/


















































































































































































Text
INSTITUT ZOOLOGIQUE DE L’ACADfeMIE PES SCIENCES DE L4JRSS
NOUVELLE SERIE n° 16
FAUNE DE L’URSS
Sous la direction de S. A. SERNOV, de I’Academic
Redige par A. A. STACKELBERG
CRUSTACES
Vol. X, П» 3
DECAPODES ANOMURES
par V. V. MAKAROV
EDITION DE 1’ACADEMIE DES SCIENCES DE L’URSS
MOSCOU 1938 • LENINGRAD
ЗООЛОГИЧЕСКИЙ ИНСТИТУТ АКАДЕМИИ ПАУК СССР
НОВАЯ СЕРИЯ № 16
ФАУНА СССР
Главный редактор акад. С. А. ЗЕРНОВ
Редактор А. А. ШТАКЕЛЬБЕРГ
РАКООБРАЗНЫЕ
Т. X, вып. 3
В. В. МАКАРОВ
ANOMURA
ИЗДАТЕЛЬСТВО АКАДЕМИИ НАУК СССР
МОСКВА . 1938 • ЛЕНИНГРАД
Ответственный редактор Директор Зоологического Института
акад. С. А. Зернов
Редактор издания А. А. Штакельберг
Технический редактор и ученый корректор Н. Г. Редько
Сдано в набор 2 марта 1937 г. — Подписано к печати 10 января 1938 г.
X -+- 324 стр. (5 таблиц)
Формат бум. 72Х1Ю см.—217/s печ. л.—24.66 уч.-авт. л.—53728 тип. зи. в л.—Тираж 1200
Ленгорлит Xs 252.РИСО № 164. —АНИ № 1430. — Заказ Хе 301
Типография Академии Наук СССР. Ленинград, В. О.; 9 линия, 12
ПРЕДИСЛОВИЕ
Настоящий выпуск охватывает большой систематический раздел
Decapoda, так называемых среднехвостовых десятиногих раков, Апотига.
В следующем выпуске будут разобраны истинные крабы, или Brachyura.
Обработка более примитивных групп десятиногих раков задерживается
из-за невыхода ряда еще неопубликованных работ, в которых описы-
вается довольно много новых видов, относящихся к нашей стране, без
учета которых составление „Фауны СССР" по этим группам страдало бы
значительной неполнотой. Этой причиной и объясняется некоторое наруше-
ние систематического порядка в выходе выпусков, посвященных десятиногим
ракам. Более ранний выход Апотига диктуется еще тем обстоятельством,
что эта группа заключает в себе ряд важных промысловых объектов.
Во введении дается краткий анатомо-морфологический очерк, а также
самые краткие сведения по биологии и зоогеографии рассматриваемой
группы. Семейству Paguridae, в виду его своеобразия, предшествует
отдельное введение, где более подробно разбирается анатомия, тогда
как в общем введении главное внимание отводится внешней морфологии
и некоторым общим вопросам (экология, хозяйственное значение, филоге-
ния). Акватории, фауна которых разбирается в настоящем выпуске,
перечисляются в зоогеографическом очерке семейства Paguridae', для
отдельных видов, типы которых хранятся в Зоологическом Институте
Академии Наук, граница по западному побережью Северной Америки
отодвинута несколько южнее острова Кадьяк, именно до Ситки.
Обработанные коллекции Зоологического Института Академии
Наук относятся главным образом к нашим дальневосточным морям; из
них главнейшие — сборы И. Г. Вознесенского (1840), Петелина (1852),
А. А. Бунге (1897—1898), П. Ю. Шмидта (1900, 1901, 1929, 1933), В. К.
Бражникова (1899, 1908—1910), Н. Смирнова и А. Бегак (1907—1910),
Ф. А. Дербека (1908—1913), Б. А. Гейнемана (1909—1910), В. К. Ар-
сеньева (1910—1911), Л. М. Старокадомского (1911—1913, 1934), Э. Э.
Арнольда (1911), Ф. П. Рябушинского (1911), Е. К. Суворова (1911),
М. Н. Павленко и В. К. Солдатова (1911—1915), Мясковского (1912),
сборы Гидрографической экспедиции Восточного океана (1914—1925)
и Рошковского (1917). По Черному морю сборы К. П. Ягодовского (1908)
и С. А. Зернова (1908—1911). t
Рлсуякя. исполнены художником Н. Кондаковым, фотографии —
автором.
В. Макаров.
СИСТЕМАТИЧЕСКИМ УКАЗАТЕЛЬ ВИДОВ
Стр.
Надсем. Thalassinidea
1. Сем. Axiidae
1. Род Axiopsis Borradaile
1. A. princeps (Boas)................................................... 48
2. Сем. Callianassidae Bate
1 Род Upogebia Leach
1. U. littoralis (Risso) ............................................... 52
2. U. major (de Haan)................................................... 54
3. U. yokoyai nom. nov................................................. 57
4. U. issaeffi (Balss).................................................. 59
2. Род Callianassa Leach
1. C. subterranea (Montagu)............................................. 63
2. C. harmandi Bouvier , ........................... 66
3. C. gigas eoa nom. ................................................... 68
4. C. japonica (Ortmann)................................................ 70
5. C. californiensis bouvieri nom. ..................................... 72
6. C. pontica (Czerniavsky)............................................. 73
3. Род Ctenocheles Kishinouye
1. C. balssi Kishinouye............,.................................... 76
Надсем. Galatheidea
3. Сем. Galatheidae
1. Род Galathea Fabricius
1. G. strigosa (Linne).................................................. 83
2. G. acanthomera Stimpson.............................................. 86
3. G. Integra Benedict.................................................. 88
4. G. pubescens Stimpson................................................ 91
2. Род Munida Leach
1. M. bamffica (Pennant)................................................ 93
2. M. japonica Stimpson................................................ 97
VIII
ANOMURA
Стр.
3. Род Munidopsis Whiteaves
1. M. beringana Benedict....................................................... 101
4. Род Cervimunida Benedict
1. C. princeps Benedict....................................................... 103
4. Сем. Porcellanidae
1. Род Porcellana Lamarck
1. P. longicornis (Pennant).................................................... 107
2. P. longimana Risso................................. ...................... 110
2. Род Pachycheles Stimpson
1. P. stevensii Stimpson...................................................... Ill
Над сем. Hippidea
5. Сем. Albuneidae
1. Род Blepharipada Randall
1. B. japonica Durufle........................................................ 115
Надсем. Paguridea
6. Сем. Pylochelidae
1. Род Mixtopagurus Milne Edwards
1. M. jeffreysii (Miers)...................................................... 120
7. Сем. Paguridae
1. Род Diogenes Dana
1. D. edwardsii (de Haan)..................................................... 158
2. D. penicillatus Stimpson................................................... 159
3. D. varians (Costa) . ...................................................... 160
2. Род Clibanarius Dana
1. C. misanthropus (Risso)................................................... 162
2. C. japonicus Rathbun..................................................... 163
3. Род Pagaristes Dana
1. P. digitalis Stimpson...................................................... 166
2. P. setosus (Milne Edwards)................................................. 167
4. Род Dardanus Paulson
1. D. arrosor (Herbst)....................................................... 168
5. Род Pagurus Fabrici us
1. P. middendorffii Brandt.................................................... 174
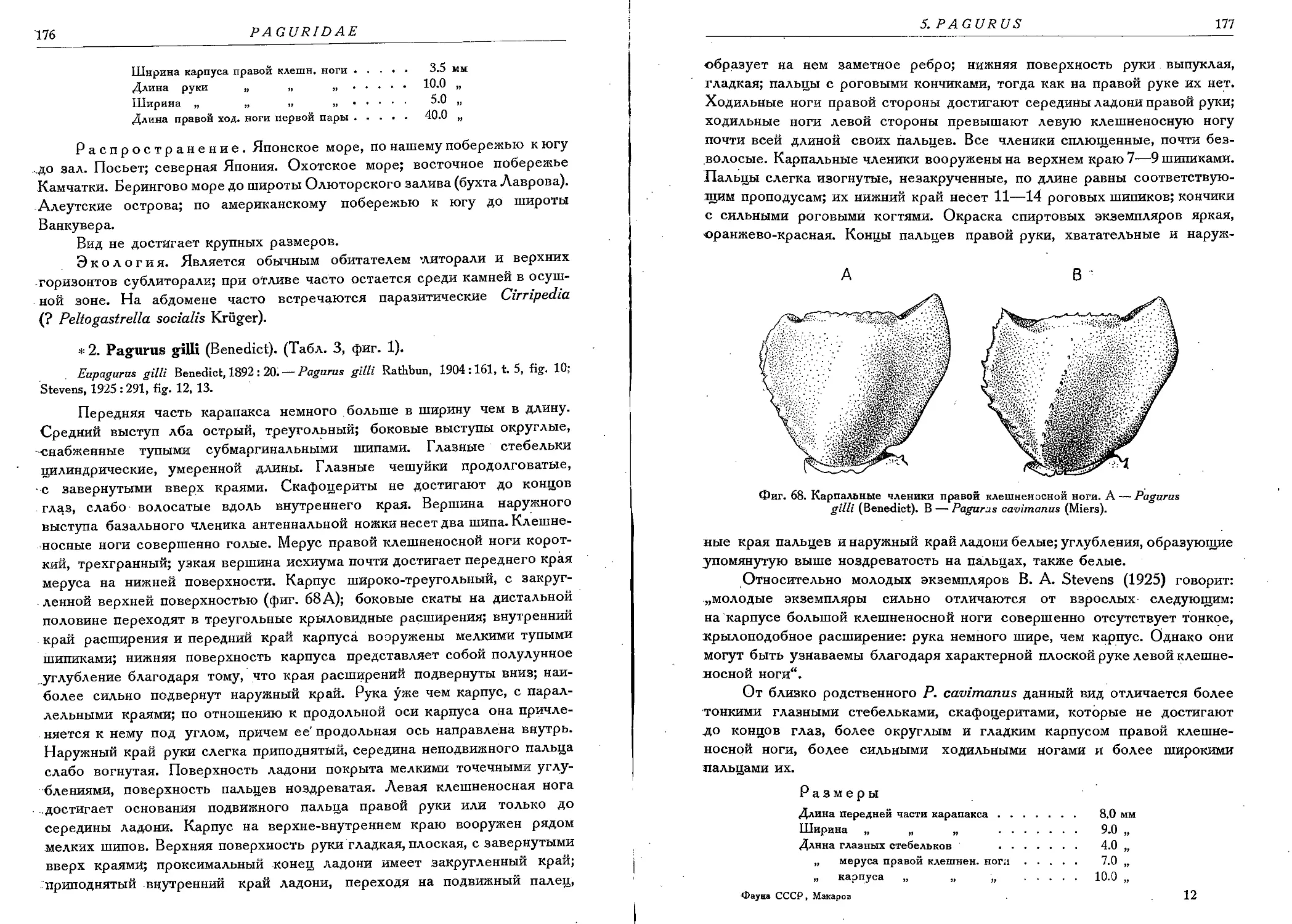
2. P. gilli (Benedict)....................................................... 176
3. P. cavimanus (Miers)...................................................... 178
СИСТЕМАТИЧЕСКИМ УКАЗАТЕЛЬ
IX
Стр.
4. Р. tenuimanus (Dana).......................................................... 179
5. Р. hirsutiusculus (Dana)..................................................... 181
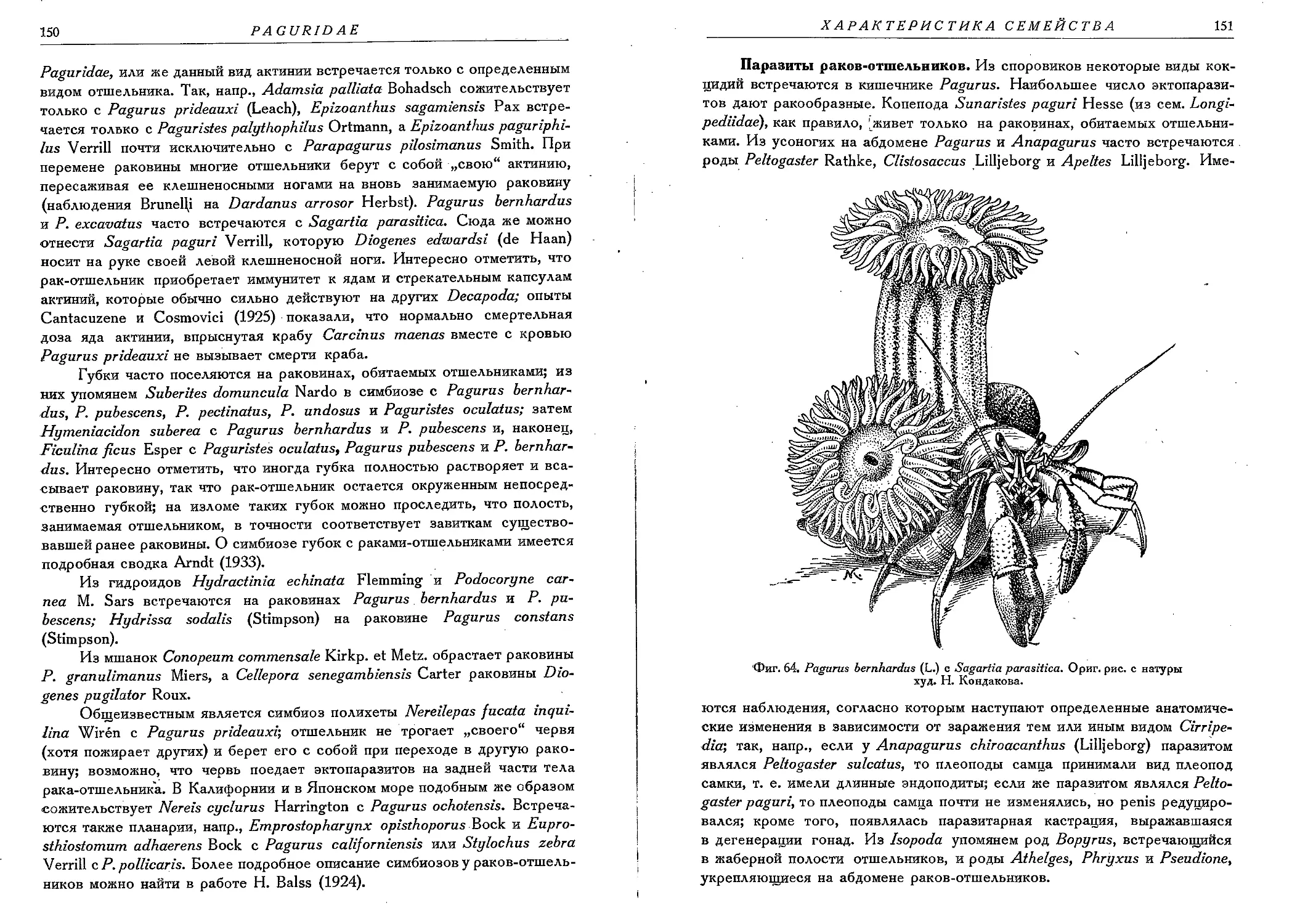
6. Р. bernhardus (Linne)......................................................... 183
7. P. gracilipes (Stimpson)...................................................... 185
8. P. beringanus (Benedict) ................................................... 186
9. P. dubius (Ortmann)........................................................- 188
10. P. samuelis (Stimpson)......................................................... 189
11. P. cornutus (Benedict)......................................................... 191
12. P. undosus (Benedict).......................................................... 192
13. P. tanneri (Benedict).......................................................... 194
14. P. anomalus (Balss)........................................................... 196
15. P. splendescens Owen........................................................... 197
16. P. ochotensis Brandt...................................................... 199
16a.P.ochotensis aleuticus (Benedict)............................................... 202
17. P. rathbuni (Benedict)....................................................... 203
18. P. granosimanus (Stimpson)..................................................... 205
19. P. dalli (Benedict) ........................................................... 206
20. P. pubescens Kroyer......................................................... 208
21. P. brachiomastus (Thallwitz)................................................... 211
22. P. lanuginosus de Haan ........................................................ 212
23. P. armatus (Dana).............................................................. 213
24. P. pectinatus (Stimpson)..................................................... 214
25. P. obtusifrons (Ortmann)....................................................... 216
26. P. setosus (Benedict).......................................................... 216
27. P. kennerlyi (Stimpson)............................................... • .218
28. P. capillatus (Benedict)....................................................... 219
29. P. constans (Stimpson)......................................................... 221
6. Род Parapagurus Smith
1. P. pilosimanus Smith............................................................ 223
2. P. mertensii Holmes............................................................ 226
7. Род Orthopaguras Stevens
1. O. minimus (Holmes)......................................................... 227
2. O. schmitti (Stevens)........................................................ 229
8. Род Spiropaguras Stimpson
1. S. spiriger (de Haan)........................................................... 231
8. Сем. Lithodidae
1. Род Hapalogaster Brandt
1. H. grebnitzkii Schalfeew....................................................... 236
2. H. mertensii Brandt............................................................. 238
3. H. dentata (de Haan)........................................................... 239
2. Род Dermaturus Brandt
1. D. mandtii Brandt............................................................. 242
3. Род Oedignathus Benedict
1. O. inermis (Stimpson)........................................................ 244
X
ANO MURA
Стр.
4. Род Placetron Schalfeew
1. P. wosnessenskii Schalfeew......................................................... 246
5. Род Phyllolithod.es Brandt
1. P. papillosus Brandt............................................................... 251
6. Род Paralithodes Brandt
1. P. camtschatica (Tilesius)......................................................... 254
2. P. platypus Brandt................................................................ 257
3. P. brevipes (Milne Edwards et Lucas)............................................... 262
7. Род Lithod.es Latreille
1. L. aequispina Benedict............................................................. 264
2. L. maja (Linne) ................................................................... 267
3. L. couesi Benedict . .............................................................. 270
8. Род Paralomis White
1. P. multispina (Benedict) .......................................................... 272
2. P. verrilli (Benedict) .......................................................... 273
9. Род Rhinolithodes Brandt
1. R. wosnessenskii Brandt . ........................................................ 275
10. Род Sculptolithodes Makarov
1. S. derjugini Makarov............................................................... 277
11. Род Lopholithodes Brandt
1. L. mandtii Brandt.................................................................. 280
12. Род Cryptolithodes Brandt
1. C. sitchensis Brandt............................................................... 284
2. C. typicus Brandt................................................................. 286
3. C. expansus Miers.................................................................. 287
ВВЕДЕНИЕ
По внешнему виду, разбираемые в настоящем выпуске Decapoda
распадаются на две группы: представители одной группы имеют гарнеле-
видную или шримсообразную (макрурообразную) форму тела, представи-
тели второй группы имеют крабообразную форму тела.1 Этот внешний
habitus в основном определяется формой карапакса и формой и положе-
нием абдомена. У представителей первой группы карапакс продолговатый
и более или менее сжатый с боков, абдомен также удлиненный, вытянутый
и не подогнутый под головогрудь. У представителей второй группы кара-
пакс широкий, сплющенный в дорзо-вентральном направлении, а абдомен
укорочен и подогнут под головогрудь. Верхняя поверхность карапакса
в редких случаях совершенно гладкая, чаще всего она покрыта различного
рода украшениями: то это поперечные борозды или гребни, то тупые бугорки
или острые шипы, то чешуеобразные возвышения, то, наконец, 'крупные,
самой разнообразной формы, выступы и углубления. На верхней поверх-
ности карапакса, в особенности при его крабообразной форме, разли-
чаются следующие области (фиг. 1): по средней линии расположены
непарные лобная, или фронтальная, гастральная, кардиальная и интести-
нальная области; парными областями будут орбитальные, хепатические
и бранхиальные области. Гастральная область в свою очередь распа-
дается на непарные мезо- и урогастральную подобласти и на парные эпи-,
прото- и метагастральные подобласти. Каждая бранхиальная область также
распадается на мезо-, эпи- и метабранхиальные подобласти. Дугообраз-
ная цервикальная борозда разграничивает хепатические и бранхиальные
области и гастральную и кардиальную области. Парные бранхио-кар-
диальные или бранхиальные борозды отграничивают кардиальную область
от бранхиальных областей. В контуре крабообразной формы карапакса
различается передний фронтальный, или лобный, край, который обычно
несет рострум, затем передне-боковые края, которые иногда встречаются
друг с другом под явственным углом, иногда же плавно переходят друг
в друга, и, наконец, задний край. Боковые стороны карапакса носят наз-
вание грудных плевр или бранхиостегитов. Важно отметить, что боковой
1 Здесь мы не будем останавливаться на обзоре своеобразного семейства Paguridae,
жоторый дается во введении к этому семейству.
Фауна СССР, Макаров
1
2
ANOMURA
край карапакса здесь не соответствует границе' между notum и плеврами?
эта граница помещается ниже бокового края, имеет вид перепончатого-
шца и известна у Апотига как linea anomurica или linea thalassinica;,
шов начинается от переднего угла ротового отверстия и тянется назад на
порядочном расстоянии от бокового края карапакса, постепенно прибли-
жаясь к нему, пока, наконец, не достигнет заднего края карапакса над
основанием пятой пары перейопод. Таким образом верхняя половина боко-
вой поверхности карапакса, лежащая между боковым краем карапакса
и linea anomurica, представляет собой подвернутый вниз край notum’a,
тогда как плеврами являются лишь те боковые части, которые находятся
ниже linea anomurica. Передний отдел грудных плевр обозначается как.
птеригостомиальная область, задний отдел — как субхепатическая область.
Linea anomurica особенно резко выражена у рода Pachycheles из сем. Рог-
cellanidae. Карапакс не сливается с эпистомом. Рострум имеет самую
различную форму: иногда он совершенно отсутствует или рудиментарный
(Callianassinae), иногда имеет форму выступа, которым оканчивается при-
поднятая гастральная область (Upogebiinae), иногда он выглядит в виде
широкого цельного треугольника (Hapalogastrinae, Pachycheles) или рас-
сечен на несколько долей (Porcellana)', то он узко треугольной формы
с зубчатыми краями (Galat he а), то в виде длинного, острого, гладкого-
шипа (Munida), то в виде шипа, вооруженного различным числом дополни-
тельных шипов (Lithodes, Paralithodes); в редких случаях рострум имеет
полупластинчатую форму (Cryptolithodes), наконец, у некоторых родов
Lithodinae он имеет вид выступа самой различной формы (Phyllolithodes,.
Lopholithodes, Rhinolithodes, Sculptolithodes). Лобный край карапакса не
образует замкнутых орбит и поэтому глаза имеют свободную подвижность.,
Стернальные пластинки узкие, сливающиеся друг с другом, за исключе-
нием последнего торакального сегмента, стернальная пластинка которого
свободная.
Глазные стебельки чаще всего стебельчатой формы с терминально
расположенными роговицами. Обычно стебелек состоит из двух сегмен-
тов, короткого базального и более длинного терминального. В редких,
случаях, как, напр., у Emerita из Hippidea, базальный сегмент вторично
раздваивается, так что общее число члеников составляет три. Свое-
образное изменение претерпевают стебельки у родов Callianassa-
и А1Ъипеа‘. они сплющены в дорзо-вентральном направлении, имеют
форму треугольной пластинки, причем роговицы находятся на дорзальной
стороне в центре этих пластинок, иногда смещаясь несколько вперед,
или назад.
Антеннулы не обнаруживают каких-либо особенностей: ножка,
состоит из трех члеников, терминальный членик несет два жгута. У рода
Albunea (из Hippidea), ведущего роющий образ жизни, антеннулярные
жгуты, будучи сложены вместе, образуют своего рода трубку для дыха-
тельного тока воды.
ВВЕДЕНИЕ
3
1*
4
ANOMURA
Антеннальная ножка обычно четырехчленистая, в редких случаях
пятичленистая, жгут не такой длинный, как у представителей Natantia.
У рода Emerita опушенные длинными волосками антеннальные жгуты
употребляются для улавливания мелких планктонных водорослей и живот-
ных, служащих в качестве пищи. На втором членике ножки у большинства
представителей Апотига имеется скафоцерит; его форма очень разно-
образна, начиная от маленькой пластинки (Callianassidae) и простого шипа,
до ветвистых шипов, изогнутых зубчатых пластинок (Cryptolithodes) или
длинных шипов, усаженных в виде елочки многочисленными шипами
(Lopholithodes). В общем, скафоцерит заметно редуцирован у группы
Апотига по сравнению с группой Natantia, и утрачивает свою функцию
статического органа в связи с преобладанием ползающего образа жизни;
у шримсообразных форм Апотига, как, напр., у Galatheidae, отсутствие
скафоцеритов компенсируется чрезмерно удлиненными передними перейо-
подами, которые при движении животного назад с помощью сгибательных
и разгибательных движений абдомена вытягиваются в горизонтальной
плоскости и таким образом выполняют статическую функцию.
Ротовые конечности. Жевательная часть мандибул распадается
на головной отдел и апофиз; головной отдел у Апотига не расчленяется
на молярную и инцизорную части; пальп обычно двучленистый. Максил-
лулы имеют две жевательных доли и обычно одночленистый или двучле-
нистый (Thalassinidea) пальп. Максиллы несут четырехдольчатую жеватель-
ную часть, одночленистый пальп (эндоподит) и пластинчатый скафогнат
(экзоподит). Соха и basis первой максиллярной ножки имеют по одной
жевательной доле (у Axius коксальная доля раздвоенная); пальп одночле-
нистый (у Axius двучленистый); экзоподит широкий у основания, с тонким,
иногда многочленистым жгутом. Вторая максиллярная ножка состоит из дву-
членистого протоподита, не несущего никаких жевательных долей,
пятичленистого эндоподита (некоторые членики иногда сливаются) и экзо-
подита с жгутом. По своему строению третьи максиллярные ножки подобны
вторым; исхиум иногда вооружен шиповатой лопастью, употребляющейся
для размельчения пищи (Galatheidea)-, у Galatheidea эндоподитам третьих
максиллярных ножек Zimmermann (1913) приписывает следующие функции:
1) собирание мелких частиц пищи сметающими движениями; 2) очищение
антеннул и антенн; 3) защита при согнутом положении выходных каналов
жаберной полости. Очищение антеннул и антенн у Galatheidae происходит
следующим образом: антеннулы и антенны резко наклоняются вниз,
эндоподиты третьих максиллярных ножек расправляются, после чего
антеннулы и антенны медленно тянутся вверх и таким образом очищаются
волосками трех последних члеников эндоподитов, которые имеют форму
двусторонних пил или гребней. Волоски, приспособленные для сметания
пищевых частиц, длинные и тонкие и несут два ряда мелких ветвей.
У Porcellanidae, которые имеют крабообразный habitus и антенны, кото-
рых в основном утратили свою функцию .органа осязания, наблюдается
ВВЕДЕНИЕ
5
редукция вычесывающих волосков и расширение и уплощение члеников
эндоподита, благодаря чему их третьи максиллярные ножки приближаются
к типу Brachyura (напр., Cancer). У некоторых примитивных Paguridea
третья максиллярная ножка окан-
чивается маленькой клешней.
Экзоподит иногда отсутствует
(Callianassa). По своей форме
третьи максиллярные ножки мо-
гут быть ногообразными или
крышкообразными (фиг. 2); в пер-
вом случае исхиум и мерус та-
кой же ширины или лишь немного
шире следующих члеников, во
втором случае исхиум и мерус
расширены, целиком закрывая
ротовое отверстие*, в то время
как три следующих членика (кар-
пус, проподус и палец), узкие, ма-
ленькие и обозначаются обычно
как пальп. Соотношение рото-
вых частей показывает фиг. 3.
Фиг. 2. Два типа третьих максиллярных ножекг
а—крышкообразный тип (Callianassa harmandi),
b — ногообразный тип (Upogebia major).
Перейоподы. Каждая перейопода состоит из семи члеников; однако»
у рассматриваемой группы basis и ischium сливаются в один членик, так
Фиг. 3. Сагиттальный разрез через переднюю часть голово-
груди Galathea dispersa, показывающий расположение ротовых
частей. А—labrum (верхняя губа); В — мандибулярный пальп;
С — инцизорный отросток мандибулы; D — вторая максилляр-
ная ножка; Е — пищевод; F — молярный отросток мандибулы;
G — максиллула; Н — максилла; I — первая максиллярная
ножка; К — третья максиллярная ножка (из Nicol, 1932).
6
ANOMURA
что получается шестичленистая конечность. Первая пара перейопод почти
всегда снабжена клешнями и в систематических описаниях ноги этой пары
называются клешненосными, в то время как четыре следующих пары
называются ходильными ногами, хотя некоторые из них иногда снабжены
клешнями. Первые перейоподы бывают одинаковой или почти одинаковой
длины и формы {Galatheidea,Upogebiinae), или же обычно правая нога силь-
нее и длиннее левой (большинство Lithodidae)} особенно заметная разница
в длине и форме передних ног наблюдается у Callianassinae. По строению
различают истинную и ложную клешню. У истинной клешни неподвижный
палец (pollex или index) такой же длины, как подвижный палец (dactylus),
при смыкании внутренний край подвижного пальца прилегает к внутреннему
краю неподвижного пальца, или же, когда пальцы изогнутые, сходятся
только кончики пальцев. У ложной клешни неподвижный палец предста-
вляет собой короткий шипообразный отросток проподуса, отходящий от
одного из углов его прямого или несколько скошенного переднего края;
к противоположному углу причленяется подвижный палец, который при
сгибании прилегает своим внутренним краем к переднему краю проподуса
и лишь кончиком касается неподвижного пальца или частично скрещи-
вается с его основанием. Первые перейоподы с ложными клешнями
имеются у Albuneidae и Upogebiinae. Hippidea, в виде исключения, имеют
простые прямые перейоподы, т. е. терминально оканчивающиеся одним
пальцем, причем у рода Mastigochirus этот палец жгутообразно удлинен
и является сегментированным. Передние перейоподы служат для схваты-
вания пищи, а также как орудие нападения и защиты; функция ног только
что упомянутого строения у рода Mastigochirus не выяснена. Вторые
перейоподы простые {Hippidea, Galatheidea, Upogebiinae, Lithodidae) или
клешненосные {Axiidae, Callianassidae). Третья и четвертая ноги простые.
Пятая нога. простая {Hippidea, Axiidae, у последних иногда с ложной
клешней Upogebiinae), или клешненосная {Galatheidea, Callianassinae,
Lithodidae). У форм, ведущих роющий образ жизни, ходильные ноги при-
способлены для рытья или за счет расширения проподусов трех послед-
них перейопод {Callianassidae), или за счет расширения и уплощения паль-
цев трех ходильных ног {Hippidea), как это видно на фиг. 41. Пятые
перейоподы у Hippidea, Galatheidea и Lithodidae редуцированы и служат
для очищения жаберной полости (Putzfiisse); при этом у представителей двух
первых надсемейств они обычно только частично спрятаны в жаберной
полости, так что карпус и мерус, согнутые в своем сочленении, обычно
остаются снаружи; у Lithodidae пятые перейоподы обычно целиком скрыты
под карапаксом и снаружи не видны. У форм, живущих под камнями, как,
напр., у рода Hapalogaster, все перейоподы, а также и карапакс заметно
сплющены. У рода Cryptolithodes и в особенности у Lopholithodes бросается
в глаза уплощенная конгруентная форма боковых поверхностей перейо-
под, бладаря чему все они могут быть плотно прижаты друг к другу
и к карапаксу и образуют прекрасную защиту для всей вентральной
ВВЕДЕНИЕ
7
Фиг. 4. Два типа жабр (схематично): а — филлобранхии;
b — трихобранхии.
поверхности тела у Lopholithodes и совершенно скрываются под карапак-
сом у Cryptholithodes. Замечательное образование имеется у калифорний-
ского Lopholithodes foraminatus (Stimpson): наружный край карпуса
клешненосной ноги имеет глубокую полукруглую выемку, а внутренний
край карпуса первой ходильной ноги имеет такую же, но более мелкую
выемку; когда все ноги плотно прижаты друг к другу, обе выемки оказы-
ваются расположенными друг против друга, так что получается довольно
широкое круглое отверстие (откуда и видовое название), обеспечивающее
свободную циркуля-
цию дыхательного
тока воды.
Жабры. По строе-
нию различают три
типа жабр: дендро-
бранхии (имеются
только у Penaeided),
трихобранхии и фил-
лобранхии. Трихо-
бранхии (фиг. 4Ь) со-
стоят из централь-
ного ствола, усажен-
ного многочислен-
ными тонкими нитями
илитрубочками;по на-
правлению к дисталь-
ному концу ствола
нити утончаются; каждая нить имеет сосудистый канал, сообщающийся
с каналом центрального ствола. Филлобранхии (фиг. 4а) состоят из цен-
трального ствола, несущего два ряда плотно прилегающих друг к другу
листочков или пластинок, которые уменыцаются по направлению к ди-
стальному концу центрального ствола; оба ряда пластинок расположены
на противоположных сторонах ствола. Трихобранхиями обладают Tha-
lassinidea, род Aeglea из Galatheidea и Pylochelidae\ филлобранхии имеют
Hippidea, Galatheidea (за исключением Aeglea) и Lithodidae.
По положению (месту прикрепления) различают три вида жабр:
1) подобранхии — расположены на коксальных члениках максилляр-
ных ножек и перейопод; 2) артробранхии — расположены на сочленовной
мембране между коксальным члеником и телом; 3) плевробранхии — поме-
щаются на боковой стенке тела.
При полной жаберной формуле каждый сегмент груди должен обла-
дать одной подобранхией, двумя артробранхиями (передней и задней) и одной
плевробранхией. Однако в подавляющем большинстве случаев число жабр
меньше., так как те или другие из них обычно отсутствуют. Мастигобран-
хии могут находиться на, коксальных члениках конечностей, начиная от
8
ANOMURA
первой максиллярной ножки до четвертой перейоподы включительно»
У рода Callianidea помимо нормальных, скрытых под карапаксом жабр,
описаны многочисленные жаброобразные нити, подобные трихобранхиям,
которые находятся на краях и наружных поверхностях плеопод второго-
пятого абдоминальных сегментов; они отличаются от трихобранхий нали-
чием отчасти действительной, отчасти намечающейся благодаря перехватам
членистости, а также попарным слиянием у основания в один утолщенный
ствол.
Бранхиостегит неплотно прилегает к коксальным членикам конеч-
ностей, так что имеется Свободный доступ воды к жабрам, обеспечивае-
мый движениями скафогнатов. У Galatheidae и Lithodidae благодаря ре-
дукции пятых перейопод задний край карапакса может приподниматься
и тем самым еще больше облегчать доступ воде. Для очищения жабр,
помимо упомянутых пятых перейопод, используется также изменение
направления дыхательного потока, вызываемое обратными ударами ска-
фогната.
К. Zimmermann (1913) останавливается в своей работе на адаптив-
ных изменениях в группе Galatheidea, связанных с переходом от более
глубинных мест обитания (Galathea squamifera) к жизни на илистых фа-
циях приливо-отливной зоны (Porcellana platycheles); мы отметим особен-
ности, связанные с защитой жаберной полости от засорения илом.
У Galathea squamifera, обитающей в чистой воДе открытой части моря-,
один бранхиостегит является уже достаточной защитой жаберной полости;
волоски, окаймляющие входные отверстия жаберной полости (у заднего
конца карапакса) имеют простое строение, как и волоски на краях бран-
хиостегита; для очистки жабр, именно артробранхий, служат мастиго-
бранхии, находящиеся у G. squamifera на третьих максиллярных ножках
и трех первых перейоподах. У Galathea strigosa в связи с некоторой
редукцией скафогната поддерживание необходимого тока воды вызы-
вается также движениями самого карапакса; эти движения повели
к отсутствию, в виду возросшей опасности отрыва при таких движениях,
эпиподитов на перейоподах; защита жабр от засорения обеспечивается
возрастанием сложности строения волосков у входных отверстий жабер-
ной полости. Примерно то же самое наблюдается и у Galathea intermedia.
У Porcellana, живущей в прибрежной полосе, отсутствуют мастигобранхии
на третьих максиллярных ножках и всех перейоподах, а также отсут-
ствуют и подобранхии, но хорошо развиты артробранхии и плевробранхии;
Zimmermann ставит это в связь с привычкой данного рода цепляться за
различные предметы и растопыривать при этом ноги, а при таких движе-
ниях эпиподиты и подобранхии подвергались бы опасности повреждения
и отрыва или, в лучшем случае, терлись бы друг о друга и о края бран-
хиостегита. В соответствии с этим у Porcellana platycheles,, обитающей
в наиболее заиленных участках, волоски на краях бранхиостегита и на
коксальных члениках перейопод имеют ветвистое и двуперистое строе-
ВВЕДЕНИЕ 9’-
пие, обеспечивающее надежную фильтрацию; в то же время у Р. longicor-
nis, обитающей на каменистых фациях при отсутствии процессов заиле-
ния, волоски на тех же местах имеют простое строение, как у Galathea
squamifera.
Нарост волосков на базальных члениках третьих максиллярных но-
жек, найденный у всех упомянутых видов, защищает выходное отверстие
жаберной полости от посторонних токов воды, которые могли бы задер-
.живать и замедлять дыхательный поток. Для того, чтобы дыхательный
поток шел как можно ближе к жабрам, на внутренней поверхности бран-
хиостегита у Galathea, Munida и в особенности у Porcellana имеется гре-
бень, напоминающий такой же гребень, описанный Pearson’oM у рода..
Cancer. Zimmermann отмечает у Porcellana замечательную привычку чере-~
дования дыхательной функции на обеих сторонах тела; в состоянии покоя,
Porcellana всегда держит одну клешненосную ногу дальше от тела, чем
другую; выносящий дыхательный поток наблюдается только на той сто-
роне тела, где клешненосная нога отодвинута, что заметно по движению
жгута второй максиллярной ножки, по движению взвешенных в воде
частиц и по движению антеннул, которые направлены в сторону дыха-
тельного потока; на другой стороне дыхательный поток неощутим; при-
мерно через полчаса начинается дыхательная деятельность на другой
стороне, прекращаясь на первой. Этим чередованием обеспечивается
отдых мускулам скафогната и очистка жаберной полости, что у таких,
форм, как Carcinus, осуществляется полной приостановкой дыхательной
деятельности на некоторый период на обеих сторонах. Сказанное подтвер-
ждается также и поведением животных при помещении их в ядовитый
раствор, как, напр., в экстракт из красных водорослей; при этом у такой
формы, как Carcinus наступает более длительная приостановка дыха-,
тельного потока, тогда как у Porcellana появляется более быстрое и не-
правильное чередование дыхательной деятельности на обеих сторонах;
Galathea в таких условиях обнаруживала явственную приостановку дыха-
тельной деятельности. Последний членик пятых перейопод, служащих
для очистки как жабр, так и фильтрующих волосков, покрыт волосками,
каждый из которых представляет собой серповидный крючок, усаженный
на внутренней стороне короткими остроконечными зубчиками, и является
таким образом прекрасным скребком. У Porcellana, лишенной эпиподитов,
эта конечность засовывается также и внутрь жаберной полости, что оче-
видно, согласно Zimmermann’y, несвойственно роду Galathea, зато у нее
и у Munida пятые перейоподы служат для очищения также и верхней
поверхности карапакса, пересеченной поперечными бороздами и гребнями.
У форм, ведущих амфибиотический или иногда почти наземный
образ жизни, помимо жабр существуют дополнительные образования,
обеспечивающие газообмен. Так, у Birgus latro (Linne) у наружной стенки
жаберной полости находятся кожистые складки, обильно снабженные кро-
веносными сосудами, приносящими венозную кровь и идущую затем к пет
10
ANOMURA
рикардию; таким образом здесь налицо как бы прототип легочного дыха-
ния, т. к. газообмен осуществляется в воздушной среде. У близкого рода
Coenobita тонкая дорзальная стенка абдомена снабжена богатой капил-
лярной сетью, где несомненно также осуществляется газообмен. Однако
замечено, что, напр., Birgus latro раз в сутки должен заходить в воду,
чтобы смочить жабры. Coenobita в засушливый период впадает в своего
рода спячку, прячась при этом под камнями,
Абдомен. Форма и положение абдомена в основном определяют
вышеупомянутое разделение Апотига на две группы; при этом можно
переход от гарнелевидного habitus’a к habitus’y типичного
'проследить
Фиг. 5. Абдомен самца Pachy-
cheles stevensii (выпрямленный).
краба, что и придает всей группе ее проме-
жуточный характер. Абдомен состоит из семи
сегментов, считая и тельсон за самостоятель-
ный сегмент. У Galatheidae абдомен наиболее
близок к гарнелевидной форме: эпимеры
(плевры) еще хорошо развиты, точно так же
хорошо развит хвостовый веер; отличие за-
ключается в том, что сегменты сплющены
в дорзовентральном направлении и изгиб
абдомена приходится не.на третий сегмент,
как у Natantia, а на четвертый. У Porcella-
nidae, целиком утративших плавающий образ
жизни, абдомен подогнут под торакс, благо-
даря чему приобретается habitus типичного
краба, однако абдомен еще несет черты, ука-
зывающие на сравнительно недавний переход
к ползающему образу жизни (фиг. 5): именно
на каждом сегменте еще имеются эпимеры,
•которые, правда, довольно маленькие и направлены горизонтально, т. е.
•иначе говоря находятся в одной плоскости с тергальной частью сегмен-
тов (или их спинкой); кроме того шестой сегмент еще несет уроподы,
ветви которых имеют форму небольших узких пластинок обычно скры-
тых под тельсоном. Тельсон Galatheidea имеет своеобразную форму: он
плоский и состоит из пяти треугольных частей, соединенных швами.
Среди Thalassinidea у представителей семейства Axiidae абдоминальные
-эпимеры еще хорошо развиты, тогда как у Thalassinidae бросается в глаза
их редукция; в общем абдомен имеет макрурообразную форму. У Hippidea,
как и у Porcellanidae, абдомен подогнут под торакс. Особенного разно-
образия достигает форма абдомена в надсемействе Paguridea в связи
’С различным образом жизни его представителей. У наиболее примитивных
форм, как сем. Pylochelidae, имеются черты как „макрурного" типа, так
и типа отшельника: абдомен вытянутый в длину, явственно сегментиро-
ванный, сегменты сравнительно хорошо объизвествленные, но имеют реду-
цированные эпимеры и хвостовый веер; общий вид напоминает отшель-
ВВЕДЕНИЕ
11
ника (см. фиг. 42), это, так сказать, рак отшельник in statu nascendi;
образ жизни полускрытый — животные, очевидно, часто покидают свои
убежища и свободно ползают по дну. Своеобразные черты строения аб-
домена Paguridae, связанные с постоянным обитанием в полых ино-
родных телах, разбираются в вводной части к этому семейству и здесь
«а них мы останавливаться не будем. Если Pylochelidae являются отшель-
никами in statu nascendi, то Lithodidae являются отшельниками, оконча-
тельно покинувшими свои убежища, но еще не утратившими некоторых
своеобразных черт. Hapalogastrinae, более примитивные среди Lithodidae,
имеют кожистый мягкий мешкообразный, но сильно укороченный абдомен,
сравнительно мало подогнутый под торакс;
некоторые из Hapalogastrinae, как род Нара-
logaster, еще сохранили привычку вести полу-
скрытый образ жизни, прячась под камнями,
благодаря чему форма их тела и конечностей
уплощенные и покровы слабо объизвествлен-
ные; остальные роды Hapalogastrinae имеют
хорошо обьизвествленные карапакс и конеч-
ности. У Lithodinae наблюдается различная
степень объизвествленности абдомена, обычно
в виде отдельных пластинок. Первый сегмент
маленький, узкий и цельный, часто бывает
скрыт под задним краем карапакса, поэтому
у некоторых авторов второй сегмент непра-
вильно называется базальным. Второй сегмент
самый крупный, состоит из непарной медианной
пластинки и парных латеральных и маргиналь-
ных пластинок; пластинки иногда могут сли-
ваться, так что весь сегмент оказывается цель-
ным. После второго сегмента абдомен резко подогнут под торакс и имеет
пластинчатую форму. Три следующих сегмента состоят из медианных
пластинок, на месте которых иногда бывают перепончатые участки,
покрытые отдельными объизвествленными узелками; первые латеральные
пластинки почти всегда имеются; маргинальные пластинки имеются или
отсутствуют, у самки, если имеются, то только на правой стороне. Свое-
образное строение имеют эти три сегмента у рода Neolithodes". вся эта
часть абдомена перепончатая, густо усаженная отдельными мелкими
бугорчатыми пластинками. Шестой сегмент состоит из одной медиан-
ной пластинки. Тельсон маленький, редуцированный. Форма абдоми-
нальных пластинок и характер их расположения, описываемые в спе-
циальной части в родовых диагнозах, имеют систематическое значение.
Как Hapalogastrinae, так и Lithodinae утратили уроподы; самки сохра-
нили еще асимметрию абдомена, тогда как у самцов он вполне сим-
метричный.
Фиг. 6. Вторая пара плеопод
Callianassa californiensis bou-
vieri. ex — экзоподит; en —
эндоподит.
12
ANOMURA
Абдоминальные конечности. Плеоподы состоят из двух коротких
основных члеников (соха и basis) и двух ветвей (экзоподит и эндоподит)>
или одной ветви (эндоподит); второй тип встречается у Galatheidea,
Hippidea и Lithodidae. Первые или вторые плеоподы иногда преобразованы,
в совокупительные органы (Galatheidea, Pglochelidae). На эндоподите;
иногда имеется appendix interna или stylamblys (Thalassinidea, фиг. 6).
На приводимой таблице дается расположение абдоминальных конечностей:
на пяти первых сегментах; левая графа в рубрике пола соответствует
левой стороне абдомена, правая — правой. Об абдоминальных конеч-
ностях шестого сегмента — уроподах — говорилось выше.
Питание. Ротовые части были описаны выше. Желудок распадается
на две части: cardia, функция которой заключается в размельчении пищи
с помощью желудочной мельницы, и pylorus, который выполняет функцию
фильтра; оба отдела выстланы эктодермой. Средняя энтодермальная часть
кишечника сравнительно короткая; вентрально в нее открываются протоки
печени, и дорвально — слепые отростки (соеса); в этом участке кишеч-
ника происходит всасывание. Эктодермальная концевая кишка длинная,
открывается задним проходом на вентральной поверхности тельсона.
В настоящем разделе мы рассмотрим главным образом характер пищи
и способы ее приема.
Способам питания у Galatheidea посвящена работа A. Nicol (1932).
В качестве представителя Galatheidea автор берет Galathea dispersa,
а в качестве типичного представителя Porcellanidae взята Porcellana
longicomis. У обеих способы питания несколько различаются, что связано
и со строением ротовых частей.. Пища Galathea бывает двух сортов: или
это крупные куски животного или растительного происхождения, или же
органические остатки и микроорганизмы из осадков морского дна. Пита-
ние детритом является более обычным. При этом способе питания глав-
ную роль играют третьи максиллярные ножки, густо опушенные на тер-
минальных члениках и производящие по субстрату движения подметания;
обе ножки могут действовать как совместно, так и врозь. Застрявший
между пучками волос детрит счищается вторыми максиллярными ножками,
ВВЕДЕНИЕ
13
при этом последние действуют всегда попеременно; терминальные ще-
тинки вторых максиллярных ножек препровождают захваченные с третьих
максиллярных ножек пищевые частицы к внутренним ротовым частям,
которые и производят сортировку пищи; пригодные частицы направляются
далее в рот, а неподходящие частицы выталкиваются обратно и попадают
в выносящий дыхательный поток воды, которым и уносятся прочь. При
питании крупными кусками, если последние достаточно крупны, то они
.сперва схватываются клешненосными ногами, а затем максиллярными
ножками, или же, при незначительной величине кусков, сразу максилляр-
ными ножками. Пучки волос третьих максиллярных ножек удерживают
выпадающую пищу, а вторые максиллярные' ножки вращают пищевой
кусок, растрепывая его; при этом третьи максиллярные ножки прижимают
пищевой кусок к мандибулам, а вторые максиллярные ножки, вонзая
в пищевой кусок свои терминальные щетинки, могут оттягивать его от
мандибул. Достаточно растрепанный кусок вдавливается третьими максил-
лярными ножками между раздвинутыми мандибулами, которые ватем
смыкаются и инциворными отростками перерезают пищевой кусок, по-
ступающий далее в пищевод и желудок. В желудке Galathea находили
детрит, мелкий песок, мелкие куски красных и зеленых водорослей, диато-
меи, части мелких ракообразных, мелких Gastropoda и полихет, иногда
.крупные куски водорослей и куски мышц.
Porcellana питается исключительно детритом и микроорганивмами
и способ принятия пищи вначительно отличается от такового Galathea.
По сути дела происходит отфильтровывание пищевых частиц черпаком,
образованным густыми двуперистыми волосками третьих максиллярных
ложек; сложными движениями конечностей вокруг животного поддержи-
вается ток воды, несущий взвешенные частицы. Третья максиллярная ножка
вытягивается, при этом волоски ее расправляются, получается большая
ложкообразная сеть, стенки которой образованы волосками и их боко-
выми веточками. Затем максиллярная ножка сгибается и при этом дви-
жении отфильтровываются пищевые частички из значительного объема
воды. Обычно третьи максиллярные ножки двигаются попеременно. Дви-
жение второй максиллярной ножки точно сЪгласовано с движениями
третьей максиллярной ножки; как только последняя сгибается, терми-
нальные пучки волос второй максиллярной ножки вставляются между осно-
ваниями волос третьей максиллярной ножки, при этом вторая максил-
лярная ножка начинает двигаться ко рту, а третья максиллярная ножка раз-
гибается; таким образом осуществляется вычесывание третьей максилляр-
ной ножки, которое может происходить несколько раз подряд при значи-
тельном количестве пищи; затем вторая максиллярная ножка всовывается-
между следующими ротовыми частями и вытаскивается назад, в то время
.как ножка противоположной стороны приближается; таким образом вторые
.максиллярные ножки вычесываются друг другом и следующими ротовыми
•частями; освобожденные пищевые частицы поступают далее в рот. Только
14
A NOMURA
однажды наблюдалась у Porcellana longicornis попытка захватить малень-
кого моллюска, но последний был быстро выброшен обратно в поток,
дыхательной воды. Указание Dalyell’a о питании Porcellana двустворча-
тыми моллюсками и Potts’a о нахождении в желудке крупных кусков,
водорослей наблюдениями Nicol’a не подтверждаются.
Callianassidae, живущие в вырываемых ими норах, являются детритояд-
ными животными; способ питания их описан G. Mac Ginitie (1930) для Upo-
gebia pugettensis (Dana). Животное подползает к выходу из норы, расто-
пыривает первые и вторые перейоподы, волоски которых образуют боль-
шую фильтрационную камеру („корзину", как выражается автор); энер-
гичные движения плеопод вызывают усиленный ток воды, направляющийся,
через волоски перейопод, где и задерживаются взвешенные пищевые
частицы; карапакс плотно прижимается к верхней стенке норы, так что.
весь поток устремляется через фильтрующий аппарат; промежутки между
боками рострума и карпусами клешненосных ног заполняются волосатыми
кончиками третьих максиллярных ножек, чем достигается наибольшее
удерживание пищевых частиц. Через определенные промежутки времени
третьи максиллярные ножки сметают собранный пищевой материал, кото-
рый затем вторыми максиллярными ножками переносится к первым максил-
лярным ножкам, максиллам, а этими последними препровождается в рот.
Крупные час'тицы пищи (напр., куски моллюска в опытах Mac Ginetie}
некоторое время переворачиваются в фильтрационной камере, но затем
выталкиваются обратно током воды, вызываемым резким движением
хвостового плавника. В. A. Stevens (1928) находила в желудках Upogebia
и Callianassa растительные остатки, немногочисленные диатомеи, мелкие
песчинки. По данным Pesta, Upogebia питается мелкими частями моллю-
сков (Modiola, Venus), предварительно раскрывая их раковины.
Среди Lithodidae наиболее изучены в отношении питания промыс-
ловые объекты, главным образом, Paralithodes camtschatica. Tilesius при
описании этого вида указывает, что животное питается головоногими,,
морскими звездами и моллюсками. По данным Marukawa (1933), пищей
Р. camtschatica являются Cynthia superba, Cucumaria japonica, моллюски,
морские ежи и трупы рыб; при этом автор отмечает преобладание в< пище
Cucumaria japonica. Навозов-Лавров (1927) характеризует пищу про-
мыслового камчатского краба как разнообразную: в желудке были нахо-
димы остатки водорослей, позвонки и мелкие кости рыб и чаще всего
остатки морских звезд, морских ежей и моллюсков; известковые части
этих животных используются,- по мнению автора, крабом для образования
панцыря. И. Закс (1936) в качестве основной пищи краба приводит
• мелких моллюсков (Yoldia, Nucula, Venus), баланид, морских ежей, поли-
хет и эхиурид; попутно автор замечает, что по данным В. Фенюк пища
краба чрезвычайно разнообразна, а не ограничена животными, снабжен-
ными известковыми образованиями, хотя при просмотре желудков остатки
таких животных несомненно бросаются в глаза в первую очередь и бла-
ВВЕДЕНИЕ
15
годаря этому создают ложное впечатление превалирования именно этого
сорта пищи. Анализ содержимого желудков нескольких сотен крабов,
произведенный в лаборатории Тихоокеанского Института Рыбного Хозяй-
ства и Океанографии, не подтверждает выводов Marukawa относительно
преобладания в пище краба голотурий. Наблюдали, что молодь Paraiitho-
des camtschatica питается гидрантами Obelia longissima. В условиях аква-
риума мальки, по наблюдениям И. Закса, А. Казаева и Д. Логинович,.
употребляли в пищу литторин, акмей, икру морских ежей, Caprella и кома--
Фиг. 7. Абдомен самца Lopholithodes mandtii, вскрытый сверху. 1о — на-
ружное отверстие левого семепровода; Ivd — левый семепровод; fl — пятая
пара ног; rvd — правый семепровод; го — наружное отверстие правого
семепровода; It — левый семенник; rt — правый семенник; int — кишечник
(из Fasten, 1917).
ров, случайно попадавших в аквариум. Таким образом, Lithodidae очевидно,
являются всеядными, хотя по сравнению с предыдущими группами несо-
мненно преобладает питание животными организмами.
Размножение. Семенники расположены или в полости головогруди,
частично вдаваясь в абдомен (у Upogebia, напр., доходят даже до тельсона),.,
или же целиком расположены в полости абдомена (фиг. 7); они имеют
форму трубок и обычно соединены друг с другом одной или двумя по-
перечными анастомозами. Выходящий неподалеку от заднего конца
семенника семепровод представляет собой или простую трубку, или же
эта трубка на некотором своем протяжении, как это наблюдается у Gala-
theidean Paguridea, образует несколько спиралевидных оборотов; концевая
16
ANOMURA
мускулистая часть семепровода, так называемый ductus ejaculatorius, откры-
вается на коксальных члениках пятых перейопод. Спермин имеют раз-
личную форму и снабжены длинными протоплазматическими отростками,
чаще всего в числе трех, отходящими от шейки спермин. Спермин соеди-
няются обычно в сперматофоры, имеющие очень разнообразную форму.
Яичники представляют собой простые трубки, имеющие такое же
.расположение, как и семенники; обе трубки связаны друг с другом анасто-
мозами, за исключением Thalassinidea. Яйцеводы, отходящие по бокам яич-
ников, представляют собой простые мускулистые трубки, открывающиеся
на коксальных члениках третьих перейопод; receptaculum seminis не имеется.
Фиг. 8. Первая и вторая плео-
поды самца Munidopsis antoni
(из Milne-Edwards et Bou-
vier, 1894).
Преобразование абдоминальных конечно-
стей самцов в половые придатки наблюдается
только у примитивных Paguridea и у Galatheidea.
У Galatheidae это преобразование захватывает
обычно две первых пары плеопод, которые по
своему строению очень напоминают такие же
плеоподы примитивных Paguridea. Каждый при-
даток состоит из трех члеников (фиг. 8): первый
членик иногда бывает рудиментарный, следую-
щий соответствует ножке, а последний—эндо-
подиту, или внутренней ветви; этот последний
членик листовидно расширенный, лодкообразно
выемчатый и более или менее спирально скру-
чен; у плеопод второй пары он снабжен у осно-
вания коротким отростком, который предста-
вляет собой наружную ветвь плеоподы. Эти две пары половых придатков
•встречаются у самцов всех Galatheidae', половые придатки первой пары
отсутствуют у большинства Porcellanidae.
Все Decapoda раздельнополы; лишь в редких случаях наблюдается
гермафродитизм. Так, у рода Calocaris из семейства Axiidae имеется
протандрический гермафродитизм: на втором году жизни развиваются
семенники, которые затем дегенерируют, а яичники развиваются только
на третьем-четвертом году; семепровод, наполненный сперматофорами,
сохраняется в течение долгого времени. Однако такие гермафродитные
формы ведут себя как гонохористы, так как благодаря протандрии обе-
спечивается перекрестное оплодотворение. В семенниках Upogebia major
иногда находили яйцевые клетки, которые позже облитерировались.
В процессе размножения можно наметить четыре следующих стадии:
1) спаривание; 2) икрометание с одновременным оплодотворением; 3) вына-
шивание яиц самками на плеоподах, во время которого проходятся эмбрио-
нальные стадии развития, и 4) вылупление личинок и их дальнейшее
личиночное развитие.
Спариванию обычно предшествует линька самки. Как уже упомина-
лось выше, у Апотига, в отличие от Brachyura, не имеется receptaculum
ВВЕДЕНИЕ
17
seminis и поэтому процесс оплодотворения происходит на наружной
поверхности вентральных покровов самки. Процесс спаривания наблю-
дался лишь у немногих видов. У Galatheidae (Brandes, 1897) самец, лежа
на спине и вытянув абдомен, держит над собой самку таким образом,
что ее брюшная поверхность прилегает к его брюшной поверхности;
первыми ложкообразно расширенными на концах плеоподами самец достает
из своих половых отверстий сперматофоры, которые затем пятыми перейо-
• подами переносятся к половым отверстиям самки и к ее абдоминальным
конечностям. Весь акт продолжается приблизительно около одного часа.
У Lithodidae процесс
•спаривания наблюдался Ма-
rukawa, который описывает
его следующим образом (ци-
тируется по И. Заксу, 1936):
„В садках самцы схватывали
клешнями самок за верхнюю
часть мерусов их клешненос-
ных ног. В течение этого
„рукопожатия" оба пола не
принимали пищи. „Рукопожа-
тие" продолжалось от 3 до
7 дней, причем самка за это
время слиняла, после чего са-
мец снова схватил слинявшую
самку. Затем последовало фиг. 9. Плеоподы первого абдоминального сегмента
самки: а — Hapalogaster grebnitzkii; b — Dermatarus
откладывание икры И ОПЛО- mandtii.
дотворение. Следовательно,
линька самки совпадает со спариванием. „Рукопожатие", сопровождающее
линьку, не является для нее необходимым, так как одиночные самки
линяли нормально и без помощи самца. При откладывании икры самка
„садится" задним краем панцыря на дно и опускает абдомен до земли.
В этом положении яйца, выходят из яичников через половые отверстия
на брюшной стороне коксоподитов третьих перейопод. В это время самец
оплодотворяет икру, выпуская ленту, содержащую сперматофоры, через
отверстие на вершине базиподита. Один самец может оплодотворить за
короткое время несколько самок". По наблюдениям И. Закса и Д. Логи-
нович „рукопожатие" может быть более продолжительным (до двух месяцев
в одном из наблюдавшихся случаев у Paralithodes platypus), хотя обычно
сроки совпадают с таковыми, указанными Marukawa. Согласно тем же
авторам во время „рукопожатия" самец и самка стоят на пальцах перейо-
под, самка не принимает „сидячей позы" и сперматофорная лента выпу-
скается самцами за 1—4 суток до икрометания; после выпуска икры самец
быстро отпускает самку и пара расходится; икрометание продолжается
и после расхождения пары. Н. Навозов-Лавров (1927) несколько иначе
Фауна СССР, Макаров 2
18
ANOMURA
описывает спаривание у Paralithodes camtschaticcr. „самец схватывает
самку за обе клешни и подгибает под себя так, что последняя, пере-
вертываясь вниз спинкой, прилегает грудкой и абдоменом к грудке и
абдомену самца. Самец в это время может свободно передвигаться вместе
с прильнувшей к нему снизу самкой". Число икринок у промысловых Litho-
didae в среднем достигает 200 000 штук. Икрометание у самок с шириной
панциря в 10—12 см (при этой ширине оба пола достигают половой
зрелости) происходит один раз в год. После оплодотворения икры начи-
нается эмбриональное развитие, длящееся 10—11 месяцев, после чего
наступает массовый выход личинок в воду, где они продолжают свое
личиночное развитие до стадии §1апсоЛоё. Сроки икрометания и про-
должительность развития зависят от температурных условий. Так, напр.,
у Сахалина массовый выход личинок наблюдается уже в первых числах
мая, а не в апреле, как это наблюдается в Японском море несколько
южнее; чем дальше на север, тем сроки все больше и больше запаздывают.
Такое явление отмечено и для видов, встречающихся, напр., в Средизем-
ном и Немецком морях: в первом все процессы протекают раньше, во
втором они запаздывают. Икрометание происходит обычно на мелко-
водья, поэтому можно наблюдать нерестовые миграции к берегам. Инте-
ресно отметить, что наземные формы, как, напр., Coenobitidae, или даже:
некоторые пресноводные формы, как Callianassa turnerana, встречающаяся
в пресных водах Камеруна, в период икрометания и вынашивания икры
мигрируют в море.
При развитии Decapoda могут встречаться следующие стадии,
вкратце характеризуемые ниже по Balss’y (1927); развитие, однако,
никогда не проходит через все эти стадии: некоторые из них обычно
выпадают.
1. Nauplius. Тело несегментированное, без панцыря, простой лобный
глаз. Имеется три пары конечностей: одноветвистые антеннулы, двуветви-
стые антенны и двуветвистые мандибулы без жвал; все конечности усажены
плавательными щетинками и служат для гребли. Рядом с замкнутым задним
проходом с каждой стороны находится фуркальная щетинка.
2. Metanauplius. Позади конечностей науплиуса (мандибулы теперь,
с коксальными жвалами) появляются зачатки трех-четырех следующих
конечностей (две пары максилл и одна-две пары максиллярных ножек).
На туловище начинают обособляться некоторые сегменты. Конец тела
с двумя фуркальными отростками;
3. Protozoea. Тело делится на головогрудь (cephalon Ч~ первый тора-
кальный сегмент) и тораксч-абдомен (второй-пятый торакальные сег-
ментыч-абдомен). Сложные глаза помещаются под образовавшимся теперь
панцырем. Антенны все еще служат для плавания, но мандибула уже не
является расщепленной конечностью. Максиллы двуветвистые, более или
менее ногообразные. Перейоподы еще отсутствуют. Абдомен сегмен-
тированный, без плеопод; тельсон раздвоенный.
ВВЕДЕНИЕ
19
4. Zoea (типична у Brachyura). Закладываются перейоподы, которые
развиваются последовательно в направлении от рострума к заднему концу
тела. У Caridea имеется три максиллярных ножки, вообще же имеется
только две первых пары, которые служат для гребли (Апотига и Brachy-
ига). Торакс сперва еще несегментированный, глаза развиваются позже,
чем глазные стебельки. Абдомен расчленяется на сегменты, однако шестой
сегмент на первых стадиях еще сидит с седьмым сегментом.
5. Мизидиая стадия (mysis). Заканчивается сегментация абдомена.
Имеются все перейоподы, именно большей частью в виде расщепленных
конечностей, с экзо- и эндоподитами, которые теперь вместо антенн слу-
жат для гребли.
5а. Metazoea. Если, как у большинства Апотига и Brachyura (за
исключением Dromia), экзоподиты перейопод уже не закладываются, то
говорят о стадии Metazoea.
6. Стадия Decapoda. Ларвальные придатки головогруди и абдомена
исчезают, экзоподиты перейопод редуцируются и теперь для гребли слу-
жат плеоподы. У Paguridea эта стадия носит название £1аисо1Ьоё, у Bra-
chyura— megalopa.
В кратких чертах остановимся на развитии отдельных групп.
У Hippidea личинки напоминают личинок Galatheidea и Paguridea"
карапакс с задне-боковыми краевыми шипами, перейоподы без экзоподи-
тов; в качестве характерного признака они имеют лопастеобразный тель-
сон с многочисленными зубчиками на заднем краю.
Среди Thalassinidea развитие в отдельных семействах характери-
зуется следующими чертами. У Axiidae экзоподиты закладываются на всех
перейоподах, на пятой паре они остаются рудиментарными. Рострум длин-
ный, широкий, с зубчатыми краями. Плеоподы отсутствуют на первом
сегменте. Абдоминальные сегменты на дорзальной стороне без килей.
Тельсон с, многочисленными щетинками на краю.
Callianassinae". рострум как у Axiidae", последние перейоподы боль-
шей частью без экзоподитов. Первые плеоподы и большей частью вторые
и пятые отсутствуют. Имеется длинный шип на втором абдоминальном
сегменте, следующие сегменты с дорзальным гребнем. Тельсон с 17 шипами
(8-ь1+8).
Upogebiinae". рострум маленький, не сплющенный в горизонтальной
плоскости. Экзоподиты имеются только на первых трех перейоподах.
Абдоминальные сегменты без дорзальных или боковых шипов. Плеоподы
на первом сегменте отсутствуют. Webb (1921) следующим образом харак-
теризует отличительные черты четырех личиночных стадий рода Upogebia-.
1) каудальная пластинка простая, несущая 12 шипов; уроподы отсутствуют;
2) каудальная пластинка простая, несущая 15 шипов; уроподы отсутствуют;
3) тельсон и наружные уроподы хорошо развиты и вооружены шипами;
внутренние уроподы имеются, но они маленькие и лишены шипов; 4) обе
пары уропод хорошо развиты и вооружены шипами.
2:
20
ANOMURA
Личинки Galatheidea и Paguridea очень похожи друг на друга; они
имеют очень длинный заостренный рострум; боковой край карапакса
у Galatheidea с каждой стороны оканчивается длинным шипом.
У Galatheidea этот шип на одной из своих сторон или на обеих
сторонах усажен рядом мелких шипиков. Четыре личиночных стадии нор-
мальны, первая с экзоподитами только на двух первых максиллярных
ножках, вторая стадия с экзоподитами также и на третьих ножках (отличие
от Brachyura). Уроподы появляются только на третьей стадии. Перейоподы
уже не имеют экзоподитов, поэтому нет стадии mysis, но имеется стадия
metazoea. У metazoea заметны плеоподы в виде зачатков на втором-
четвертом или на втором-пятом сегментах. Отличительные признаки четы-
рех личиночных стадий по Webb’y следующие: 1) каудальная пластинка
простая, несущая 12 шипов; уроподы отсутствуют; 2) каудальная пластинка
простая, несущая 14 шипов; уроподы отсутствуют. Остальные две стадии,
как у Upogebiinae.
Porcellanidae имеют рострум, который в два раза длиннее тела,
а боковой край карапакса сзади и с каждой стороны также оканчивается
длинным шипом; напротив, шипы на заднем краю карапакса отсутствуют.
Последняя личиночная стадия, как и первая, имеет только две максилляр-
ных ножки с экзоподитами, уроподы отсутствуют, но имеются уже три
или четыре пары плеопод.
У Lithodidae процесс развития прослежен на промысловых крабах,
развитие которых происходит по типу Paguridea. Личинка вылупляется
на стадии рго1огоёа. Рострум подогнут вниз; тельсон и антенны усажены
длинными перистыми щетинками. Продолжительность этой стадии очень
кратковременна, после чего наступает переход в стадию zoea. Zo6a имеет
широкую головогрудь, вооруженную спереди выпрямившимся ростраль-
ным шипом; задне-боковые края карапакса оканчиваются длинными шипами;
задний край карапакса имеет небольшой выступ; поверхность карапакса
покрыта мелкими шипиками; имеются три пары ногочелюстей; абдомен
узкий, оканчивающийся широким тельсоном. В стадии гоёа личинка претер-
певает четыре линьки, после которых она переходит в стадию glaucotlum.
При последующей линьке выходит малек, уже совершенно напоминающий
взрослое животное.
Филогения Anomura. Исходной для всех Anomura является группа
Astacura, от которой развитие Anomura пошло, как нам представляется, по
четырем направлениям (см. ниже схему). Hippidea образуют своеобразную
обособленную группу, ближе всего стоящую к Galatheidea (эпимеры абдо-
минальных сегментов еще сравнительно хорошо развитые, жабры типа
филлобранхий, абдоминальные конечности самки простые); переход
к роющему образу жизни (в песке) наложил характерный отпечаток на
всех представителей группы. Другую *ветвь образуют Galatheidea, ведущие
свободный образ жизни; из них Galatheidae наиболее близки к Astacura
((абдомен с хорошо развитыми эпимерами, карапакс с хорошо развитым
ВВЕДЕНИЕ
21
рострумом); перейдя к ползающему образу жизни, Galatheidae дали в конце
концов Porcellanidae, которые приобрели внешний вид крабов, сохранив,
однако, „аномурные" признаки (карапакс не сливается с эпистомом, пятые
Porcellanidae 1 Lithodinae Hapalogastrinae
Galatheidae Caliianassidae 1
1 Paguridae
Axiidae 1
Galatheidea I Pylochelidae
Hippidea 1 Thalassinidea Astacura । Paguridea I
перейоподы редуцированы, еще имеются уроподы). Третью ветвь образуют
Thalassinidea, сохранившие более примитивные черты, чем обе предыду-
щие группы (жабры типа трихобранхий); представители группы ведут рою-
щий образ жизни, строя норы в илу и песке. От общего ствола с Thalas-
sinidea отделились Paguridea, которые вначале перешли к полускрытному
образу жизни; семейство Pylochelidae имеет еще ряд признаков, сближаю-
щих его именно с Thalassinidea: жабры типа трихобранхий, в особенности же-
форма абдомена, который еще сегментирован и имеет редуцированные-
плевры; правда, уроподы уже приспособлены для удерживания тела в полых,
инородных предметах. Дальнейший переход к постоянному пребыванию
в инородных предметах приводит к типичным Paguridae, которые затем,
покинув свои убежища и перейдя снова к свободному образу жизни, обра-
зовали Lithodidae, в своих наиболее развитых формах принявших анало-
гично Porcellanidae внешний вид крабов. На филогенезе этих двух се-
мейств — Paguridae и Lithodidae, наиболее богато представленных в нашей
фауне, мы остановимся несколько подробнее.
Еще Н. Milne-Edwards (1832), A. Milne-Edwards е^ Lucas (1841), Brandt
(1849, 1850, 1851) указывали на происхождение Lithodes от Pagurus. Boas
(1880,1880а) в своих работах уделяет много внимания этому вопросу; указав
на ошибочность сближения de Наап’ом рода Lithodes с Galatheidea, с одной
стороны, и с Birgus, с другой, он на основании изучения строения, главным
образом, конечностей и абдомена приходит к заключению о близком род-
стве рода Lithodes2 с родом Pagurus (или Eupagurus) или с формой, близко
родственной последнему роду. У обоих родов правая клешненосная нога
больше левой; третьи максиллярные ножки имеют точно такое же строе-
ние, в особенности характерно наличие шипа на исхиуме под зубчатым:
гребнем, который у всех других Decapoda отсутствует. Число жабр оди-
наковое. Самки имеют по одной конечности на левой стороне второго-
пятого абдоминальных сегментов. Стернальная часть первого, абдоминаль-
1 В род Lithodes у Boas’a входит и род Paralithodes.
'22
ANOMURA
ного сегмента плотно соединяется с последним грудным сегментом; стер-
нальные части следующих сегментов мягкие. Спинка первого сегмента
абдомена цельная, как у Pagurus", спинка второго сегмента видоизменена,
т. к. между латеральными пластинками имеется не кожистый участок,
а непарная пластинка, однако спинки следующих сегментов прерваны на
середине перепончатым участком, как и у Pagurus, но в отличие от послед-
него усажены здесь известковыми узелками; кроме того имеются марги-
нальные пластинки, аналогов которых нет у Pagurus. Все пластинки, кото-
рые у Pagurus только хитинизированы, у Lithod.es хорошо объизвествлены.
Абдомен асимметричный, главным образом у самок, так же, как и у Pagurus,
левая сторона всегда более сильно развита. Род Hapalogaster, согласно
Boas, занимает промежуточное положение между Pagurus и Lithodes-, самец,
как и у Lithodes, не имеет абдоминальных конечностей; самки имеют
некоторые конечности на втором-пятом сегментах, но не имеют их на
первом сегменте; вследствие этого парные абдоминальные конечности
первого сегмента самки Lithodes рассматриваются Воаз’ом, как явление
атавизма. Здесь же отметим ошибочность вывода Boas’a: самки всех про-
смотренных видов Hapalogaster, а также Dermaturus и Oedignathus имеют,
правда, сильно редуцированные, парные конечности на первом абдоминаль-
ном сегменте (фиг. 9, стр. 17); таким образом, Hapalogastrinae не предста-
вляют в этом отношении исключения и объяснение наличия парных плео-
под Lithodidae атавизмом, следовательно, отпадает. Спинки второго-пятого
абдоминальных сегментов у Hapalogaster прерваны на середине перепон-
чатым промежутком. Общий вывод Boas’a следующий: Lithodes — это
Pagurus, приспособившийся к свободному образу жизни, точно так же
как Birgus — видоизмененная Coenobita, „отказавшаяся" от раковины. Неко-
торые признаки Lithodes, как то: направление осей сочленения последних
члеников клешненосных ног, наличие парных конечностей на первом абдо-
минальном сегменте, указывают, что форма, от которой произошел Lithodes,
была, может- быть, более близка к Paguristes, чем к Pagurus", однако по
ряду признаков Paguristes стоит дальше от Lithodes, чем Pagurus.
Pagurus, согласно Boas’y, отличается от других отшельников тремя
следующими признаками: 1) стернальная часть, первого абдоминального
сегмента соединяется с последним грудным сегментом, так что подчас
кажется, что последние перейоподы отходят от первого абдоминального
сегмента; у всех остальных отшельников между последним грудным и пер-
вым абдоминальным сегментами имеется сочленовная мембрана; 2) спин-
ные пластинки второго-пятого абдоминальных сегментов разделены на
пары отдельных пластинок; 3) на исхиуме третьей максиллярной ножки
под зубчатым гребнем имеется шип. Во всех этих признаках Lithodes
сходен с Pagurus", единственный пункт расхождения заключается в парных
конечностях первого абдоминального сегмента, которые отсутствуют
у Pagurus. Boas (1924) указывает форму, которая обладает как тремя выше-
упомянутыми признаками, сближающими ее с Pagurus, так и парными
ВВЕДЕНИЕ
23
плеоподами на первом абдоминальном сегменте; это, прежде всего, род
Nematopagurus, который детально сравнивается Воаз’ом с Lithodes, а также
род Руlopagurus’, из этих или близких к ним форм Boas выводит род
Lithodes. Мы не можем останавливаться на всех деталях сравнения Pagu-
rus, Nematopagurus и Lithodes, производимого Воаэ’ом, которые касаются
вооружения пальцев клешненосных ног, относительной длины члеников
двух первых перейопод, формы клешни пятых перейопод, формы ротовых
конечностей (где особенно бросается в глаза постепенная редукция при-
датка на пальпе первой максиллы в направлении Pagurus — Nematopagu-
rus— Lithodes) и редукция щетинок на том же пальпе в обратном напра-
влении, а остановимся лишь на сравнении абдомена; перед тем отметим
еще, что Nematopagurus отличается от Pagurus и Lithodes наличием пар-
ных трубкообразных половых придатков у самца, подобных непарному
придатку рода Spiropagurus, причем левая трубка короче правой.
Абдомен типичных раков отшельников характеризуется следующими
чертами: стернальная или брюшная пластинка первого сегмента образует
сплошную пластинку; на втором-пятом сегментах брюшные пластинки
частично сохраняются лишь у краев, будучи прерваны на середине пере-
пончатым участком; на втором сегменте эти части брюшных пластинок
имеются с обеих сторон, на третьем-пятом сегментах — только на левой
стороне; к этим остаткам брюшных пластинок прикрепляются плеоподы;
брюшные пластинки или непосредственно примыкают к соответствующим
спинным пластинкам, или же разграничены кожистым промежутком; на
границе обеих пластинок могут находиться волоски. Для тергальных, или
спинных, пластинок характерна их разобщенность, только на первом и
втором сегментах они тесно прилегают друг к другу. Для абдомена
Pagurus и Nematopagurus характерно срастание брюшной стороны первого
сегмента с последним торакальным сегментом, разделение спинных пласти-
нок второго-пятого сегментов на отдельные, разобщенные кожистыми
промежутками пластинки (это же Boas наблюдал у одного вида Paguristes),
которые, однако, иногда соединяются узкой поперечной полоской (напр.,
на третьем сегменте у Pagurus bernhardus).
Фиг. 10 представляет собой вскрытые по средней линии вентральной
поверхности и расправленные абдомены самок Nematopagurus и Lithodes
maja, перепончатые части которых удалены. Первый сегмент ничем не
отличается у обоих родов. Второй сегмент, наоборот, сильно видоизме-
нен; у рода Paralithodes медианная пластинка соответствует перепончатой
средней части второго сегмента Pagurus (или Nematopagurus), латераль-
ные пластинки соответствуют спинным пластинкам Pagurus, а маргиналь-
ные пластинки являются новообразованиями, развившимися в перепонча-
тых участках, разобщающих у Pagurus спинные и брюшные пластинки;
в случае рода Lithodes все пять пластинок сливаются в одну. Брюшные
пластинки второго сегмента менее видоизменены, чем спинные. Три
•следующих сегмента сходны у обоих родов, только у Lithodes пластинки
24
ANOMURA
утолстились и теснее прилегают друг к другу в особенности на левой
стороне; маргинальные пластинки и узелки центральной части являются
новообразованиями; вентральные части ничем не отличаются у обоих
родов. Шестой и седьмой сегменты совершенно сходные. Абдомен Pylo-
pagurus почти такой же, как у Nematopagurus, только правые половинки
спинных пластинок третьего-пятого сегментов здесь почти совсем не-
заметны.
В заключение Boas говорит: „Lithodes развился из представи-
теля отшельников, который очевидно был весьма близко родственен1
современным родам Nematopagurus и Pylopagurus (может быть даже
относился к одному из этих родов). Освободясь от раковины, Lithodes
сохранил очень явственные следы своего происхождения; хотя абдомен
уже не помещается в раковину, он развивается, однако, до некоторой
степени так, как если бы он находился в раковине: асимметрия абдоми-
нальных сегментов и самого абдомена, размягчение средних частей
спинных пластинок третьего-пятого сегментов, мягкость почти всей
вентральной поверхности абдомена — все это наследие от отшельников"..
Уроподы (функция удерживания тела в раковине) за ненадобностью
исчезли, четвертые перейоподы вновь удлинились и превратились в ходиль-
ные ноги, а пятые перейоподы — в органы для очищения жабр; карапакс
стал твердым во всех своих частях.
Е. L. Bouvier (1895, 1896), в общем, придерживаясь тех же взглядов
на филогению Lithodidae, в деталях несколько расходится с мнением
Boas’a. На примере Hapalogaster cavicauda автор показывает близкое
сходство их с отшельником: маленький треугольный рострум без субтер-
минального выступа, сердцевидная форма карапакса, узкая передняя часть
суббранхиальных областей, форма кардиальной области, наличие пере-
пончатых линий на боках под linea anomurica, форма ротовых конечностей,
пластинчатый невооруженный скафоцерит. Что касается абдомена, то
Bouvier согласен с Воаэ’ом лишь частично: дорзальная пластинка первого
сегмента является цельной у обоих родов, четвертый и пятый сегменты
образованы у обоих родов парами отдельных пластинок, шестой и седьмой
сегменты образованы непарными пластинками, вся остальная дорзальная
поверхность мягкая, за исключением маленькой пластинки на третьем:
сегменте у Hapalogaster cavicauda, которая соответствует левой тергальной
пластинке Pagurus. Однако строение второго сегмента, по мнению Bouvier,
иное, чем у Pagurus'. обе тергальные пластинки у Hapalogaster состоят
каждая из двух, латеральной и маргинальной, четко разделенных швом;:
в середине между ними на перепончатом участке находятся объизвествлен-
ные узелки; у Pagurus эти пластинки простые, разделенные сплошной
перепончатой областью. Bouvier считает, что тергальные пластинки
Hapalogaster образуются из слияния узелков, подтверждение чему он
усматривает в существовании на краях пластинок мелких бугорков, как бы
еще не совсем слившихся и сгладившихся узелков. Bouvier говорит:
ВВЕДЕНИЕ
25
Фиг. 10. Абдомены еамок Nematopagurus (елева) и Lithodes maja (справа); перепончатые части удалены,
а — acetabulum для последней торакальной ноги; с!-6 — абдоминальные конечности; d1-7 — спинные пластинки
абдоминальных сегментов; f — отросток первой спинной пластинки; h — опушка волос, соответствующаг опушке
на краях эпимеров у Reptantia-, S1, S2 и т. д. — брюшные пластинки; S — край обреза самой передней отрезан-
ной брюшной пластинки; S2 — верхняя часть второй брюшной пластинки (из Boas, 1924).
26
ANOMURA
„следовательно, нельзя сомневаться во вторичном происхождении тергаль-
ных пластинок второго сегмента; главный признак абдомена Н. cavicauda
можно, таким образом, охарактеризовать, сказав, что все тергальные
пластинки Pagurus снова имеются и у вида, за исключением пластинок
третьего сегмента, которые частично исчезают, и пластинок второго
сегмента, которые исчезают полностью; впрочем, эти последние заменяются
известковыми узелками, которые постепенно сливаются и образуют с каждой
стороны две новых тергальных пластинки". Правильность такого вывода
Bouvier усматривает в строении абдомена Phyllolithodes papillosa, у кото-
рого слияние узелков началось с периферии и благодаря этому образовались
пластинки с центральными перепончатыми участками, покрытыми еще
неслившимися узелками; интенсивно идет слияние и на средних перепон-
чатых участках третьего-пятого сегментов, где слившиеся узелки обра-
зуют поперечные пластинки. У рода Neolithodes поражает развитие тер-
гальных пластинок в ущерб известковым узелкам; эти тергальные
пластинки своим положением напоминают соответствующие пластинки
Hapalogaster и отшельников, но отнюдь не гомологичны этим последним
и представляют собой как бы тергальные пластинки второго сегмента
Hapalogastrinae и Phyllolithodes papillosa, новые образования, характерные
для Lithodidae. Род Neolithodes Bouvier считает связывающим Hapalo-
gastrinae с Lithodinae, в то время как Phyllolithodes образует самостоятель-
ную боковую ветвь в филогении Lithodidae. У рода Paralithodes развитие
вторичных пластинок пошло еще дальше, чем у Neolithodes. Объяснение
наличия у Lithodinae парных плеопод на первом сегменте самки атавиз-
мом Bouvier избегает, производя Lithodinae через Neolithodes от Oedigna-
thus и Dermaturus, у последних из которых он, правда, лишь предполагал
наличие этих парных придатков. Как мы видим, предположение Bouvier
оправдалось, но мы можем и Dermaturus, как более „литодиновую" форму
среди Hapalogastrinae, связать с отшельниками через более примитивный
род Hapalogaster, так как все Hapalogastrinae имеют у самок парные
плеоподы на первом сегменте. У рода Acantholithus медианные узелки
третьего-пятого сегментов сливаются в пластинки, между которыми еще
остаются свободные узелки. У Lopholithod.es слияние зашло еще дальше,
так как промежуточные медианные узелки соединяются друг с другом;
кроме того наблюдается слияние маргинальных пластинок третьего сег-
мента с латеральными пластинками не только на левой стороне, но и на
правой. У Paralomis маргинальные пластинки третьего сегмента сливаются
с латеральными на обеих сторонах, маргинальные пластинки двух следую-
щих сегментов часто сливаются между собой. У Rhinolithodes все марги-
нальные пластинки сливаются с соответствующими латеральными пла-
стинками; то же самое наблюдается у Cryptolithodes. У двух последних
родов абдомен самки мало асимметричен, что указывает на тенденцию
абдомена у Lithodinae вновь приобрести симметричную форму, хотя
плеоподы второго-пятого сегментов самки имеются только на левой сто-
ВВЕДЕНИЕ
27
роне и выдают еще „пагуриновый" характер абдомена Lithodidae. В итоге
Bouvier приходит к выводу, „что абдоминальные пластинки второго-пятого
сегментов Lithodidae, хотя и аналогичные по своему положению соответ-
ствующим пластинкам абдомена Paguridae, не представляют с ними ника-
кой действительной гомологии. Чтобы превратиться в типичных Lithodidae,
Paguridae должны были сперва утратить все абдоминальные пластинки
второго-пятого сегментов, затем на обширной перепончатой поверхности
абдомена должны были появиться известковые узелки, через прогрессивное
слияние которых образовались тергальные пластинки". Как видим, вывод,
совершенно противоположный мнению Boas’a. Однако связь брюшных
пластинок, несущих плеоподы, с соответствующими спинными пластинками
подтверждает их гомологию у Paguridae и Lithodidae и, невидимому,
взгляд Boas’a следует признать более правильным.
Мы могли лишь вкратце остановиться на взглядах Bouvier на эволю-
,цию Lithodidae с точки зрения развития абдомена, опустив его исследования
того же вопроса с точки зрения развития рострума и скафоцеритов.
Отметим еще, что исходная форма Lithodidae, согласно Bouvier, прошла
через стадии Tomopagurus, Pylopagurus и Pagurus. В заключение приво-
дим цитату из Bouvier (1895:197), подытоживающую его взгляд на взаимо-
отношение отдельных родов семейства Lithodidae'. „покинув свои раковины,
Paguridae, которые явились исходными формами Lithodidae, вероятно
скрывались под камнями и приобрели до некоторой степени сходство
с Porcellanidae.1 У Hapalogaster сохранилась эта форма приспособления, но
она была только переходной к Dermaturus, который отказался от своего
убежища и стал свободно блуждать в море, как только его карапакс
достаточно объизвествился и вооружился шипами. Здесь находится исход-
ный пункт для всей большой трибы Ostracogastrica (т. е. подсемейства
Lithodinae, В. М.); Hapalogaster и Dermaturus, характеризующиеся отсут-
ствием новых тергальных пластинок на третьем-пятом сегментах, образо-
вали трибу Hapalogastrica (т. е. подсемейство Hapalogastrinae, В. М.).
Phyllolithodes, вероятно, сохранил тот же образ жизни, как и Dermaturus',
во всяком случае, он произошел прямо от этого последнего и отличается
от всех других Ostracogastrica неполным и очень причудливым слиянием
известковых узелков своего абдомена. Phyllolithodes образует в трибе
Ostracogastrica особое подразделение, признаки которого являются укло-
няющимися.1 2 3 Все нормальные представители Lithodinae произошли от
форм, аналогичных Dermaturus hispidus, через более интенсивное объизве-
ствление панцыря и через прогрессивное слияние узелков и твердых
1 В своей следующей работе (1896) Bouvier разделяет Hapalogastrinae на порцел-
ланообразных Hapalogastrica и крабообразных Hapalogastrica. Первые ведут еще полускрыт-
ный образ жизни, прячась под камнями (виды Hapalogaster), вторые свободно бродят
по дну (роды Dermaturus и Oedignathus).
2 Среди Hapalogastrinae подобной уклоняющейся формой является род Placetron
«(Bouvier, 1896).
28
ANOMURA
пластинок абдомена. К Dermaturus hispidus непосредственно примыкает
Neolithodes, у которого начинается слияние известковых узелков, а к Neoli-
thodes приближается Paralithodes; этот последний через посредство
Paralithodes camtschatica служит исходной точкой для ветви Lithodes,
а через посредство Paralithodes brevipes—для всех следующих родов.
Acantholithus происходит от Paralithodes brevipes и еще обнаруживает,
подобно Lithodes и Paralithodes, богатое вооружение шипами; через
редукцию шипов и слияние некоторых маргинальных пластинок он при-
водит к Paralomis, который в свою очередь через этот же процесс ведет
к Rhinolithodes. К Paralithodes brevipes примыкают другие формы,,
характеризующиеся прогрессивной редукцией карапакса, слиянием абдо-
минальных пластинок и, главным образом, расширением карапакса, который
все больше и больше начинает защищать основание перейопод. У Echid-
nocerus (т. е. Lopholithodes, В. М.) эти адаптивные видоизменения еще
мало выражены, но они чрезвычайно развиты у Cryptolithodes, ноги кото-
рого совершенно скрыты под карапаксом и который представляет собой,
конец эволюционной ветви Lithodidae“.
ЗООГЕОГРАФИЧЕСКИЙ ОЧЕРК
По своему распространению фауна Апотига распадается на две
крупные группы: фауну бореальной области и фауну тропической области.
К первой относятся Lithodidae (географический обзор для Paguridae дан:
отдельно), ко второй — все остальные рассматриваемые в настоящем
выпуске группы. И в той и в другой группе имеются выходцы в смежные
области; так как большинство наших морей приурочено к бореальной
области, то коренную, здесь сложившуюся, фауну образуют Lithodidae,
тогда как все остальные представители являются пришельцами тропической
области, находящими в наших или сопредельных водах северную границу
своего распространения. Арктическая область не содержит в своей фауне
ни одного вида обеих групп.
В Черном море встречается только два вида из семейства Callianassidae
(Upogebia littoralisvs. Callianassa pontica) и два вида из семейства Porcellani-
dae (Porcellana longicornis и P. longimana), которые идентичны с соответ-
ствующими видами Средиземного моря и проникли несомненно оттуда.
В Баренцовом море у побережья западного Мурмана находят восточ-
ную границу своего распространения Munida bamffica, Galathea strigosa
(редко) и Lithodes maja; в качестве случайного заноса зарегистрирована
Porcellana longicornis. Первые два вида встречаются довольно редко,
проникая в Баренцово море с теплыми струями Гольфстрима. Бедность
фауны Lithodidae в Баренцовом море, по своему характеру вполне при-
годному для обитания этой группы, объясняется большой отдаленностью
этого моря от центра происхождения и распространения Lithodidae^.
о котором скажем несколько слов ниже.
ВВЕДЕНИЕ
29
ДАЛЬНЕВОСТОЧНЫЕ МОРЯ
ЕВРОПЕЙСКИЕ МОРЯ
Axiidae
Фиг. 11. Количественные соотношения числа видов
по семействам в европейских и дальневосточных морях.
Таким образом, в наших европейских морях насчитывается всего
11 видов Апотига (включая сюда и четырех отшельников), что соста-
вляет примерно 29% общего числа представителей этой группы, известных
для наших вод; остальные 71% или 41 вид составляют фауну дальне-
восточных морей (включая в это число и отшельников). 38 видов, или
примерно 42%, из приводимых в настоящем выпуске видов приходится
на фауну сопредельных вод, главным образом, западного побережья
.Японии и западного побережья Северной Америки. В нижеследующей
табличке даются количе-
ственные соотношения ви-
дов по семействам в наших
европейских и дальневос-
точных морях:
Европейские моря
Thalassinidea . . 2 (18 0/q)
Galatheidae ... 2 (18%)
Porcellanidae . . 2 (18%)
Pagaridae ... 4 (37%)
Lithodidae . . . 1 ( 9%)
Дальневосточные моря
Axiidae .... 1 ( 2%)
Thalassinidea . 6 (15%)
Galatheidae . . 1 ( 2%)
Porcellanidae . 1 ( 2%)
Albuneidae . . 1 ( 2°|o)
Pagaridae ... 21 (52%)
Lithodidae . . 10 (25%)
Те же соотношения
представлены на фиг. 11
в виде диаграмм, где пло-
щадь круга пропорциональна числу видов, а секторы показывают про-
центное соотношение числа видов по семействам. Как видно из диа-
граммы и как упоминалось выше, основное ядро фауны дальневосточных
морей, количественно почти в 4 раза превышающей фауну наших евро-
пейских морей, составляют Paguridea; среди последних отшельники вдвое
превышают численность Lithodidae, а последние по числу видов как раз
равны числу видов всех остальных семейств вместе взятых. На диаграммах
также хорошо видно, что фауна наших европейских морей не имеет
в группе Апотига основного ядра и состоит из довольно равномерного
числа пришлых элементов. Преобладание в фауне дальневосточных морей
Pagaridae, этого в основном тропического семейства, происходит главным
образом за счет рода Pagurus, который в своем распространении приуро-
чен преимущественно к бореальной области. Не принимая в дальнейшем
изложении во внимание семейство Paguridae, рассмотрим фауну Апотига
дальневосточных морей.
30 ANO MURA
В фауне Апотига дальневосточных морей можно наметить четыре-
следующих группы:
1) Виды, свойственные всем трем дальневосточным морям: Hapalo-
gaster grebnitzkii, Dertnaturus mandtii, Paralithodes camtschatica, P. platy~
pus и P. brcvipes.
2) Виды, общие Охотскому и Японскому морям: сюда относится
только один вид — Sculptolithodes derjugini.
3) Виды, общие Охотскому и Берингову морям: Munidopsis beringana
и Lithodes aequispina.
4) Группа тождественных или близких видов с прерывчатым распро-
странением, т. е. встречающихся у противоположных побережий северного
Тихого океана и отсутствующих в промежуточных областях (амфипацифи-
ческие виды, как предложил называть их А. Андрияшев):
Азиатское побережье Американское побережье
Callianassa californiensis bouvieri Callianassa californiensis
Callianassa gigas eoa Callianassa gigas
Oedignathus inermis
Cryptolithodes expansus Cryptolithodes typicus
Перейдем к рассмотрению фауны по отдельным морям.
Японское море. Двадцать пять видов, известных для Японского
моря, можно разбить на три следующие группы:
1) Виды, общие нашему побережью и побережью Японии: Upogebia
major, Callianassa harmandi, C. japonica, C. californiensis bouvieri,
Pachgcheles stevensi, Blepharipoda japonica, Hapalogaster dentata, Oedi-
gnathus inermis, Paralithodes camtschatica и P. brevipes.
2) Виды, встречающиеся только у нашего побережья Японского
моря: Axiopsis princeps, Upogebia issaeffi, Callianassa gigas eoa, Hapaloga-
ster grebnitzkii, Dermaturus mandti, Paralithodes platypus, Sculptolithodes
derjugini (последний вид, правда, обнаружен у острова Ришири, но южнее
по японскому побережью онне известен).
3) Виды, свойственные только японскому побережью: Upogebia
yokoyai, Ctenocheles balssi, Galathea acanthomera, G. Integra, G. pubescens,
Munida japonica, Cervimunida princeps, Mixtopagurus jeffreysii и Cryptoli-
thodes expansus.
В качестве эндемиков Японского моря можно назвать: Axiopsis
princeps, Upogebia issaeffi, Callianassa gigas eoa и Ctenocheles balssi.
3. И. Кобяковой (Труды Лен. Общ. Ест., LXV, 2, 1936) отмечен новый
подвид Axiopsis spinulicauda amurensis для Амурского залива, приведен-
ный под родовым названием Axius; в виду того, что описание подвида
еще не опубликовано, в систематической части настоящей работы он не
упоминается. Виды, общие с японским побережьем, и виды, свойственные
только японскому побережью, в общем, являются довольно сильно лока-
лизованными формами, не заходящими на юг дальше южной оконечности
ВВЕДЕНИЕ
31
острова Ниппон; только Callianassa harmandi, С. japonica и С. californien-
sis bouvieri известны также из Желтого моря, a Galathea acanthomera
доходит до Бонинских островов. Бросается в глаза полное отсутствие
представителей семейства Galatheidae у нашего побережья Японского,
моря.
Охотское. море. Фауна Апотига Охотского моря состоит из
9 следующих видов: Calastacus quinqueseriatusf Munidopsis beringana,
Hapalogaster grebnitzkiii, Dermaturus mandtii, Paralithodes camtschatica,
P. platypus, P. brevipes, Lithodes aequispina и Sculptolithodes derjugini.
Таким образом, более 50% фауны Апотига Охотского моря состоит из.
видов, общих всем трем дальневосточным морям. Большинство видов
проникает в Охотское море из северной части Тихого океана. Sculptolitho-.
des derjuginiявляется, очевидно, охотоморской формой (Шантарское море),
проникающей отсюда лишь в самую северную часть Японского моря. Ни
один из видов япономорского происхождения не проникает в Охотское -
море.
Восточное побережье Камчатки. За исключением Sculptolithodes
derjugini, все виды, приведенные для Охотского моря, могут быть отне-.
сены и к фауне восточного побережья Камчатки. Правда, Calastacus.
quinqueseriatus, Munidopsis beringana и Lithodes aequispina еще не были
зарегистрированы у этого побережья Камчатки, однако, в виду близкого,
подхода глубин к побережью в этом районе, нахождение здесь этих:
глубоководных форм весьма вероятно. Здесь же отмечен Paralomis
multispina — глубоководная форма северной части Тихого океана.
Берингово море. Фауна Апотига Берингова моря насчитывает
13 видов, которые могут быть разбиты на две группы:
1) Виды, встречающиеся как в западной, так и в восточной полович
нах моря: Hapalogaster grebnitzkii, Dermaturus mandtii, Paralithodes,
camtschatica, P. platypus и P. brevipes.
2) Виды, встречающиеся только в восточной половине моря: Muni-,
dopsis beringana, Hapalagaster mertensii, Oedignathus inermis, Placetron
wosnessenskii, Lithodes aequispina, L. couesi, Paralomis verrilli и Crypto-,
lithodes typicus.
Весьма возможно, что Munidopsis beringana и Lithodes aequispina
при более детальных исследованиях глубин Берингова моря войдут-
в первую группу. Нет ни одного вида, который встречался бы только,
в западной половине Берингова моря; правда, Paralithodes platypus как
будто бы подпадает под эту категорию видов, так как он не упоминается,
в американской литературе для восточной половины Берингова моря,,
однако экземпляры Richters’a с Прибыловых островов относятся к этому
виду и поэтому мы включаем его в первую группу.
1 Отмечен 3. Кобяковой (1936) для Охотского моря без указания более точ-..
ного местонахождения.
3'2
ANOMURA
Если обратить внимание па распространение всех видов Берингова
моря в широтном направлении, то бросается в глаза приуроченность
большинства их к южной части моря, главным образом, к району Алеут-
ской гряды, и быстрое выпадение их по мере продвижения на север.
Так, напр., только в районе Алеутской гряды встречаются Hapalogaster
mertensii, Oedignathus inermis, Placetron wosnessenskii, Paralithod.es brevi-
pes, Lithodes aequispina, L. couesi и Cryptolithodes typicus, т. e. примерно
54% всего числа видов. В районе мыса Олюторского по нашему побе-
режью и у Прибыловых островов по американскому побережью выпадают:
Munidopsis beringana, Dermaturus mandtii, Paralithodes camtschatica (no
американскому побережью до залива Нортон), Lithodes aequispina, Para-
lomis verrilli. Только два вида, Hapalogaster grebnitzkii и Paralithodes
platypus, доходят до Берингова пролива.
Чукотское море. За исключением Paguridae в Чукотском море не
встречается ни одного представителя группы Anomura.
Как уже упоминалось выше, основное ядро фауны Anomura дальне-
восточных морей составляют Paguridea', так как Paguridae разбираются
в отдельном географическом обзоре, то мы остановимся несколько по-
дробнее на распространении семейства Lithodidae, используя при этом
в первую очередь прекрасный очерк Bouvier (1896), дополнив его новыми
данными. Прежде всего выясним количественные соотношения Lithodidae
IB различных областях мирового океана; при этом мы не будем загромо-
ждать изложение перечислением названий видов, а ограничимся приведе-
нием лишь процентных соотношений, которые будут наиболее показа-
тельны. Из всего количества известных видов Lithodidae 82% приходится
на Тихий океан и 18% на Атлантический океан. В Атлантическом
океане неизвестно ни одного рода, отличного от родов Тихого океана,
а в количественном отношении роды, представленные в Атлантике,
составляют 28% от общего числа родов Тихого океана. Если мы
посмотрим на распределение Lithodidae в широтном направлении в обоих
океанах, то на северное полушарие падает 87% всего числа видов,
а на южное 11%; в южном полушарии неизвестно ни одного рода,
отличного от родов северного полушария, а в количественном отно-
шении они составляют 21% от общего числа родов северного полу-
шария. Из всего сказанного вытекает, что Lithodidae в своей большей
массе приурочены, во-первых, к северному полушарию, а, во-вторых,—
к Тихому океану.
Рассмотрим теперь распределение Lithodidae в Тихом и Атлан-
тическом океанах отдельно. В Тихом океане на северное полушарие при-
ходится 89% числа видов, а на южное 11%. В Атлантическом океане на
северное полушарие приходится 80% числа видов, а на южное 20%.
Таким образом, соотношение примерно одинаковое как в обоих океанах,
так и при сравнении с вышеприведенным соотношением в мировом океане.
Наибольший интерес представляет распределение в каждом океане в его
ВВЕДЕНИЕ
33
от ГриНича iso
Фиг. 12. Центр происхождения н пути распространения Lithodidae.
Фауна СССР, Макаров
3
34 A NOM URA J
западной и восточной половинах. В Тихом океане количественные соотно-
шения видов будут следующие:
виды, свойственные западной половине..........2О°/о
„ „ восточной „ ..........62%
„ общие обеим половинам ...... 18%
Для Атлантического океана те же цифры будут: для западной
половины 50%, для восточной половины 30% и общие обеим поло-
винам 20%.
Из приведенных цифр получается второй вывод, что в Тихом океане
основная масса видов приурочена к восточной половине северной части
океана, а в Атлантическом значительно меньшее преобладание наблюдается
в западной половине северной части океана.
Если теперь учесть, что на ряду с количественным преобладанием
Lithodidae в северо-восточной части Тихого океана только к этой же части
приурочены Hapalogastrinae, т. е. наиболее примитивные представители
Lithodidae, то станет вполне очевидным, что эта часть океана могла быть
центром возникновения Lithodidae, откуда они начали свое расселение по,
мировому океану. В этой же части океана они, невидимому, проделали
и всю свою эволюцию, так как на ряду с Hapalogastrinae здесь находятся
почти все роды семейства, включая и такие специализованные формы
как Cryptolithodes. Все Hapalogastrinae являются литоральными или
сублиторальными видами; распространяясь по Алеутской гряде, восточ-
ному побережью Камчатки и по Курильской гряде, они проникли в Япон-
ское море и будучи формами холодолюбивыми,, нашли здесь южный
предел своего распространения {Hapalogaster dentata в южной Японии,,
как последний представитель этого подсемейства, Hapalogaster grebnitzkii
в заливе Гумбольдта, как наиболее южный представитель на западном
побережьи Северной Америки). Среди Lithodidae часть видов осталась
литоральными или сублиторальными {Phyllolithodes,. Cryptolithodes, неко-
торые виды Rhinolithodes и Lopholithodes) и как раз эти же виды приуро-
чены, главным образом, к северо-восточной части Тихого океана, тогда
как другая часть видов ушла на глубины и за счет этих-то видов и про-
изошло расселение Lithodidae в мировом океане; именно, наиболее широка
распространенным является абиссальный род Paralomis', в умеренной зоне
южного полушария некоторые виды вторично поднимаются в сублитораль,
как, напр., Paralomis verrucosa, однако большинство видов, проникших
в Атлантику, осталось глубоководными (500—2300 м). При своем рас-
пространении в Тихом океане на юг Lithodidae придерживались западного,
побережья Америки, т. е. того побережья, где наблюдается подъем холод-
ных глубинных вод; большинство видов, найденных в тропической зоне
Тихого океана, были встречены именно у этого побережья, тогда как
у азиатского побережья даже глубоководные-виды не пересекают экватор;
ВВЕДЕНИЕ
35
Bouvier (1896) объясняет отсутствие Lithodidae в южных широтах азиат-
ского побережья Тихого океана существованием некогда здесь китайско-
австралийского континента, разделившегося впоследствии на отдельные
острова; наличие этой суши и разыгрывавшиеся здесь орогенические
процессы препятствовали, по мнению Bouvier, расселению Lithodidae.
Нам кажется, однако, более вероятным объяснение распространения
вдоль западного побережья Америки температурными условиями, которые
для холодолюбивых Lithodidae были здесь более благоприятны благодаря
нагону холодных глубинных вод. Проникнув в южное прлушарие, одни
формы вторично поднялись в сублитораль {Lithodes antarctica и Paralomis
verrucosa Южной Америки), другие, оставшись абиссальными, обогнули
Южную Америку и проникли отсюда в южную Атлантику {Paralomis formosa
до широты Рио де ла Платы) и в Индийский океан {Lithodes murrayi
и Paralomis aculeata). Lithodidae северной Атлантики проникли туда иным
путем; наиболее ясен он в отношении Lithodes maja, который из Берингова
моря проник через Берингов пролив на север, обогнул Северную Америку
и распространился по всей северной Атлантике; близкое сходство era
с беринговоморским Lithodes couesl, от которого он очевидно и произошел,
подтверждает этот путь проникновения. Что касается остальных видов,
северной Атлантики, которые все являются глубоководными, то здесь
возможны два пути. Bouvier полагал, что они шли тем же путем, что.
и Lithodes maja-, мне кажется, однако, более вероятным путь вокруг Южной
Америки и подъем на север по западному побережью Африки, где также:
наблюдается нагон холодных глубинных вод. Для рода Lithodes весьма
возможно, что общая исходная форма, выйдя с одной стороны на субли-
тораль Южной Америки, дала здесь L. antarctica, а с другой стороны,,
оставшись глубоководной и пройдя указанным путем, дала у побе-
режья северной Африки L. ferox и L. tropicalis. Этот же путь наибо-
лее вероятен и для видов глубоководного рода Paralomis, каковыми
в северной Атлантике являются Р. bouvieri и Р. spectabilis. Несколько^
труднее установить путь проникновения Rhinolithodes biscayensis, найден-
ного в Гасконском заливе на глубине 1400 м; вероятнее всего, что он
шел также южным путем, огибая Южную Америку, однако, в настоящее
время ареал рода разорван и Rhinolithodes является типичным амфибо-
реальным родом. Наиболее труден вопрос о происхождении рода Neolitho-
des, два вида которого встречаются в северной части Атлантики.. По
строению своего абдомена род является наиболее примитивным среди
Lithodinae и как бы связывает последних с Hapalogastrinae. Процесс
вторичного возврата какого-либо атлантического представителя Lithodinae
к более примитивной форме трудно допустим; с другой стороны, в Атлан-
тике, как уже говорилось выше, нет ни одного представителя Hapalo-
gastr'nae, от которого мог бы произойти Neolithodes и в то же время
в Тихом океане этот род отсутствует, что еще больше осложняет вопрос,
если Lithodes diomedae, описанный Benedict’oM у Чилийских берегов
3*
36 ANOMURA
Америки, окажется, как предполагал Bouvier, принадлежащим к роду
Neolithodes, то для проникновения в Атлантику наиболее вероятным будет
также южный путь: Hapadogastrinae северного Тихого океана дали
в южном полушарии род Neolithodes, который проник в северную Атлан-
тику и ареал которого в настоящее время разорван. На фиг. 12 указаны
описанные выше пути распространения Lithodidae, причем для Атлантики
даны два возможных варианта, хотя наиболее вероятным, за исключением
Lithodes maja, мне представляется южный путь с проникновением на север
вдоль западного побережья Африки. Будущие, более детальные глубоко-
водные работы в тропической и умеренных областях как Тихого, так
и Атлантического океанов пополнят наши знания о составе фауны этих
•областей и тем самым внесут ясность в затронутые здесь вопросы, пока же
мы должны ограничиться вышеизложенной схемой, основанной на сов-
ременных, сравнительно отрывочных данных.
ХОЗЯЙСТВЕННОЕ ЗНАЧЕНИЕ
Промысловыми объектами нашей фауны Decapoda являются так
называемые промысловые „крабы"1 дальневосточных морей: Paralithodes
camtschatica, Р. platypus и Р. brevipes. Наибольшее значение в промысле
принадлежит первому виду. Самый интенсивный промысел приурочен
к западному побережью Камчатки, затем идет побережье Приморья
и, наконец, восточное побережье Камчатки; намечаются возможности
промысла Р. platypus в Беринговом море в районе Олюторского залива
и по побережью от Мыса Олюторского до мыса Наварина. Пионерами
крабового промысла были японцы; интенсивность промысла заметно воз-
росла с момента введения в практику консервирования крабов на бере-
говых консервных заводах и в особенности с 1920 года, когда на ряду
с береговыми заводами вступили в строй пловучие заводы, так .называе-
мые краболовы. Наш советский крабовый промысел начал интенсивно раз-
виваться лишь с 1923 года; в 1932 году появляются первые советские
краболовы. Мы не располагаем цифровыми материалами вылова крабов
и поэтому ограничимся приведением лишь немногих цифр из скудной
промысловой литературы; к сожалению, материалы, накопленные крабовой
бригадой Тихоокеанского Института Рыбного Хозяйства и Океано-
графии руководимой И. Г. Заксом, остаются в виде неопубликованных
рукописей.
Попов (1930) приводит следующую таблицу уловов крабов на запад-
ном побережьи Камчатки (в штуках):
1 Л. Г. Виноградовым (1931) была сделана попытка ввести термин „крабоид" с целью
подчеркнуть систематическое положение рассматриваемых промысловых объектов; однако
термин не привился и в промысловых кругах сохранилось название „крабы", с точки
зрения систематики, конечно, неверное.
ВВЕДЕНИЕ
37
1923 1924 1925 1926 1927 1928 1929
696 630 964430 2 963100 4454 090 5121 ПО 6 759 050 7 845360
Мы не даем приводимых автором в этой же таблице уловов на участ-
ках, арендованных японцами, так как при перерасчете на ящики получается
значительное расхождение с цифрами японской статистики.
Вылов краба японскими краболовами у западного побережья Кам-
чатки выражается следующими цифрами в штуках (по Попову, 1930):
1923 1924 1925 1926 1927 1928 1929
2391 230 282 070 7 353 853 15 360822 2254279 20562568 24 033503
Советскими краболовами в 1920 году в тех же районах добыто
4345925 штук крабов.
Продукция береговых заводов Приморья выражается следующими
цифрами (в ящиках, из расчета 55—60 крабов на ящик) (по Заксу, 1936):
1928 1929 1930 1931 1932 1933
2746 8405 6500 6512 5678 5333
По данным Marukawa общая продукция Японии по крабовым кон-
сервам в 1930 г. составила 58 000 ящиков.
Сроки лова краба связаны с его миграциями к берегам. Весенний
нерестовый подход начинается в Приморье в марте; сперва подходят
икряные самки, а за ними с некоторым опозданием подходят самцы. Послё'
периода размножения в июне самцы откочевывают на глубины, где линяют
в своей большей массе и с конца сентября снова начинают подходить
к берегу. У западного побережья Камчатки весенний подход падает на.
конец апреля. Обратное откочевывание самцов тесно связано с темпера-
турным режимом прибрежной полосы; при повышении температуры выше
4° самец обычно уходит на глубины, что для восточного побережья Кам-
чатки падает на июнь; осенний подход начинается обычно в сентябре.
Относительно того, как ведет себя краб в зимние месяцы, в литературе
имеются разногласия. У японцев существует такой взгляд, что зимой'
у краба наступает спячка, при которой он зарывается в ил. С другой
стороны, имеются указания о попадании крабов в уловы оттертрала
в зимние месяцы. По данным Л. Виноградова (1933) зимой в заливе Петра
Великого краб держится на глубинах 100—160 метров и вполне возможен
38
ANOMURA
его зимний промысел; правда, отмечается его незначительная подвижность,
но для промысла это является только плюсом; кроме того и по качеству
в этот период краб бывает наиболее полноценным, т. к. наиболее удален
от периода линьки.
Замечено, что при своих миграциях краб придерживается ложбинок,
пониженных участков дна. Скорость передвижения составляет примерно
1.5—2.0 км в сутки; по данным Marukawa она равна 2—4 милям в сутки.
Лов краба производится ставными сетями. Высыпка сетей производится
с таким расчетом, чтобы пересечь путь миграции краба; изучение этих
путей в каждом промысловом районе составляет поэтому один из важней-
ших вопросов. Для лова крабов употребляют два сорта ставных сетей:
корейский тип имеет крупную ячею, 20—28 см от узла до узла и высоту
имеет обычно 2’/2—З1^ ячеи; японский тип сетей при том же диаметре
Фиг. 13. Схема установки крабовых сетей (из Навозова-Лаврова, 1927).
ячеи имеет большую высоту — 7^2 ячей; иногда встречается промежуточный
тип: ячея несколько меньше, чем у корейского типа, при высоте в 4т/2 ячеи.
Через каждые три ячеи сеть причаливается к верхней подборе; к этой же
подборе подвязываются наплава, (балберы), придающие ей пловучесть,
а к нижней подборе — грузила. Длина каждой сети составляет 38—47
метров. При высыпке сети связываются в порядки (до 100 сетей
в порядке), при этом оба свободных конца с каждой стороны загружаются
якорем и имеют веху с буйком; на фиг. 13 дана схема установленной
крабовой сети. Сети ставятся обычно на глубине 50—120 метров и оста-
вляются в море на 3—4 дня (корейский тип) или на 7 дней (японский тип),
по истечении этого срока сети выбираются прямо через борт кунгаса или
кавасаки, или же на специальный ворот; если кавасаки обслуживают бере-
говой завод, то при выборке сетей сейчас же производится и отцепка
краба; на кавасаки, входящих в флотилию краболова, сеть без выпутки
поднимается на борт и доставляется на краболов, где уже и производится
отцепка краба. Затем краба сортируют и после приемки пускают в обра-
ботку.
Интенсивное развитие промысла, в особенности в Японии, повело
уже к некоторому истощению естественных запасов краба. Для охраны
сырьевой базы предложены следующие меры: запрет вылова самок
ВВЕДЕНИЕ
39
и недомерков, широкое применение разведки, выявляющей промысловые
скопления крабов и состав косяков, запрещение производства промысла
ставными сетями с ячеей ниже установленного размера, искусственное
разведение, охрана естественных нерестилищ и другие. Попытки аккли-
матизации камчатского промыслового краба в Баренцевом море пока,
кажется, не увенчались успехом.
Вред. Роющие формы, куда прежде всего следует отнести Callianas-
sidae, наносят крупный ущерб устричному хозяйству. Норы этих животных,
занимающие иногда всю приливо-
отливную зону, достигают в глубину
до одного метра и имеют цилиндри-
ческую форму; исследованием нор
Calllanassidae занимались В. A. Ste-
vens (1928) и Mac Ginitie (1930). Пер-
вый автор при изучении формы нор
пользовался следующей методикой:
во время отлива в нору вливался
жидкий гипс; при следующем отливе
слепок выкапывался отдельными
фрагментами и склеивался; полу-
ченные таким образом фигуры
(см. Stevens 1928, фиг. 20) имели
U-образную форму с одним или
несколькими слепыми ответвлениями.
Mac Ginitie описывает несколько
иную форму норы: имеется главный
ход, начинающийся от поверхности,
который пройдя на некоторой глубине
в горизонтальном направлении, снова
открывается наружу несколькими
Фнг. 14. Промысловые термины, употреб-
ляющиеся при разделке конечностей про-
мысловых крабов: 1 — розочка; 2 — толстая
ножка; 3 — коленце; 4 — тонкая ножка
(из Навозова-Лаврова, 1927).
разветвлениями; в нескольких местах имеются расширения, где животные
могут поворачиваться. Процесс рытья подробно изучен Mac Ginitie в спе-
циально сконструированных стеклянных сосудах. Рытье производится
двумя первыми перейоподами, вырытый материал выносится на поверхность
<с помощью тех же ног. Хождение по норе совершается с помощью третьих
и пятых перейопод, тогда как четвертые, упираясь в стенки норы, выпол-
няют статическую функцию. Тельсон, будучи расправлен, плотно загора-
живает вход в нору и служит надежной защитой; однако, если потревожить
животное, сзади, то оно доходит до ближайшего расширения норы, пово-
рачивается там и возвращается назад так, чтобы быть обращенным клешне-
носными ногами в сторону опасности. Аэрация поддерживается движениями
плеопод. В одной норе обычно находится пара животных — самец и самка.
Нанесение вреда устричному хозяйству идет по двум направлениям.
Во-первых, выносимым на поверхность илом и песком Callianassidae засы-
40
ANOMURA
пают устричную икру и молодь. Во-вторых, они подкапываются под
искусственно сооружаемые для выведения устриц хранилища. Так как
устричные банки при отливе часто обнажаются, то для предотвращения
этого определенные участки, где развивается главным образом молодь,
обносятся невысокой цементной дамбой, задерживающей воду над таким
участком при отливе; если Callianassidae изобильны в этом же районе,
то значительная часть нор оказывается расположенной таким образом,
что их входные отверстия находятся ниже участка, огороженного дамбой,
а выходные — под этим участком; при отливе вода из огороженного участка
стекает по норам и устричник обнажается. Многие устричные хозяйства
западного побережья Северной Америки были разрушены таким образом..
Засыпка участка слоем гравия или ракушняка не составляет препятствия
для Callianassidae. Цементирование или мощение крупными кусками камня
слишком дороги. Наиболее распространенной в Америке мерой защиты
является настил двойного досчатого пола: на огораживаемом участке
снимается слой грунта толщиной в 10—12 см, затем настилается один,
слой досок и сверх него под прямым углом кладутся доски второго ряда;;
вынутым ранее грунтом настил засыпается. При отсутствии трещин
подобный настил непроницаем для Callianassidae.
Враги. Врагами рассматриваемой группы прежде всего являются
рыбы. В Немецком море скаты, камбалы, акулы и осетры часто выкапы-
вают Thalassinidae из их нор. У экземпляров Hemilepidotus gilberti и
Myoxocephalus stelleri, добытых у южной оконечности острова Сахалина,,
желудки, вскрытые А. Андрияшевым, оказались сплошь набитыми экзем-
плярами Dermaturus mandtii. В особенности после линьки многие раки
поедаются рыбами. Среди головоногих осьминог часто нападает на про-
мысловых дальневосточных крабов. В период личиночной жизни промы-
словые крабы служат пищей кете, горбуше и анчоусам. Интересно отметить,
что такая полуназемная форма, как Birgus latro, на многих островах
была истреблена крысами.
Паразиты. Из червей немертина Cephalothrix galatheae Dieck пара-
зитирует на жабрах Galatheidae. Из паразитических усоногих чаще всего
встречаются Rhizocephala-, так, на Callianassidae паразитирует род Parthe-
пореа Kossman, на Galatheidea роды Lernaeodiscus Muller и Triangulus
Smith, на Lithodidae — род Peltogaster Rathke. Из паразитических Isopoda
встречаются преимущественно роды из семейства Bopyridae, как, напр.,
род Gyge на Upogebia и Galathea, род Jone на Callianassa и Upogebia,
род Pleurocrypta на Galathea и Porcellana и род Pseudione на Callianassa,.
Galathea и Munida.
Комменсализм и симбиозы. За исключением Pagaridae, эти явления
сравнительно редки у рассматриваемой группы. В норах Upogebia stellata.
на побережьи Девоншира, по наблюдениям Norman’a, всегда живет моллюск
Lepton squamosum (Montagu). В норах Upogebia pugettensis, обитающей
у западного побережья Северной Америки, живет краб Scleroplaxgranulata.
ВВЕДЕНИЕ
4Т
Rathbun из семейства Pinnotheridae и один вид полихеты Harmothoe°}-
в норах Callianassa живет тот же краб и другой вид полихеты того же
рода Harmothoe. Иногда в норах Upogebia, как отмечает Mac Ginitie,
встречается маленький бычок Clevelandia ios Jordan et Gilbert, который
скрывается там от опасности или от обнажения при отливе. Itiro
Miyazaki1 недавно описал комменсального двустворчатого моллюска,
найденного прикрепленным на вентральной поверхности головогруди
Upogebia major, систематическое положение моллюска еще не выяс-
нено» и он приводится в работе провизорно под названием Erycina sp.
Н. Навозов-Лавров (1927) отмечает на карапаксе Paralithodes camtscha-.
tica в качестве симбионта Eumicrotremus pacificus, указывая на чрезвы-
чайное сходство внешнего вида этой рыбки с поверхностью панцыря рака,
обеспечивающее ей надежную защиту; к сожалению, автор не говорит
о частоте встречаемости этого'явления, требующего поэтому дальнейших,
наблюдений.
ЛИТЕРАТУРА
А 1 с о с к, А., 1905. Catalogue of the Indian Decapod Crustacea in the collection of the Indian ,
Museum. Part. II: Anomura, fasc. I: Pagurides.
Arndt, W., 1933. Die biologischen Beziehungen zwischen Schwammen und Krebsen. Mitt. aus
dem Zoolog. Museum in Berlin, XIX.
Balss, H., 1913. Ostasiatischen Decapoden. I. Die Galatheiden und Paguriden. AbhandL.
der math.-phys. Klasse der K. Bayer. Akademie der Wissenschaften, 2. Supl.-Bd., 9.
---- 1913a. Diagnosen neuer ostasiatischen Macruren. Zoolog. Anzeiger, XLII, 5.
—— 1924. Ueber Anpassungen und Symbiose der Paguriden, eine zusammenfassende
Uebersicht. Zeitschr. fur Morph, und Oekol. der Tiere, I.
---- 1926. Decapoda. Die Tierwelt der Nord- und Ostsee, VI, 10 h2.
Bate, Sp., 1888. Report on the Crustacea Macrura (Anomura) dredged by H. M. S. „Challen-
ger" during the years 1873—1876. Challenger Rep., Zoology, XXIV.
Bell, T„ 1853. A history of the British stalk-eyed Crustacea.
Benedict, J. E., 1892. Preliminary descriptions of thirty-seven new species of hermit
crabs of the genus Eupagurus in the U. S. Nat. Museum. Proc. U. St. Nat. Mus., XV..
---- 1894. Description of new genera and species of crabs of the family Lithodidae with
notes on the young of Lithodes camtschaticus and Lithodes brevipes. Ibidem, XVII.
----1901. The hermit crabs of the Pagurus bernhardus type. Ibidem, XXIII.
---- 1903. Description of a new genus and forty-six new species of Crustaceans of the family
Galatheidae, with a list of the known marine species. Ibidem, XXVI.
(B i r u 1 а, А. А.) Б и p у л я, A. A., 1897. Материалы для биологии и зоогеографии пре-
имущественно русских морей. III. Очерк фауны Crustacea-Decapoda морей Мур-,
манского и Белого. Ежегодн. Зоол. Музея Акад. Наук, II.
---- 1906. Zoologische Ergebnisse der russischen Expeditionen nach Spitzbergen. Crustacea-
Decapoda. Ibidem, XL
Boas, J. E., 1880. Studier over Decapodemes Slaegtskabsforhold. Danske vidensk. selsk..
Skrifter. (6), I.
---- 1880a. Lithodes und Pagurus. Zool. Anzeiger, III.
1 J. Miyazaki. On the development of some marine bivalves with special reference-,
to the shelled larvae. Journal Imperial Fisheries Institute, vol. 31, № 2, 1936.
42
AN О MUR A
В о a s, J. E., 1924. Die verwmidtxclinftliche Stellung der Gattung Lithodes. Biol. Meddel. Kgl.
Danske Vidensk. Яо1икн1>., IV, 4.
---- 1926. Zur Konnlnia uymmetrischer Paguriden. Ibidem, V, 6.
Borradniio, L, A., 1903. On the classification of the Thalassinidae. Ann. and Mag. of
Nnt. Hintory, (7), XII.
----19(17. On the classification of the Decapod Crustaceans. Ibidem, (7), XIX.
Bouvier, E. L., 1894. Recherches sur les affinites des Lithodes et des Lomis avec les Pagu-
rides. Ann. Scien. Natur., Zoologie, (7), XVIII.
----1895. Sur la distribution geographique des Crustaces de la sousfamille des Lithodines.
Bull, du Museum d’hist, natur., 2.
---- 1896. Sur la classification des Lithodines et sur leur distribution dans les oceans. Ann.
Scien. Natur., Zoologie, (8), I.
---- 1898. Observations nouvelles sur les Blepharipoda Randall (Albunhippa Edw.). Ann. de
la Societe entomol. de France, LXVII.
Brandt, F., (1848) 1849. Die Gattung Lithodes Latreille nebst vier neuen ihr verwandten
von Wosnessenski entdeckten, als Typen einer besondern Unterabtheilung (Tribus Litho-
dea) der Edwards’schen Anomuren. Bull, de la classe phys.-math. de 1’Acad. Sciences
de St. Petersbourg, VII.
----(1849) 1850. Vorlaufige Bemerkungen uber eine neue aus zwei noch unbeschriebenen
Gattungen und Arten gebildete Unterabtheilung (Hapalogastrica) der Tribus Lithodina,
begleitet von einer Charakteristik der eben genannten Tribus der Anomuren. Ibidem, VIII.
---- 1851. Krebse, in: Dr. A. Th. v. Middendorff’s Reise in den aussersten Norden und Osten
Sibiriens. II, Zoologie, I.
---- 1853. Ueber eine neue Art der Gattung Cryptolithodes (C. sitchensis). Bull, de la classe
phys.-math. de 1’Acad. Sciences de St. Petersbourg, XI.
(Brazhnikov, W.) Бражников, В., 1907. Материалы по фауне русских восточных
морей, собранные шхуною „Сторож" в 1899—1902 гг. Зап. Акад. Наук, XX, 6.
Caiman, W. Т., 1898. On a collection of Crustacea from Puget Sound. Ann. of the New
York Acad. Scien., XI, 13.
(Czerniavsky, V.) Чернявский, В., 1884. Прибрежные десятиногие ракообразные
Понта. Харьков.
Dana, J. D., 1852. Crustacea. United States Exploring Expedition during the years 1838—
1842, XIII, I (Atlas 1855).
D о f 1 e i n, F., 1900. Die dekapoden Krebse der arktischen Meere. Fauna arctica, I, 2.
---- 1902. Ostasiatische Decapoden. Abhandl. math. phys.-Klass. K. Bayer. Akad. Wissen-
schaften, XXI, 1.
Faxon, W., 1895. The stalk-eyed Crustacea of the „Albatross". Mem. ot the Mus. Compar.
Zoology Harvard College, XVIII.
Grieg, J. A., 1927. Decapod Crustacea from the west coast of Norway and the North
Atlantic. Bergens Mus. Aarb. Naturv., 7.
Haan, W. d e, 1850. Crustacea. Fauna Japonica.
H a n s e n, H. J., 1908. Crustacea Malacostraca I. The Danish Ingolf Expedition, III, 2.
Heller, C., 1863. Crustaceen des siidlichen Europa. Crustacea podophthalmia. Wien.
Henderson, J. R., 1888. Report on the Anomura collected by H. M. S. „Challenger"
during the years 1873—1876. Challenger Rep., Zoology, XXVII.
Holmes, S. J., 1900. Synopsis of California stalk-eyed Crustacea. Occas. Papers of the
Calif. Acad. Scien., VII.
Jackson, H. G., 1913. Memoir on Eupagurus. Proc, and Trans, of the Liverp. Biolog.
Society, XXVII.
Jarzynsky, Th. (Яржинский, Ф.), 1870. Praemissus catalogus Crustaceorum decapodum,
inventorum in mari albo et in mari glaciali ad litus murmanica anno 1869 et 1870.
Kishinouye, K., 1926. Two rare and remarkable forms of macrurous Crustacea from
Japan. Annot. Zool. Japon., XI, 1.
ВВЕДЕНИЕ
43
Leach, W. E., 1815. Malacostraca podophthalmata Britannia. London.
Lenz, H., 1901. Ergebnisse einer Reise nach dem Pacific (Schauninsland). Crustaceen. Zool.
Jahrb., System., XI /, 6.
Mac Ginitie, G. E., 1930. The natural history of the mud shrimp Upogebia pugettensis
(Dana). Ann. and Mag. of Natur. History, VI, 31.
Makarov, W., 1934. Eine neue Gattung der Familie Lithodidae aus dem Japanischen
Meer. Zoolog. Anzeiger, CVIII, 9—10.
---- 1935. Beschreibung neuer Dekapoden-Formen aus den Meeren des Fernen Ostens.
Ibidem, CIX, 11—12.
Man, J. G. de, 1927. A contribution to the knowledge of twenty-one species of the genus
Upogebia Leach. Capita Zoologica, II, 5.
---- 1928. A contribution to the knowledge of twenty-two species and three varietites of the
Genus Callianassa Leach. Ibidem, II, 6.
Miers, E. J., 1879. On a collection of Crustacea made by Capt. H. C. St. John., R. N.,
in the Corean and Japanese seas. Proc. Zoolog. Society London for 1879.
Milne-Edwards, H., 1837. Histoire naturelle des Crustaces, comprenant 1’anatomie,
la physiologie et la classification de ces animaux, II. Paris.
—— 1848. Note sur quelques nouvelles especes du genre Pagure. Ann. des Scien. Natur., (3), X.
M i 1 n e-E d w a r d s, A. e t Bouvier, E. L., 1893. Description des Crustaces de la
familie des Paguriens recueillis pendant I’expedition du Blake. Mem. Mus. Compar.
Zool. Harvard College, XIV, 3.
---- 1894. Considerations generales sur la familie des Galatheidae. Ann. des Scien. Natur.,
Zoologie, (7), XVI.
M u r d о c h, J. A. M., 1885. Expedition to Point Barrow, Alaska. V. Marine Invertebrates.
Crustacea. Report International Polar Expedition to Point Barrow, Alaska.
(N a v о z o v -Lavrov, N.) H а в о з о в-Л а в р о в, Н., 1927. Материалы к этологии и про-
мыслу краба в заливе Петра Великого. Производит. Силы Дальнего Востока, 4.
---- 1929. Новый вид среднехвостых раков из Охотского моря, Paralithodes longirostris,
n. sp. Русский Гидробиол. жури., VIII, 6—7.
Nicol, Е. А. Т., 1932. The feeding habits of the Galatheidea. Jour. Mar. Biol. Assoc., XVIII, 1.
'Ortmann, A., 1892. Die Decapoden-Krebse des Strassburger Museums. Zool. Jahrb.,
System., VI, 2.
Orton, J. H., 1927. On the mode of feeding of the hermit crab, Eupagurus bernhardus
and some other Decapoda. Jour. Mar. Biol. Assoc., XIV, 4.
(Paulson, О.) Паулисон, О., 1875. Исследование ракообразных Красного моря,
с заметками относительно ракообразных других морей. Ч. I: Podophthalmata и Edrio-
phthalmata (Cumacea). Киев.
(Popo v, V. К.) Попов, В. К., 1930. Материалы по крабовому промыслу в водах западной
Камчатки. Социал. Реконструкция Рыбн. Хозяйства Дальнего Востока, 5—6.
Queen, J., 1930. Additional Brachyura and crab-like Anomura from Friday harbor,
Washington. Public. Puget Sound Biol. Station, VII.
.Randall, J. W., 1839. Catalogue of the Crustacea brought by Th. Nuttal and J. K.
Townsend from the west coast of North America and the Sandwich Islands and descrip-
tion of such species as are apparently new among which are included several species
of different localities previously existing in the collection of the Academy. Jour. Acad.
Nat. Scien. Philadelphia, VIII, 1.
Rathbun, M., 1902. Japanese stalk-eyed Crustaceans. Proc. U. St. Nat. Museum, XXVI.
---- 1904. Decapod Crustaceans of the North-west coast of North America. Harriman
Alaska Expedition, X-
---- 1919. The Decapod Crustaceans of the Canadian Arctic Expedition 1913—1918. Rep.
of the Canad. Arctic Exped., VII: Crustacea, A. Decapod Crustaceans.
Richters, F., 1883. Beitrag zur Kenntniss der Crustaceenfauna des Behringsmeeres.
Abh. der Senckenberg. Naturforsch. Gesellschaft, XIII.
44
ANOMURA
R i н a о, A., 1826. Histoire niiturollo dos principales productions de 1’Europe meridionale
et particulierement do Col low dos environs de Nice et des Alpes maritimes, 5. Paris.
Sc half ее w, P., (1892) IH’M. Carcinologische Bemerkungen. Mel. biol. Acad. Scien..
St. Petersb., XIII, 3.
Schmitt, W. I... 1921. The marine Decapod Crustacea of California. Univ. Calif. Public,
in Zoology, XXIII.
S i v e r t m о n, E., 1932. Crustacea Decapoda and Mysidacea from the east Siberian and Chu-
kotuk seas. The Norwegian North Polar Expedition with the ,,Maud“ 1918—1925. Scien-
tific. Results, V, 13.
Smith, S. I., 1880. The stalk-eyed Crustaceans of the atlantic coast of North America north'
of Cape Cod. Trans. Connecticut Acad, of Arts and Sciences, V, 1.
---- 1882. Blake, report on the Crustacea. Bull. Museum Comparat. Zoology Harv. College, X.
Stevens, B. A., 1925. Hermit crabs of Friday Harbor, Washington. Public. Puget Sound
Bio). Station, HI.
---- 1927. Orthopagurus, a new genus of Paguridae from Pacific coast. Ibidem,. V.
---- 1928. Callianassidae from the west coast of North America. Ibidem, VI.
Stimpson, W., 1858. Prodromus discriptionis animalium evertebratorum in expeditione
ad Oceanum Pacificum Septentrionalem missa, C. Ringgold et Johanne Rodgers duci-
bus, observatorum et descriptorum. VII: Crustacea Anomura. Proc. Acad. Nat..
Scien. Philadelphia, (2), X.
---- 1854. Descriptions of new species of marine invertebrata from Puget Sound. Ibidem, XVI.
---- 1907. Report on the Crustacea (Brachyura and Anomura) collected by the North Pacific
Exploring Expedition 1853—1856. Smiths. Miscell. Collections, XLIX..
Terao, A., 1913. A catalogue of hermit crabs found in Japan (Paguridea excluding Litho-
didae). Annot. Zool. Japon., VIII, 2.
Thallwitz, J., 1892. Decapoden-Studien insbesondere basirt auf A. B..Meyer’s Sammlun-
gen in Ostindischen Archipel, nebst einer Aufzahlung der Decapoden und Stomatopo-
den des Dresdener Museums. Abhandl. und Berichte Konig. zoolog. und anthrop.-
ethnogr. Museums zu Dresden, 1890—1891, 3.
T i 1 e s i u s, W. G., 1815. De cancris camtschaticis, Oniscis, Entomostracis et Cancellus
marinis microscopicis noctilucentibus cum tabulis IV aenaeis et appendice adnexa
de Acaris et Ricinis camtschaticis. Mem. Acad. Scien. St. Petersbourg, V.
Walker A. O., 1898. Crustacea collected by W. A. Herdman in Puget Sound. Proc, and
Trans, of the Liverp. Biol. Society, XII.
Way, E., 1917. Brachyura and crab-like Anomura of Friday Harbor, Washington. Puget
Sound Marine Station Publication, I.
Webb, G. E., 1921. The . larvae of the Decapoda Macrura and Anomura of Plymouth.
Jour. Mar. Biol. Assoc., XII.
White, A., 1856. Some remarks on Crustacea of the genus Lithodes, with a brief descrip-
tion of a species apparently hitherto unrecorded. Proc. Zoolog. Society London, XXIV-
Y о к о у a, Y., 1928. Report of the biological survey of Mutsu Bay. 10. Brachyura and crab-
shaped Anomura. Science Reports of the Tohoku Imperial University (4), III, 4, 2.
---- 1930. Report of the biological survey of Mutsu Bay. 16. Macrura of Mutsu Bay. Ibidem, V, 3...
---- 1933. On the distribution of Decapod Crustaceans inhabiting the continental shelf around
Japan chiefly based upon the materials collected by S. S. Soyo Maru, during the
years 1923—1930. Jour. College Agric. Tokyo Imper. Univ. XII, I.
Y u, Sh. Ch., 1931. On some species of shrimp-shaped Anomura from North China. Bull-
Fan Memorial Inst, of Biology, II, 6.
(Zachs, J.) Закс, И. Г., 1935. Биология и промысел краба (Paralithodes) в Приморье.
Вести. Д. В. Филиала Акад. Наук СССР, XVIII.
Zimmermann. К., 1913. Habit and habitat in the Galatheidea. Journ. Mar. BioL
Assoc., X.
СПЕЦИАЛЬНАЯ ЧАСТЬ
Карапакс обычно не сливается с эпистомом. Последний торакаль-
сный сегмент свободный (стернальная пластинка этого сегмента не сли-
вается с предыдущей). Скафоцериты обычно имеются. Наружные максил-
•лярные ножки в большинстве случаев узкие, ногообразные. Третья пара
перейопод всегда без клешней. Пятая пара перейопод по величине и по
положению всегда отличается от третьей пары. Абдомен макрурообразный,
т. е. вытянутый в длину, или же уплощенный и подогнутый под карапакс,
иногда мешкообразный и перепончатый; шестой сегмент обычно имеет
парные конечности-уроподы (отсутствуют только у Lithodidae). Поло-
вые отверстия расположены на коксальных члениках.
ОПРЕДЕЛИТЕЛЬНАЯ ТАБЛИЦА НАДСЕМЕЙСТВ
1 (2). Палец вторых-четвертых перейопод изогнутый и сплющенный.
Первая пара перейопод иглообразная или с ложными клешнями.
Хвостовой веер не приспособлен для плавания. Абдомен подогнут
под торакс. Рострум маленький или отсутствует. Наружные максил-
лярные ножки без мастигобранхий......................III. Hippidea.
2 (1). Палец вторых-четвертых перейопод не изогнутый и не сплющен-
ный. Первая пара перейопод не иглообразная, редко с ложными
клешнями.
3 (6). Шестые абдоминальные конечности приспособлены для плавания
(за исключением рода Thalassina, где они иглообразные). Абдомен
симметричный; абдоминальные плевры обычно хорошо развиты.
4 (5). Тело сплющенное. Имеются плевробранхии на последних перейо-
подах. Часто имеется поперечный шов на тельсоне. Абдомен более
или менее согнутый.................................II. Galatheidea.
5 (4). Тело сжатое с боков. Без плевробранхий на последних перейоподах.
Без поперечного шва на тельсоне. Абдомен прямой . I. Thalassinidea.
6 (3). Шестые абдоминальные конечности, если они имеются имеют не
широкие и не иглообразные ветви, но приспособлены для удержи-
вания тела в полых предметах. Абдоминальные плевры имеются
в редких случаях. Абдомен почти всегда асимметричный и либо
мягкий и закрученный, либо подогнут под торакс . . IV. Paguridea.
46
ANOMURA
I. Надсем. THALASSINIDEA
Тело большей частью цилиндрическое, отчасти креветкообразное,,
с хорошо развитым длинным абдоменом. Карапакс обычно сжат с боков,
фронтальная часть его не сливается с эпистомом. Пятый грудной сегмент
свободный. Покровы часто плохо объизвествленные. Linea thalassinica
имеется или отсутствует. Рострум широкий плоский или довольно сильно,
редуцированный. Антеннальная ножка шестичленистая; скафоцериты име-
ются или отсутствуют. Наружные максиллярные ножки узкие (ногообраз-
ные). Первые перейоподы клешненосные, клешни иногда устроены по типу
ложной клешни; вторые перейоподы часто также клешненосные; третьи
перейоподы всегда без клешней. Жабры типа трихобранхий, однако иногда
они своеобразно расширены, а у наиболее уклоняющихся форм имеется
намек на образование жабр типа филлобранхий; число жабр изменчивое,
но всегда меньше 17; плевро.бранхии почти всегда отсутствуют (на послед-
них перейоподах они всегда отсутствуют). Половые отверстия расположены
на коксоподитах третьих или пятых перейопод. Абдомен длинный, симме-
тричный, эпимеры сегментов часто редуцированы; эндоподиты плеопод
часто несут appendix interna. Хвостовой веер широкий, тельсон без попе-
речного шва.
В некоторых отношениях Thalassinidea имеют, согласно Ortmann’y,
сходство с Homaridea, с другой стороны, согласно Borradaile— с Pagu-
ridea. С первыми они сходны формой хвостового веера, а также первых,
и часто вторых перейопод. Они отличаются от Homaridia тем, что их
третьи перейоподы никогда не бывают клешненосными, часто уменьше-
нием числа жабр и тенденцией абдомена становиться мягким и утрачи-
вать свои плевры. Они сходны с Paguridea шипообразной формой скафо-
церита у тех из них, у которых он хорошо развит, свободным положе-
нием последнего грудного сегмента и своеобразным способом ношения
последней пары перейопод — довольно далеко от последнего грудного
сегмента и над ним. Они отличаются от отшельников тем, что 1) пятые
перейоподы имеют такую же форму, как и остальные, 2) их вторая пара
перейопод обычно клешненосная, и 3) их абдомен симметричный и с ши-
роким хвостовым веером.
Образ жизни обычно роющий, в илу или песке; иногда живут внутри
губок и т. п. Из четырех семейств, образующих надсемейство Thalas-
sinidea, в нашей фауне имеются представители двух семейств.
ОПРЕДЕЛИТЕЛЬНАЯ ТАБЛИЦА СЕМЕЙСТВ
1 (2). Linea thalassinica отсутствует. Скафоцериты всегда имеются, хотя
иногда и маленькие. Эпимеры абдоминальных сегментов большие.
Экзоподиты уропод со швом (diaresis) или без него. . . 1. Axiidae..
1. AXIOPSIS 47
2 (1). Linea thalassinica имеется, Скафоцериты редуцированы до степени
небольшого сплющенного придатка или отсутствуют. Эпимеры абдо- .
минальных сегментов маленькие. Экзоподиты уропод всегда без
шва .. ............................................... 2. Callianassidae.
1. Сем. AXIIDAE
Axiidae Bate, 1888 : 36; Ortmann, 1891: 46; 1901 :141; Borradaile, 1903 : 536; Schmitt, ,
1921:110.
По форме тела напоминают представителей Macrura (тело макруро-
образное). Рострум больше'й частью плоский, треугольный, порядочной
величины. Карапакс без linea thalassinica. Антеннулярные жгуты порядоч-
ной величины. Скафоцерит в виде подвижного шипообразного придатка .
между вторым и третьим члениками антеннального стебля; кроме того на
втором членике снаружи от скафоцерита имеется неподвижный шип
(антеннальный шип). Первая пара перейопод клешненосная, большая, не- .
равная. Вторая пара перейопод оканчивается маленькими одинаковыми
клешнями. Следующие три пары перейопод простые; пятая иногда бы-
вает с ложными клешнями, обычно же оканчивается маленьким шипооб-.
разным пальцем. Без жабр на первых максиллярных ножках; с мастиго-
бранхиями на первых четырех парах перейопод и с подобранхиями на
первых трех (по Bate — на четырех) парах перейопод. Жабры типа трихо-
бранхий. Абдомен длинный, эпимеры хорошо или умеренно развитые.
Экзоподиты и эндоподиты плеопод обычно узкие или довольно широкие;.
эндоподиты имеют stylamblys (= appendix interna). Хвостовой веер хо-
рошо развит; экзоподит (наружная пластинка) уропод со швом (diaresis)
или без него.
В нашей фауне семейство представлено одним родом Axiopsis Bor-
radaile.
1. Род AXIOPSIS BORR.
Axiopsis Borradaile, 1903; 538; Schmitt, 1921:110.
Тип рода: Axiopsis affinis (de Man).
Карапакс впереди цервикальной борозды сжат с боков, образуя на.
дорзальной поверхности хорошо заметную плоскую площадку, продол-
жающуюся в рострум; без медианного дорзального киля на карапаксе
позади цервикальной борозды, за исключением иногда лишь слабого ,
намека на киль как раз перед задним краем карапакса Глаза хорошо
пигментированные. Антеннальный шип на втором членике ножки антенн
длинный или средней величины. Ноги второй пары перейопод клешне-
носные. Экзоподит уропод со швом (diaresis). Указание Borradaile (1903)
относительно жабр, а именно, что имеется артробранхия и подобранхия
на вторых максиллярных ножках и что плевробранхии отсутствуют, отно-
'48
AXIIDAE
сятся только к A. affinis (de Man). Имея под руками только единствен-
ный сухой экземпляр A. princeps (Boas), я не мог установить, насколько
верны эти указания по отношению к данному виду. Кроме приводимого
3. И. Кобяковой отмечен новый подвид
ниже вида, для нашей фауны
Фиг. 15. Axiopsis princeps (Boas).
Между гребнями и на боковых
американского A. spinulicauda (Rath-
bun) из залива Петра Великого, именно
A. spinulicauda amurensis", к моменту
написания настоящей работы описание
подвида еще не было опубликовано.
* 1. Axiopsis princeps (Boas).
(Фиг. 15).
Axius princeps Boas 1880: 98, t. 2, fig. 52;
t. 3, fig. 87; t. 4, fig. 137, 137a; t. 5, fig. 174; t. 7,
fig. 214—217; Balss, 1914:88.
Мандибула имеет зубец на режущей
части. Последний членик пальпа первой
максиллы длинный. Экзогнат второй и
третьей пар максиллярных ножек снаб-
жен короткой ножкой, а экзогнат первой
пары большим эпигнатом. Карапакс
длинный, сильно сжатый с боков и очень
высокий. Рострум треугольный, прости-
рающийся дальше глаз; его вершина
увенчана тупым зубцом и четыре таких же
зубца имеются на каждой боковой сто-
роне рострума. Передняя половина кара-
пакса впереди цервикальной борозды
несет на своей верхней, слегка упло-
щенной, поверхности пять продольных
гребней, которые не доходят до церви-
кальной борозды. Средний гребень,
частично продолжающийся на рострум,
и два промежуточных гребня усажены
крупными гранулами; крайние боковые
гребни гладкие и начинаются от основа-
ния базальных боковых зубцов рострума.
поверхностях карапакса имеются пучки
коротких волос; задняя половина карапакса слегка морщинистая и также
покрыта короткими волосками. Антенны длинные; скафоцериты короткие,
шипообразные. Первая пара перейопод клешненосная, левая нога значи-
тельно больше правой. Мерус длинный, трехгранный; карпус, напротив,
очень короткий, сильно расширящийся наружу. Левая клешня очень
сильная; выпуклые поверхности ладони, равномерно переходящие друг
1. UPOGEBIA
49
в друга, покрыты пучками волос; у основания подвижного пальца, как
снаружи, так и изнутри, имеются крупные, круглые гранулы; пальцы силь-
ные, примерно такой же длины, как ладонь, слегка изогнутые на кончи-
ках, покрытые пучками длинных волос; хватательные края с крупными
закругленными буграми, в особенности на проксимальной половине. Мень-
шая клешненосная нога почти такой же формы, только ладонь более тон-
кая и узкая, а пальцы значительно длиннее ладони, более прямые, без
бугров на хватательных краях и плотно смыкающиеся. Вторая пара перейо-
под клешненосная. Три следующих пары ног простые; их проподусы на
нижне-задней стороне несут 5—6 коротких поперечных гребеночек из ро-
говых шипиков; пальцы короткие, изогнутые, выпуклые сверху и вогну-
тые снизу; передний край вогнутой нижней поверхности несет мелкие
шипики, тогда как задний край усажен более крупными шипиками. Абдо-
мен длинный, с сравнительно хорошо развитыми эпимерами. Плеоподы
третьей-пятой пар имеют appendix interna, который снабжен на своей
вершине крючковатыми щетинками; пластинки плеопод узкие. Тельсон
довольно широкий, почти прямоугольный, Шиповатый; внутренняя пла-
стинка (эндоподит) уропод с шиповатым продольным срединным гребнем;
наружная пластинка (экзоподит) уропод со слабым продольным шиповатым
гребнем, который расположен ближе к наружному шиповатому краю;
дистальная часть пластинки отделена явственным швом (diaresis), над
которым имеется ряд острых шипов.
Окраска: сухой экземпляр бледнолилового цвета, волоски грязно-
рыжего цвета.
Размеры
Длина карапакса............................. 29.0 мм
Ширина „ .............•...............13.0 „
Высота „ 16.0 „
Длина карпуса большей передней ноги.......... 9.0 „
„ ладони „ „ „ .............19.0 „
Ширина „ „ „ „ ...........14.0 „
Толщина „ „ „ 10.0 „
Длина подвижного пальца большей передней ноги . . 18.0 „
„ ладони меньшей передней ноги.............11.0 „
Ширина „ ,, „ ............... 7.0 „
Толщина „ „ „ ............... 4.6 „
Длина подвижного пальца меньшей передней ноги . 15.0 „
Распространение. Владивосток. Форма, повйдимому, роющая.
2. Сем. CALLIANASSIDAE
Callianassidae Bate, 1888:27; Ortmann, 1891:48; 1901:1142; Borradaile, 1903:541;
Schmitt, 1921 :114. — Thalassinidae Heller, 1863 : 200.
Рострум большой, плоский, или же маленький, почти совсем отсут-
ствующий. Linea thalassinica имеется. Антеннулярные жгуты короткие
или умеренно длинные. Скафоцерит маленький, в виде плоской чешуики.
Фауна СССР, Макаров
50
CALLIANASSIDAE
Первые перейоподы неравные или равные, клешненосные или с ложными
клешнями (иногда простые — род Gebicula)\ вторые перейоподы простые
или клешненосные; третьи и четвертые перейоподы простые; пятые перейо-
поды простые, клешненосные или с ложными клешнями. Без подобранхий
и плевробранхий и обычно без мастигобранхий. Жабры типа трихобранхий
или с нитями, расширяющимися в различной степени. Плеоподы тре-
тьего-пятого абдоминальных сегментов с широкими ветвями, их эндо-
подиты с appendix interna или без него. Эпимеры абдоминальных сег-
ментов почти отсутствуют. Экзоподиты уропод без шва. Все представи-
тели семейства ведут роющий образ жизни.
Семейство Callianassidae распадается на два подсемейства.
'ОПРЕДЕЛИТЕЛЬНАЯ ТАБЛИЦА ПОДСЕМЕЙСТВ
1 (2). Рострум большой. Перейоподы первой пары одинаковые. Проподус
третьих перейопод обычной ширины. Эндоподиты плеопод третьего-
пятого абдоминальных сегментов без appendix interna..................
......................................... . 1. Upogebiinae Borr.
2 (1). Рострум маленький, иногда почти совсем отсутствующий. Перейо-
поды первой пары неодинаковые. Проподус третьих перейопод очень
широкий, поперечно-овальный. Эндоподиты плеопод третьего-пятого
абдоминальных сегментов имеют appendix interna....................
..........................................2. Callianassinae Borr.
1. Подсем. UPOGEBIINAE
Upogebiinae Borradaile, 1903 : 542.
Рострум большой. Перейоподы первой пары клешненосные, с лож-
ными клешнями или простые, но одинаковые и не с очень широким кар-
пусом и ладонью. Перейоподы второй пары одинаковые и простые (за
исключением рода Bigea). Проподус третьих перейопод обычной ширины,,
не имеющий поперечно-овальной формы. Без мастигобранхий (иногда
имеются их следы на первых двух парах максиллярных ножек). Вторая
пара плеопод имеет такие же широкие ветви, как и третья-пятая пары;
эти последние не имеют на своих эндоподитах appendix interna.
Из этого подсемейства в нашей фауне имеются представители
только рода Upogebia Leach.
1. Род UPOGEBIA LEACH.
Upogebia Leach, 1814:386, 400; Holmes, 1900:157; Borradaile, 1903:542.— Gebia
Leach, 1815 : 335, 342; Milne-Edwards, 1837:312; Bell, 1853: 2z2; Heller, 1863 : .04.
Тип рода: Upogebia stellata (Montagu).
Карапакс спереди довольно сильно сжат с боков и продолжается
в большой треугольный рострум, который почти целиком закрывает глаза;
верхняя поверхность рострума обычно густо волосистая. По бокам рост-
1. UPOGEBIA
51
рума имеются боковые зубцы, от которых назад до цервикальной борозды
идут гребни, ограничивающие плоскую гастральную область. Сзади
карапакс широкий, закругленный на заднебоковых углах, бранхиальные
области очень большие. Лобный край карапакса несет над основанием
антеннальной ножки маленький шип, или же этот шип отсутствует. Антен-
нулярные ножки короткие; антеннальные ножки пяти членистые, с выдаю-
щимся слуховым бугорком на вентральной поверхности первого членика.
Скафоцерит маленький, рудиментарный, часто в виде короткого листовид-
ного придатка.1 Глазные стебельки цилиндрические, несколько расши-
ренные у основания; роговицы терминальные. Наружные максиллярные
ножки узкие, ногообразные. Передние перейоподы сильные, одинаково
развитые с обеих сторон, клешненосные или с ложными клешнями. Вто-
рая-четвертая пары перейопод простые. Проподус второй пары широкий
и сплющенный. Пятые перейоподы обычно не клешненосные. Абдомен
длинный, средние сегменты наиболее широкие. Вторая пара плеопод та-
кая же, как следующие три. Первая пара плеопод у -самца отсутствует;
у самки она маленькая, тонкая, нитеобразная. Эндоподиты вторых-пятых
плеопод без appendix interna. Тельсон широкий, почти квадратный; уро-
поды широкие и толстые, не длиннее тельсона.
Род Upogebia распадается на два подрода: Upogebia Leach и Gebi-
opsis A. Milne-Edwards. Последний подрод отличается от первого отсут-
ствием шипов на лобном краю карапакса над основаниями антеннальных
ножек и кроме того строением клешней первых перейопод; у Gebiopsis
неподвижный палец, который у Upogebia имеет вид шипообразного при-
датка на нижнем дистальном конце проподуса, достигает почти такой же
длины, как и подвижный палец, благодаря чему образуется настоящая
клешня. В нашей фауне представлен только подрод Upogebia.
ОПРЕДЕЛИТЕЛЬНАЯ ТАБЛИЦА ВИДОВ
1 (2). Первый членик ножки антеннул с шипом на дистальном конце ниж-
него края. Скафоцерит треугольный, оканчивающийся острым шипом.
Верхний край проподуса передних перейопод с сильным шипом
близ дистального конца. Верхняя поверхность подвижного пальца
гладкая, плоская . .......................... * 1. U. littoralis (Risso).
2 (1). Первый членик ножки антеннул невооруженный. Скафоцерит попе-
речно-овальный, с закругленным передним краем. Верхний край
проподуса передних перейопод без заметного шипа. Верхняя поверх-
ность подвижного пальца с бороздой или возвышением.
3 (6). Неподвижный палец клешни передних перейопод без крупного зубца
на середине хватательного края.
1 Целый ряд авторов (Bell, Heller, Milne-Edwards, Holmes) в родовых диагнозах
отмечают отсутствие скафоцеритов.
4*
52
CALLIANASS1 DAE
4 (5). Неподвижный палец клешни передних перейопод с 4—5 шипиками
у основания наружного края; имеется маленький шипик у основания
хватательного края. Без зубцов на проксимальном и дистальном
концах среднего гребня верхнего края проподуса передних перейо-
под ........................................ . * 2. U. major (de Haan).
5 (4). Неподвижный палец клешни передних перейопод без шипиков
у основания наружного края; имеется сильный зубец у основания
хватательного края. Средний гребень верхнего края проподуса
передних перейопод вооружен на проксимальном и дистальном
концах сильным зубцом ......... ................3. U. yokoyai, nom. nov.
6 (3). Неподвижный палец клешни передних перейопод с крупным зубцом
на середине хватательного края....................*4. U. issaeffi (Balss).
*1. Upogebia (s. str.) littoralis (Risso).
Thalassina littoralis Risso, 1816:76, t. 3, fig. 2. — Gebios littoralis Risso, 1826 : 51. —
Gebia littoralis H. Milne-Edwards, 1837 :313; Heller, 1863: 205, t. 6, fig. 12—15; Czerniavsky,
1884:85.— Upogebia littoralis de Man, 1927 : 29, t. 3, fig. 11—11b.
Рострум выступает вперед до середины или немного за середину
предпоследнего членика антеннулярной ножки; кончик закругленный, боко-
вой край несет 4—6 шипиков; верхняя поверхность с продольной без-
волосой бороздой, которая продолжается и на гастральную область,
заметно расширяясь по направлению к основанию, так что на задней
части гастральной области образуется отчетливый треугольный голый
участок, ограниченный сзади явственной цервикальной бороздой. Боковые
стороны гастральной области и рострума покрыты небольшими возвыше-
ниями с отходящими от них пучками волос. Боковые лобные зубцы
очень короткие, боковые гастральные гребни несут 13—14 шипиков. Шип
на лобном краю над основанием антеннальной ножки довольно длинный
и острый. Гранулы на заднем краю цервикальной борозды на linea thalas-
sinica и под ней выражены очень слабо. Глазные стебельки простираются
немного за середину рострума. Первый членик антеннулярной ножки
вооружен на дистальном конце нижнего края шипом, третий членик
антеннулярной ножки несет пластинчатый треугольный скафоцерит,
оканчивающийся острым шипом. Наружные максиллярные ножки узкие,
ногообразные, волосатые. Передние перейоподы одинаковые, умеренно
длинные; нижний край исхиума с 3—5 шипами; мерус длинный, сплющенный,
с шипом на внутренней стороне треугольной верхней площадки у дисталь-
ного конца; нижний край шиповатый и волосатый. Карпус с шипом на
дистальном конце верхнего края, который скрыт пучком длинных волос;
снаружи от этого шипа передний край несет два маленьких, едва замет-
ных шипа; внутренний дистальный угол нижней поверхности также несет
шип. Верхний закругленный край проподуса несет на внутренней
стороне продольный гранулированный волосатый гребень, затем по напра-
влению наружу гладкий, параллельный первому, гребень с отдельными
/. UPOGEBIA
53
пучками волос и, наконец, мелкую борозду с очень редкими пучками
волос; между гранулированным и гладким гребнем верхний край про-
подуса несет близ дистального конца довольно сильный, направлен-
ный вперед, шип; наружная сторона проподуса выпуклая и гладкая; ниж-
ний край на проксимальной половине образован треугольным уплощенным
участком, боковые стороны которого несут мелкие шипы и длинные
волоски; неподвижный палец острый, с совершенно гладким наруж-
ным краем; хватательный край на своих проксимальных двух третях
мелко зазубрен, иногда несет четыре довольно явственных шипика;
передний край проподуса на верхне-внутреннем углу несет у основания
подвижного пальца небольшой шип. Подвижный палец слегка изогнутый;
его верхняя поверхность гладкая, плоская, лишь на проксимальной трети
наблюдаются по бокам слабые гладкие гребни; наружная и внутренняя
стороны покрыты густыми волосами; нижне-наружный (хватательный)
край несет ближе к основанию очень маленький зубец и позади него три
круглых гранулы; нижняя сторона ладони безволосая, гладкая, за исклю-
чением гладкого продольного гребня близ внутреннего края, благодаря
чему между гребнем и этим краем образуется борозда, усаженная воло-
сами; ряд волос тянется и вдоль наружной стороны нижней поверхности.
Строение подвижного пальца одинаковое у обоих полов. Мерус вторых
перейопод с шипом близ дистального конца верхнего края; нижний край
с 1—2 шипами близ проксимального конца; карпус с шипом близ дисталь-
ного конца верхнего и нижнего краев. Мерус третьих перейопод с тремя
шипами на нижнем краю: один близ проксимального конца, второй на
середине и третий, совсем маленький, у дистального конца; верхний край
невооруженный; карпус с маленьким шипом на нижнем дистальном конце
и с пучком волос на верхнем краю. Тельсон почти квадратный, его боко-
вые края на протяжении проксимальной трети слегка расходятся, а затем
опять сходятся; задний край прямой, без вырезки; срединная борозда
верхней поверхности явственная.
Окраска: бледная беловатая или зеленоватая, абдомен часто про-
зрачный (Pesta); зеленовато-серая (Heller, Webb). Чернявский отмечает
способность животного быстро изменять окраску. Risso описывает окраску
как грязнозеленую, карапакс красноватый; кроме того он отмечает
карминово-красный цветовой вариетет с перламутрово-белым абдоменом,
который встречается на побережьях после сильных штормов; послед-
ний случай можно объяснить способностью животного изменять окраску,
а может быть и посмертным изменением окраски (Risso не указывает
в каком состоянии находились такие животные).
Размеры
Общая длина.............................. 34.0 мм
Длина меруса первой перейоподы............. 6.0 „
„ карпуса „ „ ...............3.0 „
» проподуса „ „ ............... 5.5 „
54
CALLIANASSIDAE
Длина подвижного пальца первой перейоподы .... 4.5 мм
Длина тельсона.................................. 3.5 „
Ширина „ . ............................... 4.5 „
Распространение. Черное море, Адриатическое и Средиземное
моря; побережье Португалии и юго-западное побережье Франции. Опи-
санная из Норвегии G. О. Загсом и отнесенная им к U. littoralis стадия
развития принадлежит, несомненно, другому виду (U. stellata?), так как
U. littoralis неизвестна севернее Ла-Манша.
В Черном море живет на глубинах до 3 м, зарываясь в ил или
песок на глубину нескольких десятков см; ночью животные покидают
норы и плавают на глубине 1—3 м, поднимаясь иногда и до самой поверх-
ности. Массовое появление их у поверхности после штормовых погод
объясняется механическим выносом свободно бродивших животных из
придонных слоев, или же размывом их нор; застигнутые врасплох живот-
ные являются совершенно беспомощными и становятся игрушкой в руках
волн; этим же объясняется и появление Callianassa у поверхности
во время крупной зыби, которое наблюдал В. Чернявский (см. ниже).
*2. Upogebia (s. str.) major (de Haan). (Фиг. 16, 17).
Gebia major de Haan 1850:165, t. 35, fig. 7; Miers, 1879 : 52; Ortmann, 1891 : 54, t. 1,
fig. 7; Balss, 1916:90; Yokoya, 1930:543, fig. 4.— Upogebia major de Man, 1927 :47, t. 6,
fig. 18.
Рострум треугольный, кончик закругленный, довольно тупой; боко-
вой край несет четыре маленьких шипика; боковые края основания рост-
рума несут 5 шипиков, которые частично переходят на гастральную
область; верхняя поверхность рострума густо волосатая, за исключением
срединной борозды; передние две пятых уплощенной гастральной области
также волосатые; задняя пятая часть гастральной области и узкая средняя
продольная часть безволосые. Боковые лобные зубцы короткие, перехо-
дящие в боковые гастральные гребни, отделенные от поверхности осно-
вания рострума и гастральной области глубокими гладкими безволосыми
бороздами и несущие 12—14 маленьких шипиков,которые скрыты волосами.
Задний край цервикальной борозды несет 3 гранулы над linea thalassinica
и 3 гранулы под ней, за которыми следует ряд более мелких гранул; боко-
вые стороны гастральной области также усажены мелкими гранулами.
Шип на лобном краю над основанием антеннальных ножек явственный.
Глазные стебельки достигают приблизительно до середины рострума.
Первый членик антеннулярной ножки невооруженный; третий членик
более чем в 4 раза длиннее второго. Антеннальная ножка несет на втором
членике маленький листовидный поперечно-овальный скафоцерит. Ма-
ксиллярные ножки узкие, ногообразные, густо опушенные волосами на ниж-
них краях члеников. Передние перейоподы довольно сильные, одинаковые.
Нижний край исхиума близ дистального конца несет острый шип. Мерус
сплющенный, слегка расширяющийся дистально; верхний край перед
/. UPOGEBIA
55
дистальным концом переходит в пло-
скую покатую треугольную площадку,
внутренняя сторона которой несет
шип близ дистального конца; боковые
стороны меруса гладкие; нижний край
слегка расширяется дистально, несет
6 острых зубцов, которые переходят
на внутренний край расширения, в то
время как наружный край несет ряд
мелких круглых гранул; внутренний
край несет ряд длинных мягких во-
лосков; другой ряд волосков нахо-
дится над ним на внутренней поверх-
ности меруса. Карпус короткий; его
верхне-внутренний край несет 6—7 ма-
леньких шипов; дистальный верхне-
внутренний угол занят сильным ши-
пом, верхний передний край несет
4—6 зубчиков, которые уменьшаются
по направлению наружу; с внутренней
стороны на переднем краю имеется
такой же сильный шип, как и на
передне-внутреннем углу; верхне-
наружная поверхность гладкая, пере-
сеченная сходящимися дистально под
углом бороздами, верхняя из кото-
рых заполнена густыми волосками;
треугольная нижняя поверхность
гладкая, наружный край несет 13—
15 шиповатых гранул, уменьшаю-
щихся в проксимальном направлении;
внутренний дистальный угол несет
сильный шип, под которым имеется
более маленький шип. Проподус про-
долговато-овальный, со слабо выпук-
лыми боковыми сторонами; верхний
и нижний края широко закругленные.
Верхний край снабжен, начиная из-
нутри, продольным рядом шиповатых
гранул и зазубренным гребнем; и тот
и другой несут, кроме того, длинные
Фиг. 16. Upogebia major (de Haan).
волоски, а промежуток между ними
гладкий и безволосый; параллельно зазубренному гребню и немного
отступя от него тянется по наружной поверхности гладкий гребень;
56
CALLIANASSIDAE
ближе к нижнему краю проподуса тянется продольная борозда, заполнен-
ная волосками; нижний край проподуса имеет диагонально идущий ряд
шипов, которые наиболее крупны у проксимального конца; дистальные
две трети несут мелкие гранулки с поднимающимися от них пучками длин-
ных мягких волос. Зубцеобразный выступ на нижне-внутреннем углу
(неподвижный палец) имеет 4—5 тупых шипов позади своего основания
как бы на гребне, продолжающем его наружный гладкий край; вогнутый
гладкий хватательный край несет маленький шипик близ основания;
передне-наружный край проподуса несет небольшой шип, который распо-
ложен ближе к основанию подвижного пальца. Вооружение подвижного
пальца отличается у обоих полов. У самца верхняя поверхность несет
9—10 плоских косых ногтевидных возвышений; внутренний край имеет
Фиг. 17. Подвижный палец клешни Upogebia major (de Haan),
a — вид сверху, <5; b — вид снизу, S; с — вид сверху, g; d — вид
снизу, g.
форму мелко зазубренного гребня; между этим гребнем и рядами описан-
ных плоских возвышений имеется еще продольный ряд очень мелких
гранул, из которых три проксимальных более крупные, чем остальные;
эти гранулы скрыты длинными волосами, занимающими пространство
между указанными выше гребнем и рядом гранул; верхне-наружный край
несет гладкий, опушенный снизу волосами гребень, который только,
у проксимального конца несет 3—4 гранулы; нижне-наружный край глад-
кий, несущий на середине круглый бугорок; над этим краем наружная
поверхность несет ряд волос; нижняя уплощенная поверхность пальца
несет 5 косых гребней, из которых средний самый длинный, а два
дистальных часто имеют форму простых гранул (в особенности у мелких
экземпляров); с обеих сторон имеется два ряда волосков. У самки верх-
няя поверхность подвижного пальца имеет глубокую продольную гладкую
борозду, приподнятые боковые края которой мелко гранулированы;
у молодых самок эта борозда отсутствует; боковые стороны густо воло-
сатые; нижне-наружный край примерно на проксимальной трети более
приподнят, чем остальная часть края; нижняя поверхность несет
посредине продольный ряд из косо-поперечных коротких гребней: этот ряд.
1. UPOGEBIA
57
у основания пальца отклоняется от середины к внутреннему углу. Вторые,
перейоподы сплющенные; верхний край меруса гладкий, с шипом близ
дистального края; нижний край волосатый, с сильным шипом у прокси-
мального конца и двумя более маленькими шипами впереди него; карпус
с шипом на дистальном конце верхнего и нижнего краев; проподус
широко-овальный, густо волосатый; палец особенно волосатый на верхнем,
краю. Мерус третьих перейопод с гладким верхним и шиповатым ниж-
ним краями. Пятые перейоподы с ложными клешнями, нижняя и дисталь-
ная части проподуса густо волосатые. Экзоподиты вторых-пятых плео-.
под заостренные, широкие, значительно длиннее закругленных эндопо-
дитов. Боковые края шестого абдоминального сегмента с низкой усечен-
ной долей. Тельсон с закругленными задне-боковыми углами; гребни
на тельсоне плоские, слабо заметные; задние края. уропод усажены
сверху мелкими шипиками.
Размеры
Общая длина..................................90.0 мм
Длина меруса первой перейоподы...............13.0 „
„ карпуса „ „ 8.0 „
„ проподуса „ „ 13.0 „
„ подвижного пальца „ 10.0 „
Длина тельсона ............................ 10.0 „
Ширина „ 13.5 „
Распространение. Южная Япония. В наших водах известен,
из района Владивостока (бухта Золотой Рог, бухта Патрокл) и из залива-
Ольги.
3. Upogebia (s. str.) yokoyai, nom. nov. (Фиг. 18).
Gebia affinis Yokoya, 1930:544, fig. 5 (nom. praeoccup.)1
„Из экземпляров из Asadokoro один имел 42.8 мм длины от кончика
рострума до кончика тельсона, другой — 33.5 мм. Я имел возможность-
исследовать один экземпляр самки этого вида 45.8 мм длины, кото-
рый был найден Mr. Hiroaki Aikawo на побережьи Haneda близ Токио.
Вид очень близок к G. issaeffi Balss из Владивостока и к G. major;
de Haan, но явственно отличается от них.
Лобный край карапакса подобен таковому G. major, состоит из трех,
направленных вперед отростков, средний отросток при этом крупнее
и длиннее, чем отростки с каждой стороны. У данного вида не только
средний отросток отчасти более узкий и более резко заостренный, но
и боковые отростки дальше отстоят от среднего отростка при рассматри-
вании сверху, чем таковые у G. major. При рассматривании сбоку средний.,
отросток обнаруживает в профиль непрерывную линию с верхним краем
1 Gebia affinis Say, Journ. Nat. Acad. Scien. Philad., I, 1818:241.
58
CALLIANASSIDAE
карапакса, а нижний край косо восходит до заостренного кончика; в то же
время у G. major линия верхнего края сначала непрерывна с линией кара-
пакса, но внезапно опускается вниз у кончика, а нижний край у более
мелких экземпляров представляет собой почти горизонтальную линию.
У совершенно взрослых экземпляров нижний край слегка восходящий
по направлению к кончику, где, однако, он не так остро оканчивается, как
у данного вида. Перейоподы одинаковые у обоих полов и по форме
подобны таковым G. is-
saeffi. Мерус толще чем
таковой у G. major
у экземпляров одинако-
вой величины, прибли-
зительно в 272 раза
больше в ширину чем
в длину,1 его наружная
поверхность гладкая и
лишенная волос, в то
время как зубцы на
Фиг. 18. Upogebia yokoyai nom. nov. a — общий вид живот-
ного с правой стороны; Ь — карапакс, вид сверху; с — пе-
редняя половина карапакса, вид сбоку; d — терминальная
половина первой перейоподы, вид с наружной стороны
(из Yokoya, 1930).
нижнем краю резко за-
остренные. Карпус во-
оружен рядом мелких
остроконечных зубцов
около верхнего края;
этот ряд зубцов более
выдающийся, чем тако-
вой у G. issaeffi и тер-
минальный зубец го-
раздо сильнее, чем все
остальные. Карпус во-
оружен двумя сильными
зубцами на дистальном
краю, один более силь-
ный находится на ниж-
нем краю, другой — на верхнем. Ладонь клешни приблизительно вдвое
больше в ширину чем в длину и ее поверхность почти гладкая и
сверху снабжена тремя тупыми продольными гребнями, которые опушены
длинными волосами. Из этих гребней средний наиболее выдающийся
и снабженный сильным зубцом на проксимальном и дистальном концах.
Между этим и наружным гребнем имеется продольная борозда. Ладонь
' также снабжена двумя рядами пучков волос на наружной поверхности;
эти два ряда встречаются позади сустава пальца, где волоса рассеяны
iмногочисленными пучками. Нижний край слабо гребенчатый и снабжен
1 Явная описка у автора: следует понимать больше в длину чем в ширину. В. М.
7. UPOGEBIA
59
длинными волосами. Неподвижный палец вооружен сильным треугольным
зубцом у основания внутреннего края. Подвижный палец снабжен тремя
рядами бугорков, по одному ряду на верхнем и нижнем краях и одним
рядом на наружной поверхности; у последнего бугорки выдающиеся, в то
время как у первых, за исключением одного или двух проксимальных
бугорков, они значительно выдаются. На ряду с этим
На верхнем краю и на каждой стороне нижнего края
Волоски собраны в ряд, а другой ряд волос нахо-
дится как раз под упомянутым бугорчатым рядом на
наружной поверхности. Волоски на верхнем краю
выдающиеся и наиболее длинные у основания, умень-
шаясь в длину в терминальном направлении. В других
отношениях вид совпадает с G. major" (Yokoya).
Распространение. Япония: 6VII1926 Asa-
dokoro, 2 3; 15 VIII 1926, Nonai 1 g juv.
* 4. Upogebia • (s. str.) issaeffi (Balss).
(Фиг. 19, 20).
Gebia (Upogebia) issaeffi Balss, 1913: 239; 1914:18, fig. 89,
49; Upogebia issaeffi de Man 1927 : 27.
Фиг. 19. Upogebia is-
saeffi (Balss). Клешня
левой первой перейо-
поды, вид с внутренней
стороны.
Близок к U. major, однако отличается явствен-
ными признаками главным образом первых перейопод,
почему мы и коснемся только их описания. Первые
перейоподы относительно не такие длинные, как
у U. major. Мерус и карпус в общем имеют такое жеворужение, как и у t/.
.major. У самца проподус несет главный волосатый гребень, который только
на дистальном конце несет ряд из 7 постепенно возрастающих в величине
шипов; на внутренней стороне имеется параллельный ряд очень мелких
Фиг. 20. Подвижный палец клешни Upogebia issaeffi (Balss). а — вид
сверху, <5; Ь — вид снизу, 3; с — вид сверху, g; d — вид снизу, g.
острых гранул, также усаженный волосками; на наружной стороне едва
намечается второй, гладкий, слабо волосатый гребень; ближе к нижнему
краю по наружной поверхности тянется волосатая борозда; нижний край
япроподуса с 3 шипами у основания, от которых тянется вперед ряд густых
волос, который, не доходя до неподвижного пальца, сворачивает наружу
60
C ALLI AN A SS IDA К
и сливается с вышеупомянутой волосатой бороздой; на этом повороте
волосы скрывают группу коротких шипов; передне-наружный край несет
шип между основаниями подвижного и неподвижного пальцев. Неподвиж-
ный палец с крупным зубцом на середине хватательного края и с совер-
шенно гладким основанием наружного края. У самки верхний край пропо-
дуса несет зазубренный волосатый гребень, а не гладкий; в остальном
проподус совершенно такой же, как у самца. Подвижный палец различается
по своему строению у обоих полов (Balss и de Man имели только самцов
данного вида и поэтому эти отличия не были отмечены). У самца верхняя
поверхность подвижного пальца несет гладкую продольную борозду,
боковые края которой приподняты; наружный край этой борозды усажен
мелкими гранулами; очень крупная круглая гранула находится на прокси-
мальном конце; внутренний край этой борозды несет ряд мелких попереч-
ных валиков (по подсчету de Мап’а число их приблизительно 100); нижне-
внутренний край усажен довольно крупными круглыми гранулами; поверх-
ность между этим краем и рядом поперечных валиков несет гладкий
гребень, густо усаженный волосами и с 1—2 гранулами на проксимальном
конце; нижне-наружный край несет два тупых зубца, вернее сказать, широ-
кий низкий усеченный зубец с вздутыми закругленными боковыми углами^
наружно-нижняя поверхность несет ряд крупных круглых гранул, который
не достигает ни до проксимального, ни до дистального концов пальца
и усажен с обеих сторон рядом волос, причем верхний ряд волос доходит
до дистального и проксимального концов пальца, а нижний имеет такое же
протяжение, как и ряд гранул.
Нижняя поверхность пальца с 9—13 косыми поперечными гребнями,
которые расположены ближе к внутреннему краю; имеются прямые ряды
волосков на наружном и внутреннем краях нижней поверхности. При сги-
бании середина широкого усеченного зубца на нижне-наружном краю
приходится как раз против зубца на хватательном краю неподвижного
пальца. У самки верхняя поверхность подвижного пальца напоминает
таковую у U. major, имеется гладкая продольная борозда, оба приподня-
тых края которой усажены крупными гранулами; в остальном палец,
похож на палец самца, только поперечные гребни на нижней поверхности
более короткие, толстые и выпуклые и имеют .благодаря этому скорее
вид крупных круглых гранул. Следующие пары перейопод и абдомен, как
у U. major.
Размеры
Общая длина...........................62.0 мм
Длина меруса первой перейоподы .... 10.0 „
„ карпуса „ „ .... 5.5 „
„ проподуса „ „ .... 11.0 „
„ подв. пальца „ . ... 8.5 „
Длина тельсона......................... 7.0 „
Ширина „ . ....................... 9.0 „
2. CALLIANASSA
61
Распространение. Вид является сравнительно узко локальной
«формой, так как все известные экземпляры найдены в районе Владиво-
стока.
2. Подсем. CALLIANASSINAE
Callianassinae Borradaile, 1903 : 544.
Рострум маленький, иногда почти совсем отсутствующий. Первые
перейоподы клешненосные, неодинаковые, у большей ноги карпус и ладонь
обычно довольно широкие. Перейоподы второй пары одинаковые, клешне-
носные. Проподус перейопод третьей пары широкий. Мастигобранхий
имеются или отсутствуют на задних перейоподах, но всегда имеются на
первых максиллярных ножках. Вторая пара плеопод такая же или иная, чем
третья-пятая пары, эндоподиты которых имеют appendix interna.
ОПРЕДЕЛИТЕЛЬНАЯ ТАБЛИЦА РОДОВ
1 (2). Плеоподы второго абдоминального сегмента не похожи на плеоподы
трех следующих сегментов.....................2. Callianassa Leach.
2 (1). Плеоподы второго абдоминального сегмента такие же, как и на трех
k следующих сегментах.....................3. Ctenocheles Kishinouye.
.2. Род CALLIANASSA LEACH
Callianassa Leach, 1814 : 386; Milne-Edwards, 1837 : 307; Bell, 1853 : 217; Heller,
1863 : 201; Bate, 1888 : 28; Holmes, 1900 :159; Borradaile, 1903 : 544; Balss, 1914 : 91; Schmitt,
1921:116.
Тип рода: Callianassa subterranea (Montagu).
Карапакс короткий по сравнению с общей длиной тела. Все покровы
более или менее мягкие, за исключением покровов первых перейопод,
которые, напротив, очень твердые. Рострум короткий, треугольный, руди-
ментарный или совсем отсутствующий. Глазные стебельки пластинчатые,
треугольные, роговицы помещаются на их дорзальной поверхности,
маленькие или отсутствуют; реже глазные стебельки закругленные с тер-
минальными роговицами (подрод Scallasis Bate). Наружные максиллярные
яожки. узкие, ногообразные^ или же широкие, крышкообразные; экзогнат
отсутствует. Передние перейоподы клешненосные, неравные, то левая, то
правая нога значительно больше другой; карпус и рука широкие по сравне-
нию с остальными члениками; мерус с зубцеобразной долей на прокси-
мальном конце наружного края; карпус сочленяется с мерусом при помощи
узкой шейки. Вторые перейоподы клешненосные, одинаковые; клешня
сплющенная, треугольная или сердцевидная. Третья и четвертая пары
перейопод простые; проподус третьей пары широкий, поперечно-овальный,
его задний край либо выступает за задний край карпуса (проподус с долей),
либо находится на одном уровне с ним» Пятая пара перейопод с ложной
62
CALLIANASSIDAE
клешней. Жабры по своему типу занимают промежуточное положение
между филлобранхиями и трихобранхиями: они состоят из двух рядов,
длинных тонких нитей, которые настолько плотно прижаты друг к другу,
что образуют как бы пластинки. По своему положению они являются
артробранхиями; имеется по две артробранхии с каждой стороны на третьих
максиллярных ножках и на четырех первых перейоподах; имеется крупная
мастигобранхия на первых максиллярных ножках и маленькая мастиго-
бранхия в виде луковицы на вторых максиллярных ножках. Абдомен длин-
ный; две первые пары плеопод отличаются по своей форме от следующих
пар; у самца плеоподы первого сегмента маленькие; на втором сегменте
у самца и у самки экзоподит и эндоподит плеопод узкие, нитевидные;,
на третьем-пятом сегментах у обоих полов ветви плеопод пластинчатые
и не имеют жаброобразных нитей; их эндоподиты несут appendix interna.
Уроподы длиннее или такой же длины как тельсон.
Благодаря большой прочности передних перейопод известны довольно-
многочисленные ископаемые формы из юрских и до-третичных отложений;
таковы, например, Call, maxima A. Milne-Eawards из аллювиальных отло-
жений, найденная при прорытии канала в Сиаме, Call, crassa A.. Milne-
Edwards из миоценовых песков St. Paul близ Dax, наконец, несколько,
видов из известняков de Parmes в окрестностях Gisors, в песках Blanchamp.
и меловых отложениях Богемии и многие другие.
Род Callianassa распадается на 5 подродов, из которых в нашей
фауне представлены три подрода, а подроды CalEactites Borradaile и Seal-
lasis Bate отсутствуют. В основу деления на подроды положены признаки
наружных максиллярных ножек, форма проподуса третьих перейопод,.
хвостового веера и глазных стебельков. Представленные в нашей фауне
подроды различаются формой наружных максиллярных ножек и хвостового,
веера.
ОПРЕДЕЛИТЕЛЬНАЯ ТАБЛИЦА ПОДРОДОВ
1 (4). Тельсон такой же длины, как уроподы.
2 (3). Наружные максиллярные ножки узкие, ногообразные..........
...........................................1. Cheramus Bate..
3 (2). Наружные максиллярные ножки широкие, крышкообразные ....
...........................................2. Trypsea Dana.
4 (1). Тельсон короче чем уроподы...........3. Callichirus Stimpson.,
ОПРЕДЕЛИТЕЛЬНАЯ ТАБЛИЦА ВИДОВА
1 (10). Тельсон такой же длины, как уроподы.
2 ( 3). Наружные максиллярные ножки узкие, ногообразные.........
. . *.............................1. С. subterranea (Montagu).
3 ( 2). Наружные максиллярные ножки широкие, крышкообразные.
1 Определитель дается без расчленения рода на подроды.
2. CALLIANASSA
63 ,
4 ( 5). Лобный край карапакса лишен рострума. Хватательный край
подвижного пальца большей руки имеет у основания широкий усе-.
ченный зубец и близ кончика другой, довольно крупный зубец.
Задний край тельсона близ боковых углов несет по паре шипиков..
.............................. ... *2. С. harmandi Bouvier.
5 (4). Рострум имеется, хотя иногда и очень маленький. Задний край
тельсона без шипиков близ боковых углов.
6 (7). Карпус большей клешненосной ноги в V/2 раза больше в длину чем
в ширину и длиннее чем мерус. Хватательный край подвижного
пальца большей руки вооружен зубцами, напоминающими таковые
С. harmandi ............. *3. С. gigas еоа, nom. nov.
7 (6). Карпус большей клешненосной ноги чуть-чуть больше в длину, чем.
в ширину и такой же длины, как мерус. Хватательный край подвиж-
ного пальца большей руки без выдающихся зубцов.
8 (9). Проксимальная доля на наружном краю меруса большей клешне-
носной ноги узкая, с острой вершиной; длина внутреннего края
карпуса равна ширине карпуса или несколько больше ее; карпус не
длиннее ладони; при смыкании пальцы клешни сходятся довольно
плотно; подвйжный палец короче, чем внутренний край ладони.
*4. С. japonica (Ortmann)..
9 (8). Проксимальная доля на наружном краю меруса большей клешне-
носной ноги широкая, с закругленной вершиной; длина внутрен-
него края карпуса короче, чем ширина карпуса; карпус длиннее чем
ладонь; при смыкании между пальцами клешни остается широкая
щель; подвижный палец длиннее, чем внутренний край ладони.
*5. С. califomiensis bouvieri, nom. nov.
10 (1). Тельсон короче, чем уроподы . . . . *6. С. pontica (Czerniavsky)...
1. Подрод Cheramus Bate
Cheramus Bate, 1888 : 30; Borradaile, 1903 ; 545.
Наружные максиллярные ножки узкие, ногообразные. Проподус
третьих перейопод с долей на заднем краю. Внутренние пластинки уропод .
(эндоподиты) средней ширины или широкие. Тельсон длинный.
1. Call'anassa (Cheramus) subterranea (Montagu). (Фиг. 21).
Cancer (Astacus) subterraneus Montagu, 1803:89, t. 3, fig. 1, 2. — Callianassa sub- .
ierranea Leach, 1815, t. 32; Bell, 1853 : 217; de Man, 1928: 6, t. 1, fig. 1—Ih (литература).
До сравнительно недавнего времени в европейских водах, включая ;
сюда Средиземгйэе и Черное моря, считали существующим один вид
Callianassa, именно С. subterranea (Montagu). Callianassa laticauda
и C. stebbingi считались идентичными с этим видом. В свое время В. Чер-
нявский (1884) различал у G. subterranea три формы и два вариетета,
именно: forma typica (Средиземное и Черное моря, северные европейские
воды) с вариететом laticauda (Средиземное море), forma intermedia (Среди- .
'64
CALLIANASSIDAE
земное море) с вариететом major (Callianassa major Say, восточное по-
бережье Америки) и forma pontica (Черное море). Определение последней
интересующей нас формы дано Чернявским неполно. De Man (1928) в своей
монографии пересмотрел материал по средиземноморским и северно-евро-
пейским формам и пришел к выводу, что в Средиземном море существуют
два вида Callianassa'. С. laticauda Otto (С. stebbingi Borradaile) и С. pe-
stae de Man; Callianassa subierranea свойственна северным европейским
водам (побережья Франции, Англии), однако экземпляр Colosi, добытый
в Средиземном море у северного побережья Африки, de Man относит
. к С. subierranea, так что вопрос о распространении этого вида в Среди-
земном море ещё не вполне ясен. Что касается Черного моря, то согласно
просмотренному материалу все экземпляры относятся к описанному
"de Мап’ом виду Callianassa pestae', сам de Man, правда, подвопросом, ставил
f. pontica Чернявского в синонимы своего С. pestae, однако идентичность
обеих форм несомненна и, следовательно, придерживаясь правил приори-
тета, этот вид следует называть Callianassa pontica (Czerniavsky). Таким
-образом, С. subierranea в Черном море не встречается (смешать ее с С. pon-
tica трудно, так как оба вида относятся к разным подродам); однако мы
считаем не лишним привести сокращенное описание этого вида по de Мап’у.
Рострум короткий, острый, широко-треугольный; боковые выступы
лба между глазным стебельком и антеннальной ножкой тупые. Цервикаль-
ная борозда глубокая, linea thalassinica явственная. Внутренние края
глазных стебельков прямые, незаметно расходящиеся кпереди и почти
соприкасающиеся; наружные края дугообразные; роговицы расположены
на внутренней половине дорзальной поверхности стебелька (у самки
немного более сзади чем у самца). Наружные максйллярные ножки почти
ногообразные. Исхиум большей клешненосной ноги (у самца) с маленьким
острым шипом на проксимальном конце выемчатого внутреннего края,1
наружный край с рядом из 5 маленьких одинаковых зубцов на проксималь-
ной половине. Мерус немного длиннее чем исхиум, внутренний край слегка
изогнутый, с маленьким полуострым зубцом на проксимальном конце, за
которым следует более крупный двувершинный выступ и затем через
короткий промежуток три полуострых одинаковых зубца, которые так же
малы, как первый зубец; проксимальная четверть наружного края про-
должается в большую плоскую долю, которая дистально загнута вверх, ее
1 Как здесь, так и в дальнейших описаниях видов Callianassa, принятая мною ориен-
тировка конечностей несколько отличается от таковой de Man’а; правда, при этом конеч-
ности оказываются как бы несколько вывихнутыми, потому что при нормальном положении
Они расположены широкими плоскостями друг к другу, так что то, что^у меня обозначается
верхней поверхностью является наружной, то что нижней — внутренней; внутренний край
в моих описаниях соответствует верхнему краю в описаниях de Man а и наружный край —
нижнему. Так как при определении сплющенная конечность помещается, всегда своей наи-
большей плоскостью в горизонтальном положении (под лупой, бинокуляром), то принятые
мною обозначения в этом случае являются более удобными. Во избежание путаницы
:я считал необходимым сделать эту оговорку.
2. CALLIANASSA
65
дистальный край оканчивается резким острием; перед долей наружный
край меруса несет 12 зубчиков, между которыми находятся волоски. Кар-
пус немного короче, чем мерус, имеет такую же ширину, как и длину,
края слабо гребенчатые и волосатые. Ладонь слегка суживается дистально,
края гребенчатые; неподвижный палец немного короче чем ладонь, его
кончик изогнутый; подвижный палец немного длиннее чем неподвижный,
его кончик сильно изогнутый, хватательный край зазубренный. Исхиум
меньшей клешненосной ноги (у самки) тонкий, отчасти расширяющийся
дистально; мерус немного короче чем исхиум, овальной формы, усеченный
на обоих концах, с маленьким шипом на задней трети наружного края.
Карпус тонкий, в 12/3 раза длиннее, чем мерус, расширяющийся дистально.
Фиг. 21. Callianassa subierranea (Montagu), а — лобный край карапакса
и глазные стебельки; b — шестой абдоминальный сегмент и хвостовый веер;
с — задний край тельсона (из de Man, 1928).
Длина клешни составляет 2/3 длины карпуса. Края члеников этой ноги
волосатые, особенно наружный край карпуса и клешня снабжены длинными
волосами, а пальцы также несут много пучков волосков. Тельсон одинаков
как в длину, так и в ширину, и кажется трапециевидным; задний край
имеет слабую выемку на середине; в этой выемке находится микроскопи-
ческий зубец; на некотором расстоянии от этого зубца, где боковые края
тельсона переходят в задний край, с каждой стороны имеется два микро-
скопических шипика одинаковой величины и почти такие же маленькие,
как срединный зубец. Направленная назад внутренняя уропода едва высту-
пает за тельсон, наружная уропода выступает очень мало; базальный
членик уропод невооруженный; наружная уропода с двумя почти смежными
сильными гребнями, почти круглая; дополнительный ряд коротких щетинок
простирается почти до середины заднего края и немного отклоняется
от него; внутренняя уропода такой же длины, как и ширины, закругленная
на заднем краю, с единственным явственным гребнем. У самки уроподы
простираются немного дальше за тельсон.
Распространение. Побережье Англии, Ирландии, Франции,
Средиземное море у северного побережья Африки близ Bengasi.
Фауна СССР, Макаров .
66
CALLIANASSIDAE
2. Подрод Trypaea Dana
Trypaea Dana, 1852 : 513; Borradaile, 1903 : 546.
Фиг. 22. Callianassa harmandi
Bouvier.
Наружные максиллярные ножки широкие, крышкообразные. Проподус
третьих перейопод с долей на заднем краю. Эндоподиты уропод широкие,
с прямоугольными заднебоковыми углами. Тель-
сон такой же длины, как и ширины, или обычно
больше в длину чем в ширину, такой же длины
как эндоподиты уропод.
*2. Callianassa (Trypaea) harmandi Bou-
vier. (Фиг 22, 23).
Callianassa harmandi Bouvier, 1901:332; de Man,
1928 :13, t. 3, fig-. 6—6j; Yii, 1931:92, fig. 3.
Лобный край карапакса лишен рострума,
но на уровне наружного края глазных сте-
бельков имеет короткие треугольные выступы
(боковые выступы); на месте рострума также
обнаруживаются с каждой стороны от средней
линии очень слабые, едва заметные закруг-
ленные выступы, между которыми образуется
небольшая выемка. Глазные стебельки bI1^ раза
больше в длину чем в ширину, внутренние пря-
мые края соприкасаются друг с другом почти
вплоть до закругленных кончиков, наружные
края в передней трети слегка вогнутые.
Роговицы находятся в центре стебельков и
заметно возвышаются над их поверхностью;
внутренние края кончиков стебельков заметно
приподнятые. Первый сегмент антеннулярной
ножки едва выступает за глазные стебельки;
второй сегмент немного короче, чем первый;
третий сегмент вдвое длиннее, чем второй,,
и несет два жгута, которые короче чем
ножка; ножка и более тонкий жгут опушены
длинными волосками. Антеннальная ножка немного короче антенну-
лярной, с маленьким овальным скафоцеритом; жгут составляет прибли-
зительно половину длины тела. Наружные максиллярные ножки, как
у С. japonica. Исхиум большей клешненосной ноги заметно расширяется
дистально, с зубцами на наружном краю и с маленьким зубцом на внут-
реннем проксимальном конце. Мерус такой же длины, как исхиум, слабо
выпуклый внутренний край несет ряд мелких тупых зубцов, верхняя
поверхность с явственным гребнем; проксимальная доля наружного края
напоминает таковую С. ja onica, ее задний и передний края зазубрены;
2. CALLIANASSA
67
впереди этой доли наружный край меруса мелко зазубрен; снизу под этим
краем имеется ряд гранул. Внутренний край карпуса мелко зазубрен
и слегка загнут вниз; то же самое относится и к наружному краю; длина
внутреннего края карпуса меньше, чем ширина карпуса. При сгибании
закругленный нижне-наружный край карпуса почти целиком закрывает
проксимальную долю наружного края меруса. Клешня приблизительно
в 1:/2 раза длиннее карпуса, ладонь немного больше в длину чем в ширину,
края ладони мелко зазубренные и волосатые; доля на переднем краю
ладони у основания подвижного пальца широкая, имеет гладкий незазуб-
ренный край и на верхне-наружном углу несет малень-
кий шипообразный выступ; изогнутый неподвижный
палец гладкий, с гладким хватательным краем, только
нижне-внутренний край обнаруживает слабую гранули-
рованность. Наиболее характерным для вида является
форма подвижного пальца, в частности вооружение его
хватательного края. Весь палец немного длиннее непо-
движного пальца, с сильно искривленным кончиком;
тотчас за кончиком хватательный край несет крупный
зубец, в точности копирующий форму кончика пальца;
за этим зубцом идет резкая выемка, отделяющая от упо-
мянутого зубца широкий проксимальный зубец, который
не доходит до проксимального конца пальца и имеет
почти прямоугольную форму, представляя собой как бы
основание усеченного широкого треугольного зубца;
свободный край этого зубца мелко зазубрен. При смы-
Фиг. 23. Callianassa
harmandi Bouvier,
большая клешне-
носная нога.
кании кончик подвижного пальца заходит за кончик
неподвижного пальца, так что верхний край второго
зубца хватательного края упирается в хватательный край
неподвижного пальца; при этом остается щель весьма
характерной формы. Исхиум меньшей клешненосной ноги тонкий, с 1—
2 маленькими зубцами на наружном краю. Мерус с шипом на наружном
краю. Карпус резко суживается проксимально. Клешня немного короче,
чем карпус, пальцы такой же длины как ладонь; хватательный край
неподвижного- пальца мелко зазубрен на середине; хватательный край
подвижного пальца гладкий. В общем, эта нога очень похожа на таковую
С. japonica, но только клешня немного короче, чем карпус. Следующие
перейоподы и абдомен с плеоподами такие же, как у С. japonica. Зад-
ний край тельсона несет шип на середине и по паре шипиков близ боко-
вых углов.
Размеры
Длина исхиума большей клешней, ноги...............
„ меруса
„ карпуса
Ширина „
9.0 мм
9.0 „
11.0 „
10.5 „
5*
68
CALLIANASSIDAE
Длина клешни большей клешней, ноги..............16.0 мм
Ширина ладони „ „ „ .............10.0 „
Длина подвижн. пальца ......................... 7.5 „
Распространение. Япония: провинция Bingo; Nemuro. Китай:
полуостров Shantung, бухта Kiaochow. В наших водах в бухте Патрокл,
залив Петр Великий.
*3. Callianassa (Trypaea) gigas var. еоа, nom. nov. (Фиг. 24).
С. gigas var. japonica Makarov, 1935:323, fig-. 4 (пот. praeoccup.).
Фиг. 24. Callianassa
gigas еоа, nom. nov.
большая клешненос-
ная нога.
Рострум более тупой, чем у типичной формы.
Кончики глазных стебельков также более тупые, чем
у типичной С. gigas-, пигментированные роговицы рас-
положены в центре стебельков или несколько впереди
их середины. Клешненосные ноги очень неравные,
большая значительно длиннее меньшей. Поверхность
исхиума большей клешненосной ноги ’гладкая и бле-
стящая; наружный край зубчатый на двух проксималь-
ных третях, внутренний край зубчатый примерно на
протяжении одной проксимальной трети; у С. gigas
наружный край зубчатый почти на всем своем протя-
жении, а внутренний край гладкий. Внутренний край
меруса почти гладкий, лишь на проксимальном конце
имеется группа мелких гранул; у С. gigas этот край
явственно зазубрен. Проксимальная доля на наружном
краю меруса более остроконечная и не так сильно
выступает наружу, как у С. gigas; при сгибании эта
доля наполовину закрывается закругленным наружно-
задним краем карпуса, тогда как у С. gigas она
в этом случае остается совершенно незакрытой. Кар-
пус длиннее меруса, его края гладкие, слабо волосатые
и слабо утонченные; у С. gigas эти края заметно утон-
ченные и явственно зазубренные. Клешня примерно
как карпус, и в l1^ раза длиннее его; наружный край
такой же ширины,
ладони несколько утончен и зазубрен, внутренний край гладкий; доля у осно-
вания подвижного пальца широко-закругленная, с гладким краем и с едва
заметным выступом на наружном углу; у С. gigas на наружном углу доли
имеется заметный выступ, усаженный шиповатыми гранулами. Пальцы
покрыты пучками длинных мягких волосков; хватательный край непо-
движного пальца слабо выпуклый на середине, с слабо изогнутым кончи-
ком, весьма характерно вооружение хватательного края подвижного пальца,
отчасти напоминающее таковое у С. harmandi; немного отступя от осно-
вания имеется длинный низкий усеченный зубец, свободный край которого
мелко зазубрен; затем следует короткий зубец и тотчас за ним сильно
2. CALLIANASSA
69
изогнутый кончик; при смыкании кончик подвижного пальца Заходит за
кончик неподвижного пальца, выпуклость хватательного края неподвиж-
ного пальца накрывает выемку между усеченным и приконцевым зубцами
хватательного края подвижного пальца, так что остается узкая треуголь-
ная щель, доходящая лишь до половины длины пальцев и своей формой
ничуть не напоминающая щель у С. harmandi. Пальцы совершенно гладкие,
как на поверхностях, так и на краях. У С. gigas хватательный край непо-
движного пальца имеет два низких зубца на проксимальной половине;
хватательный край подвижного пальца лишь со слабой выпуклостью
у основания, на остальном протяжении он не имеет никаких зубцов и мелко
зазубрен; при смыкании кончики пальцев сильно скрещиваются, а края
плотно соприкасаются, не оставляя никакой щели; наружный край подвиж-
ного пальца и его нижняя поверхность с рядами из круглых гранул.
Меньшая клешненосная нога ничем не отличается от таковой С. tigas.
Нижний край руки второй перейоподы совершенно не выпуклый и почти
прямой; оба пальца одинаковой ширины у основания; у С. gigas нижний
край заметно выпуклый и неподвижный палец значительно шире у осно-
вания, чем подвижный. Остальные перейоподы, как у С. gigas. У вариетета
тельсон несколько-короче по отношению к уроподам, чем у типичного
вида. Все описанные отличия полностью подтвердились после сравнения
данного вариетета с экземпляром типичного вида, полученным из Вашинг-
тонского музея.
Размеры
Общая длина ...................................
Длина большей клешиен. ноги.................
Длина исхиума большей клешней.' ноги...........
» меруса „
„ карпуса „ „
Ширина „
Длина клешни „ „
Ширина „
Длина меньшей клешней, ноги
46.0 мм
44.0 „
9.0 „
19.0 „
11.0 ,,
7-5 „
15-0 „
7.0 „
25.0 „
У четырех более маленьких самцов имеется отличие от описанного
крупного самца в вооружении хватательных краев пальцев большей
клешни: именно, оба края имеют лишь мелкую зазубренность, плотно
сходятся при смыкании, скрещиваясь кончиками. В остальных признаках
они совпадают с вышеописанным экземпляром. Этот факт еще раз
наводит на мысль о значительных возрастных изменениях в форме
передних перейопод у Callianassa, что, в свою очередь, косвенно под-
тверждает сомнительность С. californiensis bouvieri, как самостоятель-
ного вида (см. ниже).
Распространение. Бухта Патрокл; залив Петр Великий.
70
CALLIANASSIDAE
Фиг. 25. Callianassa
japonica (Ortmann),
большая клешне-
носная нога.
* 4. Callianassa (Trypaea) japonica (Ortmann). (Фиг. 25).
Callianassa subterranea (Montagu) var. japonica Ortmann, 1891 :56, t. 1, fig. 10a;
Balss, 1914 : 91; Yokoya, 1930 : 543; 1933 : 52. — Callianassa japonica Balss, 1924 : 52; de Man,
1928:19, t. 5, fig. 10—10c; Yu, 1931: 95, fig. 5.
Рострум короткий, широко-треугольный, несущий на вершине малень-
кое шипообразное острие. Передний край карапакса между наружным краем
глазного стебелька и антеннальной ножкой несет треугольный выступ,
который составляет примерно половину длины рострума. Глазные стебельки
достигают почти до дистального конца базального членика антенну-
лярной ножки, их ширина составляет немного больше половины их длины;
их внутренние края соприкасаются и только перед
самыми кончиками расходятся; наружные края
косо-закругленные, очень слабо вогнутые близ
кончиков. Роговицы несколько овальные, пигменти-
рованные, с ясно заметной сетчатой структурой,
находятся на середине верхней поверхности сте-
бельков или лишь чуть-чуть смещены вперед.
Второй членик антеннулярной ножки почти такой же
длины, как первый; третий членик тонкий, вдвое
длиннее чем второй; жгуты довольно длинные,
почти одинаковой длины; ножка и более тонкий
жгут опушены снизу длинными волосами, причем
волоса ножки перистые. Антеннальная ножка
немного короче антеннулярной; скафоцерит малень-
кий, овальный, налегающий на дорзальную поверх-
ность следующего членика; четвертый и пятый сег-
менты почти одинаковой длины, жгут составляет
примерно половину длины тела. Наружные максил-
лярные ножки широкие, крышкообразные; исхиум
треугольный, сильно суживающийся кзади; перед-
ний край, сочленяющийся с мерусом, косо срезанный; внутренний край
закругленного меруса длиннее чем наружный; проподус не расширяющийся.
Исхиум большей клешненосной ноги несет 5—7 зубцов на наружном краю.
Внутренний край меруса мелко зазубрен'на своей проксимальной поло-
вине, имея приблизительно 12 зубчиков, которые становятся мельче
в дистальном направлении и, наконец, совсем исчезают; наружный край
на проксимальной половине несет острую долю или зубец, нижний
изогнутый край которого несет 7—8 острых зубчиков; верхне-внутренний
край с 2—3 неотчетливыми зубцами; впереди доли наружный край меруса
мелко зазубрен; выпуклая верхняя поверхность имеет явственный гребень.
Карпус немного больше в длину чем в ширину, с гладкой выпуклой верхней
поверхностью и плоской нижней поверхностью; края карпуса острые, уса-
жены мелкими ресничками, внутренний край мелко зазубрен; длина внутрен-
него края карпуса равна ширине карпуса или бывает даже несколько больше
2. CALLIANASSA
71
ее. Клешня длиннее чем карпус, у основания такой же ширины, как послед-
ний, а дистально несколько суживается; неясно зубчатый и покрытый реснич-
ками наружный край ладони почти прямой, переходящий в прямой наружный
край неподвижного пальца, кончик которого слегка загнут. Верхняя поверх-
ность ладони выпуклая, нижняя уплощенная. Подвижный палец немного
длиннее неподвижного, с заметно изогнутым кончиком; хватательные края
обоих пальцев равномерно зазубрены, при этом зубчики на подвижном
пальце более крупные, чем на неподвижном. При смыкании кончики паль-
цев скрещиваются и остается короткая треугольная щель только у осно-
вания. Закругленный выступ переднего края ладони у щели между паль-
цами имеет зазубренный край (у самки этот край почти гладкий). Исхиум
меньшей клешненосной ноги длинный, узкий, его наружный край почти
гладкий. Мерус короче, чем исхиум, овальной формы, наружный край на
середине несет маленький шип. Карпус в 2*/2 раза больше в длину чем
в ширину, заметно длиннее чем мерус. Клешня чуть-чуть короче чем кар-
пус, плоские пальцы немного длиннее ладони, волосатые; хватательный
край подвижного пальца гладкий, тогда как хватательный край неподвиж-
ного пальца мелко зазубрен, в особенности на проксимальных двух третях.
Клешня второй перейоподы плоская, правильной конусообразной формы.
Проподус третьей перейоподы в виде поперечно-овальной пластинки,
верхний закругленный край которой несет короткий палец; нижний край
проподуса имеет долю, иначе говоря, выступает за нижний край карпуса.
Тельсон почти квадратный, с закругленными задними углами; задний край
слегка вогнутый и несет на середине маленький шип. Плеоподы третьего-
пятого сегментов листовидные.
Размеры.
Длина исхиума большей клешней, ноги......5.0 мм
„ меруса „ „ „ 5.0 „
» карпуса „ „ „ ........ 5.0 „
Ширина „ „ „ ...............4.5 „
Длина клешни „ „ „ 8.0 „
Ширина ладони „ „ „ 5.0 „
Длина подвижного пальца..................4.0 „
По внешнему виду С. japonica напоминает европейскую С. subter-
ranea, что и послужило поводом для выделения данного вида Ortmann’oM
как вариетета европейского вида. Borradaile (1903), а позднее и Balss (1924)
отметили, что правильнее считать японскую форму самостоятельным
видом, а не вариететом, так как. он относится к другому подроду, чем
европейский вид. Balss (1914, 1924) считает идентичными с данным видом
Callianassa petalura Stimpson и С. calif orniensis japonica Bouvier.
Распространение. Япония: Хакодатэ, Токийская бухта; Simoda;
Bingo; около устья реки Tanabe; Moura; Kagosima; Akita; Китай: Chefoo,
Chinwangtao, Hainan. В наших водах вид известен из бухты Патрокл,
залив Петр Великий. На глубинах от 9 до 192 м.
72
CALLIANASSIDAE
* 5. Callianassa (Trypaea) californiensis Dana var. bouvieri nom. nov.
(Фиг. 26).
Callianassa californiensis var. japonica Bouvier, 1901: 332 (nom. praeoccup.); de Man,.
1928:18, t. 4, fig. 9—9e; Yu, 1931: 94, fig. 4.
Очень близок к C. japonica, отличаясь от него только перейоподами^
Исхиум большей клешненосной ноги заметно расширяется в дистальном
направлении, его внутренний край вогнутый и имеет на проксимальном
конце маленький зубец, верхняя поверхность у основания несет две
маленьких гранулы, расположенных друг за другом; наружный край во-
Фиг.26. Callianassa
californiensis bou-
vieri nom. nov. боль-
шая клешненосная
нога.
оружен* зубцами на проксимальных двух третях своей
длины. Мерус немного длиннее чем исхиум, с почти пря-
мым зазубренным внутренним краем; гребень на верхней
поверхности очень явственный, резкая выемка отделяет
его от другого поперечного гребня, который переходит
на проксимальную долю наружного края; эта доля более
широкая, чем у С. japonica, с сильно закругленным
задним краем и не такая остроконечная; оба ее края,,
а также и наружный край меруса впереди доли мелко
зазубренные. Карпус более широкий по отношению
к своей длине, чем у С. japonica, и его задне-наружный
край гораздо дальше выступает назад за уровень
сочленения с мерусом, тогда как у С. japonica закруг-
ление этого края находится на одном уровне с мораль-
ным сочленением. Клешня имеет совершенно другую,
форму; подвижный палец немного длиннее чем внутрен-
ний край ладони (у С. japonica он короче), острый кон-
чик резко изогнутый; вырезка на дистальном краю
ладони у основания неподвижного пальца очень глубо-
кая, доля с зазубренным краем большая; неподвижный
палец относительно тоньше, чем у С. japonica, более сильно изогнутый,.
как весь, так и в особенности на конце, благодаря чему при смыкании
остается очень широкая щель, так как пальцы соприкасаются только
скрещивающимися кончиками. Вооружение хватательных краев как
у С. japonica. Меньшая клешненосная нога отличается от таковой С. japo-
nica тем, что клешня немного длиннее по отношению к карпусу, а пальцы
немного длиннее по сравнению с ладонью.
Размеры
Длина исхиума большей клешней, ноги .... . . . . 8.0 ММ
„ меруса 11 11 11 * * * * . . . . 9.0 11
„ карпуса 11 11 11 • • • ' . . . . 11.0 »
Ширина „ 11 11 11 • • • • . . . . 9.5 11-
Длина клешни п 11 »>•••• . . . . 14.0 »
„ ладони 11 11 11 * • • • .... 8.0 11
Ширина „ 11 11 11 • • • • . . . . 9.5 »•
Длина подвижного пальца . . . . 8.5
2. callianassa
73'
Самостоятельность данного вида подвергается сомнению. Как уже
говорилось выше, Н. Balss считает его идентичным с С. japonica', сходство
несомненно большое и, как справедливо отмечает de Man (1928), разница
наблюдается только в форме передних перейопод. Весьма возможно, что
данный вид является С. bouvieri, обнаруживающей возрастные и половые
изменения, так как, действительно, до сих пор известны только крупные
самцы С. californiensis bouvieri (к сожалению Yu, 1931, не указывает пола
своих экземпляров). Если и в дальнейшем будут попадаться только самцы
данного вида, то принадлежность его к С. japonica станет очевидной,
пока же недостаточность материала заставляет воздержаться от подоб-
ного вывода.
Распространение. Япония: окрестности Токио. Китай: Peitaiho.
В наших водах найден 1 $ в бухте Патрокл, залив Петр Великий.
3. Подрод Callichirus Stimpson
Callichirus Stimpson, 1866 : 47; Borradaile, 1903 : 546.
Наружные максиллярные ножки узкие по сравнению с таковыми
подрода Trypaea. Проподус третьих перейопод с долей на заднем краю.
Эндоподиты уропод узкие и заостренные или закругленные на заднем;
конце. Тельсон такой же ширины, как и длины, или больше в ширину чем
в длину и явственно короче, чем эндоподиты уропод.
* 6. Callianassa (Callichirus) pontica (Czerniavsky). (Фиг. 27, 28).
Callianassa subterranea (Montagu) forma pontica CzerniavsKy, 1884 : 81.— Callianassa
subterranea Heller, 1863:202, t. 6, fig. 9—11. — Callianassa pestae de Man, 1928:34, t. 9,
fig. 16—16e (литература).
Рострум широко-треугольный, острый; боковые зубцы тупые. Вну-
тренние края глазных стебельков или соприкасающиеся почти на протяже-
нии всей своей длины и расходящиеся только у тупых кончиков стебель--
ков, или же они расходятся примерно на середине, приближаясь этим
к С. laticauda; роговицы маленькие, выпуклые, расположены на передней
половине стебельков у бокового края. Глазные стебельки такой же длины,.
как базальные членики антеннулярных ножек, или же немного превышают
их. Антеннулярная ножка такой же длины, как и антеннальная; ее третий
членик значительно длиннее, чем второй. Наружные максиллярные ножкк.
с довольно широким исхиумом и мерусом (крышкообразные). Исхиум боль-
шей клешненосной ноги заметно расширяется у дистальной трети; наруж-
ный край у начала расширения несет заметный шипик, вообще же края
неявственно зубчатые, почти гладкие. Проксимальная доля на наружном
краю меруса остроконечная, ее наружный край несет 3—5 зубчиков,
внутренний край гладкий; наружный край меруса впереди доли зазубрен-
на дистальной половине; при сгибании только основание доли меруса
остается незакрытым закругленным задне-наружным краем карпуса....
74
CALLIANASSIDAE
Карпус короче чем ладонь (у самки из Венеции по de Мап’у несколько
длиннее чем ладонь), оба края его гладкие. Ладонь такой же ширины,
Фиг. 27. Callianassa pontica (Czerniavsky). а—лобный край карапакса
и глазные стебельки; b — шестой абдоминальный сегмент и хвостовый веер
(из de Man, 1928/
как карпус (у того же экземпляра.из Венеции ладонь была уже чем карпус),
с гладкими краями; доля на переднем краю ладони широкая и низкая,
Фиг. 28. Большая клешненосная нога
Callianassa pontica (Czerniavsky), два
типа пальцев.
на широкий зубец у основания,
с небольшим выступом у основания вну-
треннего края неподвижного пальца;
пальцы короче чем ладонь; подвижный
палец чуть-чуть длиннее чем неподвиж-
ный; наружные края пальцев гладкие,
кончик подвижного пальца изогнут и
при смыкании заходит за более прямой
кончик неподвижного пальца. Вооруже-
ние хватательных краев пальцев бывает
двоякого рода (фиг. 28): на неподвиж-
ном пальце имеется слабый выступ,
который расположен либо на середине,
либо смещен ближе к кончику, благо-
даря чему между ним и кончиком обра-
зуется явственная выемка; в первом
случае палец почти прямой, во втором
слегка изогнутый; в обоих случаях
у основания имеется ряд мелких зуб-
чиков; хватательный край подвижного
пальца либо мелко зазубрен на всем
своем протяжении, хотя имеется намек
свободный край которого более слабо
зазубрен, либо у основания имеется явственный широкий усеченный
зубец, затем полукруглая вырезка, за которой хватательный край мелко
2. CALLIANASSA
75
зазубрен почти до самого кончика. Оба типа вооружения хватательных
краев пальцев большей клешненосной ноги не связаны с полом: так, у про-
смотренных экземпляров первым типом обладали 4 самца и 4 самки,
а вторым типом один самец; возможно, что это связано с возрастом, так
как последний самец был самый крупный из всех просмотренных экземпля-
ров. Карпус меньшей клешненосной ноги узкий и длинный, менее широкий
чем мерус; его ширина составляет менее 1/3 длины его внутреннего края,
или же составляет как раз одну треть; рука такой же ширины, как карпус.
Проподус третьих перейопод овальный. Тельсон короткий и широкий,
почти полукруглый, на :/8 больше в ширину чем в длину, задний
край волосатый, но не несет никаких шипов. Задний край наруж-
ной уроподы широко закругленный, приподнятая часть ее верхней
поверхности (в виде поперечного гребня на наружной половине, опу-
шенного волосками) находится на некотором расстоянии от заднего
края; на проксимальном конце около базального членика имеется малень-
кий зубец.
Окраска: белая, переходящая в голубоватую или розовато-
жрасную (Heller) или беловатая с розовым или светлоголубым оттен-
ком (Pesta).
Размеры
Общая длина (от конца рострума до конца тельсона) . 52.0
Длина исхиума большей клешней, ноги . 8.0
„ меруса я я . 9.0
„ карпуса я я . 10.0
Ширина „ я » я . 8.5
Длина клешни п я я . 15.5
„ подв. пальца большей клешней, ноги . . . . . 8.0
„ меруса меньшей клешненосной ноги . . . . . . 5.5
„ карпуса а я » • • • • . 7.5
Ширина „ я . 2.5
Длина клешни » я я . . . . . 8.0
Ширина „ я я • • . • < . 2.5
Длина тельсона . 4.0
Ширина „ . . 6.0
мм
Распространение. Средиземное море (Неаполь, Сицилия);
-Адриатическое море (Венеция, Rovigno, Pizano, Lesina); западное побе-
режье Франции (St Malo); Черное море.
Встречается до глубины 10 м. Обычно зарывается в песок, или же
держится под корнями зостеры (Зернов, 1913). Молодые экземпляры
довольно часто попадаются ночью плавающими у поверхности моря; более
крупные экземпляры встречаются у поверхности во время сильной зыби.
По наблюдениям Чернявского (1884) взрослые и полувзрослые животные
очень хищные: в условиях аквариума они нападают на других животных,
или же друг на друга.
76
CALLIANASSIDAE.
3. Род CTENOCHELES KISHINOUYE
Ctenocheles Kishinouye, 1926 : 63.
Тип рода: Ctenocheles balssi Kishinouye.
Рострум маленький. Первая пара перейопод асимметричная и нерав-
ная, с хорошо развитыми клешнями. Вторая пара перейопод маленькая;
и клешненосная. Подобранхии и мастигобранхии отсутствуют. Плеоподы
второго абдоминального сегмента такие же, как и на трех следующих,
сегментах.
Род Ctenocheles близок, с одной стороны, к роду Callianassa, от кото-
рого отличается тем, что плеоподы второго сегмента имеют такую же
форму, как и следующие; с другой стороны, он близок к роду Callianidea,.
от которого отличается отсутствием мастигобранхий. К сожалению, Kishi-
nouye (1926) не говорит, имеются ли у его рода жаброобразные нити на.
плеоподах, которыми обладает род Callianidea1, если все отличие сводится,
лишь к отсутствию мастигобранхий, то едва ли наличие столь изменчивого-
признака является достаточным для выделения особого рода.
1. Ctenocheles balssi Kishinouye. (Фиг. 29).
Pentacheles sp. Balss, 1914 : 75, fig. 43. — Ctenocheles balssi Kishinouye, 1926 : 63, fig. If
Yokoya, 1933:55.
•
„Карапакс составляет почти четверть длины всего тела, он недоста-
точно длинный для того, чтобы покрыть головогрудь на всем протяжении^
оставляя сегмент последних перейопод непокрытым. Он более или менее
сжат с боков, а его края усажены ресничками. Рострум очень короткий,,
сжатый с боков и продолжающийся назад в виде низкого срединного киля
с рядом примерно из 10 мелких зубчиков. Глазные стебельки сплющен-
ные в горизонтальной плоскости на переднем конце, роговицы отсутствуют;
стебельки почти такой же длины, как первый членик антеннул. Антеннулы.
с двумя почти равными жгутами, которые длиннее чем ножки и располо-
жены как раз под глазными стебельками. Антенны без скафоцеритов.
и шипов и расположены с наружной стороны глазных стебельков; два.
проксимальных членика антеннальной ножки почти равны по длине антен-
нулярной ножке. Мандибула имеет очень острый режущий край и несет
пальп из трех члеников, из которых последний членик самый длинный
и снабжен пучком волосков. Вторые максиллярные ножки несут ного-
образный эндоподит из пяти члеников, из- которых дистальный членик
снабжен щетинками. Третьи максиллярные ножки ногообразные и длинные,,
состоящие из шести члеников; четвертый членик очень толстый, с острым
зазубренным внутренним краем. Первая пара перейопод неодинаковая,
по величине и по форме, с хорошо развитыми клешнями. Правая клешня
больше чем левая и более или менее напоминает правую клешню Thau-
mastocheles (Т. japonicus Caiman, В. М.); но у данного вида карпус очень
3. CTENOCHELES
77
короткий и кажется как бы слившимся с проподусом. Как подвижный,
так и неподвижный пальцы вооружены длинными острыми зубцами, чере-
дующимися с некоторым числом более мелких зубцов. Подвижный палец
отгибается наружу. Мерус сильный, имеющий округлый внешний вид на
проксимальном конце. Исхиум почти такой же длины, как wtepyc, но гораздо
тоньше. Левая клешня также хорошо развита, но меньше, ее подвижный
палец составляет приблизительно половину длины пальца правой клешни;
внутренняя сторона клешни зазубрена. Проподус почти такой же длины,
как проподус правой клешни, но по ширине составляет примерно половину
его. Карпус длиннее, но как мерус, так и исхиум короче по сравнению
с таковыми правой клешненосной ноги. Первая пара перейопод очень
слабо волосатая и ее поверхность почти гладкая. Вторая пара перейопод
Фиг. 29. Ctenocheles balssi Kishinouye (из Kishinouye, 1926).
жлешненосная и сплющенная, с длинными волосами на острых краях пальца,
проподуса и карпуса и на внутренних краях меруса и исхиума. На про-
подусе и карпусе мы находим несколько групп из немногочисленных
коротких волос. Третья пара перейопод немного длиннее чем вторая;
проподус и дистальная часть карпуса широкие и их роющие стороны
усажены ресничками; на карпусе и мерусе мы находим маленькие группы
коротких волос, как и у предыдущей пары. Четвертая пара перейопод
немного длиннее и тоньше чем предыдущая пара; карпус продолговатый
и палкообразный; исхиум несколько короче, чем у третьей или пятой
перейоподы. Пятая пара перейопод без ложных клешней; проподус густо
волосатый, на задне-внутреннем краю. Коксальные членики перейопод
на обеих сторонах встречаются друг с другом на вентральной срединной
линии головогруди, за исключением двух последних пар, которые разде-
лены thelycum’aMH. Абдомен очень удлиненный; его сегменты почти глад-
кие, а эпимеры слабо развиты. Плеоподы хорошо развиты; первая пара,
однако, маленькая, узкая и состоящая из двух члеников; вторая-пятая пары
совершенно сходные друг с другом, состоящие из двух широких листо-
видных пластинок на конце короткого базального членика; внутренний
край эндоподита густо усажен ресничками, в то время как дистальный
78
GALA T HEIDE A
и наружный края экзоподита снабжены короткими волосками. Шестые
плеоподы образуют боковые пластинки хвостового веера; экзоподит шире,,
чем эндоподит, и с выемкой на середине заднего края. Тельсон также
широкий, почти такой же ширины как и длины, и имеет три радиальных
борозды, идущих от середины переднего края. Жаберный аппарат очень,
простой, развиты только артробранхии; они находятся по две на каждом
сегменте, начиная от третьих максиллярных ножек до четвертых перей-
опод. Животное было поймано, кажется, в глубинных водах, так как оно
является слепым (Kishinouye)".
Распространение. Япония; западное побережье о-ва Ниппон
около Ohsu близ Kashiwasaki, префектура Niigata (Kishinouye); бухта
Wakasa, 119 м (Yokoya). Н, Balss имел в своем распоряжении только-
правую клешненосную ногу данного вида (из сбора Doflein’a, точное
местонахождение не указано).
II. Надсем. GALATHEIDEA
Карапакс хорошо развит, большею частью с явственным боковым
краем и с явственной linea anomurica. Рострум хорошо развит. Карапакс
не сливается с эпистомом. Глазные стебельки короткие и толстые.
Базальный членик антеннул более или менее вздутый, часто с одним или
несколькими стилоцеритами. Ножка антенн четырехчленистая (второй
и третий членики сливаются), реже пятичленистая (род Aeglea); редко
имеются шипообразные скафоцериты (род Uroptychus), большею частью
они отсутствуют; жгуты длинные и тонкие. Наружные максиллярные ножки
ногообразные или несколько расширенные. Первая пара перейопод клеш-
неносная, почти симметричная. Следующие три пары хорошо развитые,,
простые. Пятая пара редуцирована, снабжена маленькой клешней и ча-
стично скрыта в жаберной полости. Торакальные sterna широкие. Жабры
типа филлобранхий (за исключением рода Aeglea, где они типа трихобран-
хий), большею частью в числе 14:10 артробранхий и 4 плевробранхии;
пятые перейоподы всегда с плевробранхиями. Иногда сохраняются масти-
гобранхии на некоторых перейоподах; подобранхии отсутствуют. Абдомен,
симметричный, хорошо развитый, состоит из семи сегментов, из которых
первый обычно скрыт под карапаксом; абдомен частично или более или.
менее полно подогнут под торакс; эпимеры абдомена хорошо развиты,
первые маленькие, а следующие направлены косо наружу и почти гори-
зонтально; уроподы всегда имеются, вместе с тельсоном они образуют
широкий хвостовой веер; тельсон часто с поперечным швом. Абдоминальные
конечности у самца большею частью имеются на первом и втором сег-
ментах, на третьем-пятом сегментах имеются или отсутствую. ; первая
пара преобразована в половые придатки; согласно Boas (1880, fig. 169)
каждый придаток имеет единственную широкую ветвь (наружная пла-
стинка), а ее передний сильно утонченный край (внутренняя пластинка)
GALA THEIDAE
79
пригнут к первой пластинке, так что образуется своего рода трубка;
у второй пары каждый придаток (ibid., fig. 170) имеет рудимент наружной
ветви, а внутренняя ветвь несет на своем конце appendix mascuiina;
три следующих пары придатков, если они имеются, такой же формы, как
у самки. У самки всегда имеются абдоминальные конечности на четвертом
и пятом сегментах и часто бывают на втором и третьем сегментах; каждый
придаток состоит из простой узкой ветви, состоящей из трех члеников;,
при этом Boas считает, что терминальный членик представляет собой
appendix interna, а предпоследний членик — настолько редуцированную
внутренюю ветвь, что appendix interna оказался отходящий от ее дисталь-
ного конца.
Boas, который установил это надсемейство (почему правильнее было
бы ставить его имя, а не Henderson’a, как это принято), усматривал
некоторое сходство представителей Galatheidea с Paguridea. Ortmann
(1892) отмечает, что „Galatheidea суть Thalassinidea, которые выделяются
своеобразной внешней формой, связанной с уплощением всего тела,
а также преобразованием последней пары перейопод; абдомен, в противо-
положность Paguridea, обнаруживает еще близкое отношение к Thalas-
sinidea11.
Представители этого.надсемейства являются морскими обитателями,
встречающимися как на литорали, так и на глубинах. Уклоняющееся
во многих отношениях семейство Aegleidae представлено единственным
родом и видом, Aeglea laevis (Latreille) в пресных водах Южной Америки.,
В нашей фауне имеются представители двух семейств.
ОПРЕДЕЛИТЕЛЬНАЯ ТАБЛИЦА СЕМЕЙСТВ
1 (2). Абдомен согнутый, но не подогнутый под торакс. Наружные максил-
лярные ножки с мастигобранхиями. Первые перейоподы обычно тон-
кие и длинные. Общая форма тела гарнелевидная (макрурообраз-
ная)..........................................3. Сем. Galatheidae.
2 (1). Абдомен подогнут под торакс. Наружные максиллярные ножки без.
мастигобранхий. Первые перейоподы обычно умеренной длины, тол-
стые. Общая форма тела крабообразная . . 4. Сем. Porcellanidae.
3. Сем. GALATHEIDAE
Galatheidae Dana, 1852:1431; Ortmann, 1892:244; 1901 :1150; Benedict, 1903:244;
Schmitt, 1921: 162.
Тело гарнелевидное, карапакс продолговатый, с хорошо выражен-
ными областями и обычн с поперечными морщинами; боковые края,
и linea anomurica явственные. Рострум хорошо развит, треугольный или
шипообразный. Антеннулы не защищенные. Антеннальная ножка четырех-,
членистая (второй и третий членики сливаются), очень редко со скафоце-.
'80
GAL A THE I DAE
ритом, большею же частью без него; жгут длинный. Наружные максилляр-
ные ножки полуногообраэныс, исхиум и мерус узкие и часто шиповатые
изнутри; эпиподиты всегда имеются. Первые перейоподы клешненосные,
обычно длинные и тонкие. Эпиподиты могут быть на трех первых парах
перейопод. Жабры типа филлобранхий в числе 14 с каждой стороны:
4 плевробранхии и 10 артробранхий, которые распределяются следующим
образом: по одной плевробранхии на второй-пятой перейоподе и по две
артробранхии на первой-четвертой перейоподе и на наружной максил-
лярной ножке. Абдомен согнут вентрально, но не подогнут под торакс:
колено сгиба приходится на четвертый сегмент, а не на третий, как
у гарнелей; хвостовой веер хорошо развит, приспособлен для плавания.
У самца имеются половые придатки на двух первых абдоминальных сег-
ментах (на первом сегменте они могут быть рудиментарными, но на вто-
ром сегменте они имеются всегда); на трех следующих сегментах имеются
двучленистые, с расширенной базальной частью, плеоподы, которые могут
быть рудиментарными. У самки имеются простые трехчленистые плео-
поды на втором-пятом сегментах (на втором-четвертом сегментах они
могут быть рудиментарными); на первом сегменте придатки всегда
отсутствуют.
Представители этого семейства свойственны преимущественно тепло-
водным морям и встречаются как на мелководьи, так и на глубинах.
Большинство Galatheidae являются донными животными и, повидимому,
не способны свободно плавать на значительные расстояния. Род Uropty-
chus хорошо приспособлен для лазания по губкам, гидроидам, кораллам
и проч. Виды, обитающие на глубинах, часто бывают слепые. Benedict
(1903) отмечает разницу в количестве яиц у мелководных и глубоководных
представителей: у первых количество яиц велико и яйца мелкие, в то
время как у вторых количество яиц незначительное и яйца крупные.
Henderson (1888) разделял роды семейства на 2 группы по степени
изогнутости абдомена: в первую группу входят роды с просто согнутым
абдоменом (эта группа в свою очередь разбивается на 2 подгруппы:
мелководные роды с пигментированными глазами и глубоководные роды
с непигментированными глазами); у родов второй группы абдомен подо-
гнут под самого себя, но не под торакс; представители этой группы
являются глубоководными.
A. Milne-Edwards et Bouvier (1894) разделяли роды этого семей-
ства на две группы Galatheens flagelles и Galatheens non flagelles. Первая,
более примитивная группа характеризуется наличием нечленистого жгута
на экзоподите' первых максиллярных ножек (откуда и название группы);
кроме того плеоподы самца на третьем-пятом сегментах широкие, пластин-
чатые. Приспособление к абиссальной жизни у этой группы выражено
еще слабо: глазные стебельки свободные и всегда оканчиваются пигмен-
тированными глазами, глазной сегмент свободный. Ресничные линии
карапакса сильно развиты, сам панцырь тонкий. Передне-боковой угол
1. GALATHEA
81
карапакса всегда имеет шип. Личинки вылупляются на стадии zoea.
У представителей второй группы экзоподит первых максиллярных ножек
не имеет жгута; задние плеоподы самца утрачивают пластинчатую форму
и становятся стержневидными; глазной сегмент часто сливается с сосед-
ними частями, глаза атрофируются; панцырь сильно объизвествляется, ста-
новится толстым и жестким; передне-боковой угол карапакса без шипа;
ресничные линии карапакса постепенно исчезают. Личинка вылупляется
на стадии metazoea. „Потеря передними максиллярными ножками жгута
экзоподита, почти постоянное отсутствие эпиподитов на перейоподах,
превращение и редукция неполовых абдоминальных конечностей — все
это доказывает нам, что с этой группой мы все больше и больше уда-
ляемся от примитивных форм семейства. В то же самое время мы при-
сутствуем при постепенном усилении у этих животных собственно адап-
тивных признаков к абиссальной жизни: карапакс утолщается, глаза
перестают быть функционирующими органами, глазной сегмент теряет
независимость и глазные стебельки, которые иногда вооружаются шипом,
становятся простыми оборонительными органами и в конце концов стано-
вятся неподвижными, сливаясь со своей дужкой и рострумом. Как и сле-
довало ожидать, антеннулярные щетинки обнаруживают сильное развитие
у этой группы, которая приурочена к глубинам" (A. Milne-Edwards et
Bouvier).
Ortmann (1901) на основании тех же признаков установил подсемей-
ство Galatheinae (=Galatheens flagelles A. Milne-Edwards et Bouvier)
и подсемейство Munidopsinae (=Galatheens non flagelles A. Milne-Edwards
et Bouvier).
ОПРЕДЕЛИТЕЛЬНАЯ ТАБЛИЦА ПОДСЕМЕЙСТВ
1 (6). Глазные стебельки свободные, с хорошо развитыми глазами. Экзо-
подит первых максиллярных ножек оканчивается простым жгутом .
.................. ..........1. Подсем. Galatheinae Ortmann.
2 (1). Глазные стебельки свободные или срастаются с глазным сегментом,
который сам часто сливается с соседними частями; глаза редуциро-
ванные. Экзоподит первых максиллярных ножек без жгута ....
• .................' .... 2. Подсем. Munidopsinae Ortmann.
ОПРЕДЕЛИТЕЛЬНАЯ ТАБЛИЦА РОДОВ
1 (2). Роговицы пигментированные. Экзоподит первых максиллярных
ножек с жгутом. (Подсем. Galatheinae).
2 (5). Рострум не изогнутый и не сжатый с боков.
3 (4). Рострум треугольный, уплощенный в дорзовентральном направлении,
с зубчатыми боковыми краями. Супраокулярные шипы остутствуют
или редуцированы........................ 1. Galathea Fabricius.
Фауна СССР, Макаров 6
82
G'/lL/l I'll EID ЛЕ
4 (3). Рострум шипообразиый, гладкий. По бокам рострума имеются силь-
ные супраокулярные шипы ... •......................2. Munida Leach..
5 (2). Рострум изогнутый и сжатый с боков; его края, в особенности
верхний, вооружены зубцами...................4. Cervimunida Benedicts
6 (1). Роговицы лишены пигмента. Экзоподит первых максиллярных
ножек без жгута (Подсем. Munidopsinae) . 3. Munidopsis Whiteaves.
1. Подсем. GALATHEINAE
Galatheinae Ortmann, 1901 :1150.
Глазные стебельки свободные, глаза хорошо развиты. Экзоподит-
первых максиллярных ножек с простым одночленистым жгутом.
1. Род GALATHEA FABRICIUS
Galathea Fabricius, 1798 : 414; Leach, 1815; Milne-Edwards, 1837 : 273; Dana, 1852 : 478;
Bell, 1853:195; Heller, 1863:188; Henderson, 1888:117; Ortmann, 1892:248; Benedict,.
1903:246; Stimpson, 1907 : 230; Balss, 1913 :1.
Тип рода: Galathea strigosa (Linne).
Карапакс больше в длину чем в ширину, слабо выпуклый, поверх-
ность покрыта многочисленными поперечными линиями с отходящими от
них волосками и обычно невооруженная, за исключением передней части
гастральной области; боковые края вооружены шипами или зубчиками;;
цервикальная борозда глубокая и явственная; кардиальная область не
выдающаяся. Рострум треугольный, сплющенный в дорзовентральном
направлении, часто с продольной бороздой на верхней поверхности; его
боковые края в большинстве случаев вооружены зубчиками; из этих,
зубчиков два базальных расположены над глазами и очевидно соответ-
ствуют супраокулярным шипам рода Munida. Первый членик ножки
антеннул большой, несущий спереди несколько шипов; два следующих
членика ножки тонкие и почти такой же длины, как первый; жгуты очень
короткие. Ножка антенны с тремя толстыми проксимальными члениками;
два первых членика вооружены сильными шипами; жгут длинный. Глаз-
ные стебельки короткие, толстые, немного расширяющиеся дистально,,
частично скрытые под рострумом; орбита ограничена снизу зубчатым
гребнем. Наружные максиллярные ножки стройные, их два терминальных,
членика не листовидные и не расширенные; второй членик мелко зазуб-
рен изнутри на заднем краю и шиповатый на переднем краю; третий чле-
ник большею частью короче чем второй и имеет на внутреннем краю
1-2 шипа. Первая клешненосная пара перейопод умеренно длинная, шипо-
ватая, пальцы на конце ложкообразно выемчатые. Вторая-четвертая
пары перейопод простые. Пятая пара перейопод маленькая, частично
подогнутая под карапакс, с маленькой клешней. Абдомен почти такой же
ширины, как карапакс, и несколько длиннее его; передние края сегментов
невооруженные; хвостовой веер широкий.
/. GALATHEA
83
Виды этого рода преимущественно мелководные, но некоторые
встречаются и на значительных глубинах. Предпочитают скалистый
и каменистый грунты, в тропической области особенно изобилуют среди
губок и кораллов.
ОПРЕДЕЛИТЕЛЬНАЯ ТАБЛИЦА ВИДОВ
1 (4). Передняя часть гастральной области несет только 2 шипа.
2 (3). Базальные шипы рострума маленькие. Верхний край' мерусов вто-
рой пары перейопод вооружен мелкими шипами ......................
....................................... 1. G. strigosa (Linne).
3 (2). Базальные шипы рострума довольно крупные. Верхний край мерусов
второй пары перейопод вооружен 11-12 сильными острыми зубцами .
..................................2. G. acanthomera Stimpson.
4 (1). Передняя часть гастральной области несет более двух шипов.
5,(6). Боковые края рострума впереди базальных шипов гладкие, невоору-
женные. При смыкании между пальцами клешней остается широкая
щель. Абдоминальные сегменты имеют каждый одну поперечную
борозду. Вид не очень волосатый...............3. G. Integra Benedict.
6 (5). Боковые края рострума впереди базальных шипов вооружены
3-4 зубцами с каждой стороны. Пальцы клешней плотно смыкаются.
Каждый абдоминальный сегмент имеет 4 поперечных борозды. Вид
сильно волосатый..................... 4. G. pubescens Stimpson.
1. Galathea strigosa (Linne). (Фиг. 30).
Cancer strigosus Linne, 1766 ; 1053. — Galathea strigosa Risso, 1826 : 47; Milne-Edwards,
1837 :273; Bell, 1853:200; Heller, 1863:189, t. 6, fig. 1—2 (литература); Ortmann, 1892:250,
t. 11, fig. 6i.
Поверхность карапакса с поперечными бороздами, из которых четыре
более глубоких находятся за цервикальной бороздой; от переднего края
каждой борозды направлены вперед короткие волоски. Рострум треуголь-
ный, с острой вершиной и тремя зубцами на каждой стороне (вернее,
четырьмя, так как за самым нижним боковым зубцом обычно имеется еще
маленький зубец); дорзальная поверхность рострума у основания со сла-
бой срединной бороздой. Гастральная область несет пару шипов тотчас
за основанием рострума и по одному шипу сбоку; хепатическая область
несет шип на своей передней половине. Боковой край карапакса несет
обычно 8 шипов; перед крупным шипом на передне-боковом углу имеется
еще очень маленький шипик; ниже его находится еще 2-3 шипика. Глаза
довольно крупные, с короткими глазными стебельками. Первый членик
ножки антеннул широкий и несет три крупных шипа; остальные членики,
отходящие между внутренним и средним шипом, тонкие и цилиндрические,
жгуты очень короткие. Второй членик антеннальной ножки с шипами на
6*
84 GALATHEIDAE
дистальном конце наружного и внутреннего краев; третий членик с шипом
на дистальном конце внутреннего края. Наружные максиллярные ножки
короткие, их мерус длиннее чем карпус, с шипом на дистальном конце ниж-
него края и с мелко зазубренным внутренним краем; карпус с двумя круп-
ными шипами на нижнем краю. Клешненосные ноги умеренно длинные, шипо-
ватые и волосатые; исхиум с 1-2 шипами на внутреннем краю заостряю-
щегося дистального конца; мерус с тремя крупными шипами на дистальной
половине внутреннего края, с двумя шипами на верхней поверхно-
сти и с рядом более мелких шипов на наружном краю; на нижне-
внутренней стороне имеется ряд шипов, начинающийся от вершины
исхиума. Карпус короче чем мерус, с двумя крупными шипами на внутрен-
нем краю и двумя рядами шипов на верхней поверхности, которые распо-
ложены ближе к краям, оставляя середину членика гладкой или с 2-3 очень
небольшими шипиками; на нижне-внутренней поверхности имеется два
шипа. Рука довольно длинная, несколько расширяющаяся дистально
и закругленная у кончиков пальцев; верхняя поверхность ладони и паль-
цев уплощенная; боковые края ладони и наружные края пальцев усажены
крупными шипами; под внутренним рядом краевых шипов на. проксималь-
ной половине ладони имеется ряд обычно из трех шипов. От проксималь-
ного наружного угла верхней поверхности ладони тянется ряд мелких
шипов несколько диагонально к щели между пальцами и ряд таких же
шипов имеется на середине верхней поверхности неподвижного пальца;
сверху и снизу у основания подвижного пальца имеется шип; нижняя
поверхность ладони выпуклая и без шипов. Оба пальца одинаковой формы
и толщины, хватательные края несут острые бугорки, которые постепенно
возрастают в дистальном направлении; кончики выемчатые, имеют ложко-
образную форму, крупные зубцы одного пальца при смыкании входят
в промежутки между такими же зубцами другого пальца. Следующие три
пары перейопод с несколько сплющенными члениками; меральные и кар-
пальные членики с шиповатыми передними краями; меральные членики
с крупным шипом на дистальном задне-нижнем конце и двумя более
маленькими шипами; передний край проподуса с мелкими шипами на
проксимальной половине и на заднем краю; пальцы короткие, широкие
у основания и почти прямые. Абдоминальные сегменты невооруженные,
с такими же поперечными волосатыми бороздками, как и карапакс. Тель-
сон суживается дистально, с глубокой выемкой на заднем краю, который
благодаря этому выглядит в виде двух закругленных долей; верхняя
поверхность с плоскими гранулами и венчиками коротких, направленных
назад ресничек. Пластинки уропод с широкими ребрами,, которые нахо-
дятся ближе к наружному краю и несут поперечные ряды коротких под-
вижных шипиков.
Окраска: красноватая, с несколькими синими поперечными линиями
и пятнами (Bell, Heller); тело темнокрасное с синими полосами и рыжими
волосками (Risso); спиртовые экземпляры бледножелтого цвета.
1. GALA THEA
85
Фиг. 30. Galathea strigosa (Linne).
86
GALATHEIDAE
Размеры
Общая длина..................................... 44.0 мм
Длина рострума................................... 7.0 „
„ меруса клешней, ноги.........................12.5 „
„ карпуса „ „ 8.0 „
„ клешни „ „ 17.0 „
Ширина „ „ .......................... 5.5 „
Длина пальцев „ „ 7.5 „
„ второй перейоподы............................31.0 „
Распространение. Адриатическое и Средиземное море; побе-
режье Англии, Ирландии, Бельгии, Дании, Швеции и Норвегии (до Варан-
гер-фиорд). Яржинский (1870) отмечает в своем списке (стр. 317) данный
вид у западного Мурмана, не приводя, однако, точного местонахождения;
после этого автора никто не упоминает о G. strigosa в наших водах,
однако не исключена возможность эпизодического заноса атлантическими
водами этого вида, почему он и включен в настоящую работу.
Встречается от границы приливо-отливной зоны до глубины при-
мерно 65 м; предпочитает каменистые и песчаные грунты. Bell (1853) при-
водит сообщение R. Couch о привычках этого вида: „он посещает лужи
в приливо-отливной зоне, где имеются отдельные камни и песок. Он,
вообще говоря, очень медлителен в своих движениях, хотя часто двигается
с очень большой активностью, в особенности при испуге. Из-за большой
длины первой пары ног его движения всегда направлены назад. При
ползании его походка медлительна, но при плавании он переносится
с места на место со скоростью стрелы. Его никогда не видно в сколько-
нибудь открытой части лужи, он всегда отыскивает убежище среди камней
или какую-нибудь расщелину в скале, так что может скрываться при
самом незначительном испуге. Это весьма хорошо доказывается той точ-
ностью, с которой животное устремляется назад, на расстоянии несколь-
ких футов, в расщелину, которая очень немногим больше, чем само
животное; я часто наблюдал, как животные делали это и всегда с безоши-
бочной точностью".
2. Galathea acanthomera Stimpson.
Galathea acanthomera Stimpson, 1858 : 252; 1907 : 232; de Man, 1907 : 402; Balss,
1913:2, fig. 1; Yokoya, 1933:55.— Galathea orientalis Ortmann, 1892 : 252, t. 11, fig. 10
(nee Stimpson).
Длина рострума и его относительная ширина вариируют. Боковые
зубцы рострума острые; первый, или базальный, зубец направлен прямо
вперед или наружу, второй зубец слегка повернут внутрь или направлен
прямо вперед; терминальный шип, составляющий 2/5 общей, длины рост-
рума, несет на своих боковых краях 4-6 микроскопических зубчиков
и несколько волосков; длина и форма ростральных зубцов отчасти вари-
ируют; непосредственно позади поперечной линии, соединяющей основа-
1. GALA THEA
87
яие вырезок между первым и вторым боковыми зубцами, на середине,
находятся два шипа, расположенные близко друг от друга; верхняя по-
верхность рострума отчасти волосатая на середине, причем короткие
волоски располагаются в изогнутые параллельные ряды с каждой стороны
;и близ средней линии; длинная щетинка прикрепляется у основания чет-
вертого бокового зубца на середине, другая почти на середине рострума
с каждой стороны от средней линии. Боковые края карапакса водружены
"9 зубцами или вернее шипами; первый шип находится на наружном углу
орбиты и направлен косо наружу; второй шип менее повернут наружу
и расположен ближе к первому шипу, чем к цервикальной борозде, третий
шип более маленький чем второй, расположен на верхней поверхности
близ цервикальной борозды, немного отступя от бокового края, а четвертый
шип расположен под боковым краем и имеет такую же величину, как второй
шип. За цервикальной бороздой боковой край несет 5 других шипов одина-
ковой величины и расположенных на одинаковом расстоянии; по величине
эти шипы такие же, как второй шип, за исключением последнего, который
•отчасти короче. Поверхность карапакса позади двух шипов имеет примерно
семь поперечных гребней; волоски гребней длинные, 0.3—0.35 мм. Верхняя
поверхность карапакса, рострума и абдомена густо и грубо изрыта мелкими
ямочками. Передние края абдоминальных сегментов волосатые и несут
сверх того немного довольно длинных щетинок, которые встречаются
в очень небольшом числе также и на боковых частях верхней поверхности
карапакса. Третий от конца членик антеннальной ножки с сильным шипом
сверху и таким же шипом снизу; предпоследний членик с одним более
маленьким шипом снизу; предпоследний членик с одним более маленьким
шипом сверху. Исхиум наружных максиллярных ножек немного длиннее чем
мерус, его наружный край дистально оканчивается резким зубцом, который
•слегка повернут внутрь, а его внутренний край оканчивается коническим
более толстым, но более коротким зубцом. Два острых зубца на внутрен-
нем краю меруса более крупные, чем зубцы наружного края; передний
шип на наружном краю отчасти изогнут внутрь и находится на дисталь-
ном конце, другой находится почти на середине края. Наружный край
карпуса вооружен 2-3 острыми шипами, впереди которых находится еще
маленький острый зубец. Тонкая ножка экзоподита простирается немного
за мерус. Клешненосные ноги не совсем равные; подвижной палец боль-
шей клешни несет умеренно сильный полуострый зубец на расстоянии
л/3 длины от сочленения, а между этим зубцом и кончиком хватательный
край снабжен 25 мелкими тупыми зубчиками; неподвижный палец, как
и пальцы меньшей клешни, несет на хватательном краю только мелкие
зубцы; у крупных экземпляров самцов пальцы зияют, когда они сомкнуты.
Мерус второй пары ног вооружен по всей длине верхнего края 11-12
сильными острыми зубцами почти одинаковой величины; нижний край
также немного зубчатый и оканчивается на дистальном конце острым
дпипом, который слегка выступает за закругленный конец меруса. Нару ж-
88
GAL A THEIDAE
ный край карпуса вооружен 4-6 острыми шипами почти такой же вели-
чины, как и шипы меруса; шип на самом конце немного длиннее чем пре-
дыдущий. Проподус на проксимальной половине верхнего края вооружен
3-4 шипами, которые немного мельче чем шипы меруса, а нижний край
несет 6 подвижных шипов, имеющих почти такую же величину, как и шипы
верхнего края. Палец составляет немного более половины длины пропо-
дуса, оканчивается изогнутым когтем, нижний край вооружен 6 подвиж-
ными шипиками, постепенно увеличивающимися в длину по направлению
к кончику. Верхний край меруса покрыт волосами, которые отчасти
являются перистыми; задняя поверхность обнаруживает поперечные ряды
коротких волосков, а близ нижнего края длинные волоски; следующие
членики также волосатые. Ноги третьей , и четвертой пары сходны с опи-
санными, но шипы на верхнем краю меруса четвертой пары сравнительно
более маленькие. Клешненосные и ходильные ноги без эпиподитов.
Окраска бледная, желтовато-красная, рострум беловатый, подобно
боковым зубцам карапакса; волосатые гребни на карапаксе и сегменты
абдомена отмечены мелкими красными пятнами; мерусы и проподусы вто-
рых-четвертых перейопод каждый с двумя винно-красными поперечными
полосами (de Man). По Stimpson’у окраска сероватая.
Размеры
Длина карапакса....................... 7.2 мм
Ширина „ 5.0 „
Длина рострума....................... 2.7 „
Ширина „ (у основания).............. 1.5
Длина исхиума наружи, макс, ножки . . 1.5 „
„ меруса „ „ „ . . 1.2 „
„ большей клешней, ноги...........20.0 „
„ меиыпей „ „ 18.0 „
Galathea orientalis Stimpson (1858), описанный из Гон-конгд, является
несомненно иным видом; его карапакс покрыт короткими волосками, боко-
вые края карапакса вооружены 6 зубцами, первый боковой зубец рост-
рума маленький, клешненосные ноги довольно толстые, подвижный палец,
с 2 зубцами, верхние края мерусов ходильных ног густо шиповатые, шипы
одинаково мелкие.
Распространение. Бонинские о-ва; южная Япония: на западном
побережьи Японии отмечен у побережья Tottori. Литораль до глубины
200 м.
3. Galathea Integra Benedict. (Фиг. 31).
Galathea Integra Benedict, 1903 : 248; Balss, 1913 : 7, fig. 4—5;.Yokoya, 1933: 55.
Рострум представляет собой широко-пластинчатый листок, края,
которого совершенно гладкие и незубчатые; спереди он заострен, у осно-
вания с каждой стороны обладает двумя маленькими шипами, которые:
занимают внутренние углы орбит; поверхность рострума гладкая и отчасти
1. GALA THEA
89
выемчатая между глазами. Наружные углы орбит вооружены шипами.
Немного позади этих шипов имеются более маленькие шипы передне-
боковых углов карапакса. Карапакс сверху несет немного поперечных
полос, которые снабжены длинными волосами; на гастральной^ области
Фиг. 31. Galathea Integra Benedict (из Balss, 1913).
имеется поперечный ряд из четырех шипов; боковые края карапакса,
вооружены 6-7 шипами, которые часто отсутствуют. Мерус наружных
максиллярных ножек много короче чем исхиум и представляет собой
широкую, почти квадратную пластинку, внутренний край которой оканчи-
вается острым зубцом, а наружный край несет шип. Клешненосные ноги
длинные, узкие; мерус негусто усажен короткими толстыми шипами;
90
GALA Til HI DAE
карпус имеет ряд из 4 шипон па своей верхней поверхности и ряд из
5-6 шипов на внутреннем краю, но самым мощным его вооружением
является один очень большой шип, расположенный немного ниже внутрен-
Фиг. 32. Galathea pubescens Stimpson (из Balss, 1913).
него ряда; ладонь вооружена тремя рядами шипов, шипы гребня самые
крупные и самые многочисленные; пальцы короче чем ладонь, мелко зуб-
чатые на хватательных краях, с широкой щелью, остающейся при смыка-
нии; все членики покрыты тонкими волосками. Меральные членики
ходильных ног вооружены на верхнем краю немногочисленными шипами.
1. GALATHEA
91
Размеры
Длина карапакса и рострума........... 7.0 мм
Ширина „ 4.0 „
Длина клешненосной ноги..............17.0 „
Распространение. Южная Япония (Yokoya, 1933). В Японском
море найден севернее Noto; отмечен также для Сангарского пролива
и Aomori. На глубинах от 71 до 307 м.
4. Galathea pubescens Stimpson. (Фиг. 32, 33).
Фиг. 33. Galathea pubescens Stimpson; наружная
максиллярная ножка (из Balss, 1913).
Galathea pubescens Stimpson,
1858:252; 1907:233; Balss, 1913:11,
fig. 11-12; Yokoya, 1933 : 57.
Карапакс правильно и рав-
номерно выпуклый, несильно
суживающийся спереди; верхняя
поверхность несет много попе-
речных борозд, из которых,
однако, лишь немногие прохо-
дят целиком от одной стороны
карапакса до другой; вся по-
верхность заметно волосатая,
при этом волоски каждого ряда
довольно длинные и достигают
до следующего ряда. Гастраль-
ная область отграничена нечетко
и вооружена близ середины
тремя маленькими шипиками,
средний шип расположен не-
много позади двух других (по
Balls’y гастральная область
имеет ряд из 6-8 шипов). Боко-
вые края карапакса вооружены 6-8 шипами. Рострум большой, треуголь-
ный, сильно заостренный, густо волосатый сверху и с 4 зубцами на
каждом краю; зубцы очень острые и направлены вперед; иногда имеется
дополнительный зубец между базальным и вторым зубцами. Базальный
членик антеннул несет два направленных вперед шипа. Мерус третьих
максиллярных ножек изнутри с тремя, а снаружи с двумя шипами. Клешне-
носные ноги длинные, тонкие, цилиндрические, с рассеянными щетинками
и шипиками; однако иногда имеется сильный шип на карпусе (по Balss’y
клешненосные ноги густо покрыты чешуйками, расположенными в прямые
линии, от которых отходят длинные тонкие волоски); пальцы руки сплю-
щенные, параллельные, отнюдь не зияющие и не такие длинные, как
ладонь. Ходильные ноги умеренно длинные, густо волосатые, верхний
край меруса вооружен 7-9 очень тонкими шипиками; пальцы составляют
92
GAL A THEIDAE
приблизительно половину длины проподуса. Эпиподиты имеются только
на клешненосных ногах. Абдомен волосатый; его сегменты несут по
4 борозды, из которых средние самые глубокие. Окраска красноватая,
иногда с более темными пятнами и средней дорзальной белой полосой.
Распространение. Япония, у восточного побережья Ousima;
около Хакодате; Uraga-Kanal; Sag-ami; западнее Muroto-zaki. На глубинах
до 234 м.
2. Род MUNIDA LEACH
Munida Leach, 1820 :52; Bell, 1853 : 206; Heller, 1863 :192; Henderson, 1888 :123;
Holmes, 1900 : 111; Benedict, 1903 : 251; Stimpson, 1907 :235; Balss, 1913 :15.
Тип рода: Munida bamffica (Pennant).
Карапакс больше в длину, чем в ширину; поперечные морщины,
передние края которых густо опушены короткими волосками, более много-
численны и резче выражены, чем у рода Galathea; цервикальная борозда
всегда отчетливая; кардиальная область явственная; суббранхиальные
области не вздутые и отделены от дорзальной поверхности хорошо выра-
женными боковыми краями, которые изогнуты менее чем у предыдущего
рода; шипы, расположенные на этих боковых краях, более длинные и более
острые, а два шипа, расположенные на передне-боковом углу, гораздо
сильнее чем остальные. Рострум длинный, тонкий, шиловидный, по бокам
которого имеются хорошо развитые супраорбитальные (или супраокуляр-
ные) шипы. Вертикальная часть лобной области не образует наружного
орбитального гребня или образует лишь рудиментарный гребень, который
всегда лишен зубчиков; на месте этого гребня иногда наблюдается
довольно маленький шип, который бывает более сильно развит у некото-
рых видов подсемейства Munidopsinae. Глазные стебельки короткие
и сильно расширяющиеся дистально, глаза большие и хорошо пигменти-
рованные. Антеннулярные ножки длиннее, чем у рода Galathea, точно
так же, как и третьи максиллярные ножки. Первые перейоподы очень длин-
ные, тонкие, шиповатые. Три следующих пары перейопод также довольно
длинные, их пальцы длинные, стройные, мало изогнутые, лишенные на
нижнем краю заметных зубчиков, но снабженные многочисленными и мел-
кими роговыми шипиками. Абдомен с поперечной исчерченностью, перед-
ние края нескольких первых сегментов вооружены шипами или невоору-
женные. Тельсон широкий, почти квадратный.
Представители этого рода почти всегда более шиповатые чем виды
рода Galathea и, согласно Milne-Edwards’y и Bouvier (1894), являются
формами более удаленными от Macrura, чем представители рода Galathea.
Кроме того род Munida отличается от Galathea тем, что его виды являются
более глубоководными: так, у этого рода совсем неизвестно прибрежных
видов и только с 20—40 м начинают встречаться некоторые его предста-
вители; наиболее высоко поднимается М. bamffica (встречается на глу-
2. MUNIDA
93
бине 20 м), но в то же время она уходит и набольшие глубины. Из видов
этого рода здесь приводится европейский М. bamffica и дальневосточный
вид, М. japonica, который отмечен для западного побережья Японии.
ОПРЕДЕЛИТЕЛЬНАЯ ТАБЛИЦА ВИДОВ
1 (2). Гастральная область карапакса без поперечного ряда шипов.
Пальцы клешней первых перейопод длиннее чем ладонь. Второй
сегмент абдомена (а у более крупных экземпляров также третий
и четвертый сегменты) вооружен шипиками на переднем краю . .
..................................... *1. М. bamffica (Pennant).
2 (1). Гастральная область спереди с поперечным рядом из 13 шипов.
Пальцы клешни первых перейопод такой же длины, как ладонь,
или короче. Абдоминальные сегменты все невооруженные................
.......................................2. М. japonica Stimpson.
* 1. Munida bamffica (Pennant). (Фиг. 34).
Astacus bamfficus Pennant, 1777: 17, t. 13, fig. 25 (цит. no Hansen, 1908). —
•Galathea rugosa Leach, 1815, tab. 29; Risso, 1826:46; Milne-Edwards, 1837:274. —
Munida rugosa Heller, 1863 :192, t. 6, fig. 5—6; Ortmann, 1892 : 253. — Munida rondeletii
Bell, 1853 :208. — Munida bamffica Ortmann, 1892: 253; Milne-Edwards et Bouvier,
1894:83, t. 7, fig. 1—7; Hansen, 1908: 32, t. 2, fig. 3a.
Рострум в виде длинного, почти прямого острого шипа; супраоку-
лярные шипы немного не достигают до середины длины рострума; за ними
на верхней поверхности карапакса имеется по одному маленькому шипу;
затем имеется по паре шипов на бранхиальных областях; боковые края
карапакса шиповатые (обычно 7 шипов), из которых передний шип,
находящийся над основанием антенн, является самым крупным. Имеется
два шипа позади цервикальной борозды. Вся поверхность карапакса
пересечена поперечными морщинами с отходящими от них волосками.
Задний край карапакса несет направленные вперед шипы: 2 на середине
и 2—5 по бокам ближе к заднебоковому углу карапакса (число этих
боковых шипов вариирует). Глаза крупные; на границе между роговицей
и глазным стебельком, главным образом на дорзальной стороне, имеется
бордюр волосков, которые стелятся по роговице. Первый членик ножки
антеннул очень длинный, достигающий До кончиков глаз или несколько
превосходящий их. Наружные максиллярные ножки в вытянутом состоя-
нии почти достигают до кончика рострума; их мерус длиннее чем карпус,
его задний край мелко зазубрен, передний с длинными волосами
и вооружен на конце длинным шипом; на внутренней стороне карпуса,
несколько впереди середины имеется один шип. Передние перейоподы
очень длинные, почти одинаковой ширины на всех члениках, не очень
густо волосатые. Крупные шипы расположены на внутренних краях
меруса, карпуса и проподуса (ладони), причем шипы, находящиеся на
94
GALATHEIDAE
дистальных концах двух первых члеников, являются более крупными;
над и под этими краевыми шипами имеются более мелкие шипы, распо-
ложенные не совсем правильным продольным рядом; верхняя поверхность,
этих же члеников несет на середине продольный ряд шипов, продолжаю-
щийся с одного членика на другой, при этом дистальные шипы ряда
на каждом членике крупнее остальных; на ладони дистальный шип этого
срединного ряда смещен в сторону и находится над основанием подвиж-
ного пальца; наружные края меруса и карпуса не несут явственных
шипов, а на ладони имеется ряд мелких шипиков. На нижней стороне
ладони над основанием подвижного пальца также имеется крупный шип.
На основании подвижного пальца как сверху, так и снизу имеются шипо-
образные выступы, которые прилегают к упомянутым шипам и при движе-
нии пальца трутся о них; кроме того на наружном краю подвижного
пальца у основания имеется направленный вперед довольно крупный шип..
Ладонь руки узкая, пальцы несколько длиннее чем ладонь, с многочислен-
ными бугорками на хватательных краях, причем более крупные бугорки
чередуются с несколькими (3-5) более мелкими; кончики пальцев изог-
нутые, при этом кончик неподвижного пальца раздвоен, так что кончик
подвижного пальца при смыкании может помещаться в этой выемке..
Следующие три пары перейопод несколько сплющенные, шиповатые
на краях, в особенности моральные и короткие карпальные членики,,
у которых дистальные шипы как верхнего, так и нижнего края являются
более крупными, и не густо волосатые. Передний край второго абдоми-
нального сегмента несет 6 направленных вперед шипов (пара медианных
и по паре боковых); передний край третьего сегмента несет 4 шипика
(пара медианных и по одному боковому), а передний край четвертого
сегмента несет 2 шипика (медианная пара). Тельсон широкий, с вырезкой
на середине заднего края. Верхняя поверхность второго-четвертого
абдоминальных сегментов имеет глубокую поперечную борозду, из кото-
рой поднимаются направленные вперед короткие волоски; такие же
волоскй отходят от передних краев этих же сегментов, остальная поверх-
ность сегментов имеет поперечную морщинистость, которая на эпимерах
образует причудливые узоры, напоминающие узоры муаровой ленты.
Верхняя поверхность тельсона и уропод покрыта плоскими возвышениями,
от которых отходят короткие направленные назад волоски, а от некоторых
и шипы. Задние края уропод и тельсона мелко зазубренные и несут два
ряда длинных волос: сверху простые, тонкие и более короткие, а снизу
длинные, более толстые и перистые волоски.
Размеры 3
Общая длина (от кончика рострума) . . 48.0 мм 76.0 мм
Длина рострума....................... 9.5 „ 15.0 „
„ супраокулярного шипа.......... 5.0 „ 9.5 „
Общая длина клешиен. ноги..........55.0 „ 133.0 „
Длина меруса „ „............20.0 „ 48.0 „
2. MUN ID A 95
Фиг. 34. Munida bamjjica (Pennant).
96
GALA THEIDAE
Длина меруса клешней, ноги......... 8.0 мм
„ клешни „ „............26.5 „
Ширина „ „ „............ 3.5 „
Длина пальцев „ „............15.0 „
Общая длина второй перейоподы . . . 39.0 „
18.0 мм
62.0 „
6.5 „
3.50 „
87.0 „
Вопрос о синонимии данного вида несколько запутан. Часть авторов
считает М. rugosa Sars самостоятельным видом. Ortmann (1892) приводит
следующие отличительные признаки М. rugosa от М. bamffica-. 1) глаза
обладают ресничным бордюром, который отсутствует у М. bamffica-,
2) супраокулярные шипы едва расходящиеся и тянутся почти парал-
лельно ростральному шипу; всё три шипа находятся примерно на одной
и той же плоскости, тогда как у М. bamffica супраокулярные шипы
расположены выше, чем ростральный шип; 3) на заднем краю карапакса
находятся 4-6 направленных вперед шипов, именно 2 у средней линии
и по 1-2 ближе к бокам; 4) три абдоминальных сегмента несут шипы
(у М. bamffica только два), а именно: первый шесть, второй четыре
и третий два шипа.’
В коллекции Зоологического Института Академии Наук имеется
крупный экземпляр самца из Ниццы, определенный как М. bamffica.
Детальное изучение этого экземпляра показало, что вооружение абдоми-
нальных сегментов почти в точности соответствует указаниям Ortmann’a:
6 шипов на втором сегменте, 4 на третьем и без шипов на четвертом;
однако на границах роговиц ясно виден бордюр, правда, более коротких
волосков. Клешненосные ноги более волосатые, чем у мелких экземпля-
ров. На заднем краю карапакса нет двух средних шипов, слева имеется
один боковой, а справа два боковых шипа. У экземпляров из западной
-Норвегии, определенных как М. rugosa, следует отметить колебания
в числе боковых шипов на заднем краю карапакса. Наконец, у семи
мелких экземпляров, оказавшихся в сборах судна „Андрей Первозван-
ный", весьма показательным оказалось вооружение передних краёв
2 — 4-го абдоминальных сегментов; так, у одного экземпляра второй
сегмент имел 4 явственных шипа, на третьем сегменте имелись только
бугорки в виде намека на будущие шипы, так как они расположены на
тех же самых местах, где должны бы быть эти шипы, наконец, на четвер-
том сегменте не было никакого намека на развитие даже бугорков.
У других экземпляров на втором сегменте было 4 шипа, на третьем
сегменте два средних шипа хорошо развиты, а две пары боковых шипов
в виде бугорков, на четвертом сегменте также никаких шипов не было.
Из всего сказанного может быть сделан вывод, что отличия
М. rugosa от М. bamffica являются чисто возрастными отличиями у одного
и того же вида. Действительно, первый пункт Ortmann’a вообще отпа-
дает: реснички есть и у М. bamffica, но с возрастом они могут быть
короче. Пункт второй также легко объясняется возрастными изменениями:
по мере роста и удлинения рострума он несколько изгибается кончиком
2. MU NIDA
97
вниз и оказывается ниже уровня супраокулярных шипов. Признак третий
вообще обнаруживает большие вариации и не может являться отличитель-
ным признаком. Наконец, пункт четвертый наиболее ясно говорит
о возрастном характере приводимых изменений; вполне очевидна последо-
вательность появления шипов сперва на втором сегменте, затем на третьем
и, наконец, у самых крупных экземпляров на четвертом сегменте. Иден-
тичность М. rondeletii Bell с М. bamffica несомненна.
Окраска: красновато-коричневая (Heller), темная красновато-желтая
с более красными пятнами (Bell), красноватая, волоски желтые (Milne-
Edwards), рыжая с фиолетовыми пятнами и черточками; шесть сегментов
абдомена пересечены фиолетовыми линиями (Risso).
Распространение. Шетландские острова, побережья Англии,
Ирландии и Франции до Мадеры и мимо Cape Boyador до 25°41' N; Среди-
земное и Адриатическое моря; побережья Швеции и Норвегии до запад-
ного Мурмана (72°34; N, 17°20' Е). В материалах Зоологического Института
Академии Наук оказалась одна банка с этим видом из сборов судна
„Андрей Первозванный": 28 VII 1899, 69°50'30'’N, 47°09/Е, 75 м; кроме
того вид отмечен К. М, Дерюгиным (1924) для Кольского залива: 70°00' N,
33°30'Е, 146 м. Наконец, Н. Танасийчук (1927) указывает на нахождение
этого вида между островами Седловатым и Большим Оленьим и на юго-
восток от острова Сального. Перечисленные местонахождения показы-
вают, что М. bamffica проникает с теплыми атлантическими водами
несколько восточнее, чем это отмечалось А. А. Бирулей (72°34' N, 17°20' Е),
но все же в наших мурманских водах это сравнительно редкий вид,
находящий здесь восточные пределы своего распространения. Встре-
чается на глубинах от 23 до 1244 м.
2. Munida japonica Stimpson. (Фиг. 35).
х Muni da japonica Stimpson, 1858 : 252; 1907 : 235; Miers, 1879 : 51; Ortmann, 1892 : 254,
t. 11, fig. 11; de Man, 1902 : 724; Balss, 1913 :15, fig. 14; Yokoya, 1933 : 58. — Munida heter-
acantha Ortmann, 1892 : 255, t. 11, fig. 12.
Карапакс продолговатый. Рострум меньше половины длины кара-
пакса и более чем вдвое длиннее супраокулярных шипов. Глаза большие,
сплющенные, с бахромками длинных волос. Боковой край карапакса на
середине с 5 шипами, на передних углах с 3 шипами, из которых перед-
ний является самым крупным. Верхняя поверхность карапакса богато
скульптирована, борозды большею частью тянутся непрерывно через всю
поверхность. Гастральная область имеет спереди поперечный ряд из
13 шипиков, из которых один находится на средней линии и немного
сдвинут назад; шипы этого ряда, находящиеся как раз позади супраоку-
лярных шипов, являются самыми крупными; снаружи от них находится
по 4 шипа, а между ними и срединным шипом имеется по одному малень-
кому шипу. Боковые части головогруди несут еще три парных шипа:
•один на гастральной области, другой на участке, ограниченном раздвое-
Фауна СССР, Макаров 7
98
GALA THEIDAE
нием цервикальной борозды, и третий тотчас позади цервикальной борозды.
Клешненосные ноги длинные, прямые, полуцилиндрические, чешуйчатые,
с несколькими рассеянными шипиками; пальцы тонкие и прямые, у самца
короче чем ладонь и со щелью у основания, у самки такой же длины»
как ладонь, и почти без щели у основания. Исхиум вторых максиллярных
ножек длиннее чем мерус, который вооружен на внутренней стороне
тремя шипами. Абдоминальные сегменты невооруженные.
Н. Balss, имевший большой материал по этому виду, считает, что
вид Ortmann’a Munida heteracantha, является лишь вариететом М. japonica,
и приводит следующую таблицу уклонения крайних форм друг от друга:
М. japonica typica Stimp.
Рострум вдвое длиннее чем суп-
раокулярные шипы. Карапакс сра-
внительно широкий (отношение дли-
ны к ширине: 11:9.5).
Верхняя поверхность карапакса
сильно скульптирована, борозды
большею частью непрерывно тя-
нутся через всю его ширину.
Боковые части карапакса с 3 ши-
пами.
Второй и третий абдоминальные
сегменты без шипов.
Каждый сегмент абдомена с 4-6
узкими, граничащими друг с дру-
гом бороздами.
Поверхность ходильных ног
сильно скульптирована, с чешуй-
ками и волосами.
Клешненосные ноги сравнительно
короткие и широкие.
М. japonica heteracantha (Ortm.).
Рострум втрое или вчетверо длин-
нее, чем супраокулярные шипы.
Карапакс длинный (отношение
длины к ширине: 12:9).
Верхняя поверхность карапакса
менее бороздчатая, борозды обычно
прерванные.
Боковые части карапакса только
с 2 шипами, причем отсутствует тот
шип, который вообще находится
на участке, ограниченном разветвле-
нием цервикальной борозды.
Второй абдоминальный сегмент
с 8-11 шипами, третий — с 3 ши-
пами.
Каждый сегмент абдомена только
с 2 бороздами.
Поверхность ходильных ног слабо
скульптирована, волосатость силь-
нее. Зубчатость на нижней стороне
более сильная.
Клешненосные ноги длинные и
узкие.
Из сравнения описаний обоих видов можно добавить еще два пункта
Глаза крупные, сплющенные,
с длинными бахромками.
Мерус вторых максиллярных но-
жек с 3 шипами на внутренней сто-
роне.
Глаза менее крупные, менее сплю-
щенные и с короткими бахромками.
Мерус вторых максиллярных ножек
с 2 шипами на внутренней стороне.
2. MUNIDA
99
Фиг. 35. Munida japonica Stimpson (из Balss, 1913).
7*
100
GALA THEIDAE
Однако H. Balss далее отмечает, что все эти признаки неопределен-
ные и существуют все переходы между этими крайними формами; так,
напр., экземпляры формы typica имели вооруженные абдоминальные сег-
менты и, наоборот, экземпляры формы heteracantha не имели шипов на
третьем сегменте. Munida sagamiensis Doflein является такой промежу-
точной формой, стоящей ближе к форме heteracantha так же, как и
М. honsuensis. Вариетет heteracantha предпочитает, согласно Balss’y, более
глубокие воды.
Распространение. Южная Япония до Сангарского пролива.
На западном побережьи Японии известен у Noto, из бухты Toyama
Niigata, у острова Sado; кроме того зарегистрирован в Сангарском проливе
и Aomori. На глубинах до 543 м.
2. Подсем. MUNIDOPSINAE
Munidopsinae Ortmann, 1901:1151.
Глазные стебельки свободные или сросшиеся с глазным сегментом.
Глазной сегмент часто сливается с прилежащими частями. Глаза редуци-
рованные. Экзоподит первых максиллярных ножек без жгута.
3. Род MUNIDOPSIS WHITEAVES
Munidopsis Whiteaves, 1874: 212; Henderson, 1888:148; Milne-Edwars et Bouvier,
1894:271; Schmitt, 1921:167.
Тип рода: Munidopsis curvirostra Whiteaves.
Карапакс более или менее уплощенный, большею частью квадрат-
ный по своему очертанию; дорзальная поверхность с поперечными морщи-
нами, но эти морщины обычно не бывают длинными, а разбиваются
на короткие участки и принимают иногда форму чешуек; иногда поверх-
ность несет шипы; иногда она бывает совершенно безволосая и покрыта
либо выдающимися, либо слабо заметными бугорками; границы областей
хорошо заметны, если на карапаксе имеются перечисленные выше укра-
шения (морщины, шипы, волоски), но они становятся неотчетливыми
по мере того, как эти украшения уменьшаются или исчезают; боковые
края карапакса дугообразно изогнутые или прямые. Рострум хорошо
развит, шипообразный; иногда бывают маленькие супраантеннальные
шипы, но никогда не бывает длинных супраокулярных шипов, как у рода
Munida. Глазные стебельки у некоторых видов еще довольно длинные,
свободные и слегка расширяющиеся дистально, но чаще всего они корот-
кие, несут шип у границы роговицы, у некоторых видов срастаются
с глазной дужкой и становятся неподвижными; роговицы непигментиро-
ванные. Антеннулярные жгуты сильно развиты и в большинстве случаев
богато снабжены волосками. Клешненосные и ходильные ноги вариирую-
3. MUNIDOPSIS
101
щей длины, большею частью шиповатые. Все покровы хорошо объизве-
ствленные. Икринки крупные и немногочисленные.
Редукция глаз и параллельное с ней развитие антеннальных и антен-
нулярных жгутов несомненно показывают, что перед нами находится
абиссальный род. Основная масса видов была описана после глубоковод-
ных работ ряда экспедиций. Род оказался широко распространенным
Фиг. 36. Munidopsis
beriogana Benedict.
в морях и океанах, главным образом, тропической области, встречаясь
чаще всего на глубинах от 1000 до 2500 м; отдельные виды были най-
дены на глубине свыше 4000 м (напр., М. reynoldsi приблизительно на
4300 м у Антильских островов). В нашей фауне род представлен одним
видом.
*1. Munidopsis beringana Benedict. (Фиг. 36).
Munidopsis beringana Benedict, 1903 : 279, fig. 23; Rathbun, 1904 :167.
Карапакс больше в длину чем в ширину, густо покрыт папиллами
с венчиками волосков; папиллы на задней части карапакса часто ели-
102
GA LA TH EID AE
ваются в короткие поперечные гребни. Передне-боковой угол карапакса
несет крупный шип, позади которого находится также шип; на середине
бокового края имеется два маленьких шипа; задний край карапакса пря-
мой. Рострум длинный, шилообразный, изогнутый и направленный вверх.
Лобный край на 2/з расстояния от рострума до шипа переднебокового
угла заметно утолщен. Гастральная область позади рострума несет два
довольно крупных шипа и группу из менее отчетливых шипов на осталь-
ной поверхности гастральной области, число которых по Benedict’y
колеблется от 20 до 15. Цервикальная борозда широкая, заметная благо-
даря отсутствию на ней волосатых гранул; треугольная кардиальная
область также ограничена гладкими бороздами. Глазной стебелек несет
длинный острый шип над верхне-внутренней поверхностью несегменти-
рованной роговицы. Клешненосные ноги сравнительно короткие, покры-
тые, как и ходильные ноги, короткими волосками; мерус с двумя крупными
шипами на верхнем переднем краю, карпус с рядом шипов (обычно 4)
на внутреннем краю и с неправильным рядом более мелких шипов на
наружном краю, из которых передний является самым крупным. Ладонь
шиповатая на внутреннем краю, базальный шип самый крупный; верхняя
поверхность и наружный край с мелкими рассеянными шипами; пальцы
одинаковой толщины, немного длиннее чем ладонь, с слабо зазубренными
хватательными краями; при смыкании пальцы плотно сходятся и подвиж-
ный палец несколько заходит своим кончиком за кончик неподвижного
пальца. Меральные членики трех следующих пар перейопод с шипом
на верхнем дистальном конце и с крупными шипами на верхнем краю;
карпусы с двумя шипами на верхнем краю; проподусы без шипов; пальцы
длинные, изогнутые, сверху гладкие, нижний край мелко зазубренный,
а боковые двугранные поверхности несут на своем ребре ряд густых
волосков. Пятая перейопода в вытянутом состоянии едва превышает
дистальный конец меруса предыдущей ноги. Верхняя поверхность абдоми-
нальных сегментов покрыта такими же гранулами и волосками, как и кара-
пакс. От близкого вида, М. antonii A. M.-Edw., отличается изогнутой фор-
мой рострума, который у последнего вида также направлен вверх, но
совершенно прямой.
Размеры
Общая длина..................................45.0 мм
Длина рострума............................... 8.0 „
„ меруса клешней, ноги ...................12.0 „
„ карпуса „ „ 5.0 „
„ руки • „ „ 16.0 „
Ширина „ „ „ 4.5 „
Длина подвижного пальца „ 9.0 „
Распространение. Три экземпляра Benedict’a добыты „Альба-
тросом" в Беринговом море к юго-западу от Прибыловых островов на глу-
бине 3188 м. В коллекции Гидробиологической лаборатории Ленинград-
4. CER VIMUNIDA
103
ского Государственного Университета имеется единственный экземпляр
этого вида, добытый в Охотском море:
(„Гагара", 1932).
4. Род CERVIMUNIDA BENEDICT
Cervimunida Benedict, 1903 : 249; Balss,
1913:18.
Тип рода: Cervimunida princeps
Benedict.
Похож на род Munida, но рост-
рум сжат с боков и изогнут настолько,
что допускает свободное движение
глаз; нижний и в особенности верх-
ний края рострума вооружены зуб-
цами. Из немногочисленных видов
этого рода.здесь приводится лишь
один вид, который был отмечен
в Японском море у западного побе-
режья Японии.
1. Cervimunida princeps Bene-
dict. (Фиг. 37).
Cervimunida princeps Benedict, 1903,
249, fig. 3; Bouvier, 1906 : 480; Balss, 1913 :18,
fig. 15, t. 1, fig. 1; Yokoya, 1933 :65.
Глаза очень крупные. Рострум
вооружен тремя острыми треуголь-
ными зубцами: двумя на верхнем
краю впереди глаз и одним на нижнем
краю и впереди верхних зубцов;
между вершиной рострума и первым
зубцом верхнего края обычно имеется
несколько маленьких зубчиков. На-
правление рострума горизонтальное,
но на уровне глаз он образует дугу,
принимая за ней снова горизонтальное
направление. На поперечном сече-
нии рострум треугольный, с короткой
стороной снизу, нижние края гребен-
46°41'5 N, 147°28' Е, 3500 м
Фиг. 37. Cervimunida princeps Benedict
(из Balss, 1913).
чатые, гребни тянутся до супраокулярных шипов. Длина рострума от кончика
до основания свободной части супраокулярного шипа равна расстоянию
от этого последнего пункта до заднего края гастральной области. Супра-
•окулярные шипы достигают до середины глаз; их свободная часть по своей
104
PORCELLANIDAE
длине равна передне-боковым шипам. Шипы гастральной пары крупные
и острые; на том же уровне снаружи имеется маленький парный шип,,
а у некоторых экземпляров и второй, значительно более маленький; выдаю-
щийся шип на гастральной области находится на пересечении первой
ресничной линии с срединной линией карапакса; обычные шипы встре-
чаются на концах ресничных линий. Имеется один парный шип на раз-
ветвлении шва и один на обычном месте, как раз позади шва. Относительно-
скульптуры карапакса следует заметить, что между прямыми линиями,
которые тянутся по всей поверхности, большей частью находятся еще
ряды чешуек, которые особенно явственны на задней половине. Поверх-
ность, в особенности на боках, иризирует. Мерус наружных максиллярных
ножек несет на внутреннем краю два крупных шипа, на наружной стороне
три шипа; исхиум имеет мелко зазубренную жевательную долю. Клешне-
носные ноги длинные й сильные, покрыты на всей поверхности волосами
и расположенными в продольные ряды шипами, причем шипы на внутрен-
них краях члеников самые сильные; пальцы длиннее чем ладонь и плотно
смыкаются друг с другом мелко зазубренными хватательными краями;
многочисленные мелкие шипы рассеяны по всей поверхности ладони,,
за исключением нижней поверхности. Ходильные ноги густо покрыты
чешуйками и волосами; верхний край меруса несет приблизительно
10 шипов; экзоподиты на ходильных ногах отсутствуют. Абдомен воору-
женный: имеется обычно 8 шипов на втором и четвертом сегментах, третий,
сегмент имеет 6—8 (чаще 6) шипов; на других сегментах число шипов,
также вариирует, но не так часто.
Размеры
Длина карапакса (без рострума)............... 40.0 мм
„ рострума .... .................... 22.0 „
„ клешненосной ноги......................190.0 „
,, меруса ,, ,, . • » • •••••• • 70.0 ,,
„ карпуса ,, ....................... 19.0 „
„ проподуса „ „ .............• - 91.0 „
Ширина ладони „ „ .................... 17.0 „
Длина пальца „ „ ..................... 51.0 „
Распространение. Южная Япония. На западном побережьи:
Японии отмечен севернее острова Sado и около Yamag-ata-ken. На глуби-
нах от 76 до 452 м.
4. Сем. PORCELLANIDAE
Porcellanidae Henderson, 1888 :103; Ortmann, 1892 : 245; 1901:1151.
Карапакс широко-овальный, гладкий, со слабо выраженными обла-
стями. Лобный край широкий, часто рассеченный на три доли, которые;
никогда не бывают большой длины; средняя доля, или рострум, обычно.
1. POR CEL LAN A
105
широкий и короткий. Боковые края карапакса отчетливые, linea anomurica
явственная. Антеннулы скрытые, базальный членик их ножки широкий.
Антеннальная ножка четырехчленистая, направлена обычно назад, без
скафоцерита. Глаза всегда пигментированные и отчасти скрытые в орбитах.
Исхиум наружных максиллярных ножек широкий, мерус снабжен на внут-
ренней стороне выдающейся долей. Клешненосные ноги широкие и часто
сплющенные; ходильные ноги мощные и умеренной длины. Жабры как
у Galatheidae, мастигобранхий на перейоподах отсутствуют. Абдомен
подогнут под торакс, благодаря чему форма тела крабообразная. Абдо-
минальные конечности у самца имеются только на втором сегменте в каче-
стве половых придатков; на третьем-пятом сегментах они редуцированы
до степени маленьких бородавкообразных выступов; у самки имеются
тонкие одноветвистые абдоминальные конечности на четвертом и пятом,,
а иногда и на третьем сегментах.
Henderson (1888) рассматривает Porcellanidae как высоко специали-
зировавшихся Galatheidae, которые благодаря иному образу жизни приоб-
рели крабообразный внешний вид (редукция абдомена и почти полная
потеря его значения, как органа передвижения). Stimpson (1858) разделял
роды этого семейства на две группы по характеру антеннальной ножки:,
у родов первой группы базальный членик антеннальной ножки короткий,
частично скрытый в глазной полости и не достигающий до верхнего края
карапакса или до заднего угла верхнего края орбиты; поэтому следующие
членики не удалены от глаз. У родов второй группы этот членик удлинен-
ный, имеет острый, отчасти сплющенный отросток, который достигает до.
заднего угла верхнего края орбиты; благодаря этому следующие членики,
расположены на некотором расстоянии от орбиты. Оба рода, представлен-
ные в нашей фауне, относятся ко второй группе.
Представители этого семейства встречаются главным образом
в тропической области, причем обитают преимущественно в приливо-
отливной зоне под камнями, на что указывает сплющенная форма их
карапакса И клешненосных ног, или же встречаются на сравнительно-
небольших глубинах, где они обитают среди кораллов, губок, камней,
и проч.
ОПРЕДЕЛИТЕЛЬНАЯ ТАБЛИЦА РОДОВ
1 (2). Лобный край карапакса треугольный и зубчатый. Суббранхиаль-
ные (боковые) области карапакса сплошные, не разделенные пере-
пончатым промежутком....................... 1. Porcellana Lamarck.
2 (1). Лобный край карапакса треугольный, но никогда не бывает зубча-
тый. Суббранхиальные области карапакса разделяются каждая пере-
пончатым промежутком на большую переднюю и меньшую заднюю*
. части............................ . 2. Pachycheles Stimpson..
106
PORCELLANIDAE
1. Род PORCELLANA LAMARCK
Porcellana Lamarck, 1801 :153; Milne-Edwards, 1837 : 247; Bell, 1853 : 188; Heller,
1863 : 181; Henderson, 1888 : 109; Stimpson, 1907: 187. — Pisidia Leach, 1820 : 53. — Porcella-
nides Czerniavsky, 1884 :109.
Тип рода: Porcellana.platycheles (Pennant).
Карапакс обычно больше в длину чем в ширину, иногда почти круглый,
обычно плоский. Лобный край треугольный и зубчатый. Орбиты глубокие,
ограниченные сверху краем карапакса, изнутри и снаружи ножками антен-
нул и антенн и снизу коротким слабо выступающим краем; от этого края
тянется наружу шиповатый гребень, между ним и краем карапакса обра-
зуется глубокая борозда. Linea anomurica явственная. Глаза маленькие.
Второй членик ножки антенн самый длинный. Буккальная область квад-
ратная. Наружные максиллярные ножки большие, плотно смыкающиеся
и простирающиеся в сложенном виде до лобного края карапакса, частично
прикрывая антеннулы; их мерус большой, расширенный на внутреннем
краю и с зубцом на передне-наружном углу; карпус маленький, неправиль-
ной треугольной формы; следующие членики постепенно уменьшаются
в величине и вместе с карпусом густо опушены длинными волосами;
экзогнат с многочленистым жгутом; эпигнат отсутствует. Передние
перейоподы довольно большие и более или менее сплющенные; мерус
короткий; карпус продолговатый с утонченным зубчатым внутренним краем;
рука узкая у основания, слегка расширяющаяся дистально. Три следую-
щие пары ног умеренной величины, цилиндрические, с короткими пальцами.
Последняя пара ног тонкая, помещающаяся над основанием предыдущей
пары ног, согнутая в мерально-карпальном сочленении и несущая на
дистальном конце маленькую клешню. Абдомен широкий, пластинчатый,
семичленистый, подогнутый под торакс, последний сегмент (тельсон)
широкий, пересеченный несколькими бороздами и поэтому кажущийся
состоящим как бы из 5 пластинок; шестой сегмент несет пару уропод,
овальные пластинчатые части которых обычно подогнуты под тельсон.
Абдомен самца несет одну пару коротких и тонких плеопод на втором
сегменте; на третьем-пятом сегментам имеются бородавкообразные
выступы (рудименты абдоминальных конечностей); абдомен самки несет
по паре плеопод на четвертом и пятом сегментах; у некоторых видов
по Milne-Edwards’y, имеется и третья пара на третьем сегменте; второй
сегмент имеет такие же бородавкообразные выступы, как и самец. Жабры
(по Milne-Edwards’y) в числе 7 с каждой стороны: пара на наружной
максиллярной ножке, пара на ноге первой пары и по одной на ногах трех
следующих пар.
В. Чернявский (1884) установил новый род Porcellanid.es, который,
по его словам, занимает промежуточное положение между родами Porcellana
Lam. и Porcellanella White. Однако автор не указал, чем же отличается
установленный им род от обоих вышеупомянутых родов; в то же время,
1. PORCELLANA
107
сравнивая описание его нового рода с описанием рода Porcellana (кстати
приводимым В. Чернявским в сноске по Stimpson’y), невозможно обнару-
жить никаких отличий и поэтому мы сводим род Porcellanides в синонимию
рода Porcellana.
Виды рода преимущественно литоральные, наиболее широко рас-
пространенные в тропической зоне. В фауне наших морей род предста-
влен двумя видами.
ОПРЕДЕЛИТЕЛЬНАЯ ТАБЛИЦА ВИДОВ.
1 (2). Ладонь большей руки более или менее сплющенная, с закругленным
продольным, килем на середине; пальцы довольно длинные. Хвата-
тельные края пальцев меньшей руки не волосатые. ....................
< ...................................* 1. Р. longicornis (Pennant).
2 (1). Ладонь большей руки вздутая, без какого либо намека на продоль-
ный киль на середине; пальцы короткие. Пальцы меньшей руки густо
волосатые на вогнутых хватательных краях (эта волосатость осо-
бенно заметна, если смотреть со стороны нижней поверхности). . .
........................................*2. Р. longimana Risso.
* 1. Porcellana longicornis (Pennant). (Фиг. 38, 39b).
Cancer longicornis Pennant, YTT1, t. I, fig. 3 (цит. no Heller, 1863). — Porcellana longi-
cornis Leach, 1815, tab. 27, fig. 4—7; Milne-Edwards, 1837:257; Bell, 1853:193; Heller,
1863 :186. — Porcellanides rissoi Czerniavsky, 1884 :112.
Карапакс почти круглый, лишь незначительно больше в длину чем
в ширину, не слишком вздутый, иногда даже уплощенный. Поверхность
гладкая, при сильном увеличении и при высыхании обнаруживает мелкие
гранулы и морщинки, последние главным образов на задней части. Боко-
вые края хорошо очерченные, слабо гребенчатые. Области карапакса
довольно хорошо выражены. Лобный край рассечен на три треугольных
доли; средняя доля с сильно зазубренными краями (7—11 зубцов)
и довольно глубокой продольной бороздой, так что ее поверхность оказы-
вается вогнутой; боковые доли чуть-чуть короче средней, их края менее
зазубренные, а вершины несут заметный направленный вперед и внутрь
шипик; боковые доли отделены от средней закругленными выемками,
которые иногда могут быть более или менее угловатыми. Расстояние
между вершинами боковых долей меньше половины ширины карапакса
в самой широкой части. Тотчас за передне-боковым углом примерно над
ножкой антенны боковой край карапакса несет шип, который у более круп-
ных экземпляров может сглаживаться до степени бугорка. Боковой край
карапакса на середине, над основанием первой ходильной ноги, вооружен
2—4 шипами; кроме того каждая хепатическая область несет два малень-
ких шипа, которые расположены на ее верхней половине и ближе к краю
108
PORCKLLANIDAE
карапакса и которые иногда могут отсутствовать. Глаза довольно круп-
ные, с небольшими темнопигментированными роговицами. Первый членик
ножек антенн выступает за край карапакса и несет изнутри изогнутый
вперед шип; второй членик продолговатый, слегка расширяющийся в дис-
тальком направлении; третий членик короткий. Первые три базальных
сегмента наружных максиллярных ножек довольно широкие; более узкие
терминальные сегменты густо опушены длинными перистыми волосками;
экзогнат (базекфиз) достигает до середины второго сегмента ножки.
Клешненосные ноги неравные, одна обычно несколько длиннее и шире
другой. Внутренний дистальный угол короткого меруса вытянут в неболь-
шую долю, которая несет 2-4, а у более крупных экземпляров один шип;
Фиг. 38. Porcella.no. longicornis (Pennant).
верхний дистальный край усажен очень мелкими шипиками; выпуклая ниж-
няя сторона несет на вершине направленный вперед шип. Карпус продолго-
ватый, вооружен на внутреннем краю 2-6 шипами; наружный край несет
5 шипов, у основания которых имеются пучки коротких волосков; нижняя
поверхность гладкая, выемчатая на внутренней стороне. Большая рука
продолговатая, узкая, гораздо длиннее чем карпус; верхняя поверхность
гладкая, с мягко закругленным продольным килем на середине; наруж-
ный край шиповатый и волосатый, внутренний край ладони гладкий; пальцы
довольно тонкие, с сильно искривленными кончиками; наружный край под-
вижного пальца более слабо шиповатый, чем наружный край неподвиж-
ного пальца; хватательный край подвижного пальца на дистальной поло-
вине с глубокой продольной бороздой, края которой мелко зазубрены; близ
основания имеется явственный бугорок. На меньшей руке пальцы более
сплющенные, их наружные края более шиповатые; хватательные края
пальцев мелко зазубрены по всей своей длине сверху и только на дисталь-
ной трети снизу; пальцы плотно смыкаются, в то время как на большей
руке остается небольшая щель между ними. Первые три пары ходильных
1. PORCELLANA
109
ног тонкие; меральные членики довольно широкие по сравнению с осталь-
ными члениками; все членики очень слабо волосатые; пальцы короткие,
почти прямые, с длинными изогнутыми когтями. Четвертые ходильные ноги
{пятая пара перейопод) тонкие и длинные, обычно подогнутые под карапакс,
но в вытянутом состоянии достигающие до проксимальной трети проподу-
сов. предыдущих ног. Расположение большей и меньшей клешненосной ноги
с той или другой стороны не является признаком полового диморфизма;
однако, на основании просмотренного материала можно сказать, что, неви-
димому, у самок большая клешненосная нога чаще находится с правой
стороны, тогда как у самцов наоборот.
Размеры
Длина карапакса..........................5.0 мм
, Ширина „ 4.5 „
Длина карпуса большей клешней, ноги......3.5 „
„ ладони „ „ „ . .........5.0 „
„ пальцев ,, ,, ,, ........ 2.5 ,,
Форма в общем очень мелкая.
Окраска: беловатая, желто-коричневая или красно-коричневая, иногда
светло красноватая с более темными неправильной формы пятнами.
Распространение. Весьма обычен в Черном море, на литорали
среди камней и под камнями; идет до глубины 1.5—3 м, часто встречается
среди групп мидий. Средиземное и Адриатическое моря; восточная
Атлантика: побережья Норвегии, Швеции, Каттегат, Немецкое море, побе-
режье Англии, Франции и Испании,/на юг до Марокко; встречается до
глубины 70 м, обычно на песчаных и каменистых грунтах; часто живет в раз-
личных ходах, в особенности Pholas, в трубках Sabellidae, в плавнике и
проч. В материалах Зоологического Института Академии Наук оказалось
2 экземпляра Р. longicornis с острова Кильдин; повидимому данное место-
нахождение носит случайный характер (занос на плавнике?), так как для
Мурманского побережья Р. longicornis до сих пор еще не была известна.
Нет никаких оснований выделять черноморскую форму в особый вид,
как это делает В. Чернявский (1884), который почему-то сравнивает его
с Р. kriczagini, описанной им же из Сингапура, но совершенно не упоми-
нает о Р. longicornis. Сравнение последнего вида с черноморской формой
показывает, что это совершенно идентичные виды. Porcellana longicornis
является несомненно очень полиморфным видом. Изменения касаются,
главным образом вооружения боковых краев карапакса, вооружения хепа-
тических областей и характера боковых краев рук передних клешненос-
ных ног. Как резко выражена эта изменчивость у средиземноморских
экземпляров, судить трудно (я не имел возможности познакомиться
с работами Pesta и Parisi, а по более ранним работам это решить невоз-
можно), но в Черном море полиморфизм выражен довольно резко, что
было отмечено еще В. Чернявским.
110
PORCELLANIDAE
Фиг. 39. Рука меньшей клеш-
ненрсной ноги: а — Porcel-
lana longimana-, b — Porcel-
lana longicornis.
a
*2. Porcellana longimana Risso. (Фиг. 39a).
Porcellana longimana Risso, 1815:68; 1826:50. — Porcellanid.es longimana Czer-
niavsky, 1884:118.
Очень близок к P. longicornis. Карапакс округлый, гладкий; его бо-
ковые края вооружены тремя острыми шипами. Лобный край разделен на
три доли, средняя доля вооружена 8 шипиками;
боковые доли острые, вооруженные мелкими
шипиками. Хепатические шипы имеются или
отсутствуют. Клешненосные ноги длинные, без-
волосые, гладкие, у самцов они более крупные.
Рука большей ноги чрезвычайно длинная, взду-
тая, края с обеих сторон гладкие, пальцы уко-
роченные. Меньшая рука умеренной величины,
сплющенная, сверху слабо гребенчатая, краевые
гребни с обеих сторон гладкие, пальцы длин-
ные, не короче чем ладонь и густо волосатые
на вогнутых хватательных краях.
Таким образом, вид очень мало отли-
чается от Р. longicornis и, может быть, не
основания сводится некоторыми авторами в
нонимы последнего.
Распространение. Черное и Средиземное моря.
без
си-
2. Род PACHYCHELES STIMPSON
Pachycheles Stimpson, 1858 : 66; 1907 : 186; Henderson, 1888 : 113,.
Тип рода: Pachycheles grossimanus (Guerin).
Карапакс круглый, овальный, по крайней мере такой же ширины, как
и длины; его боковые края отмечены приподнятой линией. Лобная часть
карапакса отчасти покатая книзу, немного выступающая на середине, об-
разует полуострый, широко треугольный рострум, который никогда не
бывает зубчатым и верхушка которого скрыта волосами. Суббранхиальные
области карапакса не сплошные, задняя меньшая часть отделена от боль-
шей передней части суббранхиальной области перепончатым или кожистым
промежутком. Базальный членик антеннальной ножки удлиненный и сопри-
касается с краем карапакса; второй членик находится на некотором рас-
стоянии от орбиты. Клешненосные ноги неравные, сильные, более или
менее шероховатые благодаря гранулам; карпус короткий. Пальцы ходиль-
ных ног обычной формы.
Виды этого рода преимущественно литоральные, встречающиеся
в умеренных и тропических зонах Тихого и Индийского океанов. В нашей
фауне род представлен одним видом.
2. P ACHY CHELES
111
* 1. Pachycheles stevensii Stimpson. (Фиг. 40).
Pachycheles stevensii Stimpson, 1858 : 242: 1907: 187, t. 23, fig. 6; Miers, 1879 : 47’
Ortmann, 1892: 267; Balss, 1913 : 32; Yokoya, 1928 : 760.
Карапакс почти круглый; лобный край почти вытянут в короткий
широко-треугольный рострум, покрытый на верхней поверхности корот-
кими, мягкими, густо стоящими волосками; кончик рострума тупой. Боко-
вые края карапакса невооруженные, слегка приподнятые, резко отделяю-
щие. верхнюю поверхность карапакса от боковых суббранхиальных об-
Фиг. 40. Pachycheles stevensii Stimpson.
ластей, поверхность каждой из которых прерывается характерным длят
рода перепончатым участком, иначе говоря, необычайно широкой linea
anomurica; этот участок начинается от основания антенны в виде узкой
полоски и постепенно расширяется по направлению назад; его верхняя
граница тянется параллельно боковому краю карапакса вплоть до заднего
края карапакса, в то время как нижняя граница резко спускается над.
основанием первой ходильной ноги, так что образуется перепончатый
треугольный участок на задней половине суббранхиальной области; перед-
не-нижний угол этого треугольника занят крупной объизвествленной пла-
стинкой, вдоль задне-верхнего края которой расположен ряд из 3-4 очень
маленьких пластинок. Верхняя поверхность карапакса на более централь-
ных частях покрыта низкими, вытянутыми в поперечном направлении
гранулами, которые становятся крупнее по направлению к боковым:
краям; от каждой гранулы отходит вперед несколько очень коротких
112
PORCELLANIDAE
волосков; на кардиальной области эти гранулы почти отсутствуют;
на заднебоковых частях они, наоборот, становятся крупными, при-
обретая вид коротких поперечных гребней, придающих этой части кара-
пакса морщинистый вид. Для невооруженного глаза карапакс представляется
почти гладким, области неотчетливые. Задний край карапакса вогнутый.
Наружные орбитальные зубцы короткие. Глазные стебельки короткие
и толстые. Исхиум наружных максиллярных ножек широкий, с выступом
на верхне-наружном углу; мерус более узкий, с крупной закругленной
долей на внутреннем краю; карпус расширяется проксимально и на внут-
ренней стороне образует своеобразное углубление, куда при сгибании
помещается проксимальный конец проподуса; палец короткий; слегка изо-
гнутый, довольно широкий экзогнат достигает до середины меруса и не-
сет на конце довольно длинный жгут.' Клешненосные ноги мощные по
сравнению с карапаксом, безволосые, заметно сплющенные, правая нога
значительно больше левой. Мерус правой клешненосной ноги с плоской
широкой поверхностью, обращенной к карапаксу, которая почти под пря-
мым углом переходит в узкую верхнюю поверхность, образующую на
внутреннем краю треугольный выступ; нижне-внутренняя сторона вогнутая,
нижне-наружная — выпуклая. Верхняя поверхность карпуса покрыта гра-
нулами, которые становятся более крупными по направлению к краям;
вокруг гранул имеются венчики коротких волосков; внутренний уплощен-
ный край несет три зубца, подчас неправильной формы и также покрытых
гранулами; вогнутая нижне-внутренняя и выпуклая нижне-наружная поверх-
ности, встречаясь внизу, образуют довольно резкое ребро. Рука, довольно
узкая у сочленения с карпусом, имеет наибольшую ширину на уровне
основания подвижного пальца, при этом наружный край ладони образует
прямую линию с наружным краем неподвижного пальца, тогда как вну-
тренний край ладони образует тупой угол с наружным краем подвижного
пальца; верхняя поверхность ладони подобно карпусу покрыта гранулами,
которые наиболее крупны у наружного края; вдоль наружного края ла-
дони тянется заметное углубление, которое становится более глубоким
у основания неподвижного пальца и частично переходит на этот палец,
становясь более мелким; на середине ладони намечается более слабая
вторая борозда и, наконец, от основания подвижного пальца начинается
совсем незначительная третья борозда, так что вся верхняя поверхность
ладони, в общем, имеет неровный вид; выпуклая нижняя поверхность
ладони густо покрыта низкими плоскими гранулами. Пальцы слегка
искривлены на концах, хватательные края невооруженные, густо опушен-
ные у основания, в особенности снизу, мягкими волосами; при смыкании
остается широкая щель (в описании Stimpson’a указывается, что пальцы
смыкаются без щели; Balss отмечает, что большая клешня часто зияет).
Меньшая клешненосная нога в общем имеет такую же форму, только
борозды на верхней поверхности ладони, пожалуй, более резко выражены,
шальцы плотно смыкаются и не имеют опушки волос у основания. Ходильные
HIPPIDEA
113
ноги сравнительно маленькие, в противоположность клешненосным ногам,
густо волосатые на карпальных и проподальных члениках; пальцы корот-
кие, с изогнутым когтем. Пятая пара перейопод тонкая, обычно согнутая
в мерально-карпальном сочленении, так что при наружном осмотре виден
только мерус с подогнутым под него длинным карпусом, тогда как про-
долговатая и волосатая маленькая клешня спрятана под краем карапакса
в жаберной полости; в вытянутом состоянии эта нога достигает до ди-
стального конца карпуса предыдущей ходильной ноги или даже несколько
заходит за него. Абдомен сравнительно широкий; тельсон имеет харак-
терное для данного семейства строение. Наружная ветвь уропод у самки
значительно длиннее и шире, чем у самца. Окраска спиртовых экземпля-
ров бледная оранжево-красноватая.
Размеры
Длина карапакса.........
Ширина „ .........
Длина меруса правой клешней.
Длина руки „ „
Ширина „ „ „
Длина первой ходильной ноги
ноги
17.0 мм
17.0 „
12.0 „
26.5 „
14.0 „
21-0 „
Распространение. Западное побережье Японии от Хокайдо до
Нагассаки; Токийская бухта. По нашему побережью Японского моря от
бухты Горшкова до залива Стрелок.
5. Надсем. HIPPIDEA
Карапакс овальный или полуквадратный, сравнительно гладкий
с хорошо выраженными боковыми краями; лобный край широкий, не
сливается с эпистомом.. Рострум маленький или отсутствует. Глаза не
снабжены явственными орбитами, роговицы маленькие. Антеннулы хорошо
развиты. Один из жгутов длинный, другой умеренной величины или отсут-
ствует. Антенны с пятичленистой ножкой и коротким жгутом; скафоцерит
чаще всего отсутствует. Наружные максиллярные ножки ногообразные
или крышкообразные, без мастигобранхий. Первая пара перейопод с лож-
ными клешнями или простая; пятая пара маленькая, скрытая в жаберной
полости; остальные пары перейопод имеют сплющенные и изогнутые
пальцы. Жабры типа филлобранхий, число их сильно редуцировано (9-10,
иногда имеются еще рудиментарные). Мастигобранхии на перейоподах
отсутствуют. Абдомен подогнут под торакс, эпимеры частично редуциро-
ваны; уроподы всегда имеются; тельсон порядочной величины, его длина
превышает ширину. Абдоминальные конечности у самцов отсутствуют (не
считая, конечно, уропод); абдоминальные конечности самок простые, трех-
членистые. Половые отверстия у самцов расположены на коксах пятых
перейопод, у самок — на коксах третьих перейопод.
Фауна СССР, Макаров 8
114
ALBUNEIDAE
Miers (1877) сближает Hippidea при посредстве Raninidae Dana
c Brachyura Oxystomata de Haan’a, учитывая общий внешний вид, форму
клешненосных ног и развитие пальцев вторых-четвертых перейопод;
однако все эти признаки являются чисто конвергентными и отнюдь не
указывают на какую либо генетическую связь; достаточно вспомнить
о наличии у Hippidea уропод и, правда, не всегда, скафоцеритов, чтобы
убедиться лишь в чисто поверхностном сходстве их с Raninidae. Ort-
mann (1892) считает Hippidea за достаточно изолированную группу
и отмечает некоторое ее родство только с Galatheidea.
-Представители Hippidea свойственны почти исключительно тропи-
ческой области, являются литоральными или мелководными обитателями
и ведут роющий образ жизни (в песке).
III. Сем. ALBUNEIDAE
Albuneidae Stimpson, 1858:230; Henderson, 1888:39; Holmes, 1900:103; Ortmann,
1892:534;1901:1153.
Карапакс сплющенный, без боковых крылообразных. расширений,
которые закрывали бы ноги. Антеннальная ножка обычно с явственным
скафоцеритом на втором членике. Наружные максиллярные ножки узкие,
с экзогнатом. Первая пара перейопод с ложными клешнями, сплющенная.
Второй-четвертый (у рода Blepharipoda также и пятый) абдоминальные
сегменты с хорошо развитыми эпимерами; тельсон овальный и пластин-
чатый. Самка с абдоминальными конечностями на втором-пятом сег-
ментах.
1. Род BLEPHARIPODA RANDALL
Blepharipoda Randall, 1839:130; Holmes, 1900 :103; Balss, 1914 : 92.
Тип рода: Blepharipoda occidentalis Randali.
Карапакс продолговато-овальный, немного выемчатый сзади. Антен-'
нулы и антенны длинные; ножка антенны имеет скафоцерит.1 Глазные
стебельки длинные, тонкие, цилиндрические, с сочленением на середине.
Наружные максиллярные ножки ногообразные, не закрывающие рот, их
исхиум очень толстый, мерус почти цилиндрический, узкий и похож на
карпус, который не имеет выступа на своем передне-наружном углу; экзо-
1 Randall (1839, р. 130) в диагнозе рода говорит: „антенн четыре, они неравные,
наружные значительно больше, с одним длинным подвижным придатком, который усажен
ресничками на наружной стороне". Ряд авторов (Holmes, 1900, Ortmann, 1901) указывают,
что антенны без скафоцеритов. У сухого экземпляра, бывшего в моем распоряжении, я не
мог обнаружить никакого скафоцерита. Bouvier (1898) говорит: „антеннальные н антенну-
лярные ножки невооруженные". Весьма возможно, что, может быть, один вид, тип рода —
В. occidentalis Randall, имеет скафоцериты, тогда как нижеприводимый вид не имеет их.
1. BLEPHARIPODA
115
гнат довольно сильный и простирается почти до кончика меруса. Перейо-
поды сильные, волосатые, их основания сближены, пальцы серповидные;
передняя пара перейопод клешненосная, клешни устроены по типу лож-
ной клешни.
1. Blepharipoda japonica Durufle. (Фиг. 41).
Blepharipoda japonica Durufle, 1889 : 92. — Blepharipoda fauriana Bouvier, 1898 : 566;
1893a : 339, fig. 1—5.
Карапакс больше в длину
чем в ширину, с выпуклой
верхней поверхностью и сла-
бым срединным гребнем; бо-
ковой край вооружен 3 ши-
пами: один крупный на пе-
редне-боковом углу, другой
маленький на заднем конце
хепатической области (иногда
он бывает сглажен, как у эк-
земпляра, имевшегося у меня
под руками), наконец малень-
кий шип занимает передне-
наружный угол бранхиальной
области. Рострум в виде ши-
рокотреугольного зубца, по
бокам которого находятся
значительно более крупные
треугольные боковые зубцы,
отделенные от рострума во-
гнутым и мелко зазубренным
лобным краем; боковые края
рострума и боковых зубцов
также зазубрены. Тотчас позади лобного края тянется поперечная вдавлен-
ная линия, а на переднем краю гастральной области менее глубокая
и более короткая дугообразная линия, которая у срединной линии кара-
пакса образует глубокий изгиб, а по бокам, делая угловатую выемку,
сливается с первой линией; поверхность карапакса между этими линиями
понижена, более темно окрашена (у спиртового экземпляра), покрыта
морщинками и узелками. Остальная поверхность карапакса, в особен-
ности гастральная область, покрыта полулунными ямками, вогнутая сторона
которых занята низкой папиллой с зазубренным передним краем. Две глубо-
ких сходящихся борозды разграничивают на каждой стороне передние бран-
хиальные области. Цервикальная борозда мало изогнутая, идет почти
поперек карапакса и не сливается с упомянутыми выше бороздами. Зад-
ние края борозд несут короткие, направленные вперед волоски, которые
8*
116 ALBUNEIDAE
могут стираться (у имевшегося у меня экземпляра они сохранились только
на цервикальной борозде, на участке соприкосновения ее с кардиальной
областью). Боковые границы кардиальной области вполне отчетливы
спереди и неясны сзади. Задний край карапакса широко выемчатый;
закругленные заднебоковые углы карапакса усажены группой плоских
папилл с зазубренным передним краем, которые часто принимают очень
неправильную форму. Глазные стебельки длинные, расширенные у своего
основания, но затем становящиеся довольно тонкими и почти не расши-
ряются у роговиц, которые занимают терминальное положение и имеют
закругленный синус на своем верхне-внутреннем краю; стебельки сплю-
щены на своей наружной поверхности и слабо выпуклые на внутренней
поверхности, которая внизу несет ряд густых длинных волосков. Глазные
стебельки сочленяются с узкими, но довольно длинными глазными чешуй-
ками, которые почти соприкасаются по срединной линии; эти чешуйки
граничат снизу с перепончатым участком, где находится маленькая тре-
угольная пластинка, представляющая собой переднюю дужку тела. Антен-
нулярные и антеннальные ножки невооруженные; первые густо опушены
снизу и сверху длинными волосками; такие же волоски находятся на верх-
ней поверхности большего антеннулярного жгута, тогда как снизу они
гораздо более редкие; меньший жгут короткий, состоящий лишь из 10 чле-
ников. Антеннальные жгуты немного длиннее. Экзоподит наружных максил-
лярных ножек несет сплющенный нечленистый бич; карпус имеет на нижней
поверхности мелкие зубчики; исхиум несет на внутреннем краю 7 довольно
сильных шипов; базиподит также имеет шип на внутреннем краю; эпипо-
дитальная пластинка на своем свободном конце несет рудиментарную
подобранхию. Передние ноги сравнительно короткие; мерус с сильным
шипом на нижне-дистальном углу; передне-верхний угол карпуса вытянут
в остроконечный зубец, края которого волосатые, а передний край
к тому же шиповатый; верхне-наружная поверхность карпуса гранулиро-
ванная; клешня широкая, плоская, неправильной сердцевидной формы;
верхне-наружная поверхность ладони гранулированная; параллельно наруж-
ному краю и несколько отступя от него тянется неправильный гребень,
переходящий и на неподвижный палец; от этого гребня отходят напра-
вленные внутрь волоски; такие же волоски рассеяны и по поверхности
ладони; нижне-внутренний угол ладони несет пучок длинных волос. Непо-
движный палец очень широкий у основания, с выпуклым хватательным
краём и острым изогнутым кончиком; хватательный край несет 4 тупых
зубца, промежутки между которыми мелко зазубрены и заполнены длин-
ными волосками. Изогнутый у своего основания подвижный палец при
смыкании прилегает к хватательному краю неподвижного пальца и своим
кончиком заходит за кончик неподвижного пальца; его верхняя и нижняя
поверхности совершенно гладкие, а наружный край несет ряд гранул.
Три следующих пары перейопод имеют сплющенные членики; карпальные
и проподальные членики с плоскими тупыми треугольными выступами на
PAGURIDEA
117
передне-дистальных концах; пальцы в виде плоских пластинок изогнутой
формы, с совершенно гладкими поверхностями и волосатыми краями.
Пятая пара перейопод слабая, тонкая, оканчивающаяся волосатой клеш-
ней. Жабры (по Bouvier): плевробранхии на вторых-четвертых перейоподах,
по две артробранхии на вторых-четвертых, иногда и на первых перейо-
подах и на третьих максиллярных ножках, эпиподиты на первой и третьей
максиллярных ножках и подобранхия на третьей максиллярной ножке. Пер-
вый сегмент абдомена длинный, перепончатый в своей центральной части
и покрытый на большем протяжении карапаксом; он лишен эпимеров
и придатков; следующие четыре сегмента узкие, постепенно уменьшаю-
щиеся в величине по направлению назад, с тупыми длинными эпимерами.
Самка несет на этих сегментах по паре двуветвистых плеопод; самец
имеет лишь следы на местах прикрепления этих придатков. Шестой абдо-
минальный сегмент почти такой же длины, как и ширины; обе ветви его
уропод овальные. Тельсон немного больше в длину чем в ширину; его
центральная часть выпуклая, сильно объизвествленная и отделена от
боковых частей углублением; на заднем краю имеется небольшая
выемка.
Относительно образа жизни В. japonica Bouvier (1898) замечает: „Эти
животные должны, повидимому, зарываться в песок, как и другие Hip-
pidea, но они менее хорошо приспособлены к такой привычке, потому
что их фильтрационный аппарат менее совершенен; в самом деле, их
антеннулярные жгуты менее длинные и менее волосатые, а бич экзопо-
дита их передних максиллярных ножек представляет собой более умень-
шенную поверхность".
Распространение. Побережье Японии. В наших водах вид
известен из района Владивостока, бухты Патрокл, из Посьетского залива.
IV. Надсем. PAGURIDEA
Головогрудь спереди не соединяется с эпистомом. Форма карапакса
либо цилиндрическая, с мягкими покровами и без резких боковых краев,
или же карапакс расширен, имеет крабообразную форму, хорошо объиз-
вествленный и с отчетливыми боковыми краями. Рострум маленький,
треугольный, иногда совсем редуцированный, или же хорошо развитый,
шипообразный или иной формы. Глаза не снабжены явственными орби-
тами, глазные стебельки цилиндрические, с глазными чешуйками или без
них. Антеннальная ножка пятичленистая, второй членик несет шипообраз-
ный скафоцерит (последний редуцирован у Coenobitidae). Жгуты экзопо-
дитов вторых и третьих максиллярных ножек членистые. Третьи максил-
лярные ножки ногообразные. Все перейоподы шестичленистые. Первая
пара клешненосная, большей частью неравная; обе последних пары корот-
кие и снабжены участками с своеобразными бородавчатыми возвыше-
118
PAGURIDAE
ниями, или же только последняя пара редуцирована и в таком случае
несет на конце маленькую клешню и обычно спрятана.в жаберной поло-
сти. У более примитивных форм жабры типа трихобранхий, в большин-
стве же случаев типа филлобранхий, максимальное число их 14 с каждой
стороны; эпиподиты на перейоподах отсутствуют. Половые отверстия рас-
положены на коксах. Абдомен либо своеобразно преобразован, мягкий,
с сравнительно хорошо или слабо развитыми дорзальными пластинками,
либо подогнут под головогрудь, как у Brachyura, и в большей или мень-
шей степени покрыт объизвествленными пластинками и узелками. У самцов
иногда имеются половые придатки; абдоминальные конечности в боль-
шинстве случаев развиты только на одной стороне. Уроподы имеются
или отсутствуют.
Представители этого надсемейства либо прячутся в раковинах
Gastropoda и прочих полых инородных предметах, либо свободно ползают
по дну; и в том и в другом случае абдомен является асимметричным;
только у наиболее примитивных форм он симметричный, хотя и прячется
в трубки червей, губки и проч. Некоторые представители приспособились
к свободному амфибиотическому образу жизни (Coenobitidae), сохранив,
однако, асиммётрию тела.
ОПРЕДЕЛИТЕЛЬНАЯ ТАБЛИЦА СЕМЕЙСТВ
1 (4). Абдомен прямой или спирально закрученный, не подогнутый под
головогрудь. Карапакс цилиндрический, жесткий главным образом
в своей передней части, в то время как задняя часть более или
менее мягкая. Четвертые перейоподы не похожи на третьи. Рострум
рудиментарный или совсем отсутствует. Уроподы имеются.
2 (3). Абдомен симметричный, с хорошо объизвествленными сегментами
на дорзальной стороне. Имеются парные абдоминальные конеч-
ности на всех сегментах. Жабры типа трихобранхий................
..........................................1. Сем. Pylochelidae
3 (2). Абдомен в большинстве случаев асимметричный, спирально закру-
ченный, мягкий, несегментированный, с небольшими, слабо объизве-
ствленными пластинками на левой стороне. В редких случаях имеются
парные придатки только на первом абдоминальном сегменте, обычно
же они находятся только на левой стороне на местах второго-пятого
сегментов. Жабры обычно типа филлобранхий.......................
..............................•........... 2. Сем. Pagaridae
4 (1). Абдомен в редких случаях мягкий, короткий, мешкообразный, чаще же
всего он пластинчатый и подогнут под головогрудь. Карапакс
широкий, хорошо объизвествленный во всех частях. Общий вид
животного крабообразный. Четвертые перейоподы подобны третьим.
Рострум в большинстве случаев хорошо развит. Уроподы отсут-
ствуют .........................................3. Сем. Lithodidae
1. MIXTOPAGURUS
119
6. Сем. PYLOCHELIDAE
Pylochelidae Bate, 1888 :11; Ortmann, 1901 :1144.
Карапакс довольно хорошо объизвествленный; его боковые стенки
сплющенные и очень высокие. Глазные стебельки умеренной длины.
Антеннулы оканчиваются двумя жгутами. Имеются маленькие скафоце-
риты. Мандибулы с пальпом. Наружные максиллярные ножки сближены
у своих оснований. Первая пара перейопод клешненосная, ноги почти
равной длины. Вторая и третья пары длинные, тонкие, оканчивающиеся
игловидным пальцем. Четвертая и пятая пары перейопод короткие, оканчи-
вающиеся маленьким рудиментарным пальцем. Жабры типа трихобранхий,
нити длинные, цилиндрические и тонкие; подобранхии и мастигобранхии
отсутствуют на всех конечностях головогруди. С каждой стороны имеется
14 жабр: по две артробранхии на третьей максиллярной ножке и четырех
первых перейоподах и по одной плевробранхии на четырех последних
перейоподах. Абдомен симметричный, с хорошо объизвествленными на
дорзальной стороне сегментами. Хвостовой веер симметричный, с жесткими
и суживающимися уроподами, наружные уроподы крупнее внутренних.
Тельсон жесткий спереди и более гибкий сзади. Парные абдоминальные
конечности имеются на всех сегментах; у самцов плеоподы первого сег-
мента преобразованы в половые придатки.
По общему habitus’y представители данного семейства несколько
напоминают таковых сем. Pagaridae, однако отличаются от них симметрией
абдомена и его конечностей, а также тем, что сегменты абдомена соеди-
няются с помощью обычных суставов, имеют жесткую дорзальную поверх-
ность, а на вентральной стороне имеют также жесткие, но прерванные
на середине пластинки. Живут в губках, были находимы в затонувших
ветвях бамбука и мангровых деревьев, в раковинах Dentalium и других
предметах, имеющих полость. Boas (1926) придерживается того взгляда,
что Pylochelidae не таскают своего жилища за собой, но в поисках пищи
покидают его, т. е. ведут полусвободный образ жизни, и этим объясняет
симметрию и все строение абдомена. Несомненно, что среди всех Pagu-
ridea представители данного семейства являются самыми примитивными.
Из пяти родов этого семейства приводится представитель одного
рода, встречающийся в водах Японского моря.
1. Род MIXTOPAGURUS A. MILNE-EDWARDS
Mixtopagurus Milne-Edwards et Bouvier, 1893 :23; Balss, 1913 : 34. — Pomatocheles
Miers, 1879: 49.
Тип рода: Mixtopagurus paradoxus A. Milne-Edwards.
Представители этого рода обнаруживают близкое родство с родом
Pylocheles. Они имеют, как и виды рода Pylocheles, 28 нитевидных жабр,
120
PYLOCHELIDAE
четырехрядных и расположенных точно таким же образом. Торакс объиз-
вествленный по крайней мере в своей дорзальной части. Глазные стебельки
у оснований удалены друг от друга. Наружные максиллярные ножки сбли-
жены у оснований. Торакальные стерны очень узкие. Вентральная поверх-
ность абдомена перепончатая, за исключением слабых объизвествленных
участков на первом-шестом сегментах, дорзальная поверхность обра-
зована смежными и сочленяющимися сегментами, объизвествленными на
всем их протяжении. Имеются парные конечности на всех абдоминаль-
ных сегментах; две первые пары у самцов и первая пара у самок пре-
образованы в половые придатки. Передние максиллярные ножки с эпипо-
дитом, экзоподит с жгутом.
Mixtopagurus отличается от Pylocheles следующими признаками..
Лобный край вооружен рострумом. Антеннулярные ножки достигают или
немного превышают кончики глаз. Второй членик антеннальной ножки
не имеет снаружи сильного зубчатого выступа. Антеннальный жгут дости-
гает по крайней мере до кончиков клешней. Клешни первых перейопод.
равные или почти равные, оканчиваются роговыми когтями и не образуют
крышечки (operculum). Наружные максиллярные ножки на конце простые,,
не клешнеобразные. Четвертая пара перейопод с ложной клешней и воору-
жена развитым шиповатым участком (,,Raspel“ немецк. авторов). Шестой
абдоминальный сегмент гораздо сильнее объизвествлен, чем другие; тельсон
одночленистый. Все абдоминальные конечности, за исключением первой,
пары, двуветвистые у обоих полов.
1. Mixtopagurus jeffreysii (Miers). (Фиг. 42).
Pomatocheles jeffreysii Miers, 1879 : 49, tab. 3, fig;. 2. — Mixtopagurus jeffreysii Balss,.
1913 : 35, f. 25, tab. 2, fig-. 1; Yokoya, 1933 : 71.
„Животное стройное и продолговатое. Карапакс отмечен явственными
постфронтальным и латеральным швами. Кроме того имеются два меньших:
• и менее явственных шва на боках в направлении к боковым краям. Медиан-
ная лобная доля широко треугольная и закругленная на вершине. Первый
постабдоминальный сегмент очень маленький, пять следующих почти
одинаковые, с прямыми боковыми краями; последний сегмент маленький,,
прозрачный и перепончатый на своей дистальной половине и усаженный,
ресничками на своих краях, терминальная медианная вырезка очень малень-
кая. Глазные стебельки немного короче, чем лобный край и снабжены,
маленькими чешуйками у основания. Роговицы красно-коричневого цвета.
Антеннулы вдвое длиннее, чем глазные стебельки, антенны приблизительно
такой же длины, как антеннулы; скафоцерит очень маленький, острый,
меньше половины длины глазного стебелька. Передние ноги такие же, как
урода Cancellus, мерусы с слегка зазубренным гребнем на своей верхней
поверхности; карпусы очень короткие и слегка зазубренные сверху; сплю-
щенная верхняя поверхность ладоней покрыта толстыми короткими воло-
сами, нижняя поверхность гладкая, прямой внутренний и дугообразный на-
1. MIXTOPAGURUS
121
ружный края очень слабо зубчатые. Тонкие и удлиненные ноги второй
и третьей пар имеют короткий карпус; проподус и палец длинные и пря-
мые, в особенности па-
лец очень длинный, тон-
кий и острый. Усечен-
ный дистальный конец
последнего членика че-
твертой ноги вооружен
рядом коротких жестких
щетинок или шипиков
и маленьким когтем или
шипом; последний чле-
ник пятой пары густо
усажен ресничками.Кле-
шни передних ног, бу-
дучи сложены над голо-
вой и плотно сомкнуты
своими плоскими вну-
тренними краями, обра-
зуют прекрасную кры-
шку (греческ. тсму-ос от-
куда и название рода),
которая соответствует
отверстию занимаемой
раковины и служит для
защиты ее обитателя
от врагов. Базальная
часть уропод короткая
и широкая и несет две
неравных пластинкооб-
разных ветви, которые
имеют зубчатое строе-
ние на наружной по-
верхности и с реснич-
ками на краях; наружная
ветвь вдвое длиннее,
чем внутренняя. Длина
5 линий (Miers)“.
„Относительно био-
логии этой формы сле-
Фиг. 42. Mixtopagurus jeffreysii (Miers) (из Balss, 1913).
дует заметить, что тель-
сон у нее совершенно клапанообразный, так что он вместе с пластинкой
шестого сегмента закрывает нижнюю сторону трубки раковины Dentalium
(Balss)“.
122 PYLOCHELIDAE
Несколько спорным вопросом является родовая принадлежность
данного вида, что явствует и из сравнения вышеприведенных диагнозов
рода и вида, где можно найти противоречия. Boas (1926) склонен относить
дацный вид к роду Pylocheles. Весьма возможно, что Boas прав, однако,
располагая только литературными данными по этому виду, мы приводим
его под тем родовым названием, под которым он фигурирует в литературе.
Распространение. Япония (Yokoya, 1933). В Японском море
отмечен у западного побережья Японии севернее Nagato, Yamaguti-ken
и близ островов Oki. На глубинах 64—350 м; экземпляры Balss’a были
в раковинах Dentalium inter со statum Boissevain, D. eburneum L. и D. usi~
Latum Smith.
Сем. PAGURIDAE
ХАРАКТЕРИСТИКА СЕМЕЙСТВА
МОРФОЛОГИЧЕСКИЙ ОЧЕРК 1
Тело рака-отшельника распадается на головогрудь (cephalothorax)
и абдомен. Оба эти отдела, в особенности последний, обладают морфо-
логическими особенностями, свойственными почти исключительно пред-
ставителям рассматриваемого семейства.
Головогрудь. Особенностью головогруди является, за очень
небольшими исключениями, слабая объизвествленность большинства ее
частей. Обильно объизвествленной.являет.ся только цефалическая область,
отделенная от остальных частей хорошо выраженной цервикальной
бороздой; в систематических описаниях эта область будет упоминаться как
передняя часть карапакса. Из остальных частей более сильно объизвествлена
передняя часть кардиальной области, тогда как весь остальной карапакс
является перепончатым. Свободно лежащие последний и предпоследний
сегменты головогруди составляют вторую особенность головогруди раков-
отшельников. Стернальные пластинки узкие, отделяющиеся за пятым
сегментом друг от друга сочленовной мембраной; стернальная пластинка
четвертого сегмента смещена влево у большинства видов, имеющих
асимметричные клешненосные ноги. Свободное сочленение двух послед-
них сегментов обеспечивает большую подвижность, необходимую ракам-
отшельникам при их образе жизни в спирально закрученных раковинах.
На поверхности карапакса, кроме цервикальной борозды (линия С по
терминологии Boas; бранхиальная борозда или линия С по терминологии
Bouvier) можно различить еще следующие борозды (фиг. 43): продольная
борозда, тянущаяся от переднего края карапакса мимо цервикальной
1 Перед изложением анатомо-морфологической части необходимо оговориться, что
все данные, приводимые в этом разделе, касаются, главным образом, одного рода Pagurus
Fabricius. Объем работы не позволяет детально остановиться на различных морфологи-
ческих изменениях у тех или иных родов. Род Pagurus удобен в том отношении, что боль-
шинство видов разбираемого семейства, обитающих в наших водах, относится именно
к этому роду; кроме того большая часть работ по морфологии Paguridae посвящена этому
роду, в том числе прекрасная монография Н. G. Jackson (1913). Те или иные отклонения
будут отмечены в систематической части при описании соответствующих родов или видов.
124
PAGURIDAE
борозды до заднего его края; Borradaile целиком называет эту борозду
linea anomurica; по терминологии Bouvier только участок этой борозды
от переднего края карапакса до цервикальной борозды называется linea
anomurica; по терминологии Boas этот же участок называется линией В,
а участок позади цервикальной борозды обозначается им как linea
anomurica. От linea anomurica на уровне цервикальной борозды отходит
слабая бороздка, направляющаяся вертикально вниз, которая рассматри-
вается Boas как часть linea anomurica. От цервикальной борозды тянется
Фиг. 43. Поверхность карапакса Pa-
gurus ochotensis subsp. aleuticus
(Benedict), c—цервикальная борозда;
la — linea anomurica; d — линия D
(no Boas); A — передняя часть кара-
пакса; г — rostrum, или средний вы-
ступ лба; b — боковые выступы лба.
вперед, параллельно linea anomurica, сла-
бая борозда, называемая Boas линией D;
вместе со своим продолжением назад за
цервикальную борозду она, повидимому,
является гомологом linea thalassinica.
Поверхность карапакса, в особенно-
сти передней части его, часто покрыта
пучками волосков, иногда бывает богато-
скульптирована. Передний или лобный край
карапакса большею частью несет посере-
дине треугольный rostrum, в системати-
ческих описаниях в дальнейшем именуемый
как средний выступ лба; по бокам от рос-
трума лобный край образует супраорби-
тальные выступы, именуемые в дальнейшем
как боковые выступы лба; их форма изме-
няется от резко выраженной треугольной
до едва намечающегося широко закруг-
ленного выступа; иногда они совершенно
отсутствуют. На переднем краю карапакса,
около боковых выступов лба, часто
имеются расположенные несколько ниже
края карапакса субмаргинальные шипы.
Объизвествленные складки кутикулы (так
наз. аподемы) внутри головогруди образуют сложную систему эндофраг-
мального скелета, служащего местом прикрепления мышц и являющегося
защитой для внутренних органов.
Гистологическая структура панцыря рака-отшельника мало отличается
от таковой других ракообразных, за исключением степени объизвествле-
ния. Можно различать следующие слои панцыря: 1) тонкий пограничный
слой; 2) хитиновый слой, распадающийся в свою очередь на 4 слоя:
а) тонкий наружный слой, состоящий из пластинок, Ь) пигментный слой,
состоящий из параллельных верхней поверхности пластинок и содержащий
зернистое вещество, с) известковый (основной) слой, также состоящий
из пластинок, пропитанный слоями кальция; папиллы этого слоя могут
вдаваться в пигментный слой, d) тонкий неизвестковый внутренний слой,
ХАРАКТЕРИСТИКА СЕМЕЙСТВА
125
образованный слоистыми пластинками; 3) эпителиальный слой (hypoderma),
продуцирующий хитин. Перечисленные слои пронизаны штопорообразно
извивающимися поровыми канальцами, которые доходят до пограничного
слоя; эти канальцы рассматриваются как продолжение гиподермальных
клеток и несут, повидимому, функцию выделения извести. Кроме того, все
слои . пронизываются канальцами,
к волоскам, щетинкам и пр.
Абдомен. Абдомен рака-от-
шельника представляет собой мяг-
кий, перепончатый, колбасообраз-
ный мешок, который большею
частью спирально закручен, но
иногда бывает и прямой, лишь не-
значительно вентрально подогну-
тый на заднем конце. Стенки абдо-
мена бывают настолько тонки и
прозрачны, что сквозь них часто
можно различить находящиеся
внутри пищеварительную железу,
гонады и почечный пузырь. Объиз-
вествленными являются только
первый сегмент, шестой сегмент
•с его придатками и тельсон (неко-
торыми авторами тельсон не счи-
тается за отдельный сегмент); на
остальном протяжении расчленение
абдомена на сегменты почти нераз-
личимо. На дорзальной стороне
можно различить остатки спинок
сегментов (tergum) в виде слабо
объизвествленных пластинок, кото-
рые на левой стороне располо-
жены над плеоподами, а на пра-
вой стороне симметрично против левых (фиг. 44); эти пластинки слабее
выражены у молодых экземпляров и у самцов, чем у самок. На вентраль-
ной стороне абдомена на уровне третьего сегмента имеется неправильной
формы выступ — так наз. колюмеллярный мускул, функция которого
не совсем ясна, но который, повидимому, имеет отношение к более
прочному удерживанию животного в обитаемой им раковине.
Относительно формы абдомена имелось одно исследование, вызвавшее
в свое время много споров; именно Przibram (1907) утверждал, что при
лишении рака-отшельника раковины, форма его абдомена как бы воз-
вращается к форме его макрурообразных предков; это утверждение
оказалось неправильным, как показал недавно Brinkmann (1926).
проводящими чувствительные нервы
Фиг. 44. Абдомен Pagurus ochotensis Brandt
(дорзальный вид).
126
PAGURIDAE
Придатки головы. Глазные стебельки (фиг. 45). Глазной
стебелек состоит из двух члеников: короткого базального и более длин-
ного дистального. Форма стебелька цилиндрическая, часто суживающаяся
посредине и расширяющаяся у дистального конца. Базальный членик на
своей внутренней стороне несет небольшой придаток —
такназ. глазную чешуйку — чаще всего овальной формы,
с заостренной вершиной; снизу под вершиной часто нахо-
дится субтерминальный шип, который иногда настолько
выдается вперед, что бывает виден сверху (фиг. 46).
Дистальный членик на конце несет роговицу, которая за-
нимает терминальное положение, имеет круглую форму
и небольшую выемку на верхне-внутренней стороне.
Фиг. 45. Глазной стебелек Pagurus
pubescens Kroyer. с — роговица;
о — глазная чешуйка.
Фиг. 48. Правая?
антеннула Pagurus
pubescens Kroyer.
ex — коксоподит;
bp — базиподит;
en — эндоподит;
ex—экзоподит, или
скафоцерит; fl —
жгут.
Фиг. 47. Правая
антеннула Pagurus
pubescens Kroyer.
ex — коксоподит;
bp — базиподит ;
en — эндоподит;
fl — жгут.
Фиг. 46. Глазная чешуйка (дорзально-
боковой вид), s — субтермннальный шип
(сильно увелич.).
Антеннулы (первые антенны) (фиг. 47). Антеннулы при-
крепляются под глазами, но благодаря изгибу члеников кажутся лежащими
между глазами. Стебель или ножка каждой антеннулы состоит из трех
сегментов: коксоподита и двучленистого базиподита;1 в коксоподите рас-
1 Коксоподит и базиподит, вместе взятые, обозначаются иногда как протоподит.
ХАРАКТЕРИСТИКА СЕМЕЙСТВА
ТМ
положен статоцист, открывающийся наружу узкой щелью на дорзальной
стороне. На конце дистального членика (сегмента) базиподита имеется
экзо- и эндоподит. Экзоподит представляет собой более длинный, толстый,
много членистый, опушенный волосками жгут, тогда как эндоподит короче,
тоньше и почти лишен волосков.
. Антенны (вторые антенны) (фиг. 48). Стебель каждой антенны
состоит из четырех члеников: коксоподита, базиподита и двучленистого
Фиг. 49. А — Левая мандибула Pagurus pubescens Kroyer. В — Правая максиллула Pagurus
pubescens Kroyer. С—Правая максилла Pagurus pubescens Kroyer (эндоподит скрыт за бази-
подитом). D—Первая левая максиллярная ножка Pagurus pubescens Kroyer. Е—Вторая правая
максиллярная ножка Pagurus pubescens Kroyer. ex — коксоподит; bp — базиподит; pt — про-
топодит; ер — эпиподит; en — эндоподит; ex — экзоподит; В — жгут; ip — исхиум; тр —
мерус; ср — карпус; рр — проподус; dp — палец; а — апофиз; с — головка; sc — скафогнатит.
эндоподита, от которого отходит длинный многочленистый жгут, в боль-
шинстве случаев почти лишенный волосков. На нижней стороне коксопо-
дита имеется бугорок, на котором помещается выводное отверстие антен-
нальной железы. Базиподит, обозначаемый в систематических описаниях
как базальный членик антеннальной ножки, несет на наружной стороне
экзоподит, называемый здесь скафоцеритом (у некоторых авторов чешуй-
кой). У раков-отшельников скафоцерит имеет вид узкого, серповидно-
изогнутого придатка, приблизительно цилиндрической формы (иногда
прямой и с резкими гранями); его внутренний край часто бывает зазубрен
и несет волоски..
128
PAGURIDAE
Мандибулы (фиг. 49А). Тело мандибулы распадается на головку
и мощный придаток—апофиз, служащий местом прикрепления жевательных
мышц. Головка зазубрена и бороздчатая изнутри, но расчленения на инци-
зорный и молярный отростки не наблюдается. Имеется трехчленистый
пальп (Taster, Synaphipod), служащий для очищения жевательного края
головки и, возможно, для проталкивания пищи. С сравнительно-анатоми-
ческой точки зрения тело мандибулы соответствует коксоподиту. Отно-
сительно пальпа существуют разногласия: Giesbrecht называет его эпипо-
диальным придатком и
считает новообразова-
нием, тогда как Han-
sen усматривает в нем
остатки эндоподита.
Мандибулы хорошо объ-
извествлены и являются
главными органами раз-
мельчения пищи. Лежат
они между верхней и
нижней губой, причем
пальп чаще всего при-
жат к верхней губе.
Максиллулы
(первые м а к с и л л ы)
(фиг. 49 В). Максиллула
состоит из перепонча-
тых коксо- и базиподита,
края которых усажены
со т о длинными щетинками.
4>иг. эи. 1ретья правая максиллярная ножка ragurus
pubescens Кгоуег. (Обозначения те же, что и на фиг. 49). Наружный Придаток КО-
ксоподитасоответствует
экзоподиту (по другим авторам эпиподиту). Эндоподит представлен одно-
членистым пальпом. Относительно сравнительно-анатомического толкова-
ния коксо- и базиподита существуют еще разногласия.
Максиллы (вторые максиллы) (фиг. 49С). Коксо- и бази-
подит максиллы каждый расчленяется у дистального конца на две нерав-
ные лопасти. Имеется одночленистый пальп (эндоподит) и широкий эпипо-
дит. Экзоподит представляет собой широкую, вогнутую спереди, густо
опушенную волосками по краям пластинку, называемую скафогнатитом
(дыхательная пластинка); своим движением скафогнатит создает ток воды,
омывающий жабры.
Придатки груди. Первые максиллярные ножки (фиг. 49D)
по внешнему виду напоминают максиллулы. Имеется коксо-, бази- и эпи-
подит. Экзоподит маленький, без жгута; эндоподит крупнее и несет на вер-
шине жгут.
ХАРАКТЕРИСТИКА СЕМЕЙСТВА
129
Фиг. 51. Правая
клешненосная нога
Pagurus pubescens
Кгоуег, сх — кокса;
ip — исхиум; тр —
мерус; ср — карпус;
рр—проподус; dp—
подвижный палец;
pl — неподвижный
палец.
Фиг. 52. Первая левая
ходильная нога Pagu-
rus pubescens Кгоуег.
(Обозначения те же,
что и на фиг. 51).
Вторые максиллярные ножки (фиг. 49 Е) построены по типу
двуветвистой конечности. Протоподит (соха ч- basis) несет пятичленистый
эндоподит (эти членики суть: ischium, merus, carpus, propodus и dactylus)
и экзоподит, состоящий из трехчленистого стебля и жгута.
Третьи максиллярные
ножки (фиг. 50) по своему строе-
нию подобны вторым максилляр-
ным ножкам, только ее экзоподит
относительно меньше. Исхиум не-
сет ряд зубцов, величина которых
возрастает в проксимальном на-
правлении. Обе последние максил-
лярные ножки активно участвуют
в процессе принятия пищи. Кроме
того, по наблюдению Brock’a, экзо-
подит третьей максиллярной ножки
своим движением создает ток воды,
направляющийся к антеннулам,
благодаря чему животное может
ориентироваться, воспринимая ан-
теннулами раздражения, вызывае-
мые различными химическими ве-
ществами, растворенными в при-
текаемой воде.
Первые перейоподы
или клешненосные ноги
{фиг. 51) состоят из шести члени-
ков; базис и исхиум сливаются
в один членик. Продолжение про-
подуса образует неподвижный па-
лец (pollex). Часть проподуса от
места сочленения с карпусом до
линии, проведенной через осно-
вание подвижного пальца, носит
название ладони (palma). Про-
подус вместе с подвижным паль-
цем (dactylus) образует руку (manus). Внутренние края пальцев, обычно
усаженные большей или меньшей величины бугорками, называются
хватательными краями. Величина и форма клешненосных ног сильно
вариируют у различных родов и видов; обе ноги могут быть одинаковой
длины и строения, или же одна нога больше другой и имеет иную
форму. Поверхность члеников может быть гладкой, чаще же несет
различного рода вооружение из гранул, шиповатых гранул, шипов,
зубцов, бугорков, волос, щетинок и проч. Величина, форма и характер
Фауна СССР, Макаров 9
130
PAGURID AE
вооружения клешненосных ног являются главнейшими систематическими
признаками раков-отшельников. При отсутствии той или иной конечности
или при начальной стадии регенерации одной из конечностей зачастую
бывает трудно, а подчас и невозможно, точно определить экземпляр, обла-
дающий упомянутыми дефектами.
Вторая и третья пара перейопод (пёрвая и втор'ая
пара ходильных ног) (фиг. 52). Следующие за клешненосными
ногами четыре пары ног называются ходильными ногами.. Две первые/тары
Фиг. 53. Третья правая ходильная
нога Pagurus pubescens Kroyer.
(Обозначения те же, что и на
фиг. 51).
Фиг. 54. Четвертая правая ходильная нога
Pagurus pubescens Kroyer. (Обозначения
те же, что и на фиг. 51).
имеют большею частью одинаковое строение и могут отличаться только
длиной. При систематическом описании под именем ходильных ног будут
иметься в виду только эти две пары ног. Каждая нога состоит также
из шести члеников; палец причленяется к дистальному концу проподуса,.
так что клешни не образуются. Палец довольно длинный, прямой или
изогнутый и часто в большей или меньшей степени закручен по своей
продольной оси; на дистальном конце он обычно несет острый роговой
коготь. Поверхность всех члеников (обычно наружная) и их края так'же,,
как на клешненосных ногах, несут то или иное вооружение.
Четвертая пара перейопод (третья пара ходильных
ног) (фиг. 53) значительно короче предыдущих и состоит из шести чле-
ников. Палец причленяется несколько на боковом краю проподуса, бла-
годаря чему получается так наз. ложная клешня. Край проподуса про-
ХАРАКТЕРИСТИКА СЕМЕЙСТВА
131
тив пальца вооружен густо сидящими шипиками, так что образуется
шероховатая поверхность, напоминающая поверхность напильника. Нога
обычно согнута под прямым углом в сочленении между мерусом и кар-
пусом.
Пятая пара перейопод (четвертая пара ходильных
ног) (фиг. 54) также короткая и шестичленистая. Проподус густо
опушен волосками; палец очень короткий, причленяющийся на дистальном
конце проподуса. Часть поверхности проподуса и пальца вооружена
площадкой густо сидящих шипиков. Этой конечностью живртное проникает
Фиг. 55. Плеоподы Pagurus pubescens Kroyer. А — плеопода самца;
В — плеопода самки.
в жаберную полость и чистит жабры. Кроме того обе последние ходильные
ноги, будучи упираемы в стенки раковины, служат животному для более
прочного удержания последней.
Придатки абдомена. Плеоподы (фиг. 55). По своему строению
плеоподы одинаковы у обоих полов. Короткий протоподит несет на дисталь-
ном конце эндо- и экзоподит. Эндоподит всех плеопод самца и четвертой
плеоподы самки маленький; на остальных плеоподах самки эндоподит
почти равен экзоподиту; как тот, так и другой, густо опушены волосками?
к этим плеоподам прикрепляются яйца. Плеоподы раков-отшельников
являются непарными придатками, т. к. на правой стороне абдомена, при- -
лежащей к columella обитаемой животными раковины, они исчезают
(исключение составляет род Paguropsis Hend., у которого плеоподы, на-
оборот, развиты на правой стороне). Число плеопод различно у обоих
полов, а именно самка имеет плеоподы на втором, третьем, четвертом
и пятом абдоминальных сегментах, а самец только на третьем, четвертом
9*
132
PAGURIDAE
Фиг. 56. Шестой абдоминальный сегмент, уроподы и тельсон
Pagurus pubescens Kroyer. n — уроподы; t — тельсон; pt —
протоподит; en — эндоподит; ex — экзоподит.
и пятом сегментах. Таким образом, наличие или отсутствие плеоподы на
втором абдоминальном сегменте и величина эндоподитов (кроме эндопо-
дита плеоподы на пятом сегменте) позволяют легко различить пол живот-
ного. О преобразованиях плеопод, встречающихся у некоторых видов
раков-отшельников, будет сказано в описании соответствующих родов.
Уроподы (фиг. 56). Хорошо объизвествленный шестой сегмент
абдомена несет парные, также хорошо объизвветвленные придатки, назы-
ваемые уроподами. У обоих полов они имеют одинаковое строение. Про-
топодит несет серповидно изогнутый эндоподит и более крупный экзопо-
дит. Наружно-верхняя поверхность, как экзо-, так и эндоподита, несет
площадку из густо сидящих шипиков. Как правило, уропода левой стороны
развита сильнее. Вме-
сте с тельсоном,кото-
рый также объизве-
ствлен и несет на ди-
стальном краю острые
когтеобразно изогну-
тые шипики, уроподы
служат для прочного
укрепления живот-
ного в раковине.
Мускулатура. За
исключением абдоми-
нальных мышц, му-
скулатура -раков-от-
шельников мало отли-
чается от таковой прочих десятиногих раков. Своеобразное изменение
абдомена, вызванное образом жизни рака-отшельника, наложило отпе-
чаток и на развитие мускулатуры абдомена. Сгибатели значительно
более сильно развиты, чем разгибатели, причем сгибатели правой
«тороны, в свою очередь, развиты сильнее, чем таковые левой сто-
роны. На стадии glaucothoe расположение мускулов мало отличается
от обычного; разгибатели еще хорошо развиты и имеют продольное
направление; сгибатели состоят из нисходящего, поперечного, продольного
и петлеобразного мускулов; плеоподы также имеют еще самостоя-
тельные сгибатели. В дальнейшем у взрослого животного поперечные
мышцы постепенно исчезают, от нисходящего и боковых продольных
мышц сохраняются небольшие остатки; разгибатели остаются слабыми;
сгибатели состоят из мощного вентрального, продольного и петлеобраз-
ного мускулов. Мышцы плеопод дегенерируют. Из вентрального сгибателя
третьего абдоминального сегмента развивается уже упоминавшийся выше
колюмеллярный мускул.
Органы пищеварения. Пищеварительный канал распадается на
три отдела: переднюю кишку (пищевод и желудок), среднюю кишку
ХАРАКТЕРИСТИКА СЕМЕЙСТВА
133
и заднюю кишку. Передняя и задняя кишка выстланы хитиновой кутику-
лой. Ротовое 'отверстие лежит позади ногочелюстей, направлено вниз
и покрыто парой мандибул. Впереди, над ртом, лежит мясистая верхняя
губа (labrum), состоящая из трех долей, а позади рта находится более
маленькая нижняя губа (metastoma). Пищевод (oesophagus), следующий
непосредственно за ртом, представляет
собой толстостенную трубку, внутрен-
няя стенка которой выпячивается в виде
трех мощных продольных складок в про-
свет пищевода. Оканчивается пищевод
на нижней стороне кардиальной части
желудка. У проксимального конца пище-
вода лежат три группы звездчатых же-
лез— две передне-боковых и одна зад-
няя; выводные протоки желез' откры-
ваются на внутренней стенке пищевода.
Подобные же железы имеются и
повидимому, смазывающим веществом,
Фиг. 57. Желудок Pagurus bernhardus
(Linne) с левой стороны, oes — пище-
вод; руа — пилорические ампулы;
mg. саес — левый передний пилориче-
ский отросток; mg — средняя кишка;
d. liv —проток печени. (Из Jackson,1913)..
служит,,,
облегчающим прохождение пищевой
в губах рта. Секрет желез
массы и не оказывает никакого ферментативного влияния на последнюю..
Желудок (фиг. 57 и 58) распадается на два отдела: большая передняя.
часть называется кардиальной, меньшая задняя — пилорической..
Фиг. 58. Сагиттальный разрез желудка Pagurus
bernhardus (Linne), вид с левой стороны, ptc —
птерокардиальная косточка; со — кардиальная
косточка: zc — зигокардиальная косточка; ис —
урокардиальнад косточка; mt—срединный зубец;
et — боковой зубец; рр — препилорическая ко-
сточка; ру — пилорическая косточка; cpv — кар-
диопилорический клапан; sv — верхний клапан;
тт — срединный клапан; iv — нижний клапан;
int — кишка. (Из Jackson, 1913).
Кутикула, выстилающая желу-
док, пропитана известью и по-
этому стенки желудка никогда
не спадаются. Мелкие „кос-
точки", являющиеся производ-
ным этой кутикулы, образуют
в задней части кардиальной
области и в пилорической обла-
сти так наз. желудочную мель-
ницу — сложный аппарат для
размельчения пищи; строение
этого аппарата незначительно
отклоняется от такового у реч-
ного рака, поэтому мы не бу-
дем останавливаться на раз-
боре его, а отошлем интересующихся к монографии Huxley (1880).
У места перехода пилорической части желудка в среднюю кишку имеется
пять клапанов: верхний срединный клапан, пара верхне-боковых и пара
более маленьких нижне-боковых клапанов; обе пары густо опушены щетин-
ками. Кроме того, на верхней стенке пилорической части желудка находится
пара гребней, усаженных щетинками, и средний гребень, также со щетин-
ками. Назначение этих клапанов и гребней — препятствовать возвращению
пищи из кишки в желудок, в то время как переходу пищи из желудка
134
PAGURIDAE
в кишку они не мешают. За пилор ической частью желудка начинается самая
длинная часть пищеварительного тракта — средняя кишка (mesenteron), ко-
торая лишена хитиновой кутикулярной выстилки. Длина этой части кишеч-
ника колеблется у различных видов. На дорзальной стенке средней кишки,
на небольшом расстоянии от пилорической части желудка,. имеются два
хорошо развитых пилорических придатка (соеса anteriora dorsalia), которые
в виде тонких трубок тянутся на коротком расстоянии по верхней стенке
> желудка, а затем резко поворачивают вниз, спускаются по его боковым
стенкам и, направляясь несколько вперед, заканчиваются под желудком,
закручиваясь в клубочки. На дорзальной стороне средней кишки, как раз
у места перехода ее в заднюю кишку, возникает непарный пилорический
отросток (соесшп dorsale posterius), который простирается вперед и окан-
чивается небольшим клубочком на уровне третьего абдоминального сег-
мента; у самцов этот отросток как будто бывает длиннее, чем у самок;
кроме того, его длина колеблется у разных видов в зависимости от места
перехода средней кишки в заднюю. Короткая задняя кишка (rectum) пред-
ставляет собой толстостенную, мускулистую трубку, выстланную изнутри
хитиновой кутикулой. Она имеет железы, подобные таковым пищевода,
и сильно складчатые изнутри стенки. Анальное отверстие помещается
иа нижней поверхности тельсона и имеет замыкательный мускул (сфинктер).
Таким образом, передняя и задняя кишка имеют эктодермальное происхо-
ждение, а средняя кишка и ее пилорические отростки — эктодермальное.
При линьке, когда рак-отшельник высвобождает сперва тело, затем
конечности и, наконец, абдомен, хитиновая кутикула передней кишки
выталкивается через рот, а кутикула задней кишки через анальное
отверстие.
Пищеварительная железа (hepatopancreas), часто называемая
также печенью, у раков-отшельников представляет собой очень длинный
парный орган, простирающийся далеко в абдомен. Каждая доля состоит
из массы ветвистых трубочек, заполняющих полость абдомена, которые
впадают в осевые протоки. Кишечник проходит или между долями печени,
или сквозь правую долю. Осевые протоки тянутся вдоль сгибательных
мускулов абдомена, но при переходе в торакс они плотно прилегают
к вентрально-боковым стенкам кишечника и, наконец, открываются
в среднюю кишку на ее нижней стенке, недалеко от пилорической части
желудка и впереди места отхождения передних пилорических отростков.
Печень содержит клетки двух сортов: высокие цилиндрические альвеоляр-
ные клетки, часто заключающие в себе жировые капельки и гликоген,
и затем ферментные клетки, содержащие большую вакуоль с заключенной
в ней коричневатой жидкостью — секретом, который попадает в просвет
трубки либо через разрыв вакуоли, либо через отбрасывание всей клетки.
Весьма возможно, что эти клетки, помимо секреторной, несут также экс-
креторную функцию. На дистальных концах железистые трубки содер-
жат клетки с крупными ядрами и гомогенной плазмой, которые митоти-
X АР АКТЕРИСТИКА СЕМЕЙСТВА
135
ческим делением образуют так наз. фибриллярные клетки, из которых
в свою очередь образуются ферментные клетки. е
Процесс принятия пищи можно представить себе следующим образом:
обычно левая клешненосная нога подает куски пищи ногочелюстям;
последние препровождают их далее к челюстям, которые производят
первое размельчение пищи. Через .пищевод размельченная пища попадает
в желудок, где при помощи мельницы подвергается окончательному раз-
мельчению. Кашицеобразная масса через пилорическую часть продвигается
в среднюю клетку, а неподдающиеся размельчению частицы выбрасы-
ваются обратно через рот. Сразу по выходе из желудка пищевая кашица
смешивается с секретом печени; возможно, что частично секрет попадает
в пилорическую часть желудка, где, следовательно, и начинается перева-
ривание, которое в основном происходит все-таки в средней кишке. По
мере продвижения по средней кишке происходит всасывание; неперева-
ренные остатки перистальтическими движениями мускулистой задней
кишки выбрасываются наружу через анальное отверстие. Не совсем ясен
способ удаления фекальных масс из раковины; вероятно они выносятся
током воды, а часть из них, возможно, уничтожается часто живущими
в раковине мелкими Amphipoda. По характеру пищи раки-отшельники
являются всеядными. По данным Blegvad (1914) они поедают мелких мол-
люсков, ракообразных, иглокожих, возможно рыб и донный детрит. Часто
наблюдали промывание песка раками-отшельниками между ротовыми час-
тями; нахождение диатомовых водорослей и корненожек в кишечном канале
отшельников М. Т. Thompson объясняет именно таким образом. По наблю-
дениям Hunt (1925) Pagurus bernhardus и Р. prideauxi питаются мелкими
пластинчатожаберными моллюсками {Venus, Cultellus), иглокожими {Echy-
nocyamus, мелкие Echinocardium, офиуры), ракообразными {Amphipoda,
Crangonidae, мелкие Paguridae) и полихетами {Poiynoidae, Nephthys,
Goniada, Terebellidae, Pectin aria). Orton (1927) констатирует, что рак-
отшельник добывает пищу, главным образом, взмучиванием или соскабли-
ванием детрита при помощи маленькой клешненоносной ноги и третьих
максиллярных ножек и последующим отсеиванием из него органических
составных частей (мелкие полихеты, ракообразные, водоросли, форамини-
феры, диатомеи и проч.). Рак-отшельник может, правда, питаться и более
крупной пищей, но все же его ротовые части менее приспособлены для
такого сорта пищи, чтобы убедиться в этом Orton (1927) давал одновре-
менно одинаковой величины куски мяса отшельнику {Pagurus bernhardus)
и крабам {Carcinus moenas и Portunus ruber)’, спустя час с четвертью
у отшельника все еще оставалось от одной четверти до одной трети
количества его пищи, тогда как крабы съедали полученное мясо в тече-
ние 4—11 минут. Таким образом, форма третьих максиллярных ножек
всецело определяется способом питания животного. На питание рака-
отшельника мелкими организмами, добываемыми из детрита, указывают
также более мелкие размеры косточек желудочной мельницы, с одной
136
PAGURIDAE
стороны, и, с другой стороны, наличие гребней на боковых зубцах желудка
и щетинок на кардиопилорических клапанах, предназначенных для отсеи-
вания мелких частиц пищевого материала. Подтверждением этого является
еще тот факт, что у планктоноядных ракообразных (каковы напр., Наро-
locarinus и Cryptochirus) редукция косточек желудочной мельницы дости-
гает еще большей степени.
Органы дыхания. Органами дыхания являются жабры, распо-
ложенные в бранхиальной полости, защищаемой снаружи боковой
стенкой карапакса, так наз. бранхиостегитом. По своему положению
жабры являются артробранхиями, т. к. сидят на суставной мембране
между коксой конечности и телом. На двенадцатом сегменте иногда
имеется одна плевробранхия — жабра, сидящая на самой стенке тела.
Обычная жаберная формула следующая: восьмой, девятый, десятый, один-
надцатый, двенадцатый и тринадцатый сегменты имеют по две артррбран-
хии (одна передняя и одна задняя); на двенадцатом сегменте, кроме того,
иногда имеется плевробранхия. По своей форме у большинства раков-
отшельников жабры относятся к типу филлобранхий, т. е. состоят из
осевого стебля с двуперисто расположенными мелкими листочками.
По наружной стороне стебля каждой жабры проходит приносящий бранхи-
альный сосуд, а по внутренней — выносящий. Жабры имеют так наз.
бранхиальные железы, выделяющие слизистый секрет. Жабры, распо-
ложенные спереди, более мелкие, чем жабры, расположенные сзади.
Жаберная полость открыта спереди, снизу и сзади, так что вода имеет
свободный доступ к жабрам. Движение скафогнатита максиллы обеспечи-
вает постоянный ток воды, идущий через жаберную полость сзади напе-
ред. Для очищения жабр служит четвертая пара ходильных ног. С сравни-
тельно-анатомической точки зрения задние артробранхии и плевробранхии
рассматриваются как видоизмененные эпиподиты прекоксы, а передние
артробранхии как видоизмененные эпиподиты коксы.
Кровеносная система. Перикардиальная полость, в которой лежит
сердце, простирается от цервикальной борозды до уровня восьмого груд-
ного сегмента. Непосредственно под перикардиальной полостью проходит
средняя кишка, а сверху перикардий плотно прилегает к карапаксу. Сердце,
таким образом, расположено под кардиальной областью карапакса, имеет
сверху неправильную пятиугольную форму, обладает сравнительно мощ-
ными мускульными стенками и тремя парами отверстий, через которые кровь
входит в сердце из перикардиальной области: одна пара отверстий распо-
ложена дорзально спереди, другая вентрально и несколько сбоку и третья
латерально сзади (фиг. 59). Из сердца выходят семь артерий: три напра-
вляются вперед (arteria mediana cephalica и парные a. laterales cephalicae),.
три идут вниз (парные a. hepaticae и a. descendens) и одна назад (aorta po-
sterior). В самых кратких чертах проследим ход этих главнейших сосудов.
1) Arteria mediana cephalica (a. ophthalmica), или срединная непар-
ная аорта, проходит над желудком, затем опускается и делится на две
ХАРАКТЕРИСТИКА СЕМЕЙСТВА
137
ветви, снабжающие кровью лобную область головы. Тотчас по выходе
из сердца аорта образует луковицеобразное расширение, так наз.
cor frontale.
2) Парные arteriae laterales cephalicae (a. antennariae) выходят из
сердца по бокам непарной срединной аорты, поворачивают наружу
и снабжают ветвями желудок, выделительные органы,, мандибулярные
мускулы, антенны, антеннулы и глазные стебельки.
3) Парные arteriae hepaticae у раков-отшельников развиты слабо
и не снабжают кровью перемещенную в абдомен пищеварительную железу
(печень), но заканчиваются маленькими веточками на средней кишке.
4) Непарная arteria descen-
dens (a. sternalis) является самым
крупным сосудом, выходящим из
сердца. Она отходит от задне-
вентрального угла сердца, повора-
чивает влево и направляется вниз;
оставив справа от себя кишечник,
она поворачивает вперед, прохо-
дит горизонтально на протяжении
7—5-го грудного сегмента, затем
Фиг. 59. Сердце Pagurus bernhardus (I.inne),
дорзально-боковой вид. opth. а — arteria me-
diana cephalica,• ant. a — arteriae laterales,
cephalicae; hep. a — arteria hepatica; d. st.,
a— arteria descendens; S. Abd —aorta poste-.
. rior. (Из Jackson, 1913).
вновь поворачивает вниз и про-
ходит через центральную ган-
глиозную массу между нервами
второй и третьей пары перейопод;
под нервной цепочкой она-раздваи-
вается на переднюю ветвь, идущую к голове, и заднюю ветвь, идущую
к хвосту; эти ветви обозначаются общим названием как a. subneuralis..
Передняя ветвь снабжает кровью клешненосные ноги и ротовые части,
а также дает веточки к нижней и передней частям желудка, пилорическим
отросткам и выделительным органам. Задняя ветвь дает веточки ко всем:
ходильным ногам.
5) Непарная aorta posterior (arteria abdominalis dorsalis) снабжает
кровью абдомен и находящиеся в нем органы. В соответствии с своеобраз-
ным изменением абдомена этот сосуд также претерпевает изменения,
характерные только для рассматриваемого семейства. Выйдя из сердца
над a. descendens, задняя аорта в виде крупного ствола тянется над кишеч-
ником до первого абдоминального сегмента; здесь она делится на две
асимметричные ветви. Одна ветвь — сегментальная артерия — направляется
вниз и вправо и тянется до вершины сгибательных мускулов; на уровне
третьего абдоминального сегмента она делится на Две ветви: субмускуляр-
ную и супрамускулярную; первая ветвь направляется по ходу нервной
цепочки и оканчивается близ последнего ганглия; супрамускулярная ветвь
дает веточки к гонадам, печени, задней кишке, уроподам и тельсону. Другая из
асимметричных ветвей отклоняется влево,, проходит по поверхности печени;
138
PAGURIDAE
и отдает веточки к гонадам и плеоподам. Интересно, что у самки каждый
яичник снабжается кровью из обеих ветвей, тогда как у самца правый
семенник снабжается кровью из левой ветви, а левый семенник — из правой.
Венозная кровь изливается в кровяные синусы, которые все сообща-
ются друг с другом, образуя в конечном итоге один общий синус. Различают
•следующие кровяные синусы: 1) дорзальный синус, лежащий впереди
и над желудком; 2) стернальный (вентральный) синус, тянущийся вен-
трально по всей длине груди; 3) абдоминальный синус, являющийся продол-
жением стернального синуса в абдомене; 4) парные инфрабранхиальные
•синусы, лежащие по обеим сторонам груди; каждый из последних синусов
пятью канальцами сообщается со стернальным синусом; кроме того,
в эти же синусы открываются кровяные полости грудных конечностей.
Из инфраорбитальных синусов берут начало приносящие бранхиальные
сосуды, которые поднимаются по наружной стороне стебля каждой жабры
и в виде выносящих бранхиальных сосудов опускаются по внутренней
стороне стебля. Выносящие сосуды собираются с каждой стороны в четыре
бранхиокардиальных канала (так наз. жаберные вены), которые откры-
ваются в полость перикардия тремя щелями с каждой стороны.
Кровь представляет собою слабо опалесцирующую, почти прозрач-
ную жидкость, содержащую в себе форменные элементы (амебоциты,
гранулоциты и тромбоциты), каратиноиды (придающие крови слабый
красноватый оттенок) и гемоциан, который связывает кислород; гемоциан
•содержит в себе не железо, а медь, и при длительном стоянии крови
на воздухе окрашивает ее в синий или скорее в фиолетовый цвет (гемо-
циан -I- каратиноиды). Перед линькой кровь заметно обогащается углекис-
лым кальцием.
Органы выделения (фиг. 60). Выделительная система раков-отшель-
ников имеет более сложное строение, чем у других Decapoda; наряду
•с парными антеннальными железами имеются сложно ветвящиеся парные
везикулярные массы, лежащие в головогруди, и непарный абдоминальный
пузырь; везикулярные массы непосредственно сообщаются с антеннальг
ными железами. Рассмотрим в отдельности каждую из частей экскретор-
ной системы.
1) Парные антеннальные железы („зеленые железы" речного рака)
лежат в последней части головы позади антенн. Каждая железа состоит
из мезодермального целомического мешочка (sacculus) и эктодермального
нефридиального канальца; проксимальная часть этого канальца сильно
ветвится, образуя так наз. лабиринт (nephrostom), а дистальная часть
расширяется в собирательный пузырек; из собирательного пузырька выходит
выстланный хитиновой кутикулой выводной проток, который, как уже
упоминалось выше, открывается выводным отверстием (nephroporus) на
нижней стороне коксоподита антенны.
2) Передняя везикулярная масса лежит в . области головогруди
и соединяется канальцами с антеннальными железами. Каждый каналец
ХАРАКТЕРИСТИКА СЕМЕЙСТВА
139
расширяется в массу ветвистых трубочек,
образующих эпигастральную долю; узкий
канал от эпигастральной доли приводит
к другой массе ветвистых трубочек — пара-
гастральной доле, расположенной по бокам
.желудка; от этой массы отходит еще неболь-
шая супрагастральная доля; под желудком
находится непарная срединная вентральная
доля, соединяющаяся передней и задней
ветвями с парагастральными долями.
3) Задняя везикулярная масса. От пара-
гастральных долей начинается пара ветвистых
трубок, которые тянутся вдоль кишечника
вплоть до абдомена, где они сливаются
в непарный тонкостенный абдоминальный
пузырь (nephrosac), который простирается
примерно на три четверти длины абдомена.
На спиртовых экземплярах редко удается
обнаружить все части описанной системы
(обычно бывают заметны только антенналь-
ные железы и нефросак); поэтому для деталь-
ного изучения рекомендуется прижизненная
окраска крепким раствором метилгрюна или
кислого фуксина в морской воде; 1—2 куб. см
такого раствора вводится шприцем в сочленов-
ную мембрану под грудными конечностями.
Примерно, через 48 часов животное можно
подвергать вскрытию; правда, антеннальные
железы не окрашиваются, но вся остальная
система выделяется довольно отчетливо.
Так как экскреторная система как бы
целиком погружена в венозные синусы, то
выделение, невидимому, происходит при по-
мощи непосредственной диффузии через
стенки. Клетки везикулярных масс высту-
пают в виде прозрачных пузырьков в про-
свет трубок; экскреторная жидкость содержит
в себе такие выпавшие пузырьки, а иногда
даже и целые клетки. Экскреторную функцию
несут также железистые клетки (нефроциты),
располагающиеся вдоль стеблей жабр (эти
клетки также окрашиваются при инъекции
метилгрюном или кислым фуксином) и затем
ферментные клетки печени.
Фиг. 60. Экскреторная система
Pagurus bernhardus (Linne). Ле-
вая эпигастральная доля, все
другие органы теля, мускулы
и абдомен удалены. (Полусхема-
тично). gg — левая антеннальная
железа; egl — эпигастральная
доля; sgl — супрагастральная
доля; pgl—парагаст; альная доля;
mvl — срединная вентральная
доля; Ыс—протоки между перед-
ней и задней везикулярными
массами; Ы — нефрссак. (Из
Jackson, 1913). •
140
PAGURID АЕ
Нервная система. Нервная система обнаруживает, пожалуй, наимень-
шие отклонения в своем строении по сравнению с другими системами
органов; главные изменения касаются, как и следует ожидать, абдоми-
нальной части системы.
Надглоточный ганглий (ganglion supraoesophageum) расположен на
средней линии карапакса позади глазных стебельков. Из надглоточного
ганглия выходят следующие пары нервов: 1) nervus opticus — начинается
от передней части ганглия и идет в глазные стебельки; 2) n. oculomotoris —
маленький нерв, идущий параллельно n. opticus, иннервирует мышцы глаз;
3) n. antennarius I (n. antennularis) начинается от нижней поверхности
задней половины ганглия и состоит, в сущности^ из двух ветвей — нерва
статоцита и n. olfactorius; последний иннервирует чувствительные волоски
и мышцы антеннул; 4) n. tegumentarius, или кожный нерв, который своими:
веточками иннервирует интегумент и другие ткани в передней части
головы; 5) п. antennarius II (n. antennalis) отходит от мозга позади n. tegu-
mentarius и иннервирует антенны.
С торакальной ганглиозной массой надглоточный ганглий связан
двумя глоточными коммиссурами, которые начинаются от задней части
мозга и тянутся по бокам пищевода. На середине своего пути каждая ком-
миссура расширяется в параглоточный ганглий, от которого берут начало
четыре веточки: две наружных и две внутренних; передняя из наружных
веточек иннервирует окружающие ткани, а задняя — мышцы мандибулы.
Две внутренние ветви образуют так наз. стоматогастральную систему;
обе ветви начинаются вместе, а затем расходятся, идут впереди пище-
вода и затем вновь сливаются, образуя* непарный стоматогастральный
нерв, который тянется вниз по передней части желудка, где он образует
стоматогастральный ганглий; от этого ганглия отходят веточки к мышцам
желудка; стоматогастральный нерв охватывает затем желудок и на его
дорзальной стороне образует другой ганглий и, наконец, раздваиваясь,
оканчивается над пилорической частью желудка. Глоточные коммиссуры
примерно на середине расстояния между параглоточными ганглиями и
торакальной ганглиозной массой соединены друг с другом задне-глоточной
коммиссурой. Торакальная ганглиозная масса, в которой оканчиваются
глоточные коммиссуры, лежит на уровне 3—6-го грудного сегмента и
состоит из трех слившихся ганглиозных масс; степень слияния этих масс
больше чем у Мастита, но меньше чем у Brachyura', во всяком случае,
довольно ясно можно различить три отдела, ограниченных друг от друга
перехватами. Каждый отдел имеет отверстие для прохода артерий. От
первого отдела, являющегося продуктом слияния семи парных ганглиев,
отходят семь пар нервов; первая пара к мандибулам, вторая и третья
к максиллам, четвертая, пятая и шестая пары к максиллярным ножкам
и, наконец, седьмая пара к клешненосным ногам. Второй, или средний,
отдел получается из слияния только одной пары ганглиев и отсылает от
себя пару нервов, иннервирующих первые ходильные ноги. От заднего
ХАРАКТЕРИСТИКА СЕМЕЙСТВА
141
отдела торакальной ганглиозной массы отхо-
дят три пары нервов ко второй, третьей
и четвертой парам ходильных ног. С послед-
ним отделом торакальной массы сливается
пара первых абдоминальных ганглиев, кото-
рые отделены от остальной массы мелкой
бороздкой.
Торакальная ганглиозная масса соеди-
нена с абдоминальной цепочкой при помощи
абдоминальной коммиссуры, которая вначале
является непарной, но после входа в абдомен
разделяется на две ветви. Абдоминальная
цепочка состоит из пяти пар ганглиев, при-
чем каждая пара почти сливается в одну
•общую массу; друг с другом ганглии соеди-
нены парными коммиссурами. Вся цепочка
располагается под массой сгибательных му-
скулов и над слоем мускулов интегумента.
Ганглии располагаются во 2—6-м сегментах.
При входе в абдомен цепочка уклоняется
влево, проходя через колюмеллярный мускул;
после четвертой пары ганглиев цепочка идет
по средней линии абдомена. От каждой пары
ганглиев отходят нервы к мышцам, плеопо-
дам и интегументу; кроме того, отходят
веточки и от межганглиозных коммиссур.
От последней пары ганглиев отходят нервы
к уроподам, тельсону и задней кишке.
Органы чувств» Глаза. Г лаз состоит
из множества отдельных клиньев, так наз. ом-
матидиев (omma, ommatidium); число клиньев
довольно значительно и возрастает по мере
роста животного. Роговица фасеточная, фа-
сетки имеют шестиугольную форму; каждая
фасетка является дистальным концом омма-
тидия, а все омматидии, благодаря выпуклости
роговицы, сходятся радиально по направлению
внутрь. В состав каждого омматидия входят
следующие части (фиг. 61): 1) роговица,
состоящая из трех слоев хитина (покровного,
пигментного и основного, соответственно
слоям кутикулы) без известковой подкладки,
выделяется двумя нижележащими • клет-
ками роговицы (по Schneider их имеется
В А
Фиг. 61. А. Продольный разрез
омматидия. В. Отдельная про-
ксимальная ретинальная клетка.
(Полусхематично, очень сильно
увеличено), пи — ядра; сот — ро-
говица; chy — клетки роговицы;
vit — хрустальное тело; dre —
дистальные, ретинальные клетки
(главные пигментные клетки);
гЬ — рабдом; р. ге — прокси-
мальные ретинальные клетки;
n. fib—нервные фибриллы; bm—
базальная мембрана; n. op. f —
волокна n. opticus. (Из Jackson,
1913).
142
PAGURIDAE
четыре). Клетки роговицы имеют форму прямоугольных треугольников,,
которые соприкасаются своими гипотенузами; последние просвечивают
в виде диагоналей, если смотреть на роговицу сверху. Ядра клеток
лежат в прямых углах этих треугольников; 2) хрустальный конус обра-
зуется из четырех конических клеток (часто называемых Семперов-
скими клетками); каждая клетка имеет прозрачное светопреломляющее
тело, которое суживается к своему проксимальному концу, образуя сте-
бель, доходящий до дистального конца рабдома. Ядра конических клеток,,
(так наз. Семперовские ядра) лежат у их дистальных концов; 3) две глав-
ные пигментные клетки помещаются на противоположных концах кони-
ческих клеток; они содержат радужный пигмент и изменяют свою вели-
чину в зависимости от силы света; 4) имеется восемь ретинальных клеток»
одна из которых может быть рудиментарной; дистальные концы этих
клеток вздутые и содержат крупное ядро, проксимальные концы сужи-
ваются, доходя до базальной мембраны. Фибриллы оптического нерва
идут по наружной стороне клеток, огибают отчасти их ядро и опускаются
по внутренней стороне, заканчиваясь в поперечных пластинках рабдома..
Рабдом (rhabdome) представляет собой сложное образование,, как бы одну
общую светочувствительную палочку, окруженную ретинальными клет-
ками. В его образовании принимают участие каждые две ретинальные-
клетки, так что образуется четыре четвертушки, известные под названием/
рабдомеров. Каждый рабдомер состоит из квадратных поперечных пласти-
нок, которые отходят от внутренних продольных сторон ретинальных,
клеток (фиг. 62) и соединяются с пластинками клеток противоположной,
стороны; т. к. эти половинчатые пластинки в следующих друг за другом,
положениях образуют угол в 90°, то на продольном разрезе получается
слоистость, состоящая из светлых (поперечный разрез фибрилл) и темных
(продольный разрез фибрилл) полос. На дистальном конце rhabdome
имеется полость. Ретинальные клетки, как и главные пигментные клетки,,
содержат пигмент; согласно Parker’y, в темноте пигмент перемещается
в ретинальные фибриллы под базальной мембраной, благодаря чему
rhabdome становится доступным для света со всех сторон. В главных,
пигментных клетках в темноте пигмент собирается у дистальных концов»
а при дневном свете — у проксимальных концов; 5) добавочные клетки:
(Tapetumzellen Hesse) слабо пигментированы, лежат у основания rhabdome,
и на обеих сторонах базальной мембраны, образуя отражательную
поверхность. Базальная мембрана состоит из переплетающихся соеди-
нительно-тканных волокон. Нервные фибриллы от каждого омматидия
проходят через базальную мембрану к четырем следующим друг-
за другом глазным ганглиям (lobus opticus); последовательно эти
ганглии обозначаются как lamina ganglionaris (1-й ганглий), medulla
externa (2-й ганглий), medulla interna (3-й ганглий) и medulla terminalis.
(4-й ганглий). Первый ганглий получает волокна непосредственно от
омматидиев. Между первым и вторым ганглиями лежит перекрест воло-
ХАРАКТЕРИСТИКА СЕМЕЙСТВА 143-
кон, так наз. chiasma externa; между вторым и третьим ганглиями лежит
второй перекрест — chiasma interna и, наконец, между третьим и четвер-
тым ганглиями также имеется перекрест, не носящий особого названия.
Четвертый ганглий при помощи pedunculus lobi optici (n. opticus) связан,
с надглоточным ганглием. О более детальном строении глаза см. работы
Parker (1890, 1895), Hesse (1901) и Trojan (1913).
Относительно фототропизма имеются наблюдения над Clibanarius
misanthropus (Risso); этот отшельник меняет знак фототропизма каждые
14 дней, применяясь к интенсивности освещения в зависимости от высоты
стояния воды: при малых приливах он проявляет отрицательный фототро-
пизм, при полных — положительный; интересно, что при пересадке в аква-
риум животное сохраняет еще в течение некоторого времени эту смену
знаков фототропизма.
Minkiewicz (1907) стремился найти у отшельников хромотропизм,
т. е. предпочтение определенной окраске при выборе грунта; им была уста-
новлена следующая последовательность цветов (предпочтительная окраска,
расположена в правой стороне ряда): черный—красный — желтый —
синий — фиолетовый — зеленый — белый. Cotte (1921), повторивший опыты
Minkiewicz’a, не мог подтвердить правильность выводов этого автора.
Что отшельники могут воспринимать и запоминать различные цвета,
показывают “опыты Михайлова (1920); Dardanus arrosor (Herbst) улавливал
связь между тактильным раздражением, при котором он прятался в свою
раковину, и освещением определенного цвета; после закрепления- ре-
флекса животное реагировало на соответствующий цвет (без тактильного,
раздражения) также уходом в раковину, причем эта реакция сохранялась
в течение 200 дней; после исчезновения реакция могла быть восстановлена
в более короткий срок, чем в первый раз.
Статоцисты. Статоцист (отоцист, „слуховой мешочек" прежних
авторов) находится в базальном членике антеннулы. Он представляет
собой простое впячивание наружного покрова в виде мешка, стенки кото?
рого, следовательно, выстланы хитиновой кутикулой. С внешней средой
он сообщается узкой продольной щелью на дорзальной поверхности чле-
ника, причем отверстие для защиты от инородных тел снабжено двумя,
бахромками щетинок. От заднего конца мешка по его внутренней стенке
наружной стороны тянется небольшой гребень, а по вентральной стороне
в бороздке проходит „слуховая" ветвь антеннулярного нерва. К кутикуле
подвижно прикреплены чувствительные волоски, которые, начиная от
заднего конца мешка, образуют два или три неявственных ряда, причем
основная масса волосков располагается на гребне наружной стороны,
мешка. Каждый волосок двуперистый, полый изнутри; каждому волоску
принадлежит чувствительная биполярная клетка, один отросток которой,
входит в полость волоска, а другой соединяется с „слуховым" (статиче-
ским) нервом. В качестве статолитов служат мелкие песчинки, которые,
лежат либо свободно, либо приклеиваются к волоскам при помощи секрета.
144
PAGURIDAE
железок, которые находятся у основания статоциста. Нерв входит в ста-
тоцист на заднем конце, где делится на две ветви: большая ветвь огибает
статоцист по наружной стороне, меньшая — по внутренней. При линьке
вся внутренняя выстилка статоциста, имеющая эктодермальное происхо-
ждение, сбрасывается и заменяется новой; точно так же заменяются
и статолиты. Статоцисты, считавшиеся прежними авторами за органы
слуха, несут исключительно статическую функцию, помогая животному
ориентироваться в пространстве. Детальное описание статоцистов и обзор
взглядов различных авторов относительно их функции можно найти
в работе С. W. Prentiss (1901).
Обоняние. Обонятельные волоски помещаются на экзоподитах
антеннул. Каждый сегмент несет несколько таких волосков, которые пред-
ставляют собой полые стебли, расчлененные приблизительно на 24 сег-
мента, причем базальный сегмент широкий и окружен пучком мелких
волосков; каждый волосок имеет нервное волоконце. Помахивая антен-
нулами, животное постоянно обследует окружающую среду. Над значением
обонятельных волосков имеются подробные наблюдения Brock’a (1926) на
Dardanus arrosor (Herbst); если животному в гнездо незаметно вносилась
приманка, то спустя некоторое время рак становился беспокойным, начи-
нал усиленно бить антеннулами и затем поворачивался и двигался в напра-
влении приманки; время начала движения пропорционально’ расстоянию
приманки от животного, и кроме того зависит от температуры (зимой оно
вдвое больше, чем летом). Большое значение имеет также поток дыха-
тельной воды, который, восходя из жаберной полости, омывает антеннулы
и тем самым приносит различные раздражающие вещества к чувствитель-
ным волоскам.
Осязание. Осязательные волоски рассеяны по всему телу. Каждый
волосок является полым выростом кутикулы; канальцы, подводящие
к волоскам нервные волокна, пронизывают все слои наружного покрова
животного. Своими вторыми и третьими перейоподами ослепленные живот-
ные, напр., могут отличать поверхность раковины моллюска от поверх-
ности гладкой стеклянной трубки. При помощи осязания может разли-
чаться, повидимому, и форма предметов; так, напр., A. Drzewina добива-
лась того, что Clibanarius misanthropus мог отличать раковину Trochus,
в которой он не поселялся, от Cerithium, которой он охотно пользовался
в качестве жилища. Особенно чувствительны к прикосновению антенны,
которыми животное обычно следит за проплывающими мимо рыбами
и другими животными.
О температурных ощущениях имеется немного данных. По отношению
к Paguridae имеются наблюдения Matisse, который действием высоких
и низких температур вызывал у животного состояние оцепенения.
Органы размножения. Парные семенники (testes) самца лежат
в промежутке между долями печени, причем левый семенник лежит более
поверхностно и является более крупным, чем правый. Каждый семенник
ХАРАКТЕРИСТИКА СЕМЕЙСТВА
145
представляет собой длинную, свернутую трубку, петли которой и образуют
плоский, вытянутый в длину орган. Концевой участок трубки, образующий
вначале несколько спиралей, а затем идущий прямо, носит название семе-
провода (vas deferens); дойдя до грудной полости, выносящий проток круто
опускается вниз и открывается на коксе последней пары ходильных ног.
Концевой участок выносящего протока окружен сильной мускулатурой
и носит название семеизвергающего канала (ductus ejaculatorius). У неко-
торых родов имеется кожистый penis, обычно только на левой стороне,
или парные половые придатки (роды Paguristes, Parapagurus), которые
будут описаны при разборе соответствующих родов. Сперматозоиды обла-
дают тремя отростками и в семепроводе соединяются в сперматофоры.
Парные яичники (ovaria) самки имеют вытянутую форму, зернистый
внешний вид и простираются почти во всю длину абдомена, причем левый
яичник у переднего конца лежит выше правого. Яйцеводы (oviductus) пред-
ставляют собой мускулистые трубки, открывающиеся на коксах третьей
пары ходильных ног (только у рода Parapagurus имеется непарное вывод-
ное отверстие на коксе левой стороны). На нижней поверхности абдомена
у самки имеются еще так наз. „цементные" железы, помещающиеся у осно-
вания плеопод, слизистые выделения которых предназначаются для при-
крепления яиц к плеоподам; возможно, что в этой же слизи происходит
оплодотворение яиц, причем слизь, вероятно, .действует растворяющим
образом на оболочки сперматофоров. Весь процесс можно представить
следующим образом: самец помещает сперматофоры в пространство между
колюмеллярным мускулом и телом рамки, в которое открываются яйце-
воды, когда абдомен самки согнут; самка подгибает абдомен, наполняет
упомянутое пространство слизью и откладывает яйца; движениями плео-
под яйца смешиваются со спермой и оплодотворенные яйца при помощи
слизи прикрепляются к плеоподам второго, третьего и четвертого сегмен-
тов. Весь процесс может, вероятно, происходить внутри раковины. По
наблюдению Brandes (1897) над Pagurus prideauxi (Leach) сперматофоры
прикрепляются не к самке, а к раковине, в которой обитает последняя,
во время копуляции самец своей левой клешненосной ногой удерживает
самку за палец ее правой второй перейоподы, причем движение редуци-
рованных четвертых и пятых перейопод облегчает поступление спермато-
форов и откладывание их в раковину. Самки некоторых родов (Paguristes,
Paguropsis) имеют на абдомене с левой стороны кожистую складку, кото-
рая нависает над прикрепленными к плеоподам яйцами и тем самым
защищает их. О наружном половом диморфизме уже упоминалось при
описании плеопод; кроме того, имеет место разное положение наружных
отверстий 'Половых протоков. Иногда у самцов правая клешня бывает
крупнее, чем таковая у самок.
Постэмбриональное развитие у Paguridae протекает следующим
образом; у Paguridae можно различить б форм личинок, которые вкратце
характеризуются следующими признаками: 1) первая zoea: первые и вторые
Фауна СССР, Макаров 19
146
PAGURIDAE
максиллярные ножки развиты и их экзоподиты несут четыре перистых
щетинки. Грудных конечностей еще нет. Можно различить только 6 абдо-
минальных сегментов, шестой сегмент сливается с тельсоном; 2) вторая
zoea: имеется три максиллярных ножки, появляются зачатки грудных
конечностей. Экзоподиты максиллярных ножек несут шесть щетинок:
имеются зачатки уропод; 3) третья zoea: экзоподиты максиллярных ножек
несут 7—8 щетинок.
Фиг. 62. Стадия glaucothoe Ра-
guristes oculaius (Herbst); абдо-
мен еще сегментированный и
симметричный. (Из Balss, 1913,
по Issel, 1910).
Шестой сегмент абдомена отчетливый. Имеются
уроподы. На конечностях появляются зачатки
жабр; 4) четвертая zoea (или metazoea):
грудные конечности явственные; пятая пе-
рейопода, подогнутая под тело, является
клешненосной; первые перейоподы неравные.
Имеются зачатки плеопод; 5) стадия glau-
cothoe (фиг. 62), соответствующая стадии
megalopa у Brachyura, является характерной
стадией развития Paguridae и долгое время
считалась особым видом животного. Кара-
пакс уже такой формы, как у взрослого
животного, но бранхиостегит не сгибается
вниз под прямым углом к головному щиту.
Четвертые и пятые перейоподы редуциро-
ваны. Абдомен семичленистый. Плеоподы
симметрично развиты на обеих сторонах.
Правая уропода развита слабее левой.
Появляются статоцисты. В начале этой ста-
дии личинка ведет свободноплавающий образ
жизни, держась днем у дна моря, а ночью
поднимаясь к поверхности, где легко может
быть улавливаема планктонными сетками.
Затем наступает переход к ползающему
образу жизни, когда впервые животное начи-
нает отыскивать убежище. Стадия обычно,
продолжается 4—5 дней и за этот период,
печень, нефросак и гонады перемещаются
в абдомен, плеоподы правой стороны исчезают, а левой дегенерируют.
Анатомические изменения, как замечает Thompson, появляющиеся в тече-
ние стадии glaucothoe, не зависят от наличия, отсутствия или формы
раковины; однако, при отсутствии раковины продолжительность стадии
увеличивается до 6—8 дней, а смертность резко возрастает. Встречаются
исключения в виде сохранения рудиментарных плеопод на правой сто-
роне, хотя типично в этод период придатки отсутствуют на этой стороне;
6) юношеская фаза представляет собой вполне типичное взрослое живот-
ное. Органы полностью развиты и по плеоподам можно точно распознать
пол животного. Thompson считает, что половая зрелость достигается
ХАРАКТЕРИСТИКА СЕМЕЙСТВА 147
по прошествии года или более после линьки от стадии glaucothoe
до стадии молодого животного.
Webb (1921) в своей работе по личинкам Decapoda Плимутской
гавани следующим образом характеризует личинок встречающихся там
Paguridae-. рострум длинный и остроконечный, простирающийся за антен-
нулы. Г лаза крупные, овальные. Карапакс относительно широкий и короткий,
выемчатый сзади и оканчивающийся на задне-боковом углу острым высту-
пом, без каймы шипов с каждой стороны (которая имеется у личинок
Galatheidae). Тельсон грубой лопаточкообразной формы, с обрывисто
кончающимся задним краем. На четвертой стадии имеется три пары
щетинистых экзоподитов (1—3 максиллярных ножки). Автор различает
только четыре личиночных стадии (четвертая стадия glaucothoe), которые
отличаются следующими чертами: 1-я стадия: каудальная пластинка
простая, несущая 12 шипов; уроподы отсутствуют. 2-я стадия: каудальная
пластинка простая, несущая 14 шипов; уроподы отсутствуют. 3-я стадия:
тельсон и наружные уроподы хорошо развиты и вооружены шипами;
внутренние уроподы представлены маленькими, лишенными шипов отрост-
ками у основания наружной пары; грудные ноги умеренно развитые.
4-я стадия: тельсон и уроподы, как на третьей стадии; грудные ноги
развиты сильнее, они простираются назад, покрывая все, кроме двух
последних пар плеопод. Описывая личинку Pagurus bernhardus, автор-
говорит, что она почти прозрачная, с красно-желтым хроматофором на
спине; ротовые части с темномалиновыми пятнами; имеется заметный
оранжевый хроматофор (наиболее заметный на последней личиночной
стадии) на развивающейся клешне первой ноги. Глаза темные, с светлым
желтым отблеском.
Следует отметить полное отсутствие монографических работ по
личинкам Paguridae (что, пожалуй, можно отнести и ко всем Decapoda).
Почти в каждой работе по личинкам видовые, а подчас и родовые опре-
деления даются провизорно. Тот же Webb отмечает, что надежно были
определены только личинки Pagurus bernhardus и Р. prideauxi. М. V. Lebour
(1934), давая подробные описания трех личинок Paguridae, затрудняется
в точном отнесении их к определенному роду (возможно, Clibanarius)..
Таким образом, эта область еще очень мало изучена и ждет своих иссле-
дователей.
Жилище раков-отшельников. Стереотропизм особенно сильно раз-
вит у Paguridae, которые стремятся укрыть свой мягкий абдомен в каком
нибудь полом предмете. Чаще всего для этой цели служат спирально-
закрученные раковины Gastropoda-, затем употребляются также трубки:
различных червей, губки, кораллы, трубки дерева или бамбука и проч.;:
в экспериментальных условиях, будучи лишены раковины, животные
пользуются подброшенными в аквариум стеклянными трубками. Интересен:
способ, которым раки-отшельники овладевают раковинами. Swammerdam,,
один из ранних исследователей по анатомии Paguridae, стремился дока-
10*
148
PAGURIDAE
зать, что раковина выделяется самим раком-отшельником. Bell придержи-
вался того взгляда, что рак-отшельник нападает на живого моллюска,
выедает его и затем использует его раковину в качестве жилища; этот
взгляд как будто бы подтверждается наблюдениями рыбаков, которые
ловили раков в момент выедания ими моллюсков. Jackson, однако, сомне-
вается в способности рака-отшельника самостоятельно одолеть живого
моллюска; поэтому, не отрицая целиком взгляда Bell, он высказывает
предположение, что более жесткие части моллюска (нога и голова)
поедаются первоначально рыбами, подкрепляя свое предположение фактом
частого нахождения operculum моллюсков в желудках трески; оставшиеся
мягкйе части выедаются уже отшельником, который и занимает затем
раковину. Кроме того, Jackson не отрицает возможности занятия и уже
пустой раковины, что, повидимому, чаще всего и имеет место. Раки-
отшельники, содержавшиеся без какого-либо прикрытия, в конце-концов
хирели и погибали. Taylor метко охарактеризовал беспомощность отшель-
ника, лишенного раковины, сравнив его с купальщиком, у которого укра-
дена одежда. Рака отшельника довольно трудно вытащить из его раковины;
помимо уропод он удерживается в раковине, растопыривая две последние
пары ходильных ног; даже если просверлить раковину и раздражать
абдомен, то отшельник еще плотнее йрижимается к раковине, но не поки-
дает ее. Крышкообразно расширенная рука одной или обеих клешненосных
ног часто служит для закрывания устья раковины; у рода Cancellus для
этой же цели служат и расширенные ходильные ноги. Наличие раковины
не сильно обременяет рака в его движениях; так Jackson наблюдал отшель-
ников, довольно свободно карабкавшихся по крутой поверхности скалы.
Аэрация внутри раковины осуществляется движениями плеопод; на доста-
точность аэрации указывает факт нахождения Anomia, Amphipoda и других
мелких животных в самых последних завитках раковин, обитаемых отшель-
никами. Gray приписывал ракам-отшельникам способность растворять
занимаемую ими раковину; однако трудно допустить такой факт — автор,
повидимому, имел дело со старыми, уже от времени разрушившимися рако-
винами. Относительно выбора определенного вида раковины не суще-
ствует детальных наблюдений, но, повидимому, такая избирательная спо-
собность и вообще не имеет места—животное пользуется тем видом,
который имеется в данной области, сообразуясь только с размерами рако-
вины. Своеобразным укрытием обладает Paguropsis typica Hendersson,
который держит над собой в виде капюшона актинию из рода Mammilifera,
для каковой цели особенно преобразованы его уроподы и последние
перейоподы. Porcellanopagurus держит над собой одну из половинок
двустворчатой раковины и, таким образом, целиком закрывает свое тело.
Симбиозы. Симбиозы раков-отшельников являются наиболее частым
случаем симбиозов среди Decapoda. Классическим является симбиоз
отшельника с актиниями, которые обычно сидят на раковине, причем один
и тот же вид актинии может сожительствовать с различными видами
фиг. 63. Pagurus pubescens Kroyer. Момент перемены раковины. Ориг. рис. с натуры худ. Н. Кондакова.
I
I
150
PAGURIDAE
Paguridae, или же данный вид актинии встречается только с определенным
видом отшельника. Так, напр., Adamsia palliata Bohadsch сожительствует
только с Pagurus prideauxi (Leach), Epizoanthus sagamiensis Pax встре-
чается только c Paguristes palythophilus Ortmann, a Epizoanthus paguriphi-
lus Verrill почти исключительно c Parapagurus pilosimanus Smith. При
перемене раковины многие отшельники берут с собой „свою" актинию,
пересаживая ее клешненосными ногами на вновь занимаемую раковину
(наблюдения ВгипеЩ на Dardanus arrosor Herbst). Pagurus bernhardus
и P. excavatus часто встречаются c Sagartia parasitica. Сюда же можно
отнести Sagartia paguri Verrill, которую Diogenes edwardsi (de Haan)
носит на руке своей левой клешненосной ноги. Интересно отметить, что
рак-отшельник приобретает иммунитет к ядам и стрекательным капсулам
актиний, которые обычно сильно действуют на других Decapoda; опыты
Cantacuzene и Cosmovici (1925) показали, что нормально смертельная
доза яда актинии, впрыснутая крабу Carcinus maenas вместе с кровью
Pagurus prideauxi не вызывает смерти краба.
Губки часто поселяются на раковинах, обитаемых отшельниками; из
них упомянем Suberites domuncula Nardo в симбиозе с Pagurus bernhar-
dus, Р. pubescens, Р. pectinatus, Р. undosus и Paguristes oculatus; затем
Hymeniacidon suberea c Pagurus bernhardus и P. pubescens и, наконец,
Ficulina ficus Esper c Paguristes oculatus, Pagurus pubescens и P. bernhar-
dus. Интересно отметить, что иногда губка полностью растворяет и вса-
сывает раковину, так что рак-отшельник остается окруженным непосред-
ственно губкой; на изломе таких губок можно проследить, что полость,
занимаемая отшельником, в точности соответствует завиткам существо-
вавшей ранее раковины. О симбиозе губок с раками-отшельниками имеется
подробная сводка Arndt (1933).
Из гидроидов Hydractinia echinata Flemming и Podocoryne cor-
nea M. Sars встречаются на раковинах Pagurus bernhardus и Р. pu-
bescens; Hydrissa sodalis (Stimpson) на раковине Pagurus constans
(Stimpson).
Из мшанок Conopeum commensale Kirkp. et Metz, обрастает раковины
P. granulimanus Miers, a Cellepora senegambiensis Carter раковины Dio-
genes pugilator Roux.
Общеизвестным является симбиоз полихеты Nereilepas fucata inqui-
lina Wiren c Pagurus prideauxi’, отшельник не трогает „своего" червя
(хотя пожирает других) и берет его с собой при переходе в другую рако-
вину; возможно, что червь поедает эктопаразитов на задней части тела
рака-отшельника. В Калифорнии и в Японском море подобным же образом
сожительствует Nereis cyclurus Harrington с Pagurus ochotensis. Встреча-
ются также планарии, напр., Emprostopharynx opisthoporus Bock и Eupro-
sthiostomum adhaerens Bock c Pagurus californiensis или Stylochus zebra
Verrill с P. pollicaris. Более подробное описание симбиозов у раков-отшель-
ников можно найти в работе Н. Balss (1924).
ХАРАКТЕРИСТИКА СЕМЕЙСТВА
151
Паразиты раков-отшельников. Из споровиков некоторые виды кок-
цидий встречаются в кишечнике Pagurus. Наибольшее число эктопарази-
тов дают ракообразные. Копепода Sunaristes paguri Hesse (из сем. Longi-
pediidae), как правило, ' живет только на раковинах, обитаемых отшельни-
ками. Из усоногих на абдомене Pagurus и Anapagurus часто встречаются
роды Peltogaster Rathke, Clistosaccus Lilljeborg и Apeltes Lilljeborg1. Име-
Фиг. 64. Pagurus bernhardus (L.) c Sagartia parasitica. Ориг. рис. с натуры
худ. Н. Кондакова.
ются наблюдения, согласно которым наступают определенные анатомиче-
ские изменения в зависимости от заражения тем или иным видом Cirripe-
dia', так, напр., если у Anapagurus chiroacanthus (Lilljeborg) паразитом
являлся Peltogaster sulcatus, то плеоподы самца принимали вид плеопод
самки, т. е. имели длинные эндоподиты; если же паразитом являлся Pelto-
gaster paguri, то плеоподы самца почти не изменялись, но penis редуциро-
вался; кроме того, появлялась паразитарная кастрация, выражавшаяся
в дегенерации гонад. Из Isopoda упомянем род Bopyrus, встречающийся
в жаберной полости отшельников, и роды Athelges, Phryxus и Pseudione,
укрепляющиеся на абдомене раков-отшельников.
152
PA GURIDAE
ЗООГЕОГРАФИЧЕСКИЙ ОЧЕРК
В большей своей части представители семейства Paguridae являются
типичными обитателями литорали и сублиторали тропической области.
Бореальная область также имеет сврих представителей этого семейства»
но число их значительно падает и, наконец, Арктика совершенно не имеет
видов, свойственных исключительно ей, хотя единичные бореальные,
в большей или меньшей степени эвритермичные, виды поднимаются до
сравнительно высоких широт Арктики. В настоящее время известно при-
мерно 450 видов сем. Paguridae. Если общее число видов всех Decapoda
принять приблизительно за 6000 (это число, безусловно, несколько выше
в настоящий момент), то отшельники составят, таким образом, 7.5% этого
общего числа. В водах СССР зарегистрировано 25 видов отшельников,
что составляет 5.5% их общего числа.
При составлении настоящей работы пришлось иметь дело со следую-
щими акваториями: Черное море; Белое море; Баренцово море, на запад,
до линии Нордкап — остров Медвежий—Шпицберген; Японское море
целиком, следовательно, включая западное побережье Японии, Сангарский
и Лаперузов проливы; Охотское море с Курильской грядой;, восточное
побережье Камчатки; все Берингово море с Алеутской грядой до о-ва
Кадьяк включительно; Чукотское море на восток до меридиана мыса
Барроу.
В соответствии с этими границами число видов, рассматриваемых
в настоящей работе, возрастает до 42; они принадлежат к 8 родам.
Перейдем к рассмотрению распространения раков-отшельников по>
отдельным акваториям.
В Черном море мы находим только два вида отшельников—Dioge-
nes varians и Clibanarius misanthropus, типичных представителей среди-
земноморской фауны; несколько, иные условия существования в Черном
море (главным образом, повидимому, низкая соленость, составляющая
17—18%, а в отдельных районах падающая до 10%) сказалась в нан-
низме этих форм, резко бросающемся в глаза при сравнении их с. экзем-
плярами из Средиземного моря. Clibanarius misanthropus встречается
значительно реже, чем Diogenes varians.
Белое и Баренцово моря имеют один вид отшельников — Pagurus
pubescens. Другой вид, Р. bernhardus, сравнительно редко встречается на
Западном Мурмане, где он имеет восточную границу своего распростра-
нения. По своему распространению Р. pubescens занимает исключительное
положение среди отшельников наших вод, являясь типичным амфибо-
реальным видом.
Моря, омывающие северное побережье Сибири, не имеют в составе
своей фауны раков-отшельников, если не считать спорадических нахо-
ждений Pagurus pubescens в Карском море; отшельники вновь появляются
ХАРАКТЕРИСТИКА СЕМЕЙСТВА 153^
только в дальневосточном секторе Арктики от устья р. Колымы по
направлению на восток.
Наиболее богатой является фауна отшельников наших дальневосточ-
ных морей, составляя 84% видов, зарегистрированных в водах СССР.
По характеру своего распространения всю массу этих дальневосточных
видов можно разбить на 4 основных группы.
Первая группа: широко-распространенные виды, свойственные
всем трем дальневосточным морям (Японское, Охотское и Берингово)
и Чукотскому морю; сюда войдут Pagurus pubescens, Р. rathbuni
и Р. capillatus.
Вторая группа: виды, свойственные почти исключительно
только Японскому морю, если не считать выхода их во внутреннее
море Японии и на юго-восточное побережье: Pagurus gracilipes, Р. dubius,.
Р. anomalus, Р. brachiomastus, Р. lanuginosus, Р. pectinatus, Diogenes,
penicillatus.
Третья группа: виды, встречающиеся в Японском и Охотском
морях, а также у западного побережья Северной Америки, причем связь
осуществляется через Алеутскую гряду почти без проникновения, за одним
исключением, в Берингово море: Pagurus hirsutiusculus, Р. middendorffi,
Р. undosus, Р. ochotensis и Р. gilli.
Четвертая группа: виды с прерывчатым распространением*,
встречающиеся в Японском море и у западного побережья Северной
Америки и отсутствующие в промежуточных областях: Pagurus samuelis,
Orthopagurus minimus, О. schmitti.
Японское море. В пределах Японского моря из представителей
первой группы только Pagurus pubescens известен также из Японии;
остальные два вида распространяются только по нашему побережью,.
достигая здесь южной границы своего распространения. Все представи-
тели второй группы известны также и из Японии, причем, за исключением
Pagurus ' anomalus и Р. brachiomastus, в основном они распространены
в южной части Японского моря и на юго-восточном побережье Японии,,
находя у нашего побережья северные границы своего распространения;
наоборот, Pagurus anomalus а Р. brachiomastus, повидимому, приурочены
к нашим частям Японского моря и в южной Японии достигают южного
предела своего распространения. Из третьей группы Pagurus undosus
известен только для наших вэд и неизвестен из Японии. Из четвертой
группы Pagurus samuelis известен также и из Японии, тогда как оба вида
Orthopagurus отмечены только для нашего побережья. Особую группу
образуют виды эндемичные, главным образом, для внутреннего моря
Японии и ее восточного побережья и частично заходящие в собст-
венно Японское море, не распространяясь на север дальше Сангар-
ского пролива, таковы: Pagurus obtusifrons л Р. constans, Clibanarius-
japonicus и Pagurisles digitalis. Наконец, идет группа тропических видов,.,
находящих северный предел своего распространения у берегов южной
154
PAGURIDAE
Японии и сравнительно редко встречающихся в самом Японском море:
Spiropagurus spiriger, Diogenes edwardsii, Dardanus arrosor и Paguristes
setosus. К сожалению, имеется очень мало данных по фауне западного
побережья Японии от Цусимского пролива примерно до Ниигаты —
одного из интереснейших районов, лежащего на главном пути про-
никновения более южных форм в Японское море; при большей его изу-
ченности видовой состав отшельников Японского моря несомненно
^возрастет.
Таков сравнительно пестрый состав фауны отшельников Японского
моря. Как мы увидим ниже, такая же пестрота, хотя и в меньшей степени,
характерна и для фауны отшельников Берингова моря, что обусловли-
вается там примесью представителей калифорнийской фауны. Охотское
и Чукотское моря, напротив, имеют очень однородную по своему составу
фауну.
Охотское море. В состав фауны отшельников Охотского моря цели-
ком входят первая и третья из приведенных выше основных групп. Кроме
того, обычной формой является Pagurus splendescens, который характерен
также для Берингова И Чукотского морей и который не выходит за пре-
делы Охотского моря, замещаясь в Японском море близко родственным
Р. anomalus. Не совсем ясен вопрос о распространении Pagurus gilli’, однако,
на основании, правда, очень скудного материала, я склонен думать, что
этот вид может быть включен в третью группу. До настоящего времени
он не известен еще из собственно Охотского моря, но весьма возможно,
что область распространения его в виде дуги захватывает и Курильскую
гряду. К восточному побережью Камчатки (до широты Командорских
островов) целиком приложимо сказанное о составе фауны отшельников
Охотского моря.
Берингово море. В состав фауны входят первая и третья группы
(главным образом, по Алеутской гряде) и Pagurus splendescens. Кроме
' того, юго-восточный угол Берингова моря (Уналашка—Прибыловы о-ва—
о-в Матвея — о-в Нунивок) имеет фауну отшельников, отличающуюся от
таковой остальной, большей части моря; этот Комплекс видов образуют
представители калифорнийской фауны, находящие в рассматриваемом
районе северный предел своего распространения (на север не дальше
широты о-ва Нунивок); таковы следующие виды Pagurus: beringanus,
tanneri, granosimanus, armatus и setosus, причем три последние вида
не идут на север дальше Алеутской гряды (Уналашка и Кадьяк). Следую-
щую группу образуют виды также западного побережья Северной Аме-
рики, но идущие на юг не дальше Британской Колумбии, Вашингтона или
Орегона; сюда относятся следующие виды рода Pagurus: dalli, kennerlyi,
ochotensis subsp. aleuticus,cavimanus (?), tenuimanus и cornutus- два послед-
них вида идут вдоль всей Алеутской гряды до Командорских о-вов вклю-
чительно, причем Р. cornutus идет, кроме того, вдоль нашего побережья
на север до широты мыса Наварина. Parapagurus mertensii не встречается
ХАРАКТЕРИСТИКА СЕМЕЙСТВА
155
в Беринговом море, являясь эндемичным калифорнийским, видом; в настоя-
щей работе он приводится в виду ошибки, вкравшейся в синонимию
этого вида.
Из всего сказанного видно, что Алеутская гряда является мостом,
через который осуществляется, в отношении раков-отшельников, правда,
незначительная, связь между фаунами обеих сторон северной части
Тихого океана. При этом можно проследить, как различные стадии
существования этой связи, так отчасти и направление обмена. Большая
часть калифорнийских видов доходит пока до восточного конца Алеут-
ской гряды и здесь останавливается. Pagurus tenuimanus и Р. cornutus
осуществили переход через Алеутскую гряду, причем последний повер-
нул затем на север. Из третьей группы видов Pagurus hirsutiusculus
и Р. middendorffi дают пример наиболее полной связи, существующей
в настоящий момент между обеими сторонами северного Тихого океана.
Относительно Pagurus undosus можно с уверенностью сказать, что рас-
пространение шло в обратном направлении; центром, из которого нача-
лось расселение этого вида, можно считать Охотское море, откуда вид
частично проник в Японское море (только по нашему побережью), а с дру-
гой стороны продвинулся вдоль восточного побережья Камчатки до Але-
утской гряды, где образовал две ветви: одна идет вдоль нашего побе-
режья Берингова моря до широты мыса Чаплина, другая заняла всю Алеут-
скую гряду, не распространяясь пока по западному побережью Северной
Америки. Распространение Pagurus ochotensis шло, повидимому, тем же
путем, причем намечается разрыв этой сплошной дуги; об этом можно
судить по сравнительно редкой встречаемости данного вида у восточного
побережья Камчатки, включая и западную часть Алеутской гряды (Коман-
дорские о-ва). Если принять такое направление распространения Pagurus
ochotensis, то становится вполне ясным положение Р. ochotensis subsp.
akuticus, как подвида, выделившегося из основного вида в восточной
половине Алеутской гряды, т. е. в наиболее удаленном от центра распро-
странения конце дуги. Та же самая картина получится и в отношении
Pagurus gilli, если только окажется, что японский Р. cavimanus идентичен
с Р. gilli', тогда мы будем иметь дугообразное распространение Р. cavima-
nus (Miers) (=/’. gilli) с подвидом Р. cavimanus subsp. munitus (Benedict)
в восточной половине Алеутской гряды; Наконец, четвертая группа видов
дает пример полного, повидимому, давно происшедшего разрыва связи,
осуществлявшейся, вероятно, когда то тем же самым путем через Алеут-
скую гряду.1
1 Нахождение Pagurus samuelis в Авачинской бухте, согласно данным Н. Balss (1913),
можно было бы истолковать, как реликт, сохранившийся от этой, когда то сплошной цепи
распространения. Однако, имея сравнительно обширный материал по Авачинской бухте,
я склонен думать, что это определение ошибочно и что оба приводимые автором
экземпляра следует отнести к Р. hirsutiusculus, который по внешнему виду очень похож
на Р. samuelis.
156
PA GURIDAE
Чукотское море. В фауне отшельников Чукотского моря мы находим
также первую группу {Pagurus pubescens, Р. rathbuni, Р. capillatus) и Pagu-
rus splendescens. В виду малой изученности района пока трудно говорить
о северной, восточной и западной границах их распространения.
В заключение упомянем широко распространненного Parapagurus
pilosimanus, который вошел в состав нашей фауны отшельников, благо-
даря нахождению его у восточного побережья Камчатки.
СПЕЦИАЛЬНАЯ ЧАСТЬ
7. Сем. PAGURIDAE
Paguridae Dana, 1852: 435; Stimpson, 1858 : 232; Henderson, 1888: 52; Stebbins, 1893:
159; Ortmann, 1892:269; 1901:1145; Alcock, 1901: 215; 1905 :21. — Parapaguridae Smith,.
1883 : 20; Henderson, 1888 : 85; Stebbing-, 1893:166.
Рострум маленький, редуцированный или совершенно отсутствующий.
Карапакс обычно с твердой передней и мягкой задней частями. Глазной
сегмент более или менее свободно лежащий. Скафоцериты хорошо раз-
витые, шипообразные. Абдомен асимметричный, мягкий, прямой или, чаще
всего, спирально закрученный, обнаруживающий едва заметную сегмен-
тацию. Абдоминальные придатки развиты частью только на левой стороне,
парные придатки — уроподы — приспособлены для удерживания животного
в раковине, имеют бородавчатые поверхности. Четвертая и пятая пары
перейопод маленькие, четвертая пара обычно снабжена ложными клешнями^.
Жабры большею частью типа филлобранхий, наибольшее число их 14,.
наименьшее —11.
Семейство Paguridae распадается на два подсемейства.1
ОПРЕДЕЛИТЕЛЬНАЯ ТАБЛИЦА РОДОВ
1 (8). Наружные максиллярные ножки удалены друг от друга у своих,
оснований (Подсем. Pagurinae).
2 (7). Абдомен согнутый и спирально закрученный. Уроподы несимме-
тричные, левая развита значительно сильнее правой.
3 (4). Не имеется никаких половых придатков на коксальных члениках
побледней пары ног или на первых двух абдоминальных сегментах .
........................................5. Pagurus Fabricius.,
1 Ortmann (1892) выделил еще подсемейство Diogeninae для рода Diogenes, как един-
ственного рода, обладающего глазным рострумом. Однако, по всем остальным признакам
род Diogenes настолько близок к представителям подсемейства Pagurinae, что последую-
щими авторами он помещался в это подсемейство.
1. DIO GENES
157
4 (3). Половые придатки имеются.
5 (6). Имеются парные придатки на первом и втором абдоминальных сег-
ментах у самца. Правая клешненосная нога развита значительно
сильнее левой.................................6. Parapagurus Smith.
6 (5). Имеется непарный, спирально закрученный половой придаток на
коксальном членике левой четвертой ходильной ноги у самца. Клеш-
неносные ноги почти равные, правая нога, если и развита сильнее,
то очень незначительно.....................8. Spiropagurus Stimpson.
7 (2). Абдомен согнутый, но нисколько не закрученный спирально. Уро-
поды симметричные или левая развита очень незначительно сильнее
правой.....................................7. Orthopagurus Stevens.
•8 (1). Наружные максиллярные ножки сближены у своих оснований.
(Подсем. Paguristinae).
9 (12). Левая клешненосная нога развита сильнее правой; пальцы дви-
гаются в вертикальной плоскости.
10 (И). Лобный край карапакса посередине с тупым зубцом. Между глаз-
ными стебельками имеется глазной рострум ... 1. Diogenes Dana.
11 (10). Лобный край карапакса посредине без зубца. Глазной рострум
отсутствует.....................................4. Dardanus Paulson.
12 (9). Клешненосные ноги равной или почти равной длины, или же редко
левая незначительно длиннее правой. Пальцы двигаются в горизон-
тальной плоскости.
13 (14). Четвертая пара перейопод с ложными клешнями. Абдомен без
парных половых придатков на первом сегменте у обоих полов . .
. ..........................................2. Clibanarius Dana.
14 (13). Четвертая пара перейопод простая, без ложных клешней. Абдомен
с парными половыми придатками на первом сегменте у обоих полов
............................................3. Paguristes Dana.
1. Подсем. PAGURISTINAE, nom. nov.
Pagurinae Ortmann, 1892 : 269. — Diogeninae Ortmann, ibid.: 270.
Наружные максиллярные ножки сближены у основания; клешненосные
моги равные или же левая нога длиннее правой.
1. Род DIOGENES DANA
Diogenes Dana, 1852 :438; Stimpson, 1858 : 232; 1907: 201; Heller, 1863 :169; Henderson,
1888:53; Stebbing, 1893:160; Alcock, 1905:59; Balss, 1913 :44; Terao, 1913:362.
Тип рода: Diogenes miles (Dana).
Лобный край посредине с тупым зубцом. Между глазными стебель-
ками имеется подвижный, острый отросток — глазной рострум. Скафоце-
риты широкие у основания и часто глубоко раздвоенные. Жгуты антенн
158
PAGURIDAE
с ресничками. Наружные максиллярные ножки сближены у своих основа-
ний, их базальные членики широкие. Клешненосные ноги неодинаковые,
левая развита значительно сильнее правой, имеет широкую руку с пока-
той наружной поверхностью; рука косо причленяется к карпусу; пальцы
двигаются в вертикальной плоскости и имеют известковые кончики..
Пальцы ходильных ног длинные. Четвертая пара перейопод с ложными
клешнями.
Данный род характерен для тропической индо-пацифической области,,
но отдельные виды встречаются в восточной половине Атлантики, в Сре-
диземном море, а также в Южной Японии. В наших водах род представлен
двумя видами (D. varians и D. penicillatus'), D. edwardsii приводится как
вид, зарегистрированный для западного побережья Японии (Ниигата).
ОПРЕДЕЛИТЕЛЬНАЯ ТАБЛИЦА ВИДОВ
1 (2). Глазной рострум очень маленький, едва заметный для невооружен-
ного глаза. На наружной поверхности левой руки имеется овальной
формы участок с приподнятыми краями, на котором. обычно поме-
щается актиния ......................... 1. Di edwardsii (de Haan).
2 (1). Глазной рострум крупнее, хотя и не достигает концов глазных:
чешуек. Овальный участок на наружной поверхности левой руки:
отсутствует.
3 (4). Скафоцериты с 6—7 шипами на внутреннем краю. Ходильные ноги:
левой стороны превосходят левую клешненосную ногу..................
............................... * 2. D. penicillatus Stimpson.
4 (3). Скафоцериты с 4—5 шипами на внутреннем краю. Ходильные ноги:
левой стороны достигают конца левой клешненосной ноги..............
..................................... *3. Di varians (Costa).
1. Diogenes edwardsii (de Haan). (Табл. 1, фиг. 3).
Pagurus edwardsii de Haan, 1850 : 211, t. 50, fig'. 1. — Diogenes edwardsii Stimpson,
1858:246; 1907: 202, t. 24, fig. 1; Ortmann, 1892 :295; Rathbun, 1902 : 36; Alcock, 1905 :165?
Balss, 1913 :44; Terao, 1913 : 362; Yokoya, 1933 : 77.
Глазной рострум маленький и очень узкий, едва заметный для нево-
оруженного глаза. Скафоцериты короткие, треугольные, с мелкими шипи-
ками. Пальцы ходильных ног длинные и тонкие, в 1х/2 раза длиннее соот-
ветствующих проподусов. Наружная поверхность левой руки имеет гладкий,
овальной формы, участок, который занимает примерно 2/3 поверхности,
и частично окружен приподнятыми краями; на этом участке почти всегда,
находится актиния (Sagartia paguri Verrill); при фиксации актиния может
отваливаться; на раковине часто находится по несколько экземпляров этого
же вида актинии. Окраска живых экземпляров: карапакс светлокрасный;:
ноги с поперечными светлыми красновато-коричневыми полосами; левая
рука белая; абдомен желтый с светлокрасными боками
7. DIOGENES
159
Ortmann (1892) считает, что Pagurus spinifrons de Haan являются
молодыми экземплярами D. edwardsii.
Распространение. Китайское море (Гонконг); Южная Япония;.
Японское море (префектура Ниигата); на глубинах до 97 м.
* 2. Diogenes penicillatus Stimpson.
Diogenes penicillatus Stimpson, 1858:245; 1907:200; Terao, 1913:364; Yokoya,
1933:77.
Передняя часть карапакса такой же ширины, как и длины, глад-,
кая посредине, к бокам шероховатая, с неправильными чешуеобразными
выступами, края которых зубчатые и усажены мелкими щетинками. Сред-
ний выступ лба короткий, тупой; боковые выступы острые, выдаются
вперед дальше среднего выступа. Глазные стебельки умеренной длины,,
немного больше половины длины передней части карапакса, цилиндри-
ческой формы. Глазные чешуйки широкие, с прямыми внутренними и полу-
круглыми наружными краями, с 2—3 шипами на вершине. Глазной рострум
узкий, острый, не достигает концов верхушечных шипов глазных чешуек...
Скафоцериты короче глазных стебельков, продолговато-треугольные,,
с б—7 сильными шипами на верхне-внутреннем краю. Наружный выступ
базального членика антеннальной ножки с сильным шипом, внутренний
выступ с более коротким шипом. Жгуты антенн короткие, опушены,
снизу длинными волосками. Клешненосные ноги неодинаковые, левая
значительно больше правой. Мерус левой клешненосной ноги трехгран—
ный, гранулированный, более крупные гранулы занимают внутренний
край нижней поверхности; верхнее ребро покрыто довольно длинными,
волосками. Карпус неправильной треугольной формы, с прямым внутрен-
ним и угловатым наружным краем; наружная половина верхней поверх-
ности с крупными гранулами и отдельными волосками; внутренний край
усажен 10—12 наклоненными вперед шипами; нижняя поверхность с пло-
скими гранулами и почти без волос. Наружный край руки почти прямой;,
рука косая, с 2 параллельными рядами мелких зубцов или шипов сверху: один
ряд на краю, другой тотчас же ниже его на наружной поверхности; наруж-
ная поверхность руки густо покрыта кисточками нежных шелковистых,
волос; у основания ладони имеется шиповатый поперечный гребень, кото-
рый отделяет волосатую поверхность от гладкой, голой борозды у сочле-
нения с карпусом. Нижняя поверхность покрыта более плоскими грану-
лами. Неподвижный палец короткий, широко треугольный; подвижный
палец изогнутый, с 3 рядами гранул на верхней поверхности; хватательный
край неподвижного пальца с одним белым бугром; кончики пальцев изве-
стковые. Правая клешненосная нога простирается немного дальше дисталь-
ного конца карпуса левой ноги. Мерус с длинными волосами на верхнем
ребре. Карпус короткий, с 2 рядами шипов на верхней поверхности, воло-
сатый. Рука овальной формы, с длинными, изогнутыми пальцами; верхняя
поверхность покрыта шиповатыми гранулами и довольно густыми длинными.
160
PAGURIDAh
волосами; концы пальцев известковые. Ходильные ноги левой стороны
немного превышают левую клешненосную ногу. Карпальные членики
сверху шиповатые; верхние края всех члеников волосатые, боковые сто-
роны почти голые. Пальцы длинные, тонкие, слабо изогнутые, с острыми
роговыми ногтями.
Размеры
Длина передней части карапакса 3.0 ММ
Ширина „ „ ,, . ........ 3.0 я
Длина глазных стебельков 2.0 я
„ меруса правой клешней, ноги 2.5 я
„ карпуса „ ,, „ ........ 2.0 ff
Ширина „ ' „ я >» •..»•••• 1.5 ft
Длина руки „ » » 3.0 я
Ширина „ „ я » 2.0 ff
Длина меруса левой »> я 3.0 ir
„ карпуса „ ,, ,, ........ 3.5 я
Ширина „ „ 2.5 ft
Длина руки „ ff ff 5.0 ff
Ширина „ „ я я 4.0 ft
Длина левой ходильной ноги первой пары 13.0 ff
Распространение. Япония (у восточного побережья о-ва Нип-
'пон, 38° N; восточнее Miyagy-ken; севернее Inuboe-zaki; пролив Bungo),
на глубинах от 33 до 88 м. Из наших вод вид известен только по одному
экземпляру, добытому в бухте Шамара, Уссурийский залив.
* 3. Diogenes varians (Costa). (Табл. I, фиг. 6).
Pagurus varians Costa, 1832—1862:9, t. 2, fig. 2.—Pagurus pugilator Roux, 1828:
t. 14, fig. 3. — Pagurus drenarius Lucas, 33, t. 3, fig. 7a. — Pagurus ponticus Kessler, 1861;
Вагнер, 1864. — Diogenes varians Heller, 1863:170, t. 5, fig. 13—14; Carus, 1884:493;
Czerniavsky, 1884:99; Ortmann, 1892:294; Калишевский, 1906. .
Передняя часть карапакса почти такой же длины, как и ширины.
Передний край с острыми, треугольными боковыми выступами. Боковые
стороны почти параллельные. Глазные чешуйки имеют форму широких
треугольников с прямыми и гладкими внутренними краями и косыми
наружными краями, усаженными шипами, которые уменьшаются в напра-
влении от вершины к основанию. Глазной рострум острый, не достигает
конца верхушечного шипа глазной чешуйки. Глазные стебельки уме-
ренно длинные, цилиндрические. Скафоцериты короткие и широкие,
с 4—5 шипами на внутреннем скошенном краю, считая и длинный верху-
шечный шип. Антенны короткие, достигающие до дистального конца
карпуса левой клешненосной ноги, усажены длинными волосками. Клешне-
носные ноги очень неравные, правая достигает только дистальной трети
карпуса левой ноги. Мерус левой клешненосной ноги с неправильным
рядом коротких шипов на верхне-внутреннем ребре; наружная поверхность
почти гладкая, нижняя гранулирована; весь членик слабо волосатый. Кар-
1. DIOGENES
161
пус с шипами на внутреннем краю, с выпуклой волосатой верхней поверх-
ностью, незаметно переходящей в наружную боковую поверхность; нижняя
поверхность гранулированная и слабо волосатая. Верхняя и нижняя
поверхность руки гранулированная, с редкими волосками; пальцы короткие,
с известковыми кончиками. Правая клешненосная нога очень маленькая
и с более длинными волосами чем левая. Карпус с продольным рядом шипов
на верхней поверхности. Ладонь короткая; подвижный палец изогнутый,
так что при смыкании остается широкая щель; нижняя поверхность ладони
и карпуса гладкая и почти безволосая. Ходильные ноги левой стороны
достигают конца левой клешненосной ноги. Все членикй сплющенные,
карпальные шиповатые на верхнем краю; пальцы довольно длинные, изо-
гнутые, не закручены, негусто волосатые, с острыми роговыми когтями.
Окраска (по Heller’y): окраска тела зеленоватая или красновато-белая
с отдельными более темными красно-коричневыми пятнами на карапаксе
или такими же продольными полосами на некоторых члениках ног. Глаз-
ные стебельки красноватые, антенны светложелтые. Окраска вообще
сильно вариирует. Чернявский (1884) описал из района Сухуми цветовой
вариетет: „головогрудь и ноги грязного серовато-желтоватого цвета,
пятна — красно-коричневые и беловатые; абдомен сверху коричневато-зеле-
новатый, с черными полосками и пятнами, с боков и снизу (как и голово-
грудь) фиолетово-синий. Глаза золотисто-коричневатые, стебельки розо-
вые". Однако, необходимо отметить, что на спиртовом материале просле-
дить все эти оттенки совершенно невозможно.
Размеры
Длина передней части карапакса 4.0 MM
Ширина „ „ 11 4.5 »
Длина глазных стебельков . . 2.0 ft
„ меруса правой клешней, ноги 3.0
„ карпуса „ 2.0
руки „ ,, . . . . • . 3.0
Ширина „ „ „ ... .... 2.0 11
Длина меруса левой 4.0 If
„ карпуса „ ft 4.5 f>
„ руки it 6.0
Ширина „ „ и » 3.0 »
Длина левой ходильной ноги первой пары . , . . 14.0
Экземпляры не достигают крупных размеров; как отметил еще Кали-
шевский (1906), наши черноморские экземпляры вдвое меньше средиземно-
морских.
Распространение. Средиземное и Адриатическое моря; Черное
море. Diogenes varians является обычной формой отшельников нашего
Черноморского побережья; держится на небольших глубинах, преимуще-
ственно на песчаных грунтах; на зимний период, по данным С. А. Зернова,
мигрирует на большие глубины.
Фауна СССР, Макаров 11
162
PAGURIDAE
2. Род CLIBANARIUS DANA
Clibanarius Dana, 1852 : 461; Stimpson, 1858 : 234; 1907 : 208; Heller, 1863 :177; Miers,
1876 : 67; Henderson, 1888 : 60; Milne-Edwards et Bouvier, 1893 :156; Stebbing, 1893 :160;
Ortmann, 1901 :1146; Alcock, 1905 : 40; Balss, 1913 : 40; Terao, 1913 :360. .
Тип рода: Clibanarius depressus Dana.
Средний выступ лба острый. Глазные стебельки длинные и тонкие.
Глазные чешуйки очень маленькие. Скафоцериты короткие. Наружные
максиллярные ножки сближены у своих оснований. Клешненосные ноги
короткие, почти одинаковые по длине и по форме; пальцы клешней дви-
гаются в горизонтальной плоскости и имеют ложкообразно выемчатые,
крупные роговые кончики Четвертая пара перейопод с ложными клешнями.
Виды этого рода распространены большею частью на литорали тро-
пической области. В наших водах род представлен одним видом.
ОПРЕДЕЛИТЕЛЬНАЯ ТАБЛИЦА ВИДОВ
1 (2). Глазные чешуйки с прямыми внутренними краями. Карпус левой
клешненосной ноги без двух рядов шипов на дорзальной поверхности.
Пальцы ходильных ног значительно короче проподусов.................
..................................* 1. С. misanthropus (Risso).
2 (1). Глазные чешуйки с полуовальными внутренними краями. Карпус
левой клешненосной ноги с двумя рядами шипов на дорзальной
поверхности. Пальцы ходильных ног длиннее проподусов...............
.......................................2. С. japonicus Rathbun.
* 1. Clibanarius misanthropus (Risso). (Табл. I, фиг. 7).
Pagurus tubularis Risso, 1815 : 56. — Pagurus misanthropus Risso, 1826 : 40; Roux, 1828^
t. 14, fig. 1; Guerin, 1836:32; Milne-Edwards, 1836:277; 1837:228.—Pagurus rigritarsis
Lucas, 1849:30, t; 3, fig. 4.— Clibanarius misanthropus Heller, 1863:177, t. 5, fig. 16—18;
Carus, 1884: 495; Czerniavsky, 1884 : 97; Ortmann, 1892 : 291.
Поперечная часть цервикальной борозды довольно широкая, вслед-
ствие чего передняя часть карапакса не так сильно суживается по напра-
влению назад. Поверхность карапакса почти голая. Средний выступ лба
острый, треугольный; позади него поднимается короткий продольный:
валик. Лобный край карапакса валикообразно утолщен. Боковые выступы:
лба тупые и значительно менее выдаются вперед, чем средний выступ.
Глазные стебельки очень длинные и тонкие. Глазные чешуйки треуголь-
ные, соприкасающиеся своими прямыми внутренними краями и зазубренные
на вершине и на наружном краю. Скафоцериты сильно волосатые, немного-
превышают четвертый сегмент антеннального стебля, с острыми зубчи-
ками на внутреннем краю. Антеннулы немного длиннее чем глаза;
антенны длиннее клешненосных ног. Клешненосные ноги почти одинако-
вой длины. Мерус простирается немного дальше глаз, сплющенный, трех-
2. CLIBANARIUS
163
гранный; острый верхний край дугообразно изогнут, внутренний край
вооружен несколькими бугроватыми зубцами, наружный край тупой. Карпус
короткий, трехгранный, с группой шипов на дистальном конце верхнего
края. Рука косо покатая наружу, сплющенная изнутри и сильно вздутая
снизу у основания; пальцы короткие, слегка изогнутые, с большими ложко-
образно выемчатыми, черными роговыми кончиками; при смыкании у осно-
вания пальцев остается широкая щель. Поверхность руки и пальцев
покрыта отдельными острыми бугорками. На всех члениках, в особенности
на краях, имеются волоски. Ходильные ноги длиннее клешненосных,",
мерус сплющенный, проподус и палец более закругленные; все членики
волосатые на краях; палец значительно короче проподуса, с острым рого-
вым когтем и желтоватыми шипиками на нижнем краю. Окраска живых
экземпляров: карапакс и клешненосные ноги зеленоватые, с коричневыми
или синеватыми пятнами; шипы на руках синеватые; пальцы клешни
с двумя верхними и двумя нижними коричневато-красными продольными
полосами, кончики черные. Глазные стебельки и антенны красноватые.
Ходильные ноги желтовато-коричневые, пальцы синеватые, с красными
продольными полосами. Абдомен коричневый, на конце красноватый.
У молодых и спиртовых экземпляров окраска ходильных ног более желто-
ватая, на пальцах ясно видны 4 красных и 4 беловатых (первоначально
синих) продольных полосы; шипы и пятна на клешненосных ногах серовато-
белые.
В. Чернявский (1884) выделил в Черном море две формы этого
вида: forma suchumica и forma jaltensis, однако, приводимые им отличия
этих форм от средиземноморского типичного вида столь ничтожны и неопре-
деленны, что как будто бы нет оснований для выделения этих форм.
Экземпляры данного вида не достигают крупных размеров. Держится
обычно близ берега, на незначительных глубинах.
Распространение. Средиземное, Адриатическое и Черное моря.
В Черном море данный вид встречается значительно реже Diogenes varians
(Costa).
2. Clibanarius japonicus Rathbun. (Фиг. 65).
Clibanarius japonicus Rathbun, 1902 : 35, fig. 2—5; Terao, 1913 : 361.
Передняя и боковые части карапакса морщинистые; имеется прибли-
зительно 19 пучков волос, 13 из которых располагаются в виде груше-
образной фигуры. Срединный зубец переднего края выдается более, чем:
боковые зубцы, и вооружен маленьким шипом, который почти скрыт под.
пучком волос; тотчас под краем бокового зубца имеется маленький шип,,
который направлен наружу. Внутренняя часть глазных чешуек полуоваль-
ная и цельная; на конце под краем имеется маленький шип. Антеннулярная;
ножка длиннее глаза; третий сегмент немного длиннее второго, дости-
гая до конца предпоследнего сегмента наружных максиллярных ножек.
Антеннальная ножка не так длинна, как глаз; скафоцериТ тонкий, серпо-
11*
J 64
PAGURIDAE
видный, доходящий до середины последнего сегмента антеннальной ножки.
Клешненосные ноги более неравные, чем это обычно у данного рода.
Левая нога больше; мерус простирается за линию глаз; его нижняя поверх-
ность окаймлена шипами изнутри и снаружи; верхний край с двумя
дистальными и двумя более маленькими субтерминальными шипами. Карпус
больше в длину чем в ширину, имеет два дорзальных ряда шипов; перед-
ний край, шиповатый; наружная поверхность с коротким .рядом шипов
на верхнем дистальном конце. Проподус шиповатый сверху, шипы распо-
ложены приблизительно в 7 неравных рядов; ладонь значительно расши-
Фиг. 65. Clibanarius japonicus Rathbun. А — передняя часть
тела; В—левая клешненосная нога; С — правая клешненосная
нога; D — первая правая ходильная нога с наружной стороны.
(Из Rathbun, 1902).
ряется дистально, ее
внутренний край со-
ставляет немного бо-
лее половины длины
пальца; каждый па-
лец имеет приблизи-
тельно три ряда шипов
сверху, края пальцев
сходятся при смыка-
нии, кончики пере-
крещиваются. Шипы
имеют роговые кон-
чики. Нога усажена
пучками волос, ко-
торые поднимаются
близ основания ши-
пов. Правая клешне-
носная нога дости-
гает конца ладони
левой руки;1 мерус
не достигает конца
глаз. Шипы мельче
и менее ясно расположены в ряды, ладонь расширяется очень немного
по направлению к своему дистальному концу; палец в l1^ раза длиннее,
чем внутренний край ладони. Первая ходильная нога простирается за левую
клешненосную ногу на половину длины своего пальца; как первая, так
и вторая пары толстые, волосатые сверху, палец длиннее чем проподус.
Нижний край меруса и верхний край карпуса первой пары имеют ряд
шипов; пальцы обеих пар вооружены на внутренней поверхности несколь-
зкими рядами темных шипов. Эти ноги не полосатые и в спирту не обнару-
живают поперечных окрашенных полос.
1 Н. В а 1 s s (1913 :41) высказывает предположение, не имела ли места здесь регене-
рация, т. к. для данного рода не характерна более короткая правая клешненосная нога.
Возможно, однако, что в этом выражается половой диморфизм; М. Rathbun имела только
•одну самку данного вида и после нее вид никем не был найден.
3. PAGURISTES
165
Размеры
Длина тела . . . . •........................ 58.0 мм
„ головогруди.....................................26.2 „
Расстояние от конца рострума до цервик. шва..........16.0 „
Ширина переднего края................................11.1 „
Длина глазных стебельков........................... 9.0 „
„ проподуса первой ходильной правой ноги . . . 11.4 ,,
,, пальца „ „ ,, . 13.4 „.
„ проподуса второй „ „ „ . . л . 13.4 „
» Пальца „ „ „ „ « . . 16.6 „
Распространение. Япония, Hokkaido (М. Rathbun).
3. Род PAGURISTES DANA
Paguristes Dana, 1851 : 269; 1852 : 436; Stimpson, 1858 : 236; 1907 : 211; Heller, 1861; 172;
Henderson, 1888:77; Milne-Edwards et Bouvier, 1893:32; Ortmann, 1901:1146;, Benedict,,
1901:143; Alcock, 1905 :30; Balss, 1913:36; Terao, 1913: 373; Schmitt, 1921:122,-Ste vens.,
1925.
Тип рода: Paguristes hirtus Dana.
Лобный край карапакса с острым средним выступом. Глазные сте-
бельки тонкие и длинные. Скафоцериты мощные. Антенны короткие;
жгуты более или менее волосатые, часто с длинными ресничками. Антен-
нулы длиннее, чем у других родов, вершина второго членика достигает
конца глаз. Наружные максиллярные ножки сближены у своих осно-
ваний. Клешненосные ноги одинакового строения, равной длины или почти
равной длины, или же редко одна нога (обычно левая) несколько длиннее,
другой. Пальцы клешней двигаются в горизонтальной плоскости. Четвертая
пара перейопод простая, без ложных клешней. Самцы имеют парные
половые придатки на двух первых абдоминальных сегментах, самки —
только на первом абдоминальном сегменте. На левой стороне абдомен
самки образует кожистую складку, служащую для защиты яиц при вына-,
шивании их.
Ни один из видов этого широко распространенного, главным образом
в тропической и субтропической областях, рода не встречается в наших
водах. Paguristes digitalis был найден Stimpson’oM у Хакодате, а Р. setosus
отмечен Yokoya для Aomori, поэтому оба вида приводятся в настоящей,
работе.
ОПРЕДЕЛИТЕЛЬНАЯ ТАБЛИЦА ВИДОВ
1 (2). Наружная сторона подвижного пальца клешни с 7 косыми рядами
коричневых гранул.............................1. Р. digitalis Stimpson.
2 (1). Наружная сторона подвижного пальца клешни без таких рядов
гранул . ...............................2. Р. setosus (Milne-Edwards).
166
PAGURIDAE
1. Paguristes digitalis Stimpson. (Фиг. 66).
Paguristes digitalis Stimpson, 1858: 247; 1907 : 212, t. 25, fig-. 1; Balss, 1913 : 37, fig. 26,
27; Terao, 1913 : 374; Yokoya, 1933 ; 73.
Карапакс почти голый около середины, суживается кпереди и отчасти
расширяется кзади. Кардиальная scutella, или твердая срединная пла-
стинка задней части карапакса, отчасти алебардообразной формы, сужи-
ваясь у середины д немного расширяясь по направлению к своему
заостренному концу. Бока передней части карапакса более глубоко выем-
чатые, чем у большинства видов. Рострумообразный выступ лба про-
долговатый, очень острый, простирающийся
Фиг. 66. Paguristes digitalis Stimpson. Наружная по-
верхность подвижного пальца клешни. (Из Balss, 1913).
немного за основание глаз-
ных чешуек и гребенча-
тый вдоль середины. Глаза
очень длинные и тонкие,
чуть-чуть длиннее чем лоб
и не вполне достигают
конца ножки антеннул; сте-
белек голый, за исключе-
нием слабого пучка щетит
нок у верхнего основания.
Вершина глазной чешуйки
короткая, острая, с одним
зубчиком на наружном
краю. Наружные максил-
лярные ножки длинные,
простирающиеся за глаза.
Клешненосные ноги рав-
( ные, очень похожие на
таковые у Paguristes hirtus, Р. turgidus и др.; карпус и рука шиповатые
и волосатые сверху, шипы довольно многочисленные, с темными роговыми
кончиками. Рука довольно широкая и плоская сверху, с вдавленной поверх-
ностью на неподвижном пальце; внутренние края пальцев роговые на про-
тяжении терминальной половины их длины; подвижный палец угловатый,
с плоской наружной поверхностью, изящно и тонко украшенной косыми
гребнями приблизительно в числе семи (фиг. 66). Задние ноги волоса-
тые; ноги второй пары шиповатые сверху, их пальцы более чем на поло-
вину длиннее предыдущего членика. Дорзальный интегумент абдомена
самца довольно жесткий, образующий на каждом сегменте тонкую гнущуюся
пластинку. Доли терминального сегмента почти симметричные, как у сам-
цов, так и у самок. Мозолистые участки, образующие „растирающий
прибор" на нижней поверхности торакса и ног, менее хорошо заметны,
чем у Р. turgidus. Окраска спиртовых экземпляров светлая, кирпично-
красная; местами беловатая; глазные стебельки с красной поперечной
полосой (Stimpson).
4. D ARD ANU S
167
Распространение. Южная Япония (бухта Сагами; около Siriya-
zaki; юго-восточнее о-ва ЦусиМа). Самым северным местонахождением
является Хакодате (о-в Иезо). На глубинах от 20 до 150 м.
2. Paguristes setosus (Milne-Edwards). (Фиг. 67).
Pagurus setosus Milne-Edwards, 1848 : 64. —Paguristes setosus
Ortmann, 1892 : 281, t. 12, fig. 9; Yokoya, 1933 : 76.
Вид очень близок к Paguristes pilosus, но имеет
гораздо менее волосатые руки и более тонкие ноги
третьей пары. Окраска желто-красная. Длина прибли-
зительно 1 дюйм (Milne-Edwards). .Глазные стебельки
короче чем стебли антеннул. Клешненосные ноги оди-
наковой величины, шипы на них однородно окрашенные,
без темных верхушек. В литературе отсутствует более
полное описание этого вида.
Распространение. Новая Гвинея, Южная
Япония; бухта Сагами; к югу от Inuboe-zaki; около
Omae-zaki, между Koti и Asizuri-zaki; к востоку от
острова Цусима. Самым северным местонахождением
является пункт к западу от Tubaki-zaki в Aomori-ken.
На глубинах от 73 до 238 м.
Фиг. 67. Голово-
грудь Paguristes se-
tosus (М. Edw.) (Из
Ortmann, 1892).
4. Род DARDANUS PAULSON
Pagurus Fabricius, 1775:410 (part.); Milne-Edwards, 1837:213; de Haan, 1850:202;
Dana, 1852:449; Stimpson, 1858:233; 1907:204; Heller, 1863:174; Henderson, 1888:55;
Ortmann, 1901 :1146; Milne-Edwards et Bouvier, 1893 :161; Alcock, 1905 : 78; Balss, 1913 : 45;
Terao, 1913 :375. — Dardanus Paulson 1875:90; Rathbun, 1902:33; Schmitt, 1921:126. —
Pagurias Benedict, 1901 :141.
Тип рода: Dardanus depressus (Heller).
Средняя часть лобного края выступает несколько вперед, но перед-
ний край этого выступа прямой или закругленный и не несет никакого
среднего выступа (рострума); боковые выступы явственные. Глазные сте-
бельки короткие и толстые; глазные чешуйки с широкими, шиповатыми
вершинами. Скафоцериты короткие и толстые. Жгуты антенн длинные
и голые. Наружные максиллярные ножки сближены у своих оснований.
Клешненосные ноги развиты не одинаково, левая значительно больше
правой. Пальцы клешни двигаются в вертикальной плоскости и имеют
короткие, отчасти выемчатые, роговые кончики. Четвертая пара перейопод
с ложными клешнями. Абдомен без половых придатков. (Указание Пауль-
сона, что глазной сегмент посередине вооружен подвижным отростком,
как у Diogenes, следует считать ошибочным).
Большая часть видов этого рода является обитателями тропической
литорали, главным образом, восточного полушария. В наших водах род
не представлен ни одним видом. Dardanus arrosor (Herbst) приводится
1 68
PAGURID AE
в настоящей работе как вид, зарегистрированный в Японском море у запад-
ного побережья Японии (префектура Ниигата), а также в Северном Япон-
ском море (Yokoya).
1. Dardanus arrosor (Herbst). (Табл. 1, фиг. 1).
Cancer arrosor Herbst, 1794:170, t. 43, fig'. 1. — Cancer strigosus Bose, 1803 : 77, t. 11,
fig. 3.— Pagurus striatus Latreille, 1803:163; Risso, 1815:54; 1826:38; Roux, 1828, t. 10;
Costa, 1836 : 7; Milne-Edwards, 1836 : 270; 1837 : 218; Lucas, 1849 : 29; de Haan, 1850 : 206,
t. 49, fig'. 1; Heller, 1863 :174; Miers, 1881:274; Studer, 1882: 23; Carus, 1884 :494; Hender-
son, 1888:56; Ortmann, 1892:283; Chevreux et Bouvier, 1892:119; Whitelegge, 1900:166;
Doflein, 1902 : 645; Stimpson, 1907: 206. — Pagurus incisus Olivier, 1811 : 641. — Pagurus;.
arrjsor Milne-Edwards et Bouvier, 1900:178; Moreira, 1901 :24, 85; Alcock, 1905 :168; Nobili,
1906:121; Stubbing, 1908:22; 1910:350; Cunning-ham, 1910:121; Balss, 1912 : 95; 1913 : 46;
Terao, 1913 : 375; Parisi, 1918 :112; Yokoya, 1933 : 78.
Передняя часть карапакса такой же длины, как й ширины. Боковые
выступы лба острые, треугольные. Глазные стебельки толстые. Глазные,
чешуйки широкие, с прямым внутренним и выпуклым наружным краем;
вершина чешуйки несет раздвоенный шип, а выпуклый наружный край
усажен острыми роговыми шипами. Скафоцериты не достигают концов,
глаз, с шипами и волосами на внутреннем краю. Наружный и внутрен-
ний выступы базального членика антеннальной ножки с мощными шипами,,
причем у основания шипов имеются маленькие шипики; шип наружного
выступа часто раздвоенный. Клешненосные ноги одинаковой формы
и строения, но левая развита сильнее правой, которая простирается
немного дальше основания подвижного пальца левой руки. Мерус трех-
гранный, с острыми крупными шипами на нижне-внутреннем краю. Карпус
с шипами на внутреннем краю и с рассеянными шипами на покатой наруж-
ной поверхности. Внутренний край ладони и толстого подвижного пальца
также несет острые короткие шипы. Верхняя поверхность карпуса и руки
(включая и пальцы) исчерчена полосами, имеющими, в общем, диагональ-
ное направление; каждая полоса представляет собой небольшое возвыше-
ние, часто усаженное, в особенности к внутренним краям члеников, грану-
лами или мелкими шипиками; от этого возвышения направляются вперед
короткие, мягкие, двуперистые волоски, которые плотно прилегают
к поверхности членика. Нижняя поверхность ног гладкая, за исключением
области у наружного края ладони и подвижного пальца, куда частично
заходят описанные выше полосы. Хватательные края пальцев с крупными
белыми буграми; концы пальцев с сильными, отчасти выемчатыми, рого-
выми кончиками. Ходильные ноги значительно превосходят клешненосные..
Карпальные членики с шипами на верхнем краю. Наружная поверхность
всех члеников, за исключением меральных, имеет такие же полосы, как
и на клешненосных ногах. Пальцы толстые, волосатые на дистальном
конце и с сильными роговыми ногтями. Описанные полосы придают живот-
ному своеобразный вид, по которому он может, легко отличаться от прочих
видов рода. Окраска коричневато-красная.
5. PA GURUS
169
Размеры
Длина передней части карапакса........... 19.0 мм
Ширина „ „ „ ............. 19.0 „
Длина глазных стебельков .............. 9.5 „
„ меруса левой клешней, ноги . . . . . . . 17.0 „
„ карпуса „ „ „ ........ 16.0 „
„ Руки „ „ „ ............. 34.0 „
Ширина „ „ „ „ . ......... 20.0 „
Длина левой ходильной ноги первой пары ....... 78.0 „
Экземпляры могут достигать крупных размеров. Встречается на глу-
бинах от 37 до 600 м.
Распространение. Средиземное и Адриатическое моря; Мадейра;.
Сенегамбия; западная Африка; о-в Св. Елены; Красное море; Австралия;
Филиппинские о-ва; Южная Япония; Японское море, по западному побе-
режью Японии на север до Ниигаты, на побережье континента известен
из Фузана (Корея). К сожалению, Yokoya (1933:78), приводя этот вид.,
для Северного Японского моря, не указывает точно пункта нахождения.
Бразилия и Вест-Индия, как места нахождения Р. arrosor, вызывают-
сомнения.
2. Подсем. PAGURINAE, nom. nov.
Eupagurinae Ortmann, 1892 : 270.
Наружные максиллярные ножки широко расставлены у своих основа-
ний. Правая клешненосная нога длиннее левой; в редких случаях обе
ноги почти равные.
5. Род PAGURUS FABRICIUS
Pagurus Fabricius, 1775 : 410; Holmes, 1900 : 132; Rathbun, 1904 : 157; Schmitt,
1921 :130, — Eupagurus Brandt, 1851:105; Stimpson, 1858: 226; 1907 : 215; Heller, 1863:158;
Henderson, 1888:62; Ortmann, 1892:297; Benedict, 1892:1; Milne-Edwards et Bouvier,
1893:139; Alcock, 1905:122; Balss, 1913:51; Terao, 1913:364; Yokoya, 1933:80. - Bern- ,
hardus Dana, 1852 : 440.
Тип рода: Pagurus bernhardus (Linne).
Лобный край карапакса имеет средний выступ (рострум). Глазные
стебельки большею частью короткие и толстые, глазные чешуйки неболь-
шие и далеко отстоящие друг от друга. Скафоцериты хорошо развиты.
Антенны длинные и почти безволосые. Наружные максиллярные ножки
широко расставлены у своих оснований; коксоподиты этих ножек малень-
кие, очень короткие, не превышающие по размеру базальные членики
ножек; в промежутке между основаниями ножек виден выступ второго
стернального сегмента, который часто несет зубец или шиповидный отро-
сток. Клешненосные ноги неравные, правая развита сильнее левой; кончики
пальцев острые; пальцы двигаются в горизонтальной плоскости. Четвертая.
170
PA GURIDAE
пара перейопод с ложными клешнями. Первый и второй абдоминальные
сегменты без парных придатков.
Один из наиболее многочисленных по числу видов род. Областью
распространения являются, главным образом, бореальные области Атлан-
тического и Тихого океанов, хотя отдельные виды заходят как в аркти-
ческую, так и в тропическую области. Большинство отшельников наших
вод относится к этому роду.
В синонимии рода Pagurus Fabricius имеется некоторая двойствен-
ность: большинство авторов (в том числе все европейские авторы) сохра-
няет для данного рода брандтовское название подрода — Eupagurus-,
большинство американских авторов, к которым примкнем и мы, вполне
правильно сохраняет для данного рода старое название — Pagurus. Род
Pagurus, установленный Fabricius’oM в 1775 году с типичным видом bernhar-
dus, первоначально вмещал в себе фактически два рода. В 1851 году Brandt
выделил подрод Eupagurus с тем же типичным видом bernhardus', впослед-
ствии подрод был возведен в положение рода, а название Pagurus, лишен-
ное своего типичного, вида, стали применять ко всем формам, не уклады-
вавшимся в понятие рода Eupagurus, т. е. к тому, если можно так выра-
зиться, безымянному роду, который еще во времена Fabricius’a содержался
в его двойном роде Pagurus. Однако этот последний безымянный род был
выделен из рода Pagurus Fabr. в 1875 году Паульсоном под названием
Dardanus. Таким образом, брандтовский род Eupagurus должен имено-
ваться как Pagurus Fabricius, a Pagurus большинства авторов является
не чем иным, как родом Dardanus Paulson. Введенные Benedict’oM под-
роды {Eupagurus, Trigonocheirus, Elassocheirus и Labidocheirus) не при-
няты в современной литературе.
ОПРЕДЕЛИТЕЛЬНАЯ ТАБЛИЦА ВИДОВ
1 (6). Верхняя поверхность ладони правой руки гладкая и совершенно
безволосая.
2 (3). Карпус правой клешненосной ноги такой же ширины, как рука.
Верхняя поверхность левой руки выпуклая, без приподнятых краев .
. '...............................* 1. Р. middendorffii Brandt,
3 (2). Карпус правой клешненосной ноги значительно шире руки, благо-
даря боковым крыловидным расширениям. Верхняя поверхность
левой руки плоская, с приподнятыми краями.
4 (5). Верхняя поверхность карпуса правой клешненосной ноги закруглен-
ная и совершенно гладкая. Скафоцериты не достигают концов глаз .
.................................... *2. Р. gilli (Benedict).
5 (4). Верхняя поверхность карпуса правой клешненосной ноги имеет
явственное ребро посредине, которое несет у своего проксимального
конца несколько шиповатых гранул. Скафоцериты достигают концов
глаз........................................3. Р. cavimanus (Miers).
5. PAGURUS
171
6 (1). Верхняя поверхность ладони правой руки покрыта круглыми гра-
нулами; гранулы иногда очень мелкие и рассеянные, так что рука
выглядит почти гладкой; волоски, если и имеются, то очень малень-
кие, часто едва заметные для невооруженного глаза. Или верхняя
поверхность ладони правой руки покрыта шиповатыми гранулами
или более или менее короткими коническими шипами; в этих случаях
волосы на ладони почти совсем отсутствуют или очень короткие,
расположенные в виде пучков при основании шипов или гранул.
Или же верхняя поверхность ладони правой руки покрыта острыми
длинными шипами и большей или меньшей длины волосами, скры-
вающими иногда всю скульптуру ладони; в редких случаях шипы
ладони короткие, но имеются длинные мягкие волоски.
7 (26). Верхняя поверхность ладони правой руки покрыта круглыми гра-
нулами; гранулы иногда очень мелкие и рассеянные, так что рука
выглядит почти гладкой; волоски, если имеются, то очень маленькие,
часто едва заметные для невооруженного глаза.
-8 (21). Верхняя поверхность ладони правой руки без каких либо возвы-
.шающихся гребней или углублений, более или менее равномерно
выпуклая.
9 (16). Мерус правой клешненосной ноги без выдающихся тупых шипов
или крупных гранул на нижней поверхности.
10 (11). Рука правой клешненосной ноги значительно шире, чем карпус.
Верхняя поверхность ладони левой руки плоская, с приподнятыми
краями....................................* 4. Р. tenuimanus (Dana).
11 (10). Рука правой клешненосной ноги не шире, чем карпус, а если и
шире, то очень незначительно. Верхняя поверхность ладони левой
руки более или менее выпуклая, без приподнятых краев.
12 (13). Лобный край карапакса без боковых зубцов. Клешненосные ноги
волосатые, за исключением правой руки . * 5. Р. hirsutiusculus (Dana).
13 (12). Лобный край карапакса имеет боковые зубцы. Клешненосные ноги
почти безволосые.
14 (15). Ладонь правой руки сильно выпуклая, усеянная тупыми гранулами,
придающими ей шагреневый вид, пальцы толстые. Скафоцериты трех-
гранные. Пальцы ходильных ног толстые . * 6. Р. bernhardus (Linne).
15 (14). Ладонь правой руки слабо выпуклая и покрыта очень мелкими
рассеянными гранулами, так что часто выглядит почти гладкой.
Скафоцериты не трехгранные. Пальцы ходильных ног тонкие . . .
...................................* 7. Р. gracilipes (Stimpson).
16 (9). Мерус правой клешненосной ноги с выдающимися тупыми шипами
или крупными гранулами на нижней поверхности.
17 (18). Нижняя поверхность меруса правой клешненосной ноги с двумя
крупными бугрообразными гранулами . . 8. Р. beringauus (Benedict).
18 (17). Нижняя поверхность меруса правой ноги с одним тупым шипом
посредине.
172
PA GURIDAE
19 (20). Рука правой клешненосной ноги в 2 раза больше в длину, чем.
в ширину, с почти параллельными наружными и внутренним краями.
Верхняя поверхность ладони левой руки с длинными мягкими воло-
сами ...................................... * 9. Р. dubius (Ortmann).
20 (19). Рука правой клешненосной ноги менее чем в 2 раза длиннее
ширины, с закругленными краями. Верхняя поверхность ладони:
левой руки почти безволосая или с очень короткими одиночными
волосками...............................* 10. Р. samuelis (Stimpson).
21 (8). Верхняя поверхность ладони правой руки с заметным гребнем,
посредине.
22 (23). Гребень на верхней поверхности ладони правой руки заканчивается:
сильно выдающимся рогом.................* 11. Р. cornutus (Benedict)..
23 (22). Гребень на верхней поверхности ладони правой руки не заканчи-
вается рогом, но довольно круто спускается к неподвижному пальцу.
24 (25). Гребень на верхней поверхности ладони правой руки тянется парал-
лельно внутреннему краю; между ним и внутренним краем образуется,
углубление с параллельными краями. Наружная поверхность ладони:
левой руки выпуклая.................... * 12. Р. undosus (Benedict)..
25 (24). Гребень на верхней поверхности ладони правой руки имеет вид:
треугольного возвышения с вершиной,, обращенной к основанию,
щели между пальцами; углубление между внутренней стороной
треугольника и внутренним краем ладони с непараллельными краями..
Наружная поверхность ладони левой руки глубоко вогнутая . .
...................................... 13. Р. tanneri (Benedict)..
26 (7). Верхняя поверхность ладони правой руки покрыта шиповатыми
гранулами или более или менее короткими коническими шипами;
в этих случаях волосы на ладони почти отсутствуют или очень,
короткие, расположенные в виде пучков при основании шипов или
гранул. Или же верхняя поверхность ладони правой руки покрыта
острыми длинными шипами и большей или меньшей длины волосками,,
скрывающими иногда всю скульптуру ладони; в редких случаях шипы,
ладони короткие, но имеются длинные мягкие волоски.
27 (42). Верхняя поверхность ладони правой руки покрыта шиповатыми
гранулами или более или менее короткими, коническими шипами.
Волоса на ладони почти совсем отсутствуют или очень короткие>.
расположенные в виде пучков у оснований шипов или гранул.
28 (31). Жесткой является не только передняя часть карапакса, но вся
его верхняя поверхность.
29 (30). Средний выступ лба узкий, длинный, поднимающийся косо вверх,
с простой вершиной. На боковом выступе карапакса позади церви-
кальной борозды имеется острый выдающийся шип.....................
......................................* 14. Р. anomalus (Balss).
30 (29). Средний выступ лба широкий, более короткий,, направленный
горизонтально вперед, с тремя или четырьмя зубчиками на вершине.
5. PAG UR US
173
Боковой выступ карапакса позади цервикальной борозды без острого
выдающегося шипа...................* 15. Р. splendescens Owen.
31 (28). Жесткой является только передняя часть карапакса.
32 (35). Передняя часть карапакса больше в ширину чем в длину. Скафо-
цериты трехгранные, с сильно иризирующей внутренней поверх-
ностью. Клешненосные ноги очень мощные по сравнению с телом;
их членики почти совершенно лишены волос.
33 (34). Пальцы ходильных ног без продольного жолоба на верхней
поверхности ............................* 16. Р. ochotensis Brandt.
34 (33). Пальцы ходильных ног с продольным жолобом на верхней поверх-
ности ..........................16а. Р. ocbotensis aleuticus (Benedict).
35 (32). Передняя часть карапакса такой же длины, как и ширины, или
больше в длину, чем в ширину. Скафоцериты не трехгранные, не
иризирующие. Клешненосные ноги умеренной величины по сравнению
с телом; их членики, за исключением рук, более или менее волосатые.
36 (37). Карпус правой клешненосной ноги с круглым пучком волос на
внутреннем дистальном углу верхней поверхности. Пальцы левой
руки длинные, сравнительно тонкие и сильно изогнутые вниз . . .
....................................* 17. Р. rathbuni (Benedict).
37 (36). Карпус правой клешненосной ноги без подобного пучка волос.
Пальцы левой .руки не очень длинные и не сильно или почти не
изогнутые вниз.
. 38 (39). Мерус правой клешненосной ноги с двумя крупными, бугрообраз-
ными гранулами на нижней поверхности. Левая рука не имеет явствен-
ного гребня на верхней поверхности . 18. Р. granosimanus (Stimpson).
39 (38). Мерус правой клешненосной ноги без таких гранул на нижней по-
верхности. Левая рука с явственным гребнем на верхней поверхности.
40 (41). Меральные членики клешненосных ног с белой полосой у дисталь-
ных .концов. Шипы на клешненосных ногах сравнительно длинные
и острые. Правая рука довольно узкая . . . 19. Р. dalli (Benedict).
.41 (40). Меральные членики клешненосных ног без белой полосы у дисталь-
ных концов. Шипы на клешненосных ногах более короткие и тупые.
Правая рука сравнительно широкая ... * 20. Р. pubescens Kroyer.
. 42 (27). Верхняя поверхность ладони правой руки покрыта острыми,
длинными шипами и большей или меньшей длины волосами, которые
иногда могут скрывать всю скульптуру ладони. В редких случаях
шипы ладони короткие, но имеются длинные, мягкие волоски.
43 (44). Мерус правой клешненосной ноги с двумя сильно выдающимися ту-
пыми шипами на нижней поверхности. * 21. Р. brachiomastus (Thallwitz).
44 (43). Мерус правой клешненосной ноги без таких шипов на нижней
поверхности.
-45 (46). Неподвижный палец правой руки в 2 раза шире подвижного.
Поверхность ладони правой руки с короткими шипами и с длинными
мягкими волосами........................* 22. Р. lanuginosus de Haan.
174
PAGURIDAE
4'6 (45). Неподвижный палец правой руки такой же ширины, как подвижный,,
или немного шире его. Поверхность ладони правой руки с длинными:
шипами и волосами.
47 (48). Скафоцериты трехгранные, с иризирующей внутренней поверх-
ностью .......................................23.. Р. armatus (Dana)..
48 (47). Скафоцериты не трехгранные, не иризирующие.
49 (56). Шипы на поверхности ладони правой руки располагаются в более
или менее правильные продольные ряды.
50 (53). Шипы на поверхности ладони правой руки образуют 8—9 про-
дольных рядов.
51 (52). Скафоцериты короче глаз. Средний выступ лба острый. Глазные^
стебельки тонкие, расширенные у основания. Наружный край ладони:
правой руки с рядом длинных крупных шипов, резко отделяющих,
верхнюю поверхность руки от нижней.................................
................................. * 24. Р. pecthiatus (Stimpson)..
52 (51). Скафоцериты длиннее глаз. Средний выступ лба тупой. Глазные
стебельки довольно толстые, расширяющиеся у роговиц. Наружный
край ладони правой руки тупой, без сильных шипов............... .
...................................25. Р. obtusifrous (Ortmann).
53 (50). Шипы на поверхности ладони правой руки образуют 7 продоль-
ных рядов.
54 (55). Скафоцериты длиннее глаз. Волосы на руках значительно длиннее
чем шипы....................................26. Р. setosus (Benedict).
55 (54). Скафоцериты короче глаз. Волосы на руках не достигают концов
шипов...................................27. Р. kennerlyi (Stimpson).
56 (49). Шипы на поверхности ладони правой руки рассеянные, не обра-
зующие правильных продольных рядов.
57 (58). Боковые выступы лба округлые. Скафоцериты достигают концов
глаз или несколько короче их. Ходильные ноги превышают правую
клешненосную ногу, их пальцы закрученные....................... .
.....................................* 28. Р. capillatus (Benedict).
58 (57). Боковые выступы лба заостренные. Скафоцериты простираются
дальше глаз. Ходильные ноги короче правой клешненосной ноги,
их пальцы не закрученные..................29. Р. constans (Stimpson).
* 1. Pagurus middendorffii Brandt. (Табл. 5, фиг. 6).
Pagarus middendorffii Brandt, 1851:1013, t. 5, fig. 1—16; Holmes, 1900 : 234; Rathbun,
1902:35; 1904:160. — Eupagurus middendorffii Stimpson, 1858: 250; 1907 : 26; Ortmann,
1892 : 301; Doflein, 1900: 340; 1902 :646; Lenz, 1901:444; Alcock, 1905:178; Balss, 1913: 58;
Terao, 1913:371.
Передняя часть карапакса почти такой же ширины, как и длины.
Средний выступ лба широко-треугольный, острый; боковые выступы
отсутствуют. Глазные стебельки короткие, умеренно толстые. Глазные
чешуйки полуовальной формы, с вогнутой верхней поверхностью и силь-
5. PAGURUS
175
ними субмаргинальными шипами. Скафоцериты достигают концов глаз,
иногда они несколько длиннее; их внутренний край слабо волосатый.
Внутренний край наружного выступа базального членика антеннальной
ножки зазубренный. Клешненосные ноги очень неравные, совершенно
безволосые, с мелко гранулированной, шагреневой поверхностью. Правая
клещненосная нога очень велика по сравнению с телом; левая, наоборот,.
очень мала, достигая только дистального конца карпуса правой клешне-
носной ноги, а у мелких экземпляров даже несколько короче. Мерус
правой клешненосной ноги трехгранный, с округлыми ребрами. Карпус
заметно длиннее меруса, с выпуклой верхней поверхностью, равномерно.
переходящей в боковые поверхности. Рука длинная, наружный край слабо
изогнутый; внутренний край образует тупой угол с наружным краем по-
движного пальца. Поверхность руки настолько мелко гранулирована, что
невооруженному глазу кажется гладкой. Пальцы короткие, гранулирован-
ность на их верхней поверхности выражена более ясно. Хватательные края
пальцев с неотчетливыми белыми буграми и с редкими пучками волосков;
роговых кончиков нет. Мерус и карпус левой клешненосной ноги сжатые,
очень слабо- волосатые на нижней поверхности; верхний дистальный конец
карпуса образует небольшой, направленный вперед выступ, который более
крупно гранулирован. Рука узкая, с выпуклой верхней поверхностью и
слабо волосатой нижней поверхностью, особенно вдоль неподвижного ,
пальца. Пальцы с роговыми концами и разбросанными пучками волос. .
Ходильные ноги слабые, почти безволосые. Ходильные ноги правой
стороны достигают середины ладони правой руки; ходильные ноги левой
стороны превышают левую клешненосную ногу всей длиной своих пальцев. .
Продольные членики обеих пар шиповатые снизу; пальцы слабо изогнутые,
не закрученные, с мелкими шипами на верхнем и более крупными шипами.
на нижнем краю и с острым роговым когтем. Окраска живых экземпляров
оливковая, спиртовых экземпляров — равномерная красновато-оранжевая..
Дистальная половина пальцев правой руки часто белая. Карпальные
членики и проксимальная половина проподальных члеников ходильных ,
ног на наружной поверхности с коричневато-красной продольной полосой,
широкой на карпальных члениках и постепенно суживающейся на пропо-.
дальных члениках.
Размеры ,
Длина передней части карапакса.............. 8.5 мм
Ширина „ „ „ ............ 8.0 „
Длина глазных стебельков............... . 4.0 „
Длина меруса правой клешней, ноги . . . . . 13.0 »
Длина карпуса „ » п. * •- • . 17.0 »
Ширина „ „ п » • • • • . 10.0 »
Длина руки „ » » • • • • . 19.5 п
Ширина „ „ м » • • • • . 10.0
Длина меруса левой П 9.0 п
„ карпуса „ ., 8.5 >>
176
PAGURIDAE
Ширина карпуса правой клешн. ноги........ 3.5 мм
Длина руки „ „ ..........10.0 „
Ширина „ „ „ „........... 5.0 „
Длина правой ход. ноги первой пары....... 40.0 „
Распространение. Японское море, по нашему побережью к югу
.до зал. Посьет; северная Япония. Охотское море; восточное побережье
Камчатки. Берингово море до широты Олюторского залива (бухта Лаврова).
Алеутские острова; по американскому побережью к югу до широты
Ванкувера.
Вид не достигает крупных размеров.
Экология. Является обычным обитателем «литорали и верхних
горизонтов сублиторали; при отливе часто остается среди камней в осуш-
ной зоне. На абдомене часто встречаются паразитические Cirripedia
(? Peltogastrella socialis Kruger).
* 2. Pagurus gilli (Benedict). (Табл. 3, фиг. 1).
Eupagurus gilli Benedict, 1892 : 20.— Pagurus gilli Rathbun, 1904 :161, t. 5, fig. 10;
Stevens, 1925 :291, fig. 12, 13.
Передняя часть карапакса немного больше в ширину чем в длину.
Средний выступ лба острый, треугольный; боковые выступы округлые,
'снабженные тупыми субмаргинальными шипами. Глазные стебельки
цилиндрические, умеренной длины. Глазные чешуйки продолговатые,
с завернутыми вверх краями. Скафоцериты не достигают до концов
глаз, слабо волосатые вдоль внутреннего края. Вершина наружного
выступа базального членика антеннальной ножки несет два шипа. Клешне-
носные ноги совершенно голые. Мерус правой клешненосной ноги корот-
кий, трехгранный; узкая вершина исхиума почти достигает переднего края
меруса на нижней поверхности. Карпус широко-треугольный, с закруг-
ленной верхней поверхностью (фиг. 68А); боковые скаты на дистальной
половине переходят в треугольные крыловидные расширения; внутренний
край расширения и передний край карпуса вооружены мелкими тупыми
шипиками; нижняя поверхность карпуса представляет собой полулунное
„углубление благодаря тому, что края расширений подвернуты вниз; наи-
более сильно подвернут наружный край. Рука уже чем карпус, с парал-
лельными краями; по отношению к продольной оси карпуса она причле-
няется к нему под углом, причем ее' продольная ось направлена внутрь.
Наружный край руки слегка приподнятый, середина неподвижного пальца
слабо вогнутая. Поверхность ладони покрыта мелкими точечными углу-
блениями, поверхность пальцев ноздреватая. Левая клешненосная нога
достигает основания подвижного пальца правой руки или только до
середины ладони. Карпус на верхне-внутреннем краю вооружен рядом
мелких шипов. Верхняя поверхность руки гладкая, плоская, с завернутыми
вверх краями; проксимальный конец ладони имеет закругленный край;
приподнятый внутренний край ладони, переходя на подвижный палец,
5. PAGURUS
177
образует на нем заметное ребро; нижняя поверхность руки выпуклая,
гладкая; пальцы с роговыми кончиками, тогда как на правой руке их нет.
Ходильные ноги правой стороны достигают середины ладони правой руки;
ходильные ноги левой стороны превышают левую клешненосную ногу
почти всей длиной своих пальцев. Все членики сплющенные, почти без-
волосые. Карпальные членики вооружены на верхнем краю 7—9 шипиками.
Пальцы слегка изогнутые, незакрученные, по длине равны соответствую-
щим проподусам; их нижний край несет 11—14 роговых шипиков; кончики
с сильными роговыми когтями. Окраска спиртовых экземпляров яркая,
оранжево-красная. Концы пальцев правой руки, хватательные и наруж-
Фиг. 68. Карпальные членики правой клешненосной ноги. А — Pagurus
gilli (Benedict). В — Pagurus cavimanus (Miers).
иые края пальцев и наружный край ладони белые; углубления, образующие
упомянутую выше ноздреватость на пальцах, также белые.
Относительно молодых экземпляров В. A. Stevens (1925) говорит:
„молодые экземпляры сильно отличаются от взрослых следующим:
на карпусе большой клешненосной ноги совершенно отсутствует тонкое,
крылоподобное расширение: рука немного шире, чем карпус. Однако они
могут быть узнаваемы благодаря характерной плоской руке левой клешне-
иосной ноги".
От близко родственного Р. cavimanus данный вид отличается более
тонкими глазными стебельками, скафоцеритами, которые не достигают
до концов глаз, более округлым и гладким карпусом правой клешне-
носной ноги, более сильными ходильными ногами и более широкими
пальцами их.
Размеры
Длина передней части карапакса........... 8.0 мм
Ширина „ „ „ ........... 9.0 „
Длина глазных стебельков ................ 4.0 „
„ меруса правой клешней, нога......... 7.0 „
„ карпуса „ „ „ ........ 10.0 „
Фауна СССР, Макаров .12
178
PA GURIDAE
Ширина карпуса правой клешн. ноги 12.0 ММ
Длина руки „ » ♦ • • • • 15.0
Ширина „ ч 9.0
Длина меруса левой п ч 5.0 i>
„ карпуса „ и if . . • • • 6.5 ч
Ширина „ „ ч » 3.5 »
Длина руки „ ч » • • • • • 10,0 if
Ширина „ „ ч '• • • • • • 4.5 »
Длина правой ходильной НОГИ первой пары . . 19.5 Ч
Распространение. Командорские о-ва, Алеутская гряда, п-ов
Аляска к югу до о-ва Revillagigedo (юго-восточная Аляска), Калифор-
ния, Камчатка у м. Лопатка, Японское море у Советской гавани. До
настоящего времени Р. gilli считался сравнительно узкой локальной
формой Алеутской гряды, возможно, подвидом более широко распро-
страненного Р. cavimanus (Miers). Однако, в результате просмотра мате-
риалов Зоологического Института Академии Наук приходится рас-
ширить область распространения Р. gilli. Экземпляры с Камчатки, из
района Советской гавани и из Японии, детально сравненные мною
с экземпляром Р. cavimanus, полученным из Национального Музея
Соединенных Штатов (п° 16656), все таки без колебаний следует отнести
к Р. gilli. Таким образом, получается обратная картина: Р. gilli, невидимому,
имеет более широкое распространение в виде дуги через Курильскую и
Алеутскую гряду, тогда как Р. cavimanus, если считать определение
экземпляров из Японии ошибочными, должен явиться подвидом Р. gilli.
Однако без просмотра более обширного материала из Японии, трудно
притти к окончательному решению этого вопроса; поэтому я сохраняю
синонимию и географическое распространение обоих видов в том виде*
как они существовали до сих пор, расширяя, конечно, область распро-
странения Р. gilli согласно вышеприведенным местам нахождения.
Экология. Как и предыдущий вид, Р. gilli обычно встречается
на литорали, предпочитая песчано-галечные и каменистые грунты; ниже
глубин в 100—110 м он, повидимому, не опускается.
3. Pagurus cavimanus (Miers).
Eupagurus cavimanus Miers, 1879 : 21,48, t. 3, fig. 1; Balss, 1913:58, fig-. 34; Terao,
1913:365; Yokoya, 1933 : 81.— Eupagurus munitus Benedict, 1892 :19.— Pagurus muniius
Holmes, 1900 :150; Rathbun, 1904 :161, t. 5, fig. 2.—Eupagurus gotoi Terao, 1913 : 366, fig. 2.
Очень близок к P. gilli, от которого отличается следующими при-
знаками. Глазные стебельки более толстые. Скафоцериты длиннее, дости-
гают концов глаз. Карпус правой клешненосной ноги (фиг. 68 В) имеет
явственное ребро посредине на верхней поверхности, которое усажено
у проксимального конца мелкими шиповатыми гранулами (у Р. gilli поверх-
ность карпуса равномерно закругленная и совершенно гладкая). Верхняя
поверхность ладони правой руки покрыта неровными сетчатыми линиями:
из мелких плоских гранул, которые образуют своеобразный узор, хорошо
5. PAGURUS
179
заметный только под лупой (у Р. gilli поверхность ладони гладкая, под
лупой видны только редкие, мелкие точечные углубления). Ходильные
ноги относительно более слабые, их пальцы более узкие. Окраска спирто-
вых экземпляров (по Benedict): меральные и карпальные членики клешне-
носных ног светлопурпурные. Руки желтые. Ходильные ноги темного,
красновато-оранжевого цвета с маленькими круглыми светлыми пятнами.
Размеры
Длина передней части карапакса 10.5 мм
Ширина „ „ 10.0 „
Длина глазных стебельков 6.0 „
„ скафоцеритов 4.5 „
„ меруса правой клешненосной ноги . . . 8.5 „
„ карпуса „ „ п.о „
Ширина „ „ „ 13.0 „
Длина руки „ „ „ 18.0 „
Ширина „ „ „ 9.0 „
Длина меруса левой „ 6.0 „
„ карпуса „ „ „ . . . 6.5 „
» РУКН ,, и.о „
Ширина „ „ „ „ . . . 5.0 „
Длина правой ход. ноги первой пары . . . 35.0 „
Распространение. Сев. Япония (Miers); Южн. Япония (Yokoya).
Берингово море (широта 56°56’N) к югу и востоку до Portlok Bank
и Juneau, Аляска; от 36 до 250 м. (Rathbun).
*4. Pagurus tenuimanus (Dana). (Табл. 1, фиг. 2).
Bernhardus tenuimanus Dana, 1851:269; 1852:447, t. 27,’fig. 7.—Eupagurus tenuimanus
Stimpson, 1857:447; 1858 : 237; Smith, 1878—79: 211; Benedict, 1892:1; Walker, 1898:
274. — Pagurus tenuimanus Holmes, 1900:148; Rathbun, 1904 :160; Stevens, 1925 :293,
fig. 14.
Передняя часть карапакса немного больше в длину, чем в ширину,
совершенно голая. Средний выступ лба широко-треугольный, боковые
выступы широко округлые, но выглядят острыми благодаря довольно
сильным субмаргинальным шипам. Глазные стебельки немного длиннее
половины передней части карапакса, почти цилиндрические. Глазные
чешуйки длинные, узкие, с завернутыми краями и с острыми субтерми-
нальными шипами. Скафоцериты достигают концов глаз, а у более
молодых экземпляров немного превышают их. Наружный выступ базаль-
ного членика антеннальной ножки оканчивается двумя шипами. Клешне-
носные ноги очень неравные, правая значительно больше левой. Мерус
правой клешненосной ноги короткий, трехгранный, совершенно гладкий;
вершина переднего края и нижне-наружный дистальный край вооружены
несколькими шипами. Карпус широко-треугольной формы, с выпуклой
верхней поверхностью, которая покрыта шиповатыми гранулами с белыми
12*
180
PA GURIDAE
вершинами; на внутренней стороне верхняя поверхность резко отграни-
чена от гладкой нижней поверхности рядом крупных, белых, направленных
вперед шипов; с наружной стороны верхняя поверхность равномерно
переходит в нижнюю, постепенно утрачивая свои гранулы. На гладкой
нижней поверхности имеется углубление на внутренней стороне и полу-
круглый ряд неправильных шипов, который направляется от середины
к наружному дистальному углу. Рука значительно шире, чем карпус,
довольно тонкая; верхняя и нижняя поверхность густо покрыты крупными
гранулами; гранулы нижней поверхности более мелкие, постепенно исче-
зающие по направлению к сочленению карпуса; наружный край ладони
равномерно закругленный по направлению к кончику неподвижного пальца
и усажен бордюром мелких гранул; внутренний край ладони, обладающий
более крупными гранулами, образует угловатый выступ у основания по-
движного пальца; пальцы широко-треугольные, слегка вогнутые на верхней
поверхности, без роговых концов. Рука настолько широка, что закры-
вает на подобие operculum устье обитаемой животным раковины, когда
последнее прячется в нее. Левая клешненосная нога достигает сере-
дины ладони правой руки. Мерус гладкий, трехгранный. Покатая верхне-
наружная поверхность карпуса покрыта крупными гранулами белого цвета;
внутренний край вооружен рядом крупных белых зубцов; нижняя поверх-
ность совершенно гладкая, выпуклая. Рука такой же ширины, как карпус,
с сравнительно плоской верхней поверхностью, покрытой редкими мел-
кими гранулами; несколько приподнятые края ладони образованы бор-
дюром из более крупных гранул; нижняя поверхность выпуклая и гладкая;
пальцы с роговьпци концами. Ходильные ноги правой стороны несколько
превышают правую клешненосную ногу. Карпальные членики вооружены
вдоль верхнего края белыми зубцами, проподальные членики на верхнем
краю с красными зубцами; все членики сплющенные' и безволосые; пальцы
слабо изогнутые, шиповатые вдоль верхнего и нижнего края, с сильным
роговым когтем на конце. Окраска живых экземпляров (по Stevens’y)
светлая розовато-коричневая до коричневой, с оранжевыми, хромовыми
и серовато-фиолетово-синими пятнами. Клешненосные ноги коричневые,
с синевато-хромовым оттенком на проксимальных и дистальных концах.
Шипы белые, карпус коричневый, с слегка каштановым оттенком. Бугорки
белые, рука .коричневая. Моральные членики ходильных ног .окрашены,
как клешненосные ноги. Первые три сегмента светло-оливкового цвета
у дистальных концов. Жгуты антенн коричневые с полоской каштанового
цвета, которая тянется по наружной и внутренней поверхности. Окраска
спиртовых экземпляров бледнорозовая. Карпальные членики ходильных
ног с тремя красными продольными полосами на наружной поверхности.
Проподальные членики с красными пятнами на наружной поверхности,
частично сливающимися в полосы. Имеется резкая красная полоса вдоль
внутреннего края верхней и нижней поверхности карпуса левой клешне-
носной ноги.
5. PAGURUS
181
Размеры
Длина передней части карапакса ....
Ширина „ „ „ . . . . ... 7.0 „
Длина глазных стебельков .... ... 4.5 „
„ меруса правой клешненосн. ноги . ... 7.0 „
„ карпуса „ . . . 10.0 „
Ширина „ . . . 10.0 „
Длина руки . . . 13.0 „
Ширина „ . . 13.0 „
Толщина „ „ „ „ ... 5.0 „
Длина меруса левой „ „ ... 6.0
„ карпуса „ „ „ ... 6.5 „
Ширина „ ... 4.0 „
Длина руки ... 7.5 „
Ширина „ „ „ „ ... 4.0 „
Длина левой ходильн. ноги первой пары . ... 2.9 „
Распространение. Алеутские о-ва на запад до Командор-
ских о-вов включительно. По американскому побережью на юг до про-
лива Фука и Puget Sound (штат Вашингтон).
Экология. Встречается на глубинах от 1 до 225 м, чаще всего
на 20—70 м, на литораль не выходит.
* 5. Pagurus hirsutiusculus (Dana). (Табл. 3, фиг. 4).
Eupagurus mertensii Brandt, 1851:112. — Bemhardus hirsutiusculus Dana, 1851 : 270;
1852:443, t. 27, fig. 3. — Eupagurus hirsutiusculus Stimpson, 1858:250; 1907 : 223; Alcock,
1905 :178; Balss, 1913 :62, t. 1, fig. 9; Terao, 1913 : 369; Yokoya, 1933 : 83. — Pagurus hirsu-
tiusculus Holmes, 1900:143; Rathbun, 1904 :159; Hilton, 1916 : 69; Schmitt, 1921:137, text-
fig. 89, t. 16, fig. 4; Stevens, 1925:281, fig. 4.
Передняя часть карапакса больше в ширину чем в длину; поверх-
ность ее, за исключением центрального участка, покрыта короткими,
нежными волосками. Средний выступ лба короткий, треугольный; боковые
выступы отсутствуют. Глазные стебельки короткие, довольно толстые,
с перехватом посредине. Глазные чешуйки полуовальные, с закрученной
вершиной и сильными субтерминальными шипами. Скафоцериты немного'
превосходят глаза, но иногда могут быть и короче их. Клешненосные
ноги волосатые, за исключением правой руки. Левая клешненосная нога
достигает только середины ладони правой руки. Верхнее ребро меруса
правой клешненосной ноги с поперечными морщинами, от которых отходят
волоски; боковые поверхности сравнительно гладкие; нижняя поверхность
густо волосатая. Выпуклая поверхность карпуса гранулирована и покрыта
мелкими волосами; нижняя поверхность гладкая. Рука довольно правиль-
ной овальной формы, равномерно покрытая гранулами, с одиночными
волосками, которые едва заметны даже под лупой; наружный край,
усаженный гранулами, резко отграничивает верхнюю поверхность от ниж-
ней, которая покрыта более мелкими и редкими гранулами; наружный
край подвижного пальца несет два продольных ряда гранул; пальцы
182
PAGURIDAE
с очень маленькими роговыми кончиками. Карпус левой клешненосной
ноги узкий, волосатый, с гранулированной верхней поверхностью, гранулы
на наружном и внутреннем краях более крупные; нижняя поверхность
менее волосатая. Рука такого же характера, как правая, но более малень-
кая, незначительно волосатая, наружный край подвижного пальца гладкий,
с отдельными пучками волос. Ходильные ноги волосатые; на правой сто-
роне они немного превышают правую клешненосную ногу; пальцы корот-
кие, шиповатые на нижнем краю, с короткими роговыми когтями. Окраска
живых экземпляров (по Stevens’y) вариирует от светлой аквамариново-
синей до оливковой, которая выглядит сероватой благодаря многочислен-
ным мелким волоскам. Клешненосные ноги темносерые с светлыми
аквамариново-синими гранулами, пальцы сверху белые, снизу иногда
синие. Проподусы ходильных ног белые у дистальных и синие у прокси-
мальных концов; пальцы светлые аквамариново-синие, с коричневыми.
продольными полосами. Антенны оливково-коричневые с золотисто-
желтыми пятнами, чередующимися через каждые 2 или 3 сегмента жгута;
у первого десятка каждый сегмент с пятнами. Молодь по окраске несколько
отличается от взрослых: клешненосные и ходильные ноги вариируют
от цвета красного дерева до темносинего, а в некоторых случаях до светло-
оливкового цвета; у суставов ноги окрашены в белый или светлосиний
цвет; на дистальном конце карпуса левой клешненосной ноги часто
имеется белое треугольное пятно; антенны окрашены, как у взрослых.
Общая окраска спиртовых экземпляров красновато-желтая; правая рука
беловатая, иногда с зеленоватым оттенком; дистальные концы проподусов
ходильных ног белые, пальцы на боках с продольными красными полосами.
Размеры
Длина передней части карапакса 7.0 мм
Ширина „ „ „ 7.5 „
Длина глазных стебельков з.о „
„ меруса правой клеши, ноги 9.0 „
„ карпуса „ „ 8.5 „
Ширина „ „ 7.0 „
Длина руки „ „ „ 12.0 „
Ширина „ „ „ „ 8.0 „
Длина меруса левой ,, „ 7.0 „
„ карпуса „ „ „ 7.0 „
Ширина „ „ „ ........ 3.0 „
Длина руки „ „ 8.0 „
Ширина „ „ „ „ 4.0 „
Длина правой ход. ноги первой пары 31.0 „
Распространение. Японское море, по нашему побережью на юг
не далее залива Петр Великий; северная Япония (до Сангарского про-
лива); Охотское море; восточное побережье Камчатки; Командорские о-ва;
Алеутская гряда; Прибыловы о-ва; по американскому побережью к югу
до San Diego (Калифорния).
5. PAG UR US
183
Экология. Один из наиболее обычных видов литорали, преиму-
щественно на каменистых фациях; его часто можно находить под камнями
в полосе отлива. Встречается также в верхних горизонтах сублиторали
примерно до 30 м. Наибольшая глубина, на которой он был найден, рав-
няется 110 м (Сангарский пролив, Yokoya 1933).
* 6. Pagurus bernhardus (Linne). (Табл. 4, фиг. б).
Cancer bernhardus Linne, 1758 : 631. — Pagurus bernhardus Fabricius, 17 75 : 411; Risso,
1828: 37; Milne-Edwards, 1836 :266; 1837 :215; 1849: t. 44, fig. 2; Bell, 1853:171; Benedict,
1901:452; Яржинскнй, 1870 : 317. — Pagurus streblonyx Leach, 1815. — Eupagurus bernhardus
Brandt, 1851:106; Heller, 1863:160; Carus, 1884 : 491; Ortmann, 1892:303; Bouvier,
1896:151; Birula, 1897 : 436; Milne-Edwards et Bouvier, 1900 : 239; Hansen, 1908:25; Balss,
1912:106; Dons, 1934:40, 113. — Pagurus bernhardus var. granulaia Brandt, 1851 :107.—
Eupagurus acadianus Benedict, 1901:454.
Передняя часть карапакса почти такой же длины, как и ширины,
или немного больше в длину чем в ширину. Средний выступ лба острый,
треугольный, выдающийся немного дальше боковых выступов, которые
снабжены сильными субмаргинальными шипами. Глазные стебельки корот-
кие, толстые, с сильным перехватом посредине. Глазные чешуйки овальные,
с сильными субтерминальными шипами. Скафоцериты простираются
немного дальше глаз; их нижние ребра отчетливые, но не острые, тогда
как верхне-внутреннее ребро закругленное, неровное и несет пучки
коротких волосков. Наружный выступ базального членика антеннальной
ножки с большим концевым шипом и крупными шипами на внутреннем
крае; внутренний выступ несет сильный шип. Мерус правой клешненосной
.ноги с почти гладкими гранями и с рядом зубцов вдоль верхнего дисталь-
ного края. Верхняя поверхность карпуса покрыта крупными, тупыми, кони-
ческими гранулами, вершины которых часто бывают раздвоены; более
крупные гранулы располагаются на внутреннем крае, от оснований гранул
поднимаются редкие волоски, которые почти незаметны для невооружен-
ного глаза; боковые поверхности несут более редкие и плоские гранулы.
Рука сильная, с выпуклой верхней поверхностью, усеянной мелкими
тупыми гранулами, придающими ей шагреневый вид; более крупные гра-
нулы образуют посередине ладони Д-образную фигуру, обращенную вер-
шиной к основанию пальца. Шиповатые гранулы окаймляют руку по наруж-
ному краю; более крупные гранулы располагаются также на валикообразно
утолщенном внутреннем крае ладони; волоски почти отсутствуют; нижняя
поверхность с рассеяными плоскими гранулами; пальцы толстые, с извест-
ковыми кончиками. Левая клешненосная нога достигает основания
подвижного пальца правой руки. Карпус с рядом шипов на внутреннем
крае; покатая наружная поверхность покрыта шиповатыми гранулами,
причем между внутренним краем и этой поверхностью остается гладкий
промежуток. Поверхность руки покрыта такими же мелкими гранулами,
как и на правой руке, и также часто имеется Д-образная фигура из более
крупных гранул; наружный край несет более крупные гранулы, а вну-
184
PA GURIDAE
тренний приподнятый край шиповатые гранулы. Ходильные ноги правой
стороны очень незначительно превышают правую клешненосную ногу.
Карпальные членики сверху шиповатые; проподальные членики с гранули-
рованной верхне-наружной поверхностью. Пальцы сильные, с мощными
роговыми когтями. Окраска коричневато-желтая, иногда с красноватыми
пятнами, на ногах часто с красновато-коричневыми продольными полосами-
Размеры
Длина передней части карапакса . . . 13.0 мм
Ширина „ „ „ . . . 13.0 „
Длина глазных стебельков ... 6.0 „
„ скафоцеритов ..... ... 5.5 „
Длина меруса правой клешнен. ноги . . . 14.0 „
„ карпуса „ „ . . . 14.0 „
Ширина „ „ „ „ . . 11.0 „
Длина руки „ „ . . . 23.0 „
Ширина „ „ „ „ . . . 13.5 „
Длина меруса левой клешнен. ноги . . . 12.0 „
„ карпуса „ „ „ . . . 12.0 „
Ширина „ „ „ „ ... 6.0 „
Длина руки „ „ „ . . . 15.0 „
Ширина „ „ . „ ... 8.5 „
Длина правой ход. ноги первой пары . . . 56.0 „
Распространение. Западный Мурман (на восток до Кольского»
залива включительно), побережье Норвегии, Немецкое море, Большой
Бельт (редко); Шетландские о-ва, побережье Шотландии, Англии и Ирлан-
дии; северное и западное побережье Франции (Гасконский залив);1
Исландия; восточное побережье Северной Америки (от Большой Нью-
фаундлендской банки до залива Chesapeake).
Экология. В наших водах на Мурмане Р. bernhardus встречается:
значительно реже чем Р. pubescens и преимущественно в западной части
(Мотка, Ара, Корабельная, Подпахта). По данным А. Бирули (1897), он не
распространялся на восток далее п-ова Рыбачьего, что подтверждалось
и данными К. М. Дерюгина (1915). При позднейших работах2 вид этот
обнаружен в Кольском заливе (у губы Волковой и у Б. Оленьего о-ва);
согласно устным сообщениям сотрудников бывш. Мурманской Биологиче-
ской Станции, за последние годы ее работы наблюдалось значительное
увеличение частоты встречаемости Р. bernhardus в Кольском заливе; таким
образом, намечается тенденция к продвижению его на восток. Держится
этот вид обычно на незначительных глубинах вблизи берега. У Исландии
он опускается до 108 м, у Фарерских о-вов до 180 м, и у залива Chesa-
peake (восточное побережье Северной Америки) до 477 м.
1 Нахождения у побережья Португалии и в Средиземном море вызывают сомнение?
повидимому, как отметил еще Heller (1863), эти местонахождения следует отнести за счет-
Р. prideauxi Leach.
2 Н. П. Танасийчук. О новых и редких для фауны Кольского залива формах
животных. Докл. Акад. Наук СССР, 1927.
5. PAGURUS
185
* 7. Pagurus gracilipes (Stimpson). (Табл. 4, фиг. 4).
Eupagurus gracilipes Stimpson, 1858:248; 1907 : 217; Doflein, 1902:647, t. 6, fig'. 6—8;:
Alcock, 1905:177; Balss, 1913:56; Terao, 1913:368. — Eupagurus nipponensis Yokoya,
ibid.; 87, fig. 32. — Non Eupagurus gracilipes Yokoya, 1933:89, fig. 33.1
Передняя часть карапакса такой же ширины, как и длины, или немного
больше в ширину, чем в длину. Средний выступ лба треугольный, высту-
пающий далее боковых выступов, явственных и снабженных острыми
субмаргинальными шипиками, направленными вперед и наружу. Поверх-
ность карапакса почти голая, с отдельными разбросанными пучками воло-
сков, главным образом на периферии. Глазные чешуйки овальные, закруг-
ленные на вершине, с сильно • вогнутой поверхностью и с довольно
короткими субтерминальными шипами. Глазные стебельки короткие
и толстые, немного расширяющиеся у роговиц. Наружный выступ базаль-
ного членика антеннальной ножки плотно прилегает к следующему сег-
менту, имеет острую вершину и зазубренный внутренний край. Скафоце-
риты слабо изогнутые, заметно превышают кончики глаз. Мерус правой
клешненосной ноги гладкий, на переднем крае зубчатый. Поверхность
карпуса шиповатая, шипы располагаются примерно в четыре продольных
ряда, между которыми остаются гладкие промежутки; шипы внутреннего
края более крупные и острые; нижняя поверхность выпуклая, мелко грану-
лированная. Ладонь слабо выпуклая, негусто покрытая мелкими грану-
лами; более крупные гранулы окаймляют сильно утонченный наружный
край ладони, переходя и на край неподвижного пальца; внутренний край
ладони валикообразно утолщен и покрыт гранулами более густо, чем:
остальная поверхность ладони; нижняя поверхность выпуклая и гладкая;,
пальцы плоские, гладкие, почти лишены гранул, за исключением наруж-
ных краев; концы известковые. Левая клешненосная нога достигает сере-
дины ладони правой руки, Карпус с двумя продольными рядами шипов
на верхней поверхности; наружная поверхность гранулирована. Рука
такого же строения, как и правая, тодько гранулы поверхности более
шиповатые; концы пальцев роговые. Ходильные ноги правой стороны
превосходят правую клешненосную ногу более чем на половину длины
своих пальцев. Карпальные и проподальные членики сверху шиповатые.
Пальцы тонкие, искривленные, длинные, значительно превышающие длину
соответствующих проподусов, 'очень слабо закрученные, с острыми рого-
выми кончиками. Все конечности, как и карапакс, очень слабо волосатые.
При подсыхании спиртовых экземпляров конечности приобретают слабый
перламутровый блеск. Карпальные и проподальные членики ходильных
1 Eupagurus yokoyai nomen nov. Eupagurus gracilipes Yokoya, 1933:89, text-fig. 33-
(nomen praeoccup.). Местонахождение: ст. 353, близ Tanabe, Wakayama-ken, 192 m. 4 VIII
1928, 3 8 3, 3 § § (1 ov.). „Близок к Pag. pectinatus (Stimpson), но отличается от него сле-
дующими признаками: глазные стебельки короче. Ножки первых н вторых антенн явственно
превышают роговицы глаз, волосатый скафоцерит достигает дистального конца глаза. Ноги
тонкие (Yokoya)". Вид более близок, скорее, к Pog. brachiomastus (Thallw.).
186
PAGURIDAE
ног с красными продольными полосками на наружной поверхности.
У молодых экземпляров скафоцериты значительно короче глаз. На верх-
ней поверхности карпуса правой клешненосной ноги имеется только два
продольных ряда шипов с гладкой поверхностью между ними. Боковые
выступы лба выдаются более резко.
Размеры
Длина передней части карапакса 5.0 мм
Ширина „ „ 5.0
Длина глазных стебельков 2,5 п
„ скафоцеритов 3.5
„ меруса правой клешненосной ноги ...... 7.0
„ карпуса „ >> » * • .... 6.0 »>
Ширина „ „ » п • 4.0
Длина руки „ „ 9.0
Ширина „ „ „ 5.0
Длина меруса левой » 4.5
„ карпуса „ » 4.0 г>
,, руки „ 6.0 »>
Ширина „ „ „ • 3.0 »
Длина правой ход. ноги первой пары 26.0
Распространение. Япония, главным образом южная; самым:
северным местонахождением в Японии является Хакодате. Для матери-
кового побережья Японского моря (бухта Сидеми, бухта Троицы) при-
водится впервые.
Экология. Вид отмечен с глубин от 30 до 600 м, хотя чаще всего
встречается на глубинах 50—150 м.
8. Pagurus beringanus (Benedict). (Табл. 5, фиг. 4).
Eupagurus beringanus Benedict, 1892:17. — Eupagurus newcombei Benedict, ibid.:
17. — Pagurus beringanus Rathbun, 1904:159, t, 5, fig. 5; Schmitt, 1921:135, fig. 87; Stevens,
1925 :283, fig. 6.
Передняя часть карапакса больше в длину, чем в ширину; поверх-
ность голая, с редкими пучками волосков на боках. Средний выступ лба
очень короткий, широкотреугольный. Боковые выступы в виде тупых
углов, образуемых передним краем карапакса, без субмаргинальных шипов.
Глазные чешуйки треугольные, с тупой вершиной и с сильными, тупыми
•субтерминальными шипами. Глазные стебельки длинные, почти цилиндри-
ческие. Наружный выступ базального членика антеннальной ножки
с прямым волосатым наружным краем, с выпуклым внутренним краем
и с шипом близ верхушечного шипа. Скафоцериты изогнутые, короче
глаз, в лучшем случае достигают только основания роговиц; наружный
край скафоцеритов гладкий, внутренний волосатый и зазубренный. Мерус
правой клешненосной ноги трехгранный, с гладкими боковыми поверх-
ностями; верхняя грань дистально расширяется в небольшую площадку,
5. PAGURUS
187
усаженную короткими шипами и редкими волосками; передний край
с 15—16 шипами; нижняя поверхность с двумя крупными бугрообразными
гранулами, наружная из которых обычно больше внутренней; полукруглая
дистальная грань покрыта шиповатыми гранулами и волосами. Верхняя
поверхность карпуса выпуклая, покрыта тупыми шиповатыми гранулами;
более крупные гранулы расположены вдоль внутреннего края; другая
линия более крупных гранул тянется от внутреннего проксимального
угла косо вверх; боковая и нижняя поверхности более или менее гладкие,
почти безволосые. Рука такой же ширины, как карпус, поверхность ладони
выпуклая, довольно густо покрытая тупыми гранулами;' более крупные
гранулы, иногда имеющие вид шипов, образуют бордюр на выпуклом
крае; кроме того, более крупные гранулы образуют посредине ладони Д-
-образную, не всегда ясно различимую фигуру; вершина ее обращена
вперед, а внутренняя ветвь продолжается на карпусе в виде уже упомя-
нутой косой линии гранул; нижняя поверхность ладони выпуклая, гладкая,
с редкими.пучками волосков; пальцы короткие, с гранулированной верхней
поверхностью и с небольшими роговыми концами. Левая клешненосная
нога достигает середины ладони правой руки, у более мелких экзем-
пляров— основания подвижного пальца. Мерус трехгранный, с воло-
сатой нижней поверхностью. Наружная покатая поверхность карпуса
гранулирована, верхний край усажен шипами; внутренняя поверхность
плоская, волосатая, с редкими гранулами. Ладонь без резкого гребня
посредине, с выпуклой наружной поверхностью, густо покрытой гранулами;
более крупные гранулы группируются у верхнего тупого гребня; наружная
поверхность образует явственное ребро с нижней поверхностью, гладкой
и покрытой пучками волос. Пальцы при смыкании оставляют небольшую
щель. Ходильные ноги не достигают конца правой клешненосной ноги.
Карпальные и проподальные членики с шипами и волосами на верхних
краях. Пальцы короткие, довольно толстые, прямые, с острыми изогну-
тыми когтями. Окраска живых экземпляров (по Stevens’y) коричневая
с светлыми пятнами. Клешненосные ноги с огненно-красными шипами,
пальцы бледные, серовато-зеленые, с красноватым оттенком; дистальный
конец меруса с красной полосой, оканчивающейся шипами такого же цвета.
Ходильные ноги более светлые, чем клешненосные, на пальцах и пропо-
дусах часто зеленоватые, с неправильными красновато-коричневыми пят-
нами. Окраска спиртовых экземпляров красновато-бурая.
Размеры
Длина передней части карапакса............10.0 мм
Ширина „ „ „ ................. 9.0 „
Длина глазных стебельков .............. 5.0 „
„ скафоцеритов ........... 4.0 „
Длина меруса правой клешненосной ноги.....12.0 „
„ карпуса „ „ ..............11.0 „
Ширина „ „ „ ........ 8.0 „
188
PAGURIDAE
Длина руки правой клешненосной ноги . . .
Ширина „ Я я я • ’ • • ... 8.0 ,»
Длниа меруса левой я я * • • ... 9.0 я
„ карпуса я я я • • ... 8.0 я
„ руки я я . . . « ... 9.0 я
Ширина „ я я » • • • • ... 5.0 я
Длина правой ход. ноги первой пары ................42.0 „
Распространение. Берингово море (широта о-ва Нунивок)
к югу вдоль Алеутских о-вов и по побережью Аляски до залива Monterey,.
Калифорния (М. Rathbun).
Экология. На глубинах от 0 до 82 м, преимущественно на каме-
нистых фациях.
* 9. Pagurus dubius (Ortmann). (Табл. 3, фиг. 5).
Eupagurus dubius Ortmann, 1892 :307, t. 12, fig. 14; Doflein, 1902:646; Balss, 1933 :55;
Yokoya, 1933:81.
Передняя часть карапакса больше в длину, чем в ширину. Средний
выступ лба широко треугольный, острый, иногда более или менее приту-
пленный; боковые выступы округлые. Глазные стебельки довольно длин-
ные, цилиндрические; глазные чешуйки полуовальные, с довольно силь-
ными субтерминальными шипами. Скафоцериты не достигают концов
глаз, но у более мелких экземпляров могут несколько превышать их.
Клешненосные ноги очень неравные. Мерус правой клешненосной ноги
с редкими волосками на боковых сторонах, зубчатый на переднем крае;
нижняя поверхность более волосатая, с заметно выдающимся тупым
шипом в центре. Верхняя поверхность карпуса покрыта шиповатыми
гранулами, которые постепенно исчезают на боковых сторонах; верхняя
и полукруглая внутренняя поверхности с длинными тонкими волосами;,
плоская наружная поверхность почти безволосая; на наружной половине
нижней поверхности имеется лишенное волос возвышение, тогда как
внутренняя половина волосатая и покрыта мелкими гранулами. Рука очень
незначительно шире, чем карпус, верхняя поверхность совершенно без-
волосая, равномерно покрытая полупрозрачными на вид гранулами; более
крупные гранулы окаймляют края руки, причем гранулы наружного края
крупнее таковых внутреннего края; наружный край подвижного пальца
несет два продольных ряда подобных же гранул; концы пальцев роговые.
Левая клешненосная нога простирается немного дальше дистального
конца карпуса правой клешненосной ноги. Мерус сплющенный, волосатый
сверху, голый на боках; нижняя поверхность волосатая и с таким же
выдающимся тупым шипом в центре, как и на мерусе правой ноги. Карпус
узкий, волосатый, с двумя рядами шипов вдоль края верхней поверхности;
нижняя поверхность волосатая на внутренней половине и голая на наруж-
ной. Рука узкая, верхняя поверхность волосатая, покрыта шиповатыми
гранулами; нижняя поверхность гладкая и безволосая. Пальцы при смыка-
5. PAGURUS
189
нии оставляют небольшую щель у основания. Ходильные ноги достигают
конца правой клешненосной ноги или немного превышают его. Боковые
поверхности члеников голые, верхние и нижние края волосатые. Карпаль-
ные членики с шипами вдоль верхнего края. Пальцы слабо изогнутые,
с шиповатыми нижними краями. Окраска спиртовых экземпляров бледная,
розовато-желтая. Карпальные и проподальные членики ходильных ног
с продольными красными полосами на наружных поверхностях; пальцы
красные, с продолговатым белым пятном посредине. Верхняя поверх-
ность ладони правой руки имеет обычно красный узор причудливого,
зю более или менее постоянного очертания.
Размеры
Длина передней части [ карапакса .
Ширина „ „ 6.0 „
Длина глазных стебельков 4.0 „
Длина меруса правой клешненоснон ноги . . ..... 9.0 „
„ карпуса „ » » • • 8.5 „
Ширина „ „ 6.0 „
Длина руки „ >> » • • 12,0 „
Ширина „ „ » » • • 6.5 „
Длина меруса левой 6.0 „
„ карпуса „ 5.5 „
Ширина „ „ » • 2.0 ,,
Длина руки „ J> » • • 8.0 „
Ширина руки „ » ft * * 3.5 „
Длина правой ход. ноги первой пары . . . . 31.0 ,,
Распространение. Южная Япония; Японское море, по нашему
побережью на север до залива Ольги.
Экология. Представители этого вида не достигают крупных раз-
меров; держатся обычно на глубинах 50—120 м, изредка встречаются
на меньших глубинах.
* 10. Pagurus samuelis (Stimpson). (Табл. 3, фиг. б).
Eupagurus samuelis Stimpson, 1857 :482; 1858: 250; 1907 : 224; Ortmann, 1892:301;
Doflein, 1902: 646; Alcock, 1905:178; Balss, 1913:61; Terao, 1913: 371. — Pagurus samuelis
Holmes, 1900 :144; Rathbun, 1904:160, t. 5, fig. 7; Hilton, 1916 : 63; Schmitt, 1921 :139, text-
fig. 90, t. 16, fig. 2, 3.
Передняя часть карапакса больше в длину, чем в ширину. Средний
выступ лба широкий, треугольный, скрытый пучком длинных волосков;
боковые выступы закругленные. Глазные стебельки длинные, цилиндри-
ческие. Глазные чешуйки широко треугольные, с тупой вершиной и
сильным субтерминальным шипом. Скафоцериты не достигают концов
глаз, с волосками на внутреннем крае. Вершина наружного выступа
'базального членика антеннальной ножки заканчивается двумя шипами.
Мерус правой клешненосной ноги с гладкими боками, с шиповатыми
гранулами и волосами на дистальном конце верхней поверхности и с силь-
190
PAGURIDAE
ным тупым шипом в центре нижней поверхности. Верхняя поверхность
карпуса покрыта шиповатыми гранулами и волосами, боковая и нижняя
поверхности с широкими низкими гранулами и менее волосатые. Выпуклая
верхняя поверхность руки в сущности безволосая, одиночные мелкие
волоски заметны только при рассматривании под лупой; гранулы в виде
бусинок равномерно покрывают всю поверхность; более крупные гранулы
окаймляют края руки; нижняя поверхность покрыта мелкими гранулами,,
особенно на нижней стороне. Пальцы с роговыми концами. Мерус
левой клешненосной ноги на нижней поверхности также с тупым шипом.
Карпус шиповатый и волосатый. Верхняя поверхность ладони шиповатая
и почти безволосая, нижняя поверхность гладкая, с одиночными волосками.
Подвижный палец совершенно голый. Карпальные членики ходильных:
ног шиповатые на верхних краях. Пальцы слабо изогнутые и закрученные,
с темными'проксимальными и дистальными концами и светлой серединой.
Окраска спиртовых экземпляров желтоватая. У живых экземпляров,
антенны красновато-коричневые; верхняя поверхность дистального членика
стебля с узкими золотистыми полосками. Дистальная треть проподусов.
ходильных ног голубая или белая, у проксимального конца эти членики
синие, на остальной поверхности с синеватыми или зеленовато-коричне-
выми полосками; пальцы синие, становящиеся белыми около конца,
а у самого когтя красновато-оранжевые, с тремя более или менее ясными
синеватыми или зеленоватоукрасновато-коричневыми полосками. Правая
рука обычно темного синевато-зеленого цвета, концы пальцев белые;;
шипы на левой руке часто бывают оранжевые. W. L. Schmitt (1921) на
основании просмотра большого материала отмечает, что правая клешне-
носная нога у самцов с возрастом становится продолговатой и сравни-
тельно гладко гранулированной; крупные гранулы на наружном крае руки
исчезают. Этими признаками крупные самцы заметно отличаются от самок
и, если пренебречь сходством в остальных признаках, могут быть приняты
даже за особый вид; на большой возрастной серии можно, однако, про-
следить все промежуточные формы.
Размеры
Длина передней части карапакса 5.0 мм
Ширина „ „ „ 4-5 „
Длина глазных стебельков з.о ,,
Длина меруса правой клешненосн. ноги 4.5 „
„ карпуса „ „ 5.0 „
Ширина „ „ 4.0 „
Длина рукн 7-0 „
Ширина „ „ „ 5.0 „
Длина меруса левой „ 4-0 „
„ карпуса „ „ 4.5 „
Ширина „ „ „ 2.0 „
Длина руки „ „ „ 5.0 „
Ширина „ „ „ „ • з.о „
Длина правой ход. ноги первой пары 18.0 „
5. PAGURUS
191
Этот вид очень близок к Р. dubius (Ortm.), от которого отличается
более короткими скафоцеритами, более тупым средним выступом лба,
более короткой ладонью правой руки, более мелкой гранулированностью ее,
отсутствием красного рисунка на ладони той же руки, столь характерного
для Р. dubius, совершенно голым подвижным пальцем левой руки и более
короткими пальцами ходильных ног.
Распространение. Япония; на нашем побережье Японского
моря известен пока только из бухты Находка и залива Ольга; Куриль-
ские о-ва (о-в Итуруп); Авачинская бухта, Камчатка (согласно Н. Balss);
по американскому побережью от Ситки до San Diego, Калифорния.
* 11. Pagurus cornutus (Benedict). (Табл. 5, фиг. 1).
Eupagurus cornutus Benedict, 1892 :12. — Pagurus- cornutus Rathbun, 1904 :158, t. 5,
fig. 3.
Передняя часть карапакса немного больше в ширину, чем в длину,
у крупных экземпляров такой же ширины, как и длины. Средний выступ
лба треугольный, острый; боковые выступы выдаются меньше, с неболь-
шими субмаргинальными шипами. Глазные стебельки умеренной длины,
толстые, несколько расширяются у роговиц. Глазные чешуйки полуоваль-
ные, с сильными субтерминальными шипами. Скафоцериты простираются
дальше глаз почти на половину своей длины, их внутренние края воло-
сатые. Наружный выступ базального членика антеннальной ножки
с шипом на вершине и с 3—4 шипиками на внутреннем крае. Клешненос-
ные ноги сильные. Мерус правой клешненосной ноги трехгранный,
с шипами на верхне-дистальном крае; тупое верхнее ребро покрыто попе-
речными морщинками, от которых отходят пучки волос; нижняя поверх-
ность с шиповатыми краями. Верхняя поверхность карпуса негусто покрыта
шипами, впереди от основания которых имеются пучки длинных волос-
ков; шипы краев, в особенности внутреннего края, более сильные. Рука
с прямым внутренним и дугообразным, наружным краем. От внутрен-
ней стороны основания ладони тянется вперед по диагонали сильный
гребень, который у основания подвижного пальца образует острый, на-
правленный косо вверх и вперед выступ на подобие рога; второй, более
слабо намечающийся, гребень тянется от наружной стороны основания
ладони и сливается с первым гребнем на наружной поверхности рога.
Таким образом, у основания ладони образуется возвышенная, ограничен-
ная сходящимися гребнями, треугольная площадка, поверхность которой
гранулирована. Остальная поверхность ладони глубоко вогнутая и также
гранулирована. Наружный край подвижного пальца и весь наружный край
руки усажены тупыми шипами, а снизу опушен густыми, мягкими, длинными
волосками. Нижняя поверхность руки выпуклая, гладкая, с рассеянными,
пучками волос. Пальцы плоские, тонкие, с мелко гранулированной верхней
поверхностью и роговыми кончиками. Левая клешненосная нога достигает
середины подвижного пальца правой руки, а у более крупных экзем-
192
PAGURIDAE
пляров только его основания. Мерус сжатый, с такими же волосатыми
морщинками, как и у правой ноги. Карпус с двумя рядами сходящихся
спереди шипов, слабо волосатый, с сильно выдающимся дисталь-
ным нижне-наружным углом. Рука с сильным гребнем, который тянется
•от основания ладони к неподвижному пальцу; он усажен сверху рядом
мелких шипов и покрыт волосами; внутренняя сторона ладони очень
узкая, не шире подвижного пальца; наружная сторона, напротив, широкая,
с глубоко вогнутой гранулированной поверхностью, наружный край снизу
густо опушен волосками; нижняя поверхность гладкая, с пучками волосков.
Неподвижный палец с мелко гранулированной верхней поверхностью;
подвижный палец более длинный, с гладкой волосатой поверхностью;
концы пальцев роговые. Ходильные ноги сильные, на правой стороне
они несколько превышают • конец правой клешненосной ноги. Моральные
членики сплющенные, у первой пары шиповатые снизу. Карпальные
членики обеи?г пар шиповатые сверху. Ноги, вообще, слабо волосатые.
Пальцы изогнутые, сильно закрученные, с тонкими острыми роговыми
когтями. Окраска спиртовых экземпляров красновато-желтая.
Размеры (крупн. экз.)
Длина передней части карапакса...............17.0 мм
Ширина „ „ „ ..................17.0 „
Длина глазных стебельков.................... 9.0 „
„ меруса правой клешнен. ноги...............24.0 „
„ карпуса „ „ „ . . •...........24.0 „
Ширина „ „ „ „ ...............15.0 „
Длина руки „ „ ...................32.0 „
Ширина „ „ „ „ ...............15.0 „
Длина меруса левой клешнен. ноги.............19.0 „
„ карпуса „ „ 18.0 „
Ширина „ „ „ „ .............. 6.0 „
Длина руки „ „ „ .............25.0 „
Ширина „ „ „ 12.0 „
Длина правой ход. ноги первой пары....94.0 „
Распространение. Алеутские о-ва на запад до Командорских о-вов
включительно. Берингово море, по нашему побережью от Командор на
север до широты мыса Наварина; по американскому побережью от широты
55°38'N на юг до Queen Charlotte Sound, Британская Колумбия.
Экология. Глубоководный вид, известен с глубин 160—830 мм.
* 12. Pagurus undosus (Benedict). (Табл. 1, фиг. 5).
Eupagurus undosus Benedict, 1892 :18. — Pagurus undosus Rathbun, 1904 : 159, t. 4,
-fig. 6. — Eupagurus trigonochirus var. paulensis Balss, 1913 : 64, fig. 38, 39.
Передняя часть карапакса незначительно больше в длину, чем
в ширину. Треугольный средний выступ лба выдается немного дальше
боковых выступов, которые округлены и снабжены субмаргинальными
.шипами. Глазные стебельки умеренной длины, цилиндрические. Глазные
5. PAGUR US
193
чешуйки треугольные, с приподнятыми краями и с сильными субтерми-
нальными шипами. Скафоцериты достигают конца глаз, с волосатыми
.внутренними краями. Внутренний край наружного выступа базального
членика антеннальной ножки с 2—4 шипами. Мерус правой клешненосной
ноги трехгранный, с гладкими боковыми поверхностями; передний край
С шипами; на верхнем ребре у дистального конца имеется несколько
одиночных гранул и морщинок, с отходящими от них волосками; нижняя
поверхность волосатая. Верхняя поверхность карпуса с мелкими шипами,
^наиболее крупные шипы занимают внутренний край; шипы наружного края
;мелкие, но более тесно сидящие, чем на остальной поверхности; боковые
поверхности покрыты шиповатыми гранулами; нижняя поверхность
выпуклая, гладкая и волосатая. Внутренний край руки почти прямой,
наружный край несколько дугообразный; от середины основания ладони
к щели между пальцами тянется не очень сильно возвышающийся гребень,
:который на дистальном конце довольно круто обрывается, образуя скат
к внутреннему краю неподвижного пальца; в профиль вершина гребня
-образует совершенно прямую линию, не делая у дистального конца какого
либо повышения в виде бугра, рога и т. п. Благодаря тому, что внутренний
край ладони валикообразно утолщен, между ним и гребнем образуется
продольное углубление с параллельными краями; проксимальный конец
углубления свободно доходит до основания ладони, дистальный конец
отделен от основания подвижного пальца возвышающимся участком.
’Поверхность ладони снаружи от среднего гребня выпуклая. Вся поверх-
ность ладони густо покрыта круглыми гранулами; более крупные гранулы
занимают средний гребень и в особенности его дистальный конец.
Наружный край ладони и подвижного пальца усажен коническими шипами.
У основания гранул имеются венчики очень коротких волосков, которые
заметны только под лупой. Нижняя поверхность руки почти гладкая, слабо
.волосатая. Пальцы со слабыми роговыми концами. Левая клешненосная
нога достигает основания подвижного пальца правой руки. Карпус
с двумя рядами шипов на верхней поверхности, шипы внутреннего ряда
значительно крупнее таковых наружного ряда; внутренняя поверхность
волосатая, наружная гранулирована, нижняя волосатая. Рука с широкой,
•треугольной, косой наружной поверхностью, выпуклой посредине и слегка
вогнутой у краев; резкий шиповатый гребень круто поднимается от вну-
тренней стороны основания ладони и тянется несколько диагонально
к неподвижному пальцу. Наружный край ладони острый, усаженный тупыми
коническими шипами. Наружная косая поверхность ладони гранулирована
подобно правой руке; внутренняя поверхность в виде узкого гранулирован-
ного участка; нижняя поверхность с одиночными гранулами и пучками
волос. Ходильные ноги правой стороны достигают конца правой клеш-
неносной ноги, а у мелких экземпляров немного превышают его: Мораль-
ные членики первой пары шиповатые снизу. Карпальные членики обеих
;пар шиповатые сверху. Пальцы довольно толстые, совершенно прямые
Фауна СССР, Макаров 13
194
P AGURID АЕ
и не закрученные, слабо волосатые сверху, шиповатые снизу и с острым
изогнутым когтем на конце. Окраска спиртовых экземпляров: клешненос-
ные ноги розовато-желтые с коричневато-красной полосой на дистальных
концах мерусов. Шипы на карпусе клешненосных ног с красными вер-
шинами. Концевые части члеников ходильных ног коричневато-красные^
средние части светлые; кончик пальцев более бледно окрашен, чем его,
проксимальная часть; коготь прозрачный, светлокоричневый.
Размеры
Длина передней части карапакса.............
Ширина „ „ ......................
Длина глазных стебельков.......... . . . . .
„ меруса правой клешней. НОГИ
„ карпуса »
Ширина „ it » »
Длина руки It и
Ширина „ » и
Длина меруса левой »
„ карпуса ч и и
Ширина „ >» »» ч
Длина руки » п
Ширина „ ч а и
Длина правой ход. ноги первой пары
8.5 ММ
8.0
4.5 IK
10.0 з>
11.0 th
8.0 »
14.5 ii-
8.0 ii
8.5 ч.
8.5 ч,
3.0 ti-
11.0 ll
6.0 if.
36.0 it
Вид не достигает крупных размеров.
Распространение. Японское море от широты о-ва Фуругельма
на север только по нашему побережью (из Японии южнее Лаперузова.
пролива не известен); Охотское море; восточное побережье Камчатки;,
Берингово море (по нашему побережью до мыса Чаплина); Алеутские о-ва.
до района Прибыловых о-вов включительно.
Экология. Встречается на глубинах от 15 до 100 м, преимуще-
ственно на каменистых грунтах. Иногда поселяется в губках.
13. Pagurus tanner! (Benedict). (Табл. 5, фиг. 5).
Eupagurus tqnneri Benedict, 1892 :10. — Pagurus tanneri Holmes, 1900 :140; Rathbun,.
1904:158, t. 4, fig. 7; Schmitt, 1921:133, fig. 86.
Передняя часть карапакса почти такой же ширины, как и длины.
Средний выступ лба острый, широко-треугольный; боковые выступы
отчетливые, с острыми субмаргинальными шипами. Глазные чешуйки
овальные, с приподнятыми краями, с очень сильными субтерминальными
шипами. Глазные стебельки толстые, сравнительно короткие, расширяю-
щиеся у роговиц; стебелек и роговица немного сплющены в дорзовен-
тральном направлении. Наружный выступ базального членика антеннальной
ножки острый, с зазубренным внутренним краем. Скафоцериты изогнутые,,
такой же длины, как глаза, или немного превышают их, волосатые на вну-
треннем крае. Мерус правой клешненосной ноги гладкий, передний край
шиповатый; ниЖняя поверхность с группами крупных гранул вдоль наруж-
5. PA GURUS
195
ного и внутреннего краев. Выпуклая поверхность карпуса шиповатая,
крупные шипы окаймляют наружный и внутренний край. Рука такой же
ширины, как карпус; в центре ладони имеется треугольное возвышение,
вершина которого обращена к основанию щели между пальцами; от вер-
шины этого треугольника слабый гребень спускается по поверхности
неподвижного пальца ближе к его внутреннему краю; поверхность руки
по бокам от возвышения вогнутая. Вся поверхность ладони и пальцев
покрыта гранулами, более крупные гранулы расположены на ребрах
треугольника; кроме того, внутри треугольника имеется линия из 5—6
более крупных гранул, идущая ближе к наружному ребру треугольника.
Наружный край руки усажен довольно крупными шипами; нижняя поверх-
ность ладони выпуклая, гладкая, с пучками волосков. Пальцы с малень-
кими роговыми концами. Левая клешненосная нога простирается немного
далее основания подвижного пальца правой руки. Мерус сплющенный,
шиповатый на переднем крае и на краях нижней поверхности. Карпус
с двумя рядами шипов на верхнем крае; у дистального конца наружный
ряд пересекает по диагонали поверхность и соединяется с внутренним
рядом шипов на внутреннем дистальном углу. Ладонь с резким гребнем,
который усажен 13—14 шипиками, наружная поверхность ладони глубоко
вогнутая, густо покрытая гранулами; выпуклый наружный край усажен
шипами, уменьшающимися при переходе на неподвижный палец; внутренняя
сторона ладони в виде гладкого, глубокого желоба; пальцы смыкаются
плотно. По форме и скульптуре левая рука очейь напоминает таковую
у Р. cornutus. Ходильные ноги правой стороны превосходят правую клешне-
носную ногу. Боковые поверхности .члеников гладкие. Карпальные чле-
ники первой пары с острыми шипами на верхнем крае. Проподальные
членики первой пары и карпальные членйки второй пары с тупыми шипами
или, вернее, шиповатыми гранулами на верхнем крае. Пальцы длинные,
сплющенные, слегка изогнутые, незакрученные; шипы на нижнем крае
постепенно возрастают в величине по направлению к сравнительно корот-
кому роговому когтю.
Размеры-
Длина передней части карапакса................. 8.0 мм
Ширина „ „ „ . . - 8.0 „
Длина глазных стебельков...................... 5.0 „
„ скафоцеритов............................. 5.0 „
„ меруса правой клешиен. ноги...............10.0 „
„ карпуса „ „ 9.0 „
Ширина „ „ „ „.............. 7.0 „
Длина руки „ „ 13.5 „
Ширина „ „ „ „ 7.5 „
Длина меруса левой „ „.............. 8.9 „
„ карпуса „ „ 7.5 „
» руки if п t> ......... . 11.0 ,,
Ширина „ . „ „ „.............. 5.5 „
Длина правой ходильной ноги первой пары.......34.0 „
13*
196
PAGURIDAE
Распространение. Берингово море в районе Прибыловых о-вов,
на глубине 87—140 м. От Уналашки до залива San Simeon, Калифорния.
Экология. Вид глубоководный, встречается на глубинах от 90
до 1000 м; в Калифорнии неизвестен на глубинах меньше 380 м.
* 14. Pagurus anomalus (Balss). (Табл. 4, фиг. 5).
Eupagurus anomalus Balss, 1913 : 53, fig-. 32; Yokoya, 1933 : 80.
Передняя часть карапакса одинакова, как в длину, так и в ширину.
Жесткой является не только передняя часть карапакса, как у большинства
Paguridae, но и вся поверхность карапакса, которая покрыта бугорками,
несущими венчики тонких волосков, направленных вперед. На передней
части карапакса, ближе к лобному краю, имеется несколько довольно
крупных, изогнутых вперед шипов. Средний выступ лба длинный, треуголь-
ный, направленный косо вверх под углом примерно в 45°; боковые выступы
представлены также торчащими вверх шипами, по два с каждой стороны
от среднего выступа. У внутренней стороны ближайших к среднему
выступу боковых выступов имеются хорошо заметные субмаргинальные
шипы. Угловой выступ карапакса тотчас же за цервикальной бороздой
несет длинный шип. Глазные стебельки толстые, короткие, со слабым
перехватом посередине. Глазные чешуйки маленькие, острые, очень
жесткие. Скафоцериты простираются несколько дальше концов глаз.
Наружный выступ базального членика антеннальной ножки оканчивается
сильным и острым шипом. Клешненосные ноги сравнительно короткие,
слабые, почти одинаковой длины, правая немного длиннее левой. Мерус
правой клешненосной ноги трехгранный, его ребра закругленные; поверх-
ность покрыта поперечными морщинками с венчиками волос; передний край
сверху с одиночным длинным шипом. Верхняя поверхность корпуса плоская и
гладкая, с продольными рядами шипов на своих краях; боковые поверх-
ности с морщинками и волосами. Рука продолговатая, пальцы довольно
длинные; ряд мелких шипов тянется вдоль внутреннего края ладони, непра-
вильный ряд более крупных шипов занимает середину ладони; пальцы
немного короче чем ладонь, хватательные края с 2—3 буграми, кончики
роговые. Левая клешненосная нога по строению подобна правой, но более
тонкая; рука значительно уже, пальцы длиннее чем ладонь и заметно
изогнуты вниз. Ходильные ноги значительно длиннее клешненосных.
Первая правая ходильная нога превышает клешненосные ноги на половину
длины своего проподуса. Все членики сплющенные, покрытые, в особен-
ности на наружных поверхностях, широкими поперечными морщинами
с отходящими от них волосками; эти морщины придают ногам чешуйчатый
вид. Верхние края карпальных члеников на дистальном конце заканчи-
ваются широко-треугольным зубцом, почти прямые и слабо закрученные.
Абдомен короткий. Характерно, что данный вид выбирает для своего
•обитания раковину такой величины, чтобы скрыть в ней только свой
мягкий абдомен, все же остальные части тела остаются незащищенными.
5. PAGURUS
197
Окраска спиртовых экземпляров желтая, с красноватыми полосками около
морщин. Конечности немного иризируют.
Размеры
Длина передней части карапакса 11.0 мм
Ширина „ „ „ 11.0 „
Длина глазных стебельков 4.0 „
„ меруса правой клешней, ноги 12.0 „
„ карпуса „ „ 8-0 „
Ширина „ „ „ з.о „
Длина руки „ „ 1410 „
Ширина „ „ „ „ 5.5 „
Длина пальцев правой руки 7.0 „
„ правой ход. иоги первой пары 53.0 „
Распространение. Южная Япония, к северу до Сангарского
пролива; Японское море, по нашему побережью от широты о-ва Фуру-
гельма на север до залива Де-Кастри.
Экология. Встречается на глубинах от 15 до 270 м, преимуще-
ственно на каменистых грунтах. У нашего приморского побережья он дер-
жится обычно на глубине 20—60 м, в южной Японии — от 100 до 270 м.
* 15. Pagurus splendescens Owen. (Табл. 4, фиг. 2).
Pagurus splendescens Owen, 1839:81, t. 25, fig. 1, la; Brandt, 1851 : 111; Richters,
1883:406; Holmes, 1900:234; Rathbun, 1904:161; 1919:8; Stevens, 1925:295, fig. 15, 16. —
Eupagurus splendescens Murdoch, 1885 :138; Caiman, 1898 : 260; Alcock, 1905:178; Balss,
1913 : 62, text-fig. 36, 37, t. 2, fig. 2.
Передняя часть карапакса больше в ширину, чем в длину. Жестким
является весь карапакс, как и у предыдущего вида. Вся поверхность
карапакса покрыта бугорками, обрамленными короткими волосками.
Средний выступ лба широко треугольный, направленный горизонтально
вперед, а не поднимающийся косо вверх, как у Р. anomalus; вершина выступа
несет 3—4 тупых зубчика. Боковые выступы лба имеют вид коротких
шипов. Снаружи от боковых выступов край карапакса усажен буграми
с раздвоенными вершинами, что особенно резко выражено у крупных
экземпляров. Выступ карапакса за цервикальной бороздой покрыт мелкими
шипами. Глазные стебельки короткие, толстые, с заметным перехватом
посредине. Глазные чешуйки полуовальные, с закругленной вершиной и
сильными субтерминальными шипами. Скафоцериты значительно превос-
ходят концы глаз, трехгранные, с гладкой и блестящей внутренней поверх-
ностью, с мелкими тупыми шипиками на наружной поверхности. Наружный
выступ базального членика антеннальной ножки имеет раздвоенную вер-
шину. Клешненосные ноги довольно сильные, правая значительно длиннее
левой. Боковые поверхности меруса правой клешненосной ноги покрыты
поперечными рядами бугорков с отходящими от них волосками; верхний
дистальный край с 2—3 шипами, а не с одним, как у Р. anomalus. Верхняя
198
PAGURIDAE
поверхность карпуса плоская, боковые края с рядами шипов; имеется
также неправильный ряд шипов посредине; у более крупных экземпляров
шипы преобразуются в многовершинные бугорки. Рука продолго-
ватая, немного шире чем карпус; края окаймлены рядами шипиков,
которые резко отграничивают верхнюю поверхность руки от нижней.
Верхняя поверхность ладони покрыта многовершинными бугорками с воло-
сками; более крупные бугорки образуют посредине два ряда, которые
сходятся по направлению к основанию неподвижного пальца. Вершины
пальцев при смыкании скрещиваются; хватательные края с 3—4 белыми
бугорками и пучками волос над ними; концы роговые. Левая клешне-
носная нога достигает основания подвижного пальца правой руки;
по строению подобна правой, только рука более узкая, пальцы более
длинные, тонкие, изогнутые вверх. Нижняя поверхность всех члеников
клешненосных ног у крупных экземпляров довольно густо покрыта воло-
сами. Ходильные ноги длинные, первая правая нога превышает правую
клешненосную ногу почти всей длиной своего пальца. Внутренняя поверх-
ность всех члеников совершенно гладкая, наружная покрыта рядами бугор-
ков, подобных таковым меруса клешненосных ног. Дистальный конец
меральных члеников сверху с рядом мелких шипиков, средний из которых
наиболее длинный; нижний край довольно густо усажен простыми шипами.
Нижние края карпальных члеников гладкие, верхние шиповатые. Пальцы
длинные, слегка изогнутые и закрученные. Абдомен так же короток, как
и у Р. anomalus, и, подобно последнему виду, описываемый вид также
прячет в раковину только абдомен. Окраска (по Stevens’y) от светло-
коричневой до светлого цвета мальвы, иризирующая, имеющая металли-
ческий блеск, отливающий розовым, зеленым, бледнолиловым и золотым
цветом; бугорки темные. Нижняя сторона клешненосных и ходильных ног
от бледной розовато-желтой окраски до бледного цвета мальвы, пальцы
темнолиловые с белыми кончиками. Внутренняя поверхность антеннул
от бронзового до золотого цвета.
Данный вид близок к Р. anomalus (Balss), от которого отличается
формой и положением среднего выступа лба, вооружением лобного края
карапакса, раздвоенной вершиной наружного выступа базального членика
антеннальной ножки, иной скульптурой карапакса, отсутствием шипа на
выступе карапакса за цервикальной бороздой, более длинными клешне-
носными ногами и т. д.
Размеры
Длина передней части карапакса................15.0 мм
Ширина „ „ „ ................17.0 „
Длина глазных стебельков...................... 7.0 „
„ меруса правой клешней, ноги.............21.0 „
„ карпуса „ „ „ 16.0 „
Ширина „ „ „ „ 10.0 „
Длина руки „ „ „ 28.0 „
5. PA GURUS
199
Ширина руки правой клешнен. ноги.................. 13.0 мм
Длина пальцев правой руки...........................13.0 „
„ правой ход. ноги первой пары...................84.0 „
Распространение. Охотское море; восточное побережье Кам-
чатки; Берингово море, по американскому побережью на юг до Вашинг-
тона; Чукотское море, на север до широты мыса Барроу, на запад до
устья р. Колымы.
Экология. Встречается на глубинах от 1 до 411 м, на различных
грунтах, но главным образом, повидимому, на мягких илистых и песча-
нисто-илистых грунтах.
* 16. Pagurus ochotensis Brandt. (Табл. 2, фиг. 2).
, Pagurus bernhardus var. spirtimana seu subsp. ochotensis Brandt, 1851 :108. — Pagurus
bernhardus granulato-denticulata Brandt, ibid.: 107; Richters, 1883 : 405. — Eupagurus alasken-
sis Benedict, 1892 : 2. — Pagurus alaskensis Holmes, 1900:135; Benedict, 1901 :456,'text-fig.;
Rathbun, 1904 :157; Stevens, 1925 :277, fig. 1. — Eupagurus ochotensis Stimpson, 1907:218;
Balss, 1913 : 60; Yokoya, 1933 : 82. — Eupagurus ortmanni Balss, 1911 : 7. — Eupagurus spini-
manus Terao, 1913:372.
Передняя часть карапакса больше в ширину, чем в длину; поверх-
ность почти голая, за исключением двух пучков волос позади боковых
выступов лба. Средний выступ лба короткий, треугольный, выступающий
немного дальше боковых выступов, широко-треугольных и снабженных
сильными субмаргинальными шипами. Глазные стебельки короткие, толстые,
с перехватом посредине. Глазные чешуйки короткие, полуовальные,
с вогнутой верхней поверхностью, и с сильными субтерминальными
шипами, которые придают чешуйкам острый внешний вид. Скафоцериты
сильные, трехгранные, превосходящие глаза половиной своей длины;
верхне-внутренний край с 12—16 тупыми, не всегда хорошо заметными
шипами, которые ближе к основанию скафоцерита обычно имеют вид
сглаженных бугорков; кроме того, этот же край волосатый, тогда как
плоские стороны совершенно гладкие^ причем внутренняя сторона сильно
иризирующая. Наружный выступ базального членика антеннальной ножки
с зубчатым внутренним краем, внутренний выступ этого членика занят
тупым шипом. Клешненосные ноги сильные, совершенно безволосые для
невооруженного глаза. Мерус правой клешноносной ноги трехгранный,
с гладкими боковыми поверхностями; округлое верхнее ребро с группами
шипов в дистальной половине, которые располагаются в короткие попе-
речные ряды; вдоль переднего края тянется вогнутое голое пространство;
передний край вооружен направленными вперед шипами; нижняя поверх-
ность гранулированная, вдоль наружного края шиповатая. Верхняя
поверхность карпуса покрыта шиповатыми гранулами или короткими
коническими шипами с волосками впереди их основания, которые видны
только под лупой; поверхность между этими шипами покрыта более мел-
кими шипиками; наиболее крупные шипы имеются на внутреннем крае;
200
PAGURIDAE!
боковые поверхности покрыты шиповатыми гранулами, нижняя поверх-
ность мелко гранулированная. Рука неправильной треугольной формы; верх-
няя поверхность покрыта такими же гранулами или коническими шипами, как
и карпус; шипы наружного края наиболее крупные; внутренний край валико-
образно утолщен; боковые поверхности гранулированные, гранулирован-
ность постепенно сглаживается при переходе на нижнюю поверхность, про-
ксимальная часть которой остается совершенно гладкой; хватательные края
пальцев с белыми буграми и известковыми кончиками. Левая клешне-
носная нога достигает середины подвижного пальца правой руки. Мерус
и карпус имеют такое же строение, как и на правой ноге. Рука с слабо
выпуклой верхней поверхностью, которая вооружена так же, как правая:
рука; края приподнятые, в особенности внутрений край; нижняя поверхность
с рассеянными пучками темных волосков; пальцы с роговыми кончиками,,
при смыкании оставляют щель. Ходильные ноги сильные, на правой сто-
роне они превосходят правую клешненосную ногу. Карпальные членики
обеих пар с шиповатыми гранулами на верхнем крае, стороны совер-
шенно гладкие; проподальные членики обеих пар с шиповатыми гранулами:
на верхнем крае и на верхней половине наружной поверхности. Пальцы
сильные, значительно длиннее проподусов, изогнутые и закрученные,,
с желобком вдоль внутренней поверхности, выпуклые на наружной поверх-
ности; верхняя сторона снабжена тремя продольными рядами тупых шипов-
или шиповатых гранул; роговой коготь сравнительно тупой. У спиртовых,
экземпляров общая окраска сверху светлопурпурная, с радужным отливом,,
причем у очень крупных экземпляров заметно преобладает фиолетовый
оттенок; снизу светлая, с слегка красноватым оттенком. Кругом хвата-
тельного края неподвижного пальца правой руки тянется красная полоса,,
которая затем идет к внутреннему краю ладони. Нижне-наружная поверх-
ность карпальных члеников ходильных ног у концов красная.
Размеры
Длина передней части карапакса.................17.0 мм
Ширина „ „ „ е.......................18.5 „
Длина глазных стебельков....................... 8.0 „
„ скафоцеритов..............................10.0 „
„ меруса правой клешней, ноги...............22.0 „
„ карпуса „ „ 21.0
Ширина „ „ „ 18.0 „
Длина руки „ „ 31.0
Ширина „ „ „ 19.0 „
Длина меруса левой „ „ ...............15.0 „
„ карпуса „ „ 16.0 „
Ширина „ „ „ „ ...............10.0 „
Длина руки „ „ 21.0 „
Ширина „ „ „ „ ...............13.0 „
Длина правой ходильной ноги первой пары........79.0 „
Экземпляры этого вида могут достигать очень крупных размеров-
5. PA GURUS
201
Распространение. Берингово море, по американскому побе-
режью к югу до Орегона, у нашего побережья сравнительно редок; Охот-
ское море; Японское море до южной Японии включительно.
Экология. Встречается на глубинах от 1 до 250 м, преимуще-
ственно на песчанистых грунтах.
Вопрос о синонимии Р. ochoiensis и Р. alaskensis оказался весьма
запутанным. На основании просмотра коллекций Зоологического Инсти-
тута Академии Наук, я пришел к убеждению, что Brandt не. имел в руках
Р. ochoiensis в том понимании этого вида, которое впоследствии было
приписано ему. Типовой экземпляр Р. bernhardus var. spinimana sen subsp.
ochoiensis (n° 1075), несомненно, должен быть отнесен к Р. alaskensis-,
точно так же во всей коллекции я не мог найти ни одного экземпляра,,
который можно было бы отнести к Р. ochoiensis. При этом я считаю важ-
ным обратить внимание на следующие два пункта: 1) характер гранули-
рованности, как клешненосных, так и ходильных ног у Р. alaskensis
сально вариирует от очень мелких тупых гранул до шиповатых гранул,
(под последними следует понимать крупные гранулы с острыми вершинами
или короткие конические шипы); 2) необходимо иметь в виду, что при
описании Р. ochoiensis Brandt сравнивал свой вид с европейским Р. bernhar-
dus, конечности которого покрыты очень плоскими, тупыми гранулами;
по сравнению с Р. bernhardus, конечности Р. alaskensis, несомненно,
являются более шиповатыми, что и дало повод Brandt’y выделить вариетег
spinimana, а также, учитывая упомянутую выше изменчивость в степени:
гранулированности, и вариетет granulato-denticulata. Оба вариетета вполне
укладываются в понятие Р. alaskensis. Описанный Dana Bernhardus arma-
tus, вид, действительно близкий к Р. alaskensis, но резко отличающийся
от него клешненосными ногами, которые покрыты длинными, тонкими,
шипами и волосами, был ошибочно сведен в синонимы к Р. ochoiensis,
причиной чего следует считать, повидимому, краткость и неточность опи-
сания Brandt’a и совпадение главного отличительного признака (шипо-
ватость рук) с названием вариетета Brandt’a (spinimana). Таким образом,
Р. alaskensis (Benedict) следует именовать Р. ochoiensis Brandt, а вид,
считавшийся за Р. ochoiensis Brandt, следует называть Р. armatus (Dana).
В правильности такого заключения я еще больше убедился после полу-
чения экземпляров Р. ochoiensis (п° 52666) и Р. alaskensis (п° 16368)
из коллекции Национального Музея Соединенных Штатов; наиболее
бросающиеся в глаза отличия, выявившиеся в результате сравнения
упомянутых экземпляров, как друг с другом, так и с экземплярами
Зоологического Института Академии Наук, сопоставляются в нижепри-
водимой таблице:
Р. ochoiensis Brandt
1. Поверхность карпуса и проподуса клеш-
неиосных ног покрыта гранулами или ши-
поватыми гранулами; пучки волос у Основа-
P. armatus (Dana)
1. Поверхность карпуса и проподуса клеш-
неносных ног густо покрыта длинными,
тонкими, острыми шипами и пучками волос.
202
PAGURIDAE
ния гранул или совершенно отсутствуют или
очень маленькие, едва заметные.
2. Правая рука широкая, наружный край
дугообразный. Пальцы руки не перекрещи-
ваются при смыкании, благодаря чему ди-
стальный конец руки кажется более тупым.
Внутренний край ладони заметно валико-
образно утолщен.
3. Внутренний край ладони левой руки
с сильным валикообразным утолщением.
4. Верхне-наружная поверхность пропо-
дальных члеников двух первых пар ходиль-
ных ног покрыта тупыми или шиповатыми
гранулами, короткие шипы образуют только
один ряд на верхнем крае; волоски, если и
имеются, то очень маленькие.
5. Верхний край пальцев ходильных ног
несет продольный ряд тупых или шиповатых
гранул.
которые поднимаются от основания шипов
и достигают верхушки последних или даже
иногда немного превышают их.
2. Правая рука сравнительно узкая, равно-
мерно суживающаяся к дистальному концу,
который кажется очень острым благодаря
сильному скрещиванию кончиков пальцев
при их смыкании. Внутренний край ладони
без заметного валикообразного утолщения.
3. Внутренний край ладони левой руки со
слабым намеком на валикообразное утолще-
ние.
4. Верхне-наружная поверхность пропо-
дальных члеников двух первых пар ходиль-
ных йог покрыта шипами, которые распо-
лагаются в три продольных ряда, причем
самые длинные шипы образуют ряд на верх-
нем краю членика; от оснований шипов под-
нимаются пучки волосков, которые дости-
гают вершины шипов.
5. Верхний край пальцев ходильных ног
вооружен продольным рядом острых шипов,
которые постепенно уменьшаются в вели-
чине по направлению к дистальному концу.
Отсутствие в нашем, достаточно обширном, материале Р. armatus
(Dana) приводит меня к убеждению, что последний вид является сравни-
тельно узко-локальной формой западного побережья Северной Америки.
Поэтому все указания Р. ochotensis для Камчатки, Охотского и Японского
моря я отношу не к Р. armatus, а к Р. ochotensis.
16а. Pagurus ochotensis aleuticus (Benedict). (Табл. 2, фиг. 3).
Eupagurus aleuticus Benedict, 1892 : 3. — Pagurus aleuticus Holmes, 1900:136; Benedict,
1901: 460, text-fig.; Rathbun, 1904 :157; Stevens, 1925 :278, fig. 2.
Все три выступа лба острее и немного более выдающиеся, чем
у Р. ochotensis. Глазные стебельки толще и немного длиннее. Глазные
чешуйки более длинные и менее заостренные, субтерминальные шипы не
видны сверху. Скафоцериты у основания шире, чем у типичной формы.
Клешненосные ноги сильные. Карпальные членики вооружены много-
численными короткими шипами или шиповатыми гранулами. Пальцы обеих
рук имеют сверху гладкую косую поверхность, плоскую или слегка
вогнутую; на левой руке эта поверхность свободна от волос и гранул,
на правой руке она окаймлена короткими шиповатыми гранулами. Гранулы
на обеих руках часто имеют раздвоенные вершины, шиповатые гранулы
;на краях более крупные и всегда простые. Пальцы правой руки имеют
роговые концы, как у старых, так и у молодых экземпляров (у типичной
формы этот признак имеется только у молодых экземпляров). Пальцы
5. PAGURUS
203
ходильных ног очень широкие и тонкие, их верхний край целиком занят
продольным желобом (бороздой), который сравнительно глубок у осно-
вания пальца и постепенно мельчает по направлению к кончику (фиг. 69).
Окраска: клешненосные и ходильные ноги
светлого охристо-желтого цвета, красно-
ватые сверху; руки и пальцы желтые.
Размеры
Длина передней части карапакса .
Ширина „ „ „ .
.Длина глазных стебельков
„ скафоцеритов
„ меруса правой клешней, ноги
„ карпуса „
Ширина „
Длина руки
Ширина „
Длина меруса левой „ „
„ карпуса „
Ширина „
-Длина руки „ „ „
Ширина „
Длина правой ходильной ноги первой пары . 51.5
• 10.0 ММ
. 10.5 „
. 6.5 „
. 6.0 „
. 13.0 „
• 12.0 „
. 10.0 „
• 19.0 „
• п.о „
. 11.0 „
• Ю.0 „
• 6.5 ,,
. 12.5 „
7.0 „
Подобно Р. armatus, данный вид
является локальной формой западного
побережья Северной Америки; морфоло-
гически Р. aleuticus настолько близок
к Р. ochotensis, что я рассматриваю его
как подвид этого последнего вида.
Фиг. 69. Пальцы первой правой
ходильной ноги (задне-дорзальный
вид). А — Pagurus ochotensis subsp.
aleuticus (Benedict). В — Pagurus
ochotensis Brandt.
Распространение. Берингово море (район Прибыловых о-вов),
по западному побережью Северной Америки к югу до Орегона; от 14 до
435 м (Rathbun).
* 17. Pagurus rathbuni (Benedict). (Табл. 3, фиг. 3).
Eupagurus rathbuni Benedict, 1892. — Pagurus rathbuni Holmes, 1900 :140; Rathbun,
1904: 158, t. 4, fig-. 2. — Eupagurus (Trigonocheirus) polaris Sivertsen, 1932: 8, t. 2, fig. 3.
Передняя часть карапакса почти такой же длины, как и ширины.
Средний выступ лба острый, треугольный, выступает заметно дальше
боковых выступов, широко-треугольных и снабженных довольно сильными
суб маргинальными шипами. Глазные чешуйки конической формы, с за-
кругленной вершиной и сильными субтерминальными шипами. Глазные
стебельки толстые, слегка суженные у базальной трети и расширяющиеся
у роговиц. Скафоцериты превосходят концы глаз, с волосатыми
внутренними краями. Внутренний край наружного выступа базального
членика антеннальной ножки вооружен 3—5 тупыми шипами, включая
и верхушечный шип; Верхнее ребро меруса правой клешненосной ноги
204
PA GURIDAE
с редкими гранулами и поперечными морщинами, от которых отходят
направленные вперед волоски. Карпус четырехсторонний, такой же длины,,
как мерус; выпуклая верхняя поверхность покрыта гранулами, от основа-
ния которых поднимаются короткие волоски; на внутреннем дистальном
углу верхней поверхности имеется углубление, заполненное пучком неж-
ных, хорошо заметных волос; у молодых экземпляров пучок может отсут-
ствовать, но углубление всегда ясно выражено; внутренняя и нижняя
поверхность волосатые. Рука такой же ширины, как карпус, но немного
длиннее его; верхняя поверхность выпуклая, равномерно покрыта грану-
лами и короткими волосками, которые располагаются впереди гранул
и .мало заметны для невооруженного глаза; наружный край подвижного
пальца с двойным рядом гранул; хватательные края пальцев с белыми
буграми и пучками волосков над ними; нижняя поверхность ладони с рас-
сеянными, низкими, широкими гранулами и с пучками волосков. Пальцы
с роговыми концами. Левая клешненосная нога простирается немного
дальше основания подвижного пальца правой руки, у крупных экземпля-
ров она немного короче. Мерус сплющенный, с волосатыми морщинами
на верхнем ребре. Карпус немного короче чем мерус, расширяющийся:
дистально; его верхняя поверхность узкая, плоская, с рядами шиповатых
гранул на краях; боковые и нижняя поверхности гранулированные и воло-
сатые. Руки примерно на 2/з длиннее, чем карпус; наружная поверхность-
узко-треугольной формы, покрыта гранулами и короткими волосками,,
подобно поверхности правой руки; средний гребень не очень резкий;
внутренняя поверхность имеет голый промежуток с одиночными гранулами,,
снаружи от которого Тянутся два неправильных ряда гранул, параллель-
ных внутреннему краю и с пучками волосков у их оснований; пальцы
сплющенные, почти в два раза длиннее ладони, сильно изогнуты вниз на
дистальных концах; у крупных экземпяров при смыкании остается широкая,
щель между пальцами, особенно хорошо заметная с нижней стороны..
Ходильные ноги правой стороны немного превышают правую клешненос-
ную ногу. Моральные членики первой пары с шиповатыми гранулами на.
нижнем крае, карпальные членики с такими же гранулами на верхнем:
крае. Пальцы длиннее, чем проподусы, изогнутые и закрученные; верхний
и нижний края изнутри опушены волосками, наружная поверхность выпу-
клая и голая, с слабо намечающимся продольным желобком, внутренняя"
поверхность более плоская, с рядом волосков ближе к верхнему краю.
Окраска спиртовых экземпляров: дистальные концы члеников всех конеч-
ностей бледнокрасные; кончики пальцев левой руки красноватые, корич-
невые, или же часто черновато-бурые, как бы обугленные.
Размеры
Длина передней части карапакса . . . ......15.0 мм
Ширина „ „ „ ................ . 16.0 „
Длина меруса правой клешнен, ноги..........23.0 „
„ карпуса „ ' „ „ . ............22.5 „
5. PAGURUS
205
Ширина карпуса правой клешнен. ноги...............15.0 ми
Длина руки „ „ „.............30.0 „
Ширина „ „ „ .............14.5 „
Длина меруса левой клешнен. ноги ................17.0 „
„ карпуса „ „ ...............15.0 „
„ Руки „ „ ...............24.0 „
„ подвижного пальца левой руки................15.5 „
„ правой ходильной иоги первой пары ..........84.0 „
Распространение. Японское море (только по нашему побережью;
из Японии неизвестен); Охотское море; восточное побережье Камчатки;
Берингово море; Чукотское море, на север до широты о-ва Врангеля, на
запад до устья р. Колымы (16 VII 1912, 10 м; ил; 1 экз.).
Экология. Встречается на глубинах от 10 до 210 м, преимуще-
ственно на илистых грунтах.
18. Pagurus granosimanus (Stimpson). (Табл. 2, фиг. 5).
Eupagurus granosimanus Stimpson, 1859:90; Smith, 1878—79 : 211.—Pagurus grano-
simanus Holmes, 1900:146; Rathbun, 1904:10, t. 5, fig. 8; Schmitt, 1921:141, fig. 91;
Stevens, 1925:282, fig. 5.
Передняя часть карапакса значительно больше в длину, чем в ширину.
Средний выступ лба короткий, широко-треугольный; боковые выступы
закругленные, без субмаргинальных шипов. Глазные стебельки довольно
толстые, умеренной длины. Глазные чешуйки с очень сильными субтер-
минальными шипами. Наружный выступ базального членика антеннальной
ножки с тремя шипами на вершине и с шиповатым внутренним краем;
внутренний выступ с одним сильным шипом. Скафоцериты изогнутые,
значительно не достигают концов глаз, с волосатыми внутренними
краями. Мерус правой клешненосной ноги гладкий, с зубчатым передним
краем; нижняя поверхность несет две крупные бугрообразные гранулы.
Поверхность карпуса покрыта коническими шипами, вершины которых
направлены вперед. Рука довольно правильной овальной формы, несколько
длиннее, чем карпус; поверхность ладони и пальцев покрыта крупными
тупыми шиповатыми гранулами; наружный край несет острые, зубцеоб-
разные шипы, резко отделяющие верхнюю поверхность ладони от нижней,
которая покрыта плоскими широкими гранулами; пальцы с острыми рого-
выми кончиками. Левая клешненосная нога простирается немного дальше
основания подвижного пальца правой руки. Верхняя поверхность карпуса
и руки покрыта такими же гранулами, как на правой ноге. Рука без явствен-
ного гребня на верхней поверхности; наружная поверхность не образует
резкого ребра с нижней поверхностью, на которую переходят гранулы,
-становящиеся редкими в центре нижней поверхности. Ходильные
ноги правой стороны немного превышают правую клешненосную ногу.
Карпальные и проподальные членики обеих пар шиповатые на верхнем
крае. Пальцы почти прямые, с длинными роговыми когтями. Как клешне-
мосные, так и ходильные ноги очень слабо волосатые. Окраска (по Ste-
206
PAGURIDAE
vens’y) от желто-оливковой до оливковой. Гранулы клешненосных ног
фарфорово-синие, становящиеся светлосиними около концов пальцев, в осо-
бенности на нижней стороне руки. Ходильные ноги с мелкими синими
пятнами, от передней части которых поднимаются маленькие пучки щети-
нок. Антенны абрикосово-оранжевые, становящиеся оливковыми на базаль-
ных сегментах. Молодые экземпляры более светлой окраски и обнару-
живают очень мало синих тонов, гранулы и пятна почти белые, ходиль-
ные ноги с оранжевыми кончиками. Окраска спиртовых экземпляров
темнокрасная, с белыми бугорками на клешнях и более темными красными
шипами на карпусе и мерусе клешненосных ног. Ходильные ноги красные,,
с мелкими белыми пятнами; пальцы с широкой белой полосой посредине-
Бугорки на нижней стороне меруса правой клешненосной ноги белые.
Размеры
Длина передней части карапакса.......................8.0 мм.
Ширина „ „ „ ...................7.0
Длина глазных стебельков.......................4.0
„ скафоцеритов . .
„ меруса правой клешней, иоги
„ карпуса „
Ширина „
Длина руки „ „ „
Ширина „ „ „ „
Длина меруса левой „ „
„ карпуса „
>, РУКИ
Ширина „
.....................2.5
.....................6.5
.....................7.0
.....................4.0
.....................8.0
. . . ...............4.5
.....................5.0
.....................5.5
.....................7.0
.....................3.5
Длина правой ходильной иоги первой пары..........23.0
Я
я
я
я
я
я
я
я
»
я
я
я
гг
Данный вид довольно близок к Р. beringanus, от которого отличаете»
следующим: гранулы на клешненосных ногах более острые; наружный
край правой руки не такой дугообразный, как у Р. beringanus', наружный
край левой руки не образует резкого ребра с нижней поверхностью
ладони, тогда как у Р. beringanus этот переход достаточно резкий; суб-
терминальные шипы глазных чешуек более длинные и тонкие; ходильные
ноги менее волосатые, чем у Р. beringanus.
Распространение: От Уналашки до Ensenada, Нижняя Кали-
форния. От мелководья до глубины 27 м (Rathbun).
20. Pagurus dalli (Benedict). (Табл. 1, фиг. 4).
Eupagurus dalli Benedict, 1892:9. — Pagurus dalli Holmes, 1900:139; Rathbun,.
1904 :158, t. 4, fig. 1; Stevens 1925 : 287, f. 8, 9.
Передняя часть карапакса очень незначительно больше в длину,
чем в ширину. Средний выступ лба острый, треугольный и выступает
значительно дальше широко-закругленных боковых выступов, снабженных
сильными субмаргинальными шипами. Глазные чешуйки овальные,,
с несколько заостренной вершиной и очень сильными субтерминальными:
5. PAGURUS
207
шипами. Глазные стебельки умеренной длины, цилиндрические. Наружный
выступ базального членика антеннальной ножки с острым шипом на
вершине и двумя шипами на внутреннем крае около верхушечного шипа;
внутренний выступ с одним острым шипом. Изогнутые скафоцериты
с пучками коротких волосков на внутреннем крае, простираются немного
дальше глаз. Мерус правой клешненосной ноги с сильными шипами на
переднем крае и с 2—3 острыми шипами на верхнем ребре около
дистального конца. Верхняя поверхность карпуса довольно густо покрыта
острыми шипами и короткими волосками, отходящими от основания
шипов. Рука более чем в два раза больше в длину чем в ширину, узко-
овальной формы, очертания обоих краев почти одинаковые. Верхняя:
поверхность ладони, так же как и поверхность карпуса, покрыта острыми
шипами и довольно густыми ' пучками коротких волосков; подвижный
палец длинный, с двумя продольными рядами шипов на наружной стороне
верхней поверхности и с гладкой внутренней стороной. Проксимальные
половины хватательных краев пальцев вооружены белыми, неправильной
формы буграми; дистальная половина хватательного края подвижного
пальца вооружена гребеночкой из густо сидящих роговых шипиков;,
дистальная половина хватательного края неподвижного пальца вооружена
3—5 коническими зубцами, а промежутки между ними заняты роговыми
шипиками. Концы пальцев крючкообразно изогнуты и скрещиваются при
смыкании; концы роговые. Левая клешненосная нога достигает основа-
ния подвижного пальца правой руки. Карпус узкий, с двумя продольными
рядами шипов на верхней поверхности. Рука сравнительно узкая,,
нерасширяющаяся у основания, с косой треугольной наружной поверх-
ностью, средний гребень вооружен очень острыми и длинными шипами,
шипы на наружной поверхности более мелкие. Ходильные ноги правой
стороны превышают правую клешненосную ногу примерно на 1/s длины,
своих пальцев. Пальцы тонкие, слабо изогнутые у дистального конца
и незакрученные. По окраске похож на Р. pubescens, но имеет резко выра-
женную белую полосу у дистального конца меральных члеников клешне-
носных ног. У формалиновых экземпляров ходильные ноги желтовато-
красные, с светлой полосой на середине члеников и красные у сочленений^
Р азмеры
Длина передней части карапакса ............... 7.0 мм
Ширина „ „ „ . 6.5 „
Длина глазных стебельков 5.0 „
„ скафоцеритов 4.0 „
„ меруса правой клешней, ноги.............. 7.5 „
„ карпуса „ „ ................ 8.0 „
Ширина „ „ „ .................. 5.5 „
Длина руки правой клешней, ноги ...... . 12.0 „
Ширина „ „ „ „ 5.5 „
Длина меруса левой „ „ 7.0 „
„ карпуса „ „ „ 7.0 „
208
PAGURIDAE
Ширина карпуса левой клешней, ноги............. 2.5 мм
Длина руки „ „ „ ............. 8.5 „
Ширина „ „ „ ,..................3.5 „
Длина правой ходильной ноги первой пары........32.0 „
Pagurus dalli очень близок к Р. pubescens, но отличается от него
более далеко выступающим средним выступом лба-, более тонкими глаз-
ными стебельками, более тонкими, длинными и острыми шипами на
клешненосных ногах, более узкой правой рукой, которая имеет правиль-
ную овальную форму, и более узкой у основания левой рукой; наконец,
более тонкими и незакрученными пальцами у дистального конца моральных
члеников клешненосных ног.
Распространение. Берингово море (широта о-ва Нунивок)
к югу до Орегона. От мелководья до 172 м (Rathbun).
* 20. Pagurus pubescens Kroyer. (Табл. 4, фиг. 1).
Pagurus pubescens Kroyer, 1838 : 314; 1838 : 251; Brandt, 1851: 111; Goes, 1863 :166;
лЯржинский: 1870:317; Hoek, 1882:6; Richters, 1833:406. — Pagurus thompsoni Bell,
1853:372.—Eupagurus trigonocheirus Stimpson, 1858:249; 1907:221, t. 26, fig-. 2;
Murdoch, 1885 :138, 1.1, fig. 1, la, lb; Benedict, 1892; Balss, 1913 : 63 (part.); Terao, 1913: 373;
Yokoya, 1933:83. — Eupagurus pubescens Miers, 1877:32; Smith, 1880 : 47; Sars, 1882:42;
1886:4; Stuxberg, 1886:51; Pfeffer, 1890:22; Ortmann, 1892:305; A. Milne-Edwards et
- Bouvier, 1894: 74; Бируля, 1897:437; 1899:36, 431; Birula, 1905 :10; Doflein, 1900: 341; Ohlin,
•1901:19; Stimpson, 1907:222; Hansen, 1908:27; Balss, 1912:106; 1926: 32; Дерюгин,
1915:478; 1928:289; Dons, 1934:40, 113. — Eupagurus kroyeri Stimpson, 1859:89; Smith,
' 1880 : 48; Miers, 1881: 340. — Eupagurus pubescens var. kroyeri Henderson, 1888: 65. —
Eupagurus brandti Benedict, 1892:9. — Pagurus brandti Holmes, 1900:139; Rathbun,
1904:157, t. 4, fig. 4; 1919: 8; Stevens, 1925: 285, fig. 7. — Pagurus trigonocheirus Holmes,
11900 :138; Rathbun, 1904 :157; 1919 : 7. — Pagurus kroyeri Rathbun, 1919 :14.
Передняя часть карапакса больше в длину, чем в ширину. Все три
; выступа лба почти одинаково выдающиеся; средний выступ треугольный,
-острый, с пучком длинных волосков; боковые выступы более тупые,
с маленькими субмаргинальными шипами. Глазные стебельки довольно
сильные, с очень слабым перехватом посредине. Глазные чешуйки
остроконечные, с плоской верхней поверхностью и сильными субтерми-
нальными шипами. Скафоцериты с густо волосатым внутренним краем,
превосходят концы глаз, но у мелких экземпляров могут достигать только
концов или быть даже несколько короче. Клешненосные ноги, сильные,
их волосатость сильно вариирует, наиболее волосатыми являются кар-
пальные членики, тогда как руки совсем безволосые. Верхнее ребро меруса
правой клешненосной ноги с поперечными морщинами и волосками
на дистальной части. Выпуклая поверхность карпуса покрыта шипами,
от основания которых поднимаются пучки волосков; шипы на внутреннем
крае наиболее крупные; нижняя поверхность выпуклая, отчасти гранулиро-
ванная и волосатая. Верхняя поверхность руки покрыта коническими
..шипами, с пучками волосков впереди их оснований; волоски не превы-
.шают длины шипов и обычно заметны только под лупой; невоору-
5. PAGUR US
209
женному глазу рука представляется почти безволосой. Шипы наруж-
ного края более длинные. Нижняя поверхность с рассеянными гранулами
и пучками волос. Пальцы с маленькими роговыми концами, которые могут
утрачиваться у более крупных экземпляров. Размеры всех члеников
сильно вариируют. Левая клешненосная нога немного превышает основа-
ние подвижного пальца правой руки у мелких экземпляров, а у крупных
только достигает его. Карпус с двумя продольными рядами шипов на
краях верхней поверхности; наружная поверхность с шипами и йолосами.
Рука с косо-треугольной наружной поверхностью, довольно густо покрытой
гранулами. Косой средний гребень очень резкий, усажен шипами. Острый
наружный край ладони расширяется у основания, причем степень этого
расширения вариирует, как в зависимости от пола, так и у одного
и того же пола. Подвижный палец гладкий, с пучками темных волосков.
Ходильные ноги правой стороны превышают правую клешненосную ногу
на половину длины своих пальцев, у крупных экземпляров несколько
меньше. Меральные членики первой пары шиповатые снизу, карпальные
членики обеих пар шиповатые сверху. Все членики волосатые на верхних
краях. Пальцы изогнутые и закрученные, с сильными роговыми когтями.
Окраска спиртовых экземпляров бледножелтая. Дистальные концы мораль-
ных члеников клешненосных ног (а у мелких экземпляров и карпальных
члеников) бледнокрасные. Дистальные и проксимальные концы всех
члеников ходильных ног красные.
Размеры
.Длина передней части карапакса 17.0 ММ
'Ширина „ м 16.0
.Длина глазных стебельков 8.5
„ меруса правой клешиен НОГИ 23.0 Гг
„ карпуса V » 24.0 »
Ширина „ п * • • 17.0
.Длина руки »> >» » ••••••••• 30.0
Ширина „ „ 14.5 »
Длина меруса левой ,, 17.0
. Длина карпуса 18.0
Ширина „ >» ,> 6.5
.Длина руки » » »> 22.0 ,,
'Ширина „ ,, ...... 10.5
Длина правой ходильной KOVIA первой пары . ... . 82.0
Экземпляры этого вида могут достигать довольно крупных размеров.
Распространение. Белое море, Баренцево море до широты
Шпицбергена; Немецкое море, включая Скагеррак и Каттегат; побережье
Шотландии и северной Англии (до широты Durham), по побережью
Ирландии до ее юго-западной оконечности; побережье Исландии; западное
побережье Гренландии (у восточного сравнительно редко). По восточному
побережью Сев. Америки от Баффинова залива (67°57' N 55°30' W) до
широты 38°08'N. Чукотское море (на восток до мыса Барроу, самое
Фауна СССР, Макаров 14
210
PAGURIDAE
западное местонахождение 70°07' N 177°17' E). Берингово море; по запад-
ному побережью Северной Америки к югу до Орегона; Алеутские о-ва;
восточное побережье Камчатки; Охотское море; Японское море, по нашему
побережью известен До широты о-ва Фуругельма, в Японии до Нагасаки.
Г. П. Горбунов любезно сообщил мне о нахождении им нескольких
экземпляров Р. pubescens в Карском море (до настоящего времени данный
вид был известен только из восточной половины Югорского и Маточкина
Шара); он был найден у западного побережья п-ва Ямал в двух пунктах:
71°54'N, 67°45'Е и 69°52' N, 65°14' Е. Однако эти нахождения являются:
спорадическими и не позволяют считать Карское море населенным данным
видом, что подтверждает и Г. П. Горбунов. Таким образом, этот вил
является типичным амфибореальным видом.
Экология. Вид встречается до глубины 900 м. В Белом море не опу-
скается ниже 100 м, в Кольском заливе распространяется до 330 м, в Барен-
цовом море — до 360 м. В наших дальневосточных морях является одним
из наиболее часто встречающихся видов, обычно на глубинах от 20 до 200 м„
на самых различных грунтах, но преимущественно на илистых и песчанистых..
Вопрос о видовой самостоятельности Р. trigonocheirus (Strap.) уже
давно поднимался в карцинологической литературе. Ряд авторов (Brandt,
Richters, Stimpson) отмечали Р. pubescens в тихоокеанском бассейне.
Doflein (1900) первой высказал, правда чисто умозрительно, предположе-
ние, не является ли Р. trigonocheirus (Strap.) вариететом атлантического
Р. pubescens Кг. Hansen (1908) категорически высказывается за идентич-
ность обоих видов. Все те небольшие различия, которые обычно при-
водятся как отличающие оба вида (отношение длины правой руки к ее
ширине, ширина левой руки у ее основания, степень волосатости и др.)'
вполне укладываются, как в рамки индивидуальной изменчивости
Р. pubescens, так и полового диморфизма у того же вида, как эта
показал А. А. Бируля (1897: 439). Отличия Р. trigonocheirus (Strap.)'
от Р. pubescens Кг., приводимые Balss’oM (1913), покоятся на неправиль-
ном отождествлении последним автором Р. trigonocheirus (St.) с Р. ca.pilla.tus
(Benedict); таким образом, все приводимые отличия относятся к Р. pube-
scens и Р. capillatus, которые, действительно, легко отличаются друг
от друга. Имея в распоряжении обширный материал как по европейским
(Белое и Баренцево моря), так и по всем дальневосточным морям, я на осно-
вании детального сравнения пришел к выводу об идентичности Р. pubescens
с Р. trigonocheirus. Имеющийся в коллекции единственный экземпляр
Р. pubescens из Гренландии (п° 1086) идентичен с европейскими и тихо-
океанскими, так что мысль Doflein’a о выделении особого вариетета
(var. krbyeri) у восточного побережья Северной Америки как будто должна
отпасть. Что касается Р. brandti (Benedict), то на основании сравнения
экземпляра этого вида, полученного из Национального Музея Соединен-
ных Штатов (п° 19097), я присоединяюсь к мнению Hansen’a и Balss’a
об идентичности этого вида с Р. pubescens.
5. PAGURUS
211
* 21. Pagurus brachiomastus (Thallwitz). (Табл. 2, фиг. 6).
Eupagurus brachiomastus Thallwitz, 1891 :35; Ortmann, 1892:312; Terao, 1913:365.
Передняя часть карапакса больше в длину, чем в ширину. Средний
выступ лба широкий, треугольный, тупой, с пучком длинных волосков;
боковые выступы намечаются слабо. Глазные стебельки тонкие, длинные,
цилиндрические, с маленькими роговицами. Глазные чешуйки полуоваль-
ные, с очень сильными субтерминальными шипами. Скафоцериты не
достигают концов глаз, с волосатым внутренним краем. Клешненос-
ные ноги волосатые. Мерус правой ноги трехгранный, с шипами на перед-
нем крае, с гладкими боковыми поверхностями; нижняя поверхность несет
два крупных тупых шипа. Верхняя поверхность карпуса покрыта шипами
и длинными волосами. Верхняя поверхность руки густо покрыта длин-
ными, мягкими волосами, которые скрывают довольно длинные шипы,
равномерно покрывающие всю поверхность ладони; более крупные шипы
образуют ряд на наружном крае, резко отграничивая верхнюю поверхность
ладони от нижней, выпуклой и почти гладкой; на внутреннем крае переход
к нижней поверхности более равномерный, шипы постепенно уменьшаются
в размерах. Пальцы с роговыми кончиками. Левая клешненосная нога
достигает середины ладони правой руки. Мерус сплющенный, с гладкими
безволосыми боковыми сторонами; на нижней поверхности против вершины
исхиума имеется небольшой тупой шип. Верхняя поверхность карпуса
с двумя продольными рядами шипов и гладким промежутком между ними;
боковые поверхности волосатые. Верхняя поверхность руки покрыта
шипами и длинными волосами, нижняя поверхность с отдельными пучками
волос. Ходильные ноги правой стороны немного превышают правую
клешненосную ногу. Все членики волосатые на верхних краях. Пальцы
слабо изогнутые, с сильными роговыми когтями. Окраска желтовато-
оранжевая, волоски бледножелтые, почти бесцветные; пальцы клешненос-
ных ног часто малиновые на концах.
Размеры
Длина передней части карапакса........
Ширина „ „ „ .........
Длина глазных стебельков .............
„ меруса правой клешиен. ноги......
„ карпуса „ „ „..........
Ширина „ . „ „
Длина руки „ „ „..........
Ширина „ „ „ „..........
Длина меруса левой клешней, ноги......
„ карпуса „ „ .............
Ширина „ „ „
Длина руки „ „ ...........
Ширина „ „ ,, .......
Длина правой ходильной ноги первой пары . .
7.0 мм
6.5 „
4.0 „
9.0 „
9.0 „
6.5 „
10.0 „
6.5 „
8.0 „
7.0 „
3.0 „
8.0 „
4.0 „
28.0 „
14*
212
PAGURIDAE
От P. lanuginosus de Haan отличается наличием двух тупых шипов
на нижней поверхности меруса правой клешненосной ноги, узким непо-
движным пальцем правой руки, наличием роговых концов на пальцах
правой руки, большей волосатостью ходильных ног и отсутствием красных
пятен на члениках конечностей.
Распространение. Япония или Китай (Thallwitz), Японское
море, по нашему побережью от широты о-ва Фуругельма на север до
залива Де-Кастри; Лаперузов пролив; залив Анива.
Экология. Встречается на литорали, опускаясь до глубины
55—160 м, преимущественно на песчаных и каменистых грунтах, довольно
часто в зарослях Corallina, Desmarestia, Zostera и др.
* 22. Pagurus lanuginosus de Haan. (Табл. 5, фиг. 2).
Pagurus lanuginosus de Haan, 1850:207, t. 49, fig. 5. — Eupagurus lanuginosus
Ortmann, 1892: 312; Doflein, 1902: 647; Balss, 1913:56, fig. 33; Terao, 1913 : 370; Yokoya,
1933:87.
Передняя часть карапакса значительно больше в длину, чем в ширину.
Средний выступ лба острый, треугольный, достигает половины длины
глазных чешуек. Боковые выступы едва намечаются. Глазные стебельки
длинные, тонкие. Глазные чешуйки короткие, треугольные. Скафоцериты
не достигают концов глаз, их внутренние края покрыты густыми
длинными волосками. Клешненосные ноги волосистые. Мерус правой
клешненосной ноги с гладкими боковыми сторонами, нижняя поверхность
почти безволосая. Верхняя поверхность карпуса несет ряд крупных
шипов на внутреннем краю и другой ряд более мелких шипов ближе
к середине; вся поверхность, за исключением промежутка между этими
рядами шипов, покрыта длинными мягкими волосками; внутренняя поверх-
ность менее волосатая, наружная и нижняя почти безволосые. Рука шире
чем карпус, ее внутренний край образует почти прямую линию, тогда как
наружный край широко и равномерно закруглен; неподвижный палец
в два раза шире подвижного; вся поверхность покрыта густыми пучками
мягких, длинных волос, внутренний и наружный края усажены шипами,
которые резко отделяют почти плоскую верхнюю поверхность ладони
от сильно выпуклой и совершенно безволосой нижней поверхности.
Концы пальцев не роговые. Левая клешненосная нога достигает осно-
вания подвижного пальца правой руки. Мерус сплющенный, волосатый
на нижней поверхности. Карпус с двумя рядами шипов на верхней поверх-
ности и с пучками длинных волосков, в особенности на внутренней
поверхности. Поверхность руки выпуклая, покрыта шипами и волосами,
как и правая рука; нижняя поверхность почти гладкая, с одиночными
пучками волос; кончики пальцев роговые. Ходильные ноги правой стороны
превышают правую клешненосную ногу, все членики волосатые, в осо-
бенности на верхних кра’ях. Карпальные членики первой пары шиповатые
сверху. Пальцы короткие, прямые и не закрученные, с сильными, изогну-
5. PAGURUS
213
тыми вниз когтями. Окраска спиртовых экземпляров желтовато-оранже-
вая, с красными пятнами на члениках всех конечностей (в особенности
на наружных поверхностях ходильных ног); волоса бледно-желтые, почти
белые; когти ходильных ног темнокоричневые.
Р азмеры
Длина передней части карапакса
Ширина „ „ „ ... 6.0 „
Длина глазных стебельков ... 4.0 „
„ меруса правой клешней, ноги .... * .’ - • .4.5 „
„ карпуса ,, „ „ . . . 5.0 „
Ширина „ „ „ „ . • . 4.0 „
Длина руки „ „ „ ... 8.0 „
Ширина „ „ „ . . ... 6.0 „
Длина меруса левой „ „ . ... ... 4.5 „
„ карпуса „ „ ... 5.0 „
Ширина карпуса левой „ „ ... 2.5 „
Длина руки „ „ „ . . . 5.0 „
Ширина „ „ „ „ . . . 2.5 „
Длина правой ходильной ноги первой пары . . . . . . 20.0 „
Распространение. Южная Япония — Нагасаки, бухта Сагами,
Токийская бухта; Сангарский пролив. На глубинах от 110 до 229 м.
В наших частях Японского моря вид встречается редко (в сборах экспед.
Гос. Гидрол. Ин-та имеется 18 экз. из района залива Петр Великий).
23. Pagurus armatus (Dana). (Табл. 2, фиг. 4).
Bernhardus armatus Dana, 1851: 270; 1852: 442, t. 27, fig. 2. —• Eupagurus armatus
Stimpson, 1857:484; Bate, 1866:287; Whiteaves, 1870:471. — Eupagurus ochotensis Stimpson,
1858:248.—- Pagurus ochotensis Holmes, 1900:137; Benedict, 1901: 463, text-fig.; Rathbun,
1904:157; Schmitt, 1921 :130, fig. 84; Stevens, 1925:279, fig. 3. — Non Pagurus (Eupagurus)
bernhardus var. spinimana seu subsp. ochotensis Brandt, 1851 :108.
Передняя часть карапакса больше в ширину, чем в длину. Все три
выступа лба выдаются почти одинаково далеко. Глазные стебельки корот-
кие и толстые. Глазные чешуйки с сильными субтерминальными шипами,
заметными при взгляде сверху. Скафоцериты не такие мощные, как
у Р. ochotensis, трехгранные, но сильно сплющенные, простираются
значительно дальше глаз, гладкие; их внутренние края волосатые и очень
неровные, но не шиповатые, как у Р. ochotensis и Р. ochotensis subsp.
aleu.ticu.s-, внутренняя поверхность не очень сильно иризирующая. Наруж-
ный выступ базального членика антеннальной ножки сильно выдающийся,
внутренний выступ шиповатый. Клешненосные ноги не длинные, шипо-
ватые и волосатые. Мерус правой клешненосной ноги простирается не-
много дальше глаз. Стороны карпуса плоские и гранулированные; верхняя
поверхность карпуса и руки покрыта тонкими длинными шипами и воло-
сами, которые не достигают концов шипов, равны им или иногда даже
несколько превышают их. Рука удлиненная, равномерно округлая сверху;
214
PAGUR1DAE
подвижный палец с рядом шипов на наружном краю и другим рядом около
середины; при смыкании концы пальцев скрещиваются. Левая клешне-
носная нога простирается немного дальше основания подвижного пальца
правой руки. Поверхность карпуса и руки покрыта шипами и волосами,
как и на правой ноге. Моральные, карпальные и проподальные членики
ходильных ног шиповатые сверху. Пальцы шиповатые, не такие широкие
и не так сильно закручены, как у Р. ochotensis, с бороздкой на наружной
и внутренней поверхности и почти такой же длины, как два предыдущих
члерика вместе взятые. Окраска от оливково-желтой до оранжево-абри-
косовой, с коричневатыми пятнами; в общем похожа на окраску Р. ocho-
tensis, но радужный отлив заменяется в значительной мере фиолетовым
оттенком.
Размеры
Длина передней части карапакса............ . . 6.0 мм
Ширина „ „ .....................7.0 „
Длина глазных стебельков . ................... 3.5 „
Длина скафоцеритов............................ 4.0 „
„ меруса правой клешнен. иоги............. 7.0 „
„ карпуса „ „ „ 6.0 „
Ширина „ „ „ „ 4.5 „
Длина руки „ „ „ 10.0 „
Ширина „ „ „ „ 6.0 „
Длина правой ходильной иоги первой пары ...... 30.0 „
Распространение. Сравнительно узко распространенный, энде-
мичный вид западного побережья Северной Америки: от Уналашки до
San Diego, Калифорния, на глубинах от 10 до 146 м. В Охотском и Япон-
ском море отсутствует.
* 24. Pagurus pectinatus (Stimpson). (Табл. 4, фиг. 3).
Eupagurus pectinatus Stimpson, 1858: 249; 1907 : 220; Balss, 1913: 60, text-fig. 35, t. 1,
fig. 8; Terao, 1913:371; Yokoya, 1933:83. — Eupagurus seriespinosus Thallwitz, 1891: 34;
Terao, 1913:372.
Передняя часть карапакса больше в длину, чем в ширину. Средний
выступ лба короткий, острый, закрытый пучком длинных волосков. Боко-
вые выступы закругленные, с острыми субмаргинальными шипами, на-
правленными косо наружу. Глазные стебельки тонкие, длинные, цилин-
дрические, булавовидно расширенные у основания. Глазные чешуйки
полуовальные, маленькие, с сильными субтерминальными шипами. Скафо-
цериты изогнутые, волосатые на внутреннем краю, не достигают концов
глаз. Вершина наружного выступа базального членика антеннальной
ножки несет несколько шипов. Клешненосные ноги шиповатые и волоса-
тые. Мерус правой клешненосной ноги с закругленными ребрами, с гладкими
боковыми сторонами и слабо волосатой нижней поверхностью; верхне-
дистальный край несет 2—5 шипов. Верхняя поверхность карпуса гладкая,
покрыта пучками длинных волос; внутренний край вооружен рядом силь-
5. PAG UR US
215
пых, изогнутых вперед шипов; на внутреннем дистальном углу имеется
группа шипов; наружная сторона гладкая и безволосая, внутренняя
и нижняя поверхности слабо волосатые. Верхняя поверхность ладони
покрыта 8 неправильными продольными рядами длинных шипов, считая
и краевые ряды; шипы наружного края самые крупные, направлены косо
вверх и наружу и резко отделяют верхнюю поверхность ладони от
гладкой выпуклой, слабо волосатой нижней поверхности. От оснований
шипов на верхней поверхности поднимаются пучки длинных волос. На
пальцах поверхность довольно резко понижается, становится вогнутой,
благодаря сравнительно высоко поднимающимся шипам Наружных краев.
Хватательные края с сильными белыми буграми, концы роговые. Мерус
.левой клешненосной ноги сплющенный, гладкий, с 1—2 шипами на верхнем
дастальном краю. Верхняя поверхность карпуса с 2 рядами шипов на краях
и с гладким промежутком между ними; нижняя поверхность волосатая.
Верхняя поверхность руки с невысоким среднем гребнем, который обра-
зован неправильным рядом круглых шипов; менее крупные шипы непра-
вильным двойным рядом окаймляют наружный край ладони. Наружная
поверхность ладони шиповатая, с пучками длинных волос, внутренняя
поверхность менее шиповатая и волосатая; нижняя поверхность гладкая,
с отдельными пучками волос. Пальцы при смыкании оставляют довольно
широкую щель у основания. Ходильные ноги правой стороны превышают
правую клешненосную ногу. Моральные членики сильно сплющенные,
•слабо волосатые на краях. Карпальные членики первой пары шиповатые
на верхнем краю, проподальные членики волосатые. Пальцы с густыми
волосами на верхнем краю, не закрученные, с сильными роговыми когтями.
Окраска спиртовых экземпляров желтовато-оранжевая; в особенности
бросается в глаза ярко-оранжевая выпуклая нижняя поверхность правой
руки. Когти ходильных ног темнокоричневые.
Размеры
Длина передней части карапакса
Ширина „ „ ff ...... . . . 13.0 ff
Длина глазных стебельков ... 6.5 ff
„ меруса правой клешнен. ноги ...... . . . 13.0 ff
„ карпуса „ ft ff . . . 13.0 ff
Ширина „ „ » ff ...... . . . 9.0 ff
Длина руки „ ff >> ...... . . . 18.5 u
Ширина „ „ ff ff ...... . . . 10.5 ff
Длина меруса левой » ff ....•• . . . 12.5 ff
„ карпуса „ ff ff ...... . . . 10.5 ff
Ширина „ ff ft ...... ... 5.0
Длина руки „ ff ff ...... . . . 14.0 ff
Ширина „ „ ff ff ... 7.0 ff
Длина правой ходильной ноги первой пары . • . . . . 50.0 ff
Данный вид отличается от Р. capillatus более короткими скафоце-
ритами; верхней поверхностью карпуса правой клешненосной ноги, которая
216
PAGURIDAE
лишена шипов, за исключением ряда на внутреннем краю; более длин-
ными волосами и шипами на ладони правой руки, особенно на ее наруж-
ном краю; а также расположением шипов в более или менее правильные
продольные ряды; очень выпуклой и почти безволосой нижней поверх-
ностью правой руки, окрашенной в оранжевый цвет; большей шириной
правой руки и заметной вогнутостью ее пальцев; пальцами ходильных
ног, которые не закручены.
Распространение. Японское море, от о-ва Цусимы на север
до залива Де-Кастри; Сангарский пролив (Aomori, Хакодате); Лаперузов-
пролив; залив Анива.
Экология. Pagurus pectinatus встречается на глубинах от 5 до
220 м, в наших водах обычно на 10—60 м, преимущественно на жестких
грунтах; довольно часто попадается в губках (Suberites domuncula).
"25. Pagurus obtusifrons (Ortmann). (Фиг. 70).
Фиг. 70. Pagurus obtusifrons
(Ortmann). A — лобный край;
В — правая клешненосная нога.
(Из Ortmann, 1892).
Eupagurus obtusifrons Ortmann, 1892 : 313, t. 12,
fig. 18; Terao, 1913: 371; Yokoya, 1933:85.
Средний выступ лобного края тупой,,
едва такой же длины, как боковые выступы,
которые снабжены тонкими субмаргиналь-
ными шипами. Глазные стебельки довольно
толстые, с немного вздутыми роговицами,
короче, чем ножки антенн, и почти такой же
длины, как скафоцериты. Карпус и ладонь
правой клешненосной ноги покрыты длин-
ными волосками и шипами. Шипы на ладони
располагаются в 8—9 продольных рядов,
которые около наружного края становятся
неотчетливыми. Наружный край ладони тупой,
без сильных шипов. Верхняя поверхность
ладони выпуклая. Левая рука волосатая и ши-
поватая. Две первые пары ходильных ног волосатые, с довольно строй-
ными пальцами, которые длиннее, чем предпоследние членики. (Ortmann).
Распространение. Южная Япония, на север не идет дальше
Сангарского пролива. На глубине от 48 до 393 м.
26. Pagurus setosus (Benedict). (Фиг. 71).
Eupagurus setosus Benedict, 1892:19. — Pagurus setosus Rathbun, 1904:159, t. 5,.
fig. 1; Schmitt, 1921 :136, fig. 88; Stevens, 1925:290, fig. 11. — Non Pagurus setosus Milne-
Edwards, 1848:64.
Передняя часть карапакса немного больше в длину, чем в ширину.
Средний выступ лба закругленный, выступает вперед немного дальше
боковых выступов, которые благодаря субмаргинальным шипам кажутся
острыми. Глазные стебельки только немного более половины длины
5. PA GUR US
217
передней части карапакса, значительно толще и короче, чем у Р. ken-
nerlyi. Глазные чешуйки очень маленькие и острые, с сильными субтер-
минальными шипами. Скафоцериты простираются дальше глаз на своей
длины. Обе руки клешненосных ног густо покрыты жесткими щетинками.
Правая рука имеет семь продольных рядов шипов, считая и краевые ряды;
шипы более короткие, чем у Р. kennerlyi. Хватательные края пальцев
с крупными буграми; кончики роговые. Мерус левой клешненосной ноги.
Фиг. 71. Pagurus setosus (Benedict). (Из Schmitt, 1921).
сплющенный. Карпус толстый, с двумя гребнями. В общем, клешненос-
ные ноги очень похожи по форме на таковые Р. kennerlyi, но .шипы
на них более короткие, а волоса более длинные. Ходильные ноги воло-
сатые, верхние края без шипов, за исключением карпальных члеников
первой пары. Пальцы незакрученные, на х/3 длиннее чем передняя часть
карапакса. Окраска розовато-желтая, с неправильными коричневыми
полосами, за исключением рук и пальцев. Шипы с абрикосово-оранжевыми
концами. Волоса коричневые у основания и желтые на концах. Антенны
коричневые сверху и желтые снизу. Молодые экземпляры, в общем, более
светлых оттенков. Окраска спиртовых экземпляров бледножелтая. У фор-
малиновых экземпляров ноги с малиново-красными полосами, волоса
грязного цвета.
218
PAGURIDAE
Размеры
Длина передней части карапакса.......14.0 мм
Ширина „ „ „ .............13.0 „
Длина правой клешней, ноги ..........48.0 „
„ руки правой клешней, ноги .......... 21.0 „
Pagurus capillatus, за исключением слегка закрученных пальцев
ходильных ног, по общему виду очень похож на Р. setosus, но правая
рука в пропорциональном отношении не так длинна и ее более много-
численные шипы не так сильны и не так явственно располагаются в про-
дольные ряды. У экземпляров одинаковой величины шипы на правой руке
у Р. capillatus составляют только половину величины шипов у Р. setosus.
Правая рука у Р. setosus почти в два раза больше в длину чем в ширину
и с более пониженными пальцами чем у Р. capillatus. Правая рука у Р. capil-
latus обычно в полтора раза больше в длину чем в ширину, иногда в 13/4 раза
больше в длину чем в ширину. Я видел только один экземпляр, у которого
рука была длиннее — почти в два раза больше в длину чем в ширину, но
характерное вооружение правой руки исключало возможность смешения.
Кроме того, треугольная наружная поверхность левой руки у Р. setosus
более округлая и не настолько четко и широко треугольна, как таковая
у Р. capillatus, будучи приблизительно в 4 раза больше в длину чем
в ширину. У Р. capillatus треугольная наружная поверхность левой руки
примерно в три раза больше в длину чем в ширину (Schmitt).
Распространение. От о-ва Кадьяк, Аляска, до о-ва Santa Cruz,
Калифорния. На глубинах от 9 до 480 м. Встречается значительно реже
Р. kennerlyi.
27. Pagurus kennerlyi (Stimpson). (Фиг. 72).
Eupagarus kennerlgi Stimpson, 1864:153; Benedict, 1892:19; Walker, 1892 : 275.—
JPagurus kennerlgi Holmes, 1900:143; Rathbun, 1904:159, t. 5, fig. 4; Stevens, 1925:289,
fig. 10.
Передняя часть карапакса немного больше в длину, чем в ширину.
Средний выступ лба короткий, треугольный, иногда почти незаметный.
Боковые выступы маленькие, округлые, с острыми субмаргинальными
шипами. Глазные стебельки умеренной длины, прямые и тонкие, не длин-
нее, чем ножки антенн; роговицы немного расширяющиеся, с пучком волос
на верхушке. Скафоцериты маленькие, тонкие, волосатые, не достигают
кончиков глаз. Все ноги очень волосатые. Клешненосные ноги очень
короткие и толстые, обе значительно короче ходильных ног, сильно,
но не очень густо вооружены короткими шипами. Мерус правой клешне-
носной ноги почти трехгранный; наружная поверхность гладкая, нижний
край шиповатый. Карпус примерно такой же длины, как ладонь руки;
внутренний край усажен 7—8 тонкими, изогнутыми шипами. Рука немного
шире чем карпус, пересечена семью довольно неправильными продольными
рядами тонких, бледно окрашенных шипов, включая и краевые ряды;
5. PAGURUS
219
Фиг. 72. Pagurus kennerlgi (Stimpson).
(Из Stevens, 1925).
волосы не густые, по длине значительно превышающие шипы. Пальцы
короче, чем ладонь; на подвижном пальце имеется два явственных ряда
острых бугорков. Хватательные края с сильными буграми; кончики рого-
вые. Левая клешненосная нога едва достигает середины подвижного
пальца правой руки. Мерус сжатый, нижние края шиповатые. Карпус
немного сплющенный, с 2 рядами шипов. Наружная треугольная поверх-
ность руки покрыта крупными шипами. Ходильные ноги толстые; пальцы
незакрученные. Окраска (по Stevens’y): от бледной розовато-желтой до
розовато-коричневой с коричне-
выми пятнами. Шипы на руках
белые, волосы коричневые. Хо-
дильные ноги менее коричне-
вые, чем клешненосные. С пер-
вого взгляда антенны кажутся
состоящими из коричневых и
белых колец, но при более
тщательном рассмотрении ока-
зывается что примерно на сере-
дине жгута, кольца состоят из
4—б коричневых сегментов,
которые чередуются с одним
белым, двумя коричневыми и
опять белым сегментами; за-
тем серия повторяется, причем
кольца становятся шире, т. к.
число коричневых сегментов
увеличивается до 8—10, по на-
правлению к кончику, а затем
целиком замещается белыми
сегментами. На базальной части
коричневых сегментов меньше
экземпляров последовательность является менее выраженной.
Распространение. Алеутские о-ва, к югу до Вашингтона; на
глубинах от 3 до 177 м.
и многие кольца неполные. У
*28. Pagurus capillatus (Benedict). (Табл. 3, фиг. 2).
Eupagurus capillatus Benedict, 1892:8.—Pagurus capillatus Holmes, 1900:138;
Rathbun, 1904:157, t. 4, fig. 3; 1919 : 7; Schmitt, 1921 :132, fig. 85. — Eupagurus trigonocheirus
Balss, 1913 : 63 (part).
Передняя часть карапакса немного больше в длину, чем в ширину.
Средний выступ лба широкотреугольный, с пучком мягких длинных воло-
сков; боковые выступы округлые, с сильными субмаргинальными шипами.
Глазные стебельки длинные, тонкие, с незначительным перехватом
посредине. Глазные чешуйки конические, с закругленной вершиной, спуч-
220
PA GURIDAE
ком волос на верхней поверхности и с сильными субтерминальными
шипами. Скафоцериты изогнутые, достигают концов глаз или немного
короче их. Наружный выступ базального членика антеннальной ножки
с шипом на вершине и 3 шипами на внутреннем краю; внутренний выступ
с одним сильным шипом. Клешненосные ноги длинные, волосатые. Волоса-
тость вообще сильно вариирует у экземпляров, как одинакового, так
и разного размера, но, как правило, крупные экземпляры более волосатые.
То же самое относится и к шипам, которые вариируют, как по величине,
так и по густоте расположения. Мерус правой клешненосной ноги с глад-
кими, голыми или волосатыми боками; верхний дистальный край с шипами;
нижняя поверхность шиповатая и волосатая. Все стороны карпуса волоса-
тые; верхняя поверхность покрыта шипами, от основания которых подни-
маются пучки волос, длина которых превышает длину шипов. Рука продол-
говатая; выпуклая верхняя поверхность покрыта негусто сидящими шипами
и волосами; наиболее крупные шипы занимают середину ладони и наруж-
ный край; верхняя поверхность подвижного пальца имеет продольный ряд
довольно сильных шипов, а наружный край этого пальца окаймлен двойным
рядом более мелких шипов. Нижняя поверхность выпуклая, редко грану-
лированная и волосатая. Пальцы довольно длинные, с роговыми кончиками.
Левая клешненосная нога достигает до основания подвижного пальца
правой руки, у крупных экземпляров немного короче. Мерус сплющенный,,
волосатый, с шиповатой нижней поверхностью. Карпус с двумя рядами
шипов на краях верхней поверхности и с гладким, безволосым промежут-
ком между ними; боковые и нижняя поверхности волосатые. Рука узко-
овальной, симметричной формы; шиповатый средний гребень невысокий
и нерезкий; наружная, равномерно выпуклая поверхность покрыта шипами
и волосами; внутренняя поверхность только волосатая. Пальцы слабо
изогнуты вниз. Ходильные ноги волосатые, в особенности на верхних,
краях; на правой стороне они превышают правую клешненосную ногу.
Моральные членики первой пары шиповатые снизу, карпальные членики
шиповатые сверху. Пальцы длинные, изогнутые и слегка закрученные,
густо волосатые на верхнем и нижнем краю, с сильными роговыми
когтями. Окраска спиртовых экземпляров бледная, розовато-желтая.
Волоса на клешненосных ногах часто удерживают остатки грунта (ил„
песок и пр.).
Размеры
Длина передней части карапакса ................. 9.0 мм
Ширина „ „ „ ................... 8.0 „
Длина глазных стебельков ....................... 5.5 „
„ меруса правой клешнен. ноги.................10.0 „
„ карпуса „ „ „ 10.5 „
Ширина „ „ „ „ 8.0 „
Длина руки „ „ „ 15.0 ,„
Ширина „ „ „ „ . .... 9.0 „
5. PAGUR US
221
Длина меруса левой клешнен. ноги............... 8.5 мм
Длина карпуса „ „ ................... 8.5 „
Ширина „ „ „ „.................. 4.0 „
Длина руки „ „ „ 11-0 „
Ширина „ „ „ „ 6.0 „
Длина правой ходильной ноги первой пары........41.0 „
Об отождествлении Baiss’oM данного вида с Р. trigonocheirus
(= Р. pubescens) уже говорилось выше. Оба вида легко отличаются друг
от друга характером верхней поверхности левой руки: у Р. pubescens
имеется резко выраженный средний гребень, наружная поверхность косо-
треугольная, весь контур руки асимметричный; у Р. capillatus средний
гребень слабо выраженный, контур руки симметричный. Кроме того,
шипы у Р. capillatus более длинные, волосы более густые и имеют
перистое строение (у Р. pubescens они простые). Скафоцериты такой же
длины, как глаза, или короче, тогда как у Р. pubescens они всегда
длиннее глаз.
Распространение. Японское море, только по нашему побе-
режью, на юг до широты бухты Чогу-Чьен-Догу (северная Корея);
из Японии неизвестен; Охотское море; восточное побережье Кам-
чатки; Берингово море, по американскому побережью на юг до Кали-
форнии (широта 36°55' N); Чукотское море, его южная часть до широты
66°40'N.
Экология. Известен с глубин от 5 до 432 м; чаще всего
встречается на глубине 10—100 м, преимущественно на илистых
грунтах.
29. Pagurus constans (Stimpson). (Фиг. 73).
Eupaguras constans Stimpson, 1858 : 248; 1907 : 218, t. 24, fig. 3: Henderson, 1888: 67,
t. 6, fig. 8; Ortmann, 1892: 310; Doflein, 1892 :647; Alcock, 1905 :177; Balss, 1913 : 55; Terao,
1913:366; Yokoya, 1933:81.
Передняя часть карапакса выпуклая, очень твердая, с двумя дуго-
образными, сходящимися рядами пучков щетинок, которые могут совсем
стираться у старых экземпляров. Средний выступ лба довольно острый
выдается дальше боковых выступов, которые также заострены. Глазные
стебельки довольно длинные, но не достигают конца антеннальных
ножек; роговицы слегка расширяющиеся. Скафоцериты длинные, волоса-
тые, простираются дальше глаз (по Ortmann’y короче или, самое большое,
такой же длины, как глаза). На краю второго стернального сегмента между
основаниями наружных максилярных ножек имеются два острых зубца
или шипа. Правая клешненосная нога очень большая, простирается значи-
тельно дальше концов ходильных ног. Исхиум с длинным, острым шипом
на внутреннем углу (Ortmann не мог найти его у своих экземпляров,
у экземпляров Doflein’a он имелся). Мерус с несколькими шипами на верх-
222
PAGURIDAE
нем переднем краю. Карпус немного больше в длину, чем в ширину, и
приблизительно такой же длины, как и ладонь, или иногда длиннее. Верх-
няя поверхность руки плоская, ее края острые. Пальцы короче, чем ладонь,
плотно смыкаются друг с другом, с рядами густо сидящих пучков щетинок
на их внутренних краях. Левая клешненосная нога простирается дальше
основания подвижного пальца правой руки. Своим вооружением обе клешне-
носные ноги похожи друг на друга. Поверхность меруса почти гладкая,
негусто покрыта короткими рядами волос, которые поднимаются от слабых
Фиг. 73. Pagurus constans (Stimpson).
(По Stimpson, 1907).
поперечных линий. Карпус и рука
с шипами на верхней поверхности
и густо покрыты волосами, которые
поднимаются от оснований шипов
и достигают до их кончиков. На
середине ладони правой руки имеется
ряд более крупных шипов; шипы
краев ладони также более крупные.
На левой руке средний ряд шипов
продолжается на неподвижный па-
лец; имеется один ряд шипов на
наружном краю и несколько рас-
сеянных шипов на внутренней сто-
роне у основания. Между шипами
имеются бугорки, которые несут
щетинки. Ходильные ноги тонкие,,
негусто волосатые; карпальные чле-
ники с одним или двумя шипами
на верхнем краю; пальцы длин-
ные и тонкие, длиннее, чем пропо-
дусы, незакрученные, с короткими,
жесткими волосками; роговые когти очень короткие. Окраска бледно-
оранжевая, с мелкими пятнами на теле. Ноги сверху с темнокрасными
полосами.
Данный вид, как отмечает Stimpson и почти все последующие авторы,
встречается всегда в симбиозе с Hydractinia sodalis Strap.; этот гидроид,
обрастает небольшую, первоначально обитаемую отшельником раковину
и затем настолько разрастается, что последний теряет необходимость,
в смене раковины, как большинство Paguridae, но остается в этом свое-
образном, созданном гидроидом, жилище в течение всей своей жизни;;
в .глубине полости, занимаемой отшельником, можно найти раковину,
в которой он первоначально обитал. Иногда встречается в губках (Doflein,
Balss), причем, невидимому, губка растворяет раковину, первоначально'
занятую отшельником.
Распространение. Южная Япония, к северу до Хакодате
(о-в Иезо). На глубинах от]5 до 150 м.
6. PARAPAGURUS
223
6. Род PARAPAGURUS SMITH
Parapagurus Smith, 1880 : 50; 1882 : 20; Henderson, 1888: 85; Milne-Edwards et Bouvier,
1893: 26; Stebbing, 1893 :166; Holmes, 1900:155; Alcock, 1905 : 98; Balss, 1912 .-96; 1913 :49;
Terao, 1913 : 382; Schmitt, 1921:145. — Sympagurus Smith, 1883 :37; Henderson, 1888:52;
Milne-Edwards et Bouvier, 1893 : 58; Stebbing, 1893 :166; Alcock, 1905 :103.
Тип рода: Parapagurus pilosimanus Smith.
Абдомен самца с парными придатками на первом и втором абдоми-
нальном сегменте. У самки имеется только один яйцевод, открывающийся
на коксальном членике третьей грудной ноги левой стороны. Наружные
максиллярные ножки широко расставлены у основания. Экзоподит первой?
пары максиллярных ножек не имеет жгута. Г лаза маленькие. Атеннулы
и антенны длинные. Стернальные пластинки между первой и второй парой
ходильных ног узкие. Жабры состоят из многочисленных цилиндрических
папилл, как у большинства Macrura, а не из пластинок, как у большин-
ства Paguridae. Правая клешненосная нога развита сильнее левой; пальцы
двигаются в горизонтальной плоскости. Четвертая пара ног с ложными
клешнями.
Виды данного рода являются большею частью абиссальными.
ОПРЕДЕЛИТЕЛЬНАЯ ТАБЛИЦА ВИДОВ
1 (2). Глазные стебельки тонкие, суживающиеся в дистальном направлении..
Пальцы левой руки не длиннее ладони. Нижний край пальцев ходиль-
ных ног с очень маленькими, едва заметными шипиками................
.......................................... *1. Р. pilosimanus Smith,
2 (1). Глазные стебельки толстые, расширяющиеся в дистальном направле-
нии. Пальцы левой руки значительно длиннее ладони. Нижний край
пальцев ходильных ног с хорошо заметными шипами...............
........................................2. Р. mertensii Holmes..
* 1. Parapagurus pilosimanus Smith. (Фиг. 74).
Parapagurus pilosimanus Smith, 1880:51; 1883: 33, t. 5, fig. 3—5, t. 6, fig. 1-a; 1882:.-
20, t. 2, fig. 4; Milne-Edwards et Bouvier, 1893 : 28; 1894: 64, t. 9, fig. 1—17; 1900 :187, t. 6,
fig. 2, t. 24, fig. 1—3; Alcock, 1905 : 99, t. 10, fig. 1; Hansen, 1908: 29; Balss, 1912 : 96, t. 11,.
fig. 1—6; 1913:50; Terao, 1913 :385; Yokoya, 1933: 79.
Передняя часть карапакса немного больше в ширину, чем в длину,,
гладкая, почти сплошь безволосая. Средний выступ лба широкий и тупой,
боковые выступы очень маленькие. Глазные стебельки тонкие, суживаются:
в дистальном направлении, с длинными волосками на верхней поверхости;
роговицы очень маленькие. Глазные чешуйки маленькие, тонкие и острые.
Ножки антеннул очень длинные и тонкие; дорзальный жгут густо опушен
снизу волосами. Скафоцериты достигают концов глаз или простираются
немного дальше, густо волосатые на верхнем краю. Жгуты антенны.
224
PAGURIDAE
тонкие и почти совершенно безволосые. Правая клешненосная нога боль-
шая, почти такой же длины, как все тело животного. Мерус с мелкими
бугорками и отчасти волосатый на наружной и нижней поверхности.
Поверхность карпуса покрыта мелкими бугорками и тонкими мягкими,
светло-окрашенными волосами. Рука равномерно выпуклая сверху и снизу,
с закругленными боковыми краями; вся поверхность, за исключением
небольшого пространства снизу близ основания, а также базальной части
обоих пальцев, бугорчатая и покрыта волосами подобно поверхности кар-
Фиг. 74. Parapagurus pilosimanus Smith.
пуса. Неподвижный палец довольно тонкий, быстро суживающийся
к роговому концу, так что его хватательный край имеет косое напра-
вление, вправо; подвижный палец примерно такой же длины, как внутрен-
ний край ладони. Хватательные края обоих пальцев почти прямые и воору-
жены несколькими низкими и тупыми бугорками. Левая клешненосная
нога очень тонкая, составляет приблизительно 3/4 длины правой ноги; кар-
пус и рука менее бугорчатые, чем на правой ноге, но опушены подобными же
волосками. Пальцы почти такой же длины, как ладонь, тонкие и слегка
изогнутые вниз у концов; кончики роговые, хватательные края острые,
поверхность пальцев закругленная и голая, за исключением рассеянных
пучков коротких щетинок, которые поднимаются из небольших углублений
на поверхности. Ходильные ноги простираются за конец правой клешне-
носной’ ноги; за исключением пальцев, они почти совершенно гладкие и
6. P AR AP AG U R US
225
голые. Вторая пара немного длиннее и несколько толще, чем первая.
У обеих пар карпальные членики составляют приблизительно половину
.длины моральных и достигают дистального конца карпуса правой клеш-
неносной ноги; продольные членики немного длиннее моральных. Пальцы
значительно длиннее, чем проподусы, тонкие, сильно изогнутые, в особен-
ности по направлению к очень тонким и острым кончикам; они сплющены
<с боков, причем стороны почти гладкие, голые и имеют мелкую продоль-
ную борозду, простирающуюся до сильно изогнутой терминальной части,
которая сплющена еще более и очень слабо закручена; нижний край
и верхний, за исключением терминальной части, закруглённые; верхний
;имеет несколько рассеянных щетинок, которые становятся более много-
численными близ конца, где край сплющенный и острый на коротком
протяжении. Первая пара абдоминальных придатков самца помещается
'близко друг от друга, почти между коксами задних грудных ног. Каждый
придаток состоит из единственной пластинки, которая слегка утолщается
:по направлению к основанию; дистальная половина расширяется в тонкую'
пластинку, которая свернута в слегка суживающуюся до половины трубку
«с полостью, обращенной внутрь и назад. Вторая пара придатков поме-
щается на боках абдомена, на значительном расстоянии от первой пары,
и поэтому широко расставлена у своих оснований. Каждый придаток
состоит из цилиндрического базального сегмента, к которому причленяется
•более длинный пластинчатый терминальный сегмент; этот терминальный
сегмент, который расположен таким образом, что, когда он прилегает
к бороздчатой поверхности соответствующего придатка первого сегмента,
•то образует вместе с ним трубку, имеет на своей передней стороне мел-
кую борозду, которая оканчивается на наружной стороне придатка неда-
леко от конца; конец придатка тонкий, заостренный и волосатый.
Размеры
Длина передней части карапакса.................13.4 мм
Ширина „ „ „ .................12.0 „
Длина глазных стебельков ...................... 6.3 „
Длина карпуса правой клешней, ноги ............18.0 „
„ проподуса „ „ .................26.0 „
Ширина „ „ „ .................13.7 „
„ первой ходильной ноги левой стороны .... 83.0 „
Длина пальца правой руки...................... 14.0 „
Распространение. Parapagurus pilosimanus является типичным
глубоководным видом (известен от 210 до 4000 м), имеющим широкое
распространение, главным образом, в тропической и субтропической обла-
стях. Атлантика: у Исландии; у юго-западной Ирландии; Гасконский залив;
у Испании и Португалии; у Марокко и Сенегала; у Азорских и Канарских
о-вов; Сиерра-Леоне, Тристань д’Акунья; у Патагонии (47°48'S); у Nova
Scotia; Саргассово море; Бермудские о-ва. Индийский океан: Аравийское
JMope и Бенгальский залив. Тихий океан: море Банда, около Пайуа; Филип-
Фауна CCCPj Макаров 15
226
PAGURIDAE
пинские о-ва; Япония у Иокогамы и Muroto-zaki; у Вальпараисо; около
Галапагосских о-вов и в Калифорнийском заливе. Северным пределом
распространения Р. pilosimanus является параллель 53° N, южным — 48° S.
В наших водах вид был добыт Беринговской экспедицией ГосударственнОго
Гидрологического института (1932) у восточного побережья Камчатки
(52° 42.5' N 159° 03.5' Е) на глубине 830 м.
. 2. Parapagurus mertensii Holmes. (Фиг. 75).
Parapagurus mertensii Holmes, 1900 :155; Rathbun, 1904:162, t. 5, fig-. 6; Schmitt, 1921:.
146, t. 16, fig. 5. — Non. Pagurus mertensii Brandt, 1851 :112.
Средний выступ лба выдающийся, отчасти продолговатый, кончик-
закругленный, стороны полупараллельные; боковые выступы лба малень-
кие. Передняя часть карапакса приблизительна
такой же ширины, как и длины. Глазные стебельки
короткие, составляют примерно половину длины,
передней части карапакса. Клешненосные ноги воло-
сатые, шиповатые, очень неравные. Правая клешне-
носная нога очень большая; карпус длинный, вну-
тренний и наружный края его шиповатые, выпуклая,
верхняя поверхность вооружена двумя рядами корот-
ких шипов. Рука длинная, узкая, сплющенная в дорзо-
вентральном направлении и согнута внутрь под,
слабым углом к карпусу; округлая верхняя поверх^
ность с мелкими, расположенными примерно в ряды»
гранулами, которые на пальцах становятся более
крупными; внутренний и наружный края острые,
зубчато-гранулированные и параллельные. Левая
клешненосная нога длинная, тонкая и слабая. Карпус
полуцилиндрический, с тремя рядами шипов на верх-
ней поверхности. Рука узкая, значительно длиннее, чем карпус; ладонь
очень короткая, пальцы длинные, узкие и изогнутые вниз. Пальцы:
ходильных ног чрезвычайно длинные и тонкие, вооруженные снизу много-
численными шипами.
Этот вид дает один из тех любопытных случаев комменсализма,
с колонией гидроидов, которые Иногда встречаются среди глубоководных
отшельников. Колония гидроидов, покрывающая раковину,, в которой оби-
тает рак, образует перепончатый нарост, который распространяется за
пределы раковины и с течением времени, согласно Benedict’y, растворяет
ее, оставляя ее обитателя в перепончатом домике вместо известкового.
У экземпляра, который имелся у меня, покрытие было образовано
отчасти кончиком сломанной раковины, но главным образом разраста-
нием перепончатого нароста, образованным колонией гидроидов. Это>
позволяет отшельнику расти без затруднительной для него смены жилища.
(Holmes).
7. ORTHOPAGURUS 227
Под названием Pagurus mertensii Brandt’oM описан Pagurus hirsutius-
culus (Dana). Все просмотренные мною экземпляры Зоологического
Института Академии Наук, в том числе и оригинальные экземпляры
Brandt’a, относятся именно к этому виду. Местами нахождения Parapagurus
mertensii, таким образом, следует считать только таковые, приводимые
Holmes’oM, которые М. Rathbun считает ошибочными: побережье Калифор-
нии между Lobos Rocks (36°25z40,z N) и пунктом юго-восточнее о-ва Нико-
лая (33°00z30" N), на глубинах от 139 до 479 м. Все остальные места
нахождения, приводимые для Parapagurus mertensii (Кадья^, Ситка, Ван-
кувер, Камчатка), относятся к Pagurus hirsutiusculus.
7. Род ORTHOPAGURUS STEVENS
Orthopagurus Stevens, 1927 : 247.
Тип рода: Orthopagurus minimus (Holmes).
Лобный край карапакса со средним выступом. Наружные максилляр-
ные ножки широко расставлены у своих оснований. Экзоподиты всех трех,
пар максиллярных ножек жгутикообразные. Жабры типа филлобранхий,
в числе 11 с каждой стороны. Клешненосные ноги неравные, правая
больше левой. Четвертая пара перейопод с ложными клешнями. Абдомен
согнутый, но нисколько не закрученный спирально, с непарными плеопо-
дами на левой стороне у обоих полов. Уроподы симметричные или очень
незначительно более развитые на левой стороне.
Виды этого рода не достигают крупных размеров, обитают чаще:
всего в трубках червей.
ОПРЕДЕЛИТЕЛЬНАЯ ТАБЛИЦА ВИДОВ (ПО STEVENS).
1 (2). Правая рука дистально расширяющаяся, основание узкое и выпук-
лое, дистальная часть покатая, с завернутыми краями. Боковые
выступы лба округлые.......................* 1. О. minimus (Holmes)-
2 (1). Правая рука почти однообразна по ширине, плоская или слегка
выпуклая. Боковые выступы лба незаметные.........................
..................................... * 2. О. schmitti (Stevens)-
* 1. Orthopagurus minimus (Holmes).
Pagurus minimus Holmes, 1900:145; Rathbun, 1904: 160. — Pylopagurus minimus
Schmitt, 1921 :144, t. 16, fig. la, lb, lc. — Orthopagurus minimus Stevens, 1927; 247, fig. 1-
Передняя часть карапакса почти такой же ширины, как и длины-
Средний выступ лба острый, треугольный, с пучком волос; боковые
выступы округлые, с явственными субмаргинальными шипами. Глазные
стебельки длинные, расширяющиеся у основания, составляют примерно
2/3 длины передней части карапакса. Глазные чешуйки треугольные, с суб-
терминальными шипами. Скафоцериты короткие, достигают половины
глазных стебельков. Наружный выступ базального членика антеннальной
15*
228
PA GURIDAE
ножки с 3 шипиками на вершине. Клешненосные ноги менее волосатые,
чем у О. schmitti. Мерус правой клешненосной ноги трехгранный, гладкий,
слабо волосатый на йижней поверхности. Карпус дистально расширяющийся,
верхняя поверхность слабо волосатая, гладкая около наружного края,
с редкими шипами у внутреннего дистального угла и с рядом довольно
крупных, изогнутых вперед шипов на внутреннем краю; нижняя поверхность
волосатая. Рука продолговатая, заметно расширяющаяся к дистальному
концу; верхняя поверхность равномерно покрыта короткими шипами
и слабо заметными волосками; нижняя поверхность гладкая, с рассеян-
ными волосами. Оба пальца одинаково широки у основания, равномерно
закругляющиеся по направлению к концам, края несколько приподнятые
и усажены довольно крупными шипами; поверхность пальцев заметно
понижающаяся в сравнении с поверхностью ладони; хватательные края
с сильными белыми буграми; кончики роговые. Левая клешненосная нога
немного не достигает основания подвижного пальца правой руки. Мерус
сплющенный. Карпус сверху с двумя рядами шипов. Рука округлая,
верхняя поверхность косая, пальцы длиннее, чем ладонь; в общем, рука
похожа на таковую О. schmitti, но пальцы менее искривленные и не обра-
зуют такой широкой щели при смыкании. Ходильные ноги тонкие, сплющен-
ные, достигают конца правой клешненосной ноги, волосатые на верхних
и нижних краях; пальцы тонкие, изогнутые, несколько длиннее, чем про-
подусы. Общая окраска красноватая, с темнокрасными пятнами. Большая
клешненосная нога более темнокрасная, чем остальное тело, в особен-
ности у дистального конца. Глазные стебельки посредине с светло окра-
шенной полосой. .
Размеры
Длина передней части карапакса 3.0 мм
Ширина „ „ „ 2.5 „
Длина глазных стебельков 2-0 „
„ меруса правой клешнен. ноги . . з.с „
„ карпуса „ „ „ 2-5 „
Ширина „ „ „ „ 2-0 „
Длина руки „ „ „ 4.0 „
Ширина „ „ „ з.о „
Длина меруса левой „ „ 2.0 „
„ карпуса „ „ 2-5 „
Ширина „ „ „ „ 1.0 ,,
Длина руки „ „ з.о „
Ширина „ „ „ „ 1.5 „
Длина правой ходильной ноги первой пары Ю.0 „
Этот вид очень близок к О. schmitti (Stevens), от которого отли-
чается более явственными боковыми выступами лба, более короткими
скафоцеритами, дистально расширяющейся правой рукой, приподнятостью
ее краев, меньшей волосатостью и менее широкой щелью между пальцами
левой руки при их смыкании.
I. ORTHOPAGURUS
229
Распространение. Skidegate, Queen Charlotte Sound, Британская
Колумбия; Сан-Франциско, Monterey Bay, Laguna Beach и San-Diego,
Калифорния; от 27.4 до 64 м. (Schmitt). В материалах Государственного
Гидрологического Института имеется единственный экземпляр этого вида,
добытый в бухте Нельма (Татарский пролив) на глубине 20 м.
Экология. Orthopagurus minimus обитает обычно в трубках чер-
вей; был находим также в раковинах Dentalium.
* 2. Orthopagurus schmitti (Stevens). (Табл. 2, фиг. 1).
Pylopagurus schmitti Stevens, 1925 : 298, fig. 17—22.— Orthopagurus schmitti Stevens,
1927 :249, fig. 1—4.
Передняя часть карапакса очень незначительно больше в длину, чем
в ширину. Средний выступ лба треугольный, острый, с пучком длинных
тонких волосков; боковые выступы едва намечающиеся, почти незамет-
ные. Глазные стебельки длинные и тонкие, немного расширяются у осно-
вания. Глазные чешуйки короткие, треугольные, с сильными субтерми-
нальными шипами. Скафоцериты короткие, достигают 2/з длины глазных
стебельков. Наружный выступ базального членика антеннальной ножки
с 2 шипами на вершине и с длинными волосками на наружно-нижнем
краю. Клешненосные ноги покрыты сравнительно длинными тонкими воло-
сами, в особенности на карпальных члениках и руках. Мерус правой
клешненосной ноги трехгранный, с слабо волосатыми боками и более
волосатой нижней поверхностью. Верхняя, внутренняя и нижняя поверх-
ности карпуса с длинными волосами, тогда как наружная поверхность менее
волосатая; верхняя поверхность покрыта мелкими красными шипами;
более крупные, наклоненные вперед шипы образуют ряд на внутреннем
краю. Рука немного шире, чем карпус, с почти параллельными краями,
не расширяется дистально; выпуклая верхняя поверхность ладони покрыта
мелкими красными шипами, которые становятся более крупными у вну-
треннего края; шипы на пальцах белые, на подвижном пальце распола-
гаются в два продольных ряда; нижняя поверхность гладкая, слабо воло-
сатая, резко отграничена от верхней поверхности рядами шипов на краях
руки; хватательные края с 4 белыми буграми на подвижном пальце и 2
на неподвижном; кончики пальцев роговые. Как отмечает В. A. Stevens,
верхняя поверхность руки сильно вариирует, как по длине, так и по степени
выпуклости; у молодых экземпляров она короче по отношению к ширине
и кажется почти плоской, тогда как у старых экземпляров она длиннее
и в большинстве случаев выпуклая. Левая клешненосная нога достигает
основания подвижного пальца правой руки. Мерус сплющенный, волосатый
на нижней поверхности. Карпус немного расширяется дистально, с одним
рядом шипов сверху. Рука продолговатая, с узким основанием; намечается
слабый гребень, который тянется от основания ладони к середине непо-
движного пальца; поверхность ладони снаружи от этого гребця покрыта
мелкими шипами, внутренняя поверхность гладкая; как верхняя, так и ниж-
230
PAGURIDAE
няя поверхности волосатые. Пальцы длинные, с роговыми концами, искри-
вленные, при смыкании оставляют широкую щель. Ходильные ноги не
простираются дальше клешненосных, тонкие, сплющенные, волосатые,
в особенности на верхних и нижних краях. Пальцы тонкие, изогнутые,
незакрученные, с длинными тонкими роговыми когтями. Окраска бледная,
розовато-желтая до белой, с неправильными пятнами и полосами оранже-
вато-коричневого цвета; пальцы абрикосово-оранжевые на концах. Волосы
серебристо-желтые.
Размеры
Длина передией части каракапса
Ширина „ „ „ ... 4.5 „
" Длина глазных стебельков . . . 3.0 „
„ меруса правой клешней, ноги ...... ... 5.0 „
„ карпуса „ „ „ ... 6.0 „
Ширина „ „ „ ... 4.0 „
Длина руки „ „ ... 8.5 „
Ширина „ „ „ „ 4.5 „
Длина меруса левой „ „ ... 4.5 „
„ карпуса „ „ „ 5.5 „
Ширина „ „ „ „ ... 2.0 „
Длина руки „ „ , „ ... 6.5 „
Ширина,, „ „ ... 3.0 „
Длина правой ходильной ноги первой пары . . . . . . 21.0 „
Распространение. Западное побережье Северной Америки,
штат Вашингтон, в окр. Пёджет-Саундской Биологической Станции (Reed
Rock, у о-ва Brown, у о-ва San Juan, Turn Rock, у о-ва Lopez и др.);
Японское море, только у нашего побережья, главным образом, в районе
заливов Владимир и Ольга. Из Японии неизвестен; не отмечен также нигде
и из промежуточных областей (Охотское море, восточное побережье Кам-
чатки, Берингово море).
Экология. Как и предыдущий вид, О. schmitti поселяется в труб-
ках червей (Sabellaria cementarium, Serpula и др.). Встречается на каме-
нистых и галечных грунтах, ракушнике, на глубинах от 6 до 220 м.
8. Род SPIROPAGURUS STIMPSON
Spiropagurus Stimpson, 1858:236; 1907:214; Henderson, 1888 : 71; Milne-Edwards
et Bouvier, 1893:110; Stebbing, 1893:165; Alcock, 1905:117; Balss, 1913:65; Terao,
1913:385.
Тип рода: Spiropagurus spiriger (de Haan).
Карапакс достаточно жесткий на передней половине. Глазные сте-
бельки короткие, с расширяющимися роговицами. Антенны длинные,
скафоцериты тонкие. Наружные максиллярные ножки удалены друг от друга
у основания, их коксальные членики маленькие. Клешненосные ноги почти
равные, нога если и больше, то очень незначительно. Пальцы двигаются
8. SPIROPAGURUS
231
в горизонтальной плоскости. На коксальном членике левой четвертой
ходильной ноги у самца имеется непарный половой придаток в виде длин-
ной, спирально закрученной, перепончатой трубки, верхний край которой
снабжен роговой полоской, придающей трубке упругость. Тельсон раз-
двоен на конце, края пластинок зубчатые.
Виды этого рода распространены на сублиторали тропиков (Панамская
область, область Вест-Индии, западное побережье Африки); в Индопаци-
фической области род представлен одним видом Spiropagurus spiriger
(de Haan), который доходит до Южной Японии, но который я помещаю
в настоящей работе в виду указания, хотя и сомнительного, на нахождение
его в северной части Японского моря.
1. Spiropagurus spiriger (de Haan). (Табл. 5, фиг. 3).
Pagurus spiriger de Haan, 1850: 206, t. 49, fig. 2. — Spiropagurus spiriger Stimpson,
1858: 248; 1907: 214; Henderson, 1888: 72; 1893: 425; 1892: 297; Rathbun, 1902 :37; Alcock,
1905: 118, t. 13, fig. 1; Nobili, 1905: 3; Southwell, 1906 : 216; Balss, 1913: 65; Terao, 1913 :386;
Yokoya, 1933:91.
Передняя часть карапакса больше в ширину, чем в длину. Поверх-
ность как передней, так и задней части карапакса покрыта плоскими
возвышениями, напоминающими по форме чешую, от передних краев кото-
рых отходят довольно длинные, мягкие, лежачие волоса. Средний и боко-
вые выступы лба широко закругленные, одинаково далеко выступающие
вперед; боковые выступы снабжены короткими коническими субмарги-
нальными шипами. Глазные чешуйки широкие, с закругленной верхушкой,
с вогнутой поверхностью, в центре которой имеется возвышение, снабжен-
ное волосками; края снизу густо опушены волосами. Глазные стебельки
^короткие, сплющенные в дорзовентральном направлении, расширяющиеся
у роговиц; верхняя поверхность стебельков несет ряды лежачих воло-
сков; выемка роговицы также окаймлена рядом таких волосков. Базальный
членик антеннальной ножки с сравнительно тупым внутренним и острым
наружным выступом. Скафоцериты превышают концы глаз, волосатые,
в особенности вдоль внутреннего края. Клешненосные ноги одинаковой
длины и одинакового строения. Мерус узкий, трехгранный, нижние грани
закругленные. Карпус узкий, с закругленными гранями, с одним рядом
шипов вдоль внутреннего края; поверхность покрыта такими же возвыше-
ниями с волосами, как и карапакс. Рука длинная, продолговатая, верхняя
и нижняя поверхность покрыта дугообразными рядами коротких лежачих
волосков; наружный и внутренний края опушены длинными волосами;
на поверхности пальцев волосы расположены пучками. Хватательный край
неподвижного пальца с 10 зубцами, которые у основания пальца стано-
вятся бугрообразными; хватательный край подвижного пальца с 3 зубцами
на проксимальной половине; промежутки между этими зубцами на обоих
пальцах и дистальная половина хватательного края подвижного пальца
усажены короткими, плотно прилегающими друг к другу роговыми пластин-
232
PAGURI DAE
ками. Острые изогнутые кончики пальцев перекрещиваются при смыкании.
Ходильные ноги превышают клешненосные на длину своих пальцев. Все
членики их сплющенные, как наружные, так и внутренние поверхности
покрыты рядами коротких волосков; верхние края волосатые. Карпальные
членики с шипами на верхнем дистальном конце. Пальцы длинные, изо-
гнутые в дистальной трети, густо опушенные волосами на верхнем краю;
желобок на наружной поверхности заполнен волосками, которые напра-
влены вверх; на внутренней поверхности имеются две линии торчащих
вверх волосков: одна на середине, другая у нижнего края. Роговой коготь
короткий и сравнительно тупой. На коксе четвертой левой ходильной ноги
у самца имеется длинный, образующий 2—3 оборота спирали, половой,
придаток. Окраска спиртовых экземпляров светлая, рыжевато-желтая,,
у живых желтовато-коричневая, иногда слегка красноватая; волосы свет-
лые и блестящие.
Размеры
Длина передней части карапакса................ 8.0 мм
Ширина „ „ „ ................ 9.5 „
Длина глазных стебельков...................... 4.5 „
„ скафоцеритов............................. 4.5 „
„ меруса клешней, ноги...................... 9.0 „
„ карпуса „ „ 7.0 „
„ РУки „ „ 13.0 ,,
Ширина „ „ „ 4.0 „
Длина первой ходильной ноги левой стороны .... 46.0 „
Распространение. Восточно-Индийский архипелаг, Бенгальский"
залив, Маледивские о-ва; Занзибар; о-ва Адмиралтейства; Торресов про-
лив; Китайское море; Япония (самое северное местонахождение — Токий-
ская бухта). Местонахождение, приводимое Yokoya (Северное Японское
море), является, как указывает и сам автор, сомнительным в виду потери,
этикетки; Цуругская бухта.
Экология. Данный вид свободно плавает, имея при себе рако-
вину, ударяя по воде своими волосатыми ходильными ногами. Известен,
до глубины 20 м.
8. Сем. LITHODIDAE
Lithodidae Dana, 1852 :1430; Ortmann, 1892 : 271; 1901:1147; Schmitt, 1921:146.
Тело крабообразное. Карапакс большею частью жесткий. Рострум
хорошо развит, треугольный, плоский, булавовидный, или же шипообразный,,
вооруженный дополнительными шипами. Linea anomurica явственная. Глаз-
ные стебельки у основания без глазных чешуек. Скафоцериты шипо-
образные, часто ветвящиеся. Наружные максиллярные ножки ногообразные.
Первые перейоподы клешненосные, правая нога обычно больше левой.
Четвертые перейоподы по величине и форме подобны третьим. .Пятые
перейоподы маленькие и подогнутые под карапакс, так что по внешнему
виду представители этого семейства имеют как бы лишь четыре пары
ног. Жабры типа филлобранхий в числе 11 с каждой стороны: одна плевро-
бранхия на четвертой перейопде и по две артробранхии на третьей
максиллярной ножке и четырех первых перейоподах. Абдомен короткий
и широкий, не спрятан в раковину, но подогнут под головогрудь, у самок
заметно асимметричный; вентральная поверхность мягкая, дорзальная
поверхность мягкая, или же покрыта тем или иным числом жестких объиз-
вествленных пластинок и узелков. Абдоминальные конечности у самцов
совершенно отсутствуют. У самок имеются парные рудиментарные плео-
поды на первом абдоминальном сегменте; на втором-пятом сегментах
только на левой стороне имеются простые плеоподы, состоящйе лишь из
одного эндоподита (одноветвистые). Уроподы отсутствуют.
Семейство Lithodidae распадается на два подсемейства: Hapaloga-
strinae (= Hapalogastrica Brandt) и Lithodinae (—Ostracogasrica Brandt).
ОПРЕДЕЛИТЕЛЬНАЯ ТАБЛИЦА ПОДСЕМЕЙСТВ
1 (2). Рострум короткий, плоский, широко-треугольный. Третий-пятый
абдоминальные сегменты не вполне объизвествленные; чаще всего
абдомен имеет вид мягкого перепончатого вздутого мешка ....
...................................... . 1. Подсем. Hapalogastrinae.
2 (1), Рострум выдающийся, тупой или шипообразный. Третий-пятый абдо-
минальные сегменты состоят из хорошо объизвествленных пластинок;
абдомен обычно плоский........................2. Подсем. Lithodinae.
‘234
LITHODIDAE
1. Подсем. HAPALOGASTRINAE
Hapalogastrinae Ortmann, 1901 :1147.
Карапакс слабо выпуклый, редко покрытый шипами, его дорзальная
поверхность обычно лишена бугорков или шипов. Лобный край широкий,
рострум более или менее плоский, треугольный, короткий, обычно не
длиннее глаз. Дорзальная поверхность второго абдоминального сегмента
имеет две маргинальных, две латеральных и одну медианную пластинку
или группу узелков вместо нее; дорзальная поверхность трех следующих
сегментов мягкая, перепончатая, с отдельными объизвествленными зернами
или мелкими пластиночками. Абдомен в целом имеет вид мягкого взду-
того мешка, за исключением рода Placetron, у которого он более или
менее пластинчатый. До настоящего времени считалось, что самки дан-
ного подсемейства не имеют плеопод на первом абдоминальном сегменте;
только Bouvier (1896) предполагал их существование у самок рода Der-
maturus. У всех просмотренных представителей этого подсемейства
я нашел у самок парные рудиментарные плеоподы на первом абдоминаль-
ном сегменте. Таким образом, род Hapalogaster не образует никакой боко-
вой ветви в филогении семейства Lithodidae, как это думал Bouvier и, сле-
довательно, все подсемейство целиком связывает Lithodinae с Pagaridae.
Роды этого подсемейства преимущественно литоральные и сублито-
ральные (верхние горизонты), распространенные исключительно в север-
ной части Тихого океана.
ОПРЕДЕЛИТЕЛЬНАЯ ТАБЛИЦА РОДОВ
1 (2). Боковые края карапакса позади цервикальной борозды вооружены
шипами. Клешненосные ноги вооружены острыми шипами или зуб-
цами. Второй абдоминальный сегмент с непарной медианной пластин-
кой (за исключением Н. cavicauda, где на ее месте имеется группа
известковых узелков). Большинство видов с более или менее сильно
волосатыми карапаксом и конечностями . .1. Hapalogaster Brandt.
'2 (1); Боковые края карапакса невооруженные. Клешненосные ноги с тупыми
или чешуеобразными выступами, или же с поперечными гребнями.
Второй абдоминальный сегмент без непарной медианной пластинки.
Виды слабо волосатые.
3 (6). Клешненосные ноги неравные, правая обычно больше левой; по
сравнению с ходильными ногами клешненосные ноги очень сильные.
Без бугорка на стернальной поверхности между основаниями клешне-
носных ног. Без трех шипов на нижнем дистальном краю мерусов
ходильных ног. Между латеральными пластинками второго абдоми-
нального сегмента остается широкий перепончатый промежуток.
Абдомен мешкообразно вздутый.
/. HAPALOGASTER
235
4 (5). Поверхность карапакса и клешненосных ног покрыта поперечными
гребнями. Скафоцериты прямые, копьевидные. Два последних чле-
ника наружных максиллярных ножек не расширяющиеся.................
........................................2. Dermaturus Brandt.
5 (4). Поверхность карапакса покрыта чешуеобразными возвышениями,
а клешненосные ноги усажены круглыми буграми. Скафоцериты
изогнутой полулунной 'формы. Два последних членика наружных
максиллярных ножек расширенные .... 3. Oedignathus Benedict.
€ (3). Клешненосные ноги равные или почти равные, не слишком развитые
по сравнению с длинными ходильными ногами. Имеется бугорок на
стернальной поверхности между основаниями клешненосных ног.
Нижний дистальный край мерусов ходильных ног вооружен тремя
тонкими острыми шипами. Латеральные пластинки второго абдоми-
нального сегмента плотно сходятся на срединной линии, составляя
узкий шов, но не перепончатый участок. Абдомен плоский............
....................................... 4. Placetron Schalfeew.
1. Род HAPALOGASTER BRANDT.
Hapalogaster Brandt, 1850 : 269; Schalfeew, 1892 : 326 (part.); Holmes, 1900 :113;
Schmitt, 1921:148.
Тип рода: Hapalogaster mertensii Brandt.
Карапакс, клешненосные и ходильные ноги довольно сильно сплю-
щенные и в большей или меньшей степени волосатые. Клешни на попереч-
ном сечении трехгранные, ось сочленения с карпусом вертикальная, благо-
даря чему клешни двигаются в горизонтальной плоскости. Правая клешне-
носная нога больше чем левая и обычно длиннее ходильных ног. Карапакс
'сравнительно плохо объизвествленный, с сеткой необъизвествленных линий
на бранхиальных областях; боковые края карапакса позади цервикальной
борозды вооружены тем или иным числом шипов. Два дистальных членика
наружных максиллярных ножек расширенные. Самки с парными плеоподами
на первом абдоминальном сегменте; все плеоподы одноветвистые. Второй
сегмент абдомена состоит из пары узких маргинальных пластинок, пары
широких латеральных пластинок и непарной медианной пластинки или же
группы известковых узелков на ее месте (у Н. cavicauda). Третий-пятый
абдоминальные сегменты у самки объизвествлены на левой стороне на
своих краевых частях, где довольно отчетливо заметно расчленение на
пластинки; справа на месте третьего и четвертого сегментов имеется
по маленькой круглой пластинке; а пластинка пятого сегмента выглядит
так же, как и на левой стороне. У самца на третьем-пятом абдоминальных
сегментах имеется по одной маленькой круглой пластинке на обеих сторо-
нах. Хорошо объизвествленная пластинка шестого сегмента находится
над так же хорошо объизвествленной пластинкой тельсона. Уплощенная
236
LITHODIDAE
форма карапакса и конечностей, мягкость покровов, движение клешней
в горизонтальной плоскости — все эти адаптивные признаки показывают,
что виды данного рода живут, скрываясь под камнями.
ОПРЕДЕЛИТЕЛЬНАЯ ТАБЛИЦА ВИДОВ
1 (4). Боковой край карапакса за цервикальной бороздой вооружен 5 ши-
пами. Поверхность члеников клешненосных ног покрыта шипами.
2 (3). Верхне-наружная поверхность ладони правой клешни имеет 3 про-
дольных ряда шипов. Середина ладони левой клешни без шипа.
...................................* 1. Н. grebnitzkii Schalfeew.
3 (2). Верхне-наружная поверхность ладони правой клешни имеет 4 про-
дольных ряда шипов. Середина ладони левой клешни имеет шип.
.....................................2. Н. mertensii Brandt.
4 (1). Боковой край карапакса за цервикальной бороздой вооружен 7 ши-
пами. Поверхность члеников клешненосных ног покрыта бугро-
образными возвышенностями...................* 3. Н. dentata (de Haan).
* 1. Hapalogaster grebnitzkii Schalfeew. (Фиг. 76, 77a).
Hapalogaster grebnitzkii Schalfeew, 1892: 329, fig. 3a, 3b; Holmes, 1900:115; Rathbun,
1904 :163; Brashnikow, 1907: 64; Schmitt, 1921: 150, fig. 96, t. 29, fig. 2.
Карапакс покрыт довольно равномерно короткими волосками. Рост-
рум треугольный, острый, выступающий дальше шипов на наружных орби-
тальных углах, которые направлены несколько наружу, и дальше сильных
зубцов на передне-боковых углах, которые направлены вперед и несколько
внутрь. Боковой край карапакса позади цервикальной борозды вооружен
пятью шипами, которые постепенно уменьшаются в величине по направле-
нию назад. Скафоцерит в виде узкой остроконечной пластинки, которая
немного превышает сильный шип на наружной стороне первого членика
антеннальной ножки; внутренний край скафоцерита волосатый. Все перейо-
поды волосатые. Исхиум правой клешненосной ноги с тремя близко друг
к другу стоящими шипами на внутреннем дистальном краю; мерус' короткий,
вздутый, с тремя сильными шипами на внутреннем краю, из которых средний
самый длинный и с еще более маленьким шипом у переднего края; снаружи
за небольшой вырезкой имеется на переднем краю маленький шипик;
верхне-наружная поверхность несет поперечный ряд из трех шипов. Карпус
с двумя шипами на внутреннем краю и тремя шипами на переднем краю,
из которых нижне-наружный самый крупный; верхне-наружная поверх-
ность имеет три низких шипа или бугорка, расположенных в ряд, парал-
лельный внутреннему краю, и такой же ряд далее снаружи, только в этом:
ряду дистальный шип довольно крупный; имеется маленький шип позади
крупного шипа на нижне-переднем углу. Рука сильная по сравнению с осталь-
ными члениками; внутренняя поверхность ладони несколько вздутая
и гладкая; верхний край несет продольный ряд из четырех небольших
7. HAPALOGASTER
237
'шипов; тотчас снаружи от него находится ряд из пяти шипов, дохо-
дящий до самого основания подвижного пальца; четыре шипа этого ряда
крупные, а пятый проксимальный маленький; нижненаружный край на
•своих проксимальных двух третях несет ряд из пяти шипов, проксималь-
ный из которых также меньше остальных; между этими рядами наружная
•поверхность имеет вид плоского гладкого участка; нижняя поверхность
.ладони и подвижного пальца уплощенная; подвижной палец с сильным
шипом на верхнем краю у основания; его хватательный край с 12—14
бугорками, которые распространяются и на частично ложкообразный
кончик; хватательный край неподвижного пальца с довольно крупным
плоским бугром на
середине, позади ко-
торого у основания
находятся два малень-
ких бугорка, а впе-
реди него край мелко
зазубрен; кончик с
двумя более круп-
ными бугорками, как
отметил еще Шал-
феев (1892), на упло-
щенной нижней сто-
роне ладони близ
основания неподвиж-
ного пальца у более
крупных экземпляров
всегда имеется оваль-
ной формы отвер-
Фиг. 76. Hapalogaster grebnitzkii Schalfeew.
стие, затянутое тол-
стой, но необъизвествленной перепонкой. Трудно приписать этому обра-
зованию какое либо физиологическое значение; создаемся впечатление
снашивания жесткого покрова под влиянием постоянного трения мас-
сивной клешни о жесткий субстрат и зарубцовывание этой раны
мембраной. То же самое наблюдается и у Н. mertensii. Мерус левой
.клешненосной ноги с двумя крупными шипами на внутреннем краю
и двумя очень маленькими шипами на проксимальном и дистальном
концах этого края; передний край также с одним, но более крупным
шипом; верхне-наружная поверхность с тремя шипами в поперечном ряду;
карпус с двумя шипами на внутреннем краю, с тремя шипами на переднем
«краю, с четырьмя шипами на верхне-наружной поверхности. Рука более
уплощенная, с заметным изгибом оси на уровне основания пальцев: верхний
«край ладони с двумя рядами шипов по три шипа в каждом; на проксимальных
концах обоих рядов находятся маленькие бугорки; нижне-наружный край
несет четыре крупных шипа; пальцы заметно сплющенные, с загнутыми вниз
238
LITHODIDAE
кончиками; верхний край подвижного пальца без бугров, с гребенкой из
мелких роговых пластинок и с гранулой на нижнем краю тотчас под кон-
чиком; хватательный край подвижного пальца с мелкими бугорками, в про-
межутках между которыми также находятся роговые пластиночки, и с до-
вольно крупным бугром на нижнем краю под кончиком. Передние края
моральных, карпальных и проподальных члеников трех следующих сплю-
щенных пар ног вооружены острыми зубцами; задний край меруса первых
ходильных .ног с одним шипом близ дистального конца, а двух следующих
пар — с большим числом шипов. Пальцы короткие, с роговыми шипиками
на нижнем краю и несколько изогнутым роговым кончиком. Все конеч-
ности покрыты волосами, которые лучше удалить при детальном осмотре
шипов; волоса особенно длинны на нижне-наружных краях клешней и на
передних краях члеников ходильных ног; более короткие волоски на кон-
цах булавовидно утолщены. Дорзальная поверхность абдомена покрыта
короткими волосками.
Окраска спиртовых экземпляров красновато-желтая; палец правой
клешни довольно ярко красный, за исключением белых шипов и краевых,
зубцов.
Размеры
Длина карапакса...............................19.0 мм
Ширина „ ..............................18.0 „
Длина меруса правой клешней, ноги......... . 8.0 „
„ карпуса „ „ ..................10.5 „
» руки ,, п ,, ..........• » 25.5 „
„ подвижного пальца „ ..................12.0 „
Распространение. Берингово море, на север не проникает
дальше Берингова пролива; по западному побережью Сев. Америки до
залива Гумбольдта, Калифорния; Алеутские острова; восточное побережье
Камчатки; Охотское море; северная часть Японского моря, по нашему
побережью до острова Сибирякова. Из Японии неизвестен (не считая
южного Сахалина). Преимущественно на каменистом грунте. Литораль
до глубины 90 м.
2. Hapalogaster mertensii Brandt. (Фиг. 77b).
Hapalogaster mertensii Brandt, 1850:50; Schalfeew, 1892:307, fig. 4, 5 a; Holmes,.
1900:115; Rathbun, 1904:162; Way, 1917: 382, t. 80, fig. 6.
Очень похож на H. grebnitzkii. Поверхность карапакса имеет пучки
более длинных волосков; волоски, как и у Н. grebnitzkii, булавовидно
расширены на конце. Все отличия касаются главным образом шиповатости
клешненосных ног. Карпус правой клешненосной ноги более шиповатый
благодаря тому, что все шипы острые и конические, в то время как
у Н. grebnitzkii часть шипов имеет форму бугорков. Наружная поверх-
ность ладони там, где у Н. grebnitzkii имеется плоский гладкий участок
между двумя рядами шипов, несет у данного вида ряд из пяти шипов;;
7. HAPALOGASTER
239-
таким образом, общее число продольных рядов шипов на ладони у Н. mer-
tensii будет четыре, а у Н. grebnitzkii—три; кроме того, нижний ряд
у Н. mertensii имеет шесть шипов, благодаря чему он ближе подходит
к основанию неподвижного пальца. Верхний
край подвижного пальца у основания несет два
шипа; этот признак, правда, непостоянен, так
как у двух из просмотренных экземпляров
оказался здесь один шип, но чаще всего их
бывает все же два. Левая клешненосная нога
отличается от таковой Н. grebnitzkii только
тем, что середина ладони примерно против
щели между пальцами занята крупным ши-
пом. В остальном вид вполне сходен с Н. greb-
nitzkii.
Окраска (по Way, 1917) коричневато-
красная или серовато-коричневая, ноги на вен-
тральной стороне красные.
Фиг. 77. Рука правой клеш-
неносной ноги (верхне-на-
ружный вид): а — Hapaloga-
ster grebnitzkii', b — Hapalo-
gaster mertensii.
Распространение. Алеутские острова, на запад до острова
Атха, группа Андреяновских островов; на юг по западному побережью
Северной Америки до Puget Sound, Вашингтон. Литораль, до 35 м.
*3. Hapalogaster dentata (de Haan). (Фиг. 78).
Lomis dentata de Haan, 1850 :219, t. 48, fig. 2. — Hapalogaster deniata (-tus) Miers,
1879 : 47; Ortmann, 1892:323; Stimpson, 1907 :198; Balss, 1913 : 71; Yokoya, 1928: 758.
Отличается от первых двух видов своей гораздо меньшей волоса-
тостью. Гастральная область заметно вздутая. Рострум узко-треугольный,
острый. Шипы на наружных орбитальных углах маленькие и направлены
несколько вниз; шипы на передне-боковых углах карапакса имеют широко-
треугольную форму. Боковой край карапакса позади цервикальной борозды
вооружен семью шипами, которые уменьшаются в величине по направле-
нию назад. Глазные стебельки заметно сплющенные сверху. Скафоцериты
плоские, копьевидные, кончики слегка загнуты вверх. Мерус правой
клешненосной ноги с 2—3 шипами на внутреннем краю, с маленьким
шипом на переднем краю и с 2—3 маленькими шипами или бугорками на
верхней поверхности. Внутренний край карпуса с крупным шипом на
проксимальном конце и с широким зубцом на нижне-дистальном углу;
верхне-наружная поверхность с крупными бугрообразными возвышениями,,,
которые в свою очередь несут более мелкие круглые гранулы. Внутрен-
ний край ладони несет пять тупых бугров; затем снаружи от него имеется
примерно десять бугрообразных возвышений, расположенных в явственные
продольные ряды, снабженных круглыми тупыми гранулами и отделенных
друг от друга гладкими промежутками; примерно против щели между
пальцами по поверхности ладони тянется узкая гладкая продольная,
борозда; за ней идет шесть круглых возвышений, аналогичных вышеопи- -
240
LITHODIDAE
санным, а против каждого из этих возвышений нижне-наружный край
несет шесть остроконечных зубцов; передний край ладони несет попереч-
ный приподнятый валик, усаженный гранулами; нижне-внутренняя поверх-
ность ладони гладкая. Верхний край подвижного пальца несет три бугорка
или крупных гранулы, уменьшающихся в дистальном направлении; внутрь
от этих бугорков имеется неправильный продольный ряд из круглых гра-
нул; хватательный край вооружен четырьмя крупными буграми и более
мелкими бугорками] на закругленном кончике; хватательный край непо-
Фиг. 78. Hapalogaster dentata (de Haan).
.движного пальца напоминает таковой Н. grebnitzkii, только бугор на сере-
. дине более крупный. Мерус левой клешненосной ноги с сильным зубцом
на внутреннем дистальном углу, который иногда несет дополнительные
шипы; карпус в общем такой же, как на правой ноге. Ладонь на внутрен-
нем краю с тремя шипами, из которых проксимальный может быть очень
маленьким; затем идет ряд из восьми возвышений, который под прямым
углом поворачивает у основания подвижного пальца; затем следует глад-
кая продольная борозда, за ней пять круглых возвышений, против которых
наружный край несет пять зубцов; эти возвышения иногда выражены
очень слабо и поэтому продольная борозда кажется более широкой. По-
.движный и неподвижный пальцы с шипом на наружном краю у основания;
: хватательные края, как у Н. grebnitzkii. Три следующих пары перейопод
. вооружены так же, как и у Н. grebnitzkii, только шипы на передних краях
2. DERMATUR US
241
члеников более широкие, подчас зубцеобразные; пальцы на концах с более
длинными и изогнутыми когтями. Поверхность карапакса и конечностей
покрыта короткими коричневыми волосками; более длинные и светлые
волоски обрамляют нижне-наружные края клешней и передние края ходиль-
ных ног.
Размеры
Длина карапакса..................................15.0 мм
Ширина „ ................................16.0 „
Длина меруса правой клешнен. ноги..........• . . 6.0 „
„ карпуса „ „ ................... 9.0 „
„ клешни „ „ „ ..................21.0 „
„ подвижного пальца „ „................... 9.5 „
Распространение. Южная Япония на север до Aomori и Хако-
дате. По нашему побережью Японского моря идет на север не дальше
залива Петр Великий, в котором встречается уже довольно редко. Лито-
раль до глубины 180 м.
Я исследовал экземпляр самки, определенный, согласно Шалфееву,
Brandt’oM как Н. dentaia (№ 844); по всем признакам этот экземпляр
должен быть отнесен к этому виду; на этикетке значится „русско-амери-
канская колония" без указания сборщика; так как с тех пор никем у аме-
риканского побережья этот вид не был найден, а если он и цитируется
(Rathbun, 1904, р. 162), то с ссылкой на упомянутый экземпляр, возникает
почти полная уверенность в ошибочности этикетки и, следовательно,
в отсутствии данного вида у западного побережья Северной Америки.
2. Род DERMATURUS BRANDT
Dermaturus Brand, 1850 : 267; Holmes, 1900 :116. — Hapalogaster Schalfeew, 1892 : 326
(part.).
Тип рода: Dermaturus mandtii Brandt.
Карапакс жесткий, довольно выпуклый; поверхность карапакса пере-
сечена крупными поперечными морщинами; боковые края невооруженные.
Рострум треугольный, простой. Скафоцериты прямые, копьевидные.
Клешненосные ноги сильные, правая нога больше чем левая; ось сочле-
нения с карпусом косая, благодаря чему клешня может двигаться в диаго-
нальном направлении (сверху вниз и изнутри наружу). Два последних чле-
ника наружных максиллярных ножек не расширенные. Ходильные ноги
не сплющенные, полуцилиндрические. Самки с парными рудиментарными
плеоподами на первом абдоминальном сегменте. Второй абдоминальный
сегмент состоит из пары узких маргинальных и пары узких латеральных
пластинок, без непарной медианной пластинки, на месте которой остается
довольно широкий перепончатый участок; в остальном абдомен как
у самца, так и у самки подобен таковому рода Hapalogaster, только
у самки объизвествление на третьем — пятом сегментах, пожалуй, менее
Фауна СССР, Макаров 16
242
LITHODIDAE
резко выражено. Хорошее объизвествлёние карапакса и неуплощенная
общая форма тела заставляет думать, что животнбе ведет свободно-пол-
зающий образ жизни.
* 1. Dermaturus mandtii Brandt. (Фиг. 79).
Dermatarus mandtii Brandt, 1850 : 50; Stimpson, 1858; 232; Holmes, 1900 :116; Rathbun,.
1904 :163; Brashnikov, 1907: 65. — Hapalogaster mandti Schalfeew, 1892: 332, fig. 2, 5c.
Поверхность карапакса слабо выпуклая; карапакс очень незначи-
тельно расширяется по направлению назад. Рострум выдающийся, тре-
угольный, с довольно острой вершиной и с группой гранул на верхней
поверхности близ вершины. Лобный край над основанием антеннул несет
крупную гранулу. Переднебоковой угол карапакса в виде довольно широ-
кого зубца с гранулами на вершине у более крупных экземпляров. По-
верхность карапакса покрыта поперечными морщинами, от которых отхо-
дят вперед короткие стелящиеся волоски. Боковые края карапакса нево-
оруженные. Глазные стебельки имеют поперечные ряды коротких волосков.
Скафоцерит в виде узкой, слабо вогнутой пластинки с довольно сильна
закругленным кончиком и волосатыми краями; шип на наружном углу вто-
рого членика антеннальной ножки широко треугольный, простирающийся
несколько дальше середины скафоцерита, с гладким внутренним и зазуб-
ренным наружным краями. Мерус правой клешненосной ноги с небольшой
зубчатой долей на внутреннем краю. Внутренний край карпуса с тремя
зубцами, из которых проксимальный наиболее крупный. Верхняя поверх-
ность меруса, карпуса и ладони покрыта поперечным и несколько диаго-
нально идущими гребнями, усаженными круглыми гранулами; эти гранулы
особенно крупны на поверхности ладони, напоминая своим видом нити
жемчуга или бус. Борозды между гребнями заполнены короткими, стеля-
щимися волосками, отходящими от передних краев основания гребней.
На основании неподвижного пальца гребни становятся более короткими,
принимая форму групп крупных гранул; на внутреннем краю ладони
гребни имеют по одной (иногда по две) более крупной грануле, после
чего они переходят на нижнюю поверхность ладони, достигая там при-
мерно до ее середины. Подвижный палец у основания имеет неправильной
формы гребни и группы гранул; кончики обоих пальцев гладкие и несут
пучки довольно длинных волосков; нижняя поверхность ладони и пальцев
также с пучками волос; хватательные края с крупными буграми, кончики
пальцев слабо выемчатые и благодаря буграм выглядят двураздельными.
Левая меньшая клешненосная нога такой же формы, как правая, но пальцы
более плоские, гладкие, плотно смыкающиеся и покрытые пучками длин-
ных мягких волос. Три следующих пары ног довольно сплющенные, нево-
оруженные, покрытые поперечными волосатыми морщинами подобно кара-
паксу; нйжний край проподусов и пальцы с пучками довольно длинных
волосков; палец плоский, с роговыми шипиками на нижнем краю
и с сильно изогнутым роговым когтем.
2. DERMATURUS
243
Размеры
Длина карапакса...................................23.0 мм
Ширина „ 23.0 „
Длина меруса правой клешней, ноги................13.5 „
» карпуса „ „ ...................17.5 „
„ клешни „ „ „ ...............33.0 „
„ подвижного пальца,, .........................18.0 „
Длина первой ходильной ноги.......................41.0 „
Фиг. 79. Dermaturus mandtii Brandt.
Распространение. Берингово море, в восточной половине на
север до Прибыловых островов; по азиатскому побережью на север до
мыса Олюторского. По западному побережью Северной Америки на юг
до Ситки; восточное побережье Камчатки; Охотское море; северная часть
Японского моря. До настоящего времени вид не был известен южнее
южной оконечности Сахалина. В материалах Зоологического Института
Академии Наук оказалось 4 экземпляра из сборов М. Павленко в 3—4
милях от устья реки Тюмень-улы. Кроме того, имеется 5 экземпляров из
сбора Рошковского из Японии (30 VII 1917, Misaki, драгировка на глубине
15—20 м). Я затрудняюсь сказать, насколько можно доверять этим эти-
кеткам, в особенности последней, так как в литературе не имеется указа-
ний о нахождении D. mandtii в Японии, а тем более еще учитывая то
обстоятельство, что Misaki является одним из наиболее детально обсле-
дованных районов.
Литораль до глубины 72 м, преимущественно на каменистых грунтах.
16*
244
LITHODIDAE
3. Род OEDIGNATHUS BENEDICT
Oedignathus Benedict, 1894:487; Holmes, 1900:117. — Hapalogaster Schaifeew,
1892: 326 (part.). — Dermaturus Balss, 1913:71 (part.).
Тип рода: Oedignathus. inermis (Stimpson).
Похож на Dermaturus. Карапакс жесткий, выпуклый, покрытый полу-
круглыми чешуеобразными возвышениями. Боковые края карапакса нево-
оруженные. Рострум треугольный, простой. Скафоцерит плоский, сильно
изогнутый, полулунной формы. Два последних членика наружных максил-
лярных ножек сильно расширенные. Клешненосные ноги сильные, нерав-
ные, правая больше левой. Ходильные ноги полуцилиндрические, самки
с парными плеоподами на первом абдоминальном сегменте. Абдомен такого
же строения, как у рода Dermaturus.
* 1. Oedignathus inermis (Stimpson). (Фиг. 80).
Hapalogaster inermis Stimpson, 1860 : 243. — Hapalogaster brandti Schalfeew, 1892 :
330, fig. 1,5b. — Oedignathus gilli Benedict, 1894:487.— Oedignathus brandti Holmes,
1900:118, t. 1, fig-. 17—20. — Oedignathus inermis Holmes, 1900:119; Rathbun, 1904:163;
Schmitt, 1921:151, fig. 97, t. 19, fig. 1; Yokoya, 1928:759. — Dermaturus inermis Balss,
1913:71.
Карапакс немного больше в длину чем в ширину, несколько сердце-
видной, усеченной спереди формы и заметно расширяющийся сзади; его
верхняя поверхность покрыта гладкими, низкими, более или менее круг-
лыми чешуевидными возвышениями, от передних краев которых отходят
короткие, стелящиеся по поверхности волоски. Цервикальная борозда
неглубокая, области вообще неотчетливые, но все же различимые. Боко-
вые края карапакса невооруженные. Рострум узко-треугольный, несколько
наклоненный вниз, простирающийся примерно до середины глазных сте-
бельков. Вместо шипа на наружных орбитальных углах находится неболь-
шая круглая гранула. Переднебоковые углы карапакса имеют вид коротких
тупых выступов с двумя гранулами на вершине. Скафоцериты в виде
широких полулунных пластинок с вогнутым внутренним и выпуклым
наружным краями, наружный край несет четыре круглых гранулы, из кото-
рых одна находится на вершине. Правая клешненосная нога мощная; ее
исхиум гладкий; внутренний край меруса несет зазубренную долю и имеет
глубокую поперечную борозду; внутренний край карпуса с выступом близ
проксимального конца и с поперечной бороздой на наружной половине
верхней поверхности близ дистального конца; ладонь с узким основанием,
но затем значительно расширяющаяся. Верхняя поверхность меруса, кар-
пуса, ладони и большей части поверхности пальцев покрыта круглыми
бугрообразными возвышениями, которые в свою очередь несут более мел-
кие гранулообразные возвышения, придающие первым общий вид тутовой
ягоды; между крупными возвышениями находятся маленькие круглые
гранулы; каждое возвышение окружено венчиком коротких стелящихся
3. OEDIGNA THUS
245
волосков, а от верхней поверхности крупных возвышений между грану-
лами отходят редкие, короткие, более темно окрашенные волоски; на
пальцах по направлению к кончикам бугорки постепенно сходят на нет.
Нижняя поверхность всех члеников безволосая, покрытая мелкими низ-
кими гранулами. Хватательные края пальцев с низкими, мало выступаю-
щими буграми, наиболее выдается бугор близ середины хватательного
края неподвижного пальца; широкие внутренние края выемчатые, в осо-
бенности близ кончиков, так что последние оказываются отчасти ложко-
о'бразными; при смыкании между пальцами остается довольно широкая
Фиг. 80. Oedignathus inermis (Stimpson).
щель. Левая клешненосная нога значительно меньше, но по строению
похожа на правую, за исключением пальцев, которые при смыкании плотно
сходятся и имеют иное вооружение хватательных краев; кроме того, воз-
вышения на верхней поверхности члеников не имеют вида тутовой ягоды,
так как обычно несут только одну гранулу в центре. Следующие три пары
ног мало сплющенные, полуцилиндрические, невооруженные (если не
считать небольшого возвышения на верхних краях мерусов ближе к ди-
стальному концу, имеющему вид маленького тупого зубца), покрытые
подобно карапаксу и левой клешненосной ноге такими же возвышениями;
пальцы толстые, с пучками довольно длинных волос, с сильными роговыми
шипиками на нижнем краю с большим изогнутым роговым когтем. У имев-
шегося у меня более мелкого, но половозрелого экземпляра самки зубцы
переднебоковых углов карапакса более закругленные, скафоцериты более
узкие, с закругленной вершиной и без гранул на выпуклом наружном краю.
246
LITHODIDAE
Размеры
Длина карапакса...............................17.0 мм
Ширина „ ...............................16.0 „
Длина меруса правой клешней, ноги............. 9.0 „
карпуса „ „ „ ..............11.0 „
„ клешни „ „ „ ...............24.0 „
„ подвижного пальца........................14.5 „
„ первой ходильной ноги....................31.0 „
Распространение. У западного побережья Северной Америки
от Уналашки до Тихоокеанского жолоба, Калифорния; Япония: от Цусим-
ского пролива до Aomori (Сангарский пролив); бухта Патрокл, залив
Петр Великий.
4. Род PLACETRON SCHALFEEW
Placetron Schalfeew, 1892 : 333. — Lepeopus Benedict, 1894 :487.
Тип рода: Placetron wosnessenskii Schalfeew.
Карапакс плоский и довольно плохо объизвествленный, покрытый,
как и все конечности, своеобразными чешуевидными возвышениями;
боковые края карапакса невооруженные. Скафоцериты короткие, плоские.
Два последних членика наружных максиллярных ножек не расширяющиеся.
Клешненосные ноги не сильно развитые по сравнению с длинными ходиль-
ными ногами, почти одинаковой длины; ось сочленения клешни с карпу-
сом горизонтальная; пальцы длинные, ложкообразно-выемчатые на вну-
тренних краях. Ходильные ноги длинные, отчасти сплющенные, проподусы
на дистальном конце снизу с тремя шипами. Стернальная поверхность
головогруди между основаниями клешненосных ног имеет круглый буго-
рок. Маргинальные пластинки второго абдоминального сегмента сли-
ваются с широкими латеральными пластинками, которые плотно сходятся
на средней линии; медианная пластинка отсутствует. У самки слева на
третьем-пятом абдоминальных сегментах находятся хорошо объизвест-
вленные, довольно широкие пластинки; справа дорзальная поверхность
абдомена перепончатая. У самца, за исключением второго, шестого сег-
ментов и тельсона, весь абдомен перепончатый. Общая форма абдомена,
в отличие от всех предыдущих родов подсемейства, сплющенная. К сожа-
лению, в виду того, что материал по данному роду очень стар и большая
Часть мягких частей не сохранилась, я не мог установить, имеют ли самки
парные плеоподы на первом абдоминальном сегменте, но, повидимому,
последние имеются.
1. Placetron wosnessenskii Schalfeew. (Фиг. 81).
Placetron wosnessenskii Schalfeew, 1892:333, fig-. 6; Rathbun, 1904:163, tab. 6,
fig. 1. — Lepeopus forcipatus Benedict, 1894 :488.
Покровы всех частей тела довольно слабо объизвествленные. Кара-
пакс сплющенный, больше в ширину чем в длину, заметно расширяющийся
Фиг. 81. Placetron wosnessenskii Schalfeew.
248
LI THODIDAE
кзади, хотя у более мелких экземпляров боковые невооруженные края
почти параллельные. Поверхность карапакса покрыта закругленными, при-
поднятыми чешуеобразными выступами, которые на гастральной области
и на задней части карапакса сливаются в более длинные поперечные
гребни; от передних краев этих возвышений отходят короткие стелящиеся
волоски. Рострум треугольный, выступающий несколько дальше глаз,
с закругленной вершиной и глубокой продольной бороздой на верхней
поверхности; при рассматривании в профиль его вершина заметно накло-
нена вниз. Передне-боковые углы карапакса имеют вид больших остроко-
нечных зубцов; лобный край над основанием антенн несет явственный
шип, который иногда отсутствует. Скафоцерит в виде короткой, довольно
толстой, закругленной на вершине пластинки. Клешненосные ноги почти
одинаковой длины, правая нога немного толще и эта разница становится
тем заметнее, чем крупнее экземпляр; они несколько короче чем ходиль-
ные ноги, цилиндрические и отнюдь не имеют того мощного вида, как
у видов других родов Hapalogastrinae- поверхность их члеников покрыта
такими же чешуйками, как и карапакс, только здесь эти чешуйки мельче.
Мерус с тремя шипами на внутреннем краю и четырьмя шипами на верх-
ней поверхности; обращает на себя внимание строение шипов: каждый .
шип представляет собой как бы крупную полукруглую чешуйку, под
которую спереди была подведена игла и затем поднята, опираясь на.
острие, до угла приблизительно в 45°; получается приподнятая в виде
шалаша пластинка, обращенная своей покатой стороной назад; вершина
этой пластинки увенчана острием, направленным вперед. Карпус с одним:
шипом на внутреннем краю. Ладонь узкая и длинная, чуть-чуть расши-
ряющаяся в дистальном направлении; пальцы длинные, одинаковые, при-
мерно такой же длины, как ладонь, с глубоко выемчатыми внутренними
краями и с широко закругленными кончиками. Хватательные края воору-
жены мелкими бугорками; выше краев имеется ряд из небольших пучков
волосков; оба пальца имеют на концах острые роговые коготки. На
левой клешне пальцы заметно длиннее, чем ладонь. Следующие три пары,
ног заметно сплющенные, поверхность их члеников покрыта более круп-
ными чешуйками, чем клешненосные ноги; передний край мерусов несет
5—8 шипов; карпусы невооруженные; задний край проподусов на дисталь-
ном конце несет два иглообразных шипа, между которыми помещается:
палец в согнутом положении; позади этих двух шипов имеется такой же
непарный третий шип. Пальцы плоские, с 10—12 роговыми шипиками на.
заднем краю, возрастающими в величине по направлению к сильному
концевому когтю. Таким образом, все животное имеет своеобразный
чешуйчатый вид. Абдомен плоский. Второй сегмент состоит из пары узких
маргинальных пластинок, сливающихся с парой широких, сходящихся на
средней линии латеральных пластинок; дорзальная поверхность пласти-
нок покрыта волосатыми гребнями подобно задней части карапакса;,
в центре каждой латеральной пластинки имеется углубление. Остальная:
4. PLACETRON
249
часть абдомена у самцов перепончатая, за исключением шестого сегмента
и тельсона, а у самок на левой стороне снабжена еще тремя довольно
хорошо объизвествленными пластинками, дорзальная поверхность которых
имеет дугообразные приподнятые волосатые линии.
Размеры (по Шалфееву)
Длина карапакса.................................. 61.5 мм
Ширина „ 73.3 „
Длина правой клешней, ноги.......................131.8 „
„ меруса „ „ 31.0 „
„ карпуса „ „ 23.0 „
„ клешни „ „ 57.7 „
„ подвижного пальца ноги...................... 31.0 „
„ левой клешненосной ......................128.0 „
„ первой ходильной „ 152.8 „
Распространение. Алеутские острова до Британской Колумбии
(остров Graham).
2. Подсем. LITHODINAE
Lithodinae Ortmann, 1901: 1147,
Карапакс выпуклый, овальный или треугольный, хорошо объизвест-
вленный, покрытый на дорзальной поверхности бугорками, более крупными
возвышениями, шипами, или же в более редких случаях совершенно
гладкий. Рострум обычно длиннее глаз, острый, шипообразный, вооружен-
ный дополнительными шипами, иногда в виде тупого выступа, реже пло-
ский и усеченный спереди. Дорзальная сторона второго абдоминального
сегмента всегда целиком закрыта двумя маргинальными, двумя латераль-
ными и одной медианной пластинками, которые бывают явственно отде-
лены друг от друга швами, или же совершенно сливаются друг с другом.
Дорзальная поверхность трех следующих сегментов покрыта жесткими
хорошо объизвествленными пластинками и наряду с этим более мелкими
объизвествленными зернами или узелками; поверхность пластинок часто
покрыта бугорками, в центре пластинок иногда имеются перепончатые
участки. Самки с парными плеоподами на первом сегменте. Абдомен
в целом имеет уплощенную форму.
Роды данного подсемейства представлены почти во всех морях,
однако, большинство их приурочено к северной части Тихого океана.
Диапазон батиметрического распространения широкий.
ОПРЕДЕЛИТЕЛЬНАЯ ТАБЛИЦА РОДОВ
1 (14). Латеральные пластинки 3-5-го абдоминальных сегментов не имеют
в центре перепончатых участков.
2 (7). Верхняя поверхность карапакса покрыта острыми или тупыми
шипами. Рострум шипообразный, вооруженный тем или иным числом
дополнительных шипов.
250
LITHODIDAE
3 (6). Абдомен хорошо объизвествленный. Медианные пластинки 3-5-го
абдоминальных сегментов заменены перепончатым промежутком,
несущим то или иное число объизвествленных узелков, подчас сли-
вающихся в мелкие пластинки. Карапакс на дорзальной поверхности
обычно вооружен острыми шипами; . гастральная область несет
несколько шипов, по крайней мере не менее четырех.
4 (5). Второй сегмент абдомена состоит из пяти явственных пластинок:
непарной медианной и парных латеральных и маргинальных пласти-
нок. Скафоцериты хорошо развиты .... 6. Paralithodes Brandt.
5 (4). Пластинки второго абдоминального сегмента сливаются полностью
или частично, т. е. либо медианная пластинка сливается с латераль-
ными, либо латеральные пластинки сливаются с маргинальными.
Скафоцериты рудиментарные......................7. Lithodes Latreille.
6 (3). Абдомен более или менее слабо объизвествленный. Медианные пла-
стинки 3-5-го абдоминальных сегментов явственные. Карапакс
обычно вооружен многочисленными короткими и тупыми шипами;
гастральная область несет один сильно выдающийся шип..............
............................................8. Paralomis White.
7 (2). Верхняя поверхность карапакса гладкая или покрыта различ-
ной формы крупными выступами и мелкими бугорками. Рострум
в виде более или менее крупного тупого выступа, или же плоский
и усеченный на вершине.
8 (13). Карапакс не образует больших боковых расширений, закрывающих
ходильные ноги; его поверхность не гладкая, но несет различной
формы выступы и углубления. Рострум не плоский и не усеченный.
9 (12). Второй сегмент абдомена цельный; маргинальные пластинки
на 3-5-м сегментах имеются. Карапакс сердцевидной или 5-б-уголь-
ной формы, его заднебоковые углы не вытянуты в виде тупых
выступов. Верхняя поверхность карапакса без глубокого полукруг-
лого углубления.
10 (11). Карапакс сердцевидной формы, его поверхность снабжена различ-
ной формы крупными выступами. Рострум с широкой, закругленной
вершиной, имеет булавовидную форму. Ходильные ноги длиннее,
чем наибольшая ширина карапакса . . 10. Sculptolithodes Makarov.
11 (10). Карапакс 5-б-угольной формы, его поверхность снабжена мел-
кими бугорками. Рострум короткий. Ходильные ноги короче, чем
наибольшая ширина карапакса.................11. Lopholithodes Brandt.
12 (9). Второй сегмент абдомена состоит из пяти явственных пластинок;
маргинальные пластинки на 3-5-м сегментах отсутствуют. Карапакс
треугольной формы, с сильно вытянутыми задне-боковыми углами.
Верхняя поверхность карапакса с глубоким полукруглым углубле-
нием .......................................9. Rhinolithodes Brandt.
13 (8). Карапакс образует широкие боковые расширения, почти целиком
закрывающие ходильные ноги; его поверхность почти гладкая.
5. PHYLLOLITHODES
251
Рострум широкий, плоский, с усеченной или более или менее закруг-
ленной вершиной....................... 12. Cryptolithodes Brandt.
14 (1). Латеральные пластинки 3-5-го абдоминальных сегментов с углу-
бленным перепончатым участком в центре . 5. Phyllolithodes Brandt.
5. Род PHYLLOLITHODES BRANDT
Phyllolithodes Brandt, 1849:175; Holmes, 1900; 121; Schmitt, 1921:153.—Petalo-
cerus, White, 1856 : 134.
Тип рода: Phyllolithodes papillosus Brandt.
Карапакс треугольной формы, поверхность покрыта крупными гра-
нулами; в центре дорзальной поверхности карапакса находится углубление,
состоящее из двух ямок; боковые края карапакса вооружены шипами.
Рострум длинный, с двумя вздутиями на вершине. Скафоцерит хорошо
развит и состоит из трех листовидных долей. Клешненосные и ходильные
ноги покрыты длинными цилиндрическими тупыми шипами и более корот-
кими, стелящимися по поверхности члеников шипами- Второй сегмент
абдомена состоит из медианной и латеральных пластинок, последние сли-
ваются с маргинальными пластинками. Латеральные пластинки трех сле-
дующих сегментов имеют в центре вдавленный перепончатый участок,
покрытый известковыми узелками; перепончатые промежутки между лате-
ральными пластинками также покрыты узелками, которые, сливаясь в той
или иной степени, даЮт начало узким несовершенным медианным пла-
стинкам.
1. Phyllolithodes papillosus Brandt. (Фиг. 82).
Phyllolithodes papillosus Brandt, 1849 :175; Holmes, 190Э: 122 (литература); Rathbun,
1904:104; Way, 1917:354; fig. 7;. Schmitt, 1921:153, fig. 99, t. 22, fig. 2.
Карапакс треугольной формы; гастральная область сильно припод-
нятая; от нее идет назад приподнятое валообразное утолщение, образую-
щее на дорзальной поверхности карапакса фигуру, параллельную его
краям и имеющую сердцевидную форму; центр этой фигуры занят двумя
глубокими выемками, соединяющимися спереди друг с другом глубокой
бороздой. Рострум довольно длинный, пальцеобразный, направленный
косо вверх, с двумя вздутиями на вершине; верхний край ближе к основа-
нию несет два бугра, из которых передний более крупный и сжатый
с боков, а задний круглый; нижний край у основания несет шипообразный
отросток, направленный вперед и загнутый вверх; сверху над основанием
этого отростка имеется пара маленьких шипов. Имеется длинный тупой
шип на передне-боковом углу карапакса и более маленький шип такой же
формы на лобном краю над основанием антенн; последний шип направлен
несколько вниз. Выпуклый боковой край карапакса позади цервикальной
борозды вооружен тремя длинными, тупыми, направленными косо вверх
и наружу шипами, из которых передний короче, чем два остальных. Задне-
252
LITHODIDAE
боковые углы карапакса утолщены и несут три тупых выступа, из кото-
рых передний, самый крупный, направлен вверх наружу и вперед, а задний
небольшой, короткий и закругленный. Задний край карапакса прямой, без
срединной выемки, валикообразно утолщен, с слабыми вздутиями по бокам
от срединной линии. Поверхность наиболее пониженных частей карапакса
покрыта плоскими, лапчатыми, по контуру напоминающими форму цветка
гранулами, а поверхность более возвышенных частей карапакса, рострума
и шипов покрыта более мелкими гранулами, имеющими форму тутовых
ягод. Глазные стебельки усажены уплощенными шипами, в особенности
сверху близ роговиц. Скафоцерит состоит из трех листовидных долей,
из которых внутренняя направлена вперед, а две остальные вперед и наружу;
края долей едва заметно зубчатые. Клешненосные ноги одинаковой формы,
правая немного больше чем левая; мерус и карпус покрыты двумя сортами
шипов: одни, более длинные, торчат вертикально вверх и имеют такую же
форму, как на боковых краях карапакса; иногда их тупые вершины раз-
двоены; другие, более многочисленные шипы имеют короткое, вертикально
вверх направленное основание, а затем изгибаются под прямым углом,
так что оказываются параллельными к поверхности членика; вершины
этих шипов всегда направлены по длинной оси членика. Внутренний край
ладони усажен длинными шипами, тогда как верхняя поверхность и часть
нижней поверхности близ внутреннего края покрыты стелящимися шипами
второго сорта, вершины которых часто несут пучки волосков. Пальцы
гладкие, с пучками рыжих волос; наружный край подвижного пальца
у основания с тупым шипом; хватательные края пальцев правой руки
с крупными белыми буграми, которые на неподвижном пальце располо-
жены в два ряда; кончики тупо закругленные, роговые; на левой руке
бугры хватательных краев более маленькие и не образуют двойного ряда,
на неподвижном пальце. Ходильные ноги также покрыты шипами двух
сортов, как и клешненосные ноги; длинные шипы особенно крупны на.
меральных и карпальных члениках. Пальцы у основания снабжены напра-
вленными по оси пальца шипами; нижний край вооружен короткими рого-
выми шипиками; кончик с сильным изогнутым роговым когтем. Второй
сегмент абдомена состоит из медианной и двух латеральных пластинок,,
которые довольно полно сливаются с маргинальными пластинками, так
что швы между ними заметить довольно трудно; центральные части
латеральных пластинок по бокам от медианной пластинки вдавленные,,
дно их образовано перепонкой, покрытой отдельными объизвествленными
узелками. Три следующих сегмента имеют крупные латеральные пластинки
также с в давлениями в центре; имеются мелкие маргинальные пластинки,
которые у самки на левой стороне сливаются с латеральными пластинками.
Между латеральными пластинками этих трех сегментов имеются узкие меди-
анные пластинки, каждая из которых разделена пополам и которые отделены
друг от друга перепончатыми промежутками; все перепончатые участки
покрыты известковыми узелками. Тельсон самца с двумя шипами на конце.
6. PARALITHODES
253
У более молодых экземпляров валообразное утолщение на дорзальной
поверхности карапакса не так резко выступает, папиллы имеют еще одно-
родную форму, шипы вооружены многочисленными дополнительными
шипиками; более мелкие шипы конечностей имеют еще вертикальное на-
правление; рострум короткий, имеющий форму выступа с тремя круглыми
вздутиями на конце; у скафоцерита развиты только две передние доли,
края которых зубчатые.
Фиг. 82. Phyllolithodes napillosus Brandt.
Размеры
Длина карапакса..............................31.0 мм
Ширина .......................................31.0 „
Длина меруса правой клешиен. ноги.............8.5 „
„ карпуса „ „ „ . . •............6.0 „
„ клешни „ „ „..................13.0 „
„ первой ходильной ноги.....................32.0 .,
Распространение. Западное побережье Северной Америки от
Уналашки до залива Monterey, Калифорния. На каменистых грунтах, от
приливо-отливной зоны до глубины 30 м.
6. Род PARALITHODES BRANDT
Paralithodes Brandt, 1849 :173.
Тип рода: Paralithodes brevipes (A. Milne-Edwards et Lucas).
Рострум обычно довольно длинный, шипообразный, с острой или реже
тупой вершиной, вооруженный тем или иным числом дополнительных
шипов. Скафоцериты хорошо развитые, шипообразные, в виде одного
простого шипа или разветвленные. Дорзальная поверхность карапакса
254
LI THODIDAE
вооружена шипами. Дорзальная поверхность второго абдоминального сег-
мента состоит из пяти пластинок, разделенных явственными швами: двух
маргинальных, двух латеральных и одной медианной. Три следующих
сегмента абдомена представлены тремя парами латеральных пластинок
и различным числом маргинальных пластинок; передние и задние края
латеральных пластинок являются смежными; центр дорзальной поверх-
ности абдомена между этими тремя парами латеральных пластинок занят
перепончатым участком, на котором находятся отдельные объизвествленные
узелки, располагающиеся в более или менее правильные поперечные ряды.
Paralithodes, установленный Brandt’oM в качестве подрода Lithodes,
был впоследствии возведен в положение рода. Виды Paralithodes свой-
ственны исключительно северной части Тихого океана.1
ОПРЕДЕЛИТЕЛЬНАЯ ТАБЛИЦА ВИДОВ
1 (2). Скафоцерит в виде простого острого шипа. Дорзальная сторона
рострума несет непарный сильный шип, самая вершина которого
обычно бывает раздвоена; кончик сильного нижнего отростка острый.
Карапакс довольно равномерно покрыт шипами; гастральная область
несет шесть более крупных шипов, расположенных в 2 продольных
ряда по 3 шипа в каждом . . . , . * 1. Р. camtschatica (Tilesius).
2 (1). Скафоцерит иной формы: в виде двуветвистого или четырехветви-
стого шипа. Рострум более короткий и без сильного непарного шипа
на дорзальной поверхности. Шипы на поверхности карапакса более
редкие.
3 (4). Скафоцерит в виде длинного двуветвистого шипа. Кончик нижнего
отростка рострума острый; между ним и парой дорзальных шипов
нет маленького шипа. Кардиальная область карапакса имеет шесть
шипов. Ходильные ноги не сильно шиповатые, их пальцы сравни-
тельно длинные.................................*2. Р. platypus Brandt.
4 (3). Скафоцерит в виде четырехветвистого шипа. Кончик нижнего от-
ростка рострума тупой, часто булавовидно вздутый; между ним
и парой дорзальных шипов обычно имеется маленький шипик. Кар-
диальная область карапакса имеет четыре шипа. Ходильные ноги
сильно шиповатые, их пальцы сравнительно короткие.....................
............................*3. Р. brevipes (М. Edwards et Lucas)..
* 1. Paralithodes camtschatica (Tilesius). (Фиг. 83, 84, 85).
Maja camtschatica Tilesius, 1815:336, t. 5, 6. — Lithodes spinosissimus Brandt,
1849 : 172. — Lithodes camtschatica (-us) de Haan, 1850 : 217, t. 47; Brandt, 1851: 94; Benedict,
1894 :483; Rathbun, 1904 : 165. — Paralithodes\camtschatica Braschnikow, 1907 : 54, fig. 1, t. 2,
fig. 3.
Карапакс у взрослых экземпляров несколько больше в ширину, чем
в длину; довольно короткие,, острые, конические шипы равномерно и не;
1 О хозяйственном значении видов этого рода см. в общей части.
6. PARALITHODES
255
Фиг. 83. Paralithodes camtschatica (Tilesius).
256
LITHODIDAE
слишком густо покрывают верхнюю поверхность карапакса; шипы боковых
краев карапакса незначительно крупнее, чем шипы остальной поверх-
ности; шипы поверхности карапакса в свою очередь распадаются на более
крупные шипы и более мелкие, рассеянные между первыми. Гастральная
область несет шесть более крупных шипов, расположенных в два продоль-
ных параллельных ряда по три шипа в каждом. Кардиальная область имеет
шесть таких же шипов, расположенных таким же образом. Рострум воору-
жен четырьмя шипами: сильный нижний отросток с острой вершиной
направлен вперед и далеко выступает за глаза; от дорзальной стороны
рострума примерно на уровне середины глазных стебельков поднимается
косо вверх и вперед непарный сильный шип, вершина которого обычно
раздвоена; однако встречаются экземпляры, у которых этот шип имеет
острую ординарную вершину; наконец, боковые края расширенной базаль-
Фиг. 84. Paralithodes camtschatica
(Tilesius) вид рострума сбоку.
Фиг. 85. Paralithodes
camtschatica (Tilesius),
правый скафоцерит.
ной части рострума несут позади непарного дорзального шипа по одному
небольшому шипу. Скафоцерит в виде простого острого шипа. Клешне-
носные и ходильные ноги покрыты шипами, подобными таковым карапакса;
нижние края пальцев ходильных ног у взрослых экземпляров часто бывают
гладкие, так как шипы, имеющиеся здесь у молодых экземпляров, со вре-
менем изнашиваются. Молодые экземпляры отличаются от взрослых
несколько иной формой карапакса, именно у них он больше в длину чем
в ширину и поэтому имеет более треугольную форму; наиболее бросаю-
щимся в глаза отличием является большая длина шипов карапакса, что
и дало повод Brandt’y выделить вид spinosissimus; однако расположение
шипов такое же, как и у взрослых экземпляров; относительная длина
рострума у молодых экземпляров также больше, чем у взрослых.
Размеры
Длина карапакса (с рострумом)....................105.0 мм
Ширина „ 92.0 „
Длина меруса прав, клешнен. ноги.................35.0 „
' „ карпуса „ „ ....................30.0 „
„ клешни „ „ „ ..................56.0 „
„ меруса первой ходильной ноги.................64.0 „
6. PARALITHODES
257
Длина карпуса первой ходильной ноги..................39.0 мм
„ проподуса,, „ „ .............45.0 „
„ пальца „ и „ ...............34.0 „
Распространение. Япония, побережье острова Хоккайдо;
в наших водах Японского моря на юг доходит до мыса Гамова; Охотское
море; восточное побережье Камчатки; южная часть Берингова моря; у ази-
атского побережья встречается, очевидно, не выше мыса Олюторского;
Алеутские острова; по западному побережью Северной Америки от залива
Нортон до островов Королевы Шарлотты, Британская Колумбия. Указание
М. Rathbun (1919) этого вида для района мыса Барроу, основанное на
нахождении лишь проподуса левой клешненосной ноги, очевидно отно-
сится к Р. platypus. На глубинах от 14 до 250 м.
*2. Paralithodes platypus Brandt. (Фиг. 86—91).
Lithodes platypus Brandt, 1850 : 236; 1851: 94. — Lithodes camtschaticus Richters,
1883 : 404, fig. 9—10. — Lithodes brevipes Benedict, 1894 : 484 (part.). — Paralithodes platypus
Brashnikow, 1907 :58, fig. 2, t. 1, fig. 2.
Карапакс сердцевидной формы, верхняя поверхность покрыта не-
многочисленными короткими и довольно тупыми шипами. Гастральная
область, как и у Р. camtschatica, несет шесть шипов по три шипа в двух
продольных рядах; у более крупных экземпляров имеется еще довольно
крупный шип позади задней пары; кардиальная область несет четыре шипа;
бранхиальные области несут по шесть шипов каждая. Поверхность кара-
пакса между шипами покрыта мелкими плоскими гранулами, придающими
ей шагреневый вид. Боковые и задний края карапакса усажены шипами,
которые выделяются в виде бордюра благодаря незначительному числу
шипов на поверхности карапакса; шипы заднего края и закругленных
задне-боковых углов мельче, чем шипы краев передней части бранхиаль-
ных областей. В числе и расположении шипов на карапаксе наблюдаются
вариации; так, у одного крупного экземпляра самки на бранхиальных
областях имеется по семь шипов, а на кардиальной области в левом ряду
имеется три шипа, а в правом два и сзади еще намечается два явственных
маленьких шипа. Рострум сравнительно короткий, направлен несколько
вниз и вооружен лишь тремя крупными остриями: непарный нижний отро-
сток короткий и имеет острую вершину; над ним на дорзальной стороне
находится пара острых конических шипов; позади них рострум расширяется
и утолщается и несет четыре маленьких шипика, которые заметны лишь
у крупных экземпляров: один шипик расположен на середине позади круп-
ной пары шипов, два на боковом краю и немного за средним шипиком
и, наконец, совсем маленький, не всегда отчетливый шипик опять на
середине позади боковых маленьких шипиков. У одного экземпляра
имеется аномальная форма рострума, именно вместо парных крупных
шипов на дорзальной стороне имеется толстый конический шип с раздвоен-
Фауна СССР, Макгрэв 17
258
LITHODIDAE
Фиг. 86. Paralithodes platypus Brandt,
6. PARALITHODES
259
Фиг. 87. Paralithodes pla-
typus Brandt, вид рост-
рума сбоку.
Фиг. 88. Paralithodes platypus Brandt,. верхне-боковой
вид рострума: а — аномальная форма; Ъ — нормальная
форма.
Фиг. 89. Paralithodes pla-
typus Brandt, правый ска-
фоцернт.
Фиг. 90. Paralithodes platypus Brandt,
аномальные формы левого скафоцерита.
Фиг. 91. Paralithodes platypus Brandt, молодой экземпляр.
17*
260
LITHODIDAE
ной вершиной. Скафоцерит имеет форму двуветвистого шипа, при этом
внутренняя ветвь немного крупнее наружной; наружный край скафоцерита
иногда несет еще один или два очень маленьких дополнительных шипика.
У одной крупной самки оба скафоцерита имеют аномальную форму: правый
скафоцерит толстый, раздваивающийся лишь у самой вершины, левый
также толстый и совершенно не раздвоенный. У другой самки правый
скафоцерит нормальный, а левый имеет нормальную внутреннюю ветвь,
а на месте наружной ветви имеется два коротких смежных шипика. Клешне-
носные и ходильные ноги мало шиповатые, шипы расположены главным
образом на верхних краях члеников; членики ходильных ног у более мелких
экземпляров заметно сплющенные. Молодь Р. platypus характеризуется
отсутствием шипов на поверхности карапакса и на конечностях, их место
занимают круглые бугорки, число и расположение которых в точности
совпадает с таковыми шипов у взрослых экземпляров; таким образом,
процесс разития шипов у Р. platypus обратен таковому у двух других
видов Paralithodes, где молодь, наоборот, имеет более длинные и острые
шипы, постепенно укорачивающиеся с возрастом и становящиеся тупыми
бугорками у наболее крупных экземпляров; у Р. platypus наиболее крупные
экземпляры имеют наиболее острые шипы. Поверхность карапакса
у молоди густо покрыта мелкими круглыми гранулами. Рострум молоди
отличается тем, что два крупных дорзальных шипа имеют форму закруг-
ленных бугров.
Размеры
Длина карапакса (с рострумом)......................90.0 мм
Ширина „ ..................................82.0 „
Длина меруса правой клешней, ноги..................33.0 „
„ карпуса „ „ „ .................28.0 „
„ клешни „ „ „ • ................55.0 „
„ меруса первой ходильной ноги.................45.0 „
„ карпуса „ „ „ 30.0 „
„ проподуса,, „ 36.0 „
„ пальца „ „ „ ........ . . . . 33.0 „
Распространение. Северная часть Японского моря, по нашему
побережью на юг до мыса Гамова; из Японии неизвестен; Охотское море;
восточное побережье Камчатки; Берингово море, на север не далее
Берингова пролива. Несколько неясен вопрос о распространении Р. pla-
typus у американского побережья Берингова моря. В американской
литературе данный вид вообще не упоминается; лишь на основании дан-
ных Richters’a (1883) можно указать Р. platypus для района Прибыловых
островов. Встречается на глубинах от 12 до 500 м. Указание И. Г. Закса
(1936) о нахождении Р. platypus на глубине 800—1000 м ошибочно:
глубже 93 м данный вид Беринговской экспедицией Государственного
Гидрологического Института не ловился.
6. PARALITHODES
261
Фиг. 92. Paralithodes brevipes (M.-Edw. et Lucas).
262
LITHODIDAE
Фиг. 93. Paralitho-
des brevipes{№..AL<hN.
et. Lucas), вид рост-
рума сбоку.
Фиг. 94. Paralithodes
brevipes (M.-Edw. et
Lucas), правый скафо-
церит.
* 3. Paralithodes brevipes (A. Milne-Edwards et Lucas). (Фиг. 92—94).
Lithodes brevipes A. Milne-Edwards et Lucas, 1841:465, t. 24, 25, 26, 27; Brandt,
1851: 99; Benedict, 1894 :484 (part.); Rathbun, 1904 :165.
Карапакс немного больше в ширину, чем в длину. Поверхность
карапакса между немногочисленными крупными шипами совершенно
гладкая. Гастральная область имеет десять шипов, расположение которых
иное, чем у Р. camtschatica и Р. platypus’, передняя часть несет две пары
шипов, за ними находится пара более широко расставленных шипов
и непарный шип посередине, затем идет пара сильно сближенных
шипов и, наконец, задний непарный средний шип. Бранхиальные области
несут обычно по 7 шипов; иногда на задней части бранхиальных областей
некоторые шипы удваиваются. Боковые края карапакса вооружены силь-
ными шипами, которые еще более резко выделяются в виде бордюра, чем
у Р. platypus. Передний край
более узкой передней части
карапакса несет по три шипа
с каждой стороны рострума,
два из которых направлены
вперед, а третий находится уже
на боковом краю; между ними
иногда находятся мелкие до-
полнительные шипы. Затем
каждый боковой край несет
четыре крупных шипа, передний
из которых находится еще на
узкой части карапакса впереди цервикальной борозды, а следующие три
находятся на краю закругленной расширяющейся части. Наконец, закруг-
ленные задне-боковые углы и задний край карапакса несут по 12—13 более
мелких и более тесно стоящих шипов с каждой стороны. Рострум довольно
короткий, состоит из нижнего непарного, наклоненного вниз и изогнутого
отростка с булавовидно утолщенным терминальным концом; дорзальная
поверхность рострума несет пару направленных вверх шипов; впереди
них на дорзальной покатой части рострума имеется маленький шип, кото-
рый иногда может отсутствовать. У мальков и молодых экземпляров
терминальный кончик рострума более остроконечный. У одного взрослого
экземпляра, добытого М. Ю. Бекман и К. А. Виноградовым в бухте
Ахомтен (восточное побережье Камчатки), вершина рострума остроко-
нечная, как у Р. platypus, хотя по всем остальным признакам он должен
быть отнесен к Р. brevipes. Скафоцерит имеет форму отростка с четырьмя
шипами: три крупных шипа направлены вперед и наружу, четвертый, более
маленький, находится на внутреннем краю. Клешненосные и ходильные
ноги усажены многочисленными крупными шипами; особенно длинные
шипы занимают внутренние края меруса и карпуса клешненосных ног.
Ходильные ноги также сильно шиповатые, шипы на проподальных члени-
7. L I T HO DES
263
ках расположены в четыре продольных ряда; пальцы ходильных ног
короткие и широкие, вооруженные острыми темно окрашенными шипами
вдоль нижнего края. Шипы на медианной пластинке второго абдоминаль-
ного сегмента расположены иначе, чем у Р. camtschatica и Р. brevipes;
у последних четыре крупных шипа образуют фигуру квадрата, при этом
два нижних шипа расположены на заднем краю пластинки; у Р. brevipes
четыре шипа образуют фигуру ромба, который расположен в центре
пластинки.
Размеры
Длина карапакса (с рострумом).................83.0 мм
Ширина ,, ...............................86.0 „
Длина меруса правой клешнен. ноги.............35.0 „
„ карпуса „ „ „ 35.0 „
„ клешни „ „ „ 62.0 „
„ меруса первой ходильной ноги ............48.0 „
„ карпуса „ „ ...............34.0 „
„ проподуса „ я ........ 32.0 „
„ пальца „ „ „ ............27.0 „
Распространение. Северная часть Японского моря, на юг не
далее мыса Поворотного; Охотское море; восточное побережье Камчатки;
южная часть Берингова моря (район Алеутской гряды). С указанием И. Г.
Закса (1936) о повсеместной встречаемости Р. brevipes в Беринговом
море согласиться нельзя: у азиатского побережья он, очевидно, не идет
на север дальше Командорских островов и у американского побережья
•не дальше Уналашки, т. е. вообще не выходит за пределы Алеутской
гряды. Литораль до глубины 50 м.
7. Род LITHODES LATREILLE
Lithodes Latreille, 1806: 39; Milne-Edwards, 1837: 184; Bell, 1853:163.
Тип рода: Lithodes maja (Linne).
Рострум длинный, шиповатый. Карапакс широкий, покрытый шипами.
Скафоцериты рудиментарные или отсутствуют (описаны два случая хорошо
развитых скафоцеритов у видов этого рода, L. antarctica и L. panamensis,
при которых имелись уклонения и в строении второго абдоминального
сегмента; оба случая Bouvier истолковывает как явление атавизма).
Медианная пластинка второго абдоминального сегмента сливается с лате-
ральными пластинками, которые в свою очередь очень часто сливаются
с маргинальными пластинками, так что весь сегмент в таком случае ока-
зывается образованным как бы одной пластинкой. Три следующие абдо-
минальных сегмента, как у рода Paralithodes, т. е. снабжены латеральными
и маргинальными пластинками, а центральная часть занята объизвествлен-
ными узелками. Виды этого рода встречаются в океанах обоих полу-
шарий.
264
LITHODIDAE
ОПРЕДЕЛИТЕЛЬНАЯ ТАБЛИЦА ВИДОВ
1 (2). Карапакс равномерно покрыт шипами приблизительно одинаковой
величины; шипы боковых краев такой же величины, как и шипы
поверхности, или лишь немного длиннее их. Рострум вооружен
9 шипами. Скафоцерит имеет форму довольно длинного двуветви-
стого шипа . . •..........................*1. L. aequispina Benedict.
2 (1). Поверхность карапакса покрыта как сравнительно крупными, так
и мелкими шипами; шнпы боковых краев заметно длиннее шипов
поверхности карапакса. Рострум вооружен 8 шипами. Скафоцерит
рудиментарный.
3 (4). Переднебоковой край карапакса вооружен длинными шипами; на
краю бранхиальных областей нет шипов, которые бы заметно выде-
лялись по длине среди других шипов. Терминальная часть рострума
сравнительно толстая, с двуветвистым кончиком. Клешненосные
и ходильные ноги покрыты сравнительно длинными шипами. Палец,
первой ходильной ноги значительно больше половины длины соот-
ветствующего проподуса. Поверхность второго сегмента абдомена
покрыта шипами. . •...............................*2. L. maja (Linne)-
4 (3). Передне-боковой край карапакса вооружен не слишком длинными
шипами; только на закругленном краю каждой бранхиальной области,
два шипа выделяются своей значительной длиной. Терминальная
часть рострума тонкая с раздвоенным кончиком. Клешненосные
и ходильные ноги покрыты короткими шипами. Палец первой ходиль-
ной ноги едва больше половины длины соответствующего проподуса.
Поверхность второго сегмента абдомена покрыта бугорками . . .
.........................................3. L. couesi Benedict.
*1. Lithodes aequispina Benedict. (Фиг. 95—97).
Lithodes aequispinus (-nd) Benedict, 1894 : 481; Rathbun, 1904:166; Yokoya, 1933 : 94. —
Paralithodes longirostris Navosov-Lavroff, 1929 :174, fig. 3—5.
Карапакс равномерно и довольно густо покрыт коническими острыми
шипами, шипы на боковых краях карапакса лишь незначительно крупнее,
чем шипы на поверхности. Области карапакса достаточно отчетливые-
Наружные орбитальные углы вооружены длинными, направленными впе-
ред шипами, которые достигают как раз до кончиков глазных стебельков-
Рострум у основания довольно широкий и вооружен 9 сильными шипами:
по два шипа имеется на боковых краях рострума, которые направлены
косо вверх, наружу и вперед; затем имеется два шипа на дорзальной
стороне рострума, которые направлены косо вверх и вперед; первый из
этих шипов расположен впереди передней пары боковых шипов рострума,
а второй как раз в промежутке между передней и задней парами боковых
шипов; на вершине терминальной части рострума имеются два шипа,
7. LITHODES
265
фиг. 95. Lithodes aequispina Benedict.
266
LITHODIDAE
которые направлены горизонтально вперед и наружу, так что дистальный
конец рострума оказывается двуветвистым; наконец, снизу от основания
рострума отходит сильный, изогнутый, направленный несколько вниз
шип. Скафоцерит в виде двуветвистого шипа, причем внутренняя ветвь
более длинная чем наружная. Клешненосные ноги относительно слабые,
правая лишь незначительно больше левой; все членики шиповатые, наи-
более длинные шипы расположены на внутреннем дистальном углу меруса
и на внутреннем краю карпуса. Ходильные ноги густо покрыты сравни-
тельно длинными острыми шипами. Медианная пластинка второго абдо-
минального сегмента сливается с латеральными пластинками; однако швы
между латеральными и маргинальными пластинками еще явственные. Три
следующих сегмента состоят из латеральных и маргинальных пластинок;
Фиг. 96. Lithodes aequispina Bene-
dict, вид рострума сбоку.
Фиг. 97. Lithodes aequi-
spina Benedict, правый ска-
фоцерит.
пространство между латеральными пластинками занято мелкими пласти-
ночками, несущими конические шипы. Поверхность второго сегмента
усажена шипами, а на латеральных пластинках шипы располагаются по
краям. У молодых экземпляров шипы поверхности карапакса относи-
тельно более длинные и тонкие, густо расположенные, так что по
внешнему виду животные несколько напоминают молодь Paralithodes
camtschatica.
Размеры
Длина карапакса..............................101.0 мм
Ширина „ 80.0 „
Длина рострума ............................... 20.0 „
„ меруса правой клешней, ноги............... 30.0 „
„ карпуса „ „ „ . . ......... 24.0 „
„ клешни „ „ „ ............. 46.0 „
„ , первой ходильной ноги . . ,...........161.0 „
Распространение. Берингово море, район Прибыловых остро-
вов; Охотское море; Япония: восточнее Siwoya-zaki. На глубинах от 315
до 730 м.
7. LITHODES
267
*2. Lithodes maja (Linne). (Фиг. 98—100).
Lithodes maja Leach, 1815, t. 24; Bell, 1858:165; Ortmann, 1892: 320; Hansen, 1908:
22. — Lithodes arctica Milne-Edwards, 1837 :186; Boas, 1880; 117; Bouvier, 1894:181, t. 10,
fig. 7, t. 12, fig. 5a—5b.
Карапакс довольно правильной грушевидной или сердцевидной формы,
с хорошо заметными областями. Поверхность покрыта небольшими грану-
лами и сравнительно короткими коническими шипами, более длинные из
которых расположены в числе 5—б на каждой бранхиальной области;
четыре шипа украшают выпуклую кардиальную область, а шесть шипов,
расположенных по три в два продольные ряда, — гастральную область.
Рострум вооружен 8 шипами и направлен вперед и слегка вверх, или же
совершенно горизонтально; на боковых краях рострума имеются две
пары шипов, из которых задняя пара маленькая по сравнению с сильными
шипами передней пары, которые соприкасаются своими широкими основа-
ниями; на дорзальной поверхности рострума между этими боковыми
парами шипов, но ближе к задней паре находится маленький непарный
шип; впереди передней пары боковых шипов рострум имеет форму глад-
кого, слегка сплющенного в дорзовентральном направлении стержня,
дистальный конец которого несет два расходящихся шипа и является
таким образом двуветвистым; наконец, снизу от основания отходит
сильный непарный, направленный вниз и вперед шип; длина гладкой
•стержневой части рострума от места дистального разветвления до перед-
ней пары боковых шипов равна длине рострума от этой пары шипов до
основания рострума. Передне-боковой край карапакса вооружен шестью
длинными, острыми, направленными вверх и наружу шипами, из которых
самыми длинными являются два задних шипа, в то время как передний
шип на наружном углу орбиты является самым маленьким; закругленный
задне-боковой и задний края несут менее сильные шипы, между которыми
имеются более слабые дополнительные шипы; медианная пара шипов на
слабо вогнутом заднем краю карапакса направлена более резко вверх,
чем шипы остального края; в середине между шипами этой пары
и несколько впереди имеется небольшой, также направленный вверх шип;
таким образом каждый задне-боковой край карапакса вооружен примерно
15 шипами, по своей величине резко выделяющимися по сравнению
с шипами остальной поверхности карапакса. Глазные стебельки маленькие.
Скафоцериты в виде короткого сплющенного шипа с двумя маленькими
шипиками на наружном краю. Правая клешненосная нога больше левой;
мерус с немногочисленными острыми шипами, самый длинный шип распо-
ложен на внутреннем краю близ дистального конца; карпус с двумя силь-
ными шипами на внутреннем краю; рука равномерно вздутая, пальцы
толстые, с крупными буграми на хватательных краях. Ходильные ноги
довольно тонкие, длинные, сплющенные и шиповатые, в особенности на
краях члеников; пальцы длинные, значительно больше половины длины
соответствующего проподуса, искривленные и очень слабо вооруженные;
268
LITHODIDAE
фиг. 98. Lh
7. LITHODES
269
имеется 4 шипа на проксимальном конце и 2—3 маленьких шипа на верх-
нем краю; коготь длинный и острый. Первый сегмент абдомена с парой
медианных шипов на заднем краю. Второй сегмент цельный, хотя иногда
довольно отчетливо заметны маргинальные пластинки; два круглых боко-
вых углубления на поверхности сегмента соединяются поперечной бороз-
дой; впереди этой борозды имеются два крупных медианных шипа;
примерно такая же пара шипов находится на заднем краю; кроме того,
задний край несет еще четыре более сильных шипа, из которых средние
расположены друг над другом; закругленный передний край близ бокового
угла несет несколько более крупных шипов, в то времй как остальная
поверхность сегмента покрыта мелкими шипиками. Три следующих сег-
мента имеют крупные латеральные пластинки и несколько маргинальных
сбоку.
Фиг. 100. Lithodes maja.
(Linne), правый скафо-
церит (s).
пластинок; поверхность всех пластинок покрыта короткими шипиками;
перепончатый средний участок занят крупными и мелкими известковыми
узелками, которые иногда сливаются в мелкие неправильной формы
пластинки.
Размеры
Длина карапакса с рострумом.................108.0 мм
Ширина „ 79.0 „
Длина рострума.............................. 30.0 „
„ меруса правой клешнен. ноги............ 35.0 „
„ карпуса „ „ „ 28.0 „
„ клешни „ „ „ 58.0 „
„ меруса первой ходильной ноги.......... 67.0 „
» карпуса „ „ ..............35.0 „
„ проподуса „ „ „ ........... 51.0 „
» пальца „ ,, ,, ....... 42.0 ,,
В коллекции Зоологического Института Академии Наук имеется
крупный экземпляр самца этого вида из Белого моря (№ 4020, сбор Гре-
винга), помеченный как вариетет, видимо, на основании формы рострума;
однако, мне кажется, это результат поломки и дальнейшей регенерации
рострума, что не дает права на выделение вариетета. От широкого осно-
ио
LITHODIDAE
вания рострума, имеющего, как и в нормальном случае, три шипа на
дорзальной поверхности, отходит короткий, направленный вперед шип,
который не несет ни непарного шипа снизу, ни двух шипов на дорзальной
стороне и вершина которого не раздвоена; во всех остальных признаках
экземпляр совпадает с типичным L. maja. Точное местонахождение этого
экземпляра не указано; возможно, это случайный занос, так как для
Белого моря вид неизвестен и никем оттуда не отмечался.
Распространение. Шетландские и Фарерские острова; побе-
режье Англии; редко у побережья Бельгии; Немецкое море; Скагеррак:
и Каттегат; побережье Норвегии; западный Мурман (до Териберки).
Самые северные местонахождения: 74°25' N 17°36' Е и у западного Шпиц-
бергена (последнее местонахождение сомнительное); Исландия; восточная
и западная Гренландия. Восточное побережье Северной Америки от
Ньюфаундлэнда до 40° N. Обычно на глубинах 40—80 м, но опускается
и до 500 м.
3. Lithodes couesi Benedict. (Фиг. 101).
Lithodes couesi Benedict, 1894:481; Rathbun, 1904:166; Schmitt, 1921:162, t. 28,
t. 29, fig. 3—5.
„Этот вид напоминает L. maja (L.). Самые крупные шипы карапакса,
расположены около края; они тонкие и острые. Самые длинные шипы
расположены на наружных орбитальных углах, антеннальных углах, хепа-
тических областях и три шипа на краю бранхиальных областей. Шипы
на промежуточных участках края более многочисленные и гораздо мельче.
Поверхность карапакса усажена короткими острыми коническими шипами..
Гастральная область вздутая и хорошо заметная. Кардиальная область едва
намечается между сливающимися бранхиальными областями. Понижение
между гастральной и кардиальной областями очень глубокое. Рострум имеет
29 мм длины и устроен так же, как у L. maja, но терминальная часть за ди-
стальными боковыми отростками тонкая и скорее раздвоенная,чем двуветви-
стая, как у L. maja; базальные отростки немного дальше выступают вперед.
Скафоцерит рудиментарный; шип на наружном углу разветвляется у осно-
вания, ветвь состоит из единственного короткого острого шипа на наруж-
ной поверхности. Абдомен без шипов; шипы L. maja заменены бугорками;
бугорки первого сегмента расположены значительно теснее друг к другу,
чем соответствующие шипы у L. maja. Бугорки на нижнем краю второго
сегмента близкие и отчасти продолговатые у основания; бугорки в центре
сегмента более крупные. Клешненосные ноги тонкие и слабые. Вооруже-
ние пальцев правой руки слабое; пальцы со щелью. Пальцы левой руки
длинные и тонкие, со щелью у основания. Шипы клешненосных и ходиль-
ных ног многочисленные и расположенные приблизительно, как у L. maja,.
но являются более короткими (Benedict)".
„Benedict отмечает три молодых экземпляра с Шумагинских островов,
Аляска, которых он „без колебаний" причисляет к этому виду. Относи-
8. PAR ALU HOPES
271
тельно их он говорит: „рострум отличается тем, что является двуветвистым,
как у L. maja. Возможно, что дополнительные экземпляры взрослых может
быть покажут, что рострум бывает скорее двуветвистым, чем раздвоен-
ным". Однако это не является верным для двух взрослых самцов из района
Сан Диего („Альбатрос", станции 4400 и 4333), имевших характерный
рострум типа. Два других молодых экземпляра также были добыты около
Сан Диего на 500—530 саж. (Schmitt)".
Длина карапакса (тип, 3) с рострумом 105 мм, ширина карапакса
81 мм.
Фиг. 101. Lithodes couesi Benedict (из Schmitt, 1921).
Распространение. Берингово море севернее Уналашки; у Шума-
гинских островов, Аляска; около Сан Диего, Калифорния. На глубинах
542—1125 м.
8. Род PARALOHIS WHITE
Parolomis White, 1856 :134; Stimpson, 1858 : 69; Henderson, 1888 : 44; Schmitt, 1921:
158. — Leptolithodes Benedict, 1894 : 484. — Pristopus Benedict, 1894 :486.
Тип рода: Paralomis aculeata (Henderson).
Карапакс грубо гранулированный, бугорчатый или вооружен много-
численными, короткими, часто тупыми шипами. Гастральная область
обычно несет сильный шип, заметно выделяющийся своей величиной
среди остальных шипов поверхности карапакса. Карапакс не образует
боковых расширений. Рострум обычно имеет три шипа, два симметричных
и один непарный медианный. Скафоцерит более или менее треугольной
формы, его наружный край обычно шиповатый. Ходильные ноги сплющен-
ные и шиповатые на переднем и заднем краях, или же они трехгранные,.
272
LITHODID AE
угловатые, с шипами на гранях. Второй сегмент абдомена цельный.
Маргинальные пластинки третьего абдоминального сегмента сливаются
с соответствующими латеральными пластинками. Маргинальные пластинки
следующих сегментов часто сливаются друг с другом. Медианные пла-
стинки третьего-пятого сегментов явственные, часто отделенные друг от
друга перепончатыми промежутками; узелки, покрывающие эти перепон-
чатые промежутки, часто сливаются друг с другом, образуя нечто вроде
маленьких пластинок.
Виды рода преимущественно абиссальные.
ОПРЕДЕЛИТЕЛЬНАЯ ТАБЛИЦА ВИДОВ
1 (2). Карапакс шиповатый, в особенности на краях, в гастральной
и ростральной областях. Ходильные ноги угловатые, нисколько
не сплющенные, с шипами, расположенными рядами на углах или
ребрах................................... *1. Р. multispina (Benedict).
2 (1). Карапакс бугорчатый, края шиповатые, единственный острый шип
находится в гастральной области. Ходильные ноги сильно сплющен-
ные, передние и задние края их усажены острыми шипами ....
.......................................2. Р. verrilli (Benedict).
* 1. Paralomis multispina (Benedict). (Фиг. 102).
Leptolithod.es multispinus (-na) Benedict, 1894:484; Rathbun, 1904:165. — Paralomis
multispina Schmitt, 1921: 159, t. 23, t. 30, fig. 7, 8.
„Карапакс приблизительно такой же ширины, как и длины; области
хорошо выражены; На срединной линии на вершине гастральной области
имеется острый шип приблизительно 4 мм длины. Боковые края воору-
жены 12—16 шипами приблизительно 3 мм длины. У молоди и у некото-
рых взрослых экземпляров имеются мелкие шипы в бранхиальной области.
Полукруглая линия из 6 или 7 шипов отмечает границы бранхиальной и
интенстинальной областей. Карапакс густо усажен тупыми шипами, каждый
из которых оканчивается плоской поверхностью, косо срезанной по отно-
шению к поверхности карапакса; эта поверхность обрамлена опушкой
коротких, жестких щетинок. Рострум состоит из простого медианного
шипа с двумя базальными шипами. Под собственно рострумом имеется
очень короткий конический шип, гомологичный субростральному шипу рода
Lithodes; позади шипа имеется один или более шипиков. Абдомен у самца
после второго сегмента состоит из нескольких рядов кожистых пластинок;
второй сегмент лучше объизвествленный и более жесткий. Абдомен самки
скошен вправо, как у Lithodes. Клешненосные ноги умеренно тонкие
и простираются почти до дистального конца карпальных члеников. Шипы
на внутреннем краю карпальных члеников самые выдающиеся. Ходиль-
ные ноги длинные и тонкие, густо усаженные шипами. Шипы меруса
ме так .явственно расположены в ряды, как на карпальных и проподальных
& PARALOMIS
273
члениках; однако имеется явственный ряд на верхнем краю. Шипы
карпуса расположены в 8 более или менее явственных рядов; на пропо-
дальном членике шипы расположены в 6 полных .рядов и два полуряда.
Имеется 4 коротких ряда шипов на проксимальном конце пальца. Пальцы
сплющенные, слегка изогнутые и немного закрученные. Экземпляры
средней величины имеют 80 мм длины, 78 мм ширины, расстояние от
кончика до кончика ходильных ног 360 мм (Benedict)".
Р аспространение. Западное побережье Северной Америки:
от Шумагинских островов до Сан Диего, Калифорния; на глубинах от
Фиг. 102. Paralomis multispina (Benedict) (из Schmitt, 1921).
1125 до 1577. Беринговской экспедицией Государственного Гидрологи-
ческого Института добыт один экземпляр этого вида (<5, juv.) у восточного
побережья Камчатки, 52°42.5' N, 159°03.5/ Е, на глубине 830 м.
2. Paralomis verrilli (Benedict). (Фиг. 103).
Pristopus verrilli Benedict, 1894 : 486; Rathbun, 1904 :165. — Paralomis verrilli Schmitt,
1921:159, t. 24, t. 30, fig. 5,6.
„Карапакс бородавчатый, области выдающиеся. Гастральная область
сильно приподнятая и увенчана маленьким шипом. С каждой стороны
на краю бранхиальной области имеется глубокая ямка. От ямок к пони-
жению между гастральной и кардиальной областями тянется борозда.
На боковом краю карапакса имеется приблизительно 12 шипов от 2 до
3 мм длины. Задняя граница интестинальной области отмечена полукруг-
лым рядом бугорков. Кардиальная область треугольная; вершина треуголь-
ника упирается в интестинальную область, где понижение, которое отмечает
ее, тянется в виде глубокой щели или продолговатого медианного пони-
жения. Лобный край широкий и прямой. Шипы передних угло§ и орби-
Фауяа СССР, Макаров 18
274
LITHODID AE
тальные шипы направлены вперед; орбитальные шипы немного длиннее.
Между шипом на углу и орбитой имеется ряд мелких шипов и одна или
две гранулы. Трехветвистый рострум состоит из двуветвистого собственно
рострума и субрострального шипа, который простирается значительна
дальше двух верхних ростральных шипов. Скафоцерит суживается в острый
кончик и имеет три острых шипа или ветви на каждой стороне. Лате-
ральные пластинки на левой стороне абдомена самки окаймлены корот-
кими, тонкими, тупыми шипами. Клешненосные ноги простираются немного
за середину проподального членика первой пары ходильных ног. Правая
Фиг. 103. Paralomis verrilli (Benedict) (из Schmitt, 1921).
клешненосная нога сильнее чем левая; хватательные края ее пальцев
сильно бугорчатые. Верхний край ладони шиповатый; имеется также
несколько мелких шипов на середине и на цижнем краю. Имеется три
длинных шипа на внутреннем краю карпуса. Левая клешненосная нога
сходная, но меньше и хватательные края пальцев острые. Ходильные ноги
широкие и сильно сплющенные. Передний и задний края вооружены
острыми шипами, причем крупные шипы чередуются с мелкими. На верхней
поверхности проксимального конца меруса четвертой пары ног имеется
ряд из мелких шипов; соответствующие шипы на третьей паре ног мельче,
а на второй паре еще мельче (Benedict)".
Длина карапакса и рострума 90 мм, длина рострума 9 мм, ширина
карапакса 82 мм.
Распространение. Берингово море, от района Прибыловых остро-
вов на jor до Cortez Bank, Калифорния. На глубинах от 1238 до 1480 м.
9. RHINOLITHODES
275
9. Род RHINOLITHODES BRANDT
Rhinolithodes Brandt, 1847 :174; Schmitt, 1921 : 157.
Тип рода: Rhinolithodes wosnessenskii Brandt.
Карапакс имеет неуклюжую треугольную форму с сильно вытянутыми
задне-боковыми углами; центральная часть дорзальной поверхности занята
глубоким полукруглым углублением, которое отделяет довольно гладкую
шарообразную кардиальную область от остальной поверхности карапакса,
покрытой плоскими и несколько выступающими гранулами. Рострум
короткий, тупой. Второй сегмент абдомена состоит из пяти явственных
Фиг. 104. Rhinolithodes wosnessenski Brandt.
пластинок. Три следующих сегмента состоят каждой из медианной и пары
латеральных пластинок; маргинальные пластинки отсутствуют, так как
целиком сливаются с соответствующими латеральными пластинками.
1. Rhinolithodes wosnessenskii Brandt. (Фиг. 104).
Rhinolithodes wosnessenskii Brandt, 1849 :174; Rathbun, 1904 :164; Way, 1917 : 354,
fig. 11; Schmitt, 1921: 158, fig. 103, t. 22, fig. 1.
Общий вид животного очень неуклюжий. Карапакс треугольной
формы, с сильно вздутой гастральной областью, позади которой находится
глубокое полукруглое углубление, обращенное выпуклой частью вперед,
а задними концами доходящее до заднего края карапакса; центр этого
углубления занят шарообразно вздутой кардиальной областью, поверхность
которой покрыта низкими плоскими' папиллами и невооруженному глазу
кажется совершенно гладкой. Рострум в виде довольно короткого тупого
выступа цилиндрической формы, с сильным зубцеобразным, направленным
18*
276
LITHODIDAE
вперед концевым шипом, бока которого покрыты мелкими шипиками;
верхушка этого шипа при взгляде сверху выступает за вздутую часть
рострума. Лобный край карапакса несет неправильные, довольно беспо-
рядочно расположенные шипы, среди которых выделяются шипы на
передне-боковых углах карапакса и над основаниями ножек антенн.
Боковой край карапакса позади цервикальной борозды вооружен семью
довольно короткими коническими шипами; задне-боковые углы карапакса
вытянуты в конические, тупые, сильно выдающиеся выступы. Поверхность
карапакса покрыта плоскими круглыми гранулами и несколько более
сильно выступающими гранулами, имеющими форму тутовой ягоды;
центральное полукруглое углубление имеет совершенно гладкую поверх-
ность. Глазные стебельки сверху усажены многочисленными шипами.
Второй членик иожки антенн имеет наружный выступ, состоящий из трех
шипов: передний сильный шип направлен вперед, второй маленький
и третий у основания направлены наружу; внутренний выступ также
несет три почти одинаковых шипа; скафоцерит представляет собой длин-
ный, слегка изогнутый, усаженный тупыми шипами отросток; третий
членик ножки вооружен одним шипом сверху на дистальном конце;
последний длинный членик ножки несет четыре шипа на верхнем краю.
Клешненосные и ходильные ноги покрыты длинными мягкими волосами;
правая клешненосная нога крупнее левой. Верхняя поверхность меруса
и карпуса правой клешненосной ноги негусто покрыта сильными кони-
ческими шипами, особенно около внутренних краев; самый длинный шип
расположен на внутреннем проксимальном углу карпуса; ладонь широкая,
шиповатая главным образом на внутреннем краю; пальцы гладкие,
с пучками волос, подвижный палец с тупым шипом на наружном краю
у основания; хватательные края с крупными белыми буграми, кончики
с тупыми роговыми когтями. Ладонь левой руки мало шиповатая, хвата-
тельные края пальцев мелко зазубренные, кончики с ложкообразными
роговыми когтями. Мерусы, карпусы и проподусы ходильных ног с та-
кими же шипами, как и клешненосные ноги; пальцы сильно сплющенные,
с четырьмя шипами у основания, с роговыми шипиками на нижнем краю
и с острым когтем. Второй сегмент абдомена состоит из явственных
медианной, латеральных и маргинальных пластинок; латеральные пла-
стинки около внутренней стороны имеют углубление. Следующие три
сегмента состоят каждый из медианной и пары латеральных пластинок;
маргинальные пластинки отсутствуют. Тельсон короткий, треугольный.
Поверхность всех абдоминальных пластинок покрыта довольно крупными
гранулами; на латеральных пластинках, в особенности третьего сегмента,
имеются слабые, едва намечающиеся вдавления.
Окраска: карапакс серовато-коричневый с оранжевыми пятнами
в углублениях на дорзальной поверхности, на бугорках задней части
карапакса, на бранхиальных областях и на абдомене. Ноги не слишком
окрашенные, но серовато-коричневые, подобно общему тону карапакса.
10. SCULPTOLITHODES
277
Размеры
Длина карапакса...................................38.0 мм
Наибольшая ширина карапакса .................47.0 „
Длина меруса правой клешней, ноги............... 13.0 „
„ карпуса „ „ 10.0 „
„ клешни „ „ „ . . . ...........23.5 „
„ первой ходильной „ • * . ...........48.0 „
Распространение. Западное побережье Северной Америки;
остров Кадьяк; порт Etches; Ситка; Crescent City, Калифорния. Преиму-
щественно на каменистых грунтах и на ракушнике, на глубинах от 20
до 72 м.
10. Род SCULPTOLITHODES MAKAROV
Sculptolithodes Makarov, 1934 : 251.
Тип рода: Sculptolithodes derjugini Makarov.
Карапакс довольно правильный, сердцевидной формы. Рострум булаво-
видный, с тупой закругленной вершиной. Дорзальная поверхность карапакса
и конечностей украшена различной формы выступами и возвышениями. Бо-
ковые края карапакса вооружены тупыми шипами. Скафоцериты рудимен-
тарные. Ходильные ноги значительно длиннее, чем наибольшая ширина
карапакса. Пластинки второго абдоминального сегмента целиком сливаются
в одну общую пластинку, занимающую всю поверхность сегмента. Три
следующих сегмента состоят каждый из медианной, латеральных и
маргинальных пластинок; латеральные пластинки сплошь объизвествлен-
ные, с заметными углублениями в центральных частях.
* 1. Sculptolithodes derjugini Makarov. (Фиг. 105).
Sculptolithodes derjugini Makarov, 1934:251, fig. 1, 2.
Карапакс имеет сердцевидную форму. Рострум массивный, булаво-
образный, с тупой закругленной вершиной, с углублением в центре верх-
ней поверхности; нижняя поверхность довольно правильной конической
формы, с вершиной, которая направлена вперед и помещается между
глазами. Верхняя поверхность карапакса сильно скульптирована; вся
гастральная область сильно приподнятая, с покатыми боковыми поверхно-
стями, с довольно ясно намечающимся бугром на своей самой высокой
части; сливаясь с рострумбм, она образует как бы седло, надетое на перед-
нюю часть карапакса. Непосредственно за рострумом край карапакса
снабжен двумя тупыми шипами. Остальная часть карапакса сильно пони-
женная, с пятью возвышающимися буграми, расположенными следующим
образом: треугольный бугор на кардиальной области, парные сердцевид-
ные бугры на бранхиальных областях и парные почкообразные бугры на
метабранхиальных областях. Боковой край карапакса за цервикальной
бороздой приподнятый и несет 5—6 тупых шипов, в промежутках между
'278
LITHODIDAE
которыми могут находиться дополнительные более мелкие шипы. На
задне-боковом углу край карапакса резко утолщается, образуя бугор,
подобный вышеописанным, который непосредственно переходит в валико-
образно утолщенный задний край карапакса, прерванный на интестиналь-
ной области хорошо выраженной выемкой. Поверхность возвышающихся
частей карапакса сравнительно гладкая, обнаруживающая лишь крупную
шагренированность, в то время как поверхность пониженной части кара-
пакса покрыта круглыми гранулами. Глазные стебельки достигают до
вершины конуса нижней поверхности рострума или несколько превосхо-
дят ее; роговицы маленькие; верхне-наружный дистальный конец стебелька
снабжен несколькими шипами. Антеннулы превосходят конец рострума
половиной длины последнего членика стебля; наружный жгут толстый,
короткий и густо опушенный снизу; внутренний жгут короче наружного
и почти совершенно голый; антенны немного длиннее антеннул; скафо-
церит имеет вид небольшого чешуеобразного придатка с шипом на конце
и с зазубренным внутренним краем. Клешненосные ноги короче ходиль-
ных, одинакового строения, только левая нога несколько меньше правой.
Мерус трехгранный, с закругленным верхним ребром и с явственной бороз-
дой на боковых сторонах, немного отступя от дистального края. Внутренний
проксимальный угол карпуса несет пластинчатый выступ, снабженный на
краю 4—5 зубцами; остальные углы несут каждый по бугру, которые могут
сливаться друг с другом и имеют более гладкую поверхность. Рука
с равномерно выпуклой верхней поверхностью, с группой шипов на вну-
треннем дистальном углу; пальцы направлены косо наружу, покрыты пуч-
ками мягких желтых волосков; хватательные края с довольно крупными
белыми буграми; кончики черные, роговые. Верхняя поверхность всех чле-
ников клешненосных ног гранулированная, нижняя гладкая. Три первых
пары ходильных ног одинакового строения и почти равной длины, вторая
чуть-чуть длиннее чем первая и третья; четвертая пара ходильных ног
рудиментарная и, как вообще у всех родов этого семейства, подогнута под
задне-боковые углы карапакса. Передний и задний края меруса трех первых
пар ходильных ног несут неправильной формы зубцы; дистальный конец
меруса на верхней поверхности снабжен гладким выдающимся бугром;
края верхней поверхности карпуса валикообразно утолщены, причем на
переднем краю имеется неполное разделение валика на три бугра; про-
подус сплющенный, с продольной бороздой вдоль переднего края на верх-
ней поверхности и с грубо зазубренным задним краем; палец короткий,
сплющенный, с продольной бороздой на передней и задней поверхностях,
с рядом острых шипиков на нижнем краю и с черным роговым когтем на
конце. Вся поверхность ног гранулирована, за исключением выступающих
частей на меральных и карпальных члениках; нижняя поверхность также
гранулированная, но без заметных выступов. Первый сегмент абдомена
скрыт под задним краем карапакса; все пластинки второго сегмента сли-
ваются, так что весь сегмент представляет собой одно целое; по бокам
11. LOPHOLITHODES
279
от средней линии имеются углубления. Три следующих сегмента имеют
медианные, латеральные и маргинальные пластинки (у самок марги-
нальные пластинки только на правой стороне); медианные пластинки
четвертого и пятого сегментов разделены пополам; между медианными
пластинками имеются отдельные мелкие пластинки и объизвествленные
узелки. Поверхность всех пластинок густо покрыта круглыми гранулами;
центральные части латеральных пластинок заметно вдавленные. Тельсон
полукруглый.
Фиг. 105. Sculptolithodes derjugini Makarov.
Окраска спиртовых экземпляров: выступающие части дорзаль-
ной поверхности карапакса, карпальных члеников клешненосных ног
и меральных и карпальных члеников ходильных ног оранжево-красные;
общий тон клешненосных ног бледнорозовый, ходильных ног — бледно-
фиолетовый. Нижняя поверхность животного более светло окрашенная.
Что касается половых отличий, то, кроме асимметрии абдомена,
характерной для самок данного семейства, следует указать на относительно
более крупные размеры клешненосных ног' у самца, более густую волоса-
тость пальцев тех же ног; с другой стороны, абдомен самки более густо опу-
шен по краям, а центральные части латеральных пластинок менее заметно
вдавленные, чем у самца. Причудливая скульптура и окраска животного
изумительно гармонируют с формой и окраской литотамнйя, в зоне кото-
рого был найден вид; таким образом, этот рак может служить прекрасным
примером покровительственной формы и окраски.
280
LITHODIDAE
P а з м ер ы
Длина карапакса...................................38.0 мм
Наибольшая ширина карапакса.......................36.0 „
Длина меруса правой клешнен. ноги.................14.0 „
,, карпуса ,, „ >, 11.0 „
„ клешни „ „ „ 22.0 „
„ первой ходильной ноги ...................51.0 „
Распространение. До настоящего времени вид известен только
из северной части Японского моря: у бухты Силантьева и в районе
бухты Нельмы. В материалах Зоологического Института Академии Наук
удалось обнаружить один экземпляр этого вида из района острова Ришири
и один экземпляр из района бухты Андреева (Уссурийский залив). Преи-
мущественно на каменистых грунтах, на глубинах 20—35 м. Вид прони-
кает в Японское море, повидимому, из юго-западной части Охотского
моря.
И. Род LOPHOLITHODES BRANDT
Lopholithodes Brandt, 1849 (1848): 174; Holmes, 1900 : 127; Schmitt, 1921:155.—
Echinoceras White, 1848 : 47. — Ctenorhinus Gibbons, 1855: 48.
Тип рода: Lopholithodes mandtii Brandt.
„Карапакс широкий, пяти или шестиугольный, выпуклый сверху;;
края и верхняя поверхность вооружены щетинистыми буграми. Рострум
короткий, шиповатый. Скафоцерит с шиповатыми краями и обычно с шипо-
ватой верхней поверхностью. Терминальный членик мандибулярного пальпа
продолговатый, сплющенный и отогнут назад так, чтобы лежать между
вогнутыми внутренними поверхностями мандибул. Клешненосные ноги
неодинаковые, бугорчатые; карпус с выдающейся долей на внутренней
стороне. Ходильные ноги короткие, бугорчатые и способны складываться
под карапаксом. Второй сегмент абдомена цельный; три следующих сегмента,
с латеральными пластинками и маленькими маргинальными пластинками
на одной или обеих сторонах; предпоследний сегмент без латеральных
пластинок; тельсон очень маленький. Клешненосные и ходильные ноги,,
когда подогнуты, точно прилажены друг к другу, создавая таким образом
надежную защиту для ротовых частей и нижней стороны тела (Holmes)"..
1. Lopholithodes mandtii Brandt. (Фиг. 106, 107).
Lopholithodes mandtii Brandt, 1849:174; Holmes, 1900:128 (литература); Rathbun,.
1904:164; Way, 1917:356, fig. 12; Schmitt, 1921:156, fig. 101, t. 21, fig. 1.
„Карапакс сильно выпуклый, больше в ширину чем в длину, вся
поверхность — крупные бугры, а также находящиеся между ними проме-
жутки— шероховатая благодаря маленьким коротким щетинистым бугор-
кам, которые очень сильно вариируют в величине от низких, почти глад-
ких вздутий до грубых полуконических выступов. Медианная область.
11. LOPHOLITHODES
281
фиг. 106. Lopholithodes mandtii Brandt.
282
LITHODIDAE
очень выдающаяся, имеющая полуконическую вершину немного впереди
середины и группу мелких округлых или полуострых бугорков близ зад-
него конца; на кардиальной области имеется большое полуконическое
возвышение, впереди которого с каждой стороны имеется углубленный
сравнительно гладкий участок; медианная и кардиальная области раз-
делены сравнительно гладкой бороздой; имеется крупное полукониче-
ское возвышение на бранхиальных областях и два более маленьких бугорка
на заднем краю карапакса. Своеобразное, гладкое бородавкообразное
возвышение находится с каждой стороны медианной области. Рострум
короткий и состоит из сильного полуконического шипа, над основанием
которого находится
Фиг. 10/. Lupholithodes
mandtii Brandt, правый
скафоцерит.
шишка, несущая два латеральных шипа или бугорка,
обычно с шипом или бугорком сверху и снизу выемки
между ними. С каждой стороны лба имеется глубокий
закругленный синус, который служит в качестве
орбиты и снаружи которого имеется острый шип; за
этим шипом имеется большой шип, который прости-
рается вперед почти так же далеко, как рострум,
а в синусе между последним шипом и посторбиталь-
ным шипом обычно имеется маленький шип или зубец.
Переднебоковой край вооружен различным числом
(приблизительно 3) выдающихся шипов и несколькими
более мелкими шипами; средняя часть выпуклая, сплю-
щенная и выступающая. На задне-боковых углах
имеется крупный шишковидный выступ, который
отделяется от последнего передне-бокового шипа за-
метным синусом. Глазные стебельки двучленистые,
густо усаженные шипами сверху и немного не достигающие до кончика рост-
рума. Скафоцерит узкий, сильно превышающий кончик стебля, снабженный
примерно 30 сильными, гладкими острыми шипами. Исхиум максиллярных
ножек вооружен изнутри темно окрашенными зубцами; имеется зубец на
наружной поверхости близ передне-внутреннего угла; экзогнат отчасти
превосходит мерус. Клешненосные ноги короткие, первый и второй членики
с неправильными бугорчатыми выступами на нижней стороне; на передне-
внутреннем углу меруса находится один или более выдающихся шипов;
доля на внутренней стороне карпуса очень большая и острая, усажен-
ная по краю резкими шипобразными буграми. Шипы на наружной поверх-
ности рук большие и многочисленные; имеется 4 или 5 крупных шипов
на верхнем краю большей руки и 3 или 4 шипа на верхнем краю меньшей
руки. Ходильные ноги почти одинаковые, сильно бугорчатые, четырех-
угольные у основания; карпусы и проподусы полуцилиндрические, но очень
неправильные; пальцы короткие и толстые. Второй абдоминальный сег-
мент сильно вогнутый сзади, особенно у самок, и находится почти под
прямым углом к карапаксу; имеется два выдающихся бугра близ середины
и несколько на краях; остальные сегменты усыпаны многочисленными
12. CRYPTOLITHODES
283
полу коническими бугорками; между медианными пластинками имеются
мелкие поперечные вторичные пластинки, которые могут быть разделены
продольными щелйми; предпоследний сегмент продолговатый и дистально
вогнутый. Абдомен самца полутреугольный за вторым сегментом; абдо-
мен самки закругленный на боках и кончике и скошен в одну сторону.
Размеры: длина 200 мм, ширина 245 мм; длина большей клешненосной
ноги 202 мм; длина первой ходильной ноги 195 мм. Некоторые экземпляры
наиболее широки между буграми на задне-боковых углах, тогда как дру-
гие — между кончиками шипов на передне-боковых краях (Holmes)“.
Распространение. От Ситки (Аляска) до Monterey (Калифорния).
12. Род CRYPTOLITHODES BRANDT
Cryptolithodes Brandt, 1849 :185; Holmes, 1900 :123; Balls, 1913: 79; Schmitt, 1921; 154.
Тип рода: Cryptolithodes typicus Brandt.
Карапакс больше в ширину чем в длину, с почти гладкой дорзальной
поверхностью, высокий в средней части и широко распростертый в сто-
роны, так что целиком в виде крышки накрывает все конечности, когда
последние подобраны под него. Рострум большею частью широкий, сплю-
щенный, усеченный на вершине й сильно наклоненный вниз. Передне-
боковые края карапакса дугообразные, вооруженные в большинстве
случаев маленькими, часто почти незаметными зубцами или бугорками,
встречающиеся с-задним краем под явственным углом, или же закруглен-
ные. Глазные стебельки тонкие на терминальном конце и сильно расширен-
ные у основания. Скафоцериты широкие, пластинчатые. Клешненосные
ноги неравные, способные складываться таким образом, что целиком
скрываются под щитом карапакса. Ходильные ноги сплющенные, с острыми
краями. Абдомен за вторым сегментом резко подогнут под головогрудь,
имеет треугольную форму и плотно прилегает к углублению, образован-
ному торакальными стернитами. Второй сегмент абдомена цельный, сплош-
ной, или же состоит из двух пластинок. Три следующих сегмента состоят
каждый из медианной и пары латеральных пластинок; дополнительные
пластинки имеются или отсутствуют; маргинальные пластинки отсутствуют.
Шестой сегмент продолговатый; тельсон короткий, треугольный. Абдомен
самки не так сильно асимметричен, как у других родов семейства.
ОПРЕДЕЛИТЕЛЬНАЯ ТАБЛИЦА ВИДОВ
1 (2). Клешни гладкие. Рострум дистально расширяющийся, с явственными
передне-боковыми углами. Пластинки абдомена гладкие...............
.......................................1. С. sitchensis Brandt.
2 (1). Клешни бугорчатые. Рострум не расширяется дистально, с довольно
сильно закругленной вершиной. Пластинки абдомена с приподнятыми
краями.
284
LITHODIDAE
3 (4). Рострум сильно наклонен вниз. Боковые доли карапакса бугорчатые.
Боковые края карапакса сходятся под явственным углом. Второй,
сегмент абдомена цельный. У самца имеются дополнительные пла-
стинки на границе между медианной и латеральной пластинками:
третьего сегмента и задним краем второго сегмента .......
.............................................2. С, typicus Brandt.
4 (3). Рострум мало наклонен вниз. Боковые доли карапакса гладкие. Боко-
вые края карапакса не сходятся под углом, но образуют широкое;
закругление. Второй сегмент абдомена состоит из двух пластинок.
Дополнительные пластинки отсутствуют . . . 3. С. expansus Miers.
1. Cryptolithodes sitchensis Brandt. (Фиг. 108„ 109).
Cryptolithodes sitchensis Brandt, 1853:254; Holmes, 1900 :125,, t. 2, fig-. 21—25; Rath-
bun, 1904:164; Schmitt, 1921:155, fig. 100, t. 20, fig. 3, 4; Queen,, 1930:393, fig. 1.
Карапакс, поперечноовальной формы; средняя часть приподнятая,
и увенчанная продольным хребтом или гребнем,, который простирается
вперед до середины рострума; хепатические области имеют более слабо
выраженные поперечные гребни; кардиальная область, вздутая. Поверх-
ность карапакса гладкая, покрытая заметными под лупой мелкими круг-
лыми ямочками; боковые края сильно расширенные, целиком закрыва-
ющие конечности. Закругленный передне-боковой край несет 8—10 тупых:
зубцов, которые очень вариируют в величине и иногда настолько малы,
что придают краю лишь волнистый вид; боковые углы карапакса отчетли-
вые; задний край карапакса закругленный, без зубцов или с 1—2 ма-
ленькими, едва заметными зубцами близ бокового угла. Рострум сильна
изогнут вниз, заметно расширяется дистально и имеет явственные передне-
боковые углы; усеченный передний край рострума на середине часто
несет маленький зубец. С каждой стороны рострума имеется глубокий
орбитальный синус. Глазные стебельки маленькие, уплощенные сверху,
сильно расширенные у основания и целиком скрытые под рострумом.
Второй членик антеннальной ножки несет на внутренней стороне плоский,
направленный вверх отросток, а на наружной стороне имеется также
пластинчатый вырост, продолжающийся как вперед, так и назад; скафо-
церит в виде очень широкой, тонкой, жесткой, почти круглой пластинки,
с закругленным наружным краем и с волнистым внутренним краем,
с слегка вогнутой верхней поверхностью; последний членик ножки имеет
на верхнем краю гребень в виде тонкой пластинки. Исхиум наружных ма-
ксиллярных ножек маленький, треугольный, наружный угол снабжен ма-
леньким зубцом, внутренний край мелко зазубренный, а наружный снабжен
двумя круглыми бугорками; мерус пластинчатый, гладкий, с закруглен-
ным выступом на внутреннем краю и с сильно выдающимся вперед высту-
пом на наружном краю; остальные членики сплющенные и гребенчатые;
на краях. Правая клешненосная нога больше левой, но строение обеих,
ног одинаковое. Внутренний край исхиума острый; внутренний край меруса.
12. CRYPTOLITHODES
285
сплющенным или полуконическим зубцом. Верхняя поверхность карпуса
имеет гребень, расположенный ближе к наружной стороне; закругленный
передний край острый; .вну-
тренний дистальный угол
1несет тупой зубец. Рука
Заметно расширяется ди-
стально, ладонь со слабым
продольным гребнем на се-
редине; внутренний край ла-
дони острый и заканчивается
на дистальном конце закруг-
ленным выступом; наружный
край ладони толстый, закруг-
ленный; пальцы короткие и
толстые; наружный край по-
движного пальца гребнеоб-
разно заострен и сильно
расширен у проксимального
конца благодаря закруглен-
ному выступу; весь палец
имеет несколько крюкооб-
разную форму; хватательные
края вогнутые, с часто сли-
вающимися буграми и окру-
жены снаружи пучками ко-
ротких волосков. Ходильные
Фиг. 108. Cryptolithodes sitchensis Brandt, вид сверху
и снизу.
ноги гладкие, голые, сплю-
щенные; верхние края чле-
ников острые; нижняя сто-
рона мерусов уплощенная; нижние края карпуса и проподуса дистально
зубцом; пальцы плоские, значительно более
оканчиваются маленьким
Фиг. 109. Cryptolithodes sitchensis
Brandt, наружная поверхность
правой клешни.
.дистально, выемчатый на
угольный, часто скрытый
узкие, чем проподусы. Второй сегмент
абдомена цельный, с углублениями на бо-
ковых частях. Три следующих сегмента
состоят каждый из медианной и двух лате-
ральных пластинок; маргинальные пластинки
отсутствуют. У самца на границе между
медианной и латеральной пластинками и
между задним краем второго сегмента
с каждой стороны находится маленькая
треугольная добавочная пластинка. Шестой
сегмент продолговатый, расширяющийся
дистальном конце. Тельсон маленький, тре-
под дистальным концом шестого сегмента.
286
LITHODID AE
Поверхность всех пластинок абдомена покрыта микроскопически малень-
кими круглыми ямочками.
У молодых экземпляров задний край карапакса прямее, чем у взрос-
лых; гребень на верхней поверхности руки более отчетливый и могут
быть замечены один или больше более или менее отчетливых дополни-
тельных гребней. У самок задняя часть карапакса сильно вздутая и зад-
ний край выпячивается назад более сильно, чем у самцов.
Окраска (по Queen) темнокрасная; некоторые экземпляры бывают
коричневатые, а один маленький экземпляр был белый с коричневато-
красными пятнами. Holmes (1900) пишет: „самцы, которых я видел в жи-
вом состоянии, были все однородного яркокрасного цвета. Две самки
были красные, но с неправильными пятнами пурпурного оттенка, с пят-
нами более светлого цвета. Этот вид часто можно видеть на отливе на.
склонах скал, где на некотором расстоянии он действительно может быть
ошибочно принят за один из видов яркокрасной корковой губки, кото-
рая встречается в подобных же местах". Спиртовые экземпляры бледно-
желтого или рыжеватого цвета, иногда красноватые.
Размеры
Длина карапакса ..........................• ... 24.0 мм.
Ширина „ ............•......................38.0 „
Длина меруса правой клешней, ноги...........' ' . 5.0 „
„ карпуса „ „ ............ ... . 6.0 „
„ ладони „ „ ....................10.0 „
„ подвижн. пальца „ „ ................ 4.0 „
первой ходильной ноги.......................18.0 „
Распространение. От Ситки, Аляска, до Тихоокеанского
жолоба, Калифорния. Литораль до глубины 13 м.
2. Cryptolithodes typicus Brandt. (Фиг. 110, 111).
Cryptolithodes typicus Brandt, 1849: 185; Holmes, 1900:124; Rathbun, 1904:164; Way,
1917: 352, fig. 5; Schmitt, 1921:154, tab. 20, fig. 1, 2.
Карапакс значительно больше в ширину, чем в длину, с тупо за-
кругленными боковыми углами; задний край не такой дугообразный, как
у С. sitchensis-, поверхность позади поперечных хепатических гребней
более заметно вогнутая; зубцы на передне-боковых и заднем краях ма-
ленькие. Рострум не расширяется дистально, с закругленными передне-
боковыми углами, наклонен вниз и выдается вперед значительно дальше
края карапакса, благодаря чему орбитальные синусы совсем неглубокие.
Поверхность карапакса в области боковых углов покрыта мелкими низ-
кими возвышениями. Скафоцериты в виде поперечных пластинок, с пря-
мым усеченным передним краем. Клешненосные ноги отличаются от
таковых С. sitchensis тем, что верхняя поверхность ладони, в особен-
ности правой руки, и отчасти карпуса покрыта довольно крупными круг-
12. CRYPTOLITHODES
287
лыми бугорками. Нижние края меральных члеников ходильных ног с двумя,
гребнями, между которыми находится довольно глубокая борозда. Края
всех пластинок абдомена приподнятые, а поверхность покрыта мелкими
гранулками. В остальном вид похож на С. sitchensis. Самцы с дополни-
тельными пластинками на границе между медианной и латеральной
пластинками третьего сегмента и между задним краем второго сегмента
абдомена; Bouvier (1895) считает, что эти добавочные пластинки харак-
терны для самцов всех видов рода.
Окраска (по Way) темновато-коричневая на дорзальной стороне,,
светлосерая на вентральной стороне. Спиртовые экземпляры бледно-
желтоватые, иногда с зеленоватым оттенком.
Фиг. 111. Cryptolithodes typicus:
Brandt, наружная поверхность пра-
вой клешни.
Фиг. 110. Cryptolithodes typicus Brandt, вид
сверху.
Р а,з м е р ы
Длина карапакса...................................14.0 мм
Ширина „ ...............................23.0
Длина меруса правой клешней, ноги................. 4.0 „
„ карпуса „ „ .................... 3.5 „
„ клешни „ „ „ ............... 8.0 „
Длина подвижн. пальца „ „ ...............4.0 „
„ первой ходильной ноги.........................12.0 „
Распространение. От Уналашки (Аляска) до залива Monterey
(Калифорния). Литораль до глубины 13 м.
3. Cryptolithodes expansus Miers. (Фиг. 112, 113).
Cryptolithodes expansus Miers, 1879:47; Rathbun, 1902:32, fig. 1; Balss, 1913:79,..
f. 55; Vokoya, 1928:759.
Карапакс поперечнопродолговатый, с широко закругленными боко-
выми крылоподобными расширениями, не образующими боковых углов;;
поверхность карапакса покрыта мелкими ямочками, из которых поднимаются
маленькие щетинки, и более крупными ямочками. Имеется выдающийся
выступ на кардиальной области и по одному выступу с каждой стороны.
288
LITHODIDAE
от него на бранхиальных областях, так что все три выступа образуют
поперечный ряд и начинаются от общего основания. Подобный же выступ
занимает гастральную область; продольный медианный гребень про-
стирается от гастрального выступа до дистальной половины рострума.
Передняя половина каждого бокового расширения занята низким высту-
Фиг. 112. Cryptolithodes ехрап-
sus Miers, контур; карапакса
(из Rathbun, 1902).
пом, который является бугорчатым (по
Miers’y боковые . расширения гладкие, без
бугорков). Край карапакса снабжен мелкими
тупыми зубцами или бугорками в числе при-
близительно 34, с неправильными и длин-
ными промежутками между ними; самые
крупные находятся на наружных углах неглу-
боких орбитальных синусов. Рострум несильно
наклоненный вниз, порядочно выступает за.
передний край карапакса, почти такой же
длины, как его ширина у основания, стороны
постепенно сходящиеся и слегка выпуклые,
кончик поперечноусеченный, с маленьким бугорком на середине. Глаза
достигают до половины длины рострума. Второй членик антеннальной
ножки имеет наружный гребень с двумя шипами, из которых один шип
направлен вперед, а другой назад. Скафоцерит значительно больше
в ширину чем в длину; его дистальный край,
имеющий косое направление, является вогну-
тым. У левой клешненосной ноги1 базис и
исхиум бугорчатые снизу; мерус с тремя греб-
нями, внутренний гребень рассечен на четыре
неправильных зубца, сходный же гребень про-
должается на исхиум; верхняя поверхность
карпуса шероховатая, внутренний край и угол
пластинчатые, наружный гребень тупой, имеется
тупой зубец на нижнем дистальном углу; ладонь
и пальцы бугорчатые изнутри и снаружи, имеется
резкий гребень на верхней поверхности ладони,
.дистально оканчивающийся острым коническим
зубцом; имеется тупой гребень на нижнем краю
Фиг. 113. Cryptolithodes ех-
pansus Miers, абдомен (из
Balss, 1913).
ладони; пальцы значительно длиннее, чем верх-
ний край ладони, почти соприкасаются при смыкании, подвижный палец
«гребенчатый сверху, гребень на проксимальном конце образует выступ.
Исхиум ходильных ног снабжен зубцом на заднем дистальном углу верхнего
края, этот зубец возрастает в величине от первой до третьей пары. Края
•следующих члеников широкопластинчатые; мерусы с одной верхней и двумя
1 Описание в основном заимствовано из М. Rathbun, которая имела экземпляр самца
с потерянной правой клешненосной ногой. Miers в своем описании говорит только, что
ч,ладони передних ног снаружи бугорчатые".
12. CRYPTOLITHODES
289
нижними пластинками, карпусы с одной верхней пластинкой, проподусы
и пальцы с одной верхней и с одной нижней пластинками. В естественном
положении ноги скрыты под карапаксом, но когда вытянуты, то последний
членик и половина предпоследнего членика выступают за край карапакса.
Абдомен немного больше в длину, чем в ширину у основания. Первый сег-
мент очень короткий, вытянутый в поперечном направлении. Второй сегмент
состоит из двух пластинок, разделенных медианным швом; края каждой
пластинки приподнятые. Медианная пластинка третьего сегмента узкая,
с поперечной бороздой, латеральные пластинки более крупные; никаких
дополнительных пластинок между медианной и латеральной пластинками
и задним краем второго сегмента нет. Медианная пластинка четвертого
сегмента широкая, латеральные пластинки пятиугольные. Пятый сегмент
подобен четвертому. Швы между латеральными пластинками чередуются
со швами между сегментами. Шестой сегмент с выемкой на дистальном
конце, в которой помещается тельсон.
Размеры
Длина карапакса...................................51.6 мй
Ширина „ 78.9 „
Длина рострума................................... 9.5 „
Длина карапакса от наружного орбитального угла до
заднего края .....................................45.0 „
Распространение. Северная Япония (Miers); Minyako, провин-
ция Rikuzen (Rathbun); Aomori (Balss); около мыса Hanaguri-zaki; на линии
между Aomori и мысом Futago-zaki; на линии между островом Benten
и мысом Kurosaki (Yokoya).
’Фауна СССР, Макаров
19
SUMMARY
This fascicle is devoted to a very large systematical division of the
Decapod Crustaceans, the so called Апотига. In addition to a short anatomo-
morphological description, the introduction also contains some brief informa-
tion on the biology and zoogeography of the group under consideration. In
view bf the peculiarities of the family Paguridae a special introduction has
been added to this fascicle where the anatomy of this family is considered
in greater detail, for the general introduction mainly discusses external mor-
phology and some general questions (physiology, economic significance, phy-
logeny, etc.). The vast collections of the Zoological Institute of the Academy
of Sciences of the USSR have supplied the principal part of the materials
for the present paper.
I am much indebted to Prof. V. M. Rylov, the Curator of the Division of
Crustacea of the above-named Zoological Institute for allowing me free
access to the materials necessary for my work, and to all those who have kindly
given me their assistance. I am most grateful to Dr. Waldo L. Schmitt
(Washington), owing to whose kindness I have received the opportunity of
studying the specimens of some species absent from the other collections
examined by me. The illustrations are by N. N. Kondakov, the photographs
by the writer.
In accordance with its distribution the Anomuran fauna maybe divided
into two large groups: that of the boreal region and that of the tropical re-
gion. The Lithodidae (a survey of the geographical distribution of the Pagu-
ridae is given separately) belong to the first group, all the other families,
examined in this work belong to the second one. In both these regions there
are immigrants from adjoining regions; as the majority of the seas of the
USSR belong to the Boreal region, the Lithodidae which are indigenous,
here constitute the main bulk of the fauna» whereas all the other forms are
immigrants from the tropical region; the waters of the USSR or adjacent
areas being the northern limit of their distribution. Not a single species of
these two groups occurs in the fauna of the Arctic region.
In the Black Sea there are only 2 species of the family Callianassidae
{Upogebia littoralis and Callianassa pontica) and 2 species of the family
Porcellanidae {Porcellana longicornis and P. longimana) which are identical
with the corresponding species of the Mediterranean Sea, from where they
had undoubtedly penetrated into the Black Sea.
GEOGRAPHICAL DISTRIBUTION
291
The Barents Sea, off the western Murman coast, is the eastern limit of
distribution of Munida bamffica, Galathea strigosa (seldom), and Lithodes
maja, Porcellana longicornis being recorded as occasional finds. The first two
species occur rather seldom, penetrating there with the warmer currents of
the Gulf-stream. The scarcity of the Lithodidae in the Barents Sea, which is,
by its character, quite suitable for the habitation of this group, is evidently
due to the remoteness of this sea from the centre of origin and distribution
of the Lithodidae.
The European seas of the USSR, therefore, contain only 11 species of
Апотига (including 4 species of hermit-crabs). This forms about 29% of the
whole number of representatives of this group known from the waters of the
USSR. The remaining 71%, or 41 species, are supplied by the Far-Eastern
seas (hermit-crabs included in this number). 38 species, or about 42% of the
species described in this paper, belong to the fauna of adjacent waters,
chiefly to the western coast of Japan and the western coast of North America.
The quantitative relationship of the species occurring in the European and
Far-Eastern seas of the USSR is given by families in the following table:
European seas
F a r-E astern seas
Thalassinidea . . ...........2
Galatheidae ........ 2
Porcellanidae ........ 2
Paguridae ...... ... 4
Lithodidae ......... 1
Axiidae ......... 1
Thalassinidea ...... 6
Galatheidae ....... 1
Porcellanidae ...... 1
Albuneidae ........ 1
Paguridae ......... 21
Lithodidae ....... . 10
The same relationship is shown in diagram form in fig. 13, where the
area of the circles is proportionate to the number of. species, and where the
sectors show in percentage the relationship of the number of species by
families. As the above table shows the main stock of the Anomuran fauna
of the Far-Eastern seas are the Paguridae', for the Paguridae twice exceed
the quantity of Lithodidae which in their turn are just equal to the number
of species of all the remaining families put together. The diagram clearly
indicates that the fauna of the European seas of the USSR contains no ori-
ginal stock as regards the group of Апотига and consists of a fairly propor-
tionate number of immigrant elements. The predominance of Paguridae in the
fauna of the Far-Eastern seas is chiefly due to the genus Pagurus which
belongs in its greater part to the Boreal region, although the whole family
chiefly belongs to the tropical region. Without considering here the family
Paguridae any further, we shall pass to an examination of the Anomuran
fauna of the Far-Eastern seas, which may be subdivided into the following
four groups of species:
1) Species proper to all the Far-Eastern seas (the Japanese, the
Okhotsk and the Bering Seas): Hapalogaster grebnitzkii, Dermaturus mand-
tii, Paralithodes camtschatica, P. platypus and P. brevipes.
19*
292
ANOMURA
2) Species common to the Okhotsk and Japanese Seas: only one species,
Scu.lptolithod.es derjugini, refers to this group.
3) Species occurring in the Okhotsk and Bering Seas: Munidopsis
beringana and Lithodes aequispina.
4) Group of identical species (or of species resembling them) with an
interrupted distribution, i. e., occurring off the opposite coasts of the North
Pacific Ocean and absent from the intermediate region (amphipacific species
•according to the term proposed by A. Andriashev):
Asiatic coast American coast
Callianassa calif orniensis bouvieri Callianassa calif orniensis
Callianassa gigas eoa Callianassa gigas
Oedignathus inermis
Cryptolithodes expansus Cryptolithodes typicus
We shall now consider the fauna of the different seas, taken separately
The Japanese Sea. The 26 species of Anomura occurring in the Japa-
nese Sea may be divided into the three following groups:
1) Species common both to our coast and to that of Japan: Upogebia
major, Callianassa harmandi, C. japonica, C. californiensis bouvieri, Pachy-
cheles stevensii, Blepharipoda japonica, Hapalogaster dentata, Oedignathus
inermis and Paralithodes camtschatica.
2) Species occurring only off our coast of the Japanese Sea: Axiopsis
princeps, Upogebia issaeffi, Callianassa gigas eoa, Hapalogaster greb-
nitzkii, Dermaturus mandtii, Paralithodes platypus and Sculptolithodes der-
jugini (the last-mentioned species is certainly recorded off the Rishiri Island,
but it is not known further south along the Japanese coast).
3) Species peculiar only to the Japanese coast: Upogebiayokoyai, Cteno-
cheles balssi, Galathea acanthomera, G. integra, G. pubescens, Munida japo-
nica, Cervimunida princeps, Mixtopagurus jeffreysii and Cryptolithodes
expansus.
As examples of endemics in the Japanese Sea the following species
may be named: Axiopsis princeps, Upogebia issaeffi, Callianassa gigas eoa
and Ctenocheles balssi. L. Kobjakova (1936) has recorded for the Amur
Gulf a new subspecies: Axiopsis spinulicauda amurensis (given under the
generic name Axius). On the whole the majority of the species of the Japa-
nese Sea are fairly localized forms, not spreading south of the southern part
of Nippon Island; only Callianassa harmandi, C. japonica and C. califor-
niensis bouvieri axe also known from the Yellow Sea, and Galathea acantho-
mera extends as far as Bonin Islands. The complete absence of represen-
tatives of the family Galatheidae off our coast of the Japanese Sea must be
noted.
GEOGRAPHICAL DISTRIBUTION
293
The Okhotsk Sea. The Anomuran fauna of the Okhotsk Sea consists
of the 9 following- species: Calastacus quinqueseriatus (recorded by Z. Kobja-
kova, 1936), Munidopsis beringana, Hapalogaster grebnitzkii, Dermaturus
mandtii, Paralithodes camtschatica, P. platypus, P. brevipes, Lithodes aequi-
spina and Sculptolithodes derjugini. Thus more than 50% of this fauna con-
sists of species common to all the three Far-Eastern Seas. The first two
species and Lithodes aequispina penetrate into the Okhotsk Sea from the
northern part of the Pacific Ocean; all the three are deep-water species.
Sculptolithodes derjugini is, evidently, a form of the Okhotsk Sea (Shantarsk
Sea) spreading- from here only to the northernmost part of the Japanese Sea.
Not a single species of Japanese Sea origin reaches to the Okhotsk Sea.
The Eastern coast of Kamchatka. Sculptolithodes derjugini excepted,
all the species listed for the Okhotsk Sea belong as well to the fauna of the
eastern coast of Kamchatka. It is true that Calastacus quinqueseriatus, Muni-
dopsis beringana and Lithodes aequispina have not yet been recorded off
this coast of Kamchatka, but their presence here is quite probable. Para'omis
multispina, a deep-water form of the northern part of the Pacific Ocean, has
been found here.
The Bering Sea. The Anomuran fauna in the Bering Sea numbers 13
species and may be subdivided into two groups:
1) Species occurring both in the western and in the eastern part of
the sea: Hapalogaster grebnitzkii, Dermaturus mandtii, Paralithodes cam-
tschatica, P. platypus and P. brevipes.
2) Species occurring only in the eastern half of the sea: Munidopsis
beringana, Hapalogaster mertensii, Oedignathus inermis, Placetron wosnes-
senskii, Lithodes aequispina, L. couesi, Paralomis verrilli and Cryptolithodes
typicus.
It is quite probable that Munidopsis beringana and Lithodes aequispina
when studied more in detail will be found to belong to the first group of
species. There is not a single species which occurs only in the western half
of the Bering Sea; it is true that Paralithodes platypus might seem to be
such a species, as there is no mentioned of it in the american literature for
the eastern half of the Bering Sea. Richters’s specimens from the Pribyloff
s ands, however, belong to this species and we, therefore, include P. platypus
in the first group.
An examination of the distribution in the latitudinal direction of all
the species of Anomura in the Bering Sea discloses the fact of the majority
of them belonging to the southern part of the sea, chiefly to the region of
the Aleutian Islands, and of their rapid disappearance northwards. For instance,
in the region of the Aleutian Islands there occur Hapalogaster mertensii,
Oedignathus inermis, Placetron wosnessenskii, Paralithodes brevipes, Litho-
des aequispina, L. couesi and Cryptolithodes typicus, i. e., about 54% of the
total number of species. In the region of Cape Olutorsky along our coast
and off the Pribyloff Islands along the American coast disappear: Munidopsis
294
1. AXIIDAE, 2. CALLIANASSIDAE
beringana, Dermaturus mandtii, Paralithodes camtschatica (along the Ame-
rican coast up to Norton Bay), Lithodes aequispina and Paralomis verrilli.
Only two species, Hapalogaster grebnitzkii and Paralithodes platypus, extend
to the Bering Strait.
The Chukotsk Sea. In the Chukotsk Sea there is not a single represen-
tative of the Anomura, the Paguridae excepted.
Fig. 14 gives the centre of origin and the paths of distribution of the
Lithodidae, It is quite evident that Lithodes maja penetrated into the Atlan-
tic Ocean through the North American Archipelago. As to the deep-water
representatives of Lithodidae in the Northern Atlantic the writer believe,
contrary to E. L. Bouvier (1896), that they reached here round South America,
ascending northwards along the western coast of Africa.
KEY TO THE SUPERFAMILIES OF THE ANOMURA
1 (2). Second to fourth legs with last joint curved and flattened. First pair
styliform or subchelate. Tail-fan not adapted for swimming. Abdomen
bent under thorax. Rostrum small or wanting. Third maxillipede have
no mastigobranchiae.........................................3. Hippidea
2 (1). Second to fourth legs with last joint not curved and flattened. First
pair not styliform, rarely subchelate.
3 (6). Sixth abdominal limbs adapted for swimming (except in Thalassina,
where they are styliform). Pleura usually well developed. Abdomen sym-
metrical.
4 (5). Body depressed. Pleurobranchiae on last legs. Often a transverse
suture on telson. Abdomen more or less bent.............................
...................................................2. Galatheidea
5 (4). Body compressed. No pleurobranchiae on last leg. No transverse suture
on telson. Abdomen straight............................ 1. Thalassinidea
6 (3). Sixth abdominal limbs, when present, with rami neither broad nor
styliform, but adapted for retaining the body in hollow objects. Pleura
very rare. Abdomen nearly always asymmetrical, and either soft and
twisted or bent under the thorax............................4. Paguridea
1. Superfam. THALASSINIDEA
1 (2). Linea thalassinica wanting. Scaphocerites always present, although
sometimes small. Epimers of abdominal segments large. Exopodites of
uropods with or without a suture (diaresis)..........1 Axiidae
2 (1). Linea thalassinica present. Scaphocerites either reduced to a small
flat appendage or wanting. Epimers of abdominal segments small. Exo-
podites of uropods always without a suture .... 2. Callianassidae
1. AXIOPSIS, 1. UPOGEBIA
295
1. Family AXIIDAE
1. Genus AXIOPSIS BORRADAILE
A single species: Axiopsis princeps (Boas). — Japanese Sea: region of
Vladivostok.
2. Family CALLIANASSIDAE
1 (2). Rostrum large. Pereopods of the first pair equal. Propodite of third
pereopods of usual width.. Endopodites of pleopods of the third to fifth
segments without appendix interna .... 1. Upogebiinae Borradaile
2 (1) Rostrum small, sometimes almost quite wanting. Pereopods of the first
pair not equal. Propodite of the third pereopods very broad, trans-
versely oval. Endopodites of pleopods of the third to fifth segments
with appendix interna . .................2. Callianassinae Borradaile.
1. Subfamily UPOGEBIINAE
1. Genus UPOGEBIA LEACH
1 (2). First joint of the antennular peduncle with a spine on the distal end
of the lower margin. Scaphocerite triangular, ending in a sharp spine.
Upper margin of the propodite of anterior pereopods with a strong
spine near the distal end. Upper surface of movable finger smooth,
flat. — Black, Adriatic and Mediterranean Seas; east shore of the
Atlantic......................................1. U. littoralis (Risso).
2 (1). First joint of the antennular peduncle unarmed. Scaphocerite transver-
sely oval with a rounded anterior margin. Upper margin of propodite
of anterior pereopods without a noticeable spine. Upper surface of
movable finger with a furrow or elevations.
3 (6). Immovable finger of anterior chela without a large tooth in the middle
of prehensile margin.
4 (5). Immovable finger of anterior chela with 4—5 small spines at the base
of the external margin; a small spine at the base of prehensile margin.
Without teeth in proximal and distal ends of median anterior pereo-
pods.— Japanese Sea..........................2. U. major (de Haan).
5 (4). Immovable finger of anterior chela without spinules at the base of
external margin; strong tooth at the base of prehensile margin. Median
ridge of the upper margin of the propodite of anterior pereopods armed
with a strong tooth both on proximal and distal end. — Japanese Sea:
coast of Japan...............................3. U. yokoyai nom. nov.
€ (3). Immovable finger of anterior chela with a large tooth in the middle of
prehensile margin. — Japanese Sea: region of Vladivostok..........
.........................................4. U. issaeffi (Balss).
296
2. CALLIANASSIDAE, 3. GALATHEIDAE
2. Subfamily CALLIANASSINAE
1 (2). Pleopods of the second abdominal segment not like the pleopods of
the third to fifth abdominal segments...........2. Callianassa Leach.
2 (1). Pleopods of second abdominal segment the same as in the three
segments following it......................3. Ctenocheles Kishinouye.
2. Genus CALLIANASSA LEACH
1 (10). Telson of the same length as the uropods.
2 (3). Outer maxillipeds narrow, pediform. — European shores of the Atlan-
tic; Mediterranean Sea (northern coast of Africa)....................
....................................1. C. subterranea (Montagu).
3 (2). Outer maxillipeds broad, operculiform.
4 (5). Frontal ‘ margin of carapace without rostrum. Prehensile margin of the
movable finger of the large hand has at its base a broad truncated
tooth, and near the end another rather large tooth. Posterior margin
of the telson bears two pairs of spinules near the lateral angles. — Paci-
fic: Japanese shore; China; Japanese Sea: region of Vladivostok. . . .
.........................................2. C. harmandi Bouvier.
5 (4). Rostrum present, though sometimes very small. Posterior margin of
telson without spinules near lateral angles.
6 (7). Carpus of large cheliped lJ/2 times longer than wide, and longer than
merus. Prehensile margin of movable finger of large hand armed with
teeth resembling those of C. harmandi. — Japanese Sea: region of
Vladivostok.................................3. C. gigas eoa nom. nov.
7 (6). Carpus of large cheliped slightly longer than wide and of the same
length as merus. Prehensile margin of (he movable finger of large
hand without prominent teeth.
8 (9). Proximal lobe on outer margin of merus of the large cheliped narrow,
with sharp apex; length of inner margin of carpus equal to or some-
what greater that width of carpus; carpus not longer than the palm; fin-
gers of the chela close rather tightly; movable finger shorter than inner
margin of palm. — Japan; China; Japanese Sea: region of Vladivostok.
.......................................4. -C. japonica (Ortmann).
9 (8). Proximal lobe on outer margin of merus of larger cheliped broad,
with a rounded apex; length of inner margin of carpus shorter than the
width of carpus; carpus longer than palm; broad gap between fingers
of the chela when closed; movable finger longer than inner margin of
palm. — Japan: Tokyo; Japanese Sea: region of Vladivostok............
.............................5. C. californiensis bouvieri nom. nov.
10 (1). Telson shorter than uropods. — Mediterranean, Adriatic and Black
Seas; west coast of France................ . 6. C. pontica (Czerniavsky).
2. CALLIANASSA, 3. CT E NO CH E L E S. —1. GALATHEA 297
Callianassa (Cheramus) subterranea (Montagu).
Does not occur in the waters of the USSR; all previous information
for the Black Sea refers to Callianassa pontica.
Callianassa (Trypaea) gigas eoa nom. nov.
The younger male specimens differ from the large male in the armament,
of the prehensile margins of the large cheliped, namely: both margins only
slightly serrated; when closed, fingers of hand tightly pressed together and
strongly crossed at their tips. In their other characters these specimens agree
with the large male.
Callianassa (Trypaea) californiensis bouvieri nom. nov.
Probably identical with C. japonica, representing variations of the
latter in age and sex. The writer had only one male specimen.
Callianassa (Callichirus) pontica Czerniavsky.
Presents two types of armament of prehensile margins of the fingers of
the large hand (fig. 28). According to the rules of priority the name of de
Man is included by the writer in the synonymy of this species.
3. Genus CTENOCHELES KISHINOUYE
A single species: Ctenocheles balssi Kishinouye. — Japanese Sea.
Japan.
2. Superfam. GALATHEIDEA
1 (2). Abdomen flexed, but not bent under the thorax. Outer maxillipeds
with mastigobranchiae. First pereopods usually slender and long. Gene-
ral shape of body shrimp-like..........................3. Galatheidae
2 (1). Abdomen bent under the thorax. Outer maxillipeds without masti-
gobranchiae. First pereopods usually of moderate length, stout. General
shape of body crab-like . . . . . . . .................4, Porcellanidae
3. Family GALATHEIDAE
1 (6). Cornea pigmented. Expodite of first maxillip ds with flagellum.
2 (5). Rostrum neither curved nor compressed.
3 (4). Rostrum triangular, flattened in dorso-ventral direction, with indented,
lateral margins. Supraocular spines wanting or reduced................
.......................................... 1. Galathea Fabricius.
4 (3). Rostrum styliform, smooth. Strong supraocular spines present on the
sides of the rostrum.............................. 2. Munida Leach.
5 (2). Rostrum curved and compressed; its margins, the upper one espe-
cially, armed with teeth......................3. Cervimunida Benedict.
6 (1). Cornea devoid of pigment. Exopodite of first maxillipeds without
flagellum. . .................................4. Munidopsis Whiteaves.
298
3. GALATHEIDAE, 4. PORCELLANIDAE
1. Genus GALATHEA FABRICIUS
1 (4). Anterior part of gastric region bears only two spines.
2 (3). Basal spines of rostrum small. Upper margin of merus of second pair
of legs armed with spinules.— Mediterranean and Adriatic Seas, Euro-
pean shores of the Atlantic up to Varanger Fjord; Murman Coast (?).
. . . •...................................1. G. strigosa (Linne).
3 (2). Basal spines of rostrum rather large. Upper margin of merus of second
pair of legs armed with 11-12 strong sharp teeth. — Bonin Isles; South
Japan; west coast of Japan...............2. G. acanthomera Stimpson.
4 (1). Anterior part of gastric region carries more than two spines.
5 (6). Lateral margins of rostrum in front of basal spines smooth, unarmed.
Wide gape between fingers of the chelipeds, when closed. Each abdo-
minal segment with one transverse furrow. Not a very hairy species.—
South Japan; Tsugaru Strait; Aomori.............3. G. Integra Benedict.
6 (5). Lateral margins of rostrum in front of basal spines armed with
3—4 teeth on each side. Fingers of cheliped close together tightly.
Each abdominal segment with 4 transverse furrows. A very hairy spe-
cies.— Japan..................................4. G. pubescens Benedict.
2. Genus MUNIDA LEACH
1 (2). Gastric region of carapace without transverse row of spines. Fingers
of chelae of first pereopods longer than palm. Second abdominal seg-
ment (in larger specimens the third and fourth segments likewise) armed
with spinules on anterior margin. — Atlantic; Mediterranean and Adriatic
Seas; Northern and Norwegian Seas; west coasts of Murman............
....................................... 1. M. bamffica (Pennant).
2 (1). Gastric region of carapace with a transverse row of 13 spines in
front. Fingers of chelae of first pereopods • of the same length as the
palm or shorter. * All the abdominal segments unarmed. — South Japan
up to Tsugaru strait; west coast of Japan; Aomori...................
........................................2. M. japonica Stimpson.
3. Genus CERVIMUNIDA BENEDICT
A single species: Cervimunida princeps Benedict. — South Japan;
west coast of Japan.
4. Genus MUNID0PS1S WHITEAVES
A single species: Mnnidopsis beringana Benedict. — Bering and Okhotsk
Seas.
4. Family PORCELLANIDAE
1 (2). Frontal margin triangular and serrated. Subbranchial regions of cara-
pace continuous, not separated by membraneous interval............•
........................................1. Porcellana Lamarck.
5. ALBUNEIDAE, 6. PYLOCHELIDAE
299
2 (1). Frontal margin triangular, but never serrated. Subbranchial regions of
carapace divided by membraneous interval into larger anterior and
smaller posterior parts.....................2. Pachycheles Stimpson.
1. Genus PORCELLANA LAMARCK
1 (2). Palm of larger hand more or less flat, with a rounded longitudinal
ridge in the middle, fingers rather long. Prehensile margins of fingers
of small hand not hairy.—Black, Mediterranean and Adriatic Seas;
East Atlantic; Northern Sea; Kildin Isle; to the south till Marocco . .
................•.............. . . 1. P. longicornis (Pennant).
2 (1). Palm of large hand swollen with no signs of a longitudinal ridge
in the middle; fingers short. Fingers of small hand thickly set with
hairs on concave prehensile margins (this pubescence is especially no-
ticeable on the underside). — Black and Mediterranean Seas ....
....................................... 2. P. longimana Risso.
2. Genus PACHYCHELES STIMPSON
A single species: Pachycheles stevensii Stimpson. — Western coast of
Japan from Hokkaido to Nagasaki; Tokyo Bay. In the region of Peter the
Great Bay is known from Gorskov Bay up to Strelok Gulf.
3. Superfam. HIPPIDEA
5. Family ALBUNEIDAE
1. Genus BLEPHARIPODA RANDALL
A single species: Blepharipoda japonica Durufle. — Coast of Japan
region of Vladivostok.
4. Superfam. PAGURIDEA
1 (4). Abdomen straight or spirally twisted. Carapace cylindrical, firm mainly
in fore part and more or less soft in hinder part. Fourth pereopods
unlike the third. Rostrum rudimentary or completely wanting. Uropods
present.
2 (3). Abdomen symmetrical, with well-calcified segments on dorsal side.
Paired abdominal limbs present on all segments. Gills of trichobranchiate
type ...............................................6. Pj lochelidae
3 (2). Abdomen mostly asymmetrical, soft, not segmented, with small slightly
calcified plates on the left side. Very rare paired appendages, only on
the first abdominal segment, usually only on the left side of second
to fifth segments. Gills of phyllobranchiate type. ... 7. Paguridae
300
7. PA GURIDAE
4 (1). Abdomen bent under cephalothorax. Carapace broad, well calcified in
all parts. General shape of animal crab-like. Fourth pereopods like the
third. Rostrum for the most part well-developed. Uropods wanting-. . .
...................................................8. Lithodidae
6. Family PYLOCHELIDAE
1. Genus MIXTOPAGURUS A. MILNE-EDWARDS
A single species: Mixtopagurus jeffreysii (Miers).— Western coast of
Japan.
7. Family PAGURIDAE
Twenty-five species of the hermit-crabs (about 5.5% of the total num-
ber known) have been recorded in the seas of the USSR.
The number of species discussed in this work is however forty-three,,
for the species from the whole of the Japanese and Bering Seas have been
included here.
Only two species of hermit-crabs (Diogenes varians and Clibanarius
misanthropus), typical representatives of the Mediterranean fauna, are found
in the Black Sea; somewhat other conditions of life in the latter (chiefly,
perhaps, its low salinity) having led to a diminution of these forms, especially
noticeable in comparison with corresponding specimens trom the Mediter-
ranean Sea. Clibanarius misanthropus occurs more seldom than Diogenes
varians.
The White and the Barents Seas contain only one species of hermit-
crabs, Pagurus pubescens. Another species, Pagurus bernhardus, occurs
comparatively seldom off the western part of the coast of Murman only,,
where it finds its eastern limit. Being a typical' amphiboreal species, Pagurus
pubescens occupies an exceptional place among the hermit-crabs of our
seas.
The seas washing the northern coasts of Siberia contain no hermit-
crabs in their fauna except the sporadic occurrence of Pagurus pubescens in
the Kara Sea; the hermit-crab reappears only in the Far-Eastern part of
the Arctic eastwards of the river Kolyma.
Of all the seas of the USSR the Far-Eastern Seas have the richest
fauna of hermit-crabs, namely 84% of the total number of species listed
for our seas. According to their distribution the whole mass of these Far-
Eastern species may be subdivided into four principal groups:
The first group of widely spread species common to the three Far-
Eastern seas (the Japanese, the Okhotsk and the Bering Seas) and the Chu-
kotsk Sea, namely Pagurus pubescens, P. rathbuni, and P. capillatus.
The second group of species belonging almost exclusively to the Japa-
nese Sea (excepting their occurrence in the Inland Sea of Japan and its.
GEOGRAPHICAL DISTRIBUTION
301
south-eastern coast): Pagurus gracilipes, P. dubius, P. anomalus, P. brachio-
mastus, P. lanuginosus, P. pectinatus, and Diogenes penicillatus.
The third group of species occurring- in the Japanese and the Okhotsk
Seas as wellas off the western coast of North America (the connection being
accomplished along- the Aleutian Islands almost without penetration into the
Bering; Sea proper): Pagurus hirsutiusculus, P. middendorffii, P. undosus,
P. ochotensis, and P. gilli (?).
The fourth group of species with an interrupted distribution (species
present both in the Japanese Sea and off the western coast of North Ame-
rica but absent from the intermediate regions): Pagurus samuelis, Orthopa-
gurus minimus and O. schmitti.
The Japanese Sea. In the Japanese Sea of the first group of hermit-
crabs only Pagurus pubescens is known from Japan proper, the other two
species being distributed only along our coast where they find their south-
ern limit. All the representatives of the second group are also known from
Japan proper; with the exception of Pagurus anomalus and P. brachiomas-
tus, they are mainly distributed in the southern part of the Japanese Sea
and off the south-eastern coast of Ja an. They find their northern limit along
our coast. On the contrary Pagurus anomalus and P. brachiomastus seem
to be peculiar to our side of the Japanese Sea, finding their limit along the
south of Japan. Of the third group, Pagurus undosus is known only from our
waters of the Japanese Sea and is not known from Japan. Of the fourth'
group, Pagurus samuelis is also known from Japan while both the species
of Orthopagurus are recorded from our coast only. The species, endemic
mostly to the Inland Sea of Japan and its eastern coast and partly penetra-
ting into the Japanese Sea proper without spreading northward farther than
the Tsugaru Strait, form a special group: Pagurus obtusifrons and P. con-
stans, Clibanarius japonicus and Paguristes digitalis. Lastly, there is a group
of tropical species having their northern limit off the coast of South Japan
and occurring comparatively seldom in the Japanese Sea proper: Spiropagu-
rus spiriger, Diogenes edwardsii, Dardanus arrosor and Paguristes setosus.
Unfortunately the data are too scant concerning the fauna of the western
coast of Japan from Tsusima Strait to Niigata — one the most interesting
regions lying along the chief route of migration of the more southern forms
into the Japanese Sea. Were this region better known, the list of species
would undoubtedly be greater.
The Japanese Sea is thus distinguished by the number and diversity of
its hermit-crab population. As will be seen later, the fauna of hermit-crabs
of the Bering Sea is also characterized, though to a lesser degree, by the
diversity and number of its species due in this case to an admixture of Cali-
fornian forms. On the contrary, the faunae of the Okhotsk and the Chukotsk
seas are of a very homogeneous character.
The Okhotsk Sea. The first and the third of the above-named main
groups are fully comprised in the fauna of hermit-crabs of the Okhotsk Sea.
302
7. PAGURIDAE
Besides these, Pagurus splendescens is a common form also characteristical
of the Bering and the Chukotsk Seas; it does not occur in the Japanese Sea
where it is replaced by the closely allied Pagurus anomalus. The problem of
the distribution of Pagurus gilli is not quite clear as yet; however, on the basis
of the really scanty material in hand I am inclined to think that this species
may be included into the third group. Up till now this species is not known
from the Okhotsk Sea proper. It is quite probable, however, that the distri-
bution of this species, forming an arc-like area, finds its limit off the Kuril
Islands.
The faunal complex of hermit-crabs of the eastern coast of Kamchatka
(up to the Commandor Island) is identical with that of the Okhotsk Sea.
The Bering Sea. The first and the third (chiefly along the Aleutian
Islands) groups and Pagurus splendescens form the fauna of the Bering
Sea. But the fauna of the south-eastern portion of the Bering Sea (Unalaska,
Pribyloff Islands, St. Matthew Island, Nunivak Island) differs from that of the
rest of that sea.
The faunal complex of this south-eastern portion of the Bering Sea
consists of those representatives of the Californian fauna that find their
northern limits in this region and do not range north of Nunivak Island. Such
are Pagurus beringanus, P. tanneri, P. granosimanus, P. armatus and
P. setosus, the last three not passing beyond the Aleutian Islands (Unalaska
and Kadiak). The species which belong to the western coast of North America
and which do not runn south of British Columbia, Washington, and Ore-
gon, form the next group, namely: Pagurus dalli, P. kennerlyi, P ochotensis
subsp. aleuiicus, P. cavimanus (?), P. ienuimanus, and P. cornutus. The
last two species range along the whole Aleutian chain of islands up to the
Commandor Islands inclusively, Pagurus cornutus continuing further north-
wards along our coast of the Bering Sea till Cape Navarin. Pagurus mer-
tensii does not occur in the Bering Sea, being an endemic which has slip-
ped into the synonymy of this species.
It may be seen from the above-given data that the Aleutian Islands
form the bridge which connects the fauna of both sides of the north part of
the Pacific. It is interesting to note that it is possible to trace both the dif-
ferent stages in the existence of this connection and, partly, the direction
of exchange. The greater part of the Californian species extends to the
eastern end of the Aleutian chain of islands and stops there. Pagurus fenuima-
nus and P. cornutus have already passed beyond the Aleutian Islands, the
latter species then turning northwards.
Pagurus hirsutiusculus and P. middendorffii of the third group of spe-
cies are examples of the most complete connection between the two bor-
ders of the North Pacific. As regards Pagurus undosus, it may be quite
definitely stated that its migration proceeded in the opposite direction, the
Okhotsk Sea being the centre from which the distribution of this species
originated; from there the species partly penetrated into the Japanese Sea.
1. PAGURUS
303
(along1 our coast only); on the other hand, it spread along1 the eastern coast
of Kamchatka up to the Aleutian Islands where it divided into two branches:
one running1 northwards along our coast of the Bering Sea up to Cape Chap-
lin, the other occupying the whole Aleutian chain of islands but without
extending as yet along the western coast of North America. In all probabi-
lity the distribution of Pagurus ochotensis proceeded along the same route,
for there are some indications of a rupture of a once continuous area, for
example, the comparatively rare occcurrence of this species off the eastern
coast of Kamchatka including the western part of the Aleutian Islands (Com-
mandor Islands). If we adopt such a direction of distribution of Pagurus
ochotensis, the taxonomic position of P. ochotensis subsp. aleuticus becomes
quite obvious. It will then be a subspecies of P. ochoiensis, which had
separated from the main stock in the eastern part of the Aleutian Islands,
i. e., at the end of the arc most remote from the centre of distribution. The
same would likewise apply to Pagurus gilli, if it would only be known for cer-
tain that the Japanese P. cavimanus may be identified with P. gilli. If it is
so, we then have an arc-like range for P. cavimanus (Miers) (= P. gilli) with
the subspecies P. cavimanus subsp. munitus (Benedict) in the eastern part
of the Aleutian Islands. And, finally, the fourth group of species is an
example of a complete break of connection, which had occurred (probably
a long time ago) in the same manner along the Aleutian Islands. The occur-
rence of Pagurus samuelis in Avacha Bay (Kamchatka), recorded by H. Balss
(1913), could be considered a relic of a once continuous range; but the com-
paratively extensive material from Avacha Bay at my disposal seems to point
to the above identification being erroneous and to both the specimens des-
cribed by that author referring to Pagurus hirsutiusculus, which is very
similar in appearance to P- samuelis.
The Chukotsk Sea. The first group, i. e., Pagurus pubescens, P. rath-
buni and P. capillatus, as well as P. splendescens, constitute the fauna of
the Chukotsk Sea. As this region is still very little known, it is difficult to
say anything about the northern, eastern and western limits of distribution
of these species.
Finally we must mention the widely ranged Parapagurus pilosimanus
which was collected off the eastern coast of Kamchatka and is therefore
included in the fauna of hermit-crabs of our seas.
KEY TO THE GENERA
I - (8). Outer maxillipeds widely separated at base.
2 (7). Abdomen flexed and spirally coiled. Uropods unequal, usually better
developed on the left side than on the right.
3 (4). With no genital appendages either on the coxal joints of the last
ambulatory legs or on the first or second abdominal segments ....
.............................................1. Pagurus Fabricius..
304
7. PA GURIDAE
4 (3). With genital appendages.
5 (6). Paired appendages on the first and second abdominal segments of
the male. Right cheliped larger than the left . 2. Parapagurus Smith.
6 (5). An unpaired spirally coiled genital appendage on the coxal joint of
the left fourth ambulatory leg of the male. Chelipeds subequal . . .
........................................3. Spiropagurus Stimpson.
7 (2). Abdomen flexed but not spirally coiled. Uropods equal or the left
uropod slightly larger than the right ... 4. Orthopagurus Stevens.
8 (1). Outer maxillipeds approximated at base.
9 (12). Left cheliped larger than the right; the fingers moving in vertical
plane.
10 (11). Frontal margin of the carapace with a blunt median tooth. An
ophthalmic rostrum present between the eye-stalks. . 5. Diogenes Dana.
11 (10). Frontal margin of the carapace without a median tooth. Ophthal-
mic rostrum absent..................................6. Dardanus Paulson.
12 (9). Chelipeds equal or subequal, or the left sometimes slightly longer
than the right. Fingers moving in a horizontal plane.
13 (14). Feet of the fourth pair subchelate. Abdomen without paired genital
appendages on the first segment of either sex . - 7. Clibanarius Dana.
14 (14). Feet of the fourth pair simple. Abdomen with paired genital appen-
dages on the first segment of either sex .... 8. Paguristes Dana.
1. Genus PAGURUS FABRICIUS
1 (6). Upper surface of palm of right hand smooth and not hairy.
2 (3). Carpus of right cheliped as broad as the hand. Upper surface of left
hand convex, its margins not upturned. — Japanese and Okhotsk Seas;
eastern coast of Kamchatka; Bering Sea; Aleutian Islands; western
coast of North America southwards to Vancouver. ................... .
........................................1. P. middendorffii Brandt.
3 (2). Carpus of right cheliped much broader than the hand owing to late-
ral wing-like expansions. Upper surface of left hand flat with upturned
margins.
4 (5). Upper surface of carpus of right cheliped rounded and perfectly
smooth. Antennal acicle not reaching tip of eye. — Kamchatka; Aleutian
Islands........................................... 2. P. gilli (Benedict).
5 (4). Upper surface of right cheliped with distinct median ridge covered
with some spiny granules at its proximal end. Antennal acicle reaching
tip of eye. — North and South Japan; Bering Sea southwards to Port-
lock and Juneau (California)...................3. P. cavimanus (Miers).
‘6 (1). Upper surface of palm of right hand covered with rounded granules,
which are sometimes very small and scattered, so that the hand seems
almost smooth. Hairs, if any, very short, often scarcely visible with the
naked eye.
1. PA GURUS
305
7 (26). Upper surface of palm of right hand covered with rounded granu-
les, which are sometimes very small and scattered so that the hand
seems almost smooth. Hairs, if any, very short often scarcely visible
with the naked eye. Or upper surface of palm of right hand covered
with spiny granules or more or less short conical spines; in this case
hairs on palm either very short and arranged in bunches at bases of
spines or granules or almost quite absent. Or the upper surface of palm
of right hand covered with acute long spines and more or less long
hairs which may sometimes conceal all the armature of the palm. Occa-
sionally spines of palm short but long soft hairs also present.
8 (21). Upper surface of palm of right hand without any prominent ridges
or depressions but more or less evenly convex.
9 (16). Merus of right cheliped without prominent blunt spines or large gra-
nules on its lower surface.
10 (11). Hand of right cheliped much broader than carpus. Upper surface of
palm of left hand flat with upturned margins. —Aleutian Islands; Com-
mandor Islands; western coast of North America southwards to
Washington ...................................4. P. tenuimanus (Dana).
11 (10). Hand of right cheliped not broader or only insignificantly broader
than carpus. Upper surface of palm of left hand more or less convex
and without upturned margins.
12 (13). Frontal margin of carapace with no lateral teeth. Chelipeds hairy
except right hand. — Northern part of the Japanese Sea; Okhotsk Sea;
eastern coast of Kamchatka; Commandor Islands; Aleutian and Priby-
loff Islands; western coast of North America southwards to San Diego
(California)................................5. P. hirsutiusculus (Dana)
13 (12). Frontal margin of carapace with lateral teeth. Chelipeds hardly hairy.
14 (15). Palm of right hand very swollen, covered with blunt granules giving
it a shagreen-like appearance; fingers stout. Antennal acicles trilateral.
Dactyls of ambulatory legs stout. — North Atlantic: coast of North Ame-
rica from New Foundland to Chesapeake Bay; European coast from
West Murman to France.......................... 6. P. bernhardus (Linne).
15 (14). Palm of right hand a little swollen, covered with very small scat-
tered granules, often looking almost smooth. Antennal acicles not trila-
teral. Dactyls of ambulatory legs slender. — Japanese Sea............
.......................................7. P. gracilipes (Simpson).
16 (9). Merus of right cheliped furnished with prominent blunt spines or
large granules on its lower surface.
17 (18). Lower surface of merus of right cheliped furnished with two large
granules. — Bering Sea, south of Aleutian Islands; western coast of
North America southwards to Monterey (California)....................
...... ............ 8. P. beringanus (Benedict).
18 (17). Lower surface of merus of right cheliped furnished with one blunt
spine in the middle.
Фауна СССР. Макаров 20
306
7.PAGURIDAE
19
20
21
22
23
24
25
26
27
28
29
(20). Hand of right cheliped twice as long as wide, its outer and internal
margins subparallel. Upper surface of palm of left hand with long soft
hairs. — South Japan; Japanese Sea along our coast northwards to
Olga Bay...........................................9. P. dubius (Ortmann).
(19). Hand of right cheliped less than twice as long as wide with arcua-
ted margins. Upper surface of palm of left hand almost without hairs
or furnished with short sparse ones. — Japan; our coast of the Japanese
Sea only in Nakhodka Bay and Olga Bay......................................
...........................................10. P. samuelis (Stimpson).
(8). Upper surface of palm of right hand with distinct ridge in the middle.
(23). Ridge on the upper surface of palm of right hand ending in a very
prominent horn. — Bering Sea along our coast from Commander Islands
northward to Cape Navarin; along western coast of North America
from 55°38’ N., southwards to Queen Charlotte Sound (British Colum-
bia)............................................11. P. cornutus (Benedict).
(22). Ridge on the upper surface of palm of right hand not ending iri
a horn but descending sharply enough towards the immovable finger.
(25). Ridge on the upper surface of palm of right hand parallel to inter-
nal margin: there is a depression with parallel margins between the
ridge and the internal margin of palm. Outer surface of left cheliped
convex. — Aleutian and Pribyloff Islands . . 12. P. undosus (Benedict).
(24). Ridge on the upper surface of palm of right hand a triangular ele-
vation in shape with its apex pointing to the gape between the fingers;
depression between the internal side of the elevation and the internal
margin of palm without parallel margins. Outer surface of palm of left
hand deeply concave. — Western coast of North America from Una-
laska southwards to San Simeon Gulf (California); Pribyloff Islands . .
...............................................13. P. tanneri (Benedict)]..
(7). Upper surface of palm of right hand covered with spiny granules or
with more or less short conical spines; in this case hairs on palm either
very short and arranged in bunches at bases of spines or granules or
almost quite absent. Or upper surface of palm of right hand covered
with acute long spines and more or less long hairs which may someti-
mes conceal all the armature of the palm. Occasionally spines of palm
short but long soft hairs also present.
(42). Upper surface of palm of right hand covered with spiny granules or
with more or less short conical spines. Hairs on palm either very short
and arranged in bunches at bases of spines or granules or almost quite
absent.
(31). The whole upper surface of carapace hard, not only its anterior
portion.
(30). Median frontal tooth narrow, long, pointed obliquely upwards, with,
simple apex. Sharp prominent spine on lateral projections of carapace
behind cervical groove. — South Japan northwards to Tugaru Strait; our
1. PAGURUS
307
coast of the Japanese Sea from Furuhelm Island northwards to de
Castries Bay................................14. P. anomalus (Balss).
30 (29). Median frontal tooth broad, shorter, pointed horizontally forward,
with three or four denticles on apex. Lateral projection of carapace
behind cervical groove without a sharp prominent spine. — Okhotsk Sea;
eastern coast of Kamchatka; Bering Sea; Chukotsk Sea westwards to
the Kolyma......................................15. P. splendescens Owen.
31 (28). Only anterior portion of carapace hard.
32 (35). Anterior portion of carapace not so long as wide. Antennal acicles
trilateral, their inner surface being very iridescent. Chelipeds very
robust in comparison with the body; their joints almost quite devoid
of hair.
33 (34). Dactyls of ambulatory legs without a longitudinal sulcus on their
upper surface. — Bering Sea; Okhotsk Sea; Japanese Sea southwards
to South Japan inclusively; western coast of North America southwards
to Oregon.........................................16. P. ochotensis Brandt.
34 (33). Dactyls of ambulatory legs with a longitudinal sulcus on their upper
surface. — Western coast of North America southwards to Oregon;
Bering Sea (region of Pribyloff Islands)....................................
........................16a. P. ochotensis subsp. aleuticus (Benedict).
35 (32). Anterior portion of carapace as long as or longer than wide. Antennal
acicles not trilateral, not iridescent. Chelipeds of moderate size in
comparison with the body; their joints more or less hairy except the:
hands.
36 (37). Carpus of right cheliped with a round bunch of hairs on inner dis-
tal corner of upper surface. Fingers of left hand long, comparatively
thin and much bent downwards. — Japanese Sea only along our coast;
Okhotsk Sea; eastern coast of Kamchatka; Bering Sea; Chukotsk Sea
northwards to Wrangel Island, westwards to the mouth of the Kolyma.
........................................... 17. P. rathbuni (Benedict).
37 (36). Carpus of right cheliped without such a bunch of hairs. Fingers
of left hand not very long and slightly if at all bent downwards.
38 (39). Merus of right cheliped with two large granules on its lower surface.
Left hand without a distinct ridge on upper surface. — Western coast
of North America from Unalaska southwards to Ensenada (Lower Cali-
fornia)...................................18. P. granosimanus (Stimpson).
39 (38). Merus of right cheliped without such granules on its lower surface.
Left hand having a distinct ridge on upper surface.
40 (41). Merus joints of chelipeds with sharply defined band of white near
distal end. Spines on chelipeds comparatively long and sharp. Right
hand fairly narrow. — Bering Sea southwards to Oregon.......................
......................................... 19. P. dalli (Benedict).
41 (40). Merus joints of chelipeds without white band near distal end.
Spines on chelipeds shorter and blunter. Right hand fairly broad.—
20*
308
7. PAGURIDAE
A typical amphiboreal species. White and Barents Sea; North Atlantic.
In the Far-East it ranges from the Chukotsk Sea to South Japan
inclusively. Western coast of North America southwards to Oregon. .
............................................20. P. pubescens Kroyer.
42 (27). Upper surface of palm of right hand covered with acute long spines
and more or less long hairs, which may sometimes conceal all the
armature of the palm. Occasionally spines of palm short, but long
soft hairs also present.
43 (44). Merus of right cheliped with two very prominent blunt spines on
its lower surface.— Japanese Sea along our coast from Furuhelm Island
northwards to de Castries Bay; La Perouse Strait; Aniva Bay ....
...................................21. P. brachiomastus (Thallwitz).
44 (43). Merus of right cheliped with no such spines on its lower surface.
45 (46). Immovable finger of right hand twice broader than the movable one.
Surface of palm of right hand with short spines and long soft hairs. —
Japan; Japanese Sea: region of Vladivostok.................................
..................................... 22. P. lanuginosus de Haan.
46 (45). Immovable finger of right hand as broad as or slightly broader than
the movable one. Surface of palm of right hand with long spines and
hairs.
47 (48). Antennal acicles trilateral, with iridescent inner surface.— A compa-
ratively limited endemic species of the western coast of North America:
from Unalaska to San Diego (California) . . . 23. P. armatus (Dana).
48 (47). Antennal acicles not trilateral; their inner surface not iridescent.
49 (56). Spines on surface of palm of right hand arranged in more or less
regular longitudinal rows.
50 (53). Spines on surface of palm of right hand forming 8-9 longitudinal rows.
51 (52). Antennal acicles shorter than eyes. Median tooth of front acute.
Eye stalks slender, broader at base. Outer margin of palm of right
hand having a row of long large spines sharply separating upper sur-
face of hand from lower one. — Japanese Sea from Tsusima Island
northwards to de Castries Bay; Tsugaru Strait; Aniva Bay . .. . . .
........................................ 24. P. pectinatus (Simpson).
52 (51). Antennal acicles longer than eyes. Median tooth of front obtuse.
Eye stalks rather stout, broader at the cornea. Outer margin of palm
of right hand blunt without strong spines. — South Japan northwards
up to Tsugaru Strait...........................25. P. obtusifrons (Ortmann).
53 (50). Spines on surface of palm of right hand forming 7 longitudinal rows.
54 (55). Antennal acicles longer than eyes. Hairs on hands much longer than
spines. — Western coast of North America from Kadiak Island (Alaska)
southwards to Santa Cruz (California) . . . 26. P. setosus (Benedict).
55 (54). Antennal acicles shorter than eyes. Hairs on hands not reahing tip
of spines. — From Aleutian Islands southwards to Washington ....
....................................... 27. P. kennerlyi (Stimpson).
1. PAGURUS
309
56 (49). Spines on surface of palm of right hand scattered, not forming regular
longitudinal rows.
57 (58). Lateral teeth of front rounded. Antennal acicles reaching tip of eyes
or slightly shorter. Ambulatory legs exceeding the tip of right cheliped;
dactyls twisted.— Japanese Sea only along continental coast southwards
to Chogu-Chien-Dogu Bay, North Korea; eastern coast of Kamchatka;
Okhotsk, Bering and Chukotsk Seas . . . 28. P. capillatus (Benedict).
58 (57). Lateral teeth of front more acute. Antennal acicles extending beyond
the eyes. Ambulatory legs shorter than right cheliped; dactyls not
twisted. — South Japan northwards to Hakodate (Hokkaido) ....
.........................................29. P. constans (Simpson).
Pagurus gilli (Benedict). Up till now Pagurus gilli (Benedict) was
known as a comparatively limited local form of the Aleutian Islands.
It could, perhaps, have been considered a subspecies of the more widely
ranged Pagurus cavimanus (Miers). An examination of the collections in the
Zoological Institute of the Academy of Sciences of the USSR has, however,
shown that the area of distribution of P. gilli must be extended. The speci-
mens collected by B. Heinemann from Kamchatka (Lopatka Cape), by
V. Arssenyev from the region of Soviet Bay, and by W. Roszkowsky from
Japan (Misaki) were compared by me in detail with the specimen of P. cavi-
manus (=-P. minutus) from the U. S. National Museum (№ 16646); and their
identity with P. gilli is indisputable. P. gilli, therefore, appears to have a more
extended range in the shape of an arc along the Aleutian Islands, whilst
P. cavimanus must be considered as a subspecies of P. gilli, if the identification
of the specimens from Japan be assumed to be erroneous. It is difficult,
however, to come to a decisive conclusion without an examination of more
abundant material from Japan. The existing synonymy and geographical range
is retained for both these species with a corresponing extention of the range
of P. gilli in accordance with the above-mentioned data.
Pagurus yokoyai nom. nov.
Syn.: Eupagurus gracilipes Yokoya, 1933 : 89, text-fig. 33 (nomen praeoccup).
„Near Eupagurus pectinatus Stimpson, but it differs from this in the
followings points: Ophthalmopods shorter. The peduncles of the first and
the second antennal distinctly exceed the ophthalmic cornea, and the hairy
acicle of the second antenna reaches the distal end of the eye. Legs slender.
(Yokoya)“. Unfortunately Yokoya gives too brief a description. This species
is rather more allied to Pagurus brachiomastus (Thallwitz).
Pagurus ochotensis Brandt.. This species is very rare off our coast
of the Bering Sea; the Okhotsk Sea; the Japanase Sea southwards to South
Japan inclusively. The problem of the synonymy of P. ochotensis and
P. alaskensis proved to be very complicated. On the basis of my examina-
310
7. PAGURIDAE
tion of the collections of the Zoological Institute of the Academy of Sciences
of the USSR, I have come to the conclusion that Brandt had not had
at his disposal a specimen of P. ochotensis in the modern understanding
of this species. The type-specimen of Brandt’s Pagurus bernhardus var.
spinimana seu subsp. ochotensis (№ 1075) must undoubtedly be referred to
Pagurus alaskensis (Benedict); I could not find a single specimen in the
whole collection which might be identified as P. ochotensis. In my considera-
tion of this problem I paid special attention to the following two points: (1)
the character of granulation both on the chelipeds and the ambulatory legs
of P. alaskensis is very variable, grading from extremely small and blunt gra-
nules to spiny granules (i. e., large granules with acute apices or conical
spines). (2) It is necessary to bear in mind that in his description of
P. ochotensis Brandt compared his species with the European P. bernhardus
(L.) whose appendages are covered with flat blunt granules; in comparison
with P. bernhardus the appendages of P. alaskensis are decidedly more
spinous; this caused Brandt to distinguish the variety spinimana as well as
the variety granulato-denticulata on the basis of the above-mentioned
difference in degree of granulation. Both varieties agree well with the con-
ception of P. alaskensis. Bernhardus armatus described by Dana, a species
really closely allied to P. alaskensis but differing from it in its cheliped
covered with long slender spines and hairs, is erroneously included in the
synonymy of P. ochotensis. The reason for this probably lies in the brevity
and inexactitude of Brandt’s description and in the coincidence of the chief
distinguishing character (the spinosity of hands) with the name of Brandt’s
variety (s: inimana). Pagurus alaskensis (Benedict) must therefore be called
Pagurus ochotensis Brandt, and the species known as Pagurus ochotensis
Brandt must be called Pagurus armatus (Dana). After receiving the speci-
mens of P. ochotensis (№ 52666) and of P. alaskensis (№ 16368) from the
U. S. National Museum’s collection I become still more convinced in the
justness of such a conclusion. The most prominent differences obtained by
the comparison of both the above-mentioned specimens with each other
and with the specimens from the collection of the Zoological Institute of the
Academy of Sciences of theUSSR are given in the following table:
Pagurus ochotensis Brandt
1. Surface of carpus and of pro-
podus of chelipeds covered with
granules or spiny granules; bunches
of hairs at bases of granules either
entirely absent or very small and
only slightly visible.
2. Right hand wide, outer margin
arcuated. Fingers of hand not cros-
Pagurus armatus (Dana)
1. Surface of carpus and of pro-
podus of chelipeds thickly set with
long slender acute spines and bun-
ches of hairs rising at bases of
spines and reaching the apex,
sometimes slightly exeeding them.
2. Right hand comparatively nar-
row, gradually tapering towards the
1. PAGUR US
311
sing when closed; therefore distal
end of hand seems more obtuse.
Inner margin of palm markedly up-
turned.
3. Inner margin of palm of left
hand much upturned.
4. Upper-outer surface of pro-
podal joints of first two pairs of
ambulatory legs covered with blunt
or spiny granules; short spines form-
ing only one row on upper mar-
gin; hairs, if any, very short.
5. Upper margin of dactyls of
ambulatory legs with a longitudinal
row of blunt or spiny granules.
distal end and appearing very acute
owing to the crossing of the finger
tips when closed. Inner margin of
palm markedly upturned.
3. Inner margin of palm of left
hand with a slight trace of upturn-
ing.
4. Upper-outer surface of propo-
dal joints of first two pairs of am-
bulatory legs set with three longi-
tudinal rows of spines, the longest
forming the row on the upper mar-
gin of joint; bunches of hairs rising
at bases of spines reaching their
apexes.
5. Upper margin of dactyls of am-
bulatory legs armed with a longitu-
dinal row of acute spines gradually
diminishing in size towards distal
end.
The absence of Pagurus armatus (Dana) from our fairly extensive
materials leads me to a deeply rooted conviction that this species is a com-
paratively limited local form of the western coast of North America. All the
authors, recording P. ochotensis from Kamchatka, the Okhotsk and the
Japanese Seas, are, therefore, placed by me in the synonymy of P. ochotensis
and not in that of P. armatus.
Pagurus ochotensis subsp. aleuticus (Benedict). This species is a local
form of the western coast of North America; morphologically P. aleuticus is
so closely related to P. ochotensis that I consider it a subspecies of the latter.
Pagurus pubescens Kroyer. A typical amphiboreal species. In the
Far East it ranges from the Chukotsk Sea to South Japan inclusively.
The most widely spread species of our Far-Eastern Seas. The problem
of P. trigonocheirus (Stimpson) as an independent species had been raised
in the carcinological literature a long time ago. Some authors (Brandt, Rich-
ters and Stimpson himself) recorded P. pubescens from the Pacific. Doflein
(1900) was the first to surmise, thought quite speculatively, that P. trigono-
cheirus (Stmp.) is a variety of the Atlantic P. pubescens Kr. Hansen (1908)
decisively upheld the identity of both these species. All the slight differences
usually given as the distinguishing characters of the two species (i. e., the
length to width ratio of the right hand, the width of the left hand at its base,
the degree of hirsuteness, etc.) really belong, as has already been shown by
A. A. Birula (1897: 439), to individual variations and sexual dimorphism of
312
7. PA GURIDAE
the species, P. pubescens. The differences between P. trigonocheirus (Stmp.)>
and P. pubescens Kr. given by H. Balss (1913) are based on his erroneous,
identification of P. trigonocheirus with P. capillatus (Benedict); thus all the
differences given by H. Balss refer to P. pubescens and P. capillatus which
are usually easily distinguishable. Having had at my disposal extensive
material from both the European (the White and the Barents) Seas and all
the Far-Eastern Seas and having compared it in detail, I came to th з conclu-
sion that P. trigonocheirus is identical with P. pubescens.
The only Greenland specimen of P. pubescens (№ 1086) existing in the
collections of the Zoological Institute of the Academy of Sciences of the
USSR is identical both with European and with Pacific specimens; thus.
Doflein’s suggestion of the existence of . a peculiar variety (var. krbyeri) off
the eastern coast of North America must apparently be declined. As regards.
P. brandti (Benedict), having compared the specimen of this species from
the U. S. National Museum (№ 19097), I agree with Hansen’s and Balss’s
opinion about the identity of this species with P. pubescens. G. P. Gorbunov
has kindly informed me about his find of some specimens of P. pubescens
in the Kara Sea off the western coast of the Jamal Peninsula; up till now
this species had been known only from the eastern part of Jugorsk Shar and.
Matochkin Shar. These finds, however, are of a sporadic nature only and
do not signify that P. pubescens is proper to the Kara Sea, such is also the.-
opinion of G. P. Gorbunov.
2. Genus PARAPAGURUS SMITH
1 (2). Eye-stalks slender, tapering distally. Fingers of left hand not longer
than the palm. Lower margins of dactyls of ambulatory legs with
scarcely noticeable spines. — Tropical and subtropical parts of the
Atlantic Ocean; Indian Ocean; Pacific Ocean; northwards to 63°
N., southwards to 48° S...........................1. P. pilosimanus Smith.
2 (1). Eye-stalks stout, broadening distally. Fingers of left hand much longer
than the palm. Lower margins of dactyls of ambulatory legs with well-
developed spines. — Coast of California . . • 2. P. mertensii Holmes.
Parapagurus mertensii Holmes. Under the name of Pagurus mertensii
Brandt (1851) described Pagurus hirsutiusculus (Dana). All the specimens
in the collection of the Zoological Institute- of the Academy of Sciences
of the USSR examined by me, Brandt’s original specimens among them,,
undoubtedly refer to this species. Thus the localities of Parapagurus
mertensii are only those given by Holmes and considered erroneous by
Rathbun; the remaining localities given for Parapagurus mertensii (Islands
of Kadiak and Atka, Vancouver Island and Kamchatka) refer to Pagurus
hirsutiusculus (Dana).
2. PARAPAGURUS—8. PAGURISTES
313
3. Genus SPIROPAGURUS STIMPSON
Spiropagurus spiriger (de Haan). — Indian Ocean; Chinese Sea; South
Japan. The occurrence of this species in the northern part of the Japanese
Sea (Yokoya, 1933) appears somewhat doubtful.
4. Gen. ORTHOPAGURUS STEVENS
1 (2). Large hand distally widened, base narrow and convex, distal portion
declivate with edges upturned; lateral teeth of front rounded. — West-
ern coast of North America; Nelma Bay (Tartar Strait)...............
..................................... 1. Orth, minimus (Holmes).
2 (1). Large hand nearly uniform in width flattened or slightly convex; late-
ral teeth practically obsolete. — Western coast of North America
(Washington); Japanese Sea only along our. coast, chiefly in the region
of Vladimir Bay and Olga Bay; Okhotsk and Bering Seas; eastern
coast of Kamchatka. This species has, obviously, an interrupted distri-
bution .....................................2. Orth, schmitti (Stevens).
5. Genus DIOGENES DANA
1 (2). Ophthalmic rostrum very small, scarcely visible to the naked eye. An
oval area with upturned margins on outer surface of left hand. A small
acidian usually seated upon this area. — Chinese Sea; South Japanese
Sea.........................................1. D. edwardsii (de Haan).
2 (1). Ophthalmic rostrum larger but not reaching tip of ophthalmic scales...
Without oval area on outer surface of left hand.
3 (4). Antennal acicles with 6-7 spines on inner margin. Ambulatory legs
of left side extending beyond the tip of left cheliped. — Japan: eastern
coast of Nippon Island; Shamara Bay (Ussurijsk Gulf)................
......................................2. D. penicillatus Stimpson.
4 (3). Antennal acicles with 4-5 spines on inner margin. Ambulatory legs,
of left side reaching the tip of left cheliped. — Mediterranean, Adriatic
and Black Seas..................................3. D. varians (Costa).
6. Genus DARDANUS PAULSON
Dardanus arrosor (Herbst). — Mediterranean and Adriatic Seas; Atlantic
Ocean; Red Sea; Australia; Philippine Islands; South Japan; Korea; northern
part of the Japanese Sea (Yokoya, 1933). Yokoy a(1933) unfortunately does,
not indicate the exact locality of this species in the North Japanese Sea..
This species is not recorded from our waters of the Japanese Sea.
314
8. LITHODIDAE
7. Genus CLIBANARIUS DANA
1 (2). Ophthalmic scales with straight inner margins. Carpus of left cheliped
without two rows of spines on dorsal surface. Dactyls of ambulatory
legs much shorter than propodites. — Mediterranean, Adriatic and
Black Seas ................................1. Cl. misanthropus (Risso).
2 (1). Ophthalmic scales with suboval inner margins. Carpus of left cheliped
with two rows of spines on dorsal surface. Dactyls of ambulatory legs
longer than propodites. — Mororan, Hokkaido (Japan).................
......................................2. Cl. japoniciis Rathbun.
8. Genus PAGURISTES DANA
1 (2). Outer side of movable finger of chela with seven oblique rows of brown
granules. — South Japan.......................1. P. digitalis Stimpson.
2 (1). Outer side of movable finger of chela without such rows. — New Guinea;
South Japan.........................2. P. setosus (Milne-Edwards).
8. Family LITHODIDAE
Next to the Paguridae the Lithodidae are the most abundantly repre-
sented family in the waters of the USSR. In all representatives of the sub-
family Hapalogastrinae the writer found paired rudimentary pleopods on
first abdominal segment in the female (fig. 9). The Hapalogastrinae thus
directly link the Lithodidae to the Paguridae.
1 (2). Rostrum short, flat, widely triangular. Third to fifth abdominal segments
not quite calcified. Abdomen usually the shape of a soft, membra-
neous swollen bag........................1. Hapalogastrinae Ortmann.
2 (1). Rostrum prominent, blunt or spine-like. Third to fifth abdominal seg-
ments with well-calcified plates. Abdomen usually flat..............
......................................... 2. Lithodinae Ortmann.
1. Subfamily HAPALOGASTRINAE
1 (2). Lateral margins of carapace beyond cervical groove armed with spines.
Chelipeds armed with sharp spines or teeth. Second abdominal segment
with unpaired median plate (H. cavicauda excepted, which has instead
a group of calcareous nodules). Most species with a more or less strong
hairy carapace and limbs......................1. Hapalogaster Brandt.
2 (1). Lateral margins of carapace unarmed. Chelipeds with blunt or squama-
like prominences, or with transverse ridges. Second abdominal segment
without unpaired median plate. Species slightly pubescent.
3 (6). Chelipeds not equal, the right usually larger than the left one; in
comparison with the ambulatory legs the chelipeds are very strong.
1. HAPALOGASTER —4. PLACETRON
315
No tubercle on sternal surface between the bases of the chelipeds.
' Without three spines on distal lower marg-in of merus of the ambula-
tory legs. Wide membraneous interspace between lateral plates of
second abdominal segments. Abdomen of a swollen, bag-like shape.
4 (5). Surface of carapace and chelipeds covered with transverse ridges.
Scaphocerites straight, spear-like. Last two segments of outer maxilli-
peds not widending...............................2. Dermaturus Brandt.
5 (4). Surface of carapace covered with squama-like elevations and chelipeds
set with round tubercles. Scaphocerite curved, of semi-lunar shape.
Last two segments of outer maxillipeds widened.........................
.......................................3. Oedignathus Benedict.
‘6 (3). Chelipeds equal or subequal in size, not too well developed as com-
pared with the long ambulatory legs. Tubercle present on sternal sur-
face between the bases of the chelipeds. Distal lower margin of merus’
of the ambulatory legs armed with three sharp spines. Lateral plates
of second abdominal segment come close together along! the median
line, leaving a narrow suture, but no membraneous interspace. Abdo-
men flat.........................................4. Placetron Schalfeew.
1. Genus HAPALOGASTER BRANDT
1 (4). Lateral margin of carapace behind cervical groove armed with 5
spines. Surface of joints of chelipeds covered with spines.
2 (3). Upper outer surface of palm of right chela with 3 longitudinal rows
of spines. Middle of palm of left cheliped without a spine. — From
Bering Strait along the Asiatic coast to the Northern part of the Japa-
nese Sea; western coast of America to Humboldt Gulf (California) • .
....................................1. H. grebnitzkii Schalfeew.
3(2). Upper outer surface of palm of right chela with 4 longitudinal rows of
spines. Middle of palm of left chela with a spine. — Aleutian Islands
westwards to Atka Isle; southwards along the western coast of America
to Puget Sound, Washington.......................2. H. mertensii Brandt.
4 (1). Lateral margin of carapace behind cervical groove armed with 7 spines.
Surface of joints of chelipeds covered with tubercular elevations.
Japanese Sea: northwards to Peter the Great Bay; Hakodate and
Aomori..................................... 3. H. dentata (de Haan).
Hapalogaster dentata (de Haan).
I have examined the female specimen determined by Brandt as H. den-
iata (Ke 844); according to all its characters that specimen must certainly be
referred to this species; the label reads: „coloniae rosso-americanae“ without
the name of the collector. As since then this species has not been recorded
off the American coast, and as all mention of its presence there (M. Rath-
bun 1904, p. 62) was exclusively based on the above-named specimen, it may
316
8. LITHODIDAE
be assumed with almost complete certainty that the above-mentioned label
is a fault and that this species is absent from the western coast of North
America.
2. Genus DERMATURUS BRANDT
A single species: Dermaturus mandtii Brandt. The Bering Sea, along
our coast northwards up to Cape Olutorsky; eastern coast of Kamchatka;;
The Okhotsk Sea; northern part of the Japanese Sea. Up to the present
this species had not been known from the northern part of the Japanese
Sea southwards of the extremity of Sakhalin. In the collections of the Zoolo-
gical Institute are 4 specimens from the region near the mouth of the
river Tjumen-Ula (Pavlenko’s collection), and also 5 specimens from Misaki,
Japan (Roszkovski’s coll.). I feel some hesitation about trusting the labels of
•these specimens, in the last case, especially, as there are no indications in
the literature about the existence of D. mandtii in Japan.
3. Genus OEDIGNATHUS BENEDICT
A single species: Oedignathus inermis (Stimpson). In our waters of
the Japanese Sea the species is known from the vicinity of Vladivostok:
(Patrocle Bay). It is also known from Japan and off the western coast of
North America. The species has an interrupted distribution.
4. Genus PLACETRON SCHALFEEW
A single species: Placetron wosnessenskii Schalfeew. Aleutian Islands;:
southwards to British Columbia.
2. Subfamily LITHODINAE
1 (14). Lateral plates of third to fifth abdominal segments without membra-
neous areas in the centre.
2(7). Upper surface of carapace covered with sharp or blunt spines. Rostrum
spine-like, armed with a varying number of additional spines.
3(6). Abdomen well-calcified. Median plates of third to fifth abdominal
segments replaced by membraneous interspace bearing a varying num-
ber of calcareous nodules, sometimes fusing into minute plates. Cara-
pace usually armed with sharp spines; gastric region bearing not less
than four spines.
4(5). Second abdominal segment covered by 5 distinct plates: unpaired
median plate, and paired lateral and marginal plates. Scaphocerites
well developed..................................6. Paralithodes Brandt.
5(4). Plates of second abdominal segment completely or partly fused, i. e.v
either median plate fuses with the lateral ones, or the lateral plates.
5. PHYLLOLITHODES, 6. PARALITHODES, 7. LITHODES 317
fuse with the marginal ones. Scaphocerites rudimentary.............
...............................................7. Lithodes Latreille.
6(3). Abdomen more or less slightly calcified. Median plates of third to
fifth abdominal segments distinct. Carapace usually armed with numer-
ous short and blunt spines; gastric region bearing one strongly promi-
nent spine...........................................8. Paralomis White.
7 (2). Upper surface of carapace smooth or covered with large prominences
of different shape and minute tubercles. Rostrum in the shape of
a more or less large blunt process or flat and truncated.
-8(13). Carapace not forming large lateral enlargement covering ambula-
tory legs; its surface not smooth but bearing prominences of different
shape and depressions. Rostrum neither flat, nor truncated.
9(12). Second abdominal segment entire. Marginal plates of third to fifth
segments present. Carapace cardiform or of pentagonal-hexagonal shape,
its posterior lateral angles without blunt prominences. Upper surfaces
of carapace without deep hemispherical cavity.
10(11). Carapace cardiform, its surface with large prominences of different
shape. Rostrum with a broad rounded apex, of a mace-like shape.
Ambulatory legs longer than the largest width of carapace .....
.......................................10. Sculptolithodes Makarov.
11 (10). Carapace of pentagonal-hexagonal shape, its surface with minute
tubercles. Rostrum short. Ambulatory legs shorter than the widest
part of carapace...............................11. Lopholithodes Brandt.
12(9). Second abdominal segment consists of 5 distinct plates. Marginal
plates of third to fifth segments wanting. Carapace of triangular shape
with highly prominent posterior lateral angles. Upper surface of cara-
pace with deep hemispherical cavity..............9. Rhinolithodes Brandt.
13(8). Carapace forms wide lateral enlargements almost completely covering
ambulatory legs; its surface almost smooth. Rostrum broad, flat; with
truncated, or more or less rounded apex . 12. Cryptolithodes Brandt.
14(1). Lateral plates of third to fifth abdominal segments with concave
membraneous areas in the centre..................5. Phyllolithodes Brandt.
5. Genus PHYLLOLITHODES BRANDT
A single species: Phyllolithodes papillosus Brandt. Western coast of
America from Unalaska to Monterey (California).
6. Genus PARALITHODES BRANDT
1 (2). Scaphocerite simple, sharp, spine-like in shape. Dorsal side of rostrum
bearing unpaired strong spine, the apex of which is usually bifurcate;
tip of strong lower process sharp. Carapace rather uniformly covered
with spines; gastric region bears six larger spines, set in two longitu-
318
8. LITHODIDAE
dinal rows with three spines in each. — Japanese and Okhotsk Seas;:
southern part of Bering Sea; Aleutian Islands; western coast of America
southwards to British Columbia . . . . 1. P. camtschatica (Tilesius)..
2(1). Scaphocerite of different shapes, bifurcate or with four branches. Rost-
rum shorter and without strong unpaired spine on dorsal surface..
Spines on surface more sparse.
3 (4). Scaphocerite of the shape of a long spine with two branches; tip of;
lower process of rostrum short; small spine between it and the pair
of dorsal spines wanting. Cardiac region of carapace with six spines.
Ambulatory legs spiny, their fingers comparatively long. —Northern part:
of the Japanese Sea; Okhotsk Sea; Bering Sea; Pribyloff Islands . .
........................................... 2. P. platypus Brandt..
4(3). Scaphocerite of the shape of a spine with four branches. Tip of the
lower process of rostrum blunt, often mace-like, swollen; between it and.
the pair of dorsal spines usually a small spine. Cardiac region of cara-
pace with four spines. Ambulatory legs highly spinous, their fingers
comparatively short. — Northern part of Japanese Sea; Okhotsk Sea;,
eastern coast of Kamchatka; southern part of Bering Sea (region of
Aleutian Islands)..................3. P. brevipes (M. Edwards et Lucas)..
Paralithodes camtschatica (Tilesius).
The indication of this species by M. Rathbun (1919, p. 8) for the region
of Cape Barrow, based on the find of only the propodite of a left cheliped,
must evidently be taken as belonging to P. platypus (brought with the cur-
rent?). This species is the principal object of fishery (western coast of
Kamchatka and the northern part of the Japanese Sea; the eastern coast of
Kamchatka, Zhupanovsky region near Petropavlovsk).
Paralithodes platypus Brandt.
The question of the distribution of P. platypus off the American coast
of the Bering Sea is not quite clear; there is no mention at all of this species
in the American literature. Only according to Richters (1883) is it pos-
sible to record this species for the region of the Pribyloff Islands. The
identification of Richters specimens with P. brevipes is incorrect; according
to all their characters they must be referred to P. platypus. This species
is of lesser importance in fisheries than the preceding one.
7. Genus LITHODES LATREILLE
1(2). Carapace uniformly cov red with spines of equal size; spines of lateral
margins of the same size as spines of surface or only slightly longer.
Rostrum armed with 9 spines. Scaphocerite has the shape of a rather
long spine with two branches. — Bering and Okhotsk Seas; Japan. . .
.................................... . 1. L. aequispina Benedict.
8. PARALOMIS— 12. CRYPTOLITHODES
319
2(1). Surface of carapace covered with spines of different size; spines of
lateral margins distinctly longer than the spines of the surface of the
carapace. Rostrum armed with 8 spines. Scaphocerite rudimentary.
3 (4). Antero-lateral margin of carapace armed with long spines; on the mar-
gin of the branchial region there are no spines conspicuous by their
length among the other spines. Terminal part of rostrum comparatively
stout. Chelipeds and ambulatory legs covered with comparatively long
spines. Finger of first ambulatory leg considerably longer than half
the length of the propodite. Surface of second abdominal segment
covered with spines. — The Atlantic from Greenland and Spitsbergen
southwards to lat. 40° N...............................2. L. maja (Linne).
4(3). Antero-lateral margin of carapace armed with not too long spines;
only on rounded margin of each branchial region are there two spines
conspicuous by their considerable length. Terminal part of rostrum
slender. Chelipeds and ambulatory legs covered with short spines..
Finger of first ambulatory leg slightly longer than half the length of
the propodite. Surface of second abdominal segment covered with
tubercles. — Bering Sea; Alaska;'California . *. .3. L. couesi Benedict.
8. Genus PARALOMIS WHITE
1 (2). Carapace spiny, particularly the marginal, gastric and rostral regions.
Ambulatory legs angular, not all compressed, with spines arranged,
in rows on angles or ridges. — Western coast of America from Shumagin
Isles to San Diego, California. Eastern coast of Kamchatka...............
.........................................1. P. multispina (Benedict).
2 (1). Carapace tuberculated, margins spiny, a single acute spine on gastric
region. Ambulatory legs much compressed; anterior and posterior
margins set with sharp spines. — Bering Sea from Pribyloff Islands.
southwards to Cortez Bank, California................P. Verrilli (Benedict).
9. Genus RHINOLITHODES BRANDT
A single species: Rhinolithodes wosnessenskii Brandt. Western coast:
of America from Kadiak Isle to Crescent City, California.
10. Genus SCULPTOLITHODES MAKAROV
A single species: Sculptolithodes derjugini Makarov. Up to the present:
the species had been known only from the northern part of the Japanese
Sea: off the Silantjev Bay, and in the region of the Nelma Bay. In the col-
lection of the Zoological Institute I succeeded in discovering one specimen
of this species from the region of Rishiri Island and one specimen from the
region of Andreev Bay (Ussuriysk Gulf). The species penetrates into the
Japanese Sea, apparently, from the south-western part of the Okhotsk Sea.
320
8. LITHODIDAE
11. Genus LOPHOLITHODES BRANDT
A single species: Lopholithodes mandtii Brandt. — Western coast of
America, from Sitka to California.
12. Genus CRYPTOLITHODES BRANDT
1 (2). Chelae smooth. Rostrum distally enlarged with distinct anterolateral
angles. Abdominal plates smooth. — Western coast of America, from
Sitka to California..............................1. C. sitchensis Brandt.
2(1). Chelae tuberculated. Rostrum not enlarged distally, with a rather
rounded apex. Abdominal plates with upturned margins.
3(4). Rostrum much bent downwards. Lateral lobes of carapace tuberculated.
Lateral margins of carapace meet at a distinct angle. Second abdominal
segment entire, with many additional plates on the boundary between
the median and lateral plates of the third segment, and on the posterior
margin of the second. — Western coast of America from Unalaska to
California..........................................2. C. typicns Brandt.
4(3). Rostrum but little bent downwards. Lateral lobes of carapace smooth.
Lateral margins of carapace do not meet at an angle, but form a wide
rounding. Second abdominal segment consists of two plates. Additional
plates wanting. — Northern Japan....................3. C. expansus Miers.
АЛФАВИТНЫЙ УКАЗАТЕЛЬ ЛАТИНСКИХ НАЗВАНИЙ
acadianus, Eupagurus 183.
Acantholithus 26.
acanthomera, Galathea 83, 86, 298.
aculeata, Paralomis 271.
Aeglea 7.
Aegleidae 79.
aeguispina, Lithodes 264.
affinis, Axiopsis 47, 48.
affinis, Gebia 57.
alaskensis, Eupagurus 199.
alaskensis, Pagurus 201.
alaskensis, Pagurus 199.
Albuneidae 6, 114, 299.
aleuticus, Eupagurus 202.
aleuticus Pagurus, ochotensis 173, 202.
Amphipoda 135, 148.
amurensis spinulicauda, Axiopsis 48.
antonii, Munidopsis 102.
anomalus, Eupaguras 196.
anomalus, Pagurus 172, 196, 197, 198, 307.
Anomura 2, 4, 10, 16, 19, 20, 28, 29, 30, 31.
antarctica, Lithodes 263.
arctica, Lithodes 267.
armatus, Bernhardus 213.
-armatus, Eupagurus 213.
armatus, Pagurus 174, 201, 202, 203, 213, 308.
arrosor, Dardanus 167, 168, 313.
arrosor, Cancer 168.
arrosor, Pagurus 168.
Axiopsis 47, 295.
Axiidae 19, 46, 47, 295.
balsii, Ctenocheles 76, 297.
bamffica, Munida 92, 93, 96, 97, 298.
bamfficus, Astacus 93.
beringana, Munidopsis 30, 31, 101, 298.
beringanus, Eupagurus 186.
beringanus, Pagurus 171, 186j 206, 305.
Bernhardus, 169.
bernhardus, Cancer 183.
-bernhardus, Pagurus 135, 169, 171, 183, 184,
201, 305.
Фауна СССР, Макаров
bernhardus Eupagurus 178.
Blepharipoda 144.
bouvieri Callinassa, californensis, 30, 63, 69, 71,
72, 73, 296.
brachiomastus, Eupagurus 211.
brachiomastus, Pagurus 173, 211, 308.
Brachyura 118, 140.
brandti, Eupagurus 208.
brandti, Hapalogaster 244.
brandti, Oedignathus 244
brandti, Pagurus 208.
brandti, Pagurus 210.
brevipes, Lithodes 262.
brevipes, Paralithodes 253, 254, 262, 263, 318.
Calocaris 16.
Callichirus 62, 73.
Calliactites 62.
Callianassa 2, 54, 61, 62, 76, 296.
Callianassidae 14, 39, 40, 47, 49, 294, 295.
Callianassinae 19, 50, 61, 296.
Callianidea 8, 76.
camtschatica, Lithodes 254.
camtschatica, Maja 254.
camtschatica, Paralithodes 14, 15, 18, 254, 257,
262, 263, 266, 318.
camtschaticus, Lithodes 257.
Cancellus 120, 148.
capillatus, Eupagurus 219.
capillatus, Pagurus 174, 210, 215, 218, 219,
221, 309.
Caprella 15.
Carcinus 9.
cavicauda, Hapalogaster 24, 26.
cavimanus, Eupagurus 178. ,
cavimanus, Pagurus 170, 177, 178, 304.
Cervimunida 82, 103, 297, 298.
Cheramus 62, 63.
Cirripedia 176.
Clibanarius 157, 162, 314.
Coenobita 10.
Coenobitidae 18.
21
322
АЛФАВИТНЫЙ указатель
Constans, Eupagurus 221.
constans, Pagurus 174, 221, 309.
cornutus, Eupagurus 191.
cornutus, Pagurus 172, 191, 195, 306.
couesi, Lithodes 264, 270, 319.
crassa, Callianassa 62.
Cryptolithodes 6, 26, 251, 283, 320.
Ctenocheles 61, 76, 297.
Ctenorhinus 280.
curvirostra, Munidopsis 100.
dalli, Eupagurus 206.
dalli, Pagurus 173, 206, 208, 307.
Dardanus 157, 167, 170, 313.
Decapoda 1, 16, 18, 36, 138, 148.
dentata, Hapalogaster 236, 239, 241, 315.
dentata, Lomis 239.
derjugini, Sculptolithodes 277, 319.
Dermaturus 22, 26, 27, 30, 31, 234, 235, 241,
244, 316.
depressus, Dardanus 167.
depressus, Clibanarius 162.
digitalis, Paguristes 165, 166, 314.
Diogenes 157, 167, 313.
Diogeninae 157.
diomedae, Lithodes 36.
drenarius, Pagurus 160.
dubius, Eupagurus 188.
dubius, Pagurus 172, 188, 306.
Echinocerus, 280.
edwardsii, Diogenes 158, 313.
edwardsii, Pagurus 158.
Emerita 2, 4.
eoa, Callianassa gigas 63, 68, 296.
Eupagurus 169, 170.
expansus, Cryptolithodes 284, 287, 320.
fauriana, Blepharipoda 115.
forcipatus, Lepeopus 246.
Galathea 9, 12, 13, 81, 82, 92, 297, 298.
Galatheidae 79, 80, 105, 297.
Galatheidea 4, 12, 15, 16, 19, 20, 45, 78, 79,
114, 297.
Galatheinae 81, 82.
Gastropoda 118.
Gebicula 50.
Gebiopsis 51.
gigas, Callianassa 69.
gilberti, Hemilepidotus 40.
gilli, Eupagurus 176.
gilli, Oedignathus 244.
gilli, Pagurus 170, 176, 178,179, 304, 309.
gotoi, Eupagurus 178.
gracilipes, Eupagurus 185.
gracilipes, Pagurus 171, 185, 305.
granosimanus, Pagurus 205.
granosimanus, Pagurus 173, 205, 307.
grebnitzkii, Hapalogaster 236, 238, 239, 240,
315.
grossimanus, Pachycheles 110.
Hapalogaster, 234, 235, 241, 315.
Hapalogastrinae 11, 34, 233, 234, 248, 314.
harmandi, Callianassa 30, 31, 63, 66, 68, 69, 296..
heteracantha, Munida 97, 98.
heteracantha Munida japonica, 98.
Hippidae 6.
Hippidea 2, 45, 113, 114, 299.
hirsutiusculus, Bernhardus 181.
hirsutiusculus, Eupagurus 181.
hirsutiusculus, Pagurus 171, 181, 227, 305.
hirtus, Paguristes 165, 166.
Homaridea 46.
incisus, Pagurus 168.
inermis, Dermaturus 244.
inermis, Hapalogaster 244.
inermis, Oedignathus 32, 244, 316.
issaeffi, Cebia 59.
issaeffi, Upogebia 52, 59, 295.
integra, Galathea 83, 88, 298.
japonica, Blepharipoda 115, 117, 299.
japonica, Callianassa 63, 67, 70, 71, 72, 73, 296.
japonica, Cucumaria 14.
japonicus, Clibanarius 16.2, 163, 314.
japonica, Munida 93, 97, 98, 298.
jeffreysii, Mixtopagurus 120, 300.
jeffrejsii, Pomatocheles 120.
kennerlyi, Eupagurus 218.
kennerlyi, Pagurus 174, 217, 218, 308.
kriczagini, Porcellana 109.
krbgeri, Eupagurus 208.
kroyeri, Pagurus 208.
laevis, Aeglea 79.
lanuginosus, Eupagurus 212.
lanuginosus, Pagurus 173, 212, 308.
laticauda, Callianassa 63, 64, 73.
latro, Birgus 40.
Lepeopus 246.
Leptolithodes 271.
Lithodes 22, 23, 24, 250, 254, 263, 272, 318.
Lithodidae 14, 17,18, 27, 28, 29, 32, 34, 35, 36„
118, 233, 234, 314.
Lithodinae, 2, 14, 26, 27, 233, 234, 249, 316..
littoralis, Gebios 52.
littoralis, Thalassina 52.
littoralis, Upogebia 51, 52, 54, 295.
longicornis, Cancer 107, 109, 110.
АЛФАВИТНЫЙ УКАЗАТЕЛЬ
323
longicornis, Porcellana 107, 299.
longimana, Porcellana 107, 110, 299.
longirostris, Paralithodes 264.
Lopholithodes 67, 250, 280, 320.
Macrura 47, 92, 140, 223.
maja, Lithodes 35,36,263, 264,267,270,271,319.
major, Gebia 54.
major, Upogebia 30, 52, 54, 59, 60, 295.
mandtii, Dermaturus 40, 241, 242, 243, 316.
mandtii, Hapalogaster 242.
mandtii, Lopholithodes 280, 320.
Mastigochirus 6.
maxima, Callianassa 62.
mertensii, Eupagurus 181.
mertensii, Hapalogaster, 235, 236, 237, 238, 239,
315.
mertensii, Pagurus 226.
mertensii, Parapagurus 220, 226, 227, 312.
middendorffii, Eupagurus 174.
middendorffi, Pagurus 170, 174, 304.
miles, Diogenes 157.
minimus, Orthopagurus 227, 229, 313.
minimus, Pagurus 227.
minitus, Pylopagurus 227.
misanthropus, Clibanarius 144, 162, 314.
misanthropus, Pagurus 160.
Mixtopagurus 119, 120, 300.
multispina, Paralomis 272, 319.
miliispinus (— naj, Leptolithodes 272.
Munida 9, 82, 92,100,103, 297, 298.
Munidopsinae 81, 92,100.
Munidopsis 82, 100, 297, 298.
munitus, Eupagurus 178.
munitus, Pagurus 178.
Natantia 10. .
Nematopagurus 23, 24.
Neolithodes 11, 26, 35.
newcombei, Eupagurus 186.
nigritarsis, Pagurus 162.
nipponensis, Eupagurus 185.
obtusifrons, Eupagurus 216.
obtusifrons, Pagurus 174, 216, 308.
ochotensis, Pagurus 173, 199, 201, 202, 213, 214,
30', 309.
ochotensis, Eupagurus 199, 213.
occidentals, Blepharipoda 114.
Oedignathus 235, 244, 316.
orientalis, Galathea 86, 88.
Orthopagurus 157, 227, 313.
ortmanni, Eupagurus 199.
Pachycheles 105, 110, 299.
Pagurus 167.
Paguridae 11, 21, 27, 28, 29, 118, 119, 144,
145, 147, 150, 152, 156, 196, 222, 223, 234,
299, 300.
Paguridea 5, 15, 16, 19, 20, 45, 46,79,117,299.
Pagurinae 157, 169.
Paguristes 165, 314.
Paguristinae 157.
Pagurus 21, 22, 23, 24, 26, 27, 156, 169, 170,
304.
Pagurus 167.
paradoxus, Mixtopagurus 119.
Paralithodes 23, 28, 250, 253, 254, 260, 263,
317.
Paralomis 26, 250, 271, 319.
Parapaguridae 156.
Parapagurus 157, 223, 312.
panamensis, Lithodes 263.
papillosus, Phyllolithodes, 26, 251, 317..
pectinatus, Eupagurus 214.
pectinatus, Pagurus 174, 214, 216, 308.
penicillatus, Diogenes 158, 159, 313.
Pentacheles 16.
pestae, Callianassa 73.
Petalocerus 251,
petalura, Callianassa 71.
Phyllolithodes 251, 317.
pilosimanus, Parapagurus 223, 225, 312.
pilosus, Paguristes 167.
Pinnotheridae 41.
Pisidia 106.
Placetron 234, 235, 246, 316.
platycheles, Porcellana 8, 106, 107.
platypus, Lithodes 257.
platypus, Paralithodes 32, 254, 257, 260, 262,
318.
polaris, Eupagurus 203.
Pomatocheles 119.
pontica, Callianassa 63, 64, 73, 296.
ponticus, Pagurus 160.
Porcellana 9, 13, 14, 105, 106, 107, 298, 299.
Porcellanella 106.
Porcellanidae 2, 4, 10, 16, 20,. 21, 79, 104, 105,
297, 298.
Porcellanides 106, 107.
princeps, Axiopsis 48.
princeps, Axius 48.
princeps, Cervimunida 103, 298
Pristopus 271.
pubescens, Eupagurus 208.
pubescens, Galathea 83, 91, 298.
pubescens, Pagurus 173,184, 207, 208, 210, 221,
308.
pugilator, Pagurus 160.
21
324
АЛФАВИТНЫЙ УКАЗАТЕЛЬ
Pylocheles 119, 120.
Pylochelidae 10, 11, 118, 119, 300.
Pylopagurus 27
Raninidea 114.
rathbuni, Eupagurus 203.
rathbuni, Pagurus 173, 203, 307.
Rhinolithodes 26, 250, 275, 319.
rondeletii, Munida 93, 97.
rugosa, Galathea 93.
rugosa, Munida 93.
Sabellidae 109.
sagamiensis, Munida 100.
samuelis, Eupagurus 189.
samuelis, Pagurus 172, 189, 306.
schmitti, Orthopagurus 227, 228, 229, 230.
schmitti, Pylopagurus 229.
Scallasis 62.
Sculptolithodes 250, 277, 319.
seriespinosus, Eupagurus 214.
setosus, Eupagurus 216.
setosus, Paguristes 165, 167, 314.
setosus, Pagurus 174, 216, 218, 308.
seZosas, Pagurus 167.
sitchensis, Cryptolithodes 283, 284, 286, 287.
spinifrons, Pagurus 159.
spinimanus, Eupagurus 199.
spinulicauda, Axiopsis 48.
spiriger, Pagurus 231.
spiriger, Spiropagurus 230, 231, 313.
Spiropagurus,23, 157, 230, 313.
splendescens, Eupagurus 197.
splendescens, Pagurus 173, 197, 307.
stellata, Upogebia 50.
stevensii, Pachycheles 111, 299.
streblonyx, Pagurus 183.
striatus, Pagurus 168.
strigosa, Galathea 82, 83, 86, 298.
strigosus, Cancer 83, 168.
subterranea, Callianassa 61, 62, 63, 64, 71, 73,
296.
sulcatus, Peltogaster 151.
superba, Cynthia 14.
Sympagurus 223.
tanneri, Eupagurus 194.
tanneri, Pagurus 172, 194, 306.
tenuimanus, Bernhardus 179.
tenuimanus, Eupagurus 179.
tenuimanus, Pagurus 171, 179, 305.
Thalassinidae 49.
Thalassinidea 10, 19, 21, 45, 46, 79.
Thaumastocheles 76.
thompsoni, Pagurus 208.
Tomopagurus 27.
trigonocheirus, Eupagurus 192, 208.
trigonocheirus, Pagurus 210.
tropicalis, Lithodes 35.
Trypaea 62, 66, 73.
tubularis, Pagurus 162.
turgidus, Paguristes 166.
turnerana, Callianassa 18.
typicus, Cryptolithodes 283, 284, 286, 320..
typica japonica Munida, 98.
undosus, Eupagurus 192.
undosus, Pagurus 172, 192, 306.
Upogebia 14, 50, 51, 295.
Upogebiinae 6, 19, 20, 50, 295.
Uroptychus 80.
varians, Diogenes 158, 160, 161, 163, 313
varians, Pagurus 160.
verrilli, Paralomis 272, 273, 319.
verrilli, Prisiopus 273.
wosnessenskii, Placetron 246.
wosnessenskii, Rhinolithodes 275.
yokoyai, Pagurus 309.
yokoyai, Upogebia 52, 57, 295.
ТАБЛИЦЫ I-V
Таблица 1
1. Dardanus arrosor (Herbst).
2. Pagurus tenuimanus (Dana).
3. Diogenes edwardsii (de Haan) (no Stimpson’y).
4. Pagurus dalli (Benedict).
5. Pagurus undosus (Benedict).
6. Diogenes varians (Costa).
7. Clibanarius misanthropus (Risso).
Табл.
Таблица 2
1. Orthopagurus schmitti (Stevens).
2. Pagurus ochotensis (Brandt).
3. Pagurus ochotensis subsp. aleuticus (Benedict).
4. Pagurus armatus (Dana).
5. Pagurus granosimanus (Stimpson).
6, Pagurus brachiomastus (Thallwitz).
Табл.
*
Таблица 3
1. Pagurus gilli (Benedict).
2. Pagurus capillatus (Benedict).
3. Pagurus rathbuni (Benedict).
4. Pagurus hirsuiiusculus (Dana).
5. Pagurus dubius (Ortmann).
6. Pagurus samuelis (Stimpson).
Таблица 4
1. Pagurus pubescens (Kroyer).
2. Pagurus splendescens (Owen).
3. Pagurus pectinatus (Stimpson).
4. Pagurus gracilipes (Stimpson).
5. Pagurus anomalus (Balss).
6. Pagurus bernhardus (Linne).
Табл.
•J •
I
Таблица 5
1. Pagurus cornutus (Benedict).
2. Pagurus lanuginosus (de Haan).
3. Spiropagurus spiriger (de Haan).
4. Pagurus beringanus (Benedict).
5. Pagurus ianneri (Benedict).
6. Pagurus middendorffii (Brandt.)
СОДЕРЖАНИЕ
Стр.
Предисловие..................................................... V
Систематический указатель видов................................. VII
Введение ......................................................... 1
Специальная часть . . . ...................................... 45
I. Надсем. Thalassinidea................................. 46
1. Сем. Axiidae........................................ W
2. Сем. Callianassidae................................ 49
II. Надсем. Galatheidea................................. 78
3. Сем. Galatheidae.............................' • 79
4. Сем. Porcellanidae................................ 104
III. Надсем. Hippidea.................................... 113
5. Сем. Albuneidae................................... 114
IV. Надсем. Paguridea................................... 117
6. Сем. Pylochelidae................................. 119
7. Сем. Paguridae............................... 123
8. Сем. Lithodidae................................. 233
Резюме (Summary)................................................ 290
Алфавитный указатель латинских названий......................... 321
ФАУНА СССР
(новая серия)
Ракообразные
Том I. Жаброногие (Branchiopoda). «Вып. 1. Phyllopoda". Введение, Апо-
siraca; «Вып. 2. Notostraca, Conchostraca; «Выи. 3—4. Cladocera.
Том II. Ракушковые (Ostracoda). Вып. 1. Ракушковые континентальных
водоемов; Вып. 2. Ракушковые морские.
Том III. Веслоногие (Copepoda) Вып. 1. Введение; «Вып. 2. Calanoida кон-
тинентальных водоемов; «Вып. 3. Cyclopoida континентальных водое-
. мов; «Вып. 4. Harpadicoida континентальных водоемов.
Том IV. Веслоногие морские. «Вып. 1. Copepoda северных морей; Вып. 2.
Copepoda Черного моря. Вып. 3. Copepoda восточных морей.
Том V. «Паразитические веслоногие. ,,
Том VI. «Усоногие (Cirripedia), Leptostraca.
Том VII. Равноногие (Isopoda). Вып. 1. Введение; Вып. 2. Сухопутные [sopoda;
Вып. 3. Isopoda Дальневосточных морей; Вып. 4. Арктические мор-
ские Isopoda; «Вып. 5. Пресноводные Isopoda; Вып. 6. Tanaidacea.
Том VIII. Бокоплавы (Amphipoda). «Вып. 1—2. Пресноводные Amphipoda;
«Вып. 3—5. Морские Amphipoda.
Том IX. Вып. 1. Ситасеа; Вып. 2. Schizopoda; Вып. 3. Euphausiacea.
Том X. «Вып. 1. Пресноводные Decapoda; «Вып. 2. Macrura; Вып. 3.
Anomura; Вып. 4. Brachyura; Вып. 5. Stomatopoda.
* Отмечены выпуски, находящиеся в работе, подчеркнуты одной чертой ---
выпуски подготовленные к печати, подчеркнуты двумя чертами =
напечатанные.
_______________________________________________________________________________
ФАУНА СССР
(новая серия)
Вышли из печати:
Млекопитающие т. III, вып. 4 — Тушканчики (Б. С. Виноградов), 1937,
цена 10 руб.
Птицы т. I, вып. 5 — Дневные хищники (Б. К. Штегман), 1937, цена 17 р.
Рыбы т. VI, вып. 1 —• Triglidae (А. Н. Световидов), 1936, цена 1 р. 25 к.
Насекомые двукрылые т. III, вып. 2 — Москиты (П. П. Перфильев), 1937,
цена 7 р.; т. III, вып. 5 — Кровососущие комары Culicinae (А. А. Шта-
кельберг), 1937, цена 12 р.; т. VII, вып. 2 — Слепни (Н. Г. Олсуфьев),
1937, цена 17 р.; т. XIX, вып. 1 — Сем. Sarcophagidae, ч. 1 (Б. Б. Ро-
дендорф), 1937, цена 20 р.
Насекомые перепончатокрылые т. II, вып. 1 — Пилильщики Tenthredinodea,
ч. 1 (В. В. Гуссаковский), 1935, цена 22 р. 75 к.; т. V, вып. 2 — Сем.
Braconidae, ч. 1 (Н. А. Теленга), 1936, цена 20 р. 75 к.
Насекомые чешуекрылые или бабочки т. XIII, вып. 3 — Совки Agrotinae
(И. В. Кожанчиков), 1937, цена 34 р. 50 к.
Насекомые жесткокрылые или жуки т. XXI — Дровосеки, ч. 1 (Н. Н. Пла-
вильщиков), 1936, цена 22 р.; т. XXVI, вып. 1 — Листоеды Galerucinae
(Д. А. Оглоблин), 1935, цена 17 р.
Насекомые кожистокрылые (Г. Я. Бей-Биенко), 1936, цена 14 р.
Ракообразные т. VII, вып. 3 — Равноногие Дальневосточных морей (Е. Ф.
Гурьянова), 1936, цена 15 р. .50 к.; т. X, вып. 3 — Апотига (В. В. Ма-
каров), 1927.
Губки т. II, вып. 2 — Пресноводные губки (П. Д. Резвой), 1936, цена б р. 60 к.
Находятся в печати:
Млекопитающие т. Ill, вып. 5 — Мыши и крысы (А. И. Аргиропуло).
Птицы т. I, вып. 2 — Географическое распространение птиц Палеарктики
(Б. К. Штегман).
Насекомые жесткокрылые или жуки т. V, вып. 3—Жуки-карапузики, ч. 1
(А. Н. Рейхардт).
Паукообразные т. 1, вып. 3 — Сольпуги (А. А. Бялыницкий-Бируля); т. V,
вып. 2 — Водные клещи Hydracarina, ч. 1 (И. И. Соколов).
Моллюски т. III, вып. 1 — Сем. Unionidae (В. И. Жадин).
Подготовлены к печати:
Млекопитающие т. 1, вып. 2 — Олени (К. К. Флеров).
Птицы т. I, вып. 3 — Определитель отрядов и семейств (Б. К. Штегман), га-
гарообразные, буревестниковые (Е. В. Козлова), аистообразные (А. Я. Ту-
гаринов).
Рыбы т. VII, вып. 4 — Трескообразные (А. Н. Световидов).
Насекомые перепончатокрылые т. II, вып. 2 — Пилильщики, ч. 2 (В. В. Гус-
саковский); т. V, вып. 3 — Braconidae, ч. 2 (Н. А. Теленга); т. XIV,
вып. 1 — Муравьи, ч. 1 (К. В. Арнольди, В. А. Караваев).
Насекомые двукрылые т. VI, вып. 6 — Мошки Simuliidae (И. А. Рубцов);
т. IX, вып. 2 —Жужжала, ч. 1 (С. Я. Парамонов).
Насекомые жесткокрылые или жуки т. XII, вып. 3 — Щелкуны Elateridae,
ч. 2 (Л. А. Денисова); т. XIV, вып. 2 — Лжекороеды Bostrychidae
(В. Н. Старк); т. XXII — Дровосеки, ч. 2 (Н. Н. Плавильщиков).
Насекомые сосущие т. VI, вып. 3 — Тли. Триба Eriosomini (А. К. Мордвилко).
Насекомые таракановые (Г. Я. Бей-Биенко).
Паукообразные т. V, вып. 3 — Водные клещи, ч. 2 (И. И. Соколов); т. VI,
вып. 1 — Тироглифоидные клещи (А. А. Захваткин).
Ракообразные т. II, вып. 1 — Ракушковые Ostracoda континентальных водое-
мов (3. С. Бронштейн); т. VI, вып. 1—Усоногие Cirripedia thoracica
(Н. И. Тарасов).