescens Kiriakov — Спирогира колючая (рис. 140, 1—
3). — Вегетативные клетки (27) 32 (37) мкм шир., по длине в 5—7.7
(15) раза больше. Поперечные стенки складчатые. Хлоропластов 2—3.
Конъюгация лестничная. Конъюгационный канал образован обеими
клетками с преобладанием отдающей клетки. Воспринимающие клетки
веретеновидно вздутые, (45) 61.5 (92) мкм шир. Клетки без конъюгацион-
ной пары невздутые. Зигоспоры преимущественно удлиненно-эллипсо-
идные с широко закругленными вершинами, (40) 51.5 (65) мкм шир.,
(82.5) 114 (140) мкм дл. Экзоспорий тонкий, 0.6—1.1 мкм толщ., бес-
цветный, гладкий. Мезоспорий толстый, 5.5—6.6 мкм толщ., коричне-
во-оранжевый, с грубыми, острыми или тупыми возвышениями, 1.1—
6.6 мкм шир., 2.2—3.3 мкм выс., соединенными между собой сетью
невысоких ребер, не всегда четко видных. Эндоспорий тонкий, 0.7—
0.8 мкм толщ., коричнево-оранжевый, гладкий.
В сильно заросшем болоте. Болгария.
Редкий вид. Обнаруживает большое сходство с кругом форм S. re-
ticulata. Возможно, идентичен одной из них (f. acanthophora или f.
regularis). Представляет большой интерес окрашенность эндоспория,
не отмеченная больше ни у одного вида спирогир.
130. Spirogyra marchica Krieger — Спирогира маркская (рис. 140,
4, 5). — Вегетативные клетки 29—32 мкм шир., по длине в 5—8 раз
314
больше. Поперечные стенки складчатые. Хлоропластов 2. Конъюгация
боковая и лестничная, в конъюгации участвуют немногие клетки нити.
Конъюгационный канал при боковой конъюгации выдается примерно
на 1/2 ширины клетки, при лестничной конъюгации канал образован
обеими клетками, часто с преобладанием отдающей клетки, нити при
обоих типах конъюгации сильно изогнутые. Воспринимающие клетки
сильно вздутые со всех сторон. Клетки без конъюгационной пары не-
вздутые. Зигоспоры эллипсоидные, 46—53 мкм шир., 86—170 мкм дл.
Экзоспорий довольно тонкий, бесцветный, гладкий. Мезоспорий толще
экзоспория, коричневый, с небольшими бородавчатыми возвышениями.
Эндоспорий тонкий, гладкий.
В стоячих водах. Германия.
Редкий вид. От сходного S. graniilata отличается несколько мень-
шей шириной вегетативных клеток и особенно строением мезоспория.
Рис. 140. 1—3 — Spirogyra spinescens: 1,2 — лестничная конъюгация, 3 — строение обо-
лочек зигоспоры (вид с поверхности и на поперечном разрезе); 4,5 — S. marchica, лестнич-
ная (4) и боковая (5) конъюгации. (По:/—3 — Киряков в: Водеиичаров, Киряков, 1970;
4, 5 — Krieger in: Kolkwitz, Krieger, 1941—1944).
315
131. Spirogyra costata Kadi. — Спирогира ребристая (рис. 138, 6—
8). — Вегетативные клетки 30.6—40 мкм шир. Поперечные стенки
складчатые. Хлоропластов 2. Конъюгация боковая, очень редко лест-
ничная. Конъюгационный канал при лестничной конъюгации образован
обеими клетками. Воспринимающие клетки невздутые или слабо взду-
тые. Клетки без конъюгационной пары невздутые. Зигоспоры удлинен-
но-эллипсоидные, 38—50 мкм шир., 9(К-120 мкм дл. Экзоспорий тон-
кий, бесцветный, гладкий. Мезоспорий желто-коричневый или красно-
вато-бурый, двухслойный (однослойный ?): внешний слой из коротких,
тонко разветвленных утолщений, размещенных приблизительно по 10
в 10 мкм, внутренний слой из. утолщенных ребер, размещенных при-
близительно по 4 в 10 мкм и образующих неправильную сеть.
В водоемах торфяных разработок. Дания.
Редкий вид. По наличию двух рядов (слоев) орнаментаций мезо-
спорий должен быть двухслойным, а не однослойным, как указано в
первоописании. Очень близок к S. costulata, отличается двухслойным
скульптурированным мезоспорием и наличием, хотя и очень редким,
лестничной конъюгации.
132. Spirogyra granulata Jao — Спирогира зернистая (рис. 141, 1,
2). — Вегетативные клетки (29) 32—40.8 мкм шир., по длине в 4—9
раз больше. Поперечные стенки складчатые. Хлоропластов 2. Конъ-
югация боковая, очень редко лестничная. Конъюгационный канал при
боковой конъюгации выдается приблизительно на 1/2 ширины клетки,
нить при этом более или менее изогнутая. Воспринимающие клетки
веретеновидно вздутые вокруг зигоспоры. Клетки без конъюгационной
пары невздутые. Зигоспоры эллипсоидные или цилиндрически-эллип-
соидные, с закругленными вершинами, (40) 45—55 мкм шир., 72.8—
153 мкм дл. Экзоспорий тонкий, бесцветный, гладкий. Мезоспорий
желтовато-коричневый, двухслойный: внешний слой грубо и неправиль-
но морщинистый, внутренний слой мелкозернистый и слабоморщинис-
тый. Эндоспорий тонкий, гладкий, морщинистый.
В стоячих водах, на рисовых полях. Нидерланды, Германия, Китай.
Редкий вид. Близок к 5. reticulata и S. marchica, отличаясь в ос-
новном иным строением оболочек зигоспор. По первоописанию зер-
нистость внутреннего слоя мезоспория не всегда отчетливая, иногда
неправильная и неясная. По данным сканирующего микроскопа, только
внутренний слой мезоспория скульптурирован: он тонко сетчато-зер-
нистый. Очень сходен с S. pseudogranulata, отличается незначительно
меньшей шириной вегетативных клеток.
133. Spirogyra pseudogranulata Ley — Спирогира ложнозернистая
(рис. 141, 3—7). — Вегетативные клетки 35—42 мкм шир., по длине
в 4—8 (10) раз больше. Ризоиды латеральные, лопастные или вильчато
разветвленные. Поперечные стенки складчатые. Хлоропластов (1) 2, в
начале конъюгации почти неизвитые, истонченные. Конъюгация боко-
вая, очень редко лестничная. Конъюгационный канал при боковой
конъюгации выдается на 1/4—1/3 ширины клетки, нить при этом не-
значительно изогнутая или прямая; при лестничной конъюгации канал
316
Рис. 141. 1,2 — Spirogyra granulata: 1 — боковая конъюгация, 2 — строение оболочек зи-
госпоры; 3—7 — S. pseudogranulata: 3 — ризоид, 4, 5 — боковая конъюгация, 6 — лест-
ничная конъюгация, 7 — строение оболочек зигоспоры. (По: 1, 2 — Jao, 1935b; 3—5 —
Рундина, 1981; 6,7— Yamagishi, 1966b).
317
образован обеими клетками. Воспринимающие клетки слабо вздутые
по ширине зигоспоры, изредка веретеновидно или цилиндрически взду-
тые до 72.5 мкм шир. Зигоспоры от правильно эллипсоидных до уд-
линенных или цилиндрически-эллипсоидных, с закругленными верши-
нами, 43—68 мкм шир., 97—145 мкм дл. Экзоспорий тонкий, бес-
цветный, гладкий. Мезоспорий толстый, от золотисто-желтого до
желто-коричневого, двухслойный: внешний слой коротко ребристо-
морщинистый у незрелых зигот и крупно волнисто-складчатый у зре-
лых зигот, внутренний слой плотнобородавчатый, хорошо заметный на
зрелых зиготах.
В стоячих водоемах, прудах, канавах, на рисовых полях. Украина
(окрестности Киева).
Редкий вид, найденный также в Китае и Японии. Требует проверки
строение всех слоев мезоспория и особенно внутреннего слоя, так как,
судя по первоописанию, он тонко (мелко) сетчатый. Близок к S. gra-
nulata, но имеет несколько более широкие вегетативные клетки. Впол-
не возможно, что является формой или синонимом последнего вида.
От круга форм 5. reticulata отличается широкими вегетативными клет-
ками и двухслойностью мезоспория.
134. Spirogyra colligata Hodgetts — Спирогира связанная (рис. 142,
Г). — Вегетативные клетки 29—40 мкм шир., по длине в 8—16 (22)
раз больше. Поперечные стенки гладкие с кольцевой накладкой, со-
единяющей соседние клетки. Хлоропластов 3—6, очень слабо извитые
или почти прямые. Конъюгация лестничная, очень редко боковая.
Конъюгационный канал при лестничной конъюгации образован обеими
клетками; при боковой конъюгации канал ушковидный, выдается при-
близительно на ширину клетки, нить при этом изогнутая или почти
прямая. Воспринимающие клетки сильно вздутые вокруг зигоспоры до
84—102 мкм. Клетки без конъюгационной пары невздутые. Зигоспоры
линзовидные, в двух положениях круглые и широкоэллиптические,
48—75 мкм толщ., 62—90 мкм диам. Экзоспорий тонкий, бесцветный,
гладкий. Мезоспорий толстый, желто-коричневый, бородавчатый.
В стоячих мелких водоемах, прудах, лужах, сырых луговинах. Ан-
глия, Дания, Нидерланды, Польша, США,
Редкий вид с широким ареалом. По-видимому, распространен шире,
чем известно по литературе. Трудность определения усугубляется лег-
кой просматриваемостью кольцевой накладки. Отличается от S. silesi-
аса только незначительно меньшей шириной вегетативных клеток. У
обрих видов часто встречается терминальная конъюгация как разно-
видность лестничной. Третий вид, существующий в секции Colligata, —
S. subcolligata Bi, найденный только в Китае, отличается от двух дру-
гих гладким мезоспорием. С 5. colligata его сближает одинаковая ши-
рина вегетативных клеток.
135. Spirogyra silesiaca Kadi. — Спирогира силезская (рис. 142,
2—5). — Вегетативные клетки 43—52 мкм шир., по длине в 3—14
раз больше. Поперечные стенки гладкие с кольцевой накладкой, со-
единяющей соседние клетки. Хлоропластов 4—6, очень слабо извитые
318
Рис. 142. 1 — Spirogyra colligata', 2—5 — S. silesiaca: 2, 3 — съемное кольцо, сбоку и
анфас, 4 — лестничная и боковая (терминальная) конъюгации в одной нити, 5 — боковая
конъюгация; 6 — Sirogonium sticticum f. sticticum. (По: 1 — Kadhibowska, 1969; 2—5 — Kad-
hibowska, 1967 ; 6 — Jao, 1935b).
или почти прямые. Конъюгация лестничная, редко боковая. Конъюга-
ционный канал при лестничной конъюгации образован обеими клет-
ками, при боковой конъюгации канал ушковидный. Воспринимающие
клетки сильно вздутые вокруг зигоспоры до 99—132 мкм. Клетки без
конъюгационной пары невздутые. Зигоспоры линзовидные, в двух поло-
жениях круглые и широкоэллиптические, 38—87 мкм толщ., 60—
91 мкм диам. Экзоспорий тонкий, бесцветный, гладкий. Мезоспорий
толстый, коричневый, бородавчатый.
В рыбоводном пруду. Польша.
Редкий вид, не имеющий, возможно, самостоятельной видовой зна-
чимости. В отличие от S. colligata имеет незначительно более широкие
вегетативные клетки. Ширина кольцевой накладки (диаметр кольца) у
обоих видов равна соответственно ширине вегетативных клеток, высота
ее у обоих видов одинаковая — 22—24 мкм.
По сообщению Кадлубовской и Лангангена (Kadhibowska, Langan-
gen, 1997), поступившему ко мне уже после сдачи монографии в из-
319
дательство, в Норвегии найдены Spirogyra reflexa Trans, и S. schmidtii
W. et G. S. West, новые для Европы. Эти виды следует принять как
потенциальные для флоры России.
СОМНИТЕЛЬНЫЕ И ОШИБОЧНЫЕ УКАЗАНИЯ
ВИДОВ РОДА SPIROGYRA
Spirogyra acidoneglecta Uspensky (nom. nudum): Успенский, 1934 :
457; 1935 : 53; 1963 : 148.
S. adnata (Vauch.) Kiitz.: Бортов, 1870 : 53. — Неполно описан-
ный вид (Kiitzing, 1843 : 279; 1855 : Taf. 25, Fig. 4). Частично может
быть отнесен к S. fluviatilis Hilse f. grossii (Schmidle) V. Poljansk.
S. archangelskaja Uspensky (nom. nudum): Успенский, 1934 : 441;
1935 : 79; 1963 : 177.
S. arcta (Ag.) Kiitz.: Бортов, 1870 : 53; Змеев, 1872 : 53; Алек-
сенко, 1893 : 28; Gutwinski, 1895a. — Неполно описанный вид (Kiit-
zing, 1845 : 223; 1855 : Taf. 21, Fig. 2).
S. borwichae Uspensky (nom. nudum): Успенский, 1934 : 458;
1935 : 96; 1963 : 173.
S. brunnea Czurda f. minor V. Poljansk. (nom. nudum): Обухова,
1959 : 353.
S. crassa Kiitz.: Рябинин, 1889 : 335. — Сомнительное определение
из-за неполного описания и отсутствия зигоспор.
? S, decimina (Miill.) Kiitz. (=S. porticalis var. decimina'): Рябинин,
1889 : 336. — Сомнительное определение из-за неполного описания
и отсутствия зигоспор.
S. decimina (Miill.) Kiitz.: Христюк, 1947 : 751. — Сомнительное оп-
ределение из-за неполного описания и отсутствия фертильной стадии.
S. decimina Kiitz. [var.] laxa (sine auct.): Бортов, 1870 : 53. —
Неполно описанный таксон [разновидность ?] из-за отсутствия зиго-
спор (Kiitzing, 1855 : Taf. 23, Fig. 3, b—d). Non S. laxa Kiitz.
S. decimina (Miill.) Kiitz. f. crassior Gutw.: Gutwinski, 1890 : 5;
1895b : 314. — Неполно описанная форма из-за отсутствия зигоспор.
Non S. decimina (Miill.) Kiitz.
S. densa Kiitz.: Козловский, 1888 : 427; 1890 : 6, 23; Gutwinski,
1892 : 13; 1895b : 314; Триполитова, 1928 : 271. — Неполно опи-
санный вид из-за отсутствия зигоспор (Kiitzing, 1855 : Taf. 24, Fig. 3).
S. dubia Kiitz. [var.] longiarticulata Kiitz.: Хмелевский, 1885 :
76. — Неполно описанный таксон [разновидность ?] из-за отсутствия
зигоспор (Kiitzing, 1855 : Taf. 25, Fig. 1).
S. fallax (Hansg.) Wille f. berolinensis (sine auct.): Воронихин, 1953 :
218. — Неполно описанная форма из-за отсутствия данных о количестве
хлоропластов, типе конъюгации и строении оболочек у зигоспор.
S. fallax (Hansg.) Wille f. bohemica (sine auct.): Воронихин, 1953 :
218. — Неполно описанная форма из-за отсутствия данных о количест-
ве хлоропластов, типе конъюгации и строении оболочек у зигоспор.
320
S. fallax (Hansg.) Wille f. scanica (Nordstedt ?): Воронихин, 1953 :
218. — Неполно описанная форма из-за отсутствия данных о коли-
честве хлоропластов, типе конъюгации и строении оболочек у зиго-
спор.
S. flavescens Kiitz.: Алексенко, 1894 : 96. — Сомнительное оп-
ределение из-за неполного описания и отсутствия зигоспор. Экзем-
пляры с шириной вегетативных клеток 12—16 мкм соответствуют
S. flavescens (Hass.) Kiitz., а с шириной 16—26 мкм — 5. gracilis
(Hass.) Kiitz.
S. frigida Gay, 1884a : 90, tab. 4, fig. 4. — Неполное описание
из-за отсутствия данных о строении оболочек зигот. Non 5. frigida
Gay sensu Krieger in: Kolkwitz, Krieger, 1941—1944 : 448, Fig. 719;
Kadhibowska, 1972 : 345, rys. 1398, 1399; 1984 : 448, Fig. 694; Jao,
1988 : 110, tab. 45, fig. 4.
S. gallica Petit f. (sine nomen): Казановский, Смирнов, 1914 : 11. —
Сомнительная форма. По ширине вегетативных клеток (40—59 мкм)
и наличию 1—3 хлоропластов в клетке более соответствует S. porticalis
(Mull.) Cleve.
S. glubokolacustris Uspensky (nom. nudum): Успенский, 1934 : 181;
1935 : 39; 1963 : 138.
S. gluchowii Uspensky (nom. nudum): Успенский, 1934 : 441; 1935 :
79; 1963 : 173.
S. gracilis (Hass.) Kiitz. [f.] longiarticulata Rabenh.: Козловский,
1890 : 33; Триполитова, 1928 : 271. — Неполно описанный таксон
[форма ?] из-за отсутствия зигоспор (Rabenhorst, 1868b : 237; 1868а :
N 1537).
S. grevilleana (Hass.) Kiitz.: Христюк, 1947 : 753. — Сомнительное
определение из-за неполного описания и отсутствия зигоспор. По ши-
рине вегетативных клеток более соответствует S. weberi Kiitz. f. weberi.
S. groenlandica Rosenv.: Hirn, 1895 : 13. — Сомнительное опре-
деление из-за неполного описания и отсутствия зигоспор.
S. hassallii (Jenner) Petit: Hirn, 1900 : 166. — Сомнительное опре-
деление из-за неполного и противоречивого описания.
S. inflata (Vauch.) Rabenh. ampl. Petit: Gutwinski, 1892 : 27. —
Сомнительное определение. По ширине вегетативных клеток и отсут-
ствию вздутия у воспринимающих клеток более соответствует S. weberi
Kiitz. f. weberi.
S. insignis Kiitz. [f.] braunii Rabenh.: Козловский, 1888 : 427. —
Неполно описанный таксон [форма ?] из-за отсутствия зигоспор (Ra-
benhorst, 1868b : 236; 1868а : N 1310, 1395, 1705).
S. insignis Kiitz. f. braunii (sine auct.): Триполитова, 1928 : 271. —
Неполно описанная форма из-за отсутствия зигоспор (Rabenhorst,
1868b : 236; 1868а : N 1310, 1395, 1705).
S. insignis Kiitz. [f.] hantzschii Rabenh.: Козловский, 1890 : 33. — Не-
полно описанный таксон [форма ?] (Rabenhorst, 1868b : 236; 1868а :
N 1291).
21 Рундина Л. А.
321
S. insignis Kiitz. f. hantzschii (sine auct.): Триполитова, 1928 :
271. — Неполно описанная форма (Rabenhorst, 1868b : 236; 1868a :
N 1291).
S. intermedia Rabenh.: Gutwinski, 1892 : 26; 1895b : 314. — Не-
полно описанный вид (Rabenhorst, 1863 : 621).
S. kolae Hajdu, 1975 : 26—27, Abb. 1. — Сомнительный и недо-
статочно описанный вид. Противоречивые сведения о вздутии воспри-
нимающих клеток, форме зигоспор, строении мезоспория в описании
и рисунке.
S. krjutschkowae Uspensky (nom. nudum): Успенский, 1934 : 441;
1935 : 79; 1963 : 171.
S. kusnetzowii Uspensky (nom. nudum): Успенский, 1934 : 180;
1935 : 81, Taf. 1, Fig. 1—4; 1963 : 137, рис. 1.
S. lacustris Uspensky (nom. nudum): Успенский, 1932 : 99; 1963 :
120. Non 5. lacustris Czurda.
S. lancincola Uspensky (nom. nudum): Успенский, 1934 : 441; 1935 :
18; 1963 : 176.
S. laxa Kiitz. f. articulus (nom. nudum, sine auct.): Козловский,
1888 : 426; Триполитова, 1928 : 271.
S. longata (Vauch.) Kiitz.: Алексенко, 1888 : 211. — Сомнительное оп-
ределение из-за неполного описания. По ширине вегетативных клеток со-
ответствует диагнозам трех видов: S. decimina (Miill.) Kiitz. f. longata
(Vauch.) V. Poljansk., 5. condensata (Vauch.) Kiitz. и 5. pratensis Trans.
S. longata Kiitz. [f.] reversa (Kiitz.) Kirchn.: Gutwinski, 1890 : 5;
1895b : 312. — Неполно описанный таксон [форма ?] (Kiitzing, 1849 :
444; Kirchner, 1878 : 123).
S. major Kiitz.: Комаренко, 1956 : 193. — Неполно описанный вид
из-за отсутствия зигоспор (Kiitzing, 1855 : Taf. 24, Fig. 2).
S. majuscula Kiitz. f. (nom. nudum, sine nomen et auct.): Успенский,
1934 : 169.
S. mirabilis (Hass.) Kiitz. f. microspora Beck-Mannagetta, 1931 :
313. — Неполно описанная форма из-за отсутствия данных о размерах
вегетативных клеток, форме поперечных перегородок и количестве
хлоропластов.
S. nitida Link: Artari, 1885 : 14. — Сомнительное определение из-
за отсутствия зигоспор.
S. nitida (Dillw.) Link f. (sine nomen et auct.) (=? 5. princeps
Vauch.): Казановский, Смирнов, 1914 : 14. — Сомнительная форма.
Экземпляры с шириной вегетативных клеток 78—90 мкм соответству-
ют S. nitida (Dillw.) Link., а с шириной 90—114 мкм — 5. setiformis
(Roth) Kiitz.
S. orthospira Nag.: Famintzin, 1866 : 4; 1868 : 31, tab. 1—3; Алек-
сенко, 1891 : 74, с авторством «Kiitz.». — Сомнительное определение
из-за неполного описания (Kiitzing, 1855 : Taf. 26, Fig. 1).
S. pseudodecimina O. et W. Bock, 1956 : 436, Taf. 10, Fig. 6. —
Неполно описанный вид из-за отсутствия числа хлоропластов.
322
S. pulchra Alexenko: Алексенко, 1891 : 74, табл. 3, рис. 4. — Не-
полно описанный вид, близкий к S. sphaerospora Hirn. Неясен тип
конъюгации, спорен вопрос о форме поперечной перегородки и числе
хлоропластов (по описанию 5—7, по рисунку 1), отсутствуют данные
о форме и строении зигоспор.
S. quinina (Kiitz.) Kirchn.: Триполитова, 1928 : 271. — Неполное опи-
сание (Kirchner, 1878 : 118). Non S. quinina Kiitz. (Kiitzing, 1845 : 223).
S. quinina Kiitz. [var.] inaequalis (sine auct.): Козловский, 1888 :
427. — Неполно описанный таксон (разновидность ?) из-за отсутствия
зигоспор (Kiitzing, 1849 : 440).
S. subaequa Kiitz.: Алексенко, 1888 : 212; 1894 : 96. — Сомни-
тельное определение из-за отсутствия данных о форме поперечной пе-
регородки, типе и особенностях конъюгации, строении зигоспор.
S. varians (Hass.) Kiitz. (incl. Rhynchonema woodsii Kiitz., R. abbre-
viatum Kiitz.): Gutwifiski, 1892 : 26. — Сомнительное определение из-
за отсутствия данных о форме поперечной перегородки и зигоспор,
типе конъюгации. По ширине вегетативных клеток более отвечает ди-
агнозу S. varians (Hass.) Kiitz. f. woodsii (Hass.) V. Poljansk. Это пред-
положение подкрепляется включением Rhynchonema woodsii в число
синонимов.
S. warwarae Uspensky (nom. nudum): Успенский, 1934 : 441; 1935 :
79; 1963 : 172; Любимов, 1935 : 260.
S. weberi var. intermedia Kiitz.: Gutwifiski, 1884 : 138. — Неполно
описанная разновидность из-за отсутствия зигоспор, ошибочное автор-
ство. Узаконено: S. weberi var. intermedia Kirchner, 1878 : 120.
Conferva lubrica Roth: Weinmann, 1837 : 153, с авторством «Dillw.». —
Синоним Spirogyra lubrica (Roth) Kiitz. — неполно описанный вид
(Roth, 1797 : Tab. 2, Fig. 2; Kiitzing, 1845 : 223).
Род SIROGONIUM Kutz. — СИРОГОНИУМ
=Spirogyra Link, pr. p.
Нити неразветвленные, свободноплавающие. Вегетативные клетки
до 108 мкм шир., по длине в 2—9 раз больше. Поперечные стенки
клеток гладкие. Продольные стенки клеток довольно толстые, целлю-
лозные, без пектинового слоя, неослизненные. Хлоропласты пристен-
ные, лентовидные, довольно узкие, почти параллельные продольной
оси клетки или слегка извитые, 2—10 в каждой клетке. Ядро в центре
клетки, на плазматических тяжах, соединяющихся с постенным слоем
протоплазмы. Конъюгация лестничная. Конъюгирующие клетки дово-
льно сильно коленчато изогнутые. Формирование конъюгирующей (га-
метангиальной) клетки происходит из прогаметангиальной (вегетатив-
ной) клетки, которая перед конъюгацией делится на две неравные
части — длинную и короткую клетки. При этом одна из них, чаще
короткая, получает ббльшую часть хлоропластов от другой клетки и
становится собственно гаметангиальной; другая, опустевшая, становится
стерильной. Иногда наблюдающееся двойное деление прогаметангиаль-
323
ной клетки в мужской нити приводит к образованию 3 клеток —
одной гаметангиальной и двух стерильных. Гамета образуется из всего
содержимого гаметангиальной и большей части стерильной клетки.
Конъюгационный канал неотчетливый или отсутствует; редко канал
очень широкий и короткий, место соприкосновения коленчато согну-
тых гаметангиальных клеток часто окружено слизистым пектиновым
веществом в форме муфты. Воспринимающие клетки обычно крупнее
отдающих, слегка вздутые. Отдающие клетки короткоцилиндрические,
невздутые. Формирование зиготы в полости одного из гаметангиев —
в воспринимающей клетке. Зигоспоры преимущественно эллипсоидные,
редко цилиндрически-эллипсоидные или линзовидные. Экзоспорий тон-
кий, бесцветный, гладкий. Мезоспорий толстый, от желтого до темно-
коричневого, одно- или двухслойный, гладкий или разнообразно скуль-
птурированный. Эндоспорий тонкий, бесцветный, гладкий, обычно
трудноразличимый.
По форме хлоропластов род Sirogonium часто идентифицировался
с родом Spirogyra, в некоторых случаях ему придавался то ранг под-
рода, то ранг секции этого рода. По принятой мной системе Sirogo-
nium — устоявшийся, четко очерченный род, который хорошо отли-
чается от других родов зигнемовых водорослей отсутствием пектино-
вого покрова у вегетативных клеток, почти прямыми лентовидными
хлоропластами, отсутствием или плохо выраженным конъюгационным
каналом и усложненным процессом образования собственно гаметан-
гиальных клеток. В роде 17 видов, распространенных преимущественно
в Азии (Индия, Китай, Мьянма, Шри-Ланка), Австралии, Южн. Африке
и Америке (Канада, США, Бразилия). В России, как и во всей Европе,
род Sirogonium представлен одним видом.
Sirogonium sticticum (Engl. Bot.) Kiitz. — Сирогониум испещрен-
ный (рис. 142, 6). — Вегетативные клетки (29) 32—56 (60) мкм шир.,
по длине в 2—9 раз больше. Хлоропластов 2—5 (6), слегка извитые
или почти прямые, редко с одним неполным оборотом по спирали.
Конъюгационный канал не выражен, нередко он очень широкий и
короткий, неявственно отграниченный. Конъюгирующие клетки значи-
тельно изогнутые. Воспринимающие клетки вздутые до 84 мкм. Зиго-
споры широкоэллипсоидные, иногда почковидные или бобовидные,
41—75 (90) мкм шир., 66—119 (128) мкм дл. Экзоспорий тонкий, от
желтого до желто-коричневого, гладкий.
F. sticticum [=Spirogyra stictica (Engl. Bot.) Wille; Sirogonium me-
gasporum (Jao) Trans.] (рис. 142, 6). — Вегетативные клетки (38) 40—
56 (60) мкм шир. Хлоропластов (2) 3—5 (6). Зигоспоры 42—75
(90) мкм шир., 66—100 (127) мкм дл.
В стоячих и текучих водах, водохранилищах, прудах, реках, в мор-
ской воде при значительном опреснении (0.12—0.98 %о). Россия: севе-
ро-запад, центр и юг европ. части, устье Волги, зап. и сев. побережье
Каспийского моря, Зап. Сибирь (окрестности Томска); Латвия, Украина.
Самый распространенный представитель рода. В Европе найден в
Дании, Нидерландах, Германии, Польше и Чехии. Характеризуется зна-
324
читальным полиморфизмом в отношении ширины вегетативных клеток
и размеров зигоспор. Узнается без труда.
F. tenuius (Trans.) Rund. [-Sirogonium tenuius (Nordst.) Trans.; Spi-
rogyra tenuior (Nordst.) Krieger; S. stictica (Engl. Bot.) Wille sensu
Gutwinski, 1892; S. stictica f. tenuior (Trans.) V. Poljansk.]. — Вегета-
тивные клетки (29) 32—38 (40) мкм шир. Хлоропластов 2 (3). Зи-
госпоры 48—62 мкм шир., 60—88 мкм дл.
В стоячих и текучих водах, старицах рек. Зап. Украина (окрест-
ности Снятына), Зап. Казахстан (Уральская обл., р. Чаган).
Довольно редкая форма с широким ареалом, встреченная также в
Сев. и Южн. Америке (США, Аргентина, Бразилия) и Мьянме. Впер-
вые приводится для Аргентины Нордштедтом (Nordstedt, 1882 : 47),
отметившим меньшую, чем у Spirogyra stictica, ширину вегетативных
клеток, но не выделившим ее в самостоятельную таксономическую еди-
ницу. В качестве формы Sirogonium sticticum указывается Борге (Borge,
1903 : 262) для Бразилии, но также без имени. Последнее было пред-
ложено Трансо (Transeau, 1934а : 420), однако придача ей этим ав-
тором ранга самостоятельного вида едва ли обоснована, поскольку от-
личия от Spirogyra stictica сводятся в сущности к несколько меньшим
размерам. Поэтому, следуя Полянскому (1966 : 92), я считаю эту во-
доросль формой, но не формой Spirogyra stictica, как он считал, а
формой Sirogonium sticticum. От типовой формы' отличается меньшей
шириной вегетативных клеток и меньшим числом хлоропластов.
РЕДКИЙ И НЕДОСТАТОЧНО ИЗУЧЕННЫЙ РОД
СЕМ. SPIROGYRACEAE
Род Temnogyra Lewis (1925) насчитывает 15 видов. Типовой вид:
Т. collinsii Lewis. Отличается от рода Spirogyra только в стадии конъ-
югации — специфическим делением вегетативной (прогаметангиаль-
ной) клетки на две неравные части. При этом короткая клетка получает
от длинной клетки большую часть хлоропласта и становится собствен-
но гаметангиальной, а длинная клетка — стерильной, почти пустой.
Однако наблюдения Скуй (Skuja, 1928 : 112) показывают, что в не-
которых случаях подобное деление вегетативных клеток перед конъ-
югацией у Spirogyra (Temnogyra) punctata отсутствует и генеративные
клетки не чередуются со стерильными. Такие же наблюдения были
сделаны Цедеркрейтцем (Cedercreutz, 1935 : 131, Fig. 1). Таким обра-
зом, вопрос о родовом значении указанного признака приходится ос-
тавить открытым до дальнейших исследований. От родов Sirogonium
и Sirocladium, также имеющих подобный тип деления вегетативных
клеток перед конъюгацией, отличается наличием отчетливого конъюга-
ционного канала. Большинство фикологов, кроме Фрича (Fritsch, 1935)
и Ямагиси (Yamagishi, 1963а, 1966b), считают род Temnogyra синони-
мом рода Spirogyra. В данной работе я придерживаюсь этой точки
зрения.
ЛИТЕРАТУРА
Абибуллаева Б. Данные о водорослях скважин и прискважинных водоемов
Жанадарьинского массива Каракалпакии // Вести. Каракалпак, фил. АН УзССР.
1968. № 3 (33). С. 81.
Алексенко М. А. Очерк водорослей Chlorosporeae (Chlorophyllophyceae) ок-
рестностей г. Харькова // Тр. О-ва испыт. природы Харьков, ун-та. (1887—1888).
1888. Т. 21. С. 141—278.
Алексенко М. А. Материалы для альгологии Полтавской губернии. I. Phycoc-
hromaceae и Chlorosporeae окрестностей г. Полтавы и отчасти Полтавского
уезда // Тр. О-ва испыт. природы Харьков, ун-та. (1890—1891). 1891. Т. 25.
С. 47—88. (Паг. по отт.: с. 3—44).
Алексенко М. А. Флора водорослей Днепровских плавней и торфяников в
пределах Полтавской губернии // Тр. О-ва испыт. природы Харьков, ун-та.
(1892—1893). 1893. Т. 27. С. 59—123. (Паг. по отт.: с. 3—62).
Алексенко М. А. К флоре водорослей Лебединского и отчасти Сумского уез-
дов Харьковской губернии И Тр. О-ва испыт. природы Харьков, ун-та. (1893—
1894). 1894. Т. 28. С. 83—132.
Борцов И. Г. Материалы для ботанической географии Арало-Каспийского
края // Зап. Имп. АН. 1865. Т. 7, прил. № 1. С. 1—190.
Борцов И. Г. Материалы для флоры водорослей Черниговской губернии //
Зап. Киев, о-ва естествоиспыт. 1870. Т. 1, № 1. С. 44—55.
Водеиичаров Д. Г. К вопросу о систематике и номенклатуре сцеплянок // Но-
вости систематики низших растений. Л., 1972. Т. 9. С. 24—26.
Воденичаров Д., Киряков И. Нови видове Mougeotia и Spirogyra от южна
България // Изв. природонауч. музей-Пловдив. 1970. Т. 1. С. 19—30.
Воденичаров Д. Г., Киряков И. К. Нов вид от род Spirogyra Link, секция
Conjugata (Zygnemales) И Фитология. 1986. Т. 31. С. 3—13.
Водоросли. Указатель к «Библиографии советской литературы по водоро-
слям. 1961—1970». Л., 1983. 460 с.
Волкова Т. Ф. Новые находки зигнемовых водорослей // Реф. информ, о за-
конч. н.-и. работах в вузах УССР. Биология. 1970. Вып. 4. С. 41—42.
Волкова Т. Ф., Данилова Л. €. Матер1али до вивчення зигнемових водоростей
Кигвського водосховища И Укр. ботан. журн. 1970. Т. 27, № 5. С. 557—562.
Воронихин Н. Н. К флоре Phycomycetes Кавказа // Вести. Тифлис, ботан.
сада. 1920. Т. 50. С. 8—14.
Воронихин Н. Н. Материалы для флоры пресноводных водорослей Кавказа.
VIII. Zygnemales // Работы Сев.-Кавказ. гидробиол. станции. 1926. Т. 1, № 2.
С. 54—58.
Воронихин Н. Н. Материалы к флоре грибов Кавказа // Тр. Ботан. музея АН
СССР. 1927. Вып. 21. С. 87—253.
326
(Воронихин Н. Н. ) Voronyihin N. N. A florisztikai kutatasok elvei a kontinentdlis
vizek algologiaja teren // A botanika problemai. Budapest, 1953. P. 200—225.
Воронихин H. H. Флора водорослей континентальных водоемов европейского
Севера СССР. Сем. Zygnemataceae И Тр. Ботан. ин-та АН СССР. Сер. 2. 1954.
Т. 9. С. 5—104.
Воронихин Н. Н., Красноперова Л. А. Зигнемовые водоросли Боровского за-
поведника (Кокчетавская область) // Новости систематики низших растений. Л.,
1970. Т. 7. С. 46—63.
Голлербах М. М., Красавина Л. К. Водоросли. Сводный указатель к отечес-
твенным библиографиям по водорослям за 1737—1960 гг. Л., 1971. 623 с.
Голлербах М. М., Полянский В. И. Общая часть // Определитель пресновод-
ных водорослей СССР. Вып. 1. М., 1951. С. 1—199.
Демин Д. 3. Материалы по планктону р. Самур // Тр. Дагест. с.-х. ин-та.
1955. Т. 6. С. 181—190.
Еленкин А. А. Синезеленые водоросли СССР. Монография пресноводных и
наземных Cyanophyceae, обнаруженных в пределах СССР. Общая часть. М.; Л.,
1936. 679 с.
Змеев Л. Несколько данных для изучения низших водорослей в Кавказских
минеральных водах И Военно.-мед. журн. 1872. Ч. 114. С. 1—62.
Иванов Л. А. Краткий отчет о деятельности Бологовской биологической
станции за лето 1898 г. // Тр. СПб. о-ва естествоиспыт. 1898. Т. 29, протокол
№ 5. С. 174—177.
Иванов Л. А. Материалы для флоры водорослей (excl. Diatomaceae) Москов-
ской губ. // Bull. Soc. Natur. Moscou. (1898). 1899. T. 4. P. 350—393.
Иванов Л. А. Наблюдения над водной растительностью озерной области //
Тр. Пресноводн. биол. станции СПб. о-ва естествоиспыт. 1901. Т. 1. С. 1—152.
(Казановский В.) Kasanowsky V. Die Chlorophyllbander und Verzweigung der-'
selben bei Spirogyra Nawaschini (sp. nov.) // Ber. Dt. Bot. Ges. 1913a. Bd 31, N 1.
S. 55—59.
(Казановский В., Смирнов C.) Kasanowsky V., Smirnoff S. Spirogyra borysthe-
nica nov. spec. // Osterr. Bot. Ztschr. 19136. Bd 62, N 4. S. 137—141.
Казановский В., Смирнов С. Материалы к флоре водорослей окрестностей
Киева. I. Spirogyra И Тр. Днепр, биол. станции. 1914. № 4. С. 133—168. (Паг.
по отт.: с. 1—-36).
Козловский В. С. Материалы для флоры водорослей Сибири. I—II // Зап.
Киев, о-ва естествоиспыт. 1888. Т. 9, № 1—2. С. 395—437; 1890. Т. 11, № 1.
С. 1—36.
Комаренко Л. Е. Характеристика флоры водорослей и зоопланктона водо-
емов бассейна среднего течения р. Лены // Тр. Ин-та биологии Якут. фил. СО
АН СССР. 1956. Вып. 2. С. 145—212.
Косинская Е. К. Конъюгаты, или сцеплянки’(1). Мезотениевые и гонатози-
говые водоросли // Флора споровых растений СССР. М.; Л., 1952. Т. 2. С. 1—•
163.
Косинская Е. К Конъюгаты, или сцеплянки (2). Десмидиевые водоросли //
Флора споровых растений СССР. М.; Л., 1960. Т. 5, № 1. С. 1—706.
Красноперова Л. А. О новой форме Spirogyra subsalsa Kiitz. // Новости сис-
тематики низших растений. Л., 1966. Т. 3. С. 94—96.
Красноперова Л. А. Представители рода Spirogyra в Северном Каспии // Но-
вости систематики низших растений. Л., 1967. Т. 4. С. 82—86.
Красноперова Л. А. Представители рода Spirogyra Link в Северном Каспии.
II // Новости систематики низших растений. Л., 1968. Т. 5. С. 67—73.
Красноперова Л. А. Новые, редкие и интересные для СССР зигнемовые во-
доросли. I. Zygnema bohemicum Czurda и Mougeotia drouetii Transeau // Новости
систематики низших растений. Л., 1970. Т. 7. С. 63—67.
327
Красноперова Л. А. Зигнемовые водоросли Казахстана: Автореф. дис. ...
канд. биол. наук. Л., 1971а. 23 с.
Красноперова Л. А. Новые, редкие и интересные для СССР зигнемовые во-
доросли. И. Spirogyra dentireticulata Jao, S. punctulata Jao, S. sinensis Li // Новос-
ти систематики низших растений. Л., 19716. Т. 8. С. 79—84.
Лобик. А. И. Список пресноводных водорослей, собранных А. А. Еленкиным
и В. П. Савичем на Черноморском побережье Кавказа в 1912 году // Изв. СПб.
ботан. сада. 1915. Т. 15, № 1. С. 23—46.
Любимов В. И. Заметки о роли нитчатых водорослей в самоочистке водоема
при их массовом развитии // Микробиология. 1935. Т. 4, № 2. С. 258—261.
{Матвиенко А. М.) Матв1енко О. М. Про знаходження пульсуючих вакуоль
у cniporipH // Тр. НД1 ботаники Харьк1в. ун-ту. 1941. Т. 4. С. 49—51. (Учен. зап.
Харкав, ун-ту; Кн. 22).
Музафаров А. М., Эргашев А. Э. Водоросли прудов Средней Азии // Флора
и значение споровых растений Средней Азии. Ташкент, 1972. С. 3—20.
Обухова В. М. Водоросли рисовых полей Талды-Курганской и Кзыл-Ордин-
ской областей // Сб. работ по ихтиологии и гидробиологии Ин-та зоологии АН
КазССР. 1959. Вып. 2. С. 323—361.
Обухова В. М. Альгофлора рисовых полей некоторых районов Казахстана //
Тр. Ин-та ботаники АН КазССР. 1961. Т. 10. С. 85—187.
Паламарь-Мордвинцева Г. М. Десмидиевые водоросли Украинской ССР.
(Морфология, систематика, филогения, пути эволюции, флора и географическое
распространение). Киев, 1982а. 238 с.
Паламарь-Мордвинцева Г. М. Зеленые водоросли. Конъюгаты. Порядок Дес-
мидиевые (Chlorophyta: Conjugatophyceae, Desmidiales) (2) И Определитель пре-
сноводных водорослей СССР. Л., 19826. Вып. 11. С. 1—620.
{Паламарь-Мордвинцева Г. М.) Паламар-Мордвинцева Г. М. Кон’югати. Ч. 1.
Мезотешевц Гонатозиговц Десмщцев! // Визначник пркноводних водоростей
Украшсько! РСР. Кшв, 1984. Вип. 8. С. 1—511.
{Паламарь-Мордвинцева Г. М.) Паламар-Мордвинцева Г. М. Кон’югати. Ч. 2.
Десмщцев! // Визначник пргсноводних водоростей Укра!нсько1 РСР. Ки1'в, 1986.
Вип. 8. С. 1—319.
Полянский В. И. Новые и интересные виды рода Spirogyra Link // Ботан. ма-
териалы Отд. спор. раст. Ботан. ин-та АН СССР. Л., 1951. Т. 7. С. 84—89.
Полянский В. И. О значении приспособительных признаков в систематике //
Ботан. журн. 1954. Т. 39, № 5. С. 694—705.
Полянский В. И. Новые для флоры СССР виды рода Spirogyra Link // Ботан.
материалы Отд. спор. раст. Ботан. ин-та АН СССР. Л., 1955. Т. 10. С. 125—134.
Полянский В. И. О виде у низших водорослей. Доложено на девятом ежегод-
ном Комаровском чтении 15 декабря 1954 г. М.; Л., 1956. 74 с. (Комаровские
чтения. 9).
Полянский В. И. Понятие о виде в альгологии И Проблема вида в ботанике.
Т. 1. М.; Л., 1958. С. 7—33.
Полянский В. И. Новые виды и формы рода Spirogyra Link И Ботан. матери-
алы Отд. спор. раст. Ботан. ин-та АН СССР. Л., 1959. Т. 12. С. 138—143.
Полянский В. И. Номенклатурные заметки по роду Spirogyra Link // Новости
систематики низших растений. Л., 1966. Т. 3. С. 85—94.
Рундина Л. А. О филогении зигнемовых водорослей И Материалы V Москов.
совещ. по филогении растений. М., 1976. С. 147—148.
Рундина Л. А. Новые, редкие и интересные для СССР зигнемовые водоросли.
III. Представители родов Zygnema, Zygnemopsis и Mougeotia из Казахстана // Но-
вости систематики низших растений. Л., 1977а. Т. 14. С. 46—63.
Рундина Л. А. Новые таксоны зигнемовых водорослей из Казахстана // Но-
вости систематики низших растений. Л., 19776. Т. 14. С. 42—46.
328
Рундина Л. А. Новые, редкие и интересные зигнемовые водоросли СССР.
IV. Новые для флоры СССР представители рода Spirogyra Link И Новости сис-
тематики низших растений. Л., 1978а. Т. 15. С. 39—47.
Рундина Л. А. Новые, редкие и интересные зигнемовые водоросли СССР.
V. Новые для флоры Казахстана представители рода Spirogyra Link И Новости
систематики низших растений. Л., 19786. Т. 15. С. 47—62.
Рундина Л. А. Принципы видовой и внутривидовой систематики зигнемовых
водорослей И Тез. докл. VI Делегат, съезда Всесоюз. ботан. о-ва (Кишинев, 12—
17 сент. 1978 г.). Л., 1978в. С. 334—335.
Рундина Л. A. Spirogyra pseudogranulata Ley — новый и S. kuusamoensis f.
skujaensis V. Poljansk. — редкий виды для флоры СССР // Новости систематики
низших растений. Л., 1981. Т. 18. С. 25—31.
Рундина Л. А. К вопросу о внутриродовой систематике зигнемовых водоро-
слей // Тез. докл. VII конф. спор. раст. Средн. Азии и Казахстана (Алма-Ата,
11—14 сент. 1984 г.). Алма-Ата, 1984. С. 271—272.
{Рундина Л. А.) Рундша Л. О. Кон’югати. Ч. 3. Зигнемов! — Zygnematales //
Визначник пркноводних водоростей Укра'Гнсько! РСР. Ки1'в, 1988. Вип. 8.
С. 1—202.
Рундина Л. А. Таксономические изменения у некоторых видов порядка Zyg-
nematales (Chlorophyta) И Ботан. журн. 1991. Т. 76, № 2. С. 294—295.
{Рундина Л. A.) Rundina L. A. Disputed topics in the taxonomy of Zygnemata-
les // Альгология. 1992. T. 2, № 4. C. 63—66.
Рундина Л. A. Zygnematales Северной Осетии // Альгология. 1993. T. 3, № 1.
С. 80—94.
Рундина Л. А. Новый вид Mougeotia rimulosa (Zygnematales, Chlorophyta) //
Ботан. журн. 1995. Т. 80, № 5. С. 89—93.
Рундина Л. А., Степняк О. Э. Первая находка в СССР Spirogyra scrobiculata
(Stockm.) Czurda // Новости систематики низших растений. Л., 1979. Т. 16. С. 26—
28.
Рябинин Д. Б. Флора водорослей окрестностей города Чугуева // Тр. О-ва
испыт. природы Харьков, ун-та. 1888. Т. 22. С. 33—63.
{Рябинин Д. Б.) Riabinine D. В. Les Chlorophycees des environs de Kharkow //
Bull. Soc. Natur. Moscou. (N. S.). (1888). 1889. T. 2. P. 289—348.
Сатина С. А. Список водорослей озера Селигер и его окрестностей //
Тр. Пресноводн. биол. станции СПб. о-ва естествоиспыт. 1912. Т. 3. С. 135—
145.
{Свиренко Д. О.) Свиренко Д. О. Альголопчне достижения щкавого купиння
коло Дншропетровська // 36. праць Дншропетр. 6ion. станцй. 1927. Ч. 2.
С. 429—473. (Тр. Ф1з.-мат. вщдшу АН УРСР; Т. 3, № 7).
{Свиренко Д. О.) Swirenko D. О. Die botanischen Ergebnisse der Siid-Bugischen
hydrobiologischen Expedition // Arch. Hydrobiol. 1941. Bd 6, № 4. S. 593—770.
Седова T. В. Кариология водорослей. СПб., 1996. 368 с.
Скворцов Б. В. Материалы по флоре водорослей Азиатской России. I. Водо-
росли из Якутской области. II. Водоросли из Закаспийской области. III. О фи-
топланктоне озера Чля Приамурской области. IV. Водоросли верховьев р. Зеи
Амурской области. V. Водоросли из Акмолинской области. VI. О фитопланкто-
не оз. Марка-куль Киргизского края // Журн. Рус. ботан. о-ва. 1917. № 2.
С. 10—20, 117—128.
Скворцов Б. В. Наблюдения над жизнью нитчатых Zygnemataceae в окрест-
ностях г. Харбина, в Северной Маньчжурии // Тр. Сунгарийск. речи. биол. ст.
Сер. В. 1927а. Т. 1, № 4. С. 1—24.
{Скворцов Б. В.) Skvortzow В. W. Freshwater algae and phytoplankton of the
lakes and rivers of the Zaisan district, Altai mountains, Siberia // J. Bot. (London).
19276. Vol. 65. P. 249—254.
329
Скворцов Б. В. Новые и мало известные виды Algae, Flagellatae, Phycomice-
tae из Азии, Америки, Африки, а также с островов Японии и Цейлона, описан-
ные в 1931—45 гг., с 18 таблицами рисунков // Зап. Харбин, о-ва естествоиспыт.
и этногр. Ботаника. 1946. № 2. С. 1—34.
(Топачевский А. В.) Топачевський О. В. Походження кон’югат i типи стате-
вого процесу у них // Ботан. журн. АН УРСР. 1952. Т. 9, № 2. С. 32—35.
Топачевский А. В. Критика жгутиковой Теории происхождения водорослей //
Тр. биол.-почв. фак. Киев, ун-та. 1953. № 9. С. 115—126.
(Топачевский А. В.) Топачевський О. В. До питания про виникнення i типи
статевого процесу у водоростей // Тр. б!ол.-грунтозн. фак. Кшв. ун-ту. 1954.
№ 11. С. 65—73.
Триполитова Т. К. Материалы к флоре споровых растений Алтая и Томской
губернии. II. Водоросли И Изв. Томск, ун-та. 1928. Т. 79, № 4. С. 271—325.
Троицкая О. В. Об особенностях полового процесса у Spirogyra maxima
(Hass.) Wittr. // Изв. Гл. ботан. сада. 1930. Т. 29, № 3—4. С. 330—338.
Успенский Е. Е. К вопросу о задачах и путях микробиологии в связи с раз-
витием городского водоснабжения и в особенности при строительстве водохра-
нилищ // Микробиология. 1932. Т. 1, № 2. С. 89—111.
Успенский Е. Е. Превращение одних наследственных форм в другие и возни-
кновение новых в роде Spirogyra [1—2] // Микробиология. 1934. Т. 3, № 2.
С. 169—203; № 4. С. 437—469.
(Успенский Е. Е.) Uspensky Е. Е. Die Umwandlung einiger erblicher Formen in
andere und die Entstehung neuer Formen im Genus Spirogyra // Probleme der theo-
retischen Biologie. M.; L., 1935. S. 17—119.
Успенский E. E. Физико-химические условия среды как основа микробиоло-
гических процессов. М., 1963. 259 с.
Федорова В. А., Рундина Л. А., Любомирова К. А., Станичникова М. С.
Морфотипы зигоспор современных конъюгат и их возможные ископаемые ана-
логи // Фитостратиграфия и морфология спор древних растений нефтегазонос-
ных провинций СССР. Л., 1989. С. 93—104.
Хмелевский В. Ф. Материалы для флоры водорослей Бессарабской губер-
нии И Зап. Новорос. о-ва естествоиспыт. 1885. Т. 10, № 1. С. 71—81.
Хмелевский В. Ф. Материалы к морфологии и физиологии полового процес-
са у низших растений И Тр. О-ва испыт. природы Харьков, ун-та. (1890—1891).
1891. Т. 25. С. 89—168.
Христюк П. М. Основные черты флоры водорослей пресных вод Крыма:
Дис. ... д-ра биол. наук. Симферополь, 1947. 1222 с.
Шелль Ю. К. Материалы для ботанической географии Уфимской и Оренбур-
гской губерний. (Споровые растения) // Тр. О-ва естествоиспыт. Казанск. ун-та.
1883. Т. 12, № 1. С. 3—93.
Ширшов П. П. Зампка про мщрофлору р. ВовчоТ. Доплив р. Самари // BicH.
Дшпропетр. пдробюл. станцп. 1937. Т. 2. С. 167—182. (Наук. зап. Дншропетр.
ун-ту; Т. 2).
Эргашев А. Э. Действие минеральных удобрений на водоросли в прудах //
Культивирование водорослей и высших водных растений в Узбекистане. Таш-
кент, 1971. С. 77—82.
Эргашев А. Э. Альгофлора искусственных водоемов Средней Азии. Ташкент,
1974. 251 с.
Эргашев А. Э. Закономерности развития и распределения альгофлоры в ис-
кусственных водоемах Средней Азии. Ташкент, 1976. 359 с.
Aboal М. New records of Zygnemataceae (Zygnematales, Chlorophyceae) from
southeastern Spain // Nova Hedwigia. 1991. Vol. 52, N 3—4. P. 365—373.
330
Aboal M., Llimona X. Aportaci6n al estudio algo!6gico del sistema de Sierras de
Ponce у Quipar (N. O. de Murcia, S. E. de Espana) // Anal. Biol. 2. (Sec. Espec. 2).
1984. P. 1—17.
Agardh C. A. Synopsis algarum Scandinaviae, adjecta dispositione universali al-
garum. Lund, 1817. 135 p.
Agardh C. A. Systema Algarum. Vol. 1. Lund, 1824. 312 p.
Ahmad A., Goldstein M. E. Lloydiella, a new genus of the Zygnemataceae // Phy-
cologia. 1971. Vol. 10, N 1. P. 1—6.
Ahmad A., Goldstein M. E. Lloydina, a new generic name for later homonym
Lloydiella Ahmad and Goldstein (Zygnemaceae) // Phycologia. 1972. Vol. 11, N 2.
P. 217.
Aleem A. A. Sur I’autoecologie d’une Spirogyre d’eau Sumatra // C. r. Acad. sci.
1952. T. 234, N 27. P. 2648—2650.
Aleem A. A. Effect of salinity on the growth and reproduction of a brackish water
Spirogyra (S. salina nov. sp.) // Vie et Milieu. 1961. T. 12, N 3. P. 497—506.
Artari A. P. Liste des algues observees dans le gouvernement de Moscou // Bull.
Soc. Natur. Moscou. 1885. T. 3. P. 124—144. (Паг. no on.: с. 1—23).
Artari A. P. Materiaux pour servir a I’etude des algues du gouvernement de
Moscou // Bull. Soc. Natur. Moscou. 1886. T. 3. P. 165—184.
Beck-Mannagetta G. Neue Griinalgen aus Karnten // Arch. Protistenk. 1926. Bd 55.
S. 173—184.
Beck-Mannagetta G. Neue Grim- und Blaualgen aus Karnten und den Sudeten //
Arch. Protistenk. 1929. Bd 66, N 1. S. 1—10.
Beck-Mannagetta G. Die Algen Karntens. Erste Grunlagen einer Algenflora von
Karnten // Beih. Bot. Centralbl. 1931. Bd 47, N 2. S. 211—342.
Berger H. Chlorophyta // A. Engler. Syllabus der Pflanzenfamilien. 12 Aufl. Ber-
lin, 1954. Bd 1, N 8. S. 86—107.
Blackman F. F„ Tansley A. G. A revision of the classification of the green algae.
London, 1902. 64 p.
Bock O., Bock W. Zygnemales aus Unterfranken // Arch. Hydrobiol. 1956.
Bd 52, N 3. S. 406—448.
Borge O. F. Die Algen der ersten Regnellschen Expedition. III. Zygnemaceen
und Mesocarpaceen // Ark. Bot. 1903. Bd 1. S. 177—285.
Borge O. F. Beitrage zur Algenflora von Schweden // Ark. Bot. 1907. Bd 6, N 1.
S. 1—88.
Borge O. F. Zygnemales. Spezieller Teil // A. Pascher. Siisswasser-Flora Deut-
schlands, Osterreichs und der Schweiz. H. 9. Jena, 1913. 51 S.
Bourrelly P. Les algues d’eau douce. T. 1. Les algues vertes. Paris, 1966. 511 p.
Cedercreutz C. Die Zygnemaceen Finnlands // Mem. Soc. Fauna et Flora Fenni-
ca, (1934—1935). 1935. Bd 11. S. 122—132.
Cedercreutz C. Algen aus Kuusamo // Mem. Soc. Fauna et Flora Fennica,
(1935—1936). 1936. Bd 12. S. 46—52.
Cleve P. Th. Forsok till en monografi ofver de Svenska Arterna af algfamiljen
Zygnemaceae // Nova Acta Reg. Soc. Sci. Upsal. Ser. 3. 1868. Bd 6. S. 1—38.
Cooke M. E. British freshwater algae, exclusive of Desmidieae and Diatomaceae.
London, 1882—1884. Vol. 1—2. 329 p.
Czurda V. Experimentelle Untersuchungen uber die Sexualitatsverhaltnisse der
Zygnemalen // Beih. Bot. Centralbl. 1930. Bd 47, N 1. S. 15—68.
Czurda V. Zur Morphologie und Systematik der Zygnemalen // Beih. Bot. Cen-
tralbl. 1931. Bd 48, N 2. S. 238—285.
Czurda V. Zygnemales // A. Pascher. Siisswasser-Flora Mitteleuropas. H. 9.
2 Aufl. Jena, 1932. 232 S.
De Bary A. Untersuchungen uber die Familie der Conjugaten (Zygnemeen und
Desmidieen). Leipzig, 1858. 91 S.
331
De Toni J. B. Sylloge algarum omnium hucusque cognitarum, I. Chlorophyceae.
Padua, 1889. 1315 p.
Famintzin A. S. Die Wirkung des Kerosin-Lampenlichtes auf Spirogyra orthospi-
ra Naeg. // Bull. Acad. St. Petersb. (1865). 1866. Bd 10. S. 4—14.
Famintzin A. S. Die Wirkung des Lichtes auf die Algen und einige andere ihnen
nahe verwandte Organismen // Pringsh. Jahrb. Wiss. Bot. (1867). 1868. Bd 6. S. 1—
49. \
Fott B. Algenkunde. Jena, 1971. 581 S.
Fowke L. C.; Pickett-Heaps J. D. Cell division in Spirogyra. I. Mitosis // J. Phy-
col. 1969. Vol. 5, N 3. P. 240—259.
Fritsch F. E, The structure and reproduction of the algae. Cambridge, 1935.
Vol. 1. 791 p.
Fritsch F. E,, Rich F. Contributions to our knowledge of the freshwater algae of
Africa. 7. Freshwater algae (exlusive of diatoms) from Griqualand West // Trans.
Roy. Soc. S. Africa. (1929). 1930. Vol. 18. P. 1—92.
Gauthier-Lievre L, Zygnemacees Africaines // Beih. Nova Hedwigia. Weinheim,
1965. H. 20. P. 1—210.
Gay F. Essai d’une monographic locale des Conjuguees // Rev. Sci. Natur.
Ser. 3. (1883—1884). 1884a. T. 3. P. 1—112.
Gay F. Note sur les Conjuguees du midi de la France // Bull. Soc. Bot. France.
1884b. T. 31. P. 331—342.
Geitler L. Sexueller Dimorphismus bei einer Konjugate (Mougeotia heterogama
ii. sp.) // Osterr. Bot. Ztschr. 1958. Bd 105, N 4. S. 301—322.
Gutwinski R. Materyjaly do flory wodorostow Galicyi // Sprawozd. Komis. Fiz.
Akad. umiej^t. w Krakowie. 1884. T. 18, N 2. S. 127—138.
Gutwinski R. Materyjaly do flory glonow Galicyi. Cz. II // Sprawozd. Komis.
Fiz. Akad. umiej^t. w Krakowie. 1890. T. 25. S. 1—43.
Gutwinski R. Flora glonow okolic Lwowa // Sprawozd. Komis. Fiz. Akad. umi-
ej?t. w Krakowie. 1892. T. 27. S. 1—124.
Gutwinski R. Materyjaly do flory glonow Galicyi. Cz. Ill // Sprawozd. Komis.
Fiz. Akad. umiej^t. w Krakowie. 1893. T. 28. S. 104—166.
Gutwinski R. Flora glonow okolic Tarnopola // Sprawozd. Komis. Fiz. Akad.
umiej^t. w Krakowie. 1895a. T. 30. S. 45—173.
Gutwinski R. Prodromus florae algarum galiciensis // Rozpr. Akad. umiej^t.
wydz. mat.-przyrodn. Ser. 2. 1895b. T. 28, N 8. S. 274—440.
Hajdu L. Neue Taxa und Kombinationen bei den Griinalgen (Chlorophyta) aus
Ungarn // Ann. Hist.-natur. Mus. Nat. Hung. 1975. Bd 67. S. 25—30.
Dallas E. Om en ny Zygnema-Art med Azygosporer // Bot. Tidsskr. 1895. Bd 20.
S. 1—16.
Hansgirg A. Prodromus der Algenflora von Bohmen. T. I, H. 1 // Arch. Natur-
wiss. Landesdurchf. Bohmen. 1886. Bd 5, Bot. Abt. S. 3—96.
Hansgirg A. Prodromus der Algenflora von Bohmen. T. I, H. 2 // Arch. Natur-
wiss. Landesdurchf. Bohmen. 1888. Bd 6, N 5. S. 3—290.
Harvey F. L. The freshwater algae of Maine. Ill // Bull. Torrey Bot. Club. 1892.
Vol. 19. P. 118—125.
Hassall A. H. Observations on the genus Mougeotia, on two new genera of fresh-
water algae, and on Tyndaridea, with descriptions of species // Ann. Mag. Natur. Hist.
1843. Vol. 12, ser. 1. P. 180—188.
Hassall A. H A history of the British freshwater algae, including descriptions of
the Desmideae and Diatomaceae. Vol. 1, 2. London, 1852. 462 p.
Hirn К. E, Die finlandischen Zygnemaceen // Acta Soc. Fauna et Flora Fennica.
1895. Bd 11, N 10. S. 3—15.
Hirn К. E. Einige Algen aus Central Asien // Ofvers. Finska Vet. Soc. Forhandl.
(1899—1900). 1900. Bd 42. S. 163—173.
332
Hirose H. General phycology. Tokyo, 1959. 506 p. (In Japanese).
Hoek C. van den. Caribbean fresh and brackish water Chlorophyta // Blumea.
1959. Vol. 9, N 2. P. 590—625.
Hoek C. van den, Mann D. G., Jahns H. M. Algae: an introduction to phycology.
Cambridge, 1995. 623 p.
Hughes E. New fresh-water Chlorophyceae from Nova Scotia // Amer. J. Bot.
1948. Vol. 35. P. 424—427.
Jao Ch.-ch. New Zygnemataceae from Woods Hole // Trans. Amer. Microsc.
Soc. 1935a. Vol. 54, N 1. P. 1—7.
Jao Ch.-ch. Studies on the freshwater algae of China. I. Zygnemataceae from
Szechwan // Sinensia. 1935b. Vol. 6, N 5. P. 551—645.
Jao Ch.-ch. New Zygnemataceae collected in China // Amer. J. Bot. 1936.
Vol. 23, N 1. P. 53—60.
Jao Ch.-ch. Studies on the freshwater algae of China. XV. Oedogoniaceae and
Zygnemataceae from Kwangsi // Bot. Bull. Acad. Sinica. 1947. Vol. 1, N 2. P. 81—
102.
Jao Ch.-ch. (ed.). Zygnemataceae // Flora algarum sinicarum aquae dulcis.
Vol. 1. Beijing, 1988. 228 p. (In Chinese).
Kadhibowska J. Z. Spirogyra silesiaca sp. n. // Fragm. Florist. Geobot. 1967.
T. 13, N 1. S. 163—164.
Kadhibowska J. Z. Structure of cell-wall of Spirogyra colligata Hodgetts (1920)
and changes of diagnosis of this species // Fragm. Florist. Geobot. 1969. T. 15, N 2.
S. 255—257.
Kadhibowska J. Z. Chlorophyta V. Conjugales. Zygnemaceae // Flora stodko-
wodna Polski. Krak6w, 1972. T. 12A. 431 s.
Kadhibowska J. Z. Conjugatophyceae I. Chlorophyta VIII. Zygnemales // Siis-
swasserflora von Mitteleuropa. Jena, 1984. Bd 16. 532 S.
Kadhibowska J. Z.t Christensen T. Some danish Zygnemataceae // Bot. Tidsskr.
1979. Vol. 74, N 2—3. P. 165—173.
Kadhibowska J. Z., Langangen A. Mougeotia Ag. and Spirogyra Link (Zygnema-
ceae) — new species for the European flora // Acta Soc. Bot. Poloniae. 1997.
Vol. 66, N 1. P. 83—86.
Kirchner O. Algen // F. Cohn. Kryptogamen-Flora von Schlesien. Breslau, 1878.
Bd 2, H. 1. 284 S.
Kniep H. Die Sexualitat der niederen Pflanzen. Jena, 1928. 544 S.
Kolkwitz R.t Krieger H. Zygnemales // L. Rabenhorst. Kryptogamen-Flora von De-
utschland, Osterreich u. d. Schweiz. Leipzig, 1941. Bd 13, Abt. 2, Lfg 1—3. S. 1—294;
1944. Lfg 4. S. 295—499.
Kutzing F. T. Phycologia generalis, Oder Anatomie, Physiologie und Systemkun-
de der Tange. Leipzig, 1843. 458 S.
Kutzing F. T. Phycologia germanica, d. i. Deutschlands Algen in biindigen Bes-
chreibungen. Nordhausen, 1845. 340 S.
Kutzing F. T. Species algarum. Leipzig, 1849. 922 S.
Kutzing F. T. Tabulae phycologicae oder Abbildungen der Tange. V. Nordhau-
sen, 1855.
Lagerheim G. Bidrag till Sveriges algflora // Oefv. Kongl. Sv. Vet,-Akad. For-
handl. 1883. Bd 40, N 2. S. 37—78.
Lakowitz K. Die Algenflora der gesamten Ostsee. Danzig, 1929. 474 S.
Langer S. Spirogyra hungarica nov. sp. // Folia Cryptogamica. 1932. Bd 1, N 9.
S. 1069—1072.
Lemmermann E. Beitrage zur Kenntnis der Planktonalgen II. Beschreibung neuer
Formen // Bot. Centralbl. 1898. Bd 76, N 5—6. S. 150—156. (Паг. по отт.: S. 1—8).
Levander К. M. Die Zusammensetzung und die biologischen Verhaltnisse des
Planktons im Obersee // Arch. Biontolog. 1908. Bd 2, N 1. S. 125—143.
333
Lewis I. F A new Conjugatae from Woods Hole // Amer. J. Bot. 1925. Vol. 12.
P. 351—357.
Li L C. New species and varieties of freshwater algae from China // Ohio J. Sci.
1933. Vol. 33. P. 151—154.
Lloyd F. E. Conjugation in Spirogyra // Trans. Roy. Canad. Inst. 1924. Vol. 15,
N 1. P. 129—134.
Mobius M. Australische Siisswasseralgen
Flora. 1892. Bd 75. S. 421-450.
Muller J. Zygnemopsis Foersteri, sp. nov. // Ber. Bayer. Bot. Ges. 1964. Bd 37.
S. 109—111.
Nipkow F Uber die Sexual- und Dauerperioden einiger Zygnemalen aus schwe-
izerischen Kleingew&ssern // Schweiz. Z. Hydrol. 1962. Bd 24, N 1. S. 1—43.
Nordstedt O. Algologiska smdsaker, III. Ueber einige Algen aus Argentinien und
Patagonien // Bot. Notiser. 1882. [N 3]. S. 46—51. (Hedwigia. 1882. T. 21. P. 181—
187).
Okada Y. A new classification of Conjugatae with special reference to desmids //
Mem. Fac. Fish. Kagoshima Univ. 1953. Vol. 3, N 1. P. 165—192.
Palla E. Ueber eine neue, pyrenoidlose Art und Gattung der Conjugaten // Ber.
Dt. Bot. Ges. 1894. Bd 12. S. 228—236.
Papenfuss G. F. Classification of the algae // A century of progress in the natural
sciences, 1853—1953. San Francisco, 1955. P. 115—224.
Petit P. Observations critiques sur les generes Spirogyra et Rhynchonema // Bull.
Soc. Bot. France. 1874. T. 21. P. 1—41.
Petit P. Les spirogyres des environs de Paris. Paris, 1880. 39 p.
Prasad B. N., Godward M. В. E, Three new species of Zygnema and redefiniti-
on of Zygnema tenue Kiitz. // Phykos. 1966. Vol. 5, N 1—2. P. 187—194.
Prescott G, W. Algae of the western Great Lakes area. Dubuque (Iowa), 1962.
977 p.
Printz H. Conjugatae // A. Engler, K. Prantl. Die natiirlichen Pflanzenfamilien.
Leipzig, 1927. Bd 3, N 2. S. 339—374.
Rabenhorst L, Kryptogamen-Flora von Sachsen, der Ober-Lausitz, Thiiringen und
Nordbohmen mit Beriicksichtigung der benachbarten Lander. Abt. 1. Leipzig, 1863.
653 S.
Rabenhorst L Algae exciccatae. Leipzig, 1868a.
Rabenhorst L. Flora europaea algarum aquae dulcis et submarinae. Sect. III. Lei-
pzig, 1868b. 461 S.
Randhawa M. S. Three new species of Zygnema from Northern India // Proc.
Indian Acad. Sci. Sect. B. 1936. Vol. 4, N 3. P. 239—246.
Randhawa M. S. Genus Zygnemopsis in Northern India // Proc. Indian Acad. Sci.
Sect. B. 1937. Vol. 5, N 6. P. 297—314.
Randhawa M. S. Observations on some Zygnemales from Nothern India, I, II //
Proc. Indian Acad. Sci. Sect. B. 1938. Vol. 8, N 3. P. 131—147; N 4. P. 336—366.
Randhawa M. S. Sirocladium, a new terrestrial member of the Zygnemales // Bot.
Gazette. 1941. N 103. P. 192—197.
Randhawa M. S, Reproduction in Zygogonium ericetorum Kiitz. // Curr. Sci.
(India). 1959a. Vol. 28, N 1. P. 24—25.
Randhawa M. S. Zygnemaceae. New Delhi, 1959b. 478 p.
Rao С. B. The Zygnemoideae of the United provinces, India — I // J. Indian
Bot. Soc. 1937. Vol. 16, N 5. P. 269—288.
Rao С. B. The Zygnemoideae of the Central provinces, India — I // J. Indian
Bot. Soc. 1938. Vol. 17, N 5—6. P. 341—353.
Rattan R. S. Zygnemaceae of N. India. I // Res. Bull. Panjab Univ. 1971.
Vol. 22, N 3—4. P. 421—428.
Rieth A. Jochalgen (Konjugaten). Zieralgen und fadige Formen. Stuttgart, 1961.
87 S.
334
Rieth A. Siisswasser-Algenarten in Einzeldarstellung. I. Uber «Plagiospermum
tenue Cleve 1868» // Kulturpflanze. 1972a. Bd 19. S. 1—35.
Rieth A. Siisswasser-Algenarten in Einzeldarstellung. VII. Uber Spirogyra mira-
bilis (Hass.) Kiitz. 1849 und ihre Zygosporenbildung // Kulturpflanze. 1972b. Bd 20.
Rieth A. Siisswasser-Algenarten in Einzeldarstellung. VI. Spirogyra gatersleben-
sis, eine neue Art der Sektion Salmacis // Kulturpflanze. 1973. Bd 21. S. 77—87.
Rieth A. Eine Form von Spirogyra fluviatilis mit bezahnten Zygotenmesospor //
Kulturpflanze. 1976. Bd 24. S. 133—142.
Rosenvinge L. K. Om Spirogyra groenlandica nov. sp. og dens Parthenospore-
dannelse // Oefvers. Kongl. Vet.-Akad. Fbrhandl. 1883. Bd 40, N 8. S. 37—43.
Rosenvinge L K. Note sur le Zygnema reticulatum E. Hallas // Rev. Algolog.
1924. T. 1. P. 209—212.
Roth A. W. Bemerkungen liber das Studium der kryptogamischen Wasserge-
wachse. Hannover, 1797. 109 p.
Schmidle W. Einige Algen aus Sumatra // Hedwigia. 1895. Bd 34. S. 293—307.
Schmidle Ж Beitrage zur Flora von Africa XXII. Schizophyceae, Conjugate,
Chlorophyceae // Engler’s Bot. Jahrb. 1901. Bd 30. S. 239—445.
Schmidle Ж Algen, insbesondere solche des Plankton, aus dem Nyassa-See und
seiner Umgebung, gesammelt von Dr. Fiilleborn // Engler’s Bot. Jahrb. (1902). 1903.
Bd 32. S. 56—88.
Schneider G. Der Obersee bei Reval // Arch. Biontolog. 1908. Вd 2, N 1. S. 1—192.
Simons J. Spirogyra en verwante draadalgen in Nederland. Utrecht, 1990. 88 S.
Skuja H. Mersraga-Ragaciema piekrastes algas // Acta Univ. Latv. 1924. Bd 10.
S. 337—392.
Skuja H. Zwei neue Zygnemaceen mit blauem Mesospor // Acta Horti Bot. Univ.
. Latv. 1926. Bd 1. S4 109—114.
Skuja H. Vorarbeiten zu einer Algenflora von Lettland. IV // Acta Horti Bot.
Univ. Latv. 1928. Bd 3, N 2—3. S. 103—218.
Skuja H. Siisswasseralgen von den westestnischen Inseln Saaremaa und Hiiu-
maa // Acta Horti Bot. Univ. Latv. 1929. Bd 4, N 1—3. S. 1—76.
Skuja H. Beitrag zur Algenflora Lettlands. I // Acta Horti Bot. Univ. Latv. 1932a.
Bd 7, N 2. S. 25—86.
Skuja H. Le genre Pleurodiscus doit-il etre maintenu? // Rev. Alg. 1932b. T. 6,
N 2. P. 137—146.
Skuja H. Siisswasseralgen aus Griechenland und Kleinasien, gesammelt von Prof.
C. Regel // Hedwigia. 1937. Bd 77. S. 15—70.
Skuja H. Zur Siisswasseralgen Flora Burmas // Nova Acta Reg. Soc. Sci. Upsal.
Ser. 4. Uppsala, 1949. Bd 14, N 5. S. 1—188.
Skuja H. Griindziige der Algenflora und Algenvegetation der Fjeldgegenden um
Abisko in Schwedisch-Lappland // Nova Acta Reg. Soc. Sci. Upsal. Ser. 4. Uppsala,
1964. Bd 18, N 3. S. 1—465.
Skvortzov В. V., Noda M. On new species of Zygnema Agardh (Zygnemaceae)
from Altai district of Siberia and N.-E. China (Manchuria) // Sci. Rep. Niigata Univ.
D. 1973. N 10. P. 37—40.
Strom К. M. Algal phylogeny. An essay in the descent of plants // Blyttia. 1944.
Vol. 2, N 1. P. 10—20.
Teodoresco E, C, Materiaux pour la Flore algologique de la Roumaine // Ann.
Sci. Nat. (N. S.). Botanique, ser. 9. 1907. T. 5, N 1—3. P. 1—155.
Tiffany L. H. The filamentous algae of the west end of lake Erie // Amer. Mid-
land Natur. 1937. Vol. 18. P. 911—951.
Tiffany L, H., Britton M. E. The algae of Illinois. Chicago, 1952. 421 p.
Transeau E. N. New species of green algae // Amer. J. Bot. 1914. Vol. 1, N 6.
P. 289—301.
335
Transeau E. N. Notes on the Zygnemales // Ohio J. Sci. 1915. Vol. 16. P. 17—
31.
Transeau E. N. The genus Debarya // Ohio J. Sci. 1925. Vol. 25. P. 193—199.
Transeau E. N. The genus Mougeotia // Ohio J. Sci. 1926. Vol. 26. P. 311—
338.
Transeau E. N. Notes on the Zygnemataceae // Ohio J. Sci. 1934a. Vol. 34.
P. 420. у
Transeau E. N. The genera of the Zygnemataceae // Trans. Amer. Microsc. Soc.
1934b. Vol. 53. P. 201—207.
Transeau E. N. Notes on Zygnemataceae // Amer. J. Bot. 1938. Vol. 25, N 7.
P. 524—528.
Transeau E. N. Notes on Zygnemataceae // Ohio J. Sci. 1944. Vol. 44, N 5.
P. 243—244.
Transeau E. N. The Zygnemataceae (freshwater conjugate algae). Columbus,
1951. 327 p.
Transeau E. N., Tiffany L. FL, Taft C. Et, Li L. C. New species of Zygnemataceae //
Trans. Amer. Microsc. Soc. 1934. Vol. 53, N 8. P. 208—230.
Wang С. C. Species of Spirogyra of Nanking // Contr. Biol. Lab. Sci. Soc.
China. Bot. Ser. (1932). 1933. Vol. 8. P. 193—218.
Weinmann J. A. Enumeratio stirpium in agro Petropolitano sponte crescentium.
Petropoli (St.-Petersburg), 1837. 320 p.
West G. S. The algae flora of Cambridgeshire // J. Bot. 1899. Vol. 37. P. 49—
299.
West G. S. A treatise on the British freshwater algae. Cambridge, 1904. 372 p.
West G. S. Algae. Vol. 1. Cambridge, 1916. 475 p.
West G. S.t Fritsch F. E. A treatise on the British freshwater algae. Cambridge,
1927. 534 p.
West W., West G. S. Welwitsch’s African freshwater algae // J. Bot. 1897. Vol. 35.
P. 1—304.
West W.t West G. S. Observations on the Conjugatae // Ann. Bot. 1898. Vol. 12.
P. 29—58.
West W.t West G. S. Freshwater algae from the Orkneys and Shetlands // Trans.
Proc. Bot. Soc. Edinburgh. 1905. Vol. 23. P. 3—41.
West W., West G. S. Freshwater algae from Burma, including a few from Bengal
and Madras // Ann. Roy. Bot. Gard. Calcutta. 1907. Vol. 6, pt 2. P. 175—260.
Wittrock V. B. Bidrag till kSnnedomen om Sveriges Zygnemaceer och Mesocar-
paceer // Bot. Notiser. 1868. N 4. S. [187]—190.
Wittrock V. B. Beitrag zur Kenntnis der Zygnemaceen und Mesocarpeen // Hed-
wigia. 1871. Bd 10. S. 88.
Wittrock V. B. Om Gotlands och Olands Sotvattens-alger // Bih. Kongl. Sv. Vet.-
Akad. Handl. 1872. Bd 1, N 1. S. 1—72.
Wittrock V. B. On the spore-formation of the Mesocarpeae and especially of the
new genus Gonatonema // Bih. Kongl. Sv. Vet.-Akad. Handl. 1878. Vol. 5, N 5.
P. 1—18.
Wittrock V. B., Nordstedt O. Algae aquae dulcis exiccatae. Index // Bot. Notiser.
1886. Fasc. 15—16. S. 130—139; 1889. Fasc. 18—20. S. 157—168.
Wolle F. Freshwater algae of the United States. Bethlehem (Pennsylvania), 1887.
364 p.
Yamagishi T Classification of the Zygnemataceae // Sci. Rep. Tokyo Kyoiku Da-
igaku. 1963a. Vol. 11. P. 191—210.
Yamagishi T. New species of the Zygnemataceae collected in Japan // Bot. Mag.
(Tokyo). 1963b. Vol. 76, N 900. P. 215—219.
Yamagishi T. Studies on the genus Mougeotia in Japan // Bot. Mag. (Tokyo).
1965a. Vol. 78, N 919. P. 28—35. (In Japanese).
336
Yamagishi T. Studies on the genus Mougeotiella in Japan // Bot. Mag. (Tokyo).
1965b. Vol. 78, N 920. P. 74—77. (In Japanese).
Yamagishi T. Studies on the genus Zygnema in Japan // Bot. Mag. (Tokyo).
1965c. Vol. 78, N 929. P. 424-^431. (In Japanese).
Yamagishi T. Genera Temnogyra, Zygnemopsis and Zygogonium in Japan // Gen.
Educ. Rev. College Agr. Vet. Med, Nihon Univ. 1966a. Vol. 2. P. 26—34. (In Ja-
panese).
Yamagishi T. Studies on the genus Spirogyra collected in Japan // Sci. Rep. Tokyo
Kyoiku Daigaku. Sect. B. 1966b. Vol. 12, N 181. P. 73—105.
22 Рундина JI. A.
УКАЗАТЕЛЬ ЛАТИНСКИХ НАЗВАНИЙ1
Ancylonema 47
Chlorophyta 44, 46
Colligata sect. 9, 21, 39, 163, 179, 318
Conferva Miill., pr. p. 101, 162
— ericetorum Kiitz. 102
— lubrica Roth 323
Conjugata sect. 163
Conjugatophyceae 44, 46
Craterospermum A. Br. 107
— laetevirens A. Br. 115
Debarya Wittr. 12, 13, 34, 40, 45, 93,
107, 157
— africana G. S. West 127
— americana Trans. 96
— calospora (Palla) West 137, 138*
— cruciata Price 93
— decussata Trans. 97, 97*
— desmidioides W. et G. S. West 93,
94*
------var. orientale Carter 93
— glyptosperma (De Bary) Wittr. 36*,
157, 158*
— immersa (W. West) Trans. 127
— laevis (Kiitz.) W. et G. S. West 136
— reticulata (Hallas) Trans. 100, 105
— spiralis (Fritsch) Trans. 94, 94*
— talyschensis Woronich. 138
Debarya Wittr., pr. p. 92, 107
Debarya Wittr. sect. Mougeotiella Skuja
157
Debarya Wittr. sect. Zygnemopsis Skuja
92, 93
Debarya Wittr. sensu W. et G. S. West,
pr. p. 92, 159
Degagnya Conard 162
Desmidiales 11, 47
Entransia Hughes 45, 106
— dichloroplastes Prescott 106
— fimbriata Hughes 106
Gonatonema Wittr. 107
— ventricosum Wittr. 150*, 153
Gonatozygales 11, 12, 47
Hallasia Rosenv. 27, 34, 41, 44, 49, 92,
100, 105
— reticulata Rosenv. 100, 105
Haplocystia Segroves 13
Lecaniella Cookson et Eisenack 13, 157
Lloydiella Ahmad et Goldstein 161
Lloydina Ahmad et Goldstein 45, 107,
161
— kankensis Ahmad et Goldstein 161
Mehlisphaeridium Segroves 13
Mesocarpus Hass. 107
— angustus Hass. 121
— depressus Hass. var. ovalis (Hass.)
Rabenh. 135
— genuflexus Ag. 113
— gotlandicus Cleve 133, 134*
— intricatus Hass. 122
— macrosporus Wolle 118*, 123
— nummuloides Hass. 133
— parvulus Hass. 121
------var. angustus (Hass.) Kirchn. 121
------var. genuinus Kirchn. 121
— pleurocarpus De Bary 113
1 Курсивом обозначены синонимы,
полужирным шрифтом — страницы с
полным описанием водоросли, звездочкой — страницы с рисунком водоросли.
338
— recurvus Hass. 121
— robustus De Bary 135
— scalaris Hass. 122
Mesogerron Brand 159
— fluitans Brand 160
Mesotaeniales 11, 12, 22, 47
Mougeotia Ag. 10, 12—14, 25, 29, 31,
34, 36*, 40, 41, 45, 107, 157, 160
— abnormis Kissel. 29*, 112
— acadiana Trans. 109, 117
----— f. (sine nom.) Skuja 117
— africana (G. S. West) Trans. 126*,
127
— americana Trans. Ill, 138*, 140, 144,
156
— angusta Hass. 121
— areolata Trans. 131
— aspera Woronich. Ill, 149
— austriaca Czurda 112, 148, 148*, 152,
153
— bicalyptrata Wittr. 110, 119*, 127, 128
— boodlei (W. et G. S. West) Collins
111, 112, 149, 150*, 155
— boodlei (W. et G. S. West) Krieger
149
— caimanii Trans. 140
— calcarea (Cleve) Wittr. 110, 111,
119*, 127, 128, 130*, 140, 148
------var. bicalyptrata Wittr. 119*, 127
— calospora (Palla) Czurda 160
— capucina (Bory) Ag. Ill, 142, 144,
145*
— caricetorum Skv. 156
— cherokeana Taft 111, 128, 138*, 139
— condensata 156
— corniculata Hansg. Ill, 141*, 141
— costata O. et W. Bock 111, 148*, 150
— crassa (Wolle) De Toni 123
— cyanea Trans. 110, 118, 124, 128, 133
— decussata Kiitz. 156
— delicata Beck 111, 141*, 142
— depressa (Hass.) Wittr. 110, 130*,
133, 135
— djalonensis Gauthier-Lievre 113
— drouetii Trans. 110, 111, 125, 126*,
127, 151
— elegans Wittr. 140
— elegantula Wittr. Ill, 140, 141*, 142
— ellipsoidea (W. et G. S. West) Czur-
da 109, 121, 124
------f. ellipsoidea 124, 124*
------f. tenuissima (De Bary) Rund. 121,
124, 124*
— elongatum Wittr. 140, 156
— faurelii Gauthier-Lievre 142
— faveolatospora Kiriakov 112,154, 155*
— floridana Trans. 113
— fossa O. et W. Bock 111, 148, 148*,
153
— gelatinosa Wittr. 109, 118, 122*, 123,
133
— genuflexa (Dillw.) Ag. 21*, 109,
113, 117, 156
------f. genuflexa 13, 113, 114*
------var. gracilis (Kiitz.) Reinsch. 114
------f. reinschii (Trans.) Rund. 114,
115*, 156
— glyptosperma De Bary 157
— gotlandica (Cleve) Wittr. 110, 130*,
133, 134*
— gracilis Kiitz. 156
— gracilis (Reinsch) Czurda 114
— gracillima (Hass.) Wittr. 12, 111,
148*, 149
— heterogama Geitler 109, 118, 119*
— hirnii Trans. 109, 123, 124*, 125, 131
— laetevirens (A. Br.) Wittr. 12, 27*,
109, 115, 116*, 117, 128
------f. (sine nom.) Skuja 117
------var. varians Wittr. 128, 129*
---------f. minor Kissel. 115
— laevis (Kiitz.) Arch. 110, 135, 136,
137*
— lamellosa Jao 131
— macrospora (Wolle) De Toni 109,
118*, 123, 125
— maltae Skuja 109, 117, 118*, 130
— megaspora Wittr. 110, 130*, 136
— miamiana Trans. 150
— microverrucosa Krieger 110, 131,
132*
— minutissima Lemm. 156
— mysorensis Iyengar 156
— notabilis Hass. 112, 155*, 155, 156
— notabilis Hass, sensu Trans., pr. p. 112
— nummuloides (Hass.) De Toni 25,
110, 129*, 133, 136, 138
— oblongata Trans. 135
— obuchoviae Rund. 110, 131, 132*
— olivacea Ling 156
— operculata Trans. 133, 135
— ornata Jao 135
— ovalis (Hass.) Nordst. 110, 130*, 133,
135
— ovalispora Krieger 109, 123, 124*,
125
— paludosa G. S. West 111, 144, 146*,
148
— parvula Hass. 27*, 109,120, 121, 122,
124
339
Mougeotia parvula f. angusta (Hass.) Rund.
121, 122*, 125
------var. angusta (Hass.) De Toni 121
------var. ellipsoidea W. et G. S. West
124*
------f. parvula 120*, 121
— pawhuskae Taft 124, 135
— pectosa Trans. 130
— poinciana Trans. 113
— porticalis 156
— producta W. et G. S. West 12, 151
— prona Trans. 150
— protecta 156
— pseudocalospora Czurda 110, 137,
138*
— pseudovarians Gauthier-Lievre 128
— pulchella (Wittr.) De Toni 135, 136
— punctata Wittr. 12, 112, 149, 151,
152, 152*, 153
— quadrangulata Hass. 112, 143*, 151,
153, 154*
------f. (sine nom.) Popova 153
— quadrata (Hass.) Wittr. 153
— quadrata Randh. 144
— quadra-undulata Kissel. 144
— rava Trans. 154
— rectangularis Krieger 149
— recurva (Hass.) De Toni 109, 120*,
121
— regelii Skuja 111, 147, 147*
— reinschii Trans. 114
— reniformis Swir. 156
— renispora Swir. 156
— rimulosa Rund. 117
— robusta (De Bary) Wittr. 110, 134*,
135, 136
— scalaris Hass. 109, 122*, 122, 123,
125, 131, 156
------var. macrospora Hirn 124*, 125
— seminoleana Tiffany 130
— sphaerocarpa Wolle 110, 111, 126*,
127, 139, 151
— sphaerospora (Borge) Czurda 128
— staurosperma Skv. 156
— subcrassa G. S. West 123
— sumatrana (Schaarsch.) Schmidle
var. rotundospora Skv. 138, 139*
— sumatrana Schmidle 110, 136, 137*,
139
— talyschensis (Woronich.) Czurda 110,
112, 137, 138, 139*
— tenerrima G. S. West 154
— tenuis (Cleve) Wittr. 112
— tenuissima (De Bary) Czurda 32*,
124*
• — thylespora Skuja 112, 147*, 148, 151
— tibestica Gauthier-Lifevre 154
— transeaui Collins 109, 112, 112*
— tumidula Trans. 152
— tunicata O. et W. Bock 111, 139*, 151
— turicensis Nipkow 112, 155
—\varians (Wittr.) Czurda 110, 111,
117, 128, 129*
— ventricosa (Wittr.) Collins 112, 150*,
153, 155
— verrucosa Wolle 131
— victoriensis G. S. West 118, 121
— virescens (Hass.) Borge 111, 142, 143*
— viridis (Kutz.) Wittr. 25, 111, 142,
143*, 144
— weberi 156
Mougeotia Ag., pr. p. 157, 159, 162
Mougeotiaceae Palla 12, 22, 24, 25, 33,
34, 45, 48, 106
Mougeotiella Yamagishi 12, 34, 45, 107,
160
— africana (G. S. West) Yamagishi 127
— drouetii (Trans.) Yamagishi 125
— sphaerocarpa (Wolle) Yamagishi 127,
160
— sumatrana (Schmidle) Yamagishi 136
— talyschensis (Woronich.) Yamagishi
138
— tunicata (O. et W. Bock) Yamagishi
139*, 151
Mougeotiopsis Palla 9, 12, 24, 39, 45,
107, 159
— calospora Palla 159*, 160
Neozygnema Yamagishi 12, 34, 44, 49,
105
Ovoidites Potonie 13
Peltacystia Balme et Segroves 13, 157
Penium 11, 12
Pleurocarpus A. Br. 107
— mirabilis A. Br. 113
Rhynchonema Kiitz. 162
— abbreviatum Kiitz. 323
— quadratum (Hass.) Kiitz. 289
— woodsii Kiitz. 204, 323
Rundinella Zumb. 12
Salmacis sect. 163, 176
Schizocystia Cookson et Eisenack 13
Serpentinaria Gray 107
Sirocladium Randh. 9, 10, 14, 22, ,34,
39, 40, 45, 107, 160, 325
340
— cubense Rieth 160
— cuttackense Sarma 161
— kumaoense Randh. 160
— maharashtrense Randh. 160
— vandalurense Randh. 160
Sirogonium Kiitz. 9, 10, 12—14, 22, 25,
33—35, 39,40,45, 161, 162, 323, 325
— megasporum (Jao) Trans. 324
— pseudofloridanum (Pres.) Trans. 25
— sticticum (Engl. Bot.) Kiitz. 14, 27*,
33*, 324, 325
— — f. sticticum 319*, 324
------f. tenuius (Trans.) Rund. 325
— tenuius (Nordst.) Trans. 325
Sphaerocarpus angustus Hass. 121
Spirogyra Link 9—15, 20, 21, 24, 25,
31, 35, 39—41, 44, 162, 324, 325
— acanthophora Skuja 310
— acidonegiecta Uspensky 320
— acumbentis Vodenic. 170, 226, 227*
— adjerensis Gauthier-Lievre 177, 297,
298*, 313
— adnata (Vauch.) Kiitz. 320
------var. obscura Petkoff 215
— adnata (Vauch.) Kiitz., pr. p. 250
— aequalis Harv. 176, 285
— aequinoctialis G. S. West 220
— affinis (Hass.) Petit 197
— africana (Fritsch) Czurda 251
— africana (G. S. West) Gauthier-Lifevre
234*, 234
— alsiophila Skuja ex Rund. 177, 297,
298*, 299, 300
— alsiophila Skuja nom. provis 297
— ampla Krieger 240
— amplectans Skuja 301
— angulata Nipkow 286, 287*
— aphanosculpta Skuja 172, 175, 237*,
238
— archangelskaja Uspensky 320
— arcta (Ag.) Kiitz, 320
------var. cateniformis (Hass.) Kirchn.
197
------var. cateniformis (Hass.) Kirchn.
sensu Gutw, 196
— areolata Lagerh. 177, 294, 295*, 296
— arta Jao 176, 205*, 278, 281
------f. maxima H. Chu et W. Chu 280
— asiatica Czurda 173, 242, 243*
— australica Czurda 177, 299
— austriaca Czurda 288
— bellis (Hass.) Cleve 175, 265, 266,
267*, 268
— bellis (Hass.) Cleve sensu auct. 264
— bicalyptrata Czurda 203
— borgeana Trans. 14, 164, 188*, 189
— borwichae Uspensky 320
— borysthenica Kasan. et Smirn. 21*,
263, 313, 314*
------f. borysthenica 178, 313, 314*
------var. echinospora Kasan. et Smirn.
313, 314*
------f. echinospora (Kasan. et Smirn.)
V. Poljansk. 178, 313, 314*
— — var. typica Kasan. et Smirn. 313,
314*
— bourrellyana Gauthier-Lievre 222
— brunnea Czurda 228, 251, 253*
------f. brunnea 174, 252, 253*
------f. minor V. Poljansk. 320
------f. pulchrifigurata (Jao) V. Po-
ljansk. 174, 252, 253*
— brunnea Czurda sensu V. Poljansk.
252
— bullata Jao 199*, 200, 230
------f. bullata 167, 196*, 199*, 200
------f. major V. Poljansk. 167, 196*,
200
------f. minor Rund. 167, 199*, 200
— calcarea Trans. 189
— calospora Cleve 294, 295*, 296
------f. calospora 177, 294, 295*
------f. major Cleve 177, 294, 295*
------f. minor Borge 177, 296, 296
------f. venusta (Jao) V. Poljansk. 177,
295*, 296
— calospora Cleve sensu Krieger, pr. p.
296
— caroliniana Dillard 284
— cateniformis (Hass.) Kiitz, 13, 14, 163,
166, 186, 196*, 197, 197*, 206, 226
— cavata Vodenic. 175, 256*, 259
— chenii Jao 164, 166, 192, 193, 194*,
204
— chuniae Jao 177, 290*, 291
— circumlineata Trans. 202*, 203
— circumscissa Czurda 183*, 183
— cleveana Trans. 294
— colligata Hodgetts 179, 318, 319*, 319
— collinsii (Lewis) Printz 233, 238, 256
------f. ampla (Jao) V. Poljansk. 172,
229, 237*, 240
------var. ampla Jao 237*, 240
------f. collinsii 172, 239*, 240
------var. (sine nom.) Skuja 240
— columbiana Czurda 206*, 208
— communis (Hass.) Kiitz. 183*, 185
— communis (Hass.) Wittr. f. mirabilis
(Kiitz.) Kirchn. 179
— commutata Skv. 203
341
Spirogyra condensata (Vauch.) Kiitz.
164, 166, 188*, 189, 242, 322
— corrugata Trans. 174, 239, 242, 254,
255*
— costata Kadi. 179, 312*, 316
— costulata Kadi. 178, 311, 312*, 313,
316
— crassa Kiitz. 14, 24, 175, 211, 259,
260*, 261, 263, 320
------f. crassoidea (Trans.) V. Poljansk.
259
------f. genuina Kirchn. 261
------var. jassiensis Teod. 268
— crassa Kiitz. sensu auct. 259
— crassoidea Trans. 175, 211, 259, 260*,
261
— croasdaleae Blum 285
— cupientis Vodenic. et Kiriakov 272*,
273
— cylindrica Czurda 25, 287*, 288
— cylindrosperma (W. et G. S. West)
Krieger 187
— cylindrospora W. et G. S. West 246
— czubinskii Kadi, ex Rund. 177, 297
— czurdae Misra 226
— czurdiana Kadi. 175, 267*, 270
— daedalea Lagerh. 232*, 233, 234*,
235, 238
— — f. africana (G. S. West) V. Po-
ljansk. 171, 234*, 234
------f. daedalea 171, 233, 234*
------f. daedaleoides (Czurda) V. Po-
ljansk. 172, 232*, 235, 238
------f. oudhensis (Randh.) V. Poljansk.
172, 232*, 235
— daedalea Lagerh. sensu Skuja 235
— daedaleoides Czurda 235
— danica Kadi. 170, 224, 225*
— decimina Kiitz. [var.] laxa 320
— decimina (Mull.) Kiitz. 181, 190, 212,
216,320
------f. circumscissa (Czurda) V. Po-
ljansk. 163, 183*, 183, 187
------f. communis (Hass.) V. Poljansk.
14, 164, 165, 183*, 185, 192, 200, 206
------f. crassior Gutw. 320
------f. decimina 164, 165, 182*, 182,
185, 187, 203
------f. elongata Rabenh. 164, 167, 185
------f. juergensii (Kiitz.) V. Poljansk.
13, 30*, 164, 183*, 184*, 185, 186,
203
------f. longata (Vauch.) V. Poljansk.
14, 33*, 163, 164, 167, 184*, 185,
186, 198, 203, 224, 238, 322
------var. oblonga Wang 211, 213*
------f. olivascen^ (Rabenh.) V. Po-
ljansk. 163, 166, 183*, 185, 189,
195, 204, 214
— decimina (Miill.) Kiitz. sensu Alexen-
ko 208
— densa Kiitz. 320
— denticulata Trans. 294
— dentireticulata Jao 178, 299,303, 305*
— desikacharyensis Rattan 216
— dictyospora Jao 214, 250, 257, 258*
------f. dictyospora 174, 257, 258*
------f. subcylindrospora (Jao) V. Po-
ljansk. 174, 258*, 259
— discoidea Trans. 181
— discreta Trans. 297, 301
— diversizygotica (V. Poljansk.) Rund.
176, 277*, 278, 279*
— dubia Kutz. 169, 213*, 215, 251, 278
------var. genuina Kirchn. 215
------[var.] Iongiarticulata Kiitz. 320
— ellipsospora Trans. 168, 211, 212*,
248, 259
— elliptica Jao 36*, 168, 210*, 211
— elliptica Skv. 248
— elongata (Berk.) Kiitz. 214
------f. elongata 169, 214
— — f. rupestris (Schmidle) V. Po-
ljansk. 169, 212*, 214, 215
— emilianensis Bonhomme 208
— endogranulata O. et W. Bock 173,
174, 241*, 244
------f. (sine nom.) O. et W. Bock 244
— esthonica (Skuja) Czurda 171, 222,
231, 232*
— fallax (Hansg.) Wille, pr. p. 292, 293,
309
------f. berolinensis 320
------f. bohemica 320
------f. scanica (Nordstedt ?) 321
------var. (sine nom.) Smirn. 310
— farlowii Trans. 284
— flavescens (Hass.) Kiitz. 192, 193,
222, 321
------f. flavescens 164—166, 191*, 193
------f. parva (Hass.) V. Poljansk. 166,
191*, 193
— fluviatilis Hilse 13, 16, 19*, 174, 207,
214, 247*, 249, 250*, 257
------f. fluviatilis 173, 175, 247*, 248,
249, 250*
------f. grossii (Schmidle) V. Poljansk.
174, 247*, 250*, 250, 251, 320
------f. ramosa (Swir.) V. Poljansk. 173,
250*, 251
342
------- f. tibestica Gauthier-Lievre 249
— —- f. (sine nom.) Rieth 247*, 250
— fluviatilis Hilse, pr. p. 207, 250
— fossa Jao 250
— foveolata Czurda 301, 302*
— fragilis Jao 203
— franconica O. et W. Bock 172, 240,
241*
— frankliniana Tiffany 264
— frigida Gay 321
------f. arta Jao 278
— frigida Gay sensu auct., pr. p. 278,
280
— fritschiana Czurda 178, 297, 299,
300*, 301, 303
— fuellebornii Schmidle 206*, 208
— fuscoatra Rabenh. 187
— gallica Petit 164, 181, 187, 188*, 189,
208
------f. (sine nom.) 321
— gaterslebensis Rieth 178, 303, 304*
— ghosei Singh 248, 254
— gibberosa Jao 190*, 192
— glabra Czurda 175, 266, 267*, 269
— glubokolacustris Uspensky 321
— gluchowii Uspensky 321
— goetzii Schmidle 178, 299, 300*
— gracilis (Hass.) Kiitz. 14, 192, 195,
203, 321
— — var. flavescens (Hass.) Rabenh.
193
------f. genuina (Hass.) Kirchn. 192
------f. gibberosa (Jao) V. Poljansk.
164, 165, 190* 191*, 192, 194
------f. gracilis 164, 19.0*, 191*, 192
------[f.] longiarticulata Rabenh. 321
— gracillima Skv. 192
— granulata Jao 179, 297, 315, 316,
317*, 318
— grevilleana (Hass.) Kiitz. 284, 321
— groenlandica Rosenv. 177, 287*, 289,
321
— grossii Schmidle 250
— hartigii (Kiitz.) De Toni 292
— hassallii (Jenner) Petit 14, 177, 290*,
293, 321
— hatillensis Trans. 211
— heeriana Nag. 14, 36*, 261, 268, 269*
------f. heeriana 175, 269*, 269
------f. ucrainica (Smirn.) V. Poljansk.
176, 269*, 270
— henanensis (Bi) Jao 167, 204, 205*
— herbipolensis O. et W. Bock 178,
179, 304*, 311
— hoehnei Borge 220, 224
— hopeiensis Jao 176, 281*, 285, 286
— hungarica Langer 164, 187, 188*, 189
— hyalina Cleve 24, 168, 169, 208, 212,
213*
— inaequalis Kiitz. 284
— inaequalis Nag.
— incerta Skv. 224
— incrassata Czurda 179, 305, 306*
— indica (Rao) Krieger 186
— inflata (Vauch.) Kutz. 14, 27*, 177,
280, 287*, 288, 289, 297, 301
------var. alsiophila Skuja 297
------var. foveolata Trans, sensu Skuja
301, 302*
— inflata (Vauch.) Rabenh. ampl. Petit
321
— insignis (Hass.) Kiitz. 291, 292*
— — f. insignis 177, 285, 292*, 292,
293, 294
------var. longispora Smirn. 292*, 292
------ f. rectangularis (Trans.) V. Po-
ljansk. 177, 292*, 293
------f. wangii (Li) V. Poljansk. 177,
292*, 293
— insignis Kiitz. f. braunii 321
------[f.] braunii Rabenh. 321
------f. hantzschii 322
------[f.] hantzschii Rabenh. 321
— intermedia Rabenh. 322
— irregularis Nag. 168, 211, 213*
— irregularis Nag. sensu Gutw. 208
— jassiensis (Teod.) Czurda 175, 261,
267*, 268
— jatobae Trans. 269
— juergensii Kiitz. 183*, 184*, 185
— jugalis (Dillw.) Kiitz. 210
— kaffirita Trans. 228
— kolae Hajdu 322
— krjutschkowae Uspensky 322
— krubergii V. Poljansk. 175, 261, 262*
— kundaensis Singh 248
— kusnetzowii Uspensky 322
— kuusamoensis Him 178, 297, 300,
302*
------f. kuusamoensis 178, 301, 302*
------f. skujaensis V. Poljansk. 178,
301, 302*
— labyrinthica Trans. 171, 235
— lacustris Czurda 187
— lacustris Uspensky 322
— lagerheimii Wittr. 222, 223*, 226
------f. lagerheimii 170, 223, 223*
------f. obovata (Jao) V. Poljansk. 170,
172, 223*, 224, 226
— lambertiana Trans. 303
343
Spirogyra lamellata (Rao) Krieger 265,
267*
— lancincola Uspensky 322
— latviensis (Skuja) Czurda 296
— laxa Kiitz. 283
------f. articulus 322
— laxistrata Jao 303
— lenticularis Trans. 176, 269, 275*,
276
— liana Trans. 191*, 193
— libyca Gauthier-Lievre 297
— lodziensis Kadi. 274
— longata (Vauch.) Kiitz. 186, 322
------[f.J reversa (Kiitz.) Kirchn. 322
------var. reversa (Hass.) Kirchn. 186
— lubrica (Roth) Kiitz. 323
— lushanensis Li 233
— luteospora Czurda 170, 224, 226,
227*, 240
— lutetiana Petit 182
— macrospora (Rao) Krieger 214
— major Kiitz. 322
— majuscula Kiitz. 30*, 175, 262*, 264,
265, 266, 270
------var. brachymeres Stitzenberger 264
------var. genuina Kirchn. 264
------f. (sine nom.) 322
— malmeana Hirn 246, 251, 252
------var. verrucosa Rao 245*, 251
— manoromae Randh. 265
— maravillosa Trans, 180
— marchica Krieger 179, 306, 314, 315*,
316
— margalefii Aboal et Llimona 208
— margaritata Wollny 169, 213*, 215,
216
— maxima (Hass.) Wittr. 14, 261, 263,
266, 269, 270, 271*, 272*, 274
------f. cupientis (VodeniS. et Kiriakov)
Rund. 176, 272*, 273
------var. livida Vail. 273
------f. livida (Vail.) V. Poljansk. 176,
271*, 273
------f. maxima 176, 271*, 273
------f. tenuior (Magnus et Wille) V. Po-
ljansk. 176, 271*, 272*, 273, 274, 276
------f. woronichinii V. Poljansk. 176,
272*, 273, 274, 276
— maxima (Hass.) Wittr. sensu auct. 273,
274
— megaspora Trans. 271*, 273, 274
— microgranulata Jao 296
— micropunctata Trans. 224
— microspora Jao 186
— mienningensis Li 174, 254, 255*
— mirabilis (Hass.) Kiitz. 14, 27, 28*,
30*, 41, 163, 165, 179, 180*, 196
------f. microspora Beck-Mannagetta
322
------f. minor V. Poljansk. 179
— miranda Kadi. 178, 312*, 312
— moebii Trans, 271*, 274
— naegelii Kiitz. 286
— nawaschinii Kasan. 309*, 310, 311
— neglecta (Hass.) Kiitz. 189, 206*, 207,
209, 244, 278
— — f. columbiana (Czurda) Rund.
168, 206*, 208, 209
------f. fuellebornii (Schmidle) V. Po-
ljansk. 168, 183, 187, 206*, 208, 212,
213, 215
------f. fuellebornii (Schmidle) V. Po-
ljansk., pr. p., sensu V. Poljansk. 208
------f. neglecta 168, 206*, 208, 209
------var. ternata (Ripart) W. et
G. S. West 276
------f. varians (Singh) V. Poljansk. 208
— nitida (Dillw.) Link 25, 208, 209,
210*, 211, 244, 246, 322
------var. densa Rabenh. 209
------f. nitida 168, 209, 210*
------f. paradoxa (Rao) V. Poljansk.
168, 209, 210*
------f. (sine nom.) 322
— nodifera O. et W. Bock 171, 203, 234*,
236, 240
— novae-angliae Trans. 228
— oblata Jao 176, 276, 277*
— obovata Jao 223*, 224
— occidentalis (Trans.) Czurda 19*, 217,
218, 242
------f. occidentalis 173, 243, 243*
------var. striata Bourr. et Mang. 244
------f. striata (Bourr. et Mang.) V. Po-
ljansk. 173, 244
— olivascens Rabenh. 183*, 185
— oltmannsii Huber-Pestalozzi 180
— orbicularis (Hass.) Kiitz. 271*, 273,
274
------f. tenuior Magnus et Wille 274
— orthospira Nag. 264, 322
— oudhensis Randh. 232*, 235
— paludosa Czurda 185
— papulata Jao 172, 224, 236, 237*
— paradoxa Rao 209, 210*
— paraquayensis Borge 205*, 248, 251
------f. paraquayensis 173, 205*, 248
------f. rhizobrachialis (Jao) V. Po-
ljansk. 173, 205*, 248
— parva (Hass.) Kiitz. 191*
344
— parvula (Trans.) Czurda 166, 192,
196*, 196, 198
— pascheriana Czurda 287*, 288
— pellucida (Hass.) G. S. West 181, 263
------f. minor (Tiffany) V. Poljansk.
175, 263, 264
------f. pellucida 175, 262*, 264
— — f. sphaerospora (Hirn) V. Po-
ljansk. 181
— pellucida Kiitz. 264
------var. minor Tiffany 264
— perforans Trans. 222
— petitiana Trans. 296
— poljanskii Kadi. 200
— polymorpha Kirchn. 30*, 165—167,
180, 191*, 194*, 195, 204
------f. (sine nom.) Petkoff 195
— polymorpha Kirchn., pr. p. 179
— polymorpha Kirchn. sensu auct. 179
— polytaeniata Strasburger 169, 215
— porticalis (Miill.) Cleve 182, 183,186,
188, 189, 203, 321
------f. cylindrosperma (W. et G. S. West)
V. Poljansk. 163,187
------var. decimina (Hass.) Cooke 182,
320
------f. porticalis 164, 165, 167, 184*,
187
— porticalis (Vauch.) Cleve var. africa-
na G. S. West 234
— pratensis Trans. 167, 193, 197, 198,
200, 204, 206*, 322
------f. fennica (Cedercr.) V. Poljansk.
204
------var. minor Woronich. 204
— — f. minor (Woronich.) V. Poljansk.
204
— princeps Vauch. 322
— propria Trans. 252
— protecta Wood 294
------var. inflata Fritsch et Rich 300*,
301
------f. (sine nom.) Skuja 296
— protecta Wood sensu MObius 299
— pseudodaedaleoides Kadi. 172, 234*,
236
— pseudodecimina O. et W. Bock 322
— pseudogranulata Ley 179, 316, 317*
— pseudomaxima Kadi. 176, 274, 275*
— pseudonaegelii Skv. 286
— pseudoneglecta Czurda 212
— pseudonodifera O. et W. Bock 172,
203, 236, 240, 241*
— pseudosahnii Kadi. 167, 196*, 198,
199
— pseudospreeiana Jao 176, 279*, 285,
286
— pseudotenuissima O. et W. Bock 286
— pseudovarians Czurda 169, 213*, 216,
231, 259
— pseudowoodsii V. Poljansk. 169, 218,
219*, 220
— pulchra Alexenko 323
— pulchrifigurata Jao 252, 253*
— punctata Cleve 220, 228, 229*, 233,
239, 240, 256, 325
------f. crassior Nordst. 171, 229, 229*
------var. esthonica Skuja 231
------f. esthonica (Skuja) V. Poljansk.
231 •
------f. major Hirn 229
------var. major Him 229, 229*
----— f. punctata 172, 228, 229*, 240
— punctata Cleve sensu Kadi., pr. p. 220
— punctata Petit 294
— punctiformis Trans. 169, 172, 220,
221*
— punctulata Jao 244
------f. cylindrospora (W. et G. S. West)
Rund. 173, 246
------var. macrospora Jao 246
------f. macrospora (Jao) Rund. 173,
246
------f. punctulata 173, 243*, 245, 245*
— quadrata (Hass.) Petit 177, 289, 290*
------f. genuina Kirchn. 289
— quadrilamin ata Jao 174, 256, 256*
— quezelii Gauthier-Lifevre 173, 239*,
242
— quinina (Ag.) Kiitz. f. juergensii
(Kiitz.) Kirchn. 185
— quinina Kiitz. 187
------var. inaequalis 323
— quinina (Kiitz.) Kirchn. 323
— ramosa Swir. 250*, 251
— randhawae (Randh.) Krieger 265
— rectangularis Trans. 292*, 293
— reflexa Trans. 320
— regularis (Cedercr.) Krieger 308*,
311
— reinhardii Chm. 174, 248, 252, 253*
— reticulata Nordst. 39, 256, 293, 306,
307*, 308*, 313, 314, 316, 318
------f. acanthophora (Skuja) V. Po-
ljansk. 178, 179, 252, 303, 307*,
308*, 309, 310, 314
------f. czurdae V. Poljansk. 178, 308*,
310
------f. nawaschinii (Kasan.) V. Po-
ljansk. 178, 309*, 310
345
Spirogyra reticulata var. regularis Ce-
dercr. 311
------f. regularis (Cedercr.) V. Poljansk.
178, 307*, 308* 311, 314
------f. reticulata 179, 309
— rhizobrachialis Jao 205*, 248
— rhizoides Randh. 214
— rhizopus Jao 257
— rhodopea Petkoff 214
— rivularis (Hass.) Rabenh. 206*, 207,
212, 251
------var. minor Hansg. 207
— — f. minor (Hansg.) V. Poljansk.
168, 207, 214
------f. rivularis 168, 207
— robusta (Nyg.) Czurda 224
— rotundispora Skv. 264
— rugosa (Trans.) Czurda 178, 299, 300*
— rugulosa Iwan. 171, 226, 227*, 235
— rugulosa Iwan. sensu Krieger, pr. p.
235
— rupestris Schmidle 214
— sahnii Randh. 167, 189, 197*, 198
— salina Aleem 13, 163, 170, 221* 222
— schmidtii W. et G. S. West 220, 320
— scrobiculata (Stockm.) Czurda 171,
217, 230, 231* 236 '
— semiornata Jao 283
— setiformis (Roth) Kiitz. 168, 209,
210*, 210, 211, 215, 244, 248, 278,
322
— — f. diversizygotica V. Poljansk.
277*, 278, 279*
------f. margaritata (Wollny) V. Po-
ljansk. 215
— shantungensis Li 256
— shanxiensis Ling et Zheng 204
------var. henanensis Bi 204
— sibirica Skv. 171, 225*, 230
— silesiaca Kadi. 179, 318, 319*
— sinensis Li 175, 262*, 263, 264
— singularis Nordst. 183
— sphaerospora Hirn 163, 181, 182*,
264, 323
— spinescens Kiriakov 179, 314, 315*
— splendida G. S. West 211
— spreeiana Rabenh. 288
— stictica (Engl. Bot.) Wille 324, 325
— — f. tenuior (Trans.) V. Poljansk.
325
— subaequa Kiitz. 266, 323
— subarta Jao 278
— subcolligata Bi 318
— subcrassa Woronich. 175, 259, 260*,
261
— subcylindrica Jao f. major Bi 248
— subcylindrica var. major Bi 248
— subcylindrospora Jao 258*, 259
— submar gar itata God ward 25, 265
— submaxima Trans. 265, 266, 267*, 274
------var. lamellata Rao 265
------f. lamellata (Rao) V. Poljansk.
175, 265, 267*
------f. submaxima 175, 265, 267*
— subpapulata Jao 170, 223*, 224, 226,
230, 236
— subpol уtaeni ata Jao 216
— subpratensis Woronich. 200, 204
— sub reticulata Fritsch 257
— subsalina Cedercr. 185
— subsalsa Kiitz. 13, 163, 180,194, 196,
198, 214, 220
------f. gracilis (Borge) V. Poljansk. 165,
167, 191*, 195
------f. punctulata Kras nop. 220
------f. subsalsa 166, 167, 191* 195
------f. uralensis V. Poljansk. 165, 191*,
195
— subsalsa Kiitz. sensu Krieger, pr. p.
195
— subsalso-punctulata Kadi. 163, 169,
219*, 220
— subvelata Krieger 217
— suecica Trans. 195
— suomiana Trans. 229
— szechwanensis Jao 209
------var. varians Singh 208
— tandae Randh. 280, 281, 299, 301
— tenuior (Nordst.) Krieger 325
— tenuispina Rund. 170, 219*, 220, 222
— tenuissima (Hass.) Kiitz. 13, 280, 286,
287*, 289, 300, 301
------f. cylindrica (Czurda) V. Poljansk.
19*, 177, 286, 287*, 288, 289
------f. inflata (Vauch.) Cleve 288
----— f. punctata Skuja 299
------f. rugosa (Trans.) V. Poljansk. 299
------f. tenuissima 13, 176, 177, 286,
287*
— teodorescoi Trans. 25, 203, 204
— ternata Ripart 176, 208, 213, 276,
277*
------f. gagrensis Lobik 215
— ternata Ripart sensu Gutw. 215
— tjibodensis Faber 285
— tolosana Comere 283
— trachycarpa Skuja 173, 174, 246, 247*,
254
— troitzkajae V. Polansk. 273
— tuberculata Lagerh. 169, 218, 222
346
— tumida Jao 176, 280, 281*
— turfosa Gay 209
------var. manschurica Skv. 215
— ucrainica Smirn. 269*, 270
— ugandense Gauthier-Lievre 264
— undulisepta Randh. 299
— unocula Chm. 276
— vagans Teod. 226
— varians (Hass.) Kiitz. 14, 200, 202*,
216, 218, 224, 231, 238, 323
------f. bicalyptrata (Czurda) V. Po-
ljansk. 167, 202*, 203, 226, 236, 240
------f. circumlineata (Trans.) V. Po-
ljansk. 167, 202*, 203
------f. gracilis Borge 195
------f. minor Teod. 167, 195, 202*,
203
------f. varians 14, 167, 201*, 202
------f. woodsii (Hass.) V. Poljansk.
30*, 167, 186, 196*, 202*, 203, 204,
218, 323
— varians (Hass.) Kiitz. sensu auct. 204
— velata Nordst. 216, 217*, 243, 259
— — f. ellipsoidea (Wang) V. Po-
ljansk. 169, 217
------var. ellipsoidea Wang 217
------f. lateralis Gauthier-Lievre 169,
218
------ f. velata 163, 169, 217
— velata Nordst. sensu Riabinine 243
— venosa Kadi. 178, 312*, 312, 313
— venusta Jao 295*, 296
— verrucosa (Rao) Krieger 173, 245*,
251
— verruculosa Jao 251
— wangii Li 292*, 293
— warwarae Uspenky 323
— weberi Kutz. 25, 280, 281, 291
------var. elongata (Spree) Rabenh. 283
------f. farlowii (Trans.) V. Poljansk.
30*, 176, 283*, 284, 286, 291, 293
------var. genuina (Kiitz.) Kirchn. 283
------f. grevilleana (Hass.) V. Poljansk.
177, 283*, 284, 289, 291, 294
------var. grevilleana (Kiitz.) Kirchn.
284
------var. intermedia Kirchn. 323
------var. intermedia Kiitz. 323
------var. punctata Langer 299
------f. tjibodensis (Faber) V. Poljansk.
176, 285
------ f. weberi 176, 282*, 283, 321
— westii Trans. 234
— willei Czurda 308*, 310
— willei Skuja 309, 310
------var. acanthophora Skuja 308*,
310
— willei Skuja sensu Czurda 310
— wittrockii Alexenko 163, 181, 182*,
189
— wollnyi De Toni 210
— woodsii (Hass.) Czurda 204
— yunnanensis Li 210
Spirogyra sp. 250, 285
Spirogyra Link, pr. p. 323
Spirogyraceae Palla 12, 22, 24, 33, 34,
44, 48, 161, 325
Staurocarpus capucinus Hass. 144, 145*
Staurospermum Kiitz. 107
— quadratum (Hass.) Kiitz. 153
— viride Kiitz. 142
Temnogametum W. et G. S. West 12,
14, 33, 38, 40, 45, 107, 160
Temnogyra Lewis 12, 33, 45, 160, 162,
325
— collinsii Lewis 238, 325
— corrugata (Trans.) Yamagishi 254
— esthonica (Skuja) Yamagishi 231
— liana (Trans.) Yamagishi 193
— punctata (Cleve) Yamagishi 228, 325
Tetrapidites Klaus 12
Tetraporina Naum. 13
Ulotrichaceae 106
Uronema Lagerh. 106
Zygnema Ag. 9, 10, 12—15, 20, 22, 25,
31, 36*, 40, 44, 49, 93, 102, 105, 106
— adpectinatum Trans. 61, 63
— affine Hass. 104
— affine Kiitz. 104
— allorgei Gauthier-Lievre 51, 72, 73*
— altaicum Skv. 104
— americanum (Trans.) Czurda 96
— anomalum (Hass.) Cooke 104
— areolatum Taft et Trans. 63
— argillarii Kadi. 51, 70, 71*
— atrocoeruleum W. et G. S. West 53,
85
— biforme Jao 52, 72, 73*, 74, 75
— bohemicum Czurda 52, 84, 84*
— borzae Krieger 53, 90*, 91
— calosporum Jao 52, 75, 76*, 80, 81,
104
— carinatum Taft 50, 54*, 55*, 56
— carinthiacum Beck 53, 86, 87*, 89
— catenatum Trans. 90
— chalybeospermum Hansg. 53, 69, 83*,
85, 89
347
Zygnema circumcarinatum Czurda 50,
57* 58
— coeruleum Czurda 51, 56, 63, 64*,
65
— compressum Czurda 83*, 86
— conspicuum (Hass.) Trans. 50, 59, 60*,
105
> — cruciatum (Vauch.) Ag. 25, 52, 71*,
74, 75, 105
— cyaneum Czurda 53, 69, 83*, 85
— cyanosphaeroidica O. et W. Bock 50,
56, 57, 57*, 63
— cyanosporum Cleve 51, 63, 64*
— cylindricum Trans. 53, 90, 90*, 91
— cylindricum Trans, sensu Czurda 90*,
91
— cylindrospermum (W. et G. S. West)
Krieger 51, 67*, 69, 75
— cylindrosporum Czurda 52, 70, 71*,
75
— czurdae Randh. 50, 54, 55*, 58
— deciminum (Mull.) Ag. 182
— decussatum (Trans.) Czurda 97
— decussatum (Vauch.) Ag. 50, 59, 60*
— emersum Skv. 72
— ericetorum (Kiitz.) Hansg. 102
------var. terrestre (Kirchn.) Hansg. 102
— excompressum Trans. 53, 58, 83*, 86,
89
— extenue Jao 52, 65, 77*, 80*, 80, 81
— fanicum Li 52, 74, 78, 81, 82*, 104
— frigidum Taft 30*, 91
— gedeanum Czurda 54, 57
— genuflexum Ag. 113
— germanicum Czurda 52, 79*, 81, 82
— globosum Czurda 51, 60*, 61, 63
— gorakhporense Singh 65
— hausmannii (De Notar.) Czurda 52,
66, 79*, 81
— hypnosporum Rich 91
— inconspicuum Czurda 75
— inconspicuum Czurda emend. Skv. 75
— indicum Misra 65
— insigne (Hass.) Kiitz. 16, 25, 51, 68,
68* 85
------f. insigne 68*, 69
------f. kasachstanicum Rund. 68*, 69
— irregulare Krieger 53, 90*, 91
— kabylicum Gauthier-Lievre 92
— laetevirens Klebs 51, 62*, 63
— laetevirens Klebs sensu Skuja 61
— laevisporum Jao 105
— lawtonianum Taft 50, 59, 60*, 61
— leiospermum De Bary 27*, 32*, 51,
67, 67*, 68, 69, 74
— luteosporum Czurda 52, 71*, 72, 74,
81
— majus Czurda 63
— margalefianum Aboal 50, 58
— melanosporum Lagerh. 19*, 53, 85,
86, 88* 88
— micropunctatum Trans. 58
— mucigenum Randh. 53, 86, 87*
— mucosum Skv. 75
— neocruciatum Trans. 75
— nitidum Hass. 209
— ornatum (Li) Trans. 89
— ovoidanum Trans. 59
— pawhuskae Taft 51, 67, 67*
— pectinatum (Vauch.) Ag. 21*, 51, 59,
61, 62*, 105
— — var. anomalum 104
------f. conspicuum 104
------var. conspicuum (Hass.) Kirchn.
59, 104
— var. conspicuum (Hass.) Kirchn. sen-
su Skv. 104
------var. decussatum (Vauch.) Kirchn.
97
------f. terrestre (Rabenh.) Hansg. 61
— peliosporum Wittr. 53, 86, 88*, 89
— perforatum Gauthier-Lievre 72
— pseudogedeanum Gauthier-Lievre 50,
56, 58, 77*
— pulcherrimum Skv. 81
— quininum (Mull.) Ag. 187
— ralfsii (Hass.) De Bary 50, 54*, 55*,
58
— randhawae Krieger 58
— reticulatum Hallas 99*, 100, 105
— reticulosporum Gauthier-Lievre 72
— rhynchonema Hansg. 50, 53, 54*
— schwabei Krieger 92
— sinense Jao 61
— skujae Czurda 51, 61
— sphaericum Misra 85
-— spirale Fritsch 94
— spontaneum Nordst. 51—53, 65, 66*,
81
— stagnale (Hass.) Kiitz. 51, 66*, 69
— stellatum 76
— stellinum (Vauch.) Ag. 25, 52, 70,
74, 75, 76, 77*, 80*, 104
------var. compressum Gayral 76
------var. genuinum Kirchn. 76, 105
------f. subtile (Kiitz.) Kirchn. 105
------f. subtile Rabenh. 105
------var. subtile (Kiitz.) Kirchn. 70
------var. subtile Rabenh. 105
------var. tenue (Kiitz.) Kirchn. 78
348
------var. vaucheri (Ag.) Kirchn. 78
— sterile Trans. 53, 90*, 91, 92
— subcylindricum Krieger 53, 90*, 91
— subcylindricum Krieger sensu Aboal
et Llimona 91
— substellinum Taft 76, 80, 81
— subtile Kiitz. 51, 66*, 70, 78
— synadelphum Skuja 50, 51, 54*, 56,
63, 65, 87
— tenue Kutz. 52, 65, 70, 78, 79*, 80*,
81, 105
— — f. compressum Gauthier-Lievre
78
— tenue Kiitz. sensu Krieger, pr. p. 70,
78
— vaginatum Klebs 52, 72, 73*, 74, 75,
78, 81, 82, 105
— variabile Skv. 105
— vaucheri Ag. 52, 78, 79*, 80, 105
— venustum Jao et Hu 87
— verrucopunctatum Prasad et Godward
52, 82, 83*
— verrucosum Jao 67
Zygnema sp. 30*
Zygnema Ag., pr. p. 92, 101, 107, 162
Zygnemataceae Palla 12, 22, 24, 25, 34,
44, 48, 106
Zygnematales 9, 11, 12, 14, 22, 24, 44,
47, 161
Zygnematophyceae 9, 10, 11, 13, 44, 46
Zygnemopsis (Skuja) Trans. 12, 13, 34,
40, 44, 49, 92, 104, 105, 157
— akfaduensis Gauthier-Lievre 101
— americana Trans. 93—95, 96, 97*,
98
— atrocoerulea Gauthier-Lievre 101
— barcense Gauthier-Lievre 101
— circumcarinata (Czurda) Krieger 58
— columbiana Trans. 94, 95, 97
— decussata Trans, 93, 95, 97, 97*
— desmidioides (W. et G. S. West)
Trans. 93, 94*, 97
— foersteri Muller 93, 95*, 99
— globosa Randh. 93, 98, 99*, 100
— iyengari Randh. 98
— lamellata Randh. 93, 98, 99*, 100
------var. globosa Randh. 98, 99*
— mediterranea Gauthier-Lievre 101
— montana Gauthier-Lievre 101
— ouarseniense Gauthier-Lievre 101
— pseudodecussata (Czurda) Krieger 98
— quadrata Jao 93, 95, 95*, 97, 98
— reticulata (Hallas) Trans. 93, 99*,
100, 105
— sinensis Trans. 95—98
— spiralis (Fritsch) Trans. 93, 94, 94*
— splendens Randh. 98, 100
— stephensiae Trans. 96
— transeauiana Randh. 101
Zygnemopsis (Skuja) Trans., pr. p. 49
Zygogonium Kiitz. 9, 12, 14, 20, 22, 24,
34, 36*, 39, 44, 49, 56, 58, 63, 72,
101
— anomalum Ralfs 104
— crassum Kiitz. 105
— crassum (Kiitz.) De Toni 105
— cyanosphaeroidicum (O. et W. Bock)
Kadi. 57
— ericetorum Kiitz. 14, 32*, 101, 102,
102*, 103*
— — f. aquaticum Kirchn. 102
------f. terrestre Kirchn. 102
— laetevirens (Klebs) Migula 63
— pectinatum (Vauch.) Kiitz. 61
Zygogonium Kiitz., pr. p. 49
Zygonematis stellatii 76
СОДЕРЖАНИЕ
Предисловие.............................................. 5
I. ОБЩАЯ ЧАСТЬ
1. Общая характеристика зигнемовых водорослей.......... 9
Основные признаки зигнемовых водорослей (9). — Проис-
хождение и родственные связи (10). — Распространение
(13). — Практическое значение (15). — Методы сбора и изу-
чения (15).'
2. Морфология зигнемовых водорослей..................... 18
Строение нитей и вегетативных клеток (18). — Оболочка ве-
гетативных клеток (20). — Поперечные перегородки клеток
(20). — Протопласт (21). — Хлоропласт (22). — Пиреноиды
(24). — Запасные вещества (24). — Ядро (24). — Митоз
(25). — Мейоз (25). — Размножение (27). — Особенности
конъюгации (32). — Зигоспора (34).
3. Таксономическая оценка основных морфологических при-
знаков ................................................. 39
4. Система зигнемовых водорослей........................ 41
И. СИСТЕМАТИЧЕСКАЯ ЧАСТЬ
Отдел Chloro phyta.........................................
Класс Zygnematophyceae (Conjugatophyceae)
46
46
Порядок Zygnematales....................................... 47
Семейство Zygnemataceae ................................ 48
Род Zygnema.......................................... 49
Род Zygnemopsis...................................... 92
Род Zygogonium .................................... 101
Сомнительные и ошибочные указания видов родов Zygnema
и Zygogonium..................................... 104
Редкие, недостаточно изученные и сомнительные роды сем.
Zygnemataceae . ................................. 105
Семейство Mougeotiaceae................................ 106
Род Mougeotia...................................... 107
Сомнительные н ошибочные указания видов рода Mougeotia 156
Род Debarya
157
Род Mougeotiopsis.......................................
Редкие, недостаточно изученные и сомнительные роды сем.
Mougeotiaceae........................................
Семейство Spirogyraceae....................................
Род Spirogyra...........................................
Сомнительные и ошибочные указания видов рода Spirogyra
Род Sirogonium..........................................
Редкий и недостаточно изученный род сем. Spirogyraceae
159
160
161
162
320
323
325
Литература
Указатель латинских названий
4
326
338
Научное издание
Рундина Людмила Александровна
ЗИГНЕМОВЫЕ ВОДОРОСЛИ РОССИИ
Утверждено к печати
Ботаническим институтом им. В. Л. Комарова РАН
Редактор издательства Е. А. Тахтаджян
Художник А. Т. Пожванов
Технический редактор Е. И. Егорова
Корректоры Ф. Я. Петрова и И. А. Тюрина
Компьютерная верстка Е. М. Сальниковой
Лицензия № 020297 от 23 нюня 1997 г..Сдано в набор 02.07.98.
Подписано к печати 02.12.98. Формат 60 х 90 I/jg. Бумага офсетная.
Гарнитура тайме. Печать офсетная. Усл. печ. л. 22.
Уч.-изд. л. 25.9. Тираж 500 экз. Тип. зак. № 3840. С 234
Санкт-Петербургская издательская фирма РАН
199034, Санкт-Петербург, Менделеевская лин., 1
Санкт-Петербургская типография «Наука» РАН
199034, Санкт-Петербург, 9 лин., 12